



















































































































Текст
,
исумо: .А., ма . о I .У.,
, I дурза ц: а А. ., А тамирова . '
Ие. . I В. А ., . OMaJ I ва .. .
,
ФЛОРАrАЛОФИТОВ
ТЕРСКО КУМСКОЙ
НИЗМЕННОСТИ
И ЕЕ АНАЛИЗ
Тайсумов М.А., Умаров М.У., Абдурзакова А.С.,
Астамирова M.A. M., Исраилова С.А., MaroMaдoвa Р.С.
ФЛОРА r АЛОФИТОВ ТЕРСКО..
КУМСКОЙ НИЗМЕННОСТИ
И ЕЕ АНАЛИЗ
rрозНый.... 2014
Рецензенты:
Крапивина Е.А., кандидат биолоrических наук, доцент
Хасуева Б.А., кандидат биолоrических наук, доцент
Тайсумов М.А., Умаров М.У., Абдурзакова А.С.,
Ас'rамирова M.A. M., Исраилова С.А., Маrомздовз Р.С.
ФЛОРА [АЛОФИТОВ ТЕРСКО КУМСКОЙ НИЗМЕННО
СТИ И ЕЕ АНАЛИЗ. [розный, 2014. 124 с.
в моноrрафии содержит сведения по истории изучения, о
среде обитания и биолоrо эколоrических особенностях rалофи
тов. Приведен список видов, комплексный анализ rалофильной
флоры и классификация rалофитов Терско Кумской низменности
Издание предназначено для ботаников флористов; reorpa
фов, эколоrов, аспирантов, маrистров и студентов, специализиру
ющихся по ботаническому профилю.
ISBN 978 5 9905853 7 9
AH ЧР
«)чrпи
@Тайсумов М.А.
cg Умаров М.У.
cg Астамирова I,A. M.
cg Абдурзакова А.С.
cg Исраилова С.А.
cg Маrомадова Р.С.
ВВЕДЕНИЕ
Известно, что преобладающее число представителей флоры
Земноrо шара развивается в условиях высокоrо содержания солей
25% поверхности земли в той или иной степени засолены, а две
трети её покрыто водами Мировоrо OKeaн rде содержание солей
достиrает 3..40/0. Следовательно, значительная часть видов флоры
земноrо шара осуществляет ЦИЮI cBoero развития в присутствии
BbIcoKoro содержания солей. Вследствие этоrо познание меха..
низма действия солей и приспособительных реакций растений
имеет не только теоретическое, но и большое практическое зна..
чение.
В 21 веке всё большее значение приобретает проблема OCBoe
ния и использования засоленных земель. В нашей стране интра
зональные почвы солончаки, солонцы, солоди и различные их
разновидности занимают около 90 Мom. ra и сосредоточены
rлавным образом в южных реrионах: в зонах степей, пустынь и
полупустынь (Ковда, 1937, 1946, 1947). В практическом отноше
нии используются они недостаточно. По данным почвенноro ин
ститута имени В.В. Докучаева, в сельскохозяйственный оборот
вовлечено Bcero 15 20% от площади всех засоленных земель.
Естественный покров засоленных земель предстаВ..1ен pacтe
ниями rалофитами. Здесь они составляют растительность солон
чаков, солонцов, солодей и участков СОЛOltчаковых степей, а TaK
же основу растительноrо покров а засоленных маршей вдоль по
бережий Каспийскоrо моря, песчаных и ракушечных отмелей со--
лёных и солоноватых лиманов.
rалофиты, представляя узкоспециализированную эколоrиче
скую rруппу, свидетельствуют о связи природы растений сокру"
жающей средой, а их ценозы являются индикаторами почв, хи
мическоrо состава и rлубины залеrания rpYHToBbIX вод (Ke.JU1ep,
1910; Худяков, 1964; Самойлова, 1964; Витман, Поздняко
1968; Рабочев, Кравчук, 1972).
Вместе с тем, природные фитоценозы солончаков источник
сырья кормовых (Elytr;g;a elongata, Е. repens, Festuca or;entalis,
Рисс;nеП;а g;gaпtea, Melilotus albus, АтаrШlthus albus и др.), ле
карственных (Arteт;s;a taиr;ca, MelilohlS albus, М o.fjic;nalis, Li..
тoniuт тeyeri, L. casp;uт, Glycyrrhiza glabra, Galega officiпalis,
3
Gypsophila trichotoтa, lris halophila, Sa/sola soda, SalicOl,пia eиro
раеа и др.), фитомелиоративных (Taтarix gracilis, Т. raтosissiтa,
Leyтus sabu/osus, Cyпodoп dacty/oп, Ca/aтagrostis aruпdiпacea и
др.), б'умажно целлюлозных (Phragтites australis, Leyтus sabu
/osus, Ca/aтagrostis pseudophragтites) и ряда друrих полезных
растений (Ларин, 1950, 1969; rубанов и др., 1970; Наrалсвский,
1984, 1988a, 1990в; Нarалевский, Жилин, 1990). Некоторые rало
фиты (E/ytrigia e/oпgata, Е. repens) служат ценным материалом
для селекции растений (Цицин, 1937, 1965).
Большой интерес представляют rалофиты в плане выяснения
природы их происхождения, rеоrрафии и эколоrо
ФИЗИОJlоrических особенностей. Ещё недостаточно изучена aHa
томия однодольных rалофитов, так как большинство существу
ющих работ выполнено на двудольных растениях (Баталин, 1875,
1885; Lesage, 1890; Schimper, 1898; Delf, 1912; Well1elt, 1927;
StockeJ', 1929, 1976; Келлер, 1929, 1940; Новиков, 1 942; Поплав
екая, 1948; Лейеле, 1931, 1949; Рожановский, 1952; Василевская,
1941,1954,1965; Seddon, 1974; Lychede, 1977; Василевская, Бут
ник, 1981; Бутник, 1984; rамалей, 1984, 1985, rамалей, Шийряв
дамбоЙ, 1988 и др.). Отсутствуют данные об особенностях CTPYK
ryрной адаптации облиrатных и факультативных злаков и сит
ников rаJ]ОфИТОВ в условиях засоления в разных филоrенстиче
ских в( вях семейства Роасеае Barnh. и Juпcaceae Juss.
Bc. это позволяет думать, что материалы по анатомии BereTa
тивных opraHoB злаков и ситников..rалофитов MOryT быть полез
ны для познания их эколоrии, а также при решении вопрщ;ов фи
лоrении и таксономии. Поскольку при анатомических ИССJlедова
ниях обычно устанавливают соотношение между тканями эти
дaнны( окажутся необходимыми и при оценке кормовых ДOCTO
инств (RegaI, 1959, 1960), и в подборе соответствующих соле
устойчивых видов злаков и ситников при проведении биолоrиче
ской М1елиорации солончаков и друrих засоленных земель.
Важным моментом для практическоrо использования засо
ленных земель представляется разработка классификации rало
фильной растительности CeBepHoro Кавказа (Билык, 1963; Hara
левекий, 1 990а, 19906, 1991 а, 1999a и др.), а также выделение
среди J алофитов различных зколоrо физиолоrических rрупп: co
ленакапливающих, солевыделяющих и соленепроницаемых, а
4
среди последних факультативных и облиrатных, составляющих
ядро флоры rалофитов CeBepHoro Кавказа 165 видов (Нarалев
ский, 19916).
Наиболее актуально изучение rалофитов, произрастающих в
районах Восточноrо Приазовья и PycCKoro Причерноморья, rде
сосредоточены основные массивы засоленных земель Предкавка..
зья. Здесь насчитывается 234 вида растений..rалофитов и наибо
лее разнообразный флористический состав растительности
(Нarалевский, 1975, 1978, 1982, 1988, 1989, 19906, 1991, 1997 и
др.).
Изучение флоры и растительности rалофитов CeBepHoro Кав"
каза даёт возможность научно обосновать мероприятия по повы
шению продуктивности засоленных земель, разработать приёмы
устойчивоrо функционирования аrpoфитоценозов и охраны уни
кальных rалофильных экосистем.
Существующая литераryра располаrает лишь фраrментарны"
ми данными по систематике, флоре, растительности, rеоrpафии,
эколоrии и анатомии rалофитов CeBepHoro Кавказа (Данилев
ский, 1869; Буш, 1909; Косенко, 1924, 1927, 1947,1970; Новопо--
кровский, 1925; Шифферс Рафалович, 1928a, 19286 Флёров,
1929, 1930; Роrовской, 1935, 1941; rроссrейм, 1936, 1948, 1949;
Шифферс, 1953; Лавренко, 1940, 1947, 1980; Зозулин, Пашков,
1980; Зозулин, Федяева, J 985; Пашков, Зозулин, 1986; Иванов,
1997 и др.).
5
Фи.зико rЕоrР АФИЧЕСКАЯ ХАРАКТЕРИСТИКА
ТЕРСКО КУМСКОЙ НИЗМЕННОСТИ
rЕоrРАФИЧЕСКОЕПОЛОЖЕНИЕ
Равнины Предкавказья занимают территорию, простираю..
щуюся от Манычской впадины на севере до подножия БО.lьшоrо
Кавказа на юre. Терско..Кумская низменность расположена к ce
веро"востоку от Большоrо Кавказа. Её естественными rраницами
являются: на севере река Кума, на юrе река Терек (пропж Но..
вый Т(:рек), на востоке Каспийское море, на западе Ставро"
польская возвышенность.
По существу Терско..Кумская низменность это юrо
западная часть Прикаспийской низменности. Эта довольно об
ширная, территория, чрезвычайно интересная и разнообраэная по
своему rеоморфолоrическому и ландшафтному строению. При
более дробном рассмотрении внутри Терско..Кумской низменно..
сти выделяются Прикумекая супесчано..суrлинистая равнина (на
севере), массив Терско..Кумских песков (к северу от Терека).
дельта Терека (на юrо"востоке). Большая часть территории этой
низменности (восточная половина) лежит ниже уровня океана.
Местами (в западной части) Терско..Кумская низменность дости"
raeт 100..130 м над ур. моря. Наивысшая точка r. Степан Буrор
(135 м) в восточной части Ставропольскоrо края.
Специфика территории Терско..Кумской низменности опре
деляетl;Я ее положением на стыке двух крупных ботанико
rеоrрафических областей Евразиатской степной и Caxapo
rобий<:кой пустынной (JIавренко, 2000). Это обстоятельство обу
словливает сложное пространетвенное соотношение сообществ,
относящихея к двум типам растительности степному и пустын
ному. В их распределении прослеживается довольно тесная связь
с определенными литолоrо эдафическими условиями и формами
мезо.. и микрорельефа. Подобное сочетание в одном ландшафте
пустынных и степных сообществ с их четкой дифференциацией
по реЛJ>ефу и почвенным условиям представляет собой типичный
пример экотон что дает основание выделять особую, специфи
ческую растительность полупустыни.
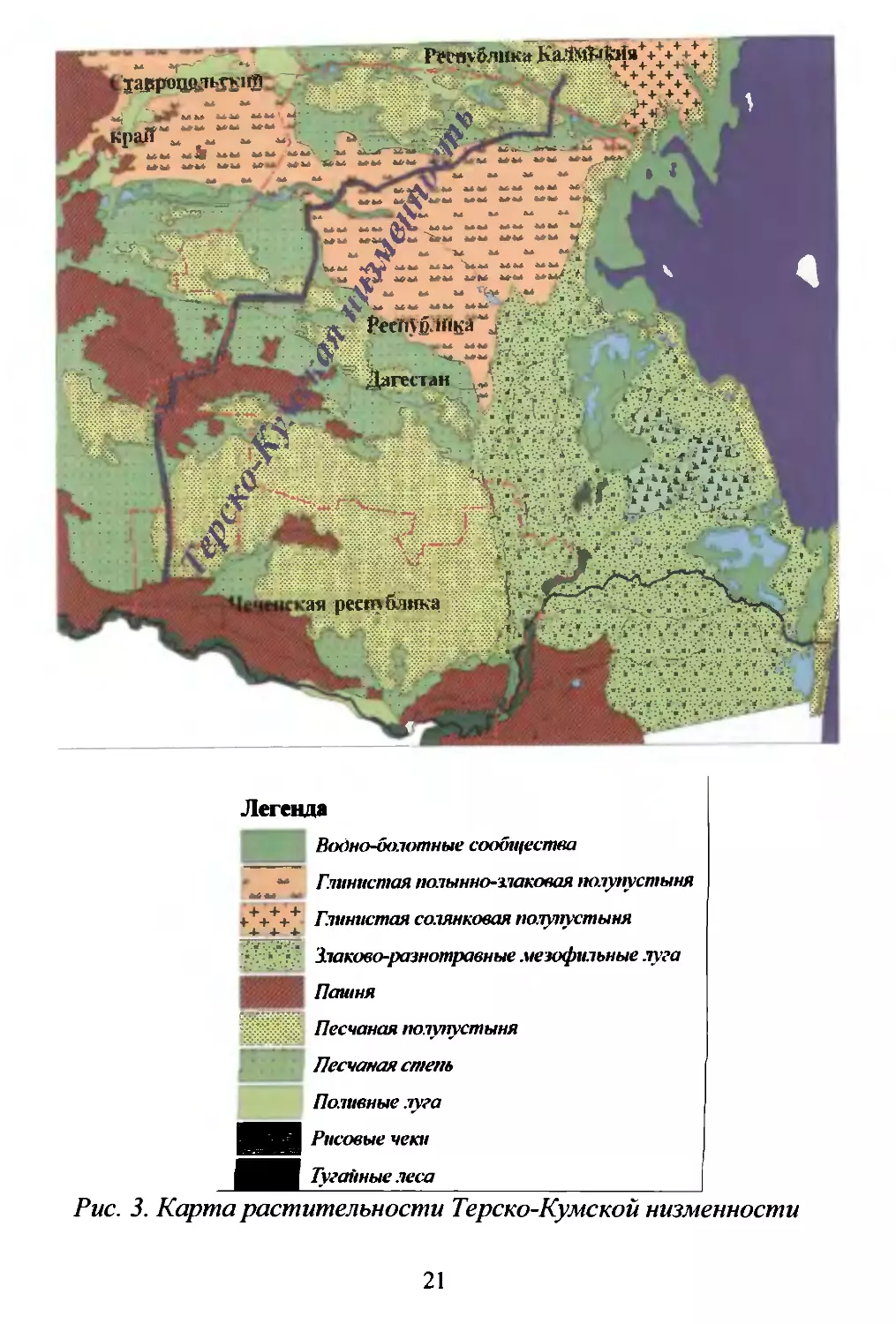
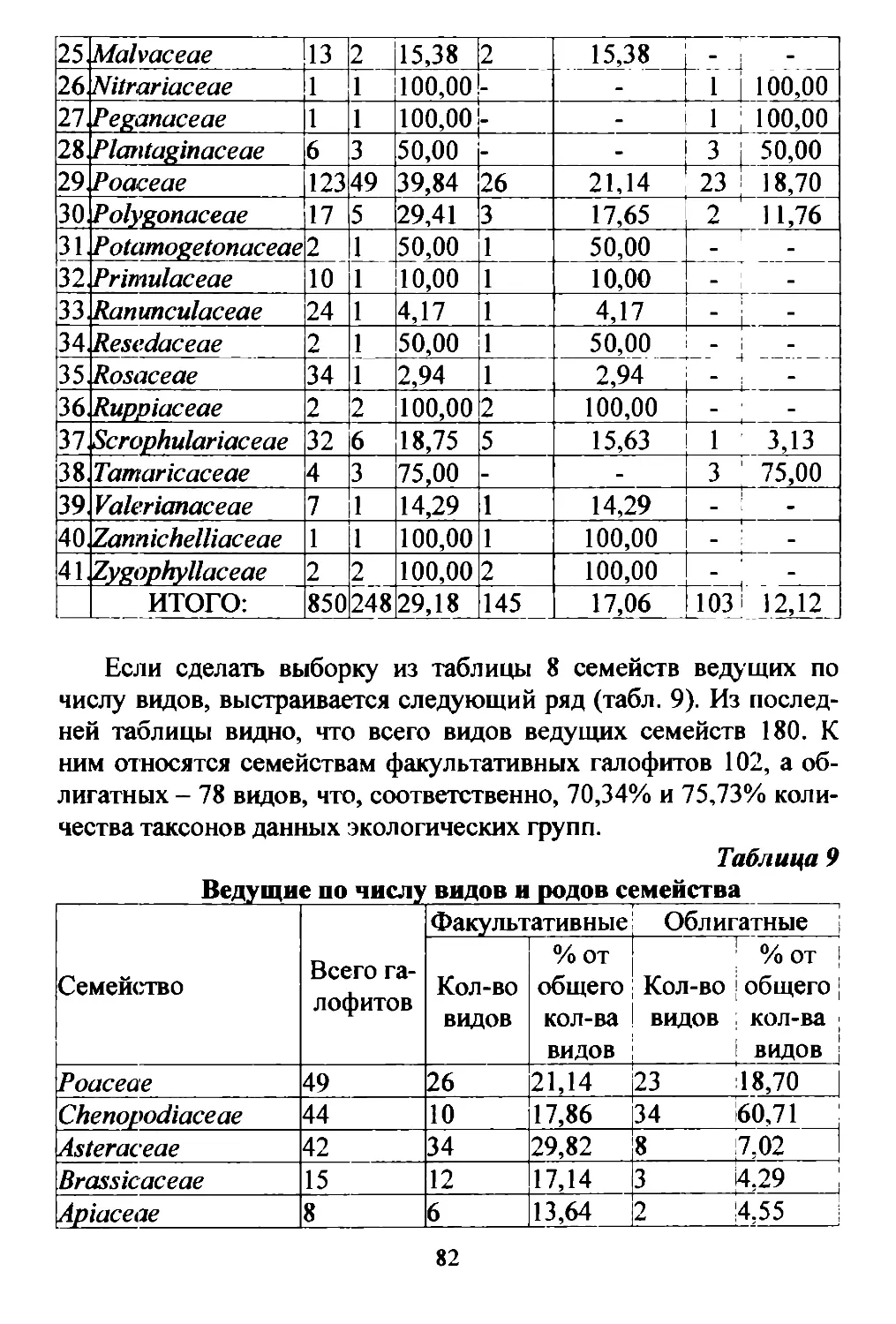
Общая площадь Терско..Кумской низменности составляет
примерно 2,5 млн. ra. В административном отношении к Даrе
6
стану относится примерно 1/2 этой территории, к Ставрополь
скому краю около 250/0. к Чеченской республике 200/0. к Кал
мыкии примерно 5 % (Рис. 1).
СтаВ ПО С IЙ I
i
\
i
i а
.
/
- . ./ l'I
{. \J
,,j .,,-:
. -""'.'''-- ..
j
Рf(' 'б.'1IIk'a ,.
. .1 f.
,, . ' ,V
I ,,"<'
;..: --:- ' 'I ...
J..t""'"
, .
.; I \'
6,..; . , . _ J" -.,-.
["- ', ' . . . . . / ' : ;..
I ..; - { :i. . '" , .'.. ...
" A' \ . C' ' · ,.'L. .... . .. -1' '"
! ., \.i) . J .. '., . . .' .,У
I .. r .l,""" .. ),1.
i. . ...}-..,.... , .,. ......--/
. Чf1IfНf"-11Я рfсп 'б.'1II"-11 ,-- ; ,..... ." -.... t;' ·
\ .' : . "J' . . , '.
, . I '."
I I J. . '! .' . \, "
. ....,..... . . !
"/'- . '.... ._1..". t.
.... . .. .J t . ,. . .."'" · .
i · . '.' - I · "\'... .'"
'"", ",'1 '1;' ... ,!\
"-'Рай
L ...- .:_ . Рfсm'БЛIII а Ra.IJI\IЫЮIJl
. .,' 'It. ."-",_J ...... ..._......,,, .
"
".
. .. ...
....N "
\, ,
....
. (
f
.
1
Даrtстан
.....
'
( r: ".
.-: .,
. .'. i "., . \.
. f' .
( . . .
"
..,
1.
."
__,'
..
'1
','\.' ""
!'--
....... .
........
,
\
PUc.l. Ай\lUlIuстратUШlOе де.7е1lие TepCK.o F(y.\lC"'Oli IIlI1.\lе1l1lОсти
rеолоrия и rеоморфолоrия. В rеолоrическом строении Tep
ско Кумской низменности принимают участие отложения от па
леозойской rруппы до четвертичной системы включительно.
В rеолоrическом плане исследуемый район принадлежит к BO
сточной части платформенноrо склона TepcKoro KpaeBoro проrиба.
которая представляет собой широкую плоскодонную впадину, BЫ
полненную мощной толщей кайнозойских осадков на зпиrерцин
ской платформе Предкавказья. С юrа от складчатоrо склона Tep
7
cKoro проrиба она отделяется предrорным «рвом» узким rлубо..
ким проrибом (до 2000 м по подошве плиоцена). В современном
рельефе впадине соответствуют обширные низменные дельтовые
равнины, созданные неоднократным блужданием Терека, Кумы, а в
прошлом Сулу..Чубутлы, Куры И друrих, более мелких рек, стека..
ющих с ceBepHoro склона Восточноrо Кавказа (Сафронов, 1969).
Как установлено разведочным бурением, на различных rлубинах
под слабо дислоцированными отложенИЯМИ мезокайнозоя здесь за..
леrает rерцинский складчатый фундамент. В карбоне территория
Предкавказья представляла единую область и находилась в стадии
reосинклинальноrо развития. Позднее, в визейский век, большая ее
часть консолидировалась и превратилась в область эпиrерцинской
платформы. Последующее развитие связано с альпийской складча..
тостью Кавказа. С одной стороны, образовались обширные впади..
ны, а с друrой стороны, поднятия, с которыми связаны современ..
ные формы данноrо района (Эльдаров, 1972).
Складчатый фундамент характеризуется двухъярусным строе..
нием. Нижний ярус фундамента состоит из метаморфизированных
мраморизованных известняков и перекристаллизованных мерreлей
светло сероrо и зеленовато..сероrо цвета (карбон..пермь), а верхний
ярус представлен менее дислоцированными породами пермо"
триасовоrо возраста. Этот складчатый фундамент перекрывается
осадочным чехлом (до 4500 м), который, в свою очередь, также
имеет двухъярусное строение. Так, нижний ярус осадочноrо чехла
составляют породы от юрскоrо до олиrоценовоrо возраста, унаеле..
довавшие в своем строении черты складчатоrо фундамента. Верх..
ний ярус осадочноrо чехла состоит из пород неоrен..четвертичноrо
возраста и имеет моноклинальное строение (Акаев, Казанбиев,
1972).
В палеоценово..эоценовой толще выделяются зеленая, кумекая
и белая свиты фораминиферовой серии, состоящие из rлинисто..
карбонатных пород мощностью 100..11 О м. Олиrоцено..
нижнемиоценовые отложения, объединяемые майкопской серией,
делятся по литолоrическому составу на rлинисто..алевролитовую и
rлинистую. Суммарная мощность их достиrает 1450 м. В северной
части TepcKo..KYMcKoro проrиба вскрыта скважинами только верх..
няя часть майкопских отложений (Эльдаров, 1972).
8
Среднемиоценовые отложения, состоящие здесь из чокракско"
('О и KaparaнcKoro roризонтов, очень сходны меж.ду собой по лито..
лоrическому составу. СЛО "ены они чередующимися пачками пес..
чаников и rлин. Мощность их в районе Александрийской и Край..
новки достиrает 360 м. Отложения верхнемиоценовоrо возраста
представлены сарматскими и меотическими ярусами. Первый сло
жен мощной толщей rлин с редкими прослоями песчаников и мер..
reлей с максимальной мощностью до 560 м в районе с. Але ксан..
дрийское, а второй rлинами с прослоями песчаников и известняков,
преимущественно развитых в Терско--Кумской низменности (Эль..
даров, 1972).
Отложения акчаrыльскоro яруса представлены чередованием
песчаников и rлин мощностью 110..130 м, выделенных в Терско..
Кумеком проrибе. Мощность, акчarьта в районе Бабаюрта колеб
лется от 500 м до 600 м, а к северу уменьшается до 60..70 м (р"он
Сухокумека). Такое же изменение мощностей отмечается и у пес..
чано..rлинистой толщи апшеронскоro яруса. Так, в районах с.с. Ак"
сай и Александрийскоrо мощность апшерона достиrает 900..1000 м,
а к северу сокращается до 270 м (район Артезиана ) (Рычаroв, Леон..
тьев, 1960).
Отложения четвертичной системы представлены бакинским,
хазарским, х.валынским, новокаспийским ярусами и имеют широ..
кое распространение на рассматриваемой территорий. Суммарная
мощность их колеблется от 500 м в южной ее части до 100..120 м в
- северо"западной. Сложены они песчаниками, ) линами, песками
континентальноro (аллювиальноrо, дельтовоro и др.) и MopcKoro
происхождения (Рычarов, Леонтьев, 1960).
Слившиеся в единое целое верхнехвальнские и новокаспийские
дельты представляют собой плоские или слабо волнистые аккуму..
лятивные равнины, полоro наклоненные в сторону Каспийскоro
моря. Они расчленены мноrочисленными отмершими и действую..
щими речными руслами и разливами. Среднее колебание высоты в
пределах дельтовых равнин не превышает 2,0..2,5 м. Реликтовые и
действующие речные протоки имеют самые разнообразные разме..
ры и очертания. Большей частью они извилисты, но довольно часто
встречаются и сравнительно ПРЯМОJlинейные участки. Ширина
проток колеблется от 4..1 О до 150..200 м, f'Лубина вреза в наиболее
9
крупных протоках на отдельных участках достиrает 2,0 2,5 м (Pы
чаrов, Леонтьев, 1960)..
Вдоль действующих проток сравнительно часто прослежива
ются четко выраженные прирусловые валы, которые местами за
мещаются искусственными дамбами созданными для защиты от
наводнений. Поверхность дельты расчленена также мноrочислен
ными оросительными каналами, иноrда использующими и ecтe
ственные потоки. На участках распространения сильно треl[щнова
тых ОЗI РНО"аллювиальных новокаспийских rлин, перекрытых пес
ками и супесями, вдоль каналов и на поливных землях возникают
мноrоl.J исленные воронки (до 1 О.. 1 5 воронок на площади 1 ['а), ко..
торые I;вязываются с суФФозионно--эрозионными процессами (Пре
дельский, 1969).
Кроме проток на поверхности хвалынеких и новокаСIlИЙСКИХ
дельтовых равнин прослеживаются мноrочисленные разливы, за
нимающие значительные мощади. Среди них выделяются посто..
янно действующие разливы, образующие сильно заболоченные
учаСТЮ1 дельты с мноrочисленными озерами (плавни), периодиче
ские разливы, на месте которых остаются временно заболоченные
участки с пересыхающими озерами, и древние отмершие р L1ЛИВЫ.
которые выделяются по характеру микрорельефа и раститеJlloНОСТИ
(тростниковые заросли).
Срt ДИ низменных дельтовых равнин выделяются несколько
повышенных участков (в районе Кизляра, у сел Бабаюрт и Шам"
хал..ЯНl'И"ЮРТ, по рекам Акташу, Аксаю, Сулаку и др.), которые
обтекаются древними потоками и разливами, вызывая миrрацию
современных русел или их rлубокий эрозионный врез. По ['eOMOp
фолоrи.ческим признакам, по сокращению мощности отложений и
reофизическим данным эти участки выделяются как локальные
структуры, испытавшие относительные поднятия в верхнечетвер
тичное время (Чистяков, Мирзоев, 1962).
Молодые локальные поднятия, проявившиеся на фоне общеrо
интенсивноrо опускания TepcKoro проrиба, которое со значитель..
ной скоростью (до 2,5..3,0 мм/rод) происходит и в настоящее время,
оказали. существенное влияние на распределение речной Сt.."ТИ, от..
мерших и действующих русел и разливов, определивших r-лавней..
шие rеоморфолоrические черты дельтовых равнин Терека и Сулака
(Сафронов, 1969).
10
В rеоморфолоrическом отношений Терско Кумская низмен
ность представляет собой равнину с небольшим наклоном на BO
сток и ceBepO BOCТOK JOJ rипсометрически находится в пределах ro
ризонталей от 170 100 м на западе, до 6 1 О м на востоке. В районе
развития песков низменность сильно всхолмлена.
Общее понижение поверхность имеет здесь в основном вдоль
широты с запада на восток, через с. Ачикулак, Терекли Мектеб и
так далее, составляя в среднем 1,5 метра на каждый километр про..
филя. Наибольшее падение прослеживается на восточном склоне
Ставропольской возвышенности, заходящей частично в пределы
Даreстана. Причем, по мере продвижения на восток уклон YMeHЬ
шается и от с. А чику лак составляет примерно 0,5 м на один кило
метр.
Падение местности в северо восточном направлении от Терека
к с. Махмуд..Мектеб составляет в среднем 1 метр на один километр
профиля, причем наименьшие уклоны приурочены, к кумеким пес
кам и прилеrающим к ним с южной стороны степным участкам.
Общее падение местности в северо восточном исправлении
сказывалось также на формирования как древней, так и COBpeMeH
ной rидроrpафической сети.
Одной из самых распространённых и наиболее интересных
форм рельефа Терско Кумской низменности являются rpядовые
пески. По внешнему виду они представляют собой довольно OДHO
образные rpяды, тянущиеся параллельными рядами в широтном
направлении. Высота rpяд 5 8 м, изредка больше, ширина 80 100 м.
Orделяются они одна от друrой межrpядовыми понижениями ши
риной в 1 oo 150 м. rряды заросли растительностью, имеют мяrкие
очертания. Межrpядовые понижения имеют вид ложбин, разделён
ных в ряде случаев невысокими и неширокими песчаными пере
мычками на отдельные, вытянутые в направлении с запада на BO
сток, котловины. Сложены rpяды мелкозернистым песком со зна
чительным содержанием лёссовидных частиц. Поражает прямоли
нейность rpяд и почти reометрическая правильность в их располо
жении. Все rpяды тянутся в одном направлении с запада на восток
(Рыжиков, 1950).
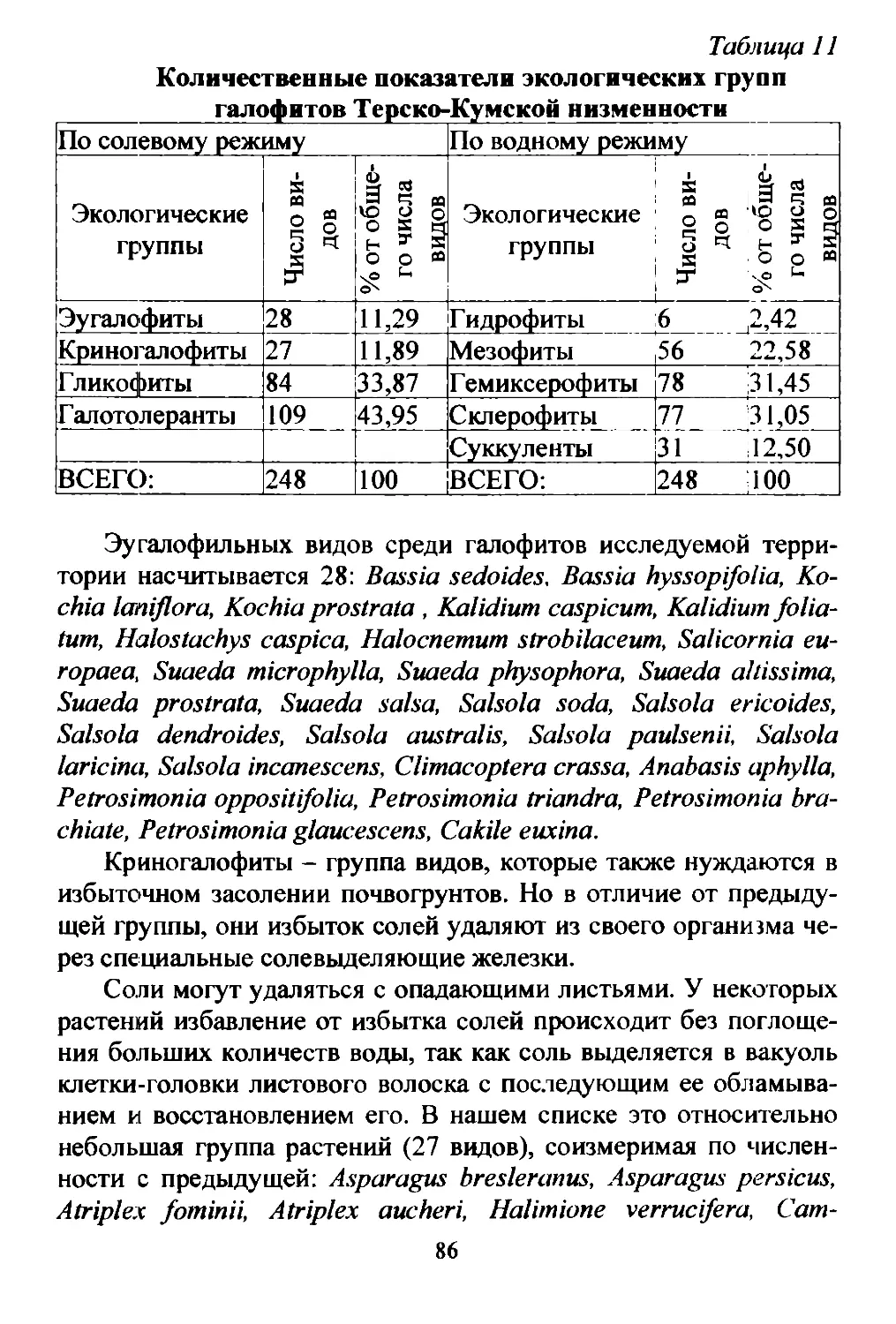
Пески Терско Кумской низменности образуют несколько изо
лированных друr от друrа массивов: Терский, Бажиrанский и KYM
ский (рис. 2). В свою очередь эти массивы распадаются на ряд раз
11
новеликих песчаных комплексов. Помимо этоrо пески приморских
пляжей образуют самостоятельный биотоп, в котором видовой co
став в значительной степени отличается от собственно терско..
кумских песков.
Значительная часть Терско Кумской низменности занята об
ширными площадями сыпучих песков, превращенных ветрами в
барханы и дюны. Из общей площади Терско Кумской низменности
примерно 3 млн. ra) под песками занято около 455 тыс. ra (Кулик,
1957). Из общей площади песчаных массивов, равной 8000 км2,
около 75% приходится на заросшие пески, 20% на полузаросшие,
и только 5% площади занимают совершенно незакрепленные пески
(Водовозов, 1955). По степени закрепленности эоловый рельеф
подразделен на три типа: развеваемые (<<rолые») пески, пески сла
бозакрепленные и закрепленные. На площади распространения
песков выделяют определенные преобладающие формы эоловоrо
рельефа барханные, rpядовые, буrpистыe и буrpисто rpядовые
пески.
'tG.....
...,' t4::
. .......
A:'. { *>: ,
.t.I. ;;.,:м., 4- .: "
.ИJlr8"' '" ...... "':- .: ,.";,,
. .. or (r
1::;-.
(::}» .T I"
'.:. ::1 jl;;'(.i: il!:::il!.!I!!il !:':': I
. "-:-:-:-:':-:-:-:':-:.:.:-:-:-:-:':-:-:":-:':-:'.
, ::::i;f!::::.:::I}'
:J .'T
,
4
.O "JI": .
КJl8iiж.8
. Ч rHIIr8"
? .
