

























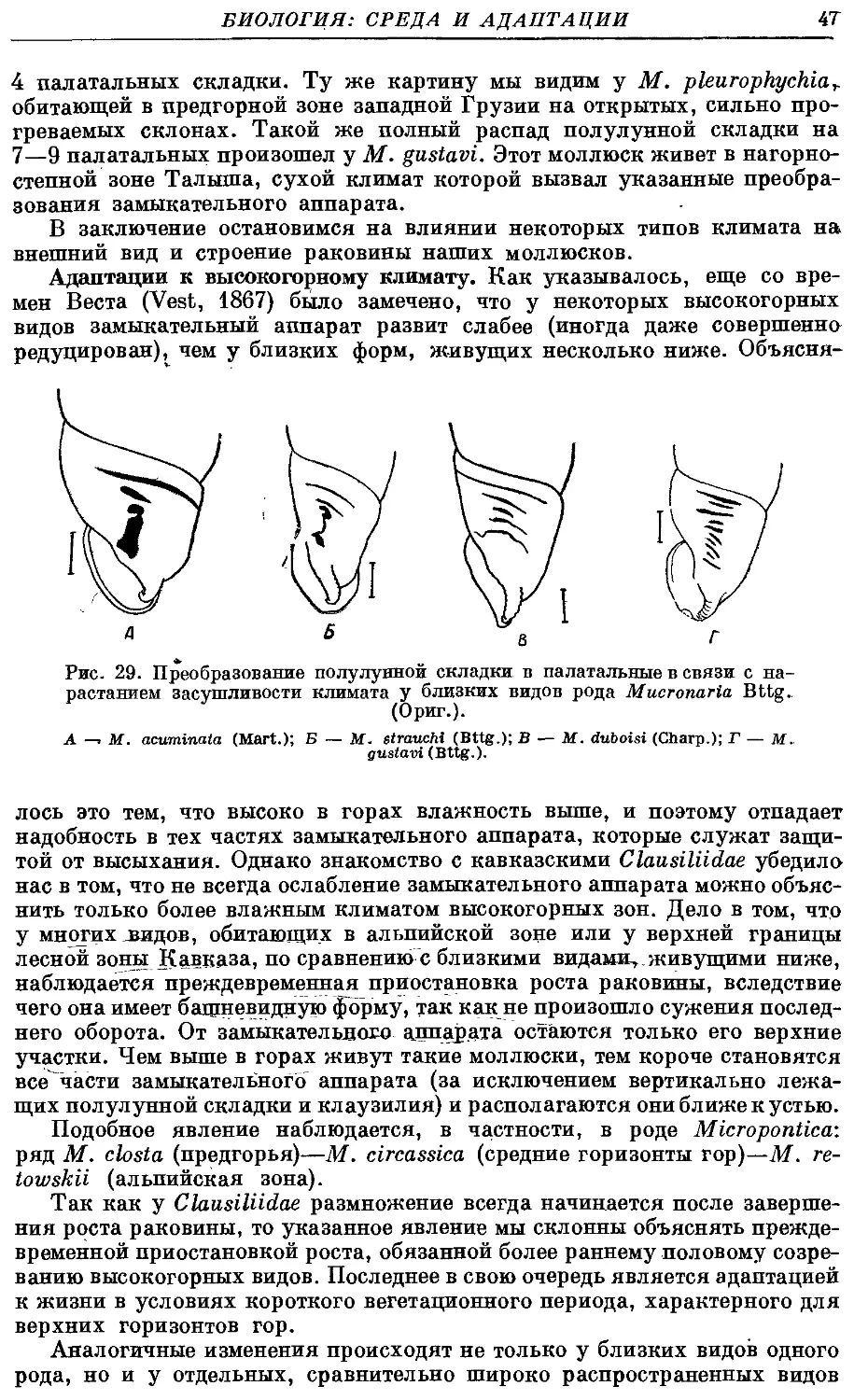





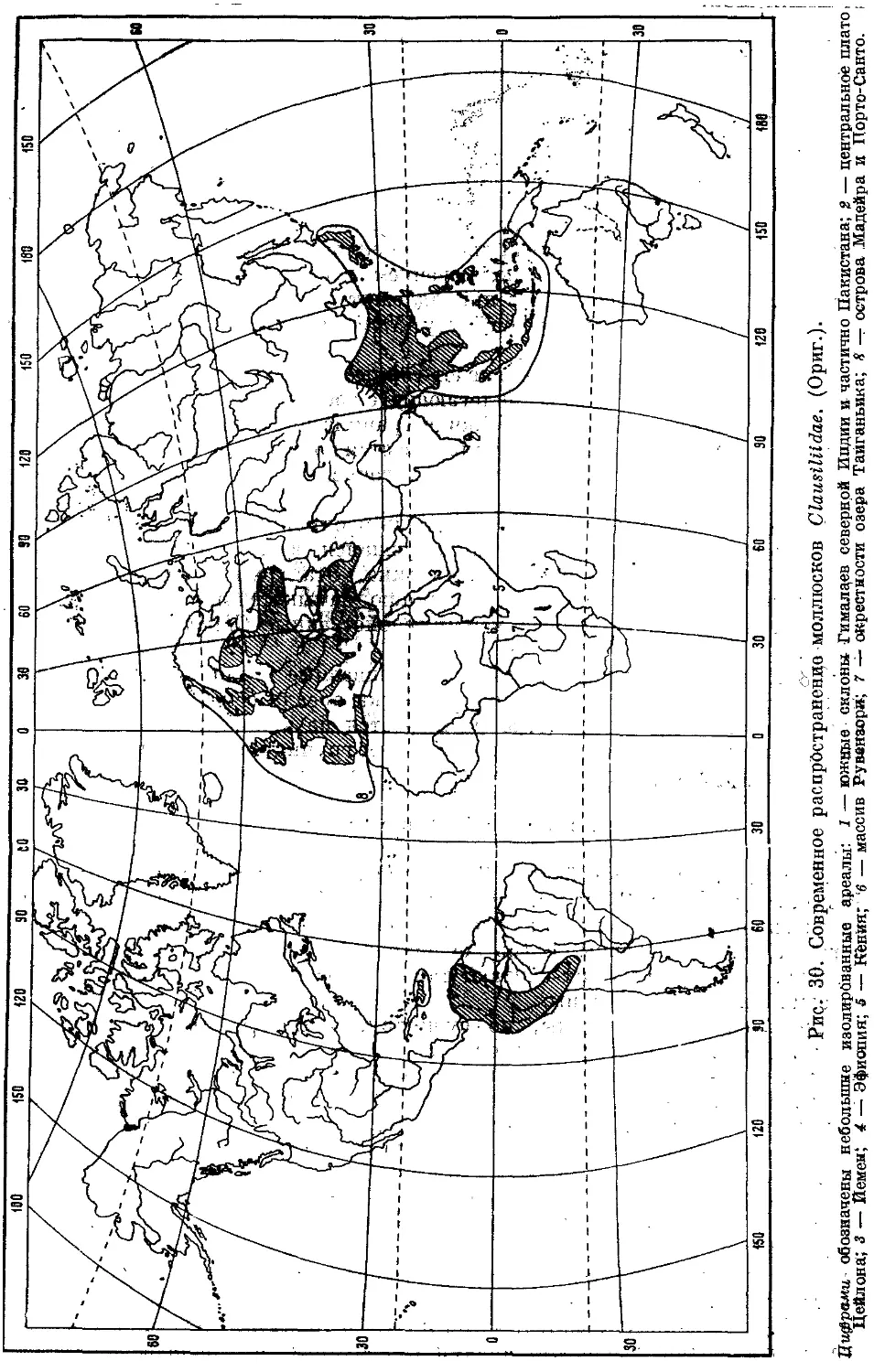


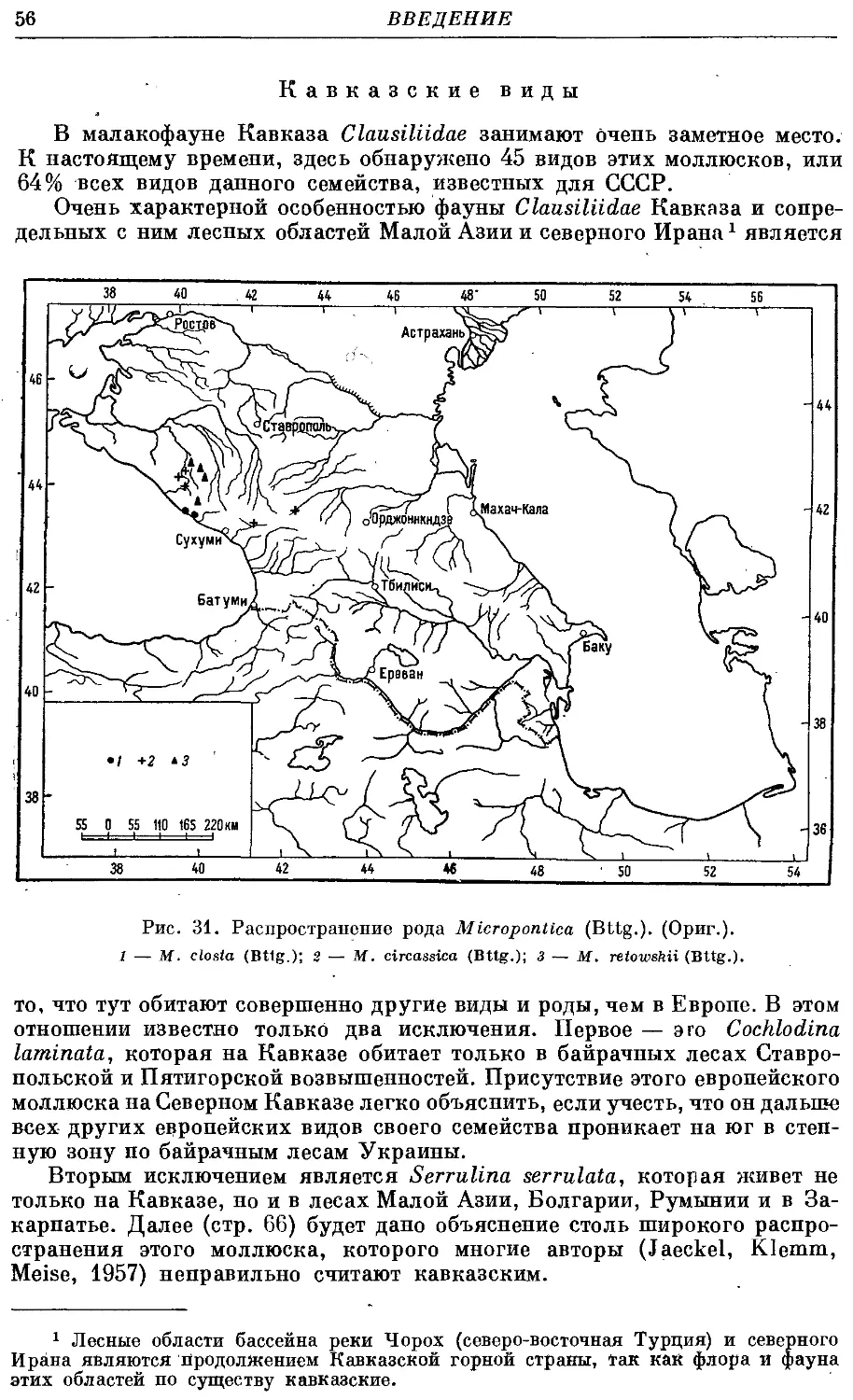


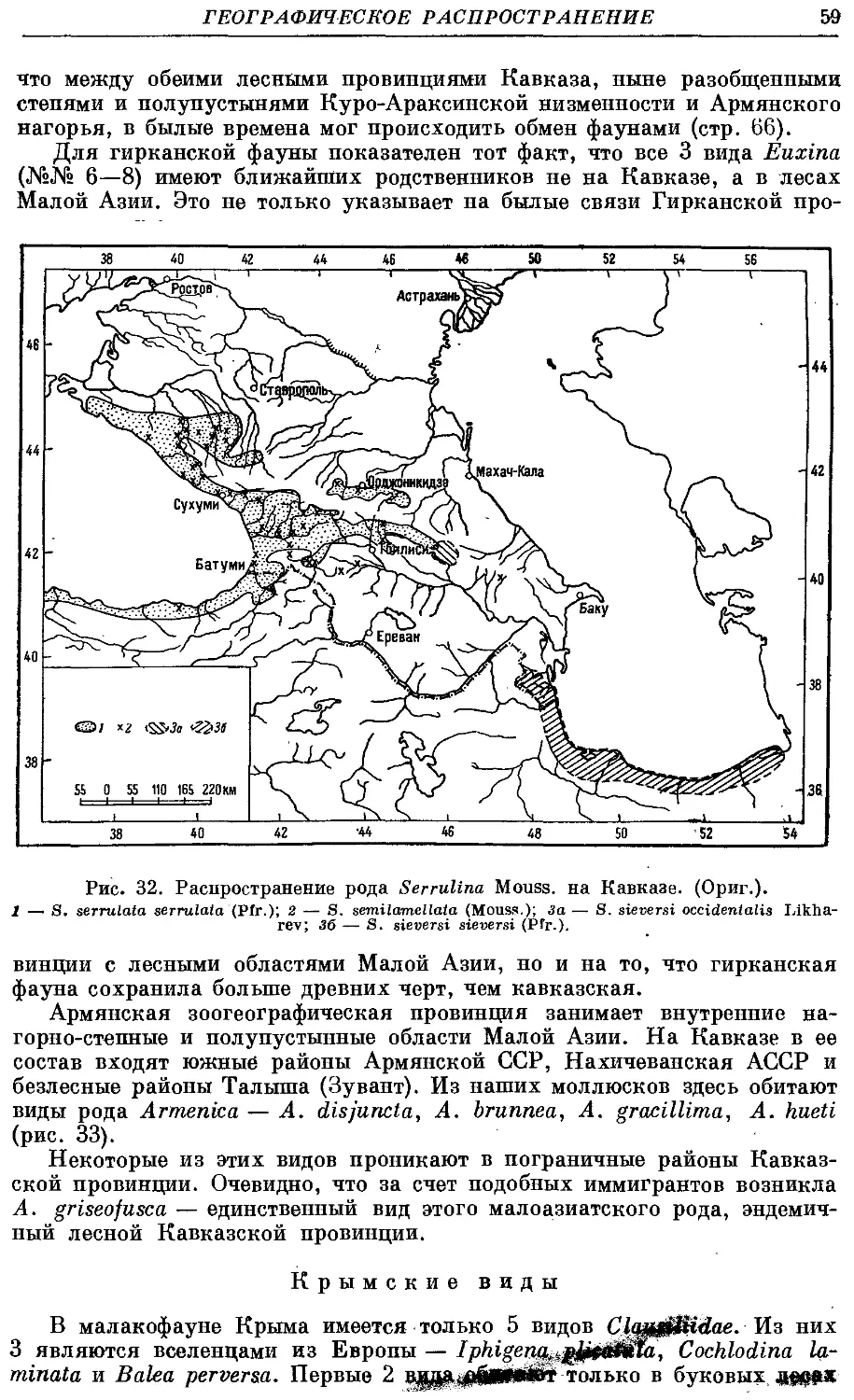































































































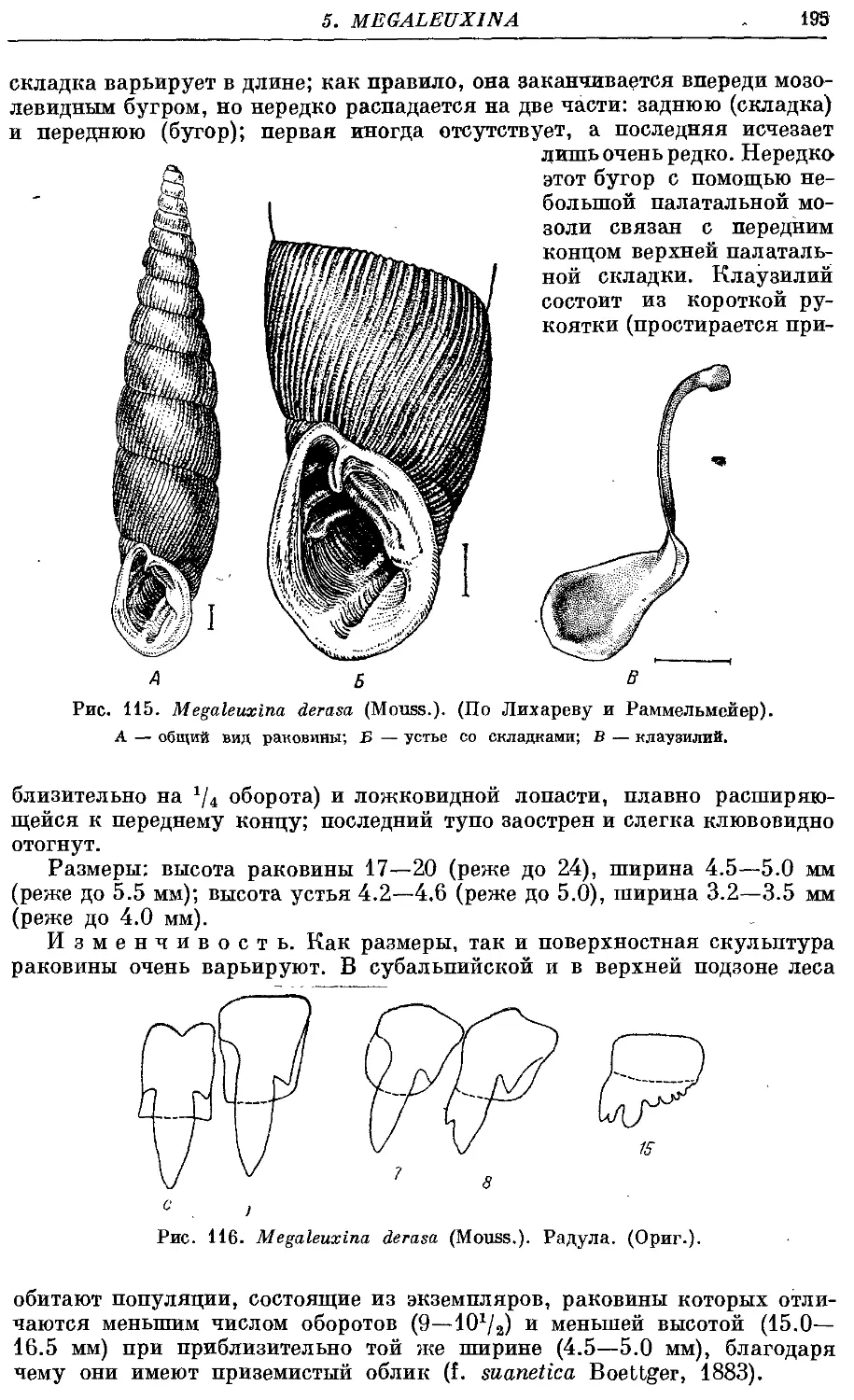
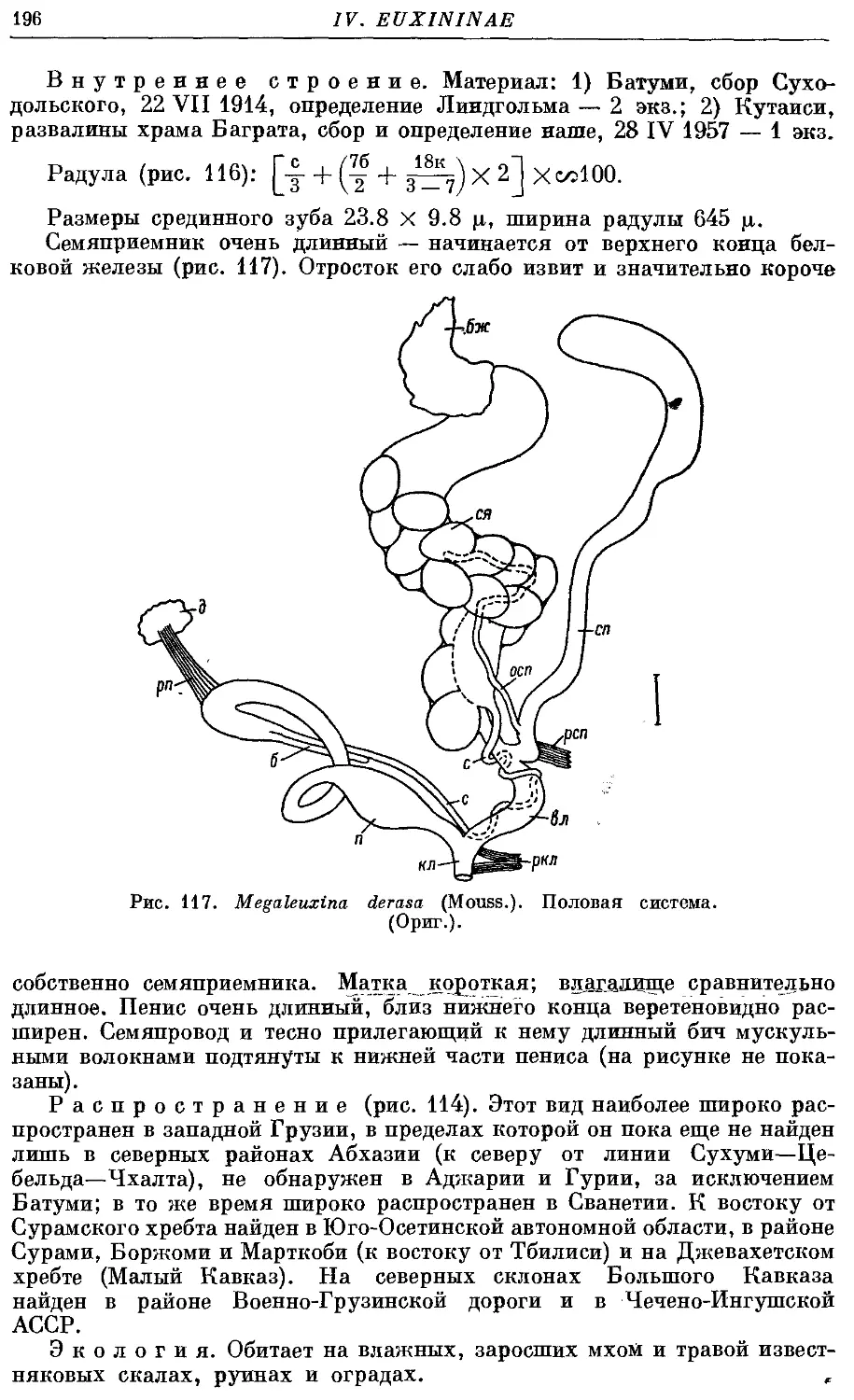
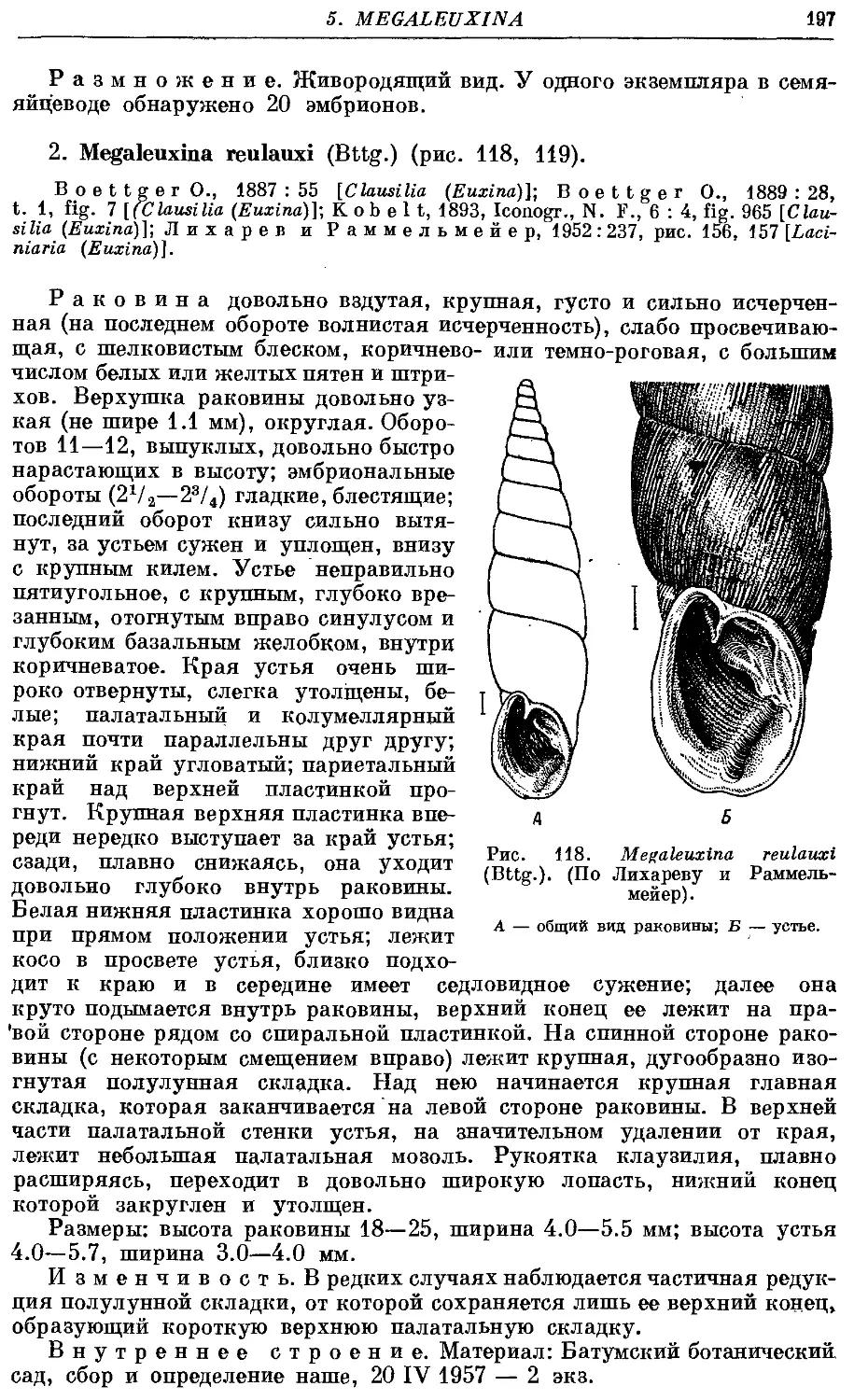
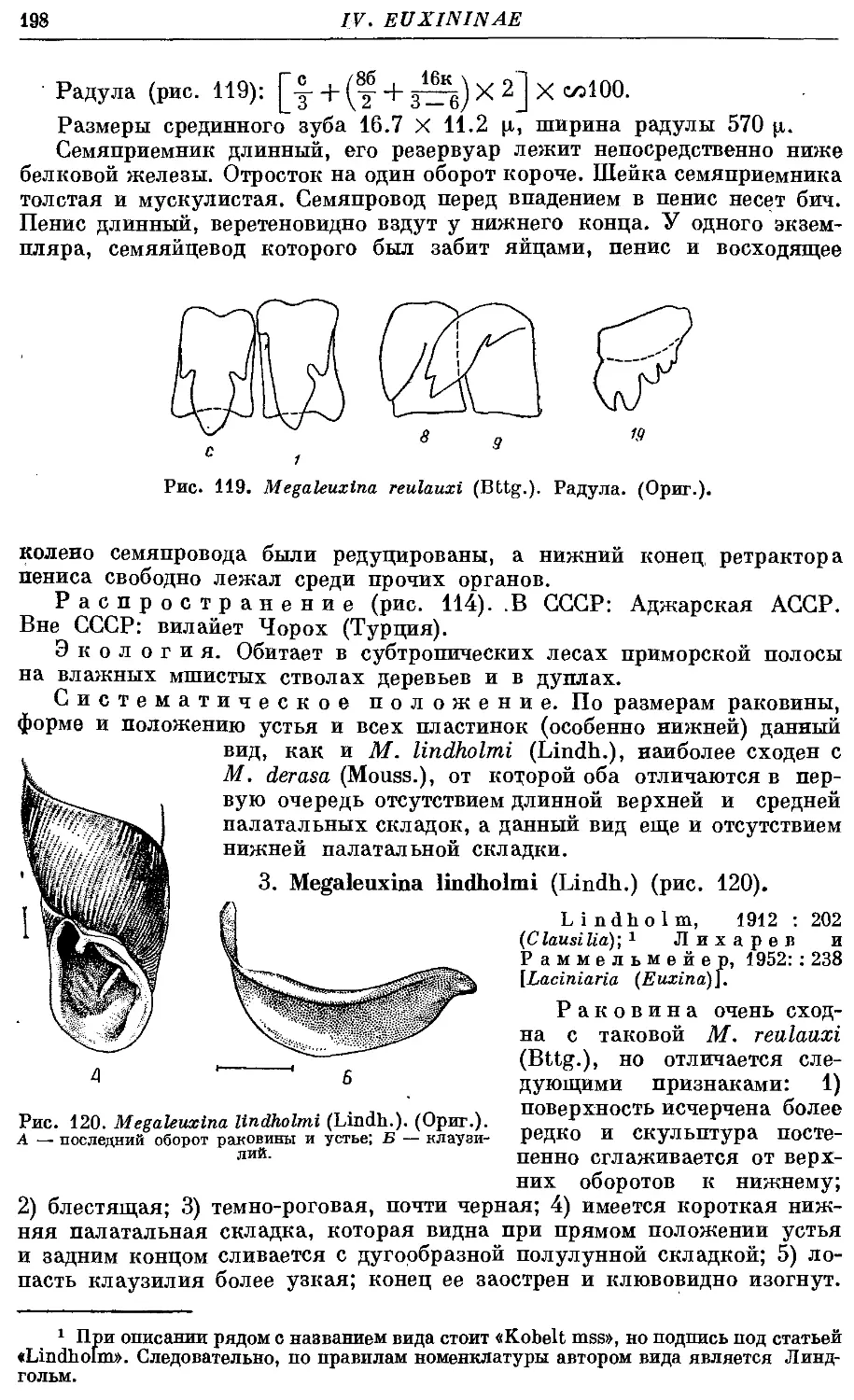




























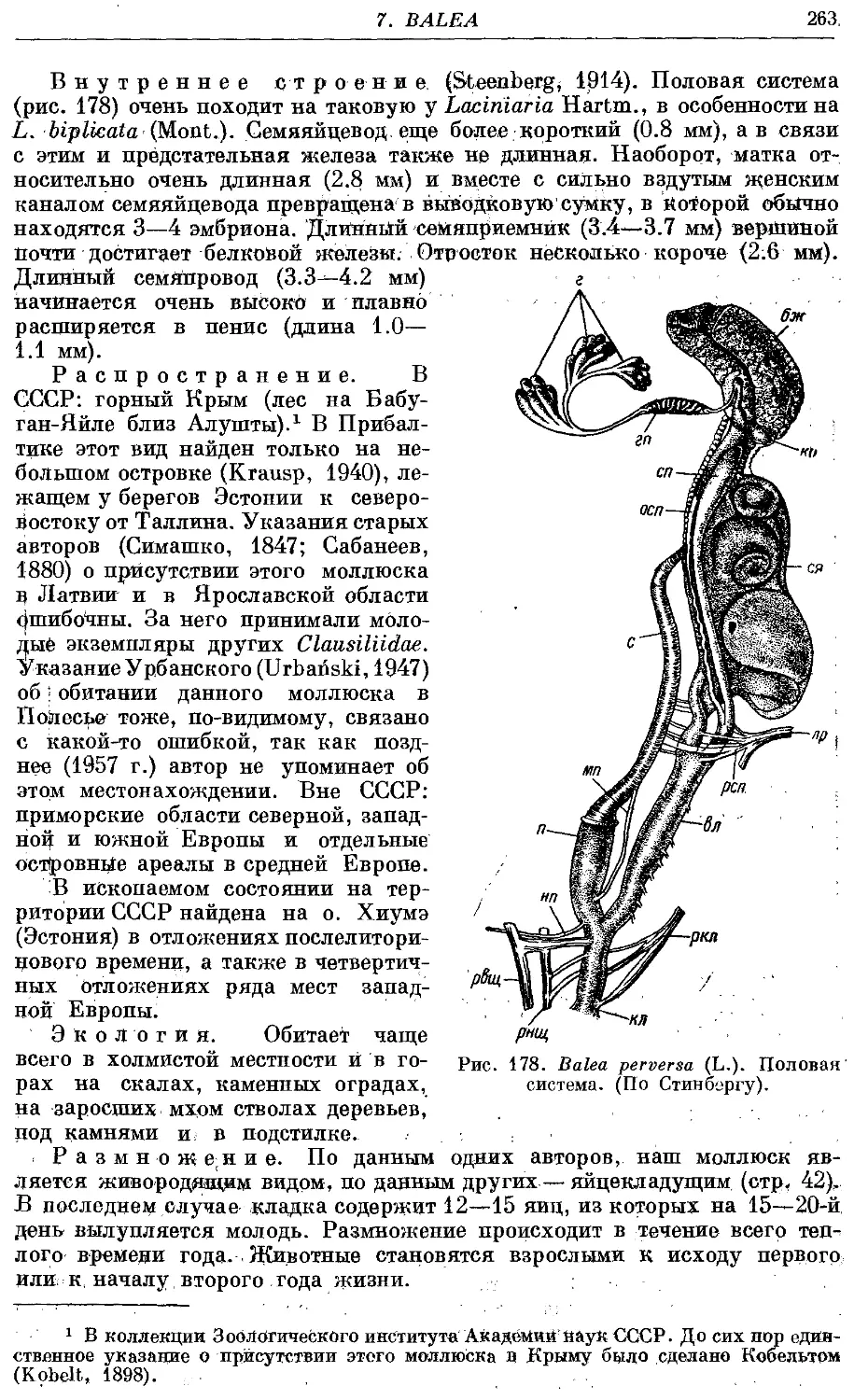
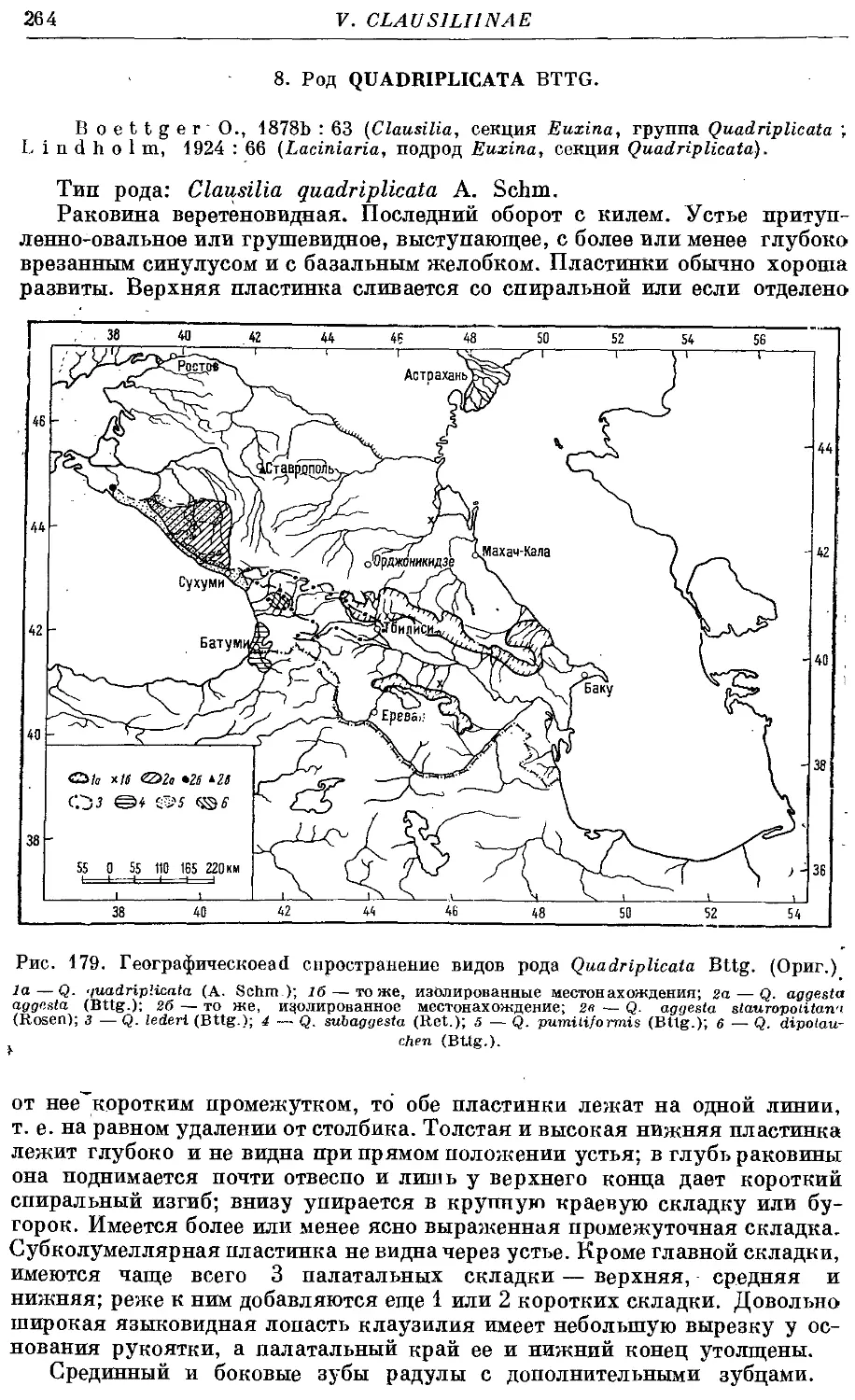


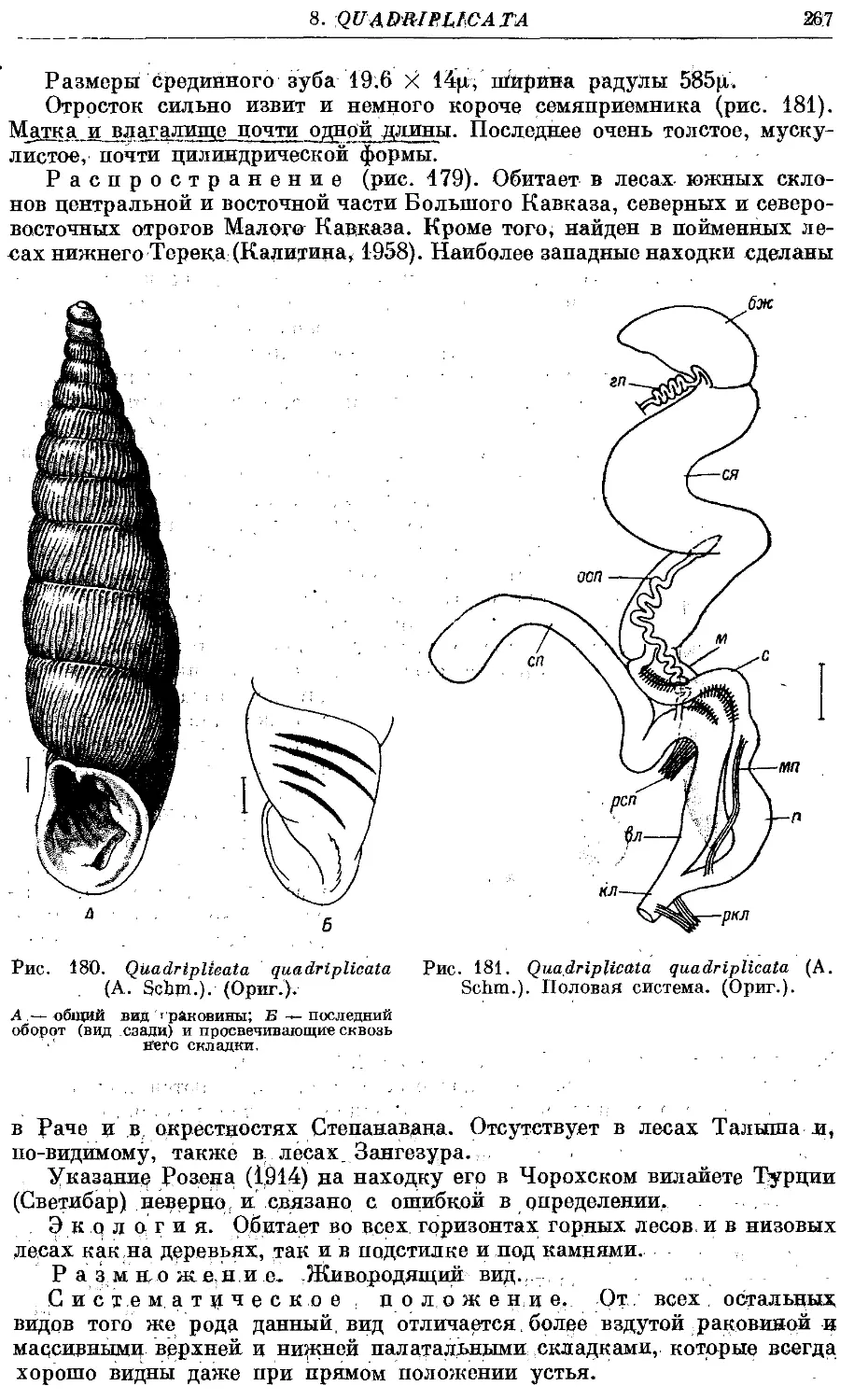
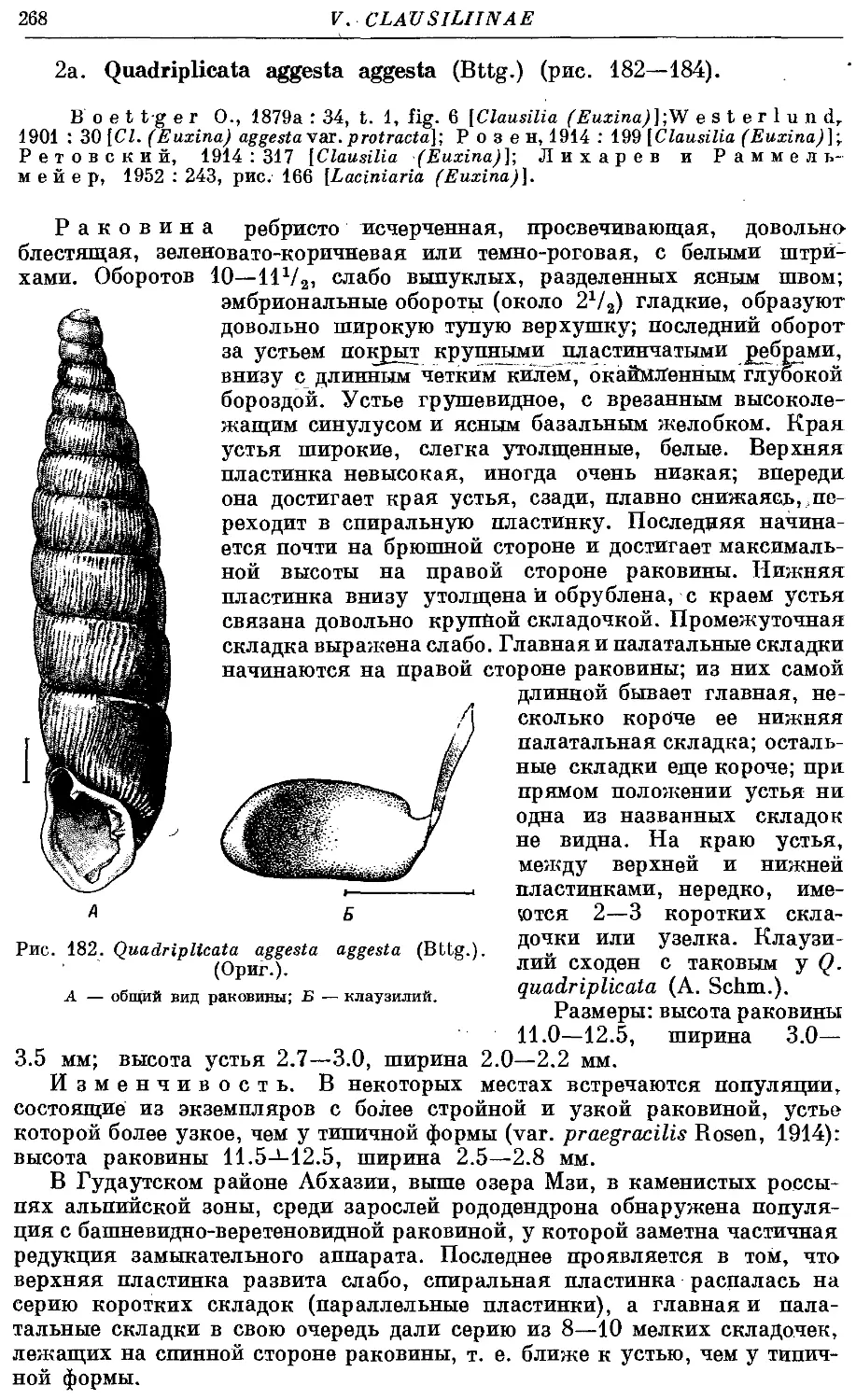




































Текст
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ, М 83
ФАУНА СССР
МОЛЛЮСКИ
Том III, вып. 4
И. М. ЛИХАРЕВ
КЛАУЗИЛИИДЫ (CLAUSILHDAE)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1962 • ЛЕНИНГРАД
Главный редактор
директор Зоологического института АН СССР
акад. Е. Н. Павловский
Редакционная коллегия
Б. Е. Быховский, И. М. Громов, А. С. Мончадский,
О. А. Скарлато, А. А. Стрелков (редактор тома),
А. А. Щтакелъберг
ПРЕДИСЛОВИЕ
Можно без преувеличения сказать, что среди наземных моллюсков
семейство Clausiliidae является одним из самых интересных. С давних
пор оно привлекало внимание многих исследователей. Тому было ряд
оснований: 1) своеобразная форма раковины, внутри которой возник
совершенно необычный замыкательный аппарат, 2) очень характерное
прерывистое распространение, 3) большая древность этой группы, о чем
свидетельствуют многочисленные находки, начиная с позднего мела
и на протяжении всего кайнозоя.
К этому можно добавить, что Clausiliidae среди наземных моллюсков
являются единственной группой тропического происхождения, которая
в умеренных широтах Европы дала пышный букет видов, родов и даже
подсемейств.
Как одну из основных задач нашего исследования мы поставили себе
изучение не только морфологии и систематического положения видов,
живущих в СССР и в сопредельных странах, но также и выяснение всей
естественной системы этого своеобразного семейства. В этом отношении
нам благоприятствовало то, что трудами наших предшественников боль-
большинство видов, населяющих западную, среднюю и южную Европу,
в настоящее время изучены довольно хорошо. Накопилось также много
новых данных о видах, обитающих в восточной Азии. Значительно хуже,
порой совершенно поверхностно, были изучены виды, живущие на Кав-
Кавказе и в Передней Азии. Поэтому основное внимание мы обратили на
исследование моллюсков этих областей, тем более, что на Кавказе
обитает более 60% всей фауны Clausiliidae СССР.
Clausiliidae по сравнению с другими группами легочных моллюсков
как будто бы дают больше возможностей для классифицирования их по
признакам раковины. В самом деле, немногие другие группы моллюсков
обладают столь сложно устроенной и богатой признаками раковиной.
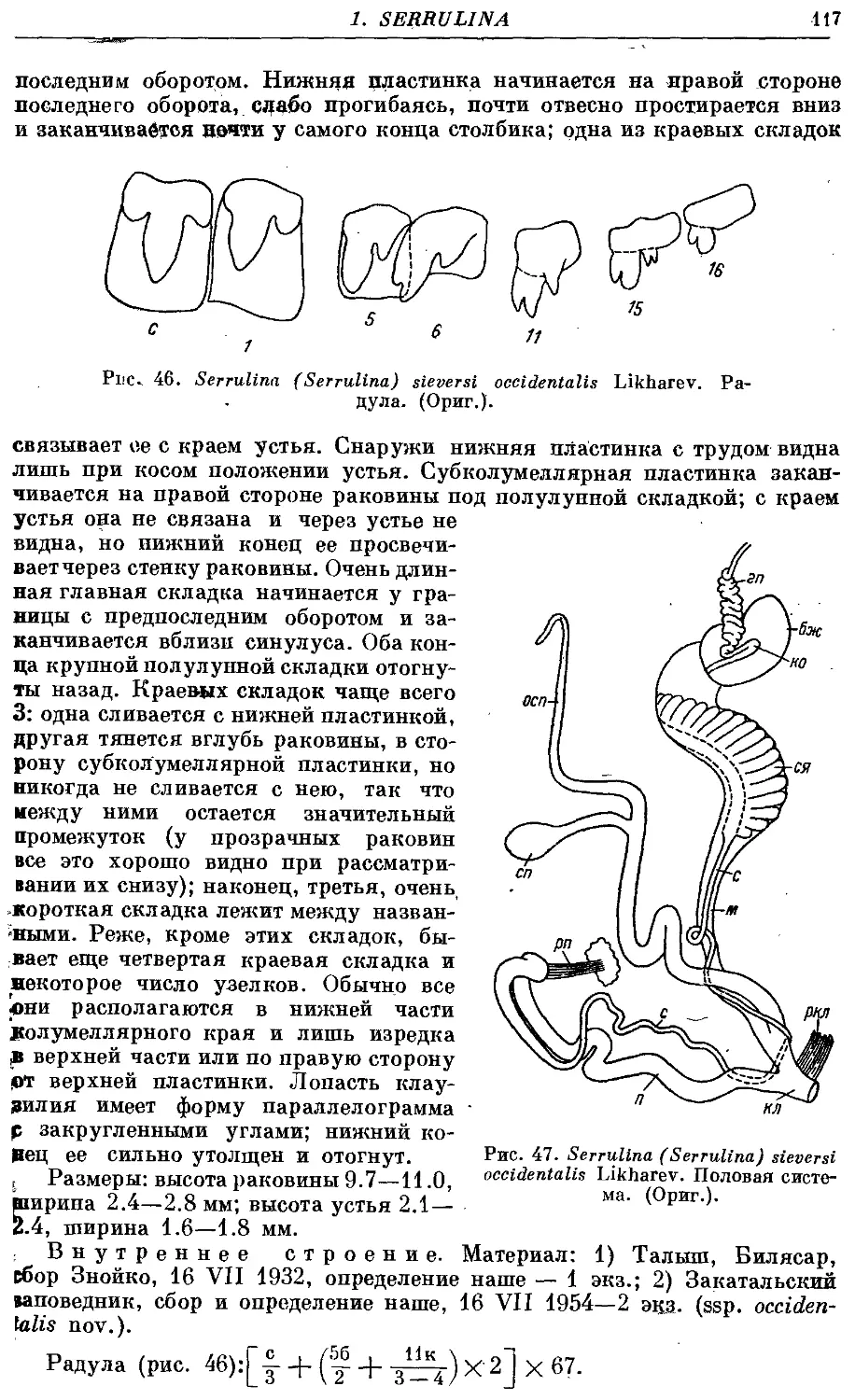
Однако в действительности даже у данных моллюсков одной раковины
недостаточно для создания естественной системы этой группы. Поэтому
предложенная в настоящей работе классификация Clausiliidae сделана
с учетом не только признаков раковины, но и внутреннего строения.
В связи с этим несколько слов о значении анатомического метода
в систематике легочных моллюсков (Pulmonata). Раковины этой группы
несравненно беднее признаками, чем таковые переднежаберных моллюс-
моллюсков (Prosobranchia). В то же время здесь очень обычны всевозможные
проявления конвергенции раковин, которые затрудняют работу система-
систематика.
Вот почему уже в середине прошлого века ряд авторов (A. Schmidt,
Moquin-Tandon, Wiegmann, Pilsbry и др.) стали прибегать к анатомии
для классификации некоторых семейств раковинных легочных моллюсков.
Это направление особенно усилилось в настоящее время (Матёкин,
Старобогатов, Watson, Hubendick, F. С. Baker, H. В. Baker, Morton,
Forcart, Riedel и др.). В результате сейчас в основу естественных систем
ПРЕДИСЛОВИЕ
многих семейств легочных моллюсков (Ellobiidae, Siphonariidae, Planor-
bidae, Lymnaeidae, Vitrinidae, Zonitidae, Helicidae, Eulotidae и др.)
наряду с признаками раковины положены признаки, характерные для
половой системы, мантийного комплекса органов, а в некоторых случаях
и радулы. Нередко анатомические признаки являются наиболее важными
для классификации некоторых групп.
Однако большинство авторов, за редкими исключениями, намечая
филогению того или иного семейства, ограничивается только сравнительно-
анатомическим анализом. В результате вместо вскрытия эволюции ор-
организма как целого дается картина нескольких рядов изменений отдель-
отдельных органов. При этом остается неясной связь между этими рядами
и адаптивное значение подмеченных различий. Нередко бывает неясным,
изменения каких органов являлись ведущими, т. е. были приспособитель-
приспособительными реакциями на определенные внешние факторы и повлекли за собой
изменения других частей тела.
Лишь сравнительно немногие исследователи (Матёкин, Morton),
прибегавшие к анатомии для изучения филогении и систематики каких-
либо семейств легочных моллюсков, не ограничивались только сравни-
сравнительно-анатомическим анализом их строения. Помимо этого, они пытались
обнаружить морфо-функциональные связи между отдельными частями
тела и представить филогению как процесс приспособления организма
к изменяющимся условиям жизни.
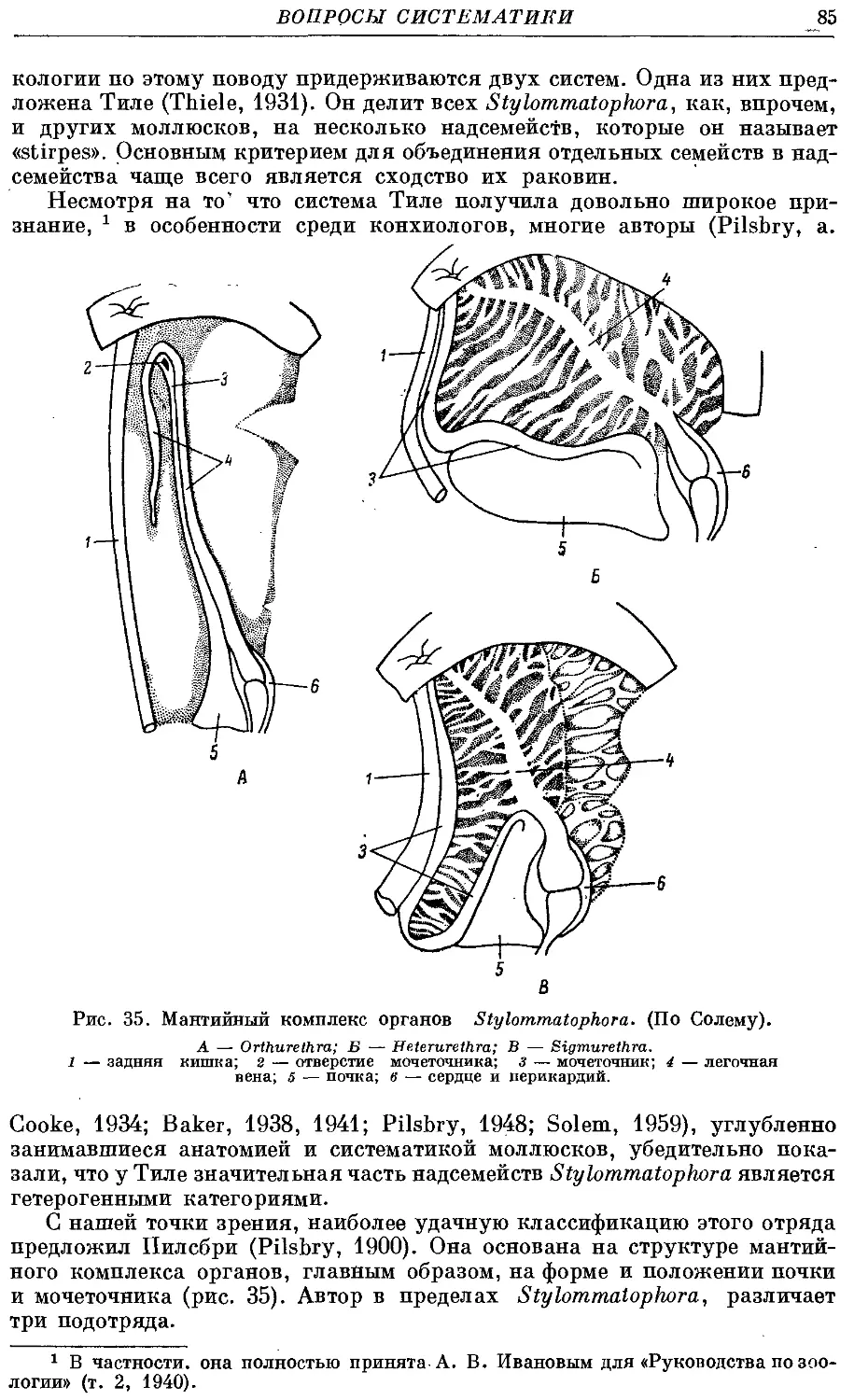
В настоящей работе, посвященной моллюскам Clauslliidae, мы пытаемся
наметить коррелятивные связи между отдельными органами, а также
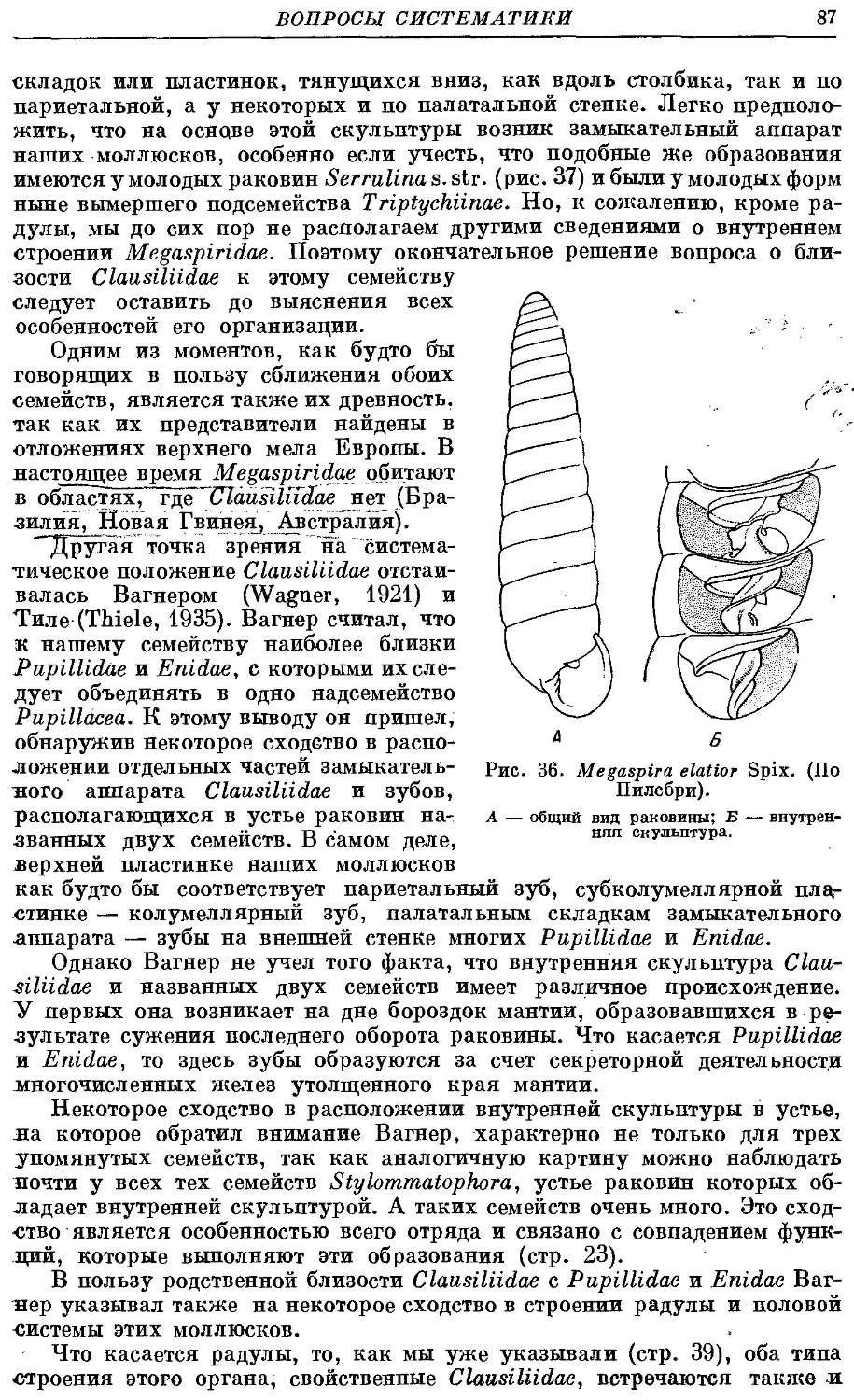
указать на зависимость строения этих органов от образа жизни мол-
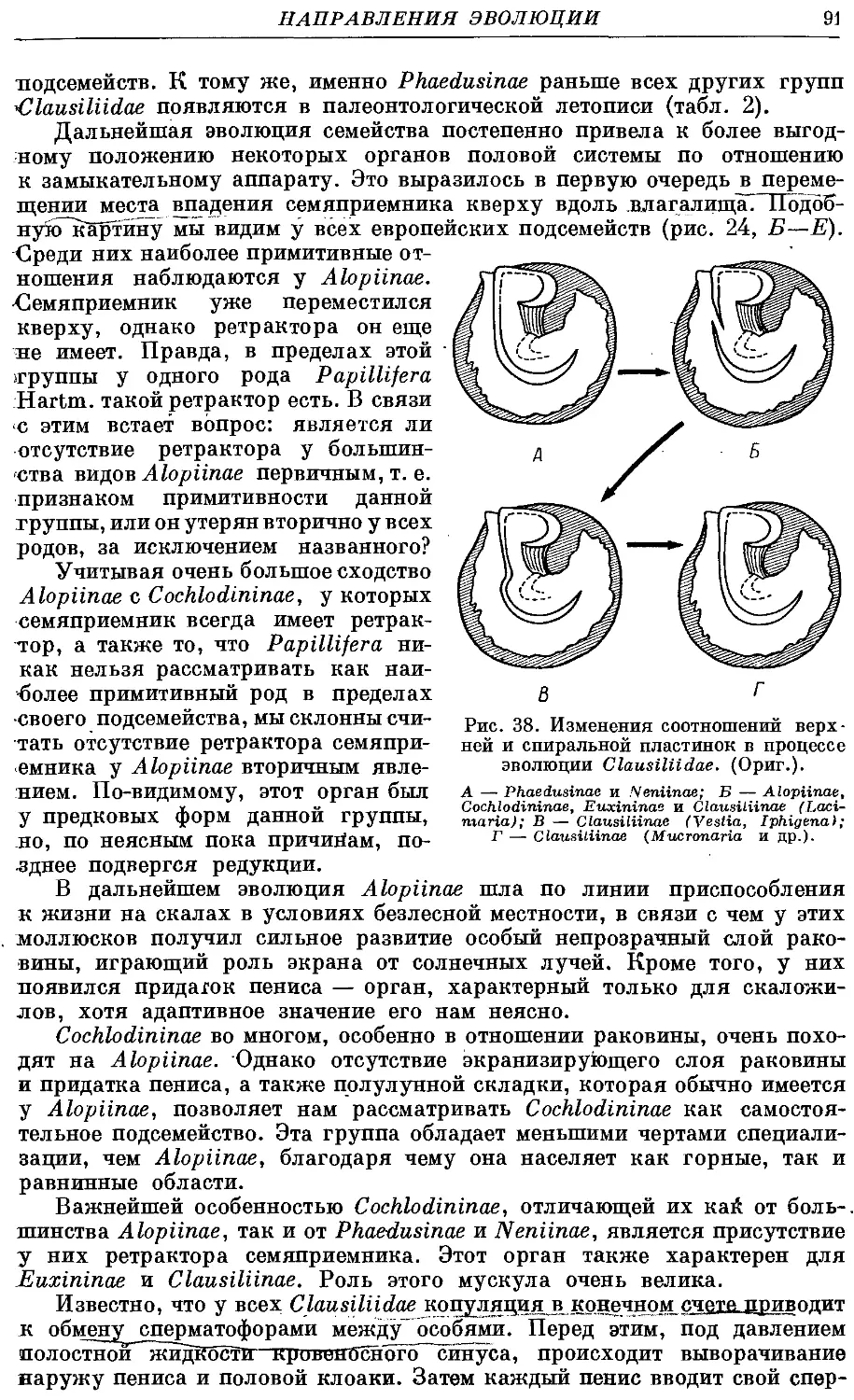
моллюсков. В разделе, посвященном эволюции данных моллюсков, мы пока-
показываем, как воздействие раковины на внутренние органы приводит к их
изменениям и к становлению основных существующих ныне подсемейств.
Одновременно мы высказываем предположение о возрасте и родственных
связях данного семейства с другими группами легочных моллюсков
и о факторах, приведших к его возникновению.
Анатомический метод помогал нам выявить конвергентные элементы
раковины. Последнее очень важно, так как дает возможность сделать
выводы о том, как различные внешние факторы влияют на строение
раковины. В данной работе мы всемерно пытаемся отметить различные
формы адаптации раковины к условиям среды.
Clausiliidae по сравнению со многими другими семействами Stylommato-
phora отличаются высокой стенобионтностью. Раковины этих моллюсков
являются очень хорошими индикаторами условий обитания. Эта особен-
особенность наших моллюсков делает их весьма ценными для стратиграфии
и палеогеографии, так как по их раковинам можно довольно уверенно
судить не только о биотопе, в котором обитал тот или иной вид, но и о кли-
климате соответствующей эпохи.
В систематической части данной работы приводятся определительные
таблицы и описания видов и родов. Изложение ведется по определенной
схеме. Все описания видов, за исключением специально оговоренных,
составлены на основании личного ознакомления с коллекционными ма-
материалами. Так как в большинстве случаев мы имели возможность по
каждому виду использовать свыше 50 экземпляров (нередко более 100),
то количество их указывается лишь в том случае, если их было немного —
менее 25. Когда мы имели дело с «оригинальными» экземплярами, т. е.
с типами и паратипами видов, то число их также приводится. В 15 слу-
случаях, используя авторские экземпляры («оригиналы»), мы выбрали из
них лектотипы, указав их размеры и места хранения.
ПРЕДИСЛОВИЕ
В разделах, посвященных изменчивости отдельных видов, мы обоз-
обозначали понятием «биологическая морфа» (forma) внутривидовые откло-
отклонения, связанные с воздействием конкретных условий биотопа или зоны
(например — альпийская зона, лес, скалы). Подобные отклонения
встречаются в пределах ареала вида всюду, где наблюдаются соответ-
соответствующие экологические условия.
В противоположность морфе, подвид (subspecies) — понятие географи-
географическое. Он населяет территорию с пестрыми физико-географическими
условиями, т. е. встречается в разных биотопах или зонах. Ареалы под-
подвидов чаще всего разобщены, однако в немногих случаях они слегка
перекрывают друг друга.
Понятие «разновидность» (varietas) сохраняется в нашей работе лишь
условно. Сюда относятся все ранее описанные и снабженные латинскими
названиями отклонения, природа которых пока неясна. Кроме того,
сюда попадают различные проявления возрастной и индивидуальной
изменчивости, вплоть до уродств (если ранее они были обозначены
специальными названиями).
В настоящей работе даются описания 102 ныне живущим видам и под-
подвидам. Внутреннее строение изучено у 80 видов, из них для 46 подобные
сведения приводятся впервые. Кроме того, описано 3 новых для наука
современных вида. Основная масса указанных здесь видов обитает на
территории нашей страны. Но кроме них, мы включили еще 32 вида, не
найденных в СССР (в систематической части они отмечены звездочкой).
При выборе некоторых из них мы руководствовались предположением,
что со временем их обнаружат и на нашей территории. Кроме того, в
данную работу мы включили несколько видов, известных из областей,
лежащих довольно далеко от границ СССР, но принадлежащих к родам,
основная масса видов которых обитает в нашей стране.
Как приложение дается описание одной коллекции третичных и чет-
четвертичных Clausiliidae Северного Кавказа. Следует учесть, что все до сих
пор известные литературные данные об ископаемых Clausiliidae Кавказа
ограничиваются очень несовершенным описанием одного вида.
Настоящая работа подготавливалась в стенах Зоологического ин-
института АН СССР, обширные коллекции которого легли в ее основу.
Нами были использованы коллекции и других учреждений — Научно-приро-
Научно-природоведческого музея АН УССР (Львов), Зоологического музея МГУ (Москва),
Музея Грузии (Тбилиси), Зоологического института АН Армянской ССР
(Ереван), Зоологического института Польской Академии наук (Варшава).
Ряд наших коллег оказал нам неоценимую помощь в получении нуж-
нужных нам коллекционных материалов и сведений о распространении от-
отдельных видов. В связи с этим мы приносим глубокую благодарность стар-
старшему научному сотруднику Зоологического института АН Армянской ССР
Н. Н. Акрамовскому, доценту Кутаисского педагогического института
Г. В. Кокочашвили, доктору А. Риделю (A. Riedel, Варшава), доктору
Л. Форкарту (L. Forcart, Базель), проф. 3. Эккелю (S. Jaeckel sen., Бер-
Берлин), доктору В. Ложеку (V. Lozek, Прага), проф. Я. Урбанскому (J. Ur-
banski, Познань).
Мы особенно благодарны А. А. Стеклову (Москва) за предоставление
нам интересной коллекции третичных и четвертичных Clausiliidae Се-
Северного Кавказа.
Изготовление рисунков для данной работы осуществлено художниками
Т. Ф. Белоцветовой, Б. Н. Толмачевым и А. И. Калининой.
Редактирование данной книги любезно взял на себя проф. А. А. Стрел-
Стрелков, за что мы приносим ему самую сердечную признательность.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ СЕМ. CLAUSILIIDAE
1. Подсемейство Phaedusinae
1. Род Serrulina Mouss.
I. Подрод Serrulina s. str. Стр.
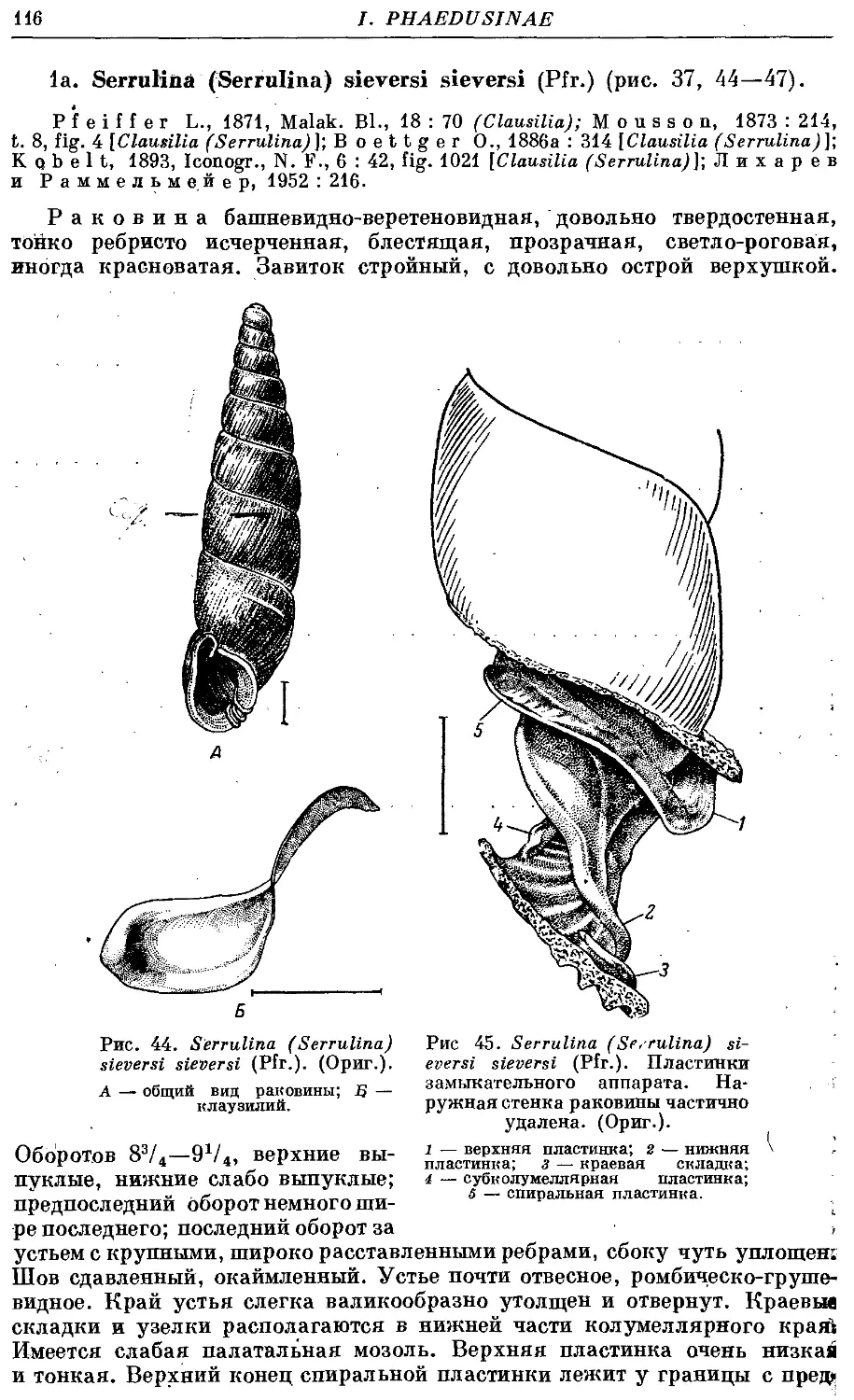
la. S. (S.) sieversi sieversi (Pfr.) 116
16. S. (S.) sieversi occidentalis Likharev, ssp. nov 118
2a. S. (S.) serrulata serrulate, (Pfr.) 118
*26. S. (S.) serrulata amanica Nagele 120
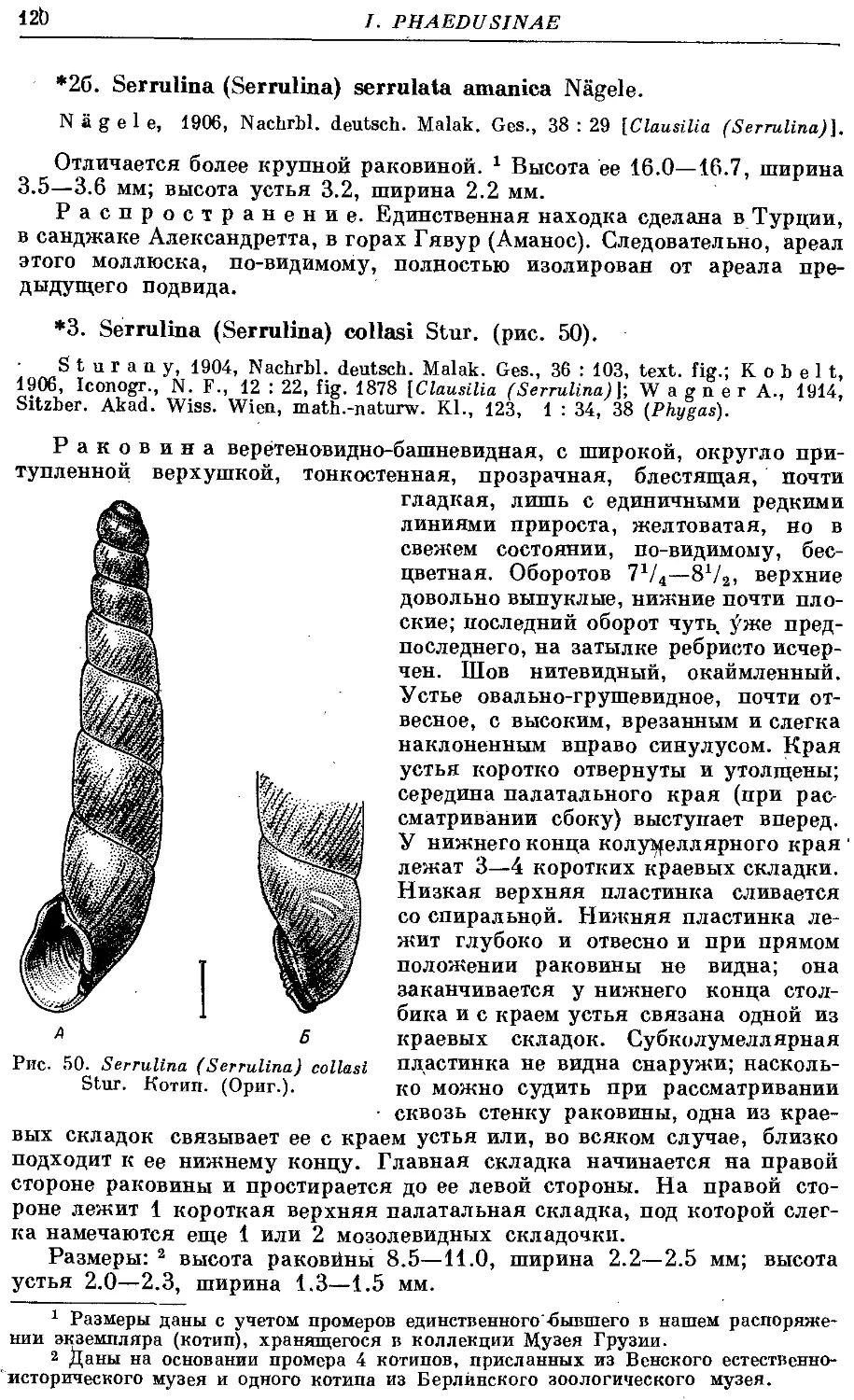
*3. S. (S.) collasi Stur 120
*4. S. (S.?J senghanensis Morgan . 121
2. Подрод Pravispira Lindh.
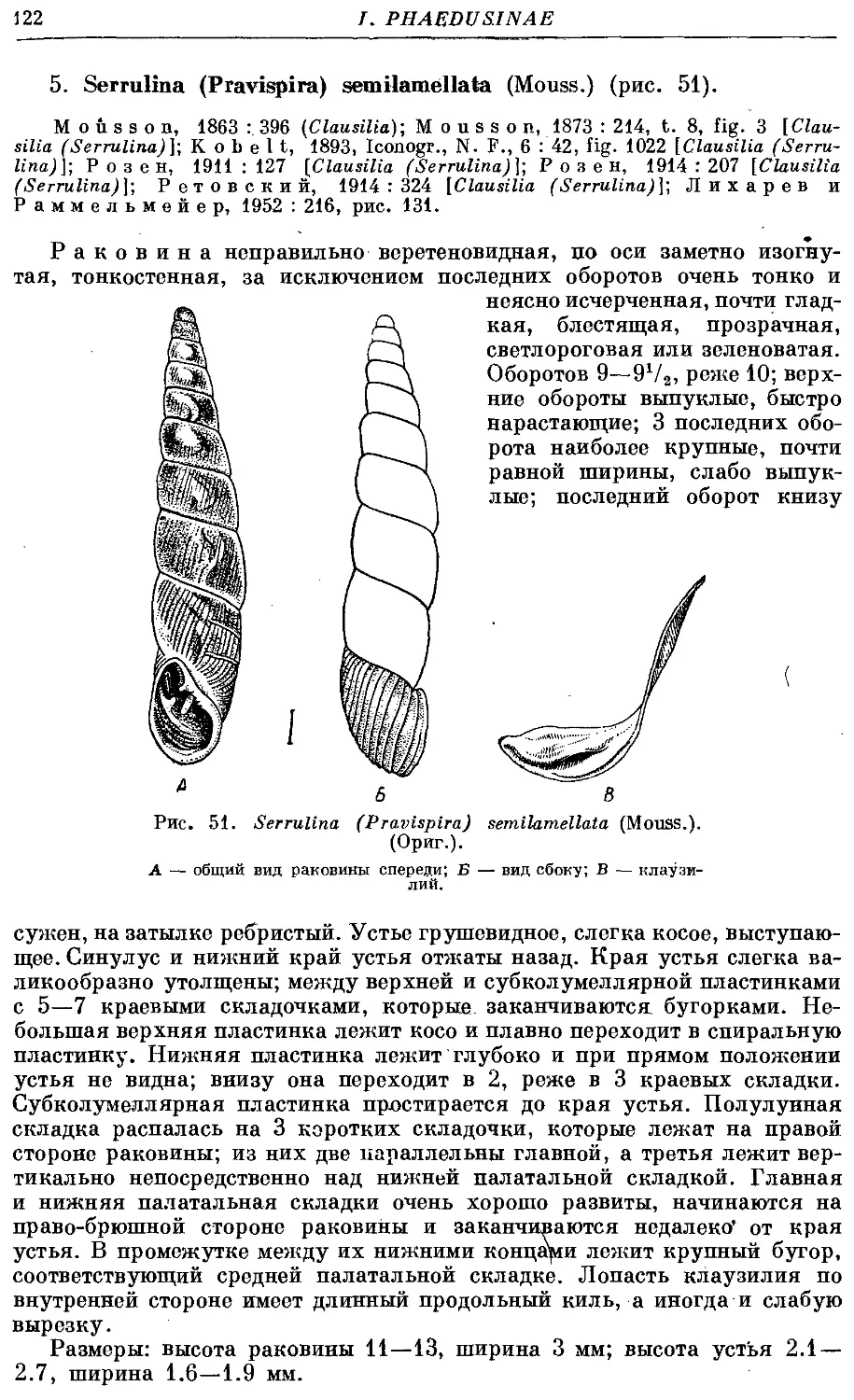
5. S. (Pr.) semilamellata (Mouss.) . . . 122
2. Род Pontophaedusa Lindh.
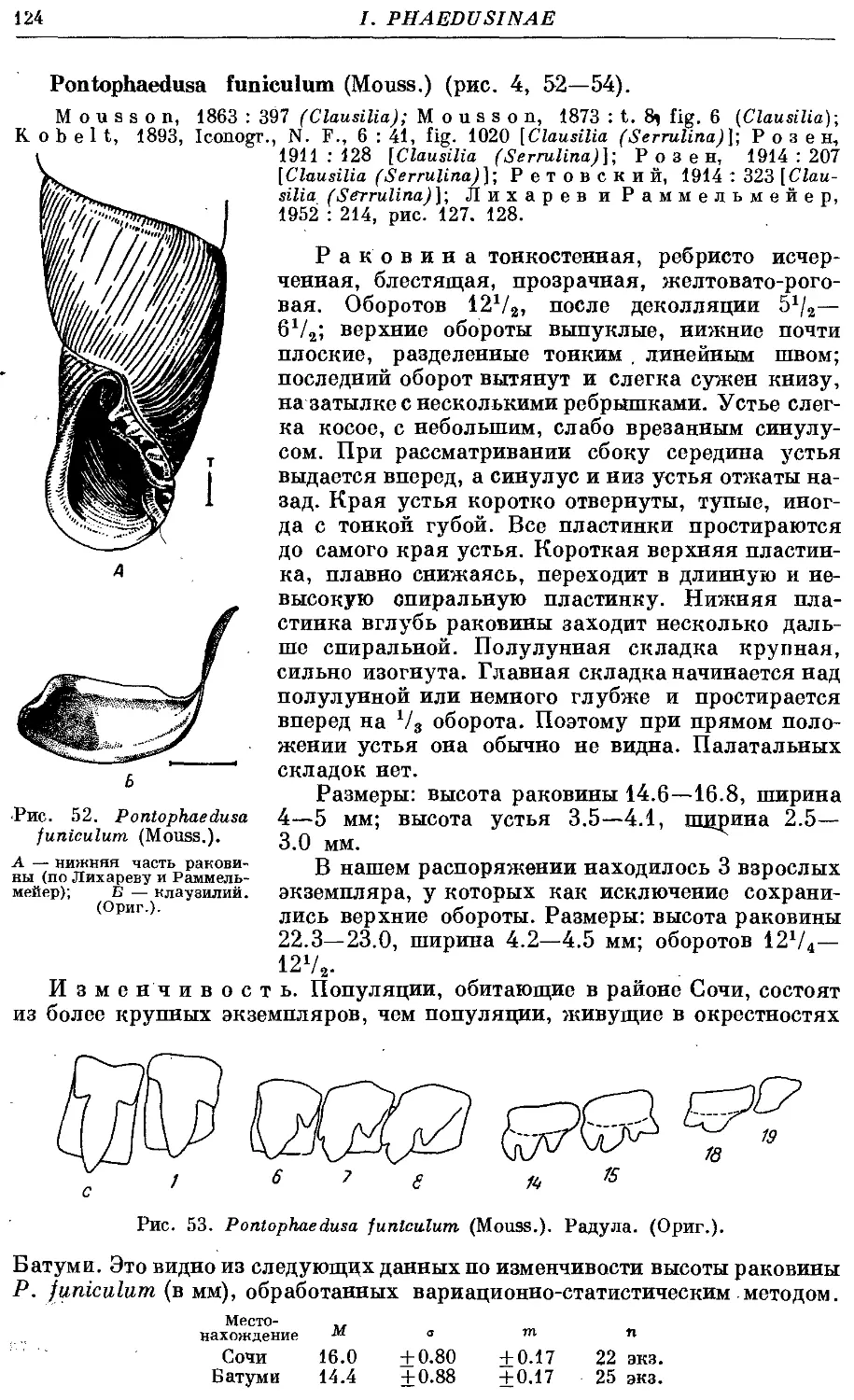
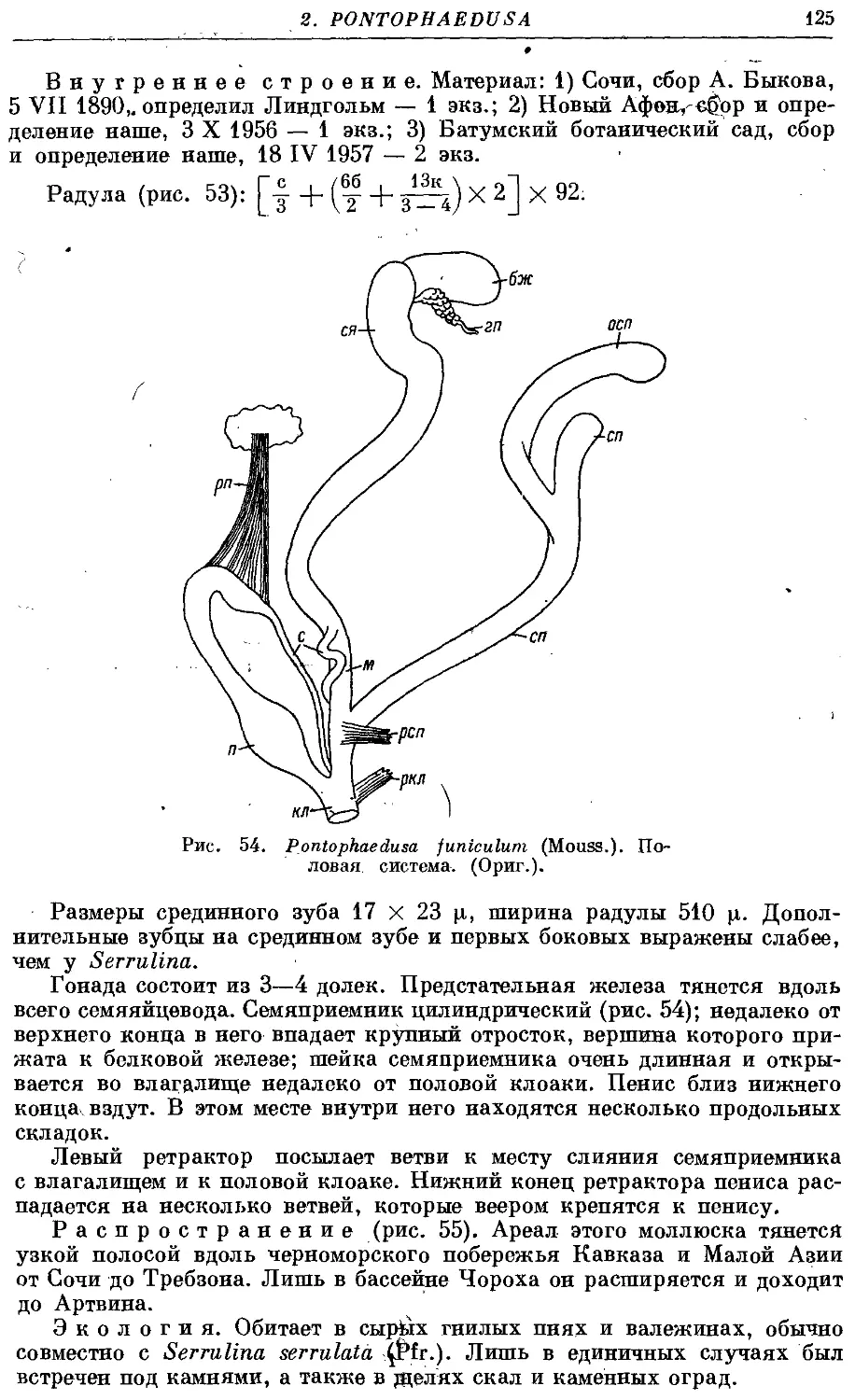
P. funiculum (Mouss.) 124
3. Род Caspiophaedusa Lindh.
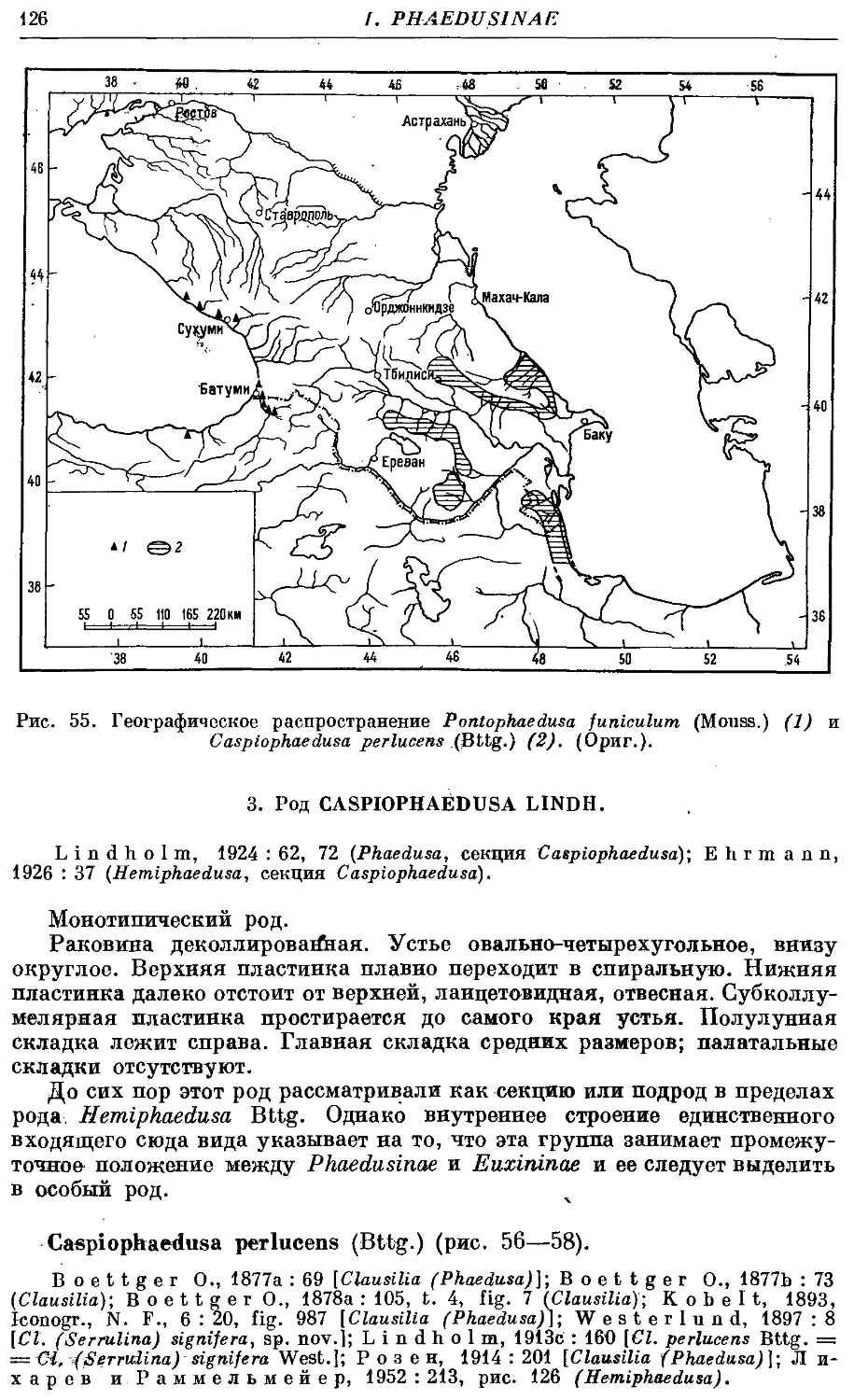
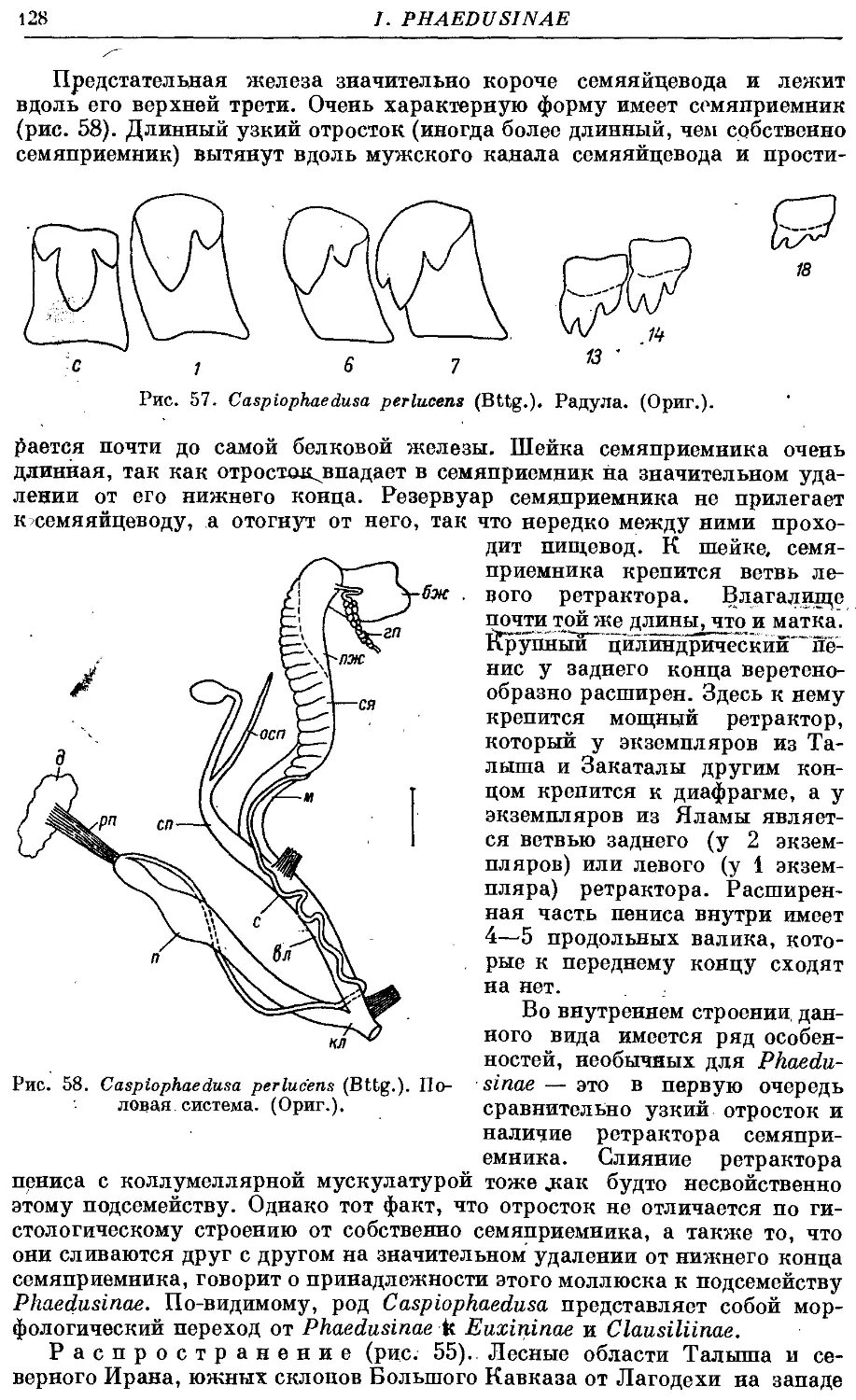
С perlucens (Bttg.) . 126
Дополнение к подсемейству Phaedusinae
* «Phaedusa» hyrcanica Morgan 129
II. Подсемейство Alopiinae
Род Alopia H. et A. Adams.
* A. glauca (E. Bielz) 131
III. Подсемейство Cochlodininae
Род Cochlodina Fer.
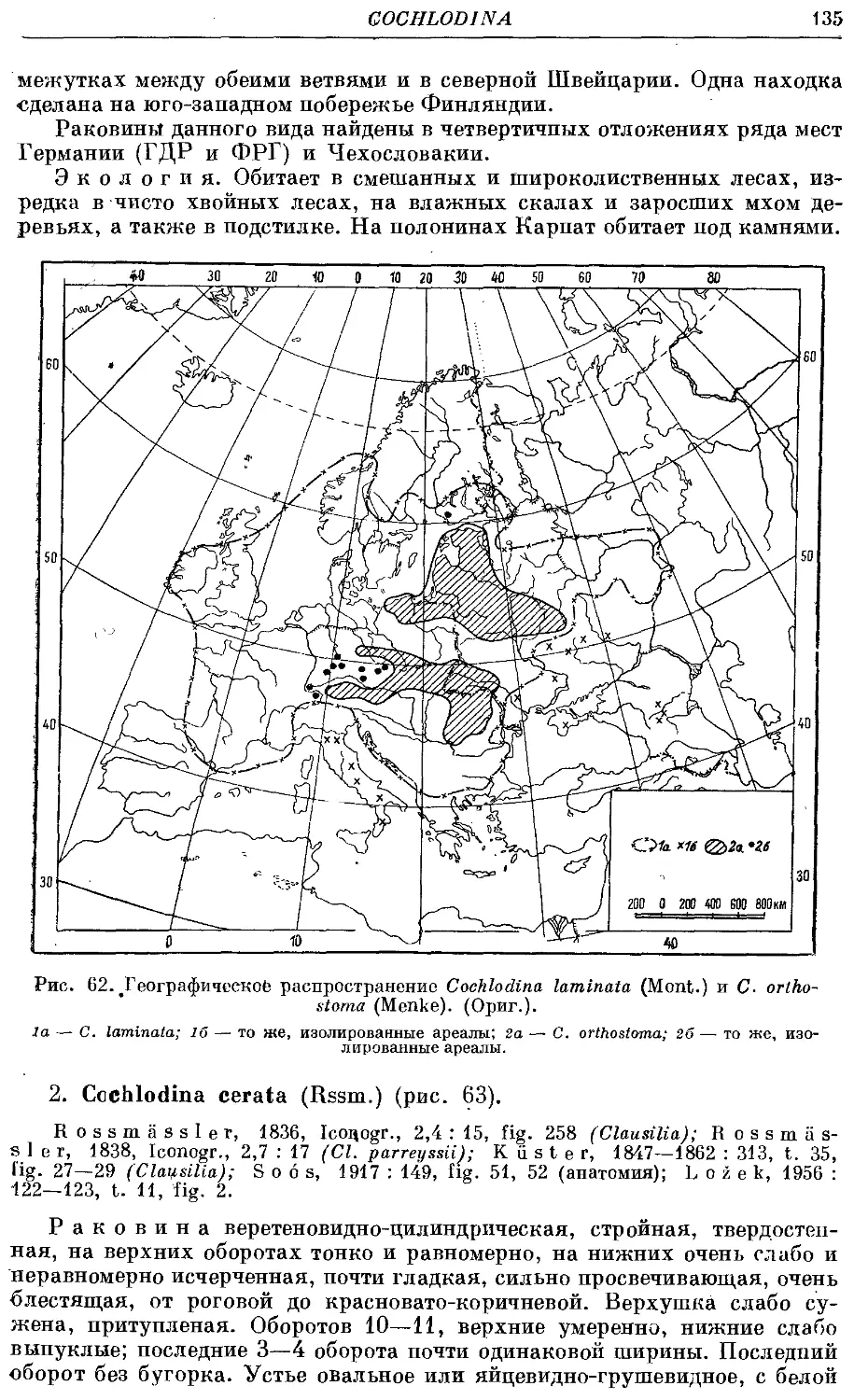
1. C. orihostoma (Menke) 134
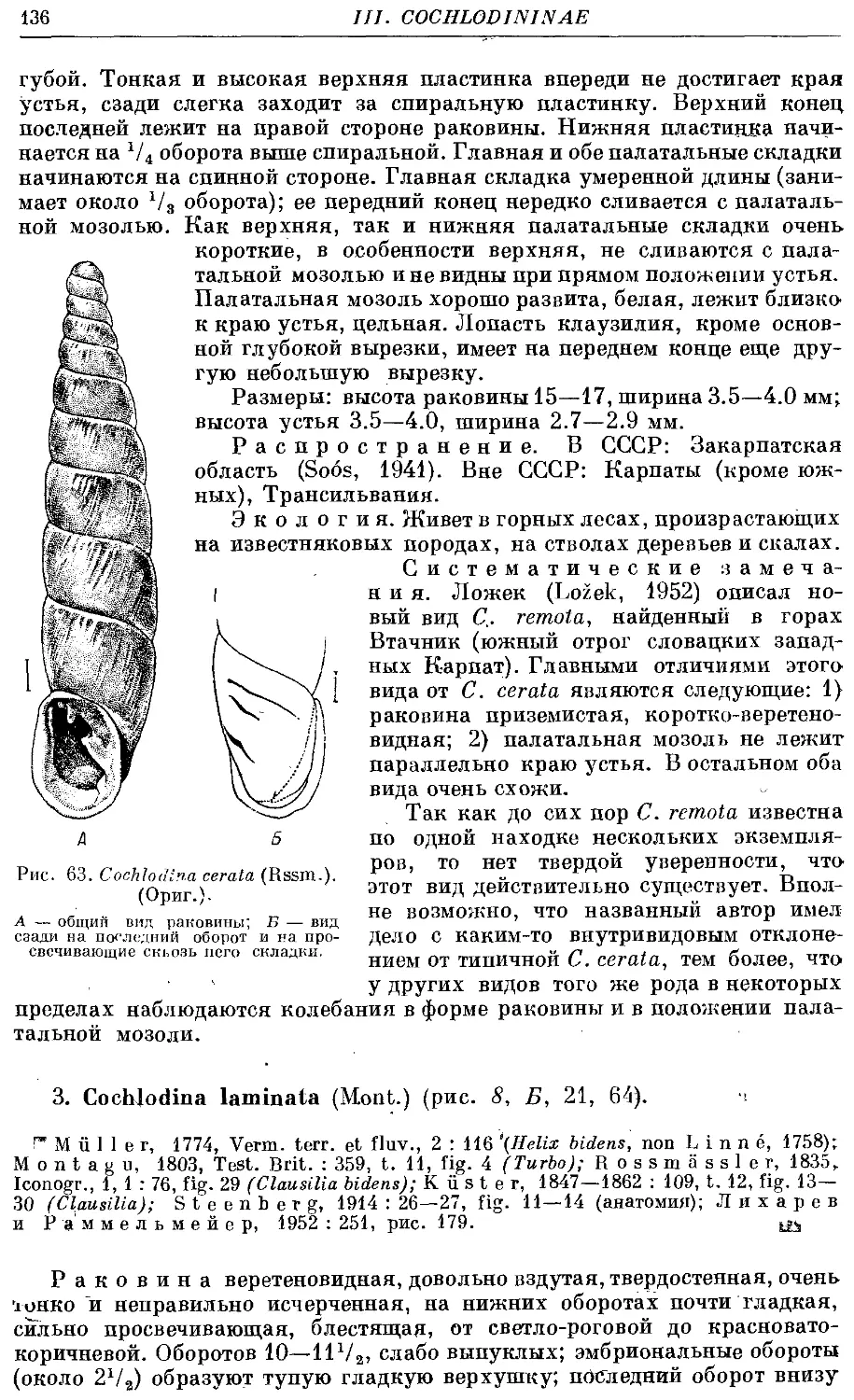
2. C. cerata (Rssm.) 135
3. С laminate. (Mont.) 136
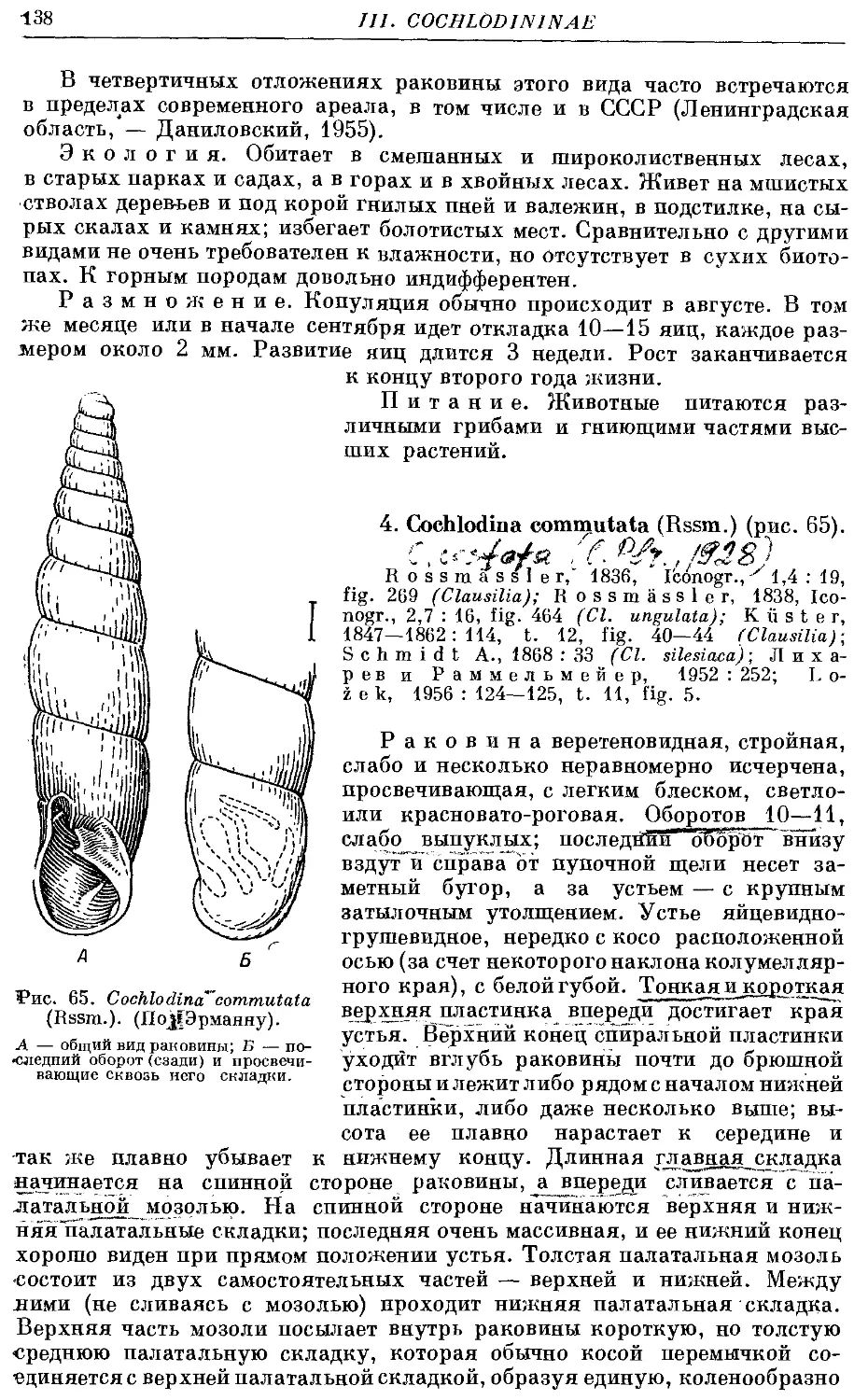
4. C. commutata (Rssm.) 138
IV. Подсемейство Enxininae subfam. nov.
Род Idyla H. et A. Adams
Подрод Scrobifera Bttg.
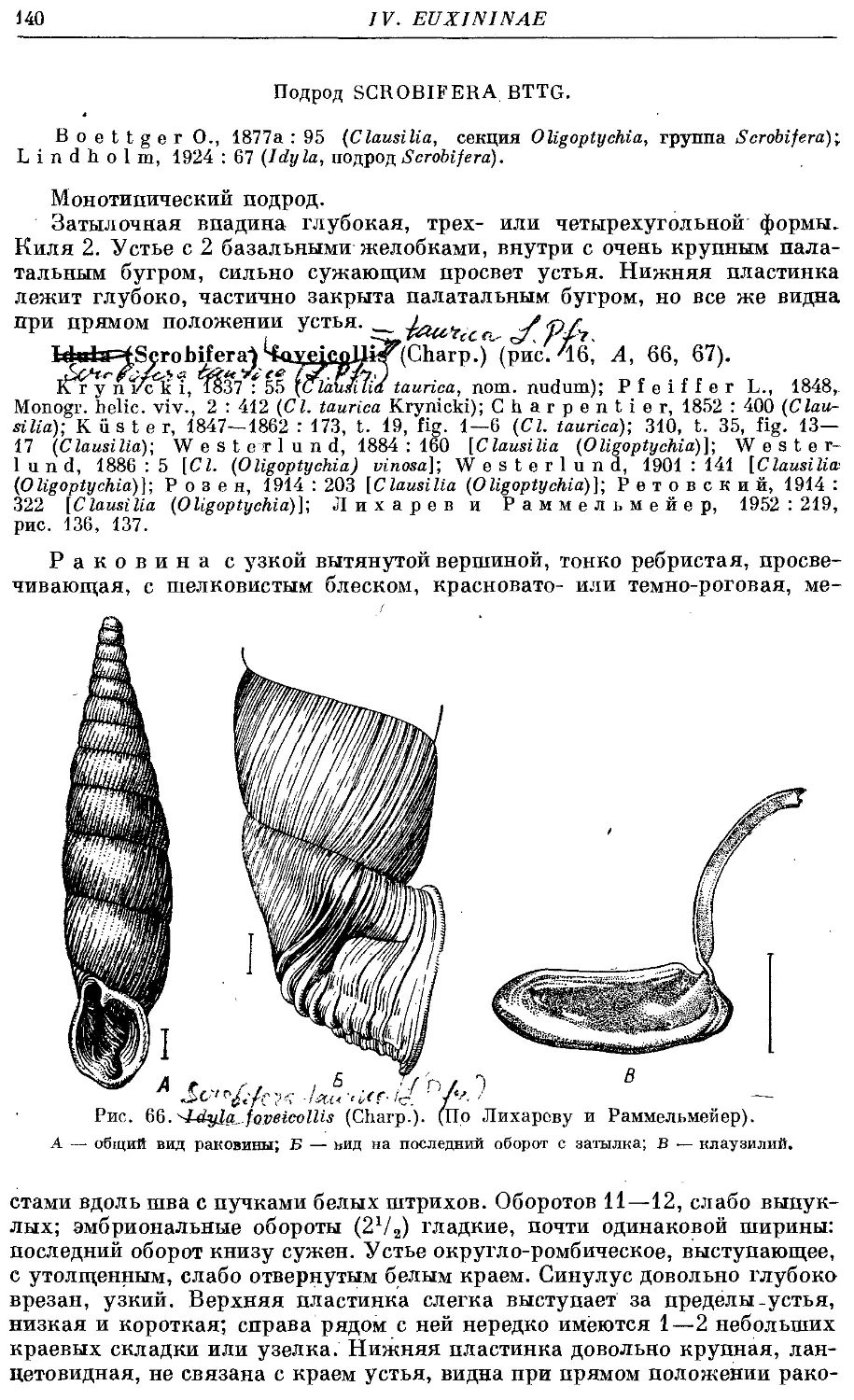
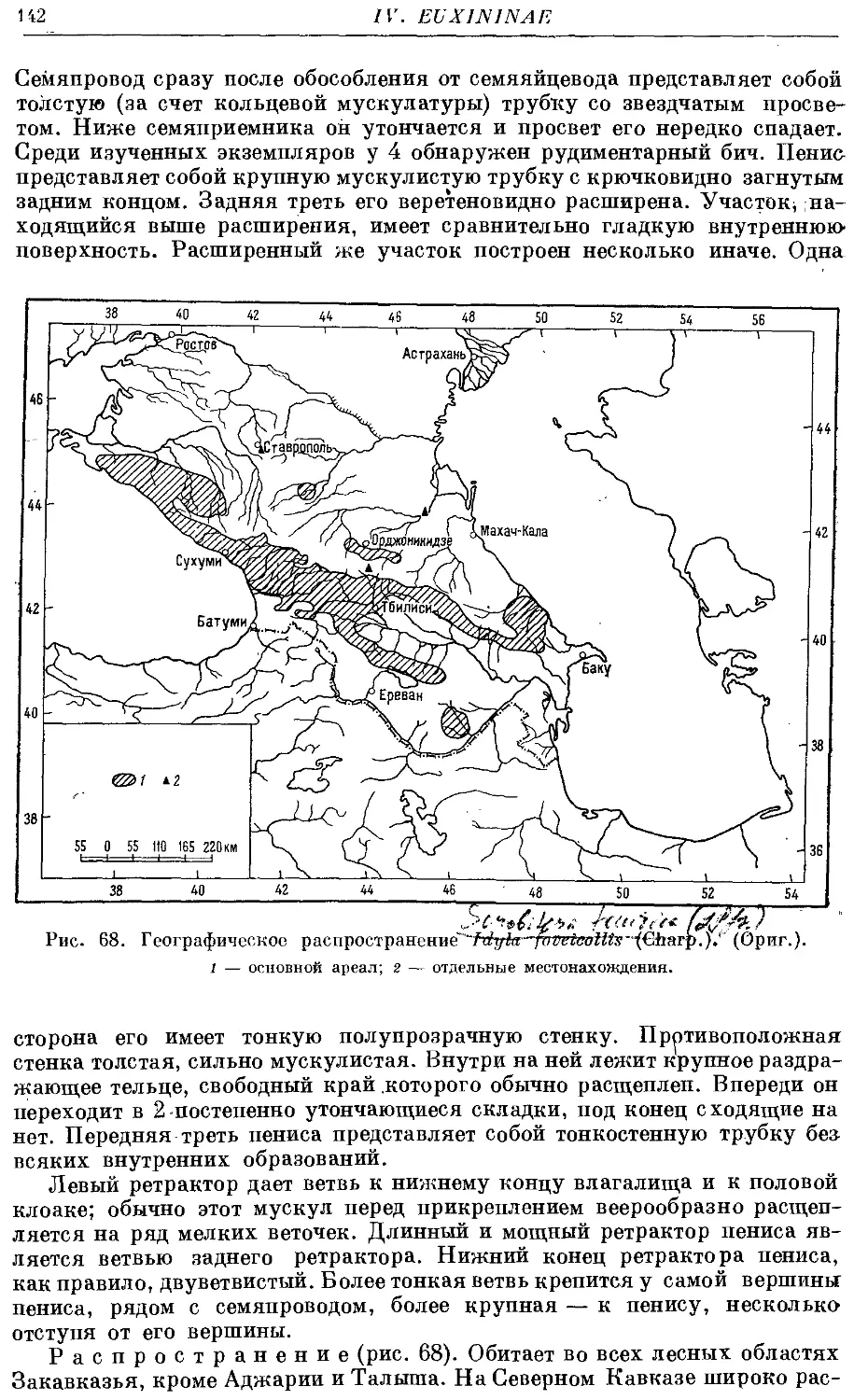
/. (S.) foveicollis (Charp.) 140
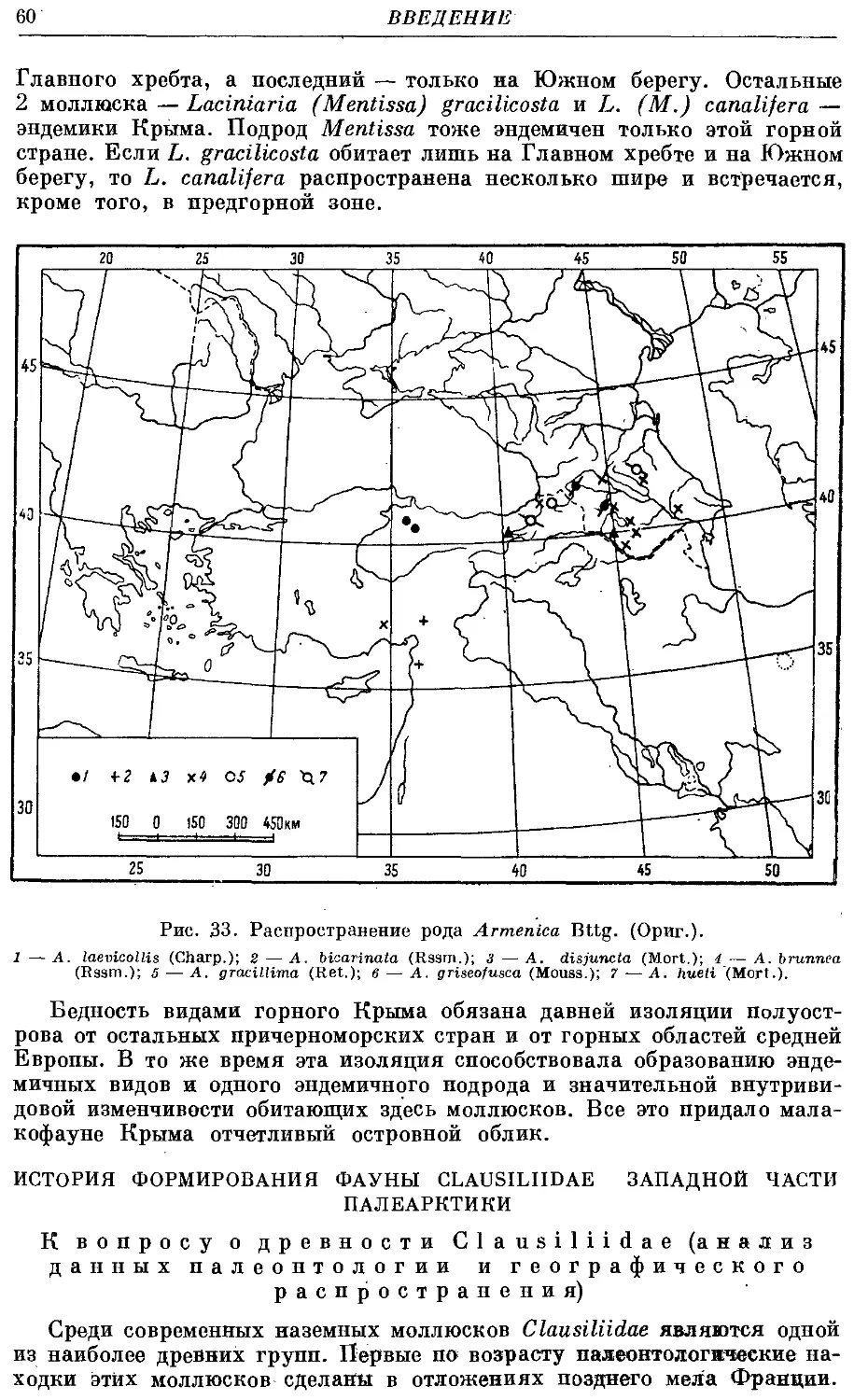
2. Род Armenica Bttg.
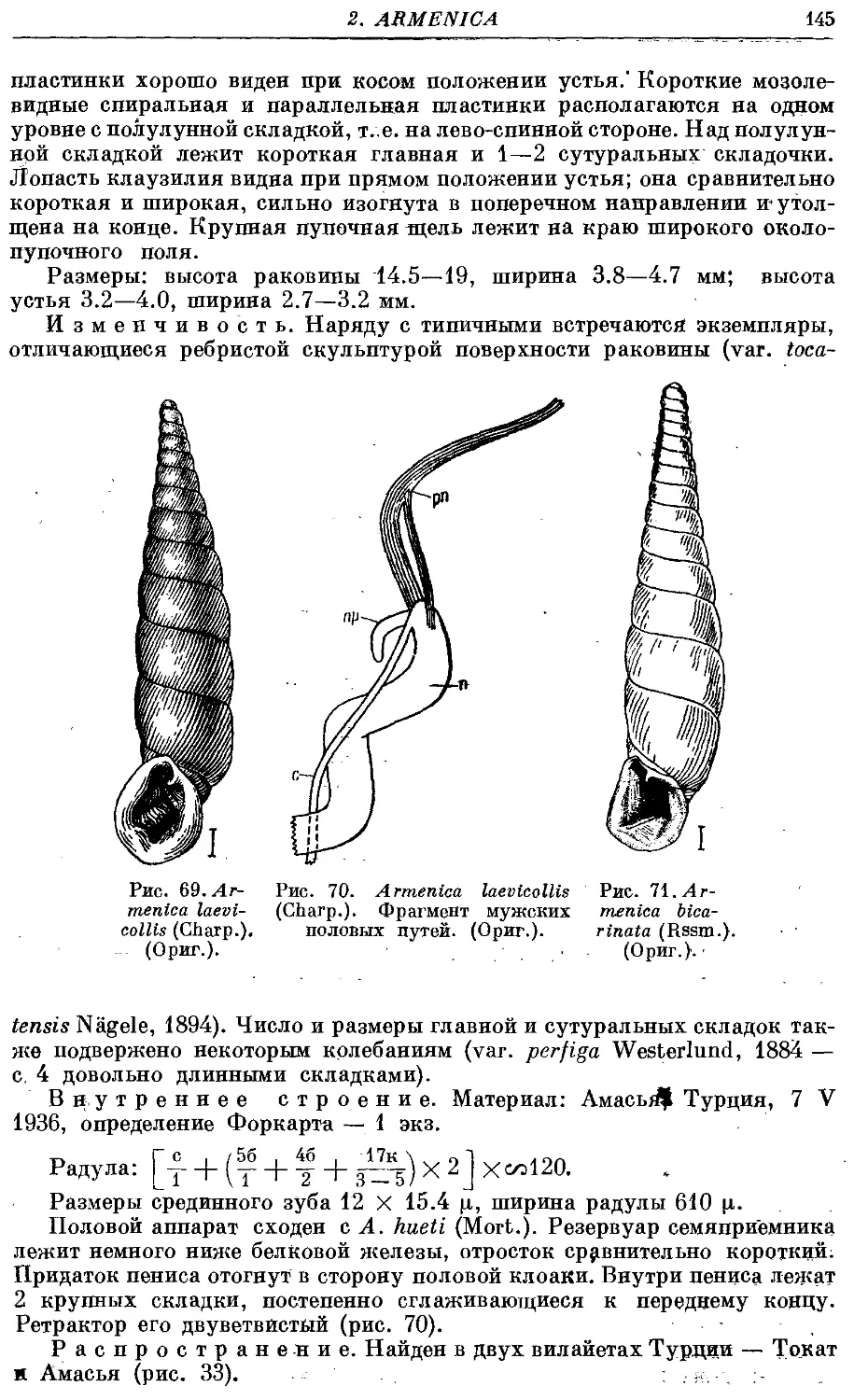
*1, A. laevicollis (Charp.) 144
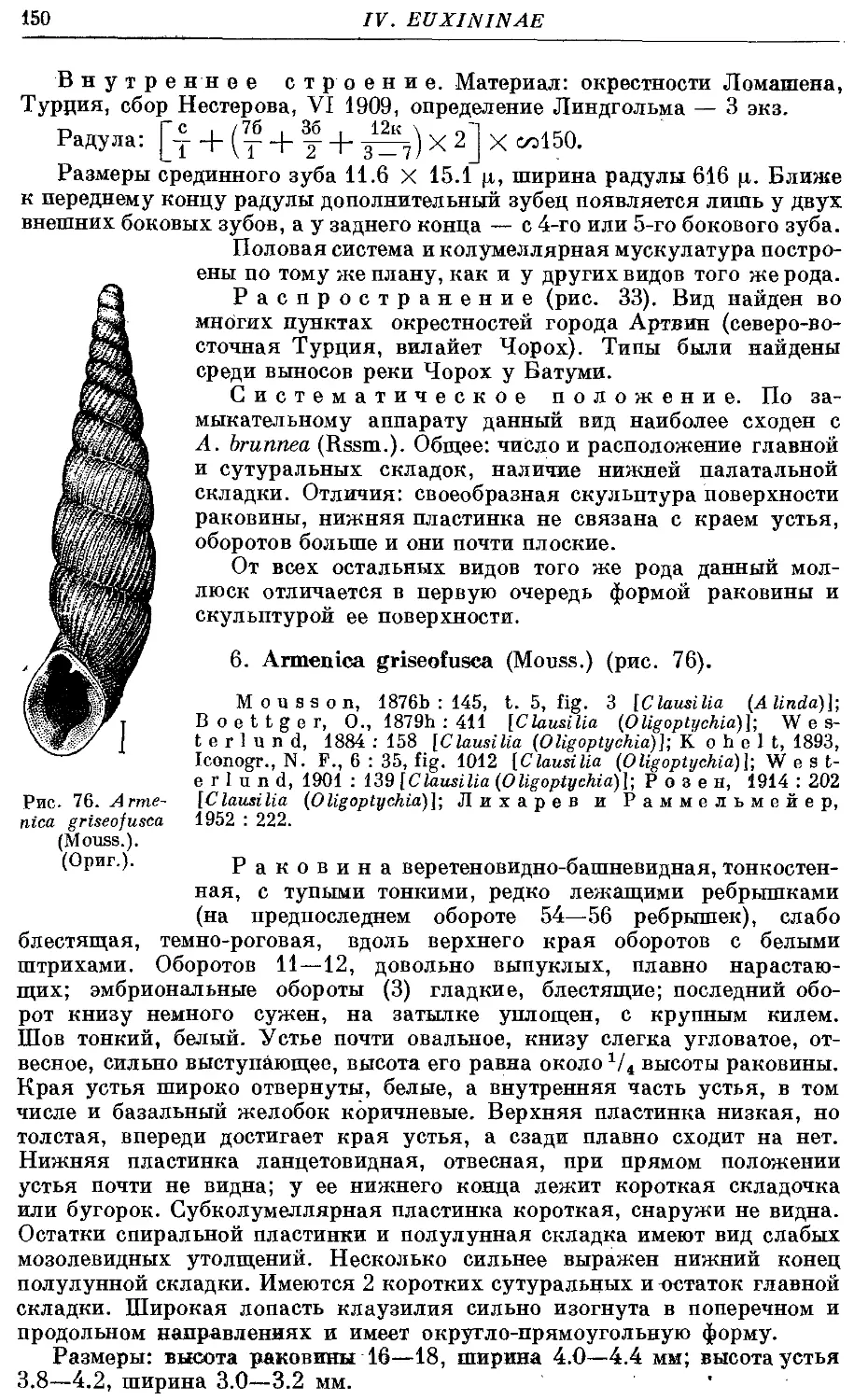
*2. A. bicarinata (Rssm.) 146
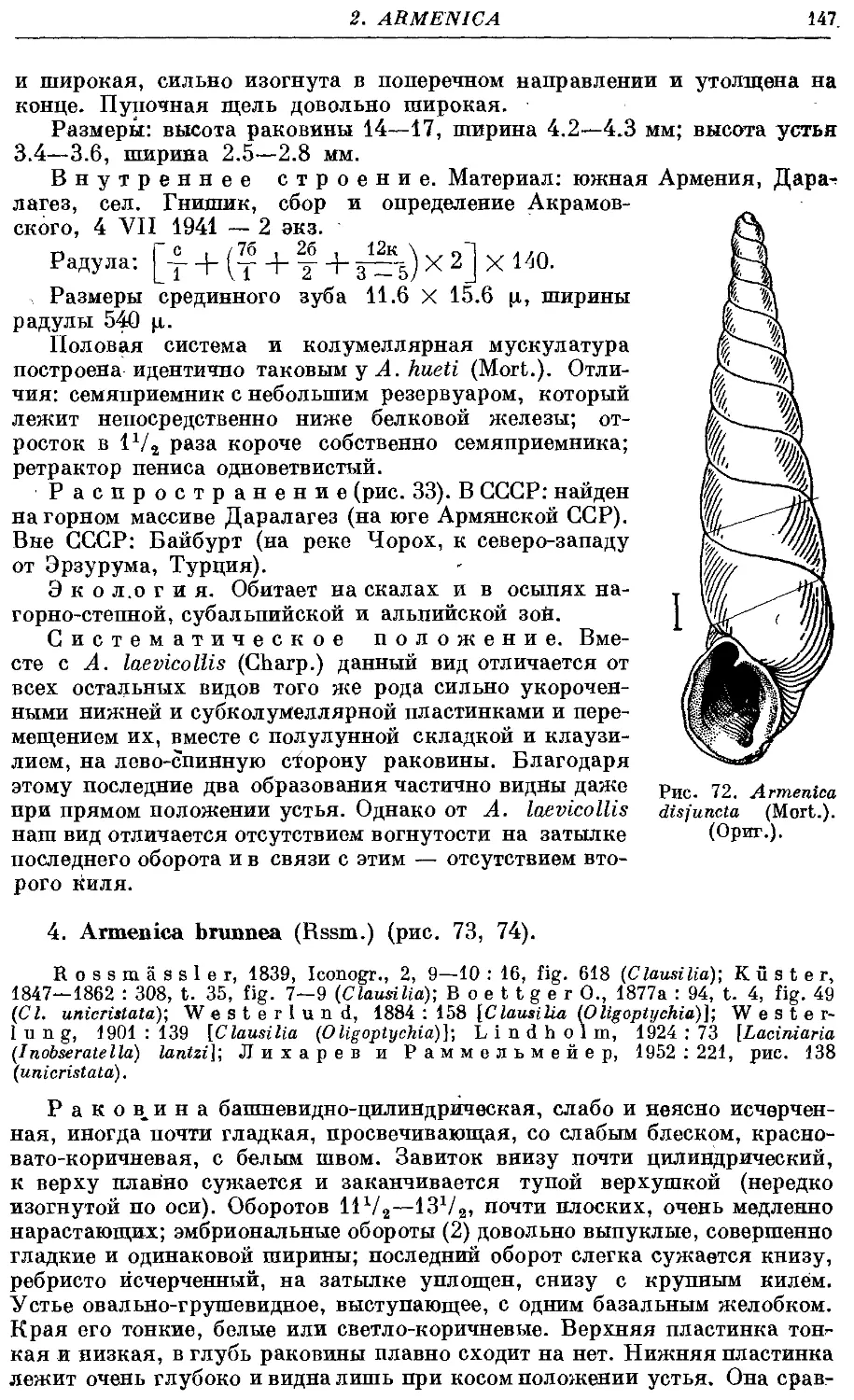
3. A. disfuncta (Mort.) 146
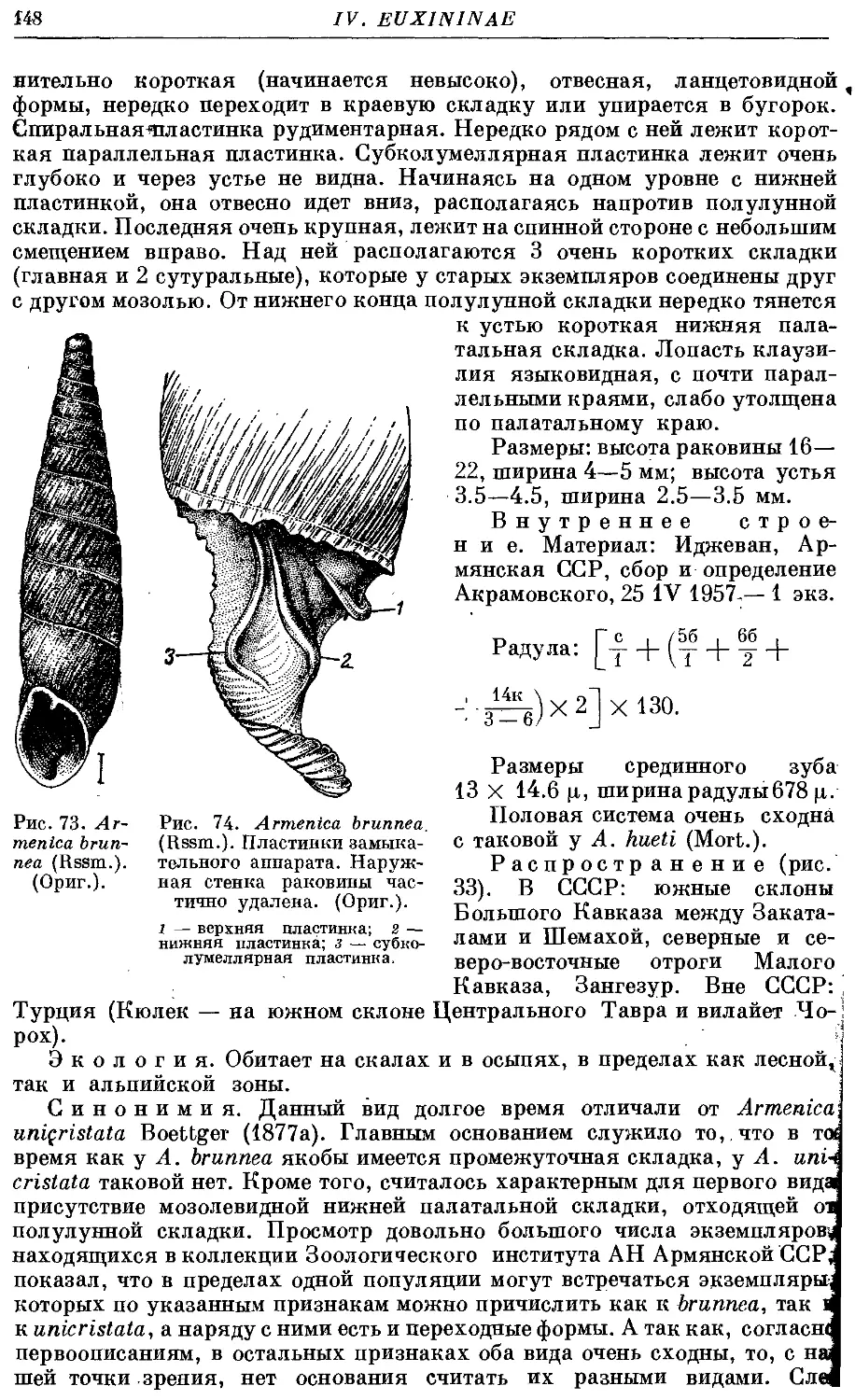
4. A. brunnea (Rssm.) 147
*5. A. gracillima (Ret.) ¦ 149
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
Стр.
6. A. griseofusca (Mouss.) 150
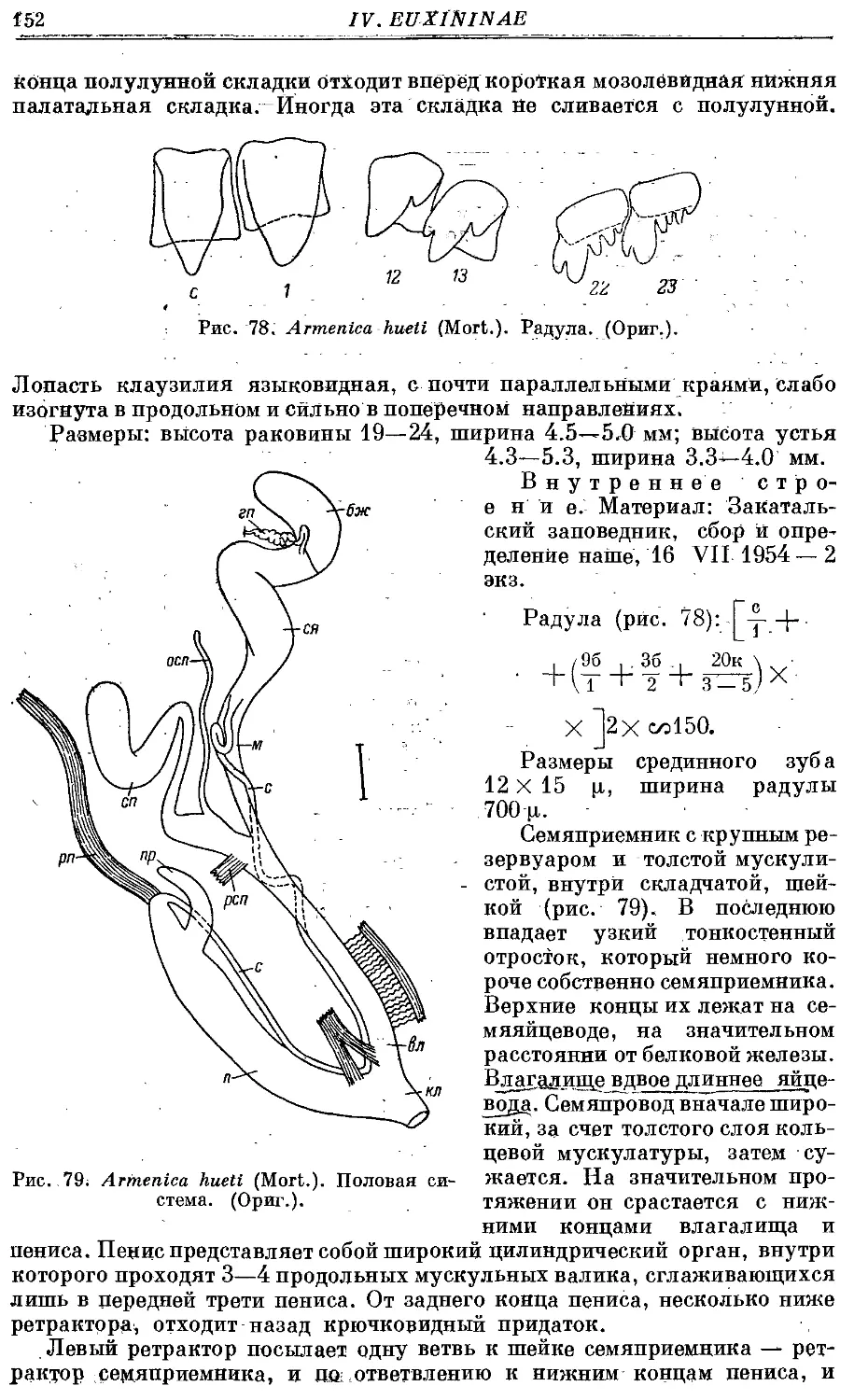
7. A. hueti (Mort.) 151
Дополнение к роду Armenica Bttg.
*8. A. rupicola (Mort.) 153
*9. A. blissi (Bttg.) 153
*10. A. euprepes Biggs 154
*11. A. mimele Biggs 154
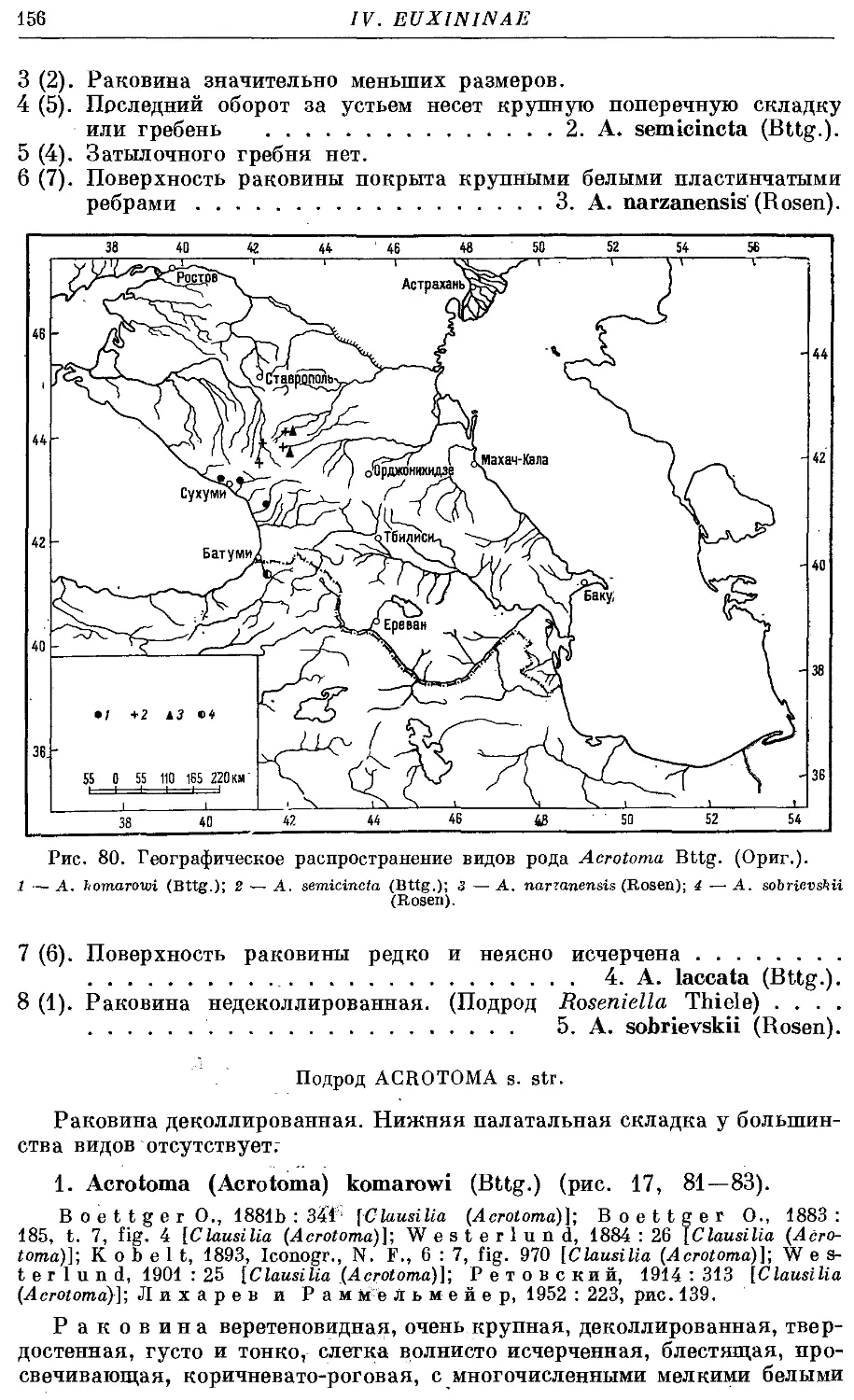
3. Род Acrotoma Bttg.
1. Подрод Acrotoma s. str.
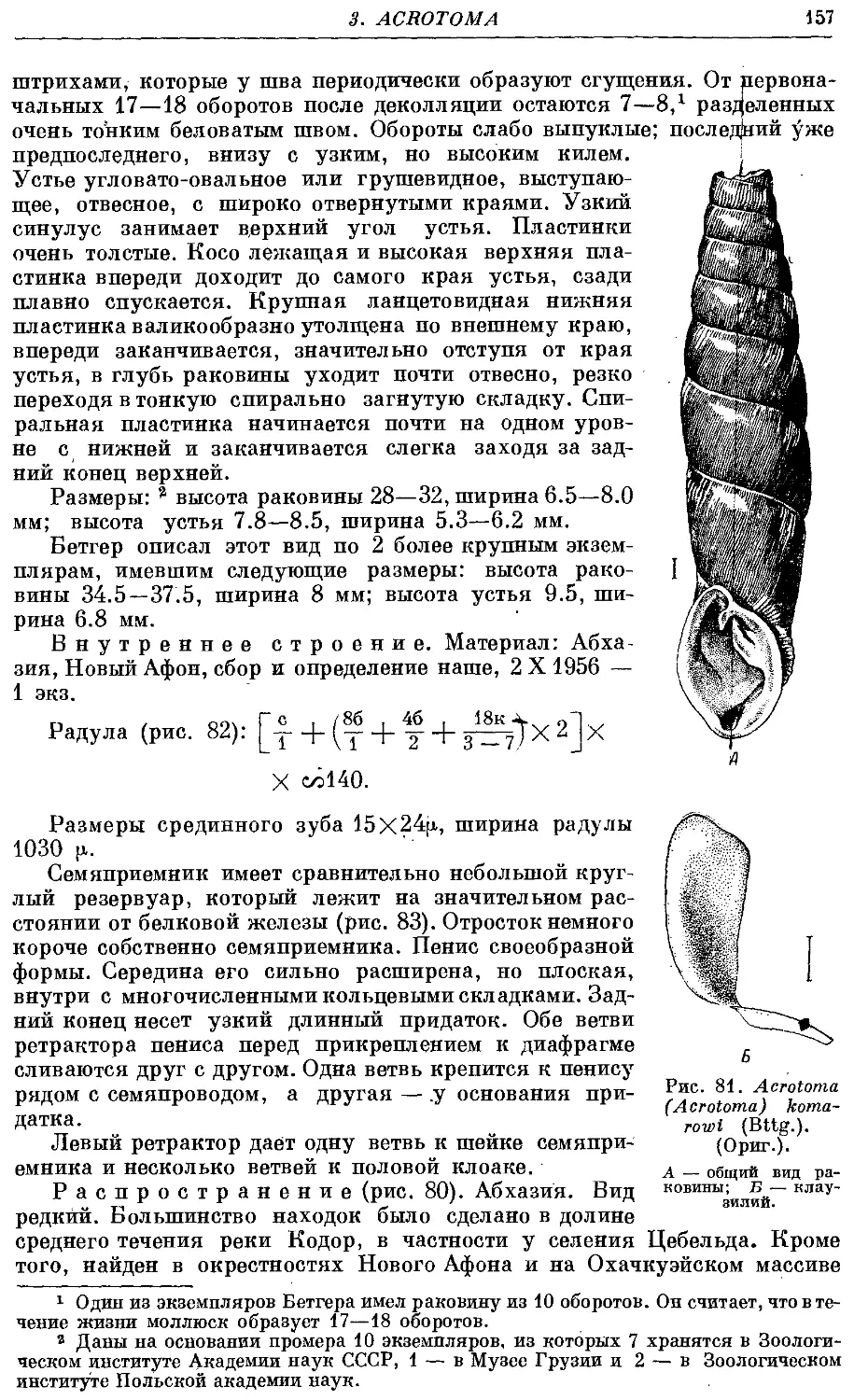
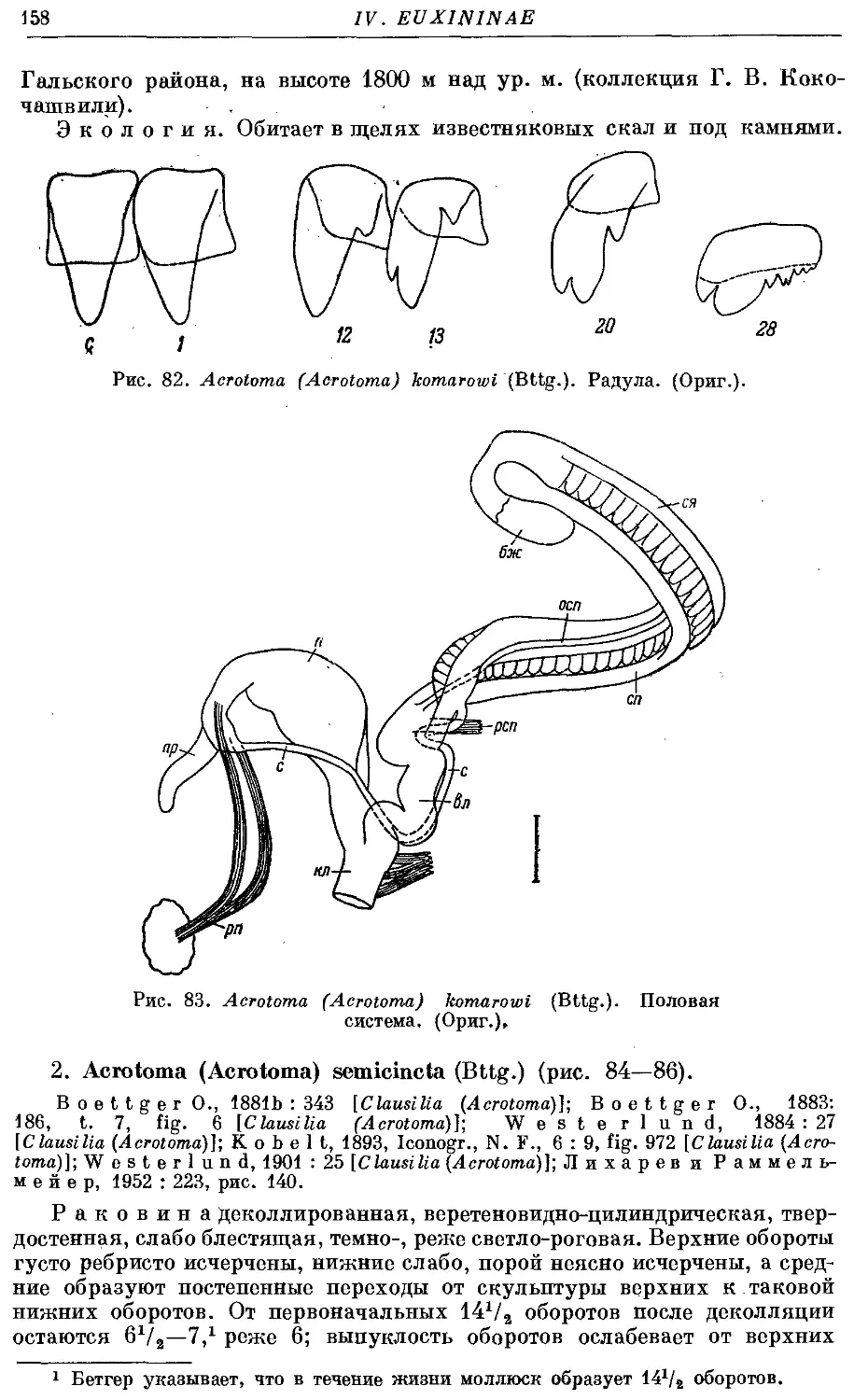
1. A. (A.) komarowi (Bttg.) 156
2. A. (A.) semicincta (Bttg.) 158
3. A. (A.) narzanensis (Rosen) 160
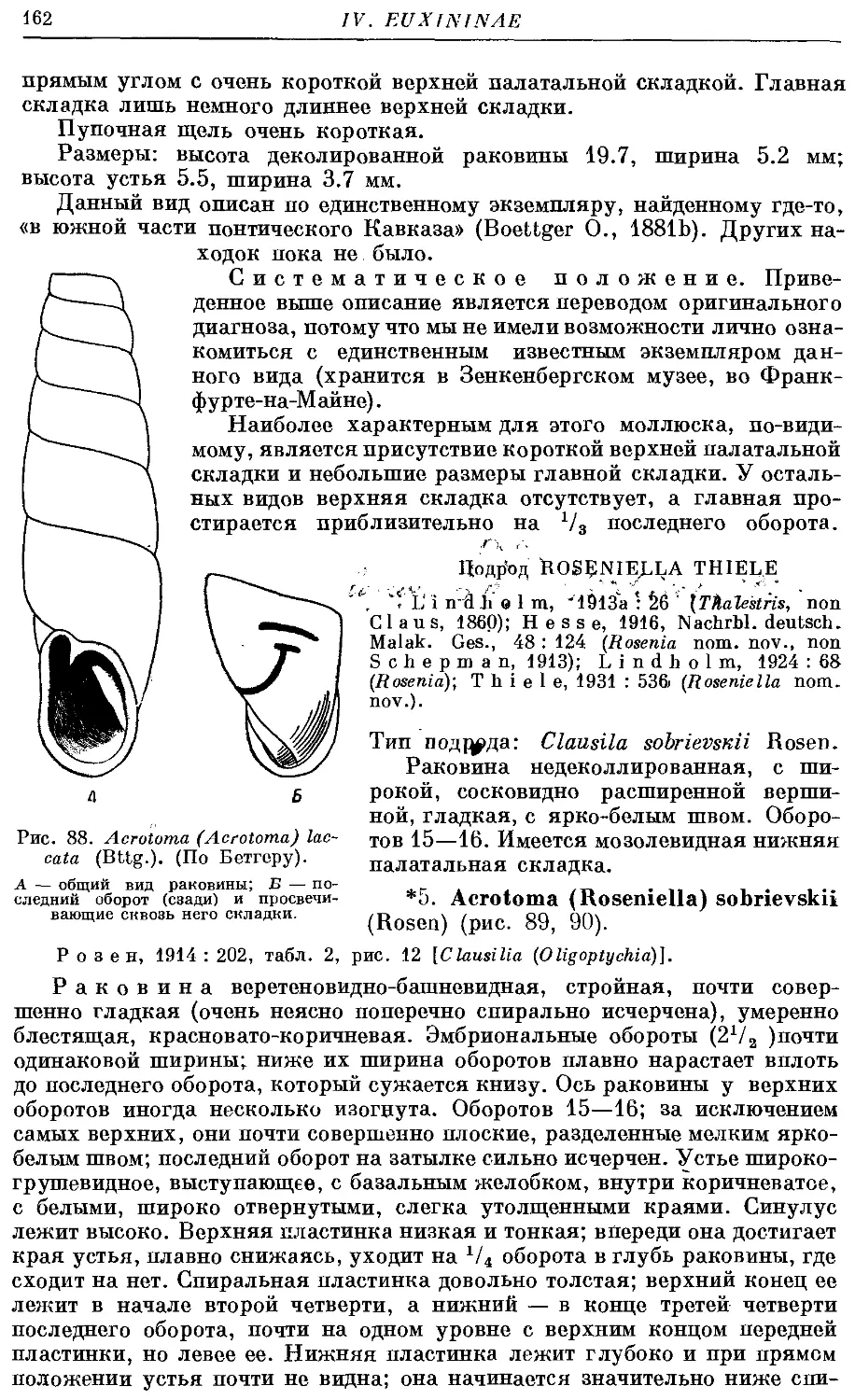
4. A. (A.) laccata (Bttg.) . 161
2. Подрод Roseniella Thiele
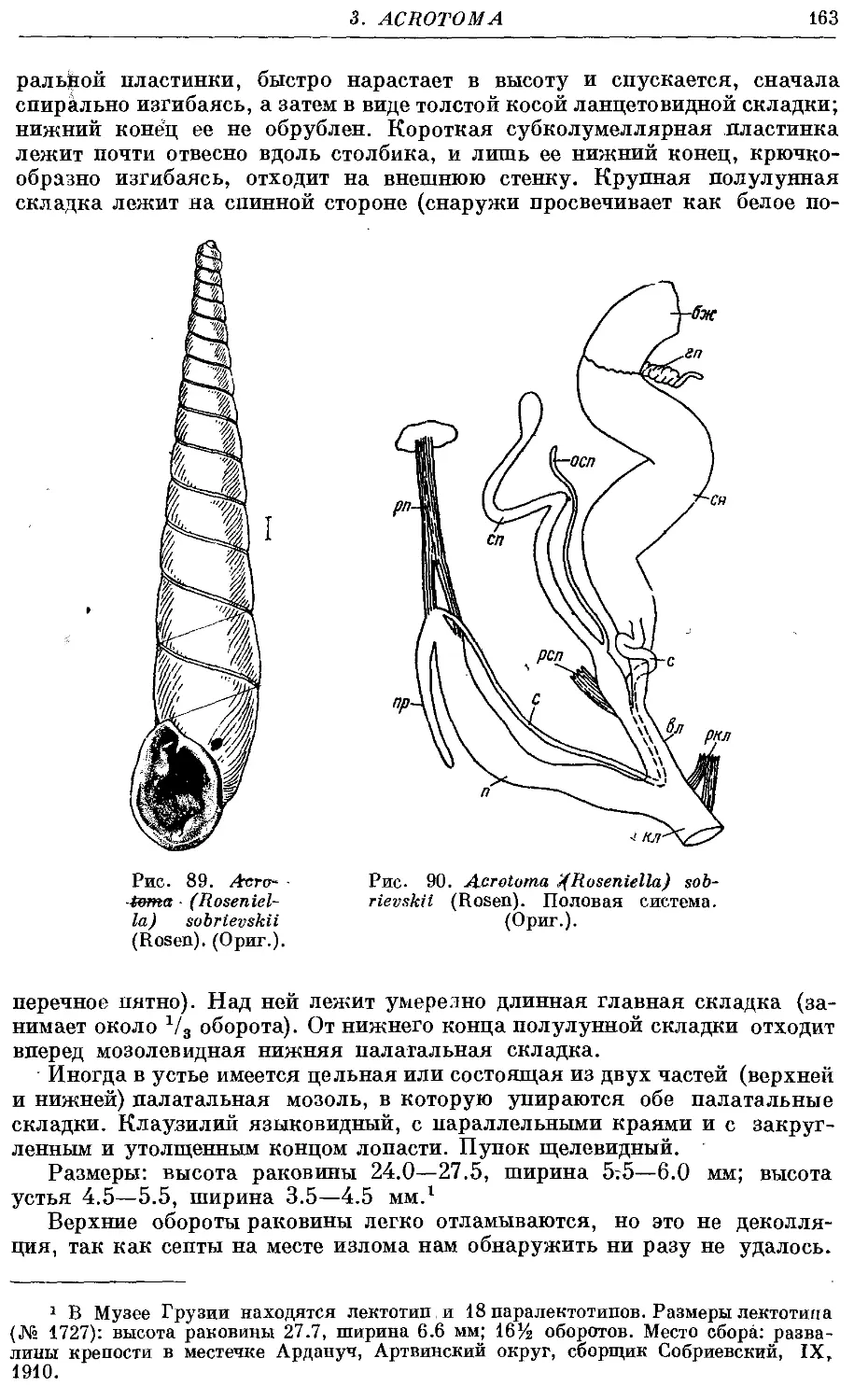
*5. A. (R.) sobrievskii (Rosen) 162
4. Род Euxina Bttg.
1. Подрод Euxina s. str.
*1. Ей. (Ей.) hetaera (Pfr.) 166
*2. Ей. (Eu.) circumdata (Pfr.) 167
*3. Ей. (Eu.) strumosa (Pfr.) 168
*4. Ей. (Ей.) difficilis (Ret.) 169
5a. Ей. (Ей.) persica persica (Bttg.) 170
*5б. Ей. (Ей.) persica paulhessei (Lindh.) 173
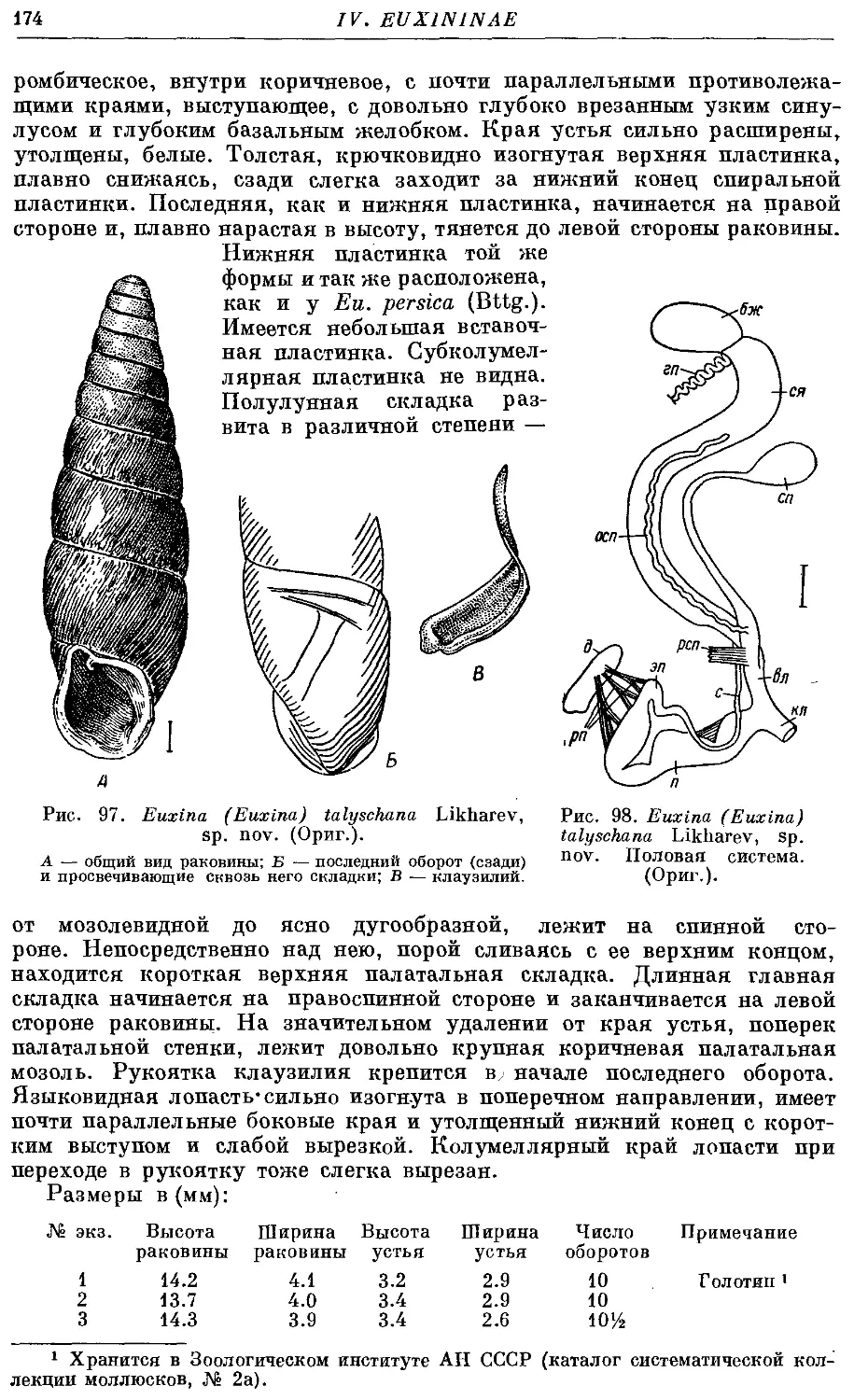
6. Ей. (Eu.) talyschana Likharev, sp. nov 173
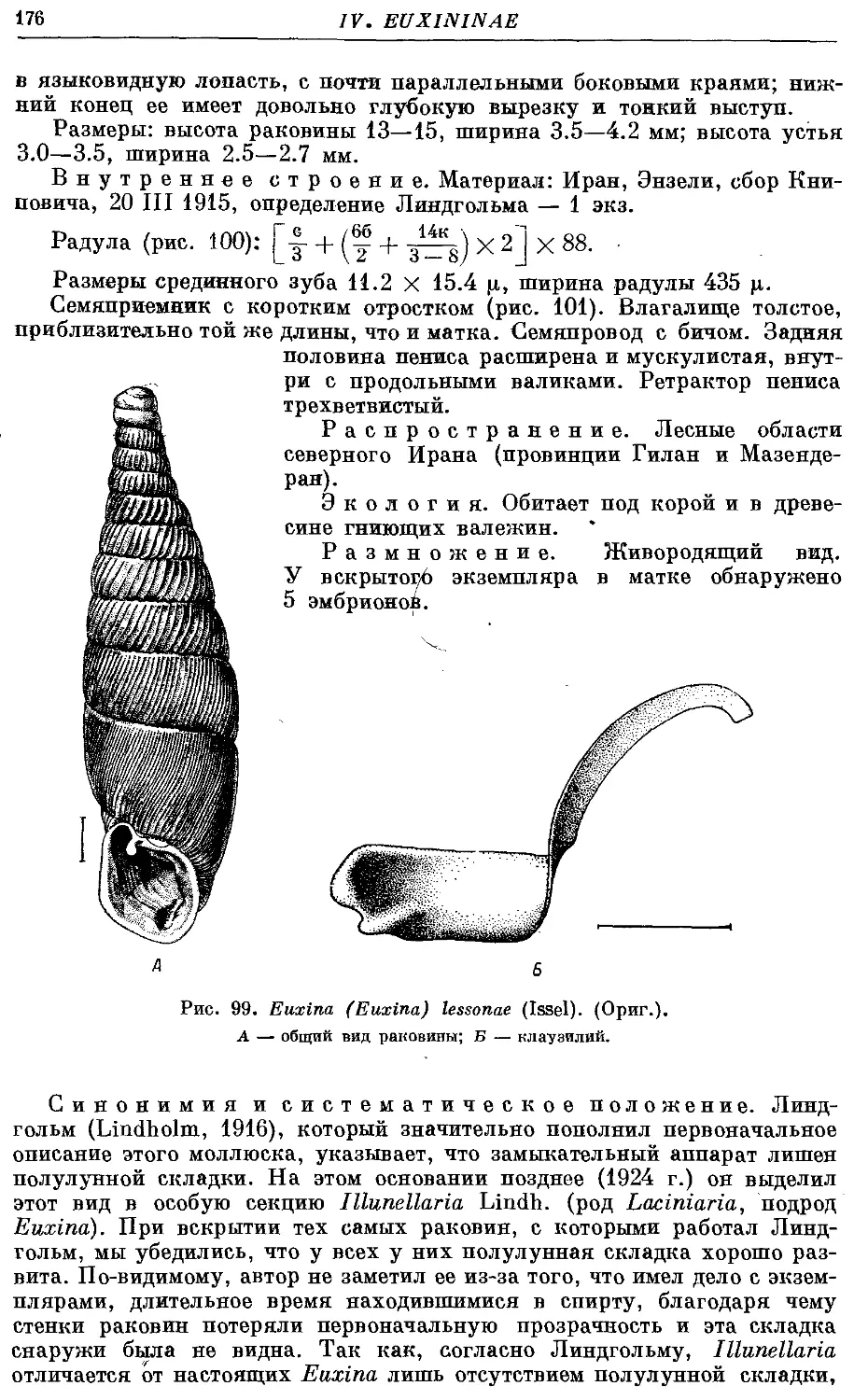
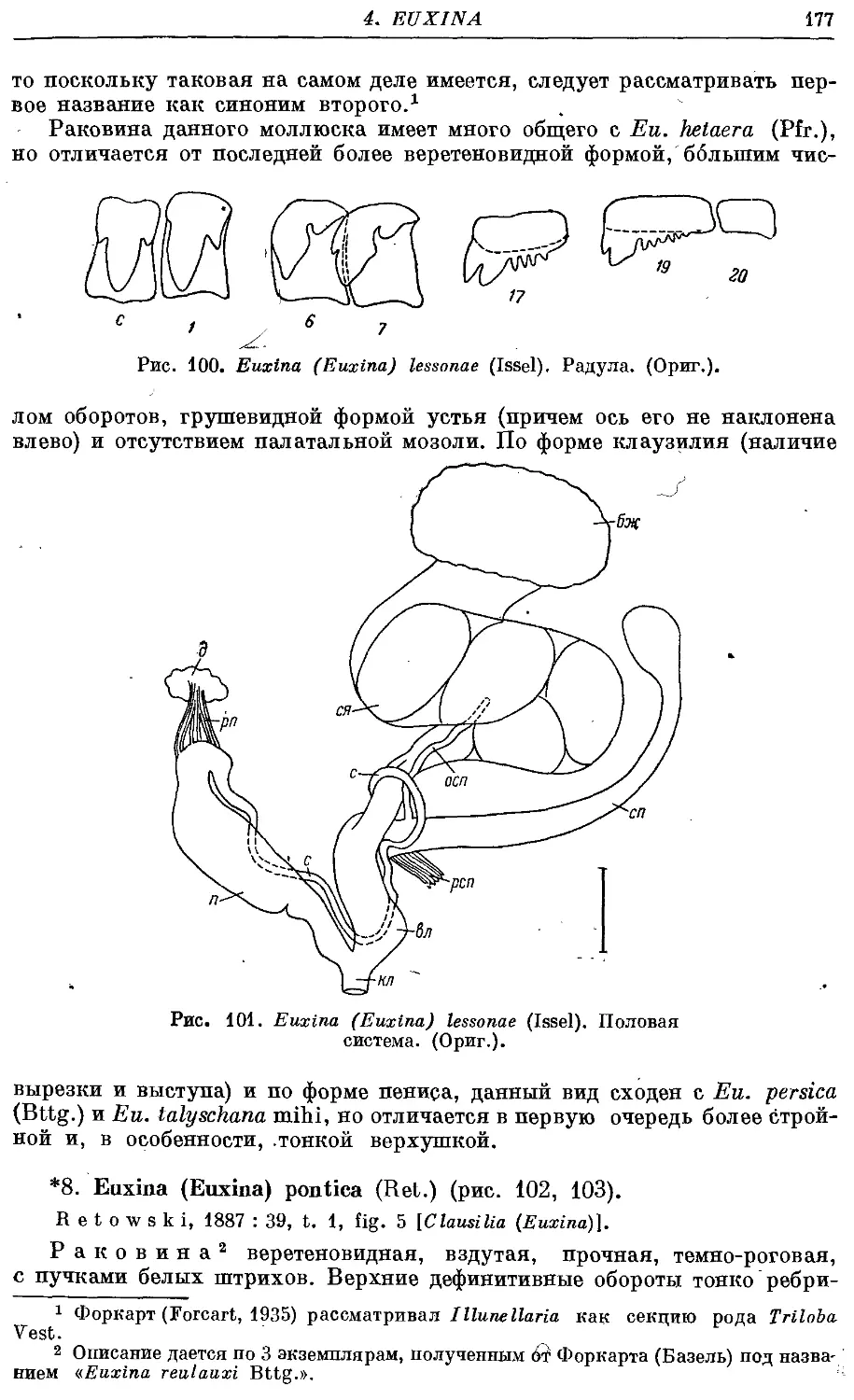
*7. Ей. (Ей.) lessonae (Issel) 175
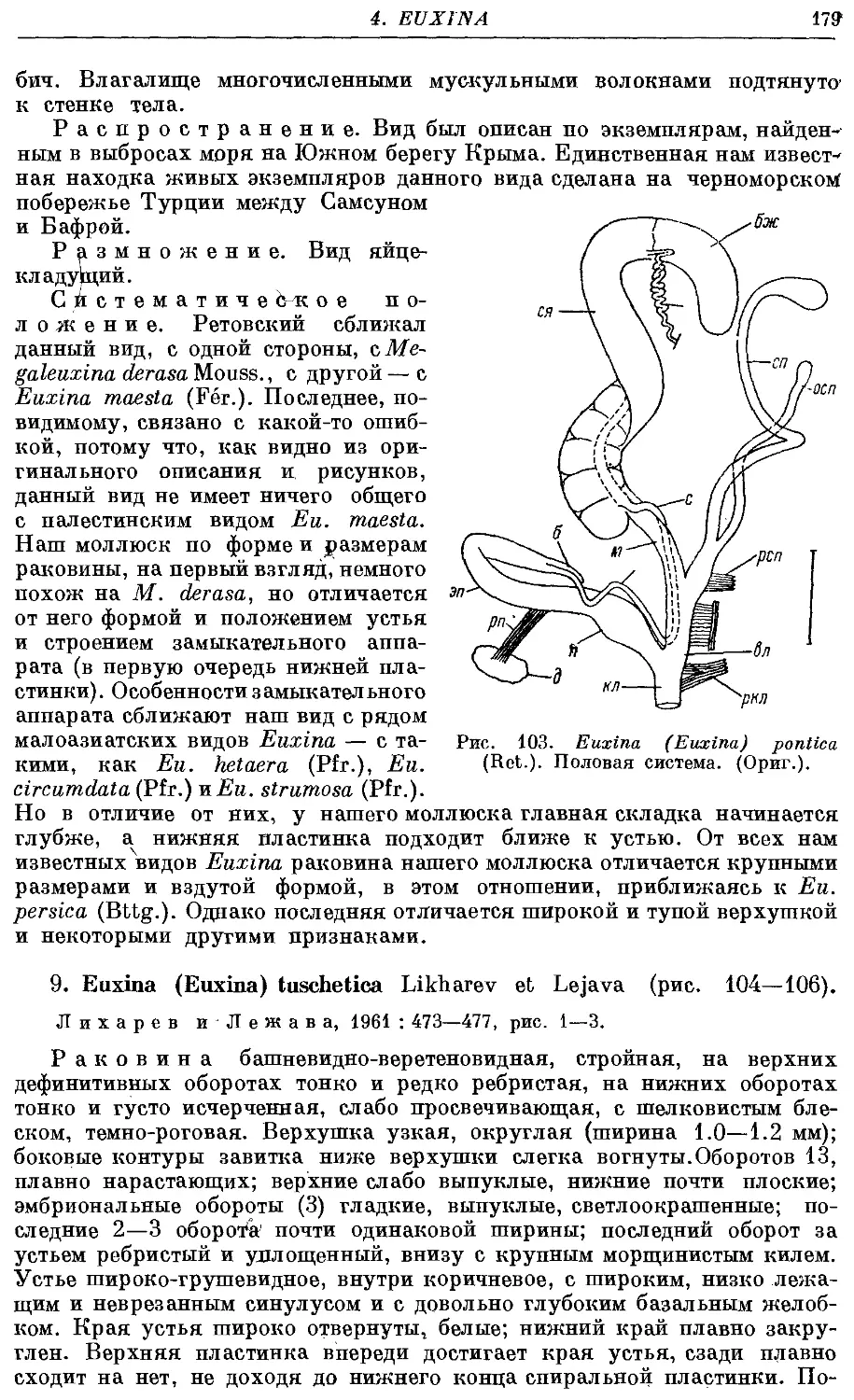
•8. Eu (Eu.) pontica1 {Ret.) 177
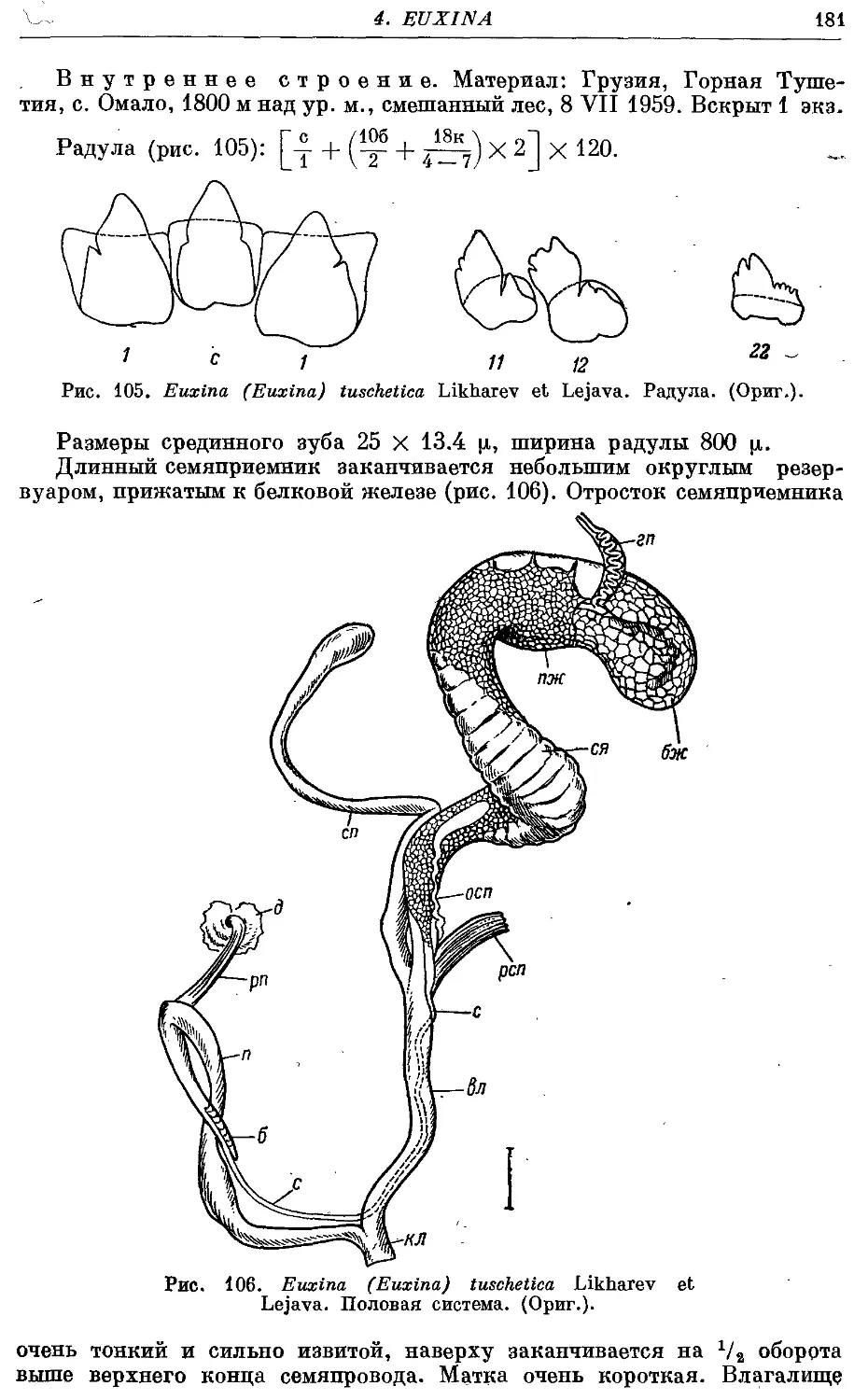
9. Eu. (Eu.) tuschetica Likharev et Lejava 179
2. Подрод Caucasica Bttg.
