




















































































































































































































































































































































Автор: Сойфер В.Н.
Теги: биологические науки в целом общая биология биология энциклопедия современное естествознание
ISBN: 5-89317-134-9
Год: 2000




Текст
щнклопедт
фтвшт
стествоЗшнт
ISSEP
МЕЖДУНАРОДНАЯ СОРОСОВСКАЯ ПРОГРАММА ОБРАЗОВАНИЯ В ОБЛАСТИ ТОЧНЫХ НАУК
э
нциклопедия
СОВРЕМЕННОЕ
ЕСТЕСТВОЗНАНИЕ
В десяти томах
МАТЕМАТИКА. МЕХАНИКА
ФИЗИКА ВОЛНОВЫХ ПРОЦЕССОВ
ФИЗИКА КОНДЕНСИРОВАННЫХ СРЕД
ФИЗИКА ЭЛЕМЕНТАРНЫХ ЧАСТИЦ. АСТРОФИЗИКА
ФИЗИЧЕСКАЯ ХИМИЯ
ОБЩАЯ ХИМИЯ
ОБЩАЯ БИОЛОГИЯ
МОЛЕКУЛЯРНЫЕ ОСНОВЫ БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
НАУКИ О ЗЕМЛЕ
СОВРЕМЕННЫЕ ТЕХНОЛОГИИ
МОСКВА
Издательский Дом
МАГИСТР-ПРЕСС
ISSEP
МЕЖДУНАРОДНАЯ СОРОСОВСКАЯ ПРОГРАММА ОБРАЗОВАНИЯ В ОБЛАСТИ ТОЧНЫХ НАУК
э
нциклопедия
СОВРЕМЕННОЕ
ЕСТЕСТВОЗНАНИЕ
Том 2
ОБЩАЯ
БИОЛОГИЯ
Главный редактор энциклопедии
В.Н. СОЙФЕР
Редактор тома
Ю.П. АЛТУХОВ
МОСКВА
Издательский Дом
МАГИСТР-ПРЕСС
2000
УДК 57
ББК 28.0 (т. 2)
ББК24.5
С 56
ТХи 658-937
Библиотеки
Конгресса США
Редактор энциклопедии Ю.А. Пашковский
Научные редакторы разделов тома
Л.И. Корочкин, О.Н. Кулаева, Е.Я. Тетушкин
Современное естествознание: Энциклопедия: В 10 т. - М.: Издательский Дом
МАГИСТР-ПРЕСС, 2000. - Т. 2. - Общая биология. - 344 с: ил.
ISBN 5-89317-134-9 (т. 2)
ISBN 5-89317-132-2
Энциклопедия «Современное естествознание» подготовлена к печати Министерством образования
Российской Федерации и Международной Соросовской Программой Образования в Области Точных Наук.
Энциклопедия знакомит читателей с достижениями в области математики, физики, химии, биологии и наук о
Земле за последнюю четверть века. Статьи написаны выдающимися учеными и преподавателями высшей
школы, большинство из которых - соросовские лауреаты. Энциклопедия рассчитана на преподавателей средних
школ, учеников старших классов, студентов и аспирантов вузов, а также на широкий круг читателей,
интересующихся естественными науками, и распространяется бесплатно по библиотекам средних школ и
высших учебных заведений России.
Издание осуществлено на средства Правительства Российской Федерации.
Copyright under International Copyright Union
All rights reserved under Universal Copyright Convention
by International Soros Science Education Program
Никакая часть данного издания не может быть воспроизведена или использована в любой форме
и любыми средствами - электронными или печатными, включая фотокопирование, ксерокопирование,
с помощью компьютерной записи и путем другого электронного воспроизведения, или любыми другими
способами хранения и распространения информации- без письменного разрешения издателя.
Налоговая льгота — общероссийский классификатор продукции
ОК-005-93, том 2; 953000 — книги, брошюры
Подписано в печать 31.01.2000 г. Формат 60x90/8. Печать офсетная.
Усл. печ. л. 43,0. Уч.-изд. л. 26,7. Тираж 5500 экз. Заказ № 4107.
ЛР № 060636 от 03.04.1997 г.
Почтовый адрес: 117292, Москва, ул. Кедрова, д. 8, корп. 3.
Оригинал-макет тома предоставлен издательством «Флинта».
Отпечатано с готовых диапозитивов в АООТ «Тверской полиграфический комбинат»
170024, г. Тверь, пр-т Ленина, 5.
Scan AAW
ISBN 5-89317-134-9
9"785893"171341
ISBN 5-89317-134-9 (т. 2)
ISBN 5-89317-132-2
© Международная Соросовская Программа
Образования в Области Точных Наук, 2000
Издательский Дом МАГИСТР-ПРЕСС, 2000
эволюция
И СИСТЕМАТИКА
И. А. Михайлова, О. Б. Бондаренко
СИСТЕМА ОРГАНИЧЕСКОГО МИРА
Построение естественной системы
органического мира является непрерывным
процессом. В настоящее время с учетом
ископаемого и современного материала
выделяют от 4 до 26 царств, от 33 до 132
типов, от 100 до 200 классов, а общее число
видов оценивается в несколько миллионов
(см.: Алтухов Ю.П. Вид и видообразование.
Наст. том). Естественно, что системы
органического мира, построенные в разное
время, существенно отличаются друг от
друга. Заранее хотим подчеркнуть, что о
некоторых, даже крупных и крупнейших,
таксонах живой природы, особенно
ископаемых, будет сказано кратко или они не
будут упомянуты вовсе.
Большинство классификаций
современных групп органического мира построены
на основе кладистического метода, или
кладистики (от греч. klados — ветвь). Кла-
дистика — один из подходов к построению
родословных древ организмов,
базирующийся на степени их родства. Полученные
таким методом родословные, основанные
на анализе молекулярно-генетических,
эмбриологических, цитологических и других
данных, в целом достаточно объективно
отражают уровни эволюции и степень
родства групп. Тем не менее без учета
палеонтологических данных, то есть
геохронологии, генеалогических связей и т. д.,
построение относительно стабильной
филогенетической системы органического
мира невозможно.
Теория и практика классификации
органических объектов получили название
таксономии (от греч. taxis — расположение,
строй, закон). Необходимо различать два
понятия: таксоны и таксономические
категории, то есть ранги таксонов. Число
таксонов как биологических объектов по мере
познания органического мира все время
возрастает.
Систематика (от греч. systematikos —
упорядоченный) представляет собой раздел
биологии, в задачи которого входят, с
одной стороны, описание всего многообразия
как современных, так и вымерших
организмов, а с другой — упорядоченное
иерархическое расположение таксономических
категорий по отношению друг к другу. (Иногда
термины «систематика», «таксономия» и
«классификация» считают синонимами,
поэтому наряду с понятием «таксономическая
категория» нередко используют понятие
«систематическая категория».) Таким
образом, систематика представляет собой прежде
всего процесс исследования, конечным
результатом которого является построение
системы.
Понятия «род» и «вид», а также
бинарное название (биномен) для обозначения
видов впервые предложил в середине XVI
века К. Геснер. В соответствии с бинарной
номенклатурой (от лат. binarius—
состоящий из двух частей и nomenclatura —
перечень имен) латинское наименование вида
состоит из двух слов: вид получает
двойное наименование — первое слово
представляет собой название рода, а второе —
вида, например Betula alba (береза белая).
Широкое применение бинарной
номенклатуры началось с работ английского
священнослужителя, ботаника-систематика,
зоолога и путешественника Дж. Рея (1628—
1705). Рей предложил разделять растения
на две большие группы, соответствующие
однодольным и двудольным.
Создателем научной таксономии и
систематики по праву является шведский
натуралист К. Линней (1707—1778). Он
разработал правила и принципы классификации
и построил иерархическую систему для
известных в то время современных и
ископаемых животных и растений. С его
работами с середины XVIII века окончательно
утвердилось применение бинарной
номенклатуры.
В настоящее время число основных
таксономических категорий возросло до
двенадцати: вид, род, триба, семейство,
отряд, когорта, класс, тип, раздел, царство,
доминион, империя. В ботанике таксонам
в ранге отряда и типа соответствуют
порядок и отдел, хотя некоторые авторы
считают, что типу в царстве животных
соответствует подотдел в царстве растений.
Благодаря систематике разнообразие
жизни предстает не как хаотическое
нагромождение организмов, а как
определенным образом упорядоченная система.
Естественно стремление построить такую
систему, которая отражала бы
филогенетическую преемственность. Исходным
может быть постулат, что более простые
организмы соответствуют предковым
состояниям, а более сложные — последующим
уровням развития.
Систему органического мира
изображают в двух основных вариантах: в виде
родословного древа, ветви которого
связаны родственными отношениями и
соответствуют определенным таксонам, или как
перечень названий таксонов в иерархичес-
8
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
кой последовательности. Излагаемая ниже
система включает два надцарства и пять
царств:
Для двух наиболее крупных царств —
растений и животных — принята
следующая иерархия высших таксонов:
Многие организмы бактериального,
растительного и животного происхождения на
одноклеточном уровне имеют ряд сходных
черт. Еще в 1866 г. Э. Геккель выделил
самостоятельное царство Protista (от греч.
protistos — самый первый). Современные
сторонники обособления царства Protista
включают в него как одноклеточных эука-
риот, так и многоклеточные водоросли.
Основу живых организмов составляет
клетка, которая функционирует как
самостоятельный организм— разнообразные
одноклеточные, либо клетки являются составной
частью многоклеточных. Основное
содержимое клетки — цитоплазма включает одно или
несколько ядер, вакуоли, митохондрии и т. д.
Наличие ядра, содержащего генетический
аппарат, или отсутствие оформленного ядра
является морфологическим признаком для
разграничения надцарства прокариот (доядер-
ные) и эукариот (ядерные).
Существует гипотеза, что на первых
этапах эволюции органического мира
широко проявлялся процесс возникновения
более сложных организмов за счет
слияния нескольких простых (симбиогенез, эн-
досимбиоз). Современная эукариотная
клетка возникла в результате длительных и
многократных эндосимбиозов. Возможно,
что такие клеточные структуры, как
реснички, жгутики, центриоли, появились за
счет серии внедрений различных бактерий
и цианобионтов (рис. 1).
Царство Животные
Царство Растения
Царство
Грибы
О
>
ее
б
ш
о
CQ
О.
