



















































































































Автор: Моисеева Н.И. Сысуев В.М.
Теги: характер изменения свойств, биологических явлений медицина биология физиология
Год: 1981




Текст
Академия наук СССР
Научный совет по проблемам прикладной
физиологии человека
Институт эволюционной физиологии и биохимии
им. И, М. Сеченова
Н. И. МОИСЕЕВА, В. М. СЫСУЕВ
ВРЕМЕННАЯ СРЕДА
И БИОЛОГИЧЕСКИЕ
РИТМЫ
И
ЛЕНИНГРАД
«НАУК А»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1981
УДК 57.034
Временная среда и биологические ритмы. Н. И. Моисеева, В. М. Сы-
суев.— Л.: Наука, 1981. — 128 с.
В монографии содержится теоретическое и экспериментальное рассмотрение
биоритмов в диапазоне колебаний с периодами от нескольких секунд и ми-
нут до циркадных и многодневных ритмов. Показано наличие циркадной
и ультрадной ритмичности многих физиологических функций, связанных
с осуществлением адаптационно-приспособительной деятельности организма,
описано большое число используемых конкретных методик оценки функцио-
нального состояния как вегетативной, так и психоэмоциональной сферы
испытуемых. Авторы ставят и решают задачу определения информативности
временных характеристик биоритмов для оценки функционального состоя-
ния организма как в ходе адаптационного процесса, при изменении'парамет-
ров внешних социальных и геофизических синхронизаторов, так и в про-
цессе лечения при неврологических заболеваниях. Книга предназначена для
физиологов, биологов, клиницистов и врачей спортивной медицины. Лит. —
190 назв., ил. — 15, табл.— 40.
Ответственный редактор
А. А. САВИЧ
М
50300-506
055(02)81
518-81 2007020000
© Издательство «Наука», 1981 г.
ПРЕДИСЛОВИЕ
Существование живых организмов в сложной и динамичной
среде обитания возможно лишь благодаря непрерывному взаимо-
действию со средой, непрекращающемуся процессу адаптации
к меняющимся внешним условиям. Ритмический характер течения
процессов в организме известен с глубокой древности — о нем
знали не только в Древней Греции, но и в Древнем Египте и
Месопотамии. Средневековые последователи Платона, например
Парацельс, уже рассматривали зависимости течения заболеваний
от экзогенных ритмов (Jovanovic, 1977). Однако учение о биоло-
гических ритмах, имеющее столь глубокие корни, возникло и
оформилось только во второй половине XIX—начале XX в.
В 1935 г. в Швейцарии было создано первое Международное об-
щество исследования биологических ритмов, преобразованное
в 1971 г. в США в Международное общество хронобиологии.
Одним из основных вопросов биоритмологии является вопрос
о происхождении биологических ритмов и способах их регуляции.
По существу никто давно не сомневается в том, что эндогенные
механизмы осуществляют как формирование, так и постоянную
регуляцию биологических ритмов и что эта регуляция осущест-
вляется под влиянием «меток времени», поступающих из внешней
среды. Однако вопросы об источниках этих экзогенных влияний
и механизмах их восприятия являются далеко не полностью
разработанными.
Если первоначально в качестве времязадающего фактора внеш-
ней среды рассматривалась преимущественно смена освещенности,
то в настоящее время внимание исследователей привлекает широ-
кий спектр гелиогеофизических и даже космических факторов.
Анализируя значение гелиогеофизических факторов в регуля-
ции биоритмов, мы обнаружили, что их влияние осуществляется
различным образом. Так, изменения магнитного поля Земли
влияют на формирование ритмичности физиологических процессов,
в то время как изменения солнечной активности (и связанные
с ней погодные факторы) преимущественно модулируют биологи-
ческие ритмы.
3
Сформированные в процессе онтогенетического развития биоло-
гические ритмы организма образуют сложную систему, в которой
ритмы с большим периодом модулируют ритмы с меньшим перио-
дом. Согласно нашим представлениям, эта система формируется
на основе индивидуальной временной шкалы данного организма.
Масштаб и вариабельность этой собственной временной шкалы
человека и соответственно его субъективного ощущения течения
времени зависит от его индивидуально-типологических особен-
ностей, от общего функционального состояния организма и от
интенсивности интеллектуально-мнестической деятельности, опре-
деляемой по объему переработанной информации и произведенной
работы.
Постоянно падающие на организм воздействия могут возникать
закономерно, согласно определенному стереотипу или, напротив,
быть непредсказуемыми (стрессорные факторы). Для того чтобы
оптимальным образом отреагировать на закономерно наступающие
изменения внешней среды, надо также обладать широким «выбором»
значений функций, но включать регулирующие механизмы, позво-
ляющие достичь этих значений, не в момент воздействий, а пред-
варительно, «упреждая» воздействие. Для того чтобы оптималь-
ным образом отреагировать на непредсказуемое воздействие, в ор-
ганизме должна быть развита способность быстро найти среди
имеющегося диапазона степени выраженности функций те значе-
ния, которые наиболее адекватны данной ситуации.
Предупреждающее реагирование осуществляется за счет пе-
риода и фазы того или иного ритма, обеспечивающих максималь-
ные функциональные возможности в определенное время суток,
и чем лучше организованы суточные кривые, тем выше адапто-
способность. Адекватная реакция на непредсказуемые воздействия
обеспечивается за счет амплитуды колебаний, и чем больше их
размах, тем большим «выбором» обладает организм и соответ-
ственно тем более адекватна его реакция.
Учитывая эти особенности, мы предложили использовать
структуру биологических ритмов в качестве критерия адапто-
’ способности и тем самым осуществлять с их помощью прогноз
как профессиональной пригодности к работе в экстремальных
условиях, так и исхода заболевания (Моисеева, 1978а, 19786).
Хорошими прогностическими признаками являются: 1) четкая
организация суточной кривой; 2) относительно высокие средние
значения и разброс этих значений в течение суток; 3) относительно
постоянное положение акрофазы при повторных исследованиях
в течение нескольких дней.
В заключение следует сказать, что изучение биологических
ритмов дает в руки врача и биолога новый инструмент для оценки
функционального состояния организма, позволяющий изучать из-
менчивость, т. е. закономерности отклонений от среднего состоя-
ния, что гораздо важнее, нежели изучение самого среднего состояния.
Глава 1
РИТМИЧЕСКИЙ ХАРАКТЕР
БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
КАК СПОСОБ ОРГАНИЗАЦИИ
ЖИВОЙ СИСТЕМЫ ВО ВРЕМЕНИ
Любые поведенческие акты и проявления биологической актив-
ности в той или иной мере отражают в своей структуре характе-
ристики внешних, в том числе периодических, воздействий среды.
Организация ритмов в процессе эволюции функций является
одним из ведущих механизмов приспособления органической
природы к неорганической среде (Войно-Ясенецкий, 1974), и соот-
ветственно от периодических режимов не свободен ни один живот-
ный и растительный организм. Постоянная работа организма
против равновесия, требуемого законами физики и химии (Бауэр,
1935), и колебательные режимы рассматриваются как фундамен-
тальное свойство всего живого (Bunning, 1964; Halberg, 1964) и
как наиболее специфическая черта обмена веществ биологических
объектов (Энгельгардт, 1960).
Более того, имеем ли мы дело с живой или неживой природой,
с увеличением размерности пространства вероятность достижения
любой произвольной траекторией движения точки «равновесия»
крайне мала. Тем самым любая перестановка частей достоверно
ведет к появлению циклов (Гончаров, 1944; Rubin, Sitgreaves,
1954; цит. по: Goodwin, 1966).
Ритмические изменения интенсивности функциональной актив-
ности любого рода (и на любом уровне функциональной органи-
зации) представляют собой универсальную форму постоянного
реагирования на воздействия внешней среды — так называемое
«опережающее реагирование». Возможность оптимальным образом
отреагировать как на предсказуемое, так и на непредсказуемое
воздействие в 'значительной мере определяется наличием в орга-
низме широкого диапазона степени выраженности функций и способ-
ностью организма найти среди этого диапазона те значения, кото-
рые наиболее адекватны данной ситуации.
Подбор оптимальных для данных условий значений параметров
физиологических функций представляет для организма немалую
функциональную нагрузку, что видно, например, из опыта при-
менения для лечения «Биотрона» — специальной изолированной
камеры, в которой созданы постоянные условия внешней среды,
т. е. поддерживаются на строго заданном уровне температура,
влажность, атмосферное давление и т. д. Помещенные на 2 нед
5
в камеру больные, страдающие гипертонической болезнью, язвой
желудка и некоторыми другими заболеваниями, оказываются
как бы «освобожденными» от необходимости осуществлять работу
по «регулированию отношений с внешней средой». Они хорошо
поправляются без применения каких-либо лечебных средств
(Панченко и др., 1964; Панченко, 1972). Таким образом, ежеднев-
ная работа по уравновешиванию во внешней среде требует значи-
тельных энергетических затрат.
Как известно, эффект любого воздействия, вызвавшего ответ
биологической системы, выражается в виде усиления распада
веществ, после чего идет интенсификация их синтеза. Но при этом
благодаря ритмическому «перемещающемуся» функционированию
каждая функциональная структура (часть органа, клетка, орга-
нелла) может восстанавливать свой трофико-пластический потен-
циал непосредственно в процессе работы (Саркисов и др., 1975;
Саркисов, 1977). Эту закономерность функционирования клеток
и тканей Н. Г. Крыжановский (1974) назвал законом «перемежаю-
щейся активности функциональных структур».
На уровне единичной клетки, как показано Гудвином (Good-
win, 1966), все виды молекул постоянно находятся в динамическом
состоянии, т. е. распадаются и синтезируются, что влечет за собой
незатухающие колебания концентрации молекул. Циркадные
ритмы обнаружены также у одноклеточных (Schweiger, 1978).
Циклическое течение процессов распространяется на нейрон-
ные сети; в них выделены два типа связей — импульсная и цик-
лическая. В последнем случае взаимодействие между двумя обра-
зованиями нервной системы осуществляется в виде правильных
ритмических колебаний потенциала (Русинов и др., 1970), кото-
рые рассматриваются М. И. Ливановым и др. (1974) в качестве
одного из основных механизмов интеграции нейронов.
Биоритмы функциональных систем связываются с процессами
квантования потоков информации, что позволяет согласовывать
процесс обработки информации с процессами обмена и трофиче-
скими функциями (Василевский, Трубачев, 1977). При нарушении
согласованности соответственной ритмической активности с внеш-
ними «датчиками времени» возникают ультраструктурные изме-
нения в виде нарушений информационных связей между ядром
и цитоплазмой (Манина, Моисеева, 1977).
Классификация биологических ритмов
О существовании биологических ритмов знали мыслители
древности по крайней мере со времен Гиппократа и Галена.
При этом известны были ритмы не только у человека и животных,
но и у растений. Суточный периодизм растений был открыт в 1729 г.
Де Мерраном и вновь описан в 1759 г. Цинни. Суточная темпера-
6
турная кривая у голубей описана в 1843 г. Шосса, у человека —
в 1887 г. Массо.
Первая диссертационная работа, касавшаяся суточных ритмов
человека, появилась в начале XIX в. (Virey, 1814). В конце XIX—
начале XX в. Вильгельм Флисс, Герман Свобода и Альфред
Тельтшер ввели представления о макроритмах, т. е. о ритмах
с многодневными периодами. Наряду с ними стали развиваться
представления о ритмах с многолетними периодами (Пэрна, 1925;
Thommen, 1968; Акрабов, 1969, 1972; Барбашова, 1972; Шапош-
никова, 1973, 1977; Lowrey, 1978).
Ритмы подразделяются по принадлежности к классу явлении
(ритмы живой и неживой природы; ритмы растений, животных,
человека и т. д.) и по признаку величины периода (микро-, мезо-,
макроритмы). И. А. Тюрин (1977) предложил различать ритмы
по признаку их функционального значения. Он выделил ритмы
«экологические», или адаптивные (суточные, приливно-отливные,
лунные, сезонные), и «функциональные» (все остальные ритмы,
обеспечивающие осуществление программы жизни и реакции на
внешние непредсказуемые воздействия).
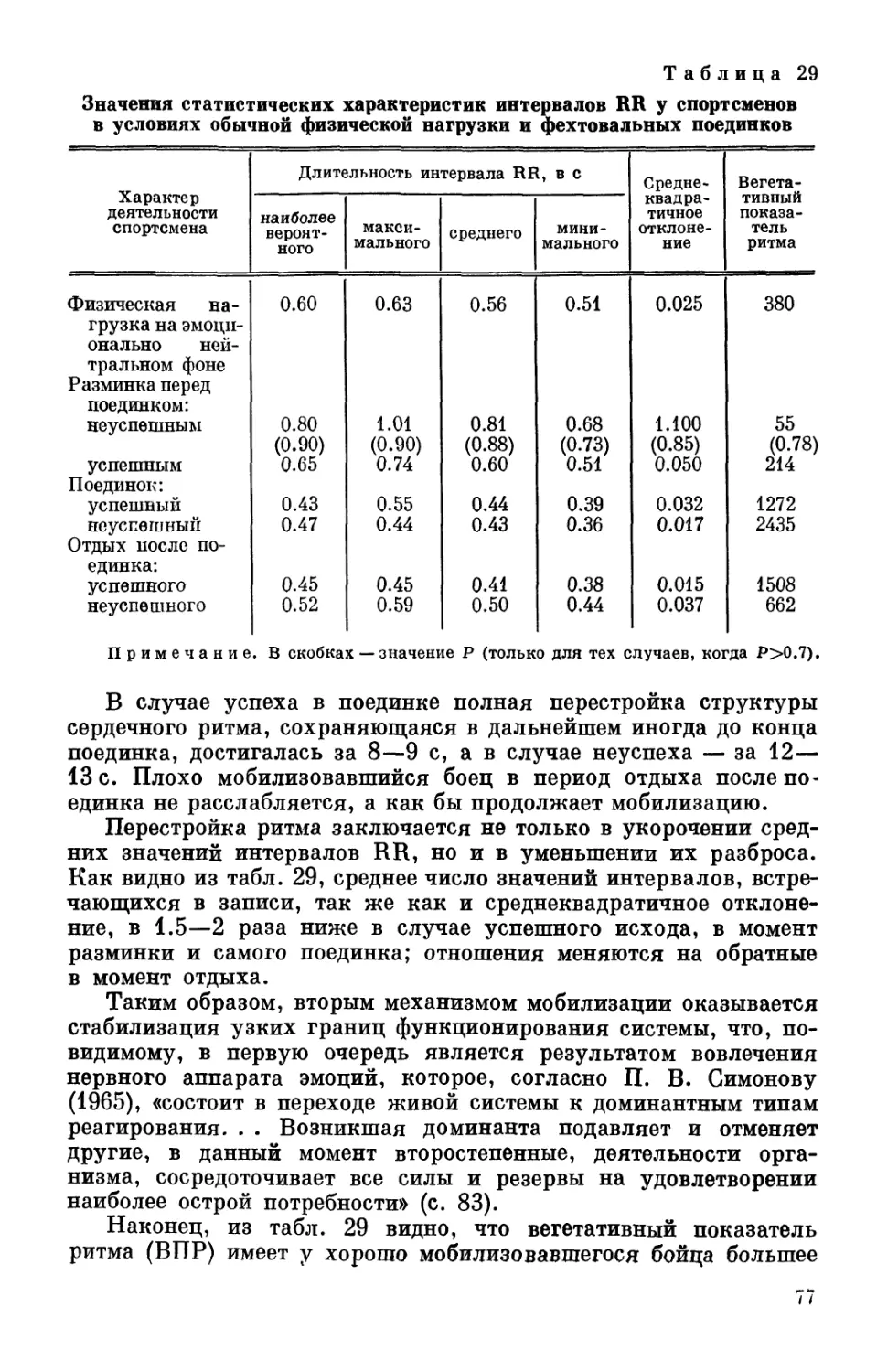
Классификация биоритмов человека по признаку принадлеж-
ности к определенному классу явлений разработана нами на ос-
нове выделенных А. Д. Слонимом (1979) уровней гомеостатических
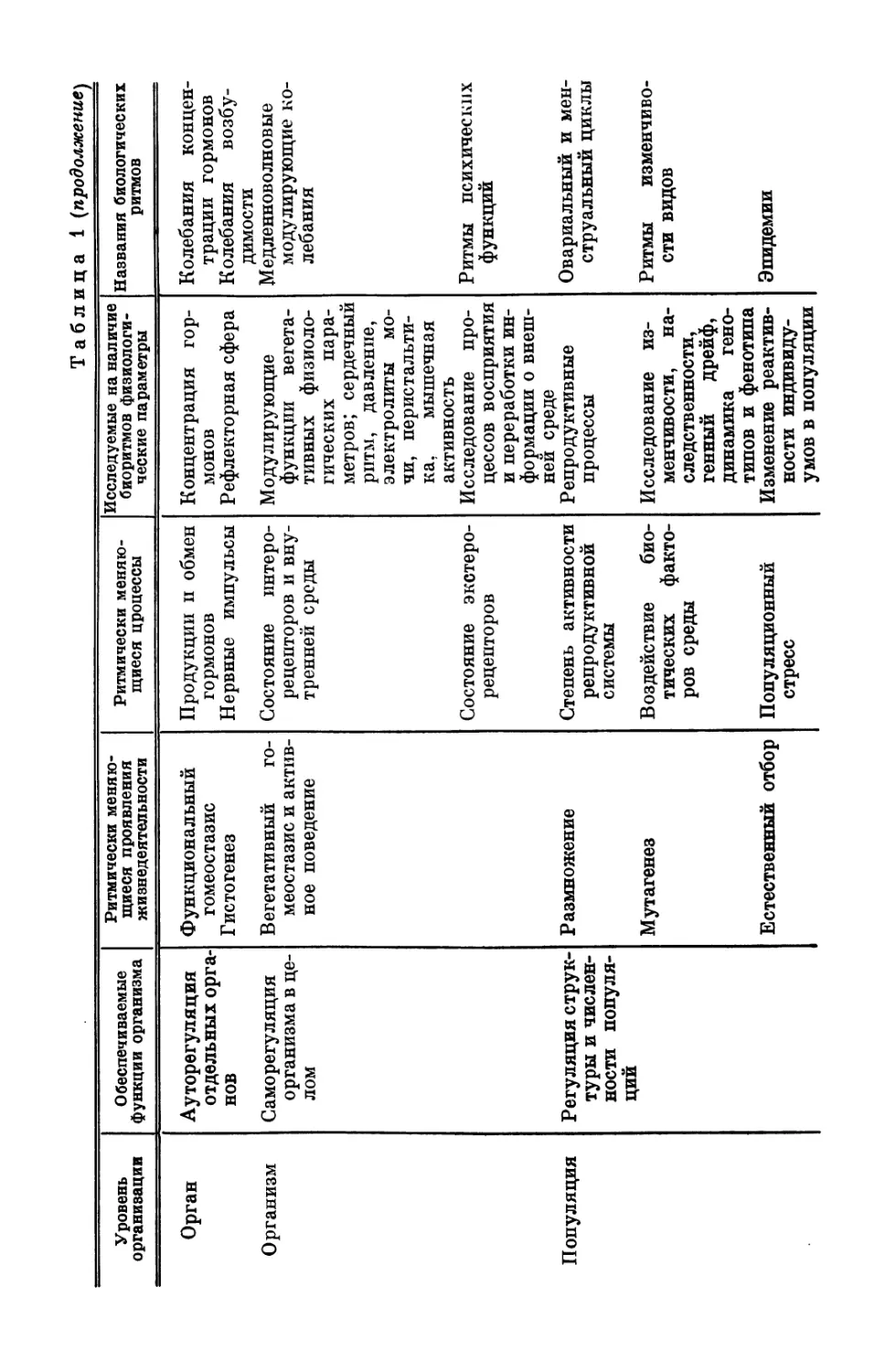
механизмов. Как видно из табл. 1, биологические ритмы присущи
практически любому процессу.
Наибольшее распространение получило деление биоритмов
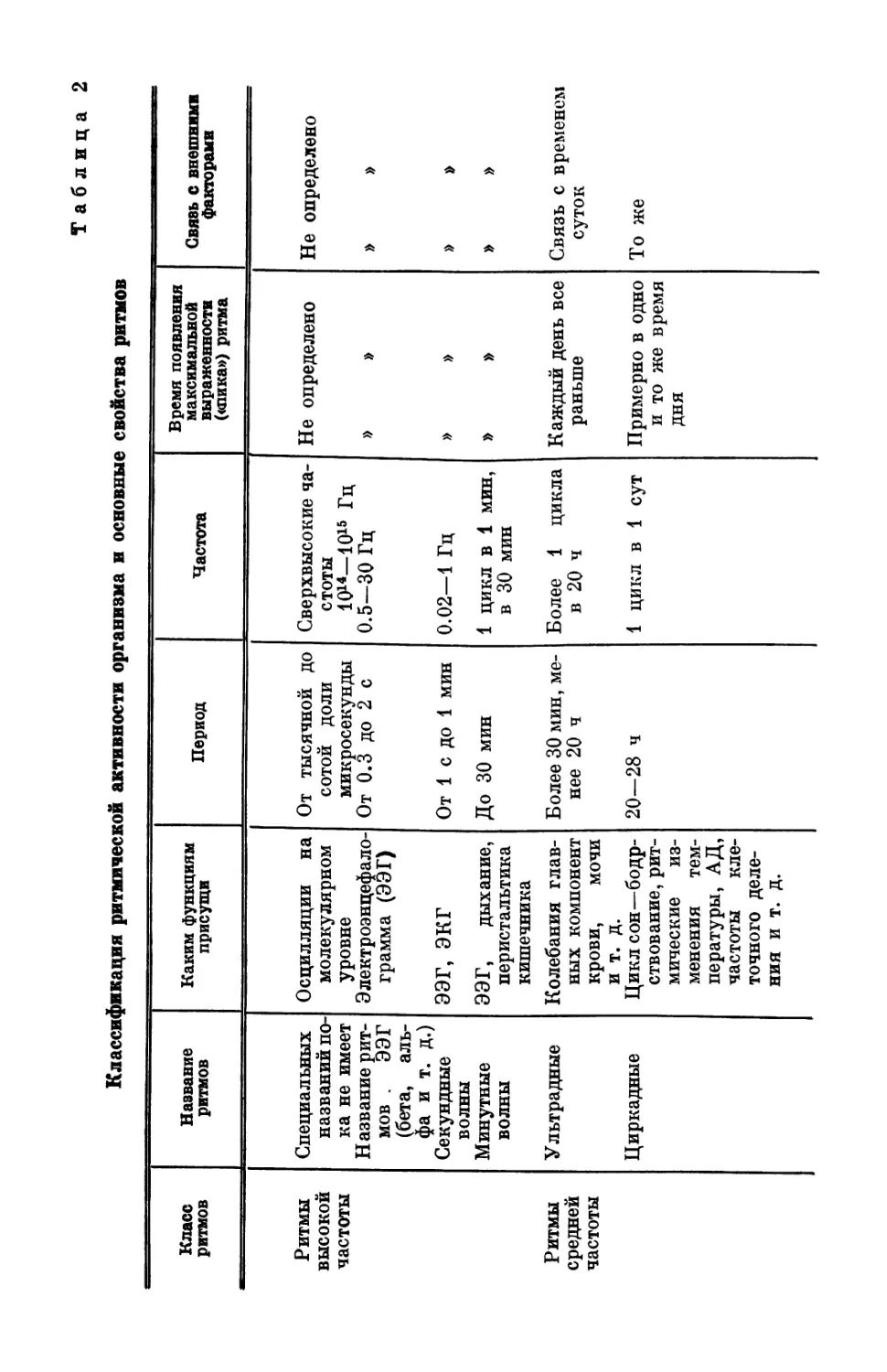
в зависимости от их периода (частоты). Крупнейший специалист
в области хронобиологии Халберг (Halberg, 1969) разделил рит-
мические процессы на три основные группы: 1) ритмы высокой
частоты с периодом до 0.5 ч; 2) ритмы средней частоты с периодом
0.5 ч—6 дней, куда входят ультрадный ритм (период 0.5—20 ч),
циркадный (период 20—28 ч), инфрадный (период 28 ч—6 дней);
3) ритмы низкой частоты, включающие циркасептанный (недель-
ный) ритм, циркавижинтанный (20-дневный); циркатригинтанный
ритм (лунный) с периодом около 30 дней и цирканный (годич-
ный) ритм.
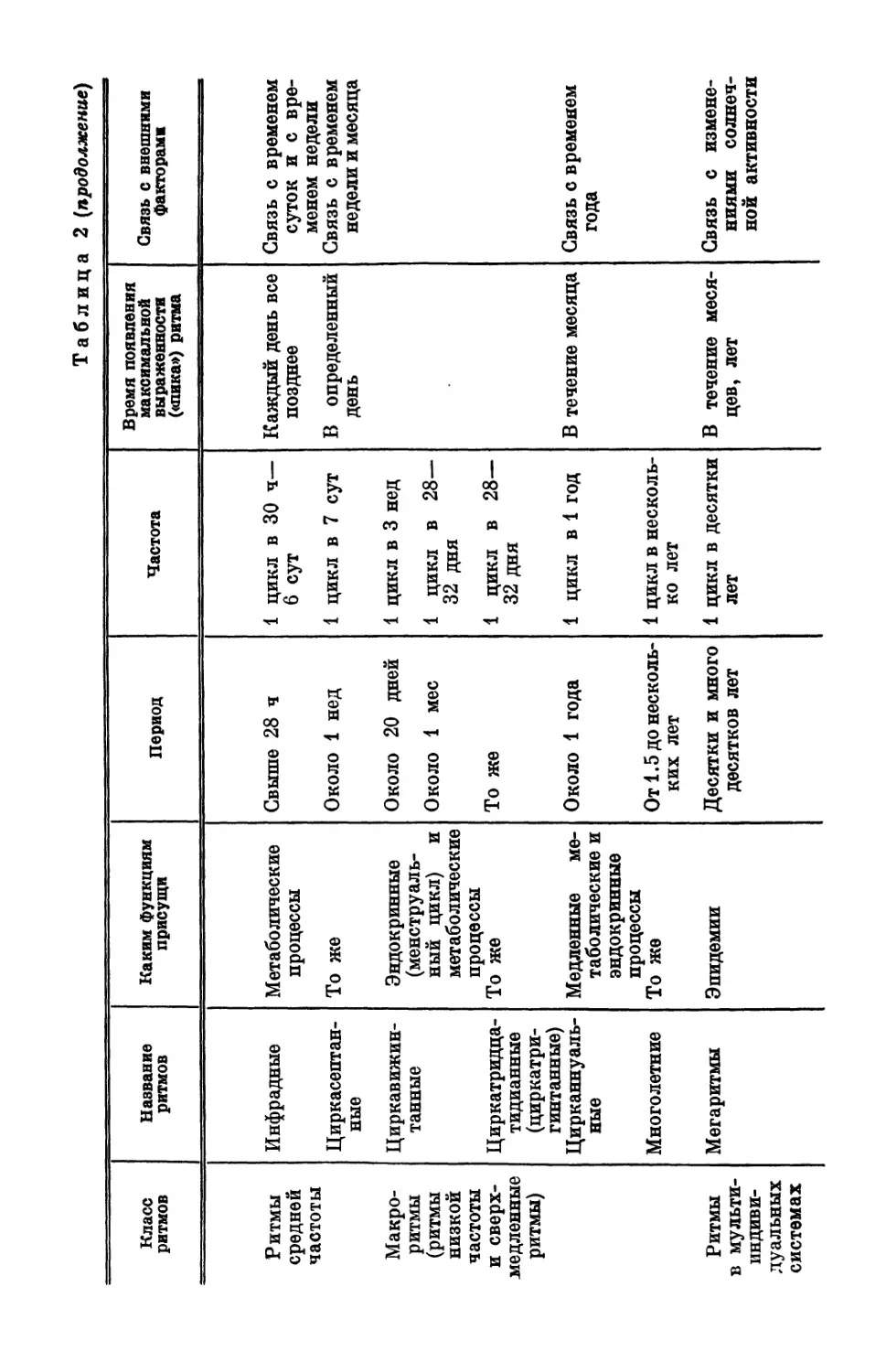
£В табл. 2 дана классификация ритмической активности орга-
низма, где кроме названия и периода ритмов приведены сведения,
каким функциям эти ритмы присущи, время появления макси-
мальной выраженности показателя (пика) и наличие или отсут-
ствие связи с внешними факторами. Видно, что живому орга-
низму присущи одновременно сосуществующие ритмы с перио-
дами, различающимися примерно на десять порядков (от 10~3 с
до 107—108 с). Эта разница в периодах колебаний имеется не
только для разных функций. Одна и та же функция изменяется
одновременно в различном диапазоне периодов. Так, например,
для колебаний времени двигательных реакций и продолжитель-
ности трудовых операций у человека наблюдаются ритмы с пе-
риодами от секунд и минут до многих часов (Смирнов, Ахметшин,
7
Т'а блица 1
Классификация биологических ритмов в зависимости от того, какие процессы, проявления жизнедеятельности
и уровни организации этими ритмами отражены
Уровень организации Обеспечиваемые функции организма Ритмически меняю- щиеся проявления жизнедеятельности Ритмически меняю- щиеся процессы Исследуемые на наличие биоритмов физиологи- ческие параметры Названия биологических ритмов
К леточное Компенсация, ре- Промежуточный об- Концентрация МО- Показатели pH, К онцентр ационные
образование, клетка, ткань гуляция тро- фических функ- ций мен Метаболизм (теп- лообразование) Возбудимость Проводимость лекул и веществ Энергия активации Мембранные про- цессы «Стратегия» фер- ментов концентрация ионов Электрическая ак- тивность мышцы, кожи, тканей Нейронная и муль- тиклеточная актив- ность Концентрация фер- ментов колебания в хи- мических си- стемах Миограмма, КГР, ЭЭГ; медленный (постоянный) по- тенциал Нейронограмма, рит- мы нейронных раз- рядов Колебания концен- трации ферментов
Орган Ауторегуляция отдельных орга- нов Температурный го- меостазис и обе- спечение тканей кислородом Химический гомео- стазис Кровоток Метаболизм Напряжение кис- лорода в тканях — оксигемометрия; параметры кро- ви, тонус сосудов; кардиограмма, реоэнцефалограм- ма Электролиты био- логических жид- костей, концент- рации химических веществ и биоло- гических компо- нент (крови, лим- фы, ликвора. . .) Сердечный ритм; со- судистые волны; колебания эле- ментов крови (лейкоциты, эри- троциты и т. д.) Концентрационные колебания
Уровень организации Обеспечиваемые функции организма Ритмически меняю- щиеся проявления жизнедеятельности
Орган Ауторегуляция отдельных орга- нов Функциональный гомеостазис Гистогенез
Организм Саморегуляция организма в це- лом Вегетативный го- меостазис и актив- ное поведение
Популяция Регуляция струк- туры и числен- ности популя- ций Размножение Мутагенез Естественный отбор
Таблица 1 (продолжение)
Ритмически меняю- щиеся цроцессы Исследуемые на наличие биоритмов физиологи- ческие параметры Названия биологических ритмов
Продукции п обмен Концентрация гор- Колебания концен-
гормонов МОНОВ трации гормонов
Нервные импульсы Рефлекторная сфера Колебания возбу- димости
Состояние интеро- Модулирующие Медленноволновые
рецепторов и вну- функции вегета- модулирующие ко-
тренней среды тивных физиоло- гических пара- метров; сердечный ритм, давление, электролиты мо- чи, перистальти- ка, мышечная активность лебания
Состояние экстеро- Исследование про- Ритмы психических
рецепторов цессов восприятия и переработки ин- формации о внеш- ней среде функций
Степень активности Репродуктивные Овариальный и мен-
репродуктивной системы процессы струальный циклы
Воздействие био- Исследование из- Ритмы изменчиво-
тических факто- ров среды менчивости, на- следственности, генный дрейф, динамика гено- типов и фенотипа сти видов
Популяционный стресс Изменение реактив- ности индивиду- умов в популяции Эпидемии
Таблица 2
Классификация ритмической активности организма и основные свойства ритмов
Класс ритмов Название ритмов Каким функциям присущи Период Частота Время появления максимальной выраженности («пика») ритма Свявь с внешними факторами
Ритмы высокой частоты Специальных названий по- ка не имеет Название рит- мов . ээг (бета, аль- фа и т. д.) Секундные волны Минутные волны Осцилляции на молекулярном уровне Электроэнцефало- грамма (ЭЭГ) ЭЭГ, ЭКГ ЭЭГ, дыхание, перистальтика кишечника От тысячной до сотой доли микросекунды От 0.3 до 2 с От 1 с до 1 мин До 30 мин Сверхвысокие ча- стоты 1Q14—1015 Гц 0.5-30 Гц 0.02—1 Гц 1 цикл в 1 мин, в 30 мин Не определено » » » » » » Не определено » » » » » »
Ритмы средней частоты Ультрадные Циркадные Колебания глав- ных компонент крови, мочи и т. д. Цикл сон—бодр- ствование, рит- мические из- менения тем- пературы, АД, частоты кле- точного деле- ния и т. д. Более 30 мин, ме- нее 20 ч 20-28 ч Более 1 цикла в 20 ч 1 цикл в 1 сут Каждый день все раньше Примерно в одно и то же время ДНЯ Связь с временем суток То же
Класс ритмов Название ритмов Каким функциям присущи Период
Ритмы средней частоты Инфрадные Циркасептан- ные Метаболические процессы То же Свыше 28 ч Около 1 нед
Макро- ритмы (ритмы низкой частоты Циркавижин- танные Эндокринные (менструаль- ный цикл) и метаболические процессы Около 20 дней Около 1 мес
и сверх- медленные ритмы) Циркатридца- тидианные (циркатри- г питанные) Цирканнуаль- ные Многолетние То же Медленные ме- таболические и эндокринные процессы То же То же Около 1 года От 1.5 до несколь- ких лет
Ритмы в мульти- индиви- дуальных системах Мегаритмы Эпидемии Десятки и много десятков лет
Таблица 2 (продолжение)
Частота - Время появления максимальной выраженности («пика») ритма Связь с внешними факторами
1 цикл в 30 ч— 6 сут Каждый день все позднее Связь с временем суток и с вре- менем недели
1 цикл в 7 сут 1 цикл в 3 нед 1 цикл в 28— 32 дня 1 цикл в 28— 32 дня В определенный день Связь с временем недели и месяца
1 цикл в 1 год 1 цикл в несколь- ко лет В течение месяца Связь с временем года
1 цикл в десятки лет В течение меся- цев, лет Связь с измене- ниями солнеч- ной активности
1976). Хотя внимание исследователей в настоящее время привле-
чено в большой мере к циркадным ритмам, вероятно, все диапа-
зоны колебаний важны для существования организма.
Механизмы формирования и поддержания
биологических ритмов
С момента начала исследования биологических ритмов законо-
мерно возник основной вопрос: генерируются ли эти ритмы са-
мим организмом, представляющим тем самым автоколебательную
систему, или являются следствием влияния импульсов («меток
времени»), постоянно поступающих из внешней среды. Для под-
тверждения этих альтернативных гипотез, носящих название
«эндогенных» и «экзогенных» часов, связанных либо с внутрен-
ними, либо с внешними синхронизаторами, проведены многочис-
ленные исследования, имеющие своей целью обнаружение влия-
ния этих синхронизаторов.
При этом высокочастотные ритмы принято считать преиму-
щественно (если не исключительно) эндогенными. Внешние воз-
действия, меняющие ритм сердца, дыхание, скорость переработки
информации и т. д., носят характер возмущений, переводящих
деятельность на другой уровень (Смирнов, 1972).
Одним из наиболее принятых приемов исследования эндоген-
ных компонентов циркадных ритмов является снятие синхрони-
зирующих воздействий окружающей среды, что влечет за собой
перестройку ритмов организма, превращая их в «свободно теку-
щие».
Такого рода перестройки достигаются путем длительного пре-
бывания испытуемых в пещерах или специальных испытательных
камерах. Условия в этих двух случаях не совсем идентичны, так
как различаются не только по микроклимату, но и по степени
ограничения двигательной активности. В пещерах люди чувство-
вали себя относительно непринужденно, а также могли свободно
выбирать режим труда и отдыха. В условиях же искусственного
ограничения подвижности нарушения нормальной ритмики от-
дельных функций и оценки времени могут возникнуть и при
неизменном суточном ритме внешних синхронизаторов (Панфе-
рова, 1967; Душков, Космолинский, 1968; Лицов, 1968; Aschoff,
1978, и др.).
Наибольшую известность получили наблюдения за состоянием
французского спелеолога Мишеля Сифра, который в 1962 г. впер-
вые провел под землей 2 мес. Связь с поверхностью поддержива-
лась лишь односторонняя. Длительность периода сон—бодрство-
вание составляла в этих условиях 24.5 ч, а период изменения
частоты сердцебиений удлинился до 24.6 ч. Время сна непрерывно
смещалось (Siffre, 1966).
В 1964 г. Жози Лор провела в пещере 3 мес, а Тони Сенни
оставался там на 1 мес дольше. Кроме прежних показателей у них
12
учитывались температура тела, частота дыхания, собирались
пробы мочи на содержание 17-гидроксикортикостероидов и 17-ке-
тостероидов. Период свободнотекущей ритмики у Жози Лор
составлял 24.6 ч, а у Тони Сенни — 24.8 ч. При 6-месячном
пребывании в пещере Жан-Пьера Марете период ритмических
колебаний температуры тела составлял 24.7 ч. За счет удлинения
периода в течение всего времени изоляции происходило смещение
акрофазы ритма (Fraisse et al., 1968; Siffre, 1968).
Изучение циркадных ритмов испытуемых проводилось в спе-
циальных камерах в условиях варьирования в широких пределах
распорядка труда и отдыха. Оказалось, что разным испытуемым
присущи различные реакции. Так, например, индивидуально
варьирует время «затягивания» ритмов при переходе к укорочен-
ным или удлиненным «суткам» и степень внутренней десинхрони-
зации в переходный период. Продолжительность перестройки
разных функций неодинакова и колеблется от 1 до 8 (Лицов, 1967)
и даже от 10 до 45 сут (Степанова, 19756; Степанова, Кукишев,
1975). При этом изменяется форма суточного хода кривой про-
цесса (Степанова, 1975а). Наиболее труднопереносимым режимом
явились регулярные 18-часовые «сутки» (Туровский и др., 1967),
а к 16- и 48-часовътм «суткам» испытуемые вообще не адаптирова-
лись (Степанова, 1973; Алякринский, 1977). Напротив, при слу-
чайном чередовании длительности периодов отдыха и работы,
когда продолжительность соответствующей фазы могла доходить
до 2 ч, через 2 нед часть функций вновь обрела 24-часовую перио-
дичность с небольшими модуляциями некоторых из них соответ-
ственно истинному режиму. Следует иметь в виду, что успешности
перестройки биоритма применительно к новым условиям может
способствовать «расшатывание» ритмики, например, посредством
длительного бодрствования (Степанова, 1976) или создания бес-
порядочного режима (Post, Gatty, 1931) и т. д.
Таким образом, при отсутствии внешних временных ориенти-
ров возникают те или иные изменения периода циркадных ритмов,
хотя сама ритмичность неизменно остается, что указывает на роль
экзогенных факторов в задании временного порядка. Такие внеш-
ние факторы носят название «синхронизаторов» или «датчиков
времени». Способами их изучения является исследование пере-
строек биоритмов под влиянием сдвинутых режимов труда и от-
дыха, климатических и сезонных колебаний внешних воздействий
и внезапного изменения временной среды, как это бывает при
перелетах с пересечением часовых поясов и космических полетах.
Если смена режима происходит часто, может возникнуть со-
стояние внутреннего десинхроноза, субъективно весьма тягостное
и даже переходящее в патологическое состояние. Известно, что
число язвенных больных и гипертоников выше среди работающих
посменно, нежели в контрольной группе. Так, при обследовании
промышленных рабочих Франции было обнаружено, что по пока-
зателям температуры тела 45% из них плохо приспосабливались
13
к сменному режиму (Руттенбург и Дэн Су-и, 1959; Алякринский,
1975; Катинас, Моисеева, 1980).
Таким образом, на создание режимов, идущих вразрез как
с внешними, так и с внутренними датчиками времени, организм
реагирует отрицательно. При этом большую роль играет мотива-
ция — в случаях очень важной и ответственной работы в сдвину-
том режиме этот сдвиг переносится во много раз лучше, чем если
работа монотонна или человек не занят целенаправленной дея-
тельностью.