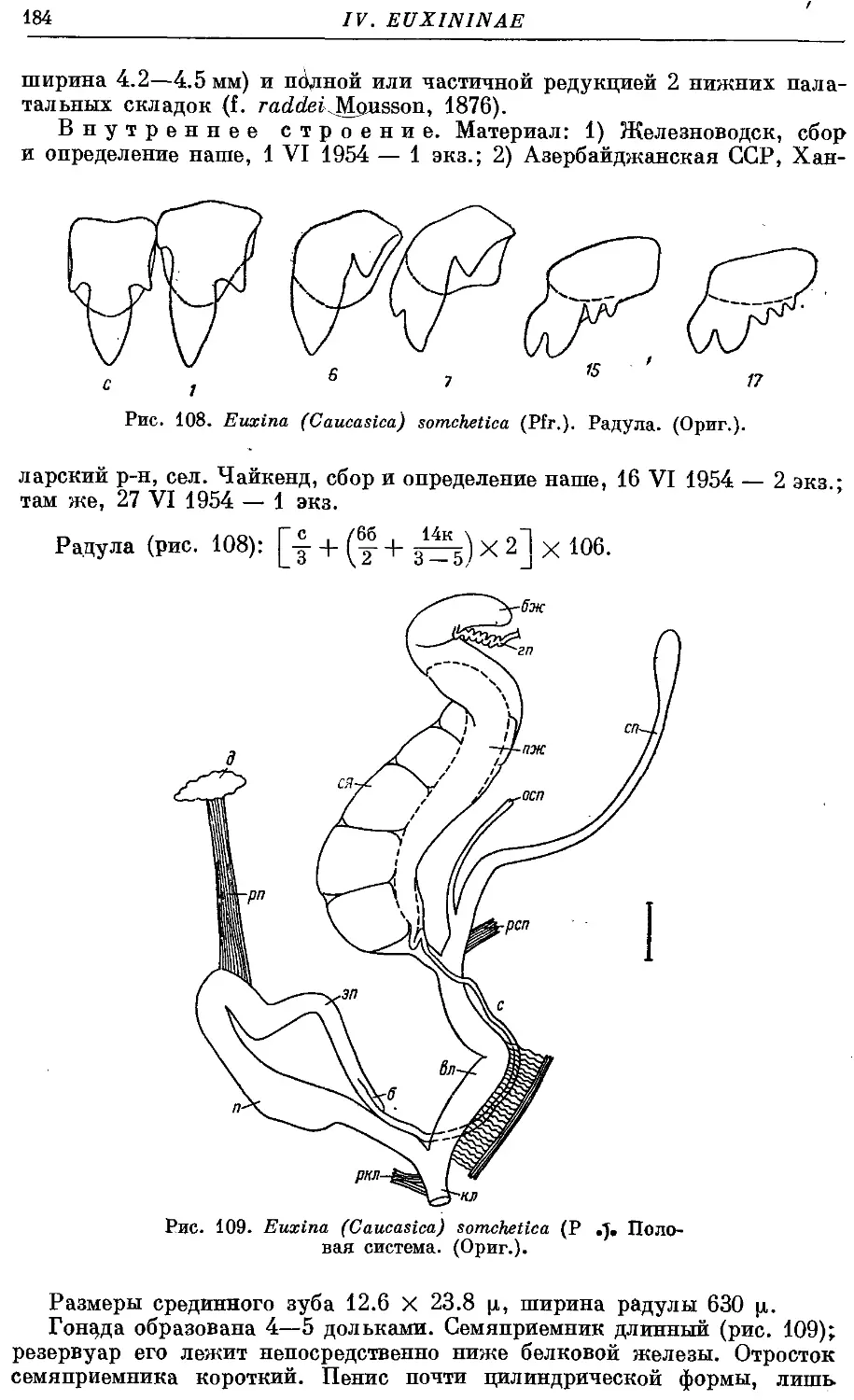
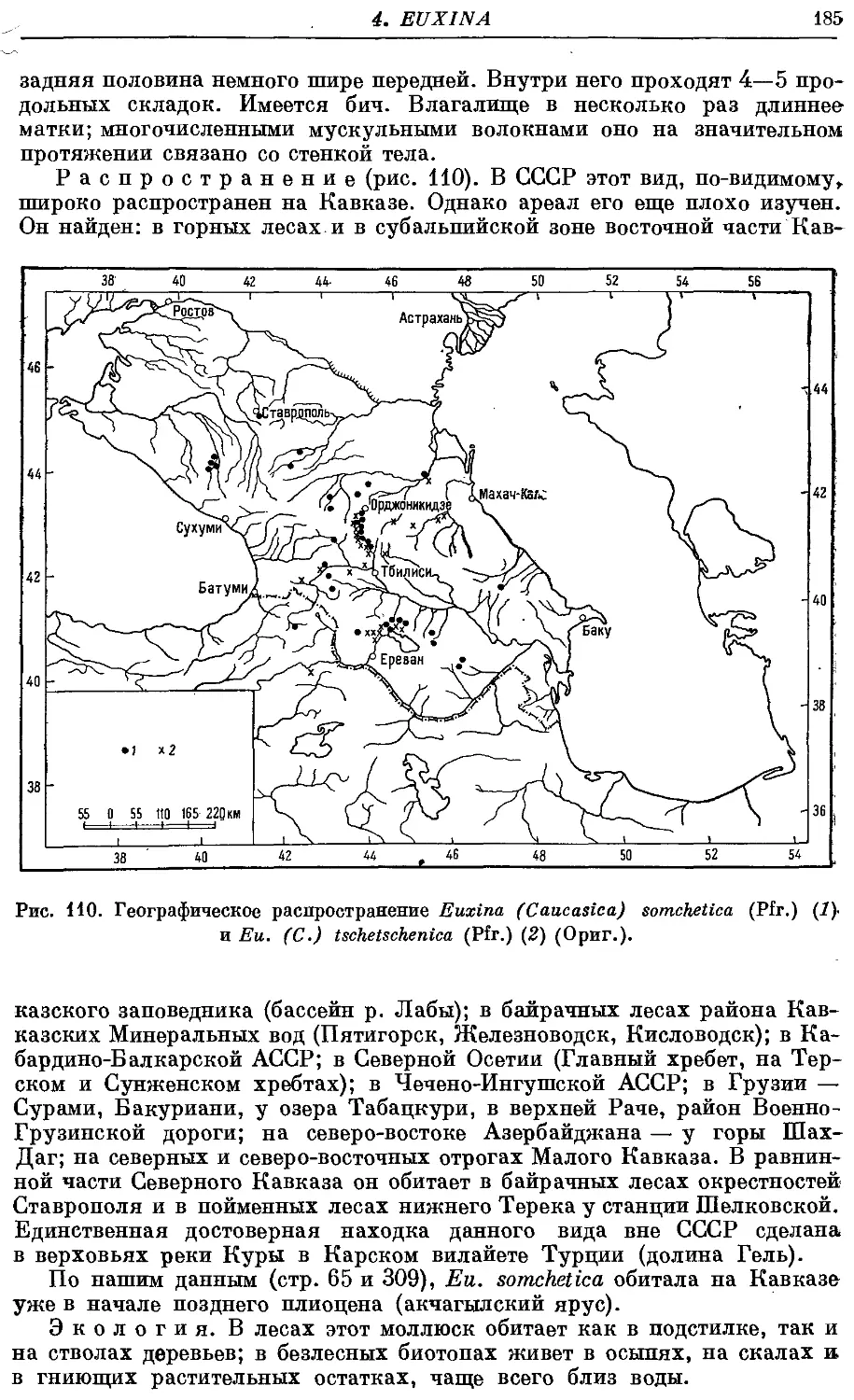
10. Eu. (C.) somchetica (Pfr.) , 182
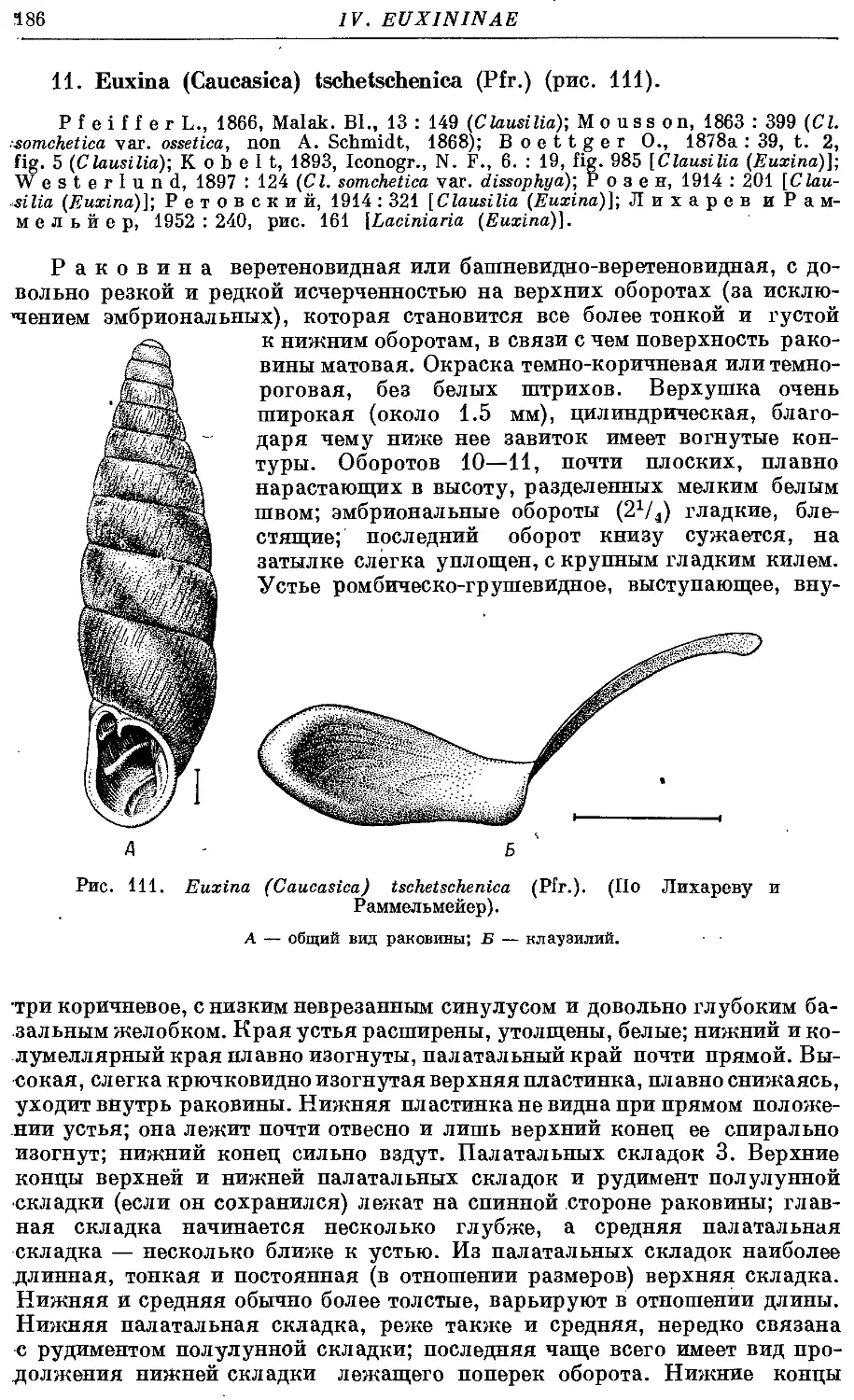
11. Ей. (С.) tschetschenica (Pfr.) 186
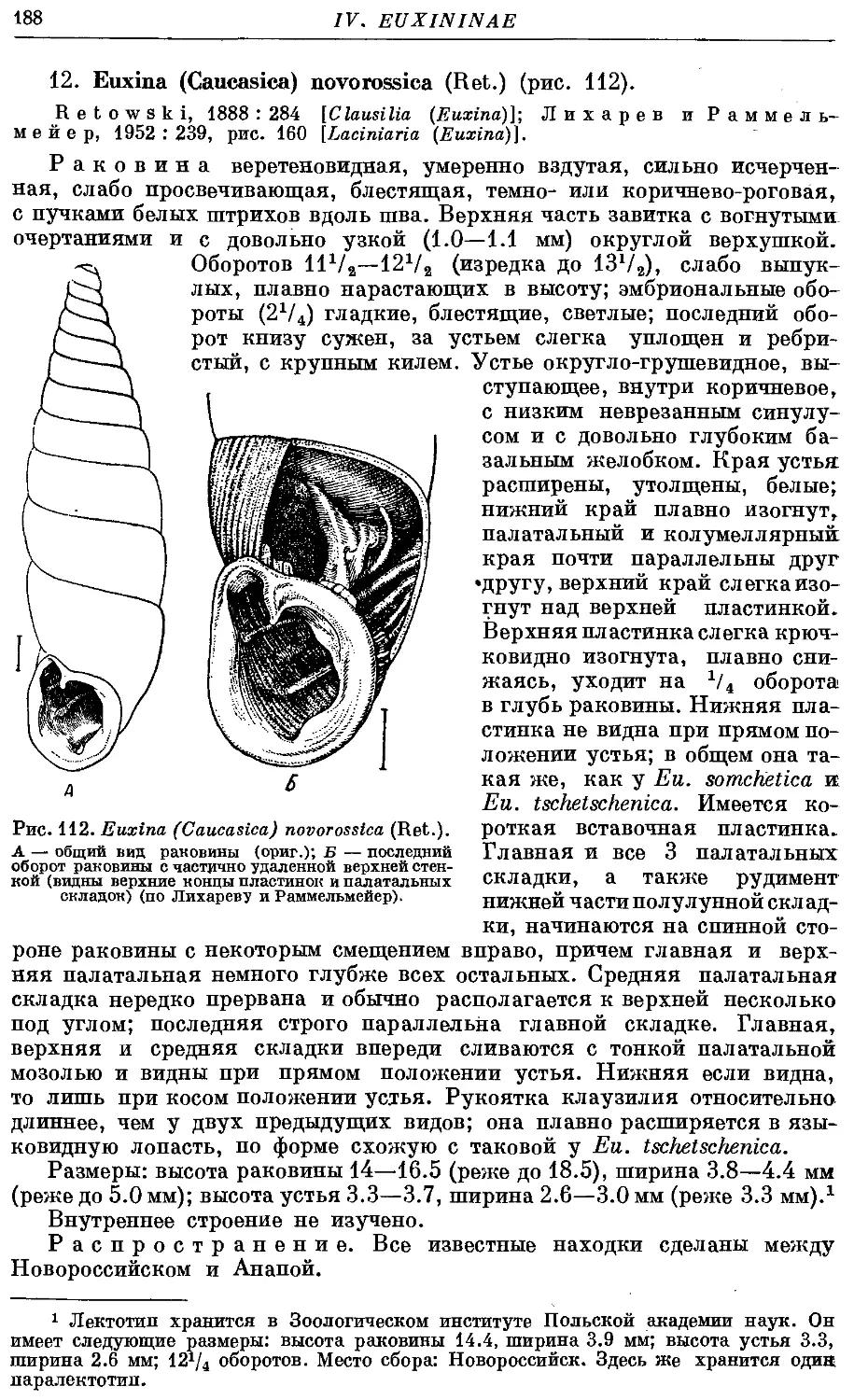
12. Ей. (C.) novorossica (Ret.) 188
3. Подрод E I i a H. et A. Adams
*13. Ей. (Е.) corpulenta (Pfr.) 189
*14. Ей. (Е.) maesta (Fer.) . 190
4. Подрод Acroeuxina Bttg.
*15. Eu. (A.) huebneri (Pfr.) 192
5. Род Megaleuxina Bttg.
1. M. derasa (Mouss.) 194
2. M. reulauxi (Bttg.) 197
3. M. lindholmi (Lindh.) 198
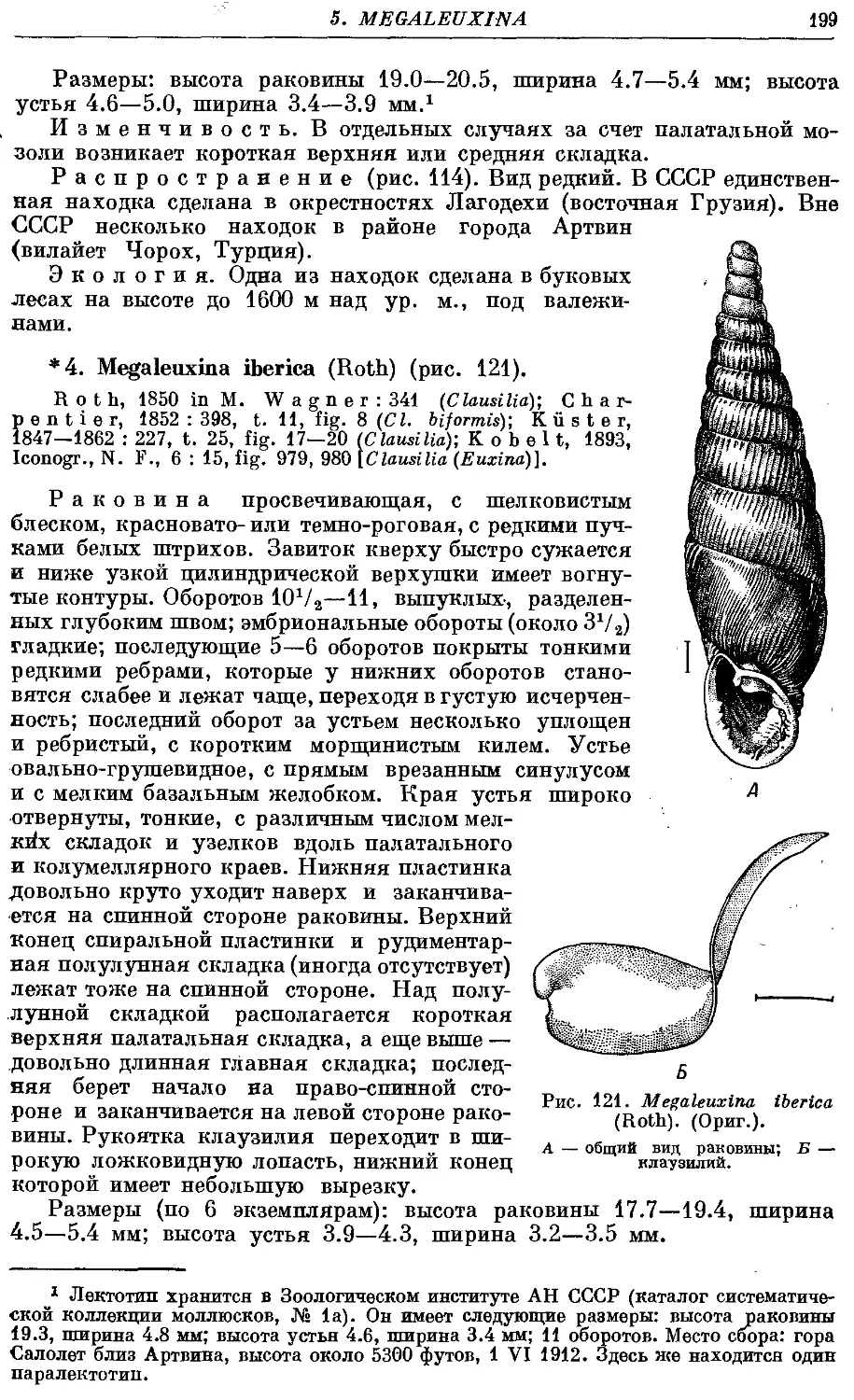
*4. M. iberica (Roth) 199
*5. M. comnena (Ret.) 200
*6. M. borisi (Hesse) 201
6. Род Euxinastra Bttg.
Eu. hamata (Bttg.) 203
7. Род Galeata Btlg.
*1. G. galeata (Rssm.) ' . . 206
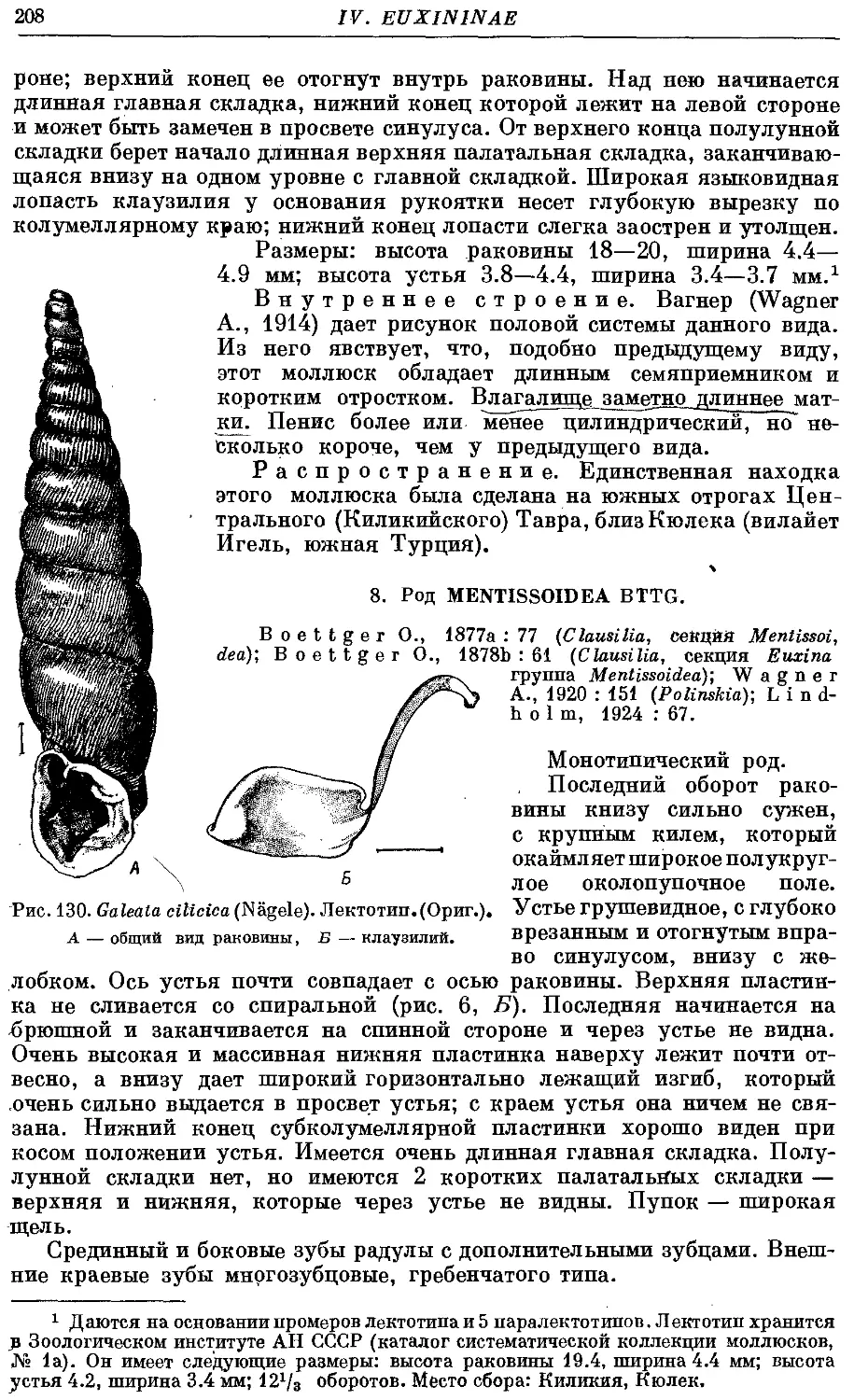
*2. G. cilicica (Nagele) 207
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
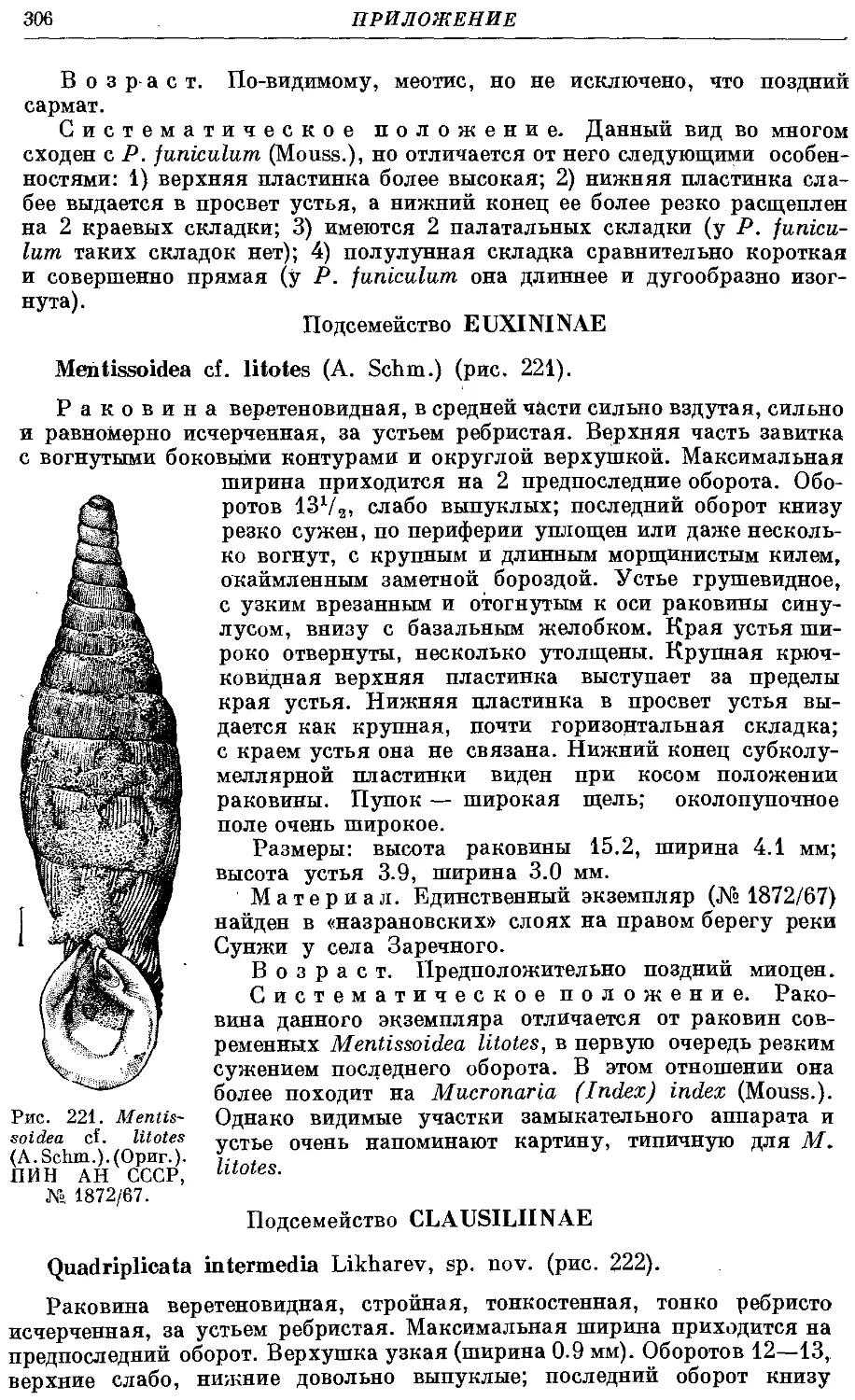
8. Род Mentissoidea Bttg. Стр.
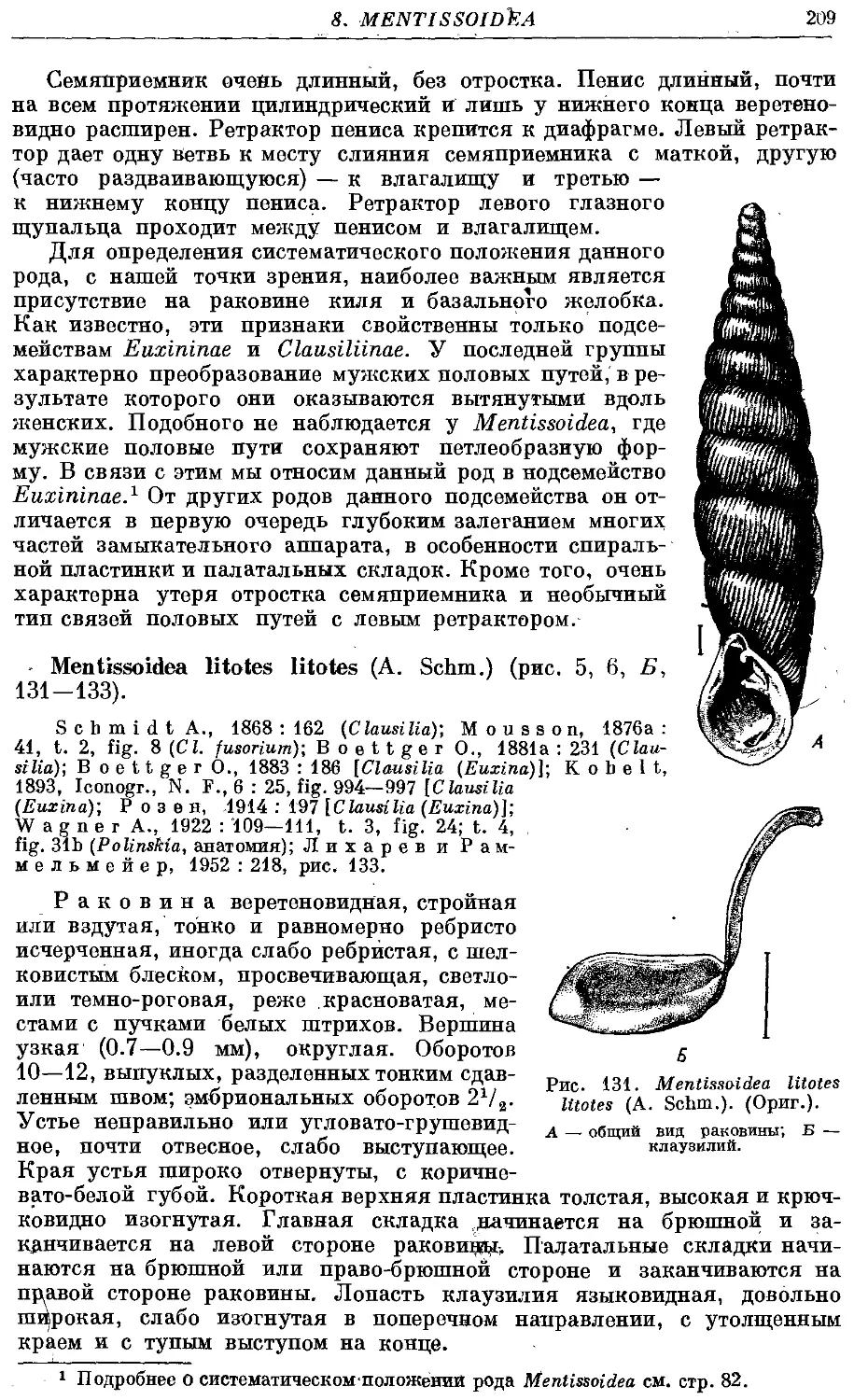
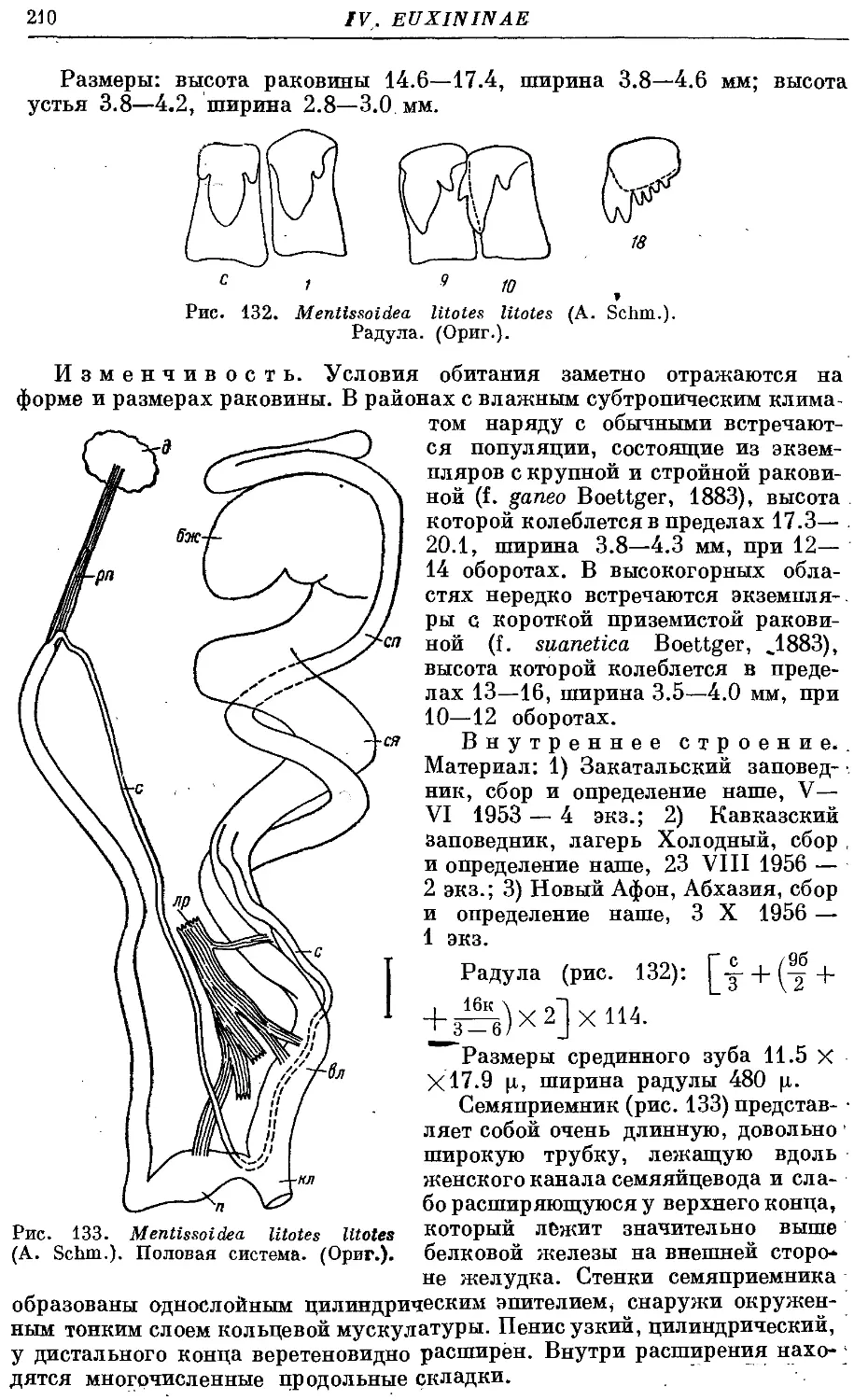
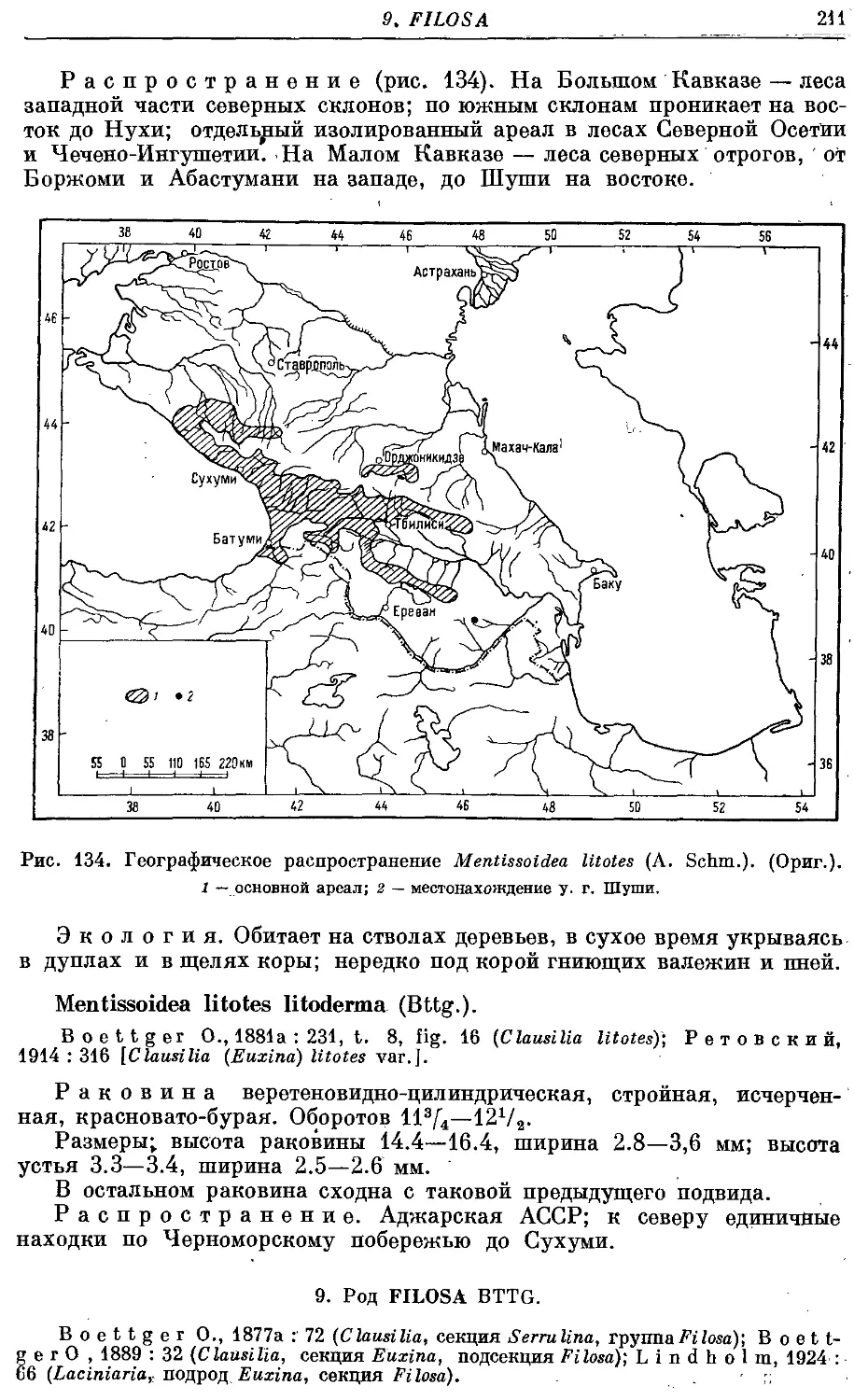
М. litotes litotes (A. Schm.) 209
М. litotes litoderma (Bttg.) 211
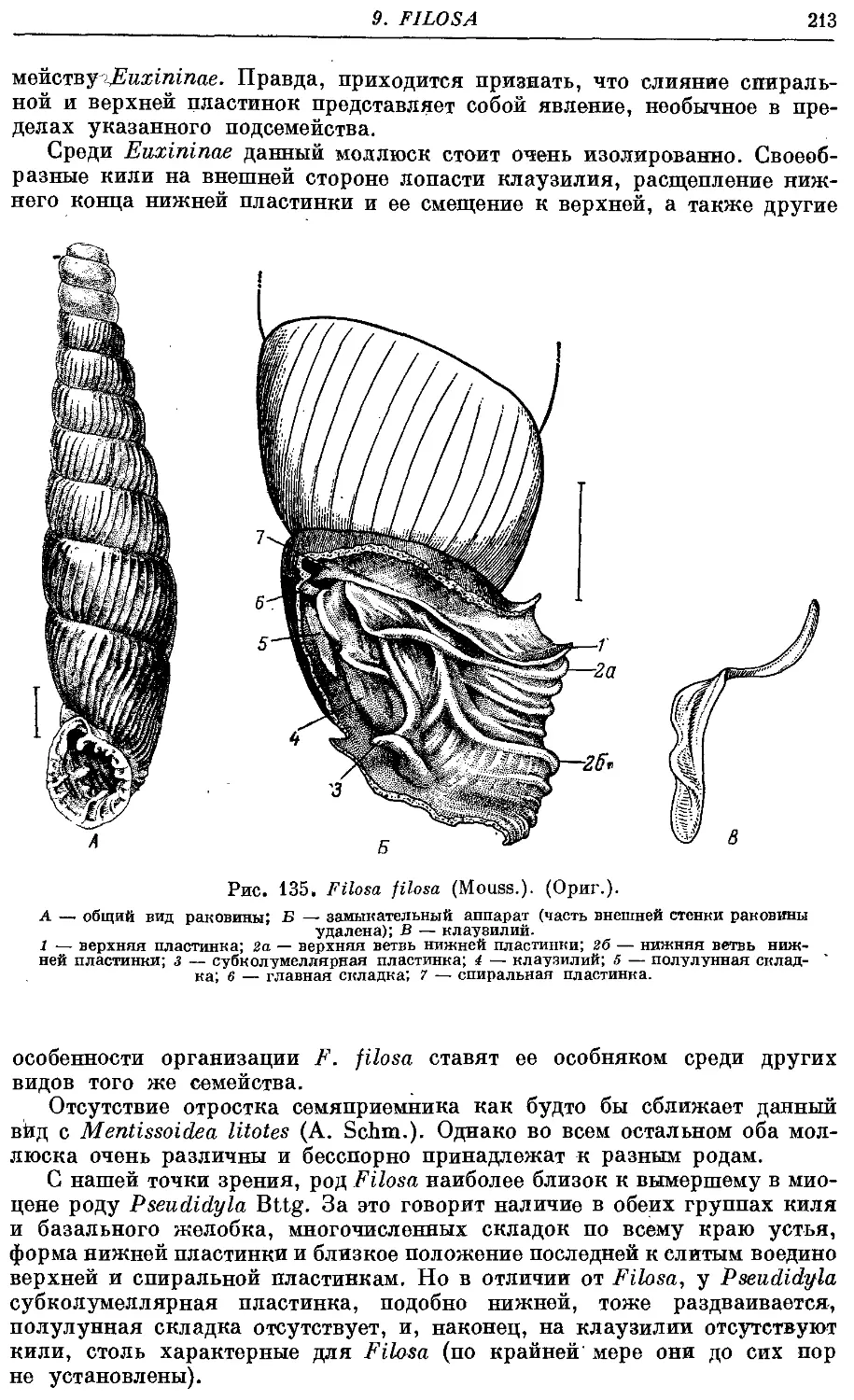
9. Род Filosa Bttg.
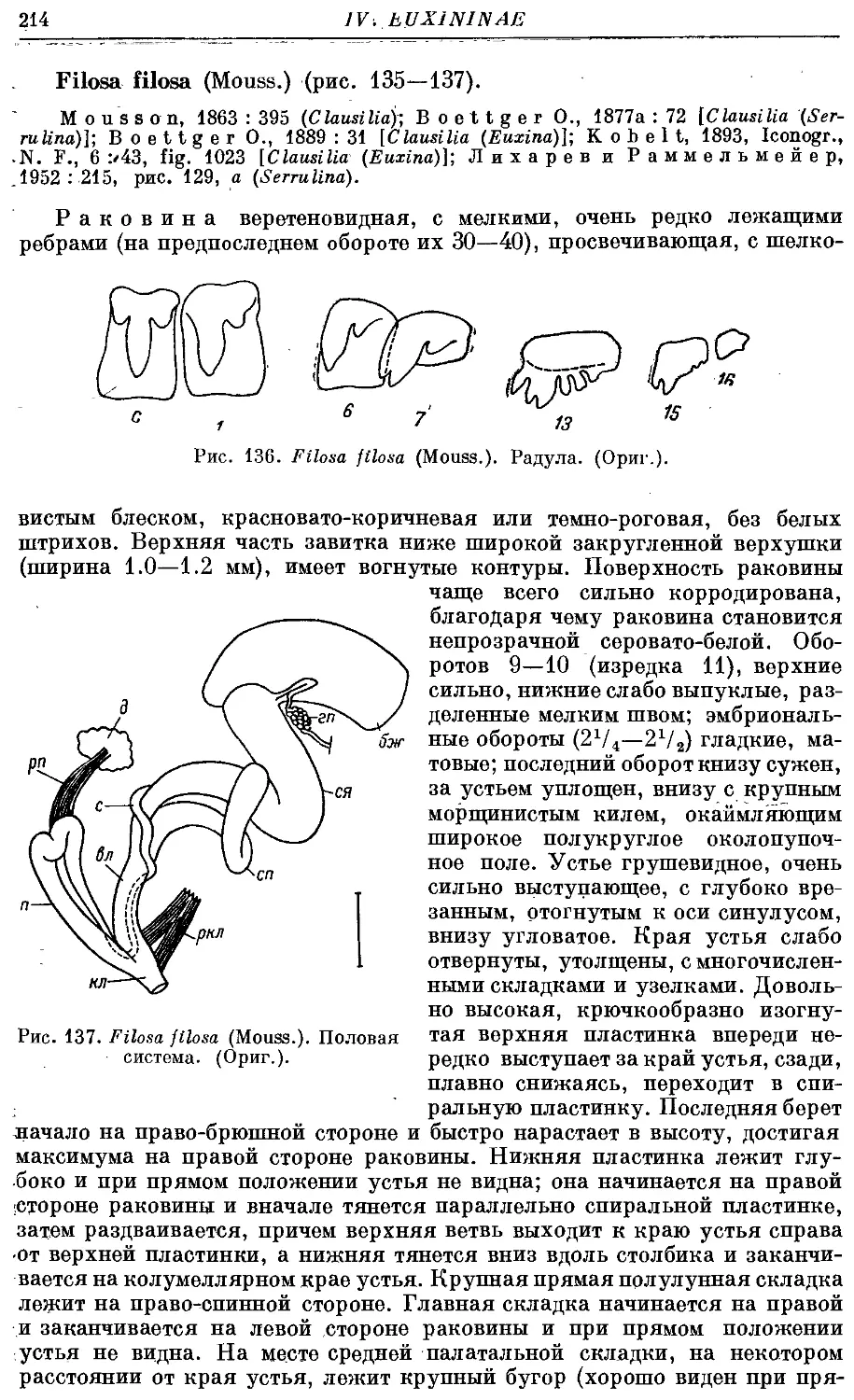
F. filosa (Mouss.) 214
V. Подсемейство Clausiliinae
1. Род Ruthenica Lindh.
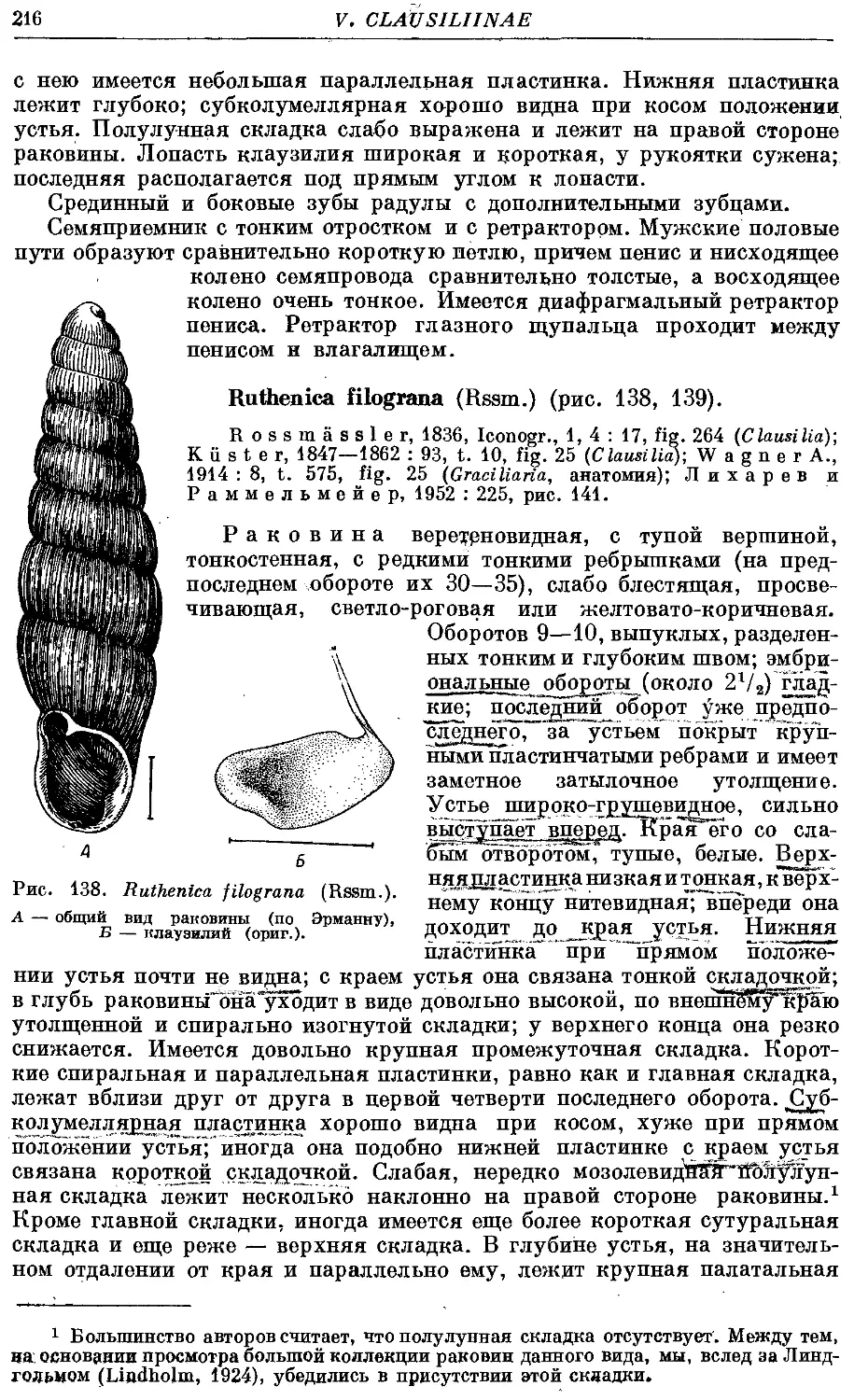
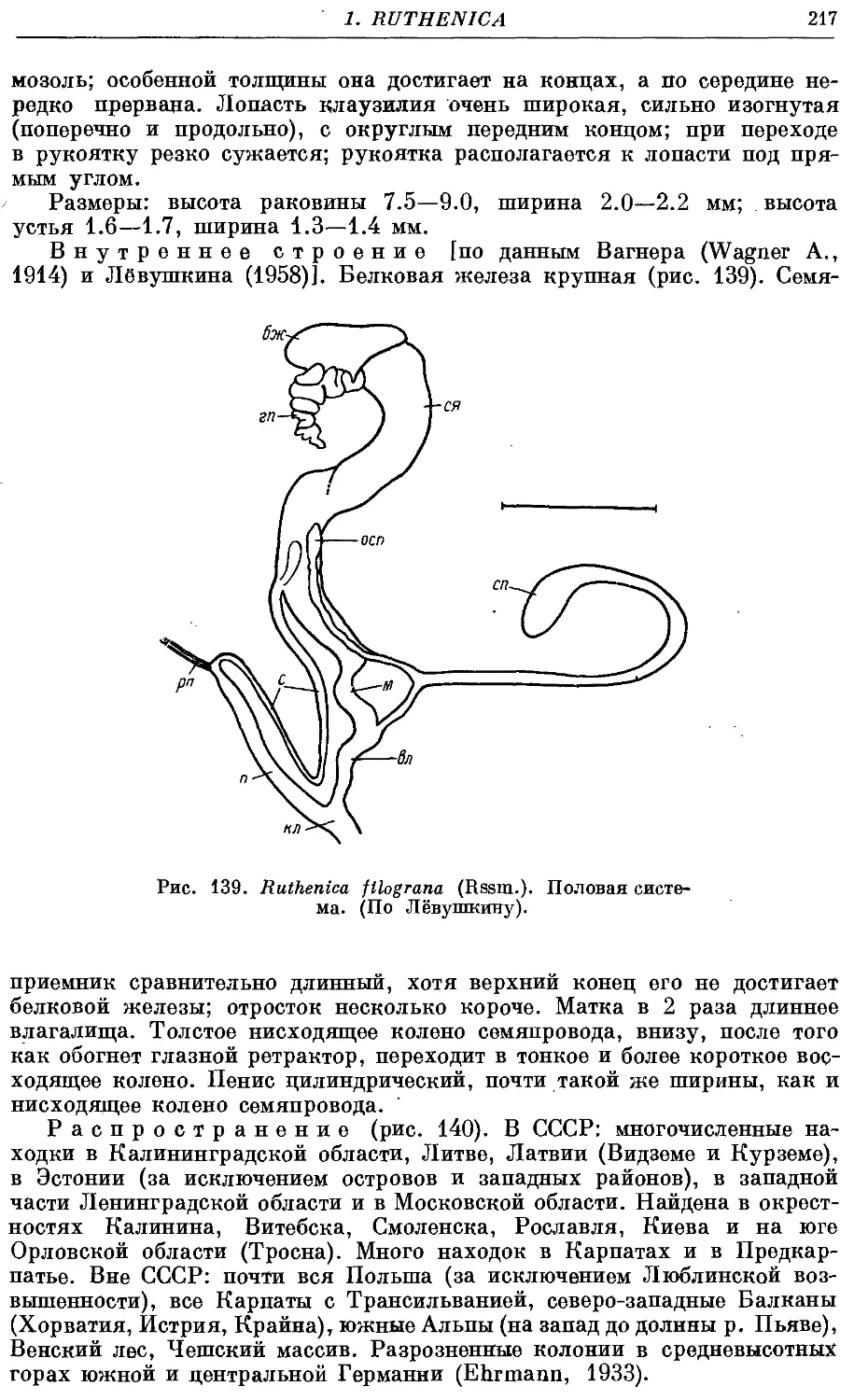
R. filograna (Rssm.) 216
2. Род Clausilia Drap.
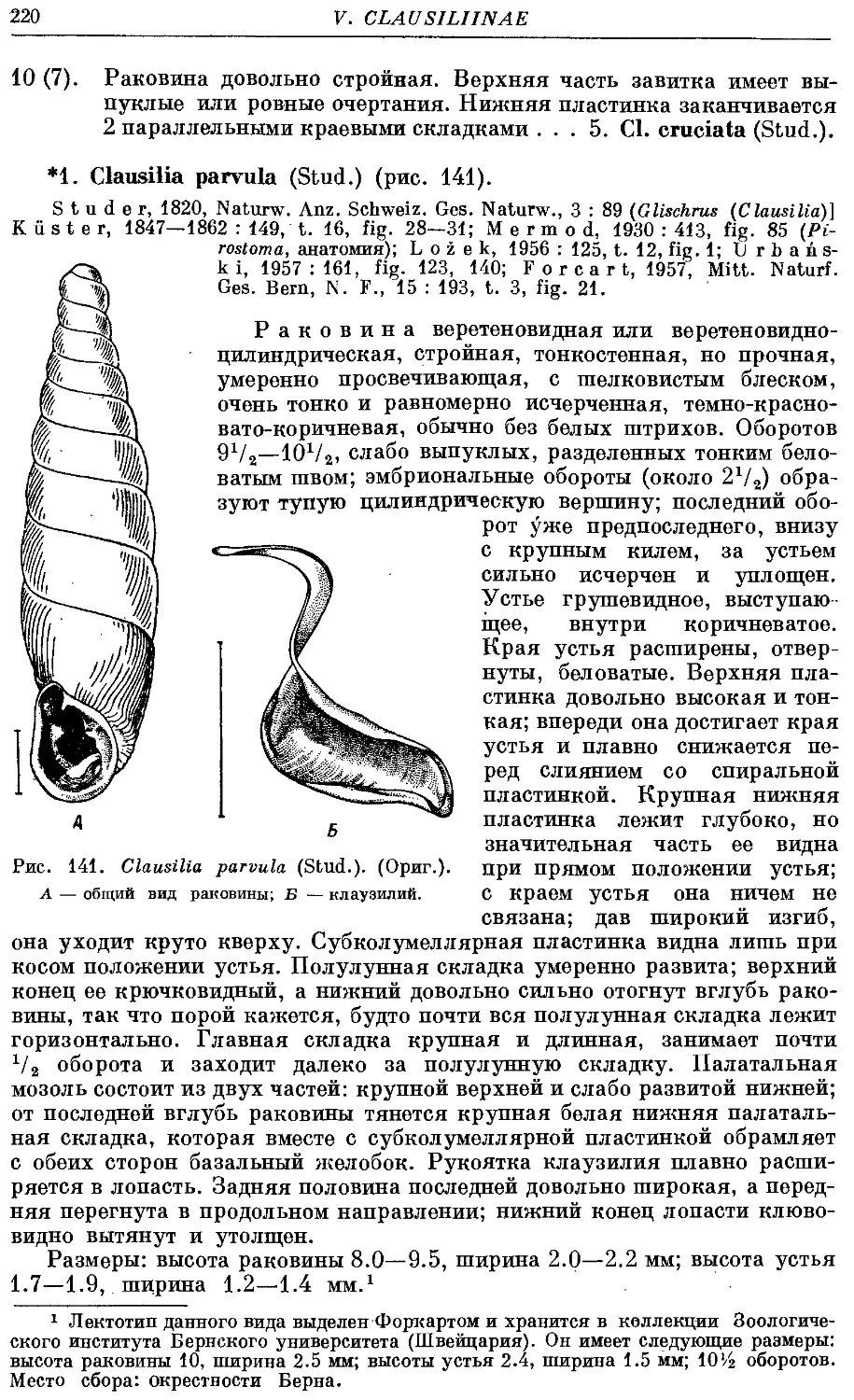
*1. Cl. parvula (Stud.) 220
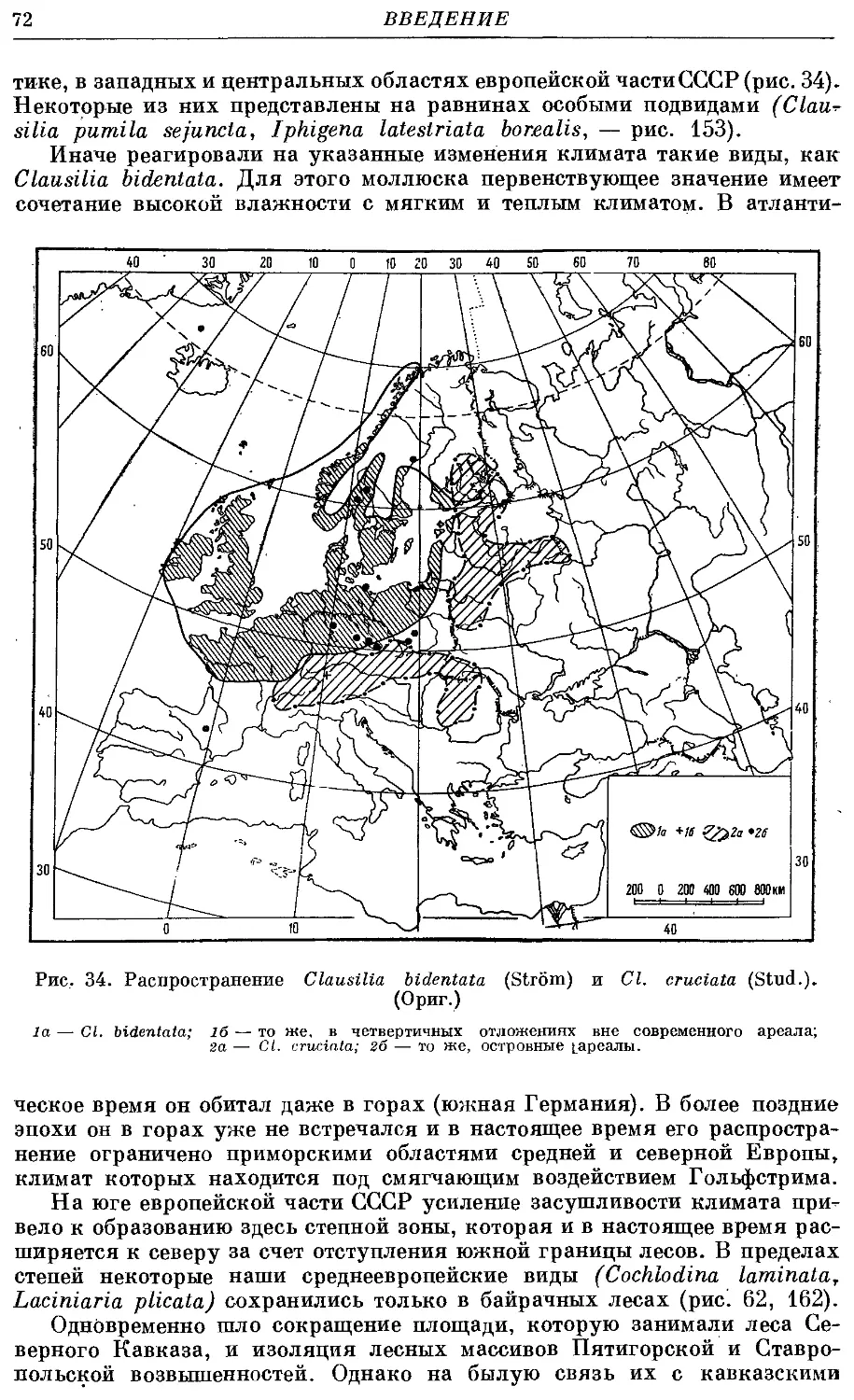
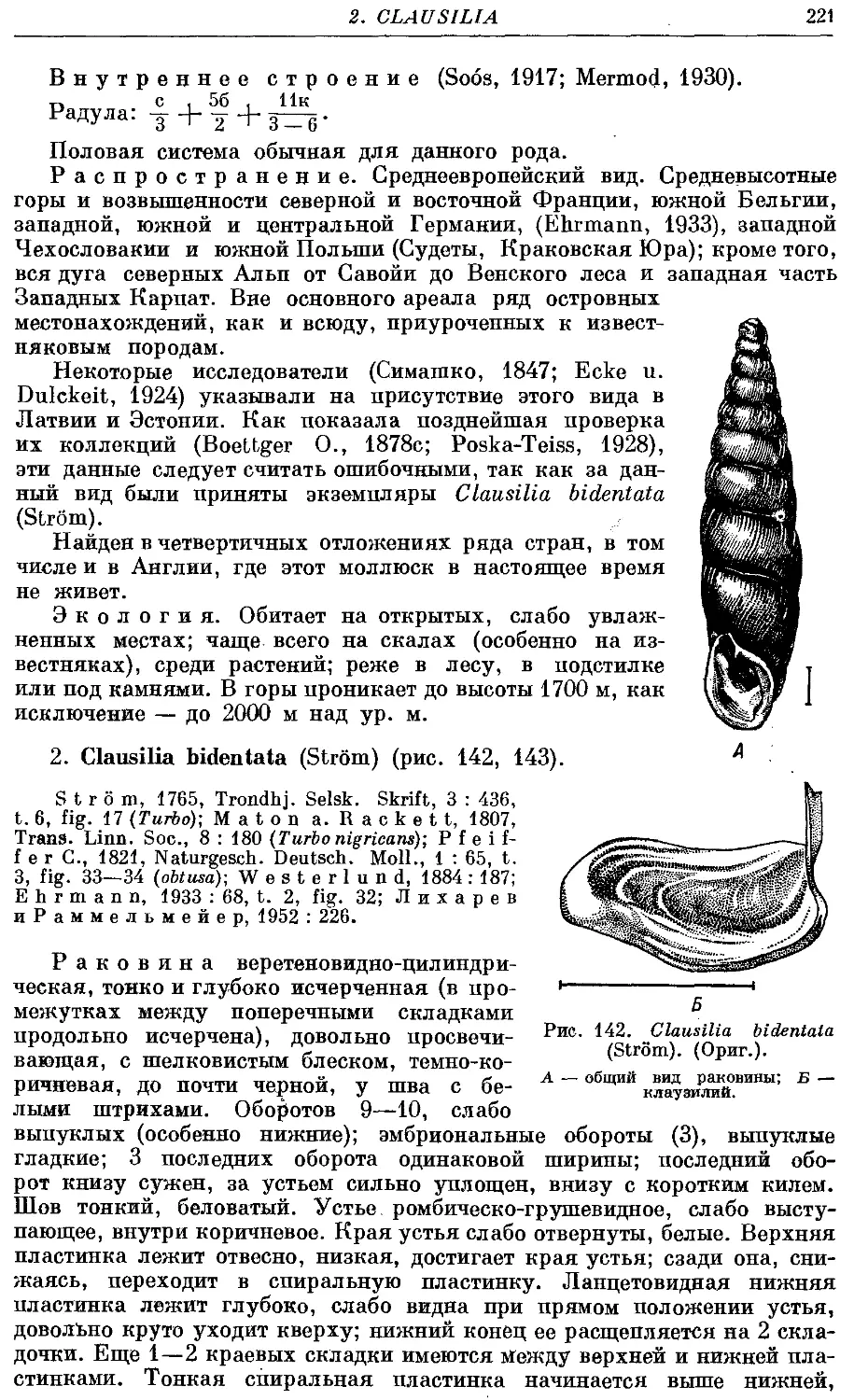
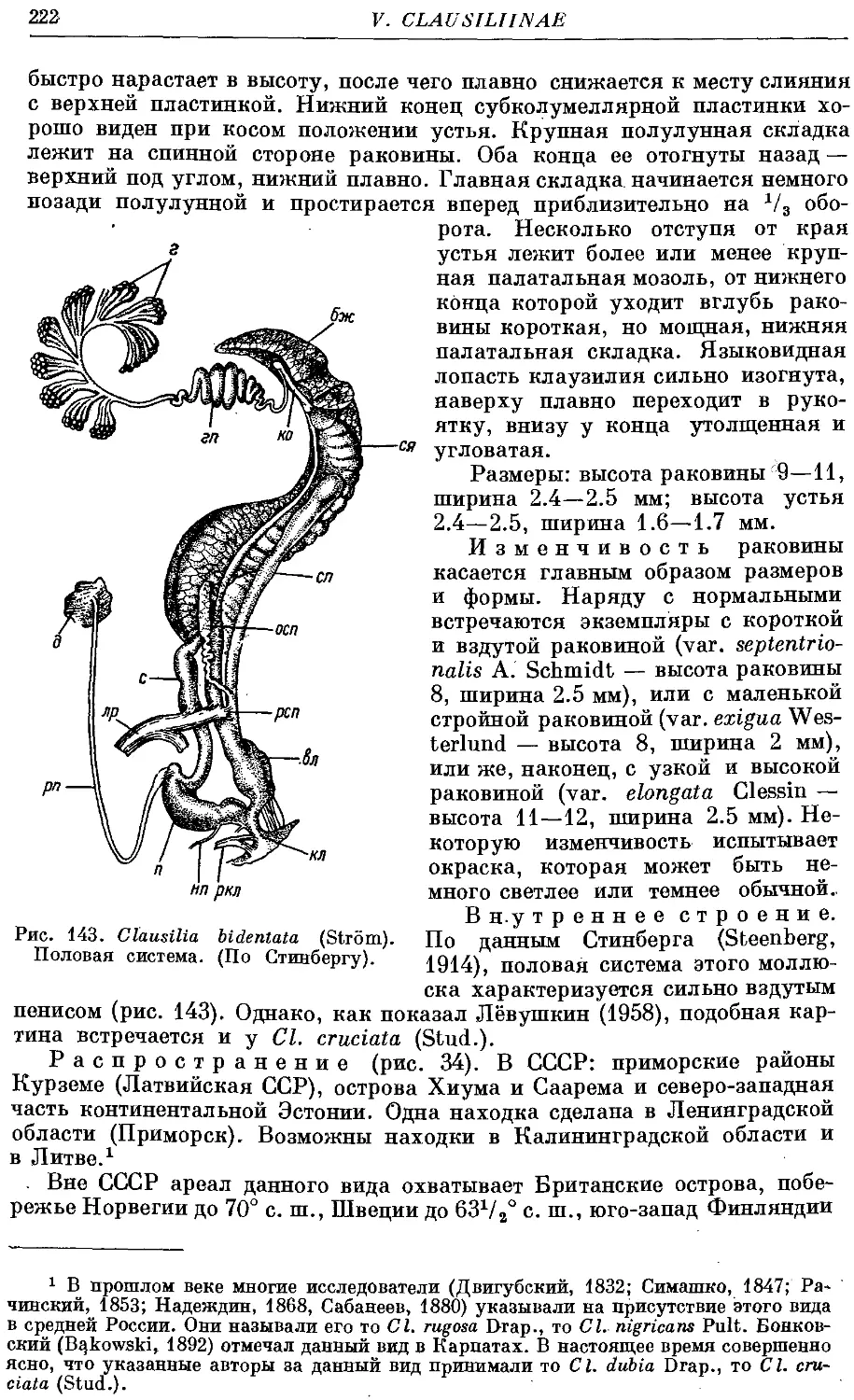
2. Cl. bidentata (Strom) 221
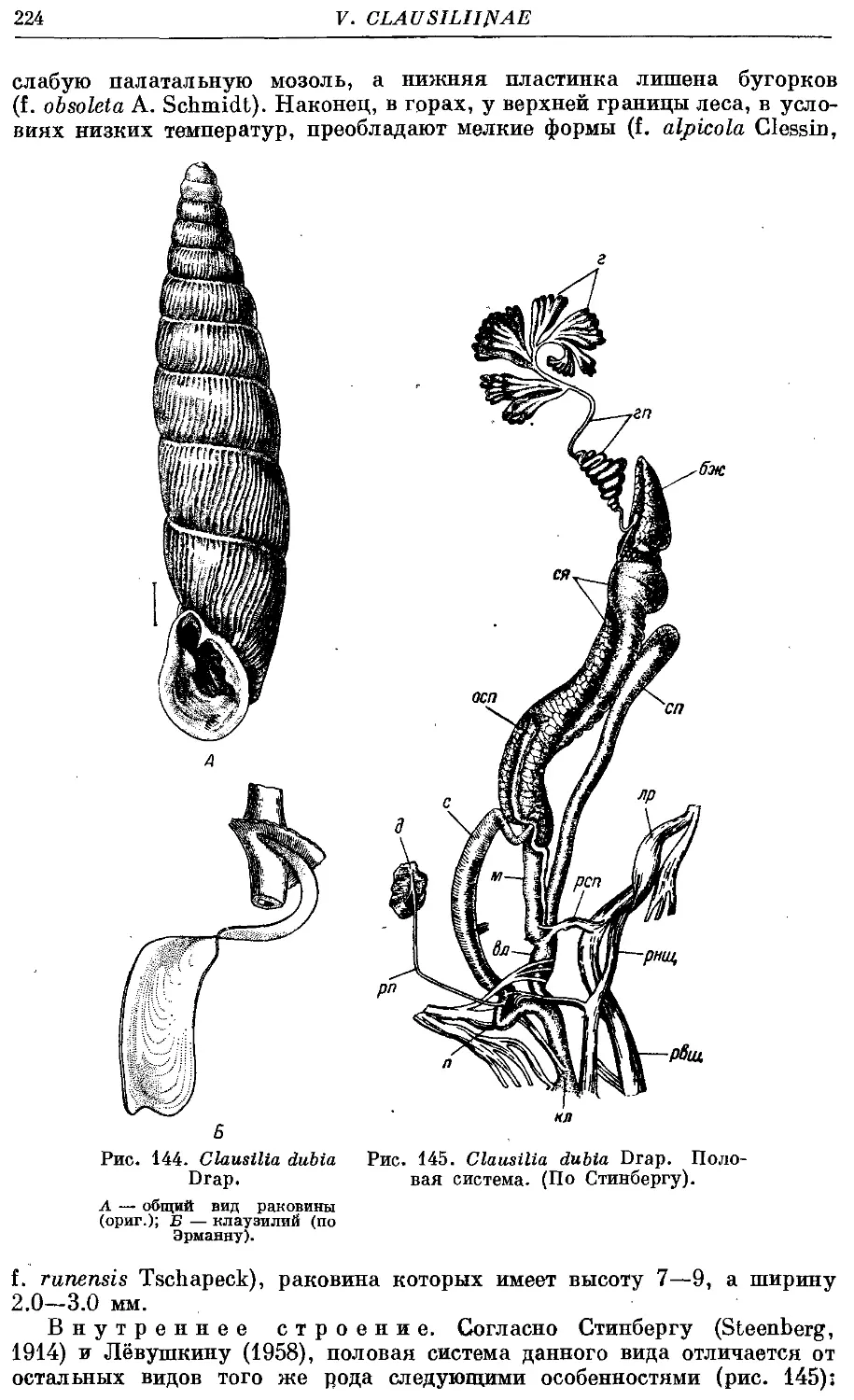
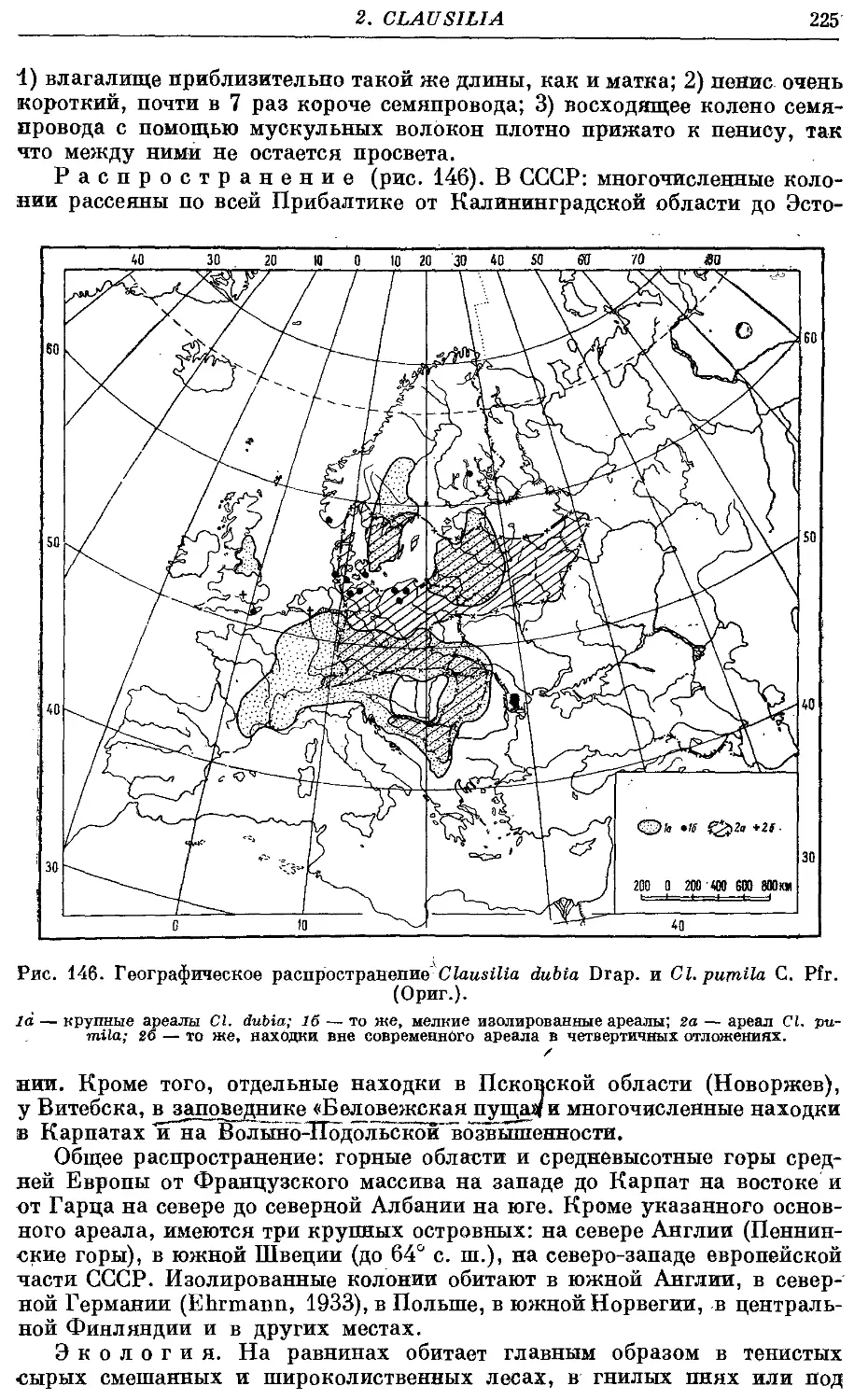
3. Cl. dubia Drap 223
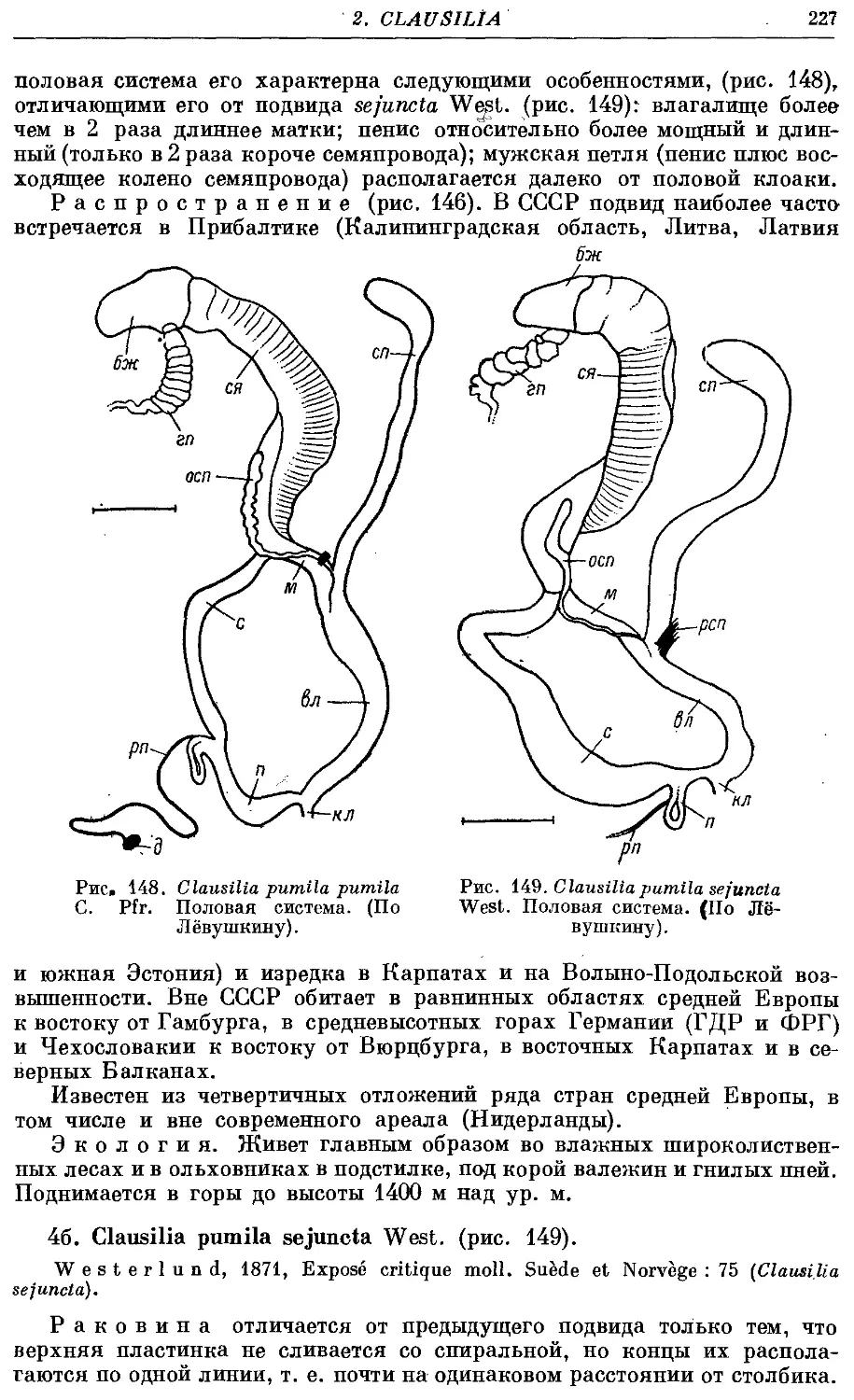
4a. Cl. pumila pumila C. Pfr 226
46. Cl. pumila sefuncta West 227
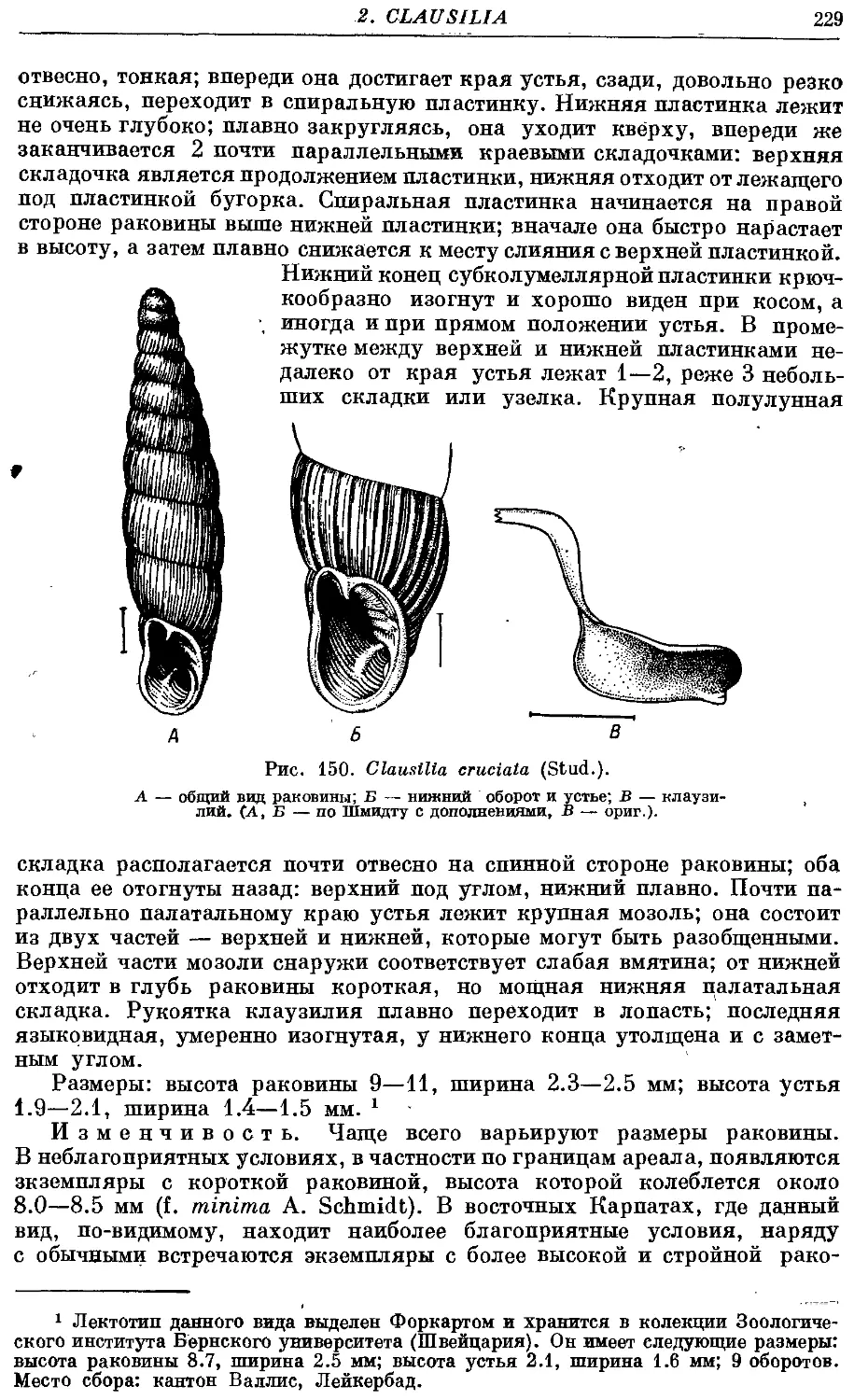
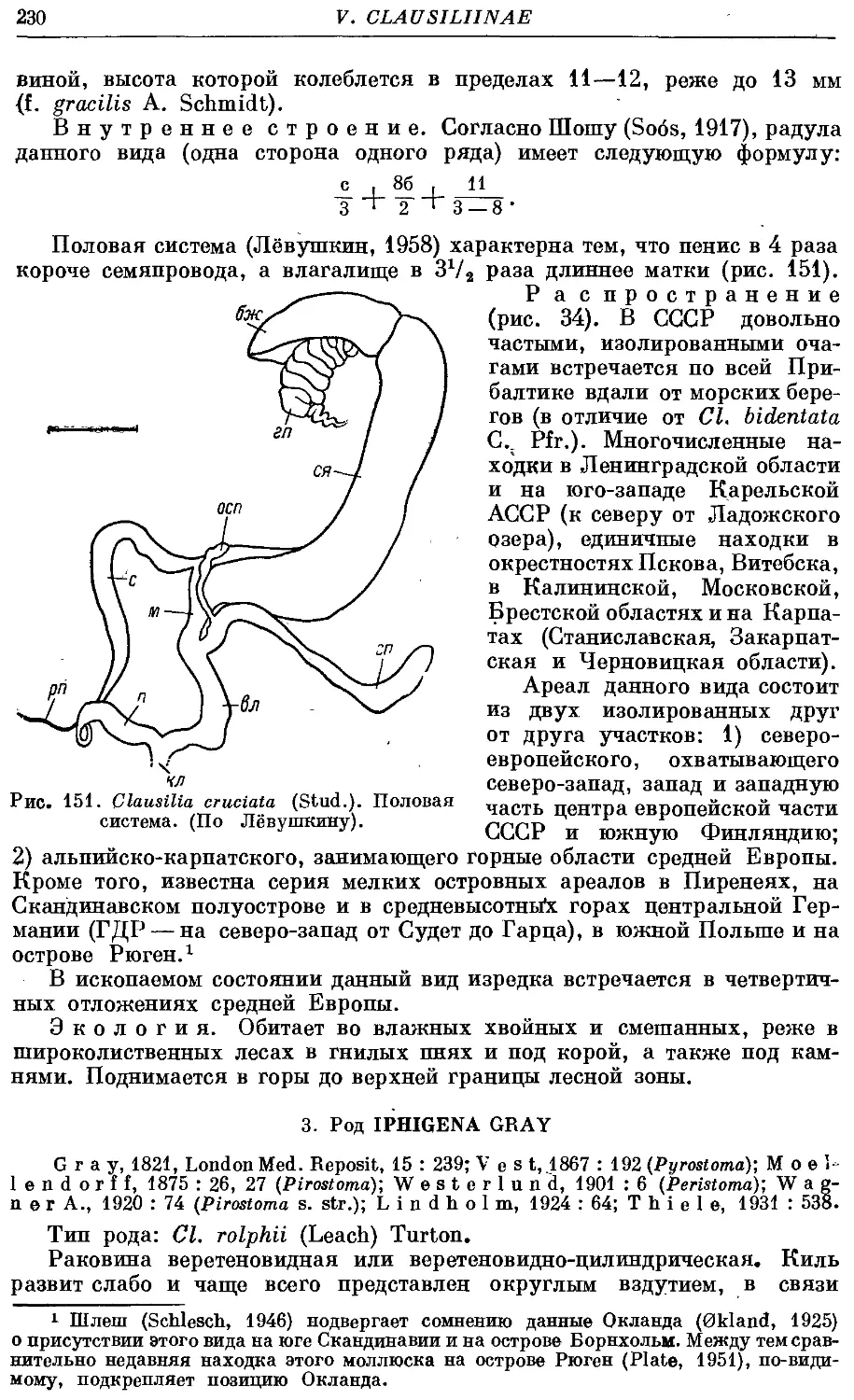
5. Cl. cruciata (Stud.) 228
3. Род Iphigena Gray
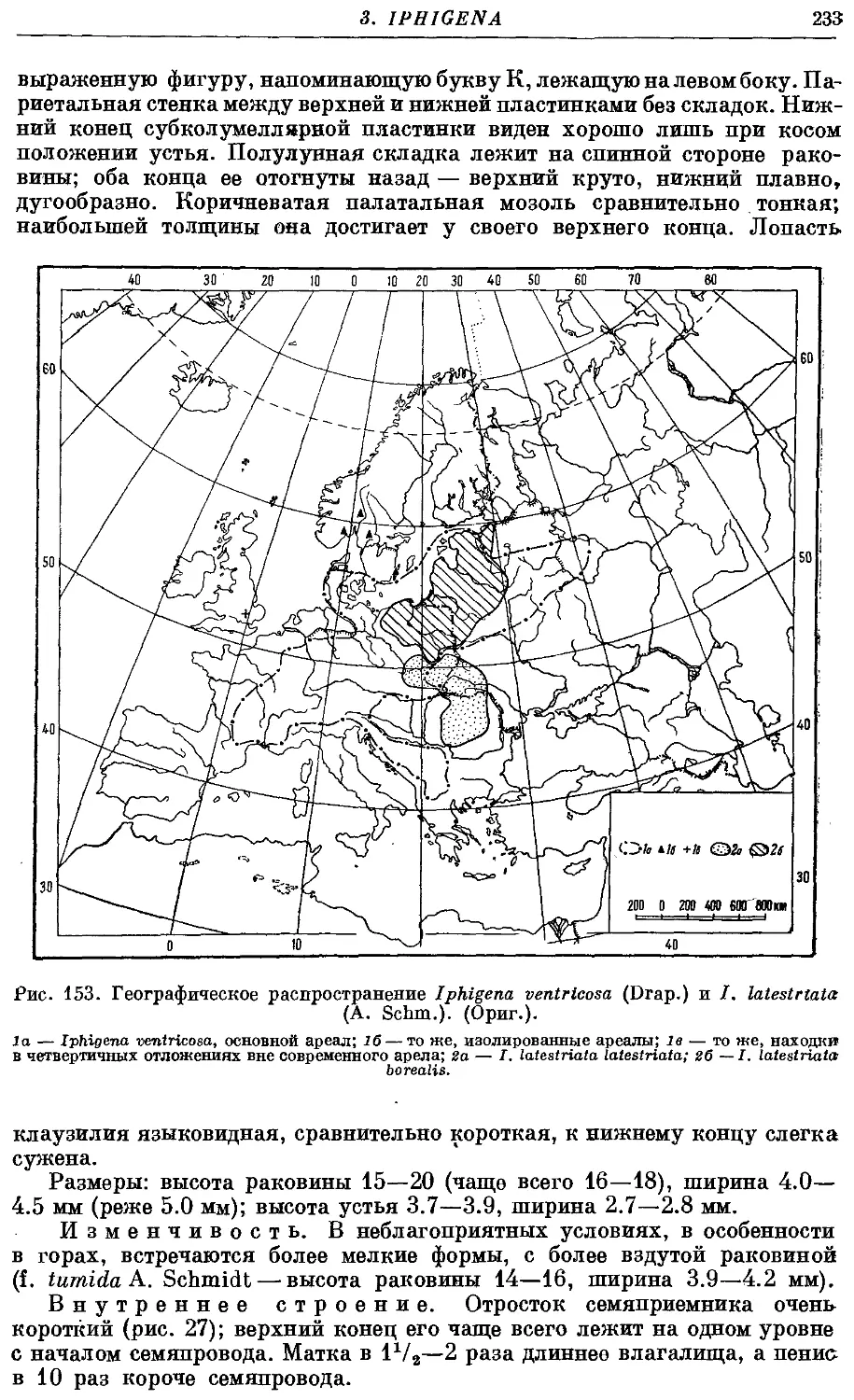
1. /. ventrieosa (Drap.) . 232
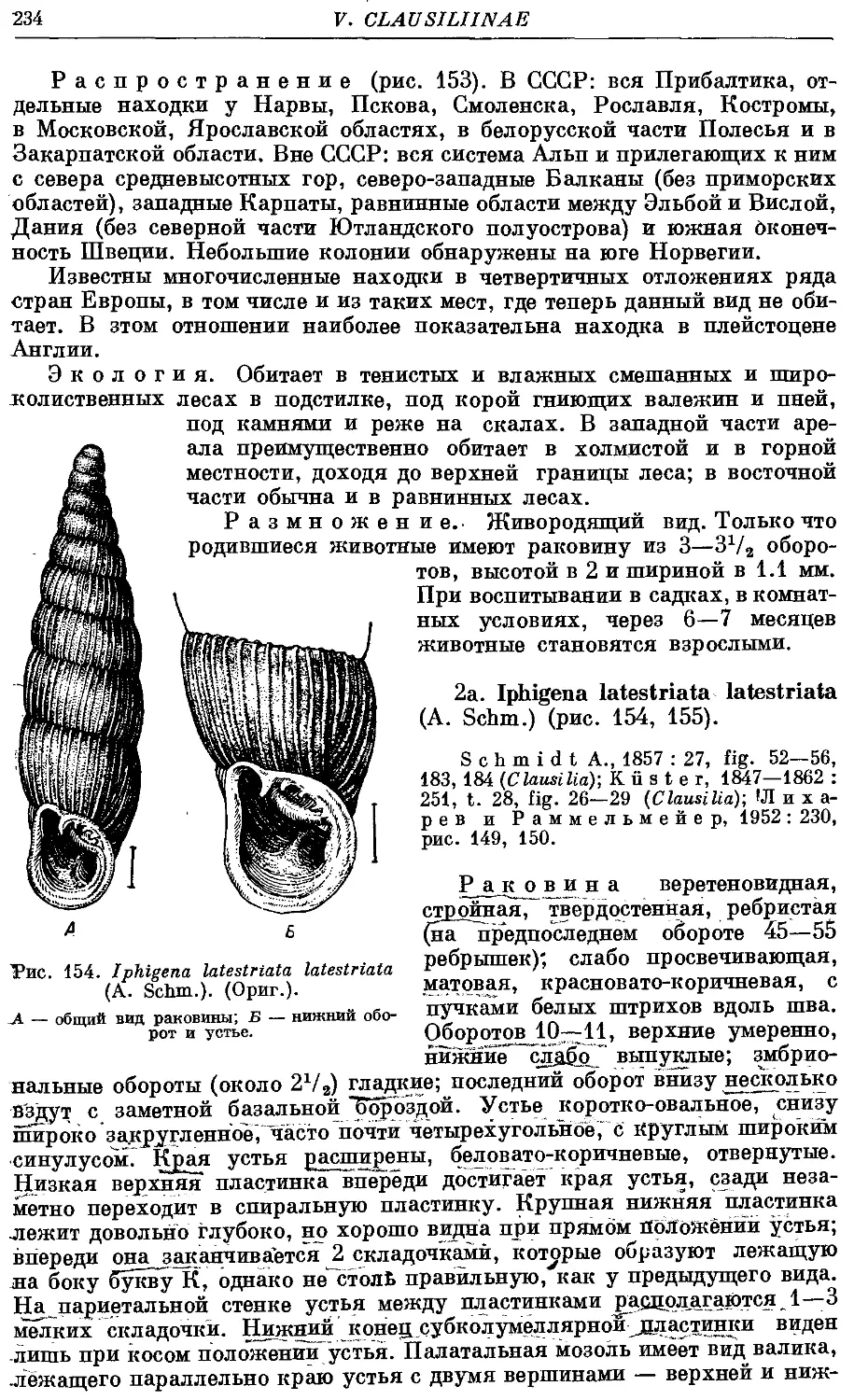
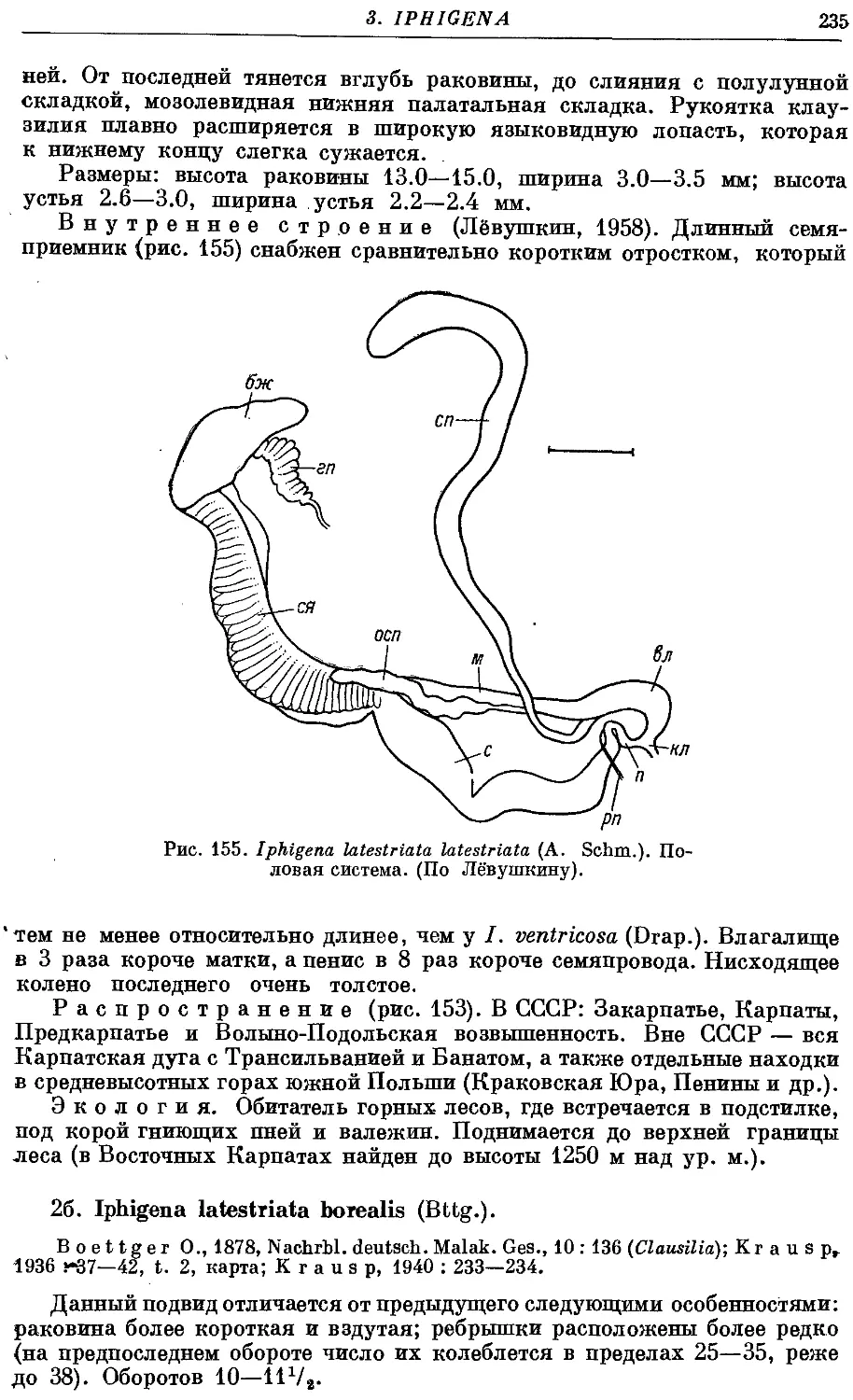
2a. /. latestriata latestriata (A. Schm.) 234
26. /. latestriata borealis (Bttg.) 235
*3. /. mucida badia (Rssm.) 236
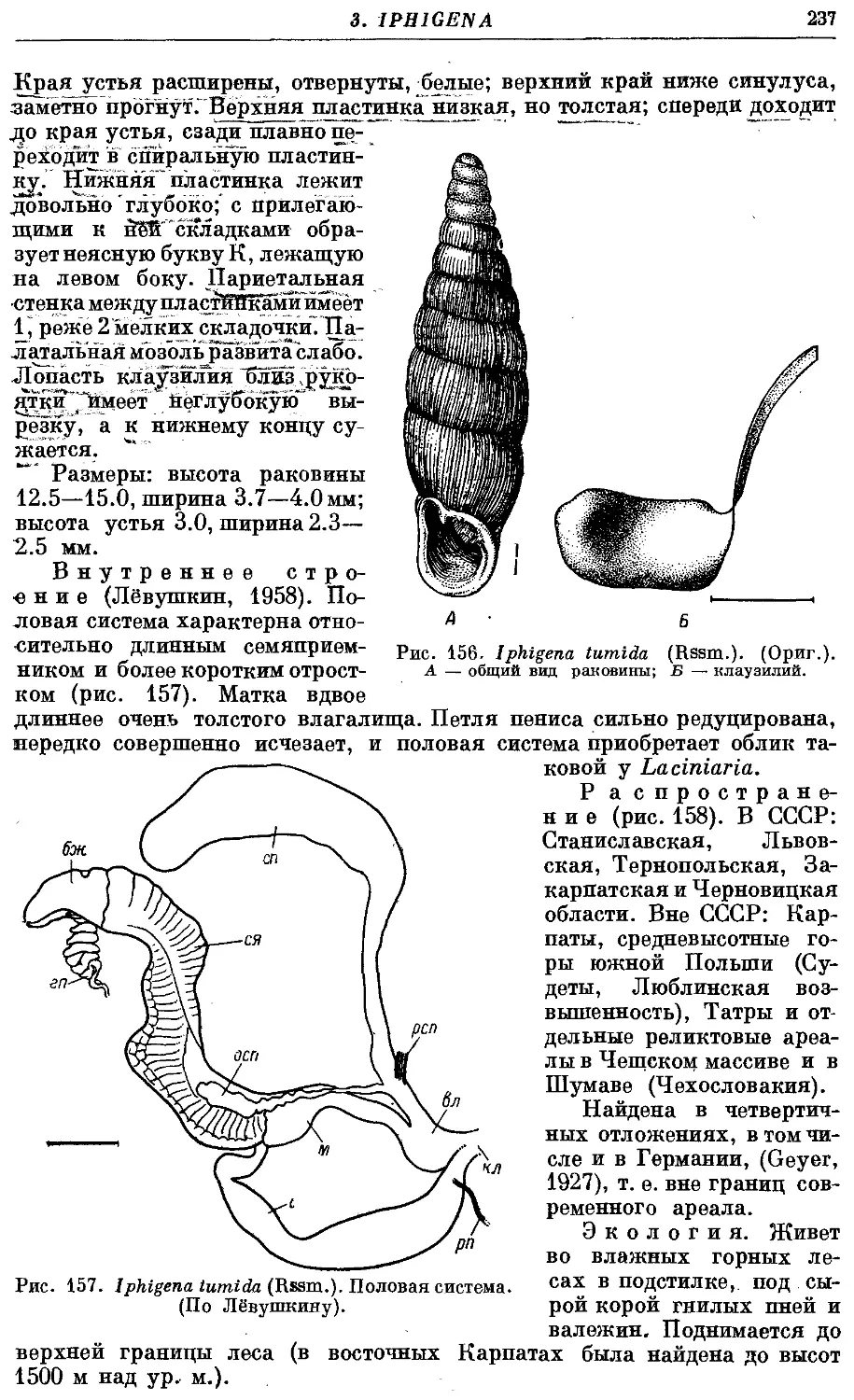
4. /. tumida (Rssm.) 236
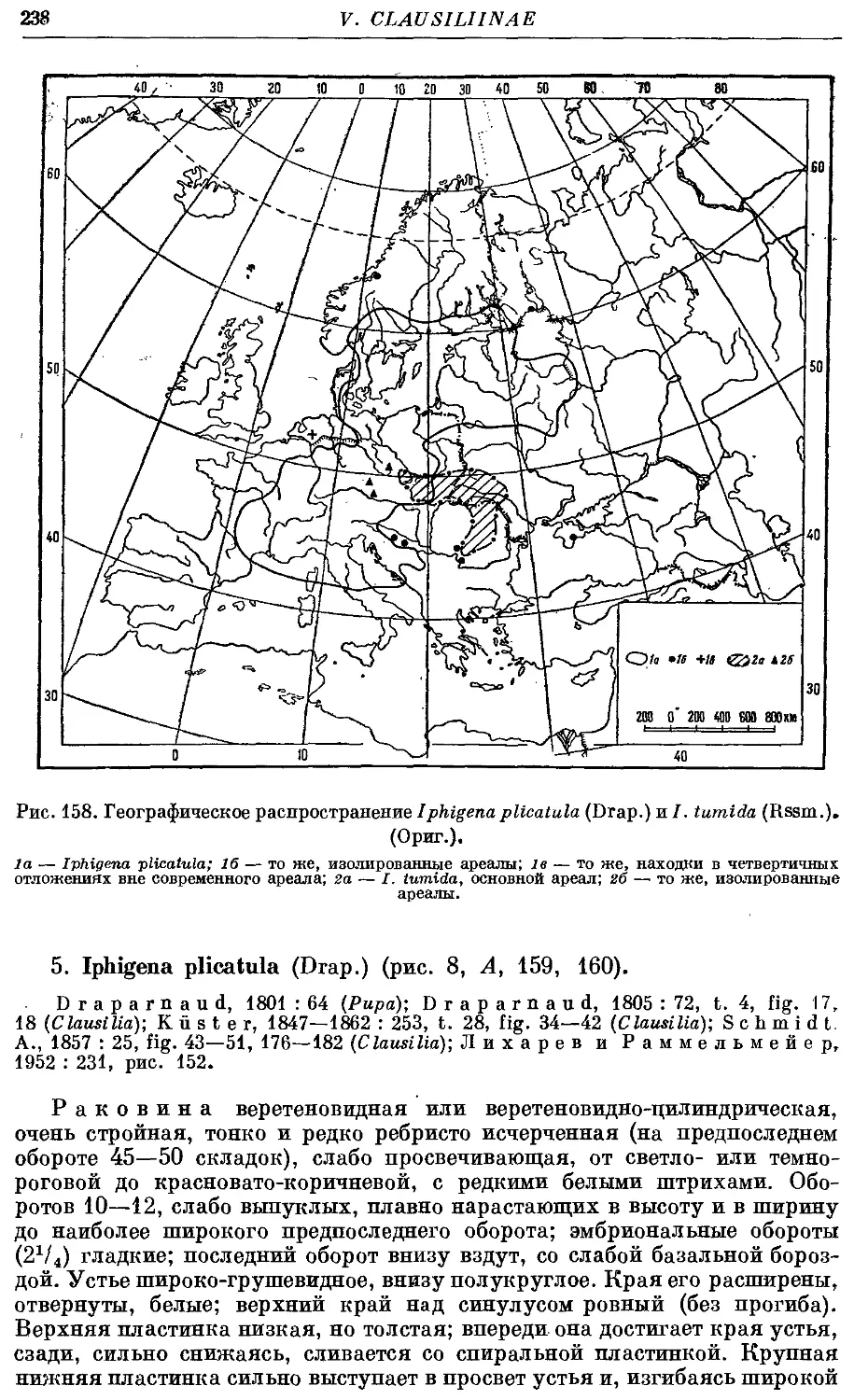
5. /. plicatula (Drap.) 238
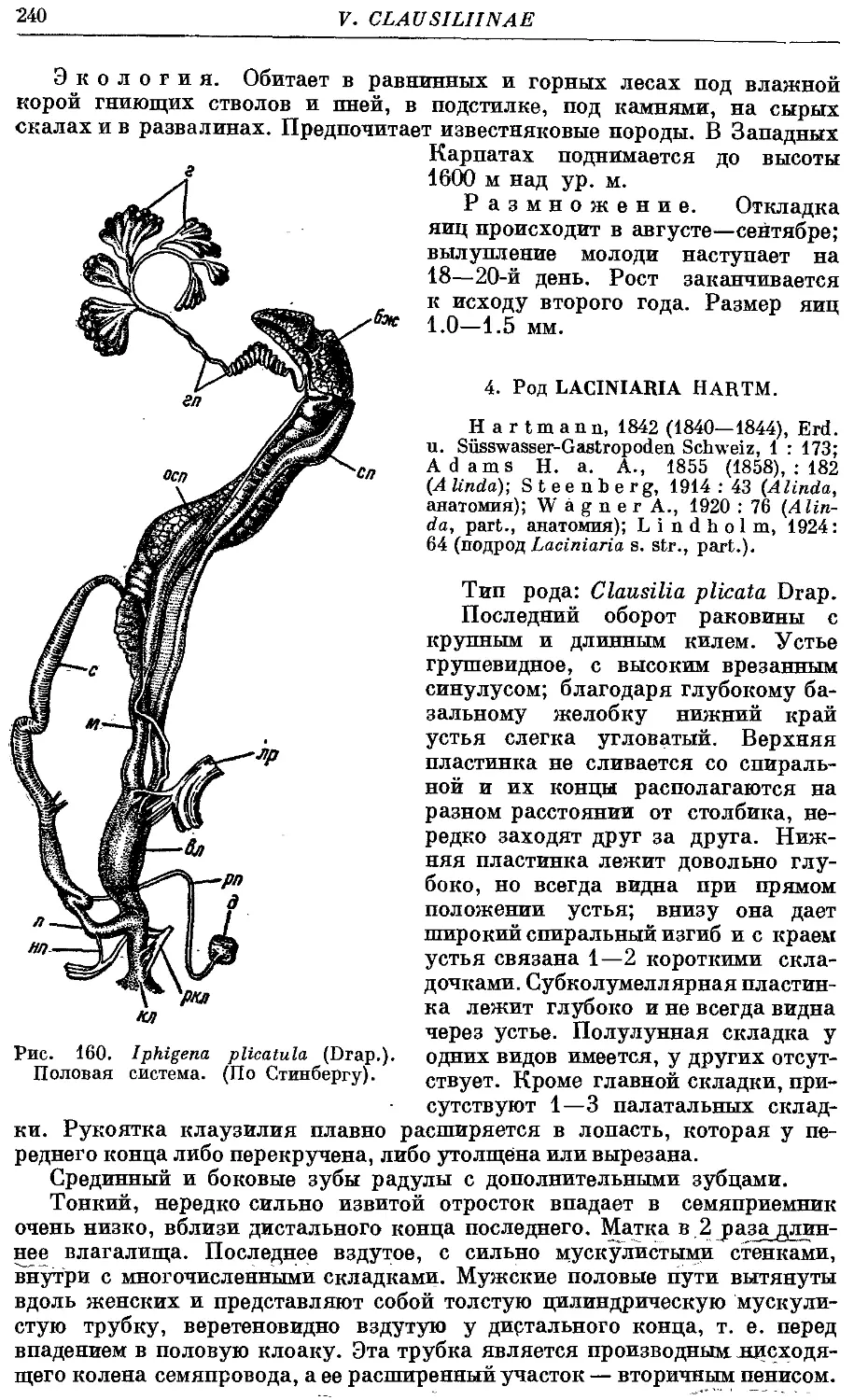
4. Род Laciniaria Hartm.
1. Подрод Laciniaria s. str.
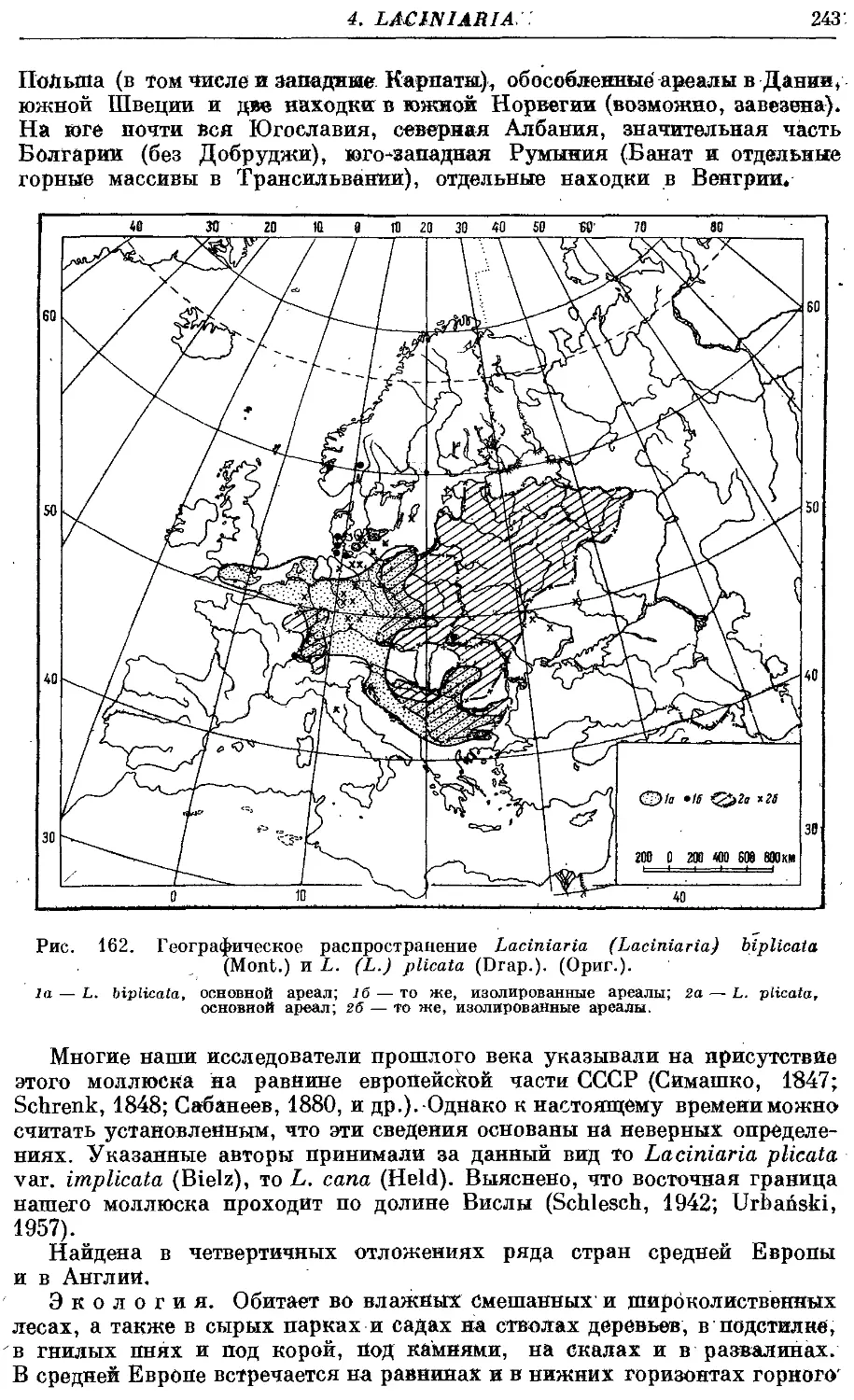
1. L. (L.) biplicata (Mont.) 242
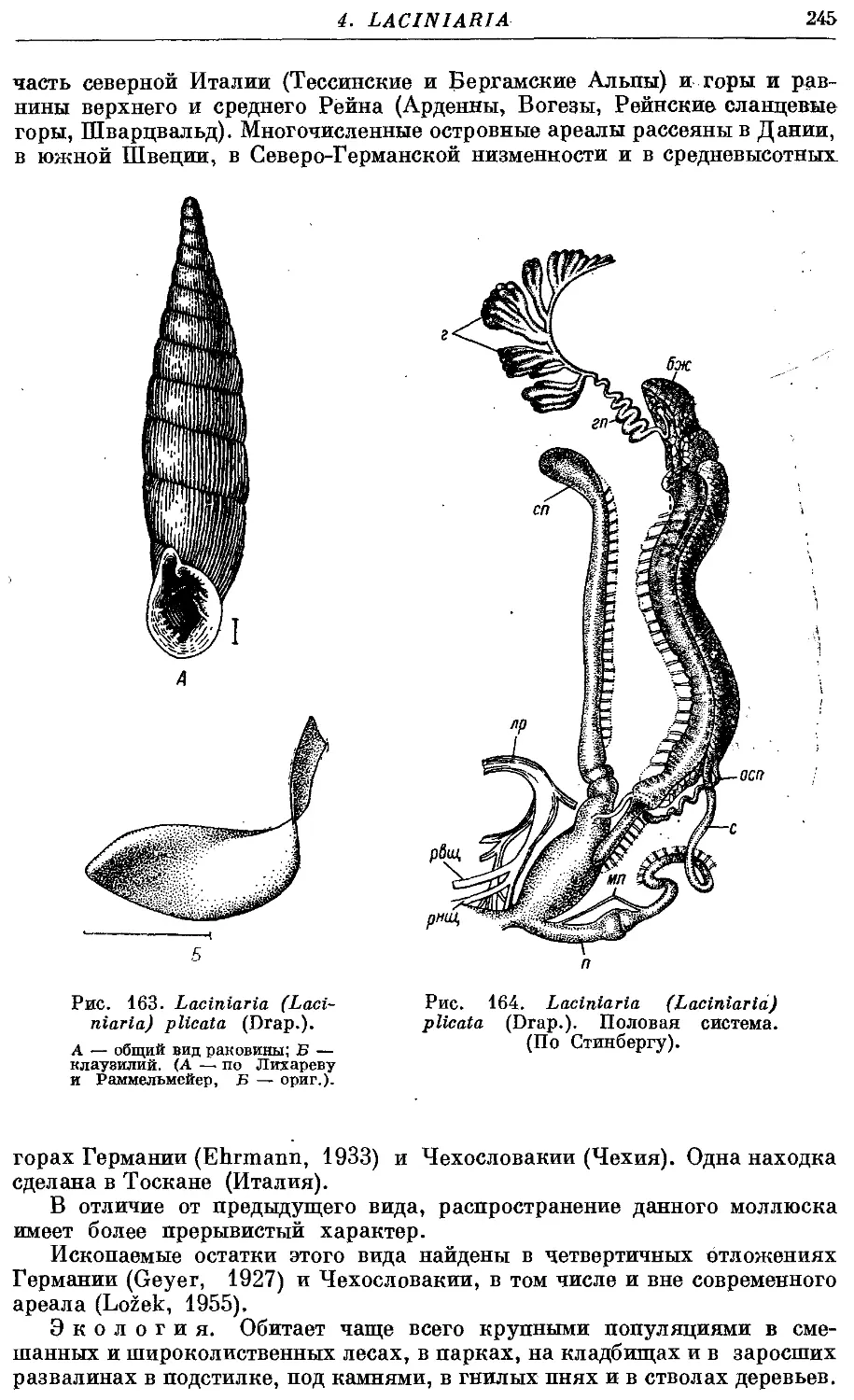
2. L. (L.) plicata (Drap.) 244
2. Подрод Strigilecula K. et W.
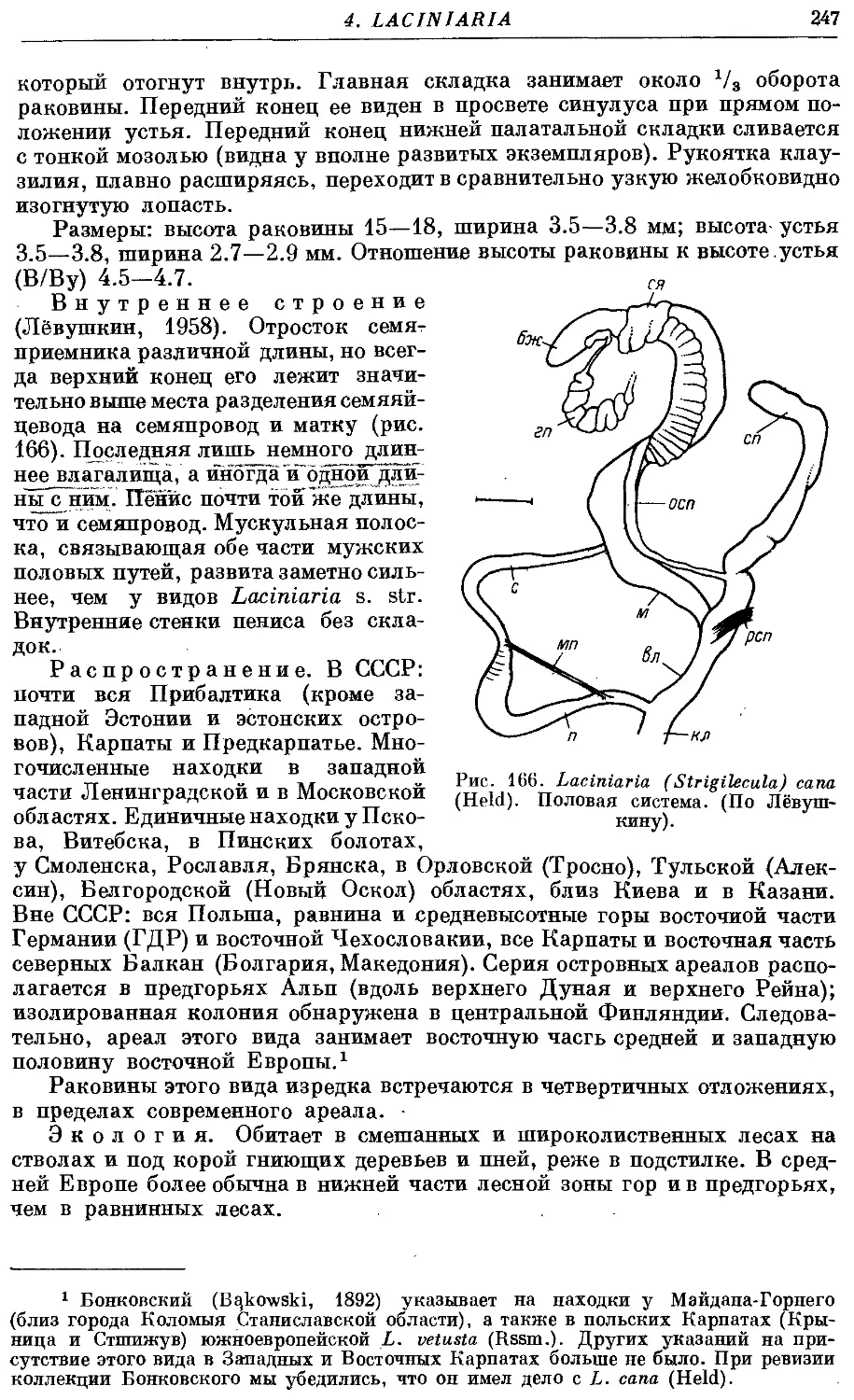
3. L. (Str.) сапа (Held) 246
*4. L. (Str.) nitidosa (Ulicny) 248
3. Подрод Mentissa H. et. A. Adams
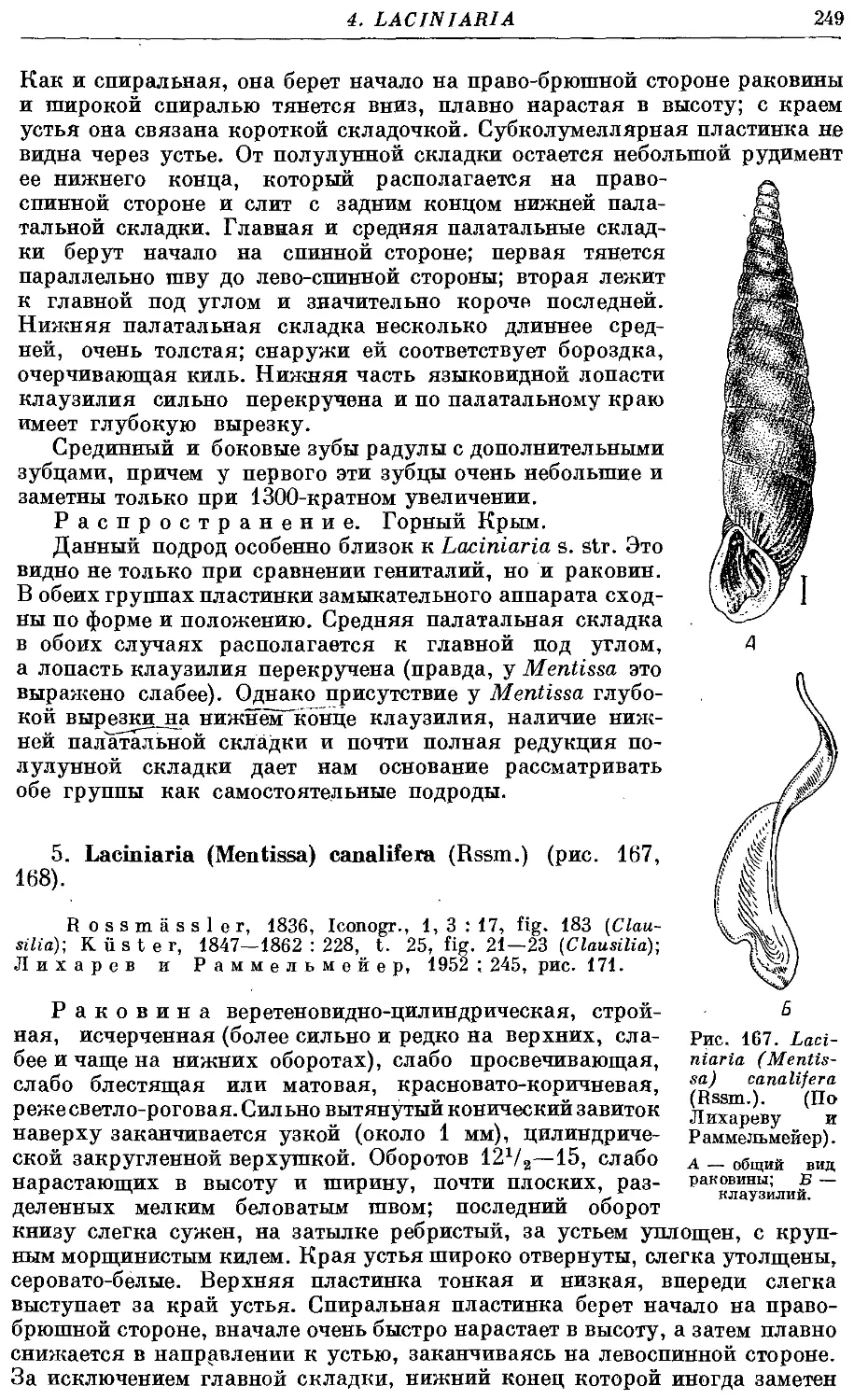
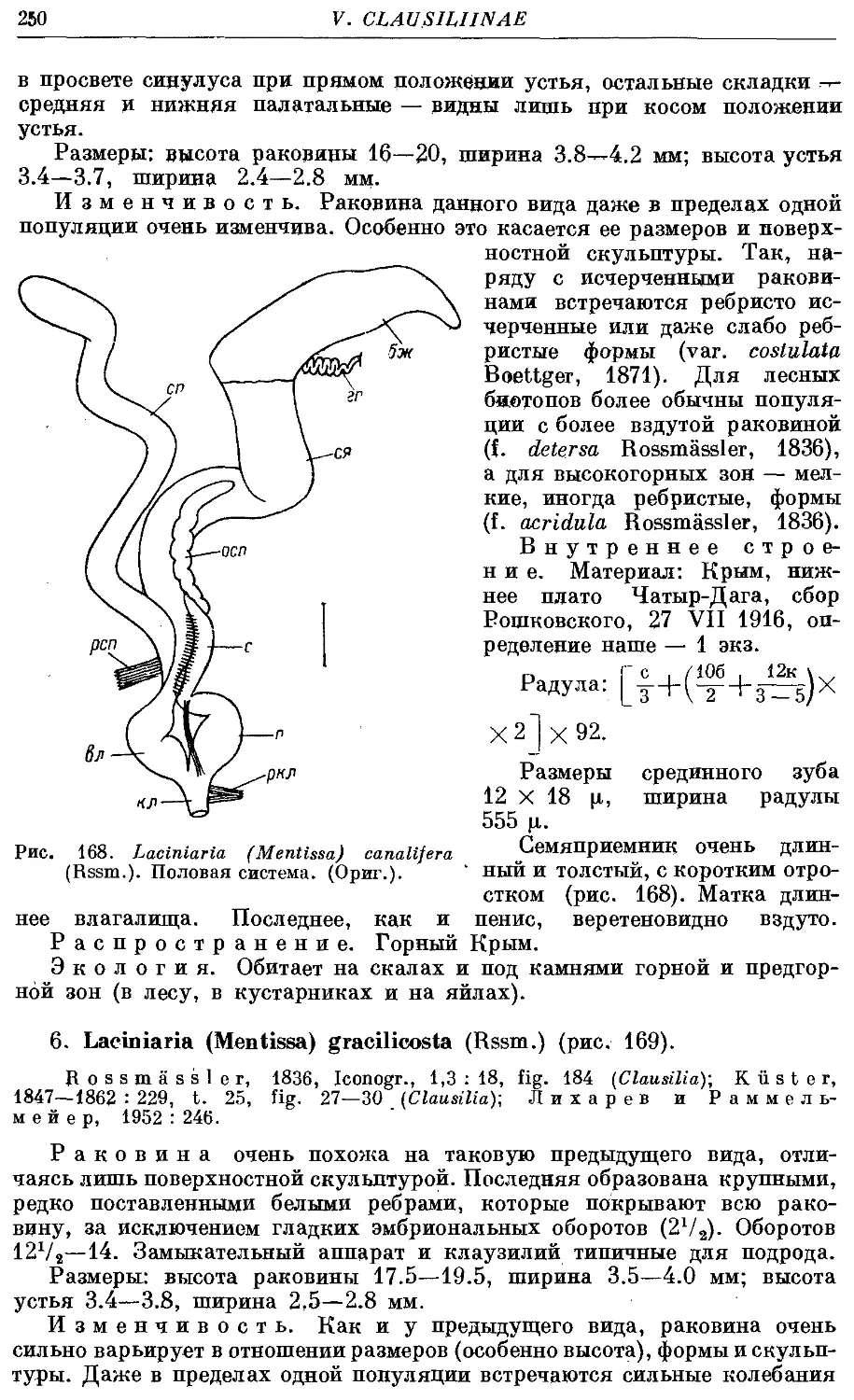
5. L. (M.) canalifera (Rssm.) 249
6. L. (M.) gracilicosta (Rssm.) 250
5. Род Pseudalinda Bttg.
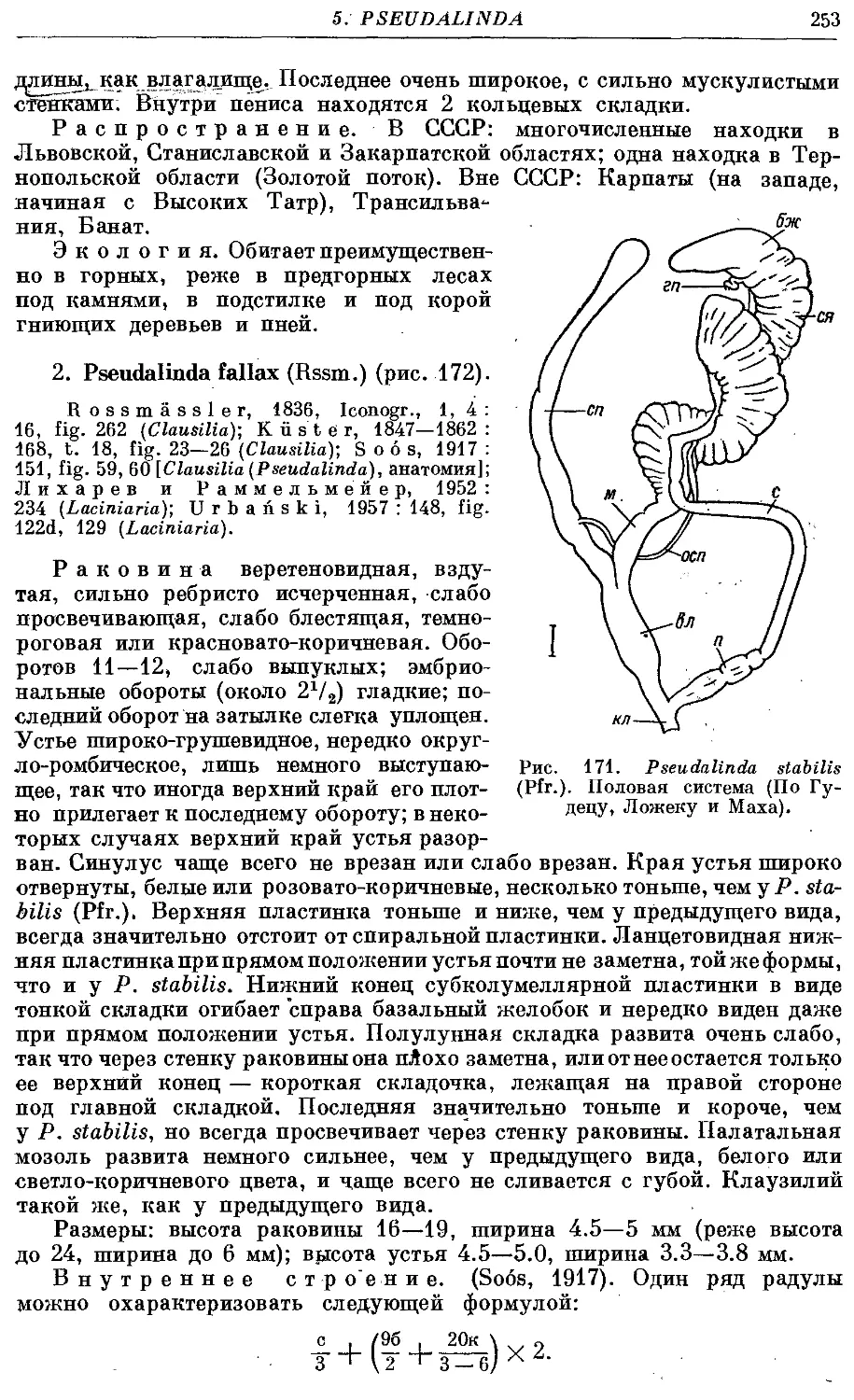
1. Ps. stabilis (Pfr.) 252
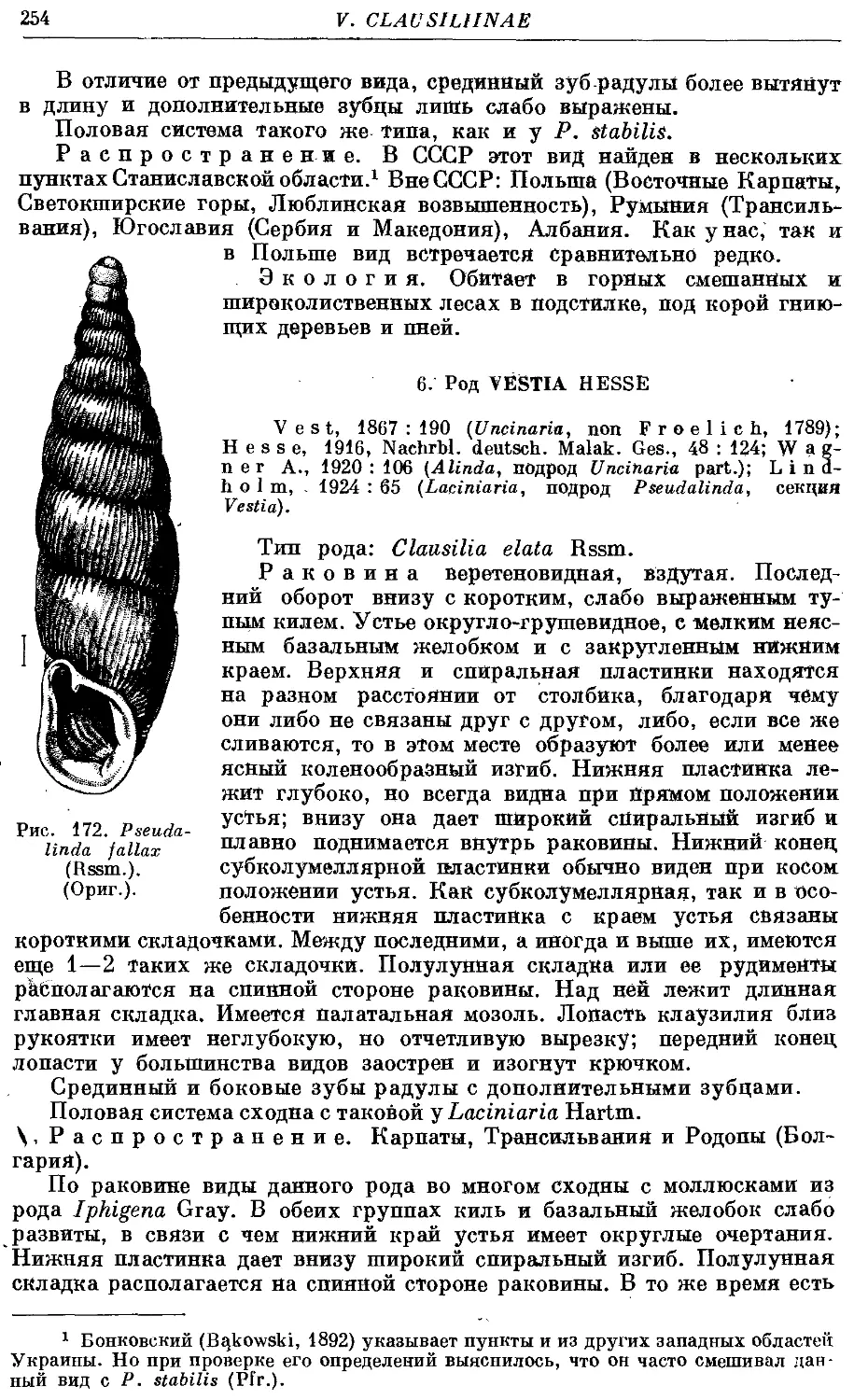
2. Ps. fallax (Rssm.) 253
6. Род Vestia Hesse
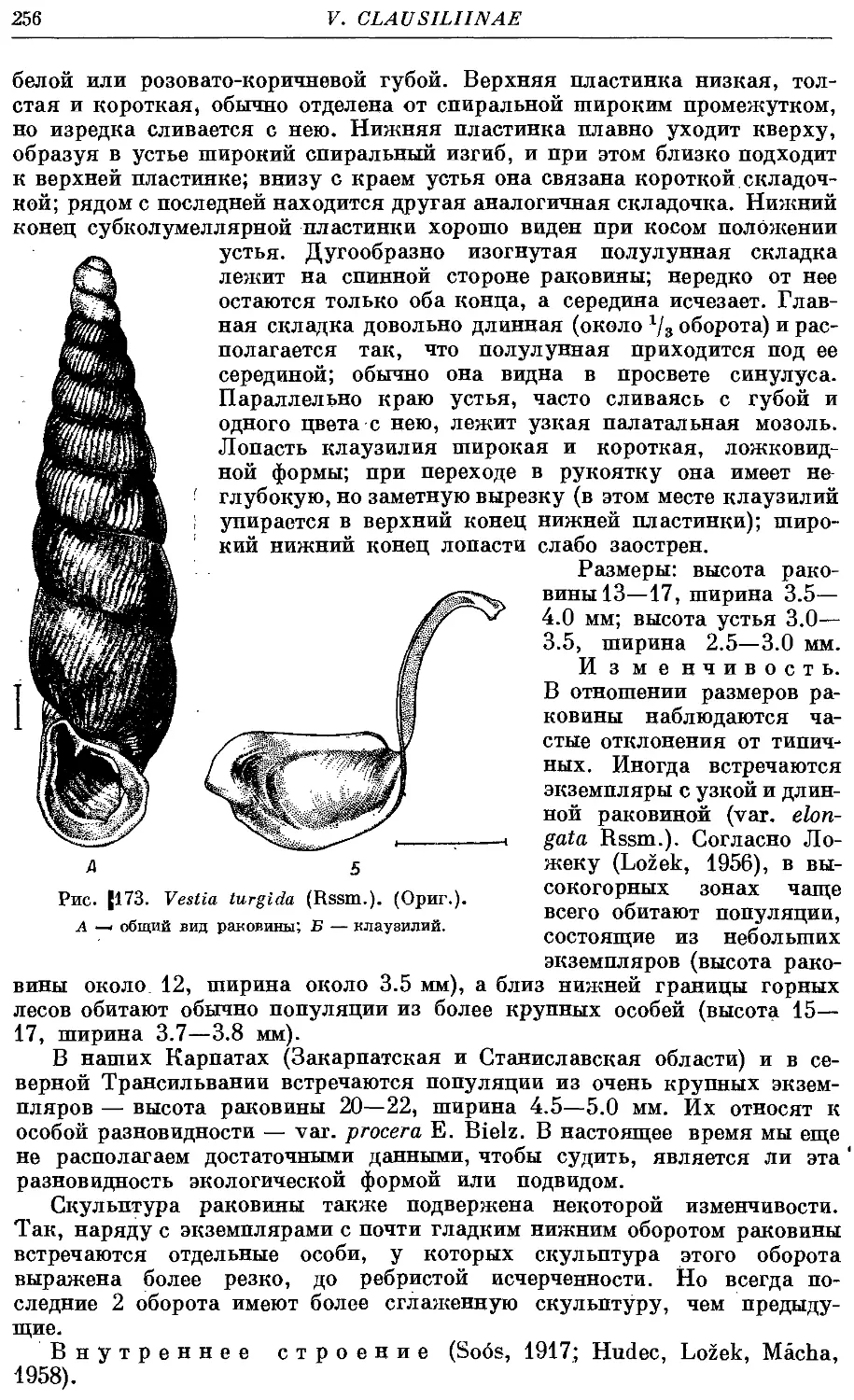
1. V. turgida (Rssm.) 255
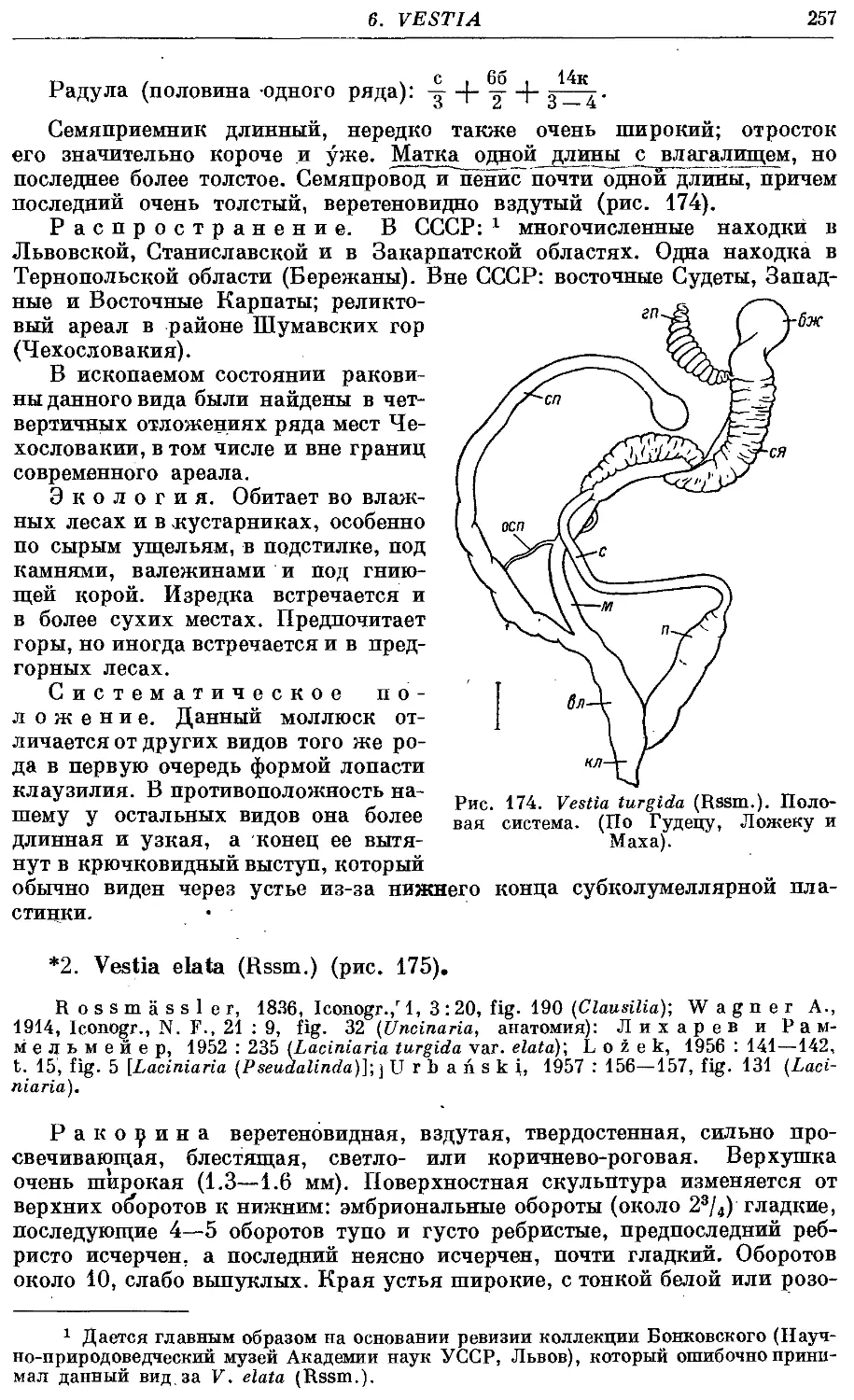
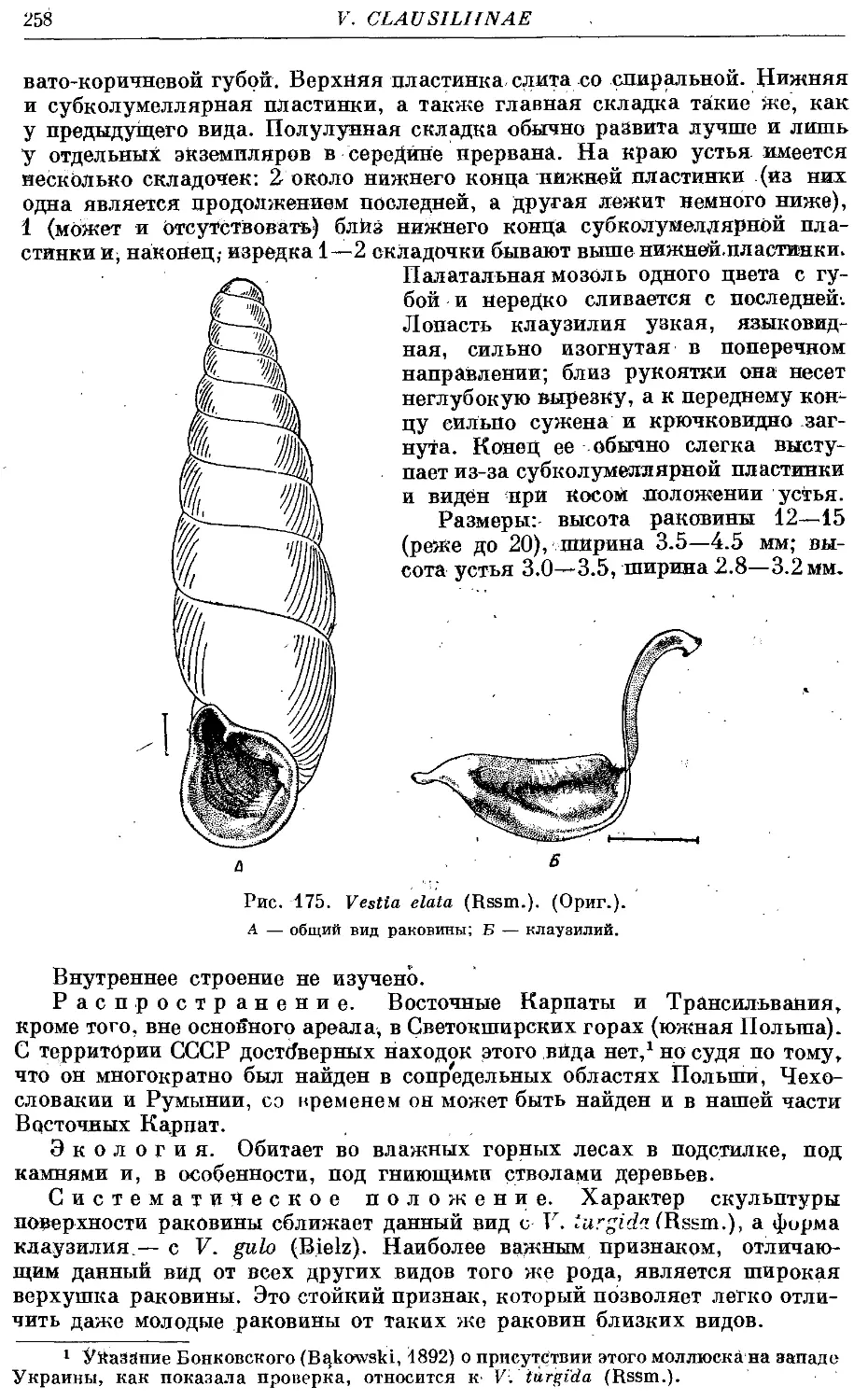
*2. V. elata (Rssm.) 257
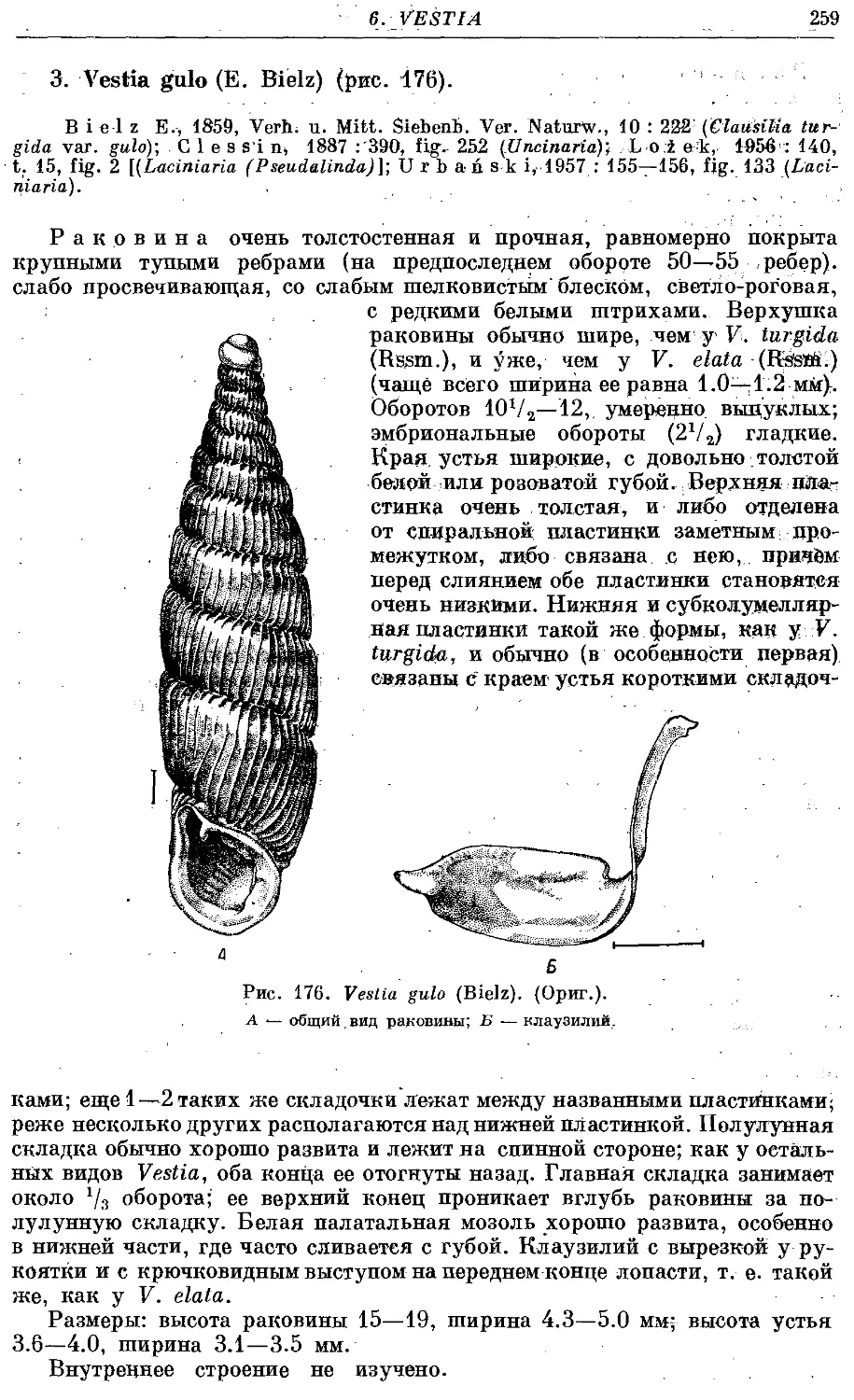
3. V. gulo (E. Bielz) 259
*4. V. riloensis (A. Wagn.) 260
7. Род Balea Gray
B. perversa (L.) 262
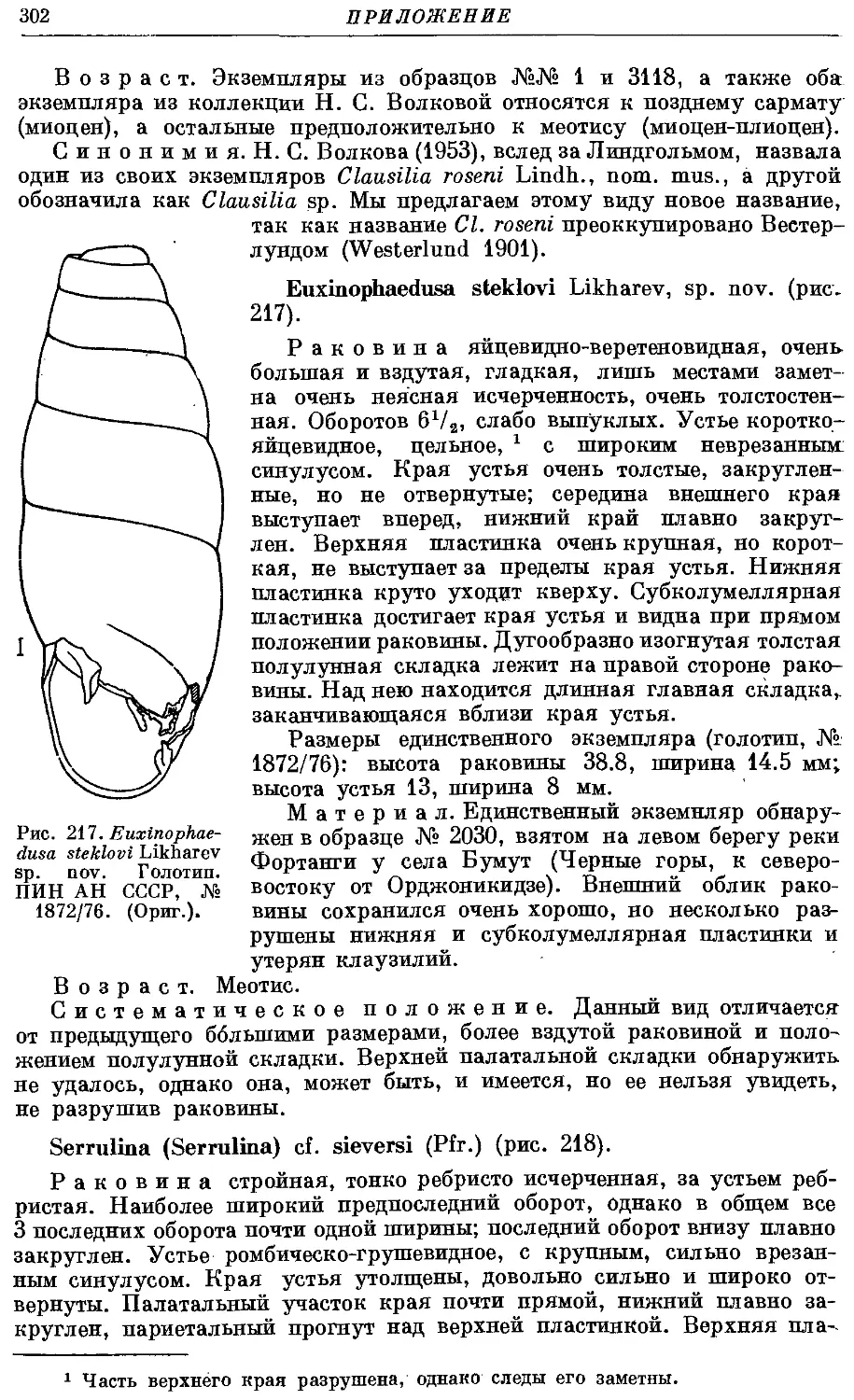
8. Род Quadriplieata Bttg.
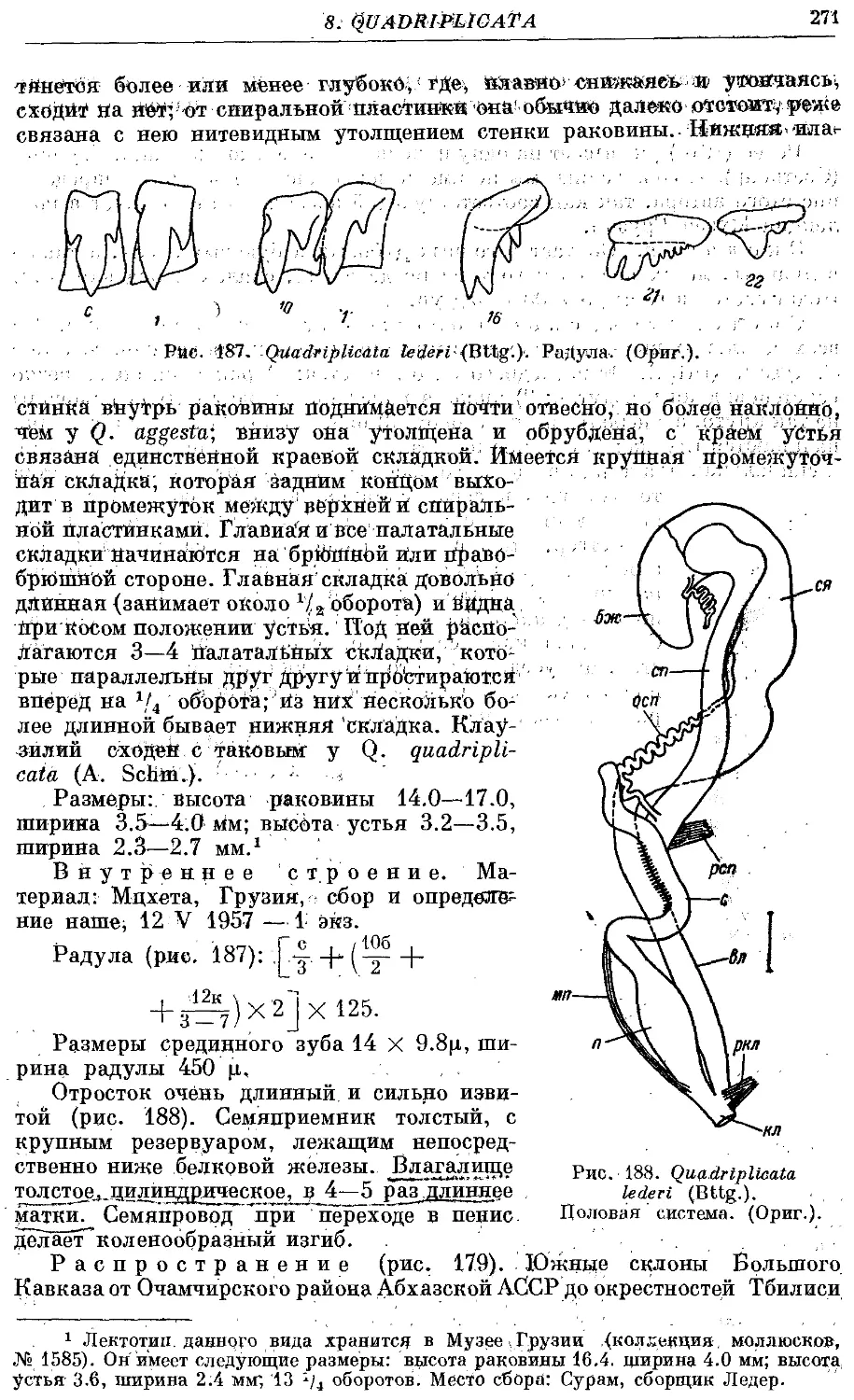
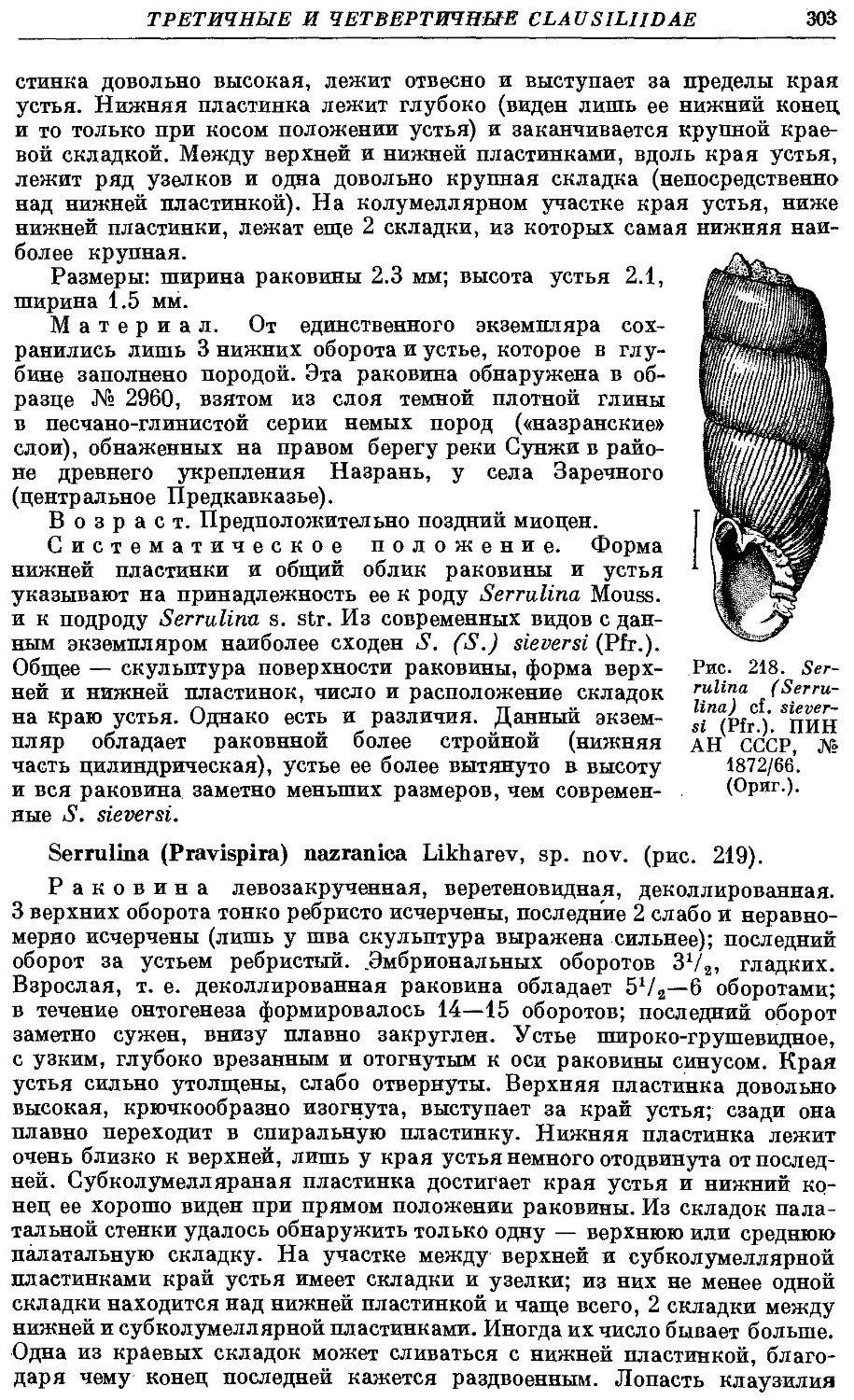
1. Q. quadriplieata (A. Schm.) 266
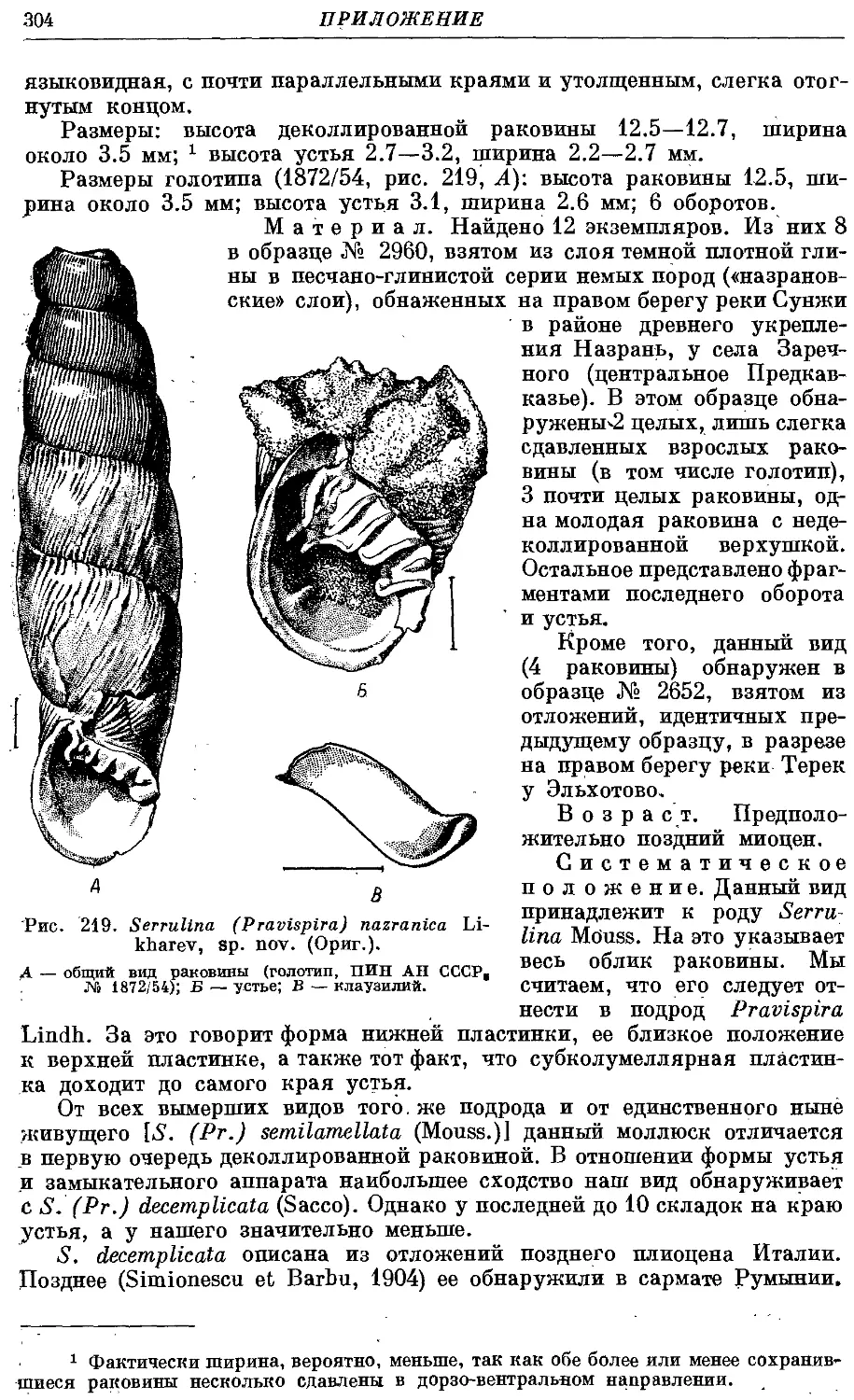
2a. Q. aggesta aggesta (Bttg.) 268
26. Q. aggesta stauropolitana (Rosen) • 269
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
Стр.