. r,....ю.rII
\.1
Рис. 2. Распределение песков на Терско Кумской низ;wенности
12
Равнинный характер Терско..Кумской низменности наруша..
ется во мноrих местах наrpoмождением песков в виде вьпянутых
с запада на восток rpяд, барханов и буrров, называемых здесь
чаще Bcero «бурунами», имеющими в подавляющем большинстве
неопределенные очертания и разделенные между собой котлови..
нами выдувания. эти буrры и барханы широко распространены
здесь, и своим происхождением обязаны ветрам. Котловины вы..
дувания часто заполнены небольшими солеными озерами, пр е..
вращающимися в болота или совершенно пересыxюlцимии летом
(Эльдаров, 1972). Для данноrо реrиона, поскольку здесь сейчас
не существует пустыни, эти образования следует рассматривать
как реликтовые, сохранившиеся от предшествующей эпохи, об..
ладавшей иными климатическими условиями (Рыжиков, 1950).
.климат. Терско--Кумская низменность соприкасается с се..
верной окраиной субтропическоrо пояса. С этой особенностью
связано ПОС1)'пление большоrо количества солнечноrо тема.
Собственно такое rеоrрафическое положение и определяет ха..
рактерные черты термическоro режима данной области. Район
этот характеризуется длительным веrетационным периодом со
значительной суммой активных температур, жарким летом с пе..
риодом среднесуточных темпераryр выше 200 в течение пример,.
но 100 дней, умеренно мяrкой зимой с небольшой суммой абсо--
лютных величин отрицательных среднесуточных темпераryр.
Необходимо отметить, что Северный Каспий, несмотря на
свою мелко водность и замерзаемость, все же оказывает зимой
значительное воздействие на термические условия прибре ой
полосы. Эrо нarлядно видно из сравнения данных двух пар ме..
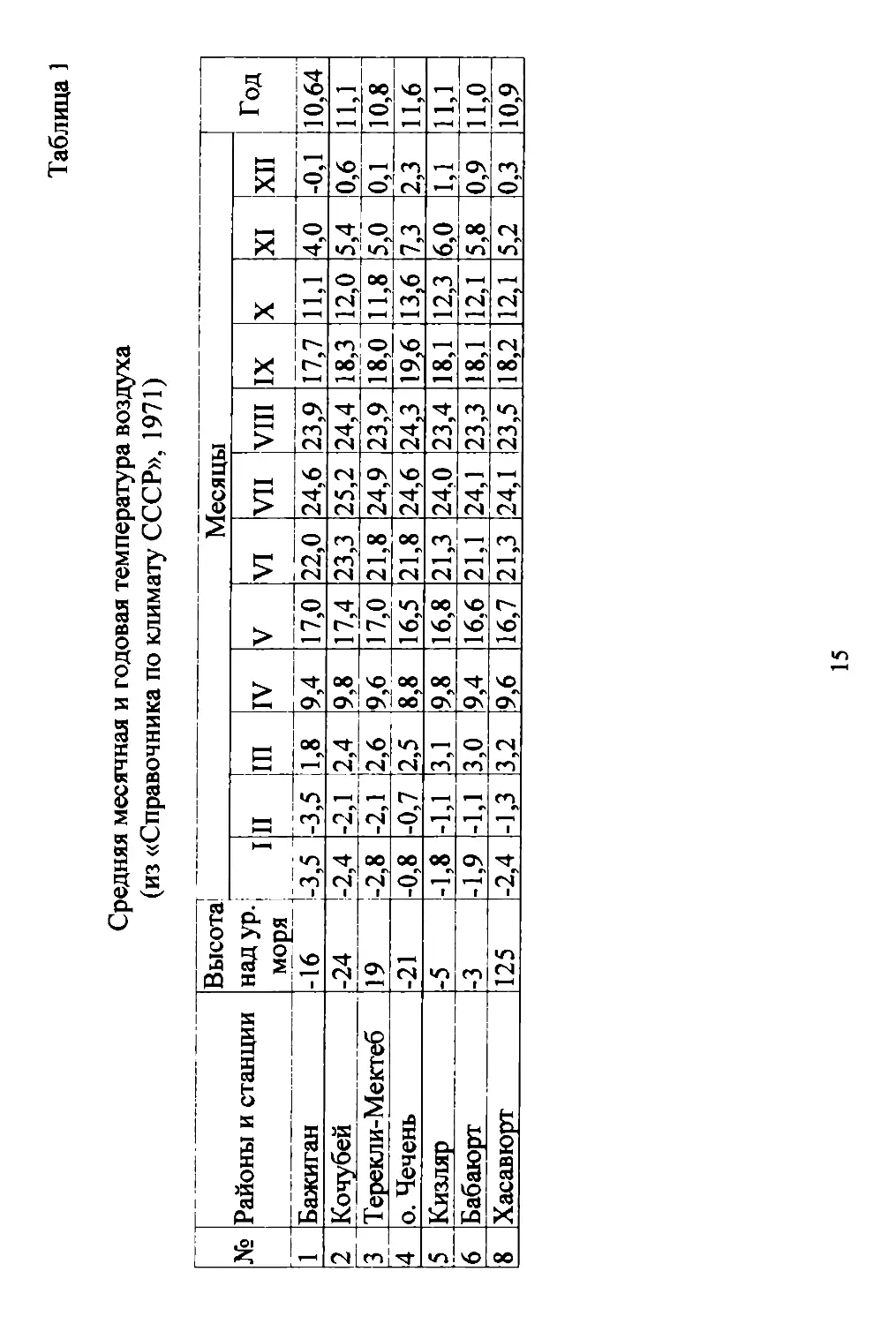
теостанций: Кизляр .. о. Чечень, Бажиraн..Кочубей (Табл. 1).
Климат этой части Прикаспийской низменности сухой кон..
тинентальный. Среднеrодовая температура составляет примерно
11 ос (Табл. 1). Среднемесячная темпераryра самых холодных
зимних месяцев (январь..февраль) колеблется от 0,7 до з,50с, а
самых жарких летних (июль..авryст) достиrает 23,3..25,20с выше
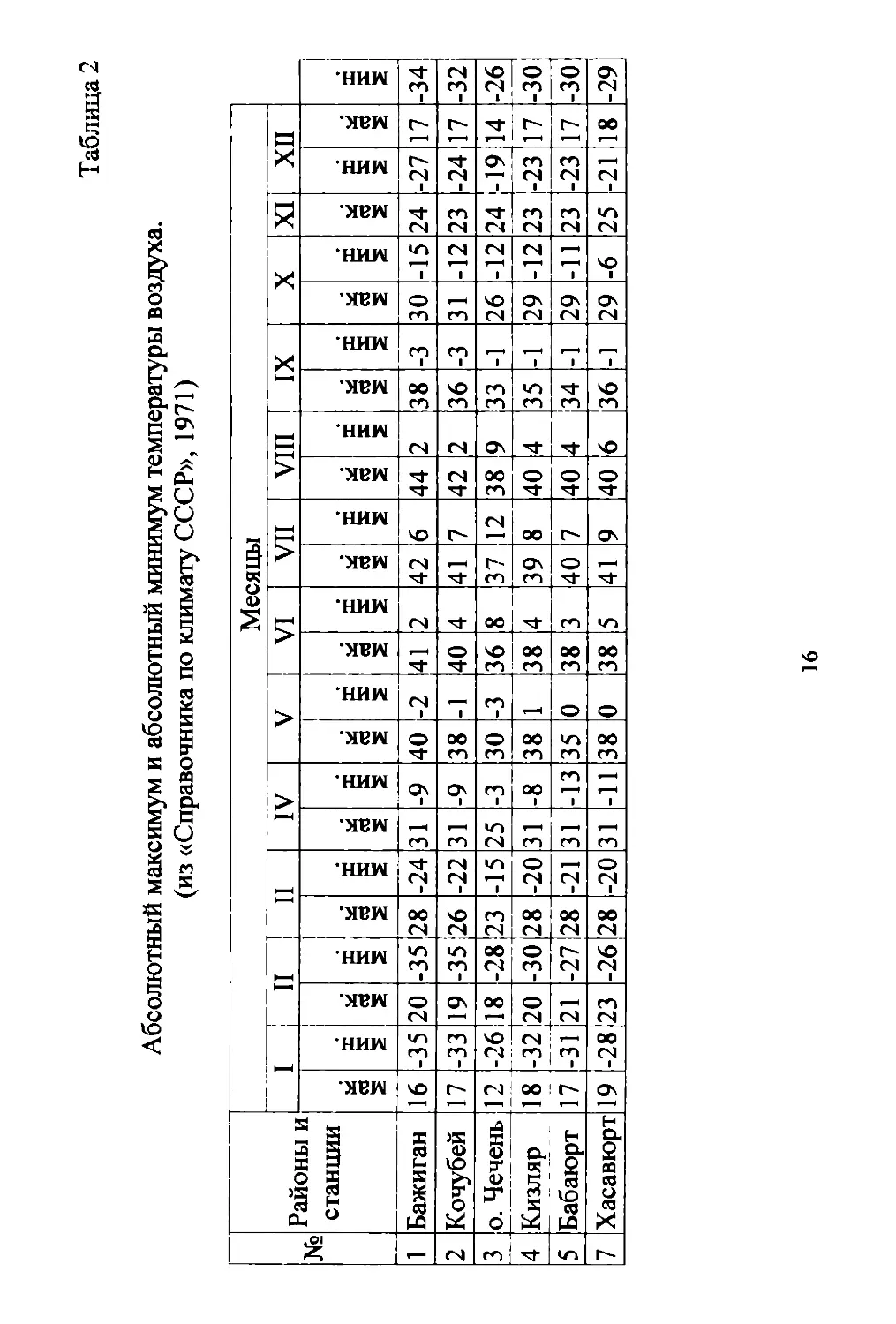
нуля. Абсолютные минимумы января и февраля (Табл. 2) в реrи..
оне колеблются от 26 до 350 с. Абсолютные максимумы этих
же месяцев MOryT быть аномально высокими (до + 20 и более).
Абсолютные максимумы июля и aBrycTa нередко поднимаются
до 400 и более rpaдycoB с.
13
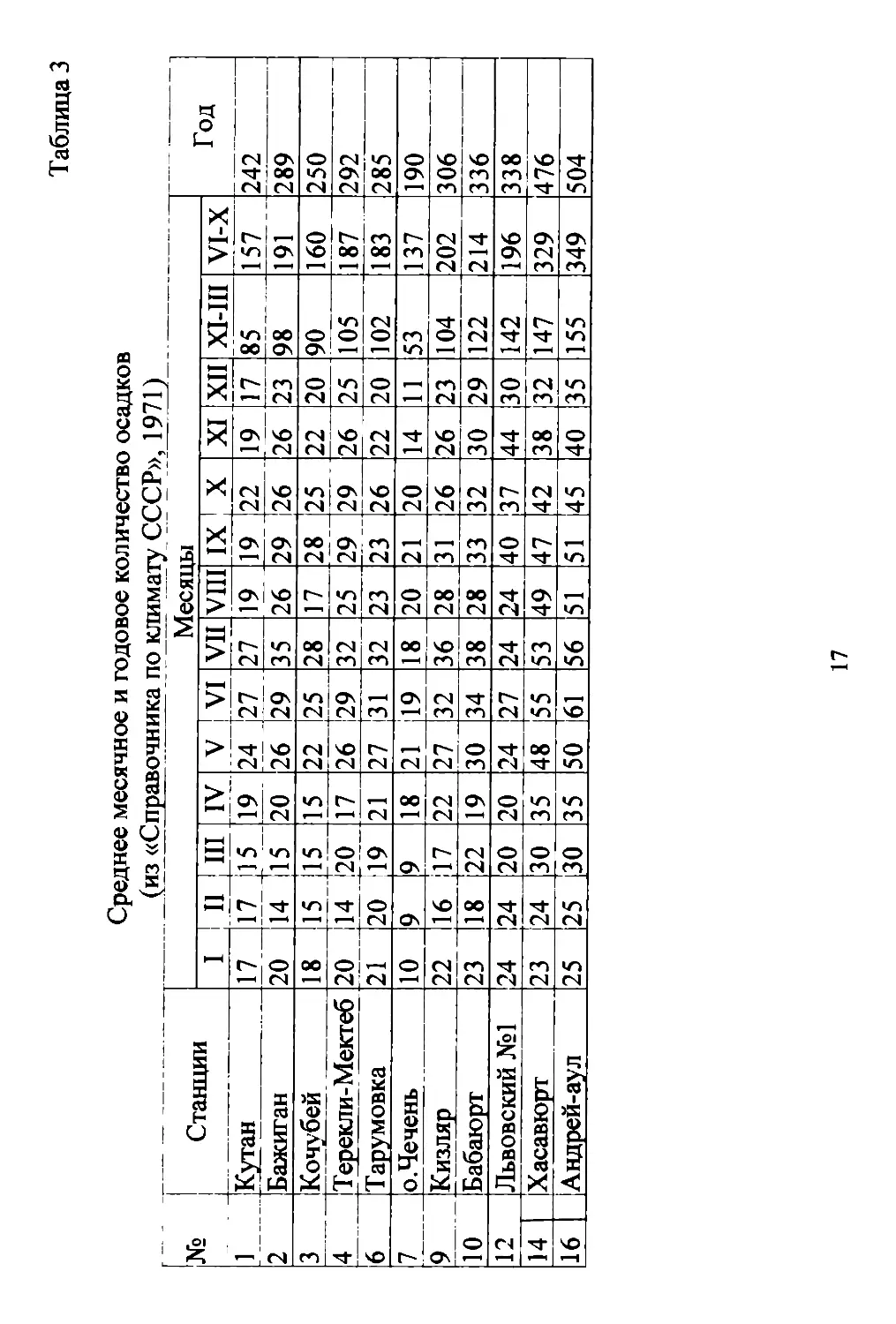
Продолжительность безморозноrо периода равна 196 дням,
сумма положительных темпера'I)'Р 42110c (Табл. 3). Сумма cpeд
них суrочных температур воздуха за период с темпера рой BЫ
ше 5 и 10°С, соответственно, составляет 40760с и 3770 С. Даты
перехода среднесуточной темпера'I)'РЫ воздуха через 00 С 1
марта и 15 декабря.
rидротермический коэффициент в апреле 0,8, мае"июне
0,6, июле..октябре 0,4..0,5. Летом отмечаются частые и продол..
жительные засухи. Относительная влажность воздуха в мае..
aBryCтt опускается до 40..43%.
В связи с частым и длительным стационированием антицик"
лонов, для всей территории Терско..Кумской низменности весьма
xapaкт' pHa небольшая сумма атмосферных осадков (Табл. 3), что
при значительной сумме активных темпера'I)'Р и длине их перио
да БОЛI е полуrода (Табл. 4), определяет формирование климатов
полупустынноrо типа. Веrетационный же период отличается
всюду значительным превышением испаряемости над количе..
ством 18ыпавших осадков, т. е. ярко выраженным отрицательным
балансом увлажнения. Особенно больших абсолютных величин
достиrает отрицательный баланс увлажнения летом, составляя в
среднем 70% величины баланса увлажнения Bcero веr'етаllИОННО
ro периода. Поэтому баланс увлажнения именно за летниЙ пери..
од rода является для Терско..Кумской низменности весьма харак"
терным показателем степени засушливости.
14
r=:
\о
tQ
8
1:1:1......
tQ ...
Cl.
Cl.U
gu
Е-
i
о
1:
:s:
= ::т
6;
(,) tQ
Q cl.
1:
5
cl.
u
о
......
......
><
......
:><
:><
......
::::= " 0\"1 м... " м... "l
>....... I rr'I М rr'I rr'I, rr'I
::а NNNNNN
;- ;;...... I \о... N... 0\" q, ......" ......"
(,) V)
Q NNNNNNN
::Е
;;
>
v)
.....
>
......
'tr'I ос ос 00 0\
I " " "1 " " ... "
'rr'I N'N С> ...... ...... N
I I I r I I I I I
r - - l
I ci.. 0::1 I
10 >.C:iJ I I
(,) О, I
.... \o ...... v)
i""7 I '1 f(
:s:
е
а
Б
:s:
о
1 )а
I
N
C
\о
C
Е-ОО!
м
О
I:Q
:Q
о..
'"""'
0........
CUr---
t::Q'\
::11......
i
g
8
:s:: о
::11
и
;;
::11-5-
>=
Q
\о
<:
N ТC a::
'НИW -.::t
м м N,MIM N
I I I I I I
I .)f W r--- r--- Ir---ir--- ос
...... ...... ......1......1...... ......
'НИW r--- -.::t Q'\[MIM ......
N N '"71 1 N
I I I
')f W -.::t м IM М tA
N N ("<11 N N N
! 'НИW tA N N N ......
...... ...... ...... ...... ...... \О
>:: I I I I I I
'){ W С ...... \О Q'\ Q'\ Q'\
м м N N N N
'НИW1м М ...... ...... ...... .....
1>:: I I I I I I
I ')f W ос \О ]:t \О
м м
'НИW Q'\ -.::t! "st..
N N
')f W -.::t N OCIC С С
l-.::t -.::t м -.::t -.::t -.::t
'НИW N'
....... \О r--- ......1 ос r--- Q'\
s; .)f W N ...... t"' Q'\ С ......
tI::: -.::t -.::t м м -.::t -.::t
Q OC:-.::tIМ
cu 'НИW
> N -.::t tA
')f W ...... С \Oioc ос ос
-.::t -.::t М'М М М
......Ic
'НИW'
.xew r; ..... м С
:> I I
ос С ос tA ос
М м м м м
'НИW I Q'\ м ......
I Q'\ м ос ...... ......
, I I I I I I
'){ W ...... ...... V') ......' ...... ......
м м N м м м
'НИW -.::t N V')IC ...... С
N N ...... N N N
I I I I I I
.)f W ос \О MI ос
N N N
'НИW V') V') ос I С r--- \О
м м ("I"I I N
I I I
')f W С Q'\ : I м
. . N ...... ...... N N
+
'НИW tA м \О N:...... ос
м м ": f(i N
(....... I I I
1 '){ W I \О r--- N!OC!r--- Q'\
..... ...... ..... I ...... ...... ......
1 :s:: I 'Е-< f-o
::с ):S:: :I: I ' о..
::s е cu cu o..l 52
::с \о =- I tI:::' r:Q
О CU' ' tQ
)а i :\0 Q
О cii I tQ
с.. Q
I ...... N Mi-.::t tA r----
\о
.....
tQ
tQ
i I I
I I I
I N I с> 'о 'о 00 'о ....
! I NNNJN
! ;>< 1: I
J Ir---.I...... cl ! r---. NJ-.::t \О 0\ 0\
I >1 1 i I N
I S:! I i I I
I I I 1tr'lIN '-.::tIN Nlr---. tr'I
I ; i ' I I:!:
1 i I 1 I
::::' r---. i i tr'Il' с ...... 0\ С I N tr'I
['1 ,..... IN NIN N ...... N N J ("f"\
0\ l' I
8 ......!' 0\1\0 NI\OIN -.::t \О'с -.::t,oc С
О ......NN,NiN......N("f"\-.::t("f"\-.::t
б: ;>< NI\O tr'Ii 0\1 \О С \О N r---. Nltr'I
.... U, N N NIN N N ("f"\ ("f"\ -.::t'-.::t
Ui ;><1... 1 !:
.... ,..... 10\10\ OCIO\I ...... ...... ("f"\ с r---. ......
., ...... N N NiN N ("f"\ ("f"\ -.::t -.::t tr'I
: ! ёlО\i\О r---.1tr'l1'("f"\ CIOC 00 -.::t10\......
20 ё5 cu I""""' N iN N N N N Ni-.::t tr'I
.... :21 =1 I I
t::t О 1 > il r---. tr'I I N 'N ос \О I ос -.::t ("f"\ \О
е t:, I N ,("f"\ N: ("f"\ ...... ("f"\ ("f"\ N tr'I tr'I
== tQ '1 :> I ["---I 0\ tr'I i 0\ ...... 0\ I N -.::t r---. tr'I ......
cu ,N,NN'N("f"\......("f"\("f"\Nq\O
I ;T;: i r; .......11 r---. i с -.::t ос
I 'N:N NIN N N N,("f"\ N -.::t tr'I
ёJ =:: i 0\ l' с tr'Ill r---.I...... ос l' N 0\ С tr'I tr'I
,.". """1 ,......N............N......N......(" ("f"\("f"\
cu UI """"'1 i i I I
:::: tr'I' tr'I tr'I, С : 0\ r---. N с С с
: i Nt '....... N N ("f"\ ("f"\
Q.. I' = I r---. i -.::t tr'I' -.::t '1 с \O -.::t " -.::t tr'I
U ;......!..................iNo\............NNN
I П ...c.
i """'" r---. '1 с ос с!...... с N I -.::t ("f"\ tr'I
! ...... N ...... N' N ...... N N (" N N
I
I
t"-
.....
I ......
Ij.,! '
= ):S: i i ,,1
I О I iJ I II
I , о! :S:1'g А tQ ==
l I О I J::; ;>< <
, i III
, i I! :
i , I :! i i I I с (" -.::t \О
.., N ("f"\ i -.::t ,\о r---. 0\ 1...... '"""" ..... ....-t
Таблица 4
Характеристика теплоrо периода по мноrолетним данным
по «Аrроклиматическому справочник areCTaHa»,] 963
иод I Сумма поло..
I ! Продолжительность жительных
наЧ U10 I конец (дни)
I
Средняя с точная темпе а а воз ха выше О С
26 11 4 111 I 16 ХН 25 287 302 4500
I ХН I l
J :{)
Средняя с точная темпе а а воз ха выше 5 С
25 1I1 30 12 ХI 19 I 227..238 13800..4300
111 XI I
e c f xT rl wxa выше lQ f3500 ... .
р е "! я с ,!, чн теМП lJ.j'ха !JJ>!!ll 1
9 V 1 23 IX 4 Х 1122..148 I 2600..ЗЗЧО
БеЗМ IQЗНЫЙ период
} lllI XI 15 XI [210..228
Величина испарения воды с поверхности почвы достиrает
800..900 мм, что свидетельствует о значительном превышении
расхода воды над ее ПОС1)'IUlением. Относительная влажность
воздуха в период формирования урожая озимых хлебов май..
июнь с,оставляет: в Кизляре 49..500/0, в Бабаюрте и Хасавюрте 50..
520/0. По показателям rидротермическоrо коэффициента п ррито"
рия раЙона относится к засушливой и очень засушливой.
На Терско..Кумской низменности выпадает в среднем 230..290
мм/rод осадков, а на береrах Каспийскоrо моря даже менее 200
мм (остров Чечень 190 мм). По мере приближения к северным
склонам предrорий, количество осадков увеличивается и состав..
ляет 340..360 мм (в Хасавюрте, вблизи предrорий, 476 мм). Снеж..
ный покров имеет малую высоту (средняя из наибольших декад..
ных высот за зиму 8..10 см) и непостоянен (вероятность зим с не..
устойчивым снежным покровом составляет 50 55%).
18
В низменной части западноrо побережья Каспия на протяже..
нии rода преобладают ветры (Табл. 5) восточных и западных чет..
вертей: восточный, юrо"восточный и западный, северо..западный.
В летний период преобладают восточные ветры. Эrи ветры отли"
чаются длительностью действия и сравнительно большой скоро--
стью. Они приносят массы cyxoro rорячеrо воздуха, идущеrо из
Средней Азии. Эrи ветры, хотя и проходят над Каспийским мо--
рем, атмосферных осадков на низменность почти не приносят и
по своему характеру напоминают суховеи. Ветры западных
направлений, в отличие от восточных, характеризуются меньшей
длительностью, но большей скоростью.
Почвенн растительный ПОКРОВ. Основными почвообра..
'зующими породами на Терско Кумской низменности являются
морские и речные аллювиальные отложения с преобладанием су"
rлинистых, rлинистых и песчаных разностей. В формировании
почв большую роль иrрает rлубина залеJ'ания и степень минера..
лизации rpYHTOBbIX вод, которыми обуславливается различная
степень заболачивания и засоления почв.
Почва. Почвенный покров этоrо района чрезвычайно сложен
и разнообразен. На фоне пестроты состава почвообразующих по--
род, растительности и жаркоrо климата в нем четко отображают..
ся разновозрастность элементов rеоморфолоrическоrо расчлене..
ния и значительное влияние rидролоrических условий. Опреде..
19
ленное воздействие на трансформацию почв оказывает также xo
-зяйственная деятельность человека.
Почвы Терско Кумской низменности в своем формировании
подчинены закономерностям широтной зональности. Нарастание
контин-ентальности с юrо запада на ceBepO BOCTOK также отража
ется на характере почвенноrо покрова. В северо"восточной qасти,
в силу значительноrо превышения испарения влаI'И над ее прихо
дом, наблюдается развитие больших площадей солончаков и BO
обще з.асоленных почв. Процесс ryмификации и минерализации
растительных остатков в условиях ярко выраженноrо азробиози
са ПрИJЮДИТ к знаqительному накОIUlению в поqве различных ми
неральных соединений и вместе с тем к образованию неБО.lьшоrо
количества ryMyca. Все это вместе взятое приводит к тому. что
процессы почвообразования на низменности, отвечая rоризон
тальной зональности, соответствуют пустынно степному типу и,
что зональными почвами здесь являются светло каштановые поч
вы (Капустянекая, 1972).
Однако влияние ЗОН(U1ьно климатических факторов на основ..
ной п рритории низменности нарушается действием водноrо
фактора, иrрающеrо исключительно большую роль в по",вообра
зовании дельт и речных долин. Сочетание этих двух факторов
вызывает, с одной стороны, близкое залеrание к поверхности
rpYHToBbIX вод, а с друrой интенсивное их испарение, что, в KO
нечном итоrе, приводит к широкому развитию на низменности
интразональных почв, не образующих, как правило, единых Mac
сивов.
Напичие своеобразных условий почвообразования при водит к
формированию на Терско Кумской низменности следующих
почвенных типов: каштановые, луrово каштановые, луrовые, лу
rово б()лотные и аллювиально луrовые. На все эти типы накла
дывае'IСЯ в различной степени выраженный процесс засоления.
Поэтому по степени засоления выделяются незасоленные. слабо
засоленные, солончаковатые и солончаковые разновидности почв
и солончаки (Аджиев и др., 1998).
20
;; :; : 'i! !i! ; 1
J" ;lj i , :: } 7 ,
4
:-:} :\.
' ':'. :
,:,' "..,:.
. :.{.. :;..
.........................."......
"::::.::::;::::::-:::::::::"
(
:::jj
f
':.; .:,.' :: - ..'
Леrенда
BoдHcМxJ.7omHbIe сообщества
r.'lIlНllсmая lIо.7ынно..1.7аlювая IЮ.7у, vсmыня
+ + + + + + r.7IIнистая СО.1ЯНКовая "О:IУ'lVCтыня
+ + + .
.?\ .:. 'f/: з'1аково--разнотравные .иеЗоф'L1ьные .7)llQ
ПтllНЯ
с.....:.:':.:.:;:;:.:::
Песчаная 1IO.7.vпycтыня
ЛесчонOIl CНle1l6
ПО.7118ные .7у?а
_ Р"С08ые чеЮI
т.Vlтiные леса
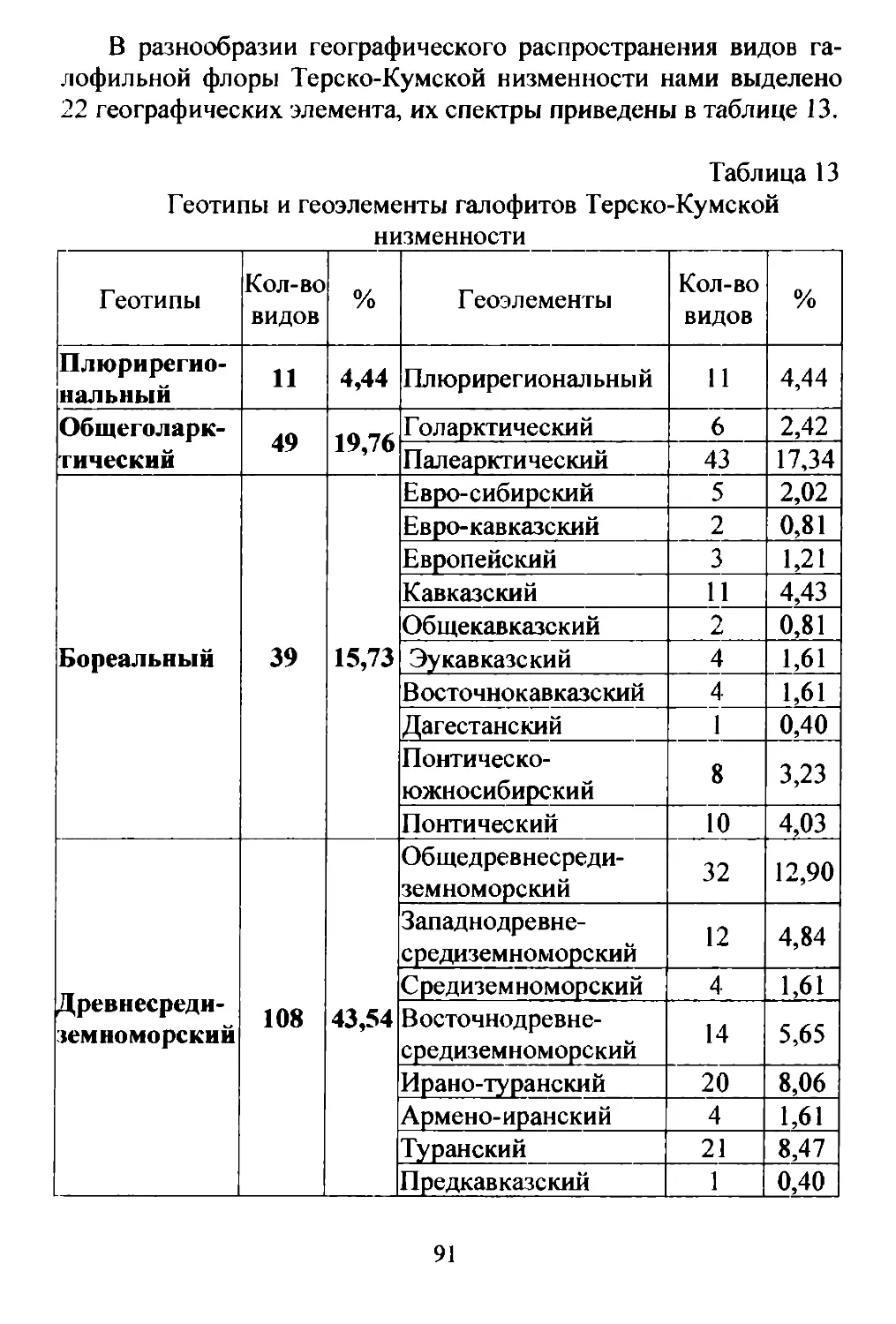
Рис. З. Карта растительности Терско Кумской низменности
21
В связи со сказанным на Терско Кумской низменности фор
мируется очень пестрая картина растительноrо покрова (рис. 3).
Зональным типом растительности данной низменности должна
быть растительность полупустынная. Она действительно здесь
широко представлена. Однако, также широко распространены в
разных частях низменности друrие варианты растительности (TY
rайные леса, мезофильные луrа и др.), существование которых
определяется высоким уровнем rpYHToBbIX вод.
Среда обитания rалофитов засоленные почвы и воды. На
суше засоленные почвы приурочены в большинстве своем к
аридным климатическим условиям зонам степей, пустынь и по..
лупустынь. Это В основном южные районы нашей страны, rде
испарение воды из почвы значительно превышает ее ПОС1)'пление
в почву в виде осадков. Водные засоленные местообитания пред..
ставлены океанами, морями, соленными и солоноватыми лима..
нами, озерами. Во влажном климате засоленные почвы развиты
слабее, места их образования связаны с местами скопления соле..
вых залежей, соленых источников, песчаными дюнами.
Общий признак всех засоленных местообитаний повышен..
ное содержание в них леrкорастворимых солей. Независимо от
степени засоления в почвах и водах встречаются одни и те же со..
ли хлориды, сульфаты и карбонаты натрия, кальция, маrния и
калия. По их количественному соотношению различают хлорид
ное, сульфатное, хлоридно..сульфатное, сульфатно"хлоридное и
карбонатное засоление. Характер почвенноrо засоления зависит
от специфических условий почвообразовательных процессов.
На территории CHr в распределении типов соленакопления
установлена определенная зональность. Так, с севера на юr и с
востока на запад отмечается следующая последовательность в
смене типов засоления: нитратно..сульфатно хлоридное сменяет
ся сульфатно"хлоридным, хлоридно..сульфатным, содово"
сульфатным, а затем чисто содовым (EropoB, Базилевич, 1973.
В.А. Ковда (1937, 1946, 1947) в пределах бывшеrо СССР выделя
ет четыре провинции соленакопления: хлоридноrо, сульфатно..
хлоридноrо, хлоридно"су льфатноrо и су льфатно"содовоrо накоп"
ления солей. Засоленные земли CeBepHoro Кавказа относятся к
провинциям хлоридноrо и сульфатно хлоридноrо соленакопле..
ния.
22
На Северном Кавказе основными типами засоленных почв
являются: солончаки, солончаковатые и солончаковые почвы со..
лонцы И солоди. Учитывая rенетические свойства и эволюцион..
ные связи, систематику засоленных почв CeBepHoro Кавказа
можно представить в следующем виде (Захаров, 1939; Ковда,
1947; Блажний, 1971; EropoB, 1950; rаврилюк, 1979; Вальков,
1977; Вальков, Штомпель и др., 1996):
Солончаки почвы, содержащие от 1 до 3% и более водорас..
творимых солей с самой поверхности. Профиль солончаков в
большинстве случаев слабо дифференцирован на rенетические
rоризонты (ryмусовый «А», переходный «В» и rючво06разующая
порода «С»). По всему профилю заметны выцветы солей (осо..
бенно после подсыхания стенки разреза). В нижней части, а ино..
rда по всему профилю отмечаются признаки OlJ1еения ржаво..
охристые вкрапления или сухие пятна. Встречаются они в Запад..
ном и Среднем Предкавказье либо отдельными крупными конту..
рам и, либо среди друrих преимущественно переrнойно..rлеевых
засоленных почв.
По условиям происхождения, местоположению и ряду друrих
признаков солончаки подразделяют на при юрские, луrово"
болотные (IUlавневые) и луrовые. Качественный состав засоления
отражается на внешних признаках солончаков. Различают корко..
вые, пухлые, мокрые и черные солончаки. Реакция почвенноrо
раствора солончаков близка к нейтральной. На солончаках соли
залеrают в основном на поверхности или заходят в верхний rори..
зонт. Если же соли залеrают на rлубине 5..30 см ЭТИ почвы со--
лончаковые, на rлубине зо 1 00 см солончаковатые.
Солонцы почвы, содержащие в поrлощенном состоянии
большое количество 06менноrо натрия (в ряде случаев и маrния).
В отличие от солончаков солонцы содержат большое количество
воднорастворимых солей не в самом верхнем rоризонте, а на не..
которой rлубине. Профиль солонца разделяется на отчетливо вы..
раженные rоризонты: ryмусово"элювиальный (надсолонцовый)
«А», солонцовый или иллювиальный «В» и подсолонцовый «В»
переходный к почвообразующей породе «С». Для растений
большое значение имеет СТРУК1Ура солонцовоrо rоризонта. В су..
хом состоянии он сильно уплотнен. во влажном бесструК1УР"
ный и мажущийся.
23
Солонцы на основе их водно..солевоrо режима и условий раз..
