





































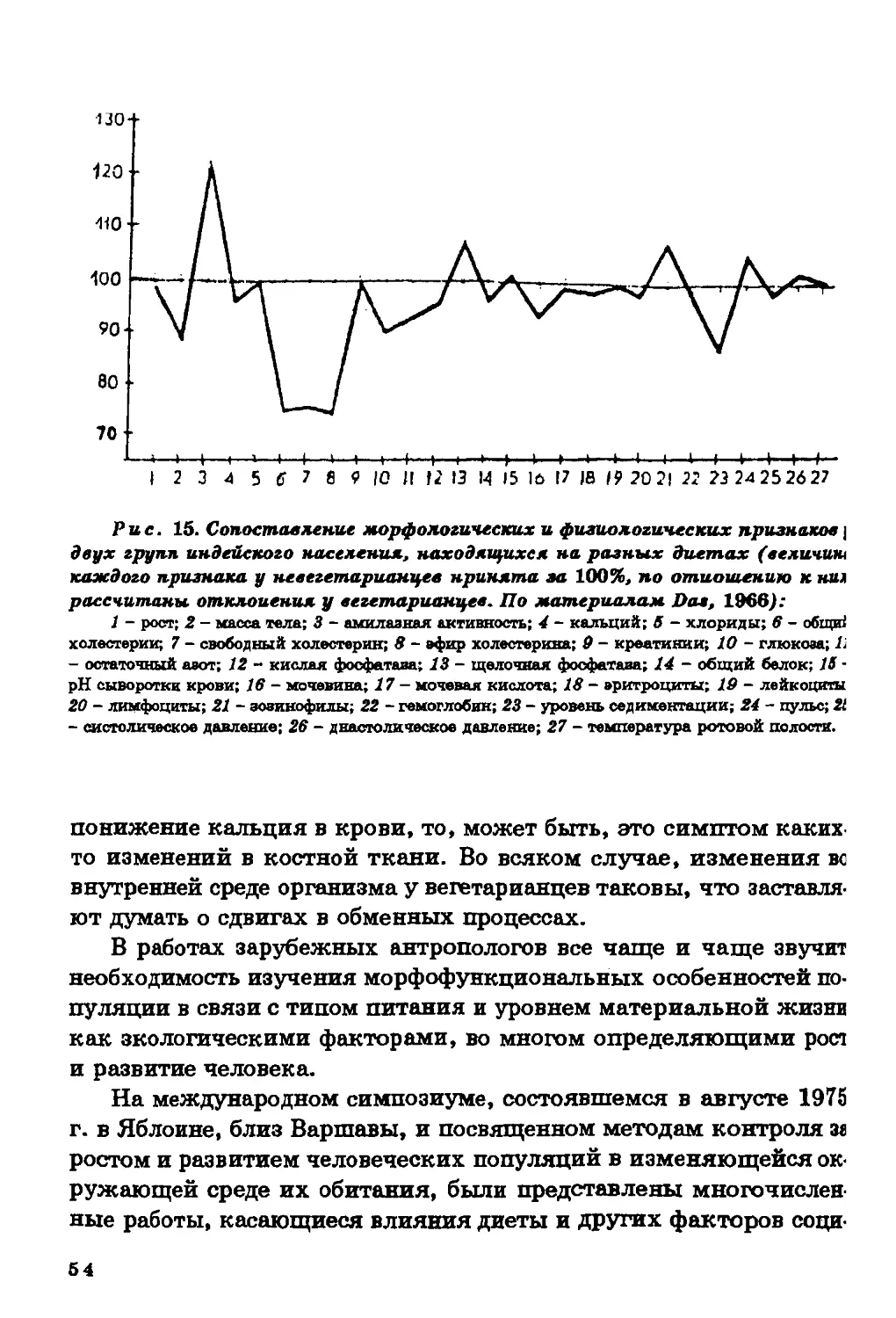
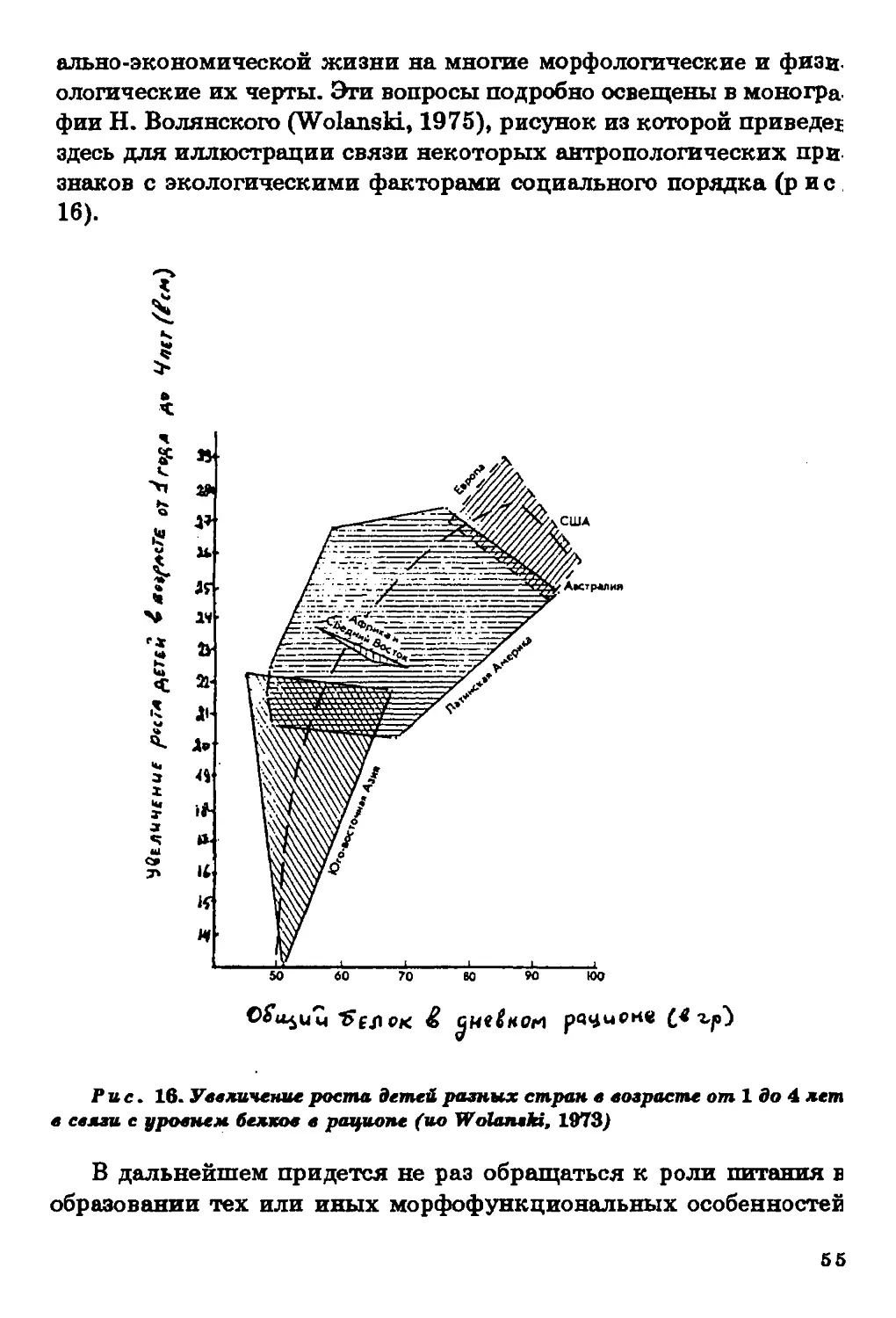






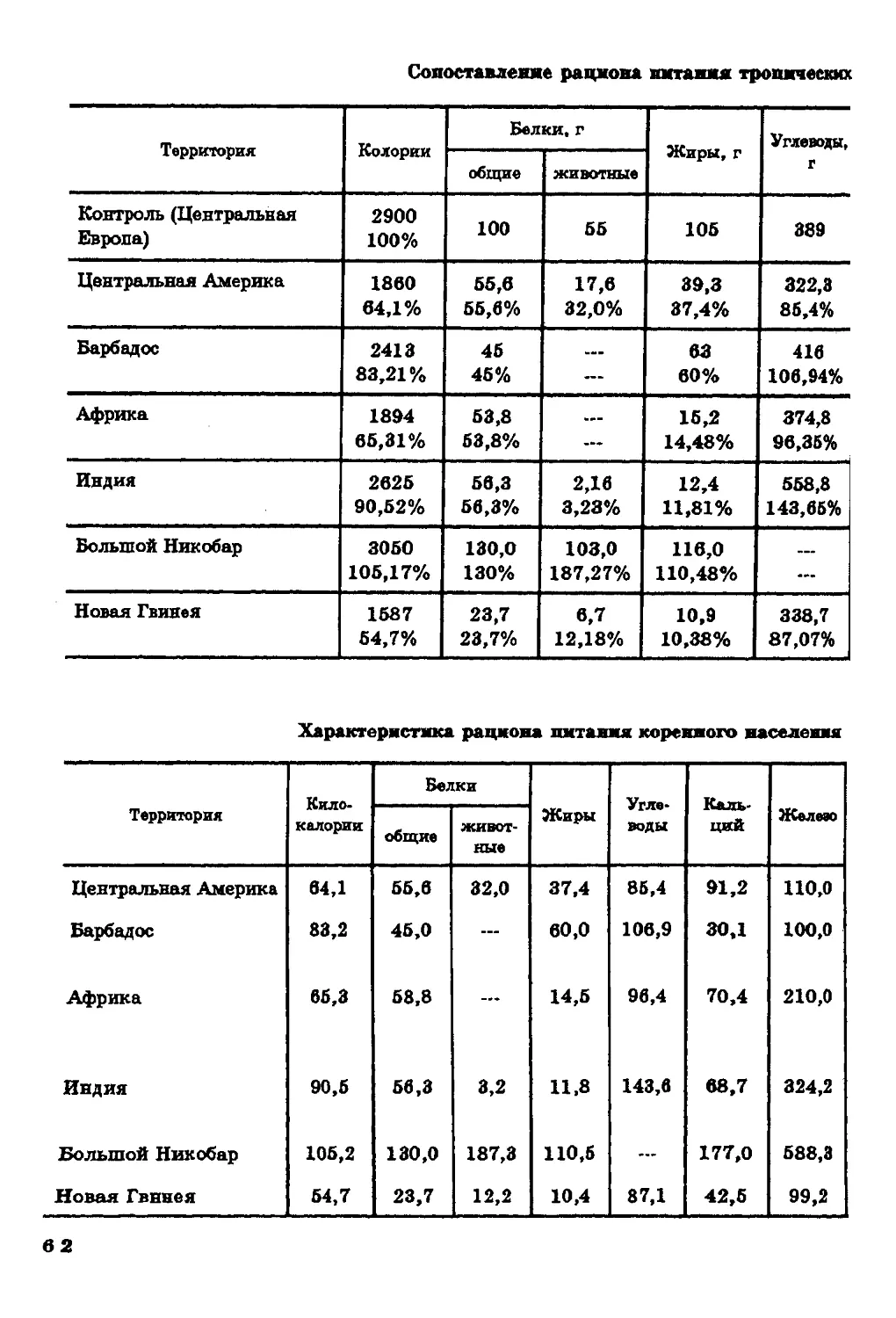
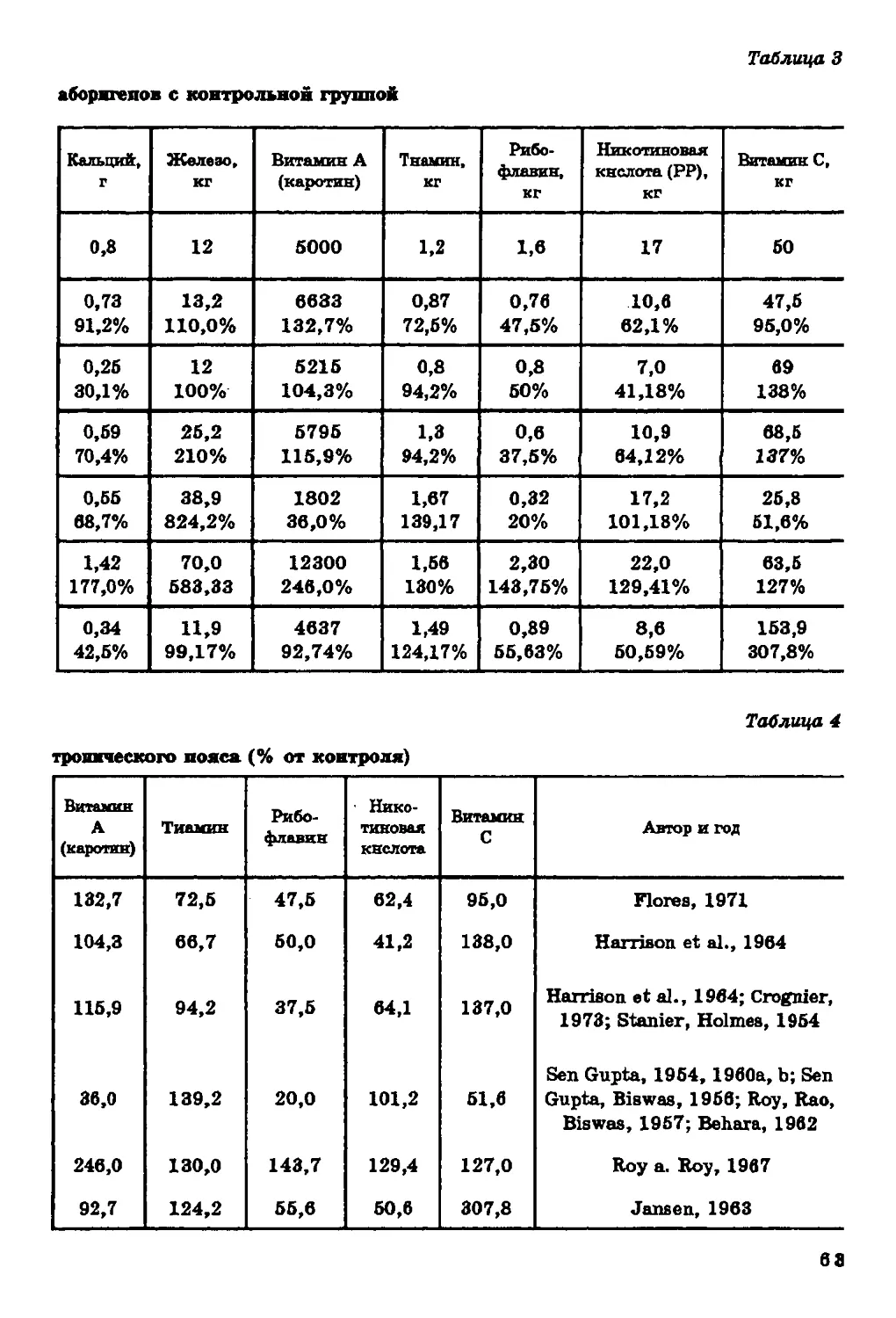



























































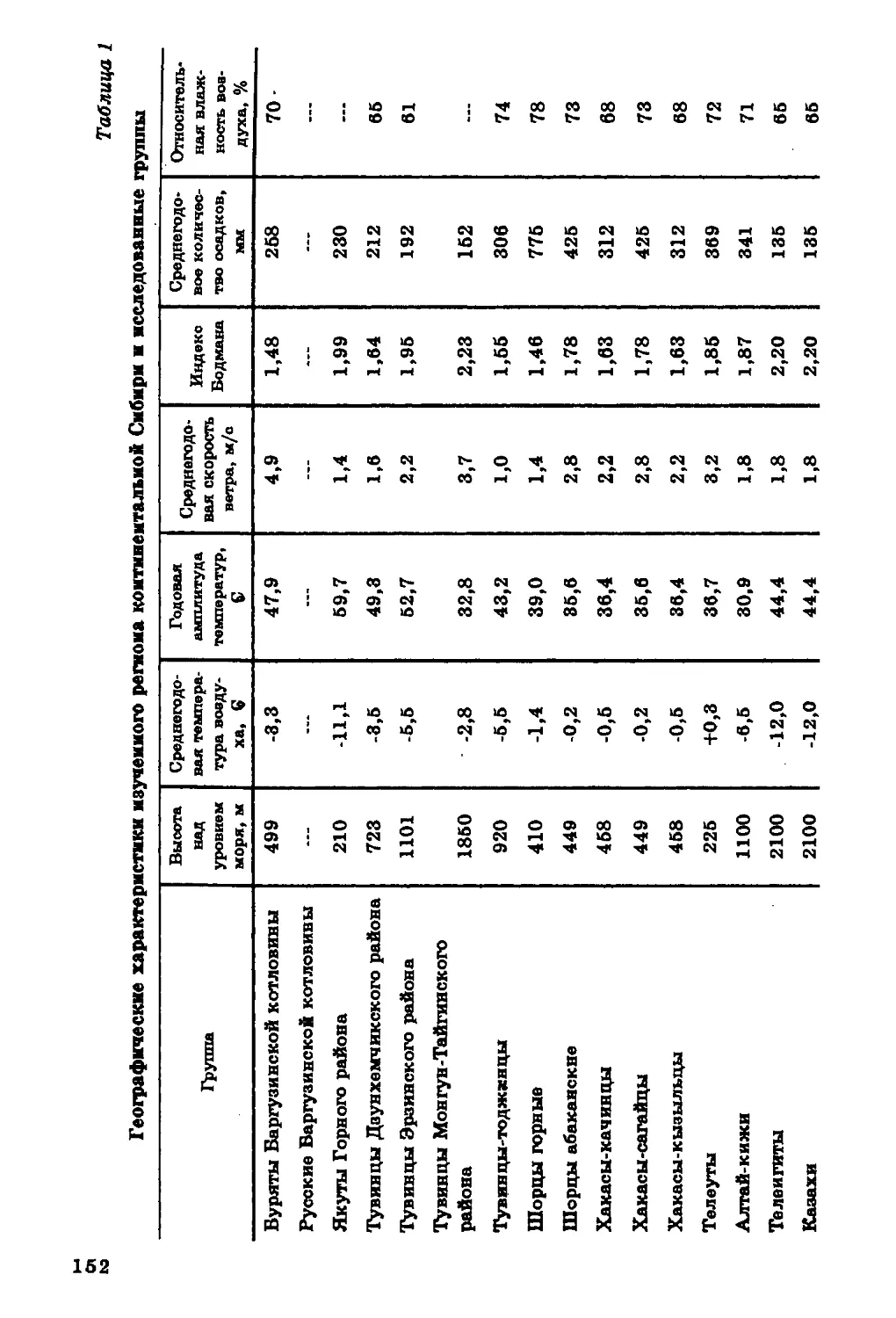

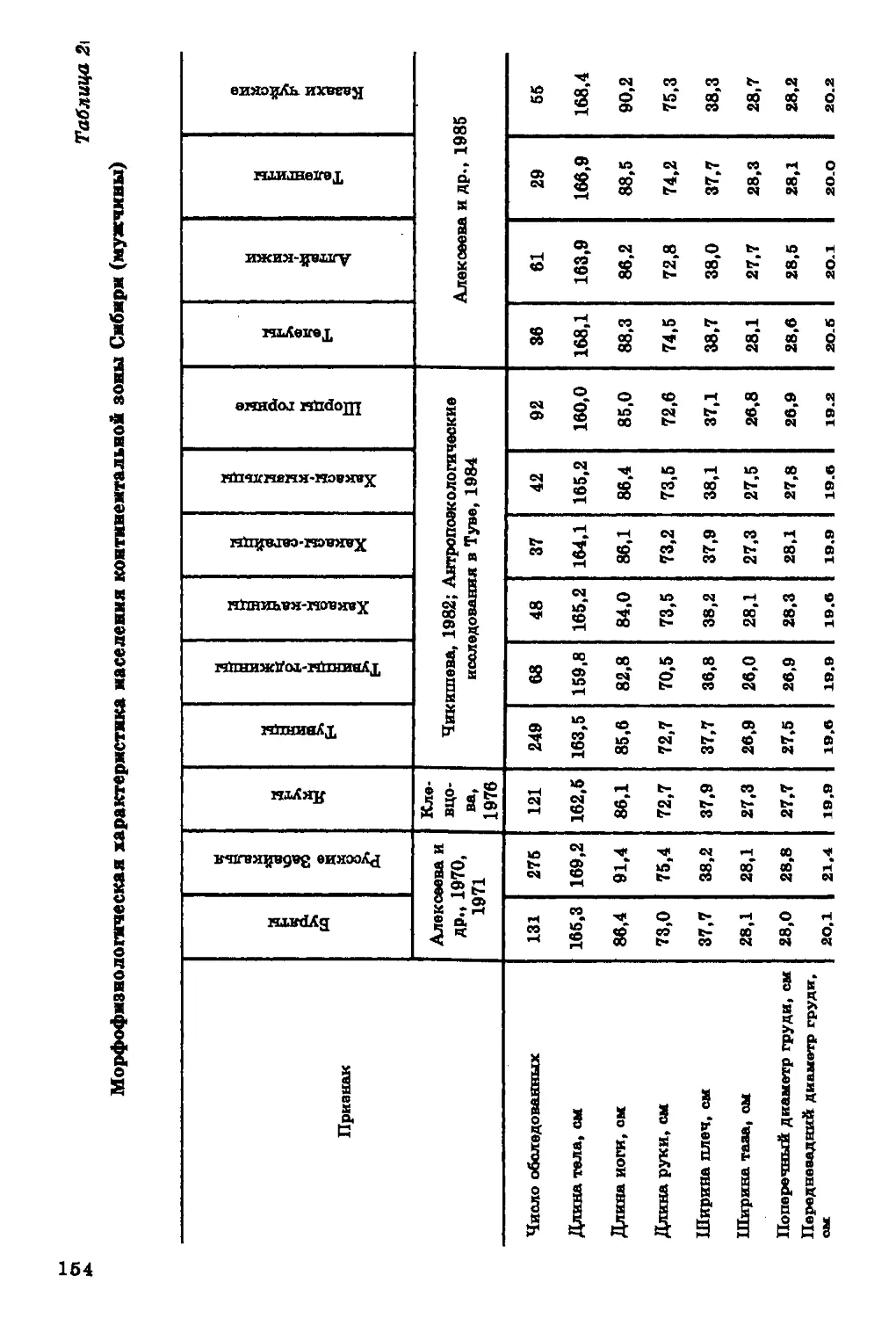
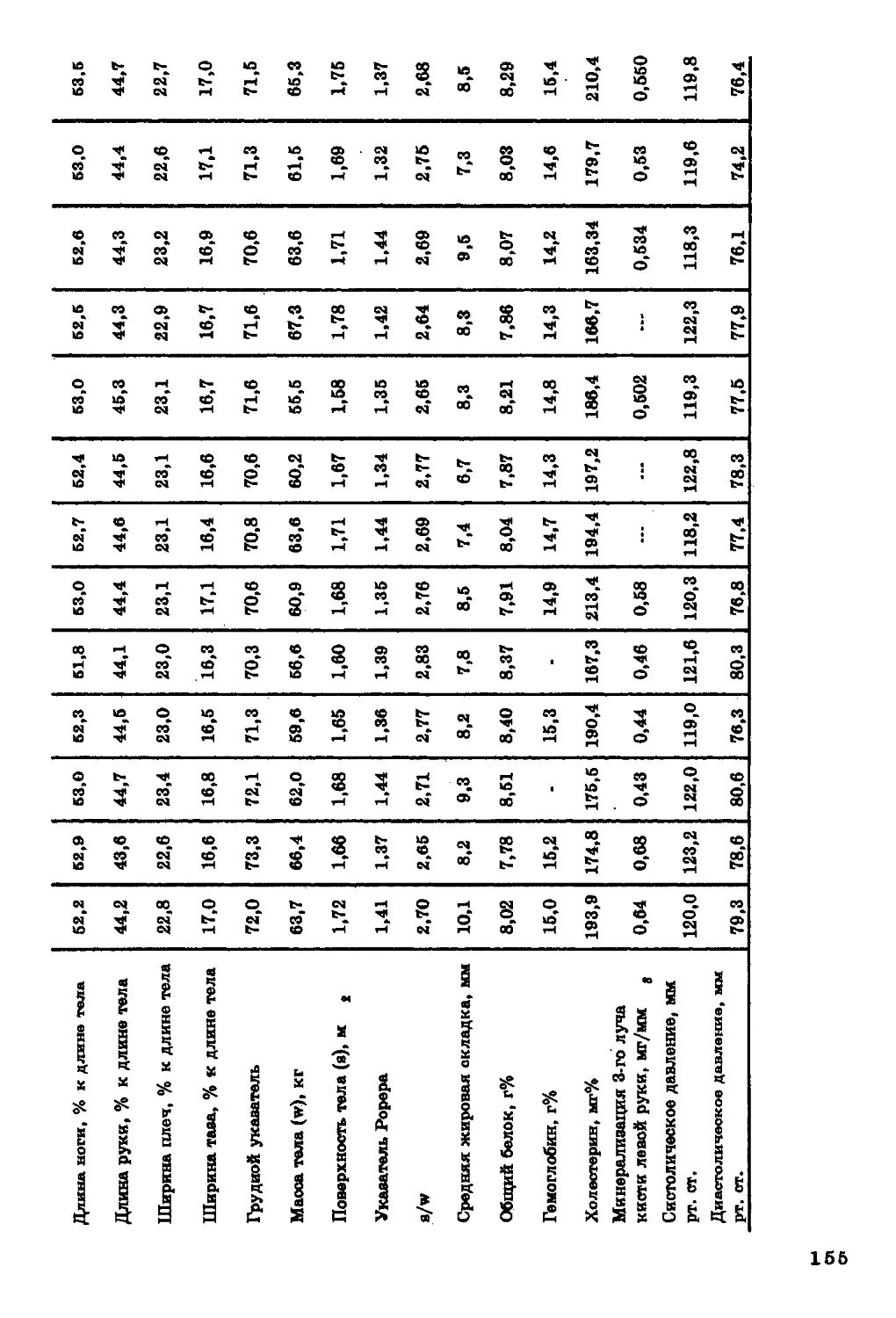
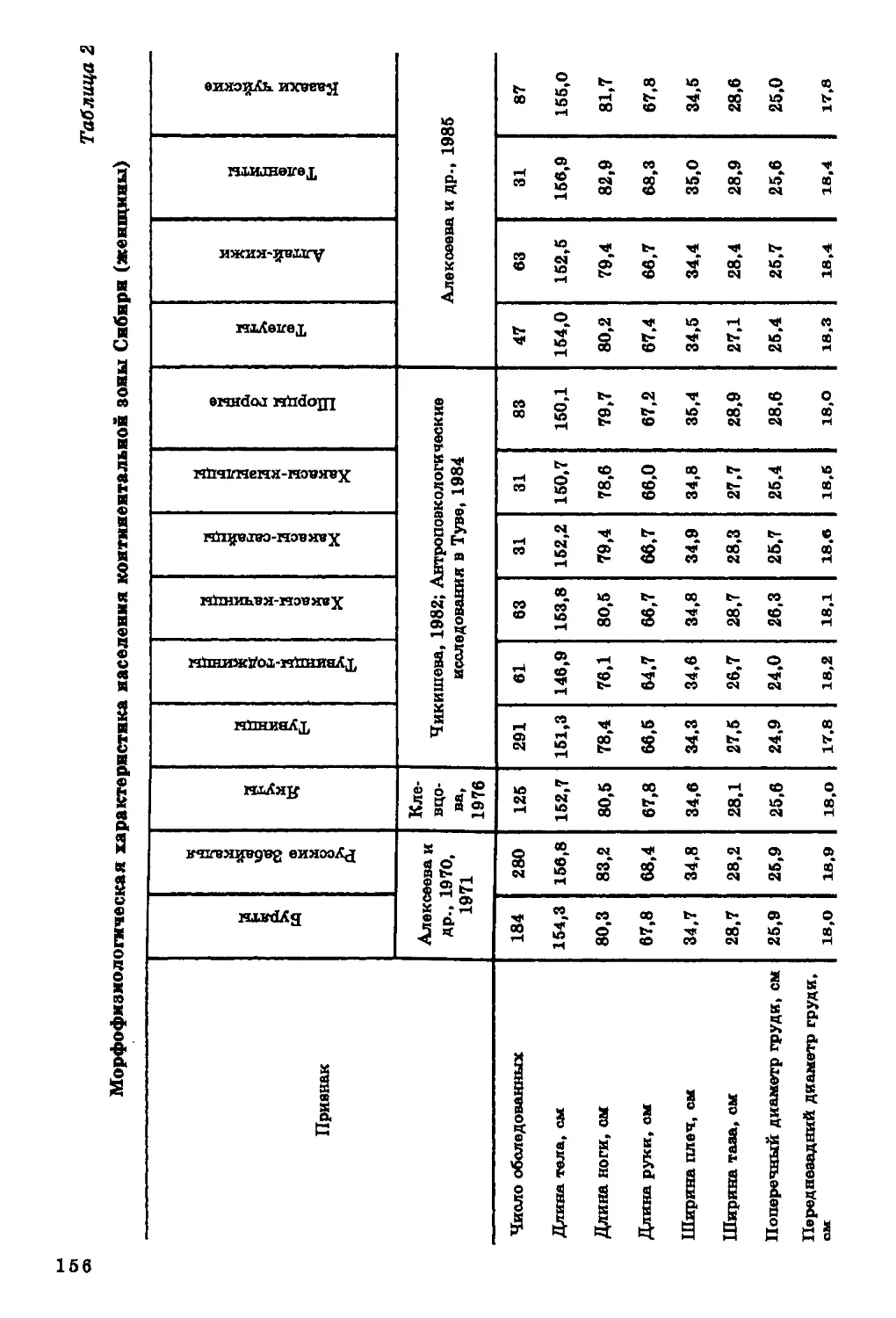
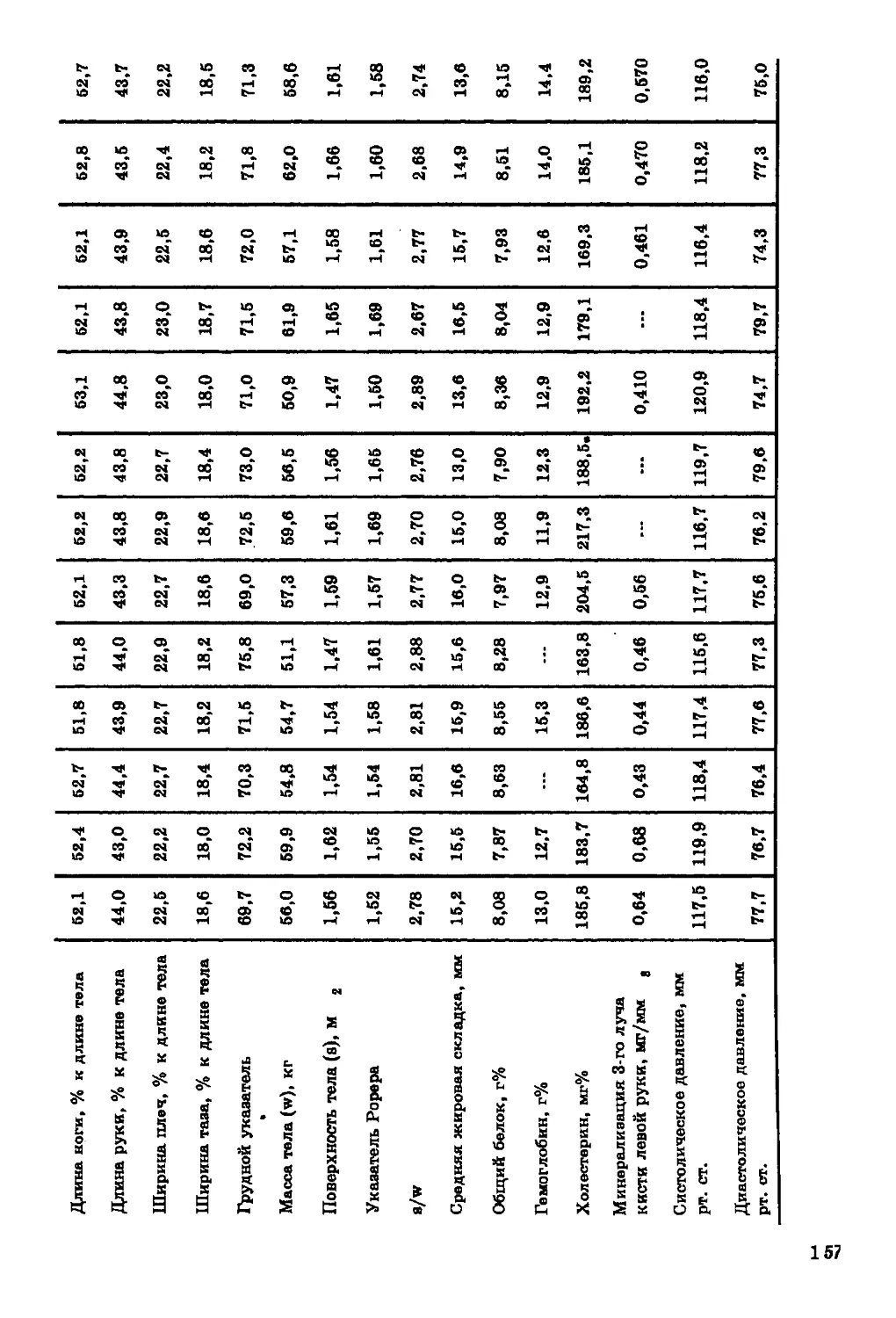

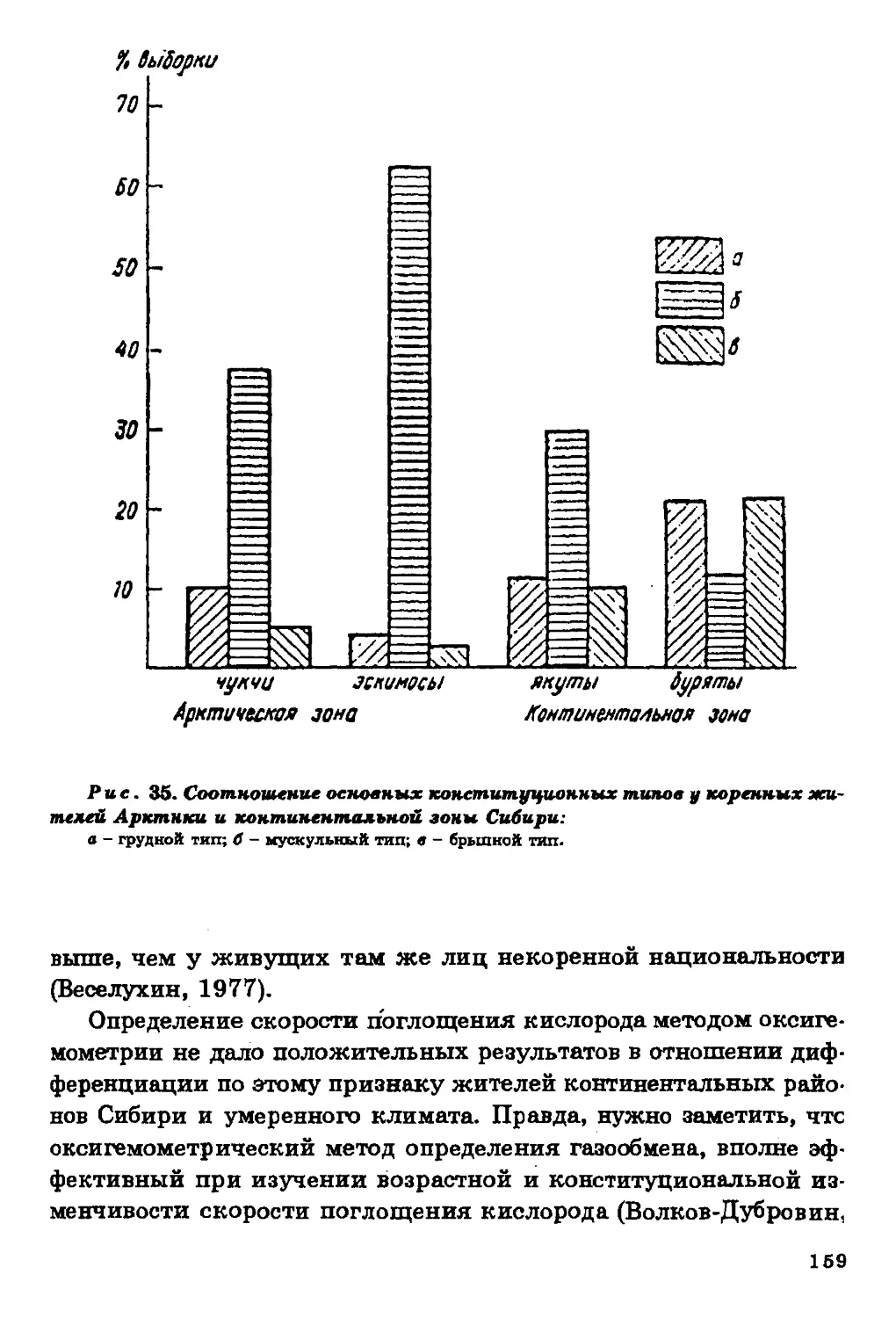


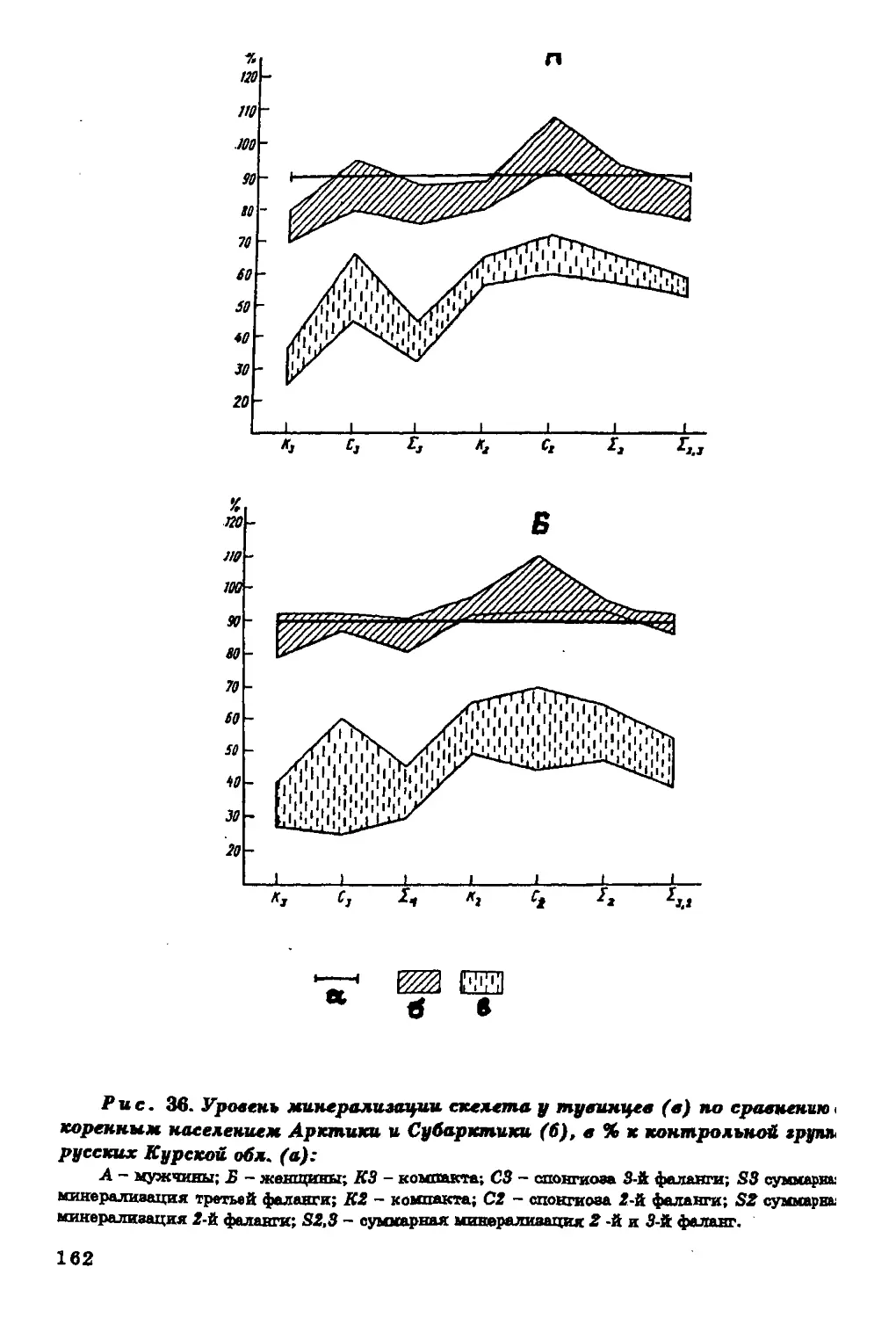


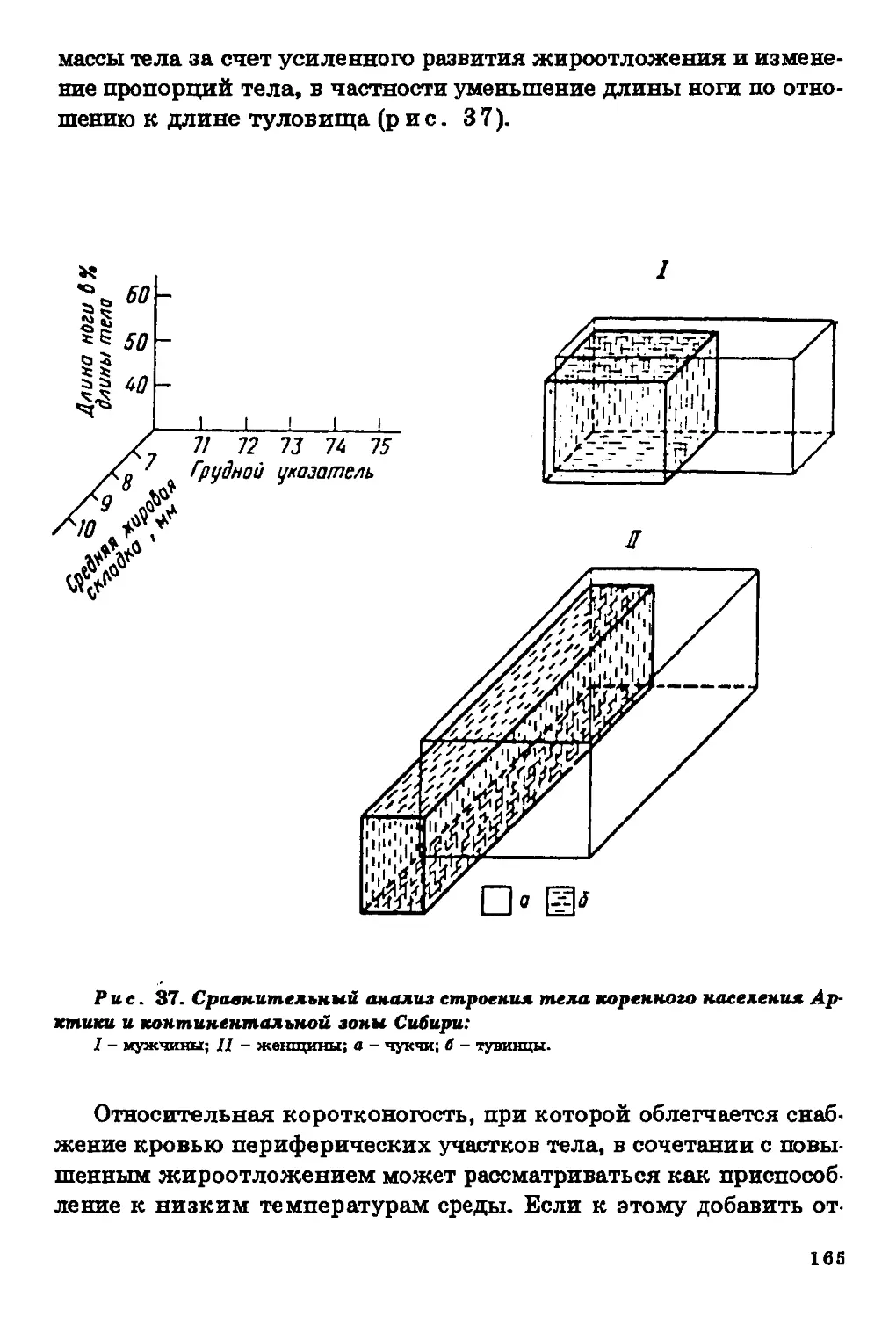





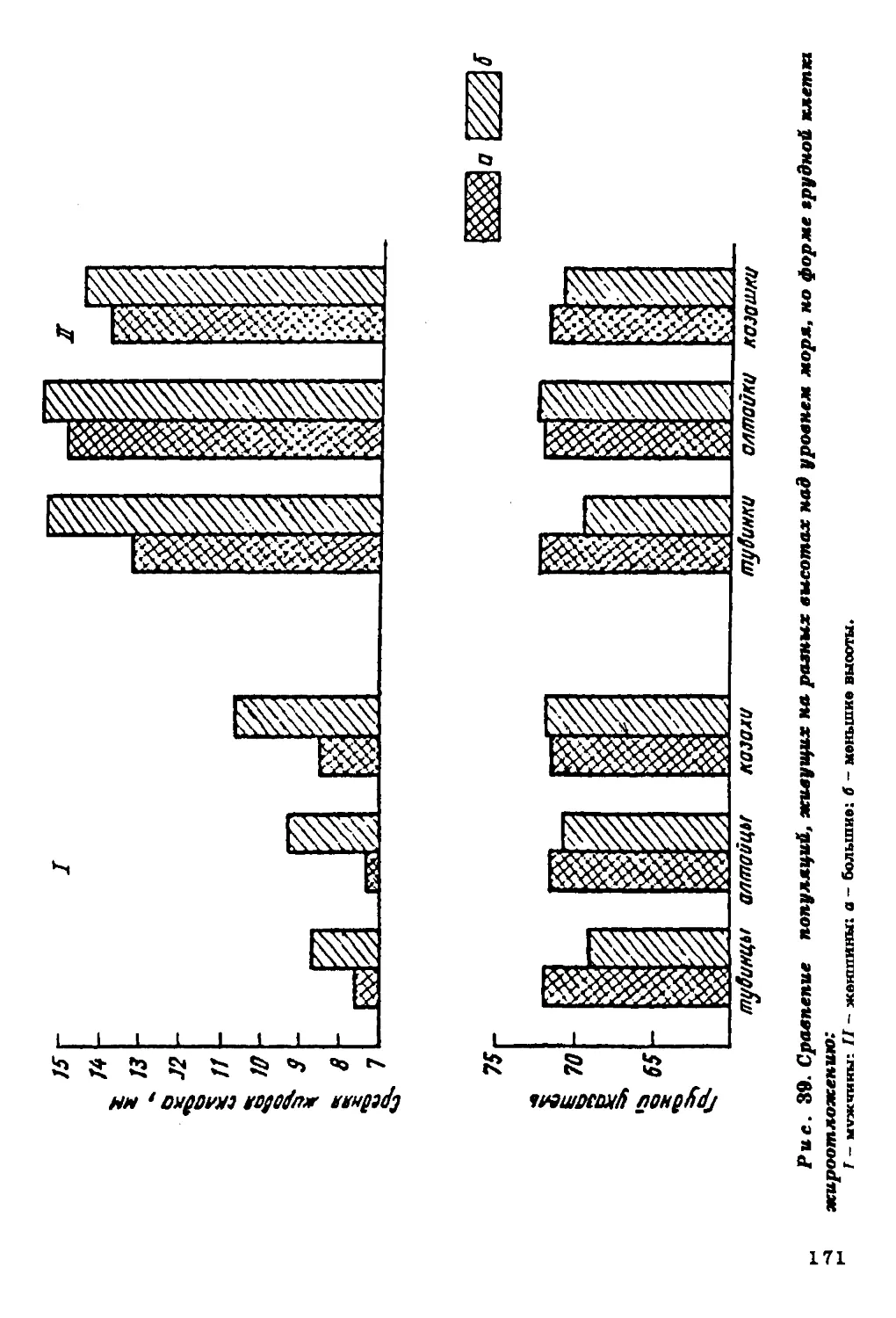


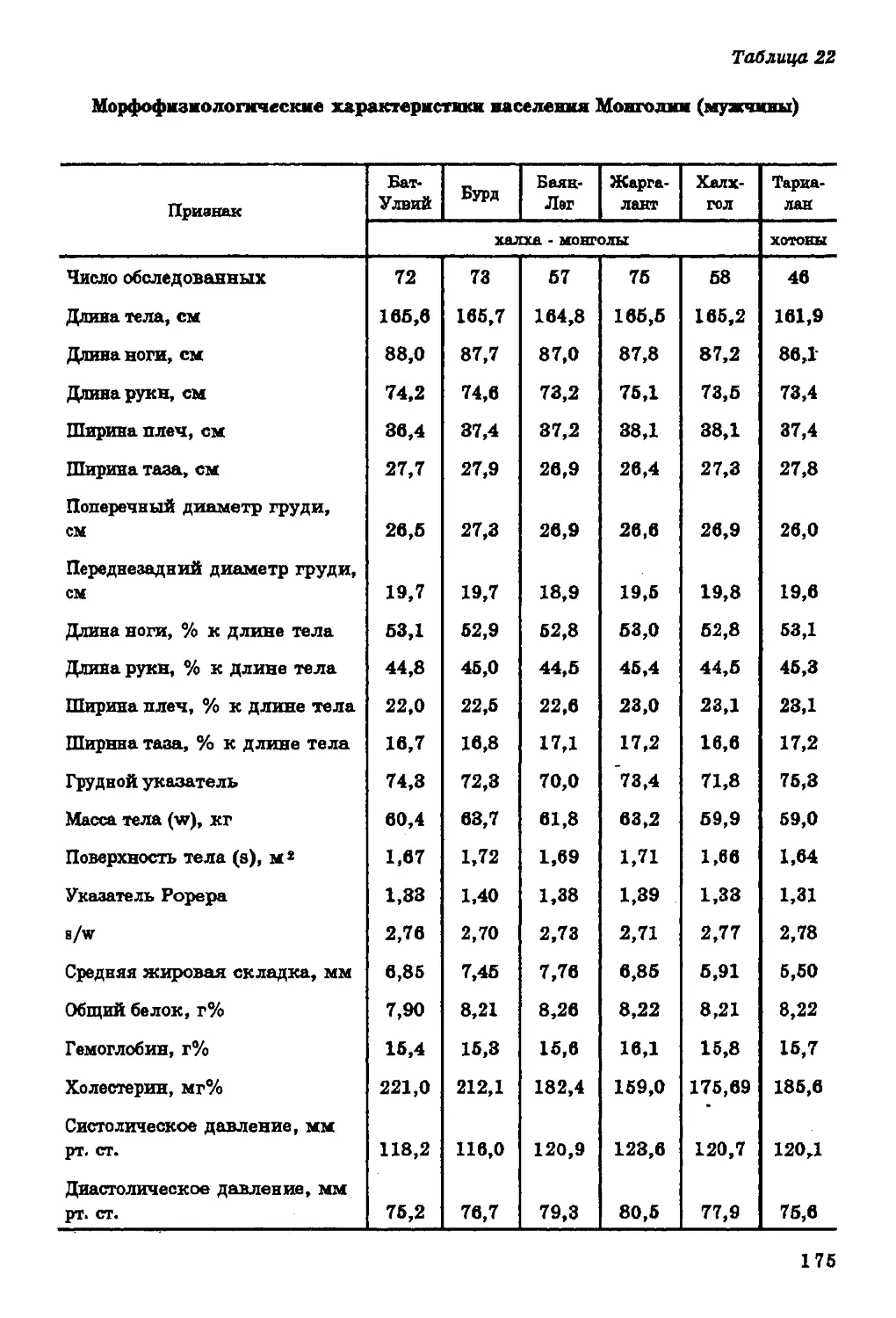
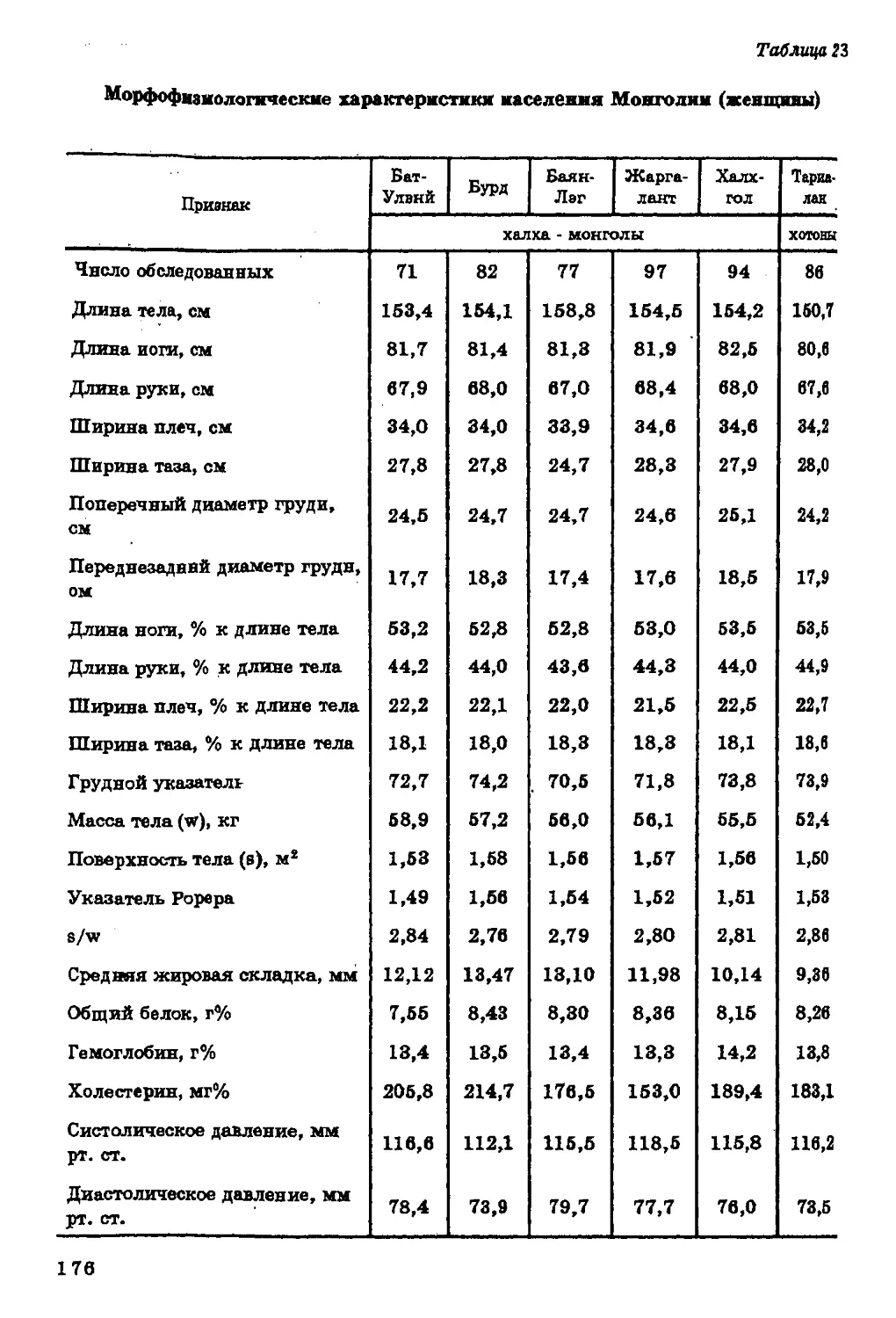




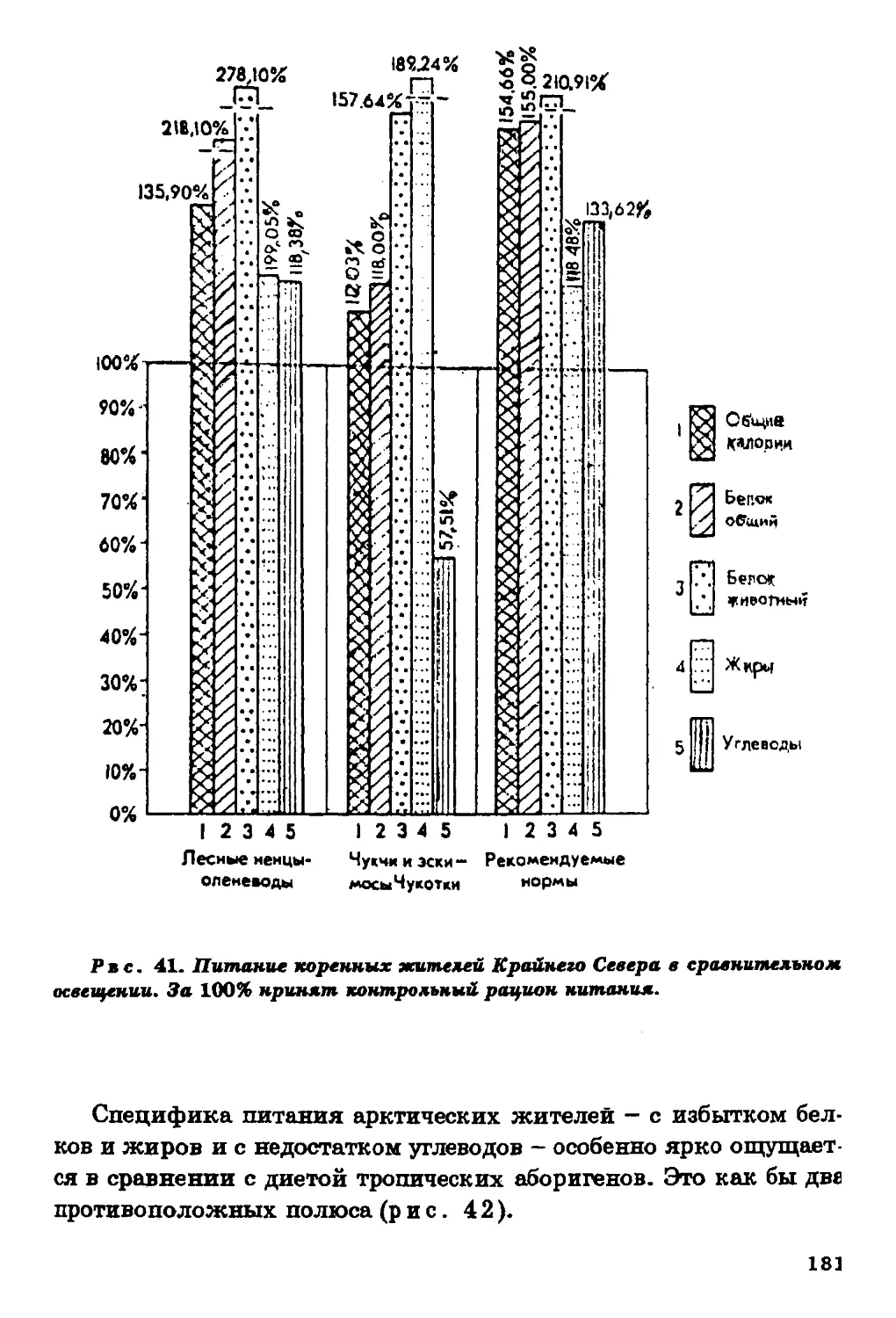
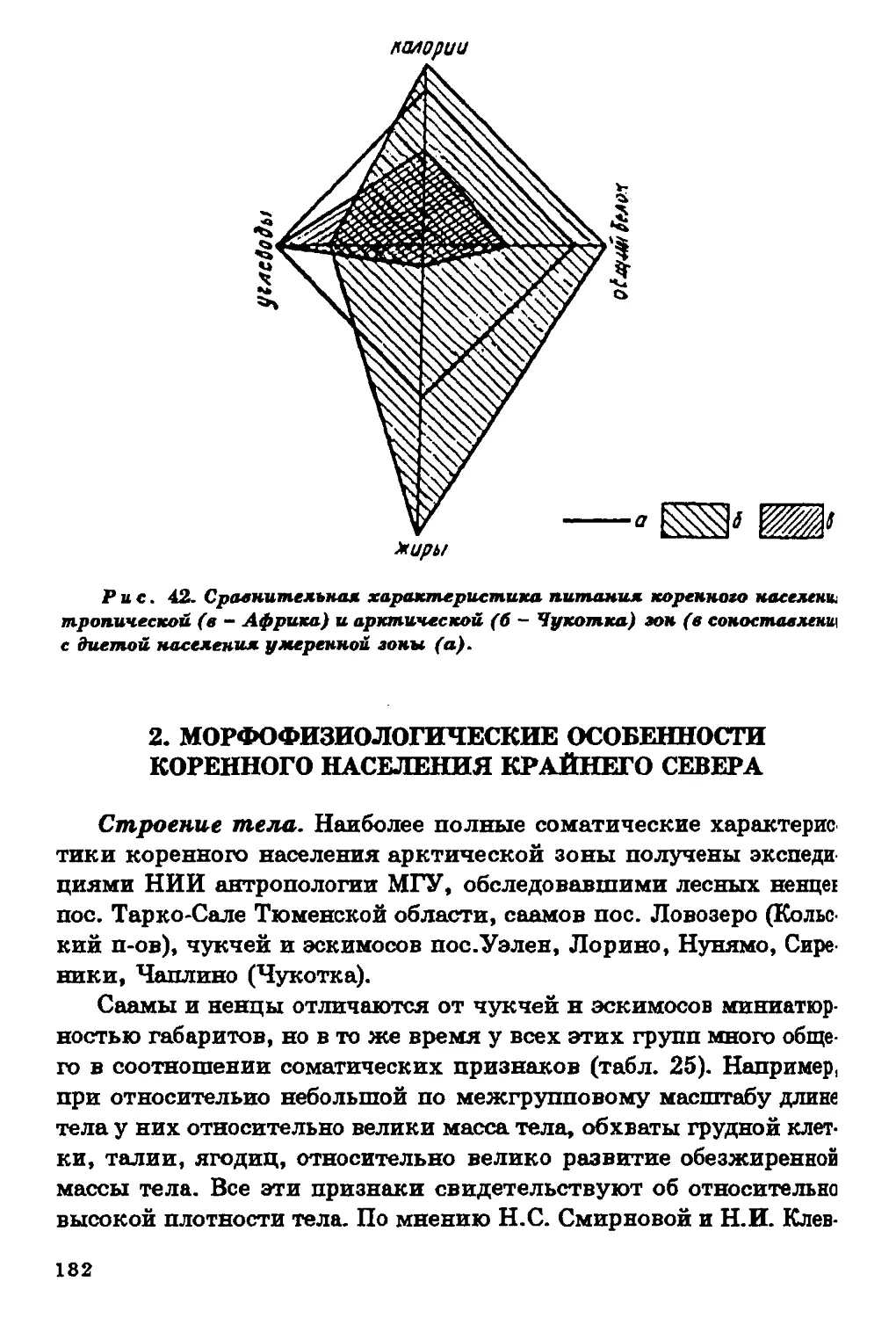
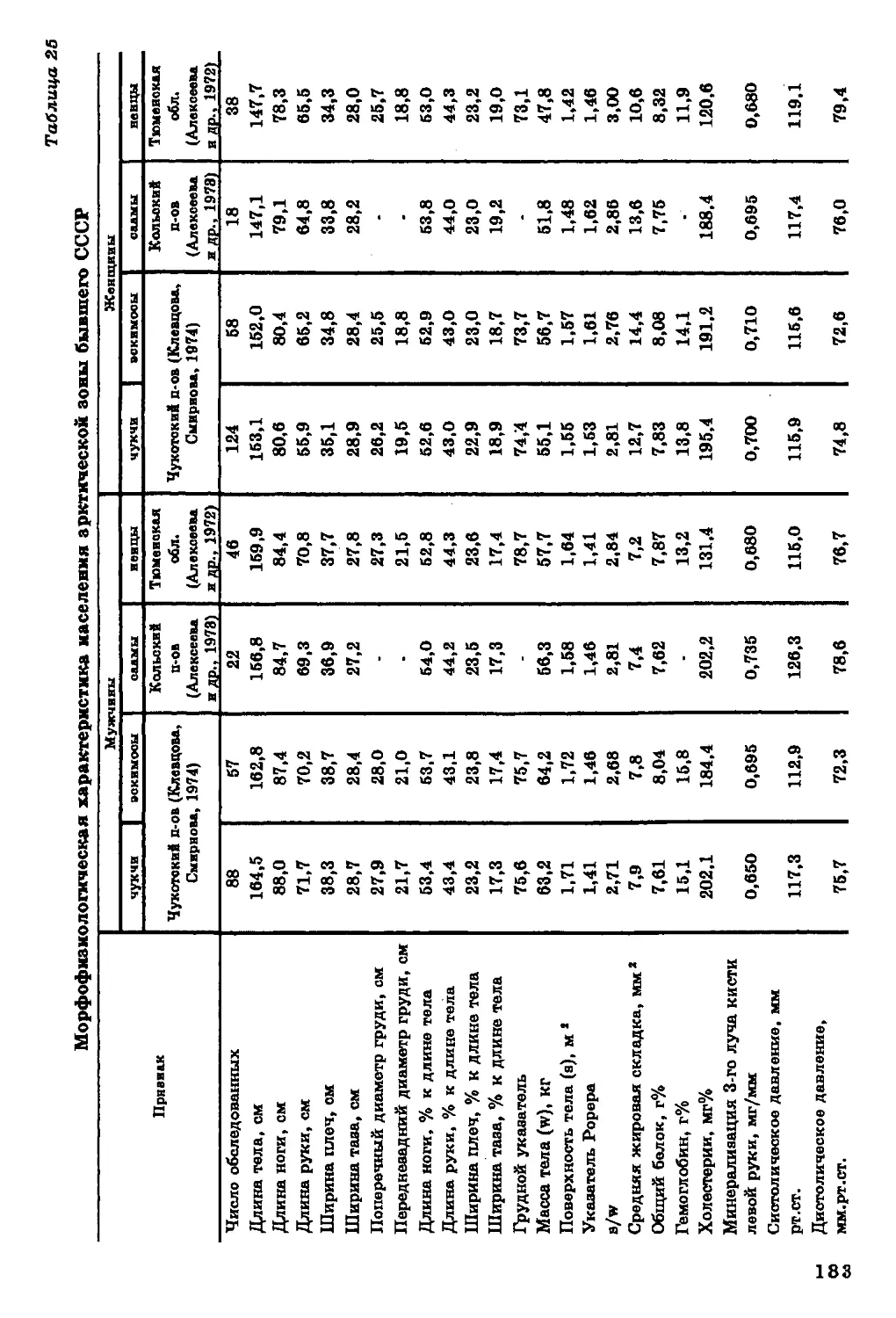






