3
м Эндосимбиоз
с различными бактериями
Эндосимбиоз
с различными бактериями и цианобионтами
СЕ
<
О
О
DC
Ql
О
00
&
а
э
X
Формирование эукариотной клетки
о?
((**
Pi***
Первичные клетки
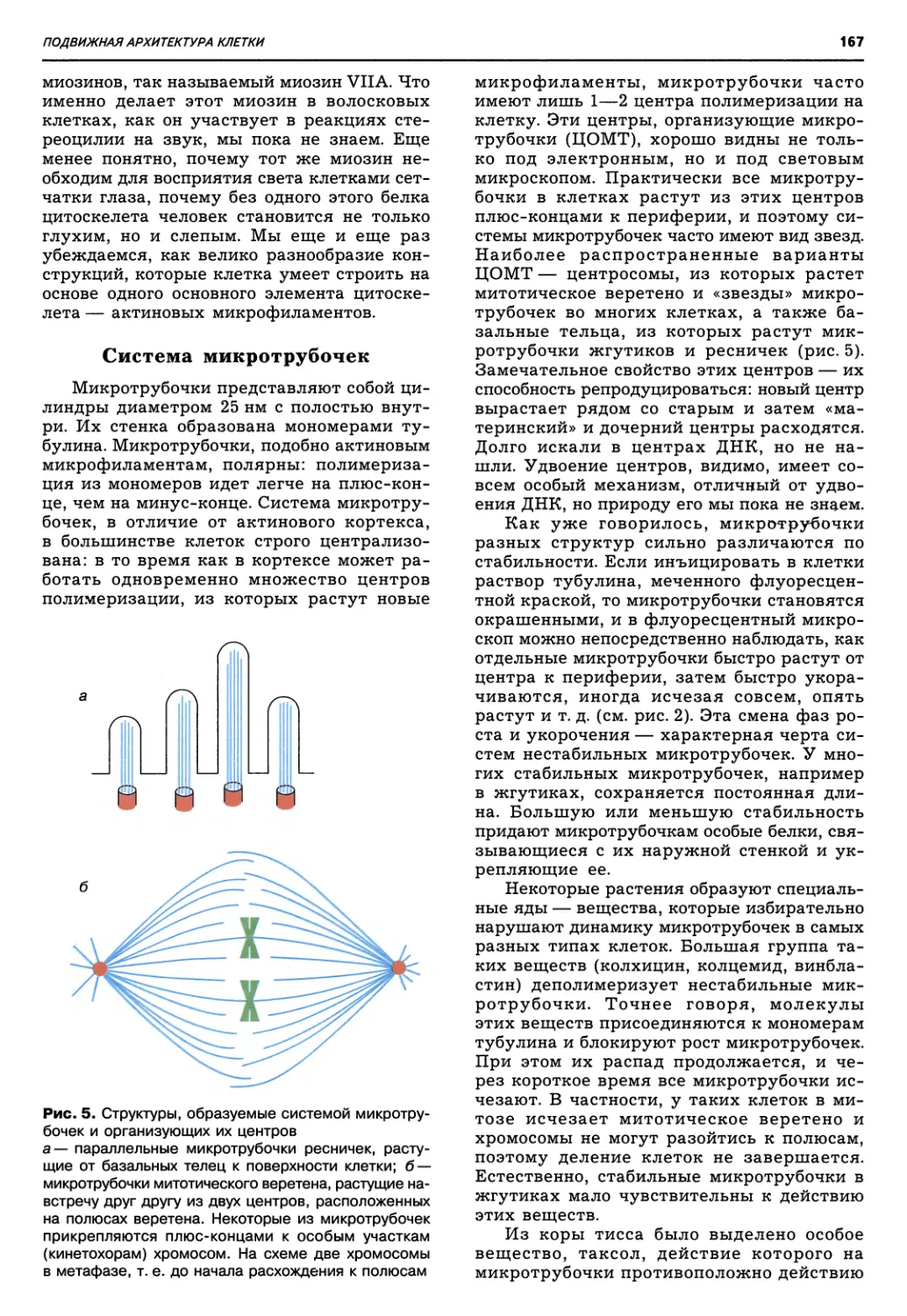
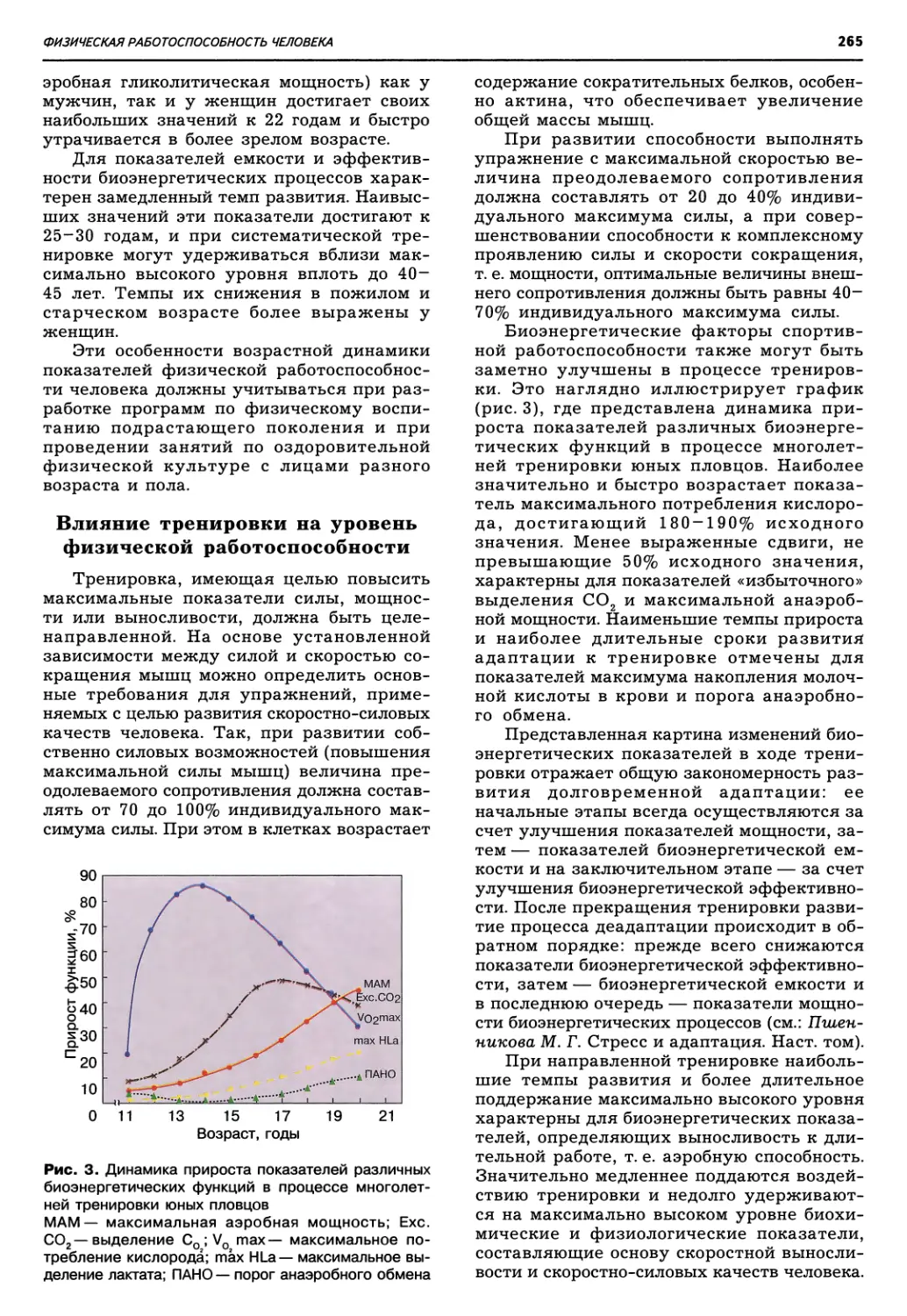
Рис. 1. Схема эволюции органического
мира согласно теории симбиогенеза
СИСТЕМА ОРГАНИЧЕСКОГО МИРА
9
НАДЦАРСТВО ДОЯДЕРНЫЕ
ОРГАНИЗМЫ.
SUPERREGNUM PROCARYOTA
Это одноклеточные и многоклеточные
(колониальные) организмы, не имеющие
обособленного ядра. Их клетки имеют стенку, а
генетическая информация сосредоточена в
единственной хромосоме. Размеры
прокариот варьируют от 0,015 мкм до 20 см
(колониальные формы). Эти организмы появились
3,8—3,1 млрд лет назад. Они разделяются на
два царства: бактерий и цианобионтов (рис. 1).
Обмен веществ осуществляется в процессе
хемосинтеза и фотосинтеза.
Царство Бактерии. Regnum Bacteria
Бактерии представляют собой
микроскопические организмы, размеры которых
составляют обычно около 1—5 мкм.
Гигантские бактерии размером более 10 мкм
обнаружены в денсалях — черных и белых
«курильщиках». Термин «денсаль», или «ден-
соабиссаль» (от лат. densum — плотный,
компактный, густой) предложен для бионо-
мической зоны, связанной преимущественно
с рифтовыми поясами, где извергаются
многочисленные гидротермальные
выбросы, струи которых отличаются по цвету от
окружающей морской воды и поэтому
напоминают струи дыма (отсюда название
«курильщики»).
Среди бактерий встречаются автотроф-
ные и гетеротрофные формы. Первые
создают органические вещества из
неорганических, вторые используют готовые
органические вещества. Большинство
бактерий являются автотрофами (литотрофами).
Процессы обмена веществ у автотрофных
бактерий идут либо (у хемолитотрофов) без
использования света (путем хемосинтеза),
либо (у фотолитотрофов) только на свету
(путем фотосинтеза). По типам обмена
веществ бактерии чрезвычайно разнообразны.
Различают серообразующие,
железисто-марганцевые, азотные, ацетатные, метано-, уг-
леродообразующие и другие группы
бактерий. Роль бактерий в геологических процессах
чрезвычайно велика. С их деятельностью
связано образование разнообразных
полезных ископаемых: железных руд
(железистых конкреций, джеспилитов), пирита, серы,
графитов, карбонатов, фосфоритов, нефти,
газа и др.
Достоверные находки бактерий
известны из кремнистых пород, имеющих возраст
около 3,5 млрд лет, проблематичные
находки датируются с уровня 3,8 млрд лет
(рис. 2). Скорее всего, бактерии появились
независимо в различных средах обитания.
В настоящее время они населяют все
водные бассейны от литорали до абиссали, а
также обитают в почве и горных породах,
в воздухе, внутри других организмов. Они
живут в горячих источниках при
температуре, превышающей 100°С, и в соленых
водах с высокой концентрацией солей.
Современная классификация царства
бактерий основана в первую очередь на
строении стенки клетки. Особую группу
представляют архебактерии, которые по
физиологическим и биохимическим свойствам
отличаются от групп истинных бактерий,
или эубактерий. Для архебактерии
характерно разнообразие по обмену веществ,
особый состав клеточной стенки, некоторым
из них свойствен своеобразный тип
фотосинтеза — свет поглощается мембранным
белком бактериородопсином, а не
хлорофиллом. Поэтому архебактерии выделяют в ранг
подцарства, а в последнее время,
особенно на основании сравнительного изучения
нуклеотидных последовательностей ДНК
эубактерий и архебактерии, возводят в ранг
самостоятельного царства.