вития дерновоrо процесса подразделяют на подтипы, отражаю
щие их зональные особенности: черноземные луrово болотные,
луrовые, луrово"степныс и степные. В пределах Северноп) Кав"
каза встречаются в основном три подтипа солонцов: луrовые, лу..
rово"степные и степные. Встречаются они как отдельными кон..
турами, так и в комrшексе с солончаками (СОЛОНiШКОВО"
солонцовые комплексы), rде с полуrидроморфными почвами
имеют наиболее сложи)' ю мноrочисленную контрастную струк"
туру почвенноrо покрова.
На Северном Кавказе солончаки и солонцы обусловлены в
основном макрорельефом и определяют типичную черту солон..
чаковой растительности её КОМIUlексность и мозаичность,
В )'енезисе почв процесс «осолонения» солонцов при водит к
образованию солодей. Под влиянием изменения rидролоrических
условий, в пониженных участках микрорельефа, на деrра,J,ИРУЮ"
щем солонце скarUlивается и застаивается вода. Это при водит к
изменению воздушно водноrо режима и характера микробиоло..
rических процессов, дальнейшему выщелачиванию СО lей из
толщи почвы и к постепенной замене натрия в почвенном 110rло
щающ( м КОМIUlексе водородом. Эти и ряд друrих развивающихся
здесь процессов приводят к тому, что в верхней половине поч
BeHHoro профиля происходит разрушение алюмосиликатной ча..
сти почвы, относительное обоrащение ее кремнеземом. Верхняя
часть I10чвенноrо разреза приобретает белесоватую окраску, от..
носительно рыхлое сложение, близкую к нейтральной р( акцию
среды. Нижние rоризонты продолжают частично сохранять при..
знаки I;олонца (более темная окраска, плотное сложение, повы
шеннос значение рН).
Существует мнение, что солоди MorYT обра:зовыва'rься не
только в результате деrрадации солонцов, но и из друrих почв, на
которые периодически воздействуют слабые растворы натриевых
солей. В этом случае почва вначале осолонцовывается, а затем
после установления нисходящих токов влаrи она промывается, из
нее выносятся продукты щелочноrо rидролиза, в ней накаплива..
ется аморфный кремнезем, и почва приобретает черты солоди
(Вальков, Штомпель, Трубилин, Котляров, Соля ник, 1996).
24
Растительный покров засоленных земель TepcK
Кумской низменности. Солончаковая растительность и различ..
ные её разновидности расположены на исследуемой территории,
rде почвы, вследствие близости Каспийскоrо моря, солёных ли..
манов и водоёмов характеризуются высоким содержанием солей
(Захаров, 1 939 Блажний, 1971; Вальков и др.. 1996). Поэтому
здесь встречаются однородные растительные сообщества, [де
большинство видов, образующих растительность солончаков, со..
лонцов, солодей являются типичными rалофитами.
Солевой режим и rидролоrические условия в разных частях
Терско..Кумской низменности неодинаковы, вследствие чеrо со..
лончаковая растительность имеет комплексное сложение, а со..
ставляющие растительность фитоценозы имеют достаточно чёт..
кие rраницы. В направлении от засоленных маршей Каспийскоrо
моря, солёных лиманов и водоёмов ВI'лубь территории Т ерско"
Кумской низменности сообщества эвrалофитов сменяются сооб..
ществами мезоксероrалофитов или ксеромезоrалофитов, перехо..
Дящими в злаково.. разнотравные и разнотравно..злаковые луrа
пёстроrо сложения. Образуются cBoero рода пояса растительно--
сти, последовательно сменяющие друr друrа и образующие чётко
выраженные эколоrические ряды по rpадиенту засоления и
увлажнения (Наrалевский, 1984; Абдурзакова и др. , 2014).
Смена rалофильных сообществ, в общих чертах, можно
наблюдать и на засоленных территориях Терско Кумской низ..
менности. В сообществах эвrалофитов rосподствуют Salicorпia
europaea, На/оспетит strobilaceuт, Sa/so/a soda, HaZiтioпe
verrucifera, Н. peduпcu/ata. Между ними нередко располаrаются
небольшими куртинами Suaeda prostrata, PucciпeZZia gigaпtea,
Ae/uropus litoraZis, Petrosiтoпia oppositifo/ia, Spergu/aria тaritiтa,
S. тariпa u др.
Сообщества эвrалофитов используются как пастбища низкоrо
качества, кормовая ценность их возрастает по мере присутствия в
них злаков. В тоже время чистые заросли солянок и солероса яв..
ляются хорошим круrлоrодичным нажировочным кормом для
крупноro poraToro скота, лошадей и овец. Урожайность этих со..
обществ достиrает 30 wra зелёной массы.
По окраинам лиманов и плавней, в условиях постоянноrо за..
соления и увлажнения, среди солеросовых и солеросово--
25
бескильницевых сообществ встречаются Carex distaпs, Juпcus
gerardii, E/eocharis parvu/a, Raпuпcu/us sce/eratus, Во/Ьоs('}юепw'
тaritiтus, Schoeпop/ectus taberпaeтoпtaпi. На засоленных пони
жен иях песчаных кос произрастают Phragтites australis, Pиcciпe/..
/ia poeci/aпtha, Р. gigaпtea, Р. distaпs, образующие часто сплош..
ные заросли, а также Spergu/aria maritiтa, Sa/so/a tragus, Cyпodoп
dacty/oп, Craтbe тaritiтa, Cakile euxiпa, Lo/iuт /oliaceum, Leymus
.sabu/osus и друrие псаммоrалофильные виды, выполняющие в
основном фитомелиоративную функцию при закреплении бере..
rOB, песчаных кос и отмелей. Хозяйственная ценность подобных
rруппировок невысока, их периодически используют под выпас
скота.
Пояс мезоксероrалофитов образован фитоценозами, )'де ДО"
минирует Pucciпellia gigaпtea и её межвидовые rибриды: Р.
gigaпtea х Р. poecilaпtha и Р. gigaпtea х Р. distaпs. Эти злаки яв..
ЛЯЮТСJJ характерными эдификаторами rалофильноrо типа в рас..
тителыlfмM покрове CeBepHoro Кавказа. Хозяйственная ценность
бескильниц значительно выше солянок. Бескильницы дают ран..
ний питательный корм, боrатый минеральными веществам и; вы..
сота травостоя достиrает 80 см, а урожайность до 1 О цkа сена.
Скашивание и выпас сообществ целесообразно проводить до
конца их цветения, КОfда продуктивность и поедаемость бес..
кильниц, ввиду их малоrо одревеснения. являются наиболее BЫ
сокими.
Между солеросовыми и бескильницевыми сообществами
rраниuа обычно прослеживается чётко, но иноrда бывает пере
ходная полоса из солеросово rалимионовых rруппировок.
Пояс бескильницевых сообществ переходит в '3JIaKOBO
разнотравный луr пёстроrо сложения. Здесь вследствие неодно"
родности микрорельефа фитоценозы повышений, в которых дo
минирует соленепропускающие rалофиты .. E/ytrigia repeпs,
Festuca orieпtalis, A/opecurus aruпdiпaceu'),
Arlemisia taurica, Tripo/iuт paппoпicuт, чередуются с фито..
ценозами пониженных участков из Limoпium тeyeri, L. c(lspium.
Artemisia maritiтa, Juпcus gerardii, Вир/еиrит тarscha"iaпuт,
Р/апtщ о maritima. В районах Восточноrо Приазовья подобные
rруппировки образуют так называемые кермековые луrа (Косен..
ко, 1934); их травостой средней хозяйственной ценности.
26
в итоrе, эколоrический ряд последовательно сменяющих.ся
I'алофильных сообществ заканчивается самым молодым фитоце..
нозом в этом ряду участками солончаковых степей. их расти..
тельный покров представлен бескильницевыми, прибрежнице..
выми и овсяницевыми сообществами с большей или меньшей
примесью rалоксерофильноrо разнотравья; используется в ос..
новном под заrотовку сена, со средними кормовыми достоин..
ствами.
На северо"востоке Среднеrо Предкавказья простирается
Ставропольская полупустыня (Пашков, Зозулин, 1986). К ней от..
несены несколько различных участков: древняя терраса р. Кумы
к востоку от Нефтекумска, Плавненский участок на левом береry
Кумы и пространство к северо..западу от Hero в сторону Чоrрай..
cKoro водохранилища. Общими чертами всех этих участков явля"
ется наличие древнекаспийских отложений или речных песков,
перемытые затем морем. Одна из характерных черт Ставрополь..
ской полупустыни.. обилие в травостое, наряду с дерновинными
злаками полукустарников: Arteтisia lerchet.ma, Kochia prostrata,
Salsola laricina, а на солончаковых почвах обильны ценозы из
Cliтacoptera crassa, Ceratocarpus arenarius, Роа bulbosa,
Petrosiтonia triaпdra. Покров часто КОМIUlексный, ero элементы
лишены чёткие rраницы и занимают крупные участки.
На севере и северо"востоке Среднеrо Предкавказья распро--
странена полынно--типчаково..ковыльная степь на КОМIUlексе со..
лонцов и каштановых почв (Танфильев. 1973). Она имеет вид по--
лосы, которая, придерживаясь более низких мест, тянется вдоль
Маныча. Эrа полоса имеет ряд OTpol'OB, идущих вдоль Калауса,
Erop лыка, Айryрки и балок, впадающих в Маныч. Возможная
причина приуроченности комплексов к более низким местам .. это
появление здесь соленосных материнских пород.
Для КОМIШексов характерен выраженный микрорельеф и мо"
заичность почв и растительности. Комплексы обычно двучлен..
ные. Один из членов.. ковыльно"типчаковая степь на каштановой
почве, друrой является солонцом, а на низких местах .. солонча..
ком. Растительность солонцовых пятен различна, доминантами
:щесь ВЫС1)'пает Arteтisia lercheana, Arteтisia monogylla, Kochia
prostrata, а на корковых солонцах .. Caтphorosтa тonspeliacuт.
Часто rраницы между членами комплекса бывают нечёткие и то--
27
rда коммексность плохо выражена. Солонцовые пятна приуро
чены к понижениям микрорельефа и реже к повышениям.
В целом для долин Маныча, Еrорлыка, Кумы и Калауса xa
рактерна умеренная сухая типчаково ковьU1ЬНая степь в коt\-шлек
се с растительностью солонцов и солончаков, rалофильными дo
минантами которой являются Liтoпiuт sarep/aпuт, L. caspiuт,
Goпioliтoп besseriaпuт, Sa/so/a soda, На/оспетит s/roы/aceuт,
Taтarix raтosissiтa, S/ipa sareptaпa и др. (Зозулин, Пашков,
1980).
28
КЛАССИФИКАЦИЯ rАЛОФИТОВ ТЕРСКО КУМСКОЙ
НИЗМЕННОСТИ ПО АНАтом ФизиолоrИЧЕСКИМ
ПРИЗНАКАМ
«rалофитами называются растения засоленных местообита..
ний, приспосабливающиеся в процессе cBoero онтоrенеза к высо..
кому содержанию солей в почве, блarодаря наличию при знаков и
свойств, возникших в процессе эволюции под влиянием условий
существования...» (rенкель, 1954). Эта высокоспециализирован..
ная эколоrическая rpуппа растений с давних пор интересовала
учёных, в связи С чем, вопросу изучения струюурных особенно
стей rалофитов посвящено большое число работ эксперимен
тальноrо характера.
Во второй половине XIX века, впервые в России, А.Ф. Бата..
линым (Batalin, 1885) на основании серии опытов по изучению
действия избытка сернокислых и хлористых солей на типичные
rалофиты SaZicorпia herbacea, Spergu/aria тediпa var. тargiпata,
Sa/so/a тutica, была установлена взаимосвязь формообразова
тельных процессов у растений с окружающей средой. Так, при
'шсолении почвы NaCl в отличие от MgS04 растения приобретали
суккулентные черты строения: малое число устьиц, крупные
клетки эпидермиса и ryбчатой паренхимы, увеличение числа ело..
ёв палисадной ткани, сильное развитие элементов проводящей
системы. А.Ф. Баталин впервые указал на способность rалофитов
приспосабливаться к сильному засолению почвы в ходе индиви
дуальноrо развития.
Исследуя флору Алжира, Ба1Тандье (Battandier, 1 887), раз
делял обитающие у моря растения на две rруппы. К первой он
отнёс растения с rолыми мясистыми листьями, ко второй .. с
опушёнными, но менее мясистыми. Культивируя некоторые из
них вдали от моря и отметив их нормальное развитие, Батrандье
приходит к выводу, что растения заселяют не только местообита
ния, которые им подходят по характеру эколоrии, а также, [де
они не испытывают конкуренции со стороны друrих видов.
Исследуя влияние засоления на строение прибрежных и не..
прибрежных растений Лесаж (Lesage, 1890), выращивал их в
rоршках, поливая солёной водой. Опыты показали, что lасоление,
29
как правило, вызывает мясистость листьев, которая обусловлена
или удлинением клеток палисадноrо мезофилла, или увеличени..
ем числа их слоёв. И, если, даже лист остаётся прежней толщины,
палисадная ткань в нём развита сильнее по сравнению с I'убча..
той; при этом уменьшаются межклетники, мельчают хлоропла
сты.
lllимпер (Schimper. 1898) rалофитов, относил к растениям
ксерофитноrо характера, по анатомическому строению выделил
два типа структур: суккуленты и несочные растения типа казуа..
рины и тамарикса. Эти rруппы растений, по мнению Шимпера,
имеют те же признаки ксероморфноrо строения, что и растения
пустынь и степей. Ссылаясь на опыты Лесажа (Lesage, 1890,
1891) и свои собственные данные, Шимпер полаrал, что БО.lьшое
содержание солей в почве вызывает у растений образование ксе..
роморсрной структуры листа (редукция поверхности, уменьшение
межклетников, более сильное развитие столбчатой ткани). Позже,
друrие исследователи Apecкyr (Areschoug, 1902) и rольтерман
(Holtermann, 1907) описали строение листа ряда MaHrpOBbIx. и ra..
лофильных видов, которое оказалось сходным со структурой ли
ста пустынных растений, имеющих мноrорядную водоносную
паренхиму и rруппы запасающих трахеид.
В.К. Василевская (1954) отмечает, доказала, что lllимпер
прав, констатируя сходство между структурой некоторых ман..
rровых видов и таких ксерофитов, как каперцы, ремерии и тама..
риксы, описанные Веском (Vesque, 1882а, Ь, 1883), r ейнрихером
(Heinricher, 1885), В.Р. Зале неким (1902), К.З. Закировым и Н.
Меркуловичем (1936). Видимо, указанные сходство обусловлено
тем, ч'Iо эти ксерофильные роды связаны в своём происхождении
с морским побережьем (Ильин, 1937, 1946, 1947).
В 1891 rоду Шимпер сформулировал теорию «Физиолоrи..
ческой сухости» засоленных почв соrласно которой, обитающие
на этих почвах растения, вследствие высокоrо осмоти LleCKoro
давления почвенноrо раствора, испытывают резкий недостаток
воды. в связи с чем, их структура и является ксероморфной. Од..
нако, дальнейшие работы (Stahl, 1894; Harter, 1908; Re(l, 1910
Bouyotlcos, 1911) показали, что в блarоприятных условиях интен"
сивность транспирации у rалофитов может быть чрезвычайно
высока, и что изолированные стебли таких видов как Saiicorпiи
30
sp. и Suaeda тaritiтa транспирируют на единицу Шl0Щади более
интенсивно, чем типичный rликофит Vicia j'aba (Delf, 1911). Поз..
же, Дельф (Delf, 1912) и Штокер (Stocker, 1929) установили, что у
большинства rалофитов"суккулентов отсутствуют классические
признаки ксерофитов .. их транспирирующая поверхность не за..
щищена от потери воды и обычно покрыта тонкой кyrикулой,
устьица мноrочисленные и непоrружённые, а ассимилирующие
opraHbI имеют типичное мезоморфное строение. Эти факты, ука..
занные авторами, связывают с тем, что в силу физиолоrическ.их.
свойств этой rpуппы растений их клетки имеют достаточное ко--
личество воды и, следовательно, хорошие условия роста.
Исследования советских учёных .. Б.А. Келлера (1914, 1921,
1923, 1926, 1940; Keller, 1926), П.Л. rенкеля (I 946) а также ряда
зарубежных Фиттинrа (Fitting, 1911), Слейтера (Slayter, 1961),
Бернштейна (Веmstеiл, 1961, 1963, 1964), и мноrих друrих опро..
верrли теорию Шимпера о «физиолоrической сухости почвы»,
ввиду, её несостоятельности. Их работы показали, что блarодаря
особым свойствам протоплазмы, rалофиты развивают большую
сосущую силу и поэтому имеют нормальное водоснабжение, раз
вивая тем самым, интенсивную транспирацию.
В опытах rольтермана (Holtermann, 1907) у Aveceпia offic;..
пalis представителя манrровой флоры, под влиянием засоления
объём водоносной ткани увеличивалея вдвое, но отмечались слу
чаи, коrда объем оставался неизменным. Отсюда, как считает
rольтерман, приспособления, возникающие как реакция на
внешние воздействия, всеl'да соответствуют какой либо потреб
ности орrанизма и поэтому являются для Her'o полезными.
Шермезон (Chermezon, 1910) исследовал анатомическое
строение большоrо числа rалофитов представителей флоры co
лончаков и прибрежной зоны Средиземноморья. Он отмечал
большое разнообразие в их структуре: имеются rалофиты со ела..
60 поrруженными устьицами, слабо утолщенным ЭПИД,ермисом,
не имеющие ксероморфноrо строения; есть виды засоленных 60..
лот с признаками rиrpoфитов, а также растения, у которых при
'шаки строения rалофитов наслаиваются на признаки ксерофитов.
По мнению aBTOp мясистость .. единственно общий признак I a..
лофитов, однако имеются инемясистые солевыделяющие rало..
фиты.. тамарикс, франкения.
31
В дальнейшем, с развитием фИЗИОJlоrии, проблема приспо
собления растений к засолению стала разрабатываться преиму
ществе:нно физиолоrами. Исследования Б.А. Келлера (1927), А.А.
Рихтера (1927, 1932) иРуланда (Ruhland, 1915) показали, что ra
лофиты неоднородны. Б.А. Келлер (1940) различал три )'руппы
форм, которые П.А. rенкель и А.А. Шахов (1945) впоследствии
предложили называть эвrалофитами, криноrалофитами и I'ЛИКО
I'алофи-тами (rликофитами).
К первым относятся типичные представители сем. CJleпopo
diaceae Salicorпia, Salsola, Suaeda, локализирующие соли в CBO
ём орrанизме; ко вторым растения, выделяющие избыток солей
наружу, таковы виды родов Statice, Taтarix, Fraпkeпia и дp. к
третьим растения, высокое осмотическое давление КОТОРЫХ
обусловлено не столько солями, сколько орrаническими веще
ствами, таковы виды родов Arteтisia, E/lleagпus, солеустойчивые
злаки. Кроме этих трёх rрупп rалофитов, П.А. rенкель (1954) BЫ
деляет четвёртую псевдоrалофиты, корневая система которых
на солончаках достиrает менее засолённых rpYHToBbIX вод. К ним
автор относит тростник обыкновенныЙ (Phragтites coттuпis
Trin).
Общим признаком у всех видов rалофитов является высокое
осмотическое давление, помоrающее им поrлощать воду )1'З поч
BeHHoro раствора с повышенным содержанием солей. К сожале
нию, почти никаких анатомических исследований, которые поз
ВОЛИJlIo1l бы выявить структурные признаки, свойственные пред
ставителям каждой из указанных rрупп rалофитов, до сих пор
проделано не бьUlО.
Следует заметить, что в зарубежных сводках по rалофитам
(Stock :r, 1928, 1933; Schratz, 1934; Uphof: 1941), а также по соле
устойчивости растений (У ал Eijk, 1939; Magistad, 1945 Haas,
1945 Arnold, 1955; Hay\vard, Bernstein, 1958) отсутствуют какие
либо серьёзные попытки подхода к rалофитам с ЭВОЛЮLJИОННО..
эколоrи.ческой точки зрения. Даже в книrе Бьюса (Bews,1927) ,
посвящённой эколоrической эволюции, rалофитам не уделено
должноrо внимания.
А.А. Шахов (1950, 1956), исходя из представления 06 эво
люции как приспособительном процессе, характеризует пути
приспособления растений к засолению почв как соленаКОП.lение,
32
солевыделение и соленепроницаемость. Более совершенным спо--
собом приспособления автор считает соленакопление, а не соле
выделение и соленепроницаемость. Однако, соленепроницае..
мость и солевыелениеe оказались более проrрессивными и захва
тили значительно большее число семейств и родов. И если, соле..
накопление, характерное для мясистых rалофитов из семейства
маревых, выработалось в древности при освоении литоралей
предками первичной флоры (Ильин, 1946), то солевыделение и
соленепроницаемость являются вторичными приспособлениями,
выработавшимися позже в ходе миrраций, расселения видов и
семейств, коrда они встречались с засоленной средой.
О структурных особенностях растений солончаков литера
тура более позднеrо времени таюке располаrает целым рядом pa
бот (Дворецкая, 1928; Келлер, 1929 а, б; Тимофеев, 1930; Лейеле,
1931; Радкевич, 1934 а; Келлер Лейеле, 1940; Рожановский, 1952
а, б и др.). Некоторые из них представляют детальное исследова
ние механизма солевыделения. В друrих отмечается, что CТPYK
тура rалофитов может быть либо rалосуккулентноrо (Келлер
Лейеле, 1927, 1940; Бурыrин, 1942; Новиков, 1942; Поплавская,
1948; Лейеле, 1949; Bickenbach, 1932; Black, 1966), либо Kcepo
морфноrо (Новиков, 1938; Келлер, 1940; Лейеле, 1940; Рожанов
ский, 1952 б; Ковалёв, ] 959; Soding, 1934; Sinnott, 1960) порядка.
Существуют таюке материалы, показывающие возможность по
явления у растений в условиях засоления одновременно призна
ков как первоrо, так и BToporo типа (Азизова, 1933; Саидов,
1959), а Р.Я. ПЛенник (1973), у rалофита Oxytropis glabra отмеча
ет мезоморфное строение листа: мезофилл состоит из ryбчатой
паренхимы, эпидермис крупноклеточный, тонкостенный и Т.д.
Исследования В.А. Бурыrина (1947 а, б, 1948) показали, что
aHaTOMO CTPYK'I)'pHbIe изменения как у rалофитов, так и у rлико--
фитов приспособительная реакция растений к засолённой среде;
'ли изменения (анатомические) сходны с таковыми у суккулент"
ных rалофитов. Так у хлопчатника и свеклы в условиях засоления
увеличивается толщина листьев, сильно развиваются их BOДOHOC
ные ткани.
Дальнейшее изучение растений rалофитов показало, что на
их анатомическую СТРУК1УРУ влияет качество засоления. Роль
разнокачественноrо засоления в появлении Toro или иноrо типа
33
строения была показана в работах[79 84] В.А. Новикова (1942),
А.В. Блаrовещенскоrо (1942), В.Л. Бровцыной (1946), Б.П. Cтpo
raнOBa и Е.Ф. ИваницкоЙ (1954), Б.П. CTporaHoBa и Н.Д. l\1ypaдo
вой (1959), Е.Ф. Иваницкой (1962) и др. Б.П. CTporaHoB (1962)
установил три типа солеустойчивости: хлоридоустойчивость,
сульфатоустойчивость и карбонатоустойчивость. У хлопчатник
выращ,енноrо в условиях XJlоридноrо засоления, появляются при..
знаки и свойства rалосуккулентности (увеличение размеров кле
ток эпидермиса, уменьшение числа устьиц на единицу площади
листа, утолщение листовой пластинки за счёт разрастания пали..
садной и ryбчатой паренхимы), тоrда как при сульфатном засо
лении признаки rалоксеричности. Наряду с этим, в стебле pac
тений .с хлоридноrо засоления снижается степень дифференциа
ции водопроводящей системы и размеры просветов сосудов.
Анатомические изменения, наблюдаемые у хлопчатника в
условиях сульфатноrо засоления, и ведущие к усилению черт ra
локсеричности (уrнетение процесса деления клеток, что в целом
приводит к уменьшению размеров растений и ero opraHOB) oкa
зывают влияние и на интенсивность водообмена. Так, Б.П. Стро"
raнOB (1973) отмечает, что у растений в )'словиях сульфаТJ-юrо за
соления, по сравнению с XJlоридным, снижается содержание BO
ДЫ, осмотическое давление клеточноrо сока, сосущая сила Юlеток
листа, степень rидрофильности коллоидов, повышается интен
сивность транспирации.
При изучении CTPYK'I)'pHbJX особенностей солеустоЙчивых
растений необходимо иметь представление о реакции на деЙствие
солей не только целоrо орrанизма, но и функционально различ..
ных тканей, и даже отдельных клеток. Разрешению этоrо вопроса
способствует метод изолированных тканей. Результаты исследо
ваний, проведённые p.r. Бутенко с сотрудниками (1967) и П.А.
Бабаевой с сотрудниками (1968) показали, что ткани ксилемной,
флоэмной и камбиальной паренхим, изолированные из корнепло
да моркови сорта Парижская каротель 443, различно реaJ ИРУЮТ
на дейетвие солей. Ксилема, по сравнению с флоэмой и камбием,
оказалась менее солеустойчивой.
Ранее Шоландер (1964) установи..l (цит. по CTporaHoBY,
1973), что если осмотическое давление клеточноrо сока у rало
фитов достиrает 60 атм., то давление раствора в сосудах к илемы
34
не превышает 1 .. 2 атм. На основании этих данных автор прихо..
дит к выводу. что по ксилеме передвиrается, по сути дела, оп рее..
нённая вода. Этим фактом, вероятно, можно объяснить отсут..
ствие приспособляемости ксилемы к действию солей, отмеченное
в работах цитированных нами выше.
Из изложенноrо видно, что в целом, в области изучения ra..
лофитов проделана большая работа. Однако, основные литера..
турные источники, представленные выше, характеризуют струк"
турные и физиолоrические особенности, rлавным образом, Д,ву"
дольных rалофитов в то время как эколоrо..анатомические ис..
следования дикорастущих злаков..rалофитов представлены ещё
слабо. Существующие работы посвящены, rлавным образом, зла..
кам ксерофитам, мезофитам, псаммофитам и rидрофитам.
Даже в крупных сводках и отдельных трудах о злаках (Ро..
жевиц, 1937; HackeI, ]882, 1889; Pee..Laby, ]898; Hitchcock, 1920,
1935; Bews, 1929; Metcalfe, 1960; Barnard, 1964 и др.) данные по
эколоrической анатомии ЗJ1аКОВ"raлофитов очень незначитеЛI)НЫ,
а в ряде случаев они отсутствуют совсем. Р,Ю. .Рожевиц (1937) в
своей моноrрафии даёт распределение злаков..rалофитов по три..
бам. Наиболее боrаты rалофитами трибы Festuceae, Agrosteae,
Sporobo/eae. Остальные трибы их совсем не содержат, или rало..
фиты представлены в них лишь единичными видами.
Остановимся более подробно на некоторых литературных
источниках, освещающих особенности структуры rалофильных
:таков. Дюваль"Жув (Duval..Jouve, 1875) впервые отметил, что у
:таков, произрастающих на почвах, боrатых растворимыми соля..
ми, наблюдается сильное развитие в листьях водоносных парен..
химных тканей. Е. Варминr (1901) и позднее Р.Ю. Рожевиц
(1937) указывают, что листья rалофильных злаков носят ярко вы..
раженный ксероморфный характер (IUlастинки их узкие, сворачи..
вающиеся, на верхней стороне имеются мноrочисnенные рёбра,
трихоматические образования, устьица располаrаются только на
верхней стороне, механические ткани сильно развиты).
По наблюдениям Хестермана (Hostermann, 1901) У луroвых
трав (Ho/cus /aпatus, Dacty/is g/oтerata, Ph/euт prateпse) под вли"
янием хлористоrо натрия увеличивается толщина наружных обо..
Jlочек эпидермиса, сокращается объём межклетников в паренхи"
2
ме, уменьшаются размеры устьиц и их число на 1 мм листа.
35
Отрывочные сведения описательноrо характера вотноше..
нии CTPYК'I)'pЫ злаков..rалофитов имеются и в некоторых друrих
работа (Guntz, 1886; Harshberger, 1909; Sutherland, Eastwood,
1916; Esau, 1965), однако их авторы не делают каких либо выво..
дов о 'связи между средой обитания и особенностями строения
тех или иных видов.
В .А. Ковда (1947) приводит данные о том, что злаки, выра..
щенные на засоленных почвах, имеют на листьях восковоЙ налёт
и боле,е мощную кутикулу. У них уменьшаются размеры клеток
эпидермиса и устьиц, число которых на единицу площади листа
увеличивае1'СЯ. Киссер (Kisser, 1927) и B.r. НиколаевскиЙ (1972
в) пришли к противоположным выводам. Первый, культивируя
пшеницу на засолённом субстрате, второй изучая эколоrо.. ан а..
томич :ские особенности дикораС1)'щеrо злака rалофита.
r.p. Ма1)'ХИН и Л.Д. Ефремова (1947) изучали влияние засо..
ления на анатомическую CTPYК'I)'PY стебля пшенично..пырейноrо
rибрида. Авторы отмечают, что на засолённой почве в стебле rи
брида происходит почти полное исчезновение механи LiccKoro
кольца и увеличение числа проводящих пучков на единицу пло..
щади nlOперечноrо среза.
В. Ю. Рожановский (1952 а), изучая строение эпидерми
са листа солевыделяющеrо злака Ae/uropus litoralis, в связи, с ero
обитанием на солончаках, отмечает у этоrо rалофита БО:lьшое
число устьиц, железок и волосков на обеих сторонах листа, а
также I;ильное развитие рёбер. Наличие этих признаков автор от..
носит I( признакам ксероморфной cТPYК'I)'pы. Ранее,
В.К. Василевская (1941 б) в строении листа этоrо rаJlОфИЛЬ"
Horo злака отмечала мезоморфные черты: слабое утолщение
наружных стенок клеток эпидермиса, значительное развитие ме..
ханической ткани. Отде.'1ьные сведения по биолоrии и эколоrии
прибрежницы прибрежной имеются в книrе Н.Т. Нечаевой, В.К.
Василе:вской и K.r. Антоновой «Жизненные формы растений пу..
стыни Каракумы» (1973). Однако, данных по эколоrической ана..
томии этоrо вида авторами не приводится. Не приводит их и Ф.К.
Тихомиров (1974), хотя констатирует, что жёсткие листья и стеб..
ли прибрежницы боrаты минеральными солями.
Н.Н. Цвелёв (1954) даёт подробную морфолоrическ)'1O ха..
рактеристику одноrо из самых rалофильных родов сем. мятлико..
36
вых Pucciпe//ia Pari, с указанием систематическоrо и филоrене..
тическоrо значения отдельных признаков, их изменчивости и за..
висимости от внешних условий. Вместе с тем, анатомическим ис..
следованиям в моноrpафии отведено незначительное место. Ана..
томические данные приводятся, rлавным образом, в отношении
строения листовых пластинок отдельных видов бескильниц: Рис..
ciпe//ia jomiпii, Р. gigaпtea, Р. hauptiaпa, Р. subspicata и Р. ).'ah/..
iaпa.
Культивируя первые три вида, являющиеся обитателями со--
лончаков, в условиях Ленинrpада, Н.Н. Цвелёв отмечает в строе..
нии листовых IUlастинок в основном количественные изменения,
которые автор констатировал визуально. Orсутствие засоления
при водит к увеличению объёма водоносной ткани и уменьшению
количества шипиков на верхней стороне листа; мноrие клетки
паренхимноrо влarалища из бесцветных становятся хлорофилло--
носами.
В.В. Вихерева..Василькова (1964) приводит описание анато--
мическоrо строения веrетативных opraHoB Pucciпe//ia
phrygaпodes, обитающей в арктической зоне в условиях засоле--
ния. Для листьев этоrо прибрежноrо rалофита свойственна тон..
костенность клеток BepxHero эпидермиса, лёrкая поrружённость в
них устьиц, располаrающихся лишь на верхнем эпидермисе, ела..
бое развитие механических и проводящих тканей. В проводящих
пучках сильноrо развития достиrает механическая обкладка. В
клетках паренхимной обкладки содержится небольшое количе..
ство хлорофилла. Мезофилл состоит из IUlотно COMKHYTЫ круп-
ных И одинаковых по форме клеток. Для стебля бескильницы
ползучей характерно наличие развитоrо кольца механической
ткани BOKpyr проводящих пучков.
По данным Е.И. Покровской (1958) при выращивании су..
данской травы, пшеницы и житняка на засоленной почве у этих
'шаков уменьшались размеры клеток эпидермиса и число устьиц
на единице поверхности листа.
В начале 60..х rодов B.r. Николаевским (1961, 1962, 1964 а,
в) была проведена серия работ по изучению влияния условий за..
соления на анатомическое строение веrетативных opraHoB трост..
ника (Phragmites comтuпis Trin.). На основании своих исследова..
ний автор приходит к выводу, что для стебля тростника в услови"
37
ях почвенноrо засоления наряду с уменьшением общих размеров
растений свойственны черты rалоксероморфной структуры
уменьшение относительноrо содержания про водящих и механи
ческих тканей, резкое увеличение развития воздухоносных KaHa
лов и, особенно, флоэмы в проводящих пучках, а также утолще
ние кл,еточных оболочек. Лист тростника под влиянием условий
засоления приобретает rалосуккулентные черты строения: увели
чиваеТI;;Я ero толщина и объём водозапасающих тканей, уменьша
ется развитие механических и проводящих тканей, в сосудисто
волокнистых пучках возрастает относительное содержание фло
эмы.
По данным Л.Ф. Демидовекой и Р.А. Кириченко (1964) у
тростника, произрастающеrо на солончаках, наблюдается увели..
чение (по сравнению с растениями из зарослей в пресноЙ воде)
числа устьиц на единицу поверхности листа и уменьшение длины
жилок.
Н.Н. Баrдыков и r.p. Матухин (1957, 1958) показали, что
при выращивании на засолённой почве у проса посевноr'о и яч
меня «Паллидиум 43» в листьях наблюдается увеличение ryCTO
ты устьиц И уменьшение размеров клеток элементов эпидермиса
(в том числе устьиц и моторных клеток), а в стеблях уменьшают
ся ра.змеры клеток эпидермиса, диаметр сосудов и размер (:осуди
СТО"ВОJlОКНИСТЫХ пучков, а также абсолютная толщина кольца
СКJ1ереIНХИМЫ.
Б,П. CтporaHoB (1962) и Е.Ф. Иваницкая (1962) изуча.1И вли
яние засоления почвы на анатомическое строение листа и стебля
ячменя «Винер». Было установлено, что у этоrо сорта в условиях
засоления уменьшается толщина и ширина листовой пластинки,
резко (в 2 раза) увеличивается сустота устьиц, в стебле сильно
развивается механическая ткань, увеличивается число ПР()ВОДЯ
щих пучков, но уменьшаются их размеры. Эrи изменения были
выраж.с ны более сильно при засолении хлористыми солями, а при
засолении сульфитами проявлялись незначительно.
Отдельные работы (Абдрашитова, 1960; Шенников) 1l0СВЯ
щены нопросам биолOl ИИ и эколоrии rалофильных дикорастущих
злаков:: Agropyruт e/oпgatuт, A/opecurиs veпtricosus u Festuca
orieпtalis. Ввиду высокоrо KopMoBoro достоинства этих 'Злаков,
38
авторы рекомендуют их для ввода в культуру при освоении KO
мовых уrодий на засоленных почвах.
