














































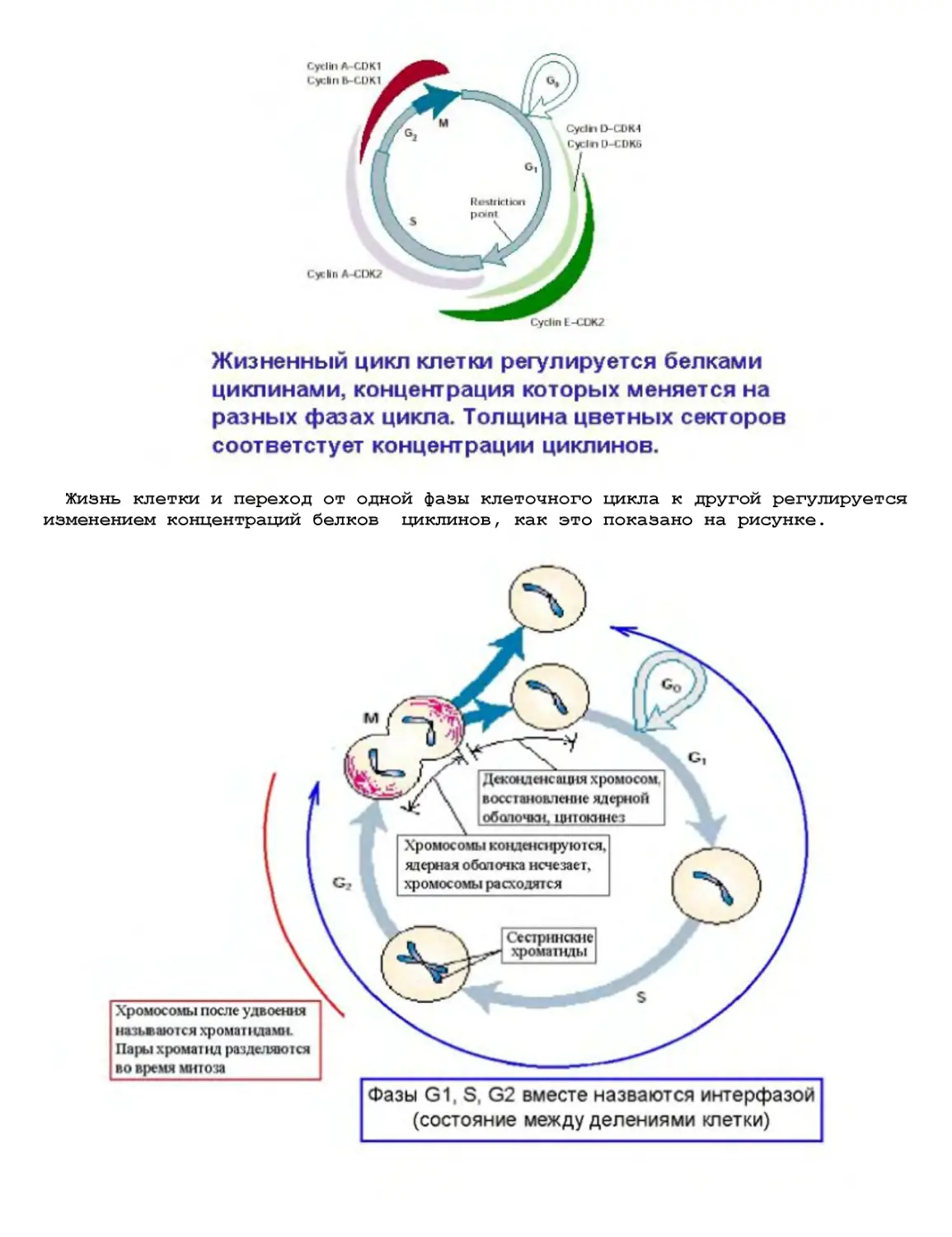

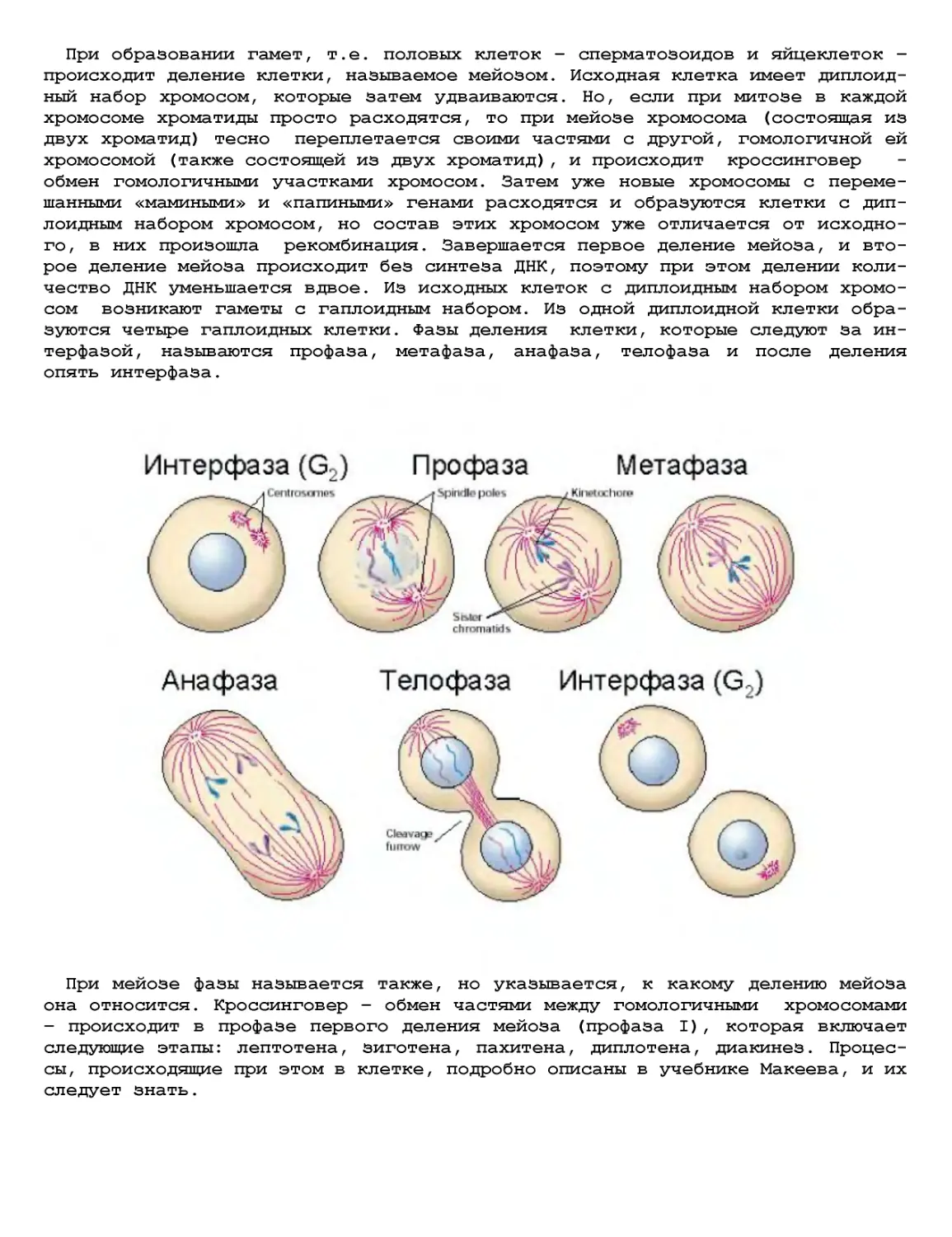
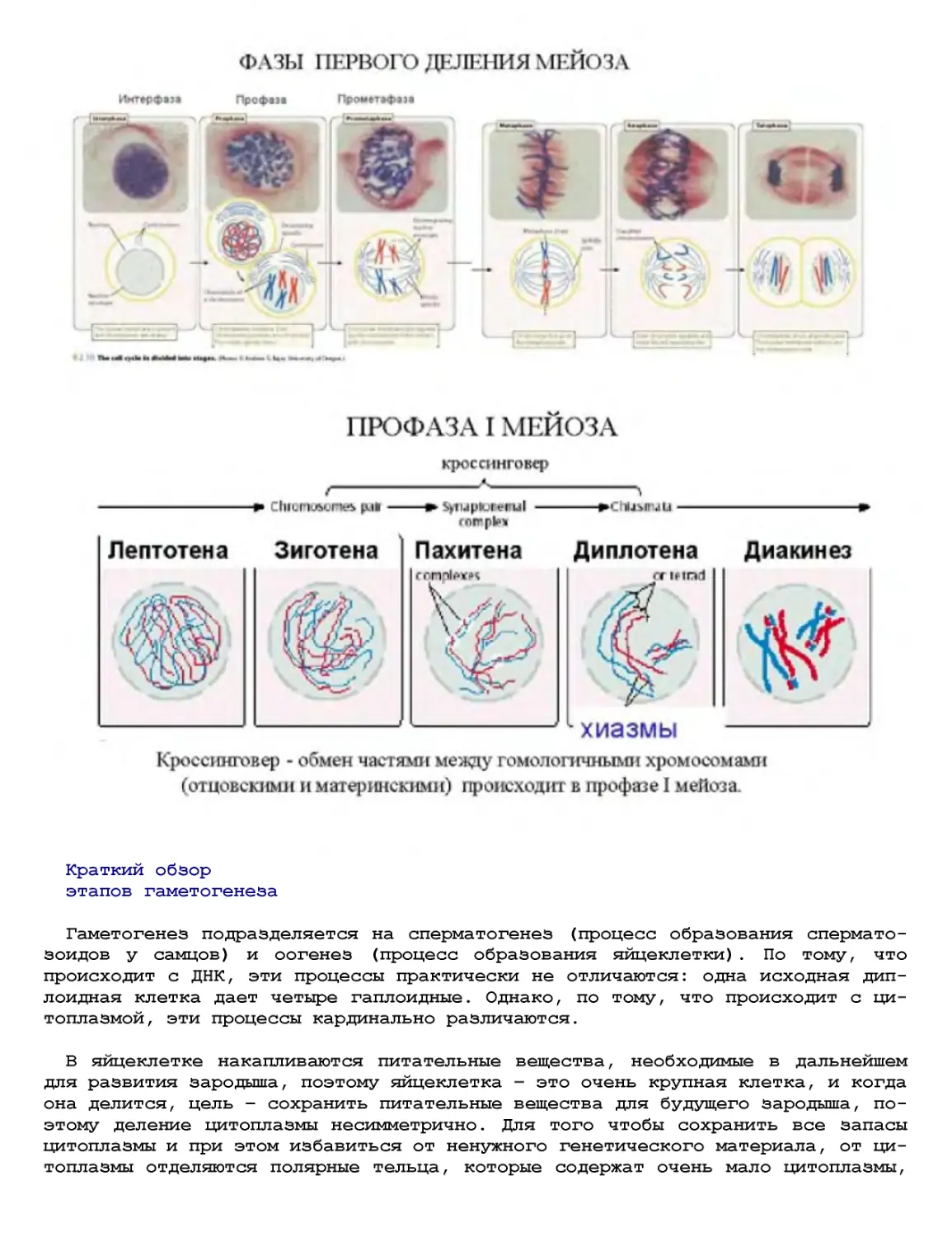
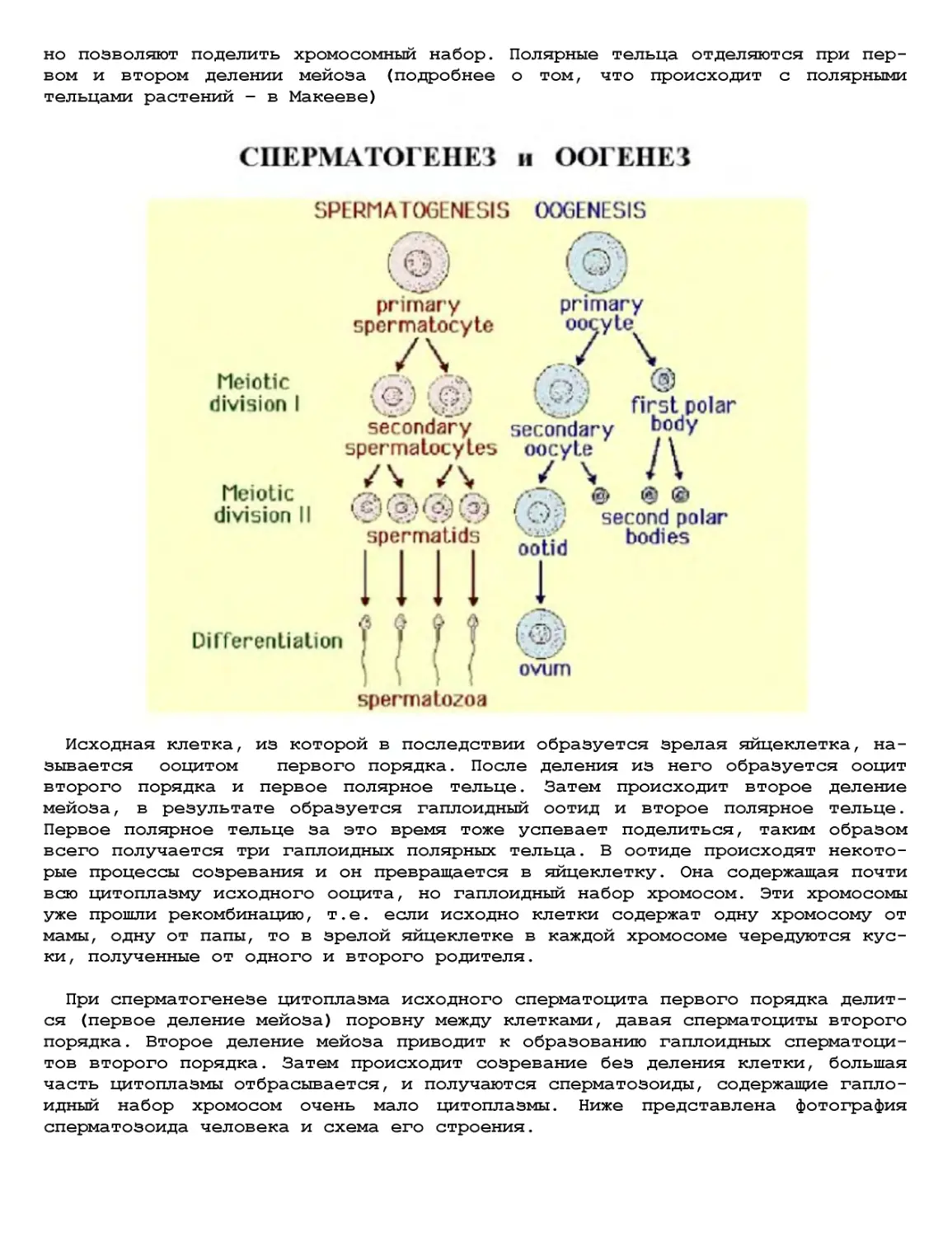

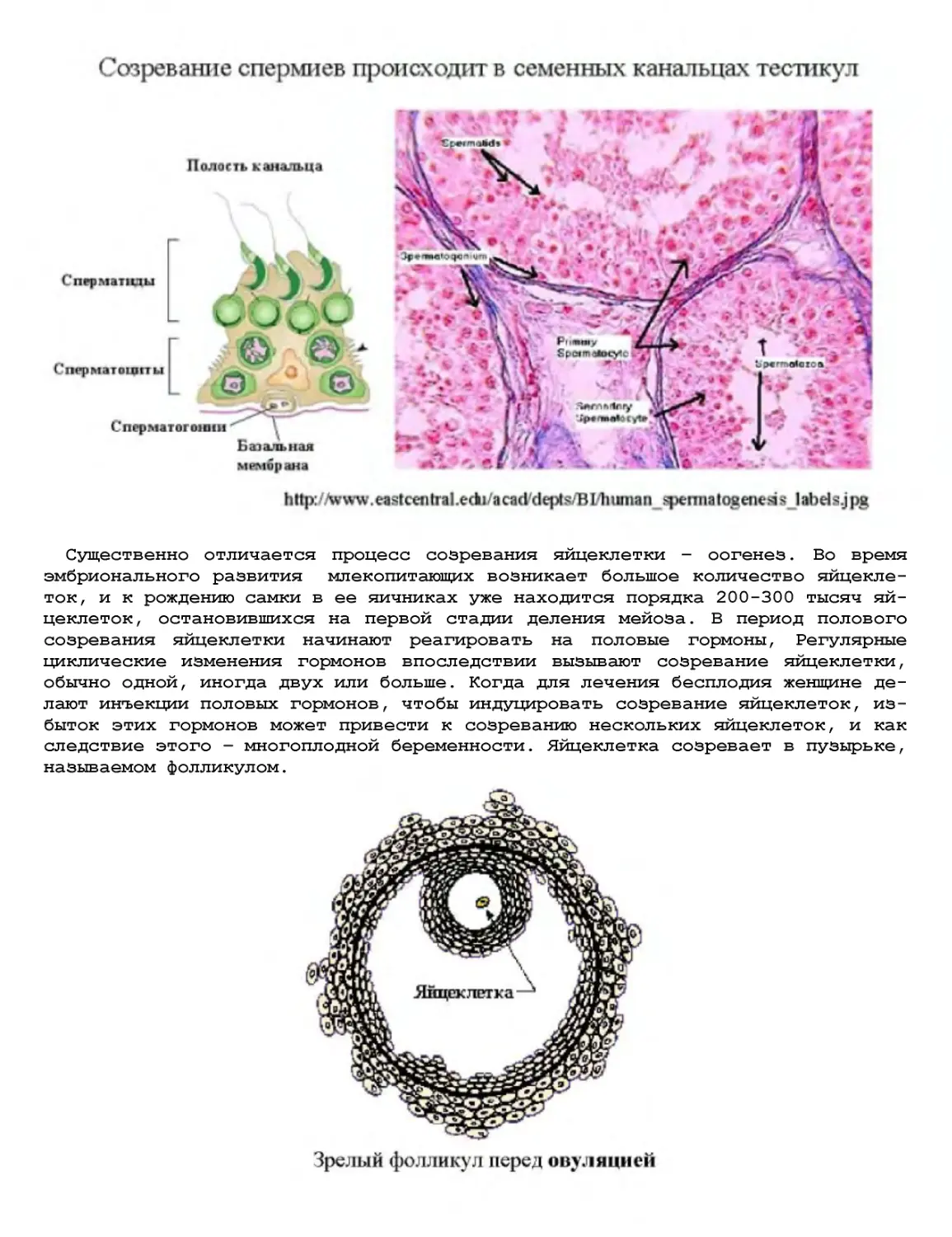

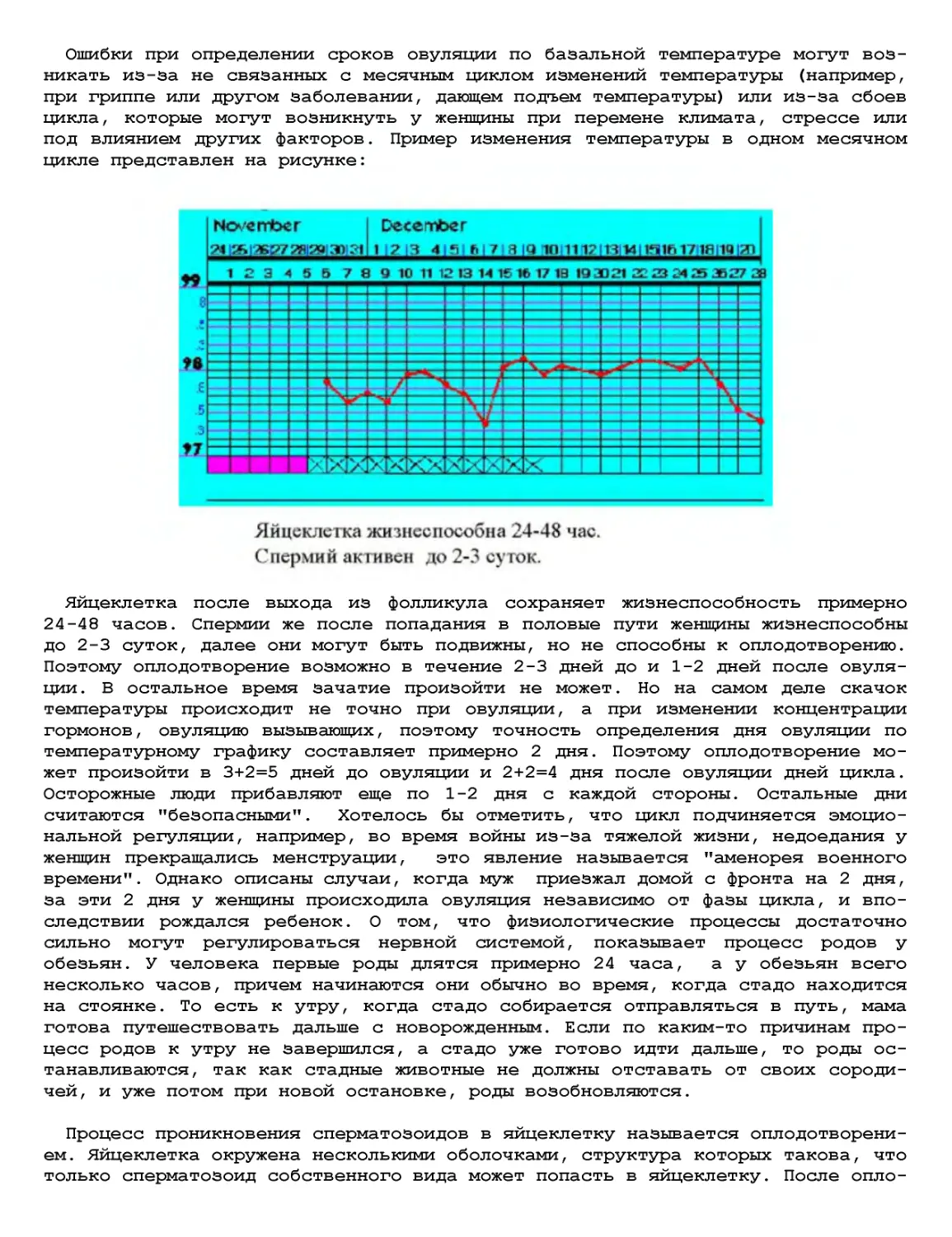


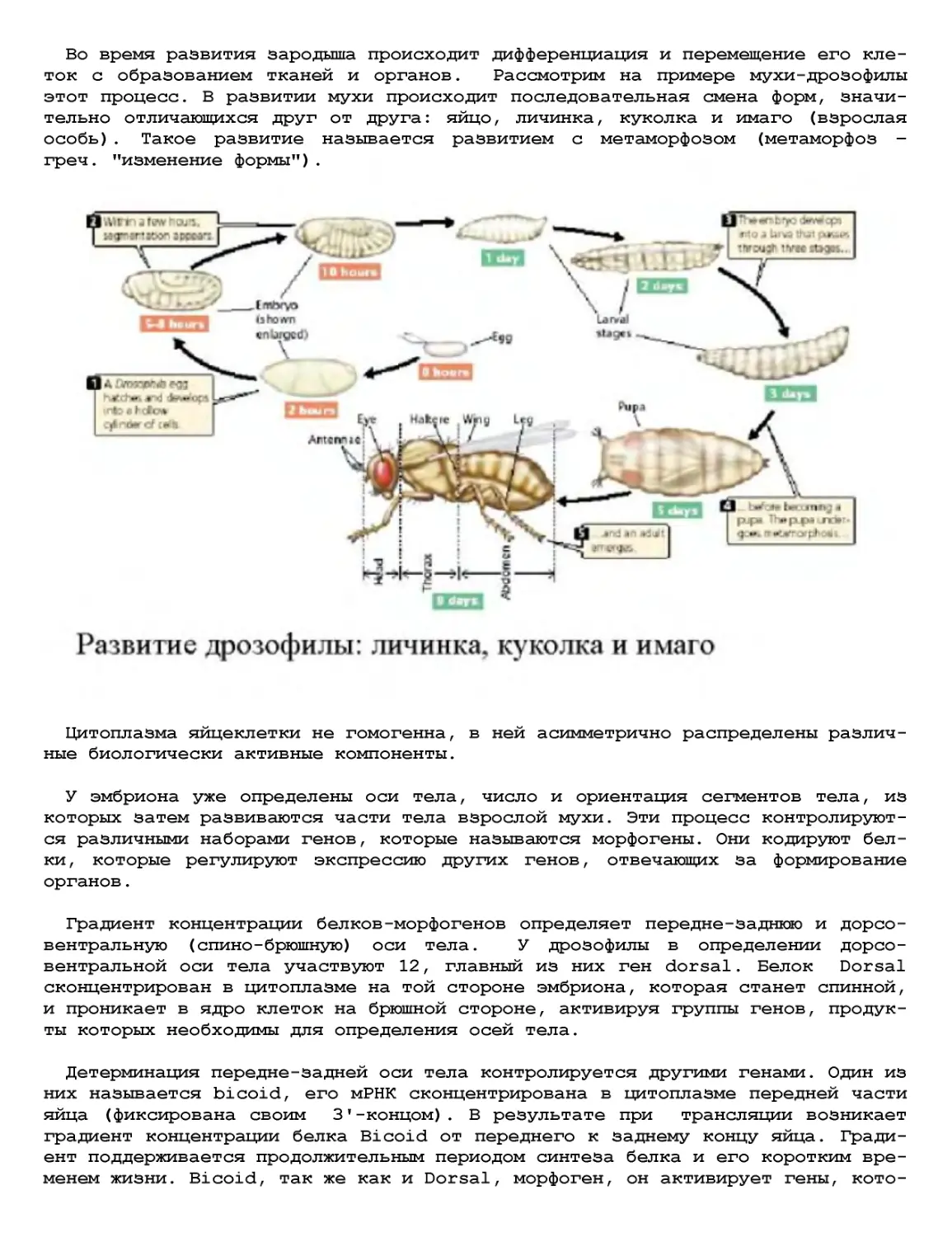

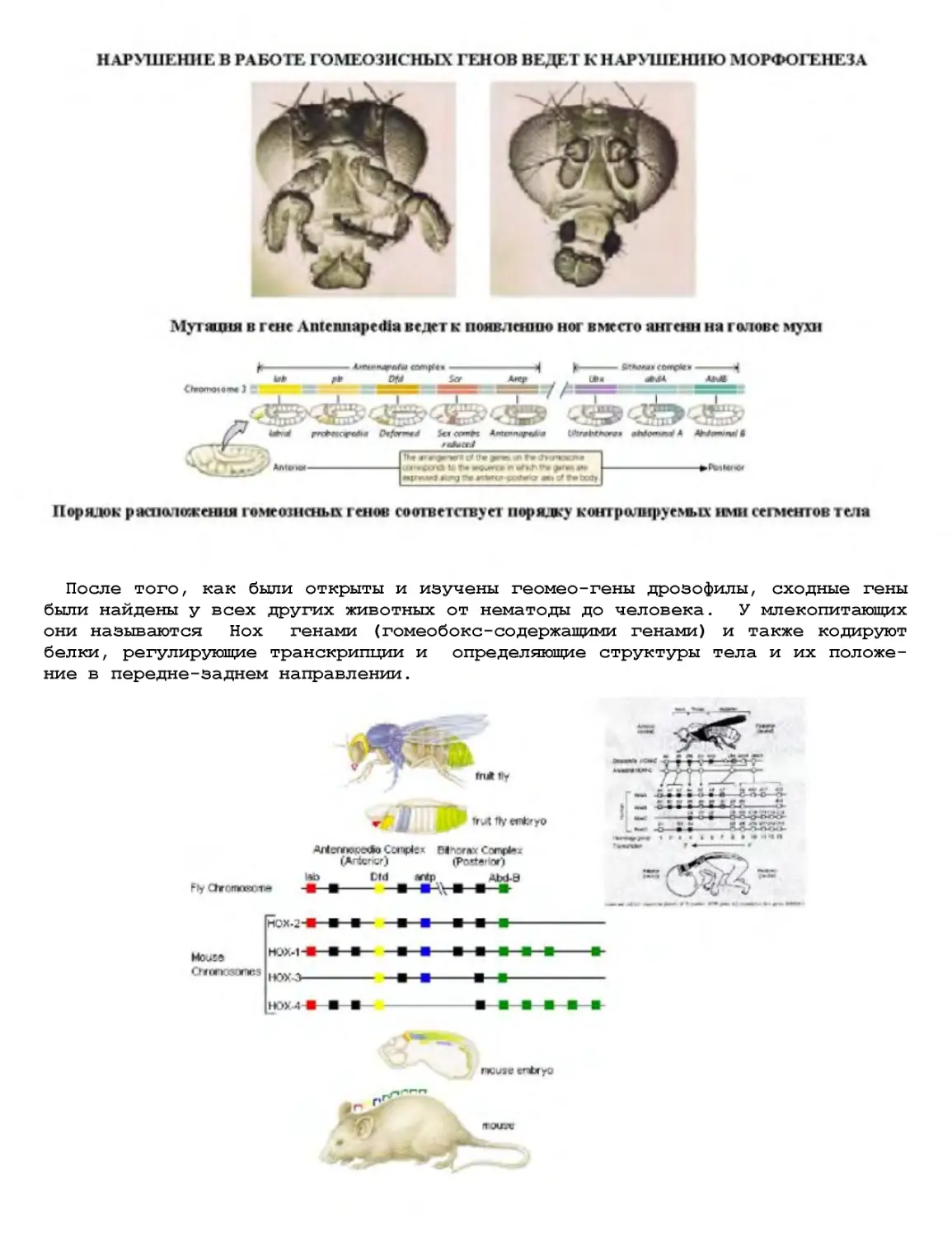








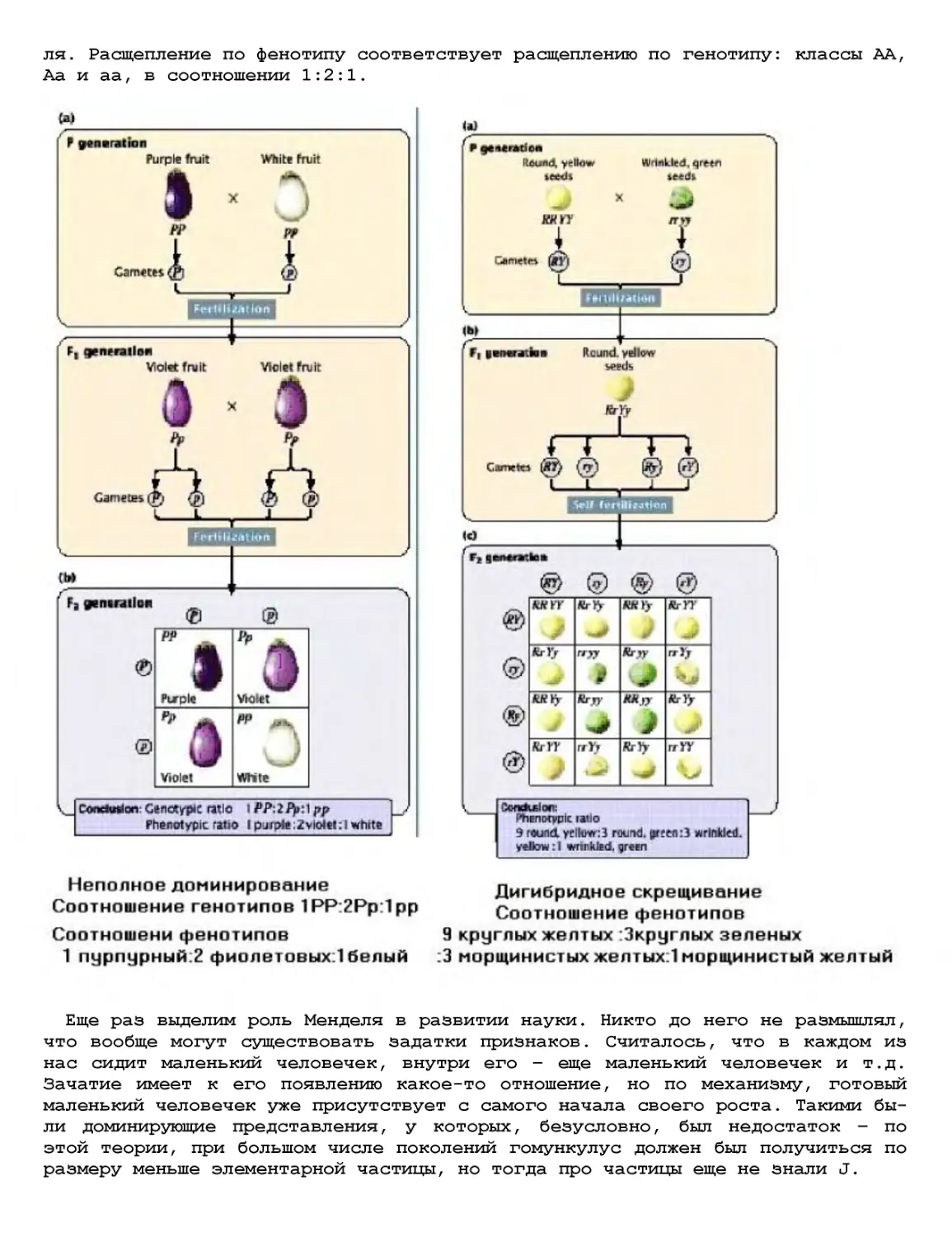

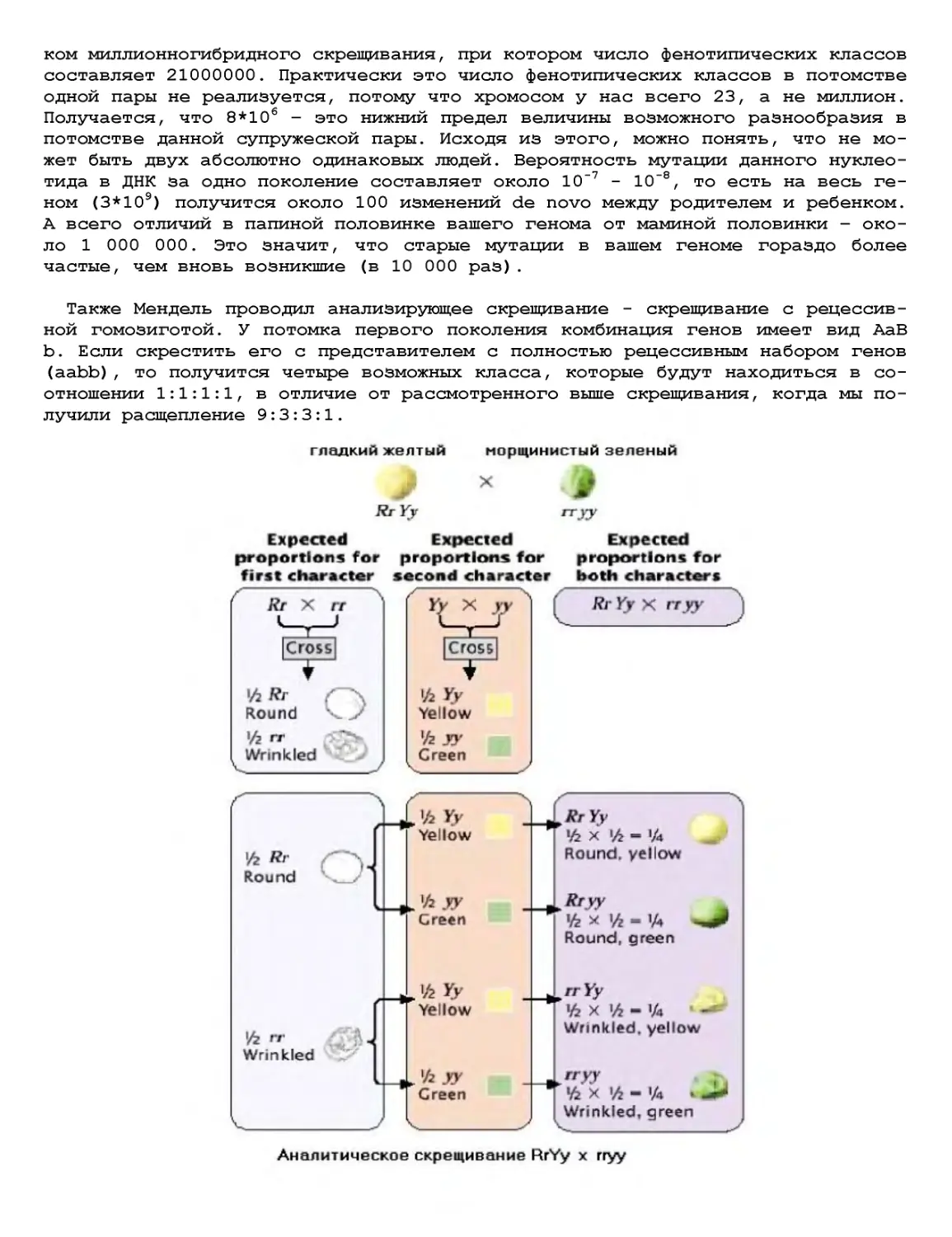



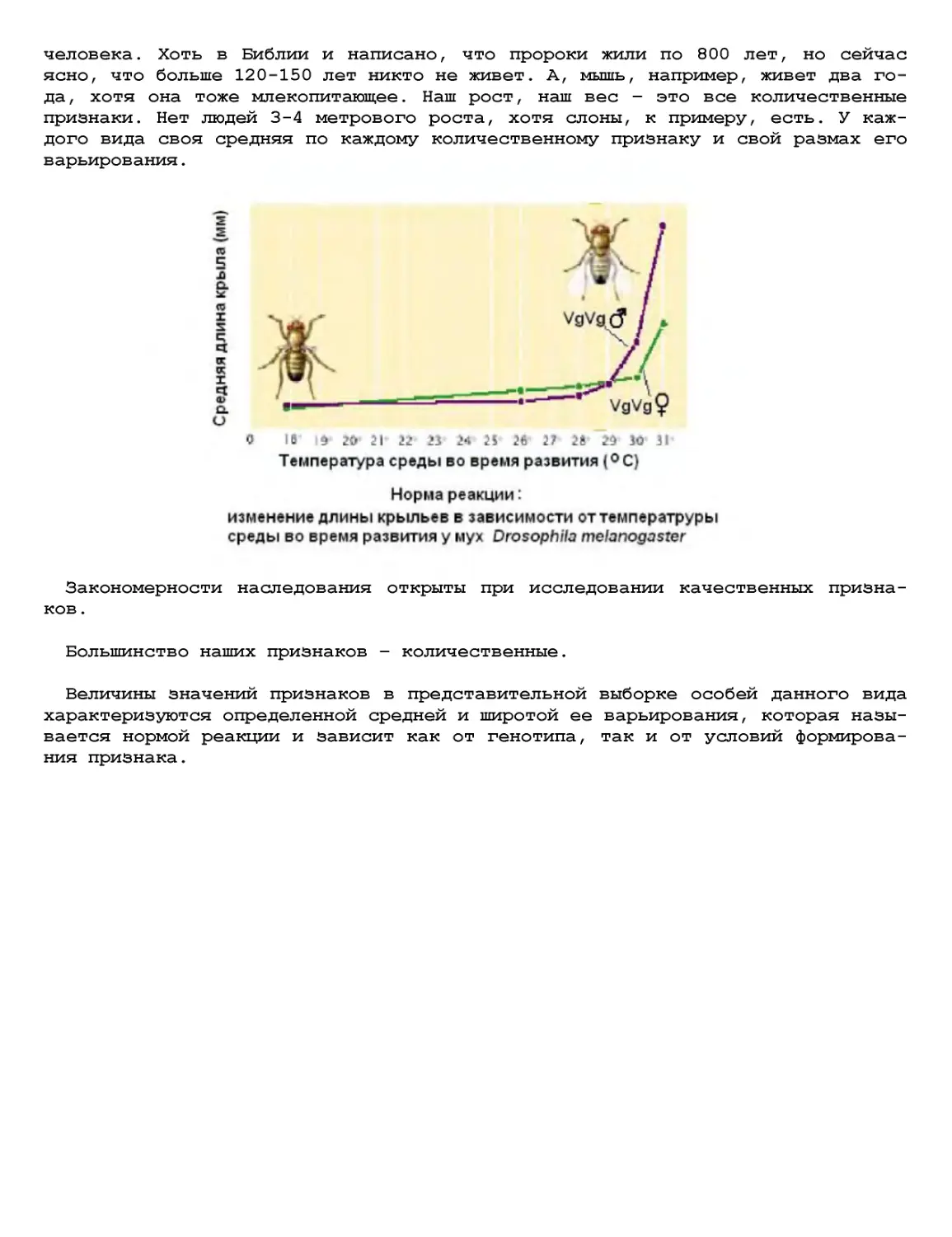





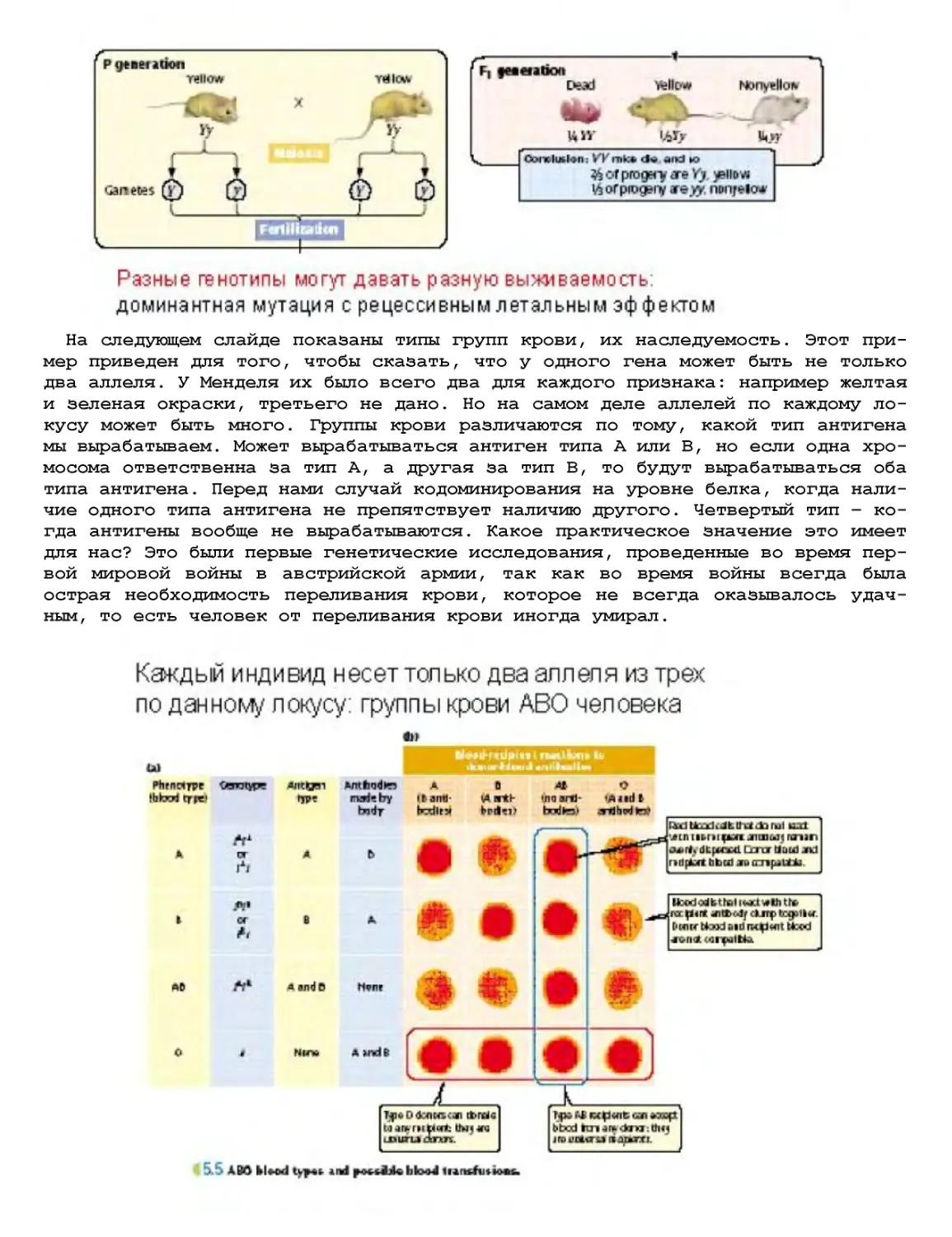

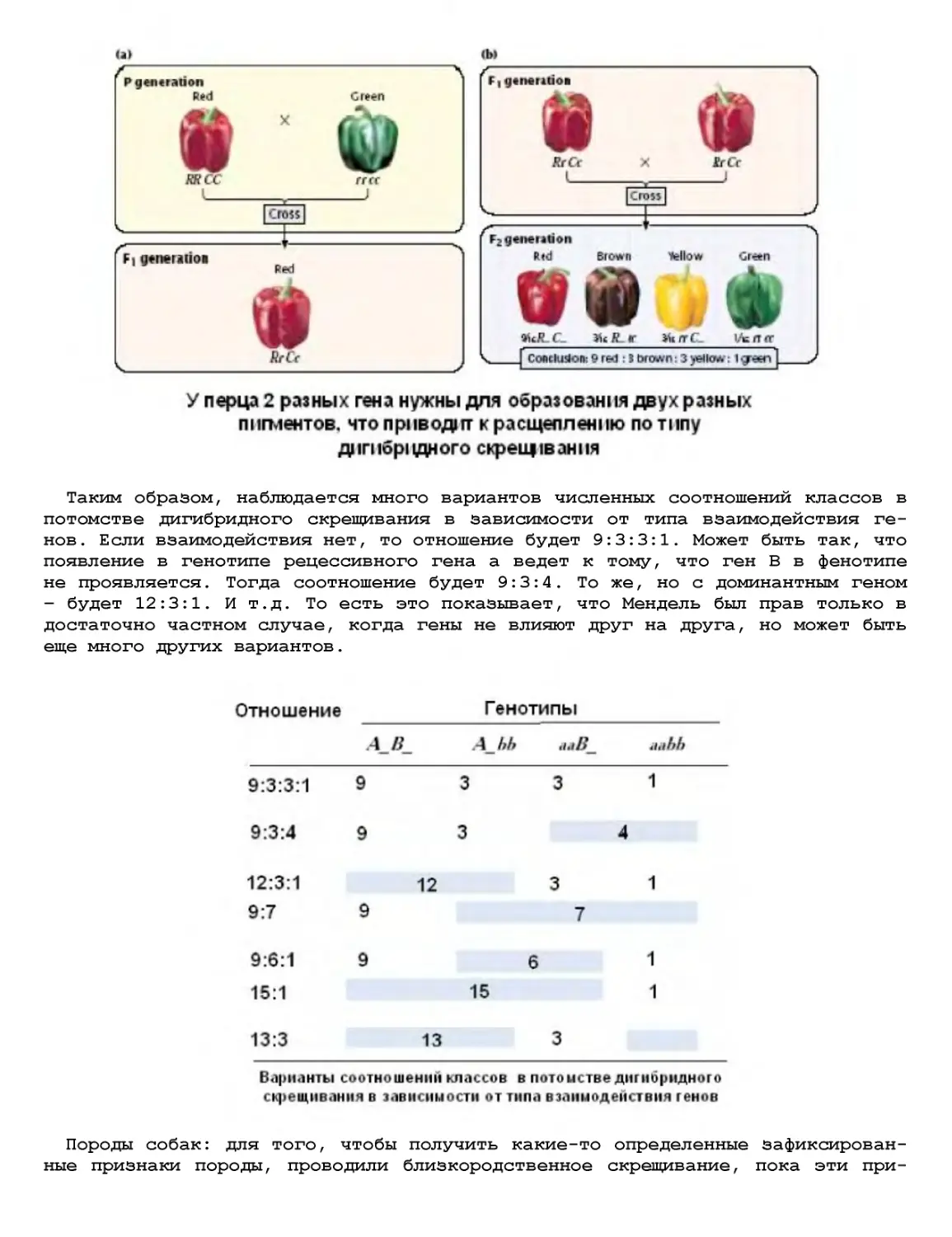





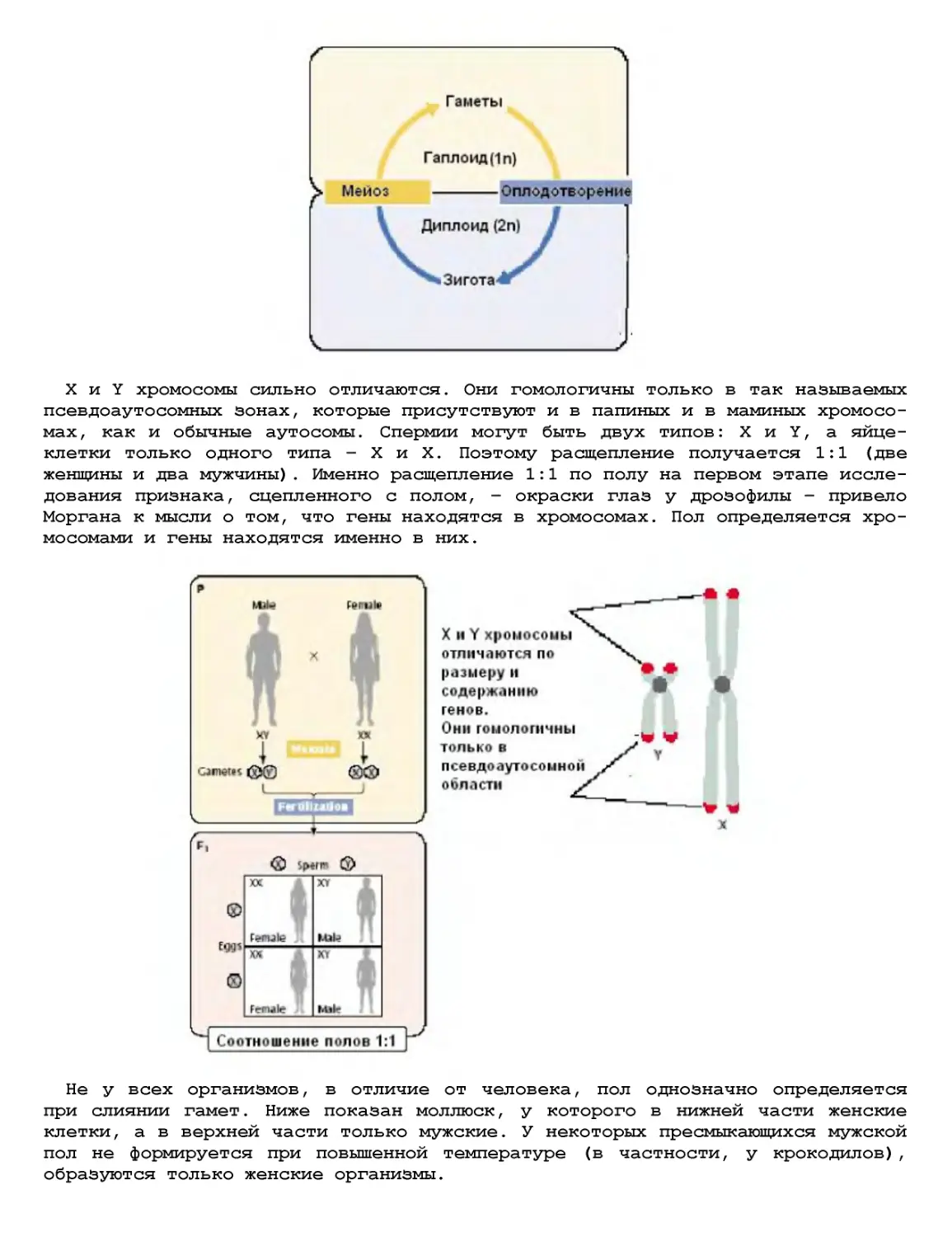

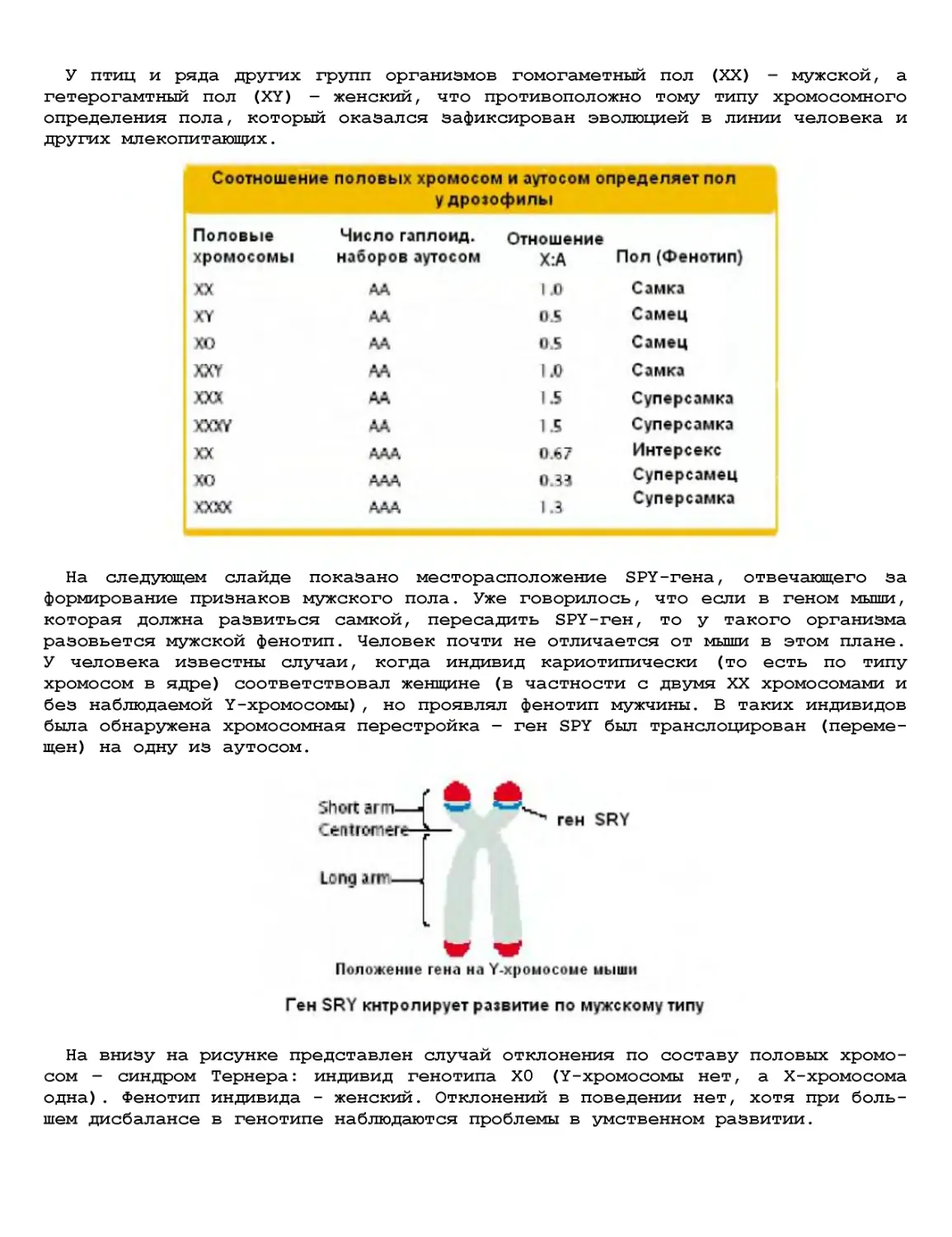



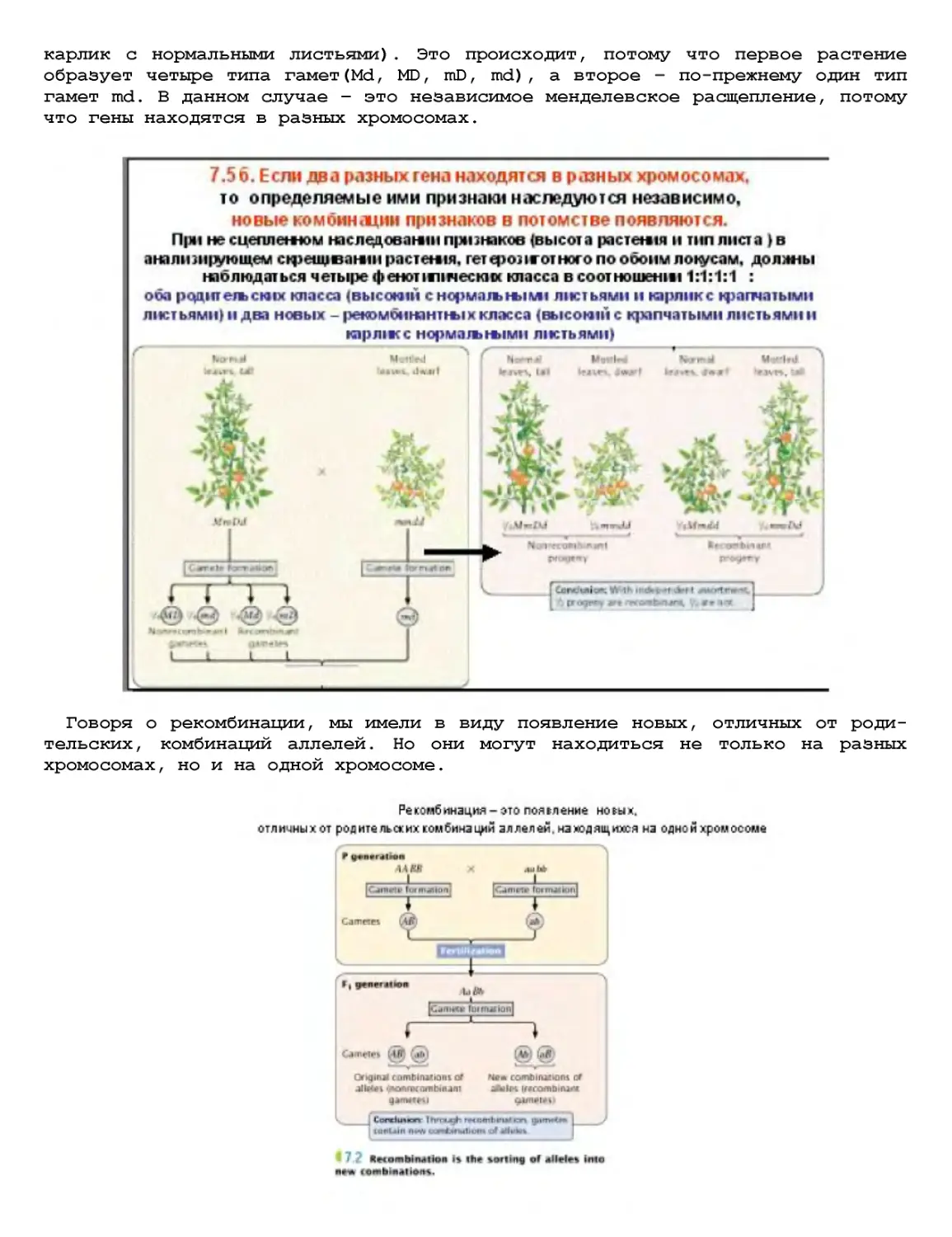













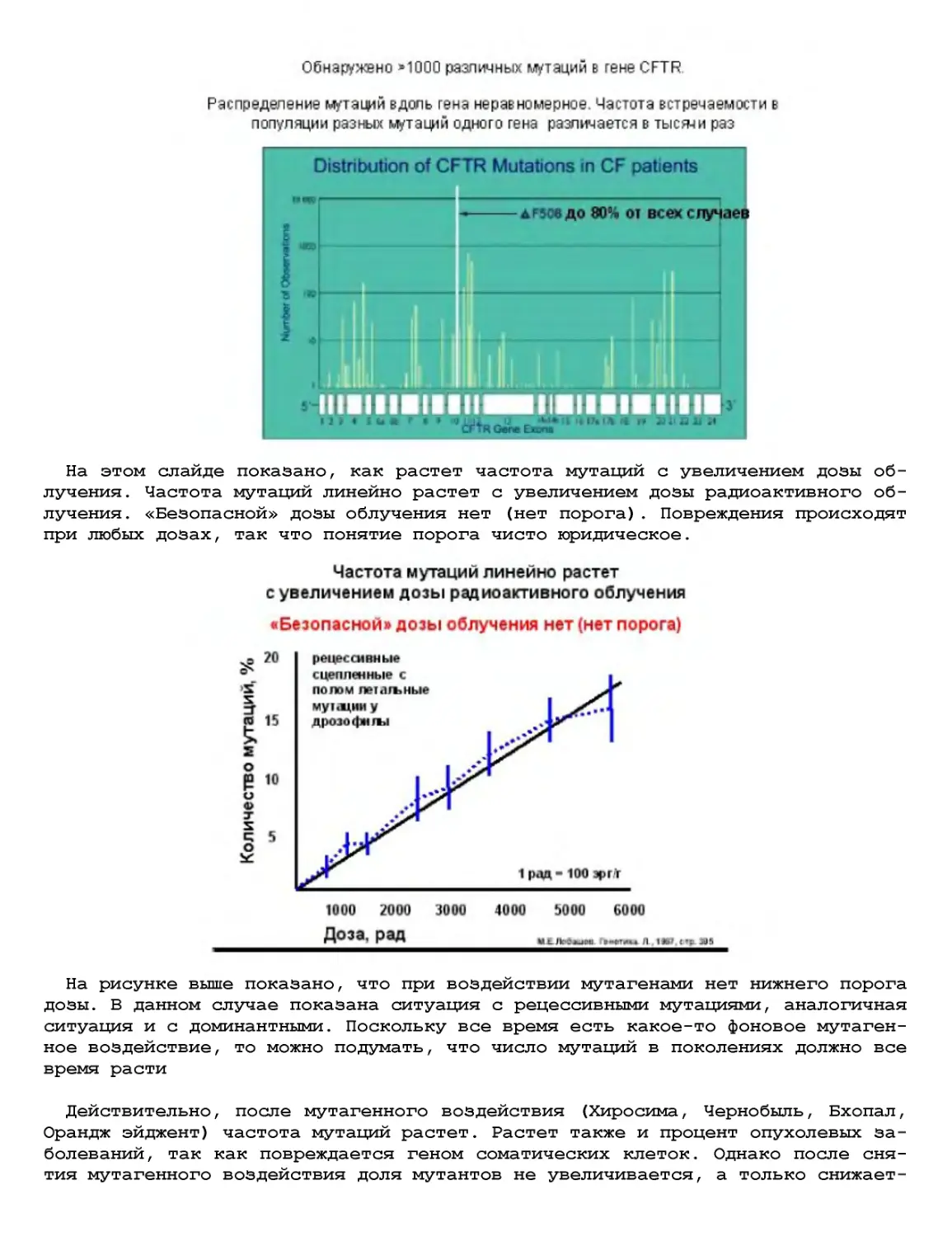


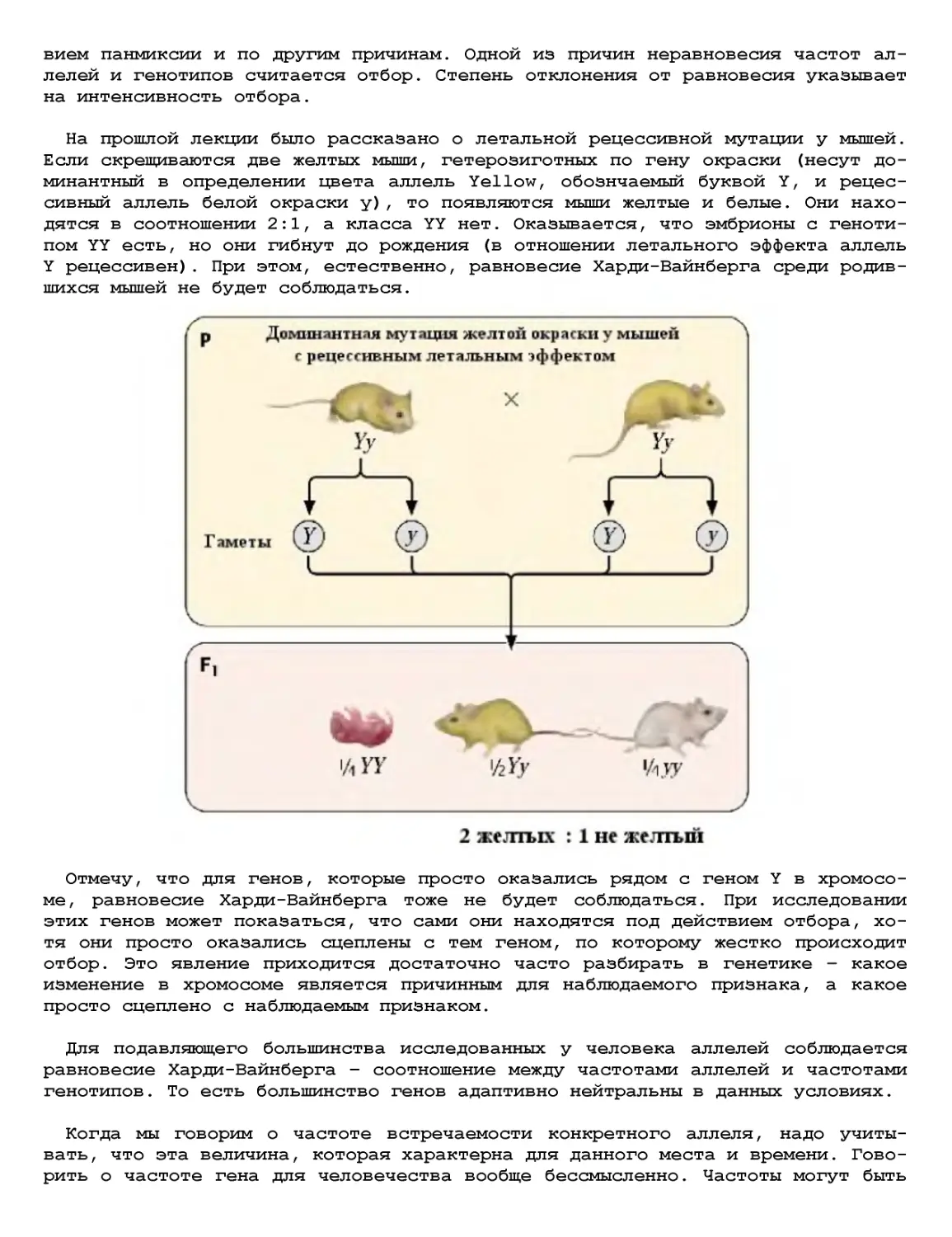
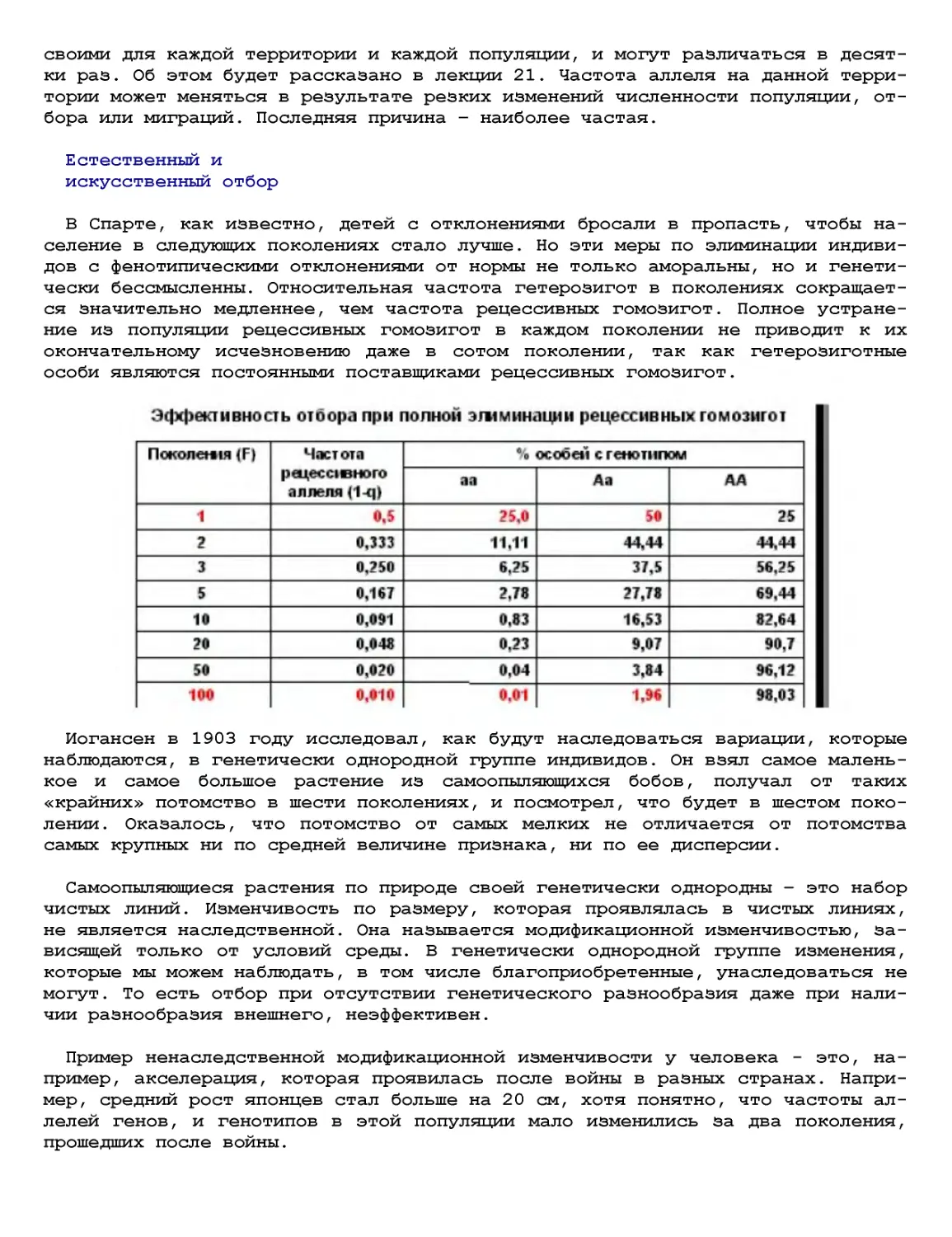
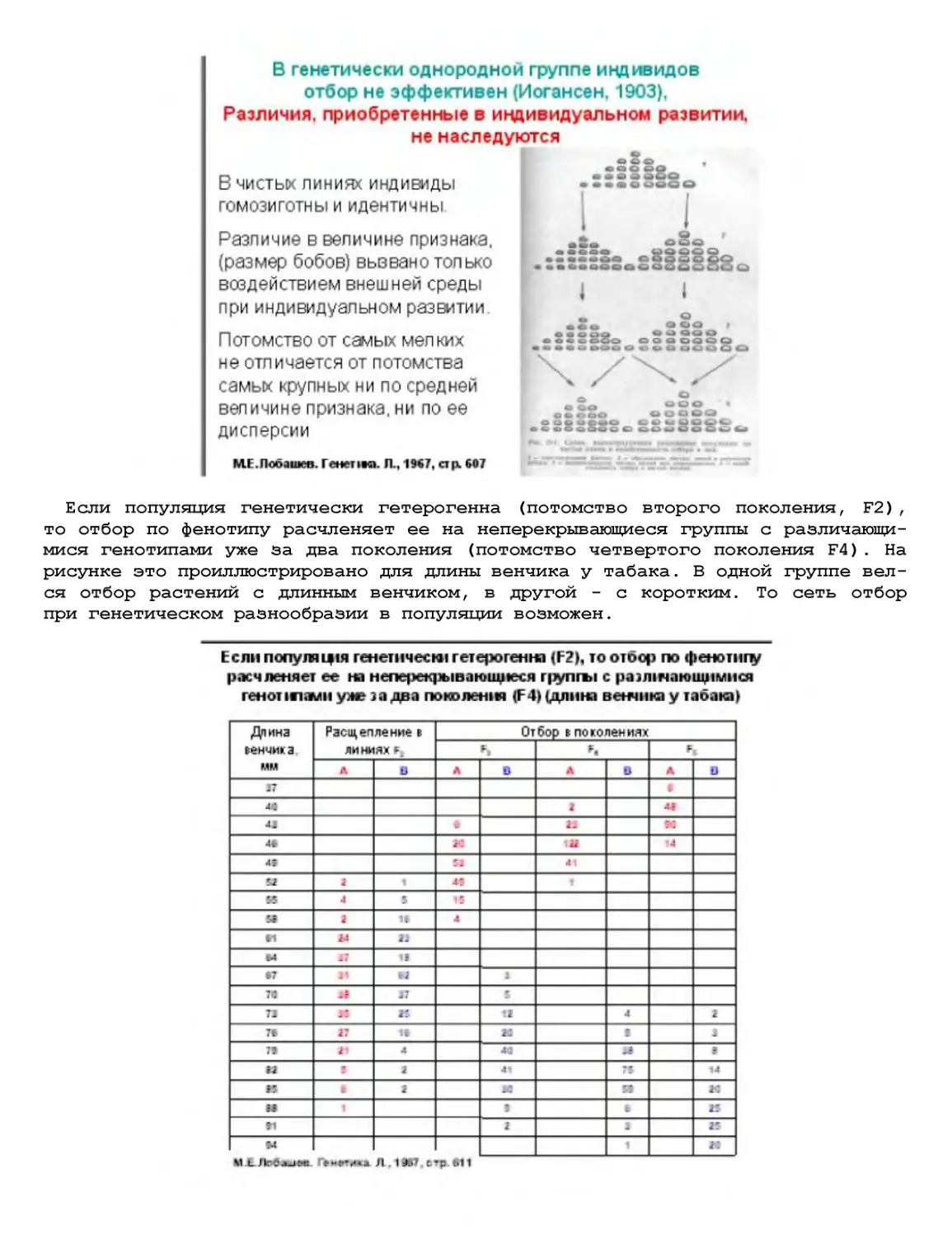
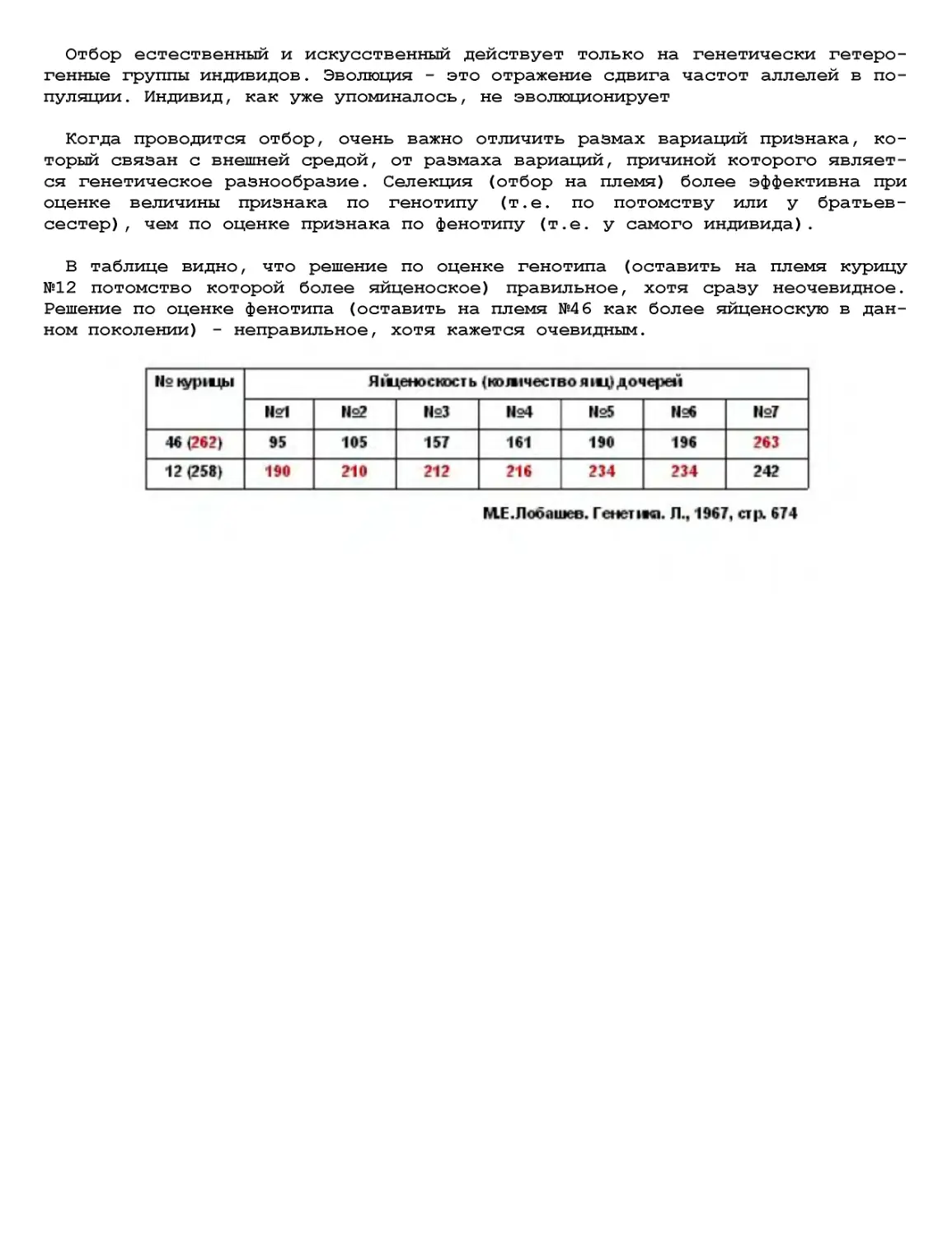





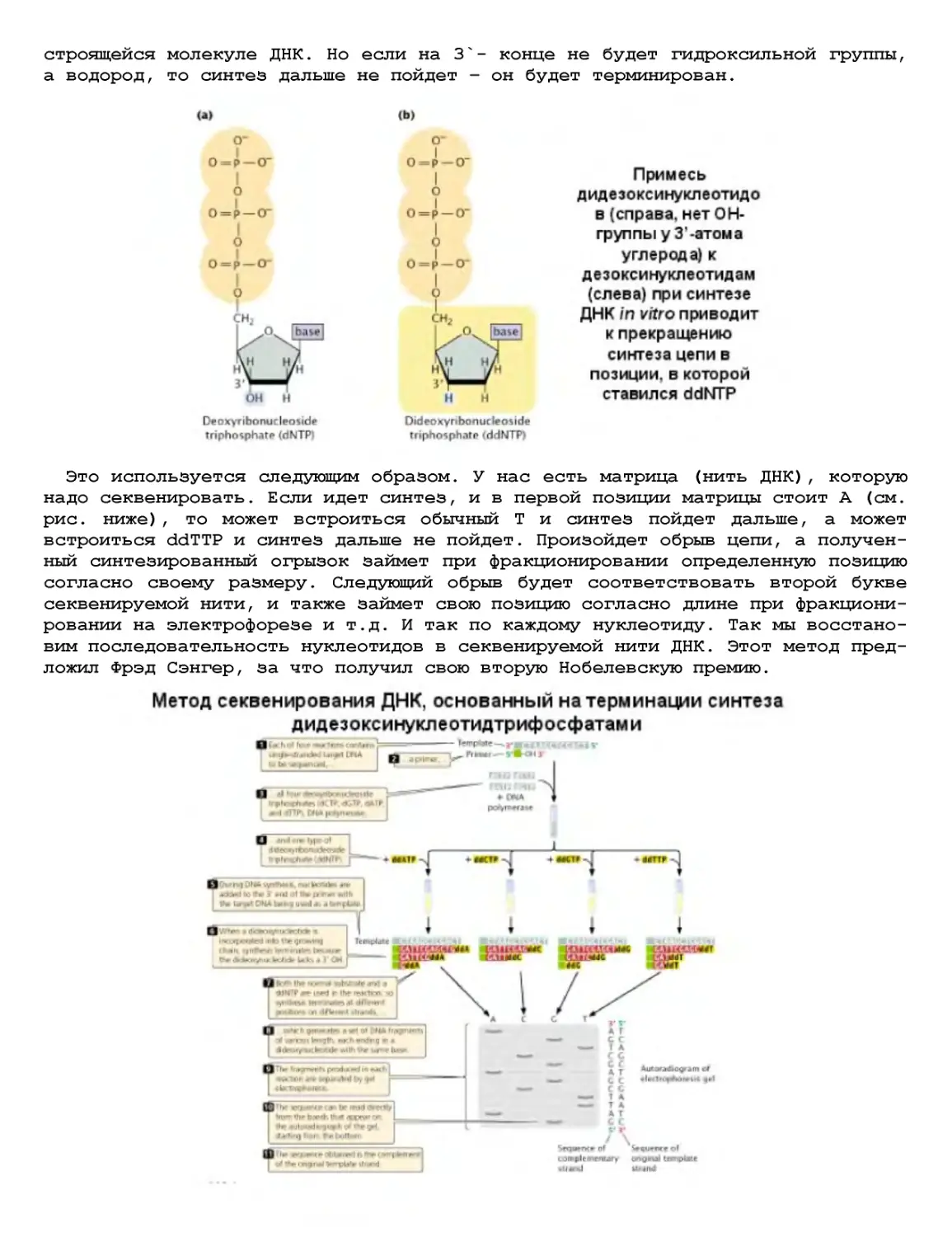


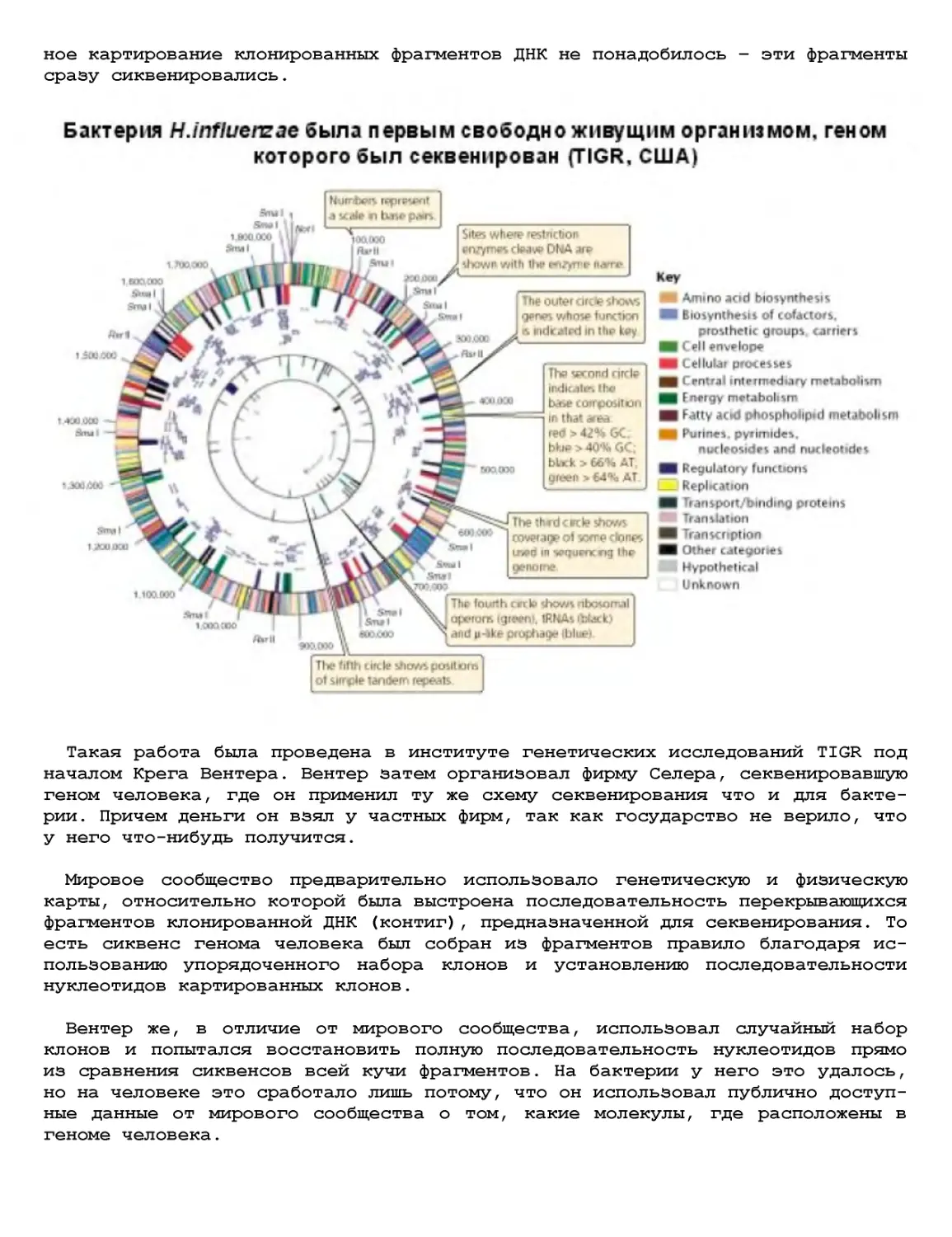










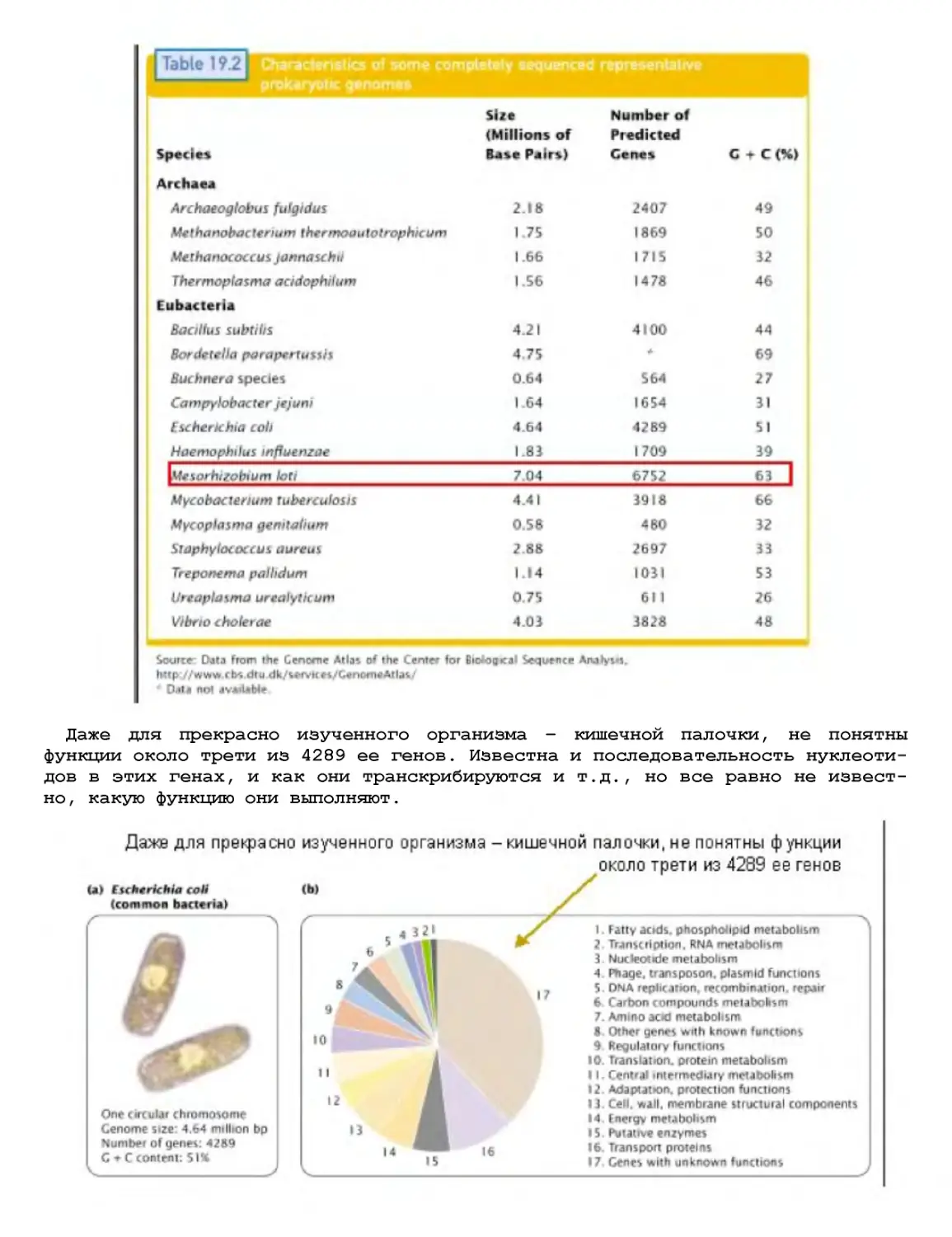
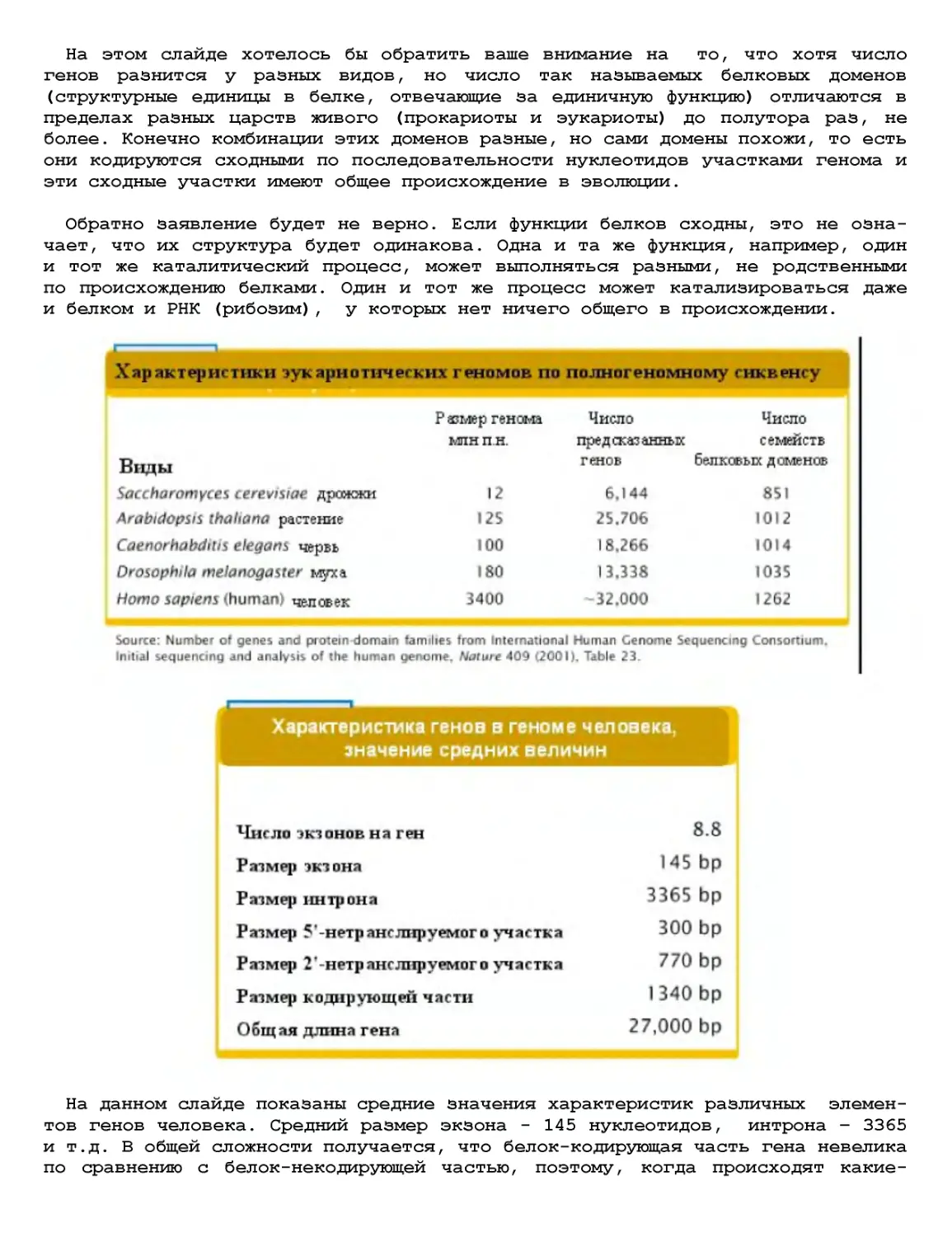






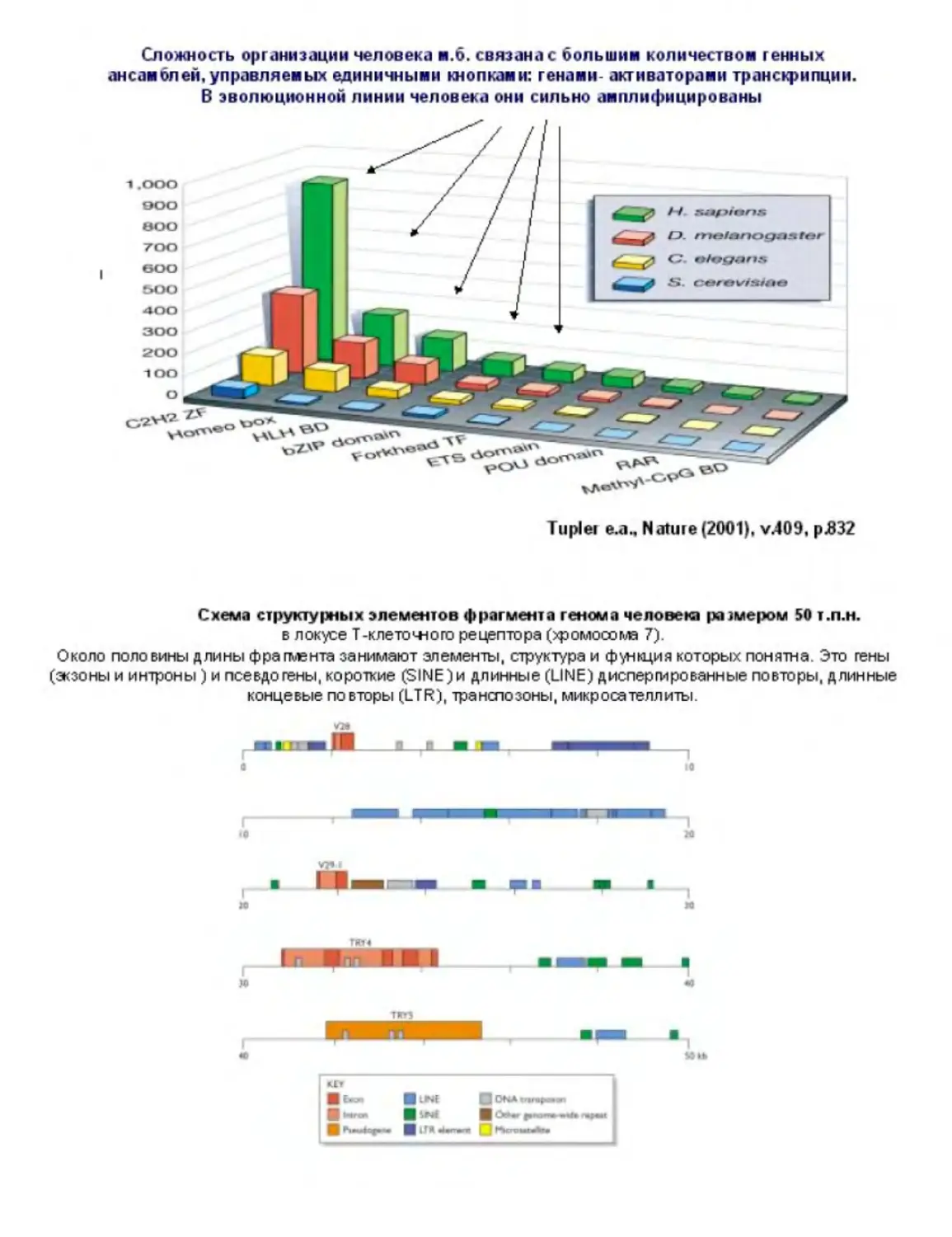


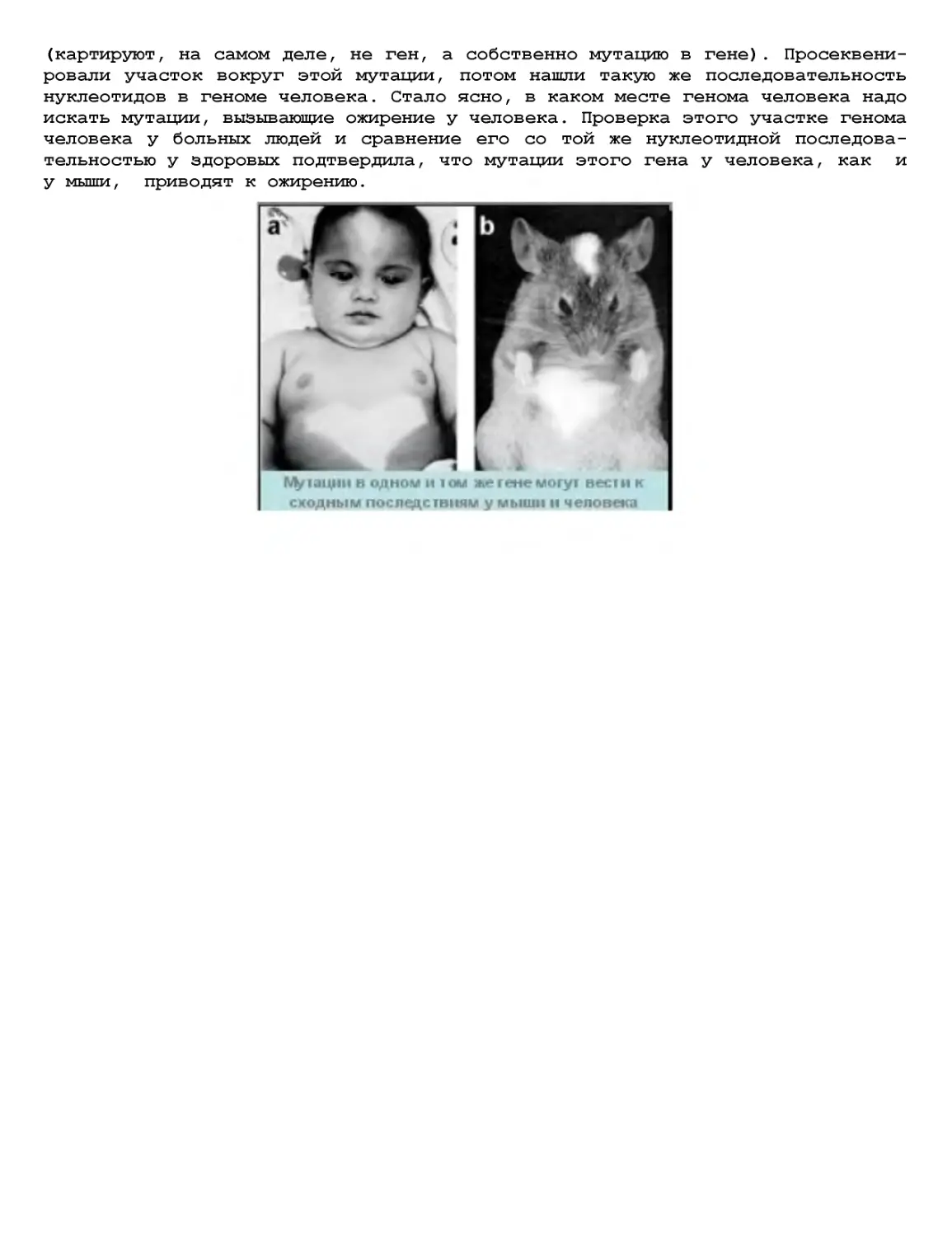






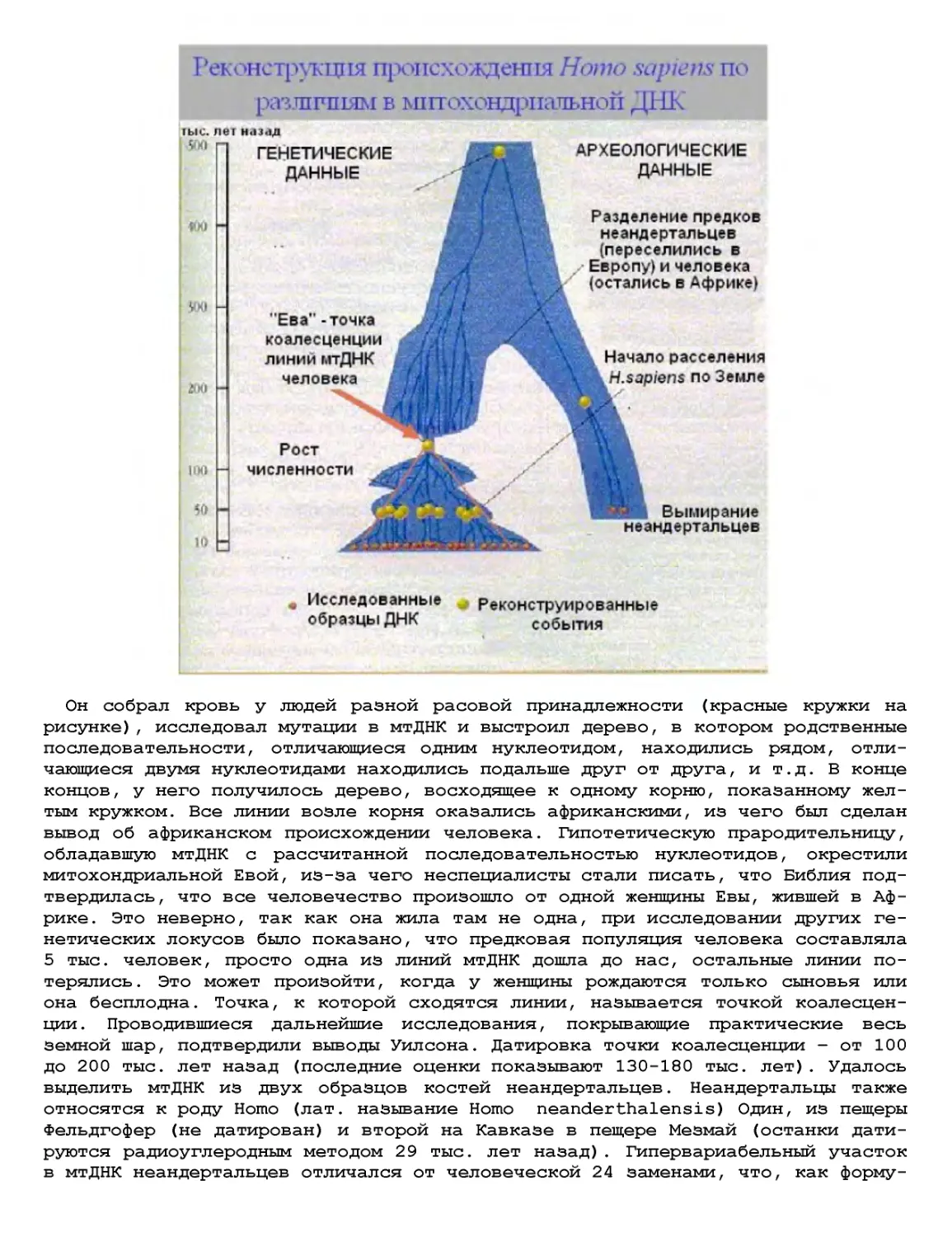

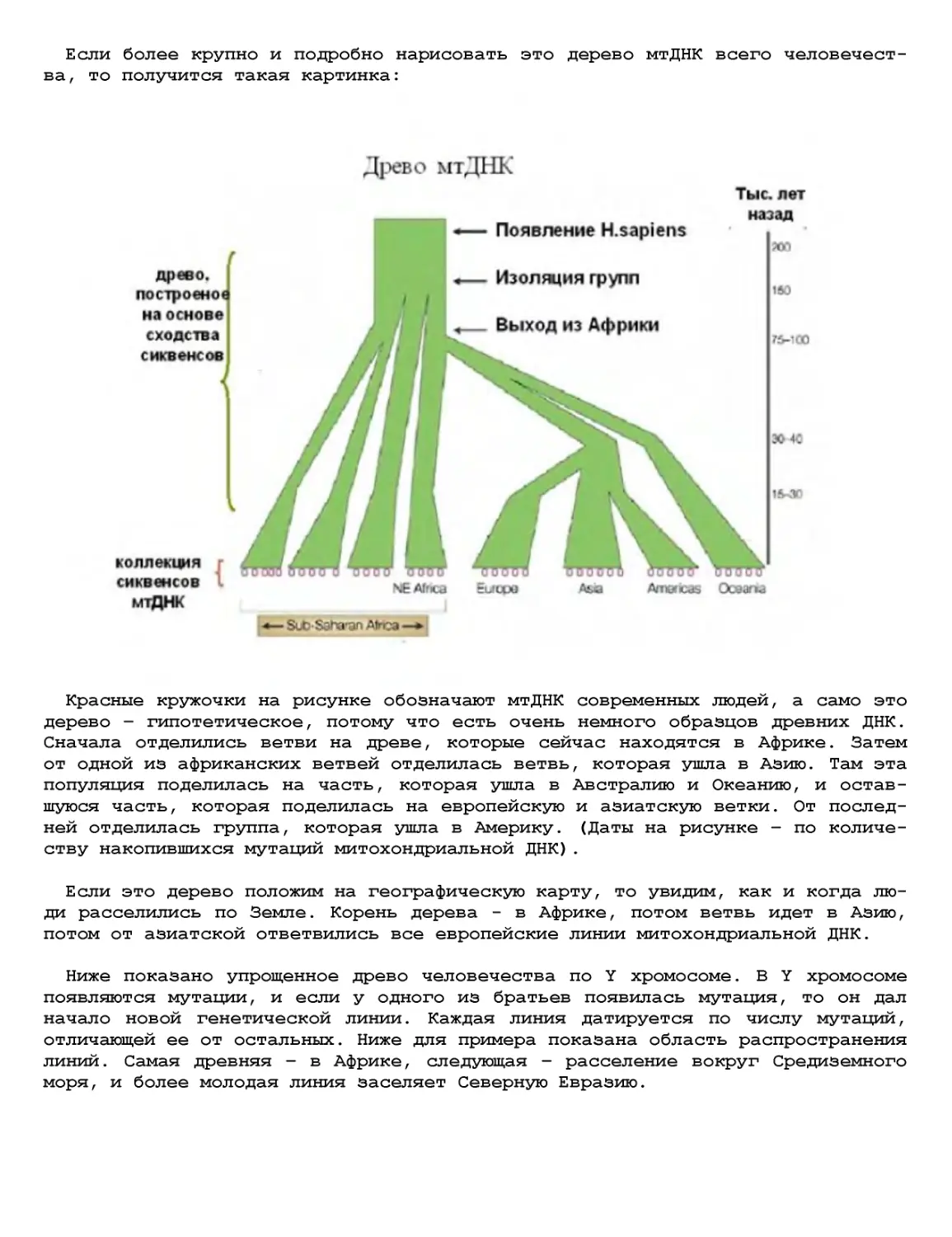

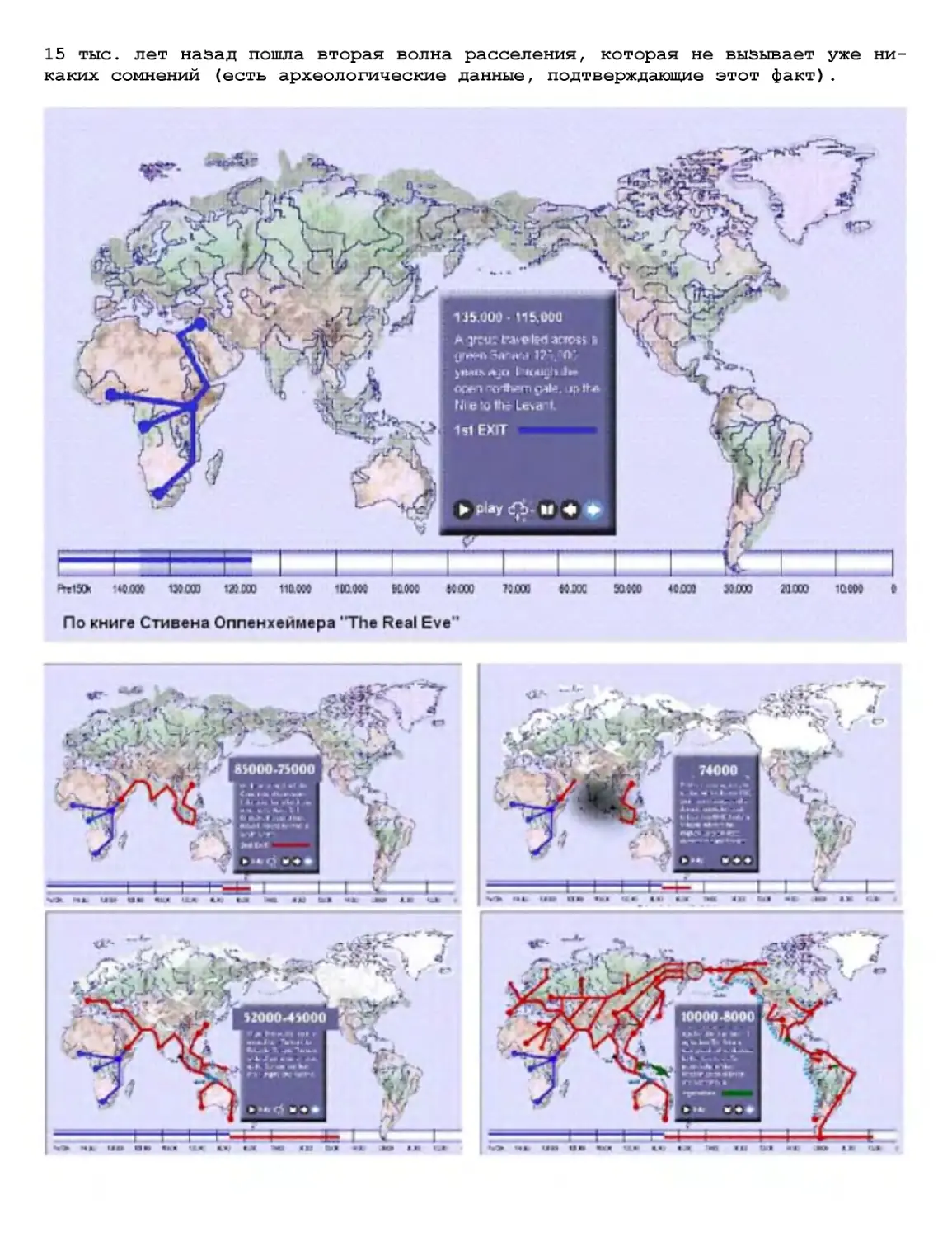






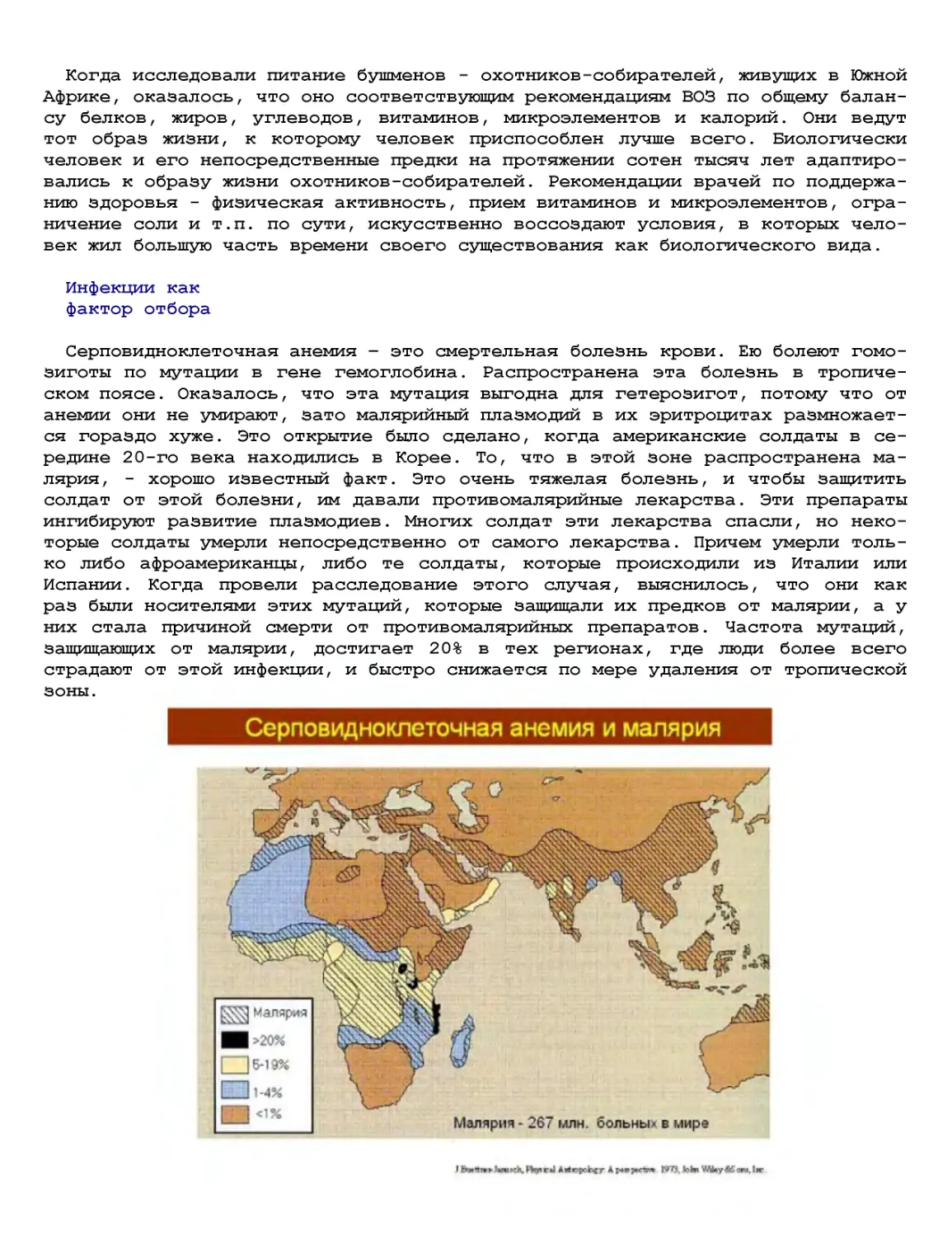



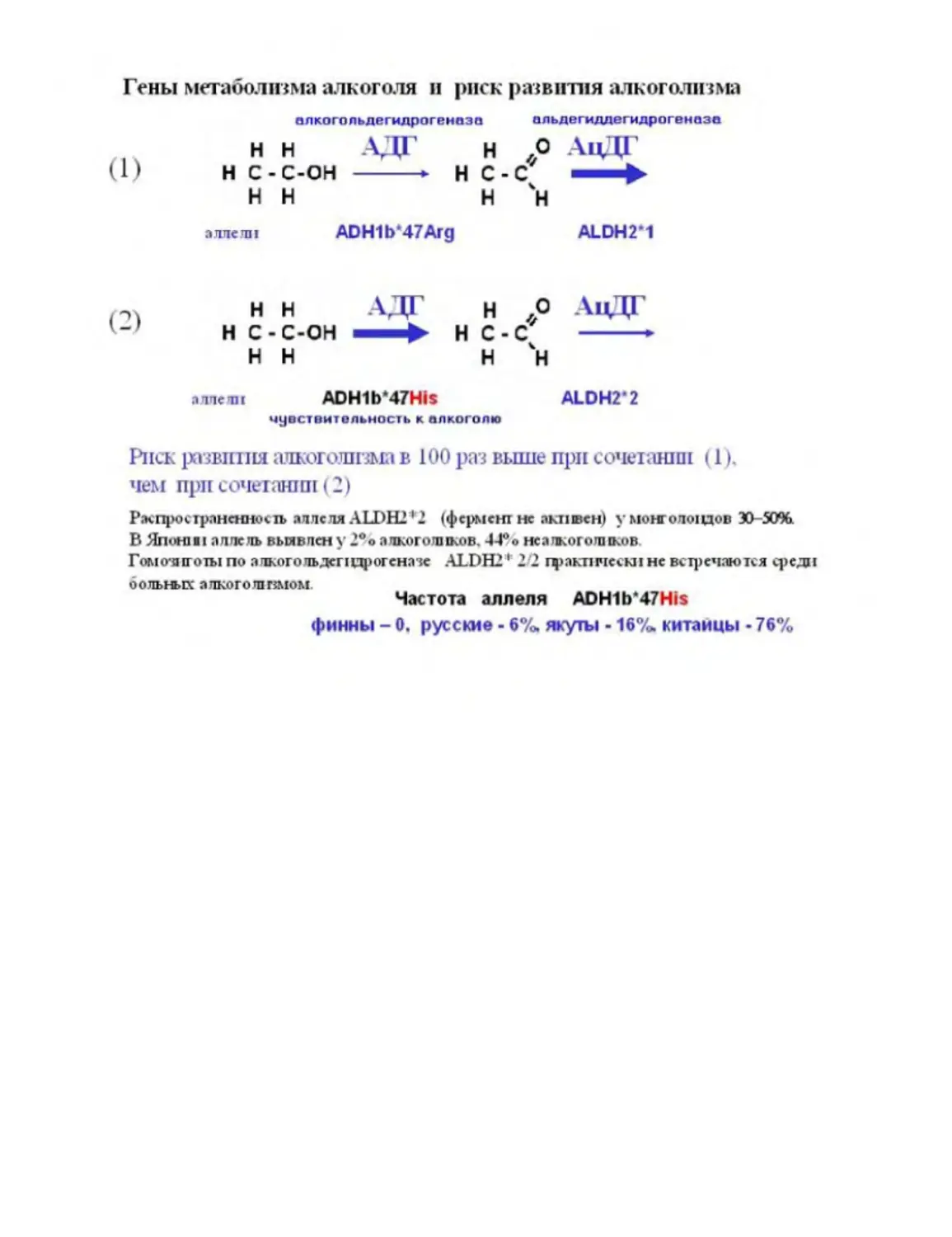









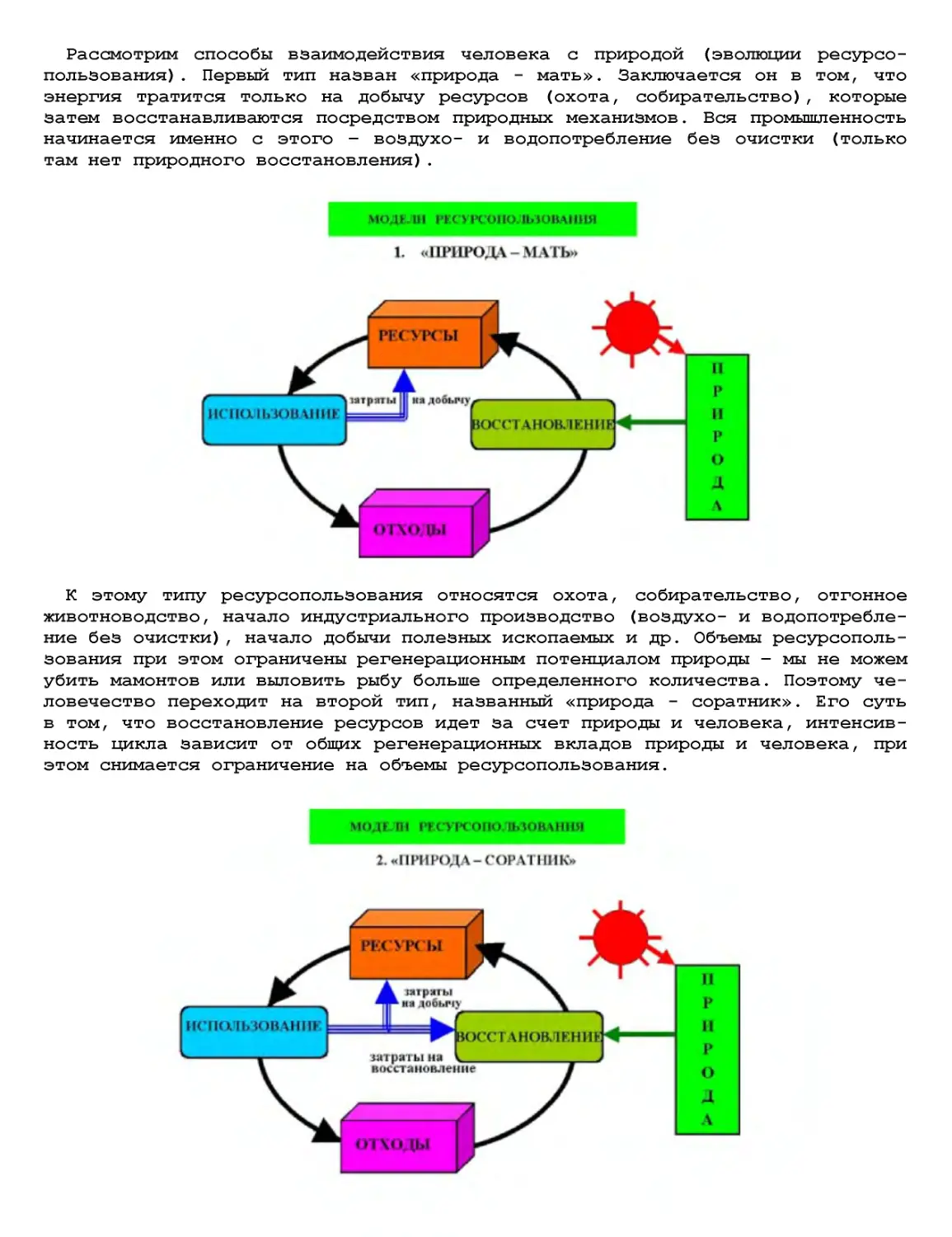
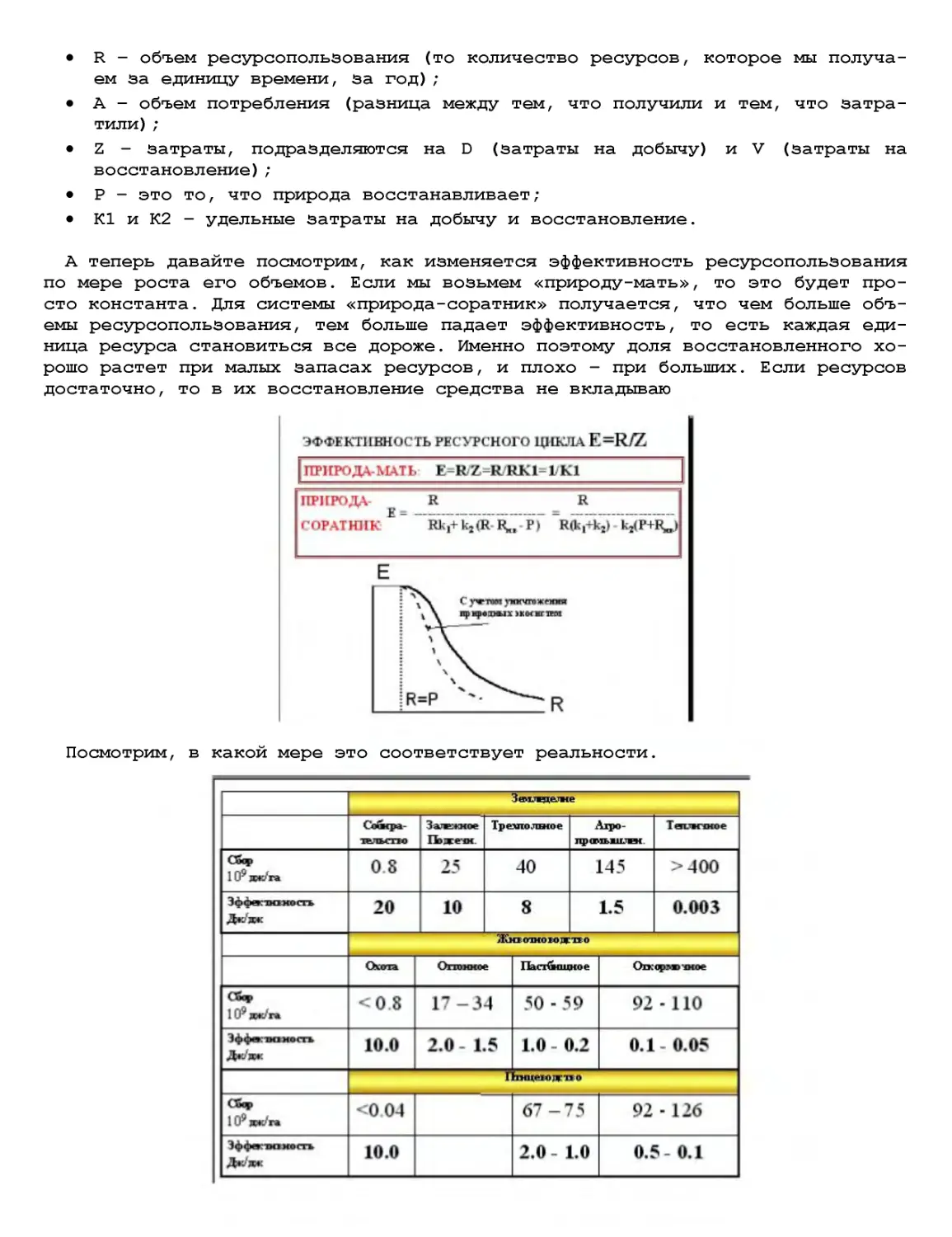




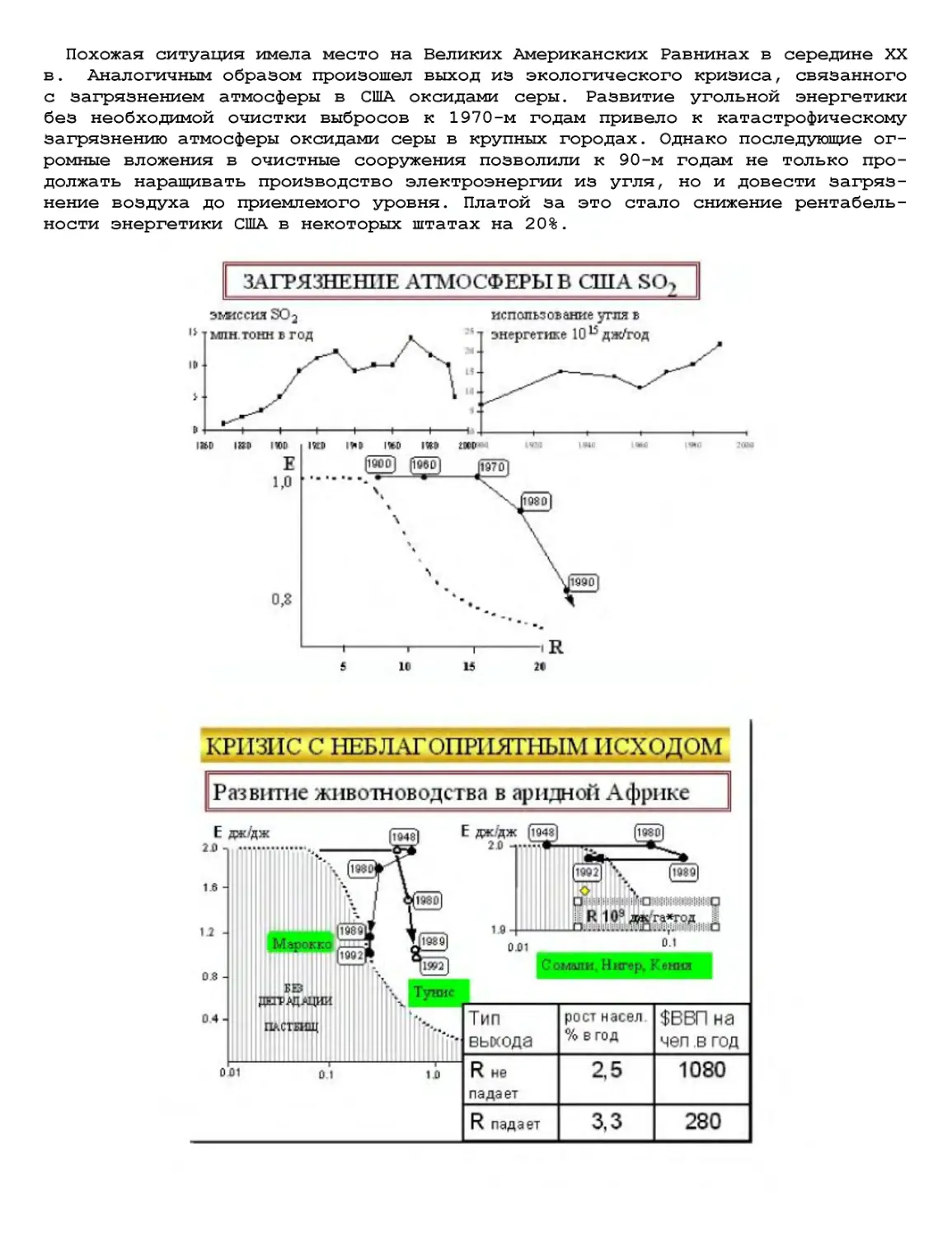

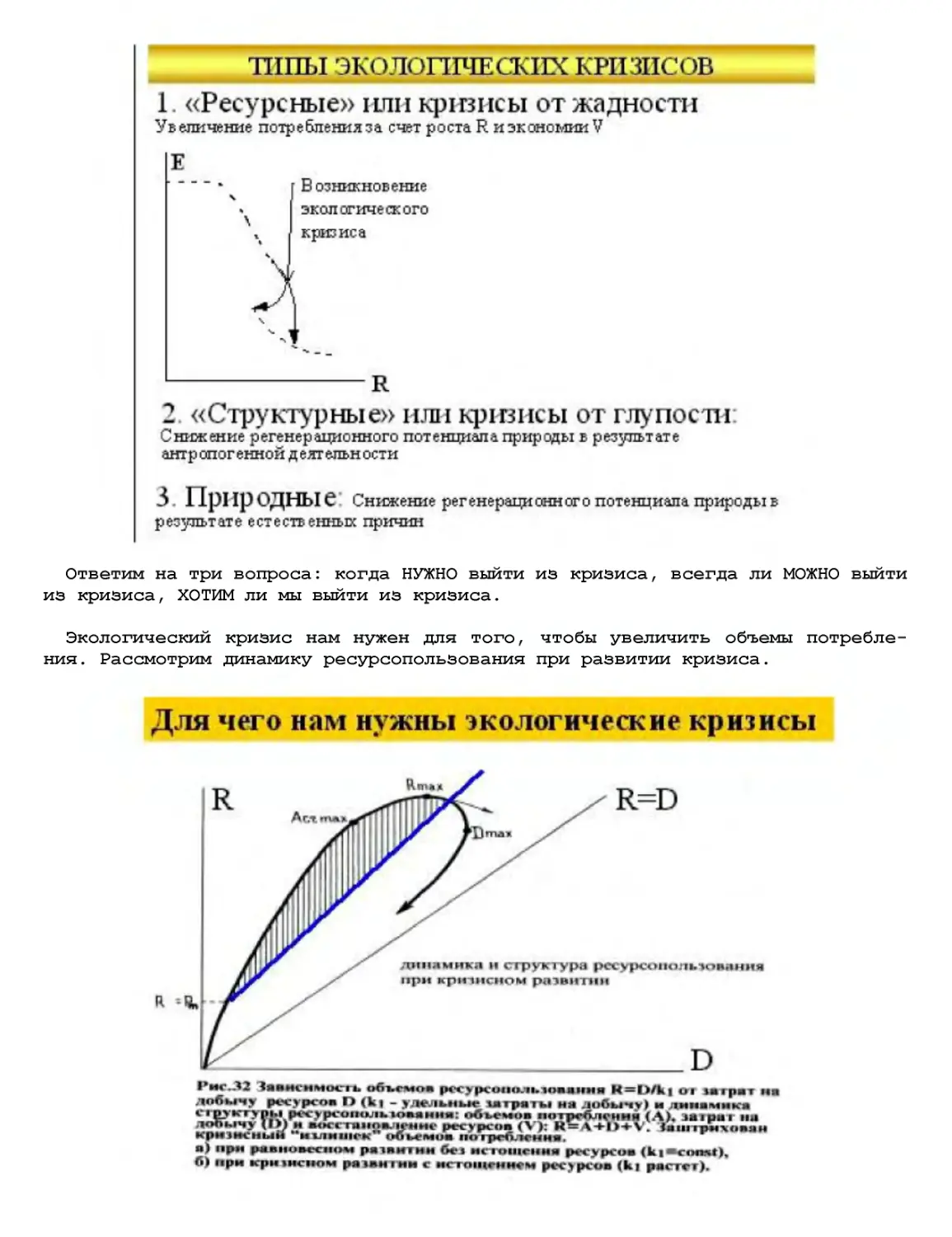

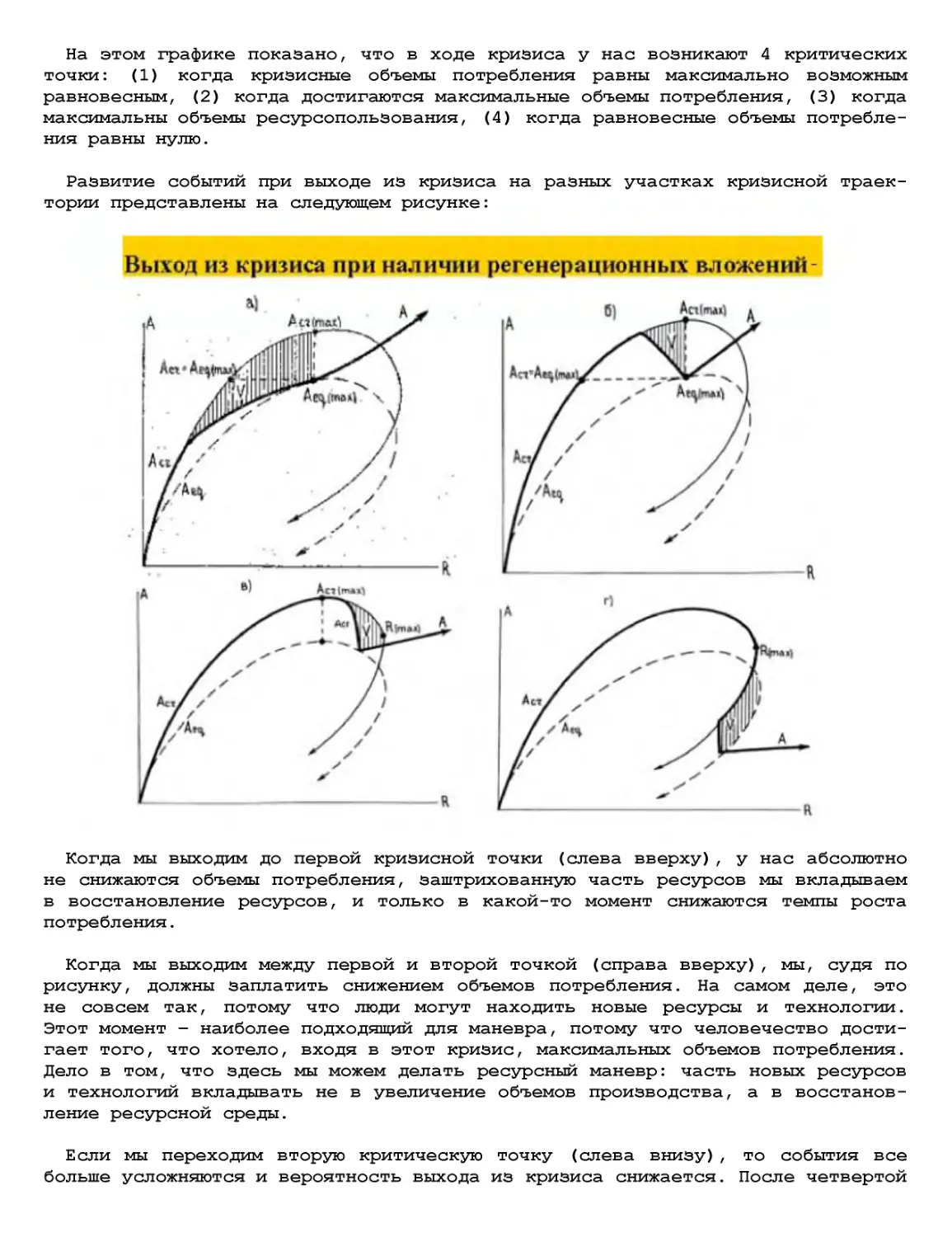
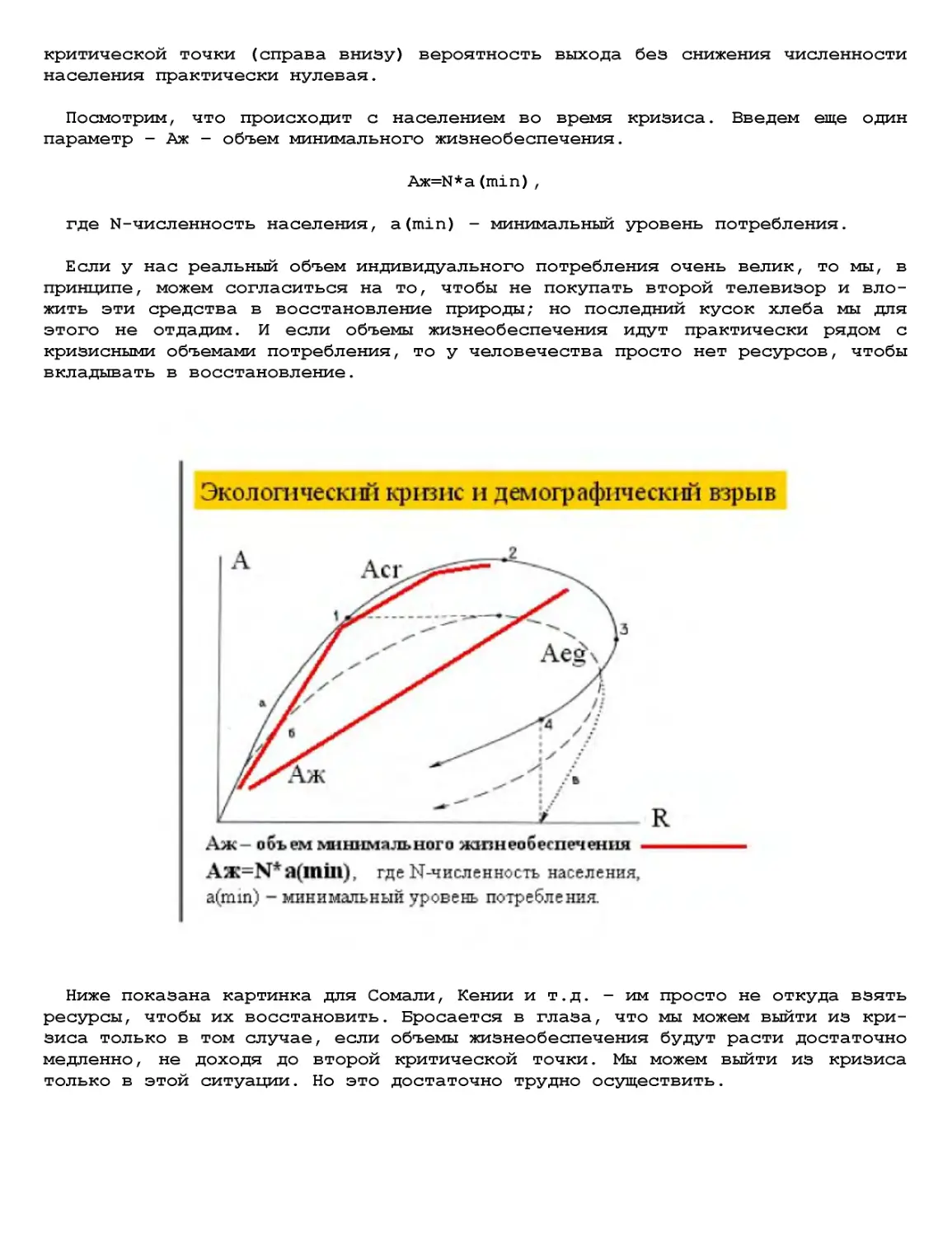

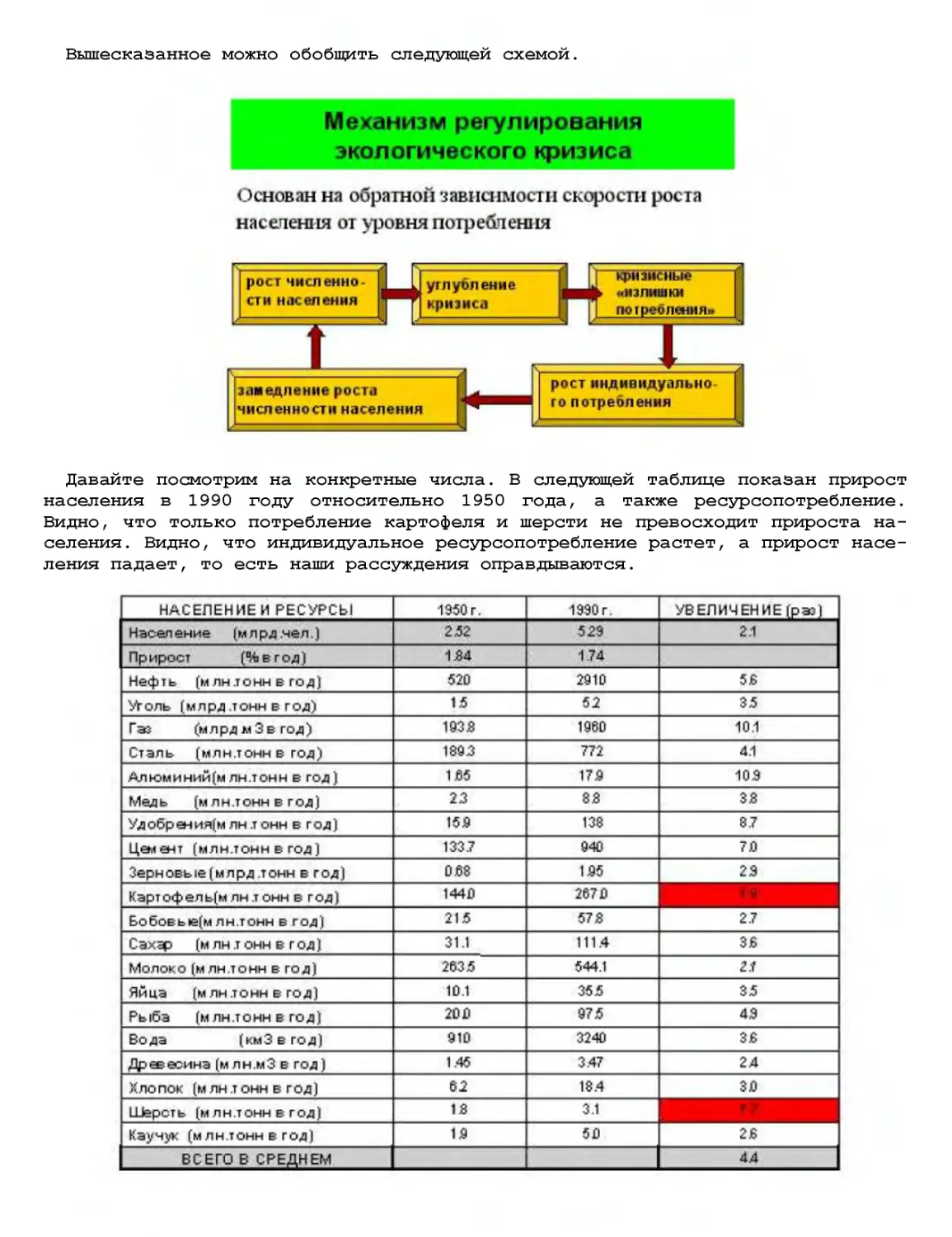




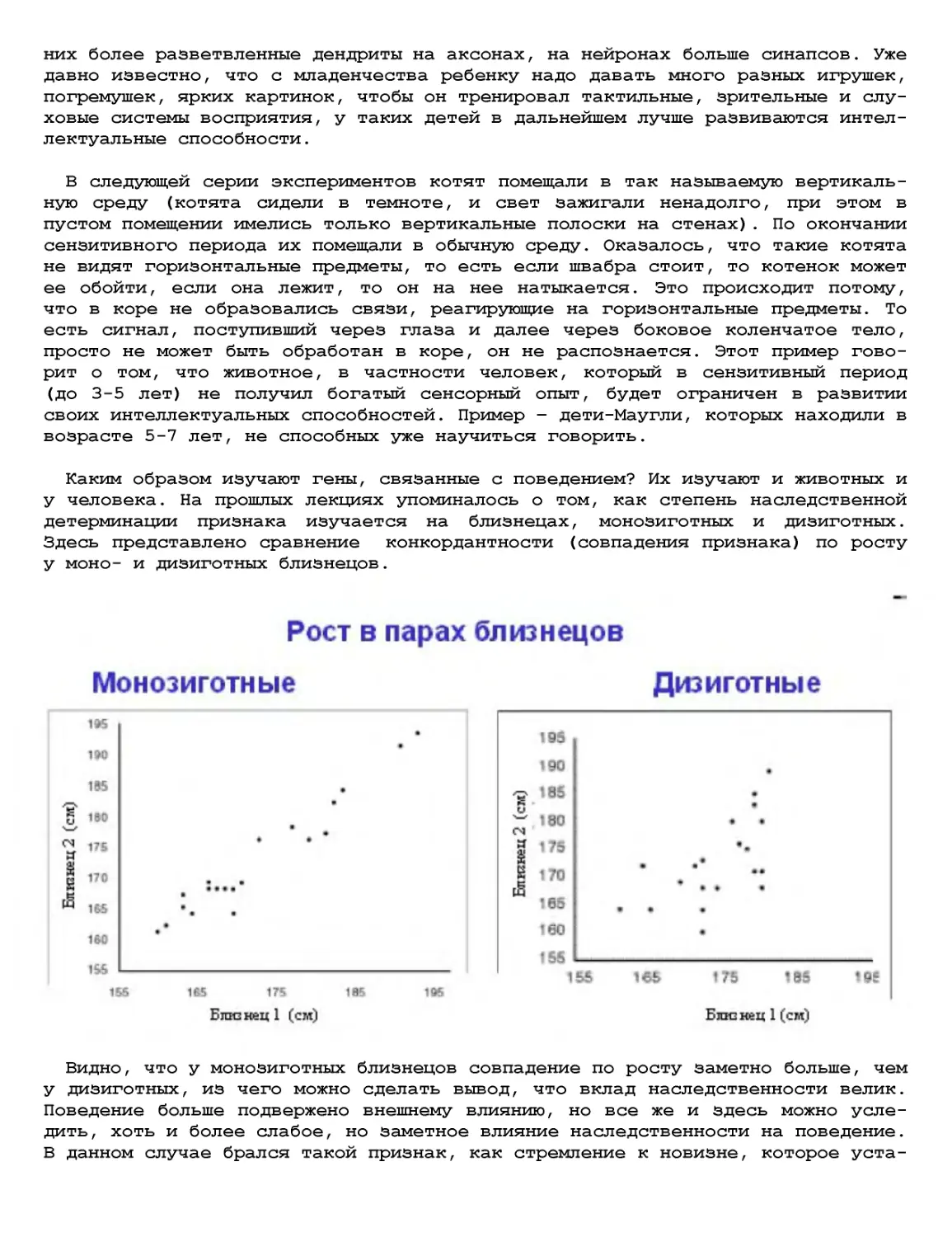
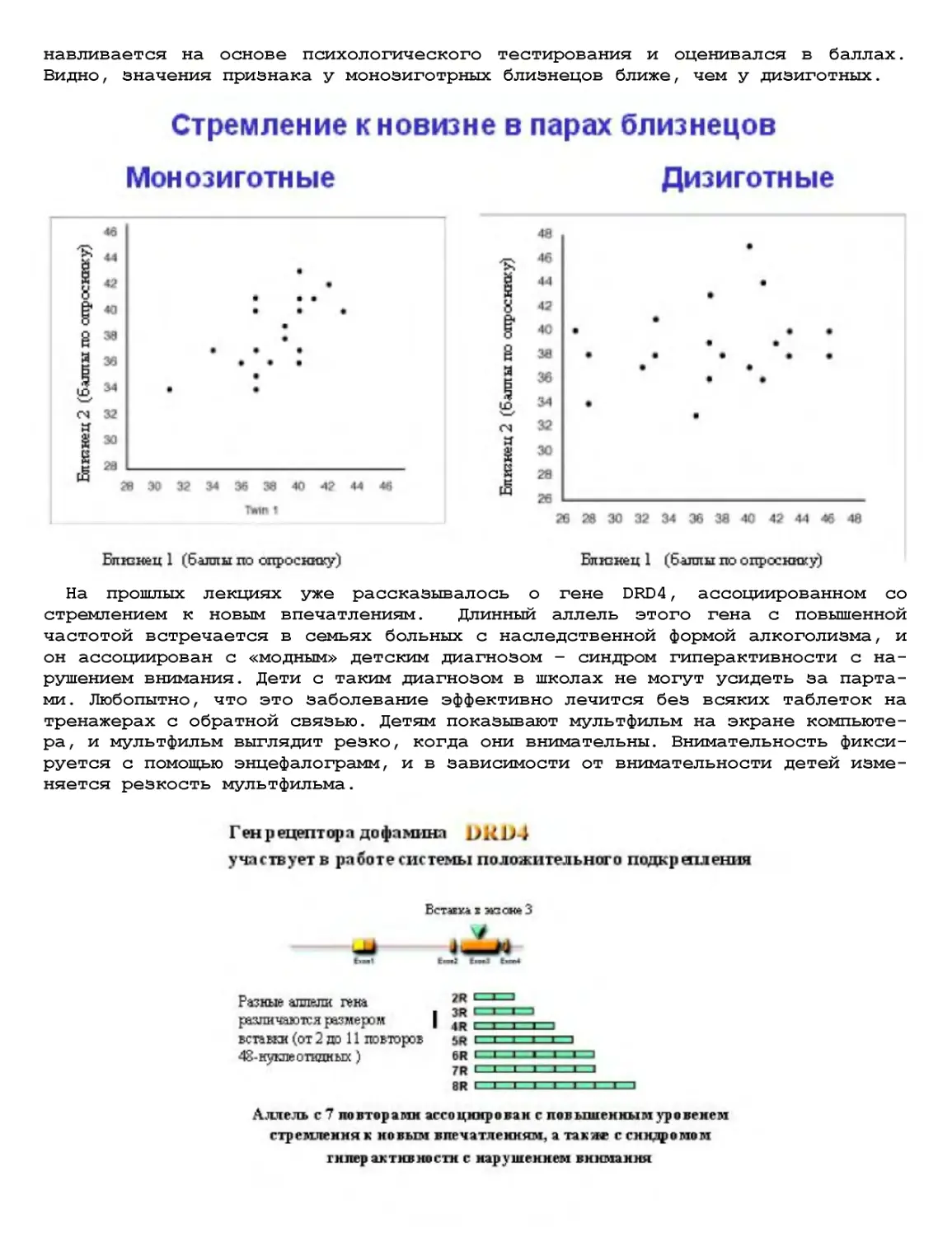










































































































































































































