Некоторые исследователи объединяют с
бактериями вирусы, полагая, что
упрощение их строения обусловлено способом
существования — внутриклеточным
паразитизмом. Другие рассматривают их как
доклеточную форму жизни и выделяют в
самостоятельное царство Virae. Вирусы в
ископаемом состоянии пока не
обнаружены. Значение вирусов в современной биоте
очень велико. Они были открыты в конце
прошлого века как возбудители болезней
(недаром их название происходит от лат.
virus — яд). В ходе столетнего изучения
болезнетворных вирусов произошло
становление и развитие науки вирусологии,
одного из разделов микробиологии. Вирусы
устроены просто, но они имеют
генетический аппарат и, подобно другим живым
организмам, обладают способностью к развитию.
Установлена вирусная природа многих
заболеваний человека и других теплокровных
позвоночных животных (их вызывают
около 500 вирусов). Более 300 вирусов живет
в клетках растений. Многочисленны
вирусы собственно бактерий, так называемые
бактериофаги. И хотя вирусы в ископаемом
состоянии неизвестны, наиболее вероятно,
что они появились на ранних этапах
развития биосферы.
Царство Цианобионты.
Regnum Cyanobionta
Одиночные и колониальные организмы
с постоянной формой клеток без
обособленного ядра. Размеры одиночных форм
микроскопические— около 10 мкм. Размеры
колоний, а особенно продуктов их
жизнедеятельности (строматолиты) могут
достигать многих сотен метров. Колониальные
формы покрыты общей слизистой оболоч-
10
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
кой. В самом организме, на его
поверхности и в слизистой оболочке может
происходить накопление карбонатов, приводящее
в дальнейшем к формированию
известняков. Известняковые слоистые образования
получили название строматолитов (рис. 2).
Цианобионты наряду с фикоцианом,
фикоэритрином, каротином имеют и
хлорофилл. Перечисленные пигменты
определяют розоватую, желтоватую, сине-зеленую,
а иногда почти черную окраску.
Цианобионты появились около 3,5 млрд лет назад.
Благодаря наличию хлорофилла они
являются первыми фотосинтезирующими
организмами, продуцирующими биогенный
молекулярный кислород. Современные
цианобионты живут и в пресных, и в
морских бассейнах; в последних в зоне
мелководья не глубже 150 м, но преимущественно
на глубине от 0 до 20 м. Цианобионты пе-
Рис. 2. Ископаемые прокариоты и
(?) эукариоты
а — древнейший организм (?) (3,8
млрд лет); б—д — прокариоты,
представленные бактериями и ци-
анобионтами (б—г— 3,5 млрд лет,
д— 2,5 млрд лет); е—к—
прокариоты и (?) эукариоты (2 млрд лет)
(а—д—по [Earth's Earliest Biosphere,
1983]; е-к— по [Крылов, 1968]); л-
ц — внешний вид и продольные
сечения строматолитов (АН-0):л, м —
пластовые строматолиты Collenla
(AR-Q); и—у— столбчатые и жел-
ваково-столбчатые строматолиты;
н, о— Kusslella (R), п — Conophyton
(PR), p—c— Baicalia (R2-3), г —
Mlnjarla (R3), у— Gymnosolen (R3);
ф—х— онколиты Osagia (AR-Q); ц —
Yakutophyton (PR), отдельные
фрагменты единой постройки отвечают
морфотипам разных родов
СИСТЕМА ОРГАНИЧЕСКОГО МИРА
11
реносят загрязнение и резкие колебания
физико-химических условий. Диапазон
температур — от минусовой в ледниках до
почти точки кипения в горячих источниках.
Среда обитания — пресные, солоновато-
водные и нормально соленые морские
бассейны, а также засоленные, обогащенные
нитратами и сульфатами. Некоторые циа-
нобионты обитают в почве и на ней, на
камнях, в пустынях и т. д.
По отсутствию ядра цианобионты
сближаются с бактериями, а по наличию
хлорофилла и способности синтезировать
биогенный молекулярный кислород — с водорослями.
Отделение синезеленых от царства растений
и перенос в надцарство прокариот привел к
их фактическому объединению с царством
бактерий и рассмотрению в качестве циано-
бактерий. Мы считаем эту группу
самостоятельным царством и поэтому используем
название цианобионты: с одной стороны, ясно,
что это «бывшие» синезеленые, а с другой —
сохраняется этимологическая дистанция от
бактерий.
Сравнительно недавно выявлена
небольшая группа ранее неизвестных прокариот-
ных организмов (род Prochloron). По составу
пигментов эта группа ближе к зеленым
водорослям, нежели к так называемым сине-
зеленым. Вопрос о месте этих организмов
в надцарстве прокариот еще ждет своего
разрешения. Возможно, их следует
причислить к цианобионтам, тем самым расширив
объем и диагноз этого царства (см. рис. 1).
В иерархии живых организмов
цианобионты находятся на более высокой ступени, чем
бактерии (имеют более сложную
структуру и пигменты), но на более низкой, чем
водоросли (отсутствует ядро).
НАДЦАРСТВО ЯДЕРНЫЕ
ОРГАНИЗМЫ.
SUPERREGNUM EUCARYOTA
Эукариоты — одноклеточные или
многоклеточные организмы, разделяющиеся на
три царства: растений, животных и грибов.
В отличие от прокариот они имеют
обособленное ядро. Размеры эукариот
изменяются в диапазоне от 10 мкм (одноклеточные)
до 33 м (длина китообразных) и 100 м
(высота некоторых гигантских хвойных).
Эукариоты появились позднее прокариот,
скорее всего 1,5—1,7 млрд лет тому назад
(ранний протерозой), хотя не исключено и
более раннее возникновение.
Не всегда можно четко разграничить
одноклеточные растения и животные. Так,
например, жгутиковые синтезируют
органические вещества, являются автотрофа-
ми и включаются в царство растений
(отдел динофитовых водорослей), а другие
питаются готовыми органическими
соединениями и причисляются к царству животных.
Некоторые формы занимают
промежуточное положение, и в этой связи выделение
самостоятельного царства Protista не
лишено основания.
Царство Растения. Regnum Phyta
Это разнообразные, преимущественно
неподвижные одноклеточные и
многоклеточные организмы, имеющие верхушечный
рост, плотные, преимущественно
целлюлозные оболочки клеток и автотрофный
способ питания. Для всех растений характерен
фотосинтез: при помощи энергии света,
поглощаемой хлорофиллом, реже другими
пигментами, они выделяют молекулярный
кислород, а из неорганических соединений
создают органические.
Клетка растений состоит из
цитоплазмы, которая содержит ядро, полости —
вакуоли и органоиды - разнообразные
пластиды. Последние ограничены мембранами
и представляют самостоятельные
внутриклеточные образования, различающиеся
формой, размерами, окраской и
функцией. Твердая целлюлозная оболочка клетки
пронизана порами, нередко
пропитывается солями и минерализуется. Достоверные
находки растений известны со времен
верхнего венда, хотя, несомненно, они возникли
раньше.
Царство растений разделяется на два
подцарства, различающиеся уровнем
организации и средой обитания: Thallophyta
(низшие растения) и Telomophyta (высшие
растения). Первые обитают в
разнообразных водных бассейнах, и для них
используется собирательное название
«водоросли», то есть растущие в воде. Высшие
растения обитают в наземных условиях,
встречаясь почти на всех широтах, лишь
небольшое их число ведет вторичноводный
образ жизни.
Подцарство Низшие растения.
Subregnum Thallophyta
Низшие растения — это одноклеточные
и многоклеточные организмы, которые
обитают в разнообразных водных бассейнах,
а изредка живут в почве. Водоросли имеют
единое тело (таллом, слоевище), в котором
не выделяются корень, стебель и листья.
Их размеры изменяются от
микроскопических (несколько микрометров) до
гигантских (свыше 50 м). Размножение
осуществляется половым и бесполым путем. У многих
групп имеются различные
минерализованные покровные образования (покровные
клетки, чехлы, оболочки). Распространение
водорослей ограничено глубиной
проникновения света (не более 200 м), среди них
имеются как донные - бентосные формы,
12
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
так и пелагические — планктонные.
Достоверные находки известны с конца венда.
В основу выделения отделов, число
которых превышает 10, положены число
клеток (одноклеточные и многоклеточные),
различный набор окрашивающих
пигментов и особенности минерального скелета.
Подцарство Высшие растения.
Subregnum Telomophyta
Подцарство высших растений отличается
от подцарства низших растений следующими
особенностями: 1) тело расчленено на
корень, стебель, листья и органы
размножения; 2) специализация клеток приводит к
образованию различных специфических
тканей, осуществляющих проводящую,
защитную, механическую и другие функции;
3) среда обитания наземная, хотя имеются
некоторые вторичноводные формы; 4)
закономерное чередование полового (гамето-
фит) и бесполого (спорофит) поколений. В
жизненном цикле всех высших растений
(исключая мохообразных) спорофит резко
преобладает над гаметофитом, особенно у
древесных форм. Основным звеном в
эволюции высших растений явилось
преобразование и усложнение органов размножения
с сокращением и последующей редукцией
гаметофита как самостоятельного растения.
Параллельно происходило усложнение
процесса полового размножения. Микроспоры
постепенно преобразовались в пыльцу, а
мегаспорангии с мегаспорами — в
семязачатки (= семяпочки). После оплодотворения
семяпочки пыльцой возникает семя.
Подлинные растения известны с середины силура
(рис. 3).
В соответствии со способом
размножения подцарство высших растений разделено
на два надотдела: Sporophyta (споровые) и
Spermatophyta (семенные).
Надотдел Споровые растения.
Superdivisio Sporophyta
Споровые растения характеризуются
следующими признаками: 1) размножение
осуществляется с помощью спор; 2) гаме-
тофит свободноживущий; 3) ксилема
состоит из трахеид — удлиненных клеток с
толстой оболочкой, которая несет
разнообразную скульптуру и поры; 4)
эволюция споровых связана с выходом растений
на сушу и формированием ствола,
листьев и корня.
К споровым растениям относится пять
отделов: моховидные, риниофиты, плауно-
видные, хвощевидные и
папоротниковидные. У моховидных спорофит не
существует как самостоятельное растение, у всех
остальных спорофит самостоятельный и
преобладает над гаметофитом. Споровые
растения появились в середине силура, они
существуют и поныне, но в современной
флоре резко уступают в численности и
разнообразии семенным растениям.
Рис. 3. Родословное древо высших растений
СИСТЕМА ОРГАНИЧЕСКОГО МИРА
13
Надотдел Семенные растения.
Superdivisio Spermatophyta
Семенные растения характеризуются
следующими признаками: 1) размножение
осуществляется при помощи семян. Общий
признак голосеменных и покрытосеменных
растений - наличие семени, но у
голосеменных отсутствует завязь, поэтому семя
считают голым; 2) мегаспоры созревают на
спорофите и не покидают его; 3) гамето-
фит не существует как самостоятельное
растение; 4) впервые появляется
сосудистая система.