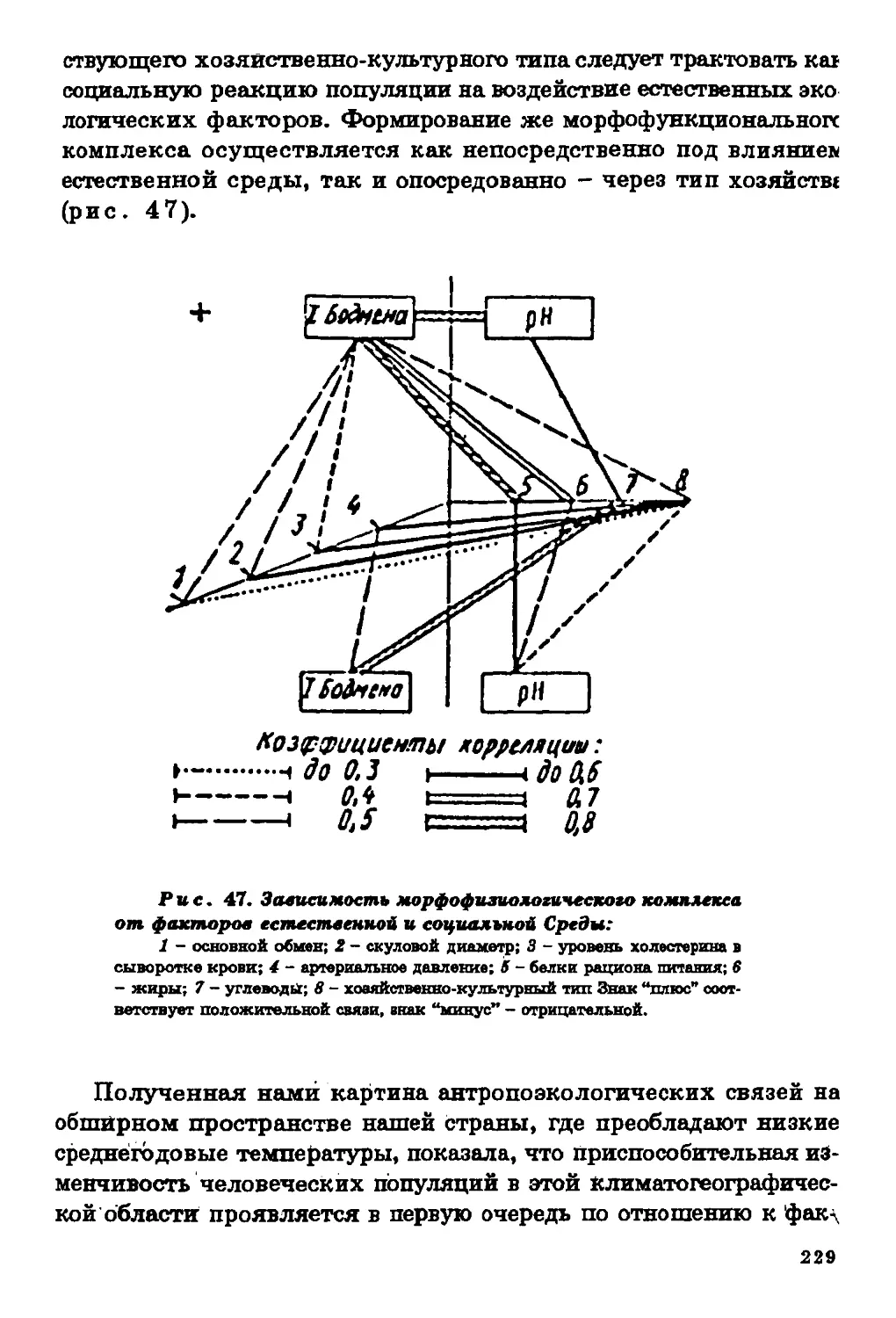









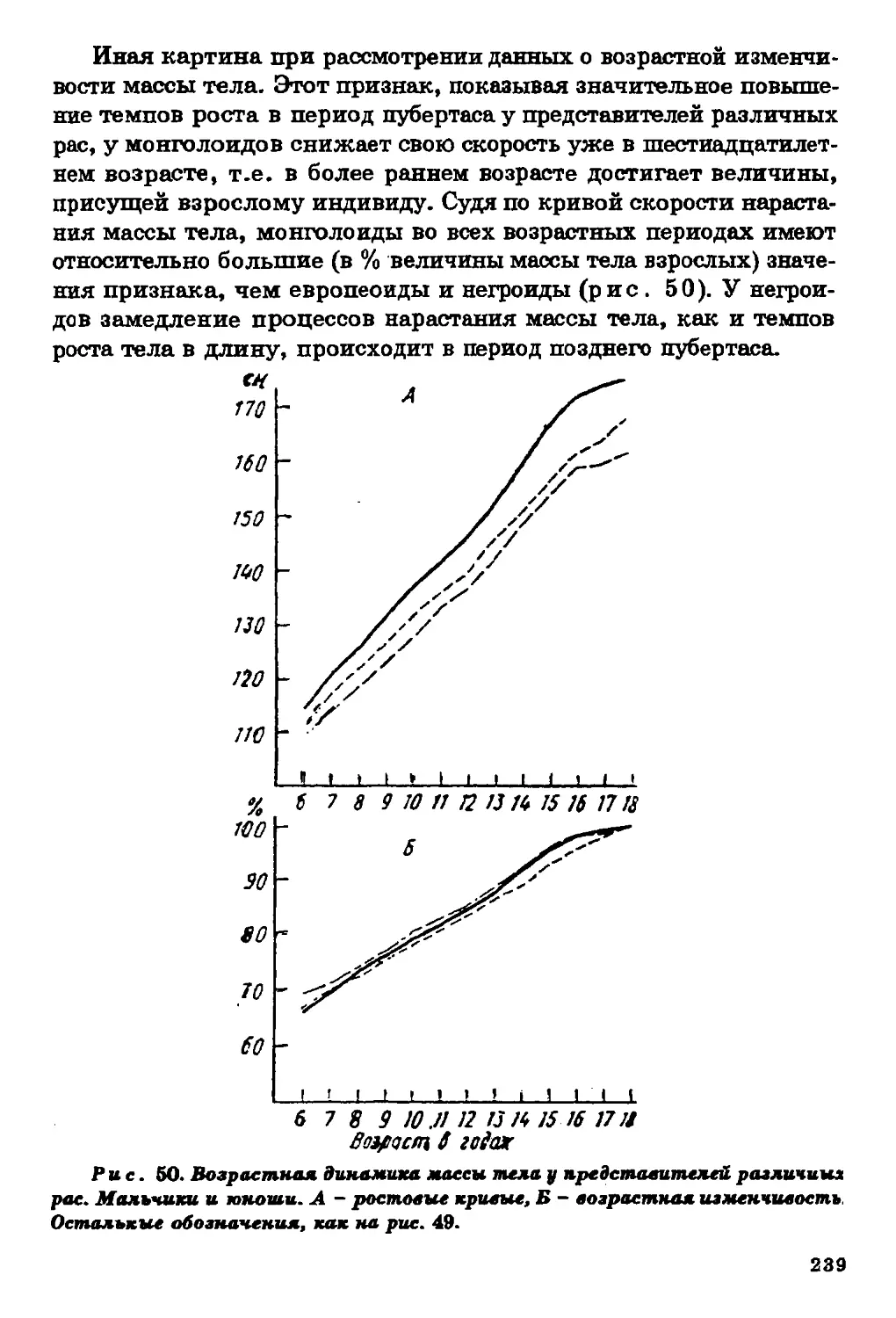

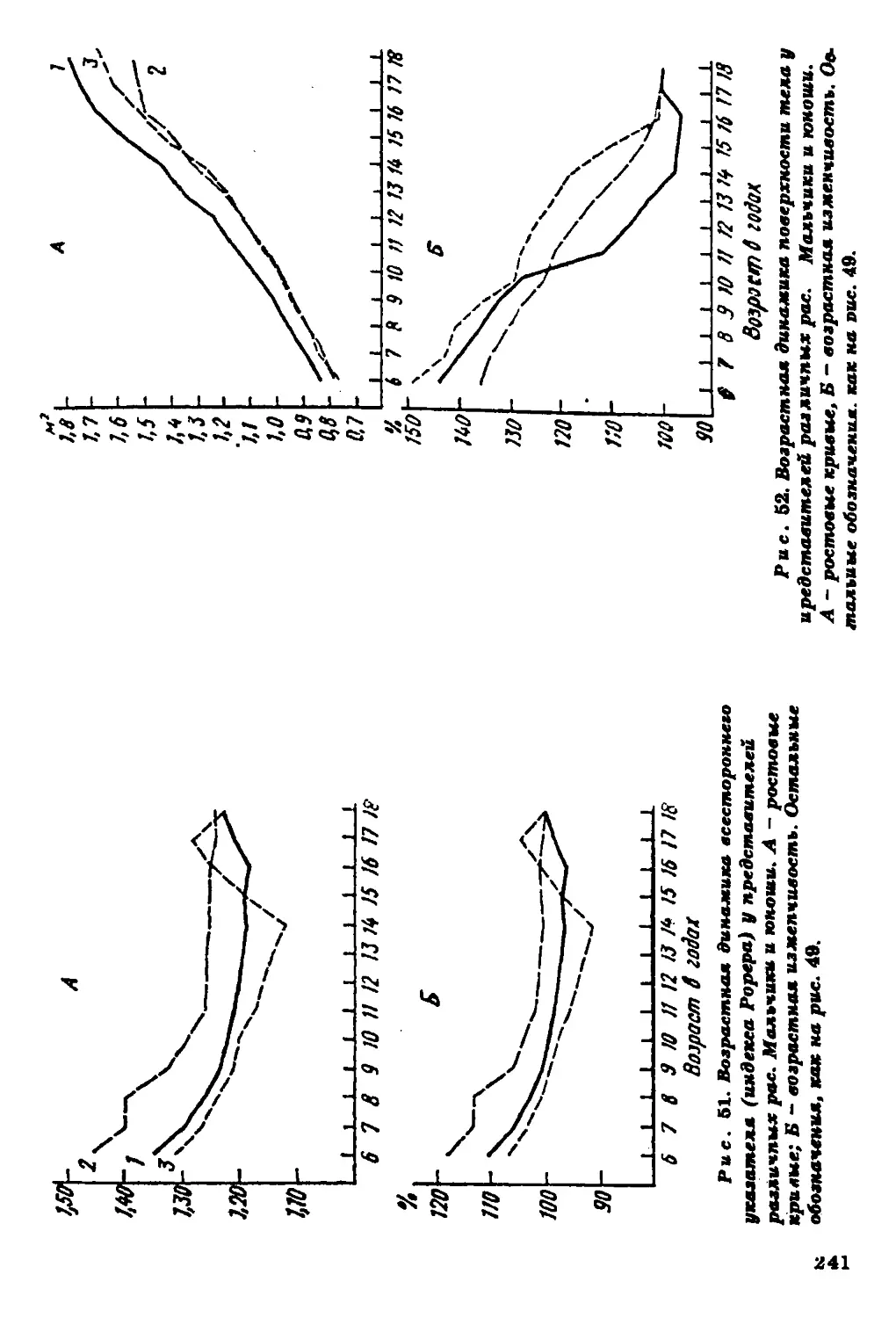

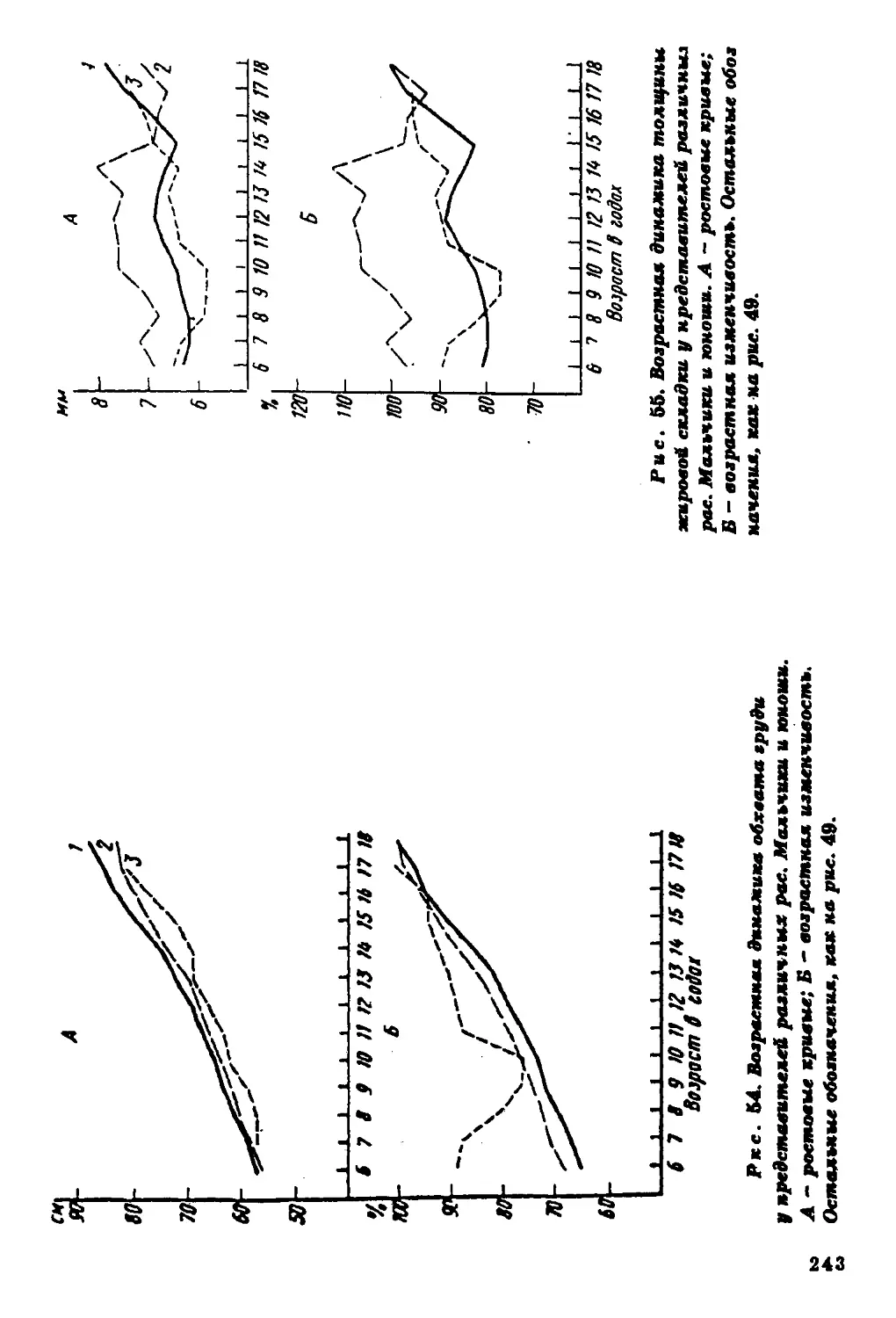


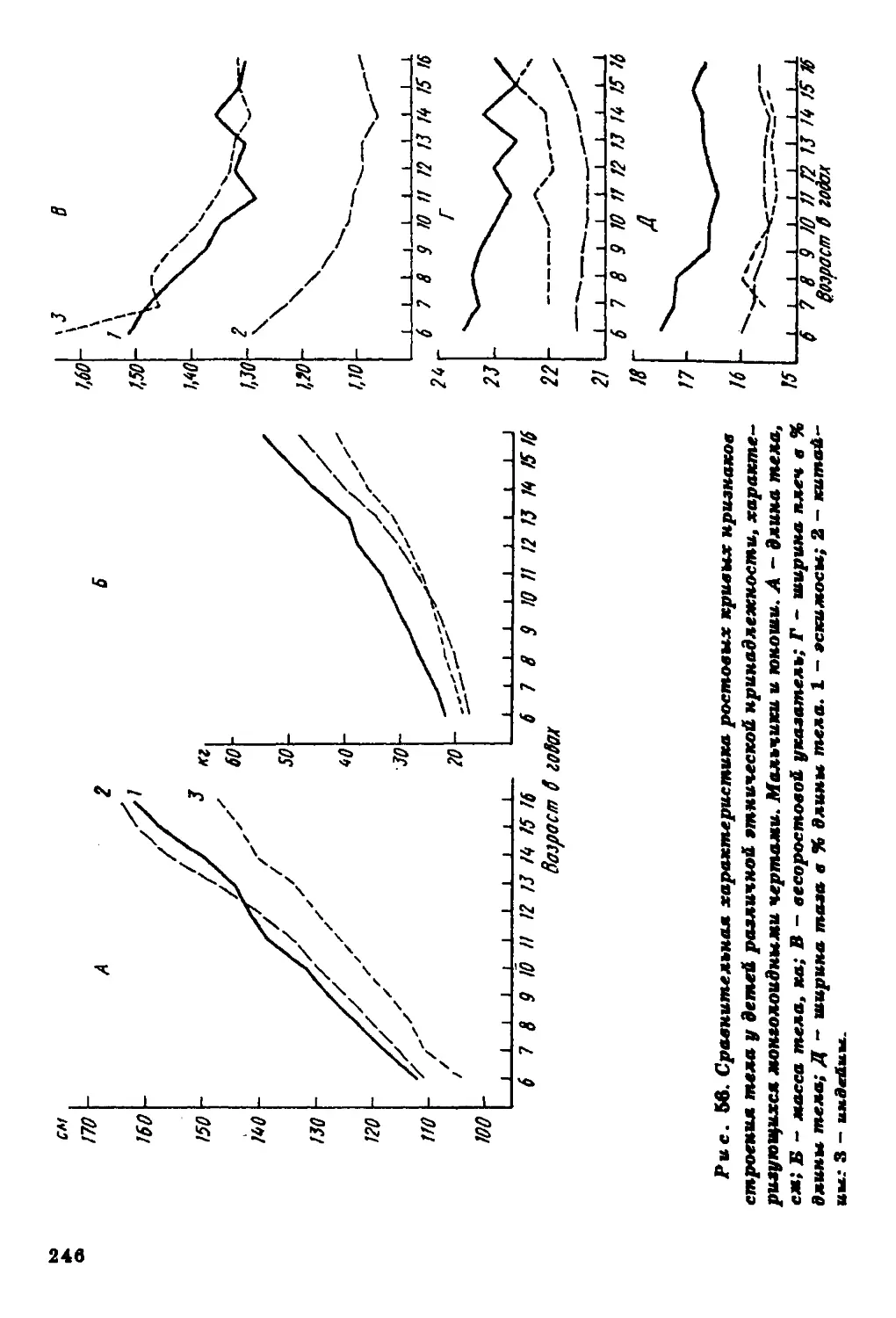

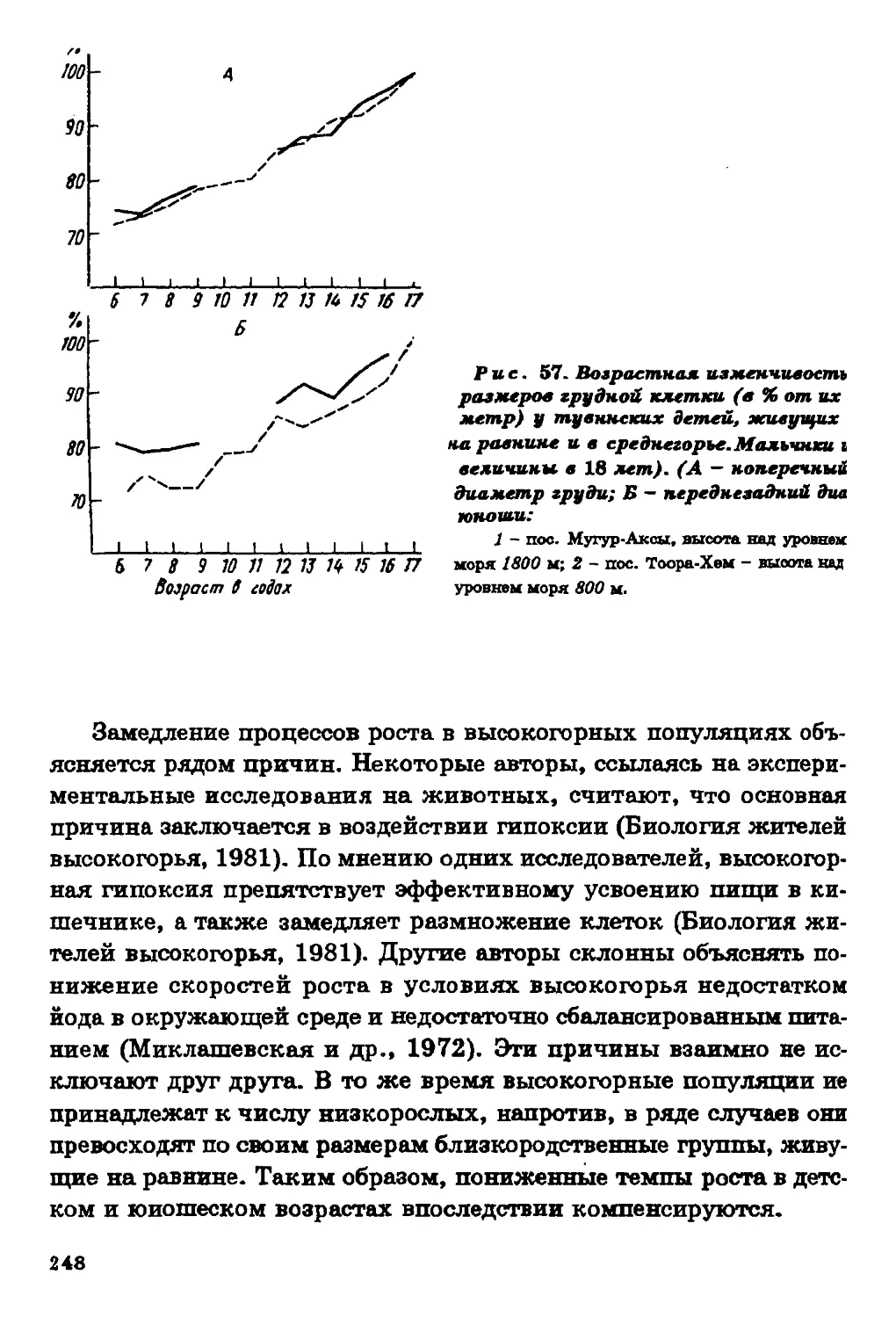




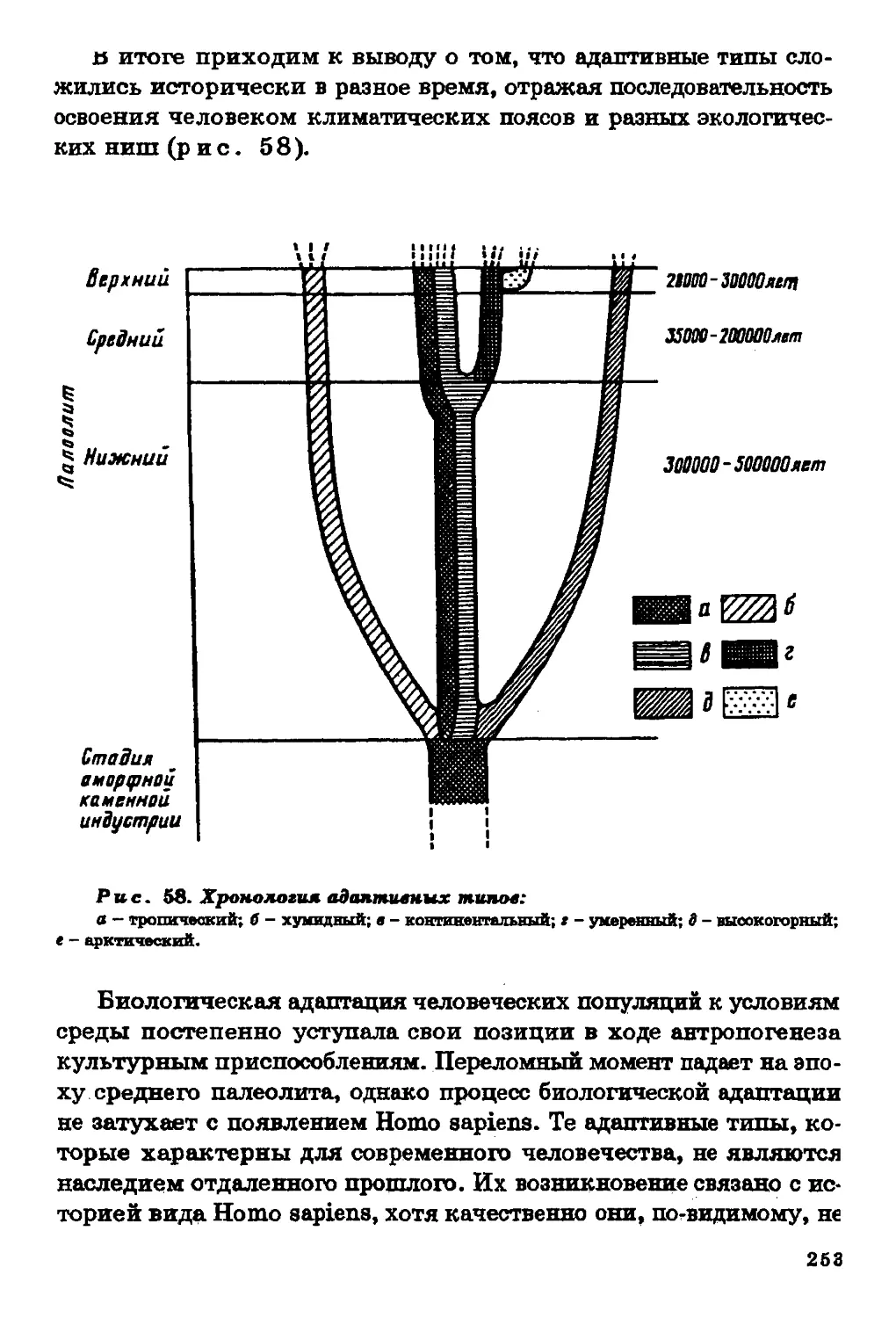

































Текст
• J&JL •
MgM МЕЖДУНАРОДНЫЙ НЕЗАВИСИМЫЙ
\Jr ЭКОЛОГО-ПОЛИТОЛОГИЧЕСКИЙ УНИВЕРСИТЕТ
Т.Е. АЛЕКСЕЕВА
АДАПТАЦИЯ ЧЕЛОВЕКА
В РАЗЛИЧНЫХ
ЭКОЛОГИЧЕСКИХ
НИШАХ ЗЕМЛИ
(биологические аспекты)
Курс лекций
Москва
Издательство МНЭПУ
1998
Алексеева Т. И. Адаптация человека в различных экологических иншаа
Земли (биологические аспекты). Курс лекций. - М.: Изд-во МНЭПУ, 1998. - 280 с.
Рецензент: д-р биол. наук, засл. проф. МГУ Е.Н. Хрисанфова
(Московский государственный университет им. М.В. Ломоносова)
В лекциях рассматривается широкий круг вопросов биологической адаптации
человека, приводятся данные по морфофизиологической изменчивости, характеру
адаптивных процессов и особенностям роста и развития коренного населения
различных регионов бывшего СССР. Составлены мировые и локальные карты
антропологических, морфофизиологических и биохимических признаков с высокой адаптивной
значимостью. Обсуждается феномен экологической дифференциации человечества во
времени и пространстве; показана зависимость ряда жизненно важных
биологических характеристик, в том числе роста и развития, от географических условий, а
также совокупное воздействие среды на процессы адаптации человеческих популяций.
Для студентов, аспирантов и специалистов - экологов, антропологов, биологов,
медиков, географов, историков и всех, интересующихся вопросами экологии человека.
The analysis is based on the material collected in expeditions for many years. The
results of the study of morpnophysiological variability as well as the character of
adaptive reactions and the peculiarities of growth and development in aboriginal populations
of the former USSR are presented. Local and world maps of traits with high adaptive
value are given, which made it possible to discuss ecological differentiation of mankind
in time and space and to reveal the relationship between vitally important biological
features and climate and geochemical environment. A new mathematical approach is
developed to study the way of adaptive reactions in human populations to both cultural
and geographical conditions.
© МНЭПУ. 1998
ПРЕДИСЛОВИЕ
Планомерные исследования адаптации человека к экстремаль
ным условиям обитания1 начались более трех десятилетий назад ]
связи с развитием физиологической антропологии, разработкоі
методик, позволяющих изучать обменные процессы на популяци
онном уровне, и внедрением их в практику полевых работ.
Интерес к социальной природе человечества, основным этапа»
его формирования, к этнической истории на долгие годы отодви
нул в тень исследования по биологии человека, в проявлении за
кономерностей которой значительная роль принадлежит влиянии
среды обитания. Между тем развитие науки о человеке подтверж
дало необходимость постановки таких исследований. В них видел
ся путь к объяснению морфологического разнообразия, специфи
ки географической изменчивости многих расово-диагностическю
черт, генетического полиморфизма. В то же время они были чре
ваты чрезмерным биологизаторством, перенесением отношение
животного мира на человека, недооценкой роли социальных явле
ний в жизни человечества. Такого рода опасения волновали уче
ных на заре исследований по биологии человека, они продолжаю':
нас волновать и сейчас. Осторожность, с которой следует относить
ся к этой проблеме, прекрасно выражена в рецензии Л. Синицко
го (1903, с. 83-84) на книгу П. Бека "Биологические корни челове
ческого общества", вышедшую в свет в 1903 г.: "Перенесение от
ношений животного мира на человека допустимо, конечно, лиш\
Под экстремальными понимаются условия, в которых для нормального труда
отдыха и поддержания высокого уровня здоровья населения требуется создание спе
циальной системы жизнеобеспечения. Экстремальность зоны оценивается по состс
янию адаптационных систем пришлого населения (ели Прохоров Б?. Медико-геогра
фическая информация при освоении новых районов Сибири. Новосибирск, 1979).
с соблюдением крайней осторожности, так как в культурную
жизнь человека привходит много новых факторов, которых нет
в жизни животных, но тем не менее небесполезно выделить из
сложного сплетения разнообразнейших факторов простейшие
силы и проследить за теми результатами, какие явились бы в
том случае, если бы налицо были одни эти силы. При этом,
конечно, необходимо не допускать смешения полученных
результатов с тем, что существует в действительности, и смотреть на
них только как на приближение к ней, которое тем ближе, чем
меньшее количество факторов мы исключили из сферы нашего
исследования9'.
Четкое оформление интереса к биологическим проблемам
человечества приходится на начало XX в. Заслуживают глубокого
внимания вопросы, волновавшие тех, кто стоял у истоков биологии
человека. Не вызывало сомнений, что при изучении влияния
климата на человеческий организм должны быть приняты во
внимание расовые особенности. Особая роль отводилась процессам
смешения с коренным населением в освоении новых территорий и
акклиматизации; отмечалась специфика некоторых этнических
заболеваний; в сравнительном аспекте рассматривалась
приспособляемость различных этносов к новым условиям обитания;
значительное внимание уделялось вопросам дифференциальной
плодовитости. Замечателен был и дифференцированный подход к
географической среде обитания с точки зрения ее воздействия на
человеческий организм, а также к некоторым социальным
влияниям, возможно нейтрализующим воздействие среды. Речь идет о
некоторых этнографических особенностях и различиях в социально-
экономических условиях жизни населения.
В отечественной антропологии среди крайне
немногочисленных работ по проблеме адаптации в 20-60-е гг. особое место
занимают работы В.В. Бунака по акклиматизации человеческих рас,
В.В. Гинзбурга - по истории и биологии горных таджиков, П.И.
Зенкевича- по формообразующим факторам скелета, Я.Я. Рогин-
ского и В.П. Волкова-Дубровина — по приспособительным
особенностям черепа в тропической зоне. Характерное для этих рабоі
внимание к биологическим процессам, происходящим в
человеческих популяциях, способствовало в дальнейшем разработке плано-
мерных исследований географической вариабельности
морфологических и физиологических признаков у человека в связи с пробле
мой приспособления к условиям обитания.
4
Было бы неверно думать, что приспособительные особенное™
человека не освещались в других работах этого периода. Им при
давалось значение, но лишь в связи с ранними этапами становле
ния человечества, и исследования эти носили в основном умозри
тельный характер (см.: Происхождение человека и древнейше
расселение человечества. Сб., 1951).
Шестидесятые годы ознаменовались бурным ростом интереса і
вопросам биологии человека, в том числе к проблеме адаптации
Первый опыт целостного и систематического обобщения разнород
ных данных, характеризующих воздействие средовых факторо]
на человеческие популяции в процессе освоения ойкумены, при
надлежит Дж. Уайнеру, одному из авторов прекрасного руковод
ства по биологии человека (Harrison, Weiner, Tanner, Barnikot
1964).
В 1977 г. вышла в свет моя книга "Географическая среда и био
логия человека", в которой на основе обобщения данных по морфо
физиологической изменчивости человеческих популяций, полу
ченных в результате систематических экспедиционных исследова
ний во многих географических регионах нашей страны, а такжі
почерпнутых из всех доступных литературных источников, был<
описано явление экологической дифференциации человечества
высказана и аргументирована гипотеза адаптивных типов как нор
мы биологической реакции человеческих популяций на комплек<
окружающих условий.
За истекшее время были проведены новые исследования мор
фофизиологических особенностей населения Центральной Азии і
Камчатки- Появилась возможность не только расширить напи
представления о характере адаптивных процессов, но и конкрети
зировать задачи изучения взаимоотношения человеческих популя
ций с естественной и социальной средой.
Выход в свет обширной сводки мировых данных о процесса:
роста и развития, выполненной в рамках Международной биоло
гической программы (IBP), позволил с достаточной полнотой рас
смотреть закономерности временной изменчивости адаптивны:
особенностей человеческих популяций (Wordwide variation in hu
man growth, 1976).
Наконец, исследования биохимического полиморфизма у чело
века, широко и плодотворно ведущиеся в последние годы, предос
тавили в наше распоряжение новые факты, подтверждающие на
следственную природу адаптивности.
В книге "Адаптивные процессы в популяциях человека", вы
шедшей малым тиражом и очень быстро исчезнувшей с книжны:
прилавков, я вновь вернулась к проблеме адаптации человечески:
популяций к окружающим условиям с целью обсуждения некото
рых фундаментальных направлений, не получивших должног*
отражения в предыдущей монографии.
Выход в свет настоящего издания обусловлен резко возросших
интересом к экологическим проблемам человечества и явным де
фицитом сведений о популяциях человека, живущих в естествен
ной среде обитания.
Интерес к проблеме биологической адаптации популяций ко
ренного населения к среде обитания в связи с усилением эколо
гического направления во всех естественных и гуманитарных на
уках не только не ослабевает, но требует дальнейшего расширени:
фактологической базы и более подробного освещения взаимоотно
шений человечества со средой обитания.
Книга значительно расширена и дополнена результатами но
вейших исследований, проведенных на территории бывшего Со
ветского Союза и в Монголии.
Охватывая в фактологическом отношении почти всю ойкуме
ну, что и отражено в названии, курс лекций рассчитан на студен
тов-экологов и представителей других родственных специальное
тей.
В основу книги, предлагаемой всем, интересующимся пробле
мами экологии человека, положены результаты многочисленны:
экспедиций, проведенных под руководством автора в различны:
экологических нишах, а также зарубежные исследования взаимо
действия человеческих популяций со средой обитания, получив
шие широкое распространение во многих странах.
Приношу глубокую благодарность моим ближайшим кол
легам и товарищам по работе: В.П. Волкову-Дубровину, ОМ
Павловскому, Н.С. Смирновой, Л.К. Гудковой, ЭЛ. Шауро, З.А
Голубчиковой, В.А. С?ицину, В.А. Бацевичу, НЯ. Клевцовой
Т.А. Чикишевой, И.Г. Клюевой, О.В. Ясиной, с чьей помощь*
были осуществлены многолетние антропологические исследи
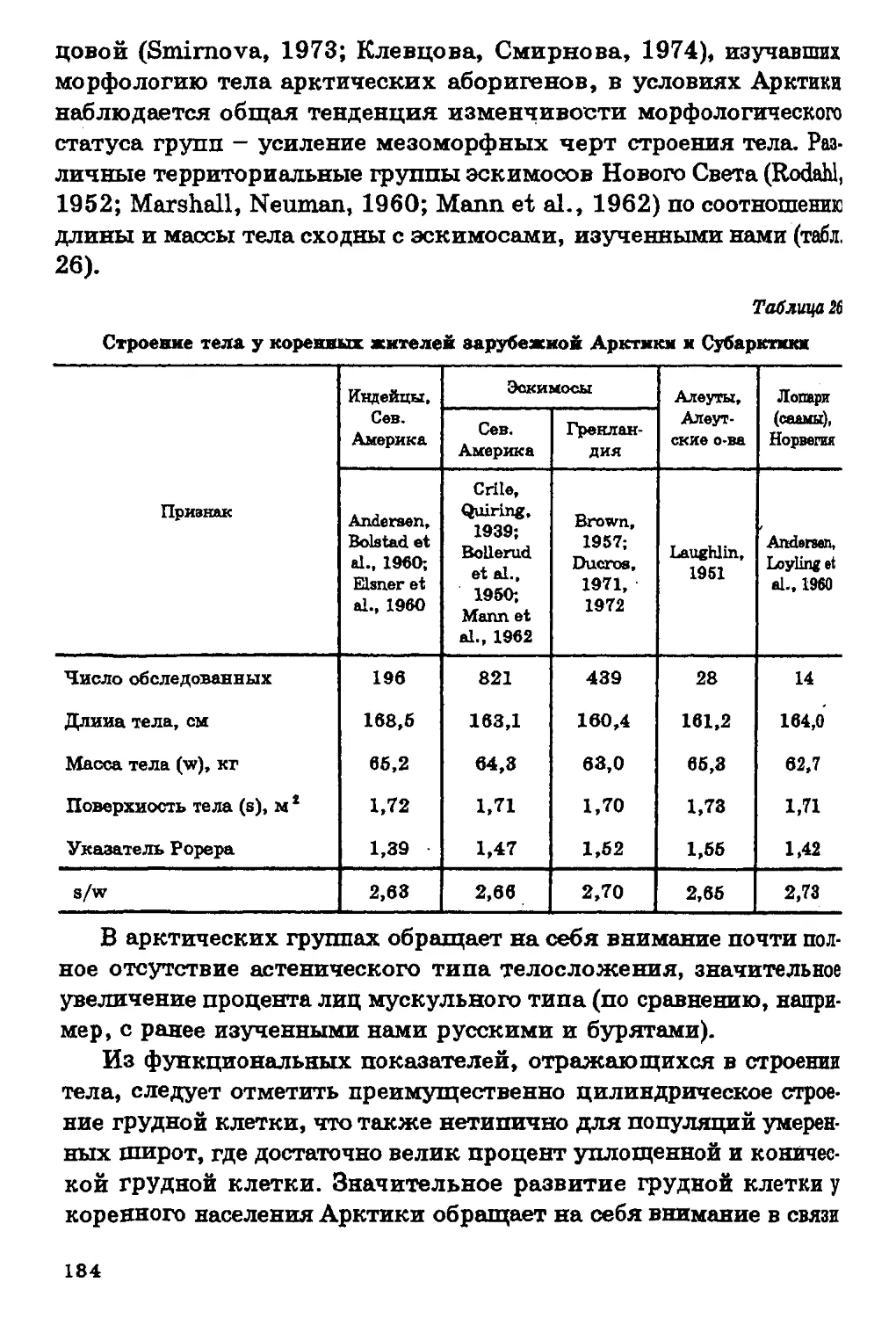
вания коренного населения различных регионов нашей странг
и зарубежья»
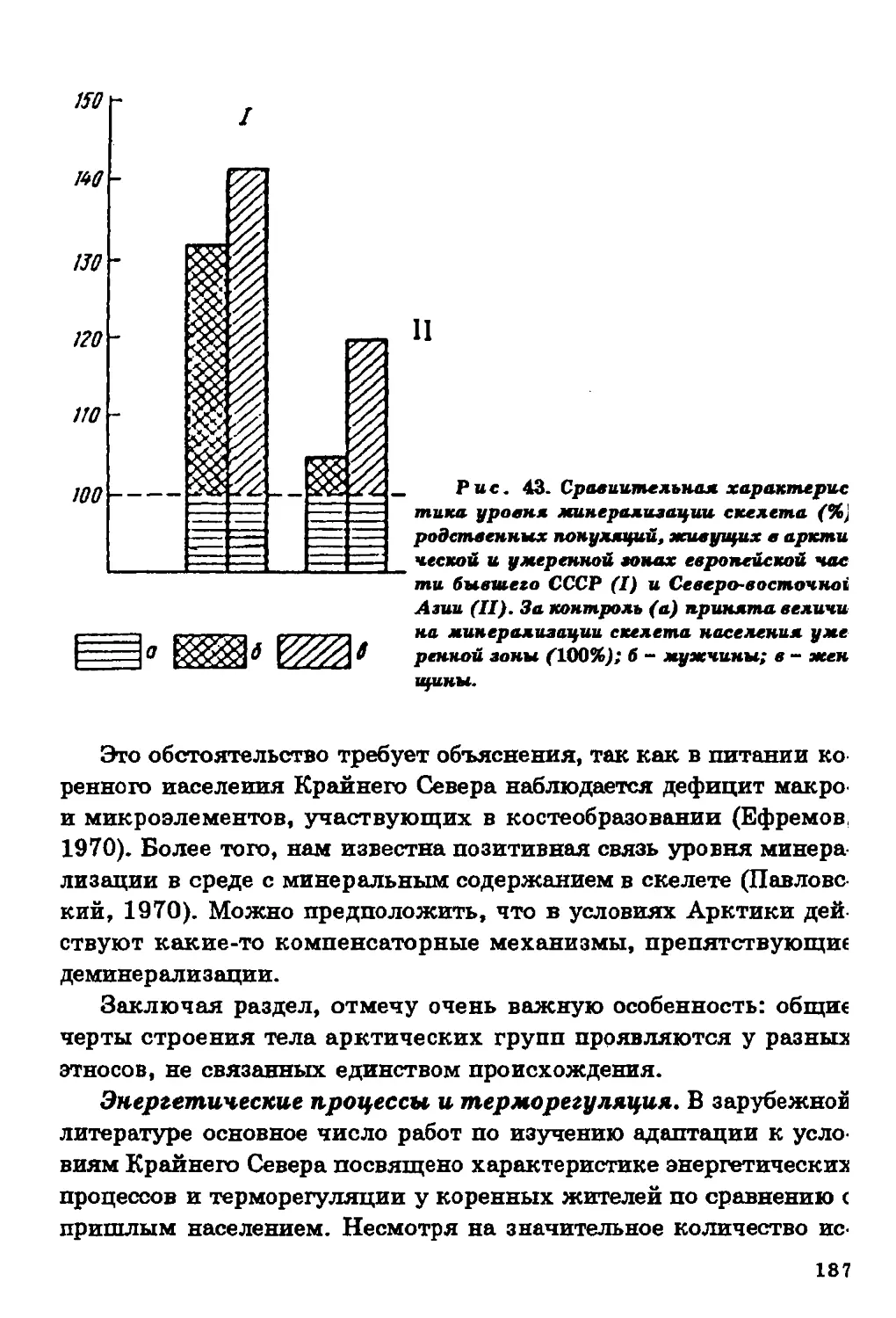
б
лекция і
ИЗУЧЕНИЕ ПРОБЛЕМЫ АДАПТАЦИИ В АНТРОПОЛОГИИ
Адаптация человека
и Международные биологические программы МВР и МАВ.
Отечественные исследования по адаптации человека.
1. КРАТКАЯ ИСТОРИЯ
АНТРОПОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
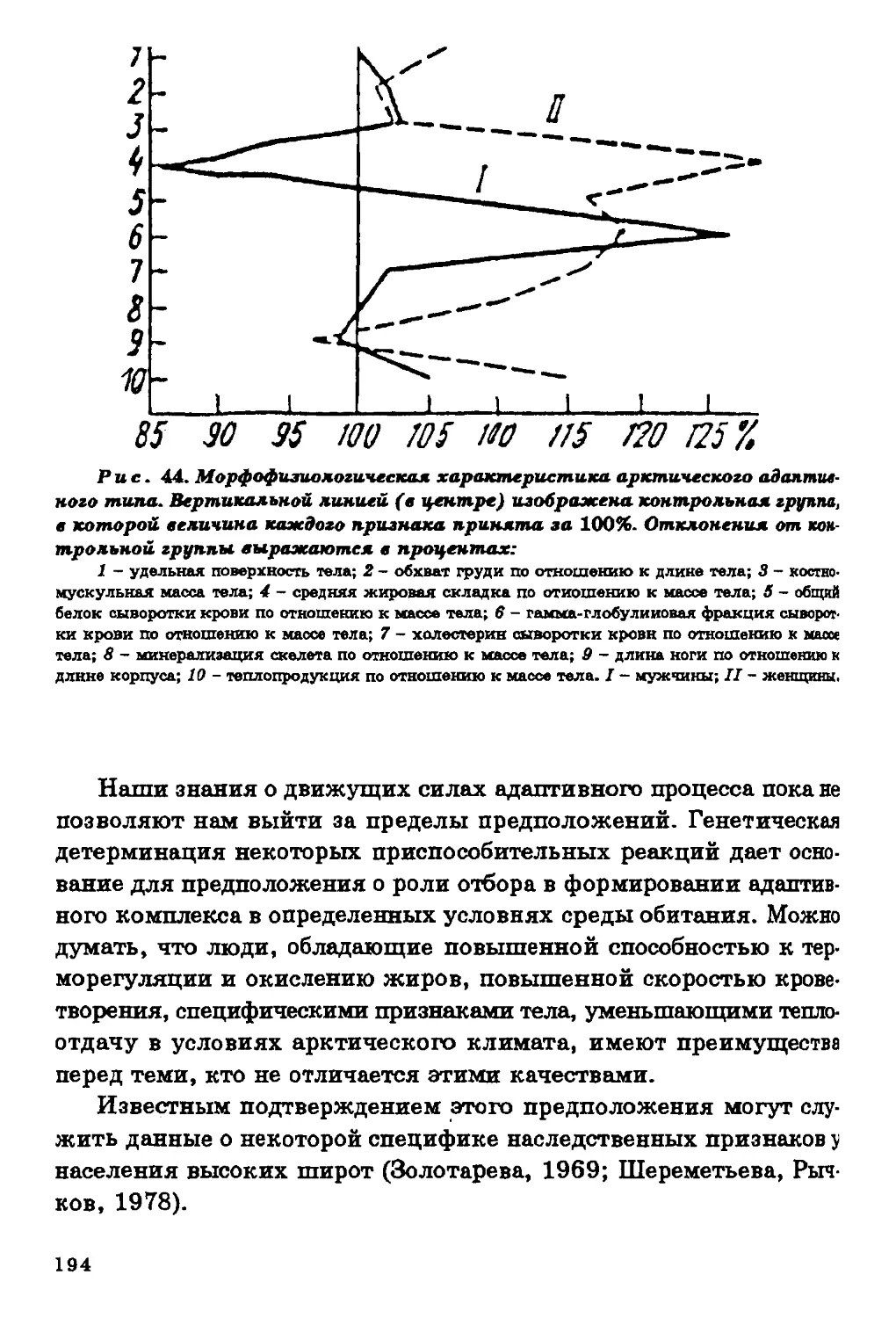
ПО АДАПТАЦИИ К ЭКСТРЕМАЛЬНЫМ УСЛОВИЯМ СРЕДЫ
На VII Международном конгрессе антропологических и этно
графических наук (МКАЭН), состоявшемся в Москве в 1964 г., по
лучила "права гражданства'9 новая отрасль антропологии - физи
ологическая антропология, основной темой которой стало изуче
ние приспособительной изменчивости у человека как вида, прояв
ляющейся не только на уровне генетических, структурных и фи
зиологических систем, но и в демографических процессах,
происходящих в популяциях.
После почти сорокалетнего перерыва в антропологической
науке вновь возник интерес к проблеме адаптации. Характерно, чтс
подобный хиатус в области изучения приспособительной
изменчивости присущ всей мировой физической антропологии, основное
интерес которой в период 20-50-х гг. сосредоточивался на
процессах антропо- и расогенеза, этнической истории, морфогенеза,
динамики ростовых процессов, прикладной антропологии. Внимание
к проблеме приспособительной изменчивости было вызвано, по-
видимому, требованиями научно-технического прогресса,
необходимостью промышленного освоения новых территорий, защиты
окружающей природной среды от последствий
научно-технической революции и, наконец, защиты человека от загрязнений
среды отходами производства.
Изучение процессов адаптации требовало широкого
комплексного подхода к оценке морфологических и физиологических осо
бенностей популяций человека в определенных условиях среды и.
соответственно, создания комплексной методики, позволяющей не
популяционном уровне производить обследование населения раз
личных экологических ниш.
Методический подход к исследованию приспособительной из
менчивости у человека был выработан на VII МКАЭН. Там же со
7
стоялся первый обмен мнениями относительно целей и задач это
го исследования, что позволило в известной мере унифицировав
программу антропологических исследований. В общем виде оні
включает изучение строения тела, различных видов обмена ве
ществ, физиологических и биохимических показателей крови
ряда генетических маркеров и демографической структуры попу
ляций. На Конгрессе определилось и основное направление иссле
дований - адаптация к экстремальным условиям обитания.
Существенную роль в разработке проблемы адаптации челове
ка на популяционном уровне сыграла Международная биологичес
кая программа (IBP, 1964-1974). Под эгидой IBP было опублико
вано несколько сводок по народам мира, заселяющим различны
в экологическом отношении территории.
В 1964 г. состоялся первый симпозиум по адаптации человека
организованный Дж. Уайнером. Итогом его был сборник докладов
представляющий собой обзор современного состояния знаний :
области изучения приспособляемости человека к условиям холода
повышенной влажности, пустыни, высокогорья и т. п. (Baker
Weiner, 1966). Большое внимание на симпозиуме было уделен<
особенностям роста и развития в экстремальных условиях, различ
ным аспектам питания и медико-географическим характеристи
кам. Накопленные ранее антропологические и генетические дан
ные были рассмотрены под углом зрения адаптивных возможное
тей человеческих популяций.
Международная биологическая программа была выполнена і
1974 г. Специальный комитет IBP представил одной из секций
работавших в рамках этой программы, а именно секции "Адап
тация человека", возможность публикации ряда сводных томов
обобщающих результаты исследований, проведенных в разны:
странах, и монографий, посвященных комплексным исследованию
коренного населения отдельных географических регионов. Пер
вый том - "Word wide variation in human growth" (1976), под ре
дакцией Дж. Таннера и Ф. Эвелеса, представляет собой системати
шрованную и весьма подробную сводку данных по росту и разви
тию детей, собранных исследователями 52 стран в рамках IBP. I
рей обобщены результаты обследования коренного населения Аме
рики, Европы, стран Ближнего Востока, Юго-Восточной Азии, Ав
етралии и Океании. Процессы роста и развития рассмотрены ]
8
непосредственной связи с особенностями климата, заболеваемости
питания и генетической конституции.
Второй том - "Population structur and human variation" (1976)
вышедший под редакцией Дж. Харрисона, охватывает серию ис
следований особенностей биологической структуры популяций
Под структурой популяции понимаются биологические взаимосвя
зи между индивидами и отдельными семьями в сообществе. Особое
внимание в этом томе уделено исследованию генетической диффе
ренциации популяций, возможность которого открылась с изуче
нием ряда полиморфных признаков с простым типом наследование
(Mourant, 1976; Rychkov, Sheremetyeva, 1976). Книга содержит об
ширные фактические данные по народностям Северо-Восточнов
Азии, островов Тихого океана, бассейна Амазонки, Африканские
саванн. Большой интерес представляют исследования
генетических особенностей человеческих популяций в свете проблемы адап
тации. Они касаются как коренного, так и пришлого населения,
причем значительное место отводится действию отбора в
современных популяциях. В этом томе опубликованы результаты
исследований генетических процессов в системе изолятов североазиатсков
части СССР (Rychkov, Sheremetyeva, 1976).
Третий том - "The biology of hightaltitude people9* (1978) издав
под редакцией П. Бейкера и переведен на русский язык в 1981 г.
Он представляет собой систематизированную сводку данных о
приспособляемости человеческих популяций к условиям высокогорья,
полученных для коренного населения Эфиопии, Перуанских Анд,
Гималаев, Тянь-Шаня и Памира. Рассмотрены особенности
генетической и физиологической адаптации к высокогорью, физическая
работоспособность коренных горцев, репродуктивная функция,
характер процессов роста и развития высокогорных популяций.
Здесь нашли отражение и результаты многолетних отечественных
исследований процесса роста и развития, проводимых Н.Н.
Миклашевской с сотрудниками в высокогорных районах Средней Азии
(см. Фрисанчо, 1981).
Жителям высокогорья посвящено также фундаментальное
многоплановое изучение индейцев кечуа в Перу, проведенное под
руководством П. Бейкера (Man in the Andes. A multidisciplinary s-
tudy of high-altitude Quechua/Ed. P. Baker, M. Little, 1976). В нем
с большой полнотой рассмотрены биологическая и социальная
адаптации человека в условиях высокогорной экосистемы. Этс
9
исследование также выполнено в рамках .международной оиологи
ческой программы, и основные его результаты включены в свод
ный том по биологии жителей высокогорья.
Региональным исследованиям эскимосов Аляски посвящена і
монография "Eskimos of Northwestern Alaska. A Biological Perspec
tive" (1978). В ней рассмотрены характерные черты популяцион
ной и демографической структуры эскимосов, особенности строе
ния тела, биохимии и физиологии. Большое внимание уделен<
физической работоспособности в условиях Арктики, а также вли
янию питания на течение метаболических процессов и физическо<
развитие.
Обращает на себя внимание то обстоятельство, что в обоих то
мах подчеркивается необходимость многодисциплинарного иссле
дования процесса приспособительной изменчивости у человека і
конкретных экологических нишах (имеется в виду не только боль
шое число морфологических, функциональных и генетически?
характеристик изученных популяций, но и многообразие их свя
зей со средой, многогранность влияния окружающей среды на че
ловека). Именно поэтому биологические характеристики эскимо
сов и кечуа рассмотрены в тесном единстве с географической і
культурной средой, определяющем своеобразие приспособитель
ных реакций в человеческих популяциях.
Новые перспективы в изучении адаптивных процессов в попу
ляциях человека были открыты Международной биологическое
программой "Человек и биосфера9' ("The Man and the Biosphen
Programm", или MAB), принятой в ноябре 1971 г. еще до оконча
ния IBP на сессии ЮНЕСКО. В рамках этой программы разработа
но 13 проектов, один из которых (№ 12) касается непосредственно
человека. На заседаниях Генеральной Ассамблеи международны}
биологических обществ в Сиэтле (США) в 1972 г. дискутировался
вопрос о преемственности исследований, проводимых IBP и про
граммой МАВ. С целесообразностью такой преемственности согла
сились все участники Ассамблеи. Проект № 12 получил название
"Взаимодействие между изменениями окружающей среды и адап
тивной, демографической и генетической структурой популя
ций" (изложение программы МАВ и Проекта № 12 см.: Барбашо
ва, 1974).
10
исновное внимание в этом проекте сосредоточено на изучении
антропогенетических, демографических и социальных изменений
в человеческом обществе и связанной с ними естественной среде
обитания в условиях научно-технического прогресса, в то время
как в IBP ведущим направлением было исследование антропогене-
тических и демографических процессов у коренного населения в
условиях естественной среды, еще не подвергшейся воздействию
человека.
Программа "Человек и биосфера" (МАВ) поставила ученых
различных стран перед необходимостью комплексных мультидисцип-
линарных исследований, решения многих проблем на стыке наук,
разработки методов и понятийного аппарата этих наук. На основе
синтеза достижений биологии человека, медицины, географии,
социологии, этнографии, философии и др. идет формирование
новой отрасли науки - экологии человека, изучающей систему
"человек - природа". Содержание понятия "экология человека" пока
остается дискуссионным (Теория и методика... 1974;
Преображенский, Райх, 1974; Географические аспекты... 1975; Казначеев,
1984). Экологию человека можно рассматривать и как
самостоятельную науку с неочерченным еще полностью комплексом
проблем взаимодействия человека с окружающей средой, и как
систему научных знаний (я склоняюсь к последнему определению).
Современный этап развития биологии, отличительной чертой
которого является поиск причинно-следственных связей, требует
внедрения в любую частную науку экологического мышления
(Шварц, 1976). В рамках экологического подхода все аспекты
изучения человека, включая и социальные стороны его
жизнедеятельности, основываются прежде всего на взаимодействии
человека со средой. Сложность экологического изучения человека
заключается в переплетении естественных и социальных факторов,
определяющих это взаимодействие, и на разных этапах
исследования мы вынуждены искусственно вычленять биологические или
социальные его стороны.
Среди антропологических работ, вызванных к жизни IBP,
особое место занимают отечественные исследования на территории
бывшего СССР, в частности изучение генетических процессов ш
Северо-Востоке Азии (Rychkov, Sheremetyeva, 1976), а также рос
та и развития высокогорных популяций Средней Азии (Микла
шевская и др., 1972, 1975).
и
Результаты отечественных исследований по проблеме адапта
ции, выполненных непосредственно в рамках Международной био
логической программы, приведены в двух сборниках: "Адаптации
человека" (1972) и "Человек и среда*9 (1975). В них представлень
антропологические, физиологические и ме дико-генетические ха
рактеристики коренного населения различных географически:
зон бывшего СССР, а также пришлых контингентов. Исследовали:
проводились в различных районах: на Крайнем Севере, в аридны:
зонах Средней Азии, на Памире, Тянь-Шане и Кавказе. К сожале
нию, материалы сборников освещены весьма фрагментарно, статы
разноплановы, не сгруппированы тематически, поэтому сборник]
носят в значительной мере информативный характер.
В Научно-исследовательском институте антропологии МГУ бы
ла разработана комплексная морфофизиологическая программ;
исследования адаптивных процессов в человеческих популяциях :
соответствии с задачами, стоявшими перед антропологами всеіч
мира с середины 60-х гг. и нашедшими отражение в Международ
ной биологической программе (Морфофизиологические исследова
ния в антропологии, 1970; Методика морфофизиологических ис
следований в антропологии, 1981). По этой программе на террито
рии бывшего СССР в течение 30 лет было обследовано коренно-
население Северо-Востока Азии (Алексеева, Клевцова, 1980; Н;
стыке Чукотки и Аляски, 1983); Центральной Азии (Алексеева ]
др., 1970,1971; Антропоэкологические исследования в Туве, 1984)
Средней Азии (Павловский, 1970; Волков-Дубровин и др., 1975
1977а, 19776; Смирнова, 1979; Архангельская и др., 1980, 1981
Алексеева и др., 1984); северных районов Западной Сибири (Алек
сеева и др., 1972); восточноевропейской части СССР (Алексеева і
др., 1973; Клевцова, 1976; Смирнова, 1977; Саливон, 1977; Тега
ко, Микулич, Саливон, 1978); Кавказа (Павловский, 1982; Волков
Дубровин и др., 1982), Русского Севера (Экологические пробле
мы... 1993), Таким образом, исследованиями были охвачены ко
ренные жители различных климатических зон - Арктики и Суб
арктики, пустыни, высокогорья, континентальных районов Сиби
ри и регионов умеренной зоны.
В моих книгах "Географическая среда и биология человека
(1977), "Адаптивные процессы в популяциях человека" (1986
были рассмотрены все накопленные к тому времени антропологи
ческие данные по различным регионам мира, характеризующи
12
морфофизиологические особенности человеческих популяций, чтс
позволило проследить закономерности экологической
изменчивости современного человечества и аргументировать гипотезу
адаптивных типов, сопряженных с различными
зонально-географическими регионами земного шара.
2. ИСТОКИ ОТЕЧЕСТВЕННЫХ ИССЛЕДОВАНИЙ
ПО АДАПТАЦИИ
Как уже отмечалось, планомерные исследования процессов
адаптации человеческих популяций к естественной среде
обитания начались в середине 60-х гг., тем не менее истоки интереса к
проблеме приспособительной изменчивости у человека восходят к
русской и зарубежной антропологии конца XIX - начала XX в.
(Бунак, 1924).
Я не имею намерения анализировать литературные источники,
относящиеся к этому времени. Они систематизированы и
проанализированы В.В. Бунаком с позиций функциональной
антропологии. В 1924 г, в "Русском антропологическом журнале9* была
опубликована его статья "Об акклиматизации человеческих рас и
сравнительном значении определяющих ее факторов'* (Этюд по
биологии человека). В статье дается определение акклиматизации
как явления, присущего человечеству в целом, и
рассматриваются различные факторы акклиматизации. Сама проблема и
характер ее освещения до сих пор сохраняют актуальность не только
для антропологов, но и представителей смежных наук.
Под акклиматизацией В.В. Бунак понимал "способность
организма подвергаться незначительным морфологическим и
функциональным изменениям, не нарушающим общий тип индивидуума
и обеспечивающим способность жить и, главное, размножаться
в не свойственной индивидууму новой обстановке" (Бунак, 1924,
с. 45).
Считая, что проблема акклиматизации представляет собой часть
проблемы приспособительной изменчивости, В.В. Бунак четко
отграничивал явление акклиматизации от глубинных изменений і
строении и функции, которые претерпевают популяции человека,
оказавшиеся в необычных для них условиях, в процессе
длительного приспособления к этим условиям. В числе механизмов при-
13
способления в данном случае В.В. Бунак назвал отбор и метиса
цию.
Среди факторов, по отношению к которым может быть осуще
ствлена акклиматизация, В.В- Бунак выделял четыре основных
метеорологический, эргологический, общебиологический и расо
вый.
Влияние метеорологического фактора рассматривается на при
мере акклиматизации европейцев к условиям тропиков и тропи
ческих аборигенов к умеренному климату.
Отмечая расстройство обмена веществ у уроженцев умеренной
зоны в условиях тропиков, понижение процессов окисления, про
дукции урины и поглотительной способности кишечника, ведущее
к катарам, гиперфункции печени, нарушению сердечной
деятельности и анемии, а также изменения в нервно-психической среде.
В.В. Бунак указывал на факт неодинаковой акклиматизационной
способности различных европейских рас. Так, например, наиболее
высокой способностью к акклиматизации в тропиках обладаю!
уроженцы южной части умеренного пояса. По аналогии можне
привести акклиматизацию русского населения в Сибири. Русские
старожилы Сибири происходят в основном из северных областей
европейской части России.
Что касается акклиматизации уроженцев тропиков в условиях
умеренных широт, в частности африканцев в Америке, то здесь
В.В. Бунак основную причину видел не столько в изменении
физиологического состояния под действием новых условий, сколько
в действии жесточайшего отбора. Ссылаясь на зарубежные данные
концаXIX в., он отмечал, что девять десятых из всех привезенных
в Америку африканцев погибли и лишь одна десятая часть
наиболее устойчивых в физическом и психическом отношениях
индивидов дала потомство.
В.В. Бунак обращал внимание на то, что акклиматизация к
холодному климату осуществляется легче. В качестве примера
приводились колонии датчан в Исландии и других регионах у
Полярного крута.
Рассматривая явление акклиматизации в пределах умеренного
пояса и метеорологические характеристики этой зоны, В.В. Бунак
ведущим фактором здесь считал не температуру, а влажность.
Акклиматизация в данном случае определяется способностью
переселенцев из влажных областей переносить холодный климат
континентальных районов.
14
Оценивая явление акклиматизации к метеорологическим фак
торам, В.В. Бунак отмечал, что успех ее зависит от достепенності
переходов во времени из одной зоны в другую1. В истории народоі
этот факт, по мнению В.В. Бунака, имел немалое значение. Освое
ние новых экологических ниш осуществлялось медленно, и на та
ком уровне явление акклиматизации уступало место адаптации і
процессе отбора.
Эргологический фактор акклиматизации заключается в способ
ности переселенцев к изменению уклада жизни и гигиенической
режима соответственно требованиям новой среды. В первую оче
редь речь здесь идет о характере быта, труда, питания.
Различия в составе пищи коренных народов разных климати
ческих областей очень велики. Тип диеты диктуется не толькс
климатическими условиями, но и особенностями обмена. Режиа
труда также во многих отношениях определяется термическим ре
жимом конкретной территории. Успех акклиматизации будет за
висеть от физиологической способности организма пришельцев в
перестройкам.
К третьему фактору акклиматизации, по мнению В.В. Бунака,
относится приспособление к биологическим условиям среды, т.е.
характер взаимоотношений человека с окружающим раститель
ным и животным миром. Особое место среди биотических
факторов среды принадлежит микроорганизмам - возбудителям
эндемических болезней. Для европейцев губительной в условиях
тропического пояса оказывается малярия, для аборигенов
тропиков в умеренных широтах -туберкулез. Акклиматизацию
осложняют не только эндемические болезни, но и отсутствие
врожденного иммунитета ко многим возбудителям болезней,
свойственного коренному населению.
Основываясь на биохимических данных о свойствах белкох
сыворотки крови у представителей разных рас, В.В. Бунак сделаі
предположение о наличии специфического расового иммунитета.
Более того, вслед за Ч. Дарвином, считавшим, что негроидная рас*
с ее своеобразными соматическими особенностями представляеп
собой продукт отбора против некоторых тропических болезней
В.В. Бунак объяснял характерные особенности европеоидной рась
В современной литературе но высокогорью это явление получило названии
ступенчатой акклиматизации (Агаджанлн, Миррахижов, 1970).
15
как результат отбора против туберкулеза. Развивая эту мысль, он
писал: "Не являются ли расовые различия результатом отбора
типов, стойких против патологических процессов?» Ведь морфо
логические расовые отличия сами по себе по большей части без
различны, и их строгая наследственная фиксация является
малопонятной. Мы можем объяснить ее,., лишь предполагая, чтс
морфологические свойства коррелятивно связаны со свойством,
существенным для жизни индивидуума, а таковым в наибольшей
степени является иммунность против различных болезней" (Бу-
нак, 1924, с. 56). Основываясь на фактах (правда, на единичных]
различия эндо- и экзопаразитов у представителей разных рас, В.В.
Бунак предполагал, что каждой расе свойствен особый эндо- и эк-
зопаразитический мир, к которому она приспособилась в
результате длительного отбора.
Наконец, четвертый фактор акклиматизации - расовый. Ов
непосредственно связан с биотическим фактором, так как
аборигенное население является носителем эндо- и экзопаразитическогс
мира. Кроме того, соприкосновение различных культурных и
психологических миров требует взаимной акклиматизации.
Последний фактор рассмотрен В.В. Бунаком лишь в общих чертах, так
как он в значительной мере обусловлен социальными причинами
и исследоваться должен методами этнографии и этнопсихологии.
В.В. Бунак отмечал, что хотя человек и заселил еще в
первобытном состоянии весь земной шар, этот процесс происходил чрез
вычайно медленно и был связан с образованием оробых расовых
типов, которые обладают ограниченным ареалом обитания и дліі
которых способность укрепиться в новых условиях достигалась
путем длительного и жесткого отбора.
Какова мера приспособительной изменчивости современного
человечества, какими ареалами она ограничивается - вот в сущно
сти вопросы, которые вытекали из самого содержания статьи В.В
Бунака и которые требовали своего разрешения.
В 1926 г. В.В. Бунак предложил программу комплексного
изучения так называемых малых народностей, справедливо полагая
что именно они, являясь коренным населением географические
регионов, не подвергшихся сильному антропогенному воздейст
вию, могут, говоря современным языком, служить "модельными'
популяциями для исследования процесса взаимодействия челове
ка со средой обитания.
16
Рассматривая коренное население как важнейшую
производительную силу страны и настаивая на необходимости исследования
хозяйственно-культурного и социального строя, семейных
отношений и гигиенических условий, В.В. Бунак подчеркивал, что "его
значение особенно велико еще потому, что оно позволяет учесть
многие своеобразные культурно-хозяйственные навыки этих
малочисленных этнических групп, основанные на совершенном
приспособлении к своеобразным условиям местной природы, - навы
ки, которые делают малые народности нередко единственными
приспособленными к эксплуатации природных богатств края и
которые грозят исчезнуть вместе с ними1* (Бунак, 1926, с. 89).
Огромное значение в программе изучения малых народностей
придавалось биологическому фактору приспособляемости к
окружающим природным условиям и взаимодействию с
соседствующим населением, т.е. приспособляемости к метеорологическим,
эргологическим и биотическим факторам. В связи с этим в
программу исследований включалось определение санитарной и
функциональной конституции, физического развития, пропорций тела,
возрастного и полового развития, некоторых биохимических
свойств (групп крови, ферментов), а также степени иммунности и
заболеваемости в отдельных семьях. Большое значение придавалось
и расово-диагностической программе.
Программа комплексного этнографо-антропологического
изучения малых народностей вскоре была воплощена в жизнь. Летом
1926 г. в Тувинскую Народную Республику (теперь Республика
Тува) отправилась первая крупная научная экспедиция,
организованная Научно-исследовательским институтом антропологии МГУ
под руководством его директора В.В. Бунака (об экспедиции см.
подробнее: Вайнштейн, 1982). Помимо В.В. Бунака в Танну-Ту-
винской экспедиции приняли участие Ф.И. Ярхо и М.Г. Левин
(публикацию материалов экспедиции см.: Bounak, 1928; Ярхо,
1947; Вайнштейн, 1982).
Комплексный антропо-этнографический подход был присуш
многим исследованиям конца 20-х - начала 30-х гг. Следует
упомянуть антропологические работы в рамках Комиссии по
изучению естественных производительных сил СССР (Материалы Бурят
Монгольской экспедиции 1931 г.); антропологическое и медико
гигиеническое исследования лопарей и изучение их быта (Золота
рев, 1927, 1928); изучение физического развития детей на Алтае
17
(Ярхо, 1929); медико-биологическое исследование горных
таджиков (Гинзбург, 1937) и многие другие комплексные исследования
башкир, якутов, ненцев.
В центре этих исследований стояли народы или народности
как производительная сила страны. Необходимость изучения этой
силы диктовалась насущными потребностями развития нашей
многонациональной страны, она стимулировала постановку многих
антропологических и этнологических работ в первое
послереволюционное десятилетие. Однако из-за недостатка антропологических
кадров, недостаточной технической оснащенности, отсутствия единой
антропологической программы столь важные в научном и
прикладном отношениях исследования не получили в свое время
должного развития.
Итак, в общей форме задачи изучения приспособительной
изменчивости у человека в советской антропологии были
сформулированы В.В. Бунаком еще в 20-х гг. Выделенные им факторы, по
отношению к которым осуществляется адаптация человеческих
популяций к среде, позволяют убедиться, что все они могут быть
включены в число географических факторов, даже в тех случаях,
когда речь идет о типе питания, режиме труда и
микроорганизмах - возбудителях эндемических болезней. Климатические, в ча
стности термические, факторы определяют и характер диеты, и
трудовые нагрузки. Что касается биотической среды, то не
случайно она стала объектом медицинской географии.
Воздействие географических факторов среды на человека
многообразно.
В литературе не раз освещалась роль гравитационного
эффекта в эволюции органического мира, в формировании строения тела
живых существ, в том числе и человека (Thompson, 1917;
Успенский, 1960, 1964; Коржуев, 1965, 1971). Работами А.Л.
Чижевского (1960,1964) открываются огромные перспективы изучения
влияния электромагнитного поля Земли на рост и функциональное
состояние человека, других представителей живой природы (Пре-
сман, 1968; Холодов, 1970). Возможность рассмотрения расогене-
тических процессов под углом зрения воздействия
электромагнитных и гравитационных сил уже обсуждалась в отечественной
антропологической литературе (Алексеев, 1974).
В этой книге основное внимание будет сосредоточено на роли
климатического фактора в формировании приспособительных осо-
18
бенностей в популяциях человека, занимающих определенные
природные регионы. А поскольку человек испытывает воздей
ствие физико- и химических агентов среды не только прямо, но в
опосредованно, через пищу, ниже рассмотрим влияние некоторых
компонентов социальной среды - хозяйственно-культурного типа
и рациона питания.
Лекция II.
ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ
ИЗМЕНЧИВОСТИ СТРОЕНИЯ ТЕЛА
И НЕКОТОРЫХ ФИЗИОЛОГИЧЕСКИХ ПОКАЗАТЕЛЕЙ
Морфологическая характеристика
различных народов мира.
Роль питания и географической среды в вариабельности черт
строения тела и состояния внутренней среды организма
1. МАТЕРИАЛ И МЕТОД ИССЛЕДОВАНИЯ
Изучение закономерностей пространственной изменчивости
антропологических признаков осуществляется главным образом с
помощью географического метода, широко применяемого в
антропологии. Введение его в исследования русской антропологической
школы связано с именем Е.М. Чепурковского (1916).
Сопряженность таких фундаментальных антропологических
характеристик, как строение тела и обмен веществ, с
особенностями географической среды теперь уже не вызывает сомнений
(Roberts, 1952, 1953; Schwidetzky, 1962; Харрисон и др., 1963;
Алексеева, 1977). Климатические правила Аллена и Бергмана,
сформулированные на заре экологической науки относительно
распределения размеров тела животных в зависимости от температурного
режима среды обитания, получили подтверждение в
географической изменчивости ряда жизненно важных признаков и у
человека. Показана зависимость длины, массы и пропорций тела, а
также основного обмена от температурного фактора; строения
грудной клетки и уровня гемоглобина - от фактора высокогорья;
минерального состава скелета - от геохимического фактора и т.п. В
19
то же время остается неясным, какой из факторов географическое
среды является ведущим в экологической дифференциации чело
века.
Для ответа на этот вопрос представлялось целесообразным
последовательное рассмотрение географической изменчивости чері
строения тела и некоторых физиологических характеристик, оп
ределяющих соматические вариации на территории ойкумены.
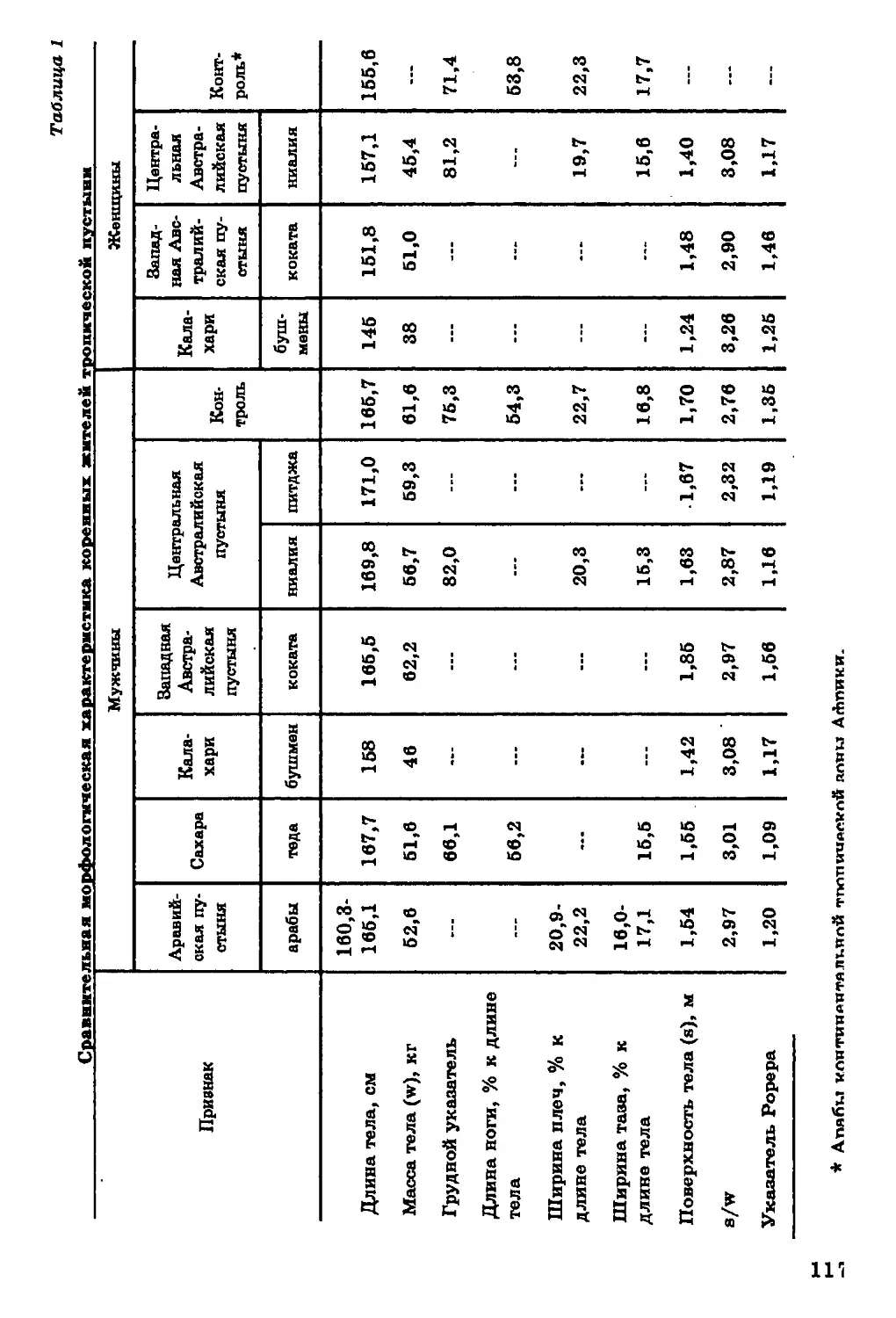
Антропологические данные, положенные в основу нашего
исследования, подробно освещены в монографии автора (Алексеева.
1977), и нет необходимости вновь обращаться к источникам, вд
которых они почерпнуты, и описанию той экологической
обстановки, в которой живут известные по морфологическим и
физиологическим характеристикам этнические группы различных
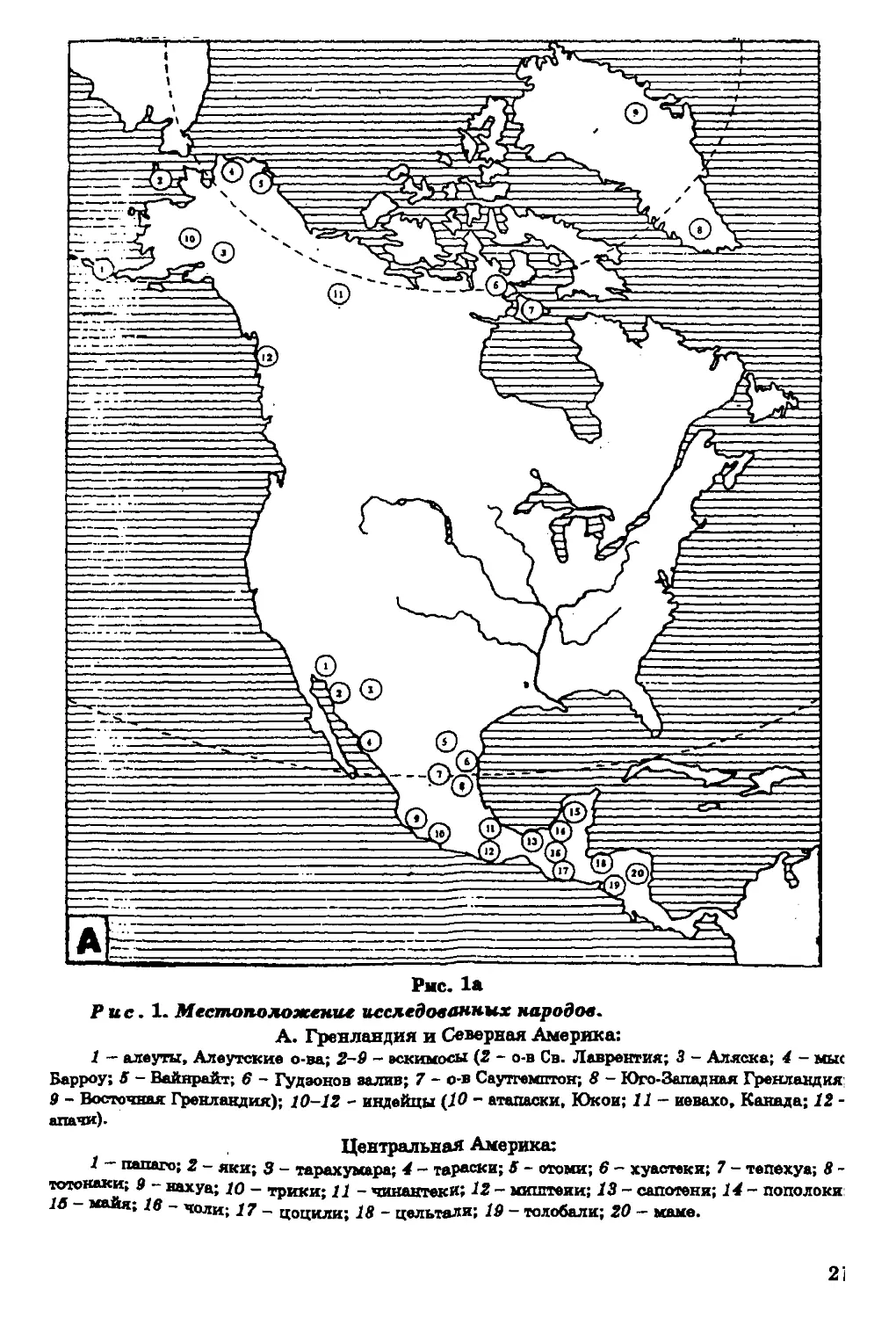
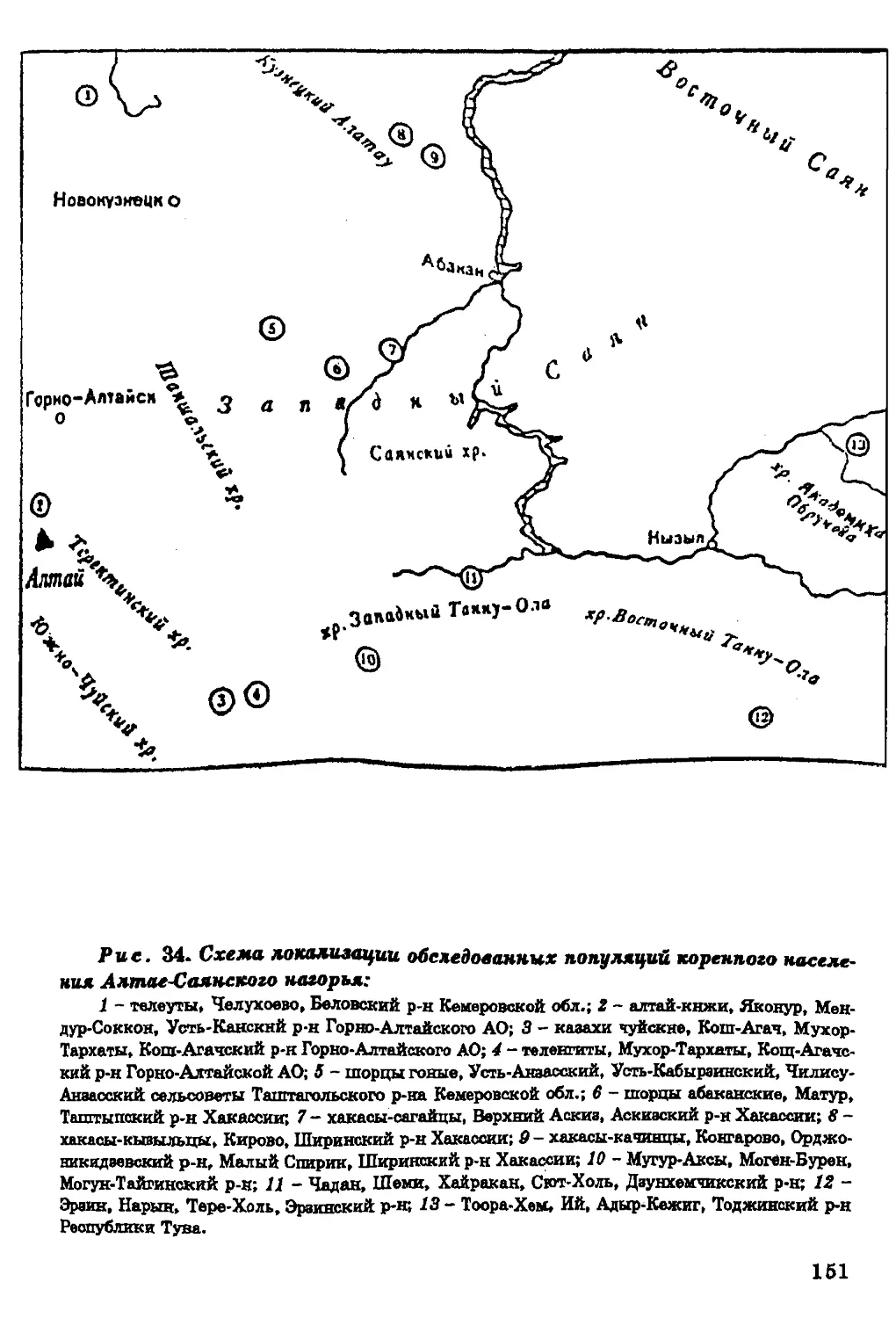
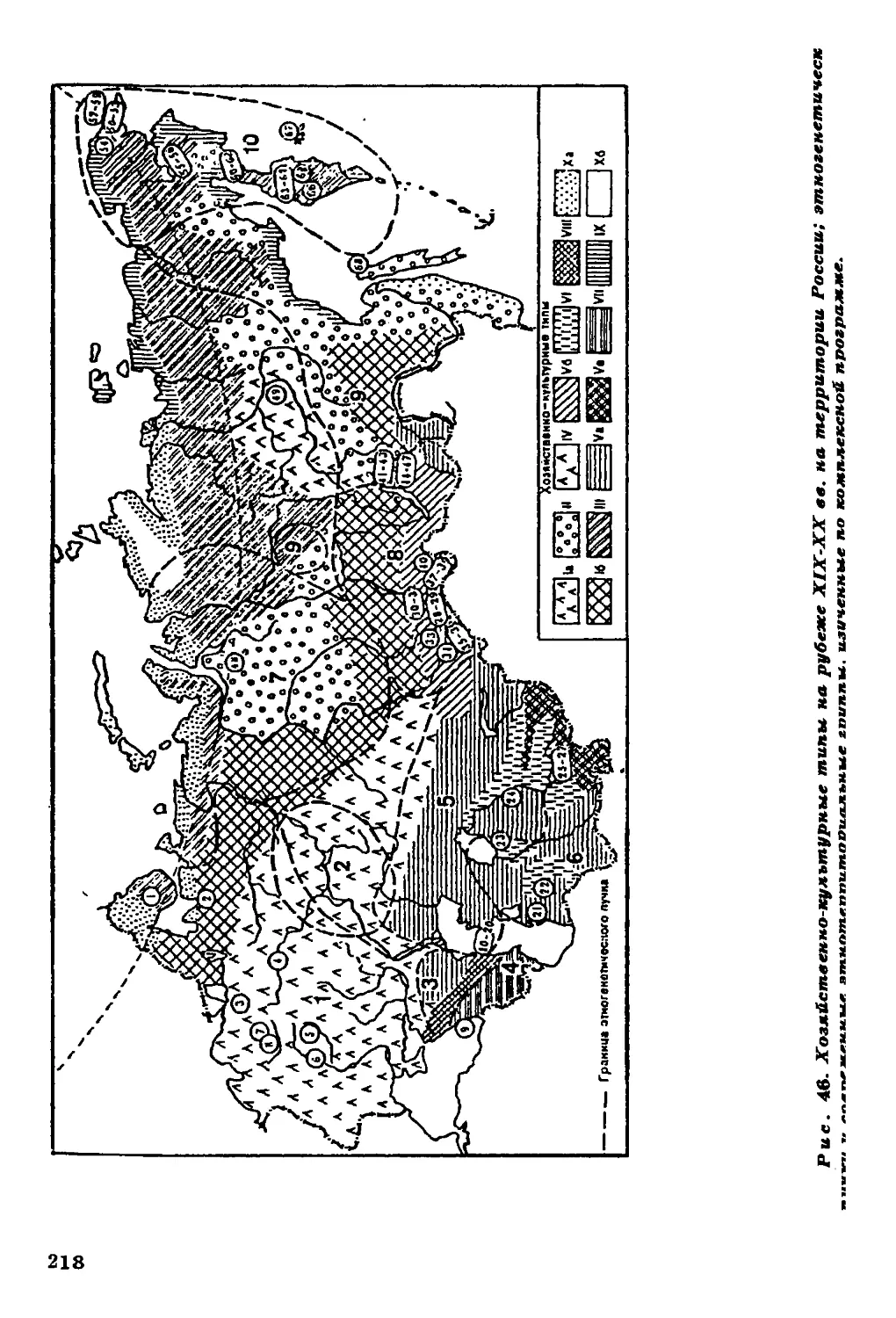
регионов мира (сведения об изученных группах см. в легенде к р и с,
1).
В дополнение к ранним материалам были рассмотрены данные
по взрослому населению различных стран, содержащиеся в
обширной сводке "Worldwide variation in human growth" (1976). Так, »
изучавшимся ранее народам Европы добавилось население
Англии, Бельгии, Болгарии, Венгрии, Греции, Дании, Италии,
Испании, ГДР и ФРГ, Норвегии, Польши, Румынии, Финляндии,
Франции, Чехословакии, Швейцарии, Швеции, Югославии.
Большинство европейских групп представлено городским населением
(студенты, служащие и рабочие). Значительный процент среди
обследованных составляют уроженцы различных, в том числе и
сельских, районов. Национальный состав не всегда точно
фиксируется в зарубежных литературных источниках, но, по-видимому,
это преимущественно доминирующая национальность страны. Вс
Франции наряду с французами обследованы баски. Баски изучены
и в Испании.
Население Индии представлено дополнительно этническими
группами из Пенджаба, Джаммы и Кашмира, Раджастана, Ассама,
среди которых также много студентов и служащих (ранее
анализируемые мной группы индийского населения преимущественнс
были представлены сельским населением). В настоящей книге
использованы новые многочисленные антропологические материалы
советско-индийской экспедиции, работавшей среди сельского
населения различных регионов Индии (Алексеев, 1983).
Пополнились новыми антропологическими данными и
характеристики строения тела коренного населения Аравийского по-
20
Рис. la
Рис. 1. Местоположение исследованных народом.
А. Гренландия и Северная Америка:
1 - алеуты, Алеутские о-ва; 2-9 - эскимосы (2 - о-в Св. Лаврентия; 3 - Аляска; 4 - мыс
Барроу; 5 - Вайнрайт; 6* - Гудаонов залив; 7 - о-в Саутгемптон; 8 - Юго-Западная Гренландия;
9 - Восточная Гренландия); 10-12 - индейцы (Ю - атапаски, Юкон; 11 - н?вахо, Канада; 12 -
апачи).
Центральная Америка:
1 - папаго; 2 - яки; 3 - тарахумара; 4 - тараски; 5 - отоми; 6* - хуастеки; 7 - тепехуа; 8 -
тотокаки; 9 - нахуа; Ю - трики; 11 - чинантекк; 12 - миштеки; 13 - сапотеки; 14 - пополоки:
16 - майя; 16 - чоли; 17' - цоцили; 18 - цельтали; 19 - толобали; 20 - маме.
2і
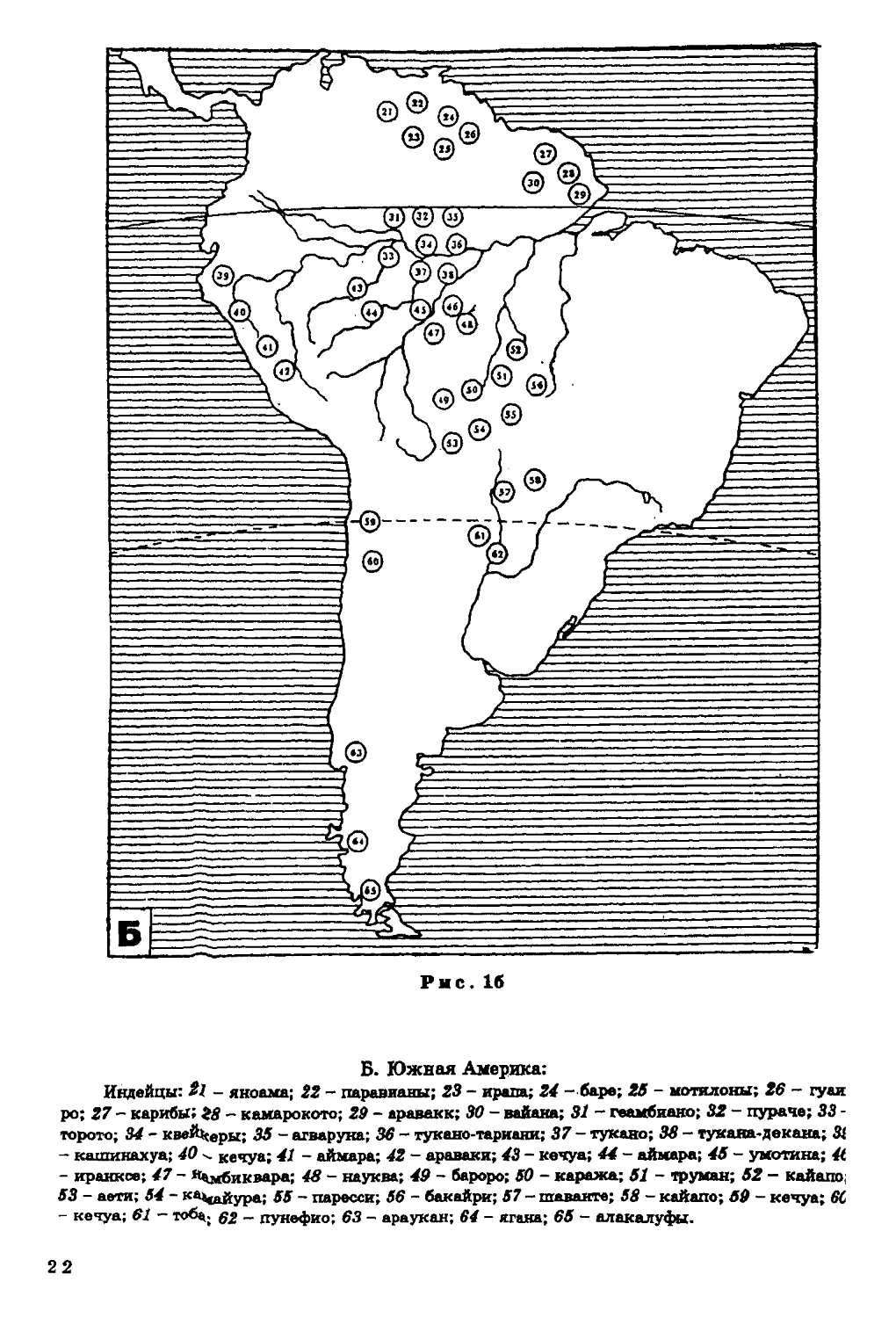
Рнс. 16
Б. Южная Америка:
Индейцы: t\ - якоама; 22 - паравианы; 23 - ирапа; 24 - баре; 26 - ыотялоны; 26 - гуаи
ро; 27 - карибы; 28 - камарокото; 29 - араваки; 30 - вайана; 31 - геамбиако; 32 - пурач?; 33 -
торото; 34 - квеЙ^еры; $$ - агваруна; 36 - тукано-тарианк; 37 - тукано; 38 - тукана-декана; Зі
- кашикахуа; 40 ^ кечуа; 41 - аймара; 42 - араваки; 43 - кечуа; 44 - аймара; 46 - умотина; 4t
- иранкое; 47 ~ *Цмбиквара; 48 - науква; 49 - бароро; 60 - каража; 61 - труман; 62 - кайапо;
53 - аети; 54 - ка^айура; 65 - паресси; 56 - бакайри; 57 - щаванте; 58 - кайапо; 60 - кечуа; 6С
- кечуа; 61 - тоба., g% _ пунефио; 63 - араукан; 64 - ягаяа; 05 - алакалуфы.
22
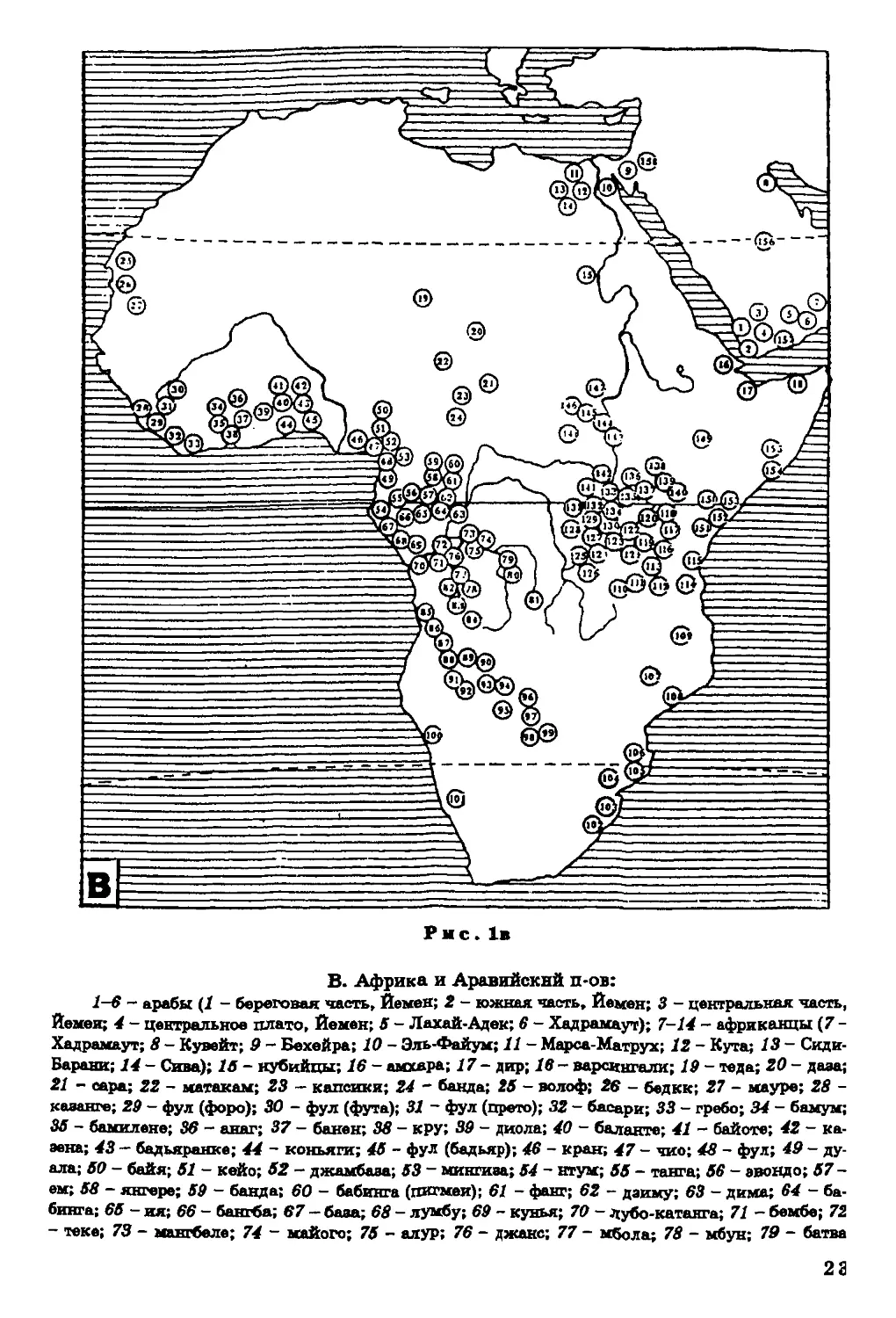
В. Африка и Аравийский п-ов:
1-6 - арабы (1 - береговая часть, Йемен; 2 - южная часть, Йемен; 3 - центральная часть,
Йемен; 4 - центральное плато, Йемен; 6 - Лахай-Аден; 6 - Хадрамауг); 7-14 - африканцы (7 -
Хадрамаут; 8 - Кувейт; 9 - Бехейра; 10 - Эль-Файум; 11 - Марса-Матрух; 12 - Куга; 13 - Сиди-
Барани; 14 - Сива); 16 - нубийцы; 16 - амхара; 17 - дир; 18 - варсивгали; 19 - теда; 20 - даэа;
21 - сера; 22 - матакам; 23 - капсики; 24 - банда; 26 - волоф; 26 ~ бедик; 27 - мауре; 28 -
казанге; 29 - фул (форо); 30 - фул (фута); 31 ~ фул (прето); 32 - басари; 33 - гребо; 34 - бамум;
36 - бамилеке; 36 - анаг; 37 - банен; 38 - кру; 39 - диола; 40 - баланте; 41 - байоте; 42 - ка-
эена; 43 - бадьяранке; 44 ~ коньяги; 46 - фул (бадьяр); 46 - кран; 47 - чио; 48 - фул; 49 - ду-
ала; 60 - байя; 61 - кейс; 52 - джамбаза; 63 - микгиаа; 64 - нтум; 66 - танга; 66 - эвондо; 07 -
ем; 68 - янгере; 69 - банда; 60 - бабинга (пигмеи); 61 - фанг; 62 - даиму; 63 - дима; 64 - ба-
бинга; 66 -яя;66- бакгба; 67 - база; 68 - лумбу; 6*0 - кунья; 70 - дубо-катанга; 71 - бембе; 72
- теке; 73 - мангбеле; 7df - какого; 76 - алур; 76 - джанс; 77 - мбола; 78 - мбун; 70 - батва
га
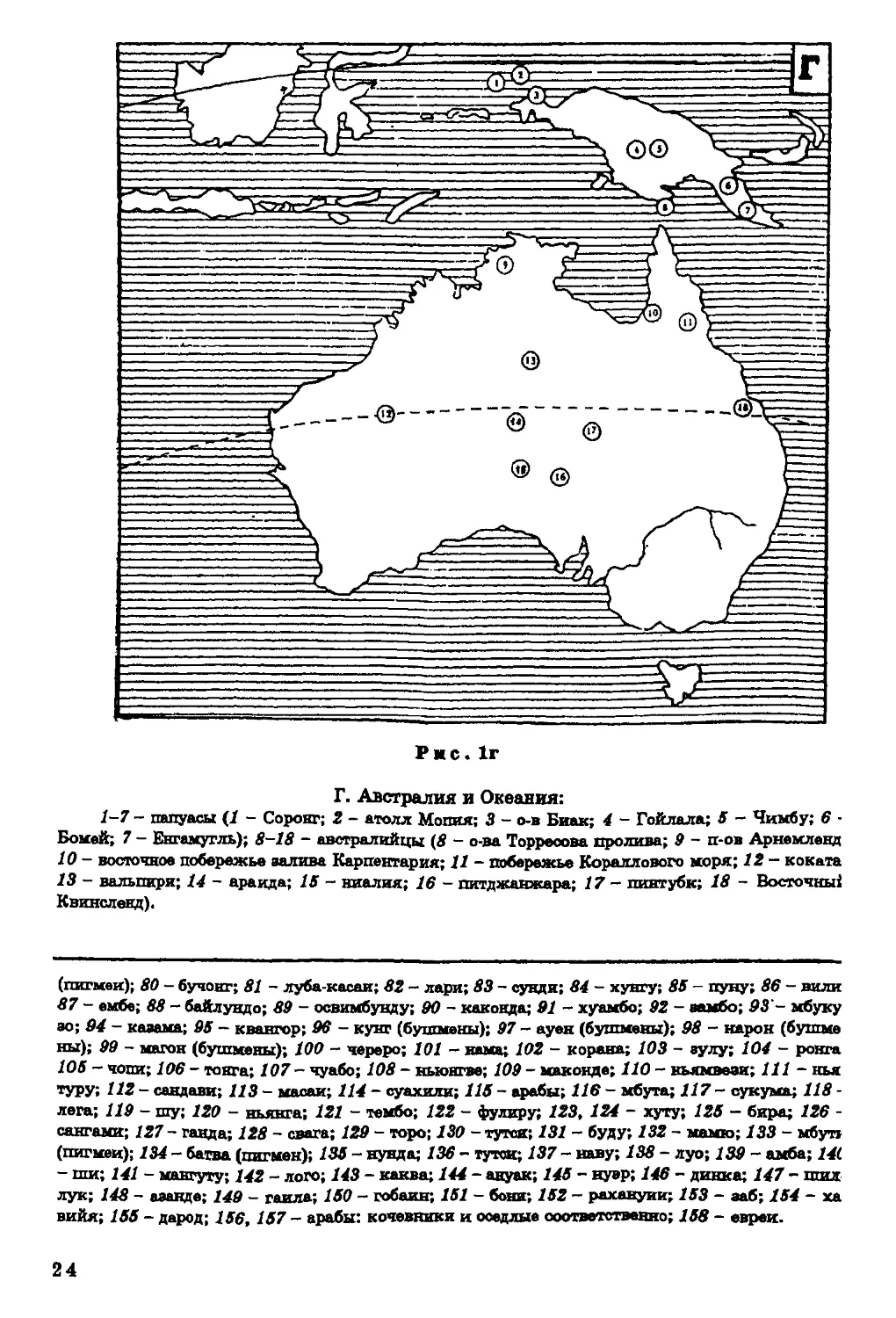
Г. Австралия и Океания:
1-7 - папуасы (/ - Соронг; 2 - атолл Мопия; 3 - о-в Биак; 4 - Гойлала; 5 - Чимбу; 6 -
Бомей; 7 - Евгамугль); 8-18 - австралийцы (8 - о-ва Торресова пролива; 9 - п-ов Арнемленд
10 - восточное побережье аалива Карпентария; 11 - побережье Кораллового моря; 12 - коката
13 - валышри; 14 - аранда; 15 - ниалия; 16 - пктджанжара; /7 - пинтуби; 18 - Восточны*
Квинсленд).
(пигмеи); 80 - бучонг; 81 - луба-касаи; 82 - лари; 83 - сунди; 84 - хунту; 85 - пуну; ДО - вили
87- ембе; 33-байлундо; 89 - освимбунду; 90 -каконда; &f -хуамбо; 92- эамбо; 03'- мбуку
зо; 94 - казана; 95 - квантор; 96 - кунг (бушмены); 97 - ауен (бушмены); 98 - карой (бушм?
ны); 99 - магон (бушмены); 100 - череро; 101 - нами; /02 - корана; ./03 - аулу; 104 - ронга
105 - чопи; /03 - тонга; 107 - чуабо; 105 - ньюнгве; 109 - ыаконде; 110 - кьямвеаи; 111 - нья
туру; 112 ~ сандави; 113 - маоаи; 114 - суахили; 115 - арабы; 116 - мбута; 117- сукума; 118 -
лета; 11$ - шу; 120 — ньянга; 121 - тембо; 122 - фулиру; 123, 124 - хуту; /25 - бира; /26* -
сайтами; 127- ганда; 128 - свата; /20 - торо; 130 ~ тутси; 131 - буду; /32 - мам»; 133 - мбута
(пигмеи); 134 - батаа (пигмеи); /35 - нунда; 136 - тутой; 137 - наву; /33 - луо; 139 - амба; /А
- ши; 141 - мавгуту; 142 - лого; /43 - каква; 144 - ануак; /45 - нувр; 146 - динка; 147 - шил
лук; /43 - ааанде; 149 - гаила; 150 - гобаин; 151 - бони; 152 - рахануин; /53 - заб; 154 - ха
вийя; 155 - дарод; /53, 167 - арабы: кочевники и оседлые соответственно; 158 - евреи.
24
Рис. 1д
Д. Индия, Пакистан, Иран:
/ - кашмирцы; 2 - пенджабцы; 3 - раджастанцы; 4 - ахиры; б - гуджары; 6 - джулахи-
чамары; 7 - джаты; 8 - раджпута; 9 - гуркхи, Непал; 10 - раджпугы; 11 - артнэаны; 12 - абор;
13 - ораоны; 14-16 - бенгальцы {IS - бенгальцы мусульмане); 17 - рканг; 18 - маратхи; 19 -
кокни; 20 - андхра; 21 - сидхи; 22 - малаяли; 23 - курумба; 24 - кота; 26 - тода; 26 - мала-
пантарам; 27 - каткари; 28 - бхилы; 29 - парсы; 30 - ирани, Ассам; 31 - пакистанцы, Лахор.
Центральная, Восточная и Юго-Восточная Азия:
1 - бирманцы; 2 - китайцы, Тайвань; 3 - китайцы, Сянган; 4 - японцы; 5 - филиппинцы;
6 - население Таиланда; 7 - население Лаоса; 8 - население Камбоджи; 9 - онг?; 10 - шоыпен;
11 - кикобарцы; 12 - индонезийцы; 13 - индонезийцы» Ява; 14 - корейцы, Южная Корея; 16 -
монголы-халха.
Западная Европа:
/ - саамы, Норвегия; 2 - норвежцы (сборная группа, далее СП; 3 - шведы (СГ); 4 - фины
(СГ); 5 - англичане (СИ; 6 - датчане (СГ); 7 - немцы (СГ); 8 - французы, Гасконь, Савойя,
Париж; 9 - баски, Франция; 10 - баски, Испания; 11 - бельгийцы (СИ; 12 - поляки, Варшава,
Краков; 18 - чехи (СП; 14 - венгры (СП; 15 - болгары, София; 16 - румыны, Мосе де Сус, Ма-
гура, Бухарест и СГ; 17 - население Югославия (СП; 18 ~ гр«кя (СП; 19 - итальянцы, Неаполь;
20 - итальянцы, Сардиния; 21 - турки (СП*
Вывший СССР:
Обозначения групп 1-88 см. на рис. 46
26
луострова, Канады, Гватемалы, Аргентины, Бразилии и Чили, раз
личных африканских государств.
Впервые могут быть привлечены для рассмотрения закономер
ностей пространственных вариаций черт строения тела антрополо
гические материалы по народам Турции, Монголии, Пакистана
Ирана и государств Юго-Восточной Азии - Бирмы, Лаоса, Камбод
жи, КНДР, Японии.
Возрастной состав вновь рассматриваемых этнических групя
относительно однородный (в соответствии с требованиями
Международной биологической программы); взрослое население
включается в возрастной диапазон до 40 лет, за редким исключением.
Ранее анализируемые нами антропологические данные относились
к возрастной категории 18-50 лет, что позволяет в целом
сопоставлять их с новыми материалами.
В общей сложности анализу было подвергнуто более 460 этно-
территориальных групп коренного населения Евразии, Африки,
Северной и Южной Америки, Австралии. Более 60 популяций
были обследованы в различных географических зонах бывшего
СССР при непосредственном участии автора и его ближайших
коллег (сведения об изученных в СССР группах см. в лекции I, п. 1 и
в легенде к рис. 1). Местоположение анализируемых групп и их
этническое наименование1 представлены на рис. 1.
2. МОРФОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ
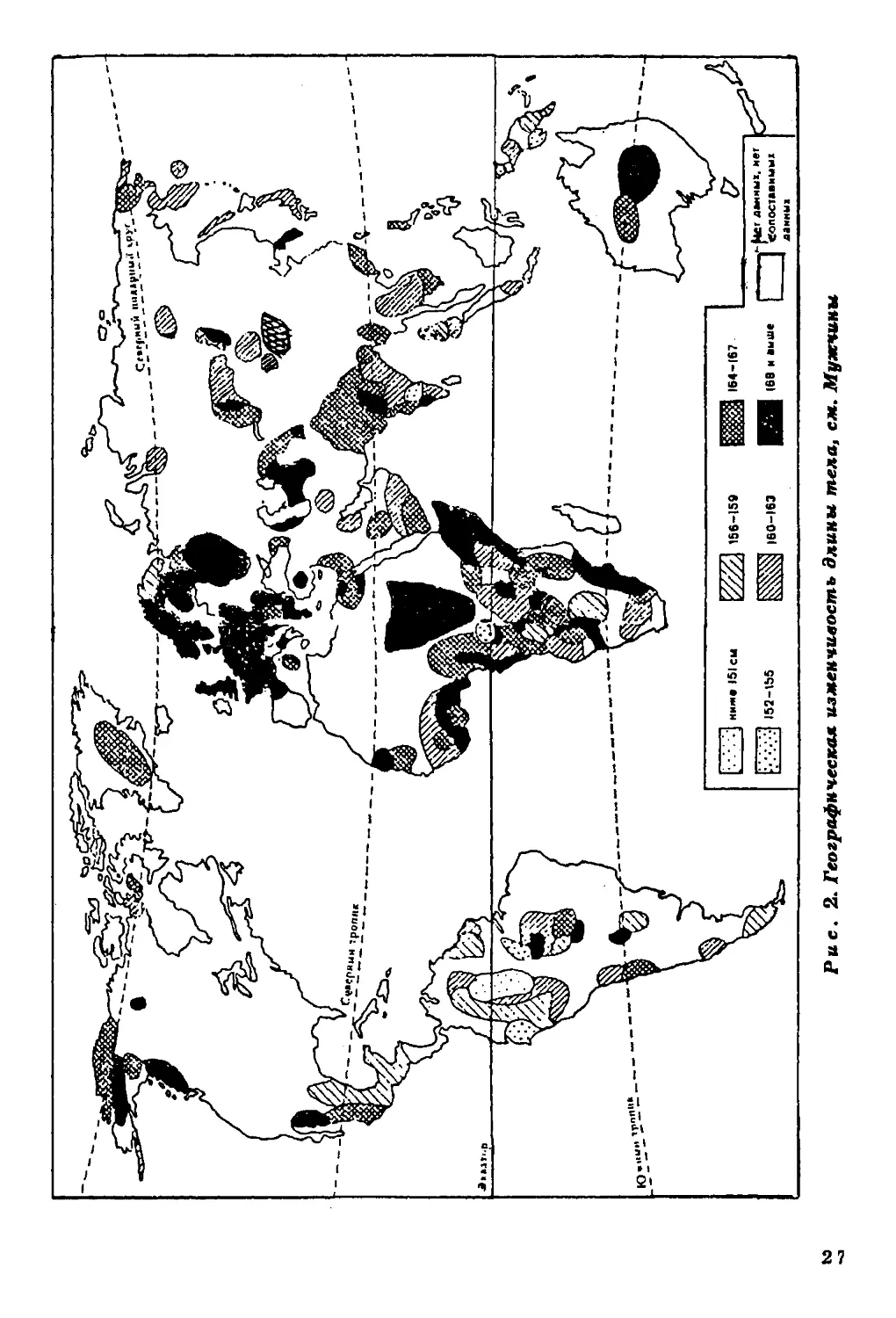
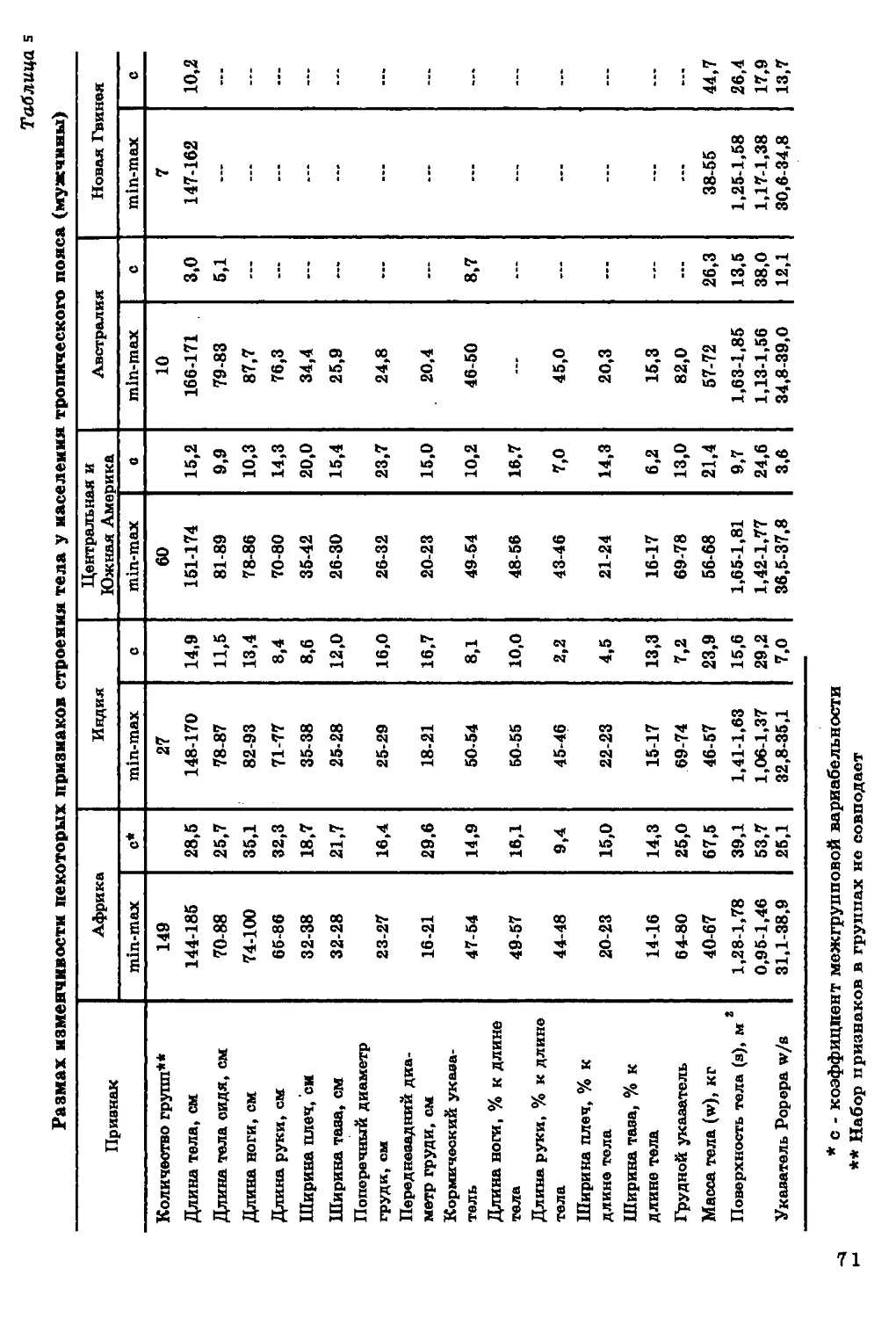
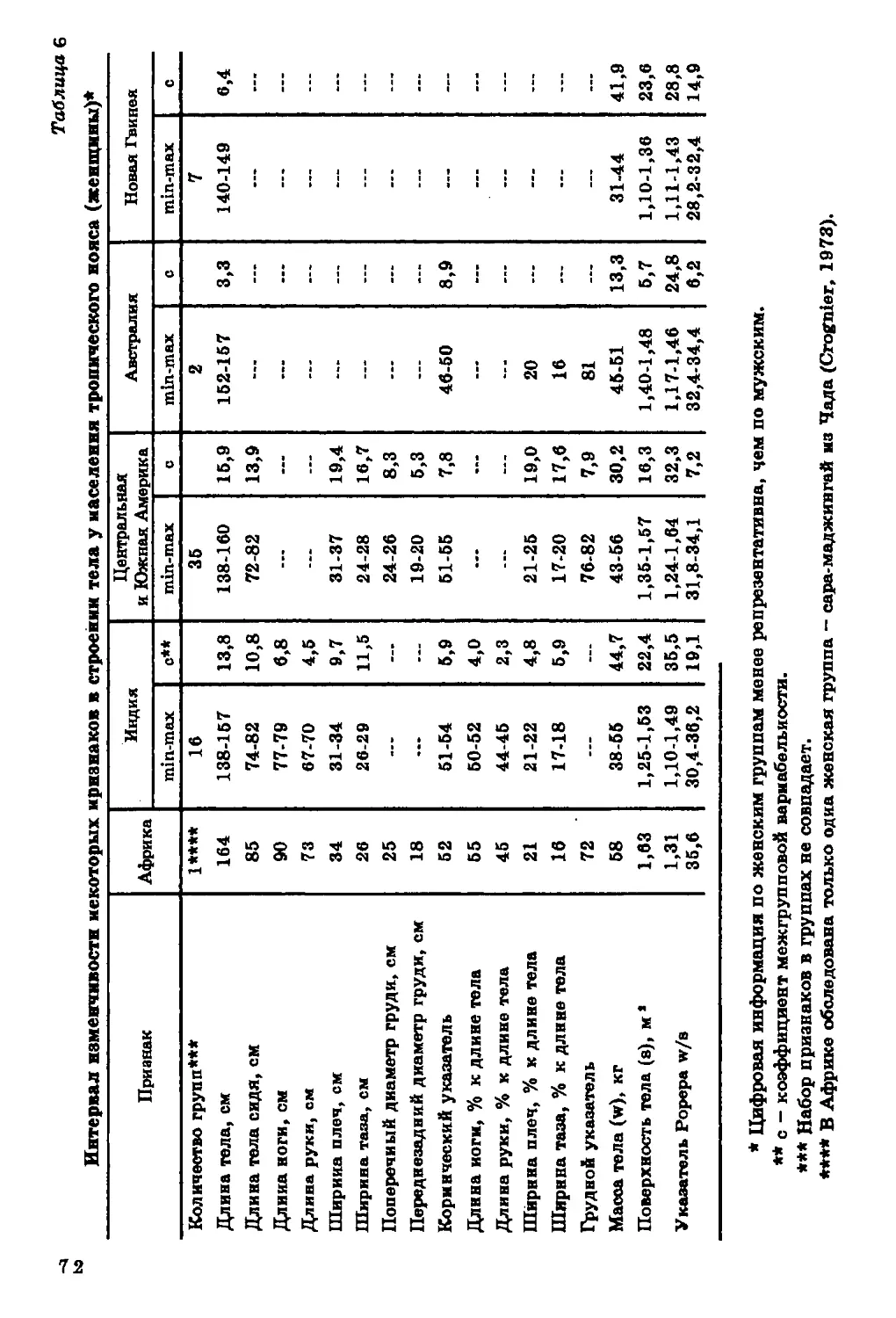
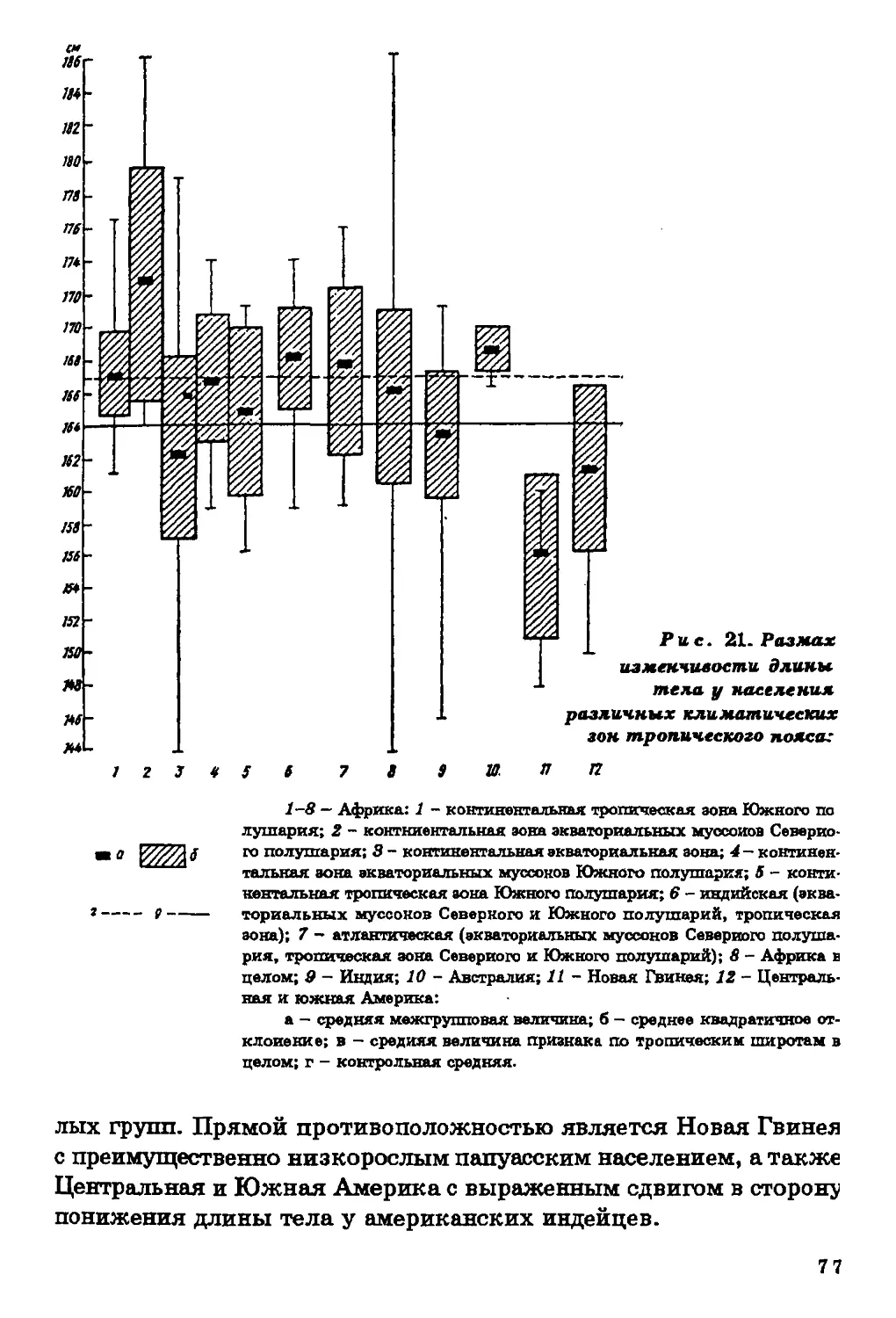
Длина тела. Диапазон географической изменчивости длины
тела (рис. 2) на территории ойкумены весьма значителен. По
имеющимся в нашем распоряжении данным, он составляет около
40 см для мужчин и 25 см для женщин. Наибольшей
дисперсностью величин длины тела характеризуется население
Африканского континента. Вариации длины тела, типичные для Африки,
превосходят размах изменчивости этого признака на территории всей
ойкумены. В то же время большинство изученных африканских
групп отличается высокорослостью, и следует признать, что ни на
одном другом континенте нет такой концентрации высоких
величин длины тела.
На рис. 1 показана локализация групп, для которых известны размеры тела.
В ряде групп Америки, Африки и Азии изучены только физиологические
характеристики (данные по этим группам см. в соответствующих разделах).
26
27
Наиболее низкорослые пигмеи мбути из Заира (длина тела у
мужчин - 144 см, у женщин - 137 см), наиболее высокорослые
народы нилотской группы из Восточного Судана - нуэр и ануак
(длина тела мужчин соответственно 185 и 177 см).
Очаг высокорослости обнаруживается на северо-западе Индии,
в штате Харьяна (длина тела мужчин 169-170 см). В остальных
частях ойкумены, за исключением Австралии и Океании,
высокорослые группы не образуют закономерных сплошных ареалов.
Прямой противоположностью Африканскому континенту по
географической изменчивости длины тела является Центральная и
Южная Америка, где очень велик процент низкорослых групп
среди индейцев Мексики, Гватемалы, Венесуэлы, Гвианы, Перу,
Боливии, Аргентины. Группы ниже среднего и низкого роста
населяют Австралию и Океанию. Островки крайней низкорослости
обнаруживаются, по имеющимся в нашем распоряжении данным, на
различных континентах, но только в экваториальном поясе.
Таким образом, наибольший размах изменчивости длины тела
характерен для тропической зоны с концентрацией высоких
величин на территории Африки, низких - в Центральной и Южной
Америке и Океании. Для населения умеренного пояса типичны
средние и вышесредние значения этого признака.
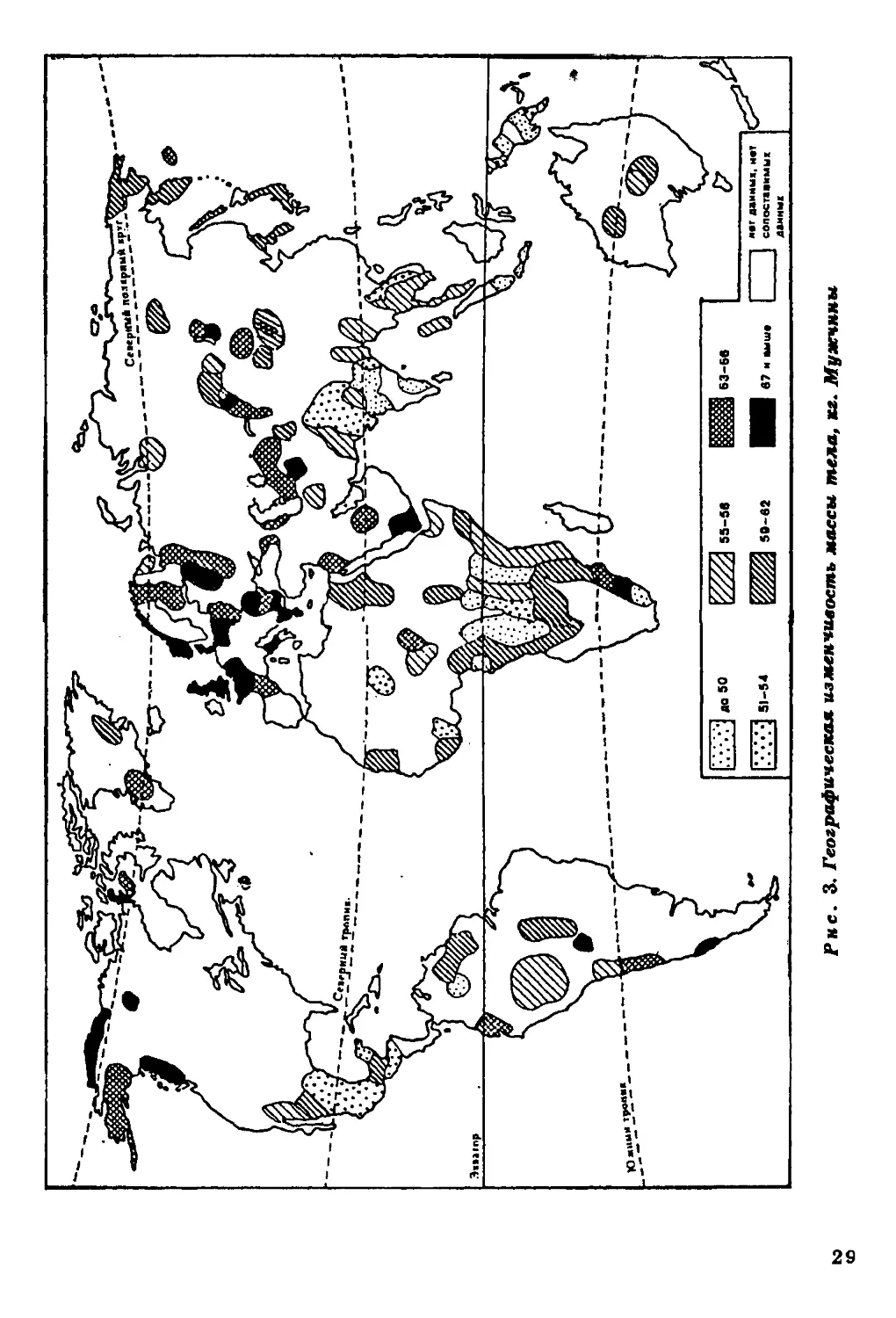
Масса тела. Данных по географической изменчивости массы
тела (рис. 3) значительно меньше, чем по вариабельности его
длины. Тем не менее и они позволяют выявить совершенно
определенную картину. Наиболее "тяжелые" формы наблюдаются за
пределами тропической зоны.
Для большинства аборигенных групп тропиков характерны
низкие значения признака. Концентрация таких групп
отмечается в Центральной Америке, Центральной Африке и Индии. Как
правило, эти области совпадают с ареалами низкорослости.
Отмечено понижение массы тела с севера на юг (Roberts, 1953).
Индекс Рорера. Значительный интерес представляет
вариабельность весоростового показателя Рорера (рис. 4), который
отражает отношение массы тела к его длине и в известной мере
характеризует плотность тела. На территории Старого Света
совершенно отчетливо проявляется горизонтальная зональность в
географическом распределении индекса Рорера. Наиболее высокие
значения индекса характерны для населения внетропических ши-
28
*
*
а
t
л
з
I
3
8
*
!
СО
а*
29
•J
0
о
»
8
о
EX
О
о
«I
л
Е
о
3
at
і
с
«4
С
в.
30
рот. На территории Африки, Индии, Австралии и Океании
распространены преимущественно низкие значения индекса. Иными
словами, в тропическом поясе сконцентрированы относительно
более облегченные формы, хотя исключения из этого правила
отмечаются на территории Африки у населения Египта, ЮАР и у
некоторых африканских народностей бассейна Конго. В
Австралии и Океании также встречаются группы с большей плотностью
тела, чем это типично для жителей тропиков, но они не
составляют большинства.
На территории Нового Света весоростовой указатель не
обнаруживает той закономерности, которая характерна для населения
Восточного полушария. Коренное население Американского
континента отличается значительной плотностью телосложения, хотя
различия в абсолютных размерах между эскимосами, алеутами и
североамериканскими индейцами, с одной стороны, и центрально-
и южноамериканскими индейцами, - с другой, достаточно
демонстративны. Можно лишь отметить, что в зоне, близкой к
экватору, наблюдается незначительное понижение плотности тела у
коренного населения.
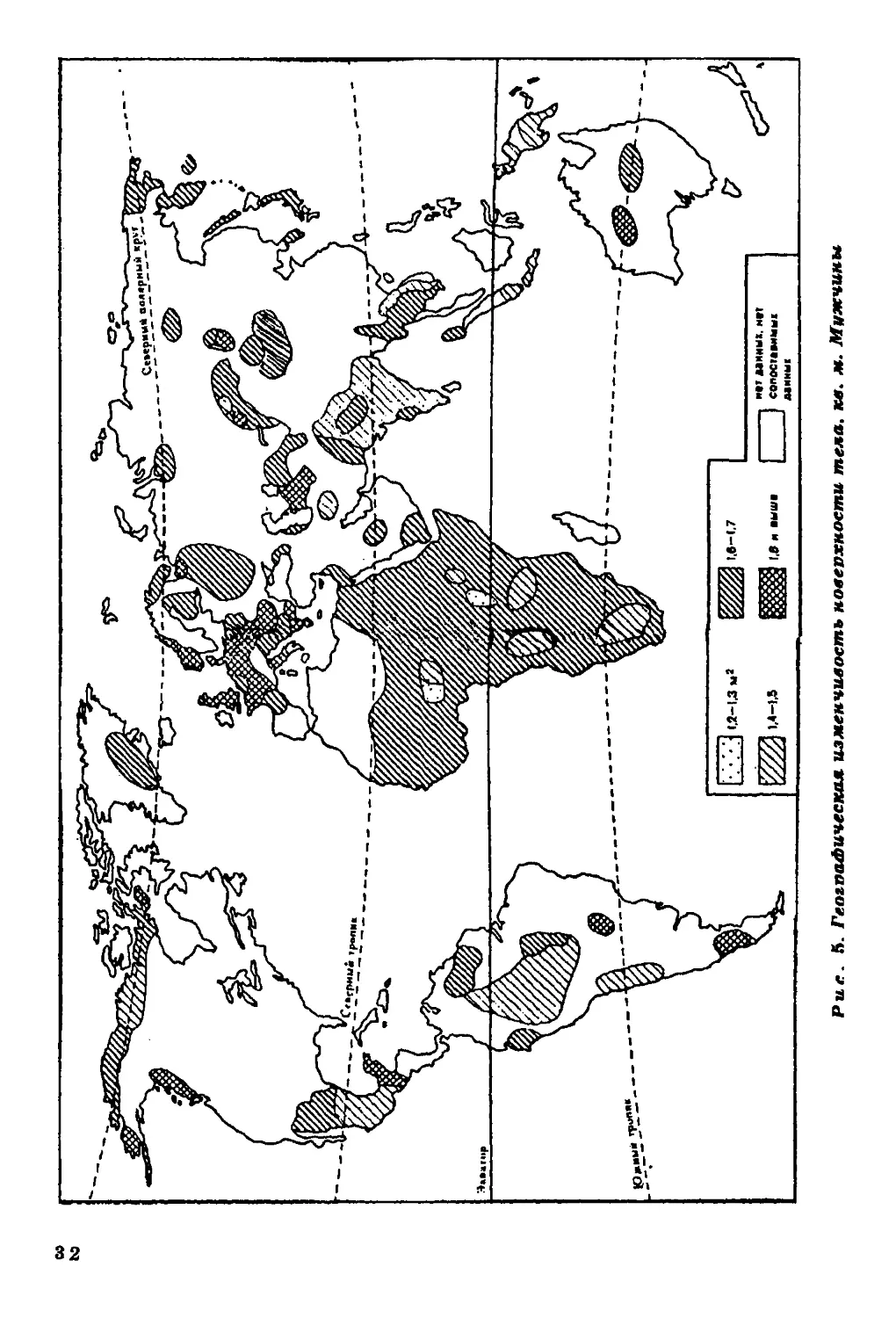
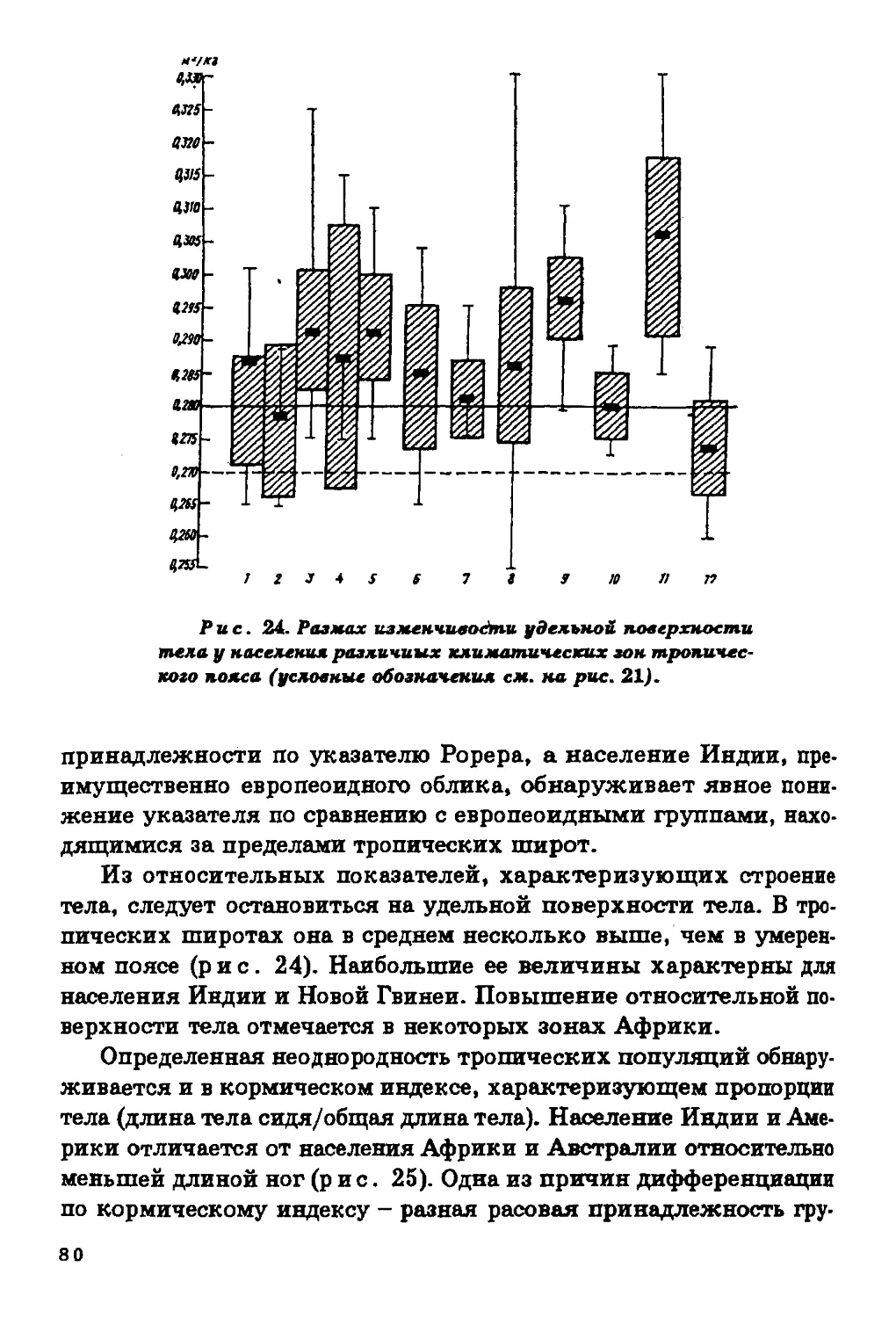
Поверхность тела. Этот показатель рассчитывается на
основе длины и массы тела, и в некотором отношении закономерности
его пространственной изменчивости сходны с теми, которые
характерны для индекса Рорера (рис. 5). Тем не менее картина его
зональной дифференциации здесь выражена четче. В Западном и
Южном полушариях сконцентрированы формы с малой
поверхностью тела. Следует заметить, однако, что относительная
поверхность тела (при расчете на 1 кг массы тела) в тропической зоне
выше, чем вне тропиков.
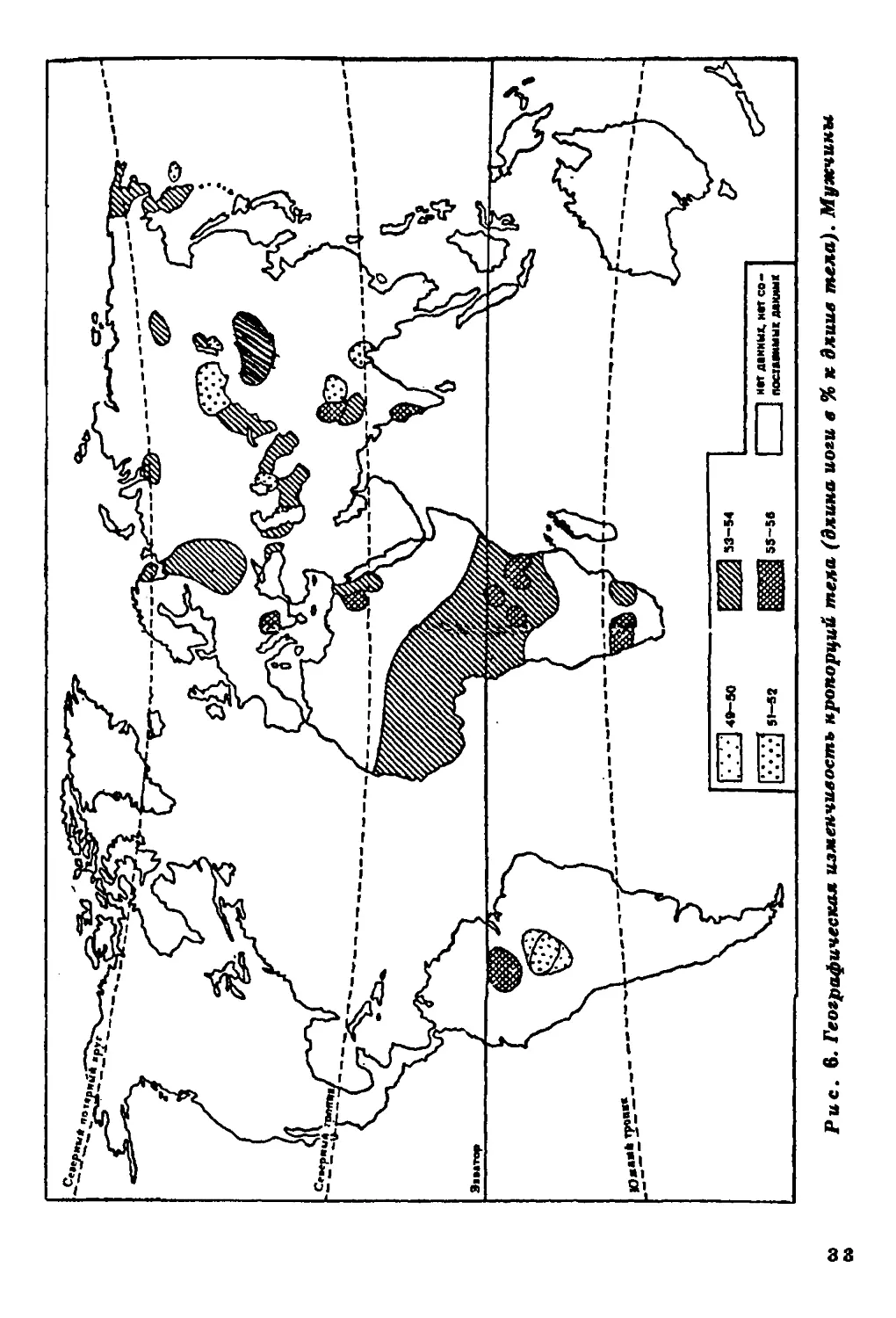
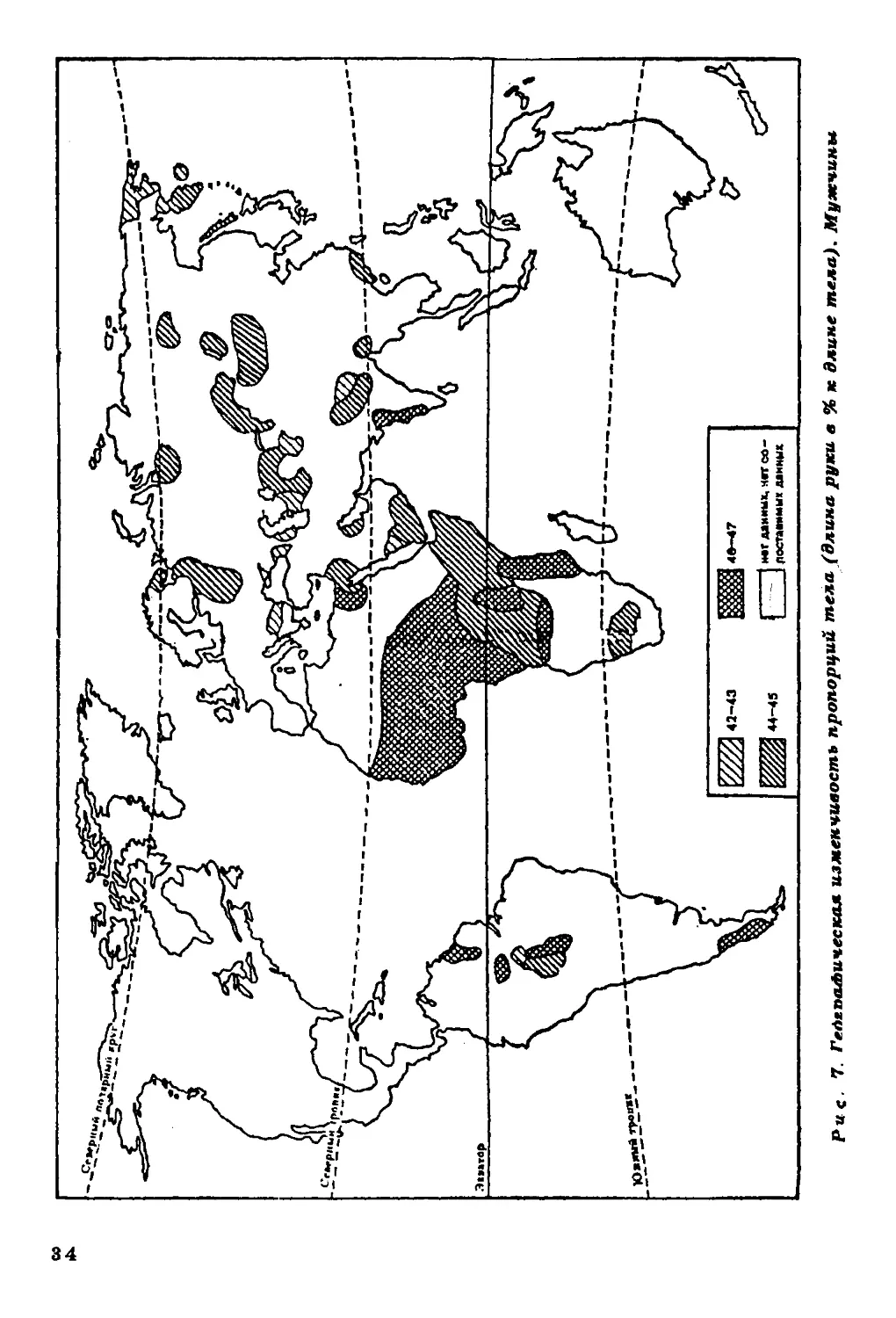
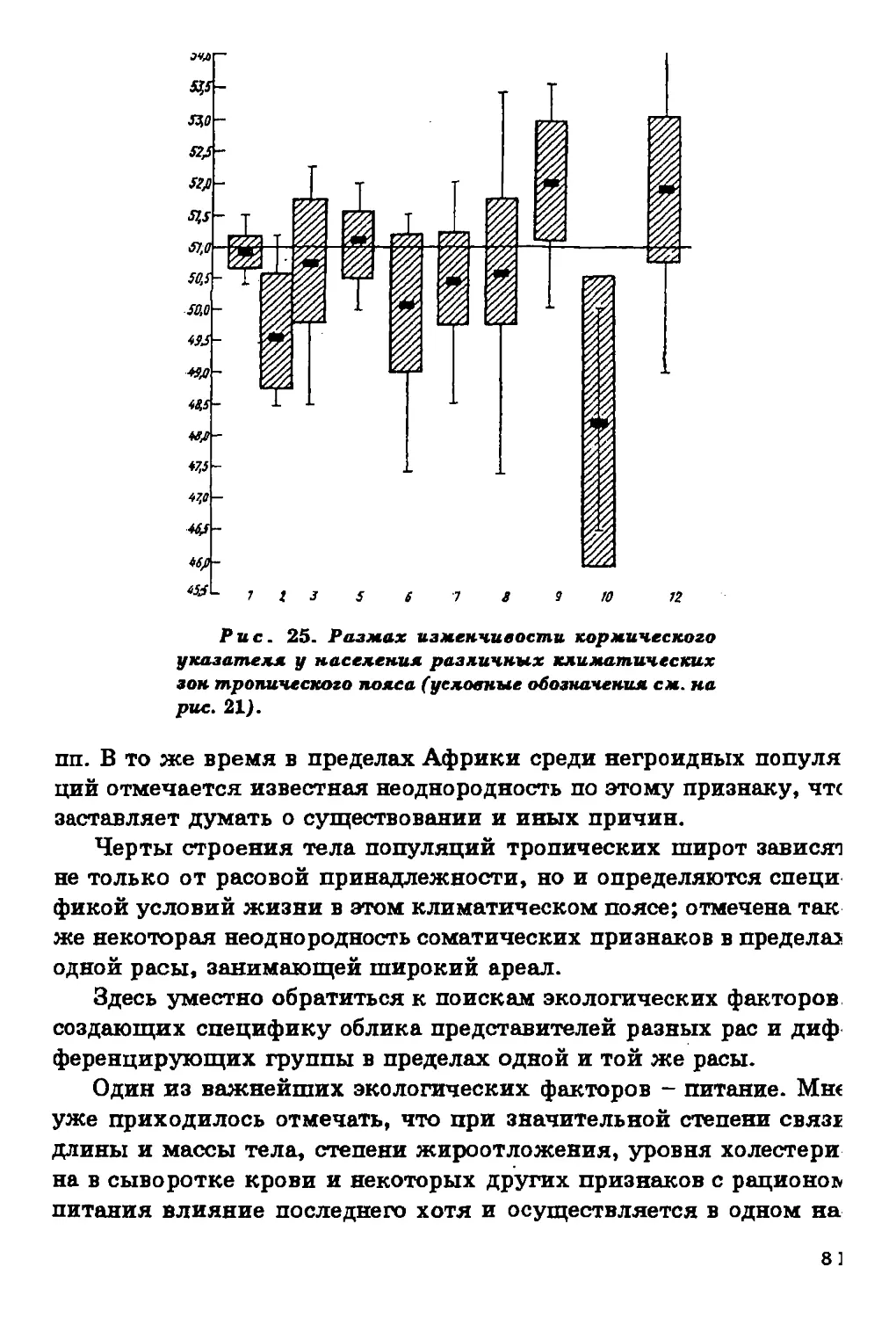
Длина ноги и длцна руки в процентах к длине тела.
Данные о продольных пропорциях тела относятся главным образом к
населению Старого Света, исключая Австралию и Океанию (рис.
6,7). Население Америки представлено весьма фрагментарно. Тем
не менее просматриваются определенные тенденции в
географической изменчивости продольных пропорций тела. Население
тропических широт характеризуется относительной длинноногостью
и длиннорукостью. На Американском континенте эти тенденции
несколько сглажены.
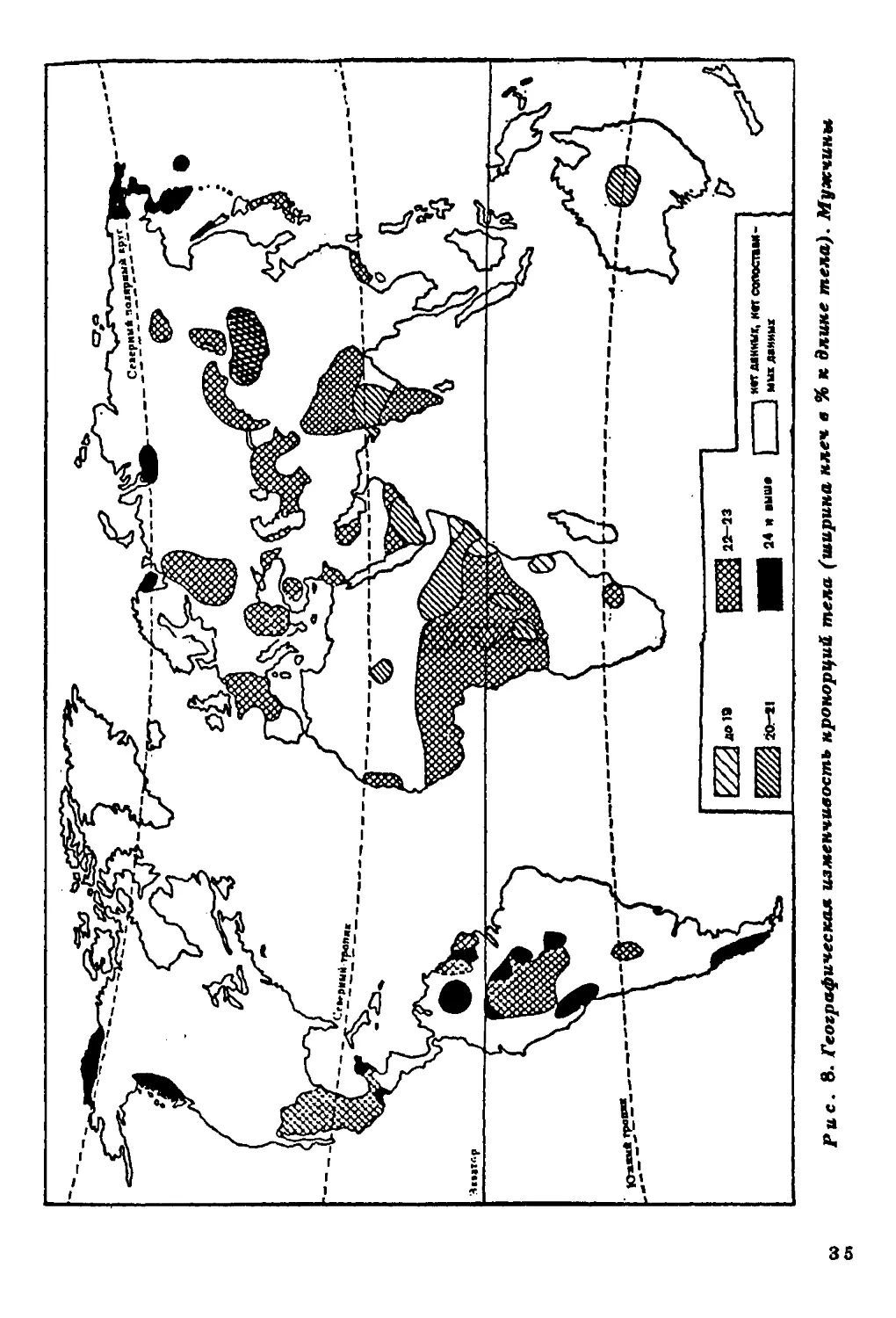
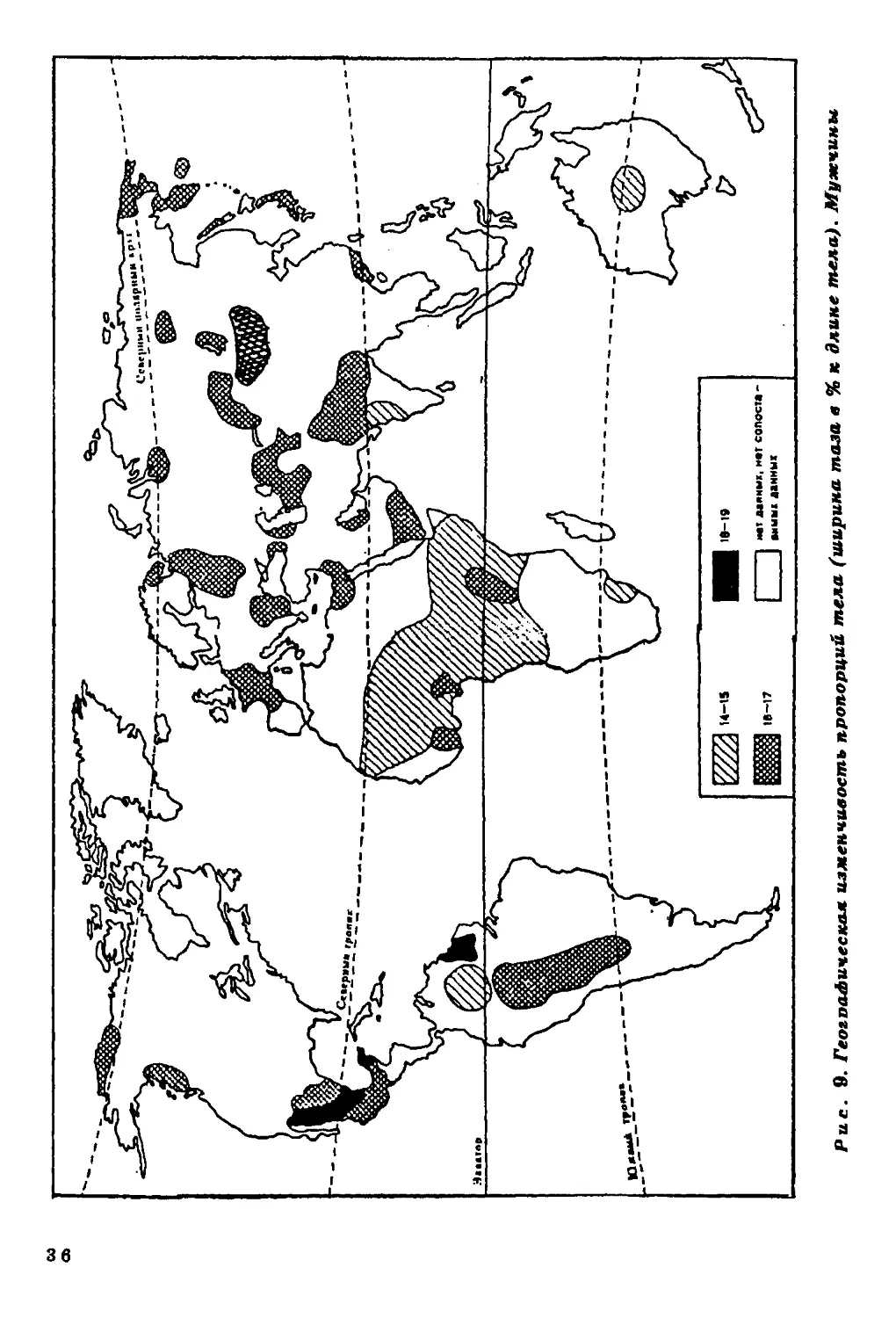
Ширина плеч и ширина таза в процентах длины тела.
Географическая дифференциация человечества по широтным
пропорциям менее отчетлива, нежели по продольным (р йс . 8, 9). В Вое-
3]
32
33
34
3
*
e
?
Я
!
a
5
»a
О
з
4
n
3
*
ft.
35
36
точном полушарии обнаруживается явная тенденция к абсолютно
му и относительному уменьшению скелета в поперечном направле
нии. Например, на территории Африки заметно увеличиваете*
число групп с относительно узкими плечами и тазом. У коренногс
населения Американского континента подобной закономерности
не обнаружено.
Строение грудной клетки. Представительные данные по
этому признаку имеются только по населению Старого Света. Продо
льный и поперечный диаметры грудной клетки находятся в пол
ном соответствии с грацилизацией скелета, типичной для больший
ства тропических групп. По направлению к тропической зоне груд
нал клетка уменьшается в поперечном направлении и становится
более плоской.
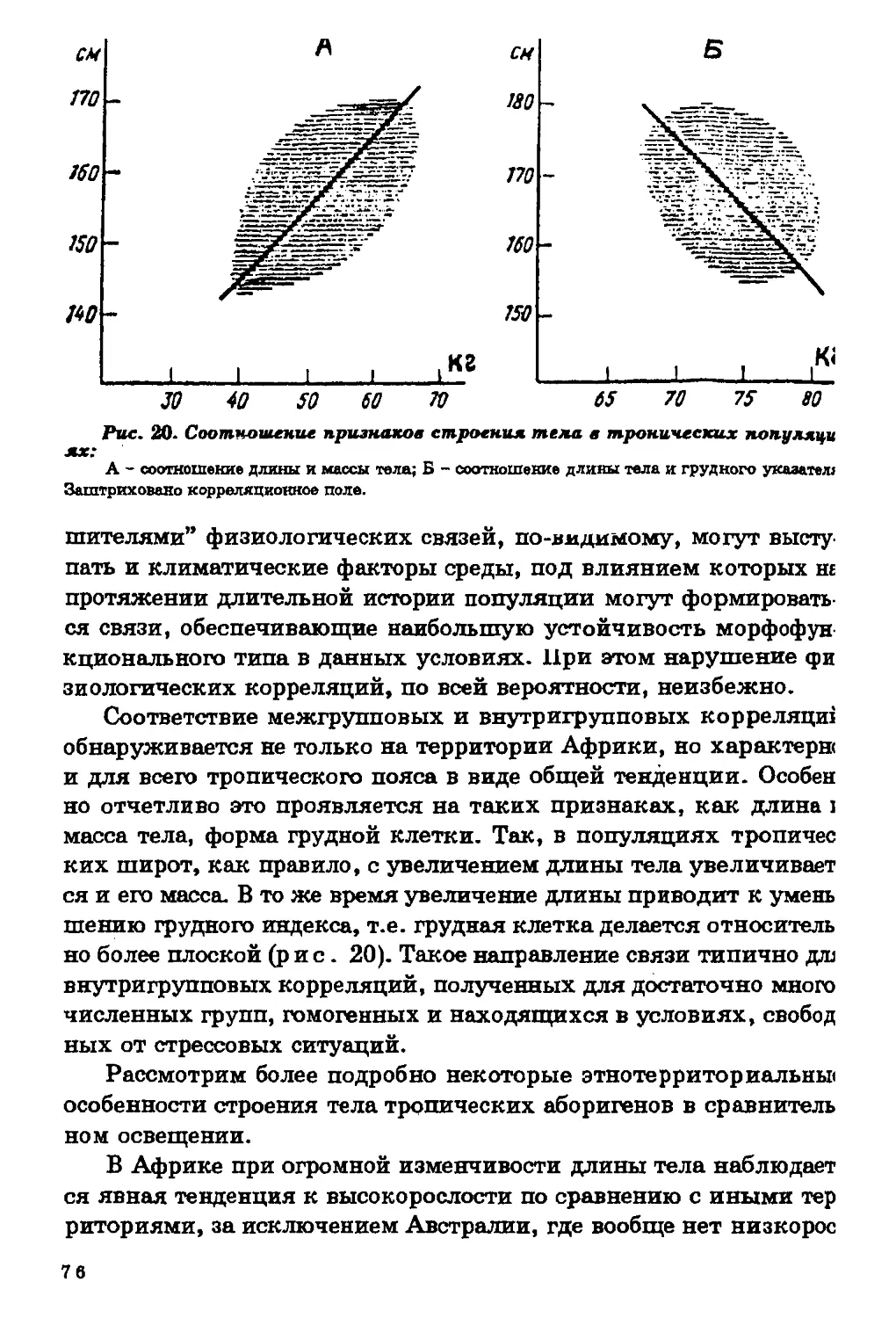
Итак, в картине пространственной изменчивости длины, мае
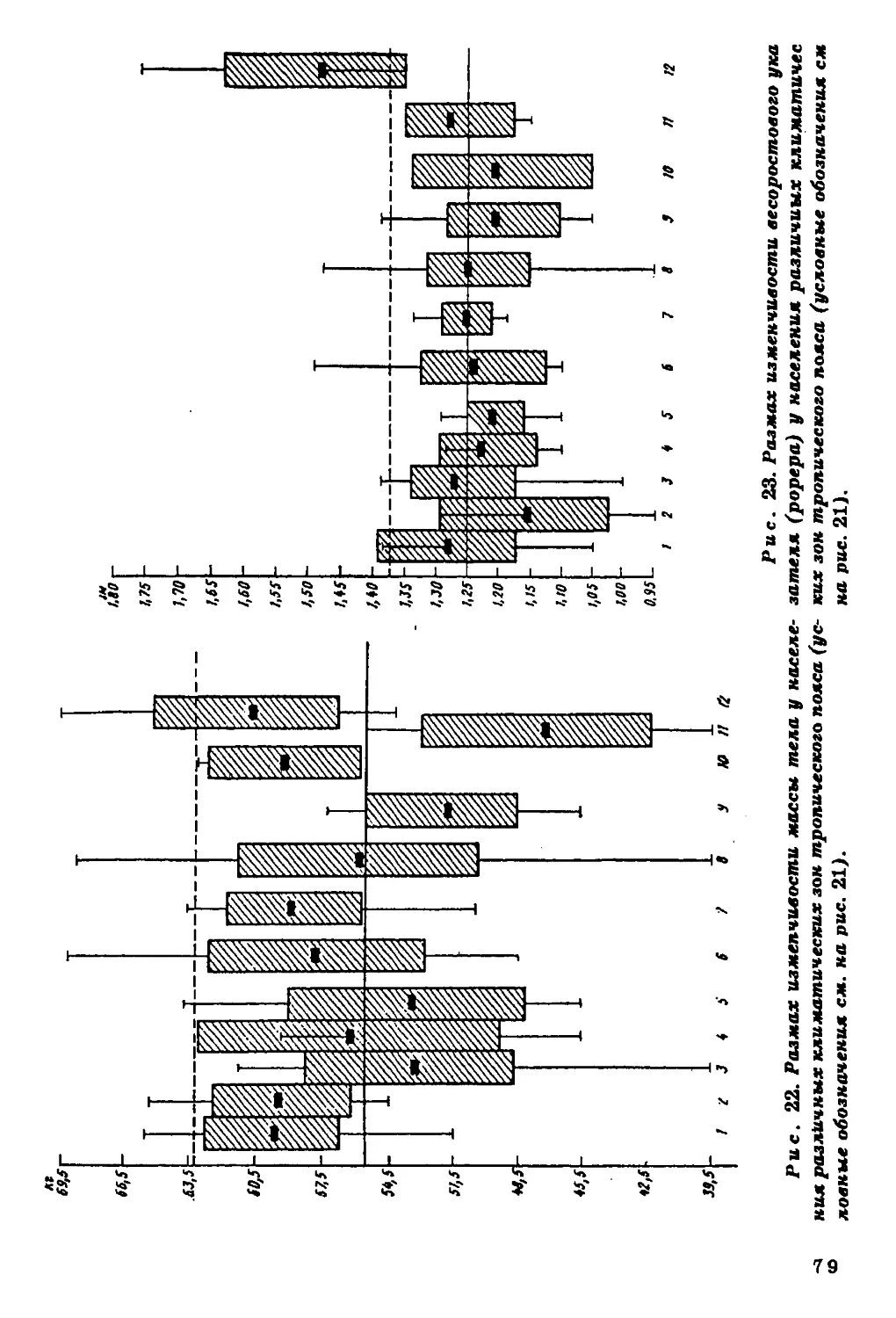
сы и поверхности тела наблюдаются следующие закономерности
1) наибольшей вариабельностью признаков характеризуется ко
ренное население тропической зоны, а в ее пределах - население
Африки; 2) большинство высокорослых групп населения сконцен
трировано в тропической зоне; 3) в тропиках сосредоточены наи
более абсолютно и относительно легкие группы, с большей относи
тельной поверхностью тела; 4) в пространственной изменчивости
пропорций тела и строения грудной клетки молено отметить одщ
закономерность - по направлению к тропической зоне возрастав1]
долихоморфия и уменьшаются поперечные оси скелета.
Таким образом, по тотальным размерам тела и их соотношени
ям все население земного шара может быть разделено на основа
нии географического признака на население тропических и вне
тропических широт.
3. ФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ
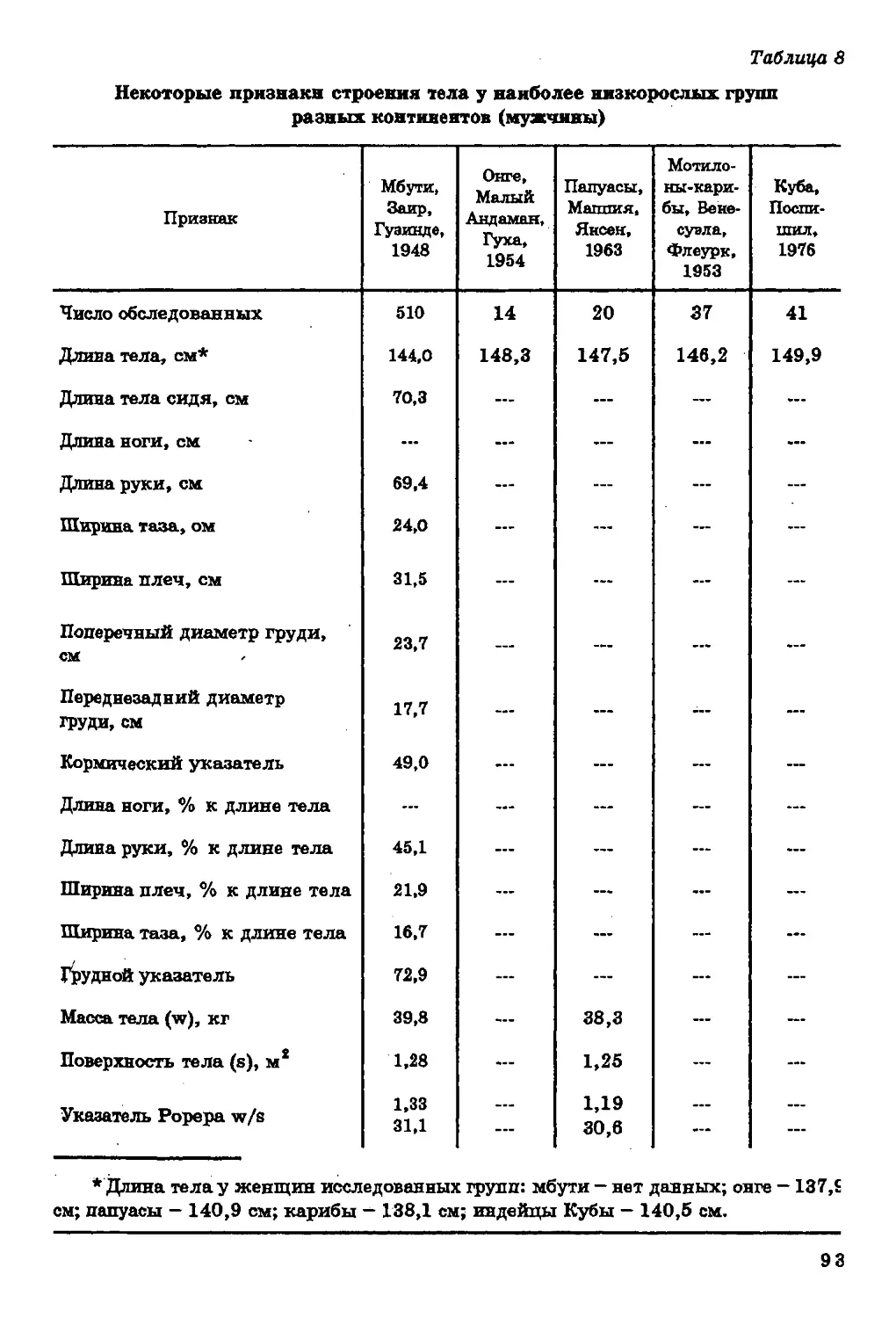
Основной обмен и терморегуляция. Географической измен
чивости основного обмена, вычисленного по формуле с учетом дли
ны и массы тела, а также температуры окружающего воздуха
посвящен© исследование Д. Робертса (Roberts, 1952), проведенное на
200 этнических группах в различных районах мира.
Определяя основной обмен (среднюю дневную
теплопродукцию, выраженную в килокалориях) по формуле: 1=28 73-4,297*-
-3,23S+19,22W, где / - средняя дневная теплопродукция, ккал; Т
37
- средняя годовая температура по Фаренгейту; S - средняя длит
тела, см; W - масса тела, кг, Д. Роберте установил, что уровень
основного обмена у человека понижается в направлении от
северных районов к экваториальным. Коэффициент корреляции
средней дневной теплопродукции и средней годовой температуры
равен -0,77.
Помимо горизонтальной зональности в изменчивости
основного обмена существует и вертикальная (Алексеева, 1974). Речь идет
о вариабельности этого признака на различных высотах над
Уровнем моря. По нашим данным, коэффициент ранговой корреляции
основного обмена с высотой над уровнем моря равен + 0,51. В
условиях высокогорья, как и в северной части ойкумены,
понижение температуры среды приводит к усилению теплопродукции;
Многими исследователями, напротив, наблюдалось понижение
газообмена у коренных жителей высокогорья. Это разногласие
обсуждалось нами в вышеназванной работе, и мы вновь вернемся к
нему в соответствующем разделе. По-видимому, в условиях
высокогорья более экономно, чем на равнине, расходуется кислород во
время работы. Причина понижения основного обмена в некоторых
высокогорных областях может заключаться в недостатке йода в
окружающей среде и связанной с этим гипофункции щитовидной
железы (Агаджанян, Миррахимов, 1970).
Исследование поглощения кислорода в лабораторных условиях
выявило повышение основного обмена у всех коренных народов
Крайнего Севера (Andersen 1960в; Irving е? аі., I960; Кандрор,
1962; Веселухин, 1977). Использование оксигемометрического
метода определения газообмена такэке дифференцирует население
внетропических и тропических широт; у последних газообмен
ниже (Алексеева и др., 1972а, 19726, 1973, 1983;
Волков-Дубровин, 1974).
Опыты с переохлаждением показывают, что у коренного
населения Севера - эскимосов, индейцев Аляски и лопарей - более
высокая температура пальцев во время охлаждения, чем в
контрольных группах (Mechan, 1955; Brown, 1957; Andersen, 1960a,
1960b; Eisner, 1960, и др.)-
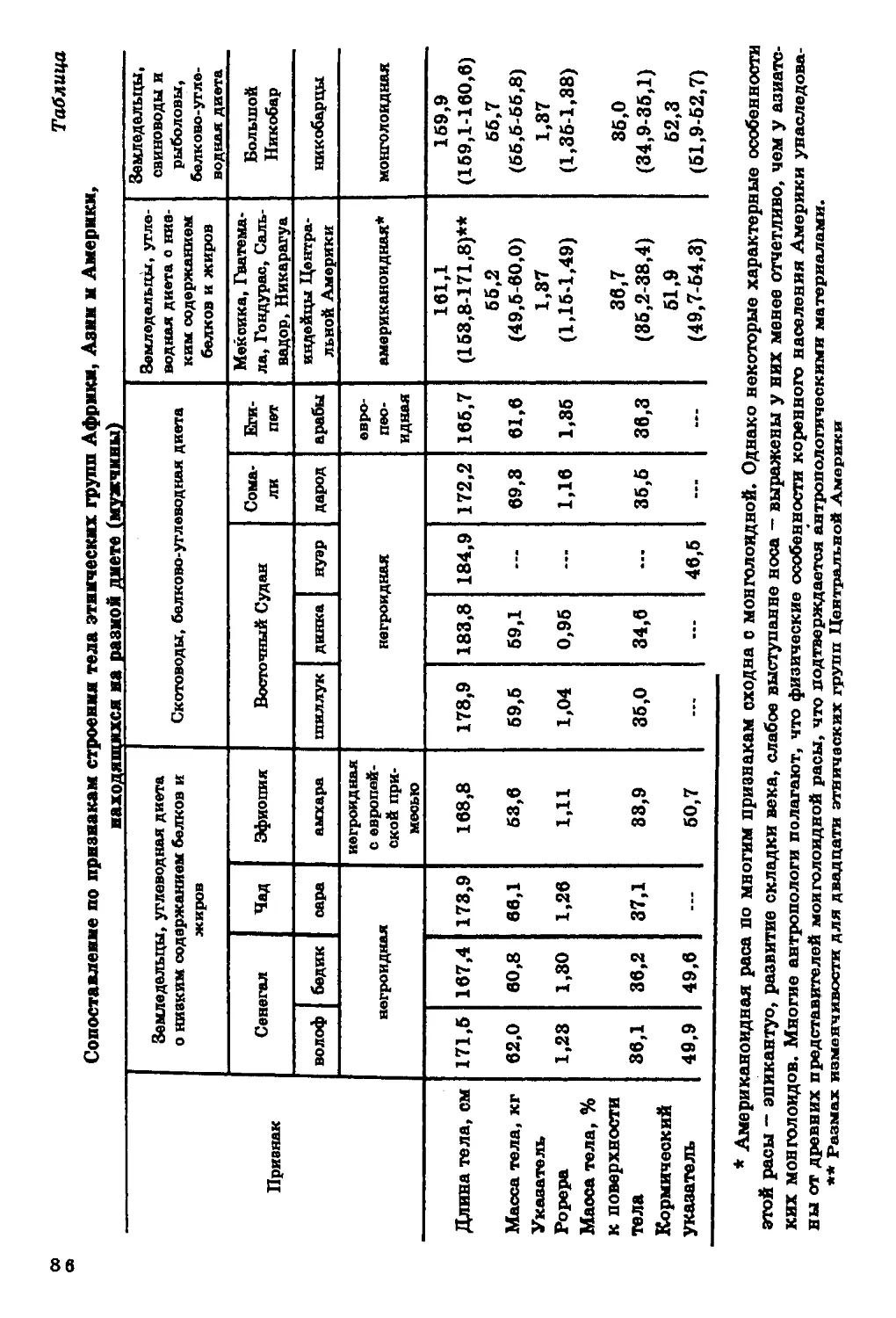
Ли?иды сыворотки крови. В антропологических
исследованиях одним из наиболее изученных физиологических признаков
является холестерин сыворотки крови.
38
При анализе географической изменчивости уровня
холестерина мы столкнулись со значительными методическими
трудностями сопоставления данных разных авторов, что обсуждалось нами
ранее (Алексеева, 1971). В настоящем разделе внимание будет
обращено лишь на общие тенденции пространственной
изменчивости этого признака.
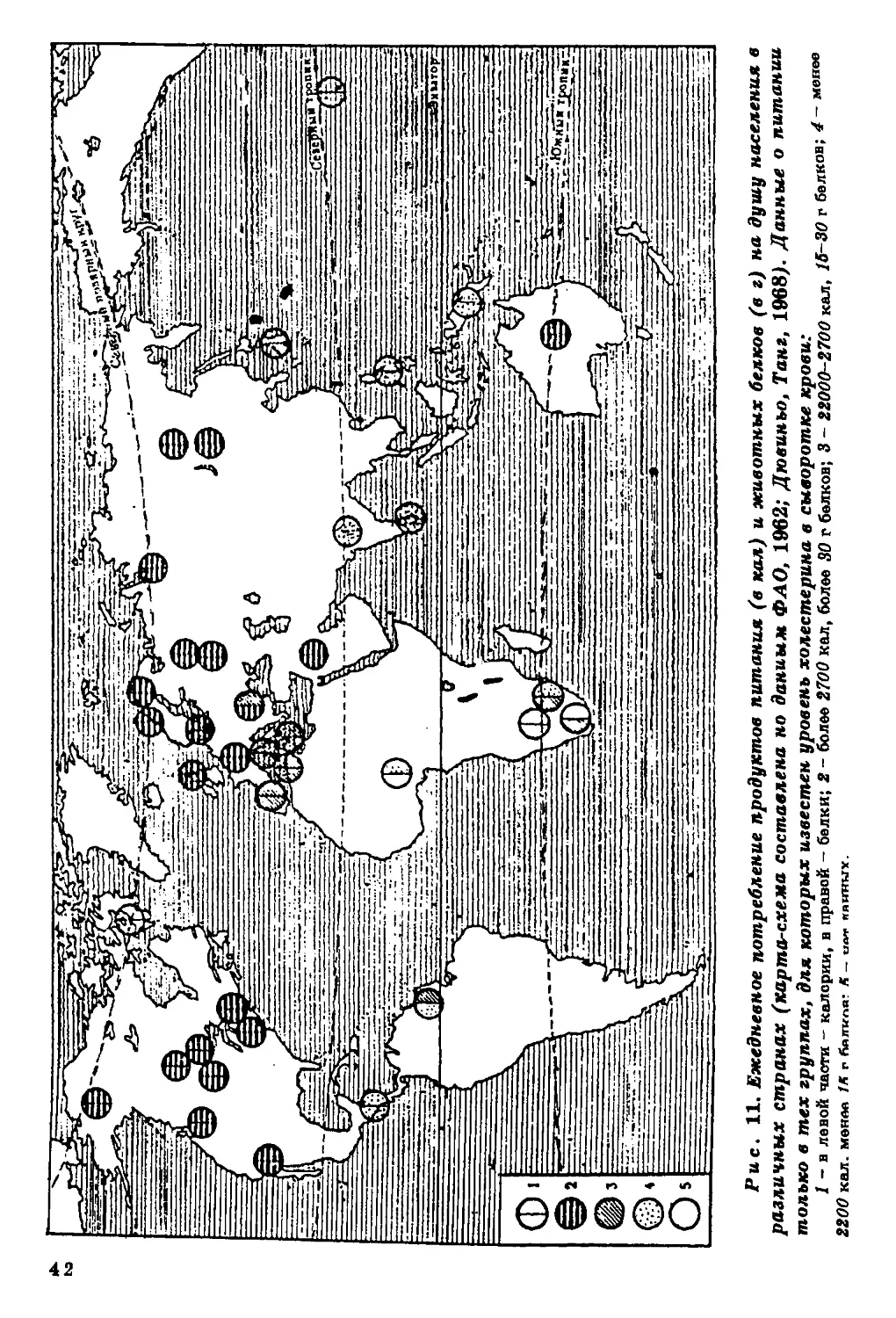
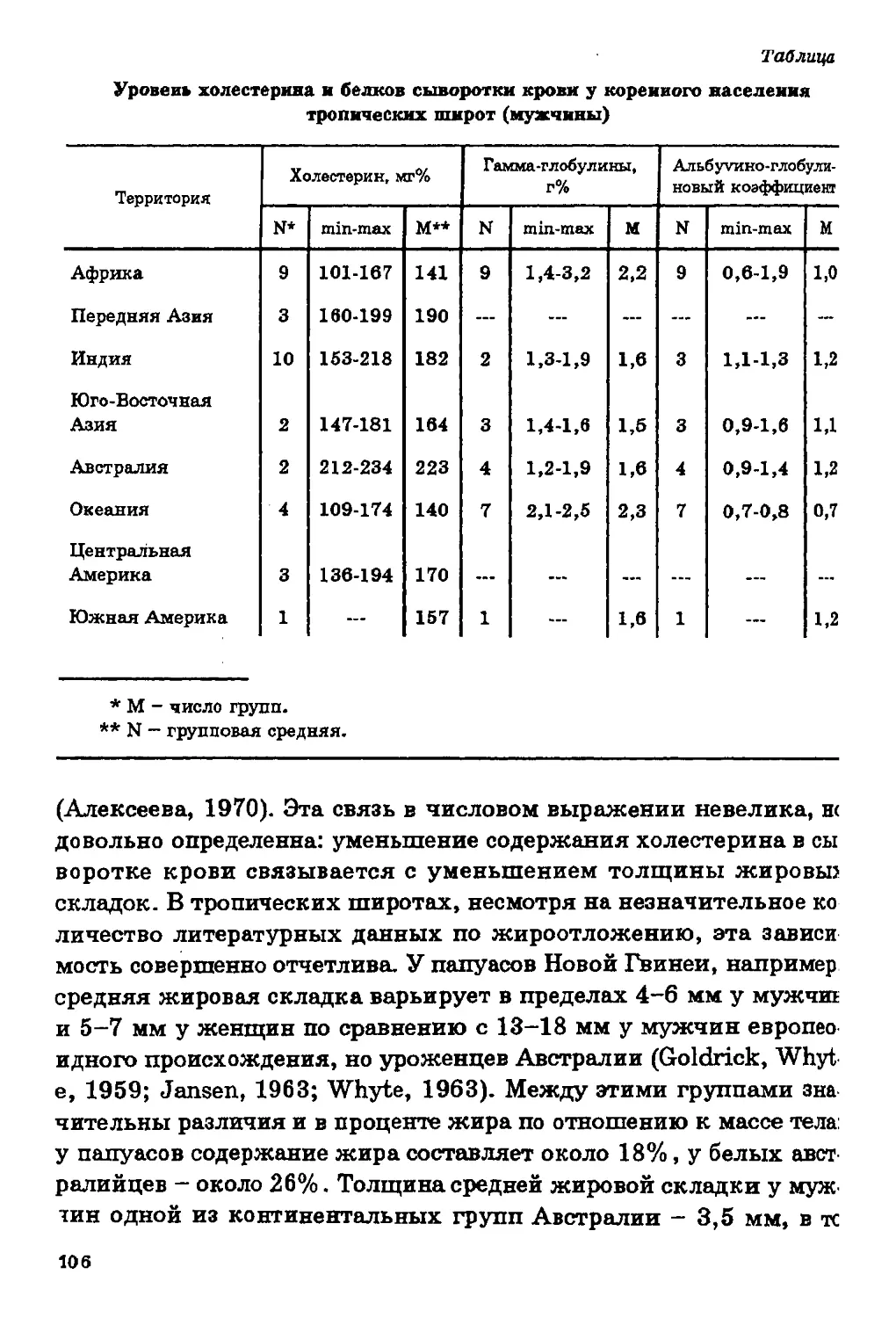
В географической изменчивости уровня холестерина
безоговорочно строгой картины не наблюдается (рис. 10). Достаточно
четко прослеживается его понижение к югу, хотя есть и группы,
где эта закономерность нарушается: эскимосы Канады и ненцы -
на севере, австралийцы и папуасы Гавайских островов - на юге.
Во многих исследованиях прослеживается значительная связь
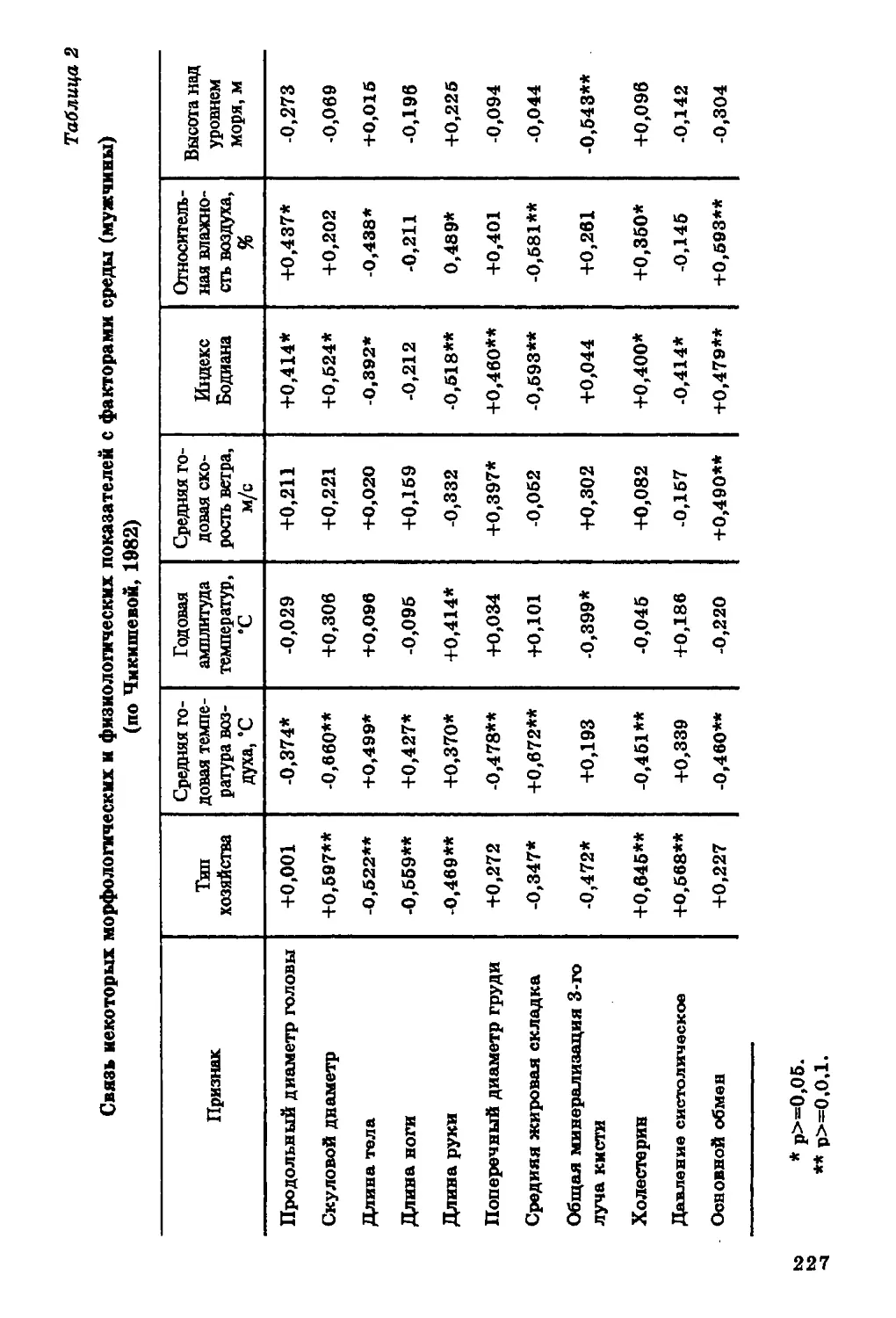
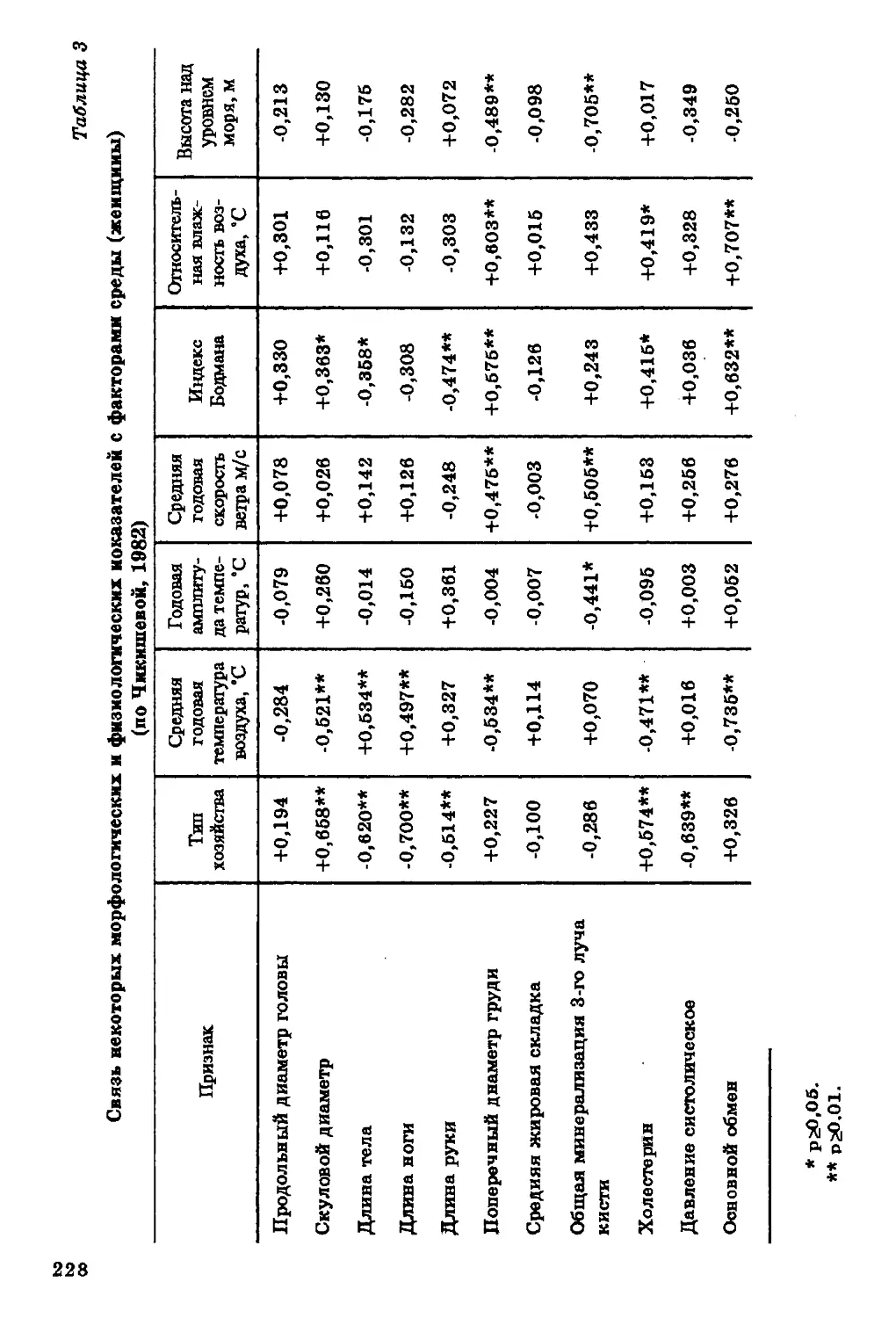
уровня холестерина в сыворотке крови с питанием: он
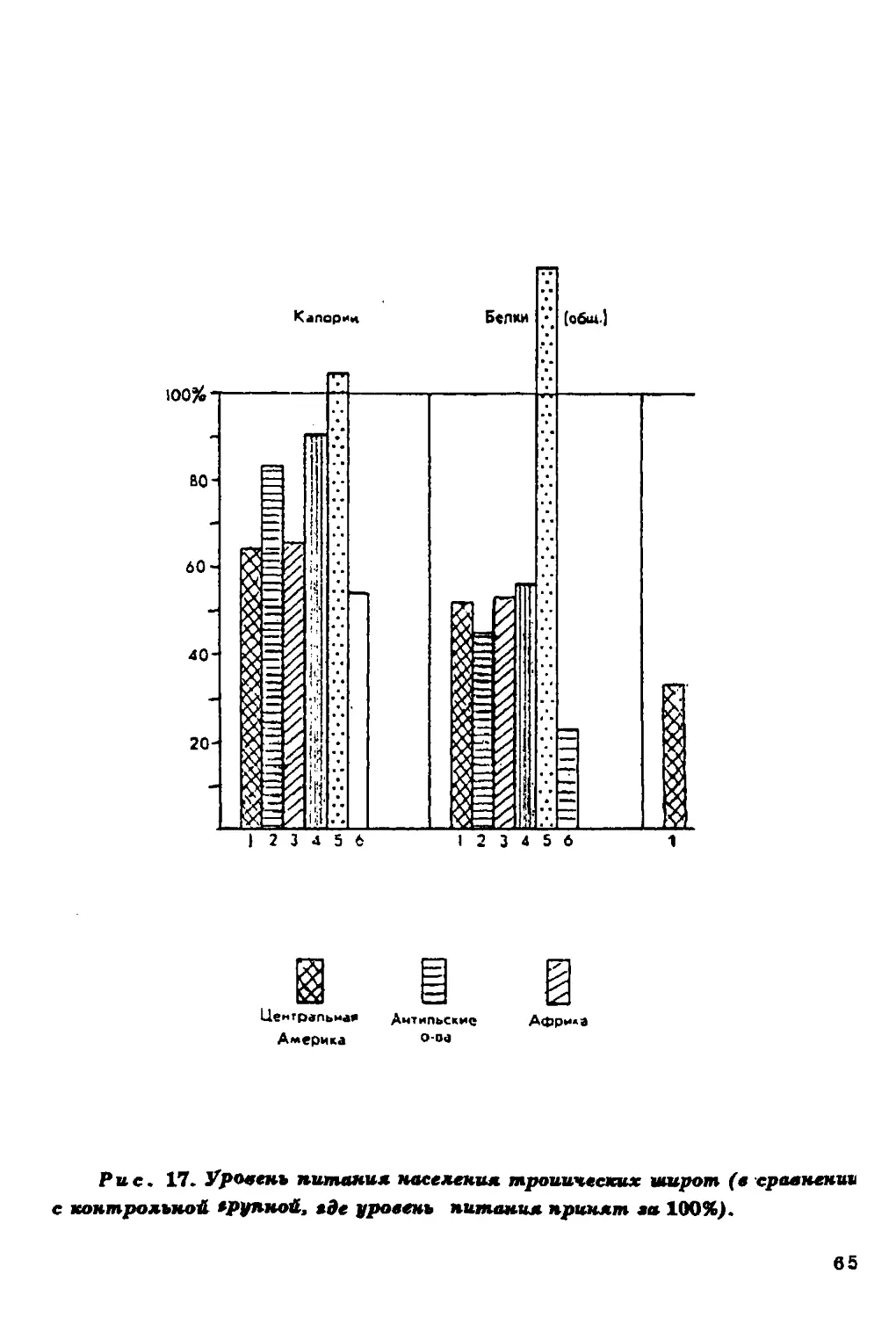
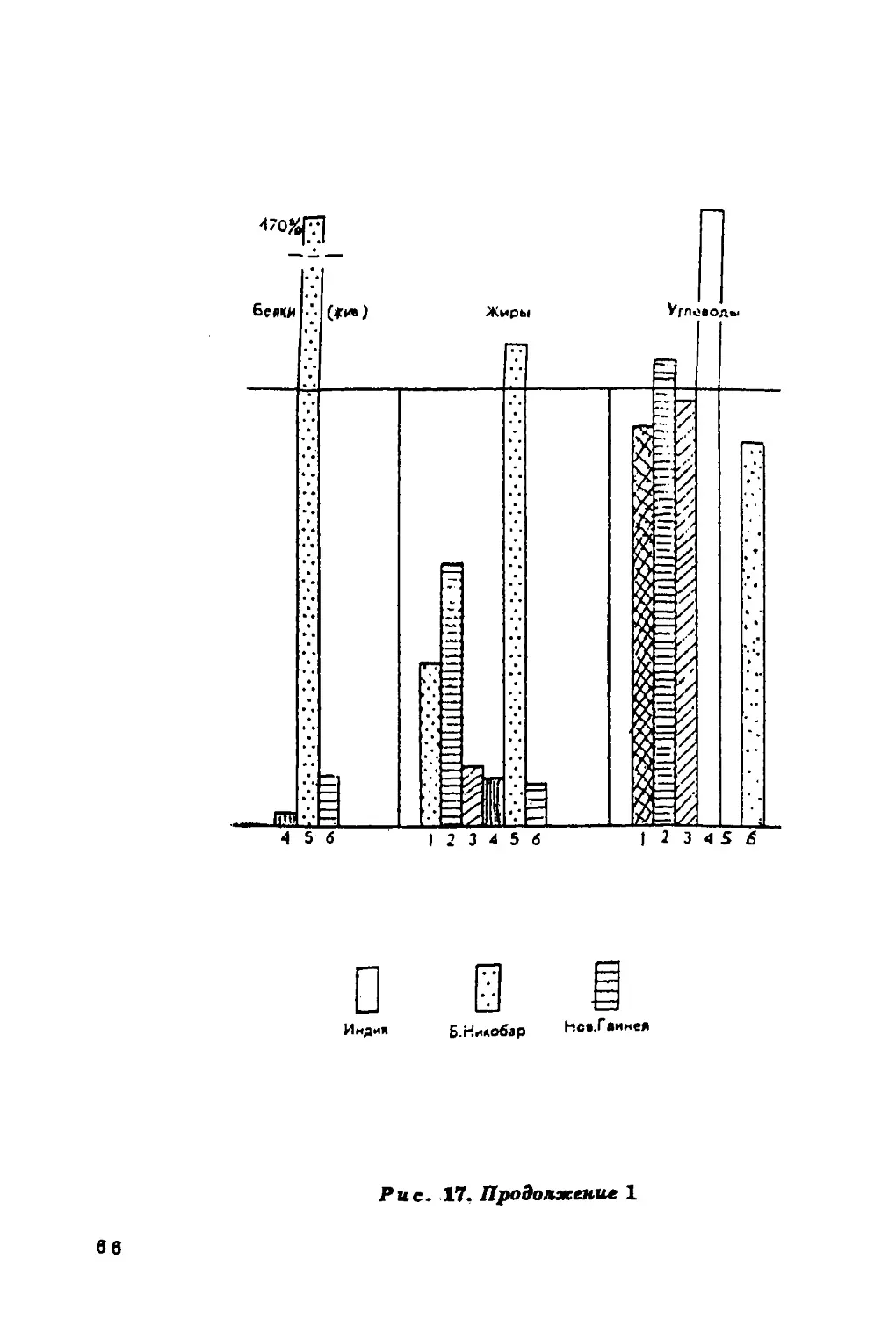
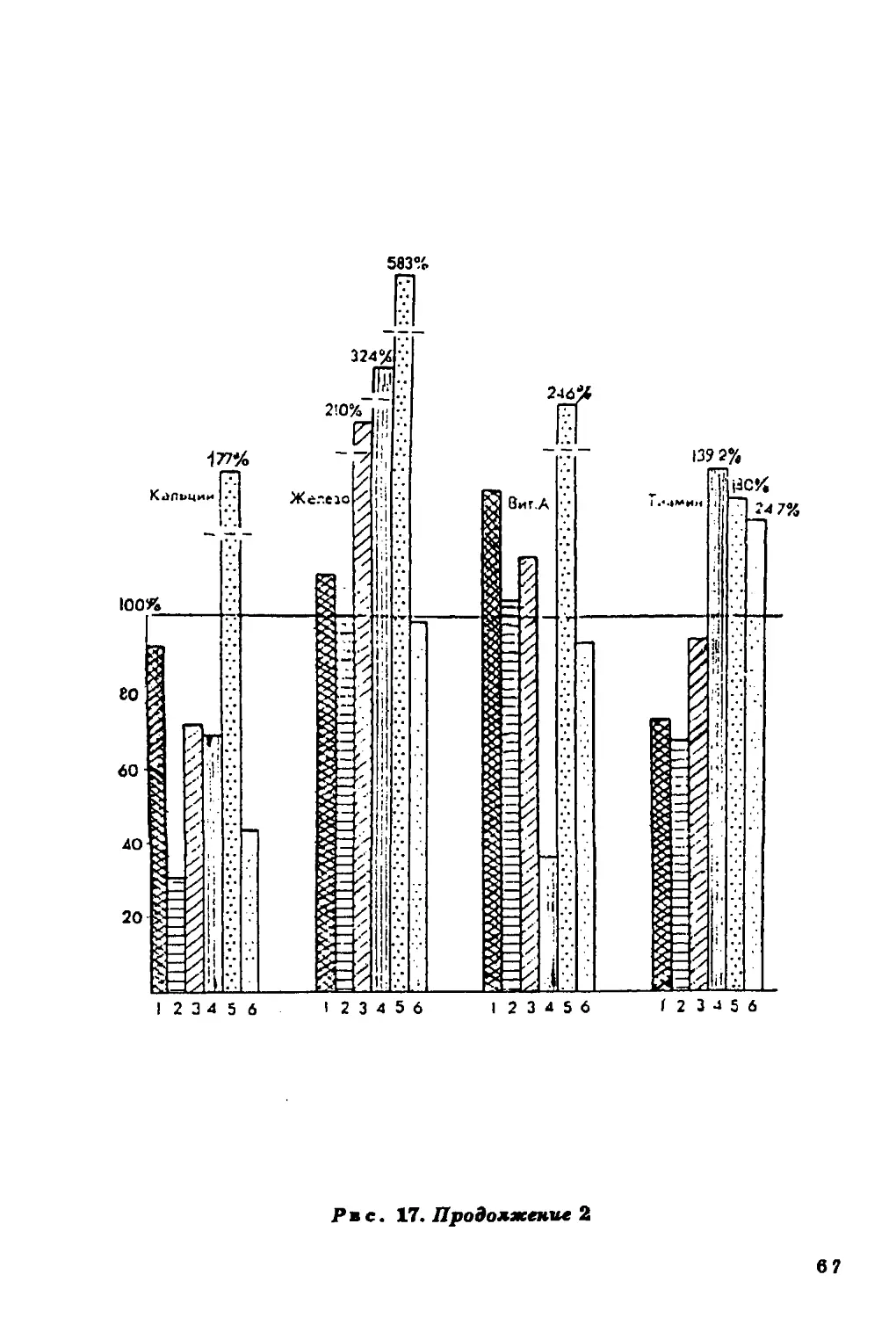
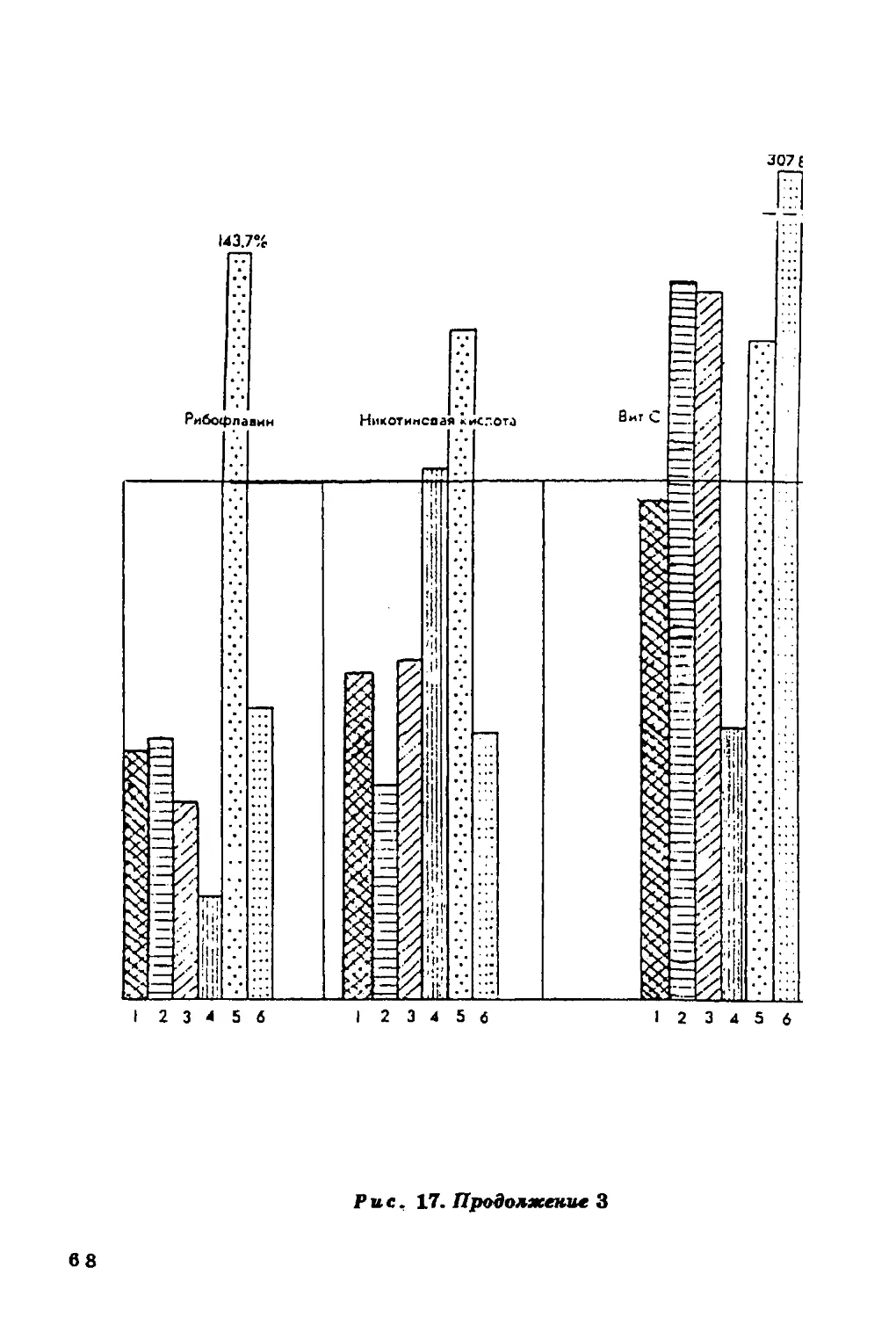
повышается при увеличении в пищевом рационе жиров и белков и
снижается при углеводной диете (та б л. 1, 2).
Таблица 1
Зависимость уровня холестерина от содержания белков,
жиров и углеводов в пище
Состав пищи
Калории
Жиры, г
Белки, г
Углеводы, г
Количество
групп
22
22
18
10
Коэффициент
корреляции Р
+0,63
+0,83
+0,48
-0,80
Пределы
достоверности
Р=0,05
Р-0,01
Р-0,05
Р=0,01
Р=0,05
Р=0,05
Р=0,01
Содержание холестерина в крови в значительной мере
определяется характером питания населения, а он, в свою очередь, если
речь идет о коренном населении, зависит во многом от
ландшафтных особенностей среды.
Связь уровня холестерина в сыворотке крови с уровнем
питания подтверждается совпадением (в общих чертах)
географической изменчивости этого признака с географическим
распределением ежедневного потребления продуктов питания (рис. 11).
39
«с
*
2
3
К
*
¦I
3
«D
О
&
S
g
*
*
©
at
«
*
и
о
в»
а?
?§
3 со
Я I Й в * — -.
SNu s IS9»* s 5*° i
i*8i 3*1 м8 I &&*
II»
¦^ S i 8c 5 Я • Мч§
5 *8 8,8.1 § ' SB ' a
I • «и • &-?>* i ?<e> a
& S ? rf I S .. 5? to л Ы f
i se
g»
I
J s в a *i Hug|
•» i a s >S i 11 и
О E M О I
«> 2 Э tr> I .^
I
'lis
¦*
ф Ф
у 2 Я — » 3 8 * V Ef
йПвч 8. * чая
? x d Э *—- Ч »Я 3 5 5
8і&8&#*«эі
gs s s«a ii «8
S W LT G
3
s
?<?§ Si
&;
41
« а
* а
I °
E. *
* .
§
*
*
i*
e*
SO
3
2 *
a «
« <*>
* 5
¦at лч
at w
* a
& i*
о
о
»
a;
4
i
H
at
о. я
a
n
о
«
ф
to
И
О со
* s
«?
* fa
*«*
SI ф
*» 5
gvg
3 к
Ho
*?
* 1
V "1
s §
is
if
8"
О ^
8-3 j
8-8 I
я •
§ i §
3 «>
с*
42
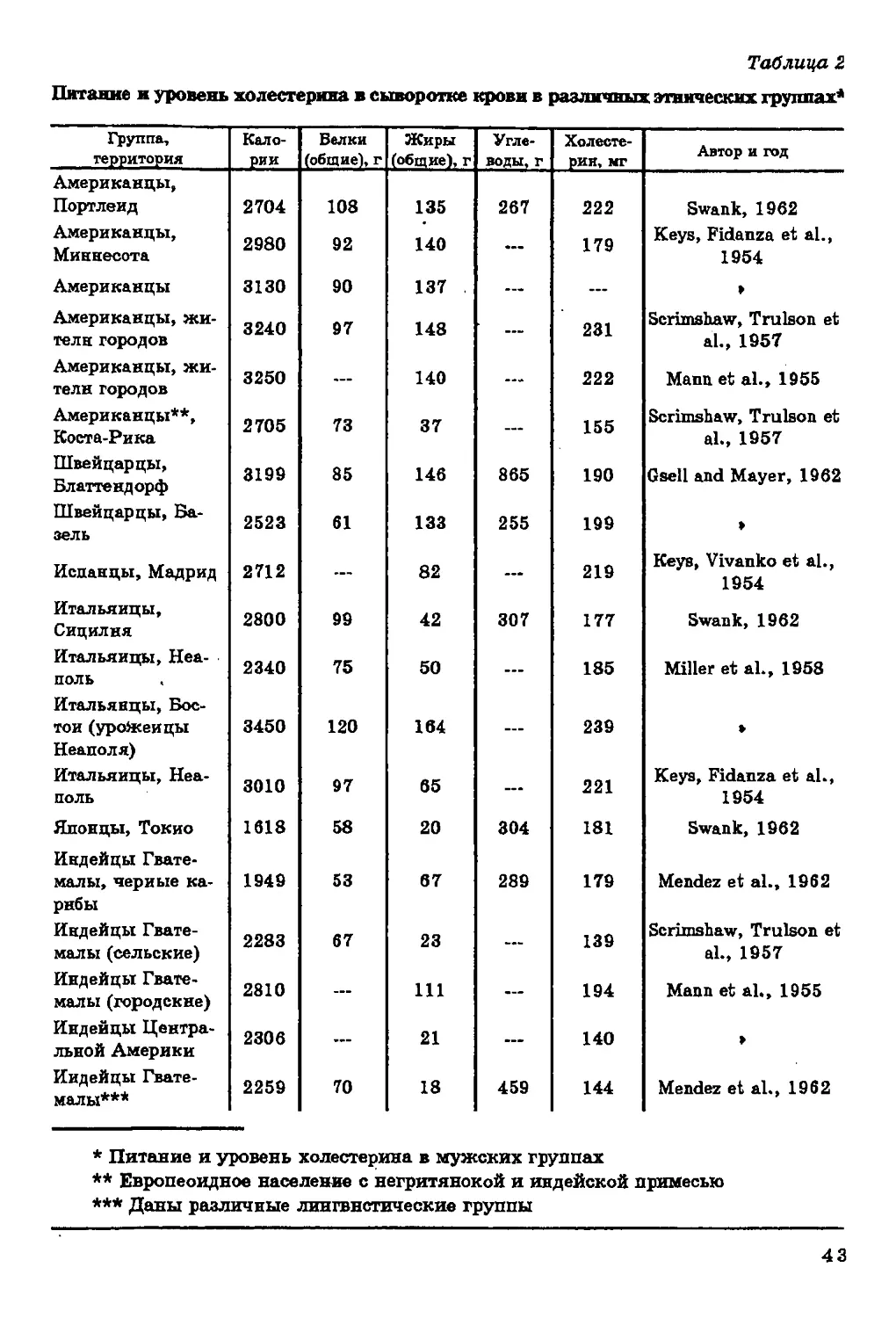
Таблица 2
Питание и уровень холестерина в сыворотке крови в различных этнических группах4
Группа,
территория
Американцы,
Портленд
Американцы,
Миннесота
Американцы
Американцы,
жители городов
Американцы,
жители городов
Американцы**,
Коста-Рика
Швейцарцы,
Блаттендорф
Швейцарцы,
Базель
Испанцы, Мадрид
Итальянцы,
Сицилия
Итальянцы,
Неаполь
Итальянцы,
Бостон (уройсенцы
Неаполя)
Итальянцы,
Неаполь
Японцы, Токио
Индейцы
Гватемалы, черные
карибы
Индейцы
Гватемалы (сельские)
Индейцы
Гватемалы (городские)
Индейцы
Центральной Америки
Индейцы
Гватемалы***
Калории
2704
2980
3130
3240
3250
2705
3199
2523
2712
2800
2340
3450
ЗОЮ
1618
1949
2283
2810
2306
2259
Белки
(общие), г
108
92
90
97
73
85
61
—
99
75
120
97
58
53
67
70
Жиры
(общие), г
135
140
137 .
148
140
37
146
133
82
42
50
164
65
20
67
23
Ш
21
18
Углеводы, г
267
—
365
255
—
307
—
304
289
459
Холестерин, мг
222
179
—
231
222
155
190
199
219
177
185
239
221
181
179
139
194
140
144
Автор и год
Swank, 1962
Keys, Fidanza et al.,
1954
»
Scrimshaw, Trulson et
al., 1957
Mann et al., 1955
Scrimshaw, Trulson et
al., 1957
Gsell and Mayer, 1962
»
Keys, Vivanko et al.,
1954
Swank, 1962
Miller et al., 1958
»
Keys, Fidanza et al.,
1954
Swank, 1962
Mendez et al., 1962
Scrimshaw, Trulson et
al., 1957
Mann et al., 1955
>
Mendez et al., 1962
* Питание и уровень холестерина в мужских группах
** Европеоидное население с негритянской и индейской примесью
*** Даны различные лингвистические группы
43
Продолжение табл. 1
Группа,
территория
Индейцы
Гватемалы
»
»
Индийцы, низкий
социально-экономический уровень
Индийцы,
высокий социально-
экономическ ий
уровень
Индийцы,
армейские офицеры
Индийцы сипаи
Калории
2144
2247
2063
2000-
2500
2500-
3000
2800-
350О
3000-
3500
Белки
(общие), г
68
74
63
30-50
70400
90-100
90-100
Жиры
(обшие), г
20
19
17
10-30
85-120
90-120
90-120
Углеводы, г
429
451
418
—
...
—
Холестерин, мг
138
127
130
153
179
212
174
Автор и год
Mendez et al., 1962
>
р
Gopalan, Ramanathan,
1957
>
»
Тип питания, правда, не единственный фактор, определяющий
уровень холестерина в сыворотке крови. Некоторые данные
заставляют думать и о существовании генетических механизмов,
регулирующих обмен липидов в организме (Алексеева, 1971; Но
Kang et al., 1971). Тем не менее влияние экзогенных факторов на
географическую изменчивость содержания холестерина очень
велико.
Белки сыворотки крови. Первая сводка данных по
географической изменчивости белков была составлена Л.К. Гудковой
(1975). В значительно дополненном виде приводим ее в настоящем
разделе (рис. 12).
В географической изменчивости сывороточных протеинов
обращает на себя внимание повышение уровня гамма-глобулинов
(иммунных белков) у коренных жителей тропических широт.
Объяснение этому явлению далеко неоднозначно. Некоторые
исследователи считают, что высокие значения гамма-глобулинов
вызываются малярийными паразитами и другими патогенными
организмами (McGregor et al., 1956, 1960); другие связывают эту
особенность с дефицитом белка в пище (Henrotte et al., 1969); третьи,
основываясь на наблюдениях уровня белков у новорожденных,
настаивают на генетической природе различий в этом признаке
между африканцами и европейцами (Edozien, 1961; Leonhardt,
1962; Lichtman et al., 1965).
44
Вопросы географической изменчивости белков сыворотки
крови будут обсуждены при характеристике морфофункциональных
особенностей населения определенных экологических зон. В
настоящем разделе важно обратить внимание на то, что высокий
уровень гамма-глобулинов характерен не только для африканцев, но
и для индейцев Америки, обитающих в тропическом поясе, для
папуасов и австралийцев, для коренного населения Индии.
В противоположность гамма-глобулинам уровень альбуминов
(структурных белков) в направлении к тропической зоне
снижается, соответственно убывает и величина альбумино-глобулинового
коэффициента. Большинство авторов склонны объяснить
пониженное содержание альбуминов в крови тропических аборигенов
белковой недостаточностью в питании, но и по отношению к ним
существуют доказательства генетической природы.
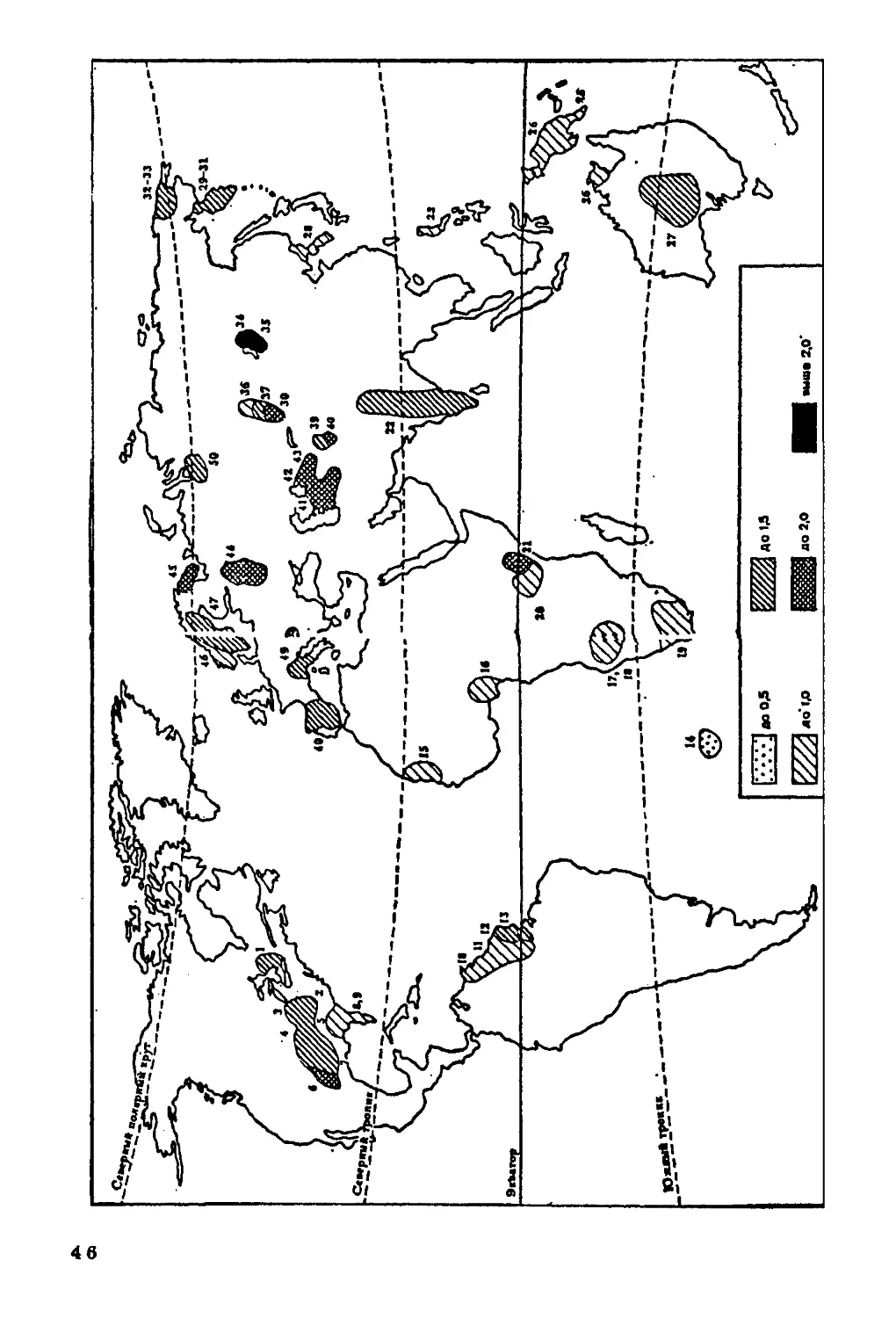
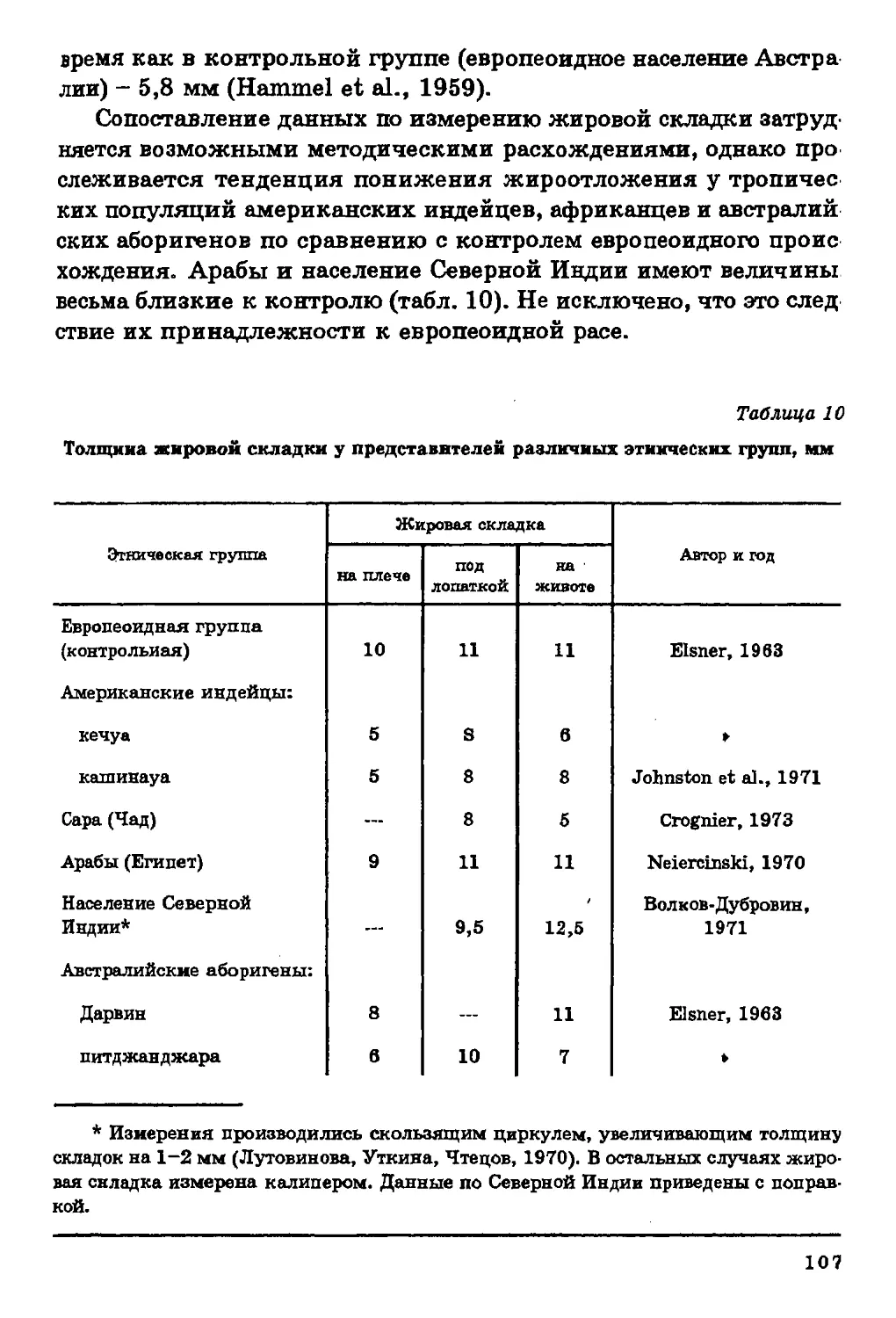
Минеральный состав костной ткани. Планомерное
исследование in vivo уровня минерализации (содержание оксиапатитов в
скелете) проводится только у нас в стране, поэтому мы не можем
судить о закономерностях пространственной изменчивости этого
показателя в мировом масштабе, но в то же время располагаем
сведениями о его территориальных вариациях.
По мнению многих исследователей, уровень минерализации
скелета в значительной степени связан с рационом питания. Так,
более низкий уровень минерализации скелета отмечен у детей
индейцев из Перу, находящихся на диете, бедной белками, жирами
и минеральными веществами, по сравнению с детьми "белых"
американцев (Schraer, Newman, 1958). Неблагоприятное воздействие
тропической диеты, бедной белками и кальцием, обнаруживается
при рентгенографическом анализе процессов окостенения у детей
из сельских районов Гватемалы (Касавина, Торбенко, 1975).
Этими же авторами отмечены минеральная недостаточность и как
следствие этого — остеопороз у сельского населения рисоводческих
районов Японии. Снижение уровня минерализации скелета
наблюдается и в районах, где для питья употребляется опресненная воде
(Хлыстов, Прибытков, 1973).
Достаточно показательна и связь уровня минерализации
скелета с геохимической средой. Напомню работу П. Вассаля (Vassal.
1957), установившего территориальную связь брахикефалии с
выходами гранитов на поверхность почв; наши данные о зависимое
ти между размерами и формой скелета и содержанием костеобра
46
46
*
и
а>
а,
а»
?
8-
Ё
#
Е
as
и
а
§
о
«1
«
о
X
а
а»
м
О
*
а»
в
л
?
о
«
а4
Я я я
ill
со
м« со а
.. .. і
в я 1
оо
t
8-
5
15
оо
© 5
^ f?Q0
f4
а
я
со
»:•
47
зующих минералов в среде обитания (Алексеева, 1979), а такж*
наблюдения, касающиеся сопряженности фосфорно-кальциевоі
недостаточности в скелете с условиями влажных тропиков (Есо
logy and anthropology, 1962). В последнее время получены исклю
чительно важные данные относительно связи микроэлементногс
состава скелета со средой обитания. Микроэлементное содержание
костной ткани отражает уровень и набор биогенных микроэлемен
тов в среде (Добровольская, 1984).
Для суждения о территориальных вариациях признака в пре
делах всей ойкумены данных пока недостаточно. Известен уровені
минерализации относительно большого количества популяций
обследованных на территории бывшего СССР, причем они могу]
рассматриваться как популяции внетропической зоны. Вместе <
тем для коренных жителей тропических широт уровень минера
лизации скелета определен рентгенофотометрически только у не
скольких индийских популяций (Волков-Дубровин, 1974). В до
полнение к этому можно обратиться к исследованиям американс
ких авторов, показавших с помощью количественных химически}
методов различия в минеральной концентрации скелета у предста
вителей европеоидной и негроидной рас (Broman, Trotter, Peters
on, 1958). Из сравнения приведенных групп видно, что как индий
ские популяции, так и группа африканского происхождения име
ют более высокий уровень минерализации, чем уроженцы умерен
ной климатической зоны. Этот результат пока несколько озадачи
вает, так как детские группы, как указывалось выше, в условияз
тропической диеты характеризуются минеральной недостаточное
тью.
К сожалению, данные о минерализации скелета детей и
взрослых относятся к разным этносам и охватывают очень небольшое
число групп. Судя по диете тропических популяций, характеризу
ющейся низким содержанием белков, жиров и кальция, уровені
минерализации костной ткани аборигенов тропиков должен был
понижен (Алексеева, 1977). В то же время известно, что грациль-
ный скелет характеризуется повышенной минерализацией
(Зенкевич, 1940; Павловский, 1970), что можно рассматривать как
компенсацию недостаточности органической основы опорной системы.
Следует признать, что пока мы не располагаем убедительными
сведениями относительно панойкуменной вариабельности уровня
минерализации костной ткани.
48
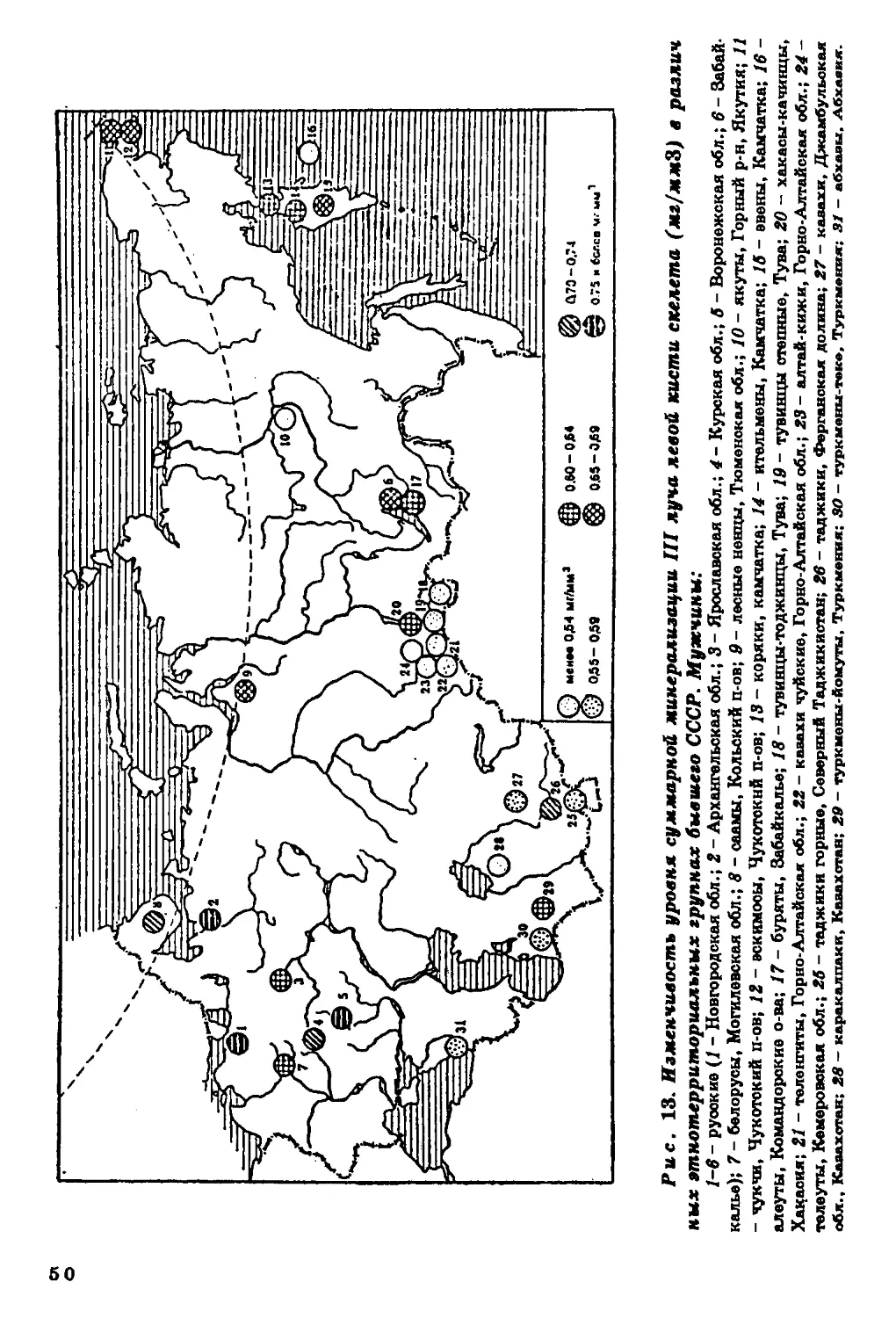
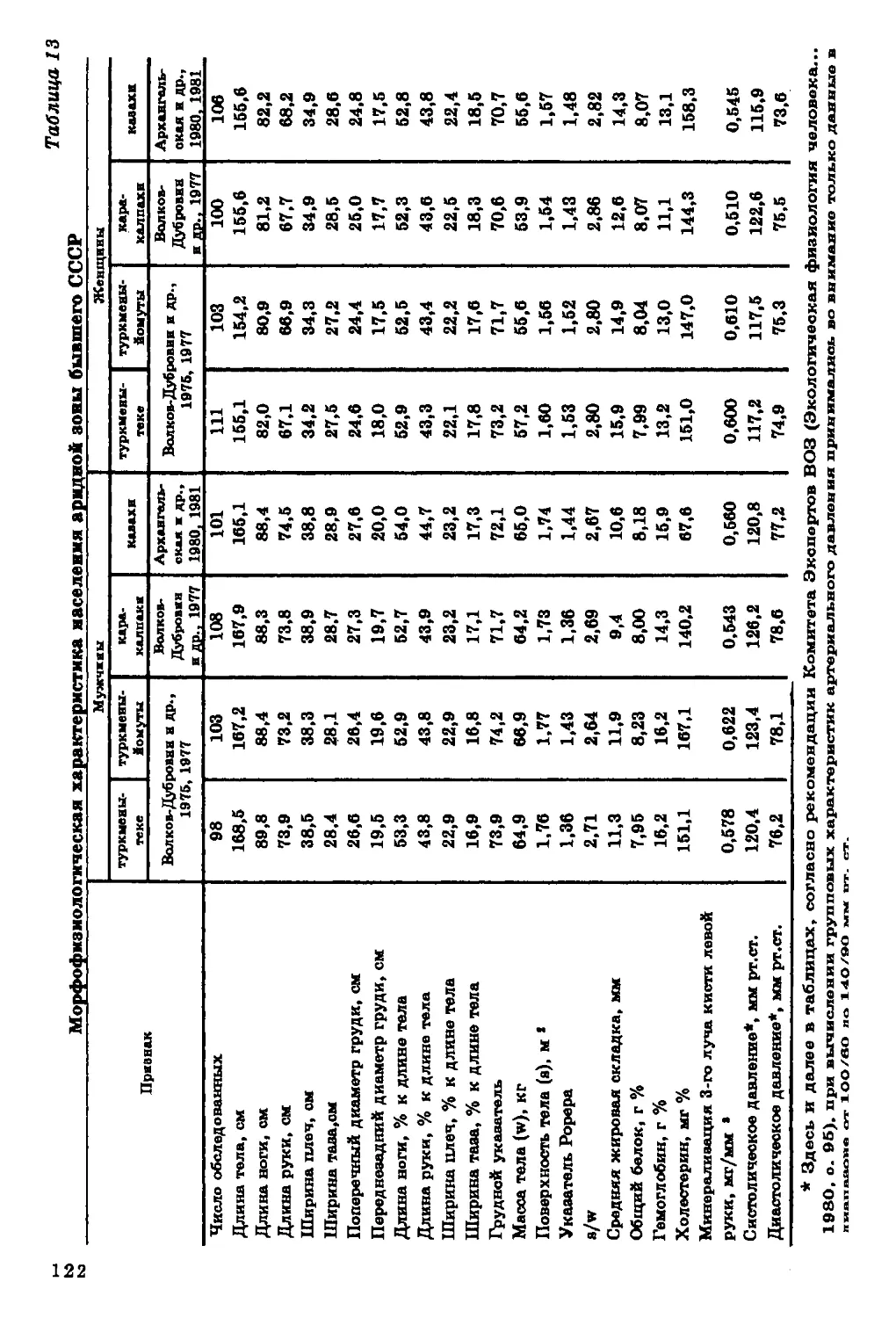
На территории бывшего СССР более чем в 60 популяциях вы
явлены некоторые географические закономерности в вариабельно
сти уровня минерализации (рис. 13). Например, у генетическі
отдаленных друг от друга, но живущих в арктической зоне этни
ческих групп наблюдается повышение уровня минерализации ске
лета, что особенно заметно при сопоставлении их с родственным!
группами умеренного климата. Так, у русских Архангельской об
ласти уровень минерализации выше, чем у русских Новгородское
области, у чукчей и эскимосов он выше, чем у народностей Кам
чатки.
Обособленное положение занимают алеуты Командорских
островов, для которых характерны наименьшие величины
показателей минерализации среди всех изученных народностей
Северо-Восточной Сибири. В то же время языковое, культурное и
антропологическое сходство их с эскимосами, коренным населением
Чукотки, несомненно. По-видимому, фактор этнической и расовой
принадлежности не играет превалирующей роли в формировании
такого важного для жизнедеятельности признака, как
минеральное содержание скелета. Причина географической
дифференциации его скорее экологическая, чем генетическая.