Текст
ИНТЕРНЕТ-ЖУРНАЛ
МАЙ 2008
ДОМАШНЯЯ
ЛАБОРАТОРИЯ
Интернет-журнал
колхозников, инженегров
и разнорабочих науки.
Адрес редакции:
domlab@inbox.com
СОДЕРЖАНИЕ
Техношок или сложность?
Май 2008
Мнения
Статьи для журнала направ-
лять , указывая в теме пись-
ма «For journal».
Журнал содержит материалы
найденные в Интернет или
написанные для Интернет.
Журнал является полностью
некоммерческим. Никакие го-
норары авторам статей не
выплачиваются и никакие оп-
латы за рекламу не принима-
ются .
Явные рекламные объявления
не принимаются, но скрытая
реклама, содержащаяся в
статьях, допускается и даже
приветствуется.
Редакция занимается только
оформительской деятельно-
стью и никакой ответствен-
ности за содержание статей
не несет.
Ретрогенетика
Литпорта^
Го стина;
Расшифровка прошлого 12
Неандертальцы снова выйдут на мамонтов 16
Поход ва разумом 19
Заразные гены... 34
...И парадоксы систематики 38
Где скрываются снежные люди 40
Ликбез
Лекции по биологии (продолжение) 46
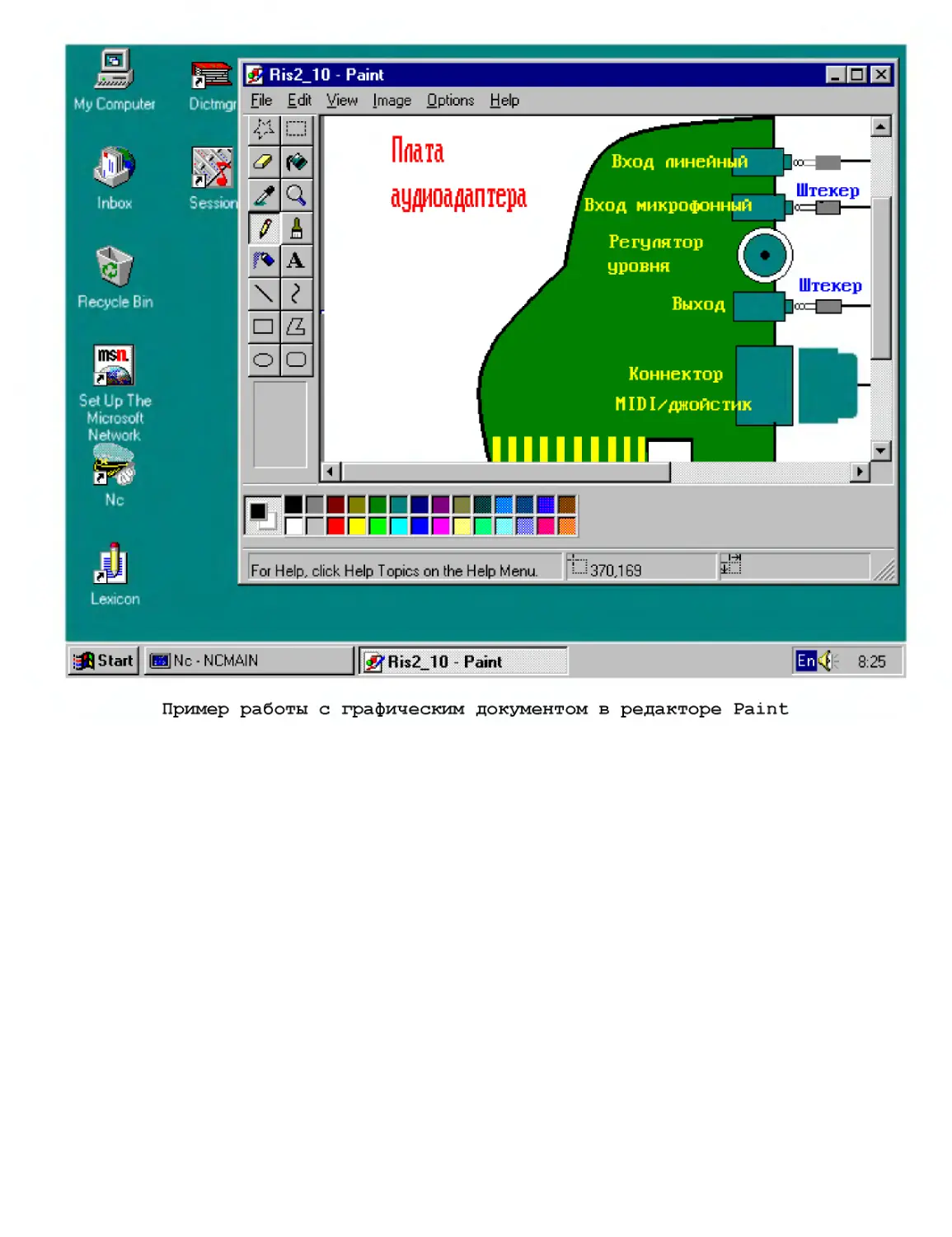
Базовый курс: Windows и Интернет
Компьюте]
294
Электроник
Термометр цифровой 416
Универсальный регулятор мощности 422
Статьи не редактируются,
орфография статей является
делом их авторов.
Технолог!
Производство грибов 426
При использовании материа-
лов этого журнала, ссылка
на него не является обяза-
тельной , но желательной.
Никакие претензии за не-
вольный ущерб авторам, за-
имствованных в Интернет
статей и произведений, не
принимаются. Произведенный
ущерб считается компенсиро-
ванным рекламой авторов и
их произведений.
Акцент
Муравьи - грибоводы 458
Юмо]
Это должен внать каждый таракан 461
По всем спорным вопросам следу-
ет обращаться лично в соответ-
ствующие учреждения провинции
Свободное государство (ЮАР).
При себе иметь, заверенные ме-
стным нотариусом, копии всех
необходимых документов на афри-
каанс, в том числе, свидетель-
ства о рождении, диплома об
образовании, справки с места
жительства, справки о здоровье
и справки об авторских правах
(в 2-х экземплярах).
НА ОБЛОЖКЕ
«Мы сами являемся организмами и окружающие нас дру-
гие системы воспринимаем по аналогии с самими собой.
Как ни естествен для нас такой подход, он не единст-
венно возможный...»
Цитата из статьи «Заразные гены»
Мнения
ТЕХНОШОК ИЛИ СЛОЖНОСТЬ?
Леонид АШКИНАЗИ
Утюг не вызывает у вас никаких эмоций - он прост; вы знаете, как он работа-
ет. Кусок металла, из коего он сделан, также не вызывает ни восторга, ни ужа-
са - хотя вы и не знаете, ни как он (металл) устроен, ни как он работает.
Компьютеры и интернет вызвали море эмоций и бурю экстаза на нем: видимо, они
лежат в каком-то смысле между утюгом и железом. В каком?
Утюг прост именно потому, что мы знаем, как он работает. Металл кажется нам
простым, потому что мы не знаем его структуры или не заморачиваемся ее нали-
чием. Увидь мы, как герой «Соляриса», все эти многоразличные атомы и бирнам-
ский лес дислокаций - возопили бы: «Почему все это не рассыпается? Как все
это передает тепло? Почему течет ток?!» Похоже, что ощущение непонимания воз-
никает, когда мы наглядно представляем себе сложность структуры, но не можем
проследить ее работу, функции, связи.
Заметим, что сложность определяется не количеством элементов. Цепь или
транспортер просты и понятны при любом количестве элементов, и даже «молния»
или веревка, состоящая из отдельных нитей, кажутся нам простыми и легкими для
понимания (хотя насчет веревки мы обманываемся). Дело в сложности структуры.
Структура радиоприемника для меня проста, и его работа мне ясна. Структура
Земли или звезды, пространственное распределение элементов в них мне не ясны,
но непонимание работы связей меня не пугает - я не вижу сложностей связей
там, где не вижу сложности структуры. Видите ли вы сложность структуры компь-
ютера или интернета? Увы, да. Компьютеры и интернет вызывают шок у людей то-
гда , когда они понимают, знают или в некоторой степени представляют себе, как
они сложны. Платы видел всякий, по крайней мере - на рекламе. Видел, но -
хоть убей - не понимает, что там и как из одной коробочки в другую бегает. И
главное - почему!
Как человек может защититься от стресса и как чаще всего он это делает?
Первый, самый простой и очевидный способ, - не заглядывать внутрь компа и не
глазеть на рекламу. Работает, потому что должно работать. Как водопровод -
работает, потому что должен. А если не работает - есть специалисты, которые
знают, как сделать так, чтобы работал. Второй, более сложный, - разобраться,
как оно работает. Третий, это который и используется чаще всего, - построение
псевдообъяснений.
Псевдообъяснение - это текст (иногда одна фраза), который содержит слова,
относящиеся к наблюдаемому явлению, и слова, обычно используемые в действи-
тельных моделях. Это слова: «все», «потому что», «всегда», «влияет», «связа-
но», «который» и многие другие. Слова эти можно разделить на несколько групп:
слова, устанавливающие категории, объединяющие объекты в группы; слова, выде-
ляющие объекты, присваивающие им какие-то свойства; слова, прямо указывающие
на связи между объектами. Стало быть, эти слова можно назвать просто и вуль-
гарно - «ученые слова». Но это не то, что обычно называют «учеными словами» -
всякие непонятные термины, имена объектов и явлений, а «на самом деле» ученые
слова. Т.е. слова, несущие научную информацию, указывающие на признаки (выде-
ляющие или объединяющие), либо на связи. Сами имена учеными словами не явля-
ются - они становятся ими лишь тогда, когда мы знаем, что за ними стоит.
«Глокая куздра» - бессмыслица, и даже утверждение «Петя - глокая куздра» -
бессмыслица, пока я не знаю, что такое эта «куздра». А вот фраза «Петя - гло-
кая куздра, и поэтому он кудланул» - уже наука.
Почему же ученые слова проливают бальзам на душу пипла? Почему вместо пони-
мания экономики пипл изрекает «все воруют», и ему делается хорошо? Почему
вместо исследования психологии конкретного человека пипл изрекает «потому что
пьет», и все понимающе кивают? Почему эти слова обладают столь сладостным
магнетическим действием? Потому что несчастный пипл учился в школе. И в том
его трепетном возрасте, когда все так безнадежно и жестоко врезается в па-
мять, уродуя нежную, только начавшую миелинизироваться, кору головного мозга,
в этом самом суперчувствительном возрасте деспот-препод со сверкающими глаза-
ми и капающим с ногтей зеленым ядом вдолбил... Сухой остаток: человек запом-
нил этот словарь, который в детстве сопровождал его попытки что-то понять.
Тогда это было отчасти настоящее понимание (не купил игрушку, потому что нет
денег; летает вокруг Земли, потому что Земля притягивает; не хочет, потому
что нашла другого), отчасти - и с течением времени все больше и больше -
псевдообъяснение.
А для расширения применимости есть еще такой прием: само слово толкуется не
совсем четко или вообще говорится одно, а понимается - в силу особенностей
психологии - другое. Вот один пример: в любом издании нынче можно прочесть
«Ученые установили, что А связано с В». Написано так, но пипл читает - «из А
следует В». Сегодня читаю: «Ученые установили, что полные женщины получают
меньшие зарплаты». Ага, - думает читатель, - мужики не любят полных и не дают
им хороших должностей. Или наоборот: они их любят, злятся, что те не любят
их, и поэтому не дают им хороших должностей. То есть можно думать любую глу-
пость или вообще никакой, но «понимание» достигнуто, связь выявлена. На самом
деле (если это вообще не простое вранье) ситуация такова: полнота иногда яв-
ляется следствием низкого уровня культуры, он коррелирует с низким образова-
тельным уровнем, а уж он имеет следствием меньшую зарплату (в некоторых стра-
нах) . Но пришлось напрягаться, и никакого тебе удовольствия. Гораздо лучше,
если пресса кормит псевдообъяснениями, а читатель радостно ими хрумкает.
Почему такая ситуация устойчива? Потому что есть система, которая по чисто
коммерческим причинам кормит потребителя «этим». Пипл хавает, отчего же его
не кормить? Система проста - она дает человеку то, чего ему хочется, за что
он готов платить. А хочется ему легкого. В некапиталистической, а еще лучше -
тоталитарной системе все может быть наоборот. Приходит диктатор и говорит:
всем ползти зигзугой, к завтрему выучить четвертый - как его? - закон Ньютона
и все объяснять посредством него. Физик радуется и прыгает. «А как насчет
русского языка?» - жалобно спрашивает гуманитарий. «А этим займется следующий
диктатор», - отвечает философ. Историк благоразумно молчит...
Не надо понимать этот пафос как призыв к диктатуре. Тем более что он беспо-
лезен - люди хотят жить хорошо, поэтому на больших временных интервалах дик-
татура обречена. А то, что прогресс науки и техники вместе с ростом произво-
дительности труда превращает большую часть человечества в грядку с овощами -
увы. Можно, конечно, предложить кое-что, но, во-первых, все это уже рассмат-
ривалось в литературе, а во-вторых, речь не о прогнозе на сотни лет, а о се-
годняшнем дне.
Вернемся к определениям. Что такое сложность? Интуитивное ощущение сложно-
сти возникает, если элементов много, структура соединений (граф, схема) слож-
на, типов элементов много. Иногда, впрочем, большие числа и не нужны. «Мол-
ния» состоит из двух типов элементов (зубья и ползунок), а можете ли вы объ-
яснить, как она работает? Ну, еще есть упругие нити ткани... Веревка состоит
из элементов двух (плетеная, с осевыми нитями) или даже одного типа (круче-
ная) . А как она работает? Поведение и «молнии», и веревки кажется простым,
но... Так что, видимо, придется ввести две сложности - сложность функциониро-
вания и сложность понимания этого функционирования. Но, о ужас, как без пони-
мания - если его нет - оценить сложность? Можно ли сказать, что сложность -
это непредсказуемость? Но как быть с бросанием монетки? Можно исключить слу-
чайное поведение, потребовав, чтобы при одинаковых исходных условиях дости-
гался одинаковый результат - но так исключаются из анализа сложные природные
системы, например, человек (поди, создай для него сто раз одинаковые усло-
вия) .
Вам не кажется, что по мере углубления в вопрос он становится все более ин-
тересным и все менее пугающим? И про техношок мы уже забыли, и сердчишко
бьется ровно... Это и есть единственная реакция на сложность этого мира, за
которую не должно быть стыдно мыслящему существу.
P.S. А началось-то все с мелочи. У Ст. Лема есть такой образ - «черная ту-
ча» . Весьма сложно ведущая себя система из элементов одного типа. Вопрос -
возможно ли такое?
Кстати - это сложный вопрос. У него, похоже, есть структура...
Литпортал
РЕТРОГЕНЕТИКА
Кир Булычев
Славный майский день завершился небольшой образцово-показательной грозой с
несколькими яркими молниями, жестяным нестрашным громом, пятиминутным ливнем
и приятной свежестью в воздухе, напоенном запахом сирени. Районный центр Ве-
ликий Гусляр нежился в этой свежести и запахах.
Пенсионер Николай Ложкин вышел на курчавый от молодой зелени, чистый и даже
кокетливый по весне двор с большой книгой в руках. По двору гулял плотный лы-
сый мужчина - профессор Лев Христофорович Минц, который приехал в тихий Гус-
ляр для поправки здоровья, подорванного напряженной научной деятельностью.
Николай Ложкин любил побеседовать с профессором на умственные темы, даже
порой поспорить, так как сам считал себя знатоком природы.
- Чем увлекаетесь? - спросил профессор. - Что за книгу вы так любовно при-
жимаете к груди?
- Увлекся антропологией, - сказал Ложкин. - Интересуюсь проблемой происхож-
дения человека от обезьяны.
- Ну и как, что-нибудь новенькое?
- Боюсь, что наука в тупике, - пожаловался Ложкин. - Сколько всего откопа-
ли, а до главного не докопались: как, где и когда обезьяна превратилась в че-
ловека .
- Да, момент этот уловить трудно, - согласился Лев Христофорович. - Может
быть, его и не было?
- Должен быть, - убежденно ответил Ложкин. - Не могло не быть такого момен-
та. Ведь что получается? Выкопают где-нибудь в Индонезии или Африке отдельный
доисторический зуб и гадают: человек его обронил или обезьяна. Один скажет -
"человек". И назовет этого человека, скажем, древнеантропом. А другой погля-
дит на тот же зуб и отвечает: "Нет, это зуб обезьяний и принадлежал он, ко-
нечно, древнепитеку". Казалось бы, какая разница, - никто не знает! А разница
в принципе!
Минц наклонил умную лысую голову, скрестил руки на тугом, обтянутом пиджа-
ком животе и спросил строго:
- И что же вы предлагаете?
- Ума не приложу, - сознался Ложкин. - Надо бы туда заглянуть. Но как? Ведь
путешествие во времени вроде бы невозможно.
- Совершенная чепуха, - ответил Минц. - Я пытался сконструировать машину
времени, забрался во вчерашний день и там остался.
- Не может быть! - воскликнул Ложкин. - Так и не вернулись?
- Так и не вернулся, - сказал Минц.
- А как же я вас наблюдаю?
- Ошибка зрения. Что для вас сегодня, для меня вчерашний день, - загадочно
ответил Минц.
- Значит, никакой надежды? Профессор глубоко задумался и ничего не ответил.
Дня через три профессор встретил Ложкина на улице.
- Послушайте, Ложкин, - сказал он. - Я вам очень благодарен.
- За что? - удивился Ложкин.
- За грандиозную идею.
- Что же, - ответил Ложкин, который не страдал излишней скромностью.
Пользуйтесь, мне не жалко.
- Вы открыли новое направление в биологии!
- Какое же? - поинтересовался Ложкин.
- Вы открыли генетику наоборот.
- Поясните, - сказал Ложкин ученым голосом.
- Помните нашу беседу о недостающем звене, о происхождении человека?
- Как же не помнить.
- И ваше желание заглянуть во мглу веков, чтобы отыскать момент превращения
обезьяны в человека?
- Помню.
- Тогда я задумался: что такое жизнь на Земле? И сам себе ответил: непре-
рывная цепь генетических изменений. Вот среди амеб появился счастливый му-
тант , он быстрее других плавал в первобытном океане или глотка у него была
шире... От него пошло прожорливое и шустрое потомство. Встретился внук этой
амебы с жуткой хищной амебихой - вот и еще шаг в эволюции. И так далее,
вплоть до человека. Улавливаете связь времен?
- Улавливаю, - ответил Ложкин и добавил: - В беседе со мной нет нужды при-
бегать к упрощениям.
- Хорошо. Мы, люди, активно вмешиваемся в этот процесс. Мы подглядели, как
это делает природа, и продолжаем за нее скрещивание, отбор, создаем новые
сорта пшеницы, продолжаем эволюцию собственными руками.
- Продолжаем, - согласился Ложкин. - Хочу на досуге вывести быстрорастущий
забор.
- Молодец. Всегда у вас свежая идея. Так вот, после беседы с вами я заду-
мался, а всегда ли правильно мы следуем за природой? Природа слепа. Она знает
лишь один путь - вперед, независимо от того, хорош он или плох.
- Путь вперед всегда прогрессивен, - заметил Ложкин.
- Тонкое наблюдение. А если нарушить порядок? Если все перевернуть? Вы ска-
зали: как бы увидеть недостающее звено? Отвечаю - распутать цепь наследствен-
ности. Прокрутить эволюцию наоборот. Углубляясь в историю, добраться до ее
истоков.
- Нам и без этого дел хватает, - возразил Ложкин.
- А перспективы? - спросил профессор, наклонив голову и прищурившись.
- Это не перспективы, а ретроспективы, - сказал Ложкин.
- Великолепно! - воскликнул Минц. - Чем пользуется генетика? Скрещиванием,
и отбором. Нашу с вами новую науку мы назовем ретрогенетикой. Ретрогенетика
будет пользоваться раскрещиванием, открещиванием и разбором. Генетика будет
выводить новую породу овец, которой еще нет, а ретрогенетика - ту породу, ко-
торой уже нет. И ученым не надо будет копаться в земле. Заказал палеонтолог в
лаборатории: выведите мне первого неандертальца, хочу поглядеть, как он вы-
глядел . Ему отвечают: будет сделано.
- Слабое место, - заявил Ложкин.
- Слабое место? У меня?
- Ваш неандерталец жил миллион лет назад. Вы что же, собираетесь миллион
лет ждать, пока его снова выведете?
- Слушайте, Ложкин. Если бы мы отдавались на милость природе, то сорта пше-
ницы, которые колосятся на колхозных полях, вывелись бы сами по себе через
миллион лет. А может, и не выведись бы, потому что природе они не нужны.
- Ну, не миллион лет, так тысячу, - не сдавался Ложкин. - Пока ваш неандер-
талец родится, да еще своих предков народит...
- Нет, нет и еще раз нет, - сказал профессор. - Зачем же нам реализовывать
все поколения? В каждой клетке закодирована ее история. Все будет, дорогой
друг, на молекулярном уровне, как учит академик Энгельгардт.
- Ну ладно, выведете вы, что было раньше. А что дальше? Какая польза от
этого народному хозяйству?
Ответ на свой вопрос Ложкин получил через три месяца, когда пожелтели липы
в городском саду и дети вернулись из пионерских лагерей.
Лев Христофорович стоял у ворот и чего-то ждал, когда Ложкин, возвращаясь
из магазина с кефиром, увидел его.
- Как успехи? - поинтересовался он. - Когда увидим живого неандертальца?
- Мы его не увидим, - отрезал профессор. Он осунулся за последние недели:
видно, много "было умственной работы. - Есть более важные проблемы.
- Какие же?
- Вы знакомы с Иваном Сидоровичем Хатой?
- Не приходилось, - сказал Ложкии.
- Достойный человек, заведующий фермой нашего пригородного хозяйства "Гус-
лярец". Зоотехник, смелый, рискованный. Большой души человек.
Тут в ворота въехал газик, из которого выскочил шустрый очкастый человечек
большой души.
- Поехали? - предложил он, поздоровавшись.
- С нами Ложкин, - сказал Минц. - Представитель общественности. Пора обще-
ственность знакомить.
- Не рано ли? - обеспокоился Хата. - Спугнут...
- Нам ли опасаться гласности? - спросил Минц. После короткого путешествия
газик достиг животноводческой фермы. Рядом с коровником стоял новый высокий
сарай.
- Ну что же, Заходите, только халат наденьте. Хата выдал Ложкину и Минцу
халаты и сам тоже облачился. Ложкин ощутил покалывание в желудке и пригото-
вился увидеть что-нибудь необычное. Может, даже страшное. Но ничего страшного
не увидел.
Под потолком горело несколько ярких ламп, освещая кучку мохнатых животных,
жевавших сено в дальнем углу.
Ложкин присмотрелся. Животные были странными, таких ему раньше видеть не
приходилось. Они были покрыты длинной рыжей шерстью, носы у них были длинные,
ноги толстые, как столбы. При виде вошедших людей животные перестали жевать и
уставились на них маленькими черными глазками. И вдруг захрюкали, заревели и
со всех ног бросились навстречу Хате и Минцу, чуть не сшибли их, ластились,
неуклюже прыгали, а профессор начал доставать из карманов халата куски сахара
и угощать животных.
- Что за звери? - спросил Ложкин, отошедший к стенке, подальше от суматохи.
- Почему не знаю?
- Не догадались? - удивился Хата. - Мамонтята. Каждому ясно.
- Мне не ясно, - сказал Ложкин, отступая перед мамонтенком, который тянул к
нему недоразвитый хоботок, требуя угощения. - Где бивни, где хоботы? Почему
мелкий размер?
- Все будет, - успокоил Ложкина Минц, оттаскивая мамонтенка За короткий
хвостик, чтобы не приставал к гостю. - Все с возрастом отрастет. Ваше удивле-
ние мне понятно, потому что вам не приходилось еще сталкиваться с юными пред-
ставителями этого славного рода.
- Я и со старыми не сталкивался, - возразил Ложкин. - И прожил, не жалуюсь.
Откуда вы их откопали?
Неужели не догадались? Они же выведены методом ретрогенетики
раскрещиванием и разбором. Из слона мы получили предка слонов и мамонтов
близкого к мастодонтам. Потом люди пошли обратно и вывели мамонта.
- Так быстро?
- На молекулярном уровне, Ложкин, на молекулярном уровне. Под электронным
микроскопом. Методом раскрещивания, открещиваиия и разбора. И вы понимаете
теперь, почему я отказался от соблазнительной идеи отыскать недостающее зве-
но, а занялся мамонтами?
- Не понимаю, - сказал Ложкин.
- Вы, товарищ, видно, далеки от проблем животноводства, - вмешался Иван Ха-
та, - Ни черта не понимаете, а критикуете. Нам мамонт совершенно необходим.
Для нашей природной зоны.
- Жили без мамонта и прожили бы еще, - упорствовал Ложкин.
- Эх, товарищ Ложкин, - в голосе Хаты звучало сострадание. - Вы когда-
нибудь думали, что мы имеем с мамонта?
- Не думал. Не было у меня мамонта.
- С мамонта мы имеем шерсть. С мамонта мы имеем питательное мясо, калорий-
ное молоко и даже мамонтовую кость...
- Но главное, - воскликнул Минц, - бесстойловое содержание! Круглый год на
открытом воздухе, ни тебе утепленных коровников, ни специальной пищи. А поду-
майте о труднодоступных районах Крайнего Севера: мамонт там - незаменимое
транспортное средство для геологов и изыскателей.
Прошло еще три месяца.
Однажды к дому № 16 по Пушкинской, где проживал Лев Христофорович, подъеха-
ла сизая "волга", из которой вышел скромный на вид человек средних лет в дуб-
ленке. Он вынул изо рта трубку, поправил массивные очки, снисходительно огля-
дел непритязательный двор, и его взгляд остановился на Ксении Удаловой, кото-
рая развешивала белье:
- Скажите, гражданка, если меня не ввели в заблуждение...
- Вы корреспондент будете? - спросила Ксения.
- Вот именно. Из Москвы. А как вы догадались?
- А чего не догадаться, - ответила Ксения. - Восемнадцатый за неделю. Под-
нимитесь на второй этаж, дверь открыта. Лев Христофорович отдыхает.
Поднимаясь по скрипучей лестнице в скромную обитель великого профессора,
журналист бормотал: "Шарлатанство. Ясно, шарлатанство. Вводят в заблуждение
общественность..."
- Заходите, - откликнулся на стук профессор Минц. Он в тот момент отдыхал,
а именно: читал "Химию и жизнь", слушал последние известия по радио, смотрел
хоккей по телевизору, гладил брюки и думал.
- Из Москвы. Журналист, - сказал гость, протягивая удостоверение. - Это вы
тут мамонтов разводите?
Журналист произнес это таким тоном, словно подразумевал: "Это вы водите за
нос общественность?"
- И мамонтов, - скромно ответил профессор, прислушиваясь к сообщениям из
Канберры и радуясь мастерству лучшего в сезоне хоккеиста.
- С помощью... - журналист извлек из замшевого кармана записную книжку, -
ретро, простите, генетики?
Доверчивый Минц не уловил иронии в голосе журналиста.
- Именно так, - подтвердил он и набрал из стакана в рот воды, чтобы обрыз-
гать брюки.
- И есть результаты?
Минц провел раскаленным утюгом по складке, поднялось облако пара.
- С этим надо что-то делать, - сказал Минц. Он имел в виду брюки и ситуацию
в Австралии.
- И все-таки, - настаивал журналист. - Можно взглянуть на ваших мамонтов?
- А почему бы и нет? Они в поле пасутся. Добывают корм из-под снега.
- Ясно. А еще каких-нибудь животных вы можете вывести?
- Будете проходить мимо речки, - сказал Минц, - поглядите в полынью. Там
бронтозавры. Думаем потом отправить их в Среднюю Азию для расчистки ирригаци-
онных сооружений.
В этот момент в окно постучала длинным, усеянным острыми зубами клювом об-
разина . Крылья у образины были перепончатые, как у летучей мыши. Образина
гаркнула так, что зазвенели стекла и форточка сама собой открылась.
- Не может быть! - сказал журналист, отступая к стене. - Это что такое? Ма-
монт?
- Мамонт? Нет, это Фомка. Фомка - птеродактиль. Когда вырастет, размахнет
свои крылья на восемь метров.
Минц отыскал под столом пакет с тресковым филе, подошел к форточке и бросил
пакет в разинутый клюв образине. Птеродактиль подхватил пакет и заглотнул, не
разворачивая.
- Зачем вам птеродактиль? - спросил журналист. - Только людей пугать.
Он был уже не так скептически настроен, как в первый момент.
- Как зачем? Птеродактили нам позарез нужны. Из их крыльев мы будем делать
плащи-болоньи, парашюты, зонтики, наконец. К тому же научим их пасти овец и
охранять стада от волков.
- От волков? Ну да, конечно... - Журналист прекратил расспросы и вскоре
удалился.
"Возможно, это, до определенной степени, и не шарлатанство, - думал он,
спускаясь по лестнице к своей машине, - но, по большому счету, это все-таки
шарлатанство".
Весь день до обеда корреспондент ездил по городу, издали наблюдал за играми
молодых мамонтов, недовольно морщился, когда на него падала тень пролетающего
птеродактиля, и вздрагивал, заслышав рев пещерного медвежонка.
- Нет, не шарлатанство, - повторял он упрямо. - Но кое в чем хуже, чем шар-
латанство .
Весной в журнале, где состоял тот корреспондент, появилась статья под суро-
вым заглавием: "ПЛОДЫ ЛЕГКОМЫСЛИЯ".
Нет смысла передавать опасения и измышления гостя. Он предупреждал, что но-
вые звери нарушат и без того неустойчивый экологический баланс, что пещерные
медведи и мамонты представляют опасность для детей и взрослых. А в заключение
журналист развернул страшную картину перспектив ретрогенетики:
"Безответственность периферийного ученого и пошедших у него на поводу прак-
тических работников гуслярского животноводства заставляет меня бить тревогу.
Эксперимент, не проверенный на мелких и безобидных тварях (жуках, кроликах и
т.д.), наверняка приведет к плачевным результатам. Где гарантия тому, что ма-
монты не взбесятся и не потопчут зеленые насаждения? Что они не убегут в ле-
са? Где гарантия тому, что бронтозавры не выползут на берег и не отправятся
на поиски новых водоемов? Представьте себе этих рептилий, ползущих по улицам,
сносящих столбы и киоски. Я убежден, что птеродактили, вместо того, чтобы
пасти овец и жертвовать крыльями на изготовление зонтиков, начнут охотиться
на домашнюю птицу, а может быть, на тех же овец. И все кончится тем, что на
ликвидацию последствий непродуманного эксперимента придется мобилизовать тру-
дящихся и тратить народные средства..."
Статья попалась на глаза профессору Минцу лишь летом. Читая ее, профессор
лукаво улыбался, а потом захватил журнал с собой на открытие межрайонной вы-
ставки .
Центром выставки, как и следовало, предполагать, был павильон "Ретро-
генетика". Именно сюда спешили люди со всех сторон, из других городов, облас-
тей и государств.
Пробившись сквозь интернациональную толпу к павильону, Лев Христофорович
оказался у вольеры, где гуляли мамонты.
Было жарко, поэтому мамонты были коротко острижены и казались поджарыми,
словно собаки породы эрдельтерьер. У некоторых уже прорезались бивни. Птеро-
дактили сидели у них на спинах и выклевывали паразитов. В круглом бассейне
посреди павильона плавали два бронтозавра. Время от времени они тяжело подни-
мались на задние лапы и, прижимая передние к блестящей груди, выпрашивали у
зрителей плюшки. У кого из зрителей не было плюшки, кидали пятаки.
Здесь, между вольерой и бассейном, Минц увидел Ложкина и Хату и прочел
друзьям скептическую статью.
Смеялись не только люди. Булькали от хохота бронтозавры, трубили мамонты, а
один птеродактиль так расхохотался, что не мог закрыть пасть, пока не прибе-
жал служитель и не стукнул весельчаку как следует деревянным молотком по ниж-
ней челюсти.
- Неужели, - сказал профессор, когда все отсмеялись, - этот наивный человек
полагает, что мы стали бы выводить вымерших чудовищ, если бы не привили им
генетически любви и уважения к человеку?
- Никогда, - отрезал Ложкин. - Ни в коем случае. Птеродактиль, все еще
вздрагивая от смеха, стуча когтями по полу, подошел к профессору, и тот уго-
стил его конфетой. Маленькие дети по очереди катались верхом на мамонтах,
подложив под попки подушечки, чтобы не колола остриженная жесткая шерсть.
Бронтозавры собирали со дна бассейна монетки и честно передавали их служите-
лям. В стороне скулил пещерный медведь, потому что его с утра никто не при-
ласкал .
...В тот день столичного журналиста, неудачливого пророка, до полусмерти
искусала его домашняя сиамская кошка.
Гостиная
У нас в гостях
журнал «Компьютерра1»
РАСШИФРОВКА ПРОШЛОГО
Юрий Ильин
"Чудища" из прошлого, которого человечество не может помнить, для большин-
ства людей не являют собой ничего более картинок в школьных учебниках, кон-
статации факта, что да, были и такие твари на Земле, но теперь их нет. И мо-
жет , хорошо, что нет: едва ли благочинного обывателя порадовало бы соседство
с тиранозаврами, - даже на одной планете.
Какими были вымершие животные, и почему они погибли? Кроме динозавров, ко-
торые исчезли задолго до появления человечества, были существа, сосущество-
вавшие с людьми. Что случилось с мамонтами или, к примеру, нашими собственны-
ми непонятными родственниками - неандертальцами? Действительно ли они оказа-
лись тупиковой ветвью, или мы соседствуем с их прямыми или не очень потомка-
ми?
Над расшифровкой геномов шерстистых мамонтов и неандертальцев учёные кро-
потливо работают и даже добились некоторых результатов. А вот с динозавра-
ми. .. В обозримом будущем материализации "Парка Юрского периода" можно не
ждать. Хотя...
1 Из той части, что Терра (т.е. Земля и, следовательно, ее обитатели, некоторые из которых в
результате эволюции научились пользоваться компьютером).
Эластичные ткани
Ранней весной 2005 года палеонтологи из Университета Северной Каролины на-
делали шуму известием, что им удалось извлечь из окаменелых, казалось бы,
костей тиранозавра, фрагменты эластичных тканей.
Палеонтолог Мэри Швайцер и её коллеги с помощью специального состава, рас-
творяющего содержащие кальций структуры, выделили фрагменты мягкой, гибкой
структуры, напоминающей кровеносные сосуды. Мало того, их строение чрезвычай-
но сильно (просто-таки до идентичности) напоминало кровеносные сосуды совре-
менных страусов, что в очередной раз доказывает высокую степень родства со-
временных птиц с древними ящерами.
Спустя несколько месяцев та же команда палеонтологов сообщила о ещё более
интересной подробности. Оказалось, что те самые недоокаменелые останки при-
надлежало молодой самке тиранозавра, погибшей в период вызревания яиц.
По мнению исследователей, та эластичная ткань представляла собой тонкий
слой так называемой медуллярной ткани - пронизанной сосудами костной ткани,
который встречается в полых костях ног у самок современных птиц в период ову-
ляции. Когда последнее вызревшее яйцо отложено, она полностью рассасывается в
организме.
По словам Мэри Швайцер, медуллярная ткань, извлечённая из окаменелых остан-
ков тиранозавра, и фрагменты такой ткани у современных страусов оказались
"практически идентичными по структуре, расположению и даже цвету". Это, веро-
ятно, отчасти отвечает на вопрос, кто сегодня на самом деле является ближай-
шими родственниками динозавров.
Когда стало известно об извлечении эластичной ткани, естественно встал во-
прос: уцелели ли в ней белки? Удастся ли их извлечь? Если это произойдёт, то
можно получить целую ДНК тиранозавра?
Похоже, что сделать это, увы, пока не удалось. По крайней мере, никаких но-
востей на этот счёт с прошлого года не поступало. И, скорее всего, это вообще
невозможно. Вероятность обнаружить ткани, где белки не замещены минералами
после миллионов лет лежания в земле, стремится к нулю.
А вот с шерстистым мамонтом всё обещало быть намного интереснее...
Дело о мамонте
В 1986 году на Чукотке из вечной мерзлоты выкопали неплохо сохранившиеся
останки шерстистого мамонта. Радиоуглеродный анализ показал, что им приблизи-
тельно 33 тысячи лет.
В начале февраля 2006 года международная группа генетиков во главе с про-
фессором Евгением Рогаевым объявила, что им удалось полностью расшифровать
митохондриальную ДНК шерстистого мамонта, основываясь как раз на том материа-
ле , который был извлечён ещё двадцать лет назад на Чукотке.
Несколькими месяцами ранее немецкие генетики отрапортовали о том, что они
смогли расшифровать фрагменты митохондриальной ДНК мамонта. Учёные использо-
вали крошечный образец мамонтовой кости (массой всего 200 мг) из кладбища ма-
монтов у реки Бёрёлёх в Якутии. Им удалось извлечь, расшифровать и затем объ-
единить 23 фрагмента митохондриальной ДНК мамонта.
Расшифровать фрагменты митохондриальной ДНК удавалось и ранее, но реконст-
руировать всю последовательность длиной 16 842 нуклеотидов - секвенсировать,
удалось только профессору Рогаеву.
С помощью митохондриальной ДНК клонировать живое существо невозможно, а вот
для эволюционных реконструкций она оказывается как нельзя кстати: митохондри-
альная ДНК передается только по материнской линии и не рекомбинирует, то есть
отцовские гены не перемешиваются с материнскими.
Благодаря расшифровке удалось установить, что популяция мамонтов, распро-
странившаяся по всей Сибири, была в генетическом плане очень однородна. Рога-
ев и его коллеги также пришли к выводу, что генетически сибирские мамонты
наиболее близки к индийским слонам, - хотя и отличаются от них довольно суще-
ственно . Общим предком был африканский слон, а разделение последующих ветвей
произошло, по мнению команды Рогаева, где-то около 4 миллионов лет назад. Его
немецкие коллеги называли несколько иной срок - 5,5 миллионов лет назад.
Между тем, из останков мамонтов удалось извлечь не только митохондриальную,
но и ядерную ДНК. Её расшифровкой активно занимались Хендрик Пойнар и Рос
Макфи - сотрудники Центра изучения ДНК древних животных при Университете Мак-
мастера в Гамильтоне (штат Онтарио, США) и Американского музея естественной
истории в Нью-Йорке. С помощью системы Genome Sequencer 20 System они секвен-
сировали 28 миллионов базовых пар из фрагментов ДНК, содержавшихся в одном
грамме костной ткани мамонта.
Когда полученные результаты сравнили с 13 миллионами базовых пар генома аф-
риканского слона, выяснилось, что совпадает около 98,55% генома. Это лишний
раз подтверждает, что разделение видов случилось приблизительно 5 миллионов
лет назад.
В октябре уже этого года Пойнар сообщил на собрании Общества палеонтологии
позвоночных в Оттаве, что его соратникам удалось провести несколько дополни-
тельных сеансов анализа с использованием Genome Sequencer 20 и проанализиро-
вать ещё 20 миллионов базовых пар.
В теории, результаты этого проекта должны были бы позволить клонировать ма-
монта и воскресить сгинувший биологический вид. Однако недавно стало извест-
но, что у проекта банально не хватает средств на "продолжение банкета". Для
того чтобы оплатить дальнейшую работу на Genome Sequencer 20 им срочно требу-
ются 500 тысяч долларов, которых у учёных нет.
Неандертальцы у ворот?
Школьные учебники биологии гласят, что на сегодняшний день Землю населяет
один вид Человека разумного. Сколь бы сильно ни отличались друг от друга
внешне представители разных рас, все шесть с лишним миллиардов людей, насе-
ляющих планету, относятся к одному и тому же биологическому виду, и генетиче-
ское разнообразие оказывается меньшим, чем у разных популяций шимпанзе, насе-
ляющих один только Африканский континент. По мнению генетиков, это означает,
что около 70 тысяч лет назад общая численность представителей Homo sapiens
составляла не более 2000 "голов".
В те времена вид Ното sapiens был ещё не единственным представителем рода
Ното. На сегодняшний день главными "конкурентами" считают неандертальцев -
Homo neanderthalis. Они соседствовали с Человеком разумным, но полностью ис-
чезли с лица Земли 24-33 тысяч лет назад.
Причины их исчезновения, как и в случае с мамонтами, неизвестны, хотя на
этот счёт есть немало гипотез. Одна из гипотез состоит в том, что Ното
sapiens оказались более приспособленными к окружающей среде, более способными
к разделению труда, и, наконец, производили более эффективный инструментарий
и оружие. В итоге люди выжили неандертальцев из пригодных для проживания
мест. Согласно другой известной гипотезе, неандертальцы и люди просто слились
в один вид.
Генетически Homo neanderthalis и Homo sapiens идентичны примерно на 99,95%,
указывает Эдвард Рубин, сотрудник Объединённого института генетических иссле-
дований при Министерстве энергетики США, возглавляющий один из проектов по
расшифровке фрагментов ДНК неандертальца.
Сходство, казалось бы, колоссальное, однако, например, и с шимпанзе у нас
весьма много общего: 98 с лишним процентов одинаковых генов. Все особенности
Человека разумного заключаются в той самой "крошечной разнице".
Полпроцента разницы - это, по всей видимости, куда как существенно. По сло-
вам Рубина, никаких признаков того, что в последние примерно 30-40 тысяч лет
имело место межвидовое скрещивание, сколько-нибудь повлиявшее на генофонд
Homo sapiens или Homo neanderthalis, увидеть пока не удаётся. "Мы не исключа-
ем этого, но признаков скрещивания не видим", - говорит он.
Коллега Рубина, Сванте Паабо из Института эволюционной антропологии имени
Макса Планка - один из первых в мире исследователей, занимавшихся расшифров-
кой ДНК неандертальцев, согласен с тем, что неандертальцы и Homo sapiens друг
с другом не скрещивались. Паабо и его сотрудники используют для расшифровки
ДНК неандертальцев примерно такой же аппарат, как тот, что использовали Хенд-
рик Пойнар и его коллеги для секвенсирования ДНК мамонта. Через два года они
рассчитывают полностью отследить геном неандертальца.
Учёных интересует, что именно сделало нас "людьми разумными"? И хотя все
отдают себе отчёт в том, что со всей степенью точности восстановить последо-
вательность исторических событий не удастся, само понимение, какие именно ге-
ны и когда изменились, и что могло стать причиной их изменения, позволит че-
ловечеству, наконец, более-менее точно установить собственное происхождение.
По крайней мере, биологическое.
Пока тайна не разгадана. И если даже это произойдёт, возникают некоторые
вопросы, как это может повлиять на самосознание человечества, и повлияет ли
вообще. Ведь, в общем и целом, западный мир более-менее свыкся с тем, что
"человек произошёл от обезьяны", даром, что ничего такого Дарвин, в сущности,
и не говорил, а лишь только называл обезьяну нашим ближайшим родичем... В то
же время растёт число тех, кто гневливо потрясая хоругвями, громогласно во-
прошает у окружающих, что им больше по душе, быть "потомками обезьяны" или
"образом и подобием божьим". И хотя речь, в сущности, совсем о другом, опре-
деление тех самых генов, которым мы обязаны наличием у нас разума, так уж ли
оно обрадует кого-то, кроме самих учёных?
НЕАНДЕРТАЛЬЦЫ СНОВА
ВЫЙДУТ НА МАМОНТОВ?
Дмитрий Шабанов
Главной идеей Николая Федорова, одного из основателей русского космизма,
была петрификация - "воскрешение отцов" (умерших поколений людей). Федоров не
знал, как естественные науки смогут решить эту задачу, но не сомневался, что
они что-то придумают. Благодаря научному чуду, человечество выполнит свое
предназначение, и Бог воплотится в нем. Не надо иронизировать над Федоровым -
им двигало острое ощущение несправедливости того, что со смертью человека
уходит целый мир.
В начале XXI века идеи наподобие федоровских реализуются синтетической ге-
номикой , которая достигла определенных промежуточных успехов. Конечно, пока
энтузиастов воскрешения интересуют не конкретные люди, а целые виды. Когда-то
распространенная вера, что сущность личности человека исчерпывается его гено-
типом, теперь стала чем-то вроде веры в плоскую Землю. Увы, представления о
том, что сущность вида можно выразить на языке четырех букв генетического ал-
фавита, пока вполне жизнеспособны (хотя имеют столь же ненадежные научные ос-
нования) .
Каковы же нынешние успехи синтетической геномики? Недавно исследовательский
центр Крейга Вентера ["КТ" уже упоминала этого мультимиллионера, одного из
лидеров расшифровки генома человека (в значительной части - своего собствен-
ного) : "Трудно быть богом" ("КТ" #599-600)] сообщил о пересадке бактериально-
го генома. Речь идет о двух видах [Надо понимать, что слово "вид" использует-
ся здесь достаточно условно. По крайней мере, вид у микоплазм - совсем не то,
что, к примеру, вид у млекопитающих] микоплазм - одних из самых простых кле-
точных организмов, известных науке. Геном Mycoplasma mycoides (возбудителя
пневмонии коров) был перенесен в культуру клеток бактерии Mycoplasma
capricolum (вызывающей полиартрит у коз) . Через некоторое время в этой куль-
туре появлялись клетки вида-донора. Вероятно, микоплазма поглощала чужую хро-
мосому (у этих бактерий нет клеточной стенки), а затем делилась и передавала
ее одной из дочерних клеток. Следующий запланированный шаг - пересадка искус-
ственной хромосомы, которую Вентер приравнивает к созданию искусственной жиз-
ни .
Что получается с бактериями, получится и с млекопитающими? Специалисты из
Института Макса Планка в Лейпциге описали, какие именно нарушения происходят
в ископаемой ДНК - например, ДНК неандертальца. Зная правила изменения гене-
тического текста, можно будет определить его исходный вид. Например, деталь-
ное сравнение наследственной информации Homo neandertalensis и Homo sapiens
должно помочь определить те белки, с изменением которых связано наше эволюци-
онное становление. Это может быть важно, например, для медицины - можно соз-
дать управляющие этими белками лекарства.
Лекарства лекарствами, но энтузиасты и в Европе, и в Америке увидели в этой
ситуации шанс на восстановление недавно вымерших видов. Соберем полные геномы
мамонтов и неандертальцев, поместим их в яйцеклетки слона и современного че-
ловека (соответственно), и, глядишь, воскрешенные неандертальцы снова смогут
охотиться на воскрешенных мамонтов! Дает ли перестановка геномов бактерий
шанс на такую перспективу?
Большинству из грамотных и культурных людей (а все прочие вообще не замора-
чиваются подобными вопросами) кажется, что механизмы развития должны быть в
главных чертах общими для всех организмов, от бактерий до млекопитающих. Ми-
фология синтетической теории эволюции предусматривает, что эволюция началась
со случайного возникновения механизма наследственности, в котором случайные
ошибки матричного копирования обеспечивают изменчивость, достаточную для то-
го, чтобы из бактерий появились секвойи, киты и люди. Но погодите, эволюция
ведь - способ выработки адаптаций (приспособлений), приведения индивидуально-
го развития в соответствие с возможностями, предоставляемыми внешней средой.
В соревновании за жизнь выигрывали те группы, которые вырабатывали требуемые
адаптации быстрее. С ходом времени количество взаимосвязей между геномом и
клеткой в целом стремительно росло. Клетка "училась" управлять своим геномом,
одной из своих многих подсистем. Наследственная информация - не причина и не
квинтэссенция клетки, а один из ее инструментов для приспособления к среде!
Эволюция эволюции привела к тому, что индивидуальное и историческое разви-
тие бактерий и людей существенно отличается. Сегодня нет никаких надежд по-
вторить на мамонтах или неандертальцах результат, полученный на микоплазмах.
Бактерия - молекулярный робот, а в наших клетках количество взаимосвязей меж-
ду элементами несоизмеримо выше. Чтобы клонировать упомянутые виды, нужны не
их геномы, а их живые клетки, которые неоткуда взять [Хотя на сегодняшнем
уровне развития биологи не смогли бы сладить даже с живой клеткой - клониро-
вание млекопитающих продолжает сталкиваться с серьезными проблемами].
Появится ли когда-то такая возможность? Помните классический пример ошибоч-
ности заявлений об ограниченности науки? В качестве примера фундаментально
недоступного знания Огюст Конт назвал химический состав звезд. Прошло не так
много времени, и был открыт спектральный анализ. Завтра сообщат о фундамен-
тальном прорыве в изучении регуляции онтогенеза? Я первый этому порадуюсь:
мне будет интересно. Впрочем, надежды на такой прорыв у меня мало, ведь слож-
ность рассматриваемых процессов многократно превосходит сложность любых за-
дач, когда бы то ни было решенных человечеством. Оно решит их не сейчас, а
позже? Когда? Путь, который нужно пройти, очень долог. Просуществует ли тре-
буемое время само человечество, будет ли оно практиковать фундаментальные на-
учные исследования?
И подумайте, какие сложности вызовет воскрешение неандертальцев. Как ре-
шить : надо будет их содержать в зоопарках или они будут получать паспорта и
водительские удостоверения?
Без комментариев.
ПОХОД ЗА РАЗУМОМ
Елена Наймарк
В конце XIX века человечество взволнованно обсуждало, действительно ли пре-
док Его Величества Человека - презренная обезьяна. Одним для смиренного при-
знания животных прародителей оказалось достаточно стройной логики Дарвина.
Другим требовалось что-то более осязаемое, нежели логика и косвенные доводы.
В 1893 году голландский антрополог Дюбуа нашел на острове Ява череп, соче-
тающий в себе обезьяньи и человеческие черты. Его обладателя так и назвали
питекантропом, то есть "обезьяночеловеком", а много позже все-таки причислили
к человеческому роду под именем "человек прямоходящий". Вскоре в Германии бы-
ли найдены неандертальцы, а во Франции - кроманьонцы. В 1925 году Раймонд
Дарт откопал в Южной Африке древнейшие останки обезьяноподобного существа с
маленькой черепной коробкой, ходившего на двух ногах и имевшего человеческие
зубы. Это существо получило имя австралопитек - "южная обезьяна". Спустя
треть века Льюис и Мэри Лики в ущелье Олдувай (Танзания) обнаружили останки
двуногого существа, изготовлявшего примитивные каменные орудия из речной
гальки, и назвали его Homo habilis, то есть человек умелый.
Таким образом, необходимые звенья цепи превращений обезьяны в человека были
собраны. Дотошные любители фактических подтверждений были удовлетворены. Все
выстроилось в простую линию: человекообразные обезьяны спустились с деревьев
на землю и стали двуногими австралопитеками. Из-за прямохождения у древних
гоминид освободились руки, что привело к эволюции руки и увеличению мозга - и
вот появился "человек умелый". Он, в свою очередь, эволюционировал, совершен-
ствуя навыки ручной работы и структуру мозга, и мало-помалу превратился в
"человека прямоходящего". Дальнейшая его эволюция привела к образованию охот-
ника-неандертальца и через него человека разумного. (Поначалу неандертальцев
считали предками Homo sapiens, но вскоре их сочли тупиковой ветвью эволюции
Ното.) Человек постепенно становился на ноги и выпрямлялся, умнел и потихонь-
ку учился разговаривать - все в рамках теории естественного отбора и нетороп-
ливого накопления полезных признаков. За столетний период изучения сформиро-
валось совершено определенное представление, кого называть человеком, то есть
относить к роду Ното. По словам Льюиса Лики, человек - это двуногое прямохо-
дящее существо с объемом черепа не меньше 600 см3. Иными словами, австралопи-
теков с когортой их обезьяньих предков оставляем за чертой Ното, а все, что
появилось после австралопитеков, причисляем к людям. Почти все новые находки,
кроме черепа человека с озера Рудольф (Homo rudolfensis), попавшие к ученым в
течение XX века, хорошо укладывались и в теорию постепенного превращения
обезьяны в человека, так что это определение человека было вполне оправдано.
Соответствие теории и фактов было до того успокаивающим, что недоверчивое от-
ношение к эволюционному превращению обезьян в людей считалось либо невежест-
вом, либо социальным эпатажем.
Лавка древностей
В 1974 году антрополог Джохансон откопал в Эфиопии остатки австралопитека,
точнее, австралопитечки, которая жила около 3 млн. лет назад, то есть раньше,
чем все ее известные сородичи. Именно эта древняя дама, названная Люси, была
признана тогда нашей древнейшей праматерью. Через двадцать лет (1995) в Кении
Мив Лики обнаружила еще более древних австралопитеков, возрастом 4 млн. лет.
Ни находка Люси, ни обнаружение ее предков не нарушили линейной человеческой
истории, только "удревнили" ее корни.
Но вот в конце XX века появились сообщения о двух новых
видах австралопитеков - Australopithecus garhi и А.
bahrelghazali, которые жили около 2,5 млн. лет назад - од-
новременно с другими видами австралопитеков. Теперь уже
невозможно было выстроить линейную цепочку от самого древ-
него и примитивного австралопитека к "самому двуногому и
рукастому" человеку. Ведь не меньше четырех видов древних
гоминид претендовали на роль предков человека умелого. Все
запуталось.
Еще большую путаницу внесли находки нескольких фрагментов костей, извлечен-
ные на свет божий тремя годами позже все тем же кланом Лики. Древние кости
лежали в слоях вулканического пепла возрастом 3,5 млн. лет. Принадлежали они
человекоподобному существу, ходившему на двух конечностях, с небольшой череп-
ной коробкой, но плоским лицом и мелкими зубами. По всем признакам это был не
австралопитек, но и не Ното. Тогда кто же он и какое отношение имеет к пре-
вращению обезьяны в человека? Лики назвали новоявленного члена человеческой
семьи кениантропом плосколицым, подыскав ему среди ранее известных загадочных
черепов близкого потомка - Homo rudolfensis. Кениантроп по ряду признаков
больше походил на представителей рода Ното, чем австралопитеки. Перед антро-
пологами встал вопрос, с каким родом следует соотносить кениантропа и от кого
вести род Ното: от австралопитеков или кениантропов? Если непосредственными
предками человеческого рода считать кениантропов, то его прямым потомком бу-
дет Homo rudolfensis; тогда в стороне от дел останутся человек умелый и позд-
ние австралопитеки, от которых раньше тянули линию людей. Если же оставить
предками человека австралопитеков, то куда девать ветвь кениантропов?
Пока ученые гадали, какие перестановки сделать в эволюционном ряду древних
гоминид, Мартин Пикфорд отправился в Кению За новым материалом. И вот среди
холмов Туджен в 2000 году экспедиция обнаруживает остатки нового, доселе не-
известного представителя гоминид. По характерным особенностям скелета ученые
установили, что он обладал небольшим мозгом, но вполне человеческими зубами и
довольно уверенно ходил на двух ногах. По этим признакам существо следовало
бы отнести к непосредственным предкам Ното. Но возраст этих существ составлял
6 млн. лет! Они жили на 2 млн. лет раньше австралопитеков и на 2,5 млн. лет
раньше кениантропа! Orrorin tugenensis - такое ученое название получило это
существо, а публика дала ему пышное прозвище - "Millennium Man" - человек ты-
сячелетия .
В том же году появились сообщения еще об одном представителе гоминид - ар-
дипитеке кадабба, найденном международной антропологической командой в Эфио-
пии, в слоях с датировками 5,8-5,7 млн. лет, то есть чуть моложе оррорина.
Его имя говорит само за себя: "кадабба" на местном афарском языке означает
"основатель рода". Как и оррорин, кадабба обитал в лесистой местности, но хо-
дил на двух ногах, и клыки у него были маленькие, не обезьяньи.
Вскоре на юге Сахары, в пустыне Дьюраб, был найден еще один древнейший пра-
человек - сахелантроп. Превосходно сохранившийся череп одной особи и фрагмен-
тарные остатки еще пяти особей позволили воспроизвести облик сахелантропа:
относительно плоское лицо, маленькая черепная коробка объемом не больше 350
см3, вытянутая в затылочной части наподобие обезьяньей, и вместе с тем непо-
хожие на обезьяньи слабые клыки. Он жил около большого озера, бродил по его
берегам на двух ногах. Это самый древний из известных гоминид, живший 6-7
млн. лет назад. Слава открытия древнейшего прародителя разделилась по крупи-
цам: сахелантропа описала целая международная команда из 38 ученых. И пра-
вильно: пусть древнейший человек принадлежит всему человечеству.
Последняя сенсационная антропологическая находка - человек с острова Фло-
рес. На этом маленьком островке на востоке Индонезии ученые обнаружили пре-
красно сохранившиеся кости крошечного существа, ростом с пятилетнего ребенка,
однако по всем анатомическим признакам вполне взрослого. Телосложением он на-
поминал уменьшенных питекантропов с объемом мозга около 380 см3, без шерсти-
стого покрова и с плоским лицом, на котором выступали вперед челюсти с круп-
ными зубами. Он пользовался отнюдь не примитивными каменными орудиями и охо-
тился на местных карликовых слонов стегодонов. Эти люди, названные Ното
floresiensis, но немедленно получившие кличку "хоббиты", жили 38-18 тысяч лет
назад. Получается, что они были современниками и неандертальцев, и кроманьон-
цев... Предполагается, что 850 тысяч лет назад одна из ветвей "человека прямо-
ходящего" - питекантропа - во время миграции в Азию попала на остров. После
этого их эволюция шла изолированно, что привело к появлению мелкого островно-
го вида. Для многих островных видов самых разных животных и растений харак-
терны более мелкие размеры, чем у их континентальных прародителей. Правда,
находятся и скептики, утверждающие, что остатки "хоббитов" принадлежат урод-
ливым или больным особям сапиенсов. Дабы установить истину, ученые продолжают
раскопки в ожидании нового материала.
Согласно данным молекулярной биологии, линии людей и человекообразных
обезьян разошлись около 6-6,5 млн. лет назад. Датировки находок сахелантропа,
оррорина и кадаббы лежат совсем близко от этого узла. Выходит, что многие че-
ловеческие признаки: двуногость, прямохождение, мелкие зубы, плоское лицо -
появились в эволюционной ветви гоминид почти сразу после размежевания с
обезьянами. И никакого постепенного превращения обезьяны в человека не было.
Были существа, получившие в дар от эволюции по кусочку от этого человеческого
пирога. И дальше они уже развивались обособленно, кто как мог.
Нужно заметить, что эволюция различных групп животных и растений, к которым
эволюционисты, да и широкая общественность относятся не так трепетно, как к
гоминидам, отнюдь не линейна. В начале развития каждой крупной группы живот-
ных и растений появляются представители с различными наборами примитивных и
продвинутых признаков. Признаки перемешиваются почти случайным образом: ран-
ний представитель группы может получить совершенный ротовой аппарат и мало-
пригодные ноги или глаза. А рядом с ним часто обнаруживается и такой, который
обладает совершенными глазами, а пережевывать пищу умеет плохо и передвигает-
ся наподобие древнего червя. Такая картина ученых не удивляет: это скорее
правило, чем исключение. Другое дело, что мозаика признаков пока не имеет
внятного научного объяснения. Судя по новейшим находкам, и человеческую исто-
рию не миновал этот необъяснимый эволюционный рок мозаики признаков.
Так или иначе, мы теперь знаем, что превращение обезьяны в человека вовсе
не было прямым и постепенным, и кандидатов в человеческие предки оказалось
много. На заре человеческой истории целый букет различных видов гоминид начал
эволюционный поход к современному разуму. Подробности этого похода нам пока
неизвестны - известны только его многочисленные и разномастные участники.
Первые пешеходы
Человек, чтобы распрощаться с животным прошлым, должен был подняться во
весь рост. То есть встать на ноги. Сразу же возникают три вопроса: скептика -
"а точно ли?", лентяя - "а зачем?" и инженера - "а как?". И антрополог вынуж-
ден изворачиваться, чтобы на них ответить.
Скептиков, не веривших, что древние люди ходили на двух ногах, а не опира-
лись на передние конечности и не висели на деревьях, убедить было проще все-
го. Тут в арсенале антрополога полно фактов. В 1976 году Мэри Лики обнаружила
на окаменевшем вулканическом пепле отпечатки ног. Возраст слоя с пеплом -
около 3,5 млн. лет, то есть по этому месту в то давнее время бродил на двух
ногах австралопитек. А следы-то были вполне человеческие - с пятками и паль-
цами ног. На некоторых следах видно, что большие пальцы прилегают к осталь-
ным, а не противопоставлены им, а это уже, несомненно, признак человеческий,
а не обезьяний. Да и походка у того австралопитека была вполне человеческая:
он ступал на пятку, потом переносил тяжесть на плюсну и отталкивался пальца-
ми .
Помимо прямых доказательств - цепочки следов - есть и ряд надежных анатоми-
ческих признаков прямохождения. Это строение костей ступни и щиколотки,
строение шейки бедра, костей таза. По костям одного лишь мизинца ученые спо-
собны заключить, как передвигался его обладатель. Также хорошо увязывается с
прямохождением положение затылочного отверстия в черепе. Оно показывает, вер-
тикально или под углом находилась голова к оси тела, а значит, определяется и
наиболее характерная поза. Кроме того, о прямохождении судят по местам при-
крепления шейных мышц и мышц плеча. Иными словами, признаков таких много, и
необязательно иметь фотографический портрет предка, чтобы восстановить его
облик. Для этого достаточно найти небольшой набор его костей. Поэтому в конце
XX века, когда у Человека разумного объявилось множество новых предков, не
составило труда выяснить, как они двигались.
От сахелантропа (6-6,5 млн. лет) нашли целый череп и несколько фрагментов
скелета. О его двуногости судят по положению затылочного гребня. К этому
гребню крепились мышцы, поддерживающие голову в нужной позиции.
От оррорина (6 млн.) сохранилось бедро. Как говорят специалисты, его строе-
ние "не исключает возможность бипедализма" (то есть двуногости), но, к сожа-
лению , и не доказывает его строго.
Ардипитек кадабба (5,2-5,8 млн.) предоставил исследователям челюсть с зуба-
ми, несколько фрагментов костей рук и ног и палец ноги. В данном случае даже
одного пальца достаточно, чтобы признать кадабба двуногим. Вдобавок кости его
конечностей очень похожи на кости Люси. А уж про эту древнюю даму, от которой
остался почти целый скелет, известно досконально - она ходила на своих двоих.
От близкого вида Ардипитек рамидус (4,4 млн.) сначала нашли только куски
черепа и челюсти, но потом откопали еще несколько костей, включая и плечевую.
Она, очевидно, не несла нагрузки, то есть не использовалась при ходьбе. Да и
затылочное отверстие черепа сдвинуто вперед.
Недавняя находка в Эфиопии явила человечеству нашего предка, жившего около
4 млн. лет назад. Руководитель раскопок Джохансон Хайле-Сайласси пока не бе-
рется судить, к какому роду причислить находку, но общий облик этого прачело-
века восстанавливается без труда. Он завещал науке скелет исключительной со-
хранности, так что имеется возможность досконально изучить анатомию и эволю-
цию прямохождения древних людей. Кстати, ученые надеются, что столь полная
информация поможет ответить на вопрос инженера - "как?". Как двигался человек
и как должны были измениться конструкция суставов и перераспределиться на-
грузки?
При исследовании этого прачеловека открылась любопытная деталь, связанная с
эволюцией прямохождения. Оказалось, что он имел весьма длинные ноги, значит,
был способен быстро ходить и бегать. У австралопитечки Люси, жившей на пол-
миллиона лет позже, ноги заметно короче. Но ведь в ходе эволюции ноги у чело-
века не укорачиваются, а удлиняются! Как же вышло, что у более древнего пред-
ставителя гоминид ноги длиннее, чем у более позднего? Этот факт удивляет тех
ученых, которые все еще пытаются уложить эволюцию гоминид в прокрустово ложе
направленных и постепенных изменений. Если же помнить о возможности мозаично-
го распределения признаков, то ничего сверхъестественного в длинных ногах
древнейшего человека нет.
Итак, антропологи доказали, что даже самые древние представители гоминид
предпочитали ходить на двух ногах. Теперь предстояло ответить лентяям: а за-
чем людям было вставать на ноги?
До недавних находок ответ представлялся простым. Обезьяны спустились с де-
ревьев для того, чтобы осваивать саванну. По плоской саванне быстрее бегать
на двух конечностях. Именно бегать, а не ходить, так как двуногие получают
преимущество в скорости только при беге, но не при ходьбе. Не случайно самые
быстрые динозавры оторвали передние лапы от земли, да и некоторые современные
ящерицы спасаются бегством на двух конечностях. (Правда, у тех и у других
важную роль в технике бега играет длинный хвост, которого с самого начала бы-
ли лишены человекообразные.) А обезьяны были к тому же предрасположены к дву-
ногости: у них ноги и руки выполняют разные функции и по-разному иннервируют-
ся . Для принятия этой простой гипотезы нужно было совсем немного: показать,
что древнейшие двуногие жили в саванне. Но факты оказались несговорчивыми.
Так, останки ардипитека и оррорина соседствуют с ископаемой лесной расти-
тельностью. Значит, эти люди жили в лесу, а вовсе не в саванне. А если вокруг
был обезьяний древесный рай, то зачем оррорину ходить на двух ногах? Получа-
лось, что древнейший предок спустился с деревьев вовсе не затем, чтобы побе-
гать по безлесным саваннам.
В связи с этим была выдвинута гипотеза, что двуногость могла поначалу сфор-
мироваться как приспособление для плавания и передвижения в воде. Например,
шимпанзе, самый близкий к человеку примат, находясь в воде, стоит и передви-
гается на двух ногах 90% времени и только 10% опирается на руки. Сторонники
"водной гипотезы" происхождения двуногости приводят и другие аргументы. У Лю-
си оказался необычно плоский таз и особенный бедренный сустав. Он устроен
так, что ноги лучше двигались не вперед-назад, а в стороны, то есть Люси и
ходила враскорячку, и бегать быстро не могла. С такими особенностями удобнее
плавать, чем ходить. Кроме того, самый древний человек сахелантроп жил на бе-
регу большого озера и питался моллюсками и рыбой. И вообще в человеческом
мозгу много незаменимых жирных кислот (организм может получить их только с
пищей, а сам синтезировать не способен), которые ближе всего по составу к
рыбьим или содержащимся в теле моллюсков. Чтобы развить мозг и поумнеть,
древний человек должен был поедать водяную живность в большом количестве. Ос-
тавим ученым судить о весомости этих аргументов.
Другие специалисты доказывали, что человек перестал висеть на ветках и на-
чал карабкаться по стволам (оррорин имеет явные признаки подобного "стволола-
за") . Третьи защищают идею об изначальном передвижении на двух ногах при
вспомогательной опоре на костяшки пальцев рук. Так могли передвигаться, на-
пример, лесные аутсайдеры - те, кого конкуренция заставила искать новое место
под солнцем, будь оно в лесу, на берегу озера или в саванне.
Гипотез много, общепринятой - ни одной. Но разнообразие мнений не должно
смущать - оно указывает лишь на то, что у науки пока нет удовлетворительного
представления об эволюции прямохождения.
На древней кухне
С появлением новых молекулярных методов исследования стали ясны детали ис-
тории и биологии некоторых из них. Например, чем питались австралопитеки, за-
чем-то поменявшие кроны деревьев на травянистые равнины? Раньше полагали, что
австралопитеки были падальщиками, доедали то, что оставляли насытившиеся гие-
ны и саблезубые тигры. По другим версиям, они унаследовали от своих обезьянь-
их предков растительноядность. Оказалось, что эти гипотезы можно проверить,
изучив изотопный состав зубной эмали австралопитеков. Австралопитеки действи-
тельно добывали пищу в саваннах, но их диета была чрезвычайно разнообразной:
от кореньев и плодов до насекомых и падали. Мясная доля в их рационе со вре-
менем возрастала: у самых древних австралопитеков больше изотопов, маркирую-
щих растительные компоненты пищи, а у поздних таковых гораздо меньше. Вероят-
но, древние гоминиды постепенно учились отгонять хищников от недоеденной до-
бычи. Жившие одновременно с австралопитеками парантропы, видимо, тоже были
всеядными (всем другим лакомствам предпочитавшие термитов). Раньше предпола-
галось , что мощные зубы и челюсти были нужны парантропам для пережевывания
жесткой волокнистой пищи - в основном кореньев и плодов. Новые данные об их
рационе заставляют пересматривать эту гипотезу: сейчас неясно, зачем паран-
тропам понадобились столь внушительные челюсти и зубы.
По изотопному составу коллагена костей определили и рацион живших после ав-
стралопитеков человека умелого и питекантропа. Они были мясоеды, и пищей им
служили остатки трапез крупных хищников. В тех случаях, когда на костях тра-
воядных жертв имеются и следы зубов хищников, и следы каменных рубил, - цара-
пины, оставленные каменными орудиями, всегда располагаются поверх зубных от-
метин . Похоже, что Человек умелый не мог добыть себе мясо охотой и добирался
до туши уже после хищника. А питекантропы умели более эффективно отгонять
хищников от жертвы - может быть, из-за того, что ростом стали повыше, а может
быть, они пользовались для этого огнем. Ведь именно питекантропы стали первы-
ми обитателями Земли, кому покорился Красный цветок.
Пищевые пристрастия потомков питекантропов не изменились: они остались вер-
ны мясной диете. Однако уже гейдельбергский человек не удовлетворился пре-
зренным занятием падальщика и стал учиться охоте. В его арсенале появилось
копье. В Германии, в Шенингене, найдены охотничьи метательные копья возрастом
400 тысяч лет - из стволов елей, без наконечников, заостренные огнем. Центр
тяжести у них находится в 1/3 длины от острия - точно как у современных мета-
тельных копий.
Виды гоминид, известные ныне, и вероятные филогенетические связи
между ними. А — по Senut В; Б — по Libermann. 40 — современные
человекообразные обезьяны (не гоминиды); Hs — Homo sapiens,
включая неандертальцев; Hh — Н.heidelbergensis; Her —
Я. ergraster; Не — Я. erectus; H/Ah — Ното (либо Australopithecus)
habilis; Кг — Kenyanthropus rudolfiensis; Kp — K. platypos; P-s
— вновь «сконструированный» французами род Praeanthropus; Ag —
Australopithecus garhi; Ab — A.bahrelghazali; Aaf — A.afarensis;
Aan - A.anamensis; Aa — A.africanus; Pb — Parantrophus boisei;
Pr — P.robustus; Pa — P. aethiopicus; Arr — Aridipithecus ramidus
ramidus; Ark — A.r. kadabba; Ot — Orrorin tugenensis; St —
Sahelantrophus tchadensis
Неандертальцы и сапиенсы тоже умели пользоваться копьями. Но копья у этих
видов были разными. Неандертальцы использовали древко с каменным наконечником
для ближнего боя, а у сапиенсов копье с наконечником служило для дальних бро-
сков. Ближний бой с хищником не сулит ничего хорошего, поэтому находят множе-
ство костей неандертальцев с характерными повреждениями, полученными в подоб-
ных схватках. Такие же повреждения встречаются и у кроманьонцев, но не столь
часто. Сапиенсы держались подальше от крупного зверя и убивали его с безопас-
ного расстояния. И неандертальцы, и первые сапиенсы были мясоеды, почти весь
их рацион состоял из животной пищи. Сапиенсы, в отличие от неандертальцев,
разнообразили свое меню рыбой и моллюсками. Переход к земледелию, разумеется,
повлиял на ход эволюции. Ведь это привело к резкому увеличению доли расти-
тельной пищи, и человек изменил своей исконной диете.
Неандертальцы и сапиенсы в течение многих тысячелетий обитали на одной тер-
ритории . Воображение рисует жестокие схватки между агрессивными приземистыми
неандертальцами и тощими хитрыми сапиенсами. Но если битвы и случались, то,
видимо, не они сыграли главную роль в вымирании неандертальцев. Неандертальцы
и сапиенсы были приспособлены к различным климатическим условиям и тяготели к
разным ландшафтам. В израильских пещерах Схул и Казеф найдены многочисленные
остатки древних людей и их орудий. Ничего удивительного в том, что древние
люди жили в пещерах, нет. Удивительно чередование обитателей этих пещер, не
раз переходивших "из рук в руки": до 130 тысяч лет назад там жили неандер-
тальцы. Между 130 и 80 тысячами лет - люди современного типа. Выше - опять
находятся кости неандертальцев возрастом 65-47 тысяч лет. Еще выше - снова
сапиенсы. Очевидно, что первая попытка сапиенсов закрепиться в этом районе
закончилась неудачей. В заманчивом жилище вновь поселились неандертальцы. Нет
никаких признаков того, что в этой борьбе сапиенсы располагали более совер-
шенной технологией, чем неандертальцы: в этом отношении они были равны. Ис-
следователи предполагают, что свою роль тут сыграли изменения климата: при
похолоданиях одерживали верх неандертальцы, при потеплениях - более теплолю-
бивые сапиенсы. Примерно 75 тысяч лет назад действительно было похолодание,
которое могло способствовать "возвращению" неандертальцев и отступлению сапи-
енсов. Кроме того, известно, что пребывание неандертальцев в этих местах со-
провождается более засушливым климатом, а сапиенсов - более влажным. Древние
сапиенсы явно предпочитали более комфортный климат, нежели неандертальцы.
Почему же сапиенсы в концов концов взяли верх над сильными, выносливыми и
неглупыми (объем их мозга не уступал нашему) неандертальцами? Некоторые спе-
циалисты считают, что главной причиной была более совершенная общественная
организация сапиенсов, их способность к согласованным коллективным действиям.
Об этом свидетельствуют более развитые лобные доли головного мозга сапиенсов.
Одна из важнейших функций лобных долей как раз и состоит в способности обуз-
дывать собственные желания, если те идут вразрез с волей коллектива.
Сапиенсы - переселенцы
или захватчики?
Человек со своей уникальной способностью существовать практически в любых
условиях ("одежда, жилища - вот что нам нужно" - говорил Страшила Мудрый)
приспособил под жилье практически всю землю. Но так было не всегда. У челове-
ка, то есть у рода Ното, была своя родина. В начале XX века таковой считалась
Юго-Восточная и Центральная Азия. Это мнение основывалось на географии самых
древних находок представителей рода человеческого, то есть питекантропов. В
середине века родину человека перенесли в Африку - там нашлись и человек уме-
лый, и питекантропы, более древние, чем азиатские. А теперь нам стало извест-
но, насколько интенсивной была эволюция гоминид в Африке 6-2 млн. лет назад,
так что африканские корни человечества на сегодня выглядят практически узако-
ненными паспортными данными.
Почти любая группа животных или растений после выхода на эволюционную арену
начинает расселяться по свету. Этот эволюционный период называется фазой рас-
цвета. Так же и род человеческий, зародившись в африканских тропиках, пустил-
ся в путь. Можно, конечно, утверждать, что человеком двигало присущее высшему
разуму любопытство или нетерпеливость, однако расселительная фаза присуща и
другим земным обитателям.
Питекантропы отправились в долгий поход из Африки в Европу и Азию около 2
млн. лет назад. Они несли с собой орудия примитивной галечной культуры. Это
обработанная галька и кремневые отщепы. В период потепления человек вместе со
слонами и носорогами преодолел преображенную аравийскую пустыню и вышел на
просторы Евразии. Самые ранние находки переселенцев датируются 1,8-1,6 млн.
лет и в Юго-Восточной Азии, и в Европе. Благодаря недавним находкам, сделан-
ным в Грузии, в Дманиси, мы знаем, что человек смог столь же рано "прописать-
ся" и в Европе. "Человек грузинский" - так назвали свою находку исследователи
этой древнейшей европейской стоянки. Они восстановили облик древнейших обита-
телей Грузии по челюстным костям и черепным крышкам четырех особей. Рядом с
костями найдены и галечные орудия. Эти люди сочетали признаки и человека уме-
лого , и питекантропа. Объяснить смешение признаков двух видов ученые пока не
берутся. Но наиболее вероятный путь европейских питекантропов проходил по за-
падному берегу Каспия, именно там найдено большинство раннепалеолитических
стоянок. В Азии питекантропы расселялись через Иран, Индию, Казахстан и далее
в Китай и на Юго-Восток.
Тем временем оставшиеся в Африке люди изобретали новые технологии. Их ноу-
хау представляло собой двухстороннее, заостренное по периметру серией мелких
сколов каменное рубило, так называемый бифас. Вооруженные новой техникой и
подбодренные хорошей погодой - пошла новая волна потепления, - африканские
питекантропы вновь пустились в путь. Распространение бифасов и других орудий,
изготовленных по той же технологии, вычерчивает на карте Евразии их путь. Это
снова Ближний Восток, а оттуда транзитом в Европу и Центральную Азию, потом в
Монголию и Сибирь. Эволюция выходцев второй волны привела к появлению неан-
дертальцев . К той эпохе относятся замечательные свидетельства человеческой
жизни. Пещерная стоянка, названная "Денисова пещера", сохранила непрерывные
следы деятельности наших предков от 300 тысяч лет назад (а возможно, даже 500
тысяч) до современности! Археологи по каменным орудиям, кухонным остаткам,
остаткам растений и животных восстанавливают образ жизни обитателей пещеры.
Охотники с каменными бифасами постепенно сменялись обладателями костяных из-
делий. Потом на их место пришли люди с копьями, металлическими орудиями, за
ними скифы, тюрки. Последний обитатель пещеры монах-отшельник Дионисий поте-
рял там свой крест. Крест достался археологам, и именем монаха была названа
эта пещера.
Еще одно удивительное свидетельство неандертальской культуры обнаружено в
конце XX века в Монголии. Там в южной Гоби располагалась Кремневая долина -
гигантская орудийная мастерская возрастом около 300 тысяч лет. На каждом мет-
ре этой мастерской - а площадь ее 20 кв. км! - в среднем откапывают шестьсот
обработанных кремневых артефактов. По предварительным подсчетам, эта мастер-
ская просуществовала примерно 200 тысяч лет. Так что есть все основания наде-
яться, что вскоре мы детально узнаем, как совершенствовались технологии ази-
атских неандертальцев.
Примерно 100 тысяч лет назад из Африки снова двинулся отряд людей. На сей
раз в поход отправились самые передовые представители человеческого рода -
Homo sapiens, которые несли с собой новые технологии и новые орудия, костяные
и каменные. На просторах Азии и Европы они встретились со своими сородичами
неандертальцами. Какой была эта встреча?
Тут к археологам подключились генетики. По изменчивости отдельных генов лю-
дей из разных частей планеты можно установить, где находится "точка сборки",
то есть очертить территорию существования "изначального" гена. Обычно для
анализа используют гены митохондрий, которые передаются строго по женской ли-
нии - от матери к детям, а также гены мужской Y-хромосомы, которая передается
строго по мужской линии - от отца к сыну. Генетики высчитали, что и митохонд-
риальная Ева - прародительница всех женщин, и Y-хромосомный Адам проживали в
Африке около 150 тысяч лет назад. И все современное человечество происходит
от этой пары. Конечно, это не значит, что данная счастливая пара жила в оди-
ночестве , размножилась и дала плодовитое потомство. Это означает, что от
большой африканской популяции в современном мире остались только митохондрии
одной женщины и Y-хромосома одного мужчины, живших 150 тысяч лет назад. Зато
другие гены человечество унаследовало от других, в том числе и более древних
сапиенсов. Так что можно говорить и о лактазных Адаме и Еве, живших 1,9 млн.
лет назад (питекантропах), гемоглобиновых Адаме и Еве, давших потомство 650
тысяч лет назад, и многих других Адамах и Евах, смотря какой ген анализиро-
вать. Так или иначе, генетики настаивают на отсутствии смешения неандерталь-
ских и сапиентных генов. Это означает, что сапиенсы, даже если и встречались
с неандертальцами, не скрещивались с ними, или от таких пар не могло быть по-
томства .
Существует гипотеза о захватнических кампаниях сапиенсов: лучше социализи-
рованные, они легко побеждали неандертальских охотников-одиночек2 и, в конце
концов, вытеснили их. Другая гипотеза утверждает, что сапиенсы и неандерталь-
цы долго жили бок о бок. Ведь в пещерах не видно никаких скачков при переходе
от неандертальской к сапиентной культуре, будто люди бережно сохраняли насле-
дие предков, кем бы те ни были. Наконец, есть предположение, что азиатская
популяция неандертальцев, в отличие от европейской, все-таки могла скрещи-
ваться с сапиенсами и, в конце концов, полностью смешалась с пришельцами. Эта
смелая гипотеза основана на некоторых находках человеческих костей древнего
возраста, имеющих смешанный комплекс неандертальских и сапиентных признаков.
Остается подождать, пока генетики не расшифруют побольше генов неандертальцев
и древнейших сапиенсов. А пока слишком мало данных, чтобы уверенно судить о
том, как вели себя наши древние предки на новом месте. Будем надеяться, что
более мирно и воспитанно, чем их потомки.
ВОПРОСЫ ПО ДЕЛУ И НЕ ОЧЕНЬ3
Зачем бесконечность?
Как могла возникнуть способность оперировать понятием бесконечности - в ча-
стности, воспринимать натуральный ряд как целое? Вряд ли эта способность мог-
ла быть фактором приспособленности?
- Эволюционным путем возник наш многофункциональный мозг, способный ко мно-
гим вещам, в том числе и к абстрагированию. Но само абстрактное мышление раз-
вилось в ходе не биологической, а культурной эволюции. В языках некоторых на-
родов хорошо видна неразвитость абстрагирования. Например, у айнов (древний
народ, живущий на Сахалине и в Японии) к началу XX века были разные числи-
тельные для предметов разной формы и разного размера. Попав в современные
школы, эти люди столкнулись с неожиданными трудностями. Если учитель задавал
вопрос: было пять деревьев, одно срубили - сколько осталось? - следовал
встречный вопрос ученика: а какие были деревья - высокие или низкие? Учитель
не понимал, в чем дело, а ученик не мог сосчитать, потому что у него для вы-
соких деревьев одна система числительных, для низких - другая.
2 И съедали их, - добавляют Злые языки. Как бы то ни было, история последних тысячелетий по-
казывает , что наш вид крайне агрессивен - его любимое Занятие война и уничтожение всех и
вся, в первую очередь представителей своего же вида.
3 Отвечает Александр Марков (мы тоже не Знаем кто это).
Где спецификации?
Возьмем вид животного, обладающий некоторым набором "умений", и его эволю-
ционного предка, этим набором не обладающего. Можно ли предъявить хотя бы ги-
потетическую последовательность жизнеспособных переходных форм? Можно ли на
генетическом уровне отследить эти переходы?
- Такие переходы часто связаны с тем, что одна и та же структура организма
может приобретать некоторую дополнительную функцию, которая потом становится
основной. Плавательный пузырь у рыб изначально был выростом пищевода. Рыба
заглатывала туда воздух, и это давало повышенную плавучесть. Но ведь в обо-
лочке пузыря есть кровеносные сосуды, там идет диффузия газов, поэтому он с
самого начала выполнял и другую функцию, второстепенную, малозначительную -
функцию газообмена. Но когда естественный отбор начал этому способствовать -
на основе пузыря возникло легкое. Мягкие ткани практически не сохраняются,
поэтому промежуточные формы сложно проследить на ископаемых рыбах. Но сравни-
тельная анатомия для современных двоякодышащих рыб (протоптеруса, например)
четко показывает гомологию (соответствие по положению относительно других ор-
ганов и по динамике индивидуального развития) плавательного пузыря и легких.
Есть и другие доказательства "эволюционной преемственности" этих органов, и
все вместе это обеспечивает достаточную строгость выводов по современным мер-
кам естественных наук. Таких примеров много.
Но можно ли дать ответ на уровне инструкции по сборке автомобиля - сначала
ставим эту гайку, потом крепим эту ось, и так далее?
- С такой подробностью отследить эти процессы невозможно в принципе! В био-
логии таких ответов не было, нет, и никогда не будет. Если принимать только
такие ответы, а все остальное считать недостоверным, то биологию надо закры-
вать . Как мы можем во всех деталях проследить, кто кого родил? Это нереально.
Как это работает у микробов?
Ограничимся бактериями - можно ли в этом случае проследить или искусственно
стимулировать появление новых видов?
- О да, есть масса работ по искусственной эволюции микробов. Их ставят в
определенные условия, и прямо на глазах исследователей начинаются эволюцион-
ные изменения. Но можно ли получить новый вид? - казуистический вопрос, пото-
му что всегда можно сказать: "а это не новый вид!" Понятие вида не определено
с математической строгостью. Тем более у бактерий, где нет четких эндогамных
кластеров - групп особей, которые предпочитают скрещиваться только внутри
группы (один из важнейших признаков вида у высших организмов). С одной сторо-
ны, скрещивание у бактерий устроено проще, чем у позвоночных, - это прямой
обмен генами, часто между совсем далекими формами. С другой стороны, есть
формы, которые вообще не обмениваются генами, а размножаются как клоны. Так
что понятие вида к микробам трудно приложимо.
Кстати, есть важные эксперименты с мутациями бактерий, проясняющие некото-
рые аспекты возникновения многоклеточности. Берутся бактерии, которые живут в
толще воды и плавают поодиночке. Когда они используют весь кислород в этой
толще, преимущество получают те, кто всплывет на поверхность. Но у этих бак-
терий иногда происходит мутация, в результате которой они выделяют порции
липкого вещества. Делясь, мутанты остаются склеенными. Важно, что склеившиеся
бактерии автоматически всплывают на поверхность - в отличие от одиночных. На
поверхности начинают образовываться круглые пятнышки - колонии бактерий, ко-
торые имеют доступ к кислороду. Тем самым мутанты получают преимущество, ко-
гда в толще кончается кислород. Но на выделение липкой слизи мутанты тратят
энергию. Если же в колонии появляется мутант-обманщик, который живет в коло-
нии , она его держит на поверхности, но сам он слизь не выделяет, - то уже он
получает эволюционное преимущество. Мутанты-обманщики начинают активно раз-
множаться - до тех пор, пока колония не разрушится, и все они не потонут. Так
процесс и идет.
Колония еще не организм, потому что естественный отбор продолжает действо-
вать на уровне отдельных бактерий. Но эксперименты можно и продолжить. Снача-
ла надо понять, что должно произойти, чтобы решить проблему обманщиков. И так
далее, по пути хотя бы к подобию настоящего многоклеточного организма.
А взять всем известный пример эволюции микробов - появление устойчивости к
антибиотикам. Прослежено, каким образом она вырабатывается и передается. У
микробов, как и в целом в природе, есть приспособления для ускоренной переда-
чи полезных признаков. Представление, что все мутации случайны, что вся эво-
люция основана только на случайных мутациях, - сильно устарело.
Мутанты и симбионты
Как оптимизируются мутации?
- Хорошо изученный пример - изменение генома в иммунной системе. Это соче-
тание целенаправленного и случайного поиска. Если в организм проник незнако-
мый микроб, клетка иммунной системы "не знает", какой нуклеотид нужно заме-
нить в своей ДНК, чтобы убивать именно этот микроб. Но клетка умеет целена-
правленно редактировать свой геном. Популяция антител формируется в организме
в два этапа. Сначала, в ходе эмбрионального и постэмбрионального развития,
определенным образом варьируются определенные участки определенных генов кле-
ток иммунной системы. Из полученного множества лимфоцитов выбраковываются те,
которые производят аутоиммунные тела (нападающие на сам организм). Остальные
размножаются, и в итоге организм получает набор лимфоцитов, которые атакуют
практически любые молекулы, кроме тех, что есть в организме. На этом этапе
набор заготовок комбинируется в нескольких миллионах вариантов, и из них в
каждом лимфоците собирается один ген антитела. А вот когда в организм попада-
ет инфекция, начинается второй этап, точная подгонка антител. Лимфоциты, ко-
торые лучше всего атакуют нового возбудителя, начинают вносить случайные, то-
чечные мутации в гены своих антител. Механизм этого процесса расшифрован. По-
сле мутаций проверяется, что получилось. Если не удалось убить нового возбу-
дителя, мутирование продолжается, и так до тех пор, пока не будет нащупан эф-
фективный вариант антитела. Тогда соответствующий лимфоцит начинает бурно
размножаться. В итоге мы получаем приобретенный иммунитет. Мутации случайны,
но конечный результат закономерен и предсказуем. Мы знаем, что дней через
пять после прививки, допустим, оспы нужные антитела появятся.
Но как возникли эти механизмы? Как определились места, где допускаются му-
тации? Как возник сам генетический код? Такие вопросы можно продолжать беско-
нечно .
- Да, и я отвечу только на первый из них: точно известно, откуда в иммунной
системе взялись белки, отвечающие за вырезание и комбинирование кусочков ге-
нома. Они были получены извне. Существуют похожие на вирусы мобильные генети-
ческие элементы - транспозоны. Они не так легко, как вирусы, передаются "по
горизонтали", но иногда это случается. Главная же особенность транспозонов в
том, что они очень хорошо умеют вырезать куски генома и переставлять их с
места на место. Они этим живут испокон веков. Вот они-то и принесли эти меха-
низмы в организм позвоночных.
Простейший такой элемент имеет только один ген - ген фермента транспозазы.
Этот фермент находит в чужом геноме нужное место и встраивает туда свой мо-
бильный элемент. Когда-то в ходе эволюции произошло важнейшее событие - по-
звоночные животные вступили в симбиоз с этими элементами, и функция, которая
эволюционно возникла для размножения транспозонов, пригодилась для создания
приобретенного иммунитета.
Это очень важный принцип эволюции - а может быть, и самый главный. Без сим-
биоза темпы эволюции были бы в миллионы раз меньше.
Сама эукариотическая (имеющая ядро) клетка возникла в результате серии по-
следовательных симбиозов безъядерных прокариотических клеток - они преврати-
лись в митохондрии и другие органеллы. Если бы этого не произошло, на Земле
до сих пор жили бы только бактерии - ведь в природе так и не появились много-
клеточные прокариотические организмы. Они не раз вплотную подходили к много-
клеточности - но не могли преодолеть некий предел сложности организации.
Симбионты и биосфера
Как работает симбиоз в эволюции на более крупных масштабах?
- Иногда он работает в масштабах всей биосферы. Современную биосферу во
многом создали растительноядные животные: насекомые, грызуны, жвачные. Они
перерабатывают биомассу растений, поддерживают соотношение лесов, степей, бо-
лот. Есть теория (которую я разделяю), что нынешняя тундра, тайга и болота в
Америке и Евразии сформировались на месте высокопродуктивных мамонтовых сте-
пей, после того как исчезли крупные травоядные, опора степной экосистемы.
Сейчас в Африке, в парке Серенгети, можно видеть гигантские стада антилоп,
стада слонов, носорогов - такая же картина была в Евразии и Северной Америке
10-12 тысяч лет назад. Роль травоядных животных в биосфере огромна - но пара-
докс в том, что они практически не умеют переваривать растительную пищу. У
них так и не выработались нужные ферменты. Тем не менее, свое главное дело
они все же делают - за счет симбиоза. У всех травоядных в пищеварительном
тракте есть симбионты, которые и переваривают растительную пищу: бактерии,
инфузории, жгутиконосцы. В некотором смысле корова служит просто термостатом
для развития инфузорий, да еще и обеспечивает их пищей!
То есть корова - устройство, которое создала инфузория для повышения своей
приспособленности?
- Здесь была совместная эволюция - если уж прибегать к метафорам, то корова
и инфузория совместно создавали друг друга. Причем заметьте, что инфузория
тоже ведь не может переварить целлюлозу! Это делают бактерии, живущие в сим-
биозе с инфузорией. Она глотает кусочки целлюлозы и отдает их своим бактериям
- а потом питается той органикой, что производят бактерии. А корова питается
тем, что производит инфузория. Если бы не симбиоз, животный мир как таковой
не получил бы столь мощного развития. Он был бы вообще не нужен биосфере.
Посмотрим и на растительный мир - сейчас доминируют высшие сосудистые на-
земные растения. Им всем нужен азот в доступной форме - нитраты или аммоний.
Но они не имеют молекулярных механизмов, чтобы эти вещества производить. Пе-
ревести азот из атмосферы в усваиваемую растениями форму умеют лишь некоторые
бактерии. Без симбиоза с азотофиксирующими бактериями не было бы современного
растительного мира. И растительный, и животный мир основаны на глобальных
симбиозах.
В эволюции участвуют симбиозы как единое целое, поэтому каждому виду не
нужно создавать свои приспособления заново, с нуля. Существует обмен новшест-
вами в пределах биосферы в широчайшем спектре: от горизонтального переноса
генов до взаимодействия и кооперации в сообществах.
Что такое "я"?
Есть ли прогнозы о дальнейшей эволюции человека?
- Прогнозы тут ненадежны, факторов слишком много, и мы заведомо знаем дале-
ко не все. Но кое-что сказать можно. Например, эволюционная судьба вида силь-
но зависит от его численности. Если численность высока, а изолированных попу-
ляций практически не осталось, эволюция резко замедляется. Новая мутация не
может быстро распространиться на все материки, на миллиарды живущих ныне лю-
дей. Даже если она дает какие-то серьезные преимущества, для этого понадобит-
ся очень много времени. Быстрые эволюционные изменения происходят в маленькой
изолированной популяции. Посадите группу людей на необитаемый остров и не
трогайте их в течение миллиона лет. Когда вы придете посмотреть, что получи-
лось, - скорее всего вас встретит новый вид.
Долгожданный новый человек!
- Вот именно. В реальности же за последние 20-30 тысяч лет строение орга-
низма, строение мозга не меняется. Разве что одни генетические варианты ста-
новятся более распространенными, нежели другие (на языке генетики - меняются
частоты аллелей). Например, лактаза, фермент, который расщепляет молочный са-
хар, образуется только у младенцев. Поэтому практически все взрослые когда-то
не могли пить молоко. Но вот появилось животноводство, и в тех районах, где
оно было особенно развито (в Северной Европе, например), мутанты, у которых
этот фермент продолжал работать во взрослом возрасте, получали преимущество -
они лучше питались, быстрее росли, производили больше детей. Сегодня в Китае,
других странах Азии, где нет традиционной культуры молочного животноводства,
70-80% взрослых людей не могут пить цельное молоко. А в Швеции - наоборот,
почти все могут.
Еще интересный пример - в районах, где высока опасность малярии, получила
распространение мутация, вызывающая серповидно-клеточную анемию. Это тяжелое
наследственное заболевание, однако, такие мутанты не болеют малярией - а ане-
мией заболевают, только если у них обе копии соответствующего гена мутантны.
Если же мутантна одна копия, то и анемии у них нет, и малярией они не болеют
- то есть получают эволюционное преимущество (хотя, по сути, мутация вред-
ная) .
Каковы перспективы искусственной эволюции?
- Это сложнейшая тема. Для иллюстрации приведу лишь один пример. Недавно
выяснилось, что у некоторых человекообразных обезьян имеется врожденный имму-
нитет к вирусу ВИЧ. У человека соответствующий белок немного другой, он защи-
щает нас от давным-давно вымершего вируса, который бушевал в Африке 2-3 млн.
лет назад. Так вот, можно сделать трансгенных людей, от рождения невосприим-
чивых к ВИЧ. Для этого вы должны отдать свою яйцеклетку, генетики над ней бу-
дут колдовать, вставлять какие-то плазмиды, и если все сработает, у ваших де-
тей , развившихся из такой яйцеклетки, будет это свойство. Но как отшлифовать
подобные технологии? Неудачные эксперименты здесь невозможны.
Сегодня уже достаточно хорошо известно, какие зоны в мозге отвечают за те
или иные интеллектуальные функции. В последние годы достигнут огромный про-
гресс и в понимании механизмов памяти. Так что, в принципе, можно представить
себе и генетические модификации, повышающие мощь интеллекта, способность к
запоминанию и анализу информации - просто за счет увеличения объема соответ-
ствующих отделов мозга.
Но вот к чему подходов пока совсем не видно, так это к пониманию биологиче-
ской основы нашего "я". Что такое "я", как реализуется самосознание, как оно
возникает в природе - об этом мы знаем сегодня несравненно меньше, чем о ме-
ханизмах эволюции в целом.
ЗАРАЗНЫЕ ГЕНЫ...
Дмитрий Шабанов
’’...Вся живая материя восстает перед
нами как одно целое, как один огром-
ный организм, заимствующий свои эле-
менты из резервуара неорганической
природы, целесообразно управляющий
всеми процессами своего прогрессивно-
го и регрессивного метаморфоза и, на-
конец, отдающий снова всё заимство-
ванное назад мертвой природе".
С. Н. Виноградский,
Лекция перед императорской
фамилией 8 декабря 1896 года
Журнал "Nature" публикует не только статьи о состоявшихся достижениях, но и
эссе, авторы которых пытаются нащупать дальнейшие пути развития науки. Одно
из последних принадлежит Нигелю Гольденфельду и Карлу Вёзе (Nigel Goldenfeld,
Carl Woese), американским ученым, которые прочат очередную революцию в биоло-
гии. С их точки зрения, новые данные об обмене генетической информацией между
представителями разных видов организмов вызовут в биологии смену парадигмы.
Речь идет о горизонтальном переносе генов - передаче наследственной инфор-
мации от организма к организму вне генеалогической (вертикальной, от предков
к потомкам) последовательности. Сам феномен известен давно: так, например,
передаются гены устойчивости к антибиотикам от одних болезнетворных микроор-
ганизмов к другим. Много лет на него смотрели как на некую оплошность приро-
ды. Энтузиасты надеялись объяснить с его помощью феномен эволюции, прагматики
искали в нем естественные технологии генетической инженерии... Лишь недавняя
лавина молекулярно-биологических данных позволила понять, насколько это явле-
ние распространено в природе.
Десятилетиями в микробиологии торжествовал принцип, возведенный в догму Ро-
бертом Кохом: изучаемый микроорганизм нужно вырастить в чистой культуре. За-
частую для культивирования используются потомки одной-единственной бактери-
альной клетки. Выращиваемые в таких условиях микроорганизмы оказываются оди-
наковыми, что считается критерием их правильного выделения. Увы, такие усло-
вия совершенно противоестественны! Естественным является именно функциониро-
вание микроорганизмов в сложной среде, информационный обмен с которой, вклю-
чает передачу фрагментов ДНК.
В таких условиях для двух основных групп "микробов" - бактерий и архей (или
архебактерий) - вообще трудно говорить о видоспецифичном геноме (наборе на-
следственной информации). Благодаря горизонтальному переносу этот геном по-
стоянно пополняется новыми фрагментами, а благодаря собственной эволюции те-
ряет многое из приобретенного. Проследив генеалогию какого-нибудь микроорга-
низма, мы увидим, что через него течет "река" разнородной по происхождению
генетической информации. В этом потоке отражается вся биосфера!
Как регистрируют горизонтальный перенос генетической информации? Иногда
удается увидеть исходный текст у одного организма и его копию у другого. Ча-
ще, однако, приходится использовать косвенные данные - вычленять участки ДНК,
которые отличаются по соотношениям разных пар нуклеотидов или по частоте ис-
пользования разных триплетов для кодирования одних и тех же аминокислот (эти
признаки, в общем, видоспецифичны). Найдя в чьем-то геноме кусок текста, от-
личающийся от окружения по частоте "букв" (нуклеотидов) или "слов" (трипле-
тов) , мы можем предположить, что имеем дело с заимствованием. Со временем хо-
зяин откорректирует текст, приведя его в соответствие с "собственным стилем".
Итак, работа современных генных инженеров, создающих генетически модифици-
рованные организмы и тем самым порождающих волну протестов и "страшилок",
оказывается для земной биосферы обыденным делом [Заодно, правда, теряет прав-
доподобие один из аргументов, которым генные инженеры пытаются успокоить
страхи растревоженного общества - что "чужие гены" никак не могут расползтись
по биосфере. Оказывается, не все так очевидно. Впрочем, вставки в геномы мо-
дифицированных организмов тоже являются продуктами развития биосферы, как и
"родные" гены культурных растений].
Что самое интересное, горизонтальный перенос свойствен не только микроорга-
низмам. Например, проведенные недавно учеными из Беркли исследования геномов
риса и проса (злаков, которые разделяет не менее 30 млн. лет независимой эво-
люции) показало, что между этими видами происходил горизонтальный перенос ге-
нетической информации. В этих растениях найдены практически идентичные транс-
позоны - участки ДНК, способные перемещаться внутри геномов и между ними с
места на место! Чаще всего транспозоны переносят информацию внутри вида, но
могут и перешагнуть видовой барьер, ведь грань между транспозонами и вирусами
весьма условна. С активностью преобразованных транспозонов может быть связана
реорганизация генома, которая происходит при видообразовании.
Горячие головы видят в горизонтальном переносе объяснение прогрессивной
эволюции. Вот так появляются новые признаки: подует ветер, принесет вирус с
куском новой информации, и - глядь! - у организма появилось новое полезное
свойство. Так, выдающийся палеоботаник В. А. Красилов считает, что в эпоху
происхождения цветковых растений именно вирусы разносили от одних голосемен-
ных к другим "блоки" генетической информации, ответственные за формирование
цветка. Увы, в это поверить нелегко. Дело в том, что конструкция любого орга-
низма - нелегкий компромисс между модульностью и монолитностью. Любое из на-
селяющих Землю существ - одновременно и целостная система, и конгломерат от-
носительно независимых признаков. Теми из свойств организма, которые связаны
со всеми остальными в тугой узел взаимосвязей, невозможно заразиться "от вет-
ру" . Именно поэтому роль горизонтального переноса в эволюции высокоорганизо-
ванных и высокоинтегрированных групп снижается. Может, когда-нибудь у генных
инженеров дойдут руки проверить идею Красилова и они перенесут "гены формиро-
вания цветов" в геномы сосен, гинкго или саговников? Скорее всего, инородные
фрагменты не смогут встроиться в систему управления развитием растения, для
которого характерен иной способ размножения.
Даже когда крупное новшество у высокоинтегрированных животных оказывается
связано с каким-то продуктом горизонтального переноса, его не следует считать
прямым следствием захвата чужеродной информации. Приведем один пример. У мле-
копитающих развитие плаценты (органа, обеспечивающего физиологическую связь
зародыша и матери) требует, кроме прочего, работы гена PeglO. Этот ген чрез-
вычайно сходен с одним из широко распространенных транспозонов. Такой факт
можно интерпретировать двояко. Наивный человек, убежденный, что всякий при-
знак - проекция определенного гена, сделает вывод, что появление плацентарных
млекопитающих (и в конечном счете нас с вами) - следствие "заражения" "геном
плаценты". Отсюда недалеко до веры в генетического Демиурга, который управля-
ет эволюцией, время от времени, запуская в оборот новые вирусы и очередные
прогрессивные качества.
Более зрелой представляется иная трактовка. Новые функции, вырабатываемые в
ходе эволюции, могут связываться не только со старыми участками генетического
текста, которые выполняют собственные задачи, а с новыми, свободными. Эти но-
вые гены могут как раз оказываться занесенными со стороны кусками. Так опыт-
ный шофер, ремонтируя сломавшуюся в дороге машину, может сделать требующуюся
деталь из какого-нибудь найденного на обочине обломка. Функция этого обломка
в отремонтированной конструкции не содержалась в нем исходно - она возникла
вследствие его определенного положения в новой системе. Такая трактовка под-
тверждается тем, что находящийся в тесном родстве с "геном плаценты" транспо-
зон в любом ином месте и окружении вовсе не вызывает формирования чего-то по-
добного .
Наоборот, горизонтальный перенос служит не двигателем, а скорее тормозом
эволюции! Существа, которые приобрели относительно изолированный геном, мор-
фологически, физиологически, поведенчески эволюционируют намного быстрее, чем
плотно вплетенные в единую сеть биосферы микроорганизмы. Эволюция направлена
от континуума геномных возможностей к отдельным генотипам. Почему же биологи
поняли это так поздно? Тому есть несколько причин. Во-первых, биология должна
была достичь технического уровня, позволяющего зарегистрировать сам факт раз-
нообразия путей передачи наследственной информации. Во-вторых, изучение на-
дорганизменных систем психологически затруднительно для исследователей, кото-
рые сами являются организмами (см. лирическое отступление ниже). В-третьих,
распространению нового понимания препятствует трактовка организма как вопло-
щения (реализации) генетической программы.
Метафора наследственной информации, как программы, настолько въелась в соз-
нание современных биологов и небиологов, что многим из них попросту трудно ее
осознать. А что же такое организм, как не воплощение хранящегося в его генах
плана? То, что использует эти гены! Наследственная информация - не истинная
суть организма, а используемая им библиотека. На протяжении большей части ис-
тории жизни эта библиотека была общей для самых разных существ, и лишь потом
некоторые из них завели себе изолированные "книжные шкафы".
Когда речь идет о людях или наших ближайших родственниках, высших животных,
мы имеем дело с относительно изолированными видами. Каждый из них характери-
зуется определенной общностью генетической программы, занимая определенное
место в экосистемах. Дискретность бактерий и архей носит иную природу. Она
скорее связана с дискретностью возможных образов жизни, экологических ниш.
Итак, есть все основания согласиться с авторами эссе в "Nature". Мы стоим
на пороге нового понимания жизни вокруг нас. Понятия организма, вида, генома,
наследственной информации нуждаются в серьезном уточнении. В то же время не
следует считать такой подход совсем уж новым. Это его нащупывал в своем "Кос-
мосе" Александр фон Гумбольдт, высказывал в лекции перед царской семьей Сер-
гей Николаевич Виноградский, воплощал в теории Геи Джеймс Лавлок, пытается
под именем "природоведческой микробиологии" развивать академик РАН Георгий
Александрович Заварзин... Но в любом случае, развитие биологии подарит нам еще
немало неожиданностей.
Лирическое отступление
Как объять необъятное
Мы сами являемся организмами и окружающие нас другие системы воспринимаем
по аналогии с самими собой. Как ни естествен для нас такой подход, он не
единственно возможный...
Позапрошлой весной автор этих строк оказался
во время нереста остромордых лягушек возле не-
большого нерестового водоема - пруда среди за-
росших дубами холмов. Были сумерки - "час меж
собакой и волком". На мелководье собралось не-
сколько тысяч самцов лягушек. В это время они
преображаются благодаря ярко-голубой брачной
окраске. Песня самца остромордой лягушки похо-
жа на клокотание кипящей крупными пузырями во-
ды. Многоголосый хор нескольких тысяч самцов
сливался в невероятный гул. Этот объединенный
голос тысяч особей разносился по окрестным
холмам, созывая неторопливых, разбухших от икры
самок...
Подходя к нерестовому котлу, самка находит себе самца и вместе с ним отме-
тывает свою порцию икры. Те самки, которые прибывают позже других, вынуждены
нереститься на сплошном поле из кладок. Часть их икринок оказывается оплодо-
творена молоками не "своего" самца, а других - тех, что были отцами соседних
кладок. Пройдет немного времени - и из икринок выйдут головастики, ферменты
вылупления которых превратят в жидкость слизистые икряные оболочки. Даже если
за время их развития уровень воды в весеннем пруду упадет, разжиженная икра
стечет под уклон к отступившей воде. Ну и пусть десятки тысяч особей (а голо-
вастик, как и икринка, - это особи!) обсохнут на окраинах пруда - сотни тысяч
попадут в воду, где продолжат развиваться. По мере роста они будут обмени-
ваться разнообразными химическими и физическими сигналами. При необходимости
те из головастиков, которые опередят прочих, притормозят рост отстающих, что-
бы снизить конкуренцию за недостающие ресурсы. В иных случаях, наоборот, рост
разновозрастных и неродственных личинок окажется синхронизирован - популяция
сама выберет стратегию своего оптимального развития.
Так вот, сумерки, пение лягушек и усталость сместили восприятие подошедшего
к нерестовому котлу человека. Он почувствовал, что стоит не перед определен-
ным числом организмов, а перед сущностью более высокого порядка - популяцией.
Потенциально бессмертный живой объект находился на одном из ключевых этапов
своего годичного цикла. Производство новых особей оправдает потери существую-
щих, ведь только через их поток и смену и осуществляется бытие популяции. Ка-
залось бы, человек не может быть замечен этой сущностью - он ведь находится
на более низком уровне бытия, чем она. Тем не менее, когда завороженный чело-
век приблизился к одному из нерестовых котлов, поющие лягушки испугались, за-
молчали и торопливо попрятались на дне. Голос популяции изменился: она заме-
тила чужака и отреагировала на его присутствие...
Вы скажете, такой опыт переживания контакта с сущностью более высокого по-
рядка относится к сфере интересов психиатра, а не биолога? Вы, конечно, пра-
вы. Но, поверите ли, такой опыт очень помогает представить себе популяцию
единым целым, а не совокупностью отдельных особей... А вы лучше попытайтесь во-
образить всепланетную общность живых организмов, связанную горизонтальным пе-
реносом генетической информации, как популяция лягушек связана совместным
размножением! Удобно ли описывать и изучать эту общность, пользуясь традици-
онными понятиями и расхожими мыслительными штампами? Новым задачам должен со-
ответствовать и новый способ мышления!
... И ПАРАДОКСЫ СИСТЕМАТИКИ
Первое дело первого человека до сих пор не доделано. Адам, по свидетельству
Библии, до грехопадения (определившего необходимость добывать хлеб насущный в
поте лица своего) придумывал имена для животных. Ныне поименовано более мил-
лиона видов, а очередь безымянных все еще скрывается за горизонтом. Классифи-
цирование названных видов позволяет разобраться в миллионе имен и сделать ин-
формацию об их сходстве и различии более компактной.
Систематики до сих пор спорят, существует ли единая, правильная классифика-
ция. Оптимисты верят, что это та классификация, которая точно отражает фило-
гению - эволюционную историю. Вот узнаем с помощью молекулярных методов, как
шла эволюция, и поставим все на свои места... Увы, эволюционные изменения
многомерны, и, проецируя их на иерархическую систему, мы неизбежно теряем
Значительную часть информации. Какую? Ту, которую сочтем менее важной. Счи-
таете, что "объективными" методами можно определить, какая часть информации
наиболее важна? Оставьте эти иллюзии!
Интерпретируйте, к примеру, такой факт. С тех пор как разошлись эволюцион-
ные пути человека и шимпанзе, человек приобрел 154 новых гена, а наши "отста-
лые" родственники шимпанзе - 233! Как измерить и сравнить эти изменения? По
числу генов? По длине последовательностей ДНК? По изменению образа жизни? Од-
на из влиятельных школ систематики говорит: если изменение видов нельзя оце-
нить "объективно", то его не следует учитывать вообще и реконструировать надо
лишь порядок ветвления эволюционного древа. Страусово решение: зачем нужна
классификация, которая не учитывает основной результат эволюции - изменение
видов?
С другой стороны, сложности в трактовке каких-то результатов не означают,
что эти результаты бесполезны. Зачастую родство (или его отсутствие) разных
групп организмов, устанавливаемое молекулярными методами, оказывается столь
явным, что приходится менять устоявшуюся классификацию.
Орнитологи давно выделили две группы грифов: Старого Света (собственно гри-
фов) и Нового Света (катартид). Хотя было ясно, что эти группы не находятся в
тесном родстве, результаты анализа ДНК оказались неожиданными. Американские
грифы - это на самом деле аистообразные. К их числу относятся и современные
кондоры, и самые крупные из когда-либо летавших птиц. Речь идет о тераторни-
сах ("ужасных птицах"), "грифах", которые в неогеновом периоде питались ос-
танками гигантских млекопитающих. У одного из них, аргентависа, размах крыль-
ев достигал 7м, а вес 120 кг! Помните, в "Детях капитана Гранта" Жюля Верна
кондор похитил и утащил по воздуху мальчика? Реальные кондоры на такие подви-
ги не способны. Но в нашем фольклоре европейские аисты носят детей в клювах
(держа за пеленочку), а американские - в когтях!
Новейшие молекулярные данные касаются примитивных групп крылатых насекомых.
Выяснилось, что термитов надо не выделять в особый отряд, а рассматривать как
семейство отряда тараканов. Неважно, что по сложности социальной организации
термиты обогнали всех животных, кроме людей. Социальное совершенство термитов
- следствие их способности расщеплять целлюлозу, основу древесины. В кишечни-
ках термитов живет сложный эндосимбиотический комплекс. Его основу составляют
высокоспециализированные жгутиконосцы (которых долго считали инфузориями, так
как под микроскопом кажется, что они покрыты множеством подвижных ресничек,
как инфузории). То, что считалось ресничками, оказалось спирохетами - удли-
ненными бактериями, способными извиваться. Населяющие кишечник термита спиро-
хеты выстраиваются на поверхности жгутиконосца, синхронизируют свои движения
и перемещаются вместе с ним, как единое целое. Жгутиконосцы сами по себе не
могут расщеплять целлюлозу, но внутри них обитают симбиотические бактерии,
овладевшие этой реакцией. Заглатывая маленькие щепочки, жгутиконосцы скармли-
вают их своим внутренним бактериям, а продуктами переработки делятся с обес-
печивающими их подвижность спирохетами. Ну а термиты питаются как продуктами
активности своего внутреннего зоопарка, так и его питомцами.
Несмотря на все чудеса эндосимбиоза, переваривание целлюлозы для термитов -
непростой процесс, в кишечнике одного насекомого ему не пройти. Поэтому одна
и та же порция пищи проходит через пищеварительные тракты многих насекомых,
поедающих фекалии друг у друга. В конечном счете, комочки из продуктов пере-
работки пищи будут использованы для возведения термитников. Эти сооружения,
иногда достигающие величественных размеров, обеспечивают защиту и благоприят-
ный климат для совместного расщепления древесины. И теперь получается, что
все это делают тараканы?! Представляете, какой победой генной инженерии и
прочих биотехнологий стало бы создание прусаков, содержащих в кишечнике сим-
биогенетический комплекс, способный расщеплять целлюлозу? Такие тараканы по-
едали бы не только пищевые остатки, но и мебель с книгами.
Сфера приложения данных молекулярной систематики растет. Последняя новость:
из остатков мягких тканей тираннозавра, найденных пару лет назад в Монтане,
извлекли фрагменты молекул белка (коллагена, основы соединительной ткани).
Анализ этих фрагментов пока принес лишь тривиальный результат: тираннозавр
состоял в более близком родстве с курицей, чем с лягушками и тритонами. Стои-
ло ли ради этого тревожить останки, ожидавшие своей судьбы 68 миллионов лет?
Стоило! Ведь лиха беда - начало. Поймем, в каких породах макромолекулы сохра-
няются лучше, - научимся их эффективнее искать и толковее анализировать, и со
временем узнаем что-то новое. Поживем - увидим...
ГДЕ СКРЫВАЮТСЯ СНЕЖНЫЕ ЛЮДИ
Кирилл Еськов
"Хочется чего-нибудь эдакого..." - при том что, помянутые классиком как ря-
доположные "севрюжина с хреном" и "конституция" совершенно уже неактуальны:
первая без ограничений продается в соседнем "Рамсторе", а вторая выложена в
открытый доступ, но так никем и не читана, ибо - многа букв... Можно на этом
месте порекомендовать для застольных бесед апробированный набор тем о загад-
ках природы: НЛО с Тунгусским метеоритом, остров Пасхи с пустыней Наска,
снежный человек с Лох-Несским чудовищем, - загадках, ответ на которые злона-
меренно скрывается от рядовых налогоплательщиков Официальной Наукой&бт;. На
некстати случившегося же за тем столом профессионального археолога, который
поведает - в простоте своей - о том, что никаких мрачных тайн в тех рисунках
из Наска не содержится, ибо это всего лишь замечательный астрономический ка-
лендарь, посмотрят с нескрываемым отвращением: испортил песню, гад!
Криптозоология определяется Википедией как лженаука: "Это связано с тем,
что криптозоологией часто занимаются люди без биологического образования, а
также тем, что ни одна находка криптозоологов, о которых они сообщали в СМИ,
не получила подтверждения". Однако при заходе на сайт Cryptozoology-ru у нор-
мального биолога (вроде меня) возникает желание всего лишь "отделить мух от
котлет". Здесь интернет-опрос "Как снежные люди связаны с НЛО?" (sic!) мирно
соседствует с отчетом о недавней зоологической экспедиции на Борнео, открыв-
шей очередные новые для науки виды мышей-и-лягушек... Тут, кстати, приходит
на память и романтическая (хотя и совершенно реальная) история мисс Латимер,
чье имя увековечено в названии открытой ею кистеперой рыбы, и с детства люби-
мый профессор Челленджер со товарищи - ну да, Как будто не все пересчитаны
звезды,// Как будто бы мир не открыт до конца!
...Ладно: с мышами-лягушками, равно как с не так давно открытыми "живыми
ископаемыми" - моллюсками-моноплакофорами и рачками-цефалокаридами (зоологи-
ческие сенсации, вовсе не замеченные широкой публикой) все ясно. А вот как
там насчет существ более корпулентных, и в силу этого более близких сердцу
той публики - Лох-Несского чудовища, гигантского кальмара, реликтового гоми-
ноида (помните: плезиозавр Лизавета, спрут Спиридон и снежный человек Федя из
Колонии необъяснимых явлений при НИИЧАВО)? Если, понятное дело, исходить из
того, что все это - нормальные зоологические объекты, по разным причинам не
попавшие пока в руки ученых, а не нечто потустороннее?
Так вот, ученый - памятуя о пренцендентах с явленными нам вживе латимерией
и метасеквойей - для начала отделит тех, чье существование в принципе допус-
тимо (хотя и не доказано) от тех, кого можно считать мифом с полной на то
уверенностью. Вот, к примеру, ящер-плезиозавр (или типа того), обитающий в
шотландском озере Лох-Несс - на радость тамошней индустрии туризма... Ну, на-
чать с того, что "мрачное и пустынное горное озеро Лох-Несс" на самом деле с
1825 года является частью пересекающего Шотландию с востока на запад Каледон-
ского канала, интенсивность судоходства по которому не в пример выше, чем,
скажем, на трассе Волго-Балта, и это не говоря уж о том, что в последние де-
сятилетия тамошние места стали культовыми для туристов - как тут спрячешься?
Еще хуже для "криптозоологов" то, что Лох-Несс, вместе со всей Шотландией,
совсем еще недавно был сплошь покрыт ледниковым щитом - условия, явно не са-
мые подходящие для выживания реликтовой рептилии...
Это, однако, косвенные доводы; самый же убийственный аргумент против суще-
ствования "нео-плезиозавра" следует из экологии. На озере, разумеется, посто-
янно работают гидробиологи; я тут имею в виду не энтузиастов, безуспешно об-
шаривающих сонарами озерные глубины, а профессионалов, ведущих рутинную рабо-
ту по изучению местной экосистемы: пробы воды и грунта, количественные учеты
фитопланктонных водорослей и зоопланктонных рачков, etc. Так вот, структура
пищевой пирамиды Лох-Несса ни на йоту не отличается от таковой соседних оли-
готрофных озер, и тамошняя экосистема просто-напросто неспособна прокормить
хищника более крупного, чем лосось (а по части такого рода энергетических
расчетов экология давно уже стала вполне точной наукой). Соответственно, ни-
какой иной "Несси", кроме той, что рисуют по всей Шотландии на стикерах и
майках (а также на эмблеме Лох-Несской биостанции Друмнадрахит - где, собст-
венно, и ведутся эти экологические исследования), нет и не было. Ну, если,
конечно, не брать в рассмотрение гипотезы о том, что Несси питается "святым
духом", либо - подобно чудо-гусыне из фантастического рассказика Азимова -
являет собою живой ядерный реактор (работающий, по доброй британской тради-
ции, на полонии-210)...
Или вот сообщения о том, что в дебрях Сибири нет-нет, да и покажется недо-
вымерший мамонт. Мамонт - это все-таки не исчезнувшие с лица Земли за десятки
миллионов лет до появления человека динозавры; на этих еще каких-то 10 тысяч
лет назад охотились наши не шибко озабоченные природоохранный идеями предки,
так что - пуркуа бы и не па?.. Увы, с симпатичным мохнатым слоном, столь лю-
бимым зрителями мультфильма "Мамонтенок ищет маму", тоже ничего не выйдет -
и, в общем-то, по той же самой причине: никак тому слону не прокормиться в
нынешней Сибири.
Тут объяснения придется начать издалека. Когда в учебниках и научно-
популярных книгах пишут, что "самой продуктивной на Земле экосистемой являет-
ся дождевой тропический лес" это, конечно, верно: суммарная масса живых орга-
низмов (представляющая собою, в конечном счете, ассимилированную экосистемой
при фотосинтезе солнечную энергию), приходящаяся на единицу площади тропиче-
ского леса, колоссальна - деревья-то вон какие, и вон их сколько! Однако есть
экосистемы, оборот вещества и энергии в которых оптимизирован по-иному: сум-
марная биомасса растений там относительно невелика, однако темпы ее прироста
и обновления очень высоки (аналогия из области экономики: важен не столько
размер капитала, сколько скорость его оборота). И вот по этому параметру вне
конкуренции "травяные биомы" - степи и саванны, в которых основу растительно-
го покрова составляют не деревья, а злаки.
Злаки с их стеблем-соломиной обладают, в отличие от большинства трав, не
верхушечным, а вставочным ростом: если отъесть у них верхушку, они начинают
лишь быстрее расти. В некотором смысле травяной биом подобен волшебному гор-
шочку из сказки Братьев Грим - чем больше ту кашу ешь, тем больше ее стано-
вится. Именно поэтому в степях растительноядные животные (копытные, грызуны,
саранчовые) могут без вредных для экосистемы последствий одномоментно изымать
до 60% растительной биомассы - цифра, совершенно немыслимая для лесных сооб-
ществ , вроде нашей тайги или того же тропического леса. Вот и выходит, что
фотосинтезирующей биомассы в степной экосистеме в каждый отдельный момент
вроде бы и немного (по сравнению с лесами) , но за счет высокой скорости ее
прироста можно прокормить гораздо больше (чем в лесах же) копытных и хищников
- вспомните знакомые всем нам по телепередачам "В мире животных" картины аф-
риканских саванн или неисчислимые стада бизонов в распаханных ныне прериях...
Так вот, одним из таких высокопродуктивных травяных биомов во времена Вели-
кого Оледенения были холодные сухие степи, окружавшие ледниковый щит. Этот
исчезнувший ныне ландшафт получил название "тундростепь". Термин возник отто-
го, что тамошняя фауна (ее так и называют - "мамонтовая фауна") представляла
собой странную смесь: часть ее сохранилась в современной тундре (северный
олень, овцебык), часть - в современной степи (сайгак, бизон), а часть (ма-
монт, шерстистый носорог, пещерный лев, саблезубый тигр) была характерна лишь
для этой экосистемы и исчезла вместе с ней.
Древние тундростепи очень похожи по структуре растительного покрова на так
называемые реликтовые степи Восточной Сибири и Аляски, существующие ныне в
виде небольших пятачков на сухих южных склонах тамошних гор. В эпоху оледене-
ния, когда колоссальные количества воды оказались заморожены в ледниковом щи-
те (создававшем к тому же устойчивый антициклон, что дополнительно иссушает
климат), эти сухие степеподобные ландшафты распространились на огромные пло-
щади. При наступившем же 10-12 тысяч лет назад потеплении, когда ледниковый
щит растаял, отступив до примерно нынешних его границ, климат стал гораздо
более влажным. Место тундростепей заняли тундра современного типа и северная
тайга, где основу растительного покрова составляют не злаки, а мхи, которые
практически несъедобны для большинства животных; изолированные пятнышки гор-
ных "реликтовых степей" просто неспособны прокормить популяции копытных и
хищников, составлявших "мамонтовую фауну". Кстати, дольше всего мамонт прожил
на арктическом острове Врангеля (открытый недавно карликовый островной подвид
- очаровательное существо высотою около полутора метров - вымер всего 5 тысяч
лет назад, против 10 тысяч лет на континенте), а там и поныне относительно
широко распространены те самые реликтовые степи.
Итак, существование мамонта в нынешние времена совершенно невозможно: ис-
чезла сама экосистема, в которую тот был встроен. И даже если в будущем ма-
монта сумеют воссоздать "в пробирке" генно-инженерными методами (благо замо-
роженных в вечной мерзлоте тканей этого существа предостаточно), жить в при-
роде ему будет все равно негде. Ну, по крайней мере, пока не закончится меж-
ледниковье, в которое мы живем, и не начнется следующее по счету оледене-
ние . . .
Иное дело - "снежный человек". Мне, по крайней мере, неизвестны законы при-
роды, налагавшие бы прямой запрет на существование в горах Центральной Азии
реликтового гоминоида - "обезьяночеловека", или просто крупной человекообраз-
ной обезьяны. С вечными снегами он, надо полагать, вопреки своему названию не
связан никак (кроме того, что иногда оставляет там следы), а обитать должен в
поясе горных лесов, где вполне достаточно и пищи, и укрытий. Ясно, что любые
сообщения о североамериканских "бигфутах" можно со спокойной совестью выкиды-
вать не читая (ибо своих видов приматов на том континенте нет и никогда не
было4, а чтобы пройти туда из Азии через приполярную Берингию, как это сдела-
ли люди, надо хотя бы обладать огнем), но вот в Гималаях или на Памире - по-
чему бы и нет? Есть даже вполне правдоподобные кандидаты на эту роль, напри-
мер мегантроп - очень крупная (около двух метров ростом) ископаемая обезьяна5
из Южной Азии, обладавшая рядом "человеческих" черт, которые сближают ее с
африканскими австралопитеками, прямыми предками гоминид...
4 Точнее говоря, в Америке нет близких человеку узконосых обезьян, широконосые обезьяны Юж-
ной Америки другая, далекая от человеческой, эволюционная линия.
5 На месте неандертальцев я бы обиделся, они в таких местах еще 30-10 тыс. лет назад обита-
ли .
Итак, допускаю ли я (как зоолог-профессионал) принципиальную возможность
существования реликтового гоминоида? - ответ: "Да". Верю ли я в его существо-
вание? - ответ: "Нет". А поскольку речь тут зашла не о "знаю/не знаю", а о
"верю/не верю", я позволю себе высказать на сей счет вполне субъективное суж-
дение , основанное на личном опыте.
...В начале 80-х мне довелось работать на плато Путорана в северной Сибири.
Место вообще глухое и безлюдное (на карте "плотности населения" оно с полным
на то основанием сохраняет девственную белизну) , а мы с напарником к тому же
сидели отдельно от всех в горах, километрах в пятидесяти от базового лагеря
экспедиции. Сам я изучал тамошнюю почвенную фауну, а напарник мой - поведение
снежных баранов и хищников, волков с росомахами. Надобно заметить, что со
спецификой работы орнитологов и "мышатников" я знаком был неплохо, а вот спе-
циалиста по крупным млекопитающим наблюдал в деле впервые - и впечатление,
прямо скажу, было ошеломляющее...
Мой спутник - зоолог-полевик старой, классической школы - умел отыскивать
(и фотографировать) свое Зверье до того лихо, что Дереу Узала с Чингачгуком
тут, что называется, нервно курят в сторонке... Замечу, что самому мне тех
шастающих по горам вокруг нашей палатки зверей углядеть не удалось просто-
таки ни разу - за изъятием случаев, когда напарник, стоя рядом, давал прямые
инструкции, куда точно направлять бинокль, да еще и разъяснял при этом (сам-
то даже в бинокль не заглядывая), что, именно сейчас вот происходит пред мои-
ми вооруженными глазами6. А уж как он читал следы... От развития тяжелого
комплекса неполноценности меня тогда уберегало лишь то детское изумление, с
каким он, в свой черед, наблюдал - сколько всякой микроскопической живности
извлекаю я из какой-нибудь неприметной моховой кочки...
Так вот, кому как, а мне тех полутора месяцев, проведенных в той палатке,
вполне хватило для обретения непоколебимой убежденности: там, где единожды
ступила нога такого вот профессионала, ни одно животное крупнее крысы не име-
ет ни единого шанса остаться "неизвестным науке". Ну, а поскольку к концу
двадцатого столетия мест, где та нога профессионала не ступала бы вовсе, по-
читай, уже не осталось (по крайней мере на суше) - выводы делайте сами... Та-
кое вот мое субъективное мнение, да; по другому говоря - "экспертная оценка".
Эпоха "бури и натиска", когда европейские путешественники что ни год, то
открывали по крупному позвоночному (горная горилла, окапи, гигантский варан и
т. д.), в которой по сию пору черпают свой энтузиазм "криптозоологи", на са-
мом деле уместилась в пару десятилетий на границе XIX и XX веков; именно то-
гда, кстати, отправился в свое путешествие за динозаврами конан-дойлевский
профессор Челленджер. С той поры технология закрашивания "белых пятен" изме-
нилась принципиально, и дело тут даже не в увеличении числа зоологов в циви-
лизованных странах. Куда важнее то, что соотечественники Челленджера, несшие
свое "бремя белого человека" (да и не только они), успешно воспитали в коло-
ниях местные кадры, способные обеспечивать не только экспедиционные, но и
стационарные исследования; а дальше, как и везде, колоритные кондотьеры усту-
пили место "большим батальонам", что "всегда правы"... Между прочим, за всю
«Снежный человек» при этом сидел в кустах и грустно размышлял о том, что нет никакой лич-
ной жизни: «Когда же, наконец, этот Еськов со своим приятелем уберутся?». Кто-то кто, а он
то, выросший в настоящих дебрях, умел прятаться и наблюдать куда лучше выросших в каменных
дебрях больших городов. Иначе хрен бы дожил до наших дней.
эпоху "бури и натиска" не бывало случая, чтобы от первых слухов о существова-
нии в неких тропических дебрях некоего загадочного зверя до триумфальной дос-
тавки в Европу его шкуры с черепом прошло больше трех-пяти лет - сравните-ка
это со снежным человеком, которого вот уж сколько десятилетий как безуспешно
"ищут прохожие, ищет милиция"...
После Первой мировой войны бурный поток зоологических открытий такого рода
(то есть достойных внимания читателя воскресных газет) резко иссяк. В 1937
году в джунглях Индокитая был открыт дикий бык коупрей; сомнения на его счет
высказывали неоднократно, и вот только что генетический анализ расставил точ-
ки над "ё": таки да, одичавший домашний скот. В конце 70-х по добыче промыш-
лявших в Антарктике советских китобоев установили было новый вид китов, но
позже его разжаловали в цветовую разновидность. В 80-90-х описали несколько
видов копытных, обитающих во Вьетнаме, - оленей и антилоп; статус большей
части этих форм столь же сомнителен, как и коупрея, хотя антилопа псевдорикс
вроде бы действительно представляет собой новый для науки род... В любом слу-
чае , результатов за три четверти столетия негусто.
По-хорошему, в сухом остатке тут - лишь история открытия в 1938 году кисте-
перой рыбы: хранительница музея мисс Латимер, заинтересовавшаяся странной до-
бычей южноафриканских рыбаков, и упрямый ихтиолог профессор Смит, после че-
тырнадцатилетних поисков обнаруживший естественное местообитание латимерий в
сотнях километров от места первоначальной, случайной как выяснилось, поимки -
у Коморских островов близ Мадагаскара, на глубине более 200 метров. А ровно
через шестьдесят лет, в 1998 году, на противоположном конце Индийского океа-
на, в Индонезии, случился почти буквальный ремейк той давней истории. Амери-
канская туристка на местном рыбном рынке; фото остатков странной рыбы; экспе-
диционный грант, тут же выданный под ту фотографию (это к вопросу об Офици-
альной Науке&tm;, злонамеренно игнорирующей-де любые сообщения с мест о зага-
дочных находках); ну и - на следующий же год - новый вид латимерии, передан-
ный американскими ихтиологами в Индонезийский национальный музей!
Обратите внимание: латимерия - редчайшая глубоководная рыба, никогда не
поднимающаяся наверх, однако профессионалы, стоило им лишь взяться за дело,
нашли ее - на раз. Гигантского кальмара архитевтиса (тоже ведь редчайшее су-
щество) то найдут на берегу в дохлом виде, и сунут в научный спирт, то снимут
целый фильм - как он себя ведет в естественных условиях, на километровой глу-
бине. А вот Морской Змей, регулярно (якобы) резвящийся на поверхности, даже и
на фото все как-то толком не попадет - прямо Маркус Вольф какой-то! Ну а как
попадет - так непременно окажется полуразложившейся акулой...
Чур-чура! - я вовсе не хочу сказать, что "эпоха великих зоологических от-
крытий" уже позади, что мир "открыт до конца", а гумилевские звезды "пересчи-
таны все". Тут достаточно вспомнить выловленного недавно в Южной Пацифике
представителя губок-археоциат, считавшихся вымершими еще в кембрийском перио-
де, и открытый в 2000 году в Намибии новый отряд ортоптероидных насекомых
(его одновременно нашли еще и в балтийском янтаре!). Новые технологии позво-
лили начать регулярные исследования древесных крон в тропических лесах, что
привело к открытию - в числе прочего - нескольких видов низших обезьян и ле-
муров. Однако все эти открытия сделаны сугубыми профессионалами, да и оцене-
ны-то по достоинству могут быть лишь ими же - это вам не мега-кальмар, питаю-
щийся яхтами непослушных олигархов...
Так вот, возвращаясь к николай-степанычевым звездам: похоже, те из них, что
можно разглядеть без соответствующей астрономической подготовки и эквипмента,
сосчитаны все ж таки полностью. Работы по пересчету остальных - еще непочатый
край, но тут уже требуется некоторое знание матчасти. Тем же, кому эту мат-
часть учить влом, осталось, боюсь, лишь обсуждать на форумах связь снежного
человека с НЛО и торсионными полями. Либо попытаться пропихнуть поиски снеж-
ного человека в Вятской губернии в число Национальных Проектов&tm;; а что -
при правильном распиле отката вполне даже может пройти!
Ликбез
ЛЕКЦИИ ПО БИОЛОГИИ
(продолжение)
Янковский Н.К. с сотоварищами
ЛЕКЦИЯ №15.
МИТОЗ, МЕЙОЗ, ГАМЕТОГЕНЕЗ,
ОПЛОДОТВОРЕНИЕ И ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Тема лекции - деление клетки (митоз, мейоз и их отличиях) и индивидуальное
развитие (образовании гамет и оплодотворения и краткий обзор эмбрионального
развития).
Клетка в своей жизни проходит разные состояния: фазу роста и фазы подготов-
ки к делению и деления. Клеточный цикл - переход от деления к синтезу ве-
ществ, составляющих клетку, а затем опять к делению - можно представить на
схеме в виде цикла, в котором выделяют несколько фаз.
Клеточный цикл
После деления клетка вступает в фазу синтеза белков и роста, эту фазу на-
зывают G1. Часть клеток из этой фазы переходит в фазу GO, эти клетки функцио-
нируют и потом погибают без деления (например, эритроциты). Но большинство
клеток, накопив необходимые вещества и восстановив свой размер, а иногда и
без изменения размеров после предыдущего деления, начинают подготовку к сле-
дующему делению. Эта фаза называется фаза S - фаза синтеза ДНК, затем, когда
хромосомы удвоились, клетка переходит в фазу G2 - фазу подготовки в митозу.
Затем происходит митоз (деление клетки), и цикл повторяется заново. Фазы G1,
G2, S вместе называются интерфазой (т.е. фазой между делениями клетки).
Жизненный цикл клетки регулируется белками
цикпинами, концентрация которых меняется на
разных фазах цикла. Толщина цветных секторов
соответстует концентрации циклинов.
Жизнь клетки и переход от одной фазы клеточного
изменением концентраций белков циклинов, как это
цикла к другой регулируется
показано на рисунке.
Фазы G1, S, G2 вместе назваются интерфазой
(состояние между делениями клетки)
При подготовке к делению происходит репликация ДНК, на каждой хромосоме
синтезируется ее копия. Пока эти хромосомы после удвоения не расходятся, ка-
ждая хромосома в этой паре называется хроматидой. После репликации ДНК кон-
денсируется, хромосомы приобретают более компактную укладку, и в таком со-
стоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы
не столь конденсированы и в большей степени расплетены. Понятно, что в кон-
денсированном состоянии им трудно функционировать. Хромосома имеет вид в виде
буквы X только во время одной из стадий митоза. Раньше считалось, что между
делениями клетки хромосомная ДНК (хроматин) находится в полностью расплетен-
ном состоянии, но сейчас выясняется, что структура хромосом достаточно слож-
ная и степень деконденсации хроматина между делениями не очень велика.
Митотические хромосомы
Митотическая
пластинка ^1
Митотическая хромосома
состоит из двух хроматид
Процесс деления, при котором исходно диплоидная клетка дает две дочерние,
также диплоидные, клетки, называется митозом. Имеющиеся в клетке хромосомы
удваиваются, выстраиваются в клетке, образуя митотическую пластинку, к ним
прикреплены нити веретена деления, которые растягиваются к полюсам клетки и
клетка делится, образуя две копии исходного набора.
МИТОЗ
МЕЙОЗ
Дочерние клетки
(2п)
Второе
мейотическое
деление клетки
Гаметы (1 п)
При образовании гамет, т.е. половых клеток - сперматозоидов и яйцеклеток -
происходит деление клетки, называемое мейозом. Исходная клетка имеет диплоид-
ный набор хромосом, которые затем удваиваются. Но, если при митозе в каждой
хромосоме хроматиды просто расходятся, то при мейозе хромосома (состоящая из
двух хроматид) тесно переплетается своими частями с другой, гомологичной ей
хромосомой (также состоящей из двух хроматид), и происходит кроссинговер
обмен гомологичными участками хромосом. Затем уже новые хромосомы с переме-
шанными «мамиными» и «папиными» генами расходятся и образуются клетки с дип-
лоидным набором хромосом, но состав этих хромосом уже отличается от исходно-
го, в них произошла рекомбинация. Завершается первое деление мейоза, и вто-
рое деление мейоза происходит без синтеза ДНК, поэтому при этом делении коли-
чество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромо-
сом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки обра-
зуются четыре гаплоидных клетки. Фазы деления клетки, которые следуют за ин-
терфазой, называются профаза, метафаза, анафаза, телофаза и после деления
опять интерфаза.
Анафаза
Телофаза Интерфаза (G2)
При мейозе фазы называется также, но указывается, к какому делению мейоза
она относится. Кроссинговер - обмен частями между гомологичными хромосомами
- происходит в профазе первого деления мейоза (профаза I) , которая включает
следующие этапы: лептотена, зиготена, пахитена, диплотена, диакинез. Процес-
сы, происходящие при этом в клетке, подробно описаны в учебнике Макеева, и их
следует знать.
ФАЗЫ ПЕРВОГО ДЕЛЕНИЯ МЕИОЗА
Лептотена
ПРОФАЗА I МЕЙОЗА
t-----
r ChiLi iJM-'nes put
Зиготена
кроссннговер
—* Sytupk-teirul yChMMTuU----------------------
rnirpki
Пахитена Диплотена Диакинез
Кроссингов ер - обмен частями между гомологичными хромосомами
(отцовскими и материнскими) происходит в профазе I меПоза.
Краткий обзор
этапов гаметогенеза
Гаметогенез подразделяется на сперматогенез (процесс образования спермато-
зоидов у самцов) и оогенез (процесс образования яйцеклетки). По тому, что
происходит с ДНК, эти процессы практически не отличаются: одна исходная дип-
лоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с ци-
топлазмой, эти процессы кардинально различаются.
В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем
для развития зародыша, поэтому яйцеклетка - это очень крупная клетка, и когда
она делится, цель - сохранить питательные вещества для будущего зародыша, по-
этому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы
цитоплазмы и при этом избавиться от ненужного генетического материала, от ци-
топлазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы,
но позволяют поделить хромосомный набор,
вом и втором делении мейоза (подробнее
тельцами растений - в Макееве)
Полярные тельца отделяются при пер-
о том, что происходит с полярными
СПЕРМАТОГЕНЕЗ и ООГЕНЕЗ
SPLRNATOGENESIS OOGLNESIS
Mei о tic
division I
Mciotic
division II
primary
spermatocyte
secondary
spermatocytes
QS I1, '„Oj
spermatids
Differentiation 1
primary
oocyte
secondary
ootid
ovum
first polar
body
second polar
bodies
spermatozoa
Исходная клетка, из которой в последствии образуется зрелая яйцеклетка, на-
зывается ооцитом первого порядка. После деления из него образуется ооцит
второго порядка и первое полярное тельце. Затем происходит второе деление
мейоза, в результате образуется гаплоидный оотид и второе полярное тельце.
Первое полярное тельце за это время тоже успевает поделиться, таким образом
всего получается три гаплоидных полярных тельца. В оотиде происходят некото-
рые процессы созревания и он превращается в яйцеклетку. Она содержащая почти
всю цитоплазму исходного ооцита, но гаплоидный набор хромосом. Эти хромосомы
уже прошли рекомбинацию, т.е. если исходно клетки содержат одну хромосому от
мамы, одну от папы, то в зрелой яйцеклетке в каждой хромосоме чередуются кус-
ки, полученные от одного и второго родителя.
При сперматогенезе цитоплазма исходного сперматоцита первого порядка делит-
ся (первое деление мейоза) поровну между клетками, давая сперматоциты второго
порядка. Второе деление мейоза приводит к образованию гаплоидных сперматоци-
тов второго порядка. Затем происходит созревание без деления клетки, большая
часть цитоплазмы отбрасывается, и получаются сперматозоиды, содержащие гапло-
идный набор хромосом очень мало цитоплазмы. Ниже представлена фотография
сперматозоида человека и схема его строения.
Сперматозоиды животных имеют одинаковое принципиальное строение, но могут
отличаться формой и размером. Сперматозоид имеет головку, в которую плотно
упакована ДНК. Головка сперматозоида окружена очень тонким слоем цитоплазмы.
На ее переднем конце находится структура, называемая акросомой. Эта структура
содержит ферменты, позволяющие сперматозоиду проникнуть через оболочку яйце-
клетки . Сперматозоид имеет хвостик. Часть хвостика, прилегающая к головке
("шейка"), окружена митохондриями. Они необходимы, чтобы обеспечить биение
хвостика и движение сперматозоида в желательном ему направлении. На спермато-
зоиде имеется для выбора направления движения хеморецепторы, сходные с обоня-
тельными клетками.
СТРОЕНИЕ СПЕРМИЯ
Созревание спермиев происходит в семенных канальцах тестикул. При превраще-
нии исходной клетки, сперматогония, в сперматоцит, сперматиды и зрелый спер-
матозоид происходит перемещение клетки от базальной мембраны семенного кана-
тика к его полости. После созревания сперматозоиды отделяются, попадая в
просвет семенных канальцев, и готовы к движению в поисках яйцеклетки и опло-
дотворению. Процесс созревания длится примерно три месяца. У млекопитающих у
особей мужского пола процесс созревания сперматозоидов - сперматогенез - на-
чинается с возраста половой зрелости и продолжается затем до глубокой старос-
ти .
Созревание спермиев происходит в семенных канальцах тестикул
Полость канальца
мембрана
http:/Avww.eastcentnl.edu/acacVdepts/BI/human tpennalogeness Jabels.jpg
Существенно отличается процесс созревания яйцеклетки - оогенез. Во время
эмбрионального развития млекопитающих возникает большое количество яйцекле-
ток, и к рождению самки в ее яичниках уже находится порядка 200-300 тысяч яй-
цеклеток , остановившихся на первой стадии деления мейоза. В период полового
созревания яйцеклетки начинают реагировать на половые гормоны, Регулярные
циклические изменения гормонов впоследствии вызывают созревание яйцеклетки,
обычно одной, иногда двух или больше. Когда для лечения бесплодия женщине де-
лают инъекции половых гормонов, чтобы индуцировать созревание яйцеклеток, из-
быток этих гормонов может привести к созреванию нескольких яйцеклеток, и как
следствие этого - многоплодной беременности. Яйцеклетка созревает в пузырьке,
называемом фолликулом.
Зрелый фолликул перед овуляцией
За всю жизнь у женщин современных индустриализованных стран созревает всего
400-500 яйцеклеток, у женщин традиционной культуры - в племенах охотников-
собирателей - менее 200 штук. Это связано с различиями в традиции деторожде-
ния: у европейских женщин рождается в среднем 1-2 ребенка, которых она кормит
в среднем 3 -5 месяцев, (а известно, что лактация тормозит восстановление ме-
сячных циклов после родов), то есть у нее больший период времени остается для
созревания яйцеклеток и прохождения менструальных циклов; в это же самое вре-
мя у бушменов женщины рожают в среднем по 5 детей, они не делают абортов, в
отличие от западных женщин, и они кормят грудью по 3-4 года, при этом овуля-
ция тормозится, поэтому месячных циклов у них в 2 раза меньше, чем у западных
женщин. Большее количество овуляторных циклов ведет к повышению риска заболе-
вания репродуктивных органов у женщин, так как каждая овуляция связана с де-
лением клеток, а чем больше делений - тем больше может возникнуть мутаций,
ведущих к появлению злокачественных образований.
Месячные циклы у женщины регулируются изменением концентрации гормонов
(верхний график на рисунке). Под действием гормонов один из покоящихся фолли-
кулов (пузырьков) с яйцеклеткой начинает развиваться. Через несколько дней
фолликул лопается и из него выходит зрелая яйцеклетка. Этот процесс называ-
ется овуляцией. Слизистая оболочка матки (эндометрий) при этом разрастается,
готовясь принять оплодотворенную яйцеклетку. Если беременность не наступает,
происходит дегенерация и отторжение верхнего слоя эндометрия, сопровождающее-
ся кровотечением. Во время овуляции у женщины происходит повышение так назы-
ваемой базальной температуры (то есть температуры, измеряемой ректально и ва-
гинально сразу после пробуждения) на несколько десятых градуса (нижний график
на рисунке), потом она может упасть или остаться слегка повышенной до начала
менструации. У каждой женщины колебания базальной температуры индивидуальны,
но более или менее постоянны при установившемся месячном цикле. Таким образом
по изменению температуры можно примерно судить, когда происходит овуляция.
гдйрмые гормоны
Базальная температура I
Изменения
Эндометрия
Фолликулостимулирующий
гормон 1
I
Гормоны яичник а
I
Эстрадиол
I 7 М 21 20
Менструация Пролиферативная Овуляция Секреторная фата
Ошибки при определении сроков овуляции по базальной температуре могут воз-
никать из-за не связанных с месячным циклом изменений температуры (например,
при гриппе или другом заболевании, дающем подъем температуры) или из-за сбоев
цикла, которые могут возникнуть у женщины при перемене климата, стрессе или
под влиянием других факторов. Пример изменения температуры в одном месячном
цикле представлен на рисунке:
Яйцеклетка жизнеспособна 24-48 час.
С перми й активен до 2-3 суток.
Яйцеклетка после выхода из фолликула сохраняет жизнеспособность примерно
24-48 часов. Спермин же после попадания в половые пути женщины жизнеспособны
до 2-3 суток, далее они могут быть подвижны, но не способны к оплодотворению.
Поэтому оплодотворение возможно в течение 2-3 дней до и 1-2 дней после овуля-
ции . В остальное время зачатие произойти не может. Но на самом деле скачок
температуры происходит не точно при овуляции, а при изменении концентрации
гормонов, овуляцию вызывающих, поэтому точность определения дня овуляции по
температурному графику составляет примерно 2 дня. Поэтому оплодотворение мо-
жет произойти в 3+2=5 дней до овуляции и 2+2=4 дня после овуляции дней цикла.
Осторожные люди прибавляют еще по 1-2 дня с каждой стороны. Остальные дни
считаются "безопасными". Хотелось бы отметить, что цикл подчиняется эмоцио-
нальной регуляции, например, во время войны из-за тяжелой жизни, недоедания у
женщин прекращались менструации, это явление называется "аменорея военного
времени". Однако описаны случаи, когда муж приезжал домой с фронта на 2 дня,
за эти 2 дня у женщины происходила овуляция независимо от фазы цикла, и впо-
следствии рождался ребенок. О том, что физиологические процессы достаточно
сильно могут регулироваться нервной системой, показывает процесс родов у
обезьян. У человека первые роды длятся примерно 24 часа, а у обезьян всего
несколько часов, причем начинаются они обычно во время, когда стадо находится
на стоянке. То есть к утру, когда стадо собирается отправляться в путь, мама
готова путешествовать дальше с новорожденным. Если по каким-то причинам про-
цесс родов к утру не завершился, а стадо уже готово идти дальше, то роды ос-
танавливаются, так как стадные животные не должны отставать от своих сороди-
чей, и уже потом при новой остановке, роды возобновляются.
Процесс проникновения сперматозоидов в яйцеклетку называется оплодотворени-
ем. Яйцеклетка окружена несколькими оболочками, структура которых такова, что
только сперматозоид собственного вида может попасть в яйцеклетку. После опло-
дотворения оболочки яйцеклетки меняются и другие сперматозоиды уже не могут в
нее проникнуть.
Яйцеклетка перед оплодотворемим
"лучистьй вснец"н одно полярное тельце.
Оплодотворяемая Яйцеклетка и сперматозоидм моллюска
Яйцеклетка иллюстрация с сайта www unr edu
У некоторых видов внутрь яйцеклетки могут проникнуть несколько сперматозои-
дов, но все равно в слиянии ядер участвует только один из них. При оплодотво-
рении в яйцеклетку проникает только ядро сперматозоида, хвостик же вместе
митохондриями отбрасывается, и в клетку не попадает. Поэтому митохондриальную
ДНК все животные наследуют только от матери. Оплодотворенное яйцо называют
зиготой (от греч. зиготос - соединенный вместе).
После оплодотворения происходит деление клетки, восстановившей диплоидный
набор хромосом. Первое и несколько последующих делений яйцеклетки происходят
без увеличения размера клеток, поэтому процесс называется дроблением яйце-
клетки .
Эмбрион (греч. "зародыш") - ранняя стадия развития живого организма от на-
чала дробления яйцеклетки до выхода из яйца или из материнского организма (в
акушерстве, в отличие от эмбриологии, термин эмбрион используют только для
первых 8 недель развития, после 8-й недели называют плодом).
Эмбриогенез (эмбриональное развитие) является частью онтогенеза (индивиду-
ального развития) - развития организма от образования зиготы до его смерти.
Эмбриогенез - это процесс, в котором презумптивные зачатки занимают свои де-
финитивные места.
Из школьного вы помните, что при развитии эмбриона ланцетника образуется
бластула (полый клеточный шарик), из которой получается двуслойная гаструла
путем инвагинации (впячивания) одной стороны бластулы внутрь.
Дробление и гаструляция яйца ланцетника
Бластула Гаструла
У млекопитающих процесс происходит несколько иным образом. Дробление яйце-
клетки у них приводит к образованию комочка клеток, называемого морулой. Мо-
рула подразделяется на внутреннюю часть, из которой потом развивается сам за-
родыш , и наружную часть, образующую полый пузырек, называемый трофобластом.
Дальнейшее развитие приводит к формированию трехслойного зародыша, состоящего
из внутреннего слоя - энтодермы, внешнего слоя - эктодермы, и третьего слоя
между ними - мезодермы. Из каждого слоя впоследствии образуются определенные
ткани и органы.
На фотографии ниже изображен червь нематода Caenorhabditis elegans (произ-
носится как "ценорабдитис элеганс"), ближайший родственник аскариды, который
интересен тем, что имеет строго фиксированное число клеток. Это дает возмож-
ность установить происхождение каждой из клеток, какая клетка из какой полу-
чилась . На рисунке показана схема происхождения разных клеток в эмбриональном
развитии С.elegans.
Клеточные линии, образующие тело нематоды c.elegans
Зигота
Во время развития зародыша происходит дифференциация и перемещение его кле-
ток с образованием тканей и органов. Рассмотрим на примере мухи-дрозофилы
этот процесс. В развитии мухи происходит последовательная смена форм, значи-
тельно отличающихся друг от друга: яйцо, личинка, куколка и имаго (взрослая
особь). Такое развитие называется развитием с метаморфозом (метаморфоз -
греч. "изменение формы").
Развитие дрозофилы: личинка, куколка и имаго
Цитоплазма яйцеклетки не гомогенна, в ней асимметрично распределены различ-
ные биологически активные компоненты.
У эмбриона уже определены оси тела, число и ориентация сегментов тела, из
которых затем развиваются части тела взрослой мухи. Эти процесс контролируют-
ся различными наборами генов, которые называются морфогены. Они кодируют бел-
ки , которые регулируют экспрессию других генов, отвечающих за формирование
органов.
Градиент концентрации белков-морфогенов определяет передне-заднюю и дорсо-
вентральную (спино-брюшную) оси тела. У дрозофилы в определении дорсо-
вентральной оси тела участвуют 12, главный из них ген dorsal. Белок Dorsal
сконцентрирован в цитоплазме на той стороне эмбриона, которая станет спинной,
и проникает в ядро клеток на брюшной стороне, активируя группы генов, продук-
ты которых необходимы для определения осей тела.
Детерминация передне-задней оси тела контролируется другими генами. Один из
них называется bicoid, его мРНК сконцентрирована в цитоплазме передней части
яйца (фиксирована своим 3’-концом). В результате при трансляции возникает
градиент концентрации белка Bicoid от переднего к заднему концу яйца. Гради-
ент поддерживается продолжительным периодом синтеза белка и его коротким вре-
менем жизни. Bicoid, так же как и Dorsal, морфоген, он активирует гены, кото-
рые необходимы для развития головы и грудных структур. Его экспрессия ингиби-
руется продуктом гена nanos, сконцентрированного на заднем конце эмбриона.
На следующем этапе включаются гены сегментации. Они контролируют дифферен-
циацию эмбриона на индивидуальные сегменты. После оплодотворения транскриби-
руется около 25 генов сегментации, их экспрессия регулируется градиентами
белков Bicoid и Nanos.
Эмбрион - 2 часа
Установлены передне-задняя и
спинно-брюшная оси тела
Эмбрион 10 часов
Определены число и ориентация
сегментов тела
Взрослая муха
Каждая часть тела взрослого
насекомого развилась из
соответствующих сегментов эмбриона
Гомеозисные гены
После сегментации и установления ориентации сегментов активируются так на-
зываемые геомеозисные гены. Различные их наборы активируются специфическими
соотношениями концентраций белков, упоминавшихся выше.
Продукты гомеозисных генов активируют другие гены, которые определяют сег-
мент -специфичные особенности. Глаза в норме возникают только на головном сег-
менте , а ноги - только на грудных сегментах.
Гомеозисные гены кодируют регуляторные белки, связывающиеся с ДНК. Каждый
из них содержит кластер нуклеотидов, называемый гомеобоксом, которые сходен
во всех гомеотических генах. Он содержит 180 нуклеотидов и кодирует 60 амино-
кислот , функционирующих как ДНК-связывающий домен.
У дрозофилы имеется два основных кластера гомеотических генов: комплекс
Antennapedia (5 генов у дрозофилы) который определяет развитие головы и пе-
редних торакальных сегментов, и комплекс Bithorax (3 гена) который контро-
лирует развитие задних торакальных и брюшных сегментов. Порядок расположения
генов тот же, что и сегментов, в которых они экспрессируются.
Впервые мутации гомеозисных генов были идентифицированы в 1894 году, когда
Уильям Бэтсон заметил, что у растений иногда части цветка появляются на не-
правильных местах. Гомеозисные гены как бы определяют адрес клетки в конкрет-
ном сегменте, сообщая клеткам, в каком районе они находятся. Когда они мути-
руют, клетки получают "ложный адрес" и ведут себя так, будто они находятся в
другом месте эмбриона
Нарушения в работе гомеозисных генов (вызванные мутациями или внешними воз-
действиями) нарушают формирование структур тела и могут привести, например, к
образованию глаз на лапках у мухи, или к тому, что вместо антенн на голову у
нее вырастут ноги (как это показано на рисунке). У человека найдены мутации в
гомеозисных генах, приводящие к недоразвитию зубов, например, и к другим, бо-
лее тяжелым нарушениям.
НАРУШЕНИЕ В РАБОТЕ ГОМЕО311СНЫХ ГЕНОВ ВЕДЕТ К НАРУШЕНИЮ МОРФОГЕНЕЗА