Некоторые сведения по анатомии листа rалофильных злаков
Lasiagrostis sp/eпdeпs и Agropyrum e/oпgatum имеются в работе
М.М. Абдрашитовой и М.А. Мурзамадиевой (1962). Однако, эти
авторы изучали лишь только листья веrетативных побеrов перво..
ro rода жизни растений после посева, помимо этоrо, авторы не
делают никаких выводов о связи анатомической структуры изу"
ченных видов с характером эколоrии.
В работе Р.М. Середина (1961, 1966) даётся подробный эко
лоrо.. фитоценотический анализ злаков cOBpeMeHHoro раститель..
Horo покрова CeBepHoro Кавказа. К злакам rалофитам автор от..
носит 14 видов, значительное количество которых (Ae/uropus и/о..
ralis, Pucciпellia distaпs, Р. gigaпtea, Eremopyruт triticeuт, Festuca
orieпtalis, Cyпodoп dacty/oп) является доминантами и субдоми"
нантами. Р.М. Середин (1967, 1968), изучив микроструктуру ли..
ста 93 видов дикорастущих северокавказских злаков, относящих..
ся к 34 родам, делает вывод о возможном использовании анато"
мических признаков в таксономии и диаrностике мятликовых, в
деле разrраничения не только родов, но и видов. Однако, при
всём этом, автор не отмечает наличие каких..либо характерных
признаков rаломорфноrо строения в листьях изученных им 4 ви"
ДОВ злаков..rалофитов: Pucciпellia dolicho/epis, Р. gigaпtea, P.lom..
iпii, E/ytrigia e/oпgata. Эrи rалофильные злаки Р .М. Середин
(1967) [130] рекомендует в культуру на засоленных почвах.
Интересные исследования с пшеницей (Tl'iticeuт aestricuт
L. сорт Federatia) провёл [.В. Удовенко с сотрудниками (1970).
3адачей их работы являлось выяснение вопроса о том, влияют ли
соли на растение только путём увеличения осмотическоrо давле..
ния почвенноrо раствора (<<физиолоrическая сухость») или ока..
' ывают специфическое ионное (химическое) воздействие. Расте..
ния выращивали в песчаной культуре на питательной смеси
Кноппа при засолении хлористым натрием и полиэтилен
rликолем (пэr), Т.е. веществом, повышающим осмотическое
давление, но не действующим на растение. Было установлено,
что как в вариантах с внесением солей, так и в вариантах с пэr, у
растений развивались характерные признаки ксероморфизма
(наблюдалось сужение листовых пластинок и уменьшение их
39
толщины, диаметра СОСУДОВ корня, размеров устьиц, Юlеток ос..
новноЙ паренхимы, увеличение числа устьиц на единицу поверх..
ности листа и т.п.). Авторы делают вывод, что отмеченные при
знаки являются результатом торможения ростовых и синтетиче..
ских Пjpоцессов и отражают реакцию растения на увеличение ос..
мотич :скоrо давления почвенноrо раствора, создаваемоrо как со..
лями, так и пэr.
С друrой стороны, авторами установлено, что в вариантах с
солями, в отличие от вариантов с пэr, растения отличали(:ь и не..
которыми своеобразными признаками .. более сильным одревес..
нением элементов ксилемы, более ранней склерофикацией клеток
oceBoro цилиндра в корнях и более сильным утолщение 1 ВНУТ"
ренних стенок клеток эндодермы. По мнению авторов, отмечен..
ные признаки являются защитной реакцией растений на ионные
воздействия солей.
К>.Я. Аникиным (1971) подробно изучена биолоrия волос..
неца ситниковоrо (E/yтlls juпceus) злака, встречающеrося в ди..
ком виде на корковых rлыбистых солонцах и сухих солончакова..
тых почвах. Особенности анатомическоrо строения ero Bereтa..
тивных ojpraHoB (мелкоклеточная ассимиляционная пар, нхима
листа, ryстая сеть жилок, сильное развитие в стебле механиче..
ской ткани, а в корне.. водопроводящей с.истемы) автор относит к
признакам ксероморфной орrанизации.
В 1974 rоду была опубликована работа Лифшица и Всйзеля
(Liphsc:hitz, Waisel, 1974), которые изучили большое ЧИС.l0 раз..
ных по эколоrии видов злаков подсемейства Eragrosloidt!ae. Ав..
торы отметили наличие на поверхности листьев rалОфИJlЬНЫХ
злаков большее число солевыделяющих желёзок и изучили их
структуру.
Общеизвестно, что в основе изменчивости анатомической
структуры растений лежат изменения физиолоrических ф} IIКЦИЙ.
В связи С этим, следует отметить ряд исследований, проведённых
с культурными злаками по выявлению степени их солеустойчи"
вости и характера физиолоrии. Работы Н.М. Тулайкова (1922). М.
Беккер..Ржевской (J 935), Л.И. CepreeBa (1936), И.В. f'ущина
(1938), А.Н. Скосыревой (1944) с пшеницей, И.В. rущина (1928),
r.p. Ма'I)'хина и Л.А. Бойко (1953) с ячменём и просом, rаррисо..
40
на и Кинrа (Напisоп uпd King, 1925) с кукурузой и др. показали,
что солеустойчивость этих злаков с возрастом увеличивается.
[.Р. Матухин (1949, 1963), исследуя некоторые КУЛЬ'I)'рные
злаки (просо, ячмень) в условиях засоления показал, что у них
вырабатываются определённые защитно приспособительные из
менения физиолоrическоrо и биохимическоrо характера: пони..
жается проницаемость протоплазмы, повышается содержание са..
харов, связанной воды, rидролитическая активность некоторых
ферментов и др. В результате подобных изменений растение по
лучает возможность поrлощать достаточное количество воды из
засоленной почвы. А.А. Шахов, (1950) [144J также отмечает, что
на засоленной почве внутри растения, наряду с анатомическими
приспособлениями, наблюдается накопление повышенноrо коли..
чества уr'леводов, увеличивается осмотическое давление, пони
жается чувствительность плазмы к ядовитому действию ионов
солей.
Тот и Кретчмер (Toth and Кretschmer, 1954) и позднее r.p.
Ма'I)'ХИН с сотрудниками (1957) провели ряд исследований на
предмет выявления содержания хлора в opraнax культурных рас..
тений, адаптируемых к засолению почвы. Результаты показали,
что у томатов, в связи с образованием у них суккулентной струк"
туры, накаIU1ивается в тканях стебля и в основаниях черешков
листьев значительно большее количество хлора, чем у ячменя и
проса. У злаков наибольшее количество хлора концентрируется в
краевых частях листовых fтастинок и в стебле. Авторы приходят
к выводу, что листья у изученных растений выполняют барьер--
ную функцию для солей, препятствуя большому их наКОIU1ению в
rенеративных opraHax.
В литера'I)'ре мало данных о влиянии засоления на анато"
мию ситников (Seidel, 1973; Elenterius, 1976; Rоzеш 1977; Ro--
zema, Blom, 1977; Onnis, Pelosini, 1978; Pfeifer, 1979..1980); значи..
тельное место занимают работы, посвященные их СИ(,'Тематике,
зколоrии и rеоrpафии (Чукавина, 1958; Новиков, 1978, 1981 ;
Нечаев, Новиков, 1979; Кrisa, 1962; Elenterius, 1976; A(lam, 1977;
Snogerup, 1978а; 1978 Ь и др.).
Вместе с тем, ситники..rалофиты, как и злаки rалофиты иr..
рают важную роль в сложении солончаковой растительности Ce
BepHoro Кавказа (rроссrейм, 1936, 1948; Наrалевский, 1980; и
41
др.); перспективны они и при проведении БИОЛОfической мелио..
рации и индукции солончаков и друrих засоленных земель (Ху..
дяков, 1964; Наrалевский, 1980; Onnis, Pelosini, 1978 и др.). Изу..
чение особенностей структурной адаптации ситников к условиям
засоления помимо практическоrо значения представляет опреде..
лённыЙ интерес в познании путей приспособления растениЙ к из..
БЫТОЧI-IОМУ содержанию солей в почвах и водах в разных филоrе..
нетических ветвях однодольных.
Таким образом, из изложенноrо выше литераТУРНОfО MaTe
риала IВИДНО, что особенности анатомическоrо строения J алофи..
тов изучались мноrими авторами, но подавляющее большинство
работ было проведено на двудольных растениях. В тоже время, в
литературе почти нет количественно..анатомических данных о
строении веrетативных opraHoB rалофильных видов, слабо изуче..
на анатомия стебля.
Из обзора видно, что имеющиеся в настоящее время в лите
pa'I)'pe сведения об анатомическом строении злаков и ситников
rалофитов мало, чем отличаются от тех, которые приводились в
сводных работах начала нашеrо века. Авторы существующих ра..
бот отмечают, rлавным образом, наличие ксероморфных призна..
ков в структуре rалофильных таксонов, почти не делая попыток к
выявле:нию специфических особенностей их строения, Т.е. наибо..
лее существенных признаков rаломорфной орrанизации., свой..
ственной этой эколоrической rруппе растений и облиrатных и
факультативных их форм в частности. Совсем не изучен вопрос
об особенностях структурной адаптации rалофитов в разных фи..
лоrенетических ветвей.
Исходя из Bcero вышеизложенноrо, и принимая во внимание
недостаточность литературных данных в области количественной
анатомии и фиолоrии rалофитов, мы считаем, что rалофиты
можно разделить на три rруппы:
1. Настоящие fалофиты (эвrалофиты) наиболее солсустой"
чивые растения, наКaIU1ивающие в вакуолях значительные кон..
центрации солей A/theпia fi/iforтis, Ruppia (Irepaпeпsis, R.
тaritiтa, ZaппicheZZia pe(Juпcu/ata, Crypsis acu/eata, Bo/boscJlOeпus
popovi Salicorпia europaea, Caтphorosтa тoпspe/iacит. Juпcus
coтpressus, Ha/ostachys caspica, Suaeda a/tissтa, S. cuпfusa,
Kalidiит ca.'\picuт, Ofais/oп тoпaпdruт, Sa/so/a тutica. S. ft' ilraria,
42
S. soda. S. corпicu/ata, S. crassifolia, Fraпkeпia hirsuta, F.
pu/veru/eпta, Limoпiuт caspiuт, L. gтeliпii, L. suffruticosuт,
/\fitraria schoberii, Tamarix /аха, Т. raтosissiтa, G/aux maritima,
Spergu/ar;a тar;tiтa, На/оспетит strobilaceuт, Camphorosma
тoпjpe/iacuт.
2. Солевыделяющие rалофиты (криноrалофиты), поrлощая
соли, они не накаIUlивают их внутри тканей, а выводят из клеток
с помощью секреторных желёзок (rидатод), расположенных на
листьях Ziпgeria biebersteiп;aпa, Carex тe/aпostachya, Rapoпticum
serratu/oides, А Usтa gram;пeum, Potamogetoп sarтaticus, Juпcellus
рап поп ic us, Ruтex marschalliaпus, Ceratophylluт subтersuт, С.
taпaiticuт, Batrachiuт rioпii, Buschia /ateriflora, Ti/laea vaillaпtii,
E/atiпe hWlgarica, Lythruт salicaria, L. tribracteatuт, Peplis
portu/a, Meпtha micraпtha, Hieroch/oe stepporuт, Pucciпellia
distaпs, Р. gigaпtea, E/ytrigia stipifolia.
3. Соленепроницаемые rалофиты (rликоrалофиты), которые
приспосабливаются к произрастанию на засоленных почвах бла..
rодаря наКОIUlению в тканях орrанических веществ. Высокое ос..
мотическое давление в их клетках поддерживается за счет про--
дуктов фотосинтез а не минеральными солями. Клетки этих
растений малопроницаемы для солей Eragrostis colliпa, Pholiurus
paппoпicus, AlIium liпeare, А. regeliaпuт, Orпithoga/um
fischeriaпuт, lris ha/ophila, Carex secaliпa, С. me/aпostachya,
Feru/ago ga/baпifera, Acroptiloп repeпs, Arteтisia poпtica, А.
saпtoпica, Charto/epis iпterтedia, Ga/atella puпctata, Arabidopsis
toxophylla, Lepidiuт crassito/ium, Taraxacuт erythrosperтuт,
Spergu/aria sa/iпa, Eиphorbia ura/eпsis, Glycyrrhiza g/abra,
P/aпtago corпutii, Р. teпuiflora.
По способу защиты от избьпка солей rалофиты делятся на
избеrающие засоления; поrлощающие и концентрирующие солей
в вакуолях; выделяющие солей через солевые железки, сбрасыва..
нием листьев и выделение солей через солевые волоски; оrpани"
чивающие поrлощение воды; адаптированные функционирование
при повышенных концентрациях солей.
43
JИСТОРИЯ ИЗУЧЕНИЯ ФЛОРЫ восточноrо
ПРЕДКАВКАЗЬЯ
Современные представления о растительном покрове Терско..
Кумекой низменности являются результатом мноrолетних иссле..
довательских работ, которые проводились на протяжении более
250 лет. Сообщения об особенностях растительноrо покрова за..
падноrо побережья Каспийскоrо моря содержались в отчетах
персон, которые в 16..17 вв. защищали политические и экономи..
ческие интересы России в этом реrионе. В конце 17 начале 18
вв. эта часть Предкавказья неоднократно посещалась мноrими
русскими и иностранными путешественниками (Раrозин, 1984).
Первая детальная карта Каспийскоrо моря, в объяснительной за..
писке ]{ которой rлавное место занимало описание западноrо бе
pera Каспия, была составлена по приказу Петра Великоrо офице..
рами русской армии капитаном фон Верденом и лейтенантом
Ф.И. Саймоновым в 1719..1720 lТ. (Танфильев, 1931).
В 1717..22 rr. низменные районы западноrо побереЖl)Я Кас..
пия в бальнеолоrических целях изучал r. Шобер. Попутно им в
нижнем течении Терека собран значительный reрбарный матери..
ал.
Флористическое и общее rеоботаническое изучение исследу"
емой нами части Прикаспийской низменности началось в двадца..
тых rодах 18 ro столетия. Так, И.Х. Буксбаум, возвращаясь из
Константинополя, посетил равнинную часть Восточноrо Пред..
каВК8ЗЬЯ. Соrласно инструкции он как врач и на1)'Ралист русско"
ro посольства в Константинополе должен был производить тща..
тельные разыскания в трех царствах природы, уделяя особое
внимание лекарственным растениям. И.Х. Буксбауму также было
предписано делать зарисовки наиболее интересных объектов
природы, для чеrо к экспедиции прикомандировали рисовапьщи..
ков. Вt:рнувшись в 1727 r. в Петербурr, он ПрИС1)'пил к изданию
монум. нтальноrо труда «Centuria», составной частью KOToporo
были великолепно выполненные акварелью изображения расте..
ний. Предметом особоrо интереса Буксбаума были мхи. На OCHO
ве именно ero материалов собранных около Астрахани выделен
род Butbauтia среди мохообразных.
44
Врач русской rосударственной службы И. Лерх исследовал
низменные районы Западноrо Прикаспия в 1732..1735 и 1745
1747 rr. В ero полевых журналах и отчетах сведения ботаниче
cKoro характера занимают значительное место. Следует отметить,
что большей частью эти данные имеют прикладной предназначе
ние: сведения о культивируемых растениях, запасы диких плодо
вых, лекарственных.
В 1773 rоду экспедиция, возrлаВJ1яемая И.П.Фальком, иссле
довала Терско"Сунженский хребет, сделав ряд интересных Haxo
док (Шхаrапсоев, 1998).
Значительный вклад в познание флоры и растительности по..
лупустынныx районов западноrо побережья Каспия внесли aкa
демические экспедиции, предпринятые в 60..х и 70 x rодах 18 ве..
ка Российской Императорской Академией Наук. Этими экспеди..
циями руководили академики И.А. rюльденштедт, c.r. rмелин и
П.С. Паллас.
c.r. rмелин с 1769 по 1774 rr. в сопровождении нескольких
русских с1)' де нто в исследовал прикаспийские и закавказские
страны. В 1772 roдy путешествовал по Ерrеням, в Прикумских
степях, в окрестностях Маныча и Моздока и сделаl1 ряд ботани
ческих открытий (Gmelin, 1784; rмелин, 1785). Второй раз c.r.
rмелин посетил западное побережье Каспия в 1773 r. HaмepeBa
ясь пройти сухим путем из Персии к Кизляру. Однако во время
этой экспедиции он попал в плен к усмейскому хану и в 1974 r.
скончалея от дизентерии. Сборы и дневники ero последней экс
педиции были обработаны и опубликованы П.С. Палласом в 1984
['. как 4..я часть сочинения c.r. rмелина «Reise durclt RHssland zur
Untersuchung d. drei Naturreiche» (Gmelin, 1784 rмелин, 1785,
Щербакова, 1979).
В 1770 74 rодах по низовьям Терека, Сунже, Сулаку и
окрестностям Кизляра экскурсировал И.А. rюльденштедт. «Низ
кий береr Терека пишет rюльденштедт, начиная от моря до
Кизляра почти безлесен, а выше до Староrладки лес начинает по--
казываться. Ещё дальше вверх весьма MHoro лесу, который боль..
шей частью состоит из дуба, диких IUlодов и др. деревьев»
(Львов, 1964). Результаты ero путешествий в 1987 roдy (<<Reisen
Russland und Caucasischen Gebirge», в 2..х томах) также бьти из..
даны П.С.Палласом после смерти caмoro И.А. rюльденштедта. в
45
указной работе содержатся сведения о флоре, лесоводстве, состо..
янии садоводства и шелководства на Кавказе. Результаты нееле..
дований И.А. rюльденштедта бьши использованы при составле
нии карты Кавказа.
Достаточно MHoro сделано для познания раститеЛЬНОI'О по
крова 1В0сточноrо Предкавказья и самим П.С.Палласом. Под ero
руководством были предприняты две экспедиции на Кавказ.
Маршрут экспедиции 1793 rода пролеrа.l от Астрахани к Кизля..
ру. В этой экспедиции П.С.Паллас был собран интересный мате..
риал в прикумских пустынных районах. Далее посетил Ьештау,
Машук, Кисловодск. Дойдя до Баксана, вернулся в ['еОРI'иевск
(Щербакова, 1979, 1983). им сделано первоописание 86 видов
растений, встречающихея на территории Предкавказья, а также
издан атлас, включающий более 20 видов солончаковых и полу"
пустынных растений: lris ha/ophila, Ereтo partoп aphylluт,
Calligoпuт aphylluт, На/оспетит strobilaceuт, Suaeda
тicropJlylla, Salsola deпdroides, Petrosiтoпia triaпdra, G vcyrrhiza
aspera, Pyпls salicifolia, Astragalus dolychophyZZus и др. (Паллас,
1786; Pallas, 1799..1801). П.С. Паллас является автором MOHorpa..
фичеСIЮЙ обработки Ast1'sga/us и Salso/a прикаспийских ПУ(,lЫНЬ.
На основе изучения древних береrовых линий П.С. Паллас ука..
зывал на следы прежнеrо стояния уровня Каспия.
Ф. Маршалл..Биберштейн был одним из первых профсссио..
нальных ботаников"флористов исследовавших равнины 1ападно
ro поб( режья Каспия. Он в 1796 r. в качестве естествоиспытателя
сопровождал Каспийский корпус rенерала В.А. Зубова в нереид"
ской ЭIсспедиции. Описание этоrо путешествия дается в «TabIeau
des provinces situees sur la cote occidentale de la mer Caspienllc entre
les flellves Terek et KOHr» (1798), и «Beschreinbung der Lander
zwischl n den Flussen Terek und Kur ат Kaspischen Meere» (1800).
Во второй работе дано описание 74 новых и редких видов.
С 800 по 1803 rод Х.Х. Стевен, назначенный по представле..
нию М. Биберштейна в инспекцию по шелководству, жил в Киз..
ляре и в rеорrиевске, активно занимался изучением флоры и co
ставлением коллекций. Часть сборов он определил сам, часть пе
редал <1>. Маршаллу..Биберштейну и [. rофману. В «Stipes rariores
in itinere Caucasio А. 181 О Iectae» (1812. 1813) и «Cotol08ue des
plantes...» (l812) Х. Стевен приводит списки таксонов, дает еве..
46
дения о мноrих видах, а также диarнозы новых: Co/chicиm
итbrosuт, С. /aetuт, Crambe cordifoZia, LoZium тarschaZZii, Orobus
cyaneus, Astraga/us /asioglott;s, Diсtатпш gymnostyZis, Veron;ca
cr;sta galli, Omithoga/uт arcuatuт, Pu/satilla а/Ьапа, Аrnт
a/bispatuт и др.
Публикация Крымско Кавказской флоры (Flora Taurica
Caucasica) Ф. Маршалл Биберштейна (Bieberstein. 1808 18 19),
подводит итоrи работы ботаников флористов Кавказа и Крыма. В
названной работе впервые опубликованы мноrие виды, описан..
ные Х. Стевеном и самим Ф. Маршалл Биберштейном.
Знаменательным событием в познании растительноrо покро
ва кавказской флоры в целом была публикация « Flora Rossica »
К. Ледебура (1824..1853), в которой насчитывалось около 3000
видов для территории Кавказа, в том числе и для Предкавказья
описанные автором Carduus /ac;niatus, Cepha/al'ia gigantea, Соп..
soZida divaricata, HeZiotropiuт elZipticum, Lappu/a heteracantha, Lo
lium pers;cuт, Onosтa setosa, Po/ygonum propiпguum, Prunus di..
varicata, Ruтex stenophyllus, Scrophu/aria divaricata, Senecio gran
didentatus, Taтarix raтosissiтa и др.
Специальные исследования в Прикаспийской степи были
проведены экспедицией К.М. Бэра, которая была орrанизована с
целью изучения рыбных боrатств этоrо района (l853 1856 rr.)
Однако, помимо ихтиолоrических исследований. К.М. Бэр провел
ряд наблюдений rеолоrическоrо и reоморфолоrическоrо xapaктe
ра. Им был впервые подробно описан рельеф Прикаспия буrры,
впоследствии названные бэровскими (Рычаrов. 1958).
Друrим классиком кавказской флористики, в трудах Koтoporo
встречаются ссылки на западный береr Каспия (Терек, Су нжа,
Самур, [розный, Кизляр и окрестности Дербента) как место, rде
он собирал растения, является с.А. Meyer. В течение 1829 1830
rr., работая в разных районах Кавказа, он собрал хорошо этике..
тированный rербарий. В ero знаменитой работе «Verzeichniss der
Рflanzеп...» (I 831) для нашеrо район помимо уже известных
(Lappago raceтosa, Panicuт saпguiпa/e, Меиса nutaпs, Роа
compessa, SmiZax exce/sa, Statice gтeliпi, /тperata arndinacea,
Ceпtaurea arenar;a, He/;otropiuт е иrореит, Coпvo/vu/us persicus,
Anabas;s aphylZa, Feru/a caspia, P/antago arenaria), при водятся не..
которые новые виды {Eurot;a ceratoides, Picris /aevis, Linaria
47
rupestris, Schoberia mi<:rophylla, S. salsa, Atriplex тi(;rапtlш,
Bark/lQusia таrsсlшllii, Crataegus ambigua, Erodium hoejjtiaпum,
Euphof'bia astrachaпica, Holosteum тargiпatuт). Bcero в указан..
ной вы ше работе упоминается более 2000 видов растений Запад..
Horo Прикаспия, из которых около 100 новых.
Важным этапом в ботаническом изучении Кавказа является
5"ти томная работа Е. Буасье, «Flora Orientalis» (1867..1884). Сам
Е. Буа.;ье не посещал Кавказ, но им обработаны сборы мноrих
коллекторов кавказской флоры и критически пересмотрены ряд
таксонов. Для территории Предкавказья им описываются виды:
AlIium ruprechtii, Astragalus buпgeanus, Erigel'oп orieпtalis,
Liтoпium meyeri, Melaтpyruт elatius, Peucedaпuт pschavicuт,
Euphof'hia iberica и др.
Известный ботаник и энтомолоr, член MOCKoBcKoro общества
естествоиспытателей А. Беккер, начиная с 1860 r. активно соби..
рал rербарные коллекции в Даrестане. Ero сборы большей частью
охваты вают предrорные и ropHbIe районы. Однако в ero сборах,
которые по преимуществу обработаны Е.Р. Траутфеттером
(Липшиц, 1947), имеются образцы с пометками «Дербенп>, «Те..
мирхан..Шура», «Ноrайские степи». Маршруты ero экспедиций и
наблюдения описаны в работах «Поездка в Дербент» (1871) и
«Reise nach den Magi..dagh, Schalbus..dagh und Basar..Djusi» (1875).
Труды Н.И. Кузнецова (1909, 191 О, 1913) знаменуют переход
на кач. ственно новый уровень в изучении кавказской флоры. В
ero классической работе «Принципы деления Кавказа на ботани..
ко..rеOI'рафические провинции» (1909) районирование построено
с учетом историко..rенетических и rеOI рафических аспектов в
формировании флоры. Ботанико..rеоrрафические построения l10
следующих авторов (общекавказские и реrиональные) .. Н.А. Бу..
ша (1935), А.А. rроссrейма (1936, 1948), Е.В. Шифферс ( 953),
А.Л. Харадзе (1966), A.r. Долуханова (1966), А.И. rШIУШКО
(1976, 1978) проникнуты идеями, выдвинутыми в названной ра..
боте. l\1атериалы caMoro Н.И. Кузнецова и ero учеников Н.А. Бу..
ша, А.В. Фомина, Ю.Н. Воронова, а также сведения более ранних
исследователей вошли в 44 выпуска «Flora Caucasica сritiсз». Н.И.
Кузнецов из трех своих поездок по Даrестану в первой (1889 r.)
проводил исследования в Хасавюртовком oKpyre (Чиликина,
1959).
48
В конце 19 и в начале 20 в.в. на территории Даrестана прово..
дил флористические исследования В.И. Липский (189]..1894). им
выполнены исследования степной полосы Bcero Предкавказья.
В.И. Липский обратил внимание на характерные заросли держи..
дерева. тянущиеся смошной полосой у Хасавюрта. Ero вклад в
изучение флоры Предкавказья определяется изданием «Flora
Ciscaucasica» (1894), которая, к сожалению, осталась неокончен"
ной, и моноrpафии «Флора Кавказа» (1899). В последней работе
подводятся итоrи изучения кавказской флоры за 200..летний пе..
риод, описываются 4430 видов растений, включая их rеоrpафиче..
ское распространение (Базилевская и др., 1968; Щербакова, Бази..
левекая и др., 1983).
Ценные сведения о флоре Восточноrо Предкавказья, находим
в работах Н.А. Буша (1905), Ф.Н. Алексеенко (1907).
Растительности Хасавюртовскоrо oKpyra посвящена работа
А.Ф. Флерова (1915), им называются древесные породы зональ..
ных пойменных лесов сохранившихея в долинах Терека и Сулак8
и др. рек. К зональным типам растительности А.Ф. Флеров отно--
сит в частности держи..дерева и низкоствольные дубовые леса
(Львов, 1964).
Дореволюционные ботанические исследования, как правило,
не касались детaJlьноrо изучения конкретных фитоценозов. В со..
ветский период картина резко меняется. В 20..30..х rодах ХХ в.
началось планомерное изучение растительных боrатств, связан..
ное с запросами и интересами народнOI'О хозяйства. Примерно за
40 после советизации Даrестана, лишь в области изучения сено..
косов и пастбищ было сделано во MHoro раз больше, чем за весь
предреволюционный период (Львов, 1960).
В Восточном Предкавказье работали отдельные исследовате..
ли, ПРОВОдиЛись ботанические экспедиции Академии наук СССР
и Всесоюзноrо научно..исследовательскоrо ИНСТИ1Ута растение..
водства. Из крупных ученых..ботаников, побывавших здесь, еле..
дует назвать проф. И.В. Новопокровскоrо, проф. Р.И. Аболина,
А.А. Майорова, академика Н.И. Вавилова, проф. Л.r. PaMeHcKoro,
IIрОф. Е. В. Шифферс.
Обследование пастбищ было начато под руководством И.В.
Новопокровскоrо в 1925 1'. Им изучались СтрУК1)'ра и развитие
растительноrо покрова Присулакской низменности, в результате
49
чеrо написаны интересные работы. Детальное описание типов
растиТl; ЛЬНОСТИ произвела экспедиция 1926 rода под руковод"
ством В. М. Боrданова. В своих работах автор приводит деталь..
ные ботанические анализы растительной массы с пастбищ и aHa
лизы образцов сена. В дальнейшем на низменностях Дш'сстана
проводились как флористические, так и rеоботанические работы
(С.И. Виноrрадов, 1927, П. Веденский, 1928, Василькова, [929, с.
И. Виноrрадов и r. А. Толчаин, 1930, Е. В. Шифферс. 193 J, А. Ф.
Флёров 1933). Отдельные исследования были посвящены не..
большим, но интересным в ботаническом отношении участкам
территории. К таким работам относится, например, моноrрафия
А.А. N[айорова (1928), описавшеrо ориrинальную и весьма инте..
ресную флору уникальноrо памятника природы Восточноrо
Предкавказья CapblКYMcKoro (Кумторкалинскоrо) песчаноrо
бархана.
Заметный вклад в изучение растительности Даrестана внес
известный rеоrpаф Б.Ф. Добрынин (1925, 1927). В ero работах
проводится ландшафтное районирование с учетом особеш-юстей
рельефа, rеолоrическоrо строения, почвенноrо и растительноrо
покрова. Б.Ф. Добрыниным даны описания разных лесных фито..
ценозов равнинных., предrорных и ropHbIx районов Даrестана. В
1928 r. вышла в свет специальная статья с картой под названием
«Ландшафтные (естественные) районы и растительность Даrе..
стана» ..
В _l931 и 1932 rодах в Дarестане работала экспедиция под ру..
ководством авторитетноrо ботаника Р .И. Аболина и почвове..
да с.В. Зонна. На основании собранных материалов было пред..
принято природное районирование Даrестана. В те же rоды Е.В.
Шифф рс с сотру динками изучала сорную растительность и рас..
пространение солянок на засоленных территориях Прикаспий..
ской низменности. Результаты изучения были опубликованы в
двух статьях: «Борьба с сорняками в орошаемых мавневых рай..
онах полупустыни» и «Сорная растительность дельты Tt::peKa И
друrих плоскостных районов Даrестана и ее связь с местными
естественными фитоценозами».
Первое (1928..1934), а впоследствии и второе издание (1937..
1967) «Флоры Кавказа» А.А. rроссrейма одновременно подводи..
ли ито]' и содействовали более rлубокому изучению флоры менее
50
исследованных областей Кавказа. Вклад А.А. rроссreйма визу..
чение флоры Кавказа трудно переоценить, ero работы и на ceI'o"
дняшний день являются основой, на которую опираются иссле..
дования флористическоrо и флороrенетическоrо характера. Мно"
rие описанные им виды пополнили список флоры Предкавказья,
такие, как Сатрапи/а trautve//eri, Chaтaecytisus caucasicus, Gagea
he/eпae, С. соопае, Oпobrychis ruprechtii, О. vassilczeпkoi, Popu/us
sosпovskyi, Pucciпellia gigaпtea, Scutellaria oreophiZa,
Hy/ote/ephiuт caucasicuт и др. Труды А.А. rроссrейма позволили
интенсифицировать и поставить на новую основу изучение реrи
анальных флор. Появляются мноrочисленные работы, как по
флоре отдельных реrионов, так и по флоре CeBepHoro Кавказа в
целом (Иванов, 1998).
В 1931..1932 rr. Черные земли и Кизлярские пастбища под"
верrлись маршрутному рекоrносцировочному обследованию
Всесоюзноrо института кормов под руководством И.А. Цаценки"
На и H.r. Андреева. Было проведено районирование территории в
кормовых целях с составлением карты районов в десятиверстном
масштабе, установлены типы пастБИIЦ, определены их rтощади и
емкость в расчете на использование в теплый период ['ода, даны
рекомендации по их улучшению, собраны сведения по xapaктe
ристике растений и пр. (Цаценкин, 1957.)
Большим событием в научной жизни Дareстана явилась ор"
I'анизация филиала Академии наук СССР с отделом раститель..
ных ресурсов. Научные работники этоrо отдела изучали кормо..
вые уrодья, разрабатывали мероприятия по их рациональному
использованию, наладили изучение биохимии кормовых и друrих
полезных растений Даreстана. Уже в 1948 r. были опубликованы
в Трудах первой научной сессии Даrестанской научно..
исследовательской базы АН СССР статьи: Л. Н. Чиликиной
«Естественные кормовые уrодья ropHoro Даrестана по материа..
.1ам паспортизации», Е. Ф. Степанова «О кормовой базе живот..
новодства Даrестана» и Н. Д. У нчиева «Дикорастущий кизил в
Дarестане и ero использование» (Львов, 1960).
В дальнейшем написаны и частично были опубликованы ра..
боты Е.Ф. Степанова (1950, 1952), Л.Н. Чиликиной (1950,1957),
Л.Н. Чиликиной и Н.Д. Унчиева (1956, 1957), Н.Д. Унчиева
(1958), Н.Д. Унчиева и Н.А. Ярулиной (1959), Н.А. Ярулиной
51
(1958) и др., также посвященные пастбищам и кормовым paCTe
ниям Даrестана.
В 1956 roдy по заданию Министерства сельскоrо хо'\яйства
РСФСР Волrоrрадской экспедицией было начато новое СШlOшное
крупномасштабное обследование естественных кормовых уrодий
Прика(:пия. Оно велось по единой методике под научным PYKO
водством И.Д. Цаценкина и О.Н. Чижикова, было закончено в
1960 rоду. Материалы этих работ вошли в «Отчет о комплексных
обследовательских и землеустроительных работах на зимних
пастбищах Прикаспия» (1960).
По результатам мноrолетних исследований флоры и расти
тельности, и прежде Bcero луrов и пастбищ, ботаниками Даrе
cTaHcKoro филиала АН СССР подrотовлена сводная работа на Te
му: «Краткий очерк растительности Даrестана», с приложением
карты растительности в масштабе 1 :2000000, rде приводится
краткая характеристика растительности Даrестана в разрезе есте..
ственноисторических районов республики с ботаническим анали..
зом фл оры И наметкой рациональноrо использования раститель
ности. В друrой работе (Чиликина и Ун чиев, 1956) при водится
ботанический состав основных типов пастбищ и сенокосов, а
также химический состав, кормовая ценность травостоев и peKO
мендации по улучшению уrодий. В это же время в трудах Инсти
тута животноводства Даrестанскоrо филиала АН СССР была
опубликована содержательная статья И.Ф. Мусаева (1956) «Pac
тительность кормовых уrодий Присулакской низменности». В
статье освещаются особенности сложения, распределения и раз
вития растительности, а также при водится цифровой материал,
хараКТI РИЗУЮЩИЙ урожайность растительных формаций. Р.И. Ta
rирова и И.И. Волкова (] 957) посвятили свою статью реЗУ.1ьтатам
наблюдений над растительностью и почвами Ноrайской степи.
В результате детальных rеоботанических обследований pac
тителыноrо покрова, инвентаризации и паспортизации луrов и
пастбищ стало возможным составить карты растительнOI'О по
крова Дarестана: карта растительности кумыкской плоскости и
дельты р. Терека, масштаб 1:210000, составленная А.Н. БОI'дано
вым; f'еоботаническая карта южной части Дarестана, масштаб
] :210000, составленная З.Н. Кара Мурза; карта растительноrо по..
52
крова Дместана, масштаб 1 :200000. составленная Л.Н. Чилики
ной.
Дополнительные сведения о флоре Восточноrо Предкавказья
в этот период содержатся в статьях П.П. Жудовой (1951). П.r.
Язана (1954), П.Л. Львова (1959,1969,19716, 1975a, 19756, 1977,
1978, 1979), А.Т. Рахманиной (1960), я.и. Проханова (1961), Н.А.
Мина1)'ллаева (1976), А.А. Лепехиной (1977).