3. Q. lederi (Bttg.) 270
4. Q. subaggesta (Ret.) 272
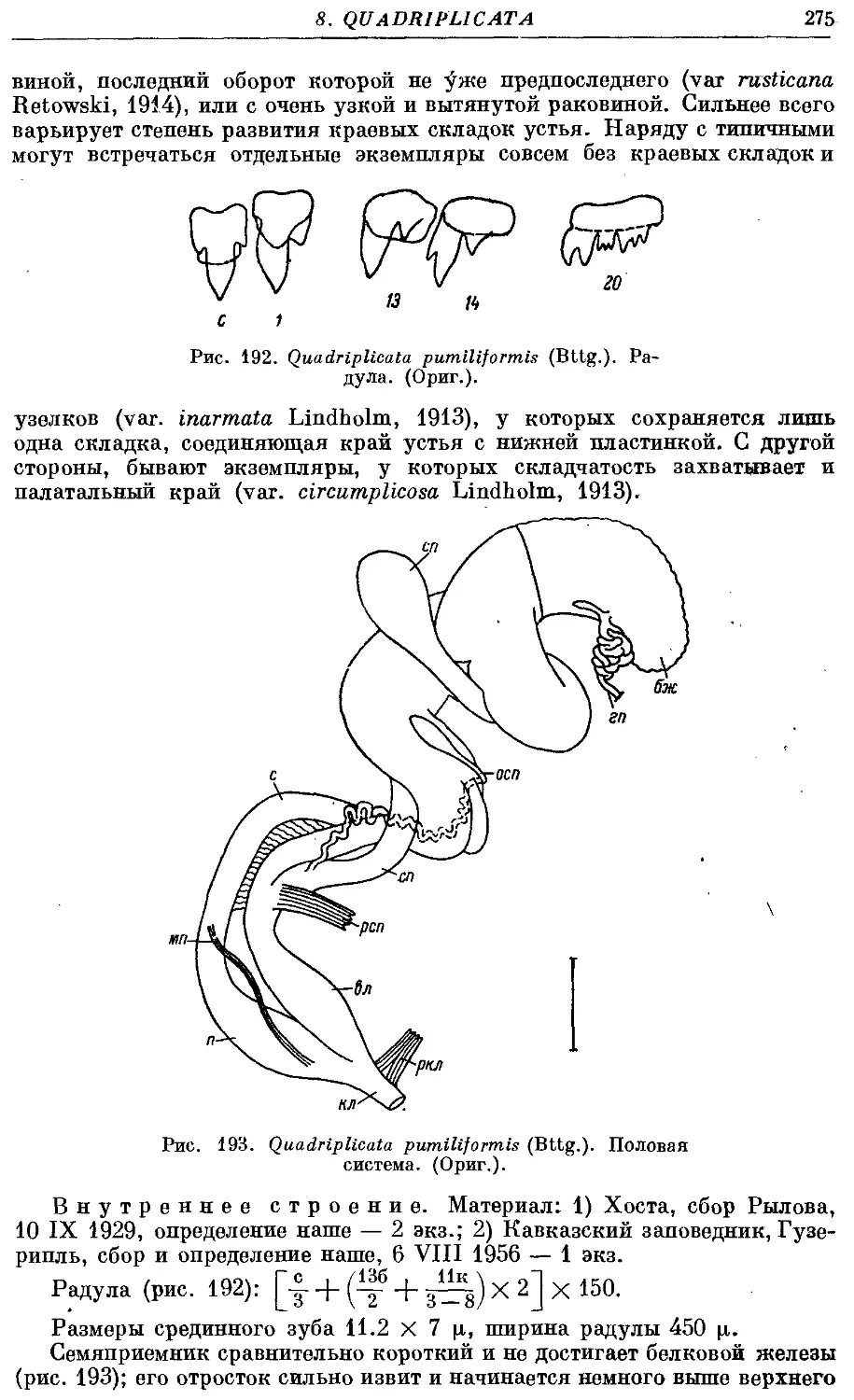
5. Q. pumiliformis (Bttg.) 274
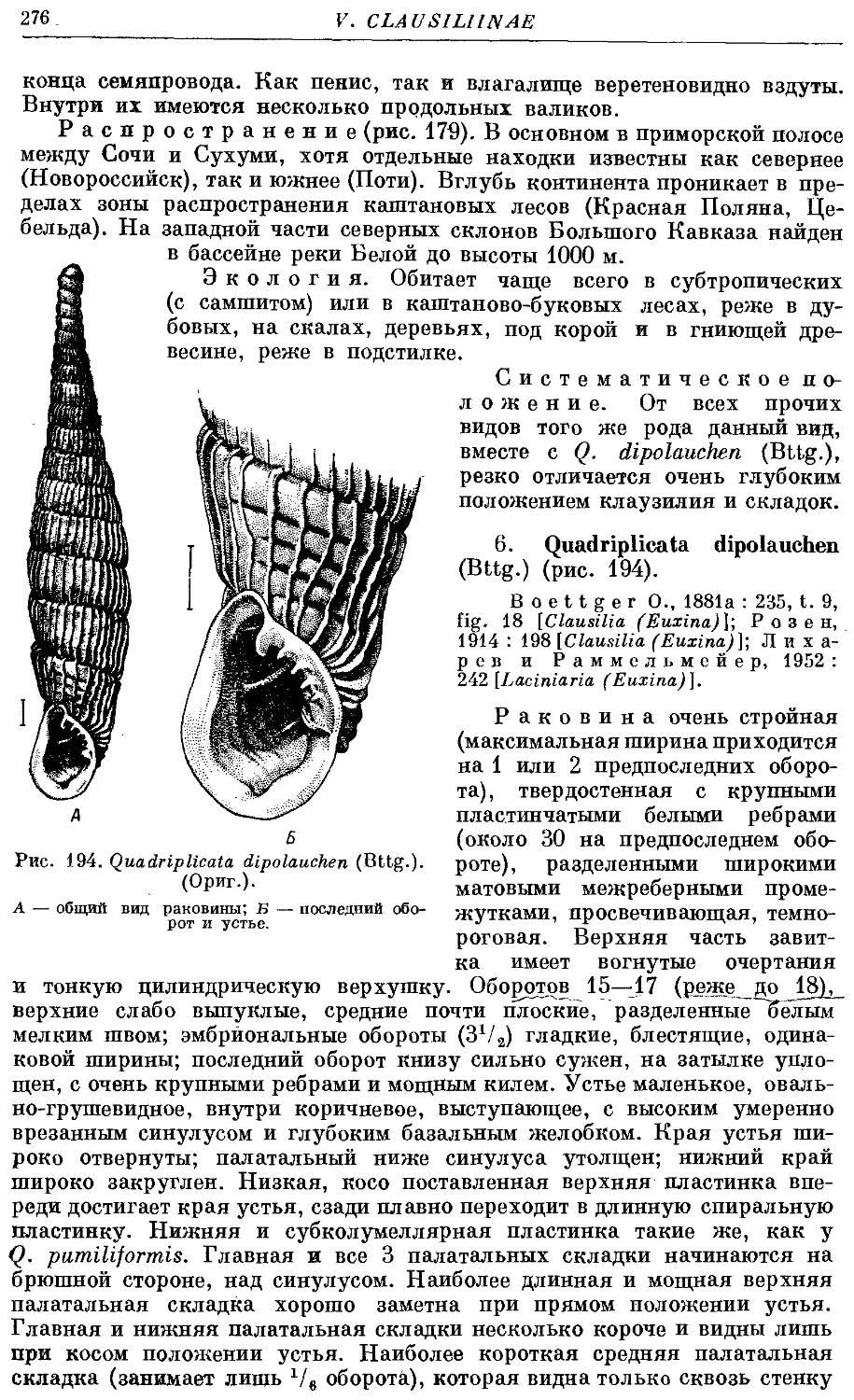
6. Q. dipolauchen (Bttg.) 276
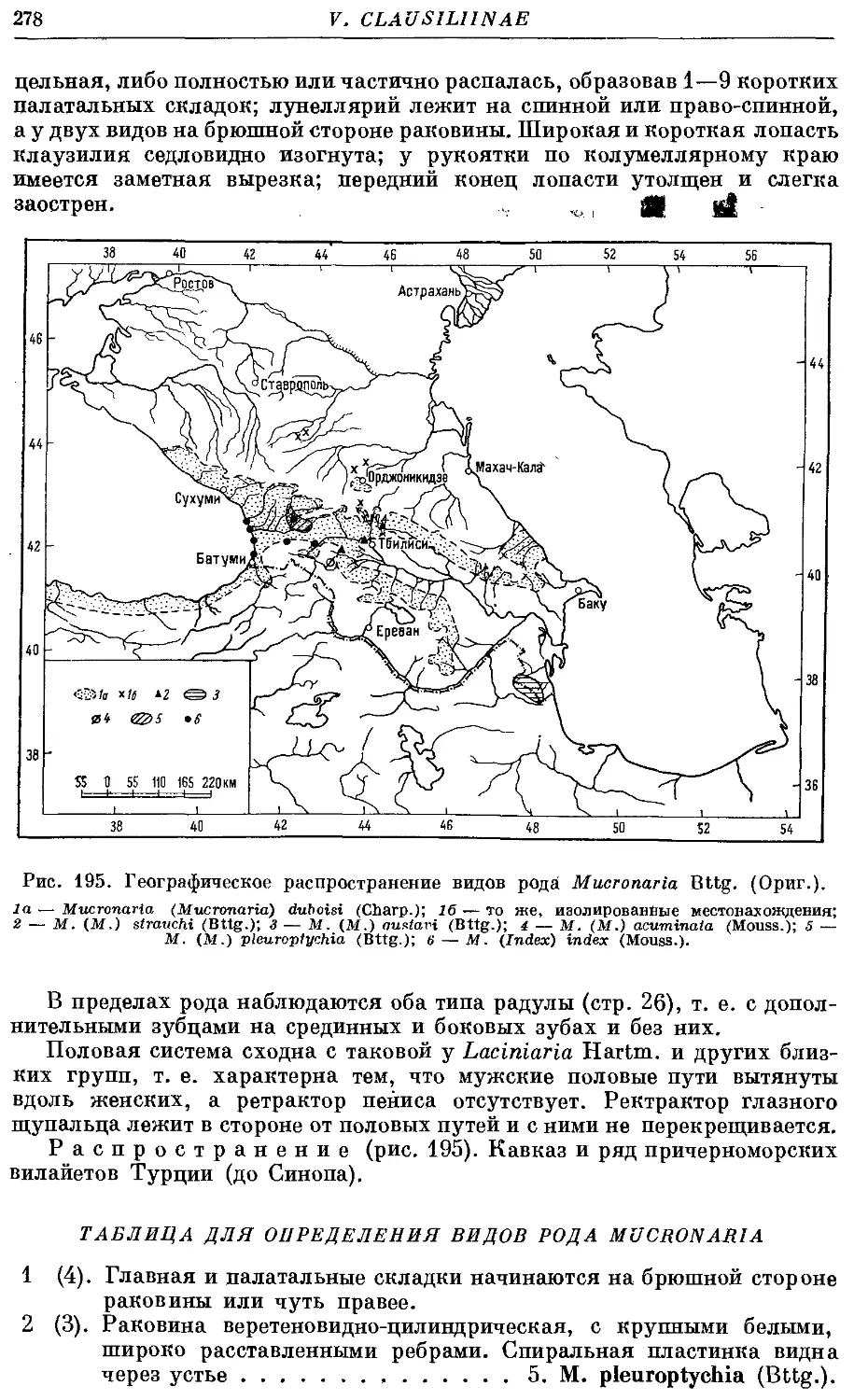
9. Род Mucronaria Bttg.
1. Подрод Mucronaria s. str.
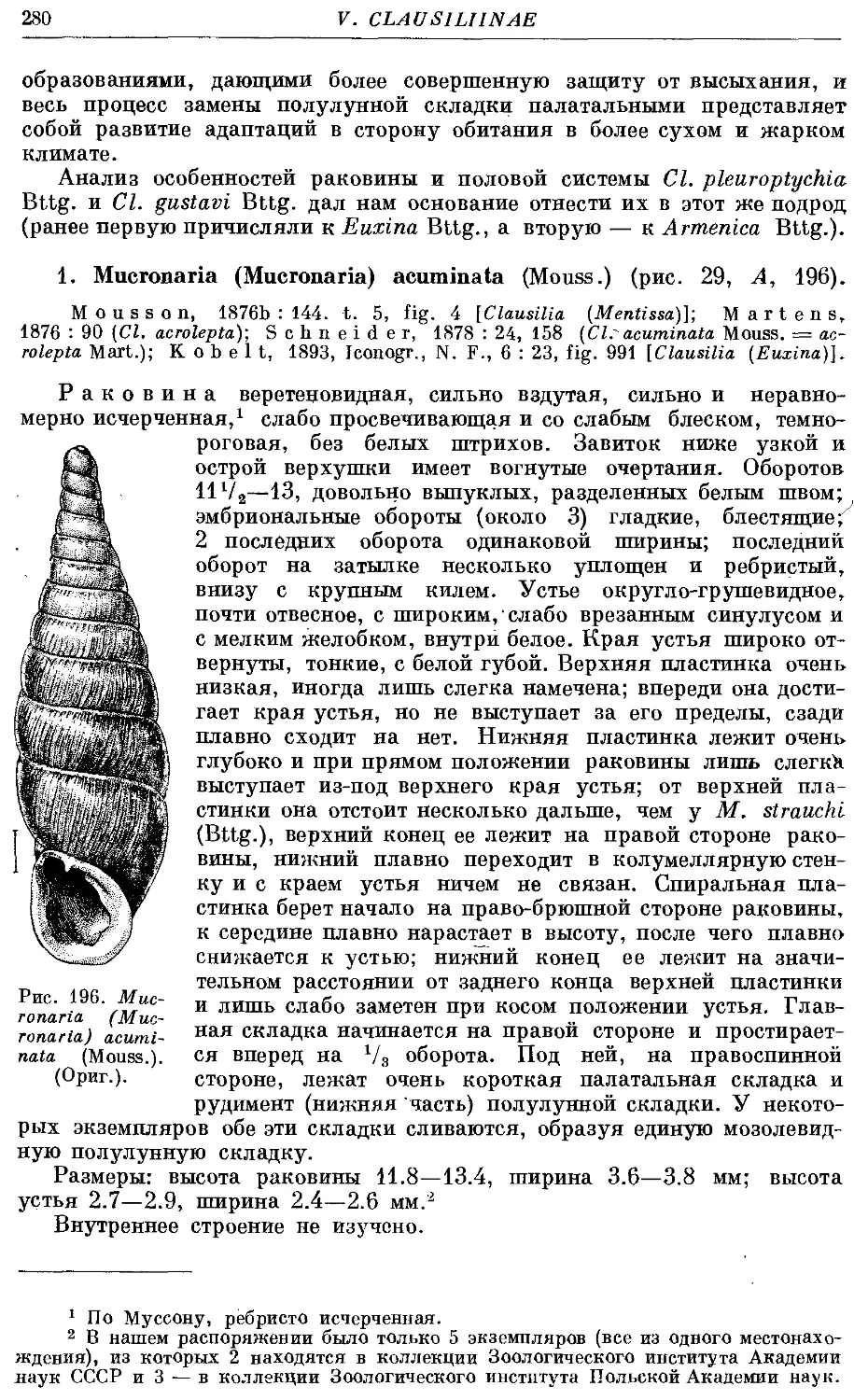
1. M. (M.) acuminata (Mouss.) 280
2. M. (M.) strauchi (Bttg.) 281
3. M. (M.) duboisi (Charp.) 282
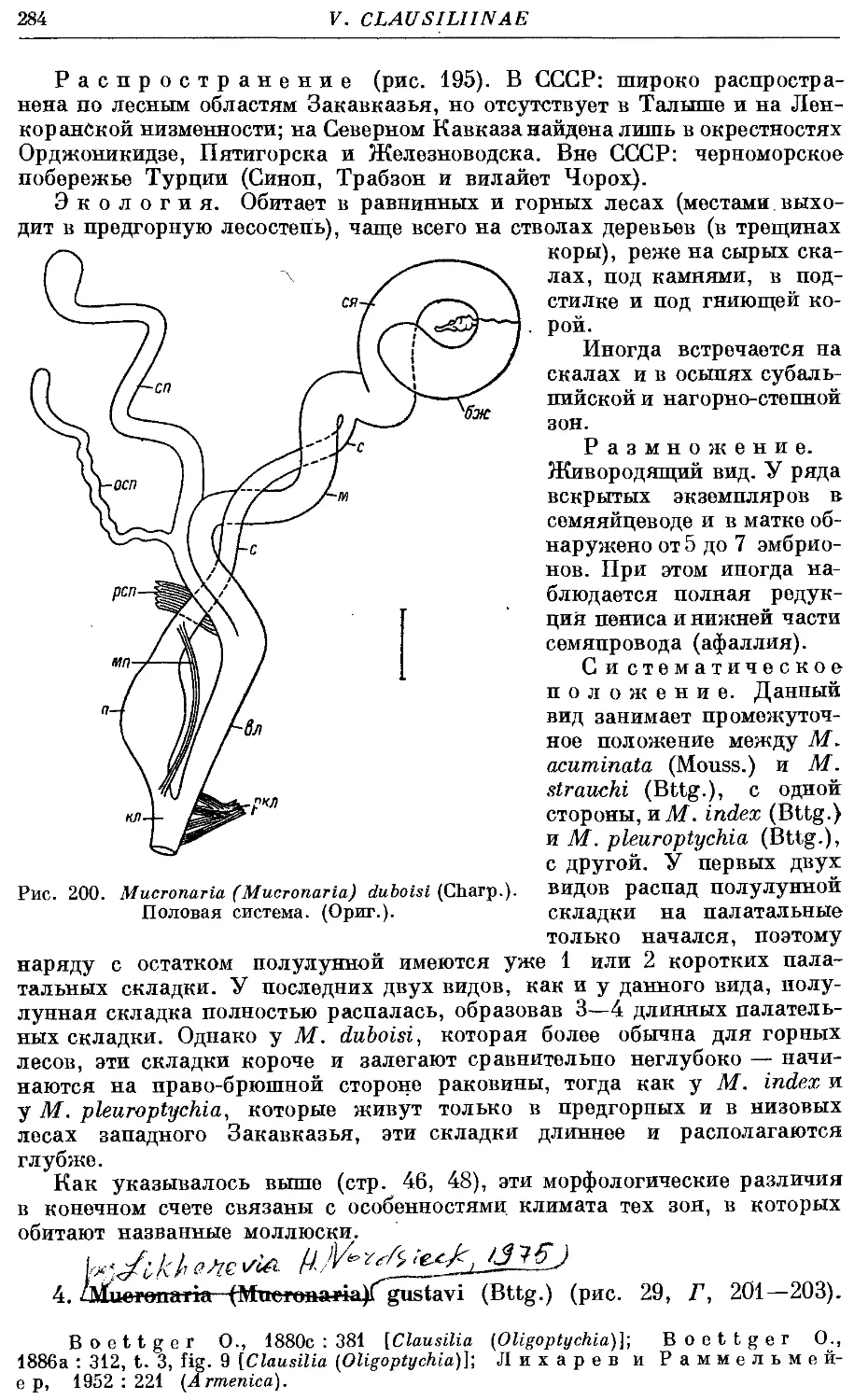
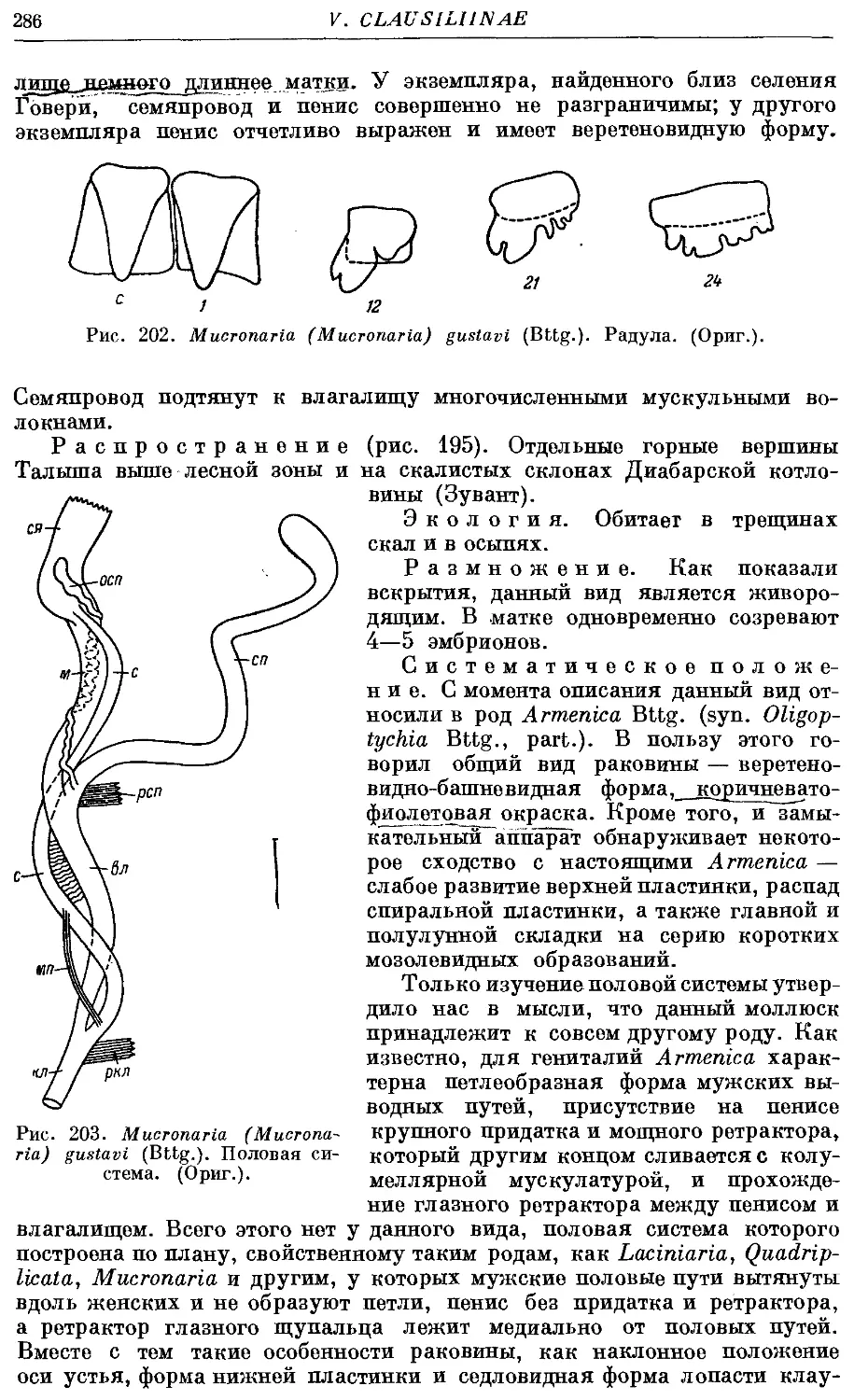
4. M. (M.) gustavi (Bttg.) 284
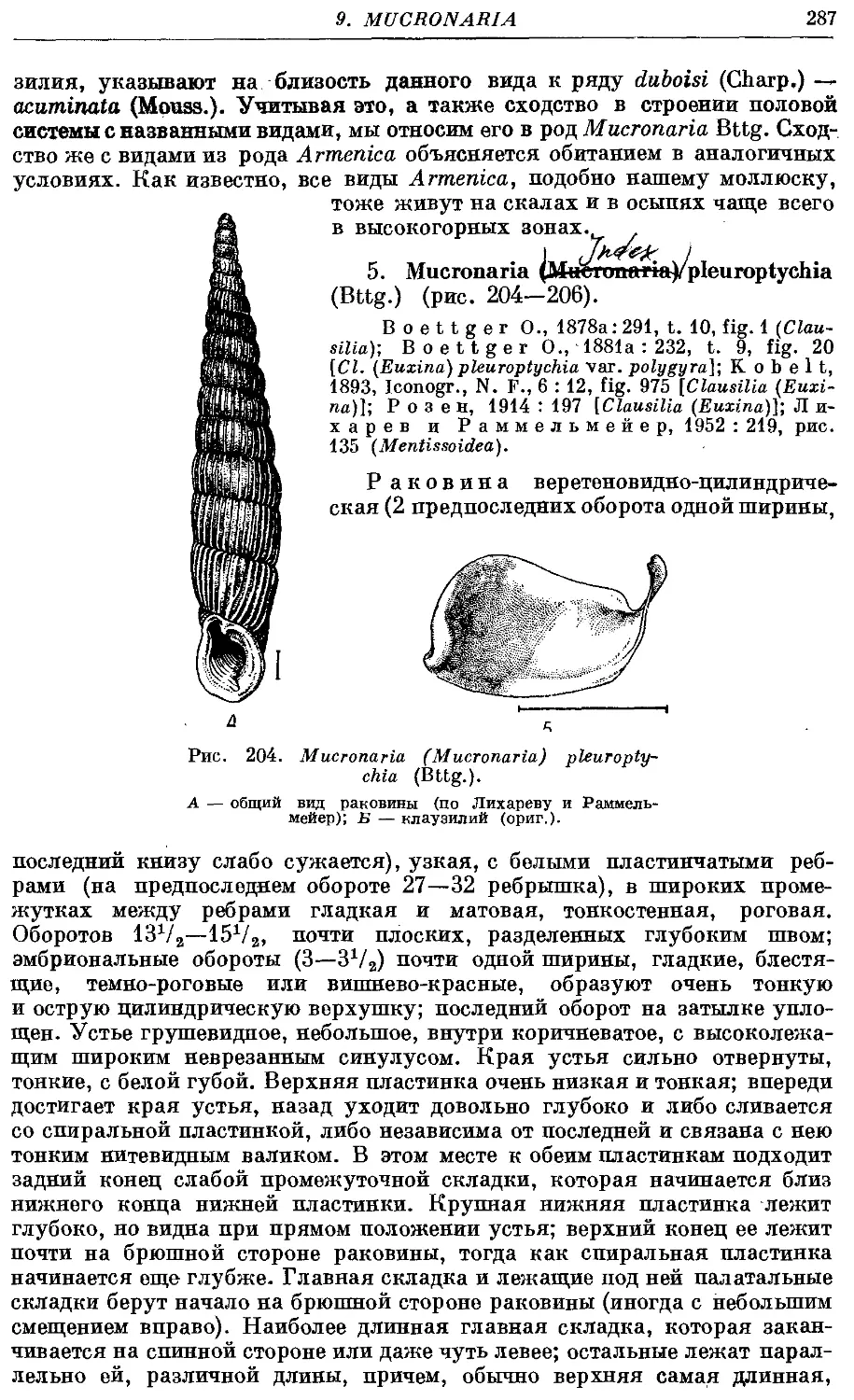
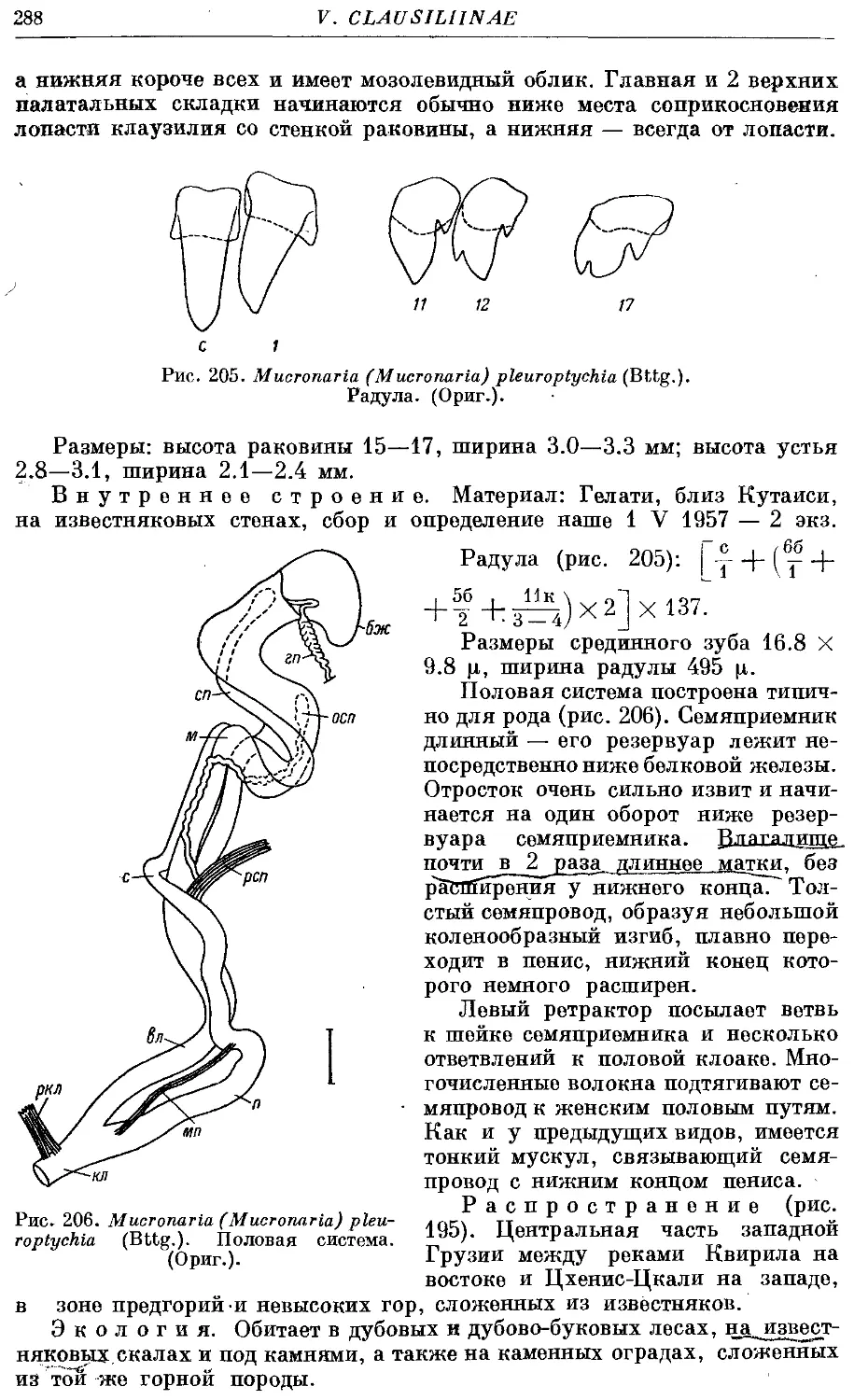
5. M. (M.) pleuroptychia (Bttg.) 287
2. Подрод Index Bttg.
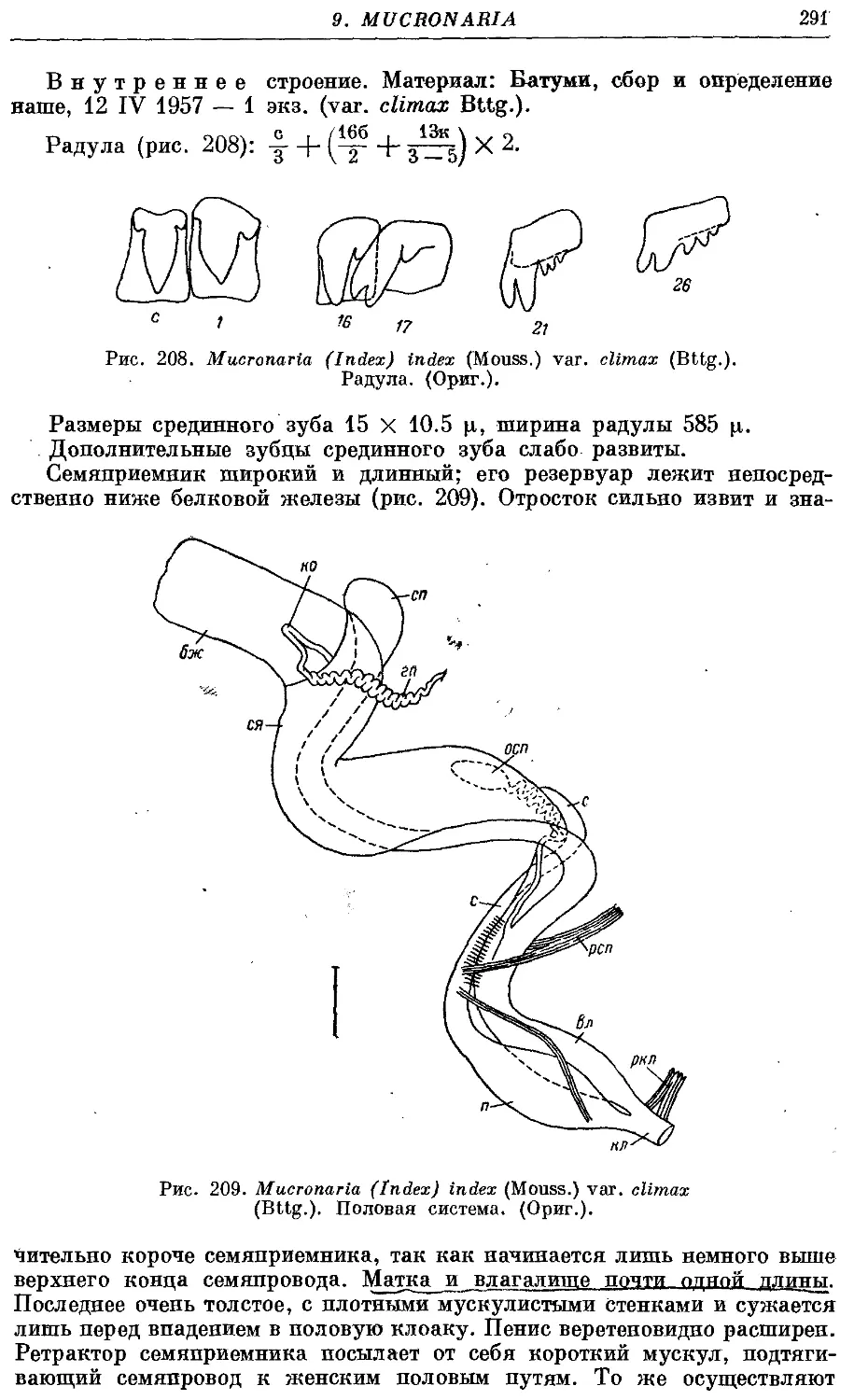
6. M. (I.) index (Mouss.) 289
10. Род Micropontica Bttg.
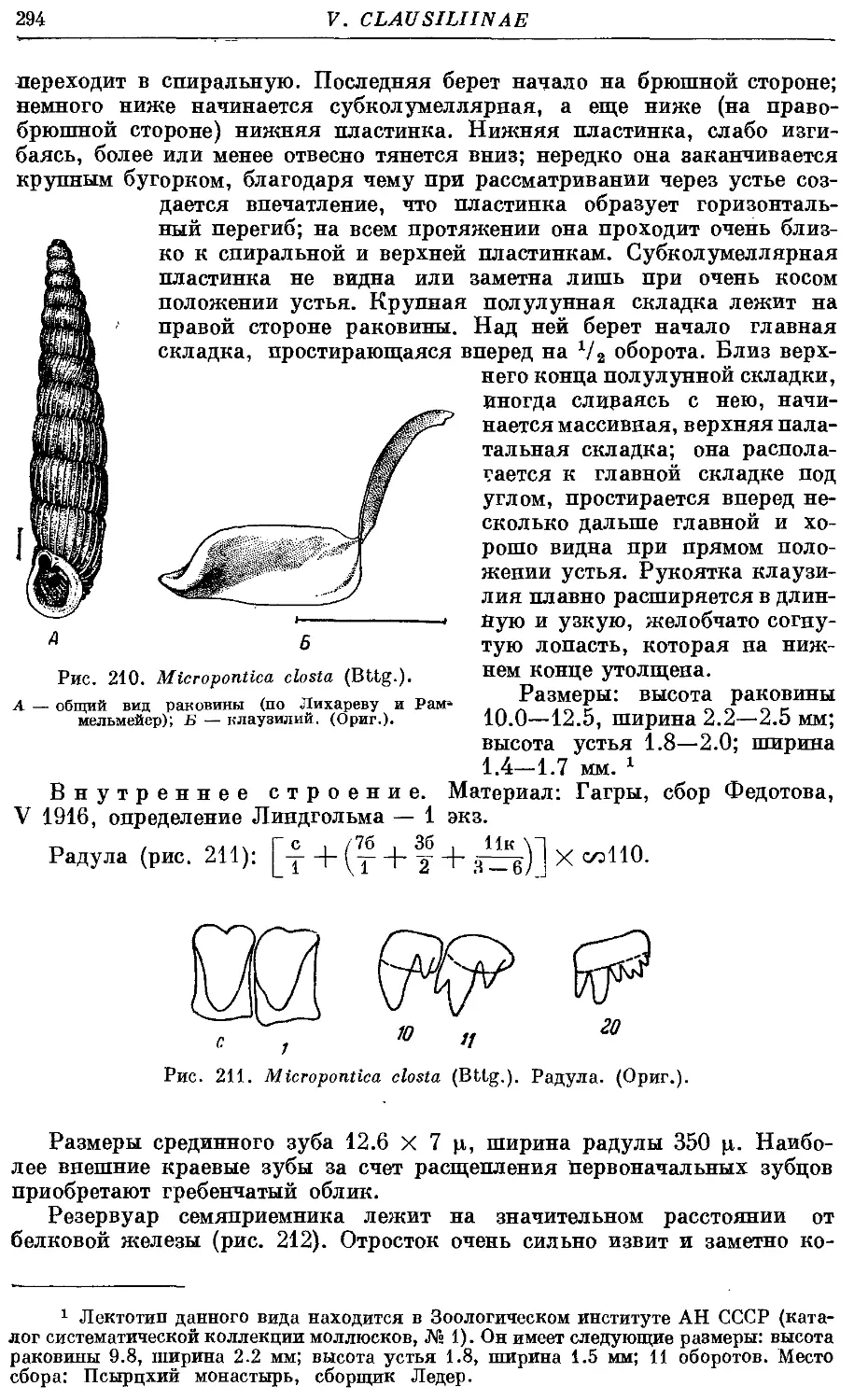
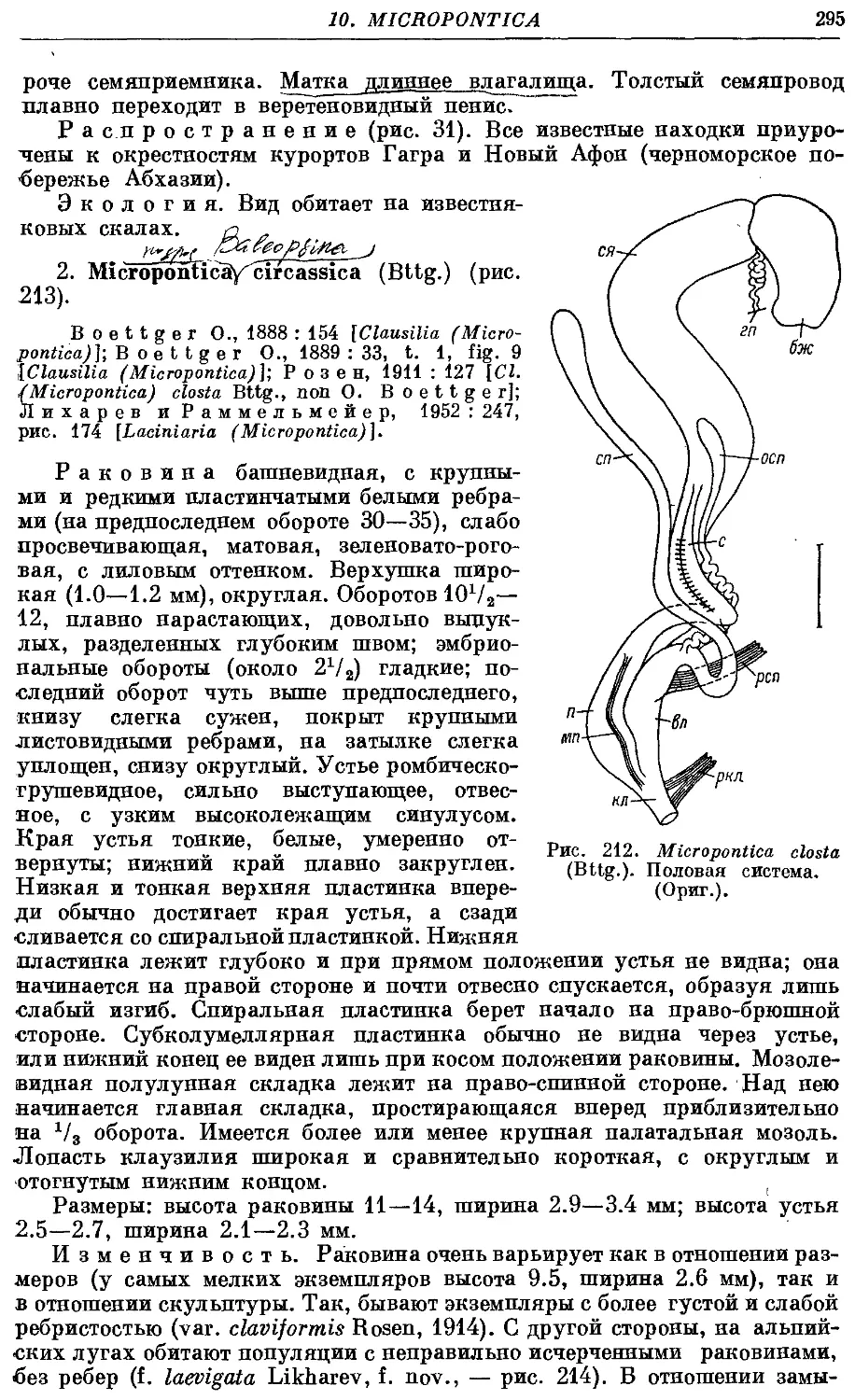
1. M. closta (Bttg.) 293
2. M. circassica (Bttg.) 295
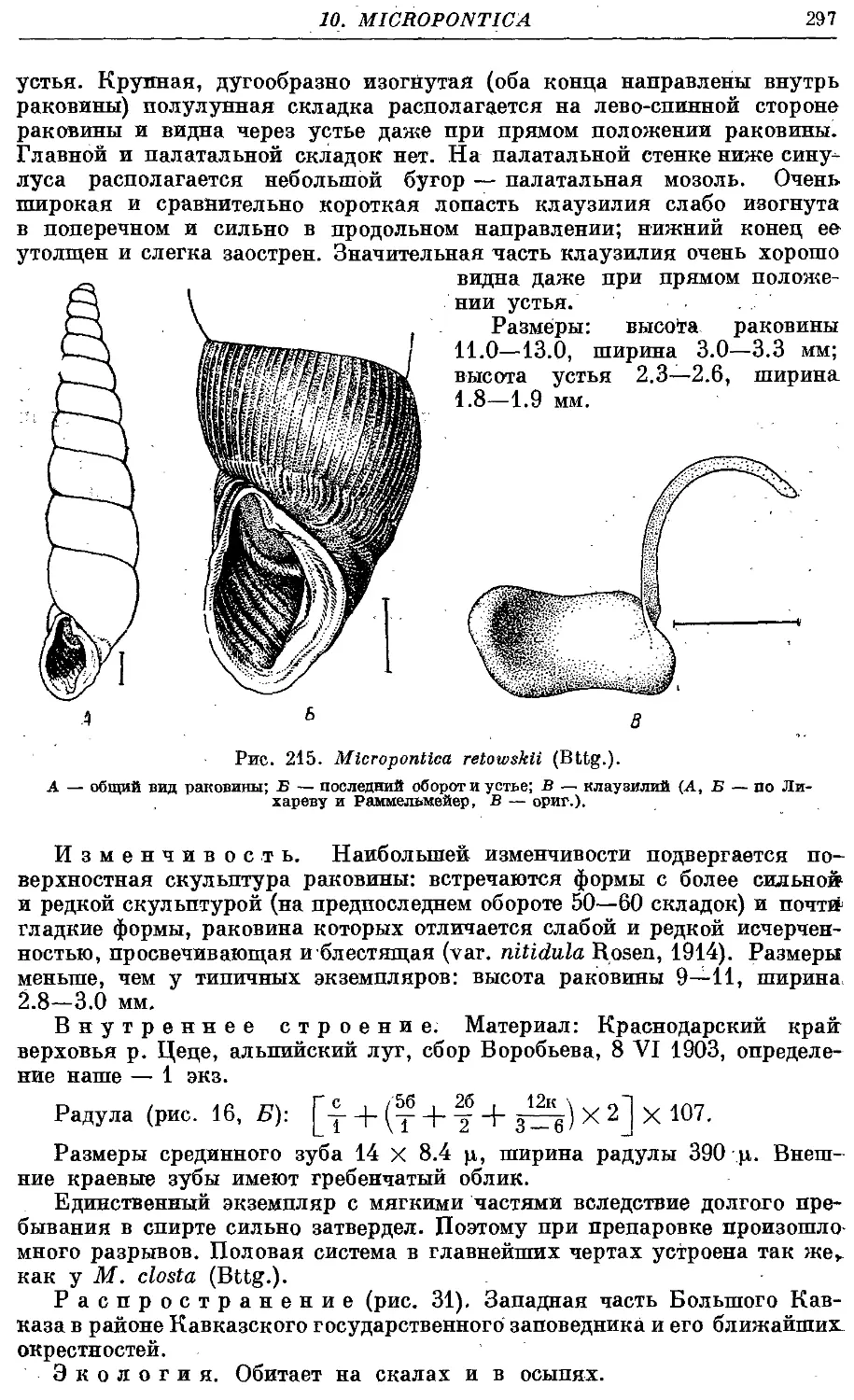
3. M. retowskii (Bttg.) 296
ВВЕДЕНИЕ
ОБЩАЯ ХАРАКТЕРИСТИКА МОЛЛЮСКОВ СЕМЕЙСТВА CLAUSILIIDAE
Раковина высокая, левозакрученная, реже правозакрученная, сложена
большим числом оборотов (до 18), веретеновидная, реже башневидная.
Устье относительно небольшое, от неправильно яйцевидного до грушевид-
грушевидного, чаще всего выступающее. Край устья цельный, реже наверху прер-
прерван, но места прикрепления к раковине соединены париетальной мозолью.
Внутри последнего оборота раковины располагается сложная скульптура—
замыкательный аппарат. Он состоит из пластинок (на париетальной стенке
и столбике), складок (на палатальной стенке) и клаузилия. Последний
представляет собой полуподвижное образование, прикрепляющееся верх-
верхним концом к столбику и свободно лежащее внутри последнего оборота.
Клаузилий служит для замыкания входа в раковину (clausilium, от латин-
латинского слова clausi — замыкать), когда моллюск втянулся внутрь ее.
Нога, спина и голова небольших размеров. Кожа с довольно резкими
морщинами. Подошва ноги гладкая, не разбита на продольные полосы.
Челюсть — дугообразно изогнутая роговая пластинка с выступом
на середине внешнего края. Радула слагается из роговой мембраны и
большого числа поперечных рядов мелких зубов.
Легкое сильно вытянуто в длину, узкое. Почка вместе с сердцем лежит
у заднего конца легкого. От почки отходит назад первичный мочеточник,
который затем переходит во вторичный мочеточник, простирающийся
вперед вдоль задней кишки вплоть до ануса, рядом с которым он откры-
открывается наружу.
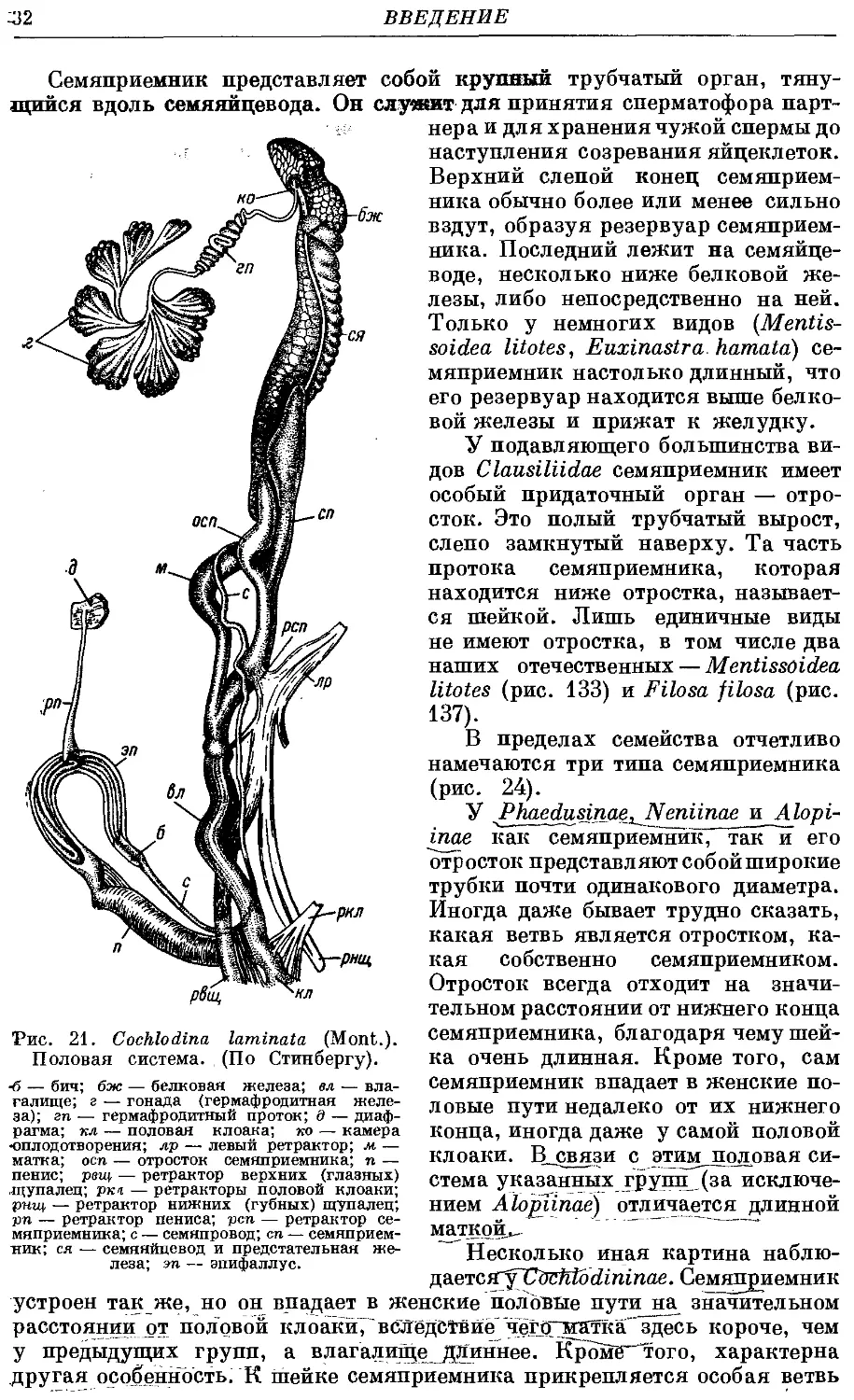
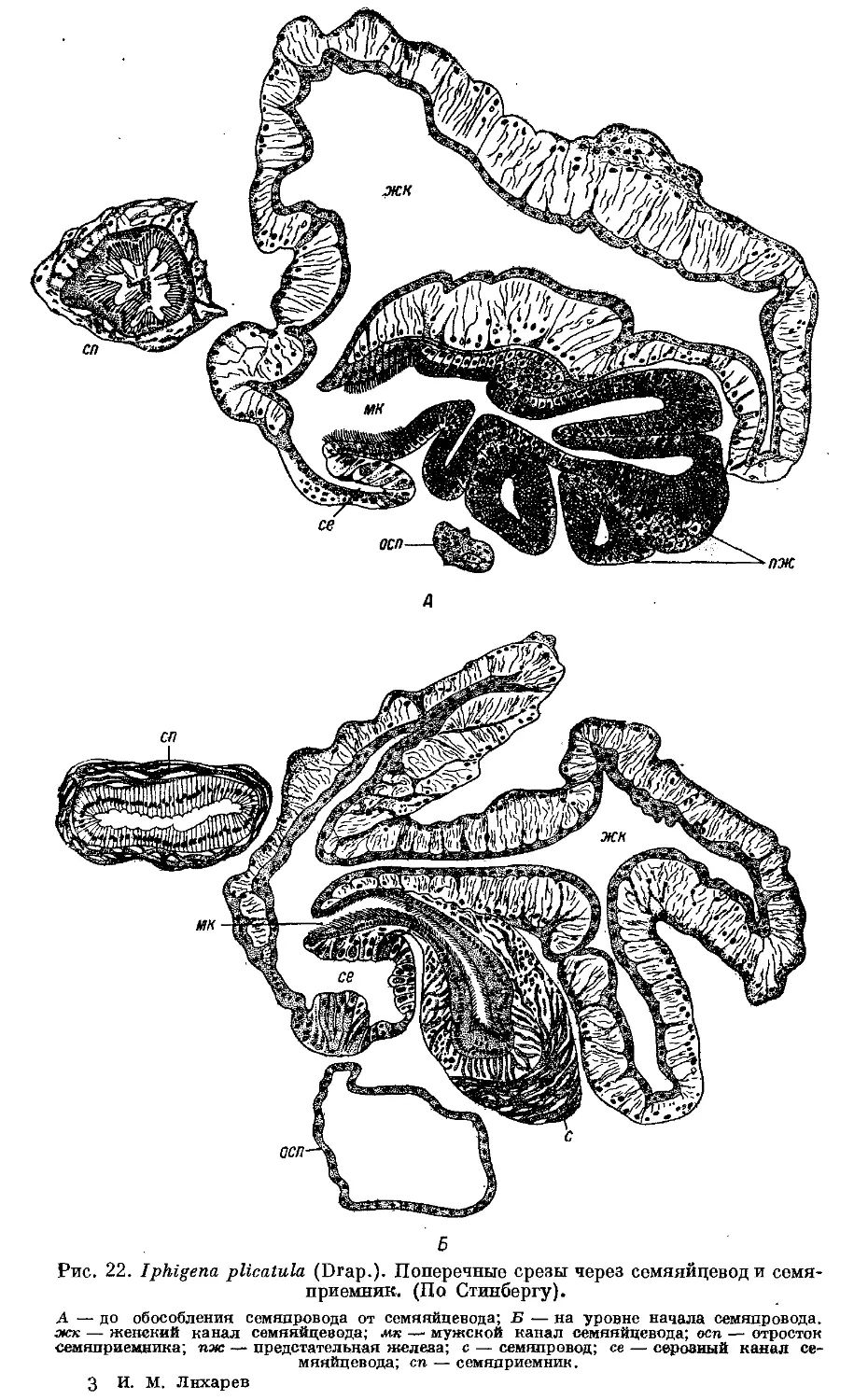
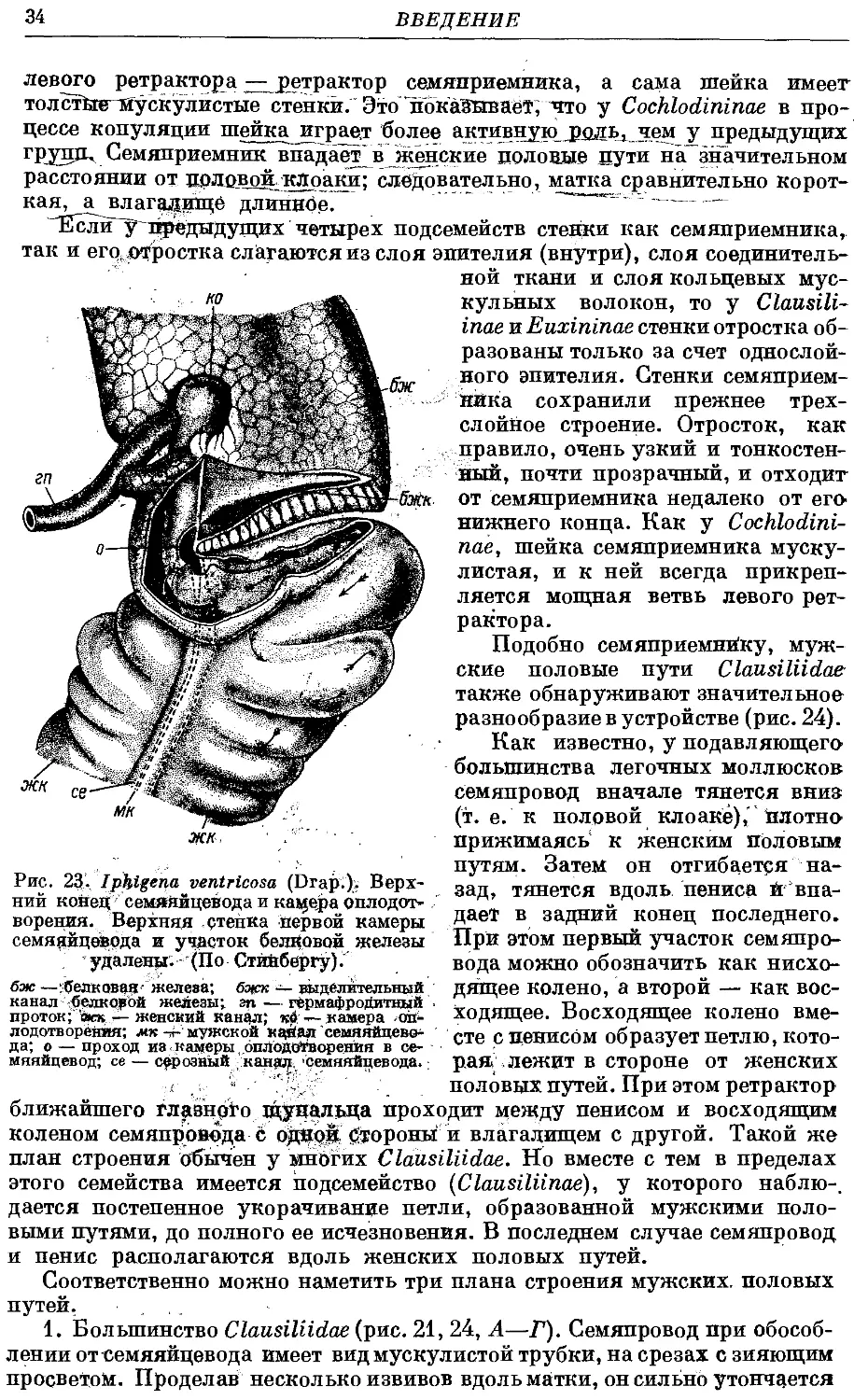
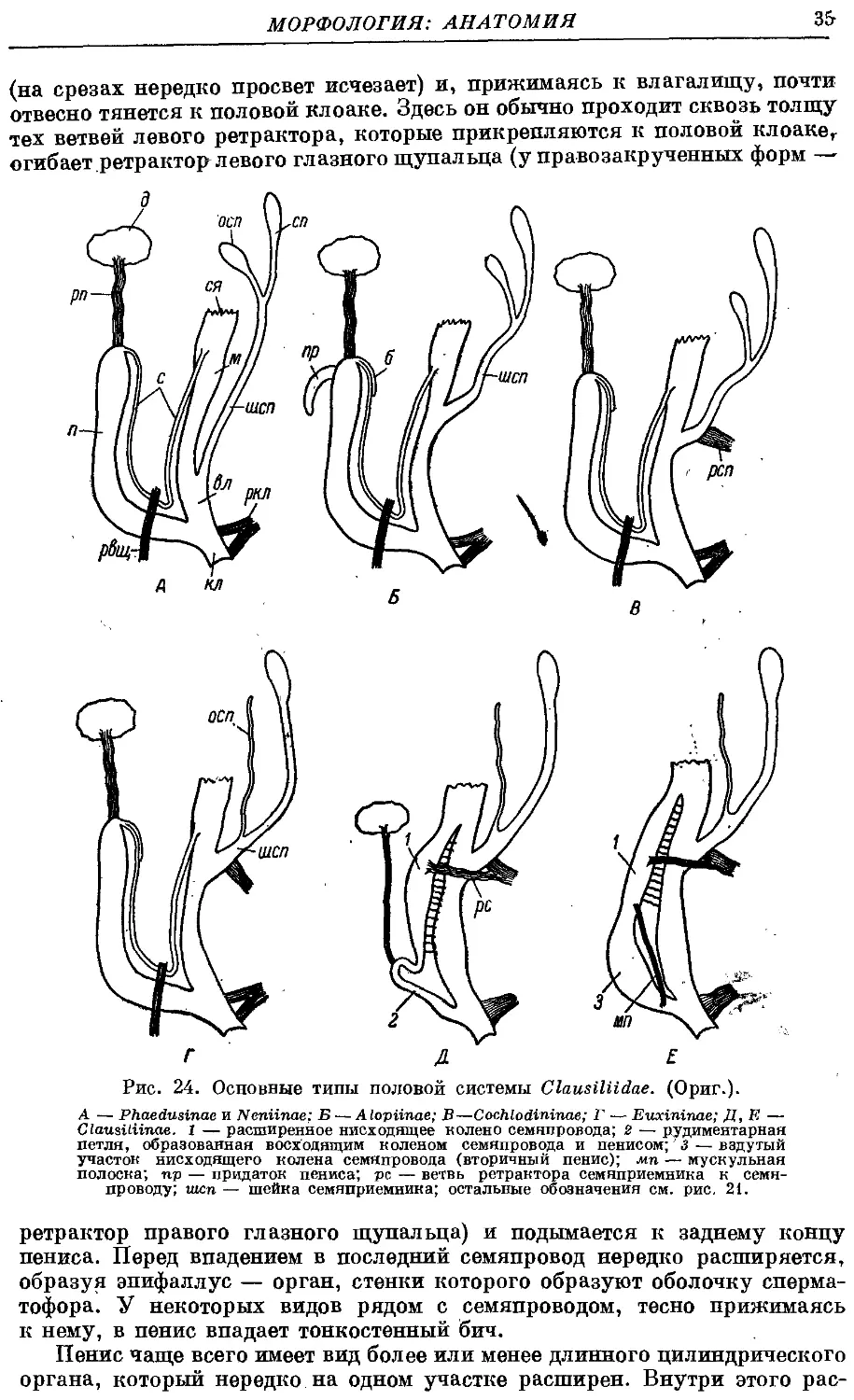
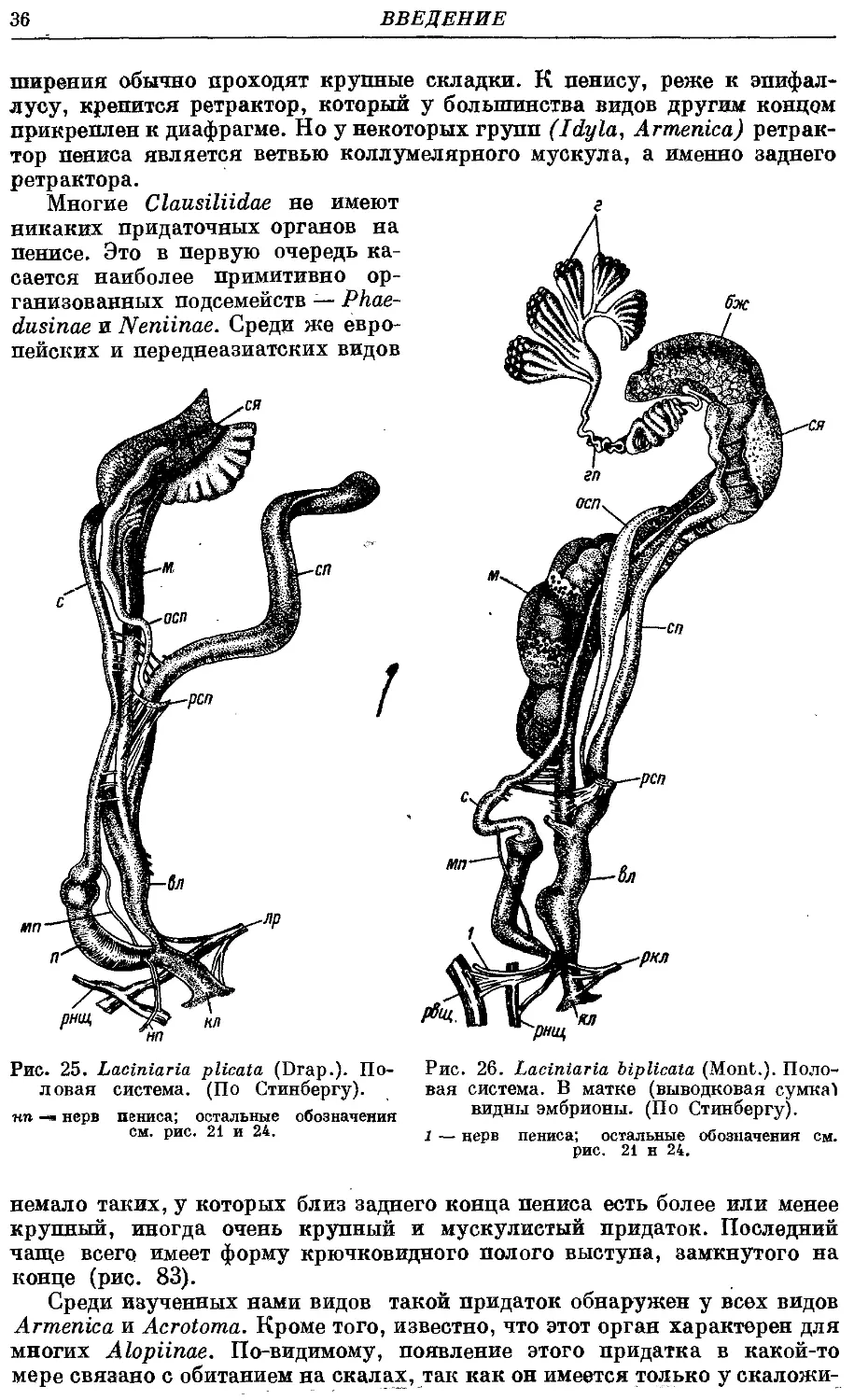
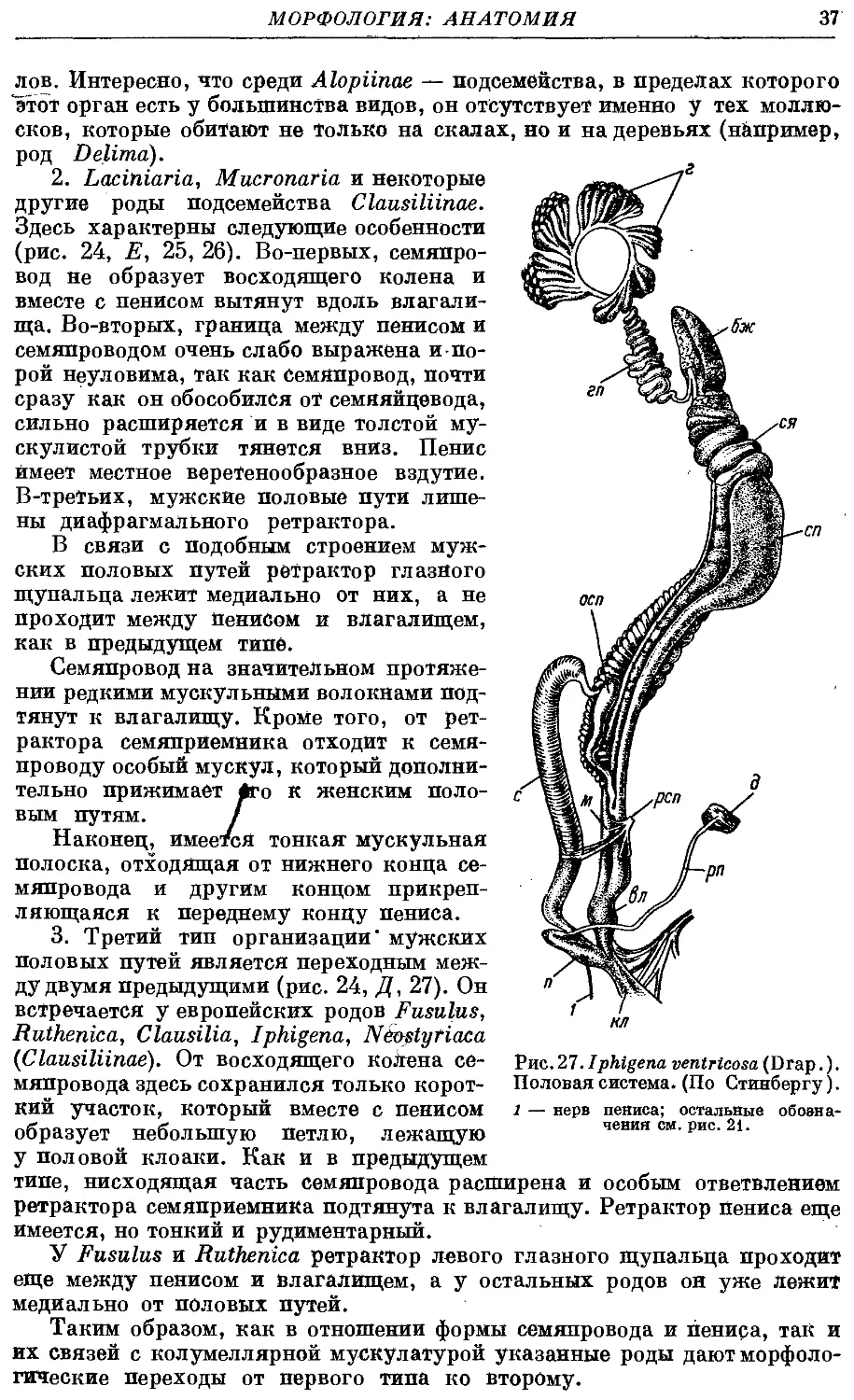
Гермафродитный половой аппарат характерен отсутствием придаточных
органов на женских половых путях. Семяприемник очень вытянут и у
подаяляющего большинства видов имеет отросток. Мужские половые пути
построены петлеобразно и имеют ретрактор пениса; однако у ряда родов
они вытянуты вдоль женских и лишены ретрактора.
Колумеллярная мускулатура образована 4 главными ретракторами
тела — левый передний, правый передний, глоточный и задний. Один
из передних ретракторов дает несколько ответвлений к женским половым
путям и к клоаке.
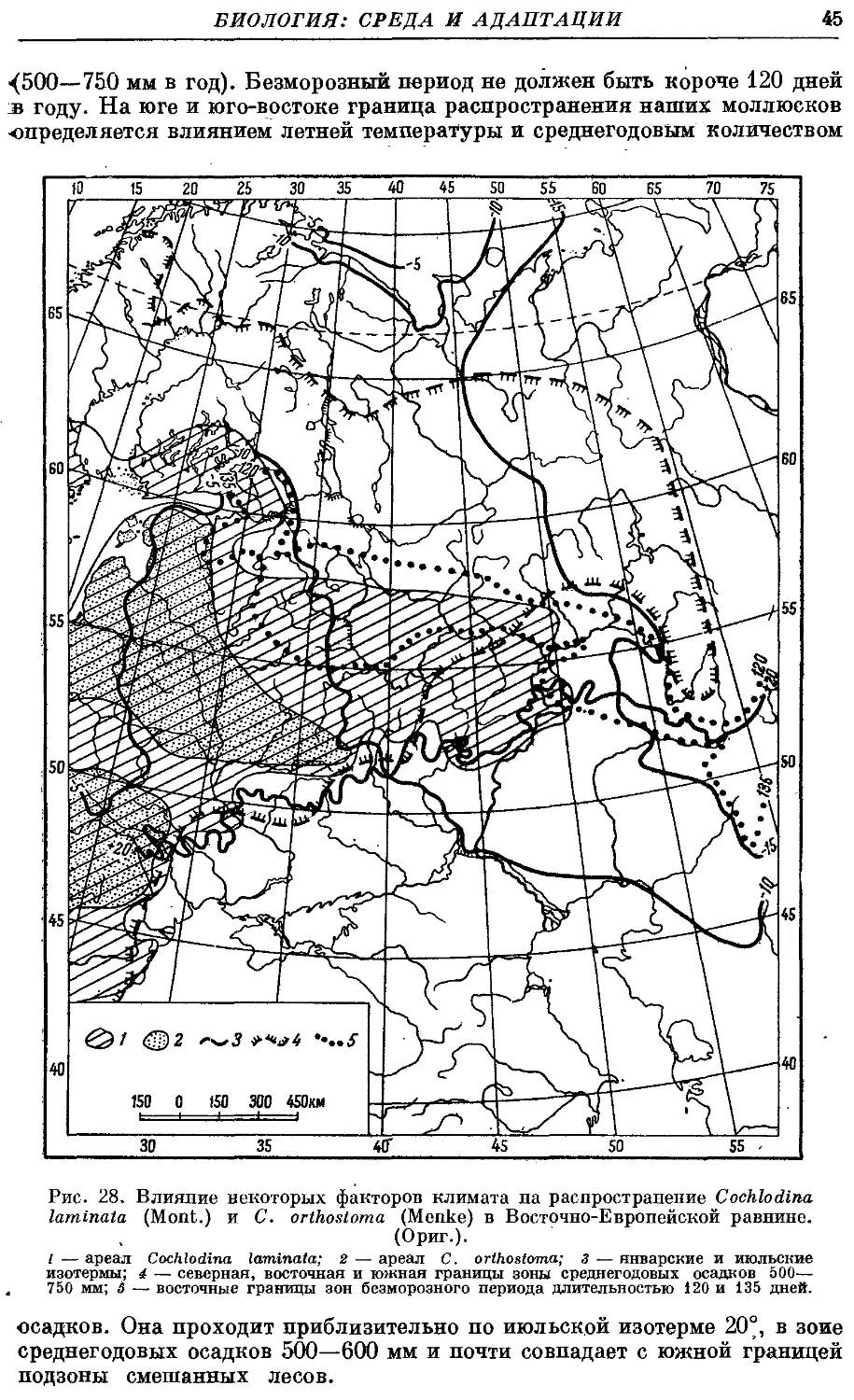
Ареал семейства состоит из трех участков, изолированных друг от
друга: 1) Европа, северная Африка и часть Передней Азии с Кавказом;
2) южная и восточная Азия (к северу, включая Японию и южную Корею);
3) северо-западные области Южной Америки.
В ископаемом состоянии раковины данной группы распространены
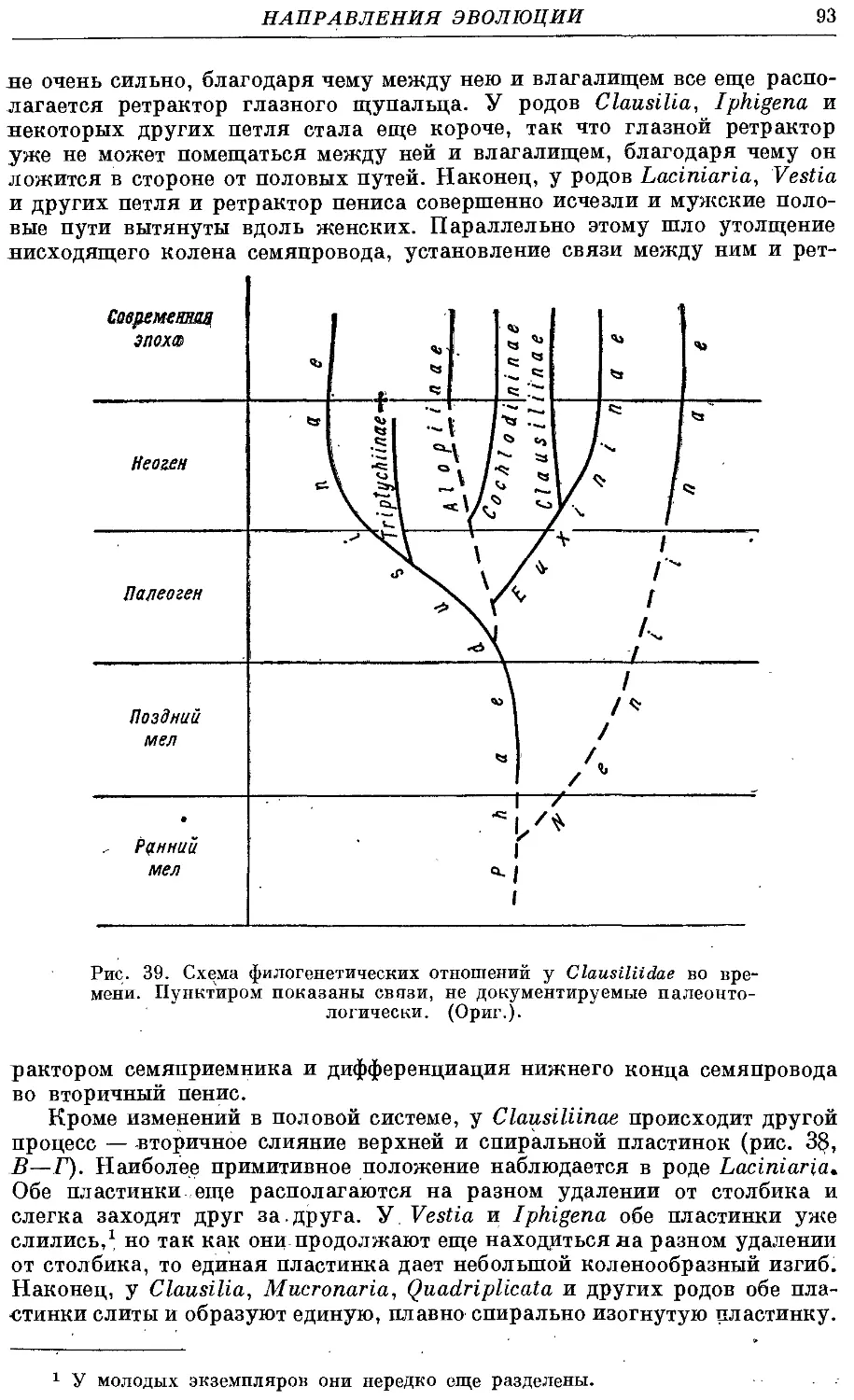
начиная с позднего мела (преимущественно в Европе).
В мировой фауне семейство содержит около 1500 видов, а в фауне
СССР — 70 видов.
Семейство Clausiliidae входит в состав отряда стебельчатоглазых
(Stylommatophora) подкласса легочных (Pulmonata) и класса брюхоногих
моллюсков (Gastropoda).
МОРФОЛОГИЯ: ВНЕШНИЙ ВИД, РАКОВИНА
11
МОРФОЛОГИЯ CLAUSILIIDAE
Внешний вид
Тело животного состоит из относительно небольшой ноги, головы и
длинного внутренностного мешка (рис. 1 и 13). Раковина всегда наружная
и хорошо развита; как правило, значительно крупнее мягких частей тела,
благодаря чему, когда животное втяги-
втягивается глубоко в раковину, нижние обо-
обороты ее остаются пустыми (рис. 2).
Голова несет пару верхних, или
глазных, щупалец, ниже и впереди ко-
которых располагается пара нижних, или
губных, щупалец. Подошва ноги глад-
гладкая и не разделена на продольные по-
полосы, как это имеет место у многих дру-
других семейств стебельчатоглазых. Верх-
Верхняя сторона ноги и голова с шеей по-
покрыты сетью поперечных и продольных
бороздок. Окраска тела серая или почти
черная; подошва, как правило, окра-
окрашена несколько светлее.
Подавляющее большинство видов,
в частности все виды,, живущие в СССР,
являются левозакрученными (синист-
ральными). Но в некоторых группах
семейства, особенно часто среди подсе-
подсемейства Alopiinae, имеются отдельные
правозакрученные (декстральные) виды
и даже роды.
У левозакрученных форм направле-
направлению спирали соответствует .и внутрен-
внутренняя асимметрия организации, внешне
выражающаяся в том, что не только
устье раковины располагается слева, но и на той же стороне
лежат дыхательное и анальное отверстия и отверстие мо-
мочеточника. Половое отверстие, напротив, располагается на
правой стороне. У право закрученных форм те же органы располагаются
соответственно наоборот.
Раковина
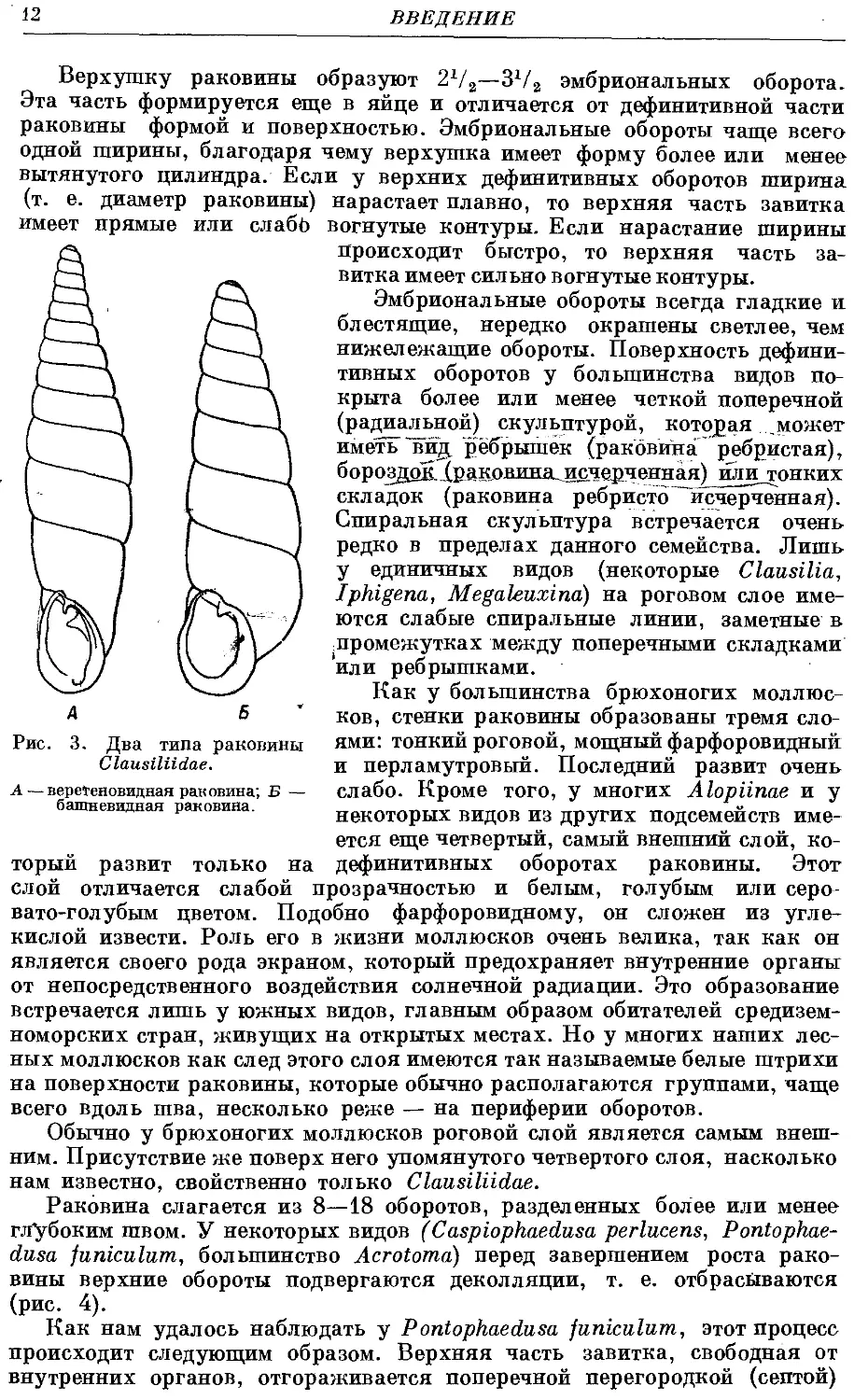
Раковина всех видов представляет собой сильно вытянутую Турбо-
спираль. По форме она бывает двух основных типов (рис. 3): 1) веретено-
видная — когда один или два последних (нижних) оборота плавно
сужаются к устью; 2) башневидная — когда раковина плавно расширяется
от верхушки до устья.
Между этими типами есть переходы. В том случае, когда раковина
более близка к первому типу, мы ее называем башневидно-веретеновидной,
когда она ближе ко второму типу — то веретеновидно-башневидной.
Если средняя часть раковины на значительном протяжении не изменяется
в ширину, то такие раковины мы обозначаем как веретеновидно-цилиндри-
ческие. Раковины бывают стройные — когда нарастание и убывание ширины
идет плавно, и вздутые — когда нижняя часть раковины очень широка
ж вся раковина имеет приземистый облик.
Рис. 1. Внешний вид
моллюска из сем.
Clausiliidae при пол-
ползании по вертикаль-
вертикальной поверхности.
(Ориг.).
Рис. 2. По-
Положение
мягких ча-
частей тела
при глубо-
глубоком втяги-
втягивании в ра-
раковину.
(Ориг.).
12
ВВЕДЕНИЕ
Верхушку раковины образуют 2V2—3V2 эмбриональных оборота.
Эта часть формируется еще в яйце и отличается от дефинитивной части
раковины формой и поверхностью. Эмбриональные обороты чаще всего
одной ширины, благодаря чему верхушка имеет форму более или менее
вытянутого цилиндра. Если у верхних дефинитивных оборотов ширина
(т. е. диаметр раковины) нарастает плавно, то верхняя часть завитка
имеет прямые или слабь вогнутые контуры. Если нарастание ширины
Происходит быстро, то верхняя часть за-
завитка имеет сильно вогнутые контуры.
Эмбриональные обороты всегда гладкие и
блестящие, нередко окрашены светлее, чем
нижележащие обороты. Поверхность дефини-
дефинитивных оборотов у большинства видов по-
покрыта более или менее четкой поперечной
(радикальной) скульптурой, которая..„может
иметь Tmjj ребрышек (раковина ребристая),
бороздо?._Хр_ак„овина.дсчерченная) или тонких
складок (раковина ребрист6~~йсчерченная).
Спиральная скульптура встречается очень
редко в пределах данного семейства. Лишь
у единичных видов (некоторые Clausilia,
Iphigena, Megaleuxina) на роговом слое име-
имеются слабые спиральные линии, заметные в
промежутках между поперечными складками
или ребрышками.
Как у большинства брюхоногих моллюс-
моллюсков, стенки раковины образованы тремя сло-
слоями: тонкий роговой, мощный фарфоровидный
и перламутровый. Последний развит очень
слабо. Кроме того, у многих Alopiinae и у
некоторых видов из других подсемейств име-
имеется еще четвертый, самый внешний слой, ко-
который развит только на дефинитивных оборотах раковины. Этот
слой отличается слабой прозрачностью и белым, голубым или серо-
серовато-голубым цветом. Подобно фарфоровидному, он сложен из угле-
углекислой извести. Роль его в жизни моллюсков очень велика, так как он
является своего рода экраном, который предохраняет внутренние органы
от непосредственного воздействия солнечной радиации. Это образование
встречается лишь у южных видов, главным образом обитателей средизем-
средиземноморских стран, живущих на открытых местах. Но у многих наших лес-
лесных моллюсков как след этого слоя имеются так называемые белые штрихи
на поверхности раковины, которые обычно располагаются группами, чаще
всего вдоль шва, несколько реже — на периферии оборотов.
Обычно у брюхоногих моллюсков роговой слой является самым внеш-
внешним. Присутствие же поверх него упомянутого четвертого слоя, насколько
нам известно, свойственно только Clausiliidae.
Раковина слагается из 8—18 оборотов, разделенных более или менее
глубоким швом. У некоторых видов (Caspiophaedusa perlucens, Pontophae-
dusa funiculum, большинство Acrotoma) перед завершением роста рако-
раковины верхние обороты подвергаются деколляции, т. е. отбрасываются
(рис. 4).
Как нам удалось наблюдать у Pontophaedusa funiculum, этот процесс
происходит следующим образом. Верхняя часть завитка, свободная от
внутренних органов, отгораживается поперечной перегородкой (септой)
Рис. 3. Два типа раковины
Clausiliidae.
А — вере!геновидная раковина; Б —
башневидная раковина.
МОРФОЛОГИЯ: РАКОВИНА
13
от остальной раковины. Одновременно, непосредственно выше перегородки,
внутренняя поверхность раковины подвергается явственной коррозии,
возможно, . за счет выделения организмом каких-то ли-
зирующих веществ. После этого достаточно небольшого
давления, чтобы отбросить верхнюю часть завитка.
Таким образом, в противоположность старым взглядам
на то, что деколляция происходит чисто механически, пу-
путем отламывания оборотов, лежащих выше септы, по-
видимому , у Clausiliidae в этом деле важную роль
играют какие-то ферменты, которые разъедают внут-
внутреннюю поверхность раковины на месте будущего излома.
У большинства наших моллюсков раковина не имеет
открытого пупка, т. е. отверстия, располагающегося на
нижней стороне ее, ведущего во внутренний канал стол-
столбика. Изредка пупок имеет вид довольно широкой щели,
но чаще всего он либо полностью закрыт отворотом колу-
меллярного края устья, либо чуть заметен, как очень уз-
узкая щель. г
Форма последнего оборота имеет важное значение при
характеристике как отдельных видов, так и целых групп.
Уже отмечалось, что у большинства Clausiliidae он более
или менее сужен в направлении к устью. У видов под- Рис- 4- Роп~
семейств Euxininae и Clausiliinae нижняя сторона послед-
последнего оборота вытянута в более или менее резкий, длинный
или короткий киль (рис. 5). У остальных подсемейств послед-
tophaedusa
funiculum
(Mouss.).
Вершина
ний оборот снизу плавно закруглен. Если киль развит раковины
хорошо, то он окружает область пупка, образуя так назы- ^ана°ЛЛИ(По
ваемое околопупочное поле. Нередко, со стороны поля, Лихареву и
вдоль киля тянется тонкая, окаймляющая его б,ороздка. Раммель-
Та часть последнего оборота, которая находится непосред- мейер).
ственно позади устья, называется затылком раковины. Этот
участок нередко уплощен иди даже вогнут внутрь (Idyla foveicollis —
рис. 66). Иногда на затылке, в непосредствен-
непосредственной близости от края устья, лежит особый
поперечный гребень, или утолщение (рис. 84).
Устье раковины чаще всего имеет груше-
грушевидную или округло-ромбическую форму.
Край устья заканчивается тонкой оторочкой,
или перистомом, расширен и отвернут. У
большинства видов устье цельное (рис. 4),
т. е. предпоследний оборот не разрывает
верхнего края устья. Сравнительно у нем-
немногих видов предпоследний оборот нару-
нарушает связь между краями устья, и обычно
в этом случае места прикрепления верх-
верхнего края устья связаны мозолью (рис.
65).
Нередко устье выступает вперед, т. е.
последний оборот под конец отходит от ос-
остальной раковины и свободно выдается впе-
вперед. В этом случае позади верхнего края устья
образуется своеобразная площадка, связанная с околопупочным полем.
Палатальный, или внешний, край устья нередко окаймлен тонкой
губой. У некоторых видов отступя от губы, т. е. глубже в устье, находится
Рис. 5. Mentissoidea litotes (A.
Schm.). Вид раковины снизу.
(Ориг.).
1 — КИЛЬ.
14 ВВЕДЕНИЕ
палатальная мозоль, имеющая вид довольно широкого валика, вытянутого
параллельно краю устья. Слева от столбика, нередко- глубоко врезаясь
в нижнюю часть палатального края устья, лежит базальный желобок.
По сути дела, его возникновение связано с образованием киля на
внешней стенке последнего оборота. Как и киль, базальный желобок
наиболее характерен для Euxininae и Clausiliinae и отсутствует у других
подсемейств.
Внутри последнего оборота раковины располагается своеобразная
скульптура, состоящая из различных складок, служащих для замыкания
входа в раковину. В связи с этим все эти образования объединяются под
понятием «замыкательный аппарат». Учитывая важность этой скульптуры
в жизнедеятельности наших моллюсков, а также для их систематики,
описание ее дано ниже.
Замыкательный аппарат
Строение. Как уже указывалось, в глубине последнего оборота рако-
раковины на различном расстоянии от устья залегает своеобразная внутренняя
скульптура, которую принято обозначать как замыкательный аппарат.
Он состоит из трех типов образований — пластинок, складок и клаузилия
(рис. 6).
Все эти образования, за исключением клаузилия, располагаются на
внутренней стенке последнего оборота раковины, Клаузилий свободно
лежит в просвете раковины. Пластинки находятся на париетальной стенке
и на столбике. Складки, за исключением промежуточной, располагаются
на палатальной стенке.
На париетальной стенке, заканчиваясь вблизи от верхнего края или
даже выступая за его пределы, лежит верхняя пластинка (lamella
superior). Чаще всего она плавно нарастает в высоту в направлении к устью
и круто снижается почти у самого края его. Левый верхний угол устья,
отграниченный верхней пластинкой, называется синулусом. Нередко он
более или менее значительно врезан вглубь париетальной стенки и как
бы отогнут вправо, к оси раковины. Позади верхней пластинки, г также
располагаясь на париетальной стенке последнего оборота, лежит другая
пластинка — спиральная (lamella spiralis). Она начинается на различной
глубине: у одних близ границы между последним и предпоследним обо-
оборотами, у других ближе к устью.
Очень важным признаком является положение спиральной пластинки
по отношению к верхней (рис. 7). Тут можно наметить три основных вари-
варианта: 1) передний конец спиральной пластинки более или менее глубоко
заходит за задний конец верхней, причем первая лежит ближе к внешней
стенке раковины, а последняя ближе к столбику; 2) спиральная пластинка
более или менее плавно переходит в верхнюю, образуя с нею единое целое;
3) обе пластинки разделены промежутком различной величины, но их
концы располагаются на одной линии, т. е. на одинаковом удалении
от столбика.
Обе названные пластинки располагаются только на париетальной
стенке последнего оборота. Нижняя же пластинка (lamella inferior) берет
начало в глубине последнего оборота. Верхний конец ее на большем или
меньшем протяжении тянется по париетальной стенке параллельно спи-
1 Здесь, как и в дальнейшем, началом пластинок и складок мы считаем их концы,
лежащие в глубине раковины. Эти концы можно также называть верхними, или зад-
задними, а противоположные им — нижними, или передними.
МОРФОЛОГИЯ: ЗАМЫКАТЕЛЬНЫЙ АППАРАТ
15
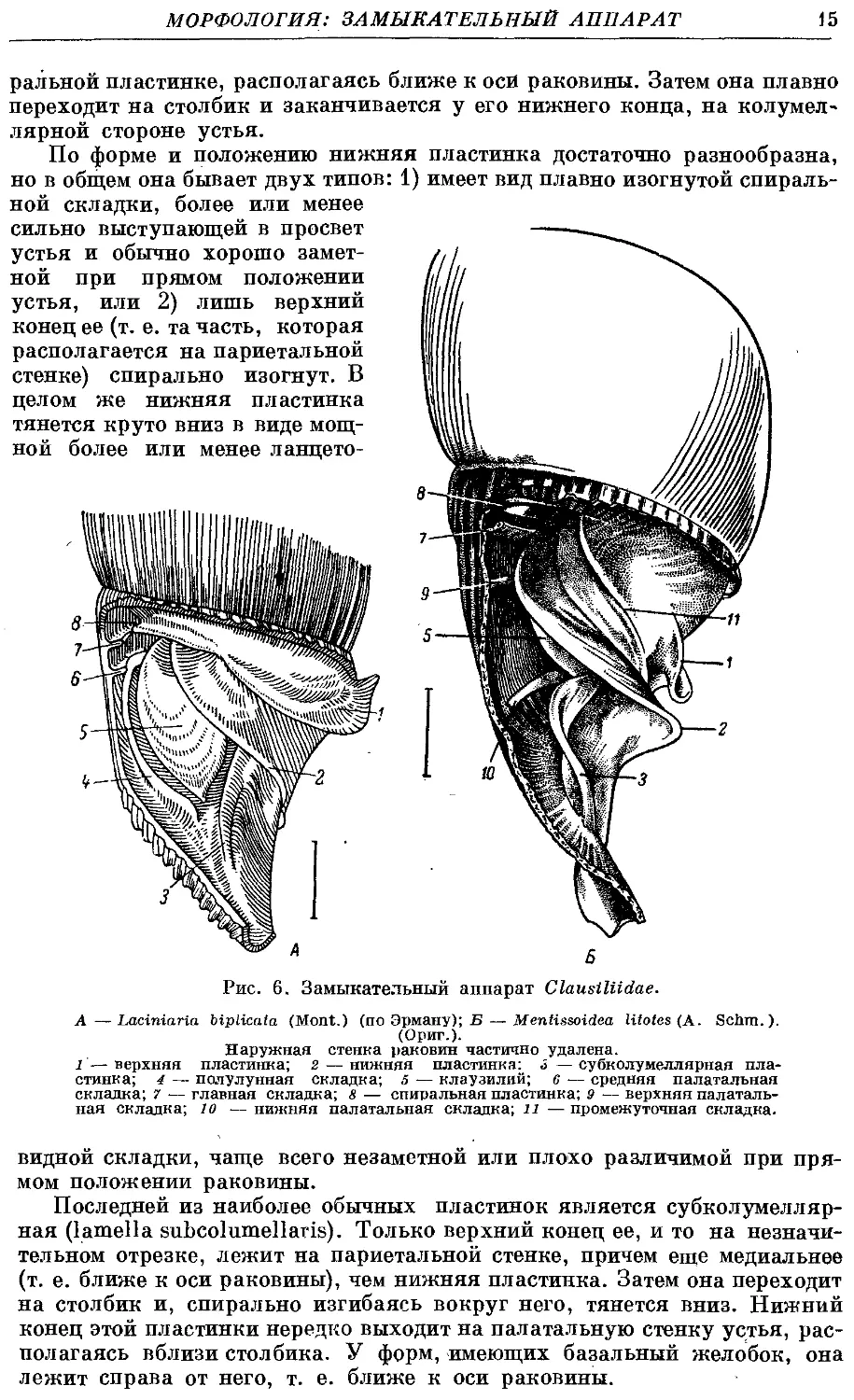
ральной пластинке, располагаясь ближе к оси раковины. Затем она плавно
переходит на столбик и заканчивается у его нижнего конца, на колумел-
лярной стороне устья.
По форме и положению нижняя пластинка достаточно разнообразна,
но в общем она бывает двух типов: 1) имеет вид плавно изогнутой спираль-
спиральной складки, более или менее
сильно выступающей в просвет
устья и обычно хорошо замет-
заметной при прямом положении
устья, или 2) лишь верхний
конец ее (т. е. та часть, которая
располагается на париетальной
стенке) спирально изогнут. В
целом же нижняя пластинка
тянется круто вниз в виде мощ-
мощной более или менее ланцето-
k-
А 6
Рис. 6. Замыкателышй аппарат Clausiliidae.
А—Laciniaria biplicata (Mont.) (по Эрману); Б — Mentissoidea litotes (A. Schm.).
(Ориг.).
Наружная стенка раковин частично удалена.
1 — верхняя пластинка; 2 — нижняя пластинка: 3 — субколумеллярная пла-
пластинка; 4 — полулунная складка; 5 — клаузилий; 6 — средняя палатальная
складка; 7 — главная складка; 8 — спиральная пластинка; 9 — верхняя палаталь-
палатальная складка; 10 — нижняя палатальная складка; и — промежуточная складка.
видной складки, чаще всего незаметной или плохо различимой при пря-
прямом положении раковины.
Последней из наиболее обычных пластинок является субколумелляр-
субколумеллярная (lamella subcolumellaris). Только верхний конец ее, и то на незначи-
незначительном отрезке, лежит на париетальной стенке, причем еще медиальнее
(т. е. ближе к оси раковины), чем нижняя пластинка. Затем она переходит
на столбик и, спирально изгибаясь вокруг него, тянется вниз. Нижний
конец этой пластинки нередко выходит на палатальную стенку устья, рас-
располагаясь вблизи столбика. У форм, имеющих базальный желобок, она
лежит справа от него, т. е. ближе к оси раковины.
JO
ВВЕДЕНИЕ
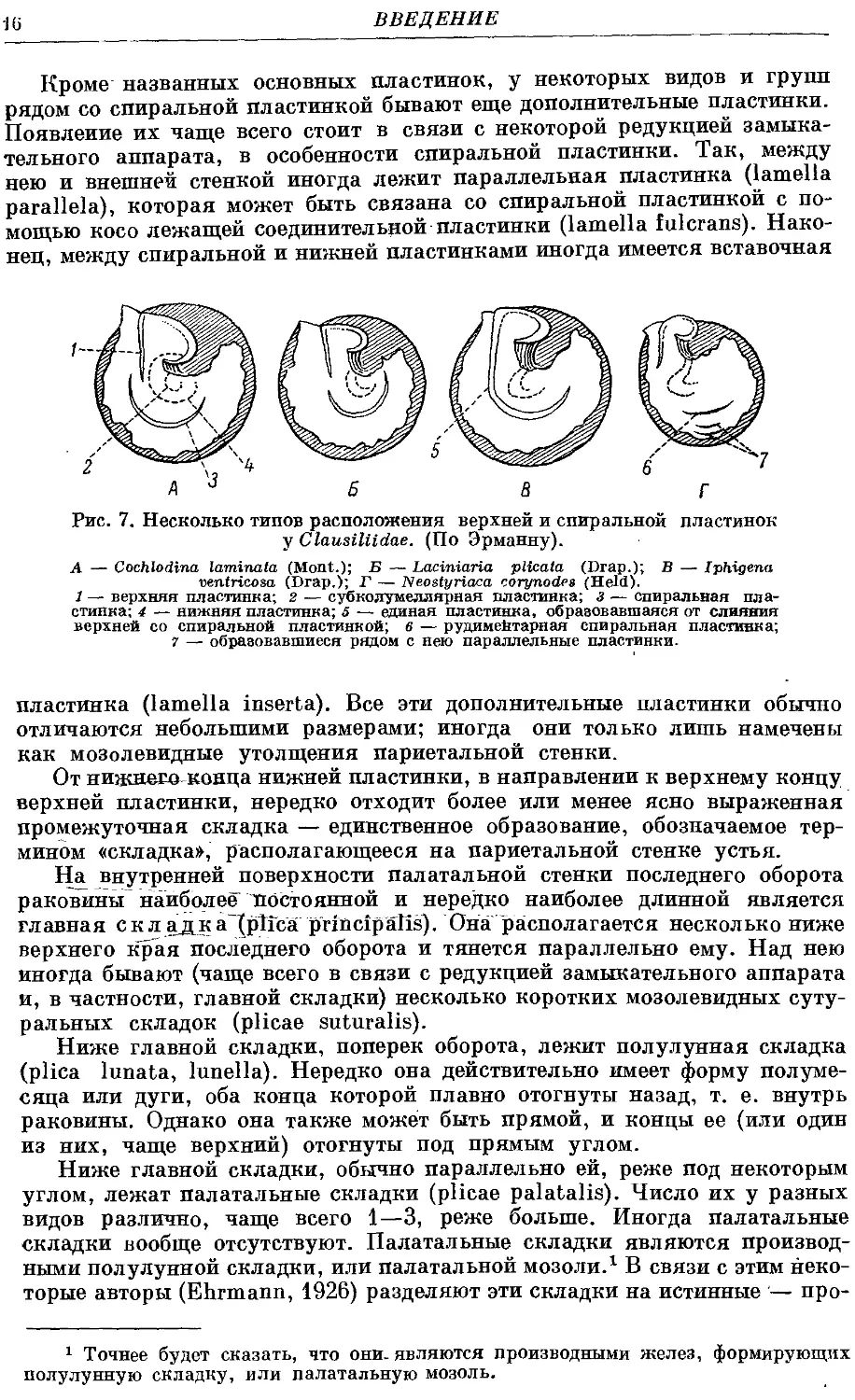
Кроме названных основных пластинок, у некоторых видов и групп
рядом со спиральной пластинкой бывают еще дополнительные пластинки.
Появление их чаще всего стоит в связи с некоторой редукцией замыка-
тельного аппарата, в особенности спиральной пластинки. Так, между
нею и внешней стенкой иногда лежит параллельная пластинка (lamella
parallela), которая может быть связана со спиральной пластинкой с по-
помощью косо лежащей соединительной пластинки (lamella fulcrans). Нако-
Наконец, между спиральной и нижней пластинками иногда имеется вставочная
Рис. 7. Несколько типов расположения верхней и спиральной пластинок
у Clausiliidae. (По Эрманну).
А — Cochlodina laminata (Mont.); Б — Laciniaria plicata (Drap.); В — Iphigena
ventricosa (Drap.); Г — Neostyriaca corynodes (Held).
1 — верхняя пластинка; г — субколумеллярная пластинка; з — спиральная пла-
пластинка; 4 — нижняя пластинка; 5 — единая пластинка, образовавшаяся от слияния
верхней со спиральной пластинкой; в — рудиментарная спиральная пластинка",
7 — образовавшиеся рядом с нею параллельные пластинки.
пластинка (lamella inserta). Все эти дополнительные пластинки обычно
отличаются небольшими размерами; иногда они только лишь намечены
как мозолевидные утолщения париетальной стенки.
От нижнего-конца нижней пластинки, в направлении к верхнему концу
верхней пластинки, нередко отходит более или менее ясно выраженная
промежуточная складка — единственное образование, обозначаемое тер-
термином «складка», располагающееся на париетальной стенке устья.
На внутренней поверхности палатальной стенки последнего оборота
раковины наиболее' постоянной и нередко наиболее длинной является
главная скл адкаП[р1Гса principaHs). Она[располагается несколько ниже
верхнего края последнего оборота и тянется параллельно ему. Над нею
иногда бывают (чаще всего в связи с редукцией замыкательного аппарата
и, в частности, главной складки) несколько коротких мозолевидных суту-
ральных складок (plicae suturalis).
Ниже главной складки, поперек оборота, лежит полулунная складка
(plica lunata, lunella). Нередко она действительно имеет форму полуме-
полумесяца или дуги, оба конца которой плавно отогнуты назад, т. е. внутрь
раковины. Однако она также может быть прямой, и концы ее (или один
из них, чаще верхний) отогнуты под прямым углом.
Ниже главной складки, обычно параллельно ей, реже под некоторым
углом, лежат палатальные складки (plicae palatalis). Число их у разных
видов различно, чаще всего 1—3, реже больше. Иногда палатальные
складки вообще отсутствуют. Палатальные складки являются производ-
производными полулунной складки, или палатальной мозоли.1 В связи с этим неко-
некоторые авторы (Ehrmann, 1926) разделяют эти складки на истинные — про-
1 Точнее будет сказать, что они. являются производными желез, формирующих
полулуннуго складку, или палатальную мозоль.
МОРФОЛОГИЯ; ЗАМЫКАТЕЛЬНЫЙ АППАРАТ
17
изводные полулунной складки и ложные — производные палатальной
мозоли. Однако это деление не всегда можно четко провести, так как порой
неясно, за счет чего возникли те или иные складки. Кроме того, совершенно
очевидно, что у ряда видов названные складки имеют смешанное происхо-
происхождение: как за счет полулунной складки, так и за счет мозоли. В связи
с этим в дальнейшем мы не придерживаемся этого деления.
Среди палатальных складок различают верхнюю, нижнюю и одну или
несколько средних. Верхняя лежит под главной складкой, чаще всего на
уровне верхнего конца полулунной складки, нередко
сливаясь с ним. Нижняя лежит слева от базального
желобка, на уровне нижнего конца полулунной скла-
складки, тоже нередко сливаясь с ним. Средние складки
занимают промежуточное положе-
положение.
Следует подчеркнуть, что если
пластинки и главная складка
•(как и клаузилий) присутствуют
у абсолютного большинства видов
Clausiliidae то полулунная или
палатальные складки нередко мо-
могут отсутствовать. При этом за-
заметна следующая закономерность:
если отсутствует полулунная скл ад-
ка, то имеются палатальные склад-
складки, (одна или больше); наоборот,
«ели отсутствуют палатальные
складки, то есть полулунная склад-
складка. Нередки однако случаи, когда
все названные складки налицо,
что бывает, когда палатальные
складки возникли за счет мозоли.
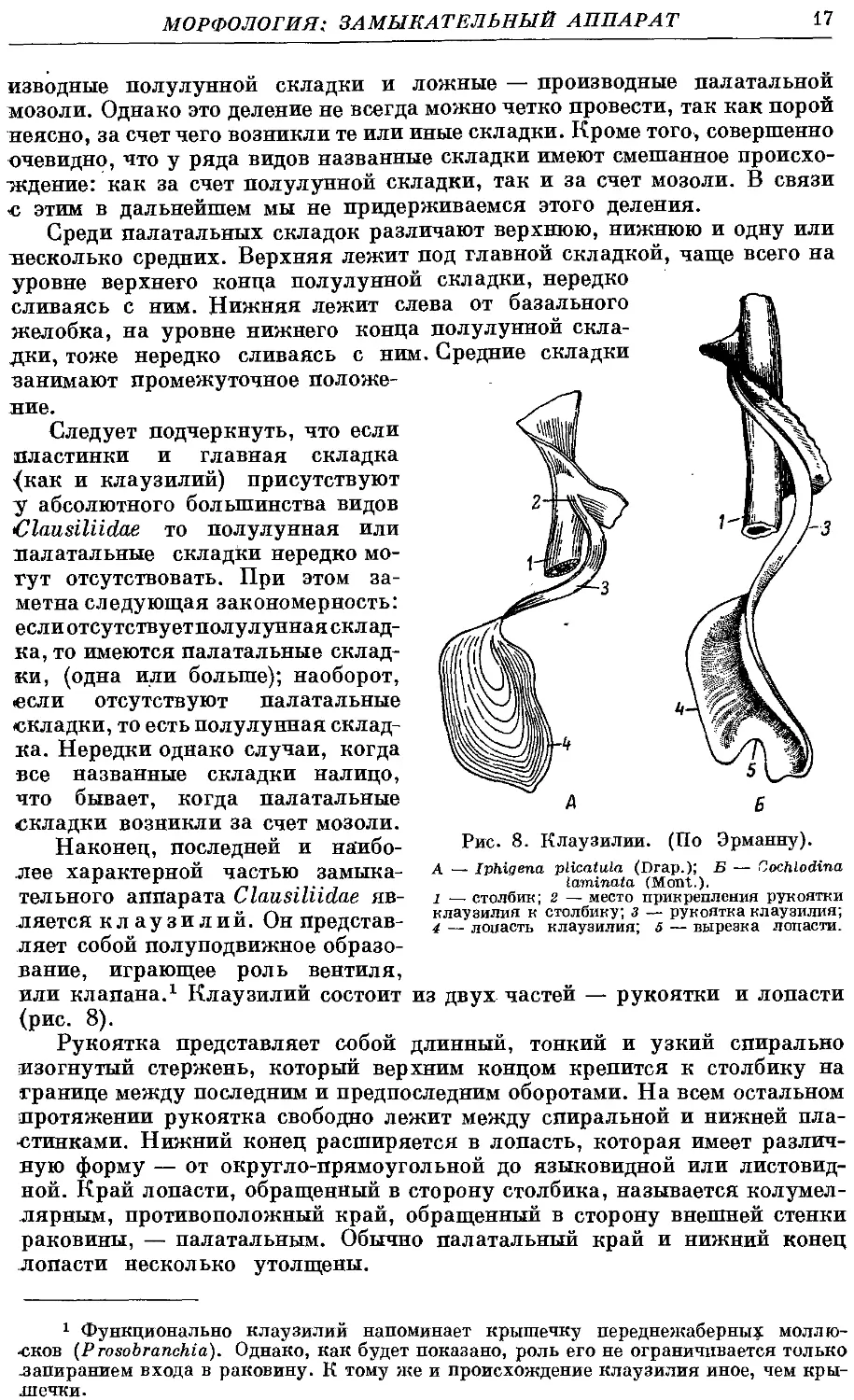
Наконец, последней и наибо-
наиболее характерной частью замыка-
тельного аппарата Clausiliidae яв-
является клаузилий. Он представ-
представляет собой полуподвижное образо-
образование, играющее роль вентиля,
или клапана.1 Клаузилий состоит из двух частей — рукоятки и лопасти
(рис. 8).
Рукоятка представляет собой длинный, тонкий и узкий спирально
изогнутый стержень, который верхним концом крепится к столбику на
границе между последним и предпоследним оборотами. На всем остальном
¦протяжении рукоятка свободно лежит между спиральной и нижней пла-
пластинками. Нижний конец расширяется в лопасть, которая имеет различ-
различную форму — от округло-прямоугольной до языковидной или листовид-
листовидной. Край лопасти, обращенный в сторону столбика, называется колумел-
лярным, противоположный край, обращенный в сторону внешней стенки
раковины, — палатальным. Обычно палатальный край и нижний конец
лопасти несколько утолщены.
Рис. 8. Клаузилий. (По Эрманну).
А — Iphigena plicatula (Drap.); Б — Cochlodina
laminafa (Mont.).
l — столбик; 2 — место прикрепления рукоятки
клаузилия к столбику; з — рукоятка клаузилия;
4 — лопасть клаузилия; 5 — вырезка лопасти.
1 Функционально клаузилий напоминает крышечку переднежаберных моллю-
моллюсков (Prosobranchia). Однако, как будет показано, роль его не ограничивается только
-запиранием входа в раковину. К тому же и происхождение клаузилия иное, чем кры-
крышечки.
18 tiv\t .'"¦'¦ ВВЕДЕНИЕ
Благодаря тонкой спирально изогнутой рукоятке клаузилий обладает
большой эластичностью, причем рукоятка играет роль пружины. Когда
моллюск втянулся вглубь раковины, лопасть клаузилия на определенной
высоте запирает вход в нее. При этом колумеллярный край лопасти упи-
упирается в субколумеллярную пластинку, а палатальный край — в полулун-
полулунную складку, или же, если ее нет, в верхние концы палатальных складок.
Когда животное расправляется, оно выталкивает лопасть немного вперед
и в сторону, и она ложится в особую щель между нижней и субколумелляр-
ной пластинками, которая называется нишей клаузилия.
Как уже отмечалось, при отсутствии полулунной складки клаузилий
опирается палатальным краем лопасти в верхние концы палатальных
складок. Однако в тех случаях, когда верхний конец нижней складки
проникает внутрь раковины дальше остальных складок, лопасть клаузи-
клаузилия в соответствующем месте имеет вырезку (Alopia, Cochlodina, Men-
tissa). Она позволяет клаузюшю при замыкании раковины занять нор-
нормальное положение, пропустив названную складку через вырезку.
Развитие. В данном случае речь идет о формировании замыкательного
аппарата в процессе онтогенеза. По этому вопросу имеются два исследо-
исследования — Кимакович-Винницки (Kimakowicz-Winnicki, 1914) и Эдлауер
(Edlauer 1941). Последняя работа, основанная на изучении Herilla bos-
niensis и некоторых других видов, отличается особой тщательностью
выполнения и глубиной анализа.
Замыкательный аппарат возникает вовремя, формирования последнего
оборота. Весь процесс проходит очень быстро — в течение 2—3 недель.
У молодых моллюсков рост раковины идет синхронно с ростом мантии,
так что та и другая представляют собой как бы два конуса, вставленные
один в другой. При этом мантия плотно прилегает к стенке раковины и не
образует складок или морщин. С началом формирования последнего обо-
оборота эта корреляция нарушается. Периметр просвета последнего оборота
раковины в большей или меньшей степени сужается по направлению
к устью (у веретеновидных раковин). В то же время поверхность мантии
продолжает увеличиваться; это приводит к тому, что она не может уже
равномерно прилегать к стенкам раковины и образует морщины и бо-
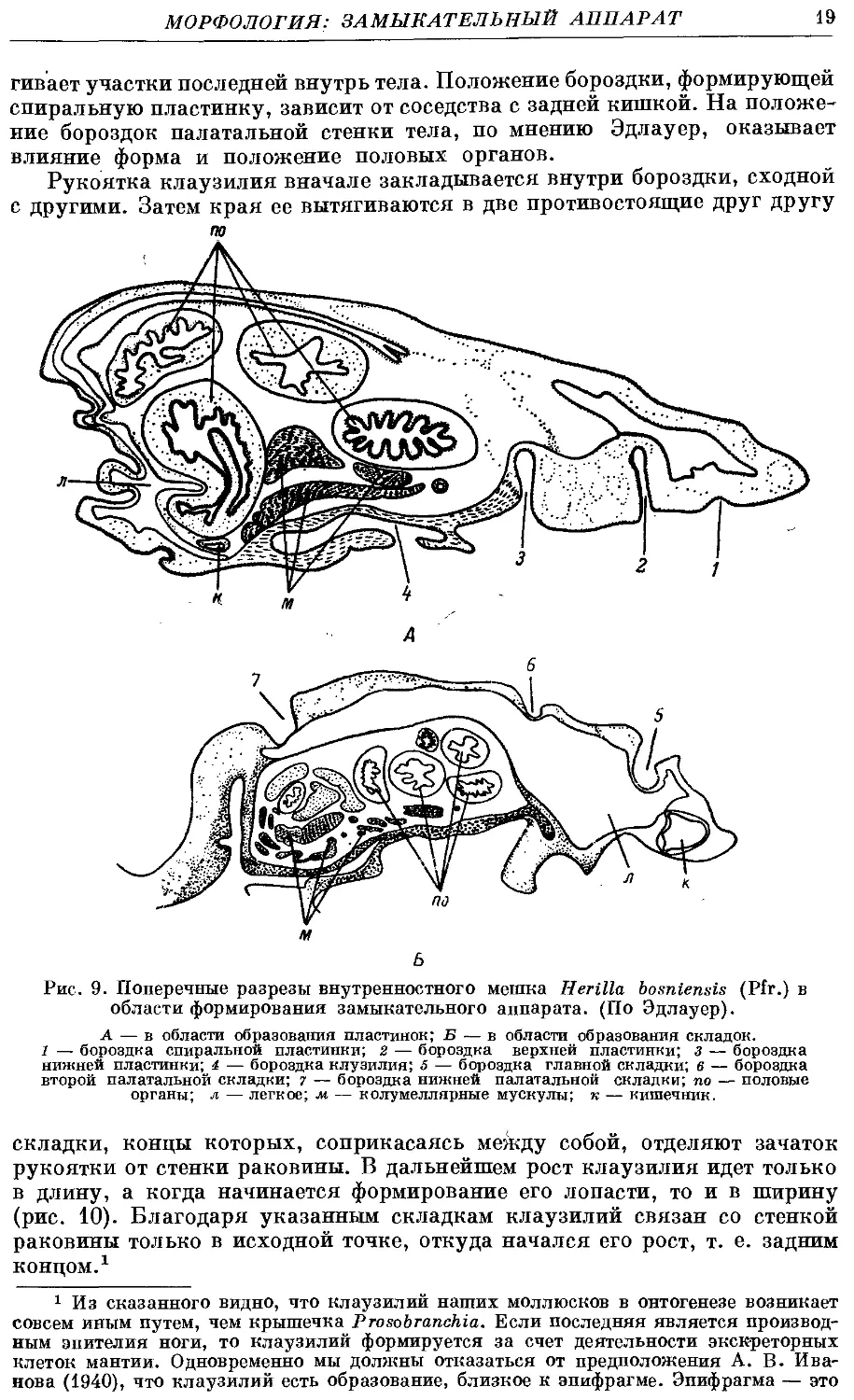
бороздки (рис. 9).
Нарастание поверхности мантии внутри последнего оборота раковины
происходит за счет увеличения размеров клеток эпителия, в особенности
на дне бороздок, где они отличаются крупными ядрами и обилием включе-
включений в протоплазме. Эти клетки, а также специальные подвижные соедини-
соединительнотканные клетки, служащие для переноса извести от внутренних
органов к мантии, участвуют в формировании всех частей замыкательного-
аппарата. Когда оно заканчивается, то вновь все эпителиальные клетки
мантии становятся низкими, почти плоскими. Одновременно уменьшаются
в размерах и их ядра.
Эдлауер убедительно показала, что бороздки мантии появляются строго
последовательно и на определенных местах. Те, внутри которых закла-
закладываются верхняя, нижняя и субколумеллярная пластинки, а также клау-
клаузилий, располагаются на колумеллярной стороне. Бороздки, внутри
которых закладываются полулунная и палатальные складки, образуются:
на палатальной стороне мантии. Спиральная пластинка закладывается
на париетальной стороне; позднее туда же смещается борозда, формирую-
формирующая верхнюю пластинку. Автор считает, что большую роль в образовании
бороздок на колумеллярной стороне оказала мускулатура диафрагмы.
При сокращении животного, а также благодаря увеличению объема!
внутренностей диафрагма в тех местах, где она переходит в мантию, затя-
МОРФОЛОГИЯ: ЗАМЫКАТЕЛЬНЫЙ АППАРАТ
19
гивает участки последней внутрь тела. Положение бороздки, формирующей
спиральную пластинку, зависит от соседства с задней кишкой. На положе-
положение бороздок палатальной стенки тела, по мнению Эдлауер, оказывает
влияние форма и положение половых органов.
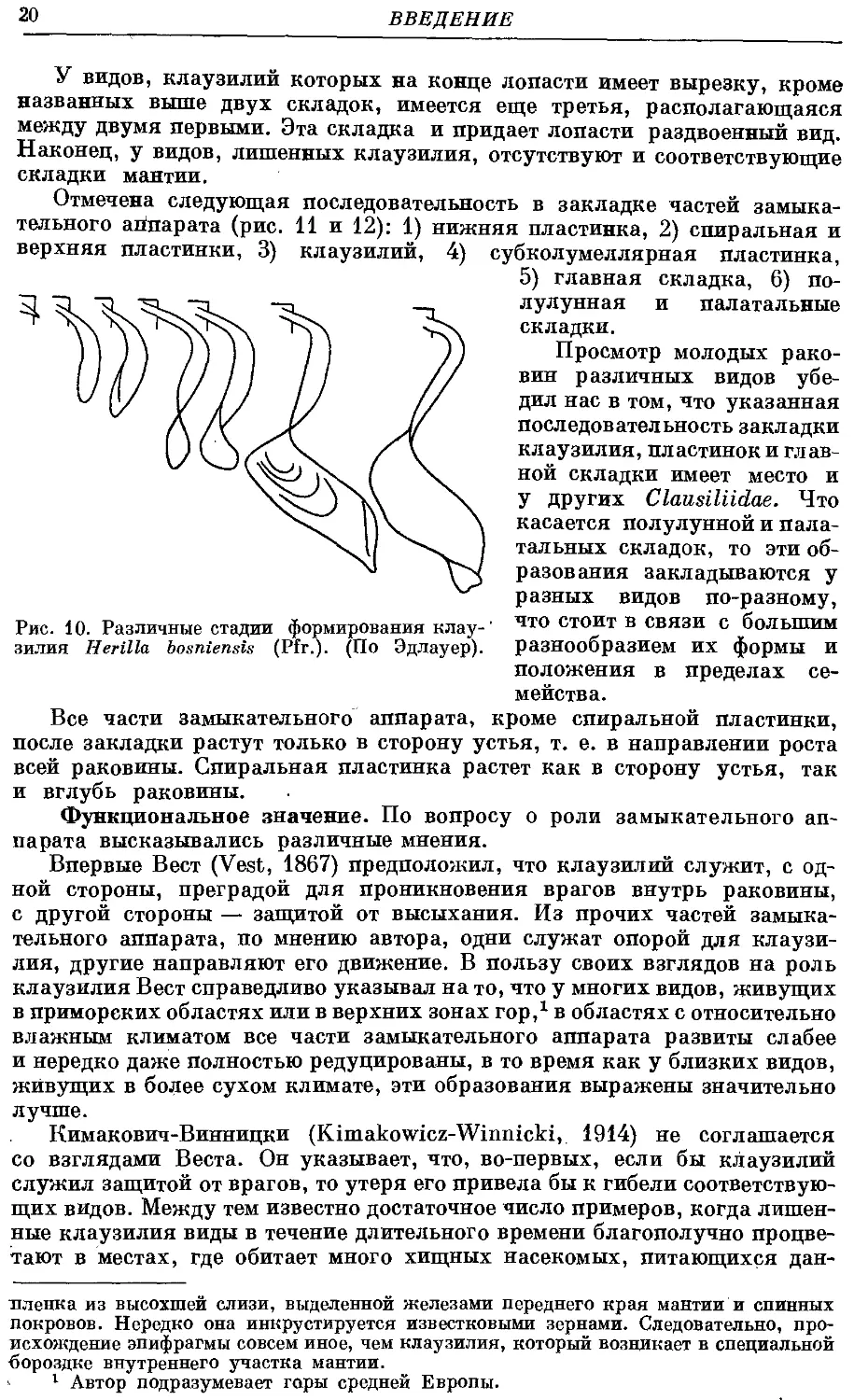
Рукоятка клаузилия вначале закладывается внутри бороздки, сходной
с другими. Затем края ее вытягиваются в две противостоящие друг другу
по
Рис. 9. Поперечные разрезы внутренностного мешка Herilla bosniensis (Pfr.) в
области формирования замыкательного аппарата. (По Эдлауер).
А — в области образования пластинок; Б — в области образования складок.
1 — бороздка спиральной пластинки; 2 — бороздка верхней пластинки; з — бороздка
нижней пластинки; 4 — бороздка клузилия; 5 — бороздка главной складки; в — бороздка
второй палатальной складки; 7 — бороздка нижней палатальной складки; по — половые
органы; л — легкое; м — колумеллярные мускулы; к — кишечник.
складки, концы которых, соприкасаясь между собой, отделяют зачаток
рукоятки от стенки раковины. В дальнейшем рост клаузилия идет только
в длину, а когда начинается формирование его лопасти, то и в ширину
(рис. 10). Благодаря указанным складкам клаузилий связан со стенкой
раковины только в исходной точке, откуда начался его рост, т. е. задним
концом.1
1 Из сказанного видно, что клаузилий наших моллюсков в онтогенезе возникает
совсем иным путем, чем крышечка Prosobranchia. Если последняя является производ-
производным эпителия ноги, то клаузилий формируется за счет деятельности экскреторных
клеток мантии. Одновременно мы должны отказаться от предположения А. В. Ива-
Иванова A940), что клаузилий есть образование, близкое к эпифрагме. Эпифрагма — это
20
ВВЕДЕНИЕ
Рис. 10. Различные стадии формирования клау-
зилия Herilla bosniensis (Pfr.). (По Эдлауер).
У видов, клаузилий которых на конце лопасти имеет вырезку, кроме
названных выше двух складок, имеется еще третья, располагающаяся
между двумя первыми. Эта складка и придает лопасти раздвоенный вид.
Наконец, у видов, лишенных клаузилия, отсутствуют и соответствующие
складки мантии.
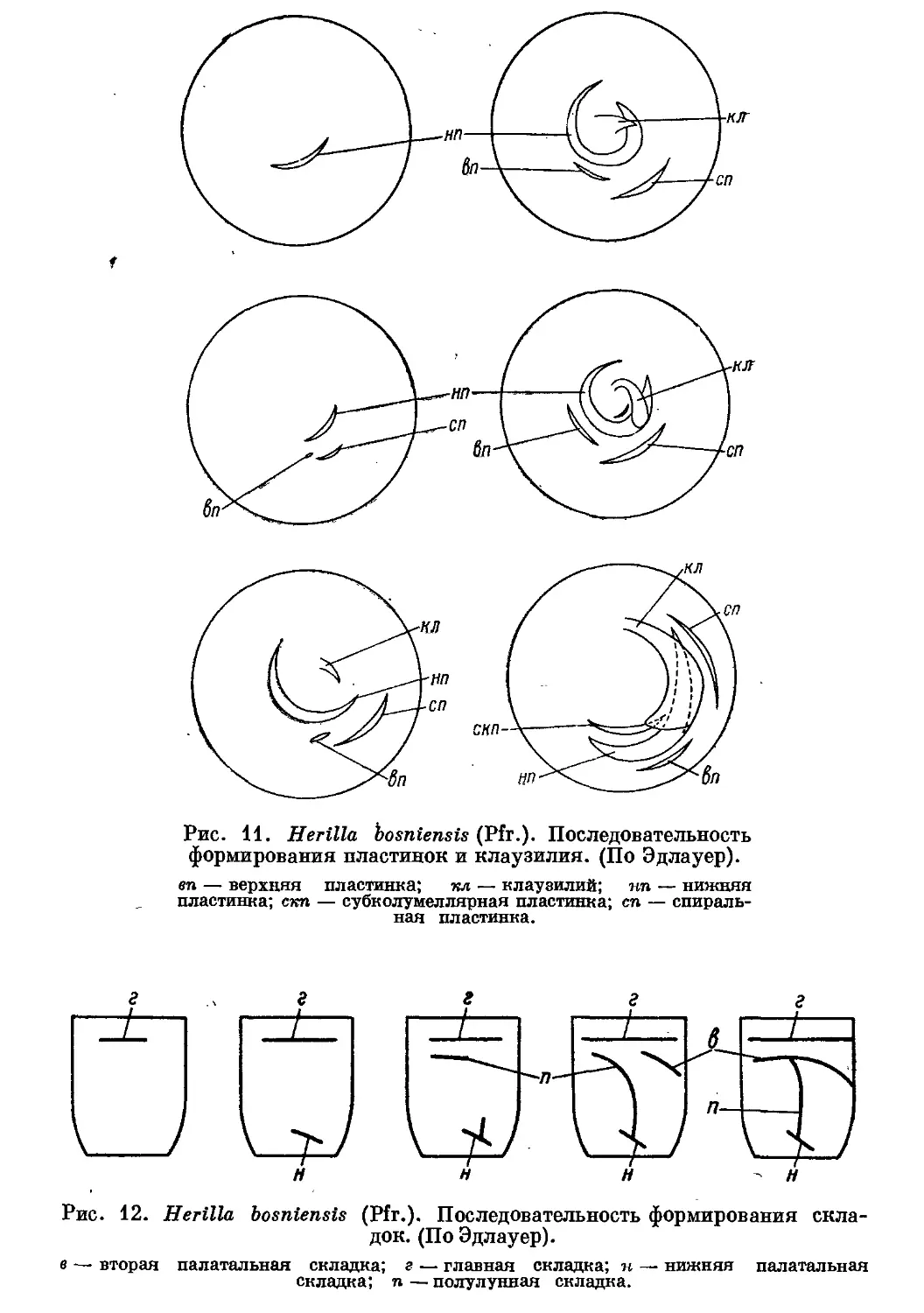
Отмечена следующая последовательность в закладке частей замыка-
тельного аппарата (рис. 11 и 12): 1) нижняя пластинка, 2) спиральная и
верхняя пластинки, 3) клаузилий, 4) субколумеллярная пластинка,
5) главная складка, 6) по-
]q -л -о -«а -^] -г;, __ лулунная и палатальные
г х\ хч >5>ч>^\ >^ч 3\ складки.
Просмотр молодых рако-
раковин различных видов убе-
убедил нас в том, что указанная
последовательность закладки
клаузилия, пластинок и глав-
главной складки имеет место и
у других Clausiliidae. Что
касается полулунной и пала-
палатальных складок, то эти об-
образования закладываются у
разных видов по-разному,
что стоит в связи с большим
разнообразием их формы и
положения в пределах се-
семейства.
Все части замыкательного аппарата, кроме спиральной пластинки,
после закладки растут только в сторону устья, т. е. в направлении роста
всей раковины. Спиральная пластинка растет как в сторону устья, так
и вглубь раковины.
Функциональное значение. По вопросу о роли замыкательного ап-
аппарата высказывались различные мнения.
Впервые Вест (Vest, 1867) предположил, что клаузилий служит, с од-
одной стороны, преградой для проникновения врагов внутрь раковины,
с другой стороны — защитой от высыхания. Из прочих частей замыка-
замыкательного аппарата, по мнению автора, одни служат опорой для клаузи-
клаузилия, другие направляют его движение. В пользу своих взглядов на роль
клаузилия Вест справедливо указывал на то, что у многих видов, живущих
в приморских областях или в верхних зонах гор,1 в областях с относительно
влажным климатом все части замыкательного аппарата развиты слабее
и нередко даже полностью редуцированы, в то время как у близких видов,
живущих в более сухом климате, эти образования выражены значительно
лучше.
Кимакович-Винницки (Kimakowicz-Winnicki, 1914) не соглашается
со взглядами Веста. Он указывает, что, во-первых, если бы клаузилий
служил защитой от врагов, то утеря его привела бы к гибели соответствую-
соответствующих видов. Между тем известно достаточное число примеров, когда лишен-
лишенные клаузилия виды в течение длительного времени благополучно процве-
процветают в местах, где обитает много хищных насекомых, питающихся дан-
пленка из высохшей слизи, выделенной железами переднего края мантии и спинных
покровов. Нередко она инкрустируется известковыми зернами. Следовательно, про-
происхождение эпифрагмы совсем иное, чем клаузилия, который возникает в специальной
бороздке внутреннего участка мантии.
<' 1 Автор подразумевает горы средней Европы.
Рис. 11. Herilla hosniensis (Pfr.). Последовательность
формирования пластинок и клаузилия. (По Эдлауер).
en — верхняя пластинка; игл — клаузилий; ип — нижняя
пластинка; скп — субколумеллярная пластинка; сп — спираль-
спиральная пластинка.
\_^J
±
т
Рис. 12. Herilla bosniensis (Pfr.). Последовательность формирования скла-
складок. (По Эдлауер).
в — вторая палатальная складка; г — главная складка; и — нижняя палатальная
складка; п — полулунная складка.
22 ВВЕДЕНИЕ
ными моллюсками. К этому от себя добавим, что известны случаи, когда
внутри раковин с хорошо развитым и исправным клаузилием обнаружи-
обнаруживали личинок хищных жуков, благополучно доедавших остатки своих
жертв. Следовательно, клаузилий не всегда является препятствием для
проникновения внутрь раковины различных хищников.
Кимакович-Винницки не согласен также с тем, что клаузилий служит
защитой от высыхания, так как считает, что способность наземных мол-
моллюсков плотно приклеиваться устьем раковины к субстрату лучше обес-
обеспечивает их изоляцию от наружного воздуха. С его точки зрения, клаузи-
клаузилий вместе с нижней пластинкой является опорой для колумеллярного
мускула, что позволяет животному приподымать свою раковину/ Осуще-
Осуществляется это следующим образом: при вытягивании из раковины мягкие
части тела отодвигают клаузилий в его нишу; давлением воздуха, находя-
находящегося в легком, между лопастью и нижней пластинкой зажимается одна
из сладок мантии.1 Тем самым колумеллярный мускул получает вторую
точку опоры (первой является столбик, к которому прикреплен верхний
конец того же мускула).
В подтверждение своих взглядов на роль клаузилия Кимакович-Вин-
Кимакович-Винницки указывает, что многие Clausiliidae, несмотря на то, что их раковина
в 1V2—2 раза тяжелее мягких частей тела, способны временами при-
приподымать ее над субстратом. Эту способность они теряют, если у них
поврежден клаузилий.
Эдлауер (Edlauer, 1941) допускает, что у некоторых видов клаузилий
действительно используется для управления раковиной. В то же время
автор вполне обоснованно указывает на то, что этой способностью обла-
обладают не все Clausiliidae и зависит она не только от веса раковины, но и от
мощности мускулатуры. Поэтому некоторые виды, обладающие относи-
относительно тяжелой раковиной, несмотря на наличие хорошо развитого клау-
клаузилия, не могут приподымать ее над субстратом и волочат ее за собой.
В то же время молодь этих же видов, несмотря на то, что клаузилий у них
еще не сформирован, обладает способностью приподымать раковину.
В итоге автор считает, что клаузилий играет роль как при защите от высы-
высыхания, так и при управлении раковиной.
Вагнер (Wagner, 1919) полагает, что, за исключением спиральной
и верхней пластинок и главной складки, все остальные части замыкатель-
ного аппарата, в том числе и клаузилий, служат для защиты от высыха-
высыхания, т. е. выполняют ту же роль, что и крышечка наземных Prosobranchia.
В пользу этого взгляда он приводит много убедительных примеров полной
или частичной редукции замыкательного аппарата у видов, живущих
в относительно влажном климате.
Мы в основном согласны с Вагнером и с теми авторами, которые рас-
рассматривали клаузилий и весь замыкательный аппарат в целом как защиту
от высыхания. Однако не только эти образования, а все строение послед-
последнего оборота раковины Clausiliidae подчинено в первую очередь той же
цели. В то же время замыкательный аппарат несет и другие функции.
Известно, что у большинства Clausiliidae последний оборот раковины
на большем или меньшем протяжении испытывает сужение, что придает
раковине веретеновидный облик. Характерно, что последний оборот не
сужен и раковина имеет башневидную форму только у видов, живущих
в достаточно влажном макро- или микроклимате (в приморских областях,
в щелях скал, в осыпях или под гниющей корой). Следовательно, сужение
1 Клаузилий, когда он отодвинут в нишу, окружен двумя складками мантии.
Как «было указано выше, сам клаузилий является производным этих складок.
МОРФОЛОГИЯ: ЗАМЫКАТЕЛЬНЫЙ АППАРАТ 23
последнего оборота и замыкание раковины в самом узком месте ее клаузи-
лием, который одной стороной упирается в полулунную складку, а другой
в субкблумеллярную пластинку, — вот главные адаптации, предупре-
предупреждающие чрезмерную потерю влаги телом моллюска.
Однако тем же целям служат и другие образования, а именно пала-
палатальные складки и мелкие складочки, развивающиеся на самом краю устья.
Но осуществляют эту же цель они иным образом. Когда животное втяги-
втягивается внутрь раковины, то эти складки, раздражая покровы спины,
вызывают обильное отделение слизи, которая скопляется у устья. Высы-
Высыхая, она образует перепонку, приклеивающую раковину устьем к суб-
субстрату. Тем самым закрывается доступ наружного воздуха к мягким
частям тела.
TonKOBaHHe^p^rajaMT^jbjiHi^ayiajroK как^^испособления к защите
от высыхания соответствует тому, что мы знаем о других группах моллюс-
моллюсков. Известно, что в таких семействах, как Pupillidae, Enidae и других,
зубы, развивающиеся внутри устья, крупнее у популяций и у видов, живу-
живущих в более засушливом климате, чем у популяций того же вида или
у близких видов, обитающих в более влажном климате (Матекин, 1950,
1959). В последнем случае зубы могут даже исчезать.
Степень развития замыкательного аппарата зависит не только от мак-
макро-, но и от микроклимата. Так, у видов, обитающих в гниющей древесине
или под корой, т. е. в условиях высокой и стабильной влажности (Caspio-
phaedusa perlucens, Pontophaedusa funiculum, Serrulina), замыкательный
аппарат развит слабее, чем у видов, живущих снаружи. Тоже наблюдается
у многих Clausiliidae, обитающих в щелях скал. Регресс замыкательного
аппарата в первую очередь проявляется в том, что клаузилий неплотно
замыкает последний оборот и между ним и внешней стенкой раковины или
столбиком остается ббльшая или меньшая щель. Кроме того, наблюдается
регресс или полное исчезновение палатальных складок, распад спираль-
спиральной пластинки на ряд коротких пластинок (параллельная и вставочная),
а также главной складки на ряд сутуральных складок. Одновременно,
как это уже отмечалось, у таких видов последний оборот раковины не
испытывает сужения.1
Наконец, как^ащитуот высыхания можно рассматривать способность
многих Clausiliidae к глубокому втягиванию в верхнюю часть раковины.
При этом происходит уменьшение поверхности тела, а следовательно,
и снижение испарения.
положения_раковины на сшгае^моллюска. ТЗ этом отношёнии~~особенно
важную роль играют ш^тяя~а'~суШ6я'^мелля.'рвйя. пластинки. Когда
моллюск расправлен, то отдельные ветви колумеллярного мускула распо-
располагаются между указанными пластинками, тем самым способствуя управ-
управлению раковиной.
И связи с сужением последнего оборота раковины у животного, когда
оно вытягивается из раковины, возникает необходимость обеспечить
свободный проход воздуха в легкое и защитить от давления окружающих
органов нижние участки задней кишки и мочеточника. Эту, третью_фу_нк^
цию замыкательного аппарата берут^^^еб^^пирал^.ная и Ъерхняя пла-
пластинки, а также "иГ'Тлавная"складка. Располагаясь друг против друга под
углом (первые две на париетальной стенке, последняя на палатальной
стенке), в целом они создают вдоль верхнего края наиболее узкой части
1 Далее будет показано, что регресс замыкательного аппарата имеет Meqro иногда
и у видов, живущих в очень сухом климате, но в таких случаях другие адаптации
защищают тело моллюска от высыхания.
24 . ВВЕДЕНИЕ
последнего оборота нечто вроде желоба, внутри которого лежат названные
органы. Соседство обеих главных ветвей легочнойвены с главной склад-
складкой также, вероятно, связащ^деоб^одимостью имейГсвчтбодное простран-
пространство для проталкивания крови.
Во всех случаях, когда в связи с обитанием во влажном климате послед-
последний оборот не сужен, наряду с редукцией остальных частей замыкательного
аппарата происходит некоторое ослабление, или даже полная редукцият
образований, слагающих этот желоб.
В двух подсемействах — Euxininae и Clausiliinae вдоль нижней части
последнего оборота спирально тянется углубление — базальный желобок.
Хотя он не имеет прямого отношения к замыкательному аппарату, роль
его аналогична главной складке, верхней и спиральной пластинкам. Он
также.BO3HiiLB_cjBH3H с,сужением выхода для мягких частей тела и служит^
для облегчения_§код1ьрйяйя ноги вдоль^стщщ1„р_аковины во время втяги-
ваана_и^вытягивания моллюска. Кроме того, так как стёЖСэтогоТКблобка
отличаются большоГмассйвностью и образуют киль, он также способст-
способствует прочности основания раковины.
В итоге отдельные части замыкательного аппарата выполняют следую-
следующие функции:
1) защита от высыхания — клаузилий, полулунная складка, субколу-
меллярная пластинка, палатальные складки и краевые складки;
2) управление раковиной — нижняя и дубкелумеллярная пластинки
и частично^шаузилий; . й
3) защита дыхательного канала, задней кишки, мочеточника и легоч-
легочных вен от давления окружающих органов — главная складка, спираль-
спиральная и верхняя пластинки.
Когда в связи с жизнью во влажном климате происходит регресс замы-
замыкательного аппарата, то первыми подвергаются редукции до лолного-
исчезновения палатальные и полулунная складки, т. е. образования, слу-
служащие только защитой от высыхания. Последними исчезают клаузилий
и нижняя пластинка. Очевидно, это связано с тем, что образования,,
обеспечивающие управление раковиной, сохраняют свое значение даже-
во влажном климате, и только когда происходит укорачивание раковиньь
(Balea), отпадает надобность и в них.
Анатомия
Легкое и мантийный комплекс органов. В связи с удлиненной формой
раковины и всего внутренностного мешка легкое наших моллюсков отли-
отличается длиной и узкой формой и у расправленного животного занимаег
2—3 нижних оборота (рис. 13). Дыхательное отверстие располагается
в синулуое устья раковины. Наружный край мантии очень толстый и He-
несет заметных придатков и лопастей.
Нижняя стенка легкого называется диафрагмой. В отличие от верхней
стенки, она обладает мускулатурой, состоящей из продольных и кольцевых
мускульных волокон. Периодические сокращения и расслабления диа-
диафрагмы изменяют объем легкого и осуществляют метаболизм.
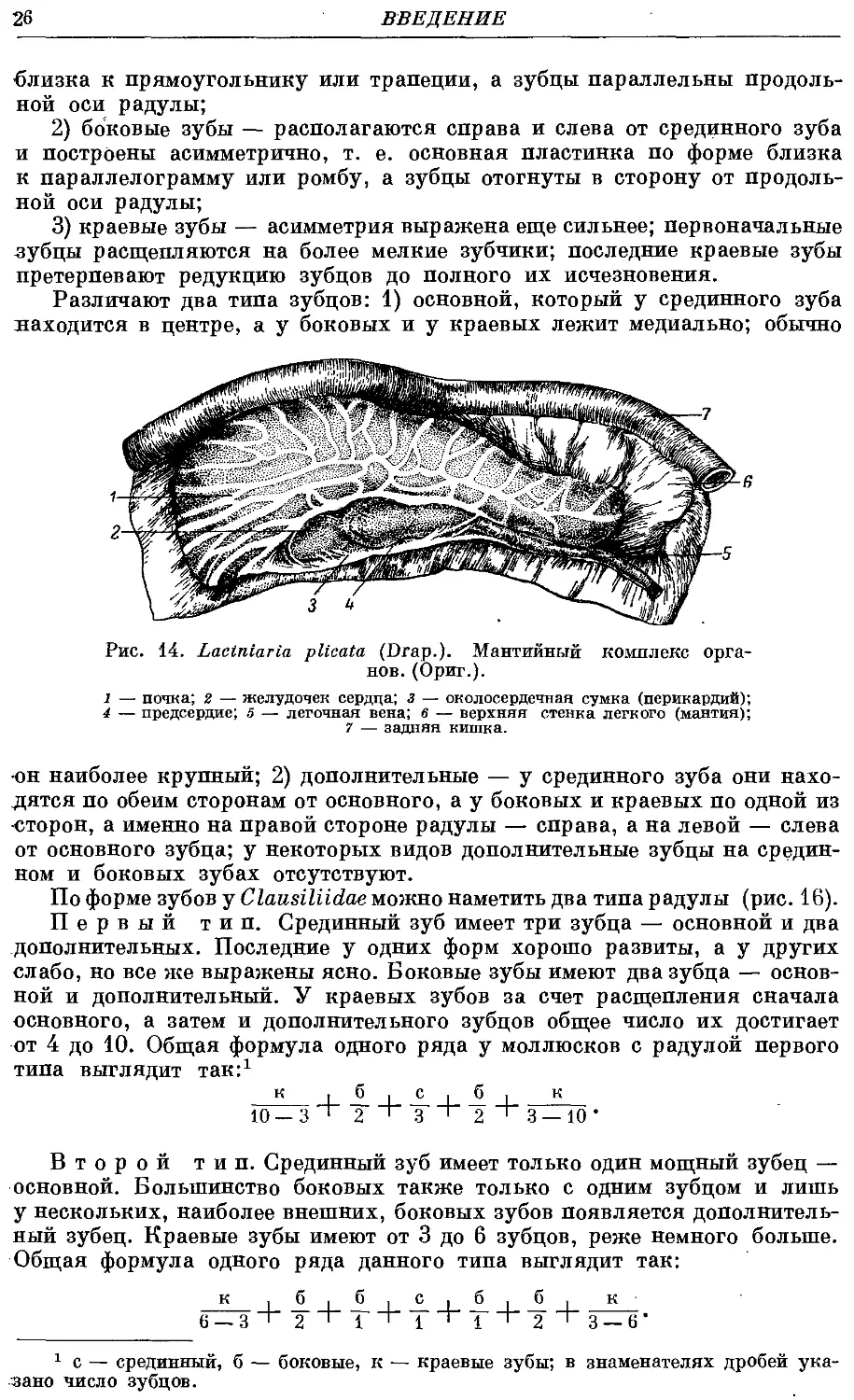
Близ заднего конца легкого, располагаясь внутри его верхней стенки,
находятся почка и сердце (рис. 14).
Почка в 4—5 раз короче легкого. От ее переднего конца отходит назад,,
тесно прилегая к ней, первичный мочеточник. Последний у заднего конца
почки переходит во вторичный мочеточник, который, тесно прижимаясь
к задней кишке, тянется вперед по направлению к дыхател-ьному отверстию
легкого и к анусу, по соседству с которыми он открывается"наружу. Таким,
МОРФОЛОГИЯ: АНАТОМИЯ
25-
$
образом, форма мочеточника, который вместе с почкой образует фигуру,:
сходную с латинской буквой S, указывает на принадлежность данного*-
семейства к подотряду Sigmurethra.
Перикардий прилегает к почке по ее нижнему краю. Они связаны
коротким рено-перикардиальным каналом. Внутри перикардия лежиг
сердце, состоящее из предсердия и же-
желудочка. От первого отходит вниз ле-
легочная вена, вскоре разветвляющаяся
на две параллельные вены. Сеть крове-
кровеносных сосудов очень тонка и без спе-
специального окрашивания слабо заметна.
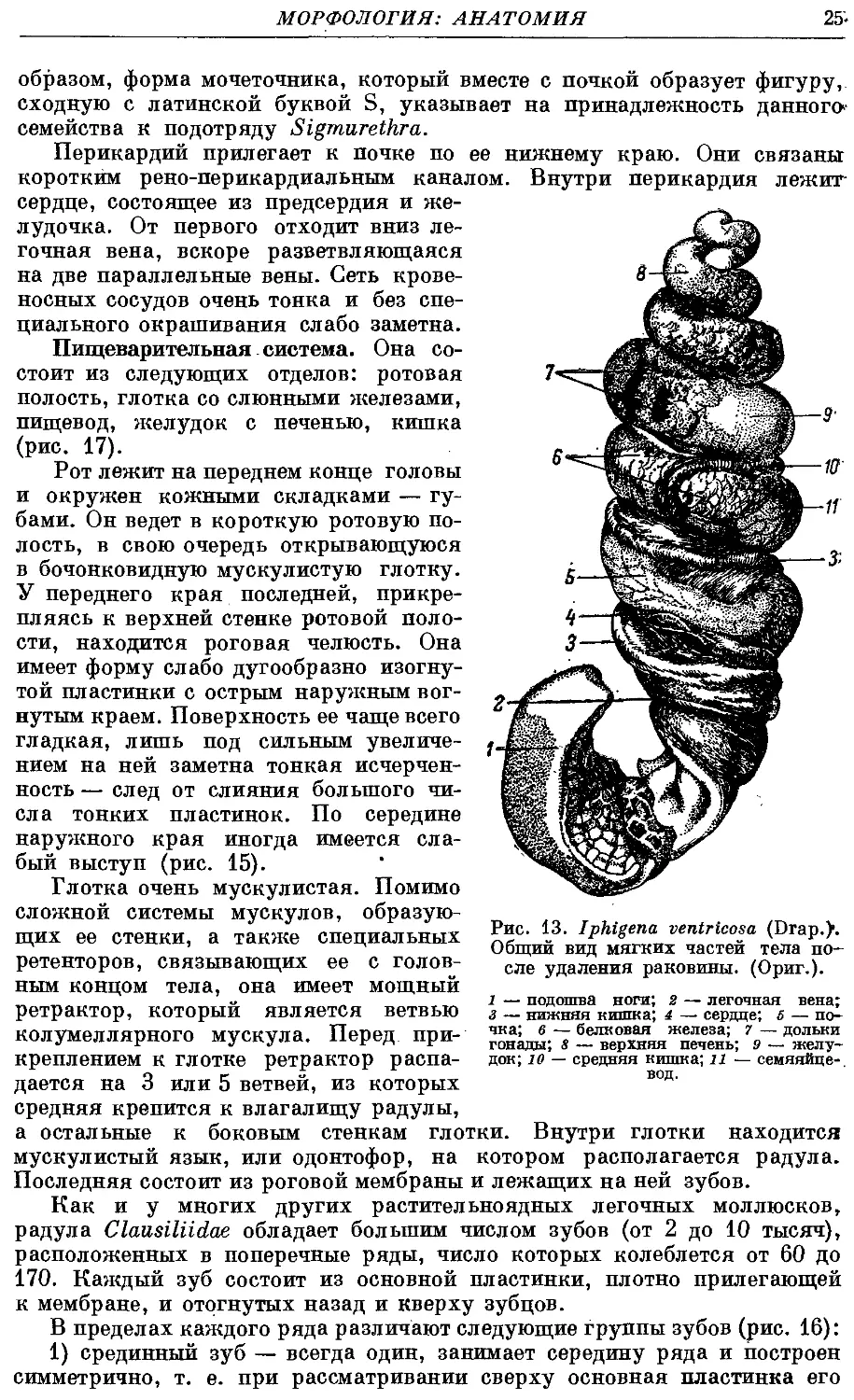
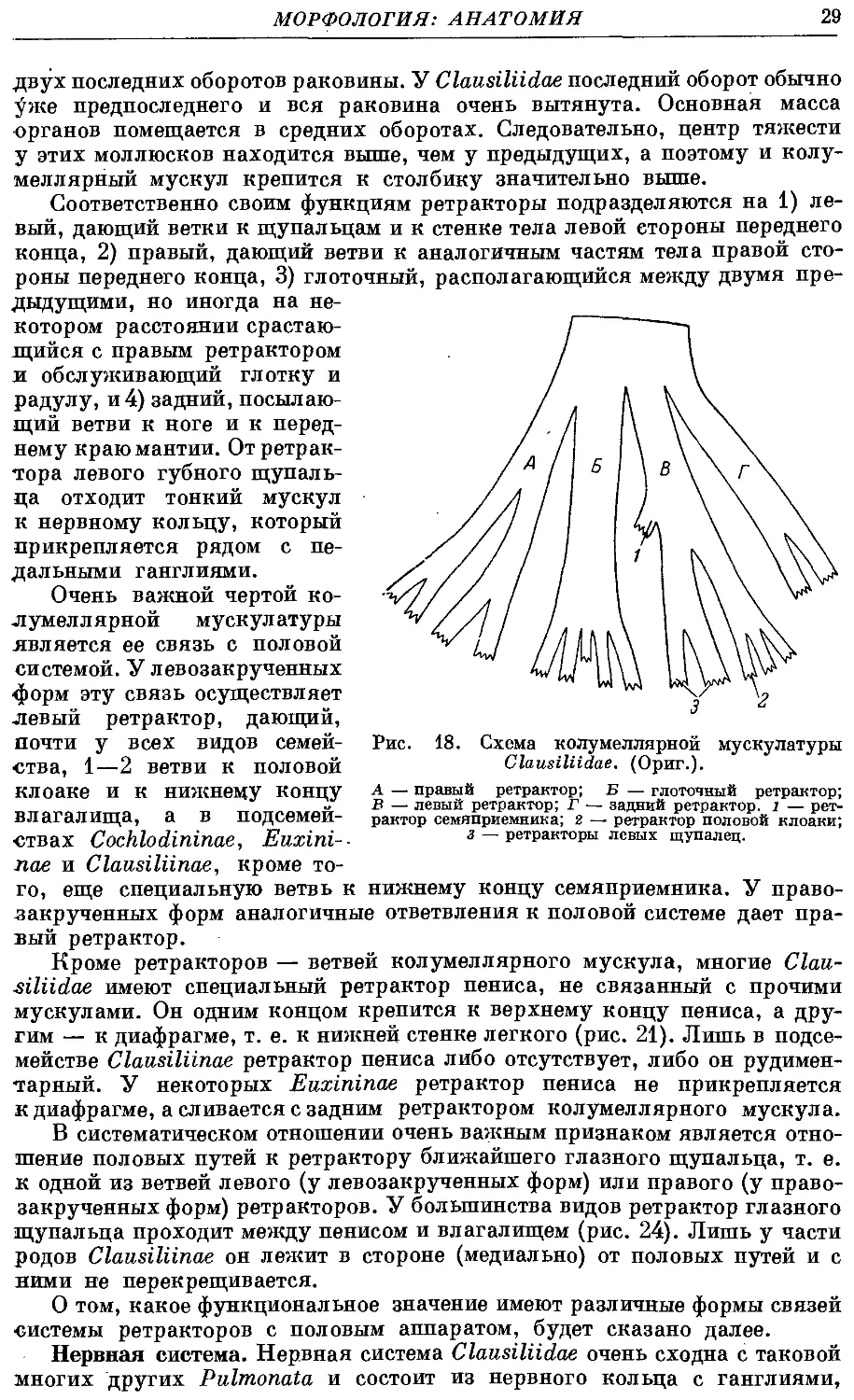
Пищеварительная система. Она со-
состоит из следующих отделов: ротовая
полость, глотка со слюнными железами,
пищевод, желудок с печенью, кишка
(рис. 17).
Рот лежит на переднем конце головы
и окружен кожными складками — гу-
губами. Он ведет в короткую ротовую по-
полость, в свою очередь открывающуюся
в бочонковидную мускулистую глотку.
У переднего края последней, прикре-
прикрепляясь к верхней стенке ротовой поло-
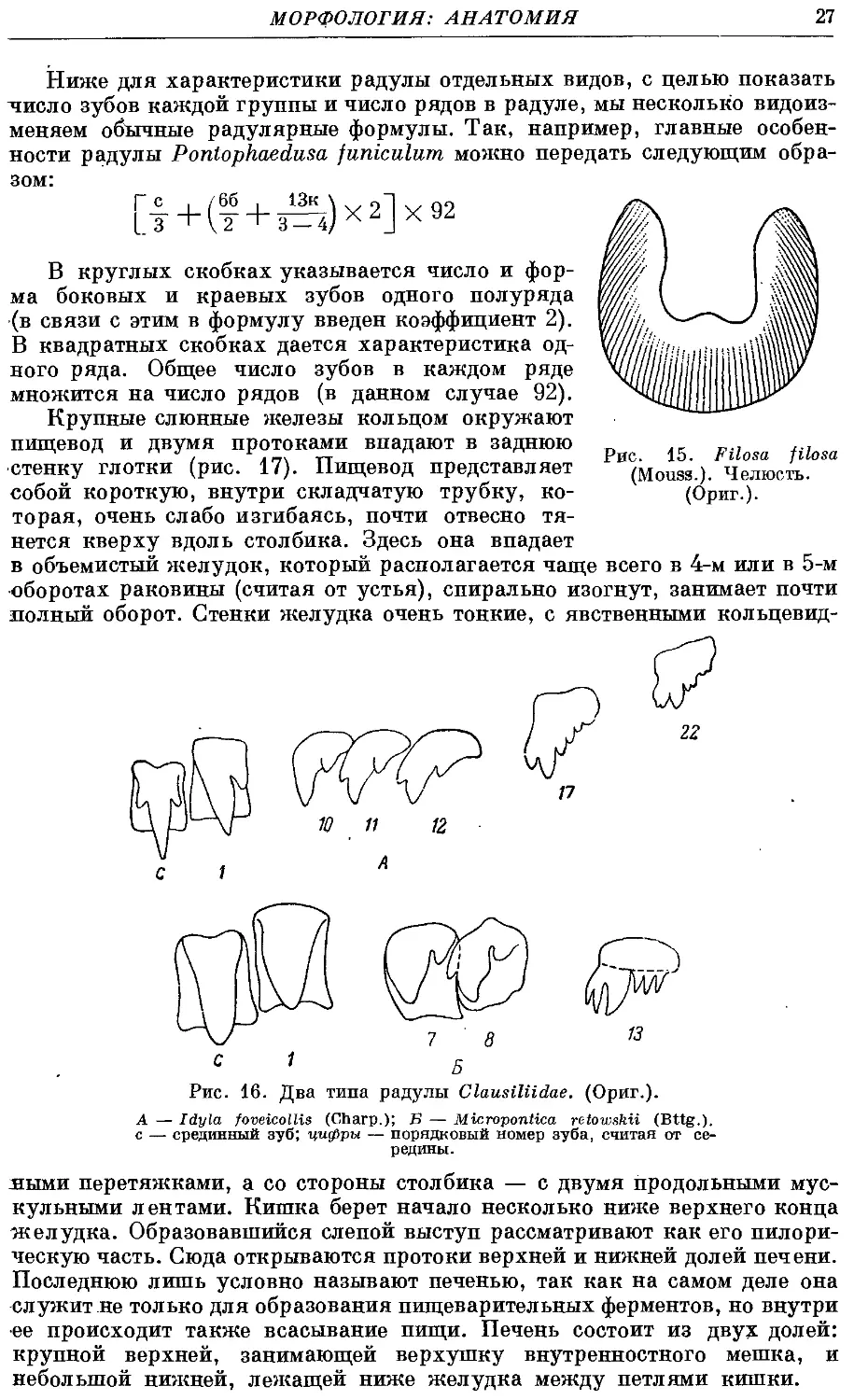
полости, находится роговая челюсть. Она
имеет форму слабо дугообразно изогну-
изогнутой пластинки с острым наружным вог-
вогнутым краем. Поверхность ее чаще всего
гладкая, лишь под сильным увеличе-
увеличением на ней заметна тонкая исчерчен-
ность — след от слияния большого чи-
числа тонких пластинок. По середине
наружного края иногда имеется сла-
слабый выступ (рис. 15).
Глотка очень мускулистая. Помимо
сложной системы мускулов, образую-
образующих ее стенки, а также специальных
ретенторов, связывающих ее с голов-
головным концом тела, она имеет мощный
ретрактор, который является ветвью
КОЛумеЛЛЯрНОГО мускула. Перед При- чка= в~ белковая железа; 7- дольки
креплением к глотке ретрактор распа-
распадается на 3 или 5 ветвей, из которых
средняя крепится к влагалищу радулы,
а остальные к боковым стенкам глотки. Внутри глотки находится
мускулистый язык, или одонтофор, на котором располагается радула.
Последняя состоит из роговой мембраны и лежащих на ней зубов.
Как и у многих других растительноядных легочных моллюсков,
радула Clausiliidae обладает большим числом зубов (от 2 до 10 тысяч),
расположенных в поперечные ряды, число которых колеблется от 60 до
170. Каждый зуб состоит из основной пластинки, плотно прилегающей
к мембране, и отогнутых назад и кверху зубцов.
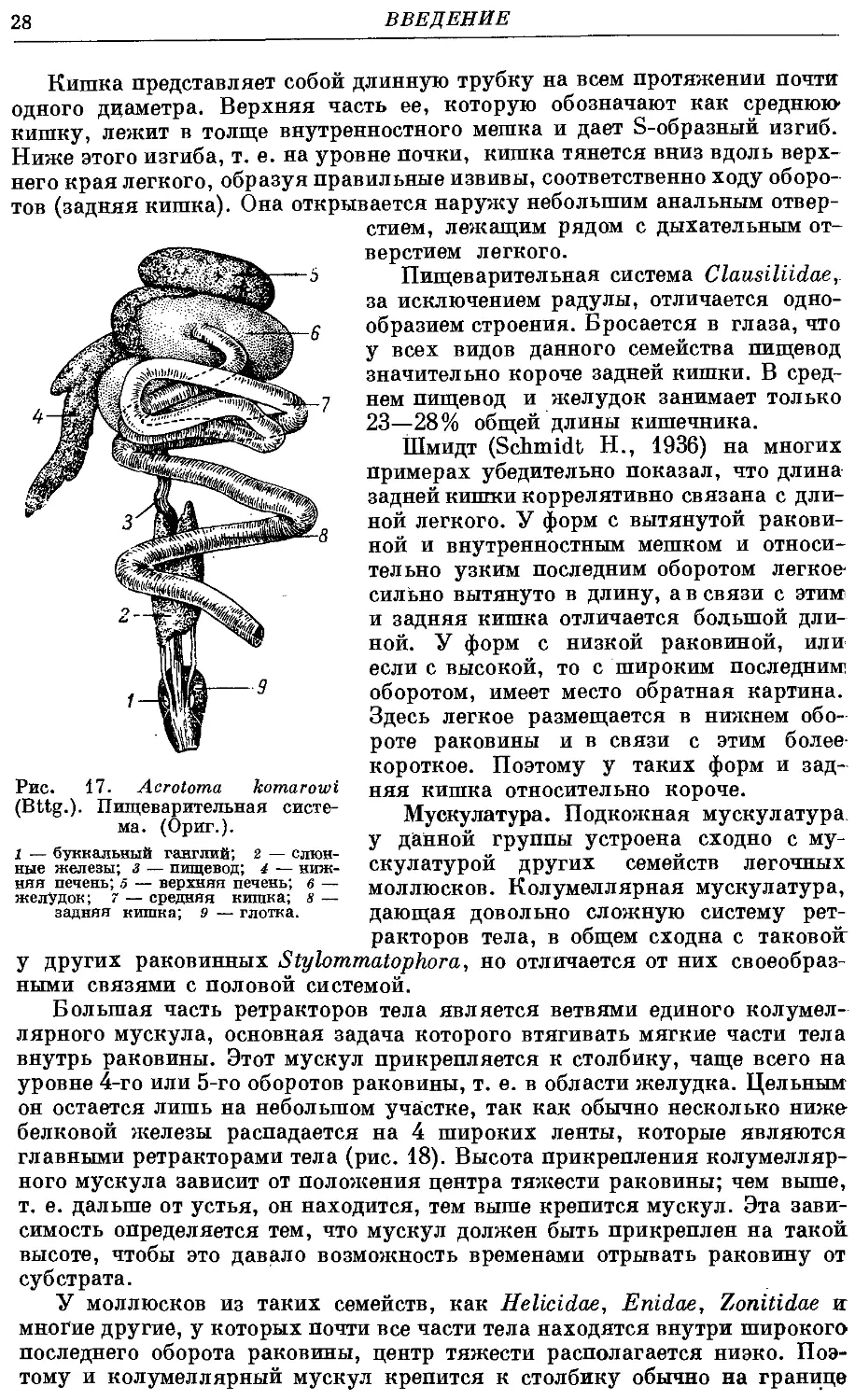
В пределах каждого ряда различают следующие группы зубов (рис. 16):
1) срединный зуб — всегда один, занимает середину ряда и построен
симметрично, т. е. при рассматривании сверху основная пластинка его
Рис. 13. Iphigena ventricosa (Drap.)".
Общий вид мягких частей тела по-
после удаления раковины. (Ориг.).
1 — подошва ноги; 2 — легочная вена;
з — нижняя кишка; 4 — сердце; 5 — по-
гонады; 8 — верхняя печень; 9 — желу-
желудок; ю — средняя кишка; 11 — семяяйпе-
вод.
26 ВВЕДЕНИЕ
близка к прямоугольнику или трапеции, а зубцы параллельны продоль-
продольной оси радулы;
2) боковые зубы — располагаются справа и слева от срединного зуба
и построены асимметрично, т. е. основная пластинка по форме близка
к параллелограмму или ромбу, а зубцы отогнуты в сторону от продоль-
продольной оси радулы;
3) краевые зубы — асимметрия выражена еще сильнее; первоначальные
зубцы расщепляются на более мелкие зубчики; последние краевые зубы
претерпевают редукцию зубцов до полного их исчезновения.
Различают два типа зубцов: 1) основной, который у срединного зуба
находится в центре, а у боковых и у краевых лежит медиально; обычно
Рис. 14. Laciniaria plicata (Drap.). Мантийный комплекс орга-
органов. (Ориг.).
1 — почка; 2 — желудочек сердца; з — околосердечная сумка (перикардий);
4 — предсердие; 5 — легочная вена; 6 — верхняя стенка легкого (мантия);
7 — задняя кишка.
•он наиболее крупный; 2) дополнительные — у срединного зуба они нахо-
находятся по обеим сторонам от основного, а у боковых и краевых по одной из
юторон, а именно на правой стороне радулы — справа, а на левой — слева
от основного зубца; у некоторых видов дополнительные зубцы на средин-
срединном и боковых зубах отсутствуют.
По форме зубов у Clausiliidae можно наметить два типа радулы (рис. 16).
Первый тип. Срединный зуб имеет три зубца — основной и два
дополнительных. Последние у одних форм хорошо развиты, а у других
слабо, но все же выражены ясно. Боковые зубы имеют два зубца — основ-
основной и дополнительный. У краевых зубов за счет расщепления сначала
основного, а затем и дополнительного зубцов общее число их достигает
от 4 до 10. Общая формула одного ряда у моллюсков с радулой первого
типа выглядит так:1
к ¦ б , с , б , к
10 — 3~+~~2~~г~1Г~т~~2+3 — 10 -
Второй тип. Срединный зуб имеет только один мощный зубец —
основной. Большинство боковых также только с одним зубцом и лишь
у нескольких, наиболее внешних, боковых зубов появляется дополнитель-
дополнительный зубец. Краевые зубы имеют от 3 до 6 зубцов, реже немного больше.
Общая формула одного ряда данного типа выглядит так:
к _|_ б _, б _, с _, б _, б _, к
1 с — срединный, б — боковые, к — краевые зубы; в знаменателях дробей ука-
указано число зубцов.
МОРФОЛОГИЯ: АНАТОМИЯ
27
Ниже для характеристики радулы отдельных видов, с целью показать
число зубов каждой группы и число рядов в радуле, мы несколько видоиз-
видоизменяем обычные радулярные формулы. Так, например, главные особен-
особенности радулы Pontophaedusa funiculum можно передать следующим обра-
образом:
[> + (? + -Й)х2]х92
В круглых скобках указывается число и фор-
форма боковых и краевых зубов одного полуряда
(в связи с этим в формулу введен коэффициент 2).
В квадратных скобках дается характеристика од-
одного ряда. Общее число зубов в каждом ряде
множится на число рядов (в данном случае 92).
Крупные слюнные железы кольцом окружают
пищевод и двумя протоками впадают в заднюю
стенку глотки (рис. 17). Пищевод представляет
собой короткую, внутри складчатую трубку, ко-
которая, очень слабо изгибаясь, почти отвесно тя-
тянется кверху вдоль столбика. Здесь она впадает
в объемистый желудок, который располагается чаще всего в 4-м или в 5-м
оборотах раковины (считая от устья), спирально изогнут, занимает почти
полный оборот. Стенки желудка очень тонкие, с явственными кольцевид-
Рис. 15. Filosa filosa
(Mouss.). Челюсть.
(Ориг.).
13
Рис. 16. Два типа радулы Clausiliidae. (Ориг.).
А — Idyla foveicollis (Charp.); В — Micropontica retoioskii (Bttg.).
с — срединный зуб; цифры — порядковый номер зуба, считая от се-
середины.
лыми перетяжками, а со стороны столбика — с двумя продольными мус-
мускульными лентами. Кишка берет начало несколько ниже верхнего конца
желудка. Образовавшийся слепой выступ рассматривают как его пилори-
ческую часть. Сюда открываются протоки верхней и нижней долей печени.
Последнюю лишь условно называют печенью, так как на самом деле она
служит не только для образования пищеварительных ферментов, но внутри
ее происходит также всасывание пищи. Печень состоит из двух долей:
крупной верхней, занимающей верхушку внутренностного мешка, и
небольшой нижней, лежащей ниже желудка между петлями кишки.
28
ВВЕДЕНИЕ
Кишка представляет собой длинную трубку на всем протяжении почти
одного диаметра. Верхняя часть ее, которую обозначают как среднюн>
кишку, лежит в толще внутренностного мешка и дает S-образный изгиб.
Ниже этого изгиба, т. е. на уровне почки, кишка тянется вниз вдоль верх-
верхнего края легкого, образуя правильные извивы, соответственно ходу оборо-
оборотов (задняя кишка). Она открывается наружу небольшим анальным отвер-
отверстием, лежащим рядом с дыхательным от-
отверстием легкого.
Пищеварительная система Clausiliidae,
за исключением радулы, отличается одно-
однообразием строения. Бросается в глаза, что
у всех видов данного семейства пищевод
значительно короче задней кишки. В сред-
среднем пищевод и желудок занимает только
23—28% общей длины кишечника.
Шмидт (Schmidt H., 1936) на многих
примерах убедительно показал, что длина
задней кишки коррелятивно связана с дли-
длиной легкого. У форм с вытянутой ракови-
раковиной и внутренностным мешком и относи-
относительно узким последним оборотом легкое-
сильно вытянуто в длину, а в связи с этим
и задняя кишка отличается большой дли-
длиной. У форм с низкой раковиной, или
если с высокой, то с широким последним!
оборотом, имеет место обратная картина.
Здесь легкое размещается в нижнем обо-
обороте раковины и в связи с этим более
короткое. Поэтому у таких форм и зад-
задняя кишка относительно короче.
Мускулатура. Подкожная мускулатура
у данной группы устроена сходно с му-
мускулатурой других семейств легочных
моллюсков. Колумеллярная мускулатура,
дающая довольно сложную систему рет-
ракторов тела, в общем сходна с таковой:
у других раковинных Stylommatophora, но отличается от них своеобраз-
своеобразными связями с половой системой.
Большая часть ретракторов тела является ветвями единого колумел-
лярного мускула, основная задача которого втягивать мягкие части тела
внутрь раковины. Этот мускул прикрепляется к столбику, чаще всего на
уровне 4-го или 5-го оборотов раковины, т. е. в области желудка. Цельным
он остается лишь на небольшом участке, так как обычно несколько ниж&
белковой железы распадается на 4 широких ленты, которые являются
главными ретракторами тела (рис. 18). Высота прикрепления колумелляр-
ного мускула зависит от положения центра тяжести раковины; чем выше,
т. е. дальше от устья, он находится, тем выше крепится мускул. Эта зави-
зависимость определяется тем, что мускул должен быть прикреплен на такой
высоте, чтобы это давало возможность временами отрывать раковину от
субстрата.
У моллюсков из таких семейств, как Helicidae, Enidae, Zonitidae и
многие другие, у которых почти все части тела находятся внутри широкого
последнего оборота раковины, центр тяжести располагается низко. Поэ-
Поэтому и колумеллярный мускул крепится к столбику обычно на границе
¦9
Рис. 17. Acrotoma komarowi
(Bttg.). Пищеварительная систе-
система. (Ориг.).
1 — буккапьный ганглий; 2 — слюн-
слюнные железы; з — пищевод; 4 — ниж-
нижняя печень; 5 — верхняя печень; 6 —
желудок; ~ — средняя кишка; 8 —
задняя кишка; в — глотка.
МОРФОЛОГИЯ: АНАТОМИЯ
29
двух последних оборотов раковины. У Clausiliidae последний оборот обычно
уже предпоследнего и вся раковина очень вытянута. Основная масса
органов помещается в средних оборотах. Следовательно, центр тяжести
у этих моллюсков находится выше, чем у предыдущих, а поэтому и колу-
меллярный мускул крепится к столбику значительно выше.
Соответственно своим функциям ретракторы подразделяются на 1) ле-
левый, дающий ветки к щупальцам и к стенке тела левой стороны переднего
конца, 2) правый, дающий ветви к аналогичным частям тела правой сто-
стороны переднего конца, 3) глоточный, располагающийся между двумя пре-
предыдущими, но иногда на не-
некотором расстоянии срастаю-
срастающийся с правым ретрактором
я обслуживающий глотку и
радулу, и 4) задний, посылаю-
посылающий ветви к ноге и к перед-
переднему краю мантии. От ретрак-
тора левого губного щупаль-
щупальца отходит тонкий мускул
к нервному кольцу, который
прикрепляется рядом с пе-
педальными ганглиями.
Очень важной чертой ко-
лумеллярной мускулатуры
является ее связь с половой
системой. У левозакрученных
форм эту связь осуществляет у ^
левый ретрактор, дающий,
почти у всех видов семей-
семейства, 1—2 ветви к половой
клоаке и к нижнему концу
влагалища, а в подсемей-
подсемействах Cochlodininae, Euxini-
пае и Clausiliinae, кроме то-
того, еще специальную ветвь к нижнему концу семяприемника. У право-
закрученных форм аналогичные ответвления к половой системе дает пра-
правый ретрактор.
Кроме ретракторов — ветвей колумеллярного мускула, многие Clau-
Clausiliidae имеют специальный ретрактор пениса, не связанный с прочими
мускулами. Он одним концом крепится к верхнему концу пениса, а дру-
другим — к диафрагме, т. е. к нижней стенке легкого (рис. 21). Лишь в подсе-
подсемействе Clausiliinae ретрактор пениса либо отсутствует, либо он рудимен-
рудиментарный. У некоторых Euxininae ретрактор пениса не прикрепляется
к диафрагме, а сливается с задним ретрактором колумеллярного мускула.
В систематическом отношении очень важным признаком является отно-
отношение половых путей к ретрактору ближайшего глазного щупальца, т. е.
к одной из ветвей левого (у левозакрученных форм) или правого (у право-
закрученных форм) ретракторов. У большинства видов ретрактор глазного
щупальца проходит между пенисом и влагалищем (рис. 24). Лишь у части
родов Clausiliinae он лежит в стороне (медиально) от половых путей и с
ними не перекрещивается.
О том, какое функциональное значение имеют различные формы связей
•системы ретракторов с половым аппаратом, будет сказано далее.
Нервная система. Нервная система Clausiliidae очень сходна с таковой
многих других Pulmonata и состоит из нервного кольца с ганглиями,
Рис. 18. Схема колумеллярной мускулатуры
Clausiliidae. (Ориг.).
А — правый ретрактор; Б — глоточный ретрактор;
В — левый ретрактор; Г — задний ретрактор. 1 — рет-
ретрактор семяприемника; 2 — ретрактор половой клоаки;
з — ретракторы левых щупалец.
ВВЕДЕНИЕ
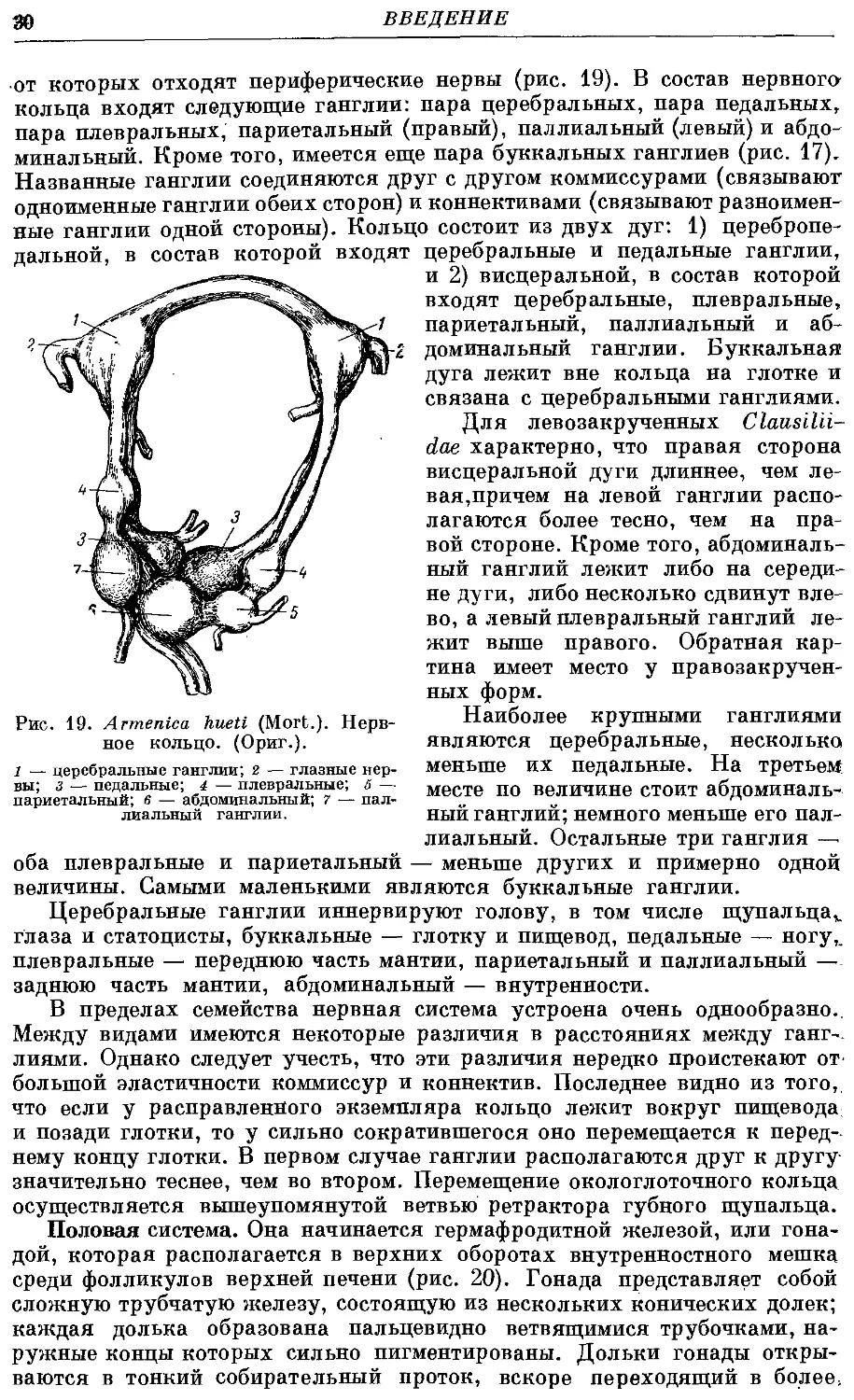
¦от которых отходят периферические нервы (рис. 19). В состав нервного-
кольца входят следующие ганглии: пара церебральных, пара педальных,
пара плевральных, париетальный (правый), паллиальный (левый) и абдо-
абдоминальный. Кроме того, имеется еще пара буккальных ганглиев (рис. 17).