Сводка данных по вопросу перестройки биоритмов под влия-
нием климатического фактора и сезонных (цирканных) колебаний
приводится в работе Г. С. Катинаса и Н. И. Моисеевой (1980).
Сезонные колебания обнаруживаются в ритмике всех исследован-
ных функций в любых климатических зонах; при этом физическая
работоспособность и двигательная активность летом выше, чем
зимой, артериальное давление снижается в весенне-летний пе-
риод и т. д., т. е. суточная динамика функций подвержена влия-
нию воздействия внешней среды, в том числе и сезонных колеба-
ний (которые имеют наряду с экзогенными и эндогенные компо-
ненты).
Наконец, изучение влияния на циркадные ритмы внезапного
и резкого изменения временной среды, проведенное многими иссле-
дователями, показало, что воздействия внешних стимулов, посту-
пающие в непривычное время, вызывают рассогласование по
времени фаз ритмов организма и среды (внешний десинхроноз),
а затем и фаз ритмов различных функций самого организма,
поскольку скорость перестроек у них различна; создается поло-
жение, когда какие-то ритмы еще протекают по «старому расписа-
нию», какие-то по новому, а какие-то беспорядочно (внутренний
десинхроноз, по О. Г. Газенко и Б. С. Алякринскому, 1970).
Десинхроноз может перейти в болезненное состояние. При этом
наблюдается последовательная смена стадий десинхронизации,
неустойчивой синхронизации и синхронизации. При пересечении
2—3 часовых поясов, по мнению ряда исследователей, десинхро-
низация не наступает (Гингст, Ярославцев, 1969; Евцихевич,
1970а, 19706), однако С. И. Степанова (1974) сдвиг на 2 ч уже
считает значимым.
По данным Фаббро (Fabbro, 1970), существен сдвиг, превы-
шающий 6—7 ч; в других наблюдениях нарушения со стороны
ритма «сон—бодрствование» проявляются после пересечения 7—
9 часовых поясов (Моисеева и др., 1972, 1978а, 19786; Моисеева,
1975в), однако изменения со стороны сердечно-сосудистой системы
возникали уже при пересечении трех часовых поясов. Как и
в опытах с изоляцией, большое влияние оказывает реальность
обстановки, в которой совершается перестройка ритма. При ими-
тации перелета Москва—Хабаровск—Москва путем изменения
распорядка дня и светового режима без осуществления реального
путешествия перестройки на новый ритм происходили медленнее,
14
а восстановление обычной ритмики быстрее, чем после совершен-
ного перелета (Парин и др., 1971).
Таким образом, изменение соотношений между внутренними и
внешними «датчиками времени» является серьезной нагрузкой на
адаптационные механизмы, и организм с этой нагрузкой легче
справляется, если все внешние датчики действуют в нужном
режиме перестройки.
Как видно из всего сказанного, накопленные факты не разре-
шают отвергнуть ни гипотезу об эндогенном, ни гипотезу об экзо-
генном происхождении биологических ритмов.
Гипотеза эндогенных часов
Гипотеза об эндогенном происхождении биоритмов сводится
к тому, что существуют собственные биологические осцилляторы
(биологические часы) и наблюдаемые периоды ритмов отражают
работу часов. При этом часов в организме много, что и объясняет
все случаи диссоциации периодов или фаз ритмов, а также сохра-
нение ритмической активности в изолированных органах и тка-
нях. В качестве источника такой основной активности было пред-
ложено рассмотреть энергию метаболизма (Pittendrigh, 1971),
однако идея эта не была подробно разработана до исследования
Е. Е. Селькова (1971, 1978), который показал, что энергетический
метаболизм, являющийся основой всего клеточного обмена, играет
роль клеточных часов. При этом важная роль принадлежит неав-
тономно регулируемым футильным циклам — циклическим после-
довательностям биохимических реакций, бесполезно рассеиваю-
щим энергию химических связей в виде тепла. Кроме того, рас-
сматривается концепция «адреналового цикла» как механизма
основной суточной ритмической деятельности организма (Halberg,
1955). Наконец, разработана концепция «хронона», основанная на
представлениях молекулярной биологии (Ehret, Trucco, 1967).
Согласно этой концепции, в полицистронном комплексе ДНК
с одного конца начинается синтез РНК, которая диффундирует
к рибосомам, программируя белковый синтез. Фактор-индикатор
диффундирует в обратном направлении и действует на другие
цистроны. Длительность всего цикла около 24 ч. Формирование
осцилляций с различным периодом, в частности колебаний, имею-
щих многоминутные периоды, по мнению Гудвина (Goodwin, 1966),
может быть следствием субгармонического резонанса в популяциях
взаимодействующих молекул. Периодичности, генерируемые инди-
видуальными химическими реакциями в молекулах, гораздо ко-
роче, поскольку во всей сети возникают функциональные колеба-
ния, базирующиеся на периодической смене скорости секреции
возбуждающих (или тормозящих) субстанций.
Кроме того, предполагается, что отсчет времени организмом
имеет непосредственно генетическую обусловленность. В одной ив
генных систем (система, контролирующая активность клукурони-
15
дазы в почках мышей) обнаружены наряду со структурными
регуляторными генами, процессинг-генами и временные гены
(Paigen, 1977).
Гипотеза экзогенных и экзогенно-эндогенных часов
Согласно этой гипотезе, ритмы организма не самостоятельны,
а возникают в связи с непрерывным поступлением импульсов (ме-
ток времени) извне. Скрытый ритмический комплекс организма
все время реагирует на геофизические ритмы воздействия света,
температуры, солнечных вспышек, влияния притяжения Солнца
и Луны, магнитных полей, электростатических полей и т. д. Наибо-
лее вероятно, что воздействия среды падают на организмы с гене-
тически закрепленными потенциальными ритмическими свой-
ствами. Их ритмы, особенно те, которые в естественных условиях
синхронны внешним циклам, при монотонных внешних воздей-
ствиях могут стать «свободнотекущими», т. е. до некоторой сте-
пени синхронизироваться вполне автономными, температурно ком-
пенсированными колебаниями в организме. Периоды этих колеба-
ний лишь приблизительно соответствуют геофизическим периодам
и в естественных условиях подвергаются непрерывной фазовой
коррекции под влиянием датчиков времени (гипотеза эндогенных
часов). Ряд авторов считают, что схемы поведения хронируются
неконтролируемыми и всепроникающими ритмическими геофизи-
ческими колебаниями (гипотеза экзогенных часов). Различные
«свободнотекущие» частоты возникают при этом в результате
взаимодействия присущего организмам естественного механизма
фазового сдвига с неизменяющимися факторами датчика времени;
это взаимодействие приводит к внутриорганизменному фазовому
сдвигу (автофазировке или повторному самовключению).
Кроме того, существует точка зрения (Hildebrandt, 1976), что
различные диапазоны биологических ритмов имеют разные меха-
низмы регуляции. Медленноволновая часть спектра (отражающая
тотальные функции организма) регулируется геофизической и
космической обстановкой. Коротковолновой диапазон зависит от
эндогенной-регуляции за счет прямого взаимодействия ритмов са-
мого организма. Гипотеза экзогенно-эндогенных часов, пожалуй,
наиболее полно объясняет как фиксированность, так и изменчи-
вость биологических ритмов, заставляющие полагать их статисти-
чески-вероятностную структуру.
Статистически-вероятностная структура
биологических ритмов
Исследования самых различных биоритмов показали, что про-
должительность периода, а также амплитуды, фазы колебаний,
форма волны меняются у разных лиц и у одного и того же лица
в разных условиях и под влиянием разных воздействий. При этом
16
существует четкая обратная зависимость между стабильностью
параметров ритма и скоростью перестроек при синхронизации или
возникновении переходных процессов. Например, непостоянные
параметры ритма двигательных функций оказываются быстро
подстраивающимися, в то время как ритмы вегетативных функций
весьма инертны.
Исследованиями закономерностей организации функций нерв-
ной системы показано, что деятельность как мозга в целом, так
и нервной клетки в отдельности подчинена статистически-вероят-
ностным закономерностям (Коган, 1964; Burns, 1969).
Те же закономерности обнаруживаются и в организации биоло-
гических ритмов. По мнению К. М. Смирнова (1980), гипотетиче-
ские генераторы любого ритма включают нервные образования
различных отделов мозга. Учитывая, что любой нервный центр
есть не что иное, как совокупность нервных клеток, деятельность
которых реализуется в эффективной импульсации и подчинена
статистическим закономерностям, флуктуация управляющих влия-
ний является естественной и обязательной. И тогда флуктуирую-
щий характер колебаний функций оказывается следствием стати-
стического характера всей регуляции в организме.
Это предположение нашло серьезное подтверждение в работах
И. П. Емельянова (1976, 1978), создавшего на основе стандартного
Косинор-анализа метод анализа косинорова тела — трехмерной
пространственной фигуры, где в полярной системе координат пред-
ставлены амплитуда и акрофаза текущей гармоники, а по оси
аппликат — период гармоники. Точка начала координат и вся
ось аппликат являются центром для «циферблатов» — кругов
с градуированной шкалой времени, периметр которых равен длине
избранного исследованного периода. Параметры косинорова тела
позволяют судить о течении во времени ритма одного определен-
ного процесса, но в случае совмещения разных косиноровых тел
(соответствующих разным функциям) в общих циферблатах (ме-
тод исследования косиноровых пещер) можно исследовать взаимо-
распределение биоритмов различных функций в пределах одного
организма.
Исследовав с помощью этих методов внутреннюю структуру
целого ряда биологических ритмов, И. П. Емельянов показал,
что они представляют собой набор гармонических колебаний
в разных комбинациях, и квазипостоянный период, характерный
для данного ритма, поддерживается за счет различных сочетаний
составляющих, т. е. что биологические ритмы вероятностные,
а не строго детерминированные события.
Это положение находит подтверждение и в некоторых клини-
ческих работах. Так, показано (Feinberg, Floyd, 1979), что дли-
тельность сна практически здоровых людей в любом возрасте
варьирует, т. е. имеется разброс показателей цикла бодрствова-
ние-сон. Отсутствие такого разброса даже рассматривается как
признак патологии.
17
Глава 2
МЕТОДИЧЕСКОЕ ОБЕСПЕЧЕНИЕ
ХРОНОБИОЛОГИЧЕСКИХ
ИССЛЕДОВАНИЙ
Наши исследования проводились на различных контингентах
здоровых лиц в обычных условиях, после перелетов с пересече-
нием часовых поясов и у больных, находившихся на стационар-
ном лечении в больницах Ленинграда. Рассматривались пара-
метры, отражающие функциональное состояние сердечно-сосуди-
стой системы, центральной нервной системы и двигательного
аппарата. Кроме того, изучался циркадный ритм сна.
Методика сбора данных
Периодичность замеров
Измерения параметров, отражающих состояние различных
физиологических функций, проводимые в клинике, в лаборатории
или в полевых условиях (например, во время туристских походов),
осуществлялись чаще всего 5 раз в сутки (в 7, 11, 15, 19, 23 ч
или в 8, 12, 16, 20, 24 ч). Реже измерения проводились 2—3 раза
в сутки (в 7, 11 и 15 ч) или каждые 2 ч (в 7, 9, 11, 13, 15, 17, 19,
23 и 1 ч ночи или в 8, 10, 12, 14, 16, 18, 20, 22, 24 ч), или каждый
час в течение дня. В группах замеры проводились 3—5 сут под-
ряд, а у отдельных лиц (и при самоизмерениях) подряд или с про-
пусками нескольких дней в течение ряда недель.
Замеры при исследовании течения циркадных ритмов в про-
цессе адаптации к изменениям воздействия внешней среды прово-
дились до воздействия в течение 3—7 дней (фон), в течение пер-
вой недели после воздействия ежедневно и затем через каждые
1—3 дня или раз в неделю вплоть до восстановления исходной
картины.
Для анализа макроритмов замеры проводились ежедневно
в 9—10 и в 16—17 ч в течение 2—3 мес.
Ночной сон регистрировался либо в течение 1—2 ночей (не
подряд), если дело касалось фоновых записей, или (после перелета
с пересечением часовых поясов) в течение 3—5 ночей (через ночь)
и затем через 1—3 нед.
18
Порядок исследования и регистрации данных
Исследование проводилось строго стандартно и начиналось
с записи точной даты (число, месяц, год) и времени (с точностью
до минуты); в условиях смены часового пояса отмечалось локаль-
ное время как места исследования, так и того места, откуда чело-
век прибыл.
Сначала, если это входило в программу, измерялась темпера-
тура, а также снимались показатели, о которых судят со слов
испытуемого (настроение, длительность ночного сна и т. п.),
и проводились пробы, не требующие физических усилий испытуе-
мого. Затем записывались ЭКГ (если это входило в программу),
изучалась капилляроскопическая картина, измерялась мышечная
сила и артериальное давление, указывалось, не произошло ли
в данный день каких-то важных событий, которые могли бы отра-
зиться на настроении, работоспособности, сне (и соответственно
на объективных показателях деятельности сердечно-сосудистой
системы). При упоминании радостных или печальных событий ни
в коем случае не выяснялись конкретные подробности, поскольку
это могло повлечь к потере контакта с испытуемым. Отмечались
только наличие события, его модальность (радостное, печальное,
устрашающее и т. д.), интенсивность и примерное время, когда
оно произошло. При учете влияния внешних воздействий во вни-
мание принималось субъективное отношение, а не объективная
значимость событий.
Исследуемые параметры и способы их оценки
1. Измерение температуры проводилось с помощью обычного
ртутного термометра в подмышечной впадине (время измерения —
5—10 мин, точность измерения — до 0.1° С).
2. Подсчет частоты дыхания за 1 мин.
3. Исследование сердечного ритма с использованием интерва-
лов RR ЭКГ или подсчета пульсовых ударов в течение 1 мин.
4. Измерение артериального давления (проводилось каждый
раз на одной и той же руке или на обеих руках) с помощью аппа-
рата Рива-Роччи.
5. Исследование капилляров ногтевого ложа четвертого пальца
руки проводилось с помощью капилляроскопа М-7 А (увеличение
в 20 раз). Кизуально оценивались окраска фона, четкость границ
капилляров, их форма, длина и ширина капилляров в пределах
трех градаций — норма, удлинение (сужение), укорочение (рас-
ширение), общее число капилляров в поле зрения (определяемого
длиной линейки окуляр-микрометра). За основной параметр при-
нималось число капилляров, остальные параметры использова-
лись при решении вопроса: имеет ли место стаз, спазм или нор-
мальная картина.
6. Измерение мышечной силы производилось с помощью дина-
мометра. Испытуемый стоял, смотря прямо перед собой, плечи
19
назад, нога слегка отставлена. Динамометр брался в правую руку,
которая отводилась в выпрямленном положении на угол 30 град,
и сжимался как можно сильнее. То же самое повторялось для
левой руки.
7. Тест исследования внимания (корректурная проба). На стан-
дартном бланке (со случайным набором букв) с помощью каждый
раз заново предложенных значков вычеркиваются три буквы.
Выполнение задания длится 5 мин, начинается по команде экспе-
риментатора, причем отмечается каждый раз разделительной
чертой, какая часть работы выполнена за 1-ю, 2-ю,. . ., 5-ю ми-
нуту. Проверяется наличие и характер ошибки — пропуск, вычер-
кивание буквы, которая не относится к числу обусловленных
в данном листе. Учитываются общее количество просмотренных
знаков и количество вычеркнутых знаков. Отношение числа верно
вычеркнутых знаков к числу секунд дает показатель, отражаю-
щий (в среднем) объем памяти. Образец бланка представлен
в табл. 3.
Таблица 3
Бланк для исследования состояния высшей нервной деятельности
по методике Анфимова (корректурная проба)
Опыт № ------ Дата----------- Время------------------
Фамилия, И. О. ------------------ Дата рождения------
3 адание-----------------------
КЕХНВАЕВХКИХВЕСКАСИЕАВЕС АЕКИАСХКНСВКИВНЕ
НСВАИХКХСАХИКНХИНЕСНВАНКЕИХСКВИНСАИХЕАИХ
АВХКАСЕНКХЕНИЕАВКСЕКИСАЕКСАВНСВКИХНИВХАВ
НКЕИВСКИВНЕХАИКХЕСАХСАХКХАСНВКАСКЕВХНСВА
КЕХКНИВХЕАВИЕХАВКНЕСИКНСЕАХНВИКХАНСАИНСЕ
НАСВЕХНВИСАВХКВАЕХНКЕСИАНКСЕХНСИКВНИЕКАХ
Параметр
Результаты за каждую минуту
исследования
Сумма
1-ю 2-ю 3-ю 4-ю 5-ю
Общее количество знаков:
просмотренных
вычеркнутых
Общее количество ошибок,
из них:
пропуск буквы
вычеркнута лишняя буква
использован не тот коррек-
турный знак
Средний коэффициент памяти
8. Тест на краткосрочную память. Проводится с помощью
листа с набором семизначных случайных чисел. В каждом тесте
изучаются скорость и точность воспроизведения девяти наборов
20
цифр. Исследуемый сам пускает секундомер, смотрит на один
набор цифр, закрывает его секундомером и под ним записывает
запомнившиеся цифры, затем процедура повторяется со следую-
щим набором цифр, и так до конца строки, после чего секундомер
останавливают. Время определяется с точностью +0.1 с. Подсчи-
тывают общее число ошибок; общее число верно воспроизведенных
чисел делится на число секунд, что дает показатель, называемый
«средняя память». Образец бланка представлен в табл. 4.
Таблица 4
Бланк для тестирования состояния краткосрочной памяти
Фамилия, И. О. ---------------------------
7410888 2228857 0740152 5704910 3501755 1475048 9683860 3628808 7873951
2230490 3147287 7173343 9283051 4911748 1210280 5804186 7177105 9621065
7395079 4431794 4056006 0478880 4325852 5890558 2163961 8498659 9326660
9. Тест определения чувства времени. С момента пуска секун-
домера испытуемый считает вслух или про себя от 1 до 60 (так,
чтобы считать приблизительно 1 мин, не смотря на секундомер).
Засекается общая длительность счета, обращается внимание на
равномерность счета.
10. Тест определения длительности заданных отрезков вре-
мени. Под контролем секундомера испытуемому предъявляются
маркированные двумя ударами молоточка (в начале и в конце)
отрезки времени длительностью от 5 до 25 с. Испытуемый немед-
ленно воспроизводит предъявленный отрезок времени также двумя
ударами молоточка по столу.
11. Счет пальцев (тест на счет и на координацию движений).
Секундомер в левой руке. Правую кисть, ладонью вверх, испы-
туемый держит перед глазами и поочередно прикасается большим
пальцем ко второму, третьему, т. е. указательному, безымянному
ит. д., и обратно, считая при этом до 25. На счет 25 большой
палец должен коснуться указательного, что покажет точность
счета. На секундомере фиксируется время счета.
12. Тест на простейшие арифметические действия (сложение).
Проводится с помощью бланка с несколькими колонками цифр.
В каждой колонке испытуемый складывает цифры попарно, про-
ставляя каждую сумму справа и между цифрами. Например, для
столбца, состоящего из цифр 7, 1, 5, 2, 9, первая сумма будет
7+1=8; следующая’"!+5=6; 5+2=7, и т. д. Если замечено, что
сделана ошибка, следует быстро исправить. В конце столбца отме-
чается время с точностью до 0.1 с. Учитывается число ошибок.
Образец бланка для проведения теста представлен в табл. 5.
13. Тест определения взаимооценок в замкнутом коллективе.
Обследовалась одновременно группа в 25—30 человек. Всем чле-
нам группы выдавались опросные листы, в которых они должны
были определить свое отношение к каждому из остальных членов
группы по пятибалльной шкале, где оценка 5 баллов соответство-
21
Таблица 5
Образец бланка теста на простейшие арифметические действия
Фамилия, И. О.-------------------------•
8 8 4 9 2 9 9 3 8 2 1 4 4 5 4 0
4 5 0 4 2 0 0 9 4 9 8 9 7 7 7 4
8 4 3 9 3 4 1 3 2 2 1 9 0 7 8 5
0 8 3 0 9 3 4 4 7 7 4 4 0 7 4 8
1 8 3 8 2 8 7 3 7 8 8 0 6 5 3 3
2 8 5 9 7 2 0 4 0 5 9 4 2 0 5 2
0 3 8 0 2 2 8 8 8 4 8 8 9 3 2 7
4 9 9 9 8 7 4 8 6 0 5 3 0 4 5 1
2 8 7 4 0 2 2 8 4 6 1 7 8 2 0 3
7 1 0 2 6 8 7 8 2 1 2 1 6 9 9 3
3 5 9 0 2 9 1 3 8 6 4 4 3 7 2 1
5 4 8 6 6 5 7 4 1 1 4 0 1 4 8 7
4 8 1 3 7 2 2 0 4 1 8 4 9 8 4 5
4 7 4 6 8 5 0 5 2 3 2 6 3 4 6 7
вала отношению «очень нравится», а оценка 1 балл — «очень не
нравится». Опрос проводился повторно 2—7 раз; на листе про-
ставлялись даты опроса и дня рождения (необходимые для состав-
ления карты биоритмов). Образец представлен в табл. 6.
Таблица 6
Образец опросного листа для определения взаимооценок в коллективе
Дата заполнения
Группа №------
Число, месяц,
год рождения
лица, заполняю-
щего лист
Поставьте оценку
№ Фамилия Оценка
1
2
3
Оценки
5 — очень нравится
4 — нравится
3 — не знаю
2 — не нравится
1 — очень не нравится
14. Оценка общего состояния проводилась согласно следую-
щим индивидуальным шкалам и описывалась по установленной
форме (табл. 7).
22
Таблица 7
Форма листа оценки общего состояния и сна
(карта суточного ритма)
Цата На£ие°е- Тонус Сон
Время
отхода
ко сну
Время
пробуж-
дения
Пробужде-
ние само-
произволь-
ное или
будили
Факторы в тече-
ние дня, которые
могли нарушить
сон и изменить
общее состояние
а. Шкала оценки настроения
1 — очень плохое настроение, злоба, раздражение;
2 — плохое настроение, депрессия;
3 — неважное настроение;
4 — обычное настроение, спокойное, ровное;
5 — хорошее настроение (слегка более веселое, чем обычно);
6 — приподнятое настроение;
7 — очень хорошее настроение.
б. Шкала оценки работоспособности (тонуса)
1 — неработоспособен («все валится из рук»);
2 — инактивен, устал, плохо работал;
3 — слегка менее активен, чем обычно; работал посредственно;
4 — обычная работоспособность;
5 — хорошая работоспособность, слегка выше обычной;
6 — отличная работоспособность;
7 — феноменальная работоспособность («горы сворачивает»).
в. Шкала оценки событий,
которые могли бы повлиять
на общее состояние испытуемого
О — событий не было;
1 — произошло очень неприятное или прямо трагическое событие (потрясе-
ние, тяжелый стресс);
2 — произошли крупные неприятности, скандал (стресс);
3 — мелкие неприятности, не грозящие серьезными последствиями, но пор-
тящие настроение;
4 — имел место прием каких-либо тонизирующих лекарств, крепкого чая,
кофе, вина (средства, влияющие на работоспособность, настроение, сон);
5 — произошло какое-то приятное событие (прогулка за город, интересная
театральная постановка и т. д.);
6 — произошло радостное событие;
7 — произошло очень радостное событие, большая удача.
Дополнительно указывалось, когда произошло событие (дата
и время суток). В той же форме дается и оценка длительности
и характера сна.
23
15. Исследование показателей ритма сна проводилось путем
записи электроэнцефалограммы (ЭЭГ), электроокулограммы (ЭОГ),
электрокардиограммы (ЭКГ) в течение ночного естественного сна,
а также в некоторых случаях во время дневного сна.
Записи проводились: 1) в специальной экранированной камере
с помощью стационарного 15-канального аппарата фирмы «Аль-
вар-электроник»; 2) в экранированной лаборатории с помощью
многоканального электро энцефалографа фирмы «Галилео».
Кроме того, в условиях клиники записывалась актограмма
с помощью барографа, герметизированный корпус которого был
соединен шлангом с резиновым кругом, находившимся под матра-
цем больного. При движении изменение давления воздуха вызы-
вало колебания пера, вычерчивавшего кривую на барабане, совер-
шавшем полный оборот за 24 ч. Получаемая таким образом кривая
позволяла судить о числе движений, совершаемых пациентом, ле-
жавшим в постели. Записи ночного сна начинались около 23—24 ч
и длились до естественного пробуждения (т. е. до 6-—9 ч).
Выделение стадий сна по ЭЭГ проводилось визуально согласно об-
щепринятой методике (Rechtschaffen, Kales, 1968) с небольшой моди-
фикацией, которая заключалась в том, что дополнительно к стадиям В,
С, D, Е по Лумису и REM-стадии учитывалась и стадия А по Лу-
мису. Стадии сна определялись на 10-секундных отрезках.
Вся запись делится на часы и для каждого часа исследуется:
длительность каждой стадии сна, в процентах; частота появления
каждой стадии сна, в процентах; среднее число смен стадий сна
в 1 мин. Для этих показателей строятся кривые, где число точек,
определяющих ход кривой, равно числу часов сна.
Соотношения между различными стадиями сна исследуются
путем определения частоты появлений каждой конкретной ста-
дии сна после каждой из остальных стадий сна; вычисления коэф-
фициента последовательности стадий сна (т. е. вычисления отно-
шения числа смен стадий сна, когда наступившая стадия была
стадией более глубокого сна, чем ей предшествовавшая, к числу
смен стадий сна в обратной последовательности).
Кроме того, для сна в целом исследуется: 1) общая длитель-
ность сна; 2) длительность каждой стадии сна за всю ночь, в про-
центах; 3) частота появления каждой стадии сна за всю ночь;
4) среднее число смен стадий сна в 1 мин; 5) коэффициент после-
довательности стадий сна (К); 6) частота появления каждой ста-
дии сна после любой другой; 7) разные значения показателей
длительности, частоты появления, среднего числа смен и стадий
сна за каждый час сна; 8) периодичность наступления сна; 9) ла-
тентный период засыпания.
Кроме того, исследуется еще и двигательная активность во
время сна (среднее число движений и средняя длительность дви-
жений в течение каждого часа и в течение всей ночи, разброс этих
показателей). Наконец, исследуется зависимость смены стадий
сна от двигательной активности.
24
Учитываемые факторы внешней среды
Исходя из предположения, что внешняя среда определенным
образом влияет на биоритмы, мы приняли во внимание следующие
параметры.
1. Метеорологические факторы: температуру воздуха, влаж-
ность, атмосферное давление. Данные были выписаны из сводок
гидрометеоцентра Ленинграда не только на день (дни) исследова-
ния, но и за 3 дня до его начала и в течение 3 дней после него.
Смысл такого подхода заключался в том, что если бы существовал
какой-то дополнительный неучтенный фактор, действующий как
на параметры биосистемы, так и на внешнюю среду, его влияние
могло быть обнаружено за счет закономерного изменения этих
параметров. Брались результаты круглосуточных изменений в 0 ч,
3, 6, 9, 12, 15, 18 и 21 ч.
Кроме того, учитывались температура с точностью до десятых
долей градуса, влажность в виде упругости водяного пара, в г/см2,
и атмосферное давление, в гектопаскалях (гПа).
2. Магнитное поле Земли с учетом всех трех составляющих
(Я, D и Z) и часовая амплитуда составляющей Н (гн), а также
магнитные бури. Данные этого рода были получены на гео-
физической станции Ленинграда, расположенной в Воейкове.
Они также были выписаны в течение всех дней исследования,
за 3 дня до и в течение 3 дней после. Учитывались результаты
круглосуточных измерений магнитного поля в 0 ч, 3, 6, 9, 12,
15, 18, 21 ч и время начала и конца, общая их длительность и вы-
раженность магнитных бурь, в баллах.
3. Солнечные хромосферные вспышки. Данные о времени
начала, максимумов, конца вспышек и их интенсивности (в бал-
лах) получены в Пулковской обсерватории по справочнику
ЮНЕСКО.
Факторы внутреннего порядка —
собственные макроритмы
и реакции на стрессорные воздействия
С целью выяснения влияния макроритмов на течение био-
ритмов с меньшим периодом на каждого испытуемого заполня-
лась карта биоритмов на текущий год, где, согласно общим пра-
вилам (Thommen, 1968), начало положительной фазы каждого
ритма задается с момента рождения и в дальнейшем предпола-
гается, что биоцикл имеет стабильную длительность.1 Вычисля-
1 Весьма логично предположить, что такого рода предпосылка доста-
точно условна и в силу различных причин могут быть некоторые отклонения
длительности цикла то в сторону ее увеличения, то в сторону уменьшения.
Однако, по-видимому, эти отклонения обычно нивелируются и, как пока-
зала практика, циклы достаточно стабильны. Механизмы, обеспечивающие
постоянство периода осцилляций колебательных систем, разбираются в спе-
циальной литературе (Alexander, Atlas, 1973).
25
ется количество прожитых дней до исследуемого момента и де-
лится на 23, 28 или 33 в зависимости от того, для какого ритма
ведется вычисление. Остаток от деления указывает, какому дню
биоцикла соответствует момент исследования.
Первоначально биоритмы рассчитывались по таблицам и по но-
мограммам (Козлов, 1976), а затем на ЭЦВМ «Минск-32» по про-
грамме «Биори», составленной С. А. Рощектаевой. После того
как карта биоритмов на год была составлена, на ней выделялись
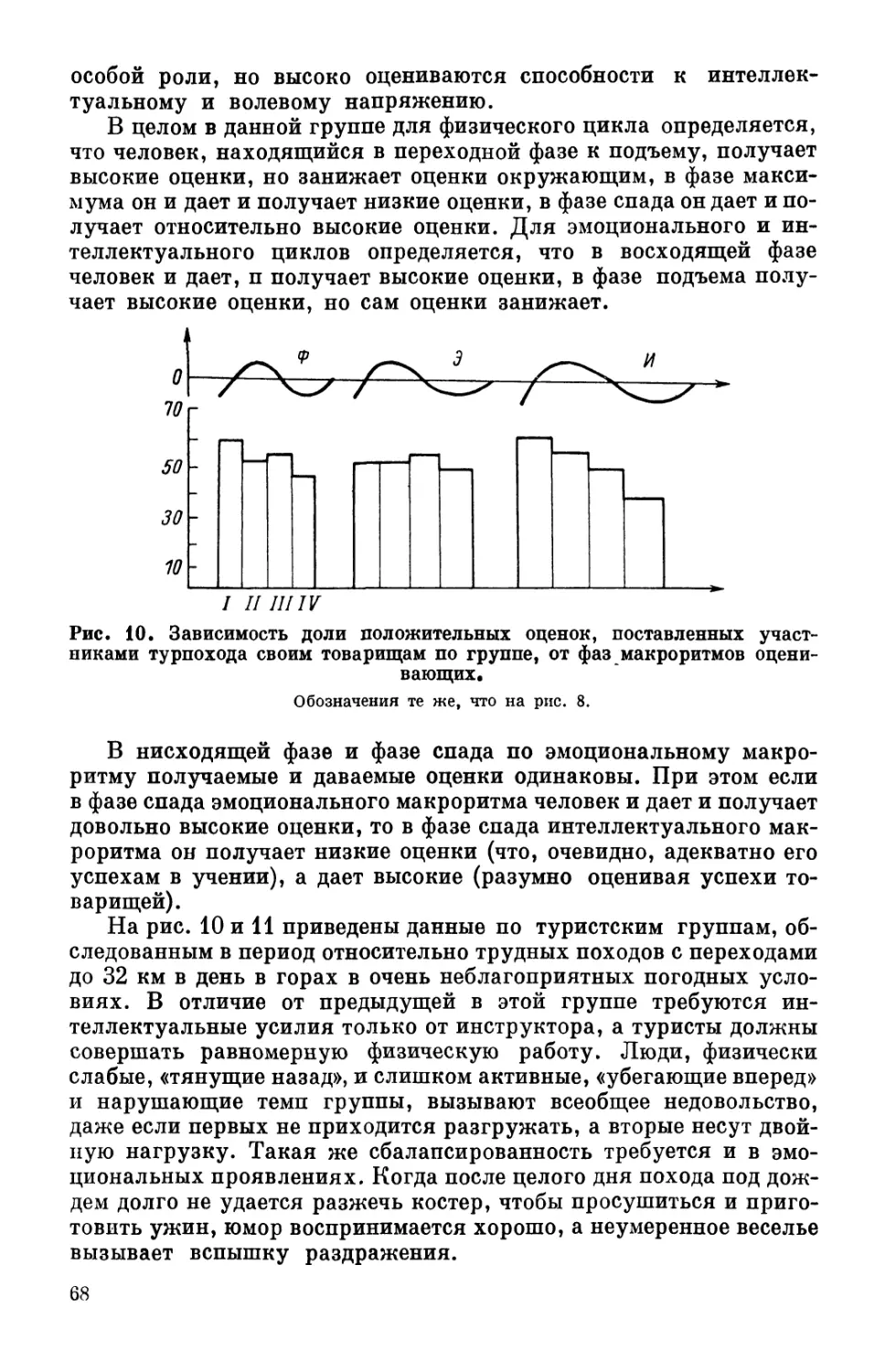
четыре фазы примерно равной длительности: I — переход от
отрицательной фазы к положительной (восходящая фаза); II —
положительная фаза (фаза подъема); III — переход от положи-
тельной фазы к отрицательной (нисходящая фаза); IV — отри-
цательная фаза (фаза спада).
Таблица 8
Таблица определения фаз (I—IV) биоритмов по дням цикла
Макроритм I (восходящая) II (подъема) III (нисходящая) IV (спада)
Физический 21-23-й, 1-3-й 4—9-Й 10-14-й 15-20-й
Эмоциональный 25—26-й, 1-3-й 4—10-й 11-17-й 18-24-й
Интеллектуальный 30-33-й, 1-4-й 5—12-й 13-21-й 22—29-й
Определение фазы биоритмов производилось согласно табл. 8,
в которой указано, какие дни цикла каждого из макроритмов
соответствуют одной из четырех фаз.
Факторы, оказывающие стрессорные воздействия, учитыва-
ются с помощью листа оценки общего состояния (табл. 7), где
есть специальная графа.
Методика обработки полученных данных
Графическое представление исходных данных
По результатам тестов № 1—12 вычерчивались циркадные
кривые для трех последовательных дней, наиболее часто с ша-
гом 4 ч (изредка с шагом 1—2 ч). В отдельных случаях при дли-
тельных измерениях (в течение недель и месяцев) кривые строи-
лись для ряда отрезков, включающих 3 дня.
Кривые вычерчивались как вручную, так и с помощью элек-
тронно-вычислительной машины. При построении графиков про-
водится нормировка, т. е. по оси откладываются не абсолютные
значения, а отклонения от среднего значения на данный кон-
кретный день. Параллельно с вычерчиванием графиков подсчи-
тывались основные статистические характеристики по 3 дням.
В целях быстрой визуальной оценки особенностей суточной
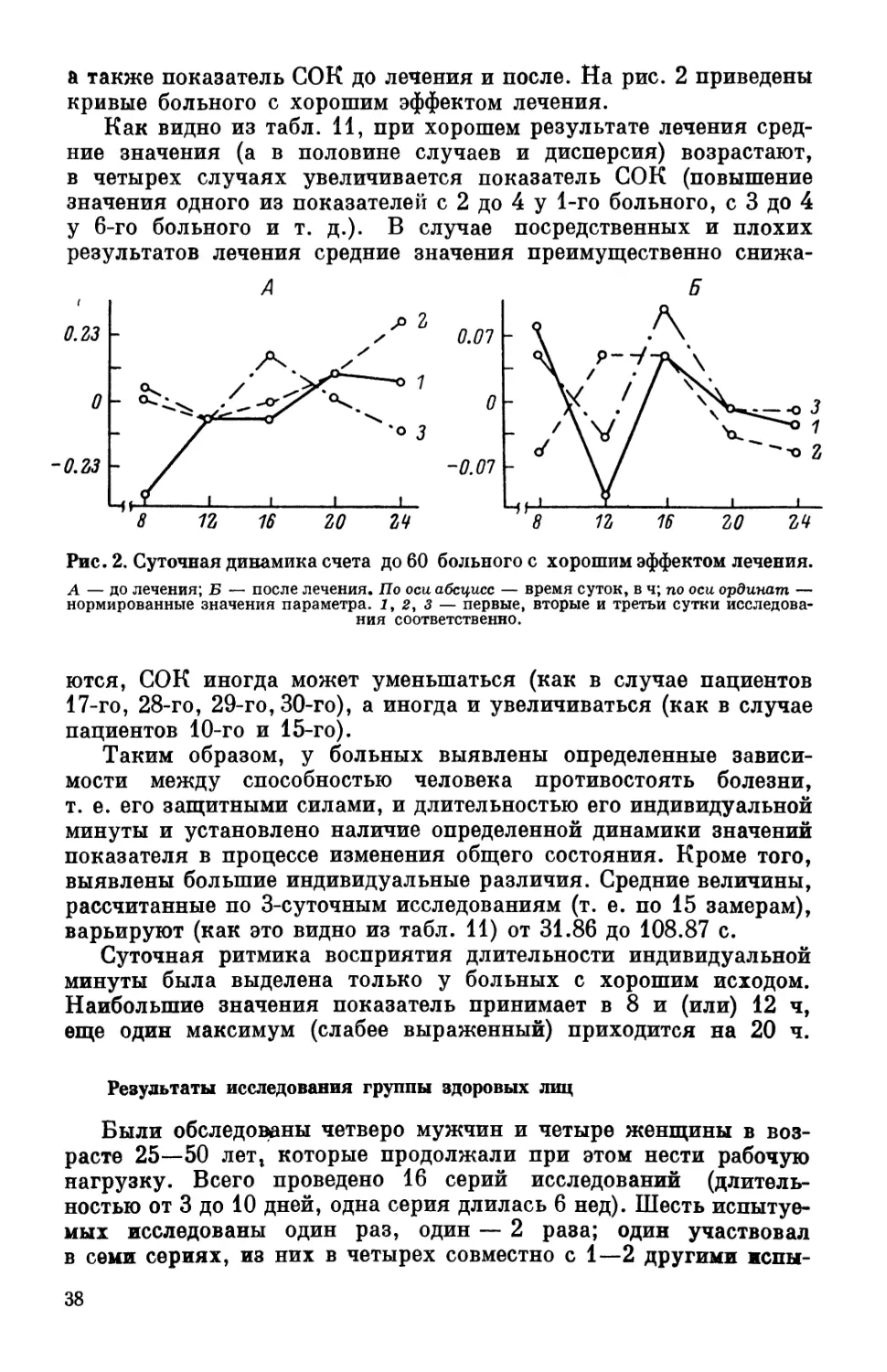
кривой был введен коэффициент степени организованности кри-
26
вой (СОК), выражаемый в баллах от 1 до 5. Коэффициент (СОК)
определяется по следующей шкале: 1) колебания отсутствуют,
вместо кривой наблюдается прямая линия с тем или иным накло-
ном; 2) кривая с одним изгибом, т. е. отражается наличие части
полуволны; 3) на кривой отчетливо вырисовывается полуволна;
4) на кривой представлена полу-
волна и еще часть полуволны;
5) на кривой представлена полная
волна (рис. 1).
Методика обработки данных сердеч-
ного ритма
Исследование временной струк-
туры сердечного ритма произ-
водилось методом периодограм-
много анализа ряда интерва-
лов RR. Вообще для анализа
экспериментальных данных при-
меняются различные варианты
периодограммных методов (Вой-
тинский, Прянишников, 1968;
Иванов-Муромский, Заславский,
Рис. 1. Степени организованности
суточных кривых.
Объяснение см. в тексте
1968; Anderson, 1976; Слауцитайс, Сысуев, 1976). Метод, исполь-
зуемый в настоящем исследовании, был основан на аппроксима-
ции ряда интервалов RR, представленных в реальной временной
шкале, тригонометрическими функциями (Сысуев, 1978).
Задавая различные значения пробных периодов тригономе-
трических функций, определяли остаточную среднеквадратич-
ную ошибку аппроксимации и ее отношение к общей дисперсии
исходного временного ряда на всем интервале наблюдения.
Полученный коэффициент (у), характеризующий по существу
долю остаточной дисперсии аппроксимации, рассматривался как
критерий близости исходного ряда интервалов RR и тригономе-
трической функции.
Минимальные значения коэффициента у при определенных
значениях пробных периодов свидетельствуют о присутствии
в исходном временном ряду колебательных компонент с соот-
ветствующими периодами. Здесь следует заметить, что обнару-
женные колебательные компоненты временной структуры сер-
дечного ритма представляют собой функции времени, модулирую-
щие длительность интервала RR, и амплитуда этих компонент
также измеряется во временных единицах —- в миллисекундах
или в долях среднего интервала RR.