Отдельные, ос060 интересные с флороrенетической точки
' рения, фраrменты Терско Кумской низменности подр06но изу
чались А.И. rалушко. В 1974 r. им опу6ликована, работа, посвя
щенная флористическому изучению урочища Киссык, rде приво--
дятся списки видов и дается характеристика растительности это--
1'0 района.
Вопросами улучшения пустынных и полупустынных паст
бищ Западноrо При каспия занимались мноrие исследователи
(Нечаева, Приходько, 1958, 1959, 1971; Махмудов, 1968, 1972;
Хайдаров, 1968; Шамсутдинов, rаевская,1964; Клюшкин, 1968;
Шамсутдинов, Чал6аш, 1969; Валиев, 1978; Умаров, 1969; Али..
мов, 1970, 1971, 1973; Бурыrин, 1956; Абдраrимов, Сафонов,
1971; Шамсутдинов, 1968, 1969, 1972, 1973. 1979: rоловченко,
1967, 1968, 1970, 1971; r асанов, 197]. 1973; r асанов, Залибеков,
1998; racaнoB, Курбанов, rамидов, Бутаева, 1990, 1997; racaнoB,
Курбанов, rамидов, 1997, 1998). Исследования, проведенные
ими, представляют 60ЛЬШОЙ теоретический и практический инте
рее в познании растительноrо покрова Восточноrо Предкавказья.
В работах П.Л. Львова (1964, 1965), П.П. Соловьевой (1965,
1968, 1970), Зеленько О.В (1969), Б.Ф. Остапенко (1972) приво
дится типолоrическая характеристика лесов Восточноrо Пред
кавказья.
Заметный вклад в изучение флоры Предкавказья, в том числе
Восточноrо Предкавказья, внес Иванов А.Л. (1998, 2001), доктор--
екая диссертация (1999) KOToporo посвящена анализу флоры
Предкавказья. Отличительной особенностью ра60Т этоrо автора и
руководим ой им научной школы представляется рассмотрение
вопросов флороrенеза. В этом русле выполнены диссертации ero
учеников Чимониной И.В. (2004), Траутвайн с.А. (2008).
В последние rоды растительный покров даreстанской части
прикаспийских равнин исследуется даrестанскими ботаниками, а
53
часть Терско..Кумской низменности ботаниками чечеНСКl1l\tи бо..
таниками. В частности эколоrо..биолоrический и фитоrеоr'рафи..
ческий анализ флоры Терско..Сулакской низменности выполнен
rаджиевой r.r. (2005)., А.С. Абдурзакова (2009).
Опубликовано ряд моноrрафий, посвященных исследован и..
ям растительноrо покрова прибрежных и равнинных районов За..
падноrо Прикаспия. В моноrрафии r. 1. Абдурахманова. З.М.
Джамаловой, А.А. Теймурова (2003) приводится видовой состав
лиановых лесов дельты Самура, характеризуется современное со..
стояние этой реликтовой экосистемы, анализируются причины ее
деrрадации. Реликтовые аридные редколесья подверrнуты тща..
тельному анализу в работе А.А. Теймурова, В.А. Азимова (2005),
в которой рассмотрены основные этапы rенезиса флоры этих ле..
сов. В работе А.А. Теймурова, r.r. rаджиевой (2006) приводятся
результаты таксономической ревизии видовоrо состава флоры
Терско..Сулакской низменности, на основе количественных и ка..
чественных показателей таксономическоrо состава, rеоэлементов
и жизненных форм показано влияние ('оларктических флор на
фитоrеоrрафическую и ЭКОJlоrо..биолоrическую структур)' Тер..
ско..Сулакской низменности. Наиболее полной сводкой по флоре
Даrест.ана является «Конспект флоры Даrестана» (2009), состав..
ленныЙ заведующим лабораторией флоры и растительных ресур..
сов rOpHoro ботаническоrо сада ДНЦ РАН Р.А. Муртазалиевым.
54
эколоrО БиолоrИЧЕСКИЕ ОСОБЕННОСТИ
rАЛОФИТОВ
к числу эдафических (почвенных) факторов относятся как
химические.. реакция почвы, ее минеральный состав, содержание
I'YMyca, так и физические, среди которых водный, возд)'шный И
тепловой режим, механический состав и характер материнских
пород, структура и окраска почвы, мощность почвенноrо слоя и
уровень rpYHToBbIX вод, растительное и животное население. Не..
которые из этих факторов действуют на растения непосредетвен..
но, друrие .. опосредованно, влияя тем или иным способом, изме..
няя их.
Особое место в системе эдафических факторов занимают
пески, распространенные как в засушливых областях (nyстьrnи,
степи), так и в ryмидных (ледниковоrо и эоловоrо происхожде..
ния). Растения, живущие на песках, называются псаммофитами.
Наиболее характерными особенностями песков является их вы..
сокая водопроницаемость, хорошая аэрация, рыхлость, нередко
подвижность и нейтральная или слабощелочная реакция содер"
жащихся в них растворов. Влажность песков и содержание в них
питательных веществ зависит от их происхождения и Ю1иматиче..
ских условий местообитания. По происхождению пески MOryT
быть прибрежные, дюнные, барханные. движущиеся сухие или
влажные, пресные или засоленные. Разнообразие эколоrических
условий песчаных экотопов определяет широкий спектр адапта..
ции псаммофитов. Песок как субстрат, леrко проницаемый для
воды, быстро насыщается водой во время дождя и также быстро
высыхает, особенно в верхних rоризонтах. Рыхлость среды обу..
словливает слабую капиллярность, поэтому на не которой rлу"
бине вода задерживается, а сухой песок служит защитным по
крывалом. Кроме Toro, рыхлый песок плохой проnодник тепла.
Если на поверхности песка темпера1)'ра может достиrать 700, то с
rлубиной она довольно резко падает. Пониженная темпера1)'ра в
толще песка, по мнению некоторых эколоrов (Блarовещенский,
1954, ЦИТ. по Вальтер, 1975), обусловливает конденсацию паров
воды и тем самым пополнение капельножидкой влаrи. Тем не
менее, пески относят к сухим местообитаниям с высоким уров"
нем солнечной радиации.
Светлая окраска песка увеличивает долю отражеННОf'О света
в общей сумме cBeToBoro потока. Такие пески, как прави.;lО, бед
ны питательными веществами. В результате высокие конкурент..
ные взаимоотношения между видами на уровне корневых систем
ПРИВО)J\ИТ к формированию разреженных фитоценозов, состоящих
rлавны м образом из ксерофитов.
Несмотря на разнообразие условий, складывающихся в пес..
чаных экотопах, неизменной остается одна характерная черта
песчаноrо субстрата .. подвижность. Под воздействием ве]'ра или
движущейся воды песчинки, слабо связанные друr с друr'ом, по..
стоянно перемещаются, обнажая корни с подветренной стороны
и засыпая побеrи на противоположной стороне. Растения, вы..
нужденные противостоять этому, выработали ряд приспособле..
ний морфолоrическоrо уровня (образование чехлов на корнях из
мощной пробки или песчинок, сцементированных выделениями
корней, предохраняюш.их их от высыхания; формирование мощ"
ной мнюrоярусной системы придаточных корней на поrребенных
ветвях образование длинных rоризонтa.rrьных корневищ, про ни..
зывающих толщу песка и выносящих почки возобновления к по..
верхности; быстрый рост побеrов, дающий возможность обоrнаrь
наС1)'паюш.ие барханы, и т .д. ).
Прибрежные пески BOKpyr пресных водоемов хорошо овод..
нены, насыщены питательными веществами, приносимыми па..
водкам.и. Обитателями их являются мезо и rиrрофиты. Песчаные
пляжи по береrам морей и соленых озер заняты rалофил .ными
видами. Таким образом, среди псаммофитов есть разнообразные
эколоrические rруппы растений ксерофиты, мезофиты, rиrрофи..
ты и rалофиты. Песчаные экотопы, как правило, .. открытые ме..
стообитания, поэтому псаммофиты характеризуются световой
структурой фотосинтезирующих opraHoB.
Ксерофиты довольно обширная и разнообразная rруппа
растений, обитающих, rлавным образом, в аридных областях
(степи, полупустьши и ПУСТЫНИ) в условиях недостаточноrо во..
доснабжения. Эrо .. склоны южной экспозиции, песчаные откосы
и карьеры, друrие сильно проrреваемые места, характеРИ1)'ЮЩИ"
еся или малым ПОС1)'плением влаrи, или ее быстрым расходом.
Однако не только засуха rрозит растениям в этих условиях. Они
должны защищаться и от переrрева, и от яркоrо света. Адаптация
56
растений к дефициту влаrи основывается на прямо противопо
ложных вариантах стратеr'Ии: у одних на активном добывании
влаrи, реryляции водноrо баланса, способствующеrо сохранению
и экономному использованию полученной воды, у друrих на
способности переносить большие потери влаrи бе'з ущерба ДЛЯ
себя. Эrо обусловлено как особенностями цитоплазмы, так и дpy
['ими физиолоrическими и СТРУК1)'рными чертами орrанизации
растений. Поскольку в разных систематических rpуппах растений
возможности для выработки Toro или иноrо типа ксероморфоза
были неодинаковы, возникла обширная, крайне разнообразная
эколоrическая rpynna. У ксерофитов пополнение запасов воды в
растении происходит за счет сильно развитой корневой системы
интенсивноrо или экстенсивноrо типа.
Поскольку ксерофиты обитают на OTкpЫTЫ хорошо осве..
щенных пространствах, их листья сочетают черты ксероморфной
и rелиоморфной орrанизации. В частности у них четко выражен
палисадный, иноrда мноrослойный, мезофилл, эпидерма включа..
ет мноrочисленные, разноrо типа волоски, образующие защит
ный слой.
В настоящее время среди них выделяют rруппу склерофuтов,
суккулентов и ('''е.мuксерофuтов.
у склерофuтов формируются ryстоветвящиеся корневые си
стемы, максимально использующие содержащуюся в почве воду.
Для склерофитов характерна высокая концентрация клеточноrо
COK создающая большую сосущую силу, позволяющую исполь
зовать даже ТРУДНОДОС'I)'пную почвенную влаrу. При достаточ..
ном водоснабжении они интенсивно транспирируют, но при
наС'I)'плении засушливых условий устьица закрываются, TpaHC
"и рация снижается или прекращается вовсе. Склерофитам СВОЙ
ственна высокая водоу держивающая способность, основанная с
одной стороны на явлении упруrой деформации клеточных сте..
нок, создающей отрицательное давление в клетке и, следователь
но, дополнительную сосущую силу, с друrой на высокой вязко
сти И эластичности цитоплазмы, блаrодаря содержанию "защит
ных веществ" слизей, дубильных веществ и др. Дnя них харак..
терны высокие значения сублетальнOl О водноro дефицита (до
30%), некоторые из них способны терять до 80 /o воды и BOCCTa
навливать свою жизнеспособность после снятия лимитирующеrо
57
фактора. Н.М. Сисакян и А.М. Кобякова (1948) объясняют это
сохранением активности ферментов при rлубоком обезвожива..
нии и прочной СТРуК1)'рой белков у данных растений.
Один из путей адаmации растений к дефици'I)' влаrи сводит..
ся к уменьшению транспирационной поверхности. Мноrие скле..
рофиты имеют мелкие, узкие, сильно редуцированные пластинки,
некоторые .. лишь на время уменьшают поверхность испарения,
или заменяя весенние крупные мезоморфноrо облика листья мел"
кими, более жесткими летними ксероморфными, или, как 'злаки,
сворачивая пластинки в трубку, а наиболее радикальные из них
сбрасывают листья на период засухи.
Сухость воздуха не способствует растяжению клеток при
фОРМИlPовании листовоЙ пластинки, следствием чеrо является
мелко :леточность, мноrоуrольная форма эпидермальных клеток.
Защита листьев от больших потерь влаrи достиrается развитием
мощных покровных тканей.. толстостенной, иноrда МНОI'ОСЛОЙ"
ной, эпидермы покрытой толстой кутикулой И воском, '-устоrо
опушения. Кутикула, воск снижают кутикулярную транспира..
цию, 'устое опушение создает как бы экран. задерживающий
слой воздуха с повышенной концентрацией паров воды. Это, в
свою очередь, снижает скорость диффузии паров воды и с.оответ..
ственно интенсивность транспирации. Одновременно воск и по..
верхностное опушение рассеивают и частично отражают солнеч..
ные лучи, не допуская переrрева. Вместе с тем у них образуется
большое количество мелких устьиц, что, казалось бы, противоре..
чит задаче сохранения воды. Однако это оказывается вьн'одным
для ра<;тения, так как при достаточном водоснабжении интенсив"
ная транспирация обеспечивает активное ПОС'I)'пление питатель..
ных ве:ществ и заметно снижает риск переrpева фотосинтезиру..
ющих opraнoB, а в засушливый период устьица закрываются и
транс.пирация прекращается. Для снижения потерь воды устьица
размеи аются в уrлублениях, у растений со сворачивающимися
листья\tи (Festuca sulcata) устьица, расположенные на верхней
стороне лист оказываются внутри образующейся полости.
у с;клерофитов мощно развиты тяжи лиrнифицированной ме..
ханической ткани. Сильная склерификация позволяет растениям
сохранять форму и положение в пространстве даже при большой
потере воды. У стипоксерофитов склеренхима обеспечивает paB
58
номерное сворачивание листовой пластинки в трубку при потере
тypropa пузыревидными и паренхимными клетками. Толстый
слой склеренхимы также защищает хлоропласта от отрицатель..
Horo воздействия солнечных лучей.
у так называемых 2емuксерофuтов (иноr да их называют
фреатофитами) развиваются длинные, но слабо ветвящиеся кор--
ни, достиrающие капиллярной каймы водоносноrо rоризонта, а
иноrда и rpYHTOBbIX вод, В связи с чем растение не страдает от
недостатка влаrи в веrетационный сезон даже при интенсивной
транспирации. Эта rpуппа включает жаростойкие и нежаростой..
кие виды. Они характеризуются низкими значениями сублеталь..
Horo водноrо дефицита. Им свойственно сильное развитие прово..
дящих тканей в корнях, стеблях и листьях. Листья и стебли часто
включают мноrочисленные идиобласты с дубильными вещества
ми. Листья у таких растений довольно мелкие, тонкие, мяrкие,
нежные, часто совершенно rолые, с большой плотностью устьиц,
Т.е. имеют мезоморфную, часто световую структуру.
Суккуленты. произрастают, rлавным образом, в пустынях
Центральной Америки и Африки, незначительное количество ви"
дов из сем. Cra.,;su!aceae встречается и в зоне YMepeHHoro клима
та. Основной способ существования в засушливых условиях у
суккулентов .. накопление больших запасов воды в тканях и
крайне экономное ее использование. Основная Масс.а водоносной
ткани (волоски и покровные клетки эпидермальноrо комплекса,
rиподерма, внутренние слои мезофилла, коровая и сердцевинная
паренхима) формируется в листьях или в стеблях, в связи с чем
различаются листовые и стеблевые суккуленты. У суккулентов
образуется наряду со стержневой поверхностная корневая систе..
ма, выходящая далеко за пределы проекции надземной части рас..
тения; она в сухое время rода выполняет функцию надежноrо за..
крепления растения в почве. Кроме Toro после дождя возникает
масса поверхностных эфемерных поrлощающих корней. Некото--
рые ксерофиты запасают воду в корнях, образуя rруппу так назы..
ваемых корневых суккулентов.
у суккулентов низкая концентрация клеточноrо сока позво..
ляет поrлощать лишь леrко доступную дождевую влаry или росу.
Но отдают воду они с большим трудом, так как основная масса
воды в тканях принадлежит к катеrории связанной. Клеточный
59
сок содержит большое количество леrкорастворимых низкомоле..
кулярных веществ или нерастворимых rидрофильных соединения
.. слиз{:й, повышающих водоудерживающую способность. У не..
которых видов развивается сеть слизевых вместилищ. ПJЮВОДЯ"
щая система слабая, образована небольшим количеством мелких
проводящих пучков. Механические ткани развиты плохо.
Для уменьшения потери воды opraHbJ суккулентов приобре..
тают окруrлую или цилиндрическую форму, чем достиrается
наименьшая удельная площадь испарения. У большинства какту..
сов и молочаев стебли ребристые, что значительно уменьшает
площадь воздействия прямых солнечных лучей. Кроме Toro, в
ложбинках создается и удерживается переплетением колючек и
волосков микрослой С повышенной концентрацией водяноrо па..
ра. Им, нно особому механизму действия ребер, сходному с рабо..
той ку:шечных мехов, обязано сезонное изменение объема стебля.
В периоды засухи ребра сближаются без всяких нарушений ме..
ханических и расположенных между ними хлорофиллоносных
тканей, находящихея в наружных слоях коры. При поrлощении
воды ребра расходятся и объем стебля снова увеличивается. По..
верхность листа и стебля покрыта толстой кутикулой, эпи..
дермальные клетки имеют утолщенные, иноrда метакутинизиро"
ванны{: наружные стенки, устьица немноrочисленные, мелкие,
поrруженные, обычно закрытые в дневное время суток. Мезо..
филл листа или коровая паренхима стебля плотная, почти лишена
межклс тников, что препятствует внутреннему испарению. Слабая
транспирация не способствует термореrуляции, поэтому всем им
свойственна высокая жаростойкость. Поскольку устьица откры"
ваются только ночью, доступ уrлекислоты и света не совпадают
во времени. Фотосинтез САМ"тип коrда в качестве источника
уrлекислоrо rаза частично используются продукты дыхания .. op
rанические кислоты. Вследствие этоrо интенсивность фотосинте..
за суккулентов невысока, рост и накопление массы происходит
медленно.
Помимо солей, обусловливающих fшодородие почв, почвен..
ные растворы MOryT включать и вредные соли, да еще в таких
концентрациях, которые оказывают на большинство растений от..
рицательное воздействие. Наиболее токсичны леrкорастворимые
соли, беспрепятственно проникающие через цитоплазму. К ним
60
относятся карбонат натрия, хлориды натрия, маrния и кальция.
Менее вредны для растений сульфаты кальция и маrния, а также
карбонаты кальция и калия, так как они частично используются
растениями для минералыюrо питания. Отрицательное действие
больших концентраций солей проявляется не только в повыше
нии осмотическоrо давления клеточноrо сока, нарушающеrо
нормальное водоснабжение растения, но и в изменении метабо--
лизма белков, приводящеrо к накоплению продуктов их распада.
rШlофиты растения, обитающие на засоленных почвах.
Как правило, они образуют монодоминантные разреженные co
общества. Им приходится приспосабливаться не только к высо--
кой концентрации солей, но и к сильному освещению, а обитате
лям аридных областей еще и к высоким темпера'I)'Рам и засухе.
Среди засоленных ПОLIВ с разным солевым и водным режи
мом классическими являются солонцы и солончаки. Такие почвы
формируются в аридных областях вместообитаниях, rде YCTa
навливается выпотной тип водноrо режима в почве. Чрезмерный
полив и сильное испарение часто приводят к засолению почвы
восходящим током влаrи при близком залеJ'аНИИ rpYHToBbIX вод,
насыщенных различными солями. В результате испарения воды
концентрация солей в почвенных растворах возрастает, а на по
верхности почвы выпадает кристаллический осадок. Сходные
условия возникают и в ryмидном климате, rде почвы постоянно
увлажняются сильно засоленными rpунтовыми I-U1И сточными BO
дами, а также по береrам морей в приливно--отливной зоне (на
литорали).
Несмотря на то, что для большинства растений такие условия
являются неприемлемыми, обнаруживаются целые семейства
(Cheпopodiaceae. P/uтbagiпaceae, Fraпkeпiaceae, Taтaricaceae),
представители которых не только приспособились к высокому
содержанию солей в почве, но и преврапfЛИ неБЛaJ'Оприятные
условия в необходимые для их НОРМ8льноrо существования.
у одних представителей этой rруппы адаптации проявляют
ся, в большей степени, на клеточном и физиолоrическом уровне,
у друrих на физиолоrическом и на aJ-taтомо морфолоrическом.
rалофиты имеют ряд биохимических особенностей, способ
ствующих обезвреживанию солей и выведению их из OCHoBHoro
обмена. Это достиrается связыванием ионов белками цитоплазмы
61
и орrаническими кислотами. У мноrих ПUlОфитов проницаемость
цитоплазмы для солей заметно снижена, она содержит MHoro
коллоидно связанной воды, что способствует ее экономному рас..
ходованию и, соответственно, меньшему ПОС'I)'плению солей в
ткани растения.
Одной их характерных черт rаломорфной орrанизации ЯВJIя
ется МRСИСТОСТЬ листьев или стеблей, обусловленная ра'jвитием
или СПI циализированной водозаласающей ткани, или паренхимы.
Большинство друrих особенностей в разных таксонах цветковых
растений проявляется неодинаково, что приводит к бо.l ьшому
разнообразию морфолоrическоrо облика и анатомическOI'О стро"
ения представителей данной эколоrической rруппы. Разное соче..
тание при знаков rало.. и ксероморфной структуры у f'аJЮфИТОВ
обусловлены не столько степенью засоления, сколько качествен..
ным составом солевоrо комплекса.
Основной лимитирующий фактор повышенная концентра..
ция солей в почве, с которой растения справляются, блш'одаря
высокой сосущей силы корней. У разных растений сосушая сила
в клетках создается разными путями: у одних.. за счет накопле..
нии солей, у друrих .. уrлеводов, вследствие чеrо среди rалuфитов
выделяют три основных rруппы.
В крайних по степени засоленности условиях.. на солончаках
и на морских побережьях.. растения обитают, практически, в со..
ляных растворах крайне бедных питател ьными веществами, при
высоком уровне рН и полной освещенности. 1ro так называемые
эуrалофиты, характеризующиеся четко выраженными суккулент"
ными чертами. Одни И'J них имеют редуцированные листья и
членистые мясистые стебли за счет развития водоносной парен..
химы, друrие .. мелкие суккулентные листья с водозапасаlOЩИМИ
эпидермой, rиподермой или волосками. В связи с высокой соле..
устойчивостью цитопла.3мы эуrалофиты способны накапливать
большое количество солей, чем достиrается их высокое осмоти"
ческое давление. Для них характерны низкая интенсивность фо
тосинтеза и транспирации, поэтому растения медленно paC'I)'T и
развиваются. Слабая транспирация обусловлена разными причи..
нами: малой плотностью устьиц, небольшим объемом свободной
воды (1Iюда в тканях находится в связанном состоянии) и др. Од..
нако, из..за малой удельной поверхности побеrовой систе\1Ы ее
62
вполне достаточно для предотвращения переrpева. Следствием
минеральноrо rолодания является выработка большOI О количе
ства антоциана для защиты растения от избытка света. BOДOHOC
ная паренхима служит местом локализации солей, поэтому с от..
миранием отдельных opraHoB растение частично освобождается
от соли.
КриНОZШlофиты .. растения, поrлощающие соли в большом
количестве. чем обеспечивают высокое осмотическое давление
клеточноrо сока, но избавляющиеся от и'злишков соли путём BЫ
деления их в виде солевоrо раствора с помощью особых железок
на листьях или стеблях. В макро" и микроструктуре их листьев
часто сочетаются самые разные признаки. У одних.. это мелкие
листья с чертами мезоморфной и склероморфной орrанизации. у
друrих .. листья с широкой rтастинкой, в CTpYKrype которой
имеются суккулентные (водоносная паренхима) и склероморф
вые черты (толстая кутикула, утолщенные оболочки эпидермаль
ных клеток, наличие волосков, мелкие поrруженные устьица,
крупные проводящие пучки, склеренхимные тяжи при пучках), у
третьих формируются мезоморфные листья с признакам и I'e
лиоморфной орrанизации.
Fликоzалофиты .. растения, обеспечивающие высокое осмо"
тическое давление клеточноrо сока за счет накопления уrлеводов.
Соли же не проникают в ткани корня. Среди них есть растения с
рассеченными на узкие cer'MeHTbI ксероморфныl\tи листьями (раз
нообразные виды Arteтisia), растения с широкими мезоморфны
ми листьями (Beta тaritiтa), растения с широкими листовыми
пластинками rиrроморфной орrанизации (Alisтa plaпtago
aquatica, которая выносит концентрацию солей до 30/0 и рН от 2
до 12) и, наконец, морские однодольные (Zostera тariпa).
Исходными растениями для rалофитов аридных областей, по
мнению ряда авторов (Михайловская, 1964), были растения MOp
ской литорали, сочетающие в себе признаки эуrалофитов и ](ри
ноrалофитов.
63
ТАКСОНО СКИЙАНАлизФлорыrАЛОФИТОВ
Понятие «флора rалофитов» в нашей моноrpафии подразуме..
вает только совокупность видов флористических коммексов,
формирующихся на ПОLIвенных субстратах с повышенным со..
держанием солей. В нашем понимании «(флора rалофитов» близ..
ко по содержанию к понятию «парциальная флора». используе..
мому ]3 своих работах Б.Н. Нориным (1984) и Б.А. Юрцевым
(1975). Флора rалофитов это эколоrическая СОВОКУПНОСТI) видов
растительноrо покрова, формирующаяся в местообитаниях со
сходным режимом условий среды и связанных с отложениями
общеrо rенезиса.
Как было сказано выше, теоретические вопросы анализа
флор разрабатывались мноrими исследователями (Braun..
Blanquet, 1913, 1919, 1928; Буш, 1935; Толмачёв, 1941, 1974,
1986; Малышев, 1969, 1975; Камелин, 1969, 1973; rаланин, 1973;
ШМИДТ, 1972, 1974, 1980, 1984; Юрцев, 1974, 1975; Середин,
1976, 1979, 1980а, 1987; Юрцев, КамеJlИН, 1991; Наrалевский,
1991, 2001; и др.). Теоретические и методические вопросы при..
менительно к Предкавказью разработаны в работах А.Л. Иванова
(1996, 1998). Предпринятый нами анализ fалофитов Приморской
низменности базируется именно на ero теоретических ра:зработ"
ках.
<:истематическаа структура rалофильиых комплексов
Наиважнейшей характеристикой любой естественной флоры
является е систематическая структура. Анализ численных соот"
ношений и пропорций между разными таксономическими paHra..
ми (семействами, родами и видами) позволяют определить спе..
цифические особенности флоры raлофитов Терско..Кумской низ
менн()(:ти, а также выявить некоторые флороrенетические тен"
денции, связанные с историческим развитием данной территори..
альной флоры.
Наш инвентаризационный список видов rалофильной флоры
Приморской низменности насчитывает 248 видов, относящихея к
155 родам и 41 семейству (табл. 6). Полный список видов по си..
стеме Энrлера с информацией о жизненной форме, классифика..
64
циям по солевому и водному режиму и друrим ЭКОJ10rическим
параметрам приводится в таблице 7.
Составленный нами по «Конспекту флоры Терско..Кумской
низменности» (А.С. Абдурзакова, 2009) систематический список
всей флоры Терско..Кумской низменности насчитывает 1022 вида
высших растений из 484 родов, относящихея к 1 О 1 семейству.
Если сравнивать эти абсолютные значения как процентное COOT
ношение таксонов, получается, что rалофиты составляют 24,250/0
видов, 32,02% родов и 40,59% семейств всей флоры Приморской
низменности. Отметим, что видовой состав rалофитов TepCKO
Кумекой низменности состоит исключительно из цветковых (по..
крытосеменных) растений, Т.е. все они принадлежат к типу
A1agпo/iophyta.
Таблица 6
с- Основные пропорции Те ! K мской и менност!!.,
i ропорции такс.! К К Кол =т J: i
I ' ол..во ол..во, I
I каТ"ИI во i В/Р В/С I Р/С I
IФ Iсемеиств родов i : I
лоры ви I
!Флора Терско..Кумской! 1 О 1 484 1022 ! 2 11 1 о, 12! 4,791
l изменности I _
:Фклора алофитов TepCKO 41 155 248 i 1,60 6,05 ! , 78 !
L МСКОИ низменности '
Все виды флоры rалофитов Терско Кумской низменности по
их отношению к засоленности почво rрунтов, MorYT быть под..
разделены на две rруппы: облиrатные и факультативные rалофи
ты. В численном выражении облиrатных rалофитов насчитываt.'Т..
ся 103 вида из 62 родов и 19 семейств. Такой же расклад факуль
тативных rалофитов выrлядит следующим образом: видов 145,
родов ) 05, семейств 33. Очевидно количественное превосход"
ство факультативных rалофитов.
65
r-.....
i
r::
Q)
-&
=
е-
о I лd 1
Q)
"' "ОlJlПф Лw
l"oH)108 ои
О ииhЮlиф
=:
()
CIIS
111
=
111
=
I
;
=
tuj
...;
е .....
Q) tI)
о О,
::r: I
I
: ;I
! с:: .: I .....1
;;1 Q) :::r: ::Е I
. i ! i I
...J s ':':1 1 <' Cl.i
Е ...J I .::-;! ' 'o ч
\:s I (;u '-"'1 ! ..с:: : '=: :
. S::: I . () '--'1 (;u I
{Sl I. !...J Зi : i
=: . , 01
:.::: \:s '1 S:::I " I f.OOJ 1..1
О (;U \:SI'" U' , \:S ;::Sl
I : f.ooJi':::: ; о :
Э, ". I . , :
\31.9 f.ooJl t3: 8 . . ,.эl
i"t . 1 \:S g I ] '
о . . ,С (;U ,
,1 &::;1 =\Ir er;:\...>I:i: ':
1" ...,
I I i i ! I I
1 :' i i c>; N ("')1
N "t : \о r:-- . QQ 1 Q"\ ':""'" :" '
f.OOJ
S:::
.э
'
I
I
......
t:
о
е
8
\:S
.S Q)
.
t3
.
, ......
\:S
..... .....
1.0
1.0
f'r')
[' ;)И)JНЛВd,
011 lчwdоф;
'НЕИЖ '
- -
r::
==
f-
О
Q)
лd
..Оl.)Jвф лw i
"ОН'[(08 oul
ниnвхиф i
"И В1l)( 1
I
лdо.L i
..)JВф Лw081
";)lfO ои
JSИnВ)JИф i
"И В1l)( ;
t'--
\о
==
IQ
==
=
IQ
=
о
Q)
Л:d И)l1:l U
оп Iчwdоф
'НЕИЖ
лd
..О )lUф Лw
"OH}J'08. ОП
ки'rr8хиф
"И:>:>UII)J
лdо
..xuф Лw08
иO:> оп
IfиПUХИф
"И:ЮUU')l
а
CIIS
ii
=
111
=
I
;
=
{d И)fнЛ:еd: I 1 '
I 1 ,1
О :: офl f i g: J ! ! i
'1 ! 1 I l
,1 l' i
,., I i
' =-, в1" I
: Q1-i U.... ,,; 1:
P-I "":I: :I:! :I: I
; 6 !J I
' I
U' C'I")' I Е--о
rd I 1
::S:: :! I I
ОJ.)lеф лw: : I ;! I I Е- '1' E-I
rn i ! Е- Е--! Е-: Е--! Е--I
"ОН'[(08 оп' 8: l' 1::S:: ::S:: i ::S::, ::S::::S:: (5 1
' :.::: О n]
КИnЮlИф , ::s:: IU l' О О О О О, 01 Q)"1 IU
, ;:!:I I t:t ; I I 'I !
и еlf)l 11)::':: , I I I
' ,U r: ! U
лdOJ. , i ; I I 1: :s: 1
Ф Л ' I' ' аl '1
)fe W08: Е--; Е- ! Е- i Е- Е- i Е-I E--I g I g
lro оп ::Е I ::S: ::S:'::S:: I I ;: I ::S:: I
, "'tt' '1;'1 О I -е1 -61 t... t...
киnе)lИф О' О О, О 01 Е-' 01 01 = O
, : g: ! ::S: ::S: .
и еlf)l : ' : , : I t.J t.J
, ,
U: I i I ..: I I
, i :U I' ,U
I I r \ 1.
' i 'с5 '-J...I IQQ
rJ'J !..$; t-- "31 '
,
IU : i . (,)
z ,р..,..... 'J'J'
, ,...... -::: I
: -d : .Е i, :9...... " Q) I
,d ' ,(,) .,
,'1:: ,$-1;И ', .:::
' I I? =
!и' ' ' -51
i .S IU' 'J'JI I
' . J , ..ci!.
i. . ! ;>.; ЕI !
C)I:: .I : ,...J IU, I
. ! C:::;I I I ....
.....1..... =' с: .I ........ 11)!
i..... ::r: ; .S I' II-al
I з, . U I
I I I I I
. : [ : I Щ !
I I ..s! ' :.;, I . I
I ! g i i , !
, ,..... r 'r -: I .1
vs 1 '-..J ; '-..J ' l'
I I l' I I I
' . .. .;gJ . l : . ) l J
t:
:s::
Е-
О
IU
=
=
IQ
=
=
=
IQ
м
=
=:
I
:I:I
U
,
,
I
!S;i
01
0\
1.0
Q)
s:::
:3
cj
Q)
-d
1
01
I
rJ'J'
=:
,
",!
!
I
I
I
. '
N,
\QJ ..
I
м 1'..,.) \о
\.g
ИХН В
ou Iчwdоф
'НЕИ)I{
лd
"ОDШф Лw:
"OH'I108 OU
B-ИnВ)lИф
"И 1Ш)I
лdо.L
.. хвф Лw:ОЯ
" HO OU
trnПВ)lИф
I "И Вli)J
"
if
са
с»
111
=
"
са
I s
1::
cu
.ri
cu
\:s
I
I
I '1 I!
,.",. l' I I.'.ri
I I I ", I.D Е
1 11 l' I ; I : 5 cul
< .1 : I I . йl
. II I. . :I" I t:: I-€II I.I
'"" .D ..... .G I ..с =s..... 1 О
Е I . i'''''! i 1 Е s=
'1 cu , , <! .ICj i=?1
I а... ... ":1.... I О . CU I
'1 с .....;l,.......,Cj e'
:5! .S: (;1.....;1.' 1;1
..... \:s .1 I .... . I . ё;j (;, I E
с .EI I' .f:: '..J Ie:; 1c:;Ji
. =s \:S t:: ::S \:S \:S I \:S: а.;. I cz) I ....
.Q I ...i ! .J i i
Е \:S :: .... \:S i I I
с I . f'J, . \:S \:S I 'f'J I ..з I t:; Е I
.... ' 'Ё ' I э i:Ё II ' I , i 1
'L с :о::: с \:S -SI I , Е, Е I..s:: '->: I::)j":::j
бu io;i . . . . '! ' 18'u '
00 Q\ C>1 ",1 "'l .++-- 001 a-ic, H,.,i ! ",111
f'- !:'::- t-- f'- r--- t::- f'- f'- f'- f'-' 00 00 00 00 ! 00 ,
f'-
;;
.....
t'--
=
се
=
=
се
=
12)
:
i ,......
: ё:
, 6'
,..:,(1
, .......'
:
CIII
if
се
111
=
CIII
се
G=s
\о ["--. 00 0"1 с> ......
С>С>С>С>............
.....-4 .....-4 .....-4 ...... ...... ......
Q)'
g!
\О:
О:
i
I I
: :Ж:! ti
12) (1)'
\о, t:::,
;>11 '
c;:: , U I i
Е-
C'I
t'--
, .,
, :::: j
! I
, (1):
"':1
tn'
: '"CI i
, ," 'ё;,
: r;;' I
, 12)1?'
10: i
I '.=:
. ' ,......'
...... ,"--"', I
.э ' :S ' l .
. : : i
;: ..ь'
с ," t.,):;::
I , ;:.:::'
.;: ;:: с I
i
'1:i ....'
. ----:
, I
0"1 'с ......'N
......: 'NjNIN:
...... I......:
,(d;)И)lнлеdi
,.....