Мутация в гене Anrcnnapcdia ведет к появлению ног вместо антенн на голове мухи
Порядок расположения го.меотненых генов соответствует поряжу контролируемо: ими се1ментовтела
После того, как были открыты и изучены геомео-гены дрозофилы, сходные гены
были найдены у всех других животных от нематоды до человека. У млекопитающих
они называются Нох генами (гомеобокс-содержащими генами) и также кодируют
белки, регулирующие транскрипции и определяющие структуры тела и их положе-
ние в передне-заднем направлении.
nuustf emU уо
гтлгл.-
Таким образом, в эмбриональном развитии исходный градиент белков и мРНК в
яйцеклетке стимулирует локальную экспрессию генов эмбриона, которая ведет
дальнейшей дифференциации генной экспрессии и определяет судьбу клеток разви-
вающегося эмбриона. Процесс, в котором формируются конечности, ткани и орга-
ны, называется морфогенезом, и определяется последовательностью переключения
экспрессии групп генов, однако эти гены пока не столь детально изучены.
В процессе эмбриогенеза осуществление записанной в генах программы развития
происходит в конкретных условиях среды. Взаимодействие генов и среды можно
описать на следующей модели. Эмбриональное развитие можно сравнить с шари-
ком, катящимся по наклонной поверхности с разными желобками. Такое представ-
ление эмбрионального развития, названное эпигенетическим ландшафтом, был
предложен Кондратом Уоддингтоном.
Эпигенетический ландшафт
Мутация
rowmcr «од р<шги>мч
Норма
paiBMrm
по момОогкс юс роят ному -
нормальному пути
Талидомид
Воздействие
среды (лечение)
может сгомгкжироаать
дпктине мут«|пм и оернути
рвлтпте к морьс
Вредное
влияние среды
жсжят примети к огпшммм»
от иор^альмого гт/гм рлтямгня
даж» при оттутегяни мутации
Самый глубокий желобок (соответствующий наиболее вероятному пути) определя-
ет нормальное развитие организма. Но у основного желобка есть много разветв-
лений, менее глубоких, соответствующих патологии, аномальному развитию орга-
низма, по ним шарик покатится с меньшей долей вероятности. Мутации меняют
соотношение вероятностей разных путей (на рисунке - меняется глубина желоб-
ков) , и увеличивают вероятность развития по "неправильному" патологическому
пути. Однако в части случаев воздействие среды (лечение) может скомпенсиро-
вать дефект и вернуть организм на нормальный путь развития. Например, фенил-
кетонурия - наследственная болезнь, которую можно лечить. Суть болезни заклю-
чается в том, что у больных отсутствует фермент фенилаланингидроксилаза, пре-
вращающий аминокислоту фенилаланин в другую аминокислоту, тирозин. При блоки-
ровании нормальных путей катаболизма фенилаланина его превращение идет други-
ми путями, обычно играющими второстепенную роль. Фенилаланин превращается в
кетокислоту фенилпируват (фенилпировиноградная кислота) и другие продукты.
Избыточные количества фенилпирувата легко определить по анализу мочи, и такой
анализ проводится всем новорожденным в нашей стране. Одним из симптомов этой
болезни является развитие умственной отсталости, которое во взрослом состоя-
нии уже необратимо. Лечить болезнь можно в детстве специальной диетой, при
которой в организме не из чего будет вырабатывать пировиноградную кислоту.
Частота заболевания около 1:10 000 новорожденных, и чем раньше начато лечение
- тем лучше результаты. Именно поэтому проводится тотальная диагностика но-
ворожденных. Интересно то, что если ребенок перестает придерживаться лечебной
диеты, то болезнь опять станет прогрессировать. Поэтому диету надо соблюдать
до остановки физиологического роста, примерно до 20 лет, когда токсичное воз-
действие будет менее опасным. Больные фенилкетонурией при беременности обяза-
тельно должны соблюдать диету, так как иначе плод будет отравлен из-за нару-
шения обмена веществ у матери.
Таким образом, при лечении, то есть полезном воздействии внешней среды,
можно вернуть развитие организма в нормальное русло. Но действие окружающей
среды может быть и вредным, то есть у организма под действием внешней среды
возникают отклонения развития при совершенно нормальных генах. Для примера
рассмотрим один случай. В 60-х годах в Германии было сильно разрекламирова-
но новое снотворное под названием талидомид. Среди принимавших новое лекар-
ство были беременные женщины. Спустя некоторое время было замечено, что в
стране стало рождаться много детей с патологией конечностей. У них отсутство-
вали длинные кости конечностей, то есть прямо от основания тела начинались
кисти или ступни.
Раньше такое заболевание встречалась один раз на несколько тысяч новорож-
денных, и вдруг такой всплеск. Начали проводиться исследования, и выяснилось,
что причина в новом лекарстве. Как оказалось, талидомид имеет большое сродст-
во к гуанину. Взаимодействуя с ДНК, он может приводить к функциональным нару-
шениям. Промотор гена, отвечающего за рост и развитие длинных конечностей,
содержит большое количество гуанина, таким образом, талидомид нарушает работу
этих генов, и зачатки костей длинных конечностей так и не начинают развивать-
ся. Многие из этих детей не выжили, часть из тех, кто выжил, ведут жизнь ин-
валидов, но есть среди них люди, которые, несмотря на инвалидность, реализо-
вали свои возможности.
После талидомидной трагедии все новые лекарства проверяют на тератогенную
(вызывающую нарушения развития плода) активность, и для каждого препарата
указано, можно ли его принимать беременным. Однако следует учитывать, что во
время беременности, особенно на ранних этапах, женщина не должна принимать
лекарства, не посоветовавшись с врачом, из-за возможных вредных воздействий
на плод.
В настоящее время уровень тяжелых врожденных уродств составляет 1-2%, из
них около трети по генетическим причинам, около трети - из-за воздействий
среды, и для трети причина неизвестна. Подбирая условия среды, соответствую-
щие индивидуальным особенностям организма, можно скомпенсировать часть врож-
денных дефектов.
Взаимодействие генов и среды в процессе индивидуального развития можно
сравнить с игрой в карты: хороший игрок может выиграть и с плохими картами.
ЛЕКЦИЯ №16.
ОСНОВНЫЕ ПРИНЦИПЫ
НАСЛЕДОВАНИЯ И НАСЛЕДСТВЕННОСТИ
Мы начнем с изложения законов Менделя, затем поговорим про Моргана, и в
конце скажем, зачем генетика нужна сегодня, чем она помогает и каковы ее ме-
тоды.
В 1860-х годах монах Мендель занялся исследованием наследования признаков.
Этим занимались и до него, и впервые об этом говорится в Библии. В Ветхом за-
вете говорится о том, что если владелец скота хотел получить определенную по-
роду, то он одних овец кормил ветками очищенными, если хотел получить потом-
ство с белой шерстью, и неочищенными, если хотел получить шкуру скота черной.
То есть, как наследуются признаки волновало людей еще до написания Библии.
Почему же до Менделя никак не могли найти законы передачи признаков в поколе-
ниях?
Дело в том, что до него исследователи выбирали совокупность признаков одно-
го индивида, с которыми было сложнее разбираться, нежели с одним признаком.
До него передача признаков рассматривалась часто как единый комплекс (типа -
у нее лицо бабушкино, хотя отдельных признаков тут очень много). А Мендель
регистрировал передачу каждого признака в отдельности, независимо от того,
как передались потомкам другие признаки.
Важно, что Мендель выбрал для исследования признаки, регистрация которых
была предельно простой. Это признаки дискретные и альтернативные:
1. дискретные (прерывистые) признаки: данный признак либо присутствует, ли-
бо отсутствует. Например, признак цвета: горошина либо зеленая, либо не
зеленая.
2. альтернативные признаки: одно состояние признака исключает наличие дру-
гого состояния. Например, состояние такого признака как цвет: горошина
либо зеленая, либо желтая. Оба состояния признака в одном организме про-
явиться не могут.
Подход к анализу потомков был у Менделя такой, который до него не применя-
ли. Это количественный, статистический метод анализа: все потомки с данным
состоянием признака (например - горошины зеленые) объединялись в одну группу
и подсчитывалось их число, которое сравнивали с числом потомков с другим со-
стоянием признака (горошины желтые).
В качестве признака Мендель выбрал цвет семян посевного гороха, состояние
которого было взаимоисключающим: цвет или желтый, или зеленый. Другой признак
- форма семян. Альтернативные состояния признака - форма или морщинистая или
гладкая. Оказалось, что эти признаки стабильно воспроизводятся в поколениях,
и проявляются либо в одном состоянии, либо в другом. В общей сложности Мен-
дель исследовал 7 пар признаков, следя за каждым по отдельности.
При скрещивании Мендель исследовал передачу признаков от родителей к их
потомкам. И вот что он получил. Один из родителей давал в череде поколений
при самоопылении только морщинистые семена, другой родитель - только гладкие
семена.
Наследственные признаки у гороха Ptsum sativum :
форма и цвет семян, форма и цвет стручка, размер растения, расположение цветков
Горох - самоопылитель. Для того, чтобы получить потомство от двух разных
родителей (гибриды), ему нужно было сделать так, чтобы растения не самоопыля-
лись. Для этого он удалял у одного родительского растения тычинки, и перено-
сил на него пыльцу с другого растения. В этом случае образовавшиеся семена
были гибридными. Все гибридные семена в первом поколении оказались одинаковы-
ми. Все они оказались гладкими. Проявившееся состояние признака мы называем
доминантным (значение корня этого слова - господствующий). Другое состояние
признака (морщинистые семена) у гибридов не обнаруживалось. Такое состояние
признака мы называем рецессивным (уступающим).
Мендель скрестил растения первого поколения внутри себя и посмотрел на фор-
му получившихся горошин (это было второе поколение потомков скрещивания). Ос-
новная часть семян оказалась гладкой. Но часть была морщинистой, точно такой
же у исходного родителя (если б мы говорили про собственную семью, то сказали
бы, что внук был точно в дедушку, хоть у папы с мамой этого состояния призна-
ка не было совсем). Он провел количественное исследование того, какая доля
потомков относится к одному классу (гладкие - доминантные), а какая к другому
классу (морщинистые - рецессивные). Оказалось, что морщинистых семян получи-
лась примерно четверть, а три четверти - гладких.
Мендель провел такие же скрещивания гибридов первого поколения по всем ос-
тальным признакам: цвету семян, окраски цветка и др. Он увидел, что соотноше-
ние 3:1 сохраняется.
Мендель провел скрещивание и в одном направлении (папа с доминантным при-
знаком, мама - с рецессивным) и в другом (папа с рецессивным признаком, мама
с доминантным). При этом качественные и количественные результаты передачи
признаков в поколениях были одинаковыми. Из этого можно сделать вывод, что и
женские и отцовские задатки признака вносят одинаковый вклад в наследование
признака у потомства.
То, что в первом поколении проявляется признак только одного родителя, мы
называем законом единообразия гибридов первого поколения или законом домини-
рования .
То, что во втором поколении вновь появляются признаки и одного родителя
(доминантный) и другого (рецессивный) позволило Менделю предположить, что на-
следуется не признак как таковой, а задаток его развития (то, что мы сейчас
называем геном). Он также предположил, что каждый организм содержит пару та-
ких задатков для каждого признака. От родителя к потомку переходит только
один из двух задатков. Задаток каждого типа (доминантный или рецессивный) пе-
реходит к потомку с равной вероятностью. При объединении у потомка двух раз-
ных задатков (доминантный и рецессивный) проявляется только один из них (до-
минантный, он обозначается большой буквой А). Рецессивный задаток (он обозна-
чается малой буквой а) у гибрида не исчезает, поскольку проявляется в виде
признака в следующем поколении
ОвиЬ— Uftnn tree wiihtau dtfcieffiTUI»— tuwri tnd
UlflUlb! UuSC Uu iltUlll 1111 IfWI utfl&C
iim .if th«i t>MXо/imo ei>iц <м а
lilltMTItlllr VBt*
hl I0UMIIO1» Mil
hjimrUln] *6r?»1fc£r*ri]
UifrvUK ©
Y
P genetMloH *9i»uftuui ~л1
"jitid к-rd-, iM-rhkd sent»
*Ih iiu« i? tn Mint» ife.WwhI
Ajihrujti t| flairs dscnir <t»e pteratvp: n( me puiirr.-
baih no мя«1 (л r. jK'Mfi'v *i 1:1 iwj<..
Схема моногрибридного
скрещивания
Явление доминирования
Так как во втором поколении появился точно такой же организм, как и роди-
тельский, Мендель решил, что задаток одного признака «не замазывается», при
объединении с другим, он остается таким же чистым. В последствии было выясне-
но то, что от данного организма передается только половина его задатков - по-
ловые клетки, они называются гаметами, несут только один из двух альтерна-
тивных признаков.
У человека насчитывается около 5 тыс. морфологических и биохимических при-
знаков , которые наследуются достаточно четко по Менделю. Судя по расщеплению
во втором поколении, альтернативные задатки одного признака комбинировались
друг с другом независимо. То есть доминантный признак мог проявиться при
комбинациях типа Аа, аА и АА, а рецессивный только в комбинации аа.
Повторим, что Мендель предположил, что наследуется не признак, а задатки
признака (гены) и что эти задатки не смешиваются, поэтому этот закон называ-
ется законом чистоты гамет. Через исследование процесса наследования можно
было сделать выводы о некоторых характеристиках наследуемого материала, то
есть что задатки стабильны в поколениях, сохраняют свои свойства, что задатки
дискретны, то есть, определяются только одно состояние признака, то, что их
два, они комбинируются случайно и т.д.
Во времена Менделя еще ничего не было известно о мейозе, хотя про ядерное
строение клетки уже знали. То, что в ядре содержится вещество, названное нук-
леином, стало известно только через пару лет после открытия законов Менделя,
причем это открытие с ним никак не было связано.
Все выводы вышеизложенного материала можно сформулировать следующим обра-
зом:
1) Каждый наследственный признак определяется отдельным наследственным фак-
тором, задатком; в современном представлении эти задатки соответствуют
генам;
2) Гены сохраняются в чистом виде в ряду поколений, не утрачивая своей ин-
дивидуальности: это явилось доказательством основного положения генети-
ки: ген относительно постоянен;
3) Оба пола в равной мере участвуют в передаче своих наследственных свойств
потомству;
4) Редупликация равного числа генов и их редукция в мужских и женских поло-
вых клетках; это положение явилось генетическим предвидением существова-
ния мейоза;
5) Наследственные задатки являются парными, один - материнский, другой -
отцовский; один из них может быть доминантным, другой - рецессивным; это
положение соответствует открытию принципа аллелизма: ген представлен ми-
нимум двумя аллелями.
К законам наследования относятся закон расщепления наследственных признаков
в потомстве гибрида и закон независимого комбинирования наследственных при-
знаков . Эти два закона отражают процесс передачи наследственной информации в
клеточных поколениях при половом размножении. Их открытие явилось первым фак-
тическим доказательством существования наследственности как явления.
Законы наследственности имеют другое содержание, и они формулируются в сле-
дующем виде:
• Первый закон - закон дискретной (генной) наследственной детерминации
признаков; он лежит в основе теории гена.
• Второй закон - закон относительного постоянства наследственной единицы -
гена.
• Третий закон - закон аллельного состояния гена (доминантность и рецес-
сивность) .
То, что законы Менделя связаны с поведением хромосом при мейозе, было обна-
ружено в начале XX века во время повторного открытия законов Менделя сразу
тремя группами ученых независимо друг от друга. Как вам уже известно, особен-
ность мейоза заключается в том, что число хромосом в клетке уменьшается
вдвое, хромосомы могут меняться своими частями при мейозе. Такая особенность
характеризует ситуацию с жизненным циклом у всех эукариот.
Для того чтобы проверить предположение о наследовании задатков в таком ви-
де, как мы уже говорили, Мендель провел также скрещивание потомков первого
поколения, имеющие желтые семена с родительскими зелеными (рецессивными).
Скрещивание на рецессивный организм он назвал анализирующим. В результате он
получил расщепление один к одному: (Аа х аа = Аа + Аа + аа + аа). Таким обра-
зом, Мендель подтвердил предположение, что в организме первого поколения есть
задатки признаков каждого из родителей в соотношении 1 к 1. Состояние, когда
оба задатка признака одинаковы, Мендель назвал гомозиготным, а когда разные -
гетерозиготным.
Поколение Р
Поколение F1
Соотношение генотипов 1Tt :1tt
Соотношение фенотипов
1 Высокий :1 Низким
Мендель учитывал результаты, полученные на тысячах семян, то есть он прово-
дил статистические исследования, которые отражают биологическую закономер-
ность. Открытые им самые законы будут действовать и на других эукариотах, на-
пример грибах. Здесь показаны грибы, у которых четыре споры, получаемые в ре-
зультате одного мейоза, остаются в общей оболочке. Анализирующее скрещивание
у таких грибов приводит к тому, в одной оболочке присутствуют 2 споры с при-
знаком одного родителя и две с признаком другого. Таким образом, расщепление
1:1 в анализирующем скрещивании отражает биологическую закономерность расщеп-
ления задатков одного признака в каждом мейозе, которая будет выглядеть как
закономерность статистическая, если все споры смешать.
То, что у родителей были разные состояния одного признака, говорит о том,
что задатки к развитию признака могут как-то меняться. Эти изменения называ-
ются мутациями. Мутации бывают нейтральными: форма волос, цвет глаз и др. Не-
которые мутации приводят к изменениям, нарушающим нормальное функционирование
организма. Это коротконогость у животных (крупный рогатый скот, овцы и др.),
безглазость и бескрылость у насекомых, бесшерстность у млекопитающих, гиган-
тизм и карликовость.
Некоторые мутации могут быть и безвредными, например бесшерстность у лю-
дей, хотя все приматы имеют волосяной покров. Но иногда встречаются изменения
интенсивности волосяного покрова на теле и у людей. Н.И.Вавилов назвал такое
явление законом гомологических рядов наследственной изменчивости: то есть
признак, типичный только для одного из двух родственных видов, может быть об-
наружен с какой-то частотой и у особей родственного вида.
На этом слайде показано то, что мутации могут быть достаточно заметными, мы
видим негритянскую семью, в которой родился белый негр - альбинос. У него де-
ти, скорее всего, будут пигментированными, поскольку мутация эта рецессивная,
а частота ее встречаемости низка.
3.3 AIMnlsm In human b«ings Is usually Inker
lead as a recessive trail. (Richard Oanuke/ss/’boto
Researchers.}'
Мы говорили до этого о признаках, которые проявляются полностью. Но это не
для всех признаков так. Например, фенотип гетерозигот может быть промежуточ-
ным между доминантным и рецессивным признаком родителей. Так, окраска плода
у баклажан в первом поколении меняется с темно-синей на менее интенсивную
фиолетовую. При этом во втором поколении расщепление по наличию окраски оста-
лось 3:1, но если учитывать интенсивность окраски расщепление стало 1:2:1
(цвет темно-синий - АА, фиолетовый - 2Аа и белый - аа, соответственно) В дан-
ном случае видно, что проявление признака зависит от дозы доминантного алле-
ля. Расщепление по фенотипу соответствует расщеплению по генотипу: классы АА,
Аа и аа, в соотношении 1:2:1.
1а»
la)
P generation
Purple fruit
X
White fruit
Г generation
Round, yellow
seeds
Gametes (ду)
Wrinkled, qreen
seeds
I »n »ll ration
Violet fruit
Gametes
Fi4 tilizat Ion
Gametes (F
L
F, generation
Violet fruit
(Ы
F, generation
Conclusion Genotypic ratio i PP: 2 P^z 1 pp
Phenotypic, ratio Ipurpie;Zviolet:i white
IM
Pi oarathe
Id
Round, yellow
seeds
Rrfj
Srlf fi’riNi
Condition:
Phenwpic tatio
9 reund. yellow: 3 round. prten:3 crinkled,
yellow: I wrinkled, green
4
x
Неполное доминирование
Соотношение генотипов 1РР:2Рр:1рр
Соотношени фенотипов
1 пурпурный?фиолетовых!белый
Дигибридное скрещивание
Соотношение фенотипов
9 круглых желтых :3круглых зеленых
:3 морщинистых желтых 1 морщинистый желтый
Еще раз выделим роль Менделя в развитии науки. Никто до него не размышлял,
что вообще могут существовать задатки признаков. Считалось, что в каждом из
нас сидит маленький человечек, внутри его - еще маленький человечек и т. д.
Зачатие имеет к его появлению какое-то отношение, но по механизму, готовый
маленький человечек уже присутствует с самого начала своего роста. Такими бы-
ли доминирующие представления, у которых, безусловно, был недостаток - по
этой теории, при большом числе поколений гомункулус должен был получиться по
размеру меньше элементарной частицы, но тогда про частицы еще не знали J.
Откуда Мендель знал, какой признак является доминантным, а какой рецессив-
ным? Ничего такого он не знал, просто взял некоторый принцип организации опы-
та. Удобно, что признаки, за которыми он наблюдал, были разными: рост, раз-
мер , цвет цветка, цвет боба и т.д. У него не было априорной модели механизма
наследования, он вывел ее из наблюдения за передачей признака в поколениях.
Еще одна особенность его метода. Он получил, что доля особей с рецессивным
признаком во втором поколении составляет четверть от всего потомства. То есть
вероятность того, что данная горошина зеленая - 1/4. Допустим, получилось в
среднем по 4 горошины в одном стручке. Будет ли в каждом стручке (это потом-
ство от двух и только от двух родителей) 1 горошина зеленая и 3 желтых? Нет.
Например, вероятность того, что там будет 2 зеленых горошины равна 1/4 х 1/4
= 1/16, а того, что все четыре зеленые - 1/256. То есть, если взять кучу бо-
бов, с четырьмя горошинами в каждом, то у каждой 256-ой все горошины будут с
рецессивными признаками, то есть зелеными. Мендель анализировал потомство
множества одинаковых пар родителей. О скрещивании было рассказано, потому что
они показывают, что законы Менделя проявляются как статистические, а в основе
имеют биологическую закономерность - 1:1. То есть гаметы разных типов в КАЖ-
ДОМ мейозе у гетерозиготы образуются в равном соотношении - 1:1, а законо-
мерности проявляется статистически, поскольку анализируются потомки сотен
мейозов - Мендель анализировал более 1000 потомков в скрещивании каждого ти-
па .
Сначала Мендель исследовал наследование одной пары признаков. Затем он за-
дался вопросом, что будет происходить, если одновременно наблюдать за двумя
парами признаков. Выше на рисунке, в правой части проиллюстрировано такое ис-
следование по двум парам признаков - цвету горошин и форме горошин.
Родители одного типа давали при самоопылении горошины желтые и круглой фор-
мы. Родители другого типа давали при самоопылении горошины зеленые и морщини-
стой формы. В первом поколении он получил все горошины желтые, а по форме -
круглые. Получившееся расщепление во втором поколении удобно рассмотреть с
помощью решетки Пенета. Получили расщепление по признакам 9:3:3:1 (желтые и
круглые: желтые и морщинистые: зеленые и круглые: зеленые и морщинистые).
Расщепление по каждой паре признаков происходит независимо друг от друга. Со-
отношение 9жк + Зжм + Ззк + 1зм соответствует независимой комбинации резуль-
татов двух скрещиваний (Зж + 1з) х (Зк + 1м). То есть и задатки признаков
этих пар (цвет и форма) комбинируются независимо.
Посчитаем, сколько разных фенотипических классов мы получили. У нас было 2
фенотипических класса: желтые и зеленые; и по другому признаку 2 фенотипиче-
ских класса: круглые и морщинистые. А всего будет 2*2=4 фенотипических клас-
са, что мы и получили выше. Если рассматривать три признака, то фенотипиче-
ских классов будет 23=8 классов. Мендель доходил до дигибридных скрещиваний.
Задатки всех признаков, к счастью Менделя, находились у гороха на разных хро-
мосомах, а всего хромосом у гороха - 7 пар. Поэтому, оказалось, что он взял
признаки, которые комбинировались независимо в потомстве.
У человека 23 пары хромосом. Если рассмотреть какой-то один гетерозиготный
признак для каждой хромосомы, может у человека может наблюдаться 223 ~ 8*106
фенотипических классов в потомстве одной супружеской пары. Как упоминалось на
первой лекции, каждый из нас содержит между папиными и мамиными хромосомами
порядка 1 различия на 1000 позиций, то есть всего порядка миллиона различий
между папиными и мамиными хромосомами. То есть каждый из нас является потом-
ком миллионногибридного скрещивания, при котором число фенотипических классов
составляет 21000000. Практически это число фенотипических классов в потомстве
одной пары не реализуется, потому что хромосом у нас всего 23, а не миллион.
Получается, что 8*106 - это нижний предел величины возможного разнообразия в
потомстве данной супружеской пары. Исходя из этого, можно понять, что не мо-
жет быть двух абсолютно одинаковых людей. Вероятность мутации данного нуклео-
тида в ДНК за одно поколение составляет около 10”7 - 10”8, то есть на весь ге-
ном (3*109) получится около 100 изменений de novo между родителем и ребенком.
А всего отличий в папиной половинке вашего генома от маминой половинки - око-
ло 1 000 000. Это значит, что старые мутации в вашем геноме гораздо более
частые, чем вновь возникшие (в 10 000 раз).
Также Мендель проводил анализирующее скрещивание - скрещивание с рецессив-
ной гомозиготой. У потомка первого поколения комбинация генов имеет вид АаВ
Ь. Если скрестить его с представителем с полностью рецессивным набором генов
(aabb), то получится четыре возможных класса, которые будут находиться в со-
отношении 1:1:1:1, в отличие от рассмотренного выше скрещивания, когда мы по-
лучили расщепление 9:3:3:1.
гладкий желтый
морщинистый зеленый
Яг Гу
Expected Expected Expected
proportions for proportions for proportions for
Аналитическое скрещивание RrYy x rryy
Ниже показаны некоторые статистические критерии - какие соотношения чисел
следует считать соответствующими ожидаемым, скажем, 3:1. Например, для 3:1 -
из четырехсот горошин вряд ли получится точно 300 к 100. Если получится,
примеру, 301 к 99, то это отношение наверное можно считать равным 3 к 1.
350 к 50 уже, наверное, не равно 3 к 1.
Статистический тест хи-квадрат (х2) используется для проверки гипотезы со-
ответствия наблюдаемого распределения ожидаемому. Произносится эта греческая
буква в русском языке как «хи», а в английском - как «чи» (chi).
Величина х2 рассчитывается как сумма квадратов отклонений наблюдаемых вели-
чин от ожидаемой, деленных на ожидаемую величину. Затем по специальной таб-
лице для данного значения х2 находят величину вероятности того, что такое
различие между наблюдаемой и ожидаемой величиной является случайным. Если ве-
роятность оказывается меньше 5% то отклонение считается не случайным (цифра в
пять процентов выбрана по договоренности.
> X
’’ Table 3.4 Критические значения Х2
Р
df .995 .975 .9 .5 .1 .05 .025 .01 .005
000 ООО 0016 0.455 2 706 3.84 1 5 024 6.635 7.879
2 0.010 0.051 0.211 1.386 4.605 5.991 7.378 9.210 10.59?
3 0.072 0216 0.584 2.366 6.251 7.815 9.348 11.345 12.838
4 0.207 0.484 1.064 3.357 7.779 9.488 11.143 13.277 I4.8G0
5 0 412 0 831 1.610 4 351 9.236 1 1.070 12.832 1 5.086 16.750
6 0.676 1.237 2.204 5 348 10.645 12.592 14 449 16.812 18.548
7 0.989 1.690 2.833 6 346 12.017 14.067 16.01 3 18.475 20.278
8 1 544 2.180 3.490 7 344 13 362 15.507 17 535 20.090 21.955
9 1.735 2.700 4.16В 8.343 14.684 16.919 19.023 21.666 23.589
10 2.1 56 3.24 7 4 865 9 342 15 987 18.307 20 48 3 23.209 25.188
2.603 3.816 5.578 10.341 17.275 19.675 21.920 24.725 26.757
12 3.074 4.404 6.304 1 1.340 18.549 21.026 23.337 26.217 28.300
13 3.56S 5.009 7.042 12.340 19.812 22.362 24.736 27.688 29.819
14 4.075 5.629 7.790 13.339 21.064 23.685 26.119 29.141 31.319
IS 4.601 6.262 8.547 14.339 22.307 24.996 27 488 30.578 32.801
Р - вероятность, df- число степеней свободы
Всегда ли будет проявляться какой-либо наследственно предопределенный при-
знак? Ведь это предположение по умолчанию лежит в основе интерпретации данных
полученных Менделем.
Оказывается, это может зависеть от многих причин. Есть такая наследуемая
черта у человека - шестипалость. Хотя у нас, как и у всех позвоночных, паль-
цев в норме пять.
Вероятность проявления задатка признака в виде наблюдаемого признака (здесь
- шестипалость) может быть меньше 100%. На фотографии у человека на обеих но-
rax по 6 пальцев. А у его близнеца этот признак не обязательно проявится.
Доля индивидов с данным генотипом, у которых проявляется соответствующий фе-
нотип, была названа пенетрантностью (этот термин ввел российский генетик Ти-
мофеев-Ресовский) .
В некоторых случаях шестой палец может быть просто обозначен некоторым кож-
ным приростом. Степень выраженности признака у индивида Тимофеев-Ресовский
предложил называть экспрессивностью.
Особенно ясно не 100% связь генотипа с фенотипом прослеживается при иссле-
довании идентичных близнецов. Генетическая конституция у них один в один, а
признаки у них совпадают в разной степени. Ниже представлена табличка, в ко-
торой представлено совпадение признаков для близнецов идентичных и неидентич-
ных. В качестве признаков в этой таблице взяты различные болезни.
Table 6.21
Trait Монозиготные конкордантны (%) Ди зиготные конкордантны (%) Ссылка
Инфаркт (мужчины) 39 26 1
Инфаркт (женщины) 44 14 1
Бронхиальная астма 47 24 2
Рак 12 15 2
Эпилепися 59 19 2
Ревматоидный артрит 32 6 3
Рассеянный склероз 28 5 4
RHc'cnces. 111 В. HavaM jnd M. Hauge, иi. Public NeJth Service PubUtanco 1101
Ipp 61 67). 166). I Havaid and M Haug*. Cmrtkt u/M thr fjrnittniciogr of Chtontc
DiitJVf U.$ Departement of Health, idiKation. .end M*lfare, 146$. (J, 5 Lawrence.
Anradi 0< ЧРшлмс/с СЪиаш 26<I97O).3$7 179. И» С. С. LLeu tt al. А/г.сгч.лп JojfMuJ
of («'nefrfS IbitOe* 44S
Признак, который присутствует у большинства особей в естественных условиях
обитания, называется диким типом. Наиболее распространенный признак часто
оказывается доминантным. Такая связь может иметь приспособительное значение,
полезное для вида. У человека доминантными признаками являются, к примеру,
черные волосы, темные глаза, кудрявые волосы. Кстати, поскольку соответствую-
щие гены находятся на разных хромосомах, то может получиться кудрявый негр,
который будет блондином - ничто это не запрещает.
Почему так получается, что в при моногибридном скрещивании трем генотипиче-
ским классам в потомстве второго поколения соответствует в некоторых случаях
три фенотипических класса (баклажаны синие, фиолетовые и белые), а в другом
случае - два класса (желтая или зеленая горошина)? Почему в одном случае
проявление доминантного признака неполное, а в другом - полное? Можно провес-
ти аналогию с фотопленкой. В зависимости от количества света, кадр может по-
лучиться совсем прозрачным, серым и совсем черным. То же самое - с генами.
Например, есть у кукурузы ген Y, который определяет образование витамина А.
Когда доза гена Y на клетку растет от одного до трех, то линейно изменяется
активность фермента, который он кодирует и, в данном случае, усиливается об-
разование витамина А и окраска зерна. (У кукурузы основная часть зерна - эн-
досперм. В каждой клетке эндосперма три генома - два от мамы и один от папы).
То есть, многие признаки зависят от дозы аллеля количественно. Чем больше ко-
пий аллеля нужного типа, тем больше будет величина контролируемого им призна-
ка. Такая связь постоянно используется в биотехнологии.
количество витамина А и его активность в эндосперме
зерна кукурузы зависят от числа (дозы) генов,
здесь - от числа доминантных аллелей гена Y в геноме.
При генотипе эндосперма У УУ 0,05
При генотипе эндосперма Yyy 2,25
При генотипе эндосперма Y Yy 5,00
При генотипе эндосперма YYY 7,50
Действие одной дозы доминантного гена
соответствует примерно 2,25 единиц активности
витамина А.
М.Е.Лобшев. Геметмка. Л. 1М7. стр. 1S4
Мендель мог благополучно свои законы и не открыть. Исследования на горохе
позволили Менделю открыть свои законы, потому что горох - самоопыляемое рас-
тение, а потому без принуждения - гомозиготный. При самоопылении доля гете-
розигот уменьшается пропорционально двум в степени номера поколения. В этом
заключалось везение Менделя - если бы доля гетерозигот была большой, то ника-
ких бы закономерностей не наблюдалось. Когда он затем взял перекрестные опы-
лители , то закономерности нарушились, что сильно расстроило Менделя, потому
что он подумал, что открыл нечто частное. Оказалось, что нет.
У самоопылителей гетерозиготное растение даст лишь половину
гетерозиготных потомков Аа х Аа => 1 АА : 2Аа : 1аа. В следующем
поколении от всех гетерозигот опять останется лишь половина.
Уже в 4-м поколении гетерозигот останется всего 0.5*=6%,
поэтому у Менделя взятые в опыт растения гороха были
гомозиготными: либо доминантными, либо рецессивными.
Законы Менделя могли бы не проявиться на перекрестниках,
где получить гомозиготную линию гораздо сложнее.
7/4. повулшис прм ибридтге ^*Nhiu соогиоаэды
гемо* а ггггроэягот ркирплнша эо одтой паре идем*).
I — кштс си *1зб)млшга.
Выше было рассказано о наследовании признаков качественных, а обычно боль-
шинство признаков - количественные. Их генетический контроль достаточно сло-
жен. Количественные признаки описываются через среднюю величину значения при-
знака и размахом варьирования, которая называется нормой реакции. И величина
средней, и норма реакции - это видоспецифические показатели, которые зависят
как от генотипа, так и от условий среды. К примеру, продолжительность жизни
человека. Хоть в Библии и написано, что пророки жили по 800 лет, но сейчас
ясно, что больше 120-150 лет никто не живет. А, мышь, например, живет два го-
да , хотя она тоже млекопитающее. Наш рост, наш вес - это все количественные
признаки. Нет людей 3-4 метрового роста, хотя слоны, к примеру, есть. У каж-
дого вида своя средняя по каждому количественному признаку и свой размах его
варьирования.
0 18 » » 21 И В М И 2« 27 2в М 50 31
Температура среды во время развития (°C)
Норма реакции:
изменение длины крыльев в зависимости оттемператруры
среды во время развития у мух Drosophila melanogaster
Закономерности наследования открыты при исследовании качественных призна-
ков .
Большинство наших признаков - количественные.
Величины значений признаков в представительной выборке особей данного вида
характеризуются определенной средней и широтой ее варьирования, которая назы-
вается нормой реакции и зависит как от генотипа, так и от условий формирова-
ния признака.
ЛЕКЦИЯ №17.
СВЯЗЬ ГЕНОТИПА И ФЕНОТИПА: ВАРИАНТЫ
ПРОЯВЛЕНИЯ МЕНДЕЛЕВСКИХ ЗАКОНОМЕРНОСТЕЙ
Мы продолжим обсуждать качественные признаки, и говорить о связи генотипа и
фенотипа, о вариантах проявления менделевских закономерностей, и что за этим
стоит с точки зрения наших представлений о работе генетического материала.
Зачем надо изучать результаты Менделя и Моргана? Прежде всего, нам интерес-
но узнать что-то о нас самих. Нам хочется быть богатыми, здоровыми и счастли-
выми (последние два признака во многом генетически контролируемые. Есть люди,
которые чувствуют себя обычно счастливыми, а есть такие, которые чаще ощущают
себя несчастными - эти ощущения связаны с определенными генетическими харак-
теристиками) .
Если мы знаем характер генетического контроля признака, то мы можем пред-
сказать с определенной вероятностью фенотип на основе генотипа (если он из-
вестен) . Если мы не знаем, как признак контролируется, то мы будем в полном
неведении, и не сможем ничего сказать относительно признаков будущих поколе-
ний. Если мы знаем связь между генотипом и фенотипом, то мы можем сделать оп-
ределенные предсказания о развитии признака (например, болезни) и, в некото-
рых случаях, предпринять действия полезные индивиду. Для этого нам надо уста-
новить генотип. Сейчас эта задача технически разрешима (поскольку секвениро-
вание генома человека проведено), хотя и непомерно дорога.
В реальности нам дано лишь наблюдать проявление признака в поколениях и на
основе этого создавать модель генетического контроля формирования признака,
которая может быть верна лишь в данных конкретных условиях. Но, тем не менее,
если мы создали такую модель, то мы можем в этом случае иметь средства для
регуляции какого-то признака, в частности, повлиять возникновение или ход ка-
кой-либо болезни. Таким образом, генетический контроль и его изучение имеет
большое практическое, в частности, медицинское, значение. И в основе всего
лежат менделевские закономерности, которые могут проявляются по-разному в за-
висимости от конкретных особенностей генотипа и среды.
Мы сейчас рассмотрим, какие могут быть условия генотипа и среды, при кото-
рых эти закономерности будут выглядеть не такими, как их наблюдал Мендель, и
почему это происходит. Мендель наблюдал, что при объединении задатков двух
признаков в одном организме может быть проявление только одного признака.
Второй задаток не проявляется. Такой тип доминирования называется полным.
Переоткрытие законов Менделя привело к выявлению других типов доминирова-
ния. Например, неполного доминирования, когда фенотип гетерозиготы является
промежуточным между двумя гомозиготами. Есть еще один тип доминирования,
очень популярный в последнее время в суде, - кодоминирование - в гетерозиготе
проявляются фенотипы каждой из гетерозигот. Это явление имеет место, в част-
ности, и у человека. Если у вас есть папина хромосома и мамина хромосома (а
это, несомненно, так), и они различаются в миллионе позиций, что можно вы-
явить разными методами, то это все случаи кодоминирования.
Первичным фенотипом организма является последовательность нуклеотидов моле-
кул его ДНК. На этом фенотипе строятся все фенотипы следующих уровней. То
есть при исследовании вашей ДНК проявляются все и папины, и мамины признаки;
каждая из молекул ДНК проявляет свой признак независимо от присутствия другой
молекулы ДНК с другим признаком: при секвенировании или при расщеплении ДНК
какими-либо ферментами видны оба состояния ДНК. Кодоминантные признаки (мар-
керы самой молекулы ДНК) характеризуют различие между хромосомами и использу-
ются для идентификации личности или установления отцовства (число таких слу-
чаев разрешаемых в суде составляет несколько сотен в год).
С от pie It
Иоппалке
д м1
la red ffcwsrv
7411 р flCWEf 5.
оиз alk-k?
Rvnotyplr iangj
Red
dririlnant
White
dnnirwnt
___________________AL—
irths hetearygote is whit
thp Л2 allele b rtmlnrrt
cwr the Л1 al lek
Типы доминирования: полное (вверху) и
неполное (внизу).
Когда мы говорим о генотипе и фенотипе - это такие крайние единого процесса
реализации наследственной информации в индивидуальном развитии. Например,
гладкая или морщинистая форма горошины, это ее фенотип. А генотип - это та
специфическая последовательность нуклеотидов, которая в данных условиях опре-
деляет, что горошина будет гладкой или морщинистой. В 1999 году с менделев-
скими линиями гороха была проведена следующая работа. Участки хромосомы, от-
вечающие за форму горошины, были клонированы, секвенированы, и было установ-
лены их особенности - различие последовательностей нуклеотидов - которые и
определяли развитие гладкой или морщинистой формы горошины.
Обратите внимание, что форма горошины является конечным признаком, а форми-
рованию признака этого уровня предшествует проявление признака на многих пре-
дыдущих уровнях. Во-первых, это наличие (аллель 1) или отсутствие (аллель 2)
олигосахарида, которое и приводит к той или иной форме горошины. Еще более
глубоким уровнем проявления фенотипа является то, что имеется соответствующий
белок (аллель 1), который необходим для синтеза олигосахарида или это тот же
белок, но альтернативной структуры (аллель 2) , при которой олигосахарид не
образуется. Еще более глубоким признаком является РНК, с которой синтезирует-
ся этот белок. Эти РНК различны по последовательности нуклеотидов (аллели 1 и
2), что и делает различными соответствующие белки. А эти РНК различны, потому
что транскрибируется с разных молекул ДНК, папиной и маминой, у которых по-
следовательности нуклеотидов в данной позиции различны (аллели 1 и 2). Все
это - проявление одного и того же фенотипа, последовательно реализуемого на
каждом из уровней.
Мы имеем право говорить о фенотипе на каждом из этих многих уровней - от
конкретных особенностей последовательности нуклеотидов ДНК до формы горошины.
При этом, как только мы продвигаемся от ДНК выше, тем больше влияние условий
окружающей среды. Например, возможность функционирования разных аллелей на
уровне ДНК (транскрипция папиной и маминой копии гена) будет мало зависеть от
температуры, а возможность функционирования тех же аллелей на уровне белка
может критически зависеть от температуры. При некоторых температурах белок
(например, аллель 1) будет работать, а другой (аллель 2) не будет работать.
Как только мы выдвигаемся на более высокий уровень в реализации фенотипа, по-
является больше возможностей для влияния окружающей среды на проявление при-
знака .
И наоборот, чем ближе мы продвигаемся к генотипу, тем предсказуемое связь
между генотипом и фенотипом. При идентификации человека в судебно-медицинской
экспертизе чаще всего используются маркеры, которые различают одни и те же
участки гомологичных (папины и мамины) хромосом. Обычно эти различия - микро-
вставки в данном месте молекулы ДНК. Принцип метода таков. Мы можем размно-
жить in vitro фрагмент молекулы ДНК в нужном месте хромосомы, например, с ко-
ординатами от позиции N до позиции N+100. Этот фрагмент мы выбрали потому,
что в нем у каждого человека в каждой хромосоме есть вставка, например, ди-
нуклеотид СА. В каждой хромосоме длина этой вставки может быть своя. Допус-
тим, в одной хромосоме этот динуклеотид повторен в данном месте 10 раз, а в
том же месте другой хромосомы - 15 раз. При размножении этого участка хромо-
сомы мы получим фрагменты двух длин 100 +2*10=120 пар нуклеотидов и
100+2*15=130 пар нуклеотидов. Фрагменты каждого из этих размеров отличимы по-
сле их фракционирования электрофорезом в геле.
Пусть и папа и мама гетерозиготны по этому локусу, подобно гибридам первого
поколения Менделевского моногибридного скрещивания. Обозначим их генотипы по
характеристической длине размножаемых фрагментов ДНК - 120/130. Тогда их дети
будут иметь такие генотипы 1 120/120 (гомозиготы по аллелю 120) : 2 120/130
(гетерозиготы): 1 130/130 (гомозиготы по аллелю 130). Естественно, присутст-
вие в образце фрагмента ДНК одной длины не влияет на возможность детекции
фрагмента ДНК другой длины, то есть, используемые маркеры ко-доминантны. Они
обе проявляются. Понятно, что здесь проявляются те же менделевские закономер-
ности передачи и расщепления признаков, хотя в качестве признака мы использо-
вали морфологию самой молекулы ДНК - различную длину фрагмента молекулы в
данном месте хромосомы.
После переоткрытия законов Менделя выяснялось, что Мендель то в одном как
бы как бы не прав, то в другом как бы не прав. Обнаруживались дополнительные
обстоятельства, которые модифицировали проявление менделевских закономерно-
стей.
Например, число классов генотипов и фенотипов зависит от плоидности, даже
если аллелей всего два. К примеру, эндосперм в зерне кукурузы триплоидный. У
него два генома маминых, а один папин (из пыльцевого зерна). Получали расте-
ния кукурузы, которые различались по числу доминантного аллеля гена Y. Этот
аллель необходим для образования витамина А. Когда в эндосперме была одна ко-
пия аллеля Y, то Если относительное количество витамина А при одной копии ал-
леля Y составляло единицу, то при двух копиях - 2,2, а при трех копиях - 3,3.
То есть выраженность конкретного признака может зависеть от дозы гена. А мо-
жет и не зависеть - как в случае полного доминирования одного аллеля над дру-
гим . Все зависит от конкретного механизма формирования признака.
То, что от дозы гена зависит количество его продукта, используется в био-
технологиях, как основа всех современных процессов, основанных на рекомби-
нантных ДНК. Во всех этих случаях в клетке увеличивают дозу гена. Увеличение
дозы гена приводит к нужным нам результатам, но клетка от этого может уме-
реть. Обычно при производстве это безразлично. Критерием эффективности в био-
технологии является получение целевого вещества с минимальными затратами.
Впрочем, если бы могли получить корову, которая бы дала сразу цистерну моло-
ка , а потом бы умерла, это было бы очень технологично, но не этично. Однако с
бактериями так и поступают: выращивают бактерии в идеальных условиях, а затем
включают синтез, который стоит бактериям жизни.
Выраженность признака может увеличиваться с ростом дозы гена в геноме
- это основа современной биотехнологии рекДНК
Количество витамина А в эндосперме зерна кукурузы зависит от числа
доминантных аллелей гена Y
Генотип-число аллелей Y Витамин А, единицы
относительные абсолютные
У У У- 0 0.02 0.05
Yyy- 1 1 2,25
YYy- 2 2.2 5.00
YYY- 3 3.3 7.50
МЕ.Лооашее. Генетта. Л.. 1967. сто. 154
Например, с помощью технологий рекомбинантных ДНК сконструированы бактерии,
превращающие сахар в аминокислоту треонин. Более половины углерода, поглощен-
ного такими бактериями превращается в единственный целевой продукт. Меньше
половины сахара превращается в тысячи других углерод-содержащих соединений
клетки, а, кроме того, расходуется еще и как источник энергии, чтобы все это
синтезировать и просто жить. Треонин накапливается в среде до концентрации в
100 грамм на литр, что близко к пределу его растворимости. Мировые объемы
производства рекомбинантных продуктов составляют десятки тысяч тонн в год
(треонин) и даже сотни тысяч тонн (лизин).
В менделевском случае каждый признак формировался под действием как бы од-
ного гена, который на другие признаки не влиял (ведь растение с гладкими се-
менами могло быть разной высоты, иметь разного цвета семена и цветы и др.) ,
На самом деле формирование каждого признака требует работы многих генов, и
большинство генов влияют более, чем на один признак. Например, относительное
содержание каждого из трех метаболитов в эндосперме кукурузы (на слайде) за-
висит от генотипа по каждому из трех исследованных генов.
Отдельный признак гложет определяется многим i генам i
(здесь признак - относительное содержание метаболита в эндосперме кукурузы)
Генотип и фенотип эндосперма кукурузы Метаболит, относительное содержание
Триптофан Никотиновая кислота Индолуксусная кислота
sud (сахаристый) 1 1 1
btj (лоький) М 2,2 1,7
sh2 (сморщенный) 2 1,5 1Д
Относительное содержание данного метаболита (признак) свое для кавдого
генотипа: количество триптофана при генотипе ЬП большее 1,4 раза, чем
для su1, а при генотипе sti2 больше в 2 раза, чем для su1
М.Е .Либашев. Генетика. Л., 1987, стр. 155
Видно, что на каждый признак влияет каждый из трех генов, и таких генов на-
верняка больше. Для формирования каждого признака нужно много генов. Объясним
это на примере слуха. Чтобы человек слышал, нужны многие структуры уха: моло-
точки, наковаленки, барабанные перепонки, и если не будет какого-то белка,
нужного, чтобы сформировать эти структуры, то человек не будет слышать. По-
этому глухота у человека может возникать из-за дефектов в десятках известных
генов (контролирующих много элементов слухового аппарата, передачу сигнала в
нервной ткани т.д.). При этом разные генотипы могут приводить к одному фено-
типу (глухота). Внешне одинаковые фенотипы, обусловленные разными генетиче-
скими причинами, называются фенокопиями. Мутации глухоты редки и обычно ре-
цессивны. Глухота проявляется обычно как моногенный аутосомный дефект с на-
следованием по менделевскому типу. Известны случаи, когда муж и жена, оба
глухие, хотели бы, чтобы и их будущий ребенок был глухим, чтобы он также при-
надлежал их сообществу. Этот пример показывает - понятие нормы и болезни от-
носительно . Оно зависит, в том числе, и от отношения индивида к своей особен-
ности и от отношения общества к этой особенности.
Менделевские закономерности при взаимодействии генов (на примере глухоты).
Допустим, что глухота определяется рецессивными мутациями в 10 разных генах и
частота этих мутаций в популяции одинакова.
Вопрос 1. Какова вероятность того, что у двух глухих родителей будет глухой
ребенок? Решение. Каждый из родителей - гомозигота по мутации в одном из ге-
нов , нужных для развития слуха. Вероятность того, что второй родитель несет
мутацию в том же гене, что и первый составляет 1/10. Следовательно, вероят-
ность того, что ребенок от такого брака будет глухим, составляет 10%, а того,
что он будет слышать - 90%. Запись генотипов будет при этом следующая: роди-
тели ааВВ х АА ЬЬ, потомство первого поколения Аа ВЬ.
Вопрос 2 . Какова будет доля слышащих потомков от скрещивания индивидов
первого поколения? (Чтобы не смущать друг друга будем считать, что этот опыт
проводится, конечно, не на людях, а на мышах - основном модельном генетиче-
ском объекте из млекопитающих). Решение. Расщепление по каждой паре аллелей
происходит независимо друг от друга. То есть скрещивание Аа ВЬ х АаВЬ можно
записать как два независимых скрещивания. По гену А: родители Аа х Аа. Расще-
пление у потомков по фенотипу составит 3 : 1 (3 слышащих к 1 глухому) . При
этом у всех слышащих есть хотя бы один доминантный аллель А. Такой генотип
Записывается в форме А- . Генотип глухих потомков - аа (гомозиготы по рецес-
сивному аллелю). Тогда генотипы соответствующие слышащим и неслышащим потом-
кам скрещивания можно представить как ЗА- : 1аа.
Таким же образом запись потомков по генотипу при расщеплении аллелей по ло-
кусу В можно представить как ЗВ- : 1ЬЬ.
Поскольку расщепление по аллелям одного локуса происходит независимо от
расщепления по аллелям другого локуса, то генотип потомков при расщеплении по
двум локусам одновременно можно записать в следующем виде:
(ЗА- : 1аа) х (ЗВ- : 1ЬЬ) = 9А-В- : ЗааВ- : 3A-bb : laabb
Чтобы индивид слышал, ему необходимо иметь хотя бы один аллель дикого типа
по каждому из локусов. Таких индивидов 9 (генотипы А-В-). Чтобы индивид не
слышал, достаточно гомозиготности по любому из двух рецессивных генов. Таких
индивидов 7 (генотипы ааВ-, A-bb и aabb)
Таким образом, при скрещивании дигетерозигот соотношение слышащих потомков
к неслышащим составит 9:7.
Отдельный ген может влиять на несколько признаков, в развитие которых он
вовлечен. Такой эффект действия гена называется плейотропным (множественным).
Например, альбинизм вызывается дефектом одного гена, контролирующего образо-
вание меланина. При этом меняются одновременно такие признаки как цвет волос
(белый), цвет кожи (белый) и цвет глаз, (он будет красным от цвета кровенос-
ных сосудов, лежащих за радужной оболочкой глаза). Множество признаков опре-
деляется геном SRY, контролирующим формирование всех признаков мужского пола
у млекопитающих. Перенос гена SRY из Y-хромосомы самца в геном самки мыши,
приводил к развитию особи с огромным количеством внешне наблюдаемых признаков
мужского пола.
У Менделя получились одинаковые численные соотношения при расщеплении алле-
лей многих пар признаков. Это в частности подразумевало одинаковую выживае-
мость индивидов всех генотипов, но это может быть и не так. Бывает, что гомо-
зигота по какому-нибудь признаку не выживает. Например, желтая окраска у мы-
шей может быть обусловлена гетерозиготностью по одной из пар аллелей (слайд).
При скрещивании таких гетерозигот друг с другом следовало бы ожидать расщеп-
ление по этому признаку соотношении 3:1. Однако наблюдается расщепление 2:1,
то есть 2 желтых к 1 белой (рецессивная гомозигота). Показано, что доминант-
ная (по окраске) гомозигота не выживает уже на эмбриональной стадии. Этот ал-
лель одновременно является рецессивной леталью (то есть рецессивной мутацией,
приводящей к гибели организма).
Разные генотипы могут давать разную выживаемость:
доминантная мутация с рецессивным летальным эф фектом
На следующем слайде показаны типы групп крови, их наследуемость. Этот при-
мер приведен для того, чтобы сказать, что у одного гена может быть не только
два аллеля. У Менделя их было всего два для каждого признака: например желтая
и зеленая окраски, третьего не дано. Но на самом деле аллелей по каждому ло-
кусу может быть много. Группы крови различаются по тому, какой тип антигена
мы вырабатываем. Может вырабатываться антиген типа А или В, но если одна хро-
мосома ответственна за тип А, а другая за тип В, то будут вырабатываться оба
типа антигена. Перед нами случай кодоминирования на уровне белка, когда нали-
чие одного типа антигена не препятствует наличию другого. Четвертый тип - ко-
гда антигены вообще не вырабатываются. Какое практическое значение это имеет
для нас? Это были первые генетические исследования, проведенные во время пер-
вой мировой войны в австрийской армии, так как во время войны всегда была
острая необходимость переливания крови, которое не всегда оказывалось удач-
ным, то есть человек от переливания крови иногда умирал.
Каждый индивид несет только два аллеля из трех
по данному локусу: группы крови АВО человека
a
lA trth
•6 Л*
it-anti
bzdlni
Amhndir»
rwl»by
hndr
О
'A I 111 I
andhod К
Al
inoarU-
trritsi
Airtlgei
a and В
пат
A >ndB
Nir»
Uxi UcoduJttiudQrui
л tn titrii fun лияьи man
шг*. dtpcacd Lcrcr Haul jnd
ridipiЛ bbcd л» апрШ1.
цк <-1 rxpait <an мвр
bled tmara-lira.tiiii
j ro voter u ti ЩГХ1.
imrntm ttNura
lduiu dona*.
IkcdodfcttBl i*act witi tte
p«rr <ntt> <4f <Mrp tcgaikw.
t'Kicr Wood aid twfltnt tk«d
avndt «irpolba
5.5 ДВО blood and poooUo blood UatttftitioM.
ш
PhenCTTPe Сятощж
fbbod trre>
О
Переливать кровь наиболее безопасно между людьми, у которых одинаковая
группа крови. Антигены групп крови у них одинаковые, антител против них нет и
агглютинация (слипание клеток крови) не произойдет.
Если донором (то есть сдающим кровь) является человек, у которого ни одного
из антигенов нет (группа крови 0), то его кровь можно переливать кому угодно
- она не будет вызывать агглютинации у реципиентов (тех, кому кровь перелива-
ют) с любым типом крови. Таким образом, люди с типом крови 0 являются универ-
сальными донорами. И наоборот, в крови людей с группой АВ не содержатся анти-
тела ни на один из этих антигенов (ни на А, ни на В) и эти люди счастливы
быть универсальными реципиентами - переливать кровь им можно от людей своей и
всех остальных групп крови. То есть человек с группой крови АВ может принять
кровь с любым типом. Самый плохой случай для реципиента - нулевая группа кро-
ви, которая будет реагировать с группами крови А, Ви АВ.
Два разных гена могут находиться в разных локусах и влиять на один и тот же
признак. Например, у дрозофилы могут быть разный цвет глаз, от коричневого
(рецессивная гомозигота по одному гену), через разные оттенки красного к бе-
лому (рецессивная гомозигота по другому гену). Красные глаза имеет муха дико-
го типа (например, гомозигота по всем аллелям дикого типа), а также гетерози-
гота по двум генам, каждый из которых нужен для пигментации глаза (см. рис.
слева). Взаимодействие двух генов в таком случае называют комплементарностью
(дополнительностью). При скрещивании дигетерозигот дает расщепление по фено-
типу в отношении 9:3:3:1. Из четырех фенотипических классов по окраске глаза
самый многочисленный - 9, это окраска дикого типа, а самый малочисленный - 1,
отсутствие окраски из за дефекта обоих генов, определяющих образование разных
пигментов.
Такая же ситуация у человека с окраской волос, у нас есть разные типы мела-
нина: эу- и фикомеланин, которые в разном сочетании дают разные оттенки, на-
пример рыжий.
Справа на рисунке приведен тот же пример, только на растениях. Гены в раз-
личных локусах могут давать вклад в развитие одной и той же фенотипической
характеристики: у перца 2 разных гена нужны для образования двух разных пиг-
ментов , что приводит к расщеплению по типу дигибридного скрещивания. Это ис-
пользуется при получении определенного цвета перца. Чтобы получит потомство с
нужным цветом плода необходимо знать генотипы родителей и правильно их подби-
рать для скрещивания.
На животных это применяется на пушных зверьках в звероводческих фермах. За
счет определенного подбора аллелей десятков известных генов, определяющих ок-
раску шерсти, можно получить большое разнообразие окрасок шкур пушных зверь-
ков , причем заранее планировать сколько и каких типов окраски получится.
Если два разных гена взаимодействуют при формировании признака
и их действие складывается, то такое действие генов называют кумулятивным
(накапливающимся). Например, степень красной окраски зерна пшеницы определя-
ется двумя разными генами, в каждом из которых есть неполностью доминантный
аллель А.
У перца 2 разных гена нужны для образования двух разных
пигментов, что привода к расщеплению по типу
д|(гибридного скрещгеания
Таким образом, наблюдается много вариантов численных соотношений классов в
потомстве дигибридного скрещивания в зависимости от типа взаимодействия ге-
нов. Если взаимодействия нет, то отношение будет 9:3:3:1. Может быть так, что
появление в генотипе рецессивного гена а ведет к тому, что ген В в фенотипе
не проявляется. Тогда соотношение будет 9:3:4. То же, но с доминантным геном
- будет 12:3:1. И т.д. То есть это показывает, что Мендель был прав только в
достаточно частном случае, когда гены не влияют друг на друга, но может быть
еще много других вариантов.
Отн ошение Ген отипы_______________
А_В_ А_ЬЬ алВ_ лаЬЬ
9:3:3:1 9331
9:3:4 9 3 4
12:3:1 12 3 1
9:7 9 7
9:6:1 9 6 1
15:1 15 1
13:3 13 3
Варианты соотношений классов в потомстве дигибридного
скрещивания в зависимости от типа взаимодействия генов
Породы собак: для того, чтобы получить какие-то определенные зафиксирован-
ные признаки породы, проводили близкородственное скрещивание, пока эти при-
знаки не закреплялись в гомозиготном состоянии, и далее при скрещивании собак
одной породы расщепление по этому признаку больше не может происходить -
признак становится стабильным, типичным для породы.
Окраска и другие признаки собак определяются
взаимодействием многих генов в большом числе локусов
«> 1Ы <с!
Каждая порода собак обладает своими признаками, и эти признаки стабильны в
поколениях, поскольку каждую породу характеризует свой набор гомозиготных ло-
кусов, определяющих эти признаки. У каждой породы собак как свой набор гомо-
зиготных локусов (у других пород они могут быть и гетерозиготны). Однако у
одних пород данный локус гомозиготен по одним аллелям, а других тот же локус
может быть гомозиготен по другим аллелям, что и определяет стабильные разли-
чия между породами (например, по окрасу). Ясно, что у каждой породы какие-то
локусы остаются гетерозиготными, и собаки одной породы по этим признакам бу-
дут различаться, но это те признаки, которые данную породу не определяют.
Если скрестить собак разных пород, то получится дворняжка. У нее по боль-
шинству генов гетерозиготное состояние, соответствующее предковым формам.
Дарвин в свое время подметил, что если скрещивать разные породы голубей, по-
лучается все время сизарь. Это происходит из-за утраты у гибрида гомозиготно-
сти характерной для каждой из родительских пород. А в гетерозиготных локусах
один из аллелей, как правило, оказывается дикого типа, поэтому и выглядят
гибриды сизарями - это дикий тип для всех пород голубей.
По генам окраски меха у норки в настоящее время установлено около 20 серий
множественных аллелей. При скрещивании гетерозиготных норок между собой во
втором поколении будет расщепление, типичное для дигибридного скрещивания в
соотношении 9:3:3:1, или 56% норок с дикой окраской меха, 19% - алеутских,
19% - платиновых и 6% с сапфировой окраской меха. Вавилов сформулировал закон
гомологических рядов: если у какого-то вида есть какой-то набор вариантов
признаков, и мы рассматриваем близкородственный вид, то у него тоже может
быть выявлен тот же набор признаков. Этот закон основан на том, что у близко-
родственных видов и набор аллелей в ДНК может быть сходен.
Комбинации генов, определяют ie окраску шерстного покрова у норок
Окраски Генотипы
Стандарт (дикий тип) А1А1 ВВ BgBg BpBp Н РР
Алеутская а1а1 ВВ BgBg BpBp Н РР
Платиновая А1А1 ВВ BgBg BpBp Н РР
Сапфир а1а1 ВВ BgBg BpBp Н РР
М.Е.Ло6ашев. Генетита. Л., 198Т, стр. 633
Гены могут находиться не только в ядре. Гены могут наследоваться однороди-
тельски, например, по материнской линии: окраска листа определяется генами
хлоропластов, а не ядра, и наследуется только от матери. То есть, какая бы ни
была мужская особь, потомство будет все однотипно - все в мать и только в
нее.
У человека тоже есть признаки, которые определяются митохондриями, то есть
наследуются по материнской линии. Митохондрии находятся в цитоплазме клетки,
которая достается потомкам только от матери. К мутациям и болезням, связанным
с митохондриями относятся, например, дефекты зрения и дефекты энергетического
обмена.
Часть ядерных генов также наследуется только однородительски. Это гены Y-
хромосомы, которая наследуется только от отца к сыну. Те гены в Y-хромосоме,
которые не имеют пары в Х-хромосоме, наследуются однородительски - от отца к
сыну. У человека один из таких генов определяет признак «волосатые уши» - у
женщин такой признак не проявляется - гена этого нет, нет и его аллеля, из-за
которого уши волосатые.
Проявление признака может зависеть также от пола индивида. Существуют при-
знаки, гены которых могут находиться в аутосомах или половых хромосомах обоих
полов, но проявляются лишь у одного из них. Такие признаки называют ограни-
ченными полом. Например, быки несут гены, определяющие молочность дочерей, но
гены свое действие у быков не проявляют. Гены казеинов - белков молока - это
случай наиболее строгого контроля транскрипции ограниченной полом. Они дейст-
вуют у особей как женского, так и мужского пола. Разница уровней экспрессии
между ними составляет 1 на 100 000. Петухи также несут гены яйценоскости и
размера яиц дочерей, хотя у петухов действие этих генов подавлено. Когда про-
водится селекция, учитывается этот факт. Здесь существенен генетический кон-
троль признаков.
Как проявление действия гена, так и тип доминирования может зависеть от по-
ла. Плешивость у человека наследуется аутосомно и моногенно. Аллель плешиво-
сти у мужчин доминирует, а у женщин рецессивен. У мужчин для облысения доста-
точно присутствия одного аллеля плешивости. При этом частота аллеля равна
частоте встречаемости фенотипа. У женщин для облысения необходимо присутствие
обоих аллелей плешивости.
Ответьте на несколько занятных вопросов.
Сколько женщин будут лысыми, если у мужчин лысый каждый десятый? Ответ.
Частота фенотипа плешивости у женщин равна квадрату частоты аллеля, следова-
тельно , одна из ста.
Может ли у неплешивых родителей быть плешивая дочь? Ответ. Нет: так как
отец неплешивый, значит у него нет такого аллеля (у мужчины аллель плешивости
доминантный и должен был проявиться) - оба аллеля определяют нормальную обво-
лосненность. Значит, дочь обязательно получит аллель волосатости от отца, а
этот аллель у женщин (в отличие от мужчин) доминантен. Плешивой она не будет
независимо от того, какой аллель придет от матери.
Может ли у неплешивых родителей быть плешивый сын? Ответ. Да: если у мамы
есть аллель плешивости он рецессивен и не проявится, он может передаться сыну
у которого этот аллель проявится как доминантный.
Если у неплешивых родителей один сын плешивый, какова вероятность того, что
другой тоже будет неплешивый? Ответ: 50%. Аллель плешивости не у отца - иначе
бы у него этот аллель проявился как доминантный. Значит аллель плешивости у
матери. Раз она волосатая, то второй аллель у нее - нормальная обволоснен-
ность, а аллель плешивости не проявляется, так как у женщин он рецессивен.
Следовательно, мать - гетерозигота - 1 аллель и нее - плешивость, 1 - норма.
Поэтому расщепление по этому признаку у ее сыновей будет 1:1 (50% плешивых)
Так генетика решает практически важные для нас задачи.
Экспрессия некоторых генотипов может зависеть от внешних условий. Ниже по-
казаны два кролика, один из которых с темными пятнами. Аллель гималайской ок-
раски у кролика температурочувствителен. При повышенной температуре белок не
функционален и необходимый пигмент не образуется, а при нормальной температу-
ре получается кролик, у которого некоторые участки шкуры окрашены.
Выращен при 20°С
Выращен при 30°С
Проявление аллеля himalayan в зависимости от температуры
Мутанты, проявление которых зависит от условий среды, сыграли очень важную
роль в биологии вообще и в молекулярной биологии в частности, в понимании то-
го, как работает генетический материал. Многие генетические феномены, были
открыты и исследованы после получения и использования условно летальных му-
тантов (условно смертельные). Наиболее распространенные мутанты такого типа -
температурочувствительные. Получены мутанты многих микроорганизмов и вирусов,
которые не способны жить при повышенной температуре (ts-мутанты). У такого
мутанта структура белка при повышенной температуре изменена, и белок не функ-
ционален (леталь), а при нормальной температуре структура обычная, и белок
функционален. При этом, возможно исследовать особенности функционирования
белка и его роли в формировании какой-либо структуры или сложной функции. Для
этого клетки растят до нужной стадии, в условиях, когда белок функционален,
затем разделяют клетки (обычно суспензию) на две части, в одной температура
остается нормальной, а в другой -поднимается, так что белок теряет функцио-
нальность .
Есть также другой тип условно летальных мутаций - нонсенс-мутации, леталь-
ные на одном генетическом фоне и нелетальные на другом. Нонсенс-мутация при-
водит к изменению кодона для какой-либо аминокислоты в данном белке на стоп-
кодон трансляции. Белок при этом образуется укороченный и нефункциональный.
Нонсенс-мутация подавляется (супрессируется), в тех клетках, где образуется
супрессорная тРНК. Супрессорная тРНК, это измененная тРНК, у которой антико-
дон по-прежнему узнает стоп-кодон в матричной РНК, но вместо терминации
(окончания) трансляции белка на этом участке происходит подстановка опреде-
ленного аминокислотного остатка. Белок с нонсенс-мутацией синтезируется пол-
норазмерным и функциональным в присутствии тРНК.
При создании модели наследования признака для выяснения его генетического
контроля начинают с проверки типа наследования по базовой, простейшей схеме -
менделевской и, если соответствия нет, то добавляют к этой схеме дополнитель-
ные условия, понятные на сегодня:
• неравную вероятность образования и сочетания гамет и выживания потомст-
ва ;
• однородительский тип наследования (Y - волосатые уши, мтДНК - болезнь
Лебера)
• взаимодействие аллелей: наличие доминирования;
• взаимодействие генов;
• сцепление генов;
• ограничения признака полом;
• вероятность проявления признака при данном генотипе;
• влияние среды (не всегда формализуемое).
Напомним, что менделевский характер наследования относится к признакам ка-
чественным. Сколько менделирующих признаков известно у человека? Они зафикси-
рованы в базе данных OMIM («Менделевское наследование у человека»). Их там
перечислено около пяти тысяч признаков. А мы у себя можем насчитать гораздо
больше признаков. Огромное количество остальных наших признаков зависит от
действия многих генов. Здесь следует отметить важный идеологический аспект.
Общей модели, по которой происходит формирование признаков под воздействием
многих генов, сейчас нет. Наверное, ее и быть не может. Чтобы понять, как
данный фенотип возникает на фоне данного генотипа, нам нужно еще узнать, как
конкретно реализуется каждый этап передачи генетической информации: что про-
исходит с модификацией ДНК, с какой из многих возможных вариантов РНК образу-
ется с данного участка генетической информации, как модифицируется белок, об-
разуемый с этой мРНК. В ДНК заложена лишь возможность всех этих вариантов
реализации генетической информации. Произойдет ли это или нет, зависит от
среды. Понятие среды мы пока толком формализовать не можем. Но если, подобно
структуре ДНК, мы их будем знать структуру и модификацию РНК и белков, то по-
нимание связи генотипа с фенотипом будет гораздо более глубоким, а предсказа-
тельная сила наших знаний будет больше. Как бы там ни было, одного знания ге-
нотипа часто недостаточно для предсказания фенотипа. Поэтому мы должны изу-
чать следующие этапы экспрессии генетической информации.
В заключительной части лекции речь пойдет об определении пола и признаках,
сцепленных с полом. Ниже показано, как выглядят половые хромосомы человека.
Х-хромосома по длине ДНК больше чем Y-хромосома приблизительно в три раза.
8»
Гаплоидныи набор яйцеклетки
млекопитающих всегда содержат X
хромосому, а спермии может нести
либо X, либо Y-хромосому.
Микрофотография Х-хромосомы
(слева) и Y хромосомы (справа).
<Biophc«o Ffcitarcht'i.l
А так выглядят клетки, транспортирующие эти хромосомы. Маленькая образована
организмом генотипа XY, а большая - генотипа XX. Такое соотношение физических
размеров имеют половые клетки.
Так выглядит человек после того, как он уже начал развиваться:
Вид человека на кончике иглы на стадии
первых делений после оплодотворения
яйцеклетки
Это игла, и на ее конце находится несколько поделившихся клеток, полученных
из оплодотворенной яйцеклетки. Такой наш размер на стадии 4-5-ого деления.
Еще раз напомню, что у нас цикл развития двухстадийный. На нижней части ри-
сунка изображена диплоидная стадия, период нашей долгой и счастливой жизни,
которая с точки зрения генетического процесса заканчивается мейозом, после
которого мы начинаем жить как бы в следующей стадии, гаплоидной. Это гаметы,
у нас они не обладают способностью к самостоятельному существованию, хотя у
многих растений именно гаплоид является основной частью жизненного цикла. Су-
ществование гамет заканчивается оплодотворением и появлением опять диплоидно-
го организма.
Мейоз
Гаметы
Гаплоид (1n)
-------Оплодотворение
Диплоид (2п) /
X и Y хромосомы сильно отличаются. Они гомологичны только в так называемых
псевдоаутосомных зонах, которые присутствуют и в папиных и в маминых хромосо-
мах, как и обычные аутосомы. Спермин могут быть двух типов: X и Y, а яйце-
клетки только одного типа - X и X. Поэтому расщепление получается 1:1 (две
женщины и два мужчины). Именно расщепление 1:1 по полу на первом этапе иссле-
дования признака, сцепленного с полом, - окраски глаз у дрозофилы - привело
Моргана к мысли о том, что гены находятся в хромосомах. Пол определяется хро-
мосомами и гены находятся именно в них.
X и Y хромосомы
отличаются по
размеру и
содержанию
генов.
Они гомологичны
только в
п се вдо ауто со м нои
области
Не у всех организмов, в отличие от человека, пол однозначно определяется
при слиянии гамет. Ниже показан моллюск, у которого в нижней части женские
клетки, а в верхней части только мужские. У некоторых пресмыкающихся мужской
пол не формируется при повышенной температуре (в частности, у крокодилов),
образуются только женские организмы.
1 ' I* Crtpulitlii fvmiiMa. the common tipper limpet, sett it
deennined by an eatiicnraeatd factor tbe Inn pet’s position in
a ttack ol linpott.
У дрозофилы всего четыре пары хромосом, три пары хромосом называются ауто-
сомы и одна пара половых хромосом. У самок набор хромосом XX, а у самца X Y
- как и у человека (у самки на конце брюшка находится яйцеклад, поэтому у нее
брюшко заостренное, а у самца оно овальное, - так их отличают).
Se*
:hr?n<i$or f-s
*ii(asam₽$
Хромосомы Drosophila melanogaster (2n=8)
состоят из трех пар аутосом (пары II. III. IV)
и одно|| пары поповых хромосом X и У
У плодовой мушки пол определяется не столько наличием той или иной половой
хромосомы, а отношением числа Х-хромосом к числу гаплоидных наборов аутосом
(А) . Это видно на схеме ниже. Набор ХХ+2А (2 Х-хромосомы и 2 набора аутосом)
дает соотношение 1,0 - такое соотношение является нормой для самок. Набор X
Y+2A (1 Х-хромосома и 2 набора аутосом) дает соотношение 0,5 - такое соотно-
шение является нормой для мужского пола. Если оказывается, что Y-хромосома
утеряна (набор 1Х+2А, соотношение 0,5), то образуется мужской организм, хоть
Y-хромосомы и нет. А если на фоне двух Х-хромосом добавилась Y-хромосома (на-
бор 2Х+2А), пол формируется женский, хоть Y-хромосома и присутствует. У чело-
века, сколько бы ни было Х-хромосом, если есть хоть одна Y-хромосома, то фе-
нотип будет мужской.
Таким образом, у человека и мухи, при внешне одинаковой хромосомной формуле
нормы (гомогаметный пол, XX, женский; гетерогаметный пол, X Y - мужской), ме-
ханизм генетического контроля образования пола различен.
У птиц и ряда других групп организмов гомогаметный пол (XX) - мужской, а
гетерогамтный пол (XY) - женский, что противоположно тому типу хромосомного
определения пола, который оказался зафиксирован эволюцией в линии человека и
других млекопитающих.
Соотношение половых хромосом и аутосом определяет пол
у дрозофилы
Половые хромосомы Число гаплоид, наборов аутосом Отношение Х:А Пол (Фенотип)
XX АД 1.0 Самка
XY АА 0.5 Самец
ХО АА 0.5 Самец
XXY АА 1.0 Самка
XXX АА 1.5 Суперсамка
XXXY АД 1.5 Суперсамка
XX АДА 0.67 Интерсекс
ХО АДА 0.33 Суперсамец
хххх ААА 1.3 Суперсамка
На следующем слайде показано месторасположение SPY-гена, отвечающего за
формирование признаков мужского пола. Уже говорилось, что если в геном мыши,
которая должна развиться самкой, пересадить SPY-ген, то у такого организма
разовьется мужской фенотип. Человек почти не отличается от мыши в этом плане.
У человека известны случаи, когда индивид кариотипически (то есть по типу
хромосом в ядре) соответствовал женщине (в частности с двумя XX хромосомами и
без наблюдаемой Y-хромосомы), но проявлял фенотип мужчины. В таких индивидов
была обнаружена хромосомная перестройка - ген SPY был транслоцирован (переме-
щен) на одну из аутосом.
Положение гена на Y-хромосоме мыши
Ген SRY кнтролирует развитие по мужскому типу
На внизу на рисунке представлен случай отклонения по составу половых хромо-
сом - синдром Тернера: индивид генотипа ХО (Y-хромосомы нет, а Х-хромосома
одна). Фенотип индивида - женский. Отклонений в поведении нет, хотя при боль-
шем дисбалансе в генотипе наблюдаются проблемы в умственном развитии.
Другой случай отклонения по составу половых хромосом - это синдром Клайн-
фельтера: генотип XXY, фенотип мужской. Хотя две Х-хромосомы присутствуют,
как в норме у женщин. То есть для определения пола у человека важно, чтобы
присутствовала Y-хромосома.
XXY. фенотип мужской. Семейные случаи
- транслокация гена SRY на аутосому.
Фенотип мужской при генотипе XX.
Справа: Синдром Тернера —
индивид генотипа ХО. фенотип женский.
». н а нм
4! «I W U !: :>
v .. V V |>1
Напомним, что гены, локализованные в Y-хромосоме и не имеющие пары в X-
хромосоме, наследуются только от отца к сыну. У человека аллель одного из та-
ких генов определяет признак «волосатые уши». Через мать к ребенку могут пе-
редаваться гены, находящиеся в митохондриях, но т.к. генов в митохондриях на
три порядка меньше чем в ядре, то и отклонений, передающихся только материн-
ской линии лишь тысячная часть от всех известных для человека.
Избыток или недостаток какой-то хромосомы в геноме приводит к очень серьез-
ным нарушениям у человека. Для примера можно привести трисомию по 21-ой хро-
мосоме - синдром Дауна. Причина заболеваний при утрате или избытке любой хро-
мосомы - дисбаланс в количестве продуктов генов этой хромосомы относительно
количества продуктов генов других хромосом (все - по две). Дисбаланс по всем
крупным хромосомам человека вообще не совместим с жизнью.
X хромосома одна из крупных хромосом человека. У женщин таких хромосом две,
а у мужчин - одна. Но оба пола живы и здоровы - хоть на себя погляди. Почему
разница в числе копий Х-хромосомы не смертельна, в отличие от ситуации с дру-
гими хромосомами?
Чтобы сбалансировать действие генов X хромосомы у мужчин (где Х-хромосома
одна) и у женщин (где Х-хромосомы две) природа предусмотрела инактивацию од-
ной из Х-хромосом в каждой клетке женского организма. У женщин одна X-
хромосома инактивирована, то есть гены в ней не работают, а работают только в
другой хромосоме - не инактивированной. Инактивиация вызвана компактизацией
хромосомы даже в интерфазе. Поэтому компактизированная хромосома видна при
окрашивании - в виде так называемого тельца Барра (слайд). Это один из диаг-
ностических признаков женского пола у млекопитающих, что у человека,
что у
слона. Некомпактизованная Х-хромосома,
как и все аутосомы,
не окрашивается и
не видна в интерфазе.
На следующем рисунке показана кошка черепаховой окраски. Поскольку это кош-
ка , а не кот, то генотип у нее XX. В каждой клетке женского организма одна из
Х-хромосом инактивирована (ее видно как тельце Барра). На Х-хромосоме у кошки
находится один из генов, определяющих окраску шерсти. У этого гена известны
разные аллели, то есть определяющие разный цвет шерсти. Кошка на рисунке ге-
терозиготна по этим аллелям. Одна из Х-хромосом в каждой клетке инактивирова-
на, поэтому только один из двух цветов может проявится в волоске, сформиро-
ванном из данной клетки. Поэтому одни волоски одного цвета, а другие - друго-
го. Какая из Х-хромосом инактивируется (папина или мамина), и, стало быть,
аллель какого цвета будет работать - дело случая. Поэтому такая кошка и имеет
черепаховую окраску. Кстати, сыновья у нее будут двух разных цветов в соотно-
шении 1:1.
Черепаховая окраска у кошки
определяется геном, находящимся
на Х-хромосоме
Еще одно доказательство того, что гены находятся в хромосомах, было получе-
но на мухах дрозофилах, называемых латеральными (боковыми, «сторонними») ги-
нандроморфами. У таких мух после первого деления яйцеклетки в одной из клеток
была утрачена Х-хромосома. Эта клетка при после-
дующих делениях дала клетки с генотипом ХО, по-
этому половина тела таких мух сформирована как у
самца и во ее всех клетках (на рисунке - правая
половина) цитологически наблюдается утрата одной
из Х-хромосом. Левая половина тела, происходящая
из клеток с нормальным генотипом XX, формируется
как самка.
В представленном на рисунке случае муха была
гетерозиготна по гену окраски глаза и по гену,
влияющему на форму крыла. Эти гены расположены в
Х-хромосоме. Поэтому правый глаз мухи неокрашен:
проявилась рецессивная мутация white, присутст-
вующая в одной из Х-хромосом. Крыло изменено по
форме. Левый глаз окрашен и крыло нормальной
формы, поскольку в нем остались обе Х-хромосомы,
в одной из которых присутствует рецессивный ген
white, а в другой - доминантный ген w+.
Латеральный гинандроморф у мухи
(вверху - схема образования)
МЕ-Побашев. Генетика. Л.. 1967. стр. 205
ЛЕКЦИЯ №18.
СЦЕПЛЕНИЕ И РЕКОМБИНАЦИЯ.
МУТАЦИИ: РОЛЬ В ЭВОЛЮЦИИ
И ИНДИВИДУАЛЬНОМ РАЗВИТИИ
Ранее было рассказано о передаче признаков, кодирующихся одним геном, и про
передачу признаков, кодирующихся разными генами с разных хромосом. В первом
случае было соотношение 3:1 в потомках (или, если скрещивать на рецессив -
1:1), а во втором - 9:3:3:1, из них два класса фенотипических родительских, а
два новых. Если же гены находятся на одной хромосоме, но все-таки достаточно
далеко друг от друга, то возникает явление рекомбинации, то есть новой комби-
нации генетических признаков, но не свободное - не 9:3:3:1. В деталях это оз-
начает следующее. Если два разных гена находятся в одной хромосоме рядом, то
определяемые ими признаки наследуются так, как если бы они определялись одним
геном; новые комбинации признаков в потомстве не появляются. Например, при
скрещивании растения, гетерозиготного по обоим локусам (MmDd, высокое и ли-
стья нормальные) с рецессивной гомозиготой (mmdd, низкое и листья крапчатые),
должны наблюдаться те же два фенотипических родительских класса в соотношении
1:1 (первое растение образует гаметы MD и md, а второе - только md, поэтому
другие классы образоваться не могут). То есть, хотя гены и разные, наследует-
ся все по Менделю.
7.5 а. Если два разных гена находятся в одной хромосоме рядом,
то определяемые ими признаки наследуются так,
как если бы они определялись одним геном -
новые комбинации признаков в потомстве не появляются.
Harpiwep прн афеицвагыи растентя,
reTqxHiroTHoro по обоим локусам {MuDd, высокое илисты» нормальные}
с рецесс1вной гомозиготой {mmdd, низкое илисты» крапчатые}, должны
наблюдаться те ле два фенотквжеских класса в соотношении 1:1 (родитегъ crater
Мама MmDd X mmdd Папа
гаметы (1 /И0+ 1mdy*. md => дети 1 MmDd : 1 mmdd
Если два разных гена находятся в разных хромосомах, то определяемые ими
признаки наследуются независимо, новые комбинации признаков в потомстве появ-
ляются. При не сцепленном наследовании признаков (высота растения и тип листа
) в анализирующем скрещивании растения, гетерозиготного по обоим локусам,
должны наблюдаться четыре фенотипических класса в соотношении 1:1:1:1 -оба
родительских класса (высокий с нормальными листьями и карлик с крапчатыми ли-
стьями) и два новых - рекомбинантных класса (высокий с крапчатыми листьями и
карлик с нормальными листьями). Это происходит, потому что первое растение
образует четыре типа гамет(Md, MD, mD, md) , а второе - по-прежнему один тип
гамет md. В данном случае - это независимое менделевское расщепление, потому
что гены находятся в разных хромосомах.
7.56. Если два разных гена находятся в разных хромосомах,
то определяемые ими признаки наследуются независимо,
новые комбинации признаков в потомстве появляются.
П|Х1 не сцепленном наследовании признаков (высота растетятя и тип листа )в
анализирующем сфеицванин растентя, гетероз нотного по обоим лотосам, должны
наблюдаться четыре ф енот iriiNecKifx класса в соотношенш :
обет родит ельеютх класса (высокий с нормать тыы! листьями и карлике крапчатыми
листьями) и два новых - ремомбинанпых класса (высокий с крапчатыми листьями и
ктрлисе нормальными листьями)
Говоря о рекомбинации, мы имели в виду появление новых, отличных от роди-
тельских, комбинаций аллелей. Но они могут находиться не только на разных
хромосомах, но и на одной хромосоме.
Ре комбинация-это поя тление нотых.
отл ионы х от род ите лыж их ком бина ций ал лел ей, на ход ящ ихся на одно й хром осоме
Огм|та1 сшяЫгшИмк of
Recombination is the * or I inf of nlleles into
combmat ions.
1 гт
lAtt l.,H
готбегшюги of
if «combi ги«ч
В профазе 1 мейоза папина и мамина хромосомы объединяются друг с другом
(конъюгация), и может происходить их кроссинговер (по-русски кроссинговер -
перекрест). То есть, когда они потом расходятся, оказывается, что они поменя-
лись своими кусочками. Гомологичные хромосомы, у которых произошел обмен ку-
сочками, называются рекомбинантными. Две из четырех гамет, образовавшихся в
мейозе 2, - рекомбинантные (две другие - родительского типа).
Перекрест хромосом происходит в профзге 1 мейоге.
<74 Crossing over takes place In m el os и and is responsible
for recombination.
Если перекреста не происходит, то все гаметы - родительского типа. В ре-
зультате одиночного перекреста половина гамет - рекомбинантная, половина -
нерекомбинантная, однако рекомбинация происходит лишь в части клеток, доля
которых зависит от расстояния между локусами (чем дальше они друг от друга,
тем больше доля кроссоверных гамет).
7.6а. Без кросстговера все гаметы родительского типа
1Ы Cresting over
ММММВ Vinretombmarit А
Kwombmant ]
ЧолгкотЬггит
7.6 Crossing over produces half noerecombinant gametes and
half recombinant gametes.
7.66. В результате од точного Kpocciitroeepa половина гамет рекомбинантная,
половт.1 - itqietcoMOitiatniiafl,
однако рекомбинация г,юисход1п лишь в части клеток, доля которых зависит от
расстояния между локусам.
Рекомбинация происходит после последнего удвоения хромосом в мейозе. Такой
вывод можно сделать из того, что наряду с рекомбинантными продуктами одного
мейоза (свидетельствующими, что рекомбинация прошла) есть и нерекомбинантные
продукты того же мейоза. А если бы рекомбинация проходила до репликации, то
все хромосомы в данной мейозе были бы рекомбинантными.
В результате крое сингов ер а
появляются и рекомбинантные и НЕ рекомбинантные гаметы
это значит, что кроссинговер происходит после синтеза ДНК
Ю% рекомбинанты
to и до репликаци/
All reconibmjj
chromoM>n>«
50% рекомбинанты
если после
3d иг к
В мейозе 1 кроссинговер между данной парой локусов происходит в части
клеток, доля которых зависит от расстояния между локусами.
В клетках, где кроссинговер произошел,
50% гамет не рекомбинантные (родительские классы, 1:1),
Гены, отвечающие за высоту куста и за тип листьев, находятся в одной хромо-
соме, но не совсем рядом. В клетке могут произойти два события. Кроссинговер
может не произойти, тогда все гаметы будут родительского типа, но может и
произойти в части клеток. Тогда половина гамет будет не кроссоверная, а поло-
вина - кроссоверная. Соотношение между такими гаметами- 1:1, а между клетка-
ми, где кроссинговер произошел и где не произошел, зависит от расстояния меж-
ду локусами. Поэтому число рекомбинантов не превосходит 50%. Возможные потом-
ки показаны на рисунке выше.
Потомков высоких с нормальными листьями получилось 55, низких с крапчатыми
листьями - 53, низких с нормальными листьями - 8, высоких с крапчатыми листь-
ями - 7. Наблюдаемая частота рекомбинации численно равна проценту рекомби-
нантного потомства, образуемого в скрещивании. Общее число рекомбинантных по-
томков составило 15, общее число всех потомков - 123. Таким образом, частота
рекомбинации равна 15/123 * 100%=12%.
Расчет частоты рекомбинации
Наблюдаемая частота рекомбинации численно равна
проценту рекомбинантного потомства, образуемого в скрещивании.
В потомстве скрещивания на рис.7.7
обнаружены растения четырех фенотипических классов:
1) 55 - высокое, листья нормальные (как родитель 1);
2) 53 - низкое, листья крапчатые (как родитель 2);
3) 8 - низкое, лисья нормальные (рекомбинант 1);
4) 7 - высокое, листья крапчатые (рекомбинант 2)
Общее число рекомбинантных потомков составило 15,
общее число всех потомков составило 123
Наблюдаемая частота рекомбинации =
число рекомбинантных потом ко в/общее число потомков х 100%
------------х 100% = х 100% = 12%
55 + 53 + 8 + 7 123
В интервале 12% рекомбинация произошла. Это событие пропорционально рас-
стоянию между участками. Если оно наблюдается с частотой 12%, то оно могло
произойти с два раза с частотой 0.12*0.12*100%=1.44%. Потомство от двух пере-
крестов между парой маркеров выглядит как НЕ рекомбинантное. Двойная (и любая
четная) рекомбинация снижает проявление каждой из одиночных рекомбинаций. Де-
тектируются только нечетные перекресты между парой маркеров, четные не детек-
тируются .
7.11. Потомство от двух кроссинговеров между парой
маркеров выглядит как НЕ рекомбинантное.
Доенная рекомбтац1я сшлоет гдоявление каедой из одиночных рекомбигаци!.
Детектируются только нечетные перекресты между парей маркеров, четные не детектируются.
Наблюдаемая доля рекомбинантов отражает итог совокупности независимых
рекомбинационных событий меду парой маркеров -одиночных, двойных,
тройных и т.д рекомбинаций.
Расчетное генетическое расстояние между парой маркеров гцмыерно |>авно
наблюдаемой частоте рекомбинацит плюс квадрат этой частоты Mityc куб и тд.
Если наблюдаемая доля рекомб1»тантов 4наблюдаемая частота рекомбинатдаО= 12°», то
Расчетное генетическое расстояние = 0.12 + 0,12x0,12 -0,12x0,12x0,12+...* 0,134= 13,4%
Наблюдаемая доля рекомбинантов отражает итог совокупности независимых ре-
комбинационных событий меду парой маркеров - одиночных, двойных, тройных и
т.д. рекомбинаций. Расчетное генетическое расстояние между парой маркеров
примерно равно наблюдаемой частоте рекомбинации плюс квадрат этой частоты -
остальные члены этого ряда будут малы. Если наблюдаемая доля рекомбинантов
(наблюдаемая частота рекомбинации) равна 12%, то расчетное генетическое рас-
стояние примерно равно 0,12 + 0,12x0,12 = 0,134 = 13,4%. Практически чтобы
величина поправки была пренебрежимо мала (меньше 5% от величины генетического
расстояния) значения наблюдаемых частот рекомбинации должны находится в пре-
делах 5%.
Группы сцепления и
хромосомная теория
наследственности
На фотографии изображен Томас
Гент Морган, который первым обна-
ружил сцепленное наследование
признаков (вначале сцепление с
полом, а затем и признаков друг с
другом) и генетическую рекомбина-
цию между ними. Это явилось гене-
тической основой хромосомной тео-
рии наследственности, за что ему
была присуждена Нобелевская пре-
мия .
Тсмас Гент Морган первым
обнаружил сцепленное
наследование признаков и
генетическую рекомбинацию
между никли. Это явилось
генетической основой
>ромосом-юй теории
наследственности, за что ему была
присуждена Нобелевская гремия.
Гены, расположенные в одной паре гомологичных хромосом наследующиеся единой
группой. Морган назвал ее группой сцепления. Совместное наследование генов,
ограничивающее свободное их комбинирование, называют сцеплением генов.
Гены в гомологичных хромосомах расположены в одном и том же порядке у всех
людей. Но аллели (альтернативные состояния этих генов) могут в различаться в
гомологичных хромосомах. Рассмотрим хромосомы вашей мамы. Обозначим аллели
генов той хромосомы, которые она получила от вашей бабушки буквами Б, а те
аллели той же хромосомы которые получила от дедушки буквами Д. Если рекомби-
нация не произошла, то у вас, как и у вашей мамы, будет присутствовать хромо-
сома с набором аллелей БББББББ (ну если дедушкина хромосома к вам попала, то
набор будет ДДДДДД ) Генетическая рекомбинация - это обмен блоками аллелей
между гомологичными хромосомами. Если рекомбинация произойдет при образовании
той яйцеклетки, из которой вам повезло родиться, то блок аллелей бабушки бу-
дет продолжен блоком аллелей дедушки. Например, если рекомбинация произойдет
между генами №4 и №5, то набор аллелей этой хромосомы будет выглядеть у вас
так: ББББДДДД.
Чем больше расстояние между генами, тем выше вероятность рекомбинации между
ними. Впервые это предположил и доказал Морган. Т.Морган предположил, что
частота кроссинговера показывает относительное расстояние между генами: чем
чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хро-
мосоме, чем реже кроссинговер, тем они ближе друг к другу.
Морган провел количественное исследование рекомбинации на дрозофилах. Он
исследовал сцепленные гены, определяющие цвет тела, цвет глаз и форму крыль-
ев. Все эти гены находятся в Х-хромосоме, то есть у самок две Х-хромосомы, а
у самцов - одна.
Самки, гетерозиготные по всем трем генам, были скрещены с самцами, несущими
рецессивные аллели этих генов. Так как самки были гетерозиготны, они обладали
признаками дикого типа. Самцы, у которых второй Х-хромосомы нет, имели рецес-
сивные признаки - желтый цвет тела (рецессивный аллель у, yellow), белые гла-
за (рецессивный аллель w, white) и расщепленные крылья (рецессивный аллель
bi, bifid),
Если бы все было «по Менделю», то маркеры должны были бы комбинироваться
независимо друг от друга. Но Морган к этому времени уже определил, что марке-
ры эти наследуются практически одним блоком - это явление и было названо ге-
нетическим сцеплением. При наследовании единым блоком самки в потомстве долж-
ны были получаться двух классов, смотря какую мамину хромосому получит дочь:
либо гетерозиготные по всем трем генам, - yY, wW, biBi (если получили от ма-
тери доминантные гены), либо гомозиготные по всем трем рецессивным генам -
уу, ww, bibi (если получили от матери хромосому с рецессивными генами). Одна-
ко иногда гены из одной группы сцепления все же наследуются раздельно, то
есть появляются также и самки с другими генотипами. Например, с набором Yy,
bibi, то есть получившие от матери сочетание аллеля Y и аллеля b i. Частота
изменения комбинации маркеров у (желтый цвет тела) и bi (расщепленные крылья)
составила 4,7%. То есть на 1000 мух таких было 47. Доля рекомбинантов между
маркерами у (желтое тело) и w (белые глаза) была равна 1,2%, а между маркера-
ми w (белые глаза) и bi (расщепленные крылья) - 3,5%.
Несложно заметить, что 3,5%+1,2%=4,7%. Если считать, что процент рекомбина-
ции отражает расстояние между генами, то это означает, что гены могут быть
расположены только линейно и никак иначе.
? О'
Х-хромосома у w bt у w bi Х-хромосома
Х-хромосома Y W Bi ... Y-хромосома
Расстояние между генами у и hi равно сумме двух одинарных
кроссинговеров между у и w, w и bi
4,7%
1,2% 3,5%
У * Ы
yellow white bifid
процент кроссинговера является функцией расстояния между генами
и их последовательного, т.е.
линейного расположения в хромосоме.
ME.Лооашев. Генет>к>. Л.. 1967, стг*. 246
На тот момент еще не было ничего известно ни про ДНК, ни про химию наслед-
ственности - были просто формальные генетические признаки. Но Морган и без
этого смог показать, что гены расположены линейно относительно друг друга,
что бы ни было носителем генов. Вывод о том, что частота кроссинговера явля-
ется функцией расстояния между генами и их линейного расположения в хромосо-
мах и принес Моргану Нобелевскую премию.
Параллельно с генетическими исследованиями Моргана шли цитологические ис-
следования. Исследовался митоз, мейоз, и было известно, что в мейозе (профаза
I) пары одинаковых (гомологичных) хромосом объединяются, а затем образуют
крестообразные фигуры (хиазмы). Предположили, из чисто цитологических данных,
что наблюдаемый под микроскопом перекрест хромосом связан с обменом сегмента-
ми между ними. Впоследствии это было подтверждено. Хромосомы маркировали -
нашли мух у которых на хромосоме были дополнительные фрагменты (транслока-
ция) , видимые под микроскопом. Было видно, что в результате перекреста изме-
ненные морфологически хромосомы родителей оказываются у потомства в новых
комбинациях.
На рисунке показано, как выглядят хиазмы - крестообразные структуры, обра-
зуемые гомологичными хромосомами в мейозе и различимые под микроскопом.
Хромосомы состоят из двух параллельных «линеечек», которые называются хро-
матидами, каждой из которых соответствует одна молекула ДНК. Под микроскопом
было видно, что одна из пар хроматид пересекается, то есть между хромосомами
происходит перекрест.
Был проведен следующий эксперимент. Х-хромосома дрозофилы была промаркиро-
вана дополнительным кусочком хромосомы, и было подтверждено соответствие ге-
нетических и цитологических данных, описывающих рекомбинацию.
MF.Лоо a inert. Генепаа. Л., 1967, ст|1 ?60
Частота видимых под микроскопом перекрестов в данной хромосоме в расчете на
1 клетку в фазе мейоза, где перекрест можно наблюдать, стремится к двукратной
частоте генетической рекомбинации между всеми маркерами этой хромосомы, по
мере роста числа маркеров. Причина этого ограничения заключается в том, что в
перекресте участвует, как правило, лишь одна из двух пар хроматид. Стало по-
нятно , что можно связать количество перекрестов наблюдаемых цитологически
(хиазмы) с частотой генетической рекомбинации, что особенно четко было пока-
зано на кукурузе в 50-х годах.
Длина генетической карты, выявляемая по анализу результатов генетической
рекомбинации, вычисляется как сумма расстояний между наиболее близкими марке-
рами, Единицей генетической карты является 1% рекомбинации и эта величина бы-
ла названа в честь Моргана 1 сантиМорган (1 сМ).
Данные о связи цитологии с генетикой оказались очень важными в последующих
генетических исследований, в частности на человеке. Они позволяют вычислить
длину генетической карты по цитологическим данным, не прибегая к получению
мутантов, чистых генетических линий и даже вообще без направленных скрещива-
ний - все это невозможно на человеке. У человека расчетная длина генетической
карты по цитологическим данным составляет около 3000 сМ. или 3000% (три тыся-
чи процентов рекомбинации).
Почему длина генетической карты человека равна 3000% (и почему нет ничего
страшного в том, что эта величина превышает 100%).
Генетическая карта - это последовательность маркеров в хромосоме и расстоя-
ния между ними, следующие из частот генетических рекомбинаций. Одна единица
карты соответствует 1% рекомбинации или одному сантиморгану (1 сМ).
Проведем следующую аналогию, чтобы было легче понять, почему длина карты
человека равна больше, чем 100%. Один и тот же термин иногда используется в
разных смыслах: 100 градусов - температура кипения воды. 90 градусов - прямой
угол. 40 градусов - крепость водки. И обычно эти смыслы никто не путает
То же и с термином процент (pro cent = на сто) который используется в раз-
ных смыслах при описании рекомбинации.
1) Генетическое расстояние между двумя маркерами (1сантиморган, сМ=1% ре-
комбинации) . Например, наблюдаемый процент рекомбинации в потомстве (12% ре-
комбинантов, как описывалось выше), где общее число потомков равно 100%. Доля
рекомбинантов по одной паре маркеров не может превышать 50% от общего числа
потомков (несцепленное наследование);
2) Общая длина генетической карты организма (N сантиморган = N %) Рассчиты-
вается как сумма минимальных экспериментально определенных генетических рас-
стояний между парами маркеров, и для каждого вида своя.
Количество последовательно расположенных пар маркеров, каждый из которых
равен, например, 12%, может быть и не ограниченным. Восемь таких отрезков со-
ставят в сумме 96%, а восемьдесят отрезков - 960%. Хотя доля рекомбинантов
между любыми двумя из этих 81 маркеров, конечно, не может превышать 50% от
общего числа потомков.
Отметим, в заключение, что длина генетической карты человека определялась
по цитологическим данным, которые были доступны уже достаточно давно. В отли-
чие от мухи, установить длину генетической карты человека на основе экспери-
ментов по скрещиванию невозможно.
Поиск или диагностика мутации, вызывающей заболевание, наследуемое по Мен-
делю, часто проводится по сонаследованию признака «болезнь» и маркера-
свидетеля, расположенного рядом с мутантным геном. Маркер-свидетель - это та-
кой маркер, который легко обнаружить при анализе. Источник ошибки диагностики
- утрата связи при рекомбинации между мутацией, вызывающей заболевание и мар-
кером-свидетелем. Точность диагностики тем выше, чем меньше это расстояние.
Вопрос. Сколько генетических маркеров нужно иметь в геноме человека, чтобы
обеспечить >95% точность диагностики для любого гена, если длина генома чело-
века - 3000%.
Точность не менее 95% означает, что не более генетическое расстояние между
мутацией вызывающей заболевание и маркером составляет не более 5% рекомбина-
ции. Следовательно до ближнего к маркеру левого и до ближнего правого маркера
должно быть не более 5%, то есть расстояние между маркерами не более 5%х2=10%
рекомбинации. В 10%-х интервалов в карте длиной 3000% будет 3000/10=300. То
есть 300 равноудаленных маркеров будет достаточно, чтобы картировать или вы-
явить мутацию с точностью >95%, даже ничего не зная о том, где находится ис-
следуемый ген. Ясно, что когда это только начали делать, примерно 10 лет на-
зад, маркеры ложились случайно, поэтому генетическую карту пришлось составить
из нескольких тысяч маркеров, чтобы большинство интервалов между маркерами не
превышало 10%. Сегодня в практической работе по общегеномному скринингу у че-
ловека используют панель из 384 равноудаленных маркеров.
Молекулярный механизм гомологичной рекомбинации, предложен Холидеем.
Рассмотрим две гомологичные хромосомы: папину и мамину. В них, как предпо-
лагается, происходит однонитевой идентичный в обеих хромосомах разрыв, после
которого эти части, перекрещиваясь, образуют так называемую структуру Холидея
(который данную схему рекомбинации первым предложил). Далее происходит пере-
нос точки надреза вдоль хромосомы, в результате чего части гомологичных хро-
мосом меняются местами. В результате получаются хромосомы, составленные из
кусков папиных и маминых хромосом. Механизм гомологичной рекомбинации - одно-
нитевой разрыв в каждой из двунитевых молекул ДНК, вытеснение и замещение ни-
ти, миграция разрыва и разрешение единичной структуры Холидея.
Гомологичная рекомбинация, механизм: однонитевой
разрыв в каждой из двунитевых молекул ДНК, вытеснение и
замещение нити, миграция разрыва и разрешение единичной
структуры Холидея
Геномные, хромосомные
и генные мутации
Теперь поговорим о мутациях: геномных, хромосомных и генных.
Пример геномной мутации - удвоение всего числа хромосом в геноме (автополи-
плоидия) , она может возникать из-за нерасхождения хромосом в митозе или мейо-
зе .
9.27.Удвоение всего числа хромосоя в геноме (автополиплоидия)
может возникать из-за нерасхождения хромосом в митозе или мейозе.
(a* Aulopotyplohty through miloti*
9 27 Aulopolyploidy can aritr through nonrliijunclion in
mitotic or tn«K>«H.
Какое это имеет отношение к нам? У прямых предков человека как вида поли-
плоидия (чаще всего удвоение) случалась неоднократно, но последний раз - бо-
лее 100 миллионов лет назад. У животных и растений она встречается часто,
особенно у культурных растений. При отборе человек просто не замечал, что от-
бирал полиплоиды. Например, культурный картофель - тетраплоид, банан - трип-
лоид, он пригоден к употреблению, так как не образует семян, в отличие от ди-
плоида, который состоит из жестких семян и почти не имеет мякоти.
Виды культурных растений часто являются полиплоидами
| Table 9 21
Plant Type of Polyploidy Ploidy Chromosome Number
Potato Autopolyploid 4л 48
Banana Autopolyploid Зл 33
Peanut Autopolyploid 4Л 40
Sweet potato Autopolyploid 6л 90
Tobacco Allopolyploid 4л 48
Cotton Allopolyploid 4л 52
Wheat Allopolyploid 6л 42
Oats Allopolyploid 6л 42
Suyat cane Allopolyploid 8л 80
Stiawbetry Allopolyploid 8л 56
Source: After R C. Dhot. Ptonf Breeder# York. MtCrav» Hill. 1956). р.
При скрещивании дикорастущих видов было ресинтезировано несколько видов
культурных растений, например, слива (Prunus domestica). Константная и гиб-
ридная форма, полностью сходная с домашней сливой получена при скрещивании
терна P.spinosa (2п=32) с алычой P.divaricata (2п=16). Это растение имело,
как и Р.domestica, 2п=48 хромосом. Вероятно, дикая слива в ходе эволюции по-
лучилась именно таким путем.
Prunus Prunus
diva ri cata spinosa (те рн )
P.domestica
(слива)
П|Н1 сфоцивашидшорастущих В1щов было
ресинт евфовано несколько вцдов нулыу]>ны>
растеши, нап|]имэд
слтва (Ргинтз domestica}.
Константная и гнб|ищная форма, полностью
сходная с домашней сливой получена
три ск]№1циванш
терна P.spinosa (2п=32)
с алычой P.divai icata (2п = 16).
Это растете млело, каки P.dorrestica, 2n=4fl
хромосом.
М .Е Лобашев. Генетика. Л., 1967, стр. 515
А триплоиды просто так не получишь, у них даже семян не образуется, но они
удобны в практических целях человека, например для получения большей урожай-
ности и плодов без семян (у арбузов, бананов и т.д.) У арбуза гаплоидный на-
бор п = 11 хромосом. Скрещивание тетраплоидного (4п = 44) и диплоидного (2п =
22) арбуза дало триплоид (Зп = 33) - бессемянный, крупноплодный, устойчивый к
заболеваниям, в отличие от родительских форм.
Триплоидность -возможный путь увеличения урожайности и получения
бессемейных форм (напр., все сорта бананов).
Гаплоидный набор арбуза п = 11 хромосом.
Скрещивание тет pan ло одного (4п = 44) и диплоидного (2п = 22) арбуза дало
триплоид (Зп = 33) - бессемянный, крупноплодный, устойчивый к
заболеваниям:
Сорт Плоидность Урожай на единицу площади
Колич.плодов Вес (в кг.)
Син-Ямато 2п 165 352,1
Син-Ям ат о 4п 115 155,6
Син-Ямато (4п) ? Отоме (2п) ЗП 215 596,2
Отоме 2п 150 253,7
М.Е.Лобашев. Генетикь Л., 1967, стрс 650
Межвидовое скрещивание можно наблюдать у близкородственных животных: осла с
лошадью - мул, или у коня с ослицей - лошак, они более выносливы, хоть и бес-
плодны. Вероятность появления потомства у мулов и лошаков (а это зависит от
того, отойдут ли при делении все хромосомы правильно к одному полюсу) равна
одной второй в степени п, где п - гаплоидное число хромосом. С такой частотой
будут образовываться яйцеклетки и сперматозоиды, соответствующие по хромосом-
ному составу родителям - ослу или лошади. Видно, что вероятность эта очень
мала.
Скрещивание между
кобылой и ослом дает мула,
скрещивание коня с ослицей
— лошака.
Такие гибриды более
вьносливы, чем родители,
но не плодовиты.
Parents
ПогЖеу
II 2л - 62
Progeny
Hmny
Теперь поговорим о хромосомных мутациях. Хромосомные мутации - изменение
числа отдельных хромосом в геноме (анеуплоидия) или целостности хромосом (пе-
рестройки) . Это одна из причин болезней человека. Моносомия (In) или трисомия
(Зп) хромосомы в оплодотворенной яйцеклетке будут смертельны для плода (кроме
половых и мелких хромосом), а в соматических клетках часто приводит к раку.
Каждая из 23 хромосом может быть покрашена в свой цвет (FISH гибридизация,
справа). Каждая хромосома прокрашивается в штрих код (G-окрашивание, слева,
полоса до 0,1% от длины генома). Это позволяет увидеть изменение числа и
структуры хромосом, в том числе при опухолевых заболеваниях. Опухолевые забо-
левания почти всегда связаны с перестройками генома. Хромосомы человека про-
нумерованы в порядке убывания размера от 1 до 22.
Хромосомы человек!
Каждая из 23 хромосом м.б. покрашена в свой цвет (FISH гибридизация, справа)
Каждая хромосома прокрашивается в штрих код (G-окраш ивание, слева , полоса до 0.1 %)
Это позволяет увидеть изменение числа и струст\ры хромосом. в том числе при раке
Хромосомы человека про номерованы в порядке убывания размера от 1 до 22
Синдром Дауна - трисомия по хромосоме 21
Подавляоцая часть случаев синдрома
Дауна вызвана нерасхождением леры
хромосом 21
в мейозе у матери
(по анализу (омкросателлитов)
Реже встречаются семейнье случаи -
транслокация гена из хр21 на другую
хромосом/
Частота рождения детей с синдромом
Дауна увеличивается с возрастом
матери
от 1 /2000 в 20 лет до 1Л 2 в 50 лет.
Изменение числа хромосом, как и всякая мутация, связано с физиологическим
состоянием организма. С возрастом организм начинает функционировать хуже, это
приводит, в частности, к повышению частоты рождения детей с синдромом Дауна.
Подавляющая часть случаев синдрома Дауна вызвана нерасхождением пары хромосом
21 в мейозе у матери. Частота рождения детей с синдромом Дауна увеличивается
с возрастом матери от 1/2000 в 20 лет до 1/12 в 50 лет. Но есть и семейные
случаи - транслокация гена из хр21 на другую хромосому.
На слайде видно, что 21-я хромосома представлена трижды. Две копии вместо
одной чаще присутствуют в еще неоплодотворенной яйцеклетке, а не в спермато-
зоиде . Это было выявлено по анализу микросателлитов, о чем будет рассказано
на следующих лекциях. Сейчас до трех месяцев беременности можно провести хро-
мосомный анализ, для этого берут клетки околоплодной жидкости и смотрят, есть
ли трисомия по 21-ой хромосоме. Во многих европейских странах после 35 лет
анализ беременной женщины на выявление у плода синдрома Дауна обязателен, в
связи с повышением частоты рождаемости больных детей при увеличении возраста
матери.
Синдром Дауна вызван присутствием
трех копий одного из генов хромосомы 21
или всей хромосомы целиком.
U)
<Ь)
Теперь поговорим о хромосомных перестройках. Основные типы хромосомных пе-
рестроек :
1. дупликация - удвоение сегмента
2. делеция - утрата сегмента,
3. инверсия - переворот сегмента,
4. транслокация - перенос сегмента на другую хромосому
Все они являются причинами многих болезней.
Перестройки хромосом в гаметах часто приводят к болезням человека - наруше-
ниям морфологии, физиологии и поведения, особенно часто встречается дуплика-
ция и делеция, как правило, все такие болезни сопровождаются заторможенным
умственным развитием, то есть чаще всего какие бы ни были хромосомные пере-
стройки, они затрагивают развитие наших умственных способностей, ну а кроме
этого болезни сопровождаются чаще всего недоразвитием каких-либо органов (на-
пример, маленькая голова).
Основные типы хромосомных перестроек:
1. дупликация - удвоение сегмента
2. делеция - утрата сегмента,
3. инверсия - переворот сегмента,
4. транслокация - перенос сегмента на другую хромосому
|b> Deletion
(о Inversion
В эволюции геномные и хромосомные мутации чаще фиксируются у растений, реже
у животных. Геномные мутации у животных обычно различают таксоны более высо-
ких порядков например отряды (сельдевые и лососевые), а у растений это могут
быть и виды (пшеница и другие культурные растения). Анеуплоидия и перестройки
хромосом в гаметах обычно приводят к болезням у животных человека - нарушени-
ям морфологии, физиологии, поведения, и резко снижают возможность оставить
потомство. Тем не менее, сотни тысяч таких случаев зафиксированы на сегодня
эволюцией - это почти любая пара родственных видов, различающиеся по структу-
ре кариотипа (набора хромосом). Ясно, что такие изменения когда-то произошли
у индивида и затем закрепились в поколениях.
Например, хромосома №2 человека образована при транслокации, произошедшей у
наших предков после ответвления от ствола всех приматов: у орангутанга, го-
риллы и даже шимпанзе (5 млн. лет расхождения). У всех этих наших родственни-
ков по две независимых хромосомы, которые лишь у человека оказались соединен-
ными. Часто, хотя и не всегда, перестройки приводят к понижению жизнеспособ-
ности, но в нашем случае получилось не так, мы получили две разных хромосомы
шимпанзе, которые соответствуют хромосоме №2 человека. Это видно при попереч-
ном окрашивании хромосом, которые выявляют идентичные в геноме фрагменты хро-
мосом. Все люди на Земле имеют общего предка, у которого произошла эта транс-
локация (меньше 5 млн. лет назад)
Число хромосом у млекопитающих может различаться в десятки раз, хотя размер
генома отличается менее чем на 20%. У человека число пар хромосом 23, а у ло-
шади - 66, у обитающего в Азии оленя-мунтжака - 6 хромосом. Исключение со-
ставляет южно-американский грызун, называемый красной вискашевой крысой (ла-
тинское название Tympanoctomys barrerae, английское - red viscacha rat), хотя
крысе оно весьма отдаленный родственник). У этого животного геном не диплоид-
ный, а тетраплоидный, содержит в два раза больше ДНК, чем у остальных млеко-
питающих, и 102 хромосомы.
Хромосома N°2 человека образована при т|>анс локации, произошедшей у наших
предков после ответвления от ствола всех приматов: у орака, гориллы и даже
шимтанзе(5 млнлет расхоедентя} две разных хромосомы соответствуют хромосоме
2 человек!. Все пводи на Земле имеют общего гр едка. у которого произошла эта
транслокащтя.
Orangutan chromosomes
Metjcentru
chromosome
fragment
Break-
points
I Robertsonian
translocation ।
Если до общего предка млекопитающих около 200 миллионов лет и около 60
транслокаций (перестроек разных хромосом), то 1 транслокация сохраняется и
дает начало новому виду не реже чем раз в 3 млн. лет. А так как недетектируе-
мых внутрихромосомных перестроек больше на 1-2 порядка, то это означает, что
носители таких перестроек выживают гораздо чаще, нежели раз в 3 млн. лет. По-
следний раз такая перестройка в линии человека произошла не более 5 млн. лет
назад.
Геномные и хромосомные мутации могут появляться и в соматических клетках
человека и животных. В этом случае они не передаются потомству, но часто свя-
заны с развитием рака. Реципрокная транслокация фрагментов между хромосомами
8 и 14 в лимфоцитах человека приводит к лимфоме Бёркита: к гену иммуноглоби-
нов присоединяется ген онкогена c-MYC, меняя его регуляцию.
Геномные и хромосомные мутащв!
в соматичесмпх клетюх человек! и mieoiHbix
часто сая заты с развтнем [мзка
Реципрокная транслокация хромосом
8 и 14 в лимф оцитах человека
приводит к лимфоме Бёркита:
К гену иммуноглобинов
присоединяется ген онкогена С-А4УС
меняя его регуляцию
14
Таким образом, перестройки, происходящие в соматических клетках, влияют
только на нас, а на следующее поколение не влияют. Те перестройки, которые
происходят в клетках зародышевого пути, могут пройти через эволюционное «си-
то» и остаться в поколениях, Это может привести к репродуктивной изоляции ин-
дивидов с перестройками от других индивидов внутри данного вида.
Хотя в клетках зародышевого пути геном остается постоянным, изменение
структуры генов и генома может быть нормальной частью жизненного цикла. У не-
которых эукариот число хромосом в соматических клетках отличается от числа
хромосом в клетках зародышевого пути. У некоторых простейших в определенной
фазе развития геном распадается по на несколько тысяч хромосом, предположи-
тельно соответствующих отдельным генам. Размер генома вегетативных клеток и
клеток зародышевого пути также может различаться. Например, у некоторых круг-
лых червей в соматических клетках (но не в клетках зародышевого пути!) подав-
ляющая часть генома утрачивается (явление называется диминуцией хроматина). У
человека перестройки генов иммуноглобулинов в лимфатических клетках - условие
образования разных антител. Только при таком условии в организме может обра-
зовываться то разнообразие антител, которое может обеспечить необходимую за-
щиту. То есть перестройки генома могут быть управляемыми и необходимыми.
Несколько слов про генные мутации. Генные мутации являются скачкообразными
изменениями отдельных локусов хромосом - генов. Мутантные гены сохраняют
свойство репродукции при делении ядра клетки, вследствие чего мутационные из-
менения наследуются. Мутации могут быть прямыми (нуклеотид Т в данной позиции
заменен на нуклеотид С) и обратными (мутантный нуклеотид С в данной позиции
заменен на нуклеотид Т, характерный для дикого типа). Частота мутирования в
обоих направления характерна для каждого локуса. Для разных типов мутаций она
варьирует от 10"6-10-8 на нуклеотид на генерацию до 3*10-1. Спонтанный мутаци-
онный процесс обусловливается свойством самого гена, системой генотипа, фи-
зиологическим состоянием организма и колебанием факторов внешней среды. Каж-
дый локус - ген может мутировать в несколько состояний, образуя серию множе-
ственных аллелей. Для примера скажем, что существует ген супрессора опухолей,
где обнаружены сотни мутаций, при каждой из которой опухоль может развиться в
разных местах.
"Горячие пятна" мутаций внутри гена распределены неравномерно. Они харак-
терны не только для спонтанного мутирования, но и при воздействии определен-
ными химическими агентами.
Например, разберем мутации в гене CFTR, которые вызывают муковисцидоз - за-
болевание , связанное с дефектом проводимости ионных каналов, проявляется в
виде заболеваний легких и др. В этом гене описано уже больше тысячи разных
мутаций. На графике представлена частота разных мутаций в разных частях гена,
видно, что распределение частоты неравномерно в разных его участках - экзо-
нах. Частота встречаемости в популяции разных мутаций одного гена различается
в тысячи раз.
Если смотреть реальное распределение мутаций среди людей, оказывается, что
каждая из них присутствует со своей частотой в группе, живущей на данной тер-
ритории. Например, для России мутация AF508 встречается у 80% больных. А ос-
тальные мутации составляют менее 20%, некоторые же не встретятся в России во-
обще .
Обнаружено »10ОО различных мутаций в гене CFTR
Распределение мутаций вдоль гена неравномерное. Частота встречаемости в
популяции разных мутаций одного гена различается в тысячи раз
На этом слайде показано, как растет частота мутаций с увеличением дозы об-
лучения. Частота мутаций линейно растет с увеличением дозы радиоактивного об-
лучения. «Безопасной» дозы облучения нет (нет порога). Повреждения происходят
при любых дозах, так что понятие порога чисто юридическое.
Частота мутаций линейно растет
с увеличением дозы радиоактивного облучения
«Безопасной» дозы облучения нет (нет порога)
На рисунке выше показано, что при воздействии мутагенами нет нижнего порога
дозы. В данном случае показана ситуация с рецессивными мутациями, аналогичная
ситуация и с доминантными. Поскольку все время есть какое-то фоновое мутаген-
ное воздействие, то можно подумать, что число мутаций в поколениях должно все
время расти
Действительно, после мутагенного воздействия (Хиросима, Чернобыль, Бхопал,
Орандж эйджент) частота мутаций растет. Растет также и процент опухолевых за-
болеваний, так как повреждается геном соматических клеток. Однако после сня-
тия мутагенного воздействия доля мутантов не увеличивается, а только снижает-
ся в поколениях из-за гибели и пониженной жизнеспособности мутантов. Если ре-
бенок родился, значит наиболее существенные для развития гены у него нормаль-
ные, ведь в противном случае он бы умер на эмбриональной стадии. Основной ле-
тальный эффект мутаций реализуется еще на клеточном уровне, а не на организ-
менном уровне. Клетка запрограммирована не пропускать мутации в следующее по-
коление. Есть специальный молекулярный механизм отслеживания повреждения в
ДНК, еще до того как поврежденный участок понадобится для реализации функции.
Если окажется, что ДНК повреждена и не может быть исправлена (репарирована),
то в такой клетке станет работать запрограммированная система самоубийства.
Вероятно, основная часть мутаций приводит к дефектам функционирования и гибе-
ли уже на стадии первых делений оплодотворенной яйцеклетки, меньшая часть
проявляется позже и приводит к спонтанному аборту, еще реже - к рождению ре-
бенка с аномалиями.
Хотя эта частота была повышенной после ядерного взрыва, уже через два поко-
ления (сейчас) у жителей с Хиросимы частота наследственных аномалий и опухо-
левых заболеваний такая же и даже ниже, чем в других городах Японии, так как
население этого города оказалось под пристальным вниманием врачей, проводи-
лось больше профилактики и т.д. Что же касается опухолевых заболеваний, то во
втором поколении частота заболеваний уже не будет больше, так как это эффект
повреждения соматических клеток, не передающееся по наследству, не учитывая
некоторые конкретные болезни.
В Чернобыле, после точки максимума, частота аномалий при рождении снизилась
в 8 раз за первые 10 лет после радиоактивного выброса. Если произошла доми-
нантная летальная мутация, то она будет устранена уже в следующем поколении.
Если же мутация рецессивная, то частота мутации будет все время падать в по-
колениях (тем быстрее чем более выражен ее вредный эффект) и это будет моно-
тонно убывающей функцией.
Генетика популяций
До этого мы говорили о том, что происходит с индивидом, но с точки зрения
эволюции больший интерес представляют изменения внутри популяций. В каждом
поколении частота каждого аллеля данного гена и частота каждого генотипа по
этому гену сохраняется постоянной. Правда при условии, если отбора нет, если
среда не меняется, если подбора пар нет (свободно скрещивающася или панмикти-
ческая популяция), если нет миграции (притока генов извне) и еще много всяких
«если». В этом случае и частоты фенотипов постоянны.
Например, мы по внешности распознаем людей с данной территории как один на-
род, хоть люди и разные. Это происходит потому, что частоты аллелей в данной
популяции постоянны в поколениях, а, следовательно, постоянны и частоты гено-
типов . Ну а среда вокруг все та же - поэтому и частоты фенотипов не меняются.
Вот эту совокупность фенотипов мы и воспринимаем либо как общую (один народ),
либо как другую (другой народ).
Если мы будем рассматривать коренных жителей достаточно удаленных регионов
(несколько тысяч километров) то принять решение, что народы, или даже расы,
разные, труда не составит. А вот если посмотреть на такие же группы, разде-
ленные расстоянием в сотни километров, не говоря уж о десятках километров, то
возникнут большие Затруднения, где же границу провести. Потому что границы-то
объективной, генетической, нет. Есть только непрерывный ряд частот огромного
количества аллелей, причем со своим направлениями и градиентами изменения в
географическом пространстве.
Приведем для наглядности такой пример, иллюстрирующий постоянство частот
аллелей в поколениях одной популяции. Для человека известен доминантный ген Т
(Taster - дегустатор). Носитель его обладает свойством определять на вкус
слабый раствор фенилтиокарбамида как горький. Обладатель рецессивной аллели
этого гена в гомозиготном состоянии (tt) тот же раствор определяет как без-
вкусный. Присутствие такого гена можно тестировать как на уровне генотипа,
так и на фенотипическом уровне: «чувствует - не чувствует» горечь. Определе-
но, что частота «дегустаторов» в данном поколении 0,36, а «не дегустаторов» -
0,64. «Не дегустаторы» несут два аллеля tt, следовательно, частота аллеля в
популяции равна корню квадратному из этой величины (0,64), то есть частота
аллеля t в популяции равна 0,8. В сумме частота двух аллелей равна 1, следо-
вательно, частота аллеля Т равна 1 - 0,8 = 0,2. Зная частоты аллелей в гаме-
тах (сперматозоидах и яйцеклетках) можно рассчитать частоты генотипов и фено-
типов которые получаться в следующем поколении.
0,2 Т 0,8 t
0,2 Т 0,04 ТТ 0,16Tt
0,8 t 0,16Tt 0,64tt
Нетрудно видеть, что частоты генотипов и фенотипов по гену Т остались неиз-
менными в следующем поколении.
Что можно ожидать в этом поколении по частотам аллелей в гаметах? Гаметы с
аллелем Т будут возникать от гомозиготы ТТ 0,04*2 + от двух гетерозигот Tt
"дегустаторов" 0,16*2 = сумма 0.4. Гаметы с аллелем t будут возникать от не-
дегустаторов tt 0,64*2 + от двух гетерозигот Tt 0,16*2 = 1.6. Соотношение
частот аллелей в гаметах 0,4:1,6= 0,2:0.8, то есть такое же, как и в предыду-
щем поколении.
Отсюда следует, что в указанной популяции в поколениях поддерживается оди-
наковое соотношение частот аллелей (0,2T:0,8t), и фенотипов (64% "недегуста-
торов" и 36% "дегустаторов").
Неизменность генетического состава свободно скрещивающейся (панмиктической)
неограниченной по численности популяции, существующей в отсутствии отбора,
называется правилом (законом) Харди-Вайнберга.
Поскольку свободное скрещивание означает случайное объединение гамет, несу-
щих аллель Т или t , то при частоте р аллеля Т и частоте q аллеля t (заметим,
что при наличии только двух аллелей, подобно рассмотренному выше случаю,
p+q=l) распределение генотипов TT, Tt и tt неизменно и соответствует уравне-
нию
р2 TT + 2pq Tt + q2 tt = 1
Если в популяции соблюдается указанное соотношение между частотами аллелей
и генотипов, то она называется равновесной по данному гену. Для каких-то ге-
нов это равновесие между частотами аллелей и генотипов может не соблюдаться.
Отклонение от равновесного распределения частот генотипов может наблюдаться
по статистическим причинам в выборках малого размера, в популяциях с отсутст-
вием панмиксии и по другим причинам. Одной из причин неравновесия частот ал-
лелей и генотипов считается отбор. Степень отклонения от равновесия указывает
на интенсивность отбора.
На прошлой лекции было рассказано о летальной рецессивной мутации у мышей.
Если скрещиваются две желтых мыши, гетерозиготных по гену окраски (несут до-
минантный в определении цвета аллель Yellow, обознчаемый буквой Y, и рецес-
сивный аллель белой окраски у), то появляются мыши желтые и белые. Они нахо-
дятся в соотношении 2:1, а класса YY нет. Оказывается, что эмбрионы с геноти-
пом YY есть, но они гибнут до рождения (в отношении летального эффекта аллель
Y рецессивен). При этом, естественно, равновесие Харди-Вайнберга среди родив-
шихся мышей не будет соблюдаться.
2 желтых : 1 не желтый
Отмечу, что для генов, которые просто оказались рядом с геном Y в хромосо-
ме, равновесие Харди-Вайнберга тоже не будет соблюдаться. При исследовании
этих генов может показаться, что сами они находятся под действием отбора, хо-
тя они просто оказались сцеплены с тем геном, по которому жестко происходит
отбор. Это явление приходится достаточно часто разбирать в генетике - какое
изменение в хромосоме является причинным для наблюдаемого признака, а какое
просто сцеплено с наблюдаемым признаком.
Для подавляющего большинства исследованных у человека аллелей соблюдается
равновесие Харди-Вайнберга - соотношение между частотами аллелей и частотами
генотипов. То есть большинство генов адаптивно нейтральны в данных условиях.
Когда мы говорим о частоте встречаемости конкретного аллеля, надо учиты-
вать, что эта величина, которая характерна для данного места и времени. Гово-
рить о частоте гена для человечества вообще бессмысленно. Частоты могут быть
своими для каждой территории и каждой популяции, и могут различаться в десят-
ки раз. Об этом будет рассказано в лекции 21. Частота аллеля на данной терри-
тории может меняться в результате резких изменений численности популяции, от-
бора или миграций. Последняя причина - наиболее частая.
Естественный и
искусственный отбор
В Спарте, как известно, детей с отклонениями бросали в пропасть, чтобы на-
селение в следующих поколениях стало лучше. Но эти меры по элиминации индиви-
дов с фенотипическими отклонениями от нормы не только аморальны, но и генети-
чески бессмысленны. Относительная частота гетерозигот в поколениях сокращает-
ся значительно медленнее, чем частота рецессивных гомозигот. Полное устране-
ние из популяции рецессивных гомозигот в каждом поколении не приводит к их
окончательному исчезновению даже в сотом поколении, так как гетерозиготные
особи являются постоянными поставщиками рецессивных гомозигот.
Э(|м|)ективность отбора при полной элиминации рецессивных гомозигот
Поколе**|я (F) Частота рецессивного аллеля (1-<|) % особей с генотипом
аа Аа АА
1 0.5 25,0 50 25
2 0,333 11,11 44,44 44,44
3 0,250 6,25 37,5 56,25
5 0,167 2,78 27,78 69,44
10 0,091 0,83 16,53 82,64
20 0,046 0,23 9,07 90,7
50 0,020 0,04 3,84 96,12
100 0,010 0,01 1,96 98,03
Иогансен в 1903 году исследовал, как будут наследоваться вариации, которые
наблюдаются, в генетически однородной группе индивидов. Он взял самое малень-
кое и самое большое растение из самоопыляющихся бобов, получал от таких
«крайних» потомство в шести поколениях, и посмотрел, что будет в шестом поко-
лении . Оказалось, что потомство от самых мелких не отличается от потомства
самых крупных ни по средней величине признака, ни по ее дисперсии.
Самоопыляющиеся растения по природе своей генетически однородны - это набор
чистых линий. Изменчивость по размеру, которая проявлялась в чистых линиях,
не является наследственной. Она называется модификационной изменчивостью, за-
висящей только от условий среды. В генетически однородной группе изменения,
которые мы можем наблюдать, в том числе благоприобретенные, унаследоваться не
могут. То есть отбор при отсутствии генетического разнообразия даже при нали-
чии разнообразия внешнего, неэффективен.
Пример ненаследственной модификационной изменчивости у человека - это, на-
пример, акселерация, которая проявилась после войны в разных странах. Напри-
мер, средний рост японцев стал больше на 20 см, хотя понятно, что частоты ал-
лелей генов, и генотипов в этой популяции мало изменились за два поколения,
прошедших после войны.
В генетически однородной группе индивидов
отбор не эффективен (Иогансен, 1903),
Различия, приобретенные в индивидуальном развитии,
не наследуются
В ЧИСТЬЕК линиях индивиды
гомозиготны и идентичны
Различие в величине признака,
(размер бобов) вызвано только
воздействием внешней среды
при индивидуальном развитии.
Потомство от самых мелких
не отличается от потомства
самых крупных ни по средней
величине признака, ни по ее
дисперсии
М.Е.Лобашев. Тенет 1Ю. Л., 19(7, стр. 607
Если популяция генетически гетерогенна (потомство второго поколения, F2) ,
то отбор по фенотипу расчленяет ее на неперекрывающиеся группы с различающи-
мися генотипами уже за два поколения (потомство четвертого поколения F4). На
рисунке это проиллюстрировано для длины венчика у табака. В одной группе вел-
ся отбор растений с длинным венчиком, в другой - с коротким. То сеть отбор
при генетическом разнообразии в популяции возможен.
Если популящя генетичесю! гете^югенна (F2), то отбор по фенотигу
расчленяет ее на неперекрывающиеся г|)упгы с раздающимися
геноипами уме за два поколения (F4) (длина венчик! у табака>
Длина венчика мм Расцепление в ЛИНИЯХ Fj Отбор в поколениях
f€
А 8 А 8 А 0 А и
37
4<3 2 Л?
41 12 - •
46 20 iH 14
49 51 41
52 2 4$
55 4 5 15
м 1 U
•п 14 21
м 11
яг S3
7в U 17 •
71 1* 15 11 4 1
76 17 19 20 2 1
7» i' 4 44 .i
91 s 1 .! 75 14
г* • 1 59 К
и < 5 к
и I 1 к
94 м
М ЬЛеблиеи 'сне’и*» Л. 1 Jt.’ егр. 611
Отбор естественный и искусственный действует только на генетически гетеро-
генные группы индивидов. Эволюция - это отражение сдвига частот аллелей в по-
пуляции . Индивид, как уже упоминалось, не эволюционирует
Когда проводится отбор, очень важно отличить размах вариаций признака, ко-
торый связан с внешней средой, от размаха вариаций, причиной которого являет-
ся генетическое разнообразие. Селекция (отбор на племя) более эффективна при
оценке величины признака по генотипу (т.е. по потомству или у братьев-
сестер), чем по оценке признака по фенотипу (т.е. у самого индивида).
В таблице видно, что решение по оценке генотипа (оставить на племя курицу
№12 потомство которой более яйценоское) правильное, хотя сразу неочевидное.
Решение по оценке фенотипа (оставить на племя №46 как более яйценоскую в дан-
ном поколении) - неправильное, хотя кажется очевидным.
№ курщы Ященосюсть (>«>Л1чество яти,) дочерей
Herl №2 №3 №4 №5 №6 №7
46 (26?) 95 105 157 161 190 196 263
12 (258) 190 210 212 216 234 234 242
МЕ.Лооашев. Генетют. Л., 1967, стр. 674
ЛЕКЦИЯ №19.
ГЕНОМИКА
Геномика - комплексная наука, изучающая геномы.
Разделы геномики:
1. структурная геномика - содержание и организация геномной информации;
2. функциональная геномика - реализация информации, записанной в геноме,
от гена - к признаку;
3. сравнительная геномика - сравнительные исследования содержания и органи-
зации геномов разных организмов;
Все эти разделы геномики вносят вклад в фундаментальную биологию (индивиду-
альное развитие, эволюция), здравоохранение, сельское хозяйство и биотехноло-
гию.
Итог структурной геномики - получение последовательности нуклеотидов (сик-
венс от англ, sequence), которая представляла бы полностью каждую из хромосом
с первого нуклеотида до последнего.
Для того, чтобы получить такой сиквенс, сегодня приходится определять по-
следовательность нуклеотидов в достаточно коротких отрезках ДНК, длиной при-
мерно 1000 позиций. В геноме человека 3 миллиарда позиций, значит, его надо
разбить на куски, которые и будут «читаться». Затем нужно восстановить единую
последовательность нуклеотидов из сравнения отдельных прочтенных отрезков
текста. Восстановление основано на сравнении определенных последовательностей
и выявлении в них перекрывающихся (идентичных) участков текста. Длина участка
перекрывания должна превышать длину последовательности, которая может встре-
титься в данном геноме по причинам случайного характера. Например, в геноме
человека 3*109 п.н. случайно может встретится последовательность длиной 15
нуклеотидов - поскольку в каждой позиции может находится один из четырех нук-
леотидов, то вероятность того, что заданные нуклеотиды окажутся в 15 позициях
подряд 415 =230 что примерно равно 109. То есть в отрезке длиной 10э позиций
заданная 15-нуклеотидная последовательность может встретиться 1 раз по причи-
нам случайного характера.
Но дело в том, что в ДНК нуклеотиды расположены не случайно, и это является
проблемой для восстановления последовательности из перекрывания отрезков. Ес-
ли две последовательности из 1000 нуклеотидов перекрываются на 20 нуклеотидов
или сто - это еще ничего не значит, так как весь этот фрагмент из 1000 нук-
леотидов может быть несколько раз повторен в геноме. Поэтому нужно было сна-
чала расставить вдоль генома фрагменты, а уже потом выявлять их перекрывание
на основе сиквенса. Таков был путь мирового сообщества при секвенировании ге-
нома человека (секвенированием в русскоязычной литературе называют процесс
определения последовательности нуклеотидов. Этот термин также является каль-
кой с английского названия).
Как это можно было сделать? Нужно было поставить какие-нибудь «буйки» в ге-
номе человека, какой участок стоит за каким. Последовательность таких участ-
ков и составляет карту генома. Первой такой картой стала карта генетическая.
Она показана на рисунке слева.
Генетическая карт a
хромосомы 1 человека
thr4------ — th<4
$UP61 —.
A6P) —
---SUP6I
---A6F?
Physical Ссппк
distance map
Физическая и генепнеская
карты хромосомы 3 дрожжей.
Относительные генетические расстояние,
уст ан селенные на основе частот рекомбинации,
отличаются от расстояний на физической карте
Рядом показана окрашенная хромосома, на которой видны поперечные полоски.
Поперечная окрашенность индивидуальна для каждой хромосомы, каждая полоска
имеет собственный номер, который представляет собой "адрес" данного участка
на хромосоме. Ясно, что в каждом таком участке миллионы пар нуклеотидов, по-
следовательность которых мы должны определить. Были получены полиморфные мар-
керы, то есть найдены такие участки хромосомы, которые у разных людей (или на
разных хромосомах одного человека) содержат неидентичные последовательности
нуклеотидов. В прошлой лекции упоминалось, что для генетической карты с ин-
тервалом в 10% рекомбинации нужно 300 равноудаленных маркеров. Эти маркеры
нужны для различения одной хромосомы от другой в данном локусе.
В основе детекции ДНК маркеров лежит метод амплификации (размножения) фраг-
ментов ДНК in vitro с точностью до нуклеотида методом полимеразной цепной
реакции (ПЦР). Методом ПЦР можно синтезировать фрагмент ДНК in vitro (в про-
бирке) и получить его как химически чистое вещество. Для синтеза используются
короткие синтетические отрезки ДНК, называемые праймерами (затравка для син-
теза) . С 3'-конца праймера начинается синтез фрагмента ДНК по матричной нити,
на которую он отжигается (прилипает при комплементарном взаимодействии между
нуклеотидами праймера и матрицы) . За один цикл достройки ДНК из двух нитей
ДНК получили 4. В следующем цикле из 4 нитей получится уже 8 и т.д. Каждый
цикл занимает несколько минут. За 30 циклов ПЦР целевой фрагмент размножится
в 1 миллиард раз, что позволяет наблюдать фрагмент (после окраски). Время
проведения каждого этапа ПЦР в будущем сократится на 2-3 порядка, таким обра-
зом, что каждый цикл будет проводиться за секунды.
Для различения папиной и маминой хромосом использовали так называемые STR-
маркеры (Short Tandem Repeat), состоящие из одинаковых звеньев, чаще всего
звено состояло из пары нуклеотидов ЦА. То есть нашли места в геноме, где по-
вторялись эти вкрапленные звенья. Допустим в папиной хромосоме в фрагменте из
100 пар нуклеотидов была вставка из 20 звеньев, а в таком же месте маминой
хромосомы было вставлено 22 звена. Этот фрагмент ДНК размножили in vitro, с
точностью до нуклеотида методом полимеразной цепной реакции (ПЦР). Длина этих
фрагментов будет у папы 100+20*2=140, а у мамы - 100+22*2=144. При фракциони-
ровании образованных фрагментов в геле под действием постоянного тока (элек-
трофорез) мы можем провести разделение фрагментов по размеру. Чем тяжелее
фрагмент, тем меньше его электрофоретическая подвижность и тем ближе к старту
он будет находиться. Если у родителей ребенка длины фрагментов составляли
(как указано в примере выше) 140 и 144 п.н., то и у ребенка будут эти полоски
присутствовать.
Описанный подход применяется не только в фундаментальных исследованиях, но
и в практике идентификации личности при судебно-медицинской экспертизе. До-
пустим данный локус в хромосоме может находиться в одном из 10 альтернативных
состояний. (Эти состояния, аллели, различимы по их электрофоретической под-
вижности) . Эти состояния различают 10 хромосом или людей с такими хромосома-
ми. Если мы возьмем в анализ еще один локус (на другой хромосоме) с такими же
характеристиками, то по этому локусу мы тоже различим 10 хромосом или людей.
А по сочетанию состояний в этих двух локусах различимы 10x10=102 хромосом.
Пять таких локусов позволят различить 105 хромосом. А поскольку хромосом у
каждого из нас по паре, то сочетания аллелей этих пяти локусов дают 105 х 105
= 1О10 вариантов. Это число вариантов больше, чем число людей на земле. На
практике при идентификации используют набор аллелей из 13 локусов, хотя и пя-
ти как мы видим, может быть волне достаточно.
Генетическая карта была первой картой генома человека, на основе которой
строилась дальнейшая работа по картированию. Эту карту соотнесли с физической
картой, показывающей порядок следования клонированных фрагментов ДНК вдоль
генома (см. рисунок 1 справа).
Физические карты генома часто представлены наборами фрагментов ДНК, клони-
рованные в векторных молекулах (рекомбинантных ДНК), упорядоченно располо-
женных относительно друг друга. Такой набор непрерывно перекрывающихся фраг-
ментов ДНК называется контиг. Для того чтобы выявить перекрывание клонирован-
ных фрагментов ДНК и понадобилась ранее установленная карта генетических мар-
керов. Перекрывание устанавливалось между «большими» молекулами ДНК, содержа-
щими примерно 106 пар нуклеотидов, которые были клонированы в искусственных
хромосомах дрожжей (YAC-клоны, сокращение от Yeast Artificial Chromosome).
Искусственные, потому что у них удалили основную часть собственно дрожжевой
ДНК и вставили человеческие фрагменты ДНК. Такие конструкции способны репли-
цироваться в клетках дрожжей. Размер хромосом дрожжей как раз примерно 1-2
миллиона пар нуклеотидов.
Как устанавливали перекрывание клонированных фрагментов ДНК? У нас есть
YAC-клон №1 с протяженным фрагментом клонированной ДНК, а в нем, предположим,
обнаружен и маркер А и маркер В, для которых из генетических данных извест-
но, что они соседние на карте. В YAC-клоне №2 уже нет маркера А, а есть мар-
керы В и С, причем также известно из генетической карты что В и С - соседи. В
клоне №3 есть маркеры С и D. Сопоставление данных по присутствию генетиче-
ских маркеров А, В, С и D в YAC-клонах показывает что они перекрываются в
последовательности YAC №1, №2, №3.
Вставки ДНК из 3000 YAC-клонов примерно равны по длине геному человека. В
анализ на перекрывание YAC-колонов взяли 30000 клонов, с тем чтобы каждая
точка генома перекрывалась несколькими клонами. Вначале неизвестно было, как
они расположены, но в среднем каждая точка генома перекрывалась 10 раз. Было
использовано порядка 3000 STR-маркеров, и посмотрели, эти как маркеры и клоны
друг с другом перекрываются. В качестве метода, выявляющего присутствие гене-
тического маркера в составе YAC-клонов, использовался ПЦР. На заключительном
этапе составления физической карты генома человека в этих 30 000 YAC-клонов,
выявлено присутствие примерно 30000 маркеров. Это один маркер на 100 000 пар
нуклеотидов. Расстояние между концами YAC-клонов также составило 100 000 п.н.
(при длине клона 1 млн. п.н.). Картирование проводили на роботизированных ма-
шинах, которые проводили приблизительно по 300 000 ПЦР-реакций в день. Позво-
лило расставить в контиг все YAC клоны. Предполагалось, что они будут непо-
средственно секвенироваться. Однако в дальнейшем была использована друга схе-
ма секвенирования клонов. Картированные YAC-клоны часто использовали для по-
иска генов, находящихся во вставке YAC, а к сиквенсу этот этап не привел.
Физические карты генома часто представлены наборами фрагментов
ДНК (в составе рекДНК), упорядоченно расположенных
относительно друг друга.
Непрерывное перекрывание фрагментов ДНК (контиг) установлено по
присутствию общего маркера - STS. выявляемого методом полимеразной
по сов нию рестрикционных карт.
цепной реакции или
YAC clones
уОХ224_Г" /\
уОХ28 —\
,-ОХ88«^—
VOX44
223—
»oX2 I CI
yOXIO —
y<JXH “
yOXj“
U4 | MUI
Ш | . IUJ
0X6,—^^^—
Часть физической карты
хромосомы Y человека,
представленная контигом YAC-
клонов. Эта карта метрическая,
ее единица - один нуклеотид,
равняется 0,34 нм
уохзг
Sequence
tagged 1441
ж=
уОХ|4,’ —
> UXI 84
да =
Перекрывание можно также посмотреть по расположению специфических рестрик-
ционных сайтов. Рассмотрим этот способ подробнее. Структура фрагмента ДНК вы-
является по положению участков расщепления специфическими ферментами - рест-
рикционными эндонуклеазами (рестриктазами). Каждая рестриктаза узнает после-
довательность нуклеотидов определенной длины и состава. Например, рестриктаза
EcoRI узнает GAATTC и никакую другую (расщеплять ДНК она будет в среднем один
раз на 46=4096 нуклеотидов), BamHI узнает GGATTC. Предположим, что у нас
есть клонированный фрагмент ДНК, длиной 13000 нуклеотидов, и мы расщепили его
рестриктазой BamHI, получив два фрагмента по 9 и 4 тысячи нуклеотидов. Затем
если мы расщепим EcoRI, получим фрагменты по 8, 3 и 2 kb. Когда мы посмотрим
двойное расщепление, получим фрагменты размерами 7, 3, 2, 1 kb. Размеры из-
вестны, потому что рядом есть дорожка, в которой идет фракционирование моле-
кул стандартного размера, что позволяет создать калибровочную кривую. Если мы
проведем второе расщепление, то увидим, что фрагмент в 9kb расщепился на
фрагменты по 7 и 2kb. Эта специфическая последовательность сайтов и специфи-
ческое расстояние между ними является портретом молекулы (см. рис. ниже). По
этим портретам мы можем сопоставлять молекулы друг с другом, независимо от
того, что они кодируют, и что в них находится. Это очень типичная процедура.
Расщепление фрагмента ДНК каждой рестриктазой по отдельности и их смесью по-
зволяет создать рестрикционную карту фрагмента.
EcoRI балиН I EcoRI + Size
a lore alone standards
^double digeUI
** I-----------------------
fcofll site BcrmHI site EcoRI site
2 kb 7 kb I kb 3 kb
Структура фрагмента ДНК выявляется по положению участков расщепления
специфическими ферментами - рестрикционными эндонуклеазами
(рестриктазами). Каждая рестриктаза узнает последовательность нуклеотидов
определенной длины и состава. Например. рестриктазаЕсоР! узнает
последовательность GAATTC, а рестриктаза Ba/nHI- GGATTC
Размер получившихся фрагментов устанавливают, разделяя их в геле под
действием электрического тока - чем меньше фрагмент, тем быстрее он
движется (слева - результат такого разделения).
Расщепление фрагмента ДНК каждой рестриктазой по отдельности и их смесью
позволяет создать рестрикционную карту фрагмента.
Итак, мы расставили молекулы методом генетического и физического картирова-
ния. Вернемся к методу секвенирования. Использовалась примесь дидезоксинукле-
отидов - ddNTP (на рисунке - справа; у них нет ОН-группы у 3'-атома углеро-
да) , которая добавлялась к обычным дезоксинуклеотидам (на рисунке слева). И
при синтезе ДНК in vitro это приводило к прекращению синтеза цепи в позиции,
в которой вставился ddNTP. Через позицию 3' идет присоединение нуклеотида к
строящейся молекуле ДНК. Но если на 3'- конце не будет гидроксильной группы,
а водород, то синтез дальше не пойдет - он будет терминирован.
(Ь)
о*
О = Р —О*
I
о
I
0 = Р —О'
I
о
Deoxyribonur leoside
triphosphate (dNTP)
Did eoxynbonuc leos ide
triphosphate (ddNTP)
Примесь
дидезоксину клеотидо
в (справа, нет 0Н-
группы у 3'-атом а
углерода) к
дезоксинуклеотидам
(слева) при синтезе
ДНК in vitro приводит
к прекращению
синтеза цепи в
позиции, в которой
ставился ddNTP
Это используется следующим образом. У нас есть матрица (нить ДНК), которую
надо секвенировать. Если идет синтез, и в первой позиции матрицы стоит А (см.
рис. ниже), то может встроиться обычный Т и синтез пойдет дальше, а может
встроиться ddTTP и синтез дальше не пойдет. Произойдет обрыв цепи, а получен-
ный синтезированный огрызок займет при фракционировании определенную позицию
согласно своему размеру. Следующий обрыв будет соответствовать второй букве
секвенируемой нити, и также займет свою позицию согласно длине при фракциони-
ровании на электрофорезе и т.д. И так по каждому нуклеотиду. Так мы восстано-
вим последовательность нуклеотидов в секвенируемой нити ДНК. Этот метод пред-
ложил Фрэд Сэнгер, за что получил свою вторую Нобелевскую премию.
Метод секвенирования ДНК, основанный на терминации синтеза
дидезоксинуклеотидтрифосфатами
Рассмотрим определение последовательности нуклеотидов в клонированном фраг-
менте ДНК. Клонированный фрагмент находится в так называемой векторной моле-
куле ДНК - молекуле, которая позволяет ввести его в клетку (обычно это клетка
бактериальная, но иногда используются и дрожжевые клетки). Все работы по сек-
венированию генома человека прошли при участии бактериальных векторных моле-
кул . Участок вектора, прилежащий к вставке, содержит последовательность нук-
леотидов , комплементарную универсальному секвенирующему праймеру. С этого
праймера инициируется синтез ДНК in vitro, который с первого нуклеотида будет
идти по матрице клонированного фрагмента ДНК человека. Универсальных прайме-
ров используется два, один к последовательности вектора прилежащей к одному
концу вставки, другой праймер к последовательности вектора прилежащей к дру-
гому концу вставки. С одного из праймеров клонированный фрагмент секвенирует-
ся с одной стороны, а с другого праймера - с другой стороны.
Участки молекулы ДНК. распознаваемые праймерами для секвенирования,
присоединены к исследуемому фрагменту ДНК путем
Исследуемый фрагмент ДНК вставляют в векторную молекулу ДНК. Участки
вектора, прилежащие к вставке, содержат последовательности нуклеотидов,
комплементарные универсальным секвенирующим праймерам -
левому и правому.
С этих праймеров инициируется синтез ДНк in vitro
Primer Primer site
'----V----P'----------v-----------K----У-----'
Vector DNA Vector
DNA I insert DNA
Primer site Primer
Вектор у нас один и тот же, а вставок - миллионы, но все они секвенирова-
лись с одной и той же пары праймеров. Основная часть генома была секвенирова-
на при клонировании фрагментов в 2 тысячи пар нуклеотидов, потому что тысяча
читалась с одной стороны и тысяча - с другой. Каждая точка генома человека
была просеквенирована несколько десятков раз в составе разных клонированных
молекул ДНК. То есть расстояние в геноме между концами клонированных и секве-
нированных фрагментов ДНК составляло меньше 200 пар нуклеотидов. От каждой
точки старта было прочитано около 1000 нуклеотидов. Из всего этого набора
«текстов» воспроизводилась структура генома человека. Но собрать эти 1000-
буквенные сиквенсы в контиги длинной в миллионы букв удалось лишь на основе
того, что большая часть фрагментов была предварительно картирована относи-
тельно хромосом человека. Без картирования сиквенс мог попасть в повторяю-
щийся участок генома, а продолжение сиквенса из такого участка имеет столько
вариантов продолжений, сколько раз повтор присутствует в геноме человека (не-
которые повторы - миллион раз). Поэтому сначала устанавливали последователь-
ность расположения клонированных фрагментов в геноме. Это было сделано для
фрагментов размером около 200 тыс пар нуклеотидов, а уже затем их секвениро-
вали.
Процесс секвенирования по методу Сенгера может быть автоматизирован. Меха-
низм представлен на следующем слайде.
Mngir-Urdnded DhlA fragment whew*
tx-f*? sequence 5 to be detenPioed i tl>?
rpcrpLatei К whltf
|)Each nf th₽ four rlcifJTTS is lagged wth л
Г'iKxuu.trI dye. ard the Sanger ^queiKing
5* - ” - 1 V
dcJCTP ddCTP ddTTP ddATP
3 □ и
\ /
f- мт
Template Printer
strand (sequence
\ I known 1
Автоматизация
секвенирования: каждый
из четырех ddhTTP имеет
флюоресцентную метку
своего цвета
H The t -H - •’ ’Ml ёП ’ in ihp r»« • к-ъй
Кър tt*> same colored dyp JttArhed
АТДАТАСТСТСТТ
ПЯПГ
? «II S'
r№
Electrophoresis
The products a/ e denatured and Пге ON A
tTAgments produced by the lour reactions
arc* ru₽d and loaded into 4 vnjle wpl an
an ebcltcphoiesfe gei The fragments
migrate through the <yM accordinq 10 sue
5* r
HATAATACTCTCTT
0 and the tluuitsKexit dye un the DHA 6
I detected by using э laser beam and detector
Q t,rh fгэдг . rt .ippcor. r. .1 p₽.>4 an trip
I Currputa printout. the сскм o4 tbe peak
•ndotes *hich base the peak represents
Терминированные
фрагменты ДНК
фракционируются
электрофоретически
ЗИатадтастстсттсссст
longest
fragment
|| The veque r.eiii'ixmaton tsread dne.’b
into the compute, which converts <1 «nto
1ho саггр4₽теп1ж>* Lrgpl -*^qiK»nce
__ Shortest
fragment
Detector
Фрагмент каждого размера имеет цвет,
соответствующий одному из четырех
ddNTP
На слайде виден праймер, синтез с которого идет влево. У нас есть дидезок-
синуклеотидфосфаты Т, А,С и G. Каждый из них занимает свою позицию во фраг-
менте синтезируемом по исследуемой матричной нити. На предыдущем слайде каж-
дой букве соответствовала отдельная дорожка геля, их всего четыре. Если каж-
дую из букв терминирующих синтез пометить в свой цвет, то все терминаторы
можно объединить в одной пробирке и фракционировать продукты в одной дорожке.
Обрыв синтеза в позиции данной буквы даст фрагмент со своим положением в ге-
ле после фракционирования. Каждое положение обрыва будет характеризоваться
цветом той- буквы терминатора, на которой произошел обрыв. В ходе фракциони-
рования терминированных фрагментов лазер будет фиксировать на детекторе по-
следовательные пики - какая прошла полоса по счету, и какого она цвета. Далее
эта последовательность пиков дешифруется в последовательность нуклеотидов в
молекуле ДНК. Точность сиквенса (установления того, какая именно буква терми-
нировала синтез в данной позиции) определяется соотношением высот пиков соот-
ветствующих разным буквам в одной и той же позиции секвенируемого фрагмента.
Между двумя пиками разных цветов в одной позиции было заданное дискриминирую-
щее значение. Техника отрабатывалась так, что буква считалась достоверно ус-
тановленной для данной позиции, если основной пик в этой позиции был выше
других в заданное количество раз.
Бактерия Н.influenzae была первым свободно живущим организмом, геном кото-
рого был полностью секвенирован. Поскольку геном бактерии маленький, около
тысячи нуклеотидов, и повторов нем мало (да и короткие они), то предваритель-
ное картирование клонированных фрагментов ДНК не понадобилось - эти фрагменты
сразу сиквенировались.
Бактерия Н. influenzae была первым свободно живущим организмом, геном
которого был секвенирован (TIGR, США)
Key
“ Amino aod biosynthesis
Biosynthesis of cofactors,
prosthetic groups carriers
IB Cell envelope
Cellular processes
Central intermediary-metabolism
i Energy metabolism
I Fatty ackJ phospholipid metabolism
в Purines, pyrimides,
nucleosides and nucleotides
Regulatory functions
Replication
BB 'ransport/binding proteins
Translation
M Transcription
BB Other categories
i Hypothetical
Unknown
Такая работа была проведена в институте генетических исследований TIGR под
началом Крега Вентера. Вентер затем организовал фирму Селера, секвенировавшую
геном человека, где он применил ту же схему секвенирования что и для бакте-
рии. Причем деньги он взял у частных фирм, так как государство не верило, что
у него что-нибудь получится.
Мировое сообщество предварительно использовало генетическую и физическую
карты, относительно которой была выстроена последовательность перекрывающихся