Названные ганглии соединяются друг с другом коммиссурами (связывают
одноименные ганглии обеих сторон) и коннективами (связывают разноимен-
разноименные ганглии одной стороны). Кольцо состоит из двух дуг: 1) церебропе-
дальной, в состав которой входят церебральные и педальные ганглии,
и 2) висцеральной, в состав которой
входят церебральные, плевральные,
париетальный, паллиальный и аб-
абдоминальный ганглии. Буккальная
дуга лежит вне кольца на глотке и
связана с церебральными ганглиями.
Для левозакрученных Clausilii-
dae характерно, что правая сторона
висцеральной дуги длиннее, чем ле-
левая,причем на левой ганглии распо-
располагаются более тесно, чем на пра-
правой стороне. Кроме того, абдоминаль-
абдоминальный ганглий лежит либо на середи-
середине дуги, либо несколько сдвинут вле-
влево, а левый плевральный ганглий ле-
лежит выше правого. Обратная кар-
картина имеет место у правозакручен-
ных форм.
Наиболее крупными ганглиями
являются церебральные, несколько
меньше их педальные. На третьем
месте по величине стоит абдоминаль-
абдоминальный ганглий; немного меньше его пал-
паллиальный. Остальные три ганглия —
оба плевральные и париетальный — меньше других и примерно одной
величины. Самыми маленькими являются буккальные ганглии.
Церебральные ганглии иннервируют голову, в том числе щупальца*,
глаза и статоцисты, буккальные — глотку и пищевод, педальные — ногу,,
плевральные — переднюю часть мантии, париетальный и паллиальный —
заднюю часть мантии, абдоминальный — внутренности.
В пределах семейства нервная система устроена очень однообразно.
Между видами имеются некоторые различия в расстояниях между ганг-
ганглиями. Однако следует учесть, что эти различия нередко проистекают от^
большой эластичности коммиссур и коннектив. Последнее видно из того,,
что если у расправленного экземпляра кольцо лежит вокруг пищевода^
и позади глотки, то у сильно сократившегося оно перемещается к перед-
переднему концу глотки. В первом случае ганглии располагаются друг к другу-
значительно теснее, чем во втором. Перемещение окологлоточного кольца
осуществляется вышеупомянутой ветвью ретрактора губного щупальца.
Половая система. Она начинается гермафродитной железой, или гона-
гонадой, которая располагается в верхних оборотах внутренностного мешка
среди фолликулов верхней печени (рис. 20). Гонада представляет собой
сложную трубчатую железу, состоящую из нескольких конических долек;
каждая долька образована пальцевидно ветвящимися трубочками, на-
наружные концы которых сильно пигментированы. Дольки гонады откры-
открываются в тонкий собирательный проток, вскоре переходящий в более,
Рис. 19. Armeniea hueti (Mort.). Нерв-
Нервное кольцо. (Ориг.).
1 — церебральные ганглии; 2 — глазные нер-
нервы; з — педальные; 4 — плевральные; 6 —
париетальный; в — абдоминальный; 7 — пал-
паллиальный ганглии.
МОРФОЛОГИЯ: АНАТОМИЯ 31
толстый гермафродитный проток. Последний тянется вниз вдоль колумел-
лярного мускула. Вблизи от белковой железы гермафродитный проток
становится толстым и образует многочисленные извивы, которые плотно
прилегают друг к другу и снаружи обтянуты соединительнотканной обо-
оболочкой. Перед впадением в семяяйцевод проток вновь становится прямым
и немного утончается (рис. 21).
Семяяйцевод представляет собой трубчатый орган, разделенный внутри
неполными перегородками на три канала: мужской, женский и серозный
(рис. 22). Узкий мужской канал служит для прохода своих спермиев.
По всей длине в него открывается лен-
лентовидная предстательная железа. Жен-
Женский канал, из всех трех наиболее ши-
широкий, служит для прохода яиц, а у
живородящих форм — для созревания
эмбрионов. Функция серозного канала
в точности неизвестна, но как, с нашей
точки зрения обоснованно, предполагает
Стинберг (Steenberg, 1914), он служит
для прохождения спермиев партнера
из семяприемника, наверх, в камеру
оплодотворения. За это говорит его
расположение — он начинается еще в
матке, у места слияния ее с семяприем-
семяприемником, и кончается у камеры оплодотво-
оплодотворения. Кроме того, секрет серозного ка-
канала как ПО своей структуре, так И ПО Рис. 20. Laciniaria plicata (Drap.).
отношению к красителям обнаруживает Верхняя печень с гонадой. (По Стин-
болыпое сходство с секретом предстатель- бергу).
ной железы. Все это позволяет предпо-
предполагать, что секрет серозного канала служит для питания спермиев парт-
партнера на пути их из семяприемника в камеру оплодотворения.
Гермафродитный проток перед слиянием с семяяйцеводом расширяется
в так называемую камеру оплодотворения (рис. 23). Она имеет вид V-об-
разно согнутой трубки, плотно прилегающей к белковой железе. Место
перегиба трубки нередко расширено и образует слепой выступ. В верхний
конец семяяйцевода, кроме камеры оплодотворения, открывается проток
белковой железы. Последняя состоит из многочисленных фолликулов,
выделяющих питательные вещества для яйцеклеток.
Верхняя часть семяяйцевода устроена довольно сложно. Здесь имеется
система камер и складок, благодаря которым яйца и спермин (свои)
направляются в соответствующие каналы. Кроме того, внутри одной из
камер формируется известковая оболочка яиц.
Внизу семяяйцевод разделяется на семяпровод и матку. Последняя,
как правило, представляет собой сравнительно с семяяйцеводом более
узкий проток различной длины. С места слияния матки с семяприемником
начинается влагалище, которое открывается в половую клоаку.
Влагалище у большинства видов заметно толще матки за счет довольно
мощной мускулатуры его стенок, особенно в верхней части.
Половая клоака, в которую открываются пенис и влагалище, невелика,
у сократившихся экземпляров не всегда даже заметна. Она сообщается
с внешним миром небольшим половым отверстием. Последнее у левозакру-
ченных форм (в том числе у всех рассматриваемых в данной работе) распо-
располагается позади левого, а у правозакрученных — за правым губным
щупальцем.
32
ВВЕДЕНИЕ
Семяприемник представляет собой крупный трубчатый орган, тяну-
тянущийся вдоль семяяйцевода. Он служит для принятия сперматофора парт-
партнера и для хранения чужой спермы до
наступления созревания яйцеклеток.
Верхний слепой конец семяприем-
семяприемника обычно более или менее сильно
вздут, образуя резервуар семяприем-
семяприемника. Последний лежит на семяйце-
воде, несколько ниже белковой же-
железы, либо непосредственно на ней.
Только у немногих видов (Mentis-
soidea litotes, Euxinastra hamata) се-
семяприемник настолько длинный, что
его резервуар находится выше белко-
белковой железы и прижат к желудку.
У подавляющего большинства ви-
видов Clausiliidae семяприемник имеет
особый придаточный орган — отро-
отросток. Это полый трубчатый вырост,
слепо замкнутый наверху. Та часть
протока семяприемника, которая
находится ниже отростка, называет-
называется шейкой. Лишь единичные виды
не имеют отростка, в том числе два
наших отечественных — Mentissoidea
litotes (рис. 133) и Filosa filosa (рис.
137).
В пределах семейства отчетливо
намечаются три типа семяприемника
(рис. 24).
У ^haedusinae^Neniinae и Alopi-
inae как семяприемнйк",~так~1н его
отросток представляют собойширокие
трубки почти одинакового диаметра.
Иногда даже бывает трудно сказать,
какая ветвь является отростком, ка-
какая собственно семяприемником.
Отросток всегда отходит на значи-
значительном расстоянии от нижнего конца
семяприемника, благодаря чему шей-
шейка очень длинная. Кроме того, сам
семяприемник впадает в женские по-
половые пути недалеко от их нижнего
конца, иногда даже у самой половой
клоаки. В_связи с этим_полрвая си-
система указанных группу (за исключе-
исключением Alojninae) отличается длинной
маткой^.
Несколько иная картина наблю-
R&eTCsrfCduMddininae. Семящгаемник
устроен так же, но он впадает в женские половые пути_на значительном
расстоянии от половой клоакйТ вследствие чейгмятка^здесь короче, чем
¦у предыдущих групп, а влагалище ^Длиннее. Кроме~"того, характерна
другая особенность. К шейке семяприемника прикрепляется особая ветвь
¦рил
¦рнщ
¦нл
Рис. 21. Cochlodina laminata (Mont.).
Половая система. (По Стинбергу).
-б — бич; бок — белковая железа; ел — вла-
влагалище; г — гонада (гермафродитная желе-
железа); гп — гермафродитный проток; д — диаф-
диафрагма; кл — половая клоака; ко — камера
¦оплодотворения; лр — левый ретрактор; м —
матка; оси — отросток семяприемника; п —
пенис; рвщ — ретрактор верхних (глазных)
щупалец; рки — ретракторы половой клоаки;
рнщ — ретрактор нижних (губных) щупалец;
рп — ретрактор пениса; реп — ретрактор се-
семяприемника; с — семяпровод; сп — семяприем-
семяприемник; ся — семяяйцевод и предстательная же-
железа; зп — эпифаллус.
се
пж
ООП
Рис. 22. Iphigena plicatula (Drap.). Поперечные срезы через семяяйцевод и семя-
семяприемник. (По Стинбергу).
А — до обособления семяпровода от семяяйпевода; Б — на уровне начала семяпровода.
жк — женский канал семяяйдевода; мк —¦ мужской канал семяяйпевода; осп — отросток
семяприемника; пж — предстательная желеаа; с — семяпровод; се — серозный канал се-
семяяйпевода; сп — семяприемник.
3 И. М. Лихарев
34
ВВЕДЕНИЕ
бж
левого ретрактора :=::_ретрактор семяприемника, а сама шейка имеет
толсйгета'ускулистые стенки. Это "йокаанвает, "что у Cochlodininae в про-
процессе копуляции шейка _играет более актиюную jiojrbi чем у предыдущих
гру_ппл Семяприемник впадаетГв женские половые пути на значительном
расстоянии от прлщой клоаки; следовательно, матка сравнительно корот-
короткая, а вдагадейце длиннбе. ----- — _=_¦- __
Т1слиГ~у~т1редыдущих четырех подсемейств стенки как семяприемника,
так и его,4игростка слагаются из слоя эпителия (внутри), слоя соединитель-
соединительной ткани и слоя кольцевых мус-
мускульных волокон, то у Clausili-
inae и Euxininae стенки отростка об-
образованы только за счет однослой-
однослойного эпителия. Стенки семяприем-
семяприемника сохранили прежнее трех-
трехслойное строение. Отросток, как
правило, очень узкий и тонкостен-
тонкостенный, почти прозрачный, и отходит
от семяприемника недалеко от его-
нижнего конца. Как у Cochlodini-
Cochlodininae, шейка семяприемника муску-
мускулистая, и к ней всегда прикреп-
прикрепляется мощная ветвь левого рет-
рактора.
Подобно семяприемнику, муж-
мужские половые пути Clausiliidae
также обнаруживают значительное
разнообразие в устройстве (рис. 24).
Как известно, у подавляющего
большинства легочных моллюсков
семяпровод вначале тянется вниз
(т. е. к половой клоаке), плотно
прижимаясь к женским половым
путям. Затем он отгибаетря на-
назад, тянется вдоль пениса й' впа-
впадает в задний конец последнего.
При этом первый участок семяпро-
семяпровода можно обозначить как нисхо-
нисходящее колено, а второй — как вос-
восходящее. Восходящее колено вме-
вместе с пенисом образует петлю, кото-
которая»; лежит в стороне от женских
i ° -^ половых путей. При этом ретрактор
ближайшего главного щуцальда проходит между пенисом и восходящим
коленом семяпровода с ОДчЩ?$оровыи влагалищем с другой. Такой же
план строения обычен у многих Clausiliidae. Но вместе с тем в пределах
этого семейства имеется подсемейство (Clausiliinae), у которого наблю-.
дается постепенное укорачивание петли, образованной мужскими поло-
половыми путями, до полного ее исчезновения. В последнем случае семяпровод
и пенис располагаются вдоль женских половых путей.
Соответственно можно наметить три плана строения мужских, половых
путей:
1. Большинство Clausiliidae(рис. 21,24, А—Г). Семяпровод при обособ-
обособлении от«емяяйцевода имеет вид мускулистой трубки, на срезах с зияющим
просветом. Проделав несколько извивов вдоль матки, он сильно утончается
жн
се
мк
ЖК;
Рис. 23., Iphigena ventricosa (Drap.). Верх-
Верхний конец семяййцевода и камера оплодот-
оплодотворения. Верхняя ртенка первой камеры
ссмяяйцевода и участок белковой железы
удалены. (По Стйнбергу).
бок —;беяковаа- желева; боцж — даделительный
канал ;белкей»ой жеяеаы; щ — гйрмафроДипщЙ
проток; окк — женский канал; кй т- камера оп-
оплодотворения; мк тК мужской иадая семяяйцево-
да; о — проход иа камеры, оплодотворения в се^-
мяяйцевод; се — сероввый ; канда, 'Семяяйревода.;
МОРФОЛОГИЯ: АНАТОМИЯ
ъъ
(на срезах нередко просвет исчезает) и, прижимаясь к влагалищу, почти
отвесно тянется к половой клоаке. Здесь он обычно проходит сквозь толщу
тех ветвей левого ретрактора, которые прикрепляются к половой клоакег
огибает ретракторлевого глазного щупальца (у правозакрученных форм —-
Г Д ?
Рис. 24. Основные типы половой системы Clausiliidae. (Ориг.).
А — Phaedusinae и Neniinae: Б — Alopiinae; В—Cochlodininae; Г — Euxininae; Д, К —
GlausUiinae. 1 — расширенное нисходящее колено семяпровода; 2 — рудиментарная
петля, образованная восходящим коленом семяпровода и пенисом; з — вздутый
участок нисходящего колена семяпровода (вторичный пенис); мп — мускульная
полоска; пр — придаток пениса; рс — ветвь ретрактора семяприемника к семя-
семяпроводу; шсп — шейка семяприемника; остальные обозначения см. рис. 21.
ретрактор правого глазного щупальца) и подымается к заднему концу
пениса. Перед впадением в последний семяпровод нередко расширяется,
образуя эпифаллус — орган, стенки которого образуют оболочку сперма-
тофора. У некоторых видов рядом с семяпроводом, тесно прижимаясь
к нему, в пенис впадает тонкостенный бич.
Пенис чаще всего имеет вид более или менее длинного цилиндрического
органа, который нередко на одном участке расширен. Внутри этого рас-
36
ВВЕДЕНИЕ
ширения обычно проходят крупные складки. К пенису, реже к эпифал-
лусу, крепится ретрактор, который у большинства видов другим концом
прикреплен к диафрагме. Но у некоторых групп (Idyla, Armenica) ретрак-
ретрактор пениса является ветвью коллумелярного мускула, а именно заднего
ретрактора.
Многие Clausiliidae не имеют г
никаких придаточных органов на
пенисе. Это в первую очередь ка-
касается наиболее примитивно ор-
организованных подсемейств — Phae- flSVWJZ/^ii 6ж
dusinae и Neniinae. Среди же евро-
европейских и переднеазиатских видов
¦ркд
рнш,
Рис. 25. Laciniaria plicata (Drap.). По-
Половая система. (По Стинбергу).
ип -т нерв пениса; остальные обозначения
см. рис. 21 и 24.
~РЩ
'/W
Рис. 26. Laciniaria biplicata (Mont.). Поло-
Половая система. В матке (выводковая сумка>
видны эмбрионы. (По Стинбергу).
1 — нерв пениса; остальные обозначения см.
рис. 21 и 24.
немало таких, у которых близ заднего конца пениса есть более или менее
крупный, иногда очень крупный и мускулистый придаток. Последний
чаще всего имеет форму крючковидного полого выступа, замкнутого на
конце (рис. 83).
Среди изученных нами видов такой придаток обнаружен у всех видов
Armenica и Acrotoma. Кроме того, известно, что этот орган характерен для
многих Alopiinae. По-видимому, появление этого придатка в какой-то
мере связано с обитанием на скалах, так как он имеется только у скаложи-
МОРФОЛОГИЯ: АНАТОМИЯ
37
ся
лов. Интересно, что среди Alopiinae — подсемейства, в пределах которого
"этот орган есть у большинства видов, он отсутствует именно у тех моллю-
моллюсков, которые обитают не только на скалах, но и на деревьях (например,
род Delima).
2. Laciniaria, Mucronaria и некоторые
другие роды подсемейства Clausiliinae.
Здесь характерны следующие особенности
(рис. 24, Е, 25, 26). Во-первых, семяпро-
семяпровод не образует восходящего колена и
вместе с пенисом вытянут вдоль влагали-
влагалища. Во-вторых, граница между пенисом и
семяпроводом очень слабо выражена и по-
порой неуловима, так как семяпровод, почти
сразу как он обособился от семяяйцевода,
сильно расширяется ив виде толстой му-
мускулистой трубки тянется вниз. Пенис
имеет местное веретенообразное вздутие.
В-третьих, мужские половые пути лише-
лишены диафрагмального ретрактора.
В связи с подобным строением муж-
мужских половых путей ретрактор глазного
щупальца лежит медиально от них, а не
проходит между Пенисом и влагалищем,
как в предыдущем типе.
Семяпровод на значительном протяже-
протяжении редкими мускульными волокнами под-
подтянут к влагалищу. Кроме того, от рет-
ретрактора семяприемника отходит к семя-
семяпроводу особый мускул, который дополни-
дополнительно прижимает Аго к женским поло-
половым путям. /
Наконец, имеется тонкая- мускульная
полоска, отходящая от нижнего конца се-
семяпровода и другим концом прикреп-
прикрепляющаяся к переднему концу пениса.
3. Третий тип организации' мужских
половых путей является переходным меж-
между двумя предыдущими (рис. 24, Д, 27). Он
встречается у европейских родов Fusulus,
Ruthenica, Clausilia, Iphigena, Ne&ptyriaca
(Clausiliinae). От восходящего колена се-
семяпровода здесь сохранился только корот-
короткий участок, который вместе с пенисом
образует небольшую петлю, лежащую
у половой клоаки. Как и в предыдущем
типе, нисходящая часть семяпровода расширена и особым ответвлением
ретрактора семяприемника подтянута к влагалищу. Ретрактор пениса еще
имеется, но тонкий и рудиментарный.
У Fusulus и Ruthenica ретрактор левого глазного щупальца проходят
еще между пенисом и влагалищем, а у остальных родов он уже лежит
медиально от половых путей.
Таким образом, как в отношении формы семяпровода и пениса, так и
их связей с колумеллярной мускулатурой указанные роды дают морфоло-
морфологические переходы от первого типа ко второму.
Рис. 27. Iphigena ventricosa (Drap.).
Половая система. (По Стинбергу).
1 — нерв пениса; остальные обозна-
обозначения см. рис. 21.
38 ВВЕДЕНИЕ
Коррелятивные связи между половой системой и мускулатурой. Суже-
Сужение последнего оборота раковины и образование внутри него замыкатель-
вого аппарата привело к тому, что органы нижней части внутренностного
мешка в отношении пространства, оставленного для них, оказались в менее
выгодных условиях, чем органы, лежащие выше. В результате намети-
наметилась определенная тенденция к преодолению этого неудобства, что при-
привело к указанному выше разнообразию в строении полового аппарата
наших моллюсков.
В этом отношении особого внимания заслуживает положение семя-
семяприемника и форма мужских половых путей. *
У наиболее примитивных подсемейств — Phaeausinae, Neniinae —
подобно большинству представителей отряда Stylommatophora (Pupilli-
dae, Enidae, Bulimulidae), семяприемник открывается в женские половые
пути рядом с половой клоакой. Однако у остальных, более высоко органи-
организованных подсемейств Clausiliidae семяприемник впадает в женские
половые пути на значительном удалении от половой клоаки (рис. 24).
Перемещение семяприемника вдоль влагалища кверху объясняется
нехваткой места для органов, расположенных в шейке внутренностного
мешка, т. е. на стыке последнего с ногой. Это перемещение повлекло за
собой другие изменения организации. Известно, что при копуляции
выворачивание женских половых путей (как и пениса) происходит за счет
давления полостной жидкости. Выворачивание идет до тех пор, пока не
откроется вход в семяприемник, куда должен быть введен сперматофор
партнера. Обратное втягивание половых органов осуществляется с по-
помощью мускулатуры. При этом у видов, семяприемник которых располо-
расположен вблизи от половой клоаки, это втягивание производят ветви одного
из передних ретракторов, крепящиеся к нижнему концу влагалища и
к клоаке. Но в тех случаях, когда семяприемник переместился кверху,
этих мускулов уже не хватает, так как они располагаются значительно
ниже его. В связи с этим у Cochlodininae, Euxininae и Clausiliinae в допол-
дополнение к указанным мускулам появляется еще особая ветвь левого ретрак-
тора — ретрактор семяприемника, который прикрепляется к шейке семя-
семяприемника и осуществляет втягивание как самой шейки, так и лежащего
ниже ее влагалища.
Отсутствие ретрактора семяприемника у Alopiinae, несмотря на то,
что семяприемник располагается высоко, указывает на то, что в этой группе
выворачивание женских половых путей ограничивается лишь нижним
концом влагалища.
Сужение последнего оборота раковины и образование в нем замыка-
тельного аппарата отразилось не только на положении семяприемника*
но и на мужских выводных путях.
В таких группах, как Helicidae, Enidae, Bulimulidae, и у многих дру-
других Stylommatophora, у которых значительная часть внутренних органов
помещается в наиболее широком, последнем обороте раковины, для втя-
втягивания пениса внутрь тела (после копуляции) вполне достаточно усилия
ретрактора пениса, который другим концом крепится к диафрагме.
Аналогичную картину мы встречаем у большинства подсемейств Clau-
Clausiliidae — Phaedusinae, Neniinae, Alopiinae, Cochlodininae и у большин-
большинства Euxininae. Однако в пределах наиболее высокоорганизованного
подсемейства — Clausiliinae мужские половые пути претерпевают ряд
последовательных изменений, которые проявляются в постепенной редук-
редукции ретрактора пениса, параллельно которой идет укорачивание петли,
составленной из пениса и восходящего колена семяпровода. В результате
мужские половые пути, вытягиваются вдоль женских и подтянуты к ним
БИОЛОГИЯ: ПИТАНИЕ 39
многочисленными мускульными волокнами и специальной ветвью ретрак-
тора семяприемника.
Тем самым втягивание пениса осуществляется уже не сравнительно
маломощным ретрактором пениса, а ретрактором семяприемника, т. е.,
в конечном счете, мощной колумеллярной мускулатурой. Если ретрактор
пениса прикрепляется к тонкой и гибкой диафрагме, то колумеллярная
мускулатура имеет более прочную опору — столбик раковины.
У двух родов Euxininae — Idyla и Armenica та же цель, т. е. передача
работы по втягиванию пениса колумеллярной мускулатуре осуществ-
осуществляется несколько иначе. Здесь ретрактор пениса становится ветвью зад-
заднего ретрактора.
Следовательно, сужение последнего оборота раковины Clausiliidae
у наиболее высоко организованных представителей этого семейства при-
приводит к тому, что втягивание пениса осуществляется колумеллярной
мускулатурой.
БИОЛОГИЯ CLAUSILIIDAE
Питание
Основными источниками питания служат гниющие растительные
остатки и гифы грибов, т. е. наши моллюски являются сапро- и микофа-
гами. Хищники среди них совершенно отсутствуют.
В связи с тем, что большинство видов обитает на деревьях и скалах,
как те, так и другие в значительной степени питаются лишайниками. Виды,
живущие в подстилке, в качестве пищи используют как гниющую листву,
так и развивающуюся на ней флору грибов. '
Среди древесных видов очень распространено питание влажной гнию-
гниющей древесиной пней и поваленных стволов. Некоторые из них (Serru-
lina, Pontophaedusa, Caspiophaedusa) только здесь и встречаются.
Как показал Фрёминг (Fromming, 1954), даже в лабораторных усло-
условиях, иогда Clausiliidae предоставлена только зеленая пища, они предпо-
предпочитаю^ гниющие части свежим. При этом обнаруживается сравнительное
безразличие к каким-нибудь определенным видам растений, а предпочте-
предпочтение оказывается растениям с мягкими тканями.
Характер пищи сказывается на форме зубов радулы. Виды, питаю-
питающиеся гниющими растительными остатками, грибами и мягкими лишай-
лишайниками, имеют радулу первого типа, т. е. с мелкими срединными и боко-
боковыми зубами, снабженными дополнительными зубцами. Это наиболее
обычный тип строения не только для данного семейства, но и для
большинства легочных моллюсков.
Скаложилы, вынужденные питаться жесткими лишайниками, произра-
произрастающими на твердом субстрате, имеют радулу второго типа, т. е. с круп-
крупными срединными и боковыми зубами, лишенными дополнительных зуб-
зубцов. Этот тип радулы следует рассматривать как вторичный, являющийся
адаптацией к питанию жесткой пищей, при котором происходит усиление
основных зубцов (они приобретают облик долота) и редукция дополнитель-
дополнительных. Подобный тип радулы свойствен не только скаложилам из нашего
«емейства, но и из других семейств. Так, Chondrina avenacea (Pupillidae)
•и Helicigona cingullela (Helicidae), которые тоже живут на скалах, имеют
такой же тип радулы.
Характер пищи отражается и на числе зубов в радуле. Как видно из
табл. 1, у скаложилов больше зубов F—10 тысяч), чем у видов, живу-
живущих на деревьях и в подстилке B—5 тысячи). Правда, здесь наблюдаются
немногие исключения. Так, например, Mucronaria gustavi (Bttg.) и М.
40
ВВЕДЕНИЕ
pleuroptychia (Bttg.), несмотря на то, что они проводят всю жизнь на ска-
скалах, обладают меньшим числом зубов D—6 тысяч), чем прочие видыт
живущие в тех же условиях. В данном случае сказываются небольшие
размеры названных моллюсков.
_, Таблица!
Зависимость числа
С I ausil iid ае от
эубов^в
радулах некоторых
характера пищи
Название вида и характер
пищи
Число
зубов
в ряде
29
33
41
33—35
43
39
41
41
37
43
41
49
41
47
49
51
47
45
51
Общее
число
зубов
в раду-
радуле
1856
2211
3198
3300
3569
3588
3608
3813
4070
4300
4346
4900
4920
4935
4998
5100
5123
5625
5712
Название вида и характер
пищи
Число
вубов
в ряде
Общее
число
вубов
в раду-
радуле
Мягкие лишайники,
растительный перегной
Serrulina semilamellata . .
S. sieversi sieversi
S. serralata serrulata . . ,
Filosa filosa
Caspiophaedusa perlucens
Pontophaedusa funiculum
Euxina lessonae
En. tscketschenica . . . .
Mucronaria duboisi . . . .
Euxina talyschana ....
Eu. somchetica ......
Megaleuxinareulauzi . . .
Quadriplicata aggesta agge-
sta
Euxinastra hamata ....
Quadriplicata quadriplicata
Megaleuxina derasa . . .
Idyla foveicollbs . . . , .
Quadriplicata lederi ....
Euxina persica persica . . .
Mentissoidea litotes . . .
Quadriplicata pumiliformis
Cochlodina Ixuninata . . .
Жесткие лишайники
LaciriimitL.canaliJe.ra ..._^
Micropontica retowskii . .
Lacir^ria_gr&cUicqsta^ ^_.
Micropontica closta . . .
Mucronaria gustavi . . .
Acrotoma semicincta . . .
Armenica disjuncta . . .
Mucronaria pleuroptychia
Armenica laevicollis . . .
A.brunnea
Euxina tuschetica . . . .
Armenica griseofusca . .
A. gracillima
Euxina difficilis
Acrotoma komarowi . . .
A. sobrievskii
Armenica hueti ......
5814
7350
7810
A1ML
4173
J.182
4730
5565
5880
602O
6165
6360
6630
6840
7420
7950
8031
8540
8850
9750
Аналогичные исключения есть и среди видов, живущих на деревьях и
в подстилке. Так, Quadriplicata pumiliformis (Bttg.) и Cochlodina laminatp
(Mont.) обладают довольно большим числом зубов G—8 тысяч).
Увеличение числа зубов у скаложилов чаще всего идет за счет нараста-
нарастания числа рядов в радуле. Однако имеет место и некоторое увеличение
числа зубов в одном ряде.
Размножение
Подобно большинству гермафродитных легочных моллюсков, у Clau-
siliidae созревание мужских половых продуктов происходит раньше жен-
женских, т. е. у них наблюдается протерандрия. Спермин передаются парт-
партнеру в сперматофорах — особых длинных пакетах с кожистыми стен-
стенками, которые формируются в заднем конце пениса или в эпифаллусе.
Копуляция в конечном счете сводится к обмену сперматофорами. Это(г
акт обычно происходит во влажную и теплую ночь или в сумерки, причем
столь скрытно, что редко когда его удавалось наблюдать. Каких-нибудь
брачных игр, обычных у других моллюсков перед копуляцией, по-види-
по-видимому, не происходит, и она завершается очень быстро. Вначале, когда
партнеры достаточно сблизились, давлением полостной жидкости вывора-
выворачиваются наружу половая клоака, пенис и частично влагалище. Пенис
через влагалище вводится в шейку семяприемника партнера и оставляет
БИОЛОГИЯ: РАЗМНОЖЕНИЕ 41i
там свой сперматофор, на чем копуляция и заканчивается. Как уже гово-
говорилось, обратное втягивание вывернутых половых органов осуществляется
с помощью мускулатуры.
Сперматофор сохраняется в семяприемнике до созревания яйцеклеток
и перемещения их по гермафродитному протоку в камеру оплодотворения.
К этому моменту стенки сперматофора под действием секрета семяприем-
семяприемника растворяются и освободившиеся спермин перемещаются по протоку
семяприемника в матку и далее поднимаются вверх по серозному каналу
семяяйцевода до камеры оплодотворения. Здесь происходит оплодотворе-
оплодотворение яйцеклеток. Формирование яиц идет несколько ниже, в первой камера
семяяйцевода. Секрет белковой железы окружает со всех сторон яйцеклетку
и образует основную массу яйца, состоящую из питательных веществ.
Далее идут три оболочки:1 1) внутренняя — тонкая* окружающая белко-
белковый слой, 2) скорлуповая, состоящая из мелких известковых зерен —
продукт выделения стенок первой камеры семяяйцевода, служащая для
защиты яйца от механических повреждений, и 3) наружная — студневид-
студневидная, довольно толстая, защищающая яйцо от высыхания.
Яйца чаще всего круглой или слегка эллипсоидальной формы, разме-
размером около 1—2 мм.
У среднеевропейских видов откладка яиц обычно происходит в конц&
лета и в начале осени, т. е. в период, когда еще довольно тепло и в то ж&
эремя достаточно осадков. Во влажных лесах Кавказа откладка яиц про-
происходит и летом. Моллюски помещают яйца под гниющую кору, в мох,
в подстилку, под камни, т. е. в места, укрытые и отличающиеся постоян-
постоянной и высокой влажностью. Время, потребное на развитие яиц, различна
у разных видов и, кроме того, зависит от погоды. В среднем, вылупление
происходит на третью—пятую неделю после откладки.
Среди наших моллюсков наряду с яйцекладущими имеется довольно
много живородящих видов. В связи с этим у них развиваются специальные-
адаптации для выведения молоди (рис. 178): женский канал семяприем-
семяприемника и матка преобразуются в обширную тонкостенную выводковую
сумку. При этом остальные части семяяйцевода — мужской и серозный
каналы, предстательная железа — занимают сравнительно небольшое
место и в виде тонкой ленты тянутся по одной из сторон выводковой сумки.
Переход к живорождению и увеличение значения матки сказывается и
на ее размерах. Так, если у близких яйцекладущих видов она короче
семяяйцевода, то у живородящих она нередко длиннее последнего-
(рис. 25 и 26). ^
Молодые улитки выходят из тела матери еще в яйцевой оболочке. Они
довольно быстро протирают ее радулой, и, по-видимому, на первое-
время эта оболочка служит им пищей.
В пределах некоторых родов могут быть как живородящие, так и яйце-
яйцекладущие виды. Так, например, Iphigena ventricosa и Quadriplicata quadri-
plicata являются единственными живородящими видами в пределах своих
родов. При изучении внутреннего строения приведенных в данной работ&
видов у некоторых нам удалось обнаружить в матке эмбрионы, что дало-
основание считать их живородящими. Ниже приводим список всех живо-
живородящих видов, упомянутых в данной работе (звездочкой отмечены мол-
моллюски, для которых живорождение отмечено впервые).
Euxina tschetschenica. * М. reulauxi. *
Ей. lessonae. * r J Filosa filosa. *
Megaleuxina derasa. Balea perversa.
1 У живородящих видов только одна оболочка — внутренняя.
42 ВВЕДЕНИЕ
Vestia riloensis. Mucronaria duboisi. *
Quadriplicata guadriplicata. * M. pleuroptychia.
'Iphigena ventricosa. M. gustavi. *
Laciniaria biplicata.
Вполне возможно, что среди изученньк нами видов есть еще несколько
живородящих, но мы могли этого и не обнаружить, если вскрытые экземп-
экземпляры находились в мужской фазе развития полового аппарата.
Живорождение встречается как у видов, живущих в умеренных широ-
широтах, так и в тропических (Loosjes, 1953а). .г
Есть основания предполагать, что имеются виды, которые1'^ зависи-
зависимости от условий обитания могут быть и яйцекладущими и живородящими.
Так, некоторые французские авторы (Moquin-Tandon,f§55; Germain,
1930) указывают, что Balea perversa является яйцекладущим видом,
и дают сведения о времени года, когда происходит откладка яиц, о их
размерах, числе и о времени, необходимом на вылупленйе молоди. В то
же время, другие авторы (Voith, 1846, — наблюдения сделаны в Баварии;
Sporleder, 1861, — в Нидерландах; Steenberg, 1914, — в Дании) считают
этого моллюска живородящим, причем последний автор пишет, что всегда
при вскрытиях он обнаруживал в матке несколько эмбрионов, иллюстри-
иллюстрируя сказанное прекрасным рисунком.
Другой пример. Анатомируя Euxina tschetschenica,мы обнаружили
внутри матки эмбрионы, что позволило нам утверщда^Ь, что данный мол-
моллюск размножается живорождением. Между тем1 Калитина A953а) ука-
указывает, что ей приходилось видеть этого моллюска рядом с яйцами, кото-
которые, как она думает, принадлежали именно ему.
Вполне возможно, что, подобно многим рептилиям (например, наша
обыкновенная живородящая ящерица — Lacerta vivipara Jacq.), некоторые
ЙЙЙюски со сравнительно широким распространением, в областях с холод-
холодным климатом, а именно на севере своего ареала и в высокогорных зонах,
становятся живородящими, хотя являются яйцекладущими в областях
и зонах с более теплым климатом. В случае с Balea perversa характерно,
что откладка яиц этим моллюском наблюдалась во Франции, в то время
как живорождение — в странах с более холодным климатом (Дания).
Рост и длительность жизни
Наблюдения за ростом некоторых Clausiliidae проводились только
в лабораториях (Degner, 1952; Fromming, 1954, — на Laciniaria biplicata),
в условиях, допускавших круглогодичную активность этих животных.
Естественно, что данные, полученные этим путем, нельзя считать пол-
полностью соответствующими отношениям, существующим в природе, где
в неблагоприятные периоды года (зима, летняя засуха) моллюски впадают
в спячку и рост их приостанавливается.
У европейских видов вылупление молоди или появление ее из тела
матери происходит осенью. За оставшийся промежуток времени до наступ-
наступления холодов и впадения в зимнюю спячку моллюски успевают лишь
очень немного подрасти. В основном рост происходит в весенне-летний
период следующего года, завершается к осени, и животное приступает
к размножению лишь на второй год жизни.
У видов, живущих во влажных субтропиках Кавказа, рост и созрева-
созревание идет быстрее. Однако точными данными на этот счет мы не распола-
располагаем.
В отношении длительности жизни Clausiliidae Дегнер (Degner, 1952)
сообщает следующее. В его опытах многочисленные экземпляры Laciniaria
БИОЛОГИЯ: СРЕДА И АДАПТАЦИИ 43
biplicata жили до 31/2 лет. Насколько точно эти цифры соответствуют
реальным отношениям, самому автору неясно. Он признает, что хотя его
моллюски не страдали от хищников, были защищены от резких колебаний
температуры и влажности и получали обильную пищу, все-таки, возможно,
они находились в худших условиях, чем в природе. Он предполагает, что
тот факт, что они не впадали в обычную для них зимнюю спячку, мог при-
привести к преждевременному старению. Кроме того, он подозревает, что его
подопытные моллюски страдали от каких-то заболеваний.
Лосьес (Loosjes, 1953a), тоже на основании наблюдений в террариумах,
пришел к выводу, что как тропические (южноазиатские), так и европей-
европейские виды этого семейства являются многолетними животными, с длитель-
длительностью жизни от 5 до 10 лет.
Средаиадаптации
В данном разделе делается попытка дать характеристику важнейшим
факторам внешней среды, влияющим на жизнедеятельность и распростра-
распространение наших моллюсков, а также наиболее обычным типам адаптации
к различным условиям жизни.
Климат
Температура и влажность. Из основных факторов, слагающих кли-
климат, наибольшое значение для наших моллюсков, как и для всех наземных
моллюсков, имеют температура и влажность. Это не значит, что другие
климатические факторы несущественны, но их воздействие сказывается
в первую очередь через изменения температуры и влажности.
Две физиологические особенности вызывают повышенную чувствитель-
чувствительность ^наземных моллюсков к этим факторам: пойкилотермность и слабая
защищенность от высыхания. Пойкилотермность ставит наших животных
в прямую зависимость от температуры внешней среды, причем это касается
как влияния на обмен веществ в целом, так и в частности прямого влияния
температуры на водный баланс организма. В последнем случае воздействие
температуры перекрещивается с влиянием влажности.
Слабая защита от высыхания связана у моллюсков с особенностями
их покровов. Тонкие слизистые покровы не могут так противостоять иссу-
иссушающему воздействию воздуха, как плотные ороговевшие покровы других
животншК Практически в период активной жизни главной защитой назем-
наземных моллюсков от высыхания становится систематическое смачивание
наружных покровов слизью, в связи с чем эти животные обладают большим
количеством слизистых желез. Однако этот способ сам по себе связан с рас-
расходом воды, которая составляет основной компонент слизи. Тем самым
еще больше увеличивается зависимость моллюсков от влажности внешней
среды.
Температура и влажность, воздействуя на моллюсков, в известных
пределах могут компенсировать друг друга. Так, например, в тех случаяху
когда температура выше оптимальной, достаточно высокая влажность
может компенсировать повышенный расход воды на испарение. Это поз-
позволяет сравнительно холодолюбивым видам проникать в области с теплым
климатом. С другой стороны, при влажности ниже оптимальной низкая
температура, замедляя испарение, способствует более экономному расходу
водных ресурсов моллюска. Однако подобная компенсация возможна лишь
на определенное время, после которого требуется восстановление утра-
утраченного запаса влаги. -
44 ВВЕДЕНИЕ
Даже по сравнению со многими другими семействами наземных мол-
моллюсков Clausiliidae обнаруживают повышенную требовательность к тем-
перАтур^и_влажности. Подавляющее большинство видов являются тепло-
теплолюбивыми и влаголюбивыми. Это хорошо видно на их распространении
как в тропиках, так и в странах умеренного пояса. Для примера рассмот-
рассмотрим распространение этих моллюсков в Азии и Европе, т. е. в областях,
где они изучены лучше всего и представлены большим числом видов н
родов.
Если придерживаться климатического районирования Алисова A950),
то в тропиках Азии наши моллюски населяют зону экваториального-
климата (Малайский архипелаг, южная половина полуострова Малакка),
зону экваториальных муссонов (Индостан, Цейлон, Бирма, страны Индо-
Индокитая, юго-восточный Китай) и часть муссонной области субтропической
зоны (южный и юго-западный Китай, южная Корея, южная и средняя Япо-
Япония). Из этого видно, что они обитают в тех тропических и субтропических
областях Азии, где в течение всего года (экваториальная зона) или только-
летом (в остальных зонах — под влиянием мусеонов) имеется высокая
влажность воздуха и обилие осадков. В то же время они отсутствуют в тро-
тропических областях, отличающихся засушливым климатом (например,
на Аравийском полуострове и в южном Иране).
Но даже в тех тропических областях, где почти круглый год стоит-
очень высокая относительная влажность воздуха и выпадает много осад-
осадков, распространение наших моллюсков зависит не только от этих факто-
факторов. Так, например, в Индонезии, по данным Лосьеса (Loesjes, 1953a) r
из 54 видов только 3 обитают на равнинах, остальные же живут в горах,
причем большинство их населяет высоты от 650 до 2500 м над уровнем
моря, т. е. зоны с умеренно-теплым или даже с прохладным климатом,,
но при минимальной температуре выше 0°. Так как в условиях экватори-
экваториального климата влажность и осадки почти не меняются по вертикали,,
то богатство видами семейства Clausiliidae в горных зонах связано с тем,
что они предпочитают умеренную температуру горного климата знойному
климату равнин.
По направлению к северу влияние обоих факторов на распространение
азиатских видов нашего семейства не ослабевает. Это видно из того, что-
они держатся областей, где годовая сумма осадков не меньше 1000 мм в год,.
а минимальная зимняя температура обычно выше 0°.
В поясе умеренного климата Clausiliidae распространены только'
в Европе, причем они тяготеют к областям с мягким морским и с умеренно-
континентальным климатом. Некоторые авторы пытались доказать связь
распространения тех или иных наземных моллюсков Европы с положением
июльских или январских изотерм^или со среднегодовым количеством осад-
осадков @kland, 1925; Krausp, 1940}. Однако в действительности северные и
северо-восточные границы ареалов" европейских Clausiliidae почти не сов-
совпадают с изотермами. Это особенно заметно на Восточно-Европейской
равнине, где ареалы наших моллюсков имеют форму клиньев, острием
направленных на восток и в большей или меньшей степени заходящих
вглубь континента. Зимние изотермы здесь проходят в основном в мериди-
меридиональном направлении, а летние — в широтном. Осадки на севере евро-
европейской части СССР, по-видимому, не лимитируют распространение наших
видов (рис. 28).
В связи с этим мы пришли к выводу, что в европейской части СССР
распространение Clausiliidae в разных местах их ареалов зависит от раз-
разных факторов. На севере и северо-востоке граница определяется длитель-
длительностью безморозного периода, так как осадков здесь вполне достаточно
БИОЛОГИЯ: СРЕДА И АДАПТАЦИИ
45
^500—750 мм в год). Безморозный период не должен быть короче 120 дней
л году. На юге и юго-востоке граница распространения наших моллюсков
определяется влиянием летней температуры и среднегодовым количеством
20 25 30 35 40 45 50 55
БО
65
70 75
Б5
60
5S
50
45
40
9
щ
г /М
150
ь=
ж
у \
у ^
I
щ
щ
ш
т
0
150 300 450км
I &
—4\
.-5 ЛЛл/f
щ
-Л
1
0
к"
30
85
35
40*
45
S0
55
50
45
40
50
55
Рис. 28. Влияние некоторых факторов климата на распространение Cochlodina
laminata (Mont.) и С. orthostoma (Menke) в Восточно-Европейской равнине.
(Ориг.).
/ — ареал Cochlodina laminata; 2 — ареал С. orthostoma; 3 — январские и июльские
изотермы; 4 — северная, восточная и южная границы зоны среднегодовых осадков 500—
, 750 мм; д —• восточные границы зон безморозного периода длительностью 120 и 135 дней.
осадков. Она проходит приблизительно по июльской изотерме 20°, в зоне
среднегодовых осадков 500—600 мм и почти совпадает с южной границей
подзоны смешанных лесов.
46 ВВЕДЕНИЕ
Таким образом, в конечном счете Clausiliidae населяют в Европе обла-
области, находящиеся под воздействием атлантических масс воздуха. С севера
и северо-востока нх распространение ограничивает влияние холодного
континентально-арктического воздуха, а на юго-востоке и юге — влия-
влияние континентально-субтропического воздуха.
Разобранные примеры показывают, какое большое воздействие на
распространение наших моллюсков оказывает климат, в особенности тем-
температура и влажность. Влияние обоих факторов перекрывает друг друга,,
и не всегда представляется возможным сказать, чье воздействие больше.
Однако на границах ареала, когда один из факторов находится в оптимуме,
а другой в пессимуме, именно последний является решающим и, в част-
частности, определяет распространение данного вида.
Основной физиологической защитой всех наземных моллюсков при
наступлении неблагоприятной температуры и влажности становится впа-
впадение в спячку. Перед этим обычно они выбирают наиболее удобные убе-
убежища, микроклимат которых чем-нибудь отличается от окружающего-
макроклимата в благоприятную для них сторону. Такими убежищами чаще-
всего являются теневые стороны и щели скал, россыпи камней, дупла,
гниющие пни и стволы деревьев, подстилка и т. д.
Что касается морфологических адаптации, то они обычно связаны
с раковиной. Сама возможность втянуться внутрь последней, закрыв вход,
в нее эпифрагмой или приклеившись устьем к субстрату, является оченв4-
эффективной защитой от большинства вредных факторов, в особенности1
от неблагоприятной температуры и влажности. Кромда того, рщшща рако-
вины^_фавма_ее_и_етр.оение замыкательного аппарата зависят от климата,
в котором обитает соответствующий моллюск.
Виды, обитающие в Тенистых и прохладных местах, как правило, обла-
обладают тонкостенной и темноокращенной раковиной. В то же время у видов,
живущих на открытых местах и при более высокой температуре, она окра-
окрашена светлее и стенки ее более толстые. ^>
Как уже отмечалось, у Clausiliidae главной защитой от высыхания
является замыкательный аппарат, образовавшийся в наиболее узкой
части последнего оборота раковины. Наибольшее значение здесь имеют-
клаузилий и те образования, которые вместе с ним служат для закрывания-
входа в раковину, т. е. полулунная складка, нижняя и субколумеллярная
пластинки. В дополнении к ним у многих видов имеются палатальные-
складки и краевые складки устья. Эти образования служат для стимули-
стимулирования выделения слизи при втягивании моллюска в раковину.
Среди многих Clausiliidae происходит процесс замены полулунной
складки палатальными. При этом задние концы последних берут на себя
функцию полулунной складки, т. е. служат опорой лопасти клаузилия,.
одновременно сохраняя свое первоначальное назначение.
Часто в пределах какой-нибудь ограниченной группы можно просле-
проследить последовательные этапы замены полулунной складки палатальными,,
причем хорошо видна связь этих изменений с различиями в климате,
в котором обитают соответствующие виды (рис. 29).
Так, например, в роде Mucronaria наиболее примитивна М. acuminata,
живущая в горах южной Грузии на больших высотах, в сравнительно-
прохладном климате. Она имеет толстую полулунную складку, однако-
ее верхний конец уже распался, дав начало одной или двум палатальным
складкам. Этот процесс еще заметнее у М. strauchi, живущей в окрестно-
окрестностях Тбилиси, т. е. в более жарком климате. У М. duboisi, широко распро-
распространенной в горных лесах Кавказа и выдерживающей большие колеба-
колебания влажности, полулунная складка полностью распалась, образовав:
БИОЛОГИЯ: СРЕДА И АДАПТАЦИИ
AT
4 палатальных складки. Ту же картину мы видим у М. pleurophychiay
обитающей в предгорной зоне западной Грузии на открытых, сильно про-
прогреваемых склонах. Такой же полный распад полулунной складки на
7—9 палатальных произошел у М. gustavi. Этот моллюск живет в нагорно-
степной зоне Талыша, сухой климат которой вызвал указанные преобра-
преобразования замыкательного аппарата.
В заключение остановимся на влиянии некоторых типов климата на
внешний вид и строение раковины наших моллюсков.
Адаптации к высокогорному климату. Как указывалось, еще со вре-
времен Веста (Vest, 1867) было замечено, что у некоторых высокогорных
видов замыкательный аппарат развит слабее (иногда даже совершенно
редуцирован), чем у близких форм, живущих несколько ниже. Объясня-
Рис. 29. Преобразование полулунной складки в палатальные в связи с на-
нарастанием засушливости климата у близких видов рода Mucronaria Bttg.
(Ориг.).
А —. М. acuminata (Mart.); Б — М. etrauchi (Bttg.); В — М. duboisi (Charp.); Г — М.
gustavi (Bttg.).
лось это тем, что высоко в горах влажность выше, и поэтому отпадает
надобность в тех частях замыкательного аппарата, которые служат защи-
защитой от высыхания. Однако знакомство с кавказскими Clausiliidae убедило
нас в том, что не всегда ослабление замыкательного аппарата можно объяс-
объяснить только более влажным климатом высокогорных зон. Дело в том, что
у многих-видов, обитающих в альпийской зоне или у верхней границы
лесной зоны Кавказа, по сравнению с близкими видами^ живущими ниже,
наблюдается преждевременная приостановка роста раковины, вследствие
чего она имеет бащневидную форму, так как не произошло сужения послед-
последнего оборота. От замыкательною аппарата остаются только его верхние
участки. Чем выше в горах живут такие моллюски, тем короче становятся
все части замыкательного аппарата (за исключением вертикально лежа-
лежащих полулунной складки и клаузилия) и располагаются они ближе к устью.
Подобное явление наблюдается, в частности, в роде Micropontica:
ряд М. closta (предгорья)—М. circassica (средние горизонты гор)—М. ге-
towskii (альпийская зона).
Так как у Clausiliidae размножение всегда начинается после заверше-
завершения роста раковины, то указанное явление мы склонны объяснять прежде-
преждевременной приостановкой роста, обязанной более раннему половому созре-
созреванию высокогорных видов. Последнее в свою очередь является адаптацией
к жизни в условиях короткого вегетационного периода, характерного для
верхних горизонтов гор.
Аналогичные изменения происходят не только у близких видов одного
рода, но и у отдельных, сравнительно широко распространенных видов
48 ВВЕДЕНИЕ
(например, Megaleuxina derasa, Quadriplicata aggesta). Как правило,
популяции, обитающие в высокогорных зонах, отличаются от живущих
в нижних зонах или на равнине меньшим числом оборотов, меньшими
размерами,1 более слабым развитием замыкательного аппарата, который
нередко лежит ближе к устью, чем у популяций, живущих ниже.
Адаптации к влажному субтропическому климату. Как уже отмечалось,
у многих моллюсков, обитающих во влажном климате, наблюдается полная
или частичная редукция замыкательного аппарата, в первую очередь тех
его частей, которые служат защитой от высыхания. Так, например, у Мис-
ronaria duboisi, которая довольно широко распространена в горных лесах
Кавказа, у популяций, живущих вдали от побережья, замыкательный
аппарат не обнаруживает признаков редукции. Напротив того, вместо
полулунной складки, обычной у близких видов, она имеет 3 хорошо разви-
развитых палатальных складки, что указывает на более совершенную защиту
от высыхания, которая нередко усиливается образованием многочис-
многочисленных мелких краевых складок устья. Но у популяций этого моллюска,
живущих на Черноморском побережье, в условиях очень влажного субтро-
субтропического климата, раковина отличается значительно меньшими размерами,
тонкостенностью и ослаблением замыкательного аппарата. Последнее
выражается в слабом развитии главной и палатальных складок, а клаузи-
лий неплотно замыкает вход в раковину. Кроме того, отсутствуют крае-
краевые складки. •
Другой пример — Euxinastra hamata. Как известно, у этого моллюска
лопасть клаузилия имеет довольно значительную вырезку (рис. 124).
У Clausiliidae эта вырезка обычно служит для пропуска крупной нижней
палатальной складки, уходящей вглубь раковины, за клаузилий. Однако
у данного вида, как редкое исключение, этой складки нет. По-видимому,
присутствие вырезки говорит за то, что предки этого вида имели нижнюю
палатальную складку, но она была утеряна в связи с обитанием во влаж-
влажных субтропиках Аджарии.
Следовательно, ослабление замыкательного аппарата под влиянием
очень влажного климата имеет место, во-первых, у видов, в основном при-
приспособленных к жизни в более сухом климате, во-вторых, у видов, предки
которых обитали при более низкой влажности. Вагнер (Wagner, 1914,1919,
1920) приводит много примеров первого рода среди Alopiinae, обитающих
ва Балканах.
Однако не у всех наших моллюсков, живущих в очень влажном кли-
климате, наблюдается ослабление или редукция замыкательного аппарата.
Так, у многих видов, обитающих в субтропиках западного Закавказья,
-он очень хорошо развит. Очевидно, это объясняется более высокими тре-
требованиями к влажности у этих моллюсков по сравнению с упомянутыми
выше видами. Однако если сравнить раковины этих моллюсков с близкими
видами, живущими в условиях более континентального или холодного
{высокогорного) климата, то бросается в глаза следующая закономер-
закономерность. У видов, живущих в субтропиках, замыкательный аппарат, в осо-
особенности палатальные складки, располагается глубже, чем у видов того же
рода, живущих в глубине континента или в высокогорных зонах. Это
заметно при сопоставлении Mucronaria index и М. pleuroptychiac осталь-
остальными видами того же рода или Quadriplicata subaggesta, Q. pumiliformis,
-$. dipolauchen с Q. quadriplicata, обитающей в лесах центрального и вос-
восточного Закавказья.
1 Уменьшение размеров касается чаще всего лишь высоты раковины, а ширина
-нередко остается прежней. Благодаря этому высокогорные популяции обычно отли-
•чаются приземистыми раковинами.
БИОЛОГИЯ: СРЕДА И АДАПТАЦИИ
В высокогорных зонах период активной жизни моллюсков приходится
pia сравнительно короткое лето. В лесах центрального и восточного Закав-
Закавказья весенне-летний период более продолжительный. Однако здесь на
время летней засухи моллюски вынуждены впадать в спячку. Тем самым
ji обоих случаях, но по разным причинам, период активности соответст-
соответствующих видов короче, чем у их родичей из влажных субтропиков запад-
западного Закавказья, которые для роста и размножения практически почти
j&e ограничены никакими фенологическими сроками. Поэтому моллюски
^высокогорных зон и областей с континентальным климатом переходят
* размножению сразу после сформирования замыкательного аппарата,
ш виды из субтропиков западного Закавказья имеют возможность продол-
i-жать рост раковины даже тогда, когда замыкательный аппарат уже сфор-
сформирован. Это имеет для них большое значение, так как более глубокое
залегание палатальных складок и клаузилия является хорошей защитой
от высыхания — моллюск может создать между клаузилием и краем
устья больше защитных пленок (эпифрагм).
Адаптации к сухому субтропическому климату. Несмотря на то, что
;большинство моллюсков данного семейства обитает в областях с влажным
климатом, некоторые виды и роды населяют области с засушливым клима-
i-том. Таких моллюсков особенно много в средиземноморских странах. Так,
ва побережье Греции и на островах Эгейского моря очень распространены
[моллюски из рода Albinaria, в Сирии и в Палестине обитают Cristataria,
[ в северной Африке (Тунис) — Barcania и Mauritanica. Все это группы из
{подсемейства Alopiinae.
Как известно, климат средиземноморских стран отличается засушливым
летом и теплой влажной зимой. В связи с этим здесь обитают исключительно
теплолюбивые моллюски, легко переносящие длительную засуху. Отсут-
Отсутствие тенистых лесов, преобладание кустарников, внутри которых мол-
моллюски не могут найти хорошей защиты от высыхания, привело к тому,
что Clausiliidae обитают преимущественно на_скдлах и в осыпях.
Раковины большинства средиземноморских Clausiliidae отличаются
значительной толщиной стенок — обычная у наземных моллюсков защита
от перегрева. Кроме того, внешний беловатый и непрозрачный слой рако-
раковины, характерный для Alopiinae, у средиземноморских родов этого под-
подсемейства/достигает особенной мощности. .Этот слой играет роль «экрана»,
так как, отражая солнечные лучи, предохраняет тело моллюски от пере-
перегрева и высыхания. В некоторых случаях, когда он достигает особенно
сильного развития (Albinaria), отпадает надобность в защитной функции
замыкательного аппарата и некоторые части его подвергаются редукции.
В других группах, напротив, замыкательный аппарат достигает большого
совершенства и плотно запирает раковину. В результате такие моллюски
обладают способностью длительное время противостоять засухе. На-
Насколько они устойчивы в этом отношении, видно из того, что однажды мы
обнаружили среди нескольких видов Barcania, присланных нам Р. Бранд-
том из Ливии, живые экземпляры, собранные почти два года назад. Ко-
Конечно, столь длительный срок существования без капельно-жидкой влаги
объясняется тем, что эти моллюски находились в помещении при умерен-
умеренной температуре, однако наши мезофильные кавказские виды в таких же
условиях не выживают более 3 месяцев.
Свет
Будучи тепло- и влаголюбивыми животными, наши моллюски соот-
соответственно реагируют на освещенность местообитания. Так как в »южных
широтах обилие света сопряжено с высокой температурой и сравнительно
50 ВВЕДЕНИЕ
низкой влажностью, то большинство видов семейства тенелюбивые, суме-
сумеречные или ночные животные, а днем они не активны и скрываются в зате-
затененных убежищах с относительно низкой температурой и высокой влаж-
влажностью. В северных широтах те же виды могут вести себя иначе — наи-
наибольшая активность их может приходиться на теплое дневное время,
а ночью из-за низкой температуры активность снижается. В связи с отри-
отрицательным в общем отношении к свету большинство видов обладает совер-
совершенно непрозрачной или лишь просвечивающей раковиной. Только те,
которые живут в пещерах, в гниющей древесине и под корой, имеют тон-
тонкую прозрачную раковину. В данном случае проявляется общая законо-
закономерность — когда наземные моллюски живут в условиях почти полной
темноты или при недостатке света, то они обладают стекловидно-прозрач-
стекловидно-прозрачной раковиной, которая тем самым не препятствует поглощению мягкими
частями тела необходимого им количества лучистой энергии.
Горные породы и почвы
Физические и химические особенности почв]ы и горных пород, на
которых обитают наши моллюски, оказывают на них заметное воздей-
воздействие, в особенности на те виды, которые всю жизнь проводят на поверх-
поверхности почвы и на скалах. i
Многиемвиды о^аруживаю^^вшю_тагр^ети^к_и^в^стн^кщ^и к почвам,
богатым кальцием, ото объясняется не только потребностью в солях
кальпия^для^построения раковины, но и некоторыми физическими свойст-
свойствами известняков. Из них главным для моллюсков является большая
пористость этих пород, обилие в них различных щелей, каверн и других
углублений, образовавшихся в результате выветривания. В этих углубле-
углублениях скопляется растительный перегной и сохраняется благоприятная
влажность и температура. К этому следует прибавить, что известняки
по сравнению с другими горными породами обладают высокой способ-
способностью акумулировать тепло. Всем этим и объясняется обилие наземных
моллюсков (в том числе и Clausiliidae) на известняках, как в видовом, так
и в количественном отношениях. ,-
Многие моллюски, обитающие на известняках, отличаются от популя-
популяций из других биотопов образованием на раковине многочисленных ребер
и мелких складок на краю устья. Бдясодаря^ебрам_раковина при сравни-
сравнительно небольшом весе обладает значительной прочностью. Краевые
складки также укрепляют хрупкий край устья и в то же время, подобно
палатальным складкам, защищают тело моллюска от высыхания.
Скалы и осыпи^~сложенные из материнских пород (граниты, гнейсы,
базальты и др.) и из пеечанников, по сравнению с известняками менее
удобны для жизни наших моллюсков. Главной причиной этого является
слабая пористость этих пород, в связи с чем моллюски здесь не могут найти
такого разнообразия микроклиматических условий, как на известняках.
Эти породы не пригодны для построения раковины и не обладают такой
способностью к аккумуляции тепла, как известняки. Кроме того, песок
и щебенка, образующиеся в результате выветривания материнских пород
и песчаников, отрицательно влияют на моллюсков, так как иссушают и
раздражают поверхность тела, забиваются внутрь легкого и перетирают
раковину.
Тяжелые глинистые почвы также малопригодны для жизни моллюс-
моллюсков, так как здесь они испытывают резкие колебания влажности, когда
обилие воды на поверхности сменяется полным высыханием почвы.
БИОЛОГИЯ: СРЕДА И АДАПТАЦИИ 51
В общем, те виды Clausiliidae, которые большую часть жизни проводят
на поверхности почвы, явно предпочитают легкие скелетные, преимуще-
преимущественно горно-лесные почвы, богатые гумусом и гниющими растительными
остатками. Благодаря хорошему дренажу здесь не происходит застоя
поверхностной влаги; в то же время обилие органических веществ делает
эти почвы достаточно влагоемкими и питательными для моллюсков.
Растительность
Отношение наших моллюсков к окружающему их растительному миру
идет в основном по линии пищевых связей и роли растительности как
среды обитания. Так как Clausiliidae- являются мико- и сапрофагами,
то узкой специфичности к каким-то определенным видам высших расте-
растений у них не обнаруживается. Значительно большее значение имеет общий
характер растительных ассоциаций, в которых они обитают. Большинство
наших моллюсков обитатели смешанных и широколиственных лесов. Это
связано не только с тем, что именно под пологом таких лесов они находят
наиболее благоприятную для них пищу, но и с тем, что здесь они обычно
встречают более подходящие условия влажности и температуры, чем на
соседних открытых местах. В хвойных лесах Clausiliidae встречаются
лишь там, где имеется хоть небольшая примесь лиственных пород или
густой травяной покров. В целом хвойные леса неблагоприятны для
наших моллюсков как в силу климатических условий — они произрастают
в областях с коротким вегетационным периодом, так и в пищевом отно-
отношении, так как хвойная подстилка, отличаясь большой смолистостью и
кислотностью, мало" пригрднТа~для питания наземных моллюсков. :
Важное значение, которое в жизни напйх моллюсков имеют смешан-
смешанные и широколиственные леса, видно, в частности, и в том, что распростра-
распространение многих видов отчетливо связано с указанными лесами. Так, наприг
мер, такие обычные виды, как Cochlodina laminata и Laciniaria plicata,
по байрачным и пойменным лесам проникают далеко на юг, в глубь степ-
степной зоны европейской части СССР, а первая доходит до Крыма и Север-
Северного Кавказа. Другим примером может быть присутствие некоторых кав-
кавказских лесных видов в байрачных лесах Ставропольской возвышен-
возвышенности — Euxina somchetica, Ей. tschetschenica, Mucronaria duboisi и другие.
Эти примеры указывают на то, что в засушливых областях юга СССР
особенно четко проявляется защитная роль лесов в жизни Clausiliidae.
Под их пологом моллюски находят достаточно высокую влажность и
прохладу. Но не меньшее значение имеют леса ив более северных обла-
областях. Это хорошо видно из того, что ареалы большинства видов, обитаю-
обитающих на Восточно-Европейской равнине, располагаются в пределах пояса
смешанных и широколиственных лесов. Но так как распространение наших
моллюсков зависит и от других факторов, в частности от длительности
безморозного периода, то северная и северо-восточная границы их ареа-
ареалов не обязательно совпадают с границей указанных лесов, которая может
проходить севернее и восточнее.
Основные типы местообитаний
Наши моллюски обитают на деревьях, ла склонах и в осыпях, в. под-
подстилке и под отдельно лежащими камнями и другими предметами, нахо-
находящимися на почве.
Многие лесные Clausilidae большую часть жизни проводят на стволах
деревьев, питаясь растущими на них лишайниками и в неблагоприятное
52 ВВЕДЕНИЕ
время; скрываясь в щелях коры и среди мха. На хвойных деревьях они
встречается редко и, по-видимому, случайно. Среди древесных видов
четко обособляется группа моллюсков, живущих под корой и в дуплах,
внутри гнилых пней и валежин, питающихся разлагающейся древесиной
(Caspiophaedusa perlucens, Pontophaedusa funiculum, Semilina).
¦Некоторые виды большую часть жизни проводят на поверхности почвы
и в подстилке (idyla foveicollis, Ruthenica filograna, многие Iphigena и
Clausilia). Правда, в период дождей, спасаясь от затопления, они заби-
забираются на деревья, но при первой возможности спускаются на землю.
Наконец, большое число видов обитает на скалах и в осыпях, причем
если эти биотопы находятся в пределах лесной зоны, то здесь также
можно встретить многих моллюсков, более свойственных деревьям. Но,
кроме них, скалы и осыпи населяют многочисленные скаложилы (Агте-
nica, Arcotomdi Mieropontica, Alopia и др.).
Скалы и осыпи' являются очень своеобразными биотопами. Моллюски
здесь встречают целую мозаику убежищ с разнообразным микроклиматом.
В особенности это касается известняковых скал.
На открытых, освещенных солнцем поверхностях обитают преиму-
преимущественно очень теплолюбивые виды, которые ради тепла способны пере-
переносить значительную сухость климата. Среди.наших видов таких сравни-
сравнительно немного. К ним, в частности, принадлежат Laciniaria biplicata и
Clausilia parvula, которые в средней Европе обитают на сухих» чаще всего
известняковых склонах. Значительно больше подобных видов в средизем-
средиземноморских странах.
Скаложилы чаще всего предпочитают теневые стороны скал, различные
ниши и щели на их поверхностях, а в осыпях живут либо в глубине их,
либо на нижних поверхностях верхнего слоя камней. В этих убежищах они
находят более умеренную температуру и более высокую влажность, чем
снаружи.
Очень велика роль скал и осыпей в южных засушливых областях,
в особенности в горных степях и полупустынях с континентальным клима-
климатом. Известно, что в этих областях вследствие сильных суточных колеба-
колебаний температуры данные биотопы являются своеобразными конденсато-
конденсаторами влаги, которая скопляется внутри щелей скал или в глубине осыпей
и обеспечивает моллюскам достаточно высокую влажность.
Большинство Clausiliidae обитает на деревьях и на скалах, т. е. над
почвой. При этом раковина свободно свисает вниз и сравнительно мало
тормозит движение моллюска. Подобный образ жизни способствует вытя-
вытягиванию раковины по оси и приобретению ею большого числа оборотов —
чаще всего 12—15, иногда до 18 оборотов (рис. 89). Жизнь на поверхности
почвы и в подстилке приводит к тому, что высокая раковина при движении
моллюска волочится за ним по субстрату и тормозит его движение. В связи
с этим у видов, почти постоянно живущих на почве, раковина приобретает
приземистую «форму, тупую верхушку, стенки ее утончаются, и сама она
слагается из сравяительн© небольшого числа оборотов — чаще всего
8—Ш; реже до 12 оборотов (рис. 138). . .
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ CLAUSILIIDAE
Прежде чем перейти к характеристике особенностей распространения
Clausiliidae СССР и некоторых сопредельных стран, кратко остановимся на
распространении всего семейства. В этом отношении, самым характерным
является то, чт© ареал его разорван на три, не связанных друг с другом
участка: европейский, азиатский и американский (рис. 30). , ,
..-':¦ • Рису 30. Современное распространенно моллюсков Clausiliidae. (Ориг.).
Дифрами обозначены небольшие изолированные ареалы: 1—южные склоны-Гималаев севернрЙ Ипдйи и частично Пакистана; g — центральное плато
Цейлона; а— Йемен! 4 — Эфиопия; 6 — Кения; '6 — массив Рувензори; 7 — окрестности озера Танганьика; Н ^-острова Мадейра и Порто-Санто.
54 ВВЕДЕНИЕ
Европейский участок занимает западную половину Палеарктики, т. е.
почти вою Европу, Кавказ, Малую Азию и средиземноморское побережье
северной Африки и Передней Азии. Эти области населяют виды из подсе-
подсемейств Clausiliinae, Euxininae, Alopiinae и Cochlodininae. Здесь же сохра-
сохранилось несколько реликтов из подсемейства Phaedusinae.
Азиатский участок занимает часть восточной (юго-западный и южный
Китай, южную Корею и среднюю и южную Японию, Филиппины) и южную
Азию (страны Индокитая, южные склоны Гималаев, за исключением Не-
Непала и Кашмира, центральный массив Цейлона), а также большую часть
Малайского архипелага (за исключением восточной половины Новой
Гвинеи). Основная масса видов принадлежит к подсемейству Phaedusinae
и небольшое число — к подсемейству Neniinae (роды Tropidauchenia и
Gamier id).
Американский участок охватывает горные области северо-запада Юж-
Южной Америки (Кордильеры и прилегающие с востока средневысотные горы,
но без Бразильского нагорья). Южная граница проходит приблизительно
по южному тропику. Близкие группы имеются на островах Гаити и Пуэрто-
Рико. Указанное области населены многочисленными видами подсемей-
подсемейства Neniinae.'
К европейскому участку тяготеет цепь изолированных ареалов плохо
изученного, рода^ Mac roptychiOj вытянувшаяся вдоль восточного берега
Африки от Эфиопии до озера Танганьика. Эта группа состоит из ви-
видов, испытавших значительную редукцию' замыкательного аппарата, но
в общем сохранивших многие черты, свойственные Clausiliinae и Euxi-
Euxininae.
Из сказанного очевидно, что рассматриваемые в данной работе виды
находятся в пределах европейского участка ареала семейства. В геогра-
географическом отношении они четко распадаются на три группы — европей-
европейские, кавказские и крымские виды. Каждая группа почти полностью изо-
изолирована друг от друга и характеризуется различным видовым составом.
Европейскиевиды
Многие наши европейские Clausiliidae, в частности виды из родов
Vestia, Pseudalinda, Clausilia, Iphigena uRuthenica, по сути дела, являются
среднеевропейскими, так как основная часть их ареалов располагается
в горах средней Европы. Здесь же находится центр их возникновения.
Правда, некоторые из них обитают также в соседних областях западной,
северной и южной Европы.
Среднеевропейские виды, по существу, являются горными моллю-
моллюсками. Лишь на востоке они спускаются на низменности, захватывая
часть Северо-Германской низменности и вклиниваясь на различную глу-
глубину в Восточно-Европейскую равнину. В этом проявляется относитель-
относительная холодостойкость этих моллюсков, позволяющая им переносить хо-
холодные зимы западных и центральных областей европейской части СССР.
Единственным видом, не живущим в горах, является Clausilia bidentata
(рис. 34). Она обитает в приморских областях северо-западной Европы и
проникает на юг не далее северных средневысотных гор Франции, Герма-
Германии, (ГДР и ФРГ) Чехословакии и Польши. Этот моллюск населяет обла-
области, находящиеся под максимальным воздействием Гольфстрима. Несмотря
на то, что Cl. bidentata — единственный вид своего семейства, проникший
вдоль берегов Норвегии за полярный круг (до 70° с. ш.), по сравнению со
всеми остальными северными видами Clausiliidae он наиболее теплолю-
теплолюбивый, адаптированный к мягкому морскому климату.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 55
Интересно отметить, что некоторые среднеевропейские виды (Clausilia
tlubia, С. cruciata, Cochlodina orthostoma) имеют ареал, состоящий из двух
несвязанных частей — горной (среднеевропейской) и равнинной (прибал-
(прибалтийско-восточноевропейской). Упомянутый разрыв приходится на тер-
территорию Полесья и восточной части Великопольской низменности, по-
покрытой широколиственно-сосновыми лесами на песчанных (зандровых)
почвах. Очевидно, такой тип почвы, а также хвойная подстилка неблаго-
неблагоприятны для названных видов.
По характеру распространения европейские виды можно разделить
на следующие группы.
A. Среднеевропейские виды в широком смысле слова, т. е. виды, на-
населяющие Альпы и Карпаты, а также прилегающие к ним области север-
северной и восточной Европы: „
1. Clausilia cruciata./ , _, ,. 5. Iphigena ventricosa. I
2. Cl. parvula. —- ^ ' ' ?' 6. /. plicatula. л
3. Cl. dubia. 7. Cochlodina orthostoma. -
4. Cl. pumila pumila. 8. Ruthenica filograna. -i
Некоторые из этих моллюсков (№№ 3—6) проникают и в северные
Балканы. Генетически с этой группой тесно связаны многие близкие
им виды из родов Clausilia и Iphigena, обитающие только в Альпах.
В данной работе они не рассматриваются.
Б. Карпатские виды. К ним принадлежат моллюски, обитающие только
в пределах Карпат и их предгорий, из которых лишь некоторые недалеко
заходят в соседние горные области.
1. Iphigena latestriata latestriata. 6. Pseudalinda stabilis.
2. /. tumida. 7. P. fallax. .
3. Vestia turgida. 8. Cochlodina cerata.
4. V. elata. • 9. Alopia glauca.
5. V. gulo.
Очень характерно, что большинство названных видов принадлежит
к родам, свойственным преимущественно Карпатам (Vestia, Pseudalinda,
Alopia). Остальные являются производными либо альпийского рода
Iphigena,'либо балканского рода Cochlodina.
B. Широко распространенные по Европе виды балканского происхо-
происхождения. Сюда относятся:
1. Laciniaria biplicata. - .. 3 Laciniaria сапа.
2. L. plicata. 4. Cochlodina laminata.
Несмотря на то, что эти моллюски очень широко распространены
в Европе, мы рассматриваем их как балканский элемент, так как боль-
большинство видов, относящихся к тем же родам, в том числе и наиболее при-
примитивные, обитают только на Балканах. Здесь же распространены и ука-
указанные моллюски.
Все перечисленные группы состоят из видов, преимущественно оби-
обитающих в горах. Но имеется несколько, которые обитают только на рав-
равнинах и лишь слегка заходят в средневысотные горы. К таким моллюскам
принадлежит упомянутая Clausilia bidentata с прибрежно атлантическим
распространением. Кроме тог.о, сюда относится подвид Iphigena latestriata
borealis, обитающий на юге и юго-востоке Прибалтики и подвид Clausilia
pumila sefuncta, свойственный части Восточно-Европейской равнины.
56
ВВЕДЕНИЕ
Кавказские виды
В малакофауне Кавказа Clausiliidae занимают очень заметное место.
К настоящему времени, здесь обнаружено 45 видов этих моллюсков, или
64% всех видов данного семейства, известных для СССР.
Очень характерной особенностью фауны Clausiliidae Кавказа и сопре-
сопредельных с ним лесных областей Малой Азии и северного Ирана1 является
38 40 .42 44 46 48" 50 52 54 56
Астрахань!
. О
44
Махач-Кала
Баку
44
42
40
36
' 50
54
Рис. 31. Распространение рода Micropontica (Bttg.). (Ориг.).
1 — и¦ closta (Bttg.); 2 — М. circassico (Bttg.); 3 — М. retowskii (Bttg.).
то, что тут обитают совершенно другие виды и роды, чем в Европе. В этом
отношении известно только два исключения. Первое — эго Cochlodina
laminata, которая на Кавказе обитает только в байрачных лесах Ставро-
Ставропольской и Пятигорской возвышенностей. Присутствие этого европейского
моллюска на Северном Кавказе легко объяснить, если учесть, что он дальше
всех других европейских видов своего семейства проникает на юг в степ-
степную зону по байрачным лесам Украины.
Вторым исключением является Serrulina serrulata, которая живет не
только на Кавказе, но и в лесах Малой Азии, Болгарии, Румынии и в За-
Закарпатье. Далее (стр. 66) будет дано объяснение столь широкого распро-
распространения этого моллюска, которого многие авторы (Jaeckel, Klemm,
Meise, 1957) неправильно считают кавказским.
1 Лесные области бассейна реки Чорох (северо-восточная Турция) и северного
Ирана являются продолжением Кавказской горной страны, так как флора я фауна
этих областей по существу кавказские.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
57
Подавляющее большинство Clausiliidae Кавказа обитатели леса.
Правда, некоторые из них проникают также в соседние альпийскую и
субальпийскую зоны, в этом случае придерживаясь скал и осыпей. По
отношению к влажности все эти виды мезофилы, т. е. отдают явное пред-
предпочтение влажным биотопам. Лишь небольшая группа из рода Armenica
населяет безлесные • ксеротермные биотопы. Однако и эти виды нельзя
рассматривать как настоящих ксерофилов, так как в условиях засушли-
засушливого ландшафта или биотопа они населяют такие складки микро-
микрорельефа, которые имеют более влажный микроклимат (щели скал, осыпи).
Правильнее считать их очень теплолюбивыми моллюсками, которые
населяют такие убежища, где они защищены от черезмерной потери влаги.
По территории Кавказа Clausiliidae распространены неравномерно.
Большинство видов обитает в западных областях перешейка, т. е. к за-
западу от Сурамского хребта и в верхней части бассейна реки Кубани. К во-
востоку от указанных районов число видов наших моллюсков заметно убы-
убывает.
Обилие нашими видами на западе Кавказа объясняется в первую
очередь более влажным и мягким климатом западных районов этой горной
страны. К востоку климат становится все более сухим и континентальным.
Кроме того, на западе Кавказа имеются обширные выходы известняковых
пород. Как указывалось (стр. 50), в известняках наши моллюски находят
больше убежищ, чем в базальтах и шиферных сланцах, обычных для
центральных и восточных районов Кавказа. Вот почему некоторые мол-
моллюски — Quadriplicata dipolaucken, Mucronaria pleuroptychia, большин-
большинство Acrotoma и Micropontica (рис. 31), обитают только на отдельных
известняковых массивах западного Кавказа.
Анализ распространения Clausiliidae на Кавказе подтверждает пра-
правильность выделения на территории этой горной страны четырех зоогео-
графических провинций — Кавказской, Гирканской, Армянской и рав-
равнинных степей (Лихарев, 1958а, 1958в). Первые две являются лесными
провинциями и поэтому наиболее богаты видами данного семейства,
третья — нагорно-степная и характеризуется скаложилами из рода Ar-
Armenica. Наконец, четвертая лишена наших моллюсков, или они имеются
только в таких интразональных формациях, как пойменные леса.
Кавказская провинция занимает лесную, субальпийскую и альпий-
альпийскую зоны Большого Кавказа, низовые леса Закавказья и леса внешних
отрогов Малого Кавказа. Только здесь живут следующие виды:
1. Serrulina semilamellata.
2. Pontophaedusa funiculum.
3. Idyla foveicollis.
4. Armenica griseofusca.
5. Acrotoma komarowi.
6. A. semicincta.
7. A. narzanensis.
8. A . laccata.
9. A. sobrievskii.
10. Euxina tuschetica.
11. Eu. somchetica.
12. Eu. tschetschenica.
13. Eu. novorossica.
14. Megaleuxina derasa.
15. M. reulauxi.
16. M. lindholmi.
17. Euxinastra hamata.
18. Mentissoidea litotes. •
19. Filosa filosa.
20. Quadriplicata quadriplicata.
21. Q. aggesta.
22. Q. lederi.
23. Q. subaggesta.
24. Q. pumiliformis.
25: Q. dipolauchen.
26. Mucronaria acuminata.
27. M. strauchi.
28. M. duboisi.
29. M. pleuroptychia.
30. M. index.
31. Micropontica closta.
32. M. circassica.
33. M. retowskii.
Из приведенного списка видно, что в Кавказской провинции обитает
подавляющее большинство видов. Это не значит, однако, что они ветре-
58 ВВЕДЕНИЕ
чаются по всей указанной территории. Мы уже отмечали, что на западе
Кавказа обитает больше видов, чем в центральных и восточных районах
этой страны.
На Северном Кавказе, приблизительно между Кубанью и Тереком,
лесной пояс, обрамляющий северные склоны Большого Кавказа, прерван,
в связи с чем здесь отсутствуют и лесные Clausiliidae. К востоку от реки
Баксан опять начинается лесная зона, которая, с небольшими разрывами,
тянется почти до побережья Каспийского моря, огибает восточные отроги
Большого Кавказа и сливается с лесами южных склонов. В связи с этим
в районе города Орджоникидзе вновь встречаются наши моллюски, хотя
число видов здесь меньше, чем на западных склонах, и значительно меньше,
чем в западном Закавказье. ,
Здесь найдены следующие 7 видов:
1. Serrulina serrulata. 5. Euxina somchetica.
2. S. semilamellata. .6. Eu. tschetschenica.
3. Idyla foveicollis. 7. Mucronaria duboisi.
4. Mentissoidea litotes.
Возможно, что многие из этих моллюсков обитают и в лесах Даге-
Дагестана. К сожалению, малакофауна этой части Кавказа слабо изучена.
Очень характерно, что некоторые виды обитают к северу от основных
лесных массивов Северного Кавказа, а именно в байрачных лесах Ставро-
Ставропольской и Пятигорской возвышенностей. Кроме того, 4 вида найдены
в пойменных лесах нижнего Терека (Шелковский лесхоз, — Калитина,
1958). Все это указывает на то, что прежде леса Кавказа и их фауна про-
простиралась значительно дальше к северу, чем в настоящее время.
В Закавказье обеднение фауны лесными видами Clausiliidae по напра-
направлению с запада на восток заметно не только на южных склонах Боль-
Большого Кавказа, но и на внешних отрогах Малого Кавказа.
Наиболее широким распространением в пределах Кавказской про-
провинции обладают 3 вида — Idyla foveicollis, Mentissoidea litotes и Mucro-
Mucronaria duboisi (рис. 68, 134, 195). Некоторые из них выходят даже за пре-
пределы СССР и вдоль черноморского побережья Малой Азии могут про-
проникать почти до Синопа.
Для Кавказской провинции характерно не только присутствие боль-
большинства эндемичных Кавказу видов, но и некоторых эндемичных данной
провинции родов и подродов — Pontophaedusa, Scrobifera, Filosa, Euxi-
nastra, Micropontica, Acrotoma. x
Гирканская провинция занимает лесные районы Талыша, Ленкоран-
ской низменности и северного Ирана. Здесь обитает значительно меньше
видов, чем в Кавказской провинции:
1. «Phaedusa» hyrcanica. 5. Mucronaria gustavi.
2. Serrulina sieveni sieversi, 6. Euxina lessonae.
3. S. senghanensis. 7. Eu.. talyschana.
4. Caspiophaedusa perlucens. 8. Eu. persica
Если ни один вид Кавказской провинции не живет на территории
Гирканской провинции, то некоторые гирканские виды обитают в восточ-
восточных районах Кавказской провинции. Так, Caspiophaedusa perlucens живет
в лесах восточной части южных склонов Большого Кавказа и северных
и восточных отрогов Малого Кавказа (рис. 55). Другой гирканский вид —
Serrulina sieversi — на южных склонах Большого Кавказа представлен
особым подвидом — occidentalis (рис. 32). Эти примеры указывают на то,
1 В настоящее время виды из рода Acrotoma обитают только на западе Кавказа,
однако в эоцене Европы эта группа была распространена более широко (стр. 64).
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
59
что между обеими лесными провинциями Кавказа, ныне разобщенными
степями и полупустынями Куро-Араксинской низменности и Армянского
нагорья, в былые времена мог происходить обмен фаунами (стр. 66).
Для гирканской фауны показателен тот факт, что все 3 вида Euxina
(№№ 6—8) имеют ближайших родственников не на Кавказе, а в лесах
Малой Азии. Это не только указывает на былые связи Гирканской про-
прозе 40
44 46 48 SO
52 54
38
55 0 55 110 165 220 км
1 I
40
38
36
40
Рис. 32. Распространение рода Serrulina Mouss. на Кавказе. (Ориг.).
1 — S. serrulate, serrulata (Pfr.); 2 — S. semilamellata (Mouss.); 3a — S. sieversi occidentalis Likha-
rev; 36 — S. sieversi sieversi (Pfr.).
винции с лесными областями Малой Азии, но и на то, что гирканская
фауна сохранила больше древних черт, чем кавказская.
Армянская зоогеографическая провинция занимает внутренние на-
горно-степные и полупустынные области Малой Азии. На Кавказе в ее
состав входят южные районы Армянской ССР, Нахичеванская АССР и
безлесные районы Талыша (Зувант). Из наших моллюсков здесь обитают
виды рода Armenica — A. disjuncta, A. brunnea, A. gracillima, A. hueti
(рис. 33).
Некоторые из этих видов проникают в пограничные районы Кавказ-
Кавказской провинции. Очевидно, что за счет подобных иммигрантов возникла
A. griseofusca — единственный вид этого малоазиатского рода, эндемич-
эндемичный лесной Кавказской провинции.
Крымские виды
В малакофауне Крыма имеется только 5 видов Clamiiiiidae. Из них
3 являются вселенцами из Европы — Iphigeiia, jjjptefiHa, Cochlodina la-
minata и Balea perversa. Первые 2 щу&и&НМй&т -только в буковых де^^х
60
ВВЕДЕНИЕ
Главного хребта, а последний — только на Южном берегу. Остальные
2 моллюска — Laciniaria (Mentissa) gracilicosta и L. (M.) canalifera —
эндемики Крыма. Подрод Mentissa тоже эндемичен только этой горной
стране. Если L. gracilicosta обитает лишь на Главном хребте и на Южном
берегу, то L. canalifera распространена несколько шире и встречается,
кроме того, в предгорной зоне.
20
25
30
30
•/ +2 кз х« os tfe ъ.7
150 0 150 300 450км
25
3D
35
«
50
Рис. 33. Распространение рода Armeriica Bttg. (Ориг.).
1 — A. iaemco/iis (Charp.); 2 — A. bicarinata (Rssm.); 3 — A. disjuncta (Mort.); 4 — A. brunnea
(Bssm.); 5 — A. graeillima (Ret.); S — A. griseofusca (Mouss.); 7 — A. hueti (Mort.).
Бедность видами горного Крыма обязана давней изоляции полуост-
полуострова от остальных причерноморских стран и от горных областей средней
Европы. В то же время эта изоляция способствовала образованию энде-
эндемичных видов и одного эндемичного подрода и значительной внутриви-
внутривидовой изменчивости обитающих здесь моллюсков. Все это придало мала-
кофауне Крыма отчетливый островной облик.
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ CLAUSILIIDAE ЗАПАДНОЙ ЧАСТИ
ПАЛЕАРКТИКИ
К вопросу о древности Clausiliidae (анализ
данных палеонтологии и географического
распространения)
Среди современных наземных моллюсков Clausiliidae являются одной
из наиболее древних групп. Первые по возрасту палеонтологические на-
находки этих моллюсков сделаны в отложениях позднего мела Франции.
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 61
Здесь жили виды из рода Proalbinaria Bttg. (Phaedusinae). Они уже об-
обладали вполне сформированным замыкательным аппаратом. Правда,
клаузилий у них пока не найден, но наличие хорошо развитой
полулунной складки указывает на то, что эти маллюски имели это обра-
образование.
К сожалению, палеонтологические находки наших моллюсков распре-
распределены очень неравномерно по территории, которую они заселяют в на-
настоящее время. Большинство находок сделано в Европе; в Южной
Америке, в неогеновых отложениях Перу, найден лишь один вид — Cirro-
basis venusta (Conrad), по-видимому, из подсемейства Neniinae; в восточной
Азии найдены в отложениях плейстоцена Японии несколько видов из
рода Melazaptyx Pils. (Phaedusinae). .
В связи со сказанным в данном разделе мы делаем попытку наметить
основные направления в формировании фауны наших моллюсков, преиму-
преимущественно для западной части Палеарктики (Европа, северная Африка,
Передняя Азия). К тому же настоящая работа посвящена главным обра-
образом видам, населяющим эту часть суши.
Но прежде чем перейти непосредственно к рассмотрению изменений,
которые фауна Clausiliidae указанной территории испытала в кайнозое,
имеет смысл вспомнить некоторые общие особенности распространения
этих моллюсков.
Известно что в настоящее время ареал Clausiliidae состоит из трех
четко изолированных друг от друга участков (стр. 54). Что разобщенность
этих участков наступила уже давно, указывает, в частности, тот факт,
что в каждом из них сформировались свои роды и даже подсемейства.
Так, в азиатском участке господствуют виды из подсемейства Phaedu-
Phaedusinae, в европейском (точнее в западнопалеарктическом) — виды из под-
подсемейств Cochlodininae, Alopiinae, Euxininae и Clausiliinae, и, наконец,
в американском — только Neniinae.
Совершенно очевидно, что в давние времена эти участки были связаны
друг с другом. За это говорит следующее: 1) несколько видов Phaedusinae
обитает в европейском участке в настоящее время {Serrulina Mouss.,
Pontophaedusa Lindh., Caspiophaedusa Liadh.); 2) с позднего мела и в те-
течение всего третичного периода в малакофауне европейского участка оби-
обитало много родов и видов Phaedusinae, из которых наиболее древние очень
походят на некоторые современные роды того же подсемейства из восточной
Азии; 3) несколько видов двух эндемичных родов Neniinae обитают в на-
настоящее время в юго-восточной Азии.
Естественно поставить вопросы: если первоначально ареал нашего
семейства был единым, то в какую эпоху и по каким причинам он распался?
Или же: откуда и какими путями шло расселение моллюсков?
В первую очередь следует учесть, что наиболее примитивные из совре-
современных групп — Phaedusinae и Neniinae — являются обитателями обла-
областей с влажным тропическим климатом, где минимальные температуры,
как правило, выше нуля. Ископаемые Phaedusinae позднего мела и палео-
палеогена Европы, по всем данным, обитали в областях со сходным климатом.
Clausiliidae возникли на юге Евразии. За это говорит присутствие
здесь как всех современных, так и ископаемых подсемейств этого семейства,
в том числе и представителей в основном южноамериканского подсемейства
Neniinae.
Отсутствие наших моллюсков в Австралийской зоогеографической об-
области, которая отделилась от всех остальных областей суши уже в юрском
периоде, свидетельствует о том, что эта группа возникла и расселилась,
по-видимому, в меловом периоде.
62 ВВЕДЕНИЕ
В это время предки Phaedusinae, подобно современным видам этой
группы^ населяли тропические прибрежные области и острова, которые
омывались водами моря Тетис. Это море, за исключением той части, ко-
которая соответствует морям, омывающим современный Индокитай и Ма-
Малайский архипелаг, в основном располагалось в аридной зоне. Поэтому
влаголюбивые виды, подобно современным тропическим Clausiliidae, оби-
обитали лишь в прибрежных областях с более влажным климатом и не про-
проникали вглубь континентов. Этим, по-видимому, и объясняется отсутствие
Clausiliidae в экваториальной Африке.1 В раннем мелу Африка имела ши-
широкую континентальную связь с Южной Америкой (Страхов, 1948; Тег-
mier H. et G., 1952), благодаря чему древние Clausiliidae проникли вдоль
южных берегов Тетиса в западное полушарие, где дали начало современ-
современным южноамериканским Neniinae.
Трудно предположить, что расселение наших моллюсков в западное
полушарие шло иным путем, т. е. через Северную Америку, хотя этот
континент неоднократно вступал в связь как с Европой, так и в особен-
особенности с Азией. Дело в том, что по данным палеогеографии (Криштофович,
1955; Марков, 1960) север указанных континентов уже с начала мезозоя
находился в зоне умеренного климата, неблагоприятного для наших тро-
тропических групп. С другой стороны, тот же вывод подкрепляет отсутствие
наших моллюсков в Северной Америке как в настоящее время, так и (по
данным палеонтологии) в предшествующие эпохи.
Третичныйпериод
В палеогене в южной и средней Европе Clausiliidae достигли пышного
расцвета. Вначале здесь господствовали только Phaedusinae (табл. 2),
многие виды и роды которых поразительно походили на современные во-
сточноазиатские группы того же подсемейства. Так, например, жившие
в палеоцене — эоцене Европы виды рода Oospiroides Wenz очень сходны
с современными видами из рода Oospira Blanf. из Бирмы.
В течение всего третичного периода климат Средиземноморской под-
подобласти изменялся в сторону нарастания засушливости (Марков, I960).
В растительном мире это привело к преобразованию влаголюбивой флоры
позднего мела и палеоцена (гелинденская флора), в так называемую пол-
полтавскую флору, отличавшуюся большей сухоустойчивостью и более вы-
высокими требованиями к теплу (Вульф, 1944; Криштофович, 1955).
Аналогичные изменения происходят и в фауне Clausiliidae. На смену
палеоценовым родам пришли другие, сильно отличавшиеся как от совре-
современных восточноазиатских Phaedusinae, так и от родов этого подсемейства,
обитающих в настоящее время в Средиземноморской подобласти. Эти
новые группы имели мощный замыкательный аппарат, что указывает на
необходимость в защите от высыхания.
Однако наряду с этими родами в эоцене появляются первые виды под-
подсемейства Triptychiinae, которое вымерло лишь в конце третичного пе-
периода. Для этой группы было характерно отсутствие клаузилия, полулун-
полулунной и главной складок. Остальные части замыкательного аппарата у наи-
наиболее древних видов были развиты нормально, но рсщ^шв^^^постерен-
ный^дегвесс кконду;9Bpjiroj^j5 этой группы. По-видимому, это подсемей-
подсемейство населяло прибрежные области с влажным климатом или какие-то
1 В настоящее время там обитают несколько видов из рода Macroptychia Bttg.,
который принадлежит к подсемейству Clausiliinae (или Euxininae) и проник сюда с се-
севера в конце третичного периода или даже позднее. ..¦•¦.
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ
63
Таблица 2
Распространение современных и ископаемых родов
и подродов Clausiliidae в третичном периоде [по Венцу
(Wenz, 1923), Венцу и Цильху (Wenz u. Zilch, 1959) и собственным данным]
Условные обозначения: р. — ранний, ср. — средний, п. — поздний;
звездочкой отмечены группы с неясным систематическим положением
Подсемейства, роды,
подроды
Phaedusinae
Proalbinaria Bttg. . . .
Neniopsis Wenz . . .
Oospiroides Wenz . . .
Palaeophaedusa Wenz .
Disjunctaria Bttg. . . .
Pachyphaedusa Wenz
Ptychophaedusa Wenz
Euclausta Oppenheim
Emarginaria Bttg. . .
Constricta Bttg. ....
Eualopia Bttg
Serrulina Mouss. S. str.
Pravispira Lindh. ...
Euxinophaedusa gen.
nov
Pontophaedusa Lindh. .
Neniinae C/>
Virrobasis ^Conrad* ,~f\
Triptychiinae
Plioptychia Bttg. . . .
Triptychia Sandn. s. str.
Milne-Edwardsia Bourg.
Cochlodininae
Cochlodina Fer
Charpentieria Stab. . .
Euxininae
Acrotoma Bttg
Pseudidyla Bttg. .
Euxina Bttg. .....
Mentissoidea Bttg. . .
Laminifera Bttg. s. str*
Baboria Cossmann* . .
Polloneria Sacco* . . .
Clausiliinae
Canalicia Bttg.* . . .
Quadriplicata Bttg. . .
Clausilia Drap
Iphigena Gray
P Мел
Третичный
палеоцен
P-
ср.
п.
эоцен
Р-
1
ср.
Г.*
п.
—
олигоцен
Р-
и
ср.
D-
п.
миоцен
Р.
¦
ср.
I
п.
плиоцен
Р-
I
I
ср.
п.
I Современный
J
-
J
очень влажные биотопы. Благодаря этому у них не было нужды в защите
мягких частей тела от высыхания.
Наиболее серьезные изменения в фауне Clausiliidae Европы произошли
в неогене. Похолодание климата в миоцене привело к гибели большинства
палеогеновых родов. На смену им пришли новые группы. К этому времени
64 ВВЕДЕНИЕ
появились современные западнопалеарктические группы Phaedunnae,
в частности род Serrulina Mouss., виды которого были широко распро-
распространены в Европе до самого конца третичного периода.
Euxininae, в лице наиболее примитивного рода Acrotoma Bttg., возникли
уже в палеогене, но более пышного расцвета достигли в неогене на тер-
территории Кавказа и Малой Азии.
К cei?ej№Hfi_T^TH4jwro_jie_pjipjtaj^nbH№niee нарастание засушливости
климата Передней Азии привело к полной изоляции ClausiTiidde, обитав-
обитавших в Европе, от их восточноазиатских родичей.
В результате, в общих чертах наметилась современная картина рас-
распространения этого семейства, т. е. его ареал оказался разорванным на
три изолированных участка.
Возвращаясь к изменениям, которые произошли с фауной Clausiliidae
в неогене в западной части Палеарктики, следует отметить, что они были
связаны со следующими основными физико-географическими факторами
того времени (Страхов, 1948; Termier H. et G., 1952; Марков, 1960): 1) даль-
дальнейшим нарастанием засушливости климата; 2) существованием на протя-
протяжении всего неогена в восточной части Средиземноморья крупного мас-
массива — Эгейской суши; 3) полной или частичной изоляцией участков
суши, соответствующих современному Крыму и Кавказу.
Аридность климата особенно сильно возросла на востоке тогдашнего
Средиземного моря, т. е. на территории Эгейской суши. В состав этой
суши входил весь современный Балканский полуостров, территория,
занятая теперь Эгейским морем и восточной частью Средиземного. По сути
дела, Эгейская суша являлась продолжением Передней Азии на северо-
запад. Длдхедьное время она соединялась с территорией современных
Альп_и Карпат.
В связи с ксерофилизацией климата на Балканах возникли совер-
совершенно новые группы Clausiliidae — Alopiinae и Cochlodininae. Будучи
значительно лучше адаптированы к сухому климату, чем Phaedusinae,
эти группы, в особенности первая, достигли здесь пышного расцвета.
По-видимому^_в_этр_2ке время где-то на севере Эгейской суши за счет
древних Euxininae возникли первые Clausiliinae. Позднее они расселились
на территорию современных Альп, Карпат, горного Крыма, а со стороны
Малой Азии^п^оникли_и_на ^авказ.
ТйшказТ "Втечение всего палеогена Большой Кавказ был островом.
Климат и растительность его носили отчетливо выраженный тропический
облик. Лишь в середине миоцена, в результате поднятий, Кавказский
остров с юга причленяется к Малой Азии и становится полуостровом
(Жижченко, Колесников, Эберзин, 1940).
Полная или частичная изоляция Кавказа от остальных учпетков суши,
становление горной зональности под влиянием горообразовательных
процессов, изменение климата в сторону похолодания привели к тому, что
первоначально тропическая фауна этого острова (а затем полуострова)
в миоцене претерпела сильные изменения.
До настоящего времени все сведения о третичных Clausiliidae Кавказа
ограничивались одним видом — «Clausilia» roseni (Lindh.) Volkova, опи-
описанным H. С. Волковой A953) из верхнесарматских отложений Армавира.
В результате изучения коллекции А. А. Стеклова (стр. 300) список верхне-
верхнемиоценовых Clausiliidae Северного Кавказа увеличился до 7 видов:
1. Euxinaphaedusa volkovae mihi 4. S. serailamellata (Mouss.)-
(=<iClausilia* roseni Volkova). 5. S. nazranica mihi.
2. Eu. steklovi mihi. 6. Pontophaedusa praefuniculum mihi.
3. Serrulina cf. sieversi (Pfr.). 7. Mentissoidea cf. litotes (A. Schm.).
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 65
Из этого списка видно, что в миоцене на Кавказе в фауне Clausiliidae
бок о бок существовали виды, принадлежавшие к современным родам
{№№. 3—7) и виды вымершего рода Euxinopkaedusa gen. nov.
По раковинам этот род напоминает некоторые группы, живущие те-
теперь в тропической Азии. По-видимому, моллюски Euxinophaedusa в мио-
миоцене Кавказа были элементом фауны палеогена. Это предположение под-
подкрепляют «гигантские» размеры раковин обоих видов, входящих в этот
род. Известно, что многие Phaedusinae палеогена Европы отличались очень
крупными размерами.
Остальные моллюски являются более молодым элементом верхнемио-
щеновой фауны Кавказа. С особой уверенностью это можно сказать о видах
из рода Serrulina Mouss., так как во многих других странах Средиземно-
Средиземноморской подобласти эта группа появилась только в миоцене.
Очень характерно присутствие одного современного вида (№ 4) и
двух форм, очень близких к современным моллюскам (№№ 3, 7). Впервые
документально подтверждается, что некоторые наземные моллюски Кав-
Кавказа, без видимых существенных изменений сохранились здесь с конца
миоцена. Наконец, в приведенном списке есть виды (№№ 5, 6) хотя и
современных родов, но вымершие в миоцене или в плиоцене.
В миоцене лесные зоны Кавказа были несколько более приподняты,
~чем в настоящее время, и флора, близкая к современной колхидской,
занимала большую территорию, в том числе и значительную часть север-
лых склонов Большого Кавказа (Вульф, 1944). Только этим можно объяс-
объяснить, что моллюски из рода Pontophaedusa Lindh. в это время оби-
обитали в районе Майкопа и Орджоникидзе. Напомним, что единст-
единственный современный вид этого рода сейчас живет лишь во влажных
субтропических лесах черноморского побережья Кавказа и Малой Азии.
В плиоцене соединение Кавказа с Малой Азией в общем продолжало
расширяться. Временами эта связь не надолго нарушалась из-за морских
трансгрессий со стороны Каспийского моря. Связь с Русской равниной
была менее постоянной, но в кимерийском веке она была очень широка
^Жижченко, Колесников и Эберзин, 1940; Страхов, 1948).
В коллекции А. А. Стеклова мы обнаружили только 3 вида из верхне-
ллиоценовых отложений Северного Кавказа (акчагыл — апшерон):
* 1. Euxina somchetica (Pfr.).
2. Ей. tschetschenica fPfr.^.
3. Quadriplicata intermedia mihi.
Совершенно ясно, что этим кратким списком далеко не исчерпывается
зся фауна Clausiliidae указанной эпохи. Однако характерно, что первые
два вида являются современными, а последний вымерший, но принадле-
принадлежащий к роду, эндемичному Кавказу и богато там теперь представленному.
•Согласно устному сообщению А. А. Стеклова, в тех же образцах, где были
найдены указанные выше виды, им обнаружены многие другие наземные
и пресноводные моллюски. Из них одни являются современными, а осталь-
остальные принадлежат к современным родам. Все это показывает, что к концу
плиоцена малакофауна Кавказа приобрела почти современный облик.
При характеристике географического распространения ныне живущих
¦Clausiliidae на Кавказе (стр. 56) обращалось внимание, что, подобно дру-
другим лесным моллюскам, в данном семействе отчетливо намечается
разделение на две географические группировки: 1) виды, населяющие
равнинные и горные леса Большого Кавказа, низовые леса Закавказья
и леса внешних отрогов Малого Кавказа, и 2) виды, населяющие леса
Талыща, Ленкоранской низменности и северных склонов Эльбурса (се-
66 ВВЕДЕНИЕ
верный Иран). Отмечалось, что подобная картина географического рас-
распространения соответствует двум основным зоогеографическим провин-
провинциям лесной малакофауны Кавказа — Кавказской и Гирканской.
В настоящее время обе лесные провинции совершенно разобщены
полупустынями и степями Куро-Араксинской низменности и Армянского
нагорья.
Для гирканских видов характерна большая родственная близость
к видам, населяющим современную Малую Азию, чем к видам тех же родов
с Большого Кавказа. С другой стороны, если виды Кавказской провинции
не проникают на территорию Гирканской, то некоторые виды, свойствен-
свойственные последней, обитают в лесах южных склонов восточной части Большого
и северных отрогов Малого Кавказа. С нашей точки зрения, обе эти осо-
особенности малакофауны Гирканской провинции связаны с историей этой
территории. Очевидно, что до соединения Кавказского острова с Малой
Азией вдоль ее северного берега и Эгейской суши вообще обитала единая
лесная фауна. Как след этого, остался ныне разорванный ареал Euxina
persica (Талыш и северный Иран, черноморское побережье Малой Азии,
южная Болгария). В середине миоцена, когда произошло соединение
Кавказа с Малой Азией, некоторые гирканские виды проникли на Боль-
Большой Кавказ. Это подтверждается находкой в верхнем миоцене Северного
Кавказа вида, очень близкого к современной Serrulina sieversi. Однако
обилие на Большом Кавказе видов, экологически сходных с гирканскими,
не дало возможности последним распространиться по всему перешейку
и они задержались лишь в некоторых восточных районах.
В то же время длительная изоляция Кавказского острова и интенсив-
интенсивное горообразование привели к формированию здесь основных родов и
подродов, характерных для Кавказской зоогеографической провинции
(Qundriplicata Bttg., Micropontica Bttg., Scrobifera Bttg., Pontophaedusa
Lindh. и др.).
Известно, что в Талыше на скалах и в осыпях высокогорной луговой
и нагорно-степной зон обитает моллюск Mucronaria gustavi. Это единствен-
единственный гирканский вид рода Mucronaria Bttg. Остальные виды этой группы
живут в лесах и в альпийской зоне Малого и Большого Кавказа, причем
один из них — Mucronaria duboisi проникает и на черноморское побе-
побережье Турции.
Вероятно, в неогене род Mucronaria Bttg. был довольно широко рас-
распространен в лесах приморских областей Малой Азии. Когда к последней
присоединился Кавказ, эта группа расселилась и туда. Позднее в связи
с нарастанием засушливости климата и возникновением безлесного Армян-
Армянского нагорья с этой территории исчезло большинство лесных видов.
Однако М. gustavi, который как скал ожил находится в более благоприят-
благоприятных микроклиматических условиях, чем обитатели открытых биотопов,
сохранился на восточной окраине Армянского нагорья, т. е. в безлесном
Талыше.
С присоединением Кавказа к Малой Азии некоторые кавказские виды
стали расселяться на запад вдоль Черного моря. Однако это не имело мас-
массового характера, так как в настоящее время сравнительно немногие виды
обитают западнее Синопа. Возможно, что это расселение произошло в бо-
более позднее время.
Известно, что один вид — Serrulina serrulata, долгое время считав-
считавшийся эндемиком Кавказа, позднее был обнаружен не только в Турции
(как на севере, так и на юге), но и в Болгарии, Румынии и в Лесистых Кар-
Карпатах Украины. Эти находки указывают на былое очень широкое рас-
распространение этого моллюска, ареал которого сократился и распался,
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 67
вероятно, уже в четвертичное время. Трудно предположить, что в неогене
этот моллюск расселился на запад из Кавказа. Скорее всего он был свой-
свойствен всем лесным областям Эгейской суши, в том числе и Малой Азии,
откуда проник на Кавказ. К тому же, как мы уже указывали, многие виды
того же рода были обычны в неогене западной Европы.
Другой моллюск — Cochloidina laminata — указывает на то, что обмен
фаунистическими элементами между Кавказом и Европой в неогене мог
происходить и другим путем — северным. В настоящее время этот моллюск
обитает в большинстве лесных областей Европы, а на Кавказе живет
только в байрачных лесах Пятигорской и Ставропольской возвышенно-
возвышенностей. В Закавказье он отсутствует. По-видимому, в указанные районы
Кавказа этот моллюск проник в конце плиоцена со стороны Русской
равнины. Позднее этот путь для него был отрезан открытыми степными
пространствами.
Крым. Мы не располагаем палеонтологическими данными о третичных
Clausiliidae данной области. Уже отмечалось, что для современной мала-
кофауны из Clausiliidae здесь наиболее характерны 2 вида эндемичного
подрода Mentissa H. et A. Adams (род Laciniaria Hartm., Clausiliinae).
Остальные 3 вида являются вселенцами из средней и южной Европы.
В течение раннего и среднего миоцена (Жижченко, Колесников, Эбер-
зин, 1940), а по некоторым авторам (Страхов, 1948) в течение всего мио-
миоцена, Крым был частью Эгейской суши, точнее ее полуостровом, ориен-
ориентированным на восток. По-видимому, в это время на севере этой суши,
т. е. на территории современных Балкан, возникли первые Clausiliinae и,
в частности, род Laciniaria Hartm., к которому принадлежат также оба
моллюска, эндемичные Крыму.
Начиная с позднего миоцена, а по другим авторам с раннего плиоцена,
обрывается прямая связь Крыма с Балканами. Вначале на месте его
находился остров, который позднее неоднократно вступал в связь с Рус-
Русской равниной.
Хотя в течение всего плиоцена юг Русской равнины был покрыт ле^-
сами, моллюски горного Крыма были практически изолированы от мала'
кофауны других горных областей. Этим и объясняется, что здесь на базе
древних Laciniaria Hartm. возник эндемичный подрод Mentissa H. et
A. Adams. Одновременно некоторые виды равнинных лесов Европы про-
проникли в Крым и обосновались там (Cochlodina laminata, Iphigena plica-
tula, Balea perversa). Когда юг Русской равнины и север полуострова
заняли степи, эти моллюски оказались отрезанными от своего основного %
ареала.
Средняя Европа. Как мы уже упоминали, с нашей точки зрения, именно
Эгейская суша, точнее та часть ее, на месте которой находятся теперь
северные области Балканского полуострова, явилась в начале неогена аре-
ареной бурного образования основных современных групп Clausiliidae
Европы. Здесь возникли Alopiinae, Cochlodininae и Clausiliinae.
Когда в миоцене, в результате поднятий и складчатых движений,
к Эгейской cjn^j^cejej^Fpnr.opj^HjwrK Альпы и Карпаты, эти терри-
территории ста'л^з^1селяться предками современных Clausiliinae и Cochlodini-
Cochlodininae. Обе горные области длительное время были разъединены морскими
проливами и Паннонским бассейном. Bflaroj^ja,^Tjo^U?a.3BHTHe фаун
Альп и Карпат шло почти^ независимо и joHkeH отдельными элементами
мог""пр6йсходить только обходным путем, т. е. черел. северные Балканы.
В Карпатах, за счет пришедших сюда балканских видов Clausiliinae,
возникли эндемичные роды Vestia Hesse и Pseudalinda Bttg., в Альпах —
Clausilia Drap. и Iphigena Gray.
68 ВВЕДЕНИЕ
Позднее, когда морские преграды исчезли, произошел некоторый
обмен фаунистическими элементами. Однако в целом известная самостоя-
самостоятельность фаун обеих горных областей прослеживается и в настоящее
время.
По-видимому, к концу плиоцена в средней Европе существовали
уже все современные виды Clausiliidae. Некоторые из них найдены в отло-
отложениях этого периода — Clausilia dubia Drap., Iphigena plicatula Drap.
(под названием I, baudoni Mich.), Cochlodina laminata Mont, (под назва-
названием С. prolaminata Sacco). Остальные обитали в горах, где условия фос-
силизации хуже, чем на равнине. Однако косвенным подтверждением
тому, что они существовали уже в плиоцене, служит массовое появление
их на равнинах в плейстоцене, куда они сместились из-за начавшегося
похолодания и разрастания горных ледников.
В итоге в третичном периоде формирование фауны Clausiliidae в запад-
западной части Палеарктики отличалось следующими особенностями:
1) Pkaedusinae, господствовавшие в палеогене, к началу миоцена вы-
вымирают или заменяются другими группами того же подсемейства;
2) Euxininae возникли еще в палеогене, но пышного расцвета достигли
лишь в неогене на востоке Средиземноморской подобласти;
3) в миоцене на севере Эгейской суши возникли новые группировки —
подсемейства Alopiinae, Cochlodininae и Clausiliinae, поэтому и в настоя-
настоящее время Балканский полуостров является местом сосредоточения боль-
большинства видов и родов этих подсемейств', в том числе и наиболее при-
примитивных;
4) фауна Clausiliidae средней Европы, Крыма и Кавказа слагалась,
с одной стороны, в результате миграций предковых форм Clausiliinae
(главным образом) и Cochlodininae (в меньшей степени) из Эгейской суши;
с другой стороны, фауна этих областей образована за счет палеоге-
палеогеновых Phaedusinae и Euxininae; полная или частичная изоляция этих
областей наряду с интенсивными горообразовательными процессами,
которые здесь происходили в течение неогена, способствовала образо-
образованию эндемичных видов и родов;
5) в миоцене произошло формирование большинства современных ро-
родов, а к концу плиоцена — современных видов Clausiliidae.
Четвертичный период
В четвертичном периоде в Европе произошли события, кардинальным
образом изменившие весь облик фауны и флоры. Важнейшие из них свя-
связаны с так называемым ледниковым периодом, т. е. с многократным
оледенением значительной части континента. При этом, за исключением
отдельных возвышенных мест, оставшихся, подобно небольшим островам
(нунатаки), среди огромных пространств, занятых ледниками, все живое
в зоне оледенения было либо уничтожено, либо оттеснено за пределы лед-
ледников (Вульф, 1944; Страхов, 1948; Termier H. et G., 1952; Шварцбах,
1955).
В упомянутых убежищах могли остаться только наиболее холодостой-
холодостойкие формы животных и растений. В связи с этим трудно предположить,
Что среди них были какие-нибудь виды Clausiliidae, так как они, по срав-
сравнению с другими наземными моллюсками умеренных широт, принадлежат
к числу наиболее теплолюбивых животных.
Наряду с образованием огромного ледникового щита на Скандинав-
Скандинавском полуострове, который, разрастаясь, распространялся на значитель-
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 69
ную часть областей, лежащих южнее, горные страны средней Европы и
Кавказа испытывали свои местные оледенения различного масштаба.
Ледниковым периодам средней и северной Европы в средиземномор-
средиземноморских странах соответствовали холодные плювиальные периоды, сопрово-
сопровождавшиеся увеличением количества осадков и высокой влажностью кли-
климата. И наоборот — в то время как на севере межледниковые фазы зна-
знаменовались теплым и влажным климатом, на юге господствовали очень
жаркие, а вместе с тем и сухие пустынные условия.
Эти события привели к почти полному исчезновению с территории
Европы влаго- и теплолюбивых третичных видов. Лишь некоторые из
них сохранились, но только в пределах Средиземноморской подобласти и
в таких местах, где в силу орографических условий климат отличается
равномерным и почти непрерывным выпадением осадков (атлантическое
побережье Пиренейского полуострова, Родопы, черноморское побережье
Болгарии и Турции, субтропики Закавказья, — Вульф, 1944).
Из Clausiliidae, в силу их тропической и субтропической природы,
почти нацело исчезли Phaedusinae. Лишь некоторые виды этого подсе-
подсемейства сохранились в лесах Кавказа, черноморского побережья Турции,
на Балканах и в Лесистых Карпатах. Показателен характер их местооби-
местообитаний. Большинство этих моллюсков живет в гниющей древесине и под
корой поваленных деревьев, т. е. в условиях постоянной и высокой влаж-
влажности. Другим убежищем явились пещеры. Как известно, несколько ви-
видов третичного рода Serrulina Mouss. живет в пещерах острова Корфу,
Далмации и западного Закавказья.
Следовательно, наиболее влаголюбивые среди Clausiliidae тропические
и субтропические реликты третичной фауны сохранились в областях с влаж-
влажным субтропическим климатом, причем в наиболее влажных убежищах.
Показательно распространение другого древнего рода — Laminifera
Bttg. В настоящее время от этой, когда-то многочисленной группы сохра-
сохранилось всего 2 вида, причем оба живут в Пиренеях.
В Средиземноморской подобласти на смену влаголюбивым тропиче-
тропическим и субтропическим видам' пришли новые, из которых большинство
принадлежит к подсемейству Alopiinae. Эта группа состоит из скаложи-
лов, адаптированных к жизни в безлесной местности. В связи с этим у них
возник экранизирующий слой на поверхности раковины (стр. 12, 49). К тем-
температуре они относятся различно. Одни живут в сравнительно холодных
высокогорных областях Балканского полуострова, другие -г— в сухих
субтропиках побережья и островов Средиземного моря.
В восточной части Средиземноморской подобласти одним из самых
важных событий четвертичного периода явилось опускание на дно моря
значительной части Эгейской суши. Эта трансгрессия произошла в рисс-
вюрмскую межледниковую фазу (Страхов, 1948; Termier H. et G., 1952).
Образование Эгейского и Мраморного морей и восточной части Среди-
Средиземного моря привело к распаду первоначально единых ареалов многих
видов и групп. В результате Alopiinae населяют не только Балканский
полуостров, но и острова и ряд прибрежных областей Эгейского моря
и восточной части Средиземного. Такие роды, как Euxina Bttg. и Mega-
leuxina Bttg., живут как в Малой Азии и на Кавказе, так и на Балканах,
а некоторые виды — Euxina persica, Ей. circumdata — обитают по обеим
сторонам Мраморного моря.
Нарастание засушливости климата Средиземноморской подобласти
в межледниковые фазы_плетстодена и в голодене способствовало расселе-
расселению засухоустойчивых моллюсков. Некоторые из них, по таким ксеротерм-
ным биотопам, как скалы и осыпи, довольно далеко проникали в области,
70
ВВЕДЕНИЕ
занятые лесами. К их числу мы относим моллюсков из рода Armenica
Bttg. Этот малоазиатский род появился на Кавказе в четвертичном пе-
периоде, когда сложились нагорно-степные и полупустынные ландшафты
Армянского нагорья. Трудно предположить более раннее появление их
на Кавказе, например в плиоцене. К тому же, за исключением двух ви-
видов — A. griseofusca и A. gracillima — в этом роде нет эндемиков кав-
кавказской фауны.
В средней и северной Европе окончание ледникового периода (плей-
(плейстоцена) знаменовалось не только постепенным освобождением ото льда
соответствующих территорий, но и изменениями их климата и ландшафта
(Вульф, 1944; Шварцбах, 1955). Некоторое время области, освободив-
освободившиеся от ледника, обладали субарктическим климатом и были заняты
тундрой. Позднее эту растительность сменили леса из березы и сосны
(время дриас). Следующая эпоха (бореальное время) отличается более
теплым и континентальным климатом. В это время указанная территория
была покрыта смешанными лесами с дубом и орешником. Наступившая
затем эпоха характеризовалась влажным и теплым океаническим кли-
климатом (атлантическое время). Она была теплее современной; это видно
из того, что снеговая линия была на 300 м выше современной. В расти-
растительности преобладал дуб.
Позднее климат стал более континентальным (суббореальное время).
В лесах преобладающее место занял бук, а потом ель. Наконец, последняя
эпоха, предшествовавшая современной (субатлантическое время), отли-
отличалась опять океаническим климатом, но несколько более холодным, "чем
в предыдущие две эпохи. Современный климат средней Европы характе-
характеризуется большей континентальностью и потеплением.
В настоящий момент мы не располагаем достаточно подробными дан-
данными, чтобы проследить, как изменялась фауна Clausiliidae в различных
областях Европы на протяжении всех стадий ледникового (плейстоцен)
и послеледникового (голоцен) времени. В этом отношении особенно скуд-
скудными данными мы располагаем с территории СССР (Даниловский, 1955).
Несколько больше материал из четвертичных отложений средней Европы,
хотя опять-таки не для всех эпох (Lozek, 1955).
Таблица 3
Распространение среднеевропейских видов Clausiliidae в
четвертичных отложениях [по Ложеку (Lozek, 1955), Гейеру
(Geyer, 1927), Даниловскому, 1955]
• Условные обозначения: 4" достоверные находки; звездочкой отмечены
находки вне современного ареала
Виды
Cochlodina orthostoma
(Menke)
С. cerata (Rssm.) ....
С. laminata (Mont.) . . .
С. commutata (Rssm.) . .
Clausilia parvula (Stud.)
Cl. bidentata (Strom) . .
Cl. dubia Drap
Cl. pumila C. Pfr
Cl. cruciata (Stud.) . . .
Iphigena ventricosa (Drap.)
/. densestriata (Rssm.) . .
/. latestriata (A. Schm.) .
/. lineolata (Held) . . • .
Плей-
Плейстоцен
+
¦i
+*
+*
+*
+
Голо-
Голоцен
+*
' +
+
+
+*
+
+ *
+*
+ *
+ *
4-
Виды
/. tumida (Rssm.) ....
/. plicatula (Drap.) . . .
Laciniaria biplicata
(Mout.) .
L. plicata (Drap.) . . .
L. сапа (Held)
L. nitidosa (Ulicny) . . .
Pseudalinda stabilis (Pfr.)
Vestia gulo (Bielz) . . .
V. turgida (Rssm.) . . .
Ruthenica filograna
(Rssm.)
Balea perversa(h.) ....
Плей-
Плейстоцен
+ *
+
+
+
+
+
+
+
Голо-
- цен
+•
+
+
+*
+
+*
+
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 71
Поэтому мы ставим задачу дать лишь общую характеристику тех
перемен, которые испытала фауна Clausiliidae, используя для этого как
данные четвертичной палеонтологии, так и зоогеографии.
Как видно из табл. 3, в отложениях плейстоцена и голоцена средней
Европы найдены почти все современные европейские виды. Если в плей-
плейстоцене они приурочены к межледниковым эпохам, то в голоцене —
главным образом к атлантическому времени.
В некоторые межледниковые фазы (датировка находок не всегда
достаточна ясна) ряд среднеевропейских видов имел более широкое
распространение, чем теперь. Так, Clausilia pumila, Cl. parvula
и Iphigena ventricosa найдены в плейстоценойых отложениях ю кной Анг-
Англии, т. е. далеко за пределами их современных ареалов (рис. 146, 153).
Возраст этих находок, по-видимому, не может быть моложе второй геж-
ледниковое^ фазы (миндель-рисс), так как в третью межледниковую фазу
(рисс-вюрм) Британские острова были уже отделены от континентальной
Европы проливом Ла-Манш, а вюрмское оледенение не достигало этой
территории и сказалось на ней незначительно (Termier H. et G., 1952).
Следовательно, можно думать, что исчезновение указанных видов с Бри-
Британских островов обязано рисскому оледенению.
В средней Европе фауна Clausiliidae достигла наиболее пышного рас-
расцвета в атлантическое время голоцена. Теплый и влажный мягкий океа-
океанический климат способствовал тому, что многие виды обитали в областях,
где теперь отсутствуют или сохранились лишь небольшие островные
ареалы в пунктах с благоприятным местным климатом.
Для фауны этого времени было характерно обитание бок о бок тепло-
теплолюбивых и относительно холодолюбивых видов (например, Clausilia
bidentata и Cl. cruciata). Подобный парадокс объясняется способностью
холодолюбивых моллюсков, при наличии высокой и постоянной влаж-
влажности, нейтрализовать влияние избытка тепла усиленным испарением
влаги.
В атлантическое время горные области средней и южной Европы (Альпы,
Карпаты, Балканы) являлись • источником расселения на север многих
наземных моллюсков, в том числе и Clausiliidae.
Примеров подобных инвазий можно привести много. Так, Шлеш
(Schlesch, 1946, 1958) отмечает, что в настоящее время в малакофауне
прибалтийских стран имеется комплекс видов карпатского и балканского
происхождения. Из Clausiliidae сюда относится карпатская Iphigena
latestriata и все 3 среднеевропейских вида рода Laciniaria Hartm. — L. Ы-
plicata, L. plicata, L. сапа. Последние З моллюска, равно как оба средне-
среднеевропейских вида из рода Cochlodina Fer. — С. laminata, С. orthostoma —
пришли с Балкан, где и поныне они обитают вместе с другими видами
тех же родов.
Сопоставив распространение этих моллюсков в отдельных, ныне изо-
изолированных областях Прибалтики с четвертичной историей Балтийского
моря, Шлеш пришел к выводу, что они расселились в этих краях в атлан-
атлантическое время, которому соответствовала литориновая фаза развития
Балтийского моря.
Окончание атлантического времени знаменовалось нарастанием кон-
тинентальности климата (суббореальное время), затем климат стал более
мягким, но холоднее предыдущих эпох (субатлантическое время).
На эти изменения климата наши среднеевропейские моллюски реаги-
реагировали по-разному. Сравнительно холодоустойчивые виды (Clausilia
dubia, Cl. cruciata, Iphigena ventricosa и др.), т. е. большинство, смести-
сместились в горы и сохранились на равнинах только на востоке — в Прибалт
72
ВВЕДЕНИЕ
тике, в западных и центральных областях европейской части СССР (рис. 34).
Некоторые из них представлены на равнинах особыми подвидами (Clau^
silia pumila sejuncta, Iphigena latestriata homalis, — рис. 153).
Иначе реагировали на указанные изменения климата такие виды, как
Clausilia bidentata. Для этого моллюска первенствующее значение имеет
сочетание высокой влажности с мягким и теплым климатом. В атланти-
30
0 200 400 600 800 км
Рис. 34. Распространение Clausilia bidentata (Strom) и Cl. cruciata (Stud.).
(Ориг.)
la — Cl. bidentata; 16 — то же, в четвертичных отложениях вне современного ареала;
га — Cl. cruciata; 26 — то же, островные [ареалы.
ческое время он обитал даже в горах (южная Германия). В более поздние
эпохи он в горах уже не встречался и в настоящее время его распростра-
распространение ограничено приморскими областями средней и северной Европы,
климат которых находится под смягчающим воздействием Гольфстрима.
На юге европейской части СССР усиление засушливости климата при-
привело к образованию здесь степной зоны, которая и в настоящее время рас-
расширяется к северу за счет отступления южной границы лесов. В пределах
степей некоторые наши среднеевропейские виды (Cochlodina laminata,
Laciniaria plicata) сохранились только в байрачных лесах (рис^ 62, 162).
Одновременно шло сокращение площади, которую занимали леса Се-
Северного Кавказа, и изоляция лесных массивов Пятигорской и Ставро-
Ставропольской возвышенностей. Однако на былую связь их с кавказскими
ИСТОРИЯ ФОРМИРОВАНИЯ ФАУНЫ 73
лесами указывает, в частности, присутствие здесь таких моллюсков,
как Euxina somchetica, Quadriplicata aggesta stauropolitana, Idyla fovei-
collis, Mucronaria duboisi.
Следует учесть, что сокращение площади, занятой лесами, как на юге
европейской части СССР, так и в западной Европе, а следовательно,
и изменения в распространении лесных моллюсков, является также ре-
результатом хозяйственной деятельности людей.
Таким образом, фауна Clausiliidae средней и северной Европы в чет-
четвертичном периоде испытала сильные изменения. Влаголюбивые тропиче-
тропические и субтропические виды третичного периода исчезли с самого начала
первого оледенения. Основной костяк как современной фауны, так и фаун
плейстоцена и голоцена образован выходцами из горных стран средней
(Карпаты) и в особенности южной Европы (Балканы и Пиренеи). Осо-
Особенно пышного расцвета здесь достигли виды из различных родов подсе-
подсемейства Clausiliinae (Clausilia Drap., Iphigena Gray, Pseudalinda Bttg.,
Vestia Hesse и др.).
Выводы
Вся история формирования фауны Clausiliidae западной части Палеарк-
тики — это процесс постепенной перестройки влаголюбивой тропической
фауны в фауну умеренного типа. Только современных Phaedusinae данной
территории можно рассматривать как реликты тропической фауны нео-
неогена. Однако эти реликты в свою очередь возникли в результате преобра-
преобразования еще более влаго- и теплолюбивых групп времен палеогена.
Только в некоторых частях Средиземноморской подобласти (Кавказ,
черноморское побережье Малой Азии, некоторые районы Балкан), где
в силу местных орографических условий климат отличается высокой влаж-
влажностью и более или менее равномерным выпадением осадков, сохранилась,
правда сильно измененная, лесная фауна неогена.
На большей же части подобласти, где в течение четвертичного пе-
периода установился новый тип климата — средиземноморский суб-
субтропический (Алисов, 1950), с жарким засушливым летом и теплой и
влажной зимой, фауна Clausiliidae приняла совершенно иной облик.
В странах восточной части Средиземного моря господствующее положение
заняли Alopiinae, в западной части — некоторые местные Clausiliinae.
Хотя Alopiinae до сих пор не обнаружены в отложениях неогена,
есть основания предполагать, что они существовали уже в ту эпоху.
Как их образ жизни (скаложилы), так и некоторые особенности строения
раковин (экранизирующий слой) говорят за то, что они возникли в усло-
условиях безлесного ландшафта. Эти черты облегчили им в четвертичном
периоде расселение на территории, ранее занимаемой тропической лесной
фауной неогена, и обеспечили им здесь пышный расцвет, несмотря на
засушливое лето. Активную жизнь они в основном ведут осенью и зимой.
Иное положение сложилось в средней и северной Европе. После окон-
окончания ледникового периода, под влиянием Гольфстрима, здесь установился
климат, отличающийся влажным умеренно теплым летом и мягкой зимой
(Алисов, 1950). Большая часть территории занята достаточно влажными
смешанными и широколиственными лесами. Благодаря такому климату
и растительности, здесь, на смену исчезнувшим третичным субтропиче-
субтропическим и тропическим видам, появились и достигли пышного расцвета со-
современные среднеевропейские Clausiliinae и некоторые Cochlodininae.
Многие виды этих подсемейств существовали уже в плиоцене, но занимали
тогда второстепенное положение, В экологическом отношении они яв-
являются более холодостойкими, чем вымершие неогеновые виды, так как
74 ВВЕДЕНИЕ
возникли в горах (Альпы, Карпаты и Балканы). В голоцене они рассе-
расселялись далеко на равнины, заняв обширные лесные пространства на се-
севере и востоке Европы. Однако все среднеевропейские виды, даже с самым
широким распространением, держатся в сфере интенсивного климатиче*-
ского воздействия атлантических масс воздуха. Это видно из того, что они
населяют территории, где безморозный период не короче 120 дней, а ко-
количество осадков не меньше 500 мм в год (рис. 28).
Тропическое происхождение европейских Clausiliidae проявляется
в том, что все они сохранили влаголюбивость своих предков и современ-
современных родственников из восточной Азии. Однако, в то время как на засуш-
засушливых пространствах Средиземноморской подобласти местные группы
(Alopiinae, некоторые Clausiliinae) приобрели ряд адаптации, защищаю-
защищающих их от потери влаги, виды средней и северной Европы (Clausiliinae,
Cochlodininae) пошли по линии приспособления к более холодному кли-
климату.
НЕКОТОРЫЕ ВОПРОСЫ СИСТЕМАТИКИ CLAUSILIIDAE
Такс он омическое значение отдельных частей
тела
При характеристике тех или иных видов и групп для правильной
оценки их систематического положения: очень важно обратить внимание
именно на те особенности строения, которые в пределах данного таксона
отличаются наибольшей стабильностью и в то же время выражают основ-
основные направления эволюции той или иной группы. В связи с этим полезно
подвести некоторые итоги и остановиться на значении отдельных частей
тела для систематики наших моллюсков.
Раковина. Сравнительно с другими, даже близкими группами назем-
наземных легочных моллюсков раковина Clausiliidae обладает относительно
малой внутривидовой изменчивостью. Особенно это касается формы и раз-
размеров. Причина этого лежит в том, что наши моллюски отличаются_боль-
шой стенобионтностью. Правда, у тех горных видов, которые обитают
в нескольких вертикальных зонах, форма и размеры раковины нередко
сильно изменяются в зависимости от того, в какой зоне обитает соответ-
соответствующая популяция. Как уже отмечалось, это связано с климатическими,
в первую очередь с фенологическими различиями между зонами.
Скульптура раковины варьирует значительно сильнее, чем форма,
причем усиление ее чаще всего связано с обитанием на известняках.
В общем можно признать, что форма, размеры и скульптура раковины
в пределах вида отличаются значительной стабильностью и нередко яв-
являются удобными признаками для характеристики видов.
Особенно важно правильно оценить систематическое значение отдель-
отдельных частей замыкательного аппарата, так как именно эти образования
резко отличают наших моллюсков от всех других групп отряда Stylom-
matoplwa и от остальных Pulmonata вообще. Кроме того, возникновение
этого аппарата наложило резкий отпечаток на всю организацию Clausi-
Clausiliidae. .
Выше мы уже отмечали троякую роль замыкательного аппарата: за-
защита от высыхания, защита одних органов от давления других в наиболее
узкой части последнего оборота раковины и создание опоры и возможности
управлять раковиной при движении моллюска. Естественно, что степень
развития тех частей, которые защищают от высыхания, находится в пря-
прямой зависимости от климата. Следовательно, и вторая функция, которая
связана с первой, в конечном счете зависит от тех же условий. Что касается
ВОПРОСЫ СИСТЕМАТИКИ 75
последней функции — механической, то она меньше всего зависит
от первых двух. Поэтому понятно, почему в замыкательном аппарате
наибольшей стабильностью .обладают именно те его части, которые обес-
обеспечивают опору и управление раковиной, т. е. нижняя и субколумелляр-
иая пластинки и клаузилий. Форма и положение этих частей мало изме-
изменяются не только в пределах одного вида, но и внутри рода.
Важное систематическое значение имеет соотношение верхней и спи-
спиральной пластинок. Многие авторы, в том числе Бетгер и Вагнер, уделяли
¦этим особенностям большое внимание, однако подходили к этому вопросу
Неверно. Их только интересовало, соединяются ли обе пластинки или нет.
Между тем главным является не тот факт, слиты ли они или разделены,
а их положение по отношению к столбику, т. е. располагаются ли они на
одном расстоянии от него или на разном. Если они лежат от столбика
на разном расстоянии, то их концы чаще всего заходят друг за друга,
и обе пластинки всегда независимы. Если они располагаются на одном
расстоянии, т. е. на одной линии, то они либо сливаются, либо концы их
разделены промежутком. Оба последних случая могут иметь место в пре-
пределах изменчивости одного вида. Этот факт давно известен для Clausilia
pumila. To же самое мы наблюдаем у кавказских Mucronaria duboisi
и Quadriplicata quadriplicata, даже в пределах одной популяции. Правда,
у первой чаще всего пластинки слиты, а у второй разделены коротким
промежутком.
Соотношение верхней и спиральной пластинок, точнее располагаются
ли они на одном или на разном расстоянии от столбика, имеет значение
не только для характеристики вида, но и родов. В пределах рода эти осо-
особенности обычно постоянны.
Что касается палатальных и полулунной складок, то их форма и число г
могут очень варьировать даже в пределах одного вида. Это объясняется
зависимостью этих образований от влажности биотопа, в котором обитает
та или иная популяция. Как правило, они развиты сильнее у популяций,
живущих при более сухом и жарком климате, чем у популяций того же
вида из относительно влажных и прохладных мест.
Бросается в глаза следующая закономерность, подмеченная еще Ло-
сьесом (Loosjes, 1953a). Названные складки могут очень варьировать
в длине, в толщине и в количестве, однако у каждого вида положение их
строго фиксировано. Это относится как к месту, которое они зани-
занимают на периферии последнего оборота, так и к тому, насколько глубоко
они проникают внутрь раковины. г Учитывая вайшость дать точную ха-
характеристику глубине залегания полулунной складки и верхних концов
палатальных складок, Эрманн (Ehrmann, 1926) ввел понятие «лунелля-
рий» (lunellarium) как условной линии, вертикально пересекающей послед-
последний оборот, на уровне которой происходит соприкосновение клаузилия
с теми образованиями палатальной стенки, в которые он упирается. По-
Положение этой условной линии относительно стабильно как в пределах
вида, так и многих родов.
Радула. Мы уже отмечали, что строение радулы зависит от характера
питания того или иного моллюска и что в общем этот орган устроен очень
однообразно —, всего два типа строения. Поэтому его очень трудно ис-
1 Последнее относится только к палатальным складкам, так как полулунная
складка всегда одна.
2 Последнее не касается видов, у которых из-за преждевременной приостановки
роста, в связи с обитанием в высокогорных зонах, сформировались лишь верхние
участки палатальных складок.
76
ВВЕДЕНИЕ
пользовать для выяснения родственных отношений между видами и ро-
родами.
Однако при характеристике групп, состоящих из видов со сходным
образом жизни, с известной осторожностью можно использовать и при-
признаки радулы. Так, например, большинство Alopiinae и Armenica
живет на скалах и в осыпях и питается лишайниками, растущими на
твердом субстрате. Поэтому им свойствен второй тип радулы, т. е. с круп-
крупными срединным и боковыми зубами, лишенными дополнительных зуб-
зубцов.
В связи со сказанным отметим, что наши предшественники склонны
были считать характерным для Phaedusinae только первый тип радулы,
т. е. с мелкими срединным и боковыми зубами, снабженными дополни-
дополнительными зубцами. Однако это вряд ли правильно, так как в это.й богатой
видами группе только у немногих видов изучено строение радулы. Можно
ожидать, что у скаложилов из названного подсемейства со временем будет
обнаружена радула второго типа.
Половая система. Такие части полового аппарата, как гонада, гер-
мафродитный проток, белковая железа и семяяйцевод, в пределах семей-
семейства обнаруживают большое однообразие строения. Различия начинаются
с того места, где семяяйцевод разделяется на мужские и женские протоки,
в которые впадает ряд придаточных органов — семяприемник, бич, при-
придаток пениса.
Выше были подробно описаны различные типы строения отдельных
органов половой системы, дана морфо-функциональная оценка их важней-
важнейшим особенностям и связям с другими системами тела, в первую очередь
с мускулатурой и замыкательным аппаратом раковины. Поэтому здесь
имеет смысл подвести только некоторые итоги, которые можно кратко-
передать в следующей таблице (табл. 4).
В пределах каждого подсемейства в отношении половой системы раз-
различия между родами чаще всего сводятся к размерам и форме пениса,
к присутствию или к отсутствию на нем придаточных органов и, наконец,
к различным типам связи пениса с колумеллярной мускулатурой.
Таблица 4
Важнейшие изменения в половой системе Clausiliidae
Положение семяприемни-
семяприемника по отношению к по-
половой клоаке.
Ретрактор семяприемника.
Положение отростка по
отношению к семяпри-
семяприемнику.
Гистологическое строение
стенок отростка.
Восходящее колено семя-
семяпровода и пенис.
Phaedu-
Phaedusinae
Nenii-
пае
Впадает во вла-
влагалище вблизи
'л от половой
клоаки.
Нет.
Alopi-
Alopiinae
Впадает
выше
СосМо-
dininae
Euxini-
пае
Clausi-
liinae
во влагалище значительно
ПОЛОВОЙ ]
I
Впадает значительно выше нижне-
нижнего конца семяприемника
Эпителий, соединительная
мускулатура.
Хорошо развиты
ткань,
клоаки.
Есть.
Впадает вблизи
нижнего конца
семяприемника.
Эпителий.
образуют крупную петлю.
Руди-
ментар-
ментарны или
отсут-
отсутствуют.
ВОПРОСЫ СИСТЕМАТИКИ 77
К вопросу о по дразделении Glausiliidae на
подсемейства
Если раковина Clausiliidae дает богатый материал для четкой харак-
характеристики видов и родов, то ее уже недостаточно для выяснения родствен-
родственных взаимоотношений между последними. Дело в том, что раковины на-
наземных моллюсков вообще и данных в частности очень чувствительны
к воздействию внешних условий и даже у далеких друг от друга видов под
влиянием сходного образа жизни появляются аналогичные образования.
Конвергенция очень распространена среди Clausiliidae. Этим и объяс-
объясняется, что многие попытки дать нашим моллюскам единую классифика-
классификацию, исходя только из их раковин, с накоплением новых данных обычно
терпели неудачу. Именно потому уже в конце прошлого века некоторые
авторы старались использовать внутреннее строение для классификации
Clausiliidae.
Но прежде чем перейти к критическому разбору старых систем и изло-
изложить собственные взгляды, мы должны остановиться на одном частном
вопросе, касающемся взаимоотношений нескольких групп, которых дли-
длительное время объединяли в один род. Подобное отступление необходимо
не только потому, что в этом случае мы наиболее сильно расходимся с на-
нашими предшественниками, так как считаем эти группы за самостоятель-
самостоятельные роды, но также и из-за того, что попутно мы приходим к выводу,
что Clausiliinae (в прежнем понимании) необходимо разделить на два само-
самостоятельных подсемейства.
' О группах Laciniaria Hart m> и Eaiina О. Bttg.
Первоначально к группе Laciniaria относили только L. plicata и L. Ы-
plicata и несколько других европейских видов, из которых большинство
в настоящее время рассматривается как разновидности первых двух.
Для раковин считалось характерным присутствие киля и базального
желобка, глубоко лежащей спирально изогнутой нижней пластинки, а
также наличие полулунной и 1—-2 палатальных складок.
Позднее (Moquin-Tandon, 1855; Schmidt A., 1855) было установлено,
что у этих моллюсков мужские половые пути не образуют петли и вытя-
вытянуты вдоль женских (рис. 25 и 26). После этого стало обычным при обна-
обнаружении у каких-либо Clausiliidae половой системы подобного типа от-
относить их в тот же род. При наличии явственных различий в раковинах
таких моллюсков помещали лишь в особые подроды и секции. Точно так
поступал и Вагнер (Wagner,-1920). На основании сходства гениталий он
отнес в род Laciniaria следующие группы, которых рассматривал как
подроды: Balea, Mentissa, Euxina, Vestia и другие.
Так как последующие авторы не занимались столь основательно ана-
анатомией Clausiliidae, как Вагнер, то в этом вопросе они придерживались
его взглядов. Только группу Balea вследствии сильных различий в рако-
раковинах, несмотря на то что по гениталиям она очень походит на Laciniaria,
все авторы единодушно рассматривают как самостоятельный род.
Группа Euxina была впервые описана О. БетгероМ (Boettger, 1877a).
Он относил сюда многие виды, обитающие в причерноморских странах.
Наиболее характерным считалось глубокое залегание нижней пластинки
и присутствие 3—4 палатальных складок. До Вагнера ничего не было
известно об анатомии этих моллюсков. В отношении конхиологической
78 ВВЕДЕНИЕ
характеристики и видового состава данной группы он, по сути дела, при-
придерживался взглядов Бетгера. Что касается их внутреннего строения, то
Вагнер изучил его лишь у части видов, а именно у duboisi, quadriplicata,
pumiliformis, subaggesta, aggesta. Обнаружив большое сходство гениталий
этих моллюсков с таковыми у европейских Laciniaria, он сделал вывод,
что Euxina является подродом последних. Кроме того, он решил, что и
все остальные виды — pleuroptychia, index, dipolauchen, strumosa, somche-
tica, derasa, tschetschenica, которых Бетгер относил к Euxina, имеют такую-
же половую систему. Линдгольм (Lindholm, 1924), в основном согласился
с Вагнером, но, заметив значительную неоднородность среди этой группы
в признаках раковины, разделил этот подрод на несколько секций.
Указание Франкенбергера (Frankenberger, 1916) на то, что половая
система Clausilia derasa устроена иначе, чем у Laciniaria, Вагнер воспри-
воспринял как ошибку.
Позднее Гессе (Hesse, 1925) в короткой заметке дал описание анатомии
нескольких Clausiliidae, в том числе somchetica и tschetschenica. Автор
отнес их в род Mentissoidea Bttg., так как не обнаружил отростка семя-
семяприемника. Между тем, как показывают наши исследования, этот орган
имеется у обоих названных видов.
Подобное расхождение с нашими данными объясняется тем, что Гессе
(как это он сам пишет в упомянутой работе) имел для анатомирования
материал, полученный размачиванием мягких частей, сохранившихся
в сухой раковине. Естественно, что такой нежный орган, как отросток
семяприемника, стенки которого образованы лишь однослойным эпите-
эпителием, мог ускользнуть от глаз автора.
При изучении половой системы кавказских, а также некоторых иран-
иранских и турецких Clausiliidae, которые, с точки зрения наших предшествен-
предшественников, принадлежали к роду Laciniaria и к подроду Euxina, мы убедились,
что половая система у этих моллюсков построена по двум типам. У одних,.
а именно у тех видов, которые исследовал еще Вагнер, а также (по нашим
данным) у gustavi, pleuroptychia, index, closta, circassica, retowskii, dipo-
dipolauchen, мужские половые пути не образуют петли и вытянуты вдоль
женских, а глазной ретрактор лежит в стороне от пениса и влагалища
(т. е. половая система такая, как у европейских Laciniaria). У других,
а именно у somchetica, tschetschenica, lessonae, talyschana, persica, ponlica,
circumdata, derasa, reulauxi, половой аппарат совершенно другого типа
(рис. 109). Мужские половые пути образуют петлю, а глазной ретрактор
проходит между пенисом и влагалищем. Иными словами, мужские поло-
половые пути построены по обычному для большинства Clausiliidae плану.
Следовательно, подрод Euxina в том объеме, как его понимали наши
предшественники, является гетерогенной, искусственной категорией. Убе-
Убедившись в этом, мы были вынуждены провести переоценку некоторых
конхиологических признаков. При этом стало ясно, что если исходить из
наиболее стабильных признаков раковины, таких, как общая форма ее,
положение верхней пластинки по отношений) к спиральной, форма нижней
пластинки, глубина залегания замыкательного аппарата и некоторых
других, то группа Euxina в старом понимании и по раковине также пред-
представляет собой комплекс далеких друг от друга видов и есть основание
разделить ее на несколько самостоятельных родов. Такие роды, как
Mucronaria, Quadriplicata и Micropontica, обладают половой системой типа
Laciniaria, а остальные — Euxina, Megaleuxina — имеют половую си-
систему, более обычную для своего семейства, т. е. с нетлей.
По тем же причинам такие группы, как Vestia и Pseudalinda, которые
тоже по половой системе сходны с Laciniaria, но по раковине сильно отли-
ВОПРОСЫ СИСТЕМА ТИКИ 79
чаются от последней, мы тоже рассматриваем как самостоятельные
роды.
К родам, у которых половая система построена по типу Laciniaria,
близко стоят также Clausilia, Iphigena, Ruthenica, Fusulus и Neostyriaca.
В этих группах преобразование мужских половых путей уже началось,
так как нисходящее колено семяпровода у них уже утолщено, а восхо-
восходящее колено, пенис и его ретрактор обнаруживают явственные следы
деградации (рис. 27). Кроме того, у части этих родов ретрактор глазного
щупальца проходит еще между пенисом и влагалищем, а у остальных он
лежит уже медиально от них, т. е. как у родов с половой системой типа
Laciniaria.
Насколько нам известно, преобразование мужских половых путей
и связанных с ними органов, которое в конечном счете приводит к половой
системе типа Laciniaria, более нигде не имеет места, не только среди
Clausiliidae, но и у брюхоногих моллюсков вообще. С нашей точки зрения,
все роды, у которых наблюдается это явление, следует выделить в особое
подсемейство, за которым, согласно правилам зоологической номенкла-
номенклатуры, надо сохранить название Clausiliinae. Эта группа, хорошо отличаясь
от остальных подсемейств по признакам половой системы, менее четко
обособлена по признакам раковины. В этом отношении особенно много об-
общих черт она обнаруживает с другим подсемейством, а именно с Euxininae.
В обеих группах у подавляющего большинства видов имеется киль и
.базальный желобок. Однако существуют различия в других частях рако-
раковины. В то время как у Euxininae верхняя и спиральная пластинки распо-
располагаются на различном удалении от столбика и их концы обычно заходят
друг за друга, у Clausiliinae обе пластинки в большинстве родов распо-
располагаются на одинаковом удалении от столбика и чаще всего их концы
слиты Друг с другом. Единственным исключением является род Lacinia-
Laciniaria, у которого концы обеих пластинок заходят друг за друга, но распо-
располагаются очень тесно один к другому.
Роды Acrotoma, Armenica, Idyla, Euxina, Megaleuxina, Euxinastra
и Galeata, подобно Clausiliinae, отличаются от всех прочих подсемейств
формой, положением и гистологическим строением отростка семяприем-
семяприемника. Однако в то время как у последних происходит преобразование
мужских половых путей, это не имеет места у указанных родов, у которых
мужские половые пути сохраняют обычный для всего семейства план
строения. При всем разнообразии в строении раковин внутри этих групп
у них, как мы уже отмечали, верхняя и спиральная пластинки всегда
располагаются на разном удалении от столбика и нередко их концы за-
заходят друг за друга. В этом отношении эти роды сходны с Alopiinae и
Cochlodininae.
Указанные группы, по своей организации занимающие промежуточ-
промежуточное положепие между Clausiliinae и остальными подсемействами, мы объ-
объединяем в особое новое подсемейство Euxininae. x
Критический обзор старых систем
Первые попытки создать единую систему для всех, к тому времени
известных видов нашего семейства были сделаны уже в середине прошлого
века. Вначале по наличию или отсутствию клаузилия все виды распреде-
1 В данной работе к подсемейству Euxininae отнесены также Mentissoidea nFilosa.
Однако последние, по ряду важных особенностей внешнего и внутреннего строения,
столь сильно отличаются от остальных родов того же подсемейства (см. стр. 209 и 212),
что объединение их с ними можно рассматривать лишь как временную меру до более
точного выяснения систематического положения названных двух родов.
80 ВВЕДЕНИЕ
ляли по двум родам — Clausilia и Balea. Позднее, Гартманн (Hartmann,
1840—1844) и Адамсы (Adams H. а. А., 1858) разделили европейские виды
Clausilia по нескольким подродам, азиатские виды собрали в подрод
Phaedusa, а южноамериканские — в подрод Nenia.
Естественно, поскольку в малакологии того времени господствовал
конхиологический метод, все авторы при разработке системы данных
моллюсков брали за основу признаки раковины. Так, Пфейфер (Pfeiffer,
1868) классифицировал по наличию и отсутствию полулунной складки,
Шмидт (Schmidt A., 1857) — по положению спиральной пластинки и по
ее связям с верхней пластинкой, Вест (Vest, 1867) — по форме клаузилия.
Позднее тот же Шмидт (Schmidt A., 1868), Кюстер (Kiister, 1847—1862)
и Бетгер (Boettger О., 1877а) использовали не один, а многие признаки
замыкательного аппарата. Наиболее совершенную по тому времени си-
систему предложил Бетгер. Впервые в ней были учтены не только ныне-
живущие, но и ископаемые виды. Однако автор хотя и знал, что строение
замыкательного аппарата сильно зависит от климата, тем не .менее не
всегда это в достаточной мере учитывал. В то же время особенности таких
более стабильных образований, как нижняя и субколумеллярная пла-
пластинки и клаузилий, не всегда им принимались во внимание.
Система Бетгера, даже с внесенными в неё позднее дополнениями Пил-
сбри (Pilsbry, 1901—1909), х в значительной степени носит чисто формаль-
формальный характер. Она не дает возможности выяснить родственные связи между
отдельными группами, так как зиждется на признаках раковины.
Первые сведения о внутреннем строении Clausiliidae появились еще
в прошлом веке (Paasch, 1845; Moquin-Tandon, 1855; Schmidt A., 1855;
Lehmann, 1873; Wiegmann, 1878a, 1878b, 1893). Однако в большинстве
этих исследований указываются только разрозненные сведения о радуле
и половой системе отдельных видов, которые не могут привести к каким-
нибудь существенным выводам в области систематики всего семейства.
Лишь последняя работа известного анатома прошлого века Вигмана
(Wiegmann, 1893) в этом отношении выгодно отличается от предыдущих.
В ней автор анализирует внутреннее строение большого числа видов из
разных групп. К сожалению, он только суммирует данные, полученные от
вскрытий и не дает ни рисунков, ни описаний по каждому из изученных
им видов. В итоге он приходит к выводу, что каждая конхиологическая
группа характеризуется определенным вну/гренним строением, и, впервые
для Clausiliidae, делает попытку классифицировать их по анатомическим
признакам. Его классификация выглядит следующим образом.
1-й ряд. Проток семяприемника лишен отростка, вместо которого
в него впадает железистый придаточный орган. % Радула первого типа,
т. е. с дополнительными зубцами как на срединном, так и на боковых
зубах.
А. Глазной ретрактор лежит в стороне от пениса и влагалища. Пение
без диафрагмального ретрактора.
а. Пенис и семяпровод не образуют петли (Balea, Strigilecula, Men-
tissa, Veslia, Pseudalinda).3
б. Пенис и семяпровод образуют петлю (Clausilia, Iphigena).
1 Серия статей, посвященных Clausiliidae Японии и опубликованных в «Nautilus»
и в «Proceedings Academia Natural Sciences Philadelphia».
2 С нашей точки зрения, этот орган есть дериват обычного отростка семяприем-
семяприемника.
3 Из родов, приводимых у Вигманна, мы здесь указываем лишь те, которые упомя-
упомянуты в нашей работе и в их современной транскрипции.
ВОПРОСЫ СИСТЕМАТИКИ 81
Б. Глазной ретрактор проходит между пенисом и влагалищем. Пенис
с диафрагмальным ретрактором.
а. Мужская петля есть.
б. Мужской петли нет (Ruthenica).
2-й ряд. Семяприемник с отростком. Пенис с диафрагмальным ретрак-
ретрактором. Глазной ретрактор лежит между пенисом и влагалищем. Радула
первого или второго типа.
А. Пенис без придатков. Шейка семяприемника связана с системой
ретракторов (Euxina, Cochlodina).
Б. Задняя часть, пениса со слепым придатком. Шейка семяприемника
не связана с системой ретракторов (Alopia).
Из приведенной схемы видно, что Вигманн наибольшее значение при-
придавал форме семяприемника, наличию или отсутствию диафрагмального
ретрактора пениса и положению глазного ретрактора по отношению к пе-
пенису и влагалищу. Несмотря на то что в его время многие авторы были
склонны преувеличивать значение радулы в систематике моллюсков, он
вполне справедливо отметил большое однообразие в структуре этого
органа, которая сводится к двум планам строения, порой встречающимся
в пределах одной и той же группы. Положительным также является то,
что автор впервые обратил внимание на связь семяприемника с колумел-
лярной мускулатурой, имеющую большое значение для систематики на-
нашего семейства. Между тем последующие авторы не придавали этому
серьезного значения.
Несмотря на то что Вигманн почти вплотную подошел к созданию
•естественной системы, он не решился дать ей окончательное оформление
в виде установления подсемейств и указания родственных связей между
отдельными группами. По-видимому, он считал, что для этого изучено
«ще недостаточное количество видов.
Сходную попытку классифицировать наших моллюсков по особенно-
особенностям внутреннего строения осуществил Стинберг (Steenberg, 1914). Он
располагал значительно меньшим материалом, чем его предшественник,
так как ограничился изучением половой системы 10 видов Clausiliidae,
обитающих в Дании. Этот материал он исследовал с большой тщатель-
тщательностью и успешно проанализировал полученные результаты. В заключе-
заключении своей работы автор дает ключ для определения исследованных им
моллюсков по внутреннему строению. При этом он пришел к выводу, что
анатомическую характеристику можно дать не только родам, но и отдель-
отдельным видам. Кроме того, он дал анатомическую характеристику двум под-
подсемействам — Cochlodininae и Clausiliinae.
Наиболее серьезную попытку использовать внутреннее строение для
систематики Clausiliidae предпринял Вагнер (Wagner, 1914, 1919, 1920,
1922). Для этого он провел серию многочисленных вскрытий самых раз-
различных видов, а также подверг глубокому анализу строение замыкатель-
ного аппарата. Многие группы видов он возвел до ранга родов, которые
в свою очередь разделил на подроды и секции. Впервые было осуществлено
разделение семейства на несколько подсемейств.
Сам автор считал, что его система основана в первую очередь на при-
признаках половой системы и радулы. При этом он отмечал, что классифика-
классификация по анатомическим признакам в общем совпадает с классификацией по
раковинам и, следовательно, подкрепляет последнюю. Однако подобная
согласованность проявляется не всюду, и в некоторых случаях он
вынужден [характеризовать ту или иную группу либо только по
-анатомическим признакам (Mentissoideinae), либо по раковине (Еи^
xina).
82 ВВЕДЕНИЕ
Главной слабостью системы Вагнера явилось то, что он не пытался
найти морфо-функциональную зависимость между различными органами.
Поэтому он не заметил, какое важное значение имеет для классификации
наших моллюсков влияние замыкательного аппарата на внутренние
органы, и не уделил внимания связям половой системы с колумеллярной
мускулатурой.
Вагнер использовал обширный материал по различным группам и»
западной и южной Европы. Значительно беднее были его сведения о ви-
видах, обитающих на Кавказе и в Передней Азии. Из многочисленных видов,
живущих в тропической Азии, автор изучил внутреннее строение лишь
единичных и совершенно не имел материала по тропической Америке.
В связи с этим его выводы в отношении европейских групп в основном:
правильны и легли в основу современной систематики семейства. В отно-
отношении кавказских и переднеазиатских видов в ряде случаев он сделал
серьезные ошибки в оценке их систематического положения. Что касается
тропических групп, то здесь его выводы по большей части ошибочны.
В итоге своих исследований Вагнер разделил Clausiliidae на 6 под-
подсемейств: х Alopiinae, «Clausiliinae» (=Cochlodininae), Phaedusinae, «5a-
leinae» (=Clausiliinae), «Priobaleinae» (z=Mentissoideinae), «Metabaleinaefr
(¦=Fusulinae).
Что касается первых четырех групп, то выделение их в ранг подсе-
подсемейств не вызывало возражений у последующих исследователей. В систе-
систематической части данной работы мы даем им краткие характеристики^
которые лишь несколько дополняют первоначальные описания Вагнера.
По поводу остальных двух подсемейств наши взгляды сильно расходятся
с Вагнером.
В подсемейство Mentissoideinae («Priobaleinae») автор включал лишь
один вид — кавказскую Mentissoidea litotes. Он справедливо считал, что-
самой характерной чертой внутреннего строения этого моллюска является
отсутствие отростка семяприемника, т. е. органа, имеющегося у пода-
подавляющего большинства видов семейства. По раковине он сравнительно
мало отличается от некоторых других родов, в особенности от Laciniaria
ш Euxina. Тем не менее, автор счел возможным выделить его в особое под-
подсемейство, которое он считал наиболее примитивным, близким к предкам
Clausiliidae.
В этом отношении Вагнер явно непоследователен, так как им самим>
аналогичное явление было обнаружено в пределах другого подсемейства —
Phaedusinae, у моллюска «Clausilia-» rudis Bav. et Dautz. (Индокитай)..
Половая система этого моллюска также лишена отростка семяприемника.
Однако автор оставил его в пределах Phaedusinae, ограничившись лишь,
тем, что выделил этот вид в особый род Synprosphyma Wagner. В данной:
работе мы указываем на третий случай подобного же рода у моллюска
Filosa filosa из Аджарии (стр. 212). По-видимому, по неясной пока причине,,
у всех трех видов утрата отростка произошла независимо, так как по всем,
остальным признакам эти моллюски сильно отличаются друг от друга.
Учитывая сказанное, нам представляется мало обоснованным выде-
выделение Mentissoidea litotes и Filosa filosa в особое подсемейство Mentissoi-
Mentissoideinae, исходя лишь из одного признака, к тому же отрицательного по-
порядка (отсутствие отростка семяприемника). Петлеобразное строение муж-
мужских половых путей и присутствие киля на раковинах обоих моллюсков:
1 Давая название подсемействам, родам и подродам, Вагнер часто отступал от
правил зоологической номенклатуры. Последующие авторы (Lindholm, 1924; Ehr-
Ehrmann, 1926; Thiele, 1931) исправили эти ошибки. В тех случаях, когда названия Ваг-
Вагнера не соответствуют общепринятым, мы берем их в кавычки.
ВОПРОСЫ СИСТЕМАТИКИ 83
как будто бы говорит за их близость к посдсемейству Euxininae. В то же
время они отличаются от последних рядом существенных особенностей
строения. Поэтому мы относим их к Euxininae лишь условно, до оконча-
окончательного выяснения их действительного положения в семействе. х
Пожалуй, еще более искусственным выглядит другое подсемейство
Вагнера — Fusulinae («Metabaleinae»). Сюда он поместил виды, значи-
значительная часть которых обнаруживает различные комбинации признаков
Alopiinae и Clausiliinae. Так, подобно первым, у части родов указанного
подсемейства (Armenica, Acrotoma) имеется придаток пениса, рудимен-
рудиментарный бич и однозубцовые срединный и боковые зубы. Подобно вторым,
у других родов (Fusulus, Ruthenica и др.) происходит укорачивание петли,
образованной пенисом и восходящим коленом семяпровода, и утолщение
нисходящего колена. Но, в отличие от Clausiliinae, в понимании Вагнера,
ретрактор глазного щупальца проходит между пенисом и влагалищем.
Когда Вагнер сравнивал Fusulinae с Alopiinae, он не учел, что одним
из наиболее важных отличий у первых является форма и гистологическое
строение отростка семяприемника. Если у Fusulinae этот орган всегда
впадает в семяприемник близ его нижнего конца и образован только одно-
однослойным эпителием, то у вторых он впадает значительно выше и имеет
более сложное гистологическое строение. Что касается формы зубов ра-
радулы и присутствия придатка на пенисе, то, как мы уже указывали, эти
особенности связаны с обитанием на скалах и не могут служить надеж-
надежным признаком для характеристики столь крупных таксонов, как подсе-
подсемейства. В итоге мы считаем, что общий план строения половой системы
и особенности раковин таких родов, как Armenica, Acrotoma и Idyla,
которых Вагнер включал в свое подсемейство «Metabaleinae» (=Fusuli-
пае), указывает на их принадлежность к подсемейству Euxininae.
Что касается родов Fusulus, Ruthenica, Neostyriaca и других, которые,
с точки зрения того же автора, принадлежали к тому же подсемейству, то
они как по строению гениталий, так и по раковине в действительности
относятся к Clausiliinae (в нашем понимании). Тот факт, что мужские
половые пути этих групп еще сохранили петлеобразную форму, а ретрак-
ретрактор глазного щупальца еще проходит между пенисом и влагалищем, от-
отнюдь не свидетельствует против их принадлежности к указанному под-
подсемейству, так как наиболее характерной особенностью Clausiliinae яв-
•ляется сокращение мужской петли, редукция пениса и вытягивание
мужских половых путей вдоль женских. Этот процесс уже отчетливо
намечен в указанных группах, которые можно рассматривать как наи-
наиболее примитивных членов своего подсемейства, связывающих его с Euxi-
Euxininae. '
В своей известной монографии, посвященной систематике моллюсков,
Тиле (Thiele, 1931), не вдаваясь в объяснения, отнес всех Fusulinae и
Mentissoideinae в подсемейство Clausiliinae, к которым он причисляет
также те группы, которые мы выделяем в особое подсемейство Euxininae.
Кроме того, он слил Alopiinae с Cochlodininae. В итоге автор делит все
семейство на следующие подсемейства: Phaedusinae, Neniinae, Clausiliinae
и Cochlodininae.
1 Как известно, XIV Международный зоологический конгресс (Копенгаген, 1953)
распространил действие закона приоритета и на семейственную группу таксонов.
Поэтому название Mentissoideinae, как более старое, как будто бы должно сохраниться
и за всей группой, названной нами Euxininae subfam. nov. Однако, поскольку род
Mentissoidea отнесен сюда лишь условно и временно И, в общем, не типичен для данной
группы, мы дали ей новое название.
84 ВВЕДЕНИЕ
Наши обоснования целесообразности выделения Euxininae приведены
уже выше. Мы также не можем согласиться со слиянием Alopiinae с Cochlo-
Cochlodininae и считаем их самостоятельными группами. Для первых харак-
характерно присутствие непрозрачного внешнего слоя раковины и отсутствие
ретрактора семяприемника, для вторых — отсутствие непрозрачного слоя
раковины и присутствие ретрактора семяприемника. Кроме того, у боль-
большинства Alopiinae имеется придаток пениса, который отсутствует у всех
Cochlodininae.
Помимо уже перечисленных подсемейств, некоторые авторы (Boett-
ger С, 1926; Ehrmann, 1926; Wenz u. Zilch, 1959) предлагают еще
два — Garnieriinae и Laminiferinae.
К первой группе относят несколько видов, живущих в юго-восточной
Азии, т. е. в области пышного расцвета подсемейства Phaedusinae. Однако
упомянутые моллюски по раковине (внутреннее строение их не изучено)
резко отличны от Phaedusinae и очень походят на южноамериканских
Neniinae. Это сходство дало основание Тиле отнести эти виды к последнему
подсемейству. Сторонники выделения их в самостоятельное подсемейство
по существу дела аппелируют к тому, что эти моллюски географически
изолированы от Neniinae. С нашей точки зрения, этот довод не выдержи-
выдерживает критики и в данном случае мы разделяем взгляды Тиле.
Что касается до Laminiferinae, то эта группа, очень многочисленная
в третичном периоде Европы, в настоящее время представлена лишь двумя
видами, живущими в Пиренеях. По раковине она очень сходна с Neniinae,
куда многие старые авторы ее и причисляли. Однако половая система
одного из современных видов — Laminifera pauli (Mabille) (Germain,
1930), сходна с таковой Clausiliinae и Euxininae, в особенности с послед-
последним подсемейством.
В настоящее время мы затрудняемся сказать, к какому из двух назван-
названных подсемейств принадлежит род Laminifera. Однако, с нашей точки
зрения, нет достаточно основания для выделения его в особое подсемей-
подсемейство.
В итоге мы ечитаем, что семейство Clausiliidae слагается из следующих
подсемейств: Phaedusinae, Neniinae, Alopiinae, Cochlodininae, Euxininae,
Clausiliinae.
К этим группам следует еще добавить вымершее подсемейство Tri-
ptychiinae, многочисленные виды которого обитали в неогене Европы. Ра-
Раковины этой группы характерны отсутствием клаузилия, главной и полу-
полулунной складок.
Положение Clausiliidae внутри
отряда стебельчатоглазых
моллюсков (Stylommatophora)
Подкласс легочных моллюсков слагается, по взглядам одних авторов,
из двух отрядов — Basommatophora и Stylommatophora, по взглядам дру-
других— из трех, уже названных и Soleolifera. В последний отряд входят
три семейства, которые сторонники первой точки зрения относят к Sty-
Stylommatophora, правда, выделяя их в особый подотряд. Среди_ указанных
отрядов наиболее крупным являются Stylommatophora, так как в состав
этой группы входят 43 семейства D6, если причислить сюда и 3 семейства
Soleolifera), т. е. более двух третей всех семейств, слагающих подкласс
легочных моллюсков.
Вопрос о систематическом положении Clausiliidae нельзя решать в от-
отрыве от общей классификации Stylommatophora. В настоящее время в мала-
ВОПРОСЫ СИСТЕМАТИКИ
85
кологии по этому поводу придерживаются двух систем. Одна из них пред-
предложена Тиле (Thiele, 1931). Он делит всех Stylommatophora, как, впрочем,
и других моллюсков, на несколько надсемейств, которые он называет
«stirpes». Основным критерием для объединения отдельных семейств в над-
семейства чаще всего является сходство их раковин.
Несмотря на то' что система Тиле получила довольно широкое при-
признание, 1 в особенности среди конхиологов, многие авторы (Pilsbry, a.
Рис. 35. Мантийный комплекс органов Stylommatophora. (По Солему).
А — Orthurethra; Б — Heterurethra; В — Sigmurethra.
1 — задняя кишка; 2 — отверстие мочеточника; з — мочеточник; 4 — легочная
вена; 5 — почка; в — сердце и перикардий.
Gooke, 1934; Baker, 1938, 1941; Pilsbry, 1948; Solem, 1959), углубленно
занимавшиеся анатомией и систематикой моллюсков, убедительно пока-
показали, что у Тиле значительная часть надсемейств Stylommatophora является
гетерогенными категориями.
С нашей точки зрения, наиболее удачную классификацию этого отряда
предложил Пилсбри (Pilsbry, 1900). Она основана на структуре мантий-
мантийного комплекса органов, главным образом, на форме и положении почки
и мочеточника (рис. 35). Автор в пределах Stylommatophora, различает
три подотряда.
1 В частности, она полностью принята А. В. Ивановым для «Руководства по зоо-
зоологии» (т. 2, 1940).
86 ВВЕДЕНИЕ
1) Orthurethra — от почки, лежащей вдоль мантийной полости и более
или менее параллельно задней кишке, отходит вперед, в направлении
к краю мантии различной длины первичный мочеточник (иногда он не
замкнут и имеет вид борозды), который открывается в полость легкого;
2) Heterurethra — почка лежит поперек широкой мантийной полости,
перпендикулярно к задней кишке; мочеточник вначале тянется вдоль
переднего края почки, а затем вдоль задней кишки, открываясь наружу
рядом с анусом;
3) Sigmurethra — почка лежит вдоль мантийной полости; от ее перед-
переднего конца тянется назад первичный мочеточник, который у заднего конца
почки отгибается вперед и проходит вдоль задней кишки (вторичный
мочеточник), открываясь наружу рядом с нею или в общую клоаку.
Позднее некоторые последователи Пилсбри (Baker, 1955; Solem, 1959),
исходя из особенностей строения, выделительной системы нескольких
аберрантных групп, предложили выделить их в еще два новых подотряда—
Tracheopulmonata и Mesurethra.
Противники классификации Пилсбри (Simroth u. Hoffmann, 1908—1928;
Thiele, 1935) отмечают, что положение мочеточника иногда очень варьи-
варьирует даже в пределах одного семейства. Однако указанные ими примеры
чаще всего относятся к тем группам, у которых раковина полностью или
частично подверглась редукции. Естественно, что утеря раковины при-
привела к изменению всего плана строения внутренностного мешка и мантий-
мантийного комплекса органов. Поэтому отклонения в строении выделительной
системы, на которые указывали противники классификация Пилсбри,
являются вторичными. 1 А так как подавляющее большинство Stylomma-
tophora сохранило хорошо развитую раковину, то классификации Пилсбри
в настоящее время является наиболее обоснованной. К таким выводам
пришли все те малакологи, которые, анализируя систематическое положе-
положение каких-нибудь групп стебельчатоглазых, учитывали не только особен-
особенности раковины, но в первую очередь также внутреннее строение моллю-
моллюсков (Watson, 1915, 1920; Steenberg, 1925; BoettgerC, 1955; Solem, 1959).
Согласно классификации Пилсбри, полностью разделяемой нами,
Clausiliidae входят в состав подотряда Sigmurethra, куда, кроме них,
относятся еще 31 семейство.
В настоящее время еще трудно окончательно решить, какое из этих
семейств является ближайшим родственником наших моллюсков. С вытя-
вытянутой раковиной имеется довольно много групп, большинство которых
распределяются по двум надсемействам — Bulimulacea и Achatinacea. Как
тут, так и там есть семейства, подобно Clausiliidae, имеющие просто уст-
устроенный половой аппарат, т. е. без придаточных органов на женских
половых путях. Семяприемник, как правило, имеет длинный проток —
признак, обычный для моллюсков с высокой раковиной. Нередко присут-
присутствует также отросток семяприемника.
В пределах надсемейства Achatinacea имеется небольшое семейство-
Megaspiridae, которое мы, вслед за другими авторами (Pilsbry, 1904;
Boettger С, 1926), склонны рассматривать как наиболее родственное
Clausiliidae.
Входящие в эту группу виды отличаются высокой раковиной (рис. 36),
сложенной из большого числа оборотов (до 20). В полости последнего из
них или ряда нижних оборотов располагается несколько спиральных
1 Характерно, что эти отклонения конвергентно повторяются у групп, в систе-
систематическом отношении далеко отстоящих друг от друга. Это еще раз подтверждает,
что эти изменения связаны с утерей раковины и с приобретением моллюсками слизне-
образного облика.
ВОПРОСЫ СИСТЕМАТИКИ
87
С
складок или пластинок, тянущихся вниз, как вдоль столбика, так и по
париетальной, а у некоторых и по палатальной стенке. Легко предполо-
предположить, что на основе этой скульптуры возник замыкательный аппарат
наших моллюсков, особенно если учесть, что подобные же образования
имеются у молодых раковин Serrulina s. str. (рис. 37) и были у молодых форм
ныне вымершего подсемейства Triptychiinae. Но, к сожалению, кроме ра-
радулы, мы до сих пор не располагаем другими сведениями о внутреннем
строении Megaspiridae. Поэтому окончательное решение вопроса о бли-
близости Clausiliidae к этому семейству
следует оставить до выяснения всех
особенностей его организации.
Одним из моментов, как будто бы
товорящих в пользу сближения обоих
семейств, является также их древность,
так как их представители найдены в
отложениях верхнего мела Европы. В
настоящее время Megaspiridae обитают
в областях, где"Т72йвшшГое_ нет (Бра-
(Бразилия, Новая Гвинея, Австралия).
""Другая точка зрения на~система-
тическое положение Clausiliidae отстаи-
отстаивалась Вагнером (Wagner, 1921) и
Тиле (Thiele, 1935). Вагнер считал, что
к нашему семейству наиболее близки
Pupillidae и Enidae, с которыми их сле-
следует объединять в одно надсемейство
РирЩасеа. К этому выводу он пришел,
обнаружив некоторое сходство в распо-
расположении отдельных частей замыкатель-
лого аппарата Clausiliidae и зубов,
располагающихся в устье раковин на-
названных двух семейств. В самом деле,
верхней пластинке наших моллюсков
как будто бы соответствует париетальный зуб, субколумеллярной плат
стинке — колумеллярный зуб, палатальным складкам замыкательного
аппарата — зубы на внешней стенке многих Pupillidae и Enidae.
Однако Вагнер не учел того факта, что внутренняя скульптура Clau-
Clausiliidae и названных двух семейств имеет различное происхождение.
У первых она возникает на дне бороздок мантии, образовавшихся в ре-
результате сужения последнего оборота раковины. Что касается Pupillidae
и Enidae, то здесь зубы образуются за счет секреторной деятельности
многочисленных желез утолщенного края мантии.
Некоторое сходство в расположении внутренней скульптуры в устье,
ла которое обратил внимание Вагнер, характерно не только для трех
упомянутых семейств, так как аналогичную картину можно наблюдать
почти у всех тех семейств Stylommatophora, устье раковин которых об-
обладает внутренней скульптурой. А таких семейств очень много. Это сход-
сходство является особенностью всего отряда и связано с совпадением функ-
функций, которые выполняют эти образования (стр. 23).
В пользу родственной близости Clausiliidae с Pupillidae и Enidae Ваг-
Вагнер указывал также на некоторое сходство в строении радулы и половой
системы этих моллюсков.
Что касается радулы, то, как мы уже указывали (стр. 39), оба типа
строения этого органа, свойственные Clausiliidae, встречаются также и
Рис. 36. Megaspira elatior Spix. (По
Пилсбри).
А — общий вид раковины; Б
няя скульптура.
• внутрен-
88
ВВЕДЕНИЕ
у видов со сходным питанием из других семейств Stylommatophora. В част-
частности, они имеются у одного из наиболее высоко организованных се-
семейств — Helicidae, хотя эта группа, по общему признанию, стоит очень
далеко от наших моллюсков.
Отмечая сходство половой системы Clausiliidae с таковой у Pupillidae
и Enidae, Вагнер обращал внимание на общую вытянутость всех органов,,
в особенности семяприемника, на отсутствие придаточных органов на
женских половых путях и на наличие отростка семя-
семяприемника (последний отсутствует у некоторых Pupil-
Pupillidae).
Известно (Матёкин, 1959), что длинные половые
пути и семяприемник — явление, свойственное видам
с высокой раковиной и длинным внутренностным
мешком. Поэтому подобную картину можно видеть у
многих моллюсков из разных семейств. Что касается
отростка семяприемника, то этот орган может быть, а
может и отсутствовать у самых различных семейств
Stylommatophora. Как известно, он отсутствует даже
у некоторых Clausiliidae, правда у очень немногих
видов.