Анализ полученных периодограмм в общем случае позволяет
выявить (в случае их наличия) одну или несколько колебатель-
ных компонент, выраженных в разной степени по коэффициенту у
и соответственно имеющих различные амплитуды. Периоды этих
27
компонент измеряются в единицах реального времени (в секундах),
что, как нам кажется, выгодно отличает использованный нами
метод от методов выделения медленных волн сердечного ритма,
где периоды этих волн измеряются количеством интервалов RR.
Методика выделения скрытой периодичности
любого из исследованных параметров
С целью выделения скрытой периодичности той или иной
функции формируются временные ряды, составленные из от-
счетов мгновенных значений данного параметра, и в этом случае,
если ряд достаточно репрезентативен, производится обработка
по программе периодограммного анализа, первоначально создан-
ной для обработки сердечного ритма. Таким образом обрабаты-
вались, например, результаты измерений артериального давле-
ния, температуры тела, частоты пульса и дыхания, мышечной
силы, тестов на память, арифметические действия и восприятия
времени, производившиеся ежедневно у 15 человек в течение
1.5—2 мес (временной ряд для каждого параметра содержит
40—60 отсчетов), а также результаты самонаблюдений с приме-
нением тех же тестов, которые один испытуемый проводил еже-
дневно каждые 2 ч в течение 1.5 мес (временной ряд для каждого
параметра содержит около 360 отсчетов).
Методика обработки данных энцефалограммы,
записанной во время сна
Детальный анализ ЭЭГ сна позволил выделить большое коли-
чество следующих параметров: длительность каждой из стадий
сна в течение 1 ч сна (шесть параметров по числу стадий: А, В,
С, D, Е, REM); частоту появления каждой из стадий сна в течение
каждого часа (шесть параметров); число смен стадий в 1 с и коэф-
фициент последовательности стадий; число и среднюю длитель-
ность движений в течение каждого часа — всего 16 показателей
для каждого часа сна.
Кроме того, для всего периода сна изучаются те же 16 пока-
зателей и их разброс в течение этого периода, т. е. всего 32 по-
казателя. Наконец, анализируются последовательность появле-
ния в течение ночи каждой из стадий сна (например, стадии В)
после всех других стадий сна (т. е. A, G, D, Е, REM) и бодрство-
вания (что составляет 36 показателей), частота изменений те-
кущей стадии сна под влиянием движения во сне и, наконец,
общая длительность сна в минутах. Всего рассматриваются
72 общих показателя течения сна в целом, показателей для от-
дельных часов сна — от 64 до 128 в зависимости от общей дли-
тельности сна (4—8 ч).
Обработка данных проводилась двумя методами.
1. Обычные статистические методы исследования средних
и среднеквадратичного отклонения в заданных массивах данных
28
и вычисление достоверности различий (Р) между группами
по ^-критерию Стьюдента. Обработка проводилась на ЭЦВМ
«Минск-32» по программе, написанной В. П. Хоптяром. Таким
образом обрабатывались, например, данные, полученные после
перелетов с пересечением часовых поясов. Проводилось сравне-
ние между картиной ЭЭГ до и после перелета, между карти-
ной ЭЭГ при перелете с востока на запад и с запада на восток;
между картиной ЭЭГ, записанных во время магнитных бурь
и в спокойной магнитной обстановке, и т. д.
2. Метод главных компонент. Стремясь выделить наиболее
информативные и не коррелирующие друг с другом признаки
процессов, мы обратились к методу главных компонент. Алго-
ритм обработки сводится к следующему. Первую главную ком-
поненту исследуемой генеральной совокупности наблюдений,
т. е. нормированную линейную комбинацию параметров, заме-
ренных на объекте исследования, которая среди прочих линей-
ных комбинаций обладает наибольшей дисперсией, мы вычис-
ляем по формуле
У1 = ^11 Х1 + ^12 х2 + • • • + hp хр = 1\Х*
где Xi, х29 . . . хр — параметры, замеренные на объекте наблюде-
ния; 1[ — первый собственный вектор ковариационной (корре-
ляционной) матрицы.
Главная компонента исследуемой генеральной совокупности —
такая линейная комбинация исходных параметров, которая
среди всех прочих линейных комбинаций, не коррелированных
с предыдущими (г—1) главными компонентами, обладает наи-
большей дисперсией, вычисляемой по формуле
Vi = + li2x2 4- • • • + lipxp = I'ix*
Собственный вектор ковариационных (или корреляционных)
матриц, построенных по исходным параметрам, определяется
с помощью методов линейной алгебры. Комплекс программ,
созданный М. А. Каганом (Каган, 1977; Каган, Симонов, 1978)
для машины «Минск-32», позволяет, кроме коэффициентов исход-
ных параметров, определить ряд сопутствующих статистических
характеристик как входных данных (среднеквадратичное от-
клонение и т. д.), так и вычисленных параметров (доверитель-
ные уровни для собственных значений, вклад в суммарную
дисперсию, ошибку автопрогноза и т. д.). С помощью данного
комплекса программ проводился, например, анализ признаков,
указывающих на степень адаптоспособности испытуемых (Мои-
сеева, Каган, 1978).
Методика обработки взаимооценок,
полученных путем опроса
Как указывалось выше, макроритмы разбиты на четыре фазы
(I—IV), примерно равные по длительности. Прежде всего опре-
делялось, в какой фазе макроритма находится обследуемый
29
в день опроса. Затем по результатам всех опросных листов для
каждого числа строится матрица, где по строкам откладываются
все оценки, которые получило данное лицо, а по вертикали —
те оценки, которые им поставлены. Для каждого столбца и каж-
дой строки подсчитывается средняя оценка а. Затем для всех
столбцов и для всех строк вместе подсчитывается общая сред-
няя А столбца (средняя оценка, которую ставит данное лицо)
и А строки (средняя оценка, которую получает данное лицо
в данном опросе).
Все матрицы суммировались по каждой строчке и по каждому
столбцу и вычислялись средние по строчкам и по столбцам всех
матриц. Средние значения по каждой строке и по каждому
столбцу сравниваются со соответствующим средним Л, и в том
случае, если а больше Л, ставится плюс, если меньше — минус,
а если они равны, то — нуль. Далее в списке, где отмечены
ранги биоритмов на сегодняшний день, проставляются отметки
плюс, минус или нуль, т. е. отмечается отклонение от средней
каждого опросного листа. Наконец, строится таблица разделен-
ных на ранги биоритмов, где для каждого ранга вписывается
(а затем подсчитывается) число отклонений от средней в ту или
иную сторону; на основании таблицы вычерчиваются соответ-
ствующие графики.
Методика поиска зависимости
течения биологических ритмов
от геофизических факторов и собственных макроритмов
Для поиска зависимости течения биологических ритмов от
различных факторов применялись следующие методы.
1. Сравнение суточных графиков течения биоритмов с графи-
ками динамики геофизических факторов в течение суток, по-
строенными на ЭЦВМ «Минск-32» тем же способом, как это
делается для биологических ритмов.
2. Обычные статистические методы, корреляционный анализ,
в частности метод Чебышева (решение корреляционных уравне-
ний п-порядка).
3. Метод главных компонент.
В процессе работы большинство использованных методов
было включено в единую программу обработки данных на ЭЦВМ,
составленную В. М. Сысуевым с помощью старших инженеров-
программистов Л. В. Шереметьевой, Е. Г. Лебедевой и Р. Е. Лю-
бицкого. Учитывая, что в отличие от других методов обработки
данных метод Чебышева, позволяющий выявить и нелинейные
зависимости, редко применяется в биологии и медицине, мы
должны дать следующие пояснения относительно интерпретации
результатов обработки.
Результаты выводятся в виде графиков, на которых по оси
абсцисс расположен упорядоченный по возрастанию ряд число-
вых значений одного из параметров, по отношению к измене-
но
ниям которого ищутся зависимости изменений второго пара-
метра (значения которого распечатаны рядом с осью ординат).
Значения в обоих случаях представлены не абсолютными циф-
рами, а отклонениями от среднего. На графике выпечатываются
как положения значений отклонений от среднего исследуемого
параметра, так и результирующая кривая аппроксимации и ука-
зание, каков порядок уравнения (от 2 до 4), на основании кото-
рого произведена аппроксимация. При анализе обращается вни-
мание прежде всего на форму кривой, позволяющей выявить ха-
рактер зависимости.
Кроме формы учитываются абсолютные значения отклонений
от средних величин изучаемого параметра, при которых возни-
кают изменения хода аппроксимирующей кривой. Использование
этого приема обработки позволяет судить о влиянии не абсолют-
ных значений того или иного геофизического параметра, а более
или менее выраженных отклонений от средних значений этого
параметра, вычисленных для настоящего отрезка времени.
Обследованные лица
Обследованы взрослые люди обоего пола в возрасте 20—
55 лет — больные, находившиеся на лечении в двух клиниках
Ленинграда: неврологическом отделении больницы № 20 Москов-
ского р-на (заведующая кандидат мед. наук Л. И. Никитина)
и нейрохирургическом отделении больницы «В память 25 Ок-
тября» (заведующий кандидат мед. наук К. К. Родионов) —
и практически здоровые лица.
Таблица 9
Нозологические формы и исходы заболеваний изученных больных
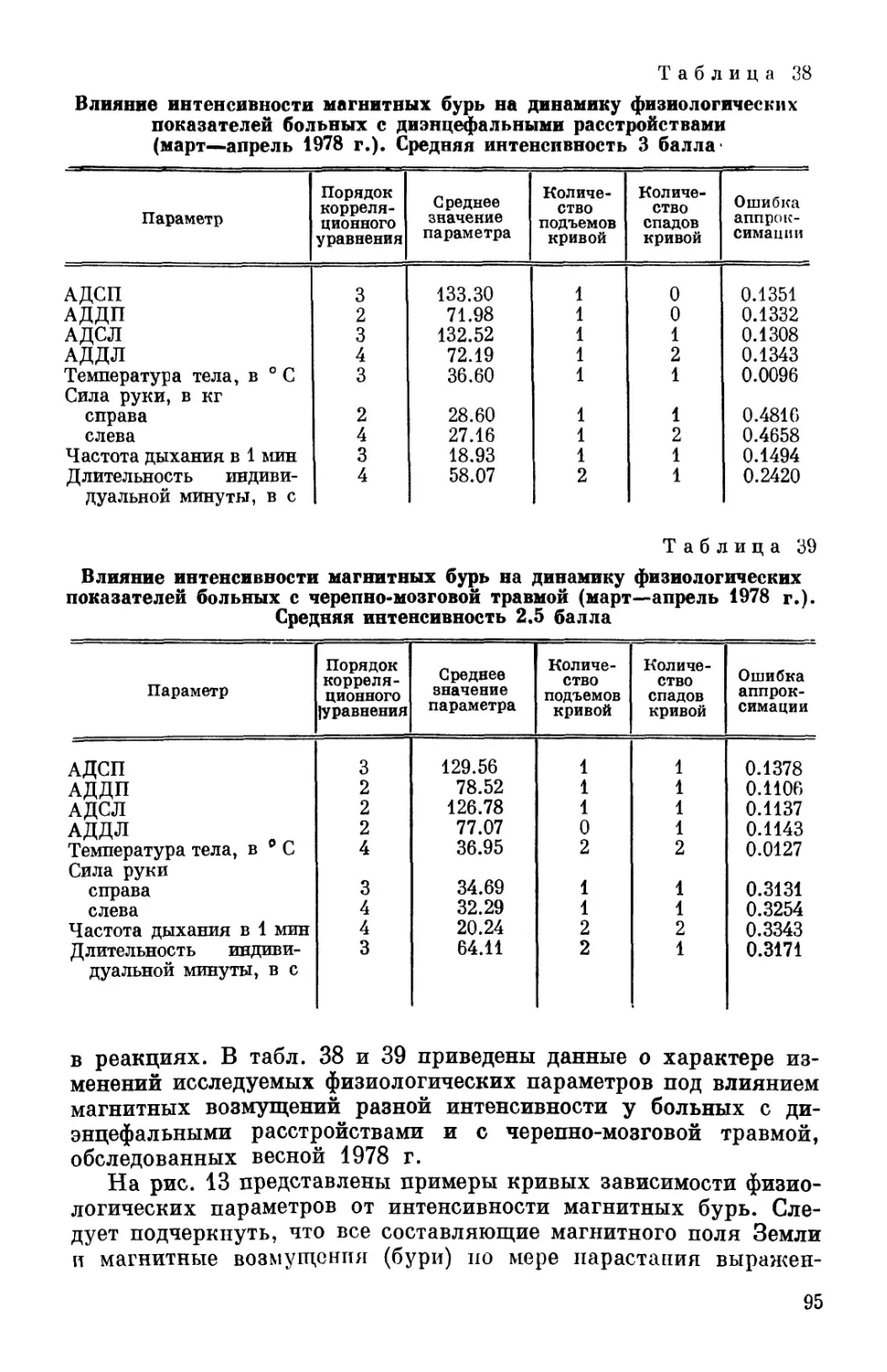
Диагноз Общее число обследован- ' ных Исход
хороший удовлет- воритель- ный плохой
Острая черепно-мозговая травма 27 9 17 1
Арахноидит головного мозга 6 4 — 2
Инфекционно-аллергический васку- лит 8 7 1 —
Вегетативно-сосудистая дистония и гипертоническая болезнь III ст. 7 4 3 —
Шейный остеохондроз 6 4 1 1
Опухоль мозга 3 — — 3
Неврастения, мигрень, митраль- ный порок 6 5 1 —
Всего: 63 33 23 7
31
Группа больных
В целях усовершенствования диагностики, прогнозирования
исхода и выбора тактики лечения в конце 1977 г. и в 1978 г.
в двух указанных клиниках проводилось биоритмологическое
обследование групп больных. В неврологическом отделении
больницы № 20 оно затем вошло в число постоянных методов
исследования больных с диэнцефальными расстройствами.
В табл. 9 приведены данные относительно нозологических
форм и исходов заболевания 63 больных, биоритмологические
данные о которых были использованы при написании этой книги.
Десять больных были исследованы в процессе лечения 2—
3 раза.
Группа здоровых
Группа испытуемых состояла из 65 практически здоровых
лиц обоего пола в возрасте 25—50 лет. 18 человек проходили
ежедневные однократные исследования в течение —2г/2 мес,
находясь в Ленинграде в обычных условиях. Из них семь чело-
век в течение длительного срока заполняли дневники, отражаю-
щие особенности их сна. 30 человек были исследованы в Ленин-
граде до и после перелетов в широтном или (и) долготном направ-
лении. У каждого человека производились 2—3-суточные замеры
различных показателей 5 раз в сутки и (или) запись ЭЭГ во
время сна. Из них пять обследованы 5 раз в сутки в течение
3 дней только в условиях Ленинграда, семь человек — в Петро-
павловске, Новосибирске, Фрунзе, Владивостоке и, по возвра-
щении из поездки, в Ленинграде. Один испытуемый в течение
45 сут подряд производил самоизмерения во время поездки по
Дальнему Востоку (Сахалин—Курильские острова—порт Ва-
нино—Комсомольск—Иркутск—Братск), он включен и в другие
серии.
Кроме того, были привлечены пять спортсменов высокого
класса (фехтовальщики), которые исследовались прямо в ходе
поединков. Отдельную группу составляли 200 человек (175 ту-
ристов и 25 студентов ЛПИ), среди которых было произведено
анкетирование. Более подробно данные об испытуемых приво-
дятся в каждом разделе при изложении материала.
Глава 3
ЦИРКАДНЫЕ КОЛЕБАНИЯ УРОВНЯ
БОДРСТВОВАНИЯ И НЕКОТОРЫХ
ПСИХИЧЕСКИХ ФУНКЦИЙ
Поскольку ритмы охватывают любые проявления живого —
от деятельности энзимов до сложных форм поведения, — они
присущи и изменениям состояния сознания как в широком,
так и в узком смысле этого слова, т. е. изменениям уровня
бодрствования, под которым понимается уровень готовности
к контакту с внеп^ней средой. Во время сна этот уровень практи-
чески является нулевым, а в бодрствовании — в значительной
мере колеблющимся, что выражается в большей или меньшей
эффективности выполнения тестов или реальной работы (Vernon,
1921; Preston et al., 1973; Биоритмы и труд, 1980).
Одним из основных факторов, влияющих на продуктивность
человека, признана не просто ритмическая смена сна и бодрство-
вания, но колебания уровня сознания, связанные с тем, что
циклы сна (выражающиеся в закономерной смене диапазона
частот биоэлектрической активности мозга) сменяют друг друга
круглосуточно (Kleitman, 1967; Webb, Agnew, Д967, 1969;
Webb, 1969). Клейтманом (Kleitman, 1969) выделен ультрадный
ритм — так называемый основной цикл активности нервной
системы (BRAC — Basic Rest-Activity Cycle) с периодом около
90 мин, обнаруживаемый в колебаниях уровня работоспособ-
ности (Огг et al., 1976; Lavie, 1977).
Показаны около-90-минутная периодичность качества выпол-
нения психологических тестов (Kripke, O’Donoghue, 1968; Orr
et al., 1974), суточная ритмика времени реакции при выполнении
сложных действий в условиях, имитирующих полет (Klein et al.,
1970), суточный ритм точности движений (Хохрякова, 1971).
Высказывается предположение (Colquhoun, 1970), что в целом
продуктивность психических процессов меняется пропорцио-
нально температуре тела.
Циркадные колебания уровня бодрствования
С целью исследования циркадных колебаний уровня бодрство-
вания мы использовали записи электроэнцефалограмм, прове-
денных на 40 практически здоровых людях в возрасте 20—35 лет
(студенты и преподаватели Ленинградского медицинского инсти-
33
тута) в дневное время. Испытуемые засыпали в звуко- и свето-
изолированной камере утром и днем. Регистрация биопотенциа-
лов проводилась с помощью 15-канального электроэнцефалографа
фирмы «Альвар». Образцы записей приведены в книге Н. Н. Де-
мина и др. (1978).
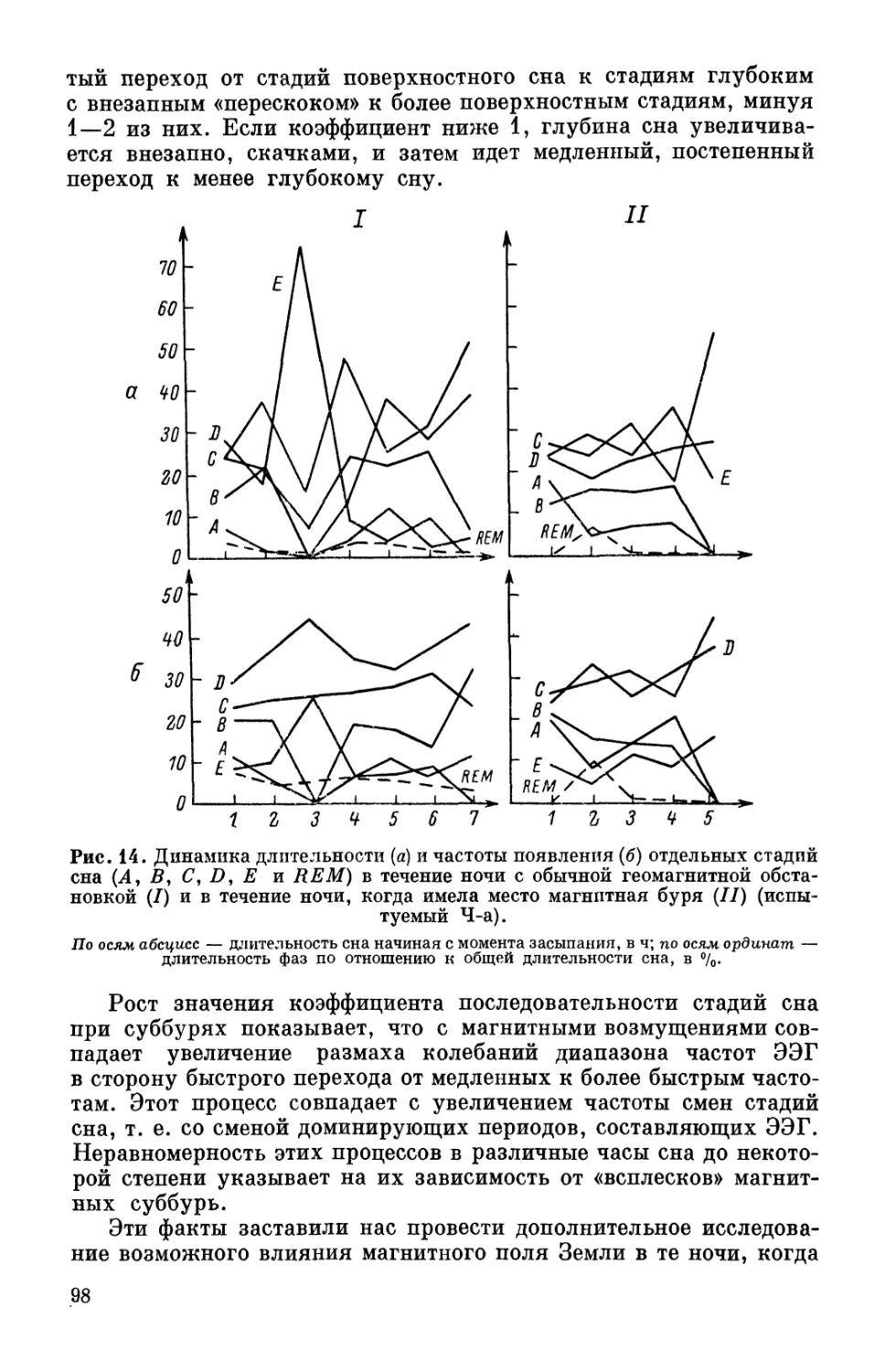
В табл. 10 дана средняя длительность стадий сна REM и Е
в течение 1—2 ч в различное время суток. Значительная раз-
Таблица 10
Средняя длительность стадии парадоксального сна (стадия REM)
и стадии глубокого сна (Е) в зависимости от времени,
когда человек засыпает, в % от общей длительности сна
Стадия Источник Приблизительное время засыпания, в ч Длительность стадий за 2 ч ночного сна
9 12 14 16 18 первых послед- них
REM Webb, Agnew, 1969 32 12 15 8 3 3
Моисеева, 1975в 7 — 0 2.3 3 3.3 8
Е Webb, Agnew, 1969 0 15 18 20 — 43 3
Моисеева, 1975в 2.5 — 17 23 0 30 9
Примечание. Нуль обозначаетнеобнаружение данной стадии, тире —отсут-
ствие исследований.
ница в длительности REM-стадии между нашими и литератур-
ными данными зависит от различной методики кодирования
этой стадии сна. Однако при различии в абсолютных цифрах
тенденция изменения представленности стадий в отрезках сна,
относящихся к разному времени суток, аналогична. Стадия глу-
бокого сна Е отчетливо прослеживается в течение двух первых
часов ночного сна, затем она почти исчезает и снова появляется
и усиливается около 14—16 ч, исчезая к 18 ч. Такого рода дина-
мика совпадает с ритмикой времени реакции на слуховые и (или)
зрительные стимулы. Самое большое время реакции приходится
на полночь и послеобеденные часы, самое малое — на раннее
утро (Aschoff et al., 1969).
Как известно, приблизительно так же меняется и работоспо-
собность независимо от вида деятельности, а также показатель
точности восприятия метрики времени.
Ритмы точности восприятия метрики времепи
Исследование восприятия времени человеком проводится
в двух основных направлениях — топологии и метрики. В ре-
зультате изучения топологии, т. е. восприятия порядка последо-
вательности стимулов, одновременности и т. д., зависящей в зна-
чительной мере от разрешающей способности анализаторов, был
34
выдвинут так называемый «сенсорный подход», согласно которому
восприятие зависит исключительно от особенностей внутренних
часов организма (Ornstein, 1970). Считается при этом, что время
квантуется с постоянной скоростью (50—100 мс) вне зависимости
от характера и содержания конкретной психической деятельности
(Kristoffersen, 1967).
Изучение метрики времени, т. е. оценки длительности про-
межутков времени (как заполненных деятельностью, так и пу-
стых), привело к созданию так называемого познавательного
подхода, согласно которому отражение времени сознанием че-
ловека — сложный процесс, зависящий от многих факторов,
в том числе от характера перцепции. Создаются комплексные
модели механизма восприятия времени, в частности модель,
согласно которой существуют два относительно независимых
блока — блок оценки времени и блок переработки информации,
заполняющей отрезок времени (Thomas, Weaves, 1975). Само
формирование в сознании настоящего времени, т. е. интервала
времени, внутри которого события присутствуют одновременно,
а не последовательно (Whitrow, 1964), обеспечивается много-
уровневой системой анализа. Экспериментально показано изме-
нение не только метрики, но и топологии восприятия времени
при изменении условий (Зинченко, Круминя, 1978).
Как любая другая функция, восприятие времени само под-
вержено колебаниям во времени, что может быть зафиксировано
в виде циркадных изменений точности оценки отрезков времени
или скорости равномерного перечисления числового ряда (На1-
berg, 1969; Моисеева и др., 1979). Однако возможны колебания
и с другими периодами — как ультрадными, так и инфрадными.
С целью изучения ритмики восприятия времени в двух сериях
были исследованы: 1) точность воспроизведения заданных отрез-
ков времени и 2) длительность «индивидуальной минуты».
Исследование точности воспроизведения
заданных отрезков времени
Практически здоровым испытуемым под контролем секундо-
мера предъявлялся отрезок времени определенной длительности
(маркируемый двумя ударами молоточком по столу). Испытуе-
мый немедленно воспроизводил заданный отрезок также двумя
ударами. Длительность отрезков от 5 до 20 с, каждый раз предъ-
являлось по десять отрезков. Двух испытуемых исследовали
4 дня подряд с 8 до 24 ч включительно через каждый час; шесть
испытуемых исследованы в течение 2—3 сут с интервалами
2-3 ч.
В протоколах фиксировалось, какой длины отрезок времени
был предъявлен, какой длины отрезок был воспроизведен, и вы-
числялось, какой процент увеличения (или уменьшения) длины
был допущен при воспроизведении. Например, при предъявле-
35
нии отрезка 7 с он был воспроизведен как 8 с, т. е. имело место
приращение на 1 с, или на 14.5%; при предъявлении отрезка
15 с он был воспроизведен как 12 с; имело место уменьшение
на 3 с, т. е. на 20%. В случае правильного воспроизведения
ошибка была равна нулю. При суммировании учитывалась
средняя ошибка в ту и другую сторону. Наиболее информатив-
ным оказался учет процента ошибок в целом (т. е. сколько раз
ошиблись, а не насколько ошиблись) в сторону увеличения или
уменьшения длины отрезка.
Определялся период наилучшей аппроксимации функций
в околосуточном диапазоне, а также выявлялась акрофаза. Кроме
того, были найдены средние значения и достоверность различий
между группами наблюдений в зависимости от фазы макроритмов
в день исследования, а также колебания тенденции психологи-
чески переоценивать и недооценивать протекшее время.
Оказалось, что наиболее точно отрезки времени распозна-
ются всеми испытуемыми около 23 ч. Кроме того, существуют
дополнительные моменты увеличения точности распознавания
около 13, 15 или 18 ч. У трех испытуемых они наблюдались
в 13 и в 15 ч, у двух — в 15 и 18 ч, у остальных — в один из
указанных сроков.
При поиске (по специальной программе) наилучшего периода
аппроксимации в околосуточном диапазоне (в часах) оказалось,
что флуктуации тенденции к переоценке времени происходят
с несколько большим периодом, чем флуктуации тенденции
к недооценке времени. Однако изучение кривых каждого испытуе-
мого по отдельности показало, что эти различия выражены
только в тех случаях, когда испытуемые почти не ошибаются.
Если же процент ошибок велик, гистограммы распределений
ошибок с плюсом и минусом являются зеркальным отражением
друг друга, что вполне естественно. Однако следует отметить,
что средняя ошибка (выявляющаяся на кривых как постоянная
составляющая) значительно чаще направлена в сторону преуве-
личения.
Наиболее точное распознавание отрезков времени в зависи-
мости от фаз макроритмов, в которых находится человек в соот-
ветствующий момент, имеет место: 1) в нисходящей фазе и фазе
спада физического цикла; 2) в восходящей фазе и фазе подъема эмо-
ционального цикла; 3) в нисходящей фазе интеллектуального цикла.
Однако статистика пока недостаточна, и различия между
группами недостоверны. Кроме того, различные сочетания фаз
разных ритмов затрудняют анализ.
Исследование особенностей восприятия
длительности «индивидуальной минуты»
В качестве теста определения чувства времени (длительность
«индивидуальной минуты») применялся равномерный счет от 1
до 60, который проводится примерно за 1 мин.
36
Изучены больные, находившиеся на лечении (результаты об-
следования группы передавались лечащему врачу и использо-
вались при определении прогноза заболевания), и группа практи-
чески здоровых лиц.
Из 50 больных 31 человек перенесли тяжелую черепно-моз-
говую травму (исследование делалось в остром периоде), 19 стра-
дали диэнцефальными расстройствами различной этиологии.
В 15 случаях показатель исследован дважды — в начале и в конце
лечения. Всего анализу подвергались 195 суточных кривых, от-
ражающих динамику внутреннего отсчета длительности проме-
жутков времени.
Оказалось, что вне зависимости от диагноза заболевания
у лиц с благополучным течением процесса счет до 60 осу-
ществлялся примерно за минутный интервал (61.81 с), в то время
как у лиц с неблагоприятным исходом отмечалась выраженная
тенденция к укорочению времени счета, которое в среднем ока-
зывается равным 53.81 с. Суточные колебания скорости отсчета
времени выражены более отчетливо у больных с благополуч-
ным исходом.
Эта же тенденция видна и при сравнении в динамике показа-
телей одного и того же больного в процессе лечения. В табл. 11
приведены средние и (дисперсия длительности счета до 60,
Таблица И
Особенности изменения параметров отсчета минутного интервала
в зависимости от исхода заболевания (средние данные за 3 дня)
Результат лечения Испы- туемый Среднее время, в с Дисперсия Показатель СОК
до лече- ния после лечения до лече- ния после лечения до лече- ния после лечения
Хороший 1-й 58.67 61.20 0.87 0.95 3; 4; 5 4; 4; 4
6-й 64.60 66.80 0.20 0.32 3; 4; 5 4; 4; 5
9-й 53.60 60.40 0.16 0.19 3; 4; 5 3; 4; 4
23-й 51.00 55.53 0.93 0.16 4; 4; 5 4; 5; 5
26-й 66.00 65.06 0.19 0.13 3; 3; 4 3; 4; 5
27-й 100.87 108.87 0.19 0.12 3; 4; 4 3; 4; 4
Посред- 10-й 47.40 48.00 0.33 0.15 3; 4; 4 3; 5; 5
ственный 15-й 63.70 62.08 0.88 0.34 2; 3; 3 3; 3; 3
17-й 66.69 65.93 0.23 0.13 4; 4; 4 2; 4; 4
28-й 35.73 31.86 0.75 0.17 4; 4; 4 2; 3; 5
29-й 61.81 59.73 0.68 0.79 4; 4; 4 2; 4; 4
30-й 60.06 60.00 0.11 0.64 4; 4; 5 4; 4; 4
31-й 66.13 64.87 0.35 0.33 3; 5; 5 4; 4; 4
Плохой 3-й 49.66 49.53 0.21 0.26 4; 4; 5 4; 4; 5
25-й 64.00 41.43 0.25 0.60 3; 4; 4 3; 4; 4
Примечание. Здесь и в табл. 15 в графе «СОК» первая цифра означает
оценку за 1-й день исследования, вторая — за 2-й, третья — за 3-й день.
37
а также показатель СОК до лечения и после. На рис. 2 приведены
кривые больного с хорошим эффектом лечения.
Как видно из табл. И, при хорошем результате лечения сред-
ние значения (а в половине случаев и дисперсия) возрастают,
в четырех случаях увеличивается показатель СОК (повышение
значения одного из показателей с 2 до 4 у 1-го больного, с 3 до 4
у 6-го больного и т. д.). В случае посредственных и плохих
результатов лечения средние значения преимущественно снижа-
Рис. 2. Суточная динамика счета до 60 больного с хорошим эффектом лечения.
А — до лечения; Б — после лечения. По оси абсцисс — время суток, в ч; по оси ординат —
нормированные значения параметра. 1, 2, 3 — первые, вторые и третьи сутки исследова-
ния соответственно.
ются, СОК иногда может уменьшаться (как в случае пациентов
17-го, 28-го, 29-го, 30-го), а иногда и увеличиваться (как в случае
пациентов 10-го и 15-го).
Таким образом, у больных выявлены определенные зависи-
мости между способностью человека противостоять болезни,
т. е. его защитными силами, и длительностью его индивидуальной
минуты и установлено наличие определенной динамики значений
показателя в процессе изменения общего состояния. Кроме того,
выявлены большие индивидуальные различия. Средние величины,
рассчитанные по 3-суточным исследованиям (т. е. по 15 замерам),
варьируют (как это видно из табл. 11) от 31.86 до 108.87 с.
Суточная ритмика восприятия длительности индивидуальной
минуты была выделена только у больных с хорошим исходом.
Наибольшие значения показатель принимает в 8 и (или) 12 ч,
еще один максимум (слабее выраженный) приходится на 20 ч.
Результаты исследования группы здоровых лиц
Были обследованы четверо мужчин и четыре женщины в воз-
расте 25—50 летг которые продолжали при этом нести рабочую
нагрузку. Всего проведено 16 серий исследований (длитель-
ностью от 3 до 10 дней, одна серия длилась 6 нед). Шесть испытуе-
мых исследованы один раз, один — 2 раза; один участвовал
в семи сериях, из них в четырех совместно с 1—2 другими испы-
38
туемыми. Данные каждой серии разбивались на 3-дневные от-
резки и анализировались последовательно, так же как клиниче-
ские серии.
Прежде всего было обнаружено, что испытуемые значительно
отличаются друг от друга по своим средним показателям, причем
для каждого конкретного лица величина его индивидуальной
минуты является относительно стойким показателем. Подобно
тем результатам, которые были получены на больных, лица
с высокими способностями к адаптации в условиях изменения
внешней среды и высоких нагрузок (эмоциональных, интеллек-
туальных и физических) отличаются более высоким показате-
лем СОК и умеют «растягивать» время. Их индивидуальная ми-
нута превышает минуту физического времени. Средние по двум
сериям исследований испытуемого с невысокими способностями
к адаптации были равны 47.06 и 46.20 с, в то время как средние
у хорошо адаптирующегося испытуемого по шести сериям варьи-
ровали от 62.90 до 69.71 с.
Оказалось далее, что различия между хорошо и слабо адапти-
рующимися лицами распространяются и на взаимозависимость
между течением различных процессов (в том числе психических
процессов) и собственным восприятием длительности времени.
Этот вопрос будет детально рассмотрен после изложения данных,
касающихся циркадных колебаний некоторых психических
функций.
Что же касается ритмики восприятия длительности индиви-
дуальной минуты — у каждого испытуемого оказывается свой
ритм. Наиболее постоянный ритм у хорошо адаптирующегося
испытуемого, проведшего шесть серий. Первый максимум при-
ходится на 8 ч утра, второй (меньший) — на 12 ч дня, в остальное
время максимумы выпадают очень редко. У других хорошо
адаптирующихся испытуемых максимальные значения функций
наблюдаются в 12 и 20 ч. У слабо адаптирующейся испытуемой
максимумы очень вариабельны, с тенденцией к 16 и к 24 ч.
Таким образом, если объединить данные, полученные на
группах больных и здоровых, то окажется, что тенденция у прак-
тически здоровых лиц, но плохо адаптирующихся, та же, что
и у больных с плохим исходом: у них нарушена ритмика, а зна-
чения средних низкие. Хорошо адаптирующиеся лица, так же
как больные с хорошим исходом, обладают адекватной ритмикой
изменений показателя восприятия времени — он имеет наиболь-
шие значения в 8 или 12 ч, т. е. тогда, когда (как известно из
обыденной жизни) большинство людей наиболее работоспособны
и когда они (как показывают многочисленные исследования
в области инженерной психологии) успешнее выполняют психо-
логические тесты. Это время совпадает с моментами, когда
в биоэлектрической активности слабо представлены (или не
представлены) медленные волны, характерные для развития
39
состояния сна, т. е. достигается самый высокий уровень бодрство-
вания. Естественно, уровень бодрствования должен отразиться
и на ритмах психической активности.
Ритмы психической активности
Ритмы психической активности исследованы с помощью пробы
проверки краткосрочной памяти тестов на способность осу-
ществлять простейшие арифметические действия, счета пальцев
и корректурной пробы.
Динамика показателей корректурной пробы
Корректурная проба (вычеркивание трех заданных букв
с помощью заданных знаков) проделывается за 5 мин; в ней учи-
тываются: а) число просмотренных знаков за каждую минуту
и за все 5 мин (что отражает работоспособность испытуемого);
б) число вычеркнутых (как правильно, так и ошибочно) знаков
за каждую минуту и все 5 мин (что отражает количественно осу-
ществление принятия решений); в) число ошибок за каждую
минуту и все 5 мин; г) коэффициент «средней памяти» — среднее
число верно вычеркнутых букв в 1 мин. Он вычисляется путем
вычитания из общего числа просмотренных знаков числа ошибок
и деления остатка на 5.
Две серии исследований проведены на десяти практически
здоровых испытуемых (четверо мужчин и шесть женщин) и двух
больных (обе женщины), страдающих диэнцефальными расстрой-
ствами. В первой серии (исследование циркадной ритмики) вы-
полнение тестов 5 раз в день шло 3 сут подряд, при этом четверо
испытуемых (из пяти) каждый раз получали новое задание, а один
выполнял все 15 раз одно и то же задание. Во второй серии —
исследование макроритмики (шесть испытуемых, один из них
исследован и в первой серии) — тесты выполнялись во вторую
половину дня 7—10 дней подряд; один испытуемый выполнял
тест в течение 3 нед.
Результаты исследования циркадных ритмов
выполнения корректурной пробы
В табл. 12 и 13 приведены сводные результаты выполнения
корректурной пробы и время максимальных и минимальных
значений параметров. Исследуемые 1—3-й—практически здоро-
вые лица, 4-й и 5-й — больные. Испытуемый 1-й имел постоян-
ное задание; 3-й, 4-й и 5-й имели одинаковые задания каждый
раз, т. е. степень трудности для всех трех была одинаковой.
Как видно из табл. 12, самые высокие показатели наблю-
даются у испытуемой 1-й, имевшей постоянное задание (рис. 3).
При этом у нее циркадная ритмика всех исследованных показа-
40
Таблица 12
Средние результаты выполнения корректурной пробы пятью испытуемыми
Параметр Испытуе- мый Результаты за каждую минуту Среднее за 5 мин
1-ю 2-ю 3-ю 4-ю 5-ю
Общее количество зна- ков: просмотренных 1-й 161 145 139 136 128 143.0
2-й 97 104 109 112 114 111.5
3-й 76 93 94 96 96 91.1
4-й 84 89 92 95 97 91.5
5-й 76 83 87 89 91 82.2
вычеркнутых 1-й 57 52 50 51 48 51.0
2-й 32 38 38 39 39 37.8
3-й 28 32 34 34 35 32.9
4-й 28 31 33 34 34 32.2
5-й 26 28 30 32 32 30.4
Число ошибок 1-й 0.2 0.5 0.4 0.4 0.3 0.36
2-й 3.3 2.0 2.0 2.7 2.3 2.46
3-й 1.1 1.7 0.4 0.6 0.4 0.85
4-й 1.9 1.7 0.5 1.4 1.0 1.30
5-й 2.6 2.1 1.6 1.1 1.9 2.50
Показатель памяти 1-й 56.8 51.5 49.6 50.6 47.7 50.6
2-й 28.7 36.0 36.0 36.3 36.7 35.3
3-й 26.9 30.3 33.6 33.4 34.6 32.1
4-й 26.1 29.3 32.5 32.6 33.0 30.9
5-й 23.4 25.9 28.4 30.9 ЗОЛ 27.9
Таблица 13
Время достижения (в ч) максимумов и минимумов средних значений
показателей корректурной пробы у пяти испытуемых
Испы- туемый Количество про- смотренных знаков Количество вычеркнутых знаков Число ошибок Показатель памяти
макси- мум минимум макси- мум минимум макси- мум минимум макси- мум мини- мум
1-й 20 (24) 8 20 (16) 8 24 (8) 20 (16) 20 (16) 8
2-й 20 (24) 8 20 (16) 12 (8) 12 (16) 8 24(16) 12
3-й 20 (8) 16 24 (8) 16 (20) 12 (16) 20(8) 20 (8) 16 (12)
4-й 8 (20) 24 (8) 8(16) 12 (24) 24 (20) 16 (20) 8 24
5-й 20 (8) 12 (16) 20 (16) 12 (24) 12 (16) 16 (20, 13) 20(8) 12 (16)
Примечание. В скобках — время дополнительных максимумов или миниму-
мов, в ч.