К сожалению, для исследования экологической зависимости
уровня минерализации скелета человека недостает конкретных
геохимических характеристик. В ряде случаев в изученных нами
регионах нет данных по важнейшим биогенным элементам.
Наиболее полно представлены геохимические характеристики
восточноевропейской части бывшего СССР, где изучено русское
население некоторых районов Архангельской, Новгородской,
Ярославской, Воронежской и Курской областей. Менее полные
геохимические характеристики имеются по Якутии, Туве, Бурятии,
Камчатке, Кольскому полуострову, северным районам Западной
Сибири, Таджикистану и Каракумам. Во всех этих районах также
исследовался уровень минерализации костной ткани у коренного
населения.
Коэффициент связи между такими биологически важными
элементами, как А1, В, Са, Со, Си, Fe, I, Mg, Mn, Mo, Si, Sr, и
содержанием оксиапатитов в скелете, определяемым рентгенофотомет-
рическим путем по уровню минерализации третьего луча левой
кисти, невелик (Бацевич, 1977). Исключение составляет высокая
(порядка 0,7 у мужчин и 0,8 у женщин) негативная связь
признака с содержанием А1. В свете биогенной роли А1 эта связь пред-
49
3
:
8 ?.- и
со
і е ° йЗ
1 II *х
'"В
S
50
ставляется вполне закономерной, так как алюминий влияет на
обмен фосфора, образуя с ним нерастворимые соединения и
выводя его из желудочно-кишечного тракта (Войнар, 1960).
Что касается других коэффициентов корреляции, то их малая
величина и нередко противоречивость с точки зрения биогенной
роли некоторых химических элементов (в частности, почти полное
отсутствие связи уровня минерализации скелета с содержанием
кальция в почвах) вызывают недоумение. Для объяснения этого
обстоятельства можно привести несколько соображений.
Во-первых, уровень минерализации скелета больше связан с
минеральным составом рационов питания, чем с содержанием минеральных
веществ в почвах; во-вторых, при корреляциях учитывались
только абсолютные значения величин содержания химических
элементов, а не их соотношение, которое нередко играет более
существенную роль во влиянии на живой организм; в-третьих, не
исключено, что химическая ситуация в большей мере воздействует на
растущий организм, и поиск антропогеохимических связей следует
перенести на детские контингенты, в которых помимо уровня
минерализации может быть изучена и скелетная зрелость.
Нарушение геохимического баланса в природной среде
особенно сильно сказывается на детях. Исследование здоровых и
больных рахитом детей в зоне нарушения стронциево-кальциевого
отношения показало, что больные дети сильно отстают от нормы по
срокам окостенения, срокам прорезывания зубов и по
физическому развитию; их физиологический возраст соответствует 12-16
месяцам при паспортном возрасте 19-21 месяц (Павловский, 1970).
Особенно велика разница между больными и контрольными
детьми по уровню минерализации. Несмотря на то, что группа
здоровых детей, взятых в качестве контроля, на 6-7 месяцев моложе
группы больных детей, минеральное содержание скелета у
последних значительно ниже (рис. 14).
В заключение следует сказать, что причины
пространственного распределения уровня минерализации недостаточно ясны.
Факторы экологические, такие, как питание и геохимические
характеристики среды обитания человеческих популяций,
бесспорно, воздействуют на процесс минерального обмена. В то же
время характер метаболизма минералов отличается большой
спецификой, свидетельствующей о наличии наследственных
регуляторов минерального обмена (Sobel, 1955).
51
Д»ГН *0Htpon*MC4 лети бЪльнме
группы рахитом
Рис, 14. Средний /*/ показатель минерализации скелета у детей г. Исфа
pat здоровых и больных рахитом- (* - минерализация определялась по ереднеі
фаланге 3-го луча кисти, плечевой и пяточной костям).
Резюмируя данные о географической изменчивости некоторых
физиологических показателей, приходим к заключению о
существовании закономерных территориальных вариаций в типе
обмена веществ. Например, отмечается понижение основного обмена и,
следовательно, теплопродукции в направлении с севера к
экватору; понижается уровень показателей липидного обмена
(холестерина) и повышается содержание иммунных белков
(гамма-глобулинов). Территориальные вариации наблюдаются и в
характеристиках минерального обмена. Строго выраженной зональности в
них нет, хотя определенные закономерности в некоторых случаях
выявляются, например повышение уровня оксиапатитов в
скелете в условиях Крайнего Севера. Закономерности пространственной
изменчивости содержания минеральных веществ в скелете
определяются геохимической ситуацией (Алексеева, 1979).
52
4, РОЛЬ УРОВНЯ ПИТАНИЯ В ГЕОГРАФИЧЕСКИХ
ВАРИАЦИЯХ АНТРОПОЛОГИЧЕСКИХ ПРИЗНАКОВ
В предшествующем изложении уже отмечалась роль питания
в географической изменчивости некоторых физиологических
признаков. Думаю, что следует более подробно остановиться на этом
вопросе.
Много внимания уделяется исследованию влияния питания на
морфофизиологические черты населения в работах индийских
антропологов и биологов. Это и понятно, так как в индийском
обществе не только велика дифференциация по уровню
социально-экономической жизни, но и значительна роль кастовых
предрассудков, запрещающих употребление в пищу тех или иных продуктов.
При этом уровень питания в Индии отнюдь не определяется
положением на иерархической лестнице общественных отношений.
Представители некоторых высокопоставленных каст имеют
гораздо больше ограничений в пище (особенно в отношении белков и
жиров), чем представители так называемых "низших" или
"средних" каст. Поэтому здесь имеет смысл проводить сопоставления не
между группами, различными в кастовом отношении, а между
вегетарианцами и невегетарианцами, как это и сделал Б. Дас (Das,
1966), обнаруживший довольно значительные различия по
комплексу морфологических и физиологических характеристик в
генетически близких группах уроженцев окрестностей Калькутты,
находящихся на разных диетах.
На рис. 15 морфофизиологические характеристики
вегетарианцев выражены в процентах от характеристик
невегетарианцев. Обе группы достаточно репрезентативны, и поэтому результат
этого сопоставления вполне достоверен. Для многих признаков
наблюдается понижение их величин у вегетарианцев; особенно
заметно падение веса, уровня общего холестерина и его
составляющих, глюкозы, остаточного азота, уровня седиментации.
Тенденцию к понижению показывает кальций, общий белок, мочевина,
кислая фосфатаза. При этом заметно увеличивается уровень ами-
ладной активности и щелочной фосфатазы. Повышение амилазной
активности, по-видимому, связано с избытком растительной пищи,
а энзим амилаза гидролизует углеводы. Что касается уровня
щелочной фосфатазы, то обычно он повышается при костных
заболеваниях (Штрауб, 1963). Поскольку у вегетарианцев наблюдается и
53
I 2 3 A 5 б 7 б 9 /О Л tf 13 И 15 16 17 Ш /9 20 ?| 22 23 24 25 26 27
Рис. 15. Сопоставление морфологических и физиологических признаков \
двух групп индейского населения, находящихся на разных диетах (величит
каждого признака у невегетарианцев принята за 100%, по отношению к нил
рассчитаны отклонения у вегетарианцев. По материалам Dae, 1966);
1 - рост; 2 - масса тела; 3 - амилазкая активность; 4 - кальций; 5 - хлориды; 6 - общиі
холестерин; 7 - свободный холестерин; 8 - эфир холестерина; 9 - креатикин; 10 - глкжоаа; 11
- остаточный авот; 12 - кислая фосфатааа; 13 - щелочная фосфатава; 14 - общий белок; 15 -
рН сыворотки крови; 16 - мочевина; 17 — мочевая кислота; 18 - эритроциты; 19 - лейкоциты
20 - лимфоциты; 21 - эовинофилы; 22 - гемоглобин; 23 - уровень седиментации; 24 - пульс; 21
- систолическое давление; 26 - дкастолич?ское давление; 27 - температура ротовой полости.
понижение кальция в крови, то, может быть, это симптом каких-
то изменений в костной ткани. Во всяком случае, изменения вс
внутренней среде организма у вегетарианцев таковы, что
заставляют думать о сдвигах в обменных процессах.
В работах зарубежных антропологов все чаще и чаще звучит
необходимость изучения морфофункциональных особенностей
популяции в связи с типом питания и уровнем материальной жизни
как экологическими факторами, во многом определяющими рост
и развитие человека.
На международном симпозиуме, состоявшемся в августе 1975
г. в Яблонне, близ Варшавы, и посвященном методам контроля зя
ростом и развитием человеческих популяций в изменяющейся
окружающей среде их обитания, были представлены
многочисленные работы, касающиеся влияния диеты и других факторов соци-
54
ально-экономической жизни на многие морфологические и
физиологические их черты. Эти вопросы подробно освещены в
монографии Н. Волянского (WolansH, 1975), рисунок из которой приведем
здесь для иллюстрации связи некоторых антропологических при
знаков с экологическими факторами социального порядка (рис
16).
OSu^uZ Ъеяок & GHtttiOrt районе (4 ър)
Рис* 16. Увеличение роста детей разных стран в возрасте от 1 до 4 лет
в связи с уровнем белков в рационе (по WolaiUki, 1973)
В дальнейшем придется не раз обращаться к роли питания в
образовании тех или иных морфофункциональных особенностей
66
популяций и их географической изменчивости. И если у населе
ния высокоразвитых стран уровень питания в основном определи
ется национальным доходом, то в странах с низким уровнем эко
номического и культурного развития характер питания завися
помимо этого от традиций, обычаев, а также ландшафтных особен
ностей среды. И поэтому связь через пищевые цепи с природное
средой у населения этих стран более тесная, чем в высокоразвитые
странах с обширными торговыми связями и, следовательно, обме
ном продуктами.
Примерно то же самое можно сказать и об этнических группах,
живущих в странах высокой цивилизации, но в экстремальны?
условиях и с натуральным типом хозяйства. Поэтому при изуче
нии связи биологических особенностей человеческих популяциі
со средой их обитания основным и непременным условием будеп
обращение именно к этим группам, которые либо, в силу каких
то исторических причин, связанных с древнейшими этапами осво
ения ойкумены, оказались обитателями экстремальных зон, либо
опять-таки в силу каких-то исторических событий, за пределам!
основных центров современной цивилизации. Это не значит, ко
нечно, что население высокоразвитых стран, а также умеренное
климатической зоны совершенно свободно от влияния фактороі
среды, тем более что примеров подобного влияния достаточно.
Возьмем хотя бы понижение уровня минерализации скелета у рус
ского населения в районе пониженного минерального статуса,
Просто в этих случаях мы сталкиваемся с гораздо большим числом
причин, нивелирующих прямую связь со средой обитания.
5. ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ
ИЗМЕНЧИВОСТИ АНТРОПОЛОГИЧЕСКИХ ПРИЗНАКОВ
Связь интенсивности обмена веществ с признаками,
характеризующими строение тела, на внутригрупповом уровне невелика,
что создает биологически оправданную независимость систем с
разным уровнем реактивности. При рассмотрении
корреляционного поля морфологических и физиологических признаков
оказывается, что особенно тесная связь существует лишь между крайними
вариантами. Это делает их наиболее динамичными показателями
по отношению к воздействию внешних факторов.
56
Примечательно, что обнаруженные внутригрупповые морфо-
физиологические связи имеют тенденцию усиливаться на
межгрупповом уровне, т.е. образуются довольно весомые исторические
корреляции между физиологическими и морфологическими
особенностями человеческих популяций (Алексеева, 1977).
На внутригрупповом уровне установлено, что с увеличением
интенсивности окислительных процессов увеличиваются длина в
масса тела, количество костно-мускульной массы и уменьшается
жироотложение (Волков-Дубровин, Смирнова, 1969). Подобногс
рода корреляции совершенно отчетливо проявляются на обшир
ной территории арктической зоны, в то время как в тропической
зоне обнаружено понижение основного обмена и уменьшение мае
сы тела. На внутригрупповом уровне фиксируется, хотя и
незначительная, связь уровня холестерина в сыворотке крови с весол
тела и жироотложением (Алексеева, 1966).
Аналогичная, но только более тесная зависимость
наблюдается на территории ойкумены: в северной ее части сосредоточенк
более ''утяжеленные" формы с тенденцией к повышению уровш
холестерина, в южной - "облегченные", с низким содержание»
холестерина.
С уменьшением уровня альбуминов и увеличением
гамма-глобулинов отмечается тенденция к ослаблению развития соматичес
ких компонентов. Эта зависимость сопряжена с тропической
зоной.
Незначительная, но вполне определенная связь наблюдается
между уровнем минерализации скелета, его размерами и формой.
Наиболее грациальные и удлиненные кости скелета
характеризуются повышенной минерализацией (Зенкевич, 1940; Алексеева,
1965).
С уровнем минерализации связана также форма черепа: брахи-
кефальные черепа характеризуются пониженной минерализацией
по сравнению с долихокефальными (Alexeeva, 1968; Алексеева,
1977).
В географическом отношении наблюдается совпадение
брахикефалии с зонами пониженного содержания костеобразующих
минеральных веществ в среде (Vassal, 1957; Alexeeva, 1968). М. Трот-
тер с коллегами (Trotter et al., 1959) показали высокую степень
минерализации скелета у африканцев, а В.П. Волков-Дубровин
67
(1974) - явное повышение уровня минерализации скелета У корен
ного населения Индии. 1?ак известно, обе группы характеризуют
ся долихоморфным типом телосложения.
Таким образом, пространственное распределение морфологи
ческих, а также некоторых физиологических признаков показы
вает, что они подчиняются экологическим факторам. Энергетичес
кие процессы во многом определяются температурным режимов
среды, минеральный обмен - геохимической ситуацией.
Пищевой рацион, как один из экологических факторов, влия
ющих на характер обмена веществ и строения тела, во многих от
ношениях определяется географическими причинами, поэтому
при обсуждении вопросов пространственной изменчивости антро
пологических признаков нет основания для противопоставления
температура среды - питание или геохимический состав среды -
питание.
Так как каждый из видов обмена веществ наследственно
обусловлен, то, по-видимому, такое соответствие экологическим уело
виям формируется путем длительной приспособительной изменчи
вости в результате действия отбора. Здесь нет альтернативы - ере
да или наследственность. Связь того или иного уровня обмена ве
ществ с расой в первую очередь определяется той экологическое
обстановкой, в какой формируется раса.
Экологические правила Бергмана, Аллена и Глогера
относительно влияния температурного режима на форму и строение тела
которые на протяжении своей длительной истории не раз подвер
гались критике, в свете имеющихся сейчас в нашем распоряженш
данных получили основательное подтверждение. Экологическая
дифференциация человечества - явление бесспорное.
Таким образом, в географической изменчивости
физиологических признаков и связанных в той или иной степени с ниме
черт строения тела наблюдается определенная зональность. Анало
гичная закономерность характерна для таких признаков, как пиг
ментация, форма волос, ширина носа, толщина губ и т.п. (Алексе
ев, 1974). Зональность морфофункциональной изменчивости под
тверждает ее приспособительное (адаптивное) значение. Адаптив
ность - черта, присущая всему виду Homo sapiens.
58
Глава III
АДАПТАЦИЯ ЧЕЛОВЕКА В ТРОПИЧЕСКИХ ШИРОТАХ
Питание населения.
Строение тела, физиологические особенности и приспособление
тропических полушарий к высоким температурам и влажности.
Морфофизиологические характеристики населения субтропиков.
1. ЭКОЛОГИЧЕСКОЕ И ЭТНИЧЕСКОЕ
РАЗНООБРАЗИЕ РЕГИОНА
Едва ли не большая часть ойкумены располагается в
тропических широтах. Разнообразие ландшафтов определяет и
разнообразие экологических условий, а также предполагает широкую
вариабельность морфофункциональных особенностей населения,
обитающего в тропиках.
Для того чтобы ответить на вопрос, существуют ли какие-либо
специфические черты в строении тела и физиологии коренногс
населения тропических широт, обязанные своим происхождением
действию тропического климата, есть ли какие-то локальные
морфологические и функциональные особенности в различных
экологических нишах этого пояса, необходимо было по возможноств
полно проанализировать морфофункциональные характеристик*
различных этнических и расовых групп тропического пояса пс
данным мировой литературы. Преобладали работы, в которых
имелись сведения о какой-либо одной системе признаков» хотя
многие из них давали информацию по нескольким этническим
группам. Данных о физиологических особенностях тропических
популяций в литературе значительно меньше. Это объясняется и
методическими сложностями, и трудностями получения
групповых физиологических характеристик. Что касается питания, то о
нем, равно как и о типе хозяйства, в этнографической литературе
сведений очень много, но количественные характеристики
рациона питания известны лишь для нескольких групп. Тем не менее,
несмотря на неравнозначность данных об изученных группах, в
целом сведения о населении тропической зоны оказываются
достаточно представительными.
Население тропического пояса разнообразно в расовом,
этническом и культурном отношениях.
В данной лекции внимание уделяется главным образом
особенностям строения тела и обмена веществ у коренного населения
59
тропической зоны в целом и населения хумидной зоны в частно
сти. Некоторые группы тропического пояса, помимо этой главы
рассматриваются в специальных разделах, посвященных морфо
физиологическим особенностям населения высокогорий и пусты
нь. Здесь также содержатся данные и о части групп умеренного по
яса Южной Америки и Австралии. Такое расположение
материала связано с желанием сохранить полноту представительности егс
по отдельным континентам.
2. КРАТКАЯ ХАРАКТЕРИСТИКА ПИТАНИЯ
НАСЕЛЕНИЯ ТРОПИКОВ
Большинство этнических групп тропического пояса
занимается земледелием и употребляет в пищу главным образом продукты
своего труда. Группы, в рационе которых мясо играет
значительную роль, перечислить нетрудно. В Африке это галла - скотоводы
Кении; нилотские народы Восточного Судана (динка, шиллук,
нуэр, ануак, луо, бари и др.); в Руанде - батутси, а также
охотничьи племена пигмеев; в Восточной Тропической Африке -
нилотские народы (масал и сукума - кочевники-скотоводы), сангаи -
полукочевники-скотоводы, немногие племена банту» у которых
наряду с земледелием развито и скотоводство. В Юго-Западной
Африке, у банту Анголы в зоне засушливых саванн и полупустынь
земледелие сочетается со скотоводством, у готтентотов
существуют многовековые скотоводческие традиции. Наконец, в Бечуана-
ленде (пустыня Калахари) основное занятие населения -
скотоводство.
Скотоводами являются кочевые арабские племена Саудовской
Аравии, употребляющие в пищу мясо и молоко верблюдов.
В Индии скотоводство развито в Кашмире и Пенджабе;
население этих районов употребляет в пищу мясо, молоко и масло.
Значительное место в питании занимает мясо в штатах Уттар-
Прадеш, Ассам, Андхра-Прадеш, Тамилнад. Надо лишь отметить,
что его едят преимущественно представители так называемых
"средних" и "низших" каст. Большая роль в рационе питания
принадлежит мясу и у никобарцев.
В основном же белки животного происхождения поступают в
пищу коренного населения Индии либо с молоком, употребление
которого значительно, либо с рыбой.
60
Среди американских индейцев тропического пояса нет групп
у которых мясо играло бы сколько-нибудь значительную роль і
рационе. То же самое относится к папуасам Новой Гвинеи и авст
ралийцам.
Количественные характеристики рациона питания населения
тропических широт достаточно убедительно свидетельствуют с
преобладании растительной пищи.
У индейцев Колумбии, например, среднедневное потребление
калорий 1796, или 79,1% рекомендуемого для Колумбии уровня.
Среднее потребление животного белка на душу в день - 18,5 г, иле
41,2% рекомендуемого уровня. Среднее потребление растительно
го белка (28,8 г в день на душу), напротив, в 1,5 раза превосходи]
рекомендуемую норму. В целом же общее потребление белков со
ставляет 73% от рекомендуемого. Колумбия, таким образом,
включается в зону белковой недостаточности.
Питание индейцев Центральной Америки характеризуется
примерно такими же чертами (Flores, 1971).
Весьма близкая ситуация наблюдается у коренного населения
острова Барбадос1 (Антильские острова, Harrison et aL, 1964).
В питании большинства африканцев много черт сходства с
питанием индейцев Америки: низкое среднесуточное содержание
калорий, белков и жиров, высокое - углеводов. Весьма,
примечательно, что чрезвычайно сходный уровень питания наблюдается і
гамбийской деревне, в горных, предгорных, равнинных, а также
городских районах Ньясаленда (Harrison et aL, 1964), в
Республике Чад (Crognier, 1973). Редким исключением является питание
масаев, употребляющих в пищу мясо зебу и очень жирное их
молоко, пьющих кровь зебу (Но Kang е? аі., 1971). Дневной рацион
масаев содержит около 3000 калорий, 66% - за счет жира, чте
превышает уровень, типичный для населения США и европейских
стран.
В целом же территория Африки относится к зоне белкового
недоедания (Stanier, Holmes, 1954; Holmes, Jones, Stanier, 1954).
Количественные характеристики рациона питания разных
этнических групп коренного населения Индии также имеют многс
общих черт: очень низкое содержание белков животного проис-
Англоязычное европейско-негрское население, в котором европейская прим?сі
составляет незначительную часть.
6]
Сопоставление рациона тетания тропических
Территория
Контроль (Центральная
Европа)
Центральная Америка
Барбадос
Африка
Индия
Большой Никобар
Новая Гвинея
Колории
2900
100%
I860
64,1%
2413
83,21%
1894
65,31%
2625
90,52%
3050
106,17%
1587
64,7%
Белки, г
общие
100
55,6
55,6%
45
45%
53,8
53,8%
56,3
56,3%
130,0
130%
23,7
23,7%
животные
55
17,6
32,0%
—
.-.
2Д6
3,23%
103,0
187,27%
6,7
12,18%
Жиры, г
105
39,3
37,4%
63
60%
15,2
14,48%
12,4
11,81%
116,0
110,48%
10,9
10,38%
Углеводы,
г
389
322,3
86,4%
416
106,94%
374,8
96,35%
558,8
143,66%
—
338,7
87,07%
Характеристика рациона литания коренного населения
Территория
Центральная Америка
Барбадос
Африка
Индия
Большой Никобар
Новая Гвинея
Килокалории
64,1
83,2
65,3
90,5
106,2
64,7
Белки
общие
55,6
45,0
53,8
56,3
130,0
23,7
животные
32,0
3,2
187,3
12,2
Жиры
37,4
60,0
14,5
11,8
110,6
10,4
Угле-
воды
86,4
106,9
96,4
143,6
87,1
Кадь-
ций
91,2
ЗОД
70,4
68,7
177,0
42,6
Железо
110,0
100,0
210,0
324,2
583,3
99,2
62
Таблица 3
аборигенов с контрольной группой
Кальций,
г
0,8
0,73
91,2%
0,25
30,1%
0,59
70,4%
0,55
68,7%
1,42
177,0%
0,34
42,5%
Железо,
кг
12
13,2
110,0%
12
100%
25,2
210%
38,9
324,2%
70,0
583,33
11,9
99,17%
Витамин А
(каротин)
5000
6633
132,7%
5215
104,3%
5796
115,9%
1802
36,0%
12300
246,0%
4637
92,74%
Тиамин,
кг
1,2
0,87
72,5%
0,8
94,2%
1,3
94,2%
1,67
139,17
1,56
130%
1,49
124,17%
Рибофлавин,
кг
1,6
0,76
47,5%
0,8
50%
0,6
37,5%
0,32
20%
2,30
143,75%
0,89
55,63%
Никотиновая
кислота (РР),
кг
17
10,6
62,1%
7,0
41,18%
10,9
64,12%
17,2
101,18%
22,0
129,41%
8,6
50,59%
ВитаминС,
кг
50
47,5
95,0%
69
138%
68,5
137%
25,8
51,6%
63,5
127%
153,9
307,8%
Таблица 4
тропического пояса. (% от контроля)
Витании
А
(каротин)
132,7
104,3
115,9
36,0
246,0
92,7
Тиамин
72,6
66,7
94,2
139,2
130,0
124,2
Рибофлавин
47,5
50,0
37,5
20,0
143,7
55,6
Никотиновая
кислота
62,4
41,2
64,1
101,2
129,4
50,6
Витамин
С
95,0
138,0
137,0
51,6
127,0
307,8
Автор и год
Flores, 1971
Harrison et al., 1964
Harrison et al., 1964; Crognier,
1973; Stanier, Holmes, 1954
Sen Gupta, 1954, 1960a, b; Sen
Gupta, Biswas, 1956; Roy, Rao,
Biswas, 1957; Behara, 1962
Roy a. Roy, 1967
Jansen, 1963
68
хождения, низкое содержание жиров и высокое - углеводов (Sen
Gupta, 1954, 1960а, б; Sen Gupta, Biswas, 1956; Roy, Rao, Biswas,
1957; Behura, 1962). Анализ уровня питания по сезонам в одной ад
каст (раджбанши), занимающихся земледелием в сочетании с
рыболовством, не выявил резких различий в соотношении белков,
жиров, углеводов, минеральных элементов и витаминов; правда, в
летние месяцы (июль) количество углеводов и витаминов,
особенно А и С, выше. Общее количество калорий также превышает то,
какое наблюдается в зимние и осенние месяцы (Roy, Rao, 1963).
Сен Гупта (Sen Gupta, 1954), рассматривая диету коренного
населения Индии, отметил, что основная масса калорий
образуется за счет злаков (98%), мясо и рыба дают лишь 0,5—0,6%.
Резко отличные от диеты тропических популяций черты
обнаруживает рацион питания никобарцев - очень высокое
содержание калорий, большое количество белка животного
происхождения и жиров. В некотором отношении питание никобарцев
напоминает питание масаев.
Диета папуасов Новой Гвинеи отражает основные черты
питания коренного населения тропических широт в форме, если
можно так сказать, максимального их проявления. Это очень низкая
общая калорийность, чрезвычайно небольшое количество
животных белков и более низкое, чем у других тропических групп,
содержание углеводов (Jansen, 1963).
Для выявления специфических особенностей диеты коренного
населения тропических широт представлялось целесообразным
сопоставление ее с диетой, типичной для населения других
территорий. В качестве контроля была выбрана диета, рекомендованная
для взрослого населения, занимающегося трудом средней тяжести
и живущего в условиях умеренного климата. Эта диета приведена
в докладе Я. Паржизковой на конференции по экологии человека,
состоявшейся в августе 1975 г. в Яблонне под Варшавой (Parizkov-
а, 1976). Величина каждого из компонентов этой диеты
принималась за 100%, и по отношению к ней рассчитывались
соответствующие значения компонентов рациона питания тропических
аборигенов (табл. 3, 4, р и с. 17).
За исключением никобарцев, все тропические группы
показали уменьшение общего количества калорий, белков и жиров в
рационе. Количество углеводов, за исключением диеты никобарцев,
находится либо на уровне, рекомендованном для популяций уме-
64
100%
во-
60-
40-
Калории
20- Л-
Белки
(о&а.
} 2 3 4 5 6
12 3 4 5 6
ііеигрвпьмар Антильские
Америка о-м
Афри»э
Рис. 17. Уровень питания населения тропических широт (в сравнении
с контрольной *Ру?пой, гЬе уровень питанил принят за 100%).
65
Л 701
бедкй
(*»>
Жиры
Vfno
^аодш
:-ЭЗ
ПЯуН
4 5 6
2 3 4 5 6
2 345 ?
D
Индия
Б.Кмобар МошТшине*
Я»е. .17. Продолжение 1
583%
Кальции
I 2 3456
I 2 3 4 5 6
12 3 4 5 6
Г 2 3 4 5 6
Рис. 17. Продолжение 2
67
143.7%
Рибоф
лавин
Никотинсоая кисг.ото.
Виг С ZT.
я-
St
=1,
.!!!
tf
V
¦Z-
У 'I'
2
У
I 2 3 4 5 6
I 2 3 4 5 6
12 3 4 5 6
Рис. 17. Продолжение 3
68
ренного климатического пояса, либо приближается к нему. У ни-
кобарцев количество углеводов в диете превышает
рекомендованный уровень. Значительно ниже, чем в контрольной группе,
уровень кальция в диете населения тропиков. У никобарцев, как и в
остальных случаях, количество кальция в пище выше, чем в
контроле.
Уровень железа в диете тропических аборигенов либо
соответствует рекомендованному, либо превышает его, как это имеет
место в Африке, Индии и на Большом Никобаре.
Более высокое, чем в контроле, содержание в диете витамина
С обнаруживается у всех групп, за исключением населения Индии.
Примерно то же самое наблюдается и в содержании витамина А.
Для диеты тропических аборигенов характерно пониженное
содержание никотиновой кислоты и рибофлавина (витамин В2).
Содержание же тиамина (витамин В1), за исключением диеты населения
Антильских островов (Барбадос) и Африки, превышает
контрольный уровень.
Из этого сопоставления можно сделать следующее заключение:
недостаток питательных веществ в диете коренного населения
тропиков в некоторой мере компенсируется избытком микро и
макроэлементов и витаминов, принимающих активное участие в
процессах обмена веществ.
В последующем изложении я коснусь вопросов питания
населения тропических широт в связи с характеристикой морфофунк-
циональных черт. В лекции II уже отмечалось, что питание, как
один из экологических факторов, оказывает существенное
влияние на строение тела и физиологические показатели организма.
Поэтому оценка морфофункциональных черт населения,
живущего в определенной климатической зоне, должна быть проведена не
только с позиции влияния климатогеографических факторов, но и
под углом зрения воздействия типа питания как фактора,
отражающего, с одной стороны, уровень экономической жизни, а с
другой - возможности, предоставляемые ландшафтными
особенностями среды.
3. СТРОЕНИЕ ТЕЛА У НАСЕЛЕНИЯ ТРОПИЧЕСКИХ ШИРОТ
Расовый и этнический состав коренного населения тропиков
весьма неоднороден, и уже одно это обстоятельство затрудняет его
соматическую оценку. В характеристике признаков тела абориге-
?9
нов тропических широт оолее всего поражает размах их
изменчивости (табл. 5, 6). Не будет преувеличением сказать, что по
некоторым признакам, например по длине и массе тела, а также по их
производным население тропиков показывает такую
вариабельность, что в ее пределы могут быть включены, по сути дела, все
известные в настоящее время человеческие популяции.
Что является причиной подобного морфологического
разнообразия? Предположение о расовой принадлежности как основной
причине соматического разнообразия населения тропических
широт не может быть принято безоговорочно. Во-первых, потому, что
нередко весьма отдаленные в территориальном отношении
различные расовые и этнические группы имеют очень близкие
морфологические характеристики (например, значения длины и массы
тела у русских Европейской равнины и чукчей и эскимосов
Северо-Восточной Азии); во-вторых, связь тех или иных соматических
характеристик с расовой принадлежностью может иметь какую-то
свою экологическую причину. Тем не менее следует рассмотреть
морфологическое разнообразие строения тела у коренного
населения тропических широт под углом зрения его расовой
принадлежности.
До сравнительно недавнего времени признаки,
характеризующие строение тела, не входили в число безусловно расово-диагно-
стических, хотя в некоторых источниках, посвященных описанию
расовых черт, фигурировали пропорции тела и с негроидами
связывалась долихоморфия (удлиненный тип строения тела), с
монголоидами - брахиморфия (укороченный тип строения). Разногласия
по этому вопросу объяснялись главным образом малым
количеством данных и, следовательно, недостаточной изученностью
признаков строения тела.
В последние годы растет интерес к территориальным
вариациям соматических особенностей коренного населения, что дало
возможность составить многочисленные и относительно полные
характеристики по различным группам земного шара.
Н.И. Клевцова (1976а, 19766), поставив целью выяснение роли
особенностей строения тела в межгрупповой дифференциации
современного человечества, показала совершенно четкие различия
по комплексу соматических признаков между негроидами
Африки, монголоидами Сибири и европеоидами, включающими пред-
70
з
к
д
n
о
О
(О* С-** СО*
О) Н rt
eg
to
ao
CO
00 00 00
Mi CO *
НИИ
I I r
w t- <o
4 *1 о
43
о *ч
со ю"
п ш он
оо
СМ
1
ч<
О
СМ
00
о
to
1 *
О
1 3
со
о
ем
j
СО
;
О
ем
оэ
to
СМ
S
со
тН
>л
ч
со
ч
»-*
00 СЯ
CO W
to о
•> Оэ
тн ео
со оо
Н СО
t- со
*ч ор
to ел
to «-
t** со ^ ел
і> to" ^Г \а
оо ь- со ем
CM
w
іН
о>
с»
со
о
.-1
со
"Ч*
»Ч
о
о
см
ч#
и»
*н
ь-
со
О)
о
ш
гН
ем
О
»н
t-
со
іН
о
t-
со
¦ч<
<"Н
°1
<о*
о -ч<
со
г. t-
tO
ем
о> CM
<о
4
Ь- Ф tO О см О СМ СО "* t& tO
О н оо оо со ^ га со ем ю ю ¦¦#
« И И СО 6 Ю в <0 О С» 00 СО
иэ оо с- с- оо ем о* ем ^ ^ «*
ем
см
r-t t- 00
с- оо op Ч Ч t-*
»Ч t>- <S гН гН СО
(О Ф Ф Ф СЦіО
fH tO Ю tO -•* уГ
НИИ
а;
л w -ч<
^ И »
Гш, ¦* tO
00 00
о
СМ*
о
to
|>
to*
00
ем
см
Ч ем « О Ч О
СО ьГ СО Ю 01 к.*
*-( *" см *н см ^
о
С— С- СО Ъ* 00 00 О
1ч. И 00 © ^ И <N ся
^COOOCMWWUO Ю
^ » СО fc- И N СМ
ем
об
«5
6
из
6
to
-ч«
1
со
см
ем
ем
СО Ъ- <Ч
с- -* ь- Ч Ч и»
Н I- Ю Н НСО
Ю CD (О И <0 ОО
r-t <© ^ Ч Ч of
н н п
к
И
А
ч
в)
*9
S
р,
§
«
о
в
0
й
е-
н
Я
5
&
s
0
о
ф
и
пах
?
S &
о
2
н
Я
8
1
ж
*&
О
X
«
Й
0
5
А
И
Р»
0
«
1
о
ю ь-
СО fc- b-
00
CM
Ю Й М ОО Н
ем со ео »ч ем
со*
со
¦
см
ч
оГ
о сэ Ч Ч *"і *і *"'
кО* V V3 t>* 05 СО* Ю*
гН Н N (О « Ю«
СП
00
3
ял w
00
6
1>
СО 00 00
оо со ем
^
и» ем
to со
ем
со
см
I
со
еч
см
«0
00 tO ф
00 СО © О Ь. *". Ч ОО
*4 СМ И 00 в гі HBD
-^ о 4 ^ 6 оо ЮН
-* см Н « ^і « в{ нв
w о* со
* В S
i з
я б
б1
ф
с*
о
J)
ч
г
«
* *
71
3
I Я
I
ш
«в
g
е
е
^ «
(О '
0} СО 00 О
К V, *> к.
И СО С0^
«rf 04 04 .н
О»
§
со со "*
* Ч X «
I
°1
«Г
w4 О »Н
С<9 Н Н
со1
о I
00 '
се
і с-
ес
°°04
ю
ем
<о
CQ
О-
а»
I
ее
я
и
«
8
2
ем
to
о
I
to
о со
04 ~Ч
СО
н Ю Н
И Л О
40 -*
to -*
t і
* 04
гН СО
°1 °і
to" со
ъ «, і
*, «, СО СО 00 і
« 2 оо « с-" J
о <о
о» t-
'* л
©4
о
со
со
«О
«04
tO
СО
о
tO CNI
i-l СО
I 1
со ээ
со і>
о
со
СО
00
1 00 Ю
* *> М
со <* а>
fc-
со
іН
со
ь-
00
С4
t*
<N
СО
©4
>С*
О)
t
о
©4
а»
іЧ
і
to
ю
.н
to
О)
US О N в
04 CM CO U3
¦ і і ¦
СО СО
04
to
со
¦н ео
^ со
N Г-
•н со
© со оо о»
•. *. *, *. *.
»0 ^* CN <4 tO
Ь- ч* Ю ^Н
С4 Ю О*
04 CO t-f
I
S
со
X
&
И
a
а
о
Ж
t-
to
I
00
СО
n а о ^ о>
00 С- С- СО ©4
і і і і і
Т|1 N N Н «О
Г> О СО СО 04
ч* 04 іО ©4 СО
1U Ю ^ N Н
•Н О Ч«
to О "4> 04
іН t-
to
to
1
00
со
CO О) ©4
- к СО
Н И(9
і і і
Ю О "ч*
N Н *
* о
г-1 »Н СО
ев
8
2«Olt-C0<NWrt»eiO^N
СО 04 CO
н с- ю
СО —I СО
И
?
3
ш
а
a
1st
ь * °
и 5 5
і в в
Sect ев
ш в
ч в вв
3 ЙЙЙЙ
В
72
ставителей северного и южного европейских стволов. Размах
изменчивости в пределах негроидных и монголоидных групп
оценивался на фоне изменчивости признаков у европеоидов. Картина
расовой дифференциации человечества по признакам строения тела
оказалась весьма отчетливой. Значения признаков у негроидов и
монголоидов располагаются диаметрально противоположно на
фоне межгруппового размаха европеоидной совокупности. Несмотря
на то что величины длины и массы тела у монголоидов попадают
в пределы вариаций этих признаков у негроидов, последние
имеют более длинные нижние конечности, более короткий корпус,
меньшие поперечные и глубинные размеры туловища.
Абсолютные величины диаметра грудной клетки, длины ноги, ширины
плеч и таза, выраженные в процентах к длине тела, у негроидов
Африки и монголоидов Сибири не трансгрессируют. Таким
образом, коренное население Африки оказывается
долихоморфным , а коренное население Сибири -брахиморфным. Более
того, эта расовая дифференциация по соматическим признакам
сохраняется и в пределах тропических широт. Так, например,
индейцы Центральной и Южной Америки, будучи представителями
американской ветви большой монголоидной расы, обладают более
длинным корпусом, более короткими ногами и относительно более
широкими плечами по сравнению с негроидами Африки, т.е.
сохраняют брахиморфный тип строения тела.
Все известные нам монголоидные группы Индии, такие, как
риаяг, гуркхи, никобарцы, имеют брахиморфный тип строения
тела.
Монголоиды тропической зоны характеризуются, как и на
других территориях, значительной массивностью по сравнению с
представителями европеоидной, негроидной и австралоидной рас.
Нередко обладая невысоким ростом, они имеют относительно
большую массу. Очень распространенный в зарубежной литературе
показатель плотности тела - индекс Рорера - у монголоидов
значительно выше, чем у представителей других рас. Европеоидное
население Передней, Юго-Западной и Южной Азии занимает
промежуточное положение между индейцами Центральной и Южной
Америки и негроидами Африки. Наиболее долихоморфными
оказываются аборигены Австралии. Итак, отвечая на вопрос о
причинах соматического разнообразия аборигенов тропических широт.
78
выскажемся в общей форме в пользу расовой дифференциации
как одной из возможных причин этого явления.
Подобное заключение, однако, не закрывает пути к поиску спе
цифики черт строения тела коренного населения тропиков, и этоі
поиск приносит определенные положительные результаты.
Сопоставление нескольких этнических групп Африки, харак
теризующихся типично негроидными чертами, и некоторых евро
пеоидных групп Индии как представителей населения тропичес
ких широт с европеоидным населением умеренного пояса и монго
лоидами Арктики и континентальной зоны показывает, что жи
тели тропиков, как европеоидного, так и негроидного облика, от
личаются от европеоидов умеренного пояса и особенно от монголо
идов арктических и континентальных районов в одном и том ж<
направлении.
Поскольку абсолютные размеры тела подвержены временны»
изменениям (например, вследствие акселерации), а в некоторой
отношении зависят и от социально-экономических причин (обе
фактора в сравниваемых группах разные), сопоставление произво
дилось по относительным величинам, т.е. по форме тела.
Европеоидное население умеренных широт (русские)
рассматривалось в качестве эталона (контроля), в котором значение каж
дого относительного признака принималось за 100% , и по отноше
нию к нему оценивались отклонения в группах тропического, ар
ктического и континентальных поясов. Оказалось, что по отноше
нию к этому эталону население тропиков и более суровых поясоі
заняло диаметрально противоположное положение (рис. 18). Пс
сравнению с эталоном население тропиков относительно боле*
длинноного, у него относительно уже плечи и таз, несколько уп
лощена грудная клетка и понижена плотность тела. Этот комплекс
у негроидов выражен ярче, чем у европеоидов.
Население арктической и континентальной территорий,
напротив, характеризуется по сравнению с контролем большим
развитием в ширину и большей плотностью тела, т.е. теми чертами, кото
рыми отличаются монголоиды внетропических широт. В то ж«
время коренные группы тропического пояса, относящиеся к мон
голоидной расе, такие, как рианг (исследовались переселенцы і
Индию из Индокитая) и гуркхи (переселенцы из Непала в Индию)
обнаруживают черты, сближающие их с населением тропически»
широт (рис. 19). Оставаясь сходными с монголоидами континен
74
Рис. 18. Сопоставление
некоторых этнических грипп тропических
широт (а) и Северо-Восточной Сибири
(б) (в % отклонений признаков
строения тела от контороля). За контроль
(в) приняты средние характеристики
четырех русских групп умеренного
пояса:
1 - отношение длины корпуса к длине
тела, %; 2 - отношение ширины плеч к длине
тела, %; 3 - отношение ширины таза к длине
тела, Уо;4- отношение перед незаднего
диаметра грудной клетки к поперечному диаметру
грудной клетки, %; 5 - указатель Рорера, %.
2 -
$
S
г
Рис. 19. Сравнение некоторые
монголоидных народностей Кк?иі
(риангов и гуркхов) (а) с негроидамъ
Африки (б)3 европеоидами Индии (в) ъ
монголоидами Сибири (г) (различал
выражены в % отклонения от
контроля, см. рис. 18);
1 - отношение длины ноги к длине тела,
%; 2 - отношение ширины плеч к длине тела,
%; 3 - отьношени? ширины таза к длине тела,
%; 4 - указатель Рорера, %.
тальной зоны по соотношению длины ног и тела, рианги и гуркхи
характеризуются более узким телосложением и значительно
меньшей плотностью тела. И хотя различия проявляются чаще всего в
виде тенденции, на них нельзя не обратить внимания, настолько
они закономерны.
В характере соотношения признаков строения тела у
населения тропического пояса, в частности на территории Африки,
обращают на себя внимание коэффициенты межгрупповых
корреляций признаков, имеющие то же направление связи, что и внутри-
групповых (Клевцова, 1976а). Как правило, такого соответствия не
наблюдается, так как на определенных территориях действия
каких-то исторических причин (изоляция, смешение и т.п.)
нарушают обычные физиологические связи между признаками. "Разру-
75
CM
no-
rn
f\
w
KO-
ch
mo
no
но
150
1
кг
Ki
лх:
JO 40 SO 60 70 65 70 75 SO
Рис. 20. Соотношение признаков строения тела а тропических популлцч
А - соотношение длины и массы тела; Б - соотношение длины тела и грудного указателе
Заштриховано корреляционное поле.
шителями" физиологических связей, по-видимому, могут
выступать и климатические факторы среды, под влиянием которых не
протяжении длительной истории популяции могут
формироваться связи, обеспечивающие наибольшую устойчивость морфофун
кционального типа в данных условиях. При атом нарушение <ри
зиологических корреляций, по всей вероятности, неизбежно.
Соответствие межгрупповых и внутригрупповых корреляциі
обнаруживается не только на территории Африки, но характера
и для всего тропического пояса в виде общей тенденции. Особен
но отчетливо это проявляется на таких признаках, как длина і
масса тела, форма грудной клетки. Так, в популяциях тропичес
ких широт, как правило, с увеличением длины тела увеличивает
ся и его масса. В то же время увеличение длины приводит к умень
шению грудного индекса, т.е. грудная клетка делается относитель
но более плоской (рис. 20). Такое направление связи типично дш
внутригрупповых корреляций, полученных для достаточно много
численных групп, гомогенных и находящихся в условиях, свобод
ных от стрессовых ситуаций.
Рассмотрим более подробно некоторые этнотерриториальнь»
особенности строения тела тропических аборигенов в сравнитель
ном освещении.
В Африке при огромной изменчивости длины тела наблюдает
ся явная тенденция к высокорослости по сравнению с иными тер
риториями, за исключением Австралии, где вообще нет низкорос
7?
Рис. 21. Размах
изменчивости длины
тела у населения
различных климатических
зон тропического пояса;
1 2 3 Ч 5 *
т п п
2 (> —
1-8 — Африка: 1 - континентальная тропическая зона Южного по
лушария; 2 - континентальная зона экваториальных муссонов
Северного полушария; 3 - континентальная экваториальная зона; 4 —
континентальная зона экваториальных муссонов Южного полушария; 5 -
континентальная тропическая зона Южного полушария; 6 - индийская
(экваториальных муссонов Северного и Южного полушарии, тропическая
зона); 7 - атлантическая (экваториальных муссонов Северного
полушария, тропическая зона Северного и Южного полушарий); 8 - Африка в
целом; 9 - Индия; 10 - Австралия; 11 - Новая Гвинея; 12 -
Центральная и южная Америка:
а - средняя межгрупповая величина; 6 - среднее квадратичное
отклонение; в - средняя величина признака по тропическим широтам в
целом; г - контрольная средняя.
лых групп. Прямой противоположностью является Новая Гвинея
с преимущественно низкорослым папуасским населением, а также
Центральная и Южная Америка с выраженным сдвигом в сторону
понижения длины тела у американских индейцев.
77
Противоположные тенденции наблюдаются в Центральной Аф
рике, в континентальной зоне экваториальных муссонов у нилот
ских народностей (высокорослость) и в континентальной эквато
риаЛьной зоне у пигмеев (низкорослость).
Несмотря на значительный размах изменчивости длины тела
большинство тропических популяций имеют среднюю или выше
среднюю длину тела (по мировому масштабу) - в пределах 160-
170 см, т.е. среднего и вышесреднего значений. Средняя величию
тела в тропических широтах несколько ниже контрольной: 164 с*
(рис. 21).
По массе тела тропические популяции также значительно раз
личаются. Население Индии, например, обнаруживает «тенденции
к понижению признака по сравнению с населением других терри
торий, за исключением Новой Гвинеи. Некоторое повышение мае
сы тела характерно для американских индейцев и некоторых гру
пп австралийцев (например, коката в Центральной Австралии)
Значения признака для большинства тропических популяций на
ходятся в пределах 49-62 кг, т.е. являются низкими и нижесред
ними (по мировому масштабу). Средняя величина - 55 кг, т.е. зна
чительно ниже контрольной - 63,5 кг (рис. 22). Понижение ма<
сы тела по направлению к югу было обнаружено Робертсом (Ro
berts, 1953) и связано с понижением основного обмена в условия»
тропиков.
Заслуживает особого внимания весоростовой указатель Рорера,
в какой-то мере характеризующий плотность тела.
Как отмечалось выше, население тропических ширсГ
характеризуется пониженной плотностью тела, за исключением
американских индейцев (рис. 23). Последние при пониженной длине
тела и относительно большой массе тела характеризуются очені
высоким указателем Рорера, намного превышающим величины,
типичные для монголоидов, у которых значения этого индексе
наибольшие. Средняя величина указателя Рорера располагается,
как видно из рис. 23, значительно ниже, чем в контрольной
группе.
Вариабельность групп тропических широт по указателю
Рорера, по-видимому, не является только следствием расовой
принадлежности, хотя представители негроидной и монголоидной рас по
этому признаку здесь бесспорно дифференцируются. В Африке,
например, существует разница между популяциями негроидной
78
3
іш-
з
*
а
і^&йіИ&Ь&^^ЬЭ!?* ^й^^й
3
79
м'/кг
Рис. 24. Размах изменчивости удельной поверхности
тела у населения различных климатических зон
тропического пояса (условные обозначения см. на рис. 21).
принадлежности по указателю Рорера, а население Индии,
преимущественно европеоидного облика, обнаруживает явное
понижение указателя по сравнению с европеоидными группами,
находящимися за пределами тропических широт.
Из относительных показателей, характеризующих строение
тела, следует остановиться на удельной поверхности тела. В
тропических широтах она в среднем несколько выше, чем в
умеренном поясе (рис. 24). Наибольшие ее величины характерны для
населения Индии и Новой Гвинеи. Повышение относительной
поверхности тела отмечается в некоторых зонах Африки.
Определенная неоднородность тропических популяций
обнаруживается и в кормическом индексе, характеризующем пропорции
тела (длина тела сидя/общая длина тела). Население Индия и
Америки отличается от населения Африки и Австралии относительно
меньшей длиной ног (рис. 25). Одна из причин дифференциации
по кормическому индексу - разная расовая принадлежность гру-
80
7 i 3 5 6 7 8 9/0 72
Рис. 25. Размах изменчивости кармического
указателя у населения различных климатических
зон тропического пояса (условные обозначения см. на
рис. 21).
пп. В то же время в пределах Африки среди негроидных популя
ций отмечается известная неоднородность по этому признаку, чт<
заставляет думать о существовании и иных причин.
Черты строения тела популяций тропических широт зависят
не только от расовой принадлежности, но и определяются специ
фикой условий жизни в этом климатическом поясе; отмечена так
же некоторая неоднородность соматических признаков в предела?
одной расы, занимающей широкий ареал.
Здесь уместно обратиться к поискам экологических факторов
создающих специфику облика представителей разных рас и диф
ференцирующих группы в пределах одной и той же расы.
Один из важнейших экологических факторов - питание. Мн«
уже приходилось отмечать, что при значительной степени связі
длины и массы тела, степени жироотложения, уровня холестери
на в сыворотке крови и некоторых других признаков с рационол
питания влияние последнего хотя и осуществляется в одном на
эчд
Sty
s&
52fi
S%5
&,Q
50,S
sao
no
47,5
Щ
Щ
6S.S.
81
правлении у представителей разных рас, но все же в предела:
уровней, типичных для каждой из них (Алексеева, 1977).
Для народов тропического пояса, за редким исключением, ти
пичен рацион, бедный белками животного происхождения и каль
цием. С недостатком белков и кальция и низким содержание)
жиров могло быть связано слабое развитие скелета, а также уме
ньшение длины и массы тела, и действительно, ни для одной гео
графической зоны земного шара не характерно такое значительно!
количество низкорослых групп, как для тропиков. В этой связі
следует напомнить о симпозиуме "Экология и антропология", со
стоявшемся в декабре 1960 г. в Нью-Йорке под председательство»
известного американского антрополога П. Бейкера. Итоги этоп
симпозиума нашли отражение как в зарубежной, так и в отече
ственной печати (Ecology and anthropology, 1962; Алексеева, 1964)
В докладе М. Ньюмена "Экология и недостаток питания у чело
века" специально рассматривается влияние недостаточности мине
ральных веществ и витаминов на рост и развитие человека. Суда
по данным Маретта (Marett, 1936), приводимым в докладе, суще
ствует большая зависимость между фосфорно-кальциевой недоста
точностью и климатическими и геохимическими характеристика
ми влажных тропиков. Маретт высказал предположение, что тел(
пигмеев представляет образец выгодной экономии при дефицит*
этих химических элементов. Другие авторы (Nickolsa, Nimalasuri
ja, 1939; Schraer, Newman, 1958) также высказывались в польз;
наследственной адаптации к недостатку кальция и фосфора, выра
жающейся в замедлении роста и развития и в уменьшении разме
ров тела взрослого человека.
О дефиците белков в питании свидетельствует очень распрост
раненная в тропическом поясе болезнь - квашиоркор. Чаще всей
она поражает младенцев, только что отнятых от груди, но иногді
наблюдается во всех возрастных группах детей и взрослых. Геогра
фическое распространение квашиоркора очень велико - Африка
Индия, Центральная и Южная Америка, Индонезия, острова Фвд
жи (Кассирский, Плотников, 1964). Болезнь вызывает снижена
массы тела, отставание детей в росте, понижение белка сыворотю
крови за счет альбуминов, увеличение гамма-глобулиновой фрак
ции. Альбумино-глобулиновый коэффициент ниже единицы, чт<
говорит об общем понижении физического развития.
Недостаточно полноценное питание, а в некоторых случаях •
избыток углеводов являются причиной некоторых болезней тро
82
пического пояса. К ним относятся: спру, характеризующаяся
угнетением функции пищеварительных желез; бери-бери, вызываемая
недостатком витамина В1 в областях, где в пищу употребляется
очищенный рис, и приводящая к тяжелым расстройствам
сердечно-сосудистой системы, нарушениям белкового, водно-солевого и
жирового обмена; тяжелые кишечные расстройства и другие
заболевания алиментарного характера.'
Влияние недоедания на строение тела и физиологию, в основе
которого лежит недостаток белков животного происхождения,
привлекает внимание многих зарубежных исследователей,
работающих среди популяций тропической зоны.
Широкие исследования питания индейцев Центральной
Америки, проведенные в течение 1965-1967 гг., выявили в связи с
нехваткой белков и низкой калорийностью пищи уменьшение
длины тела во всей этой области (Flores, 1971). Уже в возрасте
трех месяцев дети американских индейцев несколько
отклоняются от контроля. Эти отклонения в той или иной степени
проявляются на разных стадиях онтогенеза; взрослое население в возрасте
25 лет в среднем имеет длину тела 160 см и массу тела 55 кг - у
мужчин и соответственно 150 см и 50 кг - у женщин.
Здесь следует сказать, что небольшая длина тела является
отличительной чертой большинства популяций американских
индейцев как представителей монголоидной расы; в то же время
нельзя не отметить, что такие величины длины и массы тела
находятся скорее,в пределах минимума, типичного для монголоидов.
Еще большее отклонение от стандарта в сторону уменьшения дает
обхват головы. Среди этих популяций отмечено большое число
случаев маразма и квашиоркора - болезней белковой
недостаточности; около 20% детей показывают различную степень
недоедания. В крови понижены содержание витамина А и рибофлавина,
уровень гемоглобина, гематокрит. Анемия широко
распространена в Центральной Америке. Она поражает в большей мере
популяции, живущие на высоте около 1000 м над уровнем моря.
Наиболее высокий процент анемии зарегистрирован у детей до одного
года и среди беременных женщин. Наряду с дефицитом железа
имеет место низкий уровень фолиевой кислоты. Все это убеждает
в том, что питание оказывает значительное воздействие на рост,
развитие и состояние внутренней среды организма.
В связи с выяснением роли питания в процессах роста и
развития большой интерес представляют исследования, проведенные
83
среди взрослого населения Африки (Holmes, Jones, Stanier, 1954
Stanier, Holmes, 1954). В условиях белкового дефицита и высоко
го углеводного содержания в диете отмечается устойчивый угле
водный баланс- Стабильность уровня углеводов сопровождаете,
уменьшением кортизона, что, по мнению авторов эксперименте
свидетельствует о гормональном контроле углеводного обмена.
В обсуждении роли фактора питания в процессах роста, разви
тия и состояния внутренней среды организма нельзя оставить бе
внимания результаты исследований Б. Даса (Das, 1966), проведен
ных среди городского населения, находящегося на вегетарианскоі
и невегетарианской диете. Обследованные происходят из окрестно
стей Калькутты, в этническом отношении это, по-видимому, бев
гальцы. Обе группы достаточно репрезентативны- У невегетариан
цев понижаются значения многих признаков. Особенно замет»
падение массы, уровня общего холестерина и его составляющих
глюкозы, остаточного азота, уровня седиментации. Наблюдаете;
тенденция к понижению кальция, общего белка, мочевины, кис
лой фосфатазы. При этом заметно увеличивается уровень амилаз
ной активности и щелочной фосфатазы. Повышение амилазноі
активности, по-видимому, связано с избытком растительной пи
щи, а энзим амилаза гидролизует углеводы. Что касается уровн
щелочной фосфатазы, то обычно он повышается при костных за
болеваниях (Штрауб, 1963). Поскольку у вегетарианцев наблюда
ется и понижение содержания кальция в крови, то, возможно
этот симптом служит указанием на какие-то перестройки в кос?
ной ткани. Во всяком случае, изменения во внутренней среде ве
гетарианцев таковы, что позволяют предполагать сдвиги в обмев
ных процессах, которые, без сомнения, воздействуют на развита
соматических компонентов.
Дж. Вайнер в одной из глав "Биологии человека" (Harrison
Weiner, Tanner, Barnicot, 1964) ссылается на ряд исследовании
посвященных анализу зависимости физических особенностей о1
уровня питания. Из них наибольшего внимания заслуживают на
блюдения де Кастро на северо-востоке Бразилии, показавшего, чт
у населения побережья и внутренних засушливых районов длин
тела больше, чем у групп, живущих в тропических лесах. На по
бережье в рационе питания много рыбы, а у скотоводов внутрев
них областей - мяса и молока, в то время как во влажных тропи
ках основой питания служит мука из маниока.
Вайнер ссылается также на очень интересные данные Николь
са, который, изучая длину и массу тела у жителей Шри-Ланка :
84
зависимости от расовой принадлежности, а также от уровня
экономического и социального положения, пришел к выводу об
определяющем значении экономического фактора в морфологических
различиях между группами.
Такие примеры убедительно показывают, какое важное
значение в морфологической дифференциации человечества имеет
фактор питания- И в то же время в этом вопросе далеко не все так
ясно. Напомним наши данные о том, что изменение содержания
холестерина под влиянием питания происходит в пределах
уровней, типичных для негроидов и европеоидов (Алексеева, 1971).
Наблюдения пропорций тела у негроидов и японцев (Harrison,
Weiner, Tanner, Barnicot, 1964) показали, что и те и другие,
находясь на диете белых американцев, растут быстрее, чем их
сородичи в Африке и Японии, но сохраняют пропорции, типичные для
негроидной и монголоидной рас.
Материалы, находящиеся в нашем распоряжении, дают также
убедительные доказательства определенной независимости
строения тела и характера питания. Для примера возьмем, с одной
стороны, этнические группы индейцев Центральной Америки, диета
которых по сравнению с контролем менее калорийна, меньше
содержит белков и жиров, имеет пониженное количество углеводов,
кальция, витаминов группы В, никотиновой кислоты и витамина
С; с другой - никобарцев, находящихся на более калорийной
диете, с большим содержанием белков, жиров и витаминов, чем в
контроле. И индейцы Америки, и никобарцы относятся к
монголоидной расе, хотя и к разным ее ветвям.
Сопоставление американских индейцев и никобарцев дает
отрицательный результат в отношении связи строения тела с
питанием (табл. 7). В данном случае речь идет не о пропорциях тела,
как будто довольно тесно связанных с расовой принадлежностью,
а о длине и массе тела - признаках, подверженных влиянию
питания. Это тем более удивительно, что среди американских
индейцев широко распространены болезни недоедания. В какой-то мере
такая особенность может быть объяснена, во-первых, высокой
детской смертностью у аборигенов Америки от болезней недоедания
и, во-вторых, тем, что оставшиеся в живых и достигшие
взрослого состояния, по-видимому, не испытали на себе столь пагубного
воздействия пищевого дефицита. В то же время подобные
наблюдения заставляют думать, что строение тела не связано прямо с
уровнем питания, а определяется какими-то иными факторами.
86
5
s
3
s1
и
s
к «
Ml
3
о
§
?
$
а ч
і ё
I
о
і
і
I
I
2
I
I
-г оо со
О * 00
Ф Ю N «J N .
* r-f "I 10 ЕГ -Н
A i M i CO i
*° ч ? Ю и"5
" OS Ю "I
Ю Ю r-H
о g « g
8 • S •
со ю
*
*¦"» ^-4, *-%
eo^ о ев
^ t- « <o *; ¦*
to *? fi "Э * io
ih oo *° -r ^ *-*
00 4» r-t
iO w w
CO
*-.s
§4
Ю
CO
• s
S*
Oi
"*
to
to
*•
to
00
CO
¦ч
CO
CO
cs
CM
CO
at
to
to
Ю
CO
o>
184
00
183
i
i
•
tH
0Э
Ю
1
1
1
Ю
A
О
<
to
4*
CO
Ю
CO
<
OS
00
оГ
о
Ю
CO
00^
00**
to
tO -Ч
00 •>
o>
CO
CO
о
из
» -ч
CO
CO
CO
to
00
4«
CO
CO
о
со
о
CO
04
to
00
o>
¦4»
*0
о «
CO i4
-И О)
to
CO
?
ej „ 4 б
" g - ь &
a i s о л ft
° 3 и
3 д S,
n
< я 3 о в
О Й К Я Р-
Рассмотрим представителей негроидной расы различной этни
ческой принадлежности, расселенных по Африканскому конти
ненту и отличающихся типом питания (см. табл. 7). Результат со
доставления также не показывает связи размеров тела с типом ди
еты. У этнических групп Восточного Судана, занимающихся ско
товодством, в рационе питания довольно много белков и жироі
животного происхождения. Этим группам свойственна большая
длина тела, но они имеют более низкую массу тела и значительнс
меньший показатель Рорера (в какой-то мере характеризующий,
как уже отмечалось выше, плотность тела) по сравнению с этни
ческими группами Сенегала и Республики Чад, живущими на уг
леводной диете. Судя по пропорциям тела, они не только более
высокорослы, но и относительно более длинноноги, а так как про
порции тела независимы от питания, то, по-видимому, и
увеличение длины тела здесь не является результатом более
полноценного питания. В двух приведенных для сравнения группах -
европеоидной (арабы) и смешанной (негроидная с европеоидной
примесью - амхара), - также имеющих противоположную по типу ди«
ту, в строении тела проявляется их расовая принадлежность, а не
зависимость от того или иного типа, питания.
В то же время морфологические признаки у племени сара
(Республика Чад) подвержены сезонным вариациям (Crognier, 1973). В
период дождей, когда из-за гибели урожая питание населения
резко ухудшалось, а физическая активность по необходимости
увеличивалась, уменьшались толщина жировых складок и
мускульный компонент тела.
Таким образом, отрицать влияние питания на размеры тела
нет основания, но, оценивая его как один из факторов
морфологической дифференциации у населения тропических широт, не
следует придавать ему решающего значения.
Обратимся еще к одному источнику влияний на
морфологические признаки. Это комплекс природных условий, в которых
обитают популяции. Мысль о связи строения тела с климатическими
факторами не нова (Schwidetzky, 1962). Остановимся на тех
исследованиях, которые имеют непосредственное отношение к
тропическим широтам.
Представители английской биометрической школы (Thompson,
Buxton, 1923) исследовали корреляции носового указателя со
среднегодовыми температурой и влажностью воздуха в популяциях
87
своих современников и в краниологических сериях. В обоих
случаях была показана положительная связь: ширина носа в
тропическом поясе увеличивается с повышением температуры и
влажности среды. Эти данные были подтверждены на более обширном
материале А. Дэйвисом (Davies, 1932), получившим высокие
коэффициенты корреляции (порядка 0,6-0,8) для всей территории
земного шара и для отдельных материков. По материалам А. Дэйви-
са, как и предшествующих авторов, связь с температурой
оказалась выше связи с влажностью. Подобная зависимость была
обнаружена и на территории Восточной Азии (Wang Schin-chun, 1954),
где наблюдалось увеличение носового указателя по направлению к
югу. Дж. Вайнер (Weiner, 1954) показал, что величина носового
указателя в значительной степени связана не только с
температурой и влажностью воздуха, но и с атмосферным давлением. И.
Швидетская (Schwidetzky, 1962) на основании всех этих данных
сформулировала специфическое для человека экологическое
правило (правило Томпсона-Бакстона), согласно которому ширина
носа в тропической зоне больше, чем в умеренном и холодном
климате.
Такие показатели, как темный цвет кожи, курчавые волосы в
сильное развитие слизистых оболочек носа и губ, также
рассматривались под углом зрения приспособления человеческого
организма к высоким температурам, инсоляции и влажности
тропических широт (Чебоксаров, 1951).
Форма и размеры тела (соотношение массы и поверхности,
длины конечностей и длины корпуса) неоднократно служили
предметом исследования в связи с терморегуляцией (Schreider, 1950,
1952, 1954, 1964; Roberts, 1953). Выяснилось, что особенности
телосложения подчиняются климатическим правилам Бергманна
и Аллена. Согласно одному из них (Bergmaan, 1847), размеры тела
в пределах одного политипического теплокровного вида обычно
увеличиваются с понижением температуры окружающей среды;
согласно другому (Allen, 1960), у теплокровных животных,
относящихся к одному виду, имеется тенденция к увеличению
выступающих частей тела в жарком климате. В условиях тропиков
выявлено уменьшение массы тела и увеличение длины конечностей
по сравнению с жителями более высоких широт.
Заслуживают глубокого внимания экспериментальные
исследования формы головы в тропическом климате (Волков-Дубровин;
88
Рогинский, 1960), которые показали, что гипсистеноцефалия, т. е.
узкий, вытянутый в переднем направлении и высокий череп,
может рассматриваться как приспособление к повышенной
инсоляции: узкая, высокая и длинная голова, типичная для негроидов,
нагревается медленнее, чем широкая, низкая и короткая.
Е. Кронье (Crognier, 1973), исследовавший популяцию сара-
маджингай на северо-западе Республики Чад, пришел к
заключению о морфологической адаптации этой популяции к условиям
сухого тропического местообитания. Некоторые из результатов
этого исследования заслуживают особого внимания. Во-первых,
выяснено, что размеры тела у лиц, имеющих наиболее
благоприятное сочетание признаков с точки зрения приспособления к
терморегуляции в условиях сухого, жаркого климата, группируются
в пределах, близких к средним для данного местообитания.
Напомним, что средняя длина тела мужской части этой популяции ¦
173,9 см, а масса тела - 66,1 кг.
Во-вторых, наибольшее количество детей и их выживаемость
наблюдаются в семьях со средними (в масштабе данного
местообитания) размерами тела или приближающимися к ним. Это
наблюдение представляет исключительный интерес, так как допускает
мысль о селективных преимуществах определенных размеров тела
и их соотношений в конкретных условиях обитания популяции.
Вопросы климатической адаптации рассматривались и в
монографии Ж. Ерно (Hiernaux, 1968), содержащей антропологические
данные для 460 африканских популяций, живущих к югу от
Сахары. Корреляции шести климатических показателей с 23
антропологическими признаками оказались очень невысокими
(порядка 0,2-0,3), что заставило автора усомниться в универсальности
как климатических правил, так и самого процесса адаптации. В то
же время более дифференцированная оценка связи
антропологических признаков с климатическими показателями,
произведенная в определенных климатических зонах Африки с
использованием тех же антропологических данных, позволила И.И.
Крупнику прийти к заключению о том, что "в жарком климате
морфологическая изменчивость связана в первую очередь с изменчивостью
увлажнения, а не температуры; поэтому климатические
правила в их оригинальном виде не описывают точно направления
адаптивного процесса" (Крупник, 1973, с. 88).
89
Этому автору удалось показать, что в Высокой Африке в раз
ных природных условиях (саванна, влажный лес, пустыня) Климе
тическая адаптация осуществляется в разных направлениях. П
условиям теплоотдачи здесь выделяются три антропоклиматичес
кие области: 1) жаркие сухие пустыни с температурой, близко]
или выше 33°С, где идет только испарение, так как теплоотдача з
счет конвекции и излучения тепла организмом возможна лишь д
тех пор, пока температура среды не превышает температуры пс
верхности кожи человека (32-33°С); 2) влажный лес, где испарс
ние затруднено, теплоотдача осуществляется главным образом з
счет конвекции и излучения; 3) саванны, где идут испарение і
конвекция, оказываются таким образом наиболее благоприятно]
для терморегуляции климатической областью тропического пояса
Как показали исследования И.И. Крупника, население парко
вой саванны (при испаряемости 95-120 мм) наиболее высокорос
лое, в случаях же затрудненной теплоотдачи преимущество полу
чают меньшие по размерам индивиды, поскольку у них большая
относительная поверхность испарения. В областях с затрудненно]
теплоотдачей (в лесу при испаряемости 30-70 мм, в полупустыв
и пустыне - при 200-280 мм) морфологическая адаптация идет а
пути уменьшения размеров тела. Примером такого рода адаптацш
могут служить пигмеи бассейна Конго, бушмены и готтентоті
Калахари.
Морфологические особенности популяций, живущих в услови
ях крайне сухой и жаркой пустыни, где среднегодовые значена
испаряемости превышают 300 мм, будут рассмотрены в специаль
ном разделе.
О высокой степени связи морфологических признаков со ере
дой обитания свидетельствуют и полученные нами коэффициент!
корреляции между длиной и относительной поверхностью тела а
среднегодовым количеством осадков и среднегодовой температу
рой воздуха, вычисленные для 34 этнических групп субтропичес
кой, тропической и экваториальной зон; коэффициенты ранговоі
корреляции колеблются в пределах 0,6-0,9 (Алексеева, 1974). Сле
дует отметить, что направление приспособительных реакций в од
нотипных условиях среды осуществляется сходным образом ;
представителей разных рас и этносов. Эта закономерность был
90
отмечена Д. Робертсом (Roberts, 1952, 1953) в отношении массы и
длины тела, а также основного обмена в связи с температурой
среды. Единое направление адаптивных реакций у разных расовых и
этнических групп получено для территорий Арктики (Alexeeva,
1973а) и Африки (Крупник, 1973). Это положение развивается с
использованием результатов Робертса и в книге "Биология
человека" (1964).
Итак, можно сказать, что в строении тела популяций
тропических широт отражается воздействие таких экологических
факторов, как температура и влажность, испаряемость, давление и
т.п. Они в известной мере создают специфику морфологических
особенностей тропических популяций, способствуют локальной
вариабельности черт строения.
Наблюдаемые экологические закономерности позволяют еще
раз обратиться к проблеме пигмеев, которая имеет длинную
историю. Обстоятельное изложение проблемы принадлежит перу
выдающегося советского антрополога М.Г. Левина (1946), и нет
необходимости прибегать к нему вновь.
В связи с темой данной книги следует обратить внимание на
объяснение специфических черт строения тела пигмеев, данное А.
Швальбе (Schwalbe, 1910). Критикуя взгляды И. Кольмана
(Kallmann, 1894, 1905) на пигмеев как на предковую форму
современных человеческих рас, получившие широкое признание и
развитие в этнологической науке конца XIX — начала XX в., А.
Швальбе рассматривает пигмеев как локальные вариации высокорослых
рас, возникшие в результате воздействия неблагоприятных
условий и недостаточности питания, отмечая при этом роль
селективных процессов в фиксации возникающих под действием
неблагоприятной среды признаков.
Все исследователи пигмеев отмечают присущую им низкорос-
лость, грациальность скелета, прямой лоб, слабое развитие
носовых костей. Эта общность проявляется на фоне разнообразия
антропологического типа среди как африканских, так и азиатских
пигмеев (Левин, 1946). В дополнение к этому следует указать и на
тенденцию к депигментации кожи и глаз у тех и других по
сравнению с более высокорослым родственным им населением Африки
и Азии (Eickstedt, 1934).
91
Рассматривая возможные объяснения перечисленных фактов
М.Г. Левин (1946) выбирает одно как наиболее вероятное: пигме
оидные черты образовались в результате проявления законов гете
рогенного роста и условий изоляции. Географическое положена
пигаейских племен Азии и Африки способствовало их длительно!
изоляции от более высокорослых соседей.
Это объяснение нельзя снимать со счета, но оно не учитываем
всей специфики окружающих пигмеев географических условий.
В этой связи напомню отмеченную на симпозиуме "Экология г
антропология" (Нью-Йорк, 1960) недостаточность фосфорно-каль
циевых солей во влажных тропиках и высказывания в пользу на
следственной адаптации к недостатку кальция и фосфора, выра
жающейся в замедлении процессов роста и развития. Наши иссле
дования корреляций морфофизиологических признаков с факто
рами среды выявили довольно значительную связь слабого разви
тия носовых костей с пониженным содержанием костеобразующиа
минеральных веществ в среде. Напомню также и об энергетичес
ких возможностях организма в условиях влажных тропиков: прв
затрудненной теплоотдаче преимущество получают меньшие пс
размерам индивиды.
Как уже отмечалось, в антропологическом отношении пигмеи
изучались неоднократно, но данных по признакам строения тела,
на основе которых можно судить об особенностях энергетики, очень
мало, хотя и они достаточно показательны. Наиболее низкорослые
из всех, известных в настоящее время, пигмейские группы
Африки, Азии и Америки, представленные в табл. 8, по условиям
жизни чрезвычайно сходны: все они живут во влажных жарких
тропических лесах, с годовым количеством осадков в Африке от 100С
до 2000 мм, в Америке - до 3000 мм, в Азии - до 5000 мм.
В свете этих данных проблема пигмеев приобретает и
экологический аспект, а высказанные А. Швальбе в значительной мере
априорные соображения обретают фактическую основу.
В подтверждение наследственной адаптации к недостатку кос-
теобразующих минеральных веществ у пигмеев отмечу
выявленную экспериментально Д. Раймоном и сотрудниками (Rimoin et
al., 1969) пониженную чувствительность периферических тканей
к гормону роста - соматотропину.
92
Таблица 8
Некоторые признаки строения тела у наиболее низкорослых групп
разных континентов (мужчины)
Признак
Число обследованных
Длина тела, см*
Длина тела сидя, см
Длина ноги, см
Длина руки, см
Ширина таза, см
Ширина плеч, см
Поперечный диаметр груди,
см
Переднезадний диаметр
груди, см
Кормический указатель
Длина ноги, % к длине тела
Длина руки, % к длине тела
Ширина плеч, % к длине тела
Ширина таза, % к длине тела
Грудной указатель
Масса тела (w), кг
Поверхность тела (s), м2
Указатель Рорера w/s