фрагментов клонированной ДНК (контиг), предназначенной для секвенирования. То
есть сиквенс генома человека был собран из фрагментов правило благодаря ис-
пользованию упорядоченного набора клонов и установлению последовательности
нуклеотидов картированных клонов.
Вентер же, в отличие от мирового сообщества, использовал случайный набор
клонов и попытался восстановить полную последовательность нуклеотидов прямо
из сравнения сиквенсов всей кучи фрагментов. На бактерии у него это удалось,
но на человеке это сработало лишь потому, что он использовал публично доступ-
ные данные от мирового сообщества о том, какие молекулы, где расположены в
геноме человека.
Секвенирование всего генома может
быть основано на детальной
Partial nrjp-.hon ОМА г₽ч.||
In owr 1.1ДО<г<д fraqmonti th.»l
jrt then
генетической и физической карте,
относительно которой выстроена
последовательность
перекрывающихся фрагментов
клонированной ДНК (контиг),
предназначенной для
' 19 11 Map based approaches to whole genome sequencing rely
on detailed genetic and physical maps to align sequenced
fragments.
Вентер опубликовал свою работу на месяц раньше, чем мировое сообщество, по-
тому что он ничего не картировал, а использовал секвенирование совсем корот-
ких рекомбинантных молекул. Общую длина секвенированных фрагментов ДНК была у
Вентера в пять раз больше, чем сделало все мировое сообщество. Используя дан-
ные мирового сообщества о картированных фрагментах, Вентер смог восстановить
в единую последовательность нуклеотидов все то, что он насеквенировал. Если
бы данных мирового сообщества не было бы, то вся его работа была представлена
короткими отрезками, которые бы разветвлялись, из-за того, что в геноме нахо-
дятся повторы.
В результате проделанной работы вышло две статьи: статья Вентера в журнале
Science и статья Лэндера - лидера мирового сообщества - в журнале Nature.
Проект генома человека начат в 1990 г. Первая (черновая) версия последова-
тельности нуклеотидов была закончена в 2000г. Конечная версия, которая больше
не будет совершенствоваться (названная Build35) - закончена в 2004 г.
Последняя версия последовательности содержит 2,85 миллиардов пар нуклеоти-
дов с 341 брешью, то есть в этих местах по каким-то причинам секвенировать
геномную ДНК не удалось. Сиквенс покрывает около 99% той части генома челове-
ка, которая представлены в некомпактизированной форме - эухроматине. Акку-
ратность сиквенса в конечной версии - 1 ошибка на 100 тысяч позиций подряд.
Еще точнее секвенировать весь геном уже никто не будет. Напомню, что папин
геном отличается у вас от маминого генома примерно в 1 позиции на тысячу.
Genomic DNA-----
Д Ьепотк. DMA ь cut
into numerous small
overlapping fragments
, and cloned In bacteria
Each fragment
rs sequenced |
Секвенирование всего генома
может быть основано на
перекрывании
последовательности нуклеотидов,
установленной для клонированной
ДНК путем секвенирования
H Ovi t.ip hi srqi.i । '
is used to order the
clones.
Пн- Sequence uf the Human (ienonte
Регулыгпы семэегыровання генома человека были
опубликованы в двух ведущих научных журналах, в
двух статьях более 500 соавторов
Предсказанное число генов у человека теперь 20-25 тысяч, что немного мень-
ше , чем предсказывалось раньше.
Кроме данных о последовательности нуклеотидов геномной ДНК человека (рефе-
рентная последовательность) созданы также базы данных:
1) о последовательности нуклеотидов транскрибируемых участков ДНК (EST
database, EST = Expressed Sequence Tags), которая характеризует не геномную
ДНК, а то, транскрибировалось с ДНК.
2) о положении и содержании отличий (полиморфизмов, то есть нуклеотидных
замен) других известных последовательностей ДНК человека от референтной по-
следовательности (SNP database, SNP = Single Nucleotide Polymorphism)
ЛЕКЦИЯ №20.
ГЕНОМИКА
(ЧАСТЬ 2)
Геномика - недавно возникшее направление науки, объектом изучения которой
являются геномы всех организмов, не только человека. Одно из направлений ге-
номики - воссоздание суммарной карты метаболических путей живого, состоящей
из частных метаболических карт, характерных для каждого организма.
Выявление в разных геномах определенных наборов генов метаболических функ-
ций позволяет предположить, функциональную связь генов этого набора в едином
участке метаболической цепи. В частности, один из подходов такой. Исследуют
ряд видов (рисунок ниже), к примеру, бактерий. У первых трех видов есть гены
для белков 1, 3 и 6. Остальные белки у некоторых есть, а у некоторых нет.
Этот набор генов (1, 3 и 6) отсутствует у четвертого вида. Такого рода при-
сутствие-отсутствие цельного набора генов позволяет сделать предположение о
том, что кодируемые ими белки каким-то образом связаны в метаболическом цик-
ле . Гены такого набора необязательно располагаются рядом в геноме.
Е. со И
Белки Р1 Р2 РЗ Р4 Р5
Бактерия А Бактерия В Бактерия С
2 Г I Г'
Белки 1, Зи6 либо
присутствуют или
либо вое вместе
отсутствуют Из
этого можно
заключить, что они
функционально
сеязань I
Филогенетический профиль белков - основа гипотез об их
функциях: белки Р1, РЗ и Р6 присутствуют у трех разных видов
бактерий. У четвертого вида весь набор этих белков отсутствует.
Еще один критерий функциональной связи между генами, особо хорошо работаю-
щий на бактериях, основан на сохранении соседства одних и тех же (по сиквен-
су) генов у разных видов бактерий. У бактерий нередко бывает, что группа ге-
нов, расположенных вместе, отвечает за группу последовательных этапов метабо-
лизма . Такая группа генов регулируется на уровне транскрипции единым образом
и называется оперон (единица операции). Часто последовательность расположения
генов в опероне совпадает с последовательностью метаболических этапов. Для
эукариот соседнее расположение функционально связанных генов не типично, но,
хоть такие гены и разбросаны у них по геному, скоординированная регуляция
транскрипции есть и эукариот.
Genome of
species A
Genome of
species В
Genes 4 and 5 are
consistent^ linked
Genes 1 and 8 are
consistently linked
Genome of
species С
Genome of
species D
Геномы четырех бактерий.
Гены 1 и 8, а также гены 4 и 5
соседствуют в разных геномах,
хотя положение этого блока
относительно других генов в
каждом из геномов различается.
На данный момент просеквенировано несколько сотен геномов бактерий и геномы
нескольких эукариот. Теперь мы знаем, что у бактерий размеры генома не бывают
меньше 0,5 миллионов пар нуклеотидов, а максимальный размер генома около 10
миллионов п.н., у дрожжей (эукариотический организм)- порядка 12 миллионов, у
червя нематоды - 97 млн., а у человека - 3 миллиарда пар нуклеотидов. А число
генов у про- и эукариот различается уже в меньшее число раз. Минимальное ко-
личество генов у бактерии микоплазмы - 470 штук, у дрожжей - 6000, у нематоды
- 19000, а у человека около 20000, то есть от нематоды и мухи по количеству
генов мы не сильно отличаемся. Количество хромосомной ДНК, приходящейся на
один ген у бактерий - 1000 п.н. то есть гены упакованы очень плотно; у дрож-
жей - 2000 п.н., и кое-где гены разделены некоторым пространством; у нематоды
- 5000 п.н. на ген и появляются пространства внутри генов - интроны; у чело-
века - 30000 п.н. У нас в геноме большие межгенные пространства и большие
пространства внутри генов, которые не переходят в зрелую РНК.
Заметим, все эти организмы по размерам зрелых транскриптов не сильно отли-
чаются. В зрелой РНК белок-кодирующий участок занимает обычно основную часть
последовательности. Часть генов кодируют РНК, с которой белок вообще не син-
тезируется. Перед белок-кодирующей последовательностью в зрелой мРНК располо-
жены участки регуляции трансляции, а после белок кодирующей последовательно-
сти - участки определяющие стабильность (время жизни РНК). У прокариот после-
довательности перед и после белок-кодирующей части гораздо короче, чем у эу-
кариот . Так что по размерам РНК все организмы ближе, чем по размерам генов, а
по размерам белков - еще ближе.
Экспериментально проводили «выключение» каждого гена у многих бактерий, и
смотрели, выживут они в данных условиях или нет. Оказалось, что у бактерий
можно «выключить» (поочередно) около 50% генов, и они все равно будут жить. У
дрожжей можно выключить 80% генов и они все равно будут жить.
Как это было экспериментально показано? В геном клетки вставляют репортер-
ный фрагмент ДНК, который позволяет замерить скорость транскрипции и транс-
ляции в точке вставки фрагмента. Известно поэтому, что и траснкрипция и
трансляция репортерного гена через данную точку в данных условиях происходит
с регуляторных элементов гена, разорванного вставкой репортера, хотя разо-
рванный ген сам не функционален. Таким образом 80% генов дрожжей по одному
«убивали» и видели, что клетка дрожжей все равно живет.
У нематоды на 20 000 генов получено несколько десятков тысяч мутаций, ко-
торые, по-видимому, поражают около 2 000 генов (так называемых групп компле-
ментации) . Это около 10% всех генов нематоды. То есть если «выключить» около
90% генов, клетка будет продолжать жить. У человека из 20 000 генов только в
1700 (меньше 10%) известны мутации, которые связаны с болезнями, наследуемыми
по Менделю как моногенный признак.
Размеры геномов и число генов разных организмов
Организмы Геном (мпнп.н.) Число генов (тъсгми) Размер гена (Т.П.Н.) Q/щественных: генов
Бактерии 0,5-5 0,47-4,29 1-1,7 <50%
Дхжки 12 6 2 <20%
Нештсда 97 19 5 <10%
Человек 3000 30 30 <5%?
В связи с этим понятно, что количество генов, мутации в которых будут при-
водить заболеваниям человека (по крайней мере, к летальным), скорее всего,
не увеличится значительно, по сравнению с тем, что уже известно к настоящему
времени. Сейчас в интернет доступна база данных OMIM (Online Mendelian
Inheritance in Man) по генам, мутации которых приводят к заболеваниям и про-
являются как менделирующие признаки.
В геноме не все его участки транскрибируется. В связи с этим встал вопрос
экспериментального определения, где и сколько в геноме генов. Под одним геном
понимается участок ДНК, который соответствует единому транскрипту, образован-
ному с этого участка. При транскрипции участка ДНК получается так называемый
пре-мРНК, которая содержит и экзоны (участки, переходящие затем в зрелую
мРНК), и интроны (вставочные последовательности, которые удаляются из мРНК).
Интроны удаляются из пре-мРНК в результате процесса, называемого сплайсингом.
Остающиеся в результате участки пре-мРНК, называемые экзонами, соединяются в
единую нить. Она называется зрелой мРНК. ( Некоторые из РНК не кодируют бе-
лок . Называть такие РНК матричными, т.е. мРНК терминологически не верно, хотя
они соответствуют генам и имеют свои функции.)
Зрелая мРНК используется как материал для экспериментального исследования
наличия гена в геноме, его положения и интрон-экзонной структуры. Инструмен-
том для такого исследования являются биологические микрочипы.
Первый патент на микрочипы принадлежит коллективу под руководством Андрея
Дарьевича Мирзабекова, который был директором Института молекулярной биоло-
гии РАН и заведующий одной из кафедр ФМБФ МФТИ. Он предложил иммобилизовать
синтетические фрагменты ДНК на твердые матрицы, и проводить гибридизацию
этой матрицы с исследуемым образцом нуклеиновой кислоты - ДНК или РНК.
Как исследовать, действительно ли ген существует, то есть, транскрибируется
ли данный участок ДНК? Для этого ген представляют в чипе частью его последо-
вательности - олигонуклеотидом, который иммобилизован в микроплощадке с опре-
деленными координатами на этой матрице. Этот олигонуклеотид соответствует
части экзона, предсказанного компьютером на основе сиквенса геномной ДНК.
Чтобы выяснить, действительно геном в данном участке транскрибируется, берет-
ся клетка и из нее выделяется суммарная РНК. Из всех этих молекул РНК получа-
ют ДНК-копии, которые флуоресцентно метят и проводят гибридизацию с иммобили-
зованными на микрочипе олигонуклеотидами. Если в данных условиях какие-то
площадки с олигонуклеотидами «молчат» (они показаны черным), то это значит,
что участок геномной последовательности, комплементарной этому олигонуклеоти-
ду, не транскрибируется. Если же площадка матрицы «светится», значит олиго-
нуклеотиды в этой площадке прогибридизовались с флуоресцентно меченым продук-
том, то есть соответствующий участок генома транскрибировался и действитель-
но является частью какого-то гена.
Биологические микрочипы используются для одновременной
детекции экспрессии многих генов
В реальном эксперименте все участки на матрице в той или иной мере «светят-
ся». Поэтому без сравнения с некоторым стандартом, нельзя сказать, с чем
связано появление сигнала в данной площадке чипа. Чтобы определить, является
ли полученный результат ошибкой эксперимента или нет, проводится сравнение
двух объектов. Для этого берутся некие клетки А, из них получают РНК, и их
флуоресцентно метят (на слайде - красным) . То же проводят и с клетками В, но
метят РНК другим цветом (зеленым). Затем проводят гибридизацию чипа со сме-
сью этих двух препаратов РНК. Если сигнал в данной площадке на чипе получает-
ся красным, значит в клетках А транскрипция данного гена сильнее, чем в клет-
ках В. Если сигнал зеленый, то транскрипция сильнее в клетках В. Если красно-
го и зеленого поровну, то получится желтый цвет. Таким образом, возникает
возможность сравнивать уровень траснкрипции данного гена в разных клетках
В, С, D и т.д., нормируя его на уровень транскрипции этого гена в клетках А.
При этом сравнивают транскрипцию гена в разных тканях, в них гены экспресси-
руются по-разному. Можно сравнивать опухоль и норму, тогда выявляют те гены,
которые специфически более сильно транскрибируются в опухоли или в норме.
Можно посмотреть разные стадии развития, как работают гены в зародышевом раз-
витии и во взрослом состоянии. Таким образом, гибридизация на микрочипах по-
зволяет узнать, какие гены в геноме в данных условиях транскрибируются, а
именно этим он и проявляет свою жизнь.
Биологические микрочипы могут быть использованы для
установления относительного уровня экспрессии в клетках
различных типов (например, печень - почки, норма - опухоль,
зародыш - взрослый организм)
Если уровень экспрессии данного гена в образце А > В, цвет красный
Если уровень экспрессии данного гена в образце А < В, цвет зеленый
Если уровень экспрессии данного гена в образце А = В, цвет желтый
Если образец А используется как нормирующий, то можно
сравнивать экспрессию в образцах D, Е, F ... друг с другом
Гибридизация на микрочипах позволяет проверить компьютерное предсказание о
том, что данный фрагмент генома - экзон, (участок, остающийся в зрелой мРНК)
и он действительно транскрибируется. Каждый ген не обязан экспрессироваться
во всех тканях и в каждых данных условиях. Поэтому нужно исследовать много
условий и тканей, чтобы выявить все участки генома, соответствующие экзонам.
На слайде каждая гибридизация на данном чипе соответствует какому-то одному
типу ткани или условиям ее функционирования. Красным указано количество экзо-
нов в каждой из хромосом, существование которых экспериментально подтвержде-
но. На каждом чипе 1 090 408 площадок с пробами-олигонуклеотидами, соответ-
ствующими каждому из 442 785 экзонов человека, предсказанных компьютером.
Олигонуклеотиды в площадках соответствуют как транскрибируемой нити ДНК, так
комплементарной нити. В геноме человека транскрипция комплементарных нитей
ДНК, характерна для небольшой части генов. Такие гены перекрываются и, воз-
можно , взаимно регулируются на уровне транскрипции. У бактерий перекрывание
генов гораздо более частое явление, чем у эукариот.
Котранскрибируемые экзоны (границы гена) выявляются экспериментально на чи-
пе. Соседние площадки содержат олигонуклеотиды, соответствующие экзонам, со-
седним в геноме. Граница выглядит как переход от блока площадок одного цвета
(красного, олигонуклеотиды принадлежащие котранскрибируемым экзонам) к друго-
му (зеленому). Полный список данных по экспериментальному подтверждению суще-
ствования всех предсказанных компьютером экзонов пока не существует.
Границы гена
Котранскрибируемые экзоны (границы гена)
выявляются экспериментально на чипе
Shoemaker е.а., Nature (2001), v.409, р.922
Микрочипы могут быть использованы для исследования изменений уровня транс-
крипции генов, связанной с возникновением или прогрессией заболевания, (на-
пример, опухолевого или инфекционного). Предполагается, что каждая болезнь,
характеризуется своим штрих-кодом - изменением уровня транскрипции набора ге-
нов характерного именно для данной болезни. Этот анализ является очень важным
для усовершенствования функциональной диагностики в медицине.
Микрочипы могут быть использованы для
исследования экспрессии, связанной с
возникновением или прогрессией
заболевания (например, опухолевого или
инфекционного), поскольку каждая
болезнь, вероятно, характеризуется
специфичными изменениями наборов
экспрессируемых генов по сравнению
нормальным состоянием.
Провели такой опыт. Взяли образцы РНК из опухолей у двух групп больных. В
одной группе метастазы были, а в другой - нет. Метастазы - это возникновение
новых очагов опухоли в организме, пространственно отделенных от исходного
очага. На данном чипе довольно резко проходит граница между группами зеленых
и красных площадок. То есть, видны гены, изменение уровня экспрессии которых
характерно для стадии метастазирования опухоли, что можно использовать для
диагностики этой стадии. Пока этот метод диагностики недоработан. Предполага-
ется, что в будущем по штрих-коду изменения экспрессии в определенном наборе
генов можно будет диагностировать конкретные заболевания и стадии их разви-
тия , а, следовательно, и знать, как лечить.
SptXWlK tMMSt tiznours
patents years
tunoir яга <5 cm
ivmcri rxxJe negate (LNO)
Van ‘t Veer et al (2002) Nature 415: 530-536
Сделаем небольшое отступление. На прошлых лекциях было рассказано про гене-
тические карты. Такие карты были построены для многих видов. На видах с под-
робными генетическими картами проводится экспериментальный поиск мутаций свя-
занных с регистрируемыми морфологическими изменениями. На слайде показана
схема такой работы на рыбах. Вначале проводят мутагенез. После этого получают
гибриды первого поколения. Их используют для возвратных скрещиваний с мутаге-
низированными родителями. Если оказывается, что какой-то признак выявляется,
то смотрят, с какими генетическими маркерами он сонаследуется. Таким образом
исследуется, какие гены повреждены мутациями, выявляемыми фенотипически.
Гены (мутации), определяющие
морфологические или биохимические
признаки, могут быть
идентифицированы после
общегеномного мутагенеза (например,
EMS) и генетического скрининга. Для
этого проводят анализ сонаследования
исследуемого аллеля с полиморфными
маркерами ДНК, перекрывающими весь
геном в известной последовательности,
и расстояние между которыми
достаточно мало, чтобы отнести
исследуемый аллель к одному из
интервалов генетической карты.
Обобщая вышесказанное, гены (мутации), определяющие морфологические или
биохимические признаки, могут быть идентифицированы после общегеномного мута-
генеза (например, EMS) и генетического скрининга. Для этого проводят анализ
сонаследования исследуемого аллеля с полиморфными маркерами ДНК, перекрываю-
щими весь геном в известной последовательности, и расстояние между которыми
достаточно мало, чтобы отнести исследуемый аллель к одному из интервалов ге-
нетической карты.
На этом слайде показано, что существуют бактерии, у которых количество ге-
нов может быть больше, чем например у дрожжей. Мы привыкли считать, что бак-
терии устроены проще, но это не всегда так. Существуют бактерии, у которых
генов порядка 10 тысяч.
Table 19.2
Size Number of
(Millions of Predicted
Species Base Pairs) Cenes C ♦ C (%>
Archaea
Archueoglobus fulgidus 2.18 2407 49
Mcthanobactenum thcrmooutotrophicum 1.75 1869 50
Methanococcus jannaschu 1.66 1715 32
Thermopfasma acidophilum 1.56 1478 46
Eubacteria
Bacillus subtihs 4.21 4100 44
Bordetella parapertussis 4 75 * 69
Buchner a species 0.64 Sb4 27
Campylobacter jejuni 1 64 1654 31
Escherichia coll 4.64 4289 51
Haemophilus influenzae 1.83 1709 39
bfasorhrzobiurri loti 7 04 6752 63 1
Mycobacterium tuberculosis 4.41 3918 66
Mycoplasma gemtalium 0.58 4 80 32
Staphylococcus aureus 2.88 2697 33
Treponema pallidum 1.14 1031 53
Ureaplasma ureaiylit um 0.75 611 26
Vibrio c holer ae 4.03 3828 48
Source: Data from the Genome Atlas of the Center for В logical Sequence Arxatyus.
Mtp //www cta.dfu dk/tervKes/GenorneAtlas.*
0л! a no! av-A’UMe
Даже для прекрасно изученного организма - кишечной палочки, не понятны
функции около трети из 4289 ее генов. Известна и последовательность нуклеоти-
дов в этих генах, и как они транскрибируются и т.д., но все равно не извест-
но , какую функцию они выполняют.
(a> f scherrchio cob (b)
(common bacterial
One circular chromosome
Genome size: 4.64 million bp
Number of genes: 4289
С ’C content: 514
Даже для прекрасно изученного организма - кишечной палочки, не понятны ф ункции
около трети из 4289 ее генов
jC 1. Fatty acids, phospholipid metabolism
2 Transcription. RNA metabolism
3 Nucleotide metabolism
4 Phage, transposon, plasmid functions
$. DNA replication, recombination, repair
6 Carbon compounds metabolism
7. Amino acid metabolism
S Other genes with known Functions
9 Regulatory functions
10. Translation, protein metabolism
• I. Central intermediary metabolism
12. Adaptation, protection functions
U. Cell, wall, membrane structural components
14. Energy metabolism
• 5. Putative enzymes
16. Transport proteins
17. Genes with unknown functions
На этом слайде хотелось бы обратить ваше внимание на то, что хотя число
генов разнится у разных видов, но число так называемых белковых доменов
(структурные единицы в белке, отвечающие за единичную функцию) отличаются в
пределах разных царств живого (прокариоты и эукариоты) до полутора раз, не
более. Конечно комбинации этих доменов разные, но сами домены похожи, то есть
они кодируются сходными по последовательности нуклеотидов участками генома и
эти сходные участки имеют общее происхождение в эволюции.
Обратно заявление будет не верно. Если функции белков сходны, это не озна-
чает, что их структура будет одинакова. Одна и та же функция, например, один
и тот же каталитический процесс, может выполняться разными, не родственными
по происхождению белками. Один и тот же процесс может катализироваться даже
и белком и РНК (рибозим), у которых нет ничего общего в происхождении.
Характеристики эукариотических геномов по полногеномному снквенсу
P асмер генома млн п н. Число предсказанных Число семейств
Виды генов белковых доменов
Saccharomyces cerevistae дрожжи 12 6,144 8SI
Arabidopsis thahana растение I2S 25,706 1012
Caenorhabditis elegans червь 100 18,266 1014
Drosophila melanogaster муха 180 13,338 1035
Homo sapiens (human) человек 3400 -32.000 1262
Source: Number of genes and protein domain families from International Human Genome Sequencing Consortium.
Initial sequencing and analysis of the human genome. Nature 409 (2001). Table 23.
Число экзонов на ген
Размер экзона
Размер нитрона
Размер 5’-нетранслнруемого участка
Размер 2’-нетранслпруемого участка
Размер кодирующей части
Общая длина гена
8.8
145 Ьр
3365 Ьр
300 Ьр
770 Ьр
1340 Ьр
27,000 Ьр
На данном слайде показаны средние значения характеристик различных элемен-
тов генов человека. Средний размер экзона - 145 нуклеотидов, интрона - 3365
и т.д. В общей сложности получается, что белок-кодирующая часть гена невелика
по сравнению с белок-некодирующей частью, поэтому, когда происходят какие-
нибудь мутации, велика вероятность, что промутирует белок-некодирующая часть.
Такие мутации или вообще не скажутся на структуре белка, или приведут к изме-
нению его количества, но не структуры (изменения регуляторных участков ини-
циации транскрипции или стабильности РНК), или приведут к драматическим изме-
нениям структуры РНК (мутации в мишенях для сплайсинга).
Общая структура генома такова. Напомню, что размер генома человека 3200 Mb.
Гены занимают всего 1200 Mb. Основная часть этого пространства приходится на
псевдогены (нефункциональные гены, инактивированные мутациями), различные
фрагменты генов и интроны. А на экзоны функциональных генов (суммарная длина
зрелых РНК) приходится 48 Mb. Здесь есть некоторое лукавство, так как на одну
пре-мРНК в среднем приходится 1,4 зрелых РНК. А из одной зрелой мРНК в неко-
торых случаях может получиться до тысячи белков. Межгенная ДНК занимает 2000
Mb, она представлена главным образом короткими рассеянными по всему геному
повторяющимися последовательностями, которые занимают 1400 Mb. Один из таких
повторов - Alu-повтор, длиной около 300 п.н., повторен в геноме миллион раз.
Другой примечательный тип рассеянных повторов - длинные концевые повторы
(LTR, long terminal repeat). Эти элементы являются молекулярными свидетель-
ствами перескока фрагмента ДНК внутри генома. Общая протяженность таких уча-
стков на молекуле ДНК- 250 Mb.
Геном человека: размеры областей занятых элементами известной структуры
G^e»
48 Mb
H 52 Mb
Inter* pereed
repeat*
1400 Mb
~~1
Otfier
«•rgarac
region»
ЬХ Mb
Gene
fr*Xrne»>t»
Ini rent
1/TfU
LINE.
M0 Mb
LTR eiementi
250 Mb
510Mb
54NL>
420 Mb
DNA
90 Mb
Ptcudoftcfio
1
Число генов у человека оценено в 20 - 25 тысяч, (оценка 2001 г. - 35 - 40
тыс.).
Число генов у человека оценено в 20 - 25 тысяч
Основная часть генома человека занята не генами: 63-74% длины - межгенные
пространства, половина из них - повторы. Ген человека внутри «пустой»: 95%
внутригенной ДНК вырезается (интроны). Общая длина белок кодирующих областей
около 1% от геномной ДНК человека. Это лишь в 3 раза больше длины генома бак-
терий .
От 26383 до 39114 генов человека были предсказаны компьютером (в 2001г.),
но лишь менее 7000 были подтверждены на человеке. И более чем для 80% генов,
хоть в чем-то была пересмотрена структура в период с 2001 по 2003г и продол-
жает уточняться на микрочипах.
Сейчас предсказанное число генов у человека 20-25 тысяч и существование
около 19 000 из них экспериментально подтверждено - с них образуются транс-
крипты .
Имеющееся на данный момент определение гена (ген - это фрагмент геномной
ДНК с котранскрибируемыми субфрагментами) - не полное. Например, возможна
транскрипция с двух цепей. Плохо выявляются короткие гены и белок-
некодирующие гены. Их, по крайней мере, под тысячу, но точное число не из-
вестно. Такие гены - тоже гены, хоть белок они и не кодируют. Они - гены, по-
тому что с них образуется РНК. Причем РНК некоторых белк-некодирующих генов
состоит из нескольких экзонов. То есть, клетке эти РНК зачем-то нужны, но мы
пока не понимаем, зачем.
Альтернативный сплайсинг,
биологическая роль и механизмы
Предположим, образовался транскрипт зрелой мРНК, и он может содержать экзо-
ны 1, 2, 3. Это вовсе не означает, что он обязательно будет содержать их все.
У нас может появиться РНК, которая будет содержать экзоны 1 и 2 или экзоны 1
и 3, ив результате с них будут образовываться разные белки. Такой способ
процессинга (обработки) генетической информации называется альтернативный
сплайсинг.
Одному гену соответствует с среднем 1 4 зрелых мРНК
Альтернативный сплайсинг-различные комбинации экзонов остающиеся в зрелой
мРНК получаемой из одной и той же пре-мРНК
У человека есть ген slo. Он «работает» во внутреннем ухе, в частности, этот
белок присутствует в ворсинках, которые отвечают за распознавание высоты зву-
ка. Он состоит из 35 экзонов (на рисунке - прямоугольники), 8 из которых (си-
ние) могут или присутствовать, или отсутствовать в зрелой мРНК. Возможны 8! =
40 320 вариантов сплайсинга, но только около 500 из них обнаружены. Других,
может быть, и нет, то есть природа не должна, вообще говоря, реализовывать
все возможные варианты.
Биологическая роль множественного сплайсинга заключается в следующем. Раз-
ные типы волосяных клеток внутреннего уха реагируют на звуки разных частот
от 20 до 20 000 герц. Различия клеток в восприятии частоты частично определя-
ются свойствами альтернативных сплайс-форм белка Slo. Как определяется выбор
между вариантами сплайсинга неизвестно.
Ген slo человека,
Состоит из 35 экзонов (прямоугольники) .8 из которых (синие) могут или присутствовать
или отсутствовать в зрелой мРНК Возможны 8! = 40 320 вариантов сплайсинга, но только
около 500 из них обнаружены экспериментально.
» I W__ ................ ..« —
Биологическая роль множественного сплайсигта
Р азные типы волосяных клеток внутренне го уха реагирует на звуки разных частот от 20 до 20
000 герц. Различия клеток в восприятии частоты частично определяются свойствами
ал терна тивных сплайс-форм б елка Slo.
Как определяется выбор между вариантами сплайсинга неизвестно
Известны случаи, когда с одного локуса образуются тысячи разных белков. К
ним, в частности, относятся белки, которые образуются на поверхности нервных
клеток. Таким образом, они, видимо, как-то участвуют в распознавании друг
друга, и в формировании нейронных сетей. В этом случае происходит выбрасыва-
ние не только экзонов или интронов, но и может реализовываться и альтернатив-
ный участок инициации транскрипции. Такие случаи известны, в частности, для
человека, когда у разных генов есть несколько разных промоторов, каждый из
которых дает свою РНК, в которой, в зависимости от того, где он начался, бу-
дет дополнительный экзон, связанной с различной длиной транскрипта на 5' -
конце.
Механизм сплайсинга
Процесс соединения одного экзона с другим происходит в участках определен-
ной последовательности нуклеотидов. Донорный сайт сплайсинга всегда заканчи-
вается одним из двух динуклеотидов, обычно - AG.
Консенсусная последовательность
донорного и акцепторного участков сплайсинга
в интронах и экзонах эукариот
Eicxi dofv scceptc' she Exon
-------------------------------SS££££££S6SN£*0
В начале происходит нуклеофильная атака донорного экзона, затем происходит
разрезание, кусочек GU заворачивается и присоединяется к А. Затем разрезается
вторая часть, первый экзон соединяется со вторым, и образуется интрон.
(В)
Mechanism of RNA splicing (GU-AG introns).
1 Cleavage at 5'and (|)
2 Nucleophilic «tack К____)
resulting и lariat formation
II Ut SnRNP t>nds to sp-ce Oonox site
2 U2 SnRNP txnds to branch site
I. Cleavage « 3‘end
2 Lgat>on of exons
Binding of LM U5 V6
SnRNP complex. U5 SnRNP
binds to both splice donor - acceptor
Если посмотреть, какую долю гена составляют экзоны, то самый большой из-
вестный транскрипт (у гена миодистрофина) имеет длину около 2,5 миллионов
нуклеотидов. У него в зрелую часть РНК переходит 14 тыс нуклеотидов (0,6%), а
остальные 99,4% от первичного транскрипта выкидывается (интроны).
С ростом размеров гена в хромосоме его белок-кодирующая часть увеличивается
незначительно, а количество интронов в гене растет. С ростом числа интронов
растет число сайтов сплайсинга и вероятность их повреждения. Поэтому для ге-
нов с большим числом интронов потеря функции при мутации может быть связана
не с белок-кодирующей частью ДНК, а с регуляторными элементами сплайсинга.
Секвенирование генома человека показало, что некоторые экзоны многократно
повторены в геноме. Это могут повторы экзонов в составе одного гена, или
присутствие одного и того же экзона в составе нескольких разных генов. Полу-
чается, что экзоны, основные элементы структуры РНК, то есть белок-
кодирующие элементы, в процессе эволюции могут каким-то образом размножаться
в геноме и «перетасовываться» между разными генами. Такое явление получило
название exon shuffling - перетасовка экзонов. Ниже показаны разные белки, в
которых содержатся одинаковые экзоны. Таким образом, оказывается, что эволю-
ция - это нередко именно блоковые изменения генома, а не точечные изменения.
Генные дупликации и ’'тасующиеся" экзоны
Ген фибронектина содержит 12 копий экзона, кодирующего "палец" белка. Этот же модуль найдет в
продуктах других генов. Он содержит также модули, встречающихся в белках определенных типов, таких
как мембранные клеточные рецепторы и белки внеклето’+iono матрикса.
Ф иброкектин Ftxorxctn ЧШН/ нпнннннниннш н»
•эд,™ | и др
фактор роста
Тканевый активатор
плазминогена
P-ouroSirase
---------------------
I Finger modute
(Module she<e<J w<h
cell surface receptors
and other ertracelular
rnalrm pcotens
Knngle module
"1 Module found
Lj n blood dotting factors
[ Growth factor module
Проурокиназа
Классификация генов
по их функциям
На 2001 год для более 40% генов человека не было никаких предположений от-
носительно выполняемых функций. А для остальных раскладка была достаточно ус-
ловной. Принадлежность белка к одному функциональному классу не исключает его
принадлежности также и к другому классу. Например, то, что белок связывается
с ДНК, не означает, что он не может быть еще и ферментом и т.д. Это - харак-
теристика, которая была дана гену по той его части, которая связана с охарак-
теризованной функцией, но, вообще говоря, такая функция у белка может быть и
не одна.
Распределение функций 26 383 генов человека
41.8% - функция неитвестна
13 5°0- баткм, сгжывдсшяеся с ДНК
- пд> едача оахала-
10 4° о -
1 4 ° О - радод (с частотам»! >0.5% )
Больше всего генов отвечают за экспрессию, репликацию и поддержание функций
генома; около 20% - за передачу сигналов между клетками, около 17% - за то,
чтобы клетка сама по себе была здорова, и для других функции не классифициро-
ваны.
Экспрессия, репликация
Оказывается, что у человека, по сравнению с дрожжами, бактериями и т.д., в
геноме имеется больше генов регуляторов транскрипции. То есть, транскрипцион-
ные регуляторы сильно размножились в эволюционной линии млекопитающих, в ча-
стности, человека. Предполагается, что разнообразие регуляторов транскрипции
обеспечивает большую тонкость реакции генома на сигналы внешней среды. То
есть у млекопитающих больше число ансамблей координировано транскрибируемых
генов, чем в других группах.
Сложность организации человека м.б. связанас большим количеством генных
ансамблей, управляемых единичными кнопками: генами активаторами транскрипции.
В эволюционной линии человека они сильно амплифицированы
§ 8 § § 818 818
Tupler е.а., Nature (2001), v.409, р.832
Схема структурных элементов фрагмента генома человека размером 50 т.п.н.
б локусе Т-клеточного рецептора (хромосома 7).
Около половины длины фрагмента занимают элементы, структура и функция которых понятна. Это гены
(экзоны и интроны) и псевдо гены, короткие (SINE) и длинные (LINE) диспергированные повторы, длинные
концевые повторы (LTR), транспозоны, микросателлиты.
«.1т
Ц Бжх» В О ОКА rrim^Non
Mrс<* Ц fa
Ц И Itt rfanwW | t^i имГЙП
Выше показано, как выглядит кусок генома человека («50 000 п.н.). Около по-
ловины длины этого фрагмента ДНК занимают элементы, структура и функция кото-
рых понятна. Среди этих элементов есть гены (экзоны и интроны) и псевдогены -
есть гены функциональные, а есть их нефункциональные копии. Они, обычно, не
содержат интронов (считается, что после транскрипции и при преобразовании
Зрелой РНК возможен процесс встраивания ее обратно в геном в виде ДНК-копии;
тогда это будет ген, содержащий лишний «хвост» и не содержащий интронов). А
также короткие (SINE) и длинные (LINE) диспергированные повторы, длинные кон-
цевые повторы (LTR) - следы, оставшиеся после транспозиции, транспозоны, мик-
росателлиты .
К 2001 году в геноме человека было выявлено 1112 "генов болезней" (то есть
таких, мутации в которых ведут к заболеванию) и еще есть 94 "составных" гена,
образующихся при опухолевых перестройках генома. Пока, в основном, раскрыты
по механизму те заболевания, которые затрагивают белок-кодирующую часть гена.
Возможно, не меньшее количество мутаций, вызывающих болезни, будет найдено и
в участках регуляции транскрипции, сплайсинга и стабильности РНК.
В геноме человека выявлено
1112 ‘Тенов болезней”* 94 “составных" гена,
образующихся при опухолевых перестройках генома
суммарная частота мендепирующих заболеваний <1% новорожденных
(» с*о6к»< —число гемо» предрасположенности к болезням)
Мутации в кодирующей части гена -лишь часть из возможных причин
потери функции. Другая часть - дефекты процессинга генетической
информации (сплайсинга, регуляции транскрипции и трансляции)
McKusick ел, Science (20011 v291, р.1224
По представлениям на март 2005 года, у человека 24000 белок-кодирующих ге-
нов, из них 1700 генов ассоциированы с заболеваниями. Обнаружено 44,500 мута-
ций в этих 1700 генах (в среднем 26 на ген), связанных к заболеваниям. А для
остальных 10 000 000 известных мутаций подобная связь не выявлена.
20% смертей в США происходит из-за того, что прием лекарства осуществлялся
либо не того, либо не так. Но это связано не с некомпетентностью врачей, а с
тем, что мы все генетически разные. У болезней много мишеней и если бить не
по той мишени, то лекарство точно не будет полезным, а потому может быть и
вредным. У человека может быть специфическая реакция на данное лекарство
(слишком низкая или слишком высокая скорость метаболизма). А в нашей стране
ведущая пока причина смертей взрослого населения - пьянство. Эта первопричина
просто скрыта за определением «травматизм» (производственный, бытовой, дорож-
ный и т.д.), который и указывают как причину смерти. Выявлена корреляция по-
требления алкоголя в нашей стране и продолжительности жизни: когда, снижается
потребление алкоголя, продолжительность жизни идет вверх, и наоборот.
Если два белка характеризуются сходной последовательностью аминокислотных
остатков (выше критической длины, до которой совпадения могут быть чисто слу-
чайными) , то у них есть общая предковая последовательность, и соответствующий
предковый ген. Количество таких предковых структур у белков весьма ограниче-
но .
К примеру, был один ген в геноме, а потом их стало два (дупликация) . Со
временем мутации изменили эти гены каждый по-своему. А потом этот вид дал на-
чало двум новым видам (см. рис. ниже). Все эти гены являются «родственниками»
(гомологами), но по-разному называются. Гомологичные гены, которые мы рас-
сматриваем в составе разных видов, называются ортологи, а гомологичные гены
в одном геноме называют паралоги.
Гомологичные (сходные) последовательности, вероятно, являются
родственными по происхождению, если их длина выше некоторого порога.
Для генома человека этот порог равен последовательности длиной в 15
нуклеотидам
4 = 10 = длина генома чвловека в нуклеотидах.
Гомолошчные последовательности нуклеотидов в геномах разных видов
называются ортологи (А1 и В1; А2 и В2).
Гомологичные последовательности нуклеотидов в геноме одного вида
называются пар ало ги (Д1 иД2; В1 иВ2)
ПАРАЛОГИ
□РТОЛОГИ
Сравнения таких родственных генов часто используют при исследовании эволю-
ции. Эволюционная геномика (сравнительная геномика), используется очень ин-
тенсивно в медицине. Пример ее практического применения. У людей по разным
причинам бывает ожирение. В частности, есть семейные формы, менделирующие. У
человека мутации, вызывающие это заболевание, картированы не были. Сходный
фенотип ожирения наблюдали у мышей. У мышей этот ген генетически картировали
(картируют, на самом деле, не ген, а собственно мутацию в гене). Просеквени-
ровали участок вокруг этой мутации, потом нашли такую же последовательность
нуклеотидов в геноме человека. Стало ясно, в каком месте генома человека надо
искать мутации, вызывающие ожирение у человека. Проверка этого участке генома
человека у больных людей и сравнение его со той же нуклеотидной последова-
тельностью у здоровых подтвердила, что мутации этого гена у человека, как и
у мыши, приводят к ожирению.
ЛЕКЦИЯ №21.
ИЗМЕНЕНИЕ ПОПУЛЯЦИОННЫХ ЧАСТОТ
АЛЛЕЛЕЙ У ЧЕЛОВЕКА В РАЗЛИЧНОЙ
ПРИРОДНОЙ И КУЛЬТУРНОЙ СРЕДЕ
В школьном курсе биологии вы узнали, об исследованиях костных останков
предполагаемых предков человека и родственных видов, австралопитека, питекан-
тропа, неандертальцев, живших более 500 000 лет назад и исчезнувших к настоя-
щему времени. В этой лекции будет рассказано про то, как генетические иссле-
дования помогают восстановить историю народов и историю появления вида Ното
sapiens в целом.
То, что обезьяна - близкий родственник человека, известно уже давно, шим-
панзе среди всех обезьян - наш самый близкий родственник. При исследовании
ДНК происхождение человека от обезьяноподобных предков вполне подтверждается.
Генетические различия на уровне ДНК между людьми составляют в среднем 1 нук-
леотид из 1000 (то есть 0.1%) , между человеком и шимпанзе - 1 нуклеотид из
100 (т.е. 1%). По размеру генома человек и высшие приматы не отличаются друг
от друга, но отличаются по количеству хромосом - у человека на одну пару
меньше. Как было рассказано на прошлых лекциях, у человека 23 пары хромосом,
т.е. всего 46. У шимпанзе 48 хромосом, на одну пару больше. В процессе эволю-
ции у предков человека две разных хромосомы приматов объединились в одну. По-
добные изменения числа хромосом встречаются и в эволюции других видов. Они
могут быть важны для генетической изоляции группы в процессе видообразования,
так как в большинстве случаев особи с разным числом хромосом не дают потомст-
ва .
Время расхождения (дивергенции) видов, или другими словами, время существо-
вания последнего общего предка для двух видов, можно определить несколькими
способами. Первый такой: проводят датировку костных останков и определяют,
кому эти останки могли принадлежать, когда мог жить общий предок тех или иных
видов. Но костных останков предполагаемых предков человека не так много, что-
бы можно было с уверенностью восстановить и датировать полную последователь-
ность форм в процессе антропогенеза. Сейчас используют другой способ датиров-
ки времени расхождения человека и остальных приматов. Для этого подсчитывают
количество мутаций, накопившихся в одних и тех же генах в каждой из ветвей за
время их раздельной эволюции. Скорость накопления этих мутаций более менее
известна. Скорость накопления мутаций устанавливают по числу различий в ДНК
тех видов, для которых известны палеонтологические датировки расхождения ви-
дов по костным останкам. Время расхождения человека с шимпанзе по разным
оценкам варьирует от 5,4 до 7 млн. лет назад.
Вы уже знаете, что геном человека полностью прочтен (секвенирован). В про-
шлом году появилось сообщение, что прочтен также геном шимпанзе. Сравнивая
геномы человека и шимпанзе, ученые пытаются выявить те гены, которые "делают
нас людьми". Это было бы легко сделать, если бы после разделения ветвей эво-
люционировали только гены человека, но это не так, шимпанзе тоже развивались,
в их генах тоже накапливались мутации. Поэтому, чтобы понять, в какой ветви
произошла мутация - у человека или у шимпанзе - приходится сравнивать их еще
и с ДНК других видов, гориллы, орангутана, мыши. То есть то, что есть только
у шимпанзе и нет например у орангутана, это чисто «шимпанзиные» замены нук-
леотидов. Таким образом, сравнивая нуклеотидные последовательности разных ви-
дов приматов, мы можем выделить те мутации, которые произошли только в линии
наших предков. Сейчас известно около дюжины генов, которые "делают нас
людьми".
Филогенетическое древо
Генетические различия на уровне ДНК
между людьми: 1 нуклеотид из 1000
между человеком и шимпанзе : 1 нукл. из 100
Обнаружены различия между человеком и другими животными по генам обонятель-
ных рецепторов. У человека многие гены обонятельных рецепторов инактивирова-
ны. Сам фрагмент ДНК присутствует, но в нем появляются мутации, которые инак-
тивируют этот ген: либо он не транскрибируется, либо он транскрибируется, но
с него образуется нефункциональный продукт. Как только прекращается отбор на
поддержание функциональность гена, в нем начинают накапливаться мутации, сби-
вающие рамку считывания, вставляющие стоп-кодоны и т.д. То есть мутации появ-
ляются во всех генах, и скорость мутирования примерно постоянная. Удается
поддерживать ген функционирующим только за счет того, что мутации, нарушающие
важные функции, отбрасываются отбором. Такие инактивированные мутациями гены,
которые можно распознать по последовательности нуклеотидов, но накопившие му-
тации, делающие его неактивным, называются псевдогенами. Всего в геноме мле-
копитающих около 1000 последовательностей, соответствующих генам обонятельных
рецепторов. Из них у мыши 20% псевдогенов, у шимпанзе и макаки инактивирована
треть (28-26%), а у человека - более половины (54%) являются псевдогенами.
Псевдогены найдены у человека также среди генов, которые кодируют семейство
белков кератинов, входящих в состав волос. Так как волосяной покров у нас
меньше, чем у шимпанзе, то понятно, что часть таких генов могла быть инакти-
вирована .
Когда говорят об отличие человека от обезьяны, то в первую очередь выделяют
развитие умственных способностей и способность к речи. Найден ген, связанный
со способностью говорить. Этот ген выявили, изучая семью с наследственными
нарушением речи: неспособностью научиться строить фразы в соответствии с пра-
вилами грамматики, сочетавшейся с легкой степенью задержки умственного разви-
тия. На слайде представлена родословная этой семьи: кружки - это женщины,
квадратики - мужчины, закрашенные фигуры - больные члены семьи. Мутация, ас-
социированная с заболеванием, находится в гене FOXP2 (forkhead box Р2). У че-
ловека достаточно трудно исследовать функции гена, легче это делать у мышей.
Используют так называемую технику нокаута. Ген прицельно инактивируют, если
знать конкретную последовательность нуклеотидов, то это возможно, после этого
у мыши этот ген не работает. У мышей, у которых выключили ген FOXP2, наруши-
лось формирование одной из зон мозга в эмбриональный период. Видимо, у чело-
века эта зона связана с освоением речи. Кодирует этот ген фактор транскрип-
ции. Напомним, что на эмбриональной стадии развития факторы транскрипции
включают группу генов на тех или иных этапах, которые контролируют превраще-
ние клеток в то, во что они должны превратиться.
С пособность говорить
Выявлена семья с наследственными нарушенный речи
затруднениями движений. связанных с артикуляцией;
нарушением разбивания слов на фонемы
Мутация, ассоциированная с заболеванием, находится
в гене FOXP2 (forkhead box Р2)
।
«*№*413.519- 523 Q001 )Ф
A forHvead-domart gene is mut*ed in a severe speech and language dsordv
CECUAS L LATf. SIMON E FISHER*f JANE A HURST». FARANEH VARGHA KMAPEh< Л
ANTHONY P MONACO
Чтобы посмотреть, как этот ген эволюционировал, его просеквенировали у раз-
ных видов: мыши, макаки, орангутана, гориллы и шимпанзе, после этого сравнили
эти последовательности нуклеотидов с человеческой.
Молекулярная эволюция гена
on
Human
Gortla
(V5
Chimp
5.5 млн. лет
90 млн. лет
0/5
Rhesus
Mouse
Nature 418. 869 - 872 (2002): 14 August 2002
Molecular evolution of FOXP2. a gene involved in speech and
language
WOLFGANG ENARD MOLLY PRZEWORSW ,SIMON E FISHER. CECILIAS L LAI
MOTOR WIEBE TAKASHI WTANO . ANTHONYP MONACO & SVANTE PAABO
Оказалось, что этот ген очень консервативен. Среди всех приматов только у
орангутана имелась одна аминокислотная замена, и одна замена у мыши. На слай-
де у каждой линии видны две цифры, первая показывает число аминокислотных за-
мен, вторая - число так называемых молчащих (синонимических) нуклеотидных за-
мен, чаще всего это замены в третьей позиции кодона, не влияющей на кодируе-
мую аминокислоту. Видно, что молчащие замены накапливаются во всех линиях, то
есть мутации в данном локусе не запрещены, если они не ведут к аминокислотным
заменам. Это не значит, что не появлялись мутации в белок-кодирующей части,
они скорее всего появлялись, но были отсеяны отбором, поэтому мы не можем их
зафиксировать. В нижней части рисунка схематично изображена аминокислотная
последовательность белка, отмечены места, где произошли две аминокислотные
замены человека, которые, видимо, повлияли на функциональные особенности бел-
ка FOXP2.
Если белок эволюционирует с постоянной скоростью (число нуклеотидных замен
в единицу времени постоянно), то число замен в ветвях будет пропорционально
времени, в течение которого замены накапливались. Время разделения линии гры-
зунов (мыши) и приматов принимается равным 90 млн. лет, время разделения че-
ловека и шимпанзе - 5.5 млн лет. Тогда количество замен т, накопившихся, на-
копившихся суммарно в линии мыши и в линии приматов между точкой разделения с
мышью и точкой разделения человека и шимпанзе (см. рисунок), по сравнению с
числом замен h в линии человека, должно быть в 31.7 раз больше. Если же в ли-
нии человека накопилось больше замен, чем ожидается при постоянной скорости
эволюции гена, то говорят об ускорении эволюции. Во сколько раз ускорена эво-
люция , вычисляют по простой формуле:
A.I. = ( h/5.5 ) / [ m /(2 х 90 - 5.5)] = 31.7 h / m
Где A.I. (Acceleration Index) - индекс ускорения.
Теперь надо оценить, находится ли отклонение числа замен в линии человека
от в пределах случайного, или отклонение достоверно выше ожидаемого. Вероят-
ность того, что в линии человека за 5.5 млн. лет появится 2 аминокислотные
замены при том, что вероятность появления замен оценивается по линии мыши как
1/(90+84.6)=1/174.6. При этом используют биноминальное распределение B(h + m,
Th/(Th+Tm)), где h - число замен в линии человека, m-число замен в линии
мыши: Th=5.5, Тт=17 4.5.
Попробуйте самостоятельно рассчитать вероятность в приведенном примере.
Генетическое разнообразие
современного человечества
Вы знаете, что антропологи подразделяют людей на три большие расы: негрои-
ды, европеоиды и монголоиды. Представители этих рас отличаются цветом кожи,
формой тела, разрезом глаз и т.д. Но на самом деле четкие различия между раз-
ными людьми, относящимся к разным расам, имеются только если мы возьмем гео-
графически отдаленные группы. Если посмотреть на все разнообразие антропомет-
рических признаков в целом, то окажется, что четких различий нет, существует
множество переходных форм. Почему и как у людей сформировались внешние разли-
чия, где и когда зародилось человечество?. Чтобы ответить на этот вопрос в
1985 году Алан Уилсон - американский генетик - вместе со своей группой иссле-
довал митохондриальную ДНК (мтДНК), которая передается, как известно из про-
шлых лекций, только по материнской линии (обозначена красным). Y-хромосома
передается же только по отцовской линии (синяя линия), серой линией обозначе-
на передача аутосомной ДНК, то есть весь остальной геном, передающийся нам от
всех наших предков.
Отцовская линия
Y-хромосома
Материнская линия
мтДНК
Y-хромосоме не с чем рекомбинировать, поэтому она, в отличие от рекомбини-
рующих аутосом, передается из поколения в поколение от отца к сыну неизмен-
ной . Изменения происходят в ней только за счет новых мутаций. То же и с
мтДНК.
Генетическая карта мтДНК показана на рисунке, разным цветом показаны разные
гены. У человека ее размер составляет 16 500 нуклеотидов.
Генетическая карта мтДНК человека (размер • 16500 п.н.)
/казано положение мутаций, маркирующим разные линии.
Из-за того, что система репарации в митохондриях работает не так, как в яд-
ре, скорость накопления мутаций в мтДНК в 10 раз больше, чем в ядерной. В ней
есть один интересный участок примерно в 300 нуклеотидов (показан сверху на
рисунке), называемый некодирующим или гипервариабельным, в котором скорость
накопления мутаций в 10 раз выше, чем в остальной части мтДНК. Он очень ин-
тенсивно изучается. На рисунке стрелками показаны положение известных мутаций
в мтДНК и разными буквами указаны линии, маркируемые этими мутациями. Ско-
рость накопления нуклеотидных замен в гипервариабельном участке мтДНК челове-
ка: 1 нуклеотид в 18- 20 тысяч лет. Максимальные отличия между мтДНК у двух
разных людей 22 нуклеотида. Генетическое разнообразие в эволюционно родствен-
ной группе, например, среди членов одной этнической группы, меньше. Например,
максимальное число отличий в гипервариабельном участке мтДНК между индивидами
у русских - 4 нуклеотида. Скорость накопления замен у человека рассчитывается
по сравнению с числом замен, возникших после дивергенции человека и шимпанзе,
и, в зависимости от того, какая дата расхождения этих видов принята за основу
расчетов (5.5 или 7 млн. лет), полученные генетические датировки событий по-
пуляционной истории колеблются на 20-30%. Что сделал Алан Уилсон?
Реконструкция происхождения Homo sapiens по
различиям в ъпггохондрпапьноп ДНК
1ыс. ле!
УЮ п
Н<13<1Д
ЛЮ
|1Ю
50
ю
"Ева"-точка
коалесценции
линии мтДНК
человека
Рост
численности
ГЕНЕТИЧЕСКИЕ
ДАННЫЕ
АРХЕОЛОГИЧЕСКИЕ
ДАННЫЕ
Разделение предков
неандертальцев
(переселились в
Европу) и человека
(остались в Африке)
Исследованные
образцы ДНК
Реконструированные
события
Начало расселения
H.sapiens по Земле
Вымирание
неандертальцев
Он собрал кровь у людей разной расовой принадлежности (красные кружки на
рисунке), исследовал мутации в мтДНК и выстроил дерево, в котором родственные
последовательности, отличающиеся одним нуклеотидом, находились рядом, отли-
чающиеся двумя нуклеотидами находились подальше друг от друга, и т.д. В конце
концов, у него получилось дерево, восходящее к одному корню, показанному жел-
тым кружком. Все линии возле корня оказались африканскими, из чего был сделан
вывод об африканском происхождении человека. Гипотетическую прародительницу,
обладавшую мтДНК с рассчитанной последовательностью нуклеотидов, окрестили
митохондриальной Евой, из-за чего неспециалисты стали писать, что Библия под-
твердилась, что все человечество произошло от одной женщины Евы, жившей в Аф-
рике. Это неверно, так как она жила там не одна, при исследовании других ге-
нетических локусов было показано, что предковая популяция человека составляла
5 тыс. человек, просто одна из линий мтДНК дошла до нас, остальные линии по-
терялись. Это может произойти, когда у женщины рождаются только сыновья или
она бесплодна. Точка, к которой сходятся линии, называется точкой коалесцен-
ции. Проводившиеся дальнейшие исследования, покрывающие практические весь
земной шар, подтвердили выводы Уилсона. Датировка точки коалесценции - от 100
до 200 тыс. лет назад (последние оценки показывают 130-180 тыс. лет). Удалось
выделить мтДНК из двух образцов костей неандертальцев. Неандертальцы также
относятся к роду Ното (лат. называние Homo neanderthalensis) Один, из пещеры
Фельдгофер (не датирован) и второй на Кавказе в пещере Мезмай (останки дати-
руются радиоуглеродным методом 29 тыс. лет назад). Гипервариабельный участок
в мтДНК неандертальцев отличался от человеческой 24 заменами, что, как форму-
лируют генетики, выходит за пределы видового разнообразия человечества. Моле-
кулярно-генетическая датировка расхождения линий человека и неандертальца -
примерно 500 тыс. назад. По палеонтологическим данным, предки неандертальца
поселились в Европе 300 тыс. лет назад. Вымерли неандертальцы 28-30 тыс. лет
назад. Так как люди пришли в Европу 40-50 тыс. лет назад, то на протяжении 20
тыс. лет они сосуществовали с неандертальцами, а затем неандертальцы вымерли.
Предки неандертальца поселились в Европе 300 тыс. лет назад
Неандертальцы вымерли 28-30 тыс. лет назад
Пунктиром очерчен район находок останков неандертальцев
Отмечено положение пещер Фельгофер и Мез майская
Все генетические корни человека уходят в Африку, численность предковой
группы человека была 100 тыс. человек, затем был период резкого падения чис-
ленности в 10 раз, как раз это время генетики считают временем схождения раз-
ных линий - датой точки коалесценции (так называемое время прохождение через
"бутылочное горлышко"). Генетики считают, что именно в это время появился вид
Homo sapiens. У антропологов дата появления человека - 2 млн. лет назад. На
самом деле это время появления рода Ното. Можно столкнуться с тем, что неан-
дертальца также считают подвидом Homo sapiens (называя его в этом случае
Homo sapiens neanderthalensis) вместе с человеком вида (Homo sapiens
sapiens). Однако критерий принадлежности к одному виду - появление при скре-
щивании потомства, что генетики отрицают, так как генетическое разнообразие у
человека было бы гораздо больше, если бы оно накапливалось 500 тыс. лет, а не
100 или 200 тыс. лет. Раньше, в середине 20-го века предполагали, что неан-
дерталец - предок человека. Однако сейчас ясно, что он наш эволюционный
"двоюродный брат".
Если более крупно и подробно нарисовать это дерево мтДНК всего человечест-
ва , то получится такая картинка:
коллекция
сиквенсов
мтДНК
древо,
построено^
на основе
сходства
сиквенсов
Древо мтДНК
Тыс. лет
Красные кружочки на рисунке обозначают мтДНК современных людей, а само это
дерево - гипотетическое, потому что есть очень немного образцов древних ДНК.
Сначала отделились ветви на древе, которые сейчас находятся в Африке. Затем
от одной из африканских ветвей отделилась ветвь, которая ушла в Азию. Там эта
популяция поделилась на часть, которая ушла в Австралию и Океанию, и остав-
шуюся часть, которая поделилась на европейскую и азиатскую ветки. От послед-
ней отделилась группа, которая ушла в Америку. (Даты на рисунке - по количе-
ству накопившихся мутаций митохондриальной ДНК).
Если это дерево положим на географическую карту, то увидим, как и когда лю-
ди расселились по Земле. Корень дерева - в Африке, потом ветвь идет в Азию,
потом от азиатской ответвились все европейские линии митохондриальной ДНК.
Ниже показано упрощенное древо человечества по Y хромосоме. В Y хромосоме
появляются мутации, и если у одного из братьев появилась мутация, то он дал
начало новой генетической линии. Каждая линия датируется по числу мутаций,
отличающей ее от остальных. Ниже для примера показана область распространения
линий. Самая древняя - в Африке, следующая - расселение вокруг Средиземного
моря, и более молодая линия заселяет Северную Евразию.
Древо линий ¥ хромосомы человека
К;ы-дая линия соответствует появлению мутации
А>100000 BP • ancestr
По времени события развивались так. Вначале люди появились в Африке. Точное
место назвать нельзя, но наиболее древние линии встречаются у некоторых этни-
ческих групп в Танзании и Субсахарской Африке (у бушменов и готтентотов). По-
том они разделились на группы, одна из которых около 90 000 лет назад вышла в
Восточное Средиземноморье (Левант). Эта группа затем полностью вымерла. Рань-
ше считалось, что из Африки люди вышли через Левант, это на самом деле не
так: первая попытка выхода около 90 000 лет назад оказалась неудачной - люди
вымерли, и территория была заселена неандертальцами, которые потом тоже вы-
мерли. Люди выходили из Африки при потеплении, вслед За животными, на которых
они охотились, и за растениями, которые они ели. В благоприятных условиях
численность популяции росла, и они расселялись на новые территории.
Следующая попытка выхода из Африки осуществилась по южному пути, через Юж-
ную Аравию. Преимущества этого пути заключалось в получении бесконкурентного
источника питания - моллюсков (их раковину практически никто из животных не
может разбить) . Вышли они около 80 000 лет назад, а 74 000 лет назад случи-
лось извержение вулкана Тоба, которое привело к экологической катастрофе -
похолоданию по сценарию "ядерной зимы". Слой пепла, оставшегося после этого
извержения, достигает 3 метров. И, видимо, люди здесь вымерли, потому что по-
том территория Индии повторно заселялась с двух сторон. Далее люди заселили
очень благоприятную для проживания территорию Юго-Восточной Азии. Там берего-
вая линия была другой, не такой, как сейчас, водные промежутки были не очень
большими, и, скорее всего, люди уже умели плавать на плотах. Около 60 000 лет
назад они добрались до Австралии, около 50 000 они появились в Европе, а За-
тем во время потепления была заселена вся Евразия. Предполагается, что около
25 000 лет назад была первая волна расселения в Америку. Тогда туда можно бы-
ло пройти по суше. На месте Берингова пролива была суша Берингия. Эта первая
волна вызывает очень много споров - была ли она на самом деле или нет. А 12-
15 тыс. лет назад пошла вторая волна расселения, которая не вызывает уже ни-
каких сомнений (есть археологические данные, подтверждающие этот факт).
По книге Стивена Оппенхеимера "The Real Eve"
Около 10-11 тысяч лет назад появляется земледелие на Ближнем Востоке. Плот-
ность населения земледельцев была в 10 раз больше плотности населения охотни-
ков-собирателей. И эти земледельцы начинают расселяться во все стороны; идет
вторая волна заселения территорий, на которые уже живут охотники-собиратели.
Когда люди расселялись в разные стороны, мутации в географически изолиро-
ванных популяциях накапливались, давая начало новым генетическим линиям. Но
популяции не только разделялись, в какие-то периоды они смешивались.
На карте ниже обозначены народы бывшего СССР и сопредельных стран для того,
чтобы можно было представить, каковы разные этнические группы, населяющие эту
территорию. Этнический ареал русских, т.е. та область, где они сформирова-
лись, не очень большой, несмотря на то, что сейчас русские составляют около
80% населения России.
Народы бывшего СССР
Следы смешения народов можно найти в их генах. На рисунке частоты европей-
ских и азиатских линий мтДНК для некоторых народов, населяющих Россию.
Относительные частоты и
линий у народов европейской части России
ИССЛЕДОВАННЫЕ НАРОДЫ
ОТНОСЯТСЯ К РАЗНЫМ ЯЗЫКОВЫМ СЕМЬЯМ:
ИНДОЕВРОПЕЙСКОЙ Г АЛТАЙСКОЙ
На карте доля европейских линий показана синим, а азиатских - желтым. Вид-
но, что генетическая граница между Европой и Азией проходит там же, где и
географическая - по Уралу.
Расселяясь по земле, люди оставляли по земле генетический след расселения,
потому что, если какие-то мутации были частыми у африканских линий, то когда
люди уходили, накапливались другие мутации, чем дальше они уходили, тем боль-
ше их накапливалось. Есть определенный градиент частоты разных аллелей по
долготе - это и есть след расселения. Но есть и другие причины изменения час-
тот аллелей. С расселением связывают очень интересный ген, дофаминовый рецеп-
тор, DRD4, связанный с поведением человека, с такой психологической характе-
ристикой как стремление к новым впечатлениям (она тестируется по психологиче-
ским опросникам). Один из аллелей этого гена, называемый R7, ассоциирован с
более высоким уровнем стремления к новым впечатлениям. Журналисты иногда на-
зывают это ген геном «авантюризма». Частота аллеля сильно варьирует в разных
популяциях. Например, в Европе частота составляет 10-15%, на Севере Америки -
40%, а в Южной Америке доходит до 70%. Американский психолог Майкл Бертон вы-
двинул гипотезу о том, что когда народы расселялись по Земле, уходили с обжи-
того места именно люди с повышенной частотой этого аллеля.
Генетические различия между разными популяциями могут быть связаны с адап-
тацией к факторам внешней среды, таким как климат, природные условия или к
особенностям питания и типу хозяйства (производящее или присваивающее), к эн-
демичным инфекциям и некоторым другим факторам.
Адаптация к
геоклиматическим
факторам
Наиболее известный признак, связанный с адаптацией к климату - цвет кожи.
Там, где ультрафиолет наиболее интенсивен - там самая темная коже у людей. В
северных широтах кожа у людей самая светлая, если бы это было не так, у детей
был бы рахит, так как темная кожа защищает от ультрафиолета, под действием
которого вырабатывается витамин Д. Витамин Д необходим для усвоения кальция.
Интенсивность кожной пигментации связана с накоплением пигмента меланина, ко-
торое контролируется белком мелонокартиновым рецептором. Ген, кодирующий дан-
ный белок, исследован у разных народов, и было показано, что накопление мута-
ций, ведущих к ослаблению пигментации, происходило у жителей Азии и Европы.
Интересен тот факт, что хотя у народов Африки самое высокое генетическое раз-
нообразие , по этому гену мутации там отсутствуют, так как светлая кожа там
неадаптивна. На рисунке показано широтное распределение кожной пигментации.
Цвет кожи в различных регионах мира
из СЕнАс jndMEniba Aiiivpology
MC1R
ме.'ынокорт HHOP.Mii репе in op
X АЛС П. UAIOZ.lXVXDn.A TAI'A.S Щ.С AKCALI I*. rt».
Интенсивность кожной пигментации негативно коррелирует с широтой, хотя
встречаются некоторые исключения. Например, у эскимосов кожа немного темнее,
чем у других популяций, живущих на той же широте. Предполагается, что это из-
за того, что они пришли из более южных районов в недалеком прошлом и не успе-
ли адаптироваться, или из-за того, что они много едят печени морских живот-
ных, где много витамина Д.
Рост и форма тела также являются адаптацией к климатическим условиям. На
севере выгоднее всего быть низкого роста, с более короткими конечностями, так
как будут наименьшие потери тепла через кожу, на юге - наоборот, худым и вы-
соким, чтобы больше терять тепло.
Рост и форма тела - адаптация к климатическим условиям
Адаптация к
особенностям питания
и типу хозяйства
После перехода от присваивающего хозяйства многое изменилось: от кочевого
образа жизни люди перешли к оседлому, питание стало более однообразным (вме-
сто десятков диких растений и животных), возросла плотность населения и воз-
никли более благоприятные условия для распространения инфекций.
Раньше всего земледелие появилось на Ближнем Востоке в районе Месопотамии,
в так называемом «плодородном полумесяце» (здесь очень благоприятный климат),
затем в Китае. Здесь начали выращивать рис через 2000 лет после появления
земледелия в Месопотамии. И еще через несколько тысяч лет появились независи-
мые самостоятельные зоны земледелия в Америке, а только потом и в Африке.
РЕГИОНЫ И ДАТЫ <ле» до идет. времени) ПОЯВЛЕНИЯ ЗЕМЛЕДЕЛИЯ
После появления земледелия возросла частота нарушений зрения, например,
дальтонизма. Понятно, почему - альтернатива такая: либо дальтоник, либо охот-
ник. От успеха на охоте зависит количество детей; женщины предпочитают выби-
рать хороших охотников себе в мужья, так что у хорошего охотника больше шан-
сов оставить потомство. Частота нарушения цветового зрения в обществах, где
вклад охоты-собирательства 15% и выше, близка к нулю. У индустриализованных
странах частота дальтонизма может достигать 7%.
У охотников-собирателей
редко встречаются нарушения цветового зрения
Популяционная частота нарушении цветового зрения
(частоты для 19 ттнимескиа групп, по Рычтое и др , 2000)
Есть еще один интересный ген, связанный со способностью пить молоко. Все
млекопитающие кормят своих детенышей молоком, и чтобы расщеплять молочный са-
хар лактозу в кишечнике детеныша вырабатывается фермент лактаза. И у всех
нормальных млекопитающих по окончании периода грудного вскармливания лактаза
перестает синтезироваться, потому что в дикой природе им молоко больше никто
не даст.
Так же было и у людей, пока у них не появилось молочное животноводство. С
его появлением, стало выгодно пить молоко и взрослым, так как оно является
ценным пищевым ресурсом. Но дело в том, что если взрослый человек, у которого
лактаза не синтезируется, пьет молоко, ему становится очень плохо, у него
вздувается живот и начинается понос. Это происходит, потому, что сахар, не-
расщепленный лактазой в кишечнике, перерабатывается бактериями, которые на
него бурно набрасываются и расщепляют его с выделением водорода. Человек ста-
новится похожим на шарик, наполненный водородом, что порождает массу неприят-
ных симптомов. В норме синтез лактазы прекращается примерно в пять лет. Охот-
ники-собиратели кормят своих детей грудью до трех-пяти лет. Но у европейцев
распространилась мутация, которая препятствует отключению фермента в пятилет-
ием возрасте. Эта мутация найдена, она находится в регуляторном участке гена
лактазы. У 90% взрослого населения датчан синтез лактазы продолжается. У ки-
тайцев только 2% взрослого населения способны усваивать молоко, но они все
равно его не пьют, в их культуре это не принято, они молоко за еду для взрос-
лых не считают. Среди русских 30-40% взрослых не могут пить молоко. Есть кор-
реляция с широтой частоты этой мутации - на севере молоко важнее как источник
кальция, так как там меньше интенсивность ультрафиолетового излучения.
Частота переносимости/непереносимости
молочного сахара лактозы
Конвоя ЛИ, Бввщю«окая ЕВ, НурБм* С Д, Вышвохохэ^ О П
Гвколсервфпя пврх пчжов rxcxoiuxw кж х nonyweoux Старого С хата
Ганвтоа 1998,34 (4) 551-561 с разрешения А И Козлова
Еще один интересный ген - аполипопротеин Е. О нем уже шла речь на одной из
прошлых лекциях. Разные аллели этого гена связаны с разными уровнями холесте-
рина. Самый высокий уровень - у носителей аллеля е4. Уровень холестерина раз-
ный у разных этнических групп. У европейцев частота этого аллеля 5-15%, и он
ассоциирован с рядом болезней, в том числе с болезнью Альцгеймера. У гомози-
гот по аллелю е4 вероятность заболеть старческим слабоумием выше в 10 раз,
чем у тех людей, у которых этого аллеля нет. На севере уровень холестерина и
частота е4 больше, чем на юге. На экваторе частота аллеля е4 около 3-5%. У
лапландцев (саамов) - 20-30%. У бушменов он достигает 40%, но никакой болезни
Альцгеймера у них нет. И не потому, что они не доживают до такого возраста,
просто у них этот ген ни с какими рисками не ассоциирован. Когда это было об-
наружено, выдвинули предположение, что этот ген относится к так называемым
«жадным» генам. Раньше была низкохолестериновая бессолевая диета, и сахар то-
же охотникам-собирателям никто просто так не давал. И им приходилось эти цен-
ные вещества эффективно усваивать из той пищи, которая была им доступна. Те-
перь сахар, соль и масло продаются в любом магазине, а организм человека «за-
точен» на то, что это ценные вещества, и людям кажется, что они вкусные. И те
гены, которые раньше помогали человеку запастись необходимыми веществами,
стали аллелями риска. Иногда их называют «жадными» генами. Те гены, которые
обеспечивали эффективное поглощение холестерина, стали факторами риска атеро-
склероза; те, которые обеспечивали эффективное поглощение соли - факторами
риска гипертонии, сахара - факторами риска диабета.
Когда исследовали питание бушменов - охотников-собирателей, живущих в Южной
Африке, оказалось, что оно соответствующим рекомендациям ВОЗ по общему балан-
су белков, жиров, углеводов, витаминов, микроэлементов и калорий. Они ведут
тот образ жизни, к которому человек приспособлен лучше всего. Биологически
человек и его непосредственные предки на протяжении сотен тысяч лет адаптиро-
вались к образу жизни охотников-собирателей. Рекомендации врачей по поддержа-
нию здоровья - физическая активность, прием витаминов и микроэлементов, огра-
ничение соли и т.п. по сути, искусственно воссоздают условия, в которых чело-
век жил большую часть времени своего существования как биологического вида.
Инфекции как
фактор отбора
Серповидноклеточная анемия - это смертельная болезнь крови. Ею болеют гомо-
зиготы по мутации в гене гемоглобина. Распространена эта болезнь в тропиче-
ском поясе. Оказалось, что эта мутация выгодна для гетерозигот, потому что от
анемии они не умирают, зато малярийный плазмодий в их эритроцитах размножает-
ся гораздо хуже. Это открытие было сделано, когда американские солдаты в се-
редине 20-го века находились в Корее. То, что в этой зоне распространена ма-
лярия , - хорошо известный факт. Это очень тяжелая болезнь, и чтобы защитить
солдат от этой болезни, им давали противомалярийные лекарства. Эти препараты
ингибируют развитие плазмодиев. Многих солдат эти лекарства спасли, но неко-
торые солдаты умерли непосредственно от самого лекарства. Причем умерли толь-
ко либо афроамериканцы, либо те солдаты, которые происходили из Италии или
Испании. Когда провели расследование этого случая, выяснилось, что они как
раз были носителями этих мутаций, которые защищали их предков от малярии, а у
них стала причиной смерти от противомалярийных препаратов. Частота мутаций,
защищающих от малярии, достигает 20% в тех регионах, где люди более всего
страдают от этой инфекции, и быстро снижается по мере удаления от тропической
зоны.
Серповидноклеточная анемия и малярия
I Ьгакк» АйжтроЬст А 1973, lr>lw Wii»y-fi5 otv. lie
Есть мутации, защищающие от вируса иммунодефицита. На одной из предыдущих
лекций было рассказано, что ВИЧ проникает в клетку, связываясь с рецептором.
Есть люди, у которых этот рецептор отсутствует. Называется этот рецептор -
хемокиновый рецептор CCR5. Найдена мутация в кодирующем его гене, делеция 32
нуклеотидов, (мутантный аллель называется CCR5delta32) которая прерывает син-
тез белка.
ВИЧ-протекпгеные мутации CCR5A32, CCR2-64I. SDFi-3'
Эти гены кодируют рецепторы, с которыми связываются вирус.
Мутации в них нарушают связывание вируса и его проникновение в клетку
Выжпьаемость после инфицирования. Мутации увеличивают
продолжительность жизни инфицированные индивидов %
-«BeSDFl+CCR
- CCR мутации
— | - две SDF1
| | - одна SDFl
Нет мутаций O'BnenSJ, Moore JP. Immunol Rev. 2000
Есть мутации в других генах, выше приведен пример с двумя генами. На рисун-
ке выше показаны результаты обследования выборки приблизительно двух тысяч
человек. Часть из них умерла к концу исследования, длившегося 15 лет. Около
65-70% людей, у которых нет ни одной протективной мутации, при наличии инфек-
ции умерло. У людей, у которых есть три протективных мутации, симптомы СПИДа
не проявлялись на протяжении достаточно большого периода времени, и лечению
они поддавались значительно лучше. Правда, людей с тремя мутациями было всего
несколько человек.
Частота этой мутации (делеции в гене CCR5) у русских - 15%. Несложно под-
считать, что 1% русских гомозиготен по этой мутации и устойчив к заражению
СПИДом половым путем. Но, тем не менее, никто никаких экспериментов подобного
рода на них не ставит, потому что защита не абсолютная, и при инъекционном
заражении никакие мутации не помогут. На рисунке представлена карта частоты
делеции CCR5delta32 на территории Евразии. Предполагается, что в прошлом
какая-то свирепствовавшая в Европе мутация провела отбор по этой мутации.
Географическое распределение частоты
ВИЧ-протектпвноп мутации CCR5delta32
120*
МГ
150
НО*
Svetlana A brrt.orska Oleg Р BahnovskyaBena V. Balanovskaya Peter A Slominsky Mana I Schadrma Ludrrrla A Livshits S
Kravchenko MadmrM Parrpuha Eb a K. Khusnutdinovs Motor A Sptsyn Analysis of CCRjdeta32 Geographic Dis&ibution and
Correlation with Sone Clrnatic and Geoyaphic Factors// Hum htened 2О02;534в-54
Генетическая детерминация
химических зависимостей
НАСЛЕДСТВЕННОСТЬ
н
КОКАИН
Исследовали 485 монозлготных и 335 долготных близнецов
Среди них пробовали кокаин 14° о. зависимых 2.3%
Уровень конкордат ностн
Употребление Злоупотребление Зависимость
М3 54% 47% 35%
ДЗ 42% 8% 0%
» —1Ы KJ hurra СА См «а», «few* enA rtopnalaa« m« pepUcftan
Мм4»f Imib mw л rt; Oct,PJ MJ Jd
Сейчас установлено, что в развитие как химических, так и поведенческих за-
висимостей вклад наследственности составляет 40 - 60%. Вклад наследственности
определяется по близнецовым исследованиям. Если монозиготные близнецы по дан-
ному признаку больше похожи друг на друга, чем дизиготные, значит, наследст-
венность работает. Когда признаки совпадают, близнецов называют конкордантны-
ми по данному признаку. Ниже показаны результаты таких исследований на приме-
ре зависимости от кокаина. Уровень конкордантности монозиготных близнецов при
злоупотреблении кокаином достоверно выше, чем дизиготных. Вывод - развитие
злоупотребления кокаином зависит от генов, не только от среды.
Выявлены две группы генов, связанные с развитием зависимостей:
1. Гены, связанные с работой системы положительного подкрепления (преимуще-
ственно рецепторы и транспортеры нейромедиаторов). Когда вы сделали что-
нибудь хорошее для своего организма, то система подкрепления дает сигнал, что
все хорошо. То есть, в лимбической системе мозга вырабатывается дофамин, ре-
цептор воспринимает дофамин, сигнал идет дальше и организм знает, что все хо-
рошо. Когда дофаминовый рецептор слабо чувствителен, необходимо увеличить до-
зу дофамина. Наркотическая зависимость возникает, потому что наркотики дейст-
вует на эти рецепторы впрямую, обманывая организм, подавая ему ложный сигнал
«все в порядке» даже в том случае, когда человек близок к смерти.
К этой же группе генов также относится ген транспортер серотонина. Серото-
нин - это нейромедиатор, участвующий в передаче сигналов в мозге. В ряде ра-
бот показано, что различия в активности этого гена связаны со склонностью к
депрессиям.
2. Гены метаболизма алкоголя и наркотиков. В организме алкоголь окисляется
в альдегид, а потом - в кислоту. В гене алкогольдегидрогеназы ADHlb имеется
точечная мутация, ведущая к аминокислотной замене (аргинин на гистидин), от
чего сильно увеличивается скорость работы фермента. И этанол начинает быстро
перегоняться в альдегид. А альдегид - это как раз то «злобное» вещество, ко-
торое вызывает неприятные ощущения после приема спиртного, похмелье. Когда
алкоголики похмеляются, они пьют спирт, подстегивая тем самым работу фермен-
тов метаболизма алкоголя, и весь накопившийся альдегид при этом может окис-
литься дальше (до тех пор, пока ферментные системы печени не истощены регу-
лярным приемом алкоголя). Носители мутации (она называется ADHlB*47His) обла-
дают повышенной чувствительностью к алкоголю.
Частота этой мутации, которая ведет к быстрому росту концентрации альдегида
в крови, разная у разных народов. У финнов - 0, у русских - 6%, у якутов -
16%, у китайцев - 76%, у тайванцев - 86%. Фермент перерабатывающий альдегид,
также может иметь разную активность. Если он неактивен, то от очень маленьких
доз спиртного человеку становится очень плохо - концентрация альдегида в его
крови в 30 раз выше, чем у «устойчивого» индивида при тех же дозах этанола.
Частота неактивного аллеля альдегидегидрогеназы также высока в Юго-Восточной
Азии и составляет там 30-50%. В Японии аллель выявлен у 2% алкоголиков и 44%
неалкоголиков. Гомозиготы по альдегиддегидрогеназе ALDH2 практически не
встречаются среди больных алкоголизмом.
Риск развития алкоголизма в 100 раз выше при сочетании (1), чем при сочета-
нии (2) , показанным на рисунке. Ген ADHlb может быть вовлечен в адаптацию к
внешней среде: показано, что в Юго-Восточной Азии высокая частота аллеля
ADHlb*47His обусловлена действием отбора, тогда как в других регионах преоб-
ладает предковый вариант ADHlb*47Arg. Возможными факторами отбора могли быть
какие-либо особенности диеты или инфекции.
Гены метаболизма алкоголя и риск развития алкоголизма
алкогольдегидрогеназа альдегиддегидрогеназа
н н АДГ Н ,о АцЩ
(1) н с-с-он --------------► н с-с —>►
НН н чн
ЭЛЛСЛН ADH1b‘47Arg ALDH24
н н АДГ Н ,о АцДГ
н с-с-он —► Н С-С ----------------------
НН н чн
аллели ADH1b‘47His ALDH2*2
чувствительность к алкоголю
Риск развития алкоголизма в 100 раз выше при сочетании < 1).
чем при сочетании (2)
Распространенность аллеля ALDH2 *'2 (фермент нс активен) у монтопотцов 30^50%
В Японтат аллель выявлен у 2% алкоголиков. 4-Го неагвеоголнков
Гомозиготы по а лкого лъдепирогеназе ALDH2' 2 2 ^тактически не вс треплются средн
больных алкогоптгтмом
Частота аллеля ADH1b*47His
финны - 0. русские - 6%. якуты -16%, китайцы - 76%
ЛЕКЦИЯ №22.
ЭКОЛОГИЯ1
ЭКОЛОГИЯ (ойкос - «дом, жилище») - наука о взаимоотношениях между организ-
мами и средой их обитания. Такое определение дал классик современной экологии
Ю. Одум в 1975 году. Но изначально термин был предложен Э. Геккелем в 1866 г.
Как наука, экология сформировалась как раздел биологии примерно к 1900 г. Чем
же занимается экология? Есть много подразделений экологии, здесь дана упро-
щенная схема.
ПОДРАЗДЕЛЕНИЯ ЭКОЛОГИИ
БИО-ЭКОЛОГИЯ
взаимодейс!вие организмов с окружающей средой
АУТ ЭКОЛОГИЯ
взаимодействие видов с
окружающей средой
СИН-ЭКОЛОГ ИЯ
закономерности образования со общее! в
и их взаимодействие со средой
Ареал, экологическая
ниша, экологические
факторы, популяция,
адаптация
Экосистема (биогеоценоз), пищевые
цепи, трофические пирамиды, поток
энергии, круговорот веществ
1. Один из крупных блоков экологии - биоэкология, наука о взаимодействии
организмов с окружающей средой. Один из ее подразделов - аут-экология, изу-
чающая взаимодействие видов с окружающей средой. На слайде представлена фото-
графия птенцов белоголового орлана, по отношению к ним, а также и к любому
другому виду аут-экология изучает ареал распространения вида, его экологиче-
скую нишу, то есть в каких пределах температуры, влажности, скоростей ветра,
живет орлан, чем он питается, где гнездится, какие есть влияющие на его жизнь
экологические факторы - паразиты, хищники, жертвы, как происходит динамика
популяции, адаптацию к окружающей среде и т.д. Другой раздел - син-экология,
изучающая закономерности образования сообществ, а также взаимодействие этих
сообществ с окружающей средой. На слайде показана астраханская степь, в кото-
рой живет орлан. Синэкология изучает проблемы объединения разных видов в об-
щую экосистему- биогеоценоз, пищевые цепи, как по этим цепям движется энер-
гия, трофические пирамиды (как вещество и энергия движется по цепям), потоки
энергии и круговороты веществ в экосистемах. 2
2. Второй крупный блок - социоэкология, изучающая взаимодействие человече-
ского общества с окружающей средой. Она тоже делится на две части. Первая -
экология человека, изучающая влияние окружающей среды на здоровье и жизнедея-
тельность человека, как биологического вида, а конкретно комфортность среды
для человека, влияние ядовитых и токсичных веществ, заболеваемость, системы
1
Д. И. Люри
жизнеобеспечения и т.д. Вторая часть - социо- или гео- экология, изучающая
взаимодействие цивилизаций, создаваемых ею систем, с окружающей средой, а
конкретно загрязнение среды, деградация экосистем, экологические кризисы,
проблемы устойчивого развития и т.д. На слайде показано поле, на котором вы-
ращивают бахчевые культуры. Такое поле используется всего три - четыре года,
после этого оно становится непригодным.
Комфортность среды, едовитые, экосистем, экологические кризисы,
токсичные вещества, заболевав- устойчивое развитие, ресурс-
мость, системы жизнеобеспечения мененджмент, политэкология
3. Третий блок - прикладная экология, занимающаяся созданием техники, тех-
нологий и методов для:
• минимизации воздействия человека на среду,
• контроля за ее состоянием,
• управления средой,
• охраны природы и рационального природопользования,
• систем жизнеобеспечения и др.
Кроме перечисленных выше направлений есть еще псевдоэкология (экология
культуры, мышления, разума, стеклопакетов и т.д. и т.п.), служит для привле-
чения денег, клиентов и прочих благ. Экология - «несчастная» наука, стала она
несчастной лет 20 назад, когда политики и журналисты ее очень полюбили и ста-
ли использовать этот термин вне связи с его научным содержанием.
В этой лекции будет преимущественно рассказано про социоэкологию. В сферу
внимания социоэкологии входит:
• влияние природных факторов на развитие цивилизации
• влияние человеческой деятельности на окружающую среду
• деградация окружающей среды, загрязнение и др.
• закономерности ресурсопользования и управление ресурсами
• экологические угрозы
• экологические кризисы
• устойчивое развитие: правила и способы перехода
• экономическая и внеэкономическая оценка стоимости природных ресурсов
• ренатурализация и рекультивация нарушенных экосистем
• правила сохранения природных экосистем и биоразнообразия
Сейчас идут разговоры об изменении климата. Является ли это следствием дея-
тельности человека или нет, как оно подействует на человека? На этот вопрос
сейчас нет однозначного ответа. Также существует проблематика экологической
угрозы, попытки оценить экономически и внеэкономически стоимость природных
ресурсов. Возьмем, например, лес. Ясно, сколько стоит древесина, ягоды, пуш-
нина в отдельности. Но также ясно, что лес не сводится к этим ресурсам, он
еще очищает воздух, запасает углерод и т.д. Возникает вопрос, как это оце-
нить? Это огромная проблема во всем мире. В нашем современном рыночном мире
то, что не имеет стоимости, не включается в систему цивилизации, в какие-либо
программы охраны.
Можно ли выделить основную проблему геоэкологии, такую, от которой зависят
ответы на частные вопросы? Ее можно сформулировать так: является ли цивилиза-
ция составной частью системы биосферы или самостоятельной системой - биосфе-
ропользователем? В первом случае есть механизмы, регулирующие развитие циви-
лизации, направленные от биосферы к цивилизации, то есть цивилизация включена
в систему биосферных процессов, во втором случае таких механизмов нет и циви-
лизация "сидит" на биосфере как спрут.
ЯВЛЯЕТСЯ ЛИ ЦИВИЛИЗАЦИЯ СОСТАВНОЙ ЧАСТЬЮ
СИСГ ЕМЫ ЕНОС ФЕ ГЫ
ИЛИ
САМОСТ ОЯТЕ ЛЬНОЙ
СИСГ ЕМСЙЪИОСФЕРа ПОЛЬЗ ОВАТ Е ЛЕМ?
От ответа на этот вопрос зависят стратегии выживания человечества. Понятно,
что человек - потребитель ресурсов (сам он ресурсом не является, разве что
для комаров). Потребителей (называемых в экологии консументами первого поряд-
ка, консументами второго порядка) очень много, однако они никогда не смогут
«съесть» свою экосистему, потому что существуют механизмы регуляции их чис-
ленности . Это проиллюстрировано на следующем рисунке:
Б
Динамика численности
«потребителя» при
наличии (А) и отсутствии
(Б) регулирующих связей с
«ресурсом»
На верхнем графике показаны колебания численности рыси и зайца по данным о
закупках шкур этих зверей компанией Гудзонова залива. Это классическая схема
колебания численности зверей при наличии механизмов их регуляции. Рысь нико-
гда не сможет съесть всех зайцев, так как существуют механизм регуляции. В
более упрощенной схеме (справа вверху) колебания выравниваются и численность
колеблется около среднего значения.
Совершенно по-другому ведет себя система, если нет регулирующих связей
(нижний график). Есть какая-то питательная среда, туда «высевается» жертва,
после в пробирку запускают хищника, который съедает жертву и затем сам поги-
бает от голода.
Какая из этих схем соответствует отношениям цивилизации и биосферы?
Существуют два подхода к разрешению этого вопроса. Первый подход, которого,
к сожалению, до последнего придерживалось большинство ученых, представляет
человека как биосферопользователя. Этот подход представлен в ставших класси-
ческими работах супругов Даниелы и Денниса Медоузов и Й.Рандерса, выполненных
под эгидой Римского клуба (организация, созданная 100 крупнейшими промышлен-
никами, они дают заказы ученым, которые пишут книги на заказанные темы). Это
работы «Пределы роста» (1972 г.) и «За пределами роста» (1992 г.) . На схеме
из этой книги человек представлен системой, стоящей на потоке, переводящей
энергию высокого уровня и ресурсы в отходы.
Дон ЛХедоуз. Ден.Медоуз .И. Рандер с
«Пределы роста» (1972), «За пределами роста (1992)
ПЛАНЕТАРНЬЕ энергия высоко
ИСТОЧНИКИ Уровня
НАСЕЛЕНИЕ И
ЭКОНОМИКА
ИсТО'ПООЖ X стом нм® т
скчм пределы
Чеял едесп о ужепощшию
очень блмзк о к эисч
прежличн нз -за
экспсмащкалвюго роста
знс.гвзностн схоронх
Лфв1ЫЗГТ
Выход запрздехы означает
ползоенсго-щвозе
ресурсов, от-раз .то’мке
(мосферын знбель
низ м.тиз азин.
ПЛАНЕ ТАРНЬЕ СТОКИ
Теплов ые потери
Человек здесь представлен как система, стоящая на потоке, превращающая
энергию высокого уровня (солнечную энергию, нефть) и ресурсы (древесину, по-
лезные ископаемые) в энергию низкого уровня, словом, ресурсы в отходы.
Смысл работы в том, что источники ресурсов и стоки имеют свои пределы. Че-
ловечество близко подошло к этим пределам, и из-за экспоненциального роста
скоро эти пределы перейдет. Выход за эти пределы грозит катастрофой, разруше-
нием биосферы, а вместе с этим и разрушением человечества в целом. Так, как
это было представлено с моделью хищника и жертвы в пробирке.
Какие существуют ограничения по использованию ресурсов? Из 3.2 млрд, га
максимально возможных зеленых ресурсов (то есть если мы сведем все леса) мы
используем 1.5. Использовали уже почти половину доступных водных ресурсов,
треть лесных и т.д. Согласно этим расчетам, 10% стоков уже заполнено.
Сценарии будущего по модели МИР-3
население
Ни увеличение пределов,
ни развитие технологий
на результат не влияет
одолжительность жизни
СТАНДАРТНЫЙ СЦЕНАРИЙ
ресурсов
С ВЕРХ-ОПТИМИСТИг1Е СКПЙ СЦЕНАРИЙ
Указанные меры приняты
только к 2015 г. По мере
отдаления с роков оптими-
стический сценарий при-
ближается к с так парткому
2000
1900
2100
Если вложить в эту модель удвоенные значения пределов, то есть если у нас в
2 раза больше ресурсов, чем мы сейчас думаем, и если у нас будут сверхмощные,
безотходные технологии переработки, картина принципиально не изменится, толь-
ко сдвинется на 20-30 лет.
Сверху показана схема оптимистического сценария. Если в 1995 г принята про-
грамма стабилизации населения (1 семья - 2 ребенка), внедрены безотходные и
ресурсосберегающие технологии, удвоенные значения пределов. Все это приводит
к тому, что в 2005 г. ситуация стабилизируется. Но так как ничего не сделано,
Медоузы разработали модель, когда меры приняты в 2015 г. Тогда ситуация не-
сколько ухудшается, а затем стабилизируется. И чем позже приняты меры, тем
больше "оптимистический" сценарий приближается к стандартному.
Что же предлагается в социально-экономическом отношении:
• Скорейшее прекращение роста населения (к 2015 г. : 1 семья - 2 ребенка,
эффективность контроля -100%).
• Стабилизация промышленного производства на уровне 350$ на человека в год
(это примерно Южная Корея или в два раза больше, чем Бразилия в 1990
г.) .
• Внедрение «безотходных» и ресурсосберегающих технологий (снижение ресур-
сопользования и загрязнения до уровня 1975 г.).
В отношении ресурсопользования:
Темпы потребления возобновимых ресурсов не должны превышать темпов их
регенерации.
• Темпы потребления невозобновимых ресурсов не должны превышать темпов их
замены на возобновимые (очень сложно выполнить в практическом смысле,
т.е. наращивать добычу нефти так, чтобы вкладывать в лесоразведение,
чтобы количество энергии в новых лесах было таким же, как в использован-
ной нефти)
• Темпы выбросов загрязняющих веществ не должны превышать темпов их при-
родной «переработки» (очистки).
Требования очень жесткие. Но они мягкие по сравнению с другой теорией.
Вторая теория, называемая «теорией золотого миллиарда» принадлежит физику
В.Г. Горшкову, разработана в 1990-1995 гг. Она говорит о следующем:
1. Биосфера представляет собой систему, работающую по принципу Ле Шателье
(компенсация внешних воздействий внутренними механизмами).
2. Действие этих механизмов устойчивости обеспечивается «невозмущенной био-
той» , т.е. ненарушенными природными экосистемами.
3. Разрушение природных экосистем приводит к потере устойчивости биосферы,
ее разрушению и последующей гибели цивилизации
4. Современная цивилизация уже превысила пределы возмущения биоты, что при-
вело к нарушению принципа Ле Шателье (биосфера, теряющая управляемость -
об этом говорит изменение климата, нарушение/размыкание круговоротов,
загрязнение среды и др.).
Устойчивость суши, по его мнению, была нарушена в середине XVIII века, до
начала XX века устойчивость биосферы поддерживалась за счет океана, после че-
го она была нарушена глобально. Принцип, заложенный в основу работы, совер-
шенно другой, если у Медоузов были рассмотрены ресурсы, то здесь рассмотрена
термодинамическая модель биосферы.
Пределы нарушения биоты: площадь нарушенных экосистем не должна превышать
20% от площади суши, а сейчас нарушено уже 60%, доля антропогенного потребле-
ния продукции биосферы не должна превышать 1%, а сейчас она составляет 10%.
То есть здесь тоже есть пределы, но совсем другие.
ПРЕДЕЛЫ НАРУШЕНИЯ БНОТЫ
1 Площадь нарушенных экосистем не должна
превышать 20% от площади суши
СЕЙЧАС (40
ДОЛЖНО ,
БЫТЬ 1 8
2 Доля антропогенного потребления биологической
продукции биосферы не должна превышать 1®Ь
СЕЙЧАС
ДОЛЖНО
БЫТЬ
В социально-экономическом отношении предлагается сокращение численности на-
селения за несколько десятилетий в 10 раз до 0,5 - 1 млрд, человек. В отноше-
нии ресурсопользования предлагается:
1. Фактический отказ от использования невозобновимых ресурсов: уменьшение
их эксплуатации в сотни раз.
2. Прекращение роста энергопотребления (в первую очередь ГЭС и АЭС).
3. Сокращение вырубки лесов как минимум в 10 раз.
4. Прекращение экспансии на еще неосвоенные земли и сокращение уже исполь-
зуемых в 3 раза.
Как это сделать - неизвестно, в том числе и автору теории, понятно, что де-
мографическими методами это сделать не удастся (если только мерами физическо-
го воздействия)
Что общего в этих двух классических работах? Очень жесткие требования к
численности населения и ресурсопользованию. Причем если эти требования не бу-
дут выполнены в ближайшие десятилетия, нам грозит катастрофа.
Этот подход очень безрадостен. Допустим, эта модель верна. Но мы реально не
готовы не только сократить население, но даже и прекратить его рост (как по-
казывает опыт Китая). Переход только на возобновимые ресурсы - также невозмо-
жен, это другая цивилизация. Допустим, мы согласимся принять меры, а окажет-
ся, что модели неверны.
То есть в любом случае, будем ли мы принимать эти требования или нет, со-
гласно этим моделям наша цивилизация или погибнет или радикально поменяется.
Второй подход гласит, что цивилизация - часть биосферы. Основу заложили ра-
боты Вернадского, Тьер де Шардена и др. Их теория ноосферы говорит о том, что
появится некий центр, который может управлять биосферой с помощью разума.
Этот подход представлен на следующей схеме.
ЦИВИЛИЗАЦИЯ КАК ЧАСТЬ БИОСФЕРЫ
В системе
«ЦИВИЛИЗАЦИЯ -
ПРИРОДА»
существуют механизмы,
регулирующие
экспансию человечества
Основоположники
В И Вернадский
Тьер де Шарден и др
Рассмотрим с этих позиций взаимоотношения человека с ресурсами и с приро-
дой. Начнем с типов ресурсов? Ресурсы существуют возобновимые и невозобнови-
мые . Мы можем выделить 4 типа:
1. природно-возобновимые ресурсы (воздух, вода, растительная и животная
биомасса):
• они восстанавливаются после использования до исходного состояния посред-
ством природных механизмов
• производительность природных механизмов восстановления имеет свой предел
(река может переработать определенное количество отходов в год, а если
больше - то начнется загрязнение)
• человек может вложить средства в интенсификацию возобновления
2. антропогенно-возобовимые ресурсы (металлы, сера, соли, фосфаты, строи-
тельные материалы и т.д.):
• восстановления осуществляется только самим обществом за счет имеющихся у
него средств
• в принципе могут быть восстановлены после использования до исходного со-
стояния, но природные механизмы для этого отсутствуют
3. невозобновимые ресурсы (энергоресурсы углеводородные - нефть, газ,
уголь, неуглеводородные - уран, а также алмазы-брильянты т.д.). Они в принци-
пе не могут быть восстановлены после использования до исходного состояния.
4. условно неисчерпаемые ресурсы (солнечная и гравитационная энергии):
• поступают из-за границ биосферы
• за счет них функционируют природные механизмы восстановления ресурсов
Соотношение между этими группами представлено на рисунке. Видно, что возоб-
новимых ресурсов большинство, они могут вовлекаться в круговороты «ресурс -
отход - ресурс» посредством природных и антропогенных механизмов.
РЕСУРСЫ млн. т %
ПРИРОДНО-ВОЗОБНОВИМЫЕ 3655131 99.7
Вода 3240000
Воздух 400000
Растительная биомасса 14272
Животная биомасса 359
АНТ РОПОГЕ ННО-ВОЗ ОБНОВ! 1МЫЕ 3140 0.1
Металлы 1640
11р Ьчие 1500
НЕ ВОЗОБНОВИМЫЕ 6593 01
Углеводородные ископаемые “Ж
Радиоактивные материалы 0 1
ИТОГО ВСЕХ РЕ СУТСОВ 3664864 100
БОЛЬШИНСТВО РЕСУРСОВ ЯВЛЯЮТСЯ ВОЗОБНО-
ВИМЫМИ И МОГУТ ВОВЛЕКАТЬСЯ В КРУТ ОБОРОТЫ
«РЕСУРС - ОТХОД - РЕСУРС» ПОСРЕДСТВОМ
ПРИРОДНЫХ И АНТРОПОГЕННЫХ МЕХАНИЗМОВ
Рассмотрим способы взаимодействия человека с природой (эволюции ресурсо-
пользования) . Первый тип назван «природа - мать». Заключается он в том, что
энергия тратится только на добычу ресурсов (охота, собирательство), которые
затем восстанавливаются посредством природных механизмов. Вся промышленность
начинается именно с этого - воздухо- и водопотребление без очистки (только
там нет природного восстановления).
МОДЕ III РК УРСОПО. ПНОВАННЯ
1. «ПРИРОДА-МАТЬ»
К этому типу ресурсопользования относятся охота, собирательство, отгонное
животноводство, начало индустриального производство (воздухо- и водопотребле-
ние без очистки), начало добычи полезных ископаемых и др. Объемы ресурсополь-
зования при этом ограничены регенерационным потенциалом природы - мы не можем
убить мамонтов или выловить рыбу больше определенного количества. Поэтому че-
ловечество переходит на второй тип, названный «природа - соратник». Его суть
в том, что восстановление ресурсов идет за счет природы и человека, интенсив-
ность цикла зависит от общих регенерационных вкладов природы и человека, при
этом снимается ограничение на объемы ресурсопользования.
МОД Г ПI РЕ СУ Р( ОПО. ПЛОВА Н11Я
2. «ПРИРОДА-СОРАТНИК»
Человек делает ресурсные циклы все более и более интенсивными. Начало этого
этапа связано с возникновением земледелия. Сейчас преобладает почти во всей
ресурсной деятельности, т.е. человек использует ресурсы, которые требуют от
него затрат на восстановление. Приведены примеры для растительной пищевой
биомассы и для рыбных ресурсов.
Ресурсы растительной нишевой биомассы
СОБИРАТЕЛЬСТВО:
ПРИМИТИВНОЕ
ЗЕМЛЕДЕЛИЕ
Дает дж/га*год до 1 *109
Затраты на восстановление О
Дает дж/га*год 7-40Ч09
Затраты на восстановление ок. 50%
АГРОПРОМЫШЛЕННОЕ
ЗЕМЛЕДЕЛИЕ
Дает дж/га* год
10011*109
Затраты на восстановление >80%
КУКУРУЗА (США, МЕКСИКА)
Параметры Поде огнев. Агропромышленное 1945 19"0 1980
II отучено ку куру >ы 109;ск г я 28.9 35.4 84.4 1025
Затрачено всего 1(Рдж га 2.3 8.9 24.2 28.9
В тчня восстановление популяции и почв О» 52 66 81 83
Рыбные ресурсы
ПРЕСНОВОДНАЯ АКВАКУЛЬТУРА
МОРСКАЯ АКВАКУЛЬТУРА
УЛОВ кг га*год
МИРОВОЙ ОКЕАН 2
ПЕРУАНСКАЯ ЭОНА 1700
МАРИКУЛЬТУРА 2000
УЛОВ кг г я* год
ОБЫЧНЫЙ ПРУД 40 170
+ УДОБРЕНИЯ 200-550
УДОБРЕНИЯ + КОРМА >2000
Добыча tMC.tOMH Доля аквакультуры %
Проснсеод Морская
0-10 11.6 •
10-100 13Д 3.3
100 1000 13.3 4.5
1-5000 20.6 8.6
>5000 45.0
>10000 18.0
ДОЛЯ АКВАКУЛЬТУРЫ В ДОБЫЧЕ «о
18
16
14
12
10
8
б
4
2
О
1983 1988 1990 1993 1998
Видим, что чем больше добыча рыбы, тем больше доля аквакультуры. Рыболовст-
во заменяется рыбоводством. Сейчас каждая пятая рыба выращена, а не выловле-
на. Чем больше мы используем ресурсов, тем больше мы вкладываем в их восста-
новление .
Продуктивность древесных насаждений в 20 раз может превосходить продуктив-
ность естественных лесов. Использование в водных ресурсах технологии «приро-
да-соратник» - это их очистка и зацикливание. Данные по древесным и водным
ресурсам представлены ниже.
ПРОДУКТИВНОСТЬ
ИСКУССТВЕННЫХ
ЛЕСОНАСОЖДЕНИЙВ 2-20
РАЗ ВЫШЕ ЕСТЕСТВЕННЫХ
ЗАМЫКАНИЕ СИСТЕМ
ВОДОСНАБЖЕНИЯ
(т гр омы тленности США > 7044)
И ОЧИСТКА СТОКОВ
ЛЕСОПОСАДКИ тыс.га/год
ДОБЫЧА ДРЕВЕСИНЫ млн.жЗ/год
СТЕПЕНЬ ОЧИСТКИ %
ВОДОЗАБОР кмЗ/год
ЦВЕТНЫЕ МЕТАЛЛЫ И ЖЕЛЕЗО
Др.тя т тормшого мр-татги i потр<£.тасмн %
Годы 19Ы) ю ТО 1975 1980 1985 1990 2000
Cixxeu 27 26 27 30 32 34
А.'х ?химн 16 17 21 24 28 30 33
ДОЛЯ ВТОРИЧНОГО РЕСУРСА %
Перейдем к антропогенно-возобновимым ресурсам.
Выше - данные по цветным металлам. Видим, что доля вторичного металла в по-
треблении все время растет. На осях трехмерного графика отложены запасы ме-
таллов , потребление металлов, доля вторичного металла. При малых запасах при
увеличении потребления очень быстро растет доля вторичного металла, но стоит
увеличиться запасам металлов, эта зависимость ослабевает. Подобные графики
характерны для многих ресурсов.
И вполне, возможно, что если мы дальше будем развиваться по увеличению сво-
ей доли восстановления ресурсов, мы придем к третьему типу «природа-
экспонат». То есть, в рамках этой модели мы полностью тратимся на добычу и
восстановление ресурсов. В наше время этот тип встречается исключительно ред-
ко (например, в системах жизнеобеспечения на подлодках и космических станци-
ях, но и там зациклены воздух и частично вода, но не продукты питания).
3 «ПРИРОДА - ЭКСПОНАТ»
[' ВОССТАНОВЛЕНИЕ РЕ СУТСОВ ИДЕТ ТОЛЬКО ЗА СЧЕТ
ЧЕЛОВЕКА.ПРИРОДА -«МУЗЕЙНЫЙ ЭКСПОНАТ».
Мы как бы движемся к этому, но пока не подошли. Возникает вопрос: почему же
мы еще не подошли? На рисунке ниже приведены обозначения, которыми мы будем
пользоваться.
ЭФФЕКТИВНОСТЬ РЕСУРСОПОЛЬЗОВАНИЯ
И ВОССТАНОВЛЕНИЕ
• R - объем ресурсопользования (то количество ресурсов, которое мы получа-
ем за единицу времени, за год);
• А - объем потребления (разница между тем, что получили и тем, что затра-
тили) ;
• Z - затраты, подразделяются на D (затраты на добычу) и V (затраты на
восстановление);
• Р - это то, что природа восстанавливает;
• К1 и К2 - удельные затраты на добычу и восстановление.
А теперь давайте посмотрим, как изменяется эффективность ресурсопользования
по мере роста его объемов. Если мы возьмем «природу-мать», то это будет про-
сто константа. Для системы «природа-соратник» получается, что чем больше объ-
емы ресурсопользования, тем больше падает эффективность, то есть каждая еди-
ница ресурса становиться все дороже. Именно поэтому доля восстановленного хо-
рошо растет при малых запасах ресурсов, и плохо - при больших. Если ресурсов
достаточно, то в их восстановление средства не вкладываю
ЭФФЕКТИВНОСТЬ РЕСУРСНОГО ЦИКЛА E=R7Z
[ ПРИРОДА-МАТЬ E-RZ-R'RKl-l.'Kl
ПРИРОДА- R R
Е =--------------=-------------
СОРАТНИК Rk,+ k2(R R,, Р» R(krk3) k/P+I^)
Посмотрим, в какой мере это соответствует реальности.
Зсмлкделме
Со&сра- тельстто Залежное Пожечн Трелполвсое Aipo- прспышлвг Твивтосое
С&р IO9 да-.'га 0 8 25 40 145 >400
Эффа: пакость Дж/д»: 20 10 8 1.5 0.003
Жинпиоидспо
(Хота Отгонное ПасНмщное Оггорижгчное
«Ч> Ю’жжга <0 8 17-34 50 - 59 92-110
Эф(*М9.1И1МОСТЪ Дк/да. 10.0 2.0 1.5 1.0 0.2 0.1-0.05
г Ьмцеюж'П о
СЬч> 10^да?'г* <0 04 67 -75 92 -126
ЭффсгЛЮ МО ств Дк/мк 10.0 2.0 - 1.0 0.5- 0.1
Если при собирательстве, затратив 1 дж, получали 20 джоулей, то при агро-
промышленном производстве это на порядок меньше.
Рост объемов ресурсопользования сопровождается падением его эффективности.
То есть каждая единица ресурсов дается нам дороже и дороже.
Из этого следует, что у нас есть
"общество-природа", который основан
ресурсов.
первый регулирующий механизм в системе
на совместном с природой восстановлении
Регулирующая связь в системе
«Общество-Природа»
Основана на совместном
восстановлении ресурсов
по принципу «природа-
соратник»
Напомню, в системе Медоузов человек стоял на протоке, превращая ресурсы в
отходы. А здесь принципиально другая схема, человек участвует в ресурсном
цикле, восстанавливает ресурсы вместе с природой.
Итак, рост объема ресурсопользования ведет к снижению эффективности, добыча
ресурса становится все дороже и дороже, это ведет к замедлению роста объема
ресурсопользования (потом даже и к падению объема)
НАШИ ДОСТИЖЕНИЯ ИЛИ ПОЧЕМУ У НАС ТАК ПЛОХО ПОЛУЧАЕТСЯ?
ВКЛАДЫВЕМ В ВОССТАНОВЛЕНИЕ ПОЧВ ДЕГРАДАЦИЯ ЗЕМЕЛЬ ПОЧТИ ВО ВСЕХ РЕГИОНАХ МИРА
НАРАЩИВАЕМ ПЛОЩАДИ ЛЕСОПОСАДОК ОБЕЗЛЕСИВАНИЕ -ГЛОБАЛЬНАЯ ЭКОЛОГИЧЕСКАЯ ПРОБЛЕМА
РАЗВИВАЕМ АКВАКУЛЬТУРУ СОКРАЩЕНИЕ РЫБНЫХ СТАД
СТРОИМ ВО ДО- И ВОЗДУХООЧИСТНЫЕ СИСТЕМЫ ЗАГРЯЗНЕНИЕ АТМОСФЕРЫ И ГИДРОСФЕРЫ ПО ВСЕЙ ЗЕМЛЕ
УВЕЛИЧИВАЕМ РЕЦИКЛИЗАЦИЮ МИНЕРАЛЬНОГО СЫРЬЯ РОСТ ОТХОДОВ И ИСТОЩЕНИЕ МЕСТОРОЖДЕНИЙ
ИТ Д ИТ П
Получается, что, двигаясь по этой ниспадающей кривой, мы найдем где-то рав-
новесную точку, которая будет нас устраивать и по эффективности (по стоимости
добычи ресурса), и по объемам ресурсопользования.
Механизм, показанный выше, хорош всем, кроме того, что никак не соответст-
вует тому, что происходит в действительности. В таблице показано, что у нас
на самом деле получается.
Так происходит потому, что затраты на восстановление обычно гораздо ниже
необходимых, всех волнует только одно - объемы потребления. Поэтому вся наша
стратегия ресурсопользования направлена на увеличение объемов потребления.
Люди стремятся увеличить объемы ресурсопользования и снизить затраты. Снижать
затраты на добычу ресурсов почти невозможно (очевидно, если вы не посеете
зерно, то вы его и не соберете), а с восстановлением все гораздо проще - его
можно отложить на будущее. И в этом одна из основных причин экологических
кризисов.
В реальности мы не движемся по равновесной траектории. Мы сначала отходим
от нее (фаза I на рисунке), увеличивая объемы ресурсопользования без соответ-
ствующего увеличения вложений в восстановление ресурсов, и получаем в резуль-
тате излишки ресурсов.
ТРАЕКТОРИИ ЭКОЛОГ!FIECKIIX КРИЗИСОВ
1 -СОЦИАЛЬНО-БЛАГОПРИЯТНЫЙ ВЫХОД ИЗ КРИЗИСА
2 - СОЦИАЛЬНО-НЕБЛАГОПРИЯТНЫЙ ВЫХОД ИЗ КРИЗИСА
3 -КАТАСТРОФИЧЕСКИЙ «ВЫХОД ИЗ КРИЗИСА»
И все вроде бы хорошо, но затем у нас начинают истощаться ресурсы. В связи
с этим увеличиваются удельные затраты на их добычу, следовательно, падает эф-
фективность. То есть, мы сталкиваемся (фаза II) с той же проблемой, что и при
движении по равновесной траекторией, только на равновесной снижение эффектив-
ности связано с увеличением затрат на восстановление ресурсов, а на кризисной
траектории - с ростом затрат на их добычу. Причем в первом случае не происхо-
дит истощения ресурсов. Выход из сложившейся ситуации возможен тремя принци-
пиально различными путями.
(1) Первый - социально-благоприятный. Мы вкладываем средства в восстановле-
ние ресурсов, но мы платим за это падением эффективности. В результате мы
оказываемся на равновесной траектории.
(2) Второй - социально-неблагоприятный, когда у нас нет средств для восста-
новления, объем ресурсопользования сокращается до равновесного.
(3) Третий - катастрофический, когда ресурсы настолько истощены и нет меха-
низма их природного или антропогенного восстановления, что объемы ресурсо-
пользования падают до нуля, что приводит к гибели основанной на них цивилиза-
ции .
В правом верхнем углу рисунка тот же процесс показан в других координатах -
вместо эффективности (Е) здесь затраты (Z).
Таков механизм развития экологических кризисов. Под экологическими кризиса-
ми мы понимаем такой этап развития системы "общество-природа, при котором
высокоэффективное увеличение объемов ресурсопользования и потребления, дости-
гаемое за счет истощения ресурсов, тем или иным образом меняется на восста-
новление ресурсно-экологического равновесия ("вираж экологического кризиса").
Причина их заключается в стремлении человека увеличивать объемы потребления
наиболее быстрыми темпами (что характерно для большинства обществ) и, экономя
на регенерационных затратах, повышать эффективность хозяйства.
Возникновение таких этапов в ходе развития ресурсопользования является
вполне закономерным, поскольку потребительские цели главенствуют в современ-
ном обществе и поэтому постоянно толкают его к отклонению от равновесной тра-
ектории .
Некоторые характеристики кризиса: Г - глубина кризиса (максимальное удале-
ние от равновесной траектории), ср - резкость кризиса, (бкон.- бнач.) - про-
должительность кризиса (период между уходом с равновесной траектории и воз-
вращением на нее).
ХАР АКЕ ТРИСТИКИ ЭКОЛОГИЧЕСКИХ KPIDHCOB
Г - глубина кризиса
Ф - резкость кризиса
((кон.- (нам.) - продолжитепьность кризиса
Рассмотрим конкретные примеры. Ниже показана история развития сельского хо-
зяйства в центральном Черноземье России в 1785-1985 гг. В самом начале у нас
восстановление ресурсов было намного меньше, чем необходимо, за счет этого
достигалось потребление ресурсов в 2 раза большее, чем могла обеспечить рав-
новесная траектория. Эффективность была 1.4 джоуля на джоуль, тогда как надо
было бы 1.2 дж/дж, при этом шла деградация почв.
КРИЗИС С БЛАГОПРИЯТНЫМ исходом
Кризисная траектория -------------Р ав нове сная тр аекг ория
В 45-ом году луга стали настолько вытравлены, что стала падать численность
скота, вследствие этого снизилось внесение удобрений, а затем стала падать
урожайность зерновых, результатом чего стал знаменитый голод конца 19-ого ве-
ка . Здесь есть момент, похожий на то, что происходит в нашей стране сейчас.
Дело в том, что после освобождения крестьян освобожденный пахарь перестал
вкладывать средства в восстановление почв, и этот привело к кризису. Анало-
гичным образом сейчас "освобожденный бизнесмен" тоже первым делом перестает
вкладывать средства, как говорят, "в экологию", что также ведет к кризису.
В начале 20-го века кризис углубился - хотя вложения пошли, но их не хвата-
ло .
Лишь в 1950-х годах в ЦЧР началось активное увеличение вложений в регенера-
цию агроресурсов при переходе к современному агропромышленному хозяйству. Их
10-кратный рост позволил в 6 раз повысить объемы ресурсопользования и при
этом практически полностью восстановить ресурсно-экологическое равновесие -
прекратить истощение полей и ослабить пастбищную дигрессию. Платой за это
стало снижение эффективности примерно в 2 раза с 1.5 до 0.9дж/дж.
Грустно, что даже при падении объемов ресурсопользования мы не вкладываем
деньги в восстановление ресурсов, хотя, казалось бы, уже все плохо и надо
что-то делать, мы все равно пытаемся на этом экономить. В современной России,
падение объемов ресурсопользования гораздо больше, чем падение выбросов за-
грязнений .
Похожая ситуация имела место на Великих Американских Равнинах в середине XX
в. Аналогичным образом произошел выход из экологического кризиса, связанного
с загрязнением атмосферы в США оксидами серы. Развитие угольной энергетики
без необходимой очистки выбросов к 1970-м годам привело к катастрофическому
загрязнению атмосферы оксидами серы в крупных городах. Однако последующие ог-
ромные вложения в очистные сооружения позволили к 90-м годам не только про-
должать наращивать производство электроэнергии из угля, но и довести загряз-
нение воздуха до приемлемого уровня. Платой за это стало снижение рентабель-
ности энергетики США в некоторых штатах на 20%.
ЗАГРЯЗНЕНИЕ АТМОСФЕРЫ В США SO,
КРИЗИС С НЕБЛАГОПРИЯТНЫМ ИСХОДОМ
Развитие животноводства в аридной Африке
Теперь посмотрим кризис с неблагоприятным выходом. Выше проиллюстрировано
развитие животноводства в аридной Африке. График показывает зависимость де-
градации пастбищ от увеличения объемов использования мясомолочной продукции.
Некоторые страны миновали кризис, а часть стран в него вошла. Марокко и Тунис
вложили средства и достигли равновесной траектории. А Сомали, Нигеру и Кении
ничего не оставалось, только как катастрофически сократить численность скота.
Кризис с катастрофическим исходом. Ниже показано, как мы выбили китов (есть
модель того, как мы выбили мамонтов, с абсолютно идентичным графиком). Был
перейден рубеж естественного восстановления, потом мы продолжали некоторое
время истощали ресурсы, потом китов стало мало и начала падать эффективность
их добычи, а затем снизилось ресурсопользование. У нас стали слишком совер-
шенными технологии добычи, поэтому голубых китов мы добили до последнего.
КРИЗИС С КАТАСТРОФИЧЕСКИМ ИСХОДОМ
КРИЗИС КИТОБОЙНОГО ПРОМЫСЛА 1945-91 гг
Посмотрим типологию экологических кризисов. Кризисы, о которых шла речь вы-
ше - это кризисы от жадности. Представим себе другую ситуацию. Мы развиваемся
по абсолютно равновесной траектории, например, сбрасываем мало отходов в Фин-
ский залив. И в какой-то момент мы просто строим некую дамбу. В результате
изменяется вся структура потоков, механизмы восстановления начинают работать
хуже, и мы оказываемся в состоянии кризиса. Равновесная траектория сама пада-
ет, а не мы от нее отодвигаемся. Это кризисы "от глупости". И третий тип -
природные кризисы, когда снижение регенерационного потенциала природы проис-
ходит в результате естественных причин.
ТИПЫ ЭКОЛОГИЧЕСКИХ КРИЗИСОВ
1 «Ресурсные» или кризисы от жадности
Увеличение потребленияза счет роста R и экономии V
2 «Структурные» или кризисы от глупости:
Снижение регенерационного потенциала природы в результате
антр опог енной д еят ельн ости
3 . ПрИрОДНЫе Снижение per енераци
они ого потенциала природы в
результате естественных причин
Ответим на три вопроса: когда НУЖНО выйти из кризиса, всегда ли МОЖНО выйти
из кризиса, ХОТИМ ли мы выйти из кризиса.
Экологический кризис нам нужен для того, чтобы увеличить объемы потребле-
ния . Рассмотрим динамику ресурсопользования при развитии кризиса.
Для чего нам нужны экологические кризисы
лобычу ресурсов I) <к| - улелвмкм* итряш ня добычу) и диоамикя
*Р^урсопилыоминя: обм-ноя потреблении (A). JBipui ш
лобычу и ВОССТЯМОВ.1СНИС ресурсов (V): W — А + Пч-V. Ьштрмхов»
кризисный мыитек объемов потреб, зги ня.
в) при равновесном развитии бе» mcioiiichmb ресурсов <ki*comt),
б) при кризисном разам»ии с истощением ресурсов (к| растет).
Синяя линия на рисунке - это как бы росли объемы потребления, если бы мы
развивались по равновесной траектории, кривая линия показывает развитие по
кризисной траектории. Заштрихованы - кризисные излишки потребления. После
достижения точки максимального потребления и точки максимального ресурсополь-
зования кривая идет вниз, но понятно, что ниже прямой R=D она не упадет (R-
добываемые ресурсы, D - затраты на добычу).
Посмотрим, как развиваются события, если мы не вкладываем ничего в восста-
новление ресурсов. У нас начинает загибаться кривая объемов ресурсопользова-
ния, а ей навстречу падают равновесные объемы ресурсопользования. В итоге,
точка их пересечения является наиболее выгодной точкой ресурсопользования. И
чем более совершенны технологии добычи, тем глубже оказывается эта точка.
Выход из кризиса при отсутствии
регенерационных вложений
ТРАЕКТОРИИ
1 ш------------♦
2 -------* ........—
3 ------------------—
кризисные смешение точки
расктирин равновесна
Рис..Мб. Bocci ановление равновесии
в холе кризиса при отсутствии антро-
ПО1-СИИ1.1Х рсчсмсрациомиых вложений
при ратной зависимости удельных за-
трат на добычу ресурсов к) от исто-
щение их запасов Q (см.рис.33). Чем
более совершенны технологии добы-
чи, т.е. чем ме. стен нее налает ki при
истощении ресурсов (1-2-3), тем
на меньшем уровне К происходит
восстановление равновесии.
Посмотрим теперь этот же график в "более важных" координатах - объемы по-
требления (А) и объемы ресурсопользования (R).
Критические точки экологического кризиса
РасЗбДинамика объемов погр^кния Асг в холе жилсшгвхкото
крмыеса (а) и итменепме равновесных объемом потребления Хея (б).
г.е. тух. то которым нужно в каж iwfl момеш кринка еннтитъ объемы
потреб инни, чюбы павтии cpe iciwa .via рг«гперяпионныч мнвеч <и
ннй. а - И1МГМСНИС Ar*, если в холе кризиса ki превысит тманемме
и исмевмуг срелстаа дла ретгнераиноммыч ммвеггиаий.
На этом графике показано, что в ходе кризиса у нас возникают 4 критических
точки: (1) когда кризисные объемы потребления равны максимально возможным
равновесным, (2) когда достигаются максимальные объемы потребления, (3) когда
максимальны объемы ресурсопользования, (4) когда равновесные объемы потребле-
ния равны нулю.
Развитие событий при выходе из кризиса на разных участках кризисной траек-
тории представлены на следующем рисунке:
Выход из кризиса при наличии регенерационных вложений-
Когда мы выходим до первой кризисной точки (слева вверху), у нас абсолютно
не снижаются объемы потребления, заштрихованную часть ресурсов мы вкладываем
в восстановление ресурсов, и только в какой-то момент снижаются темпы роста
потребления.
Когда мы выходим между первой и второй точкой (справа вверху), мы, судя по
рисунку, должны заплатить снижением объемов потребления. На самом деле, это
не совсем так, потому что люди могут находить новые ресурсы и технологии.
Этот момент - наиболее подходящий для маневра, потому что человечество дости-
гает того, что хотело, входя в этот кризис, максимальных объемов потребления.
Дело в том, что здесь мы можем делать ресурсный маневр: часть новых ресурсов
и технологий вкладывать не в увеличение объемов производства, а в восстанов-
ление ресурсной среды.
Если мы переходим вторую критическую точку (слева внизу), то события все
больше усложняются и вероятность выхода из кризиса снижается. После четвертой
критической точки (справа внизу) вероятность выхода без снижения численности
населения практически нулевая.
Посмотрим, что происходит с населением во время кризиса. Введем еще один
параметр - Аж - объем минимального жизнеобеспечения.
Аж=Ы*а(min),
где N-численность населения, a(min) - минимальный уровень потребления.
Если у нас реальный объем индивидуального потребления очень велик, то мы, в
принципе, можем согласиться на то, чтобы не покупать второй телевизор и вло-
жить эти средства в восстановление природы; но последний кусок хлеба мы для
этого не отдадим. И если объемы жизнеобеспечения идут практически рядом с
кризисными объемами потребления, то у человечества просто нет ресурсов, чтобы
вкладывать в восстановление.
Экологический кризис и демографический взрыв
Аж- объем минимального жизнеобеспечения -----------
Аж-N* A(inill), где N-численностъ населения,
a(min) - минимальный уровень потребления.
Ниже показана картинка для Сомали, Кении и т.д. - им просто не откуда взять
ресурсы, чтобы их восстановить. Бросается в глаза, что мы можем выйти из кри-
зиса только в том случае, если объемы жизнеобеспечения будут расти достаточно
медленно, не доходя до второй критической точки. Мы можем выйти из кризиса
только в этой ситуации. Но это достаточно трудно осуществить.
Экологическим кризис гасит демографический взрыв
Падение темпов роста численности населения начинается с
700-1000 $ВВП/чел*год и 2000-2300 Кал/чел*день
Известно, что чем богаче страна, чем больше ВВП на душу населения, тем
меньше прирост населения. Падение темпов роста населения начинается в среднем
с 1000$ в год, или в других координатах с 2000 калорий в день на человека.
Экологический кризис и демографический взрыв
кто быстрее?
выход И1 кризиса с падением уровня вадивзадужлыюго потребления
не ниже aniia. «ли Аж приближайся к Aeq между первой Аог= Авалях)
и второй Асг(тах) критическими i очками.
То есть происходит следующее. Мы входим в экологический кризис, чтобы полу-
чить максимальные объемы потребления, тем самым мы увеличиваем индивидуальные
объемы потребления, и таким образом гасится демографический взрыв, являющийся
одним из основных толчков экологического кризиса. Если экологический кризис
успеет погасить демографический взрыв до второй критической точки, то есть
шанс выйти из экологического кризиса, если нет - шансы резко падают.
Вышесказанное можно обобщить следующей схемой.
Механизм регулирования
экологического кризиса
Основан на обратной зависимости скорости роста
насепенпя от уровня потребления
Давайте посмотрим на конкретные числа. В следующей таблице показан прирост
населения в 1990 году относительно 1950 года, а также ресурсопотребление.
Видно, что только потребление картофеля и шерсти не превосходит прироста на-
селения. Видно, что индивидуальное ресурсопотребление растет, а прирост насе-
ления падает, то есть наши рассуждения оправдываются.
НАСЕЛЕНИЕМ РЕСУРСЫ 1950 г. 1990 г. УВЕЛИЧЕНИЕ (раз)
Население (млрд чел.) 252 529 2.1
Прирост (% в год) 184 1.74
Нефть (м лн.тонн в год) 520 2910 56
УГоль (млрд тонн в год) 15 52 35
Газ (млрдмЗв год) 193S 1960 10.1
Сталь (млн.тонн в год) 1893 772 4.1
Алюминий(млн.тонн в год) 165 179 109
Медь (м лн.тонн в год) 23 88 38
Уд обр ен ия(м лн .т онн в г о д) 150 138 8.7
Цемент (млн.тонн в год) 133.7 940 70
Зерновые (млрд .тонн в год) 0 68 195 29
Картофель(м лн т онн в год) 1440 2670
Бобовь<е(м лн.тонн в год) 215 578 2.7
Сахар (м лн.т онн в год) 31.1 111.4 36
Молоко (м лн.тонн в год) 2835 544.1 2.1
Яйца (м лн тонн в год) 10.1 355 35
Рыба (млн.тонн в год) 200 975 49
Вода (кмЗвгод) 910 3240 36
Древесина (м лн.мЗ в год) 1.46 3.47 24
Хлопок (млн.тонн в год) 82 18.4 30
Шерсть (м лн.тонн в год) 18 3.1
Каучук (м лн.тонн в год) 19 50 26
ВСЕГО В СРЕДНЕМ 44 I
Теперь ответим на последний вопрос: хотим ли мы платить, вкладывая средства
в восстановление ресурсов? Неким образом было рассчитано для 149 стран мира
«экологичность» экономики (ВВП страны и «чистоту» производства внутри стра-
ны) , по отношению к среднему мировому значению, равному единице. На этом
слайде показана логарифмическая зависимость «экологичности» экономики от ВВП
на одного человека. Видно, что чем богаче страна, тем более высокоэкологичную
экономику она имеет, люди хотят жить в гармонии с окружающей средой и ради
этого готовы вкладывать деньги в восстановление ресурсов. Это можно назвать
неким «культурным» механизмом регулирования взаимодействия человека с приро-
дой.
Хотам ли мы платить? «Культурный» механизм
регулирования взаимодействия человека и природы
Экологичность экономики, средняя ПО КПфу= 1
В заключении хочется отметить, что сейчас происходит поворот от представле-
ния цивилизации как биосферопользователя, к представлениям о том, что все-
таки существуют некие биосферные механизмы регулирования человеческой дея-
тельности .
ЛЕКЦИЯ №23.
НЕЙРОБИОЛОГИЯ И ГЕНЕТИКА ПОВЕДЕНИЯ
Наверняка каждый бывавший в селе человек видел, как утята бегают за своей
мамой-уткой. Откуда же они знают, кто их мама и за кем надо бежать? Здесь мы
сталкиваемся с явлением импринтинга (от англ, запечатлевать), описанным лау-
реатом Нобелевской премии Конрадом Лоренцом. Дело в том, что если утенок или
цыпленок увидит в течение нескольких часов после вылупливания движущийся
объект, он будет бегать за ним всю оставшуюся жизнь. Образ движущегося объек-
та впечатывается в их зрительную память, это достаточно разумное эволюционное
приспособление, так как в обычной жизни первое, что видят тт^тплята, - это их
мать, которая обеспечивает их существование. Каким же образом формируются
нервные связи, с помощью которых мы можем что-то запомнить? Об этом и пойдет
речь.
Если вам нужно узнать время, вы можете взглянуть на часы и некоторое время
помнить, что показывали стрелки часов. Но одного мимолетного взгляда на часы
не достаточно, чтобы запомнить на всю оставшуюся жизнь, что именно в этот мо-
мент было столько-то времени, разве что в это время произойдет что-то необыч-
ное. При неоднократном повторении какого-либо действия оно запоминается и
уходит в так называемую долгосрочную память. Механизмы формирования кратко- и
долгосрочной памяти разные. Они были изучены на разных животных, от улиток до
обезьян, и знания, полученные в экспериментах, применяются сейчас в медицине,
а также помогают понять, что происходит, как происходит развитие ребенка. Ка-
ким образом формируется память? Со школы известно, что мозг состоит из нейро-
нов , у которых имеется тело, длинный отросток аксон и короткие отростки денд-
риты, которые получают сигнал от других нейронов или рецепторов, обрабатывают
его и посылают его дальше. При рождении животного часть нервных путей уже
сформирована, это то, что называют безусловными рефлексами или комплексами
фиксированных действий. Это набор реакций на какой-либо стимул, для которых
не требуется обучения. На прошлых лекциях упоминалось, что у ребенка при рож-
дении есть набор рефлексов, например хватательный и сосательный. На основе
врожденного поведения формируется дальнейшее поведение. Мы разберем формиро-
вание нервных связей на примере зрительных, то есть, как ребенок или котенок
учится различать объекты. При рождении животное не способно различать объек-
ты, хотя способно реагировать, например, на яркий свет, так как глаза у ко-
тенка устроены так же, как и у взрослой кошки, есть связи, передающие сигнал
от светочувствительных клеток к зрительной коре, но структура этих связей еще
не такая, как у кошки. После рождения, та часть коры головного мозга, которая
отвечает за зрительное и другие типы восприятия претерпевает изменения. От
глаз, светочувствительных клеток идут цепи нейронов к структуре, называемой
боковым коленчатым телом, от них сигналы поступают к зрительной коре противо-
положной части головного мозга. Сигналы, поступающие в мозг, обрабатываются
на каждой стадии поступления, то есть если в самом глазном яблоке миллионы
рецепторов, то в боковом коленчатом теле на порядок меньше нервных связей,
соответственно каждый нейрон получает сигнал от нескольких рецепторов. То,
как формируется распознавание объектов в зрительной коре, было изучено на ко-
тятах. У новорожденного котенка, зрительные нервные связи перекрываются, но
по мере накопления зрительного опыта оказывается, что глаза посылают сигналы
в не перекрывающиеся участки коры, то есть остаются только те чередующиеся
связи, которые идут только к одному глазу отдельно. Эти структуры называются
колонками глазодоминантности. На слайде они показаны на примере макаки. Они
видны, потому что макаке ввели в глаз радиоактивное вещество, которое рас-
пространилось только в тех клетках коры, которые отвечают за зрительное вос-
приятие, то есть, имеют нервные связи с этим глазом.
Колонки глазодоминантности в стриарной коре макаки
формируются в результате конкуренции
потоков нервных импульсов от левого и правого глаза
Hubei and Wiesel 1977
Исследование того, как сигнал с сетчатки глаза последовательно передается в
структуры мозга и как он обрабатывается в каждой из них, что приводит в ко-
нечном итоге к восприятию изображения, провели лауреаты Нобелевской премии
Дэвид Хьюбел и Торстен Визел. Они провели серию экспериментов по так называе-
мой монокулярной депривации у котят. Один глаз у новорожденного котенка за-
крывали на несколько недель. Сигналы от этого глаза поступали в кору. Было
показано, что соответствующие этому глазу колонки глазодоминантности были
очень узкими, в то время как от другого - очень широкими. Таким образом тот
глаз, который был закрыт, представлен в коре головного мозга небольшим числом
нервных связей. Если через месяц глаз открыть, то новые нервные связи уже не
образуются, хотя глаз и нервные структуры не повреждены. Это говорит о суще-
ствовании сензитивного (чувствительного) периода, когда клетки компетентны и
способны образовывать нервные связи, соответствующие получаемому опыту. После
этого периода такие нервные связи не образуются. До того, как были проведены
эксперименты с котятами, когда врачи лечили врожденную катаракту (помутнение
хрусталика) у детей, операцию откладывали на более поздний возраст, когда ре-
бенок легче перенесет операцию. В результате после операции зрение не вос-
станавливалось . После проведения опытов с котятами стало ясно почему - у де-
тей проходил сензитивный период, когда еще был смысл проводить операцию. Если
котятам в течение сензитивного периода закрывать глаза по очереди, то у котят
не формировалось бинокулярное зрение, то есть они видели, но не могли оценить
перспективу.
Есть концепция, основанная на ряде проведенных опытов, что процесс образо-
вания колонок глаза доминантности является конкурентным, идет конкуренция по-
токов сигналов от левого и правого глаза. Так же конкурентный характер носит
формирование связей в других участках коры, принимающих сигналы от других ре-
цепторов . Представление о конкурентном формировании нервных структур мозга
разработана Джеральдом Эдельманом (лауреат Нобелевской премии за серию работ
по структуре антител). Эдельман показал, что при поступлении сигналов между
нейронами, которые активируются одновременно, образуются нервные связи. Они
образуют группы нейронов, которые на следующем этапе обучения животного,
образуют группы более высокого порядка и так далее.
Схема различных типов связей в группах нейронов
Group 5
Edelman 1987
Таким образом, в результате приобретения жизненного опыта изначально одина-
ковые нейроны делятся на группы, которые взаимодействуют и работают вместе
при поступлении какого-либо сигнала. Например, если одна группа нейронов от-
вечает за вкус, другая - за цвет, третья - за запах, четвертая - за форму, то
при возникновении связей между этими группами можно создать целостное пред-
ставление о яблоке. На каждом этапе образование нервных связей между группами
нейронов идет за счет конкуренции, то есть если сигналы не поступают, то
нервные связи отмирают, и наоборот. Свою концепцию Эдельман назвал нейродар-
винизмом, так как процесс конкурентного отбора нервных связей очень похож на
естественный отбор, происходящий в процессе эволюции и образования видов.
Следующая серия экспериментов была проведена с совами Э.Кнудсеном. У сов
очень хорошо развита зрительная и слуховая система. На совенка надевали приз-
матические очки, в которых все образы смещались на двадцать градусов в сторо-
ну. Происходило рассогласование между слуховыми и зрительными сигналами. Че-
рез некоторое время после того, как зрительная кора сделала пересчет на два-
дцать градусов назад, слуховая система подстроилась под зрительную, и совенок
без проблем ловил мышей. Но это происходило только в том случае, если совенку
было не больше 70 дней. Если после этого очки снять, то слуховая кора может
опять подстроиться под зрительную, но только если совенку не больше 200 дней.
Однако если совенка держать в вольере, где много других сов, сензитивный пе-
риод увеличивался, они больше времени были способны к обучению. Их этих экс-
периментов был сделан вывод: богатая сенсорная среда продлевает сензитивный
период.
Похожие опыты проводились и на крысах. Оказывалось, что у крыс, у которых
была интересная, богатая среда, в коре сформировано больше нервных связей, у
них более разветвленные дендриты на аксонах, на нейронах больше синапсов. Уже
давно известно, что с младенчества ребенку надо давать много разных игрушек,
погремушек, ярких картинок, чтобы он тренировал тактильные, зрительные и слу-
ховые системы восприятия, у таких детей в дальнейшем лучше развиваются интел-
лектуальные способности.
В следующей серии экспериментов котят помещали в так называемую вертикаль-
ную среду (котята сидели в темноте, и свет зажигали ненадолго, при этом в
пустом помещении имелись только вертикальные полоски на стенах). По окончании
сензитивного периода их помещали в обычную среду. Оказалось, что такие котята
не видят горизонтальные предметы, то есть если швабра стоит, то котенок может
ее обойти, если она лежит, то он на нее натыкается. Это происходит потому,
что в коре не образовались связи, реагирующие на горизонтальные предметы. То
есть сигнал, поступивший через глаза и далее через боковое коленчатое тело,
просто не может быть обработан в коре, он не распознается. Этот пример гово-
рит о том, что животное, в частности человек, который в сензитивный период
(до 3-5 лет) не получил богатый сенсорный опыт, будет ограничен в развитии
своих интеллектуальных способностей. Пример - дети-Маугли, которых находили в
возрасте 5-7 лет, не способных уже научиться говорить.
Каким образом изучают гены, связанные с поведением? Их изучают и животных и
у человека. На прошлых лекциях упоминалось о том, как степень наследственной
детерминации признака изучается на близнецах, монозиготных и дизиготных.
Здесь представлено сравнение конкордантности (совпадения признака) по росту
у моно- и дизиготных близнецов.
Рост в парах близнецов
Монозиготные
1М
1В5 .
Т ’80
<2 175 *
S
й 170
^165 »,
180 •’
156 --------------------------------
155 165 175 185 105
Близ кец 1 (см)
Дизиготные
8
и
X—<
Г-1
Я
i
g
195
190
185
180
175
1 70
165
160
155
Близнец 1 (см)
165 175 1В5
Видно, что у монозиготных близнецов совпадение по росту заметно больше, чем
у дизиготных, из чего можно сделать вывод, что вклад наследственности велик.
Поведение больше подвержено внешнему влиянию, но все же и здесь можно усле-
дить , хоть и более слабое, но заметное влияние наследственности на поведение.
В данном случае брался такой признак, как стремление к новизне, которое уста-
навливается на основе психологического тестирования и оценивался в баллах.
Видно, значения признака у монозиготрных близнецов ближе, чем у дизиготных.
Стремление к новизне в парах близнецов
Монозиготные
Дизиготные
Близнец 1 (баллы по опроснику)
26 28 30 32 34 36 38 40 42 44 46 48
Близнец 1 (баллы по опроснику)
На прошлых лекциях уже рассказывалось о гене DRD4, ассоциированном со
стремлением к новым впечатлениям. Длинный аллель этого гена с повышенной
частотой встречается в семьях больных с наследственной формой алкоголизма, и
он ассоциирован с «модным» детским диагнозом - синдром гиперактивности с на-
рушением внимания. Дети с таким диагнозом в школах не могут усидеть за парта-
ми. Любопытно, что это заболевание эффективно лечится без всяких таблеток на
тренажерах с обратной связью. Детям показывают мультфильм на экране компьюте-
ра, и мультфильм выглядит резко, когда они внимательны. Внимательность фикси-
руется с помощью энцефалограмм, и в зависимости от внимательности детей изме-
няется резкость мультфильма.
Г ен р ецепт ор а до фл мнил [j RD 4
участвует в работе системы положительного подкрепления
Встала z экзсне 3
Разные аллели гена 2R
I3R
4R
вставки (от 2 до 11 повторов sr
4S-нуклеотидных) 6R
7R
8R
Аллель с 7 повторами ассоциирован с повьпненным уровенем
стремления к новым впечатлениям, а также с синдромом
гиперактивности с нарушением внимания
Имеется генетический вклад и в развитие наркотической зависимости - конкор-
датность монозиготных близнецов по злоупотреблению и зависимости от кокаина
выше, чем у дизиготных.
НАСЛЕДСТВЕННОСТЬ н НАРКОЗ АВИСИМОСГЬ
Исследовали 435 монозиготных и 335 дизиплных близнецов
Среди них пробовали когаин 14%, зависимых 2.3%
Уровень канкор дантгостм
Употребление Злоупотребление Зависимость
IvE 54% 47% 35%
ДЗ 42% 8% 0%
гКсаЛНм-,ККвГгакпИГ А.. ar. atanaaod ddpadacabi в
bwd iMkfic uTknadtf МииУВг J FaycHirv I Ив Ос±1Т:ЬЛ*3-Ж1
Действие кокаина было изучено на крысах. У крысы с сформированной кокаино-
вой зависимостью нейроны, опосредующие действие кокаина, имеют больше синап-
сов , чем у нормальных крыс. То есть, кокаин оказал на крыс такое же действие,
как обучение. То есть, человек или крыса, которая пользовалась наркотиком,
прошел «обучение», чтобы реагировать на наркотик, и у него сформировались па-
тологические нервные связи, которые делают для него полученный опыт легко
восстановимым, потому что нервные связи уже есть. А другие нервные связи, ко-
торые бы в норме обеспечивали ему приятные ощущения от полезных для здоровья
переживаний, из-за конкурентного формирования оказываются ослабленными. То
есть использование наркотиков, особенно в раннем возрасте, меняет морфологию
и анатомию нейронов, структуру коры головного мозга, и уклоняет развитие с
нормального пути.
Влияние кокаина на нейроны
Крысы с сформированной кокаинов ей зависимостью имеют больше
синапсов, чем нормальные в зонах коры (nucleus accumbulens and
pteftonatl cotex), связанных с действием кокаина
Как известно, сигнал в нейронах передается в два этапа. На первом этапе пе-
редается электрический импульс вдоль отростка нейрона. При этом происходит
поляризация-деполяризация мембраны, потому что поперек мембраны идет поток
ионов (К, Мд и т.д.). На участке контакта между двумя нейронами имеется
структура, которая называется синапс. Это тесный контакт меду двумя клетками,
а между ними имеется синаптическая щель. В эту щель при деполяризации мембра-
ны передающего сигнал нейрона попадают нейромедиаторы (посредники передачи
нервного импульса). На мембране воспринимающего нейрона «сидят» рецепторы,
которые воспринимают сигнал, опять происходит поляризация-деполяризация мем-
браны и сигнал передается дальше. Ниже показаны некоторые вещества, которые
выступают в роли нейромедиаторов: Глицин, ацетилхолин, у-аминомаслянная ки“
слота, гистамин, серотонин, дофамин и т.д. Структура некоторых нейро-
медиаторов представлена на рисунке.
[1' 1ирозин-3-монооксигеназа (Fe^-.THB) 1.14.16.2& дофамин-Р-монооксигеназа (Си) 1.14.17.1
2 декарбоксилаза ароматических-1-аминокис/ют m /ы»нип-»танппамин.м-мятиптпамг<ы>па-«» 9 i » од
(дофа-декарбоксилаза) (PLP) 4.1.1.28 Ш Ф®нилэтаноламин-П метилтрансфераза 2.1.1.28
К примеру, дофамин образуется из тирозина, и в результате последовательно-
сти реакций получается адреналин. Существуют последовательности реакций, в
которых происходит синтез нейромедиаторов и их распад. Рассмотрим это на
примере дофамина. В пресинаптическом нейроне синтезированный дофамин упаковы-
вается в пузырьки, которые, в ответ на поступивший сигнал, сливаются с преси-
наптической мембраной и выпускают дофамин в синаптическую щель. Поступивший в
синаптическую щель дофамин связывается с рецепторами на постсинаптической
мембране и с ауторецепторами пресинаптического нейрона. Связывание нейроме-
диатора с рецепторами постсинаптического нейрона «включает» трансмембранный
поток ионов, что приводит к изменению Заряда мембраны нейрона. Изменение За-
ряда (область переполяризации мембраны) распространяется вдоль аксона, что
собственно и представляет собой нервный импульс, и сигнал идет дальше следую-
щему нейрону. Через ауторецепторы синтез дофамина в пресинаптическом нейроне
ингибируется. Неиспользованный дофамин транспортируется обратно в пресинапти-
ческий нейрон или окисляется ферментами, работающими в синаптической щели
(катехоламин-о-метилтрансфераза, моноаминоксидаза и другие).
Другой нейромедиатор, серотонин, работает сходным образом. Есть нейрон, ко-
торый передает сигнал, и есть нейрон, который его принимает. Передающий ней-
рон синтезирует серотонин. Когда поступает сигнал, серотонин выпускается в
синаптическую щель и связывается с рецептором на мембране воспринимающего
нейрона.
ПРЕСИНАПТИЧЕСКИЙ
ПОСТСИНАПТИЧЕСКИЙ
НЕЙРОН
После того, как нейромедиатор серотонин поступил в синаптическую щель,
часть его была использована для запуска сигнала и деполяризации постсинапти-
ческой мембраны, а часть (также как и в случае с дофамином) осталась «бол-
таться» в синаптической щели. Во-первых, этот неиспользованный серотонин раз-
рушается ферментами. Но система устроена настолько экономично, что на преси-
наптической мембране (то есть на мембране подавшего сигнал нейрона) «сидит»
специальный белок - транспортер серотонина, который, как хорошая хозяйка, за-
кладывает обратно на хранение неиспользованный серотонин. Оказалось, что ге-
нетически детерминированы две формы этого белка-транспортера, одна работает
быстро, а другая медленно. Ниже показано, чем на уровне ДНК отличаются эти
две формы. Оказалось, что различие между ними вызвано различием в регулятор-
ной области гена.
tqcaqcccccccagcatctcccctgcacccccaqcatccccc
... ctgcaccccccagcatcccccc tgcagcccttccagcat...•
Вставка в регуляторном участке гена транспортера серотоюма
("длинный аллель").
Р У гомозигот по длинному аллелю уровень синтеза данного белка в 2 раза выше, |
g чем у гомозигот по короткому аллелю, что ассоциируется с повышенной
склонностью к депрессии при небл агоприягных жизненных событиях.
Если белка-транспортера много, то серотонин поступает в синаптическую щель,
и тут же начинает закачиваться обратно. Рецепторы, даже не успевают толком на
него среагировать - это случай склонности к депрессии и, возможно, повышенной
частоты самоубийств. Лекарства от депрессии блокируют работу транспортера се-
ротонина и дают возможность серотонину сделать свою работу.
Для исследования связи между генотипом по гену серотонина и склонностью к
депрессии провели следующий эксперимент. Выборку молодых людей до 26-летнего
возраста опросили о том, какие тяжелые переживания достались на их долю, и о
том, бывает ли у них депрессия, а затем определили, какие аллели он по гену
транспортера серотонина они несут. Оказалось, что существует связь между пе-
ренесенными стрессами и сообщениями о депрессии в зависимости от того, какой
вариант гена транспортера серотонина они несли. Люди с высокоактивным бел-
ком-транспортером серотонина чаще испытывают депрессию по мере накопления не-
гативных событий в их жизни, чем те, у кого транспортер обладает низкой ак-
тивностью. Если же негативных событий в жизни было мало, то различия между
людьми с разными генотипами не были обнаружены.
Связь между перенесенными стрессами и сообщениями о депрессии в
возрасте 26 лет в зависимости от генотипа
по гену транспортера серотонина
12 50 л
10.00 н
гетерозиготы
7.50 d
5 00
2
ю
5-HTTgens
SS,n»146
Ф
LL,n»264
Ф
О
О 00 +
о
2
Увеличивающееся число негативных событии
в возрасте 21-26 лет
Сейчас выявлено около 60 генов, регулирующих передачу нервного импульса в
разных системах мозга.
Гены,
влияющие на передачу нервного импульса
Дофамин
рецепторы дофамина D1-D5 (DRD1, DRD2, DRD3, DRD4, DRD5)
и транспортер дофамина DAT 1 (отрз-стЕп^пщувгг дц,
,-ч D2,14, D5 - ингИбгшувгт)
Серотонин
рецепторы серотонина (гены HTR1A, HTR1DB, HTR2A, HTR2C)
транспортер серотонинаHTT(SLC6A4)
триптофан 2,3-диоксигеназа (TD О 2)и тригл о фант идроксилаза (ТРН)
Норэпинефрин
дофамин гидроксилаза (DBH)адренергические рецепторы (ADRA2A,
ADRA2C), транспортер норэпинефрина NET (SLC6A2), фенилзтаноламин
Ы-метилтрансфераза (PNMT)
ГАМК
GABA -рецепторы (GABRA1, GABRA3, GABRB1, GABRB3)
катеиоламин-о-метилтрансфераза (СОМР)
моноамин оксидаза А (МА О А)
lTMBA-рецептор (NMDAR1),
ацетилхолиновые рецептор а4 (CHRNA4),
Выше упоминался фермент моноаминоксидаза А (МАОА), который разрушает ней-
ротрансмиттеры, оставшиеся в синаптической щели. Этот ген привлек внимание
ученых, когда была обнаружен семья с мутацией в этом гене. На прием к врачу
пришла женщина, которая хотела иметь ребенка, но боялась, что ребенок будет
болен, - у нее в семье мужчины (не все, но многие) отличались высокой агрес-
сивностью (немотивированная агрессия и легкая степень умственной отсталости).
Когда они были обследованы, оказалось, что это не просто дурной характер, а
генетически детерминированное заболевание. Родословная этой семьи представле-
на на рисунке (темные кубики - больные мужчины, кружочки с ромбиком в центре
обозначают женщин-носительниц мутации).
Однонуклеотидная замена T=>G в гене МАОА в
семье с частым агрессивным поведением мужчин
X-сцепленное наследование признака
По характеру наследования поняли, что это заболевание, сцепленное с X-
хромосомой, потому что здоровые женщины рожали больных сыновей (дочери рожда-
лись нормальными, потому что у них две Х-хромосомы, и в одной ген мутантный,
но в другой - нормальный). На Х-хромосоме нашли мутантный ген, оказалось, что
он кодирует МАОА. Мутация - единичная нуклеотидная замена, которая вела к
обрыву синтеза белка и МАОА при этом была нефункциональна. Таким образом, за-
мена одного нуклеотида (цитозина на Тимин в данном случае) приводила к изме-
нению поведения.
Этот ген стали более активно исследовать. Такая мутация, как в этой семье,
полностью инактивирующая фермент, больше нигде не была найдена, но были обна-
ружены мутации, снижающие активность МАОА. Исследовали группу людей с низкой
и нормальной активностью МАОА и сравнили количество случаев асоциального по-
ведения (нападение, нападение с целью нанесения повреждений, убийство, из-
насилование, жестокость по отношению к животным). Когда просто так исследова-
ли выборку, разницы не нашли. Но когда эту выборку разделили по условиям вос-
питания в детстве, оказалось, что разница в поведении людей, которые воспиты-
вались в благоприятных условиях, практически отсутствует, а вот если условия
были плохие, то разница становится существенной.
СВЯЗЬ АНТИСОЦИАЛЬНОГО ПОВЕДЕНИЯ.
УСЛОВИЙ ВОСПИТАНИЯ И ГЕНОТИПА ПО ГЕНУ МАОА
ИНДЕКС АНТИСОЦИАЛЬНОГО
ПОВЕДЕНИЯ
•Нападение 0 75 -
•Нападение с целью
нанесения повреждений
•Убийство
•Изнасилование
0.25 -
•Нападение на женщин)'
•Жестокость по
отношению ®
к животным
-0.25 -
Низкая активность ЫАОА
Нет Возможно Тяжелые
ПЛОХИЕ УСЛОВИЯ ВОСПИТАНИЯ
Для изучения связи моноаминоксидазы А с поведением провели серию экспери-
ментов на мышах. У мышей «нокаутировали» (то есть инактивировали) ген МАОА.
Такие мыши были очень агрессивны, они набрасывались на своих сородичей без
всякого повода, то есть вели себя также как и люди с аналогичным генетическим
дефектом. Эти исследования помогают понять, что именно в поведении детермини-
ровано генетически и в какой мере, а что подвержено влиянию среды.
Другая серия исследований была посвящена тому, как формируется память и ка-
ким образом в этом процессе участвуют гены. На мышах была проведена серия
экспериментов. Были исследованы глутаматные рецепторы. Они работают в зоне
мозга, которая связана с ориентацией на местности. Нобелевский лауреат Сусу-
ми Тонегава получил мышей, мутантных по рецептора глутамата, и исследовал их
способность к запоминанию. В разных мышиных тестах мутанты не отличались от
нормальных собратьев, но в тесте на запоминание положения предметов оказались
"двоечниками". Мышей запускали в ванну с платформой, на которую можно было
встать. Непрозрачная вода не позволяла узнать где платформа, пока мышь не на-
ткнется на нее случайно. Обычно мыши, несколько раз побывав в ванне, запоми-
нали, где находится платформа и сразу плыли к ней. Мутанты не могли запомнить
даже после десятков повторений. Их "географический кретинизм" связан с мута-
цией в рецепторе. Изменив этот рецептор, удалось получить и мышей-"отличниц".
Дело в том, что существуют как минимум два типа глутаматных рецепторов.
Один тип работает у молодых мышей и в ответный на сигнал дает сильный ионный
ток через мембрану и, следовательно, сильный и дольше действующий потенциал.
Но в определенном возрасте, по мере созревания, рецептор меняется, и вместо
белка-рецептора, который давал сильный ионный ток, в мембранах уже находится
рецептор, который дает слабый и менее продолжительный ток. Это связано с тем,
что интенсивное обучение нужно проходить в молодости, а с возрастом животное
становится более консервативным, повторяет то, что выучило в молодости. Ис-
следователи ввели в геном мышей мутацию, в результате которой синтез
"юношеского" белка усилился в несколько раз. Мутанты лучше запоминали и рас-
познавали звуки, объекты и их положение в пространстве, быстрее справлялись с
тестами. Тем самым, им как бы продлили сензитивный период. Может возникнуть
вопрос, почему способность к обучению угасает с возрастом, то это уже отдель-
ный эволюционный вопрос.
Каким же образом формируются навыки, то есть, новые синаптические связи? В
серии экспериментов на мухах и улитках морских зайцах (аплизиях - их очень
любят нейробиологи), были расшифрованы механизмы формирования кратковремен-
ной и долговременной памяти. За эти работы Эрик Кенделл получил Нобелевскую
премию
Допустим, улитке подали электрический ток на хвост. Ей нужно хвост отдер-
нуть . Каким образом это происходит.
Молекулярногенетические механизмы
формирования условного рефлекс»
СЕНСОРТНЫЙ НЕЙРОН
Кратко»
память
МОТОНЕЙРОН
Прэдажителъность реакции 1 — минуты, 2 - часы, 3 - формирование нзвых синапсов
От обиженного хвоста поступил сигнал в виде серотонина. Серотонин связыва-
ется с рецептором на мембране сенсорного нейрона. Именно здесь происходит
этап обработки информации и принятии решения. Рецептор взаимодействует с аде-
нилатциклазой, которая синтезирует циклический аденозинмонофосфат (цАМФ). По-
следний взаимодействует с киназой (киназы - это белки, которые фосфорилируют
другие белки). Киназа фосфорилирует кальциевые каналы в мембране, через них
идет ток, мембрана деполяризуется, что является сигналом к выбросу нейроме-
диаторов в синаптическую щель. Нейромедиатор связывается с рецептором на
постсинаптической мембране мотонейрона, и мотонейрон дает мышцам команду от-
дернуть хвост от неприятного раздражителя. Это - кратковременная память (ра-
ботает 3-4 минуты).
Если раздражение продолжает поступать регулярно, то эта реакция - долго-
срочная память (работает 12-24 часа). В этом случае продолжает синтезировать-
ся цАМФ, то фрагмент киназы перемещается в ядро и активирует здесь ген, моди-
фицирующий киназу - отщепляющий от нее кусочек таким образом, что она стано-
вится перманентно активной. То есть, циклический аденозинмонофосфат ей для
активации становится не нужен. Это - долговременная память.
Если сигнал продолжает поступать и дальше, то включается следующий меха-
низм. Большие количества фрагментов киназы активируют фактор транскрипции,
запускающий работу группы генов, обеспечивающей синтез белков и образование
нового синапса. Это - память на всю жизнь, именно она должна работать при
обучении.
ЛЕКЦИЯ №24.
ЭВОЛЮЦИОННАЯ ТЕОРИЯ ПОЛА.
БИОТЕХНОЛОГИИ. ИММУНОЛОГИЯ.
ПЕРЕДАЧА СИГНАЛОВ В ОРГАНИЗМЕ.
ЭВОЛЮЦИОННАЯ ТЕОРИЯ ПОЛА1
Теория полового отбора Дарвина вызывала споры, неоднократно высказывалось
мнение, что она является самым слабым местом дарвиновского учения. После Дар-
вина проблемой пола занимались крупнейшие биологи, однако современные автори-
теты продолжают говорить об отсутствии удовлетворительного объяснения того,
почему возник и сохраняется пол. Например, в книге канадского ученого Белла,
вышедшей в 1982 г. , читаем: "Пол - главный вызов современной теории эволю-
ции. . . Интуиция Дарвина и Менделя, которые осветили так много загадок, не
смогла справится с центральной загадкой полового размножения. В чем функцио-
нальное значение пола, которое приводит к его сохранению в биологических по-
пуляциях? "
Путь к решению проблемы был предложен в середине 60-х гг. в новой концепции
В.А. Геодакяна, дающей объяснение эволюционного смысла деления на два пола.
Существование двух полов чаще всего связывают с размножением. Верно ли это?
Древнейшим способом размножения является бесполое, осуществляющееся путем
простого деления клетки. Оно широко распространено у одноклеточных организмов
(бактерии, сине-зеленые водоросли, амебы, инфузории и др.). При бесполом раз-
множении не нужно искать партнера. Однако при таком способе размножения из-
менчивость достигается только за счет мутаций. Значительное увеличение измен-
чивости за счет комбинирования признаков различных особей обеспечивает поло-
вой процесс (скрещивание). Процесс скрещивания лежит в основе половых спосо-
бов размножения. Если брачным партнером особи может быть любая из остальных,
такой способ полового размножения называется гермафродитным. К гермафродитам
относятся черви, пиявки, усоногие ракообразные, многие моллюски, некоторые
рыбы. Следующим шагом в эволюции размножения стало раздельнополое размноже-
ние , при котором популяция делится на два различающихся между собой по ряду
признаков пола.
Сравним эти способы размножения по двум параметрам: количественной эффек-
тивностью и комбинаторным возможностям. В количественном отношении самая вы-
сокая эффективность принадлежит бесполым формам, которым не надо искать парт-
нера. При половом размножении эффективность в два раза меньше. Значит, с по-
явлением скрещивания количественные преимущества утрачиваются. Однако появля-
ются практически неисчерпаемые возможности быстрого обмена полезными мутация-
ми, обеспечивающими комбинаторное богатство популяции. Но по этому показателю
у половых форм явное преимущество принадлежит гермафродитам, где при скрещи-
вании нет ограничений по полу партнера.
Выходит, раздельнополое размножение уступает двум предыдущим формам или по
комбинаторному потенциалу, или по количественной эффективности. Тогда встает