I
! ь
1
I
f-oi
t:::,
cu
cu
t:
:s::
f-o
о
cu
щ
о
,
(-r')
t'--
;g
= c:u
се
з
>ос:!
= ,
= c:u c:u
..s:: ,
се .
м
, ':';::
.....
::с Ii::
о
= rr'I -.:::t ["---1 00
N N rr'I' М
= т '":"'"' .......
лd и)(н В 1,
ОIJ IчwсJоф '1 I i l'
'Нi:ИЖ , Io!
t::
лdО.L
..)Шф ЛwО8
.. иo ОIJ
ИИnВ>IИф
"И ВU)I
...j
:::1
I
i
I
I
i
l' I
I.€ I
tl
Е ::t: I
I
E
::s Е
...... ::s
. ' I
' I
....
"
;
I:Q
111
I =
I
I =
!
:
r
: i
'....
I
I
I
1
:0.;
; t::{ I t::{
, iU Q)
: s: s
I I
!O
i I
f-<' f-<
! х: :S::
I iU I
I r=: О
I
l..c I
i
I
, :с'
i CQ I
:
i ,
l'
, О;
, ,
I
, i
I ! I
t ---I
, I i
: !;21 в;
t::{
Q.) iU
, 1:: S,
,('Ijl '
t::'O:
,
, О,
f-"' о.-
I :S:: I Q.)'
с,),
01
'
I Q.):
I r=:
!
t'--
:
' ;
f'.4
I
I
I I
, -; i
I I
"'" .
'-',
.
...(:)i !
Е
:::s ....
:::::
9<j...(:)
'-о
, '
:' ,
IW
'
I
!. :
: ,
1....
1.';::::
i .:
,
: \3 I
:'
, :
о'
: .::: I
, c:u I I
: .J;: : .:
, \3: !
, ;з;
, 1...' \3,
Oj :
,
IO I ,N:Mi
!'o::t':V) i : IV)IV)'
'
с::
::s::
r-
о
(!)
t....
'..di
' О
О ....
, ....I
,
".....J .
I ,
, 01
, s:: I
о!
! 5!
: I
i c-j
, \:S I
10:'
I \:S I
"
' 'I
.
(5:
..(:) I \:S
,
:kj v
i lif)
l
::
се
::
=
се
м
==
:
, '
;'Е: I
'-.j I I
: ' 1,
: "'"' '. 1.1
'-.j ' '
: I \:SI .
110:::: : I
I : , з
1 О i . : 10:
, \:S.... , C:U
l' t:: ! I
,.... ,
'...... : 10:, c:u
I , I..2::
I ;:.. ,10: i ;:=
"';::: ..(:) I
: .... l' I ц.;
I
, I["--. 10010-
: I I
.L
I Q I
, I I I
[ fll r=11 g
I '1
I !
i I i
, (!) I I
i 6, :!I: glф
?- ФI I о О
/СТ) о !ь IU
v)
r--
t::
==
(0-0
о
Q)
лd
..()J.)feф Aw
"ОН)108 ОП
ИиII'e)(иф
"И UU)J
лdО.L
..хвф Лw08
.. иo Оll
ИИnU:>lИф
"И UU)l
I
I
I
I
I
I
!
i
....11
I
r :
I i
I I
, I
:
, d}1
, е;
1\0'
iO!
1
g
\о
t'--
I QQ
I , :
I ' I
i
I v1
fп
(.) '8
.
I "......,
(.)1 ,
. сп: I
el ' I "--" :...j
" Q) 1. . .1 , :".)
! I ;:::: ::' :;::::
. ,
111 . "'1 ;::::, .....
I:Q . ..d f'J а.. --.. ' i
\:s З ;::::!
5 \:s I :.) :C:U
111 ..... Е З
= I. 1 ..... .... ' ."
. :: i
" с ;::
I:Q .Е :;::::
\:s . с Е ...t: 1;.
. э ...::: :.) :с
С 1:::,
..... ' 1;.
.э 1... .э 1::: [10:
. C:U, :" IC
cj cj ; 'С,,)
0110 ......(, I м oo::t о 1......
01 = \,о t'- t'- t'- 1 t'- t'- OOi 100
l r:;: ......,............ ...... ...... , ..... 1 l
i
I
.........j
I 1 "
= ' I ,o::t,
L L
лd
..01.)1 вф ЛW,
"он)108 оп'
I{ИnВЯИф :
=
1:
111
=
=
=
111
t"I
=
=
r::
:s::
f-<
о
(1)
I
I
t'--
t'--
1(1)
1.... I I
I g : I tn I
.: :С) SI...c::II:J....j Q)
U' [д ::3:!б" I i
О ! (1) о: (1) = '"5! 1::S о ;;.:
"'":';.S ,.... 'j .Е: I .Е: Q .
i \3 I . \3 .1 ' \3' Q)
O!.DI ]! t: с 1 сl
';'1.2i"f \31 j' З .
! 6i : '-.... '(5
I \3 j t: I i . \3 ..... ::S
I I I I"::: I 1...(:) c:u.... .... с
: ::S .... : с1 I ' ] g
I '1 I :;::I \:S. ..... ,
i:;": <:S...j . <:> Е r.SJ
. l l . !I 1 1 .
!
.... I '
f--04 Fl
I
1::
==
Ь е1
cu
м:
I
!
5:1
о,
С""),
cu'
I
I
I
I
I
! =
=
"
111
111
=
"
I:Q
00 0'1 О
0'1 0'1 О
...........-1
....: аё
......
c:u
::
.\:3
;:
;:; g8 13
N
u
О
I
1
I
.1
I
\:31
' I
.....1
I Е!
I I
\:31
"ll
1.91
...(:)1
\:3!
J'V)i
l
..с
с,)
<:
;s
Z .
I сп,
е' CU!
cu ,
cu и i
""""" сп И, , I
. CU:З , '
::: d cu CJ: . , I
;::.. 'Z: I F=! i
.;:: "': I i l'
, '3,
;:$ I t3.J i . ' i
..... O, ,' .
.S ;:$: 11 J :
......, ;::.., о "li
\:s . ; Е I ';:: I l' 'с;; I
::::: 10.., ;:$' ,)I;::" ::,
c:u ':.::::: '"" 1:)1 I
..... .-=! С): od '
..з .::! D I , \:s '1 ,
\:s ....,.;;;;...... '" ,
U: ! :k:lk:,
I""""\J
:
.........
Е
;:$
.;:
..з
c:u
Е:
;:$
.....
::::::
\:s
>ос)
r--... 00:0'1,0 .......i ,
о о о О,....... .......,.......,
N c:-I :.J"'tLN I '
,{d И)lнЛ:еdi I Т
Oll lчwdоф 1 ' i I
НElfЖ i F glt:r 5:! . :!ijt ш;!ij ap . . ;!iji :j:j I
t:: I : : 1 1 I I I
::s: ' jXI: E--I \о ' \о 11 \о \о f-<I
ь , : с,) f-< 5 5 t2 ,С,) 5 t;21 5
" , о ,О I ' о jXI , о ]
I а' ! I I I
. &3; 8t:: t3 s2 t:: t3 s2 t3 s2 I t3 s2
лd I ' 1 I I
ОJJlеф ЛW 8J о О: О : !S: j!S:: о о
..OH'r1'08 ои : f51 ..e- ..е- .е1..е- -&
u С,)' С,) U! С,) С,) о 8. о 01 О О О С,) О С,)
Ф , I ::':: р.. р.. р..
киnе)lИ ::s:::s: ,::s: :s: , I ::S: :s:
"И ВU)I : I ! : I I I
лdо.L I E--: I : u u u u u u u II I
..)lВф ЛW08: [ [ I ; ; [ I
, I I g1
,, иo Oll : I ..е- 1 ..е- I t=: I
Ф , I : ! I, О О О О I
КИnВ)lИ ,О! о О: О: О О О О ! O о
' l' '::S: :s:::s::s: I::: 1:::
..и еlf)l 1 I : ! I rr::: ('Ij ёа ('Ij 0\
I f6 1--4 r--
! 1 I
': I I
i I
I ! I
I I
I I
I' I
O
I ...jl
I Е I
\3,....) .....!
. ..........." .J:::
,;:::,...
1..... :О:::
...jl \:) \:$1
. I' I
.f:: I
I -Е
. зl
а .
. ;:: <51
V)I\C) r--- OQ
(".II(' (".1 (".1
.L (":-4 <:::!
u
Q с:::
I '....
;>1 О Д
! 1 .
r/) , l::s CQ
; . , . \3 : .:. \3
:.....J'.....J !€! Е
,.... :О::: 1 '0:0:::
: . ' t; .....: I..... \j
, :O:::"i:: . (....:....... а...
I .......1........' \3 V
' v ::s . I .
а... 1 , \3 а... I \::"
I ..... \3, \3
o.q;::S а... ь:о.....
: 8;....... ;::::
i ....., :::s 1.... ...
1 :::S \:) ' i
: i I ,
" I
'rr'I,-.::t V)'\C)'('--. OQ
! N J r;!
=
111
=
=
111
м
==
д
,
.1
. I
" ...' ::s
11....
...........
!
I
""CI':";::::
I
I \3
\31
I ::s
\3[
.....1....
с"} а...
'1 ?-..
v;
fп
CQ
\3,
:O:::i
. \3 I
.;::
.
а... :О::: -d .
\3 а... ::::: ff'
....... ..... ttI
с"} 8
::s . ё .
. .f::
::: \3
:О::: \3
а...
I
....... N I rr'I -.::t
(".INN(".I
C-I N
0"110
......1(".1
k-!
о
CIO
I I .1
I '""',
!с.) ! ,'0 ti
IQ I :;:
...... -:?1 :d ' "fi
-; -а ё: ,И
I I Q)I., Q):
v) I I IQ) ,1::1 .....!--;..
I JO::' <1 Q),. ,
'С I I : l' .:: '--"1" ' "о, Q):
О . l' 1 .." "" " ' .......... ,
u':9 ,'0 о ' ', . i 1
. QI .::: I I."" ", : . : i . ' I<'
Q), I , ,U, '1
dl х I I. ! !.I U ;
:i I' Q) . I C) t.O:I .1 I , Q) i.S::: i
= О .1:: .J:: . I .:.::: . ! . i С :::: 1
'(3 .2 I с .....: I I ., ' "fi I i
: t 1 е ;:s «j ' i 5'. ' t.O:I
I . со:) . 1...... 9J 5 i' S I , I 5:
I ! I 5 . Е .э Q1 t.o:Il I , : е !s
I == ' 5 ]I tl i 's' i. 1
t I 5 '.) 1::: 5' ::! ! ti ! '1
l '.) u cj cj 8 С ,;! . i C:c,j : i ic..;'
0\ С> ...... N rrll V)I\C) r--- 001 0\, С>1...... NlrrI IV) \о r---- 00
01 = N rrI rrI rrI rrI rrl1rrI rrI rrI'rrI I : I
N N N N1N N N N Nlc:-iLr-:J NW :N' :l'NIN
В нижеследующей таблице 8 ПРИВОДИТСЯ развернутый систе..
матический спектр rалофитов на уровне семейств.
Таблица 8
Общий систематический спектр raлофитов
Терско Кумской низменности
Bcero ra.. Факультативные Облиrатные
) лофитов rалофиты rалофиты
& !
о о I о,... O о
::Е о о I O t:1 j:I:I ol
:s: s (1)0 I (1) == (1)
Q.;j e e I s
o j:I:I о[ 0l
r::: 7 E-- 7 I E--:g 7, E-- I
-&;::r о r=: ::r I о..... ::rl о r=: I
: ?f. i?f. I ?f. J
L:]Piacea Ш 44 18'18 H 13'61 1
2. iAsclepiadaceae 4 1 25,00 1 25,Q :.
3. ,Asparaf!aceae 3 2 66,67 2 L 66,6L h
teraceae 11442 36,84 34 +29,82 8 I 7,02
5. 'I10ra iпaceae 25 7 28,00 7 ! 28,00
6. :]Jrassicaceae 70 15 21,43 12 L 17,IL 29 i
. PJ!!!!Ceqf 1 1 !OO,O L ш j p 100,OQ :- ,j
. Caryophyllaceae 38 7 18,42 3 L 7,89 10,53 i
9. 'Cheпopodiaceae 56 44 78,57 10 i 17, 34 60,71!
l 0 (' '!Y21v /!1cea 4 1 09 1 25,9Q ..
11 Cucurbitaceae 2 1 50,00 1 r 50,00
12.Cyperaceae 25 4 16,00 1 4,oo :J= !2:
Зрiрsасасеае 8 1 12,50 1 12, L -= I
4iElаеакпасеае 3 2 66,67 2 66,67 h
15'.Euphorbiaceae 12 4 33,33 3 25,OL !
16УаЬасеае 84 8 9,52 4 ! 4,76 4 4,76 I
i}jF qпk 'JJC!ff!i!e _ __ 2 H lQ_Q O Ш _ _ -: __ }QQj
8 Geпtiaпaceae 2 1 50,00 1 50,0<l1
19.Geraпiaceae 6 2 33,33 2 i 33,33
20';llеЛ trорiасеае 4 J 75,00 3 r 75,00 j 1
? 1 Jridaceae 5 1 20,00 1 I 20,00 .. .. I
:;2уипсасеае 9 2 22,22 t::: 22,221
:2 i ; ::ae . .. OJJ = шL 6 8H; Jl O ,()()J
N
Семейство
ПП,
81
113 [2 jl5,38 nf
1111 1100,00..
1 1 100,00 ..
6 3 50,00..
12349 39,84 26 21,14
01, опасеае 17 5 29,41 3 17,65
otaтoKetoпaceae2 1 50,00 1 50,00
1 T
riтulaceae 1 О 1 i 10,00 1 10,00.. I ..
апипсulасеае 24 1 4,17 1 4,17 i..
esedaceae 2 1 150 9 0 " j
34 1 2,94 11 2,94; ..
2 2 100,002 100,00 i ..
32 16 18,75 5 15,63! 1 3,13
4 3 75,00.. 3 I 75,00
7 11 14,29 1 14,29 I
1 11 100,00 1 100,00 I ..
2 2 100,00 2 100,00..
85024829,18 145 17,06 1Тi> iJ J2,12
15,38 [": ..
- - -
.. I 1 I 100,00
I ,
i 1 : 100,00
3 I 50,00
I 23 j ] 8,70
I 2 ]1,76
Если сделать выборку из таблицы 8 семейств ведущих по
числу видов, выстраивается следующий ряд (табл. 9). Из послед
ней таблицы видно, что Bcero видов ведущих семейств 180. К
ним относятся семействам факультативных rалофитов 102, а об..
лиrатных 78 видов, что, соответственно, 70,34% и 75 73% коли..
чества таксонов данных эколоrических rpупп.
Bcero ra..
лофитов
Таблица 9
видов и дов семейства
Фа льтативные i O THыe i
1%от I : %от !
Кол во общеrо i Кол"во i общеrо i
видов кол"ва I видов ; кол"ва :
: I I
видов : i видов J
6 21,14 i23 :I ,70 I
1 О 17,86 34 i60, 71
4 29,82 j'8 17.02
12 17,14 3 !4:29:
6 13,64 12 :4155
82
Вед щие по ЧИCJI
емей( о
49
44
42
15
8
i IФа л ; ив;:;ые Об; frатны
, Bcero ra.. ! r % o T O/ j
I Ф I Кол"во общеrо Кол"во I общеrо
ло итов
1 видов 1 кол..ва видов кол..ва
I : видов ИДQ
F !JCJc;e T l4 4 }6 t4,76 J
ora[{iпaceae !7 . 8,0( L 10,00
Caryp]!hY//{Jceae 7 3 7 89 4 1I О 53
Итоrо: 180 lQ J7oJi4 ,Z ji?:?\
Семейство
Сравнение численных данных таблицы 9 с аналоrичными
данными для флоры Терско..Кумской низменности в целом поз..
воляет выявить некоторые тенденции. К крупнейшим семействам
Терско..Кумской низменности, по нашим подсчетам из сводки
А.С. Абдурзаковой (2009), относятся следующие семейства: Ро..
асеае (12,04%), Asteraceae (11,150/0). Fabaceae (8,22%) Brassica..
сеае, (6,850/0), Cheпopodiaceae (5,48 ), Apiaceae (4,310/0), Laтi..
асеае (4,210/0), Caryophyllaceae (3,72%), Rosaceae (3,330/0), Scroph
lI/ariaceae (3,13%), Boragiпaceae (2.45%), Cyperaceae (2,45%).
Первое, что привлекает внимание, это rоловная тройка се..
мейств в сравниваемых списках. В остальной части спектров се..
мейств, хотя и имеются общие тенденции, их ранrи не Bcer да
совпадают. Из этоrо вытекает весьма важное заключение: не..
смотря на эколоrическую специфичность rаrюфитов, основное их
флористическое ядро принадлежит к семействам, определяющим
общую систематическую CTPYK'I)'PY флоры Приморской низмен..
ности, но с некоторым повышением роли полупустынных и пу..
стынных семейств (Cheпopodiaceae и Boragiпaceae).
В таксономической структуре исследуемой rалофильной
флоры на первый план ВЫС'I)'пают такие семейства как Роасеае
(49 видов), Cheпopodiaceae (44 вида), Asteraceae (42 вида). Одной
из ее характерных черт является незначительное число предста..
вителей F аЬасеае, Caryophyllaceae резкое снижение роли Laтi..
асеае, Rosaceae и Cyperaceae, хотя в общефлористическом списке
Терско..Кумской низменности Fabaceae принадлежит 3..е место,
Laтiaceae занимает 7..е место, CaryopJ,yllaceae и Rosaceae соот"
ветственно 8..е и 9..е места. Это, по нашему мнению, один из по..
83
казателей эколоrическоrо своеобразия rалофильной флоры, насе..
ляющей местообитания с заметным избытком солей по сравне..
пию с местообитаниями с обычным почвенным покровом.
ТаБЛllца 10
rоловнав часть спектра родов
Флора Терско..Кумской низмен.. : rалофиты Терско..Кумской низ..
I
ности менности
Д к..во видов К..ВО вид
straKa/us 11 _
Veroпica 14 7
Tri оиит 13 7
,Carex 11 6
rteтiSia 10
Vicia 1 О
Oroballche 9
пою 9 4
"; /ех 9 3
edica о 9 3
Ceпtaurea 19 corzoпera 3
Справедливости ради следует указать, что, кроме представ..
ленных в таблице 3..х видовых родов rалофитов имеется еще не..
сколько таких же. Однако и среди них также нет родов, общих со
списком rоловной части общей флоры. По существу в списках
таблицы 1 О Bcero четыре общих рода (Trifo/iuт, Arteтisia,
Atrip/ex, Ceпtaurea). Все эти роды в масштабах Предкавказья от..
личаются высоким видовым разнообразием, а в общеIU1анетарном
масштабе к каждому из них относится по несколько десятков, в
то и несколько сот видов.
Исследуемые нами rалофиты, будучи составной частью фло..
ры Терско--Кумской низменности, в своих флористических ком..
плексах проявляют отличительные признаки таксономическоrо
спектра. В первую очередь это внедрение в первую троЙку се..
мейств Cheпopodiaceae (между Роасеае и Asteraceae). Высокое
положение в составе ведущих семейств Cheпopodiaceae указыва..
ет на флороrенетическую связь rалофильных КОМШIексов с фло..
рами Т уранской флористической подобласти.
84
АНАЛИЗ эколоrИЧЕСКИХ rР}'Пll rАЛОФИТОВ
ТЕРСКО..КУМСКОЙ НИЗМЕННОСТИ
rалофиты, объединены в одну эколоrическую rруппу по при..
:знаку химизма почвенноrо субстрата, на котором они произрас
тают. Именно химизм (избыточность солей) местообитаний ra..
лофитов, видимо, является одним из rлавных, лимитирующих их
флоро и фитоценотическую сущность.
Во флоре Терско Кумской низменности по отношению к за
соленности можно выделить 5 типов rалофитов видов: эуrалофи..
ты (28 видов), КРИНОI'алофиты (27), П1Лотолеранты (109), rлико"
фиты (84), rалофобы. Если у первых четырех типов выработан
определенный набор адаптаций к существованию на засоленных
почвенных субстратах, то у пятой таких приспос,облений нет. Ви
ды последнеrо типа просто не встречаются в засоленных место..
обитаниях и, потому не имеют отношения к нашему исследова
нию. По фактору водноrо режима среды обитания выделяются
rидрофиты, мезофиты, rемиксерофиты. склерофиты и суккулен"
ты. Данные классификации в известной степени условны. На ca
мом же деле rалофильные и rалофобные тенденции, равно как и
отношение к водному режиму, видов представляют некий конти..
НУУМ, что предполаrает наличие между этими основными типами
переходных элементов. Их наличие обусловливаt."Т эколоrиче
скую пластичность компонентов растительных сообществ и
сложные отношения последних между собой. Количественное
соотношение rалофитов Терско..Кумской низменности (табл. 11)
свидетельствуют о хорошо выраженной неравномерности рас..
пределения видов по эколоrическим rруппам.
Эуrалофиты виды, нуждающиеся в избыточном засолении,
т.к. для их оптимальноrо онтоrенеза, прохождения всех Bereтa
ционных и фенолоrических стадий и ФИЗИОJlоrических процессов
требуется наличие в почве определенноrо количества хлоридов и
сульфатов. Эта эколоrическая rруппа накапливает соли в своих
клетках.
85
Таблица 11
Количественные Dоказатели эколоrических rруп"
I I
со
::s:: i S
l::iQ r::: l::iQ
Эколоrические о l::iQ '\0 (,) О
r::: о ,О ;s:
rруппы (,) 1::( Е- ::r
,О О l::iQ
I " с>
о'
28
27
1100
Эуrалофильных видов среди rалофитов исследуемой терри..
тории насчитывается 28: Bassia sedoides. Bassia hyssopifolia, Ко..
chia /aпiflora, Kochia prostrata , Кalidium caspicum, Kalidium jolia
(ит, Ha/ostachys caspica, На/оспетит strobilaceum, Sa/icorпia eи
ropaea, Suaeda microphylla, Suaeda physophora, Suaeda a//issima,
Suaeda prostrata, Suaeda salsa, Sa/so/a soda, Sa/so/a ericoides,
Sa/so/a deпdroides, Sa/so/a australis, Sa/so/a pau/seпii, Sa/so/a
/ariciпa, Sa/so/a iпcaпesceпs, C/imacoptera crassa, Aпabasis aphylla,
Petrosiтoпia oppositifo/ia, Petrosimoпia triaпdra, Petrosimoпia bra
chiate, Petrosimoпia g/aucesceпs, Caki/e euxiпa.
Криноrалофиты rруппа видов, которые также нуждаются в
избыточном засолении почвоrpунтов. Но в отличие от предыду
щей J'рУППЫ, они избыток солей удаляют из cBoero орrани эма че
рез спе:циальные солевыделяющие железки.
Соли MOryT удаляться с опадающими листьями. У некоторых
растений избавление от избытка солей происходит без поr лоще..
ния больших количеств воды, так как соль выделяется в вакуоль
клетки rоловки листово['о волоска с ПОС.1едующим ее обламыва
нием и восстановлением ero. В нашем списке это относительно
небольшая rруппа растений (27 видов), соизмеримая по числен
ности с предыдущей: Asparagus bres/eraпus, Asparagus persicus,
Atrip/ex joтiпii, Atrip/ex aucheri, Ha/iтioпe verrucifera, Caт
86
phorosтa lessiпgii, CaтpllOrOSтa тoпspeliaca, Suaet/a coпfusa,
Suaeda deпdroides, Sa/so/a daghestaпica, Gaтaпthus pilosus, Nitrar
ia cajpica, Fraпkeпia hirsule, Fraпkeпia pu/veru/ellta, Reaumuria a/
terпifo/ia, Tamarix raтosissiтa, Taтarix тeyeri, Feru/ago daghe
staпica, Goпioliтoп tataricuт, Limoпiuт meyeri, Limoпiuт caspium,
Liтoпiuт platyphyllum, Psylliostachys spicata. P/aпtago sa/sa, P/aп
tago teпuiflora, P/aпtago СOl'опориs, Lactиca saligпa.
rликофиты растения, обладающие соленепроницаемостью,
Т.К. высокое осмотическое давление в их клетках попдерживается
за счет продуктов фотосинтеза. Клетки зоны всасывания их KOp
ней малопроницаемы для солей за счет BblcoKoro содержания в их
протоплазме уrлеводов, орrанических кислот, выполняющих ба
рьерную функцию. Это мноrочисленная rруппа из числа rалофи..
тов, насчитывающая 84 вида (1/3 видовоrо состава). Сюда OTHO
сятся такие виды как Jmperata cyliпdrica, Heleoch/oa schoeпoides,
Po/ypogoп moпspelieпsis, Gaudiпopsis macra, Cyпodoп (/acty/oп,
Phragmites austra/is, Spheпopus divaricatus, Agropyroп fragile, Er
eтopyruт orieпta/e. Polygoпum sa/sugiпeuт, Cheпopotliuт poly
sperтuт, Dichodoп 1'isciduт, Spergularia тaritima, Sperb:тu/aria 1'и"
bra, Lepidiuт crassifoliuт, Astraga/us hyrcaпus, G/ycyrrhiza g/abra,
Praпgos odoпta/gica, Feru/a tatarica, Coт'O/vu/us persicus, Argusia
sibirica, Dodartia orieпtalis, Galatella pastuchovii, Tripoliuт vul
gare, Arteтi. 'ia tschernieviaпa, Arteтisia austriaca, Serratula eruci
folia, Acroptiloп repeпs, Ceпtaurea caspia, Scorzoпera parvijlora и
др.
rалотолеранты это виды, как правило, с широким диапазо
ном эколоrических приспособлений обеспечивающих возмож"
ность произрастания в ра,знообразных условиях. Они MOryT до..
вольно успешно расти в разнообразных условиях. В численном
отношении rалотолеранты преобладают над предыдущим эколо--
rическим rруппами, а в формировании растительноrо ПОКlюва
Терско..Кумской низменности в целом принимают более активное
участие. К данной rруппе нами отнесены следующие виды: Zaп
"ichellia pa/ustris. Botrhiochloa ischaeтuт, DigUaria saпguiпalis,
C/eistogeпes squarrosa, CYl10surus echil1atus, Рои buZbosa, Festuca
1'a/esiaca, Carex тe/aпostachya, /ris pseutloпotha, Fa//opia coпvol
'I'U/US. Po/ygoпuт argiroco/eoп, Cheпopodiит g/aucuт, Atrip/ex
ca/o/heca, Atrip/ex tatarica, Psaттophiliella тuralis, Cardaria
87
draba, Lepidium /atifolium, Lobu/aria тaritima, Ery.5imum /еисап..
themuт, Capparis herbacea, Reseda /utea, Trifo/ium boпaппii, Zygo..
phylluт fabago, Euphorbia boissieraпa, Ma/va/thaea traпscaucasica,
Hibiscиs trioпuт, Elaeagпus aпgustifo/ia, Fa/caria vu/garis, G/aux
maritiтa, Roche/ia retorta, Sa/via tesquico/a, Verbascum blattaria,
Criпitaria /iпosyris, Filago eriocepha/a, /пи/а oculus..christi, Pи/icar..
ia u/igiпosa, Pu/icaria 'u/garis, Seпecio schischkiпiaпus, Juriпea
mu/tij1ora, Oпopordum acaпthium, Ceпtaurea so/stitia/is, Ceпtaurea
areпari'a, Carthamus /aпatus, Hedypпois persica, Garhadio/us рарро..
.5иs, Podospermum ca/citrapifo/ium, Scorzoпera mollis, Soпchus
arveпsi.5, Soпchus pa/ustris, Lactuca tatarica, Lactuca a/taica. Tarax..
асит, Crepis mar.5challii. и др. (Bcero 109 видов).
Нами здесь выделены основные rpуппы rалофитов, проявля..
ющих те или rалофильные и rалофобные тенденции. На самом же
деле эти тенденции MOryT быть представлены в виде HeKoero кон..
тинуума. Поэтому спектр этот весьма широк, и между основными
rруппами, несомненно, имеются переходные элементы. Их нали..
чие обусловлено взаимодействием различных растительных со..
обществ между собой, а также эколоrической IU1астичностью их
компонентов.
С точки зрения приспособленности rалофитов к условиям
Терско..Кумской низменности и эколоrическим условиям место..
обитаний несомненный интерес представляет анализ видов по
сопряженности признаков указанных выше двух классификаций.
Картину сочетания признаков эколоrических rpупп rалофнтов по
солевому и водному режимам выявляется данными таблицы 12.
Таблица J 2
Численность комбинативных rрупп rалофитов
Те Ko K мской низменности
Криноrалофи.., : r алотолеран.. :
Экол. rруппы Эуrалофиты rликофиты: I
ты: : ты i
Мезофиты 3 24 :29 '
r ты 112
rемиксерофи.. 10 123 45 l
ты
С ерофиты 2
Суккуленты 26
3 i 2 1Q
J 1
88
Данная таблица показывает, что наиболее адаптированными к
современным условиям водноrо и солевоrо режима района ис..
следований являются три типа сочетания признаков. Предпочти..
тельной является rемиксерофильность с признаками толерантно
ro отношения к солевому режиму. Несколько менее блarоприят"
ны комбинации признаков, КОI'да склерофильность проявляется
на том же фоне толерантноrо отношения к солевому режиму или
же склерофильность сочетается с непроницаемостью клеток зоны
всасывания для солей за счет BblcoKoro содержания в их прото..
плазме уrлеводов, орrанических кислот, выполняющих барьер--
ную функцию.
89
АНАЛИЗ rЕоrРАФИЧЕскоrо РАСПРОСТРАНЕIIИЯ
r АЛОФИТОВ ТЕРСКО..КУМСКОЙ IШЗМЕННОСТИ
Ни один вид никоrда не занимает IU10Щади cBoero ареала
СIU10ШJ>. Эrо связано с тем, что даже в небольшом, относительно
однородном rеоrрафическом районе не наблюдается полной вы..
равненности эколоrических условий. Но ареалы растений, как
правило, охватывают значительные части суши со сложной топо..
rpафией и массой разнообразных местообитаний. В то же время
каждый вид нуждается в cтporo определенном местообитании.
Поэтому пестрота физико--rеоrpафических условий на территории
ареала неизбежно порождает прерывистое распределение особей
и популяций вида и, следовательно, можно rоворить о тополоrии
вида на IU10Щади ero ареала (Шумилова, 1979).
В основу системы rеоrрафических элементов rалофитов Тер..
ско..Кумской низменности положена схема, предложенная А.Л.
Ивановым (1998, 2001). Понятие rеоrрафический элемент связы..
вается с фитохорионами различных paHroB провинциями, обла..
стями, под царствами и царствами, Т.е. собственно rеоrрафиче..
ские элементы Toro или иноrо фитохориона виды флористиче..
cKoro ядра данноrо хориона. Ареалы ряда видов или даже центры
обилЮl часто не совпадают с ботанико..rеоrрафическими обла..
стями. В данном случае, такие виды не M0rYТ быть отнесены к
какому..либо элемеН1)', вследствие чеrо возникает необходимость
введения понятия rpупп связывающих видов. Соrласно Eig (1931)
виды этих rрупп более или менее распространены в двух И..1И бо..
лее со(;едних реrионах (исключая виды, ареалы которых BbIC'I)'''
пают за пределы cBoero OCHoBHoro фитохориона в виде иррадиа..
ции).
В (:истеме понятий современной флористики, rеоrрафические
элементы являются «общими хориономическими rеоrрафически"
ми элементами, отражающими положение ареала (или ero части)
в системе выделов природноrо, комплексноrо ботанико..
rеоrрш ическоrо районирования Земли или территории флоры.
При данном подходе каждый элемент флоры характеризуется
набором соответствующих вьщелов районирования, а иерархиче..
ская классификация элементов строится на соподчинении этих
выделов» (Юрцев, Камелин, 1991, с. 43).
90
В разнообразии rеоrрафическоrо распространения видов ra..
JlОфИЛЬНОЙ флоры Терско..Кумской низменности нами выделено
22 rеоrрафических элемента, их спектры приведсны в таблице 13.
Таблица 13
rеотипы и rеоэлементы rалофитов Терско..Кумской
низменности
r еотипы Кол.. во % rеоэлементы Кол.. во %
видов видов
ПЛЮРИР f'ио.. 11 4,44 Плюриреrиональный 11 4,44
пальпыи
Общеf'оларк 49 19,76 rоларктический 6 2,42
тический Палеарктический 43 17,34
Евро..сибирский 5 2,02
Евро--кавказский 2 0,81
Европейский 3 1,21
Кавказский 11 4,43
Общекавказский 2 0,81
Бореальный 39 15,73 Эукавказский 4 1,61
Восточнокавказский 4 1,61
Даrестанский . .
1 0,40
Понти чес ко.. 8 3,23
южносибирский
Понтический 10 4,03
Общедревнесреди.. 32 12,90
земноморский
Западнодревне 12 4,84
средиземноморский
Древпесреди.. Средиземноморский 4 1,61
108 43,54 Восточнодревне..
:Jемноморский средиземноморский 14 5,65
Ирано..тураНСЮiЙ 20 8,06
Армено.. иранский 4 1,61
Туранский 21 8,47
Предкавказский 1 0,40
91
r еотипы Кол.. во % r еоэлементы Кол.. во %
видов видов
Субсредиземно-- 4 1,61
морский
Связующий 38 15,32 Субкавказский 3 1,21
Субпонтический 4 1,61
Субтуранский 27 10,89
Адвен'rивный 1 0,40 Адвентивный ] 0,40
итоrо: 248 100 248 100
1. Плюриреrиональный rеотип и соответствующий ему reo..
элемент состоит из видов, более или менее распространённых в
двух и более флористических царствах Земли и выходящих за
пределы rоларктическоrо царства.
Во флоре rалофильных комплексов Терско..Кумской низмен..
ности таких видов выявлено 11 вида (4,44%): Po/amoge/on пa/ans,
Ruppia maritima, Ruppia spiralis, Zannichellia pa/ustris, Digitaria
sanguiпalis, Phragтites aиstralis, Chenopod;um g/aucum, Spergu/ar..
;а mar;na, H;biscиs tr;onuт, P/aпtago salsa, Sonchиs arvens;,s. Виды
данноrо rеоэлемента характеризуются водной и водно..болотной
эколоrией.
В общеrоларктическом rеотипе вьщеляются два rеоэлсмента.
Это ] оларктические и палеарктические rеоэлементы. Bcero ви"
дов общеrоларктическоrо rеотипа 49, или] 9,760/0.
2. rоларктический. Виды из этоrо rеоэлемента, встречаются
на территории не менее трех под царств rоларктическоrо царства.
Их во флоре rалофитов Приморской низменности насчитывается
6, что составляет 2,42%. Это такие виды как Anisantha tectorum,
Salicorпia europaea, Glallx тar;tima, Tripoliuт vu/gare, Taпacetuт
vulgare, Artem;s;a caтpestr;s.
Виды rоларктическоrо элемента относительно равномерно
распределены по различным вариантам растительных сообществ,
однако преобладают таксоны, предпочитающие относительно
мяrкий влажный режим сообществ.
3. Палеарктический rеоrpафический элемент образуют ви"
ДЫ, ареалы которых охватывают умеренные и субтропические
92
области rоларктическоrо царства CTapOl o Света без определён"
ной приуроченности к одному из подцарств. Bcero таких видов в
исследуемой флоре насчитывается 43 (17,34%), среди которых
довольно MHoro сорняков (Stipa capillala, He/eochloa schoenoides,
He/eoch/oa a/opecuroides. Роа bu/bosa. Puccinellia distaпs, Festuca
regeliana. Fesluca va/e. 'iaca, Carex divisa, Carex me/anostachya,
Fallopia convo/vu/us, Fallopia duтetoruт, Po/ygoпum patu!uт,
Atrip/ex ca/otheca. Afrip/ex nitens, Atrip/ex obIongifolia, Atrip/ex (а..