Мбути,
Заир,
Гуаинде,
1948
510
144,0
70,3
—
69,4
24,0
31,5
23,7
17,7
49,0
—
45,1
21,9
16,7
72,9
39,3
1,28
1,33
31Д
Онге,
Малый
Андаман,
Гуха,
1954
14
148,3
—
—
—
—
—
—
—
—
—
—
—
—
—
—
...
...
Папуасы,
Магашя,
Янсен,
1963
20
147,6
—
—
—
—
—
—
—
—
—
—
—
—
—
38,3
1,25
1Д9
30,6
Мотило-
ны-кари-
бы,
Венесуэла,
Флеури,
1953
37
146,2
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
Куба,
Посгаг-
хпил,
1976
41
149,9
...
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
* Длина тела у женщин исследованных групп: мбути - нет данных; онге - 137,?
см; папуасы - 140,9 см; карибы - 138,1 см; индейцы Кубы - 140,5 см.
93
4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ТРОПИЧЕСКИХ ПОПУЛЯЦИЙ
Окислительно-восстановительные процессы и терморегу
ляция. Изучением основного обмена в жарком климате занима
лись многие исследователи, но их работы нередко давали противо
речивые результаты. В краткой форме они суммированы в специ
альном исследовании, посвященном энергетическому обмену (Бер
кович» 1964). Большинство авторов отмечают понижение основно
го обмена у аборигенов в условиях жаркого климата.
Де Альмейда (De Almeida, 1920,1924), К. Корлет (Corlet, 1923)
а также более поздние авторы, результаты работ которых сум
мированы О. Уилсоном (Wilson, 1956), обнаружили понижеяш
основного обмена у жителей жарких стран- Р. Гальвао (Galvao, 1948a
19486, 1950) наблюдал понижение на 7% основного обмена у бра
зильцев по сравнению с жителями США. А. Манро (Munro, 1949]
нашел понижение основного обмена у коренных жителей Индии я
англичан, Е. Хипсли (Hipsley, 1969) - у папуасов Новой Гвинеи.
Н.М- Воронин и А.Д. Слоним (1959) также склоняются к мысли а
некотором понижении метаболизма в жарком климате. Д. Роберте
(Roberts, 1952), исследовавший связь основного обмена с расой и
климатом, показал, что в разных группах (белые американцы,
европейцы, австралийцы, американские индейцы) основной обмен
падает с повышением температуры воздуха (рис. 26).
Коэффициент корреляции дневного базального метаболизма с температурой
очень высок и равен -0,773.
По мнению некоторых авторов, на основной обмен
значительно воздействует ультрафиолетовая радиация. Увеличение ее в
условиях тропических широт тормозит окислительные процессы и
снижает метаболическую активность (Kastner, Schadow, 1927;
Архангельская, Вадова, 1949; Авазбакиева, 1958).
Значительная роль в интенсивности основного обмена принадлежит
мышечной массе тела. Некоторые авторы считают, что уровень
основного обмена определяется интенсивностью обмена в мышцах
и печени в большей степени, чем в других органах (Ломов, 1975).
. Основной обмен у людей (в том числе и у детей) с развитой
мускулатурой выше, чем при слабо выраженной мышечной массе
(Cam, Clark, Portrey, 1953; Brozek, Grand, 1955).
94
ккал
2Шг
Ю 3d SO 70 JO
Рис. 26. Зависимость основной
обмена от среднегодовой тем&ерату
ры (Robert; 1952):
1 - популяция земного яара, кроме не
гроидных; 2 - американские индейцы; 3 -
монголоиды.
У коренного населения тропического пояса в основном
мышечная масса слабая, что свидетельствует о понижении у них
основного обмена в условиях тропиков.
Известно, что величина основного обмена связана с характером
питания. Высококалорийная, с большим количеством белка пища
способствует повышению основного обмена; преимущественно
углеводная диета напротив, понижает его (Слоним, 1952; Rodahl,
1952). Сопоставление нескольких групп американских индейцев
показало повышение основного обмена у чиппевеев (Канада), в
рационе которых много белков животного происхождения по
сравнению с другими индейскими группами, имеющими в рационе
преимущественно растительную пищу (Wilson, 1950).
Опыты показали, что интенсивность падения основного обмена
во время голодания связана не только с уменьшением объема
клеточной массы, но и с понижением обмена в функционирующих
тканях (Молчанова, Ежова, 1939).
Итак, понижение основного обмена в условиях тропического
климата связывают с разными причинами: высокой температурой
окружающей среды, повышенной ультрафиолетовой радиацией,
тормозящей окислительные процессы, низкой калорийностью
диеты, слабым развитием мускулатуры, понижающим уровень
энергетических процессов организма. Может создаться впечатление,
что на основной обмен воздействуют главным образом экзогенные
факторы (температура среды, недостаток белков в питании и т.п.),
96
под влиянием которых снижается степень развития мышечноі
массы тела. Однако связь эта не столь проста. Установлено (Walt
er, Bajatzadeh, 1971), 4jo все народы, у которых частота медленно
мигрирующего трансферрина TfD1 превышает 0,01, живут в тропи
ческом поясе. Такие же высокие значения Т?01отмечены иужи
вотных, обитающих в этом поясе, в частности у рогатого скота.
У жителей тропиков развито потоотделение. Авторы делаки
предположение о роли трансферрина TfD1 в терморегуляции и с
возможной селективной адаптации по этому признаку в условияз
тропиков.
Высокие частоты сывороточного протеина Нр обнаруживаются
у негров, австралийских аборигенов, полинезийцев и американс
ких индейцев тропического пояса в противоположность тому, чтс
наблюдается у лопарей, эскимосов и групп европеоидного и монго
лоидного облика, живущих за пределами тропиков (Walter, Ste
egmuller, 1969). Авторы, получившие эти результаты, считаюі
возможным объяснить их действием селективных факторов.
При изучении адаптации коренных жителей Африки к
влажному и жаркому климату, определенной с помощью
фундаментальных тестов терморегуляции, обнаружено, что негроиды имекл
пониженную температуру тела по сравнению с европеоидами,
количество выделенного пота у них выше, чем у европеоидов, а
темпы замедления потоотделения ниже (Ojikutu, 1970). Эти
физиологические особенности подтверждаются расчетами среднего
количества потовых желез у европейцев и негров на 1 кв. см
поверхности тела: у европейцев - 137,67, у негров - 148,22 (Harrison et aL,
1964).
Экспериментальное изучение территориальных вариаций
физиологических реакций на повышение температуры среды
показало, что у негров банту и белого населения Южной Африки по мере
пребывания в условиях высокой температуры резко
увеличиваются ректальная температура и частота сердечных сокращений по
сравнению с акклиматизированными банту и белыми (Wyndham,
1967, 1970а, 19706). Как в случае акклиматизации, так и при
отсутствии ее банту имеют более низкие уровень ректальной
температуры и число сердечных сокращений в 1 мин. Потоотделение,
более .интенсивное в акклиматизированных группах, понижается
начиная со второго часа эксперимента; в течение первого часа
происходит резкое возрастание потоотделения у акклиматизирован-
96
ных групп, причем как падение, так и повышение у оелых
сильнее. Создается впечатление, что в условиях высокой температуры
окружающей среды аборигенное население характеризуется
большей устойчивостью физиологических процессов, чем пришлое.
Не менее интересные результаты получены в опытах с
переохлаждением (Wyndham, 19706). Реакция на холод у белых, банту в
бушменов однотипная; она выражается в повышении температуры
пальцев ног и рук, средней температуры кожи, ректальной
температуры. В то же время интенсивность реакций в этих трех
группах разная. При акклиматизации к холоду отмечено, что
возрастание показателей метаболизма на 1 кв. м поверхности тела при
температуре до 6°С у банту сходно с таковым у белого населения;
ниже этой температуры метаболический уровень у банту
увеличивается по сравнению с европеоидами. Бушмены по показателям
метаболизма сходны с банту.
Таким образом, и в характере терморегуляции проявляется
специфика тропических популяций по сравнению с пришлым
населением; температура тела, как и ректальная, ниже,
потоотделение более интенсивное, число сердечных сокращений в минуту
уменьшено и выше устойчивость физиологических процессов,
связанных с терморегуляцией.
Белки и ли?иды крови. Интерес к характеристике состояния
внутренней среды организма, в частности уровня белков и липи-
дов в сыворотке крови, проявился в начале 60-х годов. Имеется
довольно значительное количество данных, позволяющих судить
о географической изменчивости этих признаков. Оценка
литературных и собственных данных о популяционной изменчивости
белков сделана Л.К. Гудковой (1975). В наших работах показана
географическая вариабельность липидов (Алексеева, 1971).
При изучении пространственной изменчивости белков
сыворотки крови отмечалось повышение уровня гамма-глобулина в
тропиках. Причины этого явления еще недостаточно ясны. По
мнению некоторых авторов, увеличение уровня иммунных белков
вызвано патогенными микроорганизмами, в том числе и
малярийными возбудителями. Профилактическое еженедельное введение
препарата, препятствующее развитию возбудителя малярии,
проводившееся в течение нескольких лет И, Мак-Грегором с
сотрудниками (McGregor et al., 1956, 1960) в Гамбии, снизило уровень
97
гамма-глобулинов у гамбийских детей европеоидного происхожде
ния- Между тем в Каракасе (Венесуэла), где малярия довольно ред
ка, для населения характерно увеличение гамма-глобулинов (Vera
Roche, 1956). На Новой Гвинее, например, у жителей местности
не зараженной малярией, также наблюдается высокое соде-ржа
ние гамма-глобулинов в сыворотке крови (Bakker et al., 1957).
По мнению Г. Келтца и Г. Комстока (Keltz, Coinstock, 1959)
высокие уровни гамма-глобулинов, типичные для негроидов, »
являются результатом действия какой-либо инфекции или инва
зии, а представляют собой характерную особенность коренноіч
населения тропических стран, сохраняющуюся и в более благопри
ятных условиях среды обитания. Д. Идозьен (Edozien, 1961) обна
ружил, что различия между африканцами и европейцами в уров
нях белков сыворотки крови существуют с рождения. Этот же ав
тор предположил, что индивиды с более высокими значениям)
гамма-глобулиновой фракции и, следовательно, способные к обес
печению более высоких уровней антител имеют преимущества ]
районах эпидемических заболеваний. Проявление этой особенное
ти на популяционном уровне создает большую резистентность по
пуляций по отношению к болезням, распространенным в окружа
ющей их среде. При такой трактовке повышения уровня гамма
глобулинов в сыворотке крови иммунологические и наследствен
ные факторы взаимно не исключаются.
Генетическую природу увеличения иммунных свойств организ
ма отстаивают многие авторы. М. Лихтман с сотрудниками (Li
chtman et al., 1965) установил, что у негров и белого населения пп
Джорджия (США) в одних и тех же климатических условиях
имеющих сходный высококалорийный тип питания, величин;
гамма-глобулинов резко различна: их уровень у негров на 27°Л
выше, чем у белых. Аналогичные результаты получены и для не
которых районов шт. Северная Каролина (Milam, 1946), для Чика
го (Pollack et al., 1961) и Оклахома-сити (Klein et al., 1962). Ана
лиз сывороточных белков в близнецовых исследованиях показа
более тесную связь между уровнями протеинов у монозиготны
близнецов по сравнению с дизиготными и у обоих типов близнецо:
- бо'лыпую близость, чем у людей, не связанных родством (Leonh
ardt, 1962). Близнецовые исследования подтвердили наследствен
98
ную природу сывороточных протеинов, особенно альбуминов и
гамма-глобулинов (Ггеу et al., 1968).
Очень интересны в этой связи результаты изучения
иммуноглобулина A(IgA) у масаев - одного из племен Восточной Африки
(Но Kang et al., 1971). Уровень IgA у них значительно выше, чем
у лиц европейского происхождения, особенно у детей, у которых
за период от рождения до двух лет он приближается к величине
взрослого белого населения. По мнению авторов, высокий уровень
IgA нельзя объяснить пассивной иммунизацией, скорее это
важнейший фактор генетического приспособления.
Многие исследователи, внимание которых было обращено к
пониманию причин этнотерриториальной дифференциации
сывороточных протеинов, пришли к предположению о связи высоких
уровней гамма-глобулинов с принадлежностью к африканскому
расовому стволу. Анализ географической вариабельности
сывороточных протеинов выявил скорее территориальную, чем расовую
обусловленность; высокий уровень гамма-глобулинов характерен
не только для африканцев, но и для индейцев Америки,
обитающих в тропическом поясе, для папуасов и австралийцев, для
коренного населения Индии.
Некоторые авторы объясняют вариабельность сывороточных
протеинов влиянием различных условий окружающей среды.
Например, Г. Ниб й Г. Грут (Neeb, Groot, 1963) объясняют сдвиги в
протеинограммах белковой недостаточностью в питании. Ж. Ан-
ротт с сотрудниками (Непго??е, 1961; Henrotte et al., 1963, 1964),
изучившие физиологические признаки у коренного населения
Индии, пришли к выводу о том, что содержание гамма-глобулинов
выше в сыворотке у тех людей, чье экономическое положение
ниже, и у тех, кто живет в умеренном климате (имеются в виду
уроженцы Индии, проживающие во Франции).
В отличие от гамма-глобулинов уровень альбуминов по
направлению к тропической зоне снижается, вместе с тем убывает и
величина альбумино-глобулинового коэффициента. Между
содержанием белков в пище и концентрацией альбуминов в крови
установлена тесная связь. Неоднократно отмечалось уменьшение
уровня альбуминов в связи с белковой недостаточностью в питании.
Это подтверждается экспериментальными данными на животных.
В условиях высокого белкового содержания в рационе отмечаете*
близкий уровень белков у африканцев и белых (Powell, 1958), у
99
негров Америки и белых (Lichtman. et al., 1965), у австралийских
аборигенов и европейцев (Wilkinson et al., 1958). В то же время
имеются экспериментальные данные о том, что жители Нигерии,
содержавшиеся в течение восьми недель на высококалорийной
диете, прибавили в весе, но у них осталась без каких-либо
изменений концентрация альбуминов сыворотки крови (Edozien, 1967).
Более того, упоминавшиеся выше исследования близнецов
показали наследственную детерминацию альбуминов и других белковых
фракций (Leonhardt, 1962).
Таким образом, мы сталкиваемся с очень сложным
компонентом воздействий эндогенных и экзогенных факторов на уровень
сывороточных протеинов в условиях тропиков: наследственная
обусловленность, белковая недостаточность в диете, высокая
температура среды, угнетающая энергетические процессы в
организме. Каждый из этих факторов вносит свою лепту в обеспечение
метаболического баланса тропических популяций.
Если гамма-глобулины и альбумины обнаруживают более или
менее четкую географическую локализацию, то в отношении
общего белка в сыворотке крови картина менее определенная. В
тропическом поясе наряду с низким содержанием общего белка
наблюдается и гиперпротеинемия, т.е. повышенная концентрация
белков в сыворотке крови. В основном, по-видимому, это
объясняется высоким уровнем гамма-глобулинов. Во всяком случае,
явление высокого содержания общих белков при белковой
недостаточности в питании, по мнению Л.К. Гудковой (1975), характерно для
тропических и субтропических районов.
Географическая изменчивость сывороточных протеинов
представлена на р и с. 27.
Рис* 27. Географическая изменчивость сывороточных протеинов у
населения тропических широт (% к контролю):
1 - индейцы, Оклахома; 2 - гватемальцы, Гватемала; В - панамцы, Панама; 4 -
венесуэльцы, Каракас; б - яванцы, Суринам; 6 - негры, Суринам; 7 - индейцы, Суринам; 8 - индейцы,
Мату-Гросу; 9 - чилийцы, Чили; 10 - африканцы, Сенегал; 11 - африканцы, Гамбия; 12 - аф-
риканцы, Нигерия (Илория); 13 - африканцы, Нигерия (Ибадан); 14 - африканцы, Уганда; 15
- масаи, Кения; 16 - африканцы, Заир; 17 - бушмены, Окованго, Намбия; 18 - банту, Наталц
19 ,20- банту, Кейптаун; 21-24 - индийцы (21 - Пенджаб; 22 - Нагпур; 23 - Бомбей; 24 -
Мадрас); 25 - китайцы, Юньань; 26 - филиппинцы; 27-31 - папуасы (27 - Вогелкоп; 28 - Хол-
ландия; 29 - Биак; 30 - горные области; 31 - Морсиби); 32 - меланезийцы, Бугенвиль; 33 -
меланезийцы, Соломоновы о-ва; 34-37 - австралийцы (34 - Дарвин; 55 - Центральная
Австралия; 36 - Эркабелла; 37 - Уорбертон); а - общий белок; б - гамма-глобулин; е - альбуминово-
глобулиновый коэффициент; t — данных нет.
100
101
В пространственной изменчивости уровня холестерина не
наблюдается строгой закономерности. Тем не менее в целом можно
считать более или менее определенным понижение его к югу, хотя
есть и исключения, нарушающие эту закономерность: эскимосы
Канады и ненцы - на севере, австралийцы и население Гавайских
островов - на юге. Что касается австралийцев, то высокие
значения уровня холестерина, обнаруженные у них, все-таки ниже, чем
у белого населения Австралии: 217 мг% у мужчин и 207 мг% у
женщин коренного населения по сравнению с 263 мг% у мужчин
и 284 мг% у женщин (Schwartz, 1957). Таким образом, для
тропических популяций, за редким исключением, характерно низкое
содержание холестерина в сыворотке крови (рис. 28).
Многими исследователями при оценке роли алиментарного
фактора в географической изменчивости признаков достаточно
ясно показана зависимость уровня холестерина от состава пищи:
позитивная связь отмечается с содержанием в ней жиров и белков
и с количеством калорий, а негативная - с содержанием
углеводов. Связь пониженного содержания холестерина в сыворотке
крови с диетой тропических популяций, бедной белками и жирами,
по-видимому, не должна вызывать сомнений.
В то же время существуют данные, которые заставляют
усомниться в том, что характер питания - это единственная причина,
определяющая величину липидов в крови. Прежде всего следует
обратиться к своеобразному исследованию, проведенному в
Кейптауне среди банту и европейцев (Antonis, Bersohn, Srand, 1862).
Содержание популяций на одинаковой диете показало
существование различий между ними в уровне холестерина в сыворотке
крови. Так, на диете, в которой 15% калорий определяется жирами
Рис, 28. Географическая изменчивость уровня холестерина, у населения
тропических широт (% к контролю):
1-5 индейцы (1 - киче, маме, покомам, Гватемала; 2 - "черные" карибы, Гватемала; 3 -
квква, Венесуэла; 4 - Каракас, Венесуэла; 5 - Суринам); 6 - негры, Суринам; 7 - яванцы,
Суринам; 8 - население Бразилии; 9 - население Нигерии; 10 - пигмеи, Конго; 11 - население
Эфиопии; 12 - население Кении; 13 - бушмены оседлые; 14 - банту, Батсвана; 15 - банту, Йога-
несбург; 16 - бушмены-кочевники; 17 - гагхентотско-мадайско-европекские метисы; 18 -
банту, Лесото; 19 - банту. Южная Африка; 20-23 индийцы {20 - уроженцы северных районов; 21
- уроженцы окрестностей Агры; 22 - сипаи; 23 - уроженцы южных районов); 24 -
филиппинцы; 25-27 - папуасы (25 - атолл Мапия; 26 — о-в Биак; 27 ~ уроженцы равных мест новой
Гвинеи); 28 - австралийцы, оседлая группа, Алис-Спрингс; 29 - австралийцы, кочевая группа.
102
3
¦
ft]
X
(так называемая "диета банту"), уровень холестерина у банту рав
нялся 165 мг%, у европеоидного населения -177 мг%; на диете
в которой за счет жиров образуется 40% калорий ("диета белых")
банту имели 227 мг%, европейцы - 237 мг%. Связь с питанием ]
здесь явственна, тем не менее она не уничтожает различий, суще
ствующих между этими группами.
Далее, имеются данные, свидетельствующие о том, что у раз
личных этнических групп, существующих на одной и той же дне
те, наблюдаются разные уровни холестерина (методические разли
чия в этих случаях исключены). Так, у итальянцев Неаполя (Key
et al., 1954) и сардинцев (Keys et aL, 1965) уровни холестерин
соответственно равны 229,9 и 173,2 мг%; у банту бечуанов и бав
ту басуто (Wsalker, Arvidsson, 1954а, 19546) - 149,0 и 153,0 мг%
у различных лингвистических групп индейцев Гватемалы (Scrim
shaw et al., 1957) уровень холестерина колеблется между 122 и 14-
мг%; у евреев, уроженцев Йемена и уроженцев Израиля (Вптие
et al., 1952) - 160,0 и 199,0 мг%; у коренных австралийцев и
окрестностей Алис-Спрингса и белого населения той же местносп
- 234 и 314 мг% (Charnock et al., 1959).
В дополнение к этому перечню можно привести данные о вари
ациях содержания холестерина у кочевых групп индейцев Венесу
злы, где примесь "белой" крови повышает величину этого призна
ка, в то время как диета "чистой" и смешанной групп не обнару
живает различий (Bosch, 1957): индейцы квива - 157,0 мг%, ин
дейцы с примесью "белой" крови, жители Каракаса - 197,0 мг%
Наконец, известны данные о связи уровня холестерина матери і
новорожденного ребенка (Iayalakshmi et al., 1957).
Приведенные материалы позволяют предположить, что наряд)
с экзогенными факторами на уровень холестерина в сыворотке
крови влияют и эндогенные факторы.
Обратимся к данным исключительного значения для
понимания генетической детерминации физиологических особенностей
человеческих популяций.
В первую очередь заслуживают внимания исследования
некоторых биологических характеристик у масаев Восточной Африка
(Но Kang et al., 1971). Масаи - это кочующая группа Восточной
Африки, питающаяся в основном молоком, мясом и кровью зебу.
104
Пища масаев чрезвычайно богата жирами; их дневной рацион со
ставляет 3000 ккал, причем 66% калорийности определяется со
держанием жира (ср.: с 45% в рационе белых американцев). Не
смотря на весьма значительную, неоднократно отмечаемую связі
уровня холестерина с количеством жира в пище, у масаев содер
жание холестерина приближается к мировому минимуму. Бушме
ны Калахари и пигмеи Южной Америки имеют еще более низкие
уровень холестерина, но в их диете значительно ниже, чем у ма
саев, содержание жиров и выше физическая активность.
В популяции масаев клинически не зафиксировано ни одногс
случая атеросклероза. Атеросклеротический индекс у них
наиболее низкий по сравнению с населением Индии, Японии, Ямайки.
Бостона и Лос-Анджелеса. Толщина стенок коронарной артерии >
масаев значительно меньше. Метаболизм у этой группы в
сравнения с таковым у европеоидного населения изучали путем введения
радиоактивного холестерина в дневной рацион. Отмечено
подавление синтеза эндогенного холестерина. В среднем масаи
синтезируют 650 мг холестерина в сутки, т.е. показывают 50%-ное
сокращение эндогенного холестеринового синтеза в сравнении с контролем
(1370 мг). Масаи, прожившие в США не менее 10 лет, сохраняю!
сходство с соотечественниками, что также подтверждает
наследственную природу низкого уровня холестерина.
Дополнительные доказательства генетической детерминации
уровня холестерина и триглицеридов имеются в статье Т. Бенеде-
ка и И. Зандера (Benedek, Sunder, 1970), приводящих данные
некоторых авторов о связи более высокого уровня холестерина с
группой крови А. Правда, следует отметить, что сами авторы при
обследовании негров и европейского населения США не
обнаружили такой зависимости. Ими установлено понижение уровня
триглицеридов у негров по сравнению с лицами европейского
происхождения. Краткие физиологические характеристики населения
тропической зоны представлены в табл. 9.
В соответствии с уровнем холестерина в сыворотке крови
определяются толщина жировых складок и количество общего жира у
коренного населения тропиков. Связь уровня холестерина с
развитием жирового слоя была отмечена нами ранее, при изучении
морфофизиологических корреляций на внутригрупповом уровне
105
Таблица
Уровень холестерина н белков сыворотки крови у коренного населения
тропических широт (мужчины)
Территория
Африка
Передняя Азия
Индия
Юго-Восточная
Азия
Австралия
Океания
Центральная
Америка
Южная Америка
Холестерин, мг%
N*
9
3
10
2
2
4
3
1
min-znax
101-167
160-199
163-218
147-181
212-234
109-174
136-194
—
М**
141
190
182
1?4
223
140
170
157
Гамма-глобул ины,
N
9
—
2
3
4
7
—
1
m in-max
1,4-3,2
—
1,3-1,9
1,4-1,6
1,2-1,9
2,1-2,6
—-
—
М
2,2
—
1,6
1,6
1,6
2,3
—
1,6
Аяьбууино-глобули-
новый коэффициент
N
9
—
3
3
4
7
—.
1
min-max
0,6-1,9
—
1Д-1,3
0,9-1,6
0,9-1,4
0,7-0,8
—
—
М
1,0
—
1,2
1Д
1,2
0,7
—
1,2
* М - число групп.
* N — групповая средняя.
(Алексеева, 1970). Эта связь в числовом выражении невелика, н<
довольно определенна: уменьшение содержания холестерина в сы
воротке крови связывается с уменьшением толщины жировыз
складок. В тропических широтах, несмотря на незначительное ко
личество литературных данных по жироотложению, эта завися
мость совершенно отчетлива. У папуасов Новой Гвинеи, например
средняя жировая складка варьирует в пределах 4-6 мм у мужчад
и 5-7 мм у женщин по сравнению с 13-18 мм у мужчин европео
идного происхождения, но уроженцев Австралии (Goldrick, Whyt
е, 1959; Jansen, 1963; Whyte, 1963). Между этими группами зна
чительны различия и в проценте жира по отношению к массе тела:
у папуасов содержание жира составляет около 18%, у белых авст
ралийцев - около 26%. Толщина средней жировой складки у
мужчин одной из континентальных групп Австралии - 3,5 мм, в тс
106
время как в контрольной группе (европеоидное население Австра
лии) - 5,8 мм (Haimnel et aL, 1959).
Сопоставление данных по измерению жировой складки
затрудняется возможными методическими расхождениями, однако про
слеживается тенденция понижения жироотложения у тропичес
ких популяций американских индейцев, африканцев и австралий
ских аборигенов по сравнению с контролем европеоидного проис
хождения. Арабы и население Северной Индии имеют величины
весьма близкие к контролю (табл, 10). Не исключено, что это след
ствие их принадлежности к европеоидной расе.
Таблица 10
Толщина жировой складки у представителей различных этнических групп, мм
Этническая группа
Европеоидная группа
(контрольная)
Американские индейцы:
кечуа
кашинауа
Сара (Чад)
Арабы (Египет)
Население Северной
Индии*
Австралийские аборигены:
Дарвин
литджанджара
Жировая складка
на плече
10
5
5
—
9
—
8
6
под
лопаткой
11
8
8
8
11
9,5
—
10
на
животе
11
б
8
5
11
12,5
11
7
Автор и год
Eisner, 1963
*
Johnston et ah, 1971
Crognier, 1973
Neiercinski, 1970
Волков-Дубровин,
1971
Eisner, 1963
t
* Измерения производились скользящим циркулем, увеличивающим толщину
складок на 1-2 мм (Лутовинова, Уткина, Чтецов, 1970). В остальных случаях
жировая складка измерена калипером. Данные по Северной Индии приведены с
поправкой.
107
Своеобразие коренного населения тропического пояса заключа
ется не только в характеристике терморегуляции и содержали!
белков и липидов крови.
Как уже отмечалось при изучении секреции гормона ростам
африканских пигмеев, Д. Раймон с сотрудниками (Rimoin et al.
1969) выявили генетическую детерминацию гормонального уров
ня. Пигмеи обладали пониженной чувствительностью перифери
ческих тканей к гормону роста, хотя средний уровень его близоі
к контролю (европейцы). Аналогичный результат получен дл*
бушменов Калахари (Ioffe et al., 1971). Эти факты служат эксго
риментальным подтверждением предположения о наследственно!
адаптации к дефициту белков и минеральных веществ в рацио»
(см. материалы симпозиума "Экология и антропология") (J. Amer
AnthropoL, 1962).
Пристального внимания заслуживают работы французских ис
следователей, обнаруживших непереносимость молочного сахара j
арабов, живущих в ФРГ в течение 2 лет (Sendrail, Quilici, 1970)
Подобные сведения получены и о палестинских арабах (Gilat et al.
1971). Непереносимость лактозы характерна для многих тропичес
ких популяций. В основе ее, по мнению авторов, лежит недоста
точная активность кишечной лактазы; при микроскопическое
исследовании ткани кишечника ее редукция обнаружена у 4 чело
век из 26 обследованных. Хотя некоторые авторы указывают т
адаптивный генезис лактозной непереносимости, Г. Роттхауз с со
трудниками (Rotthauwe et al., 1971) считают, что больше основа
ний для суждения в пользу генетической природы этого признака
Исключительный интерес представляют исследования концен
трации натрия и калия в клетках крови у негроидного и европео
идного населения (Munro - Faure et al.» 1971). У негроидов значи
тельно выше концентрация натрия, чем у европеоидов, у после
дних, напротив, увеличено содержание калия. У негроидов мень
ше концентрация АТФ.
Можно предположить, что изменение концентрации
электролитов и их соотношения у африканцев по сравнению с европейца
ми, по-видимому, связано со спецификой течения и регуляция
водно-солевого обмена, а уменьшение концентрации АТФ, очевид
но, коррелирует с понижением общего метаболизма у коренной
населения тропиков.
108
5. О ПРИСПОСОБИТЕЛЬНОЙ ИЗМЕНЧИВОСТИ
В ТРОПИЧЕСКИХ ШИРОТАХ
Комплекс морфофункциональных черт тропических
популяций во многих отношениях специфичен. Это проявляется как в
строении тела, так и в характеристике внутренней среды
организма, включая глубинные механизмы обмена веществ. Вряд ли
такого рода специфика может трактоваться как акклиматизационный
эффект. Скорее всего, этот морфофункциональный комплекс
возник в результате длительного, наследственно закрепленного
приспособления к высоким температурам и влажности окружающей
среды, к условиям белковой недостаточности и избытку углеводов
в диете, к некоторым эндемическим заболеваниям.
Специфическая удлиненная форма тела с повышенной
относительной поверхностью испарения, увеличение количества потовых
желез на 1 кв. см, интенсивность потоотделения, специфика
регуляции водно-солевого обмена, понижение уровня метаболизма,
достигаемое уменьшением мышечной массы тела, редукцией
синтеза эндогенных жиров и понижением концентрации АТФ, -
характерные черты тропических аборигенов. Если к этому добавить
темную пигментацию кожи, курчавоволосость, удлиненную и
высокую форму головы, то адаптивный характер морфофункцио-
нального комплекса коренного населения тропиков не будет
вызывать сомнения. Наследственная природа этого комплекса,
по-видимому, также бесспорна. Напомним концентрацию медленно
мигрирующих трансферинов, связанных с уменьшением интенсивности
основного обмена в тропической зоне, пониженную
чувствительность периферических тканей к гормону роста у пигмеев,
редукцию синтеза холестерина у масаев, лактозную непереносимость у
арабов и некоторых других групп тропического пояса.
Непереносимость лактозы, в основе которой лежит недостаточная
ферментативная активность, может быть, следует интерпретировать как
наличие избирательной способности в отношении пищи. В свете
такой интерпретации становится понятным приспособление к
диете с избытком углеводов и резким дефицитом белков.
6. МОРФОФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ
НАСЕЛЕНИЯ СУБТРОПИЧЕСКОЙ ЗОНЫ КАВКАЗА
На территории бывшего Советского Союза нет экологических
ниш тропического пояса, однако население одного из субтропичес-
109
ких регионов изучено очень подрооно, хотя некоторые признаки
в частности характеризующие белковый обмен, уже давно вошед
шие в практику антропоэкологических исследований, отсутству
ют.
Речь идет о населении Очамчирского района Абхазии - типич
ной зоны влажных субтропиков.
Средняя годовая температура воздуха здесь +13,6°С, средне*
годовое количество осадков выше 1300 мм и средняя влажності
более 80%. Село Члоу, где были проведены исследования, распо
ложено в типичной для Абхазии предгорной зоне.
Морфофизиологические исследования абхазов являлись состав
ной частью обширной советско-американской темы "Комплексно^
биолого-антропологическое и социально-этнографическое изучены
народов и этнических групп с повышенным процентом долгожи
телей" (Феномен долгожительства, 1982). Индекс долгожитель
ства, представляющий собой отношение числа лиц старше 90 лег.
к числу лиц в возрасте от 60 и старше, составляет здесь 90%, а пс
отношению ко всему населению Абхазии - 18%. Судя по "Итогал
всесоюзной переписи населения 1970 и 1972 гг.", средние значение
этих индексов по бывшему Союзу составляют соответственно 101
1,2%.
Несмотря на весьма подробные и разнообразные исследование
феномена долгожительства в Абхазии, нельзя^ с_уверенностью ска
зать, что этот феномен нашел свое окончательное решение.
Среди морфофизиологических черт, которые можно назватв
специфичными для изученного абхазского населения, отметим за
медленные темпы старения, которые у долгожителей сравнитель
но легко определяются на основе оссеографических показателе*!
остеоморфных процессов по рентгенограммам кисти, высокую эф
фективность снабжения тканей кислородом, замедленные темпь:
возрастных изменений системы кровообращения и, возможно,
существование генных комплексов, детерминирующих
долгожительство (Феномен долгожительства, 1982). Судя по
высказываниям авторов, последнее предположение безусловно нуждается в
подтверждении (Саламатина и др., 1982). Оно опирается на факт
генетической гетерогенной популяции.
Абхазы характеризуются определенными генетическими
особенностями. Некоторые из них типичны для европеоидных
популяций (системы АВО, Келл-Челлано, Даффи), другие явно тяготе-
но
ют К востоку (системы Р, резус, MNSs). Однако для
подтверждения реальности различий в генных частотах необходимы
дополнительные данные.
Из физиологических показателей» типичных для
долгожителей Абхазии, следует отметить низкий уровень окислительных
процессов организма. Низкому уровню обмена соответствует и
малая скорость кровотока. Эти характеристики дополняются
сравнительно низкими величинами частоты пульса» систолического в
диастолического давления крови, В совокупности эти показатели
свидетельствуют о слабом уровне гемодинамических процессов
(Волков-Дубровин и др., 1982). Что касается черт строения тела,
то какой-либо специфики, которая создает морфологические
особенности долгожительской популяции, не отмечается.
Это крупное, высокорослое население, с относительно малыми
размерами туловища и большими размерами рук и ног,
широкоплечее, с хорошо развитой грудной клеткой и высоким индексом
массивности (индекс Рорера) (табл. 11). По строению тела абхазы
более всего сходны с горными таджиками, дагестанцами, а также
с туркменами, каракалпаками и чуйскими казахами (рис. 29).
Сходство усугубляется довольно низкими величинами уровня
минерализации скелета, характерными для большинства изученных
нами народов, проживающих между 40 и 45 градусами северной
широты.
Абхазы, дагестанцы, таджики и казахи Чуйской степи
обитают в горных и предгорных районах, туркмены и каракалпаки - в
равнинных.
Это сходство трудно объяснить экологическими причинами.
Скорее всего, оно может быть связано с проявлением глубинного
субстрата, восходящего к европейскому скотоводческому
населению III тысячелетия до н.э., занимавшему огромное пространстве
от южнорусских степей до Западной Монголии (Медникова, 1995),
Что касается оценки морфофизиологического комплекса
абхазов с точки зрения их приспособления к субтропической зоне, тс
мы не находим здесь каких-либо специфических черт, которые
можно рассматривать как реакцию на повышенную влажность і
сочетании с высокой температурой воздуха.
Может быть, это объясняется тем, что в субтропическом
климате Кавказа повышенная влажность наблюдается только в зим
ние месяцы, а лето сухое и солнечное. Возможно, и сама субтро
113
дагестанцы
абхазы
туркмены
таджики (Чорку)
таджики (Ворух)
каракалпаки
телеуты
казахи чуйские
теленгиты
Рис. 29. строение тела абхазоё ш сравнительном освещении:
1 - длинна тела; 2 - длина ноги; 3 - длина руки; 4 - ширина плеч; 6 - ширина таза; $ -
поперечный диаметр груди; 7 - передкеаадний диаметр груди; 8 - масса тела.
112
Таблица 11
Морфофкзиолопгаеская характеристика абхазов
Признак
Число
обследованных
Длина тела, см
Длина ноги, см
Длина руки, см
Ширина плеч,
см
Ширина таза,
см
Поперечный
диаметр груди,
см
Переднезадний
диаметр груди,
см
Длина ноги, %
к длине тела
Длина руки, %
к длине тела
Ширина плеч,
% к длине тела
Ширина таза,
% к длине тела
Мужчины
123
167,3
89,5
75,4
39,2
29,3
27,3
21,0
53,6
45,1
23,4
17,6
Женщины
97
156,8
83,2
69,4
36,1
29,6
25,1
18,5
53,1
44,3
23,0
18,9
Признак
Грудной указатель
Масса тела (т), кг
Поверхность тела
(8), М2
Указатель Рорера
s/w
Средняя жировая
складка, мм
Общий белок, г%
Гемоглобин, г%
Холестерин, мг%
Минерализация 3-го
луча кисти левой
руки, мг/ммЗ
Систолическое
давление, мм рт.ст.
Диастолическое
давление, мм рт.ст.
Мужчины
76,9
65,7
1,75
1,40
2,66
8,32
191,7*
0,536
122,2
75,7
Женщины
73,8
60,3
1,63
1,56
2,70
14,20
...
0,618
119Д
76,6
* Уровень холестерина в группе 25-44 года (Даланишвили и др., 1982).
пическая зона, являясь промежуточной между тропическими в
умеренными широтами, отличается определенной комфортностью,
что не вызывает какой-либо специфической реакции со сторонк
человеческого организма.
из
К сожалению, я не располагаю достаточно представительным!
данными о населении субтропических широт, поэтому и затрудня
юсь дать сколько-нибудь определенный ответ по поводу существо
вания специфического морфофизиологического комплекса у насе
ления субтропиков.
Лекция IV
БИОЛОГИЯ НАСЕЛЕНИЯ АРИДНЫХ ЗОН
Морфофизиологические характеристики населения
тропических и внетропических пустынь
1. СПЕЦИФИКА ВОЗДЕЙСТВИЯ КЛИМАТИЧЕСКИХ
УСЛОВИЙ ПУСТЫНИ НА ОРГАНИЗМ ЧЕЛОВЕКА
Пустыня как одна из экстремальных зон обитания человека
привлекает внимание исследователей уже давно. Сочетание таких
климатических факторов, как высокая температура,
ультрафиолетовое и тепловое излучение, крайняя сухость воздуха, ветер и
пыль, оказывает нередко пагубное воздействие на организм
пришлых, неакклиматизированных людей. В то же время пустыни
обитаемы с доисторических времен, и коренное население,
особенно кочевники-скотоводы, приспособилось к жизни в такой
экстремальной ситуации, концентрируясь у источников воды.
Большинство пустынь расположено в тропическом поясе, но
климат их отличается значительным своеобразием. К. Адольф с
сотрудниками в книге "Физиология человека в пустыне" (1952)
приводят результаты сравнения климата тропиков и пустыни.
Средняя суточная температура в пустыне на 8 градусов выше, а
колебания среднесуточного цикла почти в два раза больше, чем в
тропиках. Средняя относительная влажность в тропиках
приближается к 100%, в пустыне - к 30%.
Фундаментальные исследования акклиматизации в пустыне
Колорадо показали, что на солнце термическая нагрузка для
обнаженного человека в 3, 4 раза больше, а для одетого - в 2, 4 раза
больше, чем в тропиках. Тепловая энергия, образовавшаяся в
организме в результате обмена веществ, может быть непосредственно
114
передана излучением при помощи конвекции окружающему
воздуху лишь в том случае, если его температура не превышает 33°С
(такова температура поверхности кожи). Если теплоотдачи не
происходит, то тепло остается в организме, повышая температуру
тела. При этом возможен только один путь отдачи тепла - путем
потоотделения и затем испарения содержащейся в нем воды.
Перегрев и обезвоживание - основные причины стрессовых
ситуаций в организме при акклиматизации в условиях пустыни.
Акклиматизация к жаре здесь выражается в сохранении более
низкой температуры тела, более замедленного пульса и более
низкого потоотделения. Известно, что при температуре воздуха 15,5°С
организм теряет 0,94 л из общего количества потерянной
жидкости (1,4 л/сут), тогда как при 32°С из 2,99 л на долю пота
приходится 2,4 л (Winslow et al., 1937). Таким образом, в условиях
повышенной температуры удаление тепла из организма происходи!
главным образом в результате потоотделения, в то время как в
условиях умеренного температурного режима терморегуляция
осуществляется еще и путем лучеиспускания, конвекции,
теплопроводности и др. (Ротштейн, Таубин, 1952; Гец, 1963). Что касается
приспособления к обезвоживанию, то его установить не удалось.
Привыкания к недостатку воды не происходит. Дегидратация
повышает вязкость крови, снижает скорость кровотока, приводит к
уменьшению ударного объема сердца, нарушению электролитного
равновесия и другим жизненно важным сдвигам в обмене вещесть
(обширная литература на этот счет приведена в монографии В.Г.
Воловича "Человек в экстремальных условиях природной среды",
1983).
В связи с компенсацией водопотерь, вызванных усиленным
потоотделением, уместно остановиться на исключительно
интересных опытах А.Ю. Юнусова с сотрудниками (1975), которые изучи
ли водопотребление у различных групп людей, оказавшихся в рай
онах жаркого, сухого климата: уроженцев Средней Азии (I
группа), уроженцев средней полосы (II группа) и жителей Крайнего
Севера (III группа). Выяснилось, что в такой ситуации
водопотребление возрастает тем больше, чем менее адаптирован человек в
жаркому климату. Наибольшее среднесуточное потребление водь
было отмечено для жителей Крайнего Севера, наименьшее - для
уроженцев Средней Азии. Опыты Юнусова говорят о существова
ііб
нии генетически закрепленных механизмов адаптации к условия»
дегидратации. По-видимому, есть основание думать, что коренное
население аридной зоны обнаружит адаптацию и к гипертермии.
2. МОРФОФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
НАСЕЛЕНИЯ ТРОПИЧЕСКИХ ПУСТЫНЬ
Исследования коренных жителей пустыни проводились
неоднократно, но картина адаптации к аридным условиям пока еще
недостаточно ясна.
Рассмотрим данные, которые могут быть привлечены для
характеристики черт строения тела и физиологии у пустынных
популяций. Так как большинство аридных популяций являются
коренными жителями тропиков, естественно, что те
приспособительные особенности, которые характерны в целом для тропических
аборигенов, обнаруживаются и у них. Напомним, что для
тропических жителей характерны: понижение основного обмена,
уровня холестерина и альбуминов сыворотки крови, большая
устойчивость реакций, связанных с теплообменом, понижение абсолютной
величины мускульной массы тела (что также способствует
снижению энергетических процессов) и удлиненная форма тела с
небольшим весом, приводящие к увеличению удельной поверхности тела
и, следовательно, к более интенсивной теплоотдаче путем
испарения (табл. 12). При затрудненной теплоотдаче в условиях
пустыни можно предполагать еще большую выраженность этих черт, не
исключены и какие-то специфические реакции.
И.И- Крупник, (1973), анализируя особенности климатической
адаптации в Африке, показал, что в условиях затрудненной
теплоотдачи, в сухой и жаркой пустыне, где среднегодовые значения
испаряемости превышают 300 мм, и во влажном экваториальном
лесу существует определенная морфологическая адаптация,
приводящая к увеличению относительной поверхности тела. Один и тот
же результат достигается разной комбинацией признаков
строения тела. В пустыне это сочетание высокого роста и малой массы
при крайней сухощавости (например, у теда), в экваториальном
лесу - уменьшение всех размеров тела и массы (пигмеи). У
бушменов пустыни Калахари увеличение удельной поверхности тела до-
116
5 9
I
ю 2
CD
Ю
09
00
ю
eo
»
см
см
О)
о
* о
5 3
CO
00
ем
CO
о
1-4 *M CO *H
о
4*
гЧ N
CD
см
CM
rH CO f-H
i
i
&
<d
s
I s
< s
2 3
CO
Ю
CM
CM
00
•4
CD
О
i
« b-
CN
CO
CO
о
со
eo
со
* CM
H 8
00
Ю
CO
to
CM
«J
*¦' CD
CD
CO
°1
CO*
Ю
to
Ю
kO
Ю
3
»1 CO
22 »
О* CM ер" Ь-
§ CM S ^H
CO
CO
CM pi
CO
O)
fH 04 i-4
C0fc p4
i-T CM* »H
01
CO
Ю
r-4 CM W
00 t-
O ін
CO «H
о
*H CO іЧ
Cft
О
CM
Ы C4 M
<
I
с
С
&
С
«
и
a
с
I
о
і
5 I
g§
5 2 ^
in
стигается сочетанием крайне малых размеров и массы тела. Такик
образом, на примере аборигенных африканских популяций мь
видим усиление адаптивных черт в строении тела, связанных с
увеличением поверхности теплоотдачи в условиях пустыни.
У арабов Аравийской пустыни уменьшаются размеры и массе
тела, увеличивается удельная поверхность по сравнению с араба
ми, живущими в континентальной тропической зоне Африки, не
вблизи Средиземного моря.
Для австралийских племен Центральной пустыни характерно
сочетание очень высокого роста с низкой массой, так же как у эт
нических групп Сахары. Австралийское же племя коката, обита
ющее в Западной пустыне, нарушает эту закономерность. У кока
та довольно большая масса тела (62,2 кг у мужчин и 51,0 кг j
женщин). Это сочетается с большими обхватными размерами гру
ди: у мужчин - 91,5 см, у женщин - 81,8 см (Hicks et al., 1931)
Если даже допустить методические погрешности при взвешива
нии, размеры грудной клетки говорят о достаточной массивности
тела. Так или иначе приходится признать, что коката по своин
морфологическим особенностям представляют исключение среді
тропических популяций, живущих в аридной зоне. Напомню, чт<
для австралийских групп характерен и более высокий уровені
холестерина по сравнению со всеми этническими группами тропи
ков (Алексеева, 1971).
К числу специфических физиологических особенностей абори
генов пустыни следует отнести понижение кровяного давленш
(Casley-Smith, 1959; Abbie, Schroder, 1960; Barnes, 1965), повыше
ние уровня гемоглобина и гематокрита (Davis, Pitny, 1957; Аваз
бакиева, 1958; Волович, 1983).
Такого же рода физиологические изменения наблюдаются)
пришлого населения в условиях пустыни - в результате теплово
го воздействия расширяется просвет периферических сосудов, чт<
приводит к снижению кровяного давления, а в результате обез
воживания примерно на 80% повышается вязкость крови и ні
15% увеличивается содержание гемоглобина (Бабаева и др., 1979
Волович, 1983). Все это создает дополнительную нагрузку на сер
дце, которое вынуждено сокращаться чаще для поддержания нор
мального артериального давления и минутного объема крови (Рот
штейн, Таубин, 1952; Авазбакиева, 1958).
118
Представление о специфике процессов терморегуляции у
коренного населения пустыни дают исследования бушменов
Калахари и австралийских аборигенов в условиях переохлаждения. И те
и другие практически не знают одежды и в то же время спокойно
переносят сравнительно низкие температуры ночей в пустыне.
В серии опытов с охлаждением, проведенных в 40-х годах над
австралийскими аборигенами и лицами европейского
происхождения при температуре воздуха 10-15°С, были выявлены различия,
которые позволяют предполагать большую устойчивость
процессов терморегуляции у аборигенов по сравнению с европеоидами
(Goldby et al., 1938; Hicks, O'Connor, 1938a, 1938b).
В тех случаях, когда аборигены и лица европейского
происхождения в обнаженном виде спали вблизи костров, у них не
наблюдалось каких-либо заметных метаболических отклонений.
Если же испытуемые находились вдали от костров, но в спальных
мешках, то у них изменялась температура тела. Средняя
температура, по которой судят о "силе охлаждения", у тех и других
падала до одинакового уровня, в то время как внутренняя
температура тела (в полости рта, например) у аборигенов падала, в контроле
же оставалась неизменной. Кроме того, у коренных жителей
отмечалось сужение стенок периферических сосудов в большей
степени, чем в контроле, и при более слабом действии холода, чем тот,
который вызывал подобную реакцию у лиц европеоидного
происхождения. В результате этих опытов делается предположение о
более эффективной регуляции теплообмена у аборигенов с
помощью вазомоторной деятельности (Goldby et al., 1938).
Несколько раньше (Hicks et al., 1931) была обнаружена
разница в газообмене у австралийцев коката (Западная пустыня) и
европейцев. У коката содержание кислорода по сравнению с
контролем ниже: у женщин - на 11%, у мужчин - на 13%.
Исследования с охлаждением были повторены на другой
австралийской группе - питджанджара (Центральная пустыня). При
почти одинаковом охлаждении кожи аборигены и белые
чувствовали себя по-разному. В то время как первые спали на протяжении
ночи с нормальной теплопродукцией, контрольная группа
испытывала постоянное беспокойство, и ее метаболизм резко снизился.
Авторы делают заключение о существовании у аборигенов
пустыни акклиматизационных механизмов, аналогичных тем, которые
часто обнаруживаются у млекопитающих, и представляющих со-
119
бой наследие отдаленных предков, уже утраченное европейцами]
их более цивилизованном мире (Sholander et al., 1958). Близки*
результаты получены другой группой авторов, также изучавши}
метаболизм у питджанджара (Hammel et al., 1959).
Исследовав процессы метаболизма у коренных жителей пусты
ни, австралийско-европеоидных метисов тропической зоны Авст
ралии и контрольной группы европейского происхождения, оні
показали, что при идентичных условиях экспозиции к холоду ме
тисы занимают промежуточное положение между центральным!
аборигенами и контролем. Уровень метаболизма метисов - 42,?
ккал • м2/ч, у контроля - 48,7, у аборигенов центральных районоі
Австралии - 37 ккал • м2/ч.
Различия в терморегуляции найдены также между бушменаю
Калахари и лицами европейского происхождения (Wyndham, Мог
rison, 1958; Hammel, Hildes et al., 1963).
При обследовании обнаженных бушмена и европейца во врем;
сна при температуре воздуха 12°С в течение 2,5 ч у обоих был<
обнаружено одинаковое падение температуры кожи и большее па
дение внутренней температуры тела у бушменов (так зке как ]
опытах с австралийцами). В интерпретации этих данных авторь
придерживаются иной точки зрения, нежели предыдущие. Hi
обнаружив заметных изменений в терморегуляции во время сні
вблизи костров, они склоняются к мысли об отсутствии физиоло
гической адаптации к холоду у аборигенов пустыни и о существо
вании "интеллектуальной" адаптации, создавшей определенны!
климат, близкий к термонейтральному. Подобная точка зрения (о!
отсутствии физиологической адаптации) отстаивается и в последу
ющих работах, во всяком случае одного из этих авторов (Wyndhan
et al., 1964), относительно реакции на высокую температуру ере
ды. Обнаружив большую выраженность реакции на жару, сказы
вающейся в понижении ректальной температуры, частоты сердеч
ных сокращений и повышении потоотделения у французов по ера
внению с арабами в условиях Сахары, авторы делают предположе
ние, что "недостаточная" физиологическая приспособленность ара
бов к условиям пустыни компенсируется их морфологическим!
особенностями - повышением относительной поверхности тела.
Как видим, отрицая физиологическую адаптацию к условия»
пустыни, авторы тем не менее придают значение адаптации мор
120
фологической, которая, что уже отмечалось в гл. II, не может быть
осуществлена без физиологических изменений обменных
процессов. Судя по характеру эксперимента, у французов в Сахаре резко
выражен эффект акклиматизации, в то время как арабы,
адаптированные к аридному климату, демонстрируют определенную
нейтральность физиологических процессов. Этот опыт по своим
результатам сродни приведенному выше эксперименту А.Ю. Юнусо-
ва с сотрудниками (1975), которые установили, что в условиях
пустыни водопотребление выше у уроженцев районов, наиболее
удаленных по своим климатическим условиям от пустыни.
Суммируя те данные, которые могут быть привлечены для
суждения о направлении морфофункциональной изменчивости в
аридной зоне, приходим к следующему заключению. Несмотря на
небольшое число изученных в пустыне аборигенных групп, нельзя
отрицать наличие у них определенных морфологических и
физиологических механизмов облегчения процессов теплообмена. Для
большинства групп это понижение метаболической активности и
увеличение удельной поверхности испарения. Более эффективная
сосудодвигательная регуляция потери тепла у аборигенов в
условиях резких температурных колебаний в течение суток также
может считаться более или менее типичным явлением адаптивного
характера. Этими особенностями в какой-то мере очерчивается
морфофункциональный комплекс коренного населения аридной
зоны.
3. МОРФОФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
НАСЕЛЕНИЯ ВНЕТРОПИЧЕСКИХ ПУСТЫНЬ
Климат внетропических пустынь отличается от пустынь
тропической зоны холодной бесснежной зимой. Особенно это относите*
к пустыням Каракум, Кызылкум, Мойынкум и Гоби. В Гоби, на
пример, морозы зимой достигают -40°С (Пржевальский, 1948).
В нашем распоряжении имеются весьма представительные
материалы по морфофизиологическим особенностям коренного насе
ления пустынь Каракум, Кызылкум и Мойынкум, собранные экс
педициями НИИ антропологии МГУ, работавшими в Туркмении,
Каракалпакии и Казахстане (Волков-Дубровин и др., 1975, 1977«
19776; Архангельская и др., 1980, 1981) (табл. 13, рис. 30). Эти
12]
СО
3
g
ft-
о
о
2
і
3
о
ю
¦8
ЕВ
О
К
«
ч
«
и
«в
И
3
X
в
я
а
S
О
§
і
II
г I
I
іі
1 ?
¦
S
I
§
US
Ifc-
И
в
в
а
В
: я
8<о
іЧ U3
: еч оі q
об S
04
©QOweoeofmt-et-ooweat-rt00.
-чГ да" ¦« ьГ сі" со о* оо О* ад *°. "Ч °°- V °. •«* ад
Ч ея ь- а и» о ь-
и»
со
Ф U9 СО (О О) "^ СО
^ и» n t«* ^* а" ю ь о«* со" cn" со о со" "t Ч
^ДССОЭСОСЯСЧі-іЮ'^'СМт-іЬ-Ю*-)^
s « »
те °і 0> as со ^ ^ ^ иэл ^ е^ «^ ь^ чэ^ «о ** ?| °і Я 4
*0 Ж *X мч ns ru _J irt <¦« A3 —a «s_ in —J >^ nl _l пЛ >_W ^!
lO^OHt-IOHH
Ю
«
r -r »f ^ «r ^ « «э
of b-* V t-* ^* 00 <N » N t- n >
CDttCOWMHtO^WHb-lO
^ іо л. со
О "Э
"l^wooaeooN^wH
is
f 00 00
t- та o4
о ^ ^ t- » ю а
Ъ- Tf CO
t w. О ^ Ю
©J H « H
i CO 00 CO t-
a" " " "
CO t- fc-
Mogoot-OJN»wt-H4i"
0> С* .-4 ft-
fc » *» *
t-
<N
1 « Ю О» -( О CO П
- t- со <o T о -г о
2l? др*со"с^аэ"<ооГс4'еэ*еч<о^«Г^Ч4,н^<о*"2:
Ю
CO
^ _• яГ —* _r ^* j* ^: t* со с» _r о»
ca Ж сь «* oo* ao » oT eo* eo* &f <© со" ¦*
"^ m »s_ m eo ем _^ iA -чИ ем —j is> «ft
fiSG^
122
Рис. 30. Местонахождение исследованных популяций коренного
населения внетропических пустынь:
1 - турквсены-йомугы, Узун-Су и Кульма» Кааанджикский р-н Туркмения; 2 - туркмены-
теко, Ак-Toire, Хан-Кириз, Араджа, Бахарденсккй р-н Туркмения; 3 — каракалпаки» Карауаяк,
Тахтакупырский р-н, Казахстан; 4 - казахи» Акколь, Таласский р-к Казахстан.
данные представляют большой интерес, так как относятся к
представителям двух рас, проживающих в сходных условиях
географической среды и имеющих практически один и тот же тип
хозяйства, а следовательно, и рацион питания. Традиционное занятие
населения - скотоводство; земледелие играет подчиненную роль, в
последние годы большое место в хозяйстве заняло хлопководство.
Из трех изученных народов только каракалпаки являются
сравнительно поздними пришельцами в аридную зону, однако их
формирование связано со степными и опустыненными территориями
(Рысназаров, 1972). Туркмены (теке и йомуты) относятся к
европеоидной расе, казахи и каракалпаки - к монголоидной. В
этногенезе каракалпаков и казахов просматривается участие
европеоидных компонентов, но в настоящее время монголоидные черты
выражены достаточно четко.
Характеризуя морфофизиологический статус коренного
населения внетропических пустынь, прежде всего следует обратить
123
внимание на общее укрупнение размеров тела - особенность, ти
личную для населения всей внетропической зоны. Далее, при
наличии имеющихся различий в пропорциях тела, типичных для
представителей монголоидной и европеоидной рас, население
среднеазиатской аридной зоны характеризуется комплексом общиз
морфологических и физиологических черт. Помимо увеличения
массы тела для него характерны крупные размеры скелета,
относительно слабое развитие подкожного жирового слоя и
мускулатуры, уплощенная грудная клетка, большой процент астенического
типа телосложения и несколько большая по сравнению с
контролем относительная поверхность тела.
Из физиологических признаков обращают на себя внимание
низкий уровень минерализации скелета с пониженной скоростью
возрастной перестройки кости, понижение окислительных
процессов. Понижение артериального давления и повышение
содержания гемоглобина, столь типичное для жителей тропических
пустынь, здесь проявляется лишь у туркмен и казахов. Тем не менее
эта черта наблюдается и в условиях внетропических пустынь и,
что особенно важно, у групп различного расового состава.
Аналогичные результаты были получены и другими исследователями.
Уменьшение интенсивности основного обмена, увеличение
частоты дыхания и пульса, снижение кровяного давления обнаружены
у коренных жителей аридной зоны Казахстана М.В. Авазбакиевой
(1958).
Таким образом, черты, типичные для населения тропических
пустынь, проявляются и во внетропических широтах. Различия в
морфофизиологическом статусе коренных жителей тропических и
внетропических пустынь сводятся в основном к увеличению
массы тела последних. В остальном же процесс адаптации к аридным
условиям у них сходен.
124
Лекция V.
БИОЛОГИЯ ЖИТЕЛЕЙ ВЫСОКОГОРЬЯ
Морфофизиологическая специфика высокогорных популяций
и ее природа,
1. СТРОЕНИЕ ТЕЛА И ФИЗИОЛОГИЯ
Один из интереснейших районов обитания человека -
высокогорье. Пониженное атмосферное давление, недостаток кислорода,
холод, нередко нарушение геохимического баланса, недостаток
пригодных для жизни и хозяйства земель, позволяют назвать
условия высокогорья поистине экстремальными. Исследование
физиологических реакций на больших высотах у местного населения
или у пришлых групп констатирует приспособление к основному
неблагоприятному фактору - гипоксии* т.е. пониженному
содержанию кислорода в крови. По мнению многих исследователей,
снижаются основной обмен и активность
окислительно-восстановительных ферментов, функция надпочечников и щитовидной
железы, замедляется ритм сердечных сокращений. В то же время
усиливается оксигенация крови за счет повышения уровня
гемоглобина и увеличения количества эритроцитов. Эти особенности
рассматриваются как приспособительные к более экономному
расходованию кислорода (Белекова, 1964а, 19646; Миррахимов и др.,
1968; Филатов, Каныгина, 1968; Туркменов, 1968а, 19686;
Ахмедов, 1968; Агаджанян, Миррахимов, 1970; Clegg et al., 1970).
Некоторые из физиологических сдвигов наводят на мысль об
изменениях и в морфологических признаках высокогорных
популяций. Основанием для этого служит направление морфофункци-
ональных связей (Морфофизиологические исследования в
антропологии, 1970). Много внимания уделяется изучению ростовых
процессов в высокогорье. Работы в этом направлении ведутся в
Перуанских Андах (Baker, 1963; Baket et al., 1966; Frisancho,
Baker, 1970; Baker, 1971), в горах Эфиопии (Pawson, 1971; цит. по:
Миклашевская, 1972), на Памире (Миклашевская, 1972), на Тянь-
Шане (Турусбеков, 1970) и на других территориях (Pyzuk et al.,
1967). Монография под редакцией Р. Бейкера обобщает
многолетние исследования высокогорных популяций Америки, Африки и
Азии (The biology of high-altitude peoples, 1978; переведена на
русский язык в 1981 г.).
126
Можно считать установленным, что для большинства высокс
горных популяций, независимо от их расовой и этнической про
надлежности, характерно замедление ростовых процессов и сроко
полового созревания. Значительно меньше данных о морфофунк
циональных особенностях высокогорных групп взрослого населб
ния. Обращает на себя внимание исследование Дж. Харрисона
сотрудниками (Harrison et al., 1969) популяций амхара, живущи:
на различных высотах на севере Эфиопии. Авторы показали, чт
и взрослые, и дети этой популяции имеют на высокогорье боле
крупные размеры тела, чем на равнине. Эти данные как бы протв
воречат закономерности, которая обнаружена в условиях высокс
горья, во всяком случае в детских возрастах.
Дж. Харрисон и его коллеги не случайно выбрали территорш
Эфиопии для своих исследований. Разнообразие ландшафта и кли
мата, генетическая общность групп, живущих на различных высс
тах, вполне сопоставимый характер питания - все это позволил
выявить значение геоклиматического фактора в формировани:
адаптивных особенностей у высокогорных популяций по сравне
нию с популяциями, обитающими на равнине.
Помимо более массивного телосложения в условиях высокого
рья авторы отмечали и большее развитие грудной клетки и скеле
та в целом. Последнее обстоятельство, по их мнению, можно свл
зывать с гипертрофией костного мозга, которая в свою очеред
увязывается с повышенным эритропоэзом. Крупная грудная клеі
ка горцев, сочетающаяся с более высокой жизненной емкость»
легких, также рассматривается как морфофункциональное при
способление к пониженному барометрическому давлению и сопуі
ствующему этому уменьшению парциального давления кислорода
Морфофункциональные особенности высокогорных популяциі
Эфиопии равно характерны для мужчин, женщин и детей, и во
они легко поддаются интерпретации с точки зрения приспособле
ния к гипоксии. Исключение составляет отмеченный авторам]
более низкий уровень гемоглобина по сравнению с равнинным]
популяциями. Однако при повторном исследовании были получе
ны более высокие значения гемоглобина, подтверждающие весьм;
существенную зависимость содержания гемоглобина от характер!
питания.
Несмотря на довольно высокий уровень приспособленносп
высокогорных популяций к экстремальным условиям обитания
126
проявляющейся как в строении тела, так и в физиологических
характеристиках, отмечается весьма значительный процент
выкидышей, во много раз превышающий таковой у равнинных
популяций. Авторы связывают этот факт с влиянием гипоксии на
развитие плода.
Изучение высокогорных популяций в Перуанских Андах, с
одной стороны, подтверждает результаты исследований в
Эфиопии, а с другой - противоречит им (Baker, 1971; Биология
жителей высокогорья, 1981).
В частности, в противоположность Дж. Харрисону П.Бейкер
показал не только замедление процессов роста и полового
созревания детей в высокогорных районах по сравнению с равнинными,
но также более хрупкое телосложение у взрослых. В то же время
развитие грудной клетки, особенно в продольном направлении, у
обитателей высокогорья значительно выше, чем у равнинных
жителей. Причем эта закономерность проявляется на протяжении
всего периода роста. Из физиологических особенностей;,
изученных у индейцев Перу, живущих на разных высотах, следует
остановиться на данных по метаболизму и температурной регуляции.
Пониженное давление кислорода в высокогорье оказывает
наибольшее стрессовое воздействие на энергетические процессы в
организме. Сопоставление популяций кечуа и аймара, живущих на
высотах от 3500 до 4500 м над уровнем моря, с их равнинными
сородичами, мигрировавшими в высокогорье, показывает, что
мигранты в значительно большей степени страдают от
кислородной недостаточности, чем постоянные жители больших высот. У
последних выше не только максимальная кислородная емкость, но
и легочная вентиляция, уровень гемоглобина и миоглобина, а
также число и просвет капилляров. А.Хуртадо (Hurtado, 1964) на
основании этих данных сделал предположение о существовании
разницы в усвоении кислорода на клеточном уровне у жителей
высокогорья и равнины при пониженном его напряжении в атмосфере.
Способность к более быстрому окислению гемоглобина до оксиге-
моглобина в условиях кислородной недостаточности у аборигенов
высокая по сравнению с пришлым европеоидным населением. Это
было показано и в эксперименте при оценке кривых диссоциации
кислорода. По мнению авторов, такой эффект обусловлен
модификацией молекулы гемоглобина и может рассматриваться как
феномен эволюционной адаптации в популяциях, обитающих в услови-
127
ях высокогорья в течение нескольких тысячелетий (Morpurgo e
аі., 1970).
Дж. К. Квиличи и X. Вергнес (1981), сделавшие обзор литере
туры и осуществившие сравнительный анализ гематологически:
характеристик высокогорных популяций, также отмечают рол
отбора в биологической адаптации к высокогорью, которая особев
но отчетливо проявляется в метаболических процессах, происхс
дящих в эритроцитах и обеспечивающих более эффективное ис
пользование кислорода при недостатке его в среде обитания.
Предположения о наследственной природе приспособления і
кислородной недостаточности подтверждаются данными Р. Елене
ра и А. Болстеда (Eisner, Bolstad, 1963), полученными в эксперл
менте с тестом охлаждения. У коренных жителей высокогорь,
обнаружен более высокий периферический ток крови, чем у при
шлого европейского населения и жителей равнины. Эта особен
ность, не являющаяся уникальной для населения высокогорья
рассматривается как наследственно закрепленное приспособлен
к действию низких температур (Baker, 1971).
Для характеристики высокогорных популяций Южной Амери
ки очень важными представляются демографические данные, со
держащиеся в работе П. Бейкера. Демографический анализ Боли
вии, Эквадора и Перу показал, что естественный прирост населе
ния в высокогорье ниже, чем на равнине. Однако авторы, полу
чившие эти результаты, испытывают затруднения в их интерпре
тации, так как причины биологического характера здесь не могу
быть четко отделены от причин социальных.
Мы предприняли попытку характеристики морфофункцио
нальных особенностей взрослого населения, живущего в условия:
высокогорья, с помощью антропоэкологических связей. Для сопо
ставления было взято несколько территорий, где группы моглі
быть в той или иной мере генетически связаны и в то же врем;
жили на разных высотах над уровнем моря. Так были выбрань
таджики (Гинзбург, 1937; Волкова, 1965; Рычков, 1969; Алексее
ва, 1974; Алексеева и др., 1984), население Индии (Biswas et al.
1966; Das, 1966; Henrotte, 1966), различные этнические группь
Ирана,Ирака, Саудовской Аравии, Йемена (Edholm, 1967), населе
ние Эфиопии (Harrison et al., 1969), индейцы кечуа (Frisancho, Ba
ker, 1970). Во всех этих случаях речь идет о местном (коренном
населении.
Поскольку высокогорные группы, взятые для сопоставления
живут в климатических поясах, различающихся по температур*
128
окружающей среды и влажности, необходимо учитывать эти
характеристики в связи с особенностями адаптивных реакций
населения.
Для высокогорья, независимо от того, к какому
климатическому поясу оно относится, характерно понижение температуры- В
отношении влажности картина очень разноречива. В
Таджикистане и Перу в высокогорных районах наблюдается понижение
влажности. То же самое отмечается в Иране и Ираке (в данном случае
Ирак рассматривается как равнинная территория, Иран — как
среднегорье). В Эфиопии влажность возрастает с высотой над
уровнем моря, в Индии увеличение количества среднегодовых осадков
может быть связано и с равниной, и с высокогорьем.
Население названных территорий, за исключением
Таджикистана и Эфиопии, изучено по крайне ограниченной программе, для
некоторых групп известна только длина тела.
Одной из очень важных физиологических характеристик
является основной обмен. Данные по основному обмену, полученные с
применением формулы Робертса, опубликованы в отдельной
статье (Алексеева, 1974). Имея в распоряжении 34 группы, живущие
на различных высотах в пределах субтропического, тропического
и субэкваториальных поясов, мы вычислили ряд ранговых
коэффициентов корреляции между отдельными морфологическими
признаками и высотой над уровнем моря, среднегодовой
температурой, годовым количеством осадков (табл. 14).
Таблица 14
Коэффициенты ранговой корреляции между некоторыми морфологическими
и географическими признаками
Признак
Длина тела
Масса тела
Относительная поверхность тела
Основной обмен
Среднегодовая
температура
+0,626*
+0,268***
+0,946*
-0,290***
Высота над
уровнем моря
+0,617*
+0,556*
+0,337**
+0,508*
Годовое
количество
осадков
+0,710*
+0,048***
+0,771*
+0,165
*р20,01.
** р 2: 0,06.
г** На грани достоверности.
129
Оказалось, что теплопродукция увеличивается с высотой на;
уровнем моря и уменьшается с повышением температуры. Это на
блюдение вполне увязывается с понижением температуры в высо
когорье и противоречит данным некоторых авторов о пониженш
здесь газообмена в связи с гипофункцией щитовидной железы.
Уменьшение теплопродукции с повышением температуры ок
ружающей среды было обнаружено Д. Робертсом (Roberts, 1952)
вычисленный им коэффициент корреляции был выше (-0,772)
Такое расхождение может быть объяснено тем, что обследованныі
нами группы относятся только к теплым климатическим поясам
в то время как Роберте рассматривал население гораздо более кон
трастных в климатическом отношении территорий.
Длина тела увеличивается с высотой над уровнем моря; с повы
шением среднегодовой температуры и годового количества осад
ков коэффициенты корреляции возрастают.
Масса тела достоверно увеличивается в условиях высокогорья
но с годовым количеством осадков связи нет. Очень незначитель
на (на грани достоверности) позитивная корреляция массы тела «
среднегодовой температурой. Роберте отметил негативную связі
этих признаков: г = -0,809 (коэффициент г вычислен для терри
тории с контрастными климатическими характеристиками). I
зоне теплого климата масса тела практически независима от коле
баний температуры окружающей среды. Население этой зоны от
личается очень низкой массой по сравнению с более севернымі
группами.
Обращают на себя внимание высокие положительные коэффи
циенты корреляции относительной поверхности тела со среднего
довой температурой и годовым количеством осадков. Повышена
поверхности испарения, по-видимому, является адаптивным меха
низмом, возникающим в условиях высокой влажности и темпера
туры. В высокогорье также наблюдается увеличение поверхносп
тела по отношению к массе, однако коэффициент корреляции не
велик. Это и понятно, так как для высокогорья далеко не всегді
характерно повышение влажности.
Таким образом, наши данные об усилении теплопродукции ]
высокогорье не согласуются с исследованиями, показавшими по
нижение газообмена в тех же условиях.
Для объяснения этого расхождения рассмотрим данные физи
ологических исследований. В книге Н.А. Агаджаняна и М.М. Мир
130
рахимова (1970), подводящей итог многолетним исследованиям
акклиматизации в условиях высокогорья у нас в стране и за
рубежом, приведены показатели газообмена у местного населения в
горах и на равнине, в покое и в работе.
Имеется много сообщений о понижении основного обмена э
местных жителей высокогорья. Однако редукция наблюдается
далеко не у всех обследованных. К тому же в известных нам работая
не проводились групповые измерения, так что нет основания
говорить здесь о групповых характеристиках. К.Ю. Ахмедов (1967) і
специальном обзоре, посвященном реакции на гипоксию,
отмечает, что у аборигенов высокогорья газообмен не изменяется.
X. Саха и Б. Дас (Saha, Das, 1962) зарегистрировали
повышение в покое поглощения кислорода у жителей Дарджилинга
(высота 2100 м над уровнем моря, 4,83±1,12 мл/кг-мин) и понижение
у жителей Калькутты (уровень моря, 4,61±1,21 мл/кг-мин). Н.А,
Агаджанян и М.М. Миррахимов, явно склоняясь к мнению о
редукции газообмена в высокогорье, приводят данные, которые
противоречат этому мнению. Так, у уроженцев высокогорья,
исследованных во Фрунзе (700 м), газообмен в покое был ниже, чем у лиц,
родившихся на равнине (3,91=*=0,43 и 5,1±0,38 мл/кг.мин
соответственно). При подъеме на высоту эти показатели изменились. 3
горцев в покое газообмен равнялся 5,46±0,69 мл/кг-мин, у
жителей равнины - 4,60±0,33 мл/кг-мин- Понижение у горцев
газообмена на равнине вряд ли следует рассматривать как нормальное
явление. Почти во всех работах отмечается более экономичное
расходование кислорода во время физических нагрузок у постоянных
жителей высокогорья по сравнению с жителями равнин. Мне
представляется, что о редукции газообмена можно говорить, липи
имея в виду его понижение в процессе работы. Ведь увеличение
легочной вентиляции и кроветворной активности» усиление окси-
генации крови, характерные для высокогорных популяций,
трудно было бы объяснить при общем понижении основного обмена.