К семенным растениям отнесены два
отдела: пинофиты, или голосеменные, и маг-
нолиофиты, или покрытосеменные. Семенные
растения появились в позднем девоне, в
современной флоре они резко преобладают над
споровыми.
Царство Грибы. Regnum Fungi
Царство грибов сочетает свойства как
растений, так и животных. Общие
признаки грибов и растений: неподвижность,
верхушечный рост и размножение с помощью
спор. Вместе с тем у грибов, как и у
животных, отсутствует фотосинтез, в
продуктах обмена присутствует мочевина, а в
плотных оболочках клеток имеется хитин,
поэтому оболочки клеток могут
сохраняться в ископаемом состоянии. Известно
около 100 тыс. видов грибов.
Грибы могут быть одноклеточными и
многоклеточными; клетки
преимущественно многоядерные, редко одноядерные.
Многоклеточные грибы состоят из тонких
нитевидных образований. Нити, или гифы, при
разветвлении переплетаются, образуя
грибницу, или мицелий. Размеры и строение
грибниц разнообразны — от
микроскопических скоплений до крупных шляпочных
грибов. Размножение вегетативное или
половое. Споры как подвижные со жгутиками
(=зооспоры), так и неподвижные. Последние
образуются внутри специальных полостей
или на концах гифов. Они имеют плотную
оболочку и сохраняются в ископаемом
состоянии.
По типу питания грибы являются гете-
ротрофами: сапротрофами, паразитами,
редко хищниками. Благодаря тому, что
мицелий представляет систему
ветвящихся гифов, обеспечивается большая
поверхность для осмотрофного типа питания. При
этом происходит выделение ферментов и
осмотическое всасывание органических
веществ. Грибы существуют в почве и на
ее поверхности, а также снаружи и
внутри многих объектов и организмов. Для них
характерен симбиоз с корневой системой
высших растений. Симбиоз грибов с
водорослями привел к возникновению
лишайников, известных с позднего мела.
и л
е ж з к м
а б в г д
Рис. 4. Царство Fungi (V-Q)
а—д — реконструкция вендских грибов [Бурзин, 1993]:
а—б— талломы с ризоидами, в—д— спорангии; а—
д — европейская часть России; е — низшие грибы из
карбона Англии [Криштофович, 1957]; ж, з— гифы
низших (ж) и высших (з) грибов; и—м— споры
(конидии) в различном сочетании; ж—м— неоген,
Закарпатье [Попов, Рыбакова, 1970]
В ископаемом состоянии от грибов
сохраняются преимущественно споры
(конидии), реже гифы, мицелий и еще реже
отдельные клетки (рис. 4). Максимальное
число спор встречено в бурых углях.
Достоверные остатки грибов известны с
девона. В докембрии (средний рифей)
обнаружены гифоподобные образования. Выше, при
характеристике бактерий, говорилось о
микроскопических образованиях,
встреченных в археозое на рубеже 3,8 млрд лет.
Некоторые исследователи относят их к
низшим дрожжевым грибам.
Царство Животные.
Regnum Zoa (Animalia)
Царство животных включает
одноклеточные и многоклеточные организмы, для
которых характерны следующие признаки:
1) питание осуществляется готовыми
органическими продуктами (гетеротрофы). Для
животных в отличие от грибов характерен
фаготрофный тип питания, то есть захват
(заглатывание) пищевого материала; 2)
клетки не имеют целлюлозной оболочки и
различных пигментов, свойственных
растениям; 3) на протяжении всей жизни или на
отдельных возрастных стадиях организмы
подвижные.
Размножение животных происходит
двумя способами: половым и бесполым.
Половой процесс сопровождается возникновением
половых клеток, слияние которых дает
начало новому организму. Бесполое
размножение представляет собой деление или
почкование. В результате образуются ко-
14
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
лонии, либо единый организм распадается
на несколько себе подобных особей. Коло-
ниальность характерна для многих
преимущественно прикрепленных многоклеточных
(губковых, археоциатов, книдарий, мшанок,
граптолитов), хотя колонии известны и в
подцарстве простейших (типы жгутиковых
и саркодовых). Полный жизненный цикл
нередко представляет собой чередование
полового и бесполого поколений, что
присуще как одноклеточным, так и
многоклеточным организмам.
Подцарство Простейшие,
или Одноклеточные.
Subregnum Protozoa
Это подцарство включает животных,
которые хотя и состоят из одной клетки,
но характеризуются значительным
разнообразием как по размерам, так и по
строению клетки. Простейшие многочисленны и
распространены повсюду, общее число
современных и ископаемых видов
приближается к 50 тыс. Большинство простейших
имеют микроскопические размеры (50—150
мкм), самые мелкие не достигают 10 мкм,
зато наиболее крупные превосходят 50 000
мкм (то есть 5 см), а иногда и более.
Клетка простейших является целостным
организмом, она полифункциональна, то есть
выполняет основные жизненные функции
(обмен веществ, движение, размножение).
Форма, как и строение клетки, весьма
различны. По форме тела большинство
простейших асимметрично, исключение составляют
радиально-лучистые акантарии, радиолярии
и некоторые двусторонне-симметричные фо-
раминиферы.
По способу питания простейшие
относятся к фитофагам и зоофагам: они
питаются микроорганизмами растительного и
животного происхождения. Пищеварение
внутриклеточное, то есть переваривание
пищи происходит в замкнутых вакуолях
внутри клетки, или внеклеточное
пристеночное, когда пищеварение
осуществляется в полузамкнутых полостях наружной
поверхности клетки. Размножение
простейших происходит половым и бесполым
путем, нередко наблюдается чередование
поколений.
Выполнение основных функций
производится отдельными участками клетки,
получившими название органоидов.
Особенности клетки: строение ядра, цитоплазмы,
органоидов движения, состав скелета —
основа для подразделения подцарства
простейших на типы.
Геологическая история простейших
фактически прослежена только для двух классов
типа саркодовых: фораминифер и
радиолярий. Что касается остальных типов и
классов, то можно утверждать, что
многие бесскелетные формы, несомненно,
возникли в глубокой древности, но доказать
это на палеонтологическом материале не
представляется возможным. Простейшие
ведут свое начало от каких-то прокариот-
ных организмов.
Подцарство Многоклеточные.
Subregnum Metazoa
К подцарству многоклеточных
относятся животные, тело которых состоит из
большого числа клеток, слагающих ткани и
органы и выполняющих различные
функции. По уровню строения Metazoa
подразделяются на два надраздела: Parazoa —
примитивные (?ненастоящие) и Eumetazoa —
настоящие многоклеточные. У первого из
названных надразделов нервная система
отсутствует, а у второго имеется.
Надраздел Примитивные
многоклеточные. Superdivisio Parazoa
Примитивные многоклеточные не
имеют стабильной дифференциации клеток как
по морфологии и функциям, так и по
положению в теле животного. Поэтому у них
отсутствуют ткани и органы, а в
эмбриогенезе не формируются зародышевые
листки. Это водные животные, ведущие
прикрепленный образ жизни. Они являются
фильтраторами и получают пищу вместе с
током воды. Им свойственно пристеночное
и внутриклеточное пищеварение, что
сближает этот надраздел с подцарством
простейших. К надразделу Parazoa относятся три
типа: Spongiata, Placozoa и Archaeocyathi,
третий из названных типов является
вымершим.
Надраздел Настоящие многоклеточные.
Superdivisio Eumetazoa
Настоящие многоклеточные обладают
стабильной дифференциацией клеток, у них
имеются ткани и органы, в эмбриогенезе
закладываются два или три зародышевых
листка. Для этих животных характерно
внеклеточное «резервуарное» пищеварение,
происходящее в единой пищеварительной
полости, либо в серии полостей,
образующих пищеварительную систему. При таком
типе пищеварения размер поглощаемых
пищевых частиц не зависит от размеров
клетки, что резко расширяет кормовую
базу, а отсюда и все метаболические и
физиологические процессы. Тем не менее
сохраняется внутриклеточное и
пристеночное пищеварение.
Надраздел Eumetazoa в соответствии с
типом симметрии и числом зародышевых
листков рассматривается в составе двух
разделов. К первому относятся животные,
обладающие радиальной симметрией и имеющие
в эмбриогенезе два зародышевых листка. Ко
СИСТЕМА ОРГАНИЧЕСКОГО МИРА
15
второму разделу принадлежат организмы, для
которых характерны двусторонняя
симметрия и закладка в эмбриогенезе трех
зародышевых листков. Двуслойные находятся на
более низкой ступени, чем трехслойные,
поэтому нередко говорится о низших и
высших настоящих многоклеточных.
Раздел Радиально-симметричные,
или Двуслойные. Divisio Radiata
или Diblastica
В подавляющем большинстве это
радиально-симметричные многоклеточные
животные, у которых закладывается два
зародышевых листка (эктодерма + энтодерма).
Пищеварительная система с единственным
ротовым отверстием. К этому разделу
принадлежат два типа: Стрекающие и
Гребневики, отличающиеся присутствием
стрекательных клеток у первого типа и отсутствием
таковых у второго. До недавнего времени
названные животные рассматривались в
ранге двух подтипов, входящих в единый
тип кишечнополостных.
Раздел Двусторонне-симметричные,
или Трехслойные. Divisio Bilateria
или Triblastica
К двусторонне-симметричным
относятся настоящие многоклеточные животные,
обладающие тремя зародышевыми листками
(эктодерма + энтодерма + мезодерма) и
пищеварительной системой, имеющей, как
правило, два отверстия: ротовое и
анальное. Эктодерма дает начало покровным
образованиям, включая формирование
наружного скелета, органов чувств и нервной
системы; энтодерма — прежде всего
пищеварительной системе, а за счет мезодермы
возникают внутренний скелет, кровеносная
и остальные системы.
В разделе билатерий выделяют два
подраздела: первичноротые (Protostomia) и
вторичноротые (Deuterostomia),
отличающиеся друг от друга типом дробления яйца,
способом закладки мезодермы, а также
различным положением ротового и анального
отверстий на эмбриональной и
постэмбриональной стадии развития. Достоверные
билатерий известны с вендского периода.
В последние годы возрастает число
сторонников иной концепции. Признавая, что
развитие трехслойных животных шло по
двум основным эволюционным
направлениям, некоторые исследователи считают
основополагающим признаком не положение
ротового отверстия, а тип дробления яйца.
Животные, для которых характерны
спиральное дробление яйца и телобластичес-
кий способ закладки мезодермы,
объединяются в Spiraloblastica (=Spiralia), а те,
которым свойственно радиальное
дробление яйца и чаще всего энтероцельный способ
закладки мезодермы, — в Radialoblastica
(=Radialia). Объем первичноротых и вторич-
норотых в основном совпадает с вновь
предлагаемыми эволюционными стволами.