вопрос, почему же все прогрессивные в эволюционном плане формы животных (мле-
копитающие, птицы, насекомые и другие) раздельнополы. Среди растений просле-
1
С.А.Боринская
живается та же тенденция - более поздние формы дифференцированы по полу (дву-
домные растения). Раздельнополый способ размножения появился в процессе эво-
люции в очень далеких друг от друга группах. Парадокс? Но известно, что в
эволюции ничего не бывает зря. Зачем же эволюции понадобились два пола?
Как совместить устойчивость
и изменчивость
Весь окружающий нас мир можно представить как совокупность разнообразных
систем. Каждая из них существует в определенной среде. Эволюционирует как
среда, так и система. Среда диктует системе направление эволюции. Каждая эво-
люционирующая система должна быть устойчивой, консервативной, сохраняя накоп-
ленную полезную информацию, и одновременно изменчивой, способной адаптиро-
ваться к меняющимся требованиями среды.
Как же снят этот конфликт? Возможны два решения. Первое - поддерживать
свойства системы к изменчивости в некотором промежуточном, оптимальном со-
стоянии. Однако это создает для системы довольно узкие рамки существования.
Второе решение - разделить систему на две части, подсистемы. Одна из этих
частей - консервативная - убирается подальше от среды (не в геометрическом, а
в информационном плане) для сохранения полезной информации, а другая - измен-
чивая, оперативная - воспринимает и проверяет новую информацию. Такое разде-
ление на две сопряженные подсистемы (консервативную и оперативную) повышает
устойчивость системы в целом. Но ведь популяция особей - это живая система,
эволюционирующая в изменчивой среде и нуждающаяся в повышенной устойчивости.
Логично предположить, что разделение ее на две части - два пола - и обеспечи-
вает такую устойчивость. Но тогда один из полов должен быть консервативен,
представлять собой "ядро" системы, а второй должен быть оперативной, изменчи-
вой "оболочкой". Таким образом, разделение на два пола оказывается выгодной
для популяции формой получения информации от среды и сохранения информации о
собственном строении. Возникает вопрос, кто есть кто в системе "мужской -
женский пол", какой из полов специализирован на получении информации, а какой
- на сохранении? Однако прежде чем ответить на этот вопрос, необходимо выяс-
нить, в чем суть различий между полами.
Репродуктивная стратегия
Женщина не может иметь в год более одного ребенка (если только у нее не ро-
ждаются близнецы). То есть за весь репродуктивный период своей жизни (с 13 до
45-55 лет) они способна родить примерно три десятка детей.
Возможности мужчины гораздо больше. Он может стать отцом 4-5 тысяч детей.
Рассмотрим популяцию, в которой особи разного пола скрещиваются хаотически.
Понятно, что количество потомства пропорционально количеству женских особей.
А вот качество зависит от количества мужских особей - ведь чем их больше, тем
шире возможности выбора "самых качественных".
Это значит, что при неблагоприятном воздействии среды сокращение количества
самок приведет к снижению численности потомства, а сокращение количества сам-
цов - к изменению его качества.
Мужчины отличаются друг от
друга больше, чем женщины
(дисперсия признаков)
Совокупность всех генов организм составляет его генотип. В процессе индиви-
дуального развития организма его генотип взаимодействует со средой, определяя
формирование внутренних и внешних признаков индивида. Совокупность всех этих
признаков составляет его фенотип.
Генотип - это программа, которая в разных условиях среды может реализовы-
ваться по-разному. В генотипе записано не жестко определенное значение при-
знака, а диапазон его возможных значений. В онтогенезе (процессе индивидуаль-
ного развития от зарождения до конца жизни) реализуется один самый подходящий
для данной конкретной среды фенотип. Следовательно, генотип задает диапазон,
а среда выбирает "точку" внутри него. Ширина этого диапазона называется нор-
мой реакции организма на воздействие окружающей среды.
Есть признаки, например, группа крови или цвет глаз, влияние среды на кото-
рые сводится практически к нулю. По другим признакам (например, интеллекту-
альным способностям) норма реакции очень широкая, поэтому их часто связывают
только с влиянием среды, то есть воспитанием. Третьи признаки (рост, масса)
занимают промежуточное положение. Норма реакции у женского пола значительно
шире, чем у мужского.
Возможный
диапазон
фенотипического
проявления
признака
Среднее
значение
признака,
заданное
генотипом
Рис. 1 Норма реакции
Это положение проверено при сравнении близнецов. Монозиготные братья должны
быть больше похожи друг на друга, чем монозиготные сестры. Но если близнецы
не монозиготные, то больше сходство должно быть у девочек. Иными словами,
женский пол больше поддается влияниям среды, а мужской влиянию генотипа.
Рис 2 Распределение генотипов (внизу) и фенотипов (вверху) в стабильной среде
На рисунке 2 на графике А показано распределение фенотипов у мужского и
женского пола. По горизонтальной оси отложены значения признака, а по верти-
кали - частота появления особи с данным значением признака. Так как люди со
средним значением признака - например, среднего роста, встречаются чаще, чем
карлики или великаны, кривая имеет вид колокола и у женщин, и у мужчин. Такую
же колоколообразную кривую можно получить, измеряя успеваемость учеников в
школе: выявим некоторое число отличников и двоечников, основное же количество
- среднеуспевающие. Кривые распределения признаков мужского и женского пола
отличаются - у мужского пола кривая более пологая, то есть отражает большую
дисперсию признаков. Из такого распределения можно было бы сделать вывод, что
если среди мужчин больше гениев, то среди них больше и полных идиотов.
Распределение генотипов при этом одинаково для обоих полов и показано на
графике Б. По горизонтали отложены средние значения какого-либо признака, оп-
ределяемые генотипом, а по вертикали - частота появлений генотипа с тем или
иным значением.
Из-за узкой нормы реакции (то есть низкой способности поддаваться воздейст-
виям среды) у мужского пола распределение мужских фенотипов примерно повторя-
ет распределение определяемых генотипами средних значений. Для женского же
пола картина иная. Из-за широкой нормы реакции женские фенотипы более подвер-
жены влиянию среды и ближе к среднему (оптимальному) значению в стабильной
среде. То есть самцы отличаются большей степень разнообразия признаков, чем
самки. Из-за этого первыми жертвами всех неблагоприятных изменений среды ста-
новятся мужские особи. Часть из них (обладатели крайних значений признака)
погибает, поэтому оставшиеся в живых передадут следующему поколению спектр
генотипов, измененный в соответствии с направлением воздействия среды.
Широкая норма реакции женского пола позволяет уходить от вредных воздейст-
вий среды за счет онтогенетической пластичности. Поэтому женский пол сохраня-
ет и передает потомству весь спектр исходных генотипов.
Таким образом, в каждом поколении яйцеклетки широкого разнообразия, несущие
информацию о прошлом богатстве генотипов, оплодотворяются спермиями узкого
разнообразия, генотипы которых содержат информацию только о самых подходящих
для текущих условий среды признаках. То есть, следующее поколение получает
информацию о прошлом по материнской линии, а о настоящем - по отцовской. Из
чего видно, кто есть кто в системе "мужской-женский пол". Женский пол - ста-
бильное, консервативное "ядро" популяции, а мужской - лабильная "оболочка".
Мужской пол выступает как эволюционный "авангард", принимающий на себя удары
меняющейся среды. При этом за информацию приходится платить гибелью части
мужских особей. Отсюда и повышенная смертность мужского пола.
Мы пришли к выводу, что два пола существуют не только для размножения - оно
возможно и бесполым путем. Два пола это два канала информации, один (женский)
- от предшествующих поколений, другой (мужской) - от среды. Их взаимодействие
обеспечивает системе - эволюционирующей популяции - успешное функционирова-
ние .
Природа не бережет мужчин
(соотношение полов)
Повышенная чувствительность и смертность мужского пола всегда была одной из
Загадок. Из 163 стран мира, по которым есть данные, в 152-х женщины живут в
среднем дольше мужчин. По миру в целом средняя продолжительность жизни женщин
(59,3) на 3,6 года больше, чем у мужчин (55,7).
Повышенную смертность мужчин пытались объяснить социальными причинами, та-
кими как алкоголизм, курение, опасные профессии, рискованное поведение и т.д.
Но оказалось, что аналогичная картина смертности наблюдается у большинства
животных и даже растений, при чем на всех этапах - от зачатия до самой смер-
ти. Платить за новую информацию популяции выгоднее мужскими, а не женскими
особями.
Исходя из этого, можно ожидать, что при всех неблагоприятных условиях (рез-
кие изменения климата, голод, войны, переселения) должна повышаться смерт-
ность самцов. Но чтобы популяция в экстремальных условиях не потеряла мужской
пол, это должно сопровождаться одновременным увеличением их рождаемости. То
есть увеличивается "оборачиваемость" самцов, что позволяет раздельнополой по-
пуляции быстрее приспособиться к новым условиям.
Соотношение полов потомства у позвоночных, видимо, регулируется интенсивно-
стью половой деятельности. Чем выше половая активность данного пола, тем
больше появляется потомков того же пола. Этот механизм объясняет многие факты
демографической статистики: "феномен военных лет", избыточное рождение маль-
чиков в "женских коллективах" (гаремы, текстильные города) и девочек в "муж-
ских" (экспедиции, портовые города).
У перекрестноопыляющихся растений связующим звеном является количество
пыльцы, попадающее на женский цветок. Чем больше пыльцы, тем больше женских
потомков, и наоборот.
Почему мужчины отличаются
от женщин (половой диморфизм)
У одних видов самцы почти неотличимы от самок, тогда как у других, в том
числе человека, они имеют различия по ряду признаков. Различие средних значе-
ние по любому признаку мужского пола и женского называется половым диморфиз-
мом (рис. 3).
Половой
диморфизм
Рис. 3 Распределение фенотипов в нестабильной среде
Половой диморфизм - еще одна характеристика раздельнополой популяции, кроме
соотношения дисперсий и численности полов.
Через изменение этих трех характеристик плавно регулируется степень измен-
чивости популяции. В оптимальных условиях стабильной среды, когда нет необхо-
димости что-либо менять, усиливаются консервативные тенденции: снижается рож-
даемость мальчиков, уменьшается дисперсия полов (уменьшается их разнообразие)
и уменьшается половой диморфизм новорожденных. В экстремальных условиях, ко-
гда от популяции требуется высокая пластичность, рождаемость мальчиков увели-
чивается, увеличивается дисперсия признаков (сначала у мальчиков, потом у де-
вочек) , возникает или растет половой диморфизм. Таким образом, эти три пара-
метра могут служить своеобразным индикатором состояния экологической ниши.
За последнее десятилетие в Каракалпакии рождаемость мальчиков возросла на 5
%. Печальное наблюдение. Видимо, это реакция популяции на экстремальные изме-
нения среды в связи с гибелью Аральского моря.
Мужчины - авангард эволюции
Попробуем схематически изобразить процесс эволюции какого-либо признака в
филогенезе, то есть в его историческом развитии. Допустим, что существует не-
кий стабильный признак, по которому нет полового диморфизма, то есть он оди-
наково представлен и у одного, и у другого пола. Но вот произошли какие-то
изменения в среде, и под их воздействием признак начал меняться (рис. 4). Как
мы уже выяснили, первым реагирует мужской пол. Изменения сначала происходят
только у него. Сначала увеличивается дисперсия признака, поскольку для отбора
необходимо разнообразие. Затем под давлением среды начинает меняться и сред-
нее значение признака. Женский пол остается некоторое время (может быть, мно-
го поколений) неизменным. В этот период эволюции популяции появляется и рас-
тет половой диморфизм по данному признаку. Образно выражаясь, мужской пол ве-
дет "авангардные бои" за новые эволюционные позиции, а женский остается на
месте.
Затем перемены затрагивают и женский пол. У него также начинают расти дис-
персия и скорость эволюции, достигая их значения для мужского пола. На этой
фазе эволюция идет параллельно у обоих полов. Половой диморфизм доходит до
своего максимума и остается постоянным.
Когда у мужского пола признак достигает нового стабильного значения, у жен-
ского он еще продолжает меняться. На этой стадии дисперсия и скорость эволю-
ции больше у женского пола (женский пол ведет "арьергардные бои")
признака
прнзни*а
Рисунок 4.
Таким образом, фазы эволюции признака у разных полов сдвинуты во времени: у
мужского они начинаются и заканчиваются раньше, чем у женского. При этом по-
ловой диморфизм это своего рода дистанция между полами на пути эволюции при-
знака. Смысл ее состоит в том, чтобы проверить эволюционные "новости" на муж-
ском поле прежде, чем передать их женскому. Отсюда миссия мужского пола - по-
иск и пробы. Он является той оперативной частью системы, которая делает на-
ходки, но неизбежно при этом чаще совершает ошибки. В консервативную, жен-
скую, часть системы должны попасть только находки. Поэтому миссия женского
пола - сохранение и закрепление находок. Это придает ему черты совершенства,
которые, однако, оплачиваются неизбежной эволюционной инерционностью, тогда
как новаторство и прогрессивность мужского пола сочетаются с его несовершен-
ством .
А теперь можно сделать ряд важных выводов.
Половой диморфизм существует только во время эволюции признака: появляется
с ее началом, сохраняется, пока она идет, и исчезает, как только эволюция за-
канчивается. Значит, если половой диморфизм отсутствует, то признак стабилен
и эволюция по нему не идет, а если он существует, то признак находится на
"эволюционном марше". То есть, наличие полового диморфизма может служить кри-
терием эволюции признака.
По половому диморфизму можно определить направление эволюции признака: как
правило, он меняется от женской формы к мужской.
Наконец, можно судить о фазе эволюционного процесса: только он начался или
уже завершается. На это указывает соотношение дисперсий признака у мужского и
женского пола. Таким образом, получаем своего рода хронометр эволюции.
Половой диморфизм -
"последние новости" эволюции
Все признаки можно разделить на три группы по степени различия между пола-
ми. К первой группе отнесем те признаки, по которым между мужским и женским
полом нет никакой разницы. К ним относятся качественные признаки, которые
проявляются на уровне вида - общий для обоих полов план и принципиальное
строение тела, число органов и многие другие. Половой диморфизм по этим при-
знакам в норме отсутствует. Но он наблюдается в области патологии. У девочек
чаще проявляются атавистические аномалии (возвраты или остановки развития), а
у мальчиков - футуристические (поиск новых путей). Например, среди 4000 ново-
рожденных детей с тремя почками девочек было в 2,5 раза больше, чем мальчи-
ков, а среди 2000 детей с одной почкой было примерно в 2 раза больше мальчи-
ков. Напомним, что у наших далеких предков в каждом сегменте тела имелась па-
ра выделительных органов - метанефридиев. Следовательно, три почки у девочек
- это возврат к предковому типу (атавистическое направление), а одна почка у
мальчиков - футуристическая тенденция. Такая же картина наблюдается среди де-
тей со сверхнормативным числом ребер, позвонков, зубов и т.д., то есть орга-
нов, претерпевших в процессе эволюции уменьшение числа - среди них больше де-
вочек . Среди же новорожденных с их нехваткой больше мальчиков.
Сходная картина наблюдается и в распределении врожденных пороков сердца и
магистральных сосудов. У девочек преобладают элементы, свойственные сердцу
эмбриона или филогенетическим предшественникам человека: открытое овальное
отверстие в межпредсердной перегородке, незарастающий Боталлов проток (сосуд,
соединяющий у плода легочную артерию с аортой) и др. "Мужские" пороки чаще
бывают новыми, ни у эмбрионов, ни в филогенезе они не имеют аналогий - это
разного рода стенозы (сужения) и транспозиции магистральных сосудов.
Итак, на уровне видовых признаков половые различия проявляются в области
патологии.
Теперь рассмотрим вторую группу признаков - тех, которые встречаются и у
мужского пола, и у женского, но распределены в популяции с разной частотой и
степенью выраженности. Это количественные признаки: рост, вес, размеры и про-
порции, многие морфофизиологические и этолого-психологические признаки. Поло-
вой диморфизм по ним проявляется как отношение их средний значений. Именно
этот половой диморфизм служит "компасом" эволюции признака.
Например, эволюция большинства позвоночных сопровождалась увеличением их
размеров. Многие же виды насекомых и паукообразных, наоборот, мельчали. Можно
предсказать, что у крупных позвоночных самцы должны быть крупнее самок, а у
мелких насекомых - наоборот.
Те же тенденции наблюдаются и внутри таксономических единиц низшего ранга,
скажем, в классе млекопитающих: у крупных форм чаще крупнее самцы, а у мелких
- самки. Например, самцы африканского слона весят до 6,5 т, а самки - до 3,5
т. У некоторых летучих мышей, белок-летяг, карликовой мангусты, кролика, то
есть у те формы, которые, видимо, эволюционно мельчали, самки более крупные,
чем самцы.
Здесь можно воспользоваться и обратной логикой: если мужские особи крупнее
- вид эволюционирует в сторону увеличения размеров, если мельче - наоборот.
Это правило справедливо также и для растений. Например, существует половой
диморфизм по форме листьев. У тополя женские экземпляры имеют более продолго-
ватые листья, мужские более округлые. Значит, филогенетические предшественни-
ки тополя имели узкие (как у ив) листья.
Наконец, к третьей группе относятся признаки, присущие только одному полу.
Это первичные и вторичные половые признаки: половые органы, молочные железы,
борода у человека, грива у льва, а также многие хозяйственные признаки (про-
дукция молока, яиц, икры и т.д.) Половой диморфизм по ним носит генотипиче-
ский характер, поскольку в фенотипе одного пола эти признаки отсутствуют, Но
наследственная информация об этих признаках записана в генотипе обоих полов.
Поэтому, если они эволюционируют, то по ним должен существовать генотипиче-
ский половой диморфизм. Обнаруживается он в виде реципрокных эффектов.
Исходя из авангардной роли мужского пола можно предсказать существование
реципрокного "отцовского эффекта" (доминирование отца) по всем эволюционирую-
щим, новым признакам. "Отцовский эффект" наблюдается у кур по таким хозяйст-
венно-ценным признакам, как инстинкт насиживания, скороспелость, яйценоскость
и живой вес. У крупного рогатого скота этот эффект наблюдается по удою молока
и продукции молочного жира.
У человека к эволюционно молодым признакам, видимо, можно отнести все соци-
альные, психологические характеристики, связанные с эволюцией мозга (в первую
очередь асимметрия полушарий, речь, абстрактное и логическое мышление, про-
странственное воображение, юмор и другие творческие способности).
Таким образом, по всем признакам животных и растений, по которым наблюдает-
ся половой диморфизм, его можно рассматривать, как эволюционные "последние
новости", уже попавшие к мужскому полу, но еще не дошедшие до женского. При
этом, как мы уже отметили, по видовым (качественным) признакам закономерность
проявляется в области патологии, по популяционным (количественным) - в норме,
а по половым - в виде реципрокного "отцовского эффекта".
Если по данному признаку существует половой диморфизм, то в индивидуальном
развитии (с возрастом) признак будет меняться от женской формы к мужской. То
есть для начальной, ювенильной стадии (для человека это возраст до 25 лет)
более характерна женская форма признака, а для взрослой, зрелой (старше 25
лет и до начала старческой инволюции) более характерна мужская форма.
Заключение
До сих пор мы считали, что два пола необходимы для размножения. А оказыва-
ется, что пол - это скорее способ эволюции. Теория эволюции стала основой ми-
ровоззрения современного человека. Необходимую информацию эволюционное учение
получало из трех источников: палеонтологии, эмбриологии и сравнительной ана-
томии. Теперь к ним добавляется четвертый - эволюционная теория пола, дающая
возможность извлекать ценную информацию о процессах индивидуального и истори-
ческого развития организма.
Конечно, закономерности, с которыми нас знакомит теория пола, справедливы
для всего живого. Но все же больше всего нам интересен человек, и в первую
очередь такие его признаки, как темперамент, интеллектуальные возможности,
творческие способности, юмор. Именно эти признаки, как находящиеся на "эволю-
ционном марше", теория описывает наиболее четко.
Считается, что биологическая эволюция человека завершилась 40-50 тысяч лет
назад. Теория опровергает эту точку зрения, давая возможность в перечне при-
знаков человека отличить эволюционирующие признаки от стабильных, неизменных
в данный момент.
Мужчине и женщине в эволюционном процессе отведены разные роли. Знание тео-
рии заставляет отказаться от идеи социальной одинаковости и взаимозаменяемо-
сти полов, и в противоположность ей развивать идею их различия и взаимодопол-
нительности. Не тождественность, но равноправие, не конкуренция и драматизм
неоправдавшихся притязаний, а коалиция и разумный выбор социальной ниши.
Материал подготовлен с использованием следующей литературы :
1. "Два пола: зачем и почему?" Всеросиийский институт повышения квалифика-
ции инженерно-педагогических работников профтех образования. Санкт-
Петербург, 1992. Составитель Е.И.Соколова
2. Геодакян В.А. "Эволюционная теория пола". Журнал "Природа", 1991, N 8,
с.60-69.
3. Геодакян В.А. Теория дифференциации полов в проблеме человека. В кн.:
"Человек в системе наук". М., "Наука", 1989, с. 171-179.
СИЯЮЩИЕ ВЫСОТЫ ГЕНОМНЫХ ТЕХНОЛОГИЙ2
К успехам биологии конца XX века, символизирующим прогресс науки, относятся
открытия, ставшие научными сенсациями, и разработанные на их основе техноло-
гии, которые позволяют манипулировать генным материалом всех живых существ —
от бактерий до человека. Это дает надежду на решение множества стоящих перед
человечеством проблем (разработку новых средств диагностики и лечения болез-
ней, продление жизни человека, обеспечение продовольствием) и в то же время
вызывает опасения, что внедрение биотехнологий может нарушить равновесие при-
родных экосистем и привести к катастрофическим последствиям. Современная био-
сфера есть результат миллиардов лет эволюции. Достаточно ли человеку пятиде-
сяти лет развития генных технологий, чтобы быть уверенным, что его вмешатель-
ство не вызовет глобального экологического кризиса? Чем в связи с этим пугают
людей? Утратой биологического разнообразия в результате неконтролируемой экс-
пансии созданных человеком генных конструкций. Замещением сельскохозяйствен-
ных сортов растений и пород животных немногочисленными генетически модифици-
рованными организмами в результате монополизации источников сортов и пород.
Снижением жизнеспособности будущих поколений людей в результате массового
осуществления медико-генетических мероприятий. Обоснованы ли эти опасения?
Что действительно можно ждать от этих технологий?
Как устроен геном
и как он работает
Программа развития организма записана в его генетическом коде. Геном — это
весь текст данного организма, записанный в ДНК четырьмя буквами-нуклеотидами.
Самый короткий текст из свободно живущих организмов у бактерии микоплазм —
600 тысяч знаков. В геноме человека — 3 миллиарда. Чтобы только пробежать
глазами по собственному генетическому тексту вам потребуется вся жизнь (три
миллиарда секунд).
Генетическая программа каждого организма разбита на отдельные гены — под-
программы, отвечающие за определенную часть жизни клетки. У бактерий от полу-
тысячи до почти десяти тысяч генов, а у человека, мыши и слона лишь немногим
больше — 30-40 тысяч. В каждый момент времени в клетке работает лишь неболь-
шая часть генов, необходимых для жизнедеятельности в данной конкретной ситуа-
ции, остальные «молчат».
Как у человека, так и у бактерии ген состоит из структурной и регуляторной
части. В структурной записана информация о составе синтезируемого с этого ге-
на белка, который, взаимодействуя с другими белками, участвует в построении
клеточных структур и проведении биохимических реакций. В регуляторной части
записано, когда и при каких условиях данный белок должен синтезироваться.
На структурную часть генов у бактерий приходится 80-90% ДНК, остальная ДНК
участвует в регуляции работы генов. У человека ситуация принципиально отлича-
ется. Кодирующие белок участки генома занимают меньше 3%, тогда как остальная
часть выполняет регуляторные и другие, пока неизвестные функции. Система ре-
гуляции работы генов человека (и других млекопитающих) гораздо сложнее. Ника-
ких уникальных биохимических процессов клетки человека не проводят, зато те
процессы (общие для всего живого мира), которые идут, включаются и выключают-
ся в нужное время и в нужной части тела в соответствии с генетической про-
2
С. Боринская, Н. Янковский
граммой. Например, перед человеческим геном, контролирующим переработку лак-
тозы, найдены два регуляторных участка. Один определяет место, другой — время
работы гена. Первый указывает, что ген должен работать только в клетках сли-
зистой кишечника, ведь именно здесь расщепляются поступившие с пищей сахара.
Второй полностью отключает работу гена по окончании периода грудного вскарм-
ливания (у человека в возрасте 3-5 лет), так как в естественных условиях де-
теныши млекопитающих получают лактозу только с материнским молоком, а взрос-
лым особям фермент не нужен. Однако у некоторых людей в этом регуляторном
участке имеется мутация, которая «разрешает» синтез фермента у взрослых. Та-
кие люди могут пить молоко, тогда как у носителей исходного, немутантного ва-
рианта молоко не усваивается, что приводит к расстройству пищеварения.
Записанная в генах человека программа развития реализуется в процессе роста
и деления клеток, от первого деления зародышевой клетки до последнего вздоха
на жизненном пути. Судьба каждой клетки — станет ли она клеткой эпителия или
превратится в нейрон, лейкоцит или эритроцит — определяется тем, какие группы
генов в ней работают. Постоянно работают во всех клетках только так называе-
мые гены «домашнего хозяйства» — то есть те, которые заняты синтезом клеточ-
ных структур, производством энергии, ремонтом молекулы ДНК. Большая же часть
генов обычно бездействует, и необходимы специальные сигналы для того, чтобы
они активизировались. Например, гены, контролирующие форму тела, расположены
на хромосомах несколькими блоками, причем идут один за другим в том же поряд-
ке, в каком и контролируемые ими части тела: сначала гены, которым положено
работать в голове, потом гены грудного отдела, потом те, которые определяют
развитие задней части тела. Включаются они по очереди. Причем эти свойства
генов «домашнего хозяйства» присущи и человеку, и животным. Так, в экспери-
ментах на мухах показано, что если порядок включения генов нарушен, то могут
получиться монстры, каких не придумать и Спилбергу, — с дополнительными нога-
ми вместо антенн на голове или с глазами на брюшке и крыльях. У человека из-
вестные мутации (на латыни «мутация» означает «изменение») в этих генах также
приводят к нарушениям — к изменению положения органов или, например, отсутст-
вию некоторых зубов. Более серьезные нарушения останавливают развитие плода.
Хотя прочтена последовательность нуклеотидов всего генома человека, функции
большинства генов по-прежнему неизвестны. Многие гены в нуклеотидной последо-
вательности выявлены лишь с помощью компьютерного анализа (см. «КТ» # 413), и
их существование следует подтвердить не вычислительными, а экспериментальными
методами. Мы видим текст, но не понимаем, что он означает. Кроме знания
структуры и функций генов, нужно еще представлять, чем отличается их работа в
разных клетках и на разных этапах развития. И еще — знать, как взаимодейству-
ют генные продукты. Порой утрата довольно больших фрагментов генома не приво-
дит к заметным последствиям. А в других случаях замена всего лишь одной буквы
из трех миллиардов приводит к тяжелому заболеванию.
Генные технологии
Теперь мы можем попытаться понять, каким образом генетики вмешиваются в ра-
боту наследственных программ. До появления биотехнологии и методов генной ин-
женерии генетические изменения тоже, конечно, происходили, но шли они совер-
шенно иными темпами. С очень значительными генетическими изменениями связана
вся эволюция жизни на Земле, насчитывающая более трех миллиардов лет. От вре-
мени существования общего предка обезьяны и человека прошло пять миллионов
лет, накопившиеся за это время изменения затронули 1,5% их генетических тек-
стов. Селекционная работа, которую человек вел на протяжении десяти тысячеле-
тий существования производящего хозяйства, также вызвала изменения геномов
культурных растений и одомашненных животных, являвшихся объектом отбора. Да и
сами люди были вынуждены приспосабливаться (в том числе и на генетическом
уровне) к создаваемой ими самими среде обитания.
Заставить ген одного организма работать в геноме другого можно лишь при со-
блюдении определенных условий. Во-первых, к чужеродному гену следует «под-
шить» регуляторные элементы подходящего хозяйского гена с тем, чтобы он вклю-
чился в нужное время в нужной ткани (например, чтобы его продукт секретиро-
вался в молоко у коровы), а также элементы, обеспечивающие его встраивание в
геном или самостоятельное воспроизведение в хозяйской клетке. Во-вторых, нуж-
но обеспечить систему введения генетической конструкции в клетки хозяина.
Технологии «кройки и шитья» генов для всех одинаковы, а вот системы введения
ДНК в клетки организма-хозяина сильно различаются. Сейчас такие системы отра-
ботаны и для микроорганизмов, и для растений, и для некоторых животных, при-
чем существуют методы введения ДНК в клетки, размножаемые в пробирках, и ме-
тоды, пригодные для модификации сформированного организма. Последние исполь-
зуют для генотерапии, то есть лечения наследственных болезней путем введения
человеку «здоровых» генов.
Еще одно условие — работа гена не должна вредить самому организму-хозяину.
Например, устойчивость трансгенного картофеля к колорадскому жуку обеспечена
введением в растительный геном бактериального гена, контролирующего синтез
белка, токсичного для насекомых (причем не для всех, а для определенной груп-
пы) и безвредного для растений, животных и человека. После генетической моди-
фикации полученный уникальный организм следует размножить. Для этого исполь-
зуется клонирование.
Клонирование
Клонирование (от греч. клон — ветвь, побег) — точное воспроизведение того
или иного живого объекта в некотором количестве копий. Этим термином обозна-
чают два совершенно разных процесса — клонирование (то есть получение иден-
тичных копий) фрагментов ДНК и клонирование клеток взрослого организма (то
есть получение группы клеток с одинаковым генотипом).
Клонирование фрагментов ДНК широко используется в молекулярной генетике,
так как изучать небольшой участок (размером сотни или тысячи пар нуклеотидов)
гораздо легче, чем целую хромосому. Для этого изучаемый фрагмент вводят в
клетки микроорганизмов. В частности, в биотехнологии именно с помощью клони-
рования фрагментов ДНК в бактериях получают клетки, продуцирующие нужные ме-
дицине человеческие белки.
Клонирование растений всем известно — это размножение растений черенками. А
эксперименты по клонированию животных впервые осуществили в начале 1950-х го-
дов американские эмбриологи Роберт Бриггс и Томас Кинг (Robert Briggs, Thomas
King) , пересадившие ядро зрелой клетки лягушки в яйцеклетку, собственное ядро
которой было удалено. В России такие эксперименты были проведены даже не-
сколько раньше Георгием Лопашевым, но его результаты не были опубликованы из-
за преследований генетиков в сталинское время. Английскому ученому Джону Гер-
дону (John Gurdon) удалось усовершенствовать методику и добиться того, что из
1-2% яйцеклеток с пересаженным ядром вывелись лягушата. Из остальных яйцекле-
ток или развивались дефектные эмбрионы, или не развивались вообще — слишком
велики были повреждения во время операции по пересадке ядер. Если можно кло-
нировать лягушку, то почему не попробовать сделать это и с другими животными?
В 1997 году появилось сенсационное сообщение о том, что в лаборатории Яна
Вилмута (Ian Willmut, Эдинбург, Шотландия) разработан метод клонирования мле-
копитающих . Эксперименты проводились на овце. Ядро из клетки молочной железы
взрослой особи ввели в яйцеклетку с удаленным ядром и затем активировали ее
посредством электрического удара. Развивающиеся зародыши пересадили в матку
приемной матери, где они оставались до рождения. Из 236 опытов успешным был
только один, в результате которого родилась ставшая знаменитой овечка Долли.
Позже появились сообщения о клонировании других млекопитающих — коровы, козы,
мыши, свиньи.
В принципе, технически можно клонировать и человека, но в этом случае воз-
никают моральные, этические и юридические проблемы. Но даже если удастся осу-
ществить клонирование людей, получить личность, идентичную личности донора
ядра, невозможно. Невозможно получить даже организм, полностью идентичный ис-
ходному по своим биологическим свойствам — для этого пришлось бы точно вос-
произвести условия развития плода и рождения ребенка. А предположения о мас-
совом производстве сверхгениев или сверхпослушных солдат не имеют под собой
никаких оснований. Любые возможности, которые могут быть реализованы при кло-
нировании, будут все равно лежать в границах возможностей человека как биоло-
гического вида.
Сообщения о клонировании животных и появляющиеся время от времени сообщения
о якобы успешных попытках клонирования человека (которые пока ничем не под-
тверждены) привлекают огромное внимание публики. Опросы, проведенные в евро-
пейских странах фондом Progress Educational Trust (Лондон), показали, что об
овечке Долли знают около 90% опрошенных, тогда как об имеющих гораздо большее
значение для людей и уже применяемых на практике генодиагностике и генотера-
пии слышали лишь около половины. Похоже, что клонирование ассоциируется у не-
специалистов с чем-то вроде возрождения душ и порождает страхи, подобные тем,
что были высказаны участниками студенческого митинга в Беркли, во время Меж-
дународного генетического конгресса (1973). Студенты пытались бойкотировать
ученых, обвиняя их в попытках клонировать Ленина, Гитлера, Сталина, Мао Цзэ-
дуна. Как правило, такие опасения возникают из-за недостатка информации. Для
клонирования организмов нужны живые клетки. После смерти целостность ДНК на-
рушается, и она может быть использована для уже упоминавшегося молекулярного
клонирования отдельных фрагментов, но никак не для воспроизведения генетиче-
ски идентичного организма.
Геном человека как
объект генных технологий
Геномы разных людей содержат одинаковые наборы генов, но их генетические
тексты различаются. Различия составляют в среднем один нуклеотид на тысячу
«букв» текста, то есть 0,1%. С генетическими различиями связаны видовые и ин-
дивидуальные биологические признаки каждого организма. Индивидуальные особен-
ности человека затрагивают и устойчивость к инфекциям, и адаптацию к опреде-
ленным климатическим условиям (в частности, цвет кожи является такой адапта-
цией) , и приспособленность к тому или иному типу питания. Образ жизни, к ко-
торому человек генетически и физиологически не приспособлен (включая климати-
ческие условия, уровень физической активности, диету), ведет к болезням. Не-
которые болезни можно вылечить или предотвратить, меняя среду или образ жиз-
ни. Всем известен «бег от инфаркта». Но есть наследственные заболевания, про-
являющиеся вскоре после рождения ребенка и до недавнего времени неизлечимые.
Понимание молекулярных основ развития организма в норме и при патологии по-
зволяет разработать принципиально новые подходы к лечению и профилактике за-
болеваний . Полученная при расшифровке генома человека информация уже привела
к созданию систем диагностики для нескольких сотен наследственных заболева-
ний. Еще двадцать лет назад для большинства из них в справочниках указывалось
«исходное нарушение, приводящее к развитию заболевания, неизвестно». Понима-
ние закономерностей работы генов позволяет обнаружить болезнь еще до проявле-
ния симптомов. Во многих случаях раннее начало профилактического лечения по-
зволяет предотвратить развитие заболевания или отодвинуть начало его проявле-
ния. Например, у одного из десяти тысяч новорожденных встречается серьезное
нарушение обмена веществ — фенилкетонурия. При этом заболевании недостает
фермента, превращающего аминокислоту фенилаланин в другую аминокислоту — ти-
розин. У больных накапливается промежуточный продукт обмена фенилаланина —
фенилпировиноградная кислота. Избыток ее приводит к поражению клеток мозга и
умственной отсталости. Всех младенцев проверяют на наличие этого заболевания.
Если оно выявлено, назначают специальную диету, которая позволяет избежать
или в значительной мере смягчить развитие симптомов.
Диагностика может проводиться даже еще до рождения ребенка. Для этого на
ранних сроках беременности отбирают небольшое количество околоплодной жидко-
сти, содержащей клетки плода. Затем определяют, имеются ли нарушения в гене-
тическом материале этих клеток и не содержатся ли в нем болезнетворные мута-
ции . Такая диагностика может быть проведена еще до имплантации зародыша в
матку, поэтому она называется преимплантационной.
Системы генодиагностики самых распространенных заболеваний, таких как бо-
лезнь Дауна, фенилкетонурия и др., введены в практику медико-генетического
консультирования. Для некоторых заболеваний это позволило значительно снизить
частоту рождения детей, обреченных на неизбежную мучительную смерть.
При выявлении генетических нарушений у плода врач предоставляет информацию
о возможных рисках, но только родители могут решать, прерывать беременность
или нет. Появление новых методов сделало необходимым обсуждение этических
проблем и принятие соответствующих законов, защищающих права и достоинство
человека, в том числе и в эмбриональном состоянии. В таких дискуссиях участ-
вуют медики и генетики, представители общественных организаций, религиозные
деятели, юристы, философы и специалисты по этике.
Разрабатывается новый метод лечения — генная терапия. Больным с генными на-
рушениями вводят генетический материал, который должен компенсировать исход-
ный дефект. Хотя до широкого применения метода еще далеко, однако основания
для оптимизма есть. Например, в 1990 году американский генетик Андерсон (W.F.
Anderson) успешно применил генную терапию для лечения девочки с тяжелыми вро-
жденными нарушениями иммунитета.
Большое внимание привлекают исследования по генетике рака. Рак может возни-
кать как под воздействием внешних причин (канцерогенов или вирусов), так и
при повреждении генетического аппарата клетки. Найдены гены, мутации в кото-
рых повышают риск развития злокачественных преобразований, в частности рака
груди. Это позволило проводить диагностику предрасположенности к развитию не-
которых форм рака. При лечении онкологических заболеваний может быть эффек-
тивна гемотерапия. Но разрабатываются и другие направления — например, полу-
чение вакцин против рака. В 2001 году начаты испытания вакцины, предотвращаю-
щей заражение папилломавирусом — одним из основных агентов, вызывающих рак
шейки матки (вирус передается половым путем, и формально этот вид рака можно
рассматривать как венерическое заболевание). Если они будут успешны, то в
ближайшие годы будет получена вакцина против рака шейки матки — второй по
распространенности среди 20-30-летних женщин форм рака.
Задолго до появления генотерапии люди стремились к улучшению породы. В
древней Спарте «неправильных» младенцев сбрасывали со скалы. В 30-х годах в
США с той же целью было подвергнуто принудительной стерилизации около ста ты-
сяч человек, носителей определенных, утвержденных государством признаков. Та-
кие меры бессмысленны с точки зрения генетики, так как не снижают частоты
проявления данных признаков в следующем поколении. На пороге третьего тысяче-
летия человечество стремится заплатить поменьше за свое благополучие — взять
под контроль собственные генетические процессы и вносить в них коррективы не
ценой жизни носителей неблагоприятных мутаций, а подправляя генетические тек-
сты по собственному разумению, добываемому в геномных исследованиях.
ОСНОВЫ ИММУНИТЕТА3
Введение
Иммунитет - защита организма от инфекции или, в более широком смысле, - ре-
акция организма на чужеродные макромолекулы, микроорганизмы и клетки. Защита
осуществляется с помощью двух систем - неспецифического (врожденного, естест-
венного) и специфического (приобретенного) иммунитета. Эти две системы могут
рассматриваться и как две стадии единого процесса защиты организма. Неспеци-
фический иммунитет выступает как первая линия защиты и как заключительная ее
стадия, а система приобретенного иммунитета выполняет промежуточные функции
специфического распознавания и запоминания болезнетворного агента (или чуже-
родного вещества) и подключения мощных средств врожденного иммунитета на за-
ключительном этапе процесса.
Система врожденного иммунитета действует на основе воспаления и фагоцитоза,
явлений, которые будут рассматриваться в следующей статье. Эта система реаги-
рует только на корпускулярные агенты (микроорганизмы, занозы) и на токсиче-
ские вещества, разрушающие клетки и ткани, вернее, на корпускулярные продукты
этого разрушения.
Вторая и наиболее сложная система - приобретенного иммунитета - основана на
специфических функциях лимфоцитов, клеток крови, распознающих чужеродные мак-
ромолекулы и реагирующих на них либо непосредственно, либо выработкой защит-
ных белковых молекул.
Феноменология
Рассмотрим элементарную реакцию специфического иммунитета на двух внешне
совершенно непохожих моделях - выведении чужеродного белка и отторжении чуже-
родной ткани. Если в кровь животного, например, кролика, ввести непосредст-
венно, минуя тканевые барьеры, белок крови животного другого вида (например,
альбумин лошади) в смеси с собственным альбумином, то в первые дни после вве-
3
Г. И. Абелев
дения оба белка будут вести себя неразличимо, то есть выводиться с постоянным
и довольно длительным периодом "полужизни" (рис. 1).
Днм Дни
1-3 -2-1 1 2 3 4 5 6 7 8 9101112
Дни
Рис. 1. Кривые выведения собственного и чужеродного белков из
крови кролика. САК - сывороточный альбумин кролика, САЛ - сыво-
роточный альбумин лошади, САЧ - сывороточный альбумин человека,
а - первичный ответ, б - повторный ответ, в - индукция толерант-
ности .
Чужеродный белок в нашем случае не токсичен и молекулярно-дисперсен. Он не
вызывает ни воспаления, ни фагоцитоза, то есть беспрепятственно минует первую
линию защиты и непосредственно встречается со второй. Примерно на 7-й день
кривые выведения собственного и чужеродного белка резко расходятся - первый
продолжает "двигаться" по своей кривой "полужизни", для второго наступает пе-
релом и он выделяется из кровотока с резко возросшей скоростью (рис. 1а). Ор-
ганизм, следовательно, отличает "свое" от "не своего", это первая особенность
реакции специфического иммунитета. Отличие запоминается - повторное введение
лошадиного белка приводит к сокращению латентного периода и усиленной реакции
(более крутой наклон кривой выведения). Это так называемая иммунологическая
память - вторая характерная черта реакции специфического иммунитета. Память
специфична, запоминается контакт лишь с лошадиным альбумином, но ни с каким-
либо третьим белком. Специфичность запоминания очень высока, и это третья
особенность реакции приобретенного иммунитета (рис. 16) . Иммунный ответ на
чужеродную макромолекулу можно избирательно подавить, если ввести ее в разви-
вающийся организм внутриутробно или в первые часы после рождения. Способность
отличать введенный чужеродный белок от собственного у такого животного утра-
чивается после рождения. Подавление реакции строго специфично - оно распро-
страняется только на белок, введенный в процессе развития, но не какой-либо
иной чужеродный белок (рис. 1в) . Это явление носит название толерантности
(терпимости). Оно составляет четвертую неотъемлемую особенность элементарной
реакции приобретенного, или специфического, иммунитета.
Четыре признака реакции неразделимы, они всегда вместе независимо от того,
в какой системе разыгрывается реакция иммунитета. Например, отторжение кожи
или органов у генетически разнородных животных и человека, внешне совсем не
похожее на выведение чужеродного белка, подчиняются тем же закономерностям.
Так, если взять мышей какой-либо генетически однородной линии, то все особи
этой линии будут генетически идентичны и иметь, например, белую окраску. Если
таким мышам пересадить лоскут кожи (трансплантат) другой инбредной линии,
пусть черной, а в качестве контроля - лоскут кожи от генетически идентичной
линии, то оба трансплантата сначала приживутся, но на 12 - 14-й день черный
лоскут будет окружен валом лейкоцитов, затем его кровоснабжение начнет ухуд-
шаться и через 3-4 дня он будет отторгнут, в отличие от контрольного, гене-
тически идентичного трансплантата. Очевидно, мы вновь встретились здесь со
способностью организма отличать "свое" от "не своего". Эта особенность запо-
минается: вторичная пересадка черной кожи на белую мышь ведет к усиленному
отторжению трансплантата, в более короткие сроки и более интенсивно (иммуно-
логическая память). Запоминание специфично - лоскут кожи от мыши "коричневой"
линии, пересаженный при повторной трансплантации "черного" трансплантата, от-
торгается по типу первичного, а не повторного ответа. И, наконец, толерант-
ность: при пересадке тканей она выявляется еще лучше, чем в системе с чуже-
родным белком. Введение животному живых клеток крови генетически чужеродной
линии в процессе внутриутробного развития делает его на всю жизнь восприимчи-
вым к пересадке тканей и органов линии донора крови. Именно на этой модели
толерантность и была впервые обнаружена в 1953 году.
Таким образом, в организме человека и животных (рыбы, земноводные, пресмы-
кающиеся, птицы, млекопитающие) имеется система иммунитета, способная отли-
чать "свое" от "не своего", запоминать встречу с "не своим", причем с высокой
специфичностью, отторгать "не свое" и отвечать иммунологической ареактивно-
стью (толерантностью) на контакт с чужеродным веществом, предварительно вве-
денным в процессе раннего развития. Эта система лежит в основе приобретенно-
го , или специфического, иммунитета.
Приобретенный иммунитет широко используется для вакцинации, то есть введе-
ния ослабленных или убитых микроорганизмов, или выделенных из них макромоле-
кул, вызывающих иммунологическую реакцию на эти микроорганизмы. Вакцинация
является основным способом предупреждения таких страшных заболеваний как ос-
па , туберкулез, полиомиелит, сибирская язва и многих других. Приобретенный
иммунитет составляет основное препятствие для пересадок органов (сердце, поч-
ки, печень) и тканей (кожа) от одного человека другому. Для преодоления этого
барьера несовместимости пользуются препаратами, подавляющими иммунную систе-
му.
Вещество, способное вызывать реакцию приобретенного иммунитета, носит на-
звание антигена. Антигеном может быть не всякое вещество. Оно должно быть чу-
жеродным, макромолекулярным (с мол. весом более 10 000 - 12 000) и иметь ус-
тойчивую химическую структуру. К типичным антигенам относятся белки и полиса-
хариды. В первом примере (см. рис. 1.) антигеном является сам чужеродный бе-
лок - альбумин крови, во втором - особые белки, присутствующие на мембранах
пересаженных клеток, так называемые антигены тканевой совместимости.
Антитела и антиген-распознающие
рецепторы лимфоцитов
Что же происходит в точке перелома кривых выведения и отторжения? В первом
случае, когда свободный молекулярно-дисперсный антиген циркулирует в крови, в
ответ на него появляются защитные белки - антитела, которые специфически рас-
познают его, образуют с ним комплекс, обезвреживают, если антиген токсичен
(бактериальный токсин, змеиный яд или болезнетворный вирус), и способствуют
ускоренному выведению его из организма.
При отторжении трансплантата главная роль принадлежит особой разновидности
лимфоцитов Т-лимфоцитам-киллерам, "убийцам". Эти лимфоциты несут на своей на-
ружной мембране антителоподобные рецепторы - рецепторы Т-клеток (РТК). РТК
специфически распознают антиген, находящийся на мембране чужеродных клеток, и
обеспечивают прикрепление киллера к клетке-мишени, что необходимо для осуще-
ствления киллером своей смертоносной функции. Для того чтобы киллеры убили
клетку-мишень, они должны прикрепиться к ее мембране и выделить в просвет ме-
жду киллером и мишенью особый белок, "продырявливающий" мембрану клетки-
мишени, в результате чего клетка гибнет. После этого они открепляются от ми-
шени и переходят на другую клетку, и так несколько раз. При повторной встрече
организма с антигеном образуется больше антител и киллеров, они появляются в
более короткие сроки, чем при первой встрече, и их специфичность во взаимо-
действии с антигеном возрастает. В этом и проявляется иммунологическая па-
мять, и на этом основан эффект вакцинации.
Как устроены антитела и рецепторы лимфоцитов? Очевидно, что их структура
должна быть необычной, так как они строго специфически распознают громадное
множество различных антигенов - любой чужеродный белок, полисахарид или син-
тетическую молекулу, вообще в природе не встречающуюся. При этом антитела и
Т-лимфоциты, как правило, не реагируют с белками и клетками собственного ор-
ганизма . Как природа решает эту задачу?
Принцип строения антитела и РТК представлен на рисунке 2. Антитело состоит
из двух идентичных пар полипептидных цепей: Н (Heavy - тяжелых) с молекуляр-
ным весом примерно 50 000 и L (Light - легких) - примерно 25 000. Цепи соеди-
нены друг с другом ковалентными (дисульфидными) связями (рис. 2а).
Рецептор лимфоцитов, в отличие от антител, - гетеродимер, состоящий из пары
различных цепей - а и Ь с молекулярным весом около 50 000, соединенных между
собой одной дисульфидной связью. Н-, L-, а- и b-цепи имеют сходный план
строения и относятся к одному большому суперсемейству белков - иммуноглобули-
нов. В основе их структуры лежит сходное повторяющееся звено (домен), состоя-
щее примерно из 110 аминокислот, свернутых в глобулы, напоминающие друг друга
(см. рис. 2, где эти звенья обозначены как прямоугольники). Каждый домен в
молекуле антитела или РТК выполняет свою биологическую функцию, а наиболее
важную из них - распознавание и связывание с антигеном - осуществляют конце-
вые домены (см. рис. 2). Пара концевых доменов (один из Н-цепи, другой из L-
цепи, а в РТК - один из a-цепи, другой из b-цепи) образуют активный центр,
уникальную по структуре полость, распознающую в молекуле антигена небольшие
дискретные участки, состоящие из 4-8 аминокислот. Эти участки антигена под-
ходят к структуре активного центра, "как ключ к замку", образуя прочные неко-
валентные связи антигена с антителом. Разные антитела (и РТК также) отличают-
ся друг от друга структурой активных центров. На химическом уровне отличие
активных центров определяется первичной структурой концевых доменов Н- и L-,
а- и b-цепей, то есть последовательностью аминокислот в них. Участки Н и L
цепей (равно как а и Ь), образующие "стенки" активного центра, носят название
вариабельных (или V) районов. Остальная часть молекул постоянна и носит на-
звание константного (С) района. Вариабельных районов среди молекул антител
тысячи, константных - единицы. Главная функция вариабельных районов - образо-
вание активных центров антител и РТК. Разнообразие активных центров очень ве-
лико, не менее 107 - 108, что вполне достаточно для более или менее специфи-
ческого распознавания любого произвольно взятого антигена. Таким образом,
структура антител и РТК, а точнее их активных центров, обеспечивает узнавание
любого чужеродного белка или полисахарида, попавшего в организм.
Рис. 2. Принципиальная схема строения антител и рецепторов Т-
клеток (РТК).
Н и L - тяжелая и легкая цепи антител соответственно, VL и VH -
вариабельные районы соответственно легких и тяжелых цепей; Va и
Vb - вариабельные районы а- и b-цепей; Акт. ц. - активные центры
антител и РТК. Прямоугольники - повторяющиеся сходные звенья
(домены) в структуре полипептидных цепей; М - клеточная мембра-
на .
1 D - от diversity (разнообразие) , J - от Joining (соединяющий) .
V’-фрагмент гена контролирует первые 95 аминокислот в V-доменах,
D - от 2 до 10 и J - от 5 до 15 (см. рис. 3).
Генетический контроль
синтеза антител
Как же осуществляется контроль синтеза полипептидных цепей, один из районов
которых постоянно варьирует при переходе от одного антитела (или РТК) к дру-
гому (вариабельный, V-район), а остальные строго постоянны (константные, С-
районы). Эта задача решается благодаря уникальному генетическому механизму -
независимому генетическому контролю V- и С-районов, составляющих одну поли-
пептидную цепь, и сборке гена, контролирующего вариабельный район, из фраг-
ментов в процессе созревания лимфоцита.
Гены, контролирующие структуру одной полипептидной цепи антитела или РТК,
объединены в семейства, расположенные в одном участке хромосомы. При этом ге-
ны, соответствующие V-районам, "разорваны" на 2 или 3 фрагмента - основной V’
и один или два дополнительных - D и J 1. V’-фрагментов в геноме недифференци-
рованных лимфоцитов много - от 50 до 1000 для разных цепей и все они отлича-
ются друг от друга по своей структуре. D представлены несколькими десятками
копий, a J - единичными, причем каждая из копий имеет уникальную структуру. В
процессе созревания лимфоцита фрагменты V-гена объединяются на случайной ос-
нове в V’DJ-ген (рис. 3), причем число возможных сочетаний равно произведению
числа V’-, D- и J-генов. Для различных цепей антител или РТК число вариантов
от тысячи до десятков тысяч.
С&мейстыо
С-i SUM
Семойст во V-г енои
Рис. 3. Упрощенная схема семейства антител (а) и сборки гена,
контролирующего вариабельный район антитела (б).
а - Принципиальная схема участка хромосомы, включающего семейст-
ва вариабельных (V’) и константных (С) генов. Численность V’-
генов в отдельных семействах варьирует в зависимости от типа це-
пей и вида животных от 50 до 1000, а С-генов от 1 до 9.
б - Сборка V-гена из фрагментов. V’ - фрагмент гена, контроли-
рующий первые 95 аминокислот V-района; D - короткий фрагмент,
контролирующий от 2 до 10 аминокислот, начиная с 96. (Число D -
до 20, в зависимости от типа цепи и вида животных); J - короткий
концевой фрагмент V-гена, контролирующий от 5 до 15 аминокислот
концевого района V-гена. Районы ДНК, разделяющие V’-D и D-J вы-
резаются и утрачиваются при сборке гена. Транскрибируется (то
есть переписывается в информационную РНК) лишь собранный VDJ
ген. Расстояние от VDJ до С-гена вырезается и утрачивается при
синтезе информационной РНК.
Кроме того, в процессе сборки возможны ошибки и химические модификации объ-
единяющихся фрагментов, что резко увеличивает число возможных комбинаций.
Дальнейшее разнообразие активных центров возникает за счет объединения вариа-
бельных районов пары цепей, составляющих этот центр (см. рис. 2). Таким обра-
зом, благодаря сборке V-гена из фрагментов и образования активных центров из
разных цепей, создается 107 - 108 уникальных по своей структуре участков мо-
лекулы антител и РТК, среди которых всегда найдутся способные взаимодейство-
вать с любым произвольно взятым антигеном. Все это разнообразие составляет
репертуар антител или РТК. Следует подчеркнуть, что этот репертуар возникает
до встречи с антигеном и независимо от антигена и что бЧльшая часть репертуа-
ра антител не потребуется в течение всей жизни особи.
Клеточные механизмы
иммунитета
Итак, в организме предсуществуют антитела и РТК к любому произвольно взято-
му антигену. Эти антитела и РТК присутствуют на поверхности лимфоцитов, обра-
зуя там антиген-распознающие рецепторы. Чрезвычайно важно, что на поверхности
одной клетки находятся антитела (или РТК) одной и той же специфичности. Один
лимфоцит может синтезировать антитела (или РТК) только одной специфичности,
не отличающиеся друг от друга по структуре активного центра. Это формулирует-
ся как принцип "один лимфоцит - одно антитело".
Каким же образом антиген, попадая в организм, вызывает усиленный синтез
именно тех антител, которые специфично реагируют только с ним? Ответ на этот
вопрос дала теория селекции клонов австралийского исследователя, Нобелевского
лауреата Ф.М. Бернета (1899 - 1985) . Согласно этой теории, высказанной в 1957
году и полностью подтвержденной последующими экспериментами, одна клетка син-
тезирует лишь один тип антител, которые локализуются на ее поверхности. Ре-
пертуар антител формируется до, и независимо от встречи с антигеном. Роль ан-
тигена заключается лишь в том, чтобы найти клетку, несущую на своей мембране
антитело, реагирующее именно с ним, и активировать эту клетку. Активированный
лимфоцит вступает в деление и дифференцировку. В результате из одной клетки
возникает 500 - 1000 генетически идентичных клеток (клон), синтезирующих один
и тот же тип антител, способных специфически распознавать антиген и соеди-
няться с ним. В результате дальнейшей дифференцировки лимфоцит превращается в
клетку, не только синтезирующую данное антитело, но и секретирующую его в ок-
ружающую среду. Таким образом, функции антигена - найти соответствующий ему
лимфоцит, вызвать его деление и дифференцировку в клетку, секретирующую анти-
тела . В этом и заключается суть иммунного ответа: в селекции нужных клонов и
их стимуляции к делению. Динамика первичного и повторного ответов, согласно
теории Бернета, - это отражение динамики размножения клонов клеток, продуци-
рующих антитела к данному антигену. Толерантность - утрата клона клеток
вследствие их контакта с антигеном в процессе созревания лимфоцита.
В основе образования лимфоцитов-киллеров лежит тот же принцип: селекция ан-
тигеном Т-лимфоцита, несущего на своей поверхности РТК нужной специфичности,
и стимуляция его деления и дифференцировки. В результате образуется клон од-
нотипных киллеров, несущих на своей поверхности большое количество РТК, взаи-
модействующих с антигеном, входящим в состав чужеродной клетки, и способных
убивать эти клетки.
И здесь мы встречаемся с новыми проблемами, уже выходящими за пределы кло-
нально-селекционной теории иммунитета. Первая из них: как РТК узнают антиген?
Дело в том, что киллер ничего не может сделать с растворимым антигеном, ни
обезвредить его, ни удалить из организма. Но лимфоцит-киллер очень эффективно
убивает клетки, содержащие чужеродный антиген, поэтому он проходит мимо рас-
творимого антигена, но не пропускает антиген, находящийся на поверхности чу-
жеродной клетки. Для этого существует специальный механизм, так называемое
"распознавание в контексте". Он заключается в том, что РТК не узнают соответ-
ствующий ему антиген, если он находится в свободном виде, но строго специфи-
чески реагируют с ним, если он находится в комплексе с антигеном тканевой со-
вместимости, о котором мы упоминали выше. Эти антигены всегда присутствуют на
поверхности любых клеток организма и обладают способностью комплексироваться
с чужеродными белками, вернее, с их фрагментами. Таким образом, антигены тка-
невой совместимости образуют "контекст", в котором (и только в котором!) РТК
распознают чужеродный антиген, активируя лимфоцит и стимулируя его к делению
и дифференцировке в полноценный киллер.
Вторая проблема, выходящая за пределы клонально-селекционного принципа, -
это лимфоциты-помощники. Детальное изучение реакций иммунитета показало, что
для образования клона клеток, продуцирующих антитела, или клона киллеров не-
обходимо участие специальных лимфоцитов-помощников. Сами по себе они не спо-
собны ни вырабатывать антитела, ни убивать клетки-мишени. Но, распознавая чу-
жеродный антиген, они реагируют на него выработкой ростовых и дифференциро-
вочных факторов, которые необходимы для размножения и созревания антителооб-
разующих и киллерных лимфоцитов. В этой связи интересно вспомнить о вирусе
СПИДа, вызывающем сильнейшее поражение иммунной системы (синдром приобретен-
ного иммунодефицита - СПИД). Этот вирус поражает именно лимфоциты-помощники,
делая иммунную систему не способной ни к выработке антител, ни к образованию
киллеров.
И наконец, очень важная проблема: как вырабатывается толерантность к анти-
генам собственного организма? В полном соответствии с теорией Бернета было
показано, что если незрелый лимфоцит, несущий антительный рецептор или РТК к
собственным антигенам, встречается с таким антигеном, то он инактивируется
или погибает. Таким образом, организм лишается клонов лимфоцитов, способных
реагировать с собственными антигенами, не ослабляя своего ответа на антигены
чужеродные. Важно отметить, что при некоторых заболеваниях сохраняются "за-
прещенные" клоны, отвечающие антителами или киллерами на антигены собственных
клеток. В этом случае возникают тяжелейшие заболевания, такие, например, как
красная волчанка, при которой поражаются собственные ткани организма.
Эффекторные механизмы
иммунитета
Как антитела или лимфоциты-киллеры удаляют из организма чужеродные вещества
или клетки? Очевидно, что в случае киллеров РТК выполняют лишь функцию "на-
водчика" - они распознают соответствующие мишени и прикрепляют к ним клетку-
убийцу. Так распознаются клетки, зараженные вирусом. Сам по себе РТК не опа-
сен для клетки-мишени, но "идущие за ним" Т-клетки представляют огромный раз-
рушительный потенциал. В случае антител мы встречаемся со сходной ситуацией.
Сами по себе антитела безвредны для клеток, несущих антиген, и не обладают
физиологической активностью, ведущей к разрушению антигена. Правда, они по-
давляют биологическую активность опасных антигенов, таких, как дифтерийный
или столбнячный токсины, или змеиный яд. Они также способны блокировать ви-
рус, находящийся в крови (но не в клетках). Но это лишь особые случаи дейст-
вия антител. При встрече с обычными антигенами (не токсичными), циркулирующи-
ми или входящими в состав клеточной стенки микроорганизма, к антителам под-
ключается система комплемента, резко усиливающая эффект действия антител.
Комплемент сообщает образующемуся комплексу антиген-антитело биологическую
активность: токсичность, сродство к фагоцитирующим клеткам и способность вы-
зывать воспаление.
Система комплемента включает более 10 белков, большая часть которых являет-
ся проферментами - неактивными предшественниками специфических ферментов,
действующих на белки. Первый компонент этой системы распознает комплекс анти-
ген-антитело, находящийся либо в жидкостях организма (в крови или лимфе), ли-
бо на поверхности бактериальной клетки. Распознавание комплекса антиген-
антитело ведет к активации первого компонента комплемента, в котором появля-
ется ферментативная активность к последующему компоненту. Последовательная
активация всех компонентов системы комплемента имеет ряд последствий. Во-
первых, происходит каскадное усиление реакции, почти каждый последующий этап
реакции активации комплемента является ферментативным, при котором продуктов
реакции образуется несравнимо больше, чем исходных реагирующих веществ. Во-
вторых, на поверхности бактерии фиксируются компоненты комплемента, резко
усиливающие фагоцитоз этих клеток, то есть сродство их к фагоцитирующим клет-
кам организма. В-третьих, при ферментативном расщеплении белков системы ком-
племента образуются фрагменты, обладающие мощной воспалительной активностью.
И, наконец, при включении в комплекс антиген-антитело последнего компонента
комплемента этот комплекс приобретает способность "продырявливать" клеточную
мембрану и тем самым убивать чужеродные клетки. Таким образом, система ком-
племента - важнейшее звено в защитных реакциях организма.
Однако комплемент активируется любым комплексом антиген-антитело, вредным
или безвредным для организма. В связи с этим воспалительная реакция, возни-
кающая на безвредные антигены, регулярно попадающие в организм, может вести к
аллергическим, то есть извращенным, реакциям иммунитета, тяжело переносимым
человеком или животным. Аллергия развивается при повторном, как правило, мно-
гократном попадании антигена в организм, например, при повторном введении ан-
титоксических сывороток, или у мукомолов на белки муки, или при многократной
инъекции фармацевтических препаратов, в частности, некоторых антибиотиков.
Правда, есть аллергические реакции, не требующие для своего развития компле-
мента. Борьба с аллергическими болезнями состоит в подавлении либо самой ре-
акции иммунитета, либо в нейтрализации образующихся при аллергии веществ, вы-
зывающих воспаление.
Итак, мы рассмотрели сложную и удивительно целесообразно устроенную систему
защитных реакций организма. Одной из важнейших проблем современной биологии
является вопрос о том, как и из чего она могла возникнуть в процессе эволю-
ции. Подходы к этой проблеме лишь только начинают намечаться.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
1. Ройт А.. Основы иммунологии. М.: Мир, 1991.
2. Альберте Б., Брей Д., Льюис Дж., Рафф М., Робертс К. и Уотсон Дж. Моле-
кулярная биология клетки. М.: Мир, 1994. 2 изд. Гл. 18. "Иммунная систе-
ма" .
ПЕРЕДАЧА И ТРАНСДУКЦИЯ
ГОРМОНАЛЬНОГО СИГНАЛА
В РАЗНЫЕ ЧАСТИ КЛЕТКИ4
Введение
Общим принципом передачи любой информации в клетке, как наследственной, так
и оперативно регулирующей и управляющей, является обязательность ее трансдук-
ции (преобразования) на каждом этапе. Для наследственной информации общеизве-
стны как факт трансдукции, так и ее этапы: транскрипция - процессинг про-мРНК
(созревание про-матричной РНК) - трансляция - посттрансляционная модификация
белка. Для передачи гормональной информации это во многом выяснено в послед-
ние годы.
Существуют два основных механизма трансдукции гормонального сигнала в клет-
ку (рис. 1). При первом гидрофобный гормон (стероидный, иодтиронин, активиро-
ванные витамины А и D) проникает через плазматическую мембрану, а затем через
4
Кулинекий В.И.
цитозоль (последнему, очевидно, способствуют транспортные рецепторы цитозоля)
в ядро, где образует комплекс с ядерными рецепторами и в результате изменяет
матричные синтезы. При втором гормон-рецепторный комплекс образуется на на-
ружной поверхности плазматической мембраны. Это вызывает либо быстрое откры-
тие ионного канала и вход ионов в клетку (вариант На, на рис. 1) ив резуль-
тате нервный импульс, либо включение систем вторые посредники (ВП) -
протеинкиназы (ПК), приводящее к более медленным изменениям метаболизма и
функций клеток (вариант Пб ) . Два механизма (I и Пб ) могут приводить к
поздним эффектам - изменениям процессов, которые регулируются ядром клетки.
Совокупности механизмов, осуществляющих трансдукцию межклеточных сигналов
гормонов во внутриклеточные, в том числе и во внутриорганелльные сигналы, по-
лучили название сигнал-трансдукторных систем [1]. В статье излагаются совре-
менные данные по системам ВП-ПК для разных компартментов (микроотсеков) клет-
ки. Важным достижением является выяснение того, что передача регулирующей ин-
формации в ядро и митохондрии происходит своеобразно и различно.
Рис. 1. Общая структура сигнал-трансдукторныхсистем клетки. R -реиелтор, G-G-белок, Е-фермент, образу»
ющий второй посредник. ЭПС - эндоплазматическая сеть. ПК - протеинкиназа Ras - белок Pas, HR-гормон-ре-
иегторный комплекс
Цитозоль
Четыре основные и наиболее изученные системы передачи гормонального сигнала
в цитозоль [1, 2] представлены на рис. 2. Многие гормоны (амины, пептиды,
белки, простагландины I и Е) , а также запах и вкус действуют через систему
цАМФ (циклический аденозинмонофосфат). Образование гормон-рецепторного ком-
плекса через G [ГТФ (гуанозинмонофосфат)-зависимые]-белки активирует или ин-
гибирует аденилилциклазу, которая из АТФ (аденозинтрифосфат) образует цАМФ.
Этот ВП вызывает диссоциацию зависимой от него ПК А на регуляторную и катали-
тическую субъединицы. В результате последняя активируется и фосфорилирует
многочисленные белки. Это увеличивает, например, распад гликогена и жира,
синтез катехоламинов и глюкокортикостероидов, сокращение сердца, расслабление
гладких мышц. Поэтому цАМФ часто рассматривают как сигнал голода и стресса.
Другой циклонуклеотид - цГМФ (циклический гуанозинмонофосфат) образуется
двумя гуанилилциклазами. Мембранный фермент активируется натрийуретическими
гормонами (G-белки не участвуют), а растворимый (цитозольный) - монооксидами
NOJ, СО и JOH [1-3]. Последние - новый (90-е годы) класс неорганических регу-
ляторов, проявляющих свойства иногда межклеточных, иногда внутриклеточных ре-
гуляторов . цГМФ активирует ПК G, но, кроме того, изменяет активность других
белков, включая ионные каналы (последнее важно для восприятия света [4]) .
цГМФ увеличивает выделение мочи и Na+, расслабляет гладкие мышцы; NO через
цГМФ вызывает эрекцию полового члена. Оба циклонуклеотида дезагрегируют
(разъединяют) тромбоциты.
Г орм он ы.
запах.вкус
.АТФ
—ы1цАМФ
Бо.'ки
Г ормоны
цГМФ
Белки—». Эффе
NO* орг. нитраты
(ГЦ растворимая]-—NC*. ОС.Юн
Са*’ —Г КМ
Г орм Он ы
CsT
5ело»|ФЛ С
ИФ,
ФИ
ФХ
Са-
ЭНС
Бе.-ки—•. Эс
к 2—р-Бе.'кп—* ЭфФ^.'ы
ИН. ФРК.ЦК
тк
Антигены
к.
г
Белки
Рис. 2. Основные с иг нал «тран с дукторные системы клетки. АЦ - аденил ил цикл аза, ГЦ - гуанилилииклаза. КМ -
иальмодулин. КМ-ПК - кальмодулиновая ПК, ФГ С - фосфолипаза С. ФИ - фосфоинозитиды. ФХ - фосфатидил*
холин, ИФ, - инозитол трифосфат. ДАГ — диаиилг лииерид, ИН — инсулин, ФРК - факторы роста клеток. ЦК - цито*
кины,ТК - тирозинкиназа
Многие гормоны (амины, пептиды, белки, простагландины F и тромбоксаны) че-
рез G-белок включают систему фосфолипаз С, которые из фосфатидилинозитидов
образуют два ВП: инозитолтрифосфат и диацилглицерид, а из фосфатидилхолина -
только последний. Инозитолтрифосфат увеличивает поступление в цитозоль Са2 +
- как внутриклеточного из эндоплазматической сети, так и межклеточного через
медленные кальциевые каналы. Комплекс Са2 + с его рецептором кальмодулином
активирует многие цитозольные ферменты либо прямо, либо через кальмодулиновую
ПК. Диацилглицерид при наличии Са2 + активирует ПК С. Таким образом, в этой
системе функционируют три ВП, две ПК и кальмодулин. Ее стимуляция, например,
модулирует функции ионных каналов, способствует распаду гликогена, фосфолипи-
дов и белка, активирует секрецию разных желез, выделение гормонов, вызывает
сокращение гладких мышц и агрегацию тромбоцитов [1, 2].
Тирозинкиназы (ТК) - это ПК, фосфорилирующие в белках остатки тирозина (а
не серина или треонина, как другие ПК) . Эта система отличается от остальных
отсутствием ВП. Различают рецепторные ТК, расположенные в плазматической мем-
бране и являющиеся частью рецептора или сопряженные с ним, и нерецепторные
(или клеточные). Эта система включается в действие факторов роста клеток, ци-
токинов (см. раздел "Ядро") и инсулина, опосредуя большинство его эффектов
(цитозольные). Образование гормон-рецепторного комплекса, а также антигены
активируют рецепторные ТК, и они фосфорилируют различные белки (см. рис. 2).
Это включает системы малого G-белка Ras и фосфолипаз С. В последние годы ус-
тановлено , что обе они запускают целый каскад цитозольных ПК - обычных ПК и
нерецепторных ТК [1, 2, 5].
Нет сомнений, что сигнал-трансдукторных систем больше. В 90-е годы открыты
новые ВП: церамид, фосфатидная кислота, цАДФ (циклоаденозиндифосфат)-рибоза и
даже 5 ’-АМФ (аденозинмонофосфат). Сейчас интенсивно изучаются связанные с ни-
ми сигнал-трансдукторные системы.
Ядро
Многие гормоны и другие внеклеточные факторы (некоторые витамины, сыворотка
крови) регулируют матричные синтезы, деление и дифференцировку клеток и дру-
гие процессы, определяемые ядром клетки. Однако в ядро проникает очень не-
большая группа гормонов (стероиды, иодтиронины). Все остальные гормоны регу-
лируют ядерные процессы без проникновения не только в ядро, но даже в клетку.
Это возможно только в том случае, если вместо самого гормона в ядро будет по-
ступать его сигнал. Очевидно, что для этого межклеточный сигнал, передаваемый
гормоном, должен трансдуцироваться вначале во внутриклеточный, а затем во
внутриядерный сигнал.
Первый этап преобразования гормонального сигнала нами уже разобран. Он при-
водит к накоплению в клетке ВП и / или активации ПК, то есть особой специфики
здесь нет. Отметим только, что ядерные эффекты гормонов наиболее часто связа-
ны с активацией ТК, ПК С и А [5].
Гормоны, наиболее эффективно регулирующие ядерные процессы, - это факторы
роста клеток (ФРК) и цитокины. ФРК регулируют деление и дифференцировку всех
клеток и, кроме того, поддерживают их витальность (жизнестойкость). Аналогич-
ные эффекты соматотропного гормона (гормона роста) и инсулина реализуются че-
рез ФРК. Цитокины регулируют деление и дифференцировку иммунных клеток и во-
обще процессы иммунитета и воспаления. Иммунологи часто включают ФРК в состав
цитокинов.
Теперь и начинается самое интересное - как цитозольный сигнал трансдуциру-
ется во внутриядерный. Наиболее частым механизмом регуляции транскрипции на
этапе инициации является специфическое взаимодействие белковых транскрипцион-
ных факторов с регуляторными участками ДНК. Поэтому основные усилия были на-
правлены на выявление взаимодействия этого механизма с цитозольными события-
ми. В результате в 1990-1994 годы были выявлены три основных варианта [5]
(рис. 3) . При первом, характерном для действия сыворотки крови или цАМФ-
зависимых гормонов в ядро проникают цитозольные ПК, например киназа митоген-
активированного белка (MAP-киназа) или каталитическая субъединица ПК А. В яд-
ре такие ПК фосфорилируют один (или более) из внутриядерных транскрипционных
факторов, что изменяет его сродство к ДНК и/или степень его активности. На-
пример, ПК А участвует в развитии клеток (спермы и др.) , синтезе гормонов и
поддерживании суточного ритма, то есть функции цАМФ намного шире, чем думали
недавно. При втором варианте, установленном для ряда ФРК, цитокинов и проти-
вовирусных белков интерферонов, сигнал в ядро передает не ПК, а фосфорилиро-
ванный ею белок (например, Stat). До этого он был латентным транскрипционным
фактором, но в результате фосфорилирования становится активным, проникает в
ядро и специфически связывается с ДНК. При третьем варианте, реализующем эф-
фекты ФРК, активных форм кислорода и ультрафиолета и регулирующем процессы
воспаления и иммунитета, в белковом комплексе фосфорилируется и в результате
отщепляется ингибиторная или якорная субъединица. Освобожденный от нее и
ставший активным транскрипционный фактор проникает в ядро и связывается с
ДНК. Это установлено, например, для NF-kB-ядерного фактора, первым (из очень
многих) открытым эффектом которого была стимуляция синтеза k-легкой цепи им-
муноглобулинов в В-лимфоцитах.
Таким образом, все три уже изученных варианта передачи сигнала в ядро свя-
заны с протеинкиназным фосфорилированием регуляторных белков - транскрипцион-
ных факторов или их предшественников. При всех трех вариантах связывание ак-
тивного транскрипционного фактора с регуляторным участком ДНК происходит бы-
стро и запускает или увеличивает процесс транскрипции ранних генов, то есть
генов, определяющих быстрые (в пределах ~ 15 мин) ответы клетки. Возникающие
мРНК определяют синтез белковых продуктов ранних генов, которые становятся
новыми транскрипционными факторами. Последние, как хорошо известно, стимули-
руют поздние гены (их активность реализуется в течение часов и суток) [5].
Ядро
Рис. 3. Основные пути передачи сигнала из цитозоля в ядро, тф- транскрипционный фактор. F - остаток фосфа*
та. И - ингибитор. —*-транслокация сигнальной молекулы ендро.-— другие варианты передач и сигнала
Напомним, что такие гормоны, как стероидные и иодтиронины, и активированные
витамины А (ретиноевая кислота) и D (кальцитриол) после образования комплекса
со своими ядерными рецепторами также взаимодействуют с регуляторными участка-
ми ДНК (см. рис. 1). Это означает, что конечный этап в регуляции ядерных про-
цессов разными гормонами оказывается близким - это взаимодействие гормон-
рецепторного комплекса (при механизме I на рис. 1) или - при механизме Пб -
модифицированного транскрипционного фактора (см. рис. 3) с регуляторными уча-
стками ДНК. Однако очевидны глубокие различия и в процессах трансдукции гор-
монального сигнала в ядро, и в регуляторных участках ДНК. Отметим, что изоби-
лие последних почти в каждом гене эукариот привело к термину "мозг гена", а
сам такой ген заслужил название "умного" (smart gene).
Изложенные данные не исчерпывают возможных механизмов и уровней регуляции.
Показано, что при стимуляции деления клеток в ядре увеличивается концентрация
Са2+. Следовательно, проникать в ядро могут не только ПК и модифицированные
ими транскрипционные факторы, но и ВП. Однако трансдукция сигнала Са2+ в ядре
пока не выяснена. Ясно также, что цитозольные ВП и ПК могут регулировать экс-
прессию генов и на посттранскрипционном и трансляционном уровнях. Но конкрет-
ные механизмы передачи сигналов в ядро еще не установлены.
Митохондрии
В последние годы окончательно доказано, что существует множественная регу-
ляция гормонами и ВП всех основных функций митохондрий (MX) [6] : активности
ферментов, включая цикл трикарбоновых кислот Кребса, работы дыхательной цепи,
окислительного фосфорилирования и процессов энергопотребления. Регуляция осу-
ществляется двумя группами гормонов: Са2+-мобилизующими (катехоламины через
al-рецепторы, вазопрессин, ангиотензин) и цАМФ-зависимыми (глюкагон и катехо-
ламины через Ь-рецепторы). Схема передачи гормонального сигнала в MX пред-
ставлена на рис. 4. Общее для обоих механизмов - первичный регуляторный сиг-
нал в клетке возникает в гормональных рецепторах плазматической мембраны и
затем трансдуцируется в увеличение цитозольной концентрации ВП - Са2+ и/или
цАМФ. Их влияние на MX является вторичным - в результате воздействия на на-
ружную сторону внутренней мембраны MX или проникновения через нее в матрикс
(внутренняя растворимая часть MX).
Конкретные механизмы этих процессов различны. Для Са2+ существует постоян-
ный обмен через внутреннюю мембрану MX: вход в матрикс (к минусу внутри) за
счет энергии мембранного потенциала и выход назад в гиалоплазму в обмен на
Na+ или Н+ за счет энергии, связанной с различиями pH. Расход энергии на это
невелик и оправдан: он является основой для кальциевой регуляции функций MX.
Кроме того, Са2+ активирует как минимум один фермент наружной стороны внут-
ренней мембраны MX (глицерофосфатдегидрогеназу). цАМФ взаимодействует с ре-
цепторным белком внутренней мембраны MX, что, очевидно, и приводит к актива-
ции их функций. Кроме того, цАМФ проникает во все компартменты MX, а в каждом
из них есть ПК А (но ее роль в регуляции функций MX еще не вполне доказана) .
В результате описанных процессов цитозольный сигнал - увеличение концентрации
Са+ и/или цАМФ - трансдуцируется в митохондриальный, что и вызывает множест-
венные изменения функций этих субклеточных частиц [6].
Необходимо обратить внимание на две важные особенности: 1) ионы Са2+ дейст-
вуют не через специализированные рецепторные белки типа кальмодулина, а прямо
на митохондриальные ферменты как матрикса, так и внутренней мембраны; 2) эф-
фекты цАМФ могут реализовываться не только через ПК А, но и путем связывания
с рецепторным белком внешней стороны внутренней мембраны MX, что изменяет ее
свойства и в результате приводит к изменениям функций MX. Это существенно от-
личается от классических механизмов, характерных для цитозоля эукариот. В то
же время у прокариот нет обычных рецепторных белков (кальмодулина и ПК А) для
этих вторых посредников, и цАМФ действует через внутриклеточный цАМФ-
рецепторный белок. Эту аналогию можно рассматривать как еще один важный факт
в пользу известного представления о прокариотическом происхождении MX (суть
его в том, что MX в клетке - эндосимбиоз безмитохондриального эукариота, ра-
нее жившего на гликолизе, с прокариотом, открывшим окислительное фосфорилиро-
вание) . Вместе с тем у MX есть и важные отличия от прокариот: 1) отсутствие
аденилилциклазы, 2) локализация рецепторного белка
личие в ней системы транспорта Са2+, 4) наличие
могли появиться после возникновения эндосимбиоза
клетки регулировать свои (уже свои) MX. Конечно, в
исходило разновременно: рецепторный белок цАМФ в
система транспорта Са2+ развивается только у позвоночных. Последние в регуля-
ции функций MX имеют наиболее широкие возможности для выбора: 1) выбор гормо-
на, 2) выбор одного из его рецепторов, 3) выбор ВП, 4) для
же выбор пути реализации его эффекта (рецептор внутренней
А) .
цАМФ в мембране MX, 3) на-
ПК А. Все эти особенности
как ответ на потребность
процессе эволюции это про-
МХ есть уже у дрожжей, а
цАМФ вероятен так-
мембраны MX или ПК
Рис. 4. Передача гормонального сигнала ы митохондрии. Е - фермент MX
Вероятно, регуляция функций MX не ограничивается рассмотренными четырьмя
гормонами и двумя ВП: 1) как Са2 +-мобилизующих, так и цАМФ-зависимых гормо-
нов очень много; 2) биосинтезы (особенно матричные), ионный транспорт (осо-
бенно Na+ и К+) и мышечная работа - основные пути расхода энергии, поэтому их
регуляторы должны стимулировать и энергообеспечение. Функции MX могут стиму-
лироваться и другими ВП и их рецепторными белками. Для цГМФ и ПК С такие дан-
ные уже появились, но излагать их пока преждевременно, да и регуляторные ме-
ханизмы остаются неясными.
Заключение
В каждой клетке существует комплекс сигнал-трансдукторных систем, преобра-
зующих все внешние сигналы во внутриклеточные, а затем и во внутриорганелль-
ные. Сигналы подавляющего большинства гормонов с рецепторов плазматической
мембраны в цитозоль передаются системой вторые посредники - их специфические
рецепторы (чаще всего протеинкиназы); фосфорилирование же белков изменяет их
активность. Существуют и варианты: второй посредник - неферментный рецептор-
ный белок (Са2+ - кальмодулин или цГМФ - ионный канал) и протеинкиназы (тиро-
зинкиназы), прямо активируемые гормон-рецепторным комплексом.
В ядро сигнал обычно передается путем транслокации в него цитозольной про-
теинкиназы или активированного транскрипционного фактора (фосфорилированного
ею или освобожденного из комплекса с другим белком). В митохондрии сигнал пе-
редается иначе - путем транслокации из цитозоля вторых посредников: Са2+ или
цАМФ, которые в основном действуют не через свои специфические рецепторы, а
прямо на функциональные белки митохондрий. Механизмы трансдукции в органеллах
отличаются от классических цитозольных, но они обеспечивают столь же эффек-
тивный контроль гормонами ядерных и митохондриальных процессов, как и цито-
зольных .
ЛИТЕРАТУРА
1. Кулинский В.И. Лекционные таблицы по биохимии. Иркутск: Иркут, мед. ин-
т, 1994. Вып. 4: Биохимия регуляций. 94 с.
2. Нейрохимия / Под ред. И.П. Ашмарина, П.В. Стукалова. М.: НИИ биомед. хи-
мии РАМН, 1996. С. 244-371.
3. Реутов В.П. // Успехи биол. химии. 1995. Т. 35. С. 189-228.
4. Островский М.А. // Природа. 1993. № 10. С.23-36.
5. Терентьев А.А. // Биохимия. 1995. Т. 60. С. 1923-1952.
6. Кулинский В.И. // Успехи биол. химии. 1997. Т. 37. С. 171-209.
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ
НЕЙРОЭНДОКРИННОЙ РЕГУЛЯЦИИ5
Введение
Все процессы жизнедеятельности у человека и животных находятся под контро-
лем нервных клеток, которые секретируют в синаптическую щель нейромедиаторы,
и эндокринных желез, которые выделяют в кровь гормоны. Гормоны и нейромедиа-
торы сообщают органам и тканям, что, когда и сколько они должны производить.
Когда - определяется временем секреции, сколько - количеством секретированно-
го гормона или нейромедиатора, что - наличием рецепторов к этим молекулам
только у определенной группы клеток, специализирующихся в отношении данной
функции. Среди нейроэндокринных механизмов регуляции существует своя иерар-
хия , тесно связанная со скоростями развития и гашения их сигналов, а также с
молекулярными механизмами их действия (рис. 1).
Отклонение от нормы того или иного процесса жизнедеятельности включает
нервную систему регуляции, и нейромедиаторы, изменяя активность ионных кана-
лов (являющихся одновременно рецепторами нейромедиаторов, рис. 2), вызывают
5 В. А. Ткачук
гипер- или деполяризацию мембраны. Эта регуляция клеточной активности, проис-
ходящая за счет физических процессов (перемещение ионов через мембрану),
развивается и гасится за доли секунды (рис. 1, слева).
Рис. 1. Три оснонных механи эла нойроэндокрин•
ной регуляции клеток
Если нервная система не в состоянии вернуть тот или иной фактор гомеостаза
к норме, подключаются гормоны, действующие через мембранные рецепторы и сис-
темы вторичных посредников, которые стимулируют химическую модификацию бел-
ков. Эта регуляция (рис. 3), происходящая за счет химических процессов (син-
тез и расщепление вторичного посредника, фосфорилирование и дефосфорилирова-
ние белка), развивается и гасится за минуты или десятки минут (рис. 1, в цен-
тре) .
Если же отклонения от нормы того или иного процесса достигают опасных для
организма величин или же должны произойти фенотипические изменения клеток,
подключаются стероидные и тиреоидные гормоны, которые имеют цитозольные или
ядерные рецепторы, что позволяет им взаимодействовать с хроматином и влиять
на экспрессию генов (рис. 4). Эта регуляция, развивающаяся путем индукции или
репрессии синтеза мРНК и белков, реализуется спустя 3-6 ч после появления
гормона в крови, а гасится спустя 6-12 ч (рис. 1, справа).
Промежуточное положение в этой иерархии занимают факторы роста, рецепторы
которых являются тирозиновыми киназами. Взаимодействие фактора роста с рецеп-
тором приводит сначала к фосфорилированию определенных белков по ОН-группам
тирозина, а затем к проникновению этих фосфорилированных белков или самих
факторов роста (иногда вместе с рецептором) в ядро, что может вызывать деле-
ние клеток. Следует отметить также, что многие нейромедиаторы (например, аце-
тилхолин, g-аминомасляная кислота), диффундируя из синаптической щели (кото-
рая всегда сообщается с межклеточным пространством) в кровь, приобретают
свойства гормонов, вызывающих фосфорилирование белков.
Рис. 2. Структура холинер-
гического рецептора никоти-
нового типа, формирующего
ионный канал. Субъединицы,
полипептидные цепи кото-
рых четыре раза прониаыиа-
ют липидный бис г ой, глико-
зилированы извне клетки, а
внутри взаимодействуют с
белками тубулинового и акти-
нового цитоскелета Связы-
вание ацетилхолина (АХ) с
двумя а-субъединицами хо-
лтнергичес кого рецептора вы-
эь.вает конформационные из-
менения в олигомерном ком-
плексе. в результате чего Na*
входит внутрь клетки
Фсюфопротеин-
фосфатаза
Рибосомы "Т
апологический
эффех*
Рис. 3. Система проееде-ия -ормо-альнсио сиг-
нала путем образования в-эричных гюсред-иков
и послешющей химической модификации бел-
ков Мембра-ные рецепторы. семь раз прэ-изы-
васиие липидный бнслэй i 3-Р - 5*та-адренер< и-
ческий рецептор. мг-Р - холинергический реиеп-
-эр мускариновэ-р трта) и имеющие срэдс-вэ к
□-балкам i3,-3-6ero«.стимулирующий, аЗ -3-
бегок, ин-ибирующий аденилатциклаэу), регули-
руют образование циклического А МФ щАМФ) в
цитоплазме кле-ки. циклический амф связывает-
ся с прр-еинки-зээй и треями- ее из -еачгпв-э-
го а активное состояние Фосфорил* рма-ие ряда
бамюм клетке по Он- г руппам серим или траони-
-6 наменяет их свойства и тем самым водывае-
биологический аффект данных гор'•оное Гормо-
нальное влияние -а клетку устраняется за счет
разрушен** гормонов кэв-* помшая
диссоциацию гормон-рецепторного комплекса.
Вследствие етргр происходит разэбше-ие 3-бел-
кое с аденилатииклаэой кроме того, фосфодкк-
’еразд 1ФДЭ> гидролизует циклический А МФ до
АМФ. а фэсфопротеинфэсфа-аза дефэсфоэи.-и-
рует фосфэбегки, что приводит к полному а-це-
нив г ормоналы-огэсигнала
Рис. 4. Механизм действия горч.юнэв на tipoue':-
СЫТр=н ц.-. :.---ез бегкэе Показано влия-
ние тироксина |Т*>, который проникает в клетку и
там декодируется, после чего обраэоеавшийся
трииодтиронин |Та> связь вается в ядре со своим
рецептором и. изменяя взаимодействие рецепто-
ра с гистонами и ДНК, вызывает транскрипцию
мятртьой рибонуклеиновой кислоты |мРНЮ
При трансляции мРНК на рибосомах образуется
белок, который вызывает биологические эффек-
ты гормона
Изменение мембранного
потенциала
На рис. 2 показана структура так называемого никотинового холинорецептора.
Он локализован на постсинаптической мембране клетки и при связывании ацетил-
холина изменяет свою конформацию таким образом, что через устье, сформирован-
ное субъединицами, внутрь клетки устремляются ионы Na+. Происходит деполяри-
зация, а затем и замена заряда мембраны на противоположный, что приводит к
выходу К+ из клетки. Ток ионов К+ возвращает потенциал мембраны к исходной
величине. В процессе этой перезарядки мембраны, называемой потенциалом дейст-
вия, через этот же канал-холинорецептор внутрь клетки могут входить ионы
Са2+. Следовательно, этот канал нельзя назвать избирательным в отношении ка-
тионов . В то же время это очень быстродействующая регуляторная система - по-
тенциал действия, вызываемый ацетилхолином, возникает и гасится за 1-2 милли-
секунды, благодаря чему синапс может проводить от аксона на иннервируемую
клетку до 500 имп./с. Такое быстрое развитие и гашение сигнала возможны бла-
годаря быстроте связывания ацетилхолина с рецептором, а также высоким скоро-
стям его диссоциации от рецептора и разрушения ацетилхолинэстеразой. Разуме-
ется, не менее важен и механизм открывания канала за счет конформационных пе-
реходов, происходящих за наносекунды. Продолжительное и быстрое функциониро-
вание холинергического синапса требует также большого запаса ацетилхолина,
который синтезируется впрок и накапливается в везикулах пресинаптической мем-
браны. Кроме того, в клетках должны существовать высокие градиенты ионов Na+
и К+ по обе стороны плазматической мембраны, которые создаются и поддержива-
ются Ыа+/К+-насосом (см. статью А.А. Болдырева "Na/K-АТФаЗа - свойства и био-
логическая роль": Соросовский Образовательный Журнал. 1998. № 4) .
Быстродействие ацетилхолина (как и других нейромедиаторов) определяется
также особенностями того морфологического образования, которое существует ме-
жду аксоном и иннервируемой клеткой и которое называется синапсом. Благодаря
тому что расстояние между пре- и постсинаптической мембранами составляет все-
го 300-500 Б, а холинорецепторы сконцентрированы в виде кластеров строго про-
тив мест секреции ацетилхолина, в момент разрыва секреторного пузырька нейро-
медиатор быстро оказывается в месте его рецепции. Кроме того, при раздражении
аксона происходит выброс столь большого количества ацетилхолина, что молекулы
этого нейромедиатора мгновенно насыщают все рецепторы и вызывают массирован-
ный вход Na+ в клетку (развитие потенциала действия).
Холинергические рецепторы никотинового типа имеют очень низкое сродство к
ацетилхолину - полумаксимальное насыщение рецепторов наблюдается в присутст-
вии 10- 4 М ацетилхолина, поэтому как только ацетилхолинэстераза (локализо-
ванная, кстати, рядом с холинорецептором) начинает гидролизовать ацетилхолин
и понижать его уровень в синапсе, происходят диссоциация этого нейромедиатора
от рецептора и возвращение канала в закрытое состояние. Сродство рецептора к
нейромедиатору или гормону определяется соотношением скоростей диссоциации и
ассоциации гормон-рецепторного комплекса. При константе диссоциации 10"4-10-3
М скорость диссоциации ацетилхолина от рецептора составляет доли миллисекун-
ды, что, несомненно, очень важно для быстрого (за 1-2 миллисекунды) восста-
новления синаптической передачи.
Холинорецептор-каналоформер, функционирующий на постсинаптической мембране,
получил название "никотиновый" благодаря тому, что никотин, алкалоид из ли-
стьев табака имитирует действие ацетилхолина. На рис. 2 показана структура,
которую этот рецептор имеет в электрическом органе рыб или в мышцах эмбрио-
нов. Видно, что полипептидные цепи каждой из субъединиц (две альфа, по одной
бета, гамма и дельта) четыре раза пронизывают мембрану (рис. 2, справа). В
зрелых дифференцированных мышцах взрослого животного гамма-субъединица в со-
ставе холинорецептора замещается на эпсилон-субъединицу, а в нервных клетках
этот рецептор представлен лишь тремя альфа- и двумя бета-субъединицами, что
приводит к снижению электропроводности канала для ионов Na+.
Химическая модификация белков
Совершенно иную структуру и другой механизм функционирования имеет так на-
зываемый мускариновый холинергический рецептор, который локализован преимуще-
ственно вне синапса. На этот рецептор не действует никотин, но он прекрасно
активируется мускарином (алкалоидом из ядовитых грибов), а также, разумеется,
ацетилхолином, к которому имеет сродство порядка 10”6 М. Существуют по край-
ней мере четыре типа мускариновых рецепторов, причем все они близки по струк-
туре (полипептидная цепь семь раз пронизывает мембрану) и сопряжены с G-
белками, но передают сигнал разным системам внутриклеточной сигнализации.
Так, например, М-холинорецепторы могут стимулировать фосфолипазу С, которая
гидролизует фосфоинозитиды, и могут ингибировать аденилатциклазу, синтезирую-
щую циклический АМФ (см. рис. 3) и активировать К+-канал.
Эффекты мускариновых холинорецепторов развиваются спустя минуты после взаи-
модействия с рецептором и гасятся за десятки минут. Столь разительное отличие
в скоростях проведения ацетилхолинового сигнала между никотиновым и мускари-
новым рецепторами объясняется прежде всего разной кинетикой связывания лиган-
да с соответствующими рецепторами (диссоциация ацетилхолина от мускаринового
рецептора происходит за десятки минут), сложным каскадом проведения сигнала в
случае мускариновой регуляторной системы (необходимо последовательное взаимо-
действие рецептора с соответствующим G-белком, затем G-белка с определенным
ферментом или каналом и т.д.), а также сравнительно медленно протекающими хи-
мическими реакциями синтеза вторичных посредников, фосфорилирования и дефос-
форилирования белков.
В каждой клетке функционируют обычно разные типы рецепторов к одному и тому
же гормону (например, как альфа-, так и бета-адренорецепторы). Кроме того,
клетка чувствительна обычно к 7-10 разным эндокринным регуляторам (нейроме-
диаторам, гормонам, простагландинам, факторам роста). Каждый из этих регуля-
торов имеет характерные только для него продолжительность и амплитуду регуля-
торного сигнала, для каждого характерно определенное соотношение активностей
систем генерации вторичных посредников в клетке или изменения мембранного по-
тенциала . На уровне исполнительных систем клетки может происходить как потен-
циирование, так и взаимное гашение разных регуляторных сигналов.
В каждой клетке функционирует также специальная биохимическая надстройка,
регулирующая чувствительность клеток к гормону. Проиллюстрируем ее на примере
рецептора, сопряженного с G-белками. Обычно уровень гормонов, действующих че-
рез эту систему трансмембранной сигнализации (к их числу помимо названных вы-
ше относятся простагландины, гормоны гипофиза, ангиотензин II, брадикинин,
вазопрессин, окситоцин, гистамин, дофамин, энкефалин, эндорфин, серотонин,
эндотелии, холецистокинин, гастрин, паратироидный гормон), повышается на не-
сколько минут. Этого времени достаточно, чтобы произошло образование нужного
количества вторичных посредников (циклического АМФ, Са2+, диацилглицерина),
которые вызовут активацию соответствующих протеинкиназ и последующее за этим
фосфорилирование белков. Если же уровень гормона сохраняется повышенным в те-
чение десятков минут или нескольких часов (из-за гиперфункции эндокринной же-
лезы или фармакологического вмешательства), то происходит десенсибилизация
соответствующего рецептора. Сначала протеинкиназа, которая есть в плазматиче-
ской мембране практически всех клеток, фосфорилирует рецептор, в результате
чего его сродство к гормону снижается в 2-5 раз. Эта протеинкиназа может фос-
форилировать только гормон-рецепторный комплекс, поэтому, чем дольше гормон
связан с рецептором, тем больше вероятность того, что рецептор будет фосфори-
лирован. Если такое фосфорилирование не в состоянии погасить гормональный
сигнал, то спустя 15-30 мин происходит фосфорилирование рецептора протеинки-
назой, которая активируется соответствующим вторичным посредником (например,
в случае b-адренергических рецепторов, активирующих аденилатциклазу, цАМФ-
зависимой протеинкиназой; в случае al-адренергических или Ml- и МЗ-
холинергических рецепторов, активирующих фосфолипазу С, протеинкиназой С).
Фосфорилирование рецепторов протеинкиназами, зависимыми от вторичных посред-
ников, нарушает сопряжение с G-белками, вследствие чего ослабляется активи-
рующее или ингибирующее влияние гормонов, действующих через эти рецепторы на
аденилатциклазу, фосфолипазы А, С и D, Са2+- или К+-каналы. Если высокий уро-
вень гормона сохраняется в течение нескольких часов, а перечисленные выше ме-
ханизмы десенсибилизации не в состоянии погасить регуляторный сигнал, проис-
ходит эндоцитоз гормон-рецепторных комплексов и внутри клетки появляются ре-
цепторосомы. Они могут вновь встроиться в плазматическую мембрану, если уро-
вень гормона понизится в первые 2-3 часа. Если этого не происходит, они сли-
ваются с лизосомами, после чего рецепторы разрушаются. Очевидно, что восста-
новление чувствительности клетки к этому гормону потребует нового синтеза ре-
цепторов .
Экспрессия генов
Как уже отмечалось, на определенных стадиях онтогенеза или при достижении
критического для организма отклонения от нормы того или иного фактора гомео-
стаза (гипотермия, гипогликемия, гипоксемия, потеря крови) включается медлен-
ная, но наиболее мощная система эндокринной регуляции, действующая через сте-
роидные (андрогены, эстрогены, прогестины, глюкокортикоиды и минералокорти-
коиды) и тиреоидные гормоны (тироксин и трииодтиронин). Молекулы этих регуля-
торов, имея липофильную природу, легко проникают через липидный бислой и свя-
зываются со своими рецепторами в цитоплазме или ядре (см. рис. 4). Затем гор-
мон-рецепторный комплекс, который из-за высокого сродства рецепторов к гормо-
ну (полумаксимальное насыщение рецепторов происходит в присутствии 1О-9-1О”10
М гормона) не распадается в течение 1-3 ч, связывается с ДНК и белками хрома-
тина, что стимулирует синтез матричной РНК на определенных генах. Трансляция
мРНК приводит к появлению в клетке 3-7 новых белков, которые вызывают биоло-
гический эффект этих гормонов. Стероидные и тиреоидные гормоны могут также
репрессировать некоторые гены, вызывая биологический эффект путем уменьшения
количества определенных белков в клетке. Обычно эти гормоны изменяют содержа-
ние того или иного белка не путем ускорения-замедления транскрипции функцио-
нирующих генов, а за счет включения-выключения новых генов. Так, например,
стимулирование глюкокортикоидами аминотрансферазной активности печени проис-
ходит благодаря появлению в клетках новых (индуцибельных) изоформ аминотранс-
фераз . Интересно, что и на этом этапе регуляции эндокринная система заботится
о гашении сигнала: индуцибельные изоформы белков, как правило, значительно
быстрее инактивируются эндогенными протеазами, чем конститутивные изоформы. В
процессах влияния стероидных и тиреоидных гормонов на клетки наиболее зага-
дочным представляется механизм избирательной активации только определенных
генов. По-видимому, избирательность обеспечивается тем, что рецепторы этих
гормонов (рецепторные белки) имеют специальные домены, получившие название
"цинковые пальцы", с помощью которых белки могут присоединяться только к оп-
ределенным участкам нуклеотидных последовательностей ДНК и воздействовать на
гены, несущие эти последовательности.
К числу белков, экспрессия которых в клетке контролируется гормонами, отно-
сятся не только ферменты, участвующие в метаболизме, но и протеинкиназы, фос-
фопротеинфосфатазы и другие участники систем проведения нейроэндокринного
сигнала, а также многие рецепторы-каналоформеры и рецепторы, регуляторные
белки и ферменты, участвующие в обмене вторичных посредников,. Благодаря это-
му стероидные и тиреоидные гормоны могут участвовать в формировании не только
возрастных и половых признаков, но и определять психоэмоциональный статус ор-
ганизма , а также баланс катаболических и анаболических реакций в органах и
тканях, их чувствительность к нейромедиаторам и гормонам.
Заключение
Мы рассмотрели основные механизмы, с помощью которых нейроэндокринная сис-
тема поддерживает постоянство внутренней среды организма и обеспечивает его
адаптацию к изменениям окружающей среды. Следует отметить, что у высших жи-
вотных гормоны влияют также на поведение и память, а мозг, в свою очередь,
контролирует активность эндокринных желез. Едва ли есть смысл в рассуждениях
о том, какая система - нервная или эндокринная - более эффективна или более
важна, так как эти регуляторные процессы не функционируют обособленно. У че-
ловека и животных нейроэндокринная система регуляции стоит во главе всех ре-
гуляторных процессов, обеспечивающих согласованность процессов жизнедеятель-
ности по скорости, времени и месту протекания.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
1. Розен В.Б. Основы эндокринологии. М.: Изд-во МГУ, 1994.
2. Альберте Б., Брей Д. , Льюис Дж. и др. Молекулярная биология клетки. М. :
Мир, 1994. Т. 2. С. 338-393.
3. Ткачук В.А. Введение в молекулярную эндокринологию. М. : Изд-во МГУ,
1983.
4. Авдонин П.В., Ткачук В.А. Рецепторы и внутриклеточный кальций. М.: Нау-
ка, 1994
ЛЕКЦИЯ №25.
БИОИНФОРМАТИКА1
В настоящее время слово биоинформатика стало очень модным, оно употребляет-
ся в трех разных смыслах. Первый смысл связывают с телепатией, экстрасенсори-
кой и т.д., об этом мы говорить не будем. Второй смысл связан с применением
компьютеров для изучения любого биологического объекта, но эту тему мы тоже
не будем затрагивать. Речь пойдет о биоинформатике в узком смысле слова, а
именно о применении компьютерных методов для решения задач молекулярной био-
логии, в основном анализа разных последовательностей (аминокислотных, нуклео-
тидных) . Эта наука возникла в 1976-1978 годах, окончательно оформилась в 1980
году со специальным выпуском журнала «Nucleic Acid Research» (NAR). Биоинфор-
матика включает в себя:
• базы данных, в которых хранится биологическая информация
• набор инструментов для анализа тех данных, которые лежат в таких базах
• правильное применение компьютерных методов для правильного решения био-
логических задач
На рисунке показаны соотношение этапов развития биоинформатики (справа) с
возникновением разных экспериментальных методик и полученных результатов экс-
периментальных исследований.
Технология Биоинформатика
1962 Молекулярные часы
1965 Секвенирование tRNA База данных PIR
1970 Обратная транскрипция Алгоритм выравнивания NW
1972 Клонирование
1977 Секвенирование База данных PDB
1980 Спец выпуск NAR, Базы данных нукл. Послед
1981 Алгоритм выравнивания SW
1982 Секвенирование ДНК фага лямбда
1983 PCR Алгоритм поиска по базе данных WL
1985 Секвенирование ДНК вирусов FASTA - поиск по базе данных
“1987 GeneBank . Профили
1989 Программа Теном человека" Swiss-Prot. NCBI
1991 EST
1992 Первая хромосома дрожжей BLOSSUM
1993 Автоматическое секвенирование
1995 Первый геном бактерии База данных SCOP
1996 Первый геном археиныи
1997 PSI-BLAST. Кластеры ортологичных генов
1998 Геном червя
?ПП 1 Ган ли и а пойагя
А.А. Миронов
1
В 1962 году была придумана концепция "молекулярных часов", в 1965 была сек-
венирована тРНК, определена ее вторичная структура, в это же время были соз-
даны базы данных PIR для хранения информации об аминокислотных последователь-
ностях. В 1972 году было придумано клонирование. В 1978 году были разработаны
методы секвенирования, была создана база данных пространственных структур
белков. В 1980 был выпущен спецвыпуск журнала NAR, посвященный биоинформати-
ке, затем были придуманы некоторые алгоритмы выравнивания последовательно-
стей, о которых речь пойдет дальше. Дальше был придуман метод ПЦР (полимераз-
ная цепная реакция), а в биоинформатике - алгоритмы поиска похожих фрагментов
последовательностей в базах данных. В 1987 году оформился GeneBank (коллекция
нуклеотидных последовательностей) и т.д.
Биолог в биоинформатике обычно имеет дело с базами данных и инструментами
их анализа. Теперь разберемся, какие базы данных бывают в зависимости от то-
го, что в них помещают. Первый тип - архивные базы данных, это большая свал-
ка , куда любой может поместить все, что захочет. К таким базам относятся
• GeneBank & EMBL - здесь хранятся первичные последовательности
• PDB - пространственные структуры белков, и многое другое.
В качестве курьеза могу привести пример: в архивной базе данных указано,
что в геноме археи (архебактерии) есть ген, кодирующий белок главного ком-
плекса гистосовместимости, что является полной чепухой.
Второй тип - курируемые базы данных, за достоверность которых отвечает хо-
зяева базы данных. Туда информацию никто не присылает, ее из архивных баз
данных отбирают эксперты, проверяя достоверность информации - что записано в
этих последовательностях, какие есть экпериментальные основания для того,
чтобы считать, что эти последовательности выполняют ту или иную функцию.
К базам данных такого типа относятся:
• Swiss- Prot - наиболее качественная база данных, содержащая аминокислот-
ные последовательности белков
• KEGG - информация о метаболизме (такая, которая представлена на карте
метаболических путей, которую те, кто ходит на лекции, видели на лекции
№ 2)
• FlyBase - информация о Drosophila
• COG - информация об ортологичных генах.
Поддержание базы требует работы кураторов или аннотаторов. Тем не менее,
даже в курируемых базах данных могут встречаться курьезные надписи, например
такая забавная надпись:
CAUTION: AN ORF CALLED DSDC WAS ORIGINALLY (REF.3) ASSIGNED TO THE WRONG
DNA STRAND AND THOUGHT TO BE A D- SERINE DEAMINASE ACTIVATOR, IT WAS THEN
RESEQUENCED BY REF. 2 AND STILL THOUGHT TO BE "DSDC", BUT THIS TIME TO
FUNCTION AS A D-SERINE PERMEASE. IT IS REF.l THAT SHOWED THAT DSDC IS ANOTHER
GENE AND THAT THIS SEQUENCE SHOULD BE CALLED DSDX. IT SHOULD ALSO BE NOTED
THAT THE C-TERMINAL PART OF DSDX (FROM 338 ONWARD) WAS ALSO SEQUENCED (REF.6
AND REF. 7) AND WAS THOUGHT TO BE A SEPARATE ORF (YES, DON’T WORRY, WE ALSO
HAD PROBLEMS UNDERSTANDING WHAT HAPPENED!).
По крайне мере здесь кураторы базы данных честно признаются, что не знают,
как это случилось.
Третий тип - производные базы данных. Такие базы получаются в результате
обработки данных из архивных и курируемых баз данных. Сюда входит:
• SCOP - База данных структурной классификации белков (описывается струк-
тура белков)
• РЕАМ - База данных по семействам белков
• GO (Gene Ontology) _ Классификация генов (попытка создания набора терми-
нов, упорядочивания терминологии, чтобы один ген не назывался по разно-
му, и чтобы разным генам не давали одинаковые названия)
• ProDom - белковые домены
• AsMamDB - альтернативный сплайсинг у млекопитающих
И интегрированные базы данных, в которых вся информация (курируемая, не ку-
рируемая) свалена в кучу, и введя имя гена, можно найти всю связанную с ним
информацию - в каких организмах встречается, в каком месте генома локализо-
ван , какие функции выполняет и т.д.
• NCBI Entrez - доступ к информации о нуклеотидных и аминокислотных после-
довательностях и структурах
• Есосус - все о Е. coli - гены, белки, метаболизм и пр.
Теперь перейдем к рассмотрению инструментов биоинформатике. Инструменты оп-
ределяются задачами, которые мы хотим решать.
Основу биоинформатики составляют сравнения. Если у нас есть, например, ами-
нокислотная последовательность, о которой у нас есть экспериментальные дан-
ные , и известны ее функции, и другая, похожая на нее последовательность, мы
можем предположить, что эти последовательности выполняют сходные функции. Это
задача поиска сходства последовательностей
Другая задача связана с анализом генома. Недавно было объявлено, что полно-
стью просеквенирован геном человека, но так же просеквенировали геномы и дру-
гих организмов: три генома растений, мыши, крысы, кошки, собаки, курицы, ры-
бы, лягушки завершается, шимпанзе завершается, две дрозофилы сделаны, маля-
рийный комар, червяки, дрожжи и т.д. - всего около 30 видов эукариотических
геномов. Также просеквенированы сотни бактериальных геномов. Один бактериаль-
ный геном можно просеквенировать в хорошо оборудованной лаборатории за неде-
лю . При этом получают длинную нуклеотидную последовательность нуклеотидов.
Там есть гены - белок-кодирующие участки, и участки, кодирующие тРНК и рРНК.
Возникает задача найти эти гены. Другая задача - поиск сигналов в ДНК, то
есть тех участков ДНК, которые отвечают за регуляцию - сайты связывания регу-
ляторных белков, элементы вторичной структуры мРНК, которая транскрибируется
с этого гена и др.
Есть задача предсказания вторичной структуры РНК. А также есть большой
класс задач анализа белков. Для решения этих задач надо создавать методы ана-
лиза, то есть алгоритмов (протоколов) и программ для анализа. При создании
метода надо иметь критерий того, что метод адекватен, соответствует реально-
сти .
Как оценить "правильность" метода? Геном типичной бактерии содержит около
1000 генов. Как уже упоминалось, секвенировать геном можно за неделю. Экспе-
риментальная характеристика одного белка требует как минимум 2 месяца работы
современной лаборатории.
Для того чтобы определить, насколько предложенный метод анализа хорош и
правилен, существует так называемый «золотой стандарт». Например, у нас есть
метод определения генов. Если после его применения на какой-либо последова-
тельности, в которой известно месторасположение генов, наши результаты совпа-
дают с тем, что есть на самом деле на 80-90%, значит наш метод правильный и
эффективный. В этом и заключается суть «золотого стандарта».
Или предсказание вторичной структуры РНК. Экспериментально ее определить
очень трудно, но есть РНК, структура которых хорошо известна - это рРНК и
тРНК. И если наш метод хорошо предсказывает структуру этих известных РНК, то
можно ожидать, что и для других РНК он будет давать хорошие предсказания.
Вернемся к первой задаче - сравнению последовательностей. Запишем одну по-
следовательность под другой.
attgtACcTCgTgG-AA-------
-----AC-TCaTaGcAAccag
Нам надо при сравнении найти наилучший вариант, так выровнять эту пару по-
следовательностей, чтобы количество совпадений будет максимальным (парное вы-
равнивание) . Качество выравнивания оценивают, назначая штрафы за несовпадение
букв и за наличие пробелов (когда приходится раздвигать одну последователь-
ность для того, чтобы получить наибольшее число совпадающих позиций).
Таким образом, первым делом после секвенирования последовательности ищут в
базах данных похожие последовательности, чтобы после сравнения судить о том,
какие функции несет эта последовательность. Если две буквы совпали, значит
они находятся под давлением отбора, они функционально важны. Известно, что
аминокислоты различаются по своим свойствам, поэтому если произошла аминокис-
лотная замена, это может почти никак не повлиять на работу белка, а может
сильно его изменить.
Например, если лизин (положительно заряженная аминокислота заменится на
лейцин (похожий по созвучию, но совершенно несходный по свойствам), то для
пространственной структуры и функций белка это может оказаться катастрофой. А
вот замена лизина на аргинин (также положительно заряженный) может не ска-
заться на структуре белка.
Поэтому при сравнении аминокислотных последовательностей учитывают также
матрицу сопоставления аминокислотных остатков (похожих, менее похожих и со-
всем непохожих).
Как осуществляется выравнивание? Пишем одну последовательность под другой.
Сколько есть способов написать одну последовательность S1 длиной m под
другой - S2 длиной п (со вставками)? Об этом можно доказать теорему - попро-
буйте .
(Парное) Выравнивание
• Общая задача: написать одну
последовательность под другой, чтобы
было как можно больше совпадений:
attgtAC CTCgTgG-AA-
----АС-ТС T .S АА сад
• Более аккуратная постановка
(применяется для сравнения белков)
Е M(Sa, Sfl) - Ngap*Dgap - max
Построим выборочную последовательность S длиной m + n следующим образом:
возьмем несколько символов из последовательности S1, потом несколько символов
из последовательности S2 потом опять несколько символов из S1, потом опять
несколько из S2.
• Каждой выборочной последовательности S соответствует выравнивание и по
каждому выравниванию можно построить выборочную последовательность. (До-
казать !)
• Количество выборочных последовательностей равно
N = Ст
sei ^п+т
(и + w)!
(и!)(ти!)
(Доказать!)
Таким образом количество выравниваний можно определить по формуле:
А как же найти оптимальное среди такого большого количества? Можно, конеч-
но, попробовать разные способы, но оказывается, что этот поиск сводится к за-
даче поиска оптимального пути на графе. Задача поиска оптимального пути на
графе решается методами динамического программирования следующим образом. Мы
пишем одну последовательность над другой. И у нас есть некая ячейка, в кото-
рой мы будем хранить вес наилучшего выравнивания префиксов (то фрагментов по-
следовательности от начала до данного места). И если у нас известен вес наи-
лучшего выравнивания в 3 ячейках (см. слайд ниже), то мы можем определить вес
наилучшего выравнивания в четвертой ячейке. То есть, для того, чтобы найти
вес оптимального выравнивания, нам надо просмотреть m*n ячеек (количество
ячеек в прямоугольной матрице MxN). Как принято говорить в информатике, это -
квадратичный алгоритм. Он занимает время и объем памяти, пропорциональный
квадрату длины последовательности. И вместо случайного перебора большого
числа вариантов, мы решаем задачу довольно быстро.
Динамическое программирование для оптимального выравнивания
Граф редакционного расстояния для последовательностей S’,S’: вершина v
соответствует префиксам последовательностей {S', }. (S-, J На вершине записано
редакционное расстояние между префиксами
(синие стрелки соответствуют вставкам и удалениям)
Откуда берутся матрицы замен? Мы берем некоторое количество выравниваний, в
которое по тем или иным причинам верим, и смотрим, как часто у нас происходят
такие замены. Тогда матрица замен является логарифмом отношения некоторых ве-
роятностей, которые можно оценить как частоты.
Откуда берутся параметры для
выравнивания?
• Пусть у нас есть выравнивание Если
последовательности случайные и независимые (модель
R), то вероятность увидеть букву а прошв 0
р(а, р | R) = р(а) р(/))
а вероятность выравнивания (х,у) будет равна
р(х,у | R) = II р(х,) II р(у,)
Если выравнивание не случайно (модель М), то
р(х,у |М) = II p(x„yj
Отношение правдоподобия:
Р(*.У |М)= Пр(х, ,у,)
p(x,y|R) Пр(х() Пр(у)
Логарифмируя, получаем
log( p(x,y|M)/p(x,y|R)) = Vsfx^y,);
| Матрица замен: .v(«, //) = log(p„#/рЛ pp)
Итак, у нас имеется замечательный квадратичный алгоритм поиска сходства.
Время решения задачи выравнивания пропорционально L1*L2. Мы сравниваем имею-
щуюся у нас последовательность с последовательностями в банке. L1 = размер
банка = 108, а для генома человека 3x1О9. Сравниваемая последовательность
обычно имеет размер L2=103, количество операций примерно равно 100*1011=1013.)
Обычный компьютер имеет быстродействие около 109 операций в сек. На каждый
шаг надо »102 операций. Тогда время работы равно Т»106 сек ~ 11 дней. То
есть, просеквенировав бактериальный геном из 3000 генов (приблизительно за
неделю), на то, чтобы его охарактеризовать, мы потратим 11*3000 дней, то есть
проанализировать дольше, чем секвенировать, что, конечно, не очень хорошо.
Решением является то, что мы до применения методов динамического программи-
рования сначала выбираем правильных кандидатов для сравнения. Есть такая про-
грамма BLAST (basic local alignment search tool), которую все биологи очень
любят, она почти правильная. То есть она почти всегда работает так, как тре-
бует "золотой стандарт".
Основная идея ее работы заключается в хешировании. В самом начале мы один
раз проходим по всему банку и для каждого короткого слова с заранее зафикси-
рованной длиной мы запишем список позиций, где оно встречается в банках.
BLAST
• Эвристический алгоритм позволяет
существенно сократить время поиска.
• Основная идея - Хеширование.
Проанализировав однажды банк, строим
таблицу вида:
Здесь показано для слов длиной 4, в реальности слова берут не длиной 4, как
показано на рис., а длиной 7 или 10 или 13, но принцип тот же. В каких-то
случаях "слову" соответствует три позициями, в других - 100 позиций.
Дальше мы идем вдоль последовательности "Query" (та последовательность, ко-
торую мы хотим прогнать по банку) и выбираем очередные слова. Смотрим в таб-
лице, где встречается это слово, вытягиваем найденные последовательности из
банка и строим выравнивание их с нашей исходной последовательности. Это дела-
ется быстро, так как мы сравниваем нашу последовательность не со всеми после-
довательностями из банка, а только те, которые соответствуют нашему "слову"
(tttgc в показанном случае). И выравнивание строим тоже не так аккуратно, как
это делает алгоритм динамического программирования, а используем упрощенную
схему.
Затем мы оцениваем статистическую значимость этого выравнивания - так назы-
ваемую e-value. Вообще, есть два понятия, которые очень часто встречаются в
биоинформатике: e-value и p-value. E-value - это сколько мы ожидаем увидеть
совпадений с таким весом (то есть такого качества), если бы у нас наши после-
довательность и банк были случайными. Если они случайные, то мы ожидали бы
увидеть е-2 совпадений.
e-value - это ожидаемое число событий, может быть больше единицы. Если е-
value маленькое, то, значит, совпадение значимое, и оно несет большую биоло-
гическую информацию. P-value - это вероятность встречи такого соответствия
(не может быть больше единицы) . При оценке e-value, да и вообще при любых
статистических оценках, важно, какая модель лежит в основе всего этого дела.
Модель, которая лежит в основе e-value, конечно же, неправильная, потому что
мы не знаем правильность статистических характеристик биологических последо-
вательностей. E-value просто дает нам ориентир, и реально, если мы имеем е-
value порядка 10”2, то это, как правило, мусор, незначимое соответствие.
Правда, есть некоторые специалисты с такой интуицией о структуре белков, ко-
торые могут работать с выравниваниями с e-value даже порядка 1. А обычно если
исследователи видят e-value > 10”3, они с этим не работают.
Алгоритм BLAST
• Идем вдоль Query последовательности и формируем подслова
• С помощью хеш-таблицы находим в банке подходящие последовательности
• Строим для них выравнивание
• Оцениваем статистическую значимость выравнивания (е value)
accgat ttgcgtgacgtgcagtgc
tttgc
cgatttgcgtgacgt
ct atttgcg g< cgt
e-value=1 6e-12
Есть разные модификации BLAST: BLASTp (выравнивание аминокислотных последо-
вательностей) , BLASTn (выравнивание нуклеотидных последовательностей), BLASTx
(выравнивание всех возможных транслятов нашей нуклеотидной последовательности
против банка аминокислотных последовательностей), TBLASTx (выравнивание всех
возможных транслятов нашей нуклеотидной последовательности против всех транс-
лятов банка нуклеотидных последовательностей). Еще нужно знать, что Nr Data
Base - (non redundant) - это база, против которой обычно прогоняют BLAST, в
которой нет повторяющихся последовательностей, из которой убраны дубли для
того, чтобы не гонять BLAST по одним и тем же последовательностям. И score -
это вес выравнивания.
А если на нашу последовательность при поиске налипло, например, не одна, а
двадцать последовательностей. При этом возникает задача написать все эти по-
следовательности друг под другом, чтобы увидеть, в какой мере они совпадают,
что консервативно (устойчиво повторяется), а что нет, и как устроена наша
аминокислотная последовательность. Эта задача называется
Множественное выравнивание
Множественное выравнивание - это такой способ написания нескольких последо-
вательностей друг под другом (может быть, с пропусками в каких-то позициях в
разных последовательностях), чтобы в каждом столбце стояли гомологичные пози-
ции .
Для этой задачи тоже есть «золотой стандарт». Это выравнивание, которое бы
получилось, если бы мы выровняли друг под другом последовательности, которые
имеют одинаковую пространственную структуру. То есть две экспериментально ус-
тановленные пространственные структуры белка сопоставляем и отмечаем, какие
аминокислотные остатки друг под другом встали (эти остатки соответствуют го-
мологичным позициям). Это - биологически обоснованное выравнивание. Возникает
задача - найти способ (построить алгоритм и определить параметры), который
выравнивает последовательности "золотого стандарта" (то есть последовательно-
сти, для которых пространственная структура известно) правильно. Если такой
алгоритм построен, то есть надежда, что он выровняет последовательности с не-
известной пространственной структурой тоже правильно.
Для решения задачи множественного выравнивания можно попробовать написать
многомерную матрицу и построить методом динамического программирования с про-
смотром многомерной матрицы. Тогда количество вершин будет порядка Ln , где L
- длина, а п - количество последовательностей. Так как типичное количество
последовательностей в семействе белков сотни, то 300 аминокислот дадут ЗОО100
- это очень много, этот алгоритм для множественного выравнивания не подходит.
Динамическое программирование для
множественного выравнивания
• Количество вершин равно L.= O(L^
• Количество ребер из каждой вершины = 2N-1
• Количество операций равно
Т = O(L
• Надо запоминать обратные переходы в LN вершинах.
• Если количество последовательностей > 4, то задача
практически не разрешима.
Тогда придумали метод прогрессивного выравнивания. Зная расстояния между
любой парой последовательностей, мы можем построить выравнивание, определить
вес выравнивания, и построить какое-то бинарное дерево. Затем мы обходим это
дерево, последовательно проводя парные выравнивания наиболее близких последо-
вательностей. Объединяем, получаем выравнивание. Соединяем суперпоследова-
тельности, получаем следующее выравнивание. В конце концов, получаем выравни-
вание в корне.
Такое постепенное построение выравнивание решает задачу, которую мы не мо-
жем сформулировать математически. В биоинформатике очень часто нельзя постро-
ить математическую формулировку задачи, которую мы решаем. Поэтому формули-
ровка Задачи, которую решает алгоритм BLAST, выглядит так: мы находим то, что
находит программа BLAST. Также мы не можем сказать, что мы оптимизируем при
множественном выравнивании.
Одна и та же биологическая задача может приводить к разным математическим
постановкам одной и той же задачи. Есть примеры, когда одна и та же задача
может быть построена так, что она будет математически решаемой или математи-
чески не решаемой. Есть класс задач, для которых не существует хороших алго-
ритмов . Но при построении множественных выравниваний мы решаем с помощью
данного алгоритма, без формулировки математической задачи.
Прогрессивное выравнивание
Путеводное дерево строится
приближенно -главное быстро
Обычно это кластерное дерево
• Строится бинарное дерево (guide tree, путеводное
дерево) - листья = последовательности
• Дерево обходится начиная с листьев При объединении
двух узлов строится парное выравнивание супер-
последовательностей (профилей) и получается новая
супр последовательность
Дальше идет задача
предсказания вторичной
структуры РНК
Вторичная структура РНК - структура, образуемая спаренными основаниями на
однонитевой молекуле РНК. Биологическая роль вторичной структуры: структурная
(РНК - рибосомная, тРНК), регуляция (рибопереключатели, аттенюация, мик-
роРНК), рибозимы, стабильность РНК.
На рисунке показана типичная вторичная структура РНК и разные формы пред-
ставления вторичной структуры РНК:
Вся РНК состоит из петель и спиралей (указано на рисунке). Петли бывают
следующих типов: шпилька, внутренняя, выпячивание, множественная, псевдоузел.
Так вот, возникает задача установить, кто с кем спарен. Биологическая форму-
лировка этой задачи звучит так: дана последовательность РНК, определить ее
правильную вторичную структуру. «Золотой стандарт» - тРНК и рРНК. Количество
возможных вторичных структур очень велико. Задачу можно сформулировать таким
образом (законным с точки зрения физики): надо минимизировать энергию, по-
скольку правильная вторичная структура наиболее стабильная. На самом деле, с
точки зрения биологии это не совсем верно, но формулировка очень удобная с
точки зрения физики и математики. Далее вопрос, что оптимизировать и как
оптимизировать.
Элементы вторичной структуры
Способы представления вторичных структур
Т апологическая
Массив спаренных оснований
1 2 3 4 3 i 3 4 S 10 11 12 13 14 13
30 23 28 - IS 14 13 12 - - - 8 7 6 S
t i ' i t 4 '
16 17 18 1 9 20 2 1 22 2 3 24 2 3 26 27 28 29 30
- - - - - 27 26 - - - 22 23 3 21
Предположим, что мы не будем минимизировать усилия по поиску, а все перебе-
рем. Построим такой граф, в котором вершины - потенциальные спирали, а ребра
проводятся, если две потенциальные спирали в вершинах совместимы (то есть,
если две спирали могут одновременно существовать в данной молекуле РНК).
Тогда вторичной структурой будет любой полный подграф, то есть такой граф,
в котором все вершины между собой соединены - называется "клика". Тогда зада-
ча такова: в таком графе найти клику. Клика будет соответствовать хорошей
структуре.
Но, к сожалению, задача поиска клики в графе является математически плохой
- для нее, скорее всего, не существует эффективного алгоритма ее решения
(кроме полного перебора всех вариантов).
Комбинаторный подход
а .............................................................
• Построим граф:
- верш ины-потенциальные
нуклеотидные паоы (или
потенциальные спирали)
- Ребро проводится, если
пары совместимы (не
образуют псевдоузлов и не
имеют общих оснований)
• Допустимая вторичная
структура - клика в этом
графе
Структуры без псевдоузлов
• Структура без псевдоузлов =
правильное скобочное
выражение
• Может быть представлено в
виде дерева
Оценка количества возможных
структур:
Т(1_) - 1.8 •-
(очень много)
Если мы fgh уберем, то получим клику, некую вторичную структуру. Можем по-
лучить и другую клику.
Вторичная структура может быть представлена в виде правильной скобочной
структуры, как на рисунке ниже. Левая часть - открывающая скобка, правая
часть - закрывающая скобка. Вторичная структура тоже может быть представлена
в виде дерева, но важно, что количество возможных структур порядка 1,8L (это
доказывается в теореме, которую я не буду здесь представлять). Это тоже очень
много, поэтому задача поиска клики тоже не эффективна.
Тем не менее, есть алгоритм динамического программирования, который позво-
ляет нам найти за кубичное (а не квадратичное, как раньше) время найти струк-
туру, имеющую наибольшее количество спаренных оснований. Основная идея его
(как и любого алгоритма динамического программирования) заключается в том,
что если мы знаем все решения на какой-то части, то мы можем сказать, какое
будет решение на чуть большем фрагменте.
Динамическое программирование для
количества спаренных оснований (Нуссинофф)
• Количество спаренных оснований
в оптимальной структуре
определяется как максимум: ____-
S’v>1 = max {
S’J (нет спаривания) / А
тахк(8\Л1 + 8\у)+1; |
(к спаривается с /+1) \ 7/
}; к ОС
Время работы алгоритма: 7+1
T~O(L3)
Можно минимизировать не число спаренных оснований, а минимизировать энергию
(эта задача сложнее, но ее с помощью разных ухищрений тоже можно оставить ку-
бичной) . Минимизация все равно не позволяет достигнуть большой точности пред-
сказания. Проблемы предсказания вторичной структуры РНК.
Только около 65-70% тРНК сворачиваются в правильную структуру.
Для предсказания вторичной структуры используются энергетические параметры,
а они определены не очень точно. Более того, в клетке бывают разные условия,
и, соответственно, реализуются разные параметры.
Находится единственная структура с минимальной энергией, в то время как
обычно существует несколько структур с энергией, близкой к оптимальной.
Поэтому есть предложения искать субоптимальные структуры и искать эволюци-
онно консервативные структуры (структуры тРНК и рРНК определены именно так).
То есть забыть про энергию, и если мы знаем, что эти наборы РНК выполняют од-
ну и ту же функцию, то мы можем построить такую структуру, которая была бы
общей для всех этих последовательностей.
Теперь я расскажу, как это все применяется.
Исследование консервативности
альтернативного сплайсинга, или
Почему мышь не стала человеком?
Структура генов прокариот очень проста: есть начало, есть конец, получается
мРНК, которая имеет начало и конец, идет транскрипция, трансляция и белок.
Прокариоты Структура генов
MU' 1
Транскрипция
мРНК
Трансляция
Белок
У эукариот структура гена сложнее. Из длинной мРНК удаляются (вырезаются)
интроны (insertion sequences, вставочные последовательности), а оставшиеся
экзоны сшиваются в единую нить. Из пре-мРНК получается зрелая мРНК, процесс
называется сплайсингом. Потом происходит трансляция зрелой мРНК, в результате
образуется белок. Мы будем интересоваться экзонами и интронами.
Эуксвлютъг Структура генов
Транскрипция
ДНК
Сплайсинг
пре-мРНК
Белок
Если бы мы умели правильно предсказывать интроны и экзоны, мы бы могли раз-
метить ген на белок-кодирующие и белок-некодирующие участки.
Альтернативный сплайсинг
Оказывается, ситуация еще сложнее. РНК, прочитанная с одного и того же ге-
на, может сплайсироваться по-разному, что приводит к образованию мРНК с раз-
ными наборами экзонов: какой-то экзон в один вариант мРНК попадает, а в дру-
гой - нет, и в итоге получатся две разных мРНК и, соответственно, два разных
белка. Это называется альтернативным сплайсингом. Таким образом, на уровне
созревания мРНК могут образовываться разные РНК-продукты, которые приводят к
образованию разных белков.
Сплайсинг происходит в ядре, трансляция - в цитоплазме. Для изучения того,
что же оказалось в цитоплазме (то есть того, что подвергается трансляции),
секвенируют короткие, 500-600 до 1000 нуклеотидов куски цитоплазматической
РНК. Такие сиквенсы называются EST (expresstion sequence tag - "ярлыки экс-
прессируемых последовательностей"). EST - это короткие, прочитанные однократ-
но (то есть весьма неточно), фрагменты цитоплазматической (сплайсированной,
содержащей только экзоны) РНК. Если у нас есть геном, то мы можем эти EST
картировать на геном и, тем самым, найти, где находятся интроны и экзоны.
EST - короткие се тренированные фрагменты цитоплазматиче свой
(сплайсированной) РНК
Картирование EST на геном позволяет определить интроны.
Если при картирование EST полностью, без перерывов, соответствует геномной
последовательности - это ген без интронов. Если EST ложится на геном с пере-
рывами, то мы наблюдаем результат сплайсинга. Если же разные EST демонстриру-
ют несколько вариантов расположения в одном и том же участке генома (то есть
выявляют разные сочетания экзонов), то мы наблюдаем альтернативный сплайсинг.
Экзон, который может включаться в белок, а может и не включаться, называется
кассетным экзоном. мРНК с разными наборами экзонов данного гена (то есть в
которые некий кассетный экзон или включается или не включается), называются
изоформами.
Картирование EST на геном
Если картирование разных EST показывает несколько вариантов
расположения интронов, то это-альтернативныйо сплайсинг
Частота альтернативного
сплайсинга
Сначала альтернативный сплайсинг был обнаружен у вирусов, считалось, что
это экзотика. До 1998 г. считалось, что только около 6% генов человека имеют
альтернативный сплайсинг. Рассчитали, что для того, чтобы обеспечить наблю-
даемое разнообразие белков, в геноме человека должно было быть 80 - 100 тысяч
генов. В 1998 году было показано, что около половины генов человека имеют
альтернативный сплайсинг. За счет альтернативного сплайсинга число генов мо-
жет быть меньше числа кодируемых ими белков, так как с одного гена может об-
разовываться несколько белков.
Как было написано в какой-то газете "Многолетними усилиями ученых количест-
во генов человека было сокращено со 100 тысяч до 25". Действительно, по по-
следним оценкам в геноме человека около 25-30 тысяч генов. Оценка количества
белков не изменилась - разных белков около 80-100 тысяч. Разнообразие белков
обеспечивается альтернативным сплайсингом. Например, в одних клетках белок
должен быть в цитоплазме, в других - такой же белок в мембране, в третьих -
транспортироваться наружу. И это легко делается не за счет наличия разных ге-
нов для каждого случая, а за счет альтернативного сплайсинга, который цепляет
на N-конец разные сигналы, при том что "рабочая часть" белка остается одной и
той же, и одна изоформа белка размещается в мембране, другая изоформа белка -
в цитоплазме, и т.д.
Сейчас общеизвестно, что не менее 50% генов человека альтернативно сплайси-
руется.
Альтернативный сплайсинг бывает разных типов (галочками показано, как выре-
заются экзоны):
На этом рисунке показаны кассетный экзон (вставляемый в одни изоформы и от-
сутствующий в других), альтернативный акцептор, альтернативный донор, далее
интрон может либо вырезаться, либо не вырезаться.
Теперь вернемся к вопросу о человеке и мыши. Человек и мышь биологически
очень похожи. Белки похожи - уровень сходства аминокислотных последовательно-
стей 80%, также похожа значительная часть некодирующих областей генома. Прак-
тически у всех генов одинаково устроена экзон-интронная структура, для 99%
генов экзонная структура одинакова. Только 1% генов уникален у каждого гено-
ма , остальные гены имеют гомологи в другом геноме. Интересен тот факт, что
при таком относительно невысоком уровне различий человека от мыши внешне от-
личают легко. А два вида мухи дрозофилы вряд ли кто-то различит на глаз, хотя
генетически они различаются сильнее, чем человек и мышь.
Возникает вопрос: Если геномы одинаковы, то может быть, и белки одинаковы?
Непонятно, чем же человек отличается от мыши. Одинаково ли устроен альтерна-
тивный сплайсинг у мыши и человека?
Вопрос: одинаково пи устроен альтернативный
сплайсинги мыши и человека?
Наивный подход - найти альт.сплайсинг в человеке и в мыши и сравнить
Опасность - некоторые изоформы могут не иметь представителя в базе EST
Правильный подход:
Наивный подход для ответа на этот вопрос такой: возьмем весь набор альтер-
нативного сплайсинга мыши и человека и сравним его. Этот подход неправильный,
так как при исследовании альтернативного сплайсинга мы здесь имеем дело с
EST. Если у человека EST просеквенировано несколько миллионов штук, то у мыши
сделано всего несколько тысяч, поэтому там, где мы можем увидеть альтернатив-
ный сплайсинг у человека, можем ничего не увидеть у мыши, так как базы данных
еще не совсем полные. Поэтому такое сравнение даст нам неправильный ответ.
Правильный подход в данной ситуации
имеющихся данных на человеческой ДНК
затем этот белок проецируем на мышиный
белка (или его части) нет кодирующих
заключается в следующем: мы на основе
строим мРНК, соответствующую белку, и
геном. Если оказывается, что для этого
последовательностей в мышиной ДНК, то
это значит, что тот экзон, который есть у человека, отсутствует в геноме у
мыши.
Данные:
___________ гены мыши с геномной
ASMamDB последовательностью
HASDB
гены человека с геномной blast*
последовательностью
62 пары генов
человек-мышь
116 пар генов
человек-мышь
Всего Отобрано 166 пар орюогичных генов
Геном человек мышь
Гены с альт, сплайсингом 126 _124_
Кассетные экзоны 177 123
51 _46_
Альтернативный донор . 52 53
Сохраненный интрон 12 29
Всего альтернатив 285 252 I
Возьмем человеческие и мышиные гены, происходящие от общего предкового ге-
на. Возьмем такие пары генов-ортологов, сделаем сравнение. Мы получим некото-
рую выборку, среди которым 50% генов человека имеют такие изоформы, которых
нет у мыши, то же самое и с мышью.
Сравним пары генов человек-мышь. Например, ген бета-глобина человека и мыши
- такие гены, разошедшиеся в процессе эволюционного видообразования, называ-
ются ортологами. Выборку мы взяли не очень большую, в ней присутствовали ге-
ны, имеющие альтернативный спалйсинг. И оказалось, что у 52% человеческих ге-
нов есть такие экзоны, которых нет у мыши. И половина мышиных генов имеет та-
кие изоформы, которых нет у человека.
Результаты
Геном человек мышь
Консервативность С NC С NC
Кассетные экзоны 74 26 39 9
Альт донор 16 10 17 6
Альт, акцептор 19 15 16 9
Сохр интрон 5 0 10 4
Всего альтернатив 114 82 28
69% 31% 75% 25%
генов 41 |44 30 26
48% 52% 54% 46%
Но нам могут сказать - вы использовали EST, это неточные данные. Если мы
возьмем полноразмерные мРНК (а данные по ним гораздо точнее, хотя общее коли-
чество сиквенсов по ним меньше) , и проведем с ними ту же процедуру, то ока-
жется, что примерно треть генов человека имеет изоформы, которые в геноме мы-
ши не кодируются, отсутствуют, и также в геноме человека отсутствуют мышиные
экзоны.
Контроль
Отобраны альтернативы, подтвержденные
секвенированием мРНК
Геном человек МЫШЬ
Консервативность [с Inc с NC
Кассетные экзоны 74 56 26 25 70 39 9 5
Альт, донор 16 18 10 7 24 17 6 6
Альт, акцептор 19 13 15 5 15 16 9 6
Сохр. интрон 5 4 0 3 8 10 4 7
Всего альтернатив 114 I 96 51 30 117 82 28 24
69% [76% 31% 28 83% 75% 25%
генов 41 '“4? 44 68 30 26 Г5
48% 62% 52% 76% 54% 46%
А вот конкретные примеры: сверху изображены ДНК и ее изоформы у человека, а
снизу - то же у мыши. Например, для белка р53, который участвует в регуляции
клеточных процессов (раковое перерождение, апоптоз). Видно, что у мыши есть
изоформа, которая теряет экзон, порождая стоп в другом месте.
изоформы белка Р53 у человека и мыши
rtinii llfilfi I
Представленные данные показывают, что альтернативный сплайсинг - явление
весьма распространенное, и что мышь сильно отличается от человека по альтер-
нативному сплайсингу. Можно сделать и эволюционное предположение. По-
видимому, альтернативный сплайсинг допускает большую свободу для создания но-
вых белков, или изменения функций существующих белков, и в этом и состоит его
связь с эволюцией.
ЛЕКЦИЯ № 26.
НЕКОТОРЫЕ МЕТОДЫ
МОЛЕКУЛЯРНОЙ БИОЛОГИИ1
Полимеразная цепная реакция (ПЦР) — экспериментальный метод молекулярной
биологии, позволяющий добиться значительного увеличения малых концентраций
определённых фрагментов нуклеиновой кислоты (ДНК) в биологическом материале
(пробе).
Помимо простого увеличения числа копий ДНК (этот процесс называется ампли-
фикацией) , ПЦР позволяет производить множество других манипуляций с генетиче-
ским материалом (введение мутаций, сращивание фрагментов ДНК) и широко ис-
пользуется в биологической и медицинской практике, например, для диагностики
заболеваний (наследственных, инфекционных), для установления отцовства, для
клонирования генов, выделения новых генов.
История
В начале 1970-х годов норвежскому ученому Хьеллю Клеппе (Kjell Kleppe) из
лаборатории нобелевского лауреата Хара Гобинды Хораны (Наг Gobind Khorana)
пришла в голову мысль, что можно амплифицировать ДНК с помощью пары коротких
одноцепочечных молекул ДНК — синтетических праймеров [ 1]. Однако в то время
эта идея осталась невостребованной. Полимеразная цепная реакция была вновь
открыта в 1983 году Кэри Маллисом (Kary Mullis). Его целью было создание ме-
тода, который бы позволил амплифицировать ДНК в ходе многократных последова-
тельных удвоений исходной молекулы ДНК с помощью фермента ДНК-полимераЗы. Че-
рез 7 лет после опубликования этой идеи, в 1993 г., Маллис получил за неё Но-
белевскую премию[2].
В начале использования метода после каждого цикла нагревания — охлаждения
приходилось добавлять в реакционную смесь ДНК-полимеразу, так как она быстро
инактивировалась при высокой температуре, необходимой для разделения цепей
спирали ДНК. Процедура была очень неэффективной, требовала много времени и
фермента. В 1986 г. она была существенно улучшена. Было предложено использо-
вать ДНК-полимераЗы из термофильных бактерий [3]. Эти ферменты оказались тер-
мостабильными и были способны выдерживать множество циклов реакции. Их ис-
пользование позволило упростить и автоматизировать проведение ПЦР. Одна из
первых термостабильных ДНК-полимераз была выделена из бактерий Thermus
aquaticus и названа Taq-полимеразой. Недостаток этой полимеразы заключается в
том, что вероятность внесения ошибочного нуклеотида у неё достаточно высока,
так как у этого фермента отсутствуют механизмы исправления ошибок (3’^5’ эк-
зонуклеазная активность). Полимеразы Pfu и Pwo, выделенные из архей, обладают
таким механизмом, их использование значительно уменьшает число мутаций в ДНК,
но скорость их работы (процессивность) ниже, чем у Taq. Сейчас применяют сме-
си Taq и Pfu, чтобы добиться одновременно высокой скорости полимеризации и
высокой точности копирования.
В момент изобретения метода Маллис работал в компании Цетус (Cetus), кото-
рая и запатентовала метод ПЦР. В 1992 году Цетус продала права на метод и па-
тент на использование Taq-полимеразы компании Хофман-Ла Рош (Hoffmann-La
Roche) за 300 млн. долларов. Однако оказалось, что Taq-полимераза была оха-
рактеризована русским биохимиком Алексеем Калединым в 1980 году [4], в связи
1
Д.В. Ребриков
с чем компания Промега (Promega) пыталась в судебном порядке заставить Рош
отказаться от исключительных прав на этот фермент [5]. Американский патент на
метод ПЦР истёк в марте 2005 г.
Проведение ПЦР
Метод основан на многократном избирательном копировании определённого уча-
стка ДНК при помощи ферментов в искусственных условиях (in vitro). При этом
происходит копирование только того участка, который удовлетворяет заданным
условиям, и только в том случае, если он присутствует в исследуемом образце.
В отличие от амплификации ДНК в живых организмах, (репликации), с помощью ПЦР
амплифицируются относительно короткие участки ДНК. В обычном ПЦР-процессе
длина копируемых ДНК-участков составляет не более 3000 пар оснований (3
kbp[6]). С помощью смеси различных полимераз, с использованием добавок и при
определённых условиях длина ПЦР-фрагмента может достигать 20-40 тысяч пар
нуклеотидов. Это всё равно значительно меньше длины хромосомной ДНК эукарио-
тической клетки. Например, геном человека состоит примерно из 3 млрд пар ос-
нований [ 7 ] .
Компоненты реакции
Для проведения ПЦР в простейшем случае требуются следующие компоненты:
• ДНК-матрица, содержащая тот участок ДНК, который требуется амплифициро-
вать.
• Два праймера, комплементарные концам требуемого фрагмента.
• Термостабильная ДНК-полимераза — фермент, который катализирует реакцию
полимеризации ДНК. Полимераза для использования в ПЦР должна сохранять
активность при высокой температуре длительное время, поэтому используют
ферменты, выделенные из термофилов — Thermus aquaticus (Taq-полимераза),
Pyrococcus furiosus (Pfu-полимераза), Pyrococcus woesei (Pwo-полимераза)
и другие.
• Дезоксинуклеотидтрифосфаты (dATP, dGTP, dCTP, dTTP).
• Ионы Mg2+, необходимые для работы полимеразы.
• Буферный раствор, обеспечивающий необходимые условия реакции — pH, ион-
ную силу раствора. Содержит соли, бычий сывороточный альбумин.
Чтобы избежать испарения реакционной смеси, в пробирку добавляют высококи-
пящее масло, например, вазелиновое. Если используется амплификатор с подогре-
вающейся крышкой, этого делать не требуется.
Добавление пирофософатазы может увеличить выход ПЦР-реакции. Этот фермент
катализирует гидролиз пирофосфата, побочного продукта присоединения нуклео-
тидтрифосфатов к растущей цепи ДНК, до ортофосфата. Пирофосфат может ингиби-
ровать ПЦР-реакцию [8].
Праймеры
Специфичность ПЦР основана на образовании комплементарных комплексов между
матрицей и праймерами, короткими синтетическими олигонуклеотидами длиной 18—
30 оснований. Каждый из праймеров комплементарен одной из цепей двуцепочечной
матрицы, обрамляя начало и конец амплифицируемого участка.
После гибридизации матрицы с праймером (отжиг [9]), последний служит за-
травкой для ДНК-полимеразы при синтезе комплементарной цепи матрицы (см. ни-
же) .
Важнейшая характеристика праймеров — температура плавления (Тт) комплекса
праймер-матрица. Она определяется, как температура, при которой праймер при-
соединился к половине возможных сайтов связывания. Тт можно приблизительно
определить по формуле
Тт = 2 (пА + пт) +4 (nG + пс) ,
где пх — количество нуклеотидов X в праймере. Если праймер короткий и Тт
мала, то праймер может оказаться частично комплементарен другим участкам мат-
ричной ДНК, что может привести к появлению неспецифических продуктов. Верхний
предел температуры плавления ограничен оптимумом температуры действия полиме-
разы, активность которой падает при температурах выше 80 °C.
При выборе праймеров желательно придерживаться следующих критериев:
• GC-состав ~ 40—60 %;
• близкие Тт праймеров (отличия не более чем на 5 °C);
• отсутствие неспецифических вторичных структур — шпилек[10] и диме-
ров [11] ;
• желательно, чтобы на 3'-конце был гуанин или цитозин, поскольку они об-
разуют три водородные связи с молекулой матрицы, делая гибридизацию бо-
лее стабильной.
Амплификатор
Рис. 1: Амплификатор для проведения ПЦР
ПЦР проводят в амплификаторе — приборе, обеспечивающем периодическое охлаж-
дение и нагревание пробирок, обычно с точностью не менее 0,1 °C. Современные
амплификаторы позволяют задавать сложные программы, в том числе с возможно-
стью «горячего старта», Touchdown ПЦР (см. ниже) и последующего хранения ам-
плифицированных молекул при 4 °C. Для ПЦР в реальном времени выпускают прибо-
ры, оборудованные флуоресцентным детектором. Существуют также приборы с авто-
матической крышкой и отделением для микропланшет, что позволяет встраивать их
в автоматизированные системы.
Ход реакции
Обычно при проведении ПЦР выполняется 20—35 циклов, каждый из которых со-
стоит из трех стадий (рис. 2).
Фотография геля, содержащего маркерную ДНК (1) и продукты ПЦР-
реакции (2,3). Цифрами показана длина фрагментов ДНК в парах
нуклеотидов.
Денатурация
Двухцепочечную ДНК-матрицу нагревают до 94—96°С (или до 98 °C, если исполь-
зуется особенно термостабильная полимераза) на 0,5—2 мин., чтобы цепи ДНК ра-
зошлись. Эта стадия называется денатурацией, так как разрушаются водородные
связи между двумя цепями ДНК. Иногда перед первым циклом (до добавления поли-
меразы) проводят предварительный прогрев реакционной смеси в течение 2—5 мин.
для полной денатурации матрицы и праймеров.Такой приём называется горячим
стартом, он позволяет снизить количество неспецифичных продуктов реакции.
Отжиг
Когда цепи разошлись, температуру понижают, чтобы праймеры могли связаться
с одноцепочечной матрицей. Эта стадия называется отжигом. Температура отжига
зависит от состава праймеров и обычно выбирается на 4—5°С ниже их температуры
плавления. Время стадии — 0,5—2 мин. Неправильный выбор температуры отжига
приводит либо к плохому связыванию праймеров с матрицей (при завышенной тем-
пературе) , либо к связыванию в неверном месте и появлению неспецифических
продуктов (при заниженной температуре).
Элонгация
ДНК-полимераза реплицирует матричную цепь, используя праймер в качестве за-
травки . Это — стадия элонгации. Полимераза начинает синтез второй цепи от 3 ’ -
конца праймера, который связался с матрицей, и движется вдоль матрицы. Темпе-
ратура элонгации зависит от полимеразы. Часто используемые полимеразы Taq и
Pfu наиболее активны при 72 °C. Время элонгации зависит как от типа ДНК-
полимеразы, так и от длины амплифицируемого фрагмента. Обычно время элонгации
принимают равным одной минуте на каждую тысячу пар оснований. После окончания
всех циклов часто проводят дополнительную стадию финальной элонгации, чтобы
достроить все одноцепочечные фрагменты. Эта стадия длится 7-10 мин.
Рис. 2: Схематическое изображение первого цикла ПЦР. (1) Денату-
рация при 94—96°С. (2) Отжиг при 68 °C (например). (3) Элонгация
при 72 °C (Р=полимераЗа). (4) Закончен первый цикл. Две получив-
шиеся ДНК-цепи служат матрицей для следующего цикла, поэтому ко-
личество матричной ДНК в ходе каждого цикла удваивается.
ДНК-полимераза реплицирует матричную цепь, используя праймер в качестве за-
травки . Это — стадия элонгации. Полимераза начинает синтез второй цепи от 3 ’ -
конца праймера, который связался с матрицей, и движется вдоль матрицы. Темпе-
ратура элонгации зависит от полимеразы. Часто используемые полимеразы Taq и
Pfu наиболее активны при 72 °C. Время элонгации зависит как от типа ДНК-
полимеразы, так и от длины амплифицируемого фрагмента. Обычно время элонгации
принимают равным одной минуте на каждую тысячу пар оснований. После окончания
всех циклов часто проводят дополнительную стадию финальной элонгации, чтобы
достроить все одноцепочечные фрагменты. Эта стадия длится 7-10 мин.
Количество специфического продукта реакции (ограниченного праймерами) тео-
ретически возрастает пропорционально 2п, где п — число циклов реакции. На са-
мом деле эффективность каждого цикла может быть меньше 100 %, поэтому в дей-
ствительности Р ~ (1+Е)п, где Р — количество продукта, Е — средняя эффектив-
ность цикла.
Число «длинных» копий ДНК тоже растет, но линейно, поэтому в продуктах ре-
акции доминирует специфический фрагмент.
Рост требуемого продукта в геометрической прогрессии ограничен количеством
реагентов, присутствием ингибиторов, образованием побочных продуктов. На по-
следних циклах реакции рост замедляется, это называют «эффектом плато» [12].
Разновидности ПЦР
• Вложенная» ПЦР (Nested РСИ(англ.)) — применяется для уменьшения числа
побочных продуктов реакции. Используют две пары праймеров и проводят две
последовательные реакции. Вторая пара праймеров амплифицирует участок
ДНК внутри продукта первой реакции.