М.А. Литтл и Дж. М. Ханна, изучавшие воздействие холода и
других стрессорных факторов в высокогорных районах мира,
отмечают, что, "несмотря на эффективные средства защиты от
холода, у коренных жителей высокогорья развились два физиоло
гических механизма адаптации к холоду: некоторое повышенш
основного обмена и более интенсивный приток крови к конечное
тяж при общем и периферическом охлаждении*' (1981, с. 321). По
13]
стоянно высокий уровень основного обмена у коренных жителе!
высокогорья обнаружен и другими исследователями (Petropoulos
Timiras, 1874).
Таким образом, следует признать, что увеличение основной
обмена с высотой над уровнем моря является ведущим адалтив
ным процессом, и причины отклонения от него должны рассмат
риваться специально.
В непосредственной связи с основным обменом находится ура
вень гемоглобина, обеспечивающий энергетические процессы. Уве
личение содержания гемоглобина и эритроцитов рассматривается
как реакция приспособления к гипоксии. Действительно, в боль
шинстве случаев уровень гемоглобина в высокогорье увеличивает
ся (Щекочихина, 1970).
У населения Эфиопии этой закономерности не наблюдается, не
в данном случае судить об изменении количества гемоглобина труд
но, так как у различных эфиопских групп он неизвестен, а насе
ление предгорья и высокогорья характеризуется довольно высоки
ми его величинами. К тому же, как упоминалось выше, повторно*
исследование уровня гемоглобина в эфиопском высокогорье пока
зало возрастание его по сравнению с предшествующими данными
В условиях высокогорья не только, как правило, повышаете;
уровень гемоглобина, но и изменяется морфологический состаз
крови. Известно, что у коренных жителей Перу число эритроци
тов в крови на 30% выше, чем у лиц, живущих на уровне мор*
(Harrison et al., 1964).
Обследование коренных жителей Киргизии, живущих на раз
ных высотах, также показало увеличение количества эритроци
тов, гематокритного индекса и других показателей красной крові
(Туркменов, 1968а) (рис. 31). Физиологическое значение увели
чения количества эритроцитов и уровня гемоглобина, иначе гово
ря, полиглобулии, по мнению некоторых авторов, заключается і
усилении оксигенации крови (Ахмедов, 1968). Лучшая оксигена
ция крови у коренных жителей высокогорья по сравнению с насе
лением, живущим на уровне моря, отмечалась неоднократно (Ах
медов, 1967; Туркменов, 1968а; Бускирк, 1981; Квиличи, Вергаес
1981). Таким образом, можно сделать предположение и в данной
случае об однотипности приспособительной реакции на гипоксик
независимо от этнической принадлежности з сходных условиях ее
тественной среды.
132
Нор ичсстдо jpu/npo-
Содержание
гемоглобина, г %
Средняя концентраций
пногнодина S отдель -
""* эритроцитах ,
г но 100 г *ро8и
Средний одъем
отдельны*' эритроцитов,нм3
Гк ма/покритн?ій
индекс
Цв етноО показатель
Нвцичестдо /геОкоци*
moi, тис
т°ш*
Высота над уровнем шор* , и
700 - 800*
10
О
20
10
О
100
30
80
70
60
W
0,8
0,8
0,8
0.7
0,6
7Y
6
1
і
ч
й
і
Шм
1800-2000»
У,
1
®
%
г
у
і
* *
Pi I
2 <:
№0-3400 м
у\ Щ
1
1
V
6
і
у
т
Глі
/
s
1 у
щ
¦ V
>
1
I
Рис. 31. Состав кроши у коренных жителей Киргизии на разной высоте
над уровнем моря (по материалам А1.Г. Туркменоеа, 1968);
а - мужчины, б - женщины.
133
Одной из существенных физиологических характеристик вы
сокогорных популяций оказывается довольно низкий уровень ар
териального давления. Понижение его характерно не только даь
высокогорья, но и для коренного населения среднегорья (Алексе
ева и др., 1984). П. Бейкер (1981) приводит сводку данных отно
сительно воздействия низкого парциального давления кислорода;
окружающей среде на рост и размеры капилляров и артериол ]
тканях. В период раннего онтогенеза в условиях высокогорья он<
способствует большему росту числа и размеров капилляров и арте
риол, что препятствует повышению кровяного давления. И дей
ствительно, среди жителей высокогорья очень невелик процен1
сердечно-сосудистых заболеваний (Миррахимов и др., 1980; Бей
кер, 1981).
Каковы же морфологические характеристики населения высо
когорья? Отмеченное увеличение длины тела и массы хорошо со
гласуется с увеличением основного обмена. Подробные характери
стики взрослого населения высокогорья известны по Эфиопии (На
rrisonet al., 1969), Таджикистану (Гинзбург, 1937; Волкова, 1966
Алексеева и др., 1984) и Дагестану (Кириакиди, 1973а). Посколь
ку материалы по Таджикистану были собраны в разное время
возможны расхождения в признаках физического развития, свя
занные не столько с различным происхождением исследованные
групп, сколько с разным рационом питания. Это касается в основ
ном материалов В.В. Гинзбурга. Но, во-первых, в анализе болыпо
внимание уделялось характеристикам не абсолютных размеро]
признаков, а их соотношений; во-вторых, для контроля данных п<
высокогорью, относящихся к 30-м годам, имеются материалы это
го же времени для равнины (Циммерман, 1927; Ошанин, 1927
1937). Оценка относительных величин признаков вызвана еще і
тем обстоятельством, что сравниваемые популяции в пределам
Таджикистана, несмотря на известную близость антропологичес
кого типа, не являются представителями одной и той же этничес
кой группы. Есть основание считать горцев Западного Памирі
группой, отличной в генетическом отношении от равнинных тад
жиков (Рычков, 1969а). Дж. Харрисон с сотрудниками (Har-risoi
е? а!., 1969) исследовали различные группы амхара. Относительш
антропологического облика населения Эфиопии известно, что он<
обнаруживает негроидную примесь. В Дагестане известно несколь
ко этнических групп, живущих на разных высотах и относящих
134
ся к кавкасионскому типу в пределах южной европеоидной расы
(Гаджиев, 1965; Алексеев, 19746). Следовательно, желая найти
общие закономерности у представителей разных рас и этносов, мы
должны каким-то образом нивелировать различия, обязанные
своим происхождением расовой принадлежности. В этом случае
удобны характеристики относительных размеров.
На всех указанных территориях помимо увеличения
основного обмена с высотой над уровнем моря обращают на себя внимание
изменения пропорций тела в сторону относительной длинноногос-
ти и длиннорукости и увеличение продольно-поперечного
грудного указателя (табл. 15). Последнее обстоятельство, по-видимому,
связано с усилением легочной вентиляции в условиях
высокогорья. Что касается увеличения относительной длины конечностей,
то в одной из статей я высказала предположение о вторичном
характере этого явления, возникшего в результате неравномерности
роста туловища и конечностей (Алексеева, 1974).
Не отрицая возможного действия этой причины, отмечу, что
трактовать специфику тела высокогорных популяций, имея в виду
значительное развитие их скелета, можно и с позиций
приспособления к условиям гипоксии. Большее развитие длинных костей
скелета предполагает и больший объем костно-мозгового
пространства в связи с усилением кроветворной функции. Косвенным
подтверждением этого могут служить данные о мощном развитии
рогов высокогорных архаров и козлов, которые тем больше, чем
выше над уровнем моря они живут (Коржуев, 1971а, 19716).
Закономерности изменения соматотипа с высотой над уровнем
моря подтверждаются обширными данными по морфологии
населения Дагестана (Кириакиди, 19736; табл. 16). Укрупнение
размеров тела на высоте отмечено и здесь, и хотя не все коэффициенты
корреляции отвечают критерию достоверности, общее
направление изменений здесь совершенно закономерно. Различия
касаются лишь размеров жировой массы тела, которая в Эфиопии и на
Памире уменьшается с высотой над уровнем моря, а в Дагестане
явно увеличивается. Не исключено, что это связано с различиями
в пищевом рационе; жировая складка, как правило, служит
хорошим индикатором уровня питания.
К сожалению, для анализируемых высокогорных групп
данных об уровне холестерина нет, но судя по исследованиям А.М.
135
«3
I я
a
0
I
I
©
Ю
s 1
It*
CO "
CD
CQ
CD
CD
CO
0Ofc TO i-^
со* со" *а
CN CO Ю ч*
со
о
CO
о
о
Ч СО
CD
СО
СО
со ю
00
Ю *Ч
О ~ Ю рЧ
СО М
і. со «э
N СО Ю
ю
СМ
СМ
СМ
СО
(О
ю
со
Ч со ^
о g со
ю S ю
со ч*
«5 О) .
«. * о
i-t CM СО
СО Ч< t-
о
со
см
см
ел
ч*
со
00
S
к
is
< ч»
а
о
о
«э
со
t- рч
ю
СО
ч*
ч*
со*4
ю
см
iOfc
со*
о
со
со
о
о
ю
со
» СО S
ю - со
3 3 -J
CN ^ СП СП
см со
со
ю
ч*
СМ*
ю
см
см
CD
о
со
t»
1
а,
?
*
§
>>
со
00
СП
« со
о
о
1>
і> ч< аа
t> _Г С- СО
СО
о н •* t •* оо
СО 1-4
: ю со
см со из
ч*
СП
со
со
ю
см
см
л
СО
о
СИ
CD
ю
Н со
is'
ЮСМЮі-чиЭСОСП Ю СО
см
со
рч см со
со
00
со
СО
со
см
см
о
fc-
Ч Ч н со ю
О ч* _Г СО
о со
Ю рЧ
СО рЧ СМ
*". а>
со со со см о>
см со
со
00
со
*о
рЧ О
со t-
СМ рЧ
со
°1
см
» О)
г - *- ю
°Э ^f го СО
Ю Н " 00
<* *
S?
«
s s я
I g I
ж © л
я
I & &
ч
И
& &
И
ф
я
в
А
О
И
I
8 8 S a 8 ?
136
Таблица 16
Коэффициент ранговой корреляции между морфологическими признаками
и высотой над уровнем моря на территории Дагестана (по Кнрнакндк, 19736)
Признак
Длина тела
Масса тела
Ширина плеч
Ширина таза
Обхват груди
Ширина лодыжки
Корреляция
с высотой
над уровнем
моря
0,37
0,43
0,77
0,83*
0,66
0,64
Признак
Ширина запястья
Обхват плеча
Обхват голени
Поверхность тела
Средняя жировая складка
Обезжиренная масса тела
Корреляция
с высотой
над уровней
моря
0,86*
0,60
0,66
0,46*
0,83*
0,37
Коэффициенты корреляции достоверны.
Алиева (1970) в Дагестане, Д. Гзелла иДж. Мейера (Gsejl, Mayer,
1962) в Швейцарских Альпах, А, Бака с сотрудниками (Buck et
al., 1968) в Андах и наших - в Северном Таджикистане
(Алексеева и др., 1984), концентрация холестерина понижается с высотой
над уровнем моря.
При характеристике морфофункциональных особенностей
высокогорных популяций выявляются некоторые общие для них
черты.
Из морфологических характеристик общими оказываются
форма грудной клетки и относительное увеличение длинных костей
скелета, а из физиологических особенностей - усиление
насыщения крови кислородом, обусловленное морфологическими и
функциональными факторами. И то и другое может рассматриваться
в качестве физиологического приспособления к условиям
гипоксии. По-видимому, к числу приспособительных реакций можно
отнести и общее повышение основного обмена в целом,
приводящее к увеличению длины и массы тела. Этот комплекс
морфофункциональных черт в соответствии с географическим принципом в
номенклатуре можно определить как горный. Отмеченное
некоторыми авторами (Белекова, 1968а, 19686; Миррахимов, Гринш-
137
тейн, 1968; Миррахимов, 1968) падение интенсивности основного
обмена в высокогорных областях Тянь-Шаня и Памира в
результате гипофункции щитовидной железы и связанное с ним уменыпе-
ние размеров тела, вероятно, следует рассматривать не как
правило, а как исключение, объясняемое локальными причинами.
2. О ПРИРОДЕ АДАПТИВНОГО КОМПЛЕКСА
ВЫСОКОГОРЬЯ
Р. Крус-Кок (1981), осуществивший генетический анализ
высокогорных популяций мира, пришел к выводу об отсутствии
специфической генетической адаптации человека к высокогорью и
сделал заключение о том, что адаптация в этих условиях
формируется в процессе развития и носит акклиматизационный, а не
генетический характер. В то же время он обращает внимание на
исследования Г. Морпурго с сотрудниками (Morpurgo et al., 1970)
эффекта Бора в высокогорных популяциях. Более выраженный
эффект Бора в высокогорье, по мнению Р. Крус-Кока, может иметь
адаптивное значение при низком парциальном давлении
кислорода, так как способствует более быстрому транспорту кислорода к
тканям. Сами авторы этого исследования, как уже отмечалось,
рассматривают эту особенность как феномен эволюционной
адаптации в популяциях, обитающих в условиях высокогорья в
течение нескольких тысячелетий.
Такие важные физиологические черты высокогорных
популяций, как увеличение числа и размеров капилляров и артериол,
повышенная легочная вентиляция и максимальная кислородная
емкость крови, более высокий уровень гемоглобина и миоглобина,
трактуются по-разному. А.Хуртадо (Hurtado, 1954), например,
считает это адаптацией на клеточном уровне, Р.Крус-Кок -
акклиматизацией (1981), П.Бейкер полагает, что в большинстве
случаев отличия адаптивного свойства горцев от жителей равнины
являются результатом воздействия факторов окружающей среды на
протяжении всей жизни (Baker, 1968; Бейкер, 1981).
Как бы ни трактовали природу адаптивных черт
высокогорных популяций, следует признать, что эти черты необратимы.
Так, уроженцы высокогорья Перу, проведя более 10 лет в городе
на побережье, показали те же уровни кровяного давления и
холестерина, какие характерны для жителей высокогорья.
138
Лекция VI
БИОЛОГИЯ НАСЕЛЕНИЯ УМЕРЕННОГО
КЛИМАТИЧЕСКОГО ПОЯСА
Строение тела и физиологические черты.
Антропогеографические корреляции.
1. КЛИМАТИЧЕСКОЕ И ЭТНИЧЕСКОЕ РАЗНООБРАЗИЕ
ИЗУЧЕННОГО РЕГИОНА
Население умеренного климата по своим морфофункциональ
ным особенностям не раз выступало в качестве контрольной груп
пы при характеристике жителей тропиков, Арктики, пустыни,
континентальной зоны Сибири» Оно оказалось более нейтральным
по отношению к географической среде обитания, в частности в
температурному фактору, чем коренное население других
климатических областей. Отмечу, что антропологический анализ
населения умеренного пояса в связи с адаптацией его к природным
условиям жизни затруднен тем обстоятельством, что в большей своей
части оно живет в высокоцивилизованных странах. Уровень
культуры, уклад жизни, в котором мало отражаются "требования"
естественной среды, миграции и другие проявления социальной
стороны жизни, усиливают независимость морфофункциональных
черт населения этих стран от воздействия естественной среды
обитания.
В то же время для групп, живущих в условиях умеренного
климата, получена зависимость размеров скелета и уровня его
минерализации от содержания макро- и микроэлементов в почвах
и воде, а также размеров тела от высоты местности над уровнем
моря. Следовательно, проявляя нейтральность в одних чертах,
население умеренного климата оказывается далеко не
безразличным к воздействию географических факторов в других. В связи с
этим, напрашивается вопрос: что же представляет собой население
умеренного климата в морфофизиологическом отношении? Для
характеристики черт строения тела и некоторых физиологических
показателей ограничу круг привлекаемых для анализа групп
границами бывшего СССР, обращаясь к зарубежным
антропологическим данным лишь за дополнительной информацией. Территория
СССР была выбрана не только из-за большей полноты и
сравнимости антропофизиологических характеристик, но и потому, что она
139
предоставляла возможность рассмотрения специфики строения
тела и обмена веществ у групп, генетически связанных, но живу-
щих в различных условиях среды.
Так, изученное нами население Новгородской области,
относящейся к центральной климатической области Восточной Европы (о
климатической классификации см.: Алисов, Берлин, Михель, 1954),
будучи потомками древних новгородцев, генетически увязывается
с поморским населением Архангельской области, живущим в
более экстремальной климатической ситуации; коренное население
Камчатки и Командорских островов, проживающее в
дальневосточной климатической области, родственно населению
арктической зоны Северо-Восточной Азии. Горные территории Дагестана и
Таджикистана позволяют проследить особенности адаптивных
реакций близкородственных групп, живущих на разных высотах.
Наиболее суровыми условиями отличаются дальневосточная
область, где среднегодовая температура воздуха (по данным
нескольких метеостанций) не превышает 0,3°С, а количество осадков
довольно велико (511 мм в год), и северная климатическая область
Восточной Европы со среднегодовой температурой воздуха (по
данным двух метеостанций) 1,2°С и среднегодовым количеством
осадков, достигающим 500 мм. Наиболее теплый и сухой климат в
южной климатической области Средней Азии: среднегодовая
температура - 15,0°С и среднегодовое количество осадков - 196 мм.
В антропологическом отношении анализируемые группы
умеренного пояса относятся к двум расам. Русские, абхазы, народы
Дагестана и таджики - представители различных подразделений
европеоидной расы; народы Камчатки и Командорских островов -
монголоидной. Это дает возможность оценки адаптивных реакций
в связи с расовой принадлежностью в сходных и различных
условиях географической среды.
2. СТРОЕНИЕ ТЕЛА И НЕКОТОРЫЕ ОСОБЕННОСТИ
ВНУТРЕННЕЙ СРЕДЫ ОРГАНИЗМА
Морфология тела* По абсолютным значениям признаков
строения тела (табл. 17, 18) население умеренного климата
включается в интервал изменчивости этих признаков у коренных жителей
тропиков и лишь по ширине плеч и таза выходит за ее пределы,
140
Таблица 1 ?
Вариации морфофизнолотаескнх признаков у населения
умеренной климатической зоны (мужчины)
Признак
Число обследованных
Длина тела, см
Длина ноги, см
Длина руки, см
Ширина плеч, см
Ширина таза» см
Поперечный диаметр
груди, см
Пвреднезадний
диаметр груди, см
Длина ноги,% к
длине тела
Длина руки, % к
длине тела
Ширина плеч, % к
длине тела
Ширина таза, % к
длине тела
Грудной указатель
Масса тела /w/, кг
Поверхность
тела /s/, м 2
Указатель Рорера
8/W
Средняя жировая
складка.мм
Общий белок, г%
Гемоглобин, г%
Холестерин. мг%
Минерализация 3-го
луча кисти левой
руки, мг/мм 8
Систолическое
давление, мвс.рт.ст.
Диастолическое
давление, мм.рт.ст.
Атлавтнко-
хонтинента-
львая и ков-
твнентальво-
европеЙская
французы,
румыны
Olivier, 1970;
Antoniu «t
al., 1970; Ne-
craeow, 1970;
Radu, Roeete-
anti, 1972
964
164,8-171,5
91,9
71,6
37,7-39,2
28,0-28,8
29,3
20,4
55,6
43,4
22,8-23,1
16,5-17,2
69,6
60,3-65,2
1,67-1,76
1,28-1,39
2,68-2,77
t
—
—
—
—
...
—
Центральная
климатическая
область
Восточной Европы
русские
Клевцова, 1976;
Смирнова, 1977;
Алексеева,
1971; Гудкова,
1975;
Павловский, 1970
505
166,3-168,9
89,0-90,6
73,3-74,2
37,3-38,9
27,9-28,5
27,3-28,6
20,5-21,1
53,4-53,8
44,0-44,2
22,2-23,0
16,5-17,0
73,4-77,3
63,6-69,2
1,72-1,81
1,37-1,43
2,62-2,70
8,4-10,7
7,7-8,1
14,9-15,9
143,5-185,5
0,600-0,813
123,2-124,9
78,6-80,3
Северо-
кавказская
народы
Дагестана
Кярвакядк,
1978
1941
169,2-169,9
89,6-90,6
73,1-73,6
38,7-38,9
28,5-29,0
27,5-27,8
18,6-19,6
52,9-53,5
43,0-43,5
22,8-23,0
16,8-17,1
67,6-70,0
66,3-67,4
1,76-1,78
1,35-1,39
2,64-2,65
9,3-9,8
—
—
—
—
—
—
Южная
климатическая
область Средней
Азии
таджики
Смирнова, 1979;
Алексеева я
ДР., 1984
237
164,8-167,3
87,1-89,5
71,4-74,3
37,8-38,1
28,1-28,5
26,8-27,6
19,7-20,5
52,6-53,5
43,3-44,4
22,6-23,1
16,9-17,0
74,5-74,6
61,1-64,7
1,68-1,74
1,33-1,38
2,69-2,75
8,5-11,6
7,8-7,9
12,9-15,1
141,0-157,1
0,700-0,710
116,5-127,5
—
Дальневосточная
коряки,
ительмены,
эвены, алеуты
Клевцова, 1976;
Алексеева,
1977;
Алексеева,
Клевцова, 1980
112
158,5-162,1
83,1-86,4
68,5-70,2
37,4-39,1
25,9-28,2
27,4-28,2
19,8-20,7
52,4-53,3
43,1-43,7
23,6-24,1
16,2-17,4
72,3-74,3
58,7-63,3
1,63-1,69
1,40-1,58
2,87-2,78
5,9-9,5
8,3-9,2
—
161,2-216,2
0,442-0,692
115,8-122,1
—
141
Таблица 1
Вариации морфофнзнологнческих признаков у населения
умеренной климатической зоны (женщины)
Признак
Число обследованных
Длина тела» см
Длина ноги, см
Длина руки» см
Ширина плеч, см
Ширина таза, см
Поперечный диаметр груди, см
Переднезадний диаметр груди,
см
Длина ногк,% к длине тела
Длина руки, % к длине тела
Ширина плеч, % к длине тела
Ширина таза, % к длине тела
Грудной указатель
Масса тела /w/, кг
Поверхность тела /з/, м 2
Указатель Рорера
s/w
Средняя жировая складка,мм
Общий белок, г%
Гемоглобин, г%
Холестерин, мг%
Минерализация 3-го луча кисти
левой руки, мг/мм
Систолическое давление, мм.
рт.ст.
Диастолическое давление, мм.
рт.ст.
Атлантико-
континента-
льная и кон-
тинентально-
европейская
румыны
Necraaow,
1970; Radu,
Roseteanu,
1972
132
155,0-158,9
87,4
67,8-68,8
35,5-36,2
27,5-27,9
27,4
19,4
56,3
43,7-43,8
22,9-23,1
17,5-18,0
70,8
53,7-54,0
1,54-1.55
1,41-1,43
2,87
-
-
•
¦ iii
Центральная
климатическая область
Восточной
Европы
русские
Клевцова,
1976;
Смирнова, 1977;
Алексеева,
1971; Гудко-
ва, 1976;
Павловский,
1970
587
156,0-157,4
83,0-84,3
67,7-69,2
34,9-35,8
28,2-28,9
24,8-25,7
18,2-19,2
53,1-53,4
43,4-44,1
22,2-22,8
18,0-18,5
72,7-75,1
60,7-65,9
1,64-1,72
1,55-1,70
2,61-2,70
14,2-21,1
7,6-8,1
13,0-13,7
146,0-177,6
0,600-0,853
119,9-123,4
76,7-82,2
Южная
климатическая область
Средней Азии
таджики
Смирнова,
1979;
Алексеева и
др., 1984
178
154,4-155,3
81,4-82,2
65,8-67,4
33,6-34,4
28,4-28,5
24,0-24,7
16,4-17,8
52,4-53,0
42,4-43,6
21,7-22,4
17,8-18,4
68,3-73,3
50,8-53,8
1,49-1,54
1,38-1,44
2,42-2,93
11,8-15,6
7,7-7,9
10,4-13,2
139,2-153,7
0,440-0,630
113,7-120,0
66,9-80,7
Дальневосточная
коряки,
ительмены,
эвены, алеуты
Клевцова,
197?;
Алексеева,
1977,-
Алексеева,
Клевцова,
1980
140
149,3-152,4
77,3-80,4
63,3-65,0
34,8-35,6
26,6-28,7
25,0-25,4
18,0-18,6
51,8-52,8
42,0-43,3
23,2-23,4
17,7-18,9
72,0-73,8
53,2-58,9
1,52-1,59
1,50-1.76
2,70-2,86
13,3-19,0
8,3-8,8
160,4-223,0
0,442-0,692
117,7-121,5
74,9-78,1
142
характеризуясь большими широтными размерами. Таким
образом, первое, с чем мы сталкиваемся при характеристике морфофи-
зиологических черт населения умеренного пояса, - это сужение
диапазона вариабельности морфологических признаков. Эта
особенность усиливается, как мы помним, в арктической зоне.
Второе, на что следует обратить внимание, - большая массивность,
проявляющаяся не только в увеличении массы и весоростовогс
указателя (Рорера), но и в относительном укорочении нижних
конечностей, а также увеличении обхватных размеров тела.
Подобный морфологический комплекс иногда в усиленной форме
оказывается типичным для арктических популяций, но со значительнс
меньшим диапазоном изменчивости как абсолютных, так и
относительных размеров тела.
Теплопродукция, вычисленная по формуле Робертса на
основании массы, длины тела и температуры окружающей среды, у
населения умеренного пояса выше, чем в тропиках, и ниже, чем в
Арктике. Если в тропическом поясе уровень теплопродукции
колеблется примерно от 1300 до 1700 ккал, а в Арктике - около
3000 ккал, то в умеренном поясе он варьирует в пределах 1600-
1850 ккал.
Примечательно, что в условиях умеренного климата
теплопродукция у населения повышается с увеличением экстремальности
среды: наиболее низкие величины наблюдаются в южной
климатической области Средней Азии (1602-1609 ккал), наиболее
высокие - в северной климатической области Восточной Евцопы (1817
ккал) и в дальневосточной климатической области, особенно на
Командорских островах (1856 ккал).
Отмечая как общую тенденцию увеличение весоростового
указателя и обхватных размеров тела на территории ойкумены от
тропического пояса через умеренный к Арктике, естественно задать
вопрос, существует ли эта тенденция и в частных случаях, когда
мы имеем дело с генетически близкими, но живущими в разных
климатических условиях группами. К сожалению, пока
материалов для ответа на этот вопрос мало. Однако в тех случаях, которые
сейчас можно наблюдать, эта тенденция прослеживается. Так, при
сопоставлении морфологического комплекса коряков и
ительменов, живущих на Камчатке и являющихся представителями
арктической расы, с чукчами и эскимосами оказывается, что
последние отличаются от коряков и ительменов явным укрупнением
143
72 73 74 75
[рудной указатель
Z?50
Я 60 70
Вес тела кг
а
6
Рис. 32. Сравнительный анализ размеров тела представителей аркти
ческой расы, живущих в арктической и умеренной климатических зонах. Муж
чины:
а - чукчи; б - коряки
144
72 7J 7к 75
JO Грудной указатель
4150
±
SO
эй 60 70
Л Вес тела, кг
По
Рис, 33. Сравнительный анализ размеров тела представителей
восточноевропейской расы, живущих в субарктической и умеренной зонах. Жнещины
а - руоокие Архангельской обл.; б - Новгородской обл.
145
размеров: более массивной грудной клеткой, большей массой, со
ответственно более высоким весоростовым указателем и относи
тельно большим развитием жировой массы (рис. 32).
При сопоставлении русских Новгородской и Архангельской
областей подобного рода тенденция отчетливо видна в женсков
выборке. Архангельские женщины значительно массивнее
новгородских (рис. 33). Что касается архангельских мужчин, то
выборка, которую нам удалось обследовать, по всей вероятности, не
может достоверно характеризовать мужское население северной
части Архангельской области. Дело в том, что широко
развернувшееся в последнее время городское строительство на севере
европейской части России усилило отток населения из деревень,и б
первую очередь это коснулось мужчин. Исследованные нами в
ряде деревень Ластольского сельсовета Архангельской области
мужчины оказались менее крупными, чем новгородцы, но форма
грудной клетки у них более цилиндрическая, что характерно для
жителей северных областей. Более того, у населения
Архангельской обл. выше уровень теплопродукции.
Можно обратиться еще к одному сопоставлению близких в
расовом отношении групп, живущих в разных климатических
условиях (Волков-Дубровин, 1974). Речь идет о таджиках Ферганской
долины и предгорий Памира - жителях южной климатической
области Средней Азии (умеренный пояс) и населении штата Харь-
яна в Индии (тропический пояс). Таджики по сравнению с
индийскими группами отличаются всеми морфологическими
особенностями, которые характерны для жителей более высоких широт: у
них выше весоростовой указатель, крупнее грудная клетка,
заслуживает внимания ускорение расхода кислорода в организме
таджиков, что свидетельствует об интенсификации обменных
процессов в умеренной климатической зоне.
Некоторые физиологические характеристики
Процессы метаболизма в умеренном климате по своей
интенсивности занимают промежуточное положение по сравнению с
таковыми в тропической и арктической зонах. Это обнаруживается
как при анализе скорости кислородных трат, определенных окси-
гемометрическим путем, так и при вычислении уровня
теплопродукции на основании длины и массы тела и температуры
окружающей среды.
146
В то же время другие физиологические признаки (уровень
холестерина и белков в сыворотке крови, соотношения «белковых
фракций, уровень минерализации скелета) строгой
закономерности в территориальной изменчивости в пределах умеренного пояса
не показывают (см. табл. 17,18). Согласно сводке данных о
географин холестерина на территории ойкумены, составленной автором
(Алексеева, 1971), для населения умеренного пояса характерна
высокая изменчивость этого признака, но в пределах средних в
вышесредних величин. Низкие уровни холестерина, типичные для
населения тропических широт, в условиях умеренного климата не
встречаются, так же как не встречаются и низкие величины аль-
бумин-глобулинового соотношения, которое в тропиках не
редкость (Гудкова, 1975).
В географической изменчивости содержания минералов в
скелете, по-видимому, и не следует искать строгой связи с
климатическими факторами, поскольку уровень его определяется в
значительной мере геохимической ситуацией. Правда, при анализе мор-
фофункциональных черт коренных жителей Арктики
обнаруживается явное увеличение содержания фосфорно-кальциевых солей
в их скелете по сравнению с населением умеренного пояса
(исключения очень редки). Эта закономерность проявляется и в
некоторых частных сопоставлениях: у камчатских групп уровень
минерализации ниже, чем у чукотских; у новгородцев он ниже, чем >
русских Архангельской области.
Сопоставление таджиков и населения Индии дает иную
картину: у таджиков понижено содержание фосфорно-кальциевых
солей в скелете. Не исключено, что это связано с нарушением
геохимического баланса на занимаемой ими территории. Речь идет і
данном случае о населении долины р. Исфары, стронциевой
биогеохимической провинции Таджикистана. В женской части
популяции кишлака Ворух, расположенного в верховьях Исфары на
высоте 1300-1800 м над уровнем моря, наблюдается системный осте-
опороз, рентгенологическая картина которого заключается в
истончении кортикального слоя, увеличении канала трубчатых кос
тей и характерном прозрачном рисунке кости. По-видимому, этс
отклонение вызвано увеличением содержания стронция в среде
обитания и связанным с этим нарушением стронций-кальциевогс
соотношения (Алексеева и др., 1984). При этом снижается мине
ральный уровень скелета и повышается частота скелетных дефор
147
маций. Описанное явление нетипично для населения других киш
лаков Исфаринской долины, однако минеральный дисбаланс на
блюдается и там.
В заключение следует сказать об уровне изменчивости физио
логических признаков. В характере межгрупповой изменчивости
физиологических признаков проявляется закономерность, типич
ная для морфологических признаков: уровень вариабельности
сокращается от тропического пояса через умеренный к
арктическому. Межгрупповая изменчивость уровня холестерина (с учетом
возможных методических расхождений) в тропиках почти вдвое
превышает ее величину в условиях умеренного пояса, а последняя
на 20-25% выше, чем в Арктике. Межгрупповая вариабельность
содержания общего белка в сыворотке крови в тропиках вдвое
выше, чем в умеренном климате, а в Арктике она уменьшается
почти втрое по сравнению с умеренным поясом.
Наконец, вариабельность между грутіами по уровню
минерализации в умеренной зоне более чем в пять раз превышает
таковую в Арктике. К сожалению, в нашем распоряжении нет данных
об изменчивости минерализации скелета тропических популяций
(за исключением нескольких индийских групп, территориально
очень близких).
3. РЕГИОНАЛЬНЫЕ АНТРОПОГЕОГРАФИЧЕСКИЕ СВЯЗИ
Зависимость уровня минерализации скелета человека от
содержания макро- и микроэлементов в почве и воде была впервые
описана у популяций умеренного пояса (Павловский, 1970).
Пониженный уровень минерализации скелета был обнаружен в регионе с
недостатком фосфора, кальция и некоторых других минералов в
среде (Ярославская обл.)» а также в регионе с нарушением
соотношения кальция и стронция (Северный Таджикистан).
Зависимость некоторых черт строения скелета человека от
содержания костеобразующих элементов в среде обитания также
впервые была показана на группах умеренного пояса (Alexeeva,
1968). В районах с недостатком фосфора и кальция отмечается
уменьшение длины тела, увеличение широтных размеров скелета
и относительное укорочение черепа (брахикефалия). В районах с
более благоприятной геохимической ситуацией больше длина те-
148
ла, скелет грацильнее, череп относительно удлинен
(долихокефалия).
Немаловажное значение имеют и наблюдаемые в условиях
умеренной зоны сдвиги морфофункционального статуса населения
горной местности по сравнению с равнинной. Уже говорилось о
том, что население Дагестана в своих морфологических чертах
отражает общую тенденцию высокогорных популяций - укрупнение
размеров, относительное увеличение длины ног и объема грудной
клетки (Кириакиди, 19736). Подобное направление изменений
наблюдается и в северной части Таджикистана. Таджики предгорий
Памира, живущие на высоте всего лишь 837 м над уровнем моря,
по длине тела обгоняют равнинных таджиков, они имеют также
более удлиненные пропорции тела. На высоте 1300-1800 м эти
различия уже более четкие.
Таким образом, население умеренного пояса не остается
индифферентным по отношению к среде обитания. Его морфофункцио-
нальный облик по ряду признаков занимает промежуточное
положение между морфофункциональньхми комплексами тропических
аборигенов и жителями Арктики. То же можно сказать и по
поводу межгрупповой изменчивости морфологических и некоторых
физиологических черт. Более того, население умеренного пояса
проявляет морфофункциональную реактивность и по отношению
к региональным географическим особенностям, в частности к
геохимическому статусу и высотным характеристикам местности.
Лекция VII
БИОЛОГИЯ ЖИТЕЛЕЙ КОНТИНЕНТАЛЬНОЙ ЗОНЫ
СЕВЕРНОЙ АЗИИ
Морфофизиологические особенности населения.
Антропоэкологическое разнообразие Центральной Азии.
1. СПЕЦИФИКА ГЕОГРАФИЧЕСКИХ УСЛОВИЙ
ВНУТРЕННИХ РАЙОНОВ СЕВЕРНОЙ АЗИИ
Континентальная зона Северной Азии - одна из
экстремальных зон обитания человека. Здесь располагается район "зимнегс
149
полюса холода", годовая амплитуда температур достигает наиболь
шей на земном шаре величины - 65°С, поэтому вполне естествеі
интерес к особенностям приспособительных реакций в человечес
ких популяциях, обитающих в этой зоне.
Изученный нами регион, за исключением горного района Яку
тии, располагается в южной части Сибири, наибольшее число об
следованных групп коренного населения проживает на территорш
Алтае-Саянского нагорья (рис. 34). Эта территория поражает раз
нообразием ландшафтных характеристик при выраженном клима
тическом сходстве (табл. 19). Население в основном занимается
скотоводством. На равнинных территориях подсобную роль игра
ет земледелие (Андрианов, Чебоксаров, 1972). Сходство в типехо
зяйства нивелирует различия и в рационе питания; диета в основ
ном белково-углеводная.
В антропологическом отношении якуты, буряты и тувинць
относятся к центральноазиатскому типу, алтайцы и казахи -і
южносибирскому, хакасы и шорцы - к уральскому. Все дискусси
онные вопросы расовой классификации Сибири подробно обсужде
ны в монографии В.П. Алексеева и И.И. Гохмана (1984), посвя
щенной антропологии бывшей советской Азии, и здесь нет необхо
димости касаться их. Отмечу, что различия между этими типаш
в основном сводятся к удельному весу европеоидного компонент!
в их составе. Минимум европеоидных черт обнаружен у предста
вителей центральноазиатского типа, максимум - у уральскогс
типа. Тем не менее в системе расовой классификации все они за
нимают место среди монголоидов. Европеоидный компонент т
территории степных районов Южной Сибири восходит к энеоли
ту, распространение монголоидных черт связано с эпохой монголь
ского нашествия. На границе с Монголией и в лесной таежной ча
сти Алтае-Саянского нагорья монголоидный компонент имеет бо
лее древнее происхождение.
Якуты в своем генезисе связаны с территорией Прибайкалья
В зону теперешнего пребывания они продвинулись в IX-X вв. I
процессе исторического развития якуты включили в свой состаі
эвенков, представителей байкальского антропологического типа
Есть основание думать, что байкальский антропологический тип
представителями которого являются эвенки, составлял основу древ
нейшего населения Сибири, сформировавшегося в таежной зоне і
востоку от Енисея и впоследствии проявившегося в антропологи
150
®v>
Новокузнецк о
^
Гормо-Алтайсн \
О е.
3
if
\
ч
*5
ф
*ч
?
а
?
Г"
^ ® 11
®х Г
п «/д н «УЛ,
( Саянский хр-
-—^^<5»г
почаінмй TeiiHj-
С
Оле
, *
tV
*
Ныэыл^
*¦***••*.,,
0*>
Рис. 34. Схема локализации обследованных популяций коренного
населения Алтае-Саянскаго нагорья:
1 - телеугы» Челухоево, Беловский р-н Кемеровской обл.; 2 - алтай-кижи, Яконур, Мен-
дур-Соккон, Усть-Канский р-н Горно-Алтайского АО; 3 - казахи чуйские, Кош-Агач, Мухор-
Тархаты, Кош-Агачский р-н Горно-Алтайского АО; 4 - теленгиты, Мухор-Тархаты, Кощ-Агачс-
кий р-н Горно-Алтайской АО; 5 - шорцы гоные, Усть-Анзасский, Усть-Кабырвинский, Чилису-
Анаасский сельсоветы Таштагольского р-на Кемеровской обл.; 6 - шорцы абаканские, Матур,
Таштьшский р-н Хакассии; 7 - хакасы-сагайцы, Верхний Аскиз, Аскизский р-н Хакасски; 8 -
хакасы-кызыльцы» Кирово, Ширинский р-н Хакассии; 9 - хакасы-качинцы, Кокгарово, Орджо-
никядзевский р-н, Малый Спирин, Ширинский р-н Хакассии; 10 - Мугур-Аксы, Моген-Бурен,
Могун-Тайгинский р-н; 11 - Чадан, Шеми, Хайракан, Сют-Холь, Дзунхемчикский р-н; 12 -
Эрэик, Нарын» Тбре-Холь, Эрзинский р-н; 13 - Тоора-Хем» Ий, Адыр-Кежиг, Тоджинский р-н
Республики Тува.
161
3
I
Ю r-t
to ©
f 00 W
b- b- b-
00
CO
CO
b-
00 CM iH Ю Ю
CO b- b- © ©
00
Ю
CM
О N
im ем
CM
CO Ю
о ь-
CO b-
iO CM
CM гЧ
чр СО
Ю CN
CM гЧ
4* 00
01
©
со
СО
со со
со
01 tJ< Ю
СЛ © О)
со
см
со
со
ь-
ео
СО
со
ь-
со ю ь- о о
со со со ь* ся
ем
н N М
л
V
^ *°* °1
іЧ Ы СМ*
ь^
ееГ
о ¦*
°Ч °і °Ч. °і *і Я °Ч Я
ец* csT ем" см* со" ін" гн* рн"
0>
ь-
N СО b
оГ оГ сі
ю ^« *о
С0С*О©Ч»Ю'«<Ь-СЙЧ«'«<
см
со
СО С» Ю СО
М* СО СО СО
со
со ©
со со
О 'Ч* М«
СО ^ М*
с*
2- §4 >»
о Я S
111 я
со
•ъ
со
2
ю
со
ю
*
ю
00
см
to
•ъ
ю
<«?•
і-Ц
С*
¦ъ
О
«Э
о*
О
N
О
ю » «э Я Я
©2*22
1*
о>
ел
со
N Ь- Д
О
ю
со
О О Cft 00 О»
N Н ^ «1 ^
Ф ^f ^ ^ ^
00 «Э g S g
2 S 2 2 2
см
с* о*
«3
к
о
???1
152
ческом облике многих народностей, занимавших внутренние про
странства Сибири (Рычков, 19696).
Эвенкийский субстрат имеется и в бурятском населении Баргу
зинской котловины (Золотарева, 1960), и в тувинцах-тоджинцая
(Алексеев, Гохман, 1984).
Русское население Баргузинской котловины представлено
потомками русских из северных районов европейской России,
осваивавших Сибирь в XVII-XVIII вв. (Кабузан, Троицкий, 1966).
Итак, за исключением русской группы, все исследованные і
континентальной зоне Сибири популяции коренного населения
имеют много общих черт в генофонде, что облегчает задачу их
экологической оценки в столь разнообразном географическом
регионе, как Алтае-Саянское нагорье и прилежащие к нему
территории.
2. СТРОЕНИЕ ТЕЛА
И ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НАСЕЛЕНИЯ
Поскольку данные о населении континентальной зоны и
группах, которые могут быть привлечены для сопоставления с ним.
собраны одними и теми же исследователями по единой большое
программе, мы имели возможность охарактеризовать его морфо-
функциональный комплекс более полно по сравнению с
населением какой-либо другой зоны, исключая Арктику (табл. 20, 21).
Прежде всего обратимся к морфологическим характеристикам
бурят и русских Баргузинской котловины (Алексеева и др., 1971).
Русское население в целом имеет большие длину, массу тела, об-
хваты груди, плеча, голени, более массивный скелет. У русских
больше, чем у бурят, развит мускульный компонент тела и
соответственно выше процент мускульного типа телосложения. Пс
строению тела русские Баргузинской котловины - типичные пред-
ставители русских Сибири. При сопоставлении с русским
населением Восточно-Европейской равнины баргузинцы обнаружили
наибольшую близость с новгородцами.
У бурят при относительно и абсолютно менее развитых по
сравнению с русскими мускульном и скелетном компонентах тела
более, чем у русских, развит жировой компонент. В частности,
встречаемость пикнического типа у мужчин-бурят составляет 24% вы
153
ОДЯЭ?ЛЬ. TXXBVBfi
ньшлн?ігвх
ижия-^тзигу
ічіЛ?іг?^
eiradoj нЬкЗощ
MlMffMeHH-HOFHBX
нйдвово-ик>вя«х
нішиыія-ітовявх
нйниж^ох-иіінияЛх
гйшия^х
MiXjry
K^rBHijBgug ?ияооЛ^
HxvdXg
us
00
О)
a
ч
S s
§сэ
b-
01
to
«а-
иэ
з
©4
o*
ОЭ
со
иэ*
b-
co
00
CO
00
04
00*
04
О»
s
иЭ
3
©4
fc-
b-
co
CO
00
04
OS
04
CO
со"
CO
©4
s
00
s
о
9
b»
*4
to
09
04
ш §
CO
s
us
b-
b-
00
04
00
01
04
US
00
CO
b-
b-
co
CO
CO*
CI
<0
01
3
b-
co
00
4<
s
©4
10
tH
3
.4
04
165
CO
159
"*
s
fH
CO
CO
0
3
CO
©*
CO
иэ
CO
b-
©4
09
fc-
us
CO
b-
U3
0
t-
1Ч
38,
03
fees
<M
00
CO
00
CO
CO
иэ
b-
©4
со
t-
CN
iH
00
©4
О
CO
©4
00
b-
04
И
00
04
CO
00
04
05
*
C9
04
10
0)
H
©
0)
H
e
0
H
a
0)
H
Cft
©4
"1
CO*
CO
«o
иэ
CO
CM
b-
b-
0>
CO*
©4
иэ
*
b-
04
04
u>
©4
CO
s
©4
b-
ъ*
CO
CO
b"
©4
b*
b-
04
иэ
b-
04
CO
иэ
b-
04
00
CO
CO
04
00
00*
04
04
CO
CO
иэ*
CO
00
b-
b-
co
00
04
00
04
о
C4
154
us
•к
05
US
CO
US
3
4<
еч
4
ся
О
b-
b-
09
b-
eo
о
ft
CD
UO
b-
to
b-
co
tN
CO
e*
uo
el
00
00*
4
us
to
i
со
o
uo
uo
CO
U9
00
со
us
CO
3
04
CO
©4
to*
CD
О
b-
tO
CO*
CD
3
CO
4
uo
ft
CO
fc-
о
00*
w
со
CD
00
uo
00
00*
g
US
*
us
94
b-
3
4
ъ*
to
s
C4
coe
00*
s
b-
s
°1
©Г
ем
CO
b-
»
US
сол
U9*
CO
ея
<©*
b-
W0
s
Ю
CO
uo
00
«I
ея
oo
oo*
CM
CO
00
to
eoe
ОЛ
us
uo
5
00
4
to*
4
о
2
b-
CO
CO
b-
fc-
ея
b^
to*
b-
ao^
b*
eofc
ея
ooe
©*
t>4
US
CO
3
CO
©4
4*
to
00
4
CO*
to
3
CO
to
СЯ
b*
s
CO
s
©4
00*
b-
b-
co
to
"*
^
00
©4
to
o*
b-
e>
s
US
09
to
b;
c*
us
*
00
b*
¦**
4
со*
СЯ
s
о
CO
©4
00
fH
US
¦*
^
CO
©4
CO*
°*L
o"
b-
4
to*
uo
§
CO
09
CO
CO
©4
• CO*
*¦ oo*
CO
•>
b-
CO
to
•4*
<o
ея
09
US
О
CO
us
US
uoh
t-
3
4
00*
4<
23,0
«
4
of
uo
to
.4
to*
.4
to
to
»-4
CO.
b-
1-t
¦h
b-
00
CO*
b-
4
0>
US
o*
to
8
uo
to
1-Г
1
5
b-
09
2,77
b-
*
to
to
00*
09
Cft
04
00
о
00*
t4
uo
00*
b*
CO
¦l
uo
¦
*
uo
о
CO
4
uo*
fc-
1-1
oo
г
1-C
0,44
3
о
s
о
119,0
СЦ
CO*
C4
09
8
CO
8'
to
«
of
us
СЯ
3"
00
О
ь*
tN
b-
г
b-
o
b^
c*
8.
00*
о
uo
o\
09
to
о Я
со
В
я
-г
і s.
IS &
X я S 3 Б «5S,
156
a-
g
?ИЯО9&1. ЮСЗВВЦ
ташн?ігвх
ижия-#ішгу
ніЛвіг?х
вмяйоа Khdorjl
гйгаігнеічя-новявх
пЬоезво-зчэвявх
тйтшозя-гатяех
гйшимсНоа-кйнивЛд,
MtDUie^i
міЛя#
ю
со
а
а>
в> о _.
4 Is
« в
ь-
00
со
иэ*
иэ
°1
«*
иэ
ь-
00
ем*
00
X
b*
СО
со
8
НО
со
о
со
со
со*
СМ
Cft
00*
ем
О
СМ
to
иэ
CM
17,8
?
00
H
CO
CO
Ю
Cti
b-
TO
CO
иэ
CM
b-
иэ
CM
s
fc-
CO
иэ
CO
b-
ем
иэ
CO
00
о
иэ
СП
b-
a»
<o
иэ
CO
OS
*
00
CM
4
CO*
cm
eft
ем
иэ
со
иэ
s
CO
CO
иэ
b*
CM
C7>
CM
иэ
ем
еч
to
иэ
s
b*^
CD
4*
CO
CO
CM
CO
иэ"
CM
CO
00
00
H
i4
CO
CO
CO
CO
CO
b-
150
«M
CM
иэ
00
153
03
CO
CO
b-
4*
аз
ь-
иэ
80,
W
со
b-
0
CO
CO
b-
s
b-
g
b-
s
00
¦*
CO
O)
CO
00
41
CO
CO
4*
CO
b-
b-
CM
CO
00
CM
b-
ao
01
b-
co
CM
-*•
иэ
CM
b-
иэ
ем
со
CO
см
0
см
и»
00
И
«
00
н
И
?
00
н
е*
со
1-1
со
*
Ь-
КЧ1ЛЗЯ1?В913? ?ИЯООЛ^
гшпіАд
is
а
ем
00_
СО*
иэ
со*
00
8
00
со
см
со*
см
о>
иэ
ем
00
со
иэ
со
СО
ь*
со
ев
СО
ем
иэ
ем
0>
*
со
н
И
5
&
?
I
as
156
с~
<N
U3
fc-
CO
Tf
СЧ
СЧ
СЧ
vo
00
f-4
CO
.-4
fr-
00
CO
«3
«H
«0
—«
CO
Ю
rH
4<
fc-
O)
со
CO
гЧ
U3
rH
CO
-4<
-^
i-4
oT
CO
о
b-
Ю
о
«о*
из
00
cn
из
н
ИЗ
н
о*
ю
и
со
из
сч
еч
ю
СЧ
сч
из
гЧ
04
из
00
гЧ
из
со
гН
из
feci
из
-*
еч
ИЗ
»ч
о*
ю
U3
со
^
со
со
ч*
00
со
^
00
^
чГ
?О
со
ч*
00
со
ч*
со
со
^
о
ч*
-*•
СП
со
ч*
^
ч*
ч*
о
со
"*
о
ч*
ч*
тГ
сч
ем
из
еч
еч
о
со
СЧ
о
со
©а
t-
сч
сч
а
СЧ
еч
ь-
сч
сч
оз
СЧ
СЧ
ь-
сч
СЧ
ь-
сч
СЧ
сч
СЧ
О)
«3
сч
сч
СЧ
со
»ч
со
00
гЧ
ъ-
00
гЧ
о
00
»-ч
ч*
00
іН
(0
со
гЧ
со
00
гЧ
СЧ
со
1-*
сч
00
«¦ч
ч*
00
гЧ
о
00
i-t
СО
00
»-ч
00
гЧ
с-
о
сч
ь-
иэ
с-
о
гЧ
с-
о
со
с-
из
сч
t-.
о
А
Ф
00
из
t-
ю
гЧ
t-
со
о
с-
сч
сч
ь-
fc-
«
СО
о
сч
СО
1-1
ь-
из
СП
•н
со
а
о
из
W
СО
U3
со
с»
из
со
feus
гЧ
*Ч
из
с-
ю
со
¦"ф
и»
О)
о
из
о
со
из
со
СО
гЧ
00
из
гЧ
из
со
1—1
t-
«1
гЧ
со
иэ
.-Ч
гЧ
со
гЧ
0)
из
гЧ
с-
ч*
*ч
4f
из
і-Ч
ч*
из
гЧ
сч
со
гЧ
се
ю
*ч
о
СО
гЧ
гЧ
со
гЧ
О)
со
1-f
о
из
гЧ
из
со
гЧ
05
со
ы
с-
из
гЧ
гЧ
to
*Ч
со
ю
гЧ
4*
из
гЧ
ю
из
<ч
сч
из
*ч
00
СО
сч
fc-
t-
сч
с-
СО
CN
Оі
00
сч
СО
fc-
сч
о
с-
сч
ь-
с-
сч
§8
сч
*ч
со
сч
гЧ
со
сч
о
с-
сч
00
t-
сч
с»
ч*
.Н
t-
из
гЧ
из
а»
СО
»ч
СО
со
тЧ
о
со
ы
о
из
1-і
о
со
гЧ
со
из
гЧ
0)
из
гЧ
СО
со
гН
из
из
іЧ
сч
иэ
гЧ
гЧ
из
со
со
Оі
t-
3
со
СО
со
со
о
03
ь-
S
00
с-
Оі
с-
00
сч
со
из
из
СО
ео
СО
со
t-
00
fc-
§
со
о
*
г4
СО
сч
гЧ
0>
сч
гЧ
оэ
?
?ч
со
сч
•н
CD
гЧ
А
ея
гЧ
I
со
из
гЧ
ь-
сч
гЧ
о
со
гЧ
»ч
из
00
со
со
И
гЧ
03
сч
сч
05
•
из
гЧ
со
гЧ
сч
из
6
сч
со
со
со
тЧ
со
»Ч
со
СО
гЧ
t-
со
со
гЧ
00
из
00
гЧ
470
о
гЧ
со
о
1
о
і-Ч
о
1
СО
из
о
СО
^f
о
¦*
4f
о
со
¦*
о
г
о
4J*
СО
о
еч
00
гЧ
гЧ
¦ч»
со
гЧ
рЧ
¦^
00
гЧ
гЧ
оз
о
еч
t-
^ч
гЧ
ь-
со
гЧ
t-
fc-
гЧ
гЧ
со
из
гЧ
гЧ
ч*<
»-ч
^ч
^
СО
гЧ
гЧ
03
03
гЧ
гЧ
из
гЧ
гЧ
со
ь-
ъ-
со
¦•с*
t-
fr-
05
t-
ь-
^"
t-
СО
с»
с-
сч
со
t-
со
из
t-
09
с-
t-
со
и-
с-
¦^
со
ь-
t-
со
ь-
fc-
t-
t-
ft
Си
s
№
X
ft
я
о
В
03
0)
I"
й
t
о
ЕГ
8
t
а
s
*S
ч
р-
о
Гем
№
tti
?
С)
Ф
Хол
S Е^
О
О
и
о
ВС
S
ч
О .
а
157
борки, а у русских только 9% - У женщин обеих национальности
разница в жироотложении не столь велика, хотя процент пикни
ческого типа у буряток все лее больше.
Определяя по соматическим характеристикам положение бу
рят и якутов среди коренных народностей Сибири на основаниі
большого набора морфологических признаков, характеризующие
строение тела, Н.И. Клевцова (1976а) показала, во-первых, ихзва
чительную близость друг к другу и, во-вторых, специфику по срав
нению с этническими группами Чукотки и Камчатки, относящи
мися к арктическому типу и связанными между собой генетичес
ки. Из камчатских групп только эвены обнаруживают сходство
бурятами и якутами, что не удивительно, так как эвены Камчат
ки связаны в своем происхождении с тунгусоязычными народам]
внутренних районов* Сибири, а в теперешнем месте своего обита
ния они сравнительно недавние пришельцы (40-е годы XIX в.
(Народы Сибири, 1956).
У сибирских монголоидов, изученных к настоящему времени
выявляются два определенных типа строения тела: 1) у арктичес
ких аборигенов северо-востока Азии - мезоморфный по пропорци
ям тела, с сильным развитием мускулатуры, широкоплечий, с ци
линдрической грудной клеткой и пониженным жироотлоясе
нием; 2) у коренных жителей континентальной зоны относитель
ная коротконогость и длиннорукость (в сравнении с население*
Арктики), увеличенный жировой компонент тела и более плоска:
грудная клетка (краткая морфологическая характеристика населе
ния континентальной зоны приводится в табл. 20, 21, арктичес
кой — в табл. 22). Визуальная оценка телосложения коренны:
жителей внутренних и северо-восточных районов Сибири дает чет
кое их разграничение по проценту встречаемости различных ти
нов (рис. 35).
Физиологическая характеристика населения континентальноі
зоны Сибири также имеет свою специфику (см. табл. 20, 21).
Теплопродукция, вычисленная с помощью формулы Д. Робер
тса (Roberts, 1952), в основу которой положены масса и длині
тела, а также температура окружающей среды, у населения кон
тинентальной зоны Сибири выше, чем у коренных жителей уме
ренных районов. Исследования, проведенные Институтом биоло
гических проблем Севера (Магадан) в наслегах близ Якутска, по
казали, что у якутов основной обмен не намного, но достоверні
168
% SbiBojlitu
70
60\-
SO
40
30
20 -
W
¦//,
1 1
Si
%
^
s-
\
vm°
m>
¦,W^
Ш'
чукчи эспимосы
Арктическая зона
якутьі буряты
Континентальная зона
Рис, 35. Соотношение основных конституционных типов и коренных
жителей Арктики и континентальной зоны Сибири:
а - грудкой тип; б - мускульный тип; в - брьшной тип.
выше, чем у живущих там же лиц некоренной национальности
(Веселухин, 1977).
Определение скорости поглощения кислорода методом оксиге-
мометрии не дало положительных результатов в отношении
дифференциации по этому признаку жителей континентальных
районов Сибири и умеренного климата. Правда, нужно заметить, чтс
оксигемометрический метод определения газообмена, вполне
эффективный при изучении возрастной и конституциональной
изменчивости скорости поглощения кислорода (Волков-Дубровин,
169
1966, 1970), оказался малоинформативным для суждений о геогра
фической изменчивости этого признака. Лишь при значительно]
климатической дифференциации территории мы получаем поло
жительный эффект. При обследовании коренного населения ив
дийского штата Харльяна, проведенном В.П. Волковым-Дуброви
ным (1974), было отмечено резкое понижение процессов газообме
на в условиях тропиков. Время, в течение которого кислород, со
держащийся в крови, расходуется тканями без дополнительной
доступа воздуха в организм (опыты с задержкой дыхания), у муж
ского населения Индии составляет 20,4-25,2 с, в то время как]
разнонационального населения континентальной зоны Сибири і
районов умеренного климата длительность фазы поглощения 0
колеблется от 16,2 до 17,3 с. При пересчете на массу тела разниці
еще отчетливее: в первом случае это величина порядка 0,37-0,50
во втором - 0,25-0,28.
Возвращаясь вновь к характеристикам окислительно-восстано
вительных процессов у населения континентальной Сибири, отме
чу, что здесь больше приходится опираться на данные, получен
ные экспедицией Института биологических проблем Севера, гді
основной обмен изучался методом Холдена, а расчеты теплопро
дукции производились по формуле Робертса. По этим показателя!
уровень метаболизма у населения континентальной зоны Сибирі
выше, чем у жителей умеренного климата, и очень сходен с тем
который наблюдается у коренных жителей Арктики. На повыше
ние интенсивности окислительно-восстановительных процессоі
отчасти указывает довольно высокий уровень гемоглобина и эрит
роцитов, найденный у коренных жителей континентальной Сиби
ри (Алексеева и др., 1971; Веселухин, 1977).
Уровень белков и липидов крови у населения континентальное
зоны Сибири соответствует тому, который характерен больше дои
жителей умеренного климата, чем Крайнего Севера (особенно эт(
относится к холестерину). Содержание общего белка у коренныз
народностей континентальной Сибири выше, чем у русских старо
жилов (у последних он такой же, как у русских европейской час
ти бывшего СССР). Русское население Баргузинской котловины і
целом имеет несколько большие количества альбуминов и соответ
ственно меньшие количества гамма-глобулинов по сравнению с бу
рятами: у русских мужчин альбумины составляют 70,1%, гамма
глобулины - 12,0, у мужчин-бурят - 69,3 и 14,3% соответственно
і?о
У русских женщин эти величины равны 67,9 и 14,4%, у буряток
-65,3 и 15,7%.
Повышение гамма-глобулиновой фракции отмечено не только
у коренных жителей континентальной Сибири, но и у русских
старожилов, что отличает последних от русских европейской части
бывшего СССР.
Представление о специфике белкового обмена у коренных
жителей континентальной зоны Сибири по сравнению с обитателями
Крайнего Севера можно составить и на основании данных медико-
биологического обследования, проведенного экспедицией
Института биологических проблем Севера. У якутов континентальной
зоны, как правило, выше, чем у заполярных якутов, содержание
гемоглобина и эритроцитов, меньше белка и остаточного азота (Be-
селухин, 1977). Понижение остаточного азота, по мнению этого
автора, отражает более низкое содержание белков в рационе
питания континентальных якутов по сравнению с заполярными. Это?
факт вполне согласуется с результатами исследования рационоі
питания жителей континентальной и арктической зон Сибири.
Для характеристики специфики обменных процессов у
населения континентальной зоны Сибири могут быть приведены данные
о минерализации скелета - признаке, как уже говорилось в
предшествующем изложении, обладающем большой географической
вариабельностью. Уровень минерализации, изученный рентгено-
фотометрическим путем на 2-й и 3-й фалангах среднего пальце
кисти, у жителей континентальной зоны Сибири (особенно у
тувинцев) значительно ниже, чем в контрольной группе, у
арктических аборигенов и других, изученных нами ранее групп (рис. 36).
Такое же понижение обнаружено лишь у жителей
среднеазиатских пустынь (Волков-Дубровин и др., 1975, 1977а, 19776;
Архангельская и др., 1980, 1981). Сейчас пока трудно сказать, являете*
ли подобная особенность следствием обитания в условиях
континентального климата вообще, или мы имеем дело лишь со
случайным совпадением результатов благодаря действию каких-то общих
или близких факторов в двух независимых регионах. Напомним
что содержание оксиапатитов в скелете человека обнаруживаеп
значительную связь с содержанием минералов в почвах и воде ок
ружающей среды (Alexeeva, 1968; Павловский, 1970; Хлыстов
Прибытков, 1973).
В Баргузинской котловине, где были изучены буряты, отмеча
ется нарушение геохимического баланса, в частности йодная не
16]
%
720
ЛО
JOQh
30
80
70
60
SO
40
30
20
С} Ац Л2 С} I2 2.Jti
Ш Ш
Рис. 36. Уровень минерализации скелета у тувинцев (в) по сравнению <
коренным населением Арктики и Субарктики (б), в %к контрольной групп*
русских Курской обл. (а):
А - мужчины; Б - женщины; КЗ - компакта; СЗ - спонгиоза 3-й фаланги; S3 суммарна
минерализация третьей фаланги; К2 - компакта; С2 - спонгиоза 2-й фаланги; S2 суммарна:
минерализация 2-й. фаланги; S2.3 - суммарная минерализация 2 -й и 5-Й фаланг.
162
достаточность, и этим связано широкое распространение среди
населения этой территории эндемического зоба. Не исключено, чтс
понижение содержания минералов в скелете у этих популяциі
также является отражением геохимической ситуации.
Понижение содержания минеральных веществ в организме
коренных жителей Прикаракумья авторы исследований, прове
денных в аридной зоне (Волков-Дубровин и др., 1970), склоннк
объяснять алиментарными причинами. Некоторую
недостаточность костеобразующих веществ можно предполагать и в Алтае-
Саянском нагорье, и в Якутии (см.: Карта геохимических
ландшафтов "Физико-географический атлас мира9*; Носин, 1963). В тс
же время у арктических групп, живущих в зоне вечной мерзлоты
с несбалансированным содержанием подвижных форм макро- в
микроэлементов, показатели минерализации выше, что в какой-тс
мере позволило нам аргументировать положение об известном
приспособлении арктических популяций к недостаточности
солнечной радиации и, следовательно, недостатку витамина D,
вырабатывающегося в организме под действием солнечной энергии и
стимулирующего процессы костеобразования.
На основе имеющихся в нашем распоряжении геохимических
характеристик можно сделать заключение о достаточно большом
геохимическом разнообразии континентальных регионов Сибирв
и Средней Азии (Носин, 1963; Ногина, 1964; Бен-Утяева, 1965;
Андрианова, Ковальский, 1970; Егоров и др., 1970). В то же
время по сравнению с геохимией контрольной зоны (черноземы
Курской обл.) в почвах континентального североазиатского регионе
понижено содержание некоторых костеобразующих веществ, что,
возможно, и отражается на уровне фосфорно-кальциевых солей ь
скелете коренного населения этого региона.
Мне уже неоднократно приходилось отмечать, что уровень
минерализации скелета является скорее следствием воздействия
экологических причин, чем расовой обусловленности (Алексеева,
19776, 1979). Существование генетических механизмов регуляция
минерального обмена (Sobel, 1955) отнюдь не противоречит этом}
заключению.
Высказывалось предположение о возможности воздействия
геохимических условий на рост и развитие человеческого скелете
(Alexeeva, 1968). В частности, намечается связь брахикрании,
укрупнения размеров лица и уменьшения длины тела в геохимичес-
163
ких провинциях с недостатком костеобразующих минералов. На
конец, обнаружена связь на внутригрупповом уровне между фор
мой черепа и содержанием оксиапатитов (Алексеева, 1977). Бра
хикранные черепа характеризуются понижением степени минера
лизации по сравнению с долихокранными.
Статистически далеко не всегда удается обнаружить взаимоза
висимость физиологических и морфологических признаков. В и
же время изучение внутригрупповых корреляций в четырех ту
винских группах выявило статистически достоверную связь меж
ду уровнем минерализации скелета, высотой переносья и развита
ем носовых костей. Коэффициент множественной корреляции ме
жду признаками равен 0,429 при р = 0,05.
Суммирование этих пока еще единичных фактов приводит наі
к необходимости признания роли экологического фактора в про
цессах формообразования у человека. Специфика физиологии на
селения Центральной Азии, отражающая экологические особенно
сти этого региона, позволяет по-новому взглянуть на формирова
ние антропологического типа населения. Отличительные черть
центральноазиатского типа, к которому относится болышгаств(
обследованных нами народов континентальной Сибири, - брахике
фалия, крупные размеры лица, низкое переносье, слабое выступа
ние носовых костей - могут находиться в причинно-следственноі
связи с экологической ситуацией, для которой свойственно пони
женное содержание костеобразующих минералов. Возможная рол
адаптивного фактора в формировании центральноазиатского ком
плекса отмечалась и В.П. Алексеевым (1974), обнаружившим со
впадение ареала центральноазиатской группы с ареалом макси
мального перепада зимних и летних температур.
Характер приспособительных реакций в континентальной зо
не Сибири имеет свои особенности.
Так же как и в Арктике, у жителей континентальной зонь
Сибири выражена реакция на холод. У обследованных групп по
вышены газообмен и теплопродукция по сравнению с контроль
ной группой из умеренного климата. У коренного населения кон
тинентальной Сибири, как в Арктике и Субарктике, повышен*
содержание белков в сыворотке крови, особенно гамма-глобулино
вой фракции. В то же время строение тела существенно отличает
ся от арктических аборигенов. У коренного населения континен
тальной зоны Сибири наблюдаются понижение костно-мускульноі
164
массы тела за счет усиленного развития жироотложения и
изменение пропорций тела, в частности уменьшение длины ноги по
отношению к длине туловища (рис. 37).
Рис. 37. Сравнительный анализ строения тела коренного населения
Арктики и континентальной зоны Сибири:
I - мужчины; II - женщины; а — чукчи; б - тувинцы.
Относительная коротконогость, при которой облегчается
снабжение кровью периферических участков тела, в сочетании с
повышенным жироотложением может рассматриваться как
приспособление к низким температурам среды. Если к этому добавить от-
16S
меченное усиление теплопродукции, то население
континентальной зоны Сибири представляется более адаптированным к
условиям холода, чем население Арктики. Но если учесть, что
термические условия среды определяются по температурам самого
жаркого и самого холодного месяцев, то континентальная зона Сибири
по разнице этих температур предъявляет более высокие
требования к холодовой адаптации, чем Арктика.
К сожалению, в нашем распоряжении нет данных о
механизмах липидного и белкового обмена у населения континентальной
зоны Сибири. Можно лишь предполагать, что он отличается оі
такового у жителей Арктики. В диете коренного населения
континентальной зоны много углеводов, уровень холестерина в
сыворотке крови ниже, чем у коренного населения Крайнего Севера, «
жироотложение выше. Напомню, что у него довольно значителен
и процент пикнического типа телосложения, в то время как у
населения Арктики этот тип конституции практически отсутствует
у мужчин и только в небольшом проценте случаев встречается у
женщин. Видимо, энергетические процессы у жителей
континентальной зоны Сибири проходят несколько иначе, чем в Арктике.
Можно думать, конечно, что разница в процентном отношении
типов телосложения и развитии жировой складки между
населением арктической и континентальной зон Сибири связана с
различиями в двигательной активности, тем более что в литературе
достаточно данных, показывающих связь компонентов сомы с
моторикой (Malina, 1975). И действительно, образ жизни арктических
аборигенов отличается большей активностью (охота на морского
зверя, оленеводство) по сравнению с обследованными нами
жителями континентальных районов. В то же время жители Крайнего
Севера (особенно береговые) далеко не все месяцы года ведут такой
динамичный образ жизни, как летом, во время основной охоты на
морского зверя. Поэтому различия в соматотипе коренных
жителей Арктики и континентальной зоны нельзя объяснить лишь
разницей в двигательной активности.
В связи с морфофункциональными характеристиками
аборигенов внутренних районов Сибири следует обратить внимание на
русское старожильческое население, предки которого появились в
Баргузинской котловине в середине XVII в. Хотя они
происходили из северных областей России, климат Сибири вполне мог ока-
166
зать на них воздействие. И действительно, у них повышена
теплоизоляция, они относительно более коротконоги, а у женщин прв
расчете на 1 кг массы выше и минерализация скелета. Чем же
объясняются эти особенности - воздействием экстремальных
условий континентальной зоны или метисацией с аборигенным
населением? Маловероятно, что адаптивные особенности популяции
могут возникнуть в течение двух с половиной столетий- В то же
время метисация - явление, широко распространенное в Сибири і
местах контактов русских с местным населением. И хотя процені
смешанных бурятско-русских браков на территории Баргузинской
котловины невелик (с конца XIX в. до настоящего времени он
колеблется от 7 до 8%), можно предположить, что во время
первоначального заселения котловины процесс смешения был
интенсивнее. Поэтому отличия русских старожилов от контрольной
группы и сходство их по морфофункциональному комплексу с
бурятами, по-видимому, объясняются метисацией русских с бурятами, г
не прямым влиянием климатических условий Сибири.
Остановимся подробнее на явлении метисации как на факторе,
который может оказать влияние на уровень биологической
приспособленности популяции к тем или иным условиям среды.
По таким жизненно важным признакам, как уровень
метаболизма и скорость кровотока, между бурятами и русскими нет
различий. В то же время контрольная группа, живущая в условиях
умеренного климата, показывает более низкий уровень обменных
процессов.
По пропорциям тела русские старожилы также сходны с
бурятами: отношение длины ноги к длине корпуса у русских
Забайкалья понижено по сравнению с русскими контрольной группы
европейской части бывшего СССР.
Таким образом, и по уровню метаболизма, и по строению теле
русские старожилы Сибири и буряты более адаптированы к
холодному климату по сравнению с контрольными группами. В данном
случае метисация повысила биологическую устойчивость русского
населения к низкому температурному режиму Забайкалья.
Другой пример воздействия метисации на уровень
приспособительных реакций, также взятый из исследований в БаргузинскоЁ
котловине, - способность ощущать вкус фенилтиокарбамида (РТС)
Некоторые авторы этот признак рассматривают как "экстраорди
парную группу крови" (Race, Sanger, 1958). Существует предполо
167
жение, в ряде случаев подтвержденное экспериментально, что ш
способность ощущать вкус РТС связана с большей восприимчивс
стью к действию зобообразующих веществ. Среди русских еврс
пейской части бывшего СССР процент неощущающих высокий,
то время как для коренного населения Сибири характерна явн
повышенная чувствительность к фенилтиокарбамиду. Интереснс
что в Баргузинской котловине способность к ощущению РТС
русских повышена, а у бурят понижена. Влияние метисации здес
несомненно, как очевидно в первом случае повышение резистент
ности к йодной недостаточности, а во втором ее понижение. Эта
пример с достаточной убедительностью свидетельствует о том, чъ
метисация является одним из путей выработки приспособитель
ных реакций для пришлого населения, но ее эффект неоднозна
чен.
Итак, исследования в континентальной зоне Северной Азш
выявили специфику морфофункциональных черт как местного
так и пришлого населения. Многие черты строения тела и метабо
лизма обнаруживают приспособительные особенности. Некоторьи
из характеристик очень специфичны (например, пониженный уро
вень минерализации), но пока остаются необъяснимыми с точкз
зрения полезности для организма. Так или иначе, в континенталь
ном климате Сибири у разных этнических групп наблюдается одні
и та же норма реакции, которую, имея в виду ее морфофизиоло
гическое выражение, назовем континентальным адаптивным ти
пом.
3. АНТРОПОЭКОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ
КОНТИНЕНТАЛЬНОЙ ЗОНЫ СИБИРИ
Разнообразие экологической ситуации в континентальной зоне
Сибири позволяет проследить существование региональных мор-
фофизиологических комплексов, несущих в себе все черты
адаптации к континентальному климату Сибири и в то же время
отражающих специфику воздействия локальных экологических условий.
Для иллюстрации этого наблюдения следует обратиться к
коренному населению Баргузинской котловины. Здесь выделяются
два антропологических региона: предгорно-баргузинский и цент
168
рально-северо-восточный. Для населения первого характерны
более крупные размеры головы и лица, более резко выраженная
брахикефалия, понижение уровня минерализации скелета; для
населения второго региона - уменьшение размеров головы и лица,
уменьшение головного указателя, повышение минерализации
скелета. Комплекс характерен как для бурят, так и для русских,
проживающих в этих регионах. Сочетание величин признаков в
комплексах таково, что заставляет думать об отражении в чертах этих
комплексов геохимической ситуации, характерной для регионов.
В свете данных по минерализации скелета и ее связи с формой
скелета, территорией и т.п. (Alexeeva, 1968; Павловский, 1970)
морфофункциональный комплекс предгорно-баргузинской зоны
свидетельствует о более выраженной минеральной
недостаточности почв по сравнению с центральной и северо-восточной зонами.
Сам по себе характер этих комплексов и причины их образования
на территории Баргузинской котловины имеют локальное
значение и представляют интерес главным образом для решения задач
медико-географического районирования. Нами же это наблюдение
может быть истолковано как дополнительное доказательство
независимой от расовой принадлежности нормы реакции на сходный
комплекс условий окружающей среды.
Другой пример локальной экологической дифференциации в
пределах определенного географического региона представляет
собой население Алтае-Саянского нагорья. Антропологическое
обследование тувинцев, хакасов, шорцев, телеутов, теленгитов и ал-
тай-кижи показало четкую морфофизиологическую
дифференциацию между степными и таежными группами. Таежные группы,
сохраняющие типичное для всех континентальных групп строение
тела, характеризуются уменьшением его размеров. Собственно
тувинцы, хакасы, телеуты, теленгиты и казахи, как представители
степных континентальных групп, составляют одну
морфофизиологическую совокупность, тувинцы-тоджинцы и шорцы (как
представители лесных континентальных групп) - другую (рис. 38).
Морфофизиологическая дифференциация на территории
Алтае-Саянского нагорья совпадает, как видно, с природным
разделением этой территории на таежную и степную зоны и с
дифференциацией по типу хозяйства (оленеводство и скотоводство).
Случайно ли такое совпадение? Думаю, что нет, так как в эти
совокупности включаются генетически разнородные группы. Более того, в
169
770-
}/60-
50 60 70
Вес тела, кг
ПаШі
Рис. 38. Экологическая дифференциация коренного населения Алтае-Салн
ского нагорья по признакам строения тела на степной (а) и лесной или пи
ежный (б) варианты. Мужчины.
пределах степных тувинских групп обращает на себя внимание
население горного Монгун-Тайгинского района. Здесь выделяете;
высокогорный морфофизиологический комплекс, аналогичны!
тому, который характерен для всех высокогорных территории
ойкумены, но проявляется он на фоне типично континентальные
адаптивных черт. Признаки высокогорной адаптации обнаружи
ваются и у теленгитов Горно-Алтайской области по сравнению і
населением более равнинных областей - алтай-кижи и телеутами
и у казахов Чуйской степи по сравнению с казахами равнинной
Казахстана (рис. 39).
Эти наблюдения приводят к мысли о том, что в любой специ
фической географической среде могут быть выделены главные і
второстепенные факторы, по отношению к которым осуществляет
ся адаптация. Для территории Алтае-Саянского нагорья, как и дои
Баргузинской котловины, главным фактором, по-видимому, явля
ется фактор континентальное™. Антропоэкологические исследо
вания в Алтае-Саянском нагорье позволяют ввести в понятие при
способительной изменчивости у человека принцип иерархичносп
(далее об этом см. в гл. IX, 1).
Другим (существенным, на мой взгляд) результатом исследова
ний адаптации на Алтае и в Саянах, который может быть экст
170
L_l L_J I ! L_J
ни ' ондомэ мдоіпж мндэ(1]
раполирован на другие территории, можно считать сам факт
локальной экологической дифференциации на сравнительно
ограниченной (в масштабах ойкумены) территории, разнообразие
природных условий которой делает ее ареной сложнейших
приспособительных процессов, приводящих к реальным морфофизиологи-
ческим различиям в близкородственных популяциях.
4. МОРФОФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ
НАСЕЛЕНИЯ МОНГОЛИИ
Наибольшей выраженностью континентальности, как
основной определяющей черты климата Центральной Азии,
характеризуется Монголия. Высокое гипсометрическое положение,
удаленность от морей и океанов, барьер горных хребтов делают ее самой
континентальной страной мира. Кроме того, она обладает
большим ландшафтным разнообразием.
Районирование территории Монголии по индексу суровости
погоды - показателю комплексного воздействия климатических
факторов (индекс Бодмана) на организм человека - выявило, что
в целом Монголия характеризуется более сильным Холодовым
стрессом, чем Алтае-Саянское нагорье, хотя таких высоких
величин индекса как на юге Горно-Алтайской области в районе горы
Белуха, в Монголии все же нет. По сравнению с Алтае-Саянском
нагорьем территория Монголии обладает неоспоримым
преимуществом для исследований. Здесь может быть проведен более или
менее "чистый" экологический эксперимент - одна и та же
этническая группа ( а именно халха-монголы) может быть изучена в
различной гипсиометрической, ландшафтной и климатической
обстановке на фоне чрезвычайного однообразия типа диеты
(Алексеева, 1989).
В Монголии антропологическим отрядом
Советско-монгольской экспедиции было обследовано население нескольких
экологических ниш. На территории Хангая изучены две популяции хал-
ха-монголов, проживающих на разной высоте (от 1500 до 3200 м)
над уровнем моря на восточных отрогах Гобийского Алтая -
взрослое и детское население, живущее в условиях пустыни. Дети и
взрослые обследованы также в районах горной тайги на севере
172
Монголии и в степной зоне на востоке Монголии. Единственной
группой, отличающейся по происхождению от изученных нами
халха-монголов, являются хотоны, проживающие в районе озера
Убсу-Нур. В прошлом это земледельцы, выходцы из Джунгарии, к
настоящему времени воспринявшие монгольский язык, культуру
и тип хозяйства, но сохранившие в своем облике черты памиро-
ферганской расы, характерные для тюркоязычного населения
Средней Азии. Местоположение изученных нами популяций населения
Монголии представлено на р и с. 40.
Анализ антропологических материалов, собранных на
территории Монголии, еще не завершен и поэтому результаты
приведенного здесь анализа носят предварительный характер, однако с
уверенностью можно сказать, что население Монголии несет в себе
черты высокогорной адаптации (табл. 22, 23). Прежде всего это
проявляется в строении грудной клетки, высоком уровне
гемоглобина, в понижении толщины жировой складки по сравнению с
типичными представителями континентального адаптивного типа
(Алексеева, 1989). Последний наиболее выражен у населения
пустынной зоны, обследованного нами в Гоби-Алтае и монгольским
антропологом Я. Ням-Доржем в Южной Гоби. Население
монгольских пустынь отличается более выраженной брахиморфией и
большим развитием жирового компонента теда, чем халха-монголы
других территорий.
В то же время следует признать, что некоторые черты,
типичные для населения континентальных зон, проявляются во всех
изученных монгольских группах достаточно определенно. Это
несколько пониженный уровень минерализации скелета,
пониженные темпы оссификации (скелетный возраст, определяемый in
vivo по кисти детей разного возраста, отстает от паспортного) и
замедленный темп проявления старческих изменений в скелете.
Хотоны, генетически с монголами не связанные, мало
отличаются от последних по пропорциям тела, хотя и характеризуются
меньшей его длиной, меньшей массой и более слабым развитием
жирового компонента. Черты высокогорной адаптации у хотоноі
выражены вполне определенно.
При сопоставлении с населением Алтае-Саянского нагорья,-
монголы, за исключением гобийских/при значительном сходств*
с ними в пропорциях тела отличаются большей выраженностьк