телей, кроме общего числа ошибок, на который особенно влияют
любые внешние помехи, очень четко выражена. В отличие от
остальных четырех испытуемых, у которых ясно прослежива-
ется картина тренировки во время выполнения задания, что
41
выражается в постепенном росте от минуты к минуте числа
просмотренных и вычеркнутых знаков при уменьшении числа
ошибок, у этой испытуемой наиболее высокие показатели имели
место на 1-й минуте, а затем они снижались. Таким образом,
Рис. 3. Суточная динамика показа-
телей корректурной пробы для испы-
туемой, имевшей постоянное задание.
А — суммарное количество просмотрен-
ных знаков; Б — суммарное количество
вычеркнутых знаков; В — количество оши-
бок. Остальные обозначения те же, что
на рис. 2.
в данном случае выявляется картина утомления. Новый тест
выполняется от минуты к минуте все лучше, а старый, хорошо
знакомый, — все хуже и хуже. При сравнении результатов
3 дней исследования оказалось, что во
Рис. 4. Суточная динамика среднего показа-
теля памяти больной с диэнцефальными рас-
стройствами.
Обозначения те же, что на рис. 2.
всех случаях они растут.
Как видно на рис. 3,
кривые (кроме кривых
числа ошибок) отли-
чаются более высоким
средним уровнем к 3-му
дню.
Таким образом, в
результате тренировки
повышаются показатели
во всех случаях, но
выполнение привычной,
надоевшей комбинации
имеет отрицательную
динамику внутри каж-
дой отдельной пробы,
а выполнение новой
комбинации — положи-
тельную динамику. Циркадная организация изменения иссле-
дуемых функций у здоровых испытуемых довольно четкая,
у больных ритмика нарушена. На рис. 4 приведены кривые су-
точного колебания среднего показателя памяти одной из больных
(испытуемая 5-я, см. табл. 13). Можно сказать, что в целом
исходя из данного теста, наибольшая работоспособность (о кото-
рой судят по числу просмотренных знаков) достигается примерно
42
к 20 ч, наиболее четкое принятие решения (о котором судят по
числу вычеркнутых знаков) осуществляется также в 20 ч, а наи-
меньшее число ошибок человек совершает в 20, 16 и в 8 ч.
По суммарному показателю средней памяти оптимально задание
выполняется в 20 и в 8 ч.
Результаты исследования влияния макроритмов
на выполнение корректурной пробы
Анализ зависимости результатов 83 корректурных проб
у шести практически здоровых испытуемых, от фазы их макро-
ритмов позволили обнаружить, что наиболее высокие резуль-
таты по показателю средней памяти наблюдаются: а) в нисхо-
дящую и восходящую фазы физического цикла; б) в нисходящую
фазу эмоционального цикла; в) в нисходящую фазу интеллек-
туального цикла.
Однако эта зависимость выражена незначительно (табл. 14),
и можно говорить лишь о тенденции к улучшению результатов
исследования памяти в те моменты, когда человек находится
в переходных фазах циклов. Следует напомнить, что аналогич-
Таблица 14
Значение коэффициента средней памяти (к) у шести испытуемых
в зависимости от фаз (I—IV) макроритмов (средние по 83 исследованиям)
Макроритм I II Ш IV
k п k п k п k п
Физический 32.8 25 28.1 22 32.9 11 30.0 25
Эмоциональный 31.9 15 28.63 30 34.12 24 32.5 14
Интеллектуальный 30.16 21 29.12 19 36.2 15 31.4 28
Примечание, п — число исследований, проведенных в данную фазу.
ные тенденции обнаруживались и в отношении параметра точ-
ности распознавания отрезков времени.
Динамика показателей тестов
на проверку краткосрочной памяти
и точности простейших арифметических действий
Исследования проводили 5 раз в сутки в течение 3—10 дней
на той же группе из семи человек, у которых изучалась длитель-
ность индивидуальной минуты. В тестах на кратковременную
память и сложение учитывались время выполнения теста
и число ошибок. Кроме того, для трех испытуемых учитывалось
время счета пальцев до 25.
Из-за малого числа ошибок строить суточные кривые было
нерационально, поэтому в табл. 15 приведены только данные,
43
Таблица 15
Средние показатели выполнения психологических тестов у семи
испытуемых
Испы- туемый Проба на память Счет пальцев Проба на сложение Время достиже- ния максиму- мов двух или трех тестов
сред- нее время, в с пока- затель СОК макси- мум, в ч сред- нее время, в с пока- затель СОК макси- мум, в ч сред- нее время, в с пока- затель СОК макси- мум, в ч
5-й 54.4 4; 4; 4 12; 8 40.7 3; 4; 4 12; 20 12; 8
6-й 52.0 3; 4; 4 16; 8 — — — 36.4 4; 5; 5 12; 16 16
15-й 50.2 3; 4; 4 24; 20 — — — 32.4 4; 4; 4 20; 12 20
18-й 55.0 3; 5; 5 12; 16 14.25 3; 4; 5 12; 20 57.5 4; 4; 5 12; 16 12; 16
33-й 89.0 3; 4; 4 20; 8 9.94 2; 3; 3 8; 20 69.0 3; 4; 5 12; 20 20; 8
34-й 82.0 4; 5; 5 16 13.60 3; 4; 4 8; 16 83.0 4; 4; 5 12; 24 16
35-й 45.9 3; 4; 4 20 7.15 4; 4; 4 12; 16 55.2 3; 4; 5 12; 16 12; 16
Примечание. В графе «максимум» первая цифра — время, на которое приш-
лось 2—3 максимума, вторая —один максимум.
касающиеся среднего времени выполнения каждого теста и его
суточной динамики. Среднее время выполнения проб варьирует
в широких пределах у всех испытуемых, суточные кривые пробы
на память и сложение хорошо организованы, кривые счета
пальцев (отражающего счет и координацию движения) органи-
зованы хуже. В течение трех последовательных дней совпадение
хода кривых хорошее, но у каждого испытуемого, как правило,
существуют два максимума в проявлении каждой функции.
Совпадения максимумов различных функций у всех испытуемых
имеет место около 12 ч, у пяти испытуемых второй максимум
обнаружен около 20 ч, у четырех испытуемых второй или третий
максимум — около 16 ч. Появление ошибок отмечалось чаще
во вторую половину дня, являясь, как правило, нулевым в 18
и 12 ч.
Таким образом, выполнение психологических тестов имеет
четкую организацию во времени, рисунок которой индивидуален.
Зависимость результатов психологических проб
от длительности индивидуальной минуты.
Факторы, определяющие ее продолжительность
Сравнение качес!ва выполнения психологических проб (ско-
рости выполнения и числа ошибок) у лиц с разной длительностью
индивидуальной минуты показало, что чем длиннее минута, тем
быстрее выполняется тест. Между длительностью минуты и чис-
лом ошибок отношения сложнее, но в целом они сводятся к тому,
что есть какая-то оптимальная длительность индивидуальной
минуты, при которой число ошибок минимально, при большем
затягивании число ошибок вновь растет. На табл, 16 приведены
44
Таблица 16
Зависимость результатов различных проб от длительности
индивидуальной минуты
Испытуемый Средняя длитель- ность счета до 60, в с Корректурная проба Показатель памяти Проба на память Проба на счет Время счета пальцев, в с
число просмотрен- ных знаков число вычеркну- тых знаков число ошибок (в среднем) время, в с число ошибок (в среднем) время, в с число ошибок (в среднем)
1-й 31.6 410 152 9.6 28.5 99.0 7.5 69.0 0.53 9.94
2-й 37.2 457 161 6.4 30.7 82.0 0.5 93.0 0.06 13.60
3-Й 49.5 455 164 4.3 32.0 45.9 0.5 55.2 0.13 7.15
4-й 63.3 712 255 1.8 50.6 55.5 2.8 44.6 0.21 6.68
данные четырех практически здоровых испытуемых, на табл. 17-
данные одного испытуемого, иллюстрирующие эти положения.
Используя метод Чебышева (решения корреляционных урав-
нений n-порядка), мы нашли, что на графике зависимости вероят-
ных значений времени выполнения теста на память (счет до 60)
от длительности индивидуальной минуты обнаруживается чет-
кая обратная связь. При этом кривая оказалась аппроксимиро-
ванной уравнением третьего порядка, что свидетельствует о моно-
тонно убывающей с относительно большой скоростью зависимости.
Обратной связи не обнаружено. Те же соотношения обнаружены
и для времени выполнения теста на сложение, а также счета
пальцев и длительности индивидуальной минуты (аппроксимация
уравнениями второго порядка).
Количество ошибок в тестах имеет обратную связь с индиви-
дуальной минутой, уменьшаясь по мере ее возрастания (аппро-
ксимация уравнениями второго порядка).
Нашим данным в известной мере аналогичны данные
Л. П. Гримака (1978), который исследовал влияние внушения
ускоренного или замедленного в 5 раз хода времени на выпол-
нение определенных тестов. Оказалось, что точность выдержи-
вания заданных временных промежутков (30 и 60 с) и качество
слежения за движением под влиянием ускоренного хода времени
не менялись или несколько ухудшались. Внушение замедлен-
ного хода времени повышало качество выполнения проб, их точ-
ность и стабильность результатов.
Эти изменения нашли (согласно Л. П. Гримаку) отражение
и в картине ЭЭГ — при внушении ускоренного хода времени
альфа-индекс возрастал на 10% относительно спокойного гипно-
тического состояния, амплитуда его падала. При внушении
замедленного хода времени амплитуда альфа-ритма несколько
45
Таблица 17
Показатели выполнения различных проб в зависимости от общего
состояния и уровня нагрузок (один испытуемый, исследование во время
путешествия на Дальний Восток в течение 45 сут)
Условия и время исследования Исследуемые параметры
длитель- ность индиви- дуальной минуты, в с время выпол- нения теста на память, в с число ошибок в тесте на память (среднее) время выпол- нения теста на сложе- ние, в с число ошибок в тесте на сло- жение (среднее) время теста счета пальцев, в с
Первые 3 дня после пе- релета из Ленинграда на Сахалин, нагрузок нет 67.39 50.35 0.014 — — 14.16
Легкая физическая на- грузка (поход по не- сложному маршруту) через 1 нед после пере- лета 59.79 52.60 0.021 58.67 0.0065 14.38
Тяжелый поход по Са- халину через 10 дней после перелета 66.01 50.03 0.020 62.45 0.0025 14.98
То же через 2 нед после перелета 71.52 50.38 0.026 55.74 0.0010 14.57
Перелет на Курильские острова через 3 нед после перелета на Са- халин 61.86 52.93 0.030 57.82 0.0070 14.60
Тяжелый поход на Ку- рилах через 24 дня после перелета на Са- халин 59.30 52.96 0.020 61.13 0.0040 14.84
Отъезд с Сахалина из пор- та Ванино поездом че- рез 1 мес после прибы- тия на Сахалин 58.52 50.45 0.010 56.00 0.0040 14.30
Пребывание в Иркутске, небольшие физические нагрузки (35-е сут пос- ле перелета) 61.78 50.94 0.013 54.30 0.0020 13.84
То же в Братске (39-е сут после перелета) 65.32 51.32 0.016 52.92 0.0025 13.53
Перелет Братск—Ленин- град (42-е сут путеше- ствия) 68.01 49.00 0.007 56.57 0.0060 12.86
Устройство дома 64.39 50.22 0.009 — — 12.10
Состояние недомогания (45-е сут от начала пу- тешествия) 49.50 45.90 0.530 55.20 0.1300 17.15
46
повышалась, появлялась тенденция к снижению альфа-индекса
и повышению индекса медленных волн.
Таким образом, можно сказать, что человек постоянно сущест-
вует в своем собственном временном масштабе, который меняется
под влиянием различных внешних и внутренних факторов. К по-
следним относятся изменения состояния и заболевания, к пер-
вым — гипнотические воздействия, насыщенность отрезка вре-
мени событиями и внезапная смена временной среды, возникаю-
щая при перелетах с пересечением часовых поясов, ведущих
к значительным изменениям биологических ритмов (Моисеева
и др., 1972, 1978а, 19786; Моисеева, 1975в, 1978а), в том числе
ритмов психической деятельности.
На табл. 18 приведены данные одного испытуемого, интен-
сивно работавшего непосредственно после перелета с пересече-
Таблица 18
Изменение длительности «индивидуальной минуты» (в с) в зависимости
от внешних условий и разного общего самочувствия
Условия в момент исследования Самочувствие в момент исследования
плохое хорошее
Пребывание в Ленинграде Перелет без дополнительных нагрузок с пе- ресечением часовых поясов: 49.50 62.20
одного—трех 57.70 60.30
четырех 56.15 66.01
восьми Перелет с дополнительными нагрузками с пе- ресечением часовых поясов: — 69.71
четырех (с эмоциональными нагрузками) 59.30 66.13
четырех (с физическими нагрузками) 58.52 71.52
восьми (с физическими и эмоциональ- ными нагрузками) 64.39 68.10
нием часовых поясов. Виды работы были различны: участие
в заседаниях, выступление с докладами (обычная работа) и ра-
бота в качестве инструктора в тяжелых туристских походах.
Сведены результаты серии исследований, проведенных в 1975—
1978 гг., длившихся в среднем 7—10 дней. Одно исследование
длилось 3 сут и одно — 1.5 мес. Как видно из табл. 18, длитель-
ность индивидуальной минуты растет в условиях смены внеш-
них впечатлений и больших физических и эмоциональных
нагрузок.
Нами проделано несколько исследований во время взлета,
полета и посадки самолета, которые показали, что длительность
индивидуальной минуты меняется на всех этапах. В момент
скуки и ожидания на аэродроме средние значения равны при-
47
мерно 43—45 с, во время начала взлета — 51.8—58, в середине
и конце взлета — 70—72, при полете по прямой — 53.8—55,
в момент спуска — 57—58 с. Таким образом, длительность
индивидуальной минуты увеличивается в условиях возрастания
объема поступающей извне информации. В случае перегрузки
каналов информацией внутреннего порядка (как это бывает при
заболеваниях и недомоганиях) этот механизм, по-видимому,
блокируется и происходит резкое уменьшение длительности
индивидуальной минуты.
Глава 4
ЦИРКАДНЫЕ КОЛЕБАНИЯ
ДЕЯТЕЛЬНОСТИ
СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ
Представления о циркадной ритмике показателей сердечно-
сосудистой системы и тесно с нею связанных показателей темпе-
ратуры тела и частоты дыхания разработаны в самом общем виде.
Как амплитуда колебаний, так и положение максимума функций во
времени определяются очень приблизительно. Согласно сводке
С. О. Руттенбург и А. Д. Слонима (1976), для частоты дыхания
максимум располагается в пределах 9—12 ч, для систолического
давления, частоты пульса и температуры тела — в пределах 12—
15 ч, для диастолического давления — в пределах 18—21 ч.
По данным других авторов, показателям деятельности системы
кровообращения присущ четкий суточный ритм, но при этом уро-
вень артериального давления, минутный объем крови и частота
пульса имеют максимум в 22—23 ч. Минимум этих функций отно-
сится к 2—3 ч ночи, повышение начинается утром (Ланг, 1950;
Комаров и др., 1966).
Таким образом, разница в локализации максимума функций
весьма существенна. По всей вероятности, это зависит от того,
что изучались различные контингенты лиц; эти исследования про-
водились в различных климато-географических зонах, без учета
влияния сезонной ритмики. Так, оказалось, что ритмика гемоди-
намики отчетливее выражена у лиц, физически более работоспособ-
ных (определение работоспособности делалось по степ-тесту),
причем максимумы объемной скорости выброса, мощности сер-
дечного сокращения и внутрисистолического показателя прихо-
дились на 11—23 ч, максимум общего сосудистого сопротивления —
на 3—7 ч. У лиц, физически мало работоспособных, ритмика
обнаружена только для частоты сердечных сокращений с макси-
мумом во второй половине дня (Дьячков и др., 1974).
В многочисленных исследованиях, посвященных изменениям
динамики параметров в связи со сдвинутыми режимами труда,
показано большое влияние этих режимов на амплитуду колебаний
и на положение максимумов и минимумов, т. е. существуют и про-
фессиональные особенности. Сезонные изменения ритмики показа-
телей деятельности сердечно-сосудистой системы, в первую очередь
кровяного давления, широко исследованы. Так, более чем у 2000
человек, проживающих в Заполярье, Ленинграде, Севастополе
49
и Владивостоке, выявлено статистически достоверное снижение
максимального давления у всех групп в весенне-летний период
до сравнению с осенне-зимним. При этом у лиц, проживающих во
Владивостоке, давление оказалось особенно низким (Голиков,
Голиков, 1973). Естественно, что сезонные изменения суточных
колебаний артериального давления сказываются на частоте раз-
вития гипертонических кризов и приступов коронарной недо-
статочности в различное время суток. Так, В. В. Кохановский
(1970) показал, что среди 9433 обращений в скорую помощь мак-
симум падает зимой на 8—12 ч дня (23.4%), в то время как летом
на это время приходится только 8.1% обращений.
Как показано в ряде исследований, суммированных 3. И. Бар-
башовой (1976), и сезонные, и суточные ритмы физиологических
функций неодинаковы у постоянных жителей европейской части
Советского Союза и у постоянных жителей восточных районов
(особенно горных местностей).
Исследуя сезонные изменения двигательной активности и физи-
ческой работоспособности человека в различных климатических
условиях (Чукотка, Новосибирск, Ферганская долина), К. М. Смир-
нов и др. (1974) обнаружили, что физическая работоспособность
и двигательная активность летом выше, чем зимой, во всех пунктах
обследования. Периодичность сезонных колебаний совпадает по
фазе, амплитуда колебаний больше на севере.
В исследованиях влияния высокогорного климата на суточные
ритмы Т. К. Абдылдабековым и М. Т. Туркменовым (1975а, 19756)
показано, что суточный ритм частоты дыхания и минутного объема
выдыхаемого воздуха отличается у жителей высокогорья от жите-
лей низкогорья только увеличением средней -величины без изме-
нения характера кривой и ее размаха. Для частоты сердечных
сокращений выявлены отличия по акрофазам: у жителей высоко-
горья она приходится на 17 ч, у жителей низкогорья — на 16 ч.
Минимальные значения частоты сердечных сокращений приходятся
на 5 и 4 ч соответственно. Различия акрофаз отмечаются также
и для минимального артериального давления. Его наибольшие зна-
чения у жителей высокогорья приходятся на 11 ч и 23 ч 30 мин,
у жителей низкогорья — на 9 и 21 ч. Что касается максимального
артериального давления, отличий по среднесуточным значениям,
акрофазе и величине колебаний не определялось, но у жителей
низкогорья с помощью Косинор-анализа выявлен еще и 12-часо-
вой ритм, который у жителей высокогорья не обнаруживается.
Минутное потребление кислорода, а также среднесуточная
величина энергстрат у жителей высокогорья ниже, чем у жите-
лей низкогорья. Акрофаза минутного потребления кислорода у жи-
телей высокогорья падает на 15 ч, у жителей низкогорья — на
17 ч 30 мин. Суточный ритм величины энергозатрат, температурной
кривой, пульсового давления, периферического сопротивления
и эквивалента кровотока не различаются у жителей высокогорья
и низкогорья.
50
Оказалось далее, что в условиях высокогорья, характери-
зующихся низким атмосферным давлением, высокой скоро-
стью ветра, постоянно низкой температурой и относительно
низкой влажностью, суточные ритмы ряда показателей (гемоди-
намика, легочное дыхание, газообмен, энерготраты) зависят от
метеофакторов и изменяются прямо пропорционально изменениям
температуры воздуха и скорости ветра и обратно пропорционально
изменениям атмосферного давления и относительной влажности
воздуха.
С увеличением высоты местности растет как амплитуда суточ-
ного колебания метеофакторов, так и амплитуда отклонений рит-
мов функциональной активности.
Таким образом, особенности циркадных колебаний каждого
конкретного лица обусловлены не только его индивидуальными
свойствами, но и целым рядом внешних факторов, и мы полагаем,
что правильнее оценивать не отдельные кривые, а общие свойства
циркадной организации организма, выражающиеся в трех основных
показателях: уровне средних значений, величине разброса, вре-
менной структуре суточной кривой (Моисеева, 1978а, 19786;
Моисеева и др., 1978а).
Исследование циркадных колебаний деятельности сердечно-
сосудистой системы проведено на двух группах испытуемых —
больных и здоровых.
Проанализированы данные 63 больных, находящихся на ста-
ционарном лечении в двух клиниках. Из них 27 человек перенесли
черепно-мозговую травму, у трех была опухоль мозга. 33 человека
страдали диэнцефальными расстройствами различной этиологии.
У 42 больных 3-суточное исследование было проведено однократно,
у 21 — дважды: в начале лечения или до и после лечения, из них
в 12 случаях у больных с вегетативными кризами.
В табл. 19 представлены средние значения девяти исследован-
ных Показателей у всех больных в зависимости от исхода лечения.
Таблица 19
Средние значения физиологических параметров у больных в зависимости
от результата лечения
Результат лечения
Л ® в
Е- Л S
о
ж 3
Р* ЕХ М
Сила
руки,
в кг
Очень хороший
Хороший
Посредственный
Плохой
ИЗ
112
121
121
71
72
77
81
110
112
120
118
47 43
28 27
31 29
32 28
60
61
54
49
Примечание. Здесь и в табл. 37—39 А ДСП — артериальное давление систоли-
ческое справа, АДСЛ —то же слева; АДДП — артериальное давление диастолическое
справа, АДДЛ — то же слева, в мм рт. ст.
Объединены данные обеих клиник, поскольку существенных раз-
личий по средним при раздельном анализе обнаружено не было.
Видно, что средние значения систолического и диастолического
давления, частоты пульса, дыхания и значения температуры отчет-
ливо увеличиваются от группы с очень хорошим исходом к группе
с плохим исходом, однако найденные различия недостоверны в силу
большего индивидуального разброса, и можно говорить только
об общей тенденции изменения. То же самое касается изменений
показателей физической силы, которые имеют тенденцию умень-
шаться у лиц с более плохим исходом заболевания.
Достоверны различия длительности индивидуальной минуты,
превышающей 60 с в случае хороших исходов и не достигающих
50 с в случае исходов плохих. В табл. 20 приведены данные о пара-
метре СОК у больных с очень хорошим и с плохим исходом лечения.
Таблица 20
Среднее число суточных кривых разной степени организованности
у больных в зависимости от исхода лечения, в %
Результат лечения Количество исследований Коли- чество испытуе- мых Число кривых со степенью органи- зации, выражаемой оценками
1 2 3 4 5
Очень хороший 11 9 1.7 3.1 16.1 58.4 20.7
Плохой 10 7 3.5 10.9 10.1 52.2 23.3
Количество кривых с низкой степенью организации в группе
больных с плохим исходом превышает их количество в группе
с хорошим исходом почти в 3 раза. При этом разные параметры дают,
естественно, разные картины кривых. Для артериального давле-
ния кривые у тяжелобольных чаще очень плохо организованы,
кривые температуры, напротив, показывают большие колебания.
Наши данные совпадают с результатами исследований Б. С. Фро-
лова и др. (1980), обнаруживших в случаях декомпенсации пси-
хопатий непостоянство показателей пульса и температуры и боль-
шую ригидность артериального давления.
Кроме указанных девяти показателей, характеризующих пси-
хофизиологическое состояние испытуемого, каждое однократное
исследование включало в себя также регистрацию ЭКГ в течение
2 мин с последующей обработкой соответствующих рядов интерва-
лов RR на ЭЦВМ «Минск-32» по программам В. М. Сысуева (1978).
Полученные в результате обработки показатели (среднее значение
интервалов RR, вегетативный показатель ритма,1 количество ко-
1 Вегетативный показатель ритма (ВПР), предложенный Г. И. Сидо-
ренко (1970) и Р. А. Новиковой (1970, 1971, 1973), является количественной
характеристикой формы гистограммы интервалов RR и ее местоположения
на оси длительностей интервалов. Рассчитывается по формуле ВПР =
52
лебательных компонент и их периоды) явились дополнительными
характеристиками состояния.
Оказалось, что значения вычисляемых параметров ЭКГ разли-
чаются у больных с различным исходом заболевания. В табл. 21
приведены данные, полученные у лиц с очень хорошим и с плохим
Таблица 21
Значение вычисляемых показателей электрокардиограммы у больных
с различным исходом заболевания (средние данные по 3 дням
исследования)
Показатель Результат лечения
очень хороший плохой
Среднее значение интервалов RR, в с 0.785 0.888
Среднеквадратичное отклонение, в с 0.042 0.039
Вегетативный показатель ритма 146 193
Количество колебательных компонент 6 4.3
Периоды компонент, в с:
доминирующей 25.98 33.07
первой субдоминирующей 24.35 25.32
второй » 19.58 21.55
третьей » 16.18 20.23
исходом заболевания. В случае неблагоприятного течения заболе-
вания период RR и его разброс, а также период доминирующих и
субдоминирующих компонент значительно больше. Число колеба-
тельных компонент меньше.
Как известно, учащение сердцебиений сопровождается обычно
уменьшением разброса длительности сердечных циклов, что свя-
зывается с повышением тонуса симпатической системы (Баев-
ский, 1976, и др.). Соответственно урежение сердцебиений
должно сопровождаться ростом разброса значений. По абсолютным
значениям показателей у больных с хорошим и плохим исходом
эти соотношения как будто выдерживаются, т. е. с появлением
более редких сердцебиений увеличивается разброс их значений.
Исследование суточной динамики этих показателей для двух
групп больных разрешило выявить отличия в соотношениях суточ-
ных кривых среднего значения интервалов RR и среднеквадратич-
ного отклонения их длительностей.
В группе с хорошим исходом эти кривые являются преимущест-
венно синфазными, т. е. участку роста кривой одного показателя
соответствует участок роста другого, а участку уменьшения одного
из них соответствует участок уменьшения второго (рис. 5 и 6).
Оказалось, что у больных с неудовлетворительным исходом ле-
= fMЦМХ X), где 1мх~~ объем модального класса гистограммы, в %, т. е.
доля наиболее часто встречающихся интервалов RR; Мх — значение моды
гистограммы; X — разброс значений интервалов RR.
53
чения чаще встречаются обратные соотношения между суточными
кривыми среднего интервала RR и среднеквадратичного отклоне-
ния (табл. 22).
Синфазные кривые встречаются тем реже, чем хуже исход
заболевания, при этом растет число полностью противофазных
Рис. 5. Суточная динамика среднего
интервала RR (Л) и его среднеквад-
ратичного отклонения (Б) для 3 дней
исследования (7—3) у больного с хо-
рошим исходом заболевания.
По осям — то же, что на рис. 2. «Плюс» —
случаи однонаправленного изменения обоих
параметров для каждого дня; «минус» —
случаи разнонаправленного.
Рис. 6. Суточная динамика среднего
интервала RR (А) и его среднеквад-
ратичного отклонения (Б) для
3 дней исследования (1—3) у боль-
ного с плохим исходом заболевания.
Обозначения те же, что на рис. 5.
кривых. Таким образом, динамика изменений значения параметра
определенным образом связана с динамикой среднеквадратичного
отклонения, и сдвиги в этих отношениях свидетельствуют о нару-
шениях регуляции.
Что касается времени выявления максимумов всех десяти пока-
зателей, оно весьма вариабельно и зависит, по-видимому, как от
особенностей биоритмической структуры данного больного, нередко
искаженной болезнью, так и от лечебных воздействий (иногда
весьма энергичных), получаемых еще до поступления в больницу или
в больнице. На рис. 7 представлены кривые систолического и диа-
столического давления и температуры тела больного с инфекционно-
54
Т а б л и ц а 22
Частота обнаружения синфазных и противофазных кривых среднего
интервала RR и среднеквадратичного отклонения у больных с различным
исходом заболевания, в %
Результат лечения Доля кривых
синфазных с отклонением от синфазности противофазных
Очень хороший 50 45 5
Хороший 45 45 10
Посредственный 40 40 20
Плохой 38 37 25
аллергическим цереброваскулитом до лечения и в процессе лечения,
закончившегося хорошим результатом.
Рис. 7. Суточная динамика систоли-
ческого (4) и диастолического (Б)
артериального давления, а также
температуры тела (В) больного,
лечение которого было успешным.
Обозначения те же, что на рис. 3.
Данный больной получал индивидуально рассчитанный курс
лечения дилантином (подбор дозировок и времени приема ле-
карства). Исходя из анализа структуры суточных кривых иссле-
дованных показателей, лекарство давалось дважды — в 16 и 20 ч.
Хронодиагностика и хронотерапия
Суммируя все сказанное, можно заключить, что в условиях раз-
вития патологических процессов происходят значительные изме-
нения ритмической деятельности организма.
Изменяются все три показателя циркадной организации: уро-
вень средних значений, величина разброса, временная структура
суточной кривой.
Первые два показателя зависят от характера патологического
процесса (от гипертонии или гипотонии, субфебрилитета и т. д.)
55
и поэтому теряют то диагностическое значение, которое они имеют
у здоровых при определении уровня адаптоспособности.
Показателем, уровень которого достоверно отражает функцио-
нальное состояние организма и у больных, и у здоровых, является
только длительность индивидуальной минуты. Основой для оцен-
ки состояния больного служат:
1) временная структура суточной кривой отдельного пока-
зателя;
2) соотношения этих кривых, снятых в течение 3 сут подряд;
3) соотношение кривых среднего значения показателя и сред-
неквадратичного отклонения значений того же показателя.
Используя эти три показателя, мы совместно с Л. И. Никитиной
попытались определить в каждом конкретном случае время наи-
большего отклонения функций от нормальных параметров, т. е.
провести хронодиагностику (Моисеева, 19806). Под хронодиагности-
кой мы понимаем рассмотрение не только вопроса, какие функции
поражены, но и когда (в какое время суток) функциональные из-
менения максимальны. Целью такого подхода является назначе-
ние лечения, предусматривающего воздействие на организм боль-
ного, корригирующее суточные кривые функций, а не только при-
водящее показатели к некоему среднему уровню. Так, для снижения
артериального давления у гипертоника соответствующие препа-
раты приносят пользу, если их назначать не 3 раза в день, а в мо-
менты максимального подъема, которые у каждого больного инди-
видуальны. Такой индивидуализированный подход к определению
времени лечебных воздействий мы называем хронотерапией.
Получены положительные результаты, в частности у больного,
кривые которого приведены на рис. 7. Однако рассмотрение во-
просов диагностики и лечения в нашу задачу не входит, и мы упо-
мянули об этом главным образом для демонстрации тех возможно-
стей анализа, которые открывает биоритмологический подход.
Для того чтобы проанализировать закономерности циркадных
колебаний деятельности сердечно-сосудистой системы, обратимся
к группе здоровых, у которых изучались изменения ритмики тех же
показателей, что и у больных (но в условиях перелетов с пересе-
чением большого числа часовых поясов), и дополнительно изуча-
лась капилляроскопическая картина.
Изменения циркадной ритмики
показателей деятельности сердечно-сосудистой системы
под влиянием внезапного изменения временной среды
Исследования изменений гемодинамики при широтных переле-
тах касаются обычно только пульса и артериального давления,
кривые которых выравниваются при перелете как с востока на
запад, так с запада на восток с пересечением 7—10 часовых поясов.
При этом одни авторы (Hauty, Adams, 1966; Евцихевич, 1974)
говорят о выравнивании показателей уже на 5—10-й день, другие
56
(Матюхин и др., 1976) — о выравнивании после перелета на восток
к 10—25-му дню, на запад — к 25—35-му дню.
С целью исследования особенностей адаптации сердечно-со-
судистой системы при внезапном изменении временной среды нами
было предпринято исследование на лицах, пересекающих 3—9-
часовые пояса в направлении с запада на восток и с востока на за-
пад. Учитывая стремление современной медицины к повышению
общей неспецифической сопротивляемости организма с помощью
лекарственных веществ, одна из групп обследованных принимала
в полете элеутерококк, другая группа — снотворные.
Всего обследовано девять практически здоровых лиц в возрасте
25—50 лет, совершивших перелет Ленинград—Петропавловск-на-
Камчатке (что соответствует пересечению девяти часовых поясов),
Ленинград—Новосибирск (четыре) и Ленинград—Фрунзе и об-
ратно (три пояса). Все перелеты на Камчатку происходили в ав-
густе—сентябре, во Фрунзе и Новосибирск — в июне.
Проводилось измерение артериального давления аппаратом
Рива-Роччи, счет пульса и исследование капилляров. Капилляры
исследовались на ногтевой фаланге четвертого пальца левой руки
с помощью капилляроскопа М-70-А (увеличение в 20 раз). Визу-
ально оценивались: 1) окраска фона, 2) четкость выраженности
капилляров и их форма, 3) длина и ширина, 4) число капилляров.
Исследования проводились в Ленинграде и Петропавловске
или Новосибирске в течение 4—6 сут после перелета (а также в Ле-
нинграде до перелета или через 1—2 мес после него) с промежутками
в 3—6 ч, приблизительно в одно и то же время. Пять испытуемых
во время одного из перелетов из Петропавловска принимали эле-
утерококк в виде капель официнальной настойки; шесть испытуе-
мых совершили полет в состоянии сна в результате приема сно-
творных (пять человек принимали нембутал каждые 4 ч по одной
таблетке, один — пипольфен). Всего в течение 3 лет у девяти
испытуемых проведено 18 циклов исследований (у одного — семь
циклов, у другого — три, у третьего — два цикла, у шести —
по одному циклу) и осуществлено 270 измерений пульса и арте-
риального давления и 170 исследований капилляроскопической
картины. На табл. 23 дана сводка всех данных, полученных в ре-
зультате исследования девяти испытуемых пятикратно в течение
54 сут.
Кроме того, была привлечена дополнительная группа в десять
человек." До и после перелетов у лиц, пересекающих девять часо-
вых поясов, проводились исследования и в ночное время (от 1 до
6 ч) в течение 2—3 ночей, что позволило составить полную картину
суточной ритмики. Всего в ночное время сделано 258 измерений
артериального давления, подсчетов частоты пульса и определений
капилляроскопической картины.
На табл. 24 приведена динамика состояния стенки капилляров,
среднего числа капилляров и показателей артериального давления
в разное время суток у лиц, исследованных в обычных условиях
57
Табл
Особенности циркадной ритмики показателей деятельности сердечно-сосудистой
с пересечением
Испытуемый Число пере- сечен- ных поясов Число суток иссле- дова- ния Систолическое давление Диастолическое давление
сред- нее, в мм рт. ст. раз- брос, в мм рт. ст. время достиже- ния, в ч среднее, в мм рт. ст. разброс, в мм рт. ст. время дости- жения, в ч
макси- мума мини- мума макси- мума мини- мума
М-ва 3 2 117 28 15; 23 7 68 25 15; 23 7
3 3 115 26 15 7 68 23 15 7
4 3 121 25 15 19 72 10 15 23
4 4 127 22 19 7 70 13 15 19
С-в 4 3 122 25 15 7 60 14 15 7
4 4 125 18 15; 11 19; И 72 13 15 23
Д-а 4 3 115 23 15 23 63 17 15 23
4 3 116 23 И; 15 7; 15 66 15 15 7
С-в 9 3 120 20 23 7 84 20 23 7
9 3 118 15 19 23 83 13 23 7
М-ва 9 3 124 15 11; 15; 19 И; 19 74 17 7 15
9 3 115 13 19 15 74 13 15 23
Г-а 9 2 129 20 И 19 74 10 23 И
К-в 9 1 105 25 23 19 71 25 19 15
Т-в 9 2 114 40 17 И; 23 67 24 17; 19 11; 23
И-а 9 2 101 26 23 И; 19 74 25 19; 23 И; 15
9 7 119 28 И 7 70 37 23 7
М-ва 9 3 114 17 И 15 69 25 19 7
Среднее 119 22 71 20
и на 1—3-и сутки после перелета с пересечением часовых поясов
(суммарные данные 19 испытуемых).
В табл. 25 даны суммарные значения основных показателей
деятельности сердечно-сосудистой системы в исследованиях:
1) фоновых, 2) после перелетов в различных направлениях (полет
на запад или восток) и 3) отличающихся новизной обстановки (по-
падание в новую или возвращение в привычную обстановку).
В целях выделения групп обследованных после перелета в но-
вую обстановку и после возвращения в привычную обстановку
суммированы данные полетов на запад и на восток. Оказалось, что
достоверные различия по отношению к фону имеют место только
в группе после возвращения в привычную обстановку (Р=0.9)
для разброса показателей артериального давления. Кроме того,
достоверны различия средней частоты пульса и разброса систоли-
ческого давления в группе вернувшихся в привычную обста-
новку по сравнению с группой, попавшей в обстановку новую
(Р=0.9).
58
и ц а 23
системы 9 испытуемых, исследованных в процессе адаптации после перелетов
часовых поясов
Частота пульса
Состояние капилляров
в разные часы суток
Среднее число капилляров
в поле зрения
среднее в 1 мин разброс в 1 мин время дости- жения, в ч норма спазм стаз среднее разброс
макси- мума мини- мума
72 21.0 15 19 7; И; 19 15 23 7 8
73 41.0 19 23 7; И; 15; 19 — 23 8 И
78 21.0 19 7 — — — — —
69 16.0 11 7 — — — — —
78 23.0 23 7 — — — — —
76 11.0 И; 15; 19 23 — — — — —
83 16.0 15 11 — — — — —
72 15.0 11 19; 23 — — — — —
70 15.0 И 7 23 — 7; И; 15; 19 4 8
65 13.5 И 7 15 — 7; И; 13; 19 4 8
74 16.5 15 23 — — 7; 11; 15; 19; 23 8 13
73 12.0 19 7 15 7 И; 19; 23 7 4
73 18.0 23 15 И; 19 — 15; 23 6 2
70 22.0 23 19 15 ;19 — 23 6 9
64 18.0 17 И — 19 И; 15; 23 3 6
77 27.0 23 И 15; 23 19 И 14 18
74 10.0 И 7; 23 15; 19 — 7; И; 23 5 8
75 22.0 И 7; 23 15; 19; 23 7 И 6 12
15
И
19
19
23
23
19
19 23
15 25
И; 19
11; 23
15
7
15; 19
23
И; 17; 23
И
И
7
73 17.6
6 9
Средние показателей артериального давления и частоты пульса
после перелета с пересечением девяти часовых поясов в зависимости
от условий полета (обычные условия, состояние сна, прием адап-
тогена) следующие: в фоновых исследованиях систолическое дав-
ление минимально среди ночи (4—6 ч) и имеет два максимума —
во время наиболее активной деятельности (10—15 ч) и перед сном
(22—24 ч). Диастолическое давление снижается ночью, днем его
динамика неотчетлива (табл. 26).
Перелет с запада на восток сопровождается большим общим
снижением систолического давления и разброса его значений по
часам и сохранением «ленинградского ритма» в первые дни после
перелета (минимум систолического давления в 4—9 ч по ленинград-
скому времени, что соответствует 13—18 ч камчатского).
Перелет с востока на запад (что для большинства испытуемых
было возвращением в привычную временную среду) почти не влиял
на общий уровень артериального давления, но разброс между
измерениями уменьшался. Свыше недели в редуцированном виде
59
Таблиц а 24
Динамика показателей состояния капилляров п артериального давления,
в мм рт. ст., в течение суток у лиц, исследованных в обычных условиях
и после перелета с пересечением девяти часовых поясов
Время исследова- ния, в ч Состояние капилляров, в % к общему числу исследований Среднее число капил- ляров в поле зрения Артериальное давле- ние, в мм рт. ст.