• «Инвертированная» ПЦР (Inverse РСИ(англ.)) — используется в том случае,
если известен лишь небольшой участок внутри нужной последовательности.
Этот метод особенно полезен, когда нужно определить соседние последова-
тельности после вставки ДНК в геном. Для осуществления инвертированной
ПЦР проводят ряд разрезаний ДНК рестриктазами с последующим соединением
фрагментов (лигирование). В результате известные фрагменты оказываются
на обоих концах неизвестного участка, после чего можно проводить ПЦР как
обычно.
• ПЦР с обратной транскрипцией (Reverse Transcription PCR, RT-PCR(aHrn.))
— используется для амплификации, выделения или идентификации известной
последовательности из библиотеки РНК. Перед обычной ПЦР проводят на мат-
рице мРНК синтез одноцепочечной молекулы ДНК с помощью ревертазы и полу-
чают одноцепочечную кДНК, которая используется в качестве матрицы для
ПЦР. Этим методом часто определяют, где и когда экспрессируются данные
гены.
• Ассиметричная ПЦР (англ. Asymmetric PCR) — проводится тогда, когда нужно
амплифицировать преимущественно одну из цепей исходной ДНК. Используется
в некоторых методиках секвенирования и гибридизационного анализа. ПЦР
проводится как обычно, за исключением того, что один из праймеров берет-
ся в большом избытке.
• Количественная ПЦР (Quantitative PCR, Q-PCR(англ.)) — используется для
быстрого измерения количества определенной ДНК, кДНК или РНК в пробе.
• Количественная ПЦР в реальном времени (Quantitative real-time PCR) — в
этом методе используют флуоресцентно меченые реагенты для точного изме-
рения количества продукта реакции по мере его накопления.
• Touchdown (Stepdown) ПЦР (Touchdown РСИ(англ.)) — с помощью этого метода
уменьшают влияние неспецифического связывания праймеров на образование
продукта. Первые циклы проводят при температуре выше температуры отжига,
затем каждые несколько циклов температуру снижают. При определённой тем-
пературе система пройдёт через полосу оптимальной специфичности прайме-
ров к ДНК.
• Метод молекулярных колоний (ПЦР в геле, англ. Polony - PCR Colony) — ак-
риламидный гель полимеризуют со всеми компонентами ПЦР на поверхности и
проводят ПЦР. В точках, содержащих анализируемую ДНК, происходит ампли-
фикация с образованием молекулярных колоний.
• ПЦР с быстрой амплификацией концов кДНК (англ. Rapid amplification of
cDNA ends, RACE-PCR)
• ПЦР длинных фрагментов (англ. Long-range PCR) — модификация ПЦР для ам-
плификации протяженных участков ДНК (10 тысяч оснований и больше). Ис-
пользуют две полимеразы, одна из которых — Taq-полимераза с высокой про-
цессивностью (то есть, способная за один проход синтезировать длинную
цепь ДНК), а вторая — ДНК полимераза с 3’-5 ’ эндонуклеазной активностью.
Вторая полимераза необходима для того, чтобы корректировать ошибки, вне-
сенные первой.
• RAPD PCR (англ. Random Amplification of Polymorphic DNA PCR, ПЦР co слу-
чайной амплификацией полиморфной ДНК — используется тогда, когда нужно
различить близкие по генетической последовательности организмы, напри-
мер, разные сорта культурных растений, породы собак или близкородствен-
ные микроорганизмы. В этом методе обычно используют один праймер неболь-
шого размера (20 — 25 п.н.). Этот праймер будет частично комплементарен
случайным участкам ДНК исследуемых организмов. Подбирая условия (длину
праймера, его состав, температуру и пр.), удается добиться удовлетвори-
тельного отличия картины ПЦР для двух организмов.
Если нуклеотидная последовательность матрицы известна частично или неиз-
вестна вовсе, можно использовать вырожденные праймеры, последовательность ко-
торых содержит вырожденные позиции, в которых могут располагаться любые осно-
вания. Например, последовательность праймера может быть такой: ...АТН..., где Н —
А, Т или С.
Применение ПЦР
ПЦР используется во многих областях для проведения анализов и в научных
экспериментах.
Криминалистика
ПЦР используют для сравнения так называемых «генетических отпечатков паль-
цев». Необходим образец генетического материала с места преступления — кровь,
слюна, сперма, волосы и т. п. Его сравнивают с генетическим материалом подоз-
реваемого. Достаточно совсем малого количества ДНК, теоретически — одной ко-
пии. ДНК расщепляют на фрагменты, затем амплифицируют с помощью ПЦР. Фрагмен-
ты разделяют с помощью гель-электрофореза. Полученную картину расположения
полос ДНК и называют генетическим отпечатком пальцев (англ. genetic
fingerprint).
Установление отцовства
Хотя «генетические отпечатки пальцев» уникальны (за исключением случая од-
нояйцевых близнецов), родственные связи все же можно установить, сделав не-
сколько таких отпечатков (рис. 3). Тот же метод можно применить, слегка моди-
фицировав его, для установления эволюционного родства среди организмов.
Рис. 3: Результаты электрофореза ДНК-фрагментов, амплифицирован-
ных с помощью ПЦР. (1) Отец. (2) Ребенок. (3) Мать. Ребенок
унаследовал некоторые особенности генетического отпечатка обоих
родителей, что дало новый, уникальный отпечаток.
Медицинская диагностика
ПЦР дает возможность существенно ускорить и облегчить диагностику наследст-
венных и вирусных заболеваний. Нужный ген амплифицируют с помощью ПЦР с ис-
пользованием соответствующих праймеров, а затем секвенируют для определения
мутаций. Вирусные инфекции можно обнаруживать сразу после заражения, за неде-
ли или месяцы до того, как проявятся симптомы заболевания.
Персонализированная медицина
Известно, что большинство лекарств действуют не на всех пациентов, для ко-
торых они предназначены, а лишь на 30-70 % их числа. Кроме того, многие ле-
карства оказываются токсичными или аллергенными для части пациентов. Причины
этого — отчасти в индивидуальных различиях в восприимчивости и метаболизме
лекарств и их производных. Эти различия детерминируются на генетическом уров-
не. Например, у одного пациента определенный цитохром (белок печени, отвечаю-
щий за метаболизм чужеродных веществ) может быть более активен, у другого —
менее. Для того, чтобы определить, какой разновидностью цитохрома обладает
данный пациент, предложено проводить ПЦР-анализ перед применением лекарства.
Такой анализ называют предварительным генотипированием (англ, prospective
genotyping).
Клонирование генов
Клонирование генов (не путать с клонированием организмов) — это процесс вы-
деления генов и, в результате генноинженерных манипуляций, получения большого
количества продукта данного гена. ПЦР используется для того, чтобы амплифици-
ровать ген, который затем вставляется в вектор — фрагмент ДНК, переносящий
чужеродный ген в тот же самый или другой, удобный для выращивания, организм.
В качестве векторов используют, например, плазмиды или вирусную ДНК. Вставку
генов в чужеродный организм обычно используют для получения продукта этого
гена — РНК или, чаще всего, белка. Таким образом, в промышленных количествах
получают многие белки для использования в сельском хозяйстве, медицине и др.
Рис. 4: Клонирование гена с использованием плазмиды.
(1) Хромосомная ДНК организма А. (2) ПЦР. (3) Множество копий
гена организма А. (4) Вставка гена в плазмиду. (5) Плазмида с
геном организма А. (6) Введение плазмиды в организм В. (7) Умно-
жение количества копий гена организма А в организме В.
Секвенирование ДНК
В методе секвенирования с использованием меченых флуоресцентной меткой или
радиоактивным изотопом дидезоксинуклеотидов ПЦР является неотъемлемой частью,
так как именно в ходе полимеризации в цепь ДНК встраиваются производные нук-
леотидов , меченые флуоресцентной или радиоактивной меткой. Это останавливает
реакцию, позволяя определить положения специфических нуклеотидов после разде-
ления синтезированных цепочек в геле.
Мутагенез
В настоящее время ПЦР стала основным методом проведения мутагенеза. Исполь-
зование ПЦР позволило упростить и ускорить процедуру проведения мутагенеза, а
также сделать её более надежной и воспроизводимой.
Примечания
1. Kleppe, К. et al. (1971): Studies on polynucleotides. XCVI. Repair
replications of short synthetic DNA's as catalyzed by DNA polymerases.
In: J. Mol. Biol. Bd. 56, S. 341-361. PMID 4927950
2. Нобелевские лауреаты по химии, 1993 г.(англ.)
3. R. К. Saiki, D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi, G. T.
Horn, К. B. Mullis, H. A. Erlich. Primer-Directed Enzymatic
Amplification of DNA with a Thermostable DNA Polymerase, in: Science.
239.1988, 487-491. ISSN 0036-8075 PMID: 2448875
4. Каледин А. С., Слюсаренко А. Г. , Городецкий С. И. // Биохимия.- 1980.-
T. 45.- C. 644-651.
5 . http://www.roche.com/med-cor-2005-09-12
6. 1 kbp (kilo base ра!г(англ.)) — 1 тысяча пар оснований, единица измере-
ния длины ДНК
7. Venter J, et al. (2001) . «The sequence of the human genome». Science
291 (5507): 1304-51. PMID 11181995
8 . http://www.biofidal.com/biofidal2/cat/2/pyro.php
9. Отжиг (англ, annealing) — гибридизация фрагментов ДНК
10. Шпилька — внутримолекулярная самокомплементарная структура
11. Димер — межмолекулярные структуры, образуемые праймерами друг с другом
или сами с собой
12 . [http://www.dna-technology.ru/doc/DNA-Technology_PCR-base.pdf «Теорети-
ческие основы ПЦР» (PDF)]
Литература
1. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и примене-
ние. Пер. с англ. — М.: Мир, 2002. — 589 с, илл. ISBN 5-03-003328-9
2. Щелкунов С. Н. Генетическая инженерия — Новосибирск: Сиб. унив. изд-во,
2004. - 496 с; илл. ISBN 5-94087-098-8
3. Патрушев Л. И. Искусственные генетические системы — М. : Наука, 2005 — В
2т.- ISBN 5-02-033278-Х
СЛОВАРЬ ТЕРМИНОВ
А
• Аберрация хромосомная (или хромосомная аномалия) — обобщенное название
любого из типов хромосомных мутаций: делеций, транслокаций, инверсий,
дупликаций. Иногда также обозначают и геномные мутации (анеуплодии, три-
сомии ит. д.).
• Авторадиография - способ обнаружения вещества, меченного радиоактивным
изотопом, путем наложения на чувствительную пленку.
• Акроцефалия (оксицефалия) - высокий «башенный» череп.
• Аллель — одна из двух или более альтернативных форм гена, каждая из ко-
торых характеризуется уникальной последовательностью нуклеотидов; алле-
ли , как правило, отличаются последовательностями нуклеотидов.
• Аллель дикого типа (нормальный): мутация гена, не затрагивающая его
функции.
• Аллель доминантный: аллель, одна доза которого достаточна для его фено-
типического проявления.
• Аллель мутантный: мутация гена, нарушающая его функцию.
• Аллель рецессивный: аллель, фенотипически проявляющийся только в гомози-
готном состоянии и маскирующийся в присутствии доминантного аллеля.
• Аллельные серии — моногенные наследственные заболевания, вызванные раз-
личными мутациями в одном и том же гене, но относящиеся к разным нозоло-
гическим группам по своим клиническим проявлениям.
• Алопеция - стойкое или временное, полное или частичное выпадение волос.
• Альфа-фетопротеин (АФП) - эмбриональный белок, обнаруживаемый в крови
плода, новорожденного, беременной женщины, а также в амниотической жид-
кости .
• Амниоцентез - прокол амниотического мешка с целью получения амниотиче-
ской жидкости.
• Ампликон - внехромосомная единица амплификации.
• Амплификатор ДНК (термоциклер) - прибор, необходимый для проведения по-
лимеразной цепной реакции (ПЦР); позволяет задавать нужное количество
циклов и выбирать оптимальные временные и температурные параметры для
каждой процедуры цикла.
• Амплификация — увеличение числа копий генов (количества ДНК)
• Амплификация ДНК - выборочное копирование определенного участка ДНК.
• Амфидиплоиды — эукариотические клетки, содержащие два двойных набора
хромосом в результате объединения двух геномов.
• Анеуплодия - измененный набор хромосом, в котором одна или несколько
хромосом из обычного набора или отсутствуют, или представлены дополни-
тельными копиями.
• Аниридия - отсутствие радужной оболочки.
• Анкилоблефарон - сращение краев век спайками, покрытыми слизистой обо-
лочкой .
• Анофтальмия — отсутствие одного или обоих глазных яблок.
• Антибиотик — вещество, подавляющее рост клеток или убивающее их. Обычно
антибиотики блокируют одну из стадий синтеза белков или нуклеиновых ки-
слот .
• Антиген — вещество (обычно белки, реже полисахариды), вызывающее у жи-
вотных иммунный ответ (образование антител).
• Антигенная детерминанта (эпитоп) - участок белковой или полисахаридной
молекулы, обладающей способностью вызывать образование антител данной
специфичности.
• Антикодон - последовательность из трех нуклеотидов в молекуле транспорт-
ной РНК, комплементарная кодирующему триплету в молекуле мРНК.
• Антимонголоидный разрез глаз — опущены наружные углы глазных щелей.
• Антимутагенез — процесс предотвращения закрепления (становления) мута-
ции , т. е. возврат первично поврежденной хромосомы или гена в исходное
состояние.
• Антитело — белок (иммуноглобулин), образуемый иммунной системой организ-
ма животных в ответ на введение антигена и способный вступать с ним в
специфическое взаимодействие.
• Антиципация - нарастание тяжести течения заболевания в ряду поколений.
• Анэнцефалия - полное или почти полное отсутствие головного мозга.
• Аплазия (агенезия) — полное врожденное отсутствие органа или части его.
• Арахнодактилия — необычно длинные и тонкие пальцы.
• Ассортативные браки - браки, при которых выбор брачного партнера по од-
ному или нескольким признакам неслучаен.
• Аутосома - любая неполовая хромосома. У человека имеется 22 пары ауто-
сом.
• Аутосомно-доминантное наследование — тип наследования, при котором одно-
го мутантного аллеля, локализованного в аутосоме, достаточно, чтобы бо-
лезнь (или признак) могла быть выражена.
• Аутосомно-рецессивное наследование - тип наследования признака или бо-
лезни , при котором мутантный аллель, локализованный в аутосоме, должен
быть унаследован от обоих родителей.
• Ахейрия (аподия) — недоразвитие или отсутствие кисти (стопы).
Б
• Бактериофаг — вирус бактерий: состоит из ДНК или РНК, упакованной в бел-
ковую оболочку.
• Банк (библиотека) генов — полный набор генов данного организма, получен-
ный в составе рекомбинантных ДНК.
• Белковая инженерия — создание искусственных белков с заданны ми свойст-
вами путем направленных изменений (мутаций) в генах или путем обмена ло-
кусами между гетерологичными генами.
• Биопсия хориона - процедура, осуществляемая на 7-11-й неделе беременно-
сти, с целью получения клеток для пренатальной диагностики.
• Блефарофимоз — укорочение век по горизонтали, т. е. сужение глазных ще-
лей .
• Блефарохалазия - атрофия кожи верхних век
• Блот-гибридизация по Саузерну - метод идентификации участков ДНК, содер-
жащих комплементарные ДНК-зонду последовательности, среди электрофорети-
чески разделенных фрагментов ДНК, фиксированных на твердом матриксе
(нитроцеллюлозных или нейлоновых фильтрах).
• Блотинг —перенос молекул ДНК, РНК или белка из геля, в котором шел элек-
трофорез , на нитроцеллюлозный фильтр (мембрану).
• Болезни аутосомные — обусловлены дефектами генов, локализованных в ауто-
сомах
• Болезни врожденные - присутствуют у ребенка с момента рождения
• Болезни доминантные - развиваются при наличии одного мутантного гена в
гетерозиготном состоянии
• Болезни моногенные - обусловлены дефектом одного гена
• Болезни мультифакториальные - имеющие в своей основе как генетическую,
так и средовую компоненты; генетическая компонента представляет собой
сочетание разных аллелей нескольких локусов, определяющих наследственную
предрасположенность к заболеванию при разных условиях внешней среды
• Болезни наследственные - имеющие в своей основе генетическую компоненту
• Болезни рецессивные - развиваются при наличии мутантного гена в гомози-
готном состоянии
• Болезни сцепленные с полом - обусловлены дефектом генов, локализованных
в X- или Y-хромосомах
• Болезни хромосомные - обусловлены числовыми и структурными нарушениями
кариотипа
• Брахидактилия — укорочение пальцев.
• Брахикамптодактилия — укорочение метакарпальных (метатарзальных) костей
и средних фаланг в сочетании с камптодактилией.
• Брахицефалия - увеличение поперечного размера головы при относительном
уменьшении продольного размера
В
• Вакцина — препарат ослабленного или убитого инфекционного агента (виру-
са , бактерии и т. п.) или его отдельных компонентов, несущих антигенные
детерминанты, способный вызывать образование иммунитета к данной инфек-
ции у животных (человека). Кроме того, в последнее время появились вак-
цины, произведенные методами генной инженерии (примером такой вакцины
может служить вакцина против гепатита В)
• Везикулы — мембранные пузырьки. Кроме того, везикулами в медицине назы-
вают любые элементы сыпи, представляющие собой пузырьки.
• Вектор — молекула ДНК, способная к включению чужеродной ДНК и к автоном-
ной репликации, служащая инструментом для введения генетической информа-
ции в клетку.
• Вектор для клонирования — любая небольшая плазмида, фаг или ДНК содержа-
щий вирус животных, в которые может быть встроена чужеродная вирусной
ДНК.
• Вирусы — инфекционные агенты неклеточной природы, способные в процессе
реализации генетической информации, закодирован ной в их геноме, пере-
строить метаболизм клетки, направив его в сторону синтеза вирусных час-
тиц. Вирусы могут иметь белковую оболочку, а могут и состоять только из
ДНК или РНК
• Витилиго — очаговая депигментация кожи.
• Водородная связь — образуется между электроотрицательным атомом молекулы
(кислород, азот) и электроположительным ядром водорода (протоном), кото-
рый , в свою очередь, ковалентно связан с другим электроотрицательным
атомом той же или соседней молекулы.
Врожденные болезни — болезни, имеющиеся при рождении.
Г
• р-Галактозидаза — фермент, гидролизующий - [3-галактозиды, в частности
лактозу, с образованием свободной галактозы.
• Гамета — зрелая половая клетка.
• Гаплоид — клетка, содержащая одинарный набор генов или хромосом.
• Гемизиготность — состояние организма, при котором какой-то ген представ-
лен в одной хромосоме.
• Ген — последовательность нуклеотидов в ДНК, которая обусловливает опре-
деленную функцию в организме или обеспечивает транскрипцию другого гена.
• Генетическая карта — схема расположения структурных генов и регуляторных
элементов в хромосоме.
• Генетический код — соответствие между триплетами в ДНК (или РНК) и ами-
нокислотами белков.
• Генная инженерия — совокупность приемов, методов и технологий получения
рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуще-
ствления манипуляций с генами и введения их в другие организмы.
• Генная терапия — введение генетического материала (ДНК или РНК) в клет-
ку, функцию которой он изменяет (или функцию организма).
• Геном — общая генетическая информация, содержащаяся в генах организма,
или генетический состав клетки. Термин «геном» иногда употребляется для
обозначения гаплоидного набора хромосом.
• Генотип 1) вся генетическая информация организма; 2) генетическая харак-
теристика организма по одному или нескольким изучаемым локусам.
• Ген-регулятор — ген, кодирующий регуляторный белок активирующий или по-
давляющий транскрипцию других генов.
• Ген-репортер — ген, чей продукт определяется с помощью простых и чувст-
вительных методов и чья активность в тестируемых клетках в норме отсут-
ствует . Используется в генно-инженерных конструкциях для маркирования
целевого продукта.
• Ген-усилитель (энхансер) — короткий сегмент ДНК, который влияет на уро-
вень экспрессии примыкающих к нему генов, увеличивая частоту инициации и
транскрипции.
• Гетерозигота — клетка (или организм), содержащая два различных аллеля в
конкретном локусе гомологичных хромосом.
• Гетерозиготность — наличие разных аллелей в диплоидной клетке.
• Гетерозиготный организм — организм, имеющий две различные формы данного
гена (разные аллели) в гомологичных хромосомах.
• Гетерохроматин — область хромосомы (иногда целая хромосома), имеющая
плотную компактную структуру в интерфазе.
• Гетерохромия радужки — неодинаковое окрашивание различных участков ра-
дужки .
• Гибридизация in situ — гибридизация между денатурированной ДНК клеток на
предметном стекле и меченной радиоактивными изотопами или иммунофлюорес-
центными соединениями одноцепочечной РНК или ДНК.
• Гибридизация ДНК — образование в опыте двуцепочечной ДНК или дуплексов
ДНК:РНК в результате взаимодействия комплементарных нуклеотидов.
• Гибридизация соматических клеток — слияние неполовых клеток, способ по-
лучения соматических гибридов (см.).
• Гибридный белок (полипептид) — см. Слитый белок (полипептид).
• Гибридомы — гибридные лимфоидные клетки, полученные путем слияния опухо-
левой миеломной клетки с нормальными лимфоидными клетками иммунизирован-
ного животного или человека.
• Гиперкератоз — чрезмерное утолщение рогового слоя эпидермиса.
• Гипертелоризм — увеличенное расстояние между внутренними краями глазниц.
• Гипертрихоз — избыточный рост волос.
• Гипоплазия врожденная — недоразвитие органа, проявляющееся дефицитом от-
носительной массы или размера органа.
• Гипоспадия — нижняя расщелина мочеиспускательного канала со смещением
его наружного отверстия.
• Гипотелоризм — уменьшенное расстояние между внутренними краями глазниц.
• Гирсутизм — избыточное оволосение у девочек по мужскому типу.
• Гликозилирование — присоединение к белку углеводного остатка
• Голандрическое наследование — наследование, сцепленное с Y-хромосомой.
• Голопрозэнцефалия — конечный мозг не разделен и представлен полусферой с
единственной вентрикулярной полостью свободно сообщающейся с субарахнои-
дальным пространством.
• Гомозигота — клетка (или организм), содержащая два одинаковых аллеля в
конкретном локусе гомологичных хромосом.
• Гомозиготность — наличие одинаковых аллелей в диплоидной клетке.
• Гомозиготный организм — организм, имеющий две идентичные копии данного
гена в гомологичных хромосомах.
• Гомологичные хромосомы — хромосомы, одинаковые по набору составляющих их
генов.
• Группа сцепления — все гены, локализованные в одной хромосоме.
д
• Дактилоскопия генная — выявление вариаций в числе и длине тандемных по-
второв ДНК.
• Делеция — тип хромосомной мутации, при которой утрачивается участок хро-
мосомы; тип генной мутации, при которой выпадает участок молекулы ДНК.
• Денатурация — нарушение пространственной структуры молекулы в результате
разрыва внутри- или межмолекулярных нековалентных связей.
• Дистихиаз — двойной ряд ресниц.
• ДНК-полимераза — фермент, ведущий матричный синтез ДНК.
• Долихоцефалия — преобладание продольных размеров головы над поперечными.
• Доминантность — преимущественное участие только одного аллеля в формиро-
вании признака у гетерозиготной клетки.
• Доминантный — признак или соответствующий аллель, проявляющийся у гете-
розигот .
• Дрейф генов — изменение частот генов в ряду поколений, обусловленное
случайными событиями митоза, оплодотворения и размножения.
• Дупликация — тип хромосомной мутации, при которой удвоен какой-либо уча-
сток хромосомы; тип генной мутации, при которой удвоен какой-либо уча-
сток ДНК.
3
• Зонд генетический — короткий отрезок ДИК или РНК известной структуры или
функции, меченный каким-либо радиоактивным или флуоресцентным соединени-
ем.
И
• Иммунитет — невосприимчивость организма к инфекционным агентам типа ви-
русов и микробов.
• Иммунотоксин — комплекс между антителом и каталитической субъединицей
какого-либо белкового яла (дифтерийного токсина, рицина, абрина и др.).
• Иммунофлюоресцентные Зонды — см. зонды ДНКовые, РНКовые.
• Индуктор — фактор (вещество, свет, теплота), вызывающий транскрипцию ге-
нов , находящихся в неактивном состоянии.
• Индукция профага — инициирование вегетативного развития фага в лизоген-
ных клетках.
• Интеграза — фермент, осуществляющий внедрение какого-либо генетического
элемента в геном через специфический сайт.
• Интегроны — генетические элементы, которые содержат в себе ген интегра-
зы, специфический сайт и рядом с ним промотор, что придает им способ-
ность интегрировать в себя мобильные генные кассеты и экспрессировать
присутствующие в них беспромоторные гены.
• Интерфероны — белки, синтезируемые клетками позвоночных в ответ на ви-
русную инфекцию и подавляющие их развитие.
• Интрон — некодирующий участок гена, который транскрибируется, а затем
удаляется из предшественника мРНК при сплайсинге (см. сплайсинг).
• Интронированный ген — ген, содержащий интроны.
• Итероны — повторяющиеся последовательности нуклеотидных остатков в ДНК.
К
• Каллус — масса недифференцированных клеток, образующаяся при повреждении
растения. Может образовываться из единичных клеток при их культивирова-
нии на искусственных средах.
• Кампомелия — искривление конечностей.
• Камптодактилия — сгибательная контрактура проксимальных межфаланговых
суставов пальцев кисти.
• Капсида — белковая оболочка вируса.
• Кассета экспрессионная — фрагмент ДНК, содержащий все необходимые гене-
тические элементы для экспрессии внедренного в него гена.
• кДНК — однонитевая ДНК, синтезируемая in vivo по РНКовой матрице с помо-
щью обратной транскриптазы.
• Кератоконус — коническое выпячивание роговицы.
• Клинодактилия — латеральное или медиальное искривление пальца.
• Клон — группа генетически идентичных клеток, возникших неполовым путем
от общего предка.
• Клонирование ДНК — разделение смеси рекомбинантных молекул ДНК путем их
введения в клетки методом трансформации или инфекции. Одна бактериальная
колония представляет собой клон, все клетки которого содержат одну и ту
же молекулу рекомбинантной ДНК.
• Клонирование клеток — их разделение путем рассева на питательном агаре и
получение колоний, содержащих потомство от изолированной клетки.
• Кодон — тройка расположенных подряд нуклеотидных остатков в ДНК или РНК,
кодирующая определенную аминокислоту или являющаяся сигналом окончания
трансляции.
• Компартментализация — ограничение процесса (продукта) определенной обла-
стью клетки.
• Компетентность — способность клеток к трансформации.
• Комплементарность (в генетике) — свойство азотистых оснований образовы-
вать с помощью водородных связей парные комплексы аденин—тимин (или ура-
цил) и гуанин—цитозин при взаимодействии цепей нуклеиновых кислот.
• Конкатемерная ДНК — линейная ДНК, в которой некоторый элемент (например,
фаговый геном) повторен несколько раз.
• Контиг — группа из нескольких последовательно соединенных секвенирован-
ных участков ДНК.
• Конъюгат — комплекс из нескольких ковалентно связанных молекул.
• Конъюгация — способ обмена генетической информацией у бактерий, при ко-
тором вследствие физического контакта между клетками происходит перенос
клеточной, плазмидной или транспозонной ДНК от донорной клетки в реципи-
ентную.
• Космида — вектор, содержащий cos-сайт ДНК фага Л.
• Краниосиностоз — преждевременное зарастание черепных швов, ограничиваю-
щее рост черепа и приводящее к его деформации.
• Криптофтальм — недоразвитие или отсутствие глазного яблока, век и глаз-
ной щели.
Л
• Лектины — белки, связывающие углеводы.
• Лигаза — фермент, образующий фосфодиэфирную связь между двумя полинук-
леотидами .
• Лиганд — молекула, распознаваемая специфической структурой, например,
клеточным рецептором.
• Лидерная последовательность — N-концевая последовательность секретируе-
мых белков, обеспечивающая их транспорт через мембрану и отщепляющаяся
при этом.
• Лизис — распад клетки, вызванный разрушением её оболочки.
• Лизогения — явление носительства бактериальными клетками фага в виде
профага (см. профаг).
• Линия клеток — генетически однородные клетки животных или растений, ко-
торые можно выращивать in vitro в течение неограниченно долгого времени.
• Линкер — короткий синтетический олигонуклеотид, применяемый для соедине-
ния фрагментов ДНК in vitro; обычно содержит участок узнавания опреде-
ленной рестриктазой.
• Липкие концы — комплементарные однонитевые участки ДНК, расположенные на
концах молекул ДНК.
• Липосомы — капельки жидкости, окруженные искусственной мембраной; искус-
ственные липидные везикулы (см. везикулы).
• Лиссэнцефалия (агирия) — отсутствие в больших полушариях головного мозга
борозд и извилин.
• Литическое развитие фага — фаза жизненного цикла фага, начинающаяся ин-
фекцией клетки и завершающаяся её лизисом.
• Локус — участок ДНК (хромосомы), где расположена определенная генетиче-
ская детерминанта.
М
• Макроглоссия — патологическое увеличение языка.
• Макросомия (гигантизм) — чрезмерно увеличенные размеры отдельных частей
тела или очень высокий рост.
• Макростомия — чрезмерно широкая ротовая щель.
• Макротия — увеличенные ушные раковины.
• Макроцефалия — чрезмерно большая голова.
• Маркерный ген — ген в рекомбинантной ДНК, кодирующий селективный при-
знак.
• Мегалокорнеа (макрокорнеа) — увеличение диаметра роговицы.
• Межвидовые гибриды — гибриды, полученные от слияния клеток, принадлежа-
щих к разным видам.
• Метаболизм — совокупность ферментативных процессов, обеспечивающих суще-
ствование и воспроизведение клетки.
• Метаболит — вещество, образующееся в химических реакциях живой клетки.
• Метилазы — ферменты, присоединяющие метильную группу к определенным азо-
тистым основаниям в ДНК.
• Микрогения — малые размеры нижней челюсти.
• Микрогнатия — малые размеры верхней челюсти.
• Микрокорнеа — уменьшение диаметра роговицы.
• Микростомия — чрезмерно узкая ротовая щель.
• Микротия — уменьшенные размеры ушных раковин.
• Микрофакия — малые размеры хрусталика.
• Микрофтальмия — малые размеры глазного яблока.
• Микроцефалия — малые размеры головного мозга и мозгового черепа.
• Миниклетки — клетки, не содержащие хромосомной ДНК. Модификация биополи-
мера — изменение его структуры.
• Монголоидный разрез глаз — опущены внутренние углы глазных щелей.
• Моноклональные антитела — антитела с определенной специфичностью, синте-
зируемые гибридомами (см. гибридомы).
• Морфогенез — осуществление генетической программы развития организма.
• Мугагенез — процесс индукции мутаций.
• Мутагены — физические, химические или биологические агенты, увеличиваю-
щие частоту возникновения мугаций.
• Мутация — изменение генетического материала, часто приводящее к измене-
нию свойств организма.
«Мыс вдовы» — клиновидный рост волос на лбу.
Н
• Ник — однонитевой разрыв в дуплексе ДНК с образованием 3 'ОН- и 5 'р-
концов; ликвидируется ДНК-лигаЗой (см. ДНК-лигаЗа).
• Нитрогеназа — фермент, осуществляющий фиксацию атмосферного азота.
• Нуклеазы — общее название ферментов, расщепляющих молекулы нуклеиновых
кислот.
О
• Обратная транскриптаза — фермент, катализирующий реакцию синтеза ДНК по
РНКовой матрице.
• Олигонуклеотид — цепь, состоящая из нескольких (от 2 до 20) нуклеотидных
остатков.
• Омфалоцеле — грыжа пупочного канатика.
• Онкогены — гены чьи продукты обладают способностью трансформировать эу-
кариотические клетки так, что они приобретают свойства опухолевых кле-
ток .
• Онкорнавирус — РНК-содержаший вирус, вызывающий перерождение нормальных
клеток в раковые; содержит в своем составе обратную транскриптазу.
• Оператор — регуляторный участок гена (оперона), с которым специфически
связывается репрессор (см. репрессор), предотвращая тем самым начало
транскрипции.
• Оперон — совокупность совместно транскрибируемых генов, обычно контроли-
рующих родственные биохимические функции.
П
• Пахионихия — утолщение ногтей.
• Перомелия — малая длина конечностей при нормальных размерах туловища.
• Пилонидальная ямка (сакральный синус, эпителиальный копчиковый ход) —
канал, выстланный многослойным плоским эпителием, открывающийся в межъя-
годичной складке у копчика.
• Плазмида — кольцевая или линейная молекула ДНК, реплицирующаяся автоном-
но от клеточной хромосомы.
• Полидактилия — увеличение количества пальцев на кистях и (или) стопах.
• Полилинкер — синтетический олигонуклеотид, содержащий участки узнавания
для нескольких рестриктаз (см. рестриктаза).
• Полимеразы — ферменты, ведущие матричный синтез нуклеиновых кислот.
• Полипептид — полимер, состоящий из аминокислотных остатков, связанных
пептидными связями.
• Праймер — короткая олиго- или полинуклеотидная последовательность со
свободной 3'ОН-группой, комплементарно связанная с однонитевой ДНК или
РНК; с его 3'-конца ДНК-полимераза начинает наращивать полидезоксирибо-
нуклеотидную цепь.
• Преаурикулярные папилломы — фрагменты наружного уха, расположенные впе-
реди ушной раковины.
• Преаурикулярные фистулы (преаурикулярные ямки) — слепо оканчивающиеся
ходы, наружное отверстие которых расположено у основания восходящей час-
ти завитка ушной раковины.
• Прогения — чрезмерное развитие нижней челюсти, массивный подбородок.
• Прогерия — преждевременное старение организма.
• Прогнатия — выступание верхней челюсти вперед по сравнению с нижней
вследствие её чрезмерного развития.
• Прозэнцефалия — недостаточное разделение переднего мозгового пузыря на
большие полушария.
• Прокариоты — организмы, у которых нет клеточного ядра.
• Промотор — регуляторный участок гена (оперона), к которому присоединяет-
ся РНК-полимераза с тем, чтобы начать транскрипцию.
• Протоонкогены — нормальные хромосомные гены, от которых произошли онко-
гены, содержащиеся в некоторых ретровирусах.
• Протопласт — растительная или микробная клетка, лишенная клеточной стен-
ки .
• Профаг — внутриклеточное состояние фага в условиях, когда его литические
функции подавлены.
• Процессинг — частный случай модификации (см. модификация), когда в био-
полимере уменьшается число звеньев.
• Птеригиум — крыловидные складки кожи.
Р
• Регулон — система генов, разбросанных по всему геному, но подчиняющихся
общему регуляторному белку.
• Рекомбинантная молекула ДНК (в генетической инженерии) — получается в
результате ковалентного объединения вектора и чужеродного фрагмента ДНК.
• Рекомбинантная плазмида — плазмида, содержащая фрагмент(ы) чужеродной
ДНК.
• Рекомбинантный белок — белок, часть аминокислотной последовательности
которого кодируется одним геном, а часть — другим.
• Рекомбинация in vitro — операции in vitro, приводящие к созданию реком-
бинантных молекул ДНК.
• Рекомбинация гомологическая — обмен генетическим материалом между двумя
гомологичными молекулами ДНК.
• Рекомбинация сайт-специфическая — объединение путем разрыва и слияния
двух молекул ДНК или участков одной молекулы, происходящее по определен-
ным сайтам.
• Ренатурация — восстановление исходной пространственной структуры моле-
кул .
• Репарация ДНК — исправление повреждений молекулы ДНК, восстанавливающее
её первоначальную структуру.
• Репликатор — участок ДНК, ответственный за инициацию репликации.
• Репликация — процесс удвоения молекул ДНК или геномных вирусных РНК.
• Репликон — молекула ДНК или её участок, находящиеся под контролем репли-
катора .
• Репрессия — подавление активности генов, чаще всего путем блокирования
их транскрипции.
• Репрессор — белок или антисмысловая РНК, подавляющие активность генов.
• Рестриктазы — сайт-специфические эндонуклеазы, составляющие часть систе-
мы рестрикции-модификации.
• Рестрикты — фрагменты ДНК, образовавшиеся после её гидролиза рестрикта-
зой.
• Рестрикционная карта — схема молекулы ДНК, на которой указаны места раз-
резания её различными рестриктазами.
• Рестрикционный анализ — установление мест расщепления ДНК рестриктазами.
• Ретровирусы — РНК-содержащие вирусы животных, кодирующие обратную транс-
криптазу и образующие провирус с хромосомной локализацией.
• Рецессивность — неучастие аллеля в формировании признака у гетерозигот-
ной клетки.
• Рибонуклеазы (РНКазы) — ферменты расщепляющие РНК.
С
• Сайт — участок молекулы ДНК, белка и т. п.
• Секвенирование — установление последовательности звеньев в молекулах
нуклеиновых кислот или белков (полипептидов).
• Селективные среды — питательные среды, на которых могут расти лишь клет-
ки с определенными свойствами.
• Септум — структура образующаяся в центре бактериальной клетки в конце
цикла деления и разделяющая её на две дочерние клетки.
• Симфалангия (ортодактилия) — сращение фаланг пальца.
• Синдактилия — полное или частичное сращение соседних пальцев кисти или
стопы.
• Синехии — фиброзные тяжи, соединяющие поверхности смежных органов.
• Синофриз — сросшиеся брови.
• Скафоцефалия — удлиненный череп с выступающим гребнем на месте прежде-
временно заросшего сагиттального шва.
• Скрининг — поиск в рассевах клеток или фагов тех колоний, которые содер-
жат рекомбинантные молекулы ДНК.
• Слитый белок (полипептид) — белок, образованный слиянием двух различных
полипептидов.
• Соматические гибриды — продукт слияния неполовых клеток.
• Соматические клетки — клетки тканей многоклеточных организмов, не отно-
сящиеся к половым.
• Спейсер — в ДНК или РНК — некодирующая последовательность нуклеотидов
между генами; в белках — аминокислотная последовательность, связывающая
соседние глобулярные домены.
• Сплайсинг — процесс формирования зрелой мРНК или функционального белка
путем удаления внутренних частей молекул — интронов РНК или интеинов у
белков.
• Стопа-«качалка» — стопа с провисающим сводом и выступающей кзади пяткой.
• Страбизм — косоглазие.
• Суперпродуцент — микробный штамм, нацеленный на синтез определенного
продукта в высокой концентрации.
• Сферофакия — шаровидная форма хрусталика.
т
• Телеангиоктазия — локальное чрезмерное расширение капилляров и мелких
сосудов.
• Телекант — смещение внутренних углов глазных щелей латерально при нор-
мально расположенных орбитах.
• Трансдукция — перенос фрагментов ДНК с помощью бактериофага.
• Транскриипия — синтез РНК на ДНК-матрице; осуществляется РНК-
полимеразой.
• Транскрипт — продукт транскрипции, т. е. РНК, синтезированная на данном
участке ДНК как на матрице и комплементарная одной из его нитей.
• Транскриптаза обратная — фермент, синтезирующий по РНК как по матрице
комплементарную ей однонитевую ДНК.
• Трансляция — процесс синтеза полипептида, определяемый матричной РНК.
• Транспозон — генетический элемент, реплицируемый в составе репликона и
способный к самостоятельным перемещениям (транспозиции) и интеграции в
разные участки хромосомной или внехромосомной ДНК.
• Трансфекция — трансформация клеток с помощью изолированной ДНК.
• Трансформация — изменение наследственных свойств клетки, вызванное по-
глощенной ДНК.
• Трансформация (в молекулярной генетике) — перенос генетической информа-
ции посредством изолированной ДНК.
• Трансформация (онкотрансформация) — частичная или полная дедифференци-
ровка клеток, вызванная нарушением регуляции роста клеток.
• Тригоноцефалия — расширение черепа в затылочной и сужение в лобной час-
ти .
• «Трилистник» — аномальная форма черепа, характеризующаяся высоким выбу-
хающим лбом, плоским затылком, выпячиванием височных костей, при соеди-
нении которых с теменными определяются глубокие вдавления.
У
• Умеренный фаг — бактериофаг. способный лизогенизовать клетку и в виде
профага находиться внутри бактериальной хромосомы или в плазмидном со-
стоянии .
Ф
• Фактор F (фактор фертильности, половой фактор) — коньюгативная F-
плазмида найденная в клетках Е. coli.
• Фенотип — внешнее проявление свойств организма, зависящих от его геноти-
па и факторов окружающей среды.
• Фильтр — расстояние от нижненосовой точки до красной каймы верхней губы.
• Фокомелия — отсутствие или значительное недоразвитие проксимальных отде-
лов конечностей, вследствие чего нормально раз витые стоны и (или) кисти
кажутся прикрепленными непосредственно к туловищу.
X
Химеры — лабораторные гибриды (рекомбинанты).
ц
• Центромера — локус на хромосоме, физически необходимый для распределения
гомологичных хромосом по дочерним клеткам.
Ш
• Шайн-Далгарно последовательность — участок прокариотической мРНК, необ-
ходимый для посадки на неё рибосом и её правильной трансляции. Содержит
последовательность нуклеотидов, комплементарную 3'-концу 16S рибосомной
РНК.
• Штамм — линия клеток (или вирусов), ведущая начало от одной клетки (или
вируса).
э
• Экзон — сохраняющаяся при сплайсинге часть интронированного гена.
• Экзонуклеаза — фермент, гидролизующий фосфодиэфирные связи с концов ДНК.
• Экзофтальм — смещение глазного яблока вперед, сопровождающееся расшире-
нием глазной щели.
• Эксплантат — выделенный из организма материал какой-либо ткани.
• Экспрессия гена — процесс реализации информации, закодированной в гене.
Состоит из двух основных стадий .— транскрипции и трансляции.
• Эктопия хрусталика (подвывиххрусталика, вывих хрусталика) — смещение
хрусталика из стекловидной ямки.
• Эктропион века — выворот края века.
• Электрофорез — разделение электрически заряженных полимеров в электриче-
ском поле. Обычно ведется в гелях (гель-электрофорез), чтобы зоны разде-
ляемых молекул не размывались тепловым движением.
• Эндонуклеаза — фермент, гидролизующий фосфодиэфирные связи внутри нити
ДНК.
• Энхансер — регуляторный участок ДНК, усиливающий транскрипцию с ближай-
шего к нему промотора.
• Эпибульбарный дермоид — липодермоидные разрастания на поверхности глаз-
ного яблока, чаще на границе радужки и белочной оболочки.
• Эпикант — вертикальная кожная складка у внутреннего угла глазной щели.
• Эукариоты — организмы, клетки которых содержат ядра.
Компьютер
Редактор: Иван Трус
БАЗОВЫЙ КУРС: WINDOWS И ИНТЕРНЕТ
Лекция 1
ВВЕДЕНИЕ
На протяжении всей истории человечество овладевало веществом, энергией и
информацией. Целые эпохи человеческого развития получили название по имени
наиболее передовых технологий: "каменный век" - технология обработки камня,
"бронзовый век" - технология обработки металла, "век книгопечатания" - овла-
дение технологией распространения информации, "век электричества" - техноло-
гия овладения новых видов энергии, "атомный век" - технология развития атом-
ной энергии. Потребность выразить и передать информацию привела к появлению
речи, письменности, изобразительного искусства, телефона, радио, телевидения,
почтовой связи. С появлением ЭВМ стали говорить о "веке информации" в связи с
тем, что технология информационной обработки обретает универсальный всеобщий
характер. Появление ЭВМ - универсальной машины для обработки информации озна-
чает революцию в области накопления, передачи и обработки информации. Возрас-
тает роль информационных компонент в производстве, науке, образовании, меди-
цине и т.д. (во всех сферах жизни общества). Процесс перехода к информацион-
ному обществу охватил сегодня все развитые страны. Ядро этого процесса со-
ставляет обработка информации на ЭВМ.
Отсутствие информационных сетей можно сравнить с отсутствием электричества
или бездорожьем. Компьютеризацию (компьютер - это другое название ЭВМ) - с
электрификацией или индустриализацией. Информатику - с физикой.
В современной вычислительной технике информация кодируется с помощью после-
довательности двоичных сигналов (0,1). Цифры 0,1 называются битами. Последо-
вательность из 8 нулей и единиц - байтом. Количество бит - называется инфор-
мационным объемом сообщения. Наряду с битами и байтами используются более
крупные единицы.
1 Кбит = 210 « 1000 бит = 103 бит (тысяча)
1 Мбит = 220 « 106 бит (миллион)
1 Гбит = 230 « 109 бит (миллиард)
Термин "Информация" является первичным, неопределяемым понятием (смысл и
Значимость информации в информатике теряются). Информатика изучает обработку
информации с помощью ЭВМ. Информация в информатике имеет научную (не обыден-
ную) трактовку. Скорость передачи информации измеряется в бодах 1 бод = 1
бит/сек.
Двоичная информация безлика. Поэтому используются качественные и структур-
ные характеристики. Двоичную информацию делят на управляющую (команды) и
управляемую (операнды). На множестве команд определяют структуру (пересылка,
арифметические команды, логические команды), как и на множестве операндов
(символы, числа, признаки). Команды комплектуются в блоки, операнды - в мас-
сивы. В свою очередь алгоритмы (последовательности команд) имеют свою струк-
туру рабочие алгоритмы, подпрограммы (вычисление элементарных функций, упа-
ковка, сортировка, поиск, переадресация).
Таким образом, информатика есть наука и техника, связанные с машинной обра-
боткой, хранением и передачей информации. Она занимается формализованными
способами представления информации, точными предписаниями по ее обработке, и
машинами обработки информации.
Цель информатики состоит в разработке способов решения задач информационной
обработки на компьютерах, включая синтез компьютеров и организацию связей их
в вычислительные информационные системы.
В смысле применения компьютеров информатику можно считать состоящей из 3-х
взаимосвязанных частей - технических средств (hardware), программных средств
(software) и алгоритмических средств (brainware).
В свою очередь информатику в целом и каждую ее часть обычно рассматривают с
разных позиций:
• как отрасль народного хозяйства она состоит из совокупности различных форм
хозяйствования, где занимаются производством компьютерной техники, про-
граммных продуктов и разработкой технологии переработки информации. Специ-
фика информатики, как отрасли народного хозяйства состоит в том, что от
нее во многом зависит рост производительности труда в других отраслях на-
родного хозяйства.
• Как фундаментальная наука информатика занимается разработкой методологии
создания информационного обеспечения процессов управления любыми объектами
на базе компьютерных информациооных систем. Цель фундаментальных исследо-
ваний в информатике - получение обобщенных знаний о любых информационных
ситемах, выявление общих закономерностей их построения и функционирования.
• Как прикладная дисциплина информатика занимается разработкой информацией-
них систем и технологий в различных конкретных областях и выработкой реко-
мендаций по их проектированию, производству и функционированию.
Понятие "информация" является фундаментальным для информатики. Информация в
информатике понимается как абстрактное понятие - это значение выражений, ука-
заний, высказываний, графических изображений и т.д. Только предписывание оп-
ределенного значения переводит представление в информацию. Таким образом, ин-
формация в информационной системе воспринимается как некоторая математическая
структура. Переход от представления к элементам этой математической структуры
называется интерпретацией. Информация в своем абстрактном виде не может быть
записана непосредственно, а потому всегда может быть только как-то изображе-
на . Компьютер служит универсальным средством отображения информации. Именно
поэтому персональный компьютер стал важнейшим инструментом информационного
общества.
Фактически, компьютеры подобно машинам и механизмам предшествующей эпохи,
облегчившим физический труд, облегчают и постепенно начинают видоизменять ум-
ственный труд.
Новые компьютерные технологии могут стать средством, способствующим обуче-
нию, ориентированному на конкретные возможности студента, значительно более
деликатно и точно измеряющим уровень студенческих знаний на основе стандарти-
зованного подсчета тестов, предоставляя при этом высокий уровень обучающих
инструментов. Компьютер был рожден на стыке различных отраслей науки и техни-
ки, На этом же стыке и используется, самым фактом своего существования поощ-
ряя междициплинарные исследования и подходы.
Образование, ориентированное на использование компьютеров, обогащает про-
цесс обучения. Возможности инноваций в образовании в настоящее время превыша-
ют все, что известно было в предыдущие пять веков, которые отделяют нас от
изобретения печатного станка. Это технологическое изобретение способствовало
становлению больших университетов в эпоху Возрождения, которые и до сих пор
занимают центральное место в системе образования. Грядущие технологии могут
оставить не меньший отпечаток в истории образования, обеспечивая прочные, ус-
тойчивые выгоды человечества.
Появившаяся концепция дистанционного образования, как симбиоз заочного обу-
чения и телекоммуникационного использования компьютеров, имеет особую важ-
ность . Экономическое значение дистанционного обучения состоит в интегрирова-
нии национального опыта во всех областях знаний для передачи будущим поколе-
ниям. В решении социальных задач цель дистанционного обучения состоит в пре-
доставлении гражданам равных возможностей получения образования, независимо
от географического местонахождения. Дистанционное образование обеспечивает
гибкость в выборе времени и места обучения, возможность обучения граждан в
тех географических районах страны, где нет иных возможностей получения высше-
го образования, делает процесс обучения более индивидуальным, эффектным, ув-
лекательным и интересным.
Темпы экономического развития мирового сообщества столь высоки, что струк-
тура профессиональных потребностей общества будет постоянно изменяться. Сле-
довательно, для образованного взрослого населения с особой остротой встанет
вопрос о переобучении или дополнительном высшем образования, а для России эта
проблема особенно актуальна вследствие произошедших социальных изменений. За-
дача переобучения со временем встанет перед всеми категориями специалистов, в
том числе и перед самими преподавателями. Возможность дистанционного получе-
ния образования особенно актуальна для взрослого населения, поскольку для
большинства из них затруднена возможность свободного передвижения и им важно,
чтобы университет пришел к ним, а не они к университету. Дистанционная форма
обучения решает важную психологическую проблему обучения разновозрастного
контингента. Важнейшее значение в этом процессе займет компьютерное образова-
ние и самообразования с помощью компьютерных библиотек, обучающих программ и
т.д.
Общие сведения о
персональном компьютере
Состав компьютера
Персональный компьютер (ПК) - цифровое средство для обработки информации
(информации в широком смысле слова - изображение, звук, текст и т.д.).
Предназначение ПК - следуя поступающим от человека командам, ПРИНИМАТЬ, ПЕ-
РЕРАБАТЫВАТЬ, ХРАНИТЬ И ВЫДАВАТЬ ИНФОРМАЦИЮ.
Computer = Hardware (аппаратное обеспечение)+ Software (программное обеспече-
ние)
Устройства персонального компьютера
Процессор - электронная микросхема, выполняющая действия по обработке ин-
формации. Его характеризуют фирма-производитель (Intel, AMD и др.), тип
(Celeron, Pentium III и др.), тактовая частота (например, 366МГц, 500 МГц) и
ряд других параметров.
Память компьютера подразделяется на внутреннюю и внешнюю:
Память ПК= Внутренняя память + Внешняя память
Внутренняя память находится рядом с процессором и служит для хранения ин-
формации , непосредственно участвующей в вычислительном процессе. Выполняемые
процессором программы всегда находятся во внутренней памяти.
В свою очередь внутренняя память состоит из двух частей: постоянная память
и оперативная память.
Внутренняя память ПК= Постоянная память + Оперативная память
Постоянная память обеспечивает только хранение и выдачу (чтение) информа-
ции. В ней хранятся программы, обеспечивающие запуск компьютера и поддержку
его работы.
Оперативная память служит для приема (записи), хранения и выдачи (чтения)
информации.
После выключения компьютера информация в оперативной памяти безвозвратно
теряется, поэтому для предотвращения потерь информацию необходимо размещать
(сохранять) во внешней памяти. Для этого во многих программах предусмотрена
специальная команда «Сохранить...».
Внешняя память - набор устройств для долговременного хранения информации:
жесткий магнитный диск (винчестер) - стандартное устройство для хранения
программ и данных;
дисковод для гибких магнитных дисков (дискет) обеспечивает хранение сравни-
тельно небольших объемов информации, ее конфиденциальность и возможность пе-
реноса на другие компьютеры;
дисковод для оптических дисков (CD-ROM, DVD-диски.). Оптические диски обла-
дают значительной информационной емкостью по сравнению с дискетами. Запись
информации на компакт-диски осуществляется только на специальном оборудова-
нии .
В настоящее время для хранения сверхбольших объемов информации могут ис-
пользоваться дополнительные устройства внешней памяти: стримеры, магнитооп-
тичские устройства (ZIP и другие).
Главной характеристикой памяти ПК является ее объем (емкость). Основная
единица измерения объема - байт. Упрощенно можно считать, что 1 байт хранит
информацию об одном символе. Например, для хранения в памяти компьютера слова
«информация» потребуется 10 байт. Часто используются производные единицы ем-
кости памяти:
1 Кбайт = 1024 байт
1 Мбайт = 1024 Кбайт
1 Гбайт = 1024 Мбайт.
Винчестер - это устройство, на котором чаще всего хранятся Ваши данные. Бы-
тует легенда, объясняющая, почему за жесткими дисками повелось такое причуд-
ливое название. Первый жесткий диск, выпущенный в Америке в начале 70-х го-
дов, имел емкость по 30 Мб информации на каждой рабочей поверхности. В то же
время, широко известная в той же Америке магазинная винтовка О. Ф. Винчестера
имела калибр - 0.30; может грохотал при своей работе первый винчестер как ав-
томат или порохом от него пахло - не знаю, но с той поры стали называть жест-
кие диски винчестерами.
В процессе работы компьютера случаются сбои. Вирусы, перебои энергоснабже-
ния, программные ошибки - все это может послужить причиной повреждения инфор-
мации , хранящейся на Вашем жестком диске. Повреждение информации далеко не
всегда означает ее потерю, так что полезно знать о том, как она хранится на
жестком диске, ибо тогда ее можно восстановить. Тогда, например, в случае по-
вреждения вирусом загрузочной области, вовсе не обязательно форматировать
весь диск (!), а, восстановив поврежденное место, продолжить нормальную рабо-
ту с сохранением всех своих данных.
Устройство жесткого диска Жесткий диск (HDD - Hard Disk Drive) устроен сле-
дующим образом: на шпинделе, соединенным с электромотором, расположен блок из
нескольких дисков (блинов), над поверхностью которых находятся головки для
чтения/записи информации. Форма головкам придается в виде крыла и крепятся
они на серпообразный поводок. При работе они "летят" над поверхностью дисков
в воздушном потоке, который создается при вращении этих же дисков. Очевидно,
что подъемная сила зависит от давления воздуха на головки. Оно же, в свою
очередь, зависит от внешнего атмосферного давления. Диск разбит на дорожки
(или треки), которые в свою очередь поделены на сектора. Две дорожки, равно-
удаленные от центра, но расположенные по разные стороны диска, называются ци-
линдрами .
Дорожка
разбита на
сектора
Диски
Хранение информации
Жесткий диск, как и всякое другое блочное устройство, хранит информацию
фиксированными порциями, которые называются блоками. Блок является наименьшей
порцией данных, имеющей уникальный адрес на жестком диске. Для того чтобы
прочесть или записать нужную информацию в нужное место, необходимо предста-
вить адрес блока в качестве параметра команды, выдаваемой контроллеру жестко-
го диска. Размер блока уже довольно с давних пор является стандартным для
всех жестких дисков - 512 байт. Файлы на диске записываются в свободные
кластеры, поэтому таблица постоянно изменяется. Таблица расположена на диске,
состояние которого она описывает. Таблица размещения файлов - FAT (File
Allocation Table) основной элемент организации файловых систем.
Файловая система
Часть операционной системы, обеспечивающая запись и чтение файлов на диско-
вых носителях (магнитных, магнитооптических, оптических). Файловая система
определяет логическую и физическую структуру файла, идентификацию и сопутст-
вующие данные файла. Широко известны следующие файловые системы:
- файловая система операционной системы MS-DOS, в основу которой положена
таблица размещения файлов - FAT16 (File Allocation Table). Таблица содержит
сведения о расположении всех файлов (каждый файл делится на кластеры в соот-
ветствии с наличием свободного места на диске, кластеры одного файла не обя-
зательно расположены рядом). Файловая система MS-DOS имеет значительные огра-
ничения и недостатки, например, под имя файла отводится 12 байт, работа с же-
стким диском большого объема приводит к значительной фрагментации файлов; До
появления ОС Windows95 принято было соглашение, согласно которому имя файла
состоит из двух частей: собственно имени и расширения имени, разделяемых точ-
кой . Имя файла не должно превышать 8 символов, а его расширение - 3 символов.
Имя от расширения отделяется точкой. Имя и расширение должны включать только
буквы латинского алфавита и цифры. Такой принцип именования файлов использу-
ется в MS-DOS. Данное соглашение не является стандартом, и в ряде случае слу-
чаются отклонения. Например, некоторые версии MS-DOS допускают символы рус-
ского алфавита. Недостаток коротких имен является их низкая информативность.
Трудно несколькими символами выразить характеристику файла.
- файловая система операционной системы Windows 95/98 имеет уровневую
структуру FAT 32, что позволяет поддерживать одновременно несколько файловых
систем. Старая файловая система MS-DOS поддерживается непосредственно, а фай-
ловые системы разработанные не фирмой Microsoft, поддерживаются с помощью
специальных модулей. Имеется возможность использовать длинные (до 254 симво-
лов) имена файлов.
FAT32, впервые представленная в Windows 95 SR2, была прямой наследницей
файловой системы FAT (FAT16). Отличалась она тем, что прежде всего, более эф-
фективно использовала дисковое пространство, поскольку поддерживала меньший
размер кластера на одном и том же томе. Кластер, или адресуемый блок, - это
минимальный блок дисковой памяти для записи данных на томе; он задается при
форматировании диска. Все хранящиеся на диске файлы, вне зависимости от их
реального размера, имеют размер, кратный размеру кластера. Кроме того, любой,
даже небольшой файл (например, размером 1 Кбайт) должен занимать по меньшей
мере один кластер драгоценного дискового пространства. Из-за этих ограничений
на томах FAT остается много неиспользованного пространства.
И хотя FAT32 задействует более широкий диапазон размеров кластеров, чем
FAT, и, следовательно, полнее использует дисковое пространство, NTFS справля-
ется с этой задачей еще лучше. В Таблице приведены размеры используемых FAT-
кластеров, в зависимости от размера диска. Разработчики Microsoft реализовали
в FAT32 возможности обеспечения целостности данных. В FAT32 имеется несколько
функций, обеспечивающих доступ к критичным данным о структуре системы. Напри-
мер, FAT32 может перенести корневой каталог диска из одного места в другое,
если область диска, где этот каталог располагался, случайно была испорчена.
Возможно использование резервной копии таблицы размещения файлов, если первая
копия была испорчена или недоступна. И наконец, FAT32 создает расширенную за-
грузочную запись, содержащую резервные копии структуры критичных данных. Все
эти новые особенности делают FAT32 более надежной, чем ее предшественница
FAT.
Длинные имена
В Windows95 было введено понятие "длинного" имени. Такое имя может содер-
жать до 256 символов. Длинное имя может включать любые символы, за исключени-
ем девяти специальных: \ /:*?"<> | .В имени допускаются пробелы и не-
сколько точек.
Наряду с длинным именем операционные системы Windows 95 и Windows98 создают
также и короткое имя файла - необходимое для возможности работы с данным фай-
лом на рабочих местах с устаревшими операционными системами.
Атрибуты файлов
Кроме имени и расширении файла операционная система хранит для каждого фай-
ла дату его создания (изменения) и несколько флаговых величин, называемых ат-
рибутами файла. Атрибуты - это дополнительные параметры, определяющие свойст-
ва файлов. Операционная система позволяет их контролировать и изменять; со-
стояние атрибутов учитывается при проведении автоматических операций с файла-
ми . Существует четыре основных атрибута:
• Только для чтения (Read Only)
• Скрытый (Hidden)
• Системный (System)
• Архивный (Archive)
Атрибут "Только для чтения" ограничивает возможности работы с файлом. Его
установка означает, что файл не предназначен для внесения изменений.
Атрибут "Скрытый" указывает, что данный файл не следует отображать на экра-
не при проведении файловых операций. Это мера защиты против случайного повре-
ждения файла.
Атрибут "Системный" используется для файлов, связанными с функционированием
операционной системы. Его отличительная особенность в том, что средствами
операционной системы его изменить нельзя. Как правило, большинство файлов,
имеющих установленный атрибут "Системный", имеют также установленный атрибут
"Скрытый".
Атрибут "Архивный" в прошлом использовался для работы программ резервного
копирования. Предполагалось, что любая программа, изменяющая файл, должна ав-
томатически устанавливать этот атрибут, а средство резервного копирования
должно его сбрасывать. Таким образом, очередному резервному копированию под-
лежали только те файлы, у которых этот атрибут был установлен. Современные
программы резервного копирования используют другие средства для установления
факта изменения файла, и данный атрибут во внимание не принимается, а его из-
менение вручную средствами операционной системы не имеет практического значе-
ния .
Вернемся к размещению файлов на жестком диске. К сожалению, часто происхо-
дит путаница между такими понятиями как сектор", "кластер" и "блок". Фактиче-
ски, между "блоком" и "сектором" разницы нет. Правда, одно понятие логиче-
ское, а второе топологическое. "Кластер" - это несколько секторов, рассматри-
ваемых операционной системой как одно целое. Почему не отказались от простой
работы с секторами? Переход к кластерам произошел потому, что размер таблицы
размещения файлов (FAT) был ограничен, а размер диска увеличивался. В случае
FAT16 для диска объемом 512 Мб кластер будет составлять 8 Кб, до 1 Гб - 16
Кб, до 2 Гб - 32 Кб и так далее.
Для того чтобы однозначно адресовать блок данных, необходимо указать все три числа (но-
мер цилиндра, номер сектора на дорожке, номер головки). Такой способ адресации диска был
широко распространен и получил впоследствии обозначение аббревиатурой CHS (cylinder,
head, sector). Именно этот способ был первоначально реализован в BIOS, поэтому впоследст-
вии возникли ограничения, связанные с ним. Дело в том, что BIOS определил разрядную сетку
адресов на 63 сектора, 1024 цилиндра и 255 головок. Однако развитие жестких дисков в то
время ограничилось использованием лишь 16 головок в связи со сложностью изготовления. От-
сюда появилось первое ограничение на максимально допустимую для адресации емкость жестко-
го диска: 1024*16*63*512 = 504Mb. BIOS - Базовая система ввода/вывода (от aHrn.>Basic
Input/Output System) . BIOS - это программы, предназначенные для выполнения следующих
функций:
- тестирования основных устройств компьютера;
- распознавания типов устройств, установленных в компьютере;
- вызова блока начальной загрузки операционной системы;
- обслуживания системных прерываний.
Как правило, базовую систему ввода/вывода называют BIOS. В большинстве ком-
пьютеров BIOS записывается изготовителем компьютера в постоянное запоминающее
устройство и пользователь не имеет средств изменять ее. В настоящее время вы-
пускаются компьютеры, у которых BIOS записывают во флэш-память, и у пользова-
теля появляется возможность изменять BIOS по мере необходимости. Некоторые
считают BIOS частью операционной системы
Со временем, производители стали делать HDD все большего размера. Соответ-
ственно число цилиндров на них превысило 1024, максимально допустимое число
цилиндров (с точки зрения старых BIOS). Однако, адресуемая часть диска про-
должала равняться 504 Мбайтам, при условии, что обращение к диску велось
средствами BIOS. Это ограничение со временем было снято введением так назы-
ваемого механизма трансляции адресов, о котором чуть ниже. Проблемы, возник-
шие с ограниченностью BIOS по части физической геометрии дисков, привели в
конце концов к появлению нового способа адресации блоков на диске. Этот спо-
соб довольно прост. Блоки на диске описываются одним параметром - линейным
адресом блока. Адресация диска линейно получила аббревиатуру LBA (logical
block addressing). Линейный адрес блока однозначно связан с его CHS адресом:
Iba = (cyl*HEADS + head)*SECTORS + (sector-1);
Введение поддержки линейной адресации в контроллеры жестких дисков дало
возможность BIOS’ам заняться трансляцией адресов. Суть этого метода состоит в
том, что если в приведенной выше формуле увеличить параметр HEADS (ГОЛОВКИ),
то потребуется меньше цилиндров, чтобы адресовать то же самое количество бло-
ков диска. Но зато потребуется больше головок. Однако головок-то как раз ис-
пользовалось всего 16 из 255. Поэтому ВЮЗ'ы стали переводить избыточные ци-
линдры в головки, уменьшая число одних и увеличивая число других. Это позво-
лило им использовать разрядную сетку головок целиком. Это отодвинуло границу
адресуемого BIOS’ом дискового пространства до 8Gb.
Следует упомянуть и о режиме работы с дисками большого объема (Large Mode).
Этот режим работы предназначен для работы жестких дисков объемом до 1 Гб. В
Large Mode количество логических головок увеличивается до 32, а количество
логических цилиндров уменьшается вдвое. При этом обращения к логическим го-
ловкам 0..F транслируются в четные физические цилиндры, а обращения к голов-
кам 10..1F - в нечетные. Винчестер, размеченный в режиме LBA, несовместим с
режимом Large, и наоборот.
Дальнейшее увеличение адресуемых объемов диска с использованием прежних
сервисов BIOS стало принципиально невозможным. Действительно, все параметры
задействованы по максимальной "планке" (63 сектора, 1024 цилиндра и 255 голо-
вок) . Тогда был разработан новый расширенный интерфейс BIOS, учитывающий воз-
можность очень больших адресов блоков. Однако этот интерфейс уже не совместим
с прежним, вследствие чего старые операционные системы, такие как DOS, кото-
рые пользуются старыми интерфейсами BIOS, не смогли и не смогут переступить
границы в 8GB. Практически все современные системы уже не пользуются BIOS’ом,
а используют собственные драйвера для работы с дисками. Поэтому данное огра-
ничение на них не распространяется. Но следует понимать, что прежде чем сис-
тема сможет использовать собственный драйвер, она должна как минимум его за-
грузить . Поэтому на этапе начальной загрузки любая система вынуждена пользо-
ваться BIOS’ом. Это и вызывает ограничения на размещение многих систем за
пределами 8GB, они не могут оттуда загружаться, но могут читать и писать ин-
формацию (например, DOS который работает с диском через BIOS).
Разделы, или Partitions
Обратимся теперь к размещению операционных систем на жестких дисках. Для
организации систем дисковое адресное пространство блоков разделяется на час-
ти, называемые разделами (partitions). Разделы полностью подобны целому диску
в том, что они состоят из смежных блоков. Благодаря такой организации для
описания раздела достаточно указания начала раздела и его длины в блоках. Же-
сткий диск может содержать четыре первичных раздела. Во время загрузки компь-
ютера, BIOS загружает первый сектор головного раздела (загрузочный сектор) по
адресу 0000h:7C00h и передает ему управление. В начале этого сектора располо-
жен загрузчик (загрузочный код), который прочитывает таблицу разделов и опре-
деляет загружаемый раздел (активный). А дальше все повторяется. То есть он
загружает загрузочный сектор этого раздела на этот же адрес и снова передает
ему управление. Разделы являются контейнерами всего своего содержимого. Этим
содержимым является, как правило, файловая система. Под файловой системой с
точки зрения диска понимается система разметки блоков для хранения файлов.
После того, как на разделе создана файловая система и в ней размещены файлы
операционной системы, раздел может стать загружаемым. Загружаемый раздел име-
ет в своем первом блоке небольшую программу, которая производит загрузку опе-
рационной системы. Однако для загрузки определенной системы нужно явно запус-
тить ее загрузочную программу из первого блока. Разделы с файловыми системами
не должны пересекаться. Это связано с тем, что две разные файловые системы
имеют каждая свое представление о размещении файлов, но когда это размещение
приходится на одно и то же физическое место на диске, между файловыми систе-
мами возникает конфликт. Этот конфликт возникает не сразу, а лишь по мере то-
го, как файлы начинают размещаться в том месте диска, где разделы пересекают-
ся. Поэтому следует внимательно относиться к разделению диска на разделы. Са-
мо по себе пересечение разделов не опасно. Опасно именно размещение несколь-
ких файловых систем на пересекающихся разделах. Разметка диска на разделы еще
не означает создания файловых систем. Однако, уже сама попытка создания пус-
той файловой системы (то есть форматирование), на одном из пересекающихся
разделов может привести к возникновению ошибок в файловой системе другого
раздела. Все сказанное относится в одинаковой степени ко всем операционным
системам, а не только самым популярным. Диск разбивается на разделы программ-
ным путем. То есть, Вы можете создать произвольную конфигурацию разделов. Ин-
формация о разбиении диска хранится в самом первом блоке жесткого диска, на-
зываемым главной загрузочной записью (Master Boot Record (MBR)). MBR является
основным средством загрузки с жесткого диска, поддерживаемым BIOS.
Монитор - устройство для визуального отображения информации. Отображение
информации происходит на экране монитора.
Для указания позиции экрана, в которой отображается информация, вводимая с
клавиатуры, используется КУРСОР (как правило, мигающая вертикальная черта).
Все мониторы можно классифицировать: По схеме формирования изображения. По
своим размерам. По способу воздействия на человека.
Классификация по способу формирования изображения По этому принципу монито-
ры делятся на: ЭЛТ-мониторы ЖК-мониторы Катодолюминисцентные мониторы.
ЭЛТ-мониторы (мониторы с электронно-лучевой трубкой) очень похожи на теле-
визоры. У них тот же принцип формирования сигнала - направленный электронный
пучок вызывает свечение точек на экране. Этот тип мониторов позволяет созда-
ние изображения с максимальной контрастностью, яркостью и цветностью. Их не-
достатки - высокое потребление электроэнергии и вред, наносимый здоровью.
ЖК-мониторы (жидкокристаллический) формируют изображение за счет того, что
определенные точки экрана становятся прозрачными или непрозрачными в зависи-
мости от приложенного электрического поля. Поскольку жидкокристаллические
ячейки сами не светятся, ЖК-мониторам нужна подсветка. ЖК-мониторы имеют ма-
лое потребление энергии, изображение на них приятно глазам, отсутствует ра-
диационное излучение монитора. Их недостатки - малая контрастность изображе-
ния и малые скорости регенерации (обновления изображения) экрана.
ЖК-мониторы (жидкокристаллический) формируют изображение за счет того, что
определенные точки экрана становятся прозрачными или непрозрачными в зависи-
мости от приложенного электрического поля. Поскольку жидкокристаллические
ячейки сами не светятся, ЖК-мониторам нужна подсветка. ЖК-мониторы имеют ма-
лое потребление энергии, изображение на них приятно глазам, отсутствует ра-
диационное излучение монитора. Их недостатки - малая контрастность изображе-
ния и малые скорости регенерации (обновления изображения) экрана.
Катодолюминисцентные мониторы все еще находятся в стадии разработки. Они
имеют те же характеристики изображения, что и ЭЛТ, однако в отличие от них
позволяют создавать мониторы очень большого (более одного метра) размера. Их
главный недостаток - малое время безотказной работы, после которого элементы
экрана приходится менять. Другие параметры мониторов автору не известны.
Размеры экранов
Следующим важным свойством монитора является размер его экрана. Как прави-
ло, чем больше экран, тем с большим разрешением (соответственно - меньшим
размером единицы изображения) можно на нем работать. Но при этом непропорцио-
нально высоко возрастает его цена и увеличивается требуемое место для монито-
ра на столе.
За размеры монитора считают размер его экрана по диагонали. Для ЭЛТ стан-
дартными являются размеры 14", 15", 17", 19", 21", 23" (" - обозначение дюй-
ма.) Для ЖК-мониторов - 13", 14", 15", 17", 19".
Воздействие на здоровье
Наиболее вредными для здоровья являются ЭЛТ-мониторы. Прежде всего За счет
рентгеновского излучения, возникающего из-за торможения электронов в трубке,
и паразитного ультрафиолетового излучения монитора. К тому же на глазах чело-
века отрицательно сказывается неравномерная яркость экрана, нечеткость изо-
бражения (ведущая к близорукости) и выпуклость экрана (ведущая к астигматиз-
му. )
Первым решением, которое хоть как-то ослабляло вред от мониторов, явилось
применение защитного экрана на монитор. Он увеличивал контрастность изображе-
ния, устранял солнечные блики, защищал от ультрафиолета. Однако его защита
все равно была недостаточной. В связи с этим стали выпускаться мониторы, под-
держивающие различные эргономические стандарты. Первым таким стандартом яв-
лялся шведский стандарт MPR-II. Затем за стандартизацию взялись международные
организации, и появились стандарты ТСО’92, ТСО’95 и ТСО’99. Уже для монито-
ров , удовлетворяющих стандарту ТСО’92, не требовалось защитного экрана. Стан-
дарт же ТСО’99 гарантирует непричинение вреда здоровью при 8-ми часовой рабо-
те за экраном монитора, удовлетворяющего данному стандарту.
В отличие от ЭЛТ-мониторов ЖК-мониторы не приносят вреда Здоровью.
Мышь - устройство для ввода в компьютер управляющих воздействий. Манипуля-
тор мышь значительно моложе клавиатуры - его коммерческое использование нача-
лось в 80-х годах XX века. Однако, по сравнению с клавиатурой, он имеет го-
раздо больше модификаций. Первоначально манипулятор представлял собой некий
предмет с прямоугольным основанием 7*10 см,максимальной высотой 4 см, имеющий
одну кнопку и подсоединенный к компьютеру через COM-порт длинным (1 м) прово-
дом. Именно за сходство этого манипулятора со взрослой мышью (те же размеры и
длинный хвост) эти манипуляторы также стали называть мышами.
Современные мыши обязательно содержат две кнопки - левую и правую. При пе-
редвижении мыши также передвигается курсор (стрелка и прямоугольник) на экра-
не. Поскольку большинство людей "правши" и держат мышь правой рукой, то ос-
новной клавишей мыши является левая, находящаяся под указательным пальцем.
Если же Вы - левша и держите мышь левой рукой, для Вас желательно сделать ос-
новной правую клавишу мыши. Попросите Вашего специалиста, чтобы он настроил
мышь под Вас.
Назначение клавиш меняется от программы к программе. Грубо можно считать,
что левая (основная) клавиша мыши соответствует клавише Enter на клавиатуре,
а правая - клавише Context.
Кроме двух клавиш мышь может содержать третью клавишу, колесико, боковую
клавишу и т.п. Третья клавиша мыши расположена по середине и иногда выполняет
функцию клавиши Esc на клавиатуре. Колесико служит для прокрутки текста (или
изображения) в окне Windows.
Остальные элементы используются еще реже.
В основном все выпускаемые в настоящее время мыши делятся на опто-
механические и оптические. В опто-механических мышах при движении ее по по-
верхности вращается шарик, находящийся в нижней части мыши. Вращение шарика
передается двум перпендикулярно расположенным колесикам, с которых с помощью
оптронов считывается относительное перемещение мыши. Колесико делается из ме-
талла и обтягивается резиной. Поэтому они плохо двигаются на полированной
или, наоборот, загрязненной поверхности. Лучше всего подкладывать под опто-
механическую мышь специальный коврик, увеличивающий трение между шариком и
поверхностью, по которой перемещается мышь.
Оптические мыши не содержат движущихся частей. Их работа основана на том,
что при движении мыши по поверхности меняется отражение луча света, которое
испускает мышь. Поэтому этот тип мыши меньше чувствителен к шероховатости и
чистоте поверхности, однако более чувствителен к ее оптическим свойствам. С
этой мышью также лучше использовать коврики, однако теперь они служат другой
цели - обеспечить оптическую неоднородность поверхности. Поэтому иногда про-
стой поворот коврика на 90 градусов может перевести мышь в нерабочее состоя-
ние .
Клавиатура
Клавиатура - устройство для ввода информации в ПК. Она представляет собой
"доску", на которой в 5 или в 6 рядов расположены клавиши. Все клавиши делят-
ся на 2 группы - белые и серые.
Белые клавиши служат для ввода алфавитно-цифровых знаков в компьютер. За
каждой клавишей закреплены 2 знака. Соответствие знаков клавишам называется
"раскладкой" клавиатуры. О раскладке будет сказано ниже.
К белым клавишам относят и так называемые функциональные клавиши. Всего их
двенадцать (на старых клавиатурах - десять) штук. Они служат для подачи про-
граммам, выполняемых на компьютере, определенных команд. Они отличаются от
серых клавиш тем, что имеют собственные функции, работают самостоятельно и не
управляют курсором. Серые клавиши предназначены для изменения режима ввода
символов и управлением курсором. Рассмотрим их по-подробнее:
1. Клавиша Esc (называется "Эскейп".) Обычно служит для отмены какого-либо
действия программы, выхода из чего-либо и т.д.
2. Клавиша Enter (читается "Энтр".) Служит для завершения ввода строки и пе-
рехода на следующую строку, принятия положительного ответа на вопрос, осу-
ществление действия по-умолчанию, закрытия окна с подтверждением изменений
и т.д. Это как бы антипод клавиши "Эскейп" (Esc.)
3. Клавиша Tab (читается "Таб"). Предназначена для формирования отступов в
тексте, перехода из одного пункта диалогового окна к другому и т.п.
4. Клавиша CapsLock (читается "капслок".) Предназначена для ввода прописных
букв.
5. Две клавиши Shift (читается "Шифт"). Меняет регистр клавиатуры. Дело в
том, что все клавиши на клавиатуре могут вводить два символа. На клавиату-
ре обычно нарисовано два знака, расположенные один над другим. Простое на-
жатие клавиши вводит один символ, указанный на клавише снизу. Это - нижний
регистр. Для того, чтобы ввести символ, изображенный на клавише сверху,
надо перейти в верхний регистр, т.е. нажать клавишу Shift и, не отпуская
ее, нужную клавишу.
6. Две клавиши Ctrl (читается "Контрол"). Служит для подачи компьютеру ко-
манд. Команда подается одновременным нажатием клавиши Ctrl и алфавитно-
цифрового символа (далее обозначается Сбг1+<буква>).
7. Две клавиши Alt (читается "Альт"). Их назначение сходно с клавишей Ctrl.
8. Клавиша Ins (читается "Инсерт"). Служит для переключения режима встав-
ки/замены символов при вводе, а также для выделения элементов в списке,
добавления элемента в список и т.д.
9. Backspace (читается "Бэкспейс"). Предназначена для удаления символа слева
от курсора.
10. Клавиша Del (читается "Делит"). Удаляет символ, на котором стоит курсор
(или символ справа от него). Она также предназначена для удаления элемента
графического интерфейса.
11. Клавиша Ноте (читается "Хоум"). Предназначена для перехода в начало
(строки, списка и т.п.)
12. Клавиша End (читается "Энд"). Предназначена для перехода в конец (стро-
ки , списка и т.п.)
13. Клавиша PgUp (читается "Пэйдж ап"). Предназначена для перемещения (про-
крутки) изображения на экране вниз.
14. Клавиша PgDn (читается "Пэйдж даун"). Предназначена для перемещения изо-
бражения на экране вверх.
15. Стрелка вверх. Служит для перемещения курсора на одну строку вверх.
16. Стрелка вниз. Служит для перемещения курсора на одну строку вниз.
17. Стрелка влево. Служит для перемещения курсора на одну позицию влево.
18. Стрелка вправо. Служит для перемещения курсора на одну позицию вправо.
19. Scroll Lock (читается "Скролл лок").
20. PrScr (читается "Принт скрин").
21. Pause/Break (читается "Пауз" или "Брейк").
Клавиши 19-21 используются в специальных целях и здесь не рассматриваются.
Существует также дополнительная цифровая клавиатура, расположенная справа.
Она предназначена как для ввода цифр (0 - 9 и разделитель десятичных знаков;
горит индикатор "Num Lock"), так и для для дублирования серых клавиш 8, 10 -
18 (смотри выше.) Кроме того, эта клавиатура может дублировать мышь. Управля-
ется эта клавиатура клавишей "Num Lock" (читается "Нум лок"): когда она нажа-
та (горит указатель) вводятся цифры. При повторном нажатии на эту клавишу ин-
дикатор гасится, и дополнительная клавиатура дублирует серые клавиши управле-
ния курсором.
Кроме этого, на дополнительной клавиатуре продублированы клавиши /,*,-,+ и
Enter, которые стали "серыми". Кроме ввода указанных знаков, они еще применя-
ются для других целей.
Специально для работы с Microsoft Windows 95/98/2000/NT4 на клавиатуре поя-
вились две клавиши Win (читается "Вин") с изображение логотипа Windows, и
клавиша Context (читается "контекст") с изображением пункта меню. Об этих
клавишах будет сказано при описании работы Microsoft Windows.
Так расположены клавиши на стандартной клавиатуре. Однако у обычной клавиа-
туре есть два полярных недостатка: Она занимает много места. Она неудобна эр-
гономически .
Первый недостаток существен в носимых компьютерах, и там она изменена. У
эргономичных моделей клавиатура приняла "волнистую" форму, в которой алфавит-
но-цифровая клавиатура разделена на две части, которые расположены под углом
~120 градусов. Соответственно перенесен блок клавиш управления курсором.
Раскладки клавиатуры латинский/русский.
X С V В N И < > ?
х I с v b n m « I • I /
На верхнем регистре вводятся прописные (заглавные) буквы, а на нижнем —
строчные (маленькие). На русском регистре вводятся символы кириллицы, а на
латинском — латиницы.
Мы вам советуем сразу начать осваивать так называемый слепой десятипальце-
вый метод работы на клавиатуре. Это не так уж и сложно, самое главное - не
торопиться и набраться терпения. Посмотрите на рисунок.
На нем указаны зоны ответственности каждого пальца:
• Левый мизинец постоянно находится над буквой Ф, а нажимает еще и буквы Й и
Я.
• Левый безымянный постоянно находится над буквой Ы, а нажимает еще и буквы
Ц и Ч.
• Левый средний постоянно находится над буквой В, а нажимает еще и буквы У и
С.
• Левый указательный постоянно находится над буквой А, а нажимает еще и бук-
вы К, Е, П, М и И. На большинстве клавиатур буква А отмечена риской.
• Правый указательный постоянно находится над буквой О, а нажимает еще и бу-
квы Н, Г, Р, Т и Ь. На большинстве клавиатур буква О тоже отмечена риской.
• Правый средний постоянно находится над буквой Л, а нажимает еще и буквы Ш
и Б.
• Правый безымянный постоянно находится над буквой Д, а нажимает еще и буквы
Щ и Ю.
• Правый мизинец постоянно находится над буквой Ж, а нажимает еще и буквы 3,
X, Ъ и Э.
• Большие пальцы рук нажимают на длинную клавишу пробела.
Лекция 2
Включение компьютера
Перед началом работы с компьютером необходимо включить его в электросеть.
Процесс включения компьютера следующий:
Подготовительные операции
Проверьте нет ли повреждения корпуса у компьютера и периферийных устройств.
Устройства со сломанными корпусами использовать нельзя! Заправьте принтер бу-
магой (если Вы его используете). Проверьте, нет ли каких-нибудь "висящих"
проводов, не подключенных к компьютеру. Если Вы используете компьютерную
сеть, проверьте подключения кабелей к адаптеру сети.
Порядок включения
Компьютерные устройства вообще включаются "от периферии к центру". Это зна-
чит , что необходимо включать устройства в следующей последовательности:
Включите все USB- и SCSI-устройства (USB - универсальная последовательная
шина, SCSI - стандартный интерфейс малых компьютеров.)
К USB-устройствам, как правило, относятся недорогие сканеры, цифровые фото-
аппараты и видеокамеры, иногда мониторы, клавиатура, принтер и мышь. К SCSI-
устройствам относят различные сканеры и дополнительные дисководы.
Включите приборы, подключенные к последовательным и параллельным портам
компьютера (принтеры, модемы и т.д.). Включите монитор (кнопкой на его перед-
ней панели) . Удалите 3,5" или 5,25" дискету из дисковода, если она в нем
есть. Примечание: для удаления 5.25" дискету из дисковода необходимо: а) По-
вернуть рукоятку-фиксатор дискеты на 90 градусов против часовой стрелки; Ь)
достать дискету из дисковода и положить в футляр.
Для удаления 3.5" дискеты необходимо: а) нажать кнопку выброса дискеты из
дисковода (ниже щели для вставки дискеты); Ь) вынуть дискету из дисковода и
положить в футляр.
Включите приборы, подключенные к последовательным и параллельным портам
компьютера (принтеры, модемы и т.д.) Включите кнопку "Сеть" на передней пане-
ли компьютера
Примечание: После выключения компьютера его можно повторно включать через
30 секунд.
Загрузка операционных систем
После включения компьютера начинается внутреннее тестирование компьютера. В
процессе тестирования проверяется: Источник питания. Видеокарта. Процессорный
блок. Память. Подключенные дисководы. Внешние устройства.
Если хотя бы одно из этих устройств не работает нормально, компьютер издаст
несколько длинных писков и дальше загружаться не будет. Если тест прошел ус-
пешно, то после короткого писка на экране появляется информация о конфигура-
ции компьютера, и начинает загружаться операционная система.
Далее при загрузке операционной системы на экране будет отображаться за-
ставка . Заставка может прерываться запросом на выбор конфигурации операцион-
ной системы. При этом в память загружаются различные программы. Вид и после-
довательность загрузки задается администратором. Вам важно знать, что при за-
грузке программ MS-DOS Вам предается выбрать конфигурацию компьютера. Выбери-
те конфигурацию по умолчанию (нажмите Enter.)
После загрузки программ MS-DOS появляется чистый рабочий стол Windows
95/98. Далее Вас могут попросить войти в сеть. Для этого посредине экрана
появится диалоговое окно. Вид этого окна зависит от того, в какую сеть Вы
входите. При входе в сеть у Вас могут запросить идентификационное имя (так
называемый "логин" (login) или "ник") и пароль для входа в сеть. Введите свой
логин и пароль, а если Вы его не знаете (или у Вас его нет), в поле имени на-
берите слово "guest" (без кавычек!), а в поле пароля - ничего не набирайте.
После этого Вас могут запросить выбрать пользовательские настройки. Для
этого также нужно набрать свой логин и пароль. Далее, после входа в систему,
загрузятся другие программы Microsoft Windows. Ничего на предпринимайте, пока
все программы не загрузятся (не перестанет работать винчестер.)
Сбои при загрузке Windows
Если Вы неправильно выключили компьютер, то после загрузки программ MS-DOS
система сообщит Вам об этом и начнет проверять жесткие диски. Дождитесь окон-
чания проверки. Иногда, после сбоя, Windows попросит Вас запуститься в "безо-
пасном" режиме ("Safe Mode"). При загрузке в этом режиме система устраняет
возникшие неполадки. После загрузки Windows в безопасном режиме перезагрузите
компьютер, и он запустится в нормальном режиме. Примечание: для перезагрузки
компьютера необходимо: а) выбрать пункт "Завершение работы" в главном меню;
Ь) выбрать пункт "Перезагрузить компьютер"; с) выбрать клавишу ОК и дождаться
процедуры повторного запуска компьютера.
Введение в операционную
систему Windows 95/981
Программное обеспечение (ПО) - совокупность программ, обеспечивающих функ-
ционирование ПК в интересах пользователя.
Под операционной системой (ОС) понимается набор программ, управляющих вы-
числительным процессом в компьютере и обеспечивающих диалог с пользователем.
В настоящее время в ПК широко используются операционные системы фирмы
Microsoft: Windows 95, Windows 98 и Windows 2000/ХР. Способ взаимодействия
пользователя с компьютером называется интерфейсом.. Windows 9x/2k использует
графический интерфейс. Основу его составляют окна (windows - окна, англ.).
MicrosoftWindows 98 - это высокопроизводительная, многозадачная и многопо-
токовая 32-разрядная операционная система с графическим интерфейсом и расши-
ренными сетевыми возможностями. Термин "32-разрядная" обозначает, что опера-
ционная система работает с информацией длиной 32 бита. За счет увеличения
разрядности повышается скорость выполнения программ на ПК, увеличивается на-
дежность работы компьютера, возрастают функциональные возможности самого ком-
пьютера. Термин "многозадачная" означает, что на компьютере одновременно мо-
жет работать несколько программ. Простота в работе достигается использованием
нового пользовательского интерфейса. Технология "Plug and Play" (включи и ра-
ботай) облегчает установку новых компьютерных устройств - достаточно вставить
устройство в гнездо компьютера, и далее операционная система сама найдет и
установит соответствующий драйвер. Обмен информацией и взаимосвязь между раз-
личными программами обеспечивает технология OLE - "Object Linking and
1 Windows 98 по прежнему используется, особенно на не новых, низкоскоростных компьютерах, с
не очень большими ресурсами. Ее можно порекомендовать для применения в условиях домашней ла-
боратории , там где требуется большое быстродействие, например для построения систем автома-
тизации эксперимента.
Embedding", что означает "связывание и встраивание объектов".
Запуск Windows 95/98
Запуск программы происходит автоматически после включения компьютера. После
успешного Запуска на экране появляется «Рабочий стол» Windows (см. рис.2) и
графический объект, называемый указателем (курсором) мыши. Перемещение мыши
по столу вызывает перемещение курсора по экрану, а команды передаются нажати-
ем (щелчком) кнопок.
Работа в Windows может осуществляться как с помощью клавиатуры, так и с по-
мощью мыши. На экране монитора всегда находится указатель мыши. Форма указа-
теля мыши может меняться в зависимости от того, на что он указывает. Измене-
ние формы указателя мыши «подсказывает» пользователю о тех действиях, которые
он может произвести в данных момент времени. Некоторые формы указателя мыши
приведены в табл. 1.
Таблица 1. Формы указателя мыши
Форма Возможные действия
Нормальный режим выбора объекта.
ЕЗ TF Передвигая мышь, при нажатой ле- вой клавише, можно переместить объект по экрану.
El <—► Не отпуская левую клавишу мыши, можно изменять соответствующие размеры объекта.
I Установка курсора в нужную пози- цию экрана щелчком левой клавиши мыши.
1 Ожидание завершения операции.
Можно продолжать работу, т. к. программа что-то делает в фоновом режиме.
ft) Переход по гиперссылке.
Щелчком мыши по элементу окна можно получить контекстную справ- ку.
Приемы работы с мышью
Указать на объект - подвести указатель мыши к объекту так, чтобы острие
стрелочки указывало точно на объект.
Выделить объект - указать на объект, быстро нажать и отпустить ЛЕВУЮ клави-
шу мыши.
Выбрать объект (запустить программу, открыть окно) - указать на объект, бы-
стро дважды подряд нажать и отпустить ЛЕВУЮ клавишу мыши.
Вызвать контекстное меню - указать на объект, быстро нажать и отпустить
ПРАВУЮ клавишу мыши.
Переместить объект - указать на объект, нажать ЛЕВУЮ клавишу мыши и, не от-
пуская ее, перемещать его по экрану. В нужном месте клавиша отпускается. Этот
прием называется «перетащить и опустить» (drag and drop).
Левая кнопка мыши используется:
• для запуска приложений и открытия папки (двойной щелчок). Двойной щелчок
означает выполнение двух последовательных щелчков, разделенных очень ма-
леньким
• промежутком времени (вместо двойного щелчка можно использовать клавишу
Enter);
• выбора пунктов меню и выделения отдельных объектов, таких, как файл, яр-
лык, окно (одинарный щелчок);
• выделения некоторых областей текста или рисунка;
• перемещения (перетаскивания, буксирования) объекта по экрану. Для этого
необходимо указать курсором мыши объект, нажать левую кнопку мыши и, не
отпуская левой кнопки мыши, перетащить курсор в нужное место, отпустить
левую кнопку - происходит фиксация нового положения объекта.
Правая кнопка используется не столь активно, в основном для вызова контек-
стного меню. При этом в меню будут присутствовать именно те пункты, которые
подходят для данного объекта.
На «Рабочем столе» могут находиться различные элементы (объекты), основными
из которых являются: «Панель задач», значки программ, документов, папок или
ярлыков («Мой компьютер», «Корзина», «Microsoft Word» и т.д.). В Windows 98
большое значение имеет понятие папка. Папка - это группа файлов, объединенных
по какому-либо критерию. В Windows 98 понятие папка сменило понятие каталог
(директорий), используемое в предыдущих версиях Windows и в DOS, вместо тер-
мина программа обычно используют термин приложение. В системе WINDOWS 98 зна-
чительное внимание уделено реализации документо-ориентированного подхода. Это
означает, что пользователь концентрирует внимание на своих документах, а не
на приложениях, с помощью которых эти документы обрабатываются.Документ — лю-
бой файл, обрабатываемый с помощью приложений. В Windows 98 документ может
содержать текстовую, графическую, звуковую или видеоинформацию.
Длина имени файла может состоять из 255 символов, в качестве которых можно
использовать любые символы, кроме ? \ * " < > |.
«Панель задач» содержит:
• кнопку «Пуск», которая обеспечивает доступ к «Главному меню» Windows
95/98;
• кнопки всех работающих программ (приложений) и обеспечивает переключение
между ними;
• панель индикации с кнопками для переключения алфавита, установки режимов
работы экрана (цвета, оформление, размер и т. д.) , изменение даты и време-
ни , увеличения или уменьшения громкости звука.
Вид рабочего стола
На Рабочем столе располагаются иконки (значки, пиктограммы). Иконка пред-
ставляет собой небольшой рисунок с подписью. Иконки должны отражать назначе-
ние того объекта (файла, приложения, документа), который они представляют.
Ярлык - разновидность иконки в системе Windows 95/98. Ярлык представляет со-
бой указатель на объект и помогает найти его на диске. В левом нижнем углу
ярлыка находится стрелка. Ярлык практически не занимает места на диске (всего
1 килобайт), а результат щелчка по ярлыку тот же, что и по самому объекту.
Мой компьютер
Мои документы
Microsoft Word
Корзина
Internet Explorer
Системные
Кнопки открытых
окон и приложений
Панель задач
(нопка Пуск
часы
Громкость
Индикатор текущей
раскладки клавиатуры
Мой компьютер
Двойной щелчок по ярлыку открывает объект, с которым он связан.
При стандартном виде Рабочего стола на нем располагаются иконки:
- Мой компьютер. Двойной щелчок по иконке Мой компьютер открывает папку с
одноименным названием. С помощью этой папки пользователь имеет
доступ ко всем папкам и файлам на компьютере, к дискам, различным устройст-
вам и к их настройке.
Корзина. Предназначена для удаления папок и файлов, их временного хранения.
Корзина занимает часть дискового пространства, размеры которого устанавлива-
ются пользователем. По внешнему виду Корзины видно: пуста она или нет. Инфор-
мацию о перенесенных в Корзину объектах можно получить, перейдя в режим про-
смотра корзины. Если файл удален по ошибке, то можно восстановить его при ус-
ловии, что Корзина еще не очищена. Двойной щелчок по иконке Корзины дает воз-
можность просмотреть ее содержимое.
Состав иконок, их внешний вид, расположение, форму, размеры, цвет можно ме-
нять .
Мой компьютер
Рис. Иконка папки Мой компьютер и окно просмотра содержимого
папки Мой компьютер
Пользователю постоянно доступна панель задач. Панель задач может занимать
различное положение, может находиться на рабочем столе или появляться только,
когда курсор мыши приближается к соответствующей границе экрана. Панель задач
содержит кнопку Пуск с логотипом Windows и кнопки с именами открытых приложе-
ний и папок. При установке указателя мыши на кнопке появляется подсказка с
названием приложения и документа. Кнопки можно использовать для переключения
между окнами.
Справа на панели задач расположены значки системные часы, звук и индикатор
текущей раскладки клавиатуры (буквы Ru означают выбор шрифта, соответствующе-
го кириллице). Щелчок индикатора раскладки клавиатуры вызывает окно, позво-
ляющее переключиться с одного языка на другой. После установки указателя мыши
на системных часах появляется текущая дата. Двойной щелчок системных часов
вызывает на рабочий стол большие часы с календарем. В этом режиме возможна
коррекция текущего времени и даты.
Щелчок значка звук вызывает появление значка с движком, управляющим громко-
стью Звука и опцией включение/отключения звука. Выбор в этом окне мышью Выкл
приводит к отключению звука.
Основой Windows является использование многооконного интерфейса
("интерфейс" - средство общения). Окно представляет собой обрамленную область
экрана, в которой может отображаться приложение, документ или сообщение.
На поверхности рабочего стола можно увидеть три типа окон: окна приложений,
окна документов, вспомогательные окна.
Рис. Основные элементы окна приложений
В верхней части каждого окна приложений находится строка заголовка. В ней
отображается имя открытого приложения. В левой части строки заголовка нахо-
дится иконка - это кнопка вызова системного меню. Активизируется системное
меню щелчком левой клавиши мыши по кнопке вызова системного меню. Чтобы уб-
рать его, достаточно щелкнуть той же клавишей в любом месте рабочего стола. В
правой части строки заголовка расположены три кнопки, причем на двух из них
пиктограммы постоянны, а на средней кнопке пиктограммы изменяются в зависимо-
сти от состояния окна.
Левая кнопка (со значком подчеркивания) сворачивает окно до иконки на пане-
ли задач. Средняя кнопка разворачивает окно на весь экран, или восстанавлива-
ет его первоначальные размеры. Правая кнопка (с крестиком) закрывает окно и
прекращает выполнение приложения.
Под строкой заголовка окна приложений находится строка меню. Добраться до
команд меню можно двумя способами. Во-первых, любое меню открывается при
щелчке левой клавишей мыши по названию меню. Во-вторых, получить доступ к ко-
мандам меню можно с помощью клавиатуры. В каждом пункте меню подчеркнута пер-
вая буква.
Комбинацией клавиш Alt+буква, подчеркнутая в имени меню, можно сразу от-
крыть нужное меню. Команды меню можно выбрать клавишами, соответствующими
подчеркнутым буквам в имени команды. Некоторые команды можно вызвать на вы-
полнение с помощью клавиш или комбинаций клавиш, не открывая меню. Для таких
команд справа от их имени указывается соответствующая комбинация. Например,
Alt+F4 для команды Закрыть.
У нижнего края окна находится строка состояния. Она содержит информацию о
режимах работы приложения.
Размеры окна можно легко изменять. Для этого нужно установить курсор мыши
на границу окна, он примет вид двунаправленной стрелки, показывающей направ-
ления изменения размеров окна, нажать левую клавишу мыши и установить новые
размеры окна.
Полоса прокрутки (вертикальная или горизонтальная) - это полоса вдоль ниж-
ней или правой границы окна. Она состоит из стрелок прокрутки, бегунка, соб-
ственно полосы прокрутки и служит для просмотра содержимого окна, не умещаю-
щегося в его размерах.
Окна документов всегда подчинены окнам своих приложений, не выходят за их
пределы. Окно документа может быть свернутым (при этом остается на экране
лишь заголовок с названием документа), представлено окном нормального размера
(не развернутым на все рабочее поле) или полностью развернутым в пределах ра-
бочего поля окном. Для некоторых приложений допустимо располагать на рабочем
поле несколько окон документов в различных состояниях. Если окно документа
представлено нормальным окном, то оно имеет собственную строку заголовка, со-
держащую имя документа. При увеличении размеров окна до максимального его имя
появляется в строке заголовка приложения после имени приложения. Окна доку-
ментов , также как и окна приложений, имеют кнопки для изменения вида окна, а,
установив указатель мыши на границу окна, можно изменить размеры окна.
Вспомогательные окна используются для вывода сообщений, для организации
диалога с целью запроса параметров команды. Как правило, эти окна имеют фик-
сированный размер.
Одновременно на рабочем столе могут находиться несколько окон. Чтобы упоря-
дочить окна, необходимо щелкнуть правой клавишей мыши на пустом месте панели
задач и выбрать один из способов упорядочивания: Окна каскадом (размещает от-
крытые окна "в стопку"), Окна сверху вниз, Окна слева направо. Выбор осущест-
вляется одиночным щелчком левой клавишей мыши.
Чтобы переместить окно, надо установить курсор на строку заголовка окна,
нажать левую кнопку мыши и, не отпуская ее, переместить курсор в нужное ме-
сто, отпустить левую кнопку мыши.
Щелчок мышью кнопки Пуск приводит к появлению Главного системного меню, ко-
торое позволяет запустить программу, открыть документ, вызвать панель управ-
ления для настройки компьютера, произвести поиск файла или папки, вызвать
справку и т.д.
Вид Главного системного меню может быть различным на разных компьютерах.
При стандартном виде меню на экране появятся пункты: Программы, Документы,
Настройка, Поиск, Справка, Выполнить, Завершение работы. Если у пункта есть
подменю, то справа от названия появляется значок с изображением треугольника.
Пункт меню «Программы» вызывает на экран список приложений, установленных
на компьютере. Пункт меню Документы вызывает на экран список документов, с
которыми недавно работали на данном компьютере. При выборе документа происхо-
дит запуск необходимого приложения и загрузка в него данного документа. Пункт
меню «Настройка» имеет подменю из команд Панель управления, Принтеры и Панель
задач, которые позволяют настроить Windows, внешний вид панели задач и состав
основного меню. Пункт меню «Найти» осуществляет поиск файлов или папок. Пункт
меню Справка используется для получения доступа к справочной системе. Пункт
меню «Выполнить» позволяет запустить любое приложение. Для этого в окно За-
пуск программы необходимо ввести путь и название этого приложения. Если точно
неизвестно местонахождения приложения, то можно выбрать кнопку Обзор. Пункт
меню Завершение работы позволяет выключить или перезагрузить компьютер.
Нажатие кнопки Start выводит целую систему меню, с помощью которых можно
найти и пустить заданное приложение
Для переключения шрифта с русского языка на английский и обратно с помощью
«Панели индикации» нужно:
• щелкнуть левой клавишей мыши по кнопке (индикатору) с надписью Ru или Еп ;
• в появившемся меню выбрать требуемый режим ввода букв.
My Computer
Disk Formating
3DR Version 2.0.8
[g| 3D Fl Version 2.0.8 16 bit
Inbox D Ы iskdc fg Accessories ►] ig Multimedia ►! <5 CD Player § Dictmgr
Recycle Bin 40 Cie Adobe Acrobat 2.0 ► Corel5 ► (gj IAL SW Developer CD ► _g] Maple V Release 3 ► Mathematica ► _тт] MCS Stereo ► [gj MicroCai Origin ► _—| System T ools ► @ Calculator | HyperTerminal W Notepad Paint Phone Dialer WordPad
Й MediaPlayer
Session -J Sound Recorder f | Volume Control Wave
Dodot
(gj Microsoft Office ► [gj Scankit Utility V2.2E ► (gj SPRY Mosaic ► Cg] Startup ► [gj Visual Basic 3.0 ► (gj World Atlas ► Microsoft Word Viewer MS-DOS Prompt
1 Па Programs
Documents ► Settings ► 2j Find k Help jp Run... Shut Down...
Windows Explorer
Завершение работы c Windows 95/98 и выключение ПК
Неправильный выход из Windows может привести к потере данных!
Перед выключением ПК:
• сохранить все документы;
• закрыть все работающие приложения;
• открыть «Главное меню» кнопки «Пуск» и выбрать команду «Завершение рабо-
ты» ;
• в появившемся окне выбрать пункт «Выключить компьютер»;
• отключить электропитание после того, как получите на это разрешение ПК.
Типовая структура окна
В системе Windows существуют следующие типы окон:
• программное окно - окно, в котором выполняется программа (приложение);
• окно документа - окно, связанное с конкретной прикладной программой;
• окно папки - окно, в котором можно просматривать ресурсы компьютера;
• диалоговое окно - окно, в котором происходит диалог пользователя с систе-
мой Windows (выбираются или делаются некоторые установки);
• окно сообщений - окно, в котором выдается сообщение ОС.
Строка мзню
Программное окно, окно документа и окно папки имеют общие элементы.
Элементы окна программы «Корзина»
Названия и назначение основных элементов окна приведены в табл.
Таблица. Основные элементы окна
Название Назначение
Строка заголовка В ней указано имя программы, а также имя открытого документа. Строка заголовка активного окна отличается цветом (как правило - синий) от строк заголовка всех дру- гих открытых окон. Используя строку заголовка можно перемещать окно по экрану.
Кнопка Свернуть Сворачивает окно в кнопку на «Панель задач». Содер- жимое окна при этом не уничтожается.
Кнопка Развернуть Разворачивает окно на весь экран. При развернутом окне данная кнопка превращается в кнопку «Восстановить», которая позволит вернуть ок- но в прежнее состояние.
Кнопка Закрыть Закрывает окно и завершает работу программы.
Оконное меню Осуществляет доступ к набору команд, связанных с параметрами окна.
Строка меню Содержит пункты (ключевые слова) меню. Щелчок левой клавишей мыши по пункту меню вызывает на экран ниспадающее меню данного пункта. Для завершения работы с меню можно щелкнуть мышью где-нибудь в рабочей области экрана.
Панель(и) инструментов Содержит кнопки наиболее часто используемых команд. Устанавливается в окне с помощью пункта меню «Вид».
Граница, угол Используются для плавного изменения размеров окна.
Полосы прокрутки Отсутствуют, если вся информация (документ) помеща- ется в окне. Содержат кнопки и бегунок. Кнопки вертикальной про-
Название Назначение
крутки позволяют перемещаться на строку (страницу) вверх или вниз, горизонтальной - влево или вправо. По положению бегунка можно определить, как располо- жено окно относительно документа. Бегунок можно перемещать указателем мыши по полосе прокрутки, при этом изменяется расположение доку- мента относительно окна. Щелчок по полосе прокрутки выше или ниже бегунка позволяет перемещаться постранично (вверх или вниз).
Рабочее Основная часть окна, в которой размещается доку-
поле мент.
Чтобы пустить приложение, достаточно найти его в подменю стартового меню и
щелкнуть левой клавишей мышки. На поверхности рабочего стола появится окно
работающего приложения
Окна можно открывать, закрывать, сворачивать, перемещать, изменять размеры,
упорядочивать.
Изменять представление окон можно следующим образом.
Свернуть окно в кнопку:
• или щелчком мыши по кнопке «Свернуть»,
• или щелчком мыши по значку «Оконного меню» и далее через меню.
Развернуть окно на весь экран:
• или щелчком мыши по кнопке «Развернуть»,
• или двойным щелчком мыши по ' «Строке заголовка»,
• или щелчком мыши по Значку «Оконного меню» и далее через меню
Восстановить нормальные размеры окна, свернутого в кнопку:
• щелчок мыши по кнопке на «Панели задач».
Закрыть окно:
• или щелчком мыши по кнопке «Закрыть»,
• или двойным щелчком мыши по Значку «Оконного меню»,
• или щелчком мыши по значку «Оконного меню» и далее через меню.
Для перемещения окна по экрану нужно сделать окно активным (текущим), уста-
новить указатель мыши на «Строку заголовка» и перетащить окно, используя при-
ем мыши «перетащить и отпустить».
Работа с Windows 95/98 является документно-ориентированной. Это означает,
что вам незачем вспоминать, каким приложением создан документ, в каком форма-
те он записан как файл и какие средства нужны для его модификации и просмот-
pa. Особенно отчетливо это ясно, если пустить команду Documents в стартовом
меню. Вы увидите список всех последних документов, с которыми вы работали по-
следнее время.
Для работы с любым документом просто установите курсор мышки на выбранном
документе и пару раз щелкните левой клавишей мышки - документ спустя пару-
другую секунд появится вместе с породившим его приложением
Упорядочивание открытых окон на «Рабочем столе»
Для упорядочивания открытых окон необходимо:
• установить указатель мыши на свободное место «Панели задач» и нажать ПРА-
ВУЮ кнопку мыши;
• выбрать в контекстном меню команду «Каскадом», «Сверху вниз» или «Слева
направо».
Чтобы восстановить исходные размеры и размещение окон, снова открыть кон-
текстное меню и выбрать команду «Отменить окна рядом», или «Отменить окна
каскадом».
Пример работы с графическим документом в редакторе Paint
Лекция 3
Элементы управления
Существуют три вида меню:
• строка-меню (горизонтальное меню) - находится в окне под строкой заголов-
ка ;
• контекстное меню - вызывается правой клавишей мыши на любом объекте и со-
держит команды для работы с ним;
• оконное (системное) меню - содержит команды для работы с окном.
Любое меню содержит список разных команд. Команды, доступные для выполнения
в настоящий момент времени, отображаются черным цветом, а недоступные для вы-
полнения — серым.
Если название команды заканчивается многоточием (...), то при ее выборе
появится диалоговое окно, в котором требуется сделать какие-либо дополнитель-
ные установки.
Команды, слева от названия которых находится пометка или галочка, работают
как переключатели: если галочка отображается в меню, то режим включен.
Команды, заканчивающиеся черным треугольником, вызывают еще одно меню.
Контекстное меню
Элементами управления в диалоговом окне могут быть: переключатели, регуля-
торы, флажки, поля списка, счетчики, вкладки и т.д.
[*/ Ждущий режим через
Г Отключение через
Флажки
Счетчик
мин
Некоторые элементы управления в диалоговом окне
Флажки (кнопки-переключатели) - служат для установки свойств, имеющих два
значения: да/нет, включено/выключено и т.д. «Включенный» флажок помечается
внутри черным значком. В окне может находиться несколько флажков, причем их
установки не оказывают влияния друг на друга.
Счетчики - числовые переключатели - используются для изменения числовых ха-
рактеристик в большую или меньшую стороны.
Переключатели - радиокнопки позволяют сделать выбор одного варианта из
предлагаемой группы вариантов. Признаком выбора является черная точка внутри
круга.
Регуляторы - регулятор настройки с перемещаемым движком.
Поле списка - окно, позволяющее выбрать один вариант из списка предлагае-
мых .
Поле ввода - прямоугольное окно, позволяющее ввести необходимый текст. Пе-
ред вводом текстовой информации в поле ввода щелчком мыши необходимо устано-
вить курсор.
Любое окно содержит командные кнопки «ОК» - подтвердить сделанные изменения
и «Отмена» - отменить сделанные изменения. Кроме указанных кнопок могут
встретиться и другие, назначение которых, как правило, понятно из их назва-
ния: «Справка», «Выполнить», «Печать» и т.д.
Средствами для изменения различных настроек являются «Панель управления» и
контекстное меню объектов.
Рис. Окно папки «Панель управления»
Окно «Панель управления» представляет собой основное средство для изменения
различных настроек компьютера. Это специальная папка системы Windows, которой
не соответствует никакая реальная папка на жестком диске. Открыть «Панель
управления» можно с помощью команды «Пуск / Настройка / Панель управления».
Папка «Панель управления» доступна также в окне «Мой компьютер». Имеющиеся
в папке значки вызывают открытие диалоговых окон, с помощью которых произво-
дится настройка системных часов и системного календаря, клавиатуры, звука,
мыши, шрифтов и т.д. Открытие диалоговых окон осуществляется двойным щелчком
на значке.
Настройка завершается щелчком на кнопке «Применить» (без закрытия окна) или
на кнопке «ОК» (с закрытием окна).
Ниже представлены основные утилиты контрольной панели:
Источники данных - установка опций доступности,
Установка оборудования - установка нового периферийного оборудо- вания ,
Установка/удаление программ - включение новых и уничтожение ненужных программ,
Дата и время - просмотр и установка даты и времени,
Экран - управление дисплеем,
Шрифты - установка, просмотр и удаление шрифтов,
Игровые устройства - установка опций игрового манипулятора - джойстика,
Клавиатура - установка опций клавиатуры,
Почта и факс - установка опций электронной почти и фак- са ,
Модем - установка опций телефонного модема,
Мышь - установка опций мышки,
Мультимедиа - установка средств мультимедиа и контроль за ними,
Сеть - установка сетевых средств,
Пароли - установка ключей доступа к ПК (пароля),
Языки и стандарты установка региональных параметров,
Звук - озвучивание оболочки Windows.
Каждая пиктограмма контрольной панели управляет соответствующей утилитой,
меняющей установки системы. При этом появляется то или иное диалоговое окно
для установки тех или иных опций. Многие окна имеют переключатели для групп
опций.
Файл, папка, диск
ФАЙЛОМ называется поименованная область информации на диске (гибком, жест-
ком, лазерном). Файл от англ, file - досье. Часть внешней памяти компьютера,
имеющая идентификатор (имя) и содержащая данные. В зависимости от типа храня-
щихся данных файлы разделяются на программные (выполняемые), которые управля-
ют работой компьютера, и файлы, содержащие прочие данные, например, тексты,
какие-либо сведения в виде записей. Управление файлами обеспечивает файловая
система, являющаяся частью операционной системы компьютера. На гибких и
жестких магнитных дисках файл хранится в виде наборов кластеров, как правило,
не расположены рядом.
Все файлы должны иметь УНИКАЛЬНЫЕ ИМЕНА, по которым их можно находить. Имя
файла состоит из двух частей: содержательного имени файла, длина которого ог-
раничена 256 символами (включая пробелы и русские буквы) и расширения (от 1
до 3 символов), которые разделяются между собой точкой.
Расширение определяет тип файла.
Примеры некоторых стандартных расширений:
ехе, сот - выполняемые файлы;
txt, doc - текстовые файлы;
bmp, рсх, jpg - графические файлы;
bak - резервная копия файла и др.
Каждому файлу назначается свой значок, который зависит от типа файла.
Использование длинных имен файлов в операционных системах Windows 95/98 и
Windows 2000 имеет ряд особенностей:
1. Если длинное имя включает пробелы, то в служебных операциях его надо за-
ключать в кавычки. Рекомендуется не использовать пробелы, а заменять их
символами подчеркивания.
2. В корневом каталоге (папке), т.е. верхнем уровне иерархической файловой
структуре нежелательно хранить файлы с длинными именами - в отчие от про-
стого каталога (папки) в ней ограничено количество единиц хранения, при-
чем, чем длиннее имена, тема меньше файлов можно разместить в корневом ка-
талоге (папке).
3. Кроме ограничения на длину файла (256 символов) существует гораздо более
жесткое ограничение на длину полного имени файла (в него входит путь дос-
тупа к файлу, начиная от вершины иерархической структуры). Полное имя не
может быть длиннее 260 символов.
4. Разрешается использовать символы любых алфавитов, в том числе и русского,
но если документ готовится для передачи, с потребителем этого документа
необходимо согласовать возможность воспроизведения файла с такими именами
на его оборудовании.
5. Прописные и строчные буквы не различаются операционной системой. Однако
символы разных регистров отображаются операционной системой и если для на-
глядности надо использовать прописные буквы, это можно делать.
Расширение имени файла используется для идентификации его содержимого, т.е.
нести информацию для операционной системы. Системы Windows96/98 имеют средст-
ва для регистрации свойств типов файлов по расширению их имени, поэтому во
многих случаях выбор расширения имени файла не является частным делом пользо-
вателя .
Для удобства поиска файлы, как правило, размещают в папках. Иногда папку
называют каталогом. Папкам, также как и файлам, присваиваются имена. Одни
папки могут содержать в себе другие папки (вложенные папки).
Каталоги (папки) - важные элементы иерархической структуры, необходимые для
обеспечения удобного доступа к файлам, если файлов на носителе слишком много.
Файлы объединяются в каталоги по любому общему признаку, заданному из созда-
телем (по типу, по времени создания, по имени владельца и др.). Каталоги низ-
ких уровней вкладываются в каталоги более высоких уровней и являются для них
вложенными. Верхним уровнем вложенности иерархической структуры является кор-
невой каталог диска.
В иерархической структуре данных адрес объекта задается маршрутом (путем
доступа), ведущим от вершины структуры к объекту. При записи пути доступа к
файлу, проходящего через систему вложенных каталогов, все промежуточные ката-
логи разделяются между собой определенным символом. Во многих операционных
система в качестве такого символа используются "\" (обратная черта).
Навигация по файловой системе Удобство навигации, т.е перехода от файла к
файлу или от папки к папке часто воспринимают как удобство работы с операци-
онной системой. В операционной системах, имеющих терминальный интерфейс, на-
вигация осуществляется путем ввода команд перехода с диска на диска или из
каталога (папки) в каталог (папку). Из-за неудобства такой навигации, широкое
применение нашли специальные программы, называемые файловыми оболочками. Наи-
более известной файловой оболочкой для MS-DOS является Norton Commander.
Основным инструментом для файловых операций в системе Windows 9х является
программа Проводник. Она отображает содержимое папок, позволяет копировать,
перемещать, удалять, переименовывать папки и файлы, запускать программы.
Для запуска Проводника используются следующие способы:
• команда Пуск / Программы. В списке установленных программ обязательно
находится программа Проводник;
• упрощенный метод. Состоит в том, чтобы щелкнуть правой кнопкой мыши на
кнопке Пуск и в контекстно-зависимом меню выбрать пункт Проводник;
• щелкнуть правой кнопкой мыши на любом значке папки и в открывшемся кон-
текстно-зависимом меню выбрать пункт Проводник;
Рабочая область Проводника состоит из двух панелей - левой и правой. В ле-
вой панели находится информация об иерархической структуре папок, в правой -
содержимое открытой папки. В любой момент времени только одна папка может
быть открыта. Свернутые папки имеют узел, отмеченный знаком. Если развернуть
папку щелчком на узле, то он поменяется на. Сворачивание папки выполняется
щелчком на узле . Для открытия папки необходимо щелкнуть на значке папки за-
крытой папки , папка откроется, ее значок сменится на значок открытой папки ,
и на правой панели будет отображено ее содержимое.
Запуск приложений и открытие документов выполняется двойным щелчком по
Значку приложения или документа на правой панели Проводника. При выполнении
команды запуска документа при помощи Проводника (или папки Мой компьютер) ав-
томатически запускается нужное приложение и в него загружается выбранный до-
кумент .
Панель Адрес
Значок закрытой
папки
Значок открытой
____папки______
Рис. Окно программы Проводник
С помощью пункта меню Файл можно создать новый файл или папку, удалить их,
переименовать и т.п. Через пункт меню Вид можно вывести панель Адрес, строку
состояния, просматривать содержимое папок в нескольких режимах, в том числе в
режимах Крупных значков, Мелких значков, Списка и Таблицы. Представление в
виде крупных значков удобно, если файлы имеют уникальные значки и подпись под
ними менее важна. Мелкие значки удобны в том случае, когда значки имеют стан-
дартный вид, и более важны подписи под ними. Представление в виде списка
удобно, если все документы и папке относятся к одному типу и имеют одинаковый
значок. В этом случае важна подпись под значком. При представлении в виде
таблицы приводится размер файлов, тип, дата создания или последнего измене-
ния. Если файлы отображаются в виде таблицы, их удобно сортировать по разным
признакам. Щелчок на заголовке одного из столбцов вызывает сортировку содер-
жимого по данному признаку. Уникальная возможность такого режима состоит в
том, что возможна сортировка в обратном порядке. Чтобы отсортировать файлы в
обратном порядке, надо еще раз щелкнуть на заголовке того столбца, по которо-
му выполнена сортировка.
Папка, из которой выполняется копирование или перемещение, называется ис-
точником. Папку, в которую происходит копирование или перемещение, называют
приемником. Копирование или перемещениеосуществляется нажатием левой кнопки
мыши на копируемом объекте и транспортировкой его в папку - приемник. Для
этого надо найти на правой панели Проводника копируемый объект. На левой па-
нели, разворачивая папки на узле, нужно найти папку - приемник. Если диски
файла-источника и папки-приемника не совпадают, то файл будет скопирован,
иначе он будет перенесен. Если при транспортировке файла будет нажата клавиша
Shift, то файл будет перемещен, если клавиша Ctrl, то файл будет скопирован
независимо от диска. Можно копировать сразу несколько файлов или папок, если
их предварительно выделить. Для выделения смежных файлов следует щелкнуть
около имени первого файла и растянуть появившийся прямоугольник таким обра-
зом, чтобы в нем оказалась вся группа файлов. Выделение несмежных файлов осу-
ществляется однократными щелчками по их именам при нажатой клавише Ctrl.
Можно использовать специальное перетаскивание при нажатой правой кнопке мы-
ши. В момент отпускания кнопки появляется меню, в котором надо выбрать пункт
Переместить или Копировать.
Для копирования файлов или папок можно использовать буфер обмена. Для зане-
сения информации в буфер и ее извлечения оттуда приложения Windows имеют в
своих меню команды Вырезать, Копировать, Вставить. Команда Вырезать помещает
выделенную информацию в буфер, удаляя при этом ее оригинал. Команда Копиро-
вать помещает выделенную информацию в буфер, не трогая оригинала. Команда
Вставить извлекает информацию из буфера.
Для копирования файла или папки необходимо:
• выбрать нужный файл или папку;
• выполнить команду Правка/Копировать (можно использовать контекстное меню
или панель инструментов);
• открыть диск и (или) папку, в которую предполагается переместить копию;
• выполнить команду Правка/Вставить (можно использовать контекстное меню
или панель инструментов).
Для перемещения использовать вместо пункта Копировать пункт Вырезать.
Для переименования файла или папки следует щелкнуть два раза на имени с
большим интервалом. В появившейся рамке с курсором, можно отредактировать имя
и нажать клавишу Enter. Также для переименования можно щелкнуть правой кноп-
кой мыши на имени файла или папки и в контекстном меню выбрать команду Пере-
именовать .
Для создания папки необходимо в меню Файл или в контекстном меню выбрать
пункт Создать, а затем пункт Папка. В появившейся рамке ввести имя папки и
нажать клавишу Enter.
Папки и файлы можно создавать, просматривать, переименовывать, переносить
их содержимое на новое место и удалять.
Все дисковое пространство, как правило, делится на части (логические дис-
ки) , которым также присваиваются имена:
А: или В: — обозначают гибкие магнитные диски (дискеты);
С: D: Е: F: и т.д. — обозначают логические части жесткого магнитного диска
(винчестера) или лазерного (оптического) диска (CD ROM-a).
Буфер обмена Clipboard
Буфер Clipboard - это некоторая динамически изменяемая область памяти, спо-
собная хранить информацию в типовых форматах Windows 95. Указанные выше опции
(и некоторые другие) имеют практически все приложения Windows 95 - например
упомянутая записная книжка Notepad. Сосредоточены эти опции в позиции Edit
главного меню каждого приложения, рассчитанного на обмен данными с другими
приложениями через буфер. Windows автоматически распознает и правильно приме-
няет необходимые форматы данных - текстовые, численные, графические и даже
мультимедийные (такие как звук голоса, музыка или видеоклипы).
Помимо команды Сору для заполнения буфера можно использовать нажатие клави-
ши Print Screen. Обычно при этом в буфер будет помещена полная копия экрана.
Помимо указанных команд, некоторые приложения имеют команду Cut (вырезать),
также располагаемую в подменю позиции Edit главного меню. Она переносит выде-
ленный фрагмент документа в буфер, одновременно уничтожая этот фрагмент в са-
мом документе. Возможны также специальные команды Сору, Cut и Paste, например
для работы с объемными многостраничными документами.
Папка «Мой компьютер»
После включения ПК и загрузки программы Windows на экране появляется «Рабо-
чий стол», на котором, в зависимости от выбранного типа установки, располага-
ются какие-либо значки. Значок папки «Мой компьютер» является обязательным.
Назначение этой папки - доступ к ресурсам компьютера. Открывается папка двой-
ным щелчком мыши по ее Значку. Окно папки «Мой компьютер» выглядит следующим
образом:
Ресурсы ПК
Строка меню
Панель инстру-
ментов
Окно папки «Мой компьютер»
Если подвести указатель мыши к любой кнопке «Панели инструментов» и задер-
жать его, то появиться подсказка, объясняющая назначение данной кнопки.
Используя пункт меню «Вид» можно:
• скрыть или показать некоторые элементы окна (например, «Панель инстру-
ментов») ;
• изменить вид показа ресурсов (маленькие или большие значки, таблица и
список.);
• установить параметры просмотра (команда «Параметры», вкладки «Папка» и
«Просмотр»);
• увидеть зарегистрированные в системе Windows типы файлов (команда «Пара-
метры», вкладка «Типы файлов»).
• Сводную информацию о файле, папке или диске можно получить, используя
пункт меню «Файл»/»Свойства» 1.
• Просмотр содержимого диска
• Просмотреть содержимое любого диска можно:
• двойным щелчком по значку диска, находящегося на рабочем поле папки «Мой
компьютер»;
• раскрыв список с описанием ресурсов на «Панели инструментов» и выбрав
необходимый диск из списка предлагаемых.
Просмотр содержимого папки
Для просмотра содержимого папки нужно дважды щелкнуть мышкой по ее знач-
1 Здесь и далее вместо выбора команды основного меню можно воспользоваться соответствующей
командой контекстного меню (щелчок правой клавишей мыши по объекту).
ку.
Создание папок
Для создания новой папки нужно:
• открыть диск, на котором создается новая папка;
• открыть папку, в которой создается новая папка (при вложении папок);
• выбрать команду «Создать» пункта меню «Файл»;
• выбрать пункт «Папка»;
• указать имя создаваемой папки под ее появившимся значком;
• нажать клавишу Enter или щелкнуть левой кнопкой мыши на свободном мес-
те .
Создание ярлыков
My Computet
Dictmgr
Inbox
Create Shortcut
Command line:
msn.
Set Up The
Microsoft
Network
Type the location and name of the item you want to create
a shortcut to. Or, search for the item by clicking Browse.
Recycle Bin
Nc
Lexicon
Browse...
Start
Создание ярлыка с помощью команды Create Shortcut
ЯРЛЫК - является средством быстрого доступа к информационному ресурсу
(объекту): программе, документу, папке, диску, принтеру и т.д. Ярлык «запо-
минает» адрес, по которому находится требуемый объект. Такой адрес может
содержать имя диска, имя папки (имена папок при их вложении друг в друга) и
имя файла (объекта). Ярлык может быть создан в любой папке, а также на «Ра-
бочем столе». Создать ярлык на «Рабочем столе» можно разными способами:
• или перетащить на «Рабочий стол» значок объекта, удерживая ПРАВУЮ
кнопку мыши;
• или выделить объект, для которого создается ярлык, выбрать в меню
«Файл» команду «Создать ярлык» и после создания ярлыка «перетащить»
его на «Рабочий стол»;
• или вызвать контекстное меню щелчком правой кнопкой мыши:
• выбрать в меню команду «СоЗдать»/«Ярлык»;
• используя командную кнопку «Обзор...», найти на диске объект, для
которого создается ярлык (если создается ярлык для документа, уста-
новить в окне «Тип файла»/«Все файлы»);
• указать имя создаваемого ярлыка.
Создание документов
Для создания документа необходимо:
• сделать активным папку, в которой создается документ;
• выбрать команду «Создать» пункта меню «Файл»;
• выбрать требуемый тип документа из предлагаемого списка. После этого
значок созданного документа появится в указанном месте;
• при необходимости изменить имя документа.
Двойной щелчок по значку документа запустит приложение для его редактиро-
вания .
Удаление папок, файлов и ярлыков
Для того чтобы удалить папку, файл или ярлык нужно:
• выделить удаляемый объект;
• нажать клавишу [Delete] или воспользоваться кнопкой X | «Удалить» на
«Панели инструментов»;
• подтвердить удаление.
Удаленные объекты помещаются в «Корзину»2 и остаются там до тех пор, пока
не будет дана команда «Очистить корзину». При необходимости удаленный объ-
ект из «Корзины» можно восстановить.
Копирование папок,
файлов и ярлыков
&
Вырезать
Вставить
<о пировать
Скопировать или переместить объект можно, используя пункт меню «Правка»
или кнопки «Панели инструментов»:
Для того чтобы скопировать папку, файл или ярлык нужно:
• выделить объект, который копируется;
• нажать кнопку «Копировать»;
• открыть папку, где должна появиться копия объекта;
• нажать кнопку «Вставить».
Для копирования папок, файлов и ярлыков с жесткого диска на гибкий диск
2
Кроме объектов расположенных на дискете и сетевых дисках.
удобно использовать последовательность команд: «Файл» / «Отправить» /«Диск
3,5 (А) » .
Перемещение папок,
файлов и ярлыков
Для того чтобы переместить на новое место папку, файл или ярлык нужно:
• выделить объект, который необходимо переместить;
• нажать кнопку «Вырезать»;
• открыть папку, где теперь должен быть расположен объект;
• нажать кнопку «Вставить».
Форматирование диска
Форматирование или разметка - это операция по подготовке магнитного диска
к хранению информации. При форматировании ранее записанные на диск данные
уничтожаются. Диск может быть отформатирован только в случае, если файлы,
хранящиеся на нем, не используются в данный момент другими программами
(приложениями). В частности, если содержимое диска отображается в окне «Мой
компьютер», отформатировать диск будет невозможно.
Для форматирования гибкого диска нужно:
• вставить диск в дисковод;
• в окне папки «Мой компьютер» выделить значок диска, который нужно отформа-
тировать ;
• в меню «Файл» выбрать команду «Форматировать»;
• в окне диалога установить параметры форматирования.
Дефрагментация диска
Процесс собирания файлов в одном месте на диске. Файлы при записи на диск
разбиваются на фрагменты кластеры, которые могут располагаться в разных час-
тях диска. Это связано с работой файловой подсистемы операционной системы.
Разбитые на куски файлы затрудняют доступ, увеличивают время обращения к фай-
лам. Для того, чтобы упростить доступ рекомендуется выполнять процедуру деф-
рагментации диска раз в 1-2 недели, которая собирает каждый файл в одном мес-
те (все записи подряд). Такая процедура имеется в любой операционной системе.
В MS-DOS и Windows это программа defrag.exe.
Пользование справкой
Программа Windows 95/98 снабжена удобной справочной системой, которая по-
зволяет получить пользователю вспомогательную информацию в процессе его рабо-
ты.
Вызов справочной системы Windows можно осуществить, выполнив следующие ко-
манды:
кнопка «Пуск»;
пункт «Справка».
Contents j |ndex j Find |
Disk dupe
What's This?
Iribox
New programs and accessories
Lexcon
tJ
Dodot
My Computer
Recycle Bin
Nc
Click a topic, and then click Display. Or click another tab, such as Index.
?] T our: T en minutes to using Windows
?] I f you've used Windows before
U-l Introducing Windows
(Ц Welcome
?1 Registering Windows Э5 online
?] License agreement questions and answers
lj-1 A List of What's New
?] A new look and feel
?1 Faster ways to get your work done
?] More ways to customize Windows
?] Support for portable computers
?] Improved networking
Using Windows Accessories
How To...
Display
Print...
Cancel
X
Start |
Для пуска базы данных помощи служит позиция Help (Помощь) стартового меню
Окно диалога «Справочная система Windows» состоит из трех вкладок: «Содер-
жание», «Предметный указатель» и «Поиск».
Выбрав нужный раздел помощи,
можно получить информацию о нем, которая выво-
дится в информационное окно
Help Topics: Windows Help
My Computer Diskdupe
Contents Index j Find j
Inbox
Disk Formating
Type the first few letters of the word you're looking for.
| access control
Click the index entry you want, and then click Display.
1
2
Recycle Bin
Topics Found
Click a topic., then click Display.
12-hour clock, changing to
24-hour clock, changing to
32-bit PC card
disabling support for
.-r; -,-.i rho :r=.rU;: indicator
?|X
Changing the password for a shared resource
Changing your network password
Changing your password for a NetWare server
Changing your Windows password
)r
, installing support for
d
specifying the number of
d, installing support for
effects
C.on.trgllin.q access by..u.sjnq..a.iist.of.names
Controlling access by using passwords
Controlling access to a folder or printer
Enabling others to see resource use on your computer
Protecting your files by assigning a screen-saver password
to shared resources
sssword
Display
Cancel |
Start
Display
Print...
Cancel
Выбор раздела помощи
по индексному указателю
При работе с диалоговым окном, используя кнопку со знаком вопроса в заго-
ловке окна, можно получить контекстную подсказку по любому элементу, находя-
щемуся в данном окне. Для этого необходимо:
нажать кнопку
| 9 |(указатель мыши примет форму ;
сделать щелчок по интересующему Вас элементу окна;
• после прочтения закрыть окно сообщений щелчком мыши в любом месте экрана.
ЛЕКЦИЯ 4
Текстовые файлы
Если каждому символу алфавита сопоставить определенное число (например, по-
рядковый номер), то с помощью двоичного кода можно кодировать и текстовую ин-
формацию. Восьми двоичных разрядов достаточно для кодирования 256 различных
символов. Этого хватит, чтобы выразить различными комбинациями восьми битов
все символы английского и русского алфавитов, как строчные, так и прописные,
а также знаки препинания, символы основных арифметических действий и некото-
рые общепринятые специальные символы, например
Теоретически это выглядит очень просто, однако всегда существовали доста-
точно веские организационные сложности. В первые годы развития вычислительной
техники они были связаны с отсутствием необходимых стандартов, а в настоящее
время вызваны, наоборот, изобилием одновременно действующих и противоречивых
стандртов. Для того чтобы весь мир одинаково кодировал текстовые данные, нуж-
ны единые таблицы кодирования, а это пока невозможно из-за противоречий между
символами национальных алфавитов, а также противоречий корпоративного харак-
тера .
Для английского языка, ставшего международным средством общения, противоре-
чия уже сняты. Институт стандартизации США (ANSI - American Standard
Institute) ввел в действие систему кодирования ASCII (American Standard Code
for Information Interchange - стандартный код информационного обмена США). В
системе ASCII закреплены две таблицы кодирования - базовая и расширенная. Ба-
зовая таблица закрепляет значения кодов от 0 до 127, а расширенная относится
к символам от 128 до 255.
Базовая ASCII-кодировка
При нажатии алфавитно-цифровых (символьных)
клавиш, клавиш
Tab
Esc
—Jh
Backspace х
________I, а также некоторых их комбинации с
Alt
__1или
Shift
генерируют ся
одно-
байтовые ASCII-коды или двухбайтовые расширенные коды. Исторически сложилось
так, что эта система кодировки была первой общепринятой в мире для компьюте-
ров IBM.
Название ASCII представляет собой аббревиатуру английской фразы American
Standard Code for Information Interchange (Американский Стандартный Код для
Обмена Информацией). Всего кодов 256 (с 0-го по 255-й). Первые 32 ASCII-кода
(с 0 по 31) управляющие и используются для общения с периферийными устройст-
вами или применяются для специальных целей различными программами. Им, как и
остальным, присвоены определенные символы, но на экран их можно вывести толь-
ко прямым обращением в видеопамять.
Чтобы в дальнейшем не было ошибок, запомните, что, например, ASCII 9 отно-
сится к клавише ____J, а не к цифре 9, которая имеет по этому стандарту код
ASCII 57.
Сначала в системах кодирования было много путаницы. Кодировок гуляло по ми-
ру множество. Была основная кодировка, болгарская, КОИ-7, КОИ-8, альтернатив-
ная и много других.
Многообразие способов кодирования связано с тем, что при разработке компа-
нией IBM своих первых компьютеров и создании для них компанией Microsoft опе-
рационной системы MS DOS, было заложено очень трудно обходимое техническое
ограничение на число используемых символов — не более 256. Это количество в
то время полностью перекрывали все мыслимые потребности англоязычного пользо-
вателя. Странная, на первый взгляд, цифра 256 обусловлена тем, что это макси-
мальное число (2»108) , которое может быть записано в двоичной системе в 1-ном
байте (8 бит, то есть восемь 0 и 1) . А именно 1 байт отводился в MS DOS для
хранения любого символа. Последствия этого ограничения со временем и начали
проявляться при широком распространении компьютеров не только в англоязычных
странах.
В конце концов все же пришли к разумному компромиссу:
• первые 128 кодов (с 0-го по 127-й) с управляющими символами, некоторыми
специальными знаками, цифрами и латинским алфавитом остаются в неприкосно-
венности ;
• коды со 176-го по 223-й сохранены за символами псевдографики;
• остальные же коды отданы на откуп потребителям.
У нас наибольшее распространение получила альтернативная ASCII-кодировка,
построенная именно по такому принципу: в оставшихся свободными кодах распола-
гаются буквы русского (или другого национального) алфавита. Тем самым обеспе-
чивалась преемственность программного обеспечения, разработанного в других
странах, — здесь мы были не "впереди планеты всей..
В альтернативной ASCII-кодировке:
прописные русские буквы:
• имеют номера со 128 по 159 — (с "А" по "Я"),
• буква "Ё" стоит особняком под номером 240.
строчные русские буквы (чтобы сохранить символы псевдографики) разбросаны:
• буквы с "а" по "п" имеют номера со 160 по 175,
• с "р" по "я" занимают позиции с 224 по 239,
• буква "ё" снова на отшибе и имеет номер 241.
Кодовая таблица ANSI
Появление Windows вызвало к жизни еще одну систему кодирования символов —
ANSI. Это также аббревиатура английской фразы American National Standard
Institute (Американский национальный институт стандартов). Кстати, первона-
чальное название этой кодировки было — OEM.
Система кодирования ANSI стала преемником формата ASCII, сохранив первые
128 символов на своих привычных местах. Все остальные коды также были отданы
национальным алфавитам, причем, символы псевдографики из таблицы были изъяты.
Варианты национальных кодовых таблиц получили название кодовых страниц с со-
ответствующим номером. Так, например, кодовая страница с кириллицей, исполь-
зуемая в Windows, имеет номер 1251.
На кодовой странице номер 1251 русские буквы (кириллица) размещаются сле-
дующим образом:
• Прописные буквы с "А" по "Я" (кроме "Ё") занимают места со 192 по 223.
• Строчные буквы с "а" по "я" (также кроме "ё") расположены с 224 по 255.
• Для букв "Ё" и "ё" отведены коды 168 и 184.
Ввод символов с помощью клавиши
При работе в DOS все ASCII-коды, начиная с ASCII 32, могут быть выведены на
экран с помощью клавиши
Если нажать и, удерживая нажатой клавишу
набрать на дополнительной (малой цифровой) клавиатуре необходимый ASCII-код,
Alt
то после отпускания _-1соответствующии символ появится на экране в том мес-
те, где перед этим находился текстовый курсор.
При работе в Windows (Здесь речь уже идет об ANSI-кодах) для ввода символов
с помощью клавиши _____J нужно поступать точно так же, как в DOS, но перед со-
ответствующим ANSI-кодом символа сначала надо набрать 0 (ноль) и только потом
собственно код. Причем, малая цифровая клавиатура должна быть обязательно пе-
реключена для ввода цифр (режим NumLock должен быть включен). Например, цифру
9, стоящую на 57-ом месте в таблице кодов, можно ввести с помощью клавиши
Alt
____J, набрав код 057.
Краткая информация о
других системах кодирования
Системы кодирования текстовых данных были разработаны и в других странах.
Так, например, в СССР в этой области действовала система кодирования КОИ-7
(код обмена информацией, семизначный). Однако поддержка производителей обору-
дования и программ вывела американский код ASCII на уровень международного
стандарта, и национальным системам кодирования пришлось "отступить" во вто-
рую, расширенную часть системы кодирования, определяющую значения кодов со
128 до 255. Отсутствие единого стандарта в этой области привело к множествен-
ности одновременно действующих кодировок. Только в России можно указать три
действующих стандарта кодировки и еще два устаревших.
Так, например, кодировка символов русского языка, известная как кодировка
Windows-1251, была введена "извне" - компанией Microsoft, но учитывая широкое
распространение операционных систем и других продуктов этой компании в России
она нашла широкое распространение в России. Эта кодировка используется на
большинстве локальных компьютеров, работающих на платформе Windows.
Другая распространенная кодировка носит название КОИ-8 (код обмена информа-
цией, восьмизначный) - ее происхождение относится ко времени действий Совета
Экономической Взаимопомощи государств Восточной Европы. Сегодня кодировка
КОИ-8 имеет широкое распространение в компьютерных сетях на территории России
и в российском секторе Интернета.
Международный стандарт, в котором предусмотрена кодировка символов русского
алфавита, носит название ISO (International Standard Organization - Междуна-
родный институт стандартизации). На практике данная кодировка используется
редко.
На компьютерах, работающих под операционной системой MS-DOS, могут действо-
вать еще две кодировки (кодировка ГОСТ и кодировка ГОСТ альтернативная). Пер-
вая из них считалась устаревшей даже в первые годы появления персональной вы-
числительной техники, но вторая используется и по сей день.
В связи с изобилием систем кодировки текстовых данных, действующих в России
возникает задача межсистемного преобразования данных - это одна из распро-
страненных задач информатики.
Подводя некоторый итог, можно сказать, что текстовый файл - это файл, в ко-
тором каждый байт интерпретируется как изображаемый символ в некоторой систе-
ме кодировки. Кроме кодов изображаемых символов, текстовые файлы включают
также ряд управляющих кодов, например, код возврата каретки, перевода строки,
конца файла.
Универсальная система
кодирования текстовых данных
Если проанализировать организационные трудности, связанные с созданием еди-
ной системы кодирования текстовых данных, то можно прийти к выводу, что они
вызваны ограниченным набором кодов (256). В то время, очевидно, что если, на-
пример, кодировать символы не восьмиразрядными двоичными числами, а числами с
большим количеством разрядов, то и диапазон возможных значений кодов станет
намного больше. Такая система, основанная на 16-разрядном кодировании симво-
лов, получила название универсальной Unicode. Шестнадцать разрядов позволяют
обеспечить уникальные коды для 65536 различных символов - этого поля доста-
точно для размещения в одной таблице символов большинства языков планеты.
Несмотря на тривиальную очевидность такого подхода, простой механический
переход на данную систему долгое время сдерживался из-за недостаточных ресур-
сов средств вычислительной техники (в системе кодирования UNICODE все тесто-
вые документы автоматически становятся вдвое длиннее). Во второй половине 90-
х годов технические средства достигли необходимого уровня обеспеченности ре-
сурсами, и сегодня наблюдается постепенный переход документов и программных
средств на универсальную систему кодирования.
Кодировка Unicode
В системе Unicode (другое название ISO — International Standard
Organization) для представления любого символа используется уже не один, а
два байта. Поскольку в этом случае число возможных символов равно 216 или
65536, то могут удовлетворяться практически любые потребности всех националь-
ных языков. В настоящее время система Unicode стала "de facto" мировым стан-
дартом. Тем не менее, вряд ли на этом все и остановится. Несмотря на огромное
количество возможных кодов (символов), система Unicode все же не способна
удовлетворить всем типографским требованиям. Так что, процесс пошел, но "еще
не вечер..."
ASCII-кодировка, однако, не умерла. Она осталась как базовый формат для
представления текстовой информации. Тексты, созданные в этом формате:
• не имеют различных начертаний шрифта;
• нет ни курсива, ни полужирного, ни подчеркнутого шрифта — он единый на все
случаи жизни;
• отсутствуют понятия отступов, кеглей и гарнитур;
• каждая строка представляет собой самостоятельный абзац.
Тем не менее, у ASCII-формата имеется одно очень важное достоинство — он
является стандартным, и его понимают абсолютно все программы, работающие с
текстами.
ЛЕКЦИЯ 5
Текстовые редакторы
и процессоры
Во многих видах человеческой деятельности требуется уметь обрабатывать тек-
стовую информацию: писать, компоновать результирующий текст из различных ис-
точников , переписывать текст, удалять и вставлять фрагменты текста, делать
несколько копий одного и тог же текста и т.д. Для облегчения такого традици-
онного труда для людей различных специальностей были созданы специальные ком-
пьютерные программы: текстовые редакторы и текстовые процессоры.
Первоначально появились текстовые редакторы с очень ограниченным количест-
вом функций. Сами же редакторы делились на две категории:
• Командные редакторы очень сложны для неподготовленного пользователя и
сейчас практически не находят применения.
• Экранные редакторы находят свое применение и по сей день. Типичным при-
мером экранного текстового редактора с современным интерфейсом является стан-
дартная программа Windows95/98 - Notepad, называемая в русифицированной вер-
сии - Блокнот. Ограниченное множество функций, компенсируется в этой програм-
ме абсолютно прозрачным интерфейсом, т.е. способом взаимодействия пользовате-
ля и компьютерной программы. Это дает возможность пользователям полностью
прогнозировать свои действия при работе с этой программой за очень небольшой
(1-1,5 часа) промежуток времени работы с ней.
Логическим итогом развития текстовых редакторов стало их превращение в тек-
стовые процессоры. Типичным представителем современного текстового процессора
является приложение Microsoft Word из пакета Microsoft Office. Текстовые про-
цессоры дают гораздо больше возможностей по редактированию текста, чем тек-
стовые редакторы. Поскольку эти программы значительно сложнее, они требуют
большей дисковой памяти и для их загрузки требуется больше времени. Для того,
чтобы использовать текстовый процессор, необходимо иметь его жестком диске
компьютера и постоянно пользоваться документацией по его применению (это не
обязательно печатная продукция, а, скорее всего встроенная справочная систе-
ма , обычно имеющая содержание, перечень терминов, поиск по словам).
Отличие текстового процессора от редактора состоит в том, что в файл добав-
лены специальные коды, макросы (особые программы), определяющие вид докумен-
та. "Родным" форматом файла для текстового процессора не является текстовый
файл, определение которого дано выше. Отличительной особенностью текстовых
процессоров является возможность обрабатывать файлы различных типов и конвер-
тировать их различные форматы, в том числе и обычные текстовые файлы. Приме-
нительно к Microsoft Word, если требуется сохранить созданный файл как тек-
стовый, то выбирается тип сохранения файла, как файла MS-DOS.
Текстовые процессоры включают большое количество возможностей по созданию
сложных документов содержащих текстовую информацию.
Обычно выделяют следующие возможности:
• контроль правописания;
• помощь в выборе синонимов
• средства форматирования текста
• поля
• отступы
• режимы выравнивания в строке
• атрибуты текста, повышающие наглядность и выразительность, например, выде-
ление курсивом, жирным шрифтом, подчеркиванием;
• выбор типа шрифта и его размера; задание межстрочных интервалов;
• предопределение стилей документов, определяющих вид оглавления, основного
текста, заголовков разного уровня, списков, элементов
• глоссария и других элементов оформления;
• сохранение информации в различных форматов, включая формат HTML, предна-
значенный для публикации документов в Интернет; при
• этом необходимо четко знать, какие из элементов оформления будут утрачены,
поскольку процессор при этой операции убирает
• форматирующие коды, в частности, установка полей, отступов, выравнивания и
т.д.
Текстовый редактор WordPad
Текстовый редактор WordPad Windows 95/98 призван заменить известный из
прежних версий Windows редактор Write. Редактор WordPad сильно усовершенство-
ван и является довольно продвинутым 32-разрядным приложением, которое пользо-
ватель приобретает сразу с приобретением Windows. Пользовательский интерфейс
WordPad является самым типовым из приложений Windows, так что, получив навыки
работы с ним, вы успешно справитесь с работой со многими приложениями. Конеч-
но , WordPad не заменяет полностью такие мощные редакторы как Word или Word
Perfect, которые по своим возможностям вполне отвечают требованиям к тексто-
вым процессорам настольных издательских систем.
Однако эти редакторы сложны в освоении, стоят дорого и требуют внушительных
затрат памяти. WordPad может использоваться даже для подготовки презентаций с
элементами мультимедиа-технологий, включая показ слайдов, обеспечение звуко-
вого сопровождения читаемых текстов и даже показ небольших видеофильмов. Осо-
бо следует отметить, что новый редактор позволяет задавать цвет любого симво-
ла, слова или целого фрагмента текста, что не позволял более простой редактор
Write, входящий в старые версии Windows.
При наличии струйного или лазерного принтера с цветной печатью это открыва-
ет неограниченные возможности в подготовке ярких цветных визитных карточек,
красочно оформленных бланков для деловых писем и др. Редактор позволяет им-
портировать графики из разных приложений, в том числе и из буфера промежуточ-
ного хранения Clipboard - хотя скорость построения импортируемого графика вы-
сокой не назовешь.
Для начального ввода текста используется команда New в позиции File главно-
го меню или команда Open для загрузки файла с редактируемым текстом). Пользо-
вательский интерфейс редактора WordPad наглядно свидетельствует о стремлении
его разработчиков сделать управление редактором предельно простым и нагляд-
ным. Как отмечалось, пользовательский интерфейс WordPad является своеобразным
эталоном для большинства Windows-приложений, созданных и создаваемых для опе-
рационной системы Windows.
My Computer
Desk dupe
Inbox
Recycle Bin
□
Dodot
Nc
Lexicon
Start
Is! dem° -Wo
Поскольку этот интерфейс является образцовым, рассмотрим его несколько под-
робнее. Прежде всего, заметим, что главное меню редактора содержит множество
позиций, и каждая порождает свое подменю с различными опциями и командами.
Они и определяют возможности редактора. Ниже представлены основные команды
редактора. Позиция File (работа с файлами) главного меню имеет подменю со
следующими командами:
New - очистка окна редактирования и подготовка к вводу нового текста (его файл получает имя Untitle - безымянный).
Open - открытие и загрузка файла с его поиском в диалоговом окне.
Save - запись файла на диск с текущим именем в текущую директорию.