Как у многих других групп с гермафродитным
половым аппаратом, половая система Stylommatophora
отличается большим разнообразием строения. Многие
придаточные органы, в том числе и отросток семяприем-
семяприемника, появляются в разных семействах совершенно1
независимо друг от друга. Поэтому делать какие-ни-
какие-нибудь выводы цо классификации этого отряда, исходя;
из особенностей этих органов, очень трудно, а порой
и невозможно.
Взгляды Тиле на систематическое положение Clausi-
Clausiliidae, по сути дела, совпадают с мнением Вагнера. Он
сближает наших моллюсков с одним из подсемейств
Enidae — Chondrulinae. Основание этому автор видит
в том, что у последних наряду с наличием зубов в устье
раковины нет своеобразного придатка пениса, свойст-
свойственного остальным Enidae и отсутствующего у всех
. Clausiliidae.
Наши возражения по поводу сближения отдельных семейств Stylom-
Stylommatophora, исходя только из наличия скульптурных образований в устье
раковины и некоторого сходства в половой системе, мы уже привели.
Добавим только следующее.
Chondrulinae является самой молодой группой среди Enidae. Она
возникла в плиоцене на территории Средиземноморья и Передней Азии
в результате нараставшей ксерофилизации климата и целиком состоит
из засухоустойчивых видов.
Clausiliidae значительно древнее. Их корни уходят в нижвщй мел.
В биологическом отношении эта группа состоит из значительно более
влаголюбивых видов, чем Chondrulinae.
В настоящее время доказано, что в результате длительных изменений
климата в сторону нарастания засушливости из относительно влаголюби-
влаголюбивых форм не раз воникали виды, устойчивые к высыханию. Таких при-
примеров можно привести довольно много как среди наземных моллюсков
(Helicellinae, Pupillidae, многие Enidae и др.), так и других групп жи-
животных. Однако очень сомнительно, чтобы когда-либо имел место обрат-
Рис. 37. Serrulina
sieversi (Pfr.).
(Ориг.). Раковина
молодого экзем-
экземпляра. Видна юве-
нильная внутрен-
внутренняя скульптура.
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ 89-
ный процесс, и достоверные примеры этому среди наземных моллюсков-
неизвестны.
В заключение критики взглядов Вагнера и Тиле отметим, что как
Pupillidae, так и Enidae, с которыми оба автора роднили наших моллю-
моллюсков, согласно классификации, предложенной Пилсбри, относятся к под-
подотряду Orthurethra, т. е. к самой древней группе среди стебельчатоглазых,
выделительная система которых построена по тому же плану, что и у Ва-
sommatophora. Поэтому всякое сближение указанных двух семейств
с Clausilliidae, которые, как известно, принадлежат к высшему подотряду
Sigmurethra, является искусственным.
ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ CLAUSILHDAE
Раньше, чем перейти к обсуждению путей, по которым шла филогения^
наших моллюсков, приведшая к формированию современных подсемейств,
следует остановиться на условиях, которые способствовали возникнове-
возникновению всего семейства.
В настоящее время большинство видов данной группы, в том числе
почти все виды наиболее примитивных подсемейств Phaedusinae и Neniinae,
обитают во влажных тропиках. В тропиках же обитали наиболее древни»
из уже вымерших видов и групп, так как находки их приурочены к мелу
и палеогену Европы, т. е. к тем временам, когда архипелаг на месте этой
части суши был покрыт тропическими лесами. Отсюда напрашивается
естественный вывод, что возникновение семейства произошло в условиях
очень влажного тропическото климата.
—^^Б^яъТпйнСтвсг-вядсшг Ctausiliidae — обитатели деревьев или гниющих
на земле xxBoaoJt. Сравнительно меньшее число живет только на скалах
или в лиственной подстилке. 11о^томх^?хесшшн1ш_ожидать, что основные
черты организации наших моллюсков сложились в результате жизни над
ПОЧД?Ш~
Известно, что у наземныхлегочных моллюсков подобный образ жизни
способствует вытягиванию раковины в длину и увеличению числа оборотов
(Achatinidae, Bulimulidae, Enidae, Megaspiridae и др.). Однако при этом
моллюски оказываются под большим воздействием воздушных течений,
чем при жизни на почве, что в свою очередь усиливает испарение и угрозу
для организма в результате чрезмерной потере влаги.
Как защита от высыхания произошло сужение последнего оборота;
раковины ~и за счет имевшейся на столбике внутренней скульптуры,
у предков наших моллюсков возник клаузилий, в нужный момент наглухо
закрывавший вход в раковину. Остальные части замыкательного аппарата
образовались либо как дополнение к клаузилию (полулунная и палаталь-
палатальные складки), либо для защиты дыхательного канала от давления осталь-
остальных органов (спиральная и верхняя пластинки, главная складка). Такие
образования, как нижняя и субколумеллярная пластинки, вероятно, уже-
имелись у предков наших моллюсков (см. Megaspiridae, стр. 86), т. е.
до образования клаузилия, и служили им для управления раковиной.
Следовательно, с нашей точки зрения, клаузилием обладали уже
самые первые, наиболее примитивные Clausiliidae. В этом вопросе мы ко-
коренным образом расходимся со старыми авторами (Boettger О., 1877а,
и др.), считавших представителей нашего семейства, лишенных клаузилия
(Balea, Reinia), наиболее примитивными, близкими к исходной форме,
с которой началась эволюция всей группы.
Накопленный за последние годы обильный материал о таких видах и
родах указывает на то, что отсутствие клаузилия представляет собой вто-
¦90 ВВЕДЕНИЕ
ричное явление, связанное с общей редукцией замыкательного аппарата.
У одних это произошло под влиянием очень влажного климата, у других —
в связи с обитанием в высокогорных зонах с коротким вегетационным
периодом (см. стр. 47). О том, что клаузилий имелся у самых древних из
представителей нашего семейства, свидетельствует палеонтология. Правда,
это образование отсутствовало у вымершего подсемейства Triptychiinae,
жившего преимущественно в неогене Европы и отличавшегося общим
регрессом замыкательного аппарата. Однако тот факт, что это подсемей-
подсемейство появилось уже тогда, когда существовало много групп с хорошо
развитым замыкательным аппаратом, говорит за то, что отсутствие клаузи-
лия у Triptychiinae также вторичное явление. Характерно, что более
древние представители этой группы имели менее редуцированный замы-
кательный аппарат, чем более молодые. Отсюда напрашивается вывод,
Ч1о утеря клаузилия у Triptychiinae связана с общим ослаблением замы-
замыкательного аппарата.
Возникновение клаузилия и замыкательного аппарата вместе с общим
«ужением последнего оборота раковины не прошло бесследно для всей
организации Clausiliidae. Появление этих образований вызвало нехватку
свободного пространства для органов, лежащих в шейке внутренностного
мешка. Именно этим мы объясняем большинство изменений в строении
полового аппарата, которые наблюдаются в пределах семейства.
Начались эти перемены с положения семяприемника. У двух подсе-
подсемейств — Phaedusinae и Neniinae — он открывается в женские половые
пути очень низко, почти у самой половой клоаки (рис. 24, А). Однако
уже среди Phaedusinae, * у некоторых видов семяприемник открывается
во влагалище на значительном удалении от половой клоаки. В этом отно-
отношении особенно показательна половая система Caspiophaedusa perlucens
(рис. 58). Семяприемник этого моллюска находится далеко от половой
клоаки и обладает ретрактором, который является ветвью левого перед-
переднего ретрактора тела.
Подобные отклонения встречаются лишь у очень немногих видов
Phaedusinae, у подавляющего большинства которых семяприемник впа-
впадает вблизи половой клоаки.
Раковина этого подсемейства также обнаруживает наиболее примитив-
примитивные черты. Устье лишено базального желобка, а следовательно, отсут-
отсутствует и киль на последнем обороте. Верхняя пластинка чаще всего слита
«о спиральной воедино (рис. 38, А), либо если они разделены, то во всех
случаях они лежат на одном расстоянии от столбика. Если учесть, что
у большинства других семейств Sigmurethra, как правило, соответствую-
соответствующие скульптурные образования, находящиеся на париетальной стенке
устья, представляют собой единую пластинку, более или менее глубоко
уходящую внутрь раковины, то можно считать, что положение спираль-
вой и верхней пластинок у Phaedusinae (а также и у Neniinae) является
изначальным. Следовательно, последующие изменения этой картины,
которые наблюдаются в других подсемействах Clausiliidae, представляют
собой вторичное явление.
В результате мы приходим к выводу, что Phaedusinae и Neniinae наи-
наиболее примитивные подсемейства среди остальных групп того же ранга.
Однако Neniinae более узко специализированная группа, чем Phaedusinae,
что позволяет нам считать последних родоначальниками всех остальных
1 Аналогичное, возможно, встречается и у Neniinae, но мы мало знаем о внутрен-
внутреннем строении данной группы.
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
91
подсемейств. К тому же, именно Phaedusinae раньше всех других групп
^Clausiliidae появляются в палеонтологической летописи (табл. 2).
Дальнейшая эволюция семейства постепенно привела к более выгод-
выгодному положению некоторых органов половой системы по отношению
к замыкательному аппарату. Это выразилось в первую очередь в переме-
перемещении места впадения семяприемника кверху вдоль влагалищаТ ПбдоТ)-
нукГкартину мы видим у всех европейских подсемейств (рис. 24, Б—Е).
Среди них наиболее примитивные от-
отношения наблюдаются у Alopiinae.
Семяприемник уже переместился
кверху, однако ретрактора он еще
не имеет. Правда, в пределах этой
>группы у одного рода Papillifera
Hartm. такой ретрактор есть. В связи
с этим встает вопрос: является ли
отсутствие ретрактора у большин-
большинства видов Alopiinae первичным, т. е.
признаком примитивности данной
труппы, или он утерян вторично у всех
родов, за исключением названного?
Учитывая очень большое сходство
Alopiinae с Cochlodininae, у которых
семяприемник всегда имеет ретрак-
ретрактор, а также то, что Papillifera ни-
никак нельзя рассматривать как наи-
наиболее примитивный род в пределах
•своего подсемейства, мы склонны счи-
считать отсутствие ретрактора семяпри-
семяприемника у Alopiinae вторичным явле-
явлением. По-видимому, этот орган был
у предковых форм данной группы,
но, по неясным пока причинам, по-
позднее подвергся редукции.
В дальнейшем эволюция Alopiinae шла по линии приспособления
к жизни на скалах в условиях безлесной местности, в связи с чем у этих
моллюсков получил сильное развитие особый непрозрачный слой рако-
раковины, играющий роль экрана от солнечных лучей. Кроме того, у них
появился придаток пениса — орган, характерный только для скаложи-
лов, хотя адаптивное значение его нам неясно.
Cochlodininae во многом, особенно в отношении раковины, очень похо-
походят на Alopiinae. Однако отсутствие экранизирующего слоя раковины
и придатка пениса, а также полулунной складки, которая обычно имеется
у Alopiinae, позволяет нам рассматривать Cochlodininae как самостоя-
самостоятельное подсемейство. Эта группа обладает меньшими чертами специали-
специализации, чем Alopiinae, благодаря чему она населяет как горные, так и
равнинные области.
Важнейшей особенностью Cochlodininae, отличающей их как от боль-
большинства Alopiinae, так и от Phaedusinae и Neniinae, является присутствие
у них ретрактора семяприемника. Этот орган также характерен для
Euxininae и Clausiliinae. Роль этого мускула очень велика.
Известно, что у всех Clausiliidae коп^япия^коне5н^р1^сде^се_ДЕИводит
к обменусперматофорами между особями. Перед этим, под давлением
полостнои~жидйости кровинбстого~синуса, происходит выворачивание
наружу пениса и половой клоаки. Затем каждый пенис вводит свой спер-
Рис. 38. Изменения соотношений верх-
верхней и спиральной пластинок в процессе
эволюции Clausiliidae. (Ориг.).
А —• Phaedusinae и Neniinae; Б — Alopiinae,
Cochlodininae, Euxininae и Clausitiinae (Laci-
niaria); В — Clausiliinae (Vestia, Iphigena);
Г —• Clausitiinae (Mucronaria и др.).
92 ВВЕДЕНИЕ
матофор в семяприемник партнера. Обратный процесс, т. е, втягивание
вывернутых органов после копуляции, производится различными муску-
мускулами. Пенис втягивает внутрь тела его ретрактор, половую клоаку —
ветви переднего ретрактора тела. Если у Phaedusinae и Neniinae, у которых
семяприемник находится почти у самой половой клоаки, практически
выворачивания вланалища не происходит, то перемещение семяприем-
семяприемника кверху влечет за собой при копуляции выворачивание нижней части
этого органа, а также общее перемещение женских половых путей в сто-
сторону полового отверстия. Этим мы объясняем появление ретрактора семя-
семяприемника, который служит для возврата женских половых путей в их
исходное положение.
Если у Alopiinae и Cochlodininae необходимость поставить мягкие части
тела в более выгодное положение по отношению к самому узкому месту
последнего оборота раковины привела только к перемещению семяприем-
семяприемника и появлению у него ретрактора, то у Euxininae и Clausiliinae, кроме
того, возникают другие образования, имеющие аналогичное значение.
К ним относится базальный желобок устья и связанный с ним киль. Ба-
зальный желобок облегчает скольжение мягких частей тела через устье,,
а киль служит для придания прочности этой части раковины.
Но наряду с возникновением базального желобка у предков обоих
подсемейств совсем по другим причинам шли изменения отростка семя-
семяприемника. До сих пор этот орган по форме и гистологическому строению
мало отличался от основной части семяприемника и выполнял с ним сход-
сходные функции — принятие и хранение сперматофора и растворение в нуж-
нужный момент его кожистых стенок.
У общих предков Euxininae и Clausiliinae начался процесс специализа-
специализации обеих частей семяприемника. Принятие и хранение сперматофора
осталось только за основной частью семяприемника, в то время как отро-
отросток превратился в железу, служащую для выделения лизирующего
секрета. В связи с этим стенки его утеряли мускулатуру и соединитель-
соединительнотканный слой, он стал очень узким и извитым и сместился к основанию
сем яприемника.
Euxininae, по сравнению с Clausiliinae, стоят ближе к той исходной:
форме, которая дала как это подсемейство, так и Alopiinae и Cochlodininae.
Это явствует из строения мужских половых путей, которые еще сохра-
сохранили петлеобразную форму, свойственную большинству Clausiliidae
и остальным Stylommatophora (рис. 24, Г). К тому же выводу можно
прийти, если учесть, что у Euxininae, подобно Alopiinae и Сochloidininae,
очень часто имеется бич, а верхняя и спиральная пластинки распола-
располагаются в устье так же, как у двух последних групп (рис. 38, В).
Иную картину мы видим у Clausiliinae. В этой группе под действием
отбора необходимость в преодолении отрицательного влияния замыка-
тельного аппарата на внутренние органы привела не только к переме-
перемещению семяприемника, но и к постепенному исчезновению петли, образо-
образованной пенисом и восходящим коленом семяпровода, к утере ретрактора
пениса и в конечном счете — к вытягиванию мужских половых путей
вдоль женских и к установлению связи между ними и ретрактором семя-
семяприемника (рис. 24, Д, Е). Тем самым работа по втягиванию внутрь тела
всех органов, принимавших участие в копуляции, полностью передается
наиболее мощной мускулатуре — колумеллярной, обладающей самой
прочной опорой — столбиком раковины.
Сокращение и в конечном счете полное исчезновение петли вместе
с параллельным утолщением нисходящего колена семяпровода шло по-
постепенно. Так, у родов Fusulus mRuthenica петля уже сократилась, однако
НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
93
ле очень сильно, благодаря чему между нею и влагалищем все еще распо-
располагается ретрактор глазного щупальца. У родов Clausilia, Iphigena и
некоторых других петля стала еще короче, так что глазной ретрактор
уже не может помещаться между ней и влагалищем, благодаря чему он
ложится в стороне от половых путей. Наконец, у родов Laciniaria, Vestia
и других петля и ретрактор пениса совершенно исчезли и мужские поло-
половые пути вытянуты вдоль женских. Параллельно этому шло утолщение
лисходящего колена семяпровода, установление связи между ним и рет-
Шременлщ
эпох®
Неоген
Палеоген
Поздний
мел
•
.- Рд.нний
мел
Ча
С
г
til I
(з 1 •— V
* \ \
«А
а»
?::
.* °>
¦*- 5»
¦= ,•¦
: •
/* /
А /'"
Л f
\ /
1 /*
i у
V
Рис. 39. Схема филогенетических отношений у Clausiliidae во вре-
времени. Пунктиром показаны связи, не документируемые палеонто-
палеонтологически. (Ориг.).
рактором семяприемника и дифференциация нижнего конца семяпровода
во вторичный пенис.
Кроме изменений в половой системе, у Clausiliinae происходит другой
процесс — -вторичное слияние верхней и спиральной пластинок (рис. 38,
В—Г). Наиболее примитивное положение наблюдается в роде Laciniaria*
Обе пластинки еще располагаются на разном удалении от столбика и
слегка заходят друг за,друга. У. Vestia и Iphigena обе пластинки уже
слились,1 но так как они продолжают еще находиться на разном удалении
от столбика, то единая пластинка дает небольшой коленообразный изгиб.
Наконец, у Clausilia, Mucronaria, Quadriplicata и других родов обе пла-
пластинки слиты и образуют единую, плавно спирально изогнутую пластинку.
1 У молодых экземпляров они нередко еще разделены.
94 ВВЕДЕНИЕ
Таким образом, эволюция наших моллюсков от самых примитивных,
групп P,haedusinae и Neniinae к остальным подсемействам шла по следую-
следующим направлениям:
1) перемещение семяприемника вдоль влагалища кверху;
2) дифференциация отростка семяприемника в особую железу;
3) вытягивание мужских половых путей вдоль женских;
4) передача колумеллярной мускулатуре работы по втягиванию внутрь
тела половых органов, участвовавших в копуляции;
5) изменение соотношений между верхней и спиральной пластинками
и образование базального желобка.
В заключение обзора путей, по которым шла эволюция наших моллю-
моллюсков, приведенная схема иллюстрирует филогению и родственные отноше-
отношения между подсемействами (рис. 39).
МЕТОДЫ ИССЛЕДОВАНИЯ CLAUSILIIDAE i
Сбор материала. Ввиду того что наши моллюски по сравнению с дру-
другими группами, населяющими подстилку (Pupillidae, Valloniidae и др.),
отличаются довольно крупными размерами, сбор их производится обычно
вручную, без всяких специальных приспособлений (за исключением
пинцета). Поэтому просеивание подстилки и почвы через систему сиг
имеет смысл применять лишь для сбора молодых экземпляров и немногих
мелких видов. К тому же, как уже указывалось, в лиственной подстилке
обитает сравнительно немного видов Clausiliidae. Поэтому при поисках
наших моллюсков основное внимание уделяют осмотру поверхности ство-
стволов деревьев (на высоте человеческого роста), сырых дупел, зарослей мха
и лишайника и всевозможных трещин и щелей на скалах и деревьях.
Учитывая то, что многие виды предпочитают жить под влажной корой
и в древесине гниющего валежника и пней, просмотр этих убежищ совер-
совершенно необходим. Поэтому, если видно, что на стволе поваленного дерева
кора отстает, ее надо оторвать и осмотреть как обнажившуюся поверх-
поверхность древесины, так и внутреннюю сторону коры. При обнаружении
гнилых пней, древесина которых отличается высокой влажностью и имеет
различные щели, следует, удалив отдельные куски, убедиться, нет ли
там моллюсков. Некоторые виды охотно забираются вглубь древесины
по ходам, проделанным червями и насекомыми.
Осыпи и отдельные камни, лежащие в тени деревьев и у подножья
скал, также являются убежищами для многих видов. Осмотр их, особенно-
днем и в сухую погоду, нельзя ограничивать лишь верхней поверхностью,
так как в такую пору моллюски собираются чаще всего именно на нижней
стороне, где больше влаги. Поиски в осыпях следует вести на доста-
достаточную глубину, до тех пор, пока не доберешься до влажного слоя камней.
Для сбора моллюсков следует использовать различные небольшие
коробки. Особенно удобны обыкновенные коробки от спичек, так как они
достаточно прочны и не ограничивают доступ воздуха внутрь них. Кроме
того, благодаря тому, что их можно легко приоткрывать и закрывать одной
рукой, другая рука остается свободной для сбора. Допустим также сбор
в пробирки и в стеклянные банки, закрытые пробкой; однако следует
следить за тем, чтобы за время экскурсии моллюски в них не задохлись.
1 Данная глава посвящена преимущественно некоторым методическим приемам,,
необходимым при исследовании Clausiliidae. Подробнее о методике исследования на-
наземных моллюсков см. нашу работу (Лихарев и Раммельмейер, 1952) и специальную
сводку Еккеля (Jaeckel, 1953).
МЕТОДЫ ИССЛЕДОВАНИЯ 95-
Для того чтобы во время экскурсии раковины собранных моллюсков,
не поломались от перекатывания из стороны в сторону, свободное про-
пространство внутри коробки или банки надо заполнить травой или листьями.
Собранные таким образом моллюски, если их оградить от прямого
воздействия солнца, свободно выдерживают несколько недель, а то и ме-
месяцев, не погибая. Поэтому нет необходимости их фиксировать сразу
после сбора.
Обработка сборов в полевой лаборатории. При разборе материала,
собранного во время экскурсии, следует стремиться часть его зафикси-
зафиксировать для последующих вскрытий. Однако фиксация всех собранных
моллюсков нежелательна. Во-первых, если животное расправлено (а к этому
надо стремиться при всякой фиксации мягких частей тела), то тело его
закрывает доступ ко многим частям замыкательного аппарата. Остаются
видимыми лишь те его детали, которые просвечивают сквозь внешнюю-
стенку раковины. Во-вторых, при длительном хранении в спирте раковина
теряет прозрачность, меняет окраску, а роговой слой ее нередко отстает
и даже шелушится.
. Поэтому часть материала следует сохранить в виде сухих раковин.
Для этого такие экземпляры надо помещать в коробку между слоями
гигроскопической ваты. При этом мягкие части животных быстро высы-
высыхают, не оставляя никаких следов запаха.
Вскрывать моллюсков значительно легче, когда они зафиксированы
в расправленном состоянии, так как при этом все органы располагаются
нормально и между ними остается достаточно свободных промежутков.
Поэтому всегда следует стремиться по каждому виду из любого сбора иметь
несколько экземпляров, зафиксированных в расправленном состоянии.
Однако следует учесть, что большинство видов Clausiliidae крайне
трудно поддается анестезии. При помещении их в воду г они некоторое
время интенсивно вытягиваются из раковины, но потом в большинстве
случаев втягиваются внутрь и более не высовываются. Поэтому вскры-
вскрывать такие экземпляры немногим легче, чем те, которые были сразу поме-
помещены в 70%-й спирт. Лишь немногие виды (например, Idyla foveicollis}
можно анестезировать, помещая на 1—2 суток в холодную воду.
Поэтому рекомендуется несколько видоизменять указанный метод
анестезии. Перед помещением моллюсков в воду следует, оперируя пре-
препаровальной иглой под лупой или бинокуляром, осторожно пробить
стенку раковины на границе между последним и предпоследним оборо-
оборотами, стремясь не повредить мягкие части животного. Затем отверстие
расширяют так, чтобы оно было немного больше устья раковины. При
этом следует стремиться к тому, чтобы отверстие оказывалось как можно,
ближе к устью, но так, чтобы та часть внешней стенки, на которой рас-
располагаются полулунная, главная и палатальные складки, осталась
неповрежденной (рис. 40).
Когда отверстие готово, следует, раздражая мягкие части моллюска
иглой, заставить его втянуться внутрь раковины, т. е. выше отверстия.
После этого его можно поместить в холодную кипяченую или сырую воду.
При соблюдении указанных советов, попав в воду, моллюск скоро
начнет расправляться, голова и нога выступят через пробитое отверстие
и в таком положении моллюск погибает по прошествии 1—2 суток.
1 Обычно для анестезии моллюсков рекомендуют использовать 0.1 %-й водный
раствор хлоралгидрата. Однако наш опыт с Clausiliidae показал, что присутствие?
в воде даже еще меньшего количества этого вещества влечет немедленное втягивание
моллюска в раковину, и в этом положении он так и погибает.
¦96
ВВЕДЕНИЕ
Указанный способ анестезии имеет два главных преимущества: 1) в по-
давлякнцем большинстве случаев моллюски анестезируются в расправ-
расправленном состоянии; 2) нижняя часть раковины, в которой находятся
наиболее важные для определения признаки, остается пустой, благодаря
чему перед вскрытием легко ее отделить и сохранить на случай, если
понадобится проверить правильность определения.
Нижний оборот раковины можно также сохранить, если
помещать живых моллюсков в спирт без анестезии. Тогда
животное втягивается глубоко внутрь раковины и послед-
последний оборот остается пустым. Однако вскрывать моллюсков,
зафиксированных без анестезии, значительно труднее, чем
расправленных.
Комбинируя указанные приемы, можно получить мате-
материал, пригодный впоследствии для решения различных во-
вопросов, связанных с изучением анатомии и особенностей
раковины (в том числе и конхиометрии). х
Для фиксации чаще всего используют 70%-й спирт. Од-
Однако он мало пригоден для гистологических исследований.
В этом случае следует применять какой-либо специальный
фиксатор.2 Однако следует помнить, что многие из них со-
содержат различные кислоты, растворяющие раковину. Поэ-
Поэтому лучше всего использовать фиксатор без кислоты
(например, жидкость Шаудина). Кислотные фиксаторы при-
применимы лишь для материала, надежно определенного до
вида. Из последних очень удобна жидкость Буэна.
Исследование раковины. При описании новых видов для
определения собранного материала, а также для конхио-
метрических исследований очень важно правильно ориенти-
ориентировать раковину. В этом отношении существуют общие пра-
правила для изучающих любых брюхоногих моллюсков. Ра-
Раковину следует держать так, чтобы (рис. 41): 1) завиток об-
обрисовывался максимально полно, а вершина его была бы
направлена от исследователя или кверху; 2) выполняя пре-
предыдущее указание, повернуть раковину по оси так, чтобы
устье вырисовывалось возможно полнее. Данное положение
как раковины, так и устья является «основным» или «пря-
«прямым» при характеристике формы раковины в целом, при
измерении высоты и ширины раковины и устья, при опре-
определении, насколько те или иные части замыкательного ап-
аппарата выступают в просвете устья и видны исследователю.
Кроме того, мы часто используем понятие «косое поло-
положение» устья. В этом случае дается описание тех или иных
частей не при нормальном положении устья, а когда исследователь
заглядывает внутрь его сбоку или снизу.
При характеристике замыкательного аппарата, в особенности складок,
лежащих на внешней стенке последнего оборота, очень важно точно
указать глубину залегания этих образований. С этой целью на периферии
последнего оборота намечают следующие условные линии, соответствую-"
Рис. 40. Схе-
Схема, иллюст-
иллюстрирующая
выбор участ-
участка стенки
последнего
-оборота ра-
раковины, под-
подлежащего
удалению
(обозначен
пунктиром)
и его поло-
положения по
отношению
~к палаталь-
палатальным склад-
•кам и к
устью.
(Ориг.).
1 Конхиометрия — методы математического исследования особенностей рако-
раковины, в основе которых лежат различные промеры последней.
2 См. «Большой практикум по зоологии беспозвоночных» (ч. I) А. В. Иванова,
10. И. Полянского, А. А. Стрелкова, изд. 2-е A958), а также руководства по микро-
микроскопической технике Роскина A946) и Ромейса A953).
МЕТОДЫ ИССЛЕДОВАНИЯ
97
щие основным четырем сторонам (рис. 42): брюшную, спинную, правую
и левую. Для более точной ориентации, в тех случаях, когда складки
залегают в промежутках между названными сторонами, вводятся поня-
понятия: право-брюшное, право-спинное и лево-спинное положения.
Измерение раковины ведется точным штангенциркулем, либо с по-
помощью окулярной линейки. Последний способ наиболее хорош для из-
измерений величин меньше 10 мм. Особенно удобно
измерять раковину с помощью окулярной линейки
лри микроскопе МБС-1, под объективом 1 и окуля-
окуляром X 8. Так как эта оптическая система дает увели-
увеличение в 10 раз, то результаты быстро поддаются пе-
пересчету для получения естественных величин.
Для того чтобы укрепить раковину или серию
раковин в нужном положении, рекомендуется при-
приклеивать ее к предметному стеклу, на который
нанесен слой пластилина. По окончании работы ра-
раковины легко снять со стекла, причем их поверх-
поверхность остается чистой.
Поскольку изучение наших моллюсков немыс-
немыслимо без рассмотрения замыкательного аппарата,
необходимо особо остановиться на некоторых при-
приемах, облегчающих это исследование. Естественно,
что изучение нижних участков замыкательного ап-
аппарата, видимых через устье, требует только хоро-
хорошей оптики. Лучше всего использовать микроскопы
МБС-1 и МБС-2.
Те части замыкательного аппарата, которые рас-
располагаются на внешней стенке — цолулунная, глав-
главная, сутуральные и палатальные складки—обычно
хорошо видны снаружи, так как раковины боль-
большинства наших видов обладают более или менее
прозрачными или просвечивающими стенками.
Однако иногда приходится иметь дело с совер-
совершенно непрозрачными раковинами. У некоторых
видов это объясняется большой толщиной стенок.
У других свежие раковины достаточно хорошо про-
пропускают свет, но после долгого лежания на откры-
открытом воздухе, в результате выветривания, стенки
их становятся мутными. В этих случаях для изу-
изучения внутренней скульптуры рекомендуется сма-
смачивать раковины ксилолом, толуолом или бензолом,
от чего стенки их становятся более прозрачными. Кроме того, удобно
использовать отечественные осветители для микроскопов. Они позво-
позволяют направить на раковину снизу сильный и тонкий пучок света. Для
того чтобы защитить глаза от прямого попадания света и улучшить
видимость, следует помещать исследуемую раковину на кусок кар-
картона, имеющего прорезь, размером немного меньше самой раковины.
Чтобы рассмотреть верхние концы пластинок, а также ознакомиться
•с параллельной и вставочной пластинками или вынуть клаузилий, необ-
необходимо удалить определенную часть внешней стенки последнего оборота.
Эрманн (Ehrmann, 1926) рекомендует для этих целей выпиливать нужный
участок внешней стенки раковины с помощью небольшой пилки. Этот
участок можно потом вклеить на место и раковина сохраняет вид цельной.
. Однако надо признать, что это довольно трудоемкая операция, к тому же
Рис. 41. Схема основ-
основных промеров рако-
раковины. (Ориг.).
в — высота, ш — ши-
ширина раковины; е#—вы-
е#—высота, шу — ширина
устья.
98
ВВЕДЕНИЕ
не всегда приводящая к успеху. Поэтому, если не имеешь дело с уникаль-
уникальными экземплярами, то лучше всего просто удалить нужную часть рако-
раковины иглой, соблюдая указания, сделанные в разделе, посвященном прие-
приемам анестезии моллюсков. Естественно, что надо стремиться удалить
минимальную часть стенки раковины.
Чтобы хорошо рассмотреть клаузилий, нужно его предварительно
освободить от столбика. Сделав это с помощью иглы, его вытряхивают
либо через устье, либо через пролом в раковине.
Хранить клаузилий необходимо отдельно от раковины, чтобы не
повредить это хрупкое образование, но по соседству с нею. Для этого
используют либо небольшие стеклянные пробирки, либо -желатиновые
капсулы, продающиеся в аптеках. В
последнем случае надо учитывать,
что желатина очень гигроскопична,
поэтому капсулы надо хранить в со-
совершенно сухом месте. Для предох-
предохранения капсул от нападения неко-
некоторых музейных насекомых-вредите-
насекомых-вредителей в коробки, где они находятся,
надо подсыпать небольшие порции
нафталина.
Вскрытие. Прежде чем присту-
приступать к вскрытию, полезно сделать
основные промеры раковины и их
записать. Далее, если есть возмож-
возможность, следует отделить нижний обо-
оборот и в дальнейшем хранить его в су-
сухом виде. Те обороты раковины, в ко-
которых располагается тело моллюска,
необходима разрушить и удалить все
остатки раковины. При достаточной
тренировке это легко сделать вруч-
вручную, с помощью игл. Однако есть и
более простой способ — декальцина-
декальцинация. Для этого помещают моллюска
в 10%-й водный раствор соляной кислоты, где его держат до тех пор,
пока не прекратится выделение пузырьков газа. После этого тело моллюска
промывают водой и осторожно, с помощью игл и пинцета, удаляют рого-
роговой слой раковины. В таком виде моллюск готов к анатомированию.
Вскрытие следует вести в небольшой препаровальной ванночке, дно
которой залито слоем смеси воска и парафина. Тело моллюска прижимают
в нужном положении ко дну двумя тонкими перекрещивающимися булав-
булавками (очень удобны различные номера энтомологических булавок). Самой
удобной средой для вскрышя моллюсков является 70%-й спирт. Вода
менее пригодна, так как стенки жен«кого канала семеяйцевода нередко
сильно набухают в ней и быстро разрушаются.
Вскрытие начинают с Того, что тонким ланцетом или остро заточен-
заточенными иглами х делают два надреза покровов: 1) продольный, начиная
его несколько позади полового отверстия и до края мантии; 2) от той же-
точки, что и первый, но на противоположную сторону шеи. Затем вскры-
вскрывают мантийную полость, стремясь не повредить край мантии, заднюю
ПС
Рис. 42. Положение сторон раковины
при рассматривании снизу. (По Эрману).
1 — верхняя пластинка; 2 — главная склад-
складка; з — полулунная складга; л — левая сто-
сторона: лс — лево-спинная сторона; с — спинная
сторона; пс — право-спинная сторона; п —
правая сторона; пб — право-брюшная сторо-
сторона; б — брюшная сторона.
*' Можно также использовать пинцет-ножницы, применяемые в глазной хирургии.
МЕТОДЫ ИССЛЕДОВАНИЯ 99
кишку, почку и сердце.1 После этого надрезают и отворачивают нижнюю
стенку мантийной полости. Если надо отпрепарировать всю половую
систему, то дальнейшие операции переносят в верхнюю часть внутренност-
внутренностного мешка. Так как не всегда легко отделить дольки гонады от верхней
печени, можно этого не делать, а отделить последнюю от верхнего конца
желудка, перерезав соответствующий проток. При этом следует внима-
внимательно следить, чтобы не порвать гермафродитный проток, который
тянется вдоль столбика между желудком и колумеллярным мускулом.
Поэтому необходимо осторожно удалить все пленки, связывающие эти
органы. После освобождения гермафродитного протока отделяют белко-
белковую железу от желудка, так как последняя тесно прилегает к нему по
его внешней стенке. Дальнейшая препаровка заключается в постепенном
раскручивании половой системы, кишечника и колумёллярного мускула.
Для полного отделения гениталий необходимо перерезать все ветви
колумёллярного мускула, которые к ним подходят (оставив лишь конце-
концевые участки мускулов), освободить ретрактор пениса от диафрагмы (лучше
всего отрезав с ним кусок последней), если требуется, перерезать глазной
ретрактор и под конец,, если нужно, отделить половую клоаку вместе
с участком покровов.
Для изучения особенностей радулы и челюсти необходимо отделить
глотку от остальных органов.
В первую очередь надо принять меры к тому, чтобы не повредить
окологлоточное нервное кольцо. Для этого сначала отделяют от глотки
буккальную цепочку. Затем, если нервное кольцо лежит позади глотки,
то перерезают пищевод между ним и глоткой. Если же нервное кольцо
находится впереди глотки, то, оттягивая последнюю назад, отделяют
ее от ротовой полости. При этом челюсть должна остаться с глоткой.
В заключение перерезают все ветви глоточного ретрактора.
Для освобождения радулы и челюсти от мягких тканей глотку поме-
помещают в небольшое количество горячего 10%-го раствора едкого калия
или едкого натрия. Когда растворятся мягкие части (надо внимательно
следить за этим под лупой, так как если передержать радулу в щелочи,
то она тоже может раствориться), радулу и челюсть переносят в холод-
холодную воду.2 Здесь, действуя препаровальными иглами, отделяют остатки
мягких частей, а затем, для окончательной очистки, несколько минут
оба органа держат в крутом кипятке.
Такие обычные среды для микропрепаратов, как канадский бальзам,
глицерин или жидкость Фора, не пригодны для заключения в них раду-
радулы, так как в них она становится настолько прозрачной, что не только
невозможно рассмотреть детали ее строения, но порой трудно даже найти
самый объект.
В этом отношении наилучшей средой является вода. Для того чтобы
водный препарат радулы сохранился на длительное время, рекомендуется
окантовывать его канадским бальзамом по методу Целлера. Этот метод
требует чистоты в работе и некоторую сноровку. Препарат приготовляют
следующим образом (рис. 43).
Очищенные от мягких частей радулу и челюсть в капле воды помещают
на чистое маленькое покровное стекло размером 15 X' 15 или 18 X 18 мм.
Это стекло кладут на две спички, чтобы легйо его было брать пальцами
1 Общие указания о приемах и последовательности вскрытия брюхоногих мол-
моллюсков см. в «Большом практикуме по зоологии беспозвоночных» (ч. II) А.'В. Ива-
Иванова, А. С. Мончадского, Ю. И'. Полянского, А. А. Стрелкова A946).
2 Обычно оба органа остаются связанными друг с другом тонкой перепонкой.
100
ВВЕДЕНИЕ
1 /
1
1
1
/
¦
'или пин Детом. Под бинокуляром, с помощью игл, радулу .располагают
Зубцами кверху и тщательно ее расправляют. Для этого с помощью филь-
фильтровальной бумаги иногда приходится удалить часть воды. После того
как радула расправлена, вновь добавляют воду.1 Затем сверху осторожно
накладывают другое покровное стекло, но больших размеров B0 X 20
или 24 X 24 мм) так, чтобы последнее значительно выступало за края
первого стекла. После этого быстро переворачивают препарат малым
стеклом кверху и вновь помещают его на спички. С этого момента, насту-
-таает наиболее ответственная операция, заключающаяся в том, чтобы
точно отрегулировать количество воды под малым стеклом. Для этого
¦-'¦'• ее отсасывают-фильтровальной'бумагой
или добавляют пипеткой. Вода должна
заполнять все пространство между стек-
стеклами и лишь слегка выступать за его
пределы, а объект должен быть плотно
зажат между стеклами. Если вода силь-
сильно выступает за края малого стекла или
остались капли -ее на выступах боль-
большого стекла, ее следует тщательно уда-
удалить. После этого.на чистоепредметное
стекло наносят 2---3 капли канадского
бальзама и равномерно размазывают их
на площади, несколько меньшей, чем
поверхность большого покровного стек-
стекла. Далее берут оба покровных стекла
с радулой (за свободный край большого
стекла) и вновь переворачивают препа-
препарат так, чтобы малое стекло оказалось
внизу. Оба стекла, которые в силу ка-
капиллярности промежуточной среды проч-
•но пристали друг к другу, кладут горизонтально на слой бальзама так,
чтобы поверхность его одновременно соприкоснулась со всей нижней
поверхностью покровных стекол. Таким образом, вода, в которой нахо-
находятся радула и челюсть, оказывается герметически замкнутой между по-
покровными стеклами и окружена широкой рамкой из канадского баль-
бальзама. Если количество воды было правильно отрегулировано, то смешения
обеих сред не происходит, а следовательно, не произойдет и помутнения
препарата. В таком виде препараты радул могут храниться неопреде-
неопределенно долгое -время.
Изучение отдельных зубов радулы необходимо вести под микроскопом
с иммерсионным объективом, а зарисовку — при помощи рисовального
аипарата. Для измерения зубов наиболее удобен винтовой окулярный
микрометр АМ-9-2, изготовляемый нашей промышленностью. Обычные
окулярные микрометры менее точны, и работа с ними утомительна для
глаз. ,
ЛИТЕРАТУРА
Акрамовский Н. Н. 1949. Наземные моллюски территории селенья Гнишик
в советской Армении. Зоол. сб., 4 [Инст.зоол. и фитопатол. АН Армянск. CCPI :
127-183. * - ¦
Акрамовский Н. Н. 1955. Новые и редкие наземные моллюски-из Армении.
Докл. АН Армянск. ССР, 20, 4 : 149—153.
Алисов Б. П. 1950. Климатические области зарубежных стран. М. : г— 352.
1 Для того чтобы предупредить возможность последующего загнивания препа-
препарата; в воду Ну5кно добавить немного водного раствора карболовой кислоты. '
Рис. 43. Схема водно-бальзамного
. препарата. (Ориг.).
А -=- вид сверху; Б — поперечный разрез.
1 — предметное стекло; 2 — малое покров-
покровное стекло; з — большое покровное стекло;
в — вода; к — канадский бальзам.
ЛИТЕРАТУРА 101
А с ад о в СМ. 1950. Материалы к изучению биологии Dlcrocoelium lanceatum Sti-
Stiles et Hassal и состояния дикроцелиоза в Азербайджане. Тр. Инст. зоол.
АН Азербайджанок. ССР, 14 : 76—132.
Вирштейн Я. А. 1950. Пещерная фауна западного Закавказья. Зоол. журн.,
29, 4:354-365.
Волк о в а Н. С. 1953. Фауна нижней части верхнего сармата окрестностей г. Ap-i
мавира. Сб. «Палеонтология и стратиграфия», Тр. Всесоюзн. н.-исслед. геол.
инст. (ВСЕГЕИ) : 52—76.
В у л ьф Е. В. 1944. Историческая география растений. М.—Л. : I—XIX, 1—546.'
Гитилис В. С. 1956. Фауна и распространение наземных моллюсков Буковины.
Пращ експедицп Чершвецьк. держ. утв., сер. бшл., 2 : 157—175. :
Гитилис B.C. 1957. Наземные моллюски Советской Буковины. Научн. ежегодн.
Черновицк. гос. унив. A956), 1, 2 : 142—145.
Г и т и л и с В. С. 1959. Наземные моллюски Советской Буковины. В кн.: Животный
мир Советской Буковины. Тр. экспед. .по комплексн. изуч. Карпат и Прикар-
Прикарпатья, 7, сер. биол. наук, Черновицы: 264—278.
Даниловский И. В. 1955. Опорный литолого-стратиграфический разрез отло-
отложений Скандинавского оледенения Русской равнины и руководяпще четвертич-
четвертичные моллюски. Тр. Всесоюзн. н.-иссл. геол. инст. (ВСЕГЕИ), нов. сер., 9 :
1—202.
Д в и г у б с к и й И. 1831. Опыт естественной истории всех животных Российской
империи, с изображением животных. Животные мягкие и раковинные. М. :
1—72. •
(Д в и г у б с к и й И.) Dwigubsky I. A. 1892. Primitiae Faunae Mosquensis.
Изд. 2-е, под ред. П. П. Мельгунова. М. Mollusca : 14—18.
Ельский К. М. 1862. О малакологической фауне окрестностей Киева. Изв. унив.
св. Владимира. Второй съезд естествоиспыт. в Киеве : 187—184.
(Ельский К. М.) J е 1 s k i С. 1863. Note sur la faune malacologique des envi-
environs de Kieff (Russie). Journ. Conchyl., 11 : 129—137.
Жижченко Б. П., В. А. Колесников, А. Г. Э б е р з и н. 1940. Неоген
СССР. Стратиграфия СССР, 12, Изд. АН СССР, М.—Л. : 1—677.
3 д у н В. И. 1960а. К фауне моллюсков Закарпатья. Наук. зап. наук.-природознав.
муз. АН УРСР, 8 : 83—95.
3 д у н В. И. 19606. Интересные виды моллюсков Черногоры. 36ipH. праць зоол. муз.
[АН УРСР], 29 : 45—48.
Иванов А. В. 1940. Класс брюхоногих моллюсков {Gastropoda). В кн.: Руковод-
Руководство по зоологии, 2, ред. В. А. Догель и Л. А. Зенкевич, Изд. АН СССР, М.—
Л. : 321—455.
Калитина 3. И. 1953а. К изучению наземных моллюсков Центрального Кав-
Кавказа и восточного Предкавназья (фауна, экология, роль в распространения
дикроцелиоза). Дисс. на соиск. уч. степ. канд. биол. наук (рукопись), Дзауд-
жикау : 1—247.
Калитина 3. И. 19536. К познанию фауны наземных моллюсков Грозненской
области. Изв. Грозн. обл. краеведч. муз., 5 : 139—149.
Калитина 3. И. 1958. К изучению зонального распределения наземных мол-
моллюсков северных склонов Центрального Кавказа и восточного Предкавказья.
Сб. научн. раб. Сев.-Осетинск. гос. мед. инст., 7, 1 : 157—181.
Кокочашви ли Г. В. 1940. Моллюски западной Грузии и их распределение по
Ландшафтным зонам. Дисс. на соиск. уч. степ. канд. биол. наук (рукопись),
Кутаиси : 1—361.
Кокочашвили Г. В. 1956. Определитель наземных раковинных моллюсков
окрестностей Кутаиси. Тбилиси : 1—50 (на грузинск. яз.).
Кокочашвили Г. В. 1958. Явление живорождения у клаузилиид Грузии.
Тр. Кутаисск. гос. пед. инст. им. Цулукидзе, 18 : 187—194.
(К р и н и цк и й И.) Krynicki J. 1837. Conchylia, tarn terrestria, quam fluvia-
tilia et e maribus adjacentibus Imperii Rossica indigena. Bull. Soc. Nat. Mos-
cou, 10 : 50—64.
К р и ш т о ф о в и ч А. Н. 1955. Развитие ботанико-географических областей север-
северного полушария с начала третичного периода. Вопр. геологии Азии, 2 : 824—
844. .
Круликовский Л. 1901. Зоологические заметки. III. О моллюсках Малмыж-
ского уезда. Зап. Уральск, общ. любит, естествозн., 22 : 1—4.
Круликовский Л. С. 1903. Зоологические заметки. V. Сведения о моллюсках
Уржумского уезда Вятской губернии. Зап. Уральск, общ. любит, естествозн.,
24 : 43-45. • ¦ .
К р улик о вс к и й Л; 1909. Краткий очерк фауны Вятской губернии. Календарь
и памятная книга Вятск. губ., Вятка, 30 : 36—69.
102 ВВЕДЕНИЕ
Лёвушкин СИ. 1958. Некоторые данные по анатомии и систематике Clausiliidae
европейской части СССР (без Крыма и Кавказа). Каф. зоол. беспозвон. МГУ,
рукопись дипломы, раб., М.
(ЛиндгольмВ. A.) L i n d h о 1 m W. A. 1901. Beitrage zur Kenntniss der Weich-
thierfauna Siid-Russlands. Nachrbl. deutsch. Malak. Ges., 33 : 161—186.
(Л и н д г о л ь м В. А-.) Lindholm W. A. 1902. Einige fur die Fauna des St,-
Petersburger Gouvernements neue Binnenmollusken. Nachrbl. deutsch. Malak.
Ges., 34: 208—211.
(Линдгольм В. A.) Lindholm W. A. 1908. Materialen zur Molluskenfauna
von Siidwestrussland, Polen und der Krim. Зап. Новоросс. общ. естествоиспыт.,
31 : 199—232.
(ЛиндгольмВ. A.) Lindholm W. А. 1912. Eine neue kaukasische Clausilie.
Nachrbl. deutsch. Malak. Ges., 44 : 202—203.
(Линдгольм В. A.) Lindholm W. 1913a. Ueber em neues Subgenus der Gat-
tung Clausilia Drap. Nachrbl. deutsch. Malak. Ges., 45 : 24—26.
(Линдгольм В. A.) Lindholm W. A. 1913b. Beschreibung neuer Arten und
Formen aus dem Kaukasus-Gebiete. Nachrbl. deutsch. Malak. Ges., 45 : 17—23,,
62-69.
(Линдгольм В. A.) Lindholm W. A. 1913c. Miszellen zur Malakozoologie
des Russischen Reiches, I—XIII. Ежег. Зоол. муз., 18 : 151—167.
(Линдгольм В. A.) Lindholm W. А. 1916. Redescription of a dubious Clau-
Clausilia. Ежег. Зоол. муз., 20 : XLIII—XLIV.
(Линдгольм В. A.) Lindholm W. А. 1924. A revised systematic list of the
Genera of the Clausiliidae, recent and fossil, with their subdivisions, synonymy,
and types. Proc. malac. Soc. London, 16 : 53—80.
(Линдгольм В. A.) L i n d ho 1 m W. A. 1925. A supplement to the revised
systematic list of the Genera of the Clausiliidae. Proc. malac. Soc. London, 16 :
261—266.
(Линдгольм В. A.) Lindholm W. A. 1926. Ein Beitrag zur Kenntnis der Mol-
Molluskenfauna der Krim. Arch. Molluskenk., 58 : 161—177.
Лихарев И. М. 1953. Наземные моллюски [лесной зоны СССР]. В кн.: Живот-
Животный мир СССР, 4, Изд. АН СССР, М.—Л. : 566—576.
Лихарев И. М. 1958а. Географическое распространение наземных моллюсков
Кавказа и некоторые пути происхождения этой фауны. В сб. «Проблемы зоо-
зоогеографии суши» [Львовск. гос. унив. им. Франко] : 138—149.
Лихарев И. М. 19586. Наземные моллюски [горного Крыма]. В кн.: Животный
мир СССР, 5, Изд. АН СССР, М.-Л. : 133-136.
Лихарев И. М. 1958в. Наземные моллюски [Кавказа]. В кн.: Животный мир
СССР, 5 : 464—476.
Л и х а р ев И. М. 1958г. Морфо-функциональный анализ организации Clausiliidae
и некоторые вопросы их систематики и филогении. Зоол. журн., 37, 5 : 750—764-
Лихарев И. М. 1960. Некоторые зоогеографические особенности фауны назем-
наземных моллюсков Карпат. Конф. по вивч. флори i фауни Карпат та прилегл. терр.,
тез. допов., Ки1в : 281—285.
Лихарев И.М.и Г. И.Лежав а. 1961. Новый наземный моллюск горной Туше-
тии (Gastropoda, Clausiliidae). Сообщ. АН Грузинок. ССР, 27,4:473—477.
Лихарев И.М. иЕ. С. Раммельмейер. 1952. Наземные моллюски фауны
СССР. Опред. по фауне СССР, изд. Зоол. инст. АН СССР, М—Л., 43:1—
511.
Марков К. К. 1960. Палеогеография. Изд. 2-е, М. [Московск. гос. унив.] : 1—
268.
Матёкин П. В. 1950. Фауна наземных моллюсков Нижнего Поволжья и ее зна-
значение для представления об истории современных лесов района. Зоол. журн.,
29, 3 : 193-205.
Матёкин П. В. 1959. Приспособительная изменчивость и процесс видообразова-
видообразования у среднеазиатских наземных моллюсков семейства Enidae. Зоол. журн.,
33, 10 : 1518—1536.
(Мельниченко А. Н.) Melnitschenko А. N. 1935а. Zur Landmollus-
kenfauna des westlichen Russlands. Arch. Molluskenk., 67 : 102—105.
(Мельниченко А. Н.) Melnitschenko A. N. 1935b. Zur Landmollus-
kenfauna der Schigulberge. Arch. Molluskenk., 67 : 105—107.
Мельниченко А. Н. 1936. Сравнительно-фаунистический обзор наземных
моллюсков Жигулевских гор и Западной области. В кн.: К фауне Куйбышев-
Куйбышевского края, М.—Куйбышев : 3—20. , /
(Милашевич К. О.) Milachewich С. 1881. Etudes sur la faune des mol-
lusques vivant terrestres et fluviatiles de Moseou. Bull. Soc. Nat. Moscou, 16 :
215—241.
ЛИТЕРАТУРА 103
М и н а с я н А. К. 1947. Материалы к изучению наземных моллюсков пастбищ Ах-
тинского, Севанского и Мартунинского районов Армянской ССР. Научн. тр.
[Ереванск. гос. унив.], 22 : 275—316.
Надеж дин Н. 1868. Список брюхоногих слизняков (Mollusca, Gastropoda],
водящихся в окрестностях Москвы. М. : 1—16.
Полевина А. А. 1959. К фауне наземных моллюсков Закарпатья. Докл. и сообщ.
[Ужгородск. гос. унив.], сер. биол., 3 : 65—68.
Пузанов И. И. 1925а. Наземные моллюски Симферопольского естественноисто-
рического музея. Зап, Крымск. общ. естествоиспыт., 8 : 111—120.
Пузанов И. И. 19256. Материалы к познанию наземных моллюсков Крыма, ч. I.
Моллюски горного Крыма. Бюлл. Московск. общ. испыт. прир., отд. биол.,
33 : 48—104.
Пузанов И. И. 1926. Материалы к познанию наземных моллюсков Крыма, ч. II.
Моллюски степного Крыма. Бюлл. Московск. общ. испыт. прир., отд. биол.,
35:84—101.
Пузанов И. И. 1927. Материалы к познанию наземных моллюсков Крыма, ч. III.
Состав, распределение и генезис крымской малакофауны. Бюлл. Московск.
общ. испыт. прир., отд. биол., 36 : 221—282.
Путь А. Л. 1954. Сравнительная коллекция современных моллюсков отдела палео-
палеозоологии Института зоологии АН УССР. 36ipH. праць Зоол. муз. [АН УРСР],
26:97—118.
(Рачинский С.) Ratchinsky S. 1853. Liste des mollusques gastiropodes
observes dans les gouvernements de Smolensk et de Moscou, dans les annees 1850,
51, 52. Bull. Soc. Nat. Moscou, 26,1 : 232—236.
{Ретовский O.)Retowski O. 1883a. Die Molluskenfauna der Krim. Malak.
Bl., N. F., 6 : 1—34.
{Ретовский О.) Retowski О. 1883b. Am Strande der Krim gefundene ange-
schwemmte transcaucasische (?) Binnenconchylien. Malak. BL, N. F., 6 : 53—61.
(Ретовский О.) Retowski О. 1884. Eine Sammelexcursion nach Abchasien
und Tscherkessien. Ber. Senckenb. Nat. Ges.: 126—143.
(Ретовский О.) Retowski О. 1887. Am Strande der Krim gefundene ange-
schwemmte Binnenconchylien. Malak. Bl., N. F., 9 : 22—42.
(P етовский О.) Retowski О. 1888. Beitrage zur Molluskenfauna des Kau-
kasus. Bull. Soc. Nat. Moscou, N. S., 2 : 277—288.
(Ретовский О.) Retowski О. 1889. Liste der von mir auf meiner Reise von
Konstantinopel nach Batum gesammelten Binnenmollusken. Ber. Senckenb. Nat.
Ges. : 225—265.
Ретовский О. 1899. Сухопутные раковины Кавказа. В кн.: Кавказский музей,
Тифлис, 1, зоол. : 493—510.
Ретовский О. 1914. Материалы к познанию фауны моллюсков Кавказа. Изв.
Кавказск. муз., 6 : 271—334.
Розен О. В. 1901а. К познанию фауны слизняков г. Харькова и его ближайших
окрестностей. Дневн. Зоол. отд. Общ. любит, естествозн., антропол. и этногр.,
3,2 : 6.
Розен О. В. 19016. Материалы к познанию фауны, слизняков Кавказа и Средней
Азии. Дневн. Зоол. отд. Общ. любит, естествозн., антропол. и этногр., 3,2 :
7—13.
(Розе н О.) Rosen О. 1903. Neue Arten aus dem Kaukasus und Zentralasien.
Nachrbl. deutsch. Malak. Ges., 35 : 178—182.
Розен О. В. 1905. Моллюски Московской губернии, собранные фаунистической
комиссией. Дневн. Зоол. отд. Общ. любит, естествозн., антропол. и этногр.,
3,6:8—12.
(Розен О.) Rosen О. 1905. Beitrag zur Kenntnis des Molluskenfauna des Kau-
Kaukasus. Nachrbl. deutsch. Malak. Ges., 37 : 49—64.
Розен О. В. 1907. Моллюски, собранные в Пинском и Мозырскрм уездах Минской
губернии (Полесье). Тр. студ. кружка для иссл. русск. прир., 3 : 83—93.
(Розен О.) Rosen О. 1907. Beitrag zur Kenntnis der Molluskenfauna Russlands.
Nachrbl. deutsch. Malak. Ges., 39 : 140—150.
Розен О. 1911. Моллюски Кавказа, преимущественно Кубанской области. Ежег.
Зоол. муз., 16 : 86—142.
Розен О. 1914. Каталог раковинных моллюсков Кавказа. Изв. Кавказск. муз.,
6 : 141—252.
Розен О. 1922. Материалы к познанию фауны моллюсков Кавказа. Ежег. Зоол.
муз., 23 : 367—370.
Сабанеев В. П. 1880. Список сухопутных и пресноводных слизняков, водя-
водящихся в Ярославской губернии. Тр. Общ. для иссл. Ярославск. губ., 1 : 78—
89.
104 ВВЕДЕНИЕ
(Симашко И.) Siemaschko J. 1847. Beitrag zur Kenntniss der Conchylien
Russlands. Bull. Soc. Nat. Moscou, 20: 93—131.
(Симашко H.)Simaschko J. 1839. Bemerkungen iiber einige Land- und Stis-
swasser-Mollusken Russlands. Bull. St.-Petersb. Acad. Sci., Cl. phys.-math.,
7,15 : 225—240.
G т p a x о в Н. М. 1948. Основы исторической геологии, ч. 2, М. — Л. : 1—396.
Цветков Б. Н. 1939. Новые данные о моллюсках Крыма. Сб. тр. Гос. зоол. муз.
при Московск. гос. унив., 5 : 171—182.
Цветкова Е. А. и Б. Н. Цветков. 1936. Материалы к познанию малакофауны
бывшей Калужской губернии. Тр. Зоол. инст. АН СССР, 3 : 367—392.
Шварцбах М. 1955. Климаты прошлого. Введение в палеоклиматологию. Изд.
иностр. литер., М. : 1—283.
Шивицкис П. К. 1960. Моллюски Литвы и их определение. Вильнюс : 1—352
(на литовск. яз.).
Adamowicz J. 1937. Materialy do fauny Polesia. Miqczaki (Mollusca) pow. pin-
skiego. Spraw. Poznan. Tow. Przyi. nauk, 2 : 52—55.
Adamowicz J. 1939. Materialen zur Kenntnis der Mollusken-Fauna von Polesie.
Fragm. faun. Mus. zool. Pol., Warszawa, 4,3 : 13—89.
Adams H. a. A. 1858. The genera of recent Mollusca arranged according to their or-
organization. 2. London : 1—661.
A 1 b e r s J. CL. 1860. Die Heliceen nach natiirlicher Verwanschaft systematisch geord-
net. 2-te Ausg. von E. Martens, Leipzig : I—VIII, 1—359.
Baker H. B. 1938. Zonitid snails from Pacific islands, part 1. Bernice P. Bishop
mus. Bull., 158, Honolulu : 103—201. .
Baker H. B. 1941. Zonitid snails from Pacific islands, parts 3 and 4. Bernice P. Bis-
Bishop mus. Bull., 166, Honolulu : 202—370.
Baker H. B. 1955. Heterurethrous and Aulacopod. Nautilus, 68,4:109—112.
B^kowski J. 1892. Mie,czaki (Mollusca). Wyd. Muz. Dzieduszyckich we Lwowie,
3:1—264.
Biggs H. E. J. 1931. Description of a new Clausilia from Persia. Journ. Conchol.,
19,6 : 164.
Boettger C. R. 1926. Systematic and geographical notes on Clausiliidae. Arch. Na-
turg., 91 A925), Abt. A, 5 : 1—18.
Boettger С R. 1955. Die Systematik der euthyneuren Schnecken. Zool. Anz., 18,
Suppl. Bd. : 253—280.
Boettger O. 1877a. Clausilienstudien. Palaeontographica. Beitr. zur Naturgesch.
Vorwelt, N. F., Suppl. 3, Cassel : 1—120.
Boettger O. 1877b. Diagnosen neuer Clausilien formen. Nachrbl. deutsch. Malak.
Ges., 9 : 65—76.
Boettger O. 1878a. Neue recente Clausilien. Jahrb. deutsch. Malak. Ges., 5 : 33—61,
97—107, 291—306.
Boettger O. 1878b. Systematisches Verzeichniss der lebenden Arten der Landsch-
neckengattung Clausilia Drap. Separatabdr. Ber Offenbach. Ver. Naturk., 17—
18 : 1—86.
Boettger O. 1878c. Beitrag zu einem Katalog der innerhalb der Granzen des Riis-
sischen Reiches vorkommenden Vertreter der Landschneckengattung Clausilia
Drap. Me), biol. Bull. Acad. Sci. St.-Petersb., 10 : 159—198.
Boettger O. 1879a. Kaukasische Mollusken gesammelt von Herrn, Hans Leder in
Paskau. Jahrb. deutsch. Malak. Ges., 6 : 1—42.
Boettger O. 1879b. Kaukasische Mollusken gesammelt von Herrn Dr. G. Sievors
in Tiflis. Jahrb. deutsch. Malak. Ges., 6 : 388^412.
Boettger O. 1880a. Kaukasische Mollusken gesammelt von Herrn Hans Leder,
z. Z. in Tiflis. Jahrb. deutsch. Malak. Ges., 7 : 109—150.
Boettger O. 1880b, Armenische und transkaukasische Mollusken aus einer Sen-
dung des Hrn. Dr. G. Sievers in Tiflis. Jahrb.. deutsch. Malak. Ges., 7 : 151—
161.
Boettger O. 1880c. Diagnoses molluscorum novorum ab ill. Hans Leder in regione
caspia Talysch dicta lectorum. Jahrb. deutsch. Malak. Ges., 7 : 379—383.
Boettger O." 1881a. Sechstes Verzeichniss transkaukasischer, armenischer und nord-
persischer Mollusken aus Sendungen der Herrn Hans Leder, z. Z. in Kutais und
Dr. G. Sievers in St.-Petersburg. Jahrb. deutsch. Malak. Ges., 8:167—261.
Boettger O. 1881b. Sectiones speciesque novae Clausiliarum caucasiarum. Jahrb.
deutsch. Malak. Ges., 8 : 341—346.
Boettger O. 1881c. Diagnoses molluscorum novorum Transcaucasiae, Armenian
et'Persiae. Nachrbl. deutsch. Malak. Ges., 13:117—129.
Boettger O. 1882. Triptychia Sndbg. und Serrulina Mouss. sind als Genera aufzu-
fassen. Nachrbl. deutscli. Malak. Ges., 14 : 33—35.
ЛИТЕРАТУРА 105
Boettger 0. 1883. Siebentes Verzeichniss von Mollusken der Kaukasuslander,
nach Sendungen des Hrn. Hans Leder, z. Z. in Helenendorf bei Elisabetpol (Trans-
kaukasien). Jahrb. deutsch. Malak. Ges., 10: 135—198.
Boettger O. 1884. Liste der von Herrn 0. Retowski in Abchasien gesammelten
Binnenmollusken. Ber. Senckenb. Nat. Ges. : 144—155.
Boettger O. 1886a. Die Binnenmollusken des Talysch-Gebietes. In : G. R a d d e.
Fauna und Flora des sudwestlichen Caspi-Gebietes : 256—350.
Boettger O. 1886b. Neuntes Verzeichniss (IX) von Mollusken der Kaukasuslander
nach Sendungen des Hrn. Hans Leder, z. Z. in Helenendorf bei Elisabetpol (Trans-
kaukasien). Jahrb. deutsch. Malak. Ges., 13 : 121—156.
Boettger 0. 1887. Zwei neue Formen transkaukasischer Landschnecken. Nachrbl.
deutsch. Malak. Ges., 19 : 55—57.
Boettger O. 1888. Diagnosen neuer kaukasischer Arten. Nachrbl. deutsch. Malak.
Ges., 20 : 149—155.
Boettger O. 1889. Zehntes Verzeichnis (XII) von Mollusken der Kaukasuslander.
Ber. Senckenb. Nat. Ges. : 3—37.
G h а г p e n t ierJ.de. 1852. Essai d'une classification naturelle des Clausilies. Journ
de Conchyl., 3 : 357—408.
Glessin S. 1881a. Beitrag zur Kenntniss der Molluskenfauna des Caucasus. Malak.
Bl., N. F., 3 : 129—135.
Glessin S. 1881b. Mollusken aus Taurien. Malak. Bl., N. F., 3 : 136—141.
Clessin S. 1887. Die Molluskenfauna Oesterreich-Ungarns und Schweiz. Numberg :
1—858.
Go оке А. Н. 1915. The genus Clausilia: a study of its geographical distribution,
with a few notes on the habits and general economy of certain species and groups.
Proc. malac. Soc. London, 11 : 249—269.
D egn er E. 1952. Der Erbgang der Inversion bei Laciniaria biplicata Mtg. (Gastr.,
Pulm.). Mitt. Hamburg, zool. Mus. u. Inst., 51 : 3—61.
D г а р а г n a u d J.-P.-R. 1801. Tableau des Mollusques terrestres et fluviatiles de la
France. Montpellier : 1—116.
Draparnaud J.-P.-R. 1805. Histoire naturelle des Mollusques terrestres et flu-
fluviatiles de la France. Paris : I—VIII, 1—164.
Dyrdowska M. 1928. Przyczynek do zoogeografji micjczakow Polski. Spraw. Kom.
Fizjogr. Pol. Ak. Urn., Krakow, 63 : 201—203.
Dyrdowska-Mlodzinowska M. 1930. Materiafy do fauny malakozoologi-
cznej Wilenszczyzny. Fragm. faun. Mus. zool. Pol., Warszawa, 1 : 21—56.
Ecke H. u. H. Dulckeit. 1924. Beitrag zur Schneckenkunde Osels. Korrespondbl.
Naturf.-Ver. Riga, 58 : 106—107.
EdlauerE. 1941. Die ontogenetische Entwicklung der Verschlussapparates der Clau-
siliiden, untersucht an Herillu bosniensis. Zeitschr. wiss. Zool., 155, 2 : 129—1.58.
Ehrmann P. 1926. Zur Systematik der Clausiliiden, besonders der ostasiatischen.
Sitzungsb. naturf. Ges., Leipzig, 49—52 : 18—59.
E h r m a n n P. 1933. Mollusken (Weichtiere). In: Tierwelt Mitteleuropas, 2, 1, Leip-
Leipzig : 1—264.
E i с h w a 1 d Ed. 1830. Naturhistorische Skizze von Lithauen, Volhynien und Podo-
lien in geognostisch-mineralogischer, botanischer und zoologischer Hinsicht.
Wilna. Mollusken : 203—225.
Forcart L. 1935. Die Mollusken der nordpersischen Provinz Masenderan und ihre
tiergeographische Bedeutung. Arch. Naturg., N. F., 4 : 404—447.
Frankenberger Z. 1916. Zur Anatomie und Systematik der Clausilien. Zool.
Anz., 47 : 221-236.
Fromming E. 1954. Biologie der mitteleuropaischen Landgastropoden. Berlin :
1—404.
F u с h s A. u. F. К a u f e 1. 1936. Anatomische und systematische Untersuchungen
an Land- und Siisswasserschnecken aus Griechenland und den Inseln des Agai-
schen Meeres. Arch. Naturg., N. F., 5 : 541—662.
Germain L. 1930. Mollusques terrestres et fluviatiles. Faune de France, 21, Paris :
1—477.
Germain L. 1933. Mollusques terrestres et fluviatiles de l'Asie Anterieure. Bull.
Mus. nation. Hist, nat., ser. 2, 5,5 : 389—392.
Germain L. 1936. Mollusques terrestres et fluviatiles d'Asie Mineure. Dans:
Voyage zoologique d'Henri Gadeau de Kervil J e en Asie Mineure (avril — mai
1912), 1,2 : 1—492.
Gerstfeldt G. 1859. Aufzahlung der in Ehst-, Liv.- und Kurland beobachteteri
Land- und Siisswasser-Mollusken mit besonderer Beriicksichtigung der betreflen-
den Sammlungen des Natarforschenden Vereins zu Riga. Corresp'ondbl.' Naturf.-
•Vlt. Riga. 11 : 102 — 114.
106 ВВЕДЕНИЕ
Geyer D. 1927. Unsere Land- und Suss-wasser-Mollusken. 3-te Aufl., Stuttgart : I—
XI, 1—224.
Haltenorth T. und S. Jaeckel sen. 1943. Beitrage zur Kenntnis der Land-
und Siisswassermollusken der Krim. Arch. Molluskenk., 75 : 225—240.
Hartmann J. D. W. 1840—1844. Erd- und Siisswasser-Gasteropoden der Schweiz.
St. Gallen : I—XX, 1—227.
Hasslein E.u. L. 1943. Zur Molluskenfauna des Stadtgebietes von Kauen (Litauen).
Arch. Molluskenk., 75 : 245—256.
Hensche A. 1861—1866. Preussens Molluskenfauna. Schrift. phys.-okonom. Ges.
Konigsberg, 2 : 73—96 A861), 1—9 A862), 99—101 A866).
Hesse P. 1925. On the anatomy of some Clausiliidae. Proc. malac. Soc. London,
16 : 154—162.
Icon.о graphic der Land- und Siisswasser-Mollusken. 1835—1920. Bd. 1—3 A835—
1859) von E. A. Rossmassler; fortgesetzt v. W. Kobelt: Bd. 4—7 A876—1880)
und N. F., Bd. 1—23 A884—1920) und Suppl. Bd. A895— 189Д); Bd. 21—22 A914—
1918) (Clausiliidae) v. A. Wagner. ...
I m r e L. 1941. Beitrage zur Kenntnis der Schnecken-Fauna der Nordost-Karpaten.
Fragm. Faun. Hung., 4 : 91—93.
I s s e 1 A. 1865. Catalogo dei Molluschi raccolti dalla Missione italiano in Persia. Mem.
Acad. Sci. Torino, ser 2,23 : 387—439.
Jaeckel S. jun. 1950. Zur Land- und Siisswasser-Molluskenfauna der Krim. Zool.
Anz., 145, Erganzungsb. : 343—363.
Jaeckel S. G. sen., W. Rlemm u.W. Meise. 1957. Die Land- und Siisswas-
Siisswasser-Mollusken der n6rdlichen Balkanhalbinsel. Abhandl. u. Ber. staatl. Mus.
Tierkunde, Dresden, 23,2 : 141—205.
Jaeckel S. H. 1953. Praktikum der Weichtierkunde. Jena : 1—87.
Kawall J. H. 1869. Notice sur la faune malacologique de la Courlande. Ann. Soc.
malac. Belgique, 4 : LXVIII—LXXV.
К e n n a r d A. S. a. B. B. W о о d w a r d. 1923. Note on the Nomenclature and Sys-
Systematic of the Clausiliidae. Proc. malac. Soc. London, 15 : 298—308.
Kimakowicz M. 1894. Prodromus zu einer Monographie des Clausilia—Sub-
genus Alopia H. et A. Adams. Verh. Mitt. Siebenb. Ver. Naturw., 43 : 19—58.
Kimakowicz-Winnicki M. 1914. Clausilium. Eine monograpnische-physio-
logische Studie. Zool. Jahrb., Abt. Syst., 37 : 283—328.
К lik a B. 1893. Verzeichniss der von Dr. V. Vavra wahrend seiner Kaukasus-Reise
gesammelten Binnenconchylien. Separat abdr. v. Sitzungsb. bohm. Ges. Wiss.,
math.-naturw. Cl. : 1—7.
Kobelt W. 1876—1920. См.: Iconographi e...
Kobelt W. 1897. Studien zur Zoogeographie. I. Die Mollusken der palaearktischen
Region. Wiesbaden: I—VIII, 1—344.
Kobelt W. 1898. Studien zur Zoogeographie. II. Die Fauna der meridionalen Sub-
' Region. Wiesbaden : 1-Х, 1—368.
К г a u s p C. 1936. Beitrag zur estlandischen Molluskenfauna. Arch. Molluskenk., 68 :
16-61.
Krausp C. 1940. Beitrag zur Molluskenfauna Lettlands. Ann. Soc. Reb. Natur. In-
Invest. Univ. Tartuensi, 45 : 217—270.
Krausp C. 1953. Die Clausiliiden Estlands. Mitt. Berlin. Malakol., 4 : 8—33.
Riister H. C. 1847—1862. Die Schliessschnecken und die verwandten Gattungen
(Clausilia, Balea, Cylindrella, Megaspira). In: Martini u. Chemnitz
System. Conch. Cab., Bd. I, Abt. 14 : 1—355.
Lehmann R. 1873. Die lebenden Schnecken und Muscheln der Umgegend Stettins
und in Pommern mit besonderes Berucksichtigung ihres anatomischen Baues.
Cassel : I—VI, 1—328.
Loosjes F. E. 1953a. Monograph of the indo-australian Clausiliidae (Gastropoda,
Pulmonafa, Clausiliidae, Phaedusinae). Wageningen : 1—226.
Loosjes F E. 1953b. Clausilien-Ausbeute des Herrn H. W. E. Crockewit aus der
asiatischen Ttirkei, Umgebung von Zonguldak. Basteria, 17 : 28—31.
L о i. e к V. 1952. Cochlodina remota sp. n. — novy plz z celedi Clausiliidae.
Vestn. 6eskoslov. zool. spol., 16, 1—2 : 135—142.
Lozek V. 1955. Mekkysi ceskoslovenskeho kvarteru. Praha : 1—510.
L о z e к V. 1956. Klic ceskoslovenskych mekkysu. Bratislava : 1—437.
Luther A. 1901a. Bidrag till kannedomen om land- och sotvattengastropodernas
utbredning i Finland. Acta Soc. Fauna et Flora Fennica, 20,3 : 1—125.
Luther A. 1901b. Verzeichnis der Land- und Siisswasser-Mollusken der Umgebung
Revals. Acta Soc. Fauna et Flora Fennica, 20,2 : 1—16.
Martens E. 1874. Ueber vorderasiatische Conchylien nach den Sammlungen des
Prof. Hausknecht. Cassel : I—VI, 1—127.
ЛИТЕРАТУРА 107
Martens E. 1876. Eine neue transcaucasische Clausilie. Nachrbl. deutsch. Malak.
Ges., 8 : 90—91.
Martens E. 1880. Aufzahlung der von Dr. Alex. Brandt in Russisch Armenien
gesammelten Mollusken. Bull. Acad: St.-Petersb., 26 : 142—158.
Metmod G. 1930. Gasteropodes. Catalogue Invertebr. Suisse, Geneve, 18 : I—XII,
1—583.
Moellendorff 0. 1875. Studien zur Systematik der Clausilien. Nachrbl. deutsch.
Malak. Ges., 7 : 17—28.
M о q u i n-T a n d о n A. 1855. Histoire naturelle des mollusques terrestres et flu-
viatiles de France. Paris, 2 : 315—351.
Mortillet G. 1854. Description de quelques coquilles nouvelles d'Armenie, et
considerations malacostatiques. Mem. Inst. Nat. Geneve, 2 : 1—15.
Mousson A. 1863. Coquilles terrestres et fluviatiles recueillies dans l'Orient par
M. le Dr. Alex. Schlafli. Vierteljahrsschr. Nat. Ges. Zurich, 8 : 275—320, 368—426.
Mousson A. 1873. Coquilles recueillies par M. le Dr. Sievers dans la Russie meridio-
nale et asiatique. Journ. de Conchy!., 21 : 193—230.
M о u s s о n A. 1876a. Coquilles recueillies par M. le Dr. Sievers dans les contrees Trans-
caucasiques. Journ. de Conchyl., 24 : 24—51.
Mousson A. 1876b. Coquilles recueillies par M. le Dr. Sievers dans la Russie Asia-
Asiatique. Journ. de Conchyl., 24 : 137—148.
0k land F. 1925. Die Verbreitung der Landgastropoden Norwegens. Skrift. Norsk.
Vidensk. Akad., 1, Mat.-nat. К.1., 8, Oslo : I—VIII, 1—168.
P a a s с h A. 1845. Beitrage zur genaueren Renntniss der Mollusken. Arch. Naturg.,
11,1 : 34—46.
Pfeiffer L. 1848. Diagnosen neuer von Frivaldsky gesammelter Landschnecken.
Zeitschr. Malak., 5 : 6—12.
Pfeiffer L. 1868. Monographia Heliceorum viventium. Sistens descriptiones sys-
tematicas et criticas omnium huius familiae, generum et specierum hodie cogni-
tarum. Lipsiae, 6 : 392—521.
Pilsbry H. A. 1900. On the Zoological Position of Partula and Achatinella. Proc.
Acad. Nat. Sci. Philadelphia : 561—567.
Pilsbry H. A. 1904. Manual of Conchology. Philadelphia, 16 : I—XL, 1—329.
Pilsbry H. A. 1948. The land Mollusca of North America, north of Mexico, 2,2,
Philadelphia: I—XLVII, 521—1113.
Pilsbry H. А. а. С. М. С о о к е. 1934. Partulidae of Tonga and related forms. Occ.
Pap. Bernice P. Bishop Mus., 10, 14, Honolulu : 1—22.
Plate H.-P. 1951. Clausilia cruciata Studer auf Riigen. Deutsch. Zool. Zeitung, 1 :
250—253.
Polinski W. 1917. Materialy do fauny malakozoologicznej Krolestwa Polskiego,
Litwy i Polesia. Prace Tow. Nauk. Warszawa, 3, Wyd. nauk mat. i przyr., 27 :
1—130.
P о s к a-T e i s s L. 1928. Zur Schneckenfauna Estlands. Arch. Molluskenk., 60 : 24—
31.
P г о t z A. 1903. Zur Binnenmolluskenfauna der Provinz Ostpreussen. Nachrbl.
deutsch. Malak. Ges., 35 : 1—6.
R e i b i s с h P. 1892. liber die bei Gelegenheit einer Reise nach Kaukasus gesammel-
gesammelten Gehauseschnecken. Sitzungsb. naturf. Ges., Leipzig, 17—18, 1891—1892 :
49—58.
R i к 1 i M. 1914. Natur- und Kultiir-Bilder aus den Kaukasuslandern und Hochar-
menien, von Teilnehmern der schweizerischen naturwissenschaftlichen Studien-
reise Sommer 1912. Zurich : 1—317.
Rossmassler E. A. 1835—1854. См.: Iconographi e...
Rotarides M. 1944. Schneckenaufsammlungen in Siebenbiirgen und im Nordost-
karpaten-Gebiet. Fragm. Faun. Hung., 7 : 53—55.
Rotarides M. a. H. 8 с h 1 e s с h, 1951. Regeneration of the shell and related
phenomena in the family Clausiliidae. Acta Biol. Acad. Sci. Hungar., 2,1—3 :
55—117.
Roth J. R. 1850. In: M. Wagner. Reise nach Kolchis. Leipzig : 341.
Schlesch H. 1927. Zur Kenntnis der Molluskerifauna des Ostbaltikums mit Beriick-
sichtigung der in Lettland vorkommenden Arten. Korrespondbl. Naturf.-Ver.
Riga, 59 : 92—126.
Schlesch H. 1936. Bemerkunfjen iiber die Verbreitung der Landmollusken im ost-
lichen Ostseegebietes. Tartu trlikooli Loodusuurijate Selsti Aruanded, 42 : 233—244.
Schlesch H. 1937. Zur Kenntnis der litauischen Molluskenfauna. Festschr. 60 Geb.
Prof. Em. Strand, 3 : 250—263.
Schlesch H. 1942. Die Land- und Siisswassermollusken Lettlands. Korrespondbl.
Naturf.-Ver. Riga, 74 : 246—360.
108 ВВЕДЕНИЕ
Schlesch H. 1946. On the occurence of south eastern elements in the land snaii
fauna of the eastern Baltic area. Proc. malac. Soc. London, 27 : 84—87.
Schlesch H. 1958. Uber das isolierte Vorkommen von Chondrina clienta auf Oland
und Gotland und andere siidostliche Landmollusken in der ostbaltischen Fauna.
Arch. Molluskenk., 87 : 45—52.
Schlesch H. uC. Krausp. 1938. Zur Kenntnis der Land- und Siisswasser-Mollus-
ken Litauen. Arch. Molluskenk., 70 : 73—125.
Schmidt A. 1855. Der Geschlechtsapparat der Stylommatophoren in taxonomischer
Hinsicht. Abhandl. naturw. Ver. Sachsen u. Thiiringen, Halle, 1 : 1—52.
Schmidt A. 1857. Die kritischen Gruppen der europaeischen Clausilien, I. Leipzig :
I—VI, 1—63.
Schmidt A. 1868. System der europaischen Clausilien und ihren nachsten Verwand-
ten. Cassel : 1—176.
Schmidt F. 1885. Beitrag zur Kenntnis der Molluskenfauna Ehstlands. Sitzungsb.
Naturf. Ges. Univ. Dorpat, 7 : 190.
Schmidt H. 1936. Die Abhangigkeit der Darmgestaltung der Landpulmonaten von
phylogenetischen und okologischen Faktoren sowie von der Gehauseform. Arch.
Naturg., N. F., 5 : 18—140.
Schneider О 1878. Naturwissenschaftlichen Beitrage zu der Kaukasuslandern
auf Grund seiner Sammelbeute. Dresden. Burdach'sche Hofbuchhandlung : 11—34,
158—159.
Schrenk A. G. 1848. Ubersicht der Land- und Susswasser-Mollusken Livlands.
Bull. Soc, Nat. Moscou, 21,1 : 135—185,.,
Simionescu I. et I. Z. В a r b u. 1940. La faune sarmatienne de Roumanie.
Mem. Inst. Geol. Romaniei, 3, Bucaresti : 1—194,
Simroth H. uH. Hoffmann. 1908—1928. Pulmonata. In: Bronns Klassen
u. Ordnungen des Tierreichs, III, 2,2 : 1—1354.
Sl.osarski A. 1872. Materyaiy do fauny malakologicznej Krolestwa Polskiego. War-
szawa : 1—17.
SI osarski A. 1877. Materiaux pour la faune malacologique du Royaume de Pologne.
Bull. Soc. Zool. France : 291—299.
Smith E. A. a. B. B. W о о d w а г d. 1890. On the Nomenclature of the Oral Folds
in the Shells of Clausilia. Ann. Mag. Nat. Hist., ser. 6,5 : 209—213.
S о 1 e m A. 1959. Systematics of the Land and Fresh-Water Mollusca of the New
Hebrides. Fieldiana, Zool., 43,1 : 1—238.
S о 6 s L. 1917. Zur systematischen Anatomie der ungarischen Pulmonaten. Ann. hist.-
natur. Mus. Nat. Hung., 15 : 1—165.
S о 6 s L. 1928. Az Alopia-nem. The genus Alopia. Ann. hist.-natur. Mus. Nat. Hung.,
25 : 261-426.
S о 6 s L. 1941, Further contributions to the mollusc fauna of the north-eastern Carpa-
Carpathians. Fragm. Faun. Hung., 4 : 58—60.
S о 6 s L. 1943. A Karpat-medence Mollusca-faunaja. Budapest : I—-VIII, 1—478.
Sporleder A. 1861. Fortgesetzte Beobachtungen uber die Wachstumszeit einiger
Schnecken. Malak. Bl., 7 : 115—120.
Starmiihlner F. u. A. Edlauer. 1957. Beitrage zur Kenntnis der Mollusken-
Molluskenfauna des Iran. Sitzungsb. Osterr. Akad. Wiss., math.-naturw. Kl., Abt. I, 166,
9—10 : 435—495.
Steenberg CM. 1914. Anatomie des Clausilies danoises. I. Les organes genitaux.
Mindeskrift Japetus Steenstrup, Kebenhavn : 1—46.
Steenberg CM. 1925. Etudes sur l'anatomie et la systematique des Maillots (fam.
Pupillidae). Videns. Medd. fra Dansk naturhist., 80:1—216.
T e r m i e г H. et G- 1952. Histoire geologique de la biosphere. La vie et les sediments
dans les geographies successives.. Paris : 1—721.
T h i e 1 e J. 1931. Handbuch der systematischen Weichtierkunde, 2. Jena : 461—778.
T h i e 1 e J. 1935. Handbuch der systematischen Weichtierkunde. 4, Jena : 1023—1154.
Tuomikoski T. 1958. Nilviasia sortavalasta (Lk). Luonnon tutkiia, 62,2:57—
58.
Urbanski J. 1947, Krytyczny przeglfjd mie,czakow (Mollusca) Polski. Roczn.
Uniw. Marii Curie-Sklodow., Dzial C, Nauki biol., 2 : 1—30.
U г b an ski J. .1957. Krajowe slimaki i maize. Warszawa : 1—276.
Vest W. 1867. (iber den Schliessapparat der Clausilien. Verh. Mitt. Siebenb. Ver.
Naturw., 18 : 5—18, 161—174, 188—196.
Villa A. et J, B. 1841. Dispositio; systematica conchyliarum terrestrium et fluvia-
,tilum quae adservantur in collectione fratrum Ant. et Jo. Babt. Villae. Medio-
lani : 1—63.
V о i t,h J. 1846. Zur NaturgeschichtederGattung Clausilia. Zeitschr. Malak., 3 : 55—59.
Wagner A. J. 1914. См.: I с о n о g r a p h i e... .
ЛИТЕРАТУРА 109
Wagner A. 1919. Zur Anatomie und Systematik der Clausiliiden. Nachrbl. deutsch.
Malak. Ges., 51 : 49—60, 87—104, 129—147.
Wagner A. 1920. Zur Anatomie und Systematik der Clausiliiden. Arch.
Molluskenk., 52 : 1—13, 67—78, 97—108, 145—158.
Wagner A. J. 1921. Ueber die zeitliche Entwicklung der Clausiliiden und deren
Beziehungen zu anderen Gruppen der Stylommatophoren. Arch. Molluskenk.,
53 : 98—103.
Wagner A. 1922. Erganzungen und Erlautungen zur Systematik der Clausiliiden.
Ann. Zool. Mus. Polon., 1 : 96—111.
Watson H. 1915. Studies on the carnivorous slugs of South Africa. Ann. Natal. Mus.,
3,2 : 107—267.
Watson H. 1920. The affinities of Pyramidula, Patulastra, Acanthinula and Val-
lonia Proc. malac. Soc. London, 14 : 6—30.
W e n z W. 1923. Gastropoda extramarina tertiaria, III. In: С D i e n e r (ed.). Fos-
silium Catalogue, I: Animalia, 20, Berlin : 757—831.
W e n z W. u. A. Zilch. 1959. Euthyneum. In: H*ndbuch d. Palaozoologie, herausg.
v:.0.-H. Schindewolf, Bd. 6, Gastropoda, Th. 2, Berlin : 1 — 1434.
Westerlund С A. 1884. Fauna der in der palaarktischen Region lebenden B'm-
nenconchylien. IV. Gen. Balea Prid. et Clausilia Dr. Karlskrona : I—VII, 1 —
212, 1 — 18, 1—3, 1—8.
Westerlu n d C. A. 1886. Fauna der in der palaarktischen Region lebenden Bin-
. nenconchylien. VI, 3-te Beilage, Lund : 1—16.
Westerlund С. А. 1897. Beitrage zur Molluskenfauna Russlands. Ежег.
Зоол. муз., 2 : 117—143.
Westerlund С. А. 1901. Synopsis molluscorum in regione Palaearctica viventum
ex typo Clausilia Drap. Зап. Акад. наук, СПб, сер. 8, физ.-мат. отд., И, И :
I—XXVII, 1—203.
Wiegm-.ann'F. 1878a. Bemerkungen zur Anatomie der Clausilien. Jahrb. deutscb.
Malak. Ges., 5 : 157—169.
Wiegmann F. 1878b. Anatomische Untersuchung der Claus. Reiniana Kob. Jahrb.
deutsch. Malak. Ges., 5:202—207.
W i e g:iu a n n F. 1893. Beitrage zur Anatomie der Landschnecken der Indischen Archi-
pels. In: M.-W e b e r. Zoologische Ergebnisse einer Reise in NiederlSndisch Ost-
Indien, 2. Leiden : 224—251.
УСЛОВНЫЕ ОБОЗНАЧЕНИЯ К РИСУНКАМ
Радула
С — срединный зуб;
цифры — порядковые номера зубов, считая от срединного.
Половая система
б — бич; пж — предстательная железа;
бж — белковая железа; пр — придаток пениса;
ел — влагалище; рещ — ретрактор верхних (глазных) щу-
г — гонада (гермафродитная железа); палец;
гп — гермафродитный проток; ркл — ретрактор половой клоаки;
д — диафрагма; рнщ — ретрактор нижних (губных) щу-
кл— половая клоака; палец;
ко — камера оплодотворения; рп — ретрактор пениса;
лр — левый ретрактор; реп — ретрактор семяприемника;
м — матка; с — семяпровод;
мп — мускульная полоска; сп ¦.— семяприемник;
иге — нер* пениса; ся — семяяйцевод или выводковая сум-
осп — отросток семяприемника; ка;
п — пенис; ап — эпифаллус.
Линейка при рисунках обозначает масштаб, соответствующий 1 мм.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СЕМ. CLAUSILIIDAE
ФАУНЫ СССР И СОПРЕДЕЛЬНЫХ СТРАН1
1 D2). Спиральная пластидка видна через устье.
2 A9). Передний конец спиральной пластинки заходит за задний конец
верхней, или обе пластинки лежат на разном расстоянии от
столбика.
3 ( 8). Нижний конец лопасти клаузилия по палатальному краю несет
глубокую вырезку, видимую при косом положении устья.
4E). Нижняя пластинка дает в просвете устья широкий горизонтально
лежащий изгиб. Париетальный край устья плотно прилегает
к стенке раковины (устье невыступающее) или даже разорван
и места прикрепления его связаны мозолью
Cochlodiaa Fer. (стр. 132).
5D). Нижняя пластинка не образует горизонтального изгиба. Парие-
Париетальный край устья цельный и выступает вперед.
6G). Нижняя пластинка спирально изогнута, видна при прямом по-
положении устья и с краем связана короткой складкой. Субкоду-
меллярная пластинка не видна. Имеется нижняя палатальная
складка
Laciniaria Hartm., подрод Mentissa H. et A. Adams (стр. 248).
7F). Нижняя пластинка лежит отвесно, не видна при прямом поло-
положении раковины и с краем устья не связана. Субколумеллярная
пластинка видна при косом положении устья. Нижней палаталь-
палатальной складки нет Euxinastra Bttg. (стр. 202).
8C). Клаузилий без вырезки или последняя очень мала и не видна
через устье.
9 A2). Нижняя пластинка лежит отвесно.
10 A1). Раковина деколлированная; если цельная, то высота ее 23—
30 мм, а число оборотов 15—16 . . Acrotoma Bttg. (стр. 155).
11 A0). Раковина недеколлированная; высота ее меньше 23 мм, оборотов
меньше 15 Euxina Bttg. (стр. 164).
12 ( 9). Нижняя пластинка более или менее спирально изогнута.
13 A6). Субколумеллярная пластинка видна через устье. Полулунная
складка или верхние концы палатальных складок лежат на
спинной стороне раковины.
14 A5). Раковина стройная {максимальная ширина приходится на пред-
предпоследний оборот). Устье небольшое, правильно грушевидное.
Клаузилий на внутренней стороне переднего конца лопасти несет
короткий киль
. . . Laciniaria Hartm., подрод Strigilecula К. et W. (стр. 246).
15 A4). Раковина вздутая (максимальная ширина приходится на послед-
последний оборот). Устье относительно крупное, неправильно пяти-
1 В таблицу включены также некоторые виды, которые по признакам раковины
сильно отличаются от остальных видов своих родов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ 111
угольное или овально-грушевидное. Клаузилий без киля ....
........... Megaleuxina Bttg. (стр. 192).
Субколумеллярная пластинка не видна. Полулунная складка
всегда цельная и лежит на правой стороне раковины.
Устье треугольно-грушевидное, высота его лишь немного больше
ширины. Палатальный край почти прямой и отвесный, остальные
края устья широко изогнуты. Главная складка длинная — про-
простирается на г/з—г/г оборота. Лопасть клаузилия широкая и не
перекручена Galeata Bttg. (стр. 204).
Устье грушевидное, высота его значительно больше ширины.
Палатальный край плавно изогнут. Главная складка короче
и простирается не более чем на х/4' оборота. Лопасть клаузилия
более узкая и перекручена
..... Laciniaria Hartm., подрод Laciniaria s. str. (стр. 241).
Спиральная пластинка сливается с верхней, если же обе плас-
пластинки разделены промежутком, то концы их лежат на одной ли-
линии, т. е. на равном расстоянии от столбика.1
Раковина деколлированная.
Нижняя пластинка лежит отвесно и не видна при прямом поло-
положении раковины. Край устья между верхней и нижней пластин-
пластинками гладкий, без складок. Высота раковины 12—14, ширина
3.2—3.7 мм Caspiophaedusa Lindh. (стр. 126).
Нижняя пластинка спирально изогнута и видна при прямом
положении раковины. Край устья между верхней и нижней пла-
пластинками несет несколько коротких складок. Высота раковины
15—20, ширина 4—5 мм . . . Pontophaedusa Lindh. (стр. 123).
Раковина недеколлированная.
Лопасть клаузилия на внешней стороне несет 2 крупных киля,
из них один виден через устье ...... Filosa Bttg. (стр. 211).
Лопасть клаузилия либо без килей, либо последние находятся
на внутренней стороне и через устье не видны.
Нижняя пластинка лежит совершенно отвесно.
Раковина имеет отчетливый киль.
Главная складка короткая — простирается не более чем на ]/4
оборота. Если есть полулунная складка, то нет палатальных
складок; если первая отсутствует, то имеется одна короткая верх-
верхняя палатальная складка Pseudalinda Bttg. (стр. 251).
Главная складка длинная — простирается На ХА оборота. Полу-
Полулунная складка отсутствует; вместо нее всегда имеются не менее
3 палатальных складок . ..... Quadriplicata Bttg. (стр. 264).
Раковина без киля .
Serrulina Mouss., подрод Serrulina s. str. (стр. 115).
Нижняя пластинка более или менее спирально изогнута.
Ось раковины более или менее сильно изогнута. Субколумелляр-
Субколумеллярная пластинка простирается до края устья
Serrulina Mouss., подрод Pravispira Lindh. (стр. 121).
Ось раковины прямая. Субколумеллярная пластинка не дости-
достигает края устья.
Последний оборот с крупным килем. Устье с глубоким базальным
желобком, благодаря которому нижний край имеет некоторую
угловатость.
1 Исключение: у молодых экземпляров некоторых видов Vestia пластинки иногда
не слиты, причем концы их находятся на разном расстоянии от столбика.
112 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
35 C6). Полулунная складка хорошо развита, цельная. Имеется пала-
, тальная мозоль, от которой внутрь раковины отходит короткая,
но толстая нижняя палатальная складка, видимая при прямом
положении устья ........... Clausilia Drap. (стр. 218).
36 C5). Полулунная складка полностью или частично распалась на серию
глубоколежащих палатальных складочек, которые не видны
при прямом положении устья. Палатальной мозоли нет . . .
Mucronaria Bttg. (стр. 277).
37 C4). Последний оборот без киля, или он выражен очень слабо, чаще
всего в виде небольшого вздутия. Базального желобка нет, или
он очень неглубокий, благодаря чему нижний край устья кажется
плавно закругленным.
38 D1). Нижняя пластинка изогнута широкой спиралью и связана с краем
устья короткой складкой. Полулунная складка или ее производ-
производные лежат на спинной стороне раковины. Виды живут в Европе.
39 D0). Раковина темно- или красновато-коричневая, матовая или со
слабым шелковистым блеском. Лопасть клаузилия на конце
более или менее закругленная .... Iphigena Gray (стр. 230).
40 C9). Раковина зеленовато- или коричнево-роговая, блестящая, реже
матовая. Лопасть клаузилия на конце более или менее заострена
и у большинства видов крючковидно изогнута .
. . . Vestia Hesse (стр. 254).
41 C8). Нижняя пластинка слабо спирально изогнута и не связана с краем
устья. Полулунная складка лежит на правой или правоспинной
стороне раковины. Виды живут на Кавказе . . ,
Micropontica Bttg. (стр. 292).
42 ( 1). Спиральная пластинка не видна через устье (у некоторых видов
отсутствует).
43F0). Замыкательный аппарат различно развит, но всегда имеется
субколумеллярная пластинка и клаузилий.
44 D5). Последний оборот на затылке имеет крупную и глубокую трех-
или четырехугольную вмятину, которой внутри устья соответст-
соответствует большой палатальный бугор, очень сильно сужающий про-
просвет устья Idyla H. et A. Adams (стр. 139).
45 D4). Последний оборот на затылке без глубокой вмятины, хотя и может
быть немного вдавлен; внутри устья нет указанного палатального
бугра.
46 D9). Полулунная складка и клаузилий лежат на лево-спинной сто-
стороне и заметны даже при прямом положении раковины.
47D8). Обороты выпуклые. . .Micropontica retowskii (Bttg.) (стр. 296).
48 D7). Обороты слабо выпуклые Armenica Bttg. (стр. 143).
49 D6). Полулунная складка и клаузилий лежат на спинной стороне
раковины или еще глубже и не видны при прямом положении
раковины.
50 E5). Нижний конец субколумеллярной пластинки виден через устье.
51 E4). Последний оборот с килем. Устье с базальным желобком.
52 E3). Нижпяя пластинка дает широкий горизонтальный изгиб, сильно
сужающий просвет устья, и не связана с краем устья складкой.
Спиральная пластинка очень длинная — начинается на брюшной
и кончается на спинной стороне раковины. Полулунной складки
нет. Вместо нее имеются 2 глубоко лежащих палатальных складки,
берущих начало на право-брюшной стороне и заканчивающихся
на правой стороне раковины . . Mentissoidea Bttg. (стр. 208).
53 E2). Нижняя пластинка очень слабо изогнута и почти отвесно уходит
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ 113
кверху; с краем устья она связана короткой складкой. Спиральная
пластинка очень короткая и вместе со вставочной и параллельной
лежит на спинной стороне раковины. Здесь же находится тонкая
полулунная складка с 2 короткими палатальными складками . .
Megaleuxina comnena (Ret.) (стр. 200).
54 E1). Киля и базального желобка нет . . Ruthenica Lindh. (стр. 215).
55 E0). Нижний конец субколумеллярной пластинки не виден через устье.
56 E9). Раковина башневидная или бапшевидно-веретеновидная. Главная
складка очень короткая.
57 E8). Полулунная складка цельная и располагается на спинной.стороне.
Над ней лежат 2—3 коротких складочки (изредка они отсутст-
отсутствуют Armenica Bttg. (стр. 143).
58 E7). Полулунная складка распалась на 7—9 коротких палатальных
складок (главная и сутуральные), лежащих на правоспинной
стороне раковины ... . . Mucronaria gustavi (Bttg.) (стр. 284).
59 E6). Раковина веретеновидная. Главная складка длинная, занимает
Уг оборота (хотя и не видна через устье) .
Mucronaria Bttg., подрод Index Bttg. (стр. 289).
60^D3). Замыкательный аппарат сильно редуцирован, без субколумел-
субколумеллярной пластинки и клаузилия.
61*F2). Раковина веретеновидно-цилиндрическая. Нижняя пластинка
{имеет вид легкой складки, тянущейся вдоль столбика г
Alopia H. et A. Adams (стр. 131).
62 F1). Раковина башневидная. Нижняя пластинка отсутствует ....
.'. Balea Gray (стр. 261).
I. Подсемейство PHAEDUSINAE
Раковина лево-, реже правозакрученная, нередко деколлированная*
обычно тонко исчерченная. Последний оборот снизу округлый, без киля.
Устье относительно крупное, округло-грушевидное. Края его расширены
и отвернуты. Верхняя пластинка впереди доходит до края устья, сзади
чаще всего сливается со спиральной пластинкой. Имеются полулунная
или палатальные складки, либо то и другое одновременно.
Семяприемник открывается в женские половые пути вблизи половой
клоаки, а иногда и непосредственно в нее. Отросток семяприемника
располагается очень различно, но, как правило, на значительном удале-
удалении от нижнего конца семяприемника; по гистологическому строению он
сходен с последним. Пенис крупный, чаще всего цилиндрический, с диаф-
рагмалъным ретрактором, но без придаточных органов. Глазной ретрактор
располагается между женскими и мужскими половыми путями.
Распространение. Основная масса видов обитает в южной
и восточной Азии (до Японии включительно); несколько видов живет
в юго-восточной Европе и на Кавказе. Многочисленные роды и виды насе-
населяли Европу* в течение верхнего мела и всего третичного периода.
1. Род SERRULINA MOUSS.
М о u s s о a, 18J3 : 214 (Clausilia, секция Serrulind); В oettger О., 1877а :
70—71 {Clausilia, секция Serrulina); L i n d h о 1 m, 1924 : 63, 72—73; Е h г in an n,
1926 : 45—58; Tbiele, 1931 : 532—533.
Тип рода: Clausilia sieversi Pfr.
Устье раковины выступающее, более или менее грушевидное, с высо-
высоким врезанным синулусом. Весь край устья, или только его колумелляр-
1 В данном пункте использованы признаки, характерные лишь для вида, присут-
присутствие которого возможно в пределах нашей страны или близ ее границ.
114 J. PHAEDUSINAE
пая часть, покрыт мелкими складочками или узелками. Сравнительно
короксан верхняя пластинка, плавно снижаясь, переходит в длинную
и тонкую спиральную пластинку. Нижняя пластинка различной формы,
но всегда лежит глубоко и при прямом положении раковины не видна;
с краем устья она связана 1—2 складочками, вубколумеллярная пластинка
не всегда видна через устье. Главная складка длинная или очень длин-
длинная. Полулуййая складка или образовавшиеся в результате ее распада
короткие складочки лежат на правой стороне.
Срединный и боковые зубы радулы с крупными дополнительными
зубцами. Краевые зубы с 3 зубцами, за исключением самых внешних (по
одному с каждой стороны радулярного ряда), которые имеют 4 зубца.
Семяприемник с Длинным, к концу сужающимся отростком; собственно
семяприемник короткий и резервуар его резко обогнут. Влагалище очень
короткое. Очень длинный цилиндрический пенис даже у расправленных
экземпляров дает Несколько извивов, а у сократившихся — спирально
закручен. Предстательная железа короткая и расположена вдоль верхней
части семяяйцевода.
Колумеллярный мускул сравнительно короткий; верхний конец его
находится на уровне белковой железы. Левый ретрактор дает ветвь к по-
половой клоаке.
Распространение, Лесные области Кавказа, северного Ирана
и Малой Азия. Кроме того, два вида обитают на Балканском полуострове,
а один из них найден в Румынии и в советской части восточных Карпат.
В ископаемом состоянии данный род известен начиная с нижнего
миоцена Европы.
Большинство видов обитает под корой и в гниющей древесине. Неко-
Некоторые живут только в пещерах. Несколько видов этого рода (к сожале-
сожалению, оставшихся неописанными) найдены в пещерах западного Закав-
Закавказья (Бирштейн, 1950).
Вслед за Линдгольмом (Lindholm, 1924) принято разделять данный род
на три секции: 1) Sermlina s. str., 2) Pravispira Lindh., 3) SciocochleaC. Bttg.
Мы считаем вполне обоснованным образование группы Pravispira,
которую рассматриваем как подрод, так как входящая сюда S. semila-
semilameilata (Mouss.) резко отличается от остальных видов формой нижней
и субколумеллярной пластинок (см. далее). Остальные признаки раковины
этого моллюска — распад полулунной складки, -с образованием верхней
и нижней Палатальных складок, а также искривление оси раковины,
которым Линдгольм придавал основное значение в характеристике дан-
данной группы, можно встретить не только у S. semilameilata. В самом деле,
редукция полулунной складки имеет место и у S. collasi Sturaay, а искрив-
искривление оси раковины нередко наблюдается у S. sieversi (Pfr.) и характерно
для S. senghanensis Morgan.
Что касается секции Sciocochlea, то все авторы, которые считают необ-
необходимым выделить S. collasi в эту группу, мотивируют это изолирован-
изолированностью ареала указанного моллюска от других видов того же рода.
Мы не склонны в этом Вопросе следовать за названными авторами и
относим этот вид в подрод Serrulina s. str., основанием для чего служит
большое сходство раковины данного моллюска с такими видами, как
S. sieversi и S. serrulata (Pfr.), в особенности по форме нижней и субколу-
субколумеллярной пластинок. Правда, редукция полулунной складки у S. collasi
как будто бы сближает ее с S. semilameilata. Однако следует учесть, что
у Clausiliidae, даже в пределах небольших групп, полулунная складка
варьирует значительно сильнее, чем пластинки, которые отличаются боль-
большей стабильностью.
1. SERRULINA 115
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА &ERRUL1NA
1 A2). Нижняя пластинка лежит почти отвесно и сильно удалена от
верхней. Субкояумеяляряая пластинка начинается выше нижней,
а заканчивается далеко от края устья. (Подрод Serrulina s. str.).
2 ( 7). Раковина ребристая.
3( 6). У вполне сформировавшихся экземпляров раковина серовато-
воскового цвета, слабо блестящая, непрозрачная. Край устья
на всем протяжении несет мелкие складочки и узелки.
4 ( 5). Высота раковины 11—12.5, ширина 2.6—3.2 мм
» . . 2а. S. serrulata serrulata (Pfr.).
5 ( 4). Высота раковины 16.0—16.7, ширина 3.5—3.6 мм
26. S. serrulata amanica Nagele.
6 ( 3). В любом возрасте раковина желтоватая, блестящая и прозрачная.
Складки и узелки располагаются лишь справа от верхней плас-
пластинки и по всей колумеллярной части края устья .
4. S. senghanensis Morgan.
7B). Раковина ребристо исчерченная или гладкая, лишь за устьем
ребристая или ребристо исчерченная.
8 A1). Раковина ребристо исчерченная. Вид обитает под корой и в гнию-
гниющей древесине.
9 A0). Высота раковины 9.7—11.0, ширина 2.4—2.8 мм; оборотов 83/4—
9х/4 la. S. sieversi sieversi (Pfr.).
10 ( 9). Высота раковины 11.2—12.6, ширина 2.7—3.0 мм; оборотов
9%—10 .16. S. sieversi occidentalis Likharev, ssp. nov.
11 ( 8). Раковина гладкая. Вид обитает в пещерах . , 3. S. collasi Stur.
12 ( 1). Нижняя пластинка образует широкую спираль и близко подходит
к верхней пластинке. Субколумеллярная пластинка начинается
ниже нижней, а заканчивается у края устья. (Подрод Pravispira
Lindh.) 5. S. semilamellata (Mouss.).
1. Подрод SERRULINA s. str.
Сравнительно короткая, массивная и высокая нижняя пластинка да-
еко отстоит от верхней, слабо прогнута и лежит отвесно. Субколумел-
[ярная пластинка начинается выше нижней, резко обрывается на правой
стороне раковины на значительном удалении от края устья, с которым
она ничем не связана. Имеется небольшая параллельная пластинка.
Палатальных складок нет. Лопасть клаузилия сравнительна короткая
лишена киля.
Раковины молодых экземпляров с момента вылупления из яйца ш
\о начала образования 8-го оборота имеют 1 или 2 спирально закручен-
шх колумеллярных складки (рис. 37), которые, как правило, занимаюж«2
юследних оборота (с ростом раковины верхние концы их подвергаются
|»езорбции). Когда начинается формирование 8-го оборота, эти складки
нсончательно исчезают. Вероятно, они служат для облегчения управлву
гая раковиной. ,
Бетгер (Boettger О., 1822), а вслед за ним и другие авторы считали;
гго эти складки указывают на древность данной группы и ее родственную
яизость с вымершими Triptychiinae и с ныве живущими Megaspiridae.
первых эти образования имелись лишь у молодых раковин, а у вторых
ни сохраняются на всю жизнь (стр. 86).
116
/. PHAEDUSINAE
la. Serrulina (Serrulina) sieversi sieversi (Pfr.) (рис. 37, 44—47).
P f e i f f e г L., 1871, Malak. Bl., 18 : 70 (Clausilia); M о u s s о n, 1873 : 214,
t. 8, fig. 4 [Clausilia (Serrulina)]; Boettger O., 1886a : 314 [Clausilia (Serrulina)];
К q b e 11, 1893, Iconogr., N. F., 6 : 42, fig. 1021 [Clausilia (Serrulina)]; Лихарев
и Раммельмейер, 1952 : 216.
Раковина башневидно-веретеновидная, довольно твердостенная,
тонко ребристо исчерченная, блестящая, прозрачная, светло-роговая,
иногда красноватая. Завиток стройный, с довольно острой верхушкой.
и./
Рис. 44. Serrulina (Serrulina)
sieversi sieversi (Pfr.). (Ориг.).
A — общий вид раковины; Б —
клаузилий.
Рис 45. Serrulina (Serrulina) si-
sieversi sieversi (Pfr.). Пластинки
замыкательного аппарата. На-
Наружная стенка раковины частично
удалена. (Ориг.).
1 — верхняя пластинка; 2 — нижняя
пластинка; з — краевая складка;
4 — субколумеллярная пластинка;
S — спиральная пластинка.
I
Оборотов 83/4—9V4, верхние вы-
выпуклые, нижние слабо выпуклые;
предпоследний оборот немного ши-
шире последнего; последний оборот за ;
устьем с крупными, широко расставленными ребрами, сбоку чуть уплощещ
Шов сдавленный, окаймленный. Устье почти отвесное, ромбическо-груше-
видное. Край устья слегка валикообразно утолщен и отвернут. Краевые
складки и узелки располагаются в нижней части колумеллярного краяй
Имеется слабая палатальная мозоль. Верхняя пластинка очень низкая
и тонкая. Верхний конец спиральной пластинки лежит у границы с пред*
1. SERRULINA
117
последним оборотом. Нижняя пластинка начинается на правой стороне
последнего оборота, щфо прогибаясь, почти отвесно простирается вниз
и заканчивается дачти у самого конца столбика; одна из краевых складок
Л
¦бж
осп
Рис» 46. Serrulina (Serrullna) sieversi occidentalls Likharev. Ра-
Радула. (Ориг.).
связывает ое с краем устья. Снаружи нижняя пластинка с трудом видна
лишь при косом положении устья. Субколумеллярная пластинка закан-
заканчивается на правой стороне раковины под полулунной складкой; с краем
устья она не связана и через устье не
видна, но нижний конец ее просвечи-
просвечивает через стенку раковины. Очень длин-
длинная главная складка начинается у гра-
границы с предпоследним оборотом и за-
заканчивается вблизи синулуса. Оба кон-
конца крупной полулунной складки отогну-
отогнуты назад. Краевых складок чаще всего
3: одна сливается с нижней пластинкой,
другая тянется вглубь раковины, в сто-
сторону субколумеллярной пластинки, но
никогда не сливается с нею, так что
между ними остается значительный
промежуток (у прозрачных раковин
все это хорошо видно при рассматри-
рассматривании их снизу); наконец, третья, очень
•короткая складка лежит между назван-
названными. Реже, кроме этих складок, бы-
бывает еще четвертая краевая складка и
некоторое число узелков. Обычно все
4>ни располагаются в нижней части
колумеллярного края и лишь изредка
В верхней части или по правую сторону
рт верхней пластинки. Лопасть клау-
зилия имеет форму параллелограмма
р закругленными углами; нижний ко-
корец ее сильно утолщен и отогнут.
, Размеры: высота раковины 9.7—11.0,
ширина 2.4—2.8 мм; высота устья 2.1—
2.4, ширина 1.6—1.8 мм.
: Внутреннее строение. Материал: 1) Талыш, Билясар,
сбор Знойно, 16 VII 1932, определение наше — 1 экз.; 2) Закатальский
юповедник, сбор и определение наше, 16 VII 1954—2 экз. (ssp. occiden-
lalis nov.).
Рис. 47. Serrulina (Serrulina) sieversi
occidentalis Likharev. Половая систе-
система. (Ориг.).
Радула (рис. 46):[|- + (у + ^) X 2] X 67.
¦118 /. PHAEDUSINAE
Размеры срединного зуба 17.7 X 21.4 ц, ширина радулы 360 \х.
Гонада состоит из 3 долек. Предстательная железа занимает верхнюю
половину семяяйцевода. К ней прилегает длинный, кверху плавно сужи-
суживающийся отросток семяприемника, простирающийся до конца белковой
железы (рис. 47). Собственно семяприемник значительно короче, прижат
к женскому каналу семяяйцевода; его резервуар отогнут вниз.
Распространение (рис. 32). В СССР: леса Талыша и Лен-
коранской низменности. Вне СССР: леса северного Ирана (провинции
Гил ян и Мазендеран).
Очень близкая данному виду форма (точнее определить систематиче-
систематическое положение пока трудно) найдена в отложениях позднего миоцена
центрального Предкавказья (стр. 302).
Э к о л о г и я. Живет во влажной древесине и под корой гниющих
деревьев, обычно совместно с Caspiopkaedusa perlucens (Bttg.).
16. Serrulina (Serrulina) sieversi occidentalis Likharev, ssp. nov.
От предыдущего подвида отличается более крупной раковиной и боль-
большим числом оборотов — 9V2—10.
Размеры: высота раковины 11.2—12.6, ширина 2.7—3.0 мм; высота
устья 2.3—2.7, ширина 1.8—2.0 мм.
Размеры голотипа:1 высота раковины 13, ширина 3 мм; высота устья
2.7, ширина 1.8 мм.
Распространение (рис. 32).- Обитает в горных лесах южных
склонов Большого Кавказа от Лагодехи до Закаталы и в прилегающих
низовых лесах.
Экология. Та же, что и у предыдущего подвида.
Систематическое пало жени е. Самостоятельность обоих
подвидов подтверждается не только морфологическими различиями, но
и полной разобщенностью их ареалов.
2а. Serrulina (Serrulina) serrulate serrulata (Pfr.) (рис. 48, 49).
P f e i f f e r L., 1847, Zeitschr. Malak., 4 : 71 (Clausilia); M о г t i 1 I e t, 1854 :"l4,
t. 1, fig. 8 (Clausilia); Raster, 1847—1862 : 162, t. 18, fig. 57 (Clausilia); M о u s-
s о n, 1863 : 394 (Cl. gracilior); I s s e 1, 1865 : 425, t. 3, fig. 52—54 (Cl. erivanensis);
Lindholm, 1913b : 66 [Cl. (Serrulina) serrulata var. semiserrata nov.]; Розен,
1914 : 207 [Clausilia (Serrulina)]; Ретовский, 1914 : 323 [Clausilia (Serrulina)];
Лихарев и Раммельмейер, 1952 : 215, рис. 130.
Раковина веретеновидная, довольно твердостенная, ребристая,
слабо блестящая, просвечивающая, ееровато-воскового цвета, у молодых
экземпляров — блестящая, прозрачная, зеленоватая. Оборотов 10—
111/2, умеренно выпуклых, плавно нарастающих; последний оборот книзу
сужен, у основания слегка сжат, но без киля. Шов сдавленный. Завиток
быстро сужается к верхушке; Последняя довольно острая. Устье непра-
неправильно грушевидное, узкое, выступающее, с врезанным синулусом. Края
устья слегка валикообразно утолщены, на всем протяжении с многочис-
многочисленными складками и узелками. Довольно высокая верхняя пластинка
немного выступает за пределы края устья и, почти не снижаясь, переходит
в тонкую и высокую спиральную пластинку. Последняя берет начало
в нижней части предпоследнего оборота. Нижняя пластинка начинается
на правой стороне последнего оборота; слабо изгибаясь, почти отвесно,
1 Голотип хранится в Зоологическом институте АН СССР (каталог систематиче-
систематической коллекции моллюсков, № 1а). Место сбора: Азербайджанская ССР, Закаталь-
ский р:н, р. Цилбан-Чай (приток р. Тала-Чай), 3 V 1936, сборщик Асадов.
/. SERRULINA
119
она тянется вниз и заканчивается у нижнего конца столбика, причем
одна из складок связывает ее с краем устья. Снаружи нижняя пластинка
видна лишСь при косом положении устья. Субколумеллярная пластинка
через устье не видна. Главная складка начинается у границы с предпо-
предпоследним оборотом и заканчивается недалеко от синулуса.
Оба конца крупной полулунной складки отогнуты назад.
Округло-ромбическая лопасть клаузилия на нижнем конце
несет утолщение или слабый выступ.
Размеры: высота раковины 11—12.5, ширина 2.6—
3.2 мм; высота устья 2.4—2.8, ширина 1.9—2.0 мм.
Внутреннее строение. Материал: 1) Бату-
Батуми, Зеленый мыс, сбор Рылова, 21 IX 1929, определение
наше — 1 экз.; 2) Хоста, сбор Рылова, 2 IX 1929, опре-
определение наше — 2 экз.; 3) Кавказский заповедник, Гузе-
рипль, сбор и определение наше, 2 VIII 1956 — 1 экз.; —
4) Румыния, северная Добруджа, сбор и определение
Гроссу, 16 VI 1957 — 2 экз.; 5) Закарпатская обл., Тячев-
ский р-н, окрестности села Округла, сбор и определение
Лёвушкина — 2 экз.
Радула (рис. 49): [| + G| + ^) X 2] X 78.
2; ' 3— 4/' ' J
Размеры срединного зуба 14.6 X 15.4 ц, ширина радулы
382 ц.
Половой аппарат очень схож с таковым предыдущего
вида. Отросток семяприемника еще длиннее — далеко
заходит за белковую железу и прижат к желудку. Соб-
Собственно семяприемник короткий и отвернут вниз, так что
резервуар его прилегает к почке. Пенис еще длиннее, чем
у S. sieversi; у одного экземпляра, зафиксированного в рас-
расправленном состоянии, длина пениса в 11/2 раза превы-
превышала длину внутренностного мешка.
Распространение (рис. 32). В СССР: лесные облаети Север-
Северного Кавказа (наиболее восточная находка сделана в Чечено-Ингушской
АССР), западного и центрального Закавказья (наиболее восточная на-
Рис. 48. Ser-
rulina (Ser-
rulina) ser-
rulata serru-
serrulata (Pfr.).
(Ориг.).
Рис. 49. Serrulina (Serrulina) serrulata serrulata (Pfr.). Радула.
(Ориг.).
ходка сделана в северо-западном Азербайджане в районе Белоканы).
Кроме того, недавно этот моллюск найден в Закарпатской области (Лё-
вушкин, 1958). Вне СССР: ряд пунктов вдоль черноморского побережья
Турции, Болгария (Родопы), Румыния (Добруджа, Яссы).
Экология. Обитает под гниющей корой и в гнилой древесине,
обычно большими популяциями.
12b
/. PHAEDUSINAE
*2б. Serrulina (Serrulina) serrulata amanica Nagele.
N a g e 1 e, 1906, Nachrbl. deutsch. Malak. Ges., 38:29 [Clausilia (Serrulina)].
Отличается более крупной раковиной. * Высота ее 16.0—16.7, ширина
3.5—3.6 мм; высота устья 3.2, ширина 2.2 мм.
Распространение. Единственная находка сделана в Турции,
в санджаке Александретта, в горах Гявур (Аманос). Следовательно, ареал
этого моллюска, по-видимому, полностью изолирован от ареала пре-
предыдущего подвида.
*3. Serrulina (Serrulina) collasi Stur. (рис. 50).
S t u г any, 1904, Nachrbl. deutsch. Malak. Ges., 36 : 103, text, fig.; К о b e 1 t,
1906, Iconogr., N. F., 12 : 22, fig. 1878 [Clausilia (Serrulina)]; Wagner A., 1914,
Sitzber. Akad. Wiss. Wien, math.-naturw. К.1., 123, 1 : 34, 38 (Phygas).
Раковина веретеновидно-башневидная, с широкой, округло при-
притуплённой верхушкой, тонкостенная, прозрачная, блестящая, почти
гладкая, лишь с единичными редкими
линиями прироста, желтоватая, но в
свежем состоянии, по-видимому, бес-
бесцветная. Оборотов 71/4—8V2, верхние
довольно выпуклые, нижние почти пло-
плоские; последний оборот чуть, уже пред-
предпоследнего, на затылке ребристо исчер-
исчерчен. Шов нитевидный, окаймленный.
Устье овально-грушевидное, почти от-
отвесное, с высоким, врезанным и слегка
наклоненным вправо синулусом. Края
устья коротко отвернуты и утолщены;
середина палатального края (при рас-
рассматривании сбоку) выступает вперед.
У нижнего конца колу^еллярного края '
лежат 3—4 коротких краевых складки.
Низкая верхняя пластинка сливается
со спиральной. Нижняя пластинка ле-
лежит глубоко и отвесно и при прямом
положении раковины не видна; она
заканчивается у нижнего конца стол-
столбика и с краем устья связана одной из
краевых складок. Субколумеллярная
пластинка не видна снаружи; насколь-
насколько можно судить при рассматривании
¦ сквозь стенку раковины, одна из крае-
краевых складок связывает ее с краем устья или, во всяком случае, близко
подходит к ее нижнему концу. Главная складка начинается на правой
стороне раковины и простирается до ее левой стороны. На правой сто-
стороне лежит 1 короткая верхняя палатальная складка, под которой слег-
слегка намечаются еще 1 или 2 мозолевидных складочки.
Размеры: 2 высота раковины 8.5—11.0, ширина 2.2—2.5 мм; высота
устья 2.0—2.3, ширина 1.3—1.5 мм.
1 Размеры даны с учетом промеров единственного-бывшего в нашем распоряже-
распоряжении экземпляра (котип), хранящегося в коллекции Музея Грузии.
2 Даны на основании промера 4 котипов, присланных из Венского естественно-
исторического музея и одного котипа из Берлинского зоологического музея.
Рис. 50. Serrulina (Serrulina) collasi
Stur. Котип. (Ориг.).
1. SERRULINA 121
Распространение. Вид найден в одной, из пещер о. Корфу
и в некоторых пещерах южной Далмации.
Систематическое положение. Строение замыкатель-
ного аппарата, в особенности форма нижней пластинки, указывает на
принадлежность данного вида к подроду Serrulina s. str. Следовательно,
нет основания выделять его в особую секцию или подрод, как это делал
Вагнер {Phygas Wagner, 1914, поп Treitschke, 1833) и Ц. Бетгер (Scioco-
chlea С. Boettger, 1935, пот. nov.).
*4. Serrulina (Serrulina?) senghanensis Morgan.
Morgan, dans Germain, 1933 : 391; Germain, 1936 : 315, fig. 24—25.
Раковина веретеновидно-башневидная, почти цилиндрическая,
по оси изогнутая, ребристая, желтоватая, блестящая. Оборотов 9, выпук-
выпуклых, разделенных тонким швом; первые 7 оборотов плавно нарастают,
последние 2 почти одной ширины; последний оборот книзу сужен. Устье
грушевидное, с небольшим округлым синулусом. Края устья утолщены,
слабо отвернуты, с многочисленными краевыми складками и узелками,
располагающимися по правую сторону от верхней пластинки и по всему
колумеллярному краю. Верхняя и нижняя пластинки крупные, субколу-
меллярная не видна.
Размеры: высота раковины 8.0—9.5, ширина 1.7—1.9 мм.
Р а с п р ост ранение. Северный Иран — провинция Гилан,
к востоку от Сефидруда.
Экология. Обитает во мху и среди камней, на высотах 50—280 м
над ур.< м.
Приведенное описание составлено исключительно по данным Моргана
и Жермена. Так как оба исследователя уделили внимание только внешним
особенностям раковины, то мы многого^ ле знаем о замыкательном аппа-
аппарате этого моллюска.
Судя по рисункам, раковина сильно похожа на Serrulina sieversi
(Pfr.), почему мы условно помещаем этот вид в подрод Serrulina s. str.-
2. Подрод PRAVISPIRA LINDH.
L in d"h о 1 m, 1924 : 63,73 (Serrulina, секция Pravispira).
Тип подрода: Clausilia semilamellata Mouss.
Длинная нижняя пластинка, снижаясь, образует широкую спираль
и близко подходит к верхней пластинке. Субколумеллярная пластинка
начинается ниже нижней, заканчивается у края устья и хорошо видна
при прямом положении раковины. Кроме главной и распавшейся полулун-
полулунной складки, имеется еще длинная нижняя палатальная складка, а близ
края устья лежит палатальный бугор.
Замыкательный аппарат и устье раковины входящего в этот подрод
вида имеет много общего с Pontophaedusa funiculum (Mouss.). Если бы не
важные различия между обоими моллюсками в строении половой системы,
в особенности в форме и величине пениса и предстательной железы, то,
по сути дела, не было бы оснований помещать их в разные роды. Однако
указанное сходство говорит о родственной близости обеих групп.
Распространение. Лесные области Кавказа (кроме Талыша)
и черноморского побережья Турции. Известно несколько видов из неоге-
неогеновых отложенпй Евролы и Кавказа.
122
/. PHAEDUS.INAE
5. Serrulina (Pravispira) semilamellata (Mouss.) (рис. 51).
M о u s s о a, 1863 : 396 (Clausilia); M о u s s о в, 1873 : 214, t. 8, fig. 3 [Clau-
silia (Serrulina)]; Kobelt, 1893, Iconogr., N. F., 6 : 42, fig. 1022 [Clausilia (Serru-
(Serrulina)]; Розен, 1911:127 [Clausilia (Serrulina)]; Розен, 1914:207 [Clausilia
(Serrulina)]; Ретовский, 1914 : 324 [Clausilia (Serrulina)]; Лихарев и
Раммельмейер, 1952 : 216, рис. 131.
Раковина неправильно веретеновидная, но оси заметно изогну-
изогнутая, тонкостенная, за исключением последних оборотов очень тонко и
неясно исчерченная, почти глад-
гладкая, блестящая, прозрачная,
светлороговая или зеленоватая.
Оборотов 9—91/2, реже 10; верх-
верхние обороты выпуклые, быстро
нарастающие; 3 последних обо-
оборота наиболее крупные, почти
равной ширины, слабо выпук-
выпуклые; последний оборот книзу
Рис. 51. Serrulina (Pravispira) semilamellata (Mouss.).
(Ориг.).
A — общий вид раковины спереди; В — вид сбоку; В — клаузи-
лий.
сужен, на затылке ребристый. Устье грушевидное, слегка косое, выступаю-
выступающее. Синулус и нижний край устья отжаты назад. Края устья слегка ва-
ликообразно утолщены; между верхней и субколумеллярной пластинками
с 5—7 краевыми складочками, которые заканчиваются бугорками. Не-
Небольшая верхняя пластинка лежит косо и плавно переходит в спиральную
пластинку. Нижняя пластинка лежит глубоко и при прямом положении
устья не видна; внизу она переходит в 2, реже в 3 краевых складки.
Субколумеллярная пластинка простирается до края устья. Полулунная
складка распалась на 3 коротких складочки, которые лежат на правой
стороне раковины; из них две иараллельны главной, а третья лежит вер-
вертикально непосредственно над нижней палатальной складкой. Главная
и нижняя палатальная складки очень хорошо развиты, начинаются на
право-брюшной стороне раковины и заканчиваются недалеко* от края
устья. В промежутке между их нижними концами лежит крупный бугор,
соответствующий средней палатальной складке. Лопасть клаузилия по
внутренней стороне имеет длинный продольный киль, а иногда и слабую
вырезку.
Размеры: высота раковины 11—13, ширина 3 мм; высота устья 2.1 —
2.7, ширина 1.6—1.9 мм.
2. PONTOPHAEDUSA 123
Изменчивость. Нередко палатальный бугор отсутствует. Из-
Изредка краевых складок значительно больше A0—11 на правой стороне),
и они имеются также и на левой стороне устья (var. serrulosa Retowski,
1889).
Внутреннее строение. Материал: 1) Абхазия, долина р. Ац-
гара, сбор Калишевского, 5 VIII 1905, определил Линдгольм — 1 экз.;
2) Кавказский заповедник, Гузерипль, сбор и .определение наше, 29 VII
1956 — 1 экз.; 3) Кавказский заповедник, кордон Пслух, сбор и определе-
определение наше, 1 IX 1956 — 1 экз.
РадУла:[! + F|+з^)х2]х64.
Размеры срединного зуба 18 X 21 ц, ширина радулы 405 ц.
Половой аппарат в целом очень схож с таковым S. sieversi. Отросток
семяприемника очень длинный и вытянут вдоль предстательной железы.
У одного экземпляра внутри отростка и шейки семяприемника находился
сперматофор. Собственно семяприемник очень короткий и пустой. Его
резервуар и часть протока отогнуты вниз и прилегают к почке. Пенис
приблизительно той же длины и формы, как у остальных Serrulina.
Распространение (рис. 32). Лесные области западной и цент-
центральной части Северного Кавказа, черноморского побережья от Сочи до
Трабзона (Турция), западной и южной Грузии. Наиболее восточная на-
находка сделана в ущелье р. Бум близ селения Кемерван — Куткашенский
р-н Азербайджанской ССР (коллекция Музея Грузии).
В коллекции А. А. Стеклова (стр. 308, 309) имеется один экземпляр дан-
данного вида, найденный в отложениях меотического века (миоцен-плиоцен) на
берегу р. Фортанги у села Бумут (Черные горы к северо-востоку от Орд-
Орджоникидзе). Раковина типичной формы и отличается от современных
только меньшими размерами: высота раковины 8.2, ширина 2.0 мм; 8V4 обо-
оборотов. Кроме того, нормальные раковины найдены в четвертичных отло-
отложениях у Эльхотово.
Экология. Обитает в сырых гнилых пнях и в валежинах, чаще
всего совместно с S. serrulata serrulata. Лишь изредка встречаются более
или менее крупные популяции этого моллюска, обычно же он обитает
в количестве не больше 5—8 экземпляров в одном месте.
2. Род PONTOPHAEbuSA LINDH.
L i n d h о 1 m, 1924 : 62, 72 (Phaedusa, секция Pontapha.edu.sa); Ehrmann,
1926 : 45; Лихарев и Раммельмейер, 1952 : 213.
Тип рода: Clausilia funiculum Mouss.
Раковина башневидно-веретеновидная, деколлированная. Устье гру-
грушевидное. Колумеллярный край несет несколько коротких складок и
узелков. Верхняя пластинка сливается со спиральной. Нижняя лежит
глубоко и внизу дает широкую спираль, близко подходя к верхней пла-
пластинке, а под конец расщепляется на 2 коротких складки. Субколумелляр-
ная пластинка простирается до края устья. Полулунная складка лежит
на право-спинной стороне. Лопасть клаузилия широкая, сильно изо-
изогнутая в продольном направлении.
Распространение. Черноморское побережье Кавказа и Малой
Азии.
Один вымерший вид обнаружен нами в отложениях верхнего сармата
или меотиса (стр. 305, 309).
124
/. PHAEDUSINAE
Pontophaedusa funiculum (Mouss.) (рис. 4, 52—54).
M о u s s о n, 1863 : 397 (Clausilia); M о u s s о n, 1873 : t. 8» fig. 6 (Clausilia);
Robelt, 1893, Iconogr., N. F., 6 : 41, fig. 1020 [Clausilia (Serrulina)]; Розен,
1911:128 [Clausilia (Serrulina)]; Розен, 1914:207
[Clausilia (Serrulina)); Ретовский, 1914 : 323 [Clau-
[Clausilia (Serrulina)]; Лихарев иРаммельмейер,
1952 : 214, рис. 127. 128.
Раковина тонкостенная, ребристо исчер-
исчерченная, блестящая, прозрачная, желтовато-рого-
желтовато-роговая. Оборотов 121/2, после деколляции 5г/2—
61/2; верхние обороты выпуклые, нижние почти
плоские, разделенные тонким , линейным швом;
последний оборот вытянут и слегка сужен книзу,
на затылке с несколькими ребрышками. Устье слег-
слегка косое, с небольшим, слабо врезанным синулу-
сом. При рассматривании сбоку середина устья
выдается вперед, а синулус и низ устья отжаты на-
назад. Края устья коротко отвернуты, тупые, иног-
иногда с тонкой губой. Все пластинки простираются
до самого края устья. Короткая верхняя пластин-
пластинка, плавно снижаясь, переходит в длинную и не-
невысокую ониральную пластинку. Нижняя пла-
пластинка вглубь раковины заходит несколько даль-
дальше спиральной. Полулунная складка крупная,
сильно изогнута. Главная складка начинается над
полулунной или немного глубже и простирается
вперед на V8 оборота. Поэтому при прямом поло-
положении устья она обычно не видна. Палатальных
складок нет.
Размеры: высота раковины 14.6—16.8, ширина
4—5 мм; высота устья 3.5—4.1, шщшна 2.5—
3.0 мм.
В нашем распоряжении находилось 3 взрослых
экземпляра, у которых как исключение сохрани-
сохранились верхние обороты. Размеры: высота раковины
22.3—23.0, ширина 4.2—4.5 мм; оборотов 12V4—
12V2.
И з м е н ч и в о с т ь. Популяции, обитающие в районе Сочи, состоят
из более крупных экземпляров, чем популяции, живущие в окрестностях
Рис. 52. Pontophaedusa
funiculum (Mouss.).
А — нижняя часть ракови-
раковины (по Лихареву и Раммель-
мейер); В — клаузилий.
(Ориг.).
18
19
П
Рис. 53. Pontophaedusa funiculum (Mouss.). Радула. (Ориг.).
Батуми. Это видно из следуюпщх данных по изменчивости высоты раковины
P. funiculum (в мм), обработанных вариационно-статистическим методом.
Место-
Местонахождение
Сочи
Батуми
м
16.0
14.4
а
+ 0.80
+0.88
т
+ 0.17
+0.17
22
25
п
экз.
экз.
2. PONTOPHAEDUSA
125
Внутреннее строение. Материал: 1) Сочи, сбор А. Быкова,
5 VII 1890,, определил Линдгольм — 1 экз.; 2) Новый Афож,«фор и опре-
определение наше, 3 X 1956 — 1 экз.; 3) Батумский ботанический сад, сбор
и определение наше, 18 IV 1957 — 2 экз.
Радула (рис. 53): [| + (? + ^_) X 2] X 92:
Рис. 54. Pontophaedusa juniculum (Mouss.). По-
Половая система. (Ориг.).
Размеры срединного зуба 17 X 23 |л, ширина радулы 510 (г. Допол-
Дополнительные зубцы на срединном зубе и первых боковых выражены слабее,
чем у Serrulina.
Гонада состоит из 3—4 долек. Предстательная железа тянется вдоль
всего семяяйцевода. Семяприемник цилиндрический (рис. 54); недалеко от
верхнего конца в него впадает крупный отросток, вершина которого при-
прижата к белковой железе; шейка семяприемника очень длинная и откры-
открывается во влагалище недалеко от половой клоаки. Пенис близ нижнего
KOH4av вздут. В этом месте внутри него находятся несколько продольных
складок.
Левый ретрактор посылает ветви к месту слияния семяприемника
с влагалищем и к половой клоаке. Нижний конец ретрактора пениса рас-
распадается на несколько ветвей, которые веером крепятся к пенису.
Распространение (рис. 55). Ареал этого моллюска тянется
узкой полосой вдоль черноморского побережья Кавказа и Малой Азии
от Сочи до Требзона. Лишь в бассейне Чороха он расширяется и доходит
до Артвина.
Экология. Обитает в сырых гнилых пнях и валежинах, обычно
совместно с Serrulina serrulata -^Pfr.). Лишь в единичных случаях был
встречен под камнями, а также в даелях скал и каменных оград.
126
/. PHAEDUSINAE
46
«4
42
40
38
38 •
_ r^
H
-
55 0 55
i
38
m. <*
XLT~)
_^-\ (
Батуми,
110 165 220км
i
40
44
—. г^~:\ь
'\ ///\
\ (
A
42
Ш 48
Астрахань^
Д
4 f
-' A
Ог1рджоникидзе
ЪТбилисИ^^?-
-<рЦ?ЕГУ
^Ереван ^
\ /-«ilk
44 46
50
-** 1
p
Махач-Кала
\
A
^^
^>U
\л •¦* ' rJ
Л^
~"— ,^
48
<
^Баку
50
54 56
~T [—
> ^
\ >
7
[
\
>
52
-
-
\
-
44
42
40
38
36
.54
Рис. 55. Географическое распространение Pontophaedusa funiculum (Mouss.) Gj
Caspiophaedusa perlucens (Bttg.) ^. (Ориг.).
3. Род CASPIOPHAEDUSA LINDH.
Lindholm, 1924 : 62, 72 (Phaedusa, секция Caspiophaedusa); Ehrmann,
1926 : 37 (Hemiphaedusa, секция Caspiophaedusa).
Монотипкческий род.
Раковина деколлировайкая. Устье овально-четырехугольное, внизу
округлое. Верхняя пластинка плавно переходит в спиральную. Нижняя
пластинка далеко отстоит от верхней, ланцетовидная, отвесная. Субколлу-
меляряая пластинка простирается до самого края устья. Полулунная
складка лежит справа. Главная складка средних размеров; палатальные
складки отсутствуют.
До сих нор этот род рассматривали как секцию или подрод в пределах
рода. Hemiphaedusa Bttg. Однако внутреннее строение единственного
входящего сюда вида указывает на то, что эта группа занимает промежу-
промежуточно» положение между Phaedusinae и Enxininae и ее следует выделить
в особый род.
Caepiophaedusa perlucens (Bttg.) (рис. 56—58).
Boettger О., 1877а : 69 [Clausilia (Phaedusa)]; В о е t t g е г О., 1877b : 73
(Clausilia); Boettger О., 1878а : 105, t. 4, fig. 7 (Clausilia); К о b e 1 t, 1893,
Iconogr., N. F., 6:20, fig. 987 [Clausilia (Phaedusa)]; W es t e г 1 u n d, 1897:8
[Cl. (Serrulina) signifera, sp. nov.j; Lindholm, 1913c : 160 [Cl. perlucens Bttg. =
= d. (Serrulina) signifera West.]; Розен, 1914 : 201 [Clausilia (Phaedusa)]; Л и-
харов иРаммельмейер, 1952 : 213, рис. 126 (Hemiphaedusa).
3. CASPIOPHAEDUSA
127
P а к о-в и н а башневидно-веретеновидаая, стройная, тонкостенная,
очень тонко и неясно исчерченная, прозрачная, блестящая, зеленовато-
роговая. Оборотов 11, после деколляции 5х/2—7, плавно нарастающих,
довольно выпуклых, разделенных тонким, глубоко врезанным швом;
последний оборот одной ширины с предпоследним, за устьем сильно и
равномерно исчерчен, книзу слабо сужается. Устье отвесное,
с небольшим неврезанным сйнулусом. Края устья коротко
отвернуты и утолщены, белые или слегка коричневые.
Верхняя пластинка невысокая и, лишь немного снижаясь,
переходит в спиральную пластинку. Нижняя пластинка
лежит глубоко и при прямом положении раковины обычно не
видна; внизу она резко спускается и нередко связана с краем
устья небольшой складкой; верхний конец ее лежит выше
начала спиральной пластинки. Субколумеллярная пластин-
пластинка всегда хорошо видна. Полулунная складка лежит почти
отвесно на правой стороне; верхний конец ее нередко отог-
отогнут назад под углом, а нижний плавно отогнут вперед. Глав-
Главная складка начинается немного глубже полулунной и про-
простирается вперед приблизительно на 1/2 оборота. При пря-
прямом положении устья она видна плохо или совсем не видна. Ло-
Лопасть клаузилия желобчатая, с почти параллельными края-
краями; нижний конец ее утолщен. Имеется узкая и короткая
пупочная щель.
Размеры: вы&ота деколлированной раковины 12—15,
ширина 3.6— 4.0 мм; высота устья 3.3—3.8, ширина 2.5—
2.8 мм.1 Единственная недеколлирован-
ная раковина, которой мы располагаем,
имеет следующие размеры: высота 18.7,
ширина 3.7 мм; высота устья 3.6, ширина
2.6 мм; оборотов 11.
Бетгер, а вслед за ним и другие авто-
авторы указывают на присутствие у этого
вида сутуральных складок. Однако, как
внимательно мы не просматривали все
имеющиеся в нашем распоряжении рако-
раковины этого моллюска, в том числе и не-
несколько, определенных Бетгером, указан-
указанных образований нам обнаружить не уда-
удалось.
Внутреннее строение. Ма-
Материал: 1) Талый, Лерик, сбор и опреде-
определение наше, 5 VI 1952 — 2 экз.; 2) Закаталы, сбор и определение наше,
22 VI 1953 — 2 экз.; 3) Ялама, сбор и определение наше, 14 V1955 — 3 экз.
Радула (рис. 57): [| + (f + ^i) X 2~]jX c/>85.
Размеры срединного зуба 21 X 24 ц, ширина радулы 570 ц. Средин-
Срединный и боковые зубы с довольно крупными дополнительными зубцами.
Краевые зубы с 3 зубцами, только предпоследние с 4 (по одному зубу
с каждой стороны, ряда).
Рис. 56. Caspiophaedusa perlucens
(Bttg.).
А — общий вид раковины (по Лиха-
Лихареву и Раммельмейер); Б —клауаилий.
(Ориг.у.
1 Размеры лектотина (Зенкенбергский музей, Франкфурт на Майне, № 61559а):
высота раковины 14, ширина 3.8 мм; высота устья 3.5, ширина 2.5 мм. Место сбора:
«Кавказ».
128
/. PHAEDUSINAE
Предстательная железа значительно короче семяяйцевода и лежит
вдоль его верхней трети. Очень характерную форму имеет семяприемник
(рис. 58). Длинный узкий отросток (иногда более длинный, чем собственно
семяприемник) вытянут вдоль мужского канала семяяйцевода и прости-
; 6 7
Рис. 57. Caspiophaedusa perlucens (Bttg.). Радула. (Ориг.).
Дается почти до самой белковой железы. Шейка семяприемника очень
длинная, так как отросток^впадает в семяприемник на значительном уда-
удалении от его нижнего конца. Резервуар семяприемника не прилегает
ксемяяйцеводу, а отогнут от него, так что нередко между ними прохо-
проходит пищевод. К шейке, семя-
семяприемника крепится ветвь ле-
левого ретрактора. Влагалище
почти той же длины, что и матка,
прупныи цилиндрический пе-
пенис у заднего конца веретено-
веретенообразно расширен. Здесь к нему
крепится мощный ретрактор,
который у экземпляров из Та-
лыша и Закаталы другим кон-
концом крепится к диафрагме, а у
экземпляров из Яламы являет-
является ветвью заднего (у 2 экзем-
экземпляров) или левого (у 1 экзем-
экземпляра) ретрактора. Расширен-
Расширенная часть пениса внутри имеет
4—5 продольных валика, кото-
которые к переднему концу сходят
на нет. ;
Во внутреннем строении дан-
данного вида имеется ряд особен-
особенностей, необычных для Phaedu-
sinae — это в первую очередь
сравнительно узкий отросток и
наличие ретрактора семяпри-
семяприемника. Слияние ретрактора
пениса с коллумеллярной мускулатурой тоже .как будто несвойственно
этому подсемейству. Однако тот факт, что отросток не отличается по ги-
гистологическому строению от собственно семяприемника, а также то, что
они сливаются друг с другом на значительном удалении от нижнего конца
семяприемника, говорит о принадлежности этого моллюска к подсемейству
Phaedusinae. По-видимому, род Caspiophaedusa представляет собой мор-
морфологический переход от Phaedusinae к Euxininae и Clausiliinae.
Распростр анение (рис. 55). Лесные области Талыша и се-
северного Ирана, южных склонов Большого Кавказа от Лагодехи на западе
Рис. 58. Caspiophaedusa perlucens (Bttg.). По-
Половая система. (Ориг.).
3. CASPIOPHAEDUSA 129
и.до Худата на востоке; лесные области северных и северо-восточных
отрогов Малого Кавказа; леса Зангезура.
Указание Жермена (Germain, 1936), что этот моллюск обитает и в за-
западном Закавказье, совершенно необоснованно и не соответствует дей-
действительности.
Две раковины данного вида найдены А. А. Стендовым в четвертичных
суглинках двухметровой террасы р. Ярык-Су (Дагестан).
Экология. Обитает в широколиственных лесах в дуплах, в гни-
гнилых пнях и под сырой корой гниющего валежника.
ДОПОЛНЕНИЕ К ПОДСЕМЕЙСТВУ PHAEDUSINAE
Раковина моллюска, описание которого дается ниже, бесспорно указы-
указывает на его принадлежность к подсемейству Phaednsinae. Но отсутствие
«ведений.о внутреннем строении этого моллюска затрудняет решение во-
вопроса о том, в какой род его следует отнести. Поэтому мы сохраняем за ним
первоначальное родовое название, лишь взяв его в кавычки.
*«Phaedusa» hyrcanica Morgan (рис. 59).
М organ, dans Germain, 1933 : 390 [Phaedusa (Caspiophaedusa)]; Ger-
Germain, 1936 : 310, fig. 21 [Phaedusa (Caspiophaedusa)],
P а к овина бапгаевидно-веретеновидная, стройная, тонко и неравно-
неравномерно, местами грубо, исчерченная, блестящая, просвечивающая, светло-
роговая. Верхушка острая. Оборотов 10—11, выпуклых, плавно нараста-
нарастающих, разделенных тонким и глубоким швом; последний оборот книзу
«ужен, сбоку прижат, на затылке ребристый. Устье округло-грушевидное,
сильно выдается вперед, с узким, глубоко врезанным синулусом. Края
устья слегка отвернуты и утолщены, белые. Верхняя пластинка низкая,
изогнутая, плавно снижается и переходит в спиральную. Нижняя пла-
пластинка спирально закручена, близко подходит к верхней, но при прямом
положении раковины почти не видна. Субкодумеллярная пластинка лежит
глубоко; нижний конец ее обрублен- и виден лишь сбоку. Вглубь рако-
раковины дальше всех заходят нижняя и субколумеллярная пластинки.
Полулунная складка лежит на спинной стороне раковины; верхний конец
•ее плавно отогнут назад, а нижний — к устью, где он переходит в мозоле-
видную нижнюю палатальную складку. Последней на затылке соответ-
соответствует ясная продольная борозда. Главная скла*дка начинается над полу-
полулунной и простирается вперед на V4 оборота. Рукоятка клаузилия, плавно
расширяясь, переходит в узкую желобчатую лопасть с почти параллель-
параллельными краями. Передний конец лопасти, клювовидно изогнут, заострен и
утолщен.
Размеры (по Жермену): высота раковины 15—18, ширина 3 мм. Один
из наших экземпляров имеет следующие размеры: высота раковины 14.5,
ширина 3.4 мм; высота устья 2.8, ширина 2.2 мм.
Распространение. Северный Иран (провинции Гилан и Мазен-
деран).
Экология. Обитает во влажной гниющей древесине пней и пова-
поваленных стволов деревьев.
Систематическое положение. Описания данного вида,
приведенные у Моргана и Жермена, составлены очень неполно и в основ-
основном ограничиваются внешними признаками раковины. Наше описание
«оставлено на основании изучения 2 экземпляров, полученных из Зоологи-
то
II. ALOPIINAE
ческого института Польской Академии наук (сборы Гейнриха, 1926 г.),
из которых один дефектный. п
Жермен сближал данный вид с Caspiophaedusa perlucens (Bttg.) и счи-
считал, что первый отличается от последнего целой, т. е. недеколлированной
раковиной, более овальным устьем, узким синулусом, более крупной и
изогнутой верхней пластинкой и, наконец, меньшими размерами всей
• раковины.
Между тем, кроме этих уже существенных различий,
можно добавить еще некоторые. А именно, у «Ph.» hyrca-
nica полулунная складка располагается на спинной сто-
стороне, в то время как у С. perlucens она находится на пра-
правой стороне раковины. Кроме того, у данного вида нижняя
пластинка спирально закручена и сближена с верхней,
а у С. perlucens она лежит отвесно и отстоит далеко от
верхней.
По форме замыкательного аппарата данный вид ва
многом сходен с Pontophaedusa funiculum (Mouss.) ir
с Serrulina sernilamellata (Mouss.), но отличается по дру-
другим существенным признакам.
И. Подсемейство ALOPIINAE ,
Раковина право- илилевозакрученная. Кроме 3 основ-
основных слоев, из которых периостракум окрашен в темно-
коричневый или роговой цветг
имеется еще четвертый, самый
внешний, который имеет вид свет-
светлого непрозрачного налета. Этот
слой в одних случаях есть только-
у шва, где он образует светлые-
сосочки или тонкую линию; в дру-
других случаях он образует радиаль-
радиальные полосы или ребра; наконец,.
у некоторых он почти сплошь за-
закрывает все дефинитивные оборо-
обороты раковины. Последний оборот
снизу закруглен. Устье крупное,
округло-грушевидное, невыступающее (верхний край плотно прижат к:
последнему обороту раковины). Пластинки, складки и клаузилий в од-
одних группах хорошо развиты, в других испытывают частичную или пол-
полную редукцию. Верхняя и спиральная пластинки лежат на различном рас-
расстоянии от столбика и не сливаются друг с другом. Нижняя пластинка глу-
глубоко выдается в просвет устья в виде более или менее горизонтальной склад-
складки. Полулунная складка может быть, но может и отсутствовать. В пос-
последнем случае палатальные складки обычно хорошо развиты. Клаузилий
желобчатый, с вырезкой на переднем конце, у некоторых рудиментарный:
или совсем отсутствует.
Срединный и боковые зубы радулы у большинства видов без дополни-
дополнительных зубцов, т. е. однозубцовые.
Семяприемник открывается в женские половые пути на значительном
удалении от половой клоаки, благодаря чему влагалище довольно длинное.
Отросток такой же ширины, как и семяприемник, и впадает в последний
высоко от его нижнего конца. Цилиндрический или веретеновидный Пенис
имеет небольшой рудиментарный бич и нередко еще крупный крючковидный,
1 Б
Рис. 59. «Phaedusa» hyrcanica Morgan.
(Ориг.).
А — общий вид раковины; В — клаузилий.
ALOPIA .131
придаток. У многих видов диафрагмалышй ретрактор пениса прикреп-
прикрепляется к, последнему двумя ветвями. Ретрактор глазного щупальца ледаит
между пенисом и влагалищем.
Распространение. Южные и восточные Альпы, Апеннины,
восточные и южные Карпаты, Балканский полуостров, Сицилия, Мальта,
Алжир, Тунис, Крит, острова Эгейского моря, Кипр, юго-западное, побе-
побережье Малой Азии, Сирия.
Род ALOPIA H. et A. ADAMS.
Adams Н. а. А., 1855 A858) ; 181 (Clausilia, подрод Alopia); Wagner A.r
1914, Iconogr., N. F., 21 : 5 (подрод Alopia s. str.); Wagner A., 1919 : 90 (подрод
Alopia s. str.); Lindholm, 1924 : 55 (подрод Alopia s. str.); S oi s, 1928 : 261.
Тип рода: Clausilia livida Menke.
Внешний непрозрачный слой раковины чаще всего имеет вид белова-
беловатого налета, сквозь который просвечивает темный периостракум. Замыка-
тельный аппарат у высокогорных видов и форм рудиментарный или пол-
полностью отсутствует, а у видов и форм, обитающих в средних горизонтах гор
и в предгорьях, хорошо развит. Нижний конец субколумеллярной пла-
пластинки виден через устье. Задние концы палатальных складок находятся на
спинной стороне раковины.
Распространение. Основная масса видов обитает в южных
Карпатах и в Трансильвании. Лишь единичные проникают на северо-запад
до Чехословакии {Alopia bielzi clathrata Rssm.) и в восточные Карпаты
(Alopia glauca E. Bielz). ;
Обитатели скал, сложенных из юрских, меловых и эоценовых известня-
известняков.
*Alopia glauca (E. Bielz) (рис. 60).
Bielz Е. А., 1853, Verh. Mitt. Siebenb* Ver. Naturw., IV : 120 (Balea part.);
Rossmassler, 1859, Iconogr., 3 : 123, fig. 954 (Clausilia part.); Kimakowicz,
1894 : 26 (binodis); Wagner A., 1914, Iconogr., N. F., 21 > 12, fig. 34—37 (анато-
(анатомия); S о 6 s, 1928:289,385.
Раковина левозакрученная, веретенрвидно-цилиндрическая (по-
(последние 2 оборота обычно одной ширины), тонко и редко исчерчена (линии
нарастания), реже грубо ребристая, чаще всего непросвечивающая., со
слабым блеском, красновато-коричневая или коричнево-лиловая, иногда
с тонким белым налетом. Верхняя часть завитка более темная и заканчи-
заканчивается широкой A.3—1.5 мм) округлой верхушкой. Оборотов 9—10, вы-
выпуклых, плавно нарастающих, разделенных мелким белым, иногда,!зазуб-
иногда,!зазубренным швом; последний оборот книзу слегка сужен, на затылке с более
сильной скульптурой. Устье овальное или коротко-грушевидное,*, лежит
слева от оси раковины, наверху (в области слабо выраженного" синулуса)
слегка угловатое, внизу плавно закругленное. Края устья цельные, ши-
широко отвернуты, сильно утолщены, с коричневато-белой губой. Верхняя
пластинка представлена небольшой складкой, порой распадающейся на
2 бугорка. Нижняя пластинкатожесла?д_пдздита и не всегда видна при
прямом положении устья; имеет вид легкой складки, начинающийся на
спинной стороне столбика и заканчивающейся близ его нижнего йойца,
далеко от края устья. Иногда есть намек на одну палатальную складку.
Клаузилия нет.
Размеры (по: Soos, 1943): высота раковины 13.7—16.5, ширина 3.8 —
4.0 мм.
132
///. COCHLODININAE
'Мы измерили 4 экземпляра, собранных в Молдовских горах, у Чахлэу
(Румыния, восточные Карпаты). Они имеют следующие размеры: высота
раковины 17.3—19.2, ширина 5.0—5.2 мм; высота устья 4.2—4.6, ширина
3.6—3.8 мм; оборотов 10—10У2.
•'Изменчивость. Наряду с типичными экземплярами встречаются
также такие, раковина которых крупнее и стройнее и несет многочисленные
крупные ребра, окрашенные в тот же цвет, что и вся раковина (var. latens
Pfr.); размеры: высота 14.0—18.7; ширина 4.3—4.6 мм.
Ра сп р о с т р а н е н и е. Найден в ряде пунктов
румынской части восточных Карпат в бассейнах рек
Молодова и Бистрица. Наиболее близкая к границам
СССР находка сделана близ города Кымпулунг-Молдо-
венеск.
Экология. Обитает на известняковых скалах,
на высотах 1300—1900 м над ур. м.
III. Подсемейство COCHLODININAE
Раковина левозакрученная, слабо исчерченная или
почти гладкая, от роговой до красновато-коричневой.
Последний оборот снизу округлый. Верхняя и спираль-
спиральная пластинки лежат на различном удалении от стол-
столбика и концы их нередко'заходят друг за друга. Нижняя
пластинка глубоко выдается в просвет устья в виде
горизонтальной складки со вздутым краем, широкой
спиралью уходящей вглубь раковины. Полулунная
складка всегда отсутствует. Ниже глазной складки
лежат крупные верхняя и нижняя палатальные склад-
складки. Передний конец лопасти клаузилия имеет одну боль-
большую, реже еще вторую маленькую вырезки.
Срединный и боковые зубы радулы с дополнитель-
дополнительными зубцами.
Семяприемник открывается в женские половые пути
значительно ?ыше половой клоаки. Отросток его рас-
пол агаеРгся высоко; по форме и гистологической струк-
структуре он" мало отличает^ от собственно семяприемника. Пенис верете-
' новидн6ивздут у заднего КЙЙЩа, к которому крепится диафрагмальный рет-
'рактор (рис. 21).
*: Левый ретрактор тела д&ет ответвления к шейке семяприемника, к вла-.
^галищу и к половой клоаке. Ретрактор левого глазного щупальца прохо-
проходит между пенисом и влагалищем.
' Ра с п р остранеяие. Средняя и южная Европа. Один вид —
'Cochlodiha laminata (Mont.) распространен почти по всей Европе.
Рис. 60. _, A lopia
, glauca, (BJelz).
(Ориг.).
Род COCHLODINA FER.
F e i u s s а е,
, 1821 (январь), ТаЫ. syst. Limagons : 65; Gray, 1821 (март),
iJftnd. Med. sBeyosit, 15 : 239 (Marpessa); P f e i f f e г L., 1855, Malak. BL, 2 : 180
(CtausHifiif^a)^ Wagner A., ,1920 : /- "nr'»"~'"" "«« п»„„„™„„., л <onn.
'.Liii dt/j.l.m, 1924:60 (Marpessa);
1931 : 541. " '
4 (Clausilia, non Draparnaud, 1805);
Lindholm, 1925 : 262; Thiele,
Тип рода: Clausilia bidens Drap. (= Cochlodina laminata Monfc.).
Раковина блестящая, нередко очень блестящая. Последней оборот
за Устьем с затылочным утолщением, без киля, вместо которого у многих
COCHLOPINA 133
видов имеется более или менее крупный бугорок. Устье не выступающее,
без базального желобка, с широким неглубоким и: неврезанным синулусом.
Верхний край устья плотно прилегает к раковине и либо цельный, либо
разорван; в последнем случае места прикрепления связаны более или менее,
толстой париетальной мозолью; нижний край устья широко закруглен.
Нижняя пластинка тянется вниз, нарастая в высоту и толщину; в просвете
устья она образует широкий, почти горизонтальный изгиб, после чего
вновь тянется вниз и заканчивается недалеко от края устья (но с ним не
связана). Нижний конец субколумеллярной пластинки у большинства
видов виден при прямом положении устья. Главная складка и лежащие
под ней палатальные складки начинаются на спинной стороне, -реже
несколько смещены вправо. Главная складка наиболее длинная, осй&лъ^ые
различной длины. На некотором расстоянии от края устья имеется^ обьршо
хорошо развитая, палатальная мозоль, которая у некоторых вид|>в посы-
посылает от себя внутрь раковины короткие палатальные складки (ложный).
Пупочная щель довольно широкая. Рукоятка клаузилия плавнорасщл?-
ряется в лопасть; последняя (рис. 8, Б) име&т впереди, по внешнему кра|о,
глубокую вырезку, через которую, в тех случаях, когда животное втяну-
втянулось вглубь раковины и клаузилий замкнул просвет ее, проходит нижнявг
палатальная складка. I
Тонкий семяпровод перед впадением в .пенис расширяется, образую
эпифаллус (рис. 21). Последний внутри имеет, многочисленные продольнж
складки, а от пениса обособлен перетяжкой. Пенис толще эпйфаллура,
цилиндрический. К нижнему концу семяпровода прилегает тонкий йач
(не всегда хорошо выражен). В остальном половая система такая же, к<?к
у всего подсемейства. ,'•
Р а^с пространение. Большая часть Европы, за исключением
самых северных и самых южных ее областей. Наибольшее число щДоЦ
обитает в юго-восточных Альпах и в северо-западных Балканах. * ,-?
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА COCHLODJNA /
1 B). Поверхность раковины правильно и тонко ребристо исчерчена.
Верхушка короткая, толстая и тупая. Ширина раковины 2.8—
3.2 мм 4 . 1. С. orthostoma (Menke).
2 A). Поверхность раковины тонко и неравномерно исчерчена, нередко
почти гладкая. Верхушка более вытянутая, узкая и острая. Ширина
раковины 3.2 мм и более.
3 D). Палатальная мозоль очень толстая, цельная, белая. Верхняя
и нижняя палатальные складки приблизительно одной длины и не
достигают мозоли 2. С. cerata (Rssm.),,
4C). Палатальная мозоль развита .слабее, особенно в средней части,
где она бывает нередко прервана; обычно она окрашена в цвет устья.
Нижняя палатальная складка всегда значительно длиннее верхней
и простирается до мозоли, иногда сливаясь с последней.
5 F). Раковина толстая; ширина ее около 4 мм. Нижняя палатальная
складка сливается с мозолью. Средней палатальной складки нет
3. С. laminata (Mont.)
6 E). Раковина более узкая; ширина ее 3.2—3.4 мм. Нижняя палаталь-
палатальная складка пересекает мозоль, которая в этом месте прервана
и не сливается с названной складкой^ Верхняя часть мозоли по,сыг
лает внутрь раковины среднюю палатальную складку, которая
обычно косой перемычкой соединяется с верхней палатальной
складкой ...:.. 4. С. commutat» (Rssift.),.
134
III. COCHLODININAE
1. Cochlodina orthostoma (Menke) (рис. 61).
Menke, 1830, Syn. method. Moll., ed. II : 130 (Clausilia); Rossmassler.
1836, Iconogr., 1, 3 : 21, fig. 192 (Cl. taeniata); Kiister, 1847—1862 : 123, t. 13,
fig. 23—27 (Clausilia); M e г m о d, 1930 : 385, fig. 83 (анатомия); Лихарев и
Раммельмейер, 1952 : 250, рис. 178.
Раковина веретеновидная, стройная, с короткой, цилиндрической,
толстой и очень тупой верхушкой, правильно и тонко ребристо исчерчена
(последний оборот исчерчен слабее и неправильно), про-
просвечивающая, блестящая, светло-или красновато-роговая,
нередко зеленоватая. Оборотов 91/2—Ю1/2, верхние доволь-
довольно выпуклые, нижние слабо выпуклые; эмбриональные
обороты (около 2) гладкие; последний оборот__за. устьем
имеет ясное затылочное^дтадшение" Устьеяйдамдно-
грущевидное."'Крайнего~широкии, с толстои^^етот^ли ко-
PJ55E!a!?&JCS& ' Тонкая и короткая верхняя пластинка
впереди не доходит до кра.я^у_стья. Высокая спиральная
пластинка_начжнается"Т1а пдавой^^до^ще^раковины^ й,
плавшГснижаяеь, простирается до_левой_сто?оны. Круп-
Крупная нижняя пластинка начинается выше спиральной.
Длинная главная складка "занимает около 1/2" оборота;
она начинается на спинной стороне (с некоторым смеще-
смещением вправо) и заканчивается у палатальной мозоли,
обычно сливаясь с последней. На спинной стороне рако-
раковины начинаются верхняя и нижняя палатальные склад-
^^^_я-, ки; последняя очень массивная в 2 раза длиннее первой,
?|аЯ|1|§§1 впереди доходит до палатальной мозоли, с которой не-
ЯВеИШШ редко сливается. Крупная белая палатальная мозоль
у вполне взрослых экземпляров не прервана, распола-
располагается на значительном удалении от края^устья параллель-
параллельно последнему.
Размеры: высота раковины 11—13, ширина 2.8—3.2 мм;
высота устья 2.8—3.0, ширина 2.1—2.3 мм.
Изменчивость. Раковина данного вида срав-
сравнительно мало варьирует. В горах иногда встречаются
популяции, раковины которых отличаются более слабой
исчерченностью и большей стройностью (var. filiformis
Е. Bielz, 1861) или же достаточно вздутые, и короткие — высота 10—
11 мм.
Распространение (рис. 62). В СССР: Карпаты и Предкарпатье
(Львовская, Станиславская, Тернопольская, Черновицкая, Закарпатская
области), многочисленные популяции по всей Прибалтике; отдельные
находки в Брестской (Полесье), Витебской (Полоцк, Витебск), Псковской,
Смоленской (Рославль) и Орловской (Троено) областях. По непроверен-
непроверенным данным (Ельский, 1862) обитает в окрестностях Киева.
Ареал данного моллюска состоит из двух больших изолированных
друг от друга частей: 1) равнинная, располагающаяся вдоль южного побе-
побережья Балтийского моря от Нарвы до Щецина и простирающаяся в глубь
континента на юго-восток до Рославля и Троено; 2) горная, охватывающая
всю' дугу Карпат с их предгорьями и в западной части Чехословакии рас-
распадающаяся на две ветви — судетскую (простирается на северо-запад
до Лужицких гор включительно) и северо-альпийскую (Известковые Альпы,
включая Зальцбург, северный Тироль, северную Штирию и верхнюю Ба-
Баварию). Кроме того, небольшие изолированные ареалы встречаются в про-
Рис. 61. Cochlo-
Cochlodina orthostoma
(Menke).
(Ориг.).
G0CHL0D1NA
135
межутках между обеими ветвями ив северной Швейцарии. Одна находка
сделана на юго-западном побережье Финляндии.
Раковины данного вида найдены в четвертичпых отложениях ряда мест
Германии (ГДР и ФРГ) и Чехословакии.
Экология. Обитает в смешанных и широколиственных лесах, из-
изредка в чисто хвойных лесах, на влажных скалах и заросших мхом де-
деревьях, а также в подстилке. На полонинах Карпат обитает под камнями.
«о
30
ад
Рис. 62. ^Географическое распространение Cochlodina laminata (Mont.) и С- ortho-
stoma (Menke). (Ориг.).
la — С. laminata; 16 — то же, изолированные ареалы; 2а — С. orthostoma; 26 — то же, изо-
изолированные ареалы.
2. Cochlodina cerata (Rssm.) (рис. 63).
Rossmassler, 1836, Icoqogr., 2,4 : 15, fig. 258 (Clausilia); R о s s m a s-
s 1 e r, 1838, Iconogr., 2,7 : 17 (Cl. parreyssii); Kiister, 1847—1862 : 313, t. 35,
fig. 27—29 (Claysilia); S о 6 s, 1917 : 149, fig. 51, 52 (анатомия); Lozek, 1956 :
122—123, t. 11, fig. 2.
Раковина веретеновидно-цилиндрическая, стройная, твердостеи-
ная, на верхних оборотах тонко и равномерно, на нижних очень слабо и
неравномерно исчерченная, почти гладкая, сильно просвечивающая, очень
блестящая, от роговой до красновато-коричневой. Верхушка слабо су-
сужена, притупленая. Оборотов 10—11, верхние умеренно, нижние слабо
выпуклые; последние 3—4 оборота почти одинаковой ширины. Последпий
оборот без бугорка. Устье овальное или яйцевидно-грушевидное, с белой
136
///. COCHLODININAE
Mis
губой. Тонкая и высокая верхняя пластинка впереди не достигает края
устья, сзади слегка заходит за спиральную пластинку. Верхний конец
последней лежит на правой стороне раковины. Нижняя пластинка начи-
начинается на V4 оборота выше спиральной. Главная и обе палатальные складки
начинаются на спинной стороне. Главная складка умеренной длины (зани-
(занимает около 1/3 оборота); ее передний конец нередко сливается с палаталь-
палатальной мозолью. Как верхняя, так и нижняя палатальные складки очень
короткие, в особенности верхняя, не сливаются с пала-
палатальной мозолью и не видны при прямом положении устья.
Палатальная мозоль хорошо развита, белая, лежит близко
к краю устья, цельная. Лопасть клаузилия, кроме основ-
основной глубокой вырезки, имеет на переднем конце еще дру-
другую небольшую вырезку.
Размеры: высота раковины 15—17, ширина 3.5—4.0 мм;
высота устья 3.5—4.0, ширина 2.7—2.9 мм.
Распространение. В СССР: Закарпатская
область (Soos, 1941). Вне СССР: Карпаты (кроме юж-
южных), Трансильвания.
Экология. Живет в горных лесах, произрастающих
на известняковых породах, на стволах деревьев и скалах.
Систематические замеча-
I н и я. Ложек (Lozek, 1952) описал но-
новый вид С., remota, найденный в горах
Втачник (южный отрог словацких запад-
западных Карпат). Главными отличиями этого
вида от С. cerata являются следующие: 1)
раковина приземистая, коротко-веретено-
видная; 2) палатальная мозоль не лежит
параллельно краю устья. В остальном оба
вида очень схожи.
Так как до сих пор С. remota известна
по одной находке нескольких экземпля-
экземпляров, то нет твердой уверенности, что
этот вид действительно существует. Впол-
Вполне возможно, что названный автор имел
дело с каким-то внутривидовым отклоне-
отклонением от типичной С. cerata, тем более, что
у других видов того же рода в некоторых
пределах наблюдаются колебания в форме раковины и в положении пала-
палатальной мозоли.
Ж
Рис. 63. Cochlodlna cerata (Rssm.).
(Ориг.).
А — общий вид раковины; Б — вид
сзади на последний оборот и на про-
просвечивающие скьозь него складки.
3. Cochlodina lamina la (Mont.) (рис. 8, Б, 21, 64). ¦?
!** М ti 1 1 е г, 1774, Verm. terr. et fluv., 2 : 116 '(Helix bidens, non Linne, 1758);
Montagu, 1803, Test. Brit. : 359, t. 11, fig. 4 (Turbo); Rossmiissler, 1835,.
Iconogr., 1, 1 : 76, fig. 29 (Clausilia bidens); Ruster, 1847—1862 : 109, t. 12, fig. 13—
30 (Clausilia); Steenberg, 1914 : 26—27, fig. 11—14 (анатомия); Лихарев
и Раммельме й.е р, 1952 : 251, рис. 179. ^
Раковина веретеновидная, довольно вздутая, твердостенная, очень
¦юнко "и неправильно исчерченная, на нижних оборотах почти гладкая,
сильно просвечивающая, блестящая, от светло-роговой до красновато-
коричневой. Оборотов 10—111/2, слабо выпуклых; эмбриональные обороты
(около 2V2) образуют тупую гладкую верхушку; последний оборот внизу
COCHLODINA
137
с небольшим бугорком. Устье округло-ромбическое. Края устья довольно
широкие, с белой губой. Верхняя пластинка^тонкая,впереди достигает
кр_аягхс|ья или мозоли, сзадислет^Ъаходит за передний к6н(щ"спйр1ль"Я6й
пластинки. Последняя "ввиде высокой складки начинается на правой
стороне й, плавно снижаясь, простирается до левой стороны раковины.
Крупная нижняя пластинка^ачинается выше спиральной.
Нижний конец суб^колумеллярной "пла^йгоШ^гтте^иден
или слабо заметен при косом положении устья. Длин-
Длинная главная складка занимает около V2 оборота; она на-
начинается на спинной стороне (с некоторым смещением
вправо), а заканчивается у палатальной мозоли, часто
сливаясь с последней. На спинной стороне раковины на-
начинаются верхняя и нижняя палатальные складки; по-
последняя очень массивная, в 2 раза длиннее первой, впе-
впереди сливается с палатальной мозолью. Палатальная
мозоль сравнительно слабо развита; по концам она более
или менее толстая, но в середине тонкая, нередко прерва-
прервана; снаружи ей соответствует затылочное утолщение; ниж-
нижний конец мозоли Нередко вытянут вглубь раковины в ви-
виде короткой складки. У клаузилия, кроме основной боль-
большой вырезки, нередко имеется еще другая, меньших раз-
размеров, на самом конце лопасти.
Размеры: высота раковины 14—18, ширина 3.8—4.2 мм;
высота устья 3.8—4.2, ширина 2.8—3.2 мм.
Изменчивость. Несмотря на обширный ареал
вид сравнительно мало изменчив. Наибольшим колеба-
колебаниям подвержена высота раковины, которая в неблаго-
неблагоприятных условиях (например, высоко в горах) умень-
уменьшается до 12—14 мм (f. alpestris Stoll), а в особо благо-
благоприятных условиях достигает 19—20 мм. Ширина рако-
раковины изменяется значительно меньше.
Распространение (рис. 62). В СССР: вся При-
Прибалтика, многочисленные находки в окрестностях Москвы
и Ленинграда, районы Карельской АССР к северу от Ла-
Ладожского озера, Псковская, Витебская, Брестская, Смо-
Смоленская, Калужская, Ярославская, Владимирская (Му-
(Муром), Горьковская (Васильсурск) области, юг Кировской
области (Уржум, Малмыж), Казань, Жигули, Тульская
(Алексин, Тула), Брянская, Орловская (Троена), Тамбов-
Тамбовская (Мичуринск), Белгородская (Новый Оскол), Воро-
Воронежская (Павловск), Харьковская (Харьков), Полтавская
(Гадяч), Кировоградская (Александрия), . Черкасская
(Корсунь-Шевченковский), Киевская (Киев, Ирпень),
Винницкая (Ольгополь) области; горный Крым (район
Чатыр-Дага), байрачные леса Ставрополья (Ставрополь,
Георгиевск, Железноводск, Пятигорск). Вне СССР: север-
северная граница ареала проходйт'чёрез южную Шотландию,
южную Норвегию (до 64° с. ш.), южную Швецию и Фин-
Финляндию (до 62° с. ш.). Вся средняя и значительная часть
западной Европы (Франция, Пиренеи). В южной Европе (Италия, южная
часть Балканского полуострова) ареал носит разорванный характер.
В средней Европе и в западных областях европейской части СССР
данный вид является одним из наиболее многочисленных и часто встре-
встречающихся моллюсков. ,
Рис. 64. Coch-
lodina laminata
(Mont.). (Ориг.).
A — общий вид
раковины; В —
последний оборот
(сзади) и просве-
просвечивающие сквозь
него складки.
138
///. COCHL0DIN1NAE
В четвертичных отложениях раковины этого вида часто встречаются
в пределах современного ареала, в том числе и в СССР (Ленинградская
область, — Даниловский, 1955).
Экология. Обитает в смешанных и широколиственных лесах,
в старых парках и садах, а в горах и в хвойных лесах. Живет на мшистых
стволах деревьев и под корой гнилых пней и валежин, в подстилке, на сы-
сырых скалах и камнях; избегает болотистых мест. Сравнительно с другими
видами не очень требователен к влажности, но отсутствует в сухих биото-
биотопах. К горным породам довольно индифферентен.
Размножение. Копуляция обычно происходит в августе. В том
же месяце или в начале сентября идет откладка 10—15 яиц, каждое раз-
размером около 2 мм. Развитие яиц длится 3 недели. Рост заканчивается
к концу второго года жизни.
Питание. Животные питаются раз-
различными грибами и гниющими частями выс-
высших растений.
4. Cochlodina commutata (Rssm.) (рис. 65).
с, с ^4На fc. e/r., /fJ8j
Rossmassler,' 1836, Iconogr.,-^ 1,4 : 19,
fig. 269 (Clausilia); R ossmassler, 1838, Ico-
nogr., 2,7 : 16, fig. 464 (Cl. ungulata); К ii s t e r,
1847—1862:114, t. 12, fig. 40—44 (Clausilia);
Schmidt A., 1868 : 33 (Cl. silesiaca); Лиха-
Лихарев и Раммельмейер, 1952 : 252; L o-
zek, 1956 : 124—125, t. 11, fig. 5.
Раковина веретеновидная, стройная,
слабо и несколько неравномерно исчерчена,
просвечивающая, с легким блеском, светло-
или красновато-роговая. Оборотов 10—11,
слабо выпуклых; последний" оборот"*внизу
вздут и справа 5т пупочной щели несет за-
заметный бугор, а за устьем — с крупным
затылочным утолщением. Устье яйцевидно-
грушевидное, нередко с косо расположенной
осью (за счет некоторого наклона колумелляр-
ного края), с белой губой. ToHKj*Hjnjopj3T_KaH
верхняя пластинка впереди достигает края
устья. Верхний конец спиральной пластинки
уходит вглубь раковины почти до брюшной
стороны и лежит либо рядом с началом нижней
пластинки, либо даже несколько выше; вы-
высота ее плавно нарастает к середине и
так же плавно убывает к нижнему концу. Длинная главная_скл^дка
начинается на спинной стороне раковины, а_впереди сливается с па-
палатальной^ мозолью. На спинной стороне начинаются верхняя и ниж-
нижняя "палатальные складки; последняя очень массивная, и ее нижний конец
хорошо виден при прямом положении устья. Толстая палатальная мозоль
•состоит из двух самостоятельных частей — верхней и нижней. Между
ними (не сливаясь с мозолью) проходит нижняя палатальная складка-
Верхняя часть мозоли посылает внутрь раковины короткую, но толстую
¦среднюю палатальную складку, которая обычно косой перемычкой со-
соединяется с верхней палатальной складкой, образуя единую, коленообразно
flic. 65. Cochlodina"commutata
(Rssm.). (ПоДЭрманну).