ленин- градское петропав- ловское норма спазм стаз систоли- ческое диасто- лическое
Фоновые исследования
1-3 10-12 60 0 40 6.9 110 71
4-6 13-15 66 6 28 5.3 102 64
7-9 16-18 46 15 39 6.4 116 67
10—12 19-21 65 8 27 9.8 122 67
13—15 22-24 38 12 50 8.0 122 68
16-18 1-3 66 4 30 9.0 123 70
19—21 4-6 57 8 35 9.0 112 70
22—24 7-9 26 16 58 7.0 126 70
Среднее 53 8.6 38.4 7.7 116.6 68.4
Разброс 40 16 31 4.5 24 7
П ерелет с зап ада на в о с Т О к
1-3 10-12 25 0 75 8.0 103 68
4-6 13-15 0 0 100 0.0 94 66
7-9 16-18 40 0 60 7.0 — —
10—12 19-21 0 0 0 0.0 103 65
13-15 22-24 20 0 80 10.0 107 69
16-18 1-3 40 0 60 10.0 — —
19-21 4—6 0 0 100 10.0 — —
22-24 7-9 22 0 78 8.0 100 64
Среднее 18.4 0 69 8.8 101.4 66.4
Разброс 20 0 40 3.0 13 5
П ерелет с вое тока н а запад
1-3 10-12 34 0 66 6.0 —
4-6 13-15 45 22 33 7.0 — —
7-9 16-18 12 33 55 2.0 НО 75
10-12 19-21 13 55 32 6.0 117 73
13-15 22-24 50 16 34 8.0 132 75
16—18 1-3 43 0 57 9.0 173 70
19—21 4-6 45 0 55 8.0 108 72
22-24 7-9 12 31 57 6.0 111 71
Среднее 31.6 16 48 6.5 115.2 72.7
Разброс 50 55 34 7.0 24 5
60
Таблица 25
Значения показателей деятельности сердечно-сосудистой системы
в фоновых исследованиях и после перелетов с пересечением часовых
поясов
Исследования Систолическое давление, в мм рт. ст. Диастолическое давление, в мм рт. ст. Частота пульса, в 1 мин
среднее разброс среднее разброс среднее разброс
Фоновые 118 30 ** 76 28 ** 76 18
После перелетов: с востока на запад 116 23 72 20 71 20
с запада на восток 120 23 67 18 76 19
В обстановке после пе- релета: * новой 118 26 ** 69 22 75 ** 19
привычной 118 20 72 16 ** 71 ** 19
* Данные полета на восток и запаД* суммированы без учета направления полета.
** Р = 0.9.
Таблица 26
Зависимость средних показателей деятельности
сердечно-сосудистой системы от условий перелета и времени суток
Время исследования, в ч Фоновые исследования Исследования на 2-е сут после перелета
в обычных условиях в состоянии сна при приеме элеутерококка
ленин- град- ское кам- чат- ское СД ДД ЧП
СД ДД ЧП СД ДД ЧП СД ДД ЧП
7 16 111 59 64 НО 75 80 110 60 67 115 87 66
И 20 121 62 75 117 73 76 122 70 79 120 82 71
15 24 120 81 74 130 75 79 121 70 78 117 82 67
19 4 115 73 72 110 75 73 116 69 75 112 80 72
23 8 126 67 79 104 60 72 125 75 78 НО 87 66
Среднее 119 68 73 114 72 75 119 69 75 115 84 68
за сутки
Разброс 15 22 15 26 15 8 15 15 И 10 7 6
за сутки
Примечание. СД и ДД — систолическое и диастолическое артериальное дав-
ление, в мм рт. ст.; ЧП — частота пульса , в 1 мин.
сохранялся «петропавловский ритм» в виде максимума систоли-
ческого давления в 7—9 ч утра ленинградского времени (16—18 ч
камчатского) и минимума вь22—24 ч ленинградского времени (7—9 ч
камчатского). Изменения при полете на запад сглаживаются за
10 дней — 2 нед, при полете на восток — за 1 мес и более. Неко-
61
торое общее снижение систолического давления и сглаживание
кривых наблюдались нами также и при переездах и перелетах
с юга на север и, возможно, связаны не с пересечением часовых по-
ясов, а с общим утомлением, вынужденной бездеятельностью и от-
сутствием достаточной двигательной активности.
Состояние сосудистой стенки и числа капилляров (см. табл. 23)
в фоновых исследованиях в ночные часы (в состоянии бодрствова-
ния) было преимущественно нормальным, но общее число капилля-
ров понижено по сравнению с их числом днем, что, вероятно, свя-
зано с отсутствием большой функциональной нагрузки. В 7—9 ч,
когда человек обычно приступает к активной деятельности и начи-
нает с приема пищи и соответственно ее переваривания, в большом
проценте случаев отмечается повышение порозности стенок ка-
пилляров. В 10—12 ч состояние сосудистой стенки преимущест-
венно нормальное, число капилляров максимально. В 13—15 ч (что
совпадает с обеденным временем) растет число капилляров с уве-
личенной порозностью стенок и появляются случаи спастических
капилляров. В 16—21 ч число капилляров сравнительно велико,
тонус сосудистой стенки более чем в половине наблюдений нормален.
В 22—24 ч значительно увеличивается процент наблюдений спазма
и стаза, число капилляров падает.
При перелете с запада на восток значительно растет порозность
сосудистой стенки, ритмичность изменений исчезает. При перелете
с востока на запад в обычных условиях число наблюдений, где
регистрировались стаз и нарушение проницаемости, растет по
сравнению с фоном, но не так резко, как при перелете с запада
на восток. Общая ритмика напоминает фоновую и соответствует
больше петропавловскому времени, чем ленинградскому (например,
появление спастичных капилляров в 13—24 ч по петропавловскому
времени).
Сроки развития изменений таковы: при перелете с запада на
восток они возникают на первые сутки и держатся 2—4 дня. При
перелете с востока на запад изменения возникают на вторые сутки
и начинают уменьшаться на 5—7-е сутки. При этом играет извест-
ную роль адаптированность к определенному часовому поясу,
в котором человек проживает постоянно.
Как видно на табл. 25, наибольшая разница в изменениях по-
казателей возникает в зависимости от того, переместился ли ис-
следуемый в новую обстановку или вернулся в обстановку привыч-
ную, а не от направления полета. В привычной обстановке снижа-
ются разброс показателей систолического давления и средняя ча-
стота пульса по сравнению с фоном и с перемещением в новую об-
становку. Кроме того, при возвращении к исходной временной среде
у человека сохраняется циркадная структура изменений капилля-
ров (хотя и приуроченная к другому часовому поясу), заключаю-
щихся не только в сдвигах проницаемости, но и в числе случаев
спазма (что характерно для фоновых наблюдений). При перелете
в новую временную среду ритмика полностью дезорганизуется,
62
в абсолютном большинстве наблюдений развивается стаз с наруше-
нием проницаемости капиллярной стенки.
Таким образом, при возвращении в привычную среду пере-
стройка идет быстрее и с меньшими затратами — организм не
нуждается в большом разбросе значения показателей, так как легко
отслеживает необходимый диапазон колебаний и находит оптимум.
При перелете с приемом снотворных ритмика показателей деятель-
ности сердечно-сосудистой системы уже в первые дни перестраи-
вается на ленинградское время. Ритм пульса уже на второй день
приближался к фоновому.
Прием элеутерококка до и во время перелета не способствует
облегчению процесса адаптации.
Общий процент наблюдений, в которых тонус капиллярной стен-
ки нормален, оказался ниже, чем при перелете в обычных усло-
виях, и значительно ниже, чем при перелете со снотворными. При
этом ритмика изменений неотчетлива. Нет существенного влияния
ни на ритмику артериального давления, ни на ритмику пульса
(см. табл. 26).
Неудача нашей попытки воздействовать на процесс адаптации
к новым временным условиям с помощью признанного адаптогена
элеутерококка (который, по современным воззрениям, принимает
участие в регуляции биосинтеза белка, нуклеиновых кислот и
в энергетическом обеспечении этих процессов (Дардымов, 1976))
лишний раз показала, что при всей важности обменных процессов
в данном случае на первый план выступает необходимость регуля-
ции ритмического рисунка всех функций организма, т. е. речь идет
не о гомеостазе, поддержании предсуществовавшего порядка,
а о его перестройке с целью приспособления к новым условиям —
гомеокинезе.
Положительное влияние сна во время полета на регуляцию цир-
кадных ритмов может зависеть по меньшей мере от нескольких
моментов: от выключения афферентации на то время, когда орга-
низм подвергается большому числу постоянно меняющихся воз-
действий, что упрощает регуляцию, поскольку она происходит
на конечном этапе, минуя промежуточные; от того, что вообще
регуляция начинается после сна, т. е. в момент наибольшей функ-
циональной готовности к любому роду перестроек, которая, как
известно, наблюдается в первые часы после сна. Это может зависеть
и от того, что непосредственно во время сна происходит регуляция
биоритмической структуры организма. В пользу последнего пред-
положения может свидетельствовать сама картина сна после пере-
лета в обычных условиях. Этот вопрос более подробно будет рас-
смотрен отдельно.
Таким образом, под влиянием перелетов наступают общие изме-
нения гемодинамики, что выражается как в нарушении ритма су-
точного изменения систолического давления, так и в местных
изменениях капиллярного кровообращения, причем последнее
более выражено. Так, при перелете с запада на восток ритмичность
63
изменений капиллярного кровообращения исчезает полностью.
Этот факт может быть объяснен тем, что приспособительная функ-
ция обменных процессов, реализуемая в основном за счет капил-
ляротканевых структур, имеет важное значение в адаптационных
механизмах (Ломоури, 1974), соответственно капилляры сильнее
реагируют на изменения внешней среды, чем другие отделы сер-
дечно-сосудистой системы.
Такая специфическая роль капилляротканевых структур в про-
цессах адаптации, вероятно, связана с тем, что действие на орга-
низм любого раздражителя выражается в напряжении той или иной
функции и, следовательно, в расходовании вещества, что, по мнению
Д. С. Саркисова (1973), должно вести к интенсификации синтети-
ческой активности.
Суммируя все вышеизложенное, можно сказать, что как при
заболеваниях, так и при изменении временных воздействий внеш-
ней среды ритмика колебаний всех показателей сердечно-сосуди-
стой системы меняется, причем это зависит от качества регулятор-
ных механизмов организма. Высокие потенциальные возможности
организма (хорошая адаптоспособность у здоровых, благоприятное
течение заболевания у больных) определяются у здоровых четкой
индивидуальной временной организацией суточной кривой и вы-
соким средним уровнем и значительным разбросом в течение су-
ток величины показателей деятельности сердечно-сосудистой* си-
стемы и длительности индивидуальной минуты. У больных это рас-
пространяется только на длительность индивидуальной минуты.
Глава 5
ИССЛЕДОВАНИЕ ВЛИЯНИЯ
БИОЛОГИЧЕСКИХ МАКРОРИТМОВ
НА КОЛЕБАНИЯ ЗНАЧЕНИЙ
ФИЗИОЛОГИЧЕСКИХ
И ПСИХОЛОГИЧЕСКИХ ПОКАЗАТЕЛЕЙ
Исследование влияния биологических микроритмов
на взаимооценки членов коллектива
Личностные взаимоотношения в коллективе, как правило,
подвергаются некоторым колебаниям, связанным не только с объ-
ективными причинами, но и с внешне немотивированными изме-
нениями настроения отдельных лиц, отражающимися на тех оцен-
ках, которые они при этом дают окружающим и от окружающих
получают.
Как уже говорилось, особенности ритмических процессов в жи-
вых системах (их незатухающий характер и постоянство периода
колебаний) связаны с двойным контролем — эндогенных механиз-
мов и экзогенных воздействий. При этом факторы, обусловленные
вращением Земли, регулируют циркадные ритмы, однако сущест-
вует широкий спектр повторяющихся событий (лунный цикл,
вращение Земли вокруг Солнца, солнечный цикл и т. д.) с перио-
дами значительно больше суточного. Эти явления, по-видимому, и
являются датчиками времени, регулирующими течение макро-
ритмов.
Отсчет макроритмов организма ведется от момента рождения,
т. е. от того момента, когда организм начинает функционировать
самостоятельно, не подчиняясь больше ритмам матери. На гене-
тической основе и на основе влияния воздействий внешней среды
в организме формируются свои собственные ритмы. Не имеет зна-
чения, в какой фазе геофизического или солнечного цикла проис-
ходит рождение организма; важно то, что именно эта фаза яв-
ляется условием начала какого-то биоритма и повторение этих
условий будет сигналом, поддерживающим ритмику.
Подобно биопотенциалам мозга, характеризующимся спектром
частот, макроритмы могут включать целый набор различных пе-
риодов колебаний. Например, так называемый «макроритм эмо-
циональной активности» имеет период 28 дней (что соответствует
лунному циклу), но при этом выделяются еще циклы недельные,
18-дневные, 6-недельные (Hersley, 1931; Dorland, Brinker, 1973).
Макроритм физической активности, имеющий 23-дневный период,
выражен настолько отчетливо, что этот факт уже довольно давно
65
учитывается тренерами спортивных команд при составлении ин-
дивидуальных расписаний тренировок, особенно перед соревно-
ваниями. Однако выделяется еще и 8—14-дневный ритм — «биоло-
гическая неделя» (Ткачук, Кучеров, 1965).
Для определения наличия взаимосвязи между фазами биологи-
ческих ритмов человека и оценками, которые он получает от ос-
тальных членов группы или дает остальным членам группы, был
выбран один из социометрических методов — анонимное анкети-
рование (Гончаров, Козлов, 1974; Козлов, Сысуев, 1’975). Анкета
содержала список членов группы обследуемых: фамилию и дату
Рис. 8. Зависимость доли положительных оценок, поставленных студентами
своим товарищам по группе, от фаз макроритмов физической (Ф), эмоцио-
нальной (Э) и интеллектуальной (И) активности оценивающих.
По оси абсцисс — фазы отдельных макроритмов (I—IV); по оси ординат — число поло-
жительных оценок, в %.
рождения анкетируемого, дату проведения опроса. К исследованию
были привлечены 175 спортсменов, совершающих туристические
многодневные походы по маршрутам Камчатки, и одна группа
студентов (25 человек) в процессе занятий в ЛПИ. Туристические
группы состояли из спортсменов обоего пола, ранее друг с другом
не знакомых, в возрасте от 18 до 40 лет, уроженцев разных облас-
тей СССР. Члены группы студентов ранее были знакомы друг с дру-
гом по совместному обучению.
Данные анкет каждого опроса одной группы сводились в мат-
рицу, которая обрабатывалась согласно правилам, описанным
в гл. 2. После построения графических зависимостей числа откло-
нений от фазы макроритма, в которой находится человек в момент
исследования, проводилась статистическая проверка гипотезы
о связи числа отклонений оценок и фазы макроритмов с помощью
критерия х2’
В случае исследования на маршруте анкетирование производи-
лось дважды — в начале и в конце похода (через 10—12 дней).
Обработано 250 анкет. В группе студентов, обследованных в Ленин-
граде, анкетирование проведено 7 раз с промежутками в 2—4 нед.
Обработано 125 анкет. Всего в двух группах обработано 375 анкет
66
с целью выяснения вопросов: 1) как влияют фазы макрорйтмов,
в которых человек находится в момент опроса, на те оценки, ко-
торые он ставит окружающим; 2) как влияют эти фазы на те оценки,
которые он от окружающих получает.
Результаты отражены графически на рис. 8—11, где в верхней
части показаны фазы биоритмов, а в нижней отражен процент
положительных отклонений оценок от среднего значения в каждой
из фаз макроритмов.
На рис. 8 и 9 приведены данные по студенческой группе, об-
следованной в период совместных занятий в институте. Как видно
Рис. 9. Зависимость доли положительных оценок, поставленных студентами
своим товарищам по группе, от фаз макроритмов оцениваемых.
Обозначения те же, что на рис. 8.
на рис. 8, отражающем результаты исследования отклонения
оценок, проставляемых испытуемым, от фазы его макроритмов,
наибольшее количество высоких оценок испытуемые ставили в IV
фазе собственных макроритмов, а наименьшее количество —
во II фазе, т. е. чем выше его собственные возможности, тем ниже
он оценивает других.
Следует отметить, что разница между наибольшим и наимень-
шим числом положительных оценок в разные фазы тем больше,
чем длиннее период макроритма.
На рис. 9 представлены результаты исследования отклонения
оценок, получаемых самими испытуемыми. Картина в значитель-
ной мере отличается от предыдущей. Человек получает наиболее
высокие оценки в те моменты, когда он находится в восходящей
фазе. Кроме того, в двух случаях (эмоциональный и интеллек-
туальный циклы) он получает высокие отметки (такие же, как
в переходной фазе) и в самой фазе подъема. В физическом макро-
ритме в фазе подъема оценки минимальные, в фазе спада — отно-
сительно высоки.
Такая «инверсия» может быть объяснена чисто социальными
факторами — данная группа объединена производственными от-
ношениями, где физические возможности практически не играют
67
особой роли, но высоко оцениваются способности к интеллек-
туальному и волевому напряжению.
В целом в данной группе для физического цикла определяется,
что человек, находящийся в переходной фазе к подъему, получает
высокие оценки, но занижает оценки окружающим, в фазе макси-
мума он и дает и получает низкие оценки, в фазе спада он дает и по-
лучает относительно высокие оценки. Для эмоционального и ин-
теллектуального циклов определяется, что в восходящей фазе
человек и дает, п получает высокие оценки, в фазе подъема полу-
чает высокие оценки, но сам оценки занижает.
Рис. 10. Зависимость доли положительных оценок, поставленных участ-
никами турпохода своим товарищам по группе, от фаз макроритмов оцени-
вающих.
Обозначения те же, что на рис. 8.
В нисходящей фазе и фазе спада по эмоциональному макро-
ритму получаемые и даваемые оценки одинаковы. При этом если
в фазе спада эмоционального макроритма человек и дает и получает
довольно высокие оценки, то в фазе спада интеллектуального мак-
роритма он получает низкие оценки (что, очевидно, адекватно его
успехам в учении), а дает высокие (разумно оценивая успехи то-
варищей).
На рис. 10 и 11 приведены данные по туристским группам, об-
следованным в период относительно трудных походов с переходами
до 32 км в день в горах в очень неблагоприятных погодных усло-
виях. В отличие от предыдущей в этой группе требуются ин-
теллектуальные усилия только от инструктора, а туристы должны
совершать равномерную физическую работу. Люди, физически
слабые, «тянущие назад», и слишком активные, «убегающие вперед»
и нарушающие темп группы, вызывают всеобщее недовольство,
даже если первых не приходится разгружать, а вторые несут двой-
ную нагрузку. Такая же сбалансированность требуется и в эмо-
циональных проявлениях. Когда после целого дня похода под дож-
дем долго не удается разжечь костер, чтобы просушиться и приго-
товить ужин, юмор воспринимается хорошо, а неумеренное веселье
вызывает вспышку раздражения.
68
Как видно на рис. 10, отражающем зависимость отклонения
оценок, проставляемых испытуемым, от фаз его макроритмов,
процент положительных отклонений для физического и эмоцио-
нального макроритмов максимален в обеих переходных фазах
и минимален в фазах спада. Для интеллектуального макроритма
максимальны оценки в восходящей фазе и фазе подъема и мини-
мальны в нисходящей фазе и фазе спада. Таким образом, представ-
лена картина несколько искаженной проекции собственного со-
стояния на окружающий мир.
В условиях очень большого физического утомления, но сопря-
женного с необычайно сильными новыми впечатлениями, человек
Рис. И. Зависимость доли положительных оценок, поставленных участни-
ками турпохода своим товарищам по группе, от фаз макроритмов оцениваемых.
Обозначения те же, что на рис. 8.
в целом сохраняет положительный эмоциональный настрой, и рас-
считываемые оценки у туристов в целом^выше, чем оценки, которые
ставили студенты в условиях обычной жизни и работы.
Особые условия похода, когда-человек оказывается среди мало-
знакомых людей и не имеет прочных социальных связей, создают
возможность ставить оценки исходя в значительной мере из соб-
ственного внутреннего настроя и из впечатления данного дня.
Соответственно в восходящей фазе оценки более высокие, в фазе
подъема — чуть ниже (за исключением физического макроритма),
в нисходящей фазе или выше, или еще ниже, в фазе спада — самые
низкие. Отклонения оценок, поставленных в группе туристов
испытуемому в зависимости от фазы его макроритма (рис. 11),
полностью совпадают по физическому макроритму с оценками,
поставленными студентами.
У туристов для всех трех макроритмов характерно наименьшее
количество положительных оценок в фазе подъема. Отклонения
в положительную сторону отмечаются в восходящей фазе для
всех трех ритмов и в нисходящей — для эмоционального и интел-
лектуального ритмов. Для физического и эмоционального ритмов
отклонение в положительную сторону имеется в фазе спада.
69
В целом в группе туристов (относительно изолированной от
социальных влияний) человек дает лучшие оценки окружающим
в фазах восходящей и подъема и худшие — в фазах спада. Сам же
он получает лучшие оценки в переходных фазах, а самые плохие —
в фазах подъема. Суммируя данные по обеим группам, можно ска-
зать, что в целом человек дает и получает более высокие оценки
в восходящей фазе любого ритма.
Занижение оценок окружающим в фазе подъема всех трех рит-
мов характерно при взаимооценке лиц, объединенных определен-
ного вида деятельностью, осуществление которой зависит от об-
щего напряжения всех сил и возможностей.
При отсутствии такого рода рабочих связей оценка окружаю-
щим занижается в фазе спада. Промежуточное положение биоритма
физической активности, где при оценке в группе туристов есть два
минимума — в фазе подъема и в фазе спада, — вероятно, объяс-
няется тем, что физическая работа является в данном случае
единственным объединяющим группу видом деятельности. В этой
группе, где рабочие связи слабы, наихудшие оценки человек по-
лучает каждый раз в фазе подъема.
В учебной группе студентов взаимооценки ставятся в большей
степени с учетом реального соотношения сил и менее зависят от
эмоциональных реакций членов группы. Максимумы и минимумы
оценок приблизительно соответствуют фазам подъема и спада эмо-
циональной и интеллектуальной активности. Эта закономерность
не распространяется на физический макроритм, который в учебной
деятельности студентов особой роли не играет.
Таким образом, в зависимости от фазы макроритма, в которой
находится человек, меняются и его оценка окружающих, и оценки,
которые окружающие ему дают. И, может быть, от тенденции спорт-
сменов, находящихся в фазе подъема физического макроритма,
занижать возможности противника и адекватно оценивать их в фазе
спада и зависит тот парадоксальный факт, что не менее 60% побед
в состязаниях спортсмены одержали, находясь именно в фазе спада
(Акрабов, 1972). Спортсмен, находящийся в фазе подъема, выиг-
рывая в силе и выносливости, проигрывает в тактике в силу не-
дооценки противников и, по-видимому, также переоценки собствен-
ных возможностей.
Однако это влияние макроритмов в очень большой мере корри-
гируется социальной значимостью конкретных оценок.
Чем существенней может повлиять оценка на организацию тру-
довой деятельности людей, тем меньше влияние на нее их биологи-
ческих ритмов.
Исследование влияния биологических макроритмов
на протекание процесса сна
У взрослых здоровых испытуемых с помощью электроэнцефа-
лографа фирмы «Альвар» проводилась регистрация ЭЭГ, ЭКоГ и
ЭКГ в течение ночного естественного сна в звукоизолированной
70
камере; использовались серебряные электроды, укрепляющиеся на
коже головы с помощью шлема и липкого пластыря. Отведения
биопотенциалов: лобно-теменное, теменно-височное, височно-заты-
лочное. Для записи электроокулограммы электроды крепились
пластырем на веках. ЭКГ регистрировалась с использованием
второго отведения. Записи начинались в 22—24 ч и длились всю
ночь до самопроизвольного пробуждения испытуемого. Снотвор-
ные не применялись. Всего у 25 испытуемых проведена 41 запись
ЭЭГ. Диализ записей проводился способами, описанными в гл. 2.
Значения показателей сна, достоверно различающихся (I—IV) физического макроритма Таблица 27 I в различные фазы
Показатель I II Ш IV
Общая длительность сна, в мин 400 * 329 359 354
Среднее число смен стадий сна в течение ночи за 1 с. 0.024 0.045 * 0.011 ** 0.023
Значение коэффициента последова- тельности стадий сна 1.083 1.199 * 0.945 ** 1.163
Средняя длительность стадии С, в % 22.17 17.18 28.55 * 20.01
Средняя частота появления ста- дии С, в % 22.99 21.99 28.25 * 25.25
Средняя длительность стадии Е, в % 19.629 24.13 21.39 15.49**
Средняя частота появления ста- дии Е, в % 12.45 11.56 10.33 6.594•< *
Разброс средней частоты появле- ния стадии REM в различные часы сна 7.573 ** 13.59 14.97 16.67
Примечание. Здесь и в табл. 28 значения средних, отмеченных одной звез-
дочкой., достоверно больше, двумя звездочками — достоверно меньше остальных сред- них в строке (Р = 0.95).
Для макроритма физической активности (табл. 27) было обна-
ружено, что в переходном состоянии от отрицательной к положи-
тельной фазе отмечается увеличение длительности сна. В положи-
тельной фазе макроритма наблюдалось уменьшение длительности
сна и нарастание числа смен стадий сна и коэффициента последо-
вательности стадий сна. В нисходящей фазе макроритма отмеча-
лись уменьшение числа смен стадий сна и коэффициента последо-
вательности стадий сна, нарастание средней длительности и ча-
стоты появления стадии С сна. В отрицательной фазе макроритма
отмечалось уменьшение средней длительности и частоты появления
стадии Е. Для макроритма эмоциональной активности (табл. 28)
было обнаружено уменьшение значения коэффициента последова-
тельности стадий сна и увеличение средней длительности движе-
71
Таблица 28
Значения некоторых показателей сна, исследованных в различные фазы
(I—IV) эмоционального и интеллектуального макроритма
Показатель I II Ш IV
Эмоциональная активность
Среднее число смен стадии сна 0.019 0.018 0.022 0.050 *
в течение ночи за 1 с
Значение коэффициента последова- 1.210 1.063 ♦♦ 1.177 1.185
тельности стадий сна
Длительность стадии REM, в °/0 3.926 3.566 4.414 8.550 ♦
Средняя длительность движений, в % 4.8 7.5 * 5.8 4.3
Интеллектуальная активность
Общая длительность сна, в мин 390 309 ** 358 362
Среднее число смен стадий сна в течение ночи за 1 с 0.023 0.019 0.066 ♦ 0.022
Значение коэффициента последова- тельности стадий сна 1.125 1.086 0.924 ♦♦ 1.085
Средняя длительность стадии А, в % 5.450 2.933 1.527 ** 4.561
Разброс длительности стадии А, в % 27.28 ♦ 11.92 3.166 *♦ 14.12
Разброс средней частоты появле- ния стадии А в различные часы сна 24.31 * 11.30 2.0 ♦♦ 13.17
Разброс средней частоты появле- ния стадии REM в различные ча- сы сна 10.38 * 16.03 9.622 ** 15.58
ний во сне в фазе подъема. В фазе спада нарастали среднее число
смен стадий сна и длительность стадии REM.
Для макроритма интеллектуальной активности (табл. 28) ока-
зывалось характерным наличие зависимостей почти исключительно
в переходных фазах. При переходе от отрицательной к положи-
тельной фазе отмечалось уменьшение разброса средней частоты
появления стадии REM в различные часы сна и увеличение раз-
броса длительности и средней частоты появления стадии А.
При переходе от положительной фазы к отрицательной наблюда-
лось увеличение среднего числа смен стадий сна и уменьшение
значения коэффициента последовательности стадий сна, средней
длительности стадии А (и ее разброса), а также разброса средней
частоты появления стадий А и REM в различные часы сна. В по-
ложительной фазе отмечалось уменьшение общей длительности сна.
Таким образом, можно сказать, что сложно организованный
процесс сна находится в некоторой функциональной зависимости
от ритмики медленных физиологических процессов в организме,
находящих свое выражение в периодических колебаниях физиче-
ской, эмоциональной и интеллектуальной активности.
72
Исследование влияния биологических макроритмов
на колебания значения некоторых физиологических
и психологических показателей
Обследовано И практически здоровых лиц, которые в течение
1—2х/2 мес ежедневно (кроме выходных) измеряли показатели,
приходя на работу или уходя с работы. Замеры проводились
в интервалах времени 9 ч 30 мин—11 ч или 17 ч—18 ч 30 мин.
Исследовались тесты на краткосрочную память и на сложение
цифр, а также (с помощью динамометра) измерялась сила правой
и левой руки. Всего в этой серии проведено 636 замеров шести
показателей. Кроме того, были использованы результаты само-
наблюдений одного испытуемого, который в течение 45 дней сни-
мал показатели 5—8 раз в день. (Всего 317 замеров семи показа-
телей). Выполнялись те же тесты на память и сложение, подсчи-
тывались частоты пульса и дыхания, измерялась температура.
Кроме того, проводился счет пальцев и определялась длитель-
ность «индивидуальной минуты». Данные обрабатывались по про-
грамме статистического анализа показателей по фазам макро-
ритмов.
Ни по одному из исследованных параметров (в том числе фи-
зической силе, измеряемой динамометром) не получено достовер-
ных различий (при Р=0.95) в зависимости от фаз макроритмов.
Возможно, этот результат связан с взаимоотношениями высо-
коамплитудного суточного ритма и многодневной периодики со
значительно меньшей амплитудой. Кроме того, данный материал
не был обработан с помощью методов выделения скрытой периодич-
ности, наиболее адекватных поставленной задаче и широко ис-
пользуемых нами в других случаях, поскольку длительность
временных рядов не превышала 2—2х/2 периодов, а эти методы
дают надежные результаты в условиях наличия пяти и более
периодов колебания.
Таким образом, модуляции с многодневным периодом (за счет
влияния макроритмов) выделяются статистическими методами в от-
носительно медленно меняющихся процессах (колебания оценок
и процесс сна), в которых несомненно имеются (но не доминируют)
ультрадные составляющие. В тех же процессах, в которых уль-
традные ритмы являются основными компонентами, модулирую-
щие влияния принадлежат, по-видимому, циркадным ритмам.
Макроритмы в этом случае, если и оказывают влияние, то опосре-
дованное (через циркадные ритмы), являясь при этом регулято-
ром второго порядка. Кроме того, влияние отдельных макрорит-
мов на общее состояние организма вряд ли просто суммируется.
По всей вероятности, это и обусловливает неоднозначность от-
ветов при попытках сопоставить результаты спортивных состяза-
ний с фазами многодневных биоритмов их участников.
Так, при сочетании двух ритмов в положительной фазе спор-
тивные результаты были хорошими во время тренировок у 95%
73
обследованных, но сочетание отрицательных фаз двух биоритмов
никак не сказалось на результатах (Акрабов, 1969). Поскольку
на спортивные результаты влияет не только уровень физических
возможностей и эмоциональная настройка, но и качество само-
оценки и оценки возможностей соперников, которые, как пока-
зано выше, снижаются в фазе подъема любого из ритмов, сочета-
ние двух или даже трех ритмов в положительной фазе не гаранти-
рует оптимальной стратегии не только в спортивной, но и в лю-
бых других видах деятельности.
Изучение макроритмов нужно и полезно в физиологических
исследованиях, но современное увлечение «гороскопами», когда
люди пытаются прогнозировать возможные успехи или неуспехи
своей деятельности на основании фаз макроритмов и их сочетаний,
с нашей точки зрения, просто вредны.
Повышение даже всех возможностей далеко не абсолютное
благо. В ряде случаев гораздо более оптимальны те действия,
которые совершаются в фазах упадка ритмов, когда человек более
трезво оценивает ситуацию.
Глава 6
ИЗМЕНЕНИЯ ЦИРКАДНОЙ
И УЛЬТРАДНОЙ РИТМИКИ
ПОД ВЛИЯНИЕМ ЭМОЦИОНАЛЬНОГО
СТРЕССА И ДРУГИХ СЛУЧАЙНЫХ
ВНЕШНИХ ВОЗДЕЙСТВИЙ
На воздействия внешней среды организм отвечает реакцией
адаптации — перестройки каких-то свойств в зависимости от
условий. Благодаря внутренней биоритмической структуре си-
стема способна активно исследовать качества и свойства окружаю-
щей среды и по каналам обратной афферентации в итоге каждого
элементарного акта взаимодействия получать информацию. Это
свойство живых организмов Н. Н. Василевский (1973) называет
способностью «зондировать» и считает его основой механизмов
адаптивного поведения.
Однако существуют и другие механизмы взаимодействия со сре-
дой — это те случаи, когда организм, отреагировав на воздействие,
стремится к восстановлению исходного состояния. Это реакции на
кратковременные сильные воздействия, протекающие по типу
так называемых переходных процессов. В этих случаях воздей-
ствие вызывает резкое отклонение уровня параметров от их те-
кущих значений. Как только это влияние прекращается, проис-
ходит отклонение в противоположном направлении и, переходя
из одного крайнего состояния в другое со все меньшей амплиту-
дой колебаний, система возвращается к исходному уровню. Эта
схема хороша и для биологических систем, но только в случаях
кратковременных воздействий с кратковременными переходными
процессами. Если же этот процесс происходит длительно или если
возмущающие воздействия повторны и один переходный процесс
сменяется другим, то в живом организме, обладающем сложной
биоритмической структурой функций, само «исходное состояние»
должно измениться во времени. Возврат к его былым значениям
для организма неоптимален.
С целью изучения способов перестройки ритмической актив-
ности в условиях переходных процессов мы сделали попытку их
исследования на модели ритма сердца, имеющего выраженные
ультрадные компоненты, и на модели циркадного ритма сна.
В качестве возмущающего фактора были использованы естествен-
ные стрессорные воздействия.
75
Изменение сердечного ритма
под влиянием стрессорных воздействий
Изменения сердечного ритма исследовались на спортсменах-
фехтовальщиках в период подготовки к бою, разминки, самого
боя и отдыха после боя. Передача сигналов электрокардиограммы
на чернилописец осуществлялась с помощью радиотелеметриче-
ской установки «Спорт-4» одновременно от обоих сражающихся.
Запись делалась на скорости 50 мм/с, что при ручном измерении
длительности кардиоинтервалов (RR) позволяет получить точность
не ниже + 10 мс.
Запись всего исследования (около 2 ч) разбивалась на функ-
циональные отрезки (подготовка, разминка, бой, отдых и т. д.),
а внутри каждого отрезка выделялись последовательные куски
по 130 кардиоинтервалов. В отдельных случаях (например, корот-
кий бой) их выделялось меньше (70—100). Снятые вручную после-
довательности интервалов RR вводились в ЭВМ «Минск-32» и
обрабатывались по специальным программам, включающим ме-
тоды вариационной пульсометрии и аппроксимации тригонометри-
ческими функциями.
Учитывая сложность реакций на стресс, в которых, кроме до-
минирующей мотивации, большую роль играет наличие дефи-
цита информации, вызывающее дополнительные проявления «эмо-
ции неуверенности» (Симонов, 1965), мы специально исследовали
противников, хорошо знающих друг друга. Как известно, даже
очень сильному бойцу иногда легче бороться с сильным, но знакомым
противником, чем с соперником, применяющим неожиданные
приемы, даже если он и не очень высоко квалифицирован.
Мы располагаем данными об особенностях ритма работы сердца
пяти спортсменов в течение 20 боев, т. е. в условиях высоких
эмоциональных и физических нагрузок, сопровождаемых сильной
мотивацией (желание выиграть), перед, после и во время боев,
которые были одним спортсменом выиграны, а другим — про-
играны.
Оказалось, что значения максимального, минимального, сред-
него и наиболее вероятного интервалов RR существенно разли-
чаются в разных состояниях. Кроме того, значительные отличия
обнаружены по параметрам среднеквадратичного отклонения и по
вегетативному показателю ритма (табл. 29).
Как видно из табл. 29, в случае хорошей мобилизации, обеспе-
чивающей успех в поединке, значения всех исследованных пока-
зателей достоверно ниже, чем в случае менее совершенной моби-
лизации, уже в период разминки. Эта разница сохраняется во
время поединка, а непосредственно после него отношения стано-
вятся обратными, при этом различия оказываются уже малодосто-
верными, что, по-видимому, зависит от огромного разброса значе-
ний показателей, связанного с большой динамичностью реакций.
76
Таблица 29
Значения статистических характеристик интервалов RR у спортсменов
в условиях обычной физической нагрузки и фехтовальных поединков
Характер деятельности спортсмена Длительность интервала RR, в с Средне- квадра- тичное отклоне- ние Вегета- тивный показа- тель ритма
наиболее вероят- ного макси- мального среднего мини- мального
Физическая на- грузка на эмоци- онально ней- тральном фоне Разминка перед поединком: 0.60 0.63 0.56 0.51 0.025 380
неуспешным 0.80 1.01 0.81 0.68 1.100 55
(0.90) (0.90) (0.88) (0.73) (0.85) (0.78)
успешным Поединок: 0.65 0.74 0.60 0.51 0.050 214
успешный 0.43 0.55 0.44 0.39 0.032 1272
неуспешный Отдых после по- единка: 0.47 0.44 0.43 0.36 0.017 2435
успешного 0.45 0.45 0.41 0.38 0.015 1508
неуспешного 0.52 0.59 0.50 0.44 0.037 662
Примечание. В скобках — значение Р (только для тех случаев, когда
Р>0.7).
В случае успеха в поединке полная перестройка структуры
сердечного ритма, сохраняющаяся в дальнейшем иногда до конца
поединка, достигалась за 8—9 с, а в случае неуспеха — за 12—
13 с. Плохо мобилизовавшийся боец в период отдыха после по-
единка не расслабляется, а как бы продолжает мобилизацию.
Перестройка ритма заключается не только в укорочении сред-
них значений интервалов RR, но и в уменьшении их разброса.
Как видно из табл. 29, среднее число значений интервалов, встре-
чающихся в записи, так же как и среднеквадратичное отклоне-
ние, в 1.5—2 раза ниже в случае успешного исхода, в момент
разминки и самого поединка; отношения меняются на обратные
в момент отдыха.
Таким образом, вторым механизмом мобилизации оказывается
стабилизация узких границ функционирования системы, что, по-
видимому, в первую очередь является результатом вовлечения
нервного аппарата эмоций, которое, согласно П. В. Симонову
(1965), «состоит в переходе живой системы к доминантным типам
реагирования. . . Возникшая доминанта подавляет и отменяет
другие, в данный момент второстепенные, деятельности орга-
низма, сосредоточивает все силы и резервы на удовлетворении
наиболее острой потребности» (с. 83).
Наконец, из табл. 29 видно, что вегетативный показатель
ритма (ВПР) имеет у хорошо мобилизовавшегося бойца большее
77
значение, превышая показатели слабее мобилизовавшегося в 4 раза
во время разминки и вдвое в бою. После боя, как и в предыдущих
случаях, этот показатель, напротив, оказывается у хорошо моби-
лизовавшегося в 2 раза ниже.
Можно предположить, что ВПР отражает в некоторой мере
энергетическую сторону процесса мобилизации резервов, обеспе-
чивающую возможность осуществления устойчивой эффективной
деятельности сердца в узком диапазоне частот сокращений. Во всех
случаях после успешного боя значения интервалов RR средне-
квадратичного отклонения в ВПР растут по сравнению с фоном.
Исследование временной структуры сердечного ритма обоих
бойцов на протяжении всего боя показало, что различий в дина-
мике значений интервалов RR (средних, максимальных, минималь-
ных, наиболее вероятных) не обнаруживается. Эти различия от-
мечены для среднеквадратичного отклонения и ВПР. В табл. 30
Таблица 30
Динамика статистических показателей сердечного ритма
у двух спортсменов в ходе поединка
Отрезок последова- тельной записи Среднеквадратичное отклонение у спортсмена Вегетативный показатель ритма у спортсмена
проигравшего выигравшего проигравшего выигравшего
1 0.0677 0.0706 * 119 71
2 0.0161 0.0166 1089 698 *
3 0.0110 0.0253 1846 469
4 0.0127 0.0397 * 2502 * 257
5 0.0263 * 0.0245 844 546
6 0.0223 0.0145 841 2280
7 0.0162 0.0086 981 2850 *
8 0.0138 0.0448 1040 374
9 0.0144 * 0.0477 * 1655 184
10 о.оюо 0.0154 2918 * 1749
И 0.0168 * 0.0090 1538 1945 *
12 0.0077 0.0110 * 1805 717
13 0.0104 * 0.0099 3984 1923 *
14 0.0068 0.0150 7692 * 1279
15 0.0135 * 0.0094 2514 2194 *
16 0.0075 0.0156 * 4497 * 841
17 0.0078 — 3165 —
18 0.0089 — 3053 —
19 0.0165 * — 978 —
Примечание. Звездой ко i отмечены максимальные повышения значении пока-
зателей (пики).
в качестве примера приведены результаты анализа последователь-
ных отрезков записи (разбиение по 130 кардиоинтервалов) двух
спортсменов в ходе одного длительного боя. Из-за различия в ча-
стоте сердцебиений отрезки записи соответствуют друг другу пол-
ностью в смысле последовательности, но не в смысле физического
78
времени, поскольку для того из бойцов, у которого сердцебиения
были чаще (а им оказался проигравший), число отрезков по
130 интервалов RR было за то же самое время больше. Видно, что
значения среднеквадратичного отклонения максимальны в первые
секунды боя, а в дальнейшем резко уменьшаются, причем наблю-
даются отчетливые колебания, затухающие по амплитуде, т. е.
имеется картина переходного процесса. При этом у более успешно
проводящего бой спортсмена ритмическая структура ярче выра-
жена (высокоамплитудные пики в отрезках 4, 8—9).