173
/
/
і®
\
2
1
\
1
Т
г
т.
1
і
'Ч
v
<
¦-Г
г
2
t
т
\
\
-S. і
г ^
\
х
J
I
I
V-
/ • Уі?
г
i
i
\
v
N
/і
* •-—¦ — " ч
V
\
Т
i
J
Л
; ©о
Ч.
/
V.v- ч.
V
V
\
\
\
I
Г
v*-,
N.
\?
С ¦
/
\
?-і
/
L.C
ч.
./
'Ч -ч
Г
I
Г
7
/
^
га 4>.>'\.
•
^
3*1
Таблица 22
Морфофжзнологнческне характеристнкн населенна Монголии (мужчины)
Признак
Число обследованных
Длина тела, см
Длина ноги, см
Длина руки, см
Ширина плеч, см
Ширина таза, см
Поперечный диаметр груди,
см
Переднезадний диаметр груди,
см
Длина ноги, % к длине тела
Длина руки, % к длине тела
Ширина плеч, % к длине тела
Ширина таза, % к длине тела
Грудной указатель
Масса тела (w), кг
Поверхность тела (s), м 2
Указатель Рорера
s/w
Средняя жировая складка, мм
Общий белок, г%
Гемоглобин, г%
Холестерин, мг%
Систолическое давление, мм
рт. ст.
Диастоличесхое давление, мм
рт. ст.
Бат-
Улвий
Бурд
Баян-
Л?г
Жарга-
лант
Халх-
гол
халха - монголы
72
166,б
88,0
74,2
36,4
27,7
26,5
19,7
53,1
44,8
22,0
16,7
74,3
60,4
1,67
1,33
2,76
6,85
7,90
15,4
221,0
118,2
75,2
73
165,7
87,7
74,6
37,4
27,9
27,3
19,7
52,9
45,0
22,5
16,8
72,3
63,7
1,72
1,40
2,70
7,45
8,21
15,3
212,1
116,0
76,7
67
164,8
87,0
73,2
37,2
26,9
26,9
18,9
52,8
44,5
22,6
17Д
70,0
61,8
1,69
1,38
2,73
7,76
8,26
15,6
182,4
120,9
79,3
75
165,6
87,8
75,1
38,1
26,4
26,6
19,5
53,0
45,4
23,0
17,2
73,4
63,2
1,71
1,39
2,71
6,85
8,22
16,1
159,0
123,6
80,5
58
165,2
87,2
73,6
38,1
27,3
26,9
19,3
52,8
44,5
23,1
16,6
71,8
59,9
1,66
1,33
2,77
5,91
8,21
15,8
175,69
120,7
77,9
Тариа-
лан
хотоны
46
161,9
86Д
73,4
37,4
27,8
26,0
19,6
53,1
45,3
23,1
17,2
75,3
59,0
1,64
1,31
2,78
5,50
8,22
15,7
185,6
120Д
75,6
175
Таблица 21
Морфофнзнологнческке характеристики населения Монголии (женщины)
Приэнак
Число обследованных
Длина тела, см
Длила ноги, см
Длина руки, см
Ширина плеч, см
Ширина таза, см
Поперечный диаметр груди,
см
Переднезадний диаметр груди,
см
Длина ноги, % к длине тела
Длина руки, % к длине тела
Ширина плеч, % к длине тела
Ширина тааа, % к длине тела
Грудной указатель
Масса тела (w), кг
Поверхность тела (в), м2
Указатель Рорера
s/w
Средняя жировая складка, мм
Общий белок, г%
Гемоглобин, г%
Холестерин, мг%
Систолическое давление, мм
рт. ст.
Диастолическое давление, мм
рт. ст.
Бат-
Улвий
Вурд
Баян-
Лег
Жарга-
лант
Халх-
гол
халха - монголы
71
163,4
81,7
67,9
34,0
27,8
24,5
17,7
63,2
44,2
22,2
18,1
72,7
53,9
1,53
1,49
2,84
12,12
7,55
13,4
205,8
116,6
78,4
82
154,1
81,4
68,0
34,0
27,8
24,7
18,3
52,8
44,0
22,1
18,0
74,2
57,2
1,58
1,56
2,76
13,47
8,43
13,5
214,7
112,1
73,9
77
153,8
81,3
67,0
33,9
24,7
24,7
17,4
52,8
43,6
22,0
18,3
70,5
56,0
1,56
1,54
2,79
13,10
8,30
13,4
176,5
115,5
79,7
97
154,5
81,9
68,4
34,6
28,3
24,6
17,6
53,0
44,3
21,5
18,3
71,8
56,1
1,57
1,52
2,80
11,98
8,36
13,3
153,0
118,5
77,7
94
154,2
82,5
68,0
34,6
27,9
26Д
18,5
53,5
44,0
22,5
18,1
73,8
55,5
1,56
1,51
2,81
10,14
8,15
14,2
189,4
115,8
76,0
Тариа-
лан
хотоны
86
160,7
80,6
67,6
34,2
28,0
24,2
17,9
53,5
44,9
22,7
18,6
73,9
52,4
1,50
1,53
2,86
9,36
8,26
13,8
183,1
116,2
73,5
176
черт высокогорной адаптации, особенно это проявляется в
строении грудной клетки и понижении жироотложений. По-видимому,
фактор высокогорья в воздействии на организм человека в
Монголии выражен сильнее, чем на Алтае и в Саянах, и в формировании
адаптивных особенностей монгольского населения играет ведущук
роль.
Помимо явной высокогорной адаптации у населения Монголия
обращают на себя внимание такие морфофизиологические
особенности, как поздняя скелетная зрелость и позднее появление стар
ческих изменений в скелете.
Эти особенности характерны и для некоторых групп Алтае-Са-
янского нагорья, но в Монголии они выражены значительно
определеннее. Картина скелетного созревания и инволютивных изме
нений скелета напоминает ту, которая характерна для долгожи
тельских популяций Абхазии (Павловский, 1982).
Лекция VIII.
БИОЛОГИЯ ЖИТЕЛЕЙ КРАЙНЕГО СЕВЕРА
Питание. Морфофизиологические характеристики.
Связь индекса "суровости" погоды и строения тела.
Коренное население арктической и субарктической зон
относится к различным этническим и языковым группам. В
антропологическом отношении, будучи представителем монголоидной ра
сы, оно также демонстрирует локальное разнообразие
антропологических типов. Некоторые из них - саамы (лопари), соединяют і
своем облике черты европеоидной и монголоидной рас, являясь,
по-видимому, реликтом древнего протоуральского типа, в
условиях географической изоляции приобретшим некоторые специфи
ческие черты.
В то же время хозяйство и быт аборигенов различных
этнических групп Арктики и Субарктики обнаруживают черты сходства.
На территории Крайнего Севера выделяется особая циркумполяр-
иная культура, которая во многих чертах отражает роль
географического фактора в ее создании (Circumpolar problems, 1973).
Наряду со многими чертами сходства в материальной
культуре групп Крайнего Севера выявляются общие черты и в демогра
177
фической структуре населения - характеристике, определяемой
как социальными, так и биологическими факторами.
В большинстве своем коренное население Крайнего Севера
характеризуется высоким естественным приростом. Об этом
свидетельствуют данные по аборигенам Аляски - эскимосам, индейцам
и алеутам, обследованным в 1960 г. (Lantis, Anderson, 1964), и
наши данные по ненцам, чукчам и эскимосам, собранные в 1968-
1971 гг. (Алексеева и др., 1972, 1983). Естественный прирост ко-
ренных жителей Аляски (27,5 на 1000 человек) превышает
таковой у населения США, Канады, стран Северной Европы и
некоторых других. Сходная картина наблюдается и в нашей Арктике.
Коэффициент выживаемости, вычисленный нами для ненцев,
эскимосов и чукчей, довольно высок; число родившихся в
процентах к общей численности населения вдвое превышает число
умерших, рассчитанное таким же образом.
В возрастной структуре коренного населения Крайнего Севера
обращают на себя внимание очень высокий процент детей в
возрасте до 10 лет и резкое снижение числа лиц старше шестидесяти
лет.
1. ПИТАНИЕ КОРЕННОГО НАСЕЛЕНИЯ АРКТИКИ
Будучи в основном оленеводами и охотниками на морского
зверя, жители Арктики и Субарктики имеют высококалорийный
рацион питания. Сопоставление химического состава и
калорийности питания арктических аборигенов с контрольным рационом, по
отношению к которому оценивалось питание тропических
жителей, показало явное повышение калорийности, увеличение
содержания белков и жиров в диете коренного населения Крайнего
Севера.
Трудности изучения питания коренного населения
арктической зоны отмечались неоднократно (Бартон, Эдхолм, 1957).
Большая подвижность образа жизни, резкие колебания в калорийной
ценности между разными приемами пищи, зависимость пищевого
состава от сезона значительно осложняют суждения о характере
питания аборигенов Арктики. И тем не менее довольно большое
число исследований дает возможность хотя бы ориентировочно
178
охарактеризовать их диету. Установлено, что в рационе
коренного населения Севера свыше 35-40% калорий образуется за счет
белков, тогда как для населения умеренного климата количество
"белковых" калорий составляет 12-15% (Кремер, 1965; Харрисон
и др., 1968).
Так, по данным М.Броуна (цит. по: Бартон и Эдхолм, 1967),
эскимосы Гудзонова залива в среднем потребляли в день 343 г
моржового мяса, 114 г жира и 208 г хлеба. Когда в рацион входила
свежая оленина, то потребление мяса составляло от 570 до 1110 г,
но в соответствии с этим менялось соотношение жиров и
углеводов. В среднем в период питания моржовым мясом калорийная
ценность среднесуточного пищевого рациона составляла 2500 ккал,
в период питания олениной - 3250 ккал, причем в первом случае
за счет жира образовывалось 1000 ккал, во втором - 2700 ккал.
Манн с сотрудниками (Mann et al., 1962), изучавшие питание
эскимосов Аляски в домашних условиях и в условиях
организованного питания в подразделениях Национальной гвардии,
нашли, что коренное население Аляски (эскимосы и индейцы
атапаски) питается преимущественно мясом оленя (карибу), морских
животных и рыбой. Рацион питания обеспечивает 1900-2300 ккал
со значительной долей белка и животных жиров.
Рацион питания коренного населения Чукотки изучался
Институтом питания АМН СССР (Ефремов, 1970; Зайцев, 1970; Аст-
ринский, Навасардов, 1970). Пища коренного населения Чукотки
в прошлом состояла из оленины, мяса морского зверя, рыбы,
которые часто употреблялись в сыром виде. По наблюдениям Н.В.
Слюнина (1895), чукчи употребляли в пищу 18 видов растений.
Сейчас в их рацион питания входят и привозные продукты - хлеб,
сахар и масло.
В.В. Ефремовым выявлена хроническая недостаточность
витаминов, костеобразующих минералов и микроэлементов.
Сопоставление диет чукчей тундровых и береговых выявило, что питание
тундровых жителей отличается большим содержанием белков
животного происхождения, углеводов и калорий. В целом же
рацион питания коренного населения Чукотки отличается высоким
содержанием калорий, белков и жиров - более высоким, чем то,
которое характерно для эскимосов Аляски и Гренландии, а также
арктических индейцев.
179
Оценка питания лесных ненцев, находящихся в оленеводчес
ких бригадах, произведенная нами (Алексеева и др., 1972), пока
зала высокий калорийный уровень, очень высокое содержание
белков (от 600 до 800 г оленины в день на человека) и недостатоі
животных жиров (употребляют в пищу преимущественно расти
тельное масло), кальция и витаминов. В питании ненцев, живу
щих в поселках, довольно значителен процент привозных продук
тов, оленеводы же сохраняют более или менее традиционный ра
цион питания. В рационе ненцев, так же как и у коренных
жителей Чукотки, обнаруживается недостаток витаминов
(Добронравова, Куинджи, 1962; Лукьянов, Пушкина, 1962).
Сопоставление химического состава и калорийности питания
арктических аборигенов с контрольным рационом, по отношению
к которому оценивалось питание тропических жителей, показало
явное повышение уровня калорий, белков и жиров в диете
коренного населения Крайнего Севера (табл. 24, рис. 41), хотя он все
же ниже, чем рекомендуемый Институтом питания для северных
районов.
Таблица 24
Краткая характеристика рациона коренного населения Крайнего Севера
в сравнительном освещении
Состав рациона
Калории
Белки общие, г
Белки животные, г
Жиры общие, г
Жиры животные, г
Углеводы, г
Контроль
2900
100
55
105
—
389
Ненцы-
оленеводы
3941
218
153
126
34
460
Чукчи
тундровые к
береговые
3249
118
97
199
192
224
Нормы,
рекомендованные
Ин-том
питания для
северных районов
4485
155
11?
124
100
520
L80
Лесные ненцы- Чукчи и зеки- Рекомендуемые
оленеводы мосы Ч у котки *°РМ ь*
Рис. 41. Питание коренных жителей Крайнего Севера е сравнительном
освещении. За 100% принят контрольный рацион питания.
Специфика питания арктических жителей - с избытком
белков и жиров и с недостатком углеводов - особенно ярко
ощущается в сравнении с диетой тропических аборигенов. Это как бы две
противоположных полюса (рис. 42).
183
пмории
т
ш
харь/
Рис. 42. Сравнительная характеристика питания коренного населени.
тропической (в - Африка) и арктической (б - Чукотка) зон (в сопоставленш
е диетой населения умеренной зоны (а).
2. МОРФОФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
КОРЕННОГО НАСЕЛЕНИЯ КРАЙНЕГО СЕВЕРА
Строение тела. Наиболее полные соматические характерно
тики коренного населения арктической зоны получены экспеди
циями НИИ антропологии МГУ, обследовавшими лесных ненцеі
пос. Тарко-Сале Тюменской области, саамов пос. Ловозеро
(Кольский п-ов), чукчей и эскимосов пос.Уэлен, Лорино, Нунямо, Сире-
ники, Чаплино (Чукотка).
Саамы и ненцы отличаются от чукчей и эскимосов
миниатюрностью габаритов, но в то же время у всех этих групп много
общего в соотношении соматических признаков (табл. 25). Например,
при относительно небольшой по межгрупповому масштабу длине
тела у них относительно велики масса тела, обхваты грудной
клетки, талии, ягодиц, относительно велико развитие обезжиренной
массы тела. Все эти признаки свидетельствуют об относительно
высокой плотности тела. По мнению Н.С. Смирновой и Н.И. Клев-
182
КЗ
3
3 Sw
Я Я
fits • * 2
* ^ 8
S * « Яі
в к &
Я .5 *3
f 2-й
еоюсоо^поппон
* N « .* _.
(О L 00 Ю ^<
Е- СО
ООЮООСО'ФбЭФСОЬ-
оо«чво<о«іда<0
". о
со »ч
со
00
о
о
s
о*
СП
г
о
о
О
г
I
о
л
г
ев
К
Я
ев
ЕВ
8
и
8
А
ев
А
2
«
О
14
О
ав
со
*
о
t
I
51
& $ А
2 ь-
1 Н «J 00 W
00 О О ©J
О "Ч* 0О 00
t- «с » м
СО ^1
со о
CM W
со со см to to us "Ч
.и ^ Ч °°. « Ч ¦• S
й и и « и t- 2
Л
(О
Ь-*
to
fc-
s
о
1
Ч т|і « 00 ¦* Ю 00 ffl О^ О N N N N <Н tO "¦*<?
S Я ©*в^оои?оо*вчевее?с©~ееГс©ЧЧь'1і'с*Ч
2а0<веое]с4ній^о4нь-квничнсо
1 ч*
CN
СП
0,710
т-1
72,6
и
Я4
Tf. ©аномілвОФсо^нюсон
2 S в~ W9 ю" СО СО* О* вч* СО* CN СО* ¦** UO* *Я Ч *9. <М
Ъ- СО
«* 00
00
і, ео
С»
8
©
Vft*
Яй
и
СП
to
as
<« g ^f О ь* ъ* ь* гч el* «<f со ь* оо ъ* *°- ч. -і
см
^ °° 5 °°- »
*¦ t- «н
00Ь"П«в«еіію^«ні-«нн«
ео
§
©*
to
X
і &-ж »_ о^ о*
3 S 3* оГ to* ь-*
^ *3 оо to ео е*
О ец
¦^ -^
Ю ¦-*
ьа
со
ем
со
t-
i-l
со
1 to
ю
00 <й Н - «
ш ^ со л. to
і-ч -ч е* ~" ь-
202
со
t-
со
to*
СМ
to
S
«5 оі
w>-l
?а
в S
S а
2 и
со
М5 СО
i'tfOlt-^OONHOO^t-WiNeoO
^ ь- о а со со
СО t- СО « N N Ю
- - и 2 « •
-^CM-Hb-tOiHf-tCM1^
-1 ю
СО іН
X
с»
со
со
с»
я •*
ю. 0_ ^ ео і> « N ^ ^ n и Ож n и и ін
оо Sco*»Ho6*oo*b^^cocoeob^u^eott',tfc*l
200fc-e0<NNNIO^NHfc-(OHHN
яч ^^ ^^ *
5 *. *в* я
Ь* кГ -j О
^ н «
о
из
СО
со
И
в
в
о,
???ЙЭ9ё
183
цовой (Smirnova, 1973; Клевцова, Смирнова, 1974), изучавших
морфологию тела арктических аборигенов, в условиях Арктики
наблюдается общая тенденция изменчивости морфологического
статуса групп - усиление мезоморфных черт строения тела.
Различные территориальные группы эскимосов Нового Света (Rodahl,
1952; Marshall, Neuman, 1960; Mann et aL, 1962) по соотношению
длины и массы тела сходны с эскимосами, изученными нами (табл.
26).
Таблица 26
Строение тела у коренных жителей зарубежной Арктики х Субарктккя
Признак
Число обследованных
Длина тела, см
Масса тела (w), кг
Поверхность тела (s), м *
Указатель Рорера
s/w
Индейцы,
Сев.
Америка
Andersen,
Bolstad et
аЦ I960;
Eisner et
al., 1960
196
168,6
66,2
1,72
1,39
2,63
Эскимосы
Сев.
Америка
Crlle,
Quiring,
1939;
BoUerud
et al.,
I960;
Mann et
al.f 1962
821
163,1
64,8
1,71
1,47
2,66
Гренландия
Brown,
1957;
Ducros,
1971,
1972
439
160,4
63,0
1,70
1,52
2,70
Алеуты,
Алеутские о-ва
Laughlin,
1951
28
161,2
65,3
1,73
1,65
2,66
Лопари
(саамы),
Норвегия
Andersen,
Loylinget
aL., 1960
14
164,0
62,7
1,П
1,42
2,73
В арктических группах обращает на себя внимание почти
полное отсутствие астенического типа телосложения, значительное
увеличение процента лиц мускульного типа (по сравнению,
например, с ранее изученными нами русскими и бурятами).
Из функциональных показателей, отражающихся в строении
тела, следует отметить преимущественно цилиндрическое
строение грудной клетки, что также нетипично для популяций
умеренных широт, где достаточно велик процент уплощенной и
конической грудной клетки. Значительное развитие грудной клетки у
коренного населения Арктики обращает на себя внимание в связи
184
с отмечаемым в условиях холода гипоксическим синдромом. Пс
мнению Н.П. Неверовой, Т.И. Андроновой и М.И. Мочаловой (1972).
гипоксический синдром обусловлен низкой концентрацией аскор
биновой кислоты в результате ее повышенного расхода при усиле
нии интенсивности окислительно-восстановительных процессов і
холодном климате. Изучая процессы акклиматизации у лиц,
впервые попавших в условия европейской Арктики, эти авторы
наблюдали гипервентиляцию, повышение тонуса дыхательной мускула-
туры, увеличение скорости кровотока, повышение содержание
гемоглобина крови и, что весьма существенно, кислородной
емкости крови.
На дефицит аскорбиновой кислоты и нарушение витаминногс
обмена при низких температурах указывают и другие авторы (Де-
ряпа, Рябинин, 1977).
Можно представить, что крупная, цилиндрической формы
грудная клетка арктических аборигенов является одним из
приспособлений, облегчающих процессы газообмена в условиях холода и і
некоторой степени нивелирующих действие повышенного тонусе
мускулатуры, при котором жизненная емкость легких
сокращается.
В литературе существует мнение о более высоком развитии
жироотложения у арктических жителей по сравнению с
населением умеренного климата. Г. Браун (Brown, 1957), изучавший этоі
признак наряду со многими функциональными тестами у
эскимосов о-ва Саутгемптон (Северная Америка), высказал предположе
ние о высоком функциональном значении повышения толщинь:
подкожного жирового слоя в условиях адаптации к холоду. Надо,
однако, отметить, что данные этого автора относительно
увеличения жироотложения в условиях Арктики не находят
подтверждения в абсолютных характеристиках толщины жировых складок }
других арктических популяций. В изученных нами арктических
группах толщина жировой складки ниже, чем у групп, живущих
в умеренном и континентальном климате. По данным А. Дюкрс
(Ducros, 1971), у эскимосов Ангмассалика (восточное побережье
Гренландии) толщина жировых складок меньше, чем у японцев,
белых американцев и бельгийцев. Заслуживают внимания отме
ченные А. Дюкро более высокие плотность тела и обезжиренная
масса тела, эскимосов Саутгемптона (последняя особенность харак
терна для арктических групп, изученных нами). Небольшую ве
185
личину жировых складок у эскимосов Нового Света приводит і
Р.Элснер (Eisner, 1963). Доля обезжиренной массы тела, т.е. кост
но-мускульного компонента, в величине веса тела у арктически
популяций больше, чем у популяций умеренного и континенталь
ного климата.
Большие обхваты верхних и нижних конечностей свидетель
ствуют о высоком развитии мускулатуры, особенно в верхнем от
деле тела. Довольно крупные по межгрупповому масштабу мыщел
ки говорят о значительном развитии скелета. Об этом можно су
дить и по костным материалам, полученным из современных эски
мосских могильников, датированных XVII-XX вв. Скелет эскимо
сов характеризуется некоторыми специфическими чертами: боль
шими периметрами костей при небольшой их длине, сильным раз
витием рельефа, крупным костно-мозговым каналом, относитель
но небольшой толщиной компактного слоя (Jorgensen, 1953; Алек
сеева, Коваленко, 1980). Определение уровня минеральной васы
щенности рентгенофотометрическим путем показало более вы-со
кую минерализацию скелета (по плечевой кости) эскимосов П(
сравнению с группой русского населения, взятой для контроля
При сравнительно небольшой толщине компактного слоя костеі
повышенная минерализация способствует прочности скелета. Про
чность достигается, вероятно, за счет значительного периметрі
костей (Алексеева, 1965).
В строении скелета обращает на себя внимание большой кост
но-мозговой канал трубчатых костей скелета. Такая особенност]
становится понятной, если учесть необходимость повышенной
кроветворения в условиях арктической кислородной недостаток
ности.
Из физиологических черт для населения арктической зонь
характерен высокий в целом уровень минерализации. Это наблю
дается и на костном материале, и при исследовании in vitro рент
генофотометрическим путем. Уровень минеральных веществ у ар
ктических популяций при имеющихся вариациях, как правило
выше, чем у населения умеренного и континентального климата
(рис. 43).
Исключение составляет группа русского населения, живущего в условиях ол
тимальнои геохимической ситуации, показавшая максимальные величины минера
лизации скелета.
186
150
?О
110
120
НО
100
II
а
'Ш.
Рис. 43. Сравнительна* характерис
тика уровня минерализации скелета (%}
родственных популяций, живущих в аркти
ческой и умеренной зонах европейской нас
ти бывшего СССР (I) и Сеееро-восточноі
Азии (II). За контроль (а) принята велики
на минерализации скелета населения уме
ренной зоны f 100%); б - мужчины; в - жен
щины.
Это обстоятельство требует объяснения, так как в питании ко
ренного населения Крайнего Севера наблюдается дефицит макро
и микроэлементов, участвующих в костеобразовании (Ефремов,
1970). Более того, нам известна позитивная связь уровня минера
лизации в среде с минеральным содержанием в скелете (Павлове
кий, 1970). Можно предположить, что в условиях Арктики
действуют какие-то компенсаторные механизмы, препятствующие
деминерализации.
Заключая раздел, отмечу очень важную особенность: общие
черты строения тела арктических групп проявляются у разных
этносов, не связанных единством происхождения.
Энергетические процессы и терморегуляция. В зарубежной
литературе основное число работ по изучению адаптации к
условиям Крайнего Севера посвящено характеристике энергетических
процессов и терморегуляции у коренных жителей по сравнению с
пришлым населением. Несмотря на значительное количество ис-
187
следований в этом направлении, единой точки зрения на характе
энергетических процессов в условиях Арктики нет.
Диета арктических аборигенов чрезвычайно богата белками
жирами и бедна углеводами и витаминами. А. иМ. Крог (А. Кг<
gh, М. Krogh, 1914), исследовавшие питание и процессы метаб<
лизма у гренландских эскимосов, назвали их диету "диетой плоте
ядного животного", отметив при этом, что никаких вредных ш
следствий в условиях холодного климата не наблюдается. В чает
ности, почти нет болезней, связанных с избытком мочевой кисле
ты в организме. Экспериментально показаны хорошее усвоение:
утилизация белков при низком содержании углеводов в рационе
Установленная этими авторами зависимость углеводной и водно:
экскреции от содержания в диете мяса показывает задержку угле
водов и воды в организме после обильного употребления мяса
Другой автор (Heinbecker, 1931) на основании изучения метаболиз
ма у эскимосов Баффиновой Земли отметил у них более высокуі
способность окислять жиры по сравнению с контрольной группой
У аляскинских эскимосов повышено выделение азота с мочой (Ro
dahl, 1952), что рассматривается как ферментная адаптация, деза
минирующая излишки аминокислот (Веселухин, 1977).
Таким образом, у арктических аборигенов, чья диета характе
ризуется очень высоким содержанием белков и жиров, существу
ют специфические биологические механизмы их утилизации. Эті
особенность питания отличает аборигенов от контрольных груш
умеренного пояса обитания.
Как отличительную черту коренного населения Арктики (Яма
л о-Ненецкий АО) можно отметить несколько более высокое п<
сравнению с пришлым населением содержание витамина С в орга
низме (Лукьянов, Пушкина, 1962), хотя явления гиповитаминоз
наблюдаются и у них.
Высокое содержание белков в рационе питания арктически:
народов, повышающее основной обмен, приводит некоторых авто
ров (Hoygard, 1941; Rodahl, 1952) к мысли об отсутствии расовоі
обусловленности этого признака или специфического действия в
него холода. Так, в эксперименте К. Родаля у эскимосов, переве
денных на обычный рацион американцев той же местности, вели
чина основного обмена не повысилась. По данным В. Левиноі
(Levine, 1949), основной обмен эскимосов оказывается близким і
контрольным значениям.
188
Данных о повышении основного обмена у аборигенов Севера в
литературе много. Отмечено усиление основного обмена у
эскимосов (Crile, Quiring, 1939; Brown et al., 1954; Кандрор, 1962),
якутов, чукчей (на 13-16%, Кандрор, 1962), Более высокое
потребление кислорода обнаружено у индейцев-атапасков по сравнению о
европейцами (Andersen et al., 1960b; Irwing et al., 1960); у ненцев,
саамов, чукчей и эскимосов, изученных нами оксигемометричес-
ким путем, потребление кислорода интенсивнее в сравнении о
жителями средних широт.
X. Эриксон (Erikson, 1957), исследуя эскимосов мыса Барроу и
американцев, живущих в тех же условиях, обнаружил более
высокие показатели поглощения кислорода у эскимосов (324 мл/
мин), чем у американцев европеоидного происхождения (299 мл/
мин).
Даже кратковременное пребывание в условиях холода
увеличивает потребление кислорода в 2,5 раза, что указывает на
усиление обменных процессов (Horwath et al., 1947).
Белки и липиды крови. В связи с энергетическими
процессами у коренного населения Арктики следует остановиться на
содержании белков и липидов в сыворотке крови. Больше всего
внимания уделено изучению содержания холестерина в сыворотке
крови. Анализируя географическое распределение уровня этого
признака, автор обнаружил общую тенденцию его повышения в
северных районах ойкумены (Алексеева, 1971). Большинство
арктических групп характеризуется, как правило, высоким содержанием
холестерина, хотя пределы колебания групповых величин этого
признака весьма значительны: у канадских эскимосов - от 139 до
176 мг% (Boyd, 1935; Corcoran, Rabinowitch, 1937), у эскимосов
Аляски - от 203 до 214 мг% (Wilber, Levine, 1950; Scott et al.,
1958), у эскимосов Чукотского полуострова и чукчей - от 184 до
202 мг% (Алексеева и др., 1983), в сборной группе, включающей
чукчей, эскимосов, коряков, орочей и камчадалов, обследованных
в поселках Чукотского, Берингова и Охотского морей, - от 200 до
235 мг% (Кукес, Власова, 1962), у саамов Кольского п-ова - 202
мг% (Алексеева и др., 1977), у лесных ненцев - 131 мг%
(Алексеева и др., 1972). Эти данные относятся к мужским группам. В
женских группах, по нашим материалам, уровень холестерина
несколько ниже: у эскимосов - 191 мг%, у чукчей - 195 мгУо, у
189
саамов - 188 мг%, у ненцев - 121 мг%. У эскимосов Аляски
содержание холестерина в сыворотке крови составляет 233 мг% (Wi
lber, Levine, 1950).
На общем фоне высокого содержания холестерина в сыворотке
крови среди арктических жителей исключение составляют ненцк
и эскимосы Баффиновой Земли. В рационе лесных ненцев прв
высоком белковом содержании очень мало жиров животного про
исхождения и много растительных жиров, что могло сказаться т
величине холестерина в их сыворотке KpojBH. Что касается эскимо
сов Баффиновой Земли, то они дают заниженные показатели ш
только по холестерину, но и по многим другим признакам (Cor
coran, Rabinowitch, 1937).
Высокий уровень холестерина, типичный для большинстве
арктических групп, в известной мере отражает высокое содержа
ние жиров в диете: коэффициент корреляции между уровнем хо
лестерина в сыворотке крови и содержанием жира в пище равеі
0,83 (Алексеева, 1971). В то же время есть ряд указаний на гене
тическую детерминацию этого признака. У масаев, например, жи
вущих в условиях жаркого климата Африки, обнаружено сокра
щение синтеза холестерина и снижение его содержания в сыворот
ке крови, несмотря на высокий процент животных жиров в ю
рационе (Но Kang е? аі., 1971). Есть также данные о том, что }
различных этнических групп, имеющих одну диету, уровень холе
стерина различен (Алексеева, 1971). Наконец, практически полно*
отсутствие атеросклероза в эскимосских популяциях, отмечаемо*
Ч. Вильбером и В. Левиной (Wilber, Levine, 1950), также застав
ляет думать о генетических механизмах, поддерживающих уро
вень холестерина в крови.
В европейских группах, как известно, при высоком содержа
нии жиров в диете и холестерина в крови высок процент атеро
склероза.
У арктических популяций высокий уровень холестерина в кро
ви, по-видимому, служит для обеспечения энергетических процес
сов в организме. В сочетании с повышенной способностью окисле
ния жиров, отмеченной ранее, и со сравнительно слабым жироот
ложением это предположение кажется вполне убедительным.
В изученных арктических популяциях зарегистрированы до
вольно высокие значения общего белка в сыворотке крови и явно*
190
увеличение относительного содержания гамма-глобулинов, особен
но у саамов и ненцев 0 Содержание общего белка у обследованныг
нами коренных жителей Арктики колеблется от 7,2 до 8,0 г% з
мужчин и от 7,6 до 8,3 г% у женщин. У аляскинских эскимосоі
уровень общего белка также довольно высок: 7,1-7,6 (Corcoran
Rabinowitch, 1937; Mann et al., 1962).
При сопоставлении с этническими группами, живущими в
других широтах, арктические популяции обнаруживают некоторое
повышение содержания белков в сыворотке крови, что в обще»
согласуется с их высоким энергетическим уровнем.
Далее рассмотрим данные о терморегуляции арктических
популяций. Результаты исследования реакции на переохлаждение з
эскимосов и индейцев Аляски по сравнению с неграми и белымЕ
американцами показали, что аборигены Аляски имеют более
высокую температуру пальцев в течение всего периода охлаждения
(Mechan, 1955). Подобные результаты получены для
индейцев-атапасков и другими авторами (Eisner et al., 1960). В исследованиях
с Холодовым стрессом» проведенных на оседлых и кочевых
лопарях и европеоидном населении Норвегии, более высокая
температура ног обнаружена у лопарей, у них же отмечена большая
стабильность уровня метаболизма в условиях охлаждения, чем у
европейцев (Andersen et al., 1960b; Andersen, 1963).
Более сильное наполнение тканей кровью нашел Г.М.Браув
(Brown, 1957) у эскимосов острова Саутгемптон по сравнению с
контрольной группой, состоящей из американских студентов. При
изучении циркуляции крови у лопарей во время локального
охлаждения в руке выявлена более высокая скорость кровотока, чем
у норвежских рыбаков, живущих в тех же широтах (69 град- с.ш.).
Он был гораздо выше и по сравнению с контрольной группой,
недавно поселившейся в этой местности (Krog et al., 1960). Авторы,
проводившие эксперимент, рассматривают более высокий уровень
циркуляции крови при охлаждении как общий процесс адаптация
к холоду, в основе которого лежит снижение способности сосудоі
к сужению.
Итак, большинство данных о характере терморегуляции и
газообмена дают основания утверждать об усилении энергетические
процессов у коренного населения Крайнего Севера. Не отрицал
большой роли специфического динамического действия диеты і
этих процессах, отмечу, что многие факты свидетельствуют о на
191
личии определенных, по-видимому, генетически ооусловленньг
механизмов газообмена и терморегуляции у аборигенов Севера.
Физиологическое исследование, проведенное Институтом био
логических проблем Севера в Магадане среди чукчей, эвенков ]
якутов Заполярья, во многих чертах подтвердило физиологичес
кие характеристики арктических аборигенов (Веселухин, 1977)
Дополнительно к этому было установлено повышение защитны:
свойств организма у коренного населения Крайнего Севера, прояв
ляющееся в повышении скорости оседания эритроцитов, сдвиг
лейкоцитарной формулы за счет нейтрофилов, выполняющих ;
основном фагоцитарную функцию, увеличение гамма-глобулино
вой фракции сыворотки крови. Высокое содержание гамма-глобу
линовой фракции было установлено во всех изученных нами арк
тических популяциях.
У коренного населения Арктики независимо от этническоі
принадлежности выявляется много общих черт как в строениі
тела, так и в ряде физиологических характеристик. Есть основа
ние считать эти особенности "нормой" реакции на воздействи
климатических условий Крайнего Севера (Алексеева, 19756). Эт
реакция непреходяща в сравнении с той, которая наблюдается прз
акклиматизации пришлого населения в арктической зоне (Бартов
Эдхолм, 1957; Данишевский, 1968). Кратко перечислю эти особев
ности: большая плотность тела; большое развитие костно-мускуль
ной массы; прочный скелет в сочетании с повышением гаммагло
булиновой фракции сыворотки крови, усиливающей иммунны
свойства организма; преимущественно цилиндрическая форма груд
ной клетки; почти полное отсутствие астенического типа телосло
жения; высокая вентиляционная способность легких; повышенно'
содержание гемоглобина крови; относительно большое простран
ство, занимаемое костным мозгом — органом кроветворения; высо
кое содержание белков и липидов в сыворотке крови; повышенна:
способность к окислению жиров; усиление энергетических и тер
морегуляционных процессов; более высокая стабильность уровю
метаболизма в условиях переохлаждения.
В этих характеристиках физиологические и морфологически*
черты переплетаются столь тесно и оказываются столь взаимообус
лов ленными, что позволяют рассматривать морфофункциональ
ный комплекс арктических аборигенов как продукт длительно!
истории приспособления организма человека к условиям Крайней
Севера.
192
В этой связи обращает на себя внимание сокращение
изменчивости, или диапазона индивидуальных вариаций, большинства
признаков в арктических группах по сравнению с контрольной,
живущей в условиях умеренною климата и оптимальной
геохимической ситуации1. Утрата минимальных н максимальных
признаков, по-видимому, способствует стабилизации морфофункциональ-
ного комплекса в экстремальных условиях Арктики.
Морфофункциональные особенности арктического населения
проявляются еще более отчетливо при сопоставлении с
контрольной группой (рис. 44). Для нивелирования различий в весе тела
между исследованными группами выяснялось его соотношение с
другими признаками. Затем значения признаков, характерные для
арктических групп, выражались в процентах величин признаков
в контрольной группе. Все арктические популяции показали
относительное повышение теплопродукции, увеличение костно-мус-
кульной массы тела, размеров грудной клетки, содержания
гемоглобина, количества иммунных белков в сыворотке крови, уровня
минерализации скелета.
У женщин арктических популяций все эти особенности
выражены более резко, чем у мужчин. Кроме того, женщины
коренного населения Арктики характеризуются по сравнению с контролем
относительно большим жироотложением, что показывает еще
большую степень приспособления к холодному климату Крайнего
Севера.
Удивительная взаимообусловленность морфологических и
физиологических черт, типичная для коренного населения Арктики,
предполагает длительную историю развития. В нашем
распоряжении пока еще мало данных для определения времени образования
адаптивных комплексов, характерных для той или иной
экологической ниши. Проблема хронологии адаптивных типов будет
обсуждена в специальном разделе книги. Здесь же следует отметить,
что особенности биологической адаптации коренного населения
Крайнего Севера формировались на протяжении обозримой
человеческой истории в условиях экстремального климата у
генетически независимых групп, заселявших территорию Арктики и Субар-
ктики, первоначально обитавших в разных местах.
В качестве контрольной группы взято русское население Черноземной полосы
(Курская обл., с. Рождественка).
19а
1 1 1 I I 1 I 1 I
85 90 95 WO 105 HO 115 120 125%
Рис* 44. Морфофизиологическая характеристика арктического
адаптивного типа. Вертикальной линией (в центре) изображена контрольном группа,
в которой величина каждого признака принята за 100%. Отклонения от
контрольной группы выражаются в процентах:
1 - удельная поверхность тела; 2 - обхват груди по отношению к длине тела; 3 - косгно-
мускульная масса тела; 4 - средняя жировая складка по отношению к массе тела; 5 - общий
белок сыворотки крови по отношению к массе тела; 6* - гамма-глобуликовая фракция
сыворотки крови по отношению к массе тела; 7 - холестерин сыворотки крови по отношению к массе
тела; 8 - минерализация скелета по отношению к массе тела; 9 - длина ноги по отношению к
длине корпуса; 10 - теплопродукция по отношению к массе тела. / - мужчины; II - женщины.
Наши знания о движущих силах адаптивного процесса пока не
позволяют нам выйти за пределы предположений. Генетическая
детерминация некоторых приспособительных реакций дает
основание для предположения о роли отбора в формировании
адаптивного комплекса в определенных условиях среды обитания. Можно
думать, что люди, обладающие повышенной способностью к
терморегуляции и окислению жиров, повышенной скоростью
кроветворения, специфическими признаками тела, уменьшающими
теплоотдачу в условиях арктического климата, имеют преимущества
перед теми, кто не отличается этими качествами.
Известным подтверждением этого предположения могут
служить данные о некоторой специфике наследственных признаков у
населения высоких широт (Золотарева, 1969; Шереметьева,
Рынков, 1978).
194
3. АНТРОПОЭКОЛОГИЧЕСКИЕ СВЯЗИ
НА ТЕРРИТОРИИ СЕВЕРНОЙ АЗИИ
Специфика морфофизиологического комплекса населения
Крайнего Севера особенно наглядно выявляется при оценке связи
антропологических признаков с климатическими характеристиками
обширного региона с высоким индексом "суровости" погоды. Этоі
индекс был вычислен Бодманом для оценки комплексного
влияния особенностей климата на человеческий организм (Назарова
1959). Индекс вычисляется по формуле: S = (1-0,04 t)(l+0,272 v).
где t - температура воздуха, °С; v - скорость ветра, м/с.
Недостаток индекса Бодмана состоит в том, что он несколько смягчаеі
действительную суровость погоды в условиях сильных морозоі
при безветрии.
Правомерность поиска антропоэкологических связей не
вызывает сомнения в свете результатов панойкуменного рассмотрения
приспособительных особенностей человеческих популяций
(Алексеева, 1977). Экологическая изменчивость, отражающая
зависимость многих морфологических и физиологических признаков оі
климатических и геохимических условий, - явление, типичное
для современного человечества. В то же время наличие антропо-
климатической связи еще не свидетельствует о причинной
зависимости антропологического признака от географического. Она
устанавливается в результате оценки функциональной значимости
этой связи.
Для получения большего размаха изменчивости морфофизио-
логических и географических характеристик антропоэкологичес-
кие связи изучались не только на территории арктической зоны
Северной Азии, но и в умеренной и континентальной зонах этого
обширного района. Таким образом, к анализу были .привлечены
антропологические данные по коренному населению Чукотки,
Камчатки, Командорских островов, Якутии, Бурятии и Тувы,
обследованному в течение 1970-1978 гг. экспедициями НИИ и Музея
антропологии МГУ.
В изученном нами регионе максимальные величины индекса
"суровости" погоды характерны для Чукотки, минимальные - для
территории Якутии и Тувы. Континентальные районы Камчаткв
и Забайкалья по своему воздействию на человеческий организм.
195
судя по величине индекса гюдмана, занимают положение межд
Чукоткой и береговыми районами Камчатки.
Анализ коэффициентов ранговой корреляции климатически)
показателей с морфологическими и физиологическими признака
ми показал, что с повышением индекса "суровости" погоды увели
чиваются теплопродукция (г = 0,786), весоростовой указатель Ро
рера, т.е. относительная масса тела (г = 0,754), грудной указателі
и обхват груди (г = 0,655 и 0,631), содержание холестерина в кро
ви (г — 0,809) снижается артериальное давление (для систоличес
кого г = -0,771, для диастолического - г = -0,970). Все эти коэф
фициенты корреляции достоверны при р = 0,05.
Географически усиление "суровости" погоды и влажности при
ходится на Чукотку и береговые районы Камчатки. Следователь
но, охарактеризованный выше морфофизиологический комплек<
можно связать с чукчами, эскимосами, алеутами, коряками и итель
менами.
Напротив, с понижением индекса Бодмана, уменьшением влаж
ности и увеличением амплитуды температурных колебаний, т.е. <
усилением признаков континентального климата, связывается понже
ние грудного указателя (г = -0,714) и обхвата груди (г = -0,667)
уменьшение относительной длины ноги (г — -0,857) и уровня ми
нерализации скелета (для минерализации 3-го луча левой кисти і
= -0,667), увеличение средней жировой складки (г = 0,673), отно
сительной длины руки (г = 0,929), диастолического давления (г =
0,750).
Выраженной континентальностью климата в исследование)»
регионе характеризуются территории Якутии, Бурятии, Тувы і
внутренних районов Камчатки, следовательно, отмеченный вы-ше
морфологический комплекс может быть обнаружен у якутов, бу
рят, тувинцев и эвенов Камчатки.
Картина сопряженности отдельных антропологических при
знаков с территорией может быть дополнена данными о связяз
интегральных морфофизиологических характеристик, каковымі
являются типы телосложения, с показателем холодового стресса.
Анализ коэффициентов корреляции типа телосложения с ин
дексом Бодмана на территории бывшего СССР выявил следующее
В мужских группах обнаружена достоверная позитивная связі
между индексом Бодмана и частотой встречаемости мускульногс
196
типа и достоверная негативная - с частотой грудного и брюшного
типов телосложения- Неопределенный тип практически не дает
никакой связи с индексом.
В женских группах корреляции имеют подобную
направленность. Только здесь индекс Бодмана дает достоверную связь с эв-
рипластическим и неопределенным типами телосложения.
Остальные коэффициенты корреляции, хотя и не являются
достоверными, вскрывают явную тенденцию увеличения мускульного
компонента в связи с возрастанием индекса Бодмана и уменьшения
грудного и брюшного компонентов1 (табл. 27).
Таблица 27
Коэффициенты корреляции частот встречаемости различных типов
телосложения с индексом Бодмана
Мужчины
тия телосложения
Грудной
Грудно-мускульный
Мускульно-грудной
Мускульный
Мускульно-брюшноЙ
Брюшно-мускулъный
Брюшной
Неопределенный
г
-0,62*
0,59
-0,70
0,06
Женщины
тип телосложения
Астенический
Стенопластический
Мезопластяческий
Пикнический
ЭврипластическнЙ
Неопределенный
г
0,29
0,40
0,21
0,41
0,73**
0,61
* Достоверно при р^О.Об.
** Достоверно при p«=0,01.
Атлетический и субатлетический типы у женщин в обработку не брались, таг
как во всех группах они довольно редки.
197
итак, на основании достоверных данных, полученных на муж
ских группах, и выявленных тенденций в женских группах моя
но сделать вывод о преимущественном развитии мускульного ком
понентав более суровых климатических условиях, в то время каі
грудной и брюшной чаще встречаются в районах более умеренно
го и теплого климата.
Не исключено, что при расселении человечества по ойкумев
экстремальность среды играла роль селективного фактора, физи
чески усиливающего человеческие популяции (Алексеева, Добро
ванова, 1980).
Таким образом, антропоэкологические связи, характерные дл
Северной Азии, находят реальное воплощение в морфологическоі
и физиологической специфике коренных обитателей Чукотки (ар
ктический морфофункциональный комплекс) и Якутии, Бурята
и Тувы (континентальный комплекс). Особенности каждого коми
лекса проявляются на определенных территориях у различны:
этнических групп и в значительной мере носят адаптивный харак
тер.
Население внутренних континентальных районов Камчаткі
(эвены) по комплексу морфофункциональных черт тяготеет к ко
ренным жителям континентальных районов Северной Азии, насе
ление прибрежных районов Камчатки и Командорских острово
(ительмены, коряки, алеуты) - к обитателям Чукотского полуосі
рова.
Как же интерпретировать эти связи?
Существует несколько исследований, посвященных оценке кор
реляций ряда антропологических признаков с географическим]
факторами среды. Так, показана связь носового указателя с темпе
ратурным режимом и влажностью среды (Thomson, Buxton, 1923
Davies, 1932; Weiner, 1954); основного обмена и массы тела с теля
пературой (Roberts, 1952, 1953); уровня минерализации и разме
ров скелета с содержанием макро- и микроэлементов в почва:
(Alexeeva, 1968; Павловский, 1970). Экологические связи обнару
жены не только для отдельных признаков, но и для их комплек
сов (Крупник, 1973, 1977, 1989).
Таким образом, экологическая изменчивость - явление, харак
терное для современного человечества. В то же время конкретны
связи и их интерпретация неоднократно подвергались критике
Один из аргументов в пользу отрицательного отношения к гипоте
198
зам, построенным на основе антропоэкологических связей,
заключается в том, что сопоставление антропологических признаков с
климатическими показателями должно проводиться лишь для
групп, однородных по происхождению или этнической
принадлежности (Бунак, 1972). Сложность соблюдения подобного
условия очевидна. Когда это возможно, территориальные и
климатические характеристики бывают малоизменчивы, и поэтому картина
антропоэкологических связей недостаточно убедительна.
Так, например, произошло с данными, касающимися анализа
географической изменчивости относительной массы тела в
нескольких популяциях Сибири в связи с температурным фактором
(Давыдова, 1972). Относительная масса тела - признак,
обладающий огромной географической изменчивостью на территории
ойкумены. Он разделяет популяции тропических и внетропических
широт, но оказывается неинформативным не только на
территории Сибири, но и во всем умеренном поясе (Алексеева, 1977).
Совершенно очевидно, что негативный результат исследований
получен вследствие малой дифференцированности территории по
климатическим характеристикам, а также из-за недостаточной
изученности характера панойкуменной изменчивости относительной
массы тела.
При сложности соблюдения условий для изучения
антропоэкологических связей групп, генетически близких, но живущих в
резко различных географических регионах, возможно заменить
его изучением различных по происхождению популяций,
живущих в одних и тех же географических условиях (Алексеева, Бала-
хонова, 1979).
Лекция IX
ПРИРОДА АДАПТИВНОСТИ У ЧЕЛОВЕКА
Адаптивные типы у человека. Экотипы у животнх и растений.
1. АДАПТИВНЫЕ ТИПЫ
Человек как вид характеризуется огромной изменчивостью
морфологических и физиологических признаков. Эту
изменчивость можно трактовать двояко: с одной стороны, это биологичес-
199
кая предпосылка паноикуменного существования человечества, <
другой - отражение каких-то внешних влияний по мере расселе
ния его по земному шару. Движущей силой миграционных про
цессов, сталкивающих человечество с новыми географическим*
условиями, всегда были исторические причины; приспособление
же к новым экологическим нишам осуществлялось не только (
помощью технологических усовершенствований, но и в результа
те биологических перестроек, касающихся как функциональных
так и структурных систем организма.
Можно предположить, что изменчивость в человеческих попу
ляциях нарастала по мере заселения земного шара; можно также
представить себе реализацию потенциальных приспособительные
возможностей, характерных для вида Homo sapiens в целом; мож
но, наконец, имея в виду различие ареалов у разных рас и наро
дов, высказать соображение о различных возможностях
приспособления к географической среде обитания. В любом случае мк
имеем дело с резервом адаптивных возможностей, присущих вида
в целом. И если оценить историю человечества под биологическим
углом зрения, то она представляется как процесс
аккумулирования этих возможностей.
Человеческие популяции проявляют определенную
реактивную изменчивость в соответствии с влиянием окружающей их
естественной среды. Судя по характеру морфофункциональных
особенностей популяций, живущих в определенных географических
условиях, эта реакция носит приспособительный характер, что в
дает основание для введения понятия адаптивного типа,
представляющего собой норму биологической реакции на комплекс
условий окружающей среды, обеспечивающей состояние равновесия
популяций с этой средой и находящей внешнее выражение в
морфофункциональных особенностях популяций.
Адаптивный тип независим от расовой и этнической
принадлежности. В одних и тех же геоклиматических условиях разные
по происхождению группы имеют одно и то же направление
приспособительных реакций, так же как в различных условиях
обитания близкие в генетическом отношении группы демонстрируют
морфофункциональные различия в соответствии с воздействием
окружающей среды.
По сходству морфофункциональных характеристик могут быть
выделены регионы-аналоги. Например, в адаптивном комплексе
200
арктических аборигенов проявляются черты приспособления к
гипоксии и низким температурам, так же как в высокогорье, хотя
природа гипоксии в этих регионах разная.
Адаптивные типы, проявляясь в пределах вида Homo sapiens,
не представляют собой экологически специализированных форм,
они выражаются в виде тенденции к изменению физиологических
и морфологических черт в направлении, наиболее благоприятном
для существования в определенной среде и не препятствующем
возможности существования в других экологических нишах.
Адаптивные реакции осуществляются в двух формах: общей и
специфической. Общей особенностью адаптивных типов можно,
по-видимому, считать повышение сопротивляемости организме
неблагоприятным условиям среды. Это выражается в увеличении
костно-мускульной массы тела. Специфические же реакции очен*
разнообразны. В одном случае это повышение теплопродукции, х
другом - относительной поверхности испарения, в третьем - в
того и другого признака и т.п. Именно эти специфические
реакции и дают право называть выделенные морфофункциональные
комплексы "арктическим", "высокогорным", "континентальным",
"тропическим" и т.п. Адаптивный тип - это лишь одна из форм
приспособительной реакции у человека - биологическая.
Как можно представить себе природу адаптивного типа? Он не
равноценен расе, поскольку проявляется независимо от этноса в
расы в сходных условиях существования человеческих популя
ций. Адаптивный тип - это норма реакции, конвергентно возни
кающая в сходных условиях обитания в популяциях, которые
могут быть генетически не связаны между собой, в то время кал
раса предполагает общее происхождение всех входящих в не«
групп, живущих на определенной территории.
Сходный комплекс приспособительных черт в тропической
зоне находим у европеоидного населения Индии, австралийцев е
аборигенов Африки; на Крайнем Севере единое направление
приспособительных реакций показывают саамы, ненцы, чукчи и эски
мосы. То же самое относится и к коренным жителям пустынь, раз
личающимся в этническом и расовом отношениях.
Основные расово-диагностические черты, имеющие, безуслов
но, приспособительный характер (например, цвет кожи, форме
волос, форма носа, развитие слизистой оболочки губ), сформирова
20]
лись на заре человеческой истории, в эпоху верхнего палеолита
Адаптивные же типы, судя по их географической приуроченности
формировались на протяжении всей истории человечества. Мно
гие районы ойкумены стали обитаемы в сравнительно недавнее (і
масштабе всей истории вида) время, и тем не менее адаптивны?
черты уже успели за это время оформиться вполне четко. И разно
образие человеческих популяций по морфофункциональным осо
бенностям, по-видимому, значительно возросло с расселением че
ловека по земному шару.
Наряду с этим мы сталкиваемся с сужением межгрупповой
изменчивости признаков в связи с зональностью. Тропическая зона
как мы помним, а в ее пределах Африка в особенности, поражаеп
огромной дисперсией величин морфологических признаков. Эт?
дисперсия заметно сужается к северу, т.е. в направлении, в кото
ром происходило расселение человечества.
, С точки зрения истории развития такой важной черты
человеческого вида, как адаптивность, тропическая зона представляется
колыбелью адаптивных возможностей человечества. Недаром с
тропической зоной связываются истоки вида Homo sapiens (Рогин
ский, 1969).
Если в этой связи обратиться к хронологии адаптивных типов,
то тропический Тип представляется наиболее древним, по
отношению к которому остальные (высокогорный, умеренный,
континентальный, аридный, арктический) могут рассматриваться как
дочерние. Образуется своеобразное древо, в котором ответвления
определяются историей развития вида и хронологически могуі
быть, по-видимому, с большей или меньшей точностью
датированы, а специфика каждого ответвления - соотношением
биологических и социальных закономерностей в конкретной
экологической нише.
Уместно задать вопрос: какая из зон тропического пояса можеі
рассматриваться как исходная для формирования тропического
адаптивного типа? Очевидно та, в которой термический режим
(температура, влажность) оптимален, а морфологическое и
физиологическое разнообразие достаточно для обеспечения
пластичности вида. По-видимому, этим требованиям отвечают
континентальные, но не аридные зоны тропического пояса, население которых
отличается наибольшей вариабельностью антропологических
признаков и которые наиболее густо заселены. Что касается аридной
202
зоны и зоны влажных тропических лесов, то они должны
рассматриваться по отношению к континентальным тропическим зонам
как экстремальные или, во всяком случае, менее пригодные для
жизни. И действительно, именно там обнаруживаются самые
низкорослые и самые высокорослые племена, по-видимому, несущие
черты некоторой специализации. В связи с этим хочется вновь
обратиться к интереснейшим исследованиям Е. Еронье (Crognier,
1973), показавшим, что наибольшей плодовитостью в группе сара-
маджингай (Чад) отличаются семьи, в которых родители имеют
размеры тела, приближающиеся к средним для этой группы.
Расселение человечества по земному шару привело к освоению
новых территорий и формированию новых адаптивных типов, в
которых реализовалась изменчивость, присущая исходному
адаптивному типу. Увеличение массы и плотности тела, усиление
метаболизма и изменение связанных с этим физиологических
признаков, типичные для населения внетропических широт, связаны
с более поздними этапами человеческой истории.
Сужение межгрупповой изменчивости морфофизиологических
черт во внетропических широтах сопровождается и уменьшением
диапазона вариабельности внутри популяций; особенно это
заметно в арктической зоне. Создается впечатление, что в процессе
освоения ойкумены происходила утрата крайних величин
признаков, что увеличивало фонд межгрупповой изменчивости.
Гипотеза адаптивных типов представляет собой попытку
показать экологическую изменчивость в пределах современного
человечества.
На основе конкретных данных, относящихся к аборигенному
населению различных экологических ниш, можно сделать
заключение о том, что экологическая дифференциация человечества
осуществляется на разных уровнях морфофизиологической
специфичности. В основе первого уровня лежат скорости
энергетических процессов. Замедленные темпы энергетики характерны для
населения тропических широт, повышенные - для населения
внетропических широт.
Каждый из этих типов (условно назовем их адаптивными
типами I порядка) подразделяется на более дробные единицы на
основе комплекса общих морфофизиологических черт, проявляющихся
у населения в сходных по географическим характеристикам
экологических нишах, так называемых экологических
нишах-аналогах. Это второй уровень дифференциации.
203
204
Так, в пределах адаптивного типа внетропических широт мо
гут быть выделены группы сходных адаптивных типов II порядка
Одну группу составляют арктический и горный типы, вторую -
континентальный и аридный, третью - умеренный. Они в свок
очередь подразделяются на адаптивные типы III порядка на осно
ве специфического морфофункционального комплекса. Так, выде
ляются арктический, горный, континентальный, аридный, уме
ренный типы, которые могут распадаться на еще более дробные
адаптивные типы IV порядка (рис. 45).
Внутри адаптивного типа тропических широт выделяются ху-
мидный, высокогорный, аридный и саванный адаптивные типы.
По-видимому, и в пределах тропического адаптивного типа могуі
быть выделены адаптивные типы II, III и IV порядков. Эта
условная классификационная схема должна обратить внимание на
сложный соподчиненный характер адаптивных особенностей в
человеческих популяциях, хотя очевидно, что вопросы классификация
адаптивных типов пока еще очень далеки от своего решения.
Одним из возможных направлений дальнейшего изучения
взаимодействия человеческих популяций со средой представляется
соотнесение адаптивного типа как нормы биологической реакции
на среду обитания с хозяйственно-культурным типом как нормой
социальной реакции. Во всяком случае, априори можно
предположить, что чем совершеннее хозяйственно-культурный тип, тем
большее число связей со средой осуществляется через социальные
каналы и тем нейтральнее комплекс морфофункциональных черт
населения, входящего в этот хозяйственно-культурный комплекс.
Гипотеза адаптивных типов вскрывает характер
экологической изменчивости, присущей современному человечеству. В ее
основе лежит реактивность человеческих популяций по отношению
к среде обитания. В связи с этим естественно задать вопрос: как
же формировались адаптивные типы и что они собой
представляют - обратимый акклиматизационный эффект, развитие в
онтогенезе или генетически закрепленное образование?
2. АДАПТАЦИЯ И АККЛИМАТИЗАЦИЯ
Исследуя процессы биологической адаптации в популяциях
человека, естественно обратиться к трактовке этого явления в
биологии.
205
В давней, но не устаревшей и по сей день статье В.В. Станчин
ский (1933) обратил внимание на два явления, отражающих важ
нейшие реакции организма на среду, которые, согласно принято!
в то время терминологии, он назвал феноакклиматизацией и гено
акклиматизацией.
Под феноакклиматизацией понималась непосредственная реак
ция на среду, выражающаяся в фенотипических сдвигах и ком
пенсаторных физиологических изменениях, которые позволяю*]
сохранить состояние равновесия организма с новой средой. Taieaj
акклиматизация не выходит за пределы потенциальных экологи
ческих особенностей данного вида и его генотипа и проходит бе;
генотипических изменений. По мнению В.В. Станчинского, у эко
логически пластичных видов приспособительные фенотипически*
изменения могут образовываться в первом же акклиматизирую
щемся поколении. В случае меньшей пластичности приспособле
ния развиваются в последующих поколениях. Такая изменчивое
ть, названная морфотипической, является ненаследственной реак
цией на воздействие внешней среды. К примерам морфотипичес
кой изменчивости, вызванной акклиматизацией, Станчинский от
носит более мощный шерстный покров у животных-переселенце!
из южных мест, акклиматизирующихся на севере, более высокие
уровень гемоглобина у крупного рогатого скота, переселившегося
с равнины в горную местность.
Наряду с ненаследственной акклиматизацией В.В. Станчинс
кий отмечает такой вид акклиматизации, при котором в ряду по
колений вырабатываются наследственные генотипические приспо
собительные свойства применительно к новым условиям. Этот ъщ.
приспособительной изменчивости был назван геноакклиматизаци
ей.
По мнению многих теоретиков эволюционного учения, эволю
ция всего органического мира представляет собой, по сути дела
адаптациогенез, т.е. непрерывный ряд наследственных приспособ
лений к разнообразным и изменяющимся условиям существования
(Шмальгаузен, 1938, 1940, 1946). Что касается фенотипическю
акклиматизационных сдвигов (или, как их принято называть
морфозов и модификаций), то их роль в эволюции несопоставим*
с ролью адаптивных изменений. Исследуя роль фенотипа в эволю
ции, в том числе и обратимых фенотипических изменений, М.М
Камшилов (1967) писал: "При изменении условий некоторые on
206
ределенные изменения фенотипа, будучи в принципе обратимы
ми, в силу необратимого изменения условий могут стать необра
таимыми фактически? (с. 114).
Остановимся подробнее на этом вопросе. Интерес к эволюцион
ной роли модификаций особенно усилился в последнее десятиле
тие. М.А. Шишкин (1984), оценивая значение фенотипическиэ
реакций в эволюционном процессе, обратил внимание на то, что і
биологической науке существуют два противоположных мненш
относительно роли модификаций в эволюции органического мира
Одно из них, восходящее к классической генетической теории эво
люции, полагает, что модификации представляют собой ненаслед
ственные изменения и в связи с этим не имеют эволюционногс
значения- Попадая в сферу действий отбора, они лишь замедляюі
его темпы.
В противовес этому сторонники теории стабилизирующего
отбора утверждают, что модификации являются не тормозом эволю
ции, а средством, позволяющим организмам выжить до появление
наследственных изменений, сходных с данной модификацией.
Согласно теории стабилизирующего отбора феноадаптации заменя
ются геноадаптациями лишь в относительно устойчивой внешнев
среде. В случае же изменяющихся условий приспособление орга
низма идет по пути усиления выгодных организму феноадаптацив
(Кирпичников, 1940). Более того, Г.Ф. Гаузе (1984) на основе экс
периментального экологического исследования простейших
организмов приходит к заключению об обратной связи между
величиной морфоза, характеризующей пластичность вида или расы, в
величиной генокопии, характеризующей степень их
специализации. "Можно доказать, что прямая соматическая перестройке
имеет приспособительное значение для организма. Если дальней
шее усиление соматического сдвига путем геновариации окажет
ся еще более полезным, то естественный отбор позаботится с
том, чтобы геноадаптация с течением времени стала на месте
феноадаптации"', - пишет Г.Ф. Гаузе в своем фундаментально»
труде, посвященном экологическим аспектам происхождения
видов, замечая при этом, что процесс замены морфоза
наследственным приспособлением "не идет дальше лишь самого общего сома
тического соответствия их друг другу" (1984, с. 101). В таков
трактовке модификаций их эволюционной роли не придается се
рьезного значения.
207
М.А. Шишкин обратил внимание на противоречие между об
щими положениями теории стабилизирующего отбора и ее выво
дами относительно роли модификаций в эволюции организмов. (
точки зрения этого автора, "модификация - это различные спосс
бы реализации в онтогенезе одного и того же генотипа в разны:
условиях обитания" (Шишкин, 1984, с. 199). В классах феноти
пов, которые имеются в популяциях, нет разделения на генокоши
(мутации) и фенокопии (модификации), т.е. наследственные и не
наследственные изменения. Они различаются лишь по степені
морфогенетической устойчивости, которая обусловлена генетичес
ки. Все сходные адаптивно ценные фенотипы одинаково вовлека
ются в отбор, в потомстве происходит элиминация отклоняющих
ся от наиболее приспособленного фенотипа форм, тем самым дос
тигается наибольшая устойчивость в реализации нормы. В итог
формируется новый генотип, обеспечивающий эту устойчивость. ]
этом процессе, названном Уоддингтоном генетической ассимиля
цией, М.А. Шишкин видит "единственно правильное описаны
механизма стабилизирующего отбора на генетическом уровне
позволяющее объяснить замену внешних морфогенетических фан
торов внутренними в становлении новых адаптации" (1984, с
204).
Таким образом, однотипные индивидуальные реакции (моди
фикации) являются материалом для отбора. Такова их эволюциов
ная роль.
Исходя из современных представлений о соотношении феноти
па и генотипа в эволюции, под акклиматизацией следует понимат:
те однотипные индивидуальные реакции, которые являются пер
вой и непосредственной приспособительной реакцией на изменяю
щиеся условия среды, и соответственно под адаптацией - новув
наследственно закрепленную форму, возникающую в результат
элиминации неустойчивых в данных условиях фенотипов.
Итак, что же представляет собой тот морфофизио логически)
комплекс, который получил название адаптивного типа?
Если мы вновь обратимся к данным, изложенным в главах
касающихся адаптивных особенностей населения различных кли
матических зон, то среди них найдем немало таких, которые зас
тавляют думать о наследственной природе адаптивных типов.
Вспомним увеличение концентрации медленно мигрирующи:
белков-трансферринов, связанных с понижением основного обме
208
на в тропических широтах, интенсивность потоотделения,
специфику регуляции водно-солевого обмена, понижение уровня
метаболизма, достигаемое уменьшением мышечной массы тела и
понижением концентрации АТФ (признаки, характерные для коренных
жителей тропических широт), сокращение синтеза эндогенного
холестерина в условиях тропиков, ослабление способности сосудов
к сужению в условиях переохлаждения у аборигенов Арктики,
повышенную способность к окислению жиров у них же,
способность к более быстрому переходу гемоглобина в оксигемоглобин в
условиях кислородной недостаточности у коренных жителей
высокогорья по сравнению с пришлым населением, сужение
диапазона внутригрупповой изменчивости признаков по мере освоения
ойкумены и некоторые другие особенности.
В свете этих данных адаптивные типы представляются
результатом длительной истории приспособления популяций человека к
условиям различных экологических ниш. Можно представить, что
те особи, которые обладали оптимальным для данного биотопа
соотношением морфофункциональных черт, имели больше
оснований оставить после себя потомство. Данные Е. Кронье (Crognier,
1973) об избирательной плодовитости сара-маджингай могут, как
мне кажется, служить подтверждением этого предположения.
Примеров доказательства наследственной обусловленности
многих особенностей биологии человеческих популяций немало. К их
числу относятся многочисленные данные об избирательном
характере разных заболеваний в зависимости от той или иной группы
крови (Эфроимсон, 1971; Giblett, 1969), о различной
иммунологической реактивности организмов - представителей разных рас.
Л.А. Зильбер (1958) приводит данные о высокой резистентности
негроидов к инфекционным заболеваниям, распространенным в
Африке, и восприимчивости к заболеваниям, принесенным
европейцами. Медицинская и биологическая литература начала
нашего века подна такого рода наблюдениями (Бунак, 1924).
Туберкулез, сифилис и некоторые другие заболевания нанесли огромный
урон аборигенному населению территорий, колонизованных
европейцами. Общеизвестно и отсутствие резистентности у европейцев,
впервые соприкоснувшихся с тропическими болезнями.
Наследственный характер иммунитета подтверждается
наследуемостью групп крови (Эфроимсон, 1968, 1971).
209
О наследственной природе морфологической изменчивости че
ловеческих популяций свидетельствует постоянство физическоя
типа во времени, зафиксированное многими палеоантропологичес
кими исследованиями, и промежуточное положение антропологи
ческих особенностей народов смешанного происхождения межд
исходными группами (Алексеев, 1979).
Подтверждение наследственной основы адаптивных процессов
в современных человеческих популяциях находим в исследовани
ях адаптивной значимости биохимического полиморфизма. Уста
новлена зависимость распределения фенотипических и генньп
частот ряда биохимических локусов от комплекса климатогеогра
фических параметров (Спицын, 1985).
Так, изменение частоты гена TfD1 системы трансферрина н;
территории ойкумены обусловлено динамикой суммарной солнеч
ной радиации и годовой суммой атмосферных осадков. Напомню
что высокая частота медленно мигрирующих трансферринов свя
зана с тропическими широтами. Функция трансферрина в орга
низме заключается в переносе железа из плазмы крови к клетка*
органов и тканей (Giblett, 1969). Есть данные о связи TfD1 с тер
морегуляцией (Walter, Bajatzdeh, 1971).
Установлена зависимость концентрации аллелей Ра и Р систем
ы эритроцитарной кислой фосфатазы (АсР) от амплитуды колеба
ний температуры среды и суммарной солнечной радиации. Про
порция гетерозигот АсР АВ этой системы возрастает со снижени
ем интенсивности суммарной солнечной радиации.
Определена также закономерность географического распрост
ранения щелочной фосфатазы. Частоты фенотипов Ррі и Рр2 cbj
заны с широтой местности.
Большого внимания заслуживает характер географической из
менчивости частот гена Go2 и фенотипа Gc2"1 сывороточного белк;
группоспецифического компонента, обнаруженный В.А.Спицы
ным (1985). Функциональная роль этого белка заключается в свя
зывании и переносе витамина D3 и 25-гидроксивитамина D3 ]
организме, а также в образовании меланина. Выяснено, что измен
чивость группоспецифического компонента в географическом про
странстве определяется совокупным воздействием среднегодовоі
температуры, амплитуды колебаний и интенсивности солнечно]
радиации. Выявлена связь интенсивности пигментации с особенно
стями распространения факторов системы Gc, в наибольшей степе
210
ни проявляющаяся в тропических широтах- Географическая ком
понента обнаруживается и в изменчивости концентрации аллелеі
РМ-11 и РМ-22 системы фосфоглюкомутазы-1, функционально свя
занной с углеводным обменом. Их корреляция со средней темпера
турой января наиболее тесно проявляется в циркумполярной обла
сти и на побережье Тихого океана.
Итак, особенности географической изменчивости
вышеприведенных генетических систем и их высокая функциональная ролі
в обеспечении жизнедеятельности организма позволяют сделаті
заключение о значительной доле наследственности в закономерно
стях пространственного распределения характеристик обмена ве
ществ. В данном случае речь идет об основном, минеральном, ви
таминном и углеводном обменах веществ. Что касается липидногс
и белкового обменов, которые в известной мере определяют разви
тие соматических особенностей, то их наследственная обусловлен
ность у человека доказывается такими наблюдениями, как пони
жение синтеза эндогенного холестерина у африканских популя
ций, специфика усвояемости белков у популяций Арктики, а так
же связью уровней белков и липидов с расовой принадлежностью.
Исключить генетическую компоненту в их географическом рас
пределении у нас нет основания.
При обсуждении вопросов наследственной природы адалтив
ных процессов в популяциях человека исключительную ценності
представляют результаты популяционно-генетического исследова
ния, проведенного Ю.Г. Рычковым и В.А. Шереметьевой (1978|
среди коренных народов Северо-Восточной Азии. Исследуя про
цессы генетической дифференциации популяций этого обширногс
региона, авторы обратили внимание на то обстоятельство, что прі
весьма благоприятной для случайного дрейфа генов популяцион
ной структуре народов Берингии (изолированность популяций,
малый обмен генами, малый объем популяций) генетическая диф
ференциация определяется не дрейфом генов и не интенсивностьк
генных миграций, а селективным давлением окружающей экстре
мальной среды, генетическая адаптация популяций к которому
должна стабилизировать процесс дальнейшей дифференциации е
противостоять случайному дрейфу генов, так как в противном слу
чае нарушится генетическая приспособленность популяций. Оце
нивая время формирования того генетического комплекса, кото
рый отличает народы Берингии от других народов Северной Азии
211
и соотнося результаты популяционно-генетического исследование
с историко-лингвистическими данными, авторы приходят к зак
лючению, что этот комплекс сформировался на начальных эталаз
их многотысячелетней истории.
Совершенно очевидно, что явление биологической адаптации
отмечаемое в современном человечестве, наследственно обусловле
но и представляет собой длительный исторический процесс.
Не менее очевидно также, что предпринятые в недавнем про
шлом и весьма обширные исследования процесса приспособление
пришлых контингентов людей в новых экологических условиях (і
такого рода исследования приобретают особо важное значение і
связи с освоением новых территорий) являются не чем иным, каі
исследованием процесса акклиматизации.
Литература, посвященная исследованиям процесса акклимати
зации, сейчас довольно обширна. Особый интерес представлякл