A — общий вид раковины; Б — по-
последний оборот (сзади) и просвечи-
просвечивающие сквозь него складки.
IV. EUXININAE 139
изогнутую складку, хорошо видимую с затылка.1 Нижняя часть мозоли
выпячивается вглубь раковины в виде довольно длинной и толстой складки,
располагающейся между нижней палатальной складкой и субколумелляр-
ной пластинкой.
Размеры: высота раковины 14—15, ширина 3.2—3.4 мм; высота устья
¦3.2—3.5, ширина 2.3—2.5 мм.
Распространение. В СССР: Закарпатская область УССР.2
Вне СССР: основной ареал в юго-восточных Альпах и прилегающей части
северо-западных Балкан. Островные ареалы в Венеции — Тридентина
(Италия), во Франконской Юре и в Судетах (Германия—ГДР, Чехосло-
Чехословакия и Польша).
Экология. Обитает на затененных известняковых скалах, под кам-
камнями и в руинах.
IV. Подсемейство EUXININAE, subfam. nov.
Раковина левозакрученная; скульптура от исч,ерченности до ребристо-
ребристости; окраска от роговой до красновато-коричневой, нередко с пучками бе-
белых штрихов. Последний оборот внизу с 1, реже с 2 килями. Устье цельное,
чаще всего выступающее, с широкими отвернутыми краями, внизу с желоб-
желобком, наверху с врезанным синулусом. Верхняя и спиральная пластинки
располагаются на различном удалении от столбика и, если замыкательный
аппарат развит нормально, их концы всегда заходят один за другой.
Срединный и боковые зубы радулы чаще всего с дополнительными зуб-
зубцами, но у некоторых видов без последних.
Семяприемник всегда с ретрактором; в женские половые пути он впадает
на значительном удалении от половой клоаки. Отросток семяприемника
открывается вблизи нижнего конца последнего и превращен в особый желе-
железистый орган, имеющий вид тонкостенной, сильно извитой трубки. Только
два вида лишены отростка. Пенис и восходящее колено семяпровода обра-
образуют обычную петлю. Часто пенис имеет рудиментарный бич, реже муску-
мускулистый придаток. Ретрактор ближайшего глазного щупальца лежит между
пенисом и влагалищем.
Распространение. Кавказ, значительная часть Передней
Азии; несколько видов обитает на Балканах.
1. Род IDYLA H. et A. ADAMS
Adams Н. а. А., 1855 A858) : 180 (Clausilia, подрод Idyla); Boettger О.,
1877а : 94 (Clausilia, секция О ligoptychia part.); Kennard a. Woodward,
1923 : 303 (Pleioptychia); Lindholm, 1924 : 67; T h i e 1 e, 1931 : 536.
Тип рода: Clausilia bicristata Rssm.
Раковина веретеновидная. Последний оборот за устьем вдавлен или
имеет глубокую и резкую впадину (рис. 66, Б), внизу с 1 или 2 килями.
Устье с 1 или 2 базальными желобками; на палатальной стенке более или
менее крупный бугор. Верхняя пластинка низкая и короткая. Спиральная
пластинка рудиментарная или отсутствует. Нижняя пластинка внизу
более или менее отвесная, ланцетовидной формы и видна при прямом поло-
положении устья. Полулунная складка лежит на спинной стороне. Над ней
находятся короткие главная и сутуральные складки.
Распространение. Кавказ, Малая Азия и Греция.
1 Эта перемычка между верхней и средней палатальными складками отсут-
отсутствует или слабо развита лишь у еще не вполне сформировавшихся раковин.
Реже она отсутствует у вполне взрослых особей.
2 По данным А. А. Полевиной A959) и В. И. Здуна A960а).
140
IV. EUXININAE
Подрод SCROBIFERA BTTG.
'era);
Boettger O., 1877a : 95 (Clausilia, секция Oligoptyehia, группа Scrobif
Lindholm, 1924 : 67 (Idyla, подрод Scrobifera).
Монотипический подрод.
Затылочная впадина глубокая, трех- или четырехугольной формы.
Киля 2. Устье с 2 базальными желобками, внутри с очень крупным пала-
палатальным бугром, сильно сужающим просвет устья. Нижняя пластинка
лежит глубоко, частично закрыта палатальным бугром, но все же видна
при прямом положении устья. 4гш,% У <"р?
VCharn.WTm'r, Aft
- tSerobifera)
г у n/c k i, 1837 : 5
(Charp.) (рис. /16, А, 66, 67).
rlah/i7ia taurica, nom. nudum); Pfeiffer L., 1848,
Monogr. belie, viv., 2 : 412 (Cl. taurica Krynicki); Charpentier, 1852 : 400 (Clau-
__• ?• \ _ УГ -•!_!._„ ЛОГП IDl?» . JTH x A t\ ?1~ Л О / /~< 1 ¦ i _'_\. ПЛ i\ i ОС *:_ Л О
silia); Kiister, 1847—1862 : 173, t. i9, tig. 1—6 (CI. taurica); 310, t. 35, fig. 13—
17 (Clausilia); W e s t e г 1 u n d, 1884:160 [Clausilia (Oligoptyehia)]; Wester-
1 u n d, 1886 : 5 [Cl. (Oligoptyehia) vinosa]; Westerlund, 1901 : 141 [Clausilia;
(Oligoptyehia)]; Розен, 1914 : 203 [Clausilia (Oligoptyehia)]; Ретовский, 1914 :
322 [Clausilia (Oligoptyehia)]; Лихарев и Раммельмейер, 1952:219,
рис. 136, 137.
Раковина с узкой вытянутой вершиной, тонко ребристая, просве-
просвечивающая, с шелковистым блеском, красновато- или темно-роговая, ме-
.^-dylajaveieollis (Charp.)• (По Ли?
А
Рис. 66.^Ldyla^foveieollis (Charp.). (По Лихареву и Раммельмейер).
общий вид раковины; Б — нид на последний оборот с затылка; В ¦— клаузилий.
стами вдоль шва с пучками белых штрихов. Оборотов 11—-12, слабо выпук-
выпуклых; эмбриональные обороты B1/2) гладкие, почти одинаковой ширины:
последний оборот книзу сужен. Устье округло-ромбическое, выступающее,
с утолщенным, слабо отвернутым белым краем. Синулус довольно глубоко
врезан, узкий. Верхняя пластинка слегка выступает за пределы-устья,
низкая и короткая; справа рядом с ней нередко имеются 1—2 небольших
краевых складки или узелка. Нижняя пластинка довольно крупная, лан-
ланцетовидная, не связана с краем устья, видна при прямом положении рако-
/. IDYLA
141
гп
вины, хотя ее частично закрывает палатальный бугор. Субкодумеллярная
пластинка начинается на'одном уровне с нижней и заканчивается чуть
впереди полулунной складки; нижний конец ее виден при косом положе-
положении устья, а также просвечивает сквозь стенку раковины. Крупный, косо
лежащий палатальный бугор вместе с нижней пластинкой сильно сужает
просвет устья. Очень часто вглубь раковины от бугра отходит различной
длины мозолевидная складка. Несколько реже от него отходит к краю
устья другая, более короткая складка. .
Спиральной пластинки либо нет, либо
от нее остается лишь рудимент. Нередко
рядом с последним, но ближе к шву,
лежит параллельная пластинка. На
спинной стороне лежит хорошо разви-
развитая полулунная складка. Над ней име-
имеется короткая главная складка, а выше
последней нередко еще сутуральная
складка. Лопасть клаузилия языковид-
ная, с почти параллельными боковыми
краями, у конца заострена, утолщена
и слегка клювовидно загнута. Внутрен-
Внутренность устья, в особенности палаталь-
палатальный бугор, окрашена в характерный
винно-красный цвет. Пупочная щель
узкая, окружена широким околопупоч-
околопупочным полем.
Размеры: высота раковины 13.5-—
14.5, ширина 3.3—3.7 мм; высота
устья 2.7—3.3, ширина 2.0—2.5 мм.
Изменчивость. В теплых и
влажных районах, в особенности вдоль
черноморского побережья Кавказа, оби-
обитают популяции, состоящие из экзем-
экземпляров с более крупной и стройной Sc-^'-fr-* fr > ''-/¦
раковиной, имеющей на 1—lVa обо- Рис. 67. idylLji^i^llU'{chzrp.f'
рота больше, чем типичные, с более Половая система. (Ориг.).
крупными. . ребрышками (f. brjanskii
Rosen, 1911); высота таких раковин 16.0—17.5, ширина 3.7—4.0 мм.
Популяции, обитающие в верхнем поясе блесной зоны, в более холод-
холодном климате, отличаются более низкой приземистой раковиной, со-
состоящей из 9—10 оборотов и покрытой более мелкими ребрышками
(f. schaposchnikovi Rosen, 1911); высота таких раковин 12.5—13.5, ширина
3.5—3.7 мм.
Внутреннее строение. Материал: 1) Закатальский запо-
заповедник, сбор и определение наше, V 1953 — 3 экз.; 2) Хоста, сбор и опреде-
определение наше, 13 IX 1956 — 2 экз. /
Радула (рис. 16, А): [| + (—¦ + ~) X 2] X 109..
Размеры срединного зуба 6 X 13.5 (х, ширина радулы 420 ц.
Гонада состоит из 4—5 долек черного цвета. Семяприемник очень
длинный, а отросток короткий (рис. 67). Шейка короткая, веретеновидно
вздутая за счет мощной мускулатуры, состоящей главным образом из
продольных волокон, внутри складчатая. Отросток впадает в шейку у ее
верхнего конца, несколько ниже крепится ретрактор семяприемника.
ркп
142
IV. EUX1NINAE
Семяпровод сразу после обособления от семяяйцевода представляет собой
толстую (за счет кольцевой мускулатуры) трубку со звездчатым просве-
просветом. Ниже семяприемника он утончается и просвет его нередко спадает.
Среди изученных экземпляров у 4 обнаружен рудиментарный бич. Пенис
представляет собой крупную мускулистую трубку с крючковидно загнутым
задним концом. Задняя треть его веретеновидно расширена. Участок; на-
находящийся выше расширения, имеет сравнительно гладкую внутреннюю-
поверхность. Расширенный же участок построен несколько иначе. Одна
38
36
Рис.
e~~ftiykr-foveteottts~(Gbarp;):' (Ориг.).
68. Географическое распространение*"
1 — основной ареал; 2 — отдельные местонахождения.
сторона его имеет тонкую полупрозрачную стенку. Противоположная
стенка толстая, сильно мускулистая. Внутри на ней лежит крупное раздра-
раздражающее тельце, свободный край .которого обычно расщеплен. Впереди он
переходит в 2 постепенно утончающиеся складки, под конец сходящие на
нет. Передняя треть пениса представляет собой тонкостенную трубку бе»
всяких внутренних образований.
Левый ретрактор дает ветвь к нижнему концу влагалища и к половой
клоаке; обычно этот мускул перед прикреплением веерообразно расщеп-
расщепляется на ряд мелких веточек. Длинный и мощный ретрактор пениса яв-
является ветвью заднего ретрактора. Нижний конец ретрактора пениса,
как правило, двуветвистый. Более тонкая ветвь крепится у самой вершины
пениса, рядом с семяпроводом, более крупная — к пенису, несколько
отступя от его вершины.
Распространение (рис. 68). Обитает во всех лесных областях
Закавказья, кроме Аджарии и Талыша. На Северном Кавказе широко рас-
2. ARMENICA 143
пространена в лесах западных склонов Большого Кавказа, в байрачных
лесах Ставропольской возвышенности и в Северной Осетии.
Экологи я.'В пределах своего ареала обитает во всех лесных био-
биотопах и мйкростациях с высокой влажностью и обилием растительных
остатков. По вертикали распространен от низовых равнинных и поймен-
пойменных лесов до верхней границы горного леса. Является не только самым
широкораспространенным, но и наиболее массовым видом своего семейства
на Кавказе.
Синонимия. В 1848 г. Пфейффер (Pfeiffer L., 1848) описал новый
вид Clmsilia taurica, родом из Крыма (из коллекции И. А. Криницкого).
Как видно из~онисания и рисунков (Kiister, 1847—1862), этот вид внешне
мало отличается от Idyla foveicollis (Charp.). Только затылочная впадина
на рисунке выражена слабее. Столь слабые различия между указанными
видами, вполне укладывающиеся в рамки внутривидовой изменчивости,
позволяют рассматривать их как синонимы. Ввиду того, что название
foveicollis Charp. прочно вошло в малакологическую литературу, нет осно-
ваний отдавать приоритет'па.звамжю taurica Pfr., хотя оно дано несколько
раньше. К тому же, в настоящее время твердо установлено, что данный
вид в Крыму не обитает,1 поэтому название taurica (по-латински «крым-
«крымский»), звучало бы как нонсенс.
Вестерлунд (Westerlurid, 1886) описал новый вид под названием Clau-
silia vinosa. Единственным отличием от /. foveicollis является то, что у пер-
первой отсутствует складка, которая у второй отходит от палатального бугра
к краю устья. Между тем в пределах одной популяции могут встречаться
экземпляры как с этой складкой, так и без нее. Поэтому мы считаем, чт&
Cl. vinosa является синонимом /. foveicollis.'2'
2. Род ARMENICA BTTG.
Boettger О., 1877а : 94 (Clausilia, секция Oligoptychia, группа Armenica)^
Wagner А., 1920 : 148 (Oligoptychia); L i n d h о 1 m, 1924 : 66 (Armenica); 67, 73.
(Laciniaria, подрод Inobseratella nov.).3
Тип рода: Clausilia laevicollis Charp.
Раковина башневидная или башневидно-веретеновидная. Последний
оборот с 1 или 2 килями, а устье с соответствующим числом базальных
желобков. Верхняя пластинка короткая и тонкая. Нижняя пластинка срав-
сравнительно с другими группами тоже короткая, лежит чаще вс^го глубоко-
и отвесно, реже слабо спирально изогнута и заметна при прямом положении
устья. Спиральная пластинка либо совершенно редуцировалась, либо>
распалась на 2—3 коротких складочки — спиральная, параллельная и
вставочная пластинки. Субколумеллярная пластинка короткая, чаще всего-
не видна через устье, реже видна лишь при косом положении его. Полулун-
Полулунная складка хорошо развита и видна сквозь стенку раковины. Над ней ле-
лежат 2—3 коротких складки — рудимент главной и сутуральные. Кроме
верхней пластинки, все остальные части замыкательн^цр аппарата, в том
числе и лопасть клаузилия, лежат на спинной стороне раковины, реже-
с некоторым смещением влево. Околопупочное поле полукруглой формы
и очень широкое. На краю его лежит более или менее крупная пупочная:
щель.
1 Указание И. А. Кришщкого (Krynicki, 1837), что этот вид найден в Крыму, свя
зано с какой-то ошибкой. Возможно, что оригиналы taurica были найдены в морских
выбросах Южного берега, где, как известно (Retowski, 1883, 1887), часто встречаются'
пустьгр раковины многих кавказских наземных моллюсков.
2 Бетгер (Boettger О., 1889) рассматривал vinosa как разновидность foveicrllis..
3 По поводу названия Inobseratella Lindh. см. стр. '49.
144 IV. EUXININAE
c Срединный и часть боковых зубов радулы без дополнительных зубцов,
которые' появляются только у самых внешних боковых зубов (рис. 78).
Краевые зубы чаще всего с 5—6 зубцами и лишь у немногих имеют гребен-
гребенчатый облик.
5 Пенис на заднем конце с довольно крупным придатком (рис. 79). Рет-
рактор пениса является ветвью заднего ретрактора, нередко двуветвистый.
В остальном половая система очень сходна с таковой у Idyla.
Распространение. Малая Азия, Сирия, Малый Кавказ и
южные склоны центральной и восточной части Большого Кавказа (рис. 33).
ТАБЛИ11А ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ARM E NIC A
1 D). Последний оборот с 2 килями.
2 C). Полулунная складка и клаузилий не видны при прямом цоложе-
нии устья 2. A. bicarinata (Rssm.).
-3 B). Полулунная складка и клаузилий частично видны при прямом
положении устья 1. A. laevicollis (Charp.).
4 A). Последний оборот с одним килем.
5 (8). Поверхность раковины тонко исчерчена или почти гладкая.
¦6 G). Полулунная складка и клаузилий частично видны при прямом
положении устья 3. A. disjuncta (Mort.).
7 F). Полулунная складка и клаузилий не видны при прямом положе-
положении устья 4. А. Ьгшшеа (Rssm.).
8 E). Поверхность раковины ребристая или сильно и равномерно ис-
исчерчена.
'9 A0). Раковина башневидно-цилиндрическая, тонко и отчетливо равно-
равномерно исчерчена, благодаря чему имеет шелковистый блеск.
Отношение высоты раковины к ширине (В/Ш) равно 5.0—5.2 . .
5. A. gracillima (Ret.).
10 (9). Раковина башневидная или веретеновидно-башневидная, ребрис-
ребристая или сильно ребристо исчерченная, со слабым блеском. В/Ш
4.0-4.6.
И A2). Раковина ребристая, на предпоследнем обороте 54—56 ребрышек
6- A. griseofusca (Mouss.).
12 A1). Раковина ребристо исчерчена, на предпоследнем обороте 74—
76 складок 7. A. hueti (Mort.).
*1. Armenica laevicollis (Charp.) (рис. 69, 70).
Charpentier, 1852 : 399 (Clausilia); P f e i f f e г L., 1853, Zeitschr. Malak.,
10 : 150 (Cl. fausta); Riister, 1847—1862 : 214, t. 23. fig- 24—27 (Cl. fausta); 264,
t. 30, fig. 1—3 (Clausilia); Westerlund, 1884:159 [Clausilia (Oligoptychia)];
Wagner A., 1914, Iconogr., N. F., 21 : t. 574, fig. 23 (Oligoptychia анатомия).
Раковина башневидная, стройная, прочная, очень неясно инерав-
номерно исчерченная, почти гладкая, лишь за устьем сильно ребристо
исчерченная, слабо блестящая, коричнево-роговая. Оборотов И—12, слабо
выпуклых, медленно нарастающих; эмбриональные обороты B) почти
равной ширины и образуют острую цилиндрическую верхушку; последний
•оборот с 2 крупными полукруглыми килями, на затылке несколько вдав-
вдавлен. Устье грушевидное, выступающее, ось его по отношению к оси рако-
раковины несколько отклонена вправо; имеются 2 базальных желобка, из
которых лежащий ближе к столбику достигает края устья: Края устья
широко отвернуты и утолщены, белые, с тонкой губой. Верхняя пластинка
¦очень низкая. Очень короткая нижняя пластинка лежит глубоко и отвесно,
внизу с легким спиральным изгибом. Нижний конец субколумеллярной
2. ARMENICA
145
пластинки хорошо виден при косом положении устья." Короткие мозоле-
видные спиральная и параллельная пластинки располагаются на одном
уровне с лолулунной складкой, т.е. на лево-спинной стороне. Над полулун-
вой складкой лежит короткая главная и 1—2 сутуральных складочки.
Лопасть клаузилия видна при прямом положении устья; она сравнительно
короткая и широкая, сильно изогнута в поперечном направлении и* утол-
утолщена на конце. Крупная пупочная щель лежит на краю широкого около-
околопупочного поля.
Размеры: высота раковины 14.5—19, ширина 3.8—4.7 мм; высота
устья 3.2—4.0, ширина 2.7—3.2 мм.
Изменчив ос т ь. Наряду с типичными встречаются экземпляры,
отличающиеся ребристой скульптурой поверхности раковины (var. toca-
Рис. 69. Аг-
menica laevi-
collis (Charp.).
(Ориг.).
Рис. 70. Armenica laevicollis
(Charp.). Фрагмент мужских
половых путей. (Ориг.).
I
Рис. 71. Ar-
Armenica Ыса-
rinata (Rssm.).
(Ориг.).
tensis Nagele, 1894). Число и размеры главной и сутуральных складок так-
также подвержено некоторым колебаниям (var. perfiga Westerlund, 1884^
с, 4 довольно длинными складками).
Внутреннее строение. Материал: Амасья^ Турция, 7 V
1936, определение Форкарта — 1 экз.
Радула: [± + (* + « + ^ ) X 2] Х-120.
J
Размеры срединного зуба 12 X 15.4 ц, ширина радулы 610 ц..
Половой аппарат сходен с A. hueti (Mort.). Резервуар семяприемника
лежит немного ниже белковой железы, отросток сравнительно короткий.
Придаток пениса отогнут в сторону половой клоаки. Внутри пениса лежат
2 крупных складки, постепенно сглаживающиеся к переднему концу.
Ретрактор его двуветвистый (рис. 70). ¦
Распространение. Найден в двух вилайетах Турции — Токат
Н Амасья (рис. 33). : н .:-.¦;;..„
146 IV. EUXININAE
Систематическое по л.о ж е н и е. По наличию 2 килей
данвщй вид сходен с A. bicarinata (Rssm.), но отличается от последнего
смещением замыкательного аппарата на лево-спинную сторону, что сбли-
сближает его с A. disjuncta (Mort.).
*2. Armenica bicarinata (Rssm.) (рис. 71).
Ross ma s s 1 e г, 1839, Iconogr., 2, 9—10 : 17, fig. 620 (Clausilia),
Раковина башневидно-веретеновидная, тонко, но неравномерно
исчерченная, слабо блестящая, слегка просвечивающая, светло-коричне-
светло-коричневая, с пучками белых штрихов вдоль шва. Оборотов 13—15, почти плоских,
плавно нарастающих; эмбриональные обороты B1/2) гладкие и образуют
узкую сосковидно вытянутую верхушку; последний оборот книзу сужается,
на затылке несколько вдавлен, а внизу с 2 резко очерченными килями.
Шов тонкий, белый. Устье широко-грушевидное, сильно выступающее,
отвесное, наверху с небольшим неврезанным синулусом, а внизу с 2 желоб-
желобками. Края устья широко отвернуты, белые, с тонкой губой. Верхняя пла-
пластинка достигает края устья. Нижняя пластинка лежит глубоко, на значи-
значительном удалении от верхней, имеет ланцетовидную форму, почти
отвесная и не связана с краем устья. Субколумеллярная пластинка еще
короче нижней и не видна даже при косом положении устья. Обе пластинки
располагаются на спинной стороне, напротив крупной полулунной складки.
Над последней лежит короткая главная и 1—2 сутуральных складки.
Лопасть клаузилия широкая, языковидная; конец ее слегка утолщен и
заострен.
Размеры: высота раковины 17—21, ширина 4—5 мм; высота устья 3.8—
4.5, ширина 3.3—4.0 мм.
Изменчивость. Окраска раковины сильно варьирует за счет
разной степени развития белых штрихов. В некоторых случаях они покры-
покрывают бблыную часть поверхности, придавая всей раковине пятнистую рас-
расцветку (var. tesselata Nagele, 1903).
Распространение. Первоначально была известна лишь из
Сирии, но позднее была найдена в Киликии (Турция), в западной части
Армянского Тавра (рис. 33).
3. Armenica disjuncta (Mort.) (рис, 72).
М о г t i 11 e t, 1854 : 13, t. 1, fig. 1 (Clausilia); Kiister, 1847—1862 : 160,
t. 17, fig. 41—43 (Clausilia); Акрамовский, 1949:149 (unicristata Bttg.);
Акрамовский, 1955 : 149.
Раковина башневидная, тонкостенная, слабо и неясно исчерчен-
исчерченная, просвечивающая, со слабым блеском, светло-роговая или коричнево-
роговая. Вершина острая. Оборотов 101/2—12, слабо выпуклых, плавно
нарастающих, разделенных мелким белым швом; эмбриональные обороты
B) гладкие, блестящие; последний оборот вздут, за устьем ребристо исчер-
исчерчен, с коротким килем. Устье грушевидно-ромбическое. Край его широко
отвернут, белый. Верхняя пластинка короткая, низкая, внутри устья
плавно сходит на нет. Нижняя пластинка очень короткая, почти отвесная,
но с легким спиральным изгибом, при прямом положении устья слабо
заметна, с краем устья не связана. Нижний конец субколумеллярной
пластинки виден через устье. Спиральная и параллельная пластинки
маленькие и мозолевидные. Крупная полулунная складка лежит на лево-
спинной стороне раковины. Над ней нередко видны 1—2 маленьких скла-
складочки (рудимент главной и сутуральная). Лопасть клаузилия заметна
снаружи даже при прямом положении устья; она сравнительно короткая
2. ARMENICA
147.
и широкая, сильно изогнута в поперечном направлении и утолщена на
конце. Пупочная щель довольно широкая.
Размеры: высота раковины 14—17, ширина 4.2—4.3 мм; высота устья
3.4—3.6, ширина 2.5—2.8 мм.
Внутреннее строение. Материал: южная Армения, Дара-г
лагез, сел. Гншпик, сбор и определение Акрамов-
ского, 4 VII 1941 — 2 экз.
„ Г с , /76 , 26 , 12к \ „, о~] w ¦ /п
Радула: [у + (у + у + ^П.) Х 2J X 140'
Размеры срединного зуба 11.6 X 15.6 ц, ширины
радулы 540 ц.
Половая система и колумеллярная мускулатура
построена идентично таковым у А. hueti (Mort.). Отли-
Отличия: семяприемник с небольшим резервуаром, который
лежит непосредственно ниже белковой железы; от-
отросток в 1V2 раза короче собственно семяприемника;
ретрактор пениса одноветвистый.
Распространение (рис. 33). В СССР: найден
нагорном массиве Даралагез (на юге Армянской ССР).
Вне СССР: Байбурт (на реке Чорох, к северо-западу
от Эрзурума, Турция).
Э к о л.о г и я. Обитает на скалах и в осыпях на-
горно-степной, субальпийской и альпийской зон.
Систематическое положение. Вме-
Вместе с A. laevicoliis (Gharp.) данный вид отличается от
всех остальных видов того же рода сильно укорочен-
укороченными нижней и субколумеллярной пластинками и пере-
перемещением их, вместе с полулунной складкой и клаузи-
лием, на лево-спинную сторону раковины. Благодаря
этому последние два образования частично видны даже
при прямом положении устья. Однако от A. laevicoliis
наш вид отличается отсутствием вогнутости на затылке
последнего оборота и в связи с этим — отсутствием вто-
второго киля.
4. Armenica Ьгшшеа (Rssm.) (рис. 73, 74).
Rossmassler, 1839, Iconogr., 2, 9—10 : 16, fig. 618 (Clausilia); К п s t e г,
1847—1862 : 308, t. 35, fig. 7-9 (Clausilia); В о e t t g e г О., 1877а : 94, t. 4, fig. 49
(Cl. unicristata); Westerlund, 1884 : 158 [Clausilia (Oligoptychia)]; Wester-
lung, 1901:139 [Clausilia (Oligoptychia)]; Lindholm, 1924:73 [Laciniaria
(Inobseratella) lantzi]; Лихарев и Раммельмейер, 1952:221, рис. 138
(unicristata).
Раков, ина башневидно-цилиндрическая, слабо и неясно исчерчен-
исчерченная, иногда почти гладкая, просвечивающая, со слабым блеском, красно-
красновато-коричневая, с белым швом. Завиток внизу почти цилиндрический,
к верху плавно сужается и заканчивается тупой верхушкой (нередко
изогнутой по оси). Оборотов 11V2—13V2, почти плоских, очень медленно
нарастающих; эмбриональные обороты B) довольно выпуклые, совершенно
гладкие и одинаковой ширины; последний оборот слегка сужается книзу,
ребристо исчерченный, на затылке уплощен, снизу с крупным килем.
Устье овально-грушевидное, выступающее, с одним базальным желобком.
Края его тонкие, белые или светло-коричневые. Верхняя пластинка тонт-
кая и низкая, в глубь раковины плавно сходит на нет. Нижняя пластинка
лежит очень глубоко и видна лишь при косом положении устья. Она срав-
Рис. 72. Armenica
disjuncta (Mort.).
(Ориг.).
148
IV. EUX1NINAE
нительно короткая (начинается невысоко), отвесная, ланцетовидной
формы, нередко переходит в краевую складку или упирается в бугорок.
Спиральная^ластинка рудиментарная. Нередко рядом с ней лежит корот-
короткая параллельная пластинка. Субколумеллярная пластинка лежит очень
глубоко и через устье не видна. Начинаясь на одном уровне с нижней
пластинкой, она отвесно идет вниз, располагаясь напротив полулунной
складки. Последняя очень крупная, лежит на спинной стороне с небольшим
смещением вправо. Над ней располагаются 3 очень коротких складки
(главная и 2 сутуральные), которые у старых экземпляров соединены друг
с другом мозолью. От нижнего конца полулунной складки нередко тянется
к устью короткая нижняя пала-
палатальная складка. Лопасть клаузи-
лия языковидная, с почти парал-
параллельными краями, слабо утолщена
по палатальному краю.
Размеры: высота раковины 16—
22, ширина 4—5 мм; высота устья
3.5—4.5, ширина 2.5—3.5 мм.
Внутреннее строе-
строение. Материал: Иджеван, Ар-
Армянская ССР, сбор и определение
Акрамовского, 25 IV 1957.— 1 экз.
-:-Йз)х2]х1зо.
Рис. 73. Аг-
menica Ьгип-
Ьгиппеа (Rssm.).
(Ориг.).
Рис. 74. Armenica Ьгиппеа.
(Rssm.). Пластинки замыка-
тельного аппарата. Наруж-
Наружная стенка раковины час-
частично удалена. (Ориг.).
1 — верхняя пластинка; 2 —
нижняя пластинка; з — субко-
субколумеллярная пластинка.
Размеры срединного зуба
13 X 14.6 |л, ширина радулы 678 ц.
Половая система очень сходна
с таковой у A. hueti (Mort.).
Распростр анение (рис.
33). В СССР: южные склоны
Большого Кавказа между Заката-
лами и Шемахой, северные и се-
северо-восточные отроги Малого
Кавказа, Зангезур. Вне СССР: \
Турция (Кюлек — на южном склоне Центрального Тавра и вилайет Чо-!
рох).
Экология. Обитает на скалах и в осыпях, в пределах как Лесной
так и альпийской зоны.
Синонимия. Данный вид долгое время отличали от Armenica
unifristata Boettger A877a). Главным основанием служило то, что в то
время как у А. Ьгиппеа якобы имеется промежуточная складка, у A. uni~
cristata таковой нет. Кроме того, считалось характерным для первого вида
присутствие мозолевидной нижней палатальной складки, отходящей от
полулунной складки. Просмотр довольно большого числа экземпляров^
находящихся в коллекции Зоологического института АН Армянской ССР;
показал, что в пределах одной популяции могут встречаться экземпляры
которых по указанным признакам можно причислить как к Ьгиппеа, так i
к unicristata, а наряду с ними есть и переходные формы. А так как, согласно
первоописаниям, в остальных признаках оба вида очень сходны, то, с на
шей точки зрения, нет основания считать их разными видами. Сла
2. ARMENICA
1.49
довательно, A. unicristata (Bttg.) является синонимом -Ar; ' bruhnea
(Rssm.)- ¦ !
Линдгольм (Lindholm, 1924) описал по единственному экземпляру но-
новый вид Clausilia lantzi. x Наиболее существенной особенностью этого
вида автор считал отсутствие клаузилия. На этом основании он его выде-
выделил в особый подрод Inobseratella Lindh. (род Laciniaria). Однако при
просмотре этого экземпляра под бинокуляром мы убедились, что он имел
клаузилий, так как сохранился обломок рукоятки, который Линдгольм
принял за спиральную пластинку. Несмотря на то что устье
этого экземпляра несколько повреждено, по всем остальным
признакам он походит на А. Ьгиппеа. Следовательно, Cl. lan-
lantzi Lindh. тоже является синонимом данного вида, равно как
Inobseratella Lindh. является синонимом Armenica Bttg.
*5. Armenica gracillima (Ret.) (рис. 75).
Retowski, 1889:261 [Clausilia (О ligoptychia)]; К о b e 1 t,
1893, Iconogr., N. F., 6:37, fig. 1015 [Clausilia {0ligoptychia)];
Westerlund, 1901 : 140 [Clausilia (O ligoptychia)]; Lindholm,
1913b : 66 [Clausilia (Oligoptychia)]; Розен, 1914:203, 232 [Cla-
[Clausilia (O ligoptychia)].
Раковина бапшевидно-цилиндрическая, очень строй-
стройная, очень тонко, но отчетливо и равномерно исчерчена, с
шелковистым блеском, красновато-роговая, у шва белая.
Оборотов 13—15,2 почти плоских, с 4-го по 9-й медленно
нарастающих в ширину; последние 2 оборота почти равной
ширины; эмбриональных оборотов 3, гладких и блестящих,
образующих цилиндрическую вершину; последний оборот
книзу слегка сужен, с крупным килем. Устье широко-гру-
широко-грушевидное или овальное, небольшое, с тонкими белыми, ши-
широко отвернутыми краями, с глубоким базальным желобком,
внутри коричневое, в глубине с белой палатальной мозолью.
Верхняя пластинка тонкая, впереди доходит до края устья,
назад плавно сходит на нет. Нижняя пластинка лежит на
спинной стороне и при прямом положении раковины не видна;
она имеет ланцетовидную форму, тонкая, почти отвесная, с
краем устья не связана; от нижнего конца она посылает на-
назад косо лежащую промежуточную складку. Кроме неболь-
небольшой спиральной пластинки, имеется такой же величины па-
параллельная пластинка. Лежащая на спинной стороне полу-
полулунная складка мозолевидная. Над нею находятся 3 коротких складочки:
главная и 2 сутуральных. От нижнего конца полулунной складки отхо-
отходит вперед небольшая мозолевидная нижняя палатальная складка. Лопасть
клаузилия языковидная, с почти параллельными краями, в поперечном
направлении сильно изогнута, по нижнему краю слабо утолщена.
Размеры: высота раковины 18—21, ширина 3.5—4.0 мм; высота устья
3.5—4.0, ширина 2.7—3.0 мм.
Изменчивость. Наибольшим колебаниям подвержена высота
раковины. Изредка среди нормальных экземпляров попадаются единич-
единичные альбиносы (var. albella Lindholm, 1913b).
Рис. 75. Ar-
Armenica gra-
ciiZima(Ret.).
(Ориг.).
1 Этот экземпляр был найден у горы Триал в Чорохском вилайете Турции.
2 Ретовский указывает 16 оборотов, однако среди просмотренных нами много-
многочисленных раковин из его коллекций мы не обнаружили экземпляров, имеющих больше
15 оборотов; наиболее часто встречались раковины с 14 оборотами»
150
IV. EUXININAE
В н у т р е н н ее строение. Материал: окрестности Ломашепа,
Турдия, сбор Нестерова, VI 1909, определение Линдгольма — 3 экз.
Радула: [° + (™ + f + ^) Х 2] X сл>150.
Размеры срединного зуба 11.6 X 15.1 ц,, ширина радулы 616 ц,. Ближе
к переднему концу радулы дополнительный зубец появляется лишь у двух
внешних боковых зубов, а у заднего конца — с 4-го или 5-го бокового зуба.
Половая система и колумеллярная мускулатура постро-
построены по тому же плану, как и у других видов того же рода.
Распространение (рис. 33). Вид найден во
многих пунктах окрестностей города Артвин (северо-во-
(северо-восточная Турция, вилайет Чорох). Типы были найдены
среди выносов реки Чорох у Батуми.
Систематическое положение. По за-
мыкательному аппарату данный вид наиболее сходен с
ЩШШк А. Ъгиппеа (Rssm.). Общее: число и расположение главной
ШИШИ и сутуральных складок, наличие нижней палатальной
^ШШ^ш складки. Отличия: своеобразная скульптура поверхности
щйШЩШШ раковины, нижняя пластинка не связана с краем устья,
^ШШЯт оборотов больше и они почти плоские.
Ш§Ш/Шщ От всех остальных видов того же рода данный мол-
ШЙШяиР люск отличается в первую очередь формой раковины и
скульптурой ее поверхности.
6. Armenica griseofusca (Mouss.) (рис. 76).
Mousson, 1876b : 145, t. 5, fig. 3 [Clausilia (Alinda)];
Boettger, 0., 1879b : 411 [Clausilia (Oligoptychia)]; W e s-
terlund, 1884 : 158 [Clausilia (Oligoptychia)}; К о b e 1 t, 1893,
Iconogr., N. F., 6 : 35, fig. 1012 [Clausilia {Oligoptychia)}; West-
e г 1 u n d, 1901 : 139 [Clausilia (Oligoptychia)]; Розен, 1914 : 202
[Clausilia (Oligoptychia)]; Лихарев и Раммельмейер,
1952 : 222.
Рис. 76. Arme-
Armenica griseofusca
(Mouss.).
(Ориг.).
Раковина веретеновидно-бапшевидная, тонкостен-
тонкостенная, с тупыми тонкими, редко лежащими ребрышками
(на предпоследнем обороте 54—56 ребрышек), слабо
блестящая, темно-роговая, вдоль верхнего края оборотов с белыми
штрихами. Оборотов 11—12, довольно выпуклых, плавно нарастаю-
нарастающих; эмбриональные обороты C) гладкие, блестящие; последний обо-
оборот книзу немного сужен, на затылке уплощен, с крупным килем.
Шов тонкий, белый. Устье почти овальное, книзу слегка угловатое, от-
отвесное, сильно выступающее, высота его равна около V4 высоты раковины.
Края устья широко отвернуты, белые, а внутренняя часть устья, в том
числе и базальный желобок коричневые. Верхняя пластинка низкая, но
толстая, впереди достигает края устья, а сзади плавно сходит на нет.
Нижняя пластинка ланцетовидная, отвесная, при прямом положении
устья почти не видна; у ее нижнего конца лежит короткая складочка
или бугорок. Субколумеллярная пластинка короткая, снаружи не видна.
Остатки спиральной пластинки и полулунная складка имеют вид слабых
мозолевидных утолщений. Несколько сильнее выражен нижний конец
полулунной складки. Имеются 2 коротких сутуральных игататок главной
складки. Широкая лопасть клаузилия сильно изогнута в поперечном и
продольном направлениях и имеет округло-прямоугольную форму.
Размеры: высота раковины 16—18, ширина 4.0—4.4 мм; высота устья
3.8—4.2, ширина 3.0—3.2 мм.
2. ARMENICA
т
Внутреннее строение. Материал: Иджеван, Армянская
GGP, сбор и определение Акрамовекого, 8 V 1955—1 экз.
., Г с , /56 . 66 , 14k\wO~]w ,/А
Радула: |_т + (т + У + Т=~в) Х 2J X с/э140>
Размеры срединного зуба 14.4 X 15.6 ц, ширина радулы 704 ц,.
Половой аппарат очень сходен с таковым A. hueti' (Mort.). Отличия:
семяприемник длиннее (доходит до белковой железы), с небольшим круг-
круглым резервуаром; придаток пениса отогнут вниз, к поло-
половому отверстию.
Распространение (рис. 33). Вид был описан по
экземплярам, найденным в окрестностях озера Табацкури
(южная Грузия). Кроме того, ряд находок сделан Н. Н. Акра-
мовским в Иджеванском районе Армянской ССР.
Указание Розена A914) на то, что этот моллюск обнару-
обнаружен в окрестностях Суреби, при проверке (материалы хра-
хранятся в Музее Грузии) не подтвердилось. Автор имел дело
с другим видом.
Экология. Обитает в осыпях и в трещинах скал лес-
лесной зоны.
Системат и ч еское положение. Данный
вид отличается от остальных видов того же рода в первую
очередь сильной ребристостью раковины и более выпуклыми
оборотами.
7. Armenica hueti (Mort.) (рис. 77—79).
Mortillet, 1854:12, t. 1, fig. 9 (Clausilia); Riister,
1847—1862:159, t. 17, fig. 38—40 (Clausilia); Westerlund,
1884 : 159 [Clausilia (Oligoptychia)]; Westerlund, 1901 : 140
[Clausilia {Oligoptychia)]; Розен, 1914 : 202 [Clausilia {Oligoptychia)].
Раковина башневидная, ребристо ис-
исчерчена (на предпоследнем обороте 74—76 ?
складок), слабо блестящая, темно-роговая или
темно-коричневая. Оборотов 12—14, слабо вы-
выпуклых, разделенных мелким беловатым швом;
эмбриональные обороты C) гладкие, почти
одинаковой ширины; последний оборот за
устьем уплощен, внизу с крупным килем. Устье
широко-грушевидное, слегка косое, с белыми,
широко отвернутыми краями, с базальным же-
желобком, внутри коричневое. Высота устья
обычно меньше V4 высоты раковины. Верхняя
пластинка довольно высокая и толстая, сзади
плавно сходит на нет, впереди достигает края
устья. Нижняя пластинка лежит глубоко и при
прямом положении устья почти не видна; она
отвесно уходит наверх, где заканчивается,
резко обрываясь; в устье, перед ее нижним концом, лежит более или ме-
менее четко выраженный бугорок. Отсюда тянется под углом в глубь ра-
раковины короткая промежуточная складка. Кроме небольшой спираль-
спиральной, имеется еще, тоже короткая, параллельная пластинка. Субколу-
меллярная пластинка снаружи не видна. Довольно крупная нолулунная
складка лежит на спинной стороне раковины. Над ней простираются 3
тонких и коротких складочки — 2 сутуральных и главная. От нижнего
Рис. 77. Armenica hueti
(Mort.). (Ориг.).
А — общий вид раковины; В —
последний оборот (сзади) и про-
просвечивающие сквозь него скла-
складки; В — клаузилий.
f52
IV. EUXININAE
конца полулунной складки отходит вперёд короткая мозолевиднан нижняя
палатадьная складка. Иногда эта складка йе сливается с полулунной.
; Рис. 78. Armenica hueti (Mort..). Радула. (Ориг.).
Лопасть клаузилия языковидная, g почти параллельными краями, слабо
изогнута в продольном и сильно в поперечном направлениях. :
Размеры: высота раковины 19—24, ширина 4.5—5.0 мм; высота устья
4.3—5.3, ширина 3.3*->4.0 мм.
Внутреннее с т р о-
е н и е. Материал: Закаталь-
ский заповедник, сбор и опре-
определение наше, 16 VII 1954 — 2
экз.
бж
Радула (рис. 78):,-[-f\-h-
. +(«+.»ч.1аух-.
рп-
X ]2Х -<У5150.
Размеры срединного зуба
12 X 15 (д,, ширина радулы
.70011. :
Семяприемник с крупным ре-
резервуаром и толстой мускули-
мускулистой, внутри складчатой, шей-
шейкой (рис. 79). В последнюю
впадает узкий тонкостенный
отросток, который немного ко-
короче собственно семяприемника.
Верхние концы их лежат на се-
мяяйцеводе, на значительном
расстоянии от белковой железы.
Влагадипде вдвое длиннее яйне-
вода. Семяпровод вначале широ-
широкий, за счет толстого слоя коль-
кольцевой мускулатуры, затем су-
сужается. На значительном про-
протяжении он срастается с ниж-
нижними концами влагалища и
пениса, Певис представляет собой широкий цилиндрический орган, внутри
которого проходят 3—4 продольных мускульных валика, сглаживающихся
лишь в передней трети пениса. От заднего конца пениса, несколько ниже
ретрактора:, отходит назад крючковидный придаток.
. Левый ретрактор посылает одну ветвь к шейке семяприемника — рет-
рактор семяприемника, и ГШ;,ответвлению к нижним концам пениса, и
Рис. 79; Armenica hueti (Mort.). Половая си-
система. (Ориг.).
2. ARMENICA 153
к влагалищу. Кроме того, многочисленные мускульные волокна, отходящие
от левого ретрактора к покровам, окружают влагалище и семяпровод
как бы чехлом.
У одного из вскрытых экземпляров ретрактор пениса был двуветвистым.
Распространение (рис. 33). В СССР: южные склоны Боль-
Большого Кавказа (Закатальский заповедник; Шемахинский район). Вне
СССР: Испир (Турция, к северо-западу от Эрзурума).
Экология, Обитает на скалах, в долинах рек лесной зоны.
Систематическое положение. Данный вид наиболее
близок к A. griseofusca (Mouss.), но отличается от нее более тонкой и ча-
частой ребристостью и большими размерами. По высоте раковины наш вид
сходен с A. gracillima (Ret.), но отличается от последней приземистостью
раковины (за счет большей ширины), более сильной скульптурой, более
выпуклыми оборотами и относительно более крупным устьем.
ДОПОЛНЕНИЕ К РОДУ ARMENICA BTTG.
Ниже даются переводы оригинальных описаний нескольких видов,
которые, по-видимому, принадлежат к роду Armenica Bttg. Однако непол-
неполнота этих описаний и отсутствие соответствующих коллекционных материа-
материалов затрудняет решение вопроса о том, как следует рассматривать этих
моллюсков — как самостоятельные виды или же как синонимы или внут-
внутривидовые отклонения уже описанных выше видов.
*8. Armenica rupicola (Mort.).
М о г t i 11 е Ь, 1854.- 13, t. I, fig. 7 (Clausilia).
«Раковин а стройная, тонкая, роговая, просвечивающая, равно-
равномерно исчерченная. Пупок слабо заметен. Устье овальное, слегка груше-
грушевидное, снизу закругленное, наверху сжатое. Верхняя пластинка сильно
выступает вперед. Нижняя пластинка выдается в просвет устья и направ-
направлена перпендикулярно к верхней. Край устья белый, цельный, отвернутый,
выступающий, только наверху прилегает к завитку. Полулунной складки
нет. Оборотов 12—13, довольно плоских. Шов умеренной глубины, ме-
местами окаймлен небольшими белыми сосочками».
Размеры: высота раковины 18, ширина 3 мм.
Распространение. Единственная находка сделана у Тортума
(вилайет Эрзурум, Турция).
Экология. Обитает в щелях скал.
Систематическое пол о ж е н и е. Очень неполное описание
и плохой рисунок затрудняют всякое сравнение с другими видами того же
рода. Следовательно, и вопрос о самостоятельности этого вида остается
открытым. . ,
*9. Armenica blissi (Bttg.).
В oettger О., 1899, Nacbrbl. deutsch, Malak. Ges., 31 : 167—169 [Clausilia
(Oligoptychia)].
«P а ко вин а со щелевидным пупком, башневидная, твердостенная,
темно-роговая. Завиток удлиненно-башневйдный; верхушка довольно
острая, недеколлированная. Оборотов 16, слабо выпуклых, почти плоских;
шов мелкий, окаймлен беловатой линией; поверхность оборотов почти
равномерно исчерчена; последний*- оборот исчерчен не сильнее предыду-
предыдущих, внизу с 2 крупными и изогнутыми килями; верхний киль наверху
раздвоен. Устье отвесное, овально-грушевидное, внизу с 2 желобками,
внутри бурое;: край устья белый, цельный, расширен и отвернут. Верхняя
154 IV. EUXININAE
пластинка небольшая, косолежащая почти у самого края устья. Субколу-
меллярная пластинка не видна. Широкая полулунная складка изогнута
почти под углом и лежит на спинной стороне раковины. Главная складка
и одна сутуральная складка очень короткие и слабо выражены, иногда
отсутствуют».
Размеры: высота раковины 25—26.5, ширина 5 мм; высота устья.5, ши-
ширина 4 мм.
Распространение. Единственная находка сделана в вилайете
Конья (Центральная Анатолия, Турция).
Систематическое положение. Бетгер считает, что
данный вид наиболее близок к «Clausilia» fausta Pfr. [syn.Armenica laevi-
collis (Charp.)], отличаясь от нее большим числом оборотов и более ясной
исчерченностью. С нашей точки зрения, вполне возможно, что данный мол-
моллюск является лишь разновидностью A. laevicollis.
*10. Armenica euprepes Biggs.
Biggs, 1936, Journ. Conchol., 20 : 253, text-fig.
«Раковина левозакрученная, веретеновидная, красновато-коричне-
красновато-коричневая, полупросвечивающая, матовая, без пупка. Оборотов 12V2; эмбрио-
эмбриональных оборотов 21/2, гладких, рогового цвета. Поверхность остальной
раковины равномерно покрыта параллельными тонкими и частыми реб-
ребрышками, которых около 9 штук на 1 мм? поверхности 11-го оборота. За
краем устья лежит крупный гребень. Устье грушевидное, с глубоким сину-
лусом и базальным желобком. Край его цельный, выступающий, иногда
отвернут; колумеллярная стенка плоская. Внутри устье окрашено в светло-
коричневый цвет».
Размеры голотипа (коллекция Бигса): высота раковины 19.3, ширина
3.9 мм. Средние размеры 10 паратипов: высота раковины 20.0, ширина
4.1 мм.
Распространение. Единственная находка сделана близ Мачка
(вилайет Трабзон, Турция).
Систематическое положение. Бигс склонен сближать
данный вид с A. hueti (Mort.), который, с его точки зрения, отличается рого-
роговым цветом раковины, более сильной скульптурой поверхности и более
крупными размерами. С нашей точки зрения, судя по рисунку Бигса, дан-
данный моллюск более походит на A. griseofusca (Mouss.) (общее — форма
раковины и ее ребристость). Однако за отсутствием всяких сведений о за-
мыкательном аппарате трудно решить, является ли A. euprepes синонимом
или разновидностью какого-либо уже известного вида или самостоятель-
самостоятельным видом.
*11. Armenica mimele Biggs.
Biggs, 1946, Journ. Conchol., 22 : 224, text-fig.
«Раковина левозакрученпая, веретеновидная, красновато-коричне-
красновато-коричневая, полупросвечивающая, матовая, без пупка. Оборотов 111/2—-12;
эмбриональных оборотов 2, более светлых, чем остальная раковина; де-
финтивные обороты неравномерно покрыты тонкими ребрышками, которые
заметно сглаживаются у шва; последний оборот с крупным изогнутым
килем. Устье грушевидное, с глубоким и узким базальным желобком. Край
устья цельный, выступающий и несколько отвернут. Париетальная пла-
пластинка отсутствует; верхняя колумеялярная пластинка не достигает края
устья; нижняя колумеллярная пластинка S-образной формы. На палаталь-
палатальной стенке, напротив нижнего конца нижней колумеллярной пластинки,
3. ACROTOMA 155
имеется слабая складка. Полулунная складка внизу раздваивается. • Над
нею лежал- 4 палатальных складки».
Размеры голотипа (коллекция Бигса): высота раковины 18.6, ширина
4.2 мм; высота устья 4.4, ширина 3.2 мм. Средние размеры 4 паратипов:
высота раковины 18.0, ширина 4.2 мм; высота устья 4.2, ширина 3.0 мм.
Распространение. Единственная находка сделана близ Мачка
(вилайет Трабзон, Турция).
Систематическое положение. Данный вид очень трудно
сравнивать с другими видами того же рода, так как оригинальное описание
его очень несовершенно. В частности, неясно, что Бигс подразумевает под
«париетальной пластинкой», которая якобы отсутствует. Если верхнюю
пластинку, то она хорошо видна на рисунке, приложенном к описанию.
Под «верхней колумеллярной пластинкой», по-видимому, автор подразу-
подразумевает нижнюю пластинку, а под «нижней колумеллярной пластинкой» —
колумеллярную пластинку в общепринятом понимании.
Автор сравнивает этот вид с A. euprepes Biggs, найденной в том же месте.
Последняя отличается от A. mimele более стройной формой раковины и
более сильной и редкой равномерной ребристостью.
3. Род ACROTOMA BTTG.
Boettger О., 1881Ь : 341 (Clausilia, секция Acrotoma); Wagner A., 1920 :
150; L i n d h о 1 m, 1924 : 68, T h i e 1 e, 1931 : 536. •
Тип рода: Clausilia komarowi Bttg.
Раковина средних размеров (высота около 15 мм) или очень крупная;
за исключением одного вида, у остальных верхние обороты подвергаются
деколляции. Последний оборот с крупным базальным килем. Верхняя
пластинка отделена от спиральной и ее задний конец заходит справа за
передний конец последней. Нижняя пластинка лежит более или менее
глубоко; она имеет вид толстой косо лежащей ланцетовидной складки,
довольно круто уходящей кверху. Субколумеллярная пластинка не видна
или чуть заметна при косом положении устья. Крупная полулунная
складка лежит на спинной стороне раковины; у большинства видов оба
конца ее отогнуты назад. Главная складка чаще всего довольно длинная,
занимает около V3 последнего оборота. Лопасть клаузилия языковидная,
с почти параллельными краями и с закругленным и слабо утолщенным
концом.
За исключением одного вида, у остальных срединный и большая часть
боковых зубов радулы без дополнительных зубцов. Они появляются только
у двух самых внешних боковых зубов.
В половой системе наиболее характерно устройство пениса. Он имеет
крупный придаток и двуветвистый ре.трактор. Последний другим концом
прикрепляется к диафрагме.
Распространение (рис.80). Западная часть Большого Кавказа
(подрод Acrotoma s. str.) и Чорохский вилайет Турции (подрод Roseniella
Thiele).
В ископаемом состоянии представитель этого рода найден в среднем
эоцене Италии. •
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА АСДОТОМА
1 (8). Раковина деколлированная. (Подрод Acrotoma а. str.).
2 C). Раковина очень крупная; высота ее больше 25 мм, ширина больше
5 мм . 1. A. komarowi (Bttg.).
1S6
IV. EUXININAE
3 B). Раковина значительно меньших размеров.
4 E). Последний оборот за устьем несет крупную поперечную складку
или гребень 2. A. semicincta (Bttg.).
5 D). Затылочного гребня нет.
6 G). Поверхность раковины покрыта крупными белыми пластинчатыми
ребрами 3. A. narzanensis1 (Rosen).
44
42
36,
40
44
48
Астрахань^
УЛахач-Кала
Баку;
Ереван
• ; +2 кЗ •¦
55 0 55 110 165 22G км
I 1
/ &г~,
40
42 44 46
44
42
40
36
4В
52
54
Рис. 80. Географическое распространение видов рода Acrotoma Bttg. (Ориг.).
1 — A. homarowi (Bttg.); 2 — A. semicincta (Bttg.); 3 —A. narzanensis (Rosen); 4 — A. sobriev&bii
(Rosen).
7 F). Поверхность раковины редко и неясно исчерчена
4. A. laccata (Bttg.).
8 A). Раковина недеколлированная. (Подрод Roseniella Thiele) ....
5. A. sobrievskii (Rosen).
¦ Подрод ACROTOMA s. str.
Раковина деколлированная. Нижняя палатальная складка у большин-
большинства видов отсутствует:
1. Acrotoma (Acrotoma) komarowi (Bttg.) (рис. 17, 81—83).
В о е t tge г О., 1881Ь : 341' [Clausilia (Acrotoma)]; Boettger О., 1883:
185, t. 7, fig. 4 [Clausilia (Acrotoma)]; Westerlund, 1884 : 26 [Clausilia (Acro-
(Acrotoma)]; К о b e 11, 1893, Iconogr., N. F., 6 : 7, fig. 970 [Clausilia (Acrotoma)]; W e s-
t er 1 u n d, 1901 : 25 [Clausilia (Acrotoma)]; Ретовский, 1914:313 [Clausilia
(Acrotoma}]; Лихарев и Ра м Me л ь м eie p, 1952 : 223, рис. 139.
Раковина веретеновидная, очень крупная, деколлированная, твер-
достенная, густо и тонко, слегка волнисто исчерченная, блестящая, про-
просвечивающая, коричневато-роговая, с многочисленными мелкими белыми
3. ACROTOMA
157
штрихами, которые у шва периодически образуют сгущения. От первона-
первоначальных 17—18 оборотов после деколляции остаются 7—8,1 разделенных
очень тонким беловатым швом. Обороты слабо выпуклые; последний уже
предпоследнего, внизу с узким, но высоким килем. i
Устье угловато-овальное или грушевидное, выступаю-
выступающее, отвесное, с широко отвернутыми краями. Узкий
синулус занимает верхний угол устья. Пластинки
очень толстые. Косо лежащая и высокая верхняя пла-
пластинка впереди доходит до самого края устья, сзади
плавно спускается. Крупная ланцетовидная нижняя
пластинка валикообразно утолщена по внешнему краю,
впереди заканчивается, значительно отступя от края ШкшШ
устья, в глубь раковины уходит почти отвесно, резко . НИ|
переходя в тонкую спирально загнутую складку. Спи-
Спиральная пластинка начинается почти на одном уров-
уровне с. нижней и заканчивается слегка заходя за зад-
задний конец верхней.
Размеры: % высота раковины 28—32, ширина 6.5—8.0
мм; высота устья 7.8—8.5, ширина 5.3—6.2 мм.
Бетгер описал этот вид по 2 более крупным экзем-
экземплярам, имевшим следующие размеры: высота рако-
раковины 34.5—37.5, ширина 8 мм; высота устья 9.5, ши- Jg|
рина 6.8 мм.
Внутреннее строение. Материал: Абха-
Абхазия, Новый Афон, сбор и определение наше, 2 X 1956 —
1 экз.
Радула (рис. 82): |^т + (т + у + T=VX 2JX
X сл140.
Размеры срединного зуба 15х24|л., ширина радулы
1030 [а.
Семяприемник имеет сравнительно небольшой круг-
круглый резервуар, который лежит на значительном рас-
расстоянии от белковой железы (рис. 83). Отросток немного
короче собственно семяприемника. Пенис своеобразной
формы. Середина его сильно расширена, но плоская,
внутри с многочисленными кольцевыми складками. Зад-
Задний конец несет узкий длинный придаток. Обе ветви
ретрактора пениса перед прикреплением к диафрагме
сливаются друг с другом. Одна ветвь крепится к пенису
рядом с семяпроводом, а другая — .у основания при-
придатка.
Левый ретрактор дает одну ветвь к шейке семяпри-
семяприемника и несколько ветвей к половой клоаке.
Распространение (рис. 80). Абхазия. Вид
редкий. Большинство находок было сделано в долине
среднего течения реки Кодор, в частности у селения Цебельда. Кроме
того, найден в окрестностях Нового Афона и на Охачкуэйском массиве
1 Один из экземпляров Ветгера имел раковину из 10 оборотов. Он считает, что в те-
течение жизни моллюск образует 17—18 оборотов.
2 Даны на основании промера 10 экземпляров, из которых 7 хранятся в Зоологи-
Зоологическом институте Академии наук СССР, 1 — в Музее Грузии и 2 — в Зоологическом
институте Польской академии наук.
Рис. 81. Acrotoma
(Acrotoma) koma-
rowi (Bttg.).
(Ориг.).
А — общий вид ра-
раковины; Б — клау-
зилий.
158
IV. EUXININAE
Гальского района, на высоте 1800 м над ур. м. (коллекция Г. В. Коко-
чашвили). .
Экология. Обитает в щелях известняковых скал и под камнями.
Q 1 12 13
Рис. 82. Acrotoma (Acrotoma) komarowi (Bttg.). Радула. (Ориг.).
Рис. 83. Acrotoma (Acrotoma) komarowi (Bttg.). Половая
система. (Ориг.),
2. Acrotoma (Acrotoma) scmicincta (Bttg.) (рис. 84—86).
В о e 11 ge г О., 1881b : 343 [Clausilia (Acrotoma)]; Boettger 0., 1883:
186, t. 7, fig. 6 [Clausilia (Acrotoma,)]; Weste rlund, 1884 : 27
[Clausilia (Acrotoma)]; К о b e 1 t, 1893, Iconogr., N. F., 6 : 9, fig. 972 [Clausilia (Acro-
(Acrotoma)]; Westerlund, 1901 : 25 [Clausilia (Acrotoma)]; Лихарев и Раммель-
м е й е р, 1952 : 223, рис. 140.
Раковина деколлированная, веретеновидно-цилиндрическая, твер-
достенная, слабо блестящая, темно-, реже светло-роговая. Верхние обороты
густо ребристо исчерчены, нижние слабо, порой неясно исчерчены, а сред-
средние образуют постепенные переходы от скульптуры верхних к таковой
нижних оборотов. От первоначальных 141/2 оборотов после деколляции
остаются 6V2—7,1 реже 6; выпуклость оборотов ослабевает от верхних
1 Бетгер указывает, что в течение жизни моллюск образует 14х/г оборотов.
3. ACROTOMA
159
к нижним, так что 2 последних почти плоские; последний оборот слегка
сужается книзу, на затылке ребристо исчерчен и приблизительно за 1мм
от края устья несет крупную складку или гребень. Тонкий беловатый
шов у верхних оборотов зубчатый. Устье округло-ромбическое, с почти
параллельными палатальным и колумеллярным кра-
краями, отвесное, выступающее, Синулус небольшой,
отогнут к оси раковины. Края устья широко отвер-
отвернуты, утолщены, белые; внутренность устья корич-
коричневато-лиловая. Верхняя пластинка отвесная, тон-
тонкая, впереди достигает края устья, сзади простира-
простирается почти на */4 оборота и, плавно снижаясь,
сходит на нет. Спиральная пластинка начинается
приблизительно в начале второй четверти послед-
последнего оборота и простирается вперед до начала
последней четверти; передний конец ее далеко за-
заходит за задний конец верхней пластинки. Мощ-
Мощная высокая ланцетовидная нижняя пластинка ле-
лежит глубоко в устье, валикообразно утолщена по
внешнему краю; нижний конец ее слегка обрублен
и находится на значительном удалении от края
устья; в глубь раковины она уходит почти отвесно и
заканчивается в виде низкой складки на одном уров-
уровне со спиральной пластинкой. Субколумеллярная
пластинка плохо видна даже при косом положении
устья. Довольно толстая полулунная складка лежит
на спинной стороне; оба конца ее отогнуты назад.
Главная складка начинается чуть выше полулунной
и простирается вниз на 1/3 оборота. Лопасть клаузшшя языковидная, с
почти параллельными краями и с закругленным и утолщенным концом.
Пупочная щель короткая.
Размеры: высота раковины 13.7—17.0, ширина 4.0—4.5 мм; высота
устья 3.7—4.5, ширина 2.6—3.2 мм.1 Бетгер указывает, что высота недекол-
лированной раковины 22 мм.
Изменчивость. Наиболее варьирует скульптура поверхности
раковины. Так, среди популяции с типичной скульптурой встречаются
Рис. 84. Acrotoma
(Acrotoma) semicincta
(Bttg.). Последний
оборот раковины, вид
с яатылка. (Ориг.).
23
10
1 с 1
Рис. 85. Acrotoma (Acrotoma) semicincta (Bttg.). Радула. (Ориг.
экземпляры, раковины которых покрыты крупными, широко расставлен-
расставленными ребрами [var. ciscaucasica-terensis Rosen, 1911 (syn. A. semicincta var.
narzanensis Westerlund, 1901, non Rosen, 1901)]. Кроме того, нередко
1 Лектотип данного вида хранится в Зоологическом институте Академии наук
СССР (каталог систематической коллекции моллюсков, № 1) и имеет следующие раз-
размеры: высота раковины 16.2, ширина 4.3 мм; высота устья 4.5, ширина 2.8 мм; 7 оборо-
оборотов. Место сбора: Теберда, сборщик Ледер. Один паралектотип хранится в Музее Гру-
Грузии (коллекция моллюсков, № 1489).
160
IV, EVXININAE
Вж>
встречаются экземпляры, у которых замыкательный аппарат имеет мозо-
левидную нижнюю палатальную складку, отходящую вперед от нижнего
конца полулунной складки.
Внутреннее строение. Материал: Грузия, Клухорский
район, река Доун, сбор Н. Н. Акрамовского, 19 VIII 1954, определение
наше — 1 экз.
Радула (рис. 85): [| + (^ + jZTg) X 2] Х 120.
В отличие от других видов того же рода срединный и все боковые
зубы с дополнительными зубцами. Размеры срединного зуба 21.0 X 15.2 ц,
ширина радулы 635 \i.
Половой аппарат (рис. 86) в
общем сходен с таковым у преды-
предыдущего вида. Отличия: пенис длин-
длиннее, задняя треть его расширена
(внутри с многочисленными про-
продольными складками), придаток
более короткий. Значительная
часть восходящего колена семя-
семяпровода окружена мускульным
чехлом и прижата к передней
трети пениса.
• Распространение (рис.
80). Западная часть северных скло-
склонов Большого Кавказа, в бассей-
бассейнах верхних притоков рек Кубани
и Подкумка (Теберда; Кара-Чай;
Клухори, ущелье р. Шоаны;
р. Ран-Коль у горы Бермамыт;
Кисловодск).
Экология. Обитает наиз-
вестняковых скалах и в осыпях,
как в лесу, так и на открытых
местах. «
"J?
кл
Рис. 86. Acrotoma (Acrotoma) semicincta
(Bttg.). Половая система. (Ориг.).
3. Acrotoma (Acrotoma) narza-
nensis (Rosen) (рис. 87).
Розен, 1901 : 10 [Clausilia (Ac-
(Acrotoma)}; Westerlund, 1901:174
(Cl. roseni); Лихарев и Раи-
мельмейер, 1952 : 224.
Раковина деколлированная, веретеновидно-цилиндрическая,
с крупными белыми пластинчатыми ребрами, на темно-роговом фоне, бле-
блестящая. Ребра поставлены широко, на предпоследнем обороте число их
колеблется от 30 до 40. Оборотов чаще всего 6V2—7;* верхние умеренно
выпуклые, книзу обороты становятся более плоскими; 2 последних оборота
почти одинаковой ширины; последний книзу слегка сужается. Шов тон-
тонкий, белый, благодаря ребрам зубчатый. Устье округло-ромбическое,
выступающее, с небольшим прямым синулусом и почти параллельными
палатальным и колумеллярным краями. Края устья умеренно отвернуты,
слегка утолщены, белые. Верхняя пластинка тонкая и низкая, короче,
чем у A. semicincta (Bttgr). Спиральная пластинка располагается так же.
1 В течение жизни моллюск образует 13—14 оборотов.
3. ACROTOMA
161
Крупная и высокая ланцетовидная нижняя пластинка сходна с таковой
у A. semicincta, но лежит она глубже и при прямом положении виден лишь
ее нижний конец, который плавно спускается и заканчивается на значи-
значительном расстоянии от края устья. Субколумеллярная пластинка не видна
даже при косом положении устья. Довольно толстая полулунная складка
лежит на спинной стороне раковины; оба конца ее отогнуты назад. Глав-
Главная складка начинается чуть выше полулунной и простирается вниз на
х/3 оборота. Языковидная лопасть клаузилия с почти параллельными
краями, у конца закруглена.
Размеры: высота раковины 16—19, ширина 4.0—4.7 мм,
высота устья 4.0—4.5, ширина 3.0—3.3 мм. Промерено
24 экземпляра.1
Единственная недеколлированная раковина при 13V4
оборотах имела следующие размеры: высота 19.9, ши-
ширина 4.2 мм; высота устья 4.0, ширина его 3.2 мм.
Внутреннее строение не изучено.
Распространение (рис. 80). Вид был найден
вначале лишь в окрестностях Кисловодска, позднее в ря-
ряде мест верховьев р. Малки (Кабардино-Балкарская
АССР).
Экология. Обитает на известняковых скалах.
Систематическое положение. Этот мол-
моллюск наиболее близок к A. semicincta (Bttg.). Главные
отличия описываемого вида: нижняя пластинка лежит
более глубоко; отсутствует затылочный гребень. Что ка-
касается различий в скульптуре поверхности раковины
(у A. narzanensis ребристая, у A. semicincta исчерченная),
то они отчетливо выражены только у типичных форм, потому
что, как уже указывалось, у A. semicincta иногда встре-
встречается ребристая разновидность.
• 4. Acrotoma (Acrotoma) laccata (Bttg.) (рис. 88).
Рис. 87. Acro-
Acrotoma (Acrolo-
ma) narzanensis
(Rosen). (Ориг.).
Boettger O., 1881b : 342 itlausilia (Acrotoma)]; Boettger О.Ч883 : 185,
t. 7, fig. 5 [Clausilia (Acrotoma)]; К о b e 1 t, 1893, Iconogr., N. F., 6:8, fig. 971
[Clausilia (Acrotoma)]; Лихарев и Раммельмейер, 1952 : 224.
P а к овина деколлированная, веретеновидно-цилиндрическая, редко
и неясно исчерченная, блестящая, оливково-коричневая. Оборотов 61/2;
верхние слабо, нижние умеренно выпуклые; последний оборот книзу су-
сужен, сбоку уплощен, за устьем густо и сильно исчерчен. Шов тонкий, слабо
врезанный, белый. Устье большое, ромбическое, скошенное, с почти парал-
параллельными сторонами. Края устья отстоят от стенок раковины, широко
отвернуты и слегка утолщены. Верхняя пластинка тонкая и слегка изо-
изогнутая; впереди доходит до края устья, сзади заходит за передний конец
•спиральной пластинки. Ланцетовидная нижняя пластинка косо подни-
поднимается внутрь раковины. Субколумеллярная пластинка не видна даже
при косом положении устья. Полулунная складка лежит на спинной сто-
стороне раковины; нижний конец ее отогнут назад, верхний сливается под
1 Лектотип данного вида хранится в Зоологическом институте АН СССР (каталог
систематической калл, моллюсков, № 6а) и имеет следующие размеры: высота рако-
раковины 17, ширина 4.6 мм; высота устья 4.4, ширина 3.2 мм; 7 оборотов. Место сбора:
Кисловодск, замок Коварства. Здесь же хранятся 3 паралектотипа; 2 паралектотипа на-
находятся в Музее Грузии и 6 в Зоологическом институте Польской Академии наук.
162
IV. EUXIN1NAE
прямым углом с очень короткой верхней палатальной складкой. Главная
складка лишь немного длиннее верхней складки.
Пупочная щель очень короткая.
Размеры: высота декодированной раковины 19.7, ширина 5.2 мм;
высота устья 5.5, ширина 3.7 мм.
Данный вид описан по единственному экземпляру, найденному где-то,
«в южной части понтического Кавказа» (Boettger О., 1881b). Других на-
находок пока не было.
Систематическое положение. Приве-
Приведенное выше описание является переводом оригинального
диагноза, потому что мы не имели возможности лично озна-
ознакомиться с единственным известным экземпляром дан-
данного вида (хранится в Зенкенбергском музее, во Франк-
фурте-на-Майне).
Наиболее характерным для этого моллюска, по-види-
по-видимому, является присутствие короткой верхней палатальной
складки и небольшие размеры главной складки. У осталь-
остальных видов верхняя складка отсутствует, а главная про-
простирается приблизительно на 1/3 последнего оборота.
ЦодрЪд UOSENIE^LA THIELE
V '? L' i n-Й h о 1 m, -1913a : 16 ' {ThaJestfis, ' поп
Claus, I860); Hesse, 1916, Nachrbl. deutsch.
Malak. Ges., 48 : 124 (Rosenia nom. nov., поп
Schepman, 1913); Liidholm, 1924 : 68
(Rosenia); T h i e 1 e, 1931 : 536/ (Roseniella nom.
nov.).
Тип подцеда: Clausila sobrievsnii Rosen.
Раковина недеколлированная, с ши-
широкой, сосковидно расширенной верши-
вершиной, гладкая, с ярко-белым швом. Оборо-
Оборотов 15—16. Имеется мозолевидная нижняя
палатальная складка.
*5. Acrotoma (Roseniella) sobrievskii
(Rosen) (рис. 89, 90).
Розен, 1914:202, табл. 2, рис. 12 [Clausilia (Oligoptychia)].
Раковина веретеновидно-башневидная, стройная, почти совер-
совершенно гладкая (очень неясно поперечно спирально исчерчена), умеренно
блестящая, красновато-коричневая. Эмбриональные обороты BVa )почти
одинаковой ширины; ниже их ширина оборотов плавно нарастает вплоть
до последнего оборота, который сужается книзу. Ось раковины у верхних
оборотов иногда несколько изогнута. Оборотов 15—16; за исключением
самых верхних, они почти совершенно плоские, разделенные мелким ярко-
белым швом; последний оборот на затылке сильно исчерчен. Устье широко-
широкогрушевидное, выступающее, с базальным желобком, внутри коричневатое,
с белыми, широко отвернутыми, слегка утолщенными краями. Синулус
лежит высоко. Верхняя пластинка низкая и тонкая; впереди она достигает
края устья, плавно снижаясь, уходит на V4 оборота в глубь раковины, где
сходит на нет. Спиральная пластинка довольно толстая; верхний конец ее
лежит в начале второй четверти, а нижний — в конце третей четверти
последнего оборота, почти на одном уровне с верхним концом передней
пластинки, но левее ее. Нижняя пластинка лежит глубоко и при прямом
положении устья почти не видна; она начинается значительно ниже спи-
Рис. 88. Acrotoma (Acrotoma) lac-
cata (Bttg.). (По Бетгору).
A — общий вид раковины; Б — по-
последний оборот (сзади) и просвечи-
просвечивающие сквозь него складки.
3. ACROTOMA
163
ральВой пластинки, быстро нарастает в высоту и спускается, сначала
спирально изгибаясь, а затем в виде толстой косой ланцетовидной складки;
нижний конец ее не обрублен. Короткая субколумеллярная пластинка
лежит почти отвесно вдоль столбика, и лишь ее нижний конец, крючко-
крючкообразно изгибаясь, отходит на внешнюю стенку. Крупная полулунная
складка лежит на спинной стороне (снаружи просвечивает как белое по-
Л
Рис. 89. Лсго- ¦
-tema (Roseniel-
la) sobrievskii
(Rosen). (Ориг.).
КЛ'
Рис. 90. Acrotoma j(Roseniella) sob-
sobrievskii (Rosen). Половая система.
(Ориг.).
перечное пятно). Над ней лежит умерелно длинная главная складка (за-
(занимает около V3 оборота). От нижнего конца полулунной складки отходит
вперед мозолевидная нижняя палатальная складка.
Иногда в устье имеется цельная или состоящая из двух частей (верхней
и нижней) палатальная мозоль, в которую упираются обе палатальные
складки. Клаузилий языковидный, с параллельными краями и с закруг-
закругленным и утолщенным концом лопасти. Пупок щелевидный.
Размеры: высота раковины 24.0—27.5, ширина 5т5—6.0 мм; высота
устья 4.5—5.5, ширина 3.5—4.5 мм.1
Верхние обороты раковины легко отламываются, но это не деколля-
ция, так как септы на месте излома нам обнаружить ни разу не удалось.
1 В Музее Грузии находятся лектотип и 18 паралектотипов. Размеры лектотипа
(№ 1727): высота раковины 27.7, ширина 6.6 мм; 16% оборотов. Место сбора: разва-
развалины крепости в местечке Ардануч, Артвинский округ, сборщик Собриевский, IX,
1910.
164 IV. EUXININAE
Внутреннее строение. Материал: крепость Ардануч, Тур-
Турция, сбор Андронаки, 5 VIII 1912, определение Линдгольма — 1 экз.
Радула: [| + (? + ?+ Й) * 2] * сЫ50.
Размеры срединного зуба 17 X 19.6 ц, ширина радулы 882 \i.
Как по форме, так и по размерам отдельных органов половой аппарат
(рис. 90) очень сходен с остальными видами того же рода. Некоторые
отличия обнаруживает пенис. У данного вида он более цилиндрический;
ретрактор его тоже двуветвистый, но слияние обеих ветвей происходит
раньше, а не перед самой диафрагмой.
Распространение. Пока этот вид был обнаружен лишь в ок-
окрестностях крепости Ардануч (Турция, вилайет Чорох).
4. Род EUXINA BTTG.
Boettger О., 1877а : 83 (Clausilia, секция Euxina); L i n d h о 1 m, 1924 :
65 (Laciniaria, подрод Euxina).
Тип рода: Clausilia hetaera Pfr.
Раковина от веретеновидной до башневидно- или цилиндрическо-
веретеновидной. Скульптура поверхности различная, но у большинства
видов она на верхних дефинитивных оборотах более сильная и редкая,
а к нижним оборотам становится более нежной и густой. Последний оборот
с килем. Устье с базальным желобком. Верхняя пластинка не сливается
со спипальной, и ее верхний конец более или менее глубоко заходит за
нижний конец спиральной пластинки. Верхние концы спиральной и
нижней пластинок обычно лежат на правой стороне раковины, причем
первая берет начало несколько глубже, чем последняя. Между ними у мно-
многих видов лежит небольшая вставочная пластинка. Нижняя пластинка
лежит глубоко в устье и круто уходит кверху; нижний конец ее плавно
сливается со столбиком, а с краем устья ничем не связан. Субколумелляр-
ная пластинка располагается так глубоко, что через устье не видна или
слабо заметна. Полулунная складка или верхние концы палатальных
складок лежат на спинной стороне. Главная складка начинается на право-
спинной и заканчивается на левой стороне раковины. В глубине устья
у большинства видов имеется палатальная мозоль, которая хорошо раз-
развита у видов, лишенных палатальных складок, у видов же, которые имеют
эти складки, она развита слабо или полностью уходит на их образование.
За исключением Ей. difficilis (Ret.), у всех остальных видов, внутрен-
внутреннее строение которых нам известно, срединный и боковые зубы с дополни-
дополнительными зубцами.
Отросток у большинства видов значительно короче собственно семя-
семяприемника. Восходящее колено семяпровода перед впадением в пенис рас-
расширяется, образуя эпифаллус. Здесь же нередко находится рудиментарный
бич. Пенис длинный, расширенный у переднего конца, а нередко также и
у заднего конца; у многих видов он коленообразно изогнут, причем оба
колена связаны друг с другом перепонкой. Ретрактор пениса крепится
к диафрагме. Глазной ретрактор лежит между пенисом и влагалищем.
Колумеллярная мускулатура дает ветвь к шейке семяприемника
и несколько ответвлений к половой клоаке и к влагалищу.
Распространение. Виды данного рода обитают на Кавказе,
в Малой Азии и в северном Ирате. Лишь единичные виды известны во
Фракии и в Палестине
4. EUXINA 165
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА EUXINA
1 F). Полулунная складка распалась, образовав 3—4 довольно длин-
* ных палатальных складки.
2 E). Верхушка раковины очень широкая (не меньше 1.5 мм).
3 D). Раковина довольно стройная, очень тонко и густо исчерченная
(в особенности на средних и нижних оборотах), матовая. Вер-
Верхушка цилиндрическая. Палатальные складки заканчиваются на
различном расстоянии от края устья; ближе всех подходит сред-
средняя, дальше всех — нижняя палатальная складка
11. Eu. tschetschenica (Pfr.).
4 C)., Раковина вздутая, сильно и редко исчерченная или ребристо
исчерченная, со слабым блеском. Верхушка округло-коническая.
Передние концы всех палатальных складок располагаются при-
приблизительно на одном расстоянии от края устья
•. . 10. Eu. somchetica (Pfr.).
5 B). Верхушка раковины довольно узкая A.0—1.1 мм)
12. Eu. novorossica (Ret.).
6 A). Полулунная складка цельная, хорошо, развита. В некоторых
случаях она тонкая, мозолевидная, а ее верхний конец имеет
вид короткой складочки.,
7 A2). Верхушка раковины широкая (больше 1.5 мм).
8 A1). Раковина вздутая. Высота ее больше 16, ширина больше 4.5 мм.
9 A0). Промежуточная складка слабо выражена или отсутствует. Полу-
, лунная складка мощная, просвечивает сквозь, стенку раковины.
Палатальная мозоль толстая . . 5а. Eu. persiea persica (Bttg.).
10 (9). Промежуточная складка хорошо развита. Полулунная складка
тонкая, мозолевидная, не видна сквозь стенку раковины, за ис-
исключением ее верхнего конца, который имеет вид короткой гори-
горизонтальной складки. Палатальная мозоль тонкая, слабо замет-
заметная , . 56. Ew. persica paulhessei (Lindh.).
Раковина стройная. Высота ее 13.7—14.3, ширина 3.9—4.1 мм.
6. Eu. talyschana Likharev, sp. nov.
Верхушка раковины узкая (не шире 1.3 мм).
13 B0). Под главной складкой лежит длинная, заметная через устье
верхняя палатальная складка.
14 A7). Раковина вздутая. Палатальная мозоль распалась на 4—5 пала-
палатальных складок. Края устья с многочисленными мелкими склад-
складками или узелками.
15 {16). Верхняя пластинка проникает вглубь раковины на 1/4 оборота
. 13. Eu. corpulenta (Pfr.).
16 A5). Верхняя пластинка проникает вглубь раковины на 54 оборота . .
- 14. Eu. maesta (Fer.).
17 A4). Раковина стройная. Кроме верхней палатальной складки, при-
присутствуют еще 1 или 2 складки или бугорки. Края устья без
складок, или их немного.
18 A9). Нижние обороты раковины тонко и густо исчерчены и почти
плоские. Нижняя пластинка плавно переходит в столбик . . .
9. Eu. tuschetica Likh. et Leg.
19 A8). Нижние обороты покрыты крупными, редко лежащими ребрами
и довольно выпуклые. Нижняя пластинка внизу обрублена . .
15. Eu. huebneri (Pfr.).
20 A3). Верхней палатальной складки нет, или она настолько коротка
и лежит так глубоко, что через устье не видна.
,166 IV. EUNIN1NAE
2\ B2). Последний оборот за устьем имеет мощное затылочное утолщение,
лежащее поперек и внизу сливающееся с килем
3. Eu. strumosa (Pfr.).
22 B1). Последний оборот без затылочного утолщения.
23 B4). Раковина вздутая, ширина ее 5 мм и больше. Полулунной складки
нет, либо она очень слабо развита и не видна сквозь стенку ра-
раковины 8. Eu. pontica (Ret.).
24 B3). Раковина стройная, ширина ее не больше 4.5 мм. Полулунная
складка хорошо развита и просвечивает сквозь стенку раковины.
25 C0). Палатальная мозоль хорошо развита. Нижний конец клаузилия
без вырезки и выступа.
26 B7). Раковина с плоскими оборотами. Нижняя пластинка при прямом
положении устья не видна. Вставочная пластинка отсутствует.
Высота раковины 16.5—20.5, ширина 4.0—4.5 мм .......
. 4. Eu. difficilis (Ret.).
27 B6). Обороты раковины относительно выпуклые. Нижняя пластинка
более или менее видна при прямом положении устья. Вставочная
пластинка есть. Высота раковины не больше 14, ширина не
больше 3.8 мм.
28 B9). Раковина на всем протяжении равномерно покрыта тупыми реб-
ребрышками. Устье удлиненно-ромбическое, высота его значительно
больше ширины. Ширина раковины 3.0—3.3 мм
2. Eu. circumdata (Pfr.).
29 B8). Верхние дефинитивные обороты редко ребристые, нижние тонко
и густо исчерчены. Устье округло-ромбическое, высота его не-
немногим больше ширины. Ширина раковины 3.3—3.8 мм ....
1. Eu. hetaera (Pfr.).
30 B5). Палатальная мозоль отсутствует. Нижний конец клаузилия
с небольшой вырезкой и выступом 7. Eu. lessonae (Issel).
Подрод EUXINA s. str.
Палатальная мозоль и полулунная складка у большинства видов цель-
цельные. Лишь у немногих они распались, образовав не связанные друг с дру-
другом серии коротких складок или узелков.
Распространение. Кавказ (Талыш), северный Иран, Турция,
Болгария.
*1. Ецхша (Euxina) hetaera (Pfr.) (рис. 91).
P f e i f f e г L., 1848 : 10 (Clausilia); Kuster, 1847—1862 : 167, t. 18, fig. 20—
22 (Clausilia); Rossmassler, 1856, Iconogr., 3, 3—4:71, fig. 888 (Clausilia).
Раковина веретеновиднаяиливеретеновидно-цилиндрическая про-
просвечивающая, с шелковистым блеском, темно- или коричнево-роговая, без
белых штрихов. Верхние 4—5 дефинитивных оборотов редко ребристо
исчерчены, на остальных исчерченность становится более тонкой и густой.
Верхняя часть завитка имеет слегка вогнутые контуры и коротко-цилинд-
коротко-цилиндрическую, совершенно тупую верхушку. Оборотов 11—12, слабо выпуклых,
медленно нарастающих в высоту; последние 2—3 оборота почти одинаковой
ширины; последний оборот книзу плавно сужен, на затылке ребристо
исчерчен, с крупным килем. Устье округло-ромбическое, выступающее,
внутри коричневое, ось его наклонена влево; синулус довольно глубоко
врезан, отогнут; базальный желобок очень глубокий. Края устья широко
отвернуты, утолщены; париетальный край над верхней пластинкой сильно
4. EUXINA
167
прогнут, нижний угловатый. Верхняя пластинка слегка крючковидно
изогнута, впереди немного выступает за край устья, сзади, плавно сни-
снижаясь, слегка заходит за нижний конец спиральной пластинки. Крупная
нижняя пластинка лежит глубоко, но хорошо видна при прямом положе-
положении устья; она круто уходит кверху и лишь под конец дает короткий спи-
спиральный изгиб. Имеется небольшая вставочная пластинка. Верхний конец
толстой и прямой полулунной складки либо отогнут назад под прямым
углом, либо обособлен в виде короткой складки. Главная складка начи-
начинается несколько глубже полулунной и заканчивается
на левой стороне раковины. Палатальная мозоль очень
крупная, белая; внизу она достигает базального желоб-
желобка, наверху немного не доходит до шва; иногда в сере-
середине она утончена или прервана. Рукоятка клаузилия
плавно переходит в короткую языковидную лопасть,
которая сильно изогнута в поперечном направлении,
закруглена и утолщена по нижнему краю.
Размеры: высота раковины 12.0—14.0, ширина 3.3—
3.8 мм; высота устья 2.8—3.2, ширина 2.2—2.5 мм.
Внутреннее строение не изучено.
Распространение. Западная Турция (ви-
(вилайет Бурса) и Принцевы острова (Мраморное море).
*2. Euxina (Euxina) circumdata (Pfr.) (рис. 92).
PfeiflerL, 1848 : 9 {Clausilia); Riister, 1847—1862 :
169, t. 18, fig. 27—30 (Clausilia); Rossmassler, 1856, Ico-
nogr., 3, 3—4 : 72, fig. 889 {Clausilia).
Раковина веретеновиднаяилибашневидно-вере-
теновидная, тонкостенная, густо покрыта тупыми реб-
ребрышками, со слабым блеском, темно- или коричневато-
роговая, с белыми штрихами. Верхушка довольно широ-
широкая, притуплённая. Оборотов 10У2—11, слабо выпук-
выпуклых, вначале медленно, затем быстро нарастающих в
высоту; эмбриональные обороты B—2V2) гладкие, бле-
блестящие; последний оборот выше и шире предпоследнего,
на затылке уплощен, с крупными ребрами, внизу с коротким килем. Устье
удлиненно-ромбическое, выступающее, внизу угловатое, с высоким синулу-
сом и глубоким базальным желобком. Края устья широко отвернуты и
утолщены; палатальный и колумеллярный края нередко почти параллельны
ДРУГ ДРУГУ- Верхняя пластинка довольно высокая и толстая, крючко-
крючковидно изогнутая; впереди она слегка выступает за край устья, сзади слегка
заходит за нижний конец спиральной пластинки и плавно сходит на нет.
Крупная ланцетовидная нижняя пластинка лежит глубоко (слабо видна
при прямом положении устья), кверху уходит почти отвесно. Имеется
небольшая вставочная пластинка. Полулунная складка дугообразно
изогнута. Над ней лежит главная складка, которая начинается на право-
спинной стороне и заканчивается на левой стороне раковины. Внутри
устья на палатальной стенке лежит белая мозоль. Рукоятка клаузилия,
плавно расширяясь, переходит в длинную языковидную лопасть с почти па-
параллельными боковыми краями и с округлым утолщенным нижним концом.
Размеры: высота раковины 12.5—13.5, ширина 3.0—3.3 мм; высота
устья 2.7—3.0, ширина 2.0—2.2 мм.
Изменчивость. Встречаются экземпляры, раковины которых
имеют меньшие размеры и более темно окрашены; высота раковины 11—
Рис. 91. Euxina
(Euxina) hetaera
(Pfr.). (Ориг.).
168
IV. EUX1NINAE
12, ширина 3.0—3.3 мм; высота устья 2.5—3.0, ширина 2.0—3.0 мм (var.
byzantia Nagele, 1910).
Внутреннее строение. Материал: Болгария, Малко Тыр-
ново, лиственный лес, сбор Писарского, 4 V 1958, определение наше —
1 экз.
Радула: [| + [Щ +.^L) X 2] Х с«100.
Размеры срединного зуба 20 X 13 |д,, ширина радулы 372 \и.
Резервуар семяприемника прижат к семяяйцеводу непосредственно
ниже белковой железы. Пенис булавовидно расширен у заднего конца.
Имеется сравнительно длинный бич. Диафрагмальный
ретрактор пениса распадается на серию ветвей, которые
крепятся как к пенису, так и к семяпроводу.
Распространение. Западная Турция (ви-
(вилайет Бурса и окрестности Стамбула) и южная Болга-
Болгария (Малко- Тырново, Граматиково и заповедник Ро-
потамо).1
Экология. Обитает в широколиственных лесах
и в кустарниках, в подстилке, на гниющих валежинах
и на известняковых скалах.
Систематическое положение. Рако-
Раковина данного вида наиболее сходна с таковой у Ей. he-
taera (Pfr.), от которой отличается более стройной и баш-
невидной формой, более сильной скульптурой поверх-
поверхности и удлиненным устьем.
*3. Euxina (Euxina) strumosa (Pfr.) (рис. 93).
PfeifferL., 1848:8 (С lausilia);
К ii s t e г, 1847—1862 : 178, t. 19, fig.
Rossmassler, 1856, Iconogr., 3, 3—4
silia).
11 (Cl. subulata);
27—31 (Clausilia);
66, fig. 882 (Clau-
Рис. 92. Euxina
(Euxina) circum-
dafa (Pfr.). (Ориг.).
Раковина веретеновидно-цилиндрическая, строй-
стройная, просвечивающая, блестящая, красновато-роговая,
с редкими белыми штрихами. Первые 5—6 дефинитив-
дефинитивных оборотов ребристо исчерчены, последующие тонко
и неясно исчерчены или почти гладкие. Завиток с очень
тонкой цилиндрической верхушкой. Оборотов 12—13,
почти плоских, очень медленно нарастающих в вы-
высоту; эмбриональные обороты B1/2) гладкие, блестящие;
последний оборот книзу слабо сужен, с крупным гладким килем, за устьем
с мощным поперечным затылочным утолщением, которое внизу сливается
с килем и отделено от устья глубокой перетяжкой. Устье удлиненно-
ромбическое, выступающее, внизу угловатое, с высоким синулусом и глу-
глубоким базальным желобком. Края устья широко отвернуты и утолщены,
палатальный и колумеллярный почти параллельны друг другу. В области
затылочного утолщения устье имеет поперечное углубление. Пластинки
по форме и положению сходны с Ей. hetaera (Pfr.) и Ей. circumdata (Pfr.),
но нижняя пластинка лежит так глубоко, что при прямом положении устья
не видна, а вставочная пластинка отсутствует. Полулунная складка рас-
распалась на 2—3 коротких складки или узелка. Над ними располагается
1 Сведения о распространении в Болгарии даются на основании обработки сборов
Писарского и Белявского (Зоологический институт Польской академии наук).
4. EUXINA
169
главная складка, которая берет начало на правоспинной стороне и закан-
заканчивается на левой стороне раковины. Имеется довольно длинная и толстая
нижняя палатальная складка, нередко видимая даже при прямом положе-
положении устья. Рукоятка клаузилия плавно расширяется в довольно длинную
языковидную лопасть с почти параллельными краями и закругленным и
утолщенным нижним концом.
Размеры: высота раковины 13—16, ширина 3.0—3.6 мм;
высота устья 3.0—3.5, ширина 2.0—2.4 мм.
Внутреннее строение неизвестно.
Распространение. Западная Турция (вилайет
Бурса).
Систематическое положение. Данный
вид наиболее близок к Е. hetaera (Pfr.), но отличается от
неб более стройной раковиной, острой вытянутой верхуш-
верхушкой, присутствием затылочного утолщения и формой
устья. В отношении замыкательного аппарата данный вид
отличается отсутствием вставочной пластинки, а полулун-
полулунная складка распалась на серию палатальных складок. явиме-*
*4. Euxina (Euxina) difficilis (Bet.) (рис. 94, 95).
Retowski, 1889:255 [Clausilia (Euxina)]; К о b e 1 t, f
1893, Iconogr., N. F., 6 : 26, fig. 998 [Clausilia (Euxina)]; Lind- 1
holm, 1913b : 63 [Cl. (Euxina) lasistana].
Раковина веретеновидно-цилиндрическая или ве-
ретеновидная, стройная, верхние обороты слабо, ниж-
нижние сильно и густо исчерчены, с шелковистым блеском,
коричнево-роговая, с редкими белыми штрихами. Завиток
узко-конический, с тонкой вытянутой верхушкой. Обо- рис &3. Euxi-
ротов 11—13, почти плоских, медленно нарастающих; по- па (Euxina)
следний оборот книзу сужен, за устьем уплощен, с круп- strumosa. (Pfr.).
ным морщинистым килем. Устье грушевидное, внизу (Ориг.).
угловатое, внутри коричневое, с глубоким желобком и высо-
высоким узким неврезанным синулусом. Края устья широко отвернуты, утолще-
утолщены, белые. Верхняя пластинка впереди достигает края устья, сзади плавно
сходит на нет. Вставочной пластинки нет. Крупная ланцетовидная нижняя
пластинка лежит глубоко и при прямом положении устья не видна. Почти
С / Ш и
Рис. 94. Euxina (Euxina) difficilis (Ret.). Радула. (Ориг.).
прямая полулунная складка лежит отвесно на спинной стороне раковины.
Главная складка начинается чуть глубк;е полулунной и заканчивается
на левой стороне раковины. Нижний конец полулунной складки вытянут
в толстую, но короткую нижнюю палатальную складку, видимую при
косом положении устья. В глубине устья, на внешней стенке, лежит раз-
различно развитая палатальная .мозоль, верхняя часть которой часто бывает
170
IV. IR. EUXIN1NAL
утолщенной, образуя крупный бугорок или складку. Языковидная лопасть
клаузилия желобчато согнута, с почти параллельными боковыми краями
и утолщенным нижним концом.
Размеры: высота раковины 16.5—20.5, ширина 4.0—4.5 мм; высота
устья 3.6—4.2, ширина 2.8—3.2 мм.
Внутреннее строение. Материал: Турция, окрестности
Артвина, сбор Нестеренко, 25 V 1910, определение Линдгольма (Clausilia
lasistana) — 1 экз.
п / пс\ Г с , /106 , 36 , 18к \ _ „Т . о_.
Радула (рис. 95): ^т + (— + -j + 3ТГ5) X 2J X 137.
Размеры срединного зуба 12.6 X 22.9 \i, ширина радулы 725 ц..
Единственный вскрытый экземпляр от долгого лежания в спирту
настолько высох, что при препаровке половой системы произошло много
разрывов. Тем не менее отчетливо видно, что половой аппарат в общих
чертах построен по типу, свойственному роду Euxina. Семяприемник с тон-
тонким отростком и округлым резервуаром. Бич отсутствует.
Ретрактор пениса крепится к диафрагме. Левый ретрактор
дает ветвь к шейке семяприемника и 2 ветви к половой
клоаке и к нижнему концу влагалища.
Распространение. Вилайет Чорох (северо-
(северовосточная Турция).
Экология. Обитает на скалах и среди камней.
Систематичес ко е положение. По фор-
форме раковины и устья, по форме и расположению пласти-
пластинок и по отсутствию вставочной пластинки данный вид
наиболее близок к Ей. strumosa, но отличается от послед-
последней большими размерами, отсутствием затылочного утол-
утолщения и цельной полулунной складкой. От Ей. hetaera
(Pfr.) и Ей. circumdata (Pfr.) данный вид отличается тем,
что у него ось устья не отклонена влево, нижняя пластинка
лежит глубже и совершенно не видна при прямом положе-
положении устья, а также присутствует бугор на палатальной
мозоли.
5а. Euxina (Euxina) persica persica (Bttg.) (рис. 96).
Boettger О., 1879, Jahrb. deutsch. Malak. Ges., 6 : 117,
t. 3, fig. 11 (Clausilia); F о г с а г t, 1935:431, fig. 6 [Triloba
Рис. 95. Euxina
(Euxina) diffi-
cilis (Ret.)-
(Ориг.).
(I llunellaria) lessonae Issel, non I s s el, 1865]; Starmuhlner
u. E d 1 a u e r, 1957 : 469 [Triloba (Illunellaria) lessonae].
Раковина веретеновидная или башневидно-вере-
теновидная, вздутая, с довольно резкой и редкой исчер-
ченностью на верхних оборотах (за исключением эмбрио-
эмбриональных), которая становится все более тонкой и
густой к нижним оборотам, благодаря чему поверхность имеет шелкови-
шелковистый блеск. Окраска темно-коричневая или темно-роговая, без белых
штрихов. Верхушка очень широкая A.6—1.9 мм), притуплённая и почти
цилиндрическая, в связи с чем ниже нее завиток имеет вогнутые контуры.
Оборотов 101/2—И1/2, почти плоских, плавно нарастающих в высоту,
разделенных мелким белым швом; эмбриональные обороты B1/4) гладкие,
блестящие; последний оборот книзу сужается, на затылке более сильно
исчерчен и уплощен, с крупным морщинистым килем. Устье округло-ром-
округло-ромбическое, со слегка отогнутым к оси, невысоким, слабо врезанным синулу-
сом и глубоким базальным желобком. Края устья сильно расширены, утол-
утолщены, белые; париетальный край прогнут над верхней пластинкой. Высо-
4. EUXINA 171
кая, крючковидно изогнутая верхняя пластинка, плавно снижаясь, уходит
внутрь раковины. Нижняя пластинка уходит кверху в виде высокой тол-
толстой косо лежащей складки, нижний конец которой слегка заметен при
прямом положении устья. Промежуточная складка слабо выражена или
отсутствует. Имеется короткая вставочная пластинка. Полулунная складка
лежит на спинной стороне; она крупная и хорошо просвечивает сквозь
стенку раковины; оба конца ее отогнуты назад. Главная складка начи-
начинается чуть глубже полулунной и заканчивается недалеко от края устья.
На внешней стенке перед ней располагается палатальная мозоль, особенно
толстая у своего верхнего конца, которым она иногда сливается с передним
концом главной складки. Клаузилий крепится на границе с предпоследним
оборотом. Его языковидная лопасть имеет почти параллельные края,
сильно изогнута в поперечном направлении; нижний конец ее и палаталь-
палатальный край утолщены; по колумеллярному краю, при переходе в рукоятку,
лопасть слегка сжата. Иногда нижний конец лопасти имеет 1 или 2 неболь-
небольших вырезки.
Для характеристики размеров раковины данного подвида приводим
результаты измерений, имевшихся в нашем распоряжении 4 экземпляров
из Ирана:
Высота Ширина Высота Ширина Число
раковины раковины устья устья оборотов
17.5
17.0
18.0
18.3
5.5
5.1
5.3
5.4
4.4
4.2
4.2
4.2
• —
—
3.3
3.7
40»/,
юу2
"V*
11
Первые 3 экземпляра собраны Гейнрихом в 1926 г. в провинции Гилан
(определение наше), последний — в 1899 г. Герцом у Астрабада и определен
Бетгером.
Бетгер (Boettger О., 1879) при описании этого моллюска приводит
следующие размеры: «высота раковины 16—17, ширина 4—4.5 мм; высота
устья 3V2—33/4, ширина 23/4—3 мм, при 11V2—12 оборотах».
Форкарт (Forcart, 1935) указывает, что раковина обоих экземпляров,
которых он рассматривал как Triloba lessonae (Issel) и которые в действи-
действительности являются Ей. persica (Bttg.), имеют высоту 19.3 и 20.3 мм.
Из сказанного видно, что размеры раковин данного моллюска подвер-
подвержены значительным колебаниям.
Внутреннее строение. Материал: Иран, провинция Гилан,
сбор Гейнриха, 1926 г., определение наше — 2 экз.
Радула: [| + (f+з^) X 2] X И2.
Размеры срединного зуба 12 X 18 ц, ширина радулы 690 \i.
Половая система построена по такому же плану, как у Ей. talyschana
mihi и Ей. lessonae (Issel). Семяприемник и его отросток длинные. Первый
достигает белковой железы, второй немного короче, но сильно зигзаго-
зигзагообразно извит. Пенис двухколенчатый, причем обе части соединены пере-
перепонкой. Дистальная часть пениса сильно расширена, внутри с продольными
складками. Имеется рудиментарный бич. Ретрактор пениса многоветвис-
многоветвистый и веерообразно крепится вдоль верхнего конца пениса, причем край-
крайние ветви являются наиболее крупными.
Распространение. В СССР: единственная находка сделана
Д. В. Знойко в бассейне р. Вашару-Чай у с. Торады. Вне СССР: лесные
области северного Ирана.
172
IV. EUXININAE
Лосьес (Loosjes, 1953) сообщает о находке Ей. persica (Bttg.) на черно-
черноморском побережье Турции (вилайет Зонгулдак). По форме раковины и
ее размерам, по степени развития полулунной складки турецкие экземп-
экземпляры очень сходны с экземплярами, найденными в Иране, но отличаются
более сильным развитием промежуточной складки, походя в этом отно-
отношении на болгарский подвид этого вида (см. далее).
Так как мы не имели возможности ознакомиться с турецкими экземп-
экземплярами этого вида, то в данный момент не можем принять решение, к ка-
какому из подвидов их следует отнести.
Синонимия и систематическое по-
положение. Форкарт (Forcart, 1935) принял данный
вид за Clausilia lessonae Issel (см. стр. 175). Обнару-
Обнаружив, что клаузилий этого моллюска имеет 2 небольших
вырезки (как мы уже отмечали, действительно, у неко-
некоторых экземпляров это имеет место), он отнес его в бал-
балканский род Triloba Vest (подсемейство Alopiinae).
В пользу этого вывода, с его точки зрения, говорил
тот факт, что на пенисе того же моллюска удалось об-
обнаружить подобие придатка. А как известно, этот ор-
орган свойствен многим Alopiinae, в том числе и роду
Triloba. Других частей половой си-
системы Форкарт не мог изучить из-за
плохой их сохранности.
Если провести сравнение ракови-
раковины и половой системы нашего моллюс-
моллюска с типом рода Triloba — Т. sandrii
(Kiister), то обнаружатся существен-
существенные различия. В первую очередь они
касаются формы семяприемника и его
отростка. У нашего моллюска, как у
всех Euxininae, он тонкий, извитой и
впадает в семяприемник у его основа-
основания. У Т. sandrii (см.: Wagner, 1922),
как у всех A lopiinae, он толстый и впа-
впадает в семяприемник на значительном
удалении от его основания. У нашего
моллюска семяприемник снабжен
ретрактором, у Т. sandrii его нет. Не-
Небольшой придаток, который обнару-
обнаружил у своего моллюска Форкарт, на самом деле находится не на пенисег
а на впадающем в последний семяпроводе, и является бичом. У Т. sandrii
придаток представляет собой крупный, хорошо развитый орган, сидящий
на пенисе. Радула нашего моллюска отличается наличием дополнитель-
дополнительных зубцов на срединном и боковых зубах, у Т. sandrii этих зубцов нет.
Наконец, клаузилий Т. sandrii, как и у всех Alopiinae, имеет глубокую
вырезку на конце лопасти, к которой у некоторых видов того же рода до-
добавляется еще вторая вырезка меньших размеров. Большая вырезка слу-
служит для пропуска крупной нижней палатальной складки. Наш моллюск
не имеет нижней палатальной складки, благодаря чему и вырезки клаузи-
лия очень небольшие и нередко отсутствуют. В заключение заметим, что
замыкательный аппарат нашего моллюска совершенно другого типа, чем
у рода Triloba.
Отсюда напрашивается единственный вывод, что у Форкарта не было
серьезных оснований относить моллюска, которого он неправильно опре-
Рис. 96. Euxina (Euxina) persica persi-
persica (Bttg.). (Ориг.).
A — общий вид раковины; Б — клаузилий.
4. EUXINA 173
делил как Clausilia lessonae Issel, в род Triloba. В действительности, это
настоящая Euxina, имеющая много общего с другими видами того же рода,
¦обитающими в Иране и Турции.
*5б. Euxina persica paulhessei (Lindh.).
Hesse, 1912, Nachrbl. deutsch. Malak. Ges. 44:58 (Clausilia thracica n. sp.,
non P. F i s с h e r, 1866); Liridholm, 1925 : 263 (Cl. paulhessei nom. nov.).
Отличается от предыдущего подвида: 1) скульптура поверхности рако-
раковины более резко выражена — верхние обороты ребристые, средние реб-
ребристо исчерчены, нижние тонко исч«рч«ны; 2) раковина блестящая;
3) промежуточная складка хорошо развита; 4) полулунная складка слабо
развита; середина ее тонкая, мозолевидная, оба конца имеют вид коротких
складок; 5) палатальная мозоль тонкая, слабо заметная.
Для характеристики размеров раковины приводим результаты измере-
измерений 7 экземпляров, собранных научными сотрудниками Зоологического
института Польской Академии наук Белявским и Писарским и одного из
котипов Clausilia thracica Hesse (№ 8).
№ экз. Высота Ширина Высота Ширина Число
раковины раковины устья устья оборотов
1 18.4 5.4 4.3 . 3.3 11
2 17.8 5.2 4.1 3.1 10з/4
3 16.7 4.9 3.8 3.2 10*/8
4 16.5 4.9 3.7 3.0 ЮУг
5 16.3 4.9 3.8 3.0 ЮУ2
6 16.5 5.0 3.8 3.0 10У2
7 18.0 5.0 4.4 3.6 11
8 21.4 5.7 4.7 — Ю'/з
Гессе (Hesse, 1912) для имевшихся в его распоряжении 4 экземпляров
приводит следующие размеры: высота раковины 19—21, ширина 5.5 мм.
Сопоставляя данные о размерах раковин обоих подвидов, можно сде-
сделать вывод, что в этом отношении они мало отличаются друг от друга.
Внутреннее строение. Материал: Малко Тырново (Бол-
(Болгария), сбор Писарского, 2 V 1958, определение наше — 1 экз.
Половая система данного подвида в общем мало отличается от таковой
предыдущего.
Распространение. Южная Болгария, возможно черномор-
черноморское побережье Турции. Этот моллюск первоначально был найден в южной
Болгарии у города Пловдив. Новые находки сделаны в окрестностях го-
города Малко Тырново и в заповеднике Ропотамо.
Экология. В сырых дубово-буковых лесах и в кустарниках, про-
произрастающих на известняковых породах, в гнилых валежинах и в под-
подстилке.
6. Euxina (Euxina) talyschana Likharev, sp. nov. (рис. 97, 98).
Раковина башневидно-веретеновидная, тонко и неясно исчер-
исчерченная, слабо просвечивающая, темно-роговая, без белых штрихов. Зави-
Завиток очень медленно и слабо расширяется книзу; боковые контуры его
прямые; верхушка очень широкая A.6—1.7 мм) округлая. Оборотов
10—10V2 почти плоских, разделенных очень мелким сдавленным швом;
последний оборот наверху шире предпоследнего, книзу плавно сужен,
на затылке уплощен, с крупным, почти гладким килем. Устье округло-
174
IV. EUX1NINAE
ромбическое, внутри коричневое, с почти параллельными противолежа-
противолежащими краями, выступающее, с довольно глубоко врезанным узким сину-
лусом и глубоким базальным желобком. Края устья сильно расширены,
утолщены, белые. Толстая, крючковидно изогнутая верхняя пластинка,
плавно снижаясь, сзади слегка заходит за нижний конец спиральной
пластинки. Последняя, как и нижняя пластинка, начинается на правой
стороне и, плавно нарастая в высоту, тянется до левой стороны раковины.
Нижняя пластинка той же
формы и так же расположена,
как и у Ей. persica (Bttg.). ^—~^^бж
Имеется небольшая вставоч-
вставочная пластинка. Субколумел-
лярная пластинка не видна.
Полулунная складка раз-
развита в различной степени —
ся
Рис. 97. Euxina (Euxina) talyschana Likharev,
sp. nov. (Ориг.).
A — общий вид раковины; Б — последний оборот (сзади)
и просвечивающие сквозь него складки; В — клаузилий.
Рис. 98. Euxina (Euxina)
talyschana Likharev, sp.
nov. Половая система.
(Ориг.).
от мозолевидной до ясно дугообразной, лежит на спинной сто-
стороне. Непосредственно над нею, порой сливаясь с ее верхним концом,
находится короткая верхняя палатальная складка. Длинная главная
складка начинается на правоспинной стороне и заканчивается на левой
стороне раковины. На значительном удалении от края устья, поперек
палатальной стенки, лежит довольно крупная коричневая палатальная
мозоль. Рукоятка клаузилия крепится ъ начале последнего оборота.
Языковидная лопасть* сильно изогнута в поперечном направлении, имеет
почти параллельные боковые края и утолщенный нижний конец с корот-
коротким выступом и слабой вырезкой. Колумеллярный край лопасти при
переходе в рукоятку тоже слегка вырезан.
Размеры в (мм):
№ экз.
1
2
3
Высота
раковины
14.2
13.7
14.3
Ширина Высота Ширина Число Примечание
раковины устья устья оборотов
4.1 3.2 2.9 10 Голотип1
4.0 3.4 2.9 10
3.9 3.4 2.6 Юй
1 Хранится в Зоологическом институте АН СССР (каталог систематической кол-
коллекции моллюсков, № 2а).
4. EUXINA 175
Внутреннее строение. Материал: Талыш, гора Шиндан-
Каласи, 31 V 1952, сбор и определение наше — 1 экз.
Радула: [? + (f + ^) X 2] X 100.
Размеры срединного зуба 11 X 14.3 \х, ширина радулы 524 ц.
Семяприемник и его отросток почти одной длины, и оба не достигают
белковой железы (рис. 98). Влагалище по длине почти равно матке.
Пенис состоит из трех отделов — верхнего, среднего и нижнего. Верхний
отдел, в который открываются семяпровод и бич, представляет собой
сужающуюся к концу трубку с гладкой внутренней поверхностью. К этому
отделу прикрепляется ретрактор пениса, который, вскоре после отхода
от диафрагмы, распадается на 3 ветви, из которых каждая несколько
ниже дает серию мелких мускулов. Все эти ответвления веерообразно
крепятся к пенису. Внутри среднего отдела пениса помещаются мускуль-
мускульные валики. Средний и передний отделы в месте сужения связаны пере-
перепонкой.
Распространение. Лесной Талыш. Сделано две находки:
одна Д. В. Знойко, 22 VII 1932 на горе Нюдис-Каласи (экземпляры
№№ 1 и 2), другая нами на горе Шиндан-Каласи, в лесу, на высоте
1650 м над ур. м.
Экология. Обитает на деревьях и под корой.
Систематическое положение. Данный вид наиболее
близок к Ей. persica (Bttg.), но отличается от последней стройной ракови-
раковиной, меньшими размерами и более слабым развитием верхней части пала-
палатальной мозоли и полулунной складки. По форме раковины данный вид
сходен с Ей. tschetschenica (Pfr.).
/
/
*7. Euxina (Ешрпа) lessonae (Issel) (рис. 99—101).
I s s e 1, 1865 : 426; t. 3, fig. 55—57 (Clausilia); Lindholm, 1916 : XLIII—
XLIV [Clausilia (Euxina)]; Biggs, 1931:164 (Cl. reshtensis); F о г с а г t, 1935:
430 [Laciniaria (Euxina) reshtensis Biggs].
Раковина веретеновидная, сильно просвечивающая, довольно
блестящая, от красновато-коричневой до темно-роговой. Поверхностная
скульптура сильно изменяется от верхушки к устью: эмбриональные
обороты BV2) гладкие; последующие 6 оборотов покрыты широко рас-
расставленными сильными складками (ребристо исчерчены); наконец, на
оставшихся оборотах исчерченность становится более тонкой и густой.
Верхняя часть завитка имеет очень вогнутые очертания и коротко-цилин-
коротко-цилиндрическую, совершенно тупую верхушку (последнее благодаря тому, что
первый оборот закручен почти в одной плоскости). Оборотов 12—13,
слабо выпуклых, медленно нарастающих в высоту; последний оборот
книзу плавно сужен, на затылке более сильно исчерчен, с крупным килем.
Устье широко-грушевидное, выступающее, внутри коричневое, с глубоко
врезанным синулусом и глубоким базальным желобком. Толстая, крючко-
видно изогнутая верхняя пластинка слегка выступает за край устья,
сзади, плавно снижаясь, слегка заходит за нижний конец спиральной
пластинки. Крупная нижняя пластинка уходит кверху в виде высокой
и толстой косо лежащей складки, нижний конец которой слегка заметен
при прямом положении устья. Имеется короткая вставочная пластинка.
Крупная дугообразная полулунная складка лежит на спинной стороне.
Главная складка начинается несколько глубже и простирается до левой
стороны раковины. Рукоятка клаузилия, плавно расширяясь, переходит
176
IV. EUXININAE
в языковидную лопасть, с почти параллельными боковыми краями; ниж-
нижний конец ее имеет довольно глубокую вырезку и тонкий выступ.
Размеры: высота раковины 13—15, ширина 3.5—4.2 мм; высота устья
3.0—3.5, ширина 2.5—2.7 мм.
Внутреннее строение. Материал: Иран, Энзели, сбор Кни-
повича, 20 III 1915, определение Линдгольма — 1 экз.
Радула (рис. 100): [ J + (J + ^) X 2] X 88. • ;
Размеры срединного зуба 11.2 X 15.4 ц, ширина радулы 435 р..
Семяприемник с коротким отростком (рис. 101). Влагалище толстое,
приблизительно той же длины, что и матка. Семяпровод с бичом. Задняя
половина пениса расширена и мускулистая, внут-
внутри с продольными валиками. Ретрактор пениса
трехветвистый.
Распространение. Лесные области
северного Ирана (провинции Гилан и Мазенде-
ран).
Экология. Обитает под корой и в древе-
древесине гниющих валежин.
Размножение. Живородящий вид.
У вскрытог/6 экземпляра в матке обнаружено
5 эмбрионов.
Рис. 99. Euxina (Euxina) lessonae (Issel). (Ориг.).
A — общий вид раковины; Б — клаузилий.
Синонимия и систематическое положение. Линд-
гольм (Lindholm, 1916), который значительно пополнил первоначальное
описание этого моллюска, указывает, что замыкательный аппарат лишен
полулунной складки. На этом основании позднее A924 г.) он выделил
этот вид в особую секцию Illunellaria Lindh. (род Laciniaria, подрод
Euxina). При вскрытии тех самых раковин, с которыми работал Линд-
гольм, мы убедились, что у всех у них полулунная складка хорошо раз-
развита. По-видимому, автор не заметил ее из-за того, что имел дело с экзем-
экземплярами, длительное время находившимися в спирту, благодаря чему
стенки раковин потеряли первоначальную прозрачность и эта складка
снаружи была не видна. Так как, согласно Линдгольму, Illunellaria
отличается от настоящих Euxina лишь отсутствием полулунной складки,
4. EUXINA
177
то поскольку таковая на самом деле имеется, следует рассматривать пер-
первое название как синоним второго.1
Раковина данного моллюска имеет много общего с Ей. ketaera (Pfr.),
но отличается от последней более веретеновидной формой, большим чис-
19
го
^..
Рис. 100. Euxina (Euxina) lessonae (Issel). Радула. (Ориг.).
лом оборотов, грушевидной формой устья (причем ось его не наклонена
влево) и отсутствием палатальной мозоли. По форме клаузилия (наличие
Вэк
Рис. 101. Euxina (Euxina) lessonae (Issel). Половая
система. (Ориг.).
вырезки и выступа) и по форме пениса, данный вид сходен с Ей. persica
(Bttg.) и Ей. talyschana mini, но отличается в первую очередь более строй-
стройной и, в особенности, .тонкой верхушкой.
*8. Euxina (Euxina) pontica (Ret.) (рис. 102, 103).
Retowski, 1887 : 39, t. 1, fig. 5 [Clausilia (Euxina)].
Раковина2 веретеновидная, вздутая, прочная, темно-роговая,
с пучками белых штрихов. Верхние дефинитивные обороты тонко ребри-
1 Форкарт (Forcart, 1935) рассматривал Illunellaria как секцию рода Triloba
Vest.
2 Описание дается по 3 экземплярам, полученным 6$ Форкарта (Базель) под назва-
названием «Euxina reulauxi Bttg.».
178
IV. EUXININAE
стые, на последующих скульптура становится гуще и слабее, до исчерчен-
ности. Верхняя часть завитка имеет вогнутые контуры и сравнительно
узкую A.2—1.3 мм) округло-цилиндрическую верхушку. Оборотов 11—12,
слабо выпуклых, разделенных тонким врезанным швом; эмбриональные
обороты B1/2—3) гладкие, блестящие, светлые; последний оборот на за-
затылке слегка уплощен, с крупным ^илем. Устье удлиненно-ромбическое,
внутри коричневое, с высоким, отогнутым направо врезанным синулусом.
Края/^стья широко отвернуты, утолщены,
белые; палатальный край почти прямой, ниж-
нижний плавно изогнут, париетальный сильно
прогнут над верхней пластинкой. Высокая,
слегка крючковидно изогнутая верхняя пла-
пластинка сравнительно короткая и лишь слегка
заходит за спиральную. Нижняя пластинка
лежит глубоко и чуть заметна при прямом
положении устья; она прямая и круто ухо-
уходит кверху. Имеется маленькая вставочная
пластинка. Главная складка начинается на
правой и заканчивается на левой стороне.
Ниже ее верхнего конца лежит короткая
Рис. 102. Euxina (Euxina) pontica (Ret.). (Ориг.).
A ¦• общий вид раковины; Б — клаузилий.
верхняя палатальная складка — остаток верхнего конца полулунной
складки. Последняя либо отсутствует, либо очень слабо развита и имеет
вид слабой мозоли. Палатальной мозоли нет. Рукоятка клаузилия плавно
переходит в широкую ложковидную лопасть, нижний конец которой за-
закруглен и утолщен.
Размеры: высота раковины 18.7—20.2, ширина 5.2 мм; высота устья
4.3—4.7, ширина 3.2—3.4 мм. Согласно Ретовскому A887), высота рако-
раковины 19.8—22.5, ширина 4.5—5.2 мм.
Внутреннее строение. Материал: Турция, между Самсу-
ном и Бафрой, 27 IV 1936, из коллекции Форкарта («Euxina reulauxi
Bttg.») — 1 экз.
Семяприемник длинный, его резервуар лежит непосредственно ниже
белковой железы (рис. 103). Отросток на один оборот короче. Шейка семя-
семяприемника толстая и мускулистая. Пенис цилиндрический, лишь перед
впадением в половую клоаку несколько вздут. Имеется рудиментарный
4. EUXJNA
179
ся
-осп
бич. Влагалище многочисленными мускульными волокнами подтянуто1
к стенке тела.
Распространение. Вид был описан по экземплярам, найден-
найденным в выбросах моря на Южном берегу Крыма. Единственная нам извести
ная находка живых экземпляров данного вида сделана на черноморском
побережье Турции между Самсуном
и Бафрой.
Размножение. Вид яйце-
кладуЦий.
Систематиче с~к о е по-
положение. Ретовский сближал
данный вид, с одной стороны, с Me-
galeuxina derasa Mouss., с другой — с
Euxina maesta (Fer.). Последнее, по-
видимому, связано с какой-то ошиб-
ошибкой, потому что, как видно из ори-
оригинального описания ж рисунков,
данный вид не имеет ничего общего
с палестинским видом Ей. maesta.
Наш моллюск по форме и размерам
раковины, на первый взгляд, немного
похож на М. derasa, но отличается
от него формой и положением устья
и строением замыкательного аппа-
аппарата (в первую очередь нижней пла-
пластинки). Особенности замыкательного
аппарата сближают наш вид с рядом
малоазиатских видов Euxina — с та-
такими, как Ей. hetaera (Pfr.), Ей.
circumdata (Pfr.) жЕп. strumosa (Pfr.).
Но в отличие от них, у нашего моллюска главная складка начинается
глубже, а нижняя пластинка подходит ближе к устью. От всех нам
известных видов Euxina раковина нашего моллюска отличается крупными
размерами и вздутой формой, в этом отношении, приближаясь к Ей.
persica (Bttg.). Однако последняя отличается широкой и тупой верхушкой
и некоторыми другими признаками.
9. Euxina (Euxina) tuschetica Likharev et Lejava (рис. 104—106).
Лихарев и Л е ж а в а, 1961 : 473—477, рис. 1—3.
Раковина башневидно-веретеновидная, стройная, на верхних
дефинитивных оборотах тонко и редко ребристая, на нижних оборотах
тонко и густо исчерченная, слабо просвечивающая, с шелковистым бле-
блеском, темно-роговая. Верхушка узкая, округлая (ширина 1.0—1.2 мм);
боковые контуры завитка ниже верхушки слегка вогнуты.Оборотов 13,
плавно нарастающих; верхние слабо выпуклые, нижние почти плоские;
эмбриональные обороты C) гладкие, выпуклые, светлоокрашенные; по-
последние 2—3 оборота почти одинаковой ширины; последний оборот за
устьем ребристый и уплощенный, внизу с крупным морщинистым килем.
Устье широко-грушевидное, внутри коричневое, с широким, низко лежа-
лежащим и неврезанным синулусом и с довольно глубоким базальным желоб-
желобком. Края устья широко отвернуты, белые; нижний край плавно закру-
закруглен. Верхняя пластинка впереди достигает края устья, сзади плавно
сходит на нет, не доходя до нижнего конца спиральной пластинки. По-
•ркл
Рис. 103. Euxina (Euxina) pontica
(Ret.). Половая система. (Ориг.).
180
IV. EUXINWAE
следндя лежит глубоко и через устье плохо заметна. Нижняя пластинка
лежит глубоко и видна лишь при косом положении устья; она имеет вид
толстой ланцетовидной складки с утолщенным внешним краем, которая
круто уходит в глубь раквшшы и лишь под конец дает короткий изгиб;
нижний конец ее не связан с краем устья. Между верхними концами ниж-
нижней и спиральной пластинок лежит небольшая, но хорошо выраженная
вставочная пластинка. Имеется промежуточная складка, лежащая между
нижним концом нижней пластинки и верхним
концом верхней пластинки. Субколумеллярная
пластинка вытянута почти параллельно ниж-
нижней; нижний конец ее заметен при косом поло-
положении устья. Толстая полулунная складка ле-
лежит на право-спинной стороне раковины;
нижний конец ее отогнут вперед, к устью. Не-
Несколько глубже берут начало главная и верх-
верхняя палатальные складки, простирающиеся
вперед на V4 оборота, причем первая парал-
параллельна второй и несколько длиннее ее. Имеется
короткая, но толстая средняя палатальная
Рис. 104. Euxina (Euxina) tuschetica Likharev et Lejava. (Ориг.).
A — общий вид раковины; В — последний оборот (справа) и просвечи-
просвечивающие сквозь него складки; В — клаузилий.
складка/'лежащая на левой стороне последнего оборота раковины. Сра-
Сравнительно короткая рукоятка клаузилия (крепится к столбику в на-
начале последнего оборота), плавно расширяясь, переходит в узкую, но
длинную языковидную лопасть, слегка утолщенную на конце.
Размеры (в мм):
№ экз. Высота Ширина Высота Ширина Число Примечание
раковины раковины устья устья оборотов
1
2
3
22
20.6
4.7
4.7
4.5
4.5
4.5
4.5
3.8
3.3
3.5
133/4
13У2
Голотип 1
Вершина (
1 Хранится в Зоологическом институте АН СССР (каталог систематической кол-
коллекции моллюсков, № 1). Место сбора: Грузия, Тушетия, левый берег Тушетской Ала-
зани, 1600 мнадур. м., VIII 1959, сборщик Лежава.
4. EUXINA
181
Внутреннее строение. Материал: Грузия, Горная Туше-
тия, с. Омало, 1800 м над ур. м., смешанный лес, 8 VII 1959. Вскрыт 1 экз.
Радула (рис. 105): [| + (^ + ^) X 2] X 120.
с 1 11 12
Рис. 105. Euxina (Euxina) tuschetica Likharev et Lejava. Радула. (Ориг.).
Размеры срединного зуба 25 X 13.4 ц, ширина радулы 800 \i.
Длинный семяприемник заканчивается небольшим округлым резер-
резервуаром, прижатым к белковой железе (рис. 106). Отросток семяприемника
гп
бж
Рис. 106. Euxina (Euxina) tuschetica Likharev et
Lejava. Половая система. (Ориг.).
очень тонкий и сильно извитой, наверху заканчивается на 1/2 оборота
выше верхнего конца семяпровода. Матка очень короткая. Влагалище
182 IV. EUZIN1NAE
длинное, цилиндрическое, в 4х/2 раза длиннее матки. Длинный пенис
в средней части веретенообразно расширен. Имеется сравнительно длин-
длинный (около 1 мм) бич. Ретрактор семяприемника прикреплен как к шейке
последнего, так и частично к матке.
Распространение. Горная Тушетия (юго-западная часть На-
Нагорного Дагестана, административно — северо-восточная Грузия), в бас-
бассейне Тушетской Алазани (верховья Андийского Койсу).
Экология. Обитает в смешанном лесу, на скалах и под камнями
на высотах 1600—2200 м над ур. м.
Систематическое положение. В пределах Euxina с на-
нашим видом наибольшее сходство обнаруживает Ей. difficilis (Ret.), с кото-
которым наш моллюск сближают форма и размеры раковины, форма и поло-
положение пластинок, наличие полулунной складки, ее форма и глубина зале-
залегания в раковине. Однако от Ей. difficilis наш вид отличается присут-
присутствием вставочной пластинки, верхней и средней палатальных складок,
формой клаузилия и тем, что нижний конец субколумеллярной пластинки
виден через устье, в то время как у Ей. difficilis он не виден.
Наш моллюск принадлежит к подроду Euxina s. str. За это говорит
как сходство с Ей. difficilis, которая относится к этой же группе, так и
ряд признаков, общих с другими видами того же подрода. Правда, нали-
наличие верхней палатальной складки, при сохранении цельной полулунной
складки — явление, до сих пор не отмечавшееся в пределах указанного
подрода и более свойственное подроду ЕНа Н. et A. Adams. Однако по-
последняя группа резко отличается от Euxina tuschetica в первую очередь
присутствием, кроме верхней, еще 3—4 нижележащих палатальных
складок, которые образовались за счет палатальной мозоли, а также нали-
наличием многочисленных складочек и узелков по краю устья. Поэтому мы
считаем, что нет оснований включать наш вид в подрод ЕНа.
Подрод CAUCASICA BTTG.
BoettgerO., 1877а : 85 (Clausilia, секция Euxina, группа Caucasica); L i n d-
h о 1 m, 1924 : 66 (Laciniaria, подрод Euxina, секция Caucasica).
Тип подрода: Clausilia somchetica Pfr.
Полулунная складка и палатальная мозоль распались и за счет их
возникли 3 или 4 крупных и длинных палатальных складки, заканчиваю-
заканчивающихся более или менее близко от края устья и видимых при прямом его
положении (иногда за исключением нижней складки).
Распространение. Кавказ.
10. Euxina (Caucasica) somchetica (Pfr.) (рис. 107—109).
P f e i f f e r L., 1846, Symb. Hist. Helic, 3 : 94 (Clausilia); С и м а ш к о, 1847 :
111 (Cl. Kolenatii); Rossmassler, 1856, Iconogr., 3, 3—4 : 61, fig. 877 (Clausilia);
P f e i f f e г L., 1857, Malak. Bl, 4 : 88 (Cl. colchica); Розен, 1914 : 200 [Clausilia
(Euxina)]; Ретовский, 1914 : 320 [Clausilia (Euxina)]; Лихарев и Рам-
мельмейер, 1952 : 240, рис. 162 [Laciniaria (Euxina)].
Раковина веретеновидная, очень вздутая, твердостенная, сильно
исчерченная, реже ребристо исчерченная, просвечивающая, со слабым
блеском, красновато- или зеленовато-роговая, с пучками белых штрихов.
Короткая коническая часть завитка с прямыми контурами и очень широ-
широкой A.5—1.7 мм) округлой верхушкой. Оборотов 9—10, реже 101/2,
слабо выпуклых; эмбриональные обороты BV4) гладкие, очень широкие,
одноцветные; последний оборот наверху сильно вздут, книзу плавно су-
сужается, за устьем уплощен, с длинным крупным килем, который окаймлен
4. EUXINA
183
ясной бороздой. Устье ромбическое или угловато-овальное, выступающее,
внутри коричневатое, с низким умеренно врезанным синулусом и глубо-
глубоким базальным дселобком, благодаря которому нижний край устья угло-
угловатый. Края устья белые, утолщены, слегка отвернуты; палатальный
и колумеллярные края почти параллельны, париетальный над верхней
пластинкой прогнут. Высокая, слегка крючковидно изогнутая верхняя
пластинка, плавно снижаясь, уходит внутрь раковины. Глубоко лежащая
невысокая нижняя пластинка плохо видна при
~ прямом положении устья; она лежит почти от-
отвесно, лишь верхний конец ее образует спираль-
спиральный изгиб, внизу она заканчивается далеко от
края устья. Имеется короткая вставочная пла-
пластинка. Главная, рудимент полулунной (если он
есть) и все палатальные складки начинаются на
спинной стороне раковины, причем первая не-
несколько глубже, чем остальные. Чаще всего под
главной складкой располагаются 3, реже 4 па-
палатальных складки. Однако нередко сохраняется
Рис. 107. Euxina (Caucasica) somchetica (Pfr.). (По Лихареву и Раммель-
мейер).
А — общий вид раковины; Б — клаузилий.
¦остаток нижней части полулунной складки, который, слив'аясь с
дижней палатальной складкой, создает иллюзию, будто задний ко-
конец последней отогнут кверху. Передние концы всех складок распола-
располагаются сравнительно недалеко от края устья и поэтому хорошо видны при
прямом положении раковины. Кроме главной, наиболее постоянной
является верхняя палатальная складка. Остальные при большей толщине
нередко испытывают некоторое укорачивание, которое идет в определен-
определенной последовательности: 1) у средней чаще всего исчезает средняя часть,
реже, кроме того, задний конец, и последним исчезает передний конец;
2) редукция нижней палатальной складки идет за счет постоянного уко-
укорачивания ее со стороны переднего конца. Клаузилий состоит из короткой
рукоятки (крепится к столбику в начале последнего оборота, т. е. над
синулусом), которая очень плавно расширяется в сравнительно узкую
языковидную лопасть, передний конец которой плавно закруглен и слегка
утолщен.
Размеры: высота раковины 14—16 (реже до 18), ширина 4.5—5.0 мм
(реже до 5.5 мм); высота устья 3.5—4.0 (реже до 4.5), ширина 2.7—3.0 мм.
Изменчивость. У верхней границы горного леса и в субаль-
субальпийской зоне популяции данного вида нередко состоит из экземпляров,
раковины которых отличаются меньшими размерами (высота 12.5—14.0,
184
IV. EUXININAE
ширина 4.2—4.5 мм) и йодной или частичной редукцией 2 нижних пала-
палатальных складок (f. raddeivMonsson, 1876).
Внутреннее строение. Материал: 1) Железноводск, сбор
и определение наше, 1 VI 1954 — 1 экз.; 2) Азербайджанская ССР, Хан-
с 1
Рис. 108. Euxina (Caucasica) somchetica (Pfr.). Радула. (Ориг.).
ларский р-н, сел. Чайкенд, сбор и определение наше, 16 VI 1954 — 2 экз.;
там же, 27 VI 1954 — 1 экз.
Радула (рис. 108): [| + (^ + i^) х 2] X 106.
Рис. 109. Euxina (Caucasica) somchetica (P .}. Поло-
Половая система. (Ориг.).
Размеры срединного зуба 12.6 X 23.8 ц,, ширина радулы 630 ц.
Гонада образована 4—5 дольками. Семяприемник длинный (рис. 109);
резервуар его лежит непосредственно ниже белковой железы. Отросток
семяприемника короткий. Пенис почти цилиндрической формы, лишь
4. EUXINA
185
задняя половина немного шире передней. Внутри него проходят 4—5 про-
продольных складок. Имеется бич. Влагалище в несколько раз длиннее-
матки; многочисленными мускульными волокнами оно на значительном
протяжении связано со стенкой тела.
Распространение (рис. 110). В СССР этот вид, по-видимому,,
широко распространен на Кавказе. Однако ареал его еще плохо изучен.
Он найден: в горных лесах и в субальпийской зоне восточной части Кав-
46
44
42
40
38
о.
_
Т<-
_
•г
38'
W } 1 \1 —у-
ч
с^—<
• )
55 0 55
i 1 1—
i
38
40 4
Js^PmipS4--.
\'\/;
Сухумику
Л
Батуми/
110 165 220 км
i
40
44-
I
4cTaepflnoj]bv
\ ^J,
\ V / •
\
.4
42
46 48
Астрахань^
СЖ
1 ^'Орджоникидзе
{Чч^ У \
"НЧгТбилисИт -¦¦
44 # 46
f
50
Махач-KS/jC
\
11/
~~~-—_
Щ
{
N
48
52
>—.
\
X
г^Баку
V
V
4 \
50
54 56
)
Ч Ч
\
J
44
42
40
38
36
52 54
Рис. НО. Географическое распространение Euxina (Caucasica) somchetica (Pfr.) A}
и Ей. (С.) tschetschenica (Pfr.) B) (Ориг.).
казского заповедника (бассейн р. Лабы); в байрачных лесах района Кав-
Кавказских Минеральных вод (Пятигорск, Железноводск, Кисловодск); в Ка-
Кабардино-Балкарской АССР; в Северной Осетии (Главный хребет, на Тер-
Терском и Сунженском хребтах); в Чечено-Ингушской АССР; в Грузии —
Сурами, Бакуриани, у озера Табацкури, в верхней Раче, район Военно-
Грузинской дороги; на северо-востоке Азербайджана — у горы Шах-
Даг; на северных и северо-восточных отрогах Малого Кавказа. В равнин-
равнинной части Северного Кавказа он обитает в байрачных лесах окрестностей
Ставрополя и в пойменных лесах нижнего Терека у станции Шелковской.
Единственная достоверная находка данного вида вне СССР сделана
в верховьях реки Куры в Карском вилайете Турции (долина Гель).
По нашим данным (стр. 65 и 309), Ей. somchetica обитала на Кавказе
уже в начале позднего плиоцена (акчагылский ярус).
Экология. В лесах этот моллюск обитает как в подстилке, так и
на стволах деревьев; в безлесных биотопах живет в осыпях, на скалах и.
в гниющих растительных остатках, чаще всего близ воды.
186
IV. EUXININAE
11. Euxina (Caucasica) tschetschenica (Pfr.) (рис. 111).
P f e i f f e г L., 1866, Malak. Bl., 13 : 149 (Clausilia); M о uss on, 1863 : 399 (Cl.
¦somchetica var. ossetica, non A. Schmidt, 1868); Boettger 0., 1878a : 39, t. 2,
fig. 5 (Clausilia); К о b e 1 t, 1893, Iconogr., N. F., 6. : 19, fig. 985 [Clausilia (Euxina)};
Westerlund, 1897 : 124 (Cl. somchetica var. dissophya); Розен, 1914 : 201 [Clau-
[Clausilia (Euxina)]; P e т о-в с к и й, 1914 : 321 [Clausilia (Euxina)]; Лихарев и Р а м-
иел ьйер, 1952 : 240, рис. 161 [Laciniaria (Euxina)].
Раковина веретеновидная или башневидно-веретеновидная, с до-
довольно резкой и редкой исчерченностью на верхних оборотах (за исклю-
исключением эмбриональных), которая становится все более тонкой и густой
к нижним оборотам, в связи с чем поверхность рако-
раковины матовая. Окраска темно-коричневая или темно-
роговая, без белых штрихов. Верхушка очень
широкая (около 1.5 мм), цилиндрическая, благо-
благодаря чему ниже нее завиток имеет вогнутые кон-
контуры. Оборотов 10—11, почти плоских, плавно
нарастающих в высоту, разделенных мелким белым
швом; эмбриональные обороты B1/1) гладкие, бле-
блестящие; последний оборот книзу сужается, на
затылке слегка уплощен, с крупным гладким килем.
Устье ромбическо-грушевидное, выступающее, вну-
Рис. 111. Euxina (Caucasica) tschetschenica (Pfr.). (По Лихареву и
Раммельмейер).
А — общий вид раковины; В — клауаилий.
три коричневое, с низким неврезанным синулусом и довольно глубоким ба-
зальным желобком. Края устья расширены, утолщены, белые; нижний и ко-
лумеллярный края плавно изогнуты, палатальный край почти прямой. Вы-
Высокая, слегка крючковидно изогнутая верхняя пластинка, плавно снижаясь,
уходит внутрь раковины. Нижняя пластинка не видна при прямом положе-
положении устья; она лежит почти отвесно и лишь верхний конец ее спирально
изогнут; нижний конец сильно вздут. Палатальных складок 3. Верхние
концы верхней и нижней палатальных складок и рудимент полулунной
складки (если он сохранился) лежат на спинной стороне раковины; глав-
главная складка начинается несколько глубже, а средняя палатальная
складка — несколько ближе к устью. Из палатальных складок наиболее
длинная, тонкая и постоянная (в отношении размеров) верхняя складка.
Нижняя и средняя обычно более толстые, варьируют в отношении длины.
Нижняя палатальная складка, реже также и средняя, нередко связана
45 рудиментом полулунной складки; последняя чаще всего имеет вид про-
продолжения нижней складки лежащего поперек оборота. Нижние концы
4. EUXINA 187
«кладок располагаются на различном расстоянии от края устья: наиболее
близко подходит средняя палатальная складка, несколько глубже закан-
заканчивается главная складка (обе видны при прямой положении устья); еще
глубже лежит верхняя и дальше всех от края устья заканчивается нижняя
палатальная складка. Имеется тонкая палатальная мозоль, связывающая
нижние концы складок. Клаузилий состоит из короткой рукоятки (кре-
(крепится к столбику правее синулуса), которая плавно расширяется в языко-
зидную лопасть (нижняя половина более широкая, чем у Ей. somchetica),
кон?ц которой плавно закруглен и утолщен.
Размеры: высота раковины 14—18, ширина 4.3—5.0 мм; высота устья
3.5—4.0, ширина 2.8—3.2 мм.
Изменчивость. Размеры раковины сильно варьируют в преде-
пределах: высота 12.8—19.4, ширина 3.8—5.5 мм; высота устья 3.0—4.5, ширина
2.6—3.4 мм. Мелкие экземпляры чаще всего встречаются в субальпийской
зоне, крупные — в пойменных лесах нижнего Терека. Длина средней и
нижней палатальной складок сильно варьирует, причем у первой это
связано с различным положением верхнего конца, а у второй — нижнего.
Внутреннеестроение. Материал: Чечено-Ингушская АССР,
при впадении Нувахи и Мужху в Ассу (Галахай-Лам), сбор Ананова,
11 VI 1886, определение Вестерлунда — 1 экз.
Радула: [?. + (^ + J^L) Х 2] X 93.
Размеры срединного зуба 14 X 21 |я, ширина радулы 600 \i.
Половая система не обнаруживает существенных отличий от таковой
у предыдущего вида.
Распространение (рис. 110). В СССР ряд находок в Кабар-
Кабардино-Балкарской АССР (Нальчик, Голубые озера и др.), вдоль Военно-
Грузинской дороги, в Дагестане от Ведено до Буйнакса, в Грузии (Лиси,
Манглиси, Квишхети, Абастумани, Тбатани), много находок в северной
Армении. В равнинной части Северного Кавказа этот вид найден в пой-
пойменных лесах и кустарниках нижнего Терека. Единственная досто-
достоверная находка вне СССР сделана близ Казикопаран (Карский вилййет
Турции).
По нашим данным (стр. 65 и 309), Ей. tschetschenica обитала уже на
Кавказе в конце позднего плиоцена (апшеронский ярус).
Экология. На субальпийских лугах этот моллюск обитает под
камнями и в осыпях. В лесах он живет, кроме скал и осыпей, также в под-
подстилке. Очень часто встречается совместно с Ей. somchetica (Pfr.).
Размножение. Живородящий вид.
Систематическое положение. Наш моллюск имеет много
общего с Ей. somchetica, вследствие чего старые авторы нередко их путали
или принимали за особую разновидность последнего. Однако несмотря на
некоторое сходство оба представляют два хорошо различимых вида. Так,
Ей. tschetschenica отличается от Ей. somchetica в первую очередь следую-
следующими особенностями: 1) у первой раковина более стройная, чем .у второй,
и благодаря цилиндрической верхушке боковые контуры завитка имеют
некоторую вогнутость; 2) у Ей. somchetica нижние концы всех складок
располагаются приблизительно на одном уровне и видны при прямом
положении устья, в то время как у Ей. tschetschenica ближе "всего к краю
устья подходит средняя складка, несколько глубже кончается главная,
а верхняя и в особенности нижняя палатальные складки заканчиваются
глубже всех.
188
IV. EUXININAE
12. Euxina (Caucasica) novorossica (Ret.) (рис. 112).
Retowski, 1888:284 [Clausilia (Euxina)]; Лихарев иРаммель-
иейер, 1952:239, рис. 160 [Laciniaria (Euxina)].
Раковина веретеновидная, умеренно вздутая, сильно исчерчен-
исчерченная, слабо просвечивающая, блестящая, темно- или коричнево-роговая,
с пучками белых штрихов вдоль шва. Верхняя часть завитка с вогнутыми
очертаниями и с довольно узкой A.0—1.1 мм) округлой верхушкой.
Оборотов HVa—12V2 (изредка до 13V2), слабо выпук-
выпуклых, плавно нарастающих в высоту; эмбриональные обо-
обороты BV4) гладкие, блестящие, светлые; последний обо-
оборот книзу сужен, за устьем слегка уплощен и ребри-
ребристый, с крупным килем. Устье округло-грушевидное, вы-
выступающее, внутри коричневое,
с низким неврезанным синулу-
сом и с довольно глубоким ба-
зальным желобком. Края устья;
расширены, утолщены, белые;
нижний край плавно изогнут,
палатальный и колумеллярньш
края почти параллельны друг
•другу, верхний край слегка изо-
изогнут над верхней пластинкой.
Верхняя пластинка слегка крюч-
ковидно изогнута, плавно сни-
снижаясь, уходит на х/4 оборота
в глубь раковины. Нижняя пла-
пластинка не видна при прямом по-
положении устья; в общем она та-
такая же, как у Ей. somchetica и
Ей. tschetschenica. Имеется ко-
короткая вставочная пластинка.
Главная и все 3 палатальных
складки, а также рудимент
нижней части полулунной склад-
складки, начинаются на спинной сто-
стороне раковины с некоторым смещением вправо, причем главная и верх-
верхняя палатальная немного глубже всех остальных. Средняя палатальная
складка нередко прервана и обычно располагается к верхней несколько
под углом; последняя строго параллельна главной складке. Главная,
верхняя и средняя складки впереди сливаются с тонкой палатальной
мозолью и видны при прямом положении устья. Нижняя если видна,
то лишь при косом положении устья. Рукоятка клаузилия относительно
длиннее, чем у двух предыдущих видов; она плавно расширяется в язы-
ковидную лопасть, по форме схожую с таковой у Ей. tschetschenica.
Размеры: высота раковины 14—16.5 (реже до 18.5), ширина 3.8—4.4 мм
(реже до 5.0 мм); высота устья 3.3—3.7, ширина 2.6—3.0 мм (реже 3.3 мм).1
Внутреннее строение не изучено.
Распространение. Все известные находки сделаны между
Новороссийском и Анапой.
Рис. 112. Euxina (Caucasica) novorossica (Ret.).
A — общий вид раковины (ориг.); Б — последний
оборот раковины с частично удаленной верхней стен-
стенкой (видны верхние концы пластинок и палатальных
складок) (по Лихареву и Раммельмейер).
1 Лектотип хранится в Зоологическом институте Польской академии наук. Он
имеет следующие размеры: высота раковины 14.4, ширина 3.9 мм; высота устья 3.3,
ширина 2.6 мм; 12*/4 оборотов. Место сбора: Новороссийск. Здесь же хранится один
паралектотип.
4. EUXINA 189
Подрод EIJA H. et A. ADAMS
A d a m s H. a. A., 1855 A858) : 180 (Clausilia, подрод Elia); Boettger O.,
4877a : 85 (Clausilia, секция Euxina, группа Laciniaria, non Hartmann, 1842);
t i n d h о 1 h m, 1924 : 65 (Laciniaria, подрод Laciniaria s. str., секция Elia).
Тип подрода: Clausilia maesta Fer.
Полулунная складка хорошо развита. Палатальная мозоль распалась
ва 4—5 более или менее длинных палатальных складок. Края устья
•с многочисленными краевыми складками.
Как общий характер раковины, так и те немногие сведения, которыми
jvibt располагаем о внутреннем строении данной группы [Ей. {Elia) corpu-
lenta (Pfr.)], говорят за то, что она несомненно принадлежит к роду
Euxina Bttg. С подродом Euxina s. str. ее сближает присутствие полулун-
гной складки. Однако в то время как у первой группы верхняя палаталь-
палатальная складка (если она имеется) очень коротка, а устье, как правило,
лишено краевых складок, оба вида данного подрода имеют длинную верх-
верхнюю палатальную складку и многочисленные складочки по краям устья.
О видами из подрода Caucasica Bttg. данный подрод сближает распад
палатальной мозоли на серию длинных складок, которые простираются
в глубь раковины на V5—*/\ часть оборота. Однако в то время как у Cauca-
Caucasica эти складки сливаются с соответствующими складками, образовав-
образовавшимися от распада полулунной складки, у Elia этого не происходит, так
jtaK полулунная складка остается цельной.- Следовательно, данный под-
подрод занимает промежуточное положение между двумя другими подродами
?uxina.
Распространение. Малая Азия, Сирия, Палестина. Наиболее
южное местонахождение — Порт-Саид (Euxina maesta Fer.).
*13. Euxina (Elia) corpulenta (Pfr.) (рис. 113).
P f e i f f e г L., 1848 : 7 (Clausilia); Kuster, 1847—1862 : 164, t. 18, fig. 10—
12 (Clausilia); Rossmassler, 1856, Iconogr., 3, 3—4:62, fig. 878 (Clausilia);
Retowski, 1889 : 259 ([Clausilia (Euxina)].
Раковина веретеновидная, очень вздутая, толстостенная, слабо
просвечивающая, с шелковистым блеском, красновато-коричневая. Верх-
Верхние обороты редко ребристые или ребристо исчерченные, нижние густо
ребристо исчерченные или исчерченные. Верхняя часть завитка с вогну-
вогнутыми контурами и с тонкой цилиндрической верхушкой. Оборотов 10—11,
почти плоских; эмбриональные обороты BV4—2V2) гладкие, блестящие,
•светлые; последний оборот книзу слегка сужен, на затылке вздут, с корот-
коротким морщинистым килем. Устье округло-грушевидное, с глубоко врезан-
врезанным синулусом и с довольно мелким базальным желобком. Края устья
утолщены, белые. Толстая, высокая и крючковидно изогнутая верхняя
пластинка, плавно снижаясь, уходит внутрь раковины. Крупная и тол-
толстая ланцетовидная нижняя пластинка лежит отвесно и при прямом
положении раковины не видна. Имеется небольшая вставочная пластинка.
Над полулунной складкой находится относительно длинная верхняя
палатальная складка, а еще выше — длинная главная складка. Последняя
начинается на правой стороне, а заканчивается недалеко от края устья.
Палатальная мозоль распалась на 4—5 довольно длинных складок, из
которых верхние нередко вступают в связь с передними концами главной
ж верхней палатальной складок. Короткая рукоятка клаузилия крепится
над синулусом. Языковидная лопасть имеет вырезку по колумеллярному
краю у основания рукоятки; нижний конец лопасти заострен и утолщен.
190\
IV. EUXIN1NAE
Размеры: высота раковины 11.5—13.5, ширина 3.8—4.3 мм; высота
устья 2.7—3.0, ширина 2.4—2.6 мм.
Изменчивость. Форма и размеры раковины, а также поверх-
поверхностная скульптура сильно варьируют. Наряду с более стройными фор-
формами (var. gracilior Retowski, var liostoma Retowski, 1889) обычно с глад-
гладким краем устья встречаются сильно вздутые и ребристые экземпляры:
(var. proteus Retowski, var. samsunensis Retowski, 1889, var. expansa
A. Schmidt, 1868), у которых край устья чаще всего покрыт многочислен-
многочисленными складками. Степень развития и длина палатальных складок также-
сильно варьирует.
Внутреннее строение. По
сообщению Форкарта (Базель), отросток
семяприемника очень тонкий, а ретрак-
тор пениса крепится к диафрагме.
Распространение. Черно-
Черноморское побережье Турции (Самсун, Си-
нон, Бафра), вилайет Бурса и, согласно
Бетгеру (Boettger, 1878b),Сирия, (Бейрут).
*14. Euxina (Elia) maesta (Fer.).
Ferussac, 1821, Tabl. Syst.: № 539 (Cla-
usilia); Rossmassler, 1839, Iconogr., 2,
9 : 23, fig. 634 (Cl. moesta Fer.); К ii s t e r, 1847—
1862 : 230, t.-25, fig. 31—33 (Clausilia); В о u г-
g u i g n a t, 1853, Oatal. rais. moll. Saulcy : 50v
pi. 2, fig. 7—9 (Cl. saulcyi); H a a s G., 1951,
Fieldiana, Zoology, 31, 45 : 482, fig. 90, 96A [La~
ciniaria (Elia) moesta Fer.]. •
Рис. 113. Euxina (Elia) corpulenta (Pfr.). (Ориг.).
A — общий вид раковины; Б — клаузилий.
Раковина веретеновидно-цилиндрическая, толстостенная, с шел-
шелковистым блеском, красновато-коричневая. Верхние 3—4 дефинитивных
оборота редко ребристые, последние 3 густо ребристо исчерченные. Вер-
Верхушка узкая, но тупая (ширина-1.1—1.3 мм), светлая. Оборотов 9—10
(реже 10V2), почти плоских, разделенных мелким белым швом; эмбрио-
эмбриональные обороты BV4—21/2) гладкие, светлые, блестящие; последние
2—3 оборота почти одной ширины; последний оборот на затылке ребри-
ребристый, с морщинистым килем. Устье округло-грушевидное, почти отвесное,
выступающее. Синулус широкий, слабо врезан. Края устья слабо отвер-
отвернуты, утолщены, белые. Верхняя пластинка впереди высокая, довольно
толстая и крючковидно изогнута, немного выступает за пределы устья;
в глубь раковины она уходит на 1/2 оборота, заканчиваясь на право-спин-
право-спинной стороне, на одном уровне с началом нижней пластинки. Последняя
лежит глубоко и отвесно (при прямом положении устья почти не видна),
имеет вид крупной ланцетовидной складки, которая с небольшим изгибом
4. EUXINA 19f
тянется кверху, а внизу у конца обрублена. Имеется промежуточная
складка. Спиральная пластинка начинается на право-спинной стороне и
заканчивается на левой стороне; наибольшей высоты она достигает на
спинной стороне. Субколумеллярная пластинка начинается несколько
ниже нижней и, подобно последней, круто тянется вниз, где переходит
на внешнюю стенку раковины и тянется почти до края устья, по правой
стороне базального желобка; нижний конец ее нередко виден даже при
прямом положении раковины. Крупная и прямая полулунная складка
лежит на спинной стороне и просвечивает сквозь стенку раковины. Над
ней находятся длинная главная складка (начинается на право-спинной
стороне) и несколько более короткая верхняя палатальная складка. Ниже
этих складок, располагаясь впереди полулунной, лежат 2 средних пала-
палатальных складки. Наконец, от нижнего конца полулунной складки тя-
тянется к устью нижняя палатальная складка. Обе самые нижние палаталь-
палатальные складки нередко прерваны. В некоторых случаях одна из средних
(чаще всего верхняя) палатальных складок вступает в связь с полулунной
складкой. Нижние концы главной и всех палатальных складок заканчи-
заканчиваются всегда недалеко и на одном расстоянии от края устья. На крак>
устья, между верхней и нижней пластинками имеются 3—5 (реже до 7)
мелких складочек или узелков, из которых одна нередко связывает ниж-
нижнюю пластинку с краем устья. Такие же складочки, но слабее выражен-
выраженные, могут быть и на палатальном крае устья. Лопасть клаузилия слабо
изогнута в поперечном направлении, утолщена по колумеллярному краю
и на конце с острым кяювовидным выступом; у рукоятки она немного
вырезана.
Размеры: высота раковины 11—15, ширина 3.2—4.0 мм; высота устья
2.5—3.5, ширина 2.0—3.0 мм.
Распространение. Сирия, Ливан, Палестина. По непроверен-
непроверенным данным — Малая Азия 1 и Порт-Саид.
Экология. Обитает в дубовых лесах, под камнями и на скалах,,
на высотах до 1000 м над ур. м.
Подрод AGROEUXINA BTTG.
Boettger О., 1877а : 85 (Clausilia, секция Euxina, группа Acroeuxina);.
Lindholm, 1924 : 66 (Laciniaria, подрод Euxina, секция Acroeuxina).
Монотипический подрод.
Верхушка раковины очень острая. Нижний конец нижней пластинки
резко обрублен. Полулунная складка крупная. Кроме главной складки,
имеется параллельная ей длинная верхняя палатальная складка, более
короткая нижняя и бугорковидная средняя палатальные складки.
Внутреннее строение не изучено.
Систематическое положение единственного вида, входящего в этот
подрод, нам представляется неясным. Возможно, что в дальнейшем,
с накоплением новых сведений как о раковине, так и о внутреннем строе-
строении этого моллюска, появятся основания для иной оценки его отношения,
к другим видам того же рода.
1 Указание на присутствие этого моллюска в Малой Азии, возможно, связано с тем,
что за него приняли .Ев. corpulenta (Pfr.), так как оба вида очень схожи и несомненно-
являются близкими родственниками.
192 IV. EUXININAE
*15. Euxina (Acroeuxina) huebneri (Pfr.).
Pfeiffer L., 1848, Monogr. Helic. viv., 2 : 473 (Clausilia); Rossmassler,
1856, Iconogr., 3, 3—4 : 73, fig. 890 (Clausilia); L о о s j e s, 1953b : 31 [Laciniaria
{Euxina)].
Раковина веретеновидная, внизу немного вздутая, тонкостенная,
•с крупными и редкими пластинчатыми ребрами (на предпоследнем обо-
обороте 30—40 ребрышек), просвечивающая, с шелковистым блеском, светло-
роговая. Благодаря тому, что начальная часть первого эмбрионального
¦оборота сильно приподнята, верхушка очень острая. Оборотов 93/4—101/2,
верхние выпуклые, нижние слабо выпуклые; эмбриональные обороты C)
гладкие; последний оборот значительно выше и шире предпоследнего,
внизу с коротким складчатым килем. Устье округло-грушевидное, ши-
ширина его почти равна высоте. Края его широко отвернуты, тонкие, хруп-
хрупкие. Верхняя пластинка впереди высокая и тонкая, быстро снижаясь,
-слегка заходит за спиральную пластинку. Ланцетовидная нижняя пла-
пластинка лежит очень глубоко и при прямом положении раковины чуть
заметна; нижний конец ее утолщен. Имеется короткая вставочная пла-
ютинка. Крупная полулунная складка лежит отвесно на спинной стороне;
верхний конец ее.отогнут назад. Главная складка начинается на право-
•спинной стороне. Под ней, на уровне верхнего конца полулунной складки,
.лежит верхняя палатальная складка. Обе складки простираются вперед
до левой стороны раковины. Мозолевидная нижняя палатальная складка
либо отходит от нижнего конца полулунной складки, либо только близко
подходит к последней. Средняя палатальная складка имеет вид бугорка,
лежащего на левой стороне раковины. Клаузилий неизвестен.
В нашем распоряжении имелись 2 экземпляра этого вида из коллекции
Ретовского (Зоологический институт Польской академии наук), которые
были найдены в морских выносах на Южном берегу Крыма (Retowski,
1887). Они имеют следующие размеры (в мм):
Высота Ширина Число
раковины раковины оборотов
13.5 3.1 93/4
11.8 3.2 93/4
Так как устье обоих экземпляров сильно повреждено, то размеры его
мы не приводим.
Лосьес (Loosjes, 1953) указывает следующие размеры для единствен-
единственного экземпляра, которым он располагал: высота раковины 14, ширина
-3.0 мм, число оборотов 10х/2.
Распространение. Черноморское побережье Турции (ви-
(вилайеты Зонгулдак и Самсун).
5. Род MEGALEUXINA BTTG.
Boettger О., 1877а : 85 (Clausilia, секция Euxina, группа Мegaleuxina);
L i n d h о 1 m, 1924 : 66 (Laciniaria, подрод Euxina, секция Megaleuxina).
Тип рода: Clausilia sandbergeri Mousson, 1873 (=Cl. derasa Mousson,
1863).
Раковина веретеновидная, вздутая (максимальная ширина прихо-
приходится на последний оборот), внизу с килем. Устье большое, располагается
почти на оси раковины, с базальным желобком. Высокая, слегка крючко-
крючкообразно изогнутая верхняя пластинка впереди нередко немного выступает
за край устья, а сзади, плавно снижаясь, сходит на нет, но перед этим
более или менее глубоко заходит за передний конец спиральной пла-
5. MEGALEUXINA
193
стинки.1 Последняя начинается на правой стороне раковины. Очень круп-
крупная нижняя пластинка лежит более или менее глубоко, но всегда видна
при прямом положении устья; внизу она дает спиральный изгиб и перед
слиянием со столбиком имеет заметное седловидное понижение (результат
слияния с ней крупной краевой складки). Нижний конец субколумелляр-
ной пластинки виден при косом положении устья. Полулунная складка
лежит на спинной стороне и развита различно: от крупной дугообразно
46
44
42
40
38
-
-
-
YK
\
•;
55
38
ТП
i—Хг*
<
1
"^—
+г
0
—1
1
38
—-Л
т
\
"С
¦?—¦
55 НО
—1 1—
40 42 44
Ростов""-^
<^ч. Y\° Ставрополь^--
ч '\ '/Х / /к
X4 05
165 220км
1 1
I
ли-
40 42
46 48
i ОС
Астрахань^
jr
—1
—~-~/ТЗ
оОрджоникидзе
¦^бТ6илиси5 -~
^Ереван ^
\ А -Л.
"к Ч ,
44 46
50
?
.Махач-Кала
-Ж
7\
\
48
52
(
\
X
раку
1
Г
50
54 56
}
)
ч ^
v^3
}
к
7
[
V
—^ '
1 1
44
42
40
38
36
52 54
Рис. 114. Географическое распространение видов рода Megaleuxina Bttg. на Кавказе
(Ориг.).
i —¦ Jf. ierasa (Mouss.); 2 — M. lindholmi (Lindh.); 3 — M. reulauxi (Bttg.); 4 — M. comnena (Ret.);
5 — M. iberica (Roth).
изогнутой до слабой мозоли. В некоторых случаях она распалась на
•серию палатальных складок.
Радула и половая система сходна с таковой у Euxina Bttg.
Распространение (рис. .114). Кавказ и черноморское побе-
побережье Турции. Один вид (М. borisi Hesse) живет в Болгарии.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА MEGALEUXINA
1 A0). Спиральная пластинка и главная складка длинные и видны через
устье.
2 C). Кроме длинной главной складки, имеются еще 2—3 палатальных
складки, хорошо видимые при прямом положении устья. Нижняя
1 За исключением М. comnena (Ret.), у которой спиральная пластинка и глав-
главная складка сильно редуцированы .
194 IV. EUXINIHAE
складка отделена от остальных вышележащих складок широким
промежутком 1. М. derasa (Mouss.).
3 B). На палатальной стенке либо только одна главная складка, либо,
кроме того, 1—2 коротких палатальных складки, причем верхняя
не видна через устье.
4 (9). Нижняя палатальная складка отсутствует.
5 (8). Нижняя пластинка хорошо видна при прямом положении устья.
6 G). Раковина густо и равномерно исчерчена. Края устья гладкие,
без складок и узелков 2. М. reulauxi (Bttg.) .
7 F). Верхние дефинитивные обороты покрыты тонкими редкими реб-
ребрами, которые у нижних оборотов становятся слабее и лежат
' гуще, под конец переходя в частую исчерченность. Палатальный
и колумеллярный края устья несут различное число мелких
складок и узелков 4. М. iberica (Roth).
8 E). Нижняя пластинка плохо видна при прямом положении устья.
6. М. borisi (Hesse).
9 D). Имеется короткая, но толстая и хорошо видимая при прямом
положении устья нижняя палатальная складка
3. М. lindholmi (Lindh).
10 A). Спиральная пластинка и главная складка короткие и не видны
через устье . 5. М. comnena (Ret.).
1. Megaleuxina derasa (Mouss.) (рис. 115—117).
Mouss on, 1863:400 (Clausilia); Schmidt A., 1868:150 (Cl. ossetica);
Mousson, 1873 : 216, t. 8, fig. 2 [Cl. (Laciniaria) sandbergeri]; К о b e 1 t, 1893,
Iconogr., N. F., 6 : 21, fig. 988 [(Clausilia {Euxina) ossetica]; Розен, 1914:200 [Clausilia
(Euxina)]; Ретовский, 1914 : 319 [Clausilia (Euxina)]; Лихарев иРаммель-
м е й е р, 1952:241, рис. 163, 164 [Laciniaria (Euxina)].
Раковина очень прочная, тупо ребристая (на предпоследнем
обороте 50—55 ребрышек) или ребристо исчерченная G0—90 складок),
слабо просвечивающая, со слабым блеском, темно- или зеленовато-рого-
зеленовато-роговая, с пучками белых штрихов. Верхняя часть завитка с вогнутыми кон-
контурами и с довольно узкой A.0—1.2 мм) округлой верхушкой. Оборотов.
11—12, верхние сильно, нижние слабо выпуклые; эмбриональные обороты
A1/2—2) гладкие, блестящие, одноцветные; последний оборот книзу
плавно сужен, на затылке (в области палатальных складок) вдавлен,
с крупным килем. Устье очень большое, неправильно пятиугольное, вы-
выступающее, с крупным, довольно глубоко врезанным синулусом и глубо-
глубоким базальным желобком, внутри желтовато-серое. Края устья очень
широко отвернуты, утолщены, белые. Крупная верхняя пластинка до-
довольно глубоко заходит за нижний конец спиральной пластинки. Нижняя
пластинка коричневатая, слабо выдается в просвет раковины; нижний
конец ее лежит наклонно и слегка обрублен. Верхняя часть пластинки
слабо закручена и почти отвесно подымается в глубь раковины, заканчи-
заканчиваясь рядом со спиральной пластинкой. Верхние концы главной и верх-
верхней палатальных складок, а также рудимент полулунной складки распо-
расположены на спинной стороне раковины, причем первые 2 несколько глубже
последней. От обоих концов полулунной складки отходят вперед по
одной толстой складке — средняя и нижняя палатальные. Главная, верх-
верхняя и средняя палатальные складки простираются далеко вперед, закан-
заканчиваясь близко от края устья; нижняя палатальная складка отделена от
них широким промежутком и значительно короче, так что не всегда видна
при прямом положении устья. Наиболее толстая средняя палатальная
5. MEGALEUXINA
199
складка варьирует в длине; как правило, она заканчивается впереди мозо-
левидным бугром, но нередко распадается на две части: заднюю (складка)
и переднюю (бугор); первая иногда отсутствует, а последняя исчезает
лишь очень редко. Нередко
^ этот бугор с помощью не-
Oi i большой палатальной мо-
мозоли связан с передним
концом верхней палаталь-
палатальной складки. Клаузилий
состоит из короткой ру-
рукоятки (простирается при-
I
А Б В
Рис. 115. Megaleuxina derasa (Mouss.). (По Лихареву и Раммельмейер).
А — общий вид раковины; Б — устье со складками; В — клаузилий.
близительно на г/4 оборота) и ложковидной лопасти, плавно расширяю-
расширяющейся к переднему концу; последний тупо заострен и слегка клювовидно
отогнут.
Размеры: высота раковины 17—20 (реже до 24), ширина 4.5—5.0 мм
(реже до 5.5 мм); высота устья 4.2—4.6 (реже до 5.0), ширина 3.2—3.5 мм
(реже до 4.0 мм).
Изменчивость. Как размеры, так и поверхностная скульптура
раковины очень варьируют. В субальпийской и в верхней подзоне леса
Рис. 116. Megaleuxina derasa (Mouss.). Радула. (Ориг.).
обитают популяции, состоящие из экземпляров, раковины которых отли-
отличаются меньшим числом оборотов (9—101/2) и меньшей высотой A5.0—
16.5 мм) при приблизительно той же ширине D.5—5.0 мм), благодаря
чему они имеют приземистый облик (f. suanetica Boettger, 1883).
196 IV. EUXININAE
Внутреннее строение. Материал: 1) Батуми, сбор Сухо-
дольского, 22 VII 1914, определение Линдгольма — 2 экз.; 2) Кутаиси,
развалины храма Баграта, сбор и определение наше, 28 IV 1957 — 1 экз.
Радула (рис. 116): [| + g + J^)X2] Xc^lOO.
Размеры срединного зуба 23.8 X 9.8 \i, ширина радулы 645 \i.
Семяприемник очень длинный — начинается от верхнего конца бел-
белковой железы (рис. 117). Отросток его слабо извит и значительно короче
Рис. 117. Megaleuxina derasa (Mouss.). Половая система.
(Ориг.).
собственно семяприемника. Матка короткая; влагалище сравнительно
длинное. Пенис очень длинный, близ нижнего конца веретеновидно рас-
расширен. Семяпровод и тесно прилегающий к нему длинный бич мускуль-
мускульными волокнами подтянуты к нижней части пениса (на рисунке не пока-
показаны).
Распространение (рис. 114). Этот вид наиболее широко рас-
распространен в западной Грузии, в пределах которой он пока еще не найден
лишь в северных районах Абхазии (к северу от линии Сухуми—Це-
бельда—Чхалта), не обнаружен в Аджарии и Гурии, за исключением
Батуми; в то же время широко распространен в Сванетии. К востоку от
Сурамского хребта найден в Юго-Осетинской автономной области, в районе
Сурами, Боржоми и Марткоби (к востоку от Тбилиси) и на Джевахетском
хребте (Малый Кавказ). На северных склонах Большого Кавказа
найден в районе Военно-Грузинской дороги и в Чечено-Ингушской
АССР.
Экология. Обитает на влажных, заросших мхом и травой извест-
известняковых скалах, руинах и оградах. ,
5. MEGALEUXINA
197
Размножение. Живородящий вид. У одного экземпляра в семя-
яйцеводе обнаружено 20 эмбрионов.
2. Megaleuxina reulauxi (Bttg.) (рис. 118, 119).
BoettgerO., 1887:55 [Clausilia (Euxina)]; Boettger 0., 1889:28,
t. 1, fig. 7 [(Clausilia (Euxina)]; Robelt, 1893, Iconogr., N. F., 6 : 4, fig. 965 [Clau-
[Clausilia (Euxina)]; Лихарев и Раммельмейер, 1952:237, рис. 156, 157 [Laci-
niaria (Euxina)].
Раковина довольно вздутая, крупная, густо и сильно исчерчен-
исчерченная (на последнем обороте волнистая исчерченность), слабо просвечиваю-
просвечивающая, с шелковистым блеском, коричнево- или темно-роговая, с большим
числом белых или желтых пятен и штри-
штрихов. Верхушка раковины довольно уз-
узкая (не шире 1.1 мм), округлая. Оборо-
Оборотов 11—12, выпуклых, довольно быстро
нарастающих в высоту; эмбриональные
обороты B1/2—23/4) гладкие, блестящие;
последний оборот книзу сильно вытя-
вытянут, за устьем сужен и уплощен, внизу
с крупным килем. Устье неправильно
пятиугольное, с крупным, глубоко вре-
врезанным, отогнутым вправо синулусом и
глубоким базальным желобком, внутри
коричневатое. Края устья очень ши-
широко отвернуты, слегка утолщены, бе-
белые; палатальный и колумеллярный
края почти параллельны друг другу;
нижний край угловатый; париетальный
край над верхней пластинкой про-
прогнут. Крупная верхняя пластинка впе-
впереди нередко выступает за край устья;
сзади, плавно снижаясь, она уходит
довольно глубоко внутрь раковины.
Белая нижняя пластинка хорошо видна
при прямом положении устья; лежит
косо в просвете устья, близко подхо-
подходит к краю и в середине имеет седловидное сужение; далее
круто подымается внутрь раковины, верхний конец ее лежит на
Рис. 118. Megaleuxina reulauxi
(Bttg.). (По Лихареву и Раммель-
Раммельмейер).
А — общий вид раковины; Б — устье.
она
пРа"
'вой стороне рядом со спиральной пластинкой. На спинной стороне рако-
раковины (с некоторым смещением вправо) лежит крупная, дугообразно изо-
изогнутая полулунная складка. Над нею начинается крупная главная
складка, которая заканчивается на левой стороне раковины. В верхней
части палатальной стенки устья, на значительном удалении от края,
лежит небольшая палатальная мозоль. Рукоятка клаузилия, плавно
расширяясь, переходит в довольно широкую лопасть, нижний конец
которой закруглен и утолщен.
Размеры: высота раковины 18—25, ширина 4.0—5.5 мм; высота устья
4.0—5.7, ширина 3.0—4.0 мм.
Изменчивость. В редких случаях наблюдается частичная редук-
редукция полулунной складки, от которой сохраняется лишь ее верхний конец,
образующий короткую верхнюю палатальную складку.
Внутреннее строение. Материал: Батумский ботанический,
сад, сбор и определение наше, 20 IV 1957 — 2 экз.
198
IV. EUXININAE
Радула (рис. 119): [у + (f + 3^) X 2] X слЮО.
Размеры срединного зуба 16.7 X 11.2 ц., ширина радулы 570 (х.
Семяприемник длинный, его резервуар лежит непосредственно ниже
белковой железы. Отросток на один оборот короче. Шейка семяприемника
толстая и мускулистая. Семяпровод перед впадением в пенис несет бич.
Пенис длинный, веретеновидно вздут у нижнего конца. У одного экзем-
экземпляра, семяяйцевод которого был забит яйцами, пенис и восходящее
19
Рис. 119. Megaleuxina reulauxi (Bttg.). Радула. (Ориг.).
колено семяпровода были редуцированы, а нижний конец ретрактора
пениса свободно лежал среди прочих органов.
Распространение (рис. 114). .В СССР: Аджарская АССР.
Вне СССР: вилайет Чорох (Турция).
Экология. Обитает в субтропических лесах приморской полосы
на влажных мшистых стволах деревьев и в дуплах.
Систематическое положение. По размерам раковины,
форме и положению устья и всех пластинок (особенно нижней) данный
вид, как и М. lindholmi (Lindh.), наиболее сходен с
М. derasa (Mouss.), от которой оба отличаются в пер-
первую очередь отсутствием длинной верхней и средней
палатальных складок, а данный вид еще и отсутствием
нижней палатальной складки.
3. Megaleuxina lindholmi (Lindh.) (рис. 120).
Lindholm, 1912 : 202
(Clausilia); * Лихарев и
Раммельмейер, 1952:: 238
[Laciniaria (Euxina)].
Раковина очень сход-
сходна с таковой М. reulauxi
(Bttg.), но отличается сле-
следующими признаками: 1)
поверхность исчерчена более
редко и скульптура посте-
постепенно сглаживается от верх-
верхних оборотов к нижнему;
2) блестящая; 3) темно-роговая, почти черная; 4) имеется короткая ниж-
нижняя палатальная складка, которая видна при прямом положении устья
и задним концом сливается с дугообразной полулунной складкой; 5) ло-
лопасть клаузилия более узкая; конец ее заострен и клювовидно изогнут.
4
Рис. 120. Megaleuxina lindholmi (Lmdh.). (Ориг.).
А — последний оборот раковины и устье; Б — клауви-
лий.
1 При описании рядом с названием вида стоит «Kobelt mss», но подпись под статьей
«Lindholm». Следовательно, по правилам номенклатуры автором вида является Линд-
гольм.
5. MEGALEUXINA
199
Размеры: высота раковины 19.0—20.5, ширина 4.7—5.4 мм; высота
устья 4.6—5.0, ширина 3.4—3.9 мм.1
Изменчивость. В отдельных случаях за счет палатальной мо-
мозоли возникает короткая верхняя или средняя складка.
Распространение (рис. 114). Вид редкий. В СССР единствен-
единственная находка сделана в окрестностях Лагодехи (восточная Грузия). Вне
СССР несколько находок в районе города Артвин
(вилайет Чорох, Турция).
Экология. Одна из находок сделана в буковых
лесах на высоте до 1600 м над ур. м., под валежи-
валежинами.
*4. Megaleuxina iberica (Roth) (рис. 121).
Rotb, 1850 in M. Wagner: 341 (Сlausilia); С h a r-
p e n t i e г, 1852 : 398, t. 11, fig. 8 (Cl. biformis); К ii s t e r,
1847—1862 : 227, t. 25, fig. 17—20 (Clausilia); К о b e 11, 1893,
Iconogr., N. F., 6 : 15, fig. 979, 980 [Clausilia (Euxina)].
Раковина просвечивающая, с шелковистым
блеском, красновато- или темно-роговая, с редкими пуч-
пучками белых штрихов. Завиток кверху быстро сужается
и ниже узкой цилиндрической верхушки имеет вогну-
вогнутые контуры. Оборотов 101/2—11, выпуклых-, разделен-
разделенных глубоким швом; эмбриональные обороты (около 31/2)
гладкие; последующие 5—6 оборотов покрыты тонкими
редкими ребрами, которые у нижних оборотов стано-
становятся слабее и лежат чаще, переходя в густую исчерчен-
ность; последний оборот за устьем несколько уплощен
и ребристый, с коротким морщинистым килем. Устье
овально-грушевидное, с прямым врезанным синулусом
и с мелким базальным желобком. Края устья широко
отвернуты, тонкие, с различным числом мел-
мелких складок и узелков вдоль палатального
и колумеллярного краев. Нижняя пластинка
довольно круто уходит наверх и заканчива-
заканчивается на спинной стороне раковины. Верхний
конец спиральной пластинки и рудиментар-
рудиментарная полулунная складка (иногда отсутствует)
лежат тоже на спинной стороне. Над полу-
полулунной складкой располагается короткая
верхняя палатальная складка, а еще выше —
довольно длинная главная складка; послед-
последняя берет начало на право-спинной сто-
стороне и заканчивается на левой стороне рако-
раковины. Рукоятка клаузилия переходит в ши-
широкую ложковидную лопасть, нижний конец
которой имеет небольшую вырезку.
Размеры (по 6 экземплярам): высота раковины 17.7—19.4, ширина
4.5—5.4 мм; высота устья 3.9—4.3, ширина 3.2—3.5 мм.
Рис. 121. Megaleuxina iberica
(Rotb). (Ориг.).
А — общий вид раковины; Б —
клаузилий.