Save As... - запись файла на диск с заданным именем и заданной директо- рией (они задаются в появившемся диалоговом окне).
Print - печать документа с установкой опций печати (выбор принтера и др.) в появившемся диалоговом окне.
Print Preview - просмотр страницы перед печатью (полезно для оценки распо- ложения абзацев и встроенных рисунков).
Page Setup - установка параметров страницы.
Send ... - установка связи с электронной почтой для получения по ней документа или пересылки редактируемого документа по электрон- ной почте.
Exit - завершение работы с редактором (возможно после закрытия окон со всеми документами).
Помимо этих команд в подменю позиции File главного меню имеется список ра-
нее загруженных в редактор документов. Можно сразу указать нужный документ,
не обращаясь к его более длинному поиску с помощью команды Open. Позиция Edit
(редактирование) порождает подменю со следующими командами:
Undo - отмена последней операции редактирования.
Cut - перенос выделенного фрагмента документа в буфер промежу- точного хранения Clipboard с уничтожением его в документе.
Сору - копирование выделенного фрагмента в буфер промежуточного хранения с сохранением этого фрагмента в документе.
Paste - перенос фрагмента документа из буфера в текст редакти- руемого документа с помещением фрагмента по месту располо- жения курсора.
Paste Special ... - перенос фрагмента документа из буфера с установлением специальной связи (используется при реализации механизма OLE) .
Clear - удаление выделенного фрагмента.
Select All - выделение всего текста (например, это удобно при его пе- ре форматировании).
Find - поиск заданной строки символов (задается в диалоговом окне). В случае успеха поиска курсор устанавливается на обнаруженной строке.
Find Next - поиск следующего включения заданной строки в документе.
Replace - замена заданной строки другой строкой.
Links - установление связи с объектом (используется при реализа- ции механизма OLE).
Object Properties - установление свойств объекта.
Object - экспорт объекта.
Options - задание дополнительных опций.
Позиция View (обзор) порождает подменю со следующими позициями:
ToolBar включение/выключение инструментального ящика с пикто- граммами быстрого управления редактором.
Format Bar - включение/выключение ящика со средствами работы с тек- стом (выбор шрифтов, их размера и типа, форматирования текста).
Ruler - включение/выключение линейки с делениями.
Status Bar - включение/выключение статусной линейки.
Все команды в этом подменю работают как переключатели с двумя позициями -
включено или выключено. Если задана позиция включено, против команды появля-
ется знак птички. Соответствующий ящик при этом виден на переднем панно при-
ложения. Позиция Insert (включение) порождает подменю с двумя командами:
Date and Time... - включение по месту расположения курсора строки с указа- нием текущего времени и даты
Object - включение объекта, созданного другим приложением (рисун- ка, проигрываемого звукового файла и т.д.).
Позиция Format (форматирование) дает подменю с командами:
Font - выбор и установка фонтов.
Bullet Style - выделение строк закрашенным кружком перед ними,
Paragraph... - вызов диалогового окна с опциями форматирования парагра- фа (с равнением по левому краю, по центру или по правому краю) .
Tabs... - вызов диалогового окна для установки параметров табуля- ции .
И, наконец, позиция Help (помощь) главного меню имеет подменю с позициями:
Help Topics - вызов базы данных помощи.
About WordPad - вызов окна с данными о редакторе и об использовании сис- темных ресурсов.
Важнейшие из команд редактора продублированы пиктограммами инструментально-
го ящика. Например, пиктограмма в виде чистого листа означает ввод команды
New (создание нового документа), пиктограмма в виде открывающейся папки озна-
чает команду Open (вывести диалоговое окно с именами файлов), пиктограмма в
виде дискеты означает запись на диск, пиктограмма в виде ножниц - вырезание
отмеченного фрагмента текста, пиктограмма в виде бинокля - поиск заданного
фрагмента текста и т.д.
Совершенно очевидно назначение пиктограмм с разными стилями букв и разным
видом выравнивания текстов - по правой кромке, по середине и по левой кромке.
Помимо этого многие команды имеют ввод с помощью так называемых "горячих"
клавиш. Они указаны после названия команд в соответствующих позициях подменю.
Например, команда Save имеет горячие клавиши Ctrl и S. При их одновременном
нажатии эта команда будет немедленно исполнена.
Важно отметить, что такие команды выполняются немедленно даже без активиза-
ции соответствующего подменю. В целом правила работы с редактором WordPad ос-
тались те же, что и у редактора Write, хорошо известного пользователям более
ранних версий Windows.
Из недостатков редактора WordPad можно отметить главный - медленное пере-
ключение шрифтов, их типа и размера. В этом отношении редактор заметно усту-
пает своему старшему собрату - текстовому процессору Word (особенно входящему
в состав комплекса Office для Windows).
Описанная система команд редактора WinPad характерна для многих других при-
ложений. Например команды позиций File и Edit есть в большинстве других при-
ложений, например в графических редакторах. Поэтому опыт использования команд
редактора WinPad окажется весьма полезным при изучении работы и других прило-
жений Windows. WordPad позволяет записывать файлы в следующих трех основных
форматах - тестовом ASCII (принятом в MS-DOS), текстовом RTF и в формате по-
пулярного редактора Word. В последнем случае в тексте документа могут быть
графические вставки в виде рисунков, а также пиктограммы различных объектов,
например звуковых или анимационных файлов.
WordPad позволяет также загружать файлы формата WRI текстового редактора
Write.
В общем, редактор WordPad - приложение с весьма обширными возможностями, но
ориентированное либо на не очень взыскательного пользователя, либо на начи-
нающего работать с Windows. Главный упрек в адрес редактора - его подчеркну-
тая медлительность при смене шрифтов, при изменении их размера, при построе-
нии импортируемых рисунков и т.д.
Складывается впечатление, что разработчики Windows намеренно не приняли мер
(вполне возможных) для ускорения работы этого "быстрого 32-разрядного прило-
жения" , как нередко именуются встроенные приложения в рекламах на Windows. И
сделали они это, в целом, по вполне понятным мотивам - если бы при своих воз-
можностях WordPad работал к тому же быстро, то нашлось бы мало пользователей,
покупающих более мощные редакторы, например Word под Windows.
Текстовый редактор формата
ASCII в составе Windows
В составе Windows есть текстовый редактор, работающий с файлами формата
ASCII (American Standard Code for Information Interchange - Американский
стандартный код для обмена информацией).
Напомним, что этот формат оперирует с 256 численными кодами, имеющими зна-
чения от 0 до 255. В соответствие каждому коду ставится определенный символ
(буква, цифра, знак препинания, математический символ или символ псевдографи-
ки) .
Это соответствие задается с помощью стандартных кодовых таблиц с различными
номерами (например, таблица 866 предназначена для русскоязычных пользовате-
лей) .
Принятая в Windows кодировка символов отличается от ASCII. Поэтому Windows-
редакторы не могут работать с документами, имеющими формат ASCII, без пере-
форматирования текстовых файлов. Поэтому в составе Windows 95 сохранен и су-
щественно доработан хорошо известный пользователям текстовый редактор MS-DOS
Edit.
Этот редактор не инсталлируется вместе с инсталляцией Windows 95, так что
его невозможно обнаружить среди приложений, доступных с помощью стартового
меню. Редактор представлен файлом Edit.com в поддиректории COMMAND директории
WIN95. Так что для его Запуска надо использовать команду Run и задать в диа-
логовом окне этой команды командную строку C:\WIN95\COMMAND\Edit.com Можно
также отыскать этот файл, используя опцию Browse. После этого дайте команду
Ок в диалоговом окне, и окно редактора MS-DOS Edit появится на рабочем столе.
Работа с редактором ускорена и стала более удобной благодаря наличию инст-
рументального ящика ToolBar.
Но, пожалуй, главное достоинство этого обновленного редактора - возможность
загрузки поистине огромных текстовых файлов с объемом до 4 Мбайт. Это полезно
и для разбивки больших файлов на более мелкие для последующего их редактиро-
вания иными редакторами.
Редактор MS-DOS Edit многооконный - в него можно загружать сразу до девяти
файлов и перекидывать выделяемые фрагменты текста из одного окна в другое.
Быстрое управление редактором обеспечивается с помощью клавиш управления -
полный перечень их и реализуемых команд легко найти в соответствующей позиции
подменю Help. И еще одно достоинство - редактор поддерживает длинные имена
файлов, которые можно использовать в Windows (в том числе в режиме эмуляции
MS-DOS).
Графический редактор PAINT
Понятие о компьютерной графике
Компьютерная графика - специальная область информатики, изучающая методы и
средства создания и обработки изображений с помощью программно- аппаратных
вычислительных комплексов.
Компьютерная графика охватывает все виды и формы представления изображения,
доступных для восприятия человеком на экране монитора, либо в виде копии на
внешнем носителе. Без компьютерной графики невозможно сейчас представить ма-
териальный мир. Например, медицина использует компьютерную томографию; науч-
ные исследования - визуализацию строения вещества, векторных полей и других
данных; моделирование тканей и одежды, опытно-конструкторские разработки так-
же используют компьютерную графику.
Сферы применения машинной графики:
• компьютерное моделирование;
• САПР (системы автоматизированного проектирования);
• компьютерные игры;
• обучающие программы;
• реклама и дизайн;
• мультимедиа презентации;
• Internet.
Компьютерная графика всего лишь инструмент, но ее структура и методы осно-
ваны на передовых достижениях фундаментальных и прикладных наук: математики,
физики, химии, биологии, статистики, программирования и множества других.
Компьютерная графика является одним из наиболее бурно развивающихся направле-
ний информатики.
Виды компьютерной графики
В зависимости от способа формирования изображений, компьютерную графику
принято делить на:
- растровую;
- векторную;
- фрактальную.
Отдельным предметом считается трехмерная графика, изучающая приемы и методы
построения объемных моделей объектов в виртуальном пространстве.
Эти виды отличаются принципами формирования изображения при отображении на
экране монитора или при печати на бумаге. Каждый вид используется в опреде-
ленной области.
Растровую графику применяют при разработке мультимедийных проектов. Иллюст-
рации, выполненные средствами растровой графики, чаще вводятся в компьютер с
помощью сканера, а затем обрабатываются специальными программами - графиче-
скими редакторами.
Программные средства для работы с векторной графикой наоборот предназначены
для создания иллюстраций на основе простейших геометрических элементов. В ос-
новном применение векторной графики - это оформительские работы.
Создание фрактальной художественной композиции состоит не в рисовании, а
скорее в программировании. Программные средства для работы с фрактальной гра-
фикой предназначены для автоматической генерации изображений путем математи-
ческих расчетов. Например, таким образом можно получить заставки на ТВ.
Графическими редакторами называются программы, предназначенные для создания
и редактирования графических изображений - рисунков. Существует два класса
графических редакторов - растровый и векторный. В растровой графике мельчай-
шим элементом изображения является точка, которой на экране монитора соответ-
ствует экранная точка - пиксел, мельчайшим элементом векторной графики явля-
ется линия, описываемая математическим выражением. Например, программа Corel
Draw -редактор векторной графики, программа Paint -редактор растровой графи-
ки. Основными функциями приложения Paint являются функции создания и редакти-
рования графических изображений - рисунков. Программа запускается командой
Пуск / Программы / Стандартные / Paint.
Рабочее окно программы Paint состоит из строки заголовка, строки меню, ра-
бочей области. Слева от рабочей области располагается панель инструментов
(команд графического редактора), под которой находится палитра настройки ин-
струмента. Она содержит элементы управления инструментами для рисования в ви-
де кнопок с пиктограммами. Каждый инструмент имеет подсказку, обозначающую
назначение инструмента.
Необходимый инструмент можно выбрать щелчком мыши по соответствующей кнопке
на панели инструментов, с помощью палитры настройки можно задать параметры
вызванного инструмента, например толщину линии, метод заполнения фигуры. Ниже
рабочей области располагается цветовая палитра (стандартная). Она содержит
набор цветов, который можно использовать при рисовании, а также окно слева с
двумя наложенными квадратами - окно просмотра цветов. Если нужный цвет в па-
литре отсутствует, его можно создать и заместить им любой из цветов палитры.
Элементы цветовой палитры служат для выбора основного цвета изображения
(щелчок левой кнопки), и фонового цвета (щелчок правой кнопки).
До начала создания документа задаются параметры страницы командой Файл /
Макет страницы. Здесь можно выбрать:
- формат бумаги из раскрывающегося списка Размер;
- ориентацию бумаги: Книжная или Альбомная;
- поля - интервалы между текстом и краями листа;
- колонтитулы (верхний и нижний) - в них могут проставляться номера стра-
ниц, дата, время создания документа, его название и т.д.
Щелчок по кнопке Ок закрывает окно Макет страницы с сохранением всехустано-
вок.
Перед печатью файла на принтере можно сделать его предварительный просмотр
командой Файл / Предварительный просмотр. Рисунок может занимать часть рабо-
чей области, всю ее и выходить за ее пределы. В последнем случае по краям ра-
бочей области появятся полосы прокрутки. На границах рисунка располагаются
маркеры изменения размера (темные точки в середине сторон и по углам рисун-
ка) .
Перед тем, как начать рисование, необходимо задать размер будущего рисунка,
воспользовавшись командой Рисунок / Атрибуты. По этой команде открывается
диалоговое окно Атрибуты, в котором можно выбрать палитру (черно - белую или
цветную), единицу измерения (дюймы, сантиметры или пикселы), задать ширину и
высоту рисунка. Если рисунок предназначен для воспроизведения на экране (на-
пример, в качестве фона рабочего стола), нужно выбрать единицу измерения пик-
селы, для подготовки печатных документов - дюймы или сантиметры. Следует
учесть, что преобразование рисунка в черно-белый вызовет потерю сведений о
цвете, после чего восстановление исходного рисунка будет невозможным.
Меню Paint состоит из следующих пунктов: Файл, Правка, Вид, Рисунок, Палит-
ра и Справка. Каждый из них содержит несколько команд, позволяющих выполнять
какие - либо действия.
Создание нового файла выполняется командой Файл/Создать, открытие сущест-
вующего - Файл/Открыть. Имя файла появится в поле Имя файла. Затем нужно на-
жать кнопку Открыть. Двойной щелчок по имени сразу открывает файл.
При первом сохранении файла можно выполнить команду Файл/Сохранить или
Файл/Сохранить как. В диалоговом окне выбирается папка, тип файла и в поле
Имя файла вводится его имя. Сохранение выполняется щелчком на кнопке Сохра-
нить .
Если файл уже имеет имя, то сохранить его можно командой Файл/Сохранить.
Файл запишется в предыдущий файл. Если нужно изменить имя и расположение фай-
ла, то необходимо воспользоваться командой Файл/Сохранить как. В этом случае
происходит вывод на экран окна, где необходимо указать новые координаты со-
храняемого файла.
Редактор Paint сохраняет рисунки в формате растровой графики *.bmp.
Команда Замостить рабочий стол Windows позволяет заполнить рабочий стол по-
вторяющимися копиями текущего рисунка, команда В центр рабочего стола Windows
- разместить текущий рисунок в центре рабочего стола Windows.
Меню Правка содержит команды:
• Отменить - отменяет последние действия. Отменить можно только действия,
выполненные активным в данный момент инструментом. Этой команде соответст-
вует комбинация клавиш Ctrl + Z, действующая в большинстве приложений
Windows.
• Вырезать - (соответствует комбинация клавиш Ctrl + X) - удаляет выделенный
фрагмент из рисунка и помещает его в буфер обмена.
• Копировать - (Ctrl + С) - помещает копию выделенного фрагмента в буфер об-
мена .
• Вставить - (Ctrl + V) - вставляет в рисунок содержимое буфера обмена. Изо-
бражение вставляется в левый верхний угол, после чего его можно «отбукси-
ровать» в нужное место.
• Очистить выделение - удаляет выделенную часть рисунка.
• Выделить все - выделяется весь рисунок.
• Копировать в файл ... - позволяет сохранить выделенный фрагмент рисунка в
виде самостоятельного файла.
Пункт меню Вид состоит из следующих команд:
• Набор инструментов - показать/убрать панели инструментов.
• Палитра - показать/убрать Палитру.
• Строка состояния - показать/убрать Строку состояния.
• Панель атрибутов текста - показать/убрать Панель атрибутов текста.
• Масштаб - изменение масштаба рисунка. Можно выбрать команду Обычный, Круп-
ный (увеличивает масштаб изображения в четыре раза) или Другой. Выбрав ко-
манду Другой, можно задать необходимый масштаб в процентах от 100% до
800%. Команда Вид / Масштаб / Окно масштаба 100% позволяет показать часть
изображения в небольшом окне в обычном масштабе для быстрого просмотра
проделанных изменений.
• Просмотреть рисунок - рисунок будет занимать всю область просмотра. Чтобы
вернуться в прежний режим, щелкните любое место рисунка. Этот режим не
предназначен для изменения рисунка.
Меню Рисунок содержит команды, позволяющие преобразовать любой выделенный
объект:
• Отразить/повернуть - позволяет зеркально отобразить выбранный фрагмент или
повернуть его как одно целое. Вариант преобразования выбирается в диалого-
вом окне Отражение и поворот - установкой соответствующего переключателя.
Переключатель Отразить слева направо соответствует горизонтальному отраже-
нию, Отразить сверху вниз - вертикальному. Переключатель Поворот позволяет
повернуть рисунок на угол 90°, 180° или 270°.
• Растянуть/наклонить - позволяет растянуть или сжать изображение по одной
из осей и одновременно наклонить его.
• Обратить цвета - каждый цвет будет заменен на обратный к нему. Например,
красный будет преобразован в голубой, а синий в желтый.
• Атрибуты - позволяет изменить полный размер рисунка в заданных единицах
измерения, а также указать, должен ли он быть цветным или черно - белым.
• Очистить - служит для очистки выделенного фрагмента или всего изображения
и заполнения его фоновым цветом.
• Непрозрачный фон - назначает один цвет в качестве «прозрачного». Прозрач-
ность цвета означает, что если данный рисунок будет отображаться поверх
другого, фонового, то нижний рисунок будет виден сквозь верхнийв тех точ-
ках, которые имеют цвет, назначенный прозрачным. Свойство прозрачности со-
храняется в файле рисунка в тех случаях, когда при сохранении рисунка вы-
бран формат *.gif.
• В меню Палитра находится только одна команда Изменить палитру..., позволяю-
щая создать произвольный цвет. Для этого нужно выбрать на палитре цвет,
который следует изменить, вызвать команду Изменить Палитру... открывается
диалоговое окно Изменение палитры, которое содержит расширенную палитру.
Если нужного цвета нет и там, можно щелкнуть на кнопке Определить цвет.
Диалоговое окно расширяется и позволяет определить цвет подбором по спек-
тру или заданием параметров Оттенок, Контраст, и Яркость. Щелчком кнопки
Добавить в набор вновь созданный цвет включается в палитру. Щелчком кнопки
ОК новый цвет выбирается в качестве основного.
Инструменты графического редактора Paint
По назначению инструменты графического редактора Paint можно разделить на
несколько групп:
1) Инструменты свободного рисования. Они позволяют рисовать произвольные
фигуры. Рисование с помощью этих инструментов осуществляется путем протягива-
ния мыши:
при движении указателя мыши за ним остается след. Протягивание с нажатой
левой кнопкой мыши дает изображение фигуры основным цветом, а при нажатой
правой кнопке - цветом фона. К инструментам свободного рисования относят:
9
Карандаш - предназначен для рисования произвольных линий. Толщину ли-
нии выбирают в палитре настройки. Чтобы линия была строго горизонтальной или
вертикальной, во время рисования необходимо удерживать нажатой клавишу Shift.
° Кисть - позволяет провести более широкую линию. Этот инструмент можно
использовать как для свободного рисования произвольных кривых, как Карандаш,
но чаще его используют для рисования методом набивки. Сначала выбирают форму
кисти в палитре настройки, а потом щелчками левой кнопки мыши наносят оттиски
на рисунок без протягивания мыши.
* Распылитель - создает «размытое» пятно, его используют как для свобод-
ного рисования, так и для рисования методом набивки. Форму пятна выбирают в
палитре настройки.
Ластик - стирает изображение, заменяя его фоновым цветом. Все инстру-
менты, кроме ластика, выполняют рисование основным цветом.
2) Инструменты рисования линий:
Линия - предназначена для вычерчивания прямых линий. Толщину линии вы-
бирают в палитре настройки. Чтобы линия получилась строго вертикальной, гори-
зонтальной или наклонной под углом 450, при ее вычерчивании следует держать
нажатой клавишу Shift.
Кривая - служит для рисования гладких кривых линий. Толщину предвари-
тельно выбирают в палитре настройки. Построение кривых линий производится в
три приема: сначала проводят отрезок прямой, концы которого совпадают с кон-
цами отрезка будущей кривой, затемщелчком и протягиваниемв стороне от линии
задают первый и второй радиусы кривизны.
3) Инструменты создания стандартных фигур:
Прямоугольник - применяют для рисования прямоугольных фигур. В палитре
настройки можно выбрать метод заполнения прямоугольника. Возможны три вариан-
та: без заполнения (рисуется только рамка), заполнение фоновым цветом и за-
полнение основным цветом. Если при создании прямоугольника держать нажатой
клавишу Shift, получается квадрат.
----- Скругленный прямоугольник - действует точно так же, но при этом полу-
чается прямоугольник со скругленными углами.
Многоугольник - предназначен для рисования произвольных многоугольни-
ков . Стороны многоугольника рисуют последовательно методом протягивания.
Контур замыкают протягиванием указателя от начальной точки к конечной.
Замкнутые фигуры могут автоматически заливаться краской в соответствии с цве-
том заполнения, выбранным в палитре настройки.
Эллипс - служит для изображения эллипсов и
стный случай правильного эллипса) получается при
ше Shift.
окружностей. Окружность (ча-
рисовании при нажатой клави-
4) Заполнение областей цветом:
Заливка - служит для заполнения замкнутых контуров основным или фоно-
вым цветом. Заполнение основным цветом производится щелчком левой кнопки мы-
ши, а заполнение фоновым цветом - щелчком правой кнопки. Если контур не замк-
нут, инструмент работает неправильно: краска «вытекает» за границы контураи
заполняет весь рисунок.
s Выбор цветов - позволяет точно выбрать основной или дополнительный цвет
не из палитры красок, а непосредственно из рисунка. Это важно, когда надо
обеспечить один и тот же цвет в разных областях изображения. После выбора ин-
струмента наводят указатель на участок рисунка с нужным цветом и щелкают
кнопкой мыши: левой - текущий цвет становится основным, правой - фоновым.
5) Инструменты выделения областей:
' Выделение произвольной области. При выборе этого инструмента курсор
принимает вид крестика. Нужный фрагмент обводится курсором при нажатой левой
кнопке мыши. При этом пунктирная линия отметит пройденный путь и ограничит
выделенную область. Если отпустить левую кнопку до того, как линия пересечет-
ся сама с собой, начальная и конечная точки соединятся по кратчайшему рас-
стоянию. Если нажать правую кнопку мыши до того, как отпустить левую, выделе-
ние будет отменено.
Выделение - формирует прямоугольную выделенную область. Выделенный
фрагмент можно удалить (клавиша Delete), поместить в буфер обмена, и вставить
из буфера обмена, переместить методом перетаскивания. Если при перемещении
удерживать нажатой клавишу Shift, то перетаскиваемый обьект оставляет за со-
бой след. При копировании выделенных областей возможны два режима вставки: с
сохранением фоновой графики или без нее (точки фонового цвета во вставляемой
области игнорируются). Переключение режима выполняют в палитре настройки.
6) Ввод текста:
Графический редактор Paint не предназначен для работы с текстом. Поэтому
текст после ввода становится частью рисунка и так же, как изображение, будет
состоять из точек растра. Режим работы с текстом предназначен в основном для
создания кратких и крупных заголовков.
Надпись - служит для ввода текста. После того, как выбран инструмент,
нужно выделить прямоугольную область протягиванием мыши там, где должна быть
надпись. Закончив ввод, необходимо вызвать панель атрибутов текста щелчком
внутри рамки или с помощью команды Вид / Панель атрибутов текста. С помощью
нее можно выбрать форму шрифта, его начертание и размер. В случае необходимо-
сти текстовую рамку можно сжать или растянуть. После щелчка вне рамки текст
становится частью рисунка, его нельзя будет изменить.
Изменение масштаба рисунка
Для точной корректировки рисунка иногда необходимо увеличить его масштаб.
Увеличение можно проделать двумя способами: с помощью команды Вид / Масштаб
или с помощью кнопки - Лупана панели инструментов. Начиная с четырехкратного
увеличения на рисунок можно наложить вспомогательную сетку командой ВидМас-
штаб / Показать сетку. Каждая ячейка этой сетки - увеличенная точка изображе-
ния, это позволяет редактировать рисунок по отдельным точкам.
ЛЕКЦИЯ 6
Microsoft Word для Windows
Технология WYSIWYG
Редактируя документ в операционной системе Windows, пользователь видит на
экране изображение, максимально близкое у тому, что он увидит на бумаге при
распечатке документа на принтере. Это называется What You See Is What You
Get, или технологией точного отображения макета. Благодаря WYSIWYG Вы можете
добиться максимальной выразительности документов даже не будучи профессиона-
лом. Используя масштабируемые шрифты, пользователь может не опасаться, что
изображение на бумаге "огрубеет" и "поплывет" (т.е. изменится вид шрифта при
его масштабировании), как это часто случалось с растровыми шрифтами. Эта тех-
нология используется в текстовых процессорах Word 97 и Word 2000.
Общего у Word 2000 и Word 97 очень много, но немало и различий. Microsoft
Office 2000 предоставляет самые последние технологии работы с документами,
позволяет создать принципиально новую рабочую среду, оптимизирующую обмен
данными, и резко повысить эффективность работы с информацией благодаря:
• простоте установки, настройки и использования;
• интеллектуальному интерфейсу;
• тесной интеграции приложений;
• возможностям работы в Интернете и интрасетях;
• инструментам анализа информации.
Интеллектуальный интерфейс
При запуске Word 2000 каждое меню содержит только команды, используемые на
протяжении 95% общего времени работы. Редко вызываемые команды его не загро-
мождают .
& Открыть... Ctrl+O
Сохранить как...
Печать... Ctrl+P
1 Особенности ввода текста на националы..
2 С: WlNDOWS\TEMPV\NDROID-dos
3 С: WNDOWS\TEMP\Android1
4 E:tlTML-npoeKTbiV..Vitmls\subject_10
Сокращенный вариант выпадающего меню
В нижней части каждого меню располагается кнопка расширения, выводящая на
экран полный набор вариантов данного меню и позволяющая очень легко получить
доступ ко всем командам.
Все меню в Word 2000 автоматически расширяются в соответствии с правилами
технологии IntelliSense (например, по истечении определенного промежутка вре-
мени, при задержке курсора мыши на меню и так далее).
П Создать... Ctrl+N
Открыть... Ctrl+O
Закрыть
12] Сохранить Shift+F12
Сохранить как...
jd] Сохранить как Web-страницу...
Версии...
Предварительный просмотр Web-страницы
Параметры страницы...
|~с\ Предварительный просмотр
Печать... Ctrl+P
Отправить ►
Свойства
1_Особенности ввода текста на националы..
2 C:WNDOWS\TEMP\ANDROID-dos
3 C:WlNDOWS\TEMP\Android1
4 E:\HTML-npoeKTbi\...\htmls\subject_10
Выход
Полный вариант выпадающего меню
Когда вы выбираете какую-то команду, она "получает повышение" и отображает-
ся ближе к началу меню, а неиспользуемые команды в конце концов перестают
отображаться в настраиваемых меню. После некоторого периода работы с програм-
мой меню будут содержать наиболее часто используемые команды, а неиспользуе-
мые или редко используемые пункты окажутся скрытыми. Однако, если вы захотите
познакомиться с новыми функциями, то, подведя курсор к кнопке расширения, ме-
ню можно быстро расширить и найти другие команды. В остальное время вы видите
только те средства, которыми реально пользуетесь при работе в Word 2000.
Автоматическое самовосстановление
Office 2000 (а значит и Word 2000) включает в себя функции самовосстановле-
ния. При каждом запуске система Office 2000 проверяет все важнейшие файлы и
соответствующие элементы реестра. Если что-то отсутствует или повреждено,
производится автоматическая повторная установка в фоновом режиме. Так что не
удивляйтесь, если при очередном вызове программы вы увидите окно с сообщением
об инсталляции. Не нервничайте, а подождите немного — эта небольшая пауза по-
может вам избежать значительно больших потерь времени при работе.
Ручной запуск самовосстановления
Если вам вдруг показалось, что Word 2000 что-то делает не так, то есть воз-
можность убедиться в справедливости или ошибочности своего впечатления.
1. Щелкните по меню Справка (Help)
2. Выберите в нем строку Найти и устранить (Detect and Repair)
(V) Справка по Microsoft Word F1
Показать помощника
1^? Что это такое? Shift+F 1
Office на Web
Справка по WordPerfect...
Найти и устранить...
О программе
Меню Справка (Help), команда Найти и устранить (Detect and Repair)
После щелчка на этой строке Word 2000 выполнит полную проверку состояния
всех подключенных шрифтов и шаблонов1. В случае обнаружения непорядка, Word
2000 попросит Вас вставить диск, с которого производилась установка програм-
мы. Так что, держите его наготове.
Щелкни и печатай
При работе в Word 2000 можно дважды щелкнуть мышкой в любое место как но-
вой, так и уже имеющейся страницы документа, и начать вводить текст.
Такая технология получила название Click-and-Type (Щелкни и печатай).
Теперь ввод текста не обязательно начинать в верхнем левом углу, а значит
подготовить документ с любой желаемой компоновкой стало значительно проще.
Этот же прием распространяется и на вставку рисунков или любых других объ-
ектов .
В качестве шрифта, используемого по умолчанию при первом запуске, в Word
2000 теперь установлен Times New Roman, кегль 12.
Автоматическое определение языка
В версии Word 2000 появилось любопытное новшество, на которое не было даже
и намека ни в одной из предыдущих реализаций Word.
Word 2000 автоматически определяет, на каком языке вводится текст и, ис-
пользуя соответствующие компоненты для проверки орфографии, грамматики, авто-
замены и другие средства проверки правописания, "на лету" исправляет ваши
ошибки. При этом усовершенствованная программа автозамены сама находит и ис-
правляет очевидные орфографические ошибки, которые не обязательно должны на-
ходиться в списке для автозамены.
1
Сначала наводит беспрорядок, потом наводит порядок и требует За это благодарности.
Каждому документу свое окно
В отличие от своих предшественников, Word 2000 для каждого документа созда-
ет свое отдельное окно. При этом на Панели задач (Task bar) появляется для
каждого из них отдельная кнопка.
• Во-первых, это позволяет очень быстро переходить из одного документа в
другой, причем, функции меню Окно (Window) сохраняются.
• Во-вторых, появилась возможность выводить на экран несколько документов
одновременно в разных окнах
Панель задач (Task bar) при открытии нескольких документов
Набор текста в Word’e
Для начала просто попытайтесь набрать любой текст, например, этот текст,
как если бы Вы его набирали на пишущей машинке. Однако помните, что:
1. В Word’e возврат каретки (клавиша <Enter>) нажимается не в конце строки,
а в конце абзаца. Перенос текста с одной строки на другую осуществляется
автоматически.
2. Каждое нажатие клавиши <Enter> добавляет в текст один абзац (или пустую
строку). Пустые строки удаляются клавишей <Del> (если пустая строка на-
ходится справа от курсора) или <Backspace> (если пустая строка находится
справа от курсора.
3. Чтобы разбить абзац на два, нажмите клавишу <Enter> там, где должен быть
конец абзаца. Чтобы соединить текущий абзац со следующим, необходимо
подвести курсор к концу абзаца и нажать клавишу <Del>.
4. Слова в абзаце отделяются друг от друга пробелами, вне зависимости от
присутствия знаков препинания. Это значит, что вне зависимости от того,
находится или нет между словами знаки препинания (точка, запятая, двое-
точие, или конец абзаца), после любого слова или знака препинания необ-
ходимо вставлять пробел (нажимать клавишу <Пробел>). Исключение скобки и
кавычки. Буквы, расположенные в скобках или кавычках, пишутся с ними
слитно, а пробелы делаются между скобками или кавычками и "внешним окру-
жением" .
5. Неправильно набранный символ можно удалить следующими способами:
а) Если курсор расположен правее символа, необходимо нажать <Backspace>.
b) Если курсор расположен левее символа, необходимо нажать клавишу <Del>.
6. Если удалить символ надо лишь для того, чтобы на его месте вставить дру-
гой символ, удобно перевести клавиатуру в режим замены. Переключение ме-
жду режимами вставки и замены осуществляется с помощью клавиши Insert
(<Ins>). Когда активен режим замены, в строке состояния Microsoft Word
активен индикатор "ЗАБ", забой ("OVR" - overtype). При вставке символа в
строку в режиме вставки все последующие символы сдвигаются вправо. В ре-
жиме замены старые символы исчезают, а на их месте появляются новые,
вводимые Вами с клавиатуры.
Обратите внимание, что когда Вы подводите курсор мыши к любой кнопке Панели
инструментов, у курсора мыши появляется всплывающая подсказка с названием
этой кнопки, а в строке состояния - объясняющая надпись.
Обратите внимание на несколько кнопок, расположенных в левом нижнем углу
окна, слева от горизонтальной линейки скроллинга. Это так называемые кнопки
смены оформления текста. Так, самая левая из этих кнопок позволяет отображать
текст в окне Microsoft Word не как в текстовом процессоре, а как в обычном
текстовом редакторе (типа блокнота). В этом окне правильно отображаются шриф-
ты и форматирование абзаца текста, но не показываются колонтитулы, вставки,
сноски, внедренные объекты и изображения. Однако в этом режиме отображение
текста всегда поспевает за нажатиями клавиш на клавиатуре, вследствие чего
можно предъявить более низкие требования к аппаратной части компьютера.
Вторая слева (в Microsoft Word 97 и 2000 - это третья слева) кнопка оформ-
ления показывает документ Word как будто он уже распечатан на бумаге. В этом
режиме отображаются все элементы форматирования текста, однако он наиболее
требователен к аппаратной части компьютера, а потому часто набор текста опе-
режает его появление на экране. Поэтому рекомендуется использовать этот режим
при окончательном оформлении документа.
И, наконец, последняя кнопка отображает документ в режиме "макета". Этот
режим почти ничего не имеет общего с WYSIWYG-технологией, однако он позволяет
быстро изменить оформление документа, перемещая целиком заголовки, абзацы и
иллюстрации по документу и тем самым упрощая окончательную верстку.
Если символы текста кажутся Вам слишком крупными или слишком мелкими, а
также если Вы не удовлетворены использованием ширины экрана документом, Вос-
пользуйтесь меню "Вид" -> "Масштаб" ("View" -> "Zoom") для изменения масштаба
изображения, или измените масштаб при помощи инструмента "Масштаб" панели ин-
струментов "Главная". Обычно можно выбрать либо точный масштаб отображения (в
процентах от истинного), или выбрать "плавающие" масштабы "по ширине страни-
цы" , "страница целиком", "несколько страниц". Автор книги предпочитает рабо-
тать в масштабе "по ширине страницы" и оформлении "как текстовый редактор",
поскольку он максимально использует ширину монитора и не перегружает аппарат-
ную часть компьютера.
Операции с фрагментом текста
При удалении нескольких слов или нескольких строк имеет смысл воспользо-
ваться следующим приемом:
1. Установите курсор на начало фрагмента, требующего удаления.
2. Теперь, придерживая клавишу Shift, подведите курсор к концу фрагмента
клавишей "стрелка вправо" и "стрелка вниз". Курсор потянет за собой
"подсветку" на весь фрагмент (обычно текст отображается инверсным цве-
том. )
3. По нажатию клавиши Del весь фрагмент будет удален.
Разумеется, подсветку можно "тянуть" и, наоборот, с конца фрагмента в его
начало.
Есть и другой способ удаления выделенного фрагмента - просто начать наби-
рать текст поверх него.
Выделять фрагмент можно и другим способом:
• Установить курсор в начало фрагмента (мышью или клавиатурой). Для установ-
ки курсора мышью необходимо однократно щелкнуть левой клавишей мыши в том
месте, где должен находится текстовый курсор - и он там окажется!
• Удерживая клавишу Shift, щелкнуть левой клавишей мыши в конце фрагмента.
• Можно обойтись вообще без клавиатуры. Для этого надо:
1. Установить курсор в начало фрагмента выделения.
2. Удерживая левую клавишу мыши, проведите курсор мыши от начала до
конца фрагмента.
Кроме того:
• двойной щелчок левой клавишей мыши на слове выделит это слово;
• щелчок левой клавишей мыши на предложении с нажатой клавишей Ctrl выделит
это предложение;
• щелчок левой клавишей мыши слева от строки выделит эту строку;
• тройной щелчок левой клавишей мыши на абзаце выделит этот абзац;
• щелчок мышью при нажатой клавише Ctrl левее текста выделит этот текст.
• выделить весь текст можно и нажатием Ctrl+A (Ctrl+Ф при русской раскладке
клавиатуры) или Ctrl+5 (на цифровой части клавиатуры).
Чтобы перенести фрагмент текста из одного места в другое при помощи клавиа-
туры, необходимо:
1. Выделить фрагмент.
2. Нажать Shift+Del или Ctrl+X, чтобы "вырезать" фрагмент текста с экрана.
3. Поместить курсор туда, куда необходимо перенести фрагмент.
4. Нажать клавиши Shift+Ins или Ctrl+V, чтобы вставить вырезанный фрагмент
в новое место.
Операции переноса фрагментов в Microsoft Word удобнее выполнять при помощи
мыши. Для его перемещения нужно его "взять" (направив курсор мыши на выделен-
ную часть текста и нажав левую клавишу мыши) и переместить мышиный курсор ту-
да , куда необходимо перенести текст. Для того, чтобы скопировать фрагмент на
новое место при помощи клавиатуры необходимо:
1. Выделить фрагмент.
2. Нажать клавиши Ctrl+Ins или Ctrl+C.
3. Поместить курсор туда, куда нужно скопировать фрагмент.
4. Нажать клавиши Shift+Ins или Ctrl+V, чтобы вставить на новое место копию
фрагмента текста.
На панели инструментов "Главная" Microsoft Word есть три специальные кноп-
ки, на которых изображены ножницы, листки документов и портфель с документа-
ми, предназначенные соответственно для выполнения операций удаления в карман,
копирования и вставки.
Чтобы перенести (скопировать) текст при помощи мыши необходимо:
• "взять" выделенный фрагмент текста;
• переместить курсор туда, куда необходимо перенести текст, удерживая нажа-
той клавишу Ctrl.
Текст можно перемещать не только в пределах документа, но и вне документа:
на рабочий стол, в корзину или папку. При нажатии клавиши Ctrl данные будут
перенесены, если Shift - скопированы. На самом деле они попадут в файл типа
Scarp, имеющее расширение SHS. Данные из Scarp можно вставить в любой доку-
мент в окне Word. Это позволяет иметь готовыми к вставке любое разумное коли-
чество фрагментов, держа их в буквальном смысле слова перед глазами.
Для того чтобы застраховаться от последствий ошибочных действий, в Word
предусмотрена процедура отмены предыдущих операций редактирования - отмена
редактирования. Она находится в меню "Правка" -> "Отменить" ("Edit"->"Undo").
Для той же цели служат клавиши Alt+Backspace и Ctrl+Z. Возвратить отмененную
операцию можно при помощи меню "Правка" -> "Вернуть" ("Edit" -> "Redo") или
клавишей Ctrl+Y. На панели инструментов "Главная" имеются кнопки для отмены и
возвращения операций.
Операции с документами
Чтобы сохранить документ, подвергшийся редактированию, необходимо выбрать
пункт меню "Файл" -> "Сохранить" ("File" -> "Save"), клавишами Ctrl+S или
кнопкой с изображением дискеты на панели инструментов "Главная". Если доку-
мент не был еще ни разу сохранен, на экране появится диалоговое окно "Сохра-
нить" ("Save as") . В поле "Файл" ("File name") необходимо ввести имя файла,
под которым будет сохранен документ. Когда необходимо сохранить документ в
файле с новым именем, примените пункт меню "Файл" -> "Сохранить как" ("File"
-> "Save as") . Для открытия уже готового файла следует использовать меню
"Файл" -> "Открыть" ("File" -> "Open"), кнопку с открывающейся папкой на па-
нели инструментов "Главная", или сочетание клавиш Ctrl+O (CTRL+щ в русской
раскладке клавиатуры). В диалоговом окне "Открыть" ("Open") в списке выбрать
нужное имя файла. Он загрузится в отдельное окно программы. Вы, в принципе,
можете открыть большое количество окон с документами. Переключение между ок-
нами возможно при помощи меню "Окно" ("Window").
Чтобы начать работу над новым файлом, необходимо дать команду в меню "Файл"
-> "Создать", или воспользоваться либо клавишами Ctrl+N (CTRL+т в русской
раскладке), либо выбрать мышью кнопку с чистым листом. Обратите внимание, что
при пользовании первым методом Вас запросят выбрать шаблон, на основании ко-
торого Вы бы хотели открыть файл, в то время как во втором случае сразу соз-
дается документ на основе шаблона "Обычный" ("Normal").
Меню "Файл" -> "Печать" ("File" -> "Print") позволяет распечатать на прин-
тере любую выделенную часть документа, либо отдельные страницы, либо весь
текст (появляется соответствующее диалоговое окно), а при нажатии кнопки с
принтером - сразу распечатать весь документ весь документ. При помощи пункта
меню "Файл" -> "Просмотр" ("File" -> "Print Preview") можно просмотреть, как
в точности будет выглядеть распечатка текста на принтере. В режиме просмотра
можно также изменять масштаб просматриваемого текста.
Шрифтовое оформление
Для того чтобы изменить шрифтовое выделение текущего слова, выберите в меню
команду "Формат" -> "Шрифт" ("Format" -> "Font"). Если необходимо изменить
шрифт на участке, большим или меньшем, чем одно слово, следует предварительно
выделить этот участок. В диалоговом окне находятся все характеристики шрифта,
а в правой нижней части диалогового окна отображаются примеры написания. Для
быстрой смены начертания шрифта, его размера, выделения букв "жирностью",
курсивом, подчеркиванием и цветом существуют соответствующие кнопки на панели
инструментов "Форматирование".
Чтение и восприятие документа в огромной степени зависит от того, какие
шрифты применялись для его оформления и каким стилем они поданы. Один и тот
же шрифт Word может выдавать в нескольких вариантах, то есть стилях.
Какие бывают шрифты
Очень часто Word используют при окончательной подготовке материалов, вклю-
чая верстку издания, для печати в типографии. То есть Word выполняет не свой-
ственные ему функции настольной издательской системы. Поскольку Microsoft
Corporation явно не планировала использовать свое детище для подобных целей,
то имеются разночтения в терминологии полиграфистов и той, что использована в
Word. Впрочем, возможно, что Microsoft Corporation и не причем, а то, что мы
имеем, — это творчество переводчиков. Но, как бы то ни было, для устранения
возможных неясностей сначала разберемся, что под термином шрифт имеют в виду
полиграфисты.
Шрифт с точки
зрения полиграфистов
В полиграфической терминологии шрифт — это набор символов определенного ри-
сунка и размера. Шрифты объединяются в гарнитуры.
Гарнитура — это художественное решение шрифта, которое отличает его от дру-
гих. Гарнитура имеет свое имя и состоит из нескольких шрифтов. Например, гар-
нитура Arial включает в себя шрифты Arial, Arial Black, Arial Narrow
Начертание — это один из вариантов шрифта в гарнитуре. Начертания шрифтов
включают в себя насыщенность, наклон, пропорциональность и кегль (размер).
• Насыщенность шрифта характеризует толщину линий литер и имеет стандартизи-
рованные названия — это нормальная или обычная (normal), полужирная
(semibold) и жирная (bold).
• Наклонный шрифт именуется курсив (Italic).
• Пропорциональность — это относительная ширина знаков шрифта. То есть каж-
дый тип букв (литер) шрифта имеет свою ширину, например, буквы "Ж" и "П".
Для более точного определения пропорциональности обычно выбирается соотно-
шение ширины и высоты литеры "М". По этому признаку различают сверхузкое,
узкое, нормальное и широкое начертания. Пропорциональные шрифты в докомпь-
ютерную эру применялись только в типографиях. На привычных пишущих машин-
ках абсолютно все буквы имели одинаковую ширину. Такие шрифты называются
моноширинными.
• Кегль — вертикальный размер шрифта в пунктах с учетом верхних и нижних
(или надстрочных и подстрочных) выносных элементов.
।— Верхний выносной элемент
(рХ> Г "Дубль
шрифта ---Базовая линия
।-Нижний выносном элемент
Элементы шрифта
Единицы измерения,
применяемые в полиграфии
Полиграфия ведет свою историю с 40-х годов XV века, когда Гуттенберг изо-
брел свой печатный станок. Как и любая отрасль науки и техники полиграфия об-
завелась своей собственной терминологией. Но если другие отрасли на этом чаще
останавливаются, то полиграфисты пошли дальше — они изобрели и свою собствен-
ную систему единиц измерения. Эта система может показаться странной — дейст-
вительно, где еще есть единицы, кратные 72? Однако такая система единиц есть
и приходится с ней мириться, поскольку нынешнее поколение полиграфистов рев-
ностно придерживается традиций предков. Поэтому, если мы хотим быть понятыми
полиграфистами и понимать их, то давайте разберемся немного и с этими экзоти-
ческими единицами измерения.
Несколько порадовать, однако, может тот факт, что подобная система единиц
позволяет соотнести размер шрифта с размерами полей, колонок, страниц и дру-
гими элементами публикации.
Создатели Word напрямую использовали только одну из полиграфических единиц
для обозначения размера (кегля) шрифта и отступов (отбивки) абзаца — Перед
(Before) и После (After). Эта единица называется пункт (point или pt):
1 дюйм (inch) = 25,4 мм = 72 пункта (points)
1 пункт (point) = 1/72 дюйма (inch) = 0,353 мм
Если речь идет о шрифте размером 12, то это высота (кегль) шрифта в пунк-
тах. Причем, это не величина букв, а высота строки с текстом. В понятие высо-
та строки входит высота букв плюс верхние и нижние выносные элементы — такие,
как в буквах "б" и "у".
Если набрать текст различными шрифтами одинакового кегля, то высота букв в
разных шрифтах будет разная.
AGOpus 12, Arial 12, Academy 12,
п, TimesET 12
Примеры разных шрифтов (гарнитур) одинакового кегля
Высота и остальные размеры отдельных букв задаются автором (художником)
шрифта из расчета гармоничного восприятия текста в целом. Обычно величина
кегля соответствует ширине прописной буквы М. Однако бывают и исключения.
Так, с множителем 72 и пунктами мы разобрались. Что же есть еще? А еще есть
пики (picas).
Пика равна 4,236 мм или 1/6 дюйма (точнее 1/6 дюйма равна 4,233 мм, но Зна-
чение пики чаще округляют до 1/6 дюйма).
1 дюйм (inch) = 25,4 мм = 72 пункта (points, pt) = 6 пик (picas)
1 пика = 12 пунктов
В пиках полиграфисты измеряют ширину и высоту полос и колонок.
Разумеется, этими двумя единицами дело не ограничивается. Но нам и их дос-
таточно для того, чтобы с пониманием подходить к форматированию наших доку-
ментов . О других полиграфических единицах, а их — приличное количество, необ-
ходимо знать, только если вы займетесь версткой с использованием настольных
издательских систем типа Page Maker или Ventura Publisher.
Что такое междустрочный интервал
Теперь, когда мы определились с терминологией, можно четко охарактеризовать
смысл понятия междустрочный интервал, который определяет расстояние по верти-
кали между строками текста. По умолчанию в Word используется одинарный интер-
вал . Выбранный междустрочный интервал появится между всеми строками текста в
выделенном абзаце или в абзаце, на котором стоит текстовый курсор.
Интервалы отсчитываются от базовой линии шрифта. Величина одинарного между-
строчного интервала соответствует шрифту наибольшего размера, используемому в
строке. Можно с достаточной точностью утверждать, что значение междустрочного
интервала равно расстоянию между базовыми линиями двух соседних строк.
То есть для кегля номер 10 одинарный интервал также равен 10 пунктам.
Полуторный и двойной междустрочные интервалы превышают одинарный в соответ-
ствующее число раз.
Шрифты в Word
В любой версии Word шрифты различаются по трем признакам:
• название,
• размер,
• начертание.
С названием все ясно. Под размером шрифта понимается то же самое, что в по-
лиграфии обозначается термином кегль. А вот с термином начертание в Word вы-
шла некоторая неувязка с полиграфией.
В пику полиграфистам (надо же отличаться) Word различает четыре вида начер-
тания шрифта, которые здесь называются стилями:
• нормальный (normal),
• полужирный (bold),
• курсив (italic) ,
• подчеркнутый (underline).
Именно для стиля шрифта определены специальные кнопки на инструментальной
панели Форматирование (Formatting) и введены "горячие клавиши":
- (Bold),
— (Italic) ,
— (Underline).
Все эти стили можно применять в произвольном наборе. Например, можно одно-
временно назначить шрифту три вида стиля: полужирный, курсив, подчеркнутый и
получить следующий » Стиль шрифта.
Запомните эти отличия в терминологии, и вам, при необходимости, будет легче
общаться с полиграфистами, а им с вами.
Как и в полиграфии, в Windows имеются и моноширинные и пропорциональные
шрифты, причем пропорциональных шрифтов — большинство. Какие же из них узкие,
широкие или любые другие — при работе в Word 6.0/95/97 надо было помнить, так
как там индикация вида шрифтов предусмотрена не была. Только в Word 2000 это
упущение наконец-то было исправлено. В стандартном варианте поставки Windows
(и Word, разумеется) русских шрифтов немного. Обычно присутствуют Arial,
Courier New и Times New Roman. В этих шрифтах, кроме букв латинского алфавита
(латиницы), присутствуют и русские (кириллица). Есть также специальный шрифт,
каждый символ которого представляет собой какую-либо картинку — Wingdings.
Однако сейчас уже существует огромное количество русских шрифтов, которые
вы можете подключить и использовать в своей работе.
Обратите только внимание на то, чтобы все используемые вами шрифты были
масштабируемыми. Они называются True Туре и в списках обозначены символами
Т. Такие шрифты одинаково выглядят и на экране, и на принтере. С другими
у вас могут быть проблемы.
Word 2000, как и Word 97, работают с кодировкой символов в стандарте
Unicode. Поэтому, если вам необходимо документы, созданные в Word 6.0/95, чи-
тать в Word 97 или Word 2000, а также наоборот, то все шрифты на вашем компь-
ютере должны быть обработаны конвертером TTFCONV.EXE.
т
В результате ваши
шрифты будут переведены в стандарт Unicode и проблемы уйдут.
ифты будут переведены в стандарт Unicode и проблемы уйдут.
Примеры шрифтов из стандартной поставки Windows
Шрифт Начертание Применение
Arial Нормальный Полужирный Курсив Подчеркнутый Пропорциональный. Хорошо выглядит в заголовках, но может использоваться и для не больших фрагментов текста.
Courier- New Нормал ьный Полужирный Курсив Подчеркнутый Мон оширинный. Преимущественно используется в заголовках и при наборе текстов программ (листингов).
Times New Roman Нормальный Полужирный Курсив Подчеркнутый Пропорциональный. Чаще всего им набирают основной текст. В последнее время наряду с этим шрифтом ча сто применяется шрифт Times L Г
Wing dings Для украшения и придания большей выразительности документа.
Как назначить шрифт
В Word 2000 разработчики из Microsoft Corporation, видимо ориентируясь на
конкурентов, внесли удобный сервисный элемент в процедуру выбора необходимого
шрифта. Теперь уже не надо помнить, как выглядят те или иные шрифты. При рас-
крытии списка Шрифт (Font) на панели Форматирование (Formatting) каждый шрифт
выводится в своем оригинальном виде.
Чтобы назначить какому-либо фрагменту текста тот или иной шрифт, сначала
надо его выделить. Далее есть несколько способов.
• Нажимаем клавиши
и переходим в поле Шрифт (Font) на панели
Форматирование (Formatting).
• Клавишами
ный.
Enter
___мли
раскрываем список доступных шрифтов и выбираем нуж-
- 12 ’ ж к ч
т Times New Roman
Ч Symbol А|ЗХВЕфГг|1ф
т Keystroke
ч а
Ч a CiiyNova
Ч й_Е0ИЕ@Р^Э^0Ь0^Ь
Ч a_GroTicL:
Ч a_GroticRoghXBIack
Ч a GroticRough
Ч a GroticRoughQb!
Список Шрифт (Font) на панели Форматирование (Formatting)
• Этот же список раскрывается и щелчком левой кнопки мыши на списке Шрифт
(Font) на панели Форматирование (Standard).
Если, не раскрывая список, повторно нажать клавиши
Ctrl + Shift + F
, откро-
ется диалог Шрифт (Font).
• Если в данный момент панели Форматирование (Formatting) на экране нет, то
диалог Шрифт (Font) открывается после первого же нажатия клавиш
Ctrl -|- Shift + F
Если вы уже использовали какие-либо шрифты, то пять последних из них располо-
жены в верхней части списка и отделены от остальных двойной линией.
Список и диалог можно вызвать с помощью мышки:
• Выберите в меню Формат | Шрифт (Format | Font) или щелкните правой кнопкой
мыши в любом месте текстового поля и в контекстном меню выберите Шрифт
(Font). Диалог Шрифт (Font) позволит не только установить вид шрифта, но и
его размер, стиль и другие.
• Если текст не выделен, то вновь назначенный шрифт будет действовать:
о для вновь вводимых символов, если курсор стоит вне слова;
о на все слово, внутри которого стоит курсор.
С помощью правой кнопкой мыши и клавиатуры можно менять шрифт и тогда, когда
инструментальные панели отсутствуют на экране.
По сравнению со списком на инструментальной панели Форматирование (Format)
диалог Шрифт (Font) предоставляет гораздо большие возможности для управления
шрифтами. Рассмотрим их по порядку.
Вкладка Шрифт (Font) диалога Шрифт (Font)
Подчеркивание
(Underline Style)
Если в диалоге Шрифт (Font) (Рис. 3.4) рас-
крыть список Подчеркивание (Underline Style),
то можно задать любой из пятнадцати вариантов
подчеркивания шрифта. Два из этих вариантов
подчеркивания (одинарное и двойное) можно за-
дать с помощью горячих клавиш остальные —
только через диалог Шрифт (Font). В этом же
диалоге Шрифт (Font) можно из списка Цвет
шрифта (Font Color) выбрать и цветовое реше-
ние для выводимых символов. Аналогичную воз-
AI’L тт
можность предоставляет и кнопка “ — Цвет
шрифта (Font Color) на панели Форматирование
(Formatting).
Оформление абзацев
Чаще всего требуется менять такие параметры оформления абзаца, как тип вы-
равнивания (по левому, правому краю, выравнивание по ширине страницы (выключ-
ка) , центрирование абзаца, межстрочное расстояния и отбивки абзаца. Для того,
чтобы переформатировать текущий абзац, выберите команду в меню "Формат" ->
"Абзац" ("Format" -> "Paragraph"). Если необходимо переформатировать более
одного абзаца, необходимо их предварительно выделить. Диалоговое окно "Абзац"
содержит множество настроек свойств абзаца, включая все вышеперечисленные.
Доступ к нему также возможен через контекстное меню. Некоторые настройки из
диалогового окна "Абзац" доступны на панели инструментов "Форматирование".
Например, кнопки выравнивания абзаца (по левому краю, центрирование, по пра-
вому краю, выключка), пошагового увеличения либо уменьшения левого отступа,
Абзацная линейка, которая отображается по команде в меню "Вид" -> "Линейка"
("View" -> "Ruler")позволяет менять отступы и величину красной строки. Верх-
ний левый маркер меняет величину красной строки, средний меняет абзацный от-
ступ, нижний сдвигает красную строку и абзацный отступ одновременно. Правый
маркер служит для изменения правого отступа.
Другие параметры
оформления текста
При помощи пункта меню "Файл" -> "Параметры страницы" ("File" ->
"Parameters") появляется диалоговое окно, определяющее размещение текста на
листе бумаги:
• верхнее, нижнее, левое, правое поля страницы;
• отступы под переплет;
• расстояние до верхнего или нижнего колонтитула;
• размер бумаги (один из стандартных или выборочный);
• положение бумаги - вертикальное ("Портрет", "Portrait") или горизонталь-
ное ("Ландшафт", "Landscape").
Кроме того, Word позволяет применять многие профессиональные методы оформ-
ления текста. Например, для форматирования списков, в том числе нумерованных,
можно применять пункт меню "Формат" -> "Список" ("Format" ->
"Bullets&Numbering"). Это меню предлагает множество вариантов оформления спи-
сков. На панели инструментов "Форматирование" существуют две кнопки для соз-
дания списка.
Первая кнопка, со списком в виде кружков, создает ненумерованный список.
Кнопка со списком в виде цифр создает нумерованный список, в котором элементы
пронумерованы по возрастанию. При нажатии клавиши Enter нумерованный и нену-
мерованные списки автоматически продолжаются. Для отмены продолжения нумера-
ции в конце списка нажмите два раза клавишу Enter. Кроме того, в диалоговом
окне "Списки" можно изменить вид нумерованного и ненумерованного списка, из-
менить порядок нумерации и т.п.
Понятие составного документа
При создании документа часто возникает необходимость вставить в него фраг-
менты из документов, созданных в других приложениях. Например, текст, создан-
ный в текстовом процессоре Word, может содержать графики, таблицы, подготов-
ленные в Excel, рисунки, созданные в графическом редакторе Paint и пр. Можно
создать документ, включающий в себя не только текст, графические изображения,
таблицы, но также звуковое сопровождение (музыку, речь), и даже видеоролики.
В результате появляется документ, содержащий данные разного типа, хотя внешне
это не проявляется. Его можно печатать, хранить и производить с ним любые
другие действия как с обычным документом, созданным в одной среде и содержа-
щим однотипные данные. Документ, в котором объединены данные разного типа,
созданные в разных приложениях, называется составным (интегрированным) доку-
ментом .
источник
ГиЦ и ч cl
ПРИЕМНИК
ТвсстовьЛ процессор Word
Объектом называется произвольный элемент, созданный в каком-либо приложе-
нии, который можно поместить в файл другого приложения (например, в документ
MS Word, рабочую книгу MS Excel и или базу данных MS Access).
Приложение, средствами которого создаются объекты, называется сервером или
приложением - источником. Приложение, принимающее объекты, подготовленные в
приложении - источнике, называется клиентом или приложением - приемником.
Обмен объектами
с помощью мыши
Обмен данными между приложениями возможен перетаскиванием объекта с помощью
мыши. Для этого окна приложений целесообразно расположить рядом. Предвари-
тельно объект должен быть выделен. Перемещение осуществляется нажатием левой
кнопки мыши на объекте в приложении - источнике и транспортировкой его в при-
ложение - получатель. Копирование осуществляется аналогично процедуре переме-
щения при одновременно нажатой клавише Ctrl. При этом необходимо соблюдать
следующую последовательность отпускания клавиш - сначала кнопку мыши, а затем
клавишу Ctrl. Также можно сначала объект протащить, не отпуская кнопки мыши,
а затем уже при необходимости нажать Ctrl. При копировании указатель мыши на
экране будет сопровождаться знаком +.
Обмен объектами с
помощью буфера обмена
Еще один способ обмена объектами между приложениями Windows - это использо-
вание буфера обмена. Буфер обмена (Clipboard) - это область оперативной памя-
ти для временного хранения данных. Процесс обмена данными с помощью буфера
обмена следует начинать в приложении - источнике данных. Сначала выделяется
фрагмент документа (или весь документ) и помещается на хранение в буфер обме-
на . Записанный в буфере фрагмент можно вставить: в другое место того же доку-
мента, в другой документ того же приложения, в документ другого приложения.
Например, можно переместить рисунок из редактора Paint в любое место докумен-
та WordPad.
Запуск и завершение программ сами по себе на содержимое буфера обмена никак
не влияют. Каждое последующее занесение в буфер какой бы то ни было информа-
ции уничтожает предыдущее содержимое. Один и тот же фрагмент можно вставлять
в документы несколько раз: при вставке содержимое буфера обмена не меняется.
Способы помещения
объектов в буфер обмена
Во всех приложениях Windows схема работы с буфером обмена стандартизирова-
на . Для занесения информации в буфер и ее извлечения оттуда в меню Правка
имеются команды Вырезать, Копировать, Вставить. Команда Вырезать помещает вы-
деленную информацию в буфер, удаляя при этом ее оригинал. Команда Копировать
помещает выделенную информацию в буфер, сохраняя ее в оригинале. Команда
Вставить извлекает информацию из буфера. Во многих приложениях этим командам
соответствуют кнопки на панели инструментов. Так же их можно вызвать, исполь-
зуя контекстное меню). Команда Правка/Специальная вставка позволяет выбрать
формат данных, в котором они вставятся из буфера обмена.
Клавиша Print Screen копирует графический образ всего экрана в виде растро-
вой картинки в буфер обмена. Затем можно вставить эту картинку в документ ка-
кого-либо графического редактора, например, Paint, отредактировать и сохра-
нить. Клавиша Alt+Print Screen копирует в буфер обмена только активное окно.
Технология внедрения и
связывания объектов OLE
Более гибким способом обмена данными между приложениями Windows является
технология OLE (Object Linking and Embedding - связывание и внедрение объек-
тов) . Технология OLE - это возможность использовать в одном документе объекты
разного происхождения, например, текст, фотографии, музыку и т.д. Основное
достоинство технологии OLE состоит в том, что она позволяет организовать пе-
редачу данных практически между любыми приложениями.
Технология OLE определяется правилами (протоколом) взаимодействия Windows-
приложений. Базовый вариант технологии OLE был впоследствии расширен до вер-
сии OLE 2.0, которая в настоящее время является наиболее используемой. Объек-
том OLE называется произвольный элемент, созданный средствами какого-либо
приложения Windows, который можно поместить в документ другого приложения.
Это может быть весь документ или его фрагмент.
OLE-серверы и OLE-клиенты
Приложение, средствами которого создается объект OLE, называется сервером
OLE (приложение-источник). Приложение, принимающее объект OLE, называется
клиентом OLE (приложение-приемник). Полностью технологию OLE поддерживают не
все Windows-приложения, а только те, которые предназначены для разработки со-
ставных документов. Одни приложения могут исполнять только роль OLE - серве-
ра, другие - OLE -клиента. Например, текстовый редактор WordPad может испол-
нять функции и OLE-сервера, и OLE-клиента, так как он предназначен для созда-
ния документов, в которых могут быть рисунки, звуки, таблицы, видеоклипы и
ДР-
Графический редактор Paint предназначен для подготовки несложных рисунков,
он может работать по технологии OLE только как приложение-источник. Текстовый
процессор Word представляет собой сложный комплекс и фактически включает в
себя несколько приложений: редактор рисунков, редактор формул, текстовый ре-
дактор и др. Эти приложения по-разному используют технологию OLE. Редактор
формул действует только как приложение-источник (OLE-сервер), а текстовый ре-
дактор может выступать как в роли источника, так и в роли приемника (OLE-
клиента).
Просмотреть список действующих в системе OLE-серверов можно, выполнив ко-
манду Вставка/Объект...
Особенности связывания
и внедрения объектов
Особенность обмена объектами состоит в следующем. При выполнении связывания
(linking) объект существует только в приложении-источнике, а приложение-
приемник содержит лишь указатель (ссылку) на его местоположение. При измене-
нии объекта в исходном документе соответствующим образом изменяется связанная
с ним копия в другом приложении. Например, можно связать электронную таблицу
с текстовым документом. Изменения, выполняемые в программе обработки элек-
тронных таблиц, будут автоматически воспроизводиться в текстовом документе.
Когда же выполняется внедрение (embedding), в целевой документ вставляется
копия объекта и требуется дополнительный объем памяти для ее хранения. Ника-
кой связи между копиями объекта не сохраняется: изменение одной копии никак
не влияет на остальные копии объекта.
При связывании объектов размер целевого документа практически не увеличива-
ется (в отличие от внедрения), так как указатель на местоположение объекта
занимает мало места. Но при этом надо следить за тем, чтобы все связанные
объекты хранились в тех папках, в которые они были помещены в момент создания
связи.
Способы вставки объектов
в целевой документ
1. Создание связанного или внедренного объекта на основе данных из сущест-
вующего файла с помощью команд Вставить или Специальная вставка:
• в приложении-источнике выделить объект, который необходимо внедрить или
связать и поместить его в буфер обмена;
• в приложении-приёмнике курсором указать место вставки объекта, выполнить
команду Правка/Вставить (произойдет внедрение объекта) или Прав-
ка /Специальная вставка... Также можно использовать контекстное меню;
• в случае выбора команды Правка/Специальная вставка... на экране появится
диалоговое окно Специальная вставка, в котором надо выбрать формат встав-
ляемого объекта2. Например, если был скопирован текст из документа Word,
то выбрать Объект Документ Microsoft Word;
• для отображения связанного объекта в виде значка установить флажок в виде
значка;
• для создания внедренного объекта установить переключатель Вставить, а для
создания связанного объекта - Связать;
• нажать кнопку ОК, в указанное место будет внедрён объект из приложения-
источника .
Примечание. При вставке объекта, в случае выбора формата данных, отличного от
оригинального, объект утратит связь с породившим его приложением, и, следовательно, станет
невозможно редактировать его непосредственно в документе.
Специальная вставка
ВЕЗ
Результат
Источник: Объект Документ Microsoft Word
С:\Мои.. ДПример.doc OLE_UNK2
встарить:
С1 св«реть:
Как:
Г В. виде значка
Вставка содержимого буфера обмена в
документ, так что его можно изменять,
используя Документ Microsoft Word.
Рис. Окно Специальная вставка
2. Создание внедрённого объекта из коллекции объектов или создание нового
внедрённого или связанного объекта:
• указать в составном документе место внедрения;
• выполнить команду Вставка / Объект;
• на экране появится диалоговое окно Вставка объекта с двумя вкладками Соз-
дание и Создание из файла. Вкладка Создание позволяет создать новый объект
и внедрить его. В списке Тип объекта следует выбрать приложение (OLE-
сервер), с помощью которого будет создаваться объект, который после выхода
из приложения-источника будет вставлен в приложение-приемник. В этом спи-
ске отображаются только установленные на данном компьютере программы. При
выборе вкладки Создание из файла можно вставить или связать объект, ранее
созданный и сохраненный на диске в виде отдельного файла. В списке Имя
файла нужно ввести имя файла, из которого будет создан связанный или вне-
дренный объект или нажать кнопку Обзор для выбора файла из списка. Для
создания связанного объекта установить флажок Связь с файлом. Для отобра-
жения связанного объекта в виде значка установить флажок В виде значка;
• нажать кнопку ОК.
Не разрешается применять команду Вставка/Объект для вставки графики и неко-
торых типов файлов. Для вставки графики в Word рекомендуется использовать ко-
манду Вставка/Рисунок.
Объекты, у которых связи с их приложениями незначительно увеличивают объем
создаваемого документа, воспринимаются некоторыми приложениями только как
внедренные. Например, при вставке фрагмента рисунка графического редактора
Paint, после выполнения команды Вставка или Специальная вставка произойдет
внедрение объекта.
Рис. Окно Вставка объекта
Редактирование объекта
При просмотре или распечатке составного документа связанный объект ничем не
отличается от внедрённого. Различие проявляется лишь при их редактировании.
Объект, внедрённый по технологии OLE, можно редактировать в приложении-при-
емнике следующими способами:
• вызвать контекстное меню и выбрать соответствующий инструментарий редакти-
рования ;
• дважды щелкнуть левой кнопкой мыши на внедренном объекте.
• Изменить связанный объект можно, отредактировав документ-источник, в кото-
ром он размещен. Это можно осуществить одним из двух способов:
• открыть документ-источник непосредственно в среде приложения - источника;
• открыть документ-источник из составного документа, где находится связанный
объект. Открытие можно осуществить либо с помощью контекстного меню, либо
двойным щелчком кнопки мыши на этом объекте.
Поскольку связанные объекты хранятся отдельно от составного документа в
своих документах-источниках, важно иметь сведения о связях составного доку-
мента . Просмотреть, обновить и изменить связи составного документа можно в
диалоговом окне Связи. Его можно вызвать, например, в приложении Word, коман-
дой Правка/Связи.
Если команды Связи нет в наличии, это означает, что данный файл не содержит
связанных данных. Диалоговое окно Связи содержит следующую информацию о свя-
зях данного объекта:
• путь к файлу;
• тип или формат объекта;
• режим обновления объекта в составном документе - автоматическое или по за-
просу .
Рис. Окно Вставка объекта
При автоматическом обновлении связанный объект в составном документе изме-
няется сразу, как только произведена его коррекция в документе - источнике.
При ручном обновлении необходимо явное указание со стороны пользователя. Из
списка Исходный файл выбирается источник для связанного объекта, а затем вы-
полняется команда Обновить. Для выделения нескольких связанных объектов, не
опуская клавишу Ctrl, надо выбрать каждый связанный объект.
ЛЕКЦИЯ 7
Компьютерные вирусы
и средства защиты1
Компьютерный вирус - это небольшая программа, выполняющая несанкционирован-
ные действия на компьютере. Компьютерный вирус инфицирует компьютер, встраи-
ваясь в другую программу или в документ, или в определенные области носителя
данных (винчестер) при каких-то условиях он активизируется, например, запус-
кается в определенный день.
Выделяют три типа вирусов: программные вирусы, загрузочные вирусы, макрови-
русы.
Программные вирусы внедряются в другие прикладные программы и активизирует-
ся при запуске этих программ. Программные вирусы инфицируют компьютер при за-
пуске непроверенных программ на внешнем носителе (дискета, компакт-диск).
Распространенным способом инфицирования компьютера, является чтение инфициро-
ванного письма, присланного по электронной почте. Знаменитым вирусом этого
типа, является вирус "I love you", появившейся в мае 2000 г. запуске происхо-
дит активизация имплантированного вируса.
Загрузочные вирусы отличаются от программных методом распространения. Они
поражают не программные файлы, а определенные системные области магнитных но-
сителей (дискеты, винчестеры). На включенном компьютере они могут временно
располагаться в основной памяти. Обычно заражение происходит при попытке за-
грузки компьютера с магнитного носителя, системная область которого содержит
загрузочный вирус. При загрузке компьютера с дискеты происходит сначала про-
никновение вируса в оперативную память, а затем в загрузочный сектор винче-
стера . Далее этот компьютер сам становится источником распространения загру-
зочного вируса.
Макровирусы поражают документы, выполненные в некоторых прикладных програм-
мах, имеющих средства для исполнения так называемых макрокоманд. В частности,
к таким документам относятся документы текстового процессора Microsoft Word и
Microsoft Excel. При открытии такого документа вирусы активизируются, перено-
сятся на компьютер и внедряются в шаблоне "Обычный". После этого каждый со-
храняемый документ заражается вирусом, а когда другие пользователи открывают
зараженный документ, вирус переносится на их компьютеры. Word или Excel не
выполняют проверку гибкого, жесткого или сетевого диска на предмет поиска и
уничтожения вирусов в макросах. Тем не менее, при открытии в Word любого до-
кумента, который содержит макросы и, следовательно, может содержать вирусы,
выводится предупреждение, и пользователю предлагается принять решение, как
следует открыть документ: с макросами или без них.
Если ожидается, что документ содержит полезные макросы (например, форму за-
каза, используемую в данной компании), можно открыть документ вместе с макро-
сами. Однако если считается, что в документе не должно быть макросов, или ис-
точник документа не внушает доверия, целесообразно открыть документ без мак-
росов , чтобы предотвратить возможное заражение. Например, следует проявлять
подобную осторожность при открытии содержащих макросы документов, вложенных в
сообщения электронной почты, или полученных с малознакомого узла Интернета.
Чтобы отключить обязательную проверку документов Word или Exel на наличие
макросов, которые могут содержать вирусы, снимите флажок ’’Всегда выводить это
окно’’ при открытии документа, содержащего макросы в предупреждающем диалоге-
1
Материал устарел, но по-прежнему актуален.
вом окне или выберите в меню "Сервис" команду "Параметры", вкладку "Общие" и
снимите флажок "Защита от вирусов в макросах" (Word), "Защита от макровиру-
сов" (Excel) .
Наиболее разрушительное действие вирусной атаки состоит в форматировании
винчестера, что ведет к полной потере данных. Для защиты компьютера от виру-
сов разрабатывается и постоянно обновляется специальное прикладное программ-
ное обеспечение, называемое антивирусными программное обеспечение. Основным
средством антивирусной защиты является периодическая проверка компьютера с
помощью антивирусного программного обеспечения. В случае если произошло инфи-
цирование компьютера вирусом, неизвестным антивирусному программному обеспе-
чению и если существует реальная угроза потери ценных данных или эта потеря
уже произошла, то переформатируют винчестер, устанавливают заново операцион-
ную систему и восстанавливают данные с резервной копии.
Резервные копии сохраняют на внешних носителях, хранящихся в сейфах. При
разработке организационного плана резервного копирования, как правило, созда-
ют две резервные копии. Таким образом, надежная антивирусная защита с помощью
антивирусного программного обеспечение требует его постоянного обновления.
Допустимая периодичность обновления составляет три месяца. Наиболее популяр-
ным программным обеспечением являются: Антивирусная система Касперского,
DrWEB, AidTest и др.
Архивирование данных
Для организации компьютерных архивов используется специальное программное
обеспечение, называемое архиваторами. Характерной особенностью различных фор-
матов, в которых хранятся данные на компьютере, является повторяемость неко-
торых порций информации, которую можно трактовать как некоторую избыточность.
Степень избыточность различных форматов сильно отличаются, так избыточность
графических форматов значительно выше текстовых.
Архиваторы, во-первых, позволяют сжать данные, устранить избыточность, а
во-вторых, организовать архив под одним именем из нескольких объектов, напри-
мер файлов. В целом эти возможности обеспечивают эффективное хранение и дос-
туп данным. Рекомендуется создавать архивы при пересылке данных по компьютер-
ным сетям.
Существует три основных способа устранения избыточности: изменение содержа-
ния, изменение структуры данных, изменение содержания и структуры. Если про-
исходит изменение содержания, то при восстановлении файла из архива происхо-
дит его частичная утрата.
Такие методы называются методами сжатия с регулируемой потерей информации.
Они применимы только для тех типов, для которых формальная утрата части со-
держания не приводит с значительному снижению потребительских свойств, прежде
всего, к мультимедийным данным. Такие методы нельзя применять к текстовым до-
кументам, базам данных и программам.
Если при сжатии, т.е. устранении некоторой избыточности, происходит только
изменение их структуры, то метод сжатия обратим.
Существует много форматов архивированных данных, самыми популярными на се-
годняшний день являются ZIP, ARJ, RAR. Программные средства, предназначенные
для создания и обслуживания архивов, выполненных в данных форматах, называе-
мые диспетчерами архивов следующие: WinZip, WinRAR, WinArj, ZipMagic.
Базовыми функциями диспетчеров архивов являются:
• извлечение файлов из архивов;
• создание новых архивов;
• добавление файлов в имеющийся архив;
• создание самораспаковывающихся архивов;
• создание распределенных архивов на носителях малой емкости (дискета);
• тестирование целостности архивов;
• полное или частичное восстановление поврежденных файлов;
• защита архивов от просмотра и несанкционированной модификации.
Самораспаковывающиеся архивы создают, когда имеются сомнения в наличии у
того, кто будет им пользоваться требуемого диспетчера архива.
В этом случае архив получает расширение .EXE, характерное для загрузочных
модулей программ. Пользователь архива может запустить его как программу, по-
сле чего начнется его распаковка.
Распределенные архивы создают, когда архив создается на носителях малой ем-
кости, например дискетах и он весь не умещается на одном носителе. Диспетчер
WinZip выполняет сразу разбиение на архива на дискетах, причем имена всех
фрагментов совпадают, что не позволяет по названиям файлов правильно выстро-
ить фрагменты в последовательность.
Поэтому каждую дискету надо маркировать на наклейке и аккуратно складывать
дискеты в нужном порядке. Также номер фрагмента архива можно узнать по так
называемой метке тома (дискеты), становящейся видимой при просмотре свойств
носителя в программе Explorer (Проводник).
WinRAR и WinArj позволяют выполнить предварительное разбиение архива на
фрагменты заданного размера на жестком диске, а потом их можно будет скопиро-
вать на дискеты. Эти диспетчеры создают фрагменты распределенного архива под
разными именами и поэтому не возникает проблем аналогичных с использованием
WinZIP.
Следует помнить, что гибкие носители являются ненадежными, поэтому жела-
тельно хранить две копии архива.
Защита архивов выполняется с помощью пароля, который запрашивается при по-
пытке просмотреть, распаковать или изменить архив. Защита с помощью пароля
считается ненадежной и поэтому ее не следует применять в работе с особо цен-
ной информацией.
WinZip, WinRAR и WinArj позволяют выполнить предварительное разбиение архи-
ва на фрагменты заданного размера на жестком диске, а потом их
можно будет скопировать на дискеты.
ЛЕКЦИЯ 8
Понятие компьютерной сети
Компьютерной сетью называется два и более компьютера, взаимодействующих че-
рез среду передачу данных. Под средой передачи данных будем понимать кабель-
ную систему (например, обычный телефонный провод, оптический волоконный ка-
бель) и различные типы беспроводной связи (инфракрасное излучение, лазер и
специальные виды радиопередачи).
Компьютеры, входящие в сеть, могут совместно использовать данные, принтеры,
факсимильные аппараты, модемы и другие устройства.
Этот список можно пополнять, так как возникают новые способы совместного
использования ресурсов. Компьютерные сети отличаются сложностью и сферой дея-
тельности. Вследствие этого они классифицируются различными способами. Однако
наиболее распространенный способ оценки сетей основывается на размерах гео-
графической площади, покрываемой сетью. Первоначально компьютерные сети были
небольшими и объединяли до десяти компьютеров и один принтер.
Технология ограничивала размеры сети, в том числе количество компьютеров в
сети и ее физическую длину. Например, в начале 1980-х годов наиболее популяр-
ный тип сетей состоял не более чем из 30 компьютеров, а длина ее кабеля не
превышала 185 метров. Такие сети легко располагались в пределах одного этажа
здания или небольшой организации. Для маленьких фирм подобная конфигурация
подходит и сегодня.
Эти сети называются локальными вычислительными сетями ЛВС (LAN, Local Area
Network). Самые первые типы локальных сетей не могли соответствовать потреб-
ностям крупных предприятий.
Вследствие этого возникла необходимость в расширении локальных сетей. Сего-
дня, когда географические рамки сетей раздвигаются, чтобы соединить пользова-
телей из разных городов и государств, ЛВС превращаются в глобальную вычисли-
тельную сеть ГВС (WAN, Wide Area Network) , а количество компьютеров в сети
практически не ограничено.
Основное назначение компьютерных сетей - совместное использование ресурсов
и осуществление интерактивной связи как внутри одной фирмы, так и за ее пре-
делами .
Ресурсы - это данные, приложения и периферийные устройства, такие, как
внешний дисковод, принтер, мышь, модем и джойстик. Понятие интерактивной свя-
зи компьютеров подразумевает обмен сообщениями в реальном режиме времени.
Типы сетей
Все сети имеют некоторые общие компоненты, функции и характеристики. В их
числе:
• серверы (server) - компьютеры, предоставляющие свои ресурсы сетевым поль-
зователям;
• клиенты (client) - компьютеры, осуществляющие доступ к сетевым ресурсам,
предоставляемым сервером;
• среда (media) - способ соединения компьютеров;
• совместно используемые данные - файлы, предоставляемые серверами по сети;
• совместно используемые периферийные устройства, например, принтеры, биб-
лиотеки CD-ROM и прочие ресурсы - другие элементы, используемые в сети;
Несмотря на определенные сходства, сети разделяются на два типа:
• одноранговые (peer-to-peer);
• на основе сервера (server based).
В одноранговой сети все компьютеры равноправны: нет иерархии среди компью-
теров и нет выделенного сервера. Каждый компьютер функционирует и как клиент
и как сервер. Все пользователи такой сети самостоятельно решают, какие данные
на своем компьютере сделать общедоступными по сети.
Если к сети подключено более 10 компьютеров, то одноранговая сеть может
оказаться недостаточно производительной. Поэтому большинство сетей использует
выделенные серверы. Выделенным называется такой сервер, который функционирует
только как сервер.
Серверы специально специализированы для быстрой обработки запросов от сете-
вых клиентов и для управления защитой файлов и каталогов. Сети на основе сер-
вера стали промышленным стандартом. Круг задач, которые должны выполнять сер-
веры, многообразен и сложен. Чтобы приспособиться к возрастающим потребностям
пользователей, серверы в больших сетях стали специализированными. Например, в
сети Windows NT существуют различные типы серверов.
Файл-серверы и принт-серверы
Файл-серверы и принт-серверы управляют доступом пользователей к файлам и
принтерам. Например, чтобы работать с текстовым процессором, Вы прежде всего
должны запустить его на своем компьютере. Документ текстового процессора,
хранящийся на файл-сервере, загружается в память Вашего компьютера, и, таким
образом, Вы можете работать с этим документом на своем компьютере. Другими
словами, файл-сервер предназначен для хранения файлов и данных.
Серверы приложений
На серверах приложений выполняются прикладные части клиент-серверных прило-
жений, а также находятся данные, доступные клиентам.
Например, чтобы упростить извлечение данных, серверы хранят большие объемы
информации в структурированном виде. Эти серверы отличаются от файл-серверов.
В последних файл или данные целиком копируются на запрашивающий компьютер. В
сервере приложений на запрашивающий компьютер пересылаются только результаты
запроса. То есть в место всей базы данных на Ваш компьютер с сервера загружа-
ется только результат запроса, например, Вы можете получить список студентов,
имеющих средний бал за успеваемость равный 4,5.
Почтовые серверы
Почтовые серверы управляют передачей электронных сообщений между пользова-
телями сети.
Факс серверы
Факс-серверы управляют потоком входящих и исходящих факсимильных сообщений
через один или несколько факс-модемов.
Коммуникационные серверы
Коммуникационные серверы управляют потоком данных и почтовых сообщений меж-
ду этой сетью и другими сетями или удаленными пользователями через модем и
телефонную линию.
Следует отметить, что компьютер в сети клиент/сервер может быть сервером
для одного типа приложений и клиентом для другого. Существуют также и комби-
нированные сети, обедняющие свойства и одноранговых сетей и сетей на основе
сервера.
Топология сети
Топология сети характеризует физическое расположение компьютеров, кабелей и
других компонентов сети. Топология сети обуславливает ее характеристики. В
частности, выбор той или иной топологии влияет на состав необходимого сетево-
го оборудования и на его характеристики, на возможности расширения сети, на
способ управления сетью.
Чтобы совместно использовать ресурсы или выполнять другие сетевые задачи,
компьютеры должны быть подключены друг к другу. Для этой цели в большинстве
сетей применяется кабель.
Однако просто подключить компьютер к кабелю, соединяющего другие компьюте-
ры, не достаточно. Различные типы кабелей в сочетании с различными сетевыми
платами, сетевыми операционными системами и другими компонентами требуют и
различного взаимного расположения компьютеров. Каждая топология сети налагает
ряд ограничений. Например, она может диктовать не только тип кабеля, но и
способ его прокладки.
Топология может также определять способ взаимодействия компьютеров в сети.
Различным видам топологий соответствуют различные методы взаимодействия.
Все сети строятся на основе трех базовых топологий:
• шина;
• звезда;
• кольцо.
Если компьютеры подключены вдоль одного кабеля, называемого сегментом или
магистралью, топология называется шиной.
Если компьютеры подключены к сегментам кабеля, исходящим из одной точки,
топология называется звездой. В том случае когда, кабель, к которому подклю-
чены компьютеры, замкнут в кольцо, такая топология называется кольцом.
ЛЕКЦИЯ 9
Введение в ИНТЕРНЕТ
Что такое Интернет?
Интернет - это глобальная СЕТЬ КОМПЬЮТЕРНЫХ СЕТЕЙ, соединенных КАНАЛАМИ
СВЯЗИ и передающих информацию друг другу по определенным правилам, называемых
ПРОТОКОЛАМИ. В качестве каналов связи могут использоваться обычные телефонные
линии. В этом случае для подключения компьютера к сети используется устройст-
во, называемое МОДЕМОМ (от комбинации слов модуляция/демодуляция). Предостав-
ляют доступ к сети Интернет фирмы или организации, называемые ПРОВАЙДЕРАМИ
(от английского provide - обеспечивать).
Рождение Интернет относят к 1969 году. Сеть Интернет (Internet) можно опи-
сать как огромную цифровую магистраль - систему, связывающую миллионы компью-
теров, подключенных к тысячам сетей по всему миру. Ее прошлое уходит своими
корнями в эпоху холодной войны, конец 60-х - начало 70-х годов. Первоначально
данные разработки финансировались правительством США, и сеть, ставшая предше-
ственницей Internet, была специально спроектирована таким образом, чтобы
обеспечить коммуникации между правительственными узлами в том случае, если
часть ее выйдет из строя в результате ядерной атаки. Применяемый в ней алго-
ритм управления передачей информации (межсетевой протокол) был разработан
так, чтобы компьютеры всех видов могли совместно использовать сетевые средст-
ва и непосредственно взаимодействовать друг с другом как одна эффективно ин-
тегрированная компьютерная сеть. Сегодня Internet сильно разрослась и связы-
вает уже десятки миллионов пользователей компьютеров во всем мире. Эта гло-
бальная "сеть сетей" охватывает тысячи университетских, правительственных и
корпоративных сетевых систем, связанных высокоскоростными частными и общедос-
тупными сетями.
Internet позволяет легко взаимодействовать друг с другом самым различным
видам компьютерных систем. Каким же образом это происходит? Благодаря стан-
дартам. В Internet применяются стандартизованные методы передачи данных, по-
зволяющие скрыть от пользователя все многообразие сетей и машин.
Наиболее фундаментальным стандартом, применяемым в Internet, является набор
сетевых протоколов TCP/IP (Transmission Control Protocol/Internet Protocol),
определяющих алгоритмы передачи данных. Поскольку все компьютеры взаимодейст-
вуют с Internet через TCP/IP, нет необходимости в сложном и дорогостоящем
преобразовании протоколов, что значительно упрощает операции в Internet. Про-
токол TCP/IP не зависит от конкретной операционной системы и, таким образом,
реализуется для всех типов компьютеров - IBM, APPLE и др.
TCP/IP позволяет присвоить каждому компьютеру уникальный адрес Internet.
Такой адрес называется IP-адресом или адресом TCP/IP и выполняет ту же роль,
что и обычный телефонный номер, то есть позволяет установить связь между дву-
мя пунктами сети. Например, адрес TCP/IP 137.65.1.3 идентифицирует в сети
один из основных информационных компьютеров корпорации NOVELL. Однако в боль-
шинстве случаев пользователи Internet применяют более удобную адресацию, на-
зываемую системой имен доменов (Domain Name System, DNS). DNS - это иерархи-
ческий распределенный метод организации пространства имен в Internet, который
позволяет уйти от цифровой адресации и дает ряд других преимуществ. Например,
вместо указания адреса TCP/IP 137.65.1.3 пользователи могут применять ассо-
циированное с этим адресом имя www.novell.com.
На этих базовых стандартах основываются другие стандарты, такие как прото-
кол электронной почты SMTP (Simple Mail Transfer Protocol). SMTP дает возмож-
ность подключенным к Internet пользователям обмениваться электронной почтой.
Благодаря этому и другим стандартам можно передавать электронную почту из од-
ного места в другое, причем не только сообщения, но и программы, графику,
Звук, видео и другие типы данных.
Важную функцию в Internet реализуют стандарты, позволяющие "публиковать"
информацию - размещать ее на специальных компьютерах (хост-узлах сети), где с
ней могут работать другие пользователи. Система компьютеров, публикующих та-
кую информацию, называется World Wide Web, а протокол, составляющий основу
Web - протоколом передачи гипертекста (HTTP -- Hypertext Transfer Protocol).
Если TCP/IP дает возможность пользователям обращаться к хост-узлам Internet,
то HTTP обеспечивает их доступ к документам World Wide Web (WWW).
World Wide Web - наиболее новая и самая быстро развивающаяся сегодня служба
Internet. Она имеет почти неограниченный потенциал в плане сбора, распростра-
нения и изучения информации. Обеспечиваемые ею графические межплатформенные
средства завоевывают все большую популярность у пользователей и компаний, ко-
торым необходимо собирать информацию, обмениваться своими идеями и самим
предлагать коммерческую информацию в Internet.
Система World Wide Web была впервые разработана Тимом Бернерсом-Ли из Евро-
пейской лаборатории физики элементарных частиц (CERN) в Женеве (Швейцария)
как способ организации информации для ее научных сотрудников. Информация на
Web-серверах хранится в виде набора документов. Каждый документ содержит ги-
пертекстовые ссылки, с помощью которых пользователь может обращаться к инфор-
мации в других документах по данной теме. Таким образом, выбирая выделенные
слова, изображения и графические элементы в тексте документа, пользователи
способны перемещаться в любом направлении и прозрачным образом "перескаки-
вать" на другие интересующие их документы (независимо от того, где эти доку-
менты находятся).
Такая технология позволяет наряду с текстом включать в Web-документы графи-
ку, звук и видеоизображения. Перемещаясь по огромным массивам электронных до-
кументов, хранящихся на тысячах серверов в Internet, пользователи, которые
ищут конкретную информацию, могут фактически путешествовать по всему свету.
Для этого им достаточно выбирать ссылки в документах и прозрачным образом пе-
реходить к нужной информации.
Web-документы создаются с помощью гипертекстового языка описания документов
HTML (Hypertext Markup Language). Такие документы могут содержать графику и
гипертекстовые ссылки. При щелчке на ней мышью гипертекстовая ссылка выводит
пользователю другой документ. Таким образом, эта ссылка содержит "указатель"
на документ, который становится доступным при нажатии кнопки мыши. Такой ука-
затель носит название унифицированного указателя ресурса - URL (Uniform
Resource Locator). Указатели URL обычно описывают транспортный протокол доку-
мента (например, HTTP или FTP) и имя хост-компьютера, на котором он находит-
ся . Кроме того, указатели URL могут включать в себя маршрут доступа к доку-
менту на данном компьютере. Эти маршруты указываются в конце строки URL.
Поиск и чтение информации во всемирной паутине WWW осуществляют с помощью
специальных программ. Протокол передачи гипертекста HTTP позволяет этим про-
граммам обращаться к файлам на любом Web-сервере. Например, по URL www.fss.ru
можно попасть на web-сервер Фонда Социального Страхования, где содержится
различная информация.
Существует и ряд других базовых средств, позволяющих передавать живую речь
(телефония Internet), проводить видеоконференции, осуществлять коллективную
работу над одним проектом и т.п. Однако основой технологий Internet, обеспе-
чившей этой глобальной компьютерной сети широкое распространение, популяр-
ность и эффективность, являются TCP/IP и World Wide Web (WWW) или «Всемирная
паутина».
Единое информационное пространство Интернет (Internet) не только сокращает
громадные расстояния, но и разрывает национальные и классовые границы, обес-
печивает каждому индивидууму возможность для самовыражения и удовлетворения
различных духовных потребностей.
Интернет (Internet) и интрасети (Intranet) - локальные компьютерные сети
предприятия, внутри которых используются технологии Internet предоставляют
беспрецедентные возможности повышения продуктивности работы, продажи товаров
и услуг на новых быстро расширяющихся рынках, а также реализуют недорогой
способ глобальных коммуникаций, как внутри предприятия, так и вне его. Техно-
логии Internet/intranet осваивают малые и большие предприятия, коммерческие
фирмы, банки, правительственные организации, учреждения образования, науки,
культуры, здравоохранения и других сфер человеческой деятельности. Осваивают
их и многочисленные отдельные пользователи, а также просто граждане, откры-
вающие для себя впечатляющие возможности коллективной работы и глобального
доступа к информации.
Новые информационные технологии меняют не только методы и формы деятельно-
сти. Они изменяют мышление и психологию людей, создают предпосылки для эволю-
ции человечества от общества постиндустриального к обществу информационному.
Естественно, что все эти сегодняшние и, еще большие, грядущие изменения за-
трагивают философию и психологию взаимоотношений, способы и формы деятельно-
сти людей и в профессиональных сферах.
Простота доступа к информации, ее безбумажный характер, высокая скорость
обмена данными между сотрудниками предприятия, возможность для деловых опера-
тивных коммуникаций с "внешним миром" оказывают существенное влияние, прежде
всего, на методы и формы системы управления. Суть этих изменений сводится к
демократизации процессов подготовки и принятия управленческих решений, более
высокому уровню их аналитичности и обоснованности, повышению роли каждого со-
трудника в управлении предприятием, высокой степени доступности управленче-
ской информации, сглаживанию границ субординации и т.п.
Внедрение новых информационных технологий на предприятии должно сопровож-
даться изменением его структуры и методов управления. Данное утверждение мож-
но обосновать, по крайней мере, тремя следующими факторами.
Во-первых, новые технологические средства дают весомый экономический эффект
лишь при реорганизации методов работы. Иначе они будут лишь модными игрушка-
ми .
Во-вторых, информационные технологии позволяют в существенно большей мере
использовать интеллектуальный потенциал сотрудников предприятия. Важно также
отметить, что высококвалифицированные специалисты при выборе места работы,
как показывают социологические исследования, придают большее значение исполь-
зованию на предприятиях новых технологий труда, чем даже уровню заработной
платы.
В третьих, широкое распространение новых информационных технологий меняет
методы и средства взаимодействия, формы взаимоотношений на рынках товаров и
услуг, например, в многочисленных цепочках "Заказчик-Поставщик", "Продавец-
Покупатель" и т.п. Если сотрудники предприятия не используют технологий
Internet в рекламе, маркетинге, финансовой, снабженческой и других сферах
управленческой деятельности, то скоро с ними перестанут сотрудничать даже из-
за проблем коммуникации.
Предприятие, не учитывающее эти и ряд других сопутствующих требований со-
временного общества, вряд ли будет процветать.
Кто же может работать в Internet? В настоящее время Internet - это общедос-
тупная сеть, открытая для любого пользователя, имеющего модем и/или инсталли-
рованное программное обеспечение для работы по стандартному протоколу переда-
чи данных в сети. Допуск в Internet для организации или частного лица через
постоянное сетевое соединение или коммутируемую линию (обычный телефон) пре-
доставляется так называемым провайдером услуг Internet (Internet Service
Provider - ISP). Круглосуточный доступ к Internet имеют уже порядка 100 мил-
лионов пользователей в 96 странах мира, и их количество растет лавинообразно.
Поскольку все больше людей обращаются к ней для коммуникаций друг с другом и
получения информации, компании, подключенные к этой глобальной компьютерной
сети, открывают множество новых возможностей для своего бизнеса.
Internet функционирует, не имея никакой центральной организации, которая
осуществляла бы управление или руководство ею, за исключением, разве что,
Центра сетевой информации Internet - InterNIC (Internet Network Information
Center), организации, предлагающей информационные и регистрационные услуги
для пользователей Internet.
«Всемирная паутина» сегодня это не только огромное количество разнообразной
информации, но и новые мультимедийные технологии ее передачи, позволяющие го-
ворить об Интернет как о «виртуальной» реальности - новом способе общения и
сосуществования людей.
Навигация по Интернет
После установления связи компьютера пользователя с сетью, например, через
сеансовое подключения к серверу2 провайдера по телефонному каналу, для путе-
шествия по просторам Интернет необходимо запустить специальную программу-
проводник. Эти программы часто называют БРАУЗЕРАМИ (от англ, browse - лис-
тать, просматривать) или обозревателями. Наиболее широко распространены брау-
зеры Netscape Communicator и Microsoft Internet Explorer. Эти программы, хотя
и разработаны фирмами-конкурентами, для начинающего пользователя очень похо-
жи .
БРАУЗЕРЫ позволяют просматривать гипертексты, получаемые из Интернет по
указанным пользователем адресам. ГИПЕРТЕКСТ - это текст, содержащий ССЫЛКИ.
Основным признаком нахождения пользователя в гипертексте jx является
появление на экране монитора указателя мыши в форме кисти руки
человека.
Гипертекстовое содержимое WWW создается с помощью языка разметки гипертек-
стовых документов - HTML (HyperText Markup Language).
Полезную информацию при работе с браузером пользователь может получить из
СТРОКИ СОСТОЯНИЯ, которая находится в нижней части окна. Профессиональная ра-
бота с Netscape Communicator и Internet Explorer обязательно включает в себя
умение разбираться в надписях, появляющихся на этой строке. В процессе работы
с Интернетом в строку состояния периодически выводятся сообщения об адресах
источников информации, режиме ожидания, готовности запрошенного документа и
ряд других полезных сведений.
Для того, чтобы получить документ из Интернет, можно в написать адрес этого
документа в поле ввода «Адрес» нажать на клавишу Enter, передав тем самым
указанный адрес браузеру:
Необходимые адреса либо берутся из справочников, либо конструируются поль-
зователем, исходя из понимания структуры адреса и интуиции, либо находятся с
помощью специальных средств поиска информации в Интернет.
Структура адресов WWW
В Интернет используются два типа адресов:
• адреса компьютеров (узлов Интернет);
• адреса информационных ресурсов, расположенных на этих компьютерах.
2
Сервер - компьютер общего доступа, подключенный к сети на постоянной основе и обслуживае-
мый специальной «сетевой» операционной системой.
Каждый из множества ПК, входящих в Интернет, имеет свой собственный уни-
кальный адрес IP. Это цифровой адрес, состоящий из четырех групп цифр, напри-
мер, 212.193.2.201 - адрес сервера Санкт-Петербургского отделения института
«Открытое общество». Для удобства IP-адресам поставлены в соответствие сим-
вольные или ДОМЕННЫЕ адреса: сервер СПб отделения ИОО имеет адрес
www.spb.osi.ru. Домен - это множество компьютеров, имеющих общую часть имени.
Составные части доменного адреса называются СЕГМЕНТАМИ: <сегмент>.<сегмент>.
... .<сегмент>.
Сегменты образуют иерархическую систему и самый последний (правый) называ-
ется именем доменом верхнего уровня. Среди доменов верхнего уровня различают
географические и тематические. Географические доменные имена верхнего уровня
- двухбуквенные. Они определяют принадлежность владельца имени к сети кон-
кретной страны (например www.spb.ru -) Тематические адреса дают возможность
представить сферы деятельности их владельцев: сот - коммерческие фирмы (на-
пример, www.microsoft.сот); edu - образовательные учреждения; gov - государ-
ственные, mil - военные, org - прочие организации.
Информационным ресурсам ставят в соответствие URL-адреса (от англ. Uniform
Resource Locator - универсальный адрес ресурса). URL-адрес состоит из двух
частей, разделяемых двоеточием. Первая (левая) часть указывает на то, к КАКО-
МУ ТИПУ принадлежит ресурс и КАК получить к нему доступ, т.е. определяет кон-
кретный сетевой протокол. Вторая часть URL-адреса сообщает, ГДЕ расположен
искомый ресурс. Таким образом, URL содержит информацию не только о том, ГДЕ
данный ресурс расположен, но и КАК к нему следует обращаться. Пример URL-
адреса : http://www.spb.osi.ru.
ВАЖНО: без особой необходимости нет надобности указывать префикс HTTP://.
Браузер сам добавит эти символы, резонно «полагая», что если пользователь пу-
тешествует по WWW, его интересуют скорее всего гипертекстовые ресурсы и, ста-
ло быть, к ним нужно обращаться по протоколу HTTP (hyper text transfer
protocol) - оптимизированному для работы с гипертекстовыми ресурсами Интер-
нет .
Работа со списками адресов
Если страница, на которую Вы попали, путешествуя по Интернет, представляет
интерес, ее адрес можно запомнить, используя кнопку панели инструментов «Из-
браное» или пункт меню «Избранное», которые обеспечивают доступ к формируемо-
му пользователем списку адресов. Этот список может в дальнейшем редактиро-
ваться, адреса можно систематизировать по папкам, подобно тому, как это дела-
ется с файлами, наконец, адреса или их часть можно как файлы переносить на
дискетах, пересылать по электронной почте или экспортировать в файл избранных
адресов другого браузера для использования на других компьютерах. Используйте
для этого обычные приемы работы с файловой системой: выделение группы адресов
или папок; перенос выделенных объектов в другую папку или диск, а также воз-
можности, предоставляемые меню «Файл» или локальным меню, вызваемым нажатием
на правую кнопку мыши, когда курсор находится в пределах окна избранных адре-
сов .
Перемещение в Интернет протоколируется браузером, что может быть использо-
вано как для поиска ранее посещенных адресов, которые не были внесены в спи-
сок избранных, так и для простого контроля работы пользователя. В браузере
Internet Explorer списки посещенных адресов доступны через кнопку панели ин-
струментов «Журнал». Они группируются по сайтам и времени, что удобно при ра-
боте с протоколом.
Рис. Работа с папкой «Избранное» Internet Explorer3
- Microsoft Internet Lxpfcxei ЕЮЕЗ
llp<3t»-a bm Избрл-ное Сервис £лроека
> - -» J 4 Laj'j gjr -J iff Ф__________________________________________________________________________
£мцж I®] Ntp//*♦«*¥ a*u&ru/iue/
Jjj aes s егоя ;atoo Irrez: егсег y sb
JU spb |w«m sphosirul
Jll Ной компьютер
Покупки • Обзоры • Популярные • НрЯм» сайты • Р«»:пм*-нд±
-' З'ТОшшл^ Сегодня продается... ал< у зн.тк
‘ffUd настоящее ЭХО "Я , ско|
. ЕПЗЛ IПГ!П'| 11Ш!Ш [
Сдклатк АРИЗЫЛЙ сткянлцки
Оши бдимдиц амаии Csaaacaso (Jaiss«*®Ii ietam
ВыИнк 1а>аю:н Лнминня» С в ста го Анекдот ьн» Агссииу [ща
Ф»т Наваеяептметы
Игнаты | | gflus
Навигация | В
Бизнес
Фкыансы Паедлриятия. Промышленность
Информация. Работа. Тоогоопя Тоанспост
Медицина и здоровье
Печение Лек кете а. Пег, галактика
Специалистам Псткляс-ая медици
Ден и семья
Автзмэби.-ь Путешествия Хобби. Мир
жвншима Дли Знакомства Кулинария
Нацка и оО|мн>ва»ае
ЕпЕИбаниые Метемлиа ибц,ес
Высшее образование Рефераты Э
Интерне»
Поиск Дизайн Поомйлевы. Услуги
Фооумы/Чаты Часнне страницы
Комтыргецы
Процтаммы Железо Сети Безопасность
Публикации Рынок. Игры графика
Оишссгво
т-авости СМИ. Арми». Радио Sails.
Выборы ГВ Г азоты Атентстеа ин*
Развлечения
Игры Музыка Зоотика Юмор Кин
Геймерам. Футбол, чтение он-лайн
* Ичткснм
Рис. Работа с папкой «Журнал» в Internet Explorer
3 Редакция рекомендует не использовать IE, из-за наличия многочисленных проблем с безопасно-
стью. Правительство США тоже согласно с этим мнением, и не рекомендовало использовать IE в
государственных учреждениях и бизнесе.
Проблема кодировок кириллицы
Информация в компьютере хранится и обрабатывается в цифровом виде. Истори-
чески сложилось, что имеются различные таблицы кодировок, т.е. правила сопос-
тавления символам соответствующих им цифровых кодов, для русского и других
национальных алфавитов: Windows (СР1251), K0I8, DOS (СР866) ISO-8859-5 и др.
Вследствие этого порой текст в окне браузера оказывается невозможно прочесть.
Обычно затруднение можно преодолеть, выбрав подходящую кодировку через меню
«Вид/Выбор кодировки». Если это не помогает, нужно попытаться найти гипер-
ссылку, позволяющую получить документ в «читаемой» кодировке, если, разумеет-
ся , разработчик это сайта такую возможность предусмотрел. Гиперссылки (WIN,
KOI, ENG и т.п.), позволяющие выбрать кодировку, в которой документы будут
загружаться с сайта, часто имеются на стартовой странице сайта, и это счита-
ется негласным правилом. Современные браузеры Internet Explorer и Netscape
Communicator в большинстве случаев автоматически настраиваются на правильное
соответствие кодировки загружаемого документа и кодировки, в которой этот до-
кумент отображается.
3 P'Rus Microsoft Internet Explorer
£айл Правка Еид Избранное Сервис Справка
Панели инструментов
_________________ Строка состояния
Адрес |в] ktp/Лм Панели обозревателя
Сделать дома»
Esc
Размер шрифта
Переход
Остановить
Обновить
глярные • Новые сайты
ра были проданы
20 копеек
Автоматический вьбор
• Кириллица (Windows!
Западноевропейский (ISO)
Дополнительно ►
Искать В | | ioIRus
Бизнес
Финансы, Предприятия, Промышленность,
Информация Работа. Торговля. Транспорт.
Дом и семья
Автомобиль Путешествия, Хобби Мир
женщины. Дехи. Знакомства Кулинария
Интернет
Поиск, Дизайн, Гровэйдеры, Услуги.
Форумы/Чаты. Частные страницы
Компьютеры
Программы. Железо. Сети. Безопасность.
Публикации. Рынок. Игры. Графика
1
Медицина и здоровь
Лечение, Лекарств»
Сдециэдшам. Иж
Наука и образование
Ьстественные. Мате
Высшее обоазэванг
Общество
Новости, СМИ, Арм
ВыбШ!. IB, Газету
Развлечения
Игры. Музыка. Эрст
Геймерам. Футбол.
Культура и искусство
Ро П1Д Г|ТГ IZ| « П 11 т/~> г» пти п -ст "ilTurs r»ni дгч
Справка
И > 1« Г>Г»Г|ДГ»1И I t
Арабский
Прибалтийский (ISO) Прибалтийский (Windows) ipexon Ссылки у>
А
Центра гьноеврогейский (DOS) Центрагьноевросейский (ISO) цди десять
цтличий
Цеитрагьноеерогейский (Windows)
Китайский - упрошенное письмо 1ымграй
Китайский традиционное письмо т□аппарат
Кириллица (DOS) Кириллица (ISO) Кириллица (К0И8-Р)
Кириллица (К0И8-У) (ЦИОН
Греческий (ISO) Г реческий (Wndows)
Иврит Японский ипи ПИЯМ отдых
Корейский ^1гГ
Тайский бесплатно
Турецкий (ISO) Т урецкий (Wridows) ити»иость по
недели и г
Unx;ode(UTF-8) ИЫОСТИ от
нтаци* ,,
Определенный пользователем Вьетнамский Ьк । 5 пойман? _U
Зас<цнсеьр1.г>ей<.кий (Windows)
Рис. Установка подходящей кодировки кириллицы в Internet Explorer
Работа с электронной почтой
Пользователи Интернет имеют возможность обмениваться информацией друг с
другом с помощью электронной почты - E-mail - одного из самых популярных и
дешевых сервисов. В Интернет есть много сайтов, предоставляющих почтовые ящи-
ки и услуги электронной почты бесплатно с доступом через WWW (т.е. по прото-
колу http), как и к другим гипертекстовым страницам, например, www.mail.ru,
www.hotmail.ru, www.hotmail.com.
Кроме того, многие провайдеры информационных услуг предоставляют клиентам
возможность иметь собственный почтовый ящик бесплатно, например,
www.yahoo.com, www.yandex.ru, www.atrus.ru. Почтовые услуги от различных про-
вайдеров различаются сервисом и объемом дискового пространства для хранения
корреспонденции.
Для получения собственного почтового ящика необходимо:
• выбрать сайт, предоставляющий почтовые услуги (например, www.mail.ru);
• зарегистрироваться в качестве нового пользователя, сообщив некоторые све-
дения о себе.
Результатом регистрации является создание личного почтового ящика, доступ к
которому обеспечивается через выбранные Вами Имя пользователя (Login) и Па-
роль .
ЛЕКЦИЯ 10
Настройка электронной
почты в Outlook Express
Этот раздел мы начнем с очень краткого совета, каким образом вы можете по-
лучить адрес электронной почты и соответствующую учетную запись на почтовом
сервере провайдера.
Как получить адрес
электронной почты
Согласно распространенной практике, получение адреса электронной почты яв-
ляется стандартной услугой, которую предоставляют провайдеры при заключении
договора на подключение к Интернету по телефонной линии (Dial-Up). При этом
пользователь получает свой адрес E-mail, а сотрудник провайдера заносит этот
адрес в список пользователей почтовых серверов, заведя тем самым соответст-
вующую учетную запись пользователя на этих серверах. Сейчас мы перечислим,
какие сведения необходимо иметь для начала работы с электронной почтой, ис-
пользуемой через коммутируемое соединение:
• ваш адрес электронной почты (адрес вида ваше_имя@имя_провайдера.ru),
• тип сервера входящей почты (POP или IMAP) и имя этого сервера входящей
почты,
• имя сервера исходящей почты (сервер SMTP),
• ваше имя для учетной записи почты (также называемое как имя входа в систе-
му или имя пользователя - login name или user name), - обычно то же самое,
что при подключении к Интернету,
• пароль, который вы используете, чтобы обратиться к вашей электронной почте
(обычно тот же самый, что при подключении к Интернету),
• сведения о том, поддерживает ли ваш сервер почты защищенный режим передачи
пароля Secure Password Authentication (SPA) . Обычно такой режим не поддер-
живается .
Начальная настройка
программы Outlook Express
Если вы ранее уже настраивали на своем компьютере доступ к Интернету для
работы с броузером, то программа "Мастер подключения к Интернету" уже запра-
шивала необходимые для работы Outlook Express сведения, и если вы их ввели,
то электронная почта сейчас должна быть готова к действию. В этом случае вы
можете перейти к следующему разделу, в котором мы рассмотрим пользовательский
интерфейс Outlook Express и затем далее. А если вы пропустили этап настройки
почты и новостей при прежнем запуске Мастера, то необходимые для этого дейст-
вия можно проделать прямо сейчас.
Итак, чтобы начать настройку Outlook Express, достаточно щелкнуть по значку
"Запустить Outlook Express" (Launch Outlook Express) на панели задач Windows
98. Иначе программу Outlook Express можно вызвать с помощью последовательно-
сти меню Пуск-Программы-Internet Explorer-Outlook Express.
Замечание. Возможно, по каким-либо причинам приложение Outlook Express не
установлено на вашем компьютере, но это можно легко исправить, выбрав с помо-
щью последовательности меню Пуск-Настройки-Панель управления - Установка и
удаление программ закладку Установка Windows и установив флажок у Microsoft
Outlook Express. Поскольку это - стандартная операция установки программных
модулей из состава Windows 98, мы не будем на ней останавливаться.
Собственно начальный процесс настройки Outlook Express, необходимый для
функционирования сервиса электронной почты, предельно прост, - при первом вы-
зове программы Outlook Express еще раз Запускается Мастер подключения к Ин-
тернету, который запросит у вас необходимые данные о почтовых серверах и ад-
ресе электронной почты, которые вы должны были получить от провайдера.
1. В самом первом окне надо будет ввести ваше имя как абонента электронной
почты, которое будет появляться в заголовке писем, когда они придут к адреса-
там. Обычно пользователи указывают свое настоящее имя, но здесь можно про-
явить гибкость, и если вы пока не планируете использовать свой адрес для де-
ловой корреспонденции, то можно обойтись и псевдонимом. Кроме того, можно на-
стоятельно рекомендовать записывать свое имя латинским шрифтом, чтобы как из-
бежать проблем неправильного отображения заголовков с русским шрифтом в ряде
программ электронной почты (которые могут стоять у ваших коллег по перепис-
ке) , так и не создавать лишних проблем вашим возможным зарубежным адресатам.
Впоследствии, кстати, указанное в этом окне имя можно будет легко изменить с
псевдонима на настоящее имя и наоборот, - если в этом возникнет необходи-
мость .
2. В следующем окне вам надо будет ввести ваш адрес электронной почты. Как
правило, адрес E-mail состоит из вашего имени и доменного имени провайдера,
соединенных с помощью знака @. К примеру, это может быть адрес вида
user@narod.ru. Когда вы будете называть свой адрес кому-либо вслух, то знак @
произносится как словосочетание "ат", хотя русские пользователи чаще называют
его сленговым термином "собака" (этот сленговый термин для знака @ ведет свою
историю со времен самых первых компьютерных игр).
3. Далее Мастер запросит у вас тип сервера входящей почты и доменные имена
почтовых серверов провайдера, предназначенных, соответственно, для входящей
(POP3) и исходящей почты (SMTP). Введите эти адреса. В случае нашего примера
с Peterlink это будут адреса pop.narod.ru и relay2.narod.ru, соответственно.
Наличие разных почтовых серверов связано с тем, что сообщения электронной поч-
ты пересылаются между узлами Интернета (узлами различных провайдеров) по про-
токолу SMTP (Simple Mail Transfer Protocol), а на последнем отрезке между уз-
лом вашего провайдера и вашим компьютером — по POP (Post Office Protocol). Вве-
дите в соответствующих полях диалогового окна те адреса почтовых серверов, ко-
торые получены вами от провайдера при заключении договора. Что касается типа
сервера входящей почты, то если ваш провайдер специально не указал данный пара-
метр, то по умолчанию оставьте POP3.
4. Теперь в новом окне укажите сведения об учетной записи вашего соединения -
имя пользователя и пароль. Это те же самые сведения, которые вы обычно исполь-
зуете для подключения к Интернету. В данном случае эти сведения нужны для того,
чтобы программа Outlook Express могла автоматически дозваниваться до провайдера
и устанавливать соединение для отправки и приема электронных писем. В следующем
окне, рисунок которого мы пропустим в этом пособии, вам будет предложено ввести
"дружественное имя" для данной учетной записи почты. Можете ввести что-нибудь
типа "Моя почта на Narod" или оставить ту запись, которая предлагается по умол-
чанию.
5. В очередном окне укажите тип вашего соединения, - для нашего случая это
соединение по модему, хотя если вы применяете Outlook Express в офисе, то там
можно использовать и подключение через локальную сеть. Эти соединения будут ус-
танавливаться автоматически, но можно выбрать и третье значение переключателя,
предлагающее вручную соединиться с провайдером перед каждым сеансом работы с
электронной почты (на взгляд автора, это ненужные хлопоты).
6. Теперь остались два самых последних окна - в одном надо выбрать учетную
запись (если у вас всего один провайдер, то и выбор, соответственно, будет
невелик), но сегодня нередка ситуация, когда пользователь имеет учетные запи-
си у нескольких провайдеров и плюс учетную запись для модемного доступа к ло-
кальной сети своей компании. И наконец, осталось то окно, где нужно подтвер-
дить данные, введенные в предыдущих окнах, щелкнув по кнопке Готово (Finish).
После нажатия кнопки Готово Мастер подключения к Интернету завершит процесс
настройки Outlook Express.
В принципе, уже можно приступать к обмену почтовыми сообщениями с другими
пользователями Интернета. Однако не стоит спешить: поскольку вы только что
настроили почту, то для вас вряд ли есть какие-либо сообщения (кроме стан-
дартного поздравления, обычно рассылаемого провайдерами в таких случаях). По-
этому будет лучше, если вы уделите еще некоторое время изучению интерфейса
программы и дополнительной настройке почтовой службы Outlook Express. Чуть
позже, через раздел, мы поучимся получать и отправлять письма. Далее мы ука-
жем на ряд дополнительных тем по работе с электронной почтой, с которыми вам
можно порекомендовать познакомиться самостоятельно, а завершат это пособие
небольшие разделы, в котором вы узнаете, как завести себе адрес с бесплатной
почтой и расшифровывать поврежденные сообщения e-mail.
Пользовательский интерфейс
Outlook Express
Теперь, когда программа Outlook Express настроена на вашего провайдера, по-
ра познакомиться с ее пользовательским интерфейсом. Если вы уже имеете опыт
работы с другими программами электронной почты (например, MS Exchange или
Eudora for Windows, MS Internet Mail and News и т.д.), то вы вряд ли испытае-
те какие-нибудь затруднения и в работе с Outlook Express. Если же вы новичок
в вопросе электронных коммуникаций — не беда, благодаря очень сильному сход-
ству пользовательских интерфейсов броузера Internet Explorer 4 и приложения
Outlook Express вы попадете "в знакомую обстановку" и быстро разберетесь в
различных функциях этой почтовой программы.
Несколько слов о том, как интерпретируются значки с различными изображения-
ми конверта в области просмотра:
• Открытый конверт отмечает уже прочитанное вами письмо
• Закрытый конверт плюс жирный шрифт отмечает письмо, которое вы еще не чи-
тали
• Скрепка у письма говорит о том, что в письмо вложен отдельный файл (напри-
мер, документ в формате Word, графический файл и т. д.). Если выделить та-
кое письмо, и щелкнуть по изображению скрепки в правом углу нижней части
окна, то будет показано имя файла. Двойной щелчок по имени вложенного фай-
ла позволит просмотреть его содержимое с помощью соответствующей програм-
мы.
Совет. Изначально интерфейс и область просмотра Outlook Express выглядят
очень нарядно, но для эффективной работы стоит немного пожертвовать привлека-
тельностью во имя функциональности. Во-первых, чтобы сберечь место на экране,
можно посоветовать сразу же отключить1 две панели - самую левую колонку со
значками папок (т.к. ее содержимое повторяется левее) и серую строку с надпи-
сью Outlook Express над областью просмотра (как не несущую функциональной на-
грузки) . Для этого выберите в меню Вид (View) пункт Раскладка. Повторный вы-
бор этого пункта меню восстанавливает эти панели. Что еще можно посоветовать,
1 А лучше вообще отключить ОЕ и перейти на использование почтового клиента с хорошей Защитой
от спама, например, Thunderbird.
- так это установить в нижней части области просмотра Outlook Express флажок
Переходить в папку "Входящие" при запуске (When starting, go directly to my
Inbox folder). Тогда сразу после запуска Outlook Express будет переходить к
папке «Входящие», что позволит вам быстрее приступить к чтению новых писем.
Почтовые папки Outlook Express
Хотя названия почтовых папок Outlook Express во многом говорят сами за се-
бя, но если вы не имеете опыта работы с почтовыми программами, все же стоит
познакомиться с их назначением:
• Входящие (Inbox). Сюда по умолчанию поступает вся новая почта и здесь хра-
нятся все пришедшие сообщения. Впоследствии вы можете создать дополнитель-
ные папки (например, посвященные разным проектам или письмам от постоянных
адресатов) и настроить Outlook Express таким образом, чтобы при поступле-
нии новых писем вся почта автоматически разбиралась и складывалась в от-
дельные папки.
• Исходящие (Outbox). Это папка предназначена для временного хранения от-
правляемых писем. Зачем это нужно? Представьте, что вы подготавливаете не-
сколько писем друг за другом. Чтобы не соединяться каждый раз с Интернетом
для отправки очередного письма, письма временно накапливаются в этой пап-
ке. Затем при нажатии на кнопку Доставить почту (Send and Reseive) они ра-
зом уходят на почтовый сервер провайдера и далее - к своим адресатам.
Именно такой режим отправки писем устанавливается в Outlook Express по
умолчанию.
• Отправленные (Sent Items). Здесь по умолчанию хранятся копии отправленных
сообщений, и вы всегда сможете вспомнить, - что, кому и когда вы отсылали.
• Удаленные (Deleted Items). Если вы решите удалить ненужные сообщения, то
они временно помещаются на хранение в эту папку (на случай, если вы пере-
думаете их удалять) . Если вы решите насовсем удалить сообщения из этой
папки, сделайте правый щелчок по значку папки и из выпавшего меню выберите
пункт Очистить папку (Empty folder).
• Черновики (Drafts). Если вы готовите новое письмо, но в процессе работы
над ним решите дописать письмо позже, то выберите в меню Файл (File) пункт
Сохранить (Save). Такое «недописанное» письмо временно сохраняется в папке
Черновики (Drafts). Чтобы продолжить впоследствии работу над письмом из
этой папки, просто откройте эту папку и дважды щелкните по черновику пись-
ма. Затем, если письмо готово, то его можно отправить и оно будет помещено
в папку Исходящие (Outbox). Если же письмо по-прежнему не готово к отправ-
ке , то его опять можно сохранить в папке черновиков.
Совет. Если вы получаете много почты с вложенными файлами (документами,
графическими файлами и пр.), то неплохо периодически, - например, хоть раз в
месяц проводить чистку своих папок от старых сообщений. Это способствует зна-
чительной экономии места на жестком диске машины. И не забывайте чистить не
только папку с входящими письмами, но и папку Исходящие (Outbox), где хранят-
ся копии ваших собственных писем. Ну и конечно, не забывайте опорожнять папку
Удаленные (Deleted Items) от уже ненужных писем, которые явно никогда не по-
надобятся .
Получение входящей почты
Получение входящей почты - это, наверно, самое простое действие из всего
спектра работ с Outlook Express, вся функциональность которого состоит в том,
что надо установить соединение с почтовым сервером провайдера. Поэтому доста-
точно запустить программу Outlook Express, которая по умолчанию сразу прело-
жит вам соединиться с сервером провайдера. Если соединение с провайдером уже
настроено, то достаточно, выбрав название соединения, нажать кнопку ОК. Иначе
появится дополнительное окно, где надо будет ввести ваше имя пользователя и
пароль. В любом случае итог будет один, - компьютер начнет устанавливать со-
единение с почтовым сервером провайдера.
Совет. Помните, что соединиться с почтовым сервером провайдера можно легко
и просто в любой момент времени- достаточно нажать кнопку Доставить почту
(Send and Reseive) на панели инструментов Outlook Express.
После того как ваш модем осуществит соединение, появится окно, в котором
Outlook Express будет проверять наличие пришедших писем и осуществлять их за-
грузку на ваш компьютер. После завершения этого действия почтовая программа
проверит, нет ли у вас в папке Исходящие (Outbox) готовых писем для отправки,
и если есть, то в свою очередь, перешлет их на почтовый сервер провайдера,
откуда они уже уйдут адресатам (см. рис.).
Совет. Outlook Express позволяет устанавливать сеанс связи с провайдером
только на время доставки писем (получения пришедших писем и отправки своих),
сразу же отключаясь от Интернета по завершению передачи. Чтобы включить эту
опцию, нужно установить флажок Разорвать соединение по завершению доставки
(Disconnect...) на вкладке Удаленная связь (Connection) окна свойств или пря-
мо в окне доставки (см. рисунок). Тем самым вы избежите нерациональных затрат
времени при "холостом режиме" соединения с почтовым сервером, когда почта уже
доставлена. Помните: время пересылки почты оплачивается так же, как и ваши
путешествия по Web, и здесь тоже следует быть экономными.
Совет. При поступлении новой почты в правом углу строки состояния Windows
98 (а также в самой программе Outlook Express) будет выведен значок с изобра-
жением конверта. Это служит удобным напоминанием о пришедших письмах в том
случае, когда вы работаете с почтой в фоновом режиме, и Outlook Express пе-
риодически проверяет наличие новых писем на сервере провайдера.
Чтение пришедшей почты
Как мы уже говорили, по умолчанию все пришедшая почта помещается в папку
Входящие и сортируется там по дате поступления. Письма, которые вы еще не чи-
тали, помечаются жирным шрифтом и значком закрытого конверта. Если рядом с
письмом есть значок с изображением скрепки, то это говорит о том, что в пись-
мо вложен отдельный файл (например, документ в формате Word, графический файл
и т. д.) . Кроме того, слева от заголовка письма может стоять значок в виде
восклицательного знака, который говорит, что адресат пометил данное письмо
как сообщение повышенной важности. Если же стоит значок в виде направленной
вниз синей стрелки, то наоборот, содержимое письма отправитель пометил как
второстепенной важности.
Познакомиться с содержимым письма совсем просто - нужно выделить его, сде-
лав однократный левый щелчок мышью по заголовку письма. Тогда в нижней части
окна просмотра Outlook Express появится содержимое письма. Письмо можно по-
смотреть и в отдельном окне, которое раскроется (см. рис. в этом разделе),
если сделать двойной щелчок по заголовку выбранного письма.
Совет. Чтобы быстрее разобрать почту, удобнее всего вначале раскрывать при-
шедшие сообщения одинарным щелчком. Тогда даже беглого взгляда достаточно,
достойно ли это сообщение вашего дальнейшего внимания. Если да, то лучше рас-
крыть это сообщение двойным щелчком в отдельном окне.
В окне с пришедшим письмом вы найдете ряд новых элементов интерфейса, наи-
более важными из них являются следующие кнопки:
• Ответить отправителю (Reply to Author) открывает окно для формирования от-
ветного письма. Текст исходного письма может быть автоматически помещен в
это окно. Знаки, отмечающие начало строк цитируемого текста, определяются
там же, на закладке Отправка.
• Ответить всем (Reply to All) — эта кнопка панели инструментов действует со-
вершенно аналогично предыдущей кнопке Ответить отправителю. Исключение со-
стоит в том, что ваше ответное письмо рассылается не только первоначальному
автору, но и всем иным адресатам, которым этим автором было послано анало-
гичное письмо (которые были включены в поле Копия исходного сообщения). Та-
ким образом, эта функция имеет смысл лишь в бизнес-переписке, когда вы ор-
ганизуете таким образом что-то вроде дискуссии. Однако вы должны быть уве-
рены, что все адресаты будут согласны на получения вашего письма. Добавим,
что традиция электронной переписки запрещает "веерную рассылку" посланий
незнакомым людям, особенно содержащих рекламу (так называемый "Spam",
сленговое словечко для непрошеной почты, дословно переводимое как «фарш») и
пр.
• Переслать (Forward) — эта кнопка позволяет перенаправить (переадресовать)
полученное письмо другому пользователю, если вы считаете, что ему(ей) оно
будет более интересно, чем вам. Данная функция очень часто используется в
бизнес-переписке. В разговорной речи среди пользователей почты для этой
функции часто используется транслитерированный сленговый термин "форвард-
нуть" .
• Удалить (Delete) — нажатие на эту кнопку удаляет сообщение из текущей пап-
ки и помещает его в папку Удаленные. Впоследствии вы можете очистить папку
Удаленные вручную или сообщения будут удаляться из нее автоматически.
• Печать (Print) - кнопка с изображением принтера посылает текст письма на
принтер. Обратите внимание, что при этом печатается только текст письма, -
если письмо содержит вложенные файлы, то для их печати надо открыть соот-
ветствующую программу просмотра и выполнить печать уже из данной програм-
мы.
Теперь осталось добавить, что если в письмо вложены отдельные файлы, то
двойной щелчок по ярлыку каждого вложенного файла позволит просмотреть его
содержимое с помощью соответствующей программы. Обратите внимание, что при
попытке открыть вложенный файл система безопасности Outlook Express выдаст на
экран стандартное предупреждение, и если вы не уверены в том, что пришедший
файл абсолютно безопасен для компьютера, то лучше его вначале сохранить на
дискету, затем проверить антивирусной программой, и уже затем только открыть
для просмотра. Чтобы сохранить файл на дискету (или иной носитель), сделайте
правый щелчок по значку вложенного файла в окне с открытым письмом и выберите
пункт Сохранить как (Save as).
Совет. Не следует пугаться всех пришедших с почтой файлов, т. к. далеко не
все файлы могут содержать вирусы. К примеру, графические файлы (файлы с рас-
ширениями .jpg, .bmp, . tif, .gif и т.д.) и текстовые файлы (файлы с расшире-
ниями .txt, .rtf) в общем случае совершенно безопасны, в то время как доку-
менты Word (.doc) с большой вероятностью могут содержать макро-вирусы2. Си-
туация, когда надо быть предельно осторожными - это при получении исполняемых
файлов (с расширением .ехе, .сот). Сегодня в книжных магазинах и компьютерной
периодике можно найти достаточно материалов по защите от вирусов, поэтому
здесь на этих проблемах мы подробно останавливаться не будем.
Подготовка и отправка
писем электронной почты
Давайте познакомимся теперь с порядком подготовки и отправки сообщения по
электронной почте с помощью Outlook Express. В целом эта операция совершенно
аналогична для случаев, когда вы подготавливаете собственное сообщение и от-
правляете его «с нуля», или же отвечаете на пришедшее письмо (или третий ва-
риант - пересылаете письмо далее другому человеку). Во всех этих случаях на
экране появится одно и то же окно (окно с одним и тем же интерфейсом). Конеч-
но, при ответе на чужое письмо или его переадресации другому пользователю от-
личие будет состоять в том, что Outlook Express автоматически помещает в от-
ветное письмо тот текст, который поступил к вам от адресата. При этом в нача-
ле каждой строки исходного письма ставится знак угловой скобки ">", чтобы от-
личить ее таким образом от текста вашего ответа. Если текст "пропутешество-
вал" несколько раз между вами и вашим адресатом, то количество угловых скобок
в начале каждой строки будет соответствовать количеству «ходок» вашего письма
туда и обратно.
Совет. Если вы отвечаете на деловое письмо, то принято полностью оставлять
в конце вашего ответного письма тот текст, что пришел от адресата. Таким об-
разом сохраняется нить переписки и всегда можно выяснить, кто и что написал.
Если же вы пишете друзьям, то в традициях Интернета можно оставить лишь те
строки исходного текста (цитаты), на которые вы конкретно отвечаете. Важно
помнить: если в пришедшее к вам письмо были вложены какие-либо файлы, то по
умолчанию почтовые программы их обратно адресату не пересылают.
Формирование нового сообщения
Нажмите на кнопку Создать сообщение (Compose Message) на панели инструмен-
тов Outlook Express, что вызовет отдельное окно (см. рис. ниже). Работу с но-
вым письмом следует начать с заполнения заголовка письма, который содержащий
поля: Кому: (То:), Копия: (Сс:), Скрытая: (Вес:), Тема: (Subject:). Заметим,
что по понятным причинам обязательным является заполнение только поля Кому:
(То:) , — иначе письмо просто не найдет своего адресата.
• Кому: (То:) — так же, как и на обычном конверте, в этом поле вы должны
ввести адрес — в данном случае адрес электронной почты вашего адресата. Вы
можете не вводить адрес вручную, а выбрать его из адресной книги, если за-
несли его туда заранее (см. далее раздел "Адресная книга").
• Копия: (Сс:) — если вы хотите, чтобы аналогичное письмо пришло и другим ад-
ресатам, введите нужные адреса в данном поле, разделяя их с помощью знака
";" (точка с запятой). Словосочетание "Сс" — это сокращение от английского
"канцелярского" термина Carbon Сору (по-русски — "копирка"). Все люди, к ко-
торым пришло данное письмо, легко смогут узнать из заголовка, кому вы еще
послали данное письмо.
2 Увы, jpg файлы уже могут содержать вирусы, тогда как Word хорошо Защищен от макровирусов и
их удаление не составляет особого труда.
• Скрытая: (Вес:) — если вы хотите, чтобы аналогичное письмо пришло и другим
адресатам, но они не знали, кому еще вы отослали данное письмо, то введите
нужные адреса в данном поле, разделяя их с помощью знака ";" (точка с запя-
той) . Словосочетание "Вес" — это сокращение от английского Blind Carbon Сору
(по-русски можно было бы перевести как "слепая копирка").
• Тема: (Subject:) — здесь следует вписать несколько слов, характеризующих
тему сообщения. Заголовок лучше писать по-английски, если вы не уверены,
что почтовая программа вашего адресата поддерживает 8-битовую кодировку
заголовков и на его машине инсталлированы русские шрифты.
Совет. Обратите внимание на то, что Outlook Express подсказывает вам назна-
чение каждого незаполненного поля с помощью поясняющей надписи серого цвета.
После заполнения заголовка письма вы на первых порах можете воспользоваться
кнопкой на панели инструментов окна Проверить имена (Check Names), чтобы быть
уверенным в правильном соответствии введенных адресов тому формату, который
предусмотрен в Интернете для сообщений E-mail (он должен выглядеть как
user@host.domain, где user — имя адресата, host.domain — доменное имя почто-
вого сервера адресата).
Далее уже можно приступать к самому письму, для чего щелкните курсором мыши
в поле письма и введите нужный текст. При необходимости вы можете "вложить" в
письмо файл любого формата (см. ниже подраздел о пересылке файлов по Е-
mail"). По традиции, завершает письмо вставка подписи, для чего следует на-
жать кнопку с изображением авторучки на панели инструментов (оригинальная и
остроумная подпись - одна из традиций Интернета) . Если у вас еще не создана
подпись, то создать ее можно, выбрав в меню Сервис (Tools) пункт Бланк сооб-
щений (Stationary) и далее - нажав на кнопку Подпись (Signature). Созданную
таким образом подпись можно потом многократно использовать. В принципе, "ин-
тернетовская" подпись — вещь необязательная и вы можете подписать письмо
вручную, напечатав обычные слова, подходящие к данному случаю — например,
"Искренне ваш..." и т. п.
Обратите еще внимание на изображение логотипа Internet Explorer (стилизо-
ванной буквы е) в правой верхней части нового сообщения. Лого является иден-
тификатором "важности" отправляемого сообщения. В меню Сервис (Tools) имеется
пункт выбора важности сообщений, при нажатии на который будет выведен список
возможных опций важности сообщений (Высокая, Обычная и Низкая). Важно знать:
это не влияет на скорость доставки, а всего лишь управляет наличием изображе-
ния с соответствующим значком у вашего письма по приходу письма к адресату.
Так, многие почтовые программы маркируют важные сообщения, пришедшие на ком-
пьютер, помещая восклицательный знак рядом с заголовком. Теперь дело за адре-
сатом, обратит ли он на это внимание. Поэтому можете не увлекаться установкой
этих параметров в ваших письмах.
Теперь осталось лишь нажать на кнопку отправки письма (самая левая кнопка
на панели инструментов этого окна с изображением летящего конверта и надписью
Отправить) и ваше письмо - на пути к адресату. Если установлен флажок Отправ-
лять сообщения немедленно (Send messages...) на вкладке Отправка в окне Пара-
метры (см. меню Сервис), то Outlook Express сразу же произведет соединение с
провайдером и отправит письмо. Если же флажок снят, ваши письма будут времен-
но помещаться в папку Исходящие (Outbox), где они будут накапливаться перед
отправкой. В последнем случае на экран будет выведено сообщение о помещении
письма в папку Исходящие. Когда вы завершите составление всех писем, нажмите
на кнопку Доставить почту (Send and Receive) на панели инструментов основного
окна Outlook Express, и далее процесс соединения с провайдером пойдет совер-
шенно аналогично тому, как мы рассмотрели в предыдущем разделе.
Пересылка вложенных
файлов по E-mail
Возможность отправить по электронной почте файл любого формата — одно из
самых полезных качеств E-mail. Тем самым вы можете направить своим адресатам
и документ Word, и файл с нужным изображением, звуковое или видео-приветствие
и т.д. Единственное условие — не посылайте очень большие файлы, если вы знае-
те о том, что ваш адресат не обладает выделенным каналом в Интернет, а рабо-
тает по обычной телефонной линии. Для таких случаев файл размером 200—300 Кб
считается обычно пределом "приличий". Также имейте ввиду, что многие почтовые
серверы провайдеров просто возвращают назад почту, если ее размер более опре-
деленного предела, чаще всего - более 1 Мб.
Замечание. По-английски процесс вставки файла в письмо называется
"attaching", поэтому в разговорной речи среди наших пользователей часто ис-
пользуется транслитерированный сленговый термин "приаттачить файл". Microsoft
называет такие файлы "вложенными" — и этот термин чаще всего используется в
литературе.
Итак, для вложения файла в письмо, разместив курсор в окне для ввода пись-
ма , нажмите кнопку с изображением скрепки на панели инструментов. В ответ на
экран будет выведено стандартное диалоговое окно с изображением файловой
структуры вашего компьютера. Когда вы найдете на диске нужный файл, выделите
его и нажмите кнопку Вложить (Attach). Outlook Express вставит файл в ваше
электронное письмо, и ниже тела письма появится дополнительное окно со знач-
ком вставленного файла (см. рис.).
Стоит напомнить, что аналогичным образом выглядят и пришедшие к вам письма,
которые содержат вложенные файлы. Достаточно дважды щелкнуть на значке с вло-
женным файлом, как благодаря механизму OLE (Object Linking and Embedding)
операционной системы Windows будет запущено соответствующее приложение для
просмотра этого файла.
Совет. Некоторые форматы файлов настолько "пухлые", что занимают многие
сотни килобайт (характерный пример таких файлов, - рисунки в формате .bmp).
Для снижения размера больших вложенных файлов используйте предварительно ка-
кую-нибудь программу-архиватор (например, WinZip), и вкладывайте в электрон-
ные письма уже сжатые файлы. Также, если вам надо отослать нескольких вложен-
ных файлов в одном письме, "скрутите вместе" нужные файлы программой-
архиватором и отправьте в письме уже единственный архивный файл.
Совет. Если вам надо постоянно отсылать файлы очень большого размера, кото-
рые не "проходят" через почтовые серверы (более предела, разрешенного для пи-
сем на данном почтовом сервере), используйте для этой цели FTP-сервер вашей
фирмы, университета или какой-либо иной доступный вам FTP-сервер (например,
сервер провайдера). Поместите туда свои файлы, и вышлите адресату соответст-
вующее письмо по E-mail, где укажите URL этих файлов. Используя броузер или
программу работы с FTP-серверами, ваш адресат без труда перепишет файлы на
свой компьютер. О том, как завести себе место на FTP-сервере своего провайде-
ра, вашей фирмы или университета, посоветуйтесь с персоналом соответствующей
организации.
Ответы на пришедшие письма и
их пересылка другим адресатам
Как и многие другие почтовые программы, Outlook Express позволяет ответить
на письмо, автоматически совершив такие подготовительные действия как откры-
тие нового окна, ввод адрес абонента в поле Кому: (То:) , и размещение в от-
ветном послании первоначального текста для цитирования.
Рассмотрим последовательность действий по ответу на пришедшее письмо:
1. Выберите (пометьте) письмо, на которое хотите ответить, и нажмите на
кнопку Ответить отправителю (Reply to Author) в основном окне Outlook
Express. Если это письмо уже было открыто в отдельном окне, то там также при-
сутствует кнопка Ответить отправителю. Нажатие на данную кнопку вызовет появ-
ление нового окна для ответного сообщения с заполненным полем Кому: (То:) и
текстом первоначального письма. В поле Тема: (Subject:) будет приведена тема
первоначального письма с пометкой "Re:" в начале строки. По этой пометке ваш
адресат поймет, что вы прислали ему ответ на письмо по конкретной теме.
2. Если вы пишете друзьям и хотите, чтобы ваше письмо выглядело «в традици-
ях Интернета», то отредактируйте текст первоначального сообщения, оставив
только те строки, которые важны для понимания вашего ответа (цитаты). После
обычных приветствий и вступительных слов напечатайте под каждой такой цитатой
ваш ответ на данное предложение или вопрос.
3. Если же это ответ на деловое письмо, то оставьте весь прежний текст без
изменений, а сверху напишите свой ответ.
4. Завершите письмо как обычно и нажмите на кнопку отправки письма (самая
левая кнопка на панели инструментов с изображением летящего конверта).
Если ваш адресат разослал такие же письма, как и вам, другим людям, и вы
хотите всех познакомить со своим ответом (разослать ваш ответ всем), то вме-
сто кнопки Ответить отправителю (Reply to Author) выберите на панели инстру-
ментов Outlook Express кнопку Ответить всем (Reply to All). В остальном про-
цесс ответа совершенно аналогичен.
Когда же вы считаете, что пришедшее письмо было бы интересно вашему коллеге
(например, по фирме), то вы можете переадресовать его соответствующему лицу,
нажав на кнопку Переслать (Forward). В этом случае в поле Кому: (То:) следует
ввести адрес нового получателя. Вы можете ввести адрес как вручную, так и ис-
пользовать адресную книгу (если нужный адрес внесен в книгу). Обратите внима-
ние на то, что тема первоначального письма будет приведена с пометкой "Fw:" в
начале строки. По ней ваш коллега узнает, что это письмо пришло не напрямую,
а было переадресовано от вас.
Перед пересылкой вы можете отредактировать текст первоначального письма и
добавить вначале письма свои комментарии, если считаете это необходимым. Сто-
ит заметить, что в электронной переписке следует придерживаться таких же пра-
вил приличия, как и в обычной жизни. Поэтому не принято переадресовывать
письма, которые содержат информацию, адресованную лично вам. В таких случаях
лучше пересказать текст другому лицу "своими словами".
Адресная книга Outlook Express
Адресная книга — это сборник адресов e-mail ваших коллег по электронной пе-
реписке, организованный с помощью удобной программной оболочки. Адресная кни-
га в программе Outlook Express представляет собой упрощенную копию адресной
книги из мощной корпоративной почтовой программы MS Exchange. Если вы исполь-
зуете MS Exchange в офисе, a Outlook Express — дома, то в справочной системе
Outlook Express есть пункты, рассказывающие о том, как экспортировать адреса
из MS Exchange в адресную книгу Outlook Express. Кроме экспорта адресов из MS
Exchange, можно также экспортировать адреса из целого ряда других почтовых
программ: MS Internet Mail, Eudora Pro, Eudora Light, а также из целого ряда
почтовых клиентов Netscape разных версий. Мы не будем останавливаться на этой
возможности, — просто имейте это в виду. Сведения о том, как произвести экс-
порт адресов из адресных книг этих приложений, вы найдете в справочной систе-
ме по Outlook Express.
Заполнять адресную книгу Outlook Express можно в двух режимах: во-первых,
потратить сначала некоторое время и заранее ввести данные о ваших коллегах, и
во-вторых, пополнять адресную книгу "на лету", по мере работы с почтой, —
просто копируя туда адреса пришедших писем. Ниже мы рассмотрим эти два слу-
чая .
1. Если вы хотите заполнить адресную книгу заранее, нажмите кнопку Адресная
книга (Address Book) на панели инструментов основного окна Outlook Express,
что вызовет появление главного окна этого сборника адресов и др. контактной
информации (см. рис.).
Кроме имени и адреса E-mail в адресной книге можно хранить множество раз-
личной информации — номера телефонов, пейджеров, личных и служебных страниц
Web, обычные почтовые адреса абонентов и пр. Для того, чтобы внести в адрес-
ную книгу новое имя, нажмите кнопку Создать адрес (New Contact). Введите в
соответствующих полях имя абонента и адрес его электронной почты, при желании
можете заполнить дополнительные данные на абонента на других закладках. Если
вы захотите отредактировать адрес электронной почты или иной параметр, то вы-
берите имя в адресной книге и нажав кнопку Свойства (Properties), измените
данные на закладках аналогичным образом.
Замечание. Если к вашему компьютеру имеют доступ посторонние люди, не стоит
помещать в адресную книгу конфиденциальную информацию, такую как домашние но-
мера телефонов и адреса ваших адресатов.
2. Вторая возможность: после того, как вы завели адресную книгу и наполнили
ее некоторыми адресами, в дальнейшем книгу можно пополнять за счет адресов из
вновь пришедших писем. Для этого откройте нужное письмо, отметьте имя адреса-
та в поле заголовка, нажмите правую клавишу мыши и из всплывающего контекст-
ного меню выберите пункт Добавить в адресную книгу (Add То Address Book).
Теперь, когда адресная книга содержит данные по адресам E-mail ваших коллег
по переписке, мы вкратце рассмотрим порядок работы с ней при формировании но-
вых писем и заполнении поля Кому: (То:):
• Нажмите кнопку Создать сообщение (New Message) в основном окне Outlook
Express, поместите курсор в поле Кому: (То:) в окне создания нового сооб-
щения .
• Щелкните по значку с изображением оторванного листка бумаги рядом со сло-
вом Кому:. Это действие вызовет появление диалогового окна (см. рис.), где
можно легко выбрать получателей данного письма.
• Выберите из списка абонентов нужного человека и нажмите кнопку Кому ->
(То->) в средней части окна.
• Аналогичным образом добавьте абонентов в полях Копия: (Сс:) или Скрытая:
(Вес:), если необходимо разослать это письмо еще нескольким адресатам.
• Нажмите кнопку ОК, и выбранные адреса появятся в соответствующих полях за-
головка нового письма.
• Теперь можно приступать к заполнению тела письма и его отправке, о чем мы
уже рассказали ранее.
Замечание. Имена абонентов, введенных в поля Кому: (То:), Копия: (Сс:),
Скрытая: (Вес:) с помощью адресной книги, могут быть представлены в окне
письма именами абонентов, а не их электронными адресами. Не беспокойтесь —
программа Outlook Express сама подставит адреса E-mail при отправке сообще-
ния .
Еще одна опция, которую вы можете использовать в адресной книге Outlook
Express — это групповые имена. Групповые имена представляют собой ваши личные
списки рассылки и удобны в том случае, если вам надо часто рассылать письма
постоянным группам людей (например, участникам какого-либо совместного с вами
проекта). Групповые имена в Outlook Express создаются нажатием на кнопку Соз-
дать группу (New Group) на панели значков адресной книги. Затем вы можете
указать имя группы и составить список людей в этой группе. После того как вы
создали группу, вы можете использовать имя группы в поле Кому: или Копия: от-
сылаемого сообщения — это сообщение будет направлено всем, кто входит в эту
группу. Чтобы в последствии отредактировать список членов группы и их адреса
e-mail, надо выбрать (отметить) уже существующую группу и щелкнуть по кнопке
Свойства.
Выражение эмоций в
электронной почте
В этом небольшом разделе мы коснемся еще одной традиции электронной почты —
выражения эмоций. Дело в том, что E-mail по сравнению с живой речью плохо пе-
редает эмоции и чувства автора сообщения по отношению к предмету письма. Для
того чтобы как-то компенсировать этот серьезный недостаток, были придуманы
специальные значки, среди обитателей Сети именуемые "смайликами".
Смайлик — это просто комбинация из нескольких текстовых символов, которые в
совокупности похожи на лицо человека — улыбающееся, подмигивающее, огорченное
ит. д. Обычно смайлик вставляют в конце той фразы, которая вызвала особые
чувства у автора. Чаще всего используются, к примеру, такие смайлики:
:-) - обсуждаемая тема рассмешила автора,
; -) - автор письма делает намек,
:- ( - автор письма расстроен.
Кроме "общепринятых", существуют различные экзотические смайлики, отражаю-
щие более сложные чувства авторов электронных писем. В Интернете вы встретите
достаточно много коллекций подобных значков (см., к примеру,
http://www.alp.ru/smile.htm или http://www.Igg.ru/~dima/smiles.htm). Только
не используйте смайлики в деловых письмах.
Шифрование сообщений
Достаточно важный элемент современного использования электронной почты,
особенно в сфере бизнеса, - это применение шифрования текста писем и защита
сообщений с помощью цифровой подписи. В случае шифрования вы будете уверены,
что никто, кроме вашего адресата, не сможет познакомиться с содержимым вашего
письма, а цифровая подпись гарантирует, что если кто-то по пути открывал файл
с вашим письмом и изменял содержимое, то это не останется незамеченным.
Как мы уже отмечали, сообщение e-mail по пути следования от вашего компью-
тера до конечного адресата проходит через несколько почтовых серверов и в
принципе, на любом из них оно может быть прочитано персоналом провайдеров,
отредактировано или даже подменено на ложное сообщение. Разумеется, через
почтовые серверы проходят в день тысячи, если не миллионы писем, и прочитать
их все просто невозможно. Вдобавок положение о провайдерах Интернета в любой
стране запрещает персоналу читать электронные письма клиентов. Поэтому, если
вы пишете самое обычное письмо своему приятелю, то можете быть на "на все
сто" уверены, что его вряд ли кто прочтет. Но... любители чужих секретов могут
в принципе поставить на поток проходящих через почтовый сервер писем некие