Рис. 5. Многоцарственная система органического мира [Кусакин, Дроздов, 1994]
7 — MethanobacterlobJonta, 2 — Halobacteriobionta, 3 — Thermoacidobacteriobionta, 4 — Archaetericutobacteriobion-
ta, 5— Spirochaetobionta, 6— Oxyphotobacteriobionta, 7— Anoxyphotobacteriobionta, 8— Scotobacteriobionta, 9 —
Rufirmicutobacteriobionta, 10 — Actinobacteriobionta, 71 — Tenericutobacteriobionta, 72—Rhidobionta, 73 —Crypto-
bionta, 74—Chlorobionta (a —Thallophyta, 6—Embryophyta), 15— Inferiobionta, 16— Metazoa, 77—Mycobionta,
18— Euglenobionta, 79— Dinobionta (a — Dinoflagellata, 6— Ciliaturata), 20 — Chromobionta, 27 — Microsporobionta,
а также Virae
16
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Необходимо отметить, что большинство
систем органического мира построены по
принципу монофилии и дивергенции. В
последнее время увеличивается число
сторонников концепции параллельного развития
различных ветвей (парафилии, но не по-
лифилии). Более того, рассматривается идея
о радиальном многоцарственном развитии
(рис. 5) органического мира,
насчитывающего 22 царства. О радиальном развитии
свидетельствует и схема эволюции живых
существ, уточненная с помощью геномных
исследований.
Литература
Биологический энциклопедический словарь.
М., 1986. 832 с.
Бурзин М.Б. Древнейший хитридиомицет
(Mycota, Chytridiomycetes incertae sedis) из
верхнего венда Восточно-Европейской платформы
// Фауна и экосистемы геологического прошлого.
М.: Наука, 1993. 125 с.
Криштофович А.Н. Палеоботаника. 4-е изд.
Л.: Гостоптехиздат, 1957. 650 с.
Крылов И.Н. Древнейшие следы жизни на
Земле // Природа. 1968. № 11. С. 41—54.
Кусакин О.Г., Дроздов А.Л. Филема
органического мира. СПб.: Наука, 1994. Ч. 1:
Пролегомены к построению филемы. 282 с.
Малахов В.В. Проблема основного плана
строения брахиопод и их положение в системе
животного царства // Современное состояние и
основные направления изучения брахиопод. М.:
ПИН РАН, 1995. С. 51—82.
Мейен СВ. Основы палеоботаники. М.:
Недра, 1987. 404 с.
Михайлова И.А.,Бондаренко О.Б.
Палеонтология. М.: Изд-во МГУ, 1997. 446 с.
Попов П.А., Рыбакова И.О. Грибы неогеновых
отложений Закарпатья // Докл. АН СССР. 1970.
Т. 191, № 6. С. 1393—1395.
Сойфер В.Н. Международный проект «Геном
человека» // Сорос, образоват. журн. 1998. № 12.
С. 4—11.
Earth's Earliest Biosphere: Its Origin and
Evolution / Ed. G.W. Schopf. Princeton: Princeton
Univ. Press, 1983. 543 p.
Б. В. Громов
УДИВИТЕЛЬНЫЙ МИР АРХЕЙ
Введение
Ко второй половине XX в. стало
очевидным, что все живые организмы,
имеющие клеточное строение, могут быть
отнесены к одной из двух групп: прокариотам
или эукариотам. Клетки прокариот
устроены проще клеток эукариот; они лишены
окруженного оболочкой ядра. В течение
некоторого времени считали, что
термины «прокариоты» и «бактерии» —
синонимы и что все бактерии представляют
единую самостоятельную ветвь эволюции
живых организмов. Однако постепенно
накапливались данные о том, что различия
между некоторыми бактериями могут быть
очень существенными. Это свидетельствует
о том, что они не находятся в близком
эволюционном родстве. Доказательства
отсутствия такого родства получены
главным образом в результате
сравнительного изучения рибосомных и транспортных
рибонуклеиновых кислот (РНК),
сопоставление нуклеотидных последовательностей
которых дает информацию о генеалогии
организмов. Эти данные, а также
исследования других особенностей прокариот
показали, что на самом деле их следует
разделить по крайней мере на две
группы, имеющие различное эволюционное
происхождение.
Эукариотические клетки не только
сложнее прокариотических, но и
обладают элементами, которые они не могли
получить от предков современных
прокариот. Так, например, их цитоплазмати-
ческие рибосомы отличны от рибосом
прокариот.
Принципиальное сходство
генетического кода, организации макромолекул и
биохимического аппарата синтеза белка
свидетельствует о единстве
происхождения всех живых организмов.
Предполагают, что у них был один общий предок —
«прогенот», но что он собой представлял,
неизвестно. Этот «прогенот» мог дать
начало трем самостоятельным ветвям
эволюционного древа. С точки зрения
иерархической систематики, они представляют
таксоны, имеющие ранг выше
традиционных «царств»; такие систематические
группы предложено называть доменами.
Выделяют три домена — бактерии, археи
и эукариоты. Археи, которых до
недавнего времени называли архебактериями,
разнообразны и могут быть разделены на
два царства: кренархеот и эвриархеот.
Общая характеристика архей
По форме и размерам клеток, общим
принципам их организации и характеру
деления археи сходны с бактериями, хотя
только среди них обнаружены организмы
кубической формы. Многие археи
подвижны и имеют жгутики, похожие на
бактериальные, но несколько отличающиеся
деталями организации. Однако
представители этого домена имеют и существенные
особенности.
1. Уже было отмечено, что важнейшей
особенностью архей является своеобразие
их рибосомных и транспортных РНК.
Кроме того, их рибосомы различаются по форме.
Отличительные черты обнаружены и в
других компонентах системы синтеза белка.
2. В отличие от всех других организмов
археи имеют в составе мембранных липи-
дов не жирные кислоты, а многоатомные
спирты, обычно с 20 или 40 атомами
углерода. В последнем случае липидная
пластина мембраны образована
мономолекулярным слоем, что, вероятно, придает ей особую
прочность.
3. Покровы клеток у разных архей
могут иметь различное строение и
химический состав, но им часто присуще наличие
поверхностных слоев, образованных
определенным образом структурированными и
регулярно уложенными белковыми или гли-
копротеидными молекулами правильной или
довольно причудливой формы (рис. 1).
Иногда в состав клеточных стенок архей входят
пептиды и полисахариды.
4. Некоторые археи осуществляют
биохимические процессы, не свойственные
другим организмам. Например, только
определенные представители архей в процессе
своей жизнедеятельности образуют метан.
5. Большинство архей — экстремофилы,
т. е. развиваются в экстремальных условиях:
при высокой температуре, кислотности или
в насыщенных солевых растворах.
6. Археи, видимо, неспособны к
паразитизму. По крайней мере, к настоящему
времени археи, приносящие вред каким-
либо другим организмам, неизвестны, хотя
среди них много симбионтов, извлекающих
взаимную пользу из совместной жизни с
другими организмами. Среди архей много
автотрофных форм, не нуждающихся в
органической пище и получающих
необходимую для жизни энергию за счет
окислительно-восстановительных реакций, в
которые вовлечены неорганические молекулы.
18
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
а
нм
6,3
18,0
4,5
20,5 нм
б
Рис. 1. Строение белковой оболочки Sulfolobus
а— схема поперечного сечения. Овальные
элементы встроены в цитоплазматическую мембрану и
служат якорями, через соединители они связаны с
фигурными субъединицами трехлучевой симметрии,
образующими пористый поверхностный белковый
слой; б— реконструкция поверхности на основе
данных электронной кристаллографии [из: Baumeister,
Lembcke. Bioenerget. Biomembr., 1992, vol. 24, № 6,
p. 567]
Царство кренархеот
Организмы, принадлежащие к кренар-
хеотам, образуют довольно гомогенную
группу. Все они получают энергию в процессах,
связанных с восстановлением или
окислением соединений серы, и являются
гипертермофилами, т. е. развиваются при
температуре выше 80°С. Название этого царства
происходит от греческих корней «крен», что
означает источник, ключ, и «архе» —
древний. Действительно, эти археи обитают
исключительно в горячих источниках, на
поверхности Земли или на дне океана, обычно
в зонах вулканической активности. Местом
их обитания являются, в частности,
окрестности глубоководных вулканических
источников — «черных курильщиков»,
расположенных в океане на тысячеметровых
глубинах. Температура воды в них,
благодаря высокому давлению, может достигать
200-300°С. При взаимодействии воды
источника с морской водой образуется темный
осадок, источник как бы дымит. Отсюда и
название этих источников. Около таких
источников развиваются экстремально
термофильные археи, некоторые из них
растут даже при температурах 100-110°С. К
ним относится, например, Pyrodictium
occultum— «огненная сеточка», организм,
представляющий собой сеточку,
образованную шариками-кокками, соединенными
тяжами. Лучше всего «сеточка» развивается
при температуре 105°С, используя для
жизни энергию окисления элементарной
серой молекулярного водорода,
содержащегося в вулканическом газе: Н2 + S = H2S.
Другой представитель кренархеот —
Sulfolobus acidocaldarius — растет при
температурах до 100°С, используя энергию
окисления элементарной серы
молекулярным кислородом: 2S + 02 + 2Н20 = 2H2S04.
Очевидно, в результате
жизнедеятельности Sulfolobus происходит сильное подкис-
ление среды, но это для него полезно,
поскольку он растет при рН 1-6. Sulfolobus
не имеет строго определенной формы и
легко ее изменяет, так как окружен
только структурированным слоем гликопроте-
иновых субъединиц (рис. 1). На его
поверхности присутствуют волоски — пили, при
помощи которых Sulfolobus может
прикрепляться к частичкам серы, используемой им
в качестве субстрата для дыхания.
Царство эвриархеот
Организмы, относящиеся к эвриархео-
там, распространены повсеместно.
Некоторые формы являются экстремально
термофильными и живут около «черных
курильщиков», как, например, Pyrococcus
furiosus («яростные огненные шарики»).
«Шарики» развиваются при отсутствии
молекулярного кислорода за счет
использования органического вещества при
температурах 70-103°С. Однако представители
эвриархеот обнаружены и в арктической
тундре и даже в Антарктиде.
К эвриархеотам относится обширная
группа метанобразующих архей. Как уже
было отмечено, биологическое образование
метана осуществляется только археями.
Основным путем его образования является
окисление молекулярного водорода
углекислотой— «карбонатное дыхание»: 4Н2 +
+ С02 = СН4 + 2Н20. В некоторых случаях
для этого используются соли муравьиной и
уксусной кислот, метиловый спирт и
метиламины. В результате эти археи получают
необходимую им энергию. Среди
метанобразующих архей есть формы палочковидные,
шарообразные (кокки), спиральные; их
организм может быть образован одной клеткой,
УДИВИТЕЛЬНЫЙ МИР АРХЕЙ
19
а может состоять и из многих клеток.
Строение и состав клеточных стенок этих
микроорганизмов сильно варьируют. Метаноб-
разующие археи широко распространены;
1,0—1,5% углерода, участвующего в
круговороте в биосфере, проходит через
стадию метана. При образовании метана
может быть использован водород
вулканического происхождения. Существуют
экстремально термофильные формы,
развивающиеся в зонах горячих источников, как,
например, Methanothermus fervidus,
растущий при температурах 65—97°С.
Образование метана происходит в осадках морей и
пресноводных водоемов, болотах, почвах
тундры и рисовых полей. Метанобразующие
археи входят в состав кишечной
микрофлоры, в частности, они развиваются в
отделе желудка — рубце жвачных животных.