1 Лектотип хранится в Зоологическом институте АН СССР (каталог систематиче-
систематической коллекции моллюсков, № 1а). Он имеет следующие размеры: высота раковины
19.3, ширина 4.8 мм; высота устья 4.6, ширина 3.4 мм; 11 оборотов. Место сбора: гора
Салолет близ Артвина, высота около 5300 футов, 1 VI 1912. Здесь нее находится один
паралектотип.
200
IV. EUX1NINAE
Внутреннее строение. Как нам любезно сообщил Фор-
карт (Базель), у данного вида отросток семяприемника очень тонкий,
а ретрактор лениса крепится к диафрагме.
Распространение (рис. 114). Все достоверные находки этого
вида сделаны в окрестностях города Самсун (черноморское побережье-
Турции). Указание М. Вагнера (Wagner, 1859) на то, что этот моллюск
обитает в Грузии, является ошибкой (Boettger, 1878c).
Систематическое положение. Данный вид отличается
от трех предыдущих скульптурой поверхности раковины, более груше-
грушевидной формой устья и присутствием многочисленных краевых складок
и узелков.
*5. Megaleuxina comnena (Ret.) (рис. 122).
Retowski, 1889 : 262 [Clausilia (Oligoptychia)]; К о b e 1 t, 1893, Iconogr.,
N. Г., 6 : 37, fig. 1014 [Clausilia (Oligoptychia].
Раковина сильно и густо исчерченная, просвечивающая, довольно-
блестящая, темно-роговая, с многочисленными белыми штрихами. Зави-
Завиток конический,с узкой, округлой верхушкой (ширина 0.8—0.9 мм). Обо-
Оборотов 10—12, слабо выпуклых; верхние медленно, нижние
быстро нарастают в высоту; эмбриональные обороты BV2)
гладкие, блестящие, светлые; последний оборот книзу
немного сужен, с коротким слабым килем. Шов сдавленный,
белый. Устье овально-грушевидное, внизу угловатое, ле-
лежит почти по оси раковины, с неврезанным широким,
слегка отогнутым вправо
синулусом и мелким базаль-
ным желобком. Края его
широко отвернуты, тонкие,
белые. Тонкая и низкая,
крючковидно изогнутая верх-
верхняя пластинка, плавно сни-
снижаясь, уходит в гдубь рако-
раковины, где сходит на нет.
Короткая рудиментарная
. спиральная пластинка ле-
лежит на спинной стороне
Рис. 122. Megaleuxina comnena (Ret.). Лектотип. (через устье не видна) вместе-
(ириг.). „
А - общий вид раковины; В - клаузилий. С ТаКИМИ Же ВСТаВОЧНОИ И Па-
раллельнои пластинками.
Крупная нижняя пластинка лежит глубоко, почти отвесно уходит
кверху; при прямом положении устья обычно хорошо видна только
крупная складка, соединяющая названную пластинку с краем устья.
Тонкая прямая мозолевидная полулунная складка лежит отвесно
на спинной стороне и просвечивает сквозь стенку раковины. Над
нею располагаются 3 коротких складки — главная и 2 сутуральных.
Имеется палатальная мозоль, состоящая из двух слабо связанных частей:
верхней и нижней. Клаузилий с широкой, слабо изогнутой лопастью,
нижний конец которой слегка утолщен и изогнут.
Размеры: высота раковины 12.2—16.3, ширина 3.5—3.8 мм; вьгсота
устья 3.0—4.0, ширина 2.2—3.0 мм.1
1 Размеры даны на основании промеров лектотипа и 10 паралектотипов из коллек-
коллекции Ретовского. Лектотип хранится в Зоологическом институте АН СССР (каталог
систематической коллекции моллюсков, № 1а). Он имеет следующие размеры: высота
раковины 13.5, ширина 3.8 мм; высота устья 3.2, ширина 2.6 мм; 10% оборотов. Место
сбора: Сефанос, Малая Азия.
5. MEGALEUXINA
201
Внутреннее строение не изучено.
Распространение (рис. 114). Вид был найден лишь один раз
на черноморском побережье Турции близ города Трабзона.
Экология. Обитает на стволах деревьев.
Систематическое положение. До настоящего времени
данный вид рассматривали как член рода Armenica Bttg. (=Oligoptychia
Bttg.). Главным доводом в пользу этого служило то, что у нашего мол-
моллюска спиральная пластинка и главная складка подверглись некоторой
редукции; кроме того, имеется параллельная пластин-
пластинка. Однако если внимательно сравнить данный вид
с настоящими Armenica, то легко обнаружить, что он
от них отличается рядом существенных особенностей.
К ним относятся веретеновидная форма раковины и
сильно вздутый последний оборот, окраска (белые штри-
штрихи), положение устья (по оси раковины) и присутствие
крупной краевой складки, сливающейся с нижней пла-
пластинкой и образующей в этом месте седловидное пониже-
понижение. Все эти признаки характерны для рода Megaleuxina
Bttg., куда мы относим и данный вид. Однако полная
ясность, к какому роду его надо отнести, появится толь-
только тогда, когда будет известно его внутреннее строение.
*6. Megaleuxina borisi (Hesse) (рис. 123).
Hesse, 1912, Nachrbl. deutsch. Malak. Ges., 44 : 59 [Clau-
silia (Wagneria)].
Раковина ребристо исчерченная, довольно бле-
блестящая, слабо просвечивающая, красновато-коричневая,
с многочисленными пучками белых штрихов. Оборотов
10—И, довольно выпуклых: эмбриональные обороты
B1/2) гладкие, блестящие; последний оборот книзу слегка
сужен, с крупным килем, за устьем ребристый и упло-
уплощен. Устье большое, широко-овальное, слегка угловатое,
с крупным, глубоко врезанным и отогнутым вправо сину-
лусом. Края его широко отвернуты, слегка утолщены,
белые. Внутренность устья коричневая. Верхняя пластинка крючковидно
изогнутая, впереди высокая, назад плавно снижается, слегка заходит за
спиральную пластинку и сходит на нет. Спиральная пластинка начинается
на брюшной стороне. Нижняя пластинка берет начало несколько правее;
в просвете устья лежит глубоко и при прямом его положении чуть высту-
выступает справа от верхней пластинки; седловидное сужение на ней слабо вы-
выражено. Имеется небольшая вставочная пластинка. На спинной сторона
лежит тонкая мозолевидная полулунная складка, верхний конец которой
загнут назад под прямым углом и имеет вид короткой складочки. Главная
складка начинается на правой и заканчивается на левой стороне рако-
раковины. Других образований на палатальной стенке нет. Клаузилий с длин-
длинной рукояткой, прикрепленной к столбику в пределах предпоследнее
оборота; лопасть широкая, с почти параллельными краями, изогнутая как
в поперечном, так и в продольном направлении, у рукоятки немного-
сжата; конец лопасти утолщен и несет 1—2 небольших вырезки.
Размеры: ,
~ ~ Число
оборотов
И1/*
Рис, 123. Me-
Megaleuxina borisi
(Hesse). (Ориг.)..
Высота
раковины
19.7
21.1
Ширина
раковины
4.9
5.2
Высота
устья
4.5
4.9
Ширина
устья
3.4
3.5
202 IV. EUXININAE
Гессе (Hesse, 1912) приводит следующие размеры: высота раковины
19—20, ширина 5 мм.
Распространение. Южная Болгария. Первоначально этот
моллюск был найден в окрестностях города Пловдива. Позднее он был
обнаружен в заповеднике Ропотамо (сбор Белявского, 8 VI 1959) и у Ра-
ковеца (сбор Явловского, IX 1937).
Экология. Обитает в сырых буковых лесах под кусками гнилой
древесины.
Систематическое положение. Мы имели возможность
сравнить наши экземпляры этого вида с одним типовым экземпляром,
хранящимся в Зоологическом институте Польской академии наук, и
убедились в их полной идентичности.
Данный вид отличается от всех прочих видов того же рода в первую
-очередь глубоким положением нижней пластинки. Внешне, т. е. формой
и размерами раковины, а также поверхностной скульптурой он напоми-
напоминает М. derasa (Mouss.), но отличается от последней отсутствием пала-
палатальных складок. Слабое развитие полулунной складки сближает наш
вид с М. iberica (Roth), однако форма устья и нижней пластинки, а также
¦отсутствие краевых складок, столь характерных для последнего вида,
позволяет их легко различать.
6. Род EUXINASTRA BTTG.
Boettger О., 1888 : 152 (Clausilia, секция Euxinastra); Boettger О., 1889 :
26 (Clausilia, секция Euxinastra); Lindholm, 1924 : 66 (Laciniaria, подрод Euxi-
Euxinastra).
Монотипический род.
Раковина веретеновидная, довольно вздутая. Последний оборот с ки-
килем. Устье большое, с базальным желобком. Длинная верхняя пластинка
заходит за спиральную пластинку. Крупная ланцетовидная нижняя пла-
пластинка лежит отвесно, глубоко и чуть заметна при прямом положении
устья; внизу она плавно сливается со столбиком, с краем устья не свя-
связана. Нижний конец субколумеллярной пластинки виден при косом поло-
положении устья. Имеется крупная главная складка. Полулунная складка
распалась, образовав короткую верхнюю, палатальную складку; нередко
появляются 1—2 еще более коротких мозолевидных складки или узелка.
Лопасть клаузилия близ нижнего конца, со стороны палатальной стенки,
имеет глубокую вырезку.
Половая система сходна с таковой у Euxina и Megaleuxina. Наиболее
характерным является очень длинный семяприемник и его положение по
отношению к желудку и к средней кишке (см. далее).
О. Бетгер справедливо считал, что главной отличительной особенностью
раковины этого рода является присутствие вырезки на лопасти клаузи-
клаузилия при отсутствии нижней палатальной складки. Де*ло в том, что во всех
других группах данного семейства подобная вырезка служит для про-
пропуска крупной нижней палатальной складки, которая в этом случае за-
заходит в глубь раковины выше клаузилия (многие Alopiinae, Mentissa,
Cochlodina и др.). Бетгер предполагал, что отсутствие указанной складки
связано с обитанием на субстрате, бедным кальцием в условиях влажного
субтропического климата. Дефицит кальция привел к утере нижней пала-
палатальной складки, имевшейся у предков Ей. hamata (Bttg.), а влажный
климат устранил угрозу гибели моллюска от чрезмерной потери влаги,
несмотря на то, что вырезка на клаузилии ослабляет герметичность замы-
лательного аппарата. Кобельт (Kobelt, 1893, Iconogr., N. Г., 6:4) спра-
6. EUXINASTRA
203
ведливо сомневался в том, что данный вид испытывает дефицит кальция,
так как общий облик раковины говорит о противном.
Что касается систематического положения Euxinastra, то, с нашей
точки зрения, данный род наиболее близок к роду Megaleuxina Bttg.
За это говорит величина и форма раковины и устья. В обоих родах нижний
-конец субколумеллярной пластинки виден при косом положении устья.
Но в отличие от Megaleuxina, у Euxinastra нижняя пластинка совсем
иной формы и не связана складкой с краем устья. Что касается вырезки
лопасти клаузилия, то у тех видов Mega-
Megaleuxina, у которых она имеется, она зна-
значительно меньших размеров и иной фор-
формы, чем у видов Euxinastra.
В связи со сказанным мы считаем, что
хотя оба рода имеют ряд общих черт, они
достаточно хорошо обособлены друг от
друга.
Euxinastra hamata (Bttg.) (рис. 124—
126).
BoettgerO., 1888 : 152 [Clausilia (Euxi-
(Euxinastra)]; Boettger O., 1889 : 27, t.. 1, fig. 6
{Clausilia (Euxinastra)]; К о Ь е 1 t, 1893, Ico-
nogr., N. F., 6 : 3, fig. 964 [Clausilia (Euxinast-
(Euxinastra)]; Лихарев и Раммельмейер, 1952 :
245, рис. 169, 170 [Laciniaria (Euxinastra)].
Раковина тупо ребристая, слабо
просвечивающая, матовая, зеленовато-
роговая, с отдельными белыми штриха-
штрихами. Завиток конический, наверху со слег-
слегка вогнутыми контурами и с довольно
широкой A.1—1.2 мм) округлой верхуш-
верхушкой. Оборотов 11—11х/а, слабо выпук-
выпуклых, плавно нарастающих, разделенных
врезанным, беловатым швом; эмбриональ-
эмбриональные обороты BV4) гладкие, блестящие; последний оборот книзу слабо
«ужен, на затылке уплощен, с морщинистым килем. Устье округло-ром-
округло-ромбическое, внутри белое, с широким, слегка отогнутым вправо, слабо вре-
врезанным синулусом и мелким базальным желобком. Края устья широко
отвернуты, утолщены, белые; палатальный край почти прямой, нижний
и колумеллярньш края широко закруглены, париетальный над верхней
пластинкой прогнут. Высокая, слегка крючковидно изогнутая верхняя
пластинка впереди немного выступает за край устья; в глубь раковины,
плавно снижаясь, простирается на х/4—V8 оборота и заходит за передний
конец спиральной пластинки.1 Последняя берет начало на правой сто-
стороне раковины. Нижняя пластинка наверху дает короткий спиральный
изгиб и начинается немного ниже спиральной пластинки. Имеется круп-
крупная промежуточная складка. Главная складка начинается на право-
спинной стороне и заканчивается на левой стороне. Ниже ее верхнего
конца лежит короткая верхняя палатальная складка, а под последней
нередко располагаются 1—2 мозолевидных складочки, или узелки. Ру-
I
Рис. 124. Euxinastra hamata
(Bttg.). (По Лихареву и Раммель-
Раммельмейер).
А — общий вид раковины, В — клау-
зилий.
1 О. Бетгер и последующие авторы считали, что она сливается со спиральной пла-
пластинкой.
204
IV. EUXININAE
коятка клаузилия плавно переходит в длинную языковидную лопасть,
нижний конец которой со стороны палатальной стенки несет глубокую
вырезку (видна при косом положении устья).
Размеры: высота раковины 18—21, ширина 4.8—5.2 мм; высота устья
4.2—4.7, ширина 3.4—3.8 мм.
Рис. 125. Euxinastra hamata (Bttg.). Радула. (Ориг.).
28
29
гп
Внутреннее строение. Материал: Батумский ботанический
сад, сбор и определение наше, 18—20 IV 1957 — 3 экз.
Радула (рис. 125): [| + (| + ^) Х 2] X сл105.
Размеры срединного зуба 10.5 X 17 ц, ширина радулы 490 \и.
Семяприемник очень длинный (рис. 126). Его резервуар лежит выше
белковой железы на внешней стенке желудка и хорошо виден снаружи.
При этом проток его прохо-
проходит через петлю средней киш-
кишки (при препаровке его
приходится выдергивать из
~сп этой петли). Отросток семя-
семяприемника очень короткий.
Влагалище длинное, матка
в несколько раз короче
его. Пенис цилиндрический,
лишь немного расширен впе-
впереди. Семяпровод без бича.
Ретрактор пениса крепится
к диафрагме.
Колумеллярная мускула-
мускулатура дает ветвь к шейке
семяприемника и 1—2 ветви
к половой клоаке. Влагали-
Влагалище на значительном протя-
протяжении подтянуто к стенке
тела многочисленными тонки-
тонкими мускульными волокнами.
Р асп р о странение.
В СССР — окрестности Ба-
Батуми (Аджарская АССР).
Вне СССР — вилайет Чорох
(Турция).
Экология. Обитаетва
влажных субтропических
лесах под камнями, валежи-
валежинами и на стволах деревьев.
рил
кл-
Рис.
126. Euxinastra hamata (Bttg.). Половая
система. (Ориг.).
ho
7. Род GALEATA BTTG.
Boettger О., 1877а : 84 (Clausilia, секция Euxina, группа Galeata)-t L i n d-
1 m, 1924 : 65 (Laciniaria, подродLaciniaria s. str., секция Galeata).
7, GALE AT A 205
Тип рода: Clausilia schwerzenbachi Gharp.
Раковина веретеновидная или башневидная, с узкой цилиндрической
верхушкой. Последний оборот на затылке очень уплощен или даже не-
немного вогнут, с крупным и длинным килем, окружающим большое около-
околопупочное поле. Устье треугольно-грушевидное, выступающее. Узкий
неврезанный синулус и базальный желобок сильно сдвинуты влево.
Края устья широко отвернуты, тонкие; палатальный край почти прямой
и отвесный; остальные края образуют широкую дугу. Верхний конец
верхней пластинки заходит за нижний конец спиральной. Последняя
начинается на право-брюшной стороне, быстро нарастает в высоту, до-
достигая максимума на правой стороне, и далее плавно снижается. Нижняя
пластинка располагается очень глубоко и близко от верхней пластинки;
лри прямом положении устья виден лишь ее нижний конец и крупная
складка, соединяющая ее с краем устья; сама пластинка, спирально за-
закручиваясь, круто уходит кверху и ее верхний конец лежит на спинной
стороне раковины. Субколумеллярная пластинка не видна. Имеется глав-
главная и довольно длинная верхняя палатальная складка. Передние концы
их упираются в небольшую палатальную мозоль.
Длинный семяприемник имеет тонкий и короткий отросток. Шейка
семяприемника снабжена ретрактором. Влагалище длинное. Пенис ци-
цилиндрический, с диафрагмальным ретрактором, без придатка и бича.
Ретрактор глазного шупальца проходит между пенисом и влагалищем.
Распространение. Малая Азия.
В оценке систематического положения данного рода высказывалось
много противоречивых взглядов. Ветгер (Boettger О., 1877а), по-видимому,
исходя из соображений географического порядка, относил его к Euxina.
Линдгольм (Lindholm, 1924) поместил обоих моллюсков, которых мы
относим в данный род, в разные роды. Анатомически тогда еще неизучен-
неизученную Clausilia schwerzenbachi Gharp. (с нашей точки зрения, разновидность
G. galeata Rssm.) автор отнес в род Laciniaria. По-видимому, основанием
послужило то, что у названного моллюска главная и верхняя палаталь-
палатальные складки располагаются под некоторым углом друг к другу, т. е.
как у Laciniaria biplicata (Mont.) и у L. plicata (Drap.).
Другой вид — Clausilia cilicica Nagele, половая система которого
к тому времени была изучена Вагнером (Wagner A., 1914), Линдгольм,
вслед за ним, отнес в род Idyla, хотя конхиологически он сильно отли-
отличается от остальных видов того же рода.1 В то же время ни Вагнер, ни
Линдгольм почему-то не обратили внимание на большое сходство рако-
раковин Clausilia schwerzenbachi и Cl. cilicica. Теперь, когда нам удалось озна-
ознакомится с половой системой первого моллюска, совершенно очевидно,
что оба они и в этом отношении очень сходны и нет никаких оснований
помещать их в разные группы.
Петлеобразное строение мужских выводных путей обоих видов гово-
говорит против сближения их с Laciniaria. Меньше различий в строении поло-
половой системы наших моллюсков и настоящих Idyla. Однако оба сильно
отличаются от видов указанного рода по раковине. У Idyla спиральная
пластинка претерпевает редукцию до полного исчезновения, а нижняя
пластинка имеет ланцетовидную форму и лежит отвесно. Иное положение
у обоих указанных видов.
Учитывая сказанное, мы считаем, что имеются основания для их выде-
выделения в особый род — Galeata Bttg.
1 Заметив это, Линдгольм в той же работе вынес этот вид в отдельную секцию —
Plistoptychia Lindh.
206
IV. EUXIN1NAE
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА GALE AT A
1 B). Раковина башневидная, небольшая. Вставочная пластинка и полу-
полулунная складка отсутствуют. Верхняя палатальная складка лежит
по отношению к главной складке под некоторым углом. Высота
раковины 13.5—14.7, ширина 3.1—3.2 мм . . 1. G. galeata (Rssm.).
2 A). Раковина веретеновидная, крупная. Имеются крупные вставочная
пластинка и полулунная складка. Верхняя палатальная складка
параллельна главной. Высота раковины 18—20, ширина 4.4—
4.9 мм 2. G. cilicica (Nagele).
*1. Galeata galeata (Rssm.) (рис. 127—129).
Rossmassler, 1839, Iconogr., 2, 3—4 : 17, fig. 621 {Clausilia).
Раковина башневидная, густо покрыта тонкими белыми реб-
ребрами, слабо просвечивающая, матовая, коричнево-роговая. Стройный
узкоконический завиток заканчивается тонкой, заострен-
заостренной цилиндрической верхушкой. Оборотов 12—13, верх-
верхние довольно* сильно, нижние слабо выпуклые, медлен-
медленно нарастающие; эмбриональные обороты C) темные,,
нередко вишнево-красные, иногда располагаются под не-
некоторым углом к остальной части раковины. Шов бело-
беловатый, тонкий. Низкая и короткая верхняя пластинка,,
быстро снижаясь, сходит на нет, но перед этим слегка
заходит за нижний конец спиральной пластинки. Вставоч-
Вставочной пластинки нет, так же как и полулунной складки.
Главная складка начинается на право-спинной и закан-
заканчивается на левой сдороне раковины и может быть видна
в просвете синулуса. Несколько ближе к устью начи-
начинается верхняя складка, которая располагается к глав-
главной под углом и заканчивается на одном уровне с по-
последней. Короткая ложковидная лопасть клаузилия
сильно изогнута в обоих направлениях; близ рукоятки
она слабо вырезана по колумеллярному краю; нижний:
конец ее слегка заострен и утолщен.
Размеры (по 2 экземплярам): высота раковины 13.5—
14.7, ширина 3.1—3.2 мм; высота устья 2.7—2.9, шири-
ширина 2.3—2.5 мм.1
Внутреннее строение. Материал: Турция,,
между Самсуном и Чаршамбой, 19 IV 1936, определе-
определение Л. Форкарта (Euxina schwerzenbachi Charp.) — 1 экз^
Рис. 127. Ga-
Galeata galeata
(Rssm.). (Ориг.).
Радула (рис. 128): [| + g + ^) X 2] Х 100.
Размеры срединного зуба 11.3 X 16.7 \i, ширина ра-
радулы 450 ц.
Семяприемник достигает белковой железы (рис. 129): отросток его
короткий. Пенис длинный, на границе между средней и нижней третью-
кол енообразно согнут; оба колена стянуты перепонкой. Влагалище очень
длинное, матка значительно короче.
Изменчивость. Разновидностью данного вида является так
называемая Clausilia schwerzenbachi Charpentier, 1852 (syn. Cl. belone
1 До сих пор известна была только ширина раковины данного вида, так как опи-
описание его было составлено по дефектному экземпляру.
7. GALEATA
2OT
Boettger, 1879), которая отличается от типичной формы более мягкой
скульптурой — тонкая ребристая исчерченность, и более косым положе-
положением верхней палатальной складки по отношению к главной складке.
Высота раковины 13.6—15.0, ширина 3.1—3.3 мм; высота устья 2.7—3.0,
ширина 2.1—2.5 мм; оборотов 12.
'ЯГ
Рис. 128. Galeata galeata (Rssm.) var. schwerzenbachi
. (Charp.). Радула. (Ориг.).
Распространение. Турция (Бурса, Самсун, Гулек к северо-
западу от Адана) и Сирия.
Систематическое положение. Раковина данного вид»
во многом сходна с G. cilicica (Nagele) — по форме завитка, киля и устья,,
форме и расположению пластинок,
но отличается от нее меньшими раз-
размерами, башневидной формой, отсут-
отсутствием вставочной пластинки и полу-
полулунной складки и косым положе-
положением верхней палатальной складки.
*2. Galeata cilicica (Nagele)
(рис. 130).
Nagele, 1902, Nachrbl. deutsch.
Malak. Ges., 34 : 7 [Clausilia (Euxina)};
WagnerA., 1914, Iconogr., N. F., 21: 7,
t. 574, fig. 22 (Pleioptychia, анатомия).
Раковина веретеновидная,
стройная (максимальная ширина при-
приходится на предпоследний оборот),
сильно просвечивающая, с шелкови-
шелковистым блеском, светло-роговая, с от-
отдельными белыми штрихами. Верхняя
часть завитка благодаря наличию уз-
узкой цилиндрической верхушки имеет
вогнутые контуры. Первые 5 дефини-
дефинитивных оборотов редко и сильно исчер-
исчерчены. На последующих исчерчен-
исчерченность становится все гуще и слабее.
Кроме того, на последних оборотах появляются следы спиральной исчер-
ченности. Оборотов 12, верхние слабо выпуклые, нижние почти плоские;
эмбриональные обороты C) гладкие, блестящие, темные; последний обо-
оборот книзу сильно сужен, на затылке сильно уплощен и даже несколько
вдавлен, ребристо исчерчен. Шов беловатый. Устье окрашено внутри
в темные тона. Довольно крупная верхняя пластинка впереди несколько
выступает за край устья. Имеется сравнительно длинная вставочная
пластинка, от верхнего конца которой отходит к столбику короткий мозо-
левидный отросток. Крупная полулунная складка лежит на правой сто-
¦—рял
Рис. 129. Galeata galeata (Rssm.) var.
schwerzenbachi (Charp.). Половая систе-
система. (Ориг.).
208
IV. EUXININAE
роне; верхний конец ее отогнут внутрь раковины. Над нею начинается
длинная главная складка, нижний конец которой лежит на левой стороне
и может быть замечен в просвете синулуса. От верхнего конца полулунной
складки берет начало длинная верхняя палатальная складка, заканчиваю-
заканчивающаяся внизу на одном уровне с главной складкой. Широкая языковидная
лопасть клаузилия у основания рукоятки несет глубокую вырезку по
колумеллярному краю; нижний конец лопасти слегка заострен и утолщен.
Размеры: высота раковины 18—20, ширина 4.4—
4.9 мм; высота устья 3.8—4.4, ширина 3.4—3.7 мм.1
Внутреннее строение. Вагнер (Wagner
А., 1914) дает рисунок половой системы данного вида.
Из него явствует, что, подобно предыдущему виду,
этот моллюск обладает длинным семяприемником и
коротким отростком. В^1аг^л^ще^з^метао_длиннеел1ат-
ки. Пенис более или менее цилиндрический, но не-
несколько короче, чем у предыдущего вида.
Распространение. Единственная находка
этого моллюска была сделана на южных отрогах Цен-
Центрального (Киликийскбго) Тавра, близ Кюлека (вилайет
Игель, южная Турция).
8. Род MENTISSOIDEA BTTG.
Boettger О., 1877а : 77 (Clausilia, секций Mentissoi,
dea); Boettger О., 1878b : 61 (Clausilia, секция Euxina
группа Mentissoidea); Wagner
A., 1920 : 151 (Polinskia); Lind-
holm, 1924 : 67.
Монотипический род.
. Последний оборот рако-
раковины книзу сильно сужен,
с крупным килем, который
окаймляет широкое полукруг-
s Б лое околопупочное поле.
Рис. 130. Galeata cilicica (Nagele). Лектотип.(Ориг.). Устье грушевидное, с глубоко
А — общий вид раковины, Б - клаузилий. врезаННЫМ И ОТОГНУТЫМ ВПра-
во синулусом, внизу с же-
желобком. Ось устья почти совпадает с осью раковины. Верхняя пластин-
пластинка не сливается со спиральной (рис. 6, Б). Последняя начинается на
брюшной и заканчивается на спинной стороне и через устье не видна.
Очень высокая и массивная нижняя пластинка наверху лежит почти от-
отвесно, а внизу дает широкий горизонтально лежащий изгиб, который
.очень сильно выдается в просвет устья; с краем устья она ничем не свя-
связана. Нижний конец субколумеллярной пластинки хорошо виден при
косом положении устья. Имеется очень длинная главная складка. Полу-
Полулунной складки нет, но имеются 2 коротких палатальных складки —
верхняя и нижняя, которые через устье не видны. Пупок — широкая
щель.
Срединный и боковые зубы радулы с дополнительными зубцами. Внеш-
Внешние краевые зубы мнргозубцовые, гребенчатого типа.
1 Даются на основании промеров лектотипа и 5 паралектотипов. Л ектотип хранится
р Зоологическом институте АН СССР (каталог систематической коллекции моллюсков,
№ 1а). Он имеет следующие размеры: высота раковины 19.4, ширина 4.4 мм; высота
устья 4.2, ширина 3.4 мм; 121/3 оборотов. Место сбора: Киликия, Кюлек.
8. MENTISSOID'kA
209
Семяприемник очейь длинный, без отростка. Пенис длинный, почти
на всем протяжении цилиндрический и лишь у нижнего конца веретено-
видно расширен. Ретрактор пениса крепится к диафрагме. Левый ретрак-
тор дает одну ветвь к месту слияния семяприемника с маткой, другую
(часто раздваивающуюся) — к влагалищу и третью —
к нижнему концу пениса. Ретрактор левого глазного
щупальца проходит между пенисом и влагалищем.
Для определения систематического положения данного
рода, с нашей точки зрения, наиболее важным является
присутствие на раковине киля и базального желобка.
Как известно, эти признаки свойственны только подсе-
подсемействам Euxininae и Clausiliinae. У последней группы
характерно преобразование мужских половых путей, в ре-
результате которого они оказываются вытянутыми вдоль
женских. Подобного не наблюдается у Mentissoidea, где
мужские половые пути сохраняют петлеобразную фор-
форму. В связи с этим мы относим данный род в подсемейство
Euxininae.1 От других родов данного подсемейства он от-
отличается в первую очередь глубоким залеганием многих
частей замыкательного аппарата, в особенности спираль-
спиральной пластинки и палатальных складок. Кроме того, очень
характерна утеря отростка семяприемника и необычный
тип связей половых путей с левым ретрактором.
Mentissoidea litotes litotes (A. Schm.) (рис. 5, 6, Б,
131-133).
Schmidt A., 1868 : 162 (Clausilia); М о u s s о n, 1876a : ЩР™^3/ .
41, t. 2, fig. 8 (Cl. fusorium); Boettger O., 1881a : 231 (Clau- W^*</ A
silia); Boettger 0., 1883 : 186 [Clausilia (Euxina)]; Kobelt,
1893, Iconogr., N. F.,6 : 25, fig. 994—997 [Clausilia
(Euxina); Розен, .1914 : 197 [Clausilia (Euxina)];
Wagner A., 1922 : 109—111, t. 3, fig. 24; t. 4,
fig. 31b (Polinsteia, анатомия); Лихарев и Рам-
мельмейер, 1952 : 218, рис. 133.
Раковина веретеновидная, стройная
или вздутая, тонко и равномерно ребристо
исчерченная, иногда слабо ребристая, с шел-
шелковистым блеском, просвечивающая, светло-
или темно-роговая, реже красноватая, ме-
местами с пучками белых штрихов. Вершина
узкая @.7—0.9 мм), округлая. Оборотов
10—12, выпуклых, разделенных тонким сдав-
сдавленным швом; эмбриональных оборотов 21/2.
Устье неправильно или угловато-грушевид-
угловато-грушевидное, почти отвесное, слабо выступающее.
Края устья широко отвернуты, с коричне-
коричневато-белой губой. Короткая верхняя пластинка толстая, высокая и крюч-
ковидно изогнутая. Главная складка начинается на брюшной и за-
заканчивается на левой стороне раковину» Палатальные складки начи-
начинаются на брюшной или право-брюшной стороне и заканчиваются на
ппавой стороне раковины. Лопасть клаузилия языковидная, довольно
широкая, слабо изогнутая в поперечном направлении, с утолщенным
краем и с тупым выступом на конце.
1 Подробнее о систематическомположений рода Mentissoidea см. стр. 82.
Рис. 131. Mentissoidea litotes
litotes (A. Schm.). (Ориг.).
А — общий вид раковины; Б —
клаузилий.
210
IV. EUXININAE
Размеры: высота раковины 14.6—17.4, ширина 3.8—4.6 мм; высота
устья 3.8—4.2, ширина 2.8—3.0 мм.
Рис. 132. Menttssoidea litotes litotes (A. Schm.)-
Радула. (Ориг.).
Изменчивость. Условия обитания заметно отражаются на
форме и размерах раковины. В районах с влажным субтропическим клима-
климатом наряду с обычными встречают-
встречаются популяции, состоящие из экзем-
экземпляров с крупной и стройной ракови-
раковиной (f. ganeo Boettger, 1883), высота
которой колеблется в пределах 17.3— .
20.1, ширина 3.8—4.3 мм, при 12—
14 оборотах. В высокогорных обла-
областях нередко встречаются экземпля-
экземпляры с короткой приземистой ракови-
раковиной (f. suanetica Boettger, „1883),
высота которой колеблется в преде-
пределах 13—16, ширина 3.5—4.0 мм, при
10—12 оборотах.
Внутреннее строение..
Материал: 1) Закатальский заповед- •
ник, сбор и определение наше, V—
VI 1953 — 4 экз.; 2) Кавказский
заповедник, лагерь Холодный, сбор ,
и определение наше, 23 VIII 1956 —
2 экз.; 3) Новый Афон, Абхазия, сбор
и определение наше, 3 X 1956 —
1 экз.
Г-
+d
+
Радула (рис. 132):
+ Й)Х2]ХШ.
Размеры срединного зуба 11.5 X
Х17.9 [х, ширина радулы 480 ц.
Семяприемник (рис. 133) представ-
представляет собой очень длинную, довольно
широкую трубку, лежащую вдоль
женского канала семяяйцевода и сла-
слабо расширяющуюся у верхнего конца,
который лежит значительно выше
белковой железы на внешней сторо-
стороне желудка. Стенки семяприемника
образованы однослойным цилиндрическим эпителием^ снаружи окружен-
окруженным тонким слоем кольцевой мускулатуры. Пенис узкий, цилиндрический,
у дистального конца веретен обидно расширен. Внутри расширения нахо-
находятся многочисленные продольные складки.
Рис. 133. Mentissoidea litotes litotes
(A. Schm.). Половая система. (Ориг.).
9. FJLOSA
211
Распространение (рис. 134). На Большом Кавказе — леса
западной части северных склонов; по южным склонам проникает на вос-
восток до Нухи; отдельный изолированный ареал в лесах Северной Осетии
и Чечено-Ингушетии. На Малом Кавказе — леса северных отрогов, от
Боржоми и Абастумани на западе, до Шуши на востоке.
Рис. 134. Географическое распространение Mentissoidea litotes (A. Schm.). (Ориг.).
1 — основной ареал; 2 — местонахождение у. г. Шуши.
Экология. Обитает на стволах деревьев, в сухое время укрываясь
в дуплах и в щелях коры; нередко под корой гниющих валежин и пней.
Mentissoidea litotes litodenna (Bttg.).
Boettger О., 1881a : 231, t. 8, fig. 16 (Clausilia litotes); Ретовский,
1914 : 316 [Clausilia (Euxina) litotes var.].
Раковина веретеновидно-цилиндрическая, стройная, исчерчен-
исчерченная, красновато-бурая. Оборотов ll3/—12V2.
Размеры; высота раковины 14.4—16.4, ширина 2.8—3,6 мм; высота
устья 3.3—3.4, ширина 2.5—2.6 мм.
В остальном раковина сходна с таковой предыдущего подвида.
Распространение. Аджарская АССР; к северу единичные
находки по Черноморскому побережью до Сухуми.
9. Род FILOSA BTTG.
Boettger О., 1877а : 72 (Clausilia, секция Serrulina, группа Fi losa); В о е 11-
g е г О , 1889 : 32 {Clausilia, секция Euxina, подсекция Filosa); L i n d h о 1 m, 1924 :
G6 (Laciniariat подрод Euxina, секция Fi losa). . ;;
212 IV. EUXININAE
Монотипический род.
Последний оборот раковины с крупным килем. Устье с базальным
желобком. Верхняя пластинка, плавно снижаясь, переходит в спираль-
спиральную пластинку. Одна из краевых складок париетальной стенки устья сли-
сливается с нижней пластинкой, благодаря чему создается впечатление, что
последняя внизу раздваивается (складка образует верхнюю ветвь). Ниж-
Нижний конец субколумеллярной пластинки хорошо, виден при косом поло-
положении устья. Полулунная складка лежит на правосшшной стороне.
Главная складка длинная. Языковидная лопасть клаузилия по внешней
стороне лопасти несет 2 крупных пластинчатых выступа, пересекающих ее
по диагонали (рис. 135, В).
ь, Срединный и боковые зубы радулы с дополнительными зубцами.
Внешние краевые зубы гребенчатого типа.
N> Семяприемник сравнительно короткий, лишен отростка и ретрактора.
Семяпровод, плавно расширяясь, переходит в эпифаллус, а последний
в цилиндрический пенис. Ретрактор пениса кренится к диафрагме.
Глазной ретрактор лежит между пенисом и влагалищем.
В этот род входит только один вид — F. filosa (Mouss.). Раковина его
столь своеобразна, что порой одни и те же авторы относили его то к роду
Serrulina Mouss., то к Euxina Bttg., следовательно, в разные подсемей-
подсемейства. Основанием причислять этот вид к роду Serrulina служило то, что
раковина этого моллюска на краю устья имеет многочисленные мелкие
складки, в этом отношении особенно походя на iS". serrulata (Pfr.). Кроме
того, главная складка длинная, верхняя пластинка слита со спиральной
в одно целое — тоже признаки, свойственные всем видам того же рода.
Наконец, как показали наши исследования (Лихарев и Раммельмейер,
1952), лопасть клаузилия данного вида несет 2 выступа, а как известно,
у одного из видов Serrulina — S. semilamellata (Mouss.) (рис. 51,В) кла-
узилий тоже с килеобразным выступом на лопасти.
С другой стороны, некоторые особенности раковины говорят как будто
бы в пользу принадлежности данного моллюска к Euxina. Так, присут-
присутствие киля на последнем обороте раковины и базального желобка в устье
несвойственно ни Serrulina, ни вообще подсемейству Phaedusinae, а ха-
характерный признак Clausiliinae и Euxininae. Кроме того, темно-коричне-
темно-коричневая окраска — явление, необычное для раковины Serrulina, у которых
она всегда светлая, нередко стекловидно-прозрачная.
Такие признаки, как обрамление устья раковины краевыми склад-
складками, слияние спиральной и верхней пластинок, встречаются в самых
различных группах семейства и не могут считаться свойственными только
Serrulina или Euxina. Что касается выступов на клаузилии, то у настоя-
настоящих Serrulina, если они имеются, то только на внутренней стороне
лопасти, тогда как у F. filosa эти образования находятся на внешней
стороне.
Внутреннее строение данного моллюска говорит за то, что он далеко
отстоит как от Phaedusinae вообще, так и от Serrulina в частности. Это
видно из того, что семяприемник у него впадает в женские половые пути
далеко от половой клоаки, благодаря чему влагалище значительно длин-
длиннее матки. Кроме того, пенис этого вида относительно короткий, в то
время как у Serrulina он очень длинный.
Утеря F. filosa отростка и ретрактора семяприемника затрудняет ре-
решение вопроса, к какому из подсемейств ее следует отнести. Однако нали-
наличие на раковине киля и базального желобка, свойственных только Clausi-
Clausiliinae ж Euxininae, петлеобразная форма мужских половых путей, с нашей
точки зрения, указывают на принадлежность этого моллюска к подсе-
9. FILOSA
213
мейству >Euxininae. Правда, приходится признать, что слияние спираль-
спиральной и верхней пластинок представляет собой явление, необычное в пре-
пределах указанного подсемейства.
Среди Euxininae данный моллюск стоит очень изолированно. Своеоб-
Своеобразные кили на внешней стороне лопасти клаузилия, расщепление ниж-
нижнего конца нижней пластинки и ее смещение к верхней, а также другие
-25т
Рис. 135, Filosa filosa (Mouss.). (Ориг.).
А — общий вид раковины; Б — замыкателышй аппарат (часть внешней стенки раковины
удалена); В — клаузилий.
1 — верхняя пластинка; 2а — верхняя ветвь нижней пластинки; 26 — нижняя ветвь ниж-
нижней пластинки; з — субколумеллярная пластинка; 4 — клаузилий; В — полулунная склад-
складка; в — главная складка; 7 — спиральная пластинка.
особенности организации F. filosa ставят ее особняком среди других
видов того же семейства.
Отсутствие отростка семяприемника как будто бы сближает данный
вид с Mentissoidea litotes (A. Schm.). Однако во всем остальном оба мол-
моллюска очень различны и бесспорно принадлежат к разным родам.
С нашей точки зрения, род Filosa наиболее близок к вымершему в мио-
миоцене роду Pseudidyla Bttg. За это говорит наличие в обеих группах киля
и базального желобка, многочисленных складок по всему краю устья,
форма нижней пластинки и близкое положение последней к слитым воедино
верхней и спиральной пластинкам. Но в отличии от Filosa, у Pseudidyla
субколумеллярная пластинка, подобно нижней, тоже раздваивается,
полулунная складка отсутствует, и, наконец, на клаузилий отсутствуют
кили, столь характерные для Filosa (по крайней мере они до сих пор
не установлены).
214
IV. tUXININAE
Filosa filosa (Mouss.) (рис. 135—137).
1863 : 395 (Clausilia); Boettger 0., 1877a : 72 [Clausilia (Ser-
ger 0., 1889:31 [Clausilia (Euxina)]; К о b e 11, 1893, Iconogr.,
Л i\C\ О Г /~1 1 . .•!_"—. / T7T_ - • . _\ 1 * TT —— •— A *л п -п -rr "D n if -шг л тг т ir л «¦ #л »ъ
M 0 U S S ОП,
• N. Г., 6 :/43, fig. 1023 [Clausilia (Euxina)]; Лихарев и Раммельмейер,
,1952:215, рис. 129, a (Serrulina).
Раковина веретеновидная, с мелкими, очень редко лежащими
ребрами (на предпоследнем обороте их 30—40), просвечивающая, с шелко-
1 7 13
Рис. 136. Filosa filosa (Mouss.). Радула. (Ориг.).
1S
Я
вистым блеском, красновато-коричневая или темно-роговая, без белых
штрихов. Верхняя часть завитка ниже широкой закругленной верхушки
(ширина 1.0—1.2 мм), имеет вогнутые контуры. Поверхность раковины
чаще всего сильно корродирована,
благодаря чему раковина становится
непрозрачной серовато-белой. Обо-
Оборотов 9—10 (изредка 11), верхние
сильно, нижние слабо выпуклые, раз-
разделенные мелким швом; эмбриональ-
эмбриональные обороты BV4—21/2) гладкие, ма-
матовые; последний оборот книзу сужен,
за устьем уплощен, внизу с крупным
морщинистым килем, окаймляющим
широкое полукруглое околопупоч-
околопупочное поле. Устье грушевидное, очень
сильно выступающее, с глубоко вре-
врезанным, отогнутым к оси синулусом,
внизу угловатое. Края устья слабо
отвернуты, утолщены, с многочислен-
многочисленными складками и узелками. Доволь-
Довольно высокая, крючкообразно изогну-
изогнутая верхняя пластинка впереди не-
нередко выступает за край устья, сзади,
плавно снижаясь, переходит в спи-
; ¦ • ральную пластинку. Последняя берет
лачало на право-брюшной стороне и быстро нарастает в высоту, достигая
максимума на правой стороне раковины. Нижняя пластинка лежит глу-
глубоко и при прямом положении устья не видна; она начинается на правой
«тороне раковины и вначале тянется параллельно спиральной пластинке,
затем раздваивается, причем верхняя ветвь выходит к краю устья справа
•от верхней пластинки, а нижняя тянется вниз вдоль столбика и заканчи-
заканчивается на колумеллярном крае устья. Крупная прямая полулунная складка
лежит на право-спинной стороне. Главная складка начинается на правой
и заканчивается на левой стороне раковины и при прямом положении
устья не видна. На месте средней палатальной складки, на некотором
расстоянии от края устья, лежит крупный бугор (хорошо виден при пря-
Рис. 137. Filosa filosa (Mouss.). Половая
система. (Ориг.).
V. CLAUSILIINAE 215
мом положении устья). Между ним и базальным желобком нередко нахо-
находятся еще 1—2 бугорка или короткие складки.
Размеры: высота раковины 10—11, ширина 2.5—2.7 мм; высота устья
2.1—2.3, ширина 1.6—1.8 мм.
Внутреннее строение. Материал: Батумский ботанический
сад, сбор и определение наше, 10—13 IV 1957 — 3 экз.
Радула (рис. 136): [| + (у + Й) X 2] X с^ЮО. '
Размеры срединного зуба 12.3—10 р., ширина радулы 335 ц.
Крупная белковая железа открывается в сравнительно короткий семя-
яйцевод (рис. 137). Вдоль женского канала последнего лежит короткий
семяприемник, лишенный отростка и специального ретрактора. Влага-
Влагалище приблизительно одной длины с маткой.
Левый ретрактор посылает ответвления к половой клоаке и к нижнему
концу влагалища.
Распространение. Черноморское побережье Аджарской АССР.
Экология. Обитает во влажных местах субтропического леса,
сильно заросших травой или папоротниками, под камнями, в подстилке
и под валежником.
Размножение. Живородящий вид.
V. Подсемейство CLAUSILIINAE
Раковина левозакрученная; скульптура от тонкой исчерченности до
ребристости; окраска от роговой до красновато-коричневой или почти
черной, нередко с пучками белых штрихов. У большинства родов послед-
последний оборот внизу с килем. Устье цельное, чаще всего выступающее, с ба-
базальным желобком и врезанным синулусом. Верхняя и спиральная пла-
пластинки, как правило, располагаются на одном расстоянии от столбика и
либо сливаются воедино, либо разделены более или менее коротким про-
промежутком. Лишь у одного рода (Laciniaria Hartm.) концы обеих пласти-
пластинок слегка заходят один за" другой.
Срединный и боковые зубы радулы чаще всего с дополнительными зуб-
зубцами; однако у некоторых видов они лишены этих зубцов.
Семяприемник всегда с ретрактором и открывается в женские половые
пути на значительном удалении от половой клоаки. Отросток семяприем-
семяприемника, подобно таковому у Euxininae, впадает в последний вблизи его
нижнего конца и превращен в особый железистый орган, имеющий вид
тонкостенной, сильно извитой трубки. Мужские половые пути в пределах
подсемейства испытывают определенные преобразования. Они заклю-
заключаются в том, что нисходящее колено семяпровода сильно утолщается,
а восходящее колено, пенис и его ретрактор претерпевают редукцию до
полного их исчезновения. В то же время дистальная часть нисходящего
колена в большей или меньшей степени вздувается и преобразуется во
вторичный пенис. В конечном счете мужские половые пути оказываются
вытянутыми вдоль женских, а ретрактор ближайшего глазного щупальца
располагается медиально от половых путей и с ними не перекрещивается.
Распространение. Европа, Крым, Кавказ, черноморское
побережье Малой Азии.
1. Род RUTHENICA LINDH.
Lindholm, 1924 : 68 (Graciliaria, секция Ruthenica); Ehrmann, 1933 : 77.
Монотипический род.
Раковина небольшая, ребристая. Последний оборот без киля. Устье
без базального желобка. Спиральная пластинка рудиментарная; рядом
216
V, CLAXJSILIINAE
с нею имеется небольшая параллельная пластинка. Нижняя пластинка
лежит глубоко; субколумеллярная хорошо видна при косом положении
устья. Полулунная складка слабо выражена и лежит на правой стороне
раковины. Лопасть клаузилия широкая и короткая, у рукоятки сужена;
последняя располагается под прямым углом к лопасти.
Срединный и боковые зубы радулы с дополнительными зубцами.
Семяприемник с тонким отростком и с ретрактором. Мужские половые
пути образуют сравнительно короткую петлю, причем пенис и нисходящее
колено семяпровода сравнительно толстые, а восходящее
колено очень тонкое. Имеется диафрагмальный ретрактор
пениса. Ретрактор глазного щупальца проходит между
пенисом и влагалищем.
Ruthenica filograna (Rssm.) (рис. 138, 139).
Rossmassler, 1836, Iconogr., 1, 4 : 17, fig. 264 (Clausilia);
Kuster, 1847—1862 : 93, t. 10, fig. 25 (Clausilia); W a g n e г A.,
1914 : 8, t. 575, fig. 25 (Graciliaria, анатомия); Лихарев и
P а м м е л ь м е й в р, 1952 : 225, рис. 141.
Раковина вереяеновидная, с тупой вершиной,
тонкостенная, с редкими тонкими ребрышками (на пред-
предпоследнем обороте их 30—35), слабо блестящая, просве-
просвечивающая, светло-роговая или желтовато-коричневая.
Оборотов 9—10, выпуклых, разделен-
разделенных тонким и глубоким швом; эмбри-
эмбриональные обор_оты_(около 2V2) глад-
гладкие; последниж^оборот уже предпо-
^йещшгоТ" за "устьем покрыт круп-
крупными пластинчатыми ребрами и имеет
заметное затылочное утолщение.
Устье широко-грушевидное, сильно
выст^1ает_вш|Р^^. Края его со сла-
слабым отворотом, тупые, белые. Верх-
няящастинка низкая и тонкая, квёрх-
нему концу нитевидная;~впереди она
доходит flo^jtyjaH устья. Нижняя
пластинка" при ""прямом положе-
положении устья почти не видна; с краем устья она связана тонкой складочкой;
в глубь раковины"*она"уходит в виде довольно высокой, по внешйм^Ткрйю
утолщенной и спирально изогнутой складки; у верхнего конца она резко
снижается. Имеется довольно крупная промежуточная складка. Корот-
Короткие спиральная и параллельная пластинки, равно как и главная складка,
лежат вблизи друг от друга в первой четверти последнего оборота. _Суб-
колумелляр_ная пластинка хорошо видна при косом, хуже при прямом
положении устья! иногда она подобно нижней пластинке с краем устья
связана короткой складочкой. Слабая, нередко мозолевидНТга'ТТбХуТГун-
ная складка лежит несколько наклонно на правой стороне раковины.1
Кроме главной складки, иногда имеется еще более короткая сутуральная
складка и еще реже — верхняя складка. В глубине устья, на значитель-
значительном отдалении от края и параллельно ему, лежит крупная палатальная
Рис. 138. Ruthenica filograna (Rssm.).
А — общий вид раковины (по Эрманну),
Б — клаувилий (оряг.).
1 Большинство авторов считает, что полулунная складка отсутствует'. Между тем,
вас основании просмотра большой коллекции раковин данного вида, мы, вслед за Линд-
гольмом (Lindholm, 1924), убедились в присутствии этой складки.
1. RUTHENICA 217
мозоль; особенной толщины она достигает на концах, а по середине не-
нередко прервана. Лопасть клаузилия очень широкая, сильно изогнутая
(поперечно и продольно), с округлым передним концом; при переходе
в рукоятку резко сужается; рукоятка располагается к лопасти под пря-
прямым углом.
Размеры: высота раковины 7.5—9.0, ширина 2.0—2.2 мм; высота
устья 1.6—1.7, ширина 1.3—1.4 мм.
Внутреннее строение [по данным Вагнера (Wagner A.,
1914) и Лёвушкина A958)]. Белковая железа крупная (рис. 139). Семя-
Q
Рис. 139. Ruthenica filograna (Rssm.). Половая систе-
система. (По Лёвушкину).
приемник сравнительно длинный, хотя верхний конец его не достигает
белковой железы; отросток несколько короче. Матка в 2 раза длиннее
влагалища. Толстое нисходящее колено семяпровода, внизу, после того
как обогнет глазной ретрактор, переходит в тонкое и более короткое вос-
восходящее колено. Пенис цилиндрический, почти такой же ширины, как и
нисходящее колено семяпровода.
Распространение (рис. 140). В СССР: многочисленные на-
находки в Калининградской области, Литве, Латвии (Видземе и Курземе),
в Эстонии (за исключением островов и западных районов), в западной
части Ленинградской области и в Московской области. Найдена в окрест-
окрестностях Калинина, Витебска, Смоленска, Рославля, Киева и на юге
Орловской области (Троена). Много находок в Карпатах и в Предкар-
патье. Вне СССР: почти вся Польша (за исключением Люблинской воз-
возвышенности), все Карпаты с Трансильванией, северо-западные Балканы
(Хорватия, Истрия, Крайна), южные Альпы (на запад до долины р. Пьяве),
Венский лес, Чешский массив. Разрозненные колонии в средневысотных
горах южной и центральной Германии (Ehrmann, 1933).
218
V. CLAUSILIINAE
В ископаемом состоянии найдена в четвертичных отложениях Гер-
Германии (Geyer, 1927) и Чехословакии (Lozek, 1955), а по данным Шоша
(Soos, 1943) и в плиоцене Венгрии.
40
10 20 3D 40 50
0 200 400 600 800 ки
40
Рис. 140. Географическое распространение Ruthenica filograna (Rssm.). (Ориг.).
1 —¦ основной ареал; г — изолированные ареалы.
Экология. Обитает во влажных смешанных и широколиственных
лесах, в подстилке, под кусками гниющей древесины и под камнями.
2. Род CLAUSILIA DRAP.
Draparnaud, 1805 : 24; Gray, 1840, Turton's Manual : 241 (Iphige-
nia pars, non Schumacher, 1817);'В г u s i n a, 1870, Contrib. malacol. Croatie :
24 (Kuzmicia); Moellendorff, 1875 : 27 (Cusmicia emend.); Westerlund,
1901 : XXXII, 6 (Cusmicia); W a g n e г A., 1920 : 75 (Kuzmicia); Lindholm, 1924 :
63; T h i о 1 e, 1931 : 538.
Тип рода: Clausilia rugosa Drap.
Последний оборот раковины с килем, окаймленным бороздой. Устье
высокое (высота заметно больше ширины), с хорошо выраженным базаль-
ным желобком; последний придает нижнему краю устья некоторую угло-
угловатость. Верхняя и спиральная пластинки чаще всего сливаются друг
с другом или, располагаясь на одной линии, разделены коротким проме-
промежутком. Нижняя пластинка лежит сравнительно глубоко. Нижний конец
субколумеллярной пластинки хорошо виден, нередко даже при прямом
: 2. CLAUSfLIA 219
положении устья. Полулунная складка лежит на спинной стороне; верх-
верхний конец ее обычно крючковидно загнут назад, нижний тоже отогнут
назад, но плавно, дугообразно. Главная складка хорошо развита и зани-
занимает около V3 последнего оборота; верхний конец ее лежит позади полулун-
полулунной складки. На палатальной стенке устья располагается мозоль, состоя-
состоящая чаще всего из двух частей: верхней и нижней; от последней внутрь
раковины тянется короткая, но толстая нижняя палатальная складка.
Лопасть клаузилия на нижнем конце всегда с более или менее выражен-
выраженным углом.
Срединный и боковые зубы радулы с дополнительными зубцами.
Верхняя (сортировочная) часть семяяйцевода развита слабо. Матка
значительно короче влагалища, реже одной длины с ним. Отросток семя-
семяприемника тонкий, короче самого семяприемника. Нисходящее колено
семяпровода очень толстое и мускулистое; восходящее колено очень
тонкое и вместе с коротким слабо развитым пенисом образует небольшую
петлю, к которой крепится очень тонкий диафрагмальный ретрактор.
Ретрактор глазного щупальца лежит медиально от половых путей
и с ними не перекрещивается. Левый ретрактор дает ветви к шейке семя-
семяприемника, к нисходящему колену семяпровода, к нижним концам вла-
влагалища и пениса, и к половой клоаке.
Р а с п р ос т р а н е н и е. Европа.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CLAUSILIA
1 B). Раковина маленькая (высота колеблется в пределах 7.0—10,
ширина 1.9—2.2 мм), очень тонко исчерчена (скульптура хорошо
видна лишь при 40-кратном увеличении), без белых штрихов
у шва . .' . 1. Cl. parvula (Stud.).
2 A). Раковина у большинства видов более крупная (высота 10.5—15.0,
ширина 2.5—3.5 мм); поверхностная скульптура всегда сильнее —
от глубокой исчерченностй'до ребристой; у шва обычно распола-
располагаются пучки белых штрихов.
3F). Раковина густо исчерчена или ребристо исчерчена (на предпо-
предпоследнем обороте 60—80 складок).
4 E). Нижняя пластинка заканчивается 2 небольшими краевыми
складочками. Париетальная стенка устья в промежутке между
верхней и нижней пластинками несет 1—2 складочки. Высота
раковины 9—11, ширина 2.4—2.5 мм ... 2. Cl. bidentata (Strom).
5 D). Нижняя пластинка заканчивается 2 бугорками. В промежутке
между верхней и нижней пластинками париетальная стенка
гладкая, без складок. Высота раковины 12—14, ширина 2.8—
3.0 мм1 . . . 3. Cl. dubia Drap.
6 C). Раковина более или менее редко ребристо исчерчена или ребрис-
ребристая (на предпоследнем обороте не более 50 складок или ребрышек).
7 A0). Раковина у нижних оборотов вздутая. Верхняя половина завитка
имеет вогнутые очертания. Нижняя пластинка заканчивается 2
расходящимися краевыми складками.
8 (9). Верхняя пластинка сливается со спиральной пластинкой . . .
................... 4а. Cl. pumila pumila G. Pfr.
9 (8). Верхняя пластинка не сливается со спиральной . .
...,.....,..., 46. Cl. pumila sejuncta West.
Встречаются мелкие угнетенные формы этого же вида.
220
V. CLAUSILIINAE
10 G). Раковина довольно стройная. Верхняя часть завитка имеет вы-
выпуклые или ровные очертания. Нижняя пластинка заканчивается
2 параллельными краевыми складками . . . 5. Cl. cruciata (Stud.).
*1. Clausilia parvula (Stud.) (рис. 141).
S t u d e г, 1820, Naturw. Anz. Schweiz. Ges. Naturw., 3 : 89 (Clischrus (Clausilia)]
Kiister, 1847—1862 : 149, t. 16, fig. 28—31; M e г m о d, 1930:413, fig. 85 (Pi-
rostoma, анатомия); L о z e k, 1956 : 125, t. 12, fig. 1; U r b a n s-
ki, 1957:161, fig. 123, 140; F о г с а г t, 1957, Mitt. Naturf.
Ges. Bern, N. F., 15 : 193, t. 3, fig. 21.
Раковина веретеновидная или веретеновидно-
цилиндрическая, стройная, тонкостенная, но прочная,
умеренно просвечивающая, с шелковистым блеском,
очень тонко и равномерно исчерченная, темно-красно-
темно-красновато-коричневая, обычно без белых штрихов. Оборотов
91/2—101/2, слабо выпуклых, разделенных тонким бело-
беловатым швом; эмбриональные обороты (около 2V2) обра-
образуют тупую цилиндрическую вершину; последний обо-
оборот уже предпоследнего, внизу
с крупным килем, за устьем
сильно исчерчен и уплощен.
Устье грушевидное, выступаю-
выступающее, внутри коричневатое.
Края устья расширены, отвер-
отвернуты, беловатые. Верхняя пла-
пластинка довольно высокая и тон-
тонкая; впереди она достигает края
устья и плавно снижается пе-
перед слиянием со спиральной
пластинкой. Крупная нижняя
пластинка лежит глубоко, но
значительная часть ее видна
при прямом положении устья;
с краем устья она ничем не
связана; дав широкий изгиб,
она уходит круто кверху. Субколумеллярная пластинка видна лишь при
косом положении устья. Полулунная складка умеренно развита; верхний
конец ее крючковидный, а нижний довольно сильно отогнут вглубь рако-
раковины, так что порой кажется, будто почти вся полулунная складка лежит
горизонтально. Главная складка крупная и длинная, занимает почти
V2 оборота и заходит далеко за полулунную складку. Палатальная
мозоль состоит из двух частей: крупной верхней и слабо развитой нижней;
от последней вглубь раковины тянется крупная белая нижняя палаталь-
палатальная складка, которая вместе с субколумеллярной пластинкой обрамляет
с обеих сторон базальный желобок. Рукоятка клаузилия плавно расши-
расширяется в лопасть. Задняя половина последней довольно широкая, а перед-
передняя перегнута в продольном направлении; нижний конец лопасти клюво-
клювовидно вытянут и утолщен.
Размеры: высота раковины 8.0—9.5, ширина 2.0—2.2 мм; высота устья
1.7—1.9, ширина 1.2—1.4 мм.1
1 Лектотип данного вида выделен Форкартом и хранится в коллекции Зоологиче-
Зоологического института Бернского университета (Швейцария). Он имеет следующие размеры:
высота раковины 10, ширина 2.5 мм; высоты устья 2.4, ширина 1.5 мм; 10!4 оборотов.
Место сбора: окрестности Берна.
Рис. 141. Clausilia parvula (Stud.). (Ориг.).
А — общий вид раковины; Б — клаузилий.
2. CLAUSILIA
221
Внутреннее строение (Soos, 1917; Mermod, 1930).
г, с > 56 . 11к
Радула: _ + - + ^_ё.
Половая система обычная для данного рода.
Распространение. Среднеевропейский вид. Средневысотные
горы и возвышенности северной и восточной Франции, южной Бельгии,
западной, южной и центральной Германии, (Ehrmann, 1933), западной
Чехословакии и южной Польши (Судеты, Краковская Юра); кроме того,
вся дуга северных Альп от Савойи до Венского леса и западная часть
Западных Карпат. Вне основного ареала ряд островных
местонахождений, как и всюду, приуроченных к извест-
известняковым породам.
Некоторые исследователи (Симашко, 1847; Ecke u.
Dulckeit, 1924) указывали на присутствие этого вида в
Латвии и Эстонии. Как показала позднейшая проверка
их коллекций (Boettger О., 1878с; Poska-Teiss, 1928),
эти данные следует считать ошибочными, так как за дан-
данный вид были приняты экземпляры Clausilia bidentata
(Strom).
Найден в четвертичных отложениях ряда стран, в том
числе и в Англии, где этот моллюск в настоящее время
не живет.
Экология. Обитает на открытых, слабо увлаж-
увлажненных местах; чаще всего на скалах (особенно на из-
известняках), среди растений; реже в лесу, в подстилке
или под камнями. В горы проникает до высоты 1700 м, как
исключение — до 2000 м над ур. м.
2. Clausilia bidentata (Strom) (рис. 142, 143).
Strom, 1765, Trondhj. Selsk. Skrift, 3 : 436,
t. 6, fig. 17 (Turbo); Maton a. Racket t, 1807,
Trans. Linn. Soc, 8 : 180 (Turbo nigricans); P f e i f-
ferC, 1821, Naturgesch. Deutsch. Moll., 1 : 65, t.
3, fig. 33—34 (obtusa); Westerlund, 1884 : 187;
Ehrmann, 1933 : 68, t. 2, fig. 32; Лихарев
иРаммельмейер, 1952 : 226.
Раковина веретеновидно-цилиндри-
ческая, тонко и глубоко исчерченная (в про-
промежутках между поперечными складками
продольно исчерчена), довольно просвечи-
просвечивающая, с шелковистым блеском, темно-ко-
темно-коричневая, до почти черной, у шва с бе-
белыми штрихами. Оборотов 9—10, слабо
выпуклых (особенно нижние); эмбриональные обороты C), выпуклые
гладкие; 3 последних оборота одинаковой ширины; последний обо-
оборот книзу сужен, за устьем сильно уплощен, внизу с коротким килем.
Шов тонкий, беловатый. Устье ромбическо-грушевидное, слабо высту-
выступающее, внутри коричневое. Края устья слабо отвернуты, белые. Верхняя
пластинка лежит отвесно, низкая, достигает края устья; сзади она, сни-
снижаясь, переходит в спиральную пластинку. Ланцетовидная нижняя
пластинка лежит глубоко, слабо видна при прямом положении устья,
довольно круто уходит кверху; нижний конец ее расщепляется на 2 скла-
складочки. Еще 1—2 краевых складки имеются между верхней и нижней пла-
пластинками. Тонкая спиральная пластинка начинается выше нижней,
Б
Рис. 142. Clausilia bidentata
(Strom). (Ориг.).
А — общий вид раковины; Б —
клаузилий.
222
V. CLAUSIL1INAE
кк
быстро нарастает в высоту, после чего плавно снижается к месту слияния
с верхней пластинкой. Нижний конец субколумеллярной пластинки хо-
хорошо виден при косом положении устья. Крупная полулунная складка
лежит на спинной стороне раковины. Оба конца ее отогнуты назад —
верхний под углом, нижний плавно. Главная складка начинается немного
позади полулунной и простирается вперед приблизительно на 1/3 обо-
оборота. Несколько отступя от края
устья лежит более или менее круп-
крупная палатальная мозоль, от нижнего
конца которой уходит вглубь рако-
раковины короткая, но мощная, нижняя
палатальная складка. Языковидная
лопасть клаузилия сильно изогнута,
наверху плавно переходит в руко-
рукоятку, внизу у конца утолщенная и
угловатая.
Размеры: высота раковины 9—11,
ширина 2.4—2.5 мм; высота устья
2.4—2.5, ширина 1.6—1.7 мм.
Изменчивость раковины
касается главным образом размеров
и формы. Наряду с нормальными
встречаются экземпляры с короткой
и вздутой раковиной (var. septentrio-
nalis A. Schmidt — высота раковины
8, ширина 2.5 мм), или с маленькой
стройной раковиной (var. exigua Wes-
terlund — высота 8, ширина 2 мм),
или же, наконец, с узкой и высокой
раковиной (var. elongate, Clessin —
высота 11—12, ширина 2.5 мм). Не-
Некоторую изменчивость испытывает
окраска, которая может быть не-
немного светлее или темнее обычной..
В ну треннее строение.
По данным Стинберга (Steenberg,
1914), половая система этого моллю-
моллюска характеризуется сильно вздутым
пенисом (рис. 143). Однако, как показал Лёвушкин A958), подобная кар-
картина встречается и у Cl. cruciata (Stud.).
Распространение (рис. 34). В СССР: приморские районы
Курземе (Латвийская ССР), острова Хиума и Саарема и северо-западная
часть континентальной Эстонии. Одна находка сделана в Ленинградской
области (Приморск). Возможны находки в Калининградской области и
в Литве.1
. Вне СССР ареал данного вида охватывает Британские острова, побе-
побережье Норвегии до 70° с. ш., Швеции до QS1/^ с. ш., юго-запад Финляндии
рп
кп
нп рнл
Рис. 143. Clausilia bidentata (Strom).
Половая система. (По Стинбергу).
1 В прошлом веке многие исследователи (Двигубский, 1832; Симашко, 1847; Ра-
чинский, 1853; Надеждин, 1868, Сабанеев, 1880) указывали на присутствие этого вида
в средней России. Они называли его то Cl. rugosa Г>гар., то Cl. nigricans Pult. Бонков-
ский (Bqkowski, 1892) отмечал данный вид в Карпатах. В настоящее время совершенно
ясно, что указанные авторы за данный вид принимали то Cl. dubia Drap., то С I. cru-
cruciata (Stud.). . -
2. CLAUSILIA 223
и все равнины и средневысотные горы от Бретани на западе и до прибал-
прибалтийских республик СССР на востоке.
Ареал данного вида занимает области с мягким и влажным океаниче-
океаническим климатом, т. е. находящиеся под воздействием Северо-Атлантиче-
Северо-Атлантического течения.
В ископаемом состоянии Cl. bidentata найдена в четвертичных отло-
отложениях ряда стран западной и северной Европы.
Экология. Обитает в тенистых смешанных и широколиственных
лесах и в старых парках, в лиственной подстилке, под гнилой корой и
валежниками и в гнилых пнях, а также в щелях сырых скал, каменных
оград и под камнями. К горным породам относится индифферентно.
3. Clausilia dubia Drap. (рис. 144, 145).
D raparnand, 1805:70, t. 4, fig. 10, Kuster, 1847—1862:152, t. 16,
fig. 41—53; 1.17, fig. 1—3; Лихарев иРаммельмейер, 1952 : 226, рис. 142.
P а к о в и н а веретеновидная или веретеновидно-цилиндрическая.
густо и тонко ребристо исчерчена; на последних? оборотах, в промежут-
промежутках между ребрышками, обычно с ясной спиральной исчерченностью, бле-
блестящая; окраска от коричнево-роговой до темно-красновато-коричневой
или вишневой, с многочисленными пучками белых штрихов, ^бодотов
10—12, слабо выпуклых; эмбрион^л^ды^^бедот^^^гладкде^^бра^уют
тупую вершину; последние" 2—3 оборота одинаковод,лпирины; последний
оборот книзу * сужается, за "устьем уплощён. Шов довольно глубокий,
белый. Устье грушевидное или ромбическо-грушёвидное, выступающее.
Края~устья~13елые, широкие, более или менее утолщены; внутренность
устья чаще всего коричневого цвета. Косо сидящая тонкая верхняя пла-
пластинка впереди достигает края устья, а сзади, немного снижаясь, плавно
переходит в спиральную пластинку. Ланцетовидная нижняя пластинка
лежит очень глубоко и при прямом положении устья почти не видна;
нижний конец~е«Рзаканчивается 2 более или менее крупными бугорками,
лежащими друг к другу под углом; иногда эти бугорки вытягиваются
к краю устья в виде слабых складочек; кверху нижняя пластинка уходит
почти отвесно и заканчивается приблизительно на спинной стороне рако-
раковины. Субколумеллярная пластинка видна при косом положении устья.
Стенкаустья между верхней и нижней пластинками всегда гладкая, без
складок. Палатальная мозоль развита слабее, чем у Cl. cruciata (Stud.)
и у Cl. bidentata (Strom); она состоит из двух частей: верхняя более круп-
крупная, нижняя поменьше. От последней вглубь раковины нередко отходит
более или менее ясно выраженная нижняя палатальная складка. Рукоятка
клаузилия плавно расширяется в языковидную лопасть; нижний конец
последней с острым или округленным углом на внешнем крае.
Размеры: высота раковины 10^14, ширина 2.5—3.0 мм; высота устья
2.6—3.0, ширина 1.8—2.0 мм.
Изменчивость. Раковина данного моллюска сильно варьирует
как в отношении размеров, так и в степени развития скульптуры. Большое
влияние оказывает химический состав субстрата. Так, на известняковых
скалах преобладают крупные формы (f. speciosa A. Schmidt, i.'podolica
Ba,kowski), у которых высота раковины достигает 16, а ширина 3.5 мм.
В таких же условиях встречаются формы, отличающиеся более сильной
поверхностной скульптурой, большим развитием мозоли, нижней пала-
палатальной складки и бугорков, лежащих у основания нижней пластинки
(f. vindobonensis A. Schmidt). Популяции, обитающие на бедных кальцием
субстратах, отличаются преобладанием форм, раковины которых имеют
224
V. CLAUSILIIJVAE
слабую палатальную мозоль, а нижняя пластинка лишена бугорков
(f. obsolete. A. Schmidt). Наконец, в горах, у верхней границы леса, в усло-
условиях низких температур, преобладают мелкие формы (f. alpicola Clessin,
Рис. 144. Clausilia dubia Рис. 145. Clausilia dubia Drap. Поло-
Drap. вая система. (По Стинбергу).
А — общий вид раковины
(ориг.); Б — клаузилий (по
Эрманну).
f. runensis Tschapeck), раковина которых имеет высоту 7—9, а ширину
2.0—3.0 мм.
Внутреннее строение. Согласно Стинбергу (Steenberg,
1914) и Лёвушкину A958), половая система данного вида отличается от
остальных видов того же рода следующими особенностями (рис. 145);
2. CLAUSILIA
225
1) влагалище приблизительно такой же длины, как и матка; 2) пенис очень
короткий, почти в 7 раз короче семяпровода; 3) восходящее колено семя-
семяпровода с помощью мускульных волокон плотно прижато к пенису, так
что между ними не остается просвета.
Распространение (рис. 146). В СССР: многочисленные коло-
колонии рассеяны по всей Прибалтике от Калининградской области до Эсто-
40
3D
200 0 200 '400 6Ш 800™
30
40
Рис. 146. Географическое распространение Clausilia dubia Drap. и Cl. pumila C. Pfr.
(Ориг.).
id — крупные ареалы Cl. dubia; 16 — то же, мелкие изолированные ареалы; 2а — ареал Cl. pu-
pumila; 26 — то же, находки вне современного ареала в четвертичных отложениях.
/
нии. Кроме того, отдельные находки в Псковской области (Новоржев),
у Витебска, взаповеднике «Беловежская^гущад и многочисленные находки
в Карпатах и на Вольшо-ТГодольйсоХ возвышенности.
Общее распространение: горные области и средневысотные горы сред-
лей Европы от Французского массива на западе до Карпат на востоке и
от Гарца на севере до северной Албании на юге. Кроме указанного основ-
основного ареала, имеются три крупных островных: на севере Англии (Пенния-
ские горы), в южной Швеции (до 64й с. ш.), на северо-западе европейской
части СССР. Изолированные колонии обитают в южной Англии, в север-
северной Германии (Ehrmann, 1933), в Польше, в южной Норвегии, в централь-
центральной Финляндии и в других местах.
Экология. На равнинах обитает главным образом в тенистых
«ырых смешанных и широколиственных лесах, в гнилых пнях или под
V. CLAUSILIINAE
корой. В горах встречается также и на открытых местах, чаще всего на
скалах, в особенности предпочитает известняковые скалы,,;; В этих усло-
условиях нередко поднимается в альпийскую зону (в Альпах до 2500 м над.
ур. м.).
4а. Clausilia pumila pumila С. Pfr. (рис. 147, 148).
Pfeiffer С, 1828, Naturgesch. Deutsch. Moll., 3:41, t. 7, fig. 16; Ross-
m a s s 1 e r, 1836, Iconogr., 1, 4 : 15, fig. 259; Kuster, 1847—1862 : 343, t. 17, fig.
10—12, 21—26; Лихарев и Раммельмейер, 1952 : 227, рис. 145, 146.
Раковина веретеновидная, у нижних оборотов вздутая, твердо-
стенная, сильно и равномерно ребристо исчерченная (промежутки между
ребрышками без спиральной исчерченности), просвечивающая, умеренно
блестящая, темно- или красновато-роговая, с редкими пуч-
пучками белых штрихов. Вследствие того, что первые 3—4 обо-
оборота слабо, а последующие обороты более быстро расши-
расширяются, верхняя половина завитка имеет вогнутые очер-
очертания. Оборотов 10—12, верхние умеренно, нижние слабо-
выпуклые; эмбриональные обороты (около 3) гладкие, обра-
зу^ют^^щлинддическую вершину; высота, .^первых 7—9 обо-
оборотов нарастает очень медленно, но затем начинает быстро-
увеличиваться; последний оборот уже предпоследнего, на
затылке уплощен и даже с легкой вмятиной (соответствует
палатальной мозоли). Шов_тонкий, беловатый. Устье косо-
грушевидное или ромбическо-^spy-
шевидное, выступающее, с узким
синулусом, который смещен квер-
кверху. Края устья расширены, утол-
утолщены, отвернуты, белые; верхний
край сильно опущен. Низкая, но-
довольно толстая верхняя пда-
стинка впереди достигает края*
устья, а сзади сливается со спи-
спиральной пластинкой. Крупная лан-
ланцетовидная нижняя плаотинкаТГё-
жит очень глубоко, и при прямом
положении слабо заметен лишь ее-
нижний конец, от которого обычно к краю устья расходятся ,2 коротких
складочки; последние могут сливаться, образуя небольшое треугольное воз-
возвышение;* сама пластинка по внешнему краю утолщена, красноватая, почти
о^тВесно уходит кверху, где, снижаясь, заканчивается спиральным изгибом.
Тонкая и высокая спиральная пластинка плавно снижается к обоим концам.
Нижний конец субколумеллярной пластинки с трудом виден при косом
положении раковины. Крупная полулунная складка лежит отвесно на
спинной стороне раковины. Палатальная мозоль относительно слабая,,
иногда почти незаметна. Нижняя палатальная складка невысокая^ но
всегда хорошо выражена, начинается от палатальной мозоли и уходит на
некоторое расстояние в глубь раковины, но не достигает полулунной
складки. Рукоятка клаузйлия, плавно расширяясь, переходит в широ-
широкую языковидную лопасть; последняя сильно изогнута в поперечном
направлении, с ясным углом на нижнем конце.
Размеры: высота раковины 12—13, ширина 3.0—3.4 мм; высота устья
2.6—2.7, ширина 1.9—2.0 мм.
Внутреннее строение. Лёвушкин A958) на основании
вскрытия большого числа экземпляров данного подвида установил, что.
Рис. 147. Clausilia pumila pumila С. Pfr.
А — общий вид раковины (по Шмидту); Б —
клаузилий (ориг.).
2. CLAUSILIA
227
половая система его характерна следующими особенностями, (рис. 148),
отличающими его от подвида sejuncta West. (рис. 149): влагалище более
чем в 2 раза длиннее матки; пенис относительно более мощный и длин-
длинный (только в 2 раза короче семяпровода); мужская петля (пенис плюс вос-
восходящее колено семяпровода) располагается далеко от половой клоаки.
Распространение (рис. 146). В СССР подвид наиболее часто
встречается в Прибалтике (Калининградская область, Литва, Латвия
бж
кл
Рис» 148. Clausilia pumila pumila
С. Pfr. Половая система. (По
Лёвушкину).
Рис. 149. Clausilia pumila sejuncta
West. Половая система. (По Лё-
Лёвушкину).
и южная Эстония) и изредка в Карпатах и на Волыно-Подольской воз-
возвышенности. Вне СССР обитает в равнинных областях средней Европы
к востоку от Гамбурга, в средневысотных горах Германии (ГДР и ФРГ)
и Чехословакии к востоку от Вюрцбурга, в восточных Карпатах и в се-
северных Балканах.
Известен из четвертичных отложений ряда стран средней Европы, в
том числе и вне современного ареала (Нидерланды).
Экология. Живет главным образом во влажных широколиствен-
широколиственных лесах и в ольховниках в подстилке, под корой валежин и гнилых пней.
Поднимается в горы до высоты 1400 м над ур. м.
46. Clausilia pumila sejuncta West. (рис. 149).
Westerlund, 1871, Expose critique moll. Suede et Norvege : 75 (Clausilia
sejuncta).
Раковина отличается от предыдущего подвида только тем, что
верхняя пластинка не сливается со спиральной, но концы их распола-
располагаются по одной линии, т. е. почти на одинаковом расстоянии от столбика.
228 V. CLAUSILIINAE
В более редких случаях пластинки соединены тонким нитевидным утол-
утолщением.
В пределах одной популяции иногда попадаются единичные экземпляры,
у которых обе пластинки сливаются друг с другом.
Внутреннее строение. Лёвушкин A958) на большом
числе вскрытий показал, что половая система (рис. 149) данного подвида
отличается от предыдущего рядом существенных особенностей. К их числу
относятся следующие: влагалище почти такой же длины, как и матка;
пенис рудиментарный, короткий и тонкий, значительно короче семяпро-
семяпровода (до 10 раз); мужская петля располагается близко от половой клоаки.
Распространение (рис. 146). В СССР данный подвид распро-
распространен в Московской, Ярославской, Калининской, Смоленской, Калуж-
Калужской и в Тульской областях. Наиболее восточная неходка сделана в ок-
окрестностях города Костромы, а наиболее южная — в Орловской области
(Троено). Несколько находок сделано в Прибалтике (Литва, Латвия —
Видземе). Вне СССР подвид обитает преимущественно в северной Европе
(Дания, южная Швеция и Норвегия).
Экология. Обитает в смешанных и широколиственных лесах,
в сырой подстилке или под корой гниющего валежника и гнилых пней.
Систематическое положение. Данный моллюск был
описан как самостоятельный вид, отличающийся от Cl. pumila С. Pfr.
только по раковине. Однако с самого начала большинство авторов рас-
рассматривали его как разновидность последней, в то же время, отмечая
значительную разобщенность их ареалов,с Лёвушкин, обнаружив указан-
указанные выше, довольно стабильные, различия в гениталиях обоих моллюсков,
полагает, что это дает основание считать их самостоятельными видами.
Однако, с нашей точки зрения, автор не учел следующие обстоятельства.
Во-первых, оба моллюска принадлежат к группе, у которой произошла
значительная редукция пениса и копуляция осуществляется мощным нис-
нисходящим коленом семяпровода, поэтому отмеченные выше различия в
длине пениса и в положении мужской петли, по-видимому, не могут по-
помешать скрещиванию обоих подвидов; остальные указанные различия
также вряд ли могут этому препятствовать. Во-вторых, следует учесть,
что у Clausiliidae близкие виды всегда хорошо отличаются друг ,.от друга
по признакам раковины. В данном случае это не имеет места. Поэтому,
с нашей точки зрения, Cl. sejuncta West, является лишь четким под-
подвидом Cl. pumila С. Pfr. За это говорит также то, что хотя значительная
часть ареалов обоих подвидов достаточно разобщена, частично они нахо-
находят друг на друга (например, в Прибалтике).
5. Clausilia cruciata (Stud.) (рис. 150, 151).
S t u d e r, 1820, Naturw. Anz. Schweiz. Ges. Naturw , 3:89 [Glischrus (Clausi-
(Clausilia)]; Kuster, 1847—1862:257, t.9, fig. 11—16; Westerlund, 1884:190;
West-erlund, 1901 : 158; Лихарев и Раммельмейер, 1952 : 227,
рис. 143, 144; F о г с а г t, 1957, Mitt. Naturf. Ges. Bern., N. F., 15 : 193, t. 3, fig. 20.
Раковина веретеновидная, с довольно широкой вершиной, твер-
достенная, с тупыми ребрами (около 40—50 на предпоследнем обороте),
от темно-роговой до вишнево-коричневой, с белыми штрихами. Последние
располагаются не только у шва, но и на периферии оборотов. Между реб-
ребрами иногда заметна густая спиральная исчерченность. Оборотов 9V2—
101/2, слабо выпуклых; эмбриональные обороты (около 2V2) гладкие; по-
последний оборот за устьем уплощен. Шов глубоко врезан. Устье грушевидно-
ромбическое, выдается вперед, с широким и низким синулусом. Края
устья белые, слегка отвернуты и утолщены. Верхняя пластинка лежит
2. CLAUSILIA
229
отвесно, тонкая; впереди она достигает края устья, сзади, довольно резко
снижаясь, переходит в спиральную пластинку. Нижняя пластинка лежит
не очень глубоко; плавно закругляясь, она уходит кверху, впереди же
заканчивается 2 почти параллельными краевыми складочками: верхняя
складочка является продолжением пластинки, нижняя отходит от лежащего
под пластинкой бугорка. Спиральная пластинка начинается на правой
стороне раковины выше нижней пластинки; вначале она быстро нарастает
в высоту, а затем плавно снижается к месту слияния с верхней пластинкой.
Нижний конец субколумеллярной пластинки крюч-
крючкообразно изогнут и хорошо виден при косом, а
•, иногда и при прямом положении устья. В проме-
промежутке между верхней и нижней пластинками не-
недалеко от края устья лежат 1—2, реже 3 неболь-
небольших складки или узелка. Крупная полулунная
Рис. 150. Clausilia cruciata (Stud.).
А — общий вид раковины; Б — нижний оборот и устье; В — клаузи-
лий. (А, Б — по Шмидту с дополнениями, В — ориг.).
складка располагается почти отвесно на спинной стороне раковины; оба
конца ее отогнуты назад: верхний под углом, нижний плавно. Почти па-
параллельно палатальному краю устья лежит крупная мозоль; она состоит
из двух частей — верхней и нижней, которые могут быть разобщенными.
Верхней части мозоли снаружи соответствует слабая вмятина; от нижней
отходит в глубь раковины короткая, но мощная нижняя палатальная
складка. Рукоятка клаузилия плавно переходит в лопасть; последняя
языковидная, умеренно изогнутая, у нижнего конца утолщена и с замет-
заметным углом.
Размеры: высота раковины 9—11, ширина 2.3—2.5 мм; высота устья
1.9—2.1, ширина 1.4—1.5 мм. х
Изменчивость. Чаще всего варьируют размеры раковины.
В неблагоприятных условиях, в частности по границам ареала, появляются
экземпляры с короткой раковиной, высота которой колеблется около
8.0—8.5 мм (f. minima A. Schmidt). В восточных Карпатах, где данный
вид, по-видимому, находит наиболее благоприятные условия, наряду
с обычными встречаются экземпляры с более высокой и стройной рако-
1 Лектотип данного вида выделен Форкартом и хранится в колекции Зоологиче-
Зоологического института Бернского университета (Швейцария). Он имеет следующие размеры:
высота раковины 8.7, ширина 2.5 мм; высота устья 2.1, ширина 1.6 мм; 9 оборотов.
Место сбора: кантон Валлис, Лейкербад.
230
V. CLAUSILIINAE
виной, высота которой колеблется в пределах 11—12, реже до 13 мм
{f. gracilis A. Schmidt).
Внутреннее строение. Согласно Шошу (Soos, 1917), радула
данного вида (одна сторона одного ряда) имеет следующую формулу:
.?. _!_ 8б о- и
бж.
Половая система (Лёвушкин, 1958) характерна тем, что пенис в 4 раза
короче семяпровода, а влагалище в 3V2 раза длиннее матки (рис. 151).
Р а с пространение
(рис. 34). В СССР довольно
частыми, изолированными оча-
очагами встречается по всей При-
Прибалтике вдали от морских бере-
берегов (в отличие от Cl. bidentata
С. Pfr.). Многочисленные на-
находки в Ленинградской области
и на юго-западе Карельской
АССР (к северу от Ладожского
озера), единичные находки в
окрестностях Пскова, Витебска,
в Калининской, Московской,
Брестской областях и на Карпа-
Карпатах (Станиславская, Закарпат-
Закарпатская и Черновицкая области).
Ареал данного вида состоит
из двух изолированных друг
от друга участков: 1) северо-
североевропейского, охватывающего
северо-запад, запад и западную
часть центра европейской части
СССР и южную Финляндию;
2) альпийско-карпатского, занимающего горные области средней Европы.
Кроме того, известна серия мелких островных ареалов в Пиренеях, на
Скандинавском полуострове и в средневысотных горах центральной Гер-
Германии (ГДР—-на северо-запад от Судет до Гарца), в южной Польше и на
острове Рюген.1
В ископаемом состоянии данный вид изредка встречается в четвертич-
четвертичных отложениях средней Европы.
Экология. Обитает во влажных хвойных и смешанных, реже в
широколиственных лесах в гнилых пнях и под корой, а также под кам-
камнями. Поднимается в горы до верхней границы лесной зоны.
3. Род IPHIGENA GRAY
Gray, 1821, London Med. Reposit, 15 : 239; Vest, 1867 : 192 (Pyrostoma); Moei-
lendorff, 1875 : 26, 27 (Pirostoma); Westerlund, 1901 : 6 (Peristoma); W a g-
n © r A., 1920 : 74 (Pirostoma s. str.); Lindholm, 1924 : 64; T h i e 1 e, 1931 : 538.
Тип рода: Cl. rolphii (Leach) Turton.
Раковина веретеновидная или веретеновидно-цилиндрическая. Киль
развит слабо и чаще всего представлен округлым вздутием, в связи
1 Шлеш (Schlesch, 1946) подвергает сомнению данные Окланда @kland, 1925)
о присутствии этого вида на юге Скандинавии и на острове Борнхольм. Между тем срав-
сравнительно недавняя находка этого моллюска на острове Рюген (Plate, 1951), по-види-
по-видимому, подкрепляет позицию Окланда.
Рис. 151. Clausilia cruciata (Stud.). Половая
система. (По Лёвушкину).
3. IPHIGENA 231
с чем базальный желобок устья у большинства видов отсутствует;
бороздка, окаймляющая киль, сохраняется у всех видов. Устье
широко-грушевидное или округло-ромбическое; высота его немногим
больше ширины; нижний край округлен, не угловатый. Верхняя
и спиральная пластинки всегда сливаются друг с другом. Нижняя
пластинка сильно выдается в просвет устья и всегда хорошо видна
при прямом положении раковины. К нижней пластинке, вблизи ее
нижнего конца, прилегают 2 расходящихся складки (промежуточная
и краевая), которые вместе с нею образуют более или менее ясно заметную
фигуру в виде буквы К, лежащей на левом боку. Полулунная складка
хорошо развита, располагается на спинной стороне раковины и чаще всего
имеет ту же форму, что и у рода Clausilia Drap. Главная складка прости-
простирается с правой стороны раковины до левой, т. е. занимает около 1/2 обо-
оборота. Палатальная мозоль развита слабее, чем у Clausilia, но той же формы.
Лопасть клаузилия к переднему концу плавно сужена.
Срединный и боковые зубы радулы с дополнительными зубцами.
Подобно Clausilia Drap., половая система данного рода характерна
рудиментарным состоянием пениса и восходящего колена семяпровода
и мощным развитием нисходящего колена семяпровода, который и в этой
группе играет основную роль в копуляции. Но в отличии от предыдущего,
у данного рода верхняя (сортировочная) часть семяяйцевода хорошо раз-
развита и крупная. Матка такой же длины, как влагалище, или значительно
длиннее последнего. В остальном половая система данного рода сходна
с таковой у Clausilia. Подобно последней, диафрагмальный ретрактор
пениса лежит медиально от половых путей и не перекрещивается с ними.
Распространение. Европа.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯМИ ДОВЩРОДА&РНЮЕЯА
--1 B). Раковина крупная: высота 15—20, ширина 4.0—5.0 мм; последний
оборот внизу с коротким, но ясным килем, а устье со слабым ба-
зальным желобком. Нижняя пластинка с 2 отходящими от нее
складками, образует отчетливую фигуру, напоминающую букву К,
лежащую на левом боку. Париетальная стенка устья, между
верхней и нижней пластинками, гладкая
1.1. ventricosa (Drap.).
2 A). Раковина меньших размеров: высота не больше 15.0, ширина
не больше 4.0 мм; последний оборот вместо киля имеет вздутие,
окаймленное бороздой; устье без базального желобка. Фигура К
менее отчетлива. Париетальная стенка с 1—4 складочками.
3 F). Имеется отчетливая мозолевидная нижняя палатальная складка,
которая обычно связывает полулунную складку с нижним концом
палатальной мозоли.-
4 E). Ребрышки располагаются сравнительно густо; на предпоследнем
обороте их чаще всего 45—55
2а. I. latestriata latestriata (A. Schm.).
5 D). Ребрышки располагаются значительно реже; на предпоследнем
обороте их 25—35 26. I. latestriata borealis (Bttg.).
6 C). Нижней палатальной складки нет.
7 (8). Кроме тонкой и густой поперечной исчерченности, имеется также
и продольная (спиральная). Нижняя пластинка внизу косо об-
обрублена. Промежуточная складка (левая ветвь фигуры К) лежит
почти параллельно нижней пластинке
3.1. mucida badia (Rssm.).
232
V. CLAUSILIINAE
8 G) Раковина ребристо исчерченная или ребристая; продольная ис-
черченность слабо выражена или отсутствует. Нижняя пластинка
внизу не обрублена и плавно сливается со столбиком. Промежу-
Промежуточная складка располагается к ней под ясным углом.
9A0). Раковина веретеновидная, вздутая, густо ребристая (на пред-
предпоследнем обороте 50—55 ребрышек). Устье округло-ромби-
округло-ромбическое. На париетальной стенке чаще всего 1, реже 2 складочки.
Ширина раковины 3.7—4.0 мм ...... 4. I. tumida (Rssm.).
10 (9). Раковина веретеновидная или веретеновидно-цилиндрическая,
стройная, редко ребристо исчерченная (на предпоследнем обороте-
45—50 складок). Устье широко-грушевидное. На париетальной
стенке 2—3 складочки. Ширина раковины 2.8—3.0 мм .....
5. I. plicatula (Drap.).
1. Iphigena ventricosa (Drap.) (рис. 27, 152).
Draparnaud, 1801 : 62 (Pupa); Draparnaud, 1805 : 71, t. 4, fig. 14
(Clausilia); К u s t e r, 1847—1862 : 245, t. 28, fig. 1—5 (Clausilia); Лихарев
и Раммельмейер, 1952:229, рис. 147, 148.
Раков гн а веретеновидная, вздутая, твердостенная, ребристо ис-
исчерченная или ребристая (на нижнем обороте нередко в промежутках
между ребрышками продольно исчерчена), слабо просвечивающая, с шел-
шелковистым блеском, красновато-коричневая, с ред-
редкими белыми штрихами у шва. Оборотов И—12,
слабо выпуклых; эмбриональные обороты C—3V2)
гладкие; первые 5 оборотов почти одинаковой вы-
высоты; последний оборот наверху чуть уже предпо-
предпоследнего, книзу плавно су-
сужается, с коротким, но ясным
килем. Устье округло-ромби-
округло-ромбическое, с почти параллель-
Рис. 152. Iphigena ventricosa (Drap.).
А — общий вид раковины; Б — нижний оборот и устье; В — клаузилий. (А, Б — по
Шмидту с дополнениями, В — ориг.)-
ными палатальными и колумеллярным краями, со слабым базальным желоб-
желобком, который сглаживается к краю устья. Края устья широко отвернуты и
утолщены, белые. Сравнительно невысокая, но толстая изогнутая верхняя
пластинка впереди доходит до края устья, а сзади, плавно снижаясь, слива-
сливается со спиральной. Крупная ланцетовидная нижняя пластинка лежит до-
довольно глубоко и, плавно изгибаясь, уходит кверху; от ее нижнего конца от-
отходят кверху 2 коротких складки,|которые образуют с пластинкой четко
3. IPHIGENA
233
выраженную фигуру, напоминающую букву К, лежащую на левом боку. Па-
Париетальная стенка между верхней и нижней пластинками без складок. Ниж-
Нижний конец субколумедлярной пластинки виден хорошо лишь при косом
положении устья. Полулунная складка лежит на спинной стороне рако-
раковины; оба конца ее отогнуты назад — верхний круто, нижний плавно,
дугообразно. Коричневатая палатальная мозоль сравнительно тонкая;
наибольшей толщины она достигает у своего верхнего конца. Лопасть
60
50
40
30
40 30
/ \ /
/ \ /
А/7
/\ / *
/ /^
/ / 'г/с
/ I ir
^\7 *-ч jJ
/ Ч/^
^-^ /
0
20 ID
7 /
/ /
/ S
$ 1 is.
>/ с
/ A
oh
л
"—If
r
10
0 10 го зо
/ /
/ /
' /
А г
$%¦ /
¦> /u\ /
IV4
~\
\
r >H
\ 1*л
( -
40
\
ж
1
^.!
-4^
-1—.
50 60
к \ 75
:эсГ"/ _У_^-—
ч ^
/
•_5—-3t—^
70
Цх
^V
\
к;
200
J
\
\
ч
;N
--^
0
40
во
, ЧУ
&"'
\
\\^
^\
3 \
v \
/-*
ъ
+и ©г»
200 «00 600
\
\
Y)
\ 1
1
1 1
У
\
{
А
800 ки
60
50
40
30
Рис. 153. Географическое распространение Iphigena ventricosa (Drap.) и /. latestrtata
(A. Schm.). (Ориг.).
la — Iphigena ventricosa, основной ареал; 16 — то же, изолированные ареалы; 1в — то же, находки
в четвертичных отложениях вне современного арела; 2а — /. latestriaia latestriata; 26 — /. latestriata
borealis.
клаузилия языковидная, сравнительно короткая, к нижнему концу слегка
сужена.
Размеры: высота раковины 15—20 (чаще всего 16—18), ширина 4.0—
4.5 мм (реже 5.0 мм); высота устья 3.7—3.9, ширина 2.7—2.8 мм.
Изменчивость. В неблагоприятных условиях, в особенности
в горах, встречаются более мелкие формы, с более вздутой раковиной
(f. tumida A. Schmidt — высота раковины 14—16, ширина 3.9—4.2 мм).
Внутреннее строение. Отросток семяприемника очень
короткий (рис. 27); верхний конец его чаще всего лежит на одном уровне
е началом семяпровода. Матка в 1х/2—2 раза длиннее влагалища, а пенис
в 10 раз короче семяпровода.
234
V. CLAUSILIINAE
Распространение (рис. 153). В СССР: вся Прибалтика, от-
отдельные находки у Нарвы, Пскова, Смоленска, Рославля, Костромы,
в Московской, Ярославской областях, в белорусской части Полесья и в
Закарпатской области. Вне СССР: вся система Альп и прилегающих к ним
с севера средневысотных гор, северо-западные Балканы (без приморских
областей), западные Карпаты, равнинные области между Эльбой и Вислой,
Дания (без северной части Ютландского полуострова) и южная оконеч-
оконечность Швеции. Небольшие колонии обнаружены на юге Норвегии.
Известны многочисленные находки в четвертичных отложениях ряда
стран Европы, в том числе и из таких мест, где теперь данный вид не оби-
обитает. В этом отношении наиболее показательна находка в плейстоцене
Англии.
Экология. Обитает в тенистых и влажных смешанных и широ-
жолиственных лесах в подстилке, под корой гниющих валежин и пней,
под камнями и реже на скалах. В западной части аре-
ареала преимущественно обитает в холмистой и в горной
местности, доходя до верхней границы леса; в восточной
части обычна и в равнинных лесах.
Размножение. Живородящий вид. Только что
родившиеся животные имеют раковину из 3—31/2 оборо-
оборотов, высотой в 2 и шириной в 1.1 мм.
При воспитывании в садках, в комнат-
комнатных условиях, через 6—7 месяцев
животные становятся взрослыми.
2а. Iphigena latestriata latestriata
(A. Schm.) (рис. 154, 155).
Schmidt A., 1857 : 27, fig. 52—56,
183, 184 (Clausilia); К u s t e г, 1847—1862 :
251, t. 28, fig. 26-29 (Clausilia); Ш и х a-
рев и Раммельмейер, 1952 : 230,
рис. 149, 150.
Р a kj^bji н а веретеновидная,
стройнаяТ^ твердостённая, ребристая
(напредпоследнем обороте 45—55
ребрышек); слабо просвечивающая,
матовая, красновато-коричневая, с
пучками белых штрихов вдоль шва.
Оборотот_10;^11, верхние умеренно,
нижние слабоL вьшУЗУ11116' эмбрио-
эмбриональные обороты (около 21/2) гладкие; последний оборот внизу несколько
вздут с заметной базальной Тэороздой. Устье коротко-овальное, снизу
широко закругленное, часто почти четырехугольное, с йруглым широким
синулусомТ Края устья р_асширены, беловато-коричневые, отвернутые.
Низкая верхняя пластинка впереди достигает края устья, сзади неза-
незаметно переходит в спиральную пластинку. Крупная нижняя пластинка
лежит довольно глубоко, но хорошо видна при прямом Положений устья;
впереди она заканчивается 2 складочками, которые образуют лежащую
ла боку букву К, однако не~столь правильную, как у предыдущего вида.
На париетальной стенке устья между пластинками располагаются 1—3
мелких складочки. Нижний конеп^ субколумеллярнои^гаастинки виден
лишь при косом положении устья. Палатальная мозоль имеет вид валика,
лежащего параллельно краю устья с двумя вершинами — верхней и ниж-
Рис. 154. Iphigena latestriata latestriata
(A. Schm.). (Ориг.).
_А — общий вид раковины; В — нижний обо-
оборот и устье.
3. IPHIGENA 235
ней. От последней тянется вглубь раковины, до слияния с полулунной
складкой, мозолевидная нижняя палатальная складка. Рукоятка клау-
зилия плавно расширяется в широкую языковидную лопасть, которая
к нижнему концу слегка сужается.
Размеры: высота раковины 13.0—15.0, ширина 3.0—3.5 мм; высота
устья 2.6—3.0, ширина устья 2.2—2.4 мм.
Внутреннее строение (Лёвушкин, 1958). Длинный семя-
семяприемник (рис. 155) снабжен сравнительно коротким отростком, который
Рис. 155. Iphigena latestriata latestriata (A. Schm.). По-
Половая система. (По Лёвушкину).
тем не менее относительно длинее, чем у /. ventricosa (Drap.). Влагалище
в 3 раза короче матки, а пенис в 8 раз короче семяпровода. Нисходящее
колено последнего очень толстое.
Распространение (рис. 153). В СССР: Закарпатье, Карпаты,
Предкарпатье и Волыно-Подольская возвышенность. Вне СССР — вся
Карпатская дуга с Трансильванией и Банатом, а также отдельные находки
в средневысотных горах южной Польши (Краковская Юра, Пенины и др.).
Экология. Обитатель горных лесов, где встречается в подстилке,
под корой гниющих пней и валежин. Поднимается до верхней границы
леса (в Восточных Карпатах найден до высоты 1250 м над ур. м.).
26. Iphigena latestriata borealis (Bttg.).
Boettger О., 1878, Nachrbl. deutsch. Malak. Ges., 10:136 (Clausilia); Rrausp,
1936 !-37—42, t. 2, карта; К г a u s p, 1940 : 233—234.
Данный подвид отличается от предыдущего следующими особенностями:
раковина более короткая и вздутая; ребрышки расположены более редко
(на предпоследнем обороте число их колеблется в пределах 25—35, реже
до 38). Оборотов 10—11V2.
236 V. CLAUSILIINAE
Размеры: высота раковины 12—14, ширина 3.0—3.3 мм; высота устья
2.6—2.9, ширина 2.1—2.4 мм.
Внутреннее строение не изучено.
Распространение (рис. 153). В СССР этот подвид широко-
широкораспространен в Прибалтике. Одна находка сделана в окрестностях
С.-Петербурга (Симашко, 1847, под названием «Clausilia densestriataZgl.y>).
Кроме того, найден в белорусской части Полесья (Adamowicz, 1937) и r
окрестностях Витебска (коллекция Зоологического института АН СССР).
Вне СССР разрозненные колонии обитают в равнинных лесах северной
Польши (наиболее западная находка сделана в окрестностях г. Гожув-
Велькопольски).
Экология. Обитает во влажных равнинных смешанных и широко-
широколиственных лесах, в подстилке и под валежинами, реже на скалах.
*3. Iphigena mucida badia (Rssm.).
Rossmassler, 1836, Ieonogr., 1, 4 : 20, fig. 271 (Clausilia); К ii s t e r, 1847—
1862 : 146, t. 16, fig. 9—12 (Clausilia); Lozek, 1956 : 1323 t. 13, fig. 3.
Раковина веретеновидная, твердостенная, умеренно просвечи-
просвечивающая, очень блестящая, тонко и густо, местами неправильно исчер-
исчерчена, с заметными продольными линиями, красновато-коричневая, с ред-
редкими одиночными белыми штрихами вдоль шва. Оборотов около 10, слабо-
слабовыпуклых. Последний оборот внизу заметно вздут и имеет базальную бо-
борозду. Устье округло-яйцевидное (без синулуса). Края устья умеренно
расширены и отвернуты, белые. Верхняя пластинка низкая, впереди до-
доходит до края устья, сзади незаметно переходит в спиральную пластинку.
Нижняя пластинка располагается косо, не закручена, внизу косо обруб-
обрублена. Фигура К выражена неясно; правая ветвь ее тянется к краю устья
в виде слабой короткой складочки; левая ветвь (соответствует промежуточ-
промежуточной складке) выражена сильнее, более длинная и тянется внутрь раковины
почти параллельно нижней пластинке. Субколумеллярная пластинка
видна лишь при косом положении устья. На париетальной стенке между
пластинками находятся 1—2 мелких складочки. Палатальная мозоль
слабо развита, цельная.
Размеры: высота раковины 12.0—14.0, ширина 3.0—3.3 мм.
Внутреннее строение не изучено.
Распространение. Восточные Альпы; отдельные реликтовые
ареалы в Судетах и в Шумаве (Чехословакия).
Экология. Обитает во влажных горных лесах, на стволах де-
деревьев.
4. Iphigena tumida (Rssm.) (рис. 156, 157).
Rossmassler, 1836, Ieonogr.; 1, 4:21, fig. 277 (Clausilia ventricosa var.
tumida); SchmidtA., 1857 : 14, fig. 10—14, 162, 163 (Clausilia); Kuster, 1847—
1862 : 247, t. 28, fig. 6—9 (Clausilia); Лихарев и Раммельмейер, 1952 :
230, рис. 151.
Раков и на веретеновидная, вздутая, очень твердостенная, jrycTj>
и правильно ребристая (на предпоследнем обороте 50—55, реже до 60 реб-
ребрышек), слабо просвечивающая, с шелковистым блеском, темно-роговая
или красновато-коричневая, с редкими белыми штрихами. Завиток быстро-
сужающийся, с вогнутыми очертаниями ниже цилиндрической вершины.
Оборотов 10—12, слабо выпуклых; эмбриональные обороты (около 2х/2)
гаадкие; последний оборот внизу вздут, с ясной базальной бороздой.
Устье округло-ромбическое; синулус широкий, округлый и врезан.
3. IPHIGENA
237
Рис. 156. Iphigena tumida (Rssm.). (Ориг.).
A — общий вид раковины; Б —¦ клаузилий.
Края устья расширены, отвернуты, белые; верхний край ниже синулуса,
заметно прогнут.ГВёрхняя пластинка" низкая, но толстая; спереди доходит
jjo края устья, сзади плавно пе-
реходит в;"спиральную пластин-
пластинку. Нижняя" пластинка лежит
довольно глубоко; с прилегаю-
прилегающими к ЙШ" "складками обра-
образует неясную букву К, лежащую
на левом боку. Париетальная
¦стенка между плас_й8Шйми~ймёет
lj реже 2 мелких складочкиТПа-
латальная мозоль^развита слабо.
Лопасть кла^зилия 1ши Грдк?
щки"Т(меёт неглубокую" вы-
^зку, а к нижнему концу су-
сужается.
"*" Размеры: высота раковины
12.5—15.0, ширина 3.7—4.0 мм;
высота устья 3.0, ширина 2.3—
2.5 мм.
Внутреннее стро-
строение (Лёвушкин, 1958). По-
Половая система характерна отно-
относительно длинным семяприем-
семяприемником и более коротким отрост-
отростком (рис. 157). Матка вдвое
длиннее очень толстого влагалища. Петля пениса сильно редуцирована,
нередко совершенно исчезает, и половая система приобретает облик та-
таковой у Laciniaria.
Р а спростране-
н и е (рис. 158). В СССР:
Станиславская, Львов-
Львовская, Тернопольская, За-
Закарпатская и Черновицкая
области. Вне СССР: Кар-
Карпаты, средневысотные го-
горы южной Польши (Су-
(Судеты, Люблинская воз-
возвышенность), Татры и от-
отдельные реликтовые ареа-
ареалы в Чешском массиве и в
Шумаве (Чехословакия).
Найдена в четвертич-
четвертичных отложениях, в том чи-
числе и в Германии, (Geyer,
1927), т. е. вне границ сов-
современного ареала.
Экология. Живет
во влажных горных ле-
лесах в подстилке, под сы-
сырой корой гнилых пней и
валежин. Поднимается до
бж
рп
Рис. 157. Iphigena tumida (Rssm.). Половая система.
(По Лёвушкину).
верхней границы леса (в восточных Карпатах была найдена до высот
1500 м над ур. м.).
238
V. CLAUSILIINAE
40,, ¦ зо го ;о о to га зо 40 so
~7В 80
О/о •» +11 €&га A2S
200 0* 200 «О 600 800X11
40
30
40
Рис. 158. Географическое распространение Iphigena plicatula (Drap.) и/, tumida (Rssm.).
(Ориг.).
la — Iphigena plicatula; 16 — то же, изолированные ареалы; 1в — то же, находки в четвертичных
отложениях вне современного ареала; 2а — I. tumida, основной ареал; 2б — то же, изолированные
ареалы.
5. Iphigena plicatula (Drap.) (рис. 8, А, 159, 160).
Draparnaud, 1801 : 64 (Pupa); Draparnaud, 1805 : 72, t. 4, fig. 17,
18 (Clausilia); Kustei, 1847—1862 : 253, t. 28, fig. 34—42 (Clausilia); Schmidt
A., 1857 : 25, fig. 43—51, 176—182 (Clausilia); Лихарев и Раммельмейер,
1952 : 231, рис. 152.
Раковина веретеновидная или веретеновидно-цилиндрическая,
очень стройная, тонко и редко ребристо исчерченная (на предпоследнем
обороте 45—50 складок), слабо просвечивающая, от светло- или темно-
роговой до красновато-коричневой, с редкими белыми штрихами. Обо-
Оборотов 10—12, слабо выпуклых, плавно нарастающих в высоту и в ширину
до наиболее широкого предпоследнего оборота; эмбриональные обороты
BV4) гладкие; последний оборот внизу вздут, со слабой базальной бороз-
бороздой. Устье широко-грушевидное, внизу полукруглое. Края его расширены,
отвернуты, белые; верхний край над синулусом ровный (без прогиба).
Верхняя пластинка низкая, но толстая; впереди она достигает края устья,
сзади, сильно снижаясь, сливается со спиральной пластинкой. Крупная
нижняя пластинка сильно выступает в просвет устья и, изгибаясь широкой
3. 1PH1GENA
23?
дугой, косо уходит кверху; впереди она заканчивается на краю устья 1—2
мелкими складками. Еще 2—3 таких складки лежат на париетальной
стенке между верхней и нижней пластинками. Нижний конец субколу-
меллярной пластинки виден при косом положении устья. Полулунная
складка слабая, лежит на спинной стороне. Главная складка занимает
около V3 оборота и лишь незначительно заходит за полулунную складку.
Палатальная мозоль сравнительно слабая; лишь верхняя часть ее более
сильно развита. Лопасть клаузилия близ рукоятки имеет
слабый вырез; передний конец лопасти широко закруг-
закруглен и утолщен.
Размеры: высота раковины 10.0—13.0, ширина 2.8—
3.0 мм; высота устья 2.4—2.7, ширина 2.0—2.2 мм.
Изменчивость. Сильнее всего варьируют вы-
высота раковины и поверхностная скульптура, что в свое
время послужило причиной описания большого числа
разновидностей, природа которых еще слабо изучена.
Известно лишь, что во влажных горных лесах популяции
отличаются преобладанием особей со сравнительно ко-
короткими и вздутыми раковинами — высота 9.0—11.0,
ширина 3.0 мм (var. curta A. Schmidt). В схожих усло-
условиях обитают в Судетах популяции с мелкими, но строй-
стройными раковинами — ширина 2.5 мм, которые к тому же
отличаются нежной исчерченностью и блеском (var. папа
Scholtz). Иногда среди нормальной популяции попада-
попадаются экземпляры, у которых на париетальной стенке
устья сохраняются только те краевые складки, которыми
заканчивается нижняя пластинка.
Внутреннее строение (Steenberg, 1914;
Лёвушкин, 1958), Отросток длинный, лишь немного ко-
короче семяприемника (рис. 160). Матка более чем в 2
раза длиннее короткого, но толстого влагалища. Пенис
в 51/2 раз короче семяпровода. Обе части петли пениса
плотно прижаты друг к другу, так что между ними не
остается просвета.
Распространение (рис. 158). В СССР: вся
Прибалтика и Ленинградская область (на востоке одна
находка в долине р. Свирь). Многочисленные находки в Московской
области, единичные на юго-западе Карельской АССР (к северу от Ладож-
Ладожского озера), в Псковской (Псков, Новоржев), Новгородской (Парфино),
Калининской (Нелидово, Соблаго), Ярославской (Тутаев, Ярославль)Y
Калужской, Тульской (Алексин), Орловской (Троено), Смоленской
(Смоленск, Рославль), Витебской областях. Обособленный ареал на-
находится в буковых лесах Чатыр-Дага (горный Крым). Вне СССР: вся
система Альп и прилегающих к ним с севера средне-высотных гор, север-
северная половина Италии, северо-западные области Югославии, Западны»
и западная часть Восточных Карпат, равнинные области между Эльбой
и Вислой, Восточная часть Ютландского полуострова, южная часть (до
64° с. ш.) Швеции и юго-восточные области Норвегии, южная Финляндия
(до 62° с. ш.). Отдельные колонии обитают в горах южной Венгрии, в.
Трансильванских Альпах и в средней Норвегии.
В пределах современного ареала найдена в четвертичных отложениях
ряда стран, в том числе и в рисс-вюрмских отложениях долины Западной
„Двины близ г. Суража (Даниловский, 1955). За пределами современного-
ареала найдена в голоценовых отложениях Нидерландов.
Рис. 159. Iphi-
gena plicatula
(Drap.)- (По
Эрманну
240
V. CLAVSILIINAE
Экология. Обитает в равнинных и горных лесах под влажной
корой гниющих стволов и пней, в подстилке, под камнями, на сырых
скалах и в развалинах. Предпочитает известняковые породы. В Западных
Карпатах поднимается до высоты
1600 м над ур. м.
Размножение. Откладка
яиц происходит в августе—сентябре;
вылупление молоди наступает на
18—20-й день. Рост заканчивается
к исходу второго года. Размер яиц
-% 1.0-1.5 мм.
4. Род LACINIARIA HARTM.
Hartmann, 1842 A840—1844), Erd.
u. Siisswasser-Gastropoden Schweiz, 1 : 173;
Adams H. a. A., 1855 A858), : 182
(A linda); Steenierg, 1914 : 43 (Alinda,
анатомия); Wagner A., 1920 : 76 (Alin-
(Alinda, part., анатомия); Lindholm, 1924:
64 (подрод Laciniaria s. str., part.).
Тип рода: Clausilia plicata Drap.
Последний оборот раковины с
крупным и длинным килем. Устье
грушевидное, с высоким врезанным
синулусом; благодаря глубокому ба-
зальному желобку нижний край
устья слегка угловатый. Верхняя
пластинка не сливается со спираль-
спиральной и их концы располагаются на
разном расстоянии от столбика, не-
нередко заходят друг за друга. Ниж-
Нижняя пластинка лежит довольно глу-
глубоко, но всегда видна при прямом
положении устья; внизу она дает
широкий спиральный изгиб и с краем
устья связана 1—2 короткими скла-
складочками. Субколумеллярная пластин-
пластинка лежит глубоко и не всегда видна
через устье. Полулунная складка у
одних видов имеется, у других отсут-
отсутствует. Кроме главной складки, при-
присутствуют 1—3 палатальных склад-
складки. Рукоятка клаузилия плавно расширяется в лопасть, которая у пе-
переднего конца либо перекручена, либо утолщёна или вырезана.
Срединный и боковые зубы радулы с дополнительными зубцами.
Тонкий, нередко сильно извитой отросток впадает в семяприемник
очень низко, вблизи дистального конца последнего. Матка в 2 раза_длин-
нее влагалища. Последнее вздутое, с сильно мускулистыми стенками,
внутри с многочисленными складками. Мужские половые пути вытянуты
вдоль женских и представляют собой толстую цилиндрическую мускули-
мускулистую трубку, веретеновидно вздутую у дистального конца, т. е. перед
впадением в половую клоаку. Эта трубка является производным ^исходя-
^исходящего колена семяпровода, а ее расширенный участок — вторичным пенисом.
нп
уркв
Рис. 160. Iphigena plicatula (Drap.).
Половая система. (По Стинбергу).
4. LACINIARIA 241-
Левый ретрактор дает одно ответвление к шейке семяприемника — рет-
рактор семяприемника, и 1—2 ветви к поповой клоаке. Ретрактор семя-
семяприемника в свою очередь посылает короткую ветвь к семяпроводу, тем
самым подтягивая его к матке и влагалищу. Той же цели служат многочис-
многочисленные мускульные волокна, связывающие женские и мужские половые
пути. Ретрактор пениса отсутствует. Пенис связан с нижним концом семя-
семяпровода тонкой мускульной полоской. Ретрактор глазного щупальца
лежит медиально от половых путей и с ними не перекрещивается.
Распр остранение. Европа и Крым.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА LACINIARIA
1 (8). Слева от базального желобка вглубь устья уходит крупная
нижняя палатальная складка.
2 E). От полулунной складки сохранился лишь нижний конец, слитый
воедино с нижней палатальной складкой. Лопасть клаузилия
имеет глубокую вырезку у нижнего конца, которая обычно видна
через устье. (Подрод Mentissa H. et A. Adams).
3 D). Раковина густо исчерченная или ребристо исчерченная . ...
5. L. canalifera (Rssm.).
4 C). Раковина редко ребристая, реже ребристо исчерченная . . . .
6. L. gracilicosta (Rssm.),
5 B). Полулунная складка хорошо развита, цельная. Лопасть клау-
клаузилия без вырезки, но на внутренней стороне имеет короткий
киль. (Подрод Strigilecula К. et W.).
6 G). Раковина веретеновидная, слабо блестящая, густо ребристая.
Губа коричневато-белая. Нижняя палатальная складка мозоле-
видная, коричневая - 3.--L. сапа (Held).
7 F). Раковина цилиндрическо-веретеновидная, очень блестящая, реб-
ребристо исчерченная. Губа белая. Нижняя палатальная складка
развита сильнее, белая . 4. L. nitidosa (Ulicny).
% A). Нижняя палатальная складка отсутствует. (Подрод Laciniaria
s. str.) .
¦9 A0). Средняя палатальная складка лежит под заметным углом к глав-
главной складке. Полулунная складка дугообразно изогнута, оба
конца ее отогнуты вглубь раковины. Палатальный край устья
без краевых складок 1. L. biplicata (Mont.).
40 (9). Средняя палатальная складка лежит под слабым углом к главной
складке, так что порой они кажутся почти параллельными. Полу-
Полулунная складка прямая, нижний конец ее слегка отогнут в сто-
сторону устья. Палатальный край обычно с 6—10 короткими склад-
складками или узелками1 2. L. plicata (Drap.).
1. Подрод LACINIARIA s. str.
Раковина с тонкой цилиндрической верхушкой, мелко и густо ребри-
ребристая, со слабым блеском, с пучками белых штрихов у шва. Субколумел-
лярная пластинка не видна через устье. Полулунная складка лежит на
правой стороне раковины. Кроме главной, имеется еще средняя палаталь-
палатальная складка, которая располагается к первой под некоторым углом.
Лопасть клаузилия более или менее сильно перекручена.
Распространение. Европа.
Имеется разновидность, лишенная краевых складок и узелков.
242
V. CLAUSILIINAE
1. Laciniaria (Laciniaria) biplicata (Mont.) (рис. 6, A, 26, 161).
Montagu, 1803, Test. Brit; : 361, t. 11, fig. 5 (Turbo); Rossmassler,
1835, Iconogr., I, 1 : 77, fig. 30. (Clausilia similis); К ii s t e r, 1847—1862 : 191, t. 21,
fig. 1—4 (Clausilia); Лихарев и Раммельмейер, 1952 : 233, рис. 124, 154.
Раковина веретеновидная или веретеновидно-цилиндрическая,
стройная, тонко и густо ребристая, просвечивающая, с шелковистым
блеском, светло^роговая, с белыми штрихами. Оборотов 12—^13, слабо
выпуклых; эмбриональные обороты (около 21/2) гладкие;
последние 2—3 оборота почти одинаковой ширины. Края
устья расширены, утолщены, отвернуты, беловатые. Верх-
Верхняя пластинка тонкая, впереди доходит до края устья,
вглубь раковины уходит приблизительно на V4 оборота, где
плавно сходит на нет. Нижняя пластинка сильно выдается
в просвет устья и близко подходит к верхней пластинке,
после чего широкой спиралью уходит кверху; нижний ко-
конец ее лежит вблизи края устья; верхний конец располагается
на, брюшной стороне на одном уровне с началом спиральной
пластинки. Субколумеллярная пластинка заканчивается вы-
высоко. Париетальная стенка устья между верхней и ниж-
нижней пластинками гладкая либо
с 1—2 складочками. Полулунная
складка хорошо развита, дугооб-
дугообразно изогнута и лежит на правой
стороне; нижний конец ее слегка
отогнут внутрь раковины. Глав-
Главная складка начинается несколько
глубже полулунной складки и про-
простирается вперед на 1/i оборота.
Имеется крупная средняя пала-
палатальная складка, которая начи-
начинается впереди полулунной (на
уровне ее верхнего конца) и за-
заканчивается ближе к краю устья,
чем главная складка; к последней
она располагается под заметным углом. Рукоятка клаузилия, плавно
расширяясь, переходит в широкую, сильно перекрученную лопасть.
Размеры: высота раковины 15—18, ширина 3.5—4.0 мм; высота устья
3.5—3.8, Ширина 2.6—2.8 мм.
Изменчивость. Раковина этого моллюска сильно варьирует,
особенно в отношении размеров, формы и ребристости; замыкательный
аппарат подвержен меньшим колебаниям.
Внутреннее строение (Steenberg, 1914). Пенис имеет
внутри многочисленные складки, среди которых 3 очень крупные. Семя-
яйцевод несколько короче матки (рис. 26). Матка и нижняя часть семя-
яйцевода превращены в крупную выводковую сумку с очень тонкими
стенками.
Распространение (рис. 162). Ареал данного вида состоит
из двух частей — среднеевропейской и северобалканской, связанных
друг с другом юго-восточными отрогами Альп и областью Хорватского
карста. В СССР достоверная находка сделана лишь в Закарпатской об-
области (Imre, 1941). Вне СССР: южная Англия, северо-восточная Франция,
Бельгия, Нидерланды, северные и восточные Альпы, средневысотные горы
Германии (Ehrmanfr, 1933) и западной Чехословакии, западная и северная
Рис. 161. Laoiniaria biplicata (Mont.).
А — общий вид раковины (по Эрманну); Б —
клаузилий. (Ориг.).
4. LAC INI ARIA.
243'
Пойьша (в том числе и западные Карпатш), обособленные ареалы в Дания,
южной Швеции ж две находквг в южной Норвегии (возможно, завезена).
На юге почти вся Югославия, северная Албания, значительная часть
Болгарии (без Добруджи), юго-западнал Румыния (Банат и отдельные
горные массивы в Трансильвании), отдельные находки в Венгрии»
Рис. 162. Географическое распространение Laciniaria (Laciniaria) biplicata
(Mont.) и L. (L.) plicata (Drap.). (Ориг.).
la — L. biplicata, основной ареал; 16 — то же, изолированные ареалы; 2а — L. plicata,
основной ареал; гб — то же, изолированные ареалы.
Многие наши исследователи прошлого века указывали на присутствие
этого моллюска на раваине европейской части СССР (Симашко, 1847;
Schrenk, 1848; Сабанеев, 1880, и др.). Однако к настоящему времени можно
считать установленным, что эти сведения основаны на неверных определе-
определениях. Указанные авторы принимали за данный вид то Laciniaria plicata
var. implicata (Bielz), то L. сапа (Held). Выяснено, что восточная граница
нашего моллюска проходит по долине Вислы (Schlesch, 1942; Urbanski,
1957).
Найдена в четвертичных отложениях ряда стран средней Европы
и в Англии.
Экология. Обитает во влажных смешанных и широколиственных
лесах, а также в сырых парках и садах на стволах деревьев, в подетюше,
в гнилых пнях и под корой, под камнями, на скалах и в развалинах.
В средней Европе встречается на равнинах и в нижних горизонтах горного^
244 V. CLAUSILIINAE
леса (до высоты 900 м над ур. м.). Редка на сухих скалах и склонах, где
обычно представлена мелкой экологической формой.
Размножение. Живородящее животное. Молодь появляется
на свет в августе—сентябре. Раковина только что родившихся экземпля-
экземпляров слагается из 21/2 оборотов. Высота ее 1.3—1.5, ширина 0.9 мм. Первые
2—3 дня молодь остается на поверхности почвы, после чего поднимается
на ближайшие предметы. Рост длится около 8—10 месяцев. Длительность
жизни до 31/2 лет.
2. Laciniaria (Laciniaria) plicata (Drap.) (рис. 25, 163, 164).
Draparnaud, 1801 : 63 {Pupa); Draparnaud, 1805 : 72, t. 4, fig. 15—16
(Clausilia); Rossmassler, 1835, Iconogr., I, 1 : 78, fig. 31 (Clausilia); К u s t e r^
1847—1862 : 194, t. 21, fig. 13—19 (Clausilia); Лихарев и Раммельмейер,
1952 : 232, рис. 153.
Раков и н а ве^ететовидно-цилиндЕическаЯд стройная, тонко й
густ?^Вристая, просвечивающая, со слабым блеском, красновато-роговая,
с~пучками белых штрихов. Оборотов 12—13, слабо выпуклых; 3 нижних
оборота одинаковой ширины; последний на затылке несколько вогнут.
Кдая^дстья распшрены, утолщены, отвернуты, беловатые. Пластинки по
^орме и положению сходны с таковыми у предыдущего вида, только ниж-
няя__нластинка менее выдается в просвет устья. Массивная полулунная
складка почти прямая и лежит на правой стороне, параллельно оси ра-
раковины; веюхний конец, ее Ълабо отогнут назад, а нижний слегка вперед —
к устью. Главная и средняя палатальные_с_кладки хорошо развиты и рас-
хЪдятся друг от друга под небольшим углом, так что порой даже кажутся
почти параллельными. Париетальный и в особенности палатальный края
устья несут 6—10 мелких краевых складочек или узелков. Рукоятка кла-
узилия, плавно расширяясь, переходит в лопасть, которая на конце
несколько перекручена.
Размеры: высота раковины 14—17, ширина 3—4 мм; высота устья
3.6—3.8, ширина 2.6—2.8 мм.
Изменчивость. Раковина данного вида меньше варьирует,
чем у L. biplicata. Тем не менее некоторые колебания в величине и форме
раковины довольно обычны. Нередко встречаются экземпляры с неболь-
небольшим числом краевых складок, причем последние слабо развиты (var.
implicata Bielz).
Внутреннее строение (Steenberg, 1914; Лёвушкин, 1958).
Пенис внутри имеет только одну крупную кольцевую складку. Семяяй-
цевод значительно длиннее матки (рис. 164). В остальном сходен с преды-
предыдущим видом.
Распространение (рис. 162). В СССР: вся Прибалтика, за
исключением эстонских островов и Ленинградской области; Карпаты,
Предкарпатье и Волыно-Подольская возвышенность. Отдельные находки
в Брестской (Беловежская пуща, Полесье), Витебской (Витебск), Смо-
Смоленской (Смоленск, Рославль), Брянской, Калужской, Тульской (Тула,
Алексин), Орловской (Троено), КиевскойуКировоградской (Александрия),
Харьковской,уЧеркасской (Корсунь-Шевченковский)У Винницкой (Оль-
гополь) областях. Наиболее северные находки сделаны у Нарвы, Пскова,
Парфино (Новгородская область), в Ярославской (Ярославль, Тутаев)
Костромской (Кострома), Горьковекой (Васильсурск) областях. Много-
Многочисленные находки в Московской области. Вне СССР: Польша, Карпаты
и^ббльшая часть северных Балкан (Болгария, Сербия, Македония, Бос-
Босния); на западе крупный ареал занимает большую часть Швейцарии,
4. LACINIARIA
245
часть северной Италии (Тессинские и Бергамские Альпы) и горы и рав-
равнины верхнего и среднего Рейна (Арденны, Вогезы, Рейнские сланцевые
горы, Шварцвальд). Многочисленные островные ареалы рассеяны в Дании,
в южной Швеции, в Северо-Германской низменности и в средневысотных
осп
Рис. 163. Laciniaria (Laci-
niaria) plicata (Drap.).
A — общий вид раковины; В —
клаувилий. (А —. по Лихареву
и Раммельмейер, Б — ориг.).
Рис. 164. Laciniaria (Laciniaria)
plicata (Drap.). Половая система.
(По Стинбергу).
горах Германии (Ehrmann, 1933) и Чехословакии (Чехия). Одна находка
сделана в Тоскане (Италия).
В отличие от предыдущего вида, распространение данного моллюска
имеет более прерывистый характер.
Ископаемые остатки этого вида найдены в четвертичных отложениях
Германии (Geyer, 1927) и Чехословакии, в том числе и вне современного
ареала (Lozek, 1955).
Экология. Обитает чаще всего крупными популяциями в сме-
смешанных и широколиственных лесах, в парках, на кладбищах и в заросших
развалинах в подстилке, под камнями, в гнилых пнях и в стволах деревьев.
246
V. CLAUAIUJNAE
Предпочитает $одмиетьш ландшафт или нижние горизонты гор, что осо-
особенно заметно в западной части ареала.
Размножение. Яйцекладущее животное.
2. Подрод STRIGILECULA К. et W.
V е s t, 1867 : 195 (Clausilia, секция Strigillaria, поп Rafinesque, 1815);
Kennard a. Woodward, 1923 : 304 (Laciniaria s. str., секция Strigilecula nom.
nov.); Lindholm, 1924 : 65 (Laciniaria s. str., секция Strigilecula).
Тип подрода: Clausilia vetusta Rssm.
Поверхность раковины покрыта пучками белых штрихов. Верхняя и
спиральная пластинки слегка заходят друг за друга. Нижний конеп
субколумеллярной пластинки виден при косом положении устья. Полу-
Полулунная складка лежит на спинной стороне или слегка сдви
нута правее, только верхний конец ее отогнут назад. Главная
скадка довольно длинная. От нижнего конца полулунной
складки отходит вперед сравнительно длинная нижняя
палатальная складка. У некоторых видов сверх того имеет-
имеется еще средняя складка. Палатальная мозоль более или менее
сильно развита. Передний конец лопасти клаузилия за-
заострен и на внутренней стороне утолщен в короткий киль.
Р аспростране-
н и е. Европа.
3. Laciniaria (Strigile-
(Strigilecula) сапа (Held) (рис.
165, 166).
Held, 1836, Isis, 29 :
275 (Clausilia); K ii s t e г,
1847—1862 : 193, t. 21, fig.
5—8 (Clausilia); Лихарев
иРаммельмейер, 1952 :
233, рис. 155.
Раковина верете-
новидная, толстостенная,
прочная, густо ребристая,
в промежутках между реб-
ребрами со следами продоль-
¦ ¦'' ' ! ¦ - ной исчерченности, слабо
просвечивающая, с легким блеском, темно- или коричневато-роговая,
с частыми белыми штрихами. Вершина тонкая, цилиндрическая. Оборо-
Оборотов 12—13, слабо выпуклых; 2" предпоследних оборота нередко почти
одинаковой ширины, последний оборот на затылке с легкой продольной
вмятиной. Внутренняя поверхность устья розовато-коричневая. Края
устья широкие, с коричневато-белой губой., Нижний конец спиральной
пластинки лишь слегка заходит за верхний конец верхней пластинки.
Коричневая нижняя пластинка образует широкий спиральный изгиб,
который сильно выдается в просвет устья и близко подходит к верхней
пластинке; с краем устья она иногда связана слабой складочкой. Субко-
лумеллярная пластинка может быть заметна при косом положении устья.
Париетальная стенка между верхней и нижней пластинками гладкая.
Полулунная складка слабо развита,, мозолевидная, прозрачная; хорошо
выражен и просвечивает сквозь стенку раковины лишь ее верхний конец,
А Б
Рис. 165. Laciniaria (Strigilecula) сапа (Held).
А — общий . вяд раковины; Б — клауэилий. (Л — по Ли-
хйрбву и Раммельмейер, Б— ориг.).
4. LACINIARIA
247
6ж.
который отогнут внутрь. Главная складка занимает около V3 оборота
раковины. Передний конец ее виден в просвете синулуса при прямом по-
положении устья. Передний конец нижней палатальной складки сливается
с тонкой мозолью (видна у вполне развитых экземпляров). Рукоятка клау-
зилия, плавно расширяясь, переходит в сравнительно узкую желобковидно
изогнутую лопасть.
Размеры: высота раковины 15—18, ширина 3.5—3.8 мм; высота устья
3.5—3.8, ширина 2.7—2.9 мм. Отношение высоты раковины к высоте,устья
(В/Ву) 4.5-4.7.
Внутреннее строение
(Лёвушкин, 1958). Отросток семят
приемника различной длины, но всег-
всегда верхний конец его лежит значи-
значительно выше места разделения семяяй-
цевода на семяпровод и матку (рис.
166). Последняя лишь немного длин-
длиннее влагалища, а иногда "и бднои~дм-
ны с ним. ГГёнйе почти тоиЗке длины,
что и семяпровод. Мускульная полос-
полоска, связывающая обе части мужских
половых путей, развита заметно силь-
сильнее, чем у видов Laciniaria s. str.
Внутренние стенки пениса без скла-
складок.
Распространение. В СССР:
почти вся Прибалтика (кроме за-
западной Эстонии и эстонских остро-
островов), Карпаты и Предкарпатье. Мно-
Многочисленные находки в западной
части Ленинградской и в Московской
областях. Единичные находки у Пско-
Пскова, Витебска, в Пинских болотах,
реп
нл
Рис. 166. Laciniaria (Strigilecula) сапа
(Held). Половая система. (По Лёвуш-
кину).
у Смоленска, Рославля, Брянска, в Орловской (Троено), Тульской (Алек-
(Алексин), Белгородской (Новый Оскол) областях, близ Киева и в Казани.
Вне СССР: вся Польша, равнина и средневысотные горы восточной части
Германии (ГДР) и восточной Чехословакии, все Карпаты и восточная часть
северных Балкан (Болгария, Македония). Серия островных ареалов распо-
располагается в предгорьях Альп (вдоль верхнего Дуная и верхнего Рейна);
изолированная колония обнаружена в центральной Финляндии. Следова-
Следовательно, ареал этого вида занимает восточную чаегь средней и западную
половину восточной Европы.1
Раковины этого вида изредка встречаются в четвертичных отложениях,
в пределах современного ареала.
Экология. Обитает в смешанных и широколиственных лесах на
стволах и под корой гниющих деревьев и пней, реже в подстилке. В сред-
средней Европе более обычна в нижней части лесной зоны гор и в предгорьях,
чем в равнинных лесах. .
1 Бонковский (Bcjkowski, 1892) указывает на находки у Майдана-Горнего
(близ города Коломыя Станиславской области), а также в польских Карпатах (Кры-
ница и Стпшжув) южноевропейской L. vetusta (Rssm.). Других указаний на при-
присутствие этого вида в Западных и Восточных Карпатах больше не было. При ревизии
коллекции Бонковского мы убедились, что он имел дело с L. сапа (Held).
248 V. CLAUSJLIINAE
*4. Laciniaria (Strigilecula) nitidosa (Ulicny).
Ulicny. 1893A892—1895), Mekkysi cesti : 90 (Clausilia vetusta striolata f.
nitidosa); Lozek, 1953, Bull, internet. Acad. Tcheque Sci., 53 : 119—128, t. 1—2
(nitidosa Ulicny emend.); Lozek, 1956 : 137, t. 14, fig. 4.
Раковина веретеновидно-цилиндрическая, стройная, довольно
тонкостенная, но прочная, ребристо исчерченная, сильно просвечивающая,
очень блестящая, светло-коричневато-роговая, с редкими короткими
белыми штрихами. Оборотов 12—14, слабо выпуклых; эмбриональные
обороты B3/4) образуют гладкую, почти цилиндрическую вершину; 2—3 пред-
предпоследних оборота почти одинаковой ширины; поверхностная скульп-
скульптура особенно сильно и правильно выражена на верхних (дефинитивных)
оборотах, а книзу постепенно слабеет и принимает неравномерный ха-
характер (только на затылке имеется несколько крупных ребер); последний
оборот, книзу сужается. Устье коротко- и отчасти косо-грушевидное, от-
относительно небольшое, с ясным базальным желобком и с высоким, нес-
несколько врезанным синулусом. Края устья широкие, с белой губой. Верх-
Верхняя и спиральная пластинки такие же, как у L. сапа. Нижняя пластинка
лежит глубже, белая. Субколумеллярная пластинка видна при косом
положении устья и достигает его края в виде слабой коричневой складки.
Крупная белая полулунная складка лежит на спинной стороне; верхний
конец ее круто отогнут назад. Крупная главная складка начинается на
правой стороне и кончается у палатальной мозоли. Нижняя палатальная
складка сходна с таковой у L. сапа, но развита сильнее и белая. Палаталь-
Палатальная мозоль более мощная, особенно в верхней части, где она толщей шире,
чем внизу, лежит параллельно краю устья. Лопасть клаузилия плавно
сужена к переднему концу.
Размеры: высота раковины 13.0—17.0, ширина 3.0—3.4 мм; высота
устья 2.8—3.2, ширина 2.1—2.3 мм; отношение высоты раковины к высоте
устья (В/Ву) 4.8-5.3.
Распространение. Район Чешского карста (западная Че-
Чехословакия).
Экология. Обитает в осыпях и на скалах, главным образом на
известняковых, реже на других горных породах. Больше характерна для
открытых (голые степи) равнинных мест, но встречается также в редколесье
и в рощах.
Систематическое положение. Данный вид длительное
время рассматривали как местную форму южнокарпатской L. vetusta
(Rssm.)ssp. striolata (Bielz.) Между тем, как показал Ложек (Lozek, 1953),
чешская форма L. vetusta отличается от всех прочих форм того же вида
четкой и постоянной особенностью раковины — тонкая и тупая ребристая
исчерченность. Это отличие, наряду с полной изолированностью ареала
L. nitidosa от ареалов других форм L. vetusta, в том числе и от ареала
наиболее, сходного с ней подвида striolata, позволило названному автору,
рассматривать var. nitidosa Ulicny как самостоятельный вид,
J3. Подрод MENTISSA H. et A. ADAMS.
Adams Н. а. А., 1855 A858) : 185 (Clausilia, подрод Mentissa); L i n d h о 1 m,
1924 : 66 (Laciniaria, подрод Mentissa).
Тип подрода: Clausilia canalifera Rssm.
Верхняя пластинка сравнительно короткая, благодаря чему не дохо-
доходит до уровня нижнего конца спиральной пластинки. Нижняя пластинка
лежит глубоко, близко от верхней и видна при прямом положении устья.
4. LAC IN I ARIA
249
w
Как и спиральная, она берет начало на право-брюшной стороне раковины
и широкой спиралью тянется вниз, плавно нарастая в высоту; с краем
устья она связана короткой складочкой. Субколумеллярная пластинка не
видна через устье. От полулунной складки остается небольшой рудимент
ее нижнего конца, который располагается на право-
спинной стороне и слит с задним концом нижней пала-
палатальной складки. Главная и средняя палатальные склад-
складки берут начало на спинной стороне; первая тянется
параллельно шву до лево-спинной стороны; вторая лежит
к главной под углом и значительно короче последней.
Нижняя палатальная складка несколько длиннее сред-
средней, очень толстая; снаружи ей соответствует бороздка,
очерчивающая киль. Нижняя часть языковидной лопасти
клаузилия сильно перекручена и по палатальному краю
имеет глубокую вырезку.
Срединный и боковые зубы радулы с дополнительными
зубцами, причем у первого эти зубцы очень небольшие и
заметны только при 1300-кратном увеличении.
Распространение. Горный Крым.
Данный подрод особенно близок к Laciniaria s. str. Это
видно не только при сравнении гениталий, но и раковин.
В обеих группах пластинки замыкательного аппарата сход-
сходны по форме и положению. Средняя палатальная складка
в обоих случаях располагается к главной под углом,
а лопасть клаузилия перекручена (правда, у Mentissa это
выражено слабее). Однако присутствие у Mentissa глубо-
глубокой вырезки_на нижнем"конце клаузилия, наличие ниж-
нижней палатальной складки и почти полная редукция по-
полулунной складки дает нам основание рассматривать
обе группы как самостоятельные подроды.
5. Laciniaria (Mentissa) canalifera (Rssm.) (рис. 167,
168).
Rossmassler, 1836, Iconogr., 1, 3 : 17, fig. 183 (Clau-
silia); К ii s t e r, 1847—1862 : 228, t. 25, fig. 21—23 (Clausilia);
Лихарев и Раммельмейер, 1952 ; 245, рис. 171.
Раковина веретеновидно-цилиндрическая, строй-
стройная, исчерченная (более сильно и редко на верхних, сла-
слабее и чаще на нижних оборотах), слабо просвечивающая,
слабо блестящая или матовая, красновато-коричневая,
реже светло-роговая. Сильно вытянутый конический завиток
наверху заканчивается узкой (около 1 мм), цилиндриче-
цилиндрической закругленной верхушкой. Оборотов 12V2—15, слабо
нарастающих в высоту и ширину, почти плоских, раз-
разделенных мелким беловатым швом; последний оборот
книзу слегка сужен, на затылке ребристый, за устьем уплощен, с круп-
крупным морщинистым килем. Края устья широко отвернуты, слегка утолщены,
серовато-белые. Верхняя пластинка тонкая и низкая, впереди слегка
выступает за край устья. Спиральная пластинка берет начало на право-
брюшной стороне, вначале очень быстро нарастает в высоту, а затем плавно
снижается в направлении к устью, заканчиваясь на левоспинной стороне.
За исключением главной складки, нижний конец которой иногда заметен
Рис. 167. Laci-
Laciniaria (Mentis-
(Mentissa) canalifera
(Rssm.). (По
Лихареву и
Раммельмейер).
А — общий вид
раковины; В —
клаузилий.
250
V. CLAU.SIUINAE
в просвете синулуса при прямом положении устья, остальные складки —
средняя и нижняя палатальные — видны лишь при косом положении
устья.
Размеры: высота раковины 16—20, ширина 3.8—4.2 мм; высота устья
3.4—3.7, ширина 2.4—2.8 мм.
Изменчивость. Раковина данного вида даже в пределах одной
популяции очень изменчива. Особенно это касается ее размеров и поверх-
поверхностной скульптуры. Так, на-
наряду с исчерченными ракови-
раковинами встречаются ребристо ис-
исчерченные или даже слабо реб-
ребристые формы (var. costulata
Boettger, 1871). Для лесных
бавтопов более обычны популя-
популяции с более вздутой раковиной
({. detersa Rossmassler, 1836),
а для высокогорных зон — мел-
мелкие, иногда ребристые, формы
(f. acridula Rossmassler, 1836).
Внутреннее строе-
строение. Материал: Крым, ниж-
нижнее плато Чатыр-Дага, сбор
Рошковского, 27 VII 1916, оп-
определение наше — 1 экз.
Ь
D Г с . /106 . 12к \
Радула: ^+^ + 3^5)
Х2~|х92.
X
ркл
ЛР
срединного зуба
ширина радулы
Рис. 168. Laciniaria (Mentissa) canalifera
(Rssm.). Половая система. (Ориг.).
как и
Размеры
12 х 18 [л,
555 fA.
Семяприемник очень длин-
длинный и толстый, с коротким отро-
отростком (рис. 168). Матка длин-
пенис, веретеновидно вздуто.
нее влагалища. Последнее,
Распространение. Горный Крым.
Экология. Обитает на скалах и под камнями горной и предгор-
предгорной зон (в лесу, в кустарниках и на яйлах).
6. Laciniaria (Mentissa) gracilicosta (Rssm.) (рис. 169).
Rossmassler,
1847—1862 : 229, t. 25,
м е й е р, 1952 : 246.
1836, Iconogr., 1,3:18, fig. 184 (Clausilia); Rnster,
fig. 27—30 (Clausilia); Лихарев и Раммель-
Раковина очень похожа на таковую предыдущего вида, отли-
отличаясь лишь поверхностной скульптурой. Последняя образована крупными,
редко поставленными белыми ребрами, которые покрывают всю рако-
раковину, за исключением гладких эмбриональных оборотов BVa). Оборотов
121/2—14. Замыкательный аппарат и клаузилий типичные для подрода.
Размеры: высота раковины 17.5—19.5, ширина 3.5—4.0 мм; высота
устья 3.4—3.8, ширина 2,5—2.8 мм.
Изменчивость. Как и у предыдущего вида, раковина очень
сильно варьирует в отношении размеров (особенно высота), формы и скульп-
скульптуры. Даже в пределах одной популяции встречаются сильные колебания
5. PSEUDALINDA
25i
от очень небольших — высота 14.0т-15.5, ширина 2.8—3.3 мм (var. gra-
cilior A. Schmidt, 1868), до очень крупных — высота до 22, ширина до
4,5 мм (var. maxima Westerlund, 1901, var. frater A. Schmidt, 1868). Из-
Изредка встречаются экземпляры с более широкой (вздутой), чем обычно,
раковиной (var sodalis A. Schmidt, 1868). Скульптура также подвергается
некоторой изменчивости. Так, бывают экземпляры с ребристо исчерченной
раковиной, но, в отличие от L. canalifera Rssm., складки располагаются
более редко как на верхних, так и на нижних оборо-
оборотах (var. taurica A. Schmidt, 1868).
Внутреннее строение. Материал: Крым,
северные склоны Чатыр-Дага, над Аяном, сбор Рошков-
ского, 27 VII 1916, определение наше — 1 экз.
Радула: [J + (? + з^)х 2] X 102.
Размеры срединного зуба 12 X 18 ц, ширина радулы
495 ц.
Половая система очень похожа на таковую L. canali-
canalifera.
Распространение. Горный Крым.
Экология. Обитает в сухих местах открытых гор-
горных плато и среди зарослей древовидного можжевельника
на скалах и под камнями.
5. Род PSEUDALINDA BTTG.
Boettger О., 1877а : 76 (Clausilia,, секция Р seudalinda);
L i n d h о 1 m, 1924: 65 (Laciniaria, подродРseudalinda, секция
Pseudalinda s. str.).
Рис. 169. Laci-
Laciniaria (Mentis-
sa) gracilicosta
(Rssm.).
(Ориг.).
Тип рода: Clausilia fallax Rssm.
Раковина со слабым блеском, без белых штрихов.
Последний оборот с коротким килем, окаймленным мел-
мелкой бороздой. Устье с коротким, но ясным базальным
желобком. Верхняя и спиральная пластинки распола-
располагаются на одном расстоянии от столбика и либо сливают-
сливаются, либо их концы находятся недалеко друг от друга. Почти
отвесная ланцетовидная нижняя пластинка лежит глубоко и не видна
или плохо различима при прямом положении устья. Полулунная складка,
или ее рудимент (у одного вида она совсем отсутствует), лежит на npar
вой стороне, реже несколько смещена к спинной стороне раковины;
верхний конец ее отогнут назад. Передний конец лопасти клаузилия
утолщен, близ палатального края угловатый.
Срединный и боковые зубы радулы с дополнительными зубцами.
Половая система сходна с- таковой у Laciniaria Hartm.
Распространение. Карпаты, Трансильвания, Банатские горы.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА PSEUDALINDA
1 B). Раковина сильно ребристая, толстостенная. Синулус устья глубоко
врезан. Полулунная складка крупная, хорошо просвечивает сквозь
стенку раковины 1. P. stabilis (Pfr..).
2 A). Раковина ребристо исчерченная, довольно тонкостенная. Синулус
не врезан или слабо врезан. Полулунная складка либо слабо раз-
развита и плохо просвечивает через стенку раковины, либо от нее
остается только ее верхний конец — короткая складочка, лежащая
под главной складкой . . . 2. P. fallax (Rssm.).
252
V. CLAUSILIINAE
1. Pseudaliuda stabilis (Pfr.) (рис. 170, 171).
P f e i f f e г L., 1847, Zeitschr. Malak., 4 : 70 (Clausilia); К u s t e r, 1847—
1862 : 199, t. 22, fig. 5—7 (Clausilia); Bakowski, 1892 : 116, t. 5, fig. 108 (Clau-
siliastra); S о 6 s, 1917:151, fig. 61, 62 [Clausilia (Pseudalinda), анатомия]; L o-
z e k, 1956 : 138—139, t. 14, fig. 5 [Laciniaria (Pseudalinda)]; Urbanski, 1957 :
149—150, fig. 130 (Laciniaria).
Раковина веретёновидная, довольно вздутая, толстостенная и
прочная, густо, равномерно тупо ребристая (на предпоследнем обороте
50—55 ребрышек), просвечивающая, с шелковистым
блеском, светло-, реже темно-роговая. O6oj>ojroB__12_—
13, слабо выпуклых, разделенных глубоким швом;
эмбриональные обороты B—2V2) гладкие, образуют
цилиндрическую верхушку; последующие обороты
заметно шире, благодаря чему верхняя часть завитка
имеет вогнутые контуры; последний оборот на затыл-
ке_? очень ^крупными редкими ребрами; оба последних
оборота нередко одинаковой ширины. Устье_грушевид-
ное, выступающее, с глубоко врезанным синулусом.
Края устья'широкие, с белой или розовато-коричневой
губой. Верхняя пластин-
ка впереди нередко вы-
выступает за край устья,
сзади плавно снижается
й либо сливается со спи-*
ральной, либо отделена
от нее, но располагается
народной линии с нею.
Крупная ланцетовидная
нижняя пластинка не
видна при прямом поло-
положении устья; она кр_уто
6
Рис. 170. Pseudalinda stabilis (Pfr.). (Ориг.).
А — общий вид раковины; Б — клаузилий.
уходит кверху, а внизу
нередко обрублена Jmh
же связана с краем
устья небольшой скла-
складочкой. Субкодумелляр-
ная пластинка чаще всего не видна даже при косом положении устья. Круп-
Крупная полулунная складка лежит на правой стороне и хорошо заметна через
стенку раковины; верхний коневу)е круто отогнут назад. Главная склад-
складка хорошо видна через стенкураковины; верхний конец ее лишь немного
заходит за полулунную складку, нижний располагается на спинной сто-
стороне.Палатальная мозоль сливается~с губой и достигает наибольшей
толщины близ базального желобка; окрашена она под цвет губы. Рукоят-
Рукоятка клаузилия плавно переходит в лопасть, нижний конец которой угло-
угловатый и утолщен.
Размеры: высота раковины 14.0—16.0, ширина 3.8—4.0 мм; высота
устья 3.3—3.6, ширина 2.5—2.8 мИ:
Внутреннее строение (So6s, 1917; Лёвушкин, 1958). Один
ряд радулы можно характеризовать следующей формулой:
'106 , 15к
¦+1— + -
^ \ 2 ^.
=!)Х2.
Отросток значительно короче семяприемника (рис. 171). Верхний конец
его располагается немного выше начала семяпровода. Матка такой же
5. PSEUDALINDA
253
^лины^как влагалищу. Последнее очень широкое, с сильно мускулистыми
стенками. Внутри пениса находятся 2 кольцевых складки.
Распространение. В СССР: многочисленные находки в
Львовской, Станиславской и Закарпатской областях; одна находка в Тер-
нопольской области (Золотой поток). Вне СССР: Карпаты (на западе,
начиная с Высоких Татр), Трансильва"
ния, Банат. —— ^ж
Экология. Обитает преимуществен-
преимущественно в горных, реже в предгорных лесах
под камнями, в подстилке и под корой
гниющих деревьев и пней.
2. Pseudalinda fallax (Rssm.) (рис. 172).
R о s s m a s s 1 e г, 1836, Iconogr., 1, 4 :
16, fig. 262 (Clausilia); К ii s t e r, 1847—1862 :
168, t. 18, fig. 23—26 (Clausilia); S о 6 s, 1917 :
151, fig. 59, 60 [Clausilia (Pseudalinda), анатомия];
Лихарев и Раммельмейер, 1952 :
234 (Laciniaria); Urbanski, 1957 : 148, fig.
122d, 129 (Laciniaria).
Раковина веретеновидная, взду-
вздутая, сильно ребристо исчерченная, слабо
просвечивающая, слабо блестящая, темно-
роговая или красновато-коричневая. Обо-
Оборотов 11—12, слабо выпуклых; эмбрио-
эмбриональные обороты (около 21/2) гладкие; по-
последний оборот на затылке слегка уплощен.
Устье широко-грушевидное, нередко округ-
округло-ромбическое, лишь немного выступаю-
выступающее, так что иногда верхний край его плот-
плотно прилегает к последнему обороту; в неко-
некоторых случаях верхний край устья разор-
разорван. Синулус чаще всего не врезан или слабо врезан. Края устья широко
отвернуты, белые или розовато-коричневые, несколько тоньше, чем у P. sta-
bilis (Pfr.). Верхняя пластинка тоньше и ниже, чем у предыдущего вида,
всегда значительно отстоит от спиральной пластинки. Ланцетовидная ниж-
нижняя пластинка при прямом положении устья почти не заметна, той же формы,
что и у i3. stabilis. Нижний конец субколумеллярной пластинки в виде
тонкой складки огибает справа базальный желобок и нередко виден даже
при прямом положении устья. Полулунная складка развита очень слабо,
так что через стенку раковины она плохо заметна, или от нее остается только
ее верхний конец — короткая складочка, лежащая на правой стороне
под главной складкой. Последняя значительно тоньше и короче, чем
у P. stabilis, но всегда просвечивает через стенку раковины. Палатальная
мозоль развита немного сильнее, чем у предыдущего вида, белого или
светло-коричневого цвета, и чаще всего не сливается с губой. Клаузилий
такой же, как у предыдущего вида.
Размеры: высота раковины 16—19, ширина 4.5—5 мм (реже высота
до 24, ширина до б мм); высота устья 4.5—5.0, ширина 3.3^-3.8 мм.
Внутреннее строение. (Soos, 1917). Один ряд радулы
можно охарактеризовать следующей формулой:
Рис. 171. Pseudalinda stabilis
(Pfr.). Половая система (По Гу-
децу, Ложеку и Маха).
-+(-4-
3^2^
2Ок Vv2
254
V. CLAUSIUINAE
В отличие от предыдущего вида, срединный зуб радулы более вытянут
в длину и дополнительные зубцы лишь слабо выражены.
Половая система такого же типа, как и у P. stabilis.
Распространение. В СССР этот вид найден в нескольких
пунктах Станиславской области.1 Вне СССР: Польша (Восточные Карпаты,
Светокширские горы, Люблинская возвышенность), Румыния (Трансиль-
вания), Югославия (Сербия и Македония), Албания. Как у нас, так и
в Польше вид встречается сравнительно редко.
Экология. Обитает в горных смешанных ж
широколиственных лесах в подстилке, под корой гнию-
гниющих деревьев и пней.
6. Род VESTIA HESSE
Vest, 1867:190 (Uncinaria, поп F г о е 1 i с h, 1789);
Hesse, 1916, Nachrbl. deutsch. Malak. Ges., 48 : 124; Wag-
Wagner A., 1920:106 (Alinda, подрод Uncinarm part.); Linfl-
holm, . 1924 : 65 (Laciniaria, подрод Pseudalinda, секц0Я
Vestia).
Тип рода: Clausilia elata Rssm.
Раковина веретеновидная, вздутая. Послед-
Последний оборот внизу с коротким, слабо выраженным ту-
тупым килем. Устье округло-грушевидное, с мелким неяс-
неясным базальным желобком и с закругленным нижним
краем. Верхняя и спиральная пластинки находятся
на разном расстоянии от столбика, благодаря чему
они либо не связаны друг с другом, либо, если все же
сливаются, то в этом месте образуют более или менее
ясный коленообразный изгиб. Нижняя пластинка ле-
лежит глубоко, но всегда видна при Прямом положении
устья; внизу она дает широкий спиральный изгиб и
плавно поднимается внутрь раковины. Нижний конец
субколумеллярной пластинки обычно виден при косом
положении устья. Как субколумеллярная, так и в осо-
особенности нижняя пластинка с краем устья связаны
короткими складочками. Между последними, а иногда и выше их, имеются
еще 1—2 таких же складочки. Полулунная складка или ее рудименты
располагаются на спинной стороне раковины. Над ней лежит длинная
главная складка. Имеется Палатальная мозоль. Лопасть клаузилия близ
рукоятки имеет неглубокую, но отчетливую вырезку; передний конец
лопасти у большинства видов заострен и изогнут крючком.
Срединный и боковые зубы радулы с дополнительными зубцами.
Половая система сходна с таковой у Laciniaria Hartm.
\, Распространение. Карпаты, Трансильвания и Родопы (Бол-
(Болгария).
По раковине виды данного рода во многом сходны с моллюсками из
рода Iphigena Gray. В обеих группах киль и базальный желобок слабо
развиты, в связи с чем нижний край устья имеет округлые очертания.
Нижняя пластинка Дает внизу широкий спиральный изгиб. Полулунная
складка располагается на спинной стороне раковины. В то же время есть
Рис. 172. Pseuda-
Pseudalinda fallax
(Rssm.).
(Ориг.).
1 Бонковский (Bakowski, 1892) указывает пункты и из других западных областей
Украины. Но при проверке его определений выяснилось, что он часто смешивал дан-
данный вид с P. stabilis (Pfr.).
6. VEST IA 255
заметные различия. Так, у Vestia раковина зеленовато- или коричневато-
роговая, чаще всего блестящая, реже матовая. Лопасть нЯаузилия на
конце более или менее заострена, у многих крючковидно изогнута. Напро-
Напротив того, у Iphigena раковина темно- или йрабновато-Коричневая, нередко
почти черная, матовая или со слабым шелковистым блеском. Лопасть
клаузйлия на конце более или менее закругленная й йикогда ие изогнута
крючком.
Значительно сильнее обе группы различаются по строению полового
аппарата. Если у Iphigena сохранилась еще петля из пениса и семяпро-
семяпровода и прикрепляющийся к ней ретрактор пениса, то у Vestia как петля,
так и ретрактор отсутствуют и мужские половые нуги вытянуты вдоль
женских.
Поэтому, несмотря на значительное сходство раковин, есть все осно-
основания для рассматривания обеих групп как самостоятельные роды, стоя-
стоящие в пределах подсемейства Clausiliinae сравнительно далеко друг от
друга.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА VESTIA
1 B). Широкая и короткая ложковидная лопасть клаузйлия у нижнего
конца слабо заострена1 1. V. turgida (Rssm.)
2 A). Узкая и длинная языковидная лопасть клаузйлия к переднему
концу сильно сужена и крючковидно изогнута.
3 D). Верхушка раковины очень широкая — 1.3—1.6 мм. Верхние дефи-
дефинитивные обороты ребристые, предпоследний ребристо исчерчен,
последний неясно исчерчен, почти гладкий . . . 2. V.elata (jRssm.).
4C). Верхушка раковины сравнительно узкая, ширина ее чаще веего
1.0—1.2 мм или еще меньше. Все дефинитивные обороты имеют при-
приблизительно сходную скульптуру.
5 F). Раковина очень толстостенная, с крупными и редкими тупыми
ребрами (на предпоследнем обороте их 50—55). Высота рако-
раковины 15—19, ширина 4.3—5.0 мм ... 3. V. gulo (E. Bielz).
6 E). Раковина тонкостенная, густо ребристо исчерченная (на пред-
предпоследнем обороте около 70—75 складок). Высота раковины 13.5—
15.5, ширина 3.5—4.0 мм 4. V. riloensis (A. Wagn.).
1. Vestia turgida (Rssm.) (рис. 173, 174).
Rossmassler, 1836, Iconogr., 1,3 : 20, fig. 191 (Clausilia); Wester-
1 u n d, 1884 : 150 [Cl. (Uncinaria)proceravsv. halieiaca]; S о 6 s, 1917 :.152, fig. 65, 66
[Clausilia (Vestia), анатомия]; Лихарев и Ра мм ельмейер, 1952 : 234
(Laciniaria); L о z e k, 1956:141, t. 15, fig. 3,4 [Laciniaria (P seudalinda)]; U r-
banski, 1957:154—155, fig. 132 (Laciniaria).
Раковина твердостенная, сильно просвечивающая, блестящая
(часто очень блестящая), коричневато-роговая или зеленоватая. Вер-
Верхушка довольно узкая (ширина около 1 мм). Поверхностная скульптура
изменяется от верхних оборотов к нижним: эмбриональные обороты (около 3)
гладкие, последующие 4—5 оборотов тупо и часто ребристые, последние
обороты с Постепенно сглаживающейся скульптурой, нередко почти глад-
гладкие. Оборотов 10—11, реже 12, слабо выпуклых; последний оборот на
затылке с Несколькими ребрышками. Края устья широкие, утолщены
1 В обоих случаях нижняя часть лопасти клаузйлия хорошо видна при косом
положении устья. Крючковидный конец ее (см. антитезу) обычно выступает из-за суб-
колумеллярной пластинки.
256
V. CLAUSILIINAE
белой или розовато-коричневой губой. Верхняя пластинка низкая, тол-
толстая и короткая, обычно отделена от спиральной широким промежутком,
но изредка сливается с нею. Нижняя пластинка плавно уходит кверху,
образуя в устье широкий спиральный изгиб, и при этом близко подходит
к верхней пластинке; внизу с краем устья она связана короткой складоч-
складочкой; рядом с последней находится другая аналогичная складочка. Нижний
конец субколумеллярной пластинки хорошо виден при косом положении
устья. Дугообразно изогнутая полулунная складка
лежит на спинной стороне раковины; нередко от нее
остаются только оба конца, а середина исчезает. Глав-
Главная складка довольно длинная (около х/3 оборота) и рас-
располагается так, что полулунная приходится под ее
серединой; обычно она видна в просвете синулуса.
Параллельно краю устья, часто сливаясь с губой и
одного цвета с нею, лежит узкая палатальная мозоль.
Лопасть клаузилия широкая и короткая, ложковид-
ной формы; при переходе в рукоятку она имеет не
глубокую, но заметную вырезку (в этом месте клаузилий
упирается в верхний конец нижней пластинки); широ-
широкий нижний конец лопасти слабо заострен.
Размеры: высота рако-
раковины 13—17, ширина 3.5—
4.0 мм; высота устья 3.0—
3.5, ширина 2.5—3.0 мм.
И з м енчивость.
В отношении размеров ра-
раковины наблюдаются ча-
частые отклонения от типич-
типичных. Иногда встречаются
экземпляры с узкой и длин-
длинной раковиной (var. elon-
gata Rssm.). Согласно Ло-
жеку (Lozek, 1956), в вы-
высокогорных зонах чаще
Рис. [173. Vestia turgida (Rssm.). (Ориг.).
А —> общий вид раковины; Б — клаузилий.
всего обитают популяции,
состоящие из небольших
экземпляров (высота рако-
раковины около 12, ширина около 3.5 мм), а близ нижней границы горных
лесов обитают обычно популяции из более крупных особей (высота 15—
17, ширина 3.7—3.8 мм).
В наших Карпатах (Закарпатская и Станиславская области) и в се-
северной Трансильвании встречаются популяции из очень крупных экзем-
экземпляров — высота раковины 20—22, ширина 4.5—5.0 мм. Их относят к
особой разновидности — var. procera E. Bielz. В настоящее время мы еще
не располагаем достаточными данными, чтобы судить, является ли эта '
разновидность экологической формой или подвидом.
Скульптура раковины также подвержена некоторой изменчивости.
Так, наряду с экземплярами с почти гладким нижним оборотом раковины
встречаются отдельные особи, у которых скульптура этого оборота
выражена более резко, до ребристой исчерченности. Но всегда по-
последние 2 оборота имеют более сглаженную скульптуру, чем предыду-
предыдущие.
Внутреннее строение (Soos, 1917; Hudec, Lozek, Macha,
1958).
6. VESTIA
257
гп~
r> i \ c i 66 , 14k
Радула (половина -одного ряда): ~о" ~r у + з —4"
Семяприемник длинный, нередко также очень широкий; отросток
его значительно короче и уже. Матка одной_длины^ с влагалищем, но
последнее более толстое. Семяпровод и пенисГ почти одной длины, причем
последний очень толстый, веретеновидно вздутый (рис. 174).
Распространение. В СССР: 1 многочисленные находки в
Львовской, Станиславской и в Закарпатской областях. Одна находка в
Тернопольской области (Бережаны). Вне СССР: восточные Судеты, Запад-
Западные и Восточные Карпаты; реликто-
реликтовый ареал в районе Шумавских гор
(Чехословакия).
В ископаемом состоянии ракови-
раковины данного вида были найдены в чет-
четвертичных отложениях ряда мест Че-
Чехословакии, в том числе и вне границ
современного ареала.
Экология. Обитает во влаж-
влажных лесах и в кустарниках, особенно
по сырым ущельям, в подстилке, под
камнями, валежинами и под гнию-
гниющей корой. Изредка встречается и
в более сухих местах. Предпочитает
горы, но иногда встречается и в пред-
предгорных лесах.
Систематическое по-
положение. Данный моллюск от-
отличается от других видов того же ро-
рода в первую очередь формой лопасти
клаузилия. В противоположность на-
нашему у остальных видов она более
длинная и узкая, а конец ее вытя-
вытянут в крючковидный выступ, который
обычно виден через устье из-за нижнего конца субколумеллярной пла-
стищш. •
*2. Vestia elata (Rssm.) (рис. 175).
R о s s m a s s 1 е г, 1836, Iconogr./ 1, 3 : 20, fig. 190 (Clausilia); Wagner A.,
1914, Iconogr., N. F., 21 : 9, fig. 32 (Uncinaria, анатомия): Лихарев и Рам-
медьмейер, 1952 : 235 (Laciniaria turgida var. elata); L о z e k, 1956 : 141—142,
t. 15, fig. 5 [Laciniaria (Pseudalinda)];\JJ r b a n s k i, 1957 : 156—157, fig. 131 (Laci-
(Laciniaria).
Раковина веретеновидная, вздутая, твердостенная, сильно про-
просвечивающая, блестящая, светло- или коричнево-роговая. Верхушка
очень широкая A.3—1.6 мм). Поверхностная скульптура изменяется от
верхних оборотов к нижним: эмбриональные обороты (около 23/4) гладкие,
последующие 4—5 оборотов тупо и густо ребристые, предпоследний реб-
ребристо исчерчен, а последний неясно исчерчен, почти гладкий. Оборотов
около 10, слабо выпуклых. Края устья широкие, с тонкой белой или розо-
Рис. 174. Vestia turgida (Rssm.). Поло-
Половая система. (По Гудецу, Ложеку и
Маха).
1 Дается главным образом на основании ревизии коллекции Бонковского (Науч-
(Научно-природоведческий музей Академии наук УССР, Львов), который ошибочно прини-
принимал данный вид за V. elata (Rssm.).
258
V. CLAUSILIINAE
вато-коричневой губой. Верхняя пластинка,слита со спиральной. Нижняя
и субколумеллярная пластинки, а также главная складка такие же, как
у предыдущего вида. Полулунная складка обычно развита лучше и лишь
у отдельных экземпляров в середине прервана. На краю устья имеется
несколько складочек: 2-около нижнего конца нижней пластинки (из них
одна является продолжением последней, а другая лежит немного ниже),
1 (может и отсутствовать) близ нижнего конца субколумелдярной пла-
пластинки и, наконец,- изредка 1—2 складочки бывают выше нижней.пластвнки.
Палатальная мозоль одного цвета с гу-
губой и нередко сливается с последней:
Лопасть клаузилия узкая, языковид-
ная, сильно изогнутая в поперечном
направлении; близ рукоятки она несет
неглубокую вырезку, а к переднему кон-
концу сильно сужена и крючковидно заг-
загнута. Конец ее обычно слегка высту-
, пает из-за субколумеяяярной пластинки
и виден при косом лоложении устья.
Размеры: высота раковины 12—15
(реже до 20), ширина 3.5—4.5 мм; вы-
высотаустья 3.0—3.5, ширина 2.8—3.2 мм.
Рис.175. Vestia elata (Rssm.). (Ориг.).
А — общий вид раковины; Б — клаузилий.
Внутреннее строение не изучено.
Распространение. Восточные Карпаты и Трансильвания,
кроме того, вне оснобного ареала, в Светокширских горах (южная Польша).
С территории СССР достоверных находок этого вида нет,1 но судя по тому,
что он многократно был найден в сопредельных областях Польши, Чехо-
Чехословакии и Румынии, со нременем он может быть найден и в нашей части
Восточных Карпат.
Экология. Обитает во влажных горных лесах в подстилке, иод
камнями и, в особенности, под гниющими стволами деревьев.
Систематическое положение. Характер скульптуры
поверхности раковины сближает данный вид и V. iurgidi (Rssia.), а форма
кдаузилия.— с V. gBZo(Bielz). Наиболее важным признаком, отличаю-
отличающим данный вид от всех других видов того же рода, является широкая
верхушка раковины. Это стойкий признак, который позволяет легко отли-
отличить даже молодые раковины от таких же раковин близких видов.
1 Указание Бонковского (Bakowski, 1892) о присутствии этого моллюскана западе
Украины, как показала проверка, относится к- V: iurgida (Rssm.).
6. VEST IA
259
3. Vestia gulo (E. Bielz) (рис. 176). !
Bielz E., 1859, Verb; u. Mitt. Siebenb. Ver. Naturw., 10 : 222 (ClausiUa tur-
gida var. gulo); С 1 e s si n» 1387 : 390, iig. 252 (Uncinaria);,' L о ± ark,.. 1056 : 140,
t. 15, fig. 2 [(Laciniaria (Pseudalinda)]; U I b an. s к i, 1957 : 155—156, fig. 133 (Laci-
niaria). ....... . ; ...... . ¦
Раковина очень толстостенная и прочная, равномерно покрыта
крупными тупыми ребрами (на предпоследнем обороте 50—55 ,ребер),
слабо просвечивающая, со слабым шелковистым' блеском, светло-роговая,
с редкими белыми штрихами. Верхушка
раковины обычно шире, чем у V. turgida
(Rsgm.), и уже, чем у V. elata (RsSteL)
(чаще всего ширина ее равна 1.0-—1-2 мй)г.
Оборотов 10V2—12, умеренно выпуклых;
эмбриональные обороты BV2) гладкие.
Края устья широкие, с довольно толстой
белой или розоватой ry6oHv Вердняя пла-
пластинка очень толстая, и ли)бо отделена
от спиральной: пластинки заметным про-
промежутком, либо связана с нею, прннбм
перед слиянием обе пластинки становятся
очень низкими. Нижняя и субкодумелляр-
ная пластинки такой же формы, как у V.
tiirgi4a, и обычно (в особенности первая)
связаны с краем устья короткими складоч-
Рис. 176. Vestia gulo (Bielz). (Ориг.).
А '— общий вид раковины; В — клаузилий.
ками; еще 1—2 таких же складочки лежат между названными пластинками;
реже несколько других располагаются над нижней пластинкой. Полулунная
складка обычно хорошо развита и лежит на спинной стороне; как у осталь-
остальных видов Vestia, оба конца ее отогнуты назад. Главная складка занимает
около */3 оборота; ее верхний конец проникает вглубь раковины за по-
полулунную складку. Белая палатальная мозоль хорошо развита, особенно
в нижней части, где часто сливается с губой. Клаузилий с вырезкой у ру-
рукоятки и с крючковидным выступом на переднем конце лопасти, т. е. такой
же, как у V. elata.
Размеры: высота раковины 15—19, ширина 4.3—5.0 мм; высота устья
3.6—4.0, ширина 3.1—3.5 мм.
Внутреннее строение не изучено. .... ,:..;...
260 V. CLAUSILIINAE
Распространение. В СССР: х Львовская, Станиславская, Тер-
нопольская, Закарпатская и Черновицкая области. Вне СССР: Карпаты
(начиная от северных отрогов Западных Бескид), Трансильвания.
. Найдена в плейстоценовых отложениях Трансильвании.
Экология. Обитает во влажных горных лесах под валежником;
под корой, под камнями и в подстилке. Нередко предпочитает очень сырые
биотопы, например берега горных речек.
*4. Vestia riloensis (A. Wagn.)
Wagner A., in Sturany u. Wagner, 1915, Denkschr. Akad. Wiss.
Wien, math.-naturw. Kl., 91 : 82 (Pirostoma); В r a b e n e c, 1952, Vestn. Ueskoslov.
spole6n. zool., 15, 1—2 : 23—31 (Laciniaria moravica); Lozek, 1956:139, t. 15,
fig. 1 [Laciniaria (Pseudalinda)]; Hudec, Lozek, Macha, 1958, Prace Brnen-
ske zakl. Ceskoslov. Akad. ved., 30, 8 : 335—368, t^ 1—6 [Laciniaria (Pseudalinda) rilo-
riloensis moravica, анатомия].
Раковина довольно тонкостенная, но прочная, густо и равномерно
ребристо исчерченная (на предпоследнем обороте около 70—75 складок),
умеренно просвечивающая, блестящая, красновато-коричневая, с редкими
белыми штрихами. Верхушка раковины узкая, ширина ее около 1.0 мм.
Оборотов 11—12, слабо выпуклых; эмбриональные обороты (около 23/4),
гладкие. Края устья широкие, с довольно плоской белой губой. Верхняя
пластинка толстая, слита со спиральной; перед слиянием обе пластинки
становятся очень низкими. Нижняя и субколумеллярная пластинки такие
же, как у V. turgida (Rssm.), лишь первая у нижнего конца слегка обруб-
обрублена; перед нижними концами обеих пластинок или между ними на краю
устья находятся 1—3 коротких складочки. Дугообразно изогнутая полу-
полулунная складка лежит на спинной стороне. Главная складка начинается
несколько глубже полулунной и тянется вперед до палатальной мозоли.
Белая или розоватая палатальная мозоль лежит параллельно краю устья,
в нижней части каплевидно утолщена, не сливается с губой. Клаузилий
такой же, как у V. gulo (E. Bielz) и V. elata (Rssm.), т. е. лопасть имеет
неглубокую вырезку близ рукоятки, к переднему концу сужена и заканчи-
заканчивается крючковидным выступом.
Размеры: высота раковины 13.5—15.5, ширина 3.5—4.0 мм; высота
устья 3.0—3.2, ширина 2.6—2.8 мм.
Внутреннее строение (Hudec, Lozek, Macha, 1958). Се-
Семяприемник довольно толстый и длинный; верхний конец его, прилегает
к основанию белковой железы. Отросток короткий; его верхний конец
лежит немного выше начала семяпровода. Матка в 3 раза длиннее
влагалища. Довольно толстый семяпровод впадает в мощный, но относи-
относительно короткий пенис. Половая система данного вида отличается от та-
таковой V. turgida более длинной маткой, коротким влагалищем, длинным
семяпроводом и коротким пенисом.
Распространение. Ареал данного вида состоит из двух изо-
изолированных частей: 1) Болгария и Македония; 2) Чехословакия (Мора-
(Моравия — Моравско-Силезские Бескиды и некоторые соседние горные мае-,
сивы).
Ископаемые остатки данного вида найдены близ чехословацкой части
современного ареала в отложениях атлантического века послеледниковой
эпохи.
1 Дается главным образом на основании ревизии коллекции Вонковского (Ba,kow-
ski, 1892), который ошибочно называл этот вид Uncinaria turgida (Rssm.).
7. BALEA 261
Экология. Обитает в горных, преимущественно буковых лесах,
произрастающих на сравнительно сухих склонах, на высотах 700—1000 м
над ур. м. (иногда несколько ниже). Живет на деревьях, в трещинах кбры
и в гниющей древесине пней и валежин. В подстилку скрывается лишь на
зимовку.
Размножение. Живородящий вид.
Систематическое положение. Несмотря на то, что
раковины моравских и балканских популяций данного вида мало отли-
отличаются друг от друга, чехословацкие авторы считают, что каждую чаеть
ареала заселяет свой подвид. Моравскому подвиду они дают название
moravica Brabenec. С их точки зрения главным доводом в пользу призна-
признания обоих подвидов является четкая разобщенность их ареалов. Мы не
разделяем этого взгляда, так как считаем, что подвиды должны разли-
различаться не только по распространению, но и по морфологическим и физио-
физиологическим признакам. А так как подобных различий пока что еще не
обнаружено, то нет и оснований для разделения вида на подвиды. Название
moravica Brabenec следует считать синонимом.
7. Род В ALE A GRAY
Gray, 1824, Zool. Journ., 1 : 61; P f e i f f e г L., 1856, Malak. Bl., 2 : 179
(Baleastra); L i n d h о 1 m, 1924:64.
Тип рода: Pupa fragilis Drap. [= Baled perversa (L.)].
Раковина бапшевидная, тонкая, хрупкая. Последний оборот без киля.
Устье без базального желобка. Замыкательный аппарат полностью реду-
редуцирован, или от него остался лишь небольшой рудимент верхней пластинки.
Клаузилий отсутствует.