программы-фильтры, которые отлавливают определенные адреса e-mail или слова
или сочетания символов в тексте писем (такие как 16-значные номера кредиток)
и далее отсортированная таким образом почта может подвергнуться уже ручному
просмотру.
Поэтому шифрование сообщений и применение цифровой подписи - основная мера,
которая может на многие порядки снизить вероятность прочтения электронного
письма и его подмену на письмо с другим содержанием. Разумеется, любой шифр
можно подобрать и дело лишь за той вычислительной мощью и временем, которые
надо на это потратить. Но мы не будем углубляться в проблемы криптографии,
поскольку наше пособие ориентировано на обычных пользователей, и в рядовой
ситуации шифрование писем - лишь излишняя трата времени. Тем не менее, по-
скольку даже обычному человеку надо хоть раз в год переслать куда-либо конфи-
денциальную информацию (или просто информацию, которую он или она считают та-
ковой) , мы не можем оставить данную проблему совсем без упоминания и потому
выносим ее на самостоятельное ознакомление читателей.
Замечание. В российском Интернете не стихает дискуссия о правомерности при-
менения шифрования сообщений с точки зрения действующего отечественного зако-
нодательства. Российское представительство Microsoft отвечает на этот вопрос
просто - "Никаких комментариев по этому вопросу". Большинство же ответов в
группах новостей сводится к тому, что вроде запрещено, но де-факто шифрование
все-таки используется пользователями, раз это позволяется программами. Осо-
бенно большое количество пользователей обсуждают планируемое внедрение систе-
мы автоматической перлюстрации электронной почты СОРМ, которую российская
служба ФСБ готовится установить на все почтовые серверы провайдеров. Статьи
об этом можно встретить на многих серверах Web и в периодических изданиях -
см. к примеру, "ФСБ-Онлайнс" в сетевом журнале "Inter.net".
Итак, завершая эту спорную тему, осталось заметить, что справочная система
Outlook Express содержит достаточно подробную информацию о том, как настроить
и использовать шифрование для пересылки электронной почты в Outlook Express.
Кроме того, на многих серверах Интернета, включая Web-узел Microsoft, вы най-
дете дополнительные сведения, статьи и советы о шифровании, применении цифро-
вых подписей и сертификатов. И, конечно, очень важно отметить, что благодаря
сотрудничеству Microsoft с ведущими поставщиками криптографических решений
настройка Outlook Express на работу с защищенной почтой крайне упрощена и
доступна даже для начинающих пользователей Интернета. В частности, это со-
трудничество гарантирует, что сертификат для индивидуального использования
шифрования можно получить бесплатно, без обычной в таких случаях оплаты шиф-
ровального ключа по кредитной карте.
Бесплатная электронная почта
С легкой руки М.Тэтчер крылатая фраза о том, что "бесплатный сыр бывает
только в мышеловке", овладела массами, и потому любое предложение о бесплат-
ной услуге многие люди встречают с подозрением - нет ли тут какого подвоха? К
счастью, Интернет изначально во многом задумывался как всеобщая бесплатная
сеть и поэтому неудивительно, что в Интернете существует возможность получить
бесплатный адрес электронной почты.
Разумеется, так называемый бесплатный адрес не является абсолютно бесплат-
ным - вы все равно платите провайдеру за время доступа к Интернету, и речь
идет только о том, чтобы не взимать абонентскую плату за сохранение вашей
почты и поддержку учетной записи. Эта плата в данном случае покрывается за
счет доходов от рекламы. Сегодня в Интернете существуют более десятка хорошо
известных и надежных служб бесплатной электронной почты - Yandex
(http://www.yandex.ru) , Gtnail.com (http: //gmail. com) и т.д.
Коренным отличием этих служб от той почты, которую вы получаете через сво-
его провайдера и с помощью почтовой программы (такой как Outlook Express),
является то, что работа с бесплатным почтовым ящиком происходит с помощью
броузера. Вы обращаетесь на Web-сервер службы, вводите свой идентификатор и
пароль, после чего получаете доступ к своему почтовому ящику. Теперь можно
читать пришедшую почту и отправлять свою собственную, но это происходит через
интерфейс броузера. Просто на Web-сервере созданы странички, которые имитиру-
ют интерфейс простенькой почтовой программы. Внешне все выглядит очень похоже
на работу с обычной почтовой программой, но реально все происходящее вы види-
те в окне броузера.
Универсальными качествами такой почты, за которые она и пользуется популяр-
ностью среди людей, является ее доступность из любой точки, с любого компью-
тера, на котором есть хоть какой-либо броузер Web и достаточно большой уро-
вень анонимности. К примеру, если вы находитесь в поездке, то вы, как прави-
ло, не можете обратиться на почтовый сервер своего провайдера за почтой. Нет,
конечно можно осуществить междугородний или даже международный звонок на но-
мер доступа своего провайдера, но это дорогое мероприятие. Другой известный
способ - использовать программу Telnet и с помощью команд UNIX обратиться на
сервер провайдера и получить свою почту. Однако далеко не все умеют работать
с Telnet и знают команды UNIX.
Ну и кроме того, предоставляемые адреса бесплатной почты безлики, их вла-
дельцы могут зарегистрироваться под каким угодно именем и находиться где
угодно - ведь к своему ящику они обращаются через броузер. Вот почему бес-
платные почтовые ящики с доступом через интерфейс броузера так популярны.
Кроме того, некоторые службы, например, GeoCities, предоставляют возможность
бесплатно разместить свою домашнюю страничку, что также привлекает туда поль-
зователей .
Замечание. У бесплатных адресов электронной почты есть и своя обратная сто-
рона . Это имидж "халявы". Поэтому с таких адресов не рекомендуется посылать
деловые письма и иные предложения незнакомым людям, поскольку они вряд ли бу-
дут восприняты серьезно. Кроме того, некоторые группы телеконференций, напри-
мер FIDO, отвергают регистрацию пользователей с такими анонимными адресами.
Как получить адрес в
службе бесплатной почты
Получить адрес бесплатной электронной почты может любой пользователь Интер-
нета, причем любое число раз - ведь один и тот же человек может зарегистриро-
ваться под разными именами. Например, один раз можно зарегистрироваться под
истинным именем для переписки с друзьями, а другой под псевдонимом - на слу-
чай участия в телеконференциях, где, как известно, "народ за словом в карман
не лезет", и т.д. Выбор той или иной службы в качестве "базы" вашего будущего
адреса особого значения не имеет, т.к. уровень функциональности всех подобных
служб примерно одинаков. В принципе, открытие ящика на любой из бесплатных
почтовых служб Интернета происходит по одной и той же схеме: вначале надо за-
регистрироваться (выбрать имя и пароль, ввести те или иные данные о себе, ко-
торые не обязательно д.б. истинными), а затем вам открывают учетную запись и
можно приступать к отправке и получении почты через этот ящик.
Как результат сотрудничества службы HotMail с Microsoft3 в интерфейсе
Internet Explorer 4 есть специальные элементы для быстрого вызова сервера
Hotmail. Это кнопка Бесплатная почта HotMail (Free Hotmail) в строке ссылок и
аналогичный пункт в меню Справка (Help). Щелкнув по любому из этих пунктов,
мы запускает Internet Explorer и загружаем в него заглавную страницу сервера
HotMail. Если вдруг у вас какая-либо иная версия Internet Explorer 4.x или вы
пользуетесь Netscape Navigator, где этого пункта в меню справки или строки
ссылок нет, то для открытия адреса просто загрузите страницу
http://www.hotmail.com.
Далее надо найти кнопку для вызова начальной регистрации (в случае Hotmail
она называется Sign Up Неге!) и заполнить все поля, которые обязательны при
регистрации. В конце этого процесса остается нажать кнопку отсылки ваших лич-
ных данных (обычно такая кнопка называется Submit), и если все было заполнено
правильно, то сервер откроет вам почтовый ящик под выбранным вами именем. Это
все! С этого момента уже можно получать и отсылать почту с данного адреса.
Как правило, сразу после открытия ящика в нем вас уже ожидает стандартное
приветствие от имени персонала данной службы.
Совет. Чтобы эффективно работать с почтовым сервисом той службы, в которой
вы завели бесплатный почтовый адрес, не поленитесь почитать раздел справки и
часто задаваемых вопросов (FAQ). Там вы наверняка найдете ряд полезных сове-
тов , которые помогут вам быстрее освоиться в интерфейсе службы и научиться
эффективно использовать ее в различных режимах работы. Так, например, HotMail
позволяет переадресовывать вашу почту с почтового сервера вашего провайдера в
ящик HotMail, сохраняя оригинал письма на почтовом сервере провайдера. Это
удобно, если надо познакомиться с содержимым своей почты в тот момент, когда
вы находитесь в поездке вдали от дома. У другой почтовой службы,
AltaVista4, - есть обратная функция, когда можно переадресовывать почту с
бесплатного ящика на почтовый сервер провайдера и получать с помощью Outlook
Express или иной почтовой программы.
Замечание. Работа обычных почтовых программ гораздо более эффективна, чем
имитация почтового ящика через броузеры в бесплатных почтовых службах. Во-
первых, почтовые программы обладают множеством функций, не реализованных в
бесплатной почте, и во-вторых, они более экономно используют соединение с Ин-
тернетом. В общем случае для перерисовки каждой странички в броузере, имити-
рующей интерфейс почтовой программы, требуется заметно большее время, чем для
перекачки почты в случае нормальной почтовой программы.
Поэтому в заключение этого раздела остается порекомендовать вначале оценить
вашу реальную потребность в адресе на одной из бесплатный почтовых служб и
если он действительно необходим, то зарегистрировать его там (например, авто-
ру нравится служба бесплатной почты на сервере AltaVista). Тогда вы сможете
самостоятельно и более объективно оценить все прелести и недостатки бесплат-
ной почты Интернета. И еще, последний совет: не заводите более одного-двух
адресов бесплатной электронной почты. Вы вряд ли будете пользоваться большим
количеством адресов одновременно, а если возникнет необходимость, то "дело за
малым" - получить новый электронный адрес вы всегда успеете.
3 Корпорация Microsoft вложила некоторую сумму денег в поддержку прежде самостоятельной
службы Hotmail, поэтому как результат этого сотрудничества в броузере Internet Explorer 4
есть специальный пункт в меню Справка для быстрого доступа к серверу Hotmail, - прим, автора
4
Ее уже нет.
Советы по расшифровке
почты с поврежденной
кодировкой русского текста
В практике любого отечественного пользователя электронной почты рано или
поздно встречается случай, когда пришедшее письмо хотя и состоит из символов
кириллицы, но абсолютно не читаемо. Попытки прочесть его в иной кодировке,
например, в КОИ-8, также не приносят успеха. Это значит, что при прохождении
через многочисленные почтовые узлы письмо подверглось множественным перекоди-
рованием из одной кодовой таблицы в другую и на каком-то этапе очередное пе-
рекодирование было выполнено с ошибкой.
Для того чтобы попытаться восстановить текст письма, можно применить одну
из программ восстановления и перекодирования русскоязычных текстов. Множество
таких программ можно найти в Интернете, например, скачать с сервера
http://www.download.ru. Здесь мы посоветуем вам программу TOT-RECODE II, ра-
ботающую под управлением Windows 95-98 и Windows NT 4.0 - 5.x beta. Эту про-
грамму можно бесплатно сгрузить с сервера http://www.smartline.ru/recode/.
Благодаря тому, что программа поставляется с подробным описанием (Help) на
русском языке, мы опустим рассмотрение правил работы с ней. Хотя на самом де-
ле эти правила очень просты - надо просто скопировать текст в письма в буфер
обмена Windows, a TOT-RECODE II сделает все остальное - сама проанализирует
содержимое письма и представит десятки, а то и сотни вариантов расшифровки.
Вам останется лишь выбрать нужный и прочесть.
Работа с новостями
в Outlook Express
Поскольку Outlook Express объединяет в себе клиентскую программу электрон-
ной почты и программу работы с новостями (телеконференциями UseNet), в сле-
дующем методическом пособии вы познакомитесь с правилами участия в различных
телеконференциях UseNet. Вы научитесь сами читать опубликованные там статьи и
направлять в группы свои собственные сообщения. UseNet - это очень увлека-
тельный мир, сравнимый по популярности с World Wide Web, поэтому многие поль-
зователи проводят в дискуссиях даже больше времени, чем обозревая Web.
Важно и то, что в UseNet вы найдете группы, в которых обсуждается работа
различных программ электронной почты и работы с новостями, в т.ч. и Outlook
Express. И там, посоветовавшись с опытными пользователями, вы сможете найти
ответы на многие вопросы, которые у вас возникнут в процессе работы с в
Outlook Express. Здесь вам будут полезны такие русскоязычные телеконференции
как microsoft.public.ru.ie40, microsoft.public.ru.ie40.outlookexpress,
relcom.newusers, relcom.fido.ru.modem, relcom.www.support, relcom.www.users,
fido7.ru.internet, fido7.ru.internet.www, fido 7.ru.internet.chainik и другие.
ЛЕКЦИЯ 11
Эффективная навигация по Web
В этой лекции приведены советы по приемам эффективной навигации по узлам
Web, повышению скорости загрузки страниц и другим вопросам, способствующим
повышению эффективности и скорости работы с Internet Explorer в течение сеан-
са связи с Интернет.
Правильный выбор
начальной страницы
Любой броузер позволяет вам загрузить начальную страницу не только в начале
каждого сеанса работы с броузером, но и в любой момент в течение работы. Для
инструментов Internet Explorer для этого достаточно нажать кнопку Домой
(Ноте) на панели и вы возвратитесь к начальной странице. Таким образом, вы
каждый раз будете возвращаться на начальную страницу, которая по умолчанию
расположена на сервере Microsoft.
Однако это не совсем удобно — во-первых, вы можете решить, что там нет дос-
таточного числа ссылок на ресурсы, представляющие для вас наибольший интерес,
и во-вторых, из-за загруженности линий скорость загрузки страницы может, как
говорится, "желать лучшего". Не проблема, - довольно просто изменить адрес
начальной страницы на тот, который вам более всего подходит.
В Internet Explorer 4 достаточно выбрать пункт Свойства обозревателя
(Internet Options) меню Вид (View). Вашему взору предстанет диалоговое окно,
в котором можно ввести адрес новой начальной страницы, и кроме того, броузер
дает возможность назначить таковой ту страницу, которая загружена в окне в
данный момент времени. В верхней части вкладки расположено окно Домашняя
страница (HomePage), куда можно или вручную ввести нужный адрес, или запол-
нить это окно автоматически - нажав на одну из кнопок, выбирающей адрес теку-
щей, пустой или исходной страницы (исходная страница - страница «Добро пожа-
ловать в Интернет». Нажмите кнопку ОК, чтобы подтвердить сделанные изменения.
Новые функции кнопок Вперед и Назад
Кнопки Вперед (Forward) и Назад (Back) - наиболее часто используемый инст-
румент броузера при навигации по Web. Как известно, нажатие на эти кнопки по-
следовательно возвращает вас к предыдущему или следующему просмотренному до-
кументу (внутри списка уже просмотренных документов). Как только вы дойдете
до последнего из просмотренных ранее документов, кнопка Назад станет недос-
тупна , а если дойдете вперед по списку из просмотренных ранее документов, -
то станет недоступна кнопка Вперед.
Хотя пользоваться этими кнопками умеют все, но многие забывают, что в если
надо вернуться к документу, просмотренному несколькими страницами ранее, то в
Internet Explorer версии 4 и старше можно не нажимать по несколько раз подряд
на кнопки Назад или Вперед, а «перескочить» к нужному к документу сразу через
несколько просмотренных документов.
Для этого щелкните по маленькой кнопке-стрелке справа от основной кнопки (к
слову, в Netscape Navigator 4 это также возможно, но надо сделать правой кла-
вишей мыши). Будет показан текущий список из последних просмотренных страниц
(числом не более 9 в списке), и можно выбрать именно ту страницу, куда вы же-
лаете перейти. Для быстрого перехода на страницы, посещенные еще раньше, в
том числе в предыдущие сеансы работы с броузером - используйте кнопку Журнал
(History) на панели броузера (см. ниже).
Использование кнопки Обновить
При навигации по Web может оказаться, что какая-либо страница загрузилась
не полностью, причем особенно часто это бывает с картинками. Или вдруг ско-
рость загрузки страницы неожиданно резко упала. Такое случается сплошь и ря-
дом, поскольку загрузка каналов Интернета все время изменяется, периодически
возникают сбои передачи и потеря части переданной информации. В таких случаях
помогает кнопка перезагрузки страницы Обновить (Refresh). При нажатии на эту
кнопку текущая страницы заново передается с сервера на ваш компьютер, и часто
этого бывает достаточно для разрешения проблемы.
Кроме того, у кнопки обновления есть еще одна функция, связанная с исполь-
зованием дисковой кэш-памяти. Дело в том, что переданные из Интернета страни-
цы хранятся на жестком диске вашего компьютера в виде временных файлов. Когда
вы нажимаете кнопки Вперед или Назад, то броузер может загрузить страницу с
сервера или же восстановить из временного файла с диска. Как правило, по
умолчанию броузер грузит страницу с жесткого диска (вы можете выбрать нужный
режим с помощью параметров из окна Свойства Обозревателя, вызываемого из меню
Вид) .
Нажатие кнопки Обновить заставляет броузер загрузить страницу с сервера за-
ново, что особенно важно для тех страниц, где информация часто меняется (на-
пример, с финансовой информацией по текущим торгам, новостями и т.п.).
Прекращение загрузки
ненужной страницы
Довольно часто случается, что вы щелкнули по ссылке на какую-либо страницу,
но грузится она очень долго, и уже видно, что ее содержание вам не интересно.
Нет ничего проще, чем прервать загрузку такой страницы — нажмите кнопку Оста-
новить (Stop) на панели инструментов броузера или просто клавишу Esc на кла-
виатуре ПК. Затем можно нажать кнопку Назад (Back), чтобы вернуться к преды-
дущей странице.
Использование внутренних
средств навигации узла
Как бы не были удобны универсальные кнопки Вперед и Назад, для перемещения
между страницами конкретного узла зачастую эффективнее использовать внутрен-
ние средства навигации узла (если такие средства предусмотрены создателями
данного узла). Обычно подобные средства навигации оформлены в виде графиче-
ских ссылок с изображением кнопок со стрелками или надписями. Такая графиче-
ская панель управления обычно располагается сбоку или же в верхней или нижней
части страницы. Графические кнопки управления позволяют вернуться на Заглав-
ную страницу узла, а также перейти к тем или иным его разделам. Достаточно
часто там же есть изображения кнопок, которые дублируют кнопки Вперед и Назад
на панели инструментов броузера.
Навигация по документам,
содержащим фреймы
В Web вам часто встретятся страницы, разделенные на несколько независимых
частей - кадров (или как их чаще называют, фреймов). Обычно в каждом из таких
фреймов содержится информация, логически отделенная от информации на другом
фрейме. В зависимости от того, как задуман художественный дизайн страницы, на
фреймах могут присутствовать или отсутствовать полосы прокрутки изображения,
а также возможность передвигать границу между фреймами (примерно также, как
вы передвигаете границу между панелями в Проводнике Windows).
Не всегда дизайн фреймов удачен, из-за чего многие пользователи Интернета
недолюбливают страницы с фреймами. Кроме того, на страницах с фреймами кнопки
Вперед и Назад на панели инструментов броузера работают несколько иначе - ес-
ли содержание фреймов может изменяться (чаще всего боковые фреймы - статичны,
и изменяется содержание только в основном фрейме), то кнопки действую на те-
кущий фрейм до тех пор, пока вы не вернетесь к самой первому изображению
фрейма. Таким образом при навигации по узлу с фреймами приходится чаще обыч-
ного нажимать кнопки Назад и Вперед.
И последнее: хотя научиться работать с фреймами достаточно просто, ряд уз-
лов предоставляют возможность ознакомиться с их содержанием без использования
фреймов. Если вы предпочитаете страницы без фреймов, поищите, имеется ли на
заглавной странице узла ссылка Non-frame version (Версия без фреймов).
Навигация в режиме
полного окна
Если вы работаете с монитором, имеющим небольшой экран (14" и 15"), то при
просмотре ряда узлов со сложным дизайнерским решением вам может помочь кнопка
Во весь экран (Full Screen) на панели инструментов Internet Explorer 4. В от-
личие от стандартной кнопки Windows-приложения "Раскрыть", кнопка Во весь эк-
ран не только раскрывает окно броузера на максимальный размер окна, но одно-
временно закрывает все панели Internet Explorer, кроме панели инструментов.
Таким образом вы можете получить дополнительное пространство для просмотра,
не занимаясь вручную операциями по закрытию панелей броузера. Чтобы вернуться
из полноэкранного режима к обычному виду окна, используйте стандартную кнопку
Свернуть в правом верхнем углу полноэкранного окна.
Навигация с помощью клавиатуры
Некоторые пользователи любят применять комбинации нажатия клавиш клавиатуры
вместо того, чтобы щелкать указателем мыши по кнопкам и меню броузера. Для
них в мы приведем основные комбинации клавиш.
Таблица. 1
Навигация с помощью клавиатуры
Действие по навигации Комбинация клавиш клавиатуры
Назад Alt + Левая стрелка или Backspace
Вперед Alt + Правая стрелка
Переход между кадрами вперед CTRL+Tab
Переход между кадрами назад Shift+CTRL+Tab
Переход к следующей ссылке Tab
Возврат к предыдущей ссылке Shift + Tab
Переход по данной ссылке Enter
Прокрутка страницы вниз (плавно) Стрелка вниз
Прокрутка страницы вниз (кусками) Page Down
Прокрутка страницы вверх (плавно) Стрелка вверх
Прокрутка страницы вверх Page Up
(кусками)
Переход в конец документа End
Переход в начало документа Home
Обновить данную страницу F5
Прекращение загрузки страни- цы Esc
Открыть новое окно CTRL+N
Сохранить текущую страницу CTRL+S
Напечатать текущую страницу или кадр CTRL+P
Навигация с помощью мыши
MS IntelliMouse
Если на вашем компьютере установлена мышь Microsoft IntelliMouse, то в
Internet Explorer 4 ее можно использовать для выполнения следующих действий.
• Чтобы пролистывать Web-страницы, вращайте колесико вперед или назад. Чтобы
непрерывно пролистывать текущие страницы Web, двигайте саму мышь, удержи-
вая нажатой кнопку-колесико.
• Вместо того, чтобы нажимать кнопки Вперед и Назад, можно соответственно,
выполнить «прокрутку» колесиком вперед, а чтобы вернуться к предыдущей
странице, — «прокрутку» назад. Для этого необходимо, не отпуская клавишу
SHIFT, вращать назад кнопку-колесико.
Панель Журнала броузера
Internet Explorer 4
Забыли, какие узлы вы посещали на прошлой неделе во вторник? Помните лишь,
что встретили очень интересную страницу, но адрес забыли записать в Избран-
ное? Не беспокойтесь, в броузере Internet Explorer 4 впервые появилась очень
полезная функция - панель Журнала (History). С ее помощью вы можете легко
найти адрес любой ранее посещенной страницы Web и загрузить ее снова.
Чтобы панель Журнала появилась на экране, достаточно нажать кнопку Журнал
на панели инструментов или выбрать пункт Панель обозревателя (Explorer Ваг) в
меню Вид (View), откуда уже выбрать подпункт Журнал (History). Сразу после
этого область просмотра броузера будет разделена на две части, где на левой
стороне вы увидите даты, включая сегодняшнюю. Чтобы познакомиться со списком
адресов узлов, просмотренных в конкретный день, следует щелкнуть по нужной
вам дате, и далее - по адресу конкретной страницы, которая будет загружена в
броузер.
По умолчанию, броузер ведет учет адресов страниц, просмотренных в течение
последних 20-ти дней. Чтобы изменить этот период в сторону увеличения или
уменьшения, пункт выберите пункт Свойства обозревателя (Internet Options) ме-
ню Вид (View). На закладке Общие (General) вы можете изменить продолжитель-
ность периода ведения Журнала, а также стереть все текущие записи, для чего
можно нажать кнопку Стереть (Clear History). Затем достаточно нажать ОК, что-
бы сделанные вами изменения вступили в силу.
Ввод URL в строке адреса
Ввод адреса нужной страницы в адресной строке броузера - один из наиболее
частых приемов навигации по Web. Вам совсем не обязательно вводить наименова-
ние протокола http:// перед каждым URL страницы Web, т. к. Internet Explorer
4 (и Netscape Navigator5 4) сделают это самостоятельно.
С помощью ввода в строке адреса можно обратиться не только к узлам Web, но
и к другим ресурсам Интернета. Если сам броузер не способен выполнить затре-
бованную функцию, то будет вызвано приложение, поддерживающее нужный прото-
кол. К примеру, если в строке адреса ввести news:relcom.newusers, то будет
вызвана программа Outlook Express (или иной клиент новостей, установленный по
умолчанию на вашем компьютере), и эта уже программа загрузит указанную группу
новостей. Ниже в таблице представлены виды ресурсов, которые можно ввести в
адресной строке броузера.
Дополнительный совет: если вы посещаете чью-либо домашнюю страницу, распо-
ложенную на сервере под управлением UNIX, то адрес этой страницы может содер-
жать знак тильды (~), - не забывайте правильно вводить этот знак с клавиату-
ры.
Таблица 2.
Тип файла (ресурса) Пример URL, на который показывает ссылка
Web-страница http://www.mysite.ru/my-page.htm
Файл file://С:/picture.bmp
Файл мультимедиа http://www.mysite.ru/video.avi
Адрес электронной почты mailto:user@ company.ru
FTP-сервер ftp://ftp.mysite.ru
Gopher-сервер gopher://gopher.mysite.com
Статья UseNet news:relcom.newusers
Сеанс Telnet telnet://mysite.ru
Автозавершение адреса
В броузере Internet Explorer 4 (и Netscape Navigator 4) можно использовать
функцию авто-завершения URL для тех адресов, которые вы уже когда-то посеща-
ли. Вам достаточно ввести несколько первых символов адреса, а броузер сам до-
пишет остальное. Чтобы включить или отключить эту функциональную возможность
в броузере Internet Explorer 4, следует установить или соответственно снять
флажок Использовать автозавершение (Use AutoComplete) на закладке Дополни-
тельно (Advanced) окна Свойства обозревателя (Internet Options). Чтобы вы-
звать это окно, выберите пункт Свойства обозревателя (Internet Options) в ме-
ню Вид (View).
Несмотря на явную полезность функции автозавершения, есть случаи, когда ее
приходится отключать. В основном это случается, когда вам надо посетить много
страниц внутри одного и того же узла, используя прямой ввод URL в адресной
строке, а броузер упрямо подставляет вам совсем другие адреса страниц этого
же узла.
Автоматический подбор
имени узла
Если вы знаете лишь название организации, но не уверены в полном адресе
Web-узла, Internet Explorer 4 поможет вам найти нужный сервер. Достаточно
ввести в адресной строке броузера название нужной организации (латинскими бу-
квами) , а броузер постарается подобрать адрес, последовательно подставляя
5 Сейчас его сменил популярный броузер Firefox, да и IE4 уже давно история, но методология
та же.
www. впереди набранного имени, а в конец - имя домена .com, .edu, .gov, .mil.
Вполне возможно, что нужный узел легко отыщется таким образом.
Т.е., к примеру, таким образом вы запросто найдете сервер Аэрофлота, не
указывая его полного адреса. Можно ввести в адресной строке броузера лишь од-
но слово aeroflot, a Internet Explorer подставит нужные префикс и суффикс и
загрузит узел www.aeroflot.com.
К сожалению, Internet Explorer 4 не подставляет доменные имена, образован-
ные от названия стран, поэтому найти таким образом серверы, кончающиеся на
.ru, .su, .fi, .de. il и т.д. не удастся6.
Переключение кодировок
русского языка
"Выберите кодировку..." — эта надпись, символизирующая классический русский
камень на распутье, встретит вас практически на всех отечественных серверах
Web. Исторически у нас получилось так, что локальные кодировки, принятые на
компьютерах с разными операционными системами (Windows, UNIX, DOS, Мас), про-
должают существовать и в виде сетевых кодировок. Поэтому, если загрузив какую
либо страницу Web, вы видите набор совершенно непонятных символов кириллицы,
то это значит, что кодировка на сервере не совпадает с кодировкой, установ-
ленной в броузере. В таком случае можно выбрать кодировку на сервере с помо-
щью имеющегося там меню или сменить кодировку у броузера. Второй способ - бо-
лее универсальный, т.к. ряд серверов не предоставляет возможности сменить ко-
дировку .
Вообще говоря, за стандартную кодировку русских страниц в Интернете принята
так называемая кодировка KOI8-R (которую поддерживают компьютеры под управле-
нием UNIX) . Именно она описана в RFC-1489, официальном документе комитета
InterNIC (ftp://ds.internic.net/rfc/rfc!489.txt). Однако повсеместное внедре-
ние Windows на ПК и серверов World Wide Web под управлением операционной сис-
темы Windows NT составляет "оппозицию" этому стандарту, реализуя "явочным по-
рядком" поддержку кодировки 1251, принятой в различных версиях Windows. Кроме
того, в российской части Интернета вы встретите и такие сервера, где докумен-
ты представлены всего лишь в одной кодировке. Пример одного из узлов, где
поддерживается только кодировка Windows 1251 — это узел российского предста-
вительства Microsoft (http://www.microsoft.com/rus).
Чтобы изменить кодировку русскоязычной страницы Web в броузере Internet
Explorer 4, выберите пункт Шрифты (Fonts) в меню Вид (View) , а в Netscape
Navigator 4 - пункт Encoding в меню View. Перед вами откроется список возмож-
ных кодировок, в котором вам нужно лишь выбрать нужную. Еще один способ - на-
жать на кнопку с буквой А на панели инструментов и в открывшемся меню выбрать
нужную кодировку.
Кроме того, в броузере Internet Explorer 4 достаточно просто переключить
кодировку страницы можно и с помощью выпадающего меню. Для этого, расположив
указатель мыши на текстовой части страницы, щелкните правой клавишей мыши.
Появится выпадающее меню, где вы найдете нужный вам пункт Язык (Language), а
далее уже следует выбрать нужную вам кодировку.
Примечание. В броузере Netscape Navigator 4 отсутствует возможность по пе-
реключению кодировки страницы с помощью выпадающего меню, и там следует поль-
зоваться пунктом Encoding в меню View.
6 С тех пор появились поисковые сайты, позволяющие найти любую информацию, например, ram-
bler .ru, google.ru.
ЛЕКЦИЯ 12
Документооборот в корпоративных
интра- и экстрасетях
Интрасеть (или intranet) - это частная корпоративная сеть, использующая
программные продукты и технологии Internet, например, Web-сервер. Интрасети
могут быть изолированы от внешних пользователей Internet с помощью специаль-
ных программ, называемых брандмауэрами, или просто функционировать как авто-
номные сети, не имеющие доступа извне. Обычно компании создают интрасети для
своих сотрудников, однако полномочия на доступ к ним иногда предоставляются
деловым партнерам и другим группам пользователей.
Другим способом обеспечения совместного доступа деловых партнеров к инфор-
мации, хранящейся в интрасети, является создание экстрасети (extranet). Этим
термином обычно называют часть интрасети, предназначенную для доступа извне.
Деловые партнеры часто создают экстрасети, обеспечивающие ограниченный доступ
к отдельным частям своих интрасетей. Деловым партнерам доступны только те
части интрасети, на которые они имеют соответствующие права доступа. Для кон-
курентов же любой доступ к такой интрасети закрыт.
Создать интрасеть несложно. Например, компании достаточно организовать в
своей локальной или территориально распределенной сети Web-сервер, снабдить
пользователей Web-браузерами и при необходимости предусмотреть брандмауэр.
Как и сама сеть Internet, интрасети быстро становятся ключевым элементом
корпоративных информационных систем. Фактически, большинство проданных на се-
годня Web-серверов используются именно в интрасетях. Компании пришли к пони-
манию того, что такие "внутренние" Web-узлы являются идеальным средством рас-
пространения информации среди сотрудников. Причина проста: интрасеть обладает
всеми достоинствами Web, включая возможность публикации документов, содержа-
щих графику, звук, видео и гипертекстовые ссылки. Поскольку все документы Web
создаются в одном и том же формате (HTML), они доступны любому работающему в
сети сотруднику, у которого есть Web-браузер. Если Internet изменила способ
взаимодействия коммерческих предприятий с "внешним миром", то интрасети со-
вершенно меняют характер внутренних коммуникаций.
Интрасети являются для компаний идеальным средством предоставления информа-
ции своим сотрудникам. Независимо от конкретной платформы, оснащенная интра-
сетью компания может публиковать на своих узлах важные внутренние документы.
Такие документы обычно включают в себя внутренние правила, различные процеду-
ры, связанные с кадрами, сведения о сотрудниках, руководства, данные о про-
дукции, рекламную информацию, отчеты отраслевых аналитиков, прайс-листы и ка-
талоги. Важно то, что все эти документы легко доступны для любого работающего
в сети сотрудника, оснащенного браузером.
Сотрудники получают преимущества мгновенного доступа к самой последней ин-
формации, хранящейся в центральном архиве, а компании могут сократить или со-
всем исключить затраты на печать документов. Короче говоря, предоставляя дос-
туп к разнообразной информации в масштабе всего предприятия, интрасети позво-
ляют сотрудникам работать эффективнее, а компания начинает функционировать
более слаженно и экономично.
Важную роль в корпоративной интрасети играет электронная почта. Она обеспе-
чивает своевременное взаимодействие между сотрудниками и ускоряет деловые
процедуры. Поскольку электронная почта позволяет присоединять к своим сообще-
ниям файлы, сотрудники могут распространять средствами электронной почты лю-
бую информацию - от простых отчетов до обновлений программного обеспечения и
полноценных мультимедиа-презентаций. Наиболее совершенные системы электронной
почты способны также маршрутизировать документы между отдельными пользовате-
ляни и досками объявлений, обеспечивая коллективную работу над проектами. С
учетом всех тех затрат, которые связаны с традиционными коммуникациями, стои-
мость электронных коммуникаций несравненно ниже.
Кроме WWW и E.mail интрасети предоставляют и ряд других весьма заманчивых
возможностей. Например, вместо обычных совещаний можно проводить видеоконфе-
ренции в интрасети. При этом можно не только слышать своих партнеров, но и
видеть их, просматривать документы, вносить в них необходимые коррективы.
Весьма показательны результаты опроса 445 немецких компаний самого разного
масштаба, проведенного в начале 1996г. фирмой Computerwoche. Большинство при-
нявших участие в опросе менеджеров информационных систем считает, что внедре-
ние технологий intranet позволит повысить экономическую эффективность корпо-
ративных компьютерных сетей. Более всего в реализации intranet заинтересованы
компании, число сотрудников в которых превышает 500 человек: они на 1-2 шага
опережают остальных, хотя пятая часть компаний в этой категории высказала
твердое "нет" сетевой революции. Что касается временных рамок, то 16% опро-
шенных компаний были намерены перейти к intranet еще в 1996 году, более поло-
вины планировали внедрить их в 1997 г. Менее одной пятой опрошенных организа-
ций оставляли решение этой проблемы до 1998 г. , одна десятая - до 1999 г. И
только 2% - до следующего столетия.
Экономия и удобство
Основные технологии, на которых строится интрасеть, были специально разра-
ботаны так, чтобы обеспечить независимость от платформы и общедоступность.
Такие технологии как Всемирная "паутина" (World Wide Web) были созданы для
того, чтобы дать возможность ученым, работающим на различных платформах, об-
мениваться информацией через Internet, не тратя времени и сил на подготовку
версий одного документа для нескольких платформ. В результате любой пользова-
тель интрасети может просматривать помещенные в ней документы с помощью Web-
браузера .
Даже небольшая компания, имеющая только один офис и располагающая сетью
скромных размеров, может выиграть от создания интрасети. Выгоды от использо-
вания этой технологии увеличиваются вместе с ростом количества сотрудников,
размеров сети и числа удаленных сетевых узлов.
По мере роста организации увеличиваются и ее затраты, если информация рас-
пространяется среди сотрудников посредством таких традиционных способов как
печатные меморандумы и бюллетени. Другие методы распространения информации,
такие как электронная почта и совместный доступ к файлам, тоже могут оказать-
ся достаточно трудоемкими и потребовать больших затрат времени на управление.
Публикация документов в интрасети оказывается наиболее рентабельным и быст-
рым среди всех способов электронного взаимодействия. Каждый сотрудник, кото-
рый имеет соответствующим образом настроенную рабочую станцию и браузер Web,
может читать документы, подготовленные в определенном формате и помещенные на
сервер интрасети. Если бы те же документы нужно было представлять в виде фай-
лов в каталоге общего доступа или в виде электронных сообщений, то зачастую
пришлось бы создавать несколько копий одних и тех же документов в разных фор-
матах, чтобы сделать их доступными для всех платформ рабочих станций и прило-
жений, имеющихся в организации.
Создав интрасеть, организация может по мере необходимости изменять содержа-
ние опубликованных в ней документов с очень небольшими дополнительными из-
держками. Если в организации, не имеющей интрасети, возникает необходимость
изменить и скорректировать содержание разосланного пользователям электронного
сообщения, то приходится посылать каждому из них новое сообщение. Почтовый
ящик каждого пользователя в таком случае содержит две версии электронного со-
общения.
Интрасети имеют преимущество и перед другими способами взаимодействия. В
интрасети можно создать такие документы, которые предоставляли бы пользовате-
лям информацию в реальном времени, например, текущие биржевые сводки или са-
мые последние данные о каком-либо сегменте рынка. Имея практически мгновенный
доступ к таким сведениям, сотрудники компании могут более оперативно реагиро-
вать на изменения рынка или на действия компаний-конкурентов, а это в свою
очередь сказывается на росте доходов компании.
Еще одно достоинство интрасети состоит в том, что, публикуя в ней докумен-
ты, компания получает обратную связь с пользователями. Например, если доку-
менты распространяются в печатном виде или в форме общедоступных файлов на
сервере, то невозможно определить, читают ли сотрудники эти материалы. Если
же материалы опубликованы на сервере интрасети, то всегда можно проследить,
сколько пользователей прочитало их и даже какие именно материалы пользователи
читают наиболее часто.
Пользователи находят все новые способы снижения затрат с помощью интрасе-
тей, особенно после того, как появились спецификации, поддерживающие видео-,
аудио- и графические форматы документов в Web. Например, многие компании ус-
тановили в своих интрасетях приложения, позволяющие сотрудникам получать дос-
туп к базам данных компании через браузер Web. Таким компаниям не приходится
создавать или приобретать специализированные программы для доступа к базам
данных, поскольку вся необходимая информация оказывается доступной через ин-
трасеть. Такие продукты как GroupWise Web Access даже дают возможность поль-
зователям читать электронные сообщения и работать с расписанием с помощью
браузера Web. Возможности снижения долгосрочных затрат и повышение удобства
работы ограничиваются только вашим воображением и размерами текущего бюджета
вашей организации.
Дополнительные выгоды от совместимости с Internet. Без особых затрат орга-
низация может соединить свою интрасеть с Internet. Тогда удаленные пользова-
тели (например, находящиеся в командировке сотрудники или клиенты компании)
получат возможность доступа к ее интрасети через Internet. Организация может
управлять доступом пользователей к документам, разрешая всем желающим про-
сматривать одну часть документов, а другую их часть, делая доступной только
для определенного круга пользователей. Можно также разрешить пользователям
интрасети выход в Internet, открыв им доступ к богатейшему фонду информации,
содержащему сведения практически на любую тему.
Документооборот в интрасети
Слежение за документами на предприятии может оказаться непосильной задачей.
Дело просто в том, что бумаг слишком много, а их содержание слишком разнооб-
разно. С чем только ни приходится иметь дело - от рекомендаций по методам ра-
боты со счетами клиентов до памятных записок, содержащих описание действий,
которые компания должна предпринять, чтобы удовлетворить современным требова-
ниям к пресс-релизам, сообщающим краткую сводку данных из квартального отчета
компании.
Как правило, для решения этих проблем компании используют системы управле-
ния документооборотом. Эти системы обеспечивают закрытый интерфейс клиентских
станций (proprietary front end) для доступа к центральному серверу; кроме то-
го, в системах имеются средства контекстного поиска, благодаря которым поль-
зователь может разыскивать документы по определенной теме, а также программы
преобразования файлов. Пользователи могут импортировать файлы из систем
управления документооборотом в программы обработки электронных таблиц и под-
готовки текстов.
Конечно, системы подготовки документов - это мощное средство работы. Однако
возможности таких систем возрастут многократно, если их функции соединить с
открытостью и простотой intranet. Комбинация этих двух технологий обеспечит
пользователям простой и быстрый доступ к большим объемам информации.
С появлением intranet, изготовители систем управления документооборотом
стали снабжать серверные части своих приложений интерфейсами с Web-
броузерами. Ожидается, что рост популярности intranet поможет расширить число
пользователей систем управления документооборотом.
Использование Web служит наглядной иллюстрацией возможных выгод компании от
обеспечения легкого доступа к информационным страницам. Все большее число
компаний начинают осознавать привлекательность такого пути к усовершенствова-
нию систем управления документооборотом.
Руководители информационных отделов предприятий обычно приветствуют переход
к использованию Web-технологий, поскольку это избавляет их от необходимости
поддерживать нестандартные интерфейсы систем управления документооборотом или
настраивать нестандартные средства взаимодействия таких систем на работу с
операционными системами, установленными на пользовательских рабочих станциях.
Вместо этого достаточно будет обеспечивать поддержку того же универсального
программного обеспечения, которое они и раньше рекомендовали пользователям
для работы с intranet.
Системы управления документооборотом с выходом на Web имеют еще одно важное
достоинство: они облегчают размещение документов на intranet, поскольку мно-
гие из них обеспечивают HTML-индексацию документов "на лету".
Еще одно преимущество состоит в том, что системы управления документооборо-
том работают точно так же, как Web-серверы, с которыми пользователи хорошо
знакомы и работа с которыми не доставляет им ни малейших затруднений. Пользо-
ватели уже научились разыскивать необходимые им документы на Web, поэтому им
не составит большого труда осознать, какие возможности предоставляют им новые
системы. Системы управления документооборотом, при этом, обеспечивают более
развитые средства поиска, чем те, что можно найти на обычных Web-серверах.
Как правило, стандартные средства поиска Web допускают поиск по одному или
двум словам, в то время как системы управления документооборотом позволяют
искать документы, вводя три или четыре ключевых слова.
Системы управления документооборотом имеют развитые функции защиты и позво-
ляют привилегированным пользователям редактировать документы и перемещать их
из одного подразделения в другое.
Многоуровневый характер
Intranet
Одним из важнейших достоинств intranet с точки зрения принятия решений о
целесообразности внедрения этих технологий на предприятии является многоуров-
невый характер их функциональных возможностей. К одному концу спектра функ-
циональных возможностей интрасети относятся статические Web-страницы, которые
заменяют корпоративные печатные документы или обеспечивают новый способ со-
вместного использования информации; на другом находятся сложные клиентские
интерфейсы для офисных серверных приложений. Естественно, что более высокий
уровень функций стоит дороже. Определение уровня возможностей своей интрасе-
ти, составляющего часть этого спектра, обеспечивает основу планирования, ко-
торое может использоваться компаниями для руководства принятием бизнес реше-
ний и осуществлением инвестиций в технологии.
Способность фирмы повысить уровень функциональности своей интрасети обу-
славливается рядом факторов, связанных с возможностью создания соответствую-
щей сетевой инфраструктуры. Сеть на базе TCP/IP с доступом к Internet, защи-
щенным с помощью брандмауэра, - это необходимый стартовый минимум. Наличие
соединений между ПК, рабочими станциями, серверами и мэйнфреймами создаст
массу благоприятных возможностей для интеграции знаний и опыта, накопленных в
компании. Наличие специалистов по технологиям клиент-сервер и Internet суще-
ственно влияет на качество и время выполнения работы.
Уровень 1. Имея базовую сетевую инфраструктуру, организации могут сделать
следующий шаг для построения интрасети. Буквально в течение одного-двух дней
можно установить недорогие браузеры и Web-серверы с базовым информационным
наполнением. Это самый распространенный способ создания интрасети "с нуля".
Организационное значение исходной конфигурации интрасети непосредственно
связано с тем, насколько полезно ее информационное наполнение. Значимость
этого наполнения способствует тому, что различные подразделения компании при-
соединяются к системе и добавляют информацию, и, следовательно, значимость
интрасети экспоненциально увеличивается. Когда сеть начинает широко использо-
ваться и появляется возможность доступа к "критической" массе данных, внима-
ние администратора переключается на структуру и организацию информации.
На этом этапе мы имеем интрасеть уровня 1, обеспечивающую статический дос-
туп к статическим данным. Термин "статический доступ" указывает на способ по-
иска информации пользователями. В рассматриваемом случае он осуществляется с
помощью имеющихся связей или известных адресов. Данные являются статическими:
они однажды созданы и могут быть изменены только владельцем информации.
Сети уровня 1 характерны для начального этапа совместного использования
корпоративной информации. Являясь относительно простым, этот уровень, тем не
менее, обеспечивает высокую отдачу от инвестиций. Компания Harris вычислила,
что одно приложение, предоставляющее в интерактивном режиме справочник корпо-
ративных методов и процедур, дает ежегодную экономию в 50 тыс. дол.
Уровень 2. Со временем, когда пользователи обнаруживают, что найти нужную
информацию становится все труднее и труднее, самая сильная сторона интрасети
уровня 1 становится ее самой большой слабостью. Богатую сложную информацию
трудно охватить с помощью структуры на основе ссылок. Значительный объем этой
информации приводит к тому, что поиск нужных данных начинает требовать серь-
езных затрат времени. Поэтому на следующем этапе компания вводит поисковые
средства.
Механизмы поиска информации и путеводители по Web упрощают пользователям
нахождение документов, однако приводят к усложнению системы, поскольку нужда-
ются в управлении индексами поиска. Кроме того, они заставляют администрато-
ров с повышенным вниманием относиться к системной и сетевой производительно-
сти .
Итак, компания добралась до интрасети уровня 2. Инвестиции растут, посколь-
ку многие из необходимых средств стоят намного дороже тех, которые требова-
лись для интрасети уровня 1. Но при этом пользователи получают динамический
доступ к статическим данным. Система помогает пользователю находить нужную
информацию, но за поддержку информации по-прежнему должны отвечать люди. В
общем случае интрасеть уровня 2 способствует более активному использованию
информационной базы корпорации.
Уровень 3. Когда компания реализует интерфейсы между системой и существую-
щими базами данных и приложениями, происходит переход к интрасети уровня 3.
Это позволяет связать с интрасетью финансовые, кадровые, инженерные, коммер-
ческие и другие приложения и базы данных, чтобы обеспечить к ним более широ-
кий доступ с помощью простого в использовании клиентского интерфейса.
Нередко такую систему считают интрасетью с полным набором услуг. Немногие
инструментальные средства способны помочь в разработке подобной среды. Для
того чтобы обеспечить интерфейс с внешними ресурсами, требуется специализиро-
ванный программный код. Во многих случаях необходим дополнительный код для
интеграции информации из множества источников в единое представление.
Пользователи получают средства доступа нового поколения, которые помогают
управлять все более усложняющейся информационной средой. Эти средства базиру-
ются на "выталкивающей" (push) модели предоставления информации. Вместо осу-
ществления запросов и поиска (по системе "вытягивания" информации), пользова-
тели создают свои профили по интересам, на которые ориентируется система,
принимая решение, какую информацию выдать ("вытолкнуть") данному пользовате-
лю. Пользователь получает сообщение, когда интересующие его данные поступают
в интрасеть или когда происходят определенные изменения элемента базы данных.
Теперь интрасеть обеспечивает динамический доступ к динамическим данным.
Интрасеть уровня 3 позволяет интегрировать все информационное богатство
корпорации. На этом уровне корпорация может строить приложения, обеспечиваю-
щие интеграцию данных из множества источников.
Уровень 4. Это последний этап метаморфоз интрасети. Имея доступ ко всей
корпоративной информации, организации могут настраивать его в соответствии с
задачами бизнеса, потребностями своих клиентов или отдельных служащих.
Сложность и богатое информационное наполнение интрасети сделает возможной
разработку нового поколения приложений. Данные о продажах будут взаимосвязаны
с процессами отслеживания заказов, составления счетов и оплаты. В случае за-
держки поставок торговые агенты автоматически получат сообщения, которые по-
зволят им оповестить заказчиков.
Организации будут разрабатывать приложения для настройки информации на спе-
цифические нужды пользователей. В качестве примера можно привести комплексную
систему управления, которая обеспечит автоматическую реализацию продаж при
выполнении определенных условий и составит расписание, автоматически устанав-
ливающее связь с нужной информацией о служащем или клиенте, либо сведениями
по конкретному вопросу для проведения деловой встречи или какого-нибудь меро-
приятия .
Интрасети будут вовлечены в процесс обслуживания заказчиков, интегрируя
внутренние системы с продажами на базе Web, маркетинговыми и сервисными при-
ложениями . Будет обеспечен персонифицированный доступ к персонифицированным
данным внутри и - при соответствующих условиях - вне корпорации. Интрасети
уровня 4 - это архитектура корпоративной информации будущего, обеспечивающая
базу для разработки и развертывания приложений.
Электроника
ТЕРМОМЕТР ЦИФРОВОЙ
А. Шамов, Г. Шик
Цифровой термометр предназначен для измерения температуры в диапазоне от О
до 99,9 °C. От известных конструкций, его отличает довольно широкий диапазон
измеряемых температур, простота конструкции и налаживания. Недостатком термо-
метра является невозможность измерения отрицательных температур. Термометром
можно быстро и точно измерить температуру тела человека, температуру раство-
ров, воды, воздуха, фоторастворов.
Предлагаемый цифровой термометр имеет следующие технические характеристики:
диапазон измеряемых температур 0...99.9 °C, разрешающая способность 0,1 °C;
точность измерения: в диапазоне 1О...9О°С — 0,1 °C; в диапазоне О...1О°С —
0,5 °C; в диапазоне 90...99,9 °C — 0,3 °C. Время измерения температуры 1 с;
время индикации температуры 3 с. Потребляемая мощность 1 Вт. Габариты
136X100X50 мм, масса 0,3 кг.
Функциональная схема термометра показана на рис. 1. Прибор состоит из пяти
основных блоков: преобразователя температура — частота (блок 1), генератора
прямоугольных импульсов (блок 2), счетчика импульсов с дешифратором (блок 3),
блока питания (4) и индикатора (блок 5).
Блок 1 преобразует прямое падение напряжения на датчике (диоде) в частоту.
Импульсы с выхода преобразователя-интегратора заполняют прямоугольные импуль-
сы, идущие с генератора, и далее поступают на счетчик — блок 3, который пре-
образует эти пакеты импульсов в код управления семисегментными индикаторами.
Во время счета импульсов индикаторы не горят — они заперты сигналом, приходя-
щим с генератора, который также вырабатывает сигнал сброса показаний в конце
цикла индикации. Блок питания 4 вырабатывает все необходимые напряжения для
питания блоков термометра.
Рис. 1. Функциональная схема термометра
Принципиальная схема термометра изображена на рис. 2. За основу устройства
взят преобразователь температура — частота в электронном термометре с непо-
средственным отсчетом. Температурная зависимость падения напряжения на р-п
переходе при фиксированном токе через него и малая нелинейность харак-
теристики температура — напряжение позволяют применять полупроводниковые дио-
ды в качестве датчиков температуры. С такими датчиками можно изготавливать
электронные термометры, не вводя в приборы специальные линеаризующие устрой-
ства . В преобразователе используется датчик — диод VD5, падение напряжения на
котором необходимо для работы интегратора. Интегратор собран на операционном
усилителе DA2 К574УД1Б, имеющем большую скорость нарастания выходного напря-
жения, чем обеспечивается высокая скорость отслеживания и достигается точ-
ность преобразования, равная 0,1 °C: Когда интегрирующий конденсатор СЗ заря-
жается до напряжения — 10 В, интегратор сбрасывается однопереходным транзи-
стором VT2. Опорное напряжение, задающее порог отпирания однопереходного
транзистора и стабилизирующее ток через датчик VD5, обеспечивается термоста-
билизированным стабилизатором VD3, VD4. Выходное напряжение интегратора через
дифференцирующую цепочку C4R16 поступает на токовый ключ — транзистор VT3,
формирующий пакеты импульсов. На базу VT3 приходят сигналы преобразователя и
генератора прямоугольных импульсов. Генератор собран на операционном усилите-
ле DA1 КН0УД8Б, обеспечивающем выходное напряжение прямоугольной формы с пе-
риодом 4 с. Скважность импульсов устанавливается резистором R2 так, что отно-
шение длительности импульса к паузе равно 1:3. За время длительности импуль-
са, равное 1 с, на вход счетчика поступают импульсы, количество которых про-
порционально измеряемой температуре; за время паузы, равное 3 с, эта информа-
ция высвечивается индикатором. Во время счета индикаторы заперты напряжением
— 15 В, приходящим с генератора. После подсчета количества импульсов, пропор-
ционального измеряемой температуре, ключ VT3 закрывается, лампы HL1 — HL3 в
течение 3 с высвечивают информацию, хранящуюся в счетчиках DD1 — DD3. В конце
периода индикации транзистор VT1 и дифференцирующая цепочка C2R9 формируют
импульс сброса показаний счетчиков. Для улучшения стабильности работы генера-
тора в качестве конденсатора С1 применяется конденсатор К73П-3 с малыми тока-
ми утечки и хорошей термостабильностью.
Блок питания (рис. 3) собран по распространенной схеме. Опорные напряжения
формируются стабилитронами VD2 — VD6. Сердечник трансформатора питания имеет
сечение 2,5 см2. Его первичная обмотка намотана проводом ПЭВ 0,1 и содержит
5000 витков. Вторичные обмотки II и III намотаны проводом ПЭВ 0,14 и содержат
2X400 витков; обмотка IV — 20 витков провода ПЭВ 0,31.
---13----
№ Д220
V92 Д229
--------
3Af
КМйУД85
a
52 0,921#K
№ Zk
VT2
КППГ
МД-ВОЗ НП6ИЕА
HLH1L3 ИЗ-ЗА
VT5
№3156
№315
39 IK
КЗ ЮЗн 35
RA (99k ?Sk
-tie
-L Wi КС A
JAS
VB3
Ш56А
Ki
2M
KH
Kt2
820
RtA 22 *
ifi
Ktf Юн
VU5 Д9Д 37
датчик t° I
Zk
R1 2 ft
m
crz
A
SC
0922
CT2
SC
083.2
95
s
6
г
tf
S A
Д
Д
£
i3
Wf.2
A
R
”0,853
W
Рис. 2. Принципиальная схема термометра
Я7
A
Q
ё
c
6
e
/
9-
a
3
V
a
e
/
till
18________4
8 ...... 2
7b T
f! IS
fits
[£_____£
9 2
a 1
9 №
H
Рис. 3. Принципиальная схема блока питания
Рис. 4. Принципиальная схема кварцевого генератора
Для увеличения точности измерения во всем диапазоне О...99,9 °C можно ис-
пользовать кварцевый генератор секундных импульсов, схема которого показана
на рис. 4. Задающий генератор собран на микросхеме DDI в одном корпусе с дву-
мя делителями частоты. Коэффициент деления первого делителя равен 29, а вто-
рого 26. Генератор с кварцевым резонатором Z1 формирует последовательность
импульсов частотой 215 Гц (32 768 Гц) . Эти импульсы подаются на 15-раЗрядный
делитель частоты. На выходе 5 микросхемы DD1 частота генератора понижается до
1 Гц. Для получения прямоугольных импульсов со скважностью 2 и периодом 2 с
применен делитель частоты на D-триггере (микросхеме DD2) . С выхода 1 этой
микросхемы снимается сигнал частотой 0,5 Гц. Этот сигнал подается на сетки
ламп HL1 — HL3 и резистор R5, сопротивление которого необходимо уменьшить до
10 кОм. Генератор, собранный по приведенной схеме, имеет хорошую временную и
температурную стабильность. В случае использования кварцевого генератора сле-
дует переделать печатную плату с учетом изменения схемы (удаляются детали
DAI, VD1 — VD2, R1 — R4, С1). Использование кварцевого генератора и термоком-
пенсированного конденсатора СЗ в преобразователе температура — частота позво-
ляет снизить погрешность измерения в диапазоне О...99,9°С до 0,1 °C и менее.
Время индикации показаний в этом варианте составляет 1 с.
/00 £
о4
PZ
о
О/
7о
go о
go о
о
7°
8? __ g
о pg в
6 о/7 О’
oj
о4
Ж?
о°2
О/
OJ
с 4.
О о/
II#
О
о
о
о
VT/ ?£-
б Q-CI
Л/
да
«О VM
О’ о
R2
Т-/4Я
"0,85$ з
7
6
о
9
о Ж
6
о
6
о
о
а
5
Рис. 5. Печатная плата генератора, счетчика и индикатора
Конструкция и детали. В термометре применены постоянные резисторы МЛТ
0,125, подстроечные резисторы R13, R14 — СП5-3 проволочные, многооборотные.
Применение однооборотных резисторов нежелательно, так как пороги срабатывания
интегратора должны быть выставлены очень точно. Резистор R15 — СПЗ-1Б или
СПЗ-22. Конденсатор СЗ — К10-23 или КМ4, КМ5. Его лучше составить из несколь-
ких конденсаторов, имеющих ТКЕ разных знаков, так, чтобы суммарный ТКЕ был
близок к нулю. Эти меры необходимы для обеспечения максимальной точности из-
мерения температуры. Для этой же цели в преобразователе используется ОУ
К574УД1Б. Если достаточна точность измерения не более 0,3...0,5°С, можно ис-
пользовать ОУ КН0УД8Б. Конденсатор С1 в генераторе может быть заменен другим,
имеющим изоляцию из фторопласта или тефлона, соответствующей емкости и габа-
ритов. Транзисторы блока питания VT1, VT2 могут быть КТ502, КТ503; КТ201,
КТ203. Счетчик может быть построен на И С серии К155, но тогда возрастет по-
требляемая мощность, потребуется внести изменения в блок питания и блок инди-
кации прибора. Датчик прибора — германиевый точечный диод Д9. Его выводы со-
гнуты в одну сторону, припаяны к кабелю с фторопластовой изоляцией, на по-
ловину корпуса надета трубка из полихлорвинила. Когда датчик опускается в то-
копроводящую среду, нужно следить, чтобы он не погружался более чем на поло-
вину длины корпуса. Для работы в агрессивных средах, с кислотами и щелочами,
датчик следует защитить эпоксидной смолой, обеспечивающей его изоляцию и хо-
рошую теплопроводность. Если возникает необходимость использования нескольких
датчиков, расположенных в разных местах при точности измерения не более
0,3... 0.5 °C, можно использовать датчики КД518А, предварительно отобрав их
по одинаковому падению напряжения при токе через диод 1 мА, также потребуется
установить переключатель П2К на необходимое количество датчиков. Для измере-
ния температуры фоторастворов на корпусе датчика можно закрепить кусочек
пробки или пенопласта так, чтобы подводящие концы датчика были изолированы, а
корпус касался измеряемой среды и плавал на ее поверхности.
Весь термометр собран на трех печатных платах из фольгированного стеклотек-
столита толщиной 1,5 мм. На одной из них, с габаритами 130X40 мм, из двусто-
роннего стеклотекстолита собран генератор прямоугольных импульсов со счетчи-
ком и индикаторами (рис. 5). На второй, с габаритами 80x40 мм, собран преоб-
разователь температура — частота (рис. 6) и на третьей, с габаритами 130X40
мм, собран блок питания, включая и трансформатор (рис. 7) . Платы с помощью
уголков крепятся к основанию из гетинакса толщиной 3 и размером 130X1 Х90 мм.
Все три платы размещены в корпусе размером 135X100X50 мм, спаянном из фольги-
рованного стеклотекстолита толщиной 2 мм. Окно для считывания показаний на
лицевой стороне корпуса термометра закрыто оргстеклом сине-зеленого цвета.
Кабель датчика наматывается на выступы на задней стенке термометра. Там же
выводится и кабель питания прибора. Для калибровки термометра использовались
цифровой частотомер 43-32 и цифровой промышленный термометр. При использова-
нии простых термометров и частотомеров точность настройки может достигать
0,3...0,5 °C.
Для калибровки преобразователя от базы транзистора VT3 отсоединяют генера-
тор и к выходу преобразователя (коллектор VT3) присоединяют частотомер. Пред-
варительно резистором R15 устанавливают ток через датчик VD5, равный 1,0 мА.
Затем датчик помещают в среду, имеющую температуру 100°С (кипящая вода), од-
новременно контролируя температуру термометром. Резистором R14 устанавливают
выходную частоту 1000 Гц. Затем датчик охлаждают до 0°С (тающий снег) и рези-
стором R13 срывают колебания интегратора — частота 0 Гц. Эти операции повто-
ряют 3 — 4 раза для устранения взаимного влияния резисторов R13 и R14. Затем
присоединяют генератор к базе транзистора VT3 и резистором R2 устанавливают
показания счетчика при температуре 99,9 °C, равным 99,9. После этого проверя-
ют линейность устройства во всем диапазоне. При необходимости настройку по-
вторяют .
80
80
RfZ
WRU
t к
о
90
---tVU
RIB
3® / o?
RIB 9
-----£4-
Ю4
Й-----
Рис. 6. Печатная плата преобразователя температура — частота
Рис. 7. Печатная плата блока питания
KOOS +&8
VT? &
? Oj
о
74
о ш
Ж
*7 № oj
№
универсальный регулятор мощности
В. Гребенщиков, В. Амелин
Предлагаемый вниманию радиолюбителей универсальный регулятор мощности (УРМ)
предназначен для плавной регулировки мощности в нагрузке до трех киловатт. С
его помощью можно поддерживать постоянными температуру, освещенность и другие
параметры.
Возможность регулирования больших мощностей весьма актуальна при конструи-
ровании ЦМУ или создании мощных термостабилизаторов.
\5,6К
R8 /я
'И VS ктшь
ВМП26 „
J пос®,
Ri 56к Р-1
М /5л П
R3 ЗЗК U
*+2208
±108
1/7
М26
J_ С /
+Г"5,0*
*50В
V3
'I WU26
R6 , ,
;r7x| Я7/я
V/0
Т25
R
/60
=г 02
Rit
Зк
R73
5/0
Д8&Г
£4
200,0 х
*25 В =£
т°пос1п.
V72
v/4
RI7 560
\НЗ
КТ80/А
и
КТЗ/5Г
V/5-V78
J7226
+220 В
V/&-V22 Д2^5А
+200 Т/
88&А
Рис. 1. Принципиальная электрическая схема универсального регу-
лятора мощности
Универсальный регулятор мощности состоит из генератора пилообразного напря-
жения, устройства управления тиристором, дифференциального каскада для под-
держания постоянной температуры (или освещенности) и блока питания. Принципи-
альная схема УРМ представлена на рис. 1.
Генератор пилообразного напряжения собран на транзисторе УЗ с RC цепочкой
(R3R5C1) и стабилитронах VI и V2. Принцип его работы следующий: конденсатор
С1 периодически заряжается через резисторы R1 и R5, а затем быстро разряжает-
ся через транзистор УЗ в то время, когда он открыт. Постоянная времени цепи
R3R5C1 выбрана такой, что За время одной полуволны выпрямленного напряжения
конденсатор С1 успевает зарядиться лишь до напряжения +15 В. На конденсаторе
получается напряжение, близкое к пилообразному и синхронированное с напряже-
нием сети. Синхронизация обеспечивается отрицательными импульсами, снимаемыми
с делителя напряжения R1R2 и стабилитронов VI, V2, открывающих транзистор V3
в конце каждого полупериода пульсирующего выпрямленного напряжения. На рис. 2
приведены временные диаграммы, поясняющие работу генератора пилообразного на-
пряжения и схемы управления тиристором. Стабилитроны V4, V5 предохраняют по-
следующие каскады от перегрузок по напряжению. Транзистор V6, включенный по
схеме эмиттерного повторителя, служит для согласования.
Устройство управления тиристором состоит из дифференциального каскада, соб-
ранного на транзисторах V7 и V8, и усилителя тока, выполненного на транзисто-
ре V9. Это устройство вырабатывает прямоугольные импульсы, длительность кото-
рых меняется в зависимости от напряжения на базе транзистора V8 и пилообраз-
ного напряжения, поступающего на базу транзистора V7. В течение каждой полу-
волны выпрямленного напряжения тиристор включается на большее или меньшее
время, благодаря чему и достигается плавная регулировка мощности.
Для поддержания постоянной температуры или освещенности используется диффе-
ренциальный каскад, собранный на транзисторах VII и V12, в одно плечо кото-
рого включен терморезистор R10, а в другое — переменный резистор R15, задаю-
щий нужную температуру. С повышением температуры уменьшается сопротивление
терморезистора и соответственно напряжение на базе транзистора VII. Это на-
пряжение сравнивается с напряжением, снимаемым с потенциометра R15. Разность
напряжений усиливается дифференциальным усилителем и подается на базу транзи-
стора V8, что приводит к уменьшению выделяемой на нагрузке мощности. С пони-
жением температуры на базу V8 подается меньшее напряжение и соответственно
мощность в нагрузке возрастает.
Рис. 2. Временные диаграммы напряжений:
а — генератора пилообразного напряжения; б — устройства управле-
ния тиристором при верхнем положении движка резистора R5; в —
устройства управления тиристором при нижнем положении движка ре-
зистора R5
Если необходимо поддерживать постоянную освещенность, вместо резистора R10
устанавливается фоторезистор или фотодиод, включенный в обратном направлении.
В этом случае конденсатор С2 следует исключить, а номинал резистора R15 по-
добрать экспериментально.
Рис. 3. Активный фильтр и детектор канала ЦМУ
При использовании УРМ с ЦМУ можно применять различные схемы фильтров и де-
текторов . Необходимо только учесть, что для нормальной работы детектированный
сигнал должен иметь постоянную составляющую напряжения +8 В. В качестве при-
мера предлагается активный фильтр на трех транзисторах с Т-образным мостом и
детектором для одного из каналов ЦМУ (рис. 3). Генератор пилообразного напря-
жения и блок питания будут общими для всего устройства, а каскад на транзи-
сторах Vll, V12, включая резистор R16, следует исключить. Выход детектора ка-
ждого канала необходимо соединить с базой транзистора V8 УРМ соответствующего
каскада управления тиристоров, а базы транзисторов V7 УРМ соединить вместе.
Настройку УРМ необходимо начать с тщательной проверки монтажа и только по-
сле этого подать питание. Блок питания должен вырабатывать напряжения, ука-
занные в схеме, с точностью ±20%. Далее следует установить переключатель S1 в
положение «Плавно», в качестве нагрузки включить лампу накаливания HI на 220
В, движок потенциометра R5 установить в нижнее по схеме положение. Передвиже-
нием движка подстроечного резистора R16 добиться такого положения, при ко-
тором лампа находится на пороге загорания, но не светится. При этом на базу
транзистора V8 должно быть подано напряжение около +8 В. Затем, передвигая
движок потенциометра R5, наблюдать за изменением яркости лампы: в нижнем по-
ложении движка лампа гореть не должна, а в верхнем — должна светиться полным
накалом. Если плавно регулировать яркость не удается, необходимо подобрать
сопротивление резистора R9, однако делать его менее 100 Ом не рекомендуется.
После этого можно приступать к проверке работы УРМ в режиме термостабилизации
и градуировке шкалы уставок температуры. Для этого в сосуд с водой следует
поместить какой-либо нагреватель, например кипятильник, термометр и терморе-
зистор. Выводы терморезистора не должны вступать в контакт с водой, места их
соединения с проводами можно залить эпоксидной смолой. Затем установить тумб-
лер S1 в положение «Т° пост.», а движок резистора R15 — в верхнее по схеме
положение, и опускать его до тех пор, пока не загорится лампа. При этом надо
помешивать воду и следить за показаниями термометра. По истечении некоторого
времени температура воды должна установиться и более не подниматься. На шкале
резистора R15 нанести риску, соответствующую полученной температуре, переме-
щая движок резистора R15 далее, нанести метки, образующие шкалу уставок тем-
пературы .
Для проверки работы активного фильтра ЦМУ (см. рис. 3) по постоянному току
необходимо замерить напряжение на базе транзистора VI, коллекторе V2 и эмит-
тере V3. Все они должны быть равными приблизительно половине напряжения ис-
точника питания (около + 5 В). Емкости конденсатора С для фильтра рас-
считываются исходя из формулы
£ „0Д6
I Ре*—
Так, для f=100 Гц С=0,16 мкФ, для f=300 Гц С=0,05 мкФ, для f=10 кГц С=1600
пФ. Для согласования работы детектора с регулятором мощности на выход детек-
тора следует подать постоянное смещение + 8 В с помощью подстроечного рези-
стора R12 .
УРМ собран из широко распространенных элементов. Транзисторы V3, V6, V7,
V8, указанные в схеме, можно заменить на любые другие германиевые транзисторы
с аналогичными параметрами. Все постоянные резисторы типа МЛТ, переменные —
типа СП-I. Необходимо учесть, что резисторы R1 и R3 должны быть рассчитаны на
мощность не менее 2 Вт. Электролиты К.50-6, но возможны и другие типы. Тран-
зисторы VII и V12 необходимо подобрать по коэффициенту усиления. Еще лучше
использовать вместо них микросхему К1НТ291 или КШТ591 с любым буквенным ин-
дексом. Трансформатор в УРМ — типа ТН12, можно взять и другие понижающие
трансформаторы с напряжением вторичной обмотки 13...14 В. Терморезистор СТЗ-
14, но можно и другой, подойдет также переход эмиттер-база германиевых тран-
зисторов. В качестве фотодатчика может быть использован фотодиод ФДК226 или
фоторезистор СФ2-4. В ЦМУ в качестве развязывающего трансформатора использо-
ван согласующий трансформатор от приемника «Альпинист». Если необходимо регу-
лировать мощность до 500 Вт, то вместо тиристора Т25 можно установить тири-
стор КУ201 или КУ202 с любыми буквенными индексами. Регулируемую мощность ка-
ждого канала можно увеличить от 3 до 10 кВт при использовании более мощных
диодов, например В25, В50, и тиристоров (Т50, Т100) . Схема УРМ при этом не
изменится, но может потребоваться снижение номинала резистора R9 до 80...50
Ом, при этом коммутация силовой части прибора (сетевой шнур, диодный мостик
V19...V22, цепь тиристора) должна быть выполнена проводом сечением не менее 2
мм2, например МГШВ 2,5. К конструкции устройства предъявляются следующие ос-
новные требования по безопасности: корпус устройства должен быть изолирован
от всех токоведущих частей. Все пайки и соединения должны быть выполнены ка-
чественно и надежно. При настройке и эксплуатации УРМ следует соблюдать пра-
вила техники электробезопасности, так как цепи УРМ гальванически связаны с
сетью. Нельзя производить пайку при включенном питании.
Технологии
ПРОИЗВОДСТВО ГРИБОВ
Ральф Курцман
Глава 1
ПОДГОТОВКА К ВЫРАЩИВАНИЮ ГРИБОВ
Выращивание грибов это искусство, основанное на науке и технологии. Руково-
дствуясь некоторыми фактами, мы можем поделить тех, кто планирует заниматься
выращиванием грибов, на несколько групп. От того, что находится в распоряже-
нии у производителя на первоначальном этапе, зависит его принадлежность к той
или иной группе.
1. Квалифицированный специалист, имеющий опыт работы на грибной ферме.
2. Наличие отходов
3. Отходы от собственного предприятия или фермы
4. У производителя есть здание или возможность приобрести его по низкой цене
Многие из вас могут предположить, что представитель первой группы добьется
наибольшего успеха, и вы будете удивлены, узнав, что каждый из них может до-
биться успеха. Большинство отходов пригодно для использования, но лучше всего
использовать их смесь. Многие владельцы ферм должны обратить на это свое вни-
мание . Хотя существует ряд исключений, большинство зданий, в которых планиру-
ется выращивать грибы, не предназначены для этого.
Не все отходы подходят для выращивания грибов. Самое важное на этом этапе
Знать, что им необходимо. Грибы больше похожи на свиней или скот, чем на ово-
щи. Хотя все, в чем они нуждаются, должно находиться рядом с ними, как и в
случае с овощами. А это не так уж просто. Пища для грибов является пищей для
животных и растений, впрочем, как и сами грибы. У животных и растений для за-
щиты есть шкура или кожура. У грибов же нет подобной защиты, кроме того, что
они с завидным упорством пытаются выжить. Самым важным орудием в выращивании
грибов является соблюдение санитарных норм.
Даже если у специалистов возникают проблемы в процессе выращивания грибов,
то, скорее всего их обсуждение не поможет остальным. Было бы целесообразно
начать разговор с рассмотрения вопроса о том, какие отходы лучше всего ис-
пользовать, так как это будет более полезным. Но, прежде всего, необходимо
рассмотреть вопрос о месте для выращивания грибов. В противном случае не по-
лучится собрать большой урожай. Поэтому мы начнем с обсуждения помещения, так
как именно оно является физической защитой для грибов.
Большинство зданий не подходят для выращивания грибов, так как для них не-
обходимы особые условия. Как мы будем изучать эти условия? Одним из лучших
способов изучения живых существ - это наблюдение за условиями их обитания в
живой природе.
ПРИРОДНАЯ ЭКОЛОГИЯ ГРИБОВ
Грибы «Вешенки» были обнаружены в лесах умеренного пояса, а некоторые виды
даже в тропических лесах, Рисунок 1. Обычно они растут на упавших стволах,
так же можно увидеть эти грибы на больных, но все еще живых деревьях. И хотя
вероятнее всего было бы предположить, что сухая древесина является наиболее
подходящим грунтом, было обнаружено, что солома и лигнино-целлюлоза лучше
подходят как почва для выращивания грибов. Чаще всего в лесах нет достаточно-
го количества света, и влажность там всегда повышена. Лесной мусор (листья,
ветви деревьев, продукты жизнедеятельности) накрывает упавшие деревья. Спо-
собность выжить у грибов зависит от способности выбраться из-под этого «по-
крывала» на поверхность.
Индикаторами для определения приближения к поверхности для вешенок служат
низкий уровень концентрации углекислого газа и свет. Деревья отфильтровывают
солнечные лучи таким образом, что преобладает голубой свет1. В результате
этого вешенки реагируют только на голубой свет. Это касается и других видов.
1 Видно в том лесу еще и марихуана росла. Вообще говоря, энергия кванта света линейно Зави-
сит от частоты изучения, поэтому поток синего света переносит больше энергии, чем поток
красного света. Более вероятно, что на Зернах хлорофилла поглотится квант из коротковолновой
(синей) части спектра, потому что он несет больше энергии, соответственно отражение таких
квантов будет меньше и поэтому листва имеет Зеленый оттенок. Многим грибам, например шам-
пиньонам, свет вообще не нужен, грибы ведь не растения. Для вешенки он, скорее всего, явля-
ется сигналом о направлении роста (где свет - там выход на поверхность для плодоношения, то
есть для распространения спор). Вместе с тем свет вызывает образование пигментов, окрашиваю-
Грибы, родственные вешенкам выращиваемые на фермах, в природных условиях
растут на более открытых пространствах, где в почве содержится компост и
сгнивший мусор. И хотя открытые пространства могут предполагать потребность к
свету, мы обнаружили, что Шампиньоны (Agaricus) и другие виды грибов, расту-
щие на гниющем лесном мусоре, не нуждаются в свете. Они так же не переносят и
углекислого газа. На открытых пространствах его уносит ветер. Но возможно бо-
лее благоприятным является фотосинтетический способ переработки углекислого
газа травой и другими растениями.
Рис. 1. Учитесь у природы.
а. Слева, Дикий Pleurotus (вешенка).
б. Справа, Дикий Agaricus (шампиньон).
Мы не можем ответить на все вопросы о среде обитания, путем простых наблю-
дений живой природы, но мы можем увидеть, что существует ряд требований, ко-
торые следует учитывать при выборе условий посадки и дальнейшей культивации.
ЗДАНИЕ И ОБОРУДОВАНИЕ
Как мы уже говорили ранее, большинство зданий не приспособлено для выращи-
вания грибов. Существует ряд требований, которые необходимо выполнять при вы-
боре здания для выращивания вешенок:
1. Температура от 15 до 20°C
2. Влажность от 80 до 95%
3. Вентилирование
4 . Свет
щих поверхность плодовых тел, но для чего грибам нужны эти пигменты, нужно спрашивать биохи-
миков .
5. Санитария
Температура и влажность должны сохраняться постоянно на указанном уровне,
остальные же требования зависят от особенностей той или иной разновидности.
Любые резкие перепады в температуре станут причиной изменения влажности, ко-
торое в свою очередь губительно скажется на грибах. При температуре равной 20
°C, влажность составит 90%. Если температура упадет до 19°С, то уровень влаж-
ности поднимется до 100%. Если температура поднимется до 21°С, то уровень
влажности снизится до 82%. При температуре 15°С изменение уровня влажности
составит один процент. Так при снижении температуры на 1°С, влажность будет
равна 99%, а при повышении на 1°С влажность снизится до 83%. Такое взаимодей-
ствие между температурой и влажностью создает потребность в изоляции. Для
изоляции можно использовать специальный укрывной материал. Для этой же цели
можно поместить на каменную кладку тонкий слой почвы, соломы или даже бумаги.
При 100% влажности, все вокруг намокает очень быстро. Наоборот, если уро-
вень влажности составляет 80%, грибы начинают засыхать. При высоком уровне
влажности многие строительные материалы начинают гнить или ржаветь. Таким об-
разом, условия выращивания грибов ограничивают нас в выборе строительных ма-
териалов . Бетон и пластик могут противостоять влажности с наименьшими потеря-
ми. Некоторые производители используют деревянную или металлическую конструк-
цию, покрытую пластиком. Другие же используют древесину и металл без покры-
тия, но такое строение прослужит не долго. Огромной проблемой является и то,
что паразиты и инфекция скапливаются в древесине и других пористых материа-
лах .
Полиэтилен и изоляционные материалы необходимы не только для защиты от сы-
рости , но и для поддержания температуры и влажности на необходимом уровне. На
следующем рисунке мы видим пластиковую прослойку между балками металлического
Здания, Рисунок 2. В Азии, например, для строительства домов люди вынуждены
использовать бамбук, полиэтилен, а для изоляции - солому, Рисунок 3.
Рис. 2. Здание.
Изоляция.
Хорошее вентилирование необходимо для здоровья, как грибов, так и работни-
ков. Для роста грибам необходим низкий уровень углекислого газа. Pleurotus
(Вешенки), как и другие грибы, будут расти в том случае, если уровень углеки-
слого газа будет оставаться низким. Для создания тока воздуха обычно исполь-
зуются центробежные вентиляторы. Прежде чем попасть в комнату воздух должен
быть подогрет или охлажден до нужной температуры, а так же увлажнен. Пар мо-
жет одновременно подогреть воздух и насытить его влагой. Вентиляция должна
очистить помещение от углекислого газа, образовавшегося в результате жизне-
деятельности грибов и людей. Люди могут выдержать 10%-ное содержание углеки-
слого газа в воздухе, что касается грибов, то они более чувствительны. Грибы
должны быть срезаны до того, как появятся споры, но даже при должном контроле
некоторые споры могут отделиться. Такие споры вызывают приступы астмы и сен-
ной лихорадки, а вентилирование поможет удалить их из воздуха.
Рис. 3. Здание. Изоляция "Соломой",
а. Тайский вариант
б. Тайвань2
Для вешенок свет имеет большее значение, чем для других видов. О необходи-
мом количестве света написано много, но существует всего несколько экспери-
ментов , проведенных с наибольшей точностью. Как я уже отмечал ранее, свет в
лесу голубой. Все мои попытки вырастить грибы при свете ламп накаливания были
2 Автор статьи явно тяготеет к экзотике. На самом деле самыми лучшими сооружениями для выра-
щивания грибов являются любые подвалы и Затененные теплицы.
тщетными3. Этот свет скорее желтый, чем голубой. Флуоресцентный свет распада-
ется на спектр. Наиболее известным является, так называемый «холодный свет4».
Если он достаточно яркий для того, чтобы при нем можно было читать, и лампы
включены на протяжении 8 часов, то такого света вполне достаточно. Солнечный
свет не подходит для освещения, так как от него зависит температура воздуха.
Чаще всего водоросли и другие растения так же подвергаются опасности5 из-за
воздействия солнечного света.
Agaricus (Шампиньоны), наиболее распространенный вид выращиваемых грибов,
не нуждаются в свете, а некоторые их разновидности даже меняют свой цвет под
воздействием солнечных лучей. Единственным источником света в помещении могут
быть фонари на шлемах6 рабочих.
Нет необходимости делать в помещении окна. В случае если грибы будут нуж-
даться в свете, то флуоресцентных ламп будет вполне достаточно. Если же осве-
щение понадобится людям, то можно будет использовать фонарь7.
Здание должно отвечать санитарным требованиям, которые можно выполнить,
приняв следующие меры:
1. Установить фильтры поступающего воздуха
2. Установить щиты или фильтры на выходе воздуха
3. «Воздушная пробка» в комнатах для переодевания
4. Использование ванны для ног8
5. Герметичность
Все отверстия должны быть защищены от проникновения паразитов и разного ро-
да инфекций. Все вентиляционные люки необходимо снабдить фильтрами или тонки-
ми защитными щитами. На отверстиях для впуска воздуха так же должны быть ус-
тановлены щиты. Для этого идеально подходят9 фильтры НЕРА. Рисунок 4.
Поскольку перебои в энергоснабжении могут вызвать остановку вентилирования
и насекомые могут проникнуть в здание, то такие фильтры просто необходимы.
Следует так же учитывать и то, что некоторые насекомые могут лететь против
воздушного потока, если он не очень сильный.
3 Что, впрочем, не мешает использовать обычные лампы накаливания, их и используют.
4 Автор, похоже, слишком долго находился в «синем» лесу, но возможно он просто пытался ска-
зать, что лампы дневного света (флуоресцентные), являются предпочтительными, поскольку
спектр их излучения более близок к дневному и, следовательно, его коротковолновая часть не
так обеднена по сравнению с излучением ламп накаливания. В хозяйственных магазинах многих
стран продается несколько видов ламп дневного света, самый распространенный (и дешевый) вид
примерно так и называется.
5 К счастью они об этом не Знают и предпочитают «Загорать на солнце». Вероятно, автор хотел
сказать, что открытая (не Затененная) солнцу теплица будет перегреваться, что очень нежела-
тельно для грибов. К тому же на солнце начнут активно размножаться водоросли (там же влажно)
и сорняки (посмотрите еще раз на соломенную теплицу).
6 Хмм...ммм, а Зачем рабочим шлемы? Возможно, это отголосок того, что для выращивания шампинь-
онов часто используют старые шахты, туннели и пещеры.
7 Или нанять людей из общества слепых.
8 Представили? Речь идет о поддоне с тканью пропитанной антисептиком, чтобы вытирать обувь.
9 Как-то это не очень подходит для соломенной теплицы, но на высокотехнологичных грибных
фермах так и делают, потому что всплеск плесени может полностью погубить урожай и привести
ферму к финансовому краху. Это окупается. Смотрите ниже пару добавленных снимков из Китая.
Рис. 4. Воздушные фильтры.
Помещение для людей, работающих на фабрике, не потребует больших финансовых
затрат, так как будет состоять из легкого каркаса и черной пластиковой плен-
ки. В нем должны храниться резиновые ботинки, ванна с солью или гипохлоритом
для обуви, а так же чистая одежда, защитные маски и резиновые перчатки. Наи-
более слабым источником света для грибов может стать натриевая лампа. Такие
лампы излучают свет, похожий на дневной. Они подходят и для людей. Насекомые
не видят их, поэтому они так же могут служить для создания «воздушной пробки»
в комнатах для переодевания. Для этой цели подходят и лампы накаливания, так
как они невидимы для насекомых10.
Если в здании кроме необходимых для вентиляции отверстий есть еще и другие,
то насекомые могут проникнуть внутрь. Для наибольшей герметичности, необходи-
мо закрыть отверстия при помощи изоляционной пены, силикона или другого уп-
лотняющего материала.
10
Рис. 5. Подготовка субстрата. Молотковая дробилка. Переработанная слома.
Что-то сомнение берет.
ПЕРВОНАЧАЛЬНАЯ ПОДГОТОВКА СУБСТРАТА
Для выращивания Agaricus (шампиньонов) необходим участок, на котором суб-
страт намокает и перемешивается с другими составляющими. Такой участок нахо-
дится под открытым небом, но иногда над ним помещается крыша. Всю тяжелую ра-
боту на данном этапе выполняют погрузчики или другие машины. Для дезинфекции
участок должен быть зацементирован.
После того как субстрат перемешают, он должен превратиться в компост. Для
перемешивания обычно используют техническое оборудование. Процесс происходит
под открытым небом. Для этой же работы люди могут использовать вилы. Большин-
ство фермеров ссыпают компост в силосные ямы. Этот процесс должен происходить
под открытым небом, так как поток воздуха проходит через субстрат и тот в ре-
зультате брожения превращается в компост. Так же возможно добавлять в ямы пар
или выполнять пастеризацию.
Для разновидностей, растущих в лесу, чаще всего используется субстрат кото-
рый пропускается через молотковую мельницу, Рисунок 5. Для получения лучшего
субстрата смешиваются несколько ингредиентов. Участок для перемалывания смеси
должен быть сухим, поэтому над ним необходимо построить крышу.
В некоторых случаях субстрат остается сухим до процесса пастеризации, а
иногда он предварительно увлажняется. В любом случае, в конечном итоге суб-
страт намокает и помещается в емкость, предназначенную для выращивания гри-
бов .
ПОСЛЕДНИЙ ЭТАП ПРИГОТОВЛЕНИЙ
Для ПАСТЕРИЗАЦИИ требуется оборудование, которое может поддерживать темпе-
ратуру ВСЕГО субстрата от 55° до 60° в течение 30 минут. Температура никогда
не должна быть выше11! Оборудование так же должно защищать смесь от загрязне-
ния и дать ей возможность остыть до 25° в течении 16 - 20 часов.
Для СТЕРИЛИЗАЦИИ требуется оборудование, которое способно подогреть смесь
до температуры 121° в течение 15 минут. Такая температура необходима для ка-
меры давления11 12. Смесь должна находиться в емкости, которая не допускает про-
никновения микроорганизмов. Охлаждение должно проходить медленно, чтобы раз-
ница между атмосферным давлением и давлением пара в емкости не была значи-
тельной .
Для стерилизации требуется больше оборудования и больше топлива. Так же не-
обходимо контролировать каждую стадию этого процесса13. Стерилизация рекомен-
дована для некоторых дорогостоящих разновидностей. Некоторые рекомендуют ее
11 Наверное, чем выше, тем лучше - и микроорганизмы гибнут, и стенки стеблей становятся более
доступны для грибов - подразрушаются. А вот ниже, наверное, не стоит, многие микроорганизмы
себя неплохо чувствуют при таких температурах. В некоторых технологиях соломенную резку За-
ливают 80-ти градусным кипятком.
12 Вообще-то такая температура необходима для стерилизации, большинство микроорганизмов и
спор гибнет, но кое-какие споры могут и выжить, процесс вероятностный. А вот такая темпера-
тура достигается только при повышенном давлении в камере стерилизации.
13 Просто много За раз не простерилизуешь (большой автоклав нужно долго нагревать), а Значит
требуется что-то вроде конвейера.
для Pleutorus (Вешенка). Это говорит о том, что они просто не знают, как про-
водить процедуру пастеризации правильно14.
ОБОРУДОВАНИЕ ДЛЯ ГРИБНИЦЫ
Санитарные требования для помещений, предназначенных для выращивания гри-
бов , ниже, чем для помещений, где проходят процессы пастеризации, охлаждения
или стерилизации. Здесь должны создаваться подходящие условия для образования
грибницы. Воздух должен быть пригоден для работников, хотя нет необходимости
контролировать уровень температуры и влажности. Лучше всего будет расположить
помещение для выращивания грибов на расстоянии нескольких километров от зда-
ния, предназначенного для пастеризации и стерилизации. Такое расположение по-
зволит избежать многих источников болезней и паразитов. Оборудование, приме-
няемое для выращивания грибов, является основным во всем процессе. В любом
случае это будет обсуждаться позже при выборе методов и видов оборудования.
ЗАКЛЮЧЕНИЕ
Выращивание грибов требует выполнения различных технических условий. Ни од-
но из этих требований не должно быть упущено15. В том случае если они не бу-
дут выполнены, то вас ожидает или провал, или низкая урожайность. Оборудова-
ние должно обеспечивать все необходимые условия для выращивания грибов. Оно
так же должно защищать от насекомых, паразитов и инфекций. Без создания бла-
гоприятных условий и защиты невозможно добиться урожая и сохранить его только
с помощью грамотного управления.
ГЛАВА 2
ПАСТЕРИЗАЦИЯ
Каждый может выращивать шампиньоны при условии, что компост правильно под-
готовлен. Хотя, субстрат можно приготовить разный. То же самое касается и ве-
шенки обыкновенной. Если использовать правильно подготовленный субстрат, все
остальное довольно просто. При подготовке субстрата неотъемлемой частью явля-
ется пастеризация. Важно ее правильно провести, т.к. параметры ее более огра-
ничены, чем выбор подходящих компонентов. Субстрат, безусловно, очень важен,
но чтобы получить максимальный результат, нужно соблюдать и остальные усло-
вия .
Почему не стерилизация?
Итак, разберемся с понятием пастеризация. Для начала, пастеризация не пре-
следует цель уничтожить все микроорганизмы. Процесс, во время которого унич-
тожаются все живые микроорганизмы, называется стерилизацией. И как раз наобо-
рот, именно стерилизация наносит больше вреда, чем пастеризация при выращива-
нии вешенки. Многие полагают, что если удобрять16 грибы более тщательным об-
разом, это только пойдет им на пользу. В действительности же, если превысить
четко определенную норму требуемых условий, это может нанести им серьезный
вред. При пастеризации влажная ткань нагревается до 55-60°С в течение 30 ми-
14 Сейчас они узнают эту тайну.
15 То есть надо делать хорошо, а плохо - оно само получится.
16 Возможно, нет, но вроде речь идет о пастеризации.
нут. Ни единая часть материи17 не должна быть нагрета менее 55°С или более
60°С.
Одно преимущество в пользу применения пастеризации очевидно: небольшие за-
траты. Для стерилизации требуется специальное оборудование с большим давлени-
ем. А оно довольно дорогое и более того, для коммерческого производства тре-
буется крупнотоннажное оборудование. Стерилизация также требует большего на-
гревания и более длительного периода времени. Таким образом, расход топлива
для стерилизации в несколько раз превышает расходы, затраченные при пастери-
зации .
Высокая себестоимость оборудования может окупиться в том случае, если про-
изводство более объемное и к тому же снижается риск вероятности инфицирования
грибов паразитами. Известно, что стерилизованный субстрат принесет меньше
грибного урожая, и к тому же с большей вероятностью заболеваний. Для возмож-
ного заболевания существуют несколько причин. Во-первых, невозможность содер-
жания крупномасштабных помещений для культивации, а также невозможность суб-
страта сохранять стерильность при производстве.
Во-вторых, пораженные микроорганизмы слабо сопротивляются микроорганизмам,
оставшимся в субстрате после правильной пастеризации. А так как стерилизация
уничтожает все живое в субстрате, то сохранившиеся микроорганизмы не присут-
ствуют в стерильном субстрате, а потому они не сопротивляются18. В-третьих,
эти сохранившиеся микроорганизмы мало поглощают или вообще не поглощают цел-
люлозу или лигнин, то есть, те вещества, на которых растут грибы. Но они так-
же используют гемицеллюлозу, - природный материал большинства видов субстра-
та, на которых лучше всего произрастают пораженные микроорганизмы.
В результате того, что микроорганизмы, сохранившиеся после пастеризации,
используют гемицеллюлозу, появляется возможность увеличения объема производ-
ства. Благодаря гемицеллюлозе, благотворные микроорганизмы, оставшиеся после
пастеризации удваиваются. Они служат удобрением для грибов. То есть, вешенки
«поедают» микроорганизмы, выращенные на гемицеллюлозе, но которую грибы не
могут использовать напрямую. Существуют признаки, что организмы, оставшиеся
после пастеризации, сберегают больше азота, который требуется грибам. Они мо-
гут добывать азот из атмосферы. Итак, целью пастеризации не является избавле-
ние от всех живых микроорганизмов. Но при пастеризации уничтожаются те микро-
организмы, которые соперничают с грибами, а также увеличивается количество
организмов, препятствующих возникновению болезней, потреблению гемицеллюлозы,
вырабатыванию азота и станут подкормкой грибам.
Для приготовления идеального субстрата необходимо знать, как избавиться от
пораженных микроорганизмов и как создать благоприятные условия для микроорга-
низмов, которые служат подкормкой грибам. Ясно, что если уничтожить все живые
микроорганизмы из подкормки, благотворные организмы смогут лишь появляться из
атмосферы или же люди, занимающиеся грибами, будут их туда вносить. Но микро-
17 Да с материей надо поосторожней.
18 Чтобы не сойти с ума, не будем комментировать этот абзац и два последующих. Скорее всего,
это просто ужасный перевод. Можно только предположить, что термофильные бактерии, сохраняю-
щиеся при пастеризации, помогают бороться с распространением низших грибов - плесеней, кото-
рые могут Запросто подавить рост и плодоношение вешенки. Да это так, но стопроцентной гаран-
тии нет, это все «балансировка на грани случайностей», однажды Заражение плесенью может про-
изойти , и придется дезинфицировать все помещение - от спор.
организмы, проникающие из атмосферы, с большей вероятностью, чем благотвор-
ные, могут являться возбудителями болезнетворных бактерий.
Шампиньоны в естественных условиях произрастают в травянистых местах, там,
где обычно имеются скопления экскрементов19 и мочи животных. Чтобы имитиро-
вать и улучшить естественный субстрат, производители используют тщательно
приготовленный компост. В процессе компостирования субстрат выдерживается во
влажном состоянии несколько недель. Сам процесс поглощает большое количество
субстрата и генерирует значительное количество тепла. К тому времени, когда
субстрат готов к пастеризации, он большей частью уже теряет способность про-
изводить тепло. Внутри компост нагревается, но снаружи утрачивает тепло быст-
рее, чем оно успевает генерироваться. С целью снизить риск заболеваний и за-
ражения паразитами, компост обрабатывается струей горячего пара. Сложно пред-
ставить иной способ пастеризации компоста для такого семейства грибов, как
шампиньоны.
Грибы семейства вешенка в естественных условиях произрастают на мертвых де-
ревьях или на необработанном лесоматериале. Между древесиной и экскрементами
животных, смешанных с травой очень мало сходства. Трава дает лигнин и целлю-
лозу грибам семейства шампиньоны, а необработанный лесоматериал дает лигнин и
целлюлозу вешенкам. Грибы, также как и все живые микроорганизмы, нуждаются в
воде.
Однако на этом и заканчиваются все сходства между шампиньонами и вешенками.
Как было упомянуто в предыдущей главе, для приготовления качественного суб-
страта используется много растительного сырья. Однако предпочтительней всегда
иметь сухое сырье и чтобы оно не являлось пригодным для пищи большинства жи-
вотных, и не представляло пищевой ценности для крупного рогатого скота и дру-
гих жвачных животных. Если сырье влажное, на нем могут быть пораженные микро-
организмы, и его будет гораздо сложнее пастеризовать. Все, что имеет какую-то
пищевую ценность для животных, имеет тенденцию к развитию заболеваний.
Многие производители начали свою деятельность с разведения шампиньонов, а в
последствии перешли на вешенки. Но они пытались выращивать вешенки тем же об-
разом, что и шампиньоны. Как я уже упоминал, их естественная среда абсолютно
разная, и выращивать их теми же способами - просто неразумно. Для достижения
желаемых результатов, производителям стоит задуматься над тем, что вешенки -
это абсолютно другое семейство грибов, требующее к себе иного подхода при их
разведении, чем шампиньоны.
Почему бы не применять пар?
Чтобы грибы смогли произрастать субстрат должен быть влажным. Но так как
пар не может давать достаточное количество влаги, нужна вода. Те, кто исполь-
зуют пар, обычно добавляют холодную воду в течение нескольких дней. Субстраты
из соломы, яичной скорлупы20 и некоторые другие качественные субстраты покры-
ваются природным воском21. Воск препятствует чрезмерному увлажнению субстра-
та. Через несколько дней вода впитается, но к тому времени могут уже появить-
ся патогены/болезнетворные микроорганизмы.
19 Навоза, наши экскременты не подойдут, хотя мы тоже вроде не растения.
20 Вряд ли что-то будет расти на яичной скорлупе, но возможно она подойдет для стабилизации
кислотности, вместо мела.
21 Откуда он взялся и что это такое?
Если большую часть сухого сырья, которое не представляет пищевой ценности
для животных22, просто увлажнить, оно не будет генерировать много тепла. По
этой же причине очень сложно пастеризовать паром субстрат вешенки тем же пу-
тем, что и субстрат шампиньона. При пастеризации идеальная температура суб-
страта - 55-60°С в течение 30-60 минут. Температура ни в коем случае не долж-
на превышать23 60°С. Субстрат вешенки - хороший изолятор тепла, поэтому, если
тепло поступает в виде пара или воздуха, на это потребуется много времени и
нельзя быть уверенным, что там не возникло холодных воздушных ям24. Засеките
время для приготовления/жарки целой курицы, и увидите, когда курица готова к
употреблению, ее внутренняя часть не такая горячая, как воздух в духовке.
Способы жарки курицы и паровой пастеризации субстрата во многом схожи. Прин-
цип парового нагревания в типичном контейнере заранее увлажненного субстрата
показан на Рис.5.
Рис.6.
3 Часа
Риг. 5 На рисунках показан метод использования пара при пастеризации
субстратов
На рисунках показаны
горячие потоки воды и пара при пастеризации.
22 Да похоже, ни навоз ни солома животных не интересуют.
23 Да верим, верим.
24 —
Пустот.
Пастеризацию можно провести лишь путем добавления горячей воды, нагретой до
температуры 55°-60°С. На Рис.6 показан принцип нагревания водой типичного
контейнера с субстратом. Субстрат полностью сухой до тех пор, пока не добав-
лена горячая вода. Сравните Рис.5 и 6. Теперь субстрат нагрет до 56°С, что
предпочтительнее температуры25 55°С, так как необходимо компенсировать тепло,
используемое для нагревания сухого субстрата. Желательно использовать больше
воды, но при этом ее температура не должна превышать 60°С. Если маленькая
часть субстрата подвергается нагреванию при температуре более 60°С в течение
короткого времени, и субстрат хорошо перемешан, благотворные микроорганизмы
смогут восстановиться. Но тогда может возникнуть небольшая потеря урожая. Од-
нако если какая-нибудь часть нагревается менее 55°С, или же не увлажнена, ве-
роятна опасность возникновения заболеваний и вредителей.
Малые производители грибов
Те, кто выращивает грибы в небольшом объеме, могут использовать довольно
простое оборудование для пастеризации, и могут в этом хорошо преуспеть. Все
что требуется, так это большой контейнер и, либо ресурсы горячей воды
(Рис.7), либо приспособление для нагрева воды в контейнере (Рис.8). Для этого
удобны в использовании чистые стальные барабаны с нагревательным устройством.
Также возможно впускать в воду пар для ее нагревания. Сначала вода нагревает-
ся до 56-60°С, затем туда добавляется субстрат, который находится там в тече-
ние 30-60 минут. После этого можно отвести воду или извлечь субстрат из воды.
Необходимо удалить излишек воды по прошествии 60 минут, так как в горячей во-
де очень мало кислорода, а «хорошие»/положительные микроорганизмы используют
кислород, после чего субстрат становится анаэробным. Тот же метод применяется
и в случае с контейнером. Берется контейнер с субстратом, куда добавляется
горячая вода (56-60°С) до тех пор, пока субстрат целиком не покрыт горячей
водой. Так же как и в первом случае, воду необходимо слить через 30-60 минут.
Но вода должна быть нагрета еще до соприкосновения с субстратом! Время и тем-
пература очень важны в обоих случаях, при добавлении воды в субстрат и наобо-
рот , помещения субстрата в воду.
Также важно соблюдать верные пропорции воды, иначе ее избыток может привес-
ти к потере урожая. Для получения максимального урожая субстрату требуется
достаточное насыщение воздухом. Исследования показывают, что углекислый газ
оказывает благотворное влияние на формирование урожая (грибов) в период высе-
ва . Желательно несколько раз открывать контейнер с урожаем для сохранения
влаги и естественного поступления углекислого газа. Тем не менее, в некоторых
видах субстрата могут образовываться другие газы, некоторые из которых пре-
пятствуют росту грибов. Очень важно подобрать оптимальное количество воды,
обеспечить достаточное количество вентиляции в пластиковый или другой контей-
нер для выращивания грибов. Нужно определить путем проб, сколько требуется
воды для субстрата, какие нужно проделывать отверстия для максимального роста
и урожая грибов. Рекомендуются субстраты с содержанием двух или более видов
остатков растений. В смеси обычно поступает некоторое количество воздуха, но
они с трудом поддаются консервированию.
Когда субстрат высушен, нужно позволить ему медленно охлаждаться в очищен-
ном от лишнего (или закрытом сверху) месте. Охлаждать его нужно до 25°С, а
потому на это потребуется, по меньшей мере, 16 и обычно не более 20 часов.
25
Поразительная точность.
Именно в период охлаждения происходит рост большинства благотворных микроор-
ганизмов. В конечном итоге субстрат должен источать сладко-кислый запах. Как
только субстрат охладился до 25°С, следует перемешать мицелий и упаковать его
в пластиковые или другие контейнеры. Все: воздух, помещение, рабочая одежда,
а также сами рабочие, должны быть чистыми. На рабочих должны быть резиновые
перчатки.
Рис. 7 Цистерна/бак для пастеризации соломы путем добавления за-
ранее нагретой воды. На рисунке субстрат из соломы был засеян
уже после охлаждения и помещен в пластиковые пакеты для выращи-
вания грибов. (Россия)
Рис.8. Стальные барабаны с газовыми горелками для нагрева воды.
Рис.8А. Пастеризованный субстрат извлекается из воды при помощи
цепного полиспаста. Он будет медленно охлаждаться в стерильном
контейнере. (Мексика)
Оба вышеперечисленных метода погружения субстрата в горячую воду хороши для
субстрата из соломы и других видов, которые будут иметь хорошую атмосферу по-
сле высушивания. Однако, не применимы субстраты из макулатуры, хлоп-
ка/хлопчатника, льна/парусины, опилок, или иные субстраты, которые с трудом
упаковываются в сжатом воздушном пространстве. Для субстратов, которые упако-
вываются в сжатом виде, требуются смесители/мешалки для перемешивания суб-
страта во время пастеризации.
Средние производители грибов
и некоторые виды субстратов
Мешалки/смесители позволяют даже нагревать и контролировать нужное количе-
ство воды в субстрате. Так как невозможно правильно пастеризовать субстрат в
простом контейнере путем применения пара, такие машины/смесители и повышенная
степень осторожности в управлении температурой делают даже паровую пастериза-
цию возможной.
Смеситель/мешалка может использоваться для пастеризации путем добавления
горячей воды в нее. Затем добавляется субстрат, или, наоборот, путем добавле-
ния горячей воды в субстрат. Так как в простых контейнерах необходимо исполь-
зовать излишки воды, то в смесителе вам требуется лишь количество воды, необ-
ходимое для увлажнения субстрата. Возможность контролировать количество воды
открывает несколько преимуществ. Во-первых, это единственный путь, когда мы
можем использовать субстраты, которые естественным путем не впитывают излишки
воды. Во-вторых, это значит, что нет сточных вод. Излишки воды, отведенные от
субстратов, крайне неблагоприятны, и их правильно ликвидировать довольно
трудно. Тем не менее, грибам необходимо огромное количество воды, поэтому до-
бавляйте столько воды, сколько может удержать субстрат. При этом не закрывай-
те вентиляционные отверстия26. Наверняка трудно оценивать вентиляционные от-
верстия, лишь взглянув на некоторые субстраты; единственный путь выяснить это
- провести исследование. Для того чтобы узнать процент содержания воды в пас-
теризованном субстрате, нужно взвесить сухой субстрат и измерить количество
воды. Когда будет определено оптимальное количество воды, таким же оно и
должно быть постоянно. Если же ингредиенты, используемые в субстрате, меняют-
ся, тогда, вероятно, должно измениться и оптимальное содержание воды в нем.
Если при пастеризации берется большее количество воды, чем требуется для
оптимального роста, сухой субстрат может лишь вызвать маленькое понижение
температуры. Однако при использовании достаточного количества воды для опти-
мального роста, температура увлажненного субстрата может быть гораздо мень-
ше/ниже температуры добавляемой воды. Есть опасность, что если температура
добавляемой воды более 60°С, положительные/благоприятные микроорганизмы по-
гибнут. Возможно, появится необходимость уравнять/компенсировать пониженную
температуру. Вероятно, лучшим способом добавления тепла будет поместить на-
гревательную рубашку на дно смесителя/мешалки. Температура в ней может быть
даже более 60°С, пока субстрат сухой. Во время процедуры нагревания сухого
субстрата, его нужно постоянно помешивать. До тех пор пока у микроорганизмов
не появится достаточное количество воды для роста, они способны переносить
большую жару27. Но когда добавляется вода, температура пакета не должна пре-
вышать 60°С и не следует прекращать процесс помешивания/перемешивания. После
того, как вода добавлена, субстрат должен находиться там в течение 30-60 ми-
нут при температуре 55-60°С.
Так же как и при всех методах, пастеризованный субстрат должен медленно ох-
лаждаться/остывать в течение 16-20 часов. Охлаждение осуществляется лишь пу-
тем отключения всех источников нагревания, после того, как будет отведен из-
лишек воды. Субстрат следует оставить в машине для пастеризации или перемес-
тить его в чистый контейнер. Если его все-таки переместить, тогда возможно
будет пастеризовать следующую порцию. Охлаждение зависит от ряда условий: ко-
личества субстрата, окружающей температуры (температуры помещения), любой
изоляции/обособления, любого движения субстрата во время охлаждения, и других
причин.
Хорошо, если процесс охлаждения будет происходить в смесителе/мешалке. Если
охлаждение слишком медленное, или если субстрат начинает саморазогреваться,
смеситель поможет его охладить. Когда субстрат охлажден и готов к упаковке в
контейнеры для выращивания (мешки), и т.д., смеситель может также использо-
ваться для посадки грибницы/ мицелия в субстрат.
На Рис.928 показана конечная часть двухвинтового смесителя с винтовым кон-
вейером. На Рис.1029 блок-схема двухвинтового смесителя и приспособлений, ко-
торые отводят излишки воды. Смеситель, показанный на Рис.9 заменил аналогич-
26 Можно догадаться, что воды должно быть только столько, чтобы субстрат (солома) был влаж-
ный , но вода не хлюпала в нем. Сожмите руками горсть влажной соломы, если вода потекла -
многовато будет.
27 Температуру.
28 Отсутствовал.
29 Аналогично.
ный, но меньших размеров. Оба этих аппарата довольно успешно применяются при
паровой пастеризации.
На Рис.II30 показаны блок-схемы частей двух одинарных смесителей/мешалок.
Одна, справа, используется для пастеризации, затем винтовой конвейер двигает
субстрат к смесителю слева, где он охлаждается. Как только субстрат охлажден,
в машину слева добавляется грибница/мицелий, и тогда другой винтовой конвейер
делает из зараженного мицелием субстрата «гигантские колбаски». На Рис.1231
показано внутреннее устройство машины-мешалки для пастеризации (справа на
Рис.11). На Рис.13 показан процесс заполнения/ наполнения «гигантских колба-
сок», контейнеров зараженным мицелием субстратом. На дне миксера/смесителя
для пастеризации находится нагревательная рубашка, а предмет/коробка вдалеке
слева - Высокоэффективный фильтр для охлаждения воздуха, который вдувается в
мешалку/смеситель слева, когда требуется дополнительное охлаждение. Использо-
вание этого оборудования позволит полностью завершить процесс по подготовке
субстрата без минимального участия людей и контакта с нефильтрованным возду-
хом. Нельзя быть уверенными, что вредные и болезнетворные микроорганизмы пол-
ностью уничтожены, но это оборудование снижает риск их появления.
Рис. 11. Конечные блоки двух одновинтовых смесителей, используе-
мые для пастеризации, охлаждения, засеивания и наполнения «ги-
гантских колбасок» - контейнеров для выращивания. (Россия)
30 Вместо блок-схем, показан сам смеситель, а может и не смеситель, но что из России - это
точно.
31 Повторяет снимок с Рис.11, поэтому не показан.
Рис.13. Наполнение «гигантских колбасок» - контейнеров для выра-
щивания субстратом, который был засеян в охладительном смесителе
(смеситель справа на Рис.11).
ЗАКЛЮЧЕНИЕ
Пастеризация - самая решающая ступень в процессе выращивания грибов. Тот,
кто их выращивает, должен уделять особое внимание времени и температуре. Тем-
пература ни одного из субстратов не должна быть менее 55°С в течение 30 или
более минут во время пастеризации. Она также ни в коем случае не должна пре-
вышать 60°С, когда субстрат влажный. Для отвода излишка воды с субстрата мо-
жет использоваться самое простое оборудование. Для тех, кто выращивает более
250 кг (1/4 тонны) грибов ежемесячно, наиболее выгодным и быстро окупаемым
окажется приобретение смесителя/мешалки. Оборудование, способное пастеризо-
вать субстрат, а также охлаждать, засевать грибницей и наполнять им контей-
нер, стоит дороже, но зато оно обеспечивает более надежную защиту от болезне-
творных микроорганизмов и паразитов, а также экономит огромные затраты чело-
веческого труда.
ГЛАВА 3
РАЗВЕДЕНИЕ ГРИБОВ «ВЕШЕНКА»
Людей, планирующих разводить грибы можно разделить на группы по некоторым
факторам, перечисленным ниже:
1. Квалификация/опыт - человек работал на фабрике по выращиванию грибов
2. Отходы, которые человек видит
3. Отходы на собственной ферме или другого бизнеса
4. Владеет зданием или имеет возможность купить по низкой цене
К настоящему моменту мы предполагаем, что вы понимаете процесс и готовы на-
чать выращивание грибов. Для этого вам необходимо выбрать материал, который
вы будете использовать в качестве субстрата, и изучить все этапы выращивания,
начиная с подготовки субстрата и заканчивая сбором урожая и уборкой после
сбора урожая.
ВЫБОР СУБСТРАТА
Прежде чем начать выращивание, нам необходимо определить, на чем мы будем
выращивать. В настоящее время с успехом используются следующие виды отходов:
1. компост для шампиньонов
2. бревна древесины твердых сортов
3. щепки и опилки древесины твердых сортов
4. зерновая шелуха
5. сухие отходы от сельхоз. культур после сбора урожая
6. бумага
7. хлопковые отходы
8. рисовая солома
9. пшеничная солома
10. кукурузные стебли и стержни початков
11. другие виды соломы
12. отходы стран-потребителей кофе32
13. отходы стран-производителей кофе
Бревна древесины твердых сортов являются естественным субстратом, но веро-
ятно в данном списке самым бедным по составу является компост для шампиньо-
нов. Все, на чем росли какие либо микроорганизмы, является неподходящим (уг-
рожающим) субстратом для грибов вешенок. Другие нежелательные материалы - это
клевер, люцерна, бобовые, все, что содержит простой сахар или любые «грибные
добавки». Принимая во внимание опыт и публикации по выращиванию грибов, можно
перечислить следующие желательные для субстрата качества:
32
1 .
2 .
3 .
4 .
чистый - хранился
содержит лигнин и
хорошо удерживает
удерживает воздух
сухим, и ничто
целлюлозу
много воды
не росло на нем ранее
Охренеть.
Мы упомянули о чистоте стартового материала, но необходимо подчеркнуть важ-
ность сказанного. Все растущее на субстрате будет уменьшать количество пита-
ния, предназначенное грибам. Некоторые организмы могут вырабатывать ядовитые
остатки. Другие могут представлять собой болезнетворные организмы, не полно-
стью убитые в процессе пастеризации. Намного труднее вывести организмы, кото-
рые уже укоренились, чем те, которые присутствуют в небольших количествах.
Большинство растительных отходов содержат лигнин, целлюлозу и гемицеллюло-
зу. Гемицеллюлозу можно назвать «улавливающей все», что содержит пектин и,
например, я так же включаю крахмал. Большое количество гемицеллюлозы и неко-
торых других материалов в растительных отходах является нежелательным, но не-
большие количества могут стать полезными, если правильно организовать работу.
Бумага и хлопковые отходы из нашего списка возможных субстратов могут состо-
ять из 100% целлюлозы, но в смесях это может быть преимуществом.
Нам необходимо, чтобы наш субстрат удерживал воду, это будет вода в грибах,
но какое-то количество воды так же испариться, и какое то количество выделит-
ся в процессе метаболизма. Субстрат должен хорошо удерживать воду, так как
нам нужен воздух, и если вода не задерживается и вытекает, она будет закупо-
ривать воздушные пространства и препятствовать росту.
ГРИБНИЦА
Грибница - это второй важный ингредиент. Мудро покупать грибницу самого
лучшего качества из доступных вариантов. Даже если стоимость намного выше,
это, вероятно, оправдает потраченные деньги. Крупные производители грибницы
имеют очень свежий посевной материал, тщательно стерилизуют и контролируют
процесс на каждом его этапе. Более мелкий производитель может по незнанию
ввести вирусы или продать старую грибницу.
Необходимо внимательно ознакомиться с процедурами по грибнице, предостав-
ленными производителем. Обычно компании-производители грибницы описывают луч-
шие условия и температуру для выращивания.
ПОДГОТОВКА СУБСТРАТА
Некоторые виды субстрата требуют предварительной подготовки. Солому необхо-
димо порубить. Бумагу порезать. Кукурузные стебли и стержни початков поло-
мать, Рис.1633. Лучшим оборудованием для такой работы может служить молотко-
вая дробилка с насадкой. Насадки для молотковой дробилки представляют собой
тяжелые стальные пластины с отверстиями. При помощи такой дробилки можно так
же обрабатывать небольшие куски древесины, ветки деревьев и т.д. существуют
несколько причин, по которым необходимо измельчать начальный материал. Наибо-
лее важная причина состоит в том, что измельчение позволяет увеличить поверх-
ность для прорастания мицелия, другая - сделать субстрат более компактным и
легким в использовании.
Следующее, что необходимо сделать, это увлажнение и пастеризация. Для этого
существуют два основных метода. Первый - так называемый метод горячей воды. В
соответствии с этим методом, вода нагревается в большом контейнере до темпе-
ратуры 55-60°С. НЕ БОЛЕЕ 60°С! Затем в воду добавляют материал для субстрата.
В идеале вода должна только покрывать субстрат. Оставить это на 30-60 минут.
33
Это тоже самое, что и рис.5.
НЕ БОЛЕЕ 60 МИНУТ! По истечении времени нужно слить воду и поставить субстрат
в такое место, где он будет медленно остывать. Через 16-20 часов он должен
остыть до температуры 25°С. По истечении этих 16-20 часов и при температуре
22-25° С субстрат будет засеян.
ТЕМПЕРАТУРА
ПАСТЕРИЗАЦИИ
Сравн: |
60
ПАСТЕРИЗАЦИЯ
Снижать:
Рис.17.
Метод горячей воды может быть использован для сырого субстрата, но в этом
случае требуется больше внимания, и теряются некоторые преимущества. Если
субстрат сырой, то вода должна быть ближе к 60°С, чем к 55°C. Необходимо вни-
мательно следить за температурой при добавлении субстрата и не позволять тем-
пературе опускаться ниже 55°С. При использовании сухого материала все, что
намокает при температуре воды 55-60°С, становиться пастеризованным. Если ма-
териал уже был сырым, то мы не можем быть полностью уверенными, что он равно-
мерно прогрелся. По этой причине его необходимо выдержать до полных 60 минут.
Еще одно преимущество метода горячей воды состоит в том, что в этом случае
материал легко увлажняется. Жар34 растворяет естественный растительный воск,
который мешает проникновению воды.
Второй метод - так называемый метод пропаривания. Субстрат необходимо намо-
чить за несколько дней до пастеризации. В течение этого времени инородные ор-
ганизмы могут начать прорастание, поэтому время должно быть ограничено. Затем
субстрат помещают в большой контейнер, возможно, использовать в этих целях
комнату, и подают пар со дна. В данном методе желательно перемешивание суб-
страта и контроль за тем, чтобы весь субстрат нагрелся до температуры 55-
60°С. В течение следующих четырех часов пар должен продолжать поступать и
держаться на температуре 55-60°С. НЕ БОЛЕЕ 60°С НА ПРОТЯЖЕНИИ ВСЕГО ВРЕМЕНИ И
В ЛЮБОЙ ТОЧКЕ! По истечению четырех часов субстрат оставляют остывать от 16
до 20 часов до температуры 25°С.
В методе горячей воды, тепло для нагрева воды может подаваться любым дос-
тупным способом. В методе пропаривания необходим пар.
34
Как это поэтично.
Я подчеркивал, что температура не должна превышать 60°. Это может показать-
ся странным, но опыт показывает, что более высокая температура будет способ-
ствовать развитию Триходермы, которая погубит посев. Кроме того, 55°С на про-
тяжении 30 минут убивают все вредные организмы в субстрате. Даже при условии,
что мы очень внимательны, мы не можем проследить, что пар равномерно нагреет
субстрат за такой короткий период времени, поэтому потребуется четыре часа.
Мы относим триходерму к оппортунистическим35 организмам. Существуют и другие
подобные организмы, но триходерма - наиболее опасна. Так же необходимо объяс-
нить лимит в 60 минут в методе горячей воды. Вода при этой температуре содер-
жит немного воздуха, и по истечении 60 минут в субстрате начинают расти ана-
эробные организмы, опасные для грибов.
ЗАСЕИВАНИЕ
После остывания субстрата до 25 ° С можно начинать засевание. На этом этапе
требуется соблюдение санитарных условий. Все в комнате должно быть чистым.
Все сотрудники, находящиеся в комнате, должны быть в чистой одежде и с чисты-
ми руками. Желательно наличие повязки на голове, хирургической маски и рези-
новых перчаток. Возможно, для комфорта рабочих потребуется вентиляция, воздух
должен поступать через фильтр, лучше использовать высокоэффективный сухой
воздушный фильтр (НЕРА filter). Полезно использование бактерицидной лампы в
течение ночи перед засеванием.
Субстрат перемешивают с грибницей и помещают в контейнеры для выращивания.
Точное соотношение грибницы и субстрата зависит определенным образом от типа
субстрата, но грибница должна составлять от 1 до 5% веса сухого субстрата.
Количество единовременно засеянной грибницы должно зависеть от того, как
быстро она может быть помещена в контейнеры для выращивания. Желательно, что-
бы субстрат был открытым в комнате как можно меньше времени, и чтобы контей-
неры были запечатаны как можно быстрее.
КОНТЕЙНЕРЫ ДЛЯ ВЫРАЩИВАНИЯ
Следующие виды контейнеров36 используются для выращивания грибов вешенок:
1. открытые (соломенные) тюки Рис. 18.
2. «поддоны» различных видов Рис. 19.
3. полиэтиленовые пакеты Рис. 20.
4. полиэтиленовые «рукава» Рис. 21.
Вероятно, бессмысленно пытаться выбрать из приведенного списка лучший или
худший тип контейнера. Лучший может легко сочетать в себе характеристики не-
скольких. Вероятно, правильный подход - перечислить желательные характеристи-
ки, о которых мы знаем. Контейнер должен:
А) вмещать субстрат в период разрастания грибницы
Б) препятствовать проникновению света, который может стать причиной прорас-
тания грибов в недоступных местах
В) не ломаться и не рваться в процессе работы
35 Да, с оппортунистами надо разбираться.
36 Еще удобны полиэтиленовые тазики, один снизу, другой сверху накрывает. В стенках несколько
отверстий.
Г) чрезмерно не нагреваться
Д) позволять получить максимальное количество продукции
Рис. 18. «Вешенка»
рост на полной кипе соломы.
Рис. 19. Поддон. Петля опытного образца слишком прекрасная37 для второго.
Внутри обтянут
полиэтиленом
37
Чево?
38
Мешок.
Рис. 20. Полиэтиленовая сумка38. А. На полках. В. Вешание С. Не-
легальный39 . D. Прозрачный.
Рис. 21. Полиэтиленовые «рукава» А. Основание сухой40 в отверстиях.
В. «вешенка» готовы. С. Горизонтальная стойка. D. Хорошее производ-
ство .
Получается, что контейнеры под номерами 3 и 4 соответствуют требованию А) ,
черный полиэтилен соответствует требованию Б). Требование В) зависит от того,
какой субстрат и в каком количестве используется. В этом случае полиэтилен
должен быть толщиной минимум 0,05 мм. Для соответствия требованию Г) (избежа-
ние самонагревания), контейнер не должен быть более 20 см в диаметре, незави-
симо от того располагается ли он горизонтально или вертикально. Даже соблюдая
лимит в 20 см, необходимо поместить разрастающуюся грибницу в прохладное ме-
сто с хорошей циркуляцией воздуха, таким образом, лимит в 15 см может быть
даже более практичным.
Наиболее сложным в определении и достижении является требование Д). Доволь-
но успешным является метод, в котором субстрат пакуют в полиэтиленовые «рука-
ва» и составляют на полках (развешивают на каркасах). Длинный поддон шириной
15-20 см с краями из проволочной сетки - одна из идей, которая используется и
может быть модифицирована. Если сетка достаточно грубая и покрыта черным по-
лиэтиленом изнутри, все 5 требований от А до Д могут быть соблюдены.
Другая идея состоит в том, чтобы пустить пластиковую трубку в центре суб-
страта . Трубка имеет множество отверстий для выхода тепла и углекислого газа,
и доступа кислорода. Такое устройство может иметь 30-40 см в диаметре. Если
трубка выступает по краям контейнера, то за нее можно горизонтально подвесить
контейнер, для обертывания субстрата в данном случае используются полиэтиле-
новые «рукава».
39
40
Час от часу не легче.
Чево?
Может непрозрачный?
РАЗРАСТАНИЕ ГРИБНИЦЫ
Разрастание грибницы - это период, когда мицелий растет и покрывает весь
субстрат. Во время этого периода углекислый газ является полезным, кислород
тоже необходим. Это так же период, во время которого из субстрата могут сте-
кать излишки воды. Нельзя допускать, чтобы вода аккумулировалась в контейне-
ре, поэтому необходимо небольшое отверстие для дренажа. После того, как мице-
лий начинает расти, необходимо большее отверстие для доступа кислорода. Лучше
всего расположить такое отверстие на верху.
Как отмечалось ранее, температура должна быть достаточно низкой, для избе-
жания самонагревания. Это температура между 15 и 20°С, но все зависит от суб-
страта и размера самого маленького контейнера. Необходимо наличие соответст-
вующей вентиляции для поддержания постоянной температуры. Доступ и свет необ-
ходимы только для контроля роста и температуры. Единственным источником света
в комнате может быть принесенный свет на батарейках (фонарь).
ПРОИЗВОДСТВО
Когда замечают хороший рост мицелия в субстрате, делают прорезы крест-
накрест по 2,5 см для того, чтобы грибы росли наружу. Флуоресцентные лампы и
вентиляция включены на полную мощность. Лучше всего расположить контейнеры на
высоте 10 см и более над полом. Если потребление электроэнергии ограничено, и
вентиляционная система не работает, углекислый газ будет собираться над по-
лом, и первое время не будет наносить большой вред. Расположение контейнеров
на некоторой высоте от пола так же будет препятствовать проникновению в них
некоторых более крупных вредителей.
После того как отверстия прорезаны, и особенно когда начинают формироваться
грибы, потребуется больше воды. Если в контейнере имеются большие отверстия,
можно добавлять воду через них, но так же поливать можно и там, где отверстия
предназначены для формирования грибов. Для этой процедуры лучше использовать
распылители, так как они позволяют распылять воду в небольших количествах и
снижают вероятность проникновения болезней. Что касается разбрызгивателей,
они не позволяют добавлять воду в достаточных количествах.
Признаком поспевания гриба является хорошо сформированная гимениальная пла-
стинка в то время, когда край гриба еще загнут вниз. Когда край выравнивает-
ся, освобождаются споры, гриб теряет вес. Освобожденные споры являются источ-
ником опасности, как для посева, так и для рабочих. Грибы не будут иметь хо-
роший вид, и не будут долго храниться. Грибы, собранные чуть раньше времени,
оставят больше питательных средств для следующей поросли, будут хорошо выгля-
деть максимальное кол-во времени и не приведут к проблемам со здоровьем рабо-
чих и вашего посева. Чуть более длительное ожидание может позволить получить
чуть больший урожай с этой поросли, но никак не с последующих. Если вы ждете
еще один день, можно прогнозировать уменьшение веса урожая и возникновения
множества проблем.
Необходимо обеспечить всех работающих на территории выращивания масками,
резиновыми перчатками и сапогами, которые прошли обработку раствором поварен-
ной соли или гипохлорита (отбеливатель для белья). Одежда и руки должны быть
чистыми для предотвращения переноса болезней и паразитов.
Собирая урожай, необходимо выдергивать грибы из субстрата. Если их срезают,
то срезанные поверхности, которые остаются в субстрате являются идеальным ме-
стом для развития триходермы (зеленой плесени). Необходимо очистить грибы пе-
ред отправкой на рынок.
Практика показывает, что вторая поросль (примерно через 10 дней после пер-
вого урожая) будет самой крупной. Не исключено, что и первая и третья поросль
может быть самой крупной. В общем, желательно ликвидировать оставшийся суб-
страт после третей поросли, но иногда его сохраняют и для четвертой.
Хотя грибы будут продолжать появляться после третей поросли, и обычно даже
после четвертой будет прорастать достаточное их количество, существуют неко-
торые причины, по которым рекомендуется закончить процесс. Во-первых, каждый
день дает болезням и паразитам больше и больше времени для проникновения и
укрепления. После того как они укрепились их труднее вывести и уберечь от них
следующий посев.
Для следующего посева необходимо свободное пространство.
Стоимость уборки урожая зависит в основном от площади, поэтому уборка уро-
жая на новом субстрате будет дешевле на каждый килограмм.
ОБРАБОТКА ПОСЛЕ СБОРА УРОЖАЯ
После того, как собрали урожай с последней поросли, необходимо произвести
уборку комнаты для выращивания. Традиционный метод представляет собой обра-
ботку комнаты паром при температуре 60°С в течение четырех часов. После чего
выносят и вывозят все использованные материалы. В данном случае превышение
нормы температуры и времени будет просто неэкономным, а в очень редких случа-
ях может привести к некоторым проблемам.
Вывоз использованных материалов предполагает их удаление на расстояние не-
скольких километров от территории разведения грибов. В процессе выращивания
грибов вешенок, при условии соблюдения всех оптимальных условий, на заключи-
тельном этапе остается очень маленькая часть начального субстрата. Заключи-
тельная обработка должна уничтожать все болезни и паразитов, впрочем, разме-
щение отходов на достаточном расстоянии от территории выращивания грибов
обеспечит устранение болезней и паразитов с производственной территории.
ПОДГОТОВКА К СЛЕДУЮЩЕМУ ПОСЕВУ
После вывоза всех остаточных материалов, необходимо тщательно очистить ком-
нату. Необходимо тщательно промыть раствором гипохлорита все поверхности и
предметы в комнате. Раствор должен содержать 0,525% гипохлорита натрия или
10% отбеливателя для белья. Дерево и некоторые другие материалы могут скры-
вать в порах болезни и вредителей. Наиболее легкий и безопасный способ обра-
ботки помещений с такими поверхностями - пропаривание в течение 6 и более ча-
сов. Можно использовать формальдегид, метил бромид и некоторые другие фуми-
ганты, но они представляют опасность для здоровья, поэтому те, кто будут с
ними работать должны быть хорошо обучены. Гипохлорит так же может вызвать
ожог, поэтому его так же необходимо использовать очень осторожно. При его ис-
пользовании в помещении должна работать вентиляционная система, пары гипохло-
рита могут повредить легкие.
ЗАКЛЮЧЕНИЕ
Каждый этап выращивания грибов требует предельного внимания. Производство
по максимуму возможно при наличии хорошего оборудования и соблюдении стан-
дартных процедур. Для достижения высокого уровня производства особенно необ-
ходимо соблюдать предельное внимание на этапах пастеризации, засевания и об-
работки после сбора урожая. Тем не менее, если выбран плохой субстрат или
грибница, производство так же будет низким, независимо от соблюдения всех ус-
ловий. Достаток воды так же очень важен.
От метода при сборе урожая зависит количество и качество грибов, а так же
здоровье рабочих. Грибы необходимо собрать, прежде чем они выпустят споры.
Если это сделано вовремя, грибы будут больше весить и дольше храниться, а
споры не перенесут вирусы на другие грибы, и рабочие не приобретут респира-
торных проблем (астма, сенная лихорадка) от спор в воздухе, которым они ды-
шат .
ГЛАВА 4
УПАКОВКА И ХРАНЕНИЕ
Основная причина выращивания грибов - продать их и получить прибыль. Если
мы считаем, что первый этап в выращивании - это подготовка помещения, то по-
следний, очевидно, сбор урожая. Но сбор урожая можно так же рассматривать как
первый шаг в подготовке к продаже. Тогда процесс подготовки к продаже выгля-
дит следующим образом:
1. Сбор грибов
2. Охлаждение (3 - 5 ° С)
3. Очистка
4. Упаковка
5. Продажа
При сборе урожая грибы нужно выдергивать. Нельзя их срезать. Очень важно,
чтобы при сборе край шляпки гриба был Загнут вниз. Когда края становятся ров-
ными, в воздух выпускаются споры, что влечет за собой следующие две крупные
проблемы. Споры в воздухе представляют опасность для рабочих, их попадание в
дыхательные пути может стать причиной таких опасных заболеваний, как сенная
лихорадка или астма. Так же споры представляют значительный вес продукта, ко-
торый теряется. Очень часто при сборе грибов допускаются ошибки. Даже если вы
считаете, что собираете весь урожай вовремя, то есть до освобождения спор,
необходимо надевать марлевую повязку. Еще одна причина раннего сбора состоит
в том, что после того, как выпущены споры, у гриба более нет биологической
цели, и он, таким образом, начинает быстро угасать и очень скоро выглядит
старым.
Охлаждение позволяет дольше хранить грибы, а так же сохранить их привлека-
тельный вид. Лучший способ охладить грибы - поместить их в камеру и создать
вакуум. Такой способ используется для листовых овощей. Конечно, требуется до-
рогостоящее оборудование, но этот метод позволяет остудить быстрее, чем какой
либо другой. Охлаждение - результат испарения воды. Быстрый способ охлаждения
позволяет дольше сохранить продукт привлекательным для покупателей. Другие
способы охлаждения - механическая рефрижерация или использование контейнера
со льдом. Но необходимо помнить, что после сбора грибы нельзя обрабатывать
при помощи воды, включая растаявший лед и т.д.
Грибы вешенки более ломкие, чем шампиньоны (Agaricus). Грибы принято соби-
рать в большие ящики, в отношении вешенок это не мудро. Шампиньоны (Agaricus)
можно даже пересыпать из ящика в ящик. Но вешенки при таком обращении полома-
ются и потеряют вид.
СТЕБЛИ41 И ОБРАБОТКА
Так как грибы выдернули из субстрата, он может в небольших количествах ос-
таться на грибах. Такие участки нужно обрезать. У большинства вешенок
(Pleurotus) будет стебель. Стебли обычно плохо жуются42 и поэтому не привле-
кательны для покупателей. Поэтому, если стебли обрезаны, у вас будет лучший
товар. Конечно, стебли добавляют вес, и если покупатели не осознают, что по-
купают лучший товар (если стебли обрезаны), то возможно будет мудрее стебли
оставить. Стебли можно использовать в производстве других продуктов, но это
требует дополнительного оборудования и рабочих. При наличии небольшого коли-
чества стеблей это не практично.
УПАКОВКА
Если понаблюдать за средним покупателем, то можно заметить, он обычно поку-
пает продукты, которые, по его мнению, выглядят привлекательно. Это касается
почти всех покупаемых товаров. Если вы не обрезаете стебли, нужно сделать
так, чтобы они не портили внешний вид ваших грибов. При приготовлении различ-
ных блюд мы разбиваем яйца, но как много людей купили бы в магазине разбитые
яйца? Грибы могут быть сломаны и повреждены, если их трогают и перебирают, но
привлекательная упаковка поможет продать больше грибов, чем без упаковки.
Некоторые типы продаж не требуют привлекательной упаковки. Обычно продажа
большого количества в рестораны идет без упаковки. Повара или другие работни-
ки ресторана обычно считают, что создавать привлекательный вид продукту - это
их работа. Так же они не хотят открывать много мелких упаковок. Но, тем не
менее, они не сильно отличаются от всех остальных людей, поэтому, наверное,
будет легче продать им товар, если в первый раз они видят его в привлекатель-
ной упаковке.
Наиболее распространена практика упаковывать грибы в пластиковые или бумаж-
ные поддоны и обтягивать их полиэтиленовой пленкой. Пленка защищает их от рук
покупателей и удерживает влажность. Грибы живы43, пока хорошо выглядят. Плен-
ка так же не пропускает кислород, необходимый для живых грибов, поэтому необ-
ходимо оставить в упаковке отверстие для поступления кислорода. Обтянутые
пленкой поддоны использовались для упаковки шампиньонов (Agaricus) более 30
лет, а теперь их используют практически для всех видов грибов.
Комнаты, где происходит упаковка, должны быть чистыми и удобными для рабо-
чих, но не требовать никаких сложных разработок.
41 Ножки.
42 Между прочим, в жареном виде очень вкусны.
43 Как это верно.
ПЕ РЕ П Р0ИЗВ0ДС ТВО
Иногда производитель грибов не может выполнить норму необходимую для удов-
летворения спроса на грибы несколько месяцев, потом вдруг дела могут пойти
неожиданно хорошо и урожай превосходит все ожидания. Так же бывает время, ко-
гда спрос на грибы внезапно снижается. Например, в некоторых местах в изоби-
лии появляются дикие грибы и люди начинают собирать их, или покупают их у
тех, кто собирает. В этих случаях существует вероятность, что грибы не будут
проданы.
Не зависимо от того, по какой причине предложение превышает спрос, грибовод
захочет сохранить имеющиеся излишки, чтобы продать их позже. Часто грибы кон-
сервируют , то есть запечатывают в стеклянный или металлический контейнер и
стерилизуют при высокой температуре. Некоторые грибы, включая шампиньоны
Agaricus, достаточно хорошо сохраняются в процессе такой обработки. Но вешен-
ки после остужения в воде имеют неприятный вид. Вешенки, как большинство гри-
бов, будут выглядеть хорошо, если их засушить. Может показаться странным, но
шампиньоны Agaricus - один из грибов, не приобретающих приятный вид в процес-
се высыхания. Когда шампиньон Agaricus сушат, большая часть твердого вещества
стекает с влагой.
Третий метод - замораживание. Ни один из сортов грибов не выглядит привле-
кательно в замороженном виде, но, тем не менее, цветная непрозрачная упаковка
может сделать продукт более привлекательным.
ПРОЦЕСС СУШКИ
Разработано много методов сушки продуктов питания: солнце, духовые шкафы,
вакуум и т.д. все они применимы к грибам. Самым высоким качеством обладает
процесс заморозки грибов, затем помещения их в вакуум, где они остаются замо-
роженными до полного высыхания. Этот процесс называется сушка сублимацией и
является очень дорогостоящим как по затратам энергии, так и по необходимому
оборудованию. Наиболее подходящей является туннельная сушка. В результате
туннельной сушки получается продукт высокого качества. Сушилку можно сконст-
руировать из обычных материалов, она потребляет намного меньше энергии, чем
сушилки других видов.
В некоторых странах производитель грибов имеет возможность найти компанию,
которая занимается засушкой других продуктов и заключить с ней договор.
КОНСТРУКЦИЯ ТОННЕЛЬНОЙ СУШИЛКИ
Туннельная сушилка состоит из вентилятора для циркуляции воздуха, нагрева-
теля для поддержания температуры до примерно от 40 до 50°C, мест для размеще-
ния продукта предназначенного для сушки и задвижки для контроля свежего воз-
духа и выхода влажного воздуха. Так же необходимы термометры, гидрометры и
термостаты для контроля температуры и влажности. Рециркуляция воздуха не
только экономит топливо для нагрева, но и позволяет продукту быстрее высы-
хать. Схемы двух сушилок различных размеров показаны на рисунках, Fig44. 3-7,
3-8, 3-9. на первом рисунке показано поперечное сечение (Fig. 3-7), так же
как и на скомбинированном виде, Fig. 3-8. Изучив рисунки, можно увидеть, что
44 Все рисунки, обозначенные как Fig, отсутствовали, поэтому были удалены даже упоминания о
них.
если вторая построена в зеркальном отражении первой, у них может быть общая
стена, а доступ может быть в стенах напротив.
ВЫСУШЕННЫЙ ПРОДУКТ
Как и свежие, высушенные грибы должны иметь привлекательный для покупателей
вид. Если они упакованы в прозрачные полиэтиленовые пакеты, они хорошо выгля-
дят. Но нужно помнить, что высушенные грибы так же очень хрупкие, поэтому мо-
жет быть выгоднее упаковать их как свежие - на поддоне, обернутые полиэтиле-
ном.
Если грибы высушены при низкой температуре в туннельной сушилке, у них бу-
дет несильный запах. Некоторые люди будут недовольны этим запахом. Возможно,
необходимо будет пересмотреть условия продажи таким клиентам.
ЗАКЛЮЧЕНИЕ
Производители грибов должны избегать потерь и продавать наилучший товар.
Необходимо собирать грибы в установленное время, обращаться с ними осторожно,
упаковывать и сохранять все те, которые невозможно продать в свежем виде.
Собирание влияет на количество и качество. При обработке может получиться
небольшая разница в количестве, но она будет иметь большое влияние на качест-
во . Упаковка так же очень сильно влияет на качество. Хранение может сильно
влиять на количество и последующий доход.
От редакции
Данный материал был найден вот на этом сайте:
http://www.oystermushrooms.net/Content.html. Там же было сказано:
«Копирование данного CD разрешено только в том случае, если в процессе ко-
пирования не вносятся изменения. Копии не подлежат продаже, но допускаются
небольшие сборы для погашения затрат, связанных с распространением и произ-
водством копий». Ну, насчет небольших сборов мы как-то не прониклись, в вот
пункт «в процессе копирования не вносятся изменения» мы постарались соблюсти
и дали текст, так как он есть. Ну, или почти так как он есть - снабдив кое-
где комментариями для лучшего понимания.
Акцент
МУРАВЬИ
- ГРИБОВОДЫ
Учёные разобрались, как американские
муравьи научились выращивать целые
грибные плантации. Чтобы научиться по-
могать друг другу, муравьям и грибам
понадобился общий враг и секретное ору-
жие - бактерии. Те же бактерии для за-
щиты от врагов через миллионы лет нау-
чился использовать человек.
То, что насекомые возникли за миллионы лет до человека, отнюдь не делает их
менее совершенными. Социальной организации колоний пчёл или муравьев может
позавидовать самая совершенная демократия. И эксплуатировать другие виды они
могут не хуже человека, при этом от охоты и собирательства к «скотоводству» и
сельскому хозяйству они перешли за миллионы лет до нас. Учёные разобрались,
как муравьи сделали это.
Кроме человека и муравьев выращивать что-то для пропитания умеют ещё терми-
ты и короеды, однако именно муравьи достигли в этом деле наибольшего совер-
шенства. Эта их уникальная черта была подмечена ещё в конце позапрошлого ве-
ка . И если для нас удивительно само по себе умение, то энтомологи-
эволюционисты смогли выделить пять отдельных сельскохозяйственных систем и
проследить эволюцию этих способностей.
Конечно, муравьи выращивают не злаки и не фруктовые деревья - им вполне
достаточно грибов; кстати, это одна из последних культур, освоенных челове-
ком.
Ученые полагают, что исходная форма сельского хозяйства возникла около 50
миллионов лет назад, когда «культурными» стали грибы семейства
Leucocoprineae. За последние 30 миллионов лет из этой системы возникли ещё
три, а еще несколько миллионов лет спустя одна из этих систем дала начало са-
мой современной - культивированию грибов муравьями-листорезами, появившимися
8-12 миллионов лет назад и ставшими доминирующими травоядными в тропиках Но-
вого Света.
Эти выводы ученые сделали после тщательного анализа поведения, особенностей
биохимических процессов и генетических исследований, давших им возможность
восстановить филогенетическое дерево и заполнить пробелы истории - предполо-
жить наличие общего предка.
Все семейство Attini состоит из 230 видов, которые преимущественно населяют
тропики Нового Света. Все виды этого семейства облигатно, то есть полностью,
Зависят от выращиваемых грибов. Сложившийся мутуализм настолько силен, что
новая муравьиная королева, когда организует новую колонию, обязательно пере-
носит туда частицы гриба, достаточные для того, чтобы возделать новый сад.
Самые совершенные из всего семейства - муравьи-листорезы, или зонтичные.
Если все остальные муравьи выращивают грибы так же, как человек растит свои
вешенки и шампиньоны, то есть на органическом детрите, то листорезы разбивают
целые сады, самостоятельно перерабатывая живые листья, цветы и траву, из ко-
торых и производят субстрат для своих подопечных.
При этом грибы, культивируемые зонтичными муравьями, не встречаются в при-
роде самостоятельно - они не способны жить на другом субстрате, кроме специ-
ально созданного для них. Взамен они образуют вздутия гиф (растущих нитей
гриба), богатые питательными веществами, которые собирают муравьи.
Остальные современные схемы «грибоводства» муравьев Attini отличаются вида-
ми культурных грибов и, как уже упоминалось, используемым субстратом. Авторы
работы, Тед Шульц и Шон Брэди, даже вспомнили сравнение колонии муравьев с
большим травоядным. Если учесть его биомассу, продолжительность жизни и коли-
чество поглощаемой травы, то сходству помешает лишь необычный внешний вид
этого «синтетического животного».
Происхождение сельского хозяйства даже несколько более сложное, чем может
показаться на первый взгляд.
К мутуалам относятся муравьи семейства Attine, их культурные грибы
(Leucocoprineae и Pterulaceae) и бактерии рода Pseudonocardia
(Actinomycetes), живущие в пищеварительном тракте муравьев. Паразит - гриб
рода Escovopsis (Ascomycetes) - известен только как «вредитель» в садах куль-
турных грибов.
Синтезируемые бактериями-симбионтами антибиотики помогают частично спра-
виться с вредителями, не поражая «культурные» грибы. Человечество «приручило»
бактерий этого же рода для производства стрептомицина и неомицина только во
второй половине прошлого века. Кстати, это ещё один пункт, по которому мура-
вьи существенно обогнали человека.
Шульц и Брэди считают, что, как и у термитов (и в отличие от человека и жу-
ка-короеда) , сельское хозяйство возникло у муравьев один раз - около 50 мил-
лионов лет назад, а затем уже изменялось и приобретало новые формы, притом
каждая новая форма тоже возникала однократно.
Более того, такая форма поведения вторична - ученые доказали, что 73 мил-
лиона лет назад существовал предок, питавшийся не только за счет сельского
хозяйства и давший начало всему многообразному семейству Attini. Так что му-
равьи на несколько миллионов лет обогнали человечество в эволюционной гонке.
Правда, это им удалось не за счет преимущества в числе конечностей, а благо-
даря нужным встречам, устроенным самой природой.
Юмор
ЧТО ДОЛЖЕН ЗНАТЬ КАЖДЫЙ ТАРАКАН
ОБ ОРУЖИИ МАССОВОГО ПОРАЖЕНИЯ
(ПАМЯТКА)
ВВЕДЕНИЕ
Помните, как радовались все прогрессивные тараканы, когда в нашу жизнь во-
шел человек? Казалось, он навсегда решит все бытовые проблемы - всюду будет
тепло, будет грязь, дешевая еда. Но, как выяснилось, человек может быть не
только другом, но и смертельно опасным врагом, когда выходит из-под таракань-
его контроля. И тогда в наш дом приходят болезни, разрушения, смерть.
Достаточно вспомнить хотя бы, сколько бед принесли тапки "Малыш" и "Тол-
стяк", сброшенные с ног на головы мирных жителей Кухни и Ванной в 1945 году!
Сегодня на Земле уже накоплено столько смертоносной обуви и отравы, что можно
было бы сотни раз уничтожить все тараканчество!
Кратко рассмотрим основные виды оружия массового поражения (ОМП) и правила
поведения тараканов в случае возникновения человека.
ТАПОК
Тапок (ботинок, сапог, сандаль, лапоть, костыль) - самый старый и самый
распространенный вид ОМП. Поражающее действие тапка основано на избыточном
давлении, возникающем в районе тапкового удара. Так, давление в эпицентре па-
дения тапка настолько велико, что от таракана, подвергшегося тапковому удару,
остается буквально мокрое место. Первым сигналом опасности при тапковой атаке
является световое излучение сверху, появляющееся после характерного щелчка.
Вслед за излучением раздается характерный звук "У, С-С-СВОЛОЧИ! ! ! ’’, после че-
го в воздухе появляется тапок.
Таракан, первым заметивший излучение, должен подать команду "ВСПЫШКА СВЕР-
ХУ!!!". По этой команде все тараканы обязаны прекратить любые работы по зага-
живанию кухни, немедленно покинуть объект и проследовать в укрытие!
ЗАПОМНИТЕ! Обыкновенные обои ослабляют силу тапкового удара в 3-5 раз. Ли-
нолеум - в 10 раз. Плинтус деревянный - в 100 раз. Железная сковорода полно-
стью защитит вас от тапкового удара, но не защитит от удара сковородой.
Сильный испуг удесятеряет силы. Известен случай, когда один внезапно напу-
ганный таракан перевернул тарелку и укрылся под нею. (К сожалению, горячий
борщ, находившийся в тарелке, оборвал жизнь безымянного героя.)
Если вы не успеваете покинуть объект, следует, поджав усы и лапки, притво-
риться едой противника ("изюм"). Примером подлинного мужества стал подвиг та-
раканов-героев братьев Прусаковых, которые в неравном бою сумели подбить 2
вражеских тапка. Обвязавшись спичками, они самоотверженно бросились под тапки
и подожгли их.
ДИХЛОФОС
Дихлофос ("Baygon", "Raid", "Egoiste") - химическое ОМП. Основными поражаю-
щими факторами дихлофоса являются: Звуковая волна ("ПШИК"), НАПРАВЛЕННОЕ ДИХ-
ЛОФОСНОЕ ИЗЛУЧЕНИЕ (АЭРОЗОЛЬНАЯ ВОЛНА) , ОСТАТОЧНЫЙ ЗАПАХ (ВЫСЕЛЯЮЩИЙ ГАЗ) .
Звуковая волна распространяется со скоростью звука по всей кухне, поражает
незащищенные уши тараканов, сеет болезнетворные децибелы. НАПРАВЛЕННОЕ ДИХЛО-
ФОСНОЕ ИЗЛУЧЕНИЕ и ОСТАТОЧНЫЙ ЗАПАХ вызывают у тараканов головокружение и
галлюцинации, временную нетрудоспособность. Как правило, у особей, регулярно
подвергающихся воздействию дихлофоса, возникает психофизиологическая зависи-
мость от него. Таким тараканам с каждым разом требуется все большая доза от-
равляющего вещества. Потомство таких тараканов, как правило, тоже становится
токсикоманами ("садятся на баллон"). Для защиты от дихлофоса следует исполь-
зовать складки противогаза, респиратора. Если под лапой не оказалось противо-
газа, можно воспользоваться влажной половой тряпкой или использованной туа-
летной бумагой.
Огромную роль играет правдивая и своевременная информация. Помните перво-
майскую демонстрацию на Кухне в 1986 году? Тысячи тараканов шли по полу, улы-
бались, не зная, что содержание выселяющего газа в воздухе превышает все до-
пустимые нормы. А руководители уже все знали и, тем не менее, тоже улыбались
демонстрантам, махали усами с хлебницы!..
ЧИСТОТА
ЧИСТОТА является самым подлым и грозным ОМП. Следите За тем, чтобы чистота
не проникала в места вашего постоянного пребывания. Чистота наводится на жи-
лище тараканов посредством веников, швабр, тряпок. Самым страшным носителем
чистоты является так называемая "чудо-швабра" (по умойной силе превосходит
веник в 100 раз, обычную швабру в 10 раз, тряпку половую мокрую - в 1 раз).
ЧТОБЫ УБЕРЕЧЬСЯ ОТ ЧИСТОТЫ, старайтесь не провоцировать человека - не до-
пускайте небрежности в обращении с собственными фекалиями! Никогда не остав-
ляйте их на полках, посуде, полотенцах. Для хранения фекалий вполне годятся
гречневая крупа, чай, сухари. Если ЧЕЛОВЕК разжег духовку газовой плиты, в
которой вы живете, надо помнить следующее:
1. Основной поражающий фактор духовки - повышенная температура, от которой
остаются ожоги на морде, животе, чреслах.
2. Как известно, у таракана шесть лап. Двумя верхними прикройте морду. Дву-
мя средними - живот и чресло. Убегайте на двух задних.
ЭВАКУАЦИЯ
План эвакуации надо повесить на видном месте (за плинтусом, возле помойного
ведра, на задней стенке шкафа с продуктами), но не на глазах у человека!
При эвакуации следует использовать заранее разведанные вентиляционные ходы
полного профиля. (Не загромождайте их!) В таких ходах можно идти не сгибаясь,
в полный рост. По дороге нужно обозначать небольшими темными шариками места
привалов, повороты, места отдыха и приема пищи.
Зоны, зараженные китайским карандашом, следует преодолевать, не прикасаясь
к ним ногами (перепрыгивать, перекатываться на спине, перелетать на мухах,
комарах). Ни в коем случае не пейте сырую воду из-под крана! Во-первых, она
кишит бактериями, а во-вторых, вас может смыть в канализацию! Поэтому пить,
мыться следует только в водоемах с кипяченой водой - в чайниках, кастрюлях,
тарелках. При этом на берегу должны оставаться двое дежурных - санинструктор
и спасатель. Помните, что сориентироваться в сложной ситуации, не поддаться
панике вам поможет радио и телевидение. Забравшись в телевизор или радиопри-
емник, вы сможете укрыться от воздействия любого ОМП. Особям, подвергшимся
воздействию ОМП, следует следовать в ближайший вычислительный центр. Там, в
системном блоке компьютера, удобно устроившись у вентилятора, они смогут бы-
стро поправить подорванное здоровье.
ИЗУЧИВ ЭТУ ПАМЯТКУ, ПЕРЕДАЙТЕ ЕЕ СОДЕРЖАНИЕ ДРУГИМ ТАРАКАНАМ ИЗ УС В УСА!