Ритмическая структура колебаний ВПР также выражена
у обоих спортсменов, но у более успешно проведшего бой коле-
бания более правильны. У спортсмена проигравшего отмечаются
меньшая регулярность колебаний и значительный рост общего
уровня ВПР, возможно отражающего степень напряжения про-
цессов. Таким образом, если у первого из них развилось состоя-
ние напряжения, оказывающего положительный эффект на дея-
тельность, то у второго, по-видимому, возникло состояние напря-
женности, характеризующееся понижением устойчивости психи-
ческих и двигательных функций вплоть до дезинтеграции деятель-
ности (Наенко, 1976).
В заключение можно сказать, что оптимальным режимом дея-
тельности сердца в условиях больших нагрузок оказалась стаби-
лизация длительности интервалов RR в определенном диапазоне.
Это выражается в уменьшении разброса их значений, а также
в сохранении определенного уровня функционирования системы
на протяжении всего периода активного действия с быстрым воз-
вращением к исходному уровню, как только необходимость в ста-
билизации характера функционирования системы ликвидирована.
Качество выполняемой деятельности было невысоким у тех
бойцов, сердечно-сосудистая система которых характеризовалась
некоторой инертностью. Во время активного действия их сердеч-
ные ритмы отличались относительно высоким разбросом значений
интервалов RR, отсутствием четкого сдвига в сторону высоких
частот и перестроек в период последействия.
Таким образом, подстройка сердечно-сосудистой системы чело-
века в условиях повышенных эмоциональных и физических нагру-
зок осуществляется путем быстрого выбора необходимого и доста-
точно узкого диапазона кардиоинтервалов и фиксации выбранного
режима на весь период активного действия. Исследование фоно-
вых записей показало, что основой этой успешной перестройки
являлась определенная временная структура сердечного ритма
до поединка, выражающаяся в наличии широкого спектра мед-
ленноволновых компонент кардиоинтервалов и большими откло-
нениями от их средних значений. Инерционные системы регуля-
ции сердца, перестраивающиеся плохо и медленно, подобным
временным паттерном не располагают.
Изменения ультрадиой ритмики (характерные для оптималь-
ного поведения системы) под влиянием возмущающих факторов
79
заключаются сначала в подборе и фиксации оптимального узкого
диапазона частот с уменьшением интервалов RR с постоянными
ритмическими отклонениями от этого диапазона, а затем (после
прекращения воздействия) в некотором увеличении периодов ко-
лебаний, среднеквадратичного отклонения интервалов RR и ВПР,
также ритмически меняющихся. В системе усиливаются периоди-
ческие составляющие, и она возвращается к исходному уровню,
но с некоторой коррекцией. Наши данные совпадают с данными
некоторых авторов, изучавших в отличие от нас реакцию не на
кратковременное, а на длительное напряжение.
При исследовании статистических характеристик сердечного
ритма у операторов при 4-часовой работе показано, что начало
работы совпадает с уменьшением значений среднего интервала RR
(с 0.96+0-04 до 0.87+0.05) и возрастанием индекса напряжения 2
(с 94+4 до 144+2). Постепенно средние значения периодов ра-
стут, а индекс напряжения уменьшается. Через 4 ч эти показа-
тели соответственно принимают значения 1.04+0.05 и 52 + 1 (Куд-
рявцева, 1980).
На усиление периодической составляющей в колебаниях фи-
зиологических процессов при повышении уровня деятельности
организма (органа) указывает К. М. Смирнов (1980). Это усиление
выявляется как на уровне внутриклеточных и тканевых процессов
(Саркисов и др., 1975), так и на уровне параметров физиологиче-
ских функций. Так, при выполнении психологических проб
в импульсной активности некоторых нейронов возникает отсут-
ствовавшая ранее периодичность (Раева, Ливанов, 1975).
Перестройка ритма сна
Переходя к вопросам изучения перестройки циркадного ритма
сна под влиянием стрессорных воздействий, мы прежде всего
должны разграничить две стороны этого вопроса. Ритм сна яв-
ляется частью основного жизненного цикла сон—бодрствование
и фактически не может быть исследован в отрыве от всего огром-
ного комплекса циркадных ритмов, прямо связанных и даже зави-
сящих от ритма сна. Так, например, ритм выделения гормона
роста настолько тесно связан со сном, что если инвертировать
время сна, время выделения гормона роста инвертируется точно
таким же образом.
Поэтому мы разделили наши исследования на две части: а) из-
учение изменений непосредственно самого ритма сна, связанных
с перелетами и с пересечением часовых поясов, при которых,
естественно, нарушены и другие ритмы, но ритм сна в первую
2 Индекс напряжения (ИН), предложенный Р. М. Баевским (1976),
по существу идентичен вегетативному показателю ритма и отличается лишь
множителем 1/2: ИН=Ямо/2(Л/о-АХ), где переменные Ам0, Мо и АХ имеют
тот же смысл, что и величины fM , Мх и X в формуле для вычисления ВПР.
80
очередь, и б) изучение воздействия на биологические ритмы эмо-
циональных стрессов, вызывающих значительные изменения дея-
тельности всех функциональных систем и опосредованно —
изменения сна.
Изменения циркадного ритма сна
в результате изменения временной среды
Наступление дня и ночи является одним из наиболее сильных
регуляторов циркадного ритма сна, и изменение этих моментов
после перелетов дает значительные перегрузки временного рас-
согласования между биоритмами организма и воздействиями
окружающей среды, которые не могли бы быть выдержаны чело-
веком, если бы тысячелетиями не была сформирована приспосо-
бительная реакция к постоянным и постепенным изменениям этих
воздействий в связи со сменой сезонов.
Согласно Астрономическому календарю, соотношения продол-
жительности светлого и темного времени суток (и соответствую-
щих изменений температуры, влажности, атмосферного давления
и т. п.) на широте Москвы колеблются от 17/7 до 7/17, на широте
Тбилиси — от 16/9 до 9/15, на широте Ленинграда — от 18/6 до в/18.
Таким образом, суточный размах достигает 6—12 ч. Однако
несмотря на эту постоянную тренировку, адаптация к внезапному
изменению временной среды при долготных перелетах все же
весьма несовершенна, что выражается в изменении самочувствия
и циркадных ритмов различных функций на довольно длительные
сроки.
С целью исследования особенностей перестроек ритма сна
после перелетов было проанализировано 98 электроэнцефалограмм
(ЭЭГ), записанных у 28 здоровых испытуемых во время сна.
Из них 40 записей были фоновыми, 53 — после перелетов, пять —
после длительной адаптации после перелета.
Фоновые исследования проводились в Ленинграде: записи
ЭЭГ в первую и пятую ночь после перелета с запада на восток
производились в Петропавловске-на-Камчатке. Там же изучалась
через мес после перелета адаптация лиц, прибывших из Ленин-
града. Исследования после перелетов с востока на запад (с пере-
сечением 7—9 часовых поясов), а также с юга на север (с пересе-
чением 1—2 часовых поясов) проводились в Ленинграде в первую
и седьмую ночи после перелетов и через 1—1х/2 мес адаптации лиц*
прибывших в Ленинград с Камчатки.
Было проведено как суммарное, так и раздельное исследование
лиц, хорошо и плохо адаптирующихся к внезапной смене обста-
новки, т. е. обладающих хорошими и плохими способностями
к перестройкам. Критерием адаптированности служили как рабо-
тоспособность и самочувствие испытуемого после конкретного
перелета и скорость, с которой он (субъективно) приспосабли-
вался к окружающей обстановке, так и сведения из жизни испы-
81
туемых, в частности их профессия и увлечения. Только хорошо
адаптирующиеся лица избирают профессии, связанные с постоян-
ными перелетами (геологи, инструкторы туризма, агенты по снаб-
жению и т. д.), или ежегодно совершают дальние путешествия
в сложных условиях.
Исследования показателей через 1—1х/2 мес адаптации в но-
вых условиях проводились только у представителей группы
хорошо адаптирующихся с помощью обычных статистических
методов и с помощью метода главных компонент (Каган, 1977;
Каган, Симонов, 1978; Моисеева, Каган, 1978).
Оказалось, что существенных изменений в картине сна пе
обнаруживается при перелетах с юго-востока на северо-запад
с пересечением 2—3 часовых поясов. Изменения, возникающие
в картине сна при пересечении 7—9 часовых поясов, приведены
в табл. 31. Общий рисунок сна (распределение длительности и
Т а б л и ц а 31
Значения некоторых характеристик ночного сна,
исследованных в обычных условиях и после перелетов
с пересечением девяти часовых поясов
Характеристики сна Фоновые исследования После перелета с запада на восток После перелета с востока на запад
длитель- ность стадии сна частота появле- ния длитель- ность стадии сна частота появления длитель- ность стадии сна частота появления
Стадия
А 4.259 5.298 0.740 ** 1.735 2.109 ** 2.734
В 16.160 19.800 26.900 * 24.620 16.760 ** 20.500
С 21.190 24.480 24.140 26.030 24.500 26.090
D 33.880 33.000 32.240 30.360 32.700 32.590
Е 19.070 * 8.575 10.610 ** 6.213 15.480 6.960
REM 5.020 ‘ 8.357 3.504 9.491 7.942 * 11.760 *
Общая длитель- ность сна, в мин 355 273 ** 285 , **
Коэффициент по- следовательности стадий сна 1.171 1.126 1.237
Примечание. Длительность стадий сна дана в процентах к общей длитель-
ности сна, частота появления — в процентах к общему числу смен стадий сна. Одна
звездочка — достоверное увеличение показателя по сравнению с исходным фоном, две
звездочки — достоверное уменьшение показателей.
частоты появления различных стадий сна в течение ночи) отчет-
ливо меняется при перелете на запад, что выражается в умень-
шении длительности и частоты появления стадии сна А и В и
нарастании этих показателей для стадии REM (для А и REM
82
различия достоверны). Кроме того, достоверно уменьшается раз-
брос длительности и частоты появления медленноволновых ста-
дий сна.
По-видимому, в связи с этой особенностью выявляются су-
щественные различия по сравнению с фоном в отношении распре-
деления стадии А в течение ночи, поскольку (при общем уменьше-
нии ее длительности и частоты появления) в середине ночи после
перелетов она так же выражена, как в первые и последние часы
сна, в то время как в фоне в середине ночи стадия А, как из-
вестно, почти не появляется. То же самое касается стадии REM,
которая оказывается в начале сна более представленной в записях
после перелета, чем в фоновых записях. Кроме того, возрастает
коэффициент последовательности стадий сна.
После перелета на восток изменения рисунка сна заключаются
только в достоверном уменьшении длительности и частоты появ-
ления стадии А, уменьшении длительности стадии Е и увеличении
длительности стадии В (соответствующие изменения претерпел и
разброс этих показателей). Описанные изменения после перелетов
обнаружены в течение 7—10 дней.
Анализ особенностей течения сна с использованием метода
главных компонент показал, что в первые 1—3-ю ночь после
перелета (как при перелете с запада на восток, так и в обратном
Таблица 32
Выделенные методом главных компонент показатели структуры
ночного сна лиц, хорошо адаптирующихся, после перелетов
с пересечением 7—9 часовых поясов (1—3-я ночь после перелета)
Перелет с запада на восток Перелет с востока на запад
описание компонент вклад в суммарную дисперсию, В 7о описание компонент вклад в суммарную дисперсию, В %
I. Сокращение дли- тельности сна и неустойчивость поверхностных стадий сна 24.46 I. Увеличение под- вижности сна в сторону пере- хода от поверх- ностных стадий к глубоким 17.42
II. Стабилизация коэф- фициента после- довательности стадий сна 16.71 II. Нарастание глу- бины сна (доми- нирование ста- дии Е) 16.23
III. Увеличение под- вижности про- цессов, учащение появления REM на фоне любой стадии сна 13.38 III. Нестабильность стадий А и REM 12.93
Примечание. Здесь и в табл. 33, 34 приведены главные компоненты, дающие
рклад в суммарную дисперсию не менее 5°/р,
83
направлении) имеет место нестабильность поверхностных стадий
сна и стадии REM (табл. 32). Но при перелете с запада на восток
резко сокращается длительность сна и стабилизируется смена
стадий, при перелете с востока на запад отмечается увеличение
подвижности сна в сторону перехода от поверхностных стадий
к глубоким.
Описанные особенности находят свое выражение, в частности,
в том, что распределение частоты появления и длительности ста-
дий сна в течение каждого последовательного часа сна повто-
ряется и соответствует примерно тому распределению, которое
в обычных условиях имеет место в течение всей ночи. Возникает
фрагментарная картина сна, где каждый фрагмент повторяется.
По мере перестроек картина сна постепенно меняется. В табл. 33
дано описание первых четырех компонент, выделенных у иссле-
Таблица 33
Выделенные методом главных компонент показатели структуры
ночного сна лиц, совершавших перелет с пересечением 7—9 часовых
поясов, для различных сроков адаптации
На 4—7-ю ночь после перелета: После 11/2—2 мес адаптации
описание компонент вклад в суммарную дисперсию, в °/о описание компонент вклад в суммарную дисперсию, в °/о
I. Уменьшение раз- броса длительно- сти стадий глубо- кого сна (С, Е) II. Увеличение раз- маха при пере- ходе от стадии поверхностного сна к стадиям глубокого III. Учащение появле- ния веретен сна 23.53 16.88 14.57 I. Способность бы- строго перехода от бодрствова- ния прямо к глу- бокому сну II. Увеличение ста- бильности ста- дии сна D III. Уменьшение дви- гательной ак- тивности во сне IV. Повышение под- вижности смен стадий сна в сто- рону перехода от глубоких к по- верхностным 44.58 22.32 17.83 14.06
дованных лиц на 4—7-ю ночь после перелета с востока на запад
и через ЬЧ2—2 мес адаптации к перелету (как с востока на запад,
так и в обратном направлении). После завершения перестройки
меняется ритмический рисунок сна. Если после перелетов как
на 1—3-ю, так и на 4—7-ю ночи была увеличена подвижность пере-
84
ходов от поверхностных стадий сна к глубоким, после адаптации
возникают тенденции к переходу в обратном направлении.
В табл. 34 приведено описание компонент, выделенных в фо-
новых записях лиц, хорошо адаптирующихся, и лиц, плохо
Таблица 34
Выделенные методом главных компонент показатели структуры ночного сна лиц, хорошо и плохо адаптирующихся (фоновые исследования)
Хорошо адаптирующиеся Плохо адаптирующиеся
описание компонент вклад в суммарную дисперсию, В % описание компонент вклад в суммарную дисперсию, в %
I. Повышение удель- ного веса и орга- низованности ста- дии сна II. Повышение ста- бильности и орга- низованности ста- дий Е — устой- чивость глубоко- го сна III. Повышение под- вижности стадий сна в сторону пе- рехода от глубо- ких к поверхност- ным IV. Реципрокные от- ношения раз- броса частоты появления ста- дий А (увеличи- вается) и REM (уменьшается) V. Стабилизация ста- дии D 12.52 10.03 8.25 6.18 5.02 I. Реципрокные от- ношения стадий REM и Е II. Снижение дли- тельности ста- дий А и D III. Повышение под- вижности в сто- рону перехода от бодрствова- ния и REM к глубокому сну IV. Увеличение раз- броса длитель- ности стадий С и REM V. Увеличение ста- бильности глу- бокого сна (ста- дия D) 17.68 11.21 10.03 8.50 6.20
адаптирующихся (т. е. группы лиц, практически здоровых, но не
выносящих дополнительных нагрузок). Мы условно приняли за
нормальную картину фоновые записи сна у хорошо адаптирую-
щихся лиц.
Анализ табл. 33 позволяет сказать, что в процессе перестройки
цикла сна прежде всего увеличивается размах колебаний диапа-
зона частот биопотенциалов. Это выражается в увеличении по-
движности в сторону перехода от поверхностных стадий к глубо-
ким, т. е. бодрствование, стадия А или В иногда сразу сменяются
стадиями D или Е (колебания ЭЭГ в диапазоне частот 7—12 Гц
85
резко переходят в диапазон 0.5—6 Гц). Стадия REM появляется
на фоне любой стадии сна. Это сочетается со стабилизацией распре-
деления стадий сна, в результате чего (как уже говорилось выше)
каждый последовательный час сна имеет примерно одинаковое
распределение каждой стадии сна.
По мере течения процесса адаптации (табл. 34) происходит
стабилизация и учащение появления глубоких стадий сна. Этот
эффект сохраняется к моменту завершения процесса адаптации,
когда наряду со способностью быстрого перехода от бодрствова-
ния прямо к глубокому сну, что характерно для процесса адапта-
ции, уже обнаруживается повышение частоты смен стадий сна
в сторону от глубоких к поверхностным.
Таким образом, переходный процесс в случае перестроек са-
мого цикла сна сводится, с одной стороны, к увеличению размаха
колебаний смены диапазона частот, с другой стороны — к строгой
фиксации представленности всего диапазона колебаний, от А
до REM (хотя поверхностные стадии и реже встречаются, чем
в обычном сне). По-видимому, в течение сна должны фигурировать
все стадии сна, причем они обязательно представлены в каждую
временную единицу (в качестве которой мы условно взяли 1 ч,
но возможно и более дробное деление).
Подобно тому как голографическое изображение в каждой
своей частице содержит полную картину объекта, каждый час сна
содержит в себе все необходимые стадии в пропорции, обычной
для сна в течение всей ночи
Для этого должна возрасти подвижность смены диапазонов,
т. е. увеличится размах колебаний при смене стадий сна, что и
происходит на самом деле.
Кроме того, примечательно, что процесс перестройки проте-
кает с отчетливым уменьшением общей длительности сна, так, как
если бы в момент перестроек удлинялась «индивидуальная ми-
нута». Как показано исследованиями ритмов точности восприятия
метрики времени, длительность «индивидуальной минуты» варьи-
рует у разных лиц и у одного и того же лица в зависимости от
состояния. Ее длительность растет в условиях смены впечатлений
и больших физических и эмоциональных нагрузок, что соответ-
ствует и моменту перестроек биоритмов после перелетов.
Как показано в табл. 18, после перелетов с пересечением 4—
8 часовых поясов у одного из испытуемых длительность минуты
варьировала от 67.39 (перелет на Сахалин) до 68.01 (перелет об-
ратно в Ленинград), составляя в среднем 67.70. Если встать на
ту точку зрения, что человек существует в своем собственном вре-
мени, то для данного испытуемого каждая минута удлинилась
на 7.70 с, а каждый час тем самым удлинился на 7 мин 42 с, т. е.
на 7х/2 мин.
Длительность сна, которая в фоновых исследованиях яв-
ляется обычной — 6ч при пересчете с добавкой 7г/2 мин к каждому
часу, стала бы равной 6 ч 45 мин (что уже было бы излишком сна),
86
а к тем 4 ч 45 мин, которые в среднем тратят на сон испытуемые
после перелета с востока на запад, нужно добавить еще 36 мин,
т. е. они спят (по их субъективному ощущению времени) 5 ч 21 мин.
Если встать на эту точку зрения, то укорочение длительности
сна после перелета является истинным во внешнем и мнимым во
внутреннем времени исследуемого.
Изменения ритма сна вследствие стрессорных воздействий
Исследования проводились на семи практически здоровых ис-
пытуемых методом анкетирования. Кроме того, у пяти из них
проведено 2—5 записей ЭЭГ во время сна (всего проанализиро-
вано 14 записей). Анализ протоколов времени отхода ко сну и
времени пробуждения (где указывались внешние факторы, воздей-
ствовавшие в течение дня), которые велись испытуемыми от 2 мес
до года, позволил установить, что имеется отчетливая зависимость
длительности сна от происшедших в течение дня событий в том
случае, если эти события вызвали эмоциональную реакцию. Мы не
касались вопросов о том, быстро ли засыпал обследуемый или
засыпание было затруднено, и о качестве сна (поскольку такого
рода указания обычно недостоверны) и оценивали лишь его общее
количество. Длительность сна в течение ночи после событий,
вызвавших отрицательную реакцию, которые могут быть расце-
нены как стрессорные факторы (создающие большую эмоциональ-
ную, а нередко и интеллектуальную и физическую нагрузки),
достоверно увеличивается (как это видно по табл. 35). После
Таблица 35
Средняя длительность сна за ночь (в мин) у испытуемых после событий
неприятных (24 исследования у 7 испытуемых в течение 2—5 ночей
у каждого) и приятных (28 исследований у 3 испытуемых
в течение 7—12 ночей у каждого)
Испытуемые Ночь накануне Ночь после непри- ятного события Ночь накануне Ночь после приятного события
1-я 2-я 1-я 2-я
С-а 401 543 438 525 415 412
Б-а 360 510 420 — — —
М-а 424 492 386 417 361 409
О-в 390 570 450 — — —
И-а 480 540 480 — — —
Х-р 387 372 372 395 418 426
К-а 450 487 540
Среднее 411 500 443 425 370 438
приятных событий длительность сна достоверно уменьшается
(г=0.95). При этом стрессорные воздействия, по-видимому, вы-
87
зывают более продолжительную реакцию, чем события прият-
ные, — их последствия выявляются и на вторую ночь.
Некоторые изменения структуры сна наблюдаются только
после воздействия стрессорных факторов и выражаются, кроме
увеличения длительности сна, в увеличении устойчивости стадий
сна (т. е. уменьшается число смен стадий сна) и в уменьшении ча-
стоты появления парадоксальной стадии сна.
Можно полагать, что возникновение стрессорных реакций,
вызывающих значительные эндокринно-обменные сдвиги, ставит
организм в новые условия. Продолжительность сна увеличи-
вается, как это показано в опытах на животных (Тонких и др.,
1977), а также у человека (Моисеева, 1976; Василевский и др.,
1978), возможно, за счет необходимости ликвидации метаболитов,
образующихся при стрессе. В течение одной (реже двух) ночи
эта функция успешно выполняется, и та же самая симпатоадрена-
ловая система, деятельность которой необходима для осуществле-
ния стрессорных реакций, по их окончании, по-видимому, присту-
пает к отправлению обычных адаптационно-трофических функций.
В целом реакция на нерегулярные «возмущающие» воздей-
ствия внешней среды на организм выражается в тенденции к «ста-
билизации» течения сна с увеличением его длительности и умень-
шением значения парадоксальной стадии, играющей для сна
роль собственного внутреннего «возмущающего фактора».
Положительные эмоции, не вызывающие гормональных сдвигов
и соответствующей мобилизации гомеостатических механизмов, не
оказывают влияния на структуру сна, только несколько понижая
потребность в нем.
Глава 7
ЗАВИСИМОСТЬ ТЕЧЕНИЯ ЦИРКАДНЫХ
РИТМОВ ОТ ГЕЛИОГЕОФИЗИЧЕСКИХ
ФАКТОРОВ
Под термином «гелиогеофизические факторы» понимается це-
лый комплекс физических факторов, влияющих на организм и
связанных с флуктуациями магнитных полей Земли, ее вращением,
особенностями строения атмосферы и т. п., а также с солнечной
активностью. В последние десятилетия проводятся многочислен-
ные исследования по проблеме «Солнце—биосфера» (Влияние сол-
нечной активности, 1971, и др.). Показаны четкие корреляции
между активностью Солнца, состоянием геомагнитных полей
Земли, ионосферы, атмосферных циркуляций и даже микросейс-
мических феноменов (Zatopek et al., 1976).
Особое значение придается воздействию на живые организмы
магнитного поля Земли, тесно связанного с магнитным полем
Солнца. На Солнце процессы перезамыкания силовых магнитных
линий порождают хромосферные вспышки, которые служат мощ-
ным источником космического излучения. На Земле взаимодей-
ствие солнечного ветра и магнитосферы самой Земли вызывает
полярные сияния, изменения наземного атмосферного давления,
температуры верхней атмосферы, холодные вторжения на Земле,
образование циклонов и другие изменения погодных и климати-
ческих факторов (Мустель, 1971).
Начаты исследования влияния комплекса биогеофизических
факторов на живой организм. Так, показано изменение ритма
прироста веса тела, связанного с фазами Луны, в магнитоактив-
ные дни (Василик, Галицкий, 1977).
Изучение геофизических факторов, влияющих на течение про-
цессов в живых организмах, приводит уже к созданию новой дис-
циплины — биогеофизики и, как одно из ее направлений,— гео-
магнитобиологии (Дубров, 1974).
Исследования влияния геомагнитных полей на биологические
объекты показали их значение для ориентации живых организмов
(особенно птиц) при движении и миграции и для организации
временной структуры биологических процессов. Электромагнит-
ное поле Земли в диапазоне низких и сверхнизких частот начали
уже рассматривать как существенный экологический фактор (Чер-
89
нышев, 1966; Чередниченко, 1973; Пресман, 1974; Холодов, 1976;
Brown, 1976; Владимирский, 1977).
Что касается механизма влияния геомагнитного поля на живой
организм, то предполагаются как прямое влияние на жидкие
среды или даже биологическую плазму, приводящее к конформа-
ционным изменениям макромолекул (Piccardi, 1967; Музалевская,
1969; Подшибякин, 1971; Дубров, 1974), так и опосредованное —
через изменения климатических воздействий.
На формирование всего сложного комплекса геофизических
факторов на Земле оказывают влияние не только Солнце (хотя,
конечно, его влияние доминирует), но и другие планеты, в пер-
вую очередь Луна (Markson, 1972).
Имеется довольно большое количество работ, в которых пока-
зано влияние погодных и геомагнитных факторов или изменений
солнечной активности на мозговую гемодинамику, динамику тех
или иных параметров крови, мочи и т. д., на развитие эпидемий
различных заболеваний (Колодченко и др., 1969; Реакция био-
логических систем. . ., 1971; Чижевский, 1973; Bartko et al.,
1974, и мн. др.).
Обнаружена зависимость развития тромбоэмболий и травма-
тизма на севере от гелиогеофизических факторов (Макаров и др.,
1978; Шалаев, Веселов, 1978). Однако лишь в единичных работах
исследуется влияние геофизических факторов на циркадные или
сезонные ритмы. Так, выявлена зависимость параметров гемоди-
намики не только от сезонов года (акрофазы ритмики частоты
пульса, артериального давления и вегетативного показателя
Кердо смещались зимой на более позднее время по сравнению
с осенью), но и от магнитного поля Земли.
Оказалось, что зависимость вегетативного индекса от напря-
женности магнитного поля Земли является прямой и при этом
более тесной в зимний период. Зависимость показателей гемоди-
намики осенью была обратной и слабо выраженной, зимой также
слабо выраженной, но прямой (Каталевская, Кузнецова, 1975).
Под действием слабого магнитного поля 10 Гц человек, поме-
щенный в специальную камеру, демонстрирует более короткие
циркадные циклы функции в те дни, когда поле включено, а также
некоторое время и после выключения поля. Частота 10 Гц соот-
ветствует примерно изменениям естественных полей в период
магнитных бурь (Wever, 1968, 1976).
С целью исследования влияния гелиогеофизических факторов
на течение циркадных ритмов данные, полученные в двух клини-
ках в Ленинграде на разных контингентах больных (с вегетососу-
дистыми расстройствами и перенесших черепномозговую травму),
были сопоставлены с геофизическими данными с использованием
метода Чебышева, позволяющего выявить как линейные, так и
нелинейные зависимости.
Изучалось влияние на физиологические параметры: 1) темпе-
ратуры воздуха, 2) влажности воздуха, 3) атмосферного давления,
90
4) значений горизонтальной (Н) и 5) вертикальной (Z) составляю-
щих магнитного поля Земли, 6) значений магнитного склоне-
ния (Р), 7) интенсивности магнитных бурь.
Кроме того, анализ некоторых показателей, полученных у здо-
ровых испытуемых, проводился с учетом влияния интенсивности
магнитных бурь и интенсивности хромосферных вспышек на
Солнце.
Влияние изменений температуры воздуха
на динамику изменений значений
физиологических показателей
За исследованный период средняя температура воздуха была
2.027° С; пределы изменения температуры от —7.3 до 4-1.2° С.
По отношению ко всем показателям, кроме температуры тела,
эта зависимость очень проста (аппроксимация уравнением второго
порядка).
С увеличением внешней температуры происходит увеличение
систолического и диастолического артериального давления, изме-
ренного как справа, так и слева, увеличение частоты дыхания и
уменьшение длительности индивидуальной минуты. Параллельно
с увеличением внешней температуры происходит нарастание мы-
шечной силы (табл. 36). Соотношения между температурой тела
Таблица 36
Направление изменении исследуемых физиологических параметров
под Влиянием усиления различных Внешних воздейстбий
Внешнее Воздействие Систола - ческое давление Диастоли- ческое давление Темпера- тура тела Сила РУ* Частота дыхания в 1 мин Длительность индивидуальной минуты
Температура воздуха
Атмосферное давление /\/\ ~\f\
Влажность воздуха [упругость водяных паров ) \f\ /V ^\f\
и внешней температурой гораздо сложнее (аппроксимация уравне-
нием четвертого порядка). При отклонении внешней температуры
от средней в сторону уменьшения температура тела также умень-
шалась. При умеренном увеличении внешней температуры темпе-
ратура тела увеличивалась, при значительном увеличении умень-
шалась. Таким образом, температура тела, во всяком случае
у больных, перенесших травму черепа, не является линейной
функцией от внешней температуры (рис. 12).
91
л
Рис. 12. Зависимость физиоло-
гических показателей больных
с вегетососудистыми расстрой-
ствами от температуры окру-
жающего воздуха.
Л, Б — систолическое и}' диастоли-
ческое артериальное давление;
Д, Е — то же на левой руке; В —
температура тела; Г — сила правой
руки; Ж — частота движения; 3 —
длительность индивидуальной ми-
нуты.
По осям абсцисс — значения внеш-
ней температуры, в °C; по осям
ординат — значение физиологиче-
ских показателей.
Зависимость изменений физиологических параметров
от влажности воздуха (упругости водяного пара)
За исследованный период времени средняя упругость водя-
ных паров составляла 4.86 г/см2. Пределы изменений — 3.3—
6.3 г/см2. Артериальное давление связано с влажностью воздуха
весьма сложными отношениями. Если систолическое давление
просто уменьшается с ростом влажности, то диастолическое дав-
ление уменьшается при значениях влажности ниже среднего,
увеличивается при значениях, близких к среднему, и уменьша-
ется при значениях выше среднего.
Таблица 37
Влияние влажности воздуха на динамику физиологических показателей
больных с черепно-мозговой травмой (декабрь 1977 г.,
средняя упругость водяных паров Кср = 4.86 г/см2)
Параметр Порядок корреля- ционного уравнения Среднее значение параметра Количе- ство подъемов кривой Количе- ство спадов кривой Ошибка аппроксима- ции
АДСП 3 125.27 0 1 0.1228
АДДП 4 79.86 1 2 0.1521
АДСЛ 3 123.25 0 1 0.1294
АДДЛ 4 78.21 1 2 0.1722
Температура тела, в °C Сила руки, в кг 4 36.73 2 1 0.0116
слева 2 43.39 1 1 0.2436
справа 4 41.32 1 1 0.2470
Частота дыхания в 1 мин 4 25.74 2 2 0.2476
Длительность инди- видуальной ми- нуты, в с 2 54.62 1 1 0.1741
В табл. 37 приведена сводка изменений параметров, возника-
ющих под влиянием нарастания влажности воздуха. Температура
тела и частота дыхания меняются при изменении влажности ана-
логичным образом: их значения увеличиваются при увеличении
влажности до среднего уровня, уменьшаются при средних значе-
ниях влажности и увеличиваются при повышении влажности выше
среднего.
Четкой зависимости силы рук от влажности не выявлено —
справа и слева динамика прямо противоположна.
Длительность индивидуальной минуты увеличивается с увели-
чением упругости водяных паров до определенного момента (зна-
чение упругости порядка 6.0), затем идет очень слабое уменьше-
ние и стационирование.
93
Влияние атмосферного давления
на изменение физиологических параметров
За исследованный период времени атмосферное давление в сред-
нем составляло 1026 гПа. Пределы изменений — 1007—1045 гПа.
Как систолическое, так и диастолическое давление с ростом атмос-
ферного давления увеличивается (аппроксимации уравнениями
второго и третьего порядка). Четкой зависимости мышечной силы
не обнаруживается.
Показатели температуры тела и частоты дыхания изменяются
в зависимости от увеличения атмосферного давления сходным об-
разом в виде ритмично повторяющихся увеличения и уменьшения
их значений.
Наконец, длительность индивидуальной минуты с ростом зна-
чений давления увеличивается вплоть до достижения среднего
уровня, а затем уменьшается (табл. 36). При этом температура и
атмосферное давление оказывают одинаковое влияние на физио-
логические параметры, а влажность влияет противоположным
образом.
Так, если под влиянием повышения температуры и атмосфер-
ного давления артериальное давление увеличивается, то под влия-
нием влажности уменьшается систолическое давление, а в пока-
зателях диастолического давления под влиянием влажности видны
волнообразные колебания (снижение диастолического давления,
затем повышение и вновь снижение). Два показателя — темпера-
тура тела и дыхание — реагируют на все три воздействия ритми-
ческими колебаниями. Эта тенденция минимальна при изменении
внешней температуры и максимальна при изменениях давления.
Каждый раз при наименьших значениях внешних параметров
температура тела и частота дыхания повышаются, затем (при зна-
чениях, близких к нулевым) снижаются.
По всей вероятности, такое сходство реакций зависит от того,
что дыхание наряду с основной своей функцией выполняет допол-
нительно и функцию терморегуляции. Длительность индивиду-
альной минуты растет с повышением влажности, падает с повы-
шением температуры и колеблется (сначала растет, затем падает)
с повышением атмосферного давления.
Влияние геомагнитного поля и магнитных бурь
на изменения физиологических параметров
Влияние геомагнитного поля и магнитных бурь изучалось на
обоих контингентах больных. При этом пациенты, перенесшие
травму черепа, обследованы во время двух сезонов года — зимой
(декабрь 1977 г.) и весной (24.III—30.IV 1978). Пациенты с диэн-
цефальными расстройствами обследованы только весной. По-
этому условия в этих трех сериях наблюдений были не вполне
одинаковы, что может быть одной из причин небольших различий
94
Таблица 38
Влияние интенсивности магнитных бурь на динамику физиологических
показателей больных с диэнцефальными расстройствами
(март—апрель 1978 г.). Средняя интенсивность 3 балла*
Параметр Порядок корреля- ционного уравнения Среднее значение параметра Количе- ство подъемов кривой Количе- ство спадов кривой Ошибка аппрок- симации
АДСП 3 133.30 1 0 0.1351
АДДП 2 71.98 1 0 0.1332
АДСЛ 3 132.52 1 1 0.1308
АДДЛ 4 72.19 1 2 0.1343
Температура тела, в ° С Сила руки, в кг 3 36.60 1 1 0.0096
справа 2 28.60 1 1 0.4816
слева 4 27.16 1 2 0.4658
Частота дыхания в 1 мин 3 18.93 1 1 0.1494
Длительность индиви- дуальной минуты, в с 4 58.07 2 1 0.2420
Таблица 39
Влияние интенсивности магнитных бурь на динамику физиологических
показателей больных с черепно-мозговой травмой (март—апрель 1978 г.).
Средняя интенсивность 2.5 балла
Параметр Порядок корреля- ционного {уравнения Среднее значение параметра Количе- ство подъемов кривой Количе- ство спадов кривой Ошибка аппрок- симации
АДСП 3 129.56 1 1 0.1378
АДДП 2 78.52 1 1 0.1106
АДСЛ 2 126.78 1 1 0.1137
АДДЛ 2 77.07 0 1 0.1143
Температура тела, в р С Сила руки 4 36.95 2 2 0.0127
справа 3 34.69 1 1 0.3131
слева 4 32.29 1 1 0.3254
Частота дыхания в 1 мин 4 20.24 2 2 0.3343
Длительность индиви- дуальной минуты, в с 3 64.11 2 1 0.3171
в реакциях. В табл. 38 и 39 приведены данные о характере из-
менений исследуемых физиологических параметров под влиянием
магнитных возмущений разной интенсивности у больных с ди-
энцефальными расстройствами и с черепно-мозговой травмой,
обследованных весной 1978 г.
На рис. 13 представлены примеры кривых зависимости физио-
логических параметров от интенсивности магнитных бурь. Сле-
дует подчеркнуть, что все составляющие магнитного поля Земли
и магнитные возмущения (бури) ио мере нарастания выражен-
95
Рис. 13. Зависимость физиоло-
гических показателей больных
с вегетососудистыми расстрой-
ствами от интенсивности маг-
нитных бурь (в баллах).
Обозначения те же, что на рис. 12.
ности вызывают однотипные существенно нелинейные реакции.
Из 108 проведенных исследований (восемь параметров в трех
группах) только в 19 случаях имела место простая реакция на-
растания или убывания значений физиологических параметров.
Во всех остальных случаях обнаружено либо нарастание, а затем
убывание, либо убывание, а затем нарастание, т. е. сложные не-
линейные отношения. В 55 случаях эта смена направлений из-
менений параметров происходит однократно, в 22 — двукратно,
в 12 — трижды.
Таким образом, медленные изменения постоянного поля Зем-
ли и магнитные возмущения влияют на все физиологические пара-
метры, вызывая не направленные реакции, а изменение исход-
ного фона в ту или иную сторону в зависимости от параметров
поля или состояния физиологической системы. Так, воздействия
горизонтальной составляющей вызывали в декабре 1977 г. четкую
двойную реакцию повышения и понижения диастолического дав-
ления. В марте—апреле 1978 г. у той же группы больных было
обнаружено однократное уменьшение и увеличение диастоличе-
ского давления слева и двукратное уменьшение с двукратным уве-
личением справа. У другой группы в то же время было просто
уменьшение диастолического давления справа и смена уменьше-
ния увеличением слева. Длительность индивидуальной минуты
то нарастала, то падала в 1977 г. у больных, перенесших черепно-
мозговую травму; монотонно нарастала в 1978 г. у той же группы
больных и монотонно уменьшалась в 1978 г. у больных с диэнце-
фальной патологией.
Кроме данного исследования, мы попытались выяснить влия-
ние магнитного поля Земли на ритмы мозга.
Влияние магнитного поля Земли на ритмы мозга
Сравнивая электрографическую картину сна в те ночи, когда
имели место магнитные суббури (слабые бури), со всеми осталь-
ными записями, сделанными в спокойное в отношении магнитных
возмущений время, мы пришли к выводу, что коэффициент после-
довательности стадий сна значимо выше в моменты суббурь.
При этом возрастает в 2 раза разрыв его значений в разные часы
у одного и того же испытуемого, а средняя частота смен стадий
сна — в 3 раза. Средняя длительность стадий сна, средняя частота
их появлений, распределение их в разные часы ночи существенно
не меняются (Моисеева и др., 1972).
Коэффициент последовательности стадий сна рассчитывался
как отношение числа смен стадий сна в «прямом» порядке (от бо-
лее поверхностного сна к более глубокому) к числу смен стадий
сна в «обратном» порядке. Он отражает направленность высоко-
амплитудных колебаний. Если коэффициент примерно равен 1,
переходы от стадии к стадии в обе стороны равномерны, если
значительно превышает 1 — имеет место постепенный ступенча-
97
тый переход от стадий поверхностного сна к стадиям глубоким
с внезапным «перескоком» к более поверхностным стадиям, минуя
1—2 из них. Если коэффициент ниже 1, глубина сна увеличива-
ется внезапно, скачками, и затем идет медленный, постепенный
переход к менее глубокому сну.
Рис. 14. Динамика длительности (а) и частоты появления (б) отдельных стадий
сна (Л, В, С, D, Е и REM) в течение ночи с обычной геомагнитной обста-
новкой (Z) и в течение ночи, когда имела место магнитная буря (II) (испы-
туемый Ч-а).
По осям абсцисс — длительность сна начиная с момента засыпания, в ч; по осям ординат —
длительность фаз по отношению к общей длительности сна, в %.
Рост значения коэффициента последовательности стадий сна
при суббурях показывает, что с магнитными возмущениями сов-
падает увеличение размаха колебаний диапазона частот ЭЭГ
в сторону быстрого перехода от медленных к более быстрым часто-
там. Этот процесс совпадает с увеличением частоты смен стадий
сна, т. е. со сменой доминирующих периодов, составляющих ЭЭГ.
Неравномерность этих процессов в различные часы сна до некото-
рой степени указывает на их зависимость от «всплесков» магнит-
ных суббурь.
Эти факты заставили нас провести дополнительное исследова-
ние возможного влияния магнитного поля Земли в те ночи, когда
98
бурь не было.-С этой целью была рассмотрена последовательность
стадий сна в течение ночи у восьми здоровых испытуемых и сделана
попытка с помощью кросскорреляционного анализа связать ее
изменения с изменениями амплитуды поля Н. Мгновенные значе-
ния амплитуды поля Н снимались вручную с помощью миллимет-
ровой линейки (отсчеты велись как вверх, так и вниз от средней
линии через каждые 10 с). Стадии сна кодировались условно также
каждые 10 с, что позволяло проследить корреляционные отноше-
ния по знаку первой производной между ними и компонентами
геомагнитного поля. Корреляций не обнаружено в семи случаях.