Половая система очень сходна с таковой у Laciniaria Hartm. Это сход-
сходство выражается в первую очередь в том, что мужские половые пути вы-
вытянуты вдоль женских и имеют вид толстой трубки, веретеновидно расши-
расширенной у нижнего конца (вторичный пенис). Они не образуют петли,
столь обычной у большинства Clausiliidae. В связи с этим отсутствует
ретрактор пениса, крепящийся к диафрагме, а вместо Пего, как у Laci-
Laciniaria, имеется тонкая мускульная полоска, соединяющая нижние концы
семяпровода и пениса. Ретрактор семяприемника дает ответвления к се-
семяпроводу и подтягивает его к женским путям.
Распространение. Европа.
До сих пор все наши предшественники, когда у каких-либо видов дру-
других родов обнаруживали половой аппарат подобного типа (т. е. без муж-
мужской петли), относили их на этом основании в род Laciniaria. Однако для
Balea делалось исключение. Считалось, что почти полная редукция замы-
кательного аппарата и отсутствие клаузилия у Balea достаточны, чтобы
рассматривать эту группу как самостоятельный род. В то же время пола-
полагали, что ближайшим родственником ее является род Laciniaria.
С нашей точки зрения, оба вывода недостаточно обоснованы. В самом
деле, различные степени редукции замыкательного аппарата, до полного
его исчезновения (в том числе и клаузилия), известны в пределах других
родов Clausiliidae (Alopia H. et A. Adams, Reinia Kobelt). Однако всем
было ясно, что редукция замыкательного аппарата не дает оснований
выделять отдельные виды указанных родов в особые роды.
Что касается Balea, все авторы единодушно рассматривали ее как са-
самостоятельный род. Они считали, что нет ясных переходов в строении
раковины этой группы с раковинами других европейских родов Clausi-
Clausiliidae, которые имеют хорошо развитый замыкательный аппарат. Однако
262
V. CLAUSILIINAE
в действительности такие переходы существуют-{правда, чисто морфоло-
морфологические). Так, среди рода Micropontica Bttg. имеются виды с раковиной,
по, форме напоминающей Balea, замыкательный аппарат которые испытал
сильную редукцию, хотя клаузилий сохранился. Линдгольм (Lindholm,
1924) на этом основании выделил их в особую секцию Baleopsina Lindh.
Стоит отметить, что половая система Micropontica построена по типу,
характерному для Laciniaria и Balea, ¦
-Сказанное не означает, что мы рассматриваем род Micropontica, как
ближайшего родственника Balea. Нам хочется только подчеркнуть, что
у Clausiliidae редукция замыкательного аппарата не может являться
надежным основанием для, решения вопроса о- родственной близости
видрв или родов. .•-.¦¦,-.
Что касается признания особенной близости Balea к. Laciniaria, исходя
только из сходства их половых систем, то с нашей точки зрения и этоне-
г - достаточно обоснованно. Мы неоднократно указывали
в данной работе (стр. 77), что подобный тип полового
аппарата свойстве^ многим среднеевропейским и кав-
кавказским Clausiliidae, причем по раковине многие из
них резко различны.
Решить вопрос о родстве двух родов данного семей-
семейства можно только: исхбдя как из внутреннего строе-
строения, так и из особенностей раковины и в первую
очередь замыкательного аппарата. А так как у Balea
этот аппарат подвергся почти полной редукции, то в
данный момент мы лишены возможности указать, какой
именно род наиболее близок к данному. Одно ясно, что
ближайшие родственники этой группы находятся
среда тех родов Сlausiliinae, мужские половые пути
которых подверглись описанной выше (стр. 92) своеоб-
своеобразной реорганизации. -л -
Balea perversa (L.) (рис. 177, 178).
767.. (Turbo); Dra-
R о s s m a s s 1 e г,
L i n h ё, 1758, Syst. nat., ed.. X.
patnau d, 1801 : 64 (Pupa fragilis); no . „ .,
1839, Ieonogr., 2, 3—4 : 24, fig. 636 (fragilis); S t e e n Ъ е г g,
1914 : 39, fig., 25 (анатомия); Лихареви Раммедь-
мейе р, 1952 : 249, рис. 177.
Рис. 177. Batea
perversa (L.).
(Орйг.) .
Р а к овин а тонко и равномерно исчерченная, с
шелковистым блеском, просвечивающая, светло- или
зеленовато-роговая, с отдельными белыми штрихами.
Завиток конический, со сравнительно широкой @.7—
0.8 мм) округлой верхушкой. Оборотов 9—1Q, умеренно выпуклых,
плавно нарастающих; эмбриональные обороты B1/2) гладкие, блестящие;
последний оборот внизу округло-угловатый. Устье округло-ромбическое
или грушевидное, не выступающее, с широким, слабо выраженным сину-
лусом. Края устья слегка отвернуты, с тонкой бедой губой; колумелляр-
ный край отвесный, нередко почти параллелен палатальному; нижний
край плавно закруглен. Париетальный край плотно прилегает к стенке
раковины, иногда, разорван и места прикрепления связаны мозолью.
На париетальцОй стенке находится небольшой зубец. Пупок, имеет вид
открытой щели. \ ..'...,
Размеры:, высота раковины 8-—10, ширина 2.2^2.6 мм; высота устья
1.8—2.1, ширина Д .4—1.6 мм. , f ^
7. В ALE A
263,
вж
Внутреннее строение (Steenberg, 1914). Половая система
(рис. 178) очень походит на таковую у Laciniaria Hartm., в особенности на
L. biplkata (Mont,). Семяяйцевод, еще более короткий @.8 мм), а в связи
с этим и предстательная железа также не длинная. Наоборот, матка от-
относительно очень длинная B.8 мм) и вместе с сильно вздутым женским
каналом семяяйцевода превращена в выводковую сумку, в которой обычно
находятся 3—4 эмбриона. Длинный семяприемник C.4—3.7 мм) вершиной
почти достигает белковой железы. Отросток несколько короче B.6 мм).
Длинный семяпровод C.3—4.2 мм)
начинается очень высоко и плавно
расширяется в пенис (длина 1.0—
1.1 мм).
Распространение. В
СССР: горный Крым (лес на Бабу-
ган-Яйле близ Алушты).1 В Прибал-
Прибалтике этот вид найден только на не-
небольшом островке (Krausp, 1940), ле-
лежащем у берегов Эстонии к северо-
востоку от Таллина. Указания старых
авторов (Симашко, 1847; Сабанеев,
1880) о присутствии этого моллюска
в Латвии и в Ярославской области
Ошибочны. За него принимали моло-
молодые экземпляры других Clausiliidae.
Указание Урбанского (Urbanski, 1947)
об! обитании данного моллюска в
Полесье тоже, по-видимому, связано
с какой-то ошибкой, так как позд-
позднее A957 г.) автор не упоминает об
этом местонахождении. Вне СССР:
приморские области северной, запад-
западной и южной Европы и отдельные
островные ареалы в средней Европе.
В ископаемом состоянии на тер-
территории СССР найдена на о. Хиумэ
(Эстония) в отложениях послелитори-
новогр времени, а также в четвертич-
четвертичных отложениях ряда мест запад-
западной Европы.
Экология. Обитает чаще
всего в холмистой местности й в го-
горах на скалах, каменных оградах,
на заросших мхом стволах деревьев,
под камнями и в подстилке. ; ; ,, ;
¦,¦¦¦ Размножение. По данным одних авторов, наш моллюск яв-
является живородящим видом, но данным других—яйцекладущим (стр, 42).
В последнем случае кладка содержит 12—15 яиц, из которых на 15—20-й
день вылупляется молодь. Размножение происходит в течение всего теп-*
лого времени года. Животные становятся взрослыми к исходу первого
или к( началу второго года жизни. :
¦ркя
рнщ
Рис. 178. Balea perversa (L.). Половая
система. (По Стинбургу).
1 В коллекции Зоологического института Академии наук СССР. До еих нор един-
единственное указание о присутствии этого моллюска я Крыму бвдо сделано Кобельтом
(Kobelt, 1898).
264
V. CLAUSIL11NAE
8. Род QUADRIPLICATA BTTG.
В oettger О., 1878b : 63 (Clausilia, секция Euxina, группа Quadriplicata ;
Lindholm, 1924 : 66 (Laciniaria, подрод Euxina, секция Quadriplicata).
Тип рода: Clausilia quadriplicata A. Schm.
Раковина веретеновидная. Последний оборот с килем. Устье притуп-
ленно-овальное или грушевидное, выступающее, с более или менее глубоко
врезанным синулусом и с базальным желобком. Пластинки обычно хороша
развиты. Верхняя пластинка сливается со спиральной или если отделено
46
44
42
40
38
ч
OJ
5S
38
-><
4—х
40 42 44
щ
Сухуми^?
>
Батуми
х« <ZZ>2a »2S 41
©4 &S <©?
0 55
i
38
110 165 220 км
40
4cTaBpjnoiibw^
А __У/
ч г
42
46 48
1 (о;
Астрахань]р
К»
о Орджоникидзе
^^
хР
4А 46
50
W
^Махач-Кала
4W4J
~~~— ^
\
48
52
[
-Г
'Ъку
>^
50
54
X
\
ь
^-"
-Г
—^
52
56
1
s
>
\
л
-
) -
54
"'
44
42
40
38
36
Рис. 179. Географическоеас! спространение видов рода Quadriplicata Bttg. (Ориг.)
la — Q. quadriplicata (A. Schm); 16 — тоже, изолированные местонахождения; 2а — Q. aggesta
aggvsta (Bttg.); 26 — то же, изолированное местонахождение; 2« — Q. aggesta stauropoiitan'i
(Rosen); 3 — Q. lederi (Bttg.); 4 —Q. subaggesta (Ret.); s — Q. pumiliformis (Bttg.); 6 — Q. dipotau-
chen (Bttg.).
от нее'коротким промежутком, то обе пластинки лежат на одной линии,
т. е. на равном удалении от столбика. Толстая и высокая нижняя пластинка
лежит глубоко и не видна при прямом положении устья; в глубь раковины
она поднимается почти отвеспо и лишь у верхнего конца дает короткий
спиральный изгиб; внизу упирается в крупную краевую складку или бу-
бугорок. Имеется более или менее ясно выраженная промежуточная складка.
Субколумеллярная пластинка не видна через устье. Кроме главной складки,
имеются чаще всего 3 палатальных складки — верхняя, средняя и
нижняя; реже к ним добавляются еще 1 или 2 коротких складки. Довольно
широкая языковидная лопасть клаузилия имеет небольшую вырезку у ос-
основания рукоятки, а палатальный край ее и нижний конец утолщены.
Срединный и боковые зубы радулы с дополнительными зубцами.
8. QUADRIPLICATA 265
Мужские половые пути вытянуты вдоль женских. Толстый семяпро-
семяпровод, плавно расширяясь, переходит в веретеновидно вздутый пенис,
внутри которого имеется несколько продольных складок. Тонкая мус-
мускульная полоска связывает друг с другом нижние концы пениса и семя-
семяпровода. Многочисленные мускульные волокна подтягивают семяпровод
к женским половым путям. В целом план строения сходен со строением
половой системы у Laciniaria Hartm.
Распространение. Кавказ (рис. 179).
Наиболее характерными особенностями раковины этого рода являются
форма и положение нижней пластинки и присутствие чаще всего 4 складок
(главной и 3 палатальных).
Длительное время данную группу рассматривали как секцию внутри
Euxina Bttg. Основанием этому служило то, что у некоторых кавказских
видов названного рода (подрод Caucasica Bttg.) тоже имеется 4 складки на
палатальной стенке последнего оборота, а нижняя пластинка сходной
формы. Однако, как мы убедились, половая система Quadriplicata постро-
построена совершенно иначе, чем у Euxina. В замыкательном аппарате обеих
групп также имеются существенные отличия. В данном роде верхняя и
спиральная пластинки располагаются на одном расстоянии от столбика
и чаще всего сливаются друг с другом. У Euxina обе пластинки распола-
располагаются на различном удалении от столбика и их концы обычно заходят
друг за друга и никогда не сливаются воедино.
Поэтому есть все основания считать Quadriplicata особым родом, до-
довольно далеко отстоящим от Euxina, который, с нашей точки зрения, при-
принадлежит к другому подсемейству (Euxininae).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА QUADRIPLICATA
1 ( 2). Раковина широкая — ширина 4.0—4.5 мм, с относительно боль-
большим устьем — высота 3.8—4.2, ширина 2.8—3.2 мм .....
1. Q. quadriplicata (A. Schm.).
2 ( 1). Раковина более узкая, ширина ее не превышает 4.0 мм, со значи-
значительно меньшим устьем.
3F). Главная и все палатальные складки начинаются на правой сто-
стороне раковины.
4( 5). Раковина веретеновидная. Оборотов 10—11%. Между сравни-
сравнительно длинными главной и нижней палатальной складками
лежат еще 2 коротких палатальных складки (верхняя и средняя)
2а. Q. aggesta aggesta (Bttg.).
5( 4). Раковина веретеновидно-цилиндрическая. Оборотов 11%—1274-
Между главной и нижней палатальной складками лежат 3—4
коротких палатальных складки
26. Q. aggesta stauropolitana (Rosen).
6C). Главная и все палатальные складки начинаются на брюшной или
правобрюшной стороне раковины.
7 ( 8). Наиболее длинной складкой является главная, которая тем не
менее не видна при прямом положении устья .
3. Q. lederi (Bttg.).
8G). Одна из палатальных складок по длине равна главной или больше
нее и обычно видна при прямом положении устья.
9 A0). Раковина светло- или зеленовато-роговая. Главная и палатальные
складки начинаются на право-брюшной стороне раковины. Из
краевых складок имеется только одна, соединяющая край устья
с нижней пластинкой 4. Q. subaggesta (Ret.).
266 Vi ClAUSILUNAE ¦
ДО ( 9). Ракованд окрашена /темнее. Главная и палатальные складки
Шчинаются на брюшной стороне раковины над еинулусом. Крает
;вых; сндадок чаде всего не ,меныце 2—&..- -.,.-. - . •¦, а
11 A2). Раковина,довольно вздутая, густо ребристо исчерченная или
,-..-:.¦',¦ ,-. ребристая-(межреберные промежутки:уже или такой же щирины4
¦ ,.-.-, как И; ребра) .j Оборотов J,2i—15. Наиболее^ длинной является сред?-
няя, наиболее короткой верхняя палатальная складка .,,.>
,:, ,•»'.-;.•.<.•..-.¦.'-.. . .; 5. Q, pttmili?ermiSr(Bttg.).
12 A1). Раковина очень стройная* редко.ребристая или ребристо исчер-
¦-.: ченная (межреберные промежутки значительно шире ребер),
Оборотов 15—17. Наиболее длинной является верхняя, наиболее
короткой средняя палатальная складка . t «. ^ ..,.(.;; . .
.ч ,. ....... .л ....... . . 6. Q»; dipolauehen (Bt%:),.
1. QuadripHcata quadriplicata (A. Schm.) (риС. t80, 481).
. S chip i (It A., 1868 : 163 (Clausilia); К о b e 1 t, 1893, Iconogr., N. F./6 : 24*,
fig. 993 [Cldusilia (Euiina)]; -P о з eir, 1914 : 198 [Clausilia1 (Euxina)]; Лих ар eh
И Р а м м e']f ь м ёж e p, 1952 : 243, рий:' 167 [Laciriiaria {Euxina)]. ' ! ;
,.. Р-д к о в.и,н,,а, вздузсая^ тонкр, густо ребристо исчерчена,: слабрпро-
свечивгиощая,. с шелковистым, блеском, коричневато-роговая,, с. пучками
белых штрихов. Верхняя часть завитка ниже довольно. трнкрй округлой
верхушки, имеет вогнутые очертания. Максимальная ширина приходится
на предпрсдедний оборот. Оборотов 12—13, реже 11, слабо выпуклых;
эмбрйойальйыё обороты B1/4—2V2) гладкие, блестящие; побледний оборот
кнвду заметнрi «ужен, на затылке с крупными ребрами» уплощен и даже
(в области,верхней палатальной складки) слегка вдавлен, вни^у с длинным
и^ высоки^ килем- Устье, притупленно-овальное, внутри красновато-корич-
красновато-коричневое, с широким, умеренно врезанным синулусоми со слабым .базальным
желобком. Края устья белые, широко отвернутые, утолщенные. Толстая
верхняя пластинка впереди достигает,края устья, другим концом глубоко
уходит внутрь раковины; чаще всего она не вступает ц связь,,с,-;лежащей
с, нею по одной- линии спиральной; пластинкой,, а лишь близко подходит
к,последней; ,реже пластинки соединены нитевидным утолщением или же
сливаются друг с,другом. У основания нижней пластинки, лежит складка
или; бугрррк; верхний конец згой пластинки находится на правой стороне
ракрвйн.Ы) где. он да^т крроткий спиральный изгиб. Главная и все 3 ца-
латаль,ных складки начин-аются,на, правой стороне раковины. Главная,
а также верхняя и нижняя палатальные складки длинные и видны при
прямом,положении устья; из них< самой длинной обычно бывает верхняя
палатальная, укладка. Средняя складка очень короткая и снаружи^ не
видна. Верхняя складка обычно сливается с небольшой палатальной мо-
мозолью. Палатальный драй и лижний, конец лопасти клаузилия сильно
утолщены. .,.,-, ; . ,.... , ..,,,. ....-,, ... ,.,-.,¦
Размеры;, высота раковины 14.0—17.5, ширина 4.0—4.5 мм; высота
устья ;3.,8—4.2, ширина,2.8—3.2,мм. ,', ,',. , ,'. '. , .¦;
В н у т р е нн е е строе н и е. Ма.териал; ,1) Закаталы, Азербай-
джанс^д С,СЙ,,сбор и определение наше, 15 V, 195$ — 1 экз.;, 2) Нагорный
Карабах, сбор и, определение, наще, 2 IX 1954 — 1 экз.
8. QUADWPUCATA
261
Размеры срединного зуба 19:6 X 'I4jx-;' п/ирйва радулы 585ц,.
Отросток сильно извит и немного короче семяприемника (рис. 181).
Матка и влагалище дочти одной длины. Последнее очень толстое, муску-
мускулистое, почти цилиндрической формы.
Распространение (рис. 479). Обитает в лесах южных скло-
склонов центральной и восточной части Большого Кавказа, северных и северо-
северовосточных отрогов Малого Кавказа. Кроме того, найден в пойменных ле-
«ах нижнего Терека (Кадитина* 1958). Наиболее западные находки сделаны
Рис. 180. Qiiadriplieatd quadriplicata
. (A. Sobpi.). (Ориг.>.
А — общий вид t раковины; В '— последний
оборот (вид сзади) и просвечивающие сквозь
' ' •' ; Него складки.
¦ркл
Рис. 181. Quadriplicata quadriplicata (A.
Schm.). Половая система. (Ориг.).
в Раче и в окрестностях Стецанавана- Отсутствует в лесах Тальппа л,
по-видимому, также в: лесах Зангезура. ,
Указание Розена A,914) да находку его в Чорохском вилайете Турции
(Светибар) неверно, и связано с ощибкдй в . определении. ,,
Эк q л о г и я. рбитает во дсех горизонтах горных лесов, и в низовых
лесах как на деревьях, так и в подстилке и под камнями.,
Р а з м д о жен и е« .Живородящий вид.. г
Систематическое , п о л о же ни е. От всех остальньщ
видов того же рода данный, вид отличается, более вздутой раковиной д
массивным^ верхней и нижней палатальными складками, которые всегда
хорошо видны даже при прямом положении устья.
268
V. CLAUSILIINAE
2a. Quadriplicata aggesta aggesta (Bttg.) (рис. 182—184).
Boettger 0., 1879a : 34, t. 1, fig. 6 [Clausilia (Euxina)];W esterlund,.
1901 : 30 [Cl. (Euxina) aggesta var. protracta]; Розен, 1914 : 199 [Clausilia (Euxina)]-T
Ретовский, 1914 : 317 [Clausilia (Euxina)]; Лихарев и Ра имел ь-
м е й е р, 1952:243, рис. 166 [Laciniaria (Euxina)].
Раковина ребристо исчерченная, просвечивающая, довольна
блестящая, зеленовато-коричневая или темно-роговая, с белыми штри-
штрихами. Оборотов 10—llVai слабо выпуклых, разделенных ясным швом;
эмбриональные обороты (около 21/2) гладкие, образуют
довольно широкую тупую верхушку; последний оборот
за устьем покрыт крупными пластинчатыми ребдами,
внизу с длинным четким килем, окайшгенным глубокой
бороздой. Устье грушевидное, с врезанным высоколе-
жащим синулусом и ясным базальным желобком. Края
устья широкие, слегка утолщенные, белые. Верхняя
пластинка невысокая, иногда очень низкая; впереди
она достигает края устья, сзади, плавно снижаясь, пе-
переходит в спиральную пластинку. Последняя начина-
начинается почти на брюшной стороне и достигает максималь-
максимальной высоты на правой стороне раковины. Нижняя
пластинка внизу утолщена й обрублена, с краем устья
связана довольно крупйой складочкой. Промежуточная
складка выражена слабо. Главная и палатальные складки
начинаются на правой стороне раковины; из них самой
длинной бывает главная, не-
А сколько короче ее нижняя
палатальная складка; осталь-
остальные складки еще короче; при
прямом положении устья ни
одна из названных складок
не видна. На краю устья,
между верхней и нижней
пластинками, нередко, име-
А- Б ются 2—3 коротких скла-
т> ,о, п . . ,. , , t ,D() ч дочки или узелка. Клаузи-
Рис. 182. Quadriplicata aggesta aggesta (Bttg.). tt „ J J „
(Ориг.). лии сходен с таковым у Q.
а - общий вид раковины; б - клаузилий. quadriplicata (A. Schm.).
Размеры: высота раковины
11.0—12.5, ширина 3.0—
3.5 мм; высота устья 2.7—3.0, ширина 2.0—2.2 мм.
Изменчивость. В некоторых местах встречаются популяции,
состоящие из экземпляров с более стройной и узкой раковиной, устье
которой более узкое, чем у типичной формы (var. praegracilis Rosen, 1914):
высота раковины 11.5-L-12.5, ширина 2.5—2.8 мм.
В Гудаутском районе Абхазии, выше озера Мзи, в каменистых россы-
россыпях альпийской зоны, среди зарослей рододендрона обнаружена популя-
популяция с башневидно-веретеновидной раковиной, у которой заметна частичная
редукция замыкательного аппарата. Последнее проявляется в том, что
верхняя пластинка развита слабо, спиральная пластинка распалась на
серию коротких складок (параллельные пластинки), а главная и пала-
палатальные складки в свою очередь дали серию из 8—10 мелких складочек,
лежащих на спинной стороне раковины, т. е. ближе к устью, чем у типич-
типичной формы.
8. QUADRIPLICATA
269
Возникновение этой экологической формы связано с обитанием в высо-
высокогорной зоне, в условиях короткого вегетационного периода. Последнее
привело к преждевременному половому созреванию и к раннему окон-
окончанию роста раковины (см. стр. 47). Эту форму мы назвали f. alpestris
Likharev. ?. nov. (рис. 183).
Внутреннее строение. Материал: 1) Красная Поляна,
сбор Рылова, 18 VIII 1929, определение наше — 1 экз,; 2) Кавказский за-
заповедник, пастбище Абаго, сбор и определение
наше, VIII 1956 — 2 экз.
D Г о ¦ /86 . 12к
Радула: ^_+(- + ^_^
Размеры срединного зуба
радулы 405 (д..
Сильно извитой отросток
яается лишь немного выше
него конца семяпровода (рис.
Семяприемник значительно
X 2] X 120.
14"х 9.8 р,
начи-
верх-
184).
длин-
ширина
нее — его резервуар лежит непосред-
непосредственно под белковой железой. ^Вла^
галище значительно__длиннее матки.
От ретрактора семяприемника отхо-
дит мускул, подтягивающий семяпро-
семяпровод к женским путям. Той же цели
служат многочисленные волокна, свя-
связывающие семяпровод с влагали-
влагалищем.
Распространение (рис.
179). Северные склоны западной
части Большого Кавказа от Ново-
Новороссийска до истоков реки Большая
Лаба_{бассейн Кубани). На южных
склонах: от Красной Поляны до Гу-
даутского района Абхазской АССР
(включительно).
Экология. В пределах сво-
своего ареала обитает во всех зонах
горных лесов, а также в субальпий-
субальпийской и альпийской зонах на деревьях,
под камнями и на скалах.
Рис 183.
Quadriplica-
ta aggesta f.
alpestris Li-
Likharev.
(Ориг.).
Рис. 184. Quadripli-
cata aggesta aggesta
(Bttg..). Половая си-
система. (Ориг.).
26. Quadriplicata aggesta stauropolitana (Rosen) (рис. 185).
Розен, 1901 : 11 [Clausilia (Euxina)]; Lindholm, 1913c : 160 (Clausi-
lia).
Отличается от типичного подвида следующими особенностями: раковина
веретеновидно-цилиндрическая; оборотов 111/2—^U\ между довольно
длинными главной и нижней палатальной складками лежат 3—4 коротких
складки.
Размеры: высота раковины 13.5—16.0, ширина 3.5—4.0 мм; высота
устья 3.0—3.5, ширина 2.2—2.5 мм.1
1 Лектотип данного подвида хранится в Музее Грузии (коллекция моллюсков,
№ 1578а). Он имеет следующие размеры: высота раковины 14.2, ширина 3.7 мм; высота
устья 3.4, ширина 2.3 мм; 113/4 оборотов. Место сбора: Ставрополь. Здесь же находится
один паралектотип. . :.
270
V. CLAU&ILLINAE
Внутрен&ее строений не изучено.
Ра си ростр а ни н и е. Леса и парки окрестностей города Став-:
рополя.
3. Quadriplicata lederi (Bttg.) (рис. 186—188).
Вое 11 ge r O., 1879a : 36, t. i, fig. 5 [Clausilia (Euxina)]; Boettger O.,
1879b : 409, t. 10, fig. 5 [Cl. (Euxina) gradata]; Bdettger 0., 1881a : 238, t. 9,
fig. 19 [Cl. (Euxina) lederi var. triadis]; К о b e 1 t, 1893, Iconogr., N. F., 6 : 12, %.
976 [Clausilia (Euxina)]; 13, fig. 977 [Cl. (Euxina) lederi var. gradata et var. triadis]i
Лихарев и Раммельмейер, 1952 : 244, рис. 168 [Laciniaria (Euxina) ].
Pa ков и на вздутая (максимальная ширина приходится на 1—2
предпоследних оборота), ребристо исчерченная, просвечивающая, с шел-
шелковистым блеском, коричнево- или светло-роговая, с редкими белыми
Рис. 185. Quadriplicata aggesta
stauropolitana (Rosen). (Ориг.).
A — общий вид раковины; Б — послед-
последний оборот (вид справа) и просвечиваю-
просвечивающие сквозь него складки.
Рис. 186. Quadriplicata lederi (Bttg.).
(Ориг.).
А — общий вид раковины; Б — последний
оборот (вид справа) и просвечивающие
сквозь него складки.
штрихами. Оборотов 11—121/2, слабо выпуклых, разделенных довольно
глубоким швом; эмбриональные обороты BV2) гладкие, блестящие и выпу-
выпуклые, образуют довольно широкую верхушку; последний оборот книзу
сильно сужен, ребристый, на затылке уплощен, внизу с крупным килем,
окаймленным четкой бороздой- Устье ромбическо-грушевидное, почти
отвесное, внутри коричневатое, с синулусом, глубоко врезанным, высо-
колежащим и отогнутым к оси раковины. Края устья широко отвернуты,
слегка утолщены, белые. Низкая верхняя пластинка впереди достигает
края устья и даже нередко слегка выступает за его пределы; назад она
8. $UAbRl&LlGA?A
271
'гйнетсл более или менее глубоко, где;, ййавмо> «нйжше!» ю утядаЯсь!,
схоДйа1 на He*t -от спиральной пяастшйкй она^ обычно далеко oigtohtv реже
связана с нею нитевидным утолщением стенки раковины. Нижняя;>Btaat-
6Э№
¦'¦¦¦¦¦;; • • Рйс. 187. Qaadfipliduta ЩёН(ВЩ'.}. Ра#уиа, (Ориг.). : ¦ ' --'
стйнка внутрь раковины йоднймйется поФги отвесно, но более наклбййб,
чем у Q. aggesta; внизу она утолщена:[ и обрубдейа,: с "краей у^йтья
связана единственной краевой складкой. Имеется кр*упнак J проМе?кутрч-
йая скЛа^ка; которая задним кЬйДОмвьгКо-
дит в промежуток мейду верхней й сДйраль-
ной йластйнкамй. Главная й ffce палатальные
складки начина1Ьтся на брй/ншой или tfpaBO-
брюшн'ой стороне. Главная складка довольно
длинная (занимает около ^^ оборота) и ЙйднА
йрй косом положении устья. ПоД яей |)асйЬ-
лагаются 3—4 палатальных складки, кото-
которые параллельны д#Уг другуипр!6'стира1ю1сй
вперед на а/4 оборота; из них несколько бо-
более длинной бывает нижняя складка. Клау-
аилйй с^ходей с таковым: у Q. quadripli-
cata (A. Sclitii.). ; ; -'-¦ 5
Размеры: высота раковины 14.0—17.0,
ширина 3.5^4;0 мм; высота устья 3.2—3.5,
ширина 2.3—2.7 мм.1
В н у т р « н н е е с т р о е н и е. i Ма-
Материал: Мцхета, Грузия, сбор и опредвЛ№
ние наше, 12 V 1957 —¦ 1 экз.
Радула (рие. 187):;,[-|.Ц (^ +
+Й)Х2]х125.
Размеры срединного зуба 14 х 9.8(х, ши-
ширина радулы 450 р., . , .
Отросток очень длинный, и сильно изви-
извитой (рис. 188). Семяприемник толстый, с
крупным резервуаром, лежащим непосред-
непосредственно ниже белковой железы. Влагалище
толс^о^_щ;шндщческое, в_4—5_раз ллиннее
матки. Семяпровод лри~~ переходе в пенис
дёлает"коленообразный изгиб. > .
Распространение (рис. 179). Южные склоны Большого
Кавказа от Очамчирского района Абхазской АССР до окрестностей Тбилиси
Рис. 188. Quadrlplicata
kderi (Bttg.).
Половая система. (Ориг.).
1 Лек^отип данного вида хранится в Музее Грузии ^коляекция моллюскор,
№ 1585). Он имеет следующие размеры; высота раковины 16.4. вдирина 4.0 мм; высота
устья 3.6, ширина 2.4 мм; 13 ^ оборотов. Место сбора: Сурам, сборщик Лёдер.
272
V. CLAUSILIINAE
и Мцхеты (наиболее восточная находка сделана у поселка Марткоби),
Сурамский хребет и лесистые отроги Малого Кавказа в районе Боржоми
и Абастумани.
Розен A914) указывает на одну находку в Чорохском вилайете Турции
(Светибар). К сожалению, мы не имели возможности проверить определе-
определение этого автора, так как соответствующий экземпляр отсутствует в кол-
коллекции Музея Грузии.
Экология. Обитает в горных дубовых и буковых лесах, а также
в лианных лесах колхидского типа на деревьях, скалах и под камнями.
Поднимается в горы до 1500 м над ур. м.
Систематическое положение и синонимия. Из
всех остальных видов того же рода к данному виду наиболее близко стоит
Q. aggesta (Bttg.). От последнего он отличается: 1) раковина более резко
веретеновидная и вздутая; 2) нижняя пластинка уходит вглубь раковины
не так круто, а с небольшим перегибом; 3) палатальные и главная складки
начинаются глубже; 4) средние палатальные складки длиннее; 5) верхняя
и спиральная пластинки чаще всего обособлены друг от друга; 6) имеется
только одна краевая складка, которая соединяет ниж-
нижнюю пластинку с краем устья (уQ. aggestasix несколько).
Бетгер, имея в своем распоряжении единичные экзем-
экземпляры данного вида, обнаружил среди них некоторые
различия. Это дало ему основание описать две разновид-
разновидности— gradataВoettger A879b) и triadis Boettger A881a).
Сопоставив наши сборы с оригиналами этих разновидно-
разновидностей, мы убедились, что они укладываются в пределы
индивидуальной изменчивости, в связи с чем их следует
рассматривать как синонимы.
4. Quadriplicata subag-
gesta (Ret.) (рис. 189—190).
Retoswki, 1887 : 37, t.
1, fig. 4 [Clausilia (Euxina)];
Boettger 0., 1889: 3ft [Clau-
[Clausilia (Euxina)}; Retoswki,
1889 : 257 [Clausilia (Euxina)];
К о b e 11, 1893, Iconogr., N. F.,
6 : 14, fig. 978 [Clausilia (Euxi-
(Euxina); Ретовский, 1914:318
[Clausilia (Euxina)]; Лихарев
и Раммельмейер, 1952:
243 [Laciniaria (Euxina)].
Раковина умеренно
вздутая (максимальная ши-
ширина приходится на предпо-
предпоследний оборот), довольно
твердостенная, сильно ребри-
ребристо исчерченная, просвечивающая, слабо блестящая, светло- или зеленовато-
роговая, с редкими белыми штрихами. Верхняя часть завитка с вогнутыми
очертаниями ниже тонкой цилиндрической верхушки. Оборотов 11—121/2,
слабо выпуклых, разделенных довольно глубоким швом; эмбриональные
обороты (около 2V2) гладкие, блестящие; последний оборот книзу сильно
сужен, с крупным килем, окаймленным ясной бороздой. Устье грушевид-
грушевидное, сильно выступающее, с узким, глубоко врезанным и наклоненным
к оси раковины синулусом и с неглубоким базальным желобком. Края
устья белые, слабо отвернуты и утолщены. Толстая и довольно высокая
косо лежащая верхняя пластинка впереди слегка выступает за край
А Б
Рис. 189. Quadriplicata subaggesta (Ret.). (Ориг.)
А — общий вид раковины; Б — клауадлий.
8. QUADRIPLICATA 273
устья, вглубь раковины уходит, плавно снижаясь, и либо сливается со
спиральной пластинкой, либо отделена от нее более или менее коротким
промежутком. Последняя берет начало на брюшной стороне, быстро на-
нарастает в высоту, достигая максимума на правой стороне, после чего
плавно спускается к нижнему концу. От нижней пластинки при прямом
положении устья видна лишь крупная краевая складка (иногда отсут-
отсутствует); нижний конец пластинки обрублен. Главная и все 3 палатальных
складки начинаются на право-брюшной. стороне раковины. Главная
складка сравнительно длинная, но короче верхней складки и не всегда
видна при прямом положении устья, в то время как верхняя складка
# 26
с 1
Рис. 190. Quadrlplicata subaggesta (Ret.). Радула. (Ориг.).
обычно видна. Средняя палатальная складка очень короткая и видна
лишь сквозь стенку раковины. Нижняя длиннее средней, но короче
верхней, приблизительно одной длины с главной складкой. Языковидная
лопасть клаузилия имеет неглубокую вырезку у рукоятки; волнисто
изогнутый палатальный край лопасти и нижний конец слегка утолщены.
Размеры: высота раковины 13.5—16.0, ширина 3.5—4.0 мм; высота
устья 3.0—3.3, ширина 2.0—2.5 мм.1
Внутреннее строение. Материал: 1) Цихис-Дзири, сбор
Быкова, 5 VII 1915, определение Линдгольма — 1 экз.; 2) Зеленый мыс,
Батуми, сбор Рылова, 21 IX 1929, определение йаше — 1 экз.; 3) Батум-
ский ботанический сад, сбор и определение наше, 20 IV 1957 — 1 экз.
Радула (рис. 190): | + I™ + ^) X 2.
Размеры срединного зуба 8.4 X 12.6 |л, ширина радулы 450 р.
Половая система устроена очень сходно с остальными видами этого же
рода.
Распространение (рис. 179). В СССР: субтропики примор-
приморской полосы Аджарской АССР. Вне СССР: Чорохский вилайет Турции.
Первоначально вид был описан по экземплярам, найденным на берегу
Крыма в выбросах моря.
Экология. Обитает в лесу, на стволах деревьев и под камнями.
Систематическое положение. Замыкательный аппарат
данного вида наиболее сходен с таковым у Q. quadriplicata (A. Schm.),
л о у последней он располагается ближе к устью, верхняя палатальная
складка более толстая и, как и все устье, окрашена в красновато-коричне-
красновато-коричневый цвет. По форме раковины и устья данный вид наиболее сходен с Q. 1е-
deri (Bttg.).
1 Лектотип данного вида хранится в Зоологическом институте Польской Акаде-
Академии наук. Он имеет следующие размеры: высота раковины 15, ширина 3.7 мм; высота
устья 3.0, ширина 2.2 мм; 13 V2 оборотов. Место сбора: «Крым, выносы».
274
V. CLAUSILIINAE
5. Quadriplicata pumiliformis (Bttg.) (рис. 191—193).
Boettger O., 1881a : 234, t. 9, fig. 17 [Clausilia (Euxina)]; Розен, 1914 :
198 [Clausilia (Euxina)]; Ретовский, 1914 : 316 [Clausilia (Euxina)]; Лиха-
Лихарев и Раммельмейер, 1952 : 249, рис. 165 [Laciniaria (Euxina)].
Раковина довольно вздутая (максимальная ширина приходится
на 2 предпоследних оборота), тонкостенная, ребристо исчерченная, реже
тупо ребристая, просвечивающая, с шелковистым блеском, коричнево-
роговая, с белыми штрихами. Верхняя часть завитка имеет вогнутые
очертания и тонкую цилиндрическую верхушку. Оборотов 12—15, слабо
выпуклых, разделенных мелким врезанным швом; эмбрио-
эмбриональные обороты B1/2—3) гладкие; последний оборот книзу
сильно сужен, на затылке уплощен и ребристый, с круп-
крупным килем, окаймленным ясной бороздой. Устье малень-
маленькое, грушевидное, сильно выступающее, внутри коричне-
коричневое, с неглубоким базальным желобком и округлым нижним
краем; синулус узкий, глубоко врезан и отогнут к оси
раковины. Края устья расширены, утолщены и слегка отвер-
отвернуты. Верхняя пластинка невысокая, в просвете устья ле-
лежит косо; впереди она дохо-
доходит до края устья и даже не-
нередко слегка выступает за
его пределы, сзади переходит
в спиральную пластинку
(иногда обе пластинки раз-
разделены коротким промежут-
промежутком). Спиральная и нижняя
пластинки начинаются на
брюшной стороне (первая не-
несколько глубже, чем вторая).
Высота спиральной пластин-
пластинки быстро нарастает к правой
стороне и плавно убывает по
направлению к устью. Глав-
Главная и все 3 палатальных
складки начинаются на брюш-
брюшной стороне, обычно над си-
нулусом. Наиболее длинна средняя палатальная складка, которая хо-
хорошо заметна при прямом положении устья. Главная и нижняя пала-
палатальная складки несколько короче и видны лишь при косом положении
устья. Наиболее коротка верхняя палатальная складка (простирается
меньше чем на 1/4 оборота), котора видна лишь сквозь стенку раковины.
Колумеллярный край устья имеет большее или меньшее число коротких
складок или узелков. Наиболее постоянна сравнительная крупная складка,
связывающая нижнюю пластинку с краем устья; обычны также еще
2—3 складки или узелка, лежащие между верхней и нижней пластинками;
несколько реже, но также довольно часто, имеются 2—3 складки ниже
нижней пластинки (точнее — ниже краевой складки, являющейся про-
продолжением последней). Клаузилий такой же, как у Q. subaggesta, но рас-
располагается глубже, так что лопасть лежит над синулусом, а рукоятка
крепится к столбику в глубине предпоследнего оборота.
Размеры: высота раковины 12.0—14.5 (реже до 17.0), ширина 3.0—
3.5 мм; высота устья 2.5—2.7 (реже 3.0), ширина 1.9—2.1 мм.
Изменчивость. Форма раковины довольно постоянна. Тем не
менее иногда встречаются экземпляры со сравнительно приземистой рако-
Рис. 191. Quadriplicaia pumiliformis (Bttg.).
А — общий вид раковины (по Лихареву и Раммель-
Раммельмейер); Б — клаузилий. (Ориг.)-
8. QUADR1PLICATA
275
виной, последний оборот которой не уже предпоследнего (var rusticana
Retowski, 1914), или с очень узкой и вытянутой раковиной. Сильнее всего
варьирует степень развития краевых складок устья. Наряду с типичными
могут встречаться отдельные экземпляры совсем без краевых складок и
Рис. 192. Quadruplicate, pumiliformis (Bttg.). Ра-
Радула. (Ориг.).
узелков (var. inarmata Lindholm, 1913), у которых сохраняется лишь
одна складка, соединяющая край устья с нижней пластинкой. С другой
стороны, бывают экземпляры, у которых складчатость захватывает и
палатальный край (var. circumplicosa Lindholm, 1913).
ил
Рис. 193. Quadriplicata pumiliformis (Bttg.). Половая
система. (Ориг.).
Внутреннее строение. Материал; 1) Хоста, сбор Рылова,
10 IX 1929, определение наше — 2 экз.; 2) Кавказский заповедник, Гузе-
рипль, сбор и определение наше, 6 VIII 1956 — 1 экз.
Радула (рис. 192): [| + (^ + ^) X 2] X 150.
Размеры срединного зуба 11.2 X 7 ц, ширина радулы 450 ц.
Семяприемник сравнительно короткий и не достигает белковой железы
(рис. 193); его отросток сильно извит и начинается немного выше верхнего
276
V. CLAUSIL11NAE
конца семяпровода. Как пенис, так и влагалище веретеновидно вздуты.
Внутри их имеются несколько продольных валиков.
Распространение (рис. 179). В основном в приморской полосе
между Сочи и Сухуми, хотя отдельные находки известны как севернее
(Новороссийск), так и южнее (Поти). Вглубь континента проникает в пре-
пределах зоны распространения каштановых лесов (Красная Поляна, Це-
бельда). На западной части северных склонов Большого Кавказа найден
в бассейне реки Белой до высоты 1000 м.
Экология. Обитает чаще всего в субтропических
(с самшитом) или в каштаново-буковых лесах, реже в ду-
дубовых, на скалах, деревьях, под корой и в гниющей дре-
древесине, реже в подстилке.
Систематическое по-
положение. От всех прочих
видов того же рода данный вид,
вместе с Q. dipolauchen (Bttg.),
резко отличается очень глубоким
положением клаузилия и складок.
6. Quadriplicata dipolauchen
(Bttg.) (рис. 194).
Boettger О., 1881а : 235, t. 9,
fig. 18 [Clausilia (Euxina)]; Розен,
1914 : 198 [Clausilia (Euxina)]; Лиха-
Лихарев и Раммельмейер, 1952 :
242 [Laciniaria (Euxina)].
Раковина очень стройная
(максимальная ширина приходится
на 1 или 2 предпоследних оборо-
оборота), твердостенная с крупными
пластинчатыми белыми ребрами
(около 30 на предпоследнем обо-
обороте) , разделенными широкими
матовыми межреберными проме-
промежутками, просвечивающая, темно-
роговая. Верхняя часть завит-
завитка имеет вогнутые очертания
и тонкую цилиндрическую верхушку. Оборотов 15—-17 (реже_до _18L_
верхние слабо выпуклые, средние почти плоские, разделенные~1эелым
мелким швом; эмбриональные обороты C1/2) гладкие, блестящие, одина-
одинаковой ширины; последний оборот книзу сильно сужен, на затылке упло-
уплощен, с очень крупными ребрами и мощным килем. Устье маленькое, оваль-
овально-грушевидное, внутри коричневое, выступающее, с высоким умеренно
врезанным синулусом и глубоким базальным желобком. Края устья ши-
широко отвернуты; палатальный ниже синулуса утолщен; нижний край
широко закруглен. Низкая, косо поставленная верхняя пластинка впе-
впереди достигает края устья, сзади плавно переходит в длинную спиральную
пластинку. Нижняя и субколумеллярная пластинка такие же, как у
Q. pumiliformis. Главная и все 3 палатальных складки начинаются на
брюшной стороне, над синулусом. Наиболее длинная и мощная верхняя
палатальная складка хорошо заметна при прямом положении устья.
Главная и нижняя палатальная складки несколько короче и видны лишь
при косом положении устья. Наиболее короткая средняя палатальная
складка (занимает лишь Ve оборота), которая видна только сквозь стенку
Рис. 194. Quadriplicata dipolauchen (Bttg.).
(Ориг.).
А — общий вид раковины; В — последний обо-
оборот и устье.
9. MUCRONARIA 277
раковины. Между верхней и нижней пластинками по краю устья распо-
располагаются 3—5 небольших складочек, из которых одна (самая крупная)
соединяется с нижней пластинкой. Реже бывают складки и в нижней части
колумеллярного края и очень редко — на палатальном крае. Клаузияий
такой же формы и занимает то же положение, как у Q. pumiliformis.
Размеры: высота раковины 15—18, ширина 3.3—3.7 мм; высота устья
2.9—3.3, ширина 2.1—2.4 мм. 2
Изменчивость. В пределах ареала встречаются популяции, ра-
раковины которых густо покрыты притуплёнными складками E5—60 на
предпоследнем обороте); однако промежутки между складками шире их
самих (var. multistriata Retowski, 1887).
Внутреннее строение. Материал: Грузия, Цагери, дубо-
дубовый лес, на юго-восточном склоне, сбор Н. Н. Акрамовского, 10 VIII 1957,
определение наше — 1 экз.
Радула: [| + (^ + 3=7) X 2] X c/d12°-
Размеры срединного зуба 13.3 X 10 \l, ширина радулы 424 \i.
Половой аппарат такой же, как у Q. pumiliformis (Bttg.).
Распространение (рис. 179). Предгорья (не выше 500м над
ур. м.) и отдельные возвышенности северной окраины Колхидской примор-
приморской равнины (не ниже 100 м над ур. м.) между реками Хоби иЦхалтубо-
Цхали (западная Грузия).
Экология. Обитает на известняковых скалах.
Систематическое положение. Данный вид наиболее
близок к Q. pumiliformis, хотя и отличается от него более стройной рако-
раковиной, образованной большим числом оборотов. Скульптура поверхности
раковины более резкая (за исключением \»ar. multistriata Ret.) и редкая.
В отношении замыкательного аппарата главное отличие заключается
в том, что у данного вида самой длинной является верхняя палатальная
складка, а наиболее короткой — средняя, тогда как у Q. pumiliformis
как раз обратная картина.
9. Род MUCRONARIA BTTG.
Boettger' О., 1877а : 84 (Clausilia, секция Euxina, группа Polyptychia nov.,
поп F e 1 d е г, 1874); 84 (Clausilia, секция Euxina, группа Mucronaria nov.); L i n d-
h 0 1 m, 1924 : 66 (Laciniaria, подрод Euxina, секция Multiplicaria, nom. nov., вместо
Polyptychia Bttg.).
/ •
Тип рода: Clausilia acuminata Mouss.
Раковина различной формы — от веретеновидной до башневидной.
Последний оборот с килем. Устье грушевидное, с базальным желобком;
ось устья наклонена влево. Верхняя и спиральная пластинки распола-
располагаются на одинаковом удалении от столбика и либо сливаются друг с дру-
другом, либо разделены более или менее значительным промежутком; в неко-
некоторых случаях спиральная пластинка отсутствует, а верхняя слабо раз-
развита. Нижняя пластинка лежит довольно глубоко, но более или менее
видна при прямом положении устья; внизу она образует широкий, почти
горизонтальный изгиб и близко подходит к верхней пластинке. Субколу-
меллярная пластинка лежит глубоко и через устье не видна. Главная
складка простирается на V3—V2 оборота. Полулунная складка либо
1 Лектотип данного вида хранится в Музее Грузии (коллекция моллюсков,
№ 1580а). Он имеет следующие размеры: высота раковины 16.3, ширина 3.3 мм; высота
устья 3.0, ширина 2.2 мм; 15 3/4 оборотов. Место сбора: Горди, сборщик Ледер.
278
V. CLAUSIL1INAE
цельная, либо полностью или частично распалась, образовав 1—9 коротких
палатальных складок; лунеллярий лежит на спинной или право-спинной,
а у двух видов на брюшной стороне раковины. Широкая и короткая лопасть
клаузилия седловидно изогнута; у рукоятки по колумеллярному краю
имеется заметная вырезка; передний конец лопасти утолщен и слегка
заострен. ,, „. Л й!
40
44
48
54
56
38 *
Астрахань^
Махач-Кала4
44
42
40
36
Рис. 195. Географическое распространение видов рода Mucronaria Bttg. (Ориг.).
1а — Mucronaria, (Mucronaria) duboisi (Charp.); 16 — то же, изолированные местонахождения;
2 — М. (М.) strauchi (Bttg.); 3 — М. (М.) oustavi (Bttg.); 4 — М. (М.) acuminata (Mouss.); S —
М. (М.) pleuropiychia (Bttg.); 6 — At. (Index) index (Mouss.).
В пределах рода наблюдаются оба типа радулы (стр. 26), т. е. с допол-
дополнительными зубцами на срединных и боковых зубах и без них.
Половая система сходна с таковой у Laciniaria Hartm. и других близ-
близких групп, т. е. характерна тем, что мужские половые пути вытянуты
вдоль женских, а ретрактор пениса отсутствует. Ректрактор глазного
щупальца лежит в стороне от половых путей и с ними не перекрещивается.
Распространение (рис. 195). Кавказ и ряд причерноморских
вилайетов Турции (до Синопа),
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА MUCRONARIA
D). Главная и палатальные складки начинаются на брюшной стороне
раковины или чуть правее.
C). Раковина веретеновидно-цилиндрическая, с крупными белыми,
широко расставленными ребрами. Спиральная пластинка видна
через устье 5. М. pleuroptychia (Bttg.).
9. MUCRONARIA 279
3 B). Раковина веретеновидная, густо и равномерно ребристо исчер-
исчерченная. Спиральная пластинка не видна через устье
6. М. index (Mouss.).
4 A). Главная и палатальные складки начинаются на правой или на
спинной стороне раковины.
5 F). Край устья имеет как минимум одну крупную складку, соеди-
соединяющую его с нижней пластинкой; нередко могут быть еще
краевые складочки и узелки 3. М. duboisi (Charp.).
6 E). Нижняя пластинка с краем устья не связана. Последний без
складок или узелков.
7 (8). Раковина веретеновидно-башневидная, нередко почти башне-
видная. Спиральной пластинки нет. Вместо полулунной и глав-
главной складок на право-спинной стороне располагаются друг над
другом 7—9 коротких складок 4. М. gustavi (Bttg.).
8 G). Раковина веретеновидная. Имеется более или менее длинная
спиральная пластинка, заметная через устье. Главная складка
простирается не менее чем на 1/3 оборота. Под ней располагаются
несколько коротких палатальных складок и мозолевидный руди-
рудимент нижней части полулунной складки. ^
9 A0). Раковина стройная, тонко ребристая . . 2. М. strauchi (Bttg.).
10 (9). Раковина вздутая, исчерченная ... 1. М. acuminata (Mouss.).
1. Подрод MUCRONARIA s. str.
В состав этого подрода включены две группы — Polyptychia Bttg.
и Mucronaria Bttg., которых Бетгер (Boettger О., 1877а) рассматривает
как самостоятельные группы секции Euxina Bttg. Первое название яв-
является преоккупированным (поп Felder, 1874), в связи с чем Линдгольм
(Lindholm, 1924) предложил в замен его новое название — Multiplicand
Lindh.
К первой группе Бетгер относил Clausilia duboisi Charp., Cl. litotes
A. Schm., Cl. quadriplicata A. Schm. и Cl. subtilis A. Schm. Ко второй
группе вначале он отнес только Cl. acuminata Mouss., а позднее (Boettger О.,
1878а) присоединил Cl. strauchi Bttg. Что касается Cl. litotes и Cl. quadri-
quadriplicata, то вскоре эти виды были отнесены самим Бетгером и последую-
последующими авторами в другие группы (Mentissoidea Bttg. m.Quadriplicata Bttg.),
a Cl. subtilis признан как синоним Cl. duboisi.
В результате сравнения раковин Cl. duboisi, Cl. acuminata и Cl. stra-
strauchi мы пришли к выводу, что они являются видами одного подрода.
Согласно правилам зоологической номенклатуры, за этим подродом должно
быть сохранено название Mucronaria Bttg. Наибольшее сходство обнару-
обнаруживается при сравнении самых стабильных частей замыкательного аппа-
аппарата названных моллюсков, а именно пластинок. Бетгер же, который
относил их к разным группам, главное внимание обратил на полулунную
и палатальные складки. В самом деле, у Cl. duboisi первой совсем нет,
зато имеются 3—4 довольно длинных палатальных складки; у осталь-
остальных же двух видов от полулунной складки сохранился лишь ее нижний
конец, а верхний распался, образовав 1—2 коротких палатальных
складки.
С нашей точки зрения, приведенные различия недостаточны для того,
чтобы помещать указанные виды в разные подроды или группы иного
ранга. Здесь наблюдается обычное для Clausiliidae явление постепенного
преобразования полулунной складки в серию палатальных складок.
Как уже отмечалось выше (см. стр. 46), палатальные складки являются
280
V. CLAUS1LIINAE
образованиями, дающими более совершенную защиту от высыхания, и
весь процесс замены полулунной складки палатальными представляет
собой развитие адаптации в сторону обитания в более сухом и жарком
климате.
Анализ особенностей раковины и половой системы Cl. pleuroptychia
Bttg. и Cl. gustavi Bttg. дал нам основание отнести их в этот же подрод
(ранее первую причисляли к Euxina Bttg., а вторую — к Armenica Bttg.).
1. Mucronaria (Mucronaria) acuminata (Mouss.) (рис. 29, A, 196).
M о u s s о n, 1876b : 144. t. 5, fig. 4 [Clausilia (Mentissa)]; Martens,
1876 : 90 (Cl. acrolepta); Schneider, 1878 : 24, 158 (CI.- acuminata Mouss. = ac-
rolepta Mart.); К о b e 1 t, 1893, Iconogr., N. F., 6 : 23, fig. 991 [Clausilia (Euxina)].
Раковина веретеновидная, сильно вздутая, сильно и неравно-
неравномерно исчерченная,1 слабо просвечивающая и со слабым блеском, темно-
роговая, без белых штрихов. Завиток ниже узкой и
острой верхушки имеет вогнутые очертания. Оборотов
111/2—13, довольно выпуклых, разделенных белым швом;
эмбриональные обороты (около 3) гладкие, блестящие;
2 последних оборота одинаковой ширины; последний
оборот на затылке несколько уплощен и ребристый,
внизу с крупным килем. Устье округло-грушевидное,
почти отвесное, с широким, слабо врезанным синулусом и
с мелким желобком, внутри белое. Края устья широко от-
отвернуты, тонкие, с белой губой. Верхняя пластинка очень
низкая, иногда лишь слегка намечена; впереди она дости-
достигает края устья, но не выступает за его пределы, сзади
плавно сходит на нет. Нижняя пластинка лежит очень
глубоко и при прямом положении раковины лишь слегка
выступает из-под верхнего края устья; от верхней пла-
пластинки она отстоит несколько дальше, чем у М. strauchi
(Bttg.), верхний конец ее лежит на правой стороне рако-
раковины, нижний плавно переходит в колумеллярную стен-
стенку и с краем устья ничем не связан. Спиральная пла-
пластинка берет начало на право-брюшной стороне раковины,
к середине плавно нарастает в высоту, после чего плавно
снижается к устью; нижний конец ее лежит на значи-
значительном расстоянии от заднего конца верхней пластинки
и лишь слабо заметен при косом положении устья. Глав-
Главная складка начинается на правой стороне и простирает-
простирается вперед на 1/3 оборота. Под ней, на правоспинной
стороне, лежат очень короткая палатальная складка и
рудимент (нижняя часть) полулунной складки. У некото-
некоторых экземпляров обе эти складки сливаются, образуя единую мозолевид^-
ную полулунную складку.
Размеры: высота раковины 11.8—13.4, ширина 3.6—3.8 мм; высота
устья 2.7—2.9, ширина 2.4—2.6 мм.2
Внутреннее строение не изучено.
Рис. 196. Muc-
Mucronaria (Muc-
(Mucronaria) acumi-
acuminata (Mouss.).
(Ориг.).
1 По Муссону, ребристо исчерченная.
2 В нашем распоряжении было только 5 экземпляров (все из одного местонахо-
местонахождения), из которых 2 находятся в коллекции Зоологического института Академии
наук СССР и 3 — в коллекции Зоологического института Польской Академии наук.
9. MUCRONARIA
281
Распространение (рис. 195). Вид очень редкий. Единствен-
Единственная находка была сделана в южной Грузии на горе Шав-Набад, к востоку
от оз. Табацкури. г
Экология. Обитает среди россыпей скал вулканического происхо-
происхождения на высоте 2000—2500 м над ур. м. (Schneider, 1878).
Систематическое положение. Раковина данного мол-
моллюска отличается от всех прочих близких видов вздутой формой и слабым
развитием замыкательного аппарата, в особенности верх-
верхней пластинки и палатальных складок. Последнее стоит
в связи с обитанием в высокогорных условиях.
2. Mucronaria (Mucronaria) strauchi (Bttg.) (рис. 29, Б,
197).
Boettger О., 1878с: 176 (Clausilia); Boettger О., 1878а :
301, t 10, fig. 6 (Clausilia); Boettger О., 1880а : 144, t. 4, fig. 6
[Cl. (Euxina) strauchi var. mezchetica]; Boettger O., 1880b : 159
(Clausilia); Boettger O., 1881a : 233 AClausilia (Euxina)]; К о-
b e 1 t, 1893, Iconogr., N. F., 6 : 18, fig. 984 [Clausilia (Euxina)]; Л и-
харев и Раммельмейер, 1952 : 239, рис. 158 2 [Lacinia-
ria (Euxina)].
Раковина веретеновидная, тонкоребристая (на пред-
предпоследнем обороте около 50—55 ребрышек), слабо просве-
просвечивающая, с шелковистым блеском, роговая, с отдельными
белыми штрихами на ребрах. Завиток близ верхушки имеет
вогнутые очертания. Оборотов 12—13, реже 14, слабо вы-
выпуклых, разделенных глубоким швом; эмбриональные обо-
обороты B3/4—3), гладкие, блестящие, образуют тонкую, почти
цилиндрическую верхушку; последний оборот на затылке
уплощен, внизу с крупным килем. Устье грушевидное, почти
отвесное, с высоким и узким умеренно врезанным синулу-
сом, внутри коричневое. Края устья широко отвернуты,
тонкие, с белой губой и желобком. Верхняя пластинка низ-
низкая, но довольно толстая; впереди она достигает края устья
и даже иногда слегка выступает за его пределы, а сзади
либо сливается со спиральной пластинкой, либо отделена
от нее промежутком. Нижняя пластинка лежит очень глубоко и при
прямом положении раковины лишь слегка выступает из-под верх-
верхнего края устья; верхний конец нижней пластинки лежит на правой сто-
стороне раковины; нижний конец плавно сливается со столбиком и с краем
устья ничем не связан. Довольно длинная главная складка начинается
на правой стороне, а кончается на левой. Непосредственно под нею на
спинной стороне раковины располагаются 2—3 палатальных складки,
а ниже их рудимент — нижняя часть полулунной складки. Из палаталь-
палатальных складок наиболее длинная — верхняя, которая начинается немного
глубже и кончается немного ближе к устью, чем остальные, очень короткие,
порой узелковидные.
Размеры: высота раковины 12—14, ширина 3.0—3.5 мм; высота устья
2.7—3.0, ширина 2.0—2.3 мм. Бетгер (Boettger О., 1881а) приводит для
Рже. 197.
Mucronaria
(Mucrona-
(Mucronaria) strauchi
(Bttg.). (По
Лихареву и
Раммель-
Раммельмейер).
1 Указание Розена A914) на обитание этого вида в Верхней Сванетпи, как пока-
показала проверка, ошибочно. Названный автор имел дело с Megaleuxinaderasai. suanetica
(Bttg.).
2 Подпись под рис. 158: uLaciniaria (Euxina) duboisi (Charp.)», следует читать —
«Laciniaria (Euxina) strauchi (Bttg.)».
282
V. CLAUSILIINAE
типов такие размеры: высота раковины 15—16, ширина 3.5—3.7 мм.
Однако все просмотренные нами экземпляры имели меньшие размеры.1
Внутреннее строение не изучено. ^Щ^.С^С^М,^ /""/^^^'cf;'^ fS%A
Распространение (рис. 1957ГВид редкий. Большинство нахо-
находок сделано в окрестностях Мцхета и по одной находке в окрестностях
Манглиси и Тианети. Все местонахождения приурочены к центральной
Грузии.
Систематическое положение. Данный
вид наиболее близок к М. duboisi (Charp.) и к М. аси-
minata (Mouss.). От первого он отличается скульптурой
поверхности раковины, более глубоко лежащей нижней
пластинкой, отсутствием краевых складок на устье, при-
присутствием рудимента полулунной складки и более корот-
короткими палатальными складками, от второго — скульптурой
поверхности раковины, более стройной формой и более
сильным развитием верхней пластинки и палатальных
складок. vw//'K''^
3. Mucronaria (Мтая"еваий) ~
duboisi (Charp.) (рис. 29, В, 198—
200).
Charpentier, 1852 : 402, t.
11, fig. 12 (Clausilia); Kiister, 1847—
1862 : 270, t. 30, fig. 25—27 (Clausi-
lia); Schmidt A., 1868: 163 (Cl.
subtilis); К obelt, 1893, Iconogr., N. F.,
6 : 27, fig. 999 [Clausilia (Euxina)]; P o-
з е Н, 1914 : 196 [Clausilia {Euxina)];
Ретовский, 1914 : 313 [Clausilia
(Euxina)]; Л и x_|i реви Раммель-
e й е р, 1952 : 238, рис. 159 2 [Lacinia-
ria (Euxina)].
Рис. 198. Mucronaria (Mucronaria) duboi-
duboisi (Charp.). (Ориг.).
A— общий вид раковины; В
¦ клаузилий.
Раковина веретеновидная
или веретеновидно-цилиндриче-
ская, узкая, тонко и густо ребристо
исчерчена, слабо просвечивающая,
с шелковистым блеском, красновато-роговая, с многочисленными пучками
белых штрихов. Оборотов 11—12, слабо выпуклых, разделенных очень мел-
мелким швом; эмбриональные обороты BV2) гладкие, блестящие; последние
2—3 оборота почти одинаковой ширины; последний оборот книзу сужен,
за устьем уплощен и ребристый, с острым килем. Устье грушевидное,
почти отвесное, с высоким узким, умеренно врезанным синулусом, внутри
коричневое. Края устья широко отвернуты, тонкие, с тонкой белой губой.
У старых экземпляров края устья (особенно между верхней и нижней
пластинками) с несколькими складочками и узелками. Верхняя пластинка
низкая, тонкая, располагается несколько наклонно; впереди она дости-
достигает края устья, сзади либо сливается со спиральной пластинкой, либо
(реже) отделена от нее коротким промежутком. Нижняя пластинка лежит
довольно глубоко, с краем устья она связана крупной складкой; верхний
конец нижней пластинки располагается на правой стороне раковины, зна-
1 Мы имели в своем распоряжении 15 экземпляров из коллекций Зоологического
института Академии наук СССР, из Музея Грузии и из Зоологического Института
Польской академии наук. Все они, по-видимому, определены Бетгером.
2 Подпись под рис. 159: «Laciniaria (Euxina) strauchi Bttg.»; следует читать:
«Laciniaria (Euxina) duboisi (Charp.)».
9. MUCRONARIA 283
чительно ниже начала спиральной пластинки. Длинная главная складка
начинается на право-брюшной или брюшной стороне раковины и кончается
на спинной стороне. Под ней лежат 3—4 палатальных складки (иногда
между ними бывают еще узелки). Все они параллельны друг другу, но
к главной складке лежат под углом; их задние концы располагаются на
право-спинной или правой стороне раковины, причем верхняя и нижняя
начинаются глубже средних складок; впереди все эти складки закан-
заканчиваются на спинной стороне; верхняя складка обычно самая корот-
короткая.
Размеры: высота раковины 12—14, ширина 2.8—3.0 мм; высота устья
2.7—3.0, ширина 2.1—2.3 мм.
Изменчивость. Как размеры, так и степень развития замыка-
тельного аппарата и ^кульптуры края устья сильно варьируют. Харак-
Характерно, что популяции, состоящие из экземпляров с раковиной типичных
13 IS
С 1 7
Рис. 199. Mucronaria (Mucronaria) duboisi (Charp.). Радула. (Ориг.).
или даже больших размеров (высота до 16 мм), обитают в удаленных от
моря районах, главным образом в буковых и дубовых лесах. Популяции,
состоящие из мелких экземпляров (f. minor A. Schmidt, 1868) — высота
раковины 8.5—9.5, ширина 2.2—2.3 мм, высота устья 1.9—2.0, ширина
1.5—1.6 мм при 9—10 оборотах — обычны для Черноморского побережья
Кавказа (Хостинская тиссо-самшитовая роща, Сухуми, Батуми). Кроме
меньших размеров, раковины этих популяций отличаются хрупкими тон-
тонкими, очень просвечивающими стенками; край устья тонкий, без краевых
складок и узелков (за исключением одной складки, соединяющей нижнюю
пластинку с краем устья); палатальных складок (кроме главной) 2—3; они
короткие, нередко в виде узелков.
Популяции, обитающие в горных лесах, произрастающих на известня-
известняковых породах, имеют много экземпляров, раковины которых обладают
многочисленными складками и узелками вдоль всего края устья (f. pli-
cata Boettger, 1880, syn. stomatica Westerlund, 1884).
Внутреннее строение. Материал: 1) Закатальский запо-
вщник, сбор и определение наше, V—VI 1952 — 2 экз.; 2) Ханларскийр-н
Азербайджанской ССР, окрестности оз. Гек-Гель, сбор и определение наше,
VI 1954 — 5 экз.; 3) Кутаиси, развалины храма Баграта, сбор и определе-
определение наше, 28 IV 1957 — 1 экз.
Радула (рис. 199): [-5- + (у + ^) X 2] X c/slOO.
Размеры срединного зуба 15.4 X 11.2 ц, ширина радулы 465 \i.
Семяприемник имеет небольшой округлый резервуар, который лежит
ниже белковой железы (рис. 200). Отросток умеренно извит, начинается
немного выше верхнего конца семяпровода и впадает в семяприемник на
значительном удалении от его нижнего конца. Матка значительно длиннее
влагалища. Толстый семяпровод, плавно расширяясь, переходит в верётё-
новидныи~ пенис.
284
V. CLAUS1LIINAE
Распространение (рис. 195). В СССР: широко распростра-
распространена по лесным областям Закавказья, но отсутствует в Талыше и на Лен-
коранской низменности; на Северном Кавказа найдена лишь в окрестностях
Орджоникидзе, Пятигорска и Железноводска. Вне СССР: черноморское
побережье Турции (Синоп, Трабзон и вилайет Чорох).
Экология. Обитает в равнинных и горных лесах (местами выхо-
выходит в предгорную лесостепь), чаще всего на стволах деревьев (в трещинах
коры), реже на сырых ска-
скалах, под камнями, в под-
подстилке и под гниющей ко-
корой.
Иногда встречается на
скалах и в осыпях субаль-
субальпийской и нагорно-степной
зон.
Размножение.
Живородящий вид. У ряда
вскрытых экземпляров в
семяяйцеводе и в матке об-
обнаружено от 5 до 7 эмбрио-
эмбрионов. При этом иногда на-
наблюдается полная редук-
редукция пениса и нижней части
семяпровода (афаллия).
С и стематическое
положение. Данный
вид занимает промежуточ-
промежуточное положение между М.
acuminata (Mouss.) и М.
strauchi (Bttg.), с одной
стороны, w.M. index (Bttg.)
и М. pleuroptychia (Bttg.),
с другой. У первых двух
видов распад полулунной
складки на палатальные
только начался, поэтому
наряду с остатком полулунной имеются уже 1 или 2 коротких пала-
палатальных складки. У последних двух видов, как и у данного вида, полу-
полулунная складка полностью распалась, образовав 3—4 длинных палатель-
ных складки. Однако у М. duboisi, которая более обычна для горных
лесов, эти складки короче и залегают сравнительно неглубоко — начи-
начинаются на право-брюшной стороне раковины, тогда как у М. index и
у М. pleuroptychia, которые живут только в предгорных и в низовых
лесах западного Закавказья, эти складки длиннее и располагаются
глубже.
Как указывалось выше (стр. 46, 48), эти морфологические различия
в конечном счете связаны с особенностями климата тех зон, в которых
обитают названные моллюски.
кл
Рис. 200. Mucronaria (Mucronaria) duboisi (Charp.).
Половая система. (Ориг.)-
4.М
¦щ
gustavi (Bttg.) (рис. 29, Г, 201—203).
Boettger О., 1880с : 381 [Clausilia (Oligoptychia)]; Boettger О.,
1886а : 312, t. 3, fig, 9 [Clausilia (Oligoptychia)]; Лихарев и Раммельмей-
е р, 1952:221 (Armenica).
9. MUCRONARIA
2S5
Раковина веретеновидно-башневидная, нередко почти бащне-
видная, стройная, верхние обороты тонко и сильно, последующие слабо
поперечно исчерчены, слабо просвечивающая, блестящая, коричневая,
с фиолетовым оттенком. Оборотов 111/2—121/2, верхние выпуклые, ниж-
нижние слабо выпуклые, почти плоские; эмбриональные обороты BV2—3)
гладкие, блестящие, образуют почти цилиндрическую, слегка заострен-
заостренную верхушку; на 2—3 последних оборота заметна слабая спиральная
исчерченность; последний оборот слабо сужен книзу, на затылке слегка
уплощен, с коротким килем. Устье округло-
или ромбическо-грушевидное, почти отвес-
отвесное, с широким слабо врезанным синулу-
сом и с глубоким желобком. Края устья
широко отвернуты, тонкие, с белой губой.
Верхняя пластинка очень тонкая, низкая и
короткая, нередко лишь слегка намечена;
впереди она достигает края устья, сзади
быстро сходит на нет. Нижняя пластинка
лежит очень глубоко и при прямом положе-
положении раковины лишь слегка выступает из-под
верхнего края устья; верхний конец ее ле-
лежит на право-спинной стороне, нижний
плавно переходит в колумеллярную стенку
и с краем устья не связан. Спиральная и
Ж
Рис. 201. Mucronaria (Mucronaria) gustavi
(Bttg.). (Ориг.).
A — общий вид раковины; Б — клаузилий.
лежащая рядом с нею параллельная пластинка короткие, мозолевидные;
обе они упираются в задний конец лопасти клаузилия. На право-спин-
право-спинной стороне раковины находятся 7—8, реже 9 коротких палатальных
складок, из которых верхние 2—3 (одна из них, вероятно, рудимент
главной складки) немного длиннее; остальные очень короткие, мозоле-
мозолевидные (являются продуктом распада полулунной складки).
Размеры: высота раковины 15.0—17.0, ширина 3.3—3.6 мм; высота
устья 3,0—3.3, ширина 2.6—2.7 мм.
Внутреннее строение. Материал: 1) Талыш, Люлякеран,
сбор Кириченко, 15—16 V 1909, определение Линдгольма — 1 экз.;
2) Талыш, Говери, сбор и определение наше, 11 VI 1952 — 1 экз.
Радула (рис. 202): \'± + C^ + Ц + ^) Х 2] X слЮ5.
Размеры срединного зуба 15 X 13.5 \л, ширина радулы 615 ц.
Семяприемник длинный, его резервуар лежит немного ниже белковой
железы (рис. 203). Отросток сильно извит, короче собственно семяприем-
семяприемника — начинается немного выше верхнего конца семяпровода. Влага-
286
V. CLAUSILI1NAE
л^щЁ_гдемвого__длиннее матки. У экземпляра, найденного близ селения
Говери, семяпровод и пенис совершенно не разграничимы; у другого
экземпляра пенис отчетливо выражен и имеет веретеновидную форму.
с-^~\
2Ь
Рис. 202. Mucronaria (Mucronaria) gustavi (Bttg.). Радула. (Ориг.).
ся
Семяпровод подтянут к влагалищу многочисленными мускульными во-
волокнами.
Распространение (рис. 195). Отдельные горные вершины
Талыша выше лесной зоны и на скалистых склонах Диабарской котло-
котловины (Зувант).
Экология. Обитает в трещинах
скал и в осыпях.
Размножение. Как показали
вскрытия, данный вид является живоро-
живородящим. В матке одновременно созревают
4—5 эмбрионов.
Систематическое положе-
положение. С момента описания данный вид от-
относили в род Armenica Bttg. (syn. Oligop-
tychia Bttg., part.). В пользу этого го-
говорил общий вид раковины — веретено-
видно-башневидная форма,__ко?ичневато-
фиолетовая окраска. Кроме того, и замы-
кательный аппарат обнаруживает некото-
некоторое сходство с настоящими Armenica —
слабое развитие верхней пластинки, распад
спиральной пластинки, а также главной и
полулунной складки на серию коротких
мозолевидных образований.
Только изучение половой системы утвер-
утвердило нас в мысли, что данный моллюск
принадлежит к совсем другому роду. Как
известно, для гениталий Armenica харак-
характерна петлеобразная форма мужских вы-
выводных путей, присутствие на пенисе
крупного придатка и мощного ретрактора,
который другим концом сливается с колу-
меллярной мускулатурой, и прохожде-
прохождение глазного ретрактора между пенисом и
влагалищем. Всего этого нет у данного вида, половая система которого
построена по плану, свойственному таким родам, как Laciniaria, Quadrip-
licata, Mucronaria и другим, у которых мужские половые пути вытянуты
вдоль женских и не образуют петли, пенис без придатка и ретрактора,
а ретрактор глазного щупальца лежит медиально от половых путей.
Вместе с тем такие особенности раковины, как наклонное положение
оси устья, форма нижней пластинки и седловидная форма лопасти клау-
ркл
Рис. 203. Mucronaria (Mucrona-
(Mucronaria) gustavi (Bttg.). Половая си-
система. (Ориг.).
9. MUCRONARIA
287
зилия, указывают на близость данного вида к ряду duboisi (Charp.) —
acuminata (Mouss.). Учитывая это, а также сходство в строении половой
системы с названными видами, мы относим его в род Mucronaria Bttg. Сход-
Сходство же с видами из рода Armenica объясняется обитанием в аналогичных
условиях. Как известно, все виды Armenica, подобно нашему моллюску,
тоже живут на скалах и в осыпях чаще всего
в высокогорных зонах.
j JA<mtr; /
5. Mucronaria (Mwcronaria)/pleuroptychia
(Bttg.) (рис. 204—206).
Boettger О., 1878а: 291, t. 10, fig. 1 (Clau-
silia); Boettger O., 1881a : 232, t. 9, fig. 20
[Cl. (Euxina) pleuroptyckia var. polygyra]; К о b e 1 t,
1893, Iconogr., N. F., 6 :12, fig. 975 [Clausilia (Euxi-
(Euxina)]; Розен, 1914 : 197 [Clausilia (Euxina)]; Л и-
харев и Раммельмейер, 1952 : 219, рис.
135 (Mentissoidea).
Р аковина веретеновидно-цилиндриче-
ская B предпоследних оборота одной ширины,
Рис. 204. Mucronaria (Mucronaria) pleuropty-
chia (Bttg.).
A — общий вид раковины (по Лихареву и Раммель-
Раммельмейер); Б — клаузилий (ориг.)-
последний книзу слабо сужается), узкая, с белыми пластинчатыми реб-
ребрами (на предпоследнем обороте 27—32 ребрышка), в широких проме-
промежутках между ребрами гладкая и матовая, тонкостенная, роговая.
Оборотов 13V2—15V2, почти плоских, разделенных глубоким швом;
эмбриональные обороты C—3V2) почти одной ширины, гладкие, блестя-
блестящие, темно-роговые или вишнево-красные, образуют очень тонкую
и острую цилиндрическую верхушку; последний оборот на затылке упло-
уплощен. Устье грушевидное, небольшое, внутри коричневатое, с высоколежа-
щим широким неврезанным синулусом. Края устья сильно отвернуты,
тонкие, с белой губой. Верхняя пластинка очень низкая и тонкая; впереди
достигает края устья, назад уходит довольно глубоко и либо сливается
со спиральной пластинкой, либо независима от последней и связана с нею
тонким нитевидным валиком. В этом месте к обеим пластинкам подходит
задний конец слабой промежуточной складки, которая начинается близ
нижнего конца нижней пластинки. Крупная нижняя пластинка лежит
глубоко, но видна при прямом положении устья; верхний конец ее лежит
почти на брюшной стороне раковины, тогда как спиральная пластинка
начинается еще глубже. Главная складка и лежащие под ней палатальные
складки берут начало на брюшной стороне раковины (иногда с небольшим
смещением вправо). Наиболее длинная главная складка, которая закан-
заканчивается на спинной стороне или даже чуть левее; остальные лежат парал-
параллельно ей, различной длины, причем, обычно верхняя самая длинная,
288
V. CLAUSILUNAE
а нижняя короче всех и имеет мозолевидный облик. Главная и 2 верхних
палатальных складки начинаются обычно ниже места соприкосновения
лопасти клаузилия со стенкой раковины, а нижняя — всегда от лопасти.
&
12
17
Рис. 205. Mucronaria (Mucronaria) pleuroptychia (Bttg.).
Радула. (Ориг.).
Размеры: высота раковины 15—17, ширина 3.0—3.3 мм; высота устья
2.8—3.1, ширина 2.1—2.4 мм.
Внутреннее строение. Материал: Гелати, близ Кутаиси,
на известняковых стенах, сбор и определение наше 1 V 1957 — 2 экз.
Радула (рис. 205): f-f + ( г +
+?+й)х2]хш:
Размеры срединного зуба 16.8 X
9.8 ц, ширина радулы 495 ц.
Половая система построена типич-
типично для рода (рис. 206). Семяприемник
длинный — его резервуар лежит не-
непосредственно ниже белковой железы.
Отросток очень сильно извит и начи-
начинается на один оборот ниже резер-
резервуара семяприемника, рлягадитпр
почтив_2^?аза_^5линнее_1аатки, без
р1Естпир1знТ1яунижнего конца. Тол-
Толстый семяпровод, образуя небольшой
коленообразный изгиб, плавно пере-
переходит в пенис, нижний конец кото-
которого немного расширен.
Левый ретрактор посылает ветвь
к шейке семяприемника и несколько
ответвлений к половой клоаке. Мно-
Многочисленные волокна подтягивают се-
семяпровод к женским половым путям.
Как и у предыдущих видов, имеется
тонкий мускул, связывающий семя-
семяпровод с нижним концом пениса.
Распространение (рис.
195). Центральная часть западной
Грузии между реками Квирила на
востоке и Цхенис-Цкали на западе,
в зоне предгорий-и невысоких гор, сложенных из известняков.
Экология. Обитает в дубовых и дубово-буковых лесах, на^извест-
няковых скалах и под камнями, а также на каменных оградах, сложенных
из той же горной породы.
-КЛ
Рис. 206. Mucronaria (Mucronaria) pleu-
pleuroptychia (Bttg.). Половая система.
(Ориг.).
9. MUCRONARIA 289
Раз множен и е. й?ивородящий вид. В матке одновременно раз-
развивается до, 6 эмбрионов. Период размножения приходится на апрель —
май (Кокочашвили, 1958).
Систематическое положение и с и н о н и м и я. Бет-
гер описал этот вид по экземплярам, якобы найденным в Сирии. Позднее
он получил рт Ледера несколько экземпляров, собранных в окрестностях
Кутаиси, которых он описал как var. polygyra Bttg. A881). При этом он
справедливо усомнился в том, что местонахождение оригиналов типичной
формы было указано правильно. В настоящее время можно считать уста-
установленным, что данный вид живет только на Кавказе, причем в ограни-
ограниченном районе, так как из других мест достоверных находок не имеется.
Что касается var. polygyra Bttg., то просмотр многочисленных серий
убедил щас в том, что все отличия этой разновидности от типичной формы
укладываются в пределы обычной популяционной изменчивости. Эта
разновидность встречается вместе с типичной формой в одних и тех же
популяциях и связана с нею плавными переходами. Следовательно, она,
скорее всего, является только синонимом данного вида.
Главными особенностями М. pleuroptychia, отличающими ее от боль-
большинства видов того же рода, является следующее: 1) своеобразная форма
раковины и ребристость; 2) главная и палатальные складки располагаются
более глубоко (в этом отношении она сходна только с М. index Bttg.).
Однако форма нижней пластинки, наклон оси устья, форма клаузилия,
вместе с рядом других признаков, с нашей точки зрения, дают основание
относить данный вид в род Mucronaria Bttg. :
2. Подрод INDEX BTTG.
Boettger 6., 1877а : 87 (Clausilia, секция Alinda, группа /гейея); L i n d-
h о 1 m, 1924 : 66 (Laoiniaria, подрод Euxina, секция Index).
Монотипический подрод. , . у
Раковина резко веретеновидная, довольно вздутая на середине, с вог-
вогнутыми боковыми очертаниями верхней части завитка и с притуплённой
верхушкой. Последний оборот сильно сужен книзу, за устьем уплощен,
внизу с крупным килем. Устье с высоко лежащим и отогнутым вправо
синулусом, внизу с базальным желобком. Спиральная пластинка длинная,
но лежит очень глубоко и через устье не видна. Нижняя пластинка очень
высокая, сильно выдается в просвет устья в виде почти горизонтальной
складки. Главная складка и 3 палатальных складки берут начало на брюш-
брюшной или право-брюшной стороне и через устье не видны. : •
Наиболее существенными отличиями от Mucronaria s. stir, является
своеобразная форма раковины, очень сильное развитие нижней пластинки,
а также глубокое залегание спиральной пластинки, главной и палаталь-
палатальных складок. • , . -
6. Mucronaria (Index) index (Mouss.) (рис. 207—209). •
, Mo u s so n, 1863 : 401 (Clausilia); Mousson, 1873 : 215, t. 8, fig, Ъ \Clau-
silia (Laciniaria)]; P'o з е н, 1914 : 200 [Clausilia (Euxina)]; P его в с кий,
1914 : 348 [Clausilia (Euxina)]; Лихарев и Раммельмейер, 1952 : 218,
рис. 134 (Mintissoidea). . .
Раковина густо и равномерно ребристо исчерченная, иногда со
спиральными линиями, просвечивающая, со слабым блеском, зеленовато-
роговая или коричнево-роговая, с многочисленными пучками белых
штрихов. Оборотов 13—131/2 (изредко 14), выпуклых; эмбриональные
290
V. CLAUSILIINAE
обороты (около 21/а) гладкие, блестящие. Устье грушевидное, отвесное,
выступающее, с узким синулусом, отогнутым к оси раковины. Края устья
широко отвернуты, утолщены, белые. Низкая верхняя пластинка распо-
располагается к оси раковины под углом; впереди она слегка выступает на
край устья, а сзади, плавно снижаясь, сходит на нет. Нижняя пластинка
наверху берет начало на правой стороне раковины; затем она быстро
нарастает в высоту, одновременно сильно спирально закручиваясь, и
в просвет устья выдается как крупная горизонтальная складка, близко
лежащая к верхней пластинке; по внешнему краю
она валикообразно утолщена, а с краем устья свя-
связана крупной складкой. Высокая спиральная пла-
пластинка берет начало на брюшной стороне, а заканчи-
заканчивается на спинной стороне; через устье она не вид-
видна, так как закрыта выступающей нижней пластин-
пластинкой. Крупная, косо лежащая промежуточная складка
Рис. 207. Mucronaria (Index) index (Mouss.).
A — общий вид раковины (по Лихареву и Раммельмейер); Б — последний
оборот и просвечивающие сквозь него^складки; В — клаузилий.
одним концом подходит к нижнему концу нижней пластинки, а другим
к нижнему концу спиральной пластинки. Длинная главная складка
начинается вместе с палатальными складками на брюшной или право-
брюшной стороне и простирается до спинной стороны раковины. Пала-
Палатальные складки короче главной; из них самая короткая верхняя пала-
палатальная складка. Широкая и сравнительно короткая лопасть клаузилия
седловидно изогнута, у рукоятки слегка вырезана по колумеллярному
краю, а передний конец ее утолщен и слегка заострен.
Размеры: высота раковины 16.5—18.0, ширина 4.0—4.5 мм; высота
устья 3.6—4.0, ширина 2.5—2.8 мм.
Изменчивость. Вид варьирует в размерах. Так, изредка встре-
встречаются отдельные экземпляры с более крупной раковиной (var. tenax
Westerlund, 1901): высота раковины 18.5—19.0, ширина 4.5—4.7 мм.
Кроме того, преимущественно в районе Батуми наряду с типичными
встречаются более мелкие формы (var. minor Mousson, 1873): высота рако-
раковины 14.5—16.0, ширина 3.7—4.0 мм; высота устья 3.3—3.7, ширина
2.5—2.8 мм. Таких же размеров имеется другая разновидность (var.
climax Boettger, 1888), отличающаяся, кроме того, тем, что нижняя пла-
пластинка закручена слабее и в проевете устья выступает более наклонно,
а главная и 4 палатальных складки короче и начинаются на правой сто-
стороне раковины.
9. MUCRONARIA
291
Внутреннее строение. Материал: Батуми, сбор и определение
наше, 12 IV 1957 — 1 экз. (var. climax Bttg.).
Радула (рис. 208): | + (^ + з
S)x
2.
Рис. 208. Mucronaria (Index) index (Mouss.) var. climax (Bttg.).
Радула. (Ориг.).
Размеры срединного зуба 15 X 10.5 ц, ширина радулы 585 ц.
Дополнительные зубцы срединного зуба слабо развиты.
Семяприемник широкий и длинный; его резервуар лежит непосред-
непосредственно ниже белковой железы (рис. 209). Отросток сильно извит и зна-
-сп
Рис. 209. Mucronaria (Index) index (Mouss.) var. climax
(Bttg.). Половая система. (Ориг.).
чительно короче семяприемника, так как начинается лишь немного выше
верхнего конца семяпровода. Матка_и влагалище почти одной гтлиньт.
Последнее очень толстое, с плотными мускулистыми стенками и сужается
лишь перед впадением в половую клоаку. Пенис веретеновидно расширен.
Ретрактор семяприемника посылает от себя короткий мускул:, подтяги-
подтягивающий семяпровод к женским половым путям. То же осуществляют
292 V.. CLAUStLJlINAE
многочисленные мускульные волокна, протянутые от матки к семяпро-
семяпроводу.
Распространение (рис, 195). Западная Грузия, преимущест-
преимущественно вдоль берега моря (от Сухуми до Батуми), реже вдали от побережья
(Кутаиси, курорт Саирме — северные склоны Аджаро-Имеретинского
хребта). Кроме того, одна находка в южной Грузии (Боржоми).
Систематическое положение. Раковина данного вида
во многом сходна с таковой у Mentissoidea litotes (A. Schm.). В свое время
(Лихарев и Раммельмейер, 1952) это дало нам основание поместить дан-
данный вид в тот же род. Теперь, когда известна половая система обоих
моллюсков, не может быть и речи об их объединении в один род. Учитывая,
что не всегда есть возможность рассмотреть внутреннее строение, важно
указать надежные различия между раковинами обоих Моллюсков. К тако-
таковым относятся следующие:
1) у нашего вида горизонтально лежащий изгиб нижней пластинки
выступает в просвет устья не так сильно, как у Mentissoidea litotes и, в от-
лячие от последней, связан с краем устья крупной складкой;
. , 2) у Mucronaria index субколумеллярная пластинка снаружи не видна,
тогда, как у Mentissoidea litotes нижний конец ее хорошо виден при косом
положении устья;
3) у нашего вида главная складка короче, так как заканчивается на
спинной стороне раковины, тогда как у Mentissoidea litotes она заканчи-
заканчивается на левой стороне;
4) у Mucronaria index 3 или 4 палатальных складки, а у Mentissoidea
litotes их 2.
10. Род MICROPONTICA BTTG.
Boettger О., 1881b : 344 (Clausilia, секция Micropontica); BoettgerO.,
1889:34 (Clausilia, секция Micropontica); Lindholm, 1924:66 (Laciniaria,
подрод Micropontica).
Тип рода: Clausilia closta Bttg.
Раковина небольшая, башневидная или башневидно-веретеновидная.
Последний оборот у двух видов без киля (устье соответственно без базаль-
ного желобка), у одного вида с килем (устье с желобком). Устье груше-
грушевидное, сильно выступающее, с высоким неврезанным синулусом. Пла-
Пластинки тонкие, относительно сЛ(або развиты. Спиральная пластинка
(если она имеется) сливается с верхней. Нижняя пластинка располагается
близко к верхней. Имеется хорошо развитая полулунная складка.
Срединный и значительная часть боковых зубов радулы без дополни-
дополнительных зубцов.
Половая система подобна таковой у Laciniaria Hartm., Mucronaria
Bttg. и других родов, у которых мужские половые пути вытянуты вдоль
женских, пенис без ретрактора, а ретрактор глазного щупальца лежит
медиально от половых путей и с ними не перекрещивается.
Распространение (рис. 31). Западная часть Большого Кав-
Кавказа.
Наиболее характерной особенностью данной группы является посте-
постеленное укорачивание последнего оборота раковины (в спиральном направ-
'лении, со стороны устья), которое можно наблюдать в ряде М. closta
(Bttg.) — М. circassica (Bttg.) — М. retowskii (Bttg.). Это приводит
К тому, что форма раковины изменяется от башневидно-веретеновидной
до башневидной, так как исчезает нижняя, т. е. суженная, часть послед-
последнего оборота. Одновременно идет укорачивание соответствующих частей
10. MICROPONTICA 293
замыкательного аппарата, а именно•— всех образований, кроме верти-
вертикально лежащей полулунной складки. Если у главной складки, верхней
и спиральной пластинки это проявляется только в том, что они становятся
все более короткими, то у нижней пластинки, кроме того, меняется и
облик, так как в результате исчезает ее нижний участок, лежавший почти
отвесно, и сохраняется только верхний участок, имеющий вид спирально
изогнутой складки. Одновременно создается впечатление, что в пределах
указанного ряда весь замыкательный аппарат перемещается ближе к устью.
Указанные изменения связаны с преждевременным окончанием роста
раковины из-за более раннего наступления половой зрелости. В свою
очередь последнее является адаптацией к жизни в высокогорном климате
с коротким вегетационным периодом (см. стр. 47).
~ таблица для определения видов рода micropontica
1 D). Последний оборот раковины снизу округлый, без киля. Устье без
базального желобка. Полулунная складка и лопасть клаузилия
располагаются на правой или спинной стороне раковины и через
устье не видны. Верхняя пластинка хорошо развита и слита со
спиральной. i
2 C). Раковина башневидно- или цилиндрическо-веретеновидная малень-
маленькая — высота 10.0—12.5, ширина 2.2—2.5 мм. Полулунная складка
хорошо развита и располагается на правой стороне раковины. Кроме
главной, всегда имеется еще верхняя палатальная складка . . .
1. М. closta (Bttg.).
3 B). Раковина башневидная,. более крупная — высота раковины 11—
14, ширина 2.9—3.4 мм. Полулунная складка рудиментарная,
мозолевидная и располагается на право-спинной стороне. Верхняя
палатальная складка чаще всего отсутствует
2. М. circassica (Bttg.).
4 A). Последний оборот раковины с заметным килем. Устье с базальным
желобком. Полулунная складка и лопасть клаузилия располагаются
на лево-спинной стороне раковины и хорошо видны через устье.
Верхняя пластинка слабо развита, спиральной пластинки нет . .
v 3. М. retowskii (Bttg.).
1. Microponticafcr6sta~XBn;g.) (рис. 210—212).
Boettger О., 1881b : 345 [Clausilia (Micropontica)]; Boettger 0., 1883 :
190, t. 7, fig. 7 [Clausilia (Micropontica)]; Ретовский, 1914 : 323 [Clausilia (Mic-
(Micropontica)]; Лихарев и Раммельмейер, 1952 : 247, рис. 173 [Laciniaria
(Micropontica)].
Раковина башневидно-, иногда цилиндрическо-веретеновидная,
стройная, с крупными, редко расставленными белыми пластинчатыми реб-
ребрами (на предпоследнем обороте 30—35), в промежутках между ними
гладкая матовая, слабо просвечивающая, коричневато-роговая. Оборотов
lOVa—12, разделенных глубоким швом; верхние обороты выпуклые,
нижние уплощены; эмбриональные обороты (около 21/2) гладкие; послед-?
ний оборот уже, но немного выше предпоследнего, с очень крупными и
редкими ребрами, на затылке уплощен, без киля. Устье относительно
маленькое, округло-грушевидное, сильно выступающее, с широким невре-
занным синулусом, внутри коричневатое. Края устья широко отвернуты,,
утолщены; нижний край устья округлый, без желобка. Низкая, но до-
довольно толстая верхняя пластинка впереди достигает края устья, а езади
294
V. CLAUSILIINAE
переходит в спиральную. Последняя берет начало на брюшной стороне;
немного ниже начинается субколумеллярная, а еще ниже (на право-
брюшной стороне) нижняя пластинка. Нижняя пластинка, слабо изги-
изгибаясь, более или менее отвесно тянется вниз; нередко она заканчивается
крупным бугорком, благодаря чему при рассматривании через устье соз-
создается впечатление, что пластинка образует горизонталь-
горизонтальный перегиб; на всем протяжении она проходит очень близ-
близко к спиральной и верхней пластинкам. Субколумеллярная
пластинка не видна или заметна лишь при очень косом
положении устья. Крупная полулунная складка лежит на
правой стороне раковины. Над ней берет начало главная
складка, простирающаяся вперед на V2 оборота. Близ верх-
верхнего конца полулунной складки,
иногда сдираясь с нею, начи-
начинается массивная, верхняя пала-
палатальная складка; она распола-
располагается к главной складке под
углом, простирается вперед не-
несколько дальше главной и хо-
хорошо видна при прямом поло-
положении устья. Рукоятка клаузи-
лия плавно расширяется в длин-
йую и узкую, желобчато согну-
согнутую лопасть, которая на ниж-
нижнем конце утолщена.
Размеры: высота раковины
10.0—12.5, ширина 2.2—2.5 мм;
высота устья 1.8—2.0; ширина
1.4-1.7 мм. х
Внутреннее строение. Материал: Гагры, сбор Федотова,
V 1916, определение Линдгольма — 1 экз.
Радула (рис. 211): [| + (^ + Ц + ^)] Х с/э110.
Рис. 210. Micropontica closta (Bttg.).
А — общий вид раковины (по Лихареву и Рам*
мельмейер); Б — клаузилий. (Ориг.).
Рис. 211. Micropontica closta (Bttg.). Радула. (Ориг.).
Размеры срединного зуба 12.6 X 7 |i, ширина радулы 350 \i. Наибо-
Наиболее внешние краевые зубы за счет расщепления первоначальных зубцов
приобретают гребенчатый облик.
Резервуар семяприемника лежит на значительном расстоянии от
белковой железы (рис. 212). Отросток очень сильно извит и заметно ко-
1 Лектотип данного вида находится в Зоологическом институте АН СССР (ката-
(каталог систематической коллекции моллюсков, № 1). Он имеет следующие размеры: высота
раковины 9.8, ширина 2.2 мм; высота устья 1.8, ширина 1.5 мм; 11 оборотов. Место
сбора: Псырцхий монастырь, сборщик Ледер.
10. MICROPONTICA
295
роче семяприемника. Матка_длиннее влагалища. Толстый семяпровод
плавно переходит в веретеновидный пенис.
Р а сп ространение (рис. 31). Все известные находки приуро-
приурочены к окрестностям курортов Гагра и Новый Афон (черноморское по-
побережье Абхазии).
Экология. Вид обитает на известня-
известняковых скалах.
ых скалах. п #
I. Microponficay"cifcassica
(Bttg.) (рис.
МП
¦рня
ал
213).
Boettger О., 1888 : 154 [Clausilia (Micro-
(Micropontica)]; Boettger О., 1889 : 33, t. 1, fig. 9
[Clausilia (Micropontica)]; Розен, 1911 : 127 [Cl.
{Micropontica) closta Bttg., non 0. Boettger];
Лихарев иРаммельмейер, 1952 : 247,
рис. 174 [Laciniaria (Micropontica)}.
Раковина башневидная, с крупны-
крупными и редкими пластинчатыми белыми ребра-
ребрами (на предпоследнем обороте 30—35), слабо
просвечивающая, матовая, зеленовато-рого-
зеленовато-роговая, с лиловым оттенком. Верхушка широ-
широкая A.0—1.2 мм), округлая. Оборотов 10V2—
12, плавно нарастающих, довольно выпук-
выпуклых, разделенных глубоким швом; эмбрио-
эмбриональные обороты (около 21/2) гладкие; по-
последний оборот чуть выше предпоследнего,
книзу слегка сужен, покрыт крупными
листовидными ребрами, на затылке слегка
уплощен, снизу округлый. Устье ромбическо-
грушевидное, сильно выступающее, отвес-
отвесное, с узким высоколежащим синулусом.
Края устья тонкие, белые, умеренно от-
отвернуты; нижний край плавно закруглен.
Низкая и тонкая верхняя пластинка впере-
впереди обычно достигает края устья, а сзади
сливается со спиральной пластинкой. Нижняя
пластинка лежит глубоко и при прямом положении устья не видна; она
начинается на правой стороне и почти отвесно спускается, образуя лишь
слабый изгиб. Спиральная пластинка берет начало на право-брюшной
стороне. Субколумеллярная пластинка обычно не видна через устье,
или нижний конец ее виден лишь при косом положении раковины. Мозоле-
видная полулунная складка лежит на право-спинной стороне. Над нею
начинается главная складка, простирающаяся вперед приблизительно
на Vg оборота. Имеется более или менее крупная палатальная мозоль.
Лопасть клаузилия широкая и сравнительно короткая, с округлым и
отогнутым нижним концом. (
Размеры: высота раковины 11—14, ширина 2.9—3.4 мм; высота устья
2.5—2.7, ширина 2.1—2.3 мм.
Изменчивость. Раковина очень варьирует как в отношении раз-
размеров (у самых мелких экземпляров высота 9.5, ширина 2.6 мм), так и
в отношении скульптуры. Так, бывают экземпляры с более густой и слабой
ребристостью (var. claviformis Rosen, 1914). С другой стороны, на альпий-
альпийских лугах обитают популяции с неправильно исчерченными раковинами,
без ребер (f. laevigata Likharev, f. nov., — рис. 214). В отношении замы-
Рис. 212. Micropontica closta
(Bttg.). Половая система.
(Ориг.).
296
V. CLAUS1LIINAE
кательнога аппарата у некоторых экземпляров (var. interfecla Rosen,
1914) варьирует положение полулунной складки, которая может быть
сдвинута на спинную сторону, главная складка отсутствует, но иногда
имеется верхняя палатальная складка. Известны случаи (var. saltuensis
Rosen, 1914), когда одновременно присутствуют обе складки, причем
верхняя тянется вглубь раковины от пала-
палатальной мозоли.
Внутреннее строение не изучено.
Распространение (рис. 31). За-
Западная часть Большого Кавказа — горный
массив Оштен-Фишт, верховья реки Кодор
южнее Клухорекого перевала. Наиболее во-
восточная находка сделана в верховьях реки
Малки (бассейн реки Терек) у горы Щиджа-
Тмас.
Систематическое положе-
положение. Данный вид во многом сходен с М.
closta (Bttg.), от которого отличается в первую
очередь формой раковины и более крупными
размерами. В отношении формы и располо-
расположения отдельных частей замыкательного ап-
аппарата данный вид занимает промежуточное
положение между JH. closta и М. retowskii.
3. Micropontica retowskii (Bttg.) (рис-
16,б, *W.^j^c&^**M?sfe\
В о е 11 g е г О., 1888 : 154 [Clausilia (Micw?
pontica)]; Boettger О., 1889 :.33, t. 1, fig. 10
[Clausilia (Micropontica)]; We sterlund, 1897 :
123 [Cl. (Euxina) baleopsis)]; Westerlund,
1901 : 168 [Cl. (Micropontica) retowskii var. simulia]'.
Розен, 1914 : 205 [Clausilia (Micropontica)}; Ли-
Лихарев и Раммельмейер, 1952 : 249, рис.
175, 176 [Laciniaria (Micropontica)).
Рис. 213.
Micropontica
circassica
(Bttg.). (По
Лихареву и
Раммель-
Раммельмейер).
Рис. 214.
Micropontica
circassica
(Bttg.) f.
laevigata
Likharev.
(Ориг.).
Раковина башневидная, очень тонко
ребристо исчерченная (на предпоследнем
обороте 60—80 белых складок), непросвечи-
вающая, матовая, желтовато-роговая или красновато-коричневая. Оборотов
10—И, выпуклых, плавно нарастающих, разделенных глубоким швом; эмб-
эмбриональные обороты BV4) гладкие, довольно блестящие, образуют тупую-
сосковидную вершину; последний оборот книзу не сужен, за устьем с не-
несколькими крупными пластинчатыми ребрами, внизу с крупным килемг
окаймленным бороздой. Устье ромбическо-грушевидное, сильно выступаю-
выступающее, с широким высоколежащим синулусом, внизу округло-угловатое,
с базальным желобком. Края устья чаще всего тонкие, слабо отвернуты,
хрупкие. Очень низкая и тонкая верхняя пластинка впереди достигает
края устья, сзади сравнительно быстро сходит на нет. Спиральной пла-
пластинки нет, или же она представлена коротким мозолевидным утолщением
стенки раковины выше лопасти клаузилия и снаружи не видимым. Ниж-
Нижняя пластинка очень короткая и низкая; она начинается на левоспинной:
стороне раковины и простирается вниз на небольшое расстояние, после'
чего сливается со столбиком. Субколумеллярная пластинка развита
очень хорошо; она начинается на спинной стороне раковины, а заканчи-
заканчивается недалеко от края устья и хорошо видна при косом положении
10. MICROPONTICA
297
устья. Крупная, дугообразно изогнутая (оба конца направлены внутрь
раковины) полулунная складка располагается на лево-спинной стороне
раковины и видна через устье даже при прямом положении раковины.
Главной и палатальной складок нет. На палатальной стенке ниже сину^
луса располагается небольшой бугор — палатальная мозоль. Очень
широкая и сравнительно короткая лопасть клаузилия слабо изогнута
в поперечном и сильно в продольном направлении; нижний конец е&
утолщен и слегка заострен. Значительная часть клаузилия очень хорошо
видна даже при прямом положе-
положении устья.
Размеры: высота раковины
' 11.0—13.0, ширина 3.0—3.3 мм;
высота устья 2.3—2.6, ширина
1.8—1.9 мм.
Рие. 215. Micropontica retowskii (Bttg.).
А — общий вид раковины; Б — последний оборот и устье; В — клаувилий (А, Б — по Ли-
Лихареву и Раммельмейер, В — ориг.).
Изменчивость. Наибольшей изменчивости подвергается по-
поверхностная скульптура раковины: встречаются формы с более сильной»
и редкой скульптурой (на предпоследнем обороте 50—60 складок) и почт»
гладкие формы, раковина которых отличается слабой и редкой исчерчен-
ностью, просвечивающая и блестящая (var. nitidula Rosen, 1914). Размеры
меньше, чем у типичных экземпляров: высота раковины 9—-11, mnpHHai
2.8—3.0 мм.
Внутреннее строение. Материал: Краснодарский край
верховья р. Цеце, альпийский луг, сбор Воробьева, 8 VI 1903, определе-
определение наше — 1 экз.
Радула (рис. 16, Б): [т + (т + 1 + з?|)х21х107'
Размеры срединного зуба 14 X 8.4 ц, ширина радулы 390 ц. Внеш-
Внешние краевые зубы имеют гребенчатый облик.
Единственный экземпляр с мягкими частями вследствие долгого пре-
пребывания в спирте сильно затвердел. Поэтому при препаровке произошло
много разрывов. Половая система в главнейших чертах устроена так же,,
как у М. closta (Bttg.).
Распространение (рис. 31). Западная часть Большого Кав-
Кавказа в районе Кавказского государственного заповедника и его ближайших,
окрестностей.
Экология. Обитает на скалах и в осыпях.
298 ВИДЫ С НЕЯСНЫМ СИСТЕМАТИЧЕСКИМ ПОЛОЖЕНИЕМ
Систематическое положение. Данный вид отличается
от всех прочих видов того же рода присутствием киля и соответствующего
ему желобка, а также неглубоким расположением замыкательного ап-
аппарата, благодаря чему такие части его, как полулунная складка и ло-
лопасть клаузилия, хорошо видны через устье. Кроме того, некоторые части
замыкательного аппарата полностью исчезли — спиральная пластинка,
главная складка, а верхняя и нижняя пластинки отличаются небольшими
размерами и слабым развитием.
ВИДЫ С НЕЯСНЫМ СИСТЕМАТИЧЕСКИМ ПОЛОЖЕНИЕМ ИЛИ
ПРИСУТСТВИЕ КОТОРЫХ В СССР МАЛОВЕРОЯТНО
После ревизии литературных данных середины прошлого века, посвя-
посвященных моллюскам нашей страны, обнаружилось, что имеется ряд видов,
систематическое положение которых неясно или присутствие которых
на нашей территории очень сомнительно. Эти же сомнения по отношению
к некоторым из этих моллюсков были высказаны еще Бетгером (Boet-
tger О., 1878с). Нередко указания на присутствие у нас этих видов свя-
связаны с грубейшими ошибками в определении.
Ниже приводится краткий перечень этих моллюсков со ссылками на
авторов, которые указывали на их наличие в СССР. Для того чтобы под-
подчеркнуть неясность систематического положения этих видов, родовое
дазвание берется в кавычки («Clausilia»):
1. «Clausilia» caucasica A. Schm.
Schmidt A., 1868 : 123.
Описание вида отличается большой неясностью и не снабжено рисун-
рисунком. Местонахождение также указано очень неопределенно — «Кавказ».
Бетгер (Boettger О., 1878Ь, 1878с) и Вестерлунд (Westerlund, 1901)
относили этот вид в среднеевропейскую группу Graciliaria Bielz. Розен
A914) справедливо усомнился в правильности этого вывода.
2. «Clausilia» dydima Parr.
Под этим названием братья Виллы (Villa, 1841) указывают какой-то
вид, якобы найденный в Галиции. В настоящее время не представляется
возможным решить, с чем имели дело названные авторы.
3. «Clausilia» eichwaldi Siem.
Симашко, 1847 : 114.
Симашко описал этот вид по экземпляру, полученному от Эйхвальда
из «южной России». Описание очень несовершенное. Поэтому уже Бетгер
(Boettger О., 1878с) пришел к выводу, что почти невозможно догадаться,
-с чем имел дело автор вида.
4. «Clausilia» gracilis С. Pfr.
Pfeif.f ег С., 1821, Naturgesch. deutsch. Land- u. Susswasser-Moll., 1 : 65,
t. 3, fig. 32.
Двигубский A831) указывает этот вид для «юго-западных губерний.
России, а Слюсарский (Slosarski, 1872) — для Тернопольской области.
В прошлом веке это название чаще всего давалось различным внутривидо-
внутривидовым формам Clausilia dubia Drap.
5. «Clausilia» livida Menke.
Menke, 1830, Syn. method. Moll., ed. II : 130.
Данный моллюск является одним из правозакрученных видов рода
ж подрода Alopia H. et A. Adams. Этот подрод обитает только в южных
ВИДЫ С НЕЯСНЫМ СИСТЕМАТИЧЕСКИМ ПОЛОЖЕНИЕМ 299
Карпатах и прилегающих горных массивах Трансильвании. Симашко
^Siemaschko, 1847) указывает, что он получил от Карпинского 2 экземпляра
этого вида с «берегов Каспийского моря».
Вряд ли когда-либо станет ясным, с чем имел дело этот автор, так как
неизвестно, где находится его коллекция. Однако можно быть абсолютно
уверенным, что определение его неверно.
6. «Clausilia» macilenta Rssm.
R ossmassler, 1842, Iconogr., 2, 11 : 6, fig. 704.
Этот моллюск в настоящее время рассматривают как подвид Laciniaria
plicata (Drap.), обитающий в лекоторых областях Балканского полу-
полуострова. Симашко (Siemaschko, 1847) указывает, что он был им найден
в окрестностях С.-Петербурга (берег р. Поповки). Вполне возможно,
он имел дело с L. plicata, которая действительно обитает в этих местах,
но вряд ли именно с подвидом macilenta Rssm.
7. «Clausilia» nana Parr.
Братья Виллы (Villa, 1841) указывают, что этот моллюск живет в Бу-
:ковине. С чем они имели дело в действительности, представляется неясным,
потому что подобное название давалось различным Clausiliidae. Парис
(Parreys), как известно, имел только коллекцию, а описания новых видов
яе опубликовывал. Бетгер (Boettger О., 1878b) рассматривает <tCl.» папа
Parr, как разновидность Iphigena plicatula (Drap.).
8. «Clausilia» pulverosa Z.
В коллекции братьев Вилла (Villa, 1841) были раковины с этим назва-
названием, собранные в Волыни. В настоящее время это название сохраняют
.за одной из бесчисленных разновидностей Laciniaria plicata (Drap.).
9. «Clausilia» similis (Cbarp.) Rssm.
Rossmassler, 1835, Iconogr., 1,1 : 77, fig. 30.
Это один из синонимов Laciniaria biplicata (Mont.). Указание Надеж-
дина A868) на присутствие «С7.» similis в окрестностях Москвы и Слюсар-
ского (Slosarski, 1872) — на обитание его в Тернопольской области, оче-
очевидно, являются ошибками в определении. Возможно, оба автора имели
дело с экземплярами Laciniaria plicata (Drap.), раковины которых
лишены обычных для этого вида складок на краю устья (var. implicata
Bielz).
10. «Clausilia» simplex Z.
Братья Виллы (Villa, 1841) указывают, что в их коллекции под этим
названием имеются раковины, найденные в Галиции. Этот вид, по-види-
по-видимому, является nomen nuduna. Единственное указание на его системати-
систематическое положение мы обнаружили у Вестерлунда (Westerlung. 1901),
который без всяких объяснений рассматривает его как синоним Iphigena
mucida badia (Rssm.).
Дополнение к корректуре. К тому времени, когда рукопись дан-
данной работы уже находилась в Издательстве, вышла в свет статья Р. Брандта —
fl. A. Brandt, 1961. Diagnosen neuer Ciausiliiden. Arch. Molluskenk., 90 : 1—20.
Для нас особый интерес представляют описания раковин трех новых видов, най-
найденных в Турции в буковых лесах близ озера Абант (вилайет. Волу, северо-западная
Анатолия). Первый вид — Euxina (Euxina) rackae Br., как признает автор, наиболее
близок к Е. (Е.) persica (Bttg.). Второй — Euxina (Euxina) abanti Br., во многом
сходен с Е. (Е.) lasistana Lindh. [с нашей точки зрения последняя является синонимом
Е. difficilis (Ret.)]. Наконец, третий вид — Serrulina (Serrulina) goettingi Br., во многом
сходен с S. (S.) serrulate, (Pit.), но отличается рядом существенных особенностей.
Вместе с упомянутыми моллюсками были найдены Serrulina serrulata (Pir.) и Euxina
(Thraciella) borisi (Hesse) (последняя, по нашим данным, принадлежит к роду Mega-
leuxina Bttg. — стр. 201).
ПРИЛОЖЕНИЕ
МАТЕРИАЛЫ К ФАУНЕ С L A U S IL I I I> A E
И 3 Т Р ЕТ ИЧНЫХ И ЧЕТВЕРТ И Ч Н Ы X
;. ОТЛОЖЕНИЙ СЕВЕРНОГО КАВКАЗА
До настоящего времени наши сведения о фауне наземных моллюсков;
третичного и четвертичного периодов Кавказа были очень скудными и
разрозненными. В полной мере это касается также и Clausiliidae, По сути
дела, они ограничиваются несовершенным описанием одного вида, сде-
сделанным Волковой A958). В связи с этим представляет большой интерес
описание коллекции ископаемых Clausiliidae Северного Кавказа, которую;
мы получили от А. А. Стеклова (Москва). Она состоит из 56 раковин тре-
третичного возраста и 7 раковин четвертичного возраста. Одновременно
А. А. Стеклов сообщил нам данные о возрасте этих раковин, которые полу-
получены как от сопоставления с хорошо изученными слоями с морской фауной,
так и в результате исследования других групп наземных моллюсков тех же
отложений, где были найдены интересующие нас раковины.
Ниже приводятся описания одного нового рода и нескольких новых
для науки видов и дается краткая характеристика всей коллекции иско-
ископаемых Clausiliidae. В настоящее время она хранится в Палеонтологи-
Палеонтологическом институте Академии наук СССР (Москва) под № 1872. Там же нахо-
находятся и голотипы новых видов.
Подсемейство PHAEDUSINAE
Род EUXINOPHAEDUSA LIKHABEV gen. nov.
Р а к о в и н а большая или очень большая, левозакрученная, декол-
лированная. Оборотов 51/2—7; последний внизу плавно закруглен. Верх-
Верхняя и спиральная пластинки лежат на разном удалении от столбика,
в связи с чем они не сливаются воедино, хотя и не заходят одна за другую.
Нижняя пластинка лежит глубоко и отвесно, плохо видна при прямом
положении раковины, за исключением ее нижнего конца, который, по-
подобно субколумеллярной пластинке, достигает края устья. Имеется
мощная полулунная складка и очень длинная главная складка. Иногда
присутствует еще короткая верхняя палатальная складка, которая лежит
очень глубоко и снаружи не видна. Клаузилий с языковидной лопастью»
нижний конец которой утолщен и отогнут.
Тип рода: Ей. volkovae Likharev, nom. nov.
Этот род сильно отличается от всех известных групп данного семейства,
существовавших в третичном периоде. Наибольшее сходство он обнару-т
живает с ныне живущими во Вьетнаме и в южном Китае видами из под-
рода Excussispira Lindh. (род Hemiphaedusa Bttgl, Phaedusinae). У обеих
сравниваемых групп раковина очень крупная, деколлированная, почти
гладкая. Однако имеются и существенные различия: у Excussispira верх-
ТРЕТИЧНЫЕ И ЧЕТВЕРТИЧНЫЕ CLAUSILIIDAE 301
ляя пластинка сливается со спиральной, край устья широкий и двойной,
часто со складками.
По общей форме раковины и по размерам, а также по многим призна-
признакам замыкательного аппарата Euxinopheadusa во многом напоминает
ныне живущие на Кавказе виды из рода Acrotoma Bttg., в особенности
A. komarowi (Bttg.), которая обладает крупной деколлированной ракови-
лой. Однако все Acrotoma имеют киль — признак, характерный для
Euzininae, в то время как данный род, как и все Phaedusinae, лишен этого
¦образования.
Соотношение спиральной и верхней пластинок
у Euxinophaeudusa позволяют рассматривать эту
группу как переход от Phaedusinae к Euxininae. От-
Отсутствие же киля указывает на принадлежность
к первому из названных подсемейств.
Euxinophaedusa volkovae Likharev, лот. nov.
<рис. 216).
Во л к о в а, 1953 : 75, табл. 5, фиг. 10,11 (Clausilia ro-
seni Lindh., nom. mus. — non Westerlund, 1901).
Раковина веретеновидно-цилиндрическая,
тонко и неясно исчерченная или гладкая, очень тол-
толстостенная. Оборотов 5V2—7, слабо выпуклых. Устье
коротко-яйцевидное, цельное или прерванное, с ши-
широким, слабо врезанным, прямым синулусом. Края
устья очень толстые, слабо отвернуты. Верхняя
пластинка высокая, толстая, сравнительно корот-
короткая, сзади, круто снижаясь, сходит на-нет. Ниж-
Нижняя пластинка толстая, отвесная. От ее нижнего
конца отходит кверху и глубь раковины короткая, рИс. 216. Euxino*
до толстая промежуточная складка. Нижний конец phaedusa volkovae Lik-
«убколумеллярной пластинки хорошо виден при пря- harev, nom. nov Го-
яом положении устья и доходит до его края. Мощ- №™1872/73 (Опит )'
лая дугообразно изогнутая полулунная складка лежит
на правобрюшной стороне. Несколько глубже ее начинается главная
¦складка, тянущаяся почти до самого края устья. Имеется короткая верх-
верхняя палатальная складка, которая лежит впереди полулунной и через
устье не видна.
Размеры деколлированных раковин: высота раковины 21.5—34.5,
ширина 7.5—10.2 мм; высота устья 6.3—9.0, ширина 5.5—6.4 мм.
Размеры голотипа (№ 1872/73): высота раковины 23.2, ширина 8.5 мм;
высота устья 6.3, ширина 6.0 мм; 53/4 оборотов.
Материал. В коллекции А. А. Стеклова находятся 7 экземпляров
данного вида. Один из них обнаружен в образце № 3118, взятом на левом
берегу реки Ардуган (Черные горы, между Орджоникидзе и Нальчиком),
другой — в образце № 1, взятом на правом берегу реки Белой у Майкопа.
3 экземпляра хорошей сохранности, в том числе голотип, найдены в об-
образце № 2960, взятом из «назранских» слоев в обнажениях правого бе-
берега реки Сунжи у села Заречного. Последние 2 экземпляра представлены
фрагментами и обнаружены в образце № 2030, взятом из средне- и крупно-
крупнозернистого песчаника с левого берега реки Фортанги у села Бумут (Чер-
(Черные горы, к северо-востоку от Орджоникидзе).
В коллекции Н. С. Волковой (Музей имени Чернышева, Ленинград^нахо-
дятся 2 экземпляра этого вида, найденные в окрестностях города Армавира.
302
ПРИЛОЖЕНИЕ
Возраст. Экземпляры из образцов №№ 1 и 3118, а также оба
экземпляра из коллекции Н. С. Волковой относятся к позднему сармату
(миоцен), а остальные предположительно к меотису (миоцен-плиоцен).
С и н о н и м и я. Н. С. Волкова A953), вслед за Линдгольмом, назвала
один из своих экземпляров Clausilia roseni Lindh., nom. mus., а другой
обозначила как Clausilia sp. Мы предлагаем этому виду новое название,
так как название Cl. roseni преоккупировано Веетер-
лундом (Westerlund 1901).
Euxinophaedusa steklovi Likharev, sp. nov. (рис.
217).
Раковина яйцевидно-веретеновидная, очень
большая и вздутая, гладкая, лишь местами замет-
заметна очень неясная исчерченность, очень толстостен-
толстостенная. Оборотов 61/2, слабо выпуклых. Устье коротко-
яйцевидное, цельное, 1 с широким неврезанным:
синулусом. Края устья очень толстые, закруглен-
закругленные, но не отвернутые; середина внешнего края
выступает вперед, нижний край плавно закруг-
закруглен. Верхняя пластинка очень крупная, но корот-
короткая, не выступает за пределы края устья. Нижняя
пластинка круто уходит кверху. Субколумеллярная
пластинка достигает края устья и видна при прямом
положении раковины. Дугообразно изогнутая толстая
полулунная складка лежит на правой стороне рако-
раковины. Над нею находится длинная главная складка,
заканчивающаяся вблизи края устья.
Размеры единственного экземпляра (голотип, №
1872/76): высота раковины 38.8, ширина 14.5 мм;
высота устья 13, ширина 8 мм.
Материал. Единственный экземпляр обнару-
обнаружен в образце № 2030, взятом на левом берегу реки
Фортанги у села Бумут (Черные горы, к северо-
востоку от Орджоникидзе). Внешний облик рако-
раковины сохранился очень хорошо, но несколько раз-
разрушены нижняя и субколумеллярная пластинки и
утерян клаузилий.
Возраст. Меотис.
Систематическое положение. Данный вид отличается
от предыдущего большими размерами, более вздутой раковиной и поло-
положением полулунной складки. Верхней палатальной складки обнаружить
не удалось, однако она, может быть, и имеется, но ее нельзя увидеть,
не разрушив раковины.
Serrulina (Serrulina) cf. sieversi (Pfr.) (рис. 218).
Раковина стройная, тонко ребристо исчерченная, за устьем реб-
ребристая. Наиболее широкий предпоследний оборот, однако в общем все
3 последних оборота почти одной ширины; последний оборот внизу плавно
закруглен. Устье ромбическо-грушевидное, с крупным, сильно врезан-
врезанным синулусом. Края устья утолщены, довольно сильно и широко от-
отвернуты. Палатальный участок края почти прямой, нижний плавно за-
закруглен, париетальный прогнут над верхней пластинкой. Верхняя пла-
Рис. 217. Euxinophae-
Euxinophaedusa steklovi Likharev
sp. nov. Голотип.
ПИН АН СССР, №
1872/76. (Ориг.).
1 Часть верхнего края разрушена, однако следы его заметны.
ТРЕТИЧНЫЕ И ЧЕТВЕРТИЧНЫЕ CLAUSILIIDAE
3<Й
стинка довольно высокая, лежит отвесно и выступает за пределы края
устья. Нижняя пластинка лежит глубоко (виден лишь ее нижний конец
и то только при косом положении устья) и заканчивается крупной крае-
краевой складкой. Между верхней и нижней пластинками, вдоль края устья,
лежит ряд узелков и одна довольно крупная складка (непосредственно
над нижней пластинкой). На колумеллярном участке края устья, ниже
нижней пластинки, лежат еще 2 складки, из которых самая нижняя наи-
наиболее крупная.
Размеры: ширина раковины 2.3 мм; высота устья 2.1,
ширина 1.5 мм.
Материал. От единственного экземпляра сох-
сохранились лишь 3 нижних оборота и устье, которое в глу-
глубине заполнено породой. Эта раковина обнаружена в об-
образце № 2960, взятом из слоя темной плотной глины
в песчано-глинистой серии немых пород («назранские»
слои), обнаженных на правом берегу реки Сунжи в райо-
районе древнего укрепления Назрань, у села Заречного
(центральное Предкавказье).
Возраст. Предположительно поздний миоцен.
Систематическое положение. Форма
нижней пластинки и общий облик раковины и устья
указывают на принадлежность ее к роду Serrulina Mouss.
и к подроду Serrulina s. str. Из современных видов с дан-
данным экземпляром наиболее сходен S. (S.) sieversi (Pfr.).
Общее — скульптура поверхности раковины, форма верх-
верхней и нижней пластинок, число и расположение складок
на краю устья. Однако есть и различия. Данный экзем-
экземпляр обладает раковиной более стройной (нижняя
часть цилиндрическая), устье ее более вытянуто в высоту
и вся раковина заметно меньших размеров, чем современ-
современные S. sieversi.
Рис. 218. Ser-
Serrulina (Serru-
(Serrulina) cf. siever-
sieversi (Pfr.). ПИН
АН СССР, №
1872/66.
(Ориг.).
Serrulina (Pravispira) nazranica Likharev, sp. nov. (рис. 219).
Раковина левозакрученная, веретеновидная, деколлированная.
3 верхних оборота тонко ребристо исчерчены, последние 2 слабо и неравно-
неравномерно исчерчены (лишь у шва скульптура выражена сильнее); последний
оборот за устьем ребристый. .Эмбриональных оборотов 3V2, гладких.
Взрослая, т. е. деколлированная раковина обладает 51/2—6 оборотами;
в течение онтогенеза формировалось 14—15 оборотов; последний оборот
заметно сужен, внизу плавно закруглен. Устье широко-грушевидное,
с узким, глубоко врезанным и отогнутым к оси раковины синусом. Края
устья сильно утолщены, слабо отвернуты. Верхняя пластинка довольно
высокая, крючкообразно изогнута, выступает за край устья; сзади она
плавно переходит в спиральную пластинку. Нижняя пластинка лежит
очень близко к верхней, лишь у края устья немного отодвинута от послед-
последней. Субколумелляраная пластинка достигает края устья и нижний ко-
конец ее хорошо виден при прямом положении раковины. Из складок пала-
палатальной стенки удалось обнаружить только одну — верхнюю или среднюю
палатальную складку. На участке между верхней и субколумеллярной
пластинками край устья имеет складки и узелки; из них не менее одной
складки находится над нижней пластинкой и чаще всего, 2 складки между
нижней и субколумеллярной пластинками. Иногда их число бывает больше.
Одна из краевых складок может сливаться с нижней пластинкой, благо-
благодаря чему конец последней кажется раздвоенным. Лопасть клаузилия
304
ПРИЛОЖЕНИЕ
языковиднаяу с почти параллельными краями и утолщенным, слегка отог-
отогнутым концом.
Размеры: высота деколлированной раковины 12.5—12.7, ширина
около 3.5 мм; х высота устья 2.7—3.2, ширина 2.2—2.7 мм.
Размеры голотипа A872/54, рис. 219, А): высота раковины 12.5, ши-
ширина около 3.5 мм; высота устья 3.1, ширина 2.6 мм; 6 оборотов.
Материал. Найдено 12 экземпляров. Из них 8
в образце № 2960, взятом из слоя темной плотной гли-
глины в песчано-глинистой серии немых пород («назранов-
ские» слои), обнаженных на правом берегу реки Сунжи
в районе древнего укрепле-
укрепления Назрань, у села Зареч-
Заречного (центральное Предкав-
Предкавказье). В этом образце обна-
обнаружены^ целых, лишь слегка
сдавленных взрослых рако-
раковины (в том числе голотип),
3 почти целых раковины, од-
одна молодая раковина с неде-
коллированной верхушкой.
Остальное представлено фраг-
фрагментами последнего оборота
и устья.
Кроме того, данный вид
D раковины) обнаружен в
образце № 2652, взятом из
отложений, идентичных пре-
предыдущему образцу, в разрезе
на правом берегу реки Терек
у Эльхотово,
В о з р а ст. Предполо-
Предположительно поздний миоцен.
Систематическое
положение. Данный вид
принадлежит к роду Serru:
В
Рис. 219. Serrulina (Pravispira) nazranica Li-
kharev, sp. nov. (Ориг.).
A — общий вид раковины (голотип, ПИН АН СССР^
J* 1872/54); Б — устье; В — клаузилий.
Una Mo'uss. На это указывает
весь облик раковины. Мы
считаем, что его следует от-
отнести в подрод Pravispira
Lindh. За это говорит форма нижней пластинки, ее близкое положение
к верхней пластинке, а также тот факт, что субколумеллярная пластин-
пластинка доходит до самого края устья.
От всех вымерших видов того. же подрода и от единственного ныне
живущего [S. (Pr.) semilamellata (Mouss.)] данный моллюск отличается
в первую очередь деколлированной раковиной. В отношении формы устья
и замыкательного аппарата наибольшее сходство наш вид обнаруживает
с S. (Pr.) decemplicata (Sacco). Однако у последней до 10 складок на краю
устья, а у нашего значительно меньше.
S. decemplicata описана из отложений позднего плиоцена Италии.
Позднее (Simionescu et Barbu, 1904) ее обнаружили в сармате Румынии.
1 Фактически ширина, вероятно, меньше, так как обе более или менее сохранив-
сохранившиеся раковины несколько сдавлены в дорзо-вентральвом направлении.
ТРЕТИЧНЫЕ И ЧЕТВЕРТИЧНЫЕ CLAUSILIIDAE
305
Pontophaedusa praefuniculum Likharev, sp. nov. (рис. 220).
Раковина левозакрученная, веретеновидная, тонко ребристо
исчерченная. Последний оборот внизу плавно закруглен, за устьем тупо-
ребристый. Устье грушевидное, цельное, выступающее, слегка косое,
с небольшим неврезанным синулусом. Края устья коротко отвернуты,
тупые, со слабой губой. Верхняя пластинка низкая, но выше, чем у P. fu-
niculum (Mouss.), плавно переходит в спиральную пластинку. Нижняя
пластинка спирально изогнута и лежит в глубине устья; на подходе к краю
устья она распадается на 2 почти параллель-
параллельных коротких складочки. Субколумеллярная
пластинка простирается до края устья (видна
при прямом положении раковины). Сравни-
Сравнительно короткая, совершенно прямая полулун-
полулунная складка лежит отвесно на право-спинной
стороне раковины. Над ней начинается глав-
главная складка, которая тянется вперед на х/з
оборота. Между главной и полулунной лежат
2 коротких, но толстых палатальных складки:
Рис. 220. Pontophaedusa praefuniculum Likharev, sp. nov. (Ориг.).
A •— общий вид нижней части раковины (голотип, ПИН АН СССР, Л! 1872/52); Б ¦
устье (вид сбоку); В — складки палатальной стенки.
верхняя — начинается на одном уровне с полулунной и простирается впе-
вперед не более чем на 1/4 оборота, средняя — начинается немного глубже
полулунной, проходит между нею и верхней складкой и тянется впе-
вперед с уклоном книзу. Лопасть клаузилия желобчатая; нижний конец ее
утолщен и отогнут кверху.
Размеры: ширина раковины 4.7 мм; высота устья 3.7—4.6, ширина 3.1 мм.
Размеры голотипа (№ 1872/52, рис. 220,А): высота устья 4.6, ширина
3.1 мм.
Материал. Найдено 4 экземпляра, из них 3 в образце № 2030,
который взят в средне- и крупнозернистом песчанике на левом берегу
реки Фортанги у села Бумут (Черные горы, к северо-востоку от Орджо-
Орджоникидзе). Сохранность этих экземпляров различная; один (голотип)
представлен 2V2 оборотами и устьем, остальные — фрагментами послед-
последнего оборота с устьем и клаузилием.
Четвертый экземпляр обнаружен в образце № 3044, взятом из песков
на левом берегу реки Белой, ниже Майкопа, напротив хутора Гавердов-
ского. Сохранность — нижняя часть последнего оборота с устьем.
306
ПРИЛОЖЕНИЕ
Возраст. По-видимому, меотис, но не исключено, что поздний
сармат.
Систематическое положение. Данный вид во многом
сходен с P. funiculum (Mouss.), но отличается от него следующими особен-
особенностями: 1) верхняя пластинка более высокая; 2) нижняя пластинка сла-
слабее выдается в просвет устья, а нижний конец ее более резко расщеплен
на 2 краевых складки; 3) имеются 2 палатальных складки (у P. funicu-
funiculum таких складок нет); 4) полулунная складка сравнительно короткая
и совершенно прямая (у P. funiculum она длиннее и дугообразно изог-
изогнута).
Подсемейство EUXININAE
Mentissoidea cf. litotes (A. Schm.) (рис. 221).
Раковина веретеновидная, в средней части сильно вздутая, сильно
и равномерно исчерченная, за устьем ребристая. Верхняя часть завитка
с вогнутыми боковыми контурами и округлой верхушкой. Максимальная
ширина приходится на 2 предпоследние оборота. Обо-
Оборотов 13V2, слабо выпуклых; последний оборот книзу
резко сужен, по периферии уплощен или даже несколь-
несколько вогнут, с крупным и длинным морщинистым килем,
окаймленным заметной бороздой. Устье грушевидное,
с узким врезанным и отогнутым к оси раковины сину-
лусом, внизу с базальным желобком. Края устья ши-
широко отвернуты, несколько утолщены. Крупная крюч-
ковидная верхняя пластинка выступает за пределы
края устья. Нижняя пластинка в просвет устья вы-
выдается как крупная, почти горизонтальная складка;
с краем устья она не связана. Нижний конец субколу-
меллярной пластинки виден при косом положении
раковины. Пупок — широкая щель; околопупочное
поле очень широкое.
Размеры: высота раковины 15.2, ширина 4.1 мм;
высота устья 3.9, ширина 3.0 мм.
Материал. Единственный экземпляр (№1872/67)
найден в «назрановских» слоях на правом берегу реки
Сунжи у села Заречного.
Возраст. Предположительно поздний миоцен.
Систематическое положение. Рако-
Раковина данного экземпляра отличается от раковин сов-
современных Mentissoidea litotes, в первую очередь резким
сужением последнего оборота. В этом отношении она
более походит на Mucronaria (Index) index (Mouss.).
Однако видимые участки замыкательного аппарата и
устье очень напоминают картину, типичную для М.
litotes.
Рис. 221. Mentis-
Mentissoidea cf. litotes
(A.Schm.).(Opnr.).
ПИН АН СССР,
N°. 1872/67.
Подсемейство CLAUSILIINAE
Quadriplicata intermedia Likharev, sp. nov. (рис. 222).
Раковина веретеновидная, стройная, тонкостенная, тонко ребристо
исчерченная, за устьем ребристая. Максимальная ширина приходится на
предпоследний оборот. Верхушка узкая (ширина 0.9 мм). Оборотов 12—13,
верхние слабо, нижние довольно выпуклые; последний оборот книзу
ТРЕТИЧНЫЕ И ЧЕТВЕРТИЧНЫЕ CLAUSILIIDAE
¦307
плавно сужается, внизу с крупным морщинистым килем. Устье широко-
широкогрушевидное, выступающее, с широким прямым неврезанным синулусом
и с базальным желобком. Края устья широко отвернуты, утолщены, ко-
коричневатые. Верхняя пластинка не выступает за пределы края устья,
низкая, толстая, плавно снижаясь, переходит в спиральную пластинку.
Нижняя пластинка лежит глубоко и при прямом положении устья не
видна; в глубь раковины она уходит круто, а с краем устья связана корот-
короткой складочкой; выше и ниже этой склад-
складки нередко лежат еще 1 или 2 слабо вы-
выраженных складки или бугорка. Ниж-
Нижний конец субколумеллярной пластин-
пластинки виден лишь при очень косом поло-
положении раковины. Главная и 3 палаталь-
палатальных складки начинаются на правой сто-
стороне раковины (лишь нижняя палаталь-
палатальная берет начало несколько ближе к
устью). Наиболее длинная верхняя па-
палатальная складка, нижний конец ко-
которой лежит на лево-брюшной стороне
раковины (на затылке ей соответствует
продольное углубление). Наиболее ко-
короткая средняя палатальная складка
(видна лишь сквозь стенку раковины).
Второй по длине является главная склад-
складка. Нижняя несколько короче, но ниж-
нижний конец ее виден снаружи даже при
прямом положении устья.
Размеры: высота раковины 13.5—
15.5, ширина 3.7—4.3 мм; высота устья
3.3—3.7, ширина 2.5—3.3 мм.
Размеры голотипа (№ 1872/77,—
рис. 222): высота раковины 15.2, ши-
ширина 3.7 мм; высота устья 3.4, шири-
ширина 2.5 мм.
Изменчивость. В образце
№ 2877а вместе с 8 типичными раковина-
раковинами обнаружена одна, покрытая крупными
редкими ребрами (на предпоследнем обо-
обороте 38 ребер). Этот экземпляр
(№ 1872/89, — рис. 223) мы рассматриваем
как разновидность — var. costata Lik-
harev, var. nov. Его размеры: .высота раковины 14, ширина 3.8 мм;
12V4 оборотов.
Материал. В нашем распоряжении находились 13 экземпляров
данного вида. Из них 9 (вместе с разновидностью) обнаружены в образце
№ 2877а, 1 в образце № 2884 и 2 — в образце № 2897. Эти образцы взяты
на правом берегу реки Сунжи у поселка Алды, немного выше Грозного.
Из них 1 экземпляр (голотип) превосходной сохранности.
Наконец, последний экземпляр очень плохой сохранности обнаружен
в образце № 2708 — балка Пседах, на северном склоне Сунженского
хребта.
Возраст. Образцы №№ 2877 и 2708 относятся к акчагылу, образец
№ 2897 — к апшерону, а образец № 2884 — переходною возраста, между
акчагылом и апшероном.
Рис. 222. Qua-
driplicata inter-
intermedia Likharev,
sp. nov. Голо-
Голотип. ПИН АН
СССР,
№ 1872/77.
(Ориг.).
Рис. 223. Quad-
riplicata inter-
intermedia var. cos-
costata Likharev,
var. nov. Голо-
Голотип. ПИН АН
СССР,
№ 1872/89.
(Ориг!).
308 ПРИЛОЖЕНИЕ
Систематическое положение. Типичная форма дан-
данного вида во многом представляет собой переход от Q. quadriplicata
(A. Schm.) к Q. subaggesta (Ret.). С первой ее сближает характер скульп-
скульптуры поверхности раковины и строение замыкательного аппарата. Однако
наш вид отличается от Q. quadriplicata более стройной раковиной, мень-
меньшими размерами, грушевидным устьем и присутствием нескольких крае-
краевых складок или бугорков. С Q. subaggesta наш вид сближает сходная
форма и величина раковины и скульптура ее поверхности. Но у Q. sub-
subaggesta устье более грушевидное, с отогнутым к оси раковины синусом,
а палатальные складки лежат глубже.
Хотя у всех просмотренных нами раковин клаузилия обнаружить
не удалось, тем не менее он несомненно был, на что указывает сильное
развитие палатальных складок.
Quadriplicata sp. В двух образцах — № 2030 с левого берега реки
Фортанги и № 3044 с левого берега реки Белой ниже Майкопа — най-
найдено 5 раковин очень плохой сохранности. Однако можно с уверенностью
сказать, что они принадлежат к роду Quadriplicata Bttg.
Предположительный возраст этих образцов меотис.
В итоге в коллекции А. А. Стеклова мы обнаружили 5 новых для
науки видов Clasiliidae (из них 2 принадлежат к новому роду) и 3 вида,
которые только условно определены как современные, так как имеют
заметные отличия от последних.
Остальные раковины той же коллекции, как третичные, так и четвер-
четвертичные, принадлежат к современным кавказским видам — Caspiophae-
dusa perlucens (Bttg.), Serrulina semilamellata (Mouss.), Euxina somche-
tica (Pfr.), Eu. tschetsckenica (Pfr.).
Для характеристики географического и хронологического распростране-
распространения ископаемых Clausiliidae, обнаруженных в коллекции А. А. Стеклова,
приводим следующую таблицу (табл. 5).
Таблица о
образ-
образца
1
3118
2030
3044
2960
2652
2703
2708
2877 а
2884
2890
2897
2864
996
2020
2460
Распределение Clausiliidae по
образцам
(в хронологическом порядке)
Местонахождение
Р. Белая, правый берег.
окраина Майкопа, под го-
городским садом.
Левый берег р. Аргудан,
Черные горы, между Орд-
Орджоникидзе и Нальчиком.
Левый берег р. Фортанги у
села Бумут, Черные горы
к северу-востоку от Орд-
Орджоникидзе.
Левый берег р. Белой ни-
ниже Майкопа, против ху-
хутора Гавердовского.
Правый берег р. Сунжи в
районе древнего укреп-
укрепления Назрань, у села
Заречного.
Правый берег р. Терек у
Эльхотово.
Балка Пседах на северном
склоне Сунженского хреб-
хребта.
Там же.
Правый берег р. Суяжи у
поселка Алды, выше Гроз-
Грозного.
Там же.
Там же.
Там же.
Заманкульская балка на
южном склоне Сунженско-
Сунженского хребта.
Район Нальчика.
Суглинки двухметровой тер-
террасы р. Терек у Эльхото-
Эльхотово.
Суглинки двухметровой тер-
террасы р. Ярык-Су (Даге-
(Дагестан).
Возраст
Поздний
сармат.
Поздний
сармат.
Меотис.
Меотис.
Меотис?
Меотис?
Акчагыл.
Акчагыл.
Акчагыл.
Акчагыл-
апше-
рон.
Апше-
роя.
Апше-
рон.
Апше-
рон.
Четвер-
тичныи
период.
Четвер-
Четвертичный
период.
Четвер-
Четвертичный
период.
Фауна
Euxinophaedusa volkovae.
Euxinophaedusa volkovae.
Euxinophaedusa volkovae.
Eu. steklovi.
Serrulina semilamellata.
Pontophaedusa praefunicu-
LUTfl.
Quadriplicata sp.
PontophaediCba praefunica-
lum.
Quadriplicata sp.
Euxinophaedusa volkovae.
Serrulina cf. sieversi.
S. nazranica.
Mentissoidea cf. litotes.
Serrulina nazranica.
Euxina somchetica.
Quadriplicata intermedia.
Quadriplicata intermedia.
Euxina somchetica.
Quadriplicata intermedia.
Euxina somchetica.
Quadriplicata intermedia.
Euxina tschetschenica.
Euxina somchetica.
Serrulina semilamellata.
Euxina somchetica.
Caspinphaedusa perlucens.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ *
abanti, Euxina (Euxina) 299
Achatinacea 86
Achatinidae 89
acridula, Laciniaria (Mentissa) canalifera
250
Acroeuxina 191
acrolepta, Clausilia 280
Acrotoma 12, 36, 52, 57, 58, 63, 64, 79,
83, 110, 155, 156, 301
Acrotoma s. str. 155, 156
acuminata, Clausilia 277, 279, 280
acuminata, Clausilia (Euxina) 280
acuminata, Clausilia (Mentissa) 280
acuminata, Mucronaria (Mucronaria) 46,
47*, 57, 278*, 279, 280*, 282, 284, 287
aggesta, Clausilia (Euxina) 268
aggesta, Laciniaria (Euxina) 268
aggesta, Quadriplicata 57, 78, 272
aggesta, Quadriplicata aggesta 40, 264*,
265, 268*, 269*
albella, Armenica •gracillima 149
Albinaria 49
Alinda 240
Alopia 18, 52, 55, 81, 113, 131, 261, 298
Alopiinae 11, 12, 32, 35*, 36—38, 48,
49, 54, 61, 64, 67—69, 73, 74, 76,
79, 82—84, 91*, 92, 93*, 130, 172, 202
alpestris, Cochlodina laminata 137
alpestris, Quadriplicata aggesta aggesta
269*
alpicola, Clausilia dubia 224
amanica, Clausilia (Serrulina) serrulata 120
amanica, Serrulina (Serrulina) serrulata
115, 120
Armenica 36, 39, 52, 57, 59, 60*, 70, 76,
79, 83, 112, 113, 143, 149, 153, 201,
280, 286, 287
avenacea, Chondrina 39
Baboria 63
badia, Clausilia 236
badia, Iphigena mucida 231, 236, 299
Balea 24, 77, 80, 89, 113, 261, 262
Baleastra 261
Baleinae 82
baleopsis, Clausilia (Euxina) 296
Barcania 49
Basommatophora 84, 89
baudoni, Iphigena 68
belone, Clausilia 206
bicarinata, Armenica 60*, 144, 145*, 146
bicarinata, Clausilia 146
bicristata, Clausilia 139
bidens, Clausilia 132, 136
bidens, IJelix 136
bidentata, Clausilia 54, 55, 70, 71, 72*,
219, 221*, 222*, 223
bidentatus, Turbo 221
biformis, Clausilia 199
binodis, Alopia 131
biplicata, Clausilia 242
biplicata, Laciniaria (Laciniaria) 15*, 36*,
42, 43, 52, 55, 70, 71, 77, 205, 241,
242*, 243*, 244, 263, 299
biplicatus, Turbo 242
blissi, Armenica 153
blissi, Clausilia (Oligoptychia) 153
borealis, Clausilia l^testriata 235
borealis, Iphigena latestriata 55, 72, 231,
233*, 235
borisi, Clausilia (Wagneria) 201
borisi, Euxina (Thraciella) 299
borisi, Megaleuxina 193, 194, 201*
bosniensis, Herilla 19*, 20*, 21*
brjanskii, Idyla foveicollis 141
brunnea, Armenica 40, 59, 60*, 144, 147,
148*, 149, 150
brunnea, Clausilia 147
brunnea, Clausilia (Oligoptychia) 147
Bulimulacea 86
Bulimulidae 38, 89
byzantia, Euxina circumdata 168
cana, Clausilia 246
cana, Laciniaria (Strigilecula) 55, 70, 71,
241, 243, 246*, 247*, 248
Canalicia 63
canalifera, Clausilia 248, 249
canalifera, Laciniaria (Mentissa) 40, 60,
241, 249*, 250*, 251
Caspiophaedusa 39, 61, 111, 126, 128
Caucasica 182, 189, 265
caucasica, «Clausilia» 298
cerata, Clausilia 135
cerata, Cochlodina 55, 70, 133, 135, 136*
Charpentieria 63
Chondrulinae 88
cilicica, Clausilia 205
cilicica, Clausilia (Euxina) 207
cilicica, Galeata 206, 207
cilicica, Pleioptychia 207
cingullela, Helicigona 39
circassica, Clausilia (Micropontica) 295
circassica, Laciniar-ia (Micropontica) 295
1 Курсивом выделены синонимы, жирным шрифтом — названия родов и подсе-
подсемейств Clausiliidae, жирные цифры обозначают страницы с описаниями, звездочкой
отмечены страницы с рисунками названной формы.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
311
circassica, Micropontica 47, 56*, 57, 78,
292, 293, 295, 296*
circumdata, Clausilia 167
circumdata, Euxina (Euxina) 69, 78, 166,
167, 168*, 170, 179
circumplicosa, Quadriplicata pumilifor-
mis 275
Cirrobasis 63
ciscaucasica-terensis, Acrotoma semicincta
159
climax, Mucronaria (Index) index 290,
291*
closta, Clausilia (Micropontica) 292, 293
closta, Laciniaria (Micropontica) 293
closta, Micropontica 40, 47, 56*, 57, 78,
292, 293, 294*, 295*, 296, 297
Clausilia 12, 37, 52, 54, 63, 67, 73, 79,
80, 93, 112, 218, 231
С lausiliastra 132
Clausiliinae 13, 14, 24, 29, 34, 35*, 37,
38, 54, 61—64, 67, 68, 73, 74, 76, 77, 79,
81—84, 91*, 92, 93*, 128, 209, 212,
215, 255, 262, 306
Clausiliinae 82
claviformis, Micropontica circassica 295
Cochlodina 18, 55, 63, 71, 81, 110, 132, 202
Cochlodininae 29, 32, 34, 35*, 38, 54, 61, 63,
64, 67, 68, 73, 74, 76, 79, 81—84,
91*, 92, 93*, 132
colchica, Clausilia 182
collasi, Clausilia (Serrulina) 120
collasi, Phygas 120
collasi, Serrulina (Serrulina) 114, 115, 120*
commutata, Clausilia 138
commutata, Cochlodina 70, 133, 138*
comnena, Clausilia (Oligoptychia) 200
comnena, Megaleuxina 113, 193*, 194,
200*
Constrieta 63
corpulenta, Clausilia 189
corpulenta, Clausilia (Euxina) 189
corpulenta, Euxina (Elia) 165, 189, 190*,
191
corynodes, Neostyriaca 16*
costata, Quadriplicata intermedia 307*
costulata, Laciniaria (Mentissa) canalifera
250
Cristataria 49
cruciata, Clausilia 55, 70*, 71, 72, 220,
222, 223, 228, 229*, 230*
cruciatus, Glischrus (Clausilia) 228
curta, Iphigena plicatula 239
Cusmicia 218
decemplicata, Serrulina (Pravispira) 304
Delima 37
densestriata, Clausilia 236
densestriata, Iphigena 70
derasa, Clausilia 78, 192, 194
derasa, Clausilia (Euxina) 194
derasa, Laciniaria (Euxina) 194
derasa, Megaleuxina 40, 41, 48, 57, 78,
179, 193*. 194, 195*. 196*, 198, 202
detersa, Laciniaria (Mentissa) canalifera
250
difficilis, Clausilia (Euxina) 169
difficilis, Euxina (Euxina) 40, 164, 166,
169*, 170*, 182, 299
dipolauchen, Clausilia (Euxina) 276
dipolauchen, Laciniaria (Euxina) 276
dipolauchen, Quadriplicata 48, 57, 78,
264*, 266, 276*
disjuncta, Armenica 40, 59, 60*, 144,
146, 147*
disjuncta, Clausilia 146
Disjunctaria 63
dissopkya, Clausilia somchetica 186
dubia, Clausilia 55, 68, 70, 71, 219, 222,
223, 224*. 225*, 298
duboisi, Clausilia 279, 282
duboisi, Clausilia (Euxina) 282
duboisi, Laciniaria (Euxina) 281, 283
duboisi, Mucronaria (Mucronaria) 40, 42,
46, 47*, 48, 51, 57, 58, 66, 73, 75, 78,
278*, 279, 282*, 283*, 284*, 287
dydyma, «Clausilia» 298
eichwaldi, «Clausilia» 298
elata, Clausilia 254, 257
elata, Laciniaria (Pseudalinda) 257
elata, Laciniaria turgida 257
elata, Uncinaria 257
; elata, Vestia 55, 255, 257, 258*, 259, 260
elatior, Megaspira 87*
Elia 182, 189
elongata, Clausilia bidentata 222
elongata, Vestia turgida 256
Emarginaria 63
Enidae 23, 28, 38, 87, 88, 89
erivanensis, Clausilia 118
Eualopia 63
Euclausta 63
euprepes, Armenica 154, 155
Euxina 59, 63, 69, 77, 78, 79, 81, 82, 110,
164, 170, 176, 179, 182, 193, 202, 205,
212, 265, 279, 280
Euxina s. str. 166, 182, 189
Euxinastra 58, 79, 110, 202, 203
Euxininae 13, 14, 24, 29, 34, 35*, 38,
39, 54, 61—63, 68, 76, 79, 83, 84, 91,
92, 93*, 126, 128, 139, 172, 209, 212,
213, 215, 265, 301, 306
Euxinophaedusa 63, 65, 300
Excussispira 300, 301
exigua, Clausilia bidentata 222
expansa, Euxina (Elia) corpulenta 190
fallax, Clausilia 251, 253
fallax, Clausilia (Pseudalinda) 253
fallax, Laciniaria 253
fallax, Pseudalinda 55, 251, 253, 254*
fausta, Clausilia 144, 154
filiformis, Cochlodina orthostoma 134
Filosa 58, 79, 111, 211, 213
filosa, Clausilia 214
filosa, Clausilia (Euxina) 214
filosa, Clausilia (Serrulina) 214
filosa, Filosa 27*, 32, 40, 42, 57, 82, 212,
213*, 214*
filosa, Serrulina 214
filograna, Clausilia 216
filograna, Graciliaria 216
312
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
filograna, Ruthenica 52, 55, 70, 216*,
217*, 218*
foveicollis, Clausilia 140
foveicollis, Clausilia (Oligoptychia) 140
foveieollis, Idyla (Scrobifera) 13, 27*, 40,
52, 57, 58, 73, 95, 140*. 141*, 142*, 143
fragilis, Balea 262
fragilis, Pupa 261, 262
frater, Laciniaria (Mentissa) gracilicosta 251
funiculum, Clausilia 123, 124
funiculum, Clausilia (Serrulina) 124
funiculum, Pontophaedusa 12, 13*, 23,
27, 40, 52, 57, 121, 124*, 125*, 126*,
130, 305, 306
fusorium, Clausilia 209
Fusulinae 82, 83 ,
Fusulus 37, 79, 83, 92
Galeata 79, 111, 204, 205, 206
galeata, Clausilia 206
galeata, Galeata 205, 206*
ganeo, Mentissoidea litotes litotes 210
Garnieria 54
Garnieriinae 84
Gastropoda 10
glauca, Alopia 55, 131, 132*
glauca, Balea 131
glauca, Clausilia 131
goettingi, Serrulina (Serrulina) 299
Graciliaria 215, 298
gracilicosta, Clausilia 250
gracilicosta, Laciniaria (Mentissa) 40, 60,
241, 250, 251*
gracilior, Clausilia 118
gracilior, Euxina (Elia) corpulenta 190
gracilior, .Laciniaria (Mentissa) gracili-
gracilicosta 251
gracilis, «Clausilia» 298
gracilis, Clausilia cruciata 230
gracillima, Armenica 40, 59, 60*, 70, 144,
149*, 153
gracillima, Clausilia (Oligoptychia) 149
gradata, Clausilia (Euxina) 270, 272
griseofusca, Armenica 40, 57, 59, 60*,
70, 144, 150*, 153, 154
griseo-fusca, Clausilia (Alinda) 150
griseo-fusca, Clausilia (Oligoptychia) 150
gulo, Clausilia turgida 259
gulo, Laciniaria (Pseudalinda) 259
gulo, Uncinaria 259
gulo, Vestia 55, 70, 255, 258, 259*, 260
gustavi, Armenica 284
gustavi, Clausilia (Oligoptychia) 280, 284
gustavi, Mucronaria (Mucronaria) 39, 40,
42, 47*, 58, 66, 78, 113, 278*, 279,
284, 285*, 286*
haliciaca, Clausilia (Uncinaria) procera 255
hamata, Clausilia (Euxinastra) 203
hamata, Euxinastra 32, 40, 48, 57, 202,
203*, 204*
hamata, Laciniaria (Euxinastra) 203
Helicellinae 88
Helicidae 28, 38, 39, 88
Hemiphaedusa 126, 300
hetaera, Clausilia 164, 166
hetaera, Euxina (Euxina) 166, 167*, 168—
170, 177, 179
Heterurethra 85*, 86
huebneri, Clausilia 192
huebneri, Euxina (Acroeuxina) 165, 192
huebneri, Laciniaria (Euxina) 192
hueti, Armenica 30*, 40, 59. 60*, 144,
145, 147, 148, 151*, 152*, 154
hueti, Clausilia 151
hueti, Clausilia (Oligoptychia) 151
hyrcanica, «Phaedusa» 58, 129, 130*
hyrcanica, Phaedusa (Caspiophaedusa) 1291
iberica, Clausilia 199
iberica, Clausilia (Euxina) 199
iberica, Megaleuxina 193*, 194, 199*, 202
Idyla 36, 39, 79, 83, 112, 139, 144, 205
Illunellaria 176, 177
implicata, Laciniaria (Laciniaria) plica-
ta 243, 244
inarmata, Quadriplicata pumiliformis 275
Index 113, 289
index, Clausilia 289
index, Clausilia (Euxina) 289
index, Clausilia (Laciniaria) 289
index, Mentissoidea 289
index, Mucronaria (Index) 48, 57, 78, 278* ,
279, 284, 289, 290*, 292, 306
Inobseratella 143, 149
interjecta, Micropontica circassica 296
intermedia, Quadriplicata 65, 306, 307*,
309
Iphigena 12, 37, 52, 54, 55, 63, 67, 73, 79,
80, 91*, 93, 112, 230, 255
Iphigenia 218
Kolenatii, Clausilia 182
komarowi, Acrotoma (Acrotoma) 28*, 40,
57, 155, 156*, 157*, 158*, 301
Komarowi, Clausilia (Acrotoma) 155, 156-
Kuzmicia 218
laccata, Acrotoma (Acrotoma) 57, 156*,
161, 162*
laccata, Clausilia (Acrotoma) 161
Laciniaria 37, 67, 71, 77—79, 82, 91*
93, 110, 111, 149, 176, 182, 205, 215,
237, 240, 241, 251, 254, 261, 262, 265,
278, 286, 292
Laciniaria s. str. 241, 247, 249
laevicollis, Armenica 40, 60*, 144, 145*,
147, 154
laevicollis, Clausilia 143, 144
laevicollis, Clausilia (Oligoptychia) 144
laevigata, Micropontica circassica 295,296*
laminata, Clausilia 136
laminate, Cochlodina 16*, 17*, 32*, 40,
45*, 51, 55, 56, 59, 67, 68, 70-72, 132,
133, 135*, 136. 137*
laminatus, Turbo 136
Laminifera 63, 69, 84
Laminiferinae 84
lantzi, Clausilia 148
lantzi, Laciniaria (Inobseratella} 147
lasistana, Clausilia (Euxina) 169, 170
lasistana, Euxina 299
latens, Alopia glauca 132
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
313*
latestriata, Clausilia 234
latestriata, Iphigena latestriata 55, 70,
71, 231, 233*, 234*, 235*
lederi, Clausilia (Euxina) 270
lederi, Laciniaria (Euxina) 270
lederi, Quadriplicata 40, 57, 264*, 265,
270*, 271*, 273
lessonae, Clausilia 172, 173, 175
lessonae, Clausilia (Euxina) 175
lessonae, Euxina (Euxina) 40, 41, 58, 78,
166, 171, 175, 176*, 177*
lessonae, Triloba (Illunellaria) 170, 171
lindholmi, Clausilia 198
lindholmi, Laciniaria (Euxina) 198
lindholmi, Megaleuxina 57, 193*, 194,
198*
lineolata, Iphigena 70
liostoma, Euxina (Elia) corpulenta 190
litoderma, Clausilia (Euxina) litotes 211
litoderma, Clausilia litotes 211
litoderma, Mentissoidea litotes 211
litotes, Clausilia 209, 279
litotes, Clausilia (Euxina) 209
litotes, Mentissoidea 13*, 15*, 32, 40, 57,
58, 82, 211», 213, 292, 306
litotes, Mentissoidea cf. 64, 306*
litotes, Mentissoidea litotes 209*, 210*
litotes, Polinskia 209
livida, Clausilia 131, 298
macilenta, «Clausilia», 299
Macroptychia 54, 62
maesta, Clausilia 189, 190
maesta, Euxina (Elia) 165, 179, 189, 190
Marpessa 132
Mauritanica 49
maxima, Laciniaria (Mentissa) gracilicosta
251
Megaleuxina 12, 69, 78, 79, 111, 192,
201, 202, 203, 299
Megaspiridae 86, 87, 89, 115
Mentissa 18, 67, 77, 80, 110, 202, 241, 248,
249
Mentissoidea 63, 78, 79, 83, 112, 208,
209, 279
Mentissoideinae 81—83
Mesurethra 86
Metabaleinae 82, 83
Metazaptyx 61
mezchetica, Clausilia (Euxina) strauchi 281
Micropontica 47, 52, 56*, 57, 58, 66, 78,
112, 262, 292, 293
Milne-Edwardsia 63
mimele, Armenica 154, 155
minima, Clausilia cruciata 229
minor, Mucronaria (Mucronaria) duboisi
283
minor, Mucronaria (Index) index 290
moesta, Clausilia 190
moesta, Laciniaria (Elia) 190
moravica, Laciniaria 260
moravica, Laciniaria (Pseudalinda) rilo-
ensis 260
Mucronaria 37, 46, 47*, 66, 78, 91*, 93,
112, 113, 277, 278, 286, 287, 289, 292
Mucronaria s. str. 279, 289
Multiplicaria 277, 279
multistriata, Quadriplicata dipolaucben.
277
nana, «Clausilia» 299
nana* Iphigena plicatula 239
narzanensis, Acrotoma (Acrotoma) 57; 156*,.
160, 161*
narzanensis, Acrotoma semicincta 159
narzanensis, Clausilia (Acrotoma) 160
nazranica, Serrulina (Pravispira) 64, 303,.
304*, 309
Nenia 80
Neniinae 32, 35*, 36, 38, 54, 61—63, 76_
83, 84, 89, 90, 91*, 92, 93*, 94
Neniopsis 63
Neostyriaca 37, 79, 83
nigricans, Clausilia 222
nigricans, Turbo 221
nitidosa, Clausilia vetusta striolata 248-
nitidosa, Laciniaria (Strigilecula) 70, 241,
248
nitidula, Micropontica retowskii 297
novorossica, Clausilia (Euxina) 188
novorossica, Euxina (Caucasica) 57, 165,
188*
novorossica, Laciniaria (Euxina) 188
obsoleta, Clausilia dubia 224
obtusa, Clausilia 221
occidentalis, Serrulina (Serrulina) sieversi
58, 59*, 115, 117*, 118
Oligoptychia 139, 143, 201, 286
Oospira 62
Oospiroides 62, 63
orthostoma, Clausilia 134
orthostoma, Cochlodina 45*, 55, 70, 71,.
133, 134*, 135*
Orthurethra 85*, 86, 89
ossetica, Clausilia 194
ossetica, Clausilia somchetica 186
Pachyphaedusa 63
Palaeophaedusa 63
Papillifera 91
parreyssii, Clausilia 135
parvula, Clausilia 52, 55, 70, 71, 219, 220*
parvula, Glischrus (Clausilia) 220
parvula, Pirostoma 220
paulhessei, Clausilia 173
paulhessei, Euxina (Euxina) persica 165r
173
pauli, Laminifera 84
perfiga, Armenica laevicollis 145
Peristoma 230
perlucens, Caspiophaedusa 12, 23, 40, 52,,
58, 90, 118, 126*, 127*, 128*, 130, 308,
309
perlucens, Clausilia 126
perlucens, Clausilia (Phaedusa) 126
perlucens, Hemiphaedusa 126
persica, Clausilia 170
persica, Euxina 58, 66, 69, 78, 172, 174,
175, 177, 179, 299
persica, Euxina (Euxina) persica 40, 165,
170, 171, 172*
perversa, Balea 41, 42, 59, 67, 70, 261,
262*, 263*
314
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
perversus, Turbo 262
Phaedusa 80
Phaedusinae 32, 35*, 36, 38, 54, 61—65,
68, 69, 73, 76, 82—84, 89, 90, 91*,
92—94, 113, 126, 128, 129, 212, 300
Phygas 121
J>irostoma 230
Pirostoma s. str. 230
Pleioptychia 139
pleuroptychia, Clausilia 280, 287
pleuroptychia, Clausilia (Euxina) 287
pleuroptychia, Mentissoidea 287
pleuroptychia, Mucronaria j (Mucronaria)
40, 42, 47, 48, 57, 78, 278*, 284, 287*,
288*, 289
plicata, Clausilia 240, 244
plicata, Laciniaria (Laciniaria) 16*, 26*,
31*, 36*, 51, 55, 70-72, 77, 205, 241,
243*, 244, 245*, 299
plicata, Mucronaria (Mucronaria) duboisi
283
plicata, Pupa 244
plicatula, Clausilia 238
plicatula, Iphigena 17*, 33*, 55, 59, 67,
68, 70, 232, 238*, 239*, 299
.plicatula, Pupa 238
Plioptychia 63
Plistoptychia 205
podolica, Clausilia dubia 223
J>olinskia 208
Polloneria 63
polygyra, Clausilia (Euxina) pleuroptychia
287, 289
Solyptychia 277, 279
pontica, Clausilia (Euxina) 177
pontica, Euxina (Euxina) 78, 166, 177,
178*, 179*
Pontophaedusa 39, 58, 61, 63, 65, 66, 111,
123
praefuniculum, Pontophaedusa64, 305*,309
praegracilis, Quadriplicata aggesta aggesta
268
Pravispira 63, 111, 114, 115, 121, 304
-Priobaleinae 82
Proalbinaria 61, 63
procera, Vestia turgida 256
j>rolaminata, Cochlodina 68
Prosobranchia 17, 19, 22
proteus, Euxina (Elia) corpulenta 190
Pseudalinda 54, 55, 67, 73, 78, 80, 111, 251
Tseudidyla 63, 213
Ptychophaedusa 63
Pulmonata 10, 29, 74
pulverosa, «Clausilia» 299
pumila, Clausilia 70, 71, 75, 225*, 228
pumila, Clausilia pumila 55, 219, 226*,
227*
pumiliformis, Clausilia (Euxina) 274
pumiliformis, Laciniaria (Euxina) Ilk
pumiliformis, Quadriplicata 40, 48, 57,
78, 264*, 266, 274*, 275*, 276, 277
Pupillacea 87
Pupillidae 23, 38, 39, 87—89, 94
Pyrostoma 230
Quadriplicata 63, 66, 78, 93, 111, 264,
265, 279, 286, 308, 309
quadriplicata, Clausilia 264, 266
quadriplicata, Clausilia (Euxina) 266, 279
quadriplicata, Laciniaria (Euxina) 266
quadriplicata, Quadriplicata 40, 41, 48,
57, 75, 78, 264*, 265, 266, 267*, 268,
271, 273, 308
rackae, Euxina (Euxina) 299
raddei, Euxina (Caucasica) somchetica
184
Reinia 89, 261
remote, Cochlodina 136
reshtensis, Clausilia 175
reshtensis, Laciniaria (Euxina) 175
retowskii, Clausilia (Micropontica) 296
retowskii, Laciniaria (Micropontica) 296
retowskii, Micropontica 27*, 40, 47, 56*,
57, 78, 112, 292, 293, 296, 297*
reulauxi, Clausilia (Euxina) 197
reulauxi, Euxina 177, 178
reulauxi, Laciniaria (Euxina) 197
reulauxi, Megaleuxina 40, 42, 57, 78, 193*,
194, 197*, 198*
riloensis, Laciniaria (Pseudalinda) 260
riloensis, Pirostoma 260
riloensis, Vestia 41, 255, 260
rolphii, Clausilia 230
roseni, Clausilia 64, 160, 301, 302
Rosen'ia 162
Roseniella 155, 162
rudis, Clausilia 82
rugosa, Clausilia 218, 222
runensis, Clausilia dubia 224
rupicola, Armenica 153
rupicola, Clausilia 153
rusticana, Quadriplicata pumiliformis 275
Ruthenica 37, 54, 79, 81, 83, 92, 113, 215
samsunensis, Euxina (Elia) corpulenta 190
sandbergeri, Clausilia 192
sandbergeri, Clausilia (Laciniaria) 194
sandrii, Triloba 172
saulcyi, Clausilia 190
schaposchnikovi, Idyla foveicollis 141
schwerzenbachi, Clausilia 205
schwerzenbachi, Euxina 206
schwerzenbachi, Galeata galeata 206, 207*
Sciocochlea 114, 121
Scrobifera 58, 66, 140
sejuncta, Clausilia 227, ?28
sejuncta, Clausilia ршпйа 55, 72, 219,
227*
semicincta, Acrotoma (A»otoma) 40, 57,
156*, 158, 159*, 160*. 161
semicincta, Clausilia (Acrotoma) 158
semilamellata, Clausilic 121, 122
semilamellata, Clausilia (Serrulina) 122
semilamellata, Serrulina (Pravispira) 40,
57, 58, 59*, 64, 114, 115, 122*, 130,
212, 304, 308, 30b
semiserrata, Clausilia (Serrulina) serru-
lata 118
senghanensis, Serrulina 68, 114, 115, 121
septentrionalis, Clausilia bidentata 222
serrulata, Clausilia lib
serrulata, Clausilia (Serrulina) 118
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
315
serrulata, Serrulina (Serrulina) 56, 58, 66,
114, 125, 212, 299
serrulata, Serrulina (Serrulina) serrulata
40, 59*, 115, 118, 119*, 123
Serrulina 23, 39, 52, 59*, 61, 63—65,
69, 87, 111, 113, 114, 115, 123, 125,
212, 303, 304
.Serrulina s. str. 114, 115, 121, 303
serrulosa, Serrulina (Pravispira) semila-
mellata 123
sieversi, Clausilia 113
sieversi, Serrulina (Serrulina) cf. 64, 302,
303*, 309
sieversi, Serrulina (Serrulina) 58, 66, 88*,
114, 121, 123, 303
sieversi, Serrulina (Serrulina) sieversi 40,
58, 59*, 115, 116*, 118
Sigmurethra 25, 85*, 86, 89, 90
signifera, Clausilia (Serrulina) 126
silesiaca, Clausilia 138
similis, Clausilia 242, 299
simplex, «Clausilia» 299
simulia, Clausilia (Micropontica) retowskii
296
sobrievskii, Acrotoma (Roseniella) 40,
57, 156*, 162, 163*
sobrievskii, Clausilia (Oligoptychia) 162
sodalis, Laciniaria (Mentissa) gracilicosta
251
.Soleolifera 84
somchetica, Clausilia 182
somchetica, Clausilia (Euxina) 182
somchetica, Euxina (Caucasica) 40, 51,
57, 58, 65, 73, 78, 165, 182, 183*, 184*,
185*, 187, 188, 308, 309
somchetica, Laciniaria (Euxina) 182
speciosa, Clausilia dubia 223
stabilis, Clausilia 252
stabilis, Clausilia (Pseudalinda) 252
-Stabilis, Clausiliastra 252
stabilis, Laciniaria (Pseudalinda) 252
stabilis, Pseudalinda 55, 70, 251, 252*,
253*, 254
stauropolitana, Clausilia (Euxina) 269
stauropolitana, Quadriplicata aggesta 73,
264*, 265, 269, 270*
steklovi, Euxinophaedusa 64, 302*, 309
stomatica, Clausilia duboisi 283
strauchi, Clausilia 279, 281
Strauchi, Clausilia (Euxina) 281
strauchi, Laciniaria (Euxina) 281, 283
strauchi, Mucronaria (Mucronaria) 46, 47*,
57, 278*, 279, 280, 281*, 284
Strigilecula 80, 110, 241, 246
Strigillaria 246
strumosa, Euxina (Euxina) 78, 166, 168,
169*, 170, 179
Stylommatophora 10, 28, 38, 74, 84, 85,
86, 87, 88, 92
suanetica, Megaleuxina derasa 195, 281
suanetica, Mentissoidea litotes litotes 210
subaggesta, Clausilia (Euxina) 272
subaggesta, Laciniaria (Euxina) 272
subaggesta, Quadriplicata 48, 57, 78, 264*,
265, 272*, 273*, 274, 308,.
subtilis, Clausilia 279, 282
subulata, Clausilia 168
Synprosphyma 82
taeniata, Clausilia 134
talyschana, Euxina (Euxina) 40, 58, 78,
165, 171, 173, 174*, 177
taurica, Clausilia 140, 143
taurica, Laciniaria (Mentissa) gracilicosta
251
tenax, Mucronaria (Index) index 290
tesselata, Armenica bicarinata 146
Thalestris 162
thracica, Clausilia 173
tocatensis, Armenica laevicollis 145
Tracheopulmonata 86
triadis, Clausilia (Euxina) lederi 270, 272
Triloba 172, 173, 177
Triptychia 63
Triptychiinae 62, 63, 84, 87, 90, 93*, 115
Tropidauchenia 54
tschetschenica, Clausilia 186
tschetschenica, Clausilia (Euxina) 186
tschetschenica, Euxina (Caucasica) 40—42,
51, 57, 58, 65, 78, 165, 175, 185*, 186*,
187, 188, 308, 309
tschetschenica, Laciniaria (Euxina) 186
tumida, Clausilia 236
tumida, Clausilia ventricosa 236
tumida, Iphigena 55, 70, 231, 236, 237*,
238*
tumida, Iphigena ventricosa 233
turgida, Clausilia 255
turgida, Clausilia (Vestia) 255
turgida, Laciniaria 255
turgida, Laciniaria (Pseudalinda) 255
turgida, Uncinaria 260
turgida, Vestia 55, 70, 255, 256*, 257*,
258—260
tuschetica, Euxina (Euxina) 40, 57, 165,
179, 180*, 181*, 182
Uncinaria 254
ungulata, Clausilia 138
unicristata, Armenica 146—149
unicristata, Clausilia 147
Valloniidae 94
ventricosa, Clausilia 232
ventricosa, Iphigena 16*, 25*, 34*, 37*,
41, 42, 55, 70, 71, 231, 232*, 233*,
235
ventricosa, Pupa 232
venusta, Cirrobasis 61
Vestia 54, 55, 67, 73, 77, 78, 80, 91*, 93,
112, 254, 255, 259
vetusta, Clausilia 246
vetusta, Laciniaria 247, 248
vindobonensis, Clausilia dubia 223
vinosa, Clausilia (Oligoptychia) 140, 143
volkovae, Euxinophaedusa 64, 300, 301*,
309
Zonitidae 28
СОДЕРЖАНИЕ
Стр-
Предисловие $
Систематический указатель видов семейства Clausiliidae &
Введение 10
Общая характеристика моллюсков сем. Clausiliidae 10
Морфология Clausiliidae 11
Внешний вид 11
Раковина 11
Замыкательный аппарат 14
Строение A4). — Развитие A8). — Функциональное значение B0).
Анатомия 24
Легкое и мантийный комплекс органов B4). — Пищеварительная
система B5). — Мускулатура B8). — Нервная система B9). — По-
Половая система C0). — Коррелятивные связи между половой систе-
системой и мускулатурой C8).
Биология Clausiliidae
Питание
Размножение ¦. . ¦
Рост и длительность жизни
Среда и адаптации
Климат
Температура и влажность D3). — Адаптации к высокогорному
климату D7). — Адаптации к влажному субтропическому кли-
климату D8).—Адаптации к сухому субтропическому климату D9).
Свет
Горные породы и почвы ,
Растительность
Основные типы местообитаний
Географическое распространение Clausiliidae
Европейские виды
Кавказские виды
Крымские виды
История формирования фауны Clausiliidae западной части Палеарктики . .
К вопросу о древности Clausiliidae (анализ данных палеонто-
палеонтологии и географического распространения) 601
Третичный период 62:
Кавказ F4). — Крым F7). — Средняя Европа F7).
Четвертичный период . 68-
Выводы . 73-
Некоторые вопросы систематики Clausiliidae 74
Таксономическое значение отдельных частей тела 74
Раковина G4). — Радула G5). — Половая система G6).
К вопросу о подразделении Clausiliidae на подсемейства 77
О группах Laciniaria Hartm. и Euxina Bttg 77
Критический обзор старых систем 79
Положение Clausiliidae внутри отряда стебельчатоглазых моллюсков (Sty-
lommatophora) 84
Основные направления эволюции Clausiliidae . 89
Методы исследования Clausiliidae 94
Сбор материала (94). — Обработка сборов в полевой лаборатории
(95). — Исследование раковины (96). Вскрытие (98).
Литература 100
Систематическая часть 110
Таблица для определения родов сем. Clausiliidae фауны СССР и сопредельных
стран 110
I. Подсемейство Phaedusinae 113
СОДЕРЖАНИЕ 317
Стр.
1. Род Serrulina Mouss 113
2. Род Pontophaedusa Lindh 123
3. Род Caspiophaedusa Lindh 126
Дополнение к подсемейству Phaedusinae . 129
П. Подсемейство Alopiinae 130
Род Alopia H. et A. Adams 131
III. Подсемейство Cochlodininae 132
Род Cochlodina Гёг 132
IV. Подсемейство Euxininae subfam. nov 139
1. Род Idyla H. et A. Adams 139
2. Род Armenica Bttg 143
Дополнение к роду Armenica Bttg 153
3. Род Acrotoma Bttg 155
4. Род Euxina Bttg 164
5. Род Megaleuxina Bttg. 192
6. Род Euxinastra Bttg. 202
7. Род Galeata Bttg 204
8. Род Mentissoidea Bttg 208
9. Род Filosa Bttg 211
V. Подсемейство Clausiliinae 215
1. Род Buthenica Lindh 215
2. Род Clausilia Drap 218
3. Род Iphigena Gray 230
4. Род Laciniaria Hartm 240
5. Род Pseudalinda Bttg 251
6. Род Vestia Hesse 254
7. Род Balea Gray 261
8. Род Quadriplicata Bttg 264
9. Род Mucronaria Bttg 277
10. Род Micropontica Bttg. 292
Виды с неясным систематическим положением или присутствие которых в
СССР маловероятно 298
Приложение. Материалы к фауне Clausiliidae из третичных и четвертичных
отложений Северного Кавказа 300
Алфавитный указатель латинских названий 310
ФАУНА СССР. МОЛЛЮСКИ
Том III, вып. 4.
Илья Михайлович Лихарев
КЛАУЗИЛИИДЫ (CLAUSILIIDAE)
Утверждено к печатай
Зоологическим институтом Академии наук СССР
Технический редактор М. Е. Зендель
Корректоры Г. А. Аухимоеич, Т. Н. Вогданова-
Иатькова и М. А. Брайнина
Сдано в набор 1A1 1962 г. Подписано к печати 20 VI
1962 г. РИСО АН СССР № 61-57В. Формат бумаги
70 X 108'/ie- Бум. л. 10. Печ. л. 20 = 27.4 усл. печ. л.
Уч.-изд. л. 27.69. Изд. № 1633. Тип. зак. № 533.
М-37376. Тираж 1400
Цена 2 р. 14 к.
Ленинградское отделение
Издательства Академии наук СССР
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. Издательства Академии наук СССР
Ленинград, В-34, 9 линия, д. 12