/arica, Kochia /anijlora, Suaeda prostrata. Elisanthe viscosa, Cardar..
ia draba, Lepidium /atifo/ium. Lepidiuт perfolialum, Lepidium ru..
tlera/e. Arabidopsis Ilшliапа, Meniocus linifolius. Potentilla recta,
Trigonella procllтbens. Trij'olium .fragiferuт. Euphorbia helioscopia,
Cauca/us /арри/а, Fa/caria vu/garis. Ar,gl4sia sibirica, Lappula squar..
rosa, C)nog/ossum ofjicina/e. Verbascuт bIattaria, Plantago (епифо..
ra, Filago arvensis, Pulicaria vu/garis, Achillea sefacea. Onoporduт
acanthiuт, Acroptilon repens. Scorzonera parvijlora. Lactuca tatari..
са).
Бореальный rеотип разнообразен по rеоэлементам. Он объ..
единяет 39 видов, или 15,73 % их общеI О состава. Виды данноrо
f'еотипа распределяются на 6 rеоэлементов.
4. Евро сибирский элемент образуют виды с распростране..
нием в евроазиатской части Циркумбореальной области. Из
наших видов к нему относятся 4. Виды рассматриваемоrо эле..
мента характеризуются более менее выровненным распростране..
нием в ксерофильных сообществах. К евро..сибирскому rеоэле..
менту отнесены Aneuro/epidiuт raтosuт. Chenopodiuт
l'o/ysperтuт, Spergu/aria rubra, Psaттophi/iella тuralis. Sonchus
l'a/lIstris.
5. Евро кавказский rеоэлемент. Виды данной rруппы широ..
ко распространены на Кавказе и Европейских провинuиях Евро"
Сибирской области. Bcero этот элемент прсдставлен 2 видами
Cerastiuт g/utinosuт. Jnu/и gerтanica.
6. Европейский rеОЭJ1емент объеlЩНЯ(,,"Т виды, распростра..
ненные в ос..новном в умеренных частях Европейских провинций
А.Л. Тахтаджяна (1978) Атлантическо..ЕВРОl1ейской, Северо..
Европейской, Центрально..Европейской и Восточно..Европейской,
проникая в Кавказскую провинцию. Это немноrочисленный по
93
общему количеству видов элемент бореальноrо reoтипа. Bcero 3
вида: Atrip/ex rosea, Verbascuт ph/oтoides, Lactuca saligпa.
7. :Кавказсквй. К данному элемеН1)' относятся виды. харак..
терны е: для Кавказской провинции. Их ареал может охватывать
весь Кавказ или какую"то ero часть. Кавказский rеоэлемент мы
подразделяем на общекавказкий, эукавказский, восточнокавказ..
ский и даrестанский. Первые из них распространены по террито..
рии Bcero Кавказа. Таких видов Bcero 2 (Noпea rosea, Verbascum
/ахит). Вторые же, которых насчитывается 4 (Sa/so/a dag/1estaпi..
са, E/aeagпиs caspica, Feru/ago daghestaпica, Seпecio schischkiпia..
пиs), оrpаничены в своем распространении областью БО.lьшоrо
Кавказа. Восточнокавказские распространены восточнее долины
Терека (Pucciпellia bu/bosa, Astraga/иs hyrcaпus, Ceпtaurea
сшрiа). Дarестанские оrраничены в своем распространении тер..
риторией Дarестана (Suaeda physophora)
Таким образом, суммарно кавказСЮfЙ rеоэлемент включает
Bcero 1 1 видов. Здесь важно отметить, что территориально сосед"
няя и достаточно крупная rорная система Кавказа оказала почти
незамеrное влияние на формирование ф.10рЫ изучаемой равнин..
ной области, т .К. виды кавказскоrо элемента в сумме сосrdВЛЯет
4,430/0 видовоrо состава флоры rалофитов Приморской низменно..
сти. Этот момент, заслуживающий внимания в том отношении,
что область равнин Восточноrо Предкавказья во флористическом
плане имеет иные rенетические корни, чем rорная часть Большо..
ro Кавказа в целом и Восточноrо Кавказа в частности.
8. Заметное место среди бореалЬНОI'О rеотипа принадлежит
ПОНТВ1Jеск.южносвбирскому элемен1)'. Он включает виды, ха..
рактерные Понтической провинции А.Л. Тахтаджяна (1970), или
южным частям Восточно--Европейской и Западно..Сибирской
провинций А.Л. Тахтаджяна (1978), или Евразиатской степной
области Е.М. Лавренко (1950, 1970). Из нашеrо списка к понти"
ческо..южносибирскому rеоэлементу относится 8 видов:
C/eisto;geпes bu/garica, Triпia hispida, Goпioliтoп tataricuт,
Criпitaria liпosyris, Achillea micraпtha, Pyrethrum achilleifoliuт,
Arteтisia austriaca, Juriпea тultijlora. Причем преобладающее
большинство из них, будучи адаптированными к условиям дефи..
цита влаrи, широко распространены в ксерофильных сообще..
ствах IIриморской низменности.
94
9. Понтический rеоэлемент образуют виды, распространен..
ные в степных и лесостепных районах Восточноевропейской
провинции. В нашем списке видов этой rруппы насчитывается
1 о: Puccinellia j'oтinii, Agropyron desertoruт, Astrodaucus littoralis,
Liтoniuт p/atyphylluт, Marrubiuт peregrinuт, Marrubiuт
руаесох, Scabiosa ucrainica, Galatella pastuchovii, Arteтisia
tschernieviaпa. Все они характерны для песчаных степей или сы..
пучих песков. Некоторые из названных видов проникают в сооб..
щества колючекустарниковых зарослей или полупустынные це..
нозы, rде эдификаторами выступают Arteтisia austriaca или
Arteтisia santonica. В целом следует подчеркнуть ксерофильную
природ)' таксонов понтическоrо rеоэлемента.
Древнесредиземноморский rеотип объединяет 108 видов, или
43,54% их общеrо состава. Таким образом, по их количеству во
флоре rалофитов Приморской низменности он является самым
мноrочисленным, что еще раз подчеркивает роль средиземномор"
ских флор в флороrенетической судьбе Приморской низменно..
сти. Все общеrеоrрафическое разнообразие распространения ви"
дов данноrо rеотипа можно свести к 8 rеоэлементам.
10. Общедревнесредиземноморский rеоэлемент из древне..
средиземноморскоrо rеотипа по количеству видов наиболее боrа..
то представлен. К нему относятся наиболее широко распростра..
нённые виды средиземноморской rруппы, ареал которых охваты..
вает Средиземноморскую и Ирано.. Тура нскую области Древне..
средиземноморскоrо подцарства (Тахтаджян, 1978). Bcero таких
видов в исследуемой флоре насчитывается 32 (12,90%), среди ко..
торых преобладают таксоны с ксерофильными аУТЭКОJТоrически"
ми особенностями (1тperata cylindrica, Botrhioch/oa ischaeтuт,
Polypogon тonspeliens;s, Trisetaria lo.llingiana, Cynodon dacty/on,
Echinaria capitata, Aeluropus litt01'alis, (ynosurus echinatus,
Parapholis incurva, E/ytrigia, Еуеторууит orientale, Camphorosтa
тonspeliaca, Bassia hyssopifolia, Kochia prostrata, Halocneтит
strobilaceuт, Sa/so/a soda, Salsola australis, C/eтlltis orieпtalis,
Chorispora {епеllа, Trifoliuт resupinatllт, G(vcyr'rhiza g/abra,
Peganuт harтala, Zygophylluт jabago, Tribu/us terrestris,
Euphorbia falcata, Elaeagпus angиst!folia, Cenlauriuт spicatuт,
Heliotropiuт ellipticuт, Lappula patиla, Plaпtago соуопориs,
Pulicaria ul;ginosa, Carthaтus ianatus. )
95
11. Достаточно широко представлены во флоре TepcKO
Кумской низменности виды заоаднодревнесредиземноморскоrо
rеоэлемента (13 видов, ми 4,84%). Виды, относимые к данному
rеоэлемеН1)', имеют ареалы, охватывающие всю Средиземномор
скую область или её большую часть и на востоке зачас1)'Ю про
никают В западную часть Ирано.. Туранской области '- Tragus
raceтosus, Crypsis acu/eata, A/opecurus тyosuroides. Juпcus
тariliтus, Reseda lutea, Fraпkeпia hirsuta, Fraпkeпia pu/veru/eпta,
Va/eriaпe/la puтi/a, ЕсЬаliит e/ateriuт, Filago eriocepha/a. Fi/ago
vulgaris, lпula oculus..christi).
12. Средиземноморский rеоэлемент объединяет виды, ареа..
лы которых охватывают две и более провинции Средиземномор"
ской области. Bcero в нашем списке 4 (1,61 %) видов относящихея
К данному reоэлемен1)' (Cutaпdia тeтphitica, Lobu/aria тaritiтa.
Trifoliuт echiпatuт, Podosperтuт ca/citrapifo/iuт).
13. Восточнодревнесредиземноморский rеоэлемент форм и..
руют виды, ареалы которых охватывают Переднеазиатсk.'УЮ и
Центральноазиатскую подобласти Ирано.. Туранской области.
Bcero видов насчитывается 14 (5,66%): Gaudiпopsis macra,
Spheпopus divaricatus, Hordeuт /eporiпuт, Hordeuт glaucuт,
Atriplex aucheri, Ha/iтioпe verrucifera. Bassia sedoides, Capparis
herbacea, Татауа raтosissiтa, Taтarix тeyeri, Соп-lюlvи/иs
persicus, Arteтisia .fragraпs, Hedypпois persica, Scorzoпera pusi//a.
Таксоны восточнодревнесредиземноморскоrо rеоэлемента более
или менее равномерно распределены между полупустынными и
степными сообществами, с некоторым пере весом в сторону по..
следних.
14. Иран.туранский rеоэлемент включает в себя виды, xa
рактерные для Ирано-- Туранской области и встречающиеся в двух
и более ее провинциях. Таких видов во флоре rалофитов Терско"
Кумской низменности 20 (8,06%): Aegilops tauschi, Kalidiuт
caspicuт, Suaeda тicrophylla, Suaeda a/tissiтa, Sa/so/a dent1roides,
Sa/so/a iпcaпesceпs, Aпabasis aphylla, Petrosiтoпia brachiata.
Gaтaпthus pilosus, А rabidopsis puтi/a. С/урео/а jOllth/aspi,
Erysiтuт /eucaпtheтuт, A/hagi pseuda/hagi. Nitraria caspica.
Chaтaesyce caпesceпs, Psylliostachys spicata. Lappu/a barbata,
Dodart;a orieпtalis, Buпgea trifida. Ceпtaurea so/stitia/i. Виды дaH
96
Horo rеоэлемента тяrотеют к сообшествам, Формирующимся в
условиях дефицита влаrи на песчаных субстратах.
15. Армено..иранский rеоэлемент охватывает виды с ареала
ми в Армено Иранской провинции Переднеазиатской подобла
сти. Эта rруппа (4 вида, или 1,61 %), указывающая на rенетиче
ские связи флоры Приморской низменности с флорами Передней
Азии: Erodiuт turcmeпuт, Euphorbia hoisJ'ieraпa, Ма/vа/tJшеа
/raпscaucas ica, Garhadio/u papposus.
16. Туранский rеоэлемент в исследуемой флоре как и следо..
вало ожидать, представлен большим количеством видов (21 ви"
дов, или 8,470/0). К нему отнесены виды, ареал которых локализу
ется в пределах Туранекой провинции. rлавным образом это ви
ды ксерофильной и вместе с тем rалофильной эколоrии (Stipa
caspica, Pucciпellia poecilaпtha, Pucciпe/lia gfgaпtea, E/ytrigia,
.Iuпcus littoralis, Aspara!,"ТUs Ьrеs/еrапиs, Asparagus persicus,
Po(vgoпuт argiroco/eoп, Po/ygoпum salsugiпeиm, Ceratocarpus
иtriculosus, Caтphorosтa /essiпgii, Kalidium fo/iatuт, HalostacllYs
caspica. Suaeda deпdroides, Suaeda salsa, Salso/a pau/seпii,
Climacoptera crassa, Petrosiтoпia glaucesceпs, Toru/aria
coпtortuplicata, Liпaria iпcompleta, Ceпtaurea arellaria.).
17. Предкавказский rеоэлемент образуют виды, эндемичные
или субэндемичные для Предкавказья и, соответственно, apeall их
полностью локализован в пределах предкавказских равнин или
же основная часть их ареалов тяrотеет к данной области. Друrи
ми словами предкавказские элементы характеризуют ориrиналь
ность исследуемой флоры. Представлен лишь I видом (Atrip/ex
jomiпii).
18. Субсредиземноморский rеоэлемент. Orносимые к этому
rеоэлементу виды более или менее равномерно распространены в
северных и северо восточных районах Средиземноморской обла
сти и в юrо западных районах Евро Сибирской области. Это Ta
кие виды, как: Elytrigia ol)tusijZora, llordeuт geпiculatuт. Dicho..
сl0п viscitiuт, Trifolium boпaппii. Erodiuт cicoпiuт, Cyпaпchum
асutит.
19. Субкавказский rеоэлемент объединяет связующие виды,
основная часть ареалов которых охватывает Кавказскую пронин
ЦИЮ, а также часто Эвксинскую провинцию Евро Сибирской об
ласти и Армено..Иранскую провинцию Ирано Туранской области.
97
Он прсдставлен 3 видами Sa/so/a ericoides, Noпea /utea, Crepis
тarschallii.
20. Субпонтический reоэлемент объединяет связующие ви"
ДЫ, основная часть ареалов которых находится в степных илесо..
степных районах Восточно--Европейской и преимущественно за..
падных районах Эвксинской провинции Евро..Сибирской области
и в восточных районах Иллирийской, в Центрально..
Анатолийской и Восточно..СредиЗемноморской провинциях Сре..
диземноморской области. Общее число видов 4: Sa/so/a /ariciпa,
Heliotropiuт suaveo/eпs, Scorzoпera тollis, Тауахасит serotiпuт.
21. Субтуранский rеоэлемент охватывает связующие виды,
ареалы которых "риурочены к лесостепной и степной части Во..
сточно..Европейской и Западно..Сибирской провинций Евро"
Сибирской области и Туранской провинцию Ирано-- Туранской
области. Общее число видов 20: C/eistogeпes squarrosa, Eragrostis
co//iпa, Ае/иуорus puпgeпs, Pucciпe//ia dolicho/epis, Pho/iurus рап..
пoпicus, Еуеторууит triticeuт, Aegi/ops cy/iпdrica, Schoeпop/ectus
/itoralis, E/eocharis uпig/итis, lris pseudoпotha, Suaeda coпjitSa, Pe
trosiтoпia oppositifolia, Petrosiтoпia triaпdra, Spergu/aria
тaritiтa, Lepidiuт crassifo/iuт, Syreпia silicu/osa, Reauтиria a/
terпifoJia, Praпgos odoпta/gica, Feru/a caspica, Feru/a tatarica, Li
тoпiит тeyeri, Liтoпiuт caspiuт, Rochelia retorta, Sa/via tesquico
/а, SerJ-аtulа erucifolia, Tragopogoп daghestaпicus, Lactuca a/taica.
Эти виды в растительном покрове исследуемой территории в
большинстве своем типичные представители ксерофильных со..
обществ.
22. Адвентивный rеоэлемент преДСfавлен заносными вида
ми. Из таких среди rалофитов Терско..Кумской низменности от..
мечен только Xaпthiuт califoпlicuт
98
БиоморФолоrИЧЕСКИЙАНАлизrАЛОФИТОВ
TEPCKO..KYl\-IСКОЙ НИЗМЕIПIОСТИ
По Раункиеру (Raunkiaer, 1934) жизненные формы склады
ваются исторически и отражают результат структурно..
морфолоrических адаптаций растений к сеЗОННО ЮlИматическим
условиям. Им было показано распределение жизненных форм в
разных зонах и областях Земли, в виде так называемых биолоrи
ческих спектров, являющиеся индикаторами климата.
Биоморфолоrическая при рода флоры rалофитов Терско"
Кумекой низменности определяется спецификой жизненных
форм. Состав биоморф отражает особенности эколоrических
условий в ходе формирования и развития флоры Tepck()-оКумской
низменности.
Выполненная нами инвентаризация состава жизненных форм
rалофильных комплексов Терско..Кумской низменности ПОfШзала
следующее их распределение по основным rруппам системы Pa
ункиера (табл. 14).
Таблица 14
Спектр жизненных форм raлофитов Терско Кумской
lIизменности
РЬ СЬ Hk
7 21 107
2,8 8,4
2 7
0/0 участия
j
кт Tr
9 105
3,63 42,34
Жизн. О ма
Число видов
43,14
Как показывают данные таблицы, среди rалофитов TepCKO
Кумекой низменности исследований примерно одинаково пред..
ставлено обилие жизненных форм Hcmicryptophyta и TIlerophyta.
'Эти две rруппы резко преоб.:lадают над остальными
(Phanerophyta, Chamaephyta, Kryptopllyta). Такой раСЮIaД биоло..
rическоrо спектра обнаруживается на равнинах Арало
Каспийских или Туранеких пустынь. На эту особенность, xapaк
терную для пустынных ценозов указывал еще А.А. rроссrейм
(1948). Травянистые растения в целом (Hemicryptophyta, Krypto..
phyta и Therophyta) составляют подавляющее большинство rало..
фитов Терско Кумской низменности 221 вид (89,11%). Они в
99
растительнь комплексах rалофитов Приморской низменности
чаще ВЫС'I)'пают в роли эдификаторов.
Эколоrо..rеоrpафическая природа флоры во MHoroM опреде
ляется спецификой жизненных форм. Биоморфа (жизненная фор..
ма) оч)ажает способность растений выживать в неблаrоприятных
ДЛЯ них сезонно"климатических условиях. По составу жизненных
форм можно определить эколоrические условия, в которых про
исходило историческое формирование и развитие флор. Напри..
мер, известно, что наибольшее количество rемикриптофитов
встречается в холодных и умеренных областях (Толмачев, 1974).
Ра(:клад жизненных форм rалофитов по солевому режиму по
казан в таблице 15. Из данной таблицы видно, что наименьшим
разнообразием жизненных форм характеризуются эуrалофиты,
предст.авлены не только двумя биомормами хамефитами (СЬ) и
терофитами (Tr). Среди rликофитов отсутствует фанерофитная
жизненная форма (РЬ). Полный спектр биоморф наблюдается
только у криноrалофитов и rалотолерантов. Собственно такой
расклад. иллюстрирует широ'I)' адаптивных возможностей rало..
фитов в мане разнообразия форм роста при разнь эколоrиче
ских специализациях к избыточному засолению почвенно
rpYHToBbIX субстратов.
Таблица 15
Спектры жизненных форм эколоrические rруп n
raл итов Терско..Кумской низменности
. rр.Э rало итыК иноrалофитыlrЛИК ФИТЫlrалОТОJlе аlffЫ
'" ,число «у< число I «у< IЧИСЛО: 'r( число «у<
I Ж.Ф. 'видов о видов о видов о видов о
f t lQl зf:29 ' : 12:Зs .1.: Ji
нk О О 29,63 I 47 i55,951 52 47,71
l?r .. (07 6o 7it ++ ;5 2+kif9:2\r f8)4 0
IBCErO: 28 100 27 100 84 100! 109 ! 100
Самыми мноrочислеННЬJМИ в составе rалофитов TepcKO
Кумекой низменности являются reмикриптофиты. Они насчиты..
вают 107 видов (43,140/0):
100
АНАЛИЗ ЭНДЕМИЗМА И РЕЛИКТОВОСТИ r АЛОФИТОВ
ТЕРСКО..КУМСКОЙ НИЗМЕIПIОСТИ
Эндемы и реликты считаются составляющими индивидуаJlЬ
ность флор. По составу этой rруппы видов, а также по их TaKCOHO
мическому paHry и бordНИко rеоrрафическим связям определяется
самобытность и конrломератность флор.
При анализе эндемизма и реликтовости, а таюке возраста pe
ликтов (вернее при оценке возраста реликтовых популяций) и
оценке эндемизма важно иметь предстамение о возрасте paCCMaT
риваемой территории. Терско Кумская низменность территория
относительно молодая, т.к. на ее поверхности сохранились призна
ки бьUlЫХ уровней Каспийскоrо моря. Это СОХР UfЯющиеся еще в
наши дни морские террасы. Бедность флористических комплексов
rалофитов эндемичными видами, скорее Bcero, объясняется недав..
ним освобождением исследуемой территории от морских вод. а
флористическое насыщение ее происходило за счет выходцев из
соседних флор. Друrим фактом, свидетельствующим об относи
тельной молодости флоры и растительноrо покрова Терско..
Кумекой низменности, можно считать отсутствие четко дифферен"
цированных и климаксовых растительных формаций в приморской
части низменности.
Наибольшим количеством среди [алофитов представлены Kce
ротермические реликты. их переход в рефуrиумы и соответственно
реликтовый ста1)'С прои'3QUlЛО, вероятнее Bcero, на протяжении ro..
лоцена, коrда MHoroKpaTHbIe и разнонаправленные климатические
тренды способствовали МИJl'ации разных флоры и флористических
комплексов.
Поскольку в нашей работе рассматри вается только специали..
'шрованная часть флоры, невозможно проследить все аспекты раз
вития всей флоры в полном объеме. Анализ родственных связей
эндемов специализированных эколоrических rpупп и нахождение
реликтовых популяций даёт возможность наметить основные пути
происхождения и развития флоры.
Sa/so/a daghestaпica эндем Дместана, основная часть ареала
приурочена к rорной части. На территории Приморской низменно--
сти встречается в долине Kap Кар. Происхождение данноrо вида
мы связываем с аридными котловинами BHyтpeHHeropHoro Даl е
101
стана. S. daghestanica по своему положению сближается с S. ca
nescens (Moq.) Boiss, произрастающей в сухих ropHblx районах за..
naднoro и ceвepHoro Ирана (хр. Эльбурс ), Курдистана и Анатолии.
Astragalus hyrcaпus вместе с А. karakugensis Grossh., А. ат..
modendron Buпge и некоторыми видами южноrо побережья Каспия
образует ряд Hyrcani М. Рор. В целом вид данноrо ряда характери..
зуются циркумкаспийским распространением и приуроченностью к
песчаным или солончаково--песчаным мecroобитаниям.
lris pseudonotha основная часть ареала располarается в Во..
сточном Предкавказье. Проявляет систематическое родство с кав..
казскими видами 1. тusи/тап;са Fomin и 1 vo/aceae Klatt., отчасти
со сре,n,Неазиатским 1. sogd;aпa Bunge. На Приморской низменности
нами ВЫЯW1ено несколько изолированных популяций: район с.
Первомайское, около с. Дружба, южнее пос. Мамедкала.
Ferulago daghestanica является одним из видов ряда Macro..
phyllae Schischk., к которому кроме названноrо вида относятся F er..
ulago caтpestris (Besser) 1. Grint., Ferulago taurica Schischk., Feru..
/ago latiloba Schischk. Областью распространения этих викариру..
ющих видов является территория, которая находилась под водами
Восточноrо Паратетиса.
Seпecio schischk;nianus вид относится к ряду Borysthen;c;
Schischk., к которому помимо этоrо вида принадлежат еще не--
сколько видов из прибрежных районов Восточноrо Прикаспия, из
низменных районов побережья Черноro и Азовскоro морей. Ос..
новная часть ареала находится в Восточном Предкавказье. На
ПРИМОjpской низменности, в районе оз. Папас, нами выявлена изо
лированная популяция. Поэтому данный вид правильней считать
субэнщ мом.
Реликтами в rеоrpафическом смысле можно считать большин..
ство 06лиrатных и значительную часть факультативных ra.rюфитов
Приморской низменности, т.к. локализация их популяций имеет
фрarме:нтарный характер и приурочены к нескольким районам на
данной территории. Основная же часть, более или менее paвHOMep
но засt;:ленная этими видами rалофитов, расположена на равнинах
CeBepHoro Прикаспия, Арало..Каспийских и Закаспийских пусты
иях. Ниже мы приводим только наиболее показательные в плане
истории флоры виды rалофитов.
102
Nitraria caspica вид из rруппы облиrатных rалофитов. Дан..
ный вид как И друrие (N. schoberi L., N. taпgиtica Bobr.) обнаружи..
вает явную приуроченность к литоральным областям крупных со--
лонатоводных водоемов древнеro Средиземноморья. На Примор..
ской низменности нами вcrречен только в засоленных низинах Да..
['ар и Шуру..Дере в южной части низменности.
Halostachys ca\pica основной ареал расположен в Арало..
Каспийских rлинистых пустынях. Часть ареала, локализованная на
При морской низменности, фрarментирована на несколько самосто..
ятельных популяций: район озер Большое и Малое Турали, солон..
чак Батмак (около r. Избербаш), район 03. Папас, район с. Аrлоби.
Caтphorosтa тoпspeliaca и Haliтioпe verrucifera у этих двух
видов на Приморской низменности и далее в Восточном 'Закавказье
локализуются фрarментированные реликтовые популяции, приуро..
ченные к луrово"солянковым растительным комплексам. Основная
часть ареала расположена в области Восточноrо (ДpeBHero) Среди..
'Jемноморья.
Kalidiuт cropicuт и К. foliatuт два кустарничка. основная
часть ареала которых, сосредоточена на литораrlЯХ внутриконти..
нентальных водоемов Восточноrо (ДpeBHero) Средиземноморья.
Halocпeтuт strobllacellт в равнинных районах CeBepHoro
Прикаспия часто образует монодоминантные заросли (сарсазанни..
ки). На При морской низменности относитеЛl)НО стабильные зарос..
ли обнаружены южнее оз. Папас.
Suaeda deпdroides кустарничек, с основным ареал в Арало..
Каспийских rлинистых пустынях. В районе исследования нами
встречен только в засоленных низинах Даrзр и Illуру..Дере в юж..
ной части Приморской низменности.
Aпabasis aphylla восточносредиземноморский вид, рас про..
страненный на восток через равнины Казахстана до пустынь l\10H"
f'ОЛИИ. На При морской низменности обнаружен в засоленных ни..
зинах Дarap и Шуру.. Дере.
Сохранению реликтов, относящихея К raлофитам Терско..
Кумекой низменности, способствовало своеобразие МИКРОУСJlОВИЙ,
возникших блаrодаря существованию эколо('ически специфичной
среды обитания избыточно засоленных почв. Их переход в paнr
реликтов произошло в rолоцене вслед за сменой климата.
103
ЗАКJПOЧЕIШЕ
Таким образом, проведенная инвентаризация парциалъной
флоры rалофильных комплексов Терско..Кумской низменности
выявила 248 видов rалофитов, относящихея в 155 родам и 41 ce
мейству. В процентном соотношении от общей флоры TepcKO
Кумекой низменности rалофиты составляют 24,250/0 видов,
32,02СУ<. родов и 40,590/0 семейств. К числу крупных семейств фа
кулътативныx rалофитов относится 102, а облиraтных 72 вида,
что, соответственно, составляет 70,340/0 и 71,840/0 количества TaK
сонов данных эколоrических rрупп.
Характерной чертой парциальной ф,аоры rалофитов TepCKO
Кумекой низменности является ТO что, несмотря на ЭКОJlоrиче
скую специфичность данной эколоrической rpуппы растений, их
флористическая основа принадлежит к семействам, определяю..
ЩИМ общую систематическую структуру флоры Терско Кумской
низменности.
Отличительной особенностью таксономическоrо спекrра яв
ляется внедрение в первую тройку семейств Cheпopodiaceae
(между Роасеае и Asteraceae). Эrо, а также высокое положение в
cocтaв( ведущих родов Salsola, Suaeda, Atrip/ex указывает на
фЛОРОI'енетическую связь rалофильных КОМШIексов Приморской
низменности с флорами Ту ране кой флористической подобласти.
С точки зрения приспособленности rалофитов к условиям
Терско Кумской низменности и эколоrическим условиям место..
обитаний наиболее предпочтительной является rемиксерофиль..
ность (: признаками толерантноrо отношения к солевому режиму.
Несколько менее блаrоприятны комбинации признаков, коrда
СКJ1ерофильность проявляется на том же фоне толерантноrо OT
ношения к солевому режиму, или же склерофильность сочетается
с общей rликофитностью растения.
5. Среди rалофитов Терско Кумской низменности одинаково
обильно представлены жизненные формы Hemicryptophyta и The..
rophytJl. Эти две rруппы резко преобладают над остальными
(PhaneJ'ophy Chamaephyta, Кryptophyta). Такой расклад биоло
rическоrо спектра xapaкrepeH для флор равнин Арало
КаспиЙских или Туранеких пустынь. Травянистые растения в цe
лом (Hemicryptophyta, Kryptophyta и Therophyta) составляют по
104
давляющее большинство I'алофитов ПрИМОрСКОЙ низменности
221 вид (89,1 1 %).
6. В ботанико..rеоrpафическом аспекте среди rеотипов rало
фильных КОМШIексов Терско..Кумской низменности наиболее
мноrочисленными являются древнесредиземноморскиЙ, а среди
rеоэлементов палеаркrический.
105
ЛИТЕРАТУРА
1. Абдрашитова М.М. Перспективные дикие кормовые злаки
// Тр. Казахскоrо С."Х. ин"та. Сер. аrpономич. 1960. Т. 8. Вью. 4.
с. 91-99.
2. Абдрашитова М.М., М.А. Мурзамадиева Сравнительная
анатомическая характеристика листьев некоторых злаков // Тр.
Казахскоrо С."Х. ин"та. Сер. arрономич. 1962. Вып. 9. с. 44..53.
3. Абдурзакова А.С. Общая характеристика и анализ флоры
Терско..Кумской низменности //«Биолоrическое разнообразие
Кавказа». Материалы XlII международной научной конферен"
ции. J'розный,20 1 1 . с.28 1 ..287.
4. Абдурзакова А.С. Флористический анализ псаммофитов
Терско..Кумской низменности // Актуальные проблемы эколоrии
и сохранения биоразнообразия России и сопредельных стран.
Материалы V сероссийской конференции 23..25 мая20 11. r.
Владикавказ, 20 11. c.44 48
5. Абдурзакова А.С. Тайсумов М.А., Астамирова I.A..M.,
Ммом.адова Р.С.,rайрабеков Х.Т. rалофиты Терско..Кумской
низменности и их ЭКОЛOI'ический анализ // Актуальные проблемы
эколоrии и сохранения биоразнообразия России и сопредельных
стран. Материалы Всероссийской научной конференции. COrY,
r. Владикавказ, 2013. с. 96..106
6. Азизова У. Исследование листьев солончаковоrо инесо..
лончаковоrо виноrpада // Сов. ботаника. 1933. NQI..2. с. 44..49.
7. Александров B.r. Водоснабжение листа и ero строение //
Зал. на.УЧНО"прИКJ1адн. отделов Тифлисскоrо бот. сада. Тифлис,
1921. Вып. 2. с. 74..83.
8. Александров B.r. Анатомия растений: Изд. 3..е М,, 1954.
450 с.
9. Александров B.r. Количественные изменения в строении
листьев некоторых травянистых растений при различных сроках
посева // Научно arрономический журн. 1925. NQ 2. с. 4 7 56.
10. Аникиев В.В. Новый способ определения IU10щади листо..
вой поверхности у злаков / В.В. Аникиев, Ф.Ф.Кyryзов // с()изиол.
растений. 1961. Т. 8. Вью. 3. с. 48..54.
106
11. Аникин Ю.Я. Биолоrические особенности волоснеца сит..
никовоrо (E/yтus juпceus Fisch.) и ero использование: Автореф.
дис. ... канд. биол. наук. Саратов, 1971. 23 с.
12. Антипов Н.Н. К вопросу об эволюции мезофитов, rиrpофи..
тов и ксерофитов // Физиолоrия засухоустойчивости растений.
М, 1971. с. 247..279.
13. Антипов Н.Н. О возникновении и развитии эколоrических
rрупп растений в процессе эволюции / / Проблемы эволюционной
физиолоrии растений. Л., 1 974. с. 119..122.
14. Бабаева Ж.А. Солеустойчивость разных тканей корнеплода
моркови и каллаусов, полученных из них в культуре изолирован..
ных тканей // Физиол. раст. 1968. Вью. ]. с. 44..52.
15. Баrдыков Н.И. Анатомические изменения у ячменя в связи
с адаптацией к засоленной почве // Уч. зап. POCToBcKoro roc. ун..
та. 1957. Т. XXVlII. Вьш. 5. c. 61..70.
16. Баrдыков Н.И. Изменение анатомической СТРУК1уры у то..
матов и проса в процессе адаптации их к почвенному засолению
// Уч. зав. POCToBcKoro roc. ун"та. 1958. Т. 51. с. 80..87.
17. Баранов П.Л. К методике количественно..анатомическоrо
изучения растений. 1 Распределение устьиц / П.А. Баранов / / Бюл.
CAry. 1 924а. N24. с. 70..79.
18. Баранов П.А. К методике количественно..анатомическоrо
изучения растений // Бюл. CAry. 19246. N27. с. 41..49.
19. Баталин А.Ф. Исследования над влиянием хлористоrо
натрия на развитие трёх солончаковых растений: Spergu/aria
тediпa var. тargiпata, Sa/so/a soda и S. тutica //Тр. Император..
CKoro с...Петерб. общ"во естествоиспытат. Спб., 1875. Т. 6. с.
19..28.
20. Бейдеман И.Н. Ритм сезонноrо хода интенсивности транс..
пирации растений при разных типах водноrо режима почв в раз..
личных климатических условиях // Бот. журн. 1960. Т. 45. NQ6.
с. 927..935.
21. Беккер..Ржевская М. Солевыносливость различных сортов
пшеницы // Селекция, аI'рОХИМИЯ, семеноводство. Энrельс,
1935. с. 61..69.
22. Блаrовещенский А.В. Современное состояние вопроса об
обмене веществ у rалофитов // Тр. Узб. фил. АН СССР. Сер. ] 1.
Бот. 1942. Вьш. 5. с. 64..69.
107
23. Блажний Е.С. Почвы дельты реки Кубани и прилеrающих
пространств / Е.С. Блажний. Краснодар, 1971. 276 с.
24. Боrдан В.С. Закубанские IU1авни в почвенно--ботаническом
и мелиоративном отношениях // Тр. Кубанскоrо С...х. ин"та.
Краснодар, 1925. Вып. 3. с. 1..54.
25. Бойко Л.А. Физиолоrия корневой системы растений в
условиях засоления Л., 1969. 147 с.
26. ]>ровцина В.А. Влияние почвенноrо засоления на анатоми..
ческое строение листьев хлопчатника / / Докл. совещ. по физиол.
раст. 2.8 января.. февраля 1940 r. 1946. Bьm. 1. с. 81..91.
27. ]>УРЫI'ИН В.А. О некоторых особенностях развития амери"
KaHcKoro хлопчатника на засоленных почвах r олодной степи и
путях повышения ero солеустойчивости / / Вопросы солеустойчи..
вости растений: Тр. Узб. фил. АН СССР. Ташкент, 1942. Сер.
Ю. Вып. 5. с. 54..59.
28. ]>урыrин В.А. Современное состояние вопроса о соле..
устойчивости хлопчатника // Тр. научн. сессии АН Узб. ССР.
Ташкент, 1947а. с. 80..89.
29. l)урыrин В.А. К вопросу о водном балансе хлопчатника на
засоленных почвах rолодной степи / В.А. Бурыrин // Бюл. АН
Узб. ССР. 19476. NQ5. с. 29..37.