Таким образом, мы нашли, что резкие изменения магнитного поля
Земли отчетливо влияют па рисунок биопотенциалов головного
мозга человека; что же касается постоянно присутствующего маг-
нитного поля, то его влияние весьма вероятно, но навряд ли имеют
место прямые корреляции.
Исследование структуры последовательности стадий сна в те-
чение ночи показало, что (в отличие от хромосферных вспышек)
влияние магнитных бурь выражается в уменьшении ритмической
организации сна. При этом обычно сокращается общая длитель-
ность сна, что несколько затрудняет анализ.
На рис. 14 приведены кривые ночного сна испытуемых, исследо-
ванных во время ночи с обычной геомагнитной обстановкой и
в момент магнитных бурь.
Таким образом, магнитные бури вносят элемент случайности
в ритмические колебания живой и неживой природы, являясь
не датчиками времени, а источником случайных воздействий
(шумовым фактором).
Влияние изменения солнечной активности
на течение биологических ритмов
В качестве параметра, отражающего солнечную активность,
были избраны солнечные (хромосферные) вспышки, данные о кото-
рых были выписаны из справочного бюллетеня ЮНЕСКО, находя-
щегося в Пулковской обсерватории.
Поскольку нас интересовало непосредственное воздействие
вспышек на формирование картины биоритмов здоровых и боль-
ных людей, мы отказались от принятого в астрономии вспышеч-
ного индекса Додсон—Уедемана и использовали данные о времени
начала, кульминации и конца вспышек и их выраженности. Время
в справочнике, указанное по Гринвичу, переводилось в ленин-
градское время (отличающееся на 3 ч). Выраженность вспышек
определяется баллами 1 (слабая), 2 (средней силы), 3 (сильная).
Прежде всего была исследована ЭЭГ — картина ночного сна
в те ночи, когда имели место хромосферные вспышки, и в периоды
спокойного солнца. Обращалось внимание на смену стадий сна,
отражающую ритмические изменения диапазона частот в ЭЭГ.
Оказалось, что появление хромосферных вспышек в течение дня,
99
предшествующего записи, и в течение той ночи, когда производи-
лась запись (случаев, когда вспышки отмечались исключительно
ночью, в наших наблюдениях не было), достоверно изменяет зна-
чения некоторых параметров. Если вспышки были только днем,
параметры не изменялись.
В табл. 40 приведены средние значения основных параметров
ЭЭГ записей ночного сна испытуемых, исследованных в обычных
Т а б л и ц а 40
Значения некоторых характеристик ночного сна, исследованных в период
спокойного Солнца и при хромосферных вспышках
Характеристики сна Отсутствие вспышек на Солнце (п = 12) X ромосферные вспышки в течение дня перед сном (п = 8) Хромосферные вспышки в течение дня перед сном и ночью во время сна (п = 9)
длитель- ность стадии сна частота появления длитель- ность стадии сна частота появления длитель- ность стадии сна частота появления
Стадия
А 2.77 5.31 1.12 2.80 3.87 6.47
В 19.96 20.80 15.90 19.38 16.10 18.10
С 17.68 22.40 21.41 25.10 24.21 28.55
D 32.64 31.94 27.61 32.15 28.20 31.50
Е 20.18 9.37 23.30 10.30 19.90 8.91
REM 4.88 9.46 7.71 10.40 4.68 7.23
Средняя частота 0.019 0.044 0.053
смен стадий сна
Коэффициент по- 1.241 1.237 1.170
следовательности
стадий сна
Среднее число дви- 11.98 5.33 6.60
жений во сне
Количество смен 49 42 г 58
стадий сна при
движениях, в %
условиях в период спокойного солнца (12 записей) и в периоды
хромосферных вспышек. Сон регистрировался в момент хромо-
сферных вспышек ночью и в течение предыдущего дня (всего 7 за-
писей).
В те ночи, когда наблюдались хромосферные вспышки, нара-
стает мозговая активность в диапазоне частот 14—16 в 1 с (вере-
тена сна), характерная для стадии сна С. Одновременно несколько
уменьшается частота и длительность стадии сна REM, т. е. пред-
ставленность высоких частот. Таким образом, процентное содер-
жание каждого диапазона частот (стадий сна) мало меняется,
но абсолютна я частота смен стадий сна нарастает почти в 3 раза.
100
Это повышение изменчивости диапазонов частот (подвижности
процессов) сочетается с ростом равномерности переходов от по-
верхностных к глубоким стадиям сна, и наоборот, что выражается
в уменьшении значений коэффициента смен стадий сна (К).
В те ночи, когда происходят хромосферные вспышки, обыч-
ный рисунок распределения диапазонов частот по часам сна меня-
I II
50
Рис. 15. Динамика длительности (а) и частоты появления (б) отдельных ста-
дий сна (PF, Л, В, С, Р, В, REM) в течение ночей, характеризующихся от-
сутствием (7) или наличием (II) хромосферной вспышки на Солнце (испы-
туемый Д-о).
Обозначения те же, что на рис. 14.
ется, о чем судят по кривым представленности каждой стадии
в разные часы сна.
Общим правилом является неравномерное распределение ста-
дий сна в течение ночи (большее количество поверхностных ста-
дий в начале и в самом конце; нарастание представленности глу-
боких стадий по мере увеличения глубины сна и т. д.).
В те ночи, когда имеют место хромосферные вспышки, кривые,
отражающие изменения длительности частоты появления каждой
стадии сна в течение ночи, приобретают ритмический характер.
101
На рис. 15 видно, что в ночи, сопровождавшиеся хромосфер-
ными вспышками, изменения как длительности, так и частоты
появления каждой стадии сна у двух здоровых испытуемых ста-
новятся более ритмичными. Таким образом, ультрадные ритмы
смены фаз сна в некоторой мере модулируются солнечной актив-
ностью.
Суммируя все сказанное о влиянии геомагнитных и погод-
ных факторов на изменения физиологических параметров, следует
отметить, что эти влияния можно разделить на два вида. Влияние
на физиологические параметры солнечной активности (хромо-
сферных вспышек) и погодных факторов (которые сами четко за-
висят от солнечной активности) чаще всего проявляется в виде
простых, линейных, отношений. Воздействия постоянного магнит-
ного поля и случайных магнитных возмущений нелинейны и со-
здают постоянный и «ритмический» фон, вызывая (в зависимости
от собственных параметров и функционального состояния живой
системы) то реакцию усиления, то ослабления функции. Таким
образом, магнитные поля Земли, по-видимому, поддерживают
существование колебательных контуров, в то время как солнеч-
ная активность и погодные факторы модулируют биологические
ритмы.
ЗАКЛЮЧЕНИЕ
Воздействия внешней среды (от которой организм настолько
неотделим, что И. М. Сеченов включал, как известно, понятие
среды, в понятие организма) могут возникать непредсказуемо
(стрессорные факторы) или совершенно закономерно, согласно
определенному стереотипу, обусловленному ритмичностью явле-
ний природы. Естественно, между этими двумя крайностями име-
ется целый ряд промежуточных видов воздействий, однако для
того чтобы охватить реакциями весь их диапазон, живым системам
было прежде всего необходимо развить механизмы ответа на эти
два основных вида.
Для того чтобы оптимальным образом отреагировать на не-
предсказуемое воздействие, в организме должна быть развита
способность быстро найти среди имеющегося диапазона степеней
выраженности функций те значения, которые наиболее адекватны
данной ситуации. Но для этого должен существовать и сам диапа-
зон, т. е. значения функций должны колебаться, и чем больше
размах колебаний, тем более широким «выбором» обладает орга-
низм. Чтобы оптимальным образом отреагировать на закономерно
наступающие изменения внешней среды, надо также обладать
широким «выбором» значений функций, но включать регулирую-
щие механизмы, позволяющие достичь этих значений, не в мо-
мент воздействий, а предварительно, «упреждая» воздействие.
Обеспечению этих двух путей реакции и служат биологические
ритмы. Существование циркадных, ультрадных и инфрадных ко-
лебаний создает в организме возможность «быть готовым» встре-
тить как предсказуемое, так и непредсказуемое воздействие. Меха-
низмы регулирования взаимодействий со средой полностью ос-
новываются на существующих эндогенных ритмических колеба-
ниях функций. Употребляя выражение «эндогенный», мы имеем
в виду, что эндогения развилась в далеком прошлом как следст-
вие закрепленных повторных временных воздействий среды,
в частности, и в первую очередь, воздействий геофизических фак-
торов (Кардашева, 1973).
Следует подчеркнуть, что наследуются только генетически
обусловленные возможности. Для их реализации каждый орга-
низм должен строить свою личную систему временных отношений,
103
формировать соответственные биоритмы, отличающиеся большей
или меньшей амплитудой колебаний, более или менее фиксирован*
ной структурой суточной кривой, большей или меньшей способ-
ностью к перестройкам.
Такого рода представления находят подтверждение во взгля-
дах рядаЗисследователей на формирование функций организма
в онтогенезе. Так, даже образующийся в процессе индивидуаль-
ного развития тип высшей нервной деятельности представляет
собой, по И. П. Павлову (1949), некий «сплав» из врожденного и
приобретенного в процессе взаимодействия с условиями среды.
Уильямс (Williams, 1960) считает, что организм наследует лишь
способность определенным образом реагировать на ряд факто-
ров внешней среды, а непосредственно характеризующие данный
организм свойства есть результат взаимодействия наследствен-
ности и внешних воздействий. При этом, как показано И. А. Арша-
вским (1979), даже «в момент существования организма в виде
зиготы декодирование генетической информации невозможно
без поступления информации из среды» (с. 259).
Построение собственной временной структуры подразумевает
и формирование механизма внутреннего отсчета временной дли-
тельности, формирование собственного эталона времени, назы-
ваемого нами индивидуальной минутой. Как показано в гл. 3,
средние значения длительности индивидуальной минуты оказа-
лись довольно постоянным показателем, свойственным каждому
конкретному лицу; при этом они выше у здоровых и хорошо адап-
тирующихся лиц, нежели у больных и плохо адаптирующихся.
Среди больных значения индивидуальной минуты выше у лиц
с хорошим прогнозом (около 60 с) по сравнению с лицами с пло-
хим прогнозом (не достигает 50 с).
Сравнение результатов повторных исследований показало,
что они меняются не только циркадно, но и в связи с общим со-
стоянием и самочувствием. Длительность индивидуальной минуты
уменьшается при ухудшении самочувствия и растет при его улуч-
шении. Кроме того, повышение интенсивности внешних воздей-
ствий и собственной деятельности влечет за собой увеличение дли-
тельности индивидуальной минуты, как это видно, например,
из наблюдений во время перелетов. В момент ожидания на аэро-
дроме длительность индивидуальной минуты наименьшая, при на-
боре высоты — наибольшая.
Индивидуальное восприятие временных интервалов прямо
зависит от содержания сознания и эмоционального отношения
к событиям, что хорошо известно и прекрасно сформулировано
в стихах С. Маршака:
Я знаю — время растяжимо,
Оно зависит от того,
Какого рода содержимым
Вы наполняете его.
104
Наличие такой «скользящей шкалы», по-видимому, объяс-
няется особенностями течения времени в организме в условиях
больших информационных и эмоциональных нагрузок. Когда
организм поставлен перед реальной необходимостью обработать
такой объем информации, который обработать за данное время
просто нельзя, внутреннее время организма течет замедленно.
Этот факт, так же, как и факт индивидуального убыстрения тече-
ния времени, многократно показан в исследованиях поведения
человека в условиях стресса. Выражения Мак-Каллоха (McCul-
loch, 1969) «если надо спешить — время идет медленнее для нас»,
и Томаса Манна «Из-за одного спешного дела непозволительно
отложить другое, их следует складывать вместе во времени»
(«Иосиф и его братья». М., 1968, ч. II, с. 497) по сути очень точно
отражают существующие отношения.
Вместе с тем в случае отсутствия внешних впечатлений время
ужасно тянется, ощущается как нечто бесконечное, а дни и не-
дели мелькают как серые мыши. Французский спелеолог Мишель
Сифр, находившийся один в пещере на дне пропасти Скарассон
около 2 мес, отмечал потерю чувства времени. Ему то казалось,
что время летит, то появлялось впечатление замедления и даже
полной остановки времени. В оценке протекшего времени
Сифр ошибся на 25 сут и категорически отказывался прек-
ратить эксперимент и подняться на поверхность (прошло
58 сут 17 ч).
Яркий художественный образ особенностей восприятия вре-
мени дает Томас Манн в романе «Волшебная гора» (Л., 1934, ч. 1,
с. 131—132): «В чем, однако, первопричина той вялости и притуп-
ленности, которые неизбежны при длительном однообразии
жизни? Не столько в физической и умственной усталости . . .
сколько в чем-то психическом, в переживании времени как тако-
вого; оно, это время, грозит, при непрерывном однообразии, вовсе
утратиться. . . Пустота и монотонность способны, правда, придать
мгновению или часу большую растянутость, сделать его скуч-
ным, но они же укорачивают и ускоряют ход больших и очень
больших масс времени вплоть до полного их уничтожения. И об-
ратно, богатое и интересное содержание способно, конечно, уко-
ротить один час или даже день и ускорить их ход, но в крупных
масштабах это же содержание сообщает времени пространствен-
ность, тяжеловесность и значительность, так что годы, богатые
событиями, протекают гораздо медленнее, чем скудные, пустые,
легковесные, разлетающие в вихре ветра. Таким образом, то, что
называется скукою, является скорее по существу болезненным
восприятием быстротечности времени, в результате монотонности:
огромные массы времени, при непрерывном однообразии, прет-
воряются убийственно устрашающим образом в пустоту. . . Когда
один день, как и все другие, т. е. все дни как один; и при совер-
шеннейшем однообразии самая долгая жизнь прожита была бы
как короткая и пролетела бы незаметно».
105
У наших испытуемых длительность индивидуальной минуты
тем больше, чем сложнее были условия существования, чем выше
эмоциональный накал и чем лучше они справлялись с текущей
работой. Эти данные совпадают с результатами исследования
М. В. Ермолаевой и М. В. Корецкого (1980), показавшими, что
у спортсмена после удачного выступления регистрируется пере-
оценка временного интервала на 5—7%. После поединков фехто-
вальщиков (всегда сопровождающихся некоторой недооценкой
интервала времени) у проигравших эта недооценка выраженнее
(9—12%), чем у выигравших (до 5%).
В случае значительных нарушений эмоциональной сферы,
заканчивающихся суицидной попыткой, обнаружено резкое со-
кращение длительности индивидуальной минуты, которая колеб-
лется от 22 до 37 с (Самохина, 1980).
Наконец, оказалось, что точность воспроизведения отрезков
времени прямо зависит от уровня бодрствования. Как видно
из сравнения показателей уровня бодрствования наших испытуе-
мых и воспроизведения ими длительности отрезка времени, наи-
более точно воспроизводится длительность в 13—15 и 23 ч, т. е.
тогда, когда уровень бодрствования снижается (Ashoff et al., 1969;
Webb, Agnew, 1969; Демин и др., 1978). По всей вероятности,
это зависит от описанных изменений масштаба «личного времени»,
связанных с насыщением сознания информацией в моменты вы-
сокого уровня бодрствования и соответственно с ошибкой в опре-
делении длительности отрезка в большую сторону.
Восприятию длительности интервалов времени в состоянии сна
возможно дать лишь опосредованную оценку, которая зависит
от того, длительность какой из стадий сна оценивается. В стадиях
так называемого «медленноволнового» (ортодоксального) сна время
как бы компрессируется — ночь проходит как одно мгновение.
В стадии «быстрого» (парадоксального) сна, сопровождаемого
сновидениями, время течет различно в зависимости от структуры
сновидения. При сновидениях строго логических оно течет так же,
как в состоянии бодрствования, при сложных многоплановых сно-
видениях за небольшой отрезок времени происходит огромное
количество событий — время как бы «растягивается», иногда
в десятки и даже сотни раз (Моисеева, 1975а, 19756, 1975г; Демин
и др., 1978).
Таким образом, в условиях возрастания объема поступающей
информации (при этом безразлично, поступает ли информация
извне, как это происходит в состоянии бодрствования, или про-
дуцируется самим мозгом, как при сновидениях) длительность
индивидуальной минуты увеличивается,.в условиях ограничения
поступления информации — уменьшается. Создается ощущение
«исчезновения времени», как это отмечали не только Сифр, но и
Марете, проведшие по нескольку месяцев в пещерах в условиях
полной звуковой изоляции и двигательной депривации (Siffre,
1966; Fraisse et al., 1968), или как это имеет место в обычном сне.
106
Не касаясь психических расстройств и таких исключительных
состояний, как пребывание в пещерах, или иных видов сенсорной
депривации, следует подчеркнуть, что экстремальные значения
длительности индивидуальной минуты (т. е. очень короткие или
очень длительные) сохраняются в течение конечных интервалов
времени, пока держится высокий (или крайне низкий, как в мед-
ленноволновом сне) уровень информационного потока. При этом
«крайние» состояния (такие, как значительное замедление тече-
ния времени в сновидениях) как бы уравновешиваются ускорением
(компрессией) течения времени в медленноволновом сне.
Такого рода уравновешивание может быть понято с точки
зрения соотношения между временем и энергией, которые, сог-
ласно некоторым исследователям (Weinberg, 1977), представляют
собой сопряженные явления в квантовомеханическом и других
смыслах этого слова. Одномоментное течение большого числа про-
цессов (вызывающее «расширение» индивидуального времени)
может проходить только кратковременно, и скорость каждого
процесса должна быть велика. Медленно идущие процессы, на под-
держание которых требуется большой запас энергии, не дол-
жны быть многочисленны.
Таким образом, в различных условиях индивидуальное время
становится то более, то менее «емким». Для одного и того же чело-
века один и тот же отрезок «физического времени» вмещает много
событий, то быстро текущих, то текущих медленно (может быть,
тех же самых). Как будто бы время, подобно резиновому кругу,
растягивается по ширине (термин «ширина настоящего» заимство-
ван из романа «Бойня № 5» Курта Воннегута), и тогда «длина»
резко сокращается (т. е. сокращается время совершения процес-
сов — растет их скорость). Появляется возможность одномомент-
ного осуществления целого ряда действий. По-видимому, емкость
времени увеличивается в процессе эволюционного развития,
а у отдельных индивидуумов — в процессе тренировки трудовых
навыков. Особенно резко она растет в момент стресса, когда орга-
низм мгновенно перестраивается, а человек может осуществить
в невероятно короткий срок и принятие решений, и проведение
сложной системы действий; т. е. время как бы растягивается, как
это бывает, например, при аварийных ситуациях с самолетами
или мчащимися автомашинами, когда пилот или шофер имеют
в своем распоряжении 30—40 с на принятие и претворение в жизнь
единственно верного решения, предотвращающего гибель (Зава-
лова, 1969; McCulloch, 1969).
В подобных ситуациях (если они завершились благополучно
и человек способен дать отчет в своих ощущениях) длительность
протекшего времени значительно переоценивается. Эта пере-
оценка, по-видимому, является одним из факторов, обеспечившим
осуществление необходимости объема деятельности. В других
случаях такая переоценка длительности прошедшего отрезка
времени становится фактором, деятельности препятствующим.
107
Так, например, некоторые, даже очень опытные парашютисты не
в состоянии провести затяжной прыжок. Они открывают парашют
немедленно, считая, что уже прошло положенное для затяжного
прыжка время (Леонов, Лебедев, 1968).
Так же широко известно наличие у некоторых людей феноме-
нальных счетных способностей, когда в уме производятся опера-
ции над многозначными числами. Н. П. Бехтерева (1974) так оце-
нивает возможности к подобному феноменальному счету: «В том
случае, если это никак не связано с умелым шарлатанством, это
действительно поддается одному и только одному объяснению —
счет осуществляется по обычным законам математических опера-
ций, но в другой системе времени» (с. 59).
Такого рода система может быть связана тоже с изменением
емкости времени, вследствие чего за абсолютную (физическую)
единицу времени организм успевает осуществить большее коли-
чество циклов процессов, т. е. течение времени субъективно за-
медляется.
Наконец, изредка человек способен охватить сущность явле-
ний в их развитии, когда его сознание фиксировано только на не-
скольких или на одном-единственном явлении. Все остальное
как бы исчезает, но зато это единственное явление воспринима-
ется во всей совокупности причинно-следственных связей как бы
существующим и в прошедшем, и в настоящем, и в будущем.
Время обретает «глубину» в миг порыва и озарения, взлета и от-
решенности. Именно эта сторона времени, эта особенность его
восприятия описывается Гете в «Фаусте» и Томасом Манном в «Док-
торе Фаустусе»: «Просто-напросто время? Нет, дорогой, этим
черт не торгует. За это нам не платили бы властью над концом.
Какого сорта время — вот в чем вопрос! Великое время, сумас-
шедшее время, совершенно чертовское время со взлетами и сверх-
взлетами, — конечно, и не без жалких падений. . .» («Доктор
Фаустус». М., 1959, с. 280).
Наши представления о трехмерности времени созвучны взгля-
дам Э. Канта (1915), который говорил о «совокупности» времени
и трех его «модусах» — последовательности, сосуществования и
устойчивости, т. е. рассмотрел время как последовательный ряд
значений, как совокупность всего существующего в данный мо-
мент и как величину. В такого рода описании «модусов» проскаль-
зывает отношение ко времени как к двух- или трехмерному явле-
нию. Во всяком случае, последовательность времени дает обычное
представление о линейном порядке течения процессов (бесконеч-
ная «стрела» времени, ассоциирующаяся с понятием «длины»).
«Модус» сосуществования — набор одномоментно происходящих
событий, который для конкретного живого организма является
конечным и, кроме того, переменным (поскольку существует из-
вестная последовательность развития жизненных процессов).
Пользуясь привычными геометрическими терминами, его можно
было бы назвать «шириной» времени.
108
Наконец, устойчивость (величину) времени можно представить
для живой системы как продуктивность каждого мгновения, как
значимость его в общем балансе времени или (пользуясь опять-таки
геометрическими представлениями) как его глубину (высоту).
При такой трактовке трехмерному замкнутому пространству кон-
кретной живой системы соответствует «трехмерное время», по всей
вероятности также в некотором смысле замкнутое. Предложен-
ная (Моисеева, 1980а) трехмерная модель времени проверялась
нами в первую очередь в исследованиях, где сопоставлялись
временные и пространственные характеристики биопотенциалов
головного мозга (здоровых людей и больных, леченных методом
вживленных электродов) и оценка испытуемыми длительности
событий в сновидениях. Показано, что в тех случаях, когда про-
исходила значительная переоценка длительности сновидения
(за несколько секунд в сновидении произошли события, которые,
по представлению испытуемого, заняли десятки минут), деятель-
ность структур головного мозга протекала независимо, т. е. струк-
туры действовали параллельно, а не последовательно. Увели-
чивалась «ширина» времени с сокращением «длины» (Моисеева,
19756, 1975г).
Кроме того, нами (Моисеева, 19796) были проанализированы
словесные описания случаев принятия решений в условиях ре-
альной опасности и обнаружено, что для этого, по-видимому, не-
обходима определенная психологическая структура, в частности
отставленная реакция страха. В условиях, когда, с одной стороны,
возможности мозга активированы за счет чувства тревоги и уси-
ленного мотивированного стремления найти решение, а, с другой
стороны, конечная цель четко сформулирована, т. е. создана мо-
дель идеального результата, включается механизм, обеспечиваю-
щий мгновенность решения. По всей вероятности, этот механизм
сводится не просто к одновременному рассмотрению вариантов
в настоящем (как это имеет место в сложных сновидениях, когда
человек, например, присутствует одновременно в двух или более
местах), а к одномоментному рассмотрению вариантов сразу
со всеми их возможными последствиями, т. е. в настоящем и буду-
щем. При этом в сознание не поступают ни непригодные варианты,
ни ход их рассмотрения — фиксируется только тот единственный
вариант, который совпал с моделью идеального результата.
Что касается механизма того удовлетворения, которое полу-
чает человек, если принимает удачное решение в стрессовой ситу-
ации, в качестве одного из возможных объяснений можно пред-
ложить следующее. Значительное возрастание уровня органи-
зации живой материи идет за счет возникновения более слож-
ных систем поведения и управления, сочетающихся с появлением
дополнительных степеней свободы только в очень неравновесных
системах, находящихся на грани своих возможностей существо-
вания (Kunicki'Goldfinger, 1978). Эти закономерности, кстати,
имеют место и во внутриутробном формировании человека. Как
109
показано И. А. Аршавским (1979), удельная величина плацен-
тарной поверхности, обеспечивающей поступление кислорода
к плоду, в четыре с лишним раза меньше удельной величины пло-
щади легких взрослого человека. Плод находится в состоянии
физиологической гипоксемии, которая в момент периодического
усиления стимулирует его подвижность. Двигательная реакция
индуцирует избыточный анаболизм, необходимый для роста и
развития.
Во время родов — момента перехода ребенка к самостоятель-
ному существованию — организм переживает тяжелейший стресс
и реально находится между жизнью и смертью, и даже акт рожде-
ния рассматривается сперва как смерть (при переходе из чрева
матери во внешнюю среду), а с первым вздохом — как начало
новой жизни (Barcroft, Barron, 1935). Таким образом, для того
чтобы окончательно сформировалось живое существо со многими
уровнями управления, организм должен пройти через барьеры
естественного отбора, оставляющего в живых только тех, кто спо-
собен на грани жизни и смерти сформировать соответствующие
более высоко организованные системы, например систему авто-
номного дыхания.
«Платой» природы за преодоление больших трудностей явля-
ется не только оставление в живых, но и повышение уровня орга-
низации.
Мозг человека в процессе эволюции сформирован как механизм
для создания сложных функциональных систем, системообразу-
ющим фактором которых, согласно П. К. Анохину (1980), яв-
ляется целевая функция. Такое системообразование идет про-
торенными рефлекторными путями, а по достижении цели функ-
циональная система распадается. В каких-то случаях человек
попадает в особо сложную ситуацию — это те случаи, когда имеет
место острый дефицит времени и информации. Последнее, со-
гласно П. В. Симонову (1965), включает интенсивные эмоциональ-
ные реакции, представляющие собой компенсаторный механизм,
восполняющий дефицит необходимой для достижения цели ин-
формации. Возникает доминанта, подавляющая все другие виды
деятельности организма, в данный момент второстепенные, —
все силы направлены на удовлетворение одной-единственной по-
требности. Так, ярость компенсирует недостаток сведений, не-
обходимых для организации борьбы. И чем больше расстояние
между исходным состоянием неинформированное™, сопровожда-
ющейся ожиданием самого худшего и достигнутым положитель-
ным результатом, тем сильнее торжество победившего в данной
ситуации человека.
Эта победа над событиями может осуществляться или за счет
обычных механизмов принятия решения, или в случаях, когда
дефицит времени и информации имеет место в ситуациях экстре-
мальных, за счет формирования функциональной системы более
высокого уровня. Новизна способа решения (не обязательно
ПО
принципиально нового само по себе) является той высокой награ-
дой, которую получает организм, превозмогший ограниченность
собственных возможностей, и, вероятно, именно это кратковремен-
ное состояние и называется ощущением полноты бытия.
Возвращаясь к оценке собственного материала, мы должны
сказать, что акрофаза длительности индивидуальной минуты
весьма вариабельна. У каких-то испытуемых она отмечалась
в 8 и 12 ч дня, у других — в 20 и даже в 24 ч; каждый раз имеет
место индивидуальная организация суточной кривой.
Хотя среди наших испытуемых «жаворонков» или «сов» в чи-
стом виде не обнаруживалось (все они были достаточно работо-
способны в разное время суток), все же лица, у которых максимум
по этому показателю падал на вечерние часы, чаще оказывались
работающими в вечернюю смену. По-видимому, тенденция к «удли-
нению» минуты в то или иное время суток относится к числу
индивидуальных циркадных характеристик, закладывающихся
очень рано и зависящих, по мнению некоторых исследователей
(Wendt, 1977), от самых различных факторов, в том числе от пола
и от того сезона года, когда человек родился.
При проведении психологических проб объем выполненного
задания, скорость и качество выполнения пробы зависят от дли-
тельности индивидуальной минуты: чем она больше, тем большую
работу и более точно успевает выполнить человек. Эта зависимость
(как показали расчеты по методу Чебышева) нелинейна. Структура
суточных кривых времени выполнения тестов и точности их
выполнения выглядит, как правило, в виде обратного изображе-
ния по отношению к кривой длительности индивидуальной минуты
(что вполне естественно, поскольку чем минута длиннее, тем меньше
времени тратится па выполнение теста и тем меньше в нем
ошибок).
Поскольку длительность индивидуальной минуты значительно
зависит от концентрации событий и уровня деятельности человека,
рисунок суточных кривых изменений этой длительности и суточ-
ных кривых параметров выполнения психологических проб весьма
вариабелен, что является физиологической нормой. Эти кривые
отражают по сути реакцию на случайные воздействия внешней
среды, и создается впечатление, что их ход не зависит от климато-
географических факторов и профессии.
На структуру суточных кривых вегетативных показателей
(деятельности сердечно-сосудистой системы и температуры тела)
отчетливо влияют следующие факторы: 1) климато-географи-
ческие зоны обитания; 2) сезоны года; 3) степень интенсивности
физических нагрузок; 4) профессии (особенно связанные со сдви-
нутыми режимами).
Для больных к этому списку необходимо еще добавить особен-
ности патогенеза и топики заболевания, лечебные воздействия.
В целом вегетативные показатели менее отчетливо меняются
под влиянием случайных явлений, чем показатели высшей нервной
111
деятельности, поэтому рисунок кривых изменения параметров
сердечно-сосудистой системы каждого отдельного человека хотя
индивидуален, но более постоянен, чем рисунок кривых изменения
параметров, отражающих психическую деятельность. Но зато они
оказываются весьма инертными при необходимости быстрой пере-
стройки (после пересечения часовых поясов). Так, например,
в обычных условиях систолическое давление минимально среди
ночи (7—6 ч) и имеет два максимума — днем в 10—15 ч (больший)
и перед сном в 22—24 ч. Диастолическое давление меняется ана-
логично, но динамика его менее выражена. При перелетах с за-
пада на восток с пересечением девяти часовых поясов рисунок
кривых резко нарушается и приходит в соответствие с местным
временем в течение месяца и более; при перелете на запад (что
для наших испытуемых было возвращением в привычную среду)
изменения сглаживаются за 10 дней—2 нед.
Возвращение в привычную обстановку характеризуется пере-
стройками не только более быстрыми, но и такими, которые, по-
видимому, происходят более «экономным» для организма образом.
Это следует из сравнения средних и разброса показателей дея-
тельности сердечно-сосудистой системы в фоне и при перелетах
в новую среду и возврате в среду привычную. В последнем слу-
чае падает величина разброса показателей артериального давле-
ния, что указывает на уменьшение зоны «поиска» оптимальных
значений физиологического параметра.
Инертность вегетативных показателей отражается в динамике
изменений суточных кривых у больных. К моменту установления
диагноза обычно имеются значительные нарушения их структуры,
что позволяет в известной мере устанавливать прогноз течения
заболевания. Чем более нарушена структура суточной кривой и
чем больше разброс значений акрофаз в разных исследованиях,
тем прогноз хуже. При этом сами средние значения физиологи-
ческих параметров и их разброс оказываются для прогноза мало-
информативными в отличие от их суточной динамики.
При улучшении общего состояния больного суточные кривые
становятся более организованными, но в силу большой инерцион-
ности сдвиги в их структуре происходят очень медленно. Изме-
нения суточных кривых оказываются одним из наиболее стой-
ких симптомов нарушений той или иной функциональной си-
стемы.
В известной мере высокие функциональные возможности орга-
низма (хорошая работоспособность и адаптоспособность у здоро-
вых, хорошая сопротивляемость болезнетворному агенту у боль-
ных) определяются: 1) четкой индивидуальной временной органи-
зацией суточных кривых; 2) относительным постоянством поло-
жения акрофаз на оси временп в повторных исследованиях;
3) большой (в нормальных пределах) величиной разброса значе-
ний в течение суток; 4) относительно высоким средним уровнем
(находящимся в пределах нормы), что обеспечивает наличие вы-
112
сокого разброса показателей (Моисеева, 1978а, 19786; Моисе-
ева и др., 1978а).
К числу факторов, требующих постоянной адаптации орга-
низма, относятся геогелиофизические воздействия — смена дня
и ночи, суточная смена температуры и давления и т. д. В этом
случае появление воздействующего сигнала строго предсказуемо,
возникает «опережающая» реакция, и, являясь повседневной, она
все же идет, по-видимому, со значительными затратами усилий
организма.
Как показал опыт применения биотрона — специальной ка-
меры, где поддерживаются постоянные условия атмосферного
давления, температуры, радиации и т. п., — избавление орга-
низма от необходимости производить работу по уравновешиванию
со средой создает дополнительные возможности по саморегулиро-
ванию, и у ряда больных наступает стойкий клинический эффект
без применения каких-либо лечебных средств (Панченко и др.,
1964; Панченко, 1972). Но пребывание в биотроне полезно только
относительно кратковременно (лечебный курс — 2 нед). Для под-
держания биоритмической структуры функции гелиогеофизиче-
ские воздействия необходимы.
Как показали наши исследования, проведенные на двух груп-
пах больных в разные сезоны разных лет, колебания составляю-
щих постоянного магнитного поля Земли оказывают влияние на
все исследованные физиологические параметры. Эти зависимости
носят нелинейный характер, и воздействие колебаний магнитного
поля сводится к попеременному усилению и ослаблению той или
иной функции, т. е. поддерживается колебательный контур —
общий «ритмический фон».
Иной характер имеет влияние солнечной активности (хромо-
сферных вспышек) и погодных факторов (температуры, влажности
и атмосферного давления). Следует подчеркнуть, что указанные
погодные факторы находятся в непосредственной зависимости
от той же солнечной активности.
Физиологические параметры связаны с погодными факторами
и солнечной активностью чаще простыми линейными отношени-
ями, т. е. с увеличением какого-то фактора погоды (например, тем-
пературы воздуха) происходит увеличение значений физиологи-
ческих показателей человека (например, мышечной силы рук,
артериального давления, частоты дыхания) или их уменьшение
(например, длительности индивидуальной минуты).
В некоторых случаях (зависимости температуры тела от внеш-
ней температуры, зависимости температуры тела и частоты ды-
хания от значений амтосферного давления) погодные факторы
вызывают попеременную реакцию усиления и ослабления, т. е.
поддерживают колебательное состояние значения функций по-
добно магнитному полю Земли. Таким образом, для организма
с уже сформированной биоритмической системой внешние воз-
действия играют роль «датчиков времени», поддерживая общий
ИЗ
уровень колебаний (как это можно видеть на действии параметров
магнитного поля и некоторых погодных факторов), регулируя
период (изменение ритмики в различные сезоны года в связи с из-
менением освещенности и других факторов) и амплитуду колеба-
ний (влияние атмосферного давления, влажности, температуры
вспышек на Солнце).
Известное значение для формирования ритмов имеет модуля-
ция ритмами с большим периодом собственных ритмов организма
(с меньшими периодами). В гл. 5 показано, что ритмика медленно
текущих (скорее всего обменных) физиологических процессов
в организме, находящих свое выражение в периодических колеба-
ниях физической, эмоциональной и интеллектуальной активности,
оказывает модулирующее влияние на течение процесса сна. Это
влияние выражается, например, в увеличении длительности сна
в переходном состоянии от отрицательной к положительной фазе
макроритма физической активности, в увеличении среднего числа
смен стадий сна REM во время фазы спада эмоциональной актив-
ности и т. д. Показано также, что оценка человеком окружающих
его людей и те оценки, которые он от окружающих сам получает,
в известной мере зависят от фазы его макроритмов в день исследо-
вания. Но такого рода зависимость отчетлива только в коллекти-
вах, мало связанных трудовой деятельностью. Чем сложнее вза-
имоотношения в коллективе и чем существеннее влияет оценка
на организацию совместной деятельности, тем меньше на нее
оказывают влияние биологические ритмы.
Следует отметить, что у человека к числу факторов, модули-
рующих биологические ритмы, относятся и так называемые
«социальные датчики времени». В условиях необходимости введе-
ния ритмов труда и отдыха, отличных от физиологических, чело-
век прекрасно к ним адаптируется, если у него высокая социаль-
ная мотивация.
Сама способность к адаптации так же точно тренируема, как
и способность адаптироваться к конкретным воздействиям.
Л. А. Орбели (1961) подчеркивает, что нужно тренировать не от-
дельную функцию, а «тренируемость». «Существенно важно, —
говорит он, — на ранних этапах развития, когда еще не вполне
фиксировались отношения, когда не выработалось никаких тра-
фаретов, бороться с выработкой узких и ненужных трафаретов
и создавать такие условия, при которых по возможности много-
гранно развивался бы организм и использовал все свои врожден-
ные способности для того, чтобы обладать максимальной пластич-
ностью нервной и мышечной систем» (с. 258). Такого рода пластич-
ность, в число составляющих элементов которой входят и биоло-
гические ритмы, весьма существенна в условиях, когда организм
должен быстро отреагировать на непредсказуемое событие, т. е.
действовать в состоянии эмоционального стресса.
В условиях жестокой борьбы за существование выживали,
по-видимому, организмы, способные ответить в кратчайший срок
114
изменениями деятельности определенного набора функций в опре-
деленном направлении.
Именно такая картина перестроек в стрессорной обстановке
спортивного поединка фехтовальщиков представлена в гл. 7.
Выигрывали схватку те спортсмены, которые, во-первых, об-
ладали широким спектром медленноволновых компонент кардио-
интервалов и большими отклонениями их значения от среднего
(т. е. широким диапазоном состояний). Во-вторых, в момент по-
единка у них осуществлялся выбор узкого диапазона кардио-
интервалов, и этот диапазон фиксировался на весь период дей-
ствия. Возможно, этот диапазон не был абсолютно оптимальным;
в другом поединке у того же спортсмена «выбор» из более широкого
спектра кардиоинтервалов в поисках наилучшего мог происхо-
дить дольше, но тогда спортсмен проигрывал. По-видимому,
здесь проявляется закономерность, отмеченная Кастлером
(Quastler, 1960): живой организм стремится минимизировать
ошибки, а не полностью их ликвидировать, ибо последнее потре-
бует слишком сложной и громоздкой системы контроля. В момент
успешного поединка должен быть осуществлен быстрый выбор
«тактики» (имеется в виду «тактика» реагирования сердечно-
сосудистой системы); до конца поединка она не должна меняться.
Как только надобность в таком самоограничении для системы ми-
новала, у хорошо адаптирующихся лиц усиливаются ритмиче-
ские компоненты, увеличивается разброс значений — из режима
«целенаправленного» действия организм переходит в «следящий»
режим, позволяющий приспосабливаться и к слабым внешним
воздействиям.
К числу промежуточных картин можно отнести перестройки,
возникающие при внезапном изменении характера внешних воз-
действий (в том числе «временных меток», что вызывает необхо-
димость регулирования практически всех биологических ритмов
организма) и текущие длительно. В этих случаях регуляция идет
совсем по другим путям. В отличие от поединка, когда желаемый
эффект (победа) может быть обеспечен только за счет кратко-
временной мобилизации физических, эмоциональных и интеллек-
туальных возможностей, при перелетах с пересечением часовых
поясов целью регулирования является приведение каждой функ-
ции в соответствие с требованиями внешней среды. Как показано
в гл. 6, на примере регуляции ритма сна вместо фиксирования
очень узкого диапазона колебаний происходит фиксация всего
диапазона частот ЭЭГ и резкое увеличение размаха колебаний
внутри этого диапазона при переходе от одной частоты к другой
(в виде смены фаз сна). Происходит как бы постоянное «отслежи-
вание» всех возможностей, и все процессы текут в более быст-
ром темпе, чем обычно. При очень больших рассогласованиях
времени и места вылета и времени и места прилета, что вызывает
значительные фазовые сдвиги биологических ритмов, потребная
длительность сна уменьшается.
115
Это, по-видимому, объясняется особенностями течения времени
в организме в условиях больших информационных и эмоциональ-
ных нагрузок, о чем подробно говорилось выше. Когда организм
поставлен перед реальной необходимостью обработать такой объем
информации, который обработать за данное время просто нельзя,
внутреннее время организма течет замедленно («растягивается»).
Таким образом, непосредственная реакция на воздействие
определенных факторов, направленная на выполнение конкрет-
ной узкой задачи, сводится к очень быстрой строгой фиксации
условий деятельности. Реакция на необходимость полной пере-
стройки взаимоотношений со средой сводится *к увеличению раз-
маха колебаний в самом широком диапазоне возможных состо-
яний.