Накопление метана, хотя и
незначительное, отмечено и в кишечнике человека.
Метанобразующие бактерии интенсивно
синтезируют витамин В12 и обеспечивают им
своих хозяев. Они являются
внутриклеточными симбионтами некоторых простейших,
особенно развивающихся в отсутствие
молекулярного кислорода.
Метанобразующие археи могут
приносить практическую пользу. Например, они
участвуют в утилизации органических
отходов в так называемых метантенках. В этих
емкостях при высокой температуре и в
отсутствие молекулярного кислорода
происходит сбраживание органических веществ
разнообразной микрофлорой, в результате
чего образуются водород и углекислота,
которые и используются археями при
образовании метана. Благодаря высокой
температуре процессы идут с высокой
интенсивностью. В литературе сообщалось, что
от трупа лошади, помещенного в такой
метантенк, через неделю остался один
скелет. Были сконструированы также установки
Рис. 2. Скопления соли на берегу Мертвого моря
для получения горючего газа метана из
соломы, которые, как предполагают, могут
обеспечить газом небольшие
сельскохозяйственные поселения.
Экстремально галофильные археи,
способные к росту в насыщенных солевых
растворах, образуют самостоятельную
группу весьма своеобразных организмов, к
которым относятся представители родов
Halobacterium, Halococcus, Natronobacterium,
Natronococcus и некоторых других. Они
развиваются при концентрациях солей,
превышающих 250-300 г/л. Natronobacterium
и Natronococcus, кроме того,
предпочитают щелочные водоемы с крайне высокими
значениями рН. У галофилов высока
внутриклеточная концентрация солей, главным
образом в результате накопления ионов К+.
Их ферменты работают при высоких
солевых концентрациях, когда аналогичные
ферменты других организмов теряют
активность. Галофилы существуют за счет
использования органических соединений, они
могут расти в присутствии
молекулярного кислорода и без него. При отсутствии
молекулярного кислорода и наличии
света у них происходит образование так
называемых пурпурных или фиолетовых
мембран — участков поверхностной мембраны
клетки, содержащих пигмент родопсин,
аналогичный родопсину человеческого глаза.
В пурпурных мембранах за счет энергии
света происходит синтез аденозинтрифос-
фата (АТФ), являющегося основным
«энергоносителем» в клетках живых организмов.
Эта энергия может быть использована
археями для поддержания жизни, хотя свет
не является единственным источником
потребляемой ими энергии. Клетки некоторых
галофилов содержат также другие типы
родопсина, а именно сенсорные
родопсины I и II, входящие в состав
рецептора света и обеспечивающие способность этих
организмов при движении определенным
образом ориентироваться в отношении
источника света. Клетки галофилов обычно
содержат также красные каротиноидные
пигменты; при массовом развитии этих
архей субстрат (соль, скопления органики
и т. п.) окрашивается в красный цвет.
Галофилы населяют соляные озера,
например, находящееся на территории
Израиля и Иордании Мертвое море, вода которого
насыщена солями (рис. 2). Думали, что в
нем нет никакой жизни, но оказалось, что
Мертвое море населено археями. Археи
обнаружены в соляных озерах США и Кении,
а также в различных солярнах (мелких
водоемах для выпаривания морской воды и
получения соли). Соляные озера на юге
России тоже заселены галофильными
археями. Известно, что раньше найденную
красную соль, как соль царскую, отправ-
20
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
ляли на телегах в Москву, в Кремль.
Существуют сообщения о том, что клетки
галофильных архей, замурованные в
окаменевшую соль при высыхании водоема,
могут сохраняться в жизнеспособном
состоянии в течение многих миллионов лет и,
попав в благоприятные условия, начинают
расти. Подобного рода данные, правда,
вызывают сомнения и нуждаются в проверке.
Особую группу эвриархеот составляют
кислотолюбивые архей, использующие для
жизни органические соединения. Сюда
относятся так называемые термоплазмы,
развивающиеся в горячих и кислых
вулканических источниках и лишенные клеточной
стенки. Окружающая их клетки цитоплаз-
матическая мембрана, очевидно, обладает
удивительной устойчивостью. Еще более
кислотолюбив Picrophilus (в переводе —
кислотолюб). Эта архея растет только при
значениях рН ниже 2,2 и существует даже
при рН около 0. Развивается она при
температуре 50-55°С. Ее клетки, кроме цито-
плазматической мембраны, окружены
структурированным слоем белковых субъединиц,
что, как уже было сказано, характерно для
многих архей. Нужно иметь в виду, что
раствор, в котором живут эти организмы,
попав на кожу человека, неизбежно
вызовет сильный ожог, а на платье образует
дырку. Изучение архей приносит все
новые свидетельства удивительной способности
живых организмов приспосабливаться к
существованию в условиях, казалось бы для
жизни непригодных.
Как уже отмечалось, архе — значит
древний, и на древней Земле в начальные
периоды эволюции жизни архей, видимо,
доминировали в биосфере и играли
ведущую роль в биологических процессах
трансформации элементов. В то время особое
значение имели процессы получения
энергии прокариотами за счет окисления
молекулярного водорода и использования в
окислительно-восстановительных реакциях
соединений серы. Эти вещества есть среди
продуктов вулканической деятельности, и
их использование микроорганизмами
должно было иметь особое значение в
периоды, когда органическое вещество
содержалось в биосфере еще в очень незначительном
количестве. В океанических глубинах
около вулканических источников типа «черных
курильщиков» могла развиваться жизнь,
независимая от энергии Солнца,
возможность существования которой еще недавно
трудно было себе представить. Влияние,
оказанное археями на эволюцию жизни на
Земле, пока оценить трудно. Однако
сведения об этой группе живых организмов
стремительно расширяются.
Заключение
Конец XX столетия характеризуется
бурным развитием биологии, вступившей в
эру биологии молекулярной, достижения
которой широко известны. Гораздо меньше
известно об успехах в изучении
биоразнообразия. Между тем описание разнообразия
жизненных форм имеет огромное
общенаучное значение. Одно из основных
достижений в изучении биоразнообразия —
выявление самостоятельных ветвей архей и других
организмов, сформировавшихся еще на заре
развития жизни. Архей, несомненно
сыгравшие существенную роль в эволюции
биосферы, и сейчас занимают в ней своеобразное
место, обеспечивая осуществление
определенных процессов круговорота веществ,
которые являются необходимым условием ее
существования.
Литература
Кнорре Д. Г. Сорос, образоват. журн., 1996,
№ 3, с. 11.
Woese С. R., Kandler О., Wheelis M. L. Ргос.
Natl. Acad. Sci. USA., 1990, vol. 87, p. 4576.
В. И. Агол
РАЗНООБРАЗИЕ ВИРУСОВ
Введение
Известно, что вирусы могут вызывать
заболевания и что их размеры очень малы.
Менее известно, как разнообразен мир этих
мельчайших существ, неспособных к
самостоятельной жизни вне заражаемых ими
клеток. Разнообразны состав, размеры и
форма вирусов. Еще важнее, что у
вирусов встречаются такие способы хранения и
передачи генетической информации,
которых больше нигде в природе не найти.
Можно даже сказать, что в каком-то
смысле генетические системы вирусов богаче
генетических систем других организмов.
Поэтому вирусы — один из излюбленных
объектов молекулярной биологии,
изучающей фундаментальные принципы
организации живого. Профилактика и лечение
вирусных болезней также требуют учета
разнообразия свойств и особенностей
функционирования вирусов.
Состав, размеры и форма
Схематически вирусы представляют
собой наследственный материал, упрятанный
в защитную белковую оболочку, иногда
содержащую также липидные и углеводные
компоненты. В наследственном веществе —
молекуле или нескольких молекулах РНК
или ДНК — обязательно закодировано все
минимально необходимое для
формирования вируса: фермент, осуществляющий
копирование (репликацию) этих вирусных
нуклеиновых кислот, а также белки,
входящие в состав вирусной частицы (вирио-
на). Некоторые вирусы (в первую очередь,
относительно крупные) производят и
другие белки, делающие их жизнь как бы
более комфортной, более приспособленной к
различным условиям окружающей среды и
ее изменениям.
Если у всех невирусных организмов
наследственное вещество — это двуцепочеч-
ные молекулы ДНК (цепочки которых
комплементарны, т. е. как бы дополняют друг
друга), то вирусы могут содержать не
только ДНК, но и РНК, причем оба типа
нуклеиновых кислот встречаются как в дву-
цепочечной, так и в одноцепочечной форме.
Для каждого вируса характерна
определенная форма нуклеиновой кислоты.
Молекулы вирусных РНК и ДНК — неразвет-
вленные (иногда кольцевые) полимеры,
построенные из множества звеньев-нукле-
отидов; в одной такой молекуле — от
нескольких тысяч до нескольких сотен
тысяч нуклеотидов. Вирусные нуклеиновые
кислоты представляют собой длинные нити,
более гибкие в случае одноцепочечных
молекул и более упругие в случае двуце-
почечных. В растворе их форма
напоминает рыхлый комок. Однако в составе ви-
риона РНК или ДНК упакована более или
менее компактно, и характер такой
упаковки зависит от архитектуры вирусной
частицы. Сама же эта архитектура
определяется, в первую очередь, свойствами
белков, из которых построены вирионы.
Существует несколько основных
вариантов формы вирионов (рис. 1). Вирусы,
построенные только из нуклеиновой
кислоты и белка, могут походить на жесткую
палочкообразную или гибкую нитевидную
спираль, на шар (точнее, правильный
двадцатигранник, или икосаэдр), а также на
структуру, имеющую как бы головку и
хвостовой отросток и отдаленно напоминающую
сперматозоид. Липиды, если присутствуют,
образуют внешнюю мембрану, в которую
включаются и некоторые вирусные белки
(часто соединенные с углеводами), и такая
липопротеидная оболочка обволакивает
белковую «сердцевину» с «запечатанной» в ней
нуклеиновой кислотой. В этих случаях ви-
рион может иметь шарообразную, пулевид-
ную или кирпичеобразную форму, а может
и не обладать правильными очертаниями.
Размеры вирусных частиц также
существенно варьируют. Наиболее «тонкие»
нитевидные вирусы имеют диаметр около
10 нм, а их длина у самых протяженных
достигает 2 мкм. Диаметр сферических
вирионов колеблется от - 20 до 300 нм.
Самые крупные из известных вирусов —
родственники вируса оспы; их кирпичеоб-
разные вирионы могут достигать 450 нм в
длину и 260 нм в ширину и толщину.
Объекты такой величины уже находятся на
пороге разрешающей способности обычного
(светового) микроскопа.
Распространение в природе
Есть вирусы, размножающиеся в клетках
животных (позвоночных и беспозвоночных),
другие облюбовали растения, третьи (их
называют бактериофагами, или фагами)
паразитируют в микробах. Икосаэдрическая форма
встречается у вирусов всех этих трех групп,
но существуют и некоторые предпочтения
(рис. 1). Например, вирионы с головкой и
хвостовым отростком характерны для фагов.