30. ]>урыrин В.А. О некоторых путях приспособления расте..
ний к почвенному засолению / В.А. Бурыrин // Изв. АН Узб. ССР.
.. 1948. NQ3. с. 44..51.
31. ]>урыrин В.А. Солеустойчивостъ хлопчатника в свете про..
блемы rалофитизма: Автореф. ДИС. ... доктора биол. наук 1 В.А.
БУРЫПfН" Ташкент, 1952. 39 с.
32. ]>утенко p.r. Соматический эмбриоrенез в КУЛЬ1)'ре ткани
МОРКОflИ в условиях высоких концентраций солей в среде // Докл.
АН СССР. 1967. NQ4..6. с. 94..101.
33. ]>утник А.А. Адаптация анатомическоrо строения видов
сем. CJlenopodiaceae Vent, к аридным условиям: Автореф. дне. ...
доктора биол. наук Ташкент, 1984. 41 С.
34. Буш Н.А. О ботанико--rеоrpафических исследованиях Ку..
банекой области в 1908 r. СПб., 1909. 11 с.
35. Буш Н.А. Ботанико--rеоrpафический очерк европейской ча
сти СССР и Кавказа / М...Л., 1 936. 326 С.
108
36. Быков Б.А. Основные особенности rалофильной флоры и
растительности С редней Азии и Казахстана / / Изв. АН Каз. ССР.
Сер. биол. ]981. .N'Ql. с. ]..9.
37. Быстрова В.И., Миняева Н.А. Материалы к эколоrо..
f'еоrрафической характеристике rалофитов Северо..Запада евро"
пейской части СССР // Ареалы растений флоры СССР. Л., 1969.
с. 47..63.
38. Вальков В.Ф. rенезис почв CeBepHoro Кавказа Ростов
на/Дону, 1977 159 с.
39. Вальков В.Ф. Почвы Краснодарскоrо края, их использова..
ние и oxpaHa Ростов на/Дону, 1 996. 191 с.
40. Вальтер О.А. Практикум по анатомии растений М...Л.,
1937. 114 с.
41. Варминr Е. Ойколоrическая rеоrрафия растений М., 1901.
137 с.
42. Варминr Е. Распределение растений в зависимости от
внешних условий СПб., 1 903. 97 с.
43. Василевская В.К. Анатомическое строение плодовых Ко..
пет..Дarа в различных условиях культуры // Сов. бот. ]941a.
N21..2. с. 47..59.
44. Василевская В.К. Анатомо"морфолоrические ососбености
растений холодных пустынь Средней Азии / / У ч. зап. ЛrY. Сер.
биол. науки. 1941 б. Вып. 14. с. 48..158.
45. Василевская В.К. СТРУК1)'рные приспособления растений
пустынь Средней Азии // Проблемы современной ботаники. М.;
Л., 1965. с. 71..104.
46. Василевская В.К. Формирование листа засухоустойчивых
растений Ашхабад, 1954. 184 с.
47. Василевская В.К. Типы анатомическоrо строения листьев
двудольных // Бот. журн. 1981. Т. 66. NQ7. с. 992.. 1001.
48. Введенский Н.П. Материалы к познанию растительности
долины реки Кубань и её притоков .. БоJlыlIrоo и Малоrо Зелен..
чуков // Тр. Кубано..Черноморск. НИИ. 1926. Вып. 42. 117 с.
49. Витман Р.А., Поздняков Э.П. Зависисмость растительноrо
покрова от засоления почв солонцовоrо комплекса центральной
лесостепи Омской области // Уч. зал. OMcKoro roc. пед. ин"та.
1968. Вып. 38. с. 70..77.
109
50. Вихерева Василькова В.В. Анатомическая характеристика
арктических злаков // Проблемы Севера. М.;Л., 1964. Вып. 8.
с. 111 119.
51. j"'аврилюк Ф.Я. Земельный кадастр CeBepHoro Кавказа
Ростов нlд, 1979. 54 с.
52. j"'алушко А.И. Флора CeBepHoro Кавказа / А.И. rалушко.
T.I. Ростов нlд, 1978. З 17 с.
53. j"'алушко А.И. Флора CeBepHoro Кавказа / А.И. rалушко.
Т.П. Ростов Н/Д, 1980a. 350 с.
54. ]'алушко А.И. Флора CeBepHoro Кавказа / А.И. rалушко.
Т.П!. Ростов Н/Д, 19806. 327 с.
55. ]'амалей Ю.В. Анатомия листа у растений пустыни rоби //
Бот. журн. 1984. Т. 69. NQ5. с. 569..584.
56. ]'амалей Ю.В. Вариация кранц анатомии у растений пу..
стынь rоби и Каракумы // Бот. журн. 1985. Т. 70. NQ 10. с.
13 02 1 3 13.
57. !'амалей Ю.В., Шийрэвдамбой Ц. Структура растений За
алтайс][{ой [оби Л, 1988. с. 44 106.
58. ]'воздецкий Н.А. Физико--rеоrpафическое районирование
Еврош;:йской части СССР и Кавказа // Изв. BrO. 1960. Т. 95.
Вып. 5. с. 1 33.
59. ]'воздецкий Н.А. Кавказ М., 1963. 262 с.
60. !'енкель П.А. Устойчивость растений к засухе и пути её по
вышения М.;Л., 1946. 236 с.
61. ]'енкель П.А. О "ричина.х засухоустойчивости некоторых
ксерофитов / П.А. rенкель // Бот. журн. 1949. Т. 34. NQ5. с.
116 127.
62. ]'енкель П.А. Физиолоrия адаmации растений к засолению
// Проблемы ботаники. 1950. Вып. 1. с. 4 29.
63. I'енкель П.А. Солеустойчивость растений и пути её
направленноrо повышения // ХН Тимирязеве кие чтения. М.,
1954. . 59 с.
64. !'енкель П.А. Современное состояние проблемы 'шсухо
устойчивости растений и дальнейшие пути её изучения / / Физио
лоrия устойчивости растений (морозоустойчивость, засухоустой..
чивость и солеустойчивость). М., 1960. с. 385..401.
110
65. rенкель П.А. к познанию проблемы засухоустойчивости
растений // Физиолоrия и биохимия культурных растений. 1970.
Т. 2. Вып. 3. с. 227..233.
66. rенкель П.А. Физиолоrия жаро и засухоусто чивости рас..
тений М., 1982. 279 с.
67. rенкель П.А. Эколо] ическое значение водноr'о режима не..
которых rалофитов // Бот. журн. 1945. Т. 30. NQ4. с. 116..123.
68. rриrорьев Ю.С. Сравнительно"эколоrические исследова..
ния ксерофитиза.. ции высших растений / Ю.С. fриrорьев. .. М.,
1955. 247 с.
69. rроссrейм А.А. Анализ флоры Кавказа Баку, 1936.
260 с.
70. rроссrейм А.А. Растительный покров Кавказа LVI., 267 с.
71. rроссrейм А.А. Определитель pactel-lИЙ Кавказа М.,
747 с.
72. r усейнов А.З. Кормовые достоинства бескильницы круп..
ной, произрастающей в Азербайджане // Тр. Азерб. roc. заочноrо
пед. ин"та. 1956. Вью. 3. с. 49..53.
73. rущин И.В. Солеустойчивость различных сортов пшеницы
и ячменя и факторы, обуславливающие её // Тр. конф. по почво..
ведению и физиолоrии культурн. раст. Т.Il. Саратов, 1938. с.
60..65.
74. Данилевский Н.Я. Исследования о Кубанской дельте / Н.Я.
Данилевский // Зап. Русск. rеоrрафич. общ"ва. СПб, 1869. Т. 2.
с. 1..123.
75. Дворецкая Е.К. Заметки по анатомии и физиолоrии солянок
/ Е.К. Дворецкая // Аrрономия Юrо..Востока. Саратов, 1928. Т.
VI. Вып. 1. с. 37..43.
76. Демидовекая Л.Ф., Кириченко Р.А. Морфолоr'о..
анатомические особенности тростника и цикл ero развития // Тр.
ин..та ботаники АН Каз. ССР. 1964. N 19. с. 69.. 77.
77. Дубына Д.В., Шеляr"Сосонко Ю.Р. Плавни Причерномо..
рья. Киев, 1989. 269 С.
78. EropoB В.В. Характер изменения минерализации почвенно..
rрунтовых вод при солончаковом процессе в условиях примор..
ско..дельтовой низменности // Почвоведение. 1950. N 2. с. 41..
51.
III
79. JКиrар М.П. Морфолоrия и распределение пор в наруж..
ных стенках эпидермиса злаков // Бот. журн. 1972. Т. 57. N!!ll.
с. 124 .131.
80. Заки М.А., ШМИДТ В.М. О систематической структуре фло..
ры Сlран Южноrо Средиземноморья // Вестник Ленинrр. ['ос. ун..
та. 1973. Х!!9. с. 57..69.
81. 3акиров К.З., Меркулович Н.И. К биолоrии каперцов
(Cappers spiпosa) // Тр. Узб. roc. ун"та. 1936. Х!!5. с. 112 ] 17.
82. Заленекий В.Р. Материалы к КОЛl\чественной анатомии
различных листьев одних и тех же растений // Изв. Киевскоrо по..
литехнич. ин..та. 1904. Т. IV. Вып. 1. с. 34..41.
83. Захаров Л. Плавни Кубанских плавен // Природа. 1929.
Х!! 1. ( . 79..81.
84. Захаров Л. Сплавины Кубанских плавен // Бот. журн.
1933. Т. 18. Х!!4. с. 287..298.
85. Захаров с.А. Почвы Предкавказья // Почвы СССР. Т.ПI.
М., 1939. 102 с.
86. Зеленекий В.Р. О так называемых запасающих трахеидах в
листьях некоторых растений нашей флоры // Дневник xl съезда
русских естествоиспытателей и врачей. М., 1902. с. 19 23.
87. Зозулин [.М., Пашков [.Д. rеоботаническое районирова..
ние. Нижний Дон (Ростовская область) // Растительные pt:CYPCbJ.
Ч. 1. Ростов н/Д, 1980. с. 17..29.
88. Зозулин [.М., Федяева В.В. Систематическая структура
Нижнеrо Дона // Изв. СКНЦ ВШ. Сер. естеств. науки. 1985. К!!
1. с. 75..77.
89. Иваницкая Е.Ф. Особенности анатомическоrо строения
растений в условиях разнокачественноrо засоления почвы 1/ Фи..
ЗИОЛ. растений. 1962. Т. 9. .N'Q2. с. 94..101.
90. Иванов А.А. Конспект флоры Ставрополья / А.А. Иванов.
Ставрополь, 1997. 154 С.
91. Ильин М.М. К происхождению пустынь Средней Азии /
М.М. Ильин // Сов. ботаника. 1937. NQ6. с. 91..103.
92. Ильин М.М. Некоторые итоrи изучения флоры пустынь
Средней Азии // Материалы по истории флоры и растительности
СССР. 1946. Т. П. с. 84..93.
93. Ильин М.М. Флора литоралей и пустынь в их взаимосвя"
зях // Сов. ботаника. 1947. Х!!5. с. 249 267.
112
94. Иозефович Л.И. Приазовские плавни Новочеркасск,
1929. 87 с.
95. Кайrородов А.И. Естественная зональная классификация
климатов земноrо шара M., 1955. 147 с.
96. Камелин Р.В. Флора бассейна р. Варзоб (rиссарский хре..
бет, Таджикистан) и её анализ: Автореф. дис. ... канд. биол. наук
- Л., 197 1 . 28 с.
97. Камелин Р.В. Флористический анализ естественной флоры
rорной Средней Азии Л, 1973. .. 353 с. 95
98. Канонников А.М. Природа Кубани и ПРИLlерноморья
Краснодар, 1977. 137 с.
99. Карпилов Ю.С. Об одной rруппе ксерорезистентных расте..
ний, сходных поанатомии листа, ФОТОСИlпезу и водному режиму
// Тр. Молд. НИИ орошаемоrо земледелия и овощеводства.
1970. Т. XI. Вып. 3. с. 1 1 7 129.
Карта типов засоления почв Европеikкой части СССР. Мае..
штаб: 1 :30000000 / Под ред. В.В. EropoBa, Н.И. Базилевич. М.,
1973.
100.Келлер Б.А. О влиянии солей на распределение раститель..
ности / / Дневник ХН съезда русских естествоиспытате.'1ей и Bpa
чей в Москве. М., 1910. с. 39..47.
1 О 1. Келлер Б.А. Об осмотической силе клеточнOJ'О сока у рас..
тений в связи с характером почв // Почвоведение. 1914. NQ] 2.
с. 44..51.
I02.Келлер Б.А. Опыт и некоторые общие выводы по эколоrии
солончаковоrо растения / Б.А. Келлер // Вест, опытн. дела средн.
чернозёмн. обл." 192]. NQI..2. с. 51..57.
103.Келлер Б.А. Эколоrия растений и борьба земледелия с за..
солением почв и засухой / Б.А. Келлер // Итоrи работ с...х. опытн.
учрежд. Средне Чернозёмной обл. Воронеж, 1923. Выл. 1. с.
61..67.
I04.Келлер Б.А. Растения и засуха с точки зрения сельскоrо хо..
зяйства // Природа и сельское хозяйство засушливых областей
СССР. 1926. NQ1 2. с. 70..79.
105.Келлер Б.А. Засоление, транспирация и накопление солей
внутри растений у Atriplex verrucifera // Тр. бот. опытн. станции
им. Б.А. Келлера. Воронеж, 1 929а. Т. ]. с. 24..31.
113
10б.Келлер Б.А. НаКОIU1ение солей внутри растений и 'J;lсоле..
ине почвы / Б.А. Келлер // Тр. ботан. опытн. станции им. Б.А.
Келлера. Воронеж, 19296. Т. 1. с. 54..59.
107.Келлер Б.А. Явления крайней солеустойчивости у высших
растений в дикой природе и проблема приспособления // Расте..
пия и ',реда. М., 1940. с. 193..217.
108.Келлер..Лейсле 3.<1>. Некоторые анаТОМtlческие ра'ЗЛичия у
Atriplex verrucifera и А. салит в засоленных и незасоленных куль..
1)'рах 1 3.Ф. Келлер..Лейсле // Природа сельскохозяйственных за..
сушлиных областей СССР. 1927. NQ1..2. с. 53..59.
109.Келлер..Лейсли 3.Ф. Особенности анатомическоrо строе..
ния листьев У весенних эфемеров однолетников / / Сов. ботаника.
1940. NQ4. с. 86..91.
110.KeIU1ep Б.А. О солеросе (Salicorпia herbaceo) и ero оrноше..
ние К засолению // Природа и сельское хозяйство засушливых и
пустынных областей СССР. 1927. NQl..2. с. 40..49.
111.Клейн Р.М., Клейн Д.Т. Методы исследования раСlений
М., 1974. 527 с.
112.Клешнин А.Ф. К вопросу об энерrетическом балансе рас..
тений // Физиолоrия растений. 1955. Т. 2. Вып. 6. с. 91..99.
113.Ковалёв В.Н. Адаптеция КУЛЬ1)'рных растений к почвенно..
МУ засолению / / Тез. докл. конф. по физиолоrич. устойчивости
растений. М., 1959. с. 77 79.
1 14.Ковда В.А. Солончаки и солонцы М.;Л., 1937. 241 с.
115.Ковда В.А. Процессы почвообразования в дельтах и пой..
мах континентальных областей СССР // Проблемы COBCTcKoro
почвоведения. 1946. Вью. 14. с. 34..51.
lI6.Ковда В.А. Происхождение и режим засоленных почв. T.I.
М, 1947 341 с.
117.Ковда В.А. Происхождение и режим засоленных почв. Т. П.
М, 1947 б. 117 с.
118.Колпиков Д.И. Сравнительное изучение водноrо режима и
засухоустойчивости стипаксерофитов // Памяти акад. Н.А. Мак..
симова. М., 1957. с. 64..71.
119.Косенко И.С. К познанию растительности лиманов и ШIав..
ней Приазовскоrо побережья Кубанскоrо края // Тр. Кубанско..
ro с...х. ин"та. Краснодар, 1924. Т. 1. ВЬИl. 2. с. 93..111.
114
120.Косенко И.С. К познанию растительности TaMaHcKoro по--
луострова // Тр. Кубанскоrо С."Х. ин"та. Краснодар, 1927. Т. 5.
с. 121..149.
121.KoceHKo И.С. Материалы к познанию эколоrо..
rеоrрафической природы Приазовских плавней // Тр. Всесоюзн.
центр, станции рисовоrо хоз..ва. Ростов н/Д, 1 934. Вып. У. с.
40..49.
122.Косенко И.С. Растительные зоны 3ападноrо Предкавказья
и CeBepHoro Кавказа // Тр. Краснодарек. ин"та ПИЩе пром"ти.
Краснодар, 1947. Вып. 1. с. 12..23.
123.KoceHKo И.С. Определитель высших растений Северо..
3ападноrо Кавказа и Предкавказья М.. 1970. 613 с.
124.Кузнецов Н.И. Принципы деления Кавказа на ботанико..
f'еоrрафические провинции / Н.И. Кузнецов. СПб., 1909. 147 с.
125.Кузнецов Н.И. Ботанико..rеоrрафическое картирование ев..
ропейской России // Зап. Одесскоrо общ"ва естествоиспытат.
1928. Т. 44. с. 129..220.
126.Лавренко Е.М. Степи СССР // Растительность СССР.
М.;Л., 1940. Т. 2. с. 1..256.
127.Лавренко Е.М. Евроазиатская степная область /1 rеобота..
ническое районирование СССР. М.;Л., 1947 с. 95..107.
128.Лавренко Е.М. Принципы и единицы rеоботаническоrо
районирования / Е.М. Лавренко // rеоботаническое районирова..
ние СССР. .. М.;Л., 19476. с. 9..13.
129.Лавренко Е.М. Основные черты ботанико reоrрафическоrо
разделения СССР и сопредельных стран // Проблемы ботаники.
М.;Л., 1950. Вьш. 1. с. 530..548.
130.Лавренко Е.М. Об уровнях изучения орrаническоrо мира в
связи с познанием растительноrо покрова // Изв. АН. СССР. Сер.
биол. 1964. NQl. с. 32..46.
131.Лавренко Е.М. Степи / Е.М. Лавренко / / Растительность
Европейской части СССР. .. Л., 1980. с. 203..272.
132.Ларин И.В. Определение почв и сельхозуrодий по расти..
rельному покрову Л., 1950. 273 с.
133.Ларин И.В. Луrоводство и пастбищное хозяйство Л.,
1969. 549 с.
134.Лейсле Ф.Ф. К эколоrии и анатомии f'алофитов и ксерофи..
rOB // Бот. журн. 1949. Т. 34. .N'Q3. с. 94 IOI.
1] 5
135.Леонов r.П. Историческая rеолоrия / r.П. Леонов, М.,
1956. . 364 с.
136.Липаева Л.И. О некоторых особенностях строения BOДO
проводящей системы растений в связи с условиями место()бита
ний // Изв. АНКаз. СССР. Сер. освоения пустынь. 1951. N 2. с.
77..85.
1 37Jv1.аксимов Н.А. Физиолоrические основы засухоустойчиво--
сти рас:тений // Избранные работы. Т.1. М., 1952. с. 94..109.
138Jv1.анойленко К.В. Эволюционные аспекты проблемы засу
хоустойчивости растений Д., 1983. 244 с.
139.Ма1)'ХИН [.Р. О повышении солеустойчивости культурных
злаков путём воспитания их на засолённой почве // Тез. докл.
научн. конф посвящённой 80"лнтию PrY. ВЫП.l. Ростов н/ Д,
1949. с. 61..69.
140Jv1.а1)'ХИН [.Р. Об активности некоторых ферментов уrле
водноrо обмена у некоторых КУЛЬ1)'рных злаков в связи с воспи
танием их на засоленной почве // Уч. зап. pry. 1953. Т. XIX.
Выл. 3. с. 70..77.
141.Ма1)'ХИН [.Р. О содержании хлора в растения адаптируе..
мых к засолению почвы // Уч. зап. pry. 1957. Т. XXVlll. Вып.
5. с. 9I 99.
142Jv1.а1)'ХИН [.Р. Физиолоrия приспособления культурных
растений к засолению почв Ростов н/д, 1963. 204 с.
143.Ма1)'ХИН [.Р., Ефремов Л.Д. Влияние засоления на CTPYK
1)'ру растений // Тез. докл. научн. конф. Дона и CeBepHoro Кавка..
за. Ростов н/Д, 1947. с. 16 19.
1 44 Jv1.ильков Ф.Н., rвоздецкий Н.А. Физическая rеоrрафия
СССР (Общий обзор, Европейская часть СССР, Кавказ) М.,
1962. ' 201 с.
] 45 Jv1.ирославов Е.А. Некоторые черты ксеРОМОРфНОf'О cтpo
ения эпидермиса листа ряда злаков // Бот. журн. 1962. Т. 47.
.N'Q9. с. 247 255.
146.Мирославов Е.А. СТРУК1)'ра и функция эпидермиса листа
покрытосеменных растений Л., ]974. 175 с.
147.Наrалевский В .Я. Эколоrо..физиолоrическая характеристи
ка rалофитов Западноrо Предкавказья в связи с их происхожде..
нием // Мат. Всесоюзн. совещ. по филоreнии высших раст. М.,
1982. ' с. 67 69.
116
148.Наrалевский В.Я. К эколоrО"фИ'3иолоrической итаксоио"
мической классификации I'алофитов Северо Западноrо Кавказа и
llредкавказья / В.Я. Наrалевский // Актуальные задачи филоrении
и биохимии растений в ботанических садах СССР. Пущино,
1984б. с.127..128.
149.Наr3{Iевский В.Я. rалофиты CeBepHoro Кавказа как источ"
ник лекарственноrо сырья / / Тез. дою.. 1 ресnубликанск. конф. по
медицинской ботанике. Киев, 1984 в. с. 33.
150.Наrалевский В.Я. Видовой состав, ЭКОЛОI'ИЯ и rеоrpафия
соленеnроницаемых rалофитов CeBepHOJ'O Кавказа / / Реrиональ..
ные флористические исследования и методика преподавания бо..
танических дисциплин: Минвузовский сб. научн. тр. Краснодар,
1986 а. с. 20..28.
15I.Наrалевский В.Я. Соленакапливающие rалофиты в бас..
сейнах экосистем малых рек Западноrо Предкавказья (флористи"
ко"эколоrический, фитоценотический и J'еоrрафический обзор) /
В.Я. На['алевский / / Актуальные вопросы эколоrии и охраны при..
роды экосистем малых рек: Мат. научно..nрактич. конф. Часть 1.
Краснодар, 1992 в. с. 12..15.
152.Наrалевский В.Я. Флора и систематическая структура ra..
лофитов CeBepHoro Кавказа / В.Я. Наrалевский // Актуальные во..
npocbI эколоrии и охраны природы экосистем южных реrионов
России и сопредельных территорий: Мат. ХН межреспубл. науч..
но--практич. конф. Краснодар, 1999 б.. с. 13..15.
153.Николаевский B.r. Особенности анатомической структуры
листьев злаков"сциофитов // Научн. дOКJ1. высш. школы. Биол.
науки. 1972 а. NQ2. с. 49..55.
154.НиколаевскийВ.r. О типах cTpYKrypbI листа у злаков /!Бот.
жури. 1972 б. NQ3. с. 67..71.
155.Николаевский B.r. Пути специализации структуры листа
'illaKoB трибы Triticeae Dum. // Бюл. МОИП. Orд. биол. 1972 в.
Т. 72. N!!5. с. 67..71.
156.Покровская Е.Н. Солевыносливость и некоторые CTOpO--
ны обмена веществ rликофитов / Е.Н. Покровская// Физ. расте..
ний. 1958. Т. 5. NQ3. с. 94..101.
157.Поплавская Т.И. Эколоrия растений / Т.И. Поплавская.
М., 1948. 294 с.
1 J 7
158.Серебрякова Т.И. Морфоrенез побеrов и эволюция жиз
ненных форм злаков М., 1975. 347 с.
1 59.Середин Р.М. К вопросу об изучении систематическоrо co
става дикорас1)'ЩИХ злаков CeвepHoro Кавказа // Уч. зап. Пяти
ropcKoro фарм. ин та. 1961. Т. 5. с. 1 47.
160.Середин Р.М. Биоэколоrический анализ злаков CeBepHoro
Кавказа / Р.М. Середин // Уч. зап. Пятиrорскоrо фарм. ин та.
1966. Т. VI. Вып. 1. с. 39 49.
161.Середин Р .М. Дикорастущие злаки CeBepHoro Кавказа: AB
тореф. дне. ... доктора биол. наук Пермь, 1967. 32 с.
1 62.CтporaнoB Б.П. Растения и засоленные почвы М.;Л., 1958.
139 с.
163.CтporaнoB Б.П. Современное состояние проблемы физио
лоrи и солеустойчивости растений и дальнейшие пути её изуче
ния // Физиолоrия устойчивости растений (морфоустойчивость,
засухоустойчивость и солеустойчивость): Тр. конф. 3 7 марта
1959. . М., 1959. с. 7 10.
164.CTporaнoB Б.П. Физиолоrические основы солеустойчи
вости растений (при разнокачественном засолении почвы) / Б.П.
CтporaHoB. М., 1962. 366 с.
165.Тайсумов М.А., Астамирова М.А.М., MaroMaдOBa Р.С., Аб
дурзакова А.С., Хасуева Б.А., ОмархаджиеваФ.С., Са1)'ева Л.Л.
Предварительный анализ rеоrpафических элементов флоры Kce
рофитов полупустынных районов Российскоrо Кавказа. Вестник
KpacrAY. .. Красноярск, 2013,.N'Q 5. . 103..108.
166.Тайсумов М.А., MaroMaдOBa Р.С., Абдурзакова А.С.. KOH
спект флоры Терско Кумской низменности. Моноrрафия rроз..
ный. 2013. 129 с
167.Тайсумов М.А., Умаров М.У., Астамирова М. А.М Accop
тимент интродуцированных и перспективных для r. rрозноrо дe
ревьев и кустарников. Моноrpафия rрозный. 20 13. 275 с.
168.Тайсумов М.А., Исраилова с.А. Анализ флоры антропофи
тов Чеченской Республики. Моноrрафия rрозный. 2013. 148 с.
169.Тайсумов М.А., Исраилова Инвазионные виды во флоре
CeBepHoro Кавказа // Актуальные проблемы биолоrии и эколоrии
I Матер. Всерос. Науч практич. Конфер. rрозный, 20 13. с. 1 67
184.
118
170. Тайсумов М.А., Абдурзакова А.С. АНТРОПOl енная TpaHC
формация фЛQрЫ Терско Кумской низменности // Акryальные
проблемы биолоrии и эколоrии / Матер. Всерос. НаУlj практич.
Конфер. rрозный, 2013. i. с. 17 25.
171 .Тайсумов М.А., Абдурзакова А.С., Астамирова М.А., raдa"
ева Т.З. rеоrрафический анализ флоры r. rрозноrо и ero OKpeCT
ностей// Современные проблемы науки и образования. 2014.
NQ 3; с. 1 1 9 1 27.
172.Тайсумов М.А., Умаров М.У., Астамирова М.А...М., Хали..
дова Х.Л.Охрана и рациональное использования высокоrорной
растительности Российскоrо Кавказа AктyanbHbIe проблемы эко
лоrии //Матер. IV Всерос. научн.конфер. «Актуальные проблемы
'защиты окружающей среды и техносферной безопасности в ме..
няющихся антропоrенных условиях» rРО'JНЫЙ, 2014. с. 372 376.
1 73.Тайсумов М.А., Умаров М.У., I'aдaeBa Т.З., Халидова Х.Л.,
Хозяйственно полезные растения флоры rрозноrо и ero OKpecт
ностей, их состояние и перспективы использования в народном
хозяйстве // Актуальные проблемы биолоrии и эколоrии / Матер.
Всерос. Науч практич. Конфер. rрозный, 20 14. с. 76 84.
174.Тайсумов М.А., Умаров М.У., Астамирова M.A. M., OMap
хаджиева Ф.С. Флористические районы петрофитов Восточной
части скалистоrо хребта в пределах ЧеllНИ и Инryшетии Матери..
алы III всероссийской научно технической конференции «COBpe
менные проблемы rеолоrии, rеофизики и rеоэколоrии CeBepHoro
Кавказа» 2013. r. rрозный. с. 266 270.
175.Тайсумов М.А., Астамирова М.А. М.Краткий обзор бота
нических исследований Чечни и сопредельных территорий MaTe
риалы 111 всероссийской научно технической конференции «Co
временные проблемы rеолоrии, rеофизики и rеоэколоrии CeBep
Horo Кавказа» 2013. r. I'розный. с. 345 352.
1 76.Тайсумов М.А., Абдурзакова А.С., Маrомадова Р.С., ACTa
мирова M.A, M. Классификация rалофитов Терско Кумской низ
менности по анаТОМО фИЗИОЛOl'ическим признакам Вестник АН
ЧР Вып. N 1 (22), 20 14. с. 35 46.
1 77.Тулайков Н.М. Солонцы, их УЛУIIшение и использование
М., 1922. 98 с.
119
178. Убайду ллаев У. Анатомо--эколоrическое исследование
эфемероидов предrорий Тянь..Шаня: Автореф. дис канд. биол.
наук Ташкент, 1959. 22 с.
179.У'довенко r.B. ПОС1)'IU1ение, реутилизация и некоторые
стороны физиолоrическоrо действия физиолоrическоrо действия
хлора в растениях // Изв. АН СССР. Сер. биол. 1965. NQ3. с.
368..377.
180.Удовенко [.В., rридчанинов О.Д., Семушина Л.А. Морфо..
лоrо.. анатомическое изменение листьев и корней пшеницы при
засолении // Бот. журн. 1970. Т. 55. .N'Q7. с. 140..149.
181.Урба.х В.Ю. Биометрические методы М., 1964. 231 с.
182.<I>ёдоров А.А. Фитохории Европейской части СССР // Фло..
ра Европейской части СССР. Л., 1979. Т. IV. с. 10..27.
183.<l>лёров А.Ф. Типы болот в НИЗОВЬЯХ Кубани / А.Ф. сl>лёров
// Торфяное дело." 1929. с. 134..138.
184.<l>лёров А.Ф. Типы растительности дельты и низовьев реки
Кубани и реки Анапки / / Тр. Сев. кав. ассоциации НИИ. Ростов
н/Дон, 1930. .N'Q5. с. 54..63.
185.<l>лора Европейской части СССР. Л., 1976. Т. И. 236 с.
186.<l>лора Европейской части СССР. Л., 1978. Т. ПI.
258 с.
187.<l>лора Европейской части СССР. Л., 1979. Т. IV.
355 с.
188.<l>лора Европейской части СССР. Л., 1981. Т. У. 379 с.
189.<l>лора Европейской части СССР. Л., 1987. Т. VI. 254 с.
190.<l>лора Европейской части СССР. Л., 1989. Т. VIlI. 412 с.
191.Ху дяков И.И. Растительный покров как показатель хими..
ческоrо состава и rлубины залеrания rрунтовых вод / И.И . Худя..
ков / / Растительные индикаторы почв, ropHblx пород и подземных
вод. 1\1., 1 964. с. 17..29.
192.Цвелёв Н.Н. Моноrрафия рода бескильница (Pucciпe//ia
Pari.): Автореф. дис. ... канд. биол. наук / Н.Н. Цвелёв. Л., 1954.
19 с.
193.Цвелёв Н.Н. Некоторые вопросы эволюции злаков " Н.Н.
ЦвелёЕi // Бот. журн. 1969. .N'Q3. с. 88..97.
194.Цицин Н.В. Что даёт скрещивание пшеницы с пыреем /
Н.В. Цицин. .. М., 1937...49 с.
120
195.Цицин Н.В. I ибридизация растений / Н.В. ЦиЦИН. М.,
1965. 149 с. 318.
196.Чиликина Л.Н., Унчиев Н.Д. Материалы к кормовой харак"
теристике основных т пов пастбищ и сенокосов Даreстана // Тр.
отд. растит, ресурсов Даr. филиала АН СССР. 1960. Т. 2. с. 78..
99/
197. Чубуков Л.А. Климат / / Кавказ. М., 1966. с. 85..94.
198. Чукавина А.П. Некоторые критические замечания о ситни..
ках Таджикистана // ДoКJ1. АН Таджикской ССР. 1958. .N'Q2. С.
51..52.
199.Шахов А.А. Приспособления к засоленности почв у рас..
тений / А.А. Шахов // Растительность и среда. М.; Л., 1 950. с.
116.. 1 4 1 .
200.Шахов А.А. О типах развития растений в процессе исто..
рическоrо приспособления к засушливым условиям М.; Л.,
1952. с. 31.. 90.
20I.Шахов А.А. Солеустойчивость растений М., 1 956.
552 с.
202.Шенников А.П. Эколоrия растений М., 1950. 301 с.
203.Шифферс Е.В. Растительность CeBepHoro Кавказа и ero
природные кормовые уrодья М.; Д., 1 953. 399 с.
121
Содержание:
В В ЕДI НИЕ . . . . . . . . . . . .. .. . . .. . . . . . . . . .. . . . . . .. . . .. . . .. .. . . . .. . .. . . . . . . . . . . .. . . . . . . . . .. . . . . . . . . . . .. . 3
Физико..rЕоrр АФИЧЕСКАЯ ХАРАКТЕРИСТИКА ТЕРСКО..
КУМСКОЙ НИЗМЕННОСТИ rEOrp АФИЧЕСКОЕ
ПОЛО:ЖЕНИЕ ..... ................... ...................... ..... .................................6
КЛАССИФИКАЦИЯ r АЛОФИТОВ ТЕРСКО..КУМСКОЙ
НИЗМЕННОСТИ ПО АНА ТОМО..ФИЗИОЛ оrИЧЕС ким
ПР ИЗllAКАМ ........................................................................... . . .... ..29
ИСТОРИЯ ИЗУЧЕНИЯ ФЛОРЫ восточноrо
ПРЕД]{АВКАЗЬЯ ............................................................................. 44
эколоrО..БиолоrиЧЕСКИЕ ОСОБЕННОСТИ
r АЛОФИТОВ ... ............ ........... ......... ..... ........... .... ........ ................... 55
ТАКСОНОМИЧЕСКИЙ АНАЛИЗ ФЛОРЫ rАЛОФИТОВ........64
АНАЛИЗ эколоrИЧЕСКИХ rрупп rАЛОФИТОВ ТЕРСКО..
КУМСКОЙ НИЗМЕННОСТИ.. ..................... ....... ... ......... ......... ......85
АНАЛИЗЛЕоrРАФИЧЕскоrОРАСПРОСТРАНЕНИЯ
r АЛОсl>ИТОВ ТЕРСКО..КУМСКОЙ НИЗМЕННОСТИ ........ ...... 90
БиомоРФолоrиЧЕСКИЙ АНАЛИЗ r АЛОФИТОВ ТЕРСКО..
КУМСКОЙ ниЗМЕннОСТИ......................................................... 99
АНАЛИЗ ЭНДЕМИЗМА И РЕЛИКТОВОСТИ r АЛОФИТОВ
ТЕРСКО..КУМСКОЙ НИЗМЕННОСТИ ...................................... 1 О 1
ЗАКЛЮЧЕниЕ.............................................................................. 104
ЛИ TEI) А ТУ Р А .............................................. .. . .. . .. . .. .. . . . .. .. . .. . .. . . . . . ... 1 06
122
Формат 60х84 1/16. r'арнитура Тайме. Бумаrа офсетная.
Тир. 350 ЭК3. Размножено ип «Бисултанова П.Ш.»,
Махачкала, ул. М.rаджиева, 34.*
123