исследования физиологической акклиматизации к условиям Арк
тики, высокогорья, пустыни, тропиков и других климатическиэ
регионов, отраженные в фундаментальном труде "Экологическая
физиология человека" (1980).
Обращает на себя внимание характер приспособительных реак
ций в типе обмена веществ и других физиологических признакам
у приезжих акклиматизирующихся лиц: сдвиг во многих физио
логических процессах происходит в направлении характеристик
типичных для коренного населения тех регионов, в которых ис
следуются процессы акклиматизации. Едва ли не единственное
исключение - артериальное давление, которое в большинстве слу
чаев повышается, особенно в начале акклиматизации, как непос
редственная реакция на ситуацию стресса.
Рассмотрение акклиматизационной изменчивости под утлой
зрения гипотезы адаптивных типов приводит к мысли о том, чте
оптимальный акклиматизационный эффект может быть достигну]
при наибольшем сходстве морфофизиологических особенностеі
пришлого и коренного населения.
Для антрополога, изучающего процессы адаптации человечес
ких популяций во времени и в пространстве, данные об акклима
тизации чрезвычайно важны для понимания начальных этапоі
адаптации. Надо думать, что на ранних стадиях образования стой
ких приспособительных реакций акклиматизационный эффекч
был первой и непосредственной реакцией на воздействие есте
ственной среды.
212
3. О СООТНОШЕНИИ АДАПТИВНЫХ ТИПОВ ЧЕЛОВЕКА
С ЭКОЛОГИЧЕСКИМИ ФОРМАМИ ЖИВОТНЫХ И РАСТЕНИЙ
Черты адаптивности, присущие человечеству в целом, в той
части, которая касается морфофункциональных перестроек, нахо
дят себе аналогии в закономерностях приспособления наземныэ
животных к различным ландшафтным условиям. Это формирова
ние сходных биологических особенностей у различных видов, оби
тающих в одной и той же среде; подчинение климатическим пра
вилам и отклонение от них в случае недостаточной энергетическое
обеспеченности вида; темная окраска некоторых видов, обитаю
щих в жарком климате, выполняющая защитную функцию (по
глощение избытка вредных коротковолновых лучей), в сочетание
со специфическими кожными образованиями, регулирующим!
температуру тела; сужение диапазона изменчивости признаков с
усилением экстремности гидротермического режима и т.п. Прав
да, у наземных животных адаптация к ландшафтно-зональнык
условиям представляет своеобразную экологическую специали
зацию, ограничивающую обитание в других условиях (Чернов
1974).
Закономерности, отмеченные у наземных млекопитающих,
открывают, как мне кажется, пути подхода к объяснению некото
рых отклонений от общего направления адаптивных процессов г
различных экологических нишах, например, терморегуляционные
преимущества крупных форм не только в холодном, но и в жар
ком сухом климате, так как расход воды на единицу массы для
охлаждения резко возрастает с уменьшением размеров теле
(Шмидт-Нильсен, 1972). Воздерживаясь от прямых аналогий,
отмечу, что и для человеческих популяций можно предположить
существование разных морфологических типов в одних и тех же
условиях среды, если эти типы в равной мере обеспечивают
терморегуляционные возможности. Вспомним относительно более
крупные, чем у других пустынных жителей, размеры тела у коренных
народов внетропических пустынь и австралийцев племени коката.
Из закономерностей, отмеченных для наземных
млекопитающих, в свете которых могут быть рассмотрены особенности
адаптивных реакций у человека, следует остановиться на активном в
пассивном характере адаптации (Чернов, 1974). К активным адап
тациям относятся приспособления, характеризующиеся активньта
213
изоирательным отношением вида к среде: высокая интенсивности
роста, развития и размножения, повышение уровня обмена, чет
кая локализация жизненного цикла. Пассивная адаптация, напро
тив, осуществляется при замедлении развития и энергетическиа
процессов. С.С. Шварц (1963) показал, что субарктическим попу
ляциям млекопитающих свойственны активные, энергетическі
менее экономные типы адаптации, а высокогорным видам, обита
ющим в условиях, близких к субарктическим, - пассивный тиг
адаптации.
Если применить представление о разных типах адаптации в
человеку, имея в виду комплекс его биологических особенностей
то для человечества наиболее распространенной формой приспо
собления к естественной среде будут активные морфофизиологи
ческие адаптации. В то же время в некоторых регионах наряду (
активными возможны и пассивные адаптации, повышающие рези
стентность организмов к неблагоприятным условиям среды и сто
собствующие более экономным энергетическим тратам. Мне ка
жется, что с этих позиций можно объяснить замедление процессов
роста и развития у высокогорных популяций.
При определенном сходстве приспособительных реакций чело
века и наземных животных в характере биологической адаптации
есть одно фундаментальное различие между ними: адаптация че
ловека к географическим условиям не представляет собой экологи
ческой специализации, ограничивающей существование в другю
ландшафтно-климатических зонах. Этой специализации препят
ствует, по-видимому, хозяйственно-культурная деятельность чело
века, многообразие его социальных связей с географической ере
дой обитания.
В заключение следует обратить внимание на принципиальное
сходство таких систематических категорий, как адаптивный тиі
у человека и экологические формы у растений и животных. Иссле
дуя явление экологической дифференциации у растений, Г, Турес
сон экспериментально доказал наследственный характер экологи
ческих форм (Лукин, 1940). До его работ большинство автороі
полагали, что экологические формы являются продуктом непос
редственного влияния условий среды на организм и не передают
ся по наследству. Турессон обнаружил в природе наследственны*
вариации видов, сгруппированные в различные типы, приурочен
214
ные к определенным местам обитания. Такие наследственные
формы он предложил называть экотипами.
Экотипы установлены многими исследователями у различных
видов и подвидов- растений и животных. Ряд систематических
единиц, в том числе и экотипов, находящихся в той или иной
степени близкого родства, представляет собой географический ряд
форм. По мнению Е.И. Лукина, "членами одного географического
ряда большей частью являются географические формы,
принадлежащие к одному и тому же виду, или виды одного и того же рода"
(1940, с. 277).
Географические ряды форм открыты у разнообразных
растительных и животных организмов. Адаптивный тип и экотип -
явления одного и того же порядка, следовательно правомерно
применить понятие "географический ряд форм" is. к человеку.
Экологический тип, по мнению Турессона, это генотипический ответ
популяции на определенные условия обитания. Адаптивный тип -
наследственно закрепленная норма реакции, обеспечивающая го-
меостаз популяции и среды. Описанные выше адаптивные типы
можно считать составляющими одного географического ряда форм
в пределах вида Homo sapiens.
Лекция X
АДАПТИВНЫЕ ТИПЫ И СРЕДА
Воздействие естественных и социальных условий
на биологию человека.
1. ПРЕДПОСЫЛКИ ИССЛЕДОВАНИЯ
КОМПЛЕКСНОГО ДЕЙСТВИЯ ЕСТЕСТВЕННЫХ
И СОЦИАЛЬНЫХ ФАКТОРОВ НА БИОЛОГИЮ ЧЕЛОВЕКА
В отечественной этнографии плодотворно развивается
концепция хозяйственно-культурных типов, впервые четко
сформулированная СП. Толстовым (1932). М.Г. Левин и Н.Н. Чебоксаров
(1955) развили и конкретизировали понятие
хозяйственно-культурного типа, дав ему следующее определение- Хозяйственно-ку-
216
льтурный тип - это исторически сложившийся комплекс хозяй
ства и культуры, типичный для народов, различных по происхож
дению, но обитающих в сходных географических условиях и нахо
дящихся на более или менее одинаковом уровне исторической
развития. В отечественной этнографической литературе предложе
на классификация хозяйственно-культурных типов и составлень
карты их распределения по земному шару (Андрианов, Чебокса
ров, 1972), прослежено их развитие во времени (Андрианов, 1968)
изучается структура (Алексеев, 1975).
В связи с содержанием понятия "хозяйственно-культурны!
тип" напомним определение адаптивного типа. Адаптивный тип -
это норма биологической реакции, выражающаяся в морфофизи
ологическом статусе и конвергентно возникающая у представите
лей разных рас в сходных географических условиях обитания
Совершенно очевидно, что одним из направлений дальнейшей
изучения взаимодействия человеческих популяций со средой пред
ставляется соотнесение адаптивного типа как нормы биологичес
кой реакции на среду обитания с хозяйственно-культурным типо»
как нормой социальной реакции.
В начале 70-х годов в биологии возникло направление, изуча
ющее конкретные формы связи биологической изменчивости чело
века с естественной и культурной окружающей средой с помощьк
количественных методов, в основе которых лежит моделирование
антропоэкологических связей. В качестве примера можно привес
ти исследования такого рода в Перуанских Андах, осуществляв
мые под руководством П. Бейкера (Baker, 1979). Авторским кол
лективом разработано несколько моделей, в основе которых лежи]
энергия, являющаяся основным требованием для функционирова
ния биотических систем, включая и человеческие популяции. По
казано, что такие характеристики, как строение тела, обмен ве
ществ, здоровье, генетическая и демографическая структура, зави
сят от энергетики. Положенный в основу моделей принцип энер
гетического гомеостаза популяции с успехом применяется в эколо
гии при характеристике биологических сообществ (Smith, 1979),
однако нельзя не признать, что подобный тип моделей может был
разработан применительно лишь к малым популяциям, стоящим
на низком уровне экономического развития и ограничивающий
свои потребности рамками натурального хозяйства.
Нельзя не признать также, что модели энергетического
гомеостаза не только могут прогнозировать перспективы дальнейшее
216
развития этих популяций, но и служить методом реконструкции
взаимоотношения человеческих сообществ и среды обитания на
различных этапах древней истории человечества-
Задачи, стоящие перед нами, диктовали иной тип
количественной оценки взаимоотношения человека с окружающей средой. В
сущности, задача сводилась к определению воздействия
совокупности естественных и социальных факторов на морфофизиологи-
ческий статус человеческих популяций. Для осуществления этой
задачи необходимо соблюдение нескольких условий. Первым
условием является наличие большой территории, разнообразной по
климатическим, ландшафтным и геохимическим
характеристикам, населенной многочисленными народами, различающимися по
антропологическим и хозяйственно-культурным
характеристикам. Второе условие предполагает полноценную информацию об
этногенетических процессах и биологической адаптации этого
населения.
Таким условиям вполне удовлетворяет территория бывшего
Советского Союза, предоставляющая неисчерпаемые возможности
для комплексных исследований взаимоотношения человека со
средой обитания. Она может служить прекрасной антропоэкологичес-
кой моделью. С целью получения информации об антропоэколо-
гических связях на территории страны, расположенной в
пределах нескольких климатических поясов, мы предприняли попытку
совмещения данных морфофункционаяьной изменчивости
коренного населения нескольких районов с хозяйственно-культурными
характеристиками. Естественно, речь идет о коренном населении,
с древнейших времен связанном с определенными территориями и
сформировавшемся в пределах этих территорий в тесном
контакте с соседними народами.
Один из вариантов классификации хозяйственно-культурных
типов, предложенный Б.В. Андриановым и Н.Н. Чебоксаровым
(1972), был использован в части, касающейся территории
бывшего СССР, для демонстрации хозяйственно-культурной
дифференциации народов бывшего Советского Союза. С этой
классификацией, нанесенной на карту, были сопоставлены территориальные
ареалы этногенетических пучков на территории бывшего СССР
(Алексеев, 1979; Алексеев, Гохман, 1984), выделенных на основе
обобщения результатов многолетних исследований процессов
этнической истории и антропологической динамики в разных рано
217
а
Q
I
а
а
Л
8:?
а
і
I
&
*
а «
И
С
«
&
с
Я
с
• «
:>
в. і
3
218
О CD
Ч Ф
219
нах нашей страны. По определению В.П. Алексеева, этногенетичес
кие пучки - это группы генетически родственных народов, имек
щих общие этногенетическио истоки.
Данные о географической изменчивости морфофизиологичес-
ких признаков были собраны в ходе многолетних экспедиций,
регулярно проводившихся антропологическими учреждениями
бывшего СССР. На карте (рис. 46) показано местоположение более
60 популяций, изученных или непосредственно под руководством
автора, или другими исследователями по тому же плану и
относящихся к различным этнотерриториальным группам. Большинство
из них проживает в экстремальных условиях.
2. КРАТКИЕ СВЕДЕНИЯ ОБ ЭТНОГЕНЕЗЕ
И ХОЗЯЙСТВЕННО-КУЛЬТУРНОМ ТИПЕ
ИССЛЕДОВАННЫХ ГРУПП
В пределах хозяйственно-культурного типа оседлых пашенных
земледельцев умеренного пояса (подтип лесов и лесостепей)
изучено несколько популяций русских и белорусов. В этногенетическом
отношении они включаются в восточноевропейский этногенети-
ческий пучок, который объединяет восточнославянские, западно-
финские и леттолитовские народы (Алексеев, 1979). Их тесное
родство, в частности наличие балтийского и финского субстратов
в этногенезе восточнославянских народов, демонстрируется как
всеми видами исторических источников, так и
антропологическими данными. Вопрос о древности восточноевропейского пучка не
может быть сейчас решен однозначно. Формирование его следует,
по-видимому, отнести к эпохе наиболее интенсивного славянского
этногенеза, т.е. к рубежу I тыс- н.э. В то же время
антропологическими исследованиями выявлены значительно более древние связи
субстратного населения, принявшего участие в этногенезе досла-
вянских народов, восходящие к эпохе неолита. В пределах
подтипа тайги с лесным подсобным промыслом того же хозяйственно-
культурного типа изучено несколько популяций русского народа,
восходящего в своем генезисе к восточноевропейскому этногенети-
ческому пучку, но с XVIII в. обитающего в субарктических
условиях.
На территории Северного Кавказа, где на рубеже XIX-XX вв.
был распространен хозяйственно-культурный тип полуоседлых в
220
полукочевых скотоводов-земледельцев аридной зоны, обследованы
равнинные и горные жители - коренное население Дагестана. В
Причерноморском районе обследованы коренные жители Абхазии,
по типу хозяйства относящиеся к оседлым пашенным
земледельцам и скотоводам теплого пояса с развитым огородничеством и
скотоводством.
Абхазы, как и народы Северного Кавказа, включаются в
северокавказский этногенетический пучок. Антропологическое,
языковое и этнографическое родство связывает многие
северокавказские народы с горными группами Грузии и закавказскими
осетинами, которые относятся к той же этногенетической общности. На
основании лексико-статистических сопоставлений формирование
общекавказского языкового единства предположительно
датируется IV тыс. до н.э., и, следовательно, обособление
северокавказского этногенетического пучка должно быть датировано более
поздним временем — эпохой энеолита и ранней бронзы (Алексеев,
1969).
На территории Средней Азии, в зоне былого распространения
хозяйственно-культурного типа кочевников и полукочевых
скотоводов аридной зоны, изучены туркмены, каракалпаки, казахи,
равнинные и горные таджики. Этногенетически они относятся к
разным общностям. Этногенез казахов и каракалпаков связан с
северным среднеазиатским пучком. Они тяготеют к киргизам,
узбекам и тюркским народам Западной Сибири. Их
антропологическую основу составило европеоидное население эпохи бронзы, а
постоянный приток монголоидных (преимущественно тюркоязыч-
ных) кочевников модифицировал эту основу, оказав
одновременно влияние на культуру и изменив язык. Можно предположить,
что вычленение среднеазиатского северного этногенетического
пучка было длительным процессом, продолжавшимся более двух
тысячелетий. Мы можем датировать его серединой II тыс. до н.э.
I тыс. н.э. (Алексеев, 1969).
Туркмены и таджики относятся к южному среднеазиатскому
этногенетическому пучку. Раннее население Таджикистана и
Туркмении, как это демонстрируется многими разнообразными
данными, в том числе и антропологическими, восходит к общему
этническому пласту. Раскопки обширных земледельческих
поселений эпохи неолита и бронзы в Туркмении и палеоантропологичес-
кое изучение оставившего их населения показали, что возникно-
221
вение южного среднеазиатского этногенетического пучка датиру
ется эпохой неолита и произошло оно под сильным влиянием кон
тактов с югом, в частности с территорией Передней Азии.
Близким в хозяйственно-культурном и этногенетическом отно
шении к населению Средней Азии оказывается коренное населе
ние Центральной Азии. В формировании антропологических осо
бенностей значительную роль играл древний европеоидный компо
нент, но сравнительно поздняя (в эпоху средневековья) монголиза-
ция за счет кочевых монголоидных племен резко изменила
внешний облик местного населения и его язык. Этногенетически
население Центральной Азии включается в центральноазиатский эт-
ногенетический пучок, обнаруживая связи с более южными
территориями. Антропологический состав народов, относящихся к
этому этногенетическому пучку, и преимущественно характерный
для них кочевой быт сформировались, по-видимому,
сравнительно поздно, не ранее рубежа нашей эры (Алексеев, Гохман, 1984).
На территории Центральной Азии были распространены два
хозяйственно-культурных типа - кочевники и полукочевники-
скотоводы аридной зоны (подтипы пустынь, степей высокогорий]
и таежный подтип пашенных земледельцев. В пределах этих
типов нами изучены тувинцы, тувинцы-тоджинцы, телеуты, хакасы-
сагайцы, хакасы-качинцы, хакасы-кызыльцы, алтай-кижи, телен-
гиты, казахи, буряты и якуты. В настоящее время якуты обитаки
севернее, тип их хозяйства имеет много своеобразных черт.
Довольно подробно изучено коренное население северо-востока
Азии, относящееся к трем хозяйственно-культурным типам. В
прошлом эскимосы, береговые чукчи, алеуты и береговые коряки
полуоседлые арктические охотники на морского зверя; тундровые
чукчи и коряки, ительмены и эвены - кочевники-оленеводы в
охотники тундры и лесотундры; нивхи - полуоседлые и оседлые
рыболовы и охотники. В этногенетическом отношении все эти
народы включаются в чукотско-камчатско-сахалинский этногенети-
ческий пучок. На основании антропологической информации с
территории распространения этих народов, а также
лингвистических данных о глубине дифференциации их языков время
формирования чукотско-камчатско-сахалинского пучка может быть
предположительно датироваіно II тыс. до н.э. (Алексеев, Гохман, 1984).
Наконец, нужно упомянуть еще о двух изученных народах.
Первый - западносибирские ненцы, в прошлом кочевники-олене-
222
воды тундры и лесотундры, ини связаны с народами, относящими
ся к западносибирскому зтногенетическому пучку, и в своем гене
зисе тяготеют к Алтае-Саянскому нагорью. Второй - саамы Коль
ского полуострова, до недавнего времени кочевники-оленеводь:
тундры и лесотундры. Их этногенез имеет много специфических
черт, но история становления антропологических особенностей
этой группы также имеет длительный период, восходя к неолиту.
Итак, среди изученных нами народов нет ни одного, генезис
которого не отличался бы значительной древностью. Даже самые
"молодые" из них имеют за плечами не менее тысячелетия, и
кроме того, в каждом из них в той или иной мере присутствует
субстрат, на тысячелетия удлиняющий время связи этих народов с
местом их обитания.
Данные о хозяйственно-культурных типах, этногенетических
пучках и изученных группах приведены на рис. 46 и легенде к
нему.
3. ОЦЕНКА АНТРОПОЭКОЛОГИЧЕСКИХ СВЯЗЕЙ
НА ТЕРРИТОРИИ БЫВШЕГО СССР
Территория бывшего СССР характеризуется не только
этническим и антропологическим разнообразием, но и большим
разнообразием ландшафта и климата. Она охватывает районы от 0 до 2000
м над уровнем моря, с амплитудой среднегодовых температур от
-11°С до + 16°С, с диапазоном скоростей ветра от 1 до 7 м/с, со
среднегодовым количеством осадков от 80 до 640 мм и
показателем кислотности почвы (рН) от 4,5 до 8,5.
В силу географического положения бывшего Советского Союза
изучаемое население проживает на территории, где в течение года
преобладают низкие температуры. Проведенный нами анализ сре-
довых факторов с помощью метода главных компонент позволил
выделить пять главных компонент, которые определяют 92%
изменчивости экологических параметров в обследованных областях.
Наибольшая нагрузка падает на арктический и субарктический
комплексы. Далее по своему значению следуют высокогорье,
дальневосточная климатическая область умеренного пояса,
континентальная Сибирь и, наконец, северная климатическая область
умеренного пояса. В изученных областях достаточно определенно
выражены черты климатической экстремальности.
222
Поэтому при анализе воздействия географических факторов на
популяции человека в условиях внетропических широт наряду с
такими факторами, как температура воздуха, скорость ветра,
количество осадков и влажность воздуха, мы придавали большое
значение индексу "суровости" погоды - комплексному показателю
воздействия холодового стресса на организм.
К числу геохимических факторов, воздействующих на
человеческие популяции, мы отнесли показатель кислотности почв (рН),
который отражает не столько химический состав, сколько
подвижность химических элементов, принимающих участие в
жизнедеятельности организмов, в том числе и человека.
Из многих экологических факторов социального характера
был выбран рацион питания, определяющий калорийность и
химический состав пищи. Рацион питания обусловлен хозяйственно-
культурным типом как в отношении количества и набора
продуктов питания, так и в отношении пищевых традиций и привычек.
Данные о рационах питания получены на основе качественных
характеристик перечня наиболее употребительных продуктов с
последующим расчетом калорийности и химического состава на
100 г каждого продукта (Алексеева, Шауро, 1970). Этот способ
дает относительные представления о характере питания на
большой территории-
Каждая из обследованных популяций изучалась по
комплексной программе, включающей около 100 антропологических
показателей, характеризующих морфофункциональный статус
человеческого организма. Использование всего этого комплекса для
выявления основных связей в сложной системе "популяция - среда
обитания" практически невозможно. Поэтому необходимо было
найти оптимальный путь исследования, и в первую очередь
оптимальный набор признаков, которые следовало бы ввести в
вычисление множественных корреляций. Были выбраны те
антропологические показатели, которые проявили наибольшую
интеркомпонентную связь в системе человеческого организма, и те, которые
оказываются относительно независимыми. К последним относятся
почти все основные физиологические особенности организма. ?
программу вошли 19 признаков, описывающих размеры скелета,
степень развития мягких тканей и некоторые показатели
минерального, белкового, жирового и основного обмена веществ (Чики-
шева, 1982).
224
Из анализа корреляций средовых факторов следовало, что в
условиях повышения холодового стресса и кислотности почв (как
правило, это почвы тундровые с вечной мерзлотой) формируется
тип хозяйства с высококалорийной диетой, с высоким
содержанием белков и жиров в рационе (охотники на морского зверя,
охотники и рыболовы, оленеводы); напротив, с понижением холодовой
экстремальности среды рацион питания претерпевает изменения
за счет увеличения удельного веса продуктов с высоким
содержанием углеводов. Такой рацион характерен для земледельческих
типов хозяйств (табл. 28).
Фактор холодового стресса выступает ведущим и для связей
морфофункциональных признаков со средовыми факторами.
Интерпретация этих связей показывает, что с увеличением индекса
"суровости" погоды укрупняются размеры головы и лица,
расширяется грудная клетка, уменьшаются продольные размеры тела,
понижается жироотложение, увеличивается уровень холестерина
в сыворотке крови и повышается основной обмен (табл. 29, 30),
Подобное направление изменчивости морфофункциональных
признаков характерно в целом для населения внетропических широт.
Наиболее выражена описанная комбинация признаков у северных
монголоидов, заселяющих территории с высокими значениями
показателя холодового стресса.
Направление и характер корреляций антропологических
признаков с рационом питания и типом хозяйства
полностью'совпадают с теми, которые обнаруживаются при оценке их связей с
климатическими факторами. Можно ли этот факт интерпретировать
как показатель только опосредованной (через
хозяйственно-культурный тип) связи популяций человека со средой обитания? По-
видимому, нет. Суровые климатические характеристики диктуюі
необходимость создания хозяйства с высокой калорийностью
питания, что должно способствовать сохранению гомеостаза
популяции в экстремальных условиях среды. В то же время мы
располагаем достаточным количеством данных, свидетельствующих о том.
что многие морфофизиологические черты являются
непосредственной приспособительной реакцией на воздействие низких и
высоких температур, гипоксии, влажности и т.п. факторов. Напомним,
что на анализе этих приспособительных реакций и основана
гипотеза адаптивных типов.
По-видимому, наблюдаемые нами антропоэкологические связв
должны быть интерпретированы по-иному. Становление соответ
225
СО
est
00
со
СО
СО
*
*
СО
*
*
О
СО
СО
о
*
*
со
t-
*-
о
ю
СО
CN
О
+
+
ем
ел
ок
о
+
ко
C0fc
о*
со
со
cq
О
ю
со
со
СО
ее
о
+
СО
О)
со
о
+
t- t>
o
+
о
+
? *
? 9
Ю
4c
*
0>
4f
CO
CM
CO
os
co
eo^
о
с»
О
+
*
«3
CD
О
+
СО
*
о
іОл
о"
•к
со
t>
ю
о
+
еч
Ю
СО
О
+
Я
ю
3
Д *
со
Я
и
g
и
К
со
Ь а
Е з
__ о в
Я© g Я
226
сч
I
?
"IS
О' 2. »,
СО О)
с- со
cq о
to to
? en
О ^
? 9"
о
со
СО
СП
о"
+
о
со
*
ь-
со
9k
О
+
-1С
ч*»
і-Ц
О
+
CN
О
О
+
*
¦^
сч
ю
о
+
*
со
со
о
N
О»
СО
О
-0,211
CN
тН
°1
О
4с
¦* i-t *
OS О м
со ^р со
^ о Ч
о + о
ф
СЯ
О
+
•к
О
ю
со
о"
+
*
*
со
с»
ю
О
+
со
гЧ
о"
*
*
О
СО
о*
+
*
¦к
со
о»
ю
4f
ч*
о
*
о
о
"^
о
+
ч*
о
*
*
О)
о*
+
CN
о
+
ся
о*
о ел
О і-(
о о
+¦ ¦+•
со
со
*
о>
сол
о"
+
вЯ
о
см
о
со
о
СЯ
со
о
о
+
a 8
Я о
+
О»
CN
о
со
о
о"
+
2 «э
? °
& ч* іЧ
5 О рН
? ? ?
*
с»
ф
со
10 ? о
¦ч» со ся
о 9 о
•к
со
*
о
со
со
*
л
а*
1*
*
і-
с*
^
о
+
о
-f-
* * Z
g 00 N
со ь- ?:
? ° ?
со
Ф
г-і
•>
О
+
* «ч *
* ? *
н W О
to со to
^ о ^f
О + О
о
о
о
+
*
-к
t»
а>
ю
•.
о
+
ся
*
о*
ю
to
* 5
^1 О
О +
4с
t-
СО
ся
*
Ю
ч*
СО
О
+
*
*
00
со
ю
о
+
о"
+
я
И
о
2
е
S
1
w
§
I
S
I
о
s
о
Я о
во
л р.
* *
227
со
а
3
|
со
со
СМ
О
О
со
о
СМ
00
CN
t-
О
О
+
-к
OS
СО
00
О»
о
о
о
о
+
СП
^»
СО
тЧ СО
О .-И
00 і-Ч
іН
О
со
(М
со
гН
со
о
со
о
+
о
+
* ю
СО гЧ
2 °
-Г °
? +
со
со
-^
О
+
ел
о
+
со
CN
СО
О
+
о
со
со
о
со
СО
СО
о
4-
со
ю
СО
00
о
СО
-# ? см
9 ? '
со
CN
«ч
о
•к
о
+
со
со
о
о
¦+
со to
ь- см
о о
о
о
см
о
-*-
со
о
00
¦к
с-
со
о
о
° о 9
¦к
ю
о
о
со
ю
гЧ
о"
+
со
ю
СМ
О
+
ел
о
о
о
СО
см
9,
о
4f
і-Ч
о
ю
со
со
о"
о
о
1>
о
о
о
ко 52
9 ?
СО
см
г*
см
-к
*
со
о
*
СЛ
о
ь-
см
со
о
+
*
со
о
+
о
с-
о
о"
*
*
СО
іН
о
о
ч*
СП
і-Н
о
+
4с
•К
со
ю
СО
о
+
о о
см о
СО Ь-
і- « 8
тн «N ,-,
5 2 °"
со
X
СМ
Si *
? го
? 9
03
3
со
а
№
8§
о
ю
см
¦к
*
t-
о
с-
о
¦+¦
см
СО
СО
о
+
СО
см
о
+
КО
о*
+
•К
со
fr-
СО
см
со^
о"
8 н
Я о
Р* О
* *
•к
228
ствующего хозяйственно-культурного типа следует трактовать каі
социальную реакцию популяции на воздействие естественных эко
логических факторов. Формирование же морфофункциональногс
комплекса осуществляется как непосредственно под влиянием
естественной среды, так и опосредованно - через тип хозяйстве
(рис. 47).
&е^'
]f Боднем
Коэффициенты корреляции:
*•- н до 0.Z \ ідо0,6
, , fft4 і і Д7
і 1 QJ \ | 0,8
Рис. 47. Зависимость морфофизиологического комплекса
от факторов естественной и социальной Среды:
1 - основной обмен; 2 - скуловой диаметр; 3 - уровень холестерина в
сыворотке крови; 4 - артериальное давление; 5 - белки рациона питания; $
- жиры; 7 - углеводы; 8 - хозяйственно-культурный тип Знак "плюс"
соответствует положительной связи, знак "минус** - отрицательной.
Полученная нами картина антропоэкологических связей на
обширном пространстве нашей страны, где преобладают низкие
среднегодовые температуры, показала, что приспособительная
изменчивость человеческих популяций в этой климатогеографичес-
кой области проявляется в первую очередь по отношению к фак-,
229
тору "суровости" погоды. Аналогичные наблюдения имеют место :
Для других территорий земного шара, с той лишь разницей, что :
Качестве ведущих факторов там оказываются иные климатогеогра
фические характеристики. Показано, например, что популяции
проживающие к югу от Сахары, адаптированы к комбинации вы
сокой температуры и высокой влажности воздуха (Ніепахіх, 1968
Крупник, 1973). На территории Западной Европы, Африки к севе
ру от Сахары, Передней Азии выявлена прямая связь антрополо
гических характеристик населения с максимально жарким и су
хим климатом (Crognier, 1979).
Приспособительная изменчивость человеческих популяций -
явление всеобщее, но при определенном сочетании внешнесредо
вых факторов ее направление меняется, и определяется оно н
столько средними характеристиками всех факторов, сколько соче
танием немногих ведущих компонентов внешней среды.
4. ЗАКОНОМЕРНОСТИ ЭКОЛОГИЧЕСКОЙ ИЗМЕНЧИВОСТИ
В ГОРОДСКИХ ПОПУЛЯЦИЯХ
Антропологическое изучение городских популяций имеет сво»
специфику. Прежде всего, здесь не может быть соблюдено основ
ное условие антропологических исследований - чистота выборки
Городское население складывается на протяжении длительное
времени с неоднократными миграционными потоками из иноро^
ной среды. Связанные с этим процессы смешения, меняющие гене
фонд и вызывающие явления гетерозиса, не могут не отразиться
на изменении физического облика населения. Более того, в формв
ровании городского населения принимают участие заведомо разно
этничные компоненты, сложившиеся в различных экологически:
условиях и, следовательно, обладающие различными приспосо
бительными возможностями.
Экологические факторы, воздействующие на городскую попу
ляцию, имеют в основном антропогенный характер, и уже само эт
обстоятельство предполагает иные ответные биологические реак
ции человеческих популяций, чем в условиях многовекового пре
бывания в естественных условиях среды, приспособление к коте
рой осуществлялось в течение длительной истории популяции.
230
Широко известны неблагоприятные воздействия такого
антропогенного фактора среды, как повышенная загрязненность
атмосферы города отходами производства, приводящая к ослаблению
физического развития детей в зоне влияния загрязнения, а
нередко и к врожденным порокам развития. Об этом достаточно
убедительно свидетельствуют антропологические и генетические
исследования. Обстоятельный анализ влияния загрязнения
окружающей среды на здоровье человека проведен М.П. Страдомской и
Е.Л. Райх (1979). Широко известны и материалы Американской
службы здоровья, демонстрирующие связь высокой плотности
населения с повышением частоты заболеваний сердечно-сосудистой
системы.
В то же время наблюдения антропологов убеждают нас в том,
что экологическая ситуация города вызывает и такие изменения в
морфофизиологическом статусе городских популяций, которые ни
в коей мере не могут быть истолкованы как отклонение от нормы.
Следует отметить, что столь широко наблюдаемое сейчас
явление акселерации, т.е. ускорения процессов роста и развития,
впервые было установлено при изучении детских контингентов
городов и характерно в значительной мере для городских популяций.
Поскольку явление акселерации - панойкуменный процесс, то,
естественно, он охватывает и сельские популяции, но мера его
проявления там значительно слабее, чем в условиях города (Рост в
развитие ребенка, 1973; Властовский, 1976; Worldwide variation in
human growth, 1976).
Акселерация выражается в ускорении физического, полового в
психического развития. Однако остается неясным, следует ли
рассматривать акселерацию только как положительное явление.
Последствия такой вспышки процессов роста и развития вряд ли
могут быть исчислимы сроками жизни одного-двух поколений.
Пока не ясно, как отразится этот процесс на репродуктивной
способности, психических и социально-психологических свойствах
личности, на характере приспособления к изменяющимся
условиям среды. Причины акселерации не ясны. Наиболее
распространено мнение о том, что она обусловлена специфическим
воздействием факторов городской жизни, таких, как повышение калорийно
сти питания, снижение физических нагрузок, усиление психоэмо
ционального напряжения (Карсаевская, 1973).
23]
Согласно одному из объяснений, акселерация вызывается гете
розисом, в основе которого лежит расширение круга брачных свя
зей вследствие усиления миграционных процессов (Бедный, 1976).
Некоторые исследователи склонны объяснять акселерацию
действием своеобразного "отбора", в результате которого из сельской
местности в город попадают люди, отличающиеся высокой ум
ственной реактивностью и более высоким физическим развитием
(Bennholdt-Thomsen, 1942). Подтверждение этой точки зрения на
ходим в довольно многочисленных данных о высоком физическом
развитии мигрантов (Kaplan, 1954; Beiguelman, 1963).
Вряд ли стоит противопоставлять друг другу факторы,
вызывающие акселерацию. Скорее всего, такое мощное явление можеп
быть вызвано их совокупным действием. Судя по данным после
дних лет, во многих странах наблюдаются затухание акселерации
и ее стабилизация. Это обстоятельство рождает предположение с
каких-то пока еще не познанных механизмах и причинах
акселерации.
Городское население отличается от сельского более крупными
размерами тела, более развитым жировым компонентом и
значительно большим процентом астенических и пикнических
конституциональных типов. Это характерно не только для детских
возрастов, но и для взрослого населения (см.материалы исследований
НИИ антропологии МГУ среди городского и сельского населения
России, Украины, Эстонии, Армении, Азербайджана и Грузии).
Наши исследования населения средневековых русских городов
обнаружили очень интересную закономерность. Независимо оі
того, на территории какого древнеславянского племени вырастал
город, его жители, будучи, бесспорно, связанными по своему
происхождению с сельским населением этого же племени, всегда
отличались большей шириной лица, более широкой головой
(брахикефалия) и высокорослостью (рис. 48). В данном случае речь
идет о краниологических и остеологических сериях из
средневековых древнерусских городов Витичева, Любеча, Чернигова, Киева,
Смоленска, Старой Рязани (Алексеева, 1973). Таким образом,
укрупнение размеров тела было присуще горожанам с самого начала
истории городов.
При формировании населения древнерусских городов наряду с
сельским населением принимали участие и пришлые контингенты
людей. Это были наемные воины из варягов, служилые люди -
232
so
женщины
ну л чины
J I ,1 I » ¦ ¦ » |_
т т т /?t /л т ю т т но ш т ш т т ш
С пуло 6о в ширимо t мы
Рис. 48. Сопоставление средневекового сельского (а) и городского (б)
населения по черепному указателю у скуловой ширине.
выходцы из Новгородской земли и южнорусских степей (Насонов,
1951; Соловьев, 1959; Плетнева, 1964). Однако удельный вес их,
как показал сравнительно-морфологический анализ, был невелик
и не мог внести существенных изменений в антропологический
облик горожан. Последние в антропологическом отношении
увязываются с сельским населением, проживающим вблизи
соответствующих городов.
Можно думать, что на изменение физического облика
древнерусских горожан влияли все те причины, которые действуют и в
современном городе: изменение социально-экономического уровня
жизни, увеличение психоэмоционального напряжения и
расширение круга брачных связей за счет миграции в город сельского
населения. Незначительный в то время процент иноплеменного и
иноязычного населения, не оставив ощутимого следа в
антропологическом облике, мог тем не менее отразиться в явлении
гетерозиса.
Существует еще одна особенность городского населения по
сравнению с сельским - высокая профессиональная дифференциация,
которая откладывает отпечаток на строение тела и
конституциональный габитус. Исследования НИИ антропологии МГУ, про во-
79
78
| 76
I"
^ 72
71
J L
J L
282
дившиеся в различных этнотерриториальных группах на террито
рии бывшего СССР неоднократно выявляли связь типа профессю
с соматотипом: высокие физические нагрузки способствуют увели
чению костно-мускульного компонента тела, среди служащих за
метно повышение астенического и пикнического типов телосложе
ния. Эти связи устанавливаются на фоне высокой степени наследу
емости типа телосложения.
Следует обратить внимание еще на одну очень важную особен
ность городского населения- Городское население - это популяции
коль скоро все жители городов имеют потенциальную возможност]
к вступлению в брак. Строго говоря, население крупных городо]
не представляет собой популяцию "в традиционном пониманш
этого термина. Скорее это центр панмиксии разнородного населе
ния обширной территории при непрерывном росте урбанизации і
расширении круга миграции во времени" (Курбатова, 1977, с. 20)
Однако из-за быстрого роста городского населения в нем не прояв
ляется один из важнейших факторов популяционной динамики -
генный дрейф (Курбатова (Русакова), 1975). Кроме того, городско
население по сравнению с популяциями малого объема испытыва
ет более интенсивное давление мутационного процесса (Бочков
1974). Далее, в городах возрастает и интенсивность отбора в ре
зультате возникновения и распространения эпидемий в места;
наибольшей скученности населения (Курбатова (Русакова), 1975)
Все эти процессы наряду с изменением образа жизни городское
населения не могут не влиять и на демографические процессы.
Понижение рождаемости и улучшение медицинского обслужи
вания изменяют возрастную структуру населения.
В современных городах наблюдаются замедленный естествен
ный прирост населения, увеличение численности городов осуще
ствляется в основном за счет прилива населения из сельской мест
ности.
Достаточно показательна и динамика смертности городской
населения. Так, например, смертность в европейских городах ]
XIX в. значительно превышала смертность в сельской местности
для большинства городов Европы XX в. характерно, напротив
снижение ее. В то же время высказывается мнение, что в некото
рых европейских странах к 2000 г. смертность несколько повысит
ся за счет дальнейшего увеличения в демографической структур"
234
городского населения удельного веса пожилых возрастов <алосто
лов, Мичков, 1977).
Влияние урбанизации на здоровье населения подробно освеще
но М.П- Страдомской (1979).
Оценивая особенности физического типа городского населения,
его популяционную и демографическую структуру, естественнс
задать вопрос: применимо ли к нему понятие "адаптивный тип" і
том его содержании, какое мы вкладываем в характеристику адап
тивных особенностей сельского населения различных
экологических ниш? По-видимому, нет.
Увеличение размеров и массы тела может иметь причиной как
акселерацию, так и более высокий социально-экономический
уровень жизни горожан. Что касается брахикефализации как
явления, характерного для городского населения, то вполне
убедительным представляется объяснение В.В. Бунака (1951),
рассматривающего ее как "частный случай общей проблемы ускорения рос
та", причины которой лежат в изменении социальных условий.
Под изменением социальных условий подразумевается не толькс
повышение уровня жизни, но и изменение демографических
показателей популяции.
Означает ли это, что процесс адаптации не касается городских
популяций? Исследование генетических процессов в городском
населении обнаруживает различия в приспособленности
фенотипов групп крови системы ABO, MN, Rhesus, Р и Lewis в
отношении репродуктивной способности, дифференциальной
заболеваемости и уровня патологии у новорожденных (Курбатова, 1977).
Эти наблюдения свидетельствуют о том, что и в городских
популяциях действуют факторы отбора, а следовательно, идут и
адаптивные процессы, которые могут отразиться, по-видимому, на разных
морфофизиологических признаках.
В заключение следует обратить внимание на то, что
антропология городских популяций, как проблема современной науки, поке
еще находится в стадии становления (Урбоэкология,1990). Тем не
менее уже сейчас можно надеяться, что существование города как
экологической ниши с точными хронологическими
характеристиками и с точной фиксацией исторических процессов, в нем проис
ходящих, позволит подойти к решению многих вопросов,
связанных со скоростью протекания адаптивных процессов и временем,
необходимым для образования адаптивных типов.
235
лекция лі
ЗАКОНОМЕРНОСТИ ВРЕМЕННОЙ ИЗМЕНЧИВОСТИ
АДАПТИВНЫХ ОСОБЕННОСТЕЙ У ЧЕЛОВЕКА
Ростовые процессы.
Эпохальная динамика адаптивных типов.
1. РОСТ И РАЗВИТИЕ ЧЕЛОВЕКА
В РАЗЛИЧНЫХ ЭКОЛОГИЧЕСКИХ НИШАХ
Наследственная обусловленность адаптивных черт в настоящее
время не подлежит сомнению, тем не менее нет основания думать
что они не изменяются в процессе онтогенетического развития :
результате влияния среды обитания на рост и развитие организма
стимулирующего развитие тех его систем, которые определяю1
уровень приспособления к конкретным экологическим нишам.
Так как адаптивные черты проявляются не только в физиоло
гических особенностях организма, но затрагивают и особенност]
строения тела, рассмотрим процессы роста и развития в человечес
ких популяциях, обитающих в различных географических услови
ях. Данных о физиологических реакциях детского организма н
воздействие окружающей естественной среды очень мало.
Все авторы, изучавшие ростовые процессы у детей в разны:
географических зонах, показали единство направлений возрастноі
изменчивости; различия выражаются лишь в темпах изменениі
(Roberts, 1960; Ashcroft, 1971; Миклашевская, 1972; Wolansky
1975; Worldwide variation in human growth, 1976). Черты, прису
щие взрослому населению каждой исследованной популяции, про
являются в самых ранних возрастных периодах.
Д. Роберте (Roberts, 1960) исследовал возрастные изменения;
детей и подростков - нилотов от 14 до 16 лет, нашел, что во все:
возрастных категориях африканские дети имели сравнительн
малую массу по отношению к длине тела при сопоставлении
детьми европеоидной принадлежности. На детском контингент
Роберте (Roberts, 1960) подтвердил связь низких величин массі
тела с жарким климатом, установленную им ранее на взрослы:
(Roberts, 1953). Коэффициенты корреляции между среднегодовоі
температурой и логарифмом массы имеют негативный знак и ве
личины порядка 0,6-0,7 для мальчиков и 0,4-0,6 для девочек.
Различия в пропорциях тела между африканцами и европейца
ми проявляются на 40-й неделе внутриутробной жизни (Schultz
236
1926). Пропорции тела, присущие монголоидам, также
формируются очень рано. Это показано для киргизских детей
(Миклашевская и др., 1972), японских детей (Greulich, 1957), детей
американских индейцев (Ashcroft, 1971).
Сопоставление изученных нами детей русских как
представителей умеренной зоны обитания, эвенков, генезис которых связан
с континентальными районами Сибири, и индийских детей,
уроженцев тропических широт, обследованных И. Шарма (Sharma,
1970), показало единое направление возрастных изменений при
сохранении во всех возрастных группах тех соотношений, которые
характерны для взрослых.
Рассмотрим более подробно данные о возрастной изменчивости
черт строения тела в различных экологических условиях. Для
этой цели мы использовали многочисленные материалы по
процессам роста и развития, собранные в рамках Всемирной
биологической программы (Worldwide variation... 1976), а также наши
данные о возрастной изменчивости признаков строения тела детей
горных районов Таджикистана (таджики), внутренних районов
Сибири (тувинцы, тувинцы-тоджинцы), Северо-Восточной Азии
(эвены, коряки, чукчи) и материалы Н.Н. Миклашевской с
сотрудниками о развитии детей и подростков в среднеазиатском
высокогорье (1972, 1975) и субтропической зоне (1979,1982). Были
привлечены данные о процессах роста и развития детей в
высокогорье тропической зоны (Биология жителей высокогорья, 1981) и
в Арктике (Eskimos of northwestern Alaska, 1978).
Все европеоидные группы, взятые для анализа, являются
коренным населением Европейского континента и, следовательно,
проживают в условиях умеренного климата (финны, эстонцы,
датчане, англичане, немцы, французы, венгры, итальянцы, испанцы,
население Югославии, Швейцарии, поляки, чехи, болгары,
русские). Негроидные группы представлены коренным населением
Африки (этнические группы Либерии, Мозамбика, Сенегала,
Судана, Танзании), а монголоидные по своему местообитанию
увязываются как с внетропическими, так и с тропическими широтами
(японцы, китайцы, корейцы, эскимосы, чукчи, эвены, коряки,
тувинцы, буряты, якуты).
В главе, посвященной закономерностям пространственной
изменчивости строения тела, обращалось внимание на характер
экологической дифференциации человечества, которая
осуществляется в первую очередь на основе признаков, обеспечивающих
энергетические процессы. Из признаков строения тела наибольшую
237
значимость в этой связи приооретают длина тела и масса тела, ве
соростовой показатель и поверхность тела.
Судя по характеру возрастной изменчивости признаков строе
ния тела, сходство процессов роста у представителей трех болыпю
рас очень велико, различие проявляется в абсолютных величина;
признаков и в скоростях роста. Так, например, у европеоидов і
монголоидов темпы роста тела в длину после шестнадцатилетнегс
возраста падают, в то время как у негроидов этого не наблюдает
ся, однако отмечается замедление ростовых процессов в позднепу
бертатный период (рис. 49).
А
/
6 7 8 9 Ю Г? Г2 U /4 15 /6 /7 ft
tr
6 7 ё 9 W *J2/JM/S/f/7fj
Возраст о ttdiQt
Рис. 49. Возрастная динамика длины тела у представителей различных
рас (X - европеоиды; 2 - монголоиды; 3 - негроиды). Мальчики и юноши. А -
ростовые кривые, В - возрастная изменчивость.
Здесь к на рисунках 50-55 возрастная изменчивость признака дается в процентах от его
величины в возрасте 18 лет.
238
Иная картина при рассмотрении данных о возрастной
изменчивости массы тела. Этот признак, показывая значительное
повышение темпов роста в период пубертаса у представителей различных
рас, у монголоидов снижает свою скорость уже в
шестнадцатилетнем возрасте, т.е. в более раннем возрасте достигает величины,
присущей взрослому индивиду. Судя по кривой скорости
нарастания массы тела, монголоиды во всех возрастных периодах имеют
относительно большие (в % величины массы тела взрослых)
значения признака, чем европеоиды и негроиды (рис. 50). У
негроидов замедление процессов нарастания массы тела, как и темпов
роста тела в длину, происходит в период позднего пубертаса.
€Н
170
160
750
Щ
ПО
120
ПО
%
W0
90
€0
70
60
\ 1-1
J L_l
J—J L
б 7 8 9 10 ft 12 П? 15 /6 ПИ
X f -т
) » і 1 I Ч t
6 7 8 9 ЮЛ 12 13 К 15 76 17Ji
&03fQc*n S гсіаж
Рис. 50. Возрастная динамика массы тела у представителей различных
рас. Мальчики и юноши. А - ростовые кривые, В - возрастная изменчивость,
Остальные обозначения, как на рис. 49.
239
Сравнительно небольшая длина тела в сочетании с
относительно большой массой создает высокую плотность тела у
монголоидов, которые по индексу Рорера значительно превосходят
представителей европеоидной и негроидной рас (рис. 51). Последние
отличаются понижением плотности тела на протяжении почти
всего изучаемого периода детства и особенно в пубертатный период.
Однако в юношеском возрасте у негроидов плотность тела резко
нарастает, что, по-видимому, связано с увеличением скорости
роста длины и массы тела в постпубертатный период после явного
затухания их в фазе позднего пубертаса.
Весоростовой указатель имеет иное направление возрастных
изменений, чем его составляющие: он неуклонно уменьшается с
возрастом, демонстрируя наибольшее падение в период
пубертатного развития. Самые большие расхождения по этому признаку
между представителями европеоидной, негроидной и
монголоидной рас наблюдаются в изучаемом возрастном диапазоне в период
нейтрального детства и в заключительной фазе пубертаса.
Весоростовой указатель новорожденных у монголоидов выше, чем у
европеоидов и негроидов; последние отличаются наименьшей
величиной указателя (Worldwide variation... 1976).
Абсолютные величины поверхности тела отражают различия,
существующие между представителями этих рас по длине и массе
тела. В то же время в кривой относительного роста поверхности
тела наблюдается своя специфика. Подобно весоростовому
отношению она падает с возрастом, но, кроме того, отчетливо
увеличивается скорость нарастания этого признака у негроидов по
сравнению с европеоидами и монголоидами, особенно в пубертатный
период (рис. 52).
Весьма показательны различия в пропорциях тела между
представителями трех больших рас во всем периоде детства (рис. 53).
У негроидов уже в период детства четко проявляются черты доли-
хоморфии: они более длинноноги, по отношению к длине тела у
них уже плечи и таз. Противоположные соотношения длиннотных
и широтных размеров характерны для монголоидов:
относительная коротконогость, относительно более широкие плечи и таз
(брахиморфный тип). Европеоиды по типу пропорций тела занимают
промежуточное положение. Надо отметить при этом, что
возрастная динамика пропорций тела очень невелика: тип пропорций,
характерный для взрослого населения, проявляется в очень ран-
240
J L
J 1 I 1 1 I
**<*» «^ *o ^ ¦*v»*s4*-^?l?5?4;
••* *c »¦<* *<-
cr^oa
^
§
I
§5
241
110
wo
so
17
16
15
lb
6 7 8 9 W 11 12 /J 1« 15 !6 1718
¦v
I I !
' ' \
6 7 8 9 10 U 12 U 14 15 15 17 IS
В
•^
f I Ч^Т"Ч--1-1._і--П ) \
6 7 8 9 10 11 12 IS I* 15 T5 1713
Г
6 7 8 9 Ю If 12 U 14 15 16 17 18
Возраст 8 годах
Рис. 53. Возрастная динамика широтных пропорций тела (ширина плеч
и таза в % длины тела) у представителей различных рас. Мальчики и
юноши. А - ростовые кривые относительной ширины плеч; Б - возрастная
изменчивость относительной ширины плеч; В - ростовые кривые относительной
ширины таза; Г - возрастная изменчивость относительной ширины таза.
Остальные обозначения, как на рис. 49.
них возрастах, включая и утробный период жизни (Shultz, 1926;
Рогинский, I960).
Существуют различия между европеоидными, негроидными в
монголоидными детьми в обхватных размерах и в толщине под
кожного жирового слоя (рис. 54, 55). Негроиды характеризуют
242
*о
** ^
і
ж
§
R?
*
§
s?
T-TF
"5Г^ ^ ^ fc
^
ч»
у»
і>5
-114
2
3
I
243
ся меньшим обхватом груди во всех возрастах, хотя
обнаруживают явное увеличение скоростей его роста в период нейтрального
детства и в пубертасе. Между европеоидами и монголоидами
существенных различий по этому признаку не отмечается. По развитию
подкожного жирового слоя монголоиды, значительно превосходя!
европеоидов и негроидов. Последние обнаруживают тенденцию к
понижению жироотложения почти во всех периодах детства, за
исключением нейтрального детства и постпубертатного периода.
Темпы нарастания жироотложения у монголоидов значительно
превышают те, которые характерны для негроидов и европеоидов.
Подводя итог рассмотрению возрастной изменчивости
некоторых признаков строения тела, имеющих адаптивное значение,
приходим к заключению о том, что негроиды отстают от
европеоидов и монголоидов по скоростям роста широтных пропорций
тела, массе тела и весоростовому соотношению и опережают и тех
и других по соотношению поверхности тела и массе тела, т.е.
отличаются по тем соматическим признакам, которые создают
специфику формы тела коренного населения тропиков. Таким
образом, экологическая дифференциация человечества на жителей
тропических и внетропических широт достаточно демонстративна
уже в детском возрасте. По-видимому, различия в скоростях
роста также могут иметь адаптивную природу, во всяком случае, это
относится к признакам строения тела, определяющим его массу и
форму.
Рассмотрим подробнее соматические особенности детей
монголоидной принадлежности. Средние величины признаков,
которыми мы оперировали, получены суммированием показателей для
групп, проживающих не только во внетропической, но и в
тропической зоне. Наряду с детьми коренных жителей
Северо-Восточной и Центральной Азии в суммарную группу были включены
дети южноазиатской ветви монголоидов и индейцев Центральной
и Южной Америки.
В главе, посвященной биологии жителей тропических широт,
отмечалось, что в строении тела коренного населения
тропического пояса, относящегося к монголоидной расе, обнаруживаются
признаки, сближающие его с представителями негроидной расы.
Оставаясь сходным по соматическому строению с монголоидами
более высоких широт, монголоидное население тропиков тем не
менее характеризуется более уз?йм телосложением и меньшей
244
плотностью тела. Это было показано на примере переселенцев і
Индию из Индокитая (рианг) и Непала (гурхи).
Сопоставляя особенности строения тела эскимосских (Аляска),
китайских (Сянган) и индейских (Гватемала, Перу) детей,
приходим к выводу, что по длине тела различия не имеют
географической увязки, но по широтным пропорциям и массе тела южные
группы отличаются от северных (рис. 56). Первые
характеризуются более узким телосложением и меньшей массой. Исключение
составляют индейские дети, у которых очень высокое весоростовое
соотношение, во многих возрастах превышающее весоростовое
соотношение эскимосских детей. Следует отметить, что
американские индейцы и во взрослом возрасте отличаются очнь высокой
плотностью тела, тем не менее отмечается тенденция к
некоторому понижению этого показателя по направлению к тропикам. Так,
индекс Рорера у мужчин-индейцев тропической зоны колеблется
от 1,30 до 1,49, а в субтропической и умеренной зоне - от 1,36 дс
1,58.
Повышенная плотность тела американских индейцев, как
нетипичная для коренных жителей тропической зоны, уже
отмечалась в литературе (Worldwide variation... 1976), однако авторы
приводят данные об относительной поверхности тела, которая
имеет величину, характерную для представителей африканского
расового ствола, и считают, что экологическое правило
поверхности тела имеет силу и в отношении американских индейцев,
заселивших Южную Америку около 30000 лет назад. Этого времени
вполне достаточно для селективного действия механизмов отбора.
Более того, как адаптивная черта в условиях тропического
климата рассматривается и пониженная толщина жировой складки у
южноамериканских индейцев по сравнению с северными.
Итак, возрастная динамика признаков строения тела
предоставляет Немало доказательств экологической дифференциации
человечества. Особенности телосложения и пропорций тела,
типичные для коренного населения тропиков, умеренной зоны и
Арктики (и Субарктики), проявляются на самых ранних стадиях росте
и развития.
Большое внимание в отечественной и зарубежной
антропологической литературе уделяется процессам роста и развития в
условиях высокогорья. Результаты исследования адаптации к высоте
валены особенно потому, что наряду с данными о строении тел*
24Б
1/ v "
Jf V -
/\ \ -
V /
\ i -
/y j
<<y '
i/ /
X s
1 1 1 ( 1 I
pi a «a & <й сь
•^ *<¦ *c •<• -c •<¦
5? §
5?
<5a
^
S>
«^
«О
*0
S^
«5i
§
§
g
§
s
I.
I
i
3
* 5 *
v * a
они содержат обширную информацию о состоянии внутренней
среды организма.
Для большинства изученных высокогорных популяций в
Андах, Гималаях, на Тянь-Шане характерно замедление процессов
роста и развития (Биология жителей высокогорья, 1981).
Исключение составляет коренное население Эфиопии - амхара, живущие
на высоте от 1200 до 3900 м над уровнем моря, где уроженцы
высокогорных селений превосходят равнинных жителей по
абсолютным размерам и темпам роста. По мнению авторов, изучавших
высокогорных амхара, эфиопское низкогорье характеризуется
менее благоприятными условиями окружающей среды: высокой
температурой и высокой влажностью, широким распространением
малярии и паразитарных кишечных инвазий.
Наиболее характерной особенностью в строении тела
высокогорных популяций по сравнению с жителями равнин является
больший размер грудной клетки. Интересно, что при понижении
темпов роста большинства признаков строения тела в высокогорье
отмечается усиление скорости роста грудной клетки в переднезад-
нем направлении. По-видимому, это усиление приводит к
увеличению объема, что вкупе с увеличением жизненной емкости легких
и аэробной способности является наиболее типичной адаптивной
реакцией в условиях высокогорной гипоксии.
Отличительные черты высокогорных популяций проявляются
не только на высоте более 2000 м. Наши исследования в Алтае-
Саянском нагорье среди тувинцев, проживающих на высоте 800 м
в пос. Тоора-Хем и на высоте 1800 м в пос.Мугур-Аксы, также
выявили понижение темпов роста основных признаков строения
тела и явное повышение скорости роста продольного диаметра
грудной клетки (рис. 57).
К числу специфических высокогорных адаптации относятся
также увеличение медуллярной полости костей и уменьшение
площади, занимаемой компактным веществом кости. Эта
особенность связана с увеличением гемопоэтической активности
костного мозга в условиях высокогорной гипоксии. Кости детей из
высокогорных популяций, по данным исследования перуанских детей
(Baker et aL, 1966; Frisaneho, 1976), детей шерпов, тибетцев (Ра-
wson, 1974) и тувинцев (наши данные), характеризуются таким же
сочетанием кортикального слоя и медуллярного пространства, как
и у взрослого населения этих областей.
241
/в
wo
99
80
70
J i i ' ' i
%
WO
90-
80
S 7 8 9 10 11 12 13 ? 15 16 ff
В
у
/ ^~-
/
n. /
¦ i * * ¦ * * ¦_
J L
7 8 9 W 11 12 U ? 15 16 T7
Возраст S codox
Рис. 57. Возрастная изменчивость
размеров грудной клетки, (в % от их
метр) у тувинских детей, живущих
на равнине и в среднегоръе. Мальчики г
величины в 18 лет). (А - поперечный
диаметр груди; В - переднезадний диа
юноши:
X - пос. Мугур-Аксы, высота над уровнем
моря 1800 и; 2 ~ пос. Тоора-Х?м - высота над
уровнем моря 800 м.
Замедление процессов роста в высокогорных популяциях
объясняется рядом причин. Некоторые авторы, ссылаясь на
экспериментальные исследования на животных, считают, что основная
причина заключается в воздействии гипоксии (Биология жителей
высокогорья, 1981). По мнению одних исследователей,
высокогорная гипоксия препятствует эффективному усвоению пищи в
кишечнике, а также замедляет размножение клеток (Биология
жителей высокогорья, 1981). Другие авторы склонны объяснять
понижение скоростей роста в условиях высокогорья недостатком
йода в окружающей среде и недостаточно сбалансированным
питанием (Миклашевская и др., 1972). Эти причины взаимно не
исключают друг друга. В то же время высокогорные популяции не
принадлежат к числу низкорослых, напротив, в ряде случаев они
превосходят по своим размерам близкородственные группы,
живущие на равнине. Таким образом, пониженные темпы роста в
детском и юношеском возрастах впоследствии компенсируются.
248
В связи с этим явлением нельзя не обратиться к
закономерностям адаптивных реакций, наблюдаемых у животных. Мне уже
приходилось отмечать, что в характере адаптивных процессов
человека и млекопитающих животных очень много общего. Так,
например, С.С. Шварц (1940) показал* что высокогорным видам
свойственны пассивные, энергетически наиболее экономные пути
адаптации в отличие от арктических видов животных. Не
исключено, что и у человека замедление процессов роста в условиях
высокогорья в какой-то мере предохраняет растущий организм оі
воздействия комплекса неблагоприятных условий окружающей
среды.
' Подобная аналогия не является исключением. Судя по данным
о скелетной зрелости, более высокие темпы процессов оссифика-
ции у арктических и субарктических популяций человека, чем у
жителей более южных широт (Архангельская, Павловский, 1982;
Алексеева и др., 1985), могут быть оценены как указание на
активный путь адаптации.
Процессы роста и развития в значительной степени
контролируются социально-экономическим уровнем жизни, в частности
характером питания. Многими авторами отмечается ускорение
физического развития детей и темпов роста под влиянием
высококалорийного питания, так же как отмечаются и замедленный
рост, и созревание детей в условиях недоедания и распространения
болезней (Salzano, Freire-Maia, 1970; Frisancho et'aL, 1973;
Worldwide variation... 1976).
Тем не менее нельзя не обратить внимания на сохранение
одинаковых пропорций тела у детей одной и той же расовой и
этнической принадлежности при различной по калорийности диете.
Итак, из рассмотрения возрастной динамики признаков
строения тела явствует, что адаптивные черты, обеспечивающие
оптимальные возможности обитания в той или иной экологической
нише, характерны для самых ранних стадий развития, что
подтверждает вывод об их наследственной обусловленности. В то же
время при исследовании процессов роста и развития отчетливо
видна огромная роль морфозов в процессе приспособления
организма к среде. Примером морфоза может служить увеличение
объема легких у жителей равнины, приехавших в высокогорье в
детском возрасте. Размеры грудной клетки и жизненная емкость
легких (ЖЗЛ) во взрослом состоянии у них соответствовали тем,
249
которые характерны для уроженцев высокогорья, а
противоположность этому жители равнины, приехавшие в горы взрослыми
и акклиматизировавшиеся там, имели значительно более низкие
величины ЖЗЛ, чем горцы (Frisancho et al., 1973). В опытах не
животных было показано, что длительная экспозиция к
высокогорной гипоксии вызывает у молодых крыс рост объема легких,
ускорение пролиферации альвеол и альвеолярной поверхности, і
то время как у взрослых животных изменений в легочном объеме
не происходит (Фрисанчо, 1981).
2. ХРОНОЛОГИЯ АДАПТИВНЫХ ТИПОВ
Биологическая адаптация к условиям существования
сопровождала человечество на протяжении всей его истории, идя бок с
бок с адаптациями социального характера. Устойчивые
адаптивные черты возникали по мере освоения человечеством ойкумены і
процессе отбора, метисации и развития в онтогенезе, и, возможно,
этот процесс еще не завершен.
Где его истоки, на каких стадиях, предшествующих виду Homo
'sapiens, в истории проявляются те особенности
морфофизиологического статуса, которые мы называем адаптивным типом?
Мне уже приходилось обращаться к проблеме биологического
приспособления в процессе антропогенеза (Алексеева, 1977).
Располагая фактическими данными о геоморфологической ситуации,
в которой протекала жизнь предшественников Homo sapiens, e
находках каменной индустрии, можно с определенной долей
вероятности реконструировать отдельные фазы процесса адаптации і
древности, экстраполируя данные о современных популяциях ш
древние.
Самые ранние находки предшественников человека связаны ее
стадией аморфной каменной индустрии - это австралопитеки е
сходные с ними формы. Древность австралопитековой стадии
составляет 5 млн. лет(Урысон, 1973, 1974, 1976). Геоморфологичес
кая ситуация, в которой обитали австралопитековые и морфологи
чески близкие им формы, была очень сходной, будь то Южная г
Восточная Африка или Сивалинские холмы в предгорьях Гимала
ев в Индии. Это открытое плато, расположенное на высоте от 50С
до 3000 м над уровнем моря со скальными выходами, перерезан
260
ными глуоокими ущелъяші с руслами высохших некогда рек;
растительность саванного типа, обилие млекопитающих, в том
числе и копытных (Bourliere, 1963; Butzer, 1971). Большинство
исследователей связывают освоение такой экологической ниши с
переходом от комбинированного древесно-наземного передвижения к
преимущественно наземному, выходом из тропического леса на
открытое пространство и включением в рацион мясной пищи,
начавшемся, возможно, еще в тропическом лесу, как об этом
свидетельствуют наблюдения над современными шимпанзе (Лавик-Гудол,
1974).
Р. Ньюман (Newman, 1970) попытался обосновать точку
зрения, согласно которой выход из тропического леса в открытую
саванну сопровождался перестройкой терморегуляции в связи с
резким повышением солнечной радиации, сухости воздуха и
интенсивности движения воздушных масс, усиливающих
потоотделение. По мнению автора этой гипотезы, которую сам он назвал
"спекулятивной", с этим временем связываются частичная утрата
волосяного покрова у наших предков и увеличение числа потовых
желез — особенности, облегчающие процесс терморегуляции.
Можно думать, что именно в это время возникают черты тропического
адаптивного комплекса, который сейчас преобладает у жителей
тропического пояса.
В эпоху нижнего палеолита произошло расширение ойкумены.
Находки палеолитических памятников сделаны в Африке, южных
и центральных районах Европы, в Средней, Передней, Южной,
Центральной, Восточной и Юго-Восточной Азии, включая
Большие Зондские острова (Окладников, 1955). Они охватывают
период от 500 тыс. до 150—120 тыс. лет до н. э.
Находки нижнепалеолитических гоминид, даже
немногочисленные и фрагментарные, дают возможность предполагать
освоение новых экологических ниш. Если шелльский человек из Олдо-
вея, синантропы, а также некоторые синхронные с ним формы
древних гоминид из Европы жили примерно в тех же условиях,
что и представители австралопитековых групп, то яванские
питекантропы были обитателями равнинных ландшафтов с буйной
растительностью влажного тропического леса.
Находки нижнепалеолитических стоянок на Кавказе (Любин,
1970) включают в зону обитания человека и высокогорные
районы, расположенные на высоте 2000 м и выше.
261
Таким образом, в эпоху нижнего палеолита возникают условия
для формирования хумидного и высокогорного адаптивных типов.
В эпоху среднего палеолита ойкумена значительно
расширилась в северном направлении. Были освоены южные районы
Сибири и многие области Западной и Восточной Европы (карту см.:
Окладников, 1965). Среднепалеолитический человек -
неандерталец, отличавшийся от своих предшественников не только
усложнением техники каменной индустрии, но и усовершенствованием
функций мышления. С этим связано обживание пещер,
конструирование искусственных жилищ, ритуальное погребение умерших.
Именно среднепалеолитический человек столкнулся с резкой
сменой сезонных температур и снежным покровом. Заселение
неандертальским человеком умеренной зоны Евразии создавало
предпосылки для формирования черт адаптивного типа умеренной
зоны и тех отличительных признаков энергетических процессов,
которые создают демаркационную линию между населением
тропических и внетропических широт. Воздействие таких черт
климата умеренной зоны, как сезонная смена температур и холод,
являющихся экстремальными по отношению к климату
субтропической и тропической зон, в некоторой мере лимитировалось
социальными моментами - изобретением искусственных жилищ
(Чернышев, 1960) и, возможно, первой примитивной одежды.
По-видимому, с эпохи среднего палеолита процесс биологической
адаптации начинает контролироваться системой социальных
адаптации.
Эпоха верхнего палеолита характеризуется значительным
повышением уровня развития производительных сил. Техническое
усложнение орудийной деятельности, изобретение лыж, с
помощью которых, согласно X. Ватанабе (Watanabe, 1968), возможно
освоение приледниковых областей с глубоким снежным покровом,
способствовали дальнейшему расширению ойкумены. Мы не
имеем верхнепалеолитических памятников, расположенных
непосредственно в арктической зоне, тем не менее можно считать
установленным факт освоения ее человеком до заселения Америки.
Время заселения Америки по последним данным составляет более ЗС
тыс. лет. По-видимому, оно было осуществлено человеком, уже
адаптированным к суровым условиям древней Берингии. Итак, в
эпоху верхнего палеолита складывается арктический адаптивный
тип, а с ним и вся система адаптивных типов, свойственная
современному человеку.
262
и итоге приходим к выводу о том, что адаптивные типы
сложились исторически в разное время, отражая последовательность
освоения человеком климатических поясов и разных
экологических ниш (рис. 58).
Верхний
Средний
Стадия
аморфной
катанной
индустрии
2Ш-Ш(Юл?т
JS№-200000мт
38Ш0-5ШООлет
*Ш6
е
д :
Рис. 58. Хронология адаптивных типов:
а — тропический; б — хумидный; в - континентальный; г ~ умеренный; 9 — высокогорный;
е - арктический.
Биологическая адаптация человеческих популяций к условиям
среды постепенно уступала свои позиции в ходе антропогенеза
культурным приспособлениям. Переломный момент падает на
эпоху среднего палеолита, однако процесс биологической адаптации
не затухает с появлением Homo sapiens. Те адаптивные типы,
которые характерны для современного человечества, не являются
наследием отдаленного прошлого. Их возникновение связано с
историей вида Homo sapiens, хотя качественно они, по-видимому, не
263
отличаются от тех, которые могли оыть типичны для наших пред.
ков на различных стадиях антропогенеза- Если мы находим мно
жество аналогий в адаптивных процессах у человека и животных,
то тем более эти аналогии правомерны в отношении наших
человекоподобных предков.
Многие адаптивные черты у современного человека в
количественном отношении могут быть выражены в меньшей степени,
чем у его предшественников, поскольку в процессе
приспособления человека современного вида к условиям естественной среды
обитания значительную роль играют механизмы социальной
адаптации. Что же касается механизмов биологической адаптации, тс
при явном ослаблении одного из них, а именно отбора,
продолжают играть роль адаптации в процессе роста и развития и
усиливается метисация - один из важнейших механизмов
приспособления к новым экологическим нишам.
Лекция XII
КРАТКИЕ ИТОГИ
И ВОЗМОЖНОСТЬ ИСПОЛЬЗОВАНИЯ ФЕНОМЕНА
ЭКОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ ЧЕЛОВЕЧЕСТВА
В ПРИКЛАДНЫХ ЦЕЛЯХ
Характерной особенностью современной науки о человеке
является оформление в ее составе экологического направления,
изучающего взаимодействие человеческих популяций с окружающей
средой. Антропология изучает те аспекты экологии человека,
которые связаны с воздействием окружающих условий на
популяцию человека, ее пространственную структуру, динамику
численности, генофонд, а также приспособительную изменчивость и
процессы роста и развития.
В число экологических факторов, воздействующих на
человеческие популяции, входят абиотические (климат, ландшафт,
структура и химический состав почв и т.д.) и биотические
(возбудители различных болезней). В век научно-технической
революции особенно усилилось воздействие экологических факторов
антропогенного характера. Непосредственное влияние окружаю-
254
щей среды на человека лимитируется хозяйственно-культурно*
деятельностью человеческих сообществ, поэтому к числу экологи