Наконец, существует еще одна постоянная форма воздействия
на организм. В течение дня на него действует большое количество
рутинных раздражений, в том числе, может быть, и стрессорного
характера. К моменту отхода ко сну в результате реакции на эти
факторы имеются фазовые рассогласования между различными
биоритмами организма, а в ряде случаев и между «метками вре-
мени» внешней среды и фазой биоритмов. Эти рассогласования
невелики и тем выше для организма, чем большее число воздей-
ствий имело место в течение всего дня. Согласно нашей гипотезе,
регуляция взаимоотношений биологических ритмов происходит
во время сна (Демин и др., 1978; Моисеева 1979а). При этом,
как показано в гл. 6, общая длительность сна возрастает после
событий, являющихся стрессорными, т. е. вызвавших большие
перестройки, нуждающиеся в достаточном времени перерегули-
рования. Кроме того, растет устойчивость стадий сна — рисунок
сна как бы больше фиксируется. Идет регулировка согласно стан-
дартным образцам, а не подстройка к новым условиям.
Описанные общие закономерности реализуются для каждого
организма в его собственной индивидуальной «шкале времени»,
в индивидуальном рисунке временной организации суточной кри-
вой биоритмов и индивидуальном диапазоне их колебаний. Орга-
низм с хорошо сформированными биологическими ритмами об-
ладает всем диапазоном возможных (и необходимых в каких-то
условиях) значений функций, позволяющих ему оказаться
не просто сильнее «противника» (реального врага или внешней
среды), но сильнее в нужное время.
Что касается наиболее благоприятных условий для формиро-
вания биоритмической структуры организма, которая происхо-
дит после рождения и длится достаточно долго (считается, напри-
мер, что ритм частоты дыхания и сердечный ритм начинают
формироваться после седьмого дня от момента рождения (Оку-
нева и др., 1978), а температура тела формируется после 7 лет
(Abe et al., 1978)), то наряду с четким соблюдением ритма жизни
маленькие дети время от времени должны иметь и резкие его нару-
шения.
116
Анализируя анамнез жизни хорошо адаптирующихся лиц
с четко организованными биоритмами, мы обнаружили в боль-
шинстве случаев, что они в детстве (до 12 лет) часто переезжали
или жили не в очень благоприятных условиях. Таким образом,
можно прийти к выводу, что способность адаптации должна тре-
нироваться с раннего детства. Четкая организация биологиче-
ских ритмов поддерживается соответствующими механизмами,
которые формируются и функционируют в условиях преодоле-
ния трудностей. Даже если высокий уровень организации зало-
жен генетически, он становится реальностью только в постоян-
ной борьбе. И это, по-видимому, закон для всего живого. Недаром
И. В. Мичурин (1948) писал: «Лесные растения живут вообще
вчетверо дольше, чем культурные сорта, выращиваемые в садах,
и, что всего интереснее, чем лучший применяется уход, тем короче
становится их жизнь» (с. 547).
ЛИТЕРАТУРА
Абдылдабеков Т. К., Туркменов М. Т. Суточный ритм легочного дыхания
у постоянных жителей низко- и высокогорья. — В кн.: Циркадные
ритмы человека и животных. Фрунзе. 1975а, с. 5—7.
Абдылдабеков Т. К., Туркменов М. Т. Суточный ритм гемодинамики у по-
стоянных жителей низко- и высокогорья. — В кн.: Циркадные ритмы
человека и животных. Фрунзе, 19756, с. 8—10.
Акрабов А. Бпоритмите и резултатпте в тренировки и състезания при щан-
гистите. — Въпросп на физпческата култура, 1969, № 9, с. 568—572.
Акрабов А. Исползване на бпорптъма на физическата работоспособност
за оптимизация на тренировочната и състезателната дейност на щан-
гистите. — Въпроси на физическата култура, 1972, № 5, с. 280—286.
Алякринский Б. С. Основы научной организации труда и отдыха космонав-
тов. М., 1975. 207 с.
Алякринский Б. С, Современное состояние космической биоритмологии. —
Космич. биол. и авиакосм, мед., 1977, т. И, Яг 2, с. 3—12.
Анохин П. К. Узловые вопросы теории функциональной системы. М., 1980.
198 с.
Аршавский И, А. К физиологическому анализу физического развития и его
особенностей у новорожденных детей. — Физиол. чел., 1979, т. 5,
№ 2, с. 254—269.
Баевский Р. М. Временная организация функций и адаптационно-приспосо-
бительная деятельность организма. — В кн.: Теоретические и при-
кладные аспекты анализа временной организации биосистем. М.,
1976, с. 83—111.
Барбашова 3. И. Исследование циклов человека в СССР участниками Между-
народной биологической программы. — J. Interdisc. Cycle Res.,
1972, vol. 3, N 2, p. 197—205.
Барбашова 3. И, Изучение биоритмов у человека. — В кн.: Ресурсы био-
сферы Л., 1976, с. 180—186. (Адаптация человека. Вып. 3).
Бауэр Э. С. Теоретическая биология. Л., 1935. 206 с.
Бехтерева Н. П. Нейрофизиологические аспекты психической деятельности
человека. 2-е изд. Л., 1974. 151 с.
Биоритмы и труд. Л., 1980. 141 с.
Василевский Н. Н. Адаптивная саморегуляция функций и ее связь с динами-
ческим управлением эндогенными ритмами. — ЖЭБиФ, 1973, т. 9,
№ 4, с. 374-382.
Василевский Н. Н., Сороко С. И., Богословский М. М. Психофизиологиче-
ские аспекты адаптации человека в Антарктиде. Л., 1978. 207 с.
Василевский Н. Н., Трубачев В. В. Системный анализ адаптивной саморегу-
ляции функций организма (экспериментальные и теоретические осно-
вания и перспективы). — В кн.: Адаптивная саморегуляция функций.
М., 1977, с. 11—19.
118
Василии П. В., Галицкий А. К. Влияние факторов внешней среды на ритм
роста дошкольников. — В кн.: Медицинская кибернетика. Киев,
1977, с. 29—32.
Владимирский В. М. Активные процессы на Солнце и биосфера. — Изв.
АН СССР. Сер. физ., 1977, т. 41, № 2, с. 403—410.
Влияние солнечной активности на атмосферу и биосферу Земли. М., Наука,
1971. 256 с.
Войно-Ясенецкий М. В. Первичные ритмы возбуждения в онтогенезе. Л.,
1974. 147 с.
Войтинский Е. Я., Прянишников В. А. Методы и системы для статистиче-
ского анализа электроэнцефалограмм. М., 1968. 87 с.
Газенко О. Г., Алякринский Б. С. Влияние космического полета на орга-
низм. — Вест. АН СССР, 1970, т. 15, № 11, с. 40—46.
Гингст В. П., Ярославцев В. Л. Внешнее дыхание и обмен веществ у гребцов
и велосипедистов в условиях переезда в места с многочасовой разницей
во времени. — В кн.: Сб. матер. 4-й науч. конф, физиол., биохим.,
фармакол. Западно-Сибирского объединения Всесоюз. физиол. общ.
им. И. П. Павлова. Красноярск, 1969, т. 1, с. 70—73.
Голиков А. П., Голиков П. П. Сезонные биоритмы в физиологии и патологии.
М., 1973. 167 с.
Гончаров В. Д. Из области комбинаторики. — Изв. АН СССР. Сер. мат.,
1944, № 1, с. 3—48.
Гончаров В. Д., Козлов Ю. Я. К вопросу о связи биологических макроритмов
и взаимооценок в коллективе студентов-спортсменов. — Теория
и практика физ. культуры, 1974, № 12, с. 56—59.
Гримак Л. П, Моделирование состояний человека в гипнозе. М., 1978. 272 с.
Гуровский Н. Н., Душков Б. М., Космолинский Ф. П. Изучение режимов
жизнедеятельности группы испытателей, находящихся в условиях
относительной изоляции. — Космпч. бпол. и мед., 1967, т. 1, № 2,
с. 54-59.
Дардымов Н. В. Женьшень, элеутерококк. М., 1976, 184 с.
Демин Н. И., Коган А. Б., Моисеева Н. И, Нейрофизиология и нейрохимия
сна. Л., 1978. 190 с.
Дубров А. П. Геомагнитное поле и жизнь. Л., 1974. 173 с.
Душков Б. А., Космолинский Ф. II. Оценка времени в условиях камерных
экспериментов. — Вопр. психол., 1968, № 6, с. 107—111.
Дьячков В. Я., Мошкин М. Ш., Маркель А. Л. Циркадные ритмы гемодина-
мики у людей с различной физической работоспособностью. — В кн.:
Адаптация и проблемы общей патологии. Новосибирск, 1974, т. 1,
с. 137—139.
Евцихевич А. В. Суточный ритм температуры тела при перелетах в широтном
направлении. — В кн.: Проблемы биоклиматологии п климатофизио-
логии. Новосибирск, 1970а, с. 244—248.
Евцихевич А. В. О суточном ритме частоты дыхания у здоровых людей в раз-
личных временных и климатических зонах при перелетах в широтном
направлении. — В кн.: Проблемы биоклиматологии и климатофизио-
логии. Новосибирск, 19706, с. 248—250.
Евцихевич А. В. Изменение циркадного ритма физиологических процессов
при перелетах в широтном направлении. — В кн.: Адаптация и проб-
лемы общей патологии. Новосибирск, 1974, т. 1, с. 141—145.
Емельянов И. П. Формы колебаний в биоритмологии. Новосибирск, 1976.
Емельянов И. П. Микромир электроэнцефалограммы. Новосибирск, 1978.
222 с.
Ермолаева М. В., Корецкий М. В. Воспроизведение временных интервалов
в условиях выполнения спортивного задания. — В кн.: Фактор вре-
мени в функциональной организации деятельности живых систем.
Л., 1980, с. 160—161.
[Завалова Н. Д.] Zavalova N. D. Decision-making models under stress. —
In: Biocybemetics of the central nervous system. Boston, 1969, p. 55—60.
119
Зинченко Т. П., Круминя С. К. Отражение времени и микрогенез зритель'
ного образа. — Вести. Ленингр. ун-та, 1978, № 23, с. 51—58.
Иванов-Муромский К. А., Заславский С. Я. Применение ЭВМ для анализа
электрограмм мозга. Киев, 1968. 144 с.
Каган М. А. Компонентный анализ как один из методов изучения факторов,
влияющих на течение процесса адаптации. — В кн.: Общие вопросы
экологической физиологии. Л., 1977, с. 16—17.
Каган М. А., Симонов М. Ю. Использование компонентного анализа в нейро-
физиологических исследованиях процесса адаптации. — Физиол. чел.,
1978, т. 4, № 2, с. 363-369.
Кант Э. Логика. Собр. соч. СПб., 1915, т. 3.
Кардашева А. С. Биологические ритмы в свете теории отражения. — В кн.:
Ленинская теория отражения и современные науки. Теория отражения
и естествознание. София, 1973, с. 253—279.
Каталевская Л. Я., Кузнецова С. С. Циркадные ритмы некоторых показателей
гемодинамики у здоровых людей в различные сезоны года. — В кн.:
Циркадные ритмы человека и животных. Фрунзе, 1975, с. 243—246.
Катинас Г. С., Моисеева Н. И. Биологические ритмы и их адаптационная
динамика. — В кн.: Экологическая физиология человека. Адаптация
человека к различным климато-географическим условиям. Л., 1980,
с. 468—515. (Сер. «Руководство по физиологии»).
Коган А. Б. Вероятностно-статистический принцип нейронной организации
функциональных систем мозга. — ДАН СССР, 1964, т. 154, № 5,
с. 1231—1234.
Козлов Ю. Я. Номограммная методика определения макробиоритмов. —
В кн.: Кибернетический подход к биологическим системам. М.; Л.,
1976, с. 52—56.
(Козлов Ю. Я., Сысуев В. М.) Kozlov Ju. J a., Sysuev V. М. The dependance
of the psychical state evaluation on biological macrorhythms phase. —
Abstracts of 2-nd Intern, congr. of collegium Internationale activitas
nervosae superiods. Praga, 1975, vol. 2, p. 475.
Колодченко В. П., Тихомирова Л. Д., Подшибякин А. К., Сиротский В. В.,
Шахова В. И. Элементы влияния геомагнитных возмущений на чело-
века и животных. — В кн.: Матер. 2-го Всесоюз. совещ. по изучению
влияния магнитных полей на биологический объект. М., 1969, с. 124—
127.
Комаров Ф. И., Захаров Л. В., Лисовский В. А. Суточный ритм физиологи-
ческих функций у здорового и больного человека. Л., 1966. 200 с.
Кохановский В. В. Влияние сезонов года на формирование суточного ритма
при некоторых заболеваниях сердечно-сосудистой системы. — В_кн.:
Биологические ритмы. Горький, 1970, с. 41—42.
Крыжановский Г. Я. Дистрофический процесс (некоторые аспекты проб-
лемы). — Арх. патол., 1974, т. 36, №л5, с. 3—12.
Кудрявцева В. И. Учет колебаний продолжительности сокращений ^сердца
при оценке функционального рабочего напряжения. — В кн.: Био-
ритмы и труд. Л., 1980, с. 54—57.
Ланг Г. Ф. Гипертоническая болезнь. Л., 1950. 496 с.
Леонов А. А., Лебедев В. И. Восприятие пространства и времени в космосе.
М., 1968. 116 с.
Ливанов М. И., Крылов Ю. В., Острякова М. В., Шульгина Г. И. О ритми-
ческих колебаниях потенциалов как одном из основных механизмов
интеграции нейронов. — В кн.: Механизмы объединения нейронов
в нервном центре. Л., 1974, с. 143—148.
Лицов А. Н. Экспериментальное изучение динамики некоторых физиологи-
ческих функций и работоспособности человека в условиях обычных
и измененных суточных режимов. — В кн.: Биологические ритмы
и вопросы разработки режимов труда и отдыха. М., 1967, с. 45—46.
Лицов А. Н. Суточная динамика некоторых физиологических функций и ра-
ботоспособности человека в условиях изоляции. — Космич. бпол.
и мед., 1968, т. 2, № 4, с. 83—86.
120
Ломоури А. И. К вопросу об адаптационной функции гемато-паренхима-
тозных барьеров. — В кн.: Адаптация и проблемы общей патологии.
Новосибирск, 1974, т. 2, с. 72—74.
Макаров Н. И., Гринберг Б. А., Орехов В. С.> Карасев М. Н, Гелиогеофизи-
ческие факторы и некоторые вопросы травматизма на Крайнем Се-
вере. — В кн.: Научно-технический прогресс и приполярная меди-
цина. Новосибирск, 1978, т. 1, с. 41.
Манина А. А., Моисеева Н. И. Изменения нейронов коры головного мозга
в процессе адаптации к временной среде. — В кн.: Адаптивная само-
регуляция функций. М., 1977, с. 67—77.
Матюхин В. А., Демин Д. В., Евцихевич А. В, Биоритмология перемещений
человека. Новосибирск, 1976. 104 с.
Мичурин И. В. Избранные сочинения. М., 1948. 792 с.
Моисеева Н. И. Некоторые методологические аспекты изучения понятия
времени в биологии. — В кн.: Методологические вопросы теоретиче-
ской медицины. Л., 1975а, с. 87—116.
Моисеева Н. И. Биоэлектрическая активность головного мозга человека
и субъективная оценка времени во время сновидений различной струк-
туры. — Физиол. чел., 19756, т. 1, № 1, с. 142—151.
Моисеева Н. И. Влияние внезапного изменения временной среды на некото-
рые циркадные ритмы человека. — Физиол. журн. СССР, 1975в,
т. 61, № 12, с. 1798—1804.
(Моисеева Н. И. 1975 г) Moiseeva N. I. The characteristics of EEG activity
and the subjective estimation of the time during dreams of different
structure. — EEG and Clin. Neurophysiol., 1975, vol. 38, N 6, p. 569—
577.
Моисеева H. И. Самоорганизация процесса сна в условиях внезапного изме-
нения временной среды. — В кн.: 'Саморегуляция процесса сна. Л.,
1976, с. 129—137.
Моисеева Н. И. Структура биоритмов как один из критериев возможностей
физиологической адаптации организма. — Физиол. журн. СССР,
1978а, т. 4, № 11, с. 1632—1640.
Моисеева Н. И. Возможности использования временной структуры физиоло-
гических функций в конкретизации понятия нормы и степени адапти-
рованности. — В кн.: Адаптивные реакции мозга и их прогнозирова-
ние. Л., 19786, с. 140—147.
(Моисеева Н. И. 1979а) Moiseeva N. I. The significance of different sleep sta-
ges for the regulation of electrical brain activity in man. — EEG and
Clin. Neuro physiol., 1979, v. 46, N 4, p. 371—381.
Моисеева H. И. Методологические вопросы в исследовании биологического
времени. — В кн.: Методологические вопросы теоретической меди-
цины. Л./ 19796, вып. 2, с. 28—37.
Моисеева Н. И. Свойства биологического времени. — В кн.: Фактор времени
в функциональной организации деятельности живых систем. Л., 1980а,
с. 21—25.
Моисеева Н. И. Хронобиологическая оценка состояния больного и
прогноз заболеваний нервной системы. — Сов. мед., 19806, № 6,
с. 30—33.
Моисеева Н. И., Богословский М. М., Колесова Е. К., Симонов М. Ю., Тон-
кова Н. В. К вопросу о влиянии внешних факторов, связанных с вра-
щением Земли, на циклы сна человека. —В кн.: Тез. докл. IV Между-
нар. биофиз. контр. (XVI—XXV секции). М., 1972, с. 80—82.
Моисеева Н. И., Каган М. А. О возможности прогнозирования адаптоспо-
собности по некоторым нейрофизиологическим показателям. — В кн.:
Адаптивные реакции мозга и их прогнозирование. Л., 1978, с. 44—55.
Моисеева Н. И., Симонов М. К)., Сысуев В. М, Особенности биоритмов и адап-
тационные возможности человека. — В кн.: Тез. докл. II Всесоюз.
конф, по адаптации человека к различным географическим, климати-
ческим п производственным условиям. Владивосток, 1978а т. 3, с. 12—
14.
121
Моисеева Н. И., Симонов М. Ю., Сысуев В. М. Значение сна в регуляции
циркадных ритмов. — Физиол. чел., 19786, т. 4, № 6, с. 995—1002.
Моисеева Н. И., Сысуев В. М., Савич А. А. Значение выбора типа временной
шкалы для построения адекватных биологических моделей. — В кн.:
Тез. и реф. докл. XI расширенной науч. конф, по пробл. физиол.
Кутаиси, 1979, с. 14—16.
Музалевская Н. И. О биологической активности возмущенного геомагнит-
ного поля. — В кн.: Адаптация организма при физических воздей-
ствиях. Вильнюс, 1969, с. 272—273.
Мустелъ Э. Р. Солнечная активность и тропосфера. — В кн.: Влияние сол-
нечной активности на атмосферу и биосферу Земли. М., 1971, с. 32—57.
Наенко И. И. Психическая напряженность. М., 1976, 112 с.
Новикова Р. А. Инструментальный анализ холодовой пробы. — В кн.: Тез.
докл. 2-й конф, кардиол. БССР. Минск, 1970, с. 98—99.
Новикова Р. А. Диагностическое значение вегетативного показателя сердеч-
ного ритма при холодовой пробе. — Здравоохранение Белоруссии,
1971, № 9, с. 26-29.
Новикова Р. А. Холодовая проба. — В кн.: Ранняя инструментальная ди-
агностика гипертонической болезни и атеросклероза. Минск, 1973,
с. 163-182.
Окунева, Г, Н,, Шевелева Л. Т., Мурашко Т. Н. Характеристика суточного
ритма газового состава крови и кислотно-щелочного равновесия в про-
цессе развития здорового человека. — В кн.: Тез. докл. II Всесоюз.
конф, по адаптации человека к различным географическим, климати-
ческим и производственным условиям. Новосибирск, 1978, т. 3, с. 18—
20.
Орбели Л. А. Физическое воспитание. — Избр. тр. М.; Л., 1961, т. 1, с. 251 —
263.
Павлов И. П. Полное собр. тр., 1949, т. 3. 516 с.
Панферова Н. Е. Изменение суточного ритма функций человека в условиях
ограниченной мышечной деятельности. — В кн.: Биологические ритмы
и вопросы разработки режимов труда и отдыха. М., 1967, с. 54—55.
Панченко Д. И. Влияние атмосферных перепадов и стабильного микрокли-
мата биотрона на приспособительные аппараты при гипертонической
болезни. — Врачебное дело, 1972, № 1, с. 1—6.
Панченко Д. И., Исаков Ю. А., Лукашевич Л. П. Медико-биологическое зна-
чение биотрона и его физико-технические ресурсы. Киев, 1964. 222 с.
Парин В. А., Баевский Р. М., Аматняк М. Г., Гехт Б. М., Гужавин Г. Н.,
Линденбратен В. Д., Коломина М. В., Кристи Е. М., Кудрявцева В. И.,
Ноздрачева Л. В., Петренко Б. Е., Семенова Т. Д., Газетдинов И, Г.,
Филиппова В. И., Шабадаш А. Л., Шарашкин Г. Ф, Влияние 7-часо-
вого сдвига времени на суточную периодику физиологических функ-
ций и умственную работоспособность человека. — В кн.: Матер,
конф. «Функциональное состояние человека в условиях переезда
в другие поясные зоны». Иркутск, 1971, с. 58—61.
Подшиб яки н А. К. О снижении надежности функционирования нервной
системы человека и животных в периоды гелиомагнитных возмуще-
ний. — В кн.: Первая республиканская летняя школа по нейробио-
нике. Киев, 1971, с. 62—63.
Пресман А. С. Электромагнитная сигнализация в живой природе. М., 1974.
64 с.
Пэрна Н. Я, Ритм, жизнь и творчество. М.; Л., 1925. 242 с.
Раева С. Н., Ливанов М. И. Микроэлектродное изучение нейрональных
механизмов произвольной мнестической деятельности человека. —
Физиол. чел., 1975, т. 1, № 1, с. 36—43.
Реакция биологических систем на слабые магнитные поля. М., 1971. 188 с.
Русинов В. С., Гриндель О. М., Болдырева Г. И. Исследование межцентраль-
ных отклонений в коре головного мозга человека методом кросскор-
реляции ЭЭГ. - ЖВНД, 1970, т. 20, № 2, с. 280-292.
122
Руттенбург С. О., Дэн Су-и. Материалы к физиологической характеристике
работы у пульта управления. — Тр. науч, сессии Науч.-исслед. ин-та
гигиены труда и профзаболеваний, посвящ. итогам работы за 1956 г.
Л., 1959, с. 138-144.
Руттенбург С. О., Слоним А. Д. Циркадный ритм физиологических процес-
сов и трудовая деятельность человека. Фрунзе, 1976. 188 с.
Самохина Т. В. Переживания настоящего и будущего у лиц, совершивших
суицидную попытку. — В кн.: Фактор времени в функциональной
организации деятельности живых систем. Л., 1980, с. 154—156.
Саркисов Д. С. Ультраструктурные основы биоритмов и проблема гомео-
стаза. — В кн.: Биологические ритмы в механизмах компенсации на-
рушенных функций. М., 1973, с. 35—46.
Саркисов Д. С. Очерки по структурным основам гомеостаза. М., 1977. 350 с.
Саркисов Д. С., Пальцын А. А., Втюрин Б. В. Приспособительная пере-
стройка биоритмов. М., 1975. 184 с.
Сельков Е. Е. Клеточные часы как автоколебательная система. — В кн.:
Колебательные процессы в биологических и химических системах.
Пущино-на-Оке, 1971, с. 5—9.
Сельков Е. Е. Временная организация энергетического метаболизма и кле-
точные часы. — В кн.: Регуляция энергетического обмена и физио-
логического состояния организма. М., 1978, с. 15—32.
Сидоренко Г. И. Актуальные вопросы анализа кардиологической информа-
ции. — В кн.: Тез. докл. 2-й конф, кардиол. БССР. Минск, 1970,
с. 125-126.
Симонов П. В, О роли эмоций в приспособительном поведении живых си-
стем. — Вопр. психол., 1965, № 4, с. 75—94.
Слауцитайс В. В., Сысуев В. М. Исследование возможностей периодограм-
много метода на моделях средней частоты сердечного ритма. — В кн.:
Кибернетический подход к биологическим системам. М.; Л., 1976,
с. 81—84.
Слоним А. Д. Учение о физиологических адаптациях. — В кн.: Общая эко-
логическая физиология адаптаций. Л., 1979, с. 79—182.
Смирнов К. М. Гипокинезия. — Успехи физиол. наук, 1972, т. 3, № 1,
с. 3-20.
Смирнов К. М. Общие вопросы учения о биологических ритмах. — В кн.:
Биоритмы и труд. Л., 1980, с. 6—20.
Смирнов К. М., Аникина Е. К., Дедовская Н. М., Хамзаев Ш, А., Швай-
кова Л. В. Сезонные изменения двигательной активности и физической
работоспособности человека в различных климатических условиях. —
В кн.: Адаптация и проблемы общей патологии. Новосибирск, 1974,
т. 3, с. 22—23.
Смирнов К. М., Ахметшин Р. X. Ритмы монотонного труда. — Гигиена
труда и проф. забол., 1976, № 8, с. 6—9.
Степанова С. И. Изучение возможностей адаптации человека к суткам
16-часовой продолжительности. — Космич. биол. и мед., 1973, т. 7,
№ 3, с. 68-75.
Степанова С. И. Влияние трансмеридиональных перелетов на организм че-
ловека. — Космич. биол. и авиакосмич. мед., 1974, т. 8, № 4, с. 3—12.
Степанова С. И. Динамика некоторых фаз температурного ритма в процессе
его инверсии. — Космич. биол. и авиакосмич. мед., 1975а, т. 9, № 1,
с. 70—79.
Степанова С. И. О биоритмологическом отборе космонавтов. — Космич. биол.
и авиакосмич. мед., 19756, т. 9, № 4, с. 40—46.
Степанова С. И. Попытка «расшатывания» суточной периодики экскреции
калия с мочой. — Космич. биол. и авиакосмич. мед., 1976, т. 10, № 1,
с. 76—86.
Степанова С, И., Кукишев С. П. Перестройка температурной периодики
человека после инверсии ритма жизни в изоляции. — Космич. биол.
и авиакосмич. мед., 1975, т. 9, № 5, с. 35—44.
123
Сысуев В, М. Методы анализа адаптивных перестроек мультиосцилляторных
систем. — В кн.: Адаптивные реакции мозга и их прогнозирование.
Л., 1978, с. 123—130.
Ткачук В. Г., Кучеров И. С. Планирование тренировки гимнаста в микро-
цикле в связи с ритмом трофических процессов. — В кн.: Матер,
конф. «Тренировка гимнастов». М., 1965, с. 63—69.
Тонких А. В., Борковская Ю. А., Фадеева О. И. Роль симпатоадреналовой
системы в регуляции сна при нормальных и экстремальных усло-
виях. — В кн.: Саморегуляция процесса сна. Л., 1977, с. 26—31.
Тюрин И, А. Специфика временных свойств биологических систем. — Вопр.
филос., 1977, № 6, с. 82—91.
Фролов Б, С., Воробьев В» М., Кузнецов О. Н., Чебаков В. П. Анализ суточной
периодики физиологических функций при дезадаптации высшей
нервной деятельности. — Физиол. чел., 1980, т. 6, № 3, с. 478—485.
Холодов Ю. А. Магнетизм в биологии. М., 1976. 96 с.
Хохрякова Е. В. Изучение суточного ритма у детей школьного возраста ме-
тодами кинематометрии и динамометрии. — В кн.: Матер, конф.
«Функциональные состояния человека в условиях переезда в другие
поясные зоны». Иркутск, 1971, с. 73—75.
Чередниченко Л. К. Природные факторы среды и их влияние на организм. —
В кн.: Общая экологическая физиология и физиология адаптаций.
Л., 1973, с. 36—78.
Чернышев В. Б. Влияние возмущений земного магнитного поля на актив-
ность насекомых. — В кн.: Тез. докл. на совещ. по изучению влияния
магнитных полей на биологические объекты. М., 1966, с. 80.
Чижевский А. Л. Земное эхо солнечных бурь. М., 1973. 348 с.
Шалаев М. И., Веселов В. П. Тромбоэмболические осложнения на европей-
ском Севере и их зависимость от метео- и гелиотропных факторов. —
В кн.: Научно-технический прогресс и приполярная медицина. Ново-
сибирск, 1978, т. \1, с. 132—133.
Шапошникова В. И. Значение биологических ритмов для спортивной прак-
тики. — В кн.: Вопросы повышения спортивного мастерства по лыж-
ным гонкам и гребле. Л., 1973, с. 19—31.
(Шапошникова В. И.) Schaposchntkova W. Z. Biologische Makrorhythmen
und «kritische» Perioden des Menschen. — In: Interdis. Cycle Res.,
1977, vol. 8, N 3—4, p. 269—271.
Энгельгардт В, А. Специфичность биологического обмена веществ. — Вопр.
филос., 1960, Кг 7, с. 113—123.
Abe К., Sasaki Н., Takebayashi К., Fukui S., Nambu Н. The development
of circadian rhythms of human body temperature. — J. Interdis. Cycle
Res., 1978, vol. 9, N 3, p. 211—216.
Alexander J., Atlas P. Cyclic activiy in nature causes of stability. — Intern.
J. Neurosci., 1973, vol. 6, N 1, p. 45—50.
(Anderson T.) Андерсон T, Статистический анализ временных рядов. М.,
1976. 756 с.
Aschoff J. Features of circadian rhythms relevant for the design of shift sche-
dules. — Ergonomics, 1978, vol. 21, N 10, p. 739—754.
Aschoff J. G., Ciedke C., Poppel U. Circadian periodicity of reaction-times. —
In: Recent advances in aerospace medicine. Amsterdam, 1969, p. 343—347.
(Barcroft J., Barron D, H.) Баркрофт Дж., Баррон Д. Г. Возникновение
дыхания при рождении. — В кн.: Тез. сообщ. ХУМеждунар. физиол.
конгр. Л., 1935, с. 24—25.
Bartko D., Wagnerova М., Danisova J. Problem vplyvu meteorologickych fac-
torov na vznik cirkularnych poruch mozgu. — Ceskoslov. neurol.
a neurochirurgie, 1974, vol. 37, N 5, p. 308—315.
Brown F. A. Biological clocks: endogenous cycles synchronized by subtile
geophysical rhythms. — Biosystems, 1976, vol. 8, N 1, p. 67—81.
(Bunning E.) Бюннинз Э. Биологические часы. — В кн.: Биологические
часы. М., 1964, с. 11—26.
(Burns В. D.) Бернс Б. Неопределенность в нервной системе. М., 1969. 251 с.
124
Colquhoun W. P. Circadian rhythms. Mental efficiency and shift work. — Ergo-
nomics, 1970, vol. 13, N 5, p. 558—560.
Dorland J., Brinker N. Fluctuation in human mood. — J. Interdis. Cycle Res.,
1973, vol. 4, N 1, p. 25-29.
Ehret C. F., Trucco E. Chronon concept for circadian clock. — J. Theoret.
Biol., 1967, vol. 15, N 5, p. 240-262.
Fabbro C. D. Ivoli di «lungo ressio» ed i ritmi circadiani. — Minerva med.,
1970, vol. 61, N 74, p. 3922-3928.
Feinberg J., Floyd T. C. Systematic trends across the night in human sleep
cycles. — Psychophysiology, 1979, vol. 16, N 3, p. 283—291.
Fraisse P., Siffre M., Oleron G. K., Luilli N. Le rhythme veilsomeil et 1’estima-
tion du temps. — In: Cycles biologiques et psychiatrie. Geneve; Paris,
1968, p. 257—265.
(Goodwin В. С.) Гудвин Б. Временная организация клетки. М., 1966. 252 с.
Halberg F. Experimentelles zur Physiologie des nebennieren Zyclus. — Acta
med. scand., 1955, suppl. 307, p. 117—118.
(Halberg F.) Халберг Ф, Временная координация физиологических функций. —
В кн.: Биологические часы. М., 1964, с. 475—509.
Halberg F. Chronobiology. — Ann. Rev. Physiol., 1969, vol. 31, p. 675—725.
Hauty G. T.) Adams V. Phase shifts of the human circadian system and per-
formance deficit during the periods of transition. I. East-West Flight.
Aerosp. Med., 1966, vol. 37, N 7, p. 668—674. II. West East Flight.
Ibid., N 10, p. 1027—1033. III. North-South Flight. Ibid., N 12,
p. 1257-1262.
Hersley R. Emotional cycles of man. — J. Men. Sci., 1931, vol. 77, N 316,
p. 151-169.
Hildebrandt G. Outline of chronohygiene. — Chronobiologia, 1976, vol. 3,
N_2, "p. 113—127.
Jovanovic U. S. Chronobiology and sleep-waking cycle. — Waking and Sleeping,
1977, vol. 1, N 3, p. 225—226.
Klein К. E., Bruner H.4 Holtman H., Kehme H., Stolze J., Steindorff W. D.,
Wegmann H. M. Circadian rhythm of pilots’ efficiency and effects of mul-
tiple time zone travel. — Aerosp. Med., 1970, vol. 41, N 2, p. 125—132.
Kleitman N. Phylogenetic, ontogenetic and environmental determinants in
the evolution of sleep-wakefullness cycle. — In: Sleep and altered sta-
tes of consciousness. Baltimore, 1967, p. 30—35.
Kleitman N. Basic rest-activity cycle in relation to sleep and wakefullness.
In: Sleep: physiology and pathology. Philadelphia, 1969. 70 p.
Kripke D., O'Donoghue J. Perceptual deprivation, REM sleep and an ultra-
dian biological rhythm. — Psychophysiology (abstract), 1968, vol. 5,
p. 231.
Kristoffersen A. B, Attention and psychological time. — Acta Psychol.,
1967, vol. 27, N 1, p. 92—100.1
(Kunicki-Goldfinger W.) К у ницкий-Гольд фингер В, Биологическая эволюция
как возрастание уровня организации. — В кн.: Проблема взаимо-
связи организации и эволюции в биологии. М., 1978, с. 102—128.
Lavie Р. Nonstationarity in human perceptual ultradian rhythms. — Ghrono-
biologia, 1977, vol. 4, N 1, p. 38—48.
Lowrey J. Biorhythms — not just crackpot gimmies, but possible lifesavers. —
Engineer, 1978, vol. 247, N 6403, p. 48—49.
Markson R. Geophysical influences on biological cycles. — J. Interdis. Gycl.
Res., 1972, vol. 3, N 2, p. 134.
McCulloch. Discussion. — In: Biocybernetics of the central nervous system.
Boston, 1969, p. 61—62.
Orr W. C., Hoffman H. J., Hegge F. W. Ultradian rhythms in extended per-
formance. — Aerosp. Med., 1974, vol. 45, N 9, p. 995—1000.
Orr W. C., Hoffman H. J., Hegge F. W. The assessment of time-dependend-
changes in human performance. — Chronobiologia, 1976, vol. 3,
N 4, p. 293-305.
Ornstein R. E. On the experience of time. London, 1970. 126 p.
125
Paigen К. Temporal genes and developmental program. — In: Human gene-
tic. Amsterdam, Excerpta Medica, Intern. Congr. Ser., 1977, N 411,
p. 33—42.
(Piccardi J.) Пиккарди Дж. Химические основы медицинской климатоло-
гии. Л., Гидрометеоиздат, 1967. 96 с.
Pittendrigh С. S. Circadian rhythms: a problem in general physiology. —
In: Abstracts of lectures and symposia XXV Intern, congr. of physiol,
sci. Munich, 1971, vol. 8, p. 83.
Post W.j Gatty H. Around the world in eight days. London, 1931. 305 p.
Preston F. 5., Batenvan S. C., Short R. V., Wilkinson R. T. Effects of flying
and of time changes on menstrual cycle length and on performance in
airline stewardesses. — Aerosp. Med., 1973, vol. 44, N 4, p. 438—443.
(Quastler H.) Кастлер Г. Место теории информации в биологии. — В кн.:
Теория информации в биологии. М., 1960, с. 183—192.
Rechtschaffen A., Kales В. (Eds.) A manual of standartized terminology,
techniques and scoring system for sleep stages of human subjects. Wa-
shington, 1968. 54 p.
Schweiger H. G. Circadian rhythms in monocellular organisms.— Arzneimit-
tel Forschung, 1978, vol. 28, N 10a, p. 1814—1818.
(S iff re M.) Сифр M. Один в глубинах земли. М., 1966. 263 с.
Siffre М. Desynchronisation of human biological rhythm in men during long
beyond time confinement (2—3—4—6-months) in constant and stress
environment of caves. — In: XVII Intern, congr. on aviation and space
medicine. Oslo, 1968, p. 129.
Thomas E. A., Weaves W. B. Cognitive processing and time perception. —
Percept. Psychophysiol., 1975, vol. 17, N 3, p. 363—367.
Thommen G. Is this your day? (How biorhythm helps you determine your life
cycles). New York, 1968. 160 p.
Vernon H. M. Industrial fatigue and efficiency. — London, G. Rouledge and
sons, 1921. 264 p.
Virey J. J. Ephemerides de la vie humaine ou recherches sur la revolution
journaliere et la periodicite de ses phenomenes dans la sante et les ma-
ladies. — These Facult. Med., Paris, 1814, t. 12, these 37, p. 5339.
Webb W. B. Twenty-four hour sleep cycling. — In: Sleep physiology and patho-
logy. Philadelphia; Toronto, 1969, p. 19—35.
Webb W. B., Agnew H. W. Sleep cycling within twenty-four hour periods. —
J. Exper. Psychol., 1967, vol. 74, N 2, p. 1, p. 158—160.
Webb W. В., Agnew H. W. Mesurement and characteristics of noctural sleep. —
In: Dreams and dreaming. New York, 1969, p. 2—27. (Progress in cli-
nical psychology, vol. VIII).
Weinberg A. M. Of time and the energy wars. — Nature, 1977, vol. 269,
N 5630, p. 638.
Wendt H. W. Population, sex and constitution in tipologies based on indivi-
dual circadian rhythms. — J. Interdis. Cycle Res., 1977, vol. 8, N 3—4,
p. 286-290.
Wever R. Different aspects of the studies of human circadian rhythms under
the influence of weak electric fields. — In: Chronobiology, 1976,
Tokyo, p. 694—699.
Wever R. Einflus schwacher electromagnetischer Felder auf die circadiane
Periodic der Menschen. — Naturwissenschaften, 1968, Bd 55, N 1,
S. 29-32.
(Williams R.) Уильямс P. Биохимическая индивидуальность. M., 1960.
295 с.
(Whitrow G. J.) Уитроу Дж. Естественная философия времени. М., 1964.
431 с.
Zatopek A., Krivsky L., Lastovicka J. Correlations between solar, interplane-
tary, geomagnetic, ionospheric, athmospheric circulation and micro-
seismic phenomena. — J. Interdis. Cycle Res., 1976, vol. 7, N 1, p. 9—13.
ОГЛАВЛЕНИЕ
Стр.
Предисловие ...................................................... 3
Глава 1. Ритмический характер биологических процессов как способ
организации живой системы во времени......................... 5
Глава 2. Методическое обеспечение хронобиологических исследований 18
Глава 3. Циркадные колебания уровня бодрствования и некоторых
психических функций......................................... 33
Глава 4. Циркадные колебания деятельности сердечно-сосудистой си-
стемы ...................................................... 49
Глава 5. Исследование влияния биологических макроритмов на колеба-
ния значений физиологических и психологических показателей 65
Глава 6. Изменения циркадной и ультрадной ритмики под влиянием
эмоционального стресса и других случайных внешних воздейст-
вий ............................................................. 75
Глава 7. Зависимость течения циркадных ритмов от гелиогеофизических
факторов ................................................... 89
Заключение ....................................................... 103
Литература ....................................................... 118
Наталья Ивановна Моисеева,
Владимир Михайлович Сысуев
«ВРЕМЕННАЯ СРЕДА
И БИОЛОГИЧЕСКИЕ РИТМЫ»
Утверждено к печати
Научным советом по проблемам прикладной физиоло-
гии человека Академии наук СССР
Редактор издательства А. А. Антонова
Художник В. П. Сысалов
Технический редактор Р. А. Кондратьева
Корректор Л. М. Бова
ИБ № 20218
Сдано в набор 20.10.80. Подписано к печати 10.12.80.
М-21081. Формат 60 х 90*/1в- Бумага типографская № 1.
Гарнитура обыкновенная. Печать высокая. Печ. л.
8=8 усл. печ. л. Уч.-изд. л. 8.91. Тираж 5450. Изд.
К/7878. Тип. зак. № 1880. Цена 55 к.
Ленинградское отделение издательства «Наука»
199164, Ленинград, В-164, Менделеевская линия, 1
Ордена Трудового Красного Знамени
Первая типография издательства «Наука»
199034, Ленинград, В-34, 9 линия, 12