22
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Вирусы спиральной и нитчатой форм
обычно паразитируют на растениях. Липопроте-
идная оболочка у вирусов животных
встречается чаще, чем в других группах.
Особенности строения заражаемой
клетки — один из факторов, от которых
зависит форма вириона. Так, бактерии и
растительные клетки помимо цитоплазматической
мембраны окружены довольно плотными и
прочными стенками. Преодолеть такую
стенку — серьезная проблема. Многие фаги
решают ее при помощи специального
приспособления, несколько напоминающего шприц.
Отсюда и обычная для них форма, в
которой хвостовой отросток выполняет роль иглы.
Вирусам растений пройти сквозь стенку часто
помогают механические повреждения,
наносимые, например, насекомыми или
сельскохозяйственными орудиями. Через такие
клеточные «раны» могут проникать вирусы,
не имеющие специальных приспособлений
для заражения неповрежденных клеток.
Некоторые вирусы заражают только
определенных «хозяев». Например, вирус
полиомиелита может жить и
размножаться лишь в клетках (да и то не во всех)
человека и приматов. Это, в частности,
означает, что заразиться полиомиелитом можно
только от людей. Кроме того, многие
вирусы передаются через переносчиков, роль
которых нередко выполняют членистоногие
(например, клещи). Такие вирусы могут
иметь широкий спектр хозяев,
включающий как позвоночных, так и
беспозвоночных животных.
Хотя сейчас уже известно несколько
тысяч вирусов, подавляющее число этих
агентов пока не описано, так как их чаще
всего открывают при поиске причины того
или иного заболевания человека,
животного, растения. Однако подавляющее число
живых организмов остаются в этом
отношении необследованными, и, стало быть,
есть все основания ожидать открытия
новых вирусов.
Хранение и передача
генетической информации
Как известно, синтез белков
осуществляется на рибосомах, а последовательность
аминокислот синтезируемых белков
(клеточных и вирусных) задается молекулами
информационных РНК (мРНК) (см.: Шат-
ский И. Н. Регуляция биосинтеза белков.
Том. 8). При описании способов хранения и
передачи генетической информации у
вирусов (рис. 2) молекулы мРНК удобно
обозначать как (+)РНК.
Есть обширная группа вирусов,
генетический материал которых представляет собой
как раз мРНК. Такие вирусы называют
вирусами с позитивным (положительным) РНК-
геномом (рис. 2, а). Сюда, например, относят
вирусы полиомиелита и клещевого
энцефалита, а из вирусов растений — вирус
табачной мозаики. Первое, что делает вирусная
РНК, попав в клетку, — обеспечивает
синтез вирусных белков. Лишь после этого
начинается размножение самих молекул ви-
Рис. 1. Разнообразие форм и размеров вирусов
Схематическое изображение некоторых вирусов растений (а), животных (б) и бактериофагов (в)
РАЗНООБРАЗИЕ ВИРУСОВ
23
русной РНК, которое просто невозможно
без предварительного образования
РНК-зависимой РНК-полимеразы — фермента,
способного синтезировать молекулы РНК без
участия ДНК, поскольку до заражения
клетки его в ней не было. На заключительной
стадии из накопившихся вирусных белков
и РНК происходит сборка вирионов.
Геном другой группы вирусов
представлен молекулами не мРНК, а их
комплементарной (зеркальной) копией, т. е.
молекулами (-)РНК (рис. 2, б). Среди таких вирусов с
негативным РНК-геномом — вирусы гриппа,
кори, бешенства, желтой карликовости
картофеля и др. Казалось бы, такие вирусы
просто не имеют права на существование.
Действительно, инфекционный процесс не
может начаться без синтеза белков:
инструкций, записанных в зеркальной форме,
рибосомы не понимают. Но и репликация
вирусной РНК кажется невозможной, поскольку,
как уже упоминалось, в клетке нет
собственных ферментов, способных осуществить этот
процесс. Вирусы с негативным РНК-геномом
решают эту проблему так. Они вводят в
заражаемую клетку свой геном не в «голом»
(свободном от белков) виде, как поступают
вирусы первой группы, а в виде более
сложных структур, содержащих, в частности,
РНК-зависимую РНК-полимеразу. Этот
вирусный фермент, синтезированный в
предыдущем цикле размножения, упакован в
вирионе в удобной для доставки в клетку форме.
Инфекционный процесс начинается с того, что
вирусный фермент копирует вирусный геном,
образуя комплементарные молекулы РНК,
т. е. (+)РНК. Эти молекулы уже «находят
общий язык» с рибосомами. Образуются
вирусные белки, в том числе и
РНК-зависимая РНК-полимераза, которая, с одной
стороны, обеспечивает размножение вирусного
генома в данной клетке, а с другой —
«консервируется впрок» во вновь образующихся
вирионах.
Есть вирусы (в том числе и
вызывающие тяжелые болезни человека —
геморрагические лихорадки), которые и по
строению и по многим другим свойствам —
близнецы вирусов с негативной РНК,
однако в их геноме наряду с участками,
соответствующими (-)РНК, есть
последовательности позитивной полярности. Хотя цикл
репродукции этих вирусов в некоторых
деталях и отличается от такового у
«классических» вирусов с негативной РНК —
отличия не столь принципиальны, чтобы здесь
на них останавливаться.
У третьей группы вирусов
наследственная информация хранится в виде двуцепо-
чечной (или ±) РНК (рис. 2, в). Сюда,
например, относятся ротавирусы, вызывающие
расстройства кишечника. Их размножение
осуществляется способом, близким к преды-
а б в
( )РНК ( )РНК ( )РНК
г д
( )РНК ( )ДНК ( )РНК
( )ДНК ж ( )РНК
( )ДНК ( )ДНК
Рис. 2. Основные стратегии репликации и
выражения генетической информации у вирусов
В рамку заключена форма нуклеиновой кислоты,
которая присутствует в вирионе; а—ж— объяснения в
тексте
дущему. Вместе с вирусной РНК в клетку
попадает и вирусная РНК-зависимая РНК-
полимераза, обеспечивающая синтез молекул
(+)РНК. В свою очередь, (+)РНК выполняет
две функции: обеспечивает производство
вирусных белков в рибосомах и служит
матрицей для синтеза новых (-)цепочек
вирусной РНК-полимеразой. Цепочки (+) и (-)РНК,
комплексируясь друг с другом, образуют
двунитевой (±)РНК-геном, который
упаковывается в белковую оболочку. Новое
поколение вирионов готово.
Четвертая группа — вирусы с двуцепо-
чечной ДНК (рис. 2, г). К ним принадлежат,
например, возбудители герпеса и оспы. Хотя
геном этих вирусов и можно условно
изобразить как (±)ДНК, во многих (но не во
всех) случаях в каждой из двух цепочек ДНК
имеются участки, соответствующие как
позитивной, так и негативной полярности.
В зараженной клетке фермент
ДНК-зависимая РНК-полимераза считывает
(транскрибирует) с генома этих вирусов
молекулы мРНК (т. е. (+)РНК), которые делают
свою обычную работу— направляют
синтез белков. Размножением вирусного ДНК-
генома занимается фермент ДНК-зависимая
ДНК-полимераза.
Поскольку геном клетки также
представлен молекулами двуцепочечных ДНК, в ней
еще до заражения имеются как
ДНК-зависимая РНК-полимераза, так и
ДНК-зависимая ДНК-полимераза. В некоторых
случаях производством вирусных мРНК и ДНК
занимаются клеточные ферменты. Другие
же вирусы (более сложные) используют
24
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
только собственные ферменты. Бывает и так,
что транскрипция и репликация
вирусного генома осуществляется при участии как
вирусных, так и клеточных ферментов.
Заканчивается инфекционный цикл, как
обычно, упаковкой генома в белковую оболочку
с образованием готовых вирионов.
Следующая группа — вирусы с одноце-
почечным ДНК-геномом (рис. 2, д), который
может быть представлен молекулами как
позитивной, так и негативной полярности.
Жертвами таких вирусов могут быть и
животные, и растения, и микроорганизмы. Попав
в клетку, вирусный геном сначала при
участии ДНК-полимеразы превращается в дву-
цепочную форму, а затем функционирует так
же, как и в предыдущем случае.
Шестая группа — ретровирусы (рис. 2, е),—
включает, в частности, широко известный вирус
иммунодефицита человека (ВИЧ) и некоторых
возбудителей злокачественных
новообразований. Геном ретровирусов, как и у
представителей первой из названных нами групп, — од-
ноцепочечная (+)РНК, но инфекционный
процесс развивается в данном случае по
совершенно иному сценарию. В вирусном
геноме закодирован необычный фермент (ревер-
таза), обладающий свойствами как
РНК-зависимой, так и ДНК-зависимой ДНК-полимеразы.
Этот фермент попадает в заражаемую клетку
вместе с вирусной РНК и обеспечивает синтез
ее ДНК-копии сначала в одноцепочечной
[(-)ДНК], а затем и в двуцепочечной форме
[(±)ДНК]. Далее события развиваются по
обычной схеме, включающей синтез вирусных
(+)РНК, синтез вирусных белков,
формирование вирионов, и выход их из клетки.
Наконец, седьмая группа — ретроидные
вирусы (рис. 2, ж), из которых наиболее
известен вирус гепатита В. В их состав
входит двуцепочечная ДНК, но реплицируется
она иначе, чем у вирусов четвертой
группы. Там вирусную ДНК копирует
ДНК-зависимая ДНК-полимераза. Здесь же сначала
с вирусной ДНК считывается (+)РНК (это
делает клеточная ДНК-зависимая РНК-по-
лимераза), а затем на ней как на матрице
синтезируются два компонента вириона:
белки и ДНК. Синтез ДНК осуществляет
вирусный фермент с активностью реверта-
зы по той же схеме, которая реализуется
у ретровирусов.
Конкретные способы выражения
генетической информации внутри упомянутых семи
групп вирусов также могут существенно
различаться. Например, синтез отдельных
белков у некоторых вирусов направляют
индивидуальные мРНК, а в других случаях
сначала образуется единый
высокомолекулярный полипептид-предшественник, который
затем «разрезается» на отдельные «зрелые»
белки. Осуществляется несколько разных схем
синтеза ДНК. Варьируют и другие важные
процессы. Например, репликация генома
некоторых вирусов происходит в клеточном ядре,
тогда как другие всю свою внутриклеточную
жизнь проводят в цитоплазме.
Типы взаимодействия с клеткой
Существуют два основных типа
взаимодействия вируса и клетки,
принципиальное различие между которыми
заключается в степени автономии вируса от своего
хозяина.
Несмотря на то что любой вирус
полностью зависит от энергетического
хозяйства клетки, от ее белоксинтезирующего
аппарата и других систем, многие вирусы
ведут себя в клетке довольно
самостоятельно. Самостоятельность проявляется в том,
что вирусные нуклеиновые кислоты
реплицируются, не подчиняясь клеточному
расписанию, а вирусные белки
синтезируются хотя и на клеточных структурах, но
по собственному расписанию. Такой путь
взаимодействия вируса с клеткой
называют продуктивным (имея в виду его
продуктивность с точки зрения вируса), и он
встречается как у РНК-содержащих, так и
ДНК-содержащих вирусов.
Продуктивное размножение вируса
может заканчиваться гибелью клетки. Но
некоторые вирусы ведут себя более
«расчетливо». Они не убивают клетку, от которой
зависит их жизнь, а находят способ
относительно мирного с ней сосуществования.
В результате развивается хронический
инфекционный процесс, при котором
выживают и вирусы, и клетки.
Но есть вирусы, более склонные к
подчинению клеточному контролю. Их так и
называют умеренными. Геном этих вирусов
включается в состав клеточной хромосомы
посредством ковалентного соединения
вирусной ДНК с клеточной. Вирусные гены как
бы превращаются в клеточные. Далее
события могут развиваться по-разному. В одних
случаях вирусный геном почти не
проявляет активности. Клетки и их хромосомы
делятся, а вместе с хромосомами в каждую
дочернюю клетку попадают и встроенные
вирусные гены. Но при определенных
обстоятельствах вирус активизируется, его гены
начинают работать, из разрушающейся
клетки выходит молодое поколение вирусных
частиц и начинает поиск новых жертв. В
самых общих чертах такое поведение
свойственно, например, фагу лямбда. В других
случаях вирусные гены в зараженной клетке
работают постоянно, производя новые и
новые поколения вирионов, но клетка при
этом не погибает. Приведем пример.
Ретровирусы — умеренные РНК-содер-
жащие вирусы. Казалось бы, в такой
формулировке есть внутреннее противоречие.
РАЗНООБРАЗИЕ ВИРУСОВ
25
Ведь вирусный геном может включаться в
клеточную хромосому только в форме ДНК.
Но мы уже знаем, что на РНК этих
вирусов с помощью ревертазы образуются ДНК-
копии, которые и встраиваются в
клеточную хромосому. Встроенные вирусные гены
во многом уподобляются клеточным, и в
частности активно транскрибируются
клеточной ДНК-зависимой РНК-полимеразой.
Образующаяся мРНК обеспечивает синтез
вирусных белков, которые затем формируют
вирионы, включающие вирусный
РНК-геном и ревертазу. Вновь образовавшиеся
вирусы покидают клетку, не убивая ее.
Возникает особая форма хронической
инфекции, при которой работающий
вирусный геном, включенный в состав клеточной
хромосомы, передается дочерним клеткам.
Другое дело, что такое сосуществование
не проходит бесследно для клетки.
Свойства ее могут драматически меняться под
влиянием присутствующих в ней вирусных
белков. В некоторых случаях зараженная
клетка становится раковой.
Вирусные болезни
Зная о таком разнообразии вирусов, мы
уже не должны удивляться разнообразию
болезней и симптомов, которые они
вызывают. Однако какой-либо корреляции
между характером болезни, с одной стороны, и
формой или особенностями генетической
системы вируса — с другой, не существует.
Так, причиной гепатита (воспаления
печени) могут быть самые разнообразные
вирусы. Например, вирусы гепатита А,
гепатита С и гепатита Е принадлежат к трем
разным семействам позитивных
РНК-вирусов (рис. 2, а), причем вирус гепатита С
имеет липопротеидную оболочку, а у двух
других вирусов она отсутствует. Вирус
гепатита В — ретроидный вирус (рис. 2, ж),
а генетическая система вируса гепатита дельта
столь необычна, что она вообще не попала
в нашу классификацию. Таким образом,
вирусы, ничего общего между собой не
имеющие, могут вызывать сходные заболевания.
Кроме того, сходные вирусы могут быть
причиной самых различных болезней.
Например, в списке заболеваний, вызываемых
пикорнавирусами (представителями одного
из семейств позитивных рибовирусов,
вирионы которых похожи друг на друга как
близнецы, а генетические системы практически
идентичны), — полиомиелит, миокардит,
диабет, конъюнктивит, простудные
заболевания, ящур, гепатит и другие болезни.
Когда мы думаем о способах борьбы с
вирусами как возбудителями болезней,
нужно отдавать себе отчет в разнообразии
устройства вирусов. Ясно, что к ним
необходим индивидуальный подход. И
поскольку нельзя привязать болезнетворное
действие вирусов к таким важнейшим их
особенностям, как форма, размеры и даже
«стратегия» размножения, приходится
углубленно изучать именно те свойства
вирусов, которые непосредственно связаны
с патогенностью.
Заключение
Таким образом, и форма, и внутреннее
содержание, и функционирование вирусов
разнообразны и индивидуальны. Можно
полагать, что особенности структуры и
функции вируса прямо или косвенно
связаны с такими качествами, которые дают
ему определенные преимущества. Уже
упоминалось о приспособленности формы
некоторых вирусов к заражению определенных
хозяев. А как сказываются на
функциональных возможностях вирусов особенности их
генетических систем?
Сравним между собой вирусы с
позитивным и негативным РНК-геномами. Первые
устроены много проще и представляют
собой по существу мРНК в изящной и
надежной упаковке. Чтобы начать полноценный
инфекционный процесс, нужна только эта
РНК и больше ничего. Соответственно РНК
вирусов данной группы обычно инфекцион-
на сама по себе. Заразив клетку, такие
вирусы могут незамедлительно начать синтез
собственных белков. Все это благоприятствует
их размножению. Но есть здесь и оборотная
сторона. В естественных условиях в
заражаемую клетку часто попадает только одна-
единственная вирусная частица. Ее РНК
оказывается в окружении множества
конкурирующих с ней клеточных мРНК. Поскольку
число претендентов на каждую свободную
рибосому велико, трансляция вирусной РНК
может не состояться. Поэтому такие
вирусы выработали специальные механизмы,
повышающие конкурентоспособность их РНК.
Вирусы с негативным РНК-геномом
устроены сложнее. В их вирионе упакована не
просто РНК, а РНК вместе с ферментом,
способным ее реплицировать. Негативная РНК
на первый взгляд никак не может повысить
конкурентоспособность ее носителей.
Однако это не так. Внедрение в клетку вирусной
РНК вместе с собственной РНК-полимеразой
обеспечивает наработку множества молекул
(+)РНК (в том числе и мРНК), которые
могут конкурировать с клеточными мРНК
просто благодаря своей многочисленности.
Литература
Агол В. И. В кн.: Молекулярная биология:
Структура и биосинтез нуклеиновых кислот. М.:
Высш. шк., 1990, с. 260.
Вирусология. В 3 т. Пер. с англ. М.: Мир, 1989.
И. С. Кулаев
ПРОИСХОЖДЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
Введение
Все живые организмы на Земле
обычно делят на прокариот и эукариот (от
греческого слова «карион» — ядро). Главной
особенностью прокариот является отсутствие
у них, в отличие от эукариот (эу —
по-гречески истинный), полноценного
клеточного ядра, покрытого мембраной. Кроме того,
прокариотические клетки обладают единой
мембранной системой, включающей как
цитоплазматическую мембрану, или плаз-
малемму, так и различные выросты из нее,
зачастую выполняющие специфические
функции. Клетки прокариот имеют очень
небольшие размеры, порядка 1 мкм.
Объем эукариотических клеток,
содержащих полноценное ядро, в 800—1000 раз
больше объема клеток прокариот. В связи
с этим для нормального выполнения
жизненных функций клеткам эукариот
необходим гораздо больший мембранный аппарат.
Действительно, в эукариотической клетке
выявлены разнообразные субклеточные
структуры, окруженные независимыми от
плазмалеммы и друг от друга мембранами.
Эти структуры называются органеллами.
Каждой органелле — митохондрии,
хлоропласту (у растительных клеток), ядру, ли-
зосомам, аппарату Гольджи и т. д.,
присущи свои специфические функции в жизни
клетки.
Следует сказать, что в последнее
десятилетие с помощью молекулярно-биологичес-
ких подходов получены убедительные
доказательства того, что все живые существа
должны быть разделены на 3 надцарства
(домена) — бактерии, археи и эукариоты.
Причем первые два домена — бактерии и
археи — являются прокариотами. С помощью
молекулярно-биологических подходов были
получены весомые доказательства в пользу
Эубактерии Эукариоты Археи
Общий предшественник
Происхождение жизни
Рис. 1. Филогенетическое древо, полученное на
основе данных молекулярной биологии [из: Oshima,
1994]
того, что бактерии и археи возникли на
Земле от общего предка — так
называемого прогенота, около 4 млрд лет назад, а
эукариоты имеют более позднее
происхождение. По данным палеонтологов,
эукариоты, в частности, дрожжеподобные
организмы, появились на Земле примерно через
500 млн лет после прокариот. Причем по
современным представлениям (рис. 1), в
возникновении эукариотических клеток,
скорее всего, участвовали как бактерии, так
и археи (совсем недавно их называли архе-
бактериями). Наиболее популярной и
достаточно обоснованной концепцией
происхождения эукариотических клеток в настоящее
время является идея многократного эндосим-
биоза различных прокариот (см.: Громов Б. В.
Удивительный мир архей. Наст. том).
По представлениям нобелевских
лауреатов А. Клюйвера и К. Ван Ниля, именно
у древних прокариот произошел отбор
клеток с наиболее экономичными и зачастую
полифункциональными биохимическими
реакциями, ставшими основой важнейших
путей метаболизма. Эти метаболические
пути, с одной стороны, явились
универсальной основой обмена веществ всех
живых существ, а, с другой, дали начало
формированию специфических биохимических
реакций, характерных для организмов,
обитающих в экстремальных условиях.
Первичные анаэробные прокариоты
отличались, по-видимому, гетеротрофным типом
питания за счет синтезированных абиогенно
органических веществ. Они должны были
довольно быстро истощить запасы
необходимой им пищи в среде своего обитания.
По мнению американского ученого Б. Аль-
бертса и его соавторов, решение
проблемы питания первичных протобионтов
(анаэробных прокариот) в условиях обедненной
органическими веществами окружающей
среды могло быть достигнуто по крайней
мере двумя независимыми путями.
Во-первых, за счет усвоения
имевшихся в атмосфере Земли необходимых для
жизни источников углерода (С02) и азота (N2).
Для того чтобы ввести в обмен веществ эти
очень стабильные, термодинамически
устойчивые соединения атмосферы, необходима
была энергия. Первичные анаэробные
организмы выработали механизмы улавливания
и использования для биосинтетических
целей химической и солнечной энергии. В той
форме, в которой эти процессы
существуют сейчас на Земле, они получили
названия хемосинтеза и фотосинтеза.
ПРОИСХОЖДЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
27
Большинство исследователей
предполагают, что первыми фотосинтетиками на
Земле явились предки современных циа-
нобактерий (синезеленых водорослей) (см.:
Громов Б. В. Цианобактерии в биосфере.
Наст. том). Важно отметить, что и в
настоящее время цианобактерии являются
теми организмами, в результате
жизнедеятельности которых углекислый газ и азот
атмосферы включаются в значительном
количестве в органические соединения и
поступают в биосферу. Цианобактерии
являлись и являются наиболее автономно
живущими организмами, способными
фиксировать и С02 и N2. Причем,
по-видимому, основные механизмы, с помощью
которых происходит питание этих организмов
(называемое автотрофным), существенно
не изменились на протяжении нескольких
миллиардов лет.
Очень важно отметить, что именно
примитивные фотосинтезирующие