ческих факторов, которые должны быть учтены в антропологи
ческих исследованиях, следует отнести тип хозяйства и связан
ный с ним рацион питания, а также физические нагрузки, вызы
ваемые характером труда.
В.И. Вернадский (1926,1944), сформулировавший идею един
ства всех геохимических, физических и биологических процес
сов, протекающих на Земле, а также развития человеческого об
щества, обратил внимание на геологические масштабы человечес
кой деятельности. Влияние человеческого фактора на развитие
биосферы растет со скоростями, превышающими скорости эволю
ционных процессов. Современное человечество находится в состо
янии большого напряжения во взаимоотношениях с окружающее
средой. Но у этих взаимоотношений есть своя история.
Освоение ойкумены человечеством отнюдь не безболезненный
процесс. За адаптацию к условиям тундры, тайги, пустыни,
высокогорья и других экологических ниш человечество "заплатило'
здоровьем и жизнью многих и многих поколений, пока пришлое
население не приспособилось к экстремальным условиям и не
выработало генетически устойчивых механизмов адаптации.
Таким образом, коренное население различных географических
регионов сформировалось в результате тысячелетней биологической
и социальной адаптации к окружающей среде, и поэтому их
генетический и морфофункциональный статус представляется
наиболее адекватным условием существования в определенных
экологических нишах.
Исследование закономерностей пространственной
изменчивости морфологических и физиологических признаков современных
людей, осуществляемое в антропологической науке в последнее
тридцатилетие, показало их значительное экологическое
разнообразие. Именно поэтому так необходим дифференцированный
подход к оценке взаимоотношений человеческих популяций со
средой обитания.
Географическая вариабельность черт строения тела и
функциональных признаков в популяциях человека чрезвычайно велика
Это качество человеческого вида сформировалось в результате
длительной его истории, связанной с освоением ойкумены, но і
255
основе его лежит высокая пластичность морфосрункциональны:
черт, присущая виду Homo sapiens.
Характер пространственного распределения черт строения тела
а также некоторых показателей основного, белкового, липидноп
и минерального обмена веществ свидетельствует о том, что он]
подчиняются требованиям экологических факторов. Энергетичес
кие процессы, о которых, помимо показателей основного и липид
но го обмена, мы судим и по массе, поверхности и пропорция?
тела, определяются в основном температурным режимом среды
минеральный обмен - геохимической ситуацией.
Питание как экологический фактор, воздействующий на об
мен веществ и строение тела человеческих популяций, живущи:
в естественных условиях среды, во многом определяется геогра
фическими причинами. Поэтому при обсуждении вопросов взаи
моотношения человека и среды не следует противопоставлять в<
влиянии на человеческий организм такие факторы, как темпера
турный режим, геохимический состав или питание.
Поскольку каждый из видов обмена веществ наследственні
обусловлен, то, по-видимому, такое соответствие экологически!
условиям формировалось путем длительной приспособительноі
изменчивости в результате действия отбора. В этом отношени]
здесь также нет основания для противопоставления действия ере
ды и наследственности (Алексеева, 1977, 1986).
Обращает на себя внимание зональный характер изменчивое
ти морфологических и функциональных признаков — специфик
морфофизиологического комплекса присуща населению арктичес
ких широт, умеренной зоны, тропиков и высокогорья. Это обсто
ятельство подтверждает мысль о приспособительном значены
многих морфофизиологических особенностей человеческих попу
ляций и дает основание для введения в научные исследования по
нятия адаптивного типа как нормы биологической реакции на об
ределенную окружающую среду, выражающейся в морфофункцн
ональных особенностях популяции и обеспечивающей состояни
равновесия популяции с этой средой.
Природа адаптивных типов у человека, их соотношение с экс
логическими формами животных и растений, специфика адаптив
ных и акклиматизационных процессов подробно освещены в пред
шествующем изложении. Здесь уместно обратить внимание лиш
на некоторые их черты. Проявляясь в пределах человеческог
256
вида, адаптивные типы не представляют экологически специали
зированных форм. Они могут быть рассмотрены лишь как тенден
ция к изменению морфофизиологических черт в направлении.
наиболее благоприятном для существования в определенной окру
ясающей среде и не препятствующем возможности существование
в других экологических нишах. Адаптивный тип независим оі
расовой и этнической принадлежности. Это норма биологической
реакции, конвергентно возникающая в сходных условиях
обитания у популяций, которые могут быть и не связаны родством.
Адаптивные типы, присущие населению различных
экологических регионов, отражают феномен экологической
дифференциации человечества. В связи с этим можно наметить некоторые
направления исследований в области экологии человека, которые
имеют не только фундаментальное значение для объяснения само-
го феномена экологического разнообразия человечества и
формообразующего эффекта роли среды жизни, но могут быть
использованы и в прикладных целях.
Основной задачей антропологических исследований в области
экологии человека представляется оценка территориальной и
временной динамики адаптивных процессов в популяциях коренногс
населения, коль скоро именно оно осуществляло стихийное
освоение ойкумены и на протяжении ряда поколений вырабатывало
черты приспособления к определенным условиям обитания. Поке
еще не удается определить конкретные сроки, необходимые для
образования адаптивного комплекса в тех или иных
экологических нишах. По-видимому, для этой цели необходимо исследование
не только изменчивости морфологических и физиологических чері
во времени, но и всего процесса этногенеза в различных
географических регионах.
Тем не менее уже сейчас открывается возможность
прогнозировать медико-биологическое состояние пришлого населения на
основе закономерностей изменчивости, характерных для коренногс
населения, и тем самым наметить пути формирования устойчивых
работоспособных коллективов в зонах народнохозяйственного
освоения новых территорий. В этой связи обращает на себя внима
ние характер приспособительных реакций многих
физиологических признаков у приезжих акклиматизирующихся лиц: сдвиі
происходит в направлении характеристик, типичных для
коренного населения тех регионов, в которых оказывается вновь при
267
бывшая группа. Что касается биологической адаптации пришлого
населения, то ее успех во многом зависит от степени адекватности
морфологических и физиологических черт организма человека
окружающим условиям.
Из работы "Статистические миграции населения" (1973)
явствует, что заболеваемость переселенцев возрастает по мере
увеличения медико-географической контрастности новых территорий
по сравнению с прежними местами их обитания. Например,
заболеваемость уроженцев южных территорий на Камчатке почти в 10
раз превосходит заболеваемость уроженцев северных районов
нашей страны. В связи с необходимостью формирования населения
в местах освоения новых территорий весьма показателен
исторический опыт сложения старожильческого населения Сибири,
основу которого составили уроженцы Русского Севера.
Одну из основных практических задач экологии человека
составляет разработка научно обоснованных моделей систем
жизнеобеспечения населения в связи с освоением новых территорий.
Решению этой задачи, как справедливо отмечает В.П. Казначее!
(1980), препятствует высокая подвижность пришлого населения.
Именно поэтому данные о широкой норме биологической реакции
человеческих популяций на комплекс условий окружающей ере
ды, обнаруженной при анализе закономерностей
пространственной изменчивости морфофизиологических особенностей, и специ
фика приспособительных черт коренного населения различных
экологических ниш могут служить основой для создания подоб
ных моделей, ибо коренное население представляет собой тот ад ад
тивный оптимум, который наиболее адекватен среде его обитания
ЛИТЕРАТУРА
Авазбакиева МЛ. Влияние климата Казахстана и Киргизии на организм
человека. Алма-Ата, 1958.
Агаджанян НА., Миррахимов ММ. Горы и резистентность организма. М., 1970.
Адаптация человека. Л., 1972.
Алексеев ВЛ. География человеческих рас. М., 1974а.
Алексеев ВЛ. Происхождение народов Кавказа. Краниологическое
исследование. М.у 19746.
Алексеев ВЛ. Системный анализ антропогеоценоза // Природа. 1975. № 7.
Алексеев ВЛ. Историческая антропология. М., 1979.
Алексеев ВЛ. Адаптация и наследственность // Окружающая среда и здоровье
человека. М., 1979а.
Алексеев ВЛ. Антропологические исследования на Кубе // Расы я народы. Т. 9.
М., 19796.
Алексеев ВЛ. Характеристика типа телосложения в трех контрастных но
расовому составу и происхождению группах населения Махараштры // Новые
материалы к антропологии Западной Индии. Результаты советско-индийских исследований.
М., 1982.
Алексеев ВЛ., Гохман ИЛ. Антропология азиатской части СССР. М., 1984.
Алексеева ТЛ. Прижизненная оценка степени минерализации костной ткани
рентгенофотометрическим путем в связи с весом отдельных костей и их
структурными особенностями // Архив анатом., гнстол., эмбриол. 1965. Т. 48, М б.
Алексеева ТЛ. Опыт сопоставления биохимических показателей крови с
основными соматическими компонентами тела человека // Anthropology. 1966. ГУ/1.
Алексеева Т.Н. Изменчивость основных компонентов тела человека в
зависимости от уровня липидов и белков в сыворотке крови // Морфофизиологические
исследования в антропологии. М., 1970.
Алексеева Т.Н. Географическая изменчивость содержания холестерина в
сыворотке крови человека (к влиянию среды и расы) // Вопр. антропол. 1971. Вып. 38.
Алексеева Т.И. Биологические аспекты изучения адаптации у человека.
Симпозиум "Антропология 70-х годов". М., 1972.
Алексеева Т.И. Этногенез восточных славян по данным антропологии. М., 1973.
Алексеева Т.И. Особенности адаптивных реакций в условиях высокогорья //
Вопр. антропол. 1974. Вып. 47.
Алексеева Т.И. Биологическая адаптация населения Арктики к экстремальным
условиям Крайнего Севера (антропологический аспект) // Географические аспекты
экологии человека. М., 1975.
Алексеева Т.И. Географическая среда и биология человека. М., 1977.
Алексеева Т.И. Биогеохимия и проблемы антропологии // Современные задачи
и проблемы биогеохимии. Тр. биогеохимич. лаб. Т. XVIL М., 1979.
Алексеева Т.И. Адаптивные процессы в популяциях человека. М., 1986.
Алексеева Т.И. Экология народов Центральной Азии. Антропологический
аспект // Вестник АМН СССР. М., 1989. J* 8.
Алексеева Т.Н., Балахонова Е.И. Антропоклиматические связи на территория
Центральной и Северо-Восточной Сибири // Вопр. антропол. 1979. Вып. 61.
Алексеева Т.Н., Доброванова СВ. География типов телосложения у человека //
Вопр. антропол. 1980. Вып. 66.
269
Алексеева ТЛ., Клевцова НЛ. Алеуты Командорских островов (морфофизиоло-
гическая характеристика) // Бопр. антропол. 1980. Вып. 64.
Алексеева ТЛ.» Коваленко В.Ю. Морфофункдиональная характеристика лост-
краНиального скелета азиатских эскимосов // Палеоантропология Сибири. М., 1980.
Алексеева ТЛ., Шауро ЭА. Аномалии черепа в их географической, социальной
и генетической обусловленности // Морфофизиологические исследования в
антропологии. М., 1970.
Алексеева ТЛ., Бацевич В А., Эрнандес Ф.О. П. Скелетный возраст у детей
некоторых народов азиатской части СССР // Вопр. антропол. 198?. Вып. 76.
Алексеева ТЛ., Бацевич В А., Медникова ЯМ. и др. Алтайцы. Морфофизиологи-
ческий очерк (рукопись).
Алексеева ТЛ.Р Алексеев ВЛ., С?ицын ВА. и др. Ферментные и другие белки
крови и дифференциация популяций Северо-Восточной Азии (некоторые итоги гене-
тико-антропологических исследований) // Вопр. антропол. 1978. Вып. 68.
Алексеева ТЛ., Асомиддинов И А., Вацевич В А. и др. Антропологические
исследования в Северном Таджикистане в связи с проблемами геохимической экологии //
Вопр. антропол. 1984. Вып. 73.
Алексеева ТЛ., Волков-Дубровин ВЛ., Голубчикова ЗА. и др. Антропологическое
изучение лесных ненцев (морфология, физиология и популяционная генетика) //
Вопр.антропол. 1972. Вып. 41, 42.
Алексеева ТЛ., Волков-Дубровин ВЛ., Голубчикдва ЗА. и др. Саамы. Морфофун-
кциональный очерк // Вопр. антропол. 1973. Вып. 43.
Алексеева ТЛ., Волков-Дубровин ВЛ., Павловский ОМ. и др.
Антропологические исследования в Забайкалье (морфология и популяционная генетика) //Вопр.
антропол. 1970. Вып. 36; 1971. Вып. 37.
Алиев AM. Содержание холестерина и лецитина в крови у жителей'различных
географических зон Дагестана // Материалы 38-й выездной научной сессии,
посвященной 50-летию установления Советской власти в Азербайджане. Баку, 1970. Ч. 1.
Алисов БЛ.Р Берлин И А., Михель ВМ. Курс климатологии. Ч. III. Климат
земного шара. Л., 1964.
Андрианов Б&. Хозяйственно-культурные типы и исторический процесс // Сов:
этнограф. 1968. № 2. ¦¦' <
Андрианов Б&., Чебоксаров НЛ. Хозяйственно-культурные типы и проблемы их
картографирования ¦// Сов. этнограф. 1972. >& 2.
Андрианова ГА., Ковальский BJ5. Микроэлементы в почвах СССР. М., 1970.
Антропоэкологические исследования в Туве. М., 1984.
Апостолов Еч Мичков X. Урбанизация. Тенденции и гиги?ническо-демографи-
ческие проблемы. М., 1977.
Арутюнов С А., Сергеев ДА. Древние культуры азиатских эскимосов (Уэленсккй
могильник). М., 1969.
.Арутюнов СА., Сергеев ДА* Проблемы этнической истории Берингоморья (Эк-
венский могильник). М., 1975.
Архангельская Af.C„ Павловский ОМ* Сравнение темпов костного созревания в
старения в современных сельских популяциях // Вопр. физической антропологии.
Тезисы. Тарту, 1982.
. Архангельская М.С^ Волков-Дубровин ВЛ^ Павловский ОМ. и др.
Морфофизиологические исследования населения аридной зоны. Ч. ГУ. Казахи Южных Муюнку-
мов; характеристика выборки и ее морфологические, особенности // Вопр. антропол.
1980. Вып. бб.
260
Архангельская М.С., Волков-Дубровин ВЛ., Павловский ОМ. и др. Морфофнзи
ологические исследования населения аридной зоны. Ч. У. Казахи Южных Муюнку
мов. Рентгеноантропологическая и гемодинамическая характеристика // Вопр. ані
ропол. 1981. Вып. 67..
Архангельская НА., Вадова A3. Влияние солнечного и ультрафиолетового све
та на химическую терморегуляцию // Рефераты АМН СССР. М., 1949. № 7.
Ахмедов KJO. Насыщение крови кислородом, давление альвеолярных газов и ко
личество гемоглобина у людей на равнине ив условиях высокогорья // Материал!
симп. "Высокогорье и красная кровь" и конф. "Высокогорье и лекарства". Фрунзе
1968. '
Ахмедов KJO. Дыхание человека при высокогорной гипоксии. Душанбе, 1971.
Бабаева AJC., Серебряков ЕЛ., Северова РЛ. Водно-солевой обмен у человека з
животных под действием факторов аридной зоны. Ашхабад, 1979.
Барбашева ЗЛ. Человек и биосфера // Человек и среда обитания. Л., 1974.
БартонА:, Эдхолм О. Человек в условиях холода. М., 1967.
Бедный М.С: Продолжительность жизни в городах и селах. М., 1976.
Бейкер П. Адаптивные возможности высокогорных популяций // Биология жи
телей высокогорья. М., 1981.
Белекова РЛ Функциональное состояние коры надпочечников у жителей высо
когорья Памира // Вопросы диагностики и терапии в клинике внутренних болезней
Вып. 1. Фрунзе, 1961а.
Белекова РЛ Функция щитовидной железы у жителей высокогорья Памира /у
Вопросы диагностики и терапии в клинике внутренних болезней. Вып. 1. Фрунзе
19646.
Бен-Утяева Т.С. Содержание микроэлементов в почвах и растениях песчанок
пустыни // Микроэлементы в сельском хозяйстве. Ташкент, 1965.
Беркович ЕМ. Энергетический обмен в норме и патологии. М., 1964.
Биология жителей высокогорья. М., 1981,
Бочков ЛЛ. Мутационный процесс у человека // Лекции по медицинской
генетике. М., 1974.
Будыко МЛ. Глобальная экология. М., 1977.
Бунак ВЛ Об акклиматизации человеческих рас и,сравнительном значении
определяющих ер факторов,// Русск. антропод. журн. 1924. Т* 13. .
. Бунак ВЛ. Задачи изучения вымирающих народностей, в частности изучения
этно-антропологического.// Русск. антропол. журн. 1926. Т. 14.
, Бунак ВЛ О современном состоянии и задачах антропологического изучения
человека как производительной силы // Бюл. орг. комитета 2-й Всесоюзной
конференции по изучению[.производит, сил СССР (февраль). М., 1927. № 3-4, 6-7.
Бунак ВЛ Структурные изменения черепа в процессе брахикефализации // Тр.
V Всесоюз. съезда анат. гистол., эмбриол. Д., 1961.
. Бунак, ВЛ Климато-зональные и этнические различия в строении лица и
головы у коренного населения Северной Азии (в связи с проблемой адаптации) //
Адаптация: .человека. Л., 1972. ,
Бускирк ЕЛ Работоспособность уроженцев высокогорья.// Биология жителей
высокогорья..М-,.1981.
Вайнштейн СЛ. Первая советская этяографс^антроп?логнческая экспедиция в
Туву .// Очерки истории русской этнографии, фольклористики и антропологии. М.,
1982. Вып. IX; Тр; Ин-та этнограф. Т. 110.'
261
Веселухин PJ3. Физиологическая характеристика адаптивных типов коренного
населения арктической и континентальной зон северо-востока Азии // Вопр. антро-
пол. 1977. Вып. 5?.
Властовский ВТ. Акселерация роста и развития детей (эпохальная и внутри-
групповая). М., 1978.
Войнар АЛ. Биологическая роль микроэлементов в организме животных и
человека. М., I960.
Волкова ТМ. Морфологические особенности горцев Западного Памира // Вопр.
антропол. 1966. Вып. 24.
Волков-Дубровин ВЛ. Половозрастная изменчивость показателей оксигемом?т-
рии // Вопр. антропол. 1966. Вып. 24.
Волков-Дубровин ВЛ. Уровень окислительных процессов в связи с тотальными
размерами и основными соматическими компонентами тела // Морфофизиологичес-
кие исследования в антропологии. М., 1970.
Волков-Дубровин ВЛ. Размеры тела и некоторые физиологические особенности
пяти эндогамных групп хиндиязычного населения Союзной территории Дели //
Вопр. антропол. 1974. Вып. 46.
Волков-Дубровин ВЛ., Смирнова Н.С. Опыт изучения взаимозависимости
признаков, относящихся к различным системам человеческого организма // Вопр.
антропол. 1969. Вып. 32.
Волков-Дубровин ВЛч Рогинский ЯЛ. Гипсистеноцефалия как
приспособительный признак в тропической зоне // Вопр. антропол. 1960. Вып. 1.
Волков-Дубровин ВЛч Воронов АА., Смирнова HjC. и др. Комплексные морфофи-
зиологические исследования в Абхазии // Вопр. антропол. 1982. Вып. 69.
Волков-Дубровин ВЛН Воронов АА., Смирнова Н.С. и др. Морфофизиологические
исследования населения аридной зоны Туркмены. Ч. I. Текинцы Ахала // Вопр.
антропол. 1975. Вып. 50.
Волков-Дубровин В Л., Гудкова К., Павловский ОМ. и др. Морфофизиологичес-.
кие исследования населения аридной зоны Туркмены. Ч. П. Йомуты Казанджикско-
го района // Вопр. антропол. 1977. Вып. 55.
Волков-Дубровин ВЛ., Гудкова К-, Павловский OJM. и др. Морфофизиологичес-
кие исследования населения аридной зоны. Часть III. Каракалпаки Западного При-
хызылкумья // Вопр. антропол. 1977. Вып. 57.
Воловик ВТ. Человек в экстремальных условиях природной среды. М., 1983.
Воронин НМ-, Слоним АД. Физиологическое действие климата как средство
профилактики и лечения // Основы курортологии. М., 1959.
Гаджиев АХ. Происхождение народов Дагестана (по антропологическим
данным). Махачкала, 1965.
Гаузе Г.Ф. Экология и некоторые проблемы происхождения видов // Экология
н эволюционная теория. Л., 1984.
Географические аспекты экологии человека. М., 1975.
Гец Л. Водно-солевой обмен при термической дегидратации // Гигиена труда и
профилактика заболеваний. 1968. № 1.
Гинзбург ВЛ. Горные таджики. Материалы по антропологии таджиков Карате-
гина и Дарваза. М.; Л., 1987.
Гудкова ЛЯ, Географическая изменчивость уровней протеинов в сыворотке
крови человека // Вопр. антропол. 1975. Вып. 49. *
Давыдова ГМ. Относительный вес тела в нескольких популяциях Сибири (в
связи с вопросами климатической адаптации) // Адаптация человека. Л., 1972.
262
Данишевский ТМ. Патология человека и профилактика заболеваний на Севере
М., 1968.
Дебец Г.Ф. Палеоантропология СССР // Тр. Ин-та этнографии АН СССР (нов
сер). 1948. Т. 4.
Дерлпа HJP., Рябинин Н.Ф. Адаптация человека в полярных районах Земли. Л.
1977.
Добровольская МЛ. Опыт количественного определения микроэлементов в
скелете человека (по материалам древних погребений) // Вопр. антропол. 1984. Вып.
74.
Егоров АД.» Григорьева ДЛ., Курелюк Т.Т. и др. Микроэлементы в почвах и
луговых растениях мерзлотных ландшафтов Якутии. Якутск, 1970.
Ефремов ВЛ. Проблема рационализации питания населения в северных районах
Советского Союза // Проблемы Севера. Вып. 14. М., 1970.
Зенкевич ИЛ. К вопросу о факторах формообразования длинных костей
человеческого скелета // Тр. Ин-та антропологии МГУ. 1940. Вып. 5.
Зильбер ЛА Основы иммунологии. М., 1958.
Золотарев ДА Кольские лопари. Л., 1928.
Золотарев ДА Карелы СССР. Л., 1930.
Золотарева ИМ. Этническая история бурят // Этнографический сб. Сибир. отд.
АН СССР. Выл. 1. Улан-Удэ, I960.
Золотарева ИМ. Распределение групп крови у народов Северной Сибири // VII
МКАЭН. М., 1969. Т. 1.
Кабузан ВМ., Троицкий СМ. Новые источники по истории населения Восточной
Сибири во второй половине XVIII в. // Сов. этнограф. 1966. № 3.
Казначеев BJI. Современные аспекты адаптации. Новосибирск, 1980.
Казначеев ВЛ. Очерки теории и практики экологии человека. М., 1983.
Камшилов ММ. Роль фенотипа в эволюции. Ссобщ. I. Фенотипическая форма
наследственной изменчивости // Генетика. 1967. № 12.
Кандор И.С. Очерки по физиологии и гигиене человека на Крайнем Севере //
Климат и здоровье человека на Крайнем Севере. М., 1961.
Карсаевская ТМ. Влияние урбанизации на сдвиги в физическом развитии
населения (обзор) // Здравоохранение Российской Федерации. 1973. № 5.
Касавина К.С., Торбенко ВЛ. Жизнь костной ткани. Мм 1976.
Кассирский ИА., Плотников ИЛ. Болезни жарких стран. М-, 1964.
Квиличи ДжЖ., Вергиес X. Гематологическая характеристика высокогорных
популяций // Биология жителей высокогорья. М., 1981.
Кириакиди Ф.С. Морфологические особенности телосложения мужчин
Дагестана // Вопр. антропол. 1973а. Вып. 44.
Кириакиди Ф.С. Морфологическая характеристика населения Дагестана. Авто-
реф. дис. канд. биол. наук. М., 19736.
Клевцова ИЛ. Соматические особенности сибирских монголоидов в
сравнительном освещении // Вопр. антропол. 1976а. Вып. 52.
Клевцова ИЛ. О межгрупповой изменчивости соматических особенностей
монголоидов Сибири // Вопр. антропол. 19766. Вып. 53.
Клевцова НЛ* Смирнова Н.С. Морфологические особенности тела чукчей и
эскимосов // Вопр. антропол. 1974. Выл* 48.
Коваленко ВJO. Соматическая характеристика сахалинских нивхов // Вопр.
антропол. 1977. Вып. 56.
263
Коржуев ПА. Силы гравитации и филогенез позвоночных животных // Усп
совр. биол. 1965. Т. 60. Вып. 2 (б).
Коржуев ПА О функциональных аспектах эволюции // Усп. совр. биол. 1971а
Т. 72. Вып. 3 (6).
Коржуев ПА Эволюция, гравитация, невесомость. М., 19716.
Крупник ИЛ. Антропологические признаки и особенности климатической адап
тации (на материале Высокой Африки) // Расы и народы. М., 1973. Вып. 3.
Крупник ИЛ. О методике антропоклиматического районирования // Вопр.ант
ропол. 1977. Вып. 54.
Крупник ИЛ, Арктическая этноэкология. М., 1939.
Крус-Кок Р. Генетическое описание высокогорных популяций // Биология жи
телей высокогорья. М., 1981.
Кукес BX.t Власова МЛ. Некоторые данные по изучению содержания холест?
рина в сыворотке крови и витамина С в плазме крови у коренных жителей Магадан
ской области // Проблемы Севера. 1962. Вып. 6.
Курбатова (Русакова) ОЛ. Опыт генодемографического исследования болыпиз
панмиксных популяций. Генетическая структура двух последовательных поколениі
жителей Москвы // Вопр. антропол. 1975. Вып. 50.
Курбатова ОМ. Генетические процессы в городском населении (опыт генодемог
рафического исследования популяций г. Москвы). Автореф. дне. канд. биол. наук
1977.
Лавик-Гудол ванДж. В тени человека. М., 1975.
Левин MX* Проблема пигмеев в антропологии и этнографии // Сов. этнограф
1946. № 2.
Левин MX., Чебоксаров НЛ. Хозяйственно-культурные типы и историко-этног
рафические области // Сов. этнограф. 1955. № 4.
Литтл МА-, Ханна Дж. М. Реакции высокогорных популяций на воздействие
холода и других стрессорных факторов // Биология жителей высокогорья. М., 1981
Ломов И А. О законе поверхности Рубнера // Количественные аспекты росте
организмов. М., 1976.
Лукин ЕЛ, Дарвинизм и географические закономерности в изменении организ
мов. М.; Л., 1940.
Лукьянов 1*3., Пушкина НЛ. Гиповитаминозы среди пришлого и коренноіч
населения некоторых районов Заполярья // Проблемы Севера. М., 1962. Вып. 6.
Лутовинова HJO., Уткина МЛ., Чтецов ВЛ. Методические проблемы изуче
ния вариаций подкожного жира // Вопр. антропол. 1970.
Любин ВЛ. Нижний палеолит // Каменный век на территории СССР. М., 1970
Материалы Бурят-Монгольской экспедиции 1931 года. Л., 1933.
Медникова М?. Древние скотоводы Южной Сибири: палеоэкологическая рекон
струкция по данным антропологии. М., 1995.
Меновщиков ГА. Язык эскимосов Берингова пролива. Языки и фольклор наро
дов Сибирского Севера. М.; Л., 1966.
Методика морфофизиологических исследований в антропологии. М., 1981.
Миклашевская НЛ, Влияние расовой принадлежности и географической средь
обитания на ростовые процессы у человека // Симпозиум "Антропология 70-х годов"
М., 1972.
Миклашевская НЛ., Година ЕЗ., Данилкович НМ. и др. Соматическое развита»
детей и подростков во влажном субтропическом климате // Вопр. антропол. 1979
Вып. 61.
264
Миклашевская НЛ., Соловьева В.С., Година ЕЗ. и др. Ростовые процессы у
человека в условиях высокогорья // Человек, эволюция и внутривидовая
дифференциация. М., 1972.
Миклашевская НЛ., Соловьева В.С., Година ЕЗ. и др. Рост и развитие детей в
условиях высокогорной Киргизии // Человек и среда. Л., 1975.
Миклашевская НЛ.> Соловьева В.С., Гилярова ОА. и др. Ростовые процессы у
абхазских детей и подростков в популяции с высоким процентом долгожителей //
Вопр. антропол. 1982. Вып. 70.
Миррахимов ММ. Сердечно-сосудистая система в условиях высокогорья. Л.,
1968.
Миррахимов ММ. Биологические и физиологические особенности коренных
жителей высокогорья Тянь-Шаня и Памира // Биология жителей высокогорья. М.,
1981.
Миррахимов ММ* Гринштейн БЯ.,ДжалобаевАД. Гематологические
исследования в условиях высокогорья Тянь-Шаня и Памира // Материалы симп. "Высоко
горье и красная кровь" и конф. "Высокогорье и лекарства'*. Фрунзе, 1968.
Многотомное руководство по патологической физиологии. Т. П. М., 1966.
Молчанова ОЛ., Ежова ЕЛ. Изучение длительного голодания. М., 1939.
Морфофизиологические исследования в антропологии. М., 1970.
Народы Сибири. М., 1956.
Насонов АЛ, "Русская земля" и образование территории древнерусского
государства. М., 1951.
Неверова НЛ., Андронова ТЛ., Мочалова МЛ. К вопросу о физиологических
механизмах начального периода акклиматизации в Арктике // Адаптация человека.
Л., 1972.
Ногина НА. Почвы Забайкалья. М., 1964.
Носин А. Почвы Тувы. М., 1963.
Окладников АЛ. Возникновение человеческого общества. Ранний
древнекаменный век (нижний палеолит) // Всемирная история. Т. 1. М., 1955.
Ошанин Л Л. Данные о телосложении узбеков, таджиков и евреев Бухары и Кер
мине // Мед. мысль Узбекистана. 1927 (ноябрь).
Павловский ОМ. Особенности динамики костной плотности в половозрастном х
этнотерриториальном аспектах // Морфофизиологические исследования в
антропологии. М., 1970.
Павловский ОМ. Оссеографические показатели возрастной; динамики остеомор
фных процессов в долгожительной популяции // Феномен долгожительства. М.
1982.
Плетнева СА. О юго-восточной окраине русских земель в домонгольское врем*
// Кр. сообщ. Ин-та археол. АН СССР. 1964. Вып. 39.
Преображенский В.С^ Райх ЕЛ, Проблемы экологии человека и географии // Те
ория и методика географических исследований экологии человека. М., 1964.
Прессман АС. Электромагнитные поля и живая природа. М., 1968.
Пржевальский НМ. Из Зайсана через Хами в Тибет. М», 1948.
Рогинский ЯЯ. К вопросу о возрастных изменениях расовых признаков у чело
века (в утробном периоде и в детстве) // Антропологический сб. П. М., 196Q (Тр. Ин
та этнографии, АН СССР (нов. сер.). Т. V).
Рогинский ЯЯ. Проблемы антропогенеза. М.,1969.
Рост и развитие ребенка (по данным антропологии). М., 1978.
265
Ротштейн А., Таубин Э. Кровообращение и температура тела ври обезвожива
нии, вызванном действием высокой температуры // Физиология человека в пусты
не. М., 1952.
Русские старожилы Сибири. М., 1973.
Рысназаров HJP. Антропологический состав каракалпаков в связи с вопросам!
этногенеза каракалпакского народа. Автореф. канд. дис. М-, 1972.
Рынков ЮХ. Антропология и генетика изолированных популяций // Древниі
изоляты Памира. М., 1969а.
Рынков ЮХ. Некоторые популядионно-генетические подходы к антрополопп
Сибири // Вопр. антропол. 19696. Вып. 33.
Саливон ИЛ. Морфофизиологические особенности населения геоморфологичес
ких зон БССР // Вопр. антропол. 1977, Вып. 66.
Саливон ИЛ. Особенности строения головы, лица и тела у населения Белорус
ского Полесья //Антропология Белорусского Полесья. Минск, 1978.
СлонимАД. Животная теплота и ее регуляция в организме животных. М., 1952
Смирнова Н.С. О межгрупповой дисперсии соматических признаков у шесті
групп русского населения // Вопр. антропол. 1977. Вып. 64.
Смирнова Н.С Сравнительная морфофизиологическая характеристика двух груш
населения Северного Таджикистана // Вопр. антропол. 1979. Вып. 61.
Соловьев СМ. История России с древнейших времен. М., 1959. Кн. I.
Спицын ВА. Биохимический полиморфизм человека. М., 1985.
Сшанчинский ВМ. Теоретические основы акклиматизации животных // Тр. Ин
та сельскохоз. гибридизации и акклиматизации животных в Аскании-Нова. М.; Л.
1933. Т. 1.
Страдомская HJI. Урбанизация и здоровье человека // Окружающая среда з
здоровье человека. М.г 1979.
Страдомская MJI., Райх EJI. Загрязнение окружающей среды и здоровье челе
века // Окружающая среда и здоровье человека. М., 1979.
Тегако ЛЛ.Р Микулин АЛ., Саливон ИЛ. Антропология Белорусского Полесь;
(демография, этническая история и генетика). Минск, 1978.
Теория и методика географических исследований экологии человека. М., 1974
Толстое СЛ. Очерки первоначального ислама // Сов. этнограф. 1932. N& 2.
Туркменов М.Т. Морфологический состав крови у постоянных жителей
низкосредне- и высокогорья // Материалы симп. "Высокогорье и красная кровь" и кон<|
"Высокогорье и лекарства**. Фрунзе, 1968а.
Туркменов М.Т. Оксиг?нация крови у постоянных жителей низко-, средне- и вк
сокогорья // Материалы симп. "Высокогорье и красная кровь" и конф. "Высокогс
рье и лекарства**. Фрунзе, 19686.
Турусбеков В. Особенности вегетативных функций у жителей Южной Киргизия
Фрунзе, 1970.
Урбоэкология. М., 1990.
Урысон МЛ. Люди или животные // Природа. 1973. № 1.
Урысон МЛ. Неужели человеку 3 миллиона лет? // Природа. 1974. № 6.
Урысон МЛ. Новые открытия древнейших гоминид в Эфиопии // Природа
1976. № 1.
Успенский СЛ. О типовых нормах строения черепов у гоминид и обезьян в ев*
зи с антропогенезом // Вопр. антропол. I960. Вып. 4.
Успенский СЛ. О некоторых биологических предпосылках очеловечения обезі
яны // У истоков человечества. М., 1964.
266
Феномен долгожительства. М., 1982.
Физиология человека в пустыне. M.f 1952.
Филатов JIJT., Каныгина КЛ. Характеристика некоторых показателей крови
человека на различных высотах // Материалы симп. "Высокогорье и красная кровь" и
конф. "Высокогорье и лекарства". Фрунзе, 1968.
Фрисанчо АР. Рост и развитие детей в высокогорных популяциях // Биология
жителей высокогорья. М., 1981.
Харрисон Дж., Уйнер Дж., Таннер Дж., Барникот Н. Биология человека. М.,
1968.
Хлыстов ВЛ., Прибытков ЮЛ. К вопросу о влиянии опресненной воды на
состояние фосфорно-кальциевого обмена и минерализацию костной ткани у населения
г. Шевченко //'Вопр. антропол. 1973. Вып. 43.
Холодов ЮЛ. Магнетизм в биологии. Мм 1970.
ХомичЛЛ. Ненцы. М.; Л., 1966.
Циммерман С.Э. К антропологии таджиков // Русск. антропол. жури. 1927.
Вып. 3-4.
Чебоксаров НЛ. Основные принципы антропологических классификаций.
Происхождение человека и древнее расселение человечества // Тр. Ин-та этнограф. АН
СССР (нов.сер.). М., 1951. Т. XVI.
Человек и среда. Л., 1975.
Чепурковский JSJW. Географическое распределение формы головы и цветности у
крестьянского населения Великороссии // ИОЛЕАЭ. Т. XXIV. Вып.2; ТАО. Т. XVIII.
Вып. 2. М., 1913.
Чернецов ВЛ. Древняя приморская культура на полуострове Ямал // Сов.этног-
раф. 1935. № 4-5.
Чернов ЮЛ. Некоторые закономерности приспособления наземных животных к
ландшафтно-зональиым условиям // Журн. общ. биол. 1974. XXXV. № е.
Черныш АЛ. Остатки жилища мустьерского времени на Днестре // Сов.
этнограф. I960. № 1.
Чижевский АЛ. Аэроионификация в народном хозяйстве. М., I960.
Чижевский АЛ. Об одном виде специфически-биоактивного или Z-излучения
Солнца // Земля во Вселенной. М., 19?4.
Чикишева ТА. Изучение изменчивости уровней жирового и белкового обмена на
примере шорцев // Вопр. антропол. 1982. Вып. 70.
Чикишева ТА. Изучение связи антропологических особенностей населения с
экологическими факторами (на примере Алтае-Саянского региона). Автореф. дне.
канд. биол. наук. М., 1982.
Шварц С.С. Пути приспособления наземных позвоночных животных к условиям
существования в Субарктике // Млекопитающие. Свердловск, 1963 (Тр. Ин-та биол.
Уральск, фил. АН СССР. Вып. 33. Т. 1).
Шварц С.С. Проблемы экологии человека // Вестник АН СССР. 197?. № 2.
Шереметьева BA.f Ринков ЮР. Популяционная генетика народов
Северо-Восточной Азии. М., 1978.
Шишкин МА. Фенотипические реакции и эволюционный процесс. (Еще раз об
эволюционной роли модификаций) // Экология и эволюционная теория. Л., 1984.
Шмальгаузен ИЛ. Организм как целое в индивидуальном и историческом
развитии. М.; Л., 1938.
Шмальгаузен ИЛ. Пути и закономерности эволюционного процесса. М.; Л.:
1940.
267
Шмалъгаузен ИМ. Факторы эволюции (теория стабилизирующего отбора). М,
Л., 1946.
Шмидт-Нильсен К. Животные пустынь. М., 1972.
Штрауб Ф?. Биохимия. Будапешт, 1963.
Щекочихина JIJC. Материалы к изучению половозрастных и территориальны
вариаций уровня гемоглобина в крови здорового человека // Вопр. антропол. 197С
Вып. 36.
Экологические проблемы в исследованиях средневекового населения Восточно]
Европы. М., 1993.
Экологическая физиология человека. Адаптация человека к различным клима
то-географическим условиям. Л., 1980.
ЭфроимсонВЛ. Введение в медицинскую генетику. М., 1968.
Эфроимсон ВЛ. Иммуногенетика. М., 1971.
Юнусое AJO., Махмудов Э.С., Ахмедов Р. и др. Влияние высокой температуры н
водно-солевое равновесие и газообмен людей, прибывших в условия Средней Азии и
различных климатических зон // Человек и среда. Л., 1975.
ЯрхоАЛ. Алтае-Саянские тюрки. Антропологический очерк. Абакан, 1947.
Ярхо АЛ. Антропологический тип кемчикских танну-тувинцев // Северна;
Азия. 1929. № б, 6.
* * *
АЬЫеАА*, Schroder I. Blood pressures in Amhem land aborigines // Med. J. Ausi
1960. Vol. 24. № 2.
Alexeeva TJ. Morpho-f unctional population studies in some biogeochemical USSI
provinces as viewed in the light of the adaptation problems // VIII Intern. Congress о
Anthropol. A. Ethnograph. Sciences (Tokio, 1968). Moscow, 1968.
Alexeeva TJ. Nature of the biological adaptation of the Arctic population to extr
erne environmental conditions // Studies in the Anthropology of the Finno-Ugrian Peop
les. Helsinki, 1973.
Allen J. The influence of physical conditions in the genesis of species // Ann. Rept
of the Smithsonian Institution for 1905. Washington, 1906.
Andersen K?. Comparisons of Scandinavian Lapps, Arctic Fishermen, and Canadia]
Arctic Indians // Fed.Proc. 1963. Vol. 22.
Andersen K?., Bolstad A, Loyning I. et al Physical fitness of Arctic Indians // J
Appl. Physiol. 1960a. Vol. 16. № 4.
Andersen KX„ Loyning /., Neims IJD. et aL Metabolic and thermal response to ;
moderate cold exposure in Nomadic lapps // J. Appl. Physiol. 1960b. Vol. 16. № 4.
Antonis A, Bersohn /., Srand W. The influence of diet on serum lipids in Sout]
African white and bantu prisoners // Amer. J. Clin. Nitrit. 1962. Vol. 10. № 6.
Antoniu ?., Ghiorchtu G., Stefanescu Gh. La conformation corporelle de la popula
tion de Moeciu de Sus // Ann. Roum. Anthropol. 1970. № 7.
Ashcroft M.T. Some aspects of growth and development in different ethnic group
in the Commonwealth West Indies // The ongoing evolution of Latin American Popu
lations. Springfield. Illinois. USA, 1971.
Baker PJT. Adaptation in high altitude cold in the Andes // Contract Rept. ДА-49
193-Md-2260, 1963.
268
Baker P.T., Frisancho AJL, Brooke ТЯ. A preliminary analysis of human growth и
the Peruvian Andes // Human adaptation to environments and physical fitness. New
Delhi, 1966.
Baker P.T. Human adaptation to high altitude // Science. 1969. >fe 163.
Baker P.T. Adaptation problems in Andean Human Populations // The ongoing
evolution of Latin American Populations. Springfield. Illinois. USA, 1971.
Baker P.T. The use of human ecological models in biological anthropology
examples from the Andes // Coll. Anthropol. Zagreb. 1979. Vol. 3. № 2.
Baker P.T., Frisancho AJL, Thomas RB. A preliminary analysis of human growth
in the Peruvian Andes // Human adaptability to environments and physical fitness.
Madras, 19?8.
Baker P.T., Weiner JJS. (ed.). The biology of human adaptability. Oxford, 1966.
Bakker A, Blick A., Luyken R. The serum proteins of malaria-free inhabitants oj
Central Netherlands New-Guines // Docum. Med. Geogr. Trop. 1957. Vol. 9. № 1.
Barnes R. Comparisons of blood pressures and blood cholesterol levels of New Gu-
means and Australians // Med. J. Aust. 1965. Vol. 1.
Behura NJC. Meals and food habits in rural India // Bull. Anthropol. Surv. India.
Calcutta, 1962. Vol. 11. M 2.
Beiguelman B. A somatometric study on Japanese immigrants and Japanese
unmixed descendants in Brasil // Ztschr. fur MorphoL, Anthropol. 1963. Bd. 53. H. 3.
Benedek T.G., Sunder IJf. Comparison of serum lipid and uric acid content in white
and negro men // Amer. J. Med. Sci. 1970. Vol. 260. № 6.
Bennholdt-Thomson C. Die Entwicklungsbeschleunigung der Jugend (Grundfatsach-
en. Theorien, Folgeeingen des Accelerations problems) // Ergebnisse der innoren Medsii
und Kinderhellmunde. Berlin, 1942. B. 62.
Bergmann С Uber die Verhaltnisse der Warmeokonomic der Here zur ihrer Grossc
// GSttinger Studien, I Abteilung. G6ttingen, 1947.
Biswas P.C., Bhattacharya DJC. Growth trends in Northern Indian
population-variation of four anthropometric traits from birth to maturity (cross sectional) // Humai
adaptability to environments and physical fitness. Madras, 1966.
Bollerud I., Edwards /., Blakely RA. Basal Metabolism of Eskimos //Arctic Ae-
romedical Laboratory. 21-01-020. 1950.
Bosch V. Lipoproteins in ultracentrifugal fractions of sera of nomadic South
American Indian tribe // Acta Cient. Venezolana, 1967. Vol. 18. № 6.
Boyad EM. Diurnal variations in plasma lipids // J.Biol. Chem. 1935. Vol. 110. H
1.
BourUere F. Observations on the ecology of some large African Mammals // Africai]
ecology and human evolution. Chicago, 1963.
Bounak V. Un pays de 1'Asie peu connu: Le Tanno-Touva // Inern. Arch. Ethnogra-
ph. Leiden. 1928. Bd. 29. H. 1-3.
Broman G^ Trotter M.. Peterson A The density of the human skeleton // Amer. J.
Phis. Anthropol. 1958. Vol. 16. № 2.
Brown GJVf, Vascular physiology of the Eskimo // Rev. Canad. Biol. 1957. Vol. 16,
№2.
Brown GM.t Bird GJS., Boag IM. et aL Blood volume and basal metabolic rate oi
Eskimos // Metabolism. 1954. Vol. 3. № 3.
Brazen /¦, Grand F, Body composition and basal metabolism in man; correlatioi
analysis versus physiological approach // Hum. Biol. 1955. Vol. 27. № 1.
269
Brunner D., Manelis L., Loebl K. Influence of age and race on lipid in Israel // La
ncet. 1962. Vol. 1. № 7082.
Buck AA., Sasaki T.T., Anderson RJ. Health and disease in four Peruvian villges
Baltimore, 1968.
Butzer K. Environment and archeology. An ecological approach to prehistory. Ch
icago; New York, 1971.
Casley-Smith IJt Blood pressures in Australian aborigines // Med. J. Aust. 1959
Vol. 1. № 18.
Charnock IS., Casley-Smith /., Schartz CJ. Serum magnesium-cholesterol relation
ships in the Central Australian aborigines and in Europeans with and without ischaemi
heart disease // Aust. J. Exptl. Biol. Med. Sci. 1969. Vol. 37.
Clegg EJ.f Harrison GA.t Bauer P.T. The impact of high altitudes on human pc
pulations // Hum. Biol. 1970. Vol. 42. № 3.
Coriet CJE. On some relation of climate, weather and fat covering to metabolism /
Med. J.Aust. 1923. № 1.
Corcoran A.C., Babinowitch JM. A study of the blood lipoids and blood protein i:
Canadian eastern Arctic Eskimos // Biochem. J. 1967. Vol. 31. № 3.
Circumpolar problems, habitat, economy and social relations in the Arctic. Oxford
New York; Toronto; Sydney; Braunschweig, 1973.
Crile G.W., Quiring DJP. Indian and Eskimo metabolism // J. Nutrit. 1939. № 18.
Crognter E, Adaptation morphologique d'une population af ricaine au biotope ti
opical: les Sara du Tchad // Bull. Mem. Soc. Anthropol. 1973. Vol. 10.
Das B.C. Multivariate investigation of blood chemistry and morphology // Huma:
adaptability to environments and physical fitness. Madras, 1966.
Das SJL, Mukherjee DJ>., Das L. A radiological study of skeletal maturity in hand
wrist area of Bengal boys and girls // Anthropological survey of India, Government о
India. Calcutta, 1976.
Davtes A. A re-survey of the morphology of the nose in relation to climate // J. Roy
Anthropol. Inst. Great Brit. a. Ireland. 1932. Vol. 62.
Davtes A, Pitney W. Some haematological observations on aborigines in the Wa
rburton ranges area // Med. J. Austr. 1957. Vol. 2. № 17.
De Almeida. Le metabolism minimum et le metabolism basale de l'homme tropica
de race blanche // J. Physiol. Path. Gener. 1920. № 18.
De Almeida. Le metabolism basal de Гпопшіе tropical // J. Physiol. Path. Gener
1924. № 22.
DucrosA. Plis cutanes d'ammassalimiut: resultats preliminaires // Bull. Mem. Soc
Anthropol. Paris, 1971. Vol. 8.
Ducros A. Adiposite et densite corporelle d'une population arctique (eskimo am
massalimiut) // Anthropologic (Paris). 1971 (1972). Vol. 76. № 7-8.
Ecology and anthropology: A simposium // Amer. Anthropol. 1962. Vol. 64. № 1
Edholm O.G. South-West Asia, with special reference to Israel // The biology о
human adaptability. Oxford, 1967.
Edozien /. The development of the serum protein pattern in Africans // J. Clin
Pathol. 1961. Vol. 14. № 6.
Eickstedt ET. Rassenkunde und Rassengeschichte der Menschheit. Stuttgart, 1934
Eisner RW. Skin fold thickness in primitive peoples native to cold climate. Bod;
composition // Ann. N.Y. Acad. Sci. 1963. Vol. 110.
Eisner AW., Bolstad A. Thermal and metabolic responses to cold of Peruvian Indi
ans native to high altitude // Arctic Aeromed. Lab. Kept. В AAL-TDR-62-64, For
Wainwright. Alaska, 1963.
270
Eisner R.W., Andersen K?., Hermansen L. Thermal and metabolic responses of Ar
ctic Indians to moderate cold exposure at the end of winter // J. Appl.Physiol. I960
Vol. 16. № 4.
Ertkson H. The respiratory response to acute exercise of Eskimos and whites /,
Acta Physiol. Scand. 1957. Vol. 41.
Eskimos of Northwestern Alaska. Dowden, Hutchinson Ross. Inc. Stroudsburg
1978.
Flares M. Nutritional studies in Central America and Panama // The ongoing evolu
tion of Latin American populations. Springfield; Illinois. USA, 1971.
Frey H.f Nanio V., Kulonen E. Serum proteins in Finnish twins // Acta Genet. S
tatist. Med. 1968. Vol. 18. № 1.
Frisancho AJL Growth and functional development at high altitude // Man in th<
Andes: A multidisciplinary Study of High Altitude Quechua. Stroudsberg. Pennsylvani
a, 1976.
Frisancho AJL, Baker P.T. Altitude and growth: A study of the patterns of physica
growth of a high altitude Peruvian Quechua population // Amer. J.Phys. Anthropol (n
s.). 1970. Vol. 32. № 2.
Frisancho AJL, Velasques Т., Sanchez I. Influences of developmental adaptation oi
lung function at high altitude // Hum. Biol. 1973. Vol. 45.
Frisancho AJL, Martinez C, Velasques T. et at Influence of developmental
adaptation on aerobic capacity at high altitude // J. Appl. Physiol. 1973a. Vol. 34.
Frisancho AJL, Sanchez I., PaUardel D. et al Adaptive significance of small bodj
size under poor socio-economic conditions in Southern Peru // Amer. J. Phys. Anthro
pol. 1973. Vol. 39.
Galvao PJL. Human heat production in relation to body weight and body surface. I.
Inapplicability of the surface law to lean men of tropical zone. II. Inapplicability of th«
surface law on well proportioned men of the tropical zone // J. Appl. Physiol. 1948. № 1
Galvao PJ5. Human heat production in relation to body weight and body surface. Ш
Inapplicability of surface law on fat men of the tropical zone. IV. General interpretati
on of climate influence of metabolism // J- Appl. Physiol. 1950. Vol. 3. № 1.
Gam ?Jlf., Clark L.C^ Portray It Relationship between body composition and bassa]
metabolic rate in children // J. Appl. Physiol. 1963. Vol. 6. № 3.
Giblett E. Genetic markers in human blood. Oxford; Edinburgh, 1969.
Gilat Т., Malachi E.G., Schochet SJi. Lactose tolerance in an Arab population //
Amer. J. Digest. Dis. 1971. >fe 16.
Goldby F., Hick CJS.t O'Conner WJ. et aL A comparison of the skin temperature and
skin circulation of naked whites and Australian aboriginals exposed to similar
environmental changes // Aust. J. Exptl. Biol. Med. Sci. 1938. № 16.
Goldrick RB., Whyte HM. A study of blood clotting and serum lipids in natives ot
New Guinea and Australians // Aust. Ann. Med. 1969. № 8.
Greulich WW. A comparison of the physical growth and development of American-
born and native Japanese children // Amer. J. Phys. Anthropol.(n.s.). 1967. № 16.
GseU D., Mayer 7. Low blood cholesterol associated with high satured fat intakes
in Swiss alpine village population // Amer. J. Clin. Nutrit. 1962. Vol. 10. № 6.
HammelH.T^ Eisner R.W., Le Messurier DY. et at Thermal and metabolic response
of the Australian aborigine exposed to moderate cold in summer // J. Appl. Physiol.
1959. Vol. 14.
HammelH.T., Htldes 1A^ Jackson D.C. et at Thermal and metabolic response of the
Kalahari bushmen to moderate cold expossure at night // Arctic Aeromed. Lab. Kept,
All-TDR-62-44,1963.
271
Harrison GA., Kuchemann CJ"., Moore МАЯ. et aL The effects of altitudinal va
riation in Ethiopian populations // Phil. Trans. Roy. Soc. L., 1969. Vol. 266.
Harrison GA., Weiner IJS., Tanner YJM., Barnicot NA. Human biology. Oxford hi
iversity press. New York and Oxford, 1964.
HelnbeckerP. Further studies on the metabolism of Eskimos // J. Biol., Chem
1932. Vol. 93. № 327.
Henrotte I.G. Enquete preliminaire sur les caracteristiques biologiques des Indies
de Madras // Bull. Soc. Anthropol. 1961. S. 11. Vol. 10.
Henrotte LG. A biological study of Indian populations at different levels of econo
mic growth // Human adaptability to environments and physical fitness. Madras, 1966
Henrotte I.G., Coquelet M.t Traverse P. Influence du mode de vie sur la compositio;
des proteines seriques chez les indiens de Madras // Acta Tropica. 1963. Vol. 20. >ft 3
Henrotte I.G., Coquelet M., Traverse P. et aL Proteines seriques d'etudiants indiens
en climat tropical et tempere // Bull. Soc. Pathol. Exot. 1964. Vol. 67. >h 1.
Hiernaux J. La diversite humaine en Afrique subsaharienne. Bruxelles, 1968.
Hiernaux J., Proment A. The correlation between anthropobiological and climati
variables in Sub-Saharan Africa: revised estimates // Hum.Biol. 1976. Vol. 48.
Hicks CJS., O'Connor WJ. The effect of changes of environmental temperature o]
the skin circulation of the naked Australian aboriginal, as measured by the Sahli-Jaque
volumebolograph // Aust. J. Exptl. Biol. Med. Sci. 1938a. Vol. 16.
Hicks CJS., O'Connor WJ. Skin temperature of Australian aboriginals under vary
ing atmospheric conditions // Aust. J. Exptl. Biol.Med. Sci. 1938. Vol. 16.
Hicks C?.f Matters RP.t MitcfieU MX. The standard metabolism of Australian ab
originals // Aust. J. Exptl. Biol. Med. Sci. 1931. Vol. 8.
Hlpsley EH. Metabolic studies in New Guineans. Oxygen uptake and carbon dioxid
excretion during eating-resting and exercising conditions // Technical Paper. 1969. Л
162.
Ho Kang J., Biss K., Mlkkelson B. et at The Massai of Rast Africa: some uniqui
biological characteristics // Arch. Pathol. 1971. Vol. 91. № 5.
Holmes E.G., Jones EJL, Stanier M.W. Malnutrition in African adults // Brit. J
Nutrit., 1964. Vol. 8. № 2.
Horwath SM., Friedman A., Golden H. Acclimatization to extreme cold // J. Clim
Invest., Amer. J- Physiol. 1947. V. 160.
Hoygaard A Studies on the nutrition and physiopathology of Eskimo. (Skiffer N
orske Videnskaps - Akad. i. Oslo I Mat - Natur. Klasse, 1940, 9). Oslo, 1941.
Human adaptation. Vol. 2. E&.AJSasu, КС Malhotra. Calcutta, 1984.
Hurtado A. Animals in high altitudes; resident man // Dill DJ3. et al. (Eds.) Hand
book of Physiology: Adaptation to the environment. Washington, 1964.
Iayalakshmi V.T., Bamanathan MJt., Goplan С Studies on the blood of poor Indiai
new-born babies and their mothers // Ind. J. Med. Res. 1967. Vol. 46. № 4.
Ioffe ?., Jackson WJP.U^ Thomas MJS. et aL Metabolic responses to oral glucose h
the Kalahari bushmen // Brit. Med. J. 1971. Vol. 4. № 6781.
Iorgemen J?. The eskimoekeleton. Contribution to the physical anthropology of th<
aboriginal greendlanders // Meddelelser am Gronland. Kobenhavn, 1963. Bd. 146. >J
2.
Irwing L* Andersen KX., Bolstad A* et aL Metabolism and temperature of Arctii
Indian men during a cold night // J. Appl. Physiol. 1960. Vol. 16. H 4.
Janeen AA Skinfold measurements from early childhood to adulthood in Papuam
from Western New Guinea // Ann. N.Y. Acad. Sci. Р.П. 1963. Vol. 110.
272
Johnston FJS., Gindhart РЯ., Jantz HL. et al The anthropometric determination of
body composition among the Peruvian Cashmahua // Amer. J. Phys. Anthropol. 1971.
Vol- 34. № 3.
Kaplan В A* Environment and human plasticity // Amer. Anthropol. 1954. Vol. 56.
Keltz H., Comstoch G. Serum globulin levelsin whites and Negroes // New Engl. J.
Med. 1959. Vol. 269. № 25.
Kestner O. Schadow. Strahlung, Atmung und Gaswechsel. Versuche am Jungfrau-
hoch // Pflug. Arch. Ges. Physiol. 1927. № 217.
Keys A., Fidanza F^ Scardi V. et al Studies of serum cholesterol and other
characteristics of clinically healthy men in Naples // Arch. Int. Med. 1954. Vol. 93.
Keys A*, Kumura N., KusukawaA et aL Serum cholesterol in Japanese cola miners.
A dietary experiment // Amer. J. Clin. Nutrit. 1957. Vol. 6. >ft 3.
Keys A., Vivanco F„ Minou 1ХЛ et al Studies on the diet, body fatness and serum
cholesterol in Madrid, Spain // Metabolism. 1954. Vol. 3. Nfe 3.
Klein G., Cummings M., Hammarstein I. Differences between the normal serum
protein patterns of American Indian, Negro and Caucasian subjects // Proc. See. Exptl.
Biol. Med. 1962. Vol. 3. J* 2.
KoUmann I. Pygmaen in Europa // Verhandl Anatom. Gesellschaft, Mai, 1894.
KoUmann I. Neue Gedaken uber das abstammung des Menschen // Korres. Blatt d.
Deutschen Anthrop. Gesellschaft. 1905. Bd. XXXVI.
Krog I., Folkow В., Fox RJf. et al Hand circulation in the cold of Lapps and North
Norwegian fischermen // J. Appl. Physiol. I960. Vol. 15. J* 4.
Krogh A., Krogh M. A study of the diet and metabolism of Eskimos unertaken in
1908 on an expedition to Greenland // Meddelelser om Gronland. Bd. 41. froste afdel-
ing. Kobenhavn, 1914.
Lantis M., Anderson EJ. Некоторые демографические, социальные и
экологические факторы полярных районов // Медицина и здравоохранение в Арктике.
Тетради общества здравоохранения ВОЗ. Женева. 1964. Т. 18.
Laughttn W. The Alaska gateway viewed from the Aleutian Islands // Physical
Anthropology of the American Indian. N.Y., 1951.
Leonhardt F. The quantitative variation of serum proteins. Electrophoretic studies
in twin materials // Acta Genet. (Basel), 1962. Vol. 12. J* 3-4.
Levine VJ5. Studies in physiological anthropology I. The basal metabolic rate of the
Eskimo // Amer. J. Phys. Anthropol. 1949. Vol. 7.
Lichtman M., Hames C, McDonough I. Serum protein electrophoretic fraction
among Negro and white subjects in Evans County, Georgia // Amer. J. Clin. Nutrit.
1965. Vol. 16. № 6.
MaUna RM. Anthropometric correlates of strength and motor performans //
Exercise and sport sciences reviews. N.Y., 1976.
Man in the Andes. A Multidisciplinary Study of High-Altitude Quechus/ Ed. P.T.
Baker, MA. Little. Stroudsburg. Pennsylvania, 197?.
Mann C.V., Scott J2Jlf„ Hursh LM. et al The health and nutritional status of
Alaskan Eskimoes // Amer. J. Clin. Nutrit. 19?2. Vol. 11. № 1.
Maret't J., Dela A Race, sex and environment // Hutchinson's Scientific and
Technical Publications. L., 1936.
Marshall B*, Newman МЛГ. Adaptations in the physique of American aborigines to
nutritional factors // Hum. Biol. 1960. Vol. 82. № 3.
McGregor I., GUles H„ Walters L et al Effects of heavy and repeated malarial
infections on Gambia infants and children // Brit. Med. J. 1956. Vol. 2, J* 4994.
273
McGregor I., Gilles H* Studies on the significance of high serum gammaglobulii
concentration in Gambian Africans // Ann. Trop. Med. I960. Vol. 54. № 3.
Mechan С J J*. Individual and racial variations in a vascular response to a cold st
imulus // Military Med. 1956. Vol. 116. JN& 5.
Metody kontroli rozwoju czlowieka i zmian struktury populacji ludzkich w zwiaz
ku z przemianami srodowiska. Warshawa, 1975.
Milam D. Plasma protein levels in normal individuals // J. Lab. Clin. Med. 1946
Vol. 31. № 3.
Morpurgo G., MattagUa P., Bernini L. et aL Higher Bohr effect in Indian natives o:
Peruvian highlands as compared with Europeans // Nature (Gr. Brit.). 1970. Vol. 227
№ 6256.
MourantAJZ. The genetic markers of the blood // Population structure and humai
variation. Cambridge, 1977.
Munro A~F. Basal metabolic rates and physical fitness scores in British and Indiz
males in tropics // J. Physiol. (L.). 1949. Vol. 110. >fe 3/4.
Munro-Faure AJ)., Hill DM.f Andersen L Ethnic differences in human blood eel
sodium concentration // Natures. 1971. Vol. 231. >й 5303.
Neeb H.t Groot H. The serum protein pattern in West New Guinean groups // Trop
Geogr. Med. 1963. Vol. 15. № 4.
Necrasov O. Sur la variabilite des caracteres anthropologique de la Moeciu de Sus
en fonction du sexe //Ann. Roum. Anthropol. 1970. № 7.
Newman A Why man is such sweaty and thirsty naked animal: a speculative revi
ew // Hum.Biol. 1970. Vol. 42. U 1.
Nichols L., Nimalasurija A Adaptation to a low calcium intake in reference to th<
calcium requirements of a tropical population // J. Nutrit. 1939. Vol. 18.
Ojikutu КО. Die Adaptation der Afrikaner an fenohtheises Klima nach thermo
regulatorishen Funktionstests // Homo. 1970. Bd. 21. J№ 1.
Olivier G. Anthropologie de la France // Bull. Mem. Soc. Paris, 1970. S. 12. Vol. 6
Pawson I.G. The effects of altitudian variation on growth in an Ethiopia populatioi
// A Thesis in Anthropology, 1971.
Pawson I.G. The growth and development of high altitude children with specia
emphasis on populations of Tibetan origin in Nepal. PhD thesis, the Pennsylvania Statt
University, 1974.
Parizkova I. Changes in the nutritional pattern and expected changes of the nutri
tional status // Metody kontroli rozwoju с czlowieka i zmian struktury populacji ludzk
ich w zwiazku z przemianami srodowiska. Wroclaw; Krakow; Gdansk, 1977.
Petropoulos ЕЛ^ Ttmiras PS. Biological effects of high altitude as related to in
creased solar radiation, temperature fluctuations and reduced partial pressure of oxygei
// Progress in biometeorology. Chapter ?. Section 7. Amsterdam, Swetz. Zeitinger
1974.
Pollack V.t Mandema E., Doig A. et aL Observations on electrophoresis of serun
proteins from healthy North American, Caucasian and Negro subjects and from patientf
with systemic lapus erythematosus // J. Lab. Clin. Med. 1961. Vol. 58. Nfe 3.
Population structure and human variation. Cambridge, 1977.
Pospisil M. Indian remnats from the orient province. Cuba Univerzita Komenske
ho, 1976.
Powell 5. The serum protein pattern and liver function tests in the Natal African /,
J. Lab. Clin. Med. 1958. Vol. 4. № 6.
274
Pyzuk M., Turuabehow V.T., Briancewa LA. Certain properties of the respiratory
system in school children in various altitude and climatic conditions // Hum. Biol. 19?7.
Vol. 39. №1.
Badu JEJ). Donnees somatiques concernantl'etude de Thomme integral, de l'adapta-
bilite et de la persnnalite. Kecherches portent sur trois lots de travaileurs de la zone
d'Argls // Ann. Roum. Anthropol. 1969. J* 6.
Radu E.f Roseteanu O. Consideratii preliminare asupra variabilitatii conformativ-
somatice a unei serii de femei din comunitatea Magura - culoarul Bran // Stud. Cere.
Anthropol. 1972. Vol. 9. № 1.
Race AA, Sanger B. Blood groups in man. Oxford, Blackwell, 1958.
Jumoin DX.f Merimee TJ.t Rabinowitz D. et at Peripheral subresponsiveness to
human growth hormone in the African Pygmies // New England J. Med. 1969. Vol. 28. H
25.
Roberts DJF. Basal metabolism, race and climate // Man. 1962. Vol. 52.
Roberts D.F. Body weight, race and climate // Amer. J. Phys. Anthropol. (n.8.),
1953. Vol. 11. №4.
Roberts DJF. Effects of race and climate on human growth as exemplified by studies
on African children // Human growth. Oxford, 1960.
Rodahl K. Basal metabolism of Eskimo // J. Nutr. 1952. № 48.
Rotthauwe H.W., El-Schallah M.O.f Flatz G. Lactose intolerance in Arabs // Human-
genetik. 1971. Vol. 13.
Roy IJC., Rao RJC., Biswas SJT. Diet surveys amongst the tribes of Madhya Prades-
h // Bull. Depart. Anthropol. Calcutta, 1957. Vol. 6. № 1.
Roy /JT., Roy B.C. Food sources, diet and body build of the people of Great Nicobai
// Bull. Anthropol. Survey India. 1967. Vol. 16. № 3-4.
Rychkov I.G* Sheremetyeva VIA. The genetic process in the system of ancient human
isolates in North Asia // Population structure and human variation. Cambridge, 1977.
Saha H., Das A. Symposium on problems of high altitude. New Delhi, 1962.
Salzano F.M., Freire-Maia N. Problems in human biology. A study of Brazilian
populations. Detroit, 1970.
ScHraer H.t Newman M. Quantitative roentgenography of skeleton mineralisation
in malnourisched Quechua India boys // Science. 1958. Vol. 128. >fe 3322.
Schreider E. Geographical distribution of the body-weight/body-surface ratio //
Nature. 1950. Vol. 165.
Schreider E. Race et regulation thermique // La semaine des hopitaux de Paris.
1952. Vol. 28.
Schreider E. Regulation thermique et evolution humaine // Actes du IV-e congres
international et sciences anthropologiues et ethnologiques. Vol. Wien, 1954.
Schreider E, Ecological rules, body-heat regulation, and human evolution //
Evolution. 1964. Vol. 18. № 1.
Schwalbe AJMT. Schmids Arbeit "Die Stellung der Pygmaenvoluer in der Entwick-
lungsgeschichte des Menschen" // Globus. Bd. XLCVTI. 1910.
Schwartz CJ., Day AJ.t Peters I A. et al Serum cholesterol and phospholipid levels
of Australian aborigines // Aust. J. Exptl. Biol. Med. Sci. 1957. Vol. 35.
Schwidetzhy I. Neuere Entwicklungen in der Rassenkunde dee Menschen. Die neue
Rassenkunde (Herausgegeben von I. Schwietzky). Stuttgart, 1962.
SchuUzAJI. Foetal growth of man and other primates // Quart.Rev.Biol. 1926,
Vol. 1. № 4.
275
* Scott ЕМ., Griffith I.V., Hosktns В J), et aL Serum cholesterol level and blood pres
sure of Alaskin Eskimo men // Lancet. 1968. Vol. 2. № 7048.
Schrimshaw N.S., Trulaon M., Fejada C. et aL Serum lipoprotein and cholestero
concentrations, comparison of rural Costa Rican, Guatemalan and United States po
pulation // Circulation. 1967. Vol. 16. № 6.
SendraUA., Quilici I.C. Etude des hemoglobines deshabitants du corridor Interand
in // Anthropologic (Franc). 1970. Vol. 74. № 3-4.
Sen Gupta P.N. Investigations into the dietary habits of the aboriginal tribes о
Abor Hills (North-Eastern Frontier). Part. 11. Manyong and Pangi // Bull. Depart
Anthropol. 1964. Vol. 3. № 2.
Sen Gupta PJJ. Investigations into the diet and nutrition of the Nokte tribe of Ti
par frontier division. N.E.F.A // Bull. Anthropol. Surv. India (Calcutta). 1964). Vol. 9. № 2
Sen Gupta PJsf., Biswas SJC. Studies on the diet and nutritional status of the Kani
kkar and urali tribes of Travancore // Bull. Depart. Anthropol. 1966a. Vol. 5. >6 1.
Sen Gupta JPJV., Biswas SJC. Studies on the diet and nutrional status of the Malap
antaram, Muthuvan and Ullatan tribes of Travancore // Bull. Depart. Anthropol. 1956b
Vol. 6. № 2.
Sharma I.C Physical growth and development of the Maharashtrians. Lucknow
1970.
Sholander Pf., Hammel H.T., Hart IS. et at Cold adaptation in Australian abori
gines // J.Appl.Physiol. 1958. Vol. 13.
Smirnova NJS. Bodybuild of the Arctic peoples in relation to climatic adaptation /,
Studies in the Anthropology of the Finno-Ugrian, Peoples. Helsinki, 1973.
Smith EA. Human adaptation and energetic efficiency // Hum. Ecol. 1979. Vol. 7
SobelAJS. Local factors in the mechanism of calcification // Ann. N.Y. Acad. Sci
1965. Vol. 60. №5.
Stonier M.W., Holmes E.G. Malnutrition in African adults // Brit, Nutrit. 1964
Vol. 8. № 2.
Swadesh M. XJnaalik and Proto-Eskimo // Inter. J. Amer. Ling. 1961. Vol. 17. № 4
Swadesh M. Linguistic relations across Bering Strait // Amer. Anthropol. 1962
Vol. 64. № 6.
The biology of high-altitude peoples. Cambridge-London, New York, Melbourne
1978.
Thompson D. On growth and form. Cambridge, 1917.
Thomson A.t Buxton L. Man's nasal index in relation to certain climatic condition)
// J. Roy. Anthropol. Inst. Gr. Brit. Ireland. 1923. Vol. 63.
Trotter M.r Broman G„ Peterson A Density of cervical vertebral and compariso]
with densities of other hones // Amer. J. Phys. Anthropol. 1959. Vol. 17. № 1.
Vassal P. Brachycephalie cloitre et terrain granitique // Rev. Pathol. Genet. Phys
iol. Clin. 1967. Vol. 67. № 688.
Vera /., Roche M. A note on the distribution of the serum protein fractions in ap
patently normal persons in Caracas // J. Lab. Clin. Med. 1966. Vol. 47. J* 3.
Walker AJLP.T Arvldsson UJS. Fat intake serum cholesterol concentration and athe
roscleroeis in the South African Bantu // J. Clin. Invest. 1964a. Vol. 33. № 10.
Walker AJLP., Arvldsson UJS. Fat intake, serum cholesterol concentration an<
atherosclerosis in South African Bantu. Part 1. Low fat intake and the age trend о
serum cholesterol concentration in the South African Bantu // J.Clin. Ubvest. 1954b
Vol. 83. >fc 13.
276
Walter В., Bajatzadeh M. Investigations on the geographical variability of th<
human transferrins // Humangenetik. 1971. Vol. 12. № 4.
Walter H., Steegmuller H. Studies on the Geographical and racial distribution of the
Hp and Go polymorphism // Hum. Hered. 1969. Vol. 19. № 3.
Wang Schin-chun. Human nasal index and apertures in relation to some climati<
conditions based on 494 Chinese skulls // Zsepou suebai. 1954. Vol. 1. № 2.
Watanabe H. Neanderthalers vs. H. sapiens. Behavioral adaptability to Arcti<
winter // Proc. VHI-th Intern. Congr. Anthropol. Ethnolog. Sci. Tokyo-Kyoto. Vol. 1
Anthropology, 1968.
Weiner J. Nose shape and climate // Amer. J. Phys. Anthropol. (n.s.), 1954. Vol
12. № 4.
Whyte HM. Body build and blood pressure of men in Australia and New Guinea /,
Aust. J. Exptl. Biol. Med. Sci. 1963. Vol. 41.
WiercinskiA* Some inter- and intrapopulation comparisons in anthropometry of tin
inhabitants of the Western desert, Siwa Oasis, El Fayum and El Beheira // Mater.
Prace Anthropol. Wroclaw, 1970. № 78.
WUber CKG., Levine V. Fat metabolism in Alaskan Eskimos // Exptl. Med. Surv
1950. Vol. 8.
Wilkinson G.f Day A, Peters A. et at Serum protein of some Central and South A
ustralian aborigines // Med. J. Aust. 1958. Vol. 2. № 5.
Wilson EA~ The basal metabolic rates of South American Indians // Handbook o:
South American Indians. Vol. 6. Washington, 1950.
Wilson O. Adaptation of the basal metabolic rate of man to climate — a review /,
Metabolism. 1956. Vol. 5. № 6.
Winslow CJEA., Herrington Z.J*. Temperature and human life. Princeton Univ. Pre
ss, 1949.
WolanskiN. Rozwoj hiologiczny czlowieka. Warszawa, 1975.
Worldwide variation in human growth. Cambridge, 197?.
Wyndham СЛ. Southern African ethnic adaptation to temperature and exercise /
The biology of human adaptability. Oxford, 1967.
Wyndham CJf. Man's adaptation to the physical environment in Southern Africa /
Biology of man in Africa. Mater, i Prace Anthropol. 1970. >6 78.
Wyndham CJf., Metz В., Munro A. Reactions to heat of Arabs and Caucasians /
J.Appl.Physiol. 1964. Vol.19.
Wyndham СЛ^ Morrison JJ?. Adjustment to cold of bushmen in the Kalahari Dese
rt // J.Appl. Physiol, 1958. Vol. 13. № 2.
СОДЕРЖАНИЕ
Лекция L КРАТКАЯ ИСТОРИЯ ИЗУЧЕНИЯ ПРОБЛЕМЫ АДАПТАЦИИ В
АНТРОПОЛОГИИ 7
1. Антропологические исследования адаптации человеческих популяций к
экстремальным условиям среды . 7
2. Истоки отечественных исследовании по адаптации 13
Лекция J/. ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ ИЗМЕНЧИВОСТИ
СТРОЕНИЯ ТЕЛА И НЕКОТОРЫХ ФИЗИОЛОГИЧЕСКИХ ПОКАЗАТЕЛЕЙ ... 19
1. Материал и метод исследования 19
2. Морфологические характеристики 26
3. Физиологические характеристики 37
4. Роль уровня питания в географических вариациях антропологических
признаков 54
5. Закономерности пространственной изменчивости антропологических
признаков 57
Лекция III. АДАПТАЦИЯ ЧЕЛОВЕКА В ТРОПИЧЕСКИХ ШИРОТАХ 60
1. Экологическое и этническое разнообразие региона 60
2. Краткая характеристика питания населения тропиков 61
3. Строение тела у населения тропических широт 67
4. Физиологические особенности тропических популяций 93
5. О приспособительной изменчивости в тропических широтах 108
6. Морфофизиологические характеристики населения субтропической зоны
Кавказа 103
Лекция IV. БИОЛОГИЯ НАСЕЛЕНИЯ АРИДНЫХ ЗОН 112
1. Специфика воздействия климатических условий пустыни на организм
человека 112
2. Морфологические особенности населения тропических пустынь 114
3. Морфофизиологическая характеристика населения внетропических
пустынь 119
Лекция V. БИОЛОГИЯ ЖИТЕЛЕЙ ВЫСОКОГОРЬЯ 123
1. Строение тела и физиология 123
2. О природе адаптивного комплекса высокогорья.. 13?
Лекция VI. БИОЛОГИЯ НАСЕЛЕНИЯ УМЕРЕННОГО КЛИМАТИЧЕСКОГО
ПОЯСА 137
278
1. Климатическое и этническое разнообразие изученного региона 13'
2. Строение тела и некоторые особенности внутренней среды организма 13
3. Региональные автропогеографические связи 14
Лекция VII. БИОЛОГИЯ ЖИТЕЛЕЙ КОНТИНЕНТАЛЬНОЙ ЗОНЫ СЕВЕРНО!
АЗИИ 14
1. Специфика географических условий внутренних районов Северной Азии 14
2. Строение тела и физиологические особенности населения 15
3. Антропоэкологич?ское разнообразие континентальной зоны Сибири 16'
4. Морфофизиологические характеристики населения Монголии 17'
Лекция VIII. БИОЛОГИЯ ЖИТЕЛЕЙ КРАЙНЕГО СЕВЕРА 17
1. Питание коренного населения Арктики 171
2. Морфофизиологические особенности коренного населения Крайнего
Севера 18і
3. Аятропоэкологические связи на территории Северной Азии 19:
Лекция IX. ПРИРОДА АДАПТИВНОСТИ Y ЧЕЛОВЕКА 19
1. Адаптивные типы 19'
2. Адаптация и акклиматизация 20:
3. О соотношении адаптивных типов человека с экологическими формам]
животных и растений 21
Лекция X. АДАПТИВНЫЕ ТИПЫ И СРЕДА 21:
1. Предпосылки исследования комплексного действия естественных и социальны:
факторов на биологию человека 21:
2. Краткие сведения об этногенезе и хозяйственно-культурном типе иоследованны
групп ¦ 211
3. Оценка антропоэкологических связей на территории бывшего СССР 22
4. Закономерности экологической изменчивости в городских популяциях... 22і
Лекция XI. ЗАКОНОМЕРНОСТИ ВРЕМЕННОЙ ИЗМЕНЧИВОСТИ АДАЛ
ТИВНЫХ ОСОБЕННОСТЕЙ У ЧЕЛОВЕКА 23
1. Рост и развитие человека в различных экологических нишах 23
2. Хронология адаптивных типов 24
ЗАКЛЮЧЕНИЕ 26;
ЛИТЕРАТУРА 25J
Учебное издание
АЛЕКСЕЕВА Татьяна Ивановна
АДАПТАЦИЯ ЧЕЛОВЕКА
В РАЗЛИЧНЫХ ЭКОЛОГИЧЕСКИХ НИШАХ ЗЕМЛИ
(биологические аспекты)
Редактор-корректор ГЛ. Секачева
Художественно-технический редактор Л.С. Солдатенкова
Компьютерная верстка Д-Ю* Балашов
Подписано в печать 29.01.98. Формат 60X84/16
Гарнитура Школьная. Усл. печ. л. 16,28
Тираж 1500 экз. Цена договорная. Заказ 0072
Издательство МНЭПУ, ! J1250, Москва, а/я 20
Типография "Информпресс-94" 109518, Москва, ул. Люблинская д. I
Татьяна Ивановна АЛЕКСЕЕВА
- известный в стране и за рубежом
антрополог. Автор 200 научных
работ и монографий. Стержнем
научных интересов являются -
этническая и физиологическая
антропология; фундаментальные
исследования этногенеза и экологии
народов России, Средней Азии и
Монголии.
Один из разработчиков российской (советской) школы биологических
аспектов экологии человека.
Член-корреспондент Российской академии наук, главный научный
сотрудник НИИ и Музея антропологии МГУ, председатель Музейного совета
РАН, руководитель группы физической антропологии Института археологии
РАН, Президент Российского отделения Европейской антропологической
Ассоциации, член Всемирной Ассоциации биологов Человека.
Авторитет ученого получил широкое признание в работе
Международных конгрессов по этнологическим и антропологическим проблемам в
Японии, Индии, США, Монголии, странах Центральной и Северной Европы.
Особое место в научных исканиях Т.И. Алексеевой занимают экспедиции,
результаты которых стали основой исследования проблемы адаптации
населения к экстремальным экологическим нишам. Педагогические и творческие
интересы ученого обращены к студенческой и научной среде. Лекции по
экологии человека пользуются повышенным вниманием учебных аудиторий
и кафедр антропологии МГУ, Тюбингенского университета (Германия),
Международного независимого эколого-политологического университета
(МНЭПУ).
Широкой известностью среди российских и зарубежных коллег
пользуется научная школа по проблемам экологии современного и древнего
населения мира, возглавляемая профессором Т.И. Алексеевой.
В педагогическом активе ученого - 19 кандидатов биологических и
исторических наук, I доктор биологических наук, открывший новое направление
в отечественной антропологии - «Биологические основы старения человека».
Основу научных трудов составляют книги - «Происхождение и
этническая история русского народа» (В соавторстве). - М., 1965; «Этногенез
восточных славян по данным антропологии» - М., 1973; «Географическая среда
и биология человека» - М., 1977 и «Адаптивные процессы в популяции
человека»-М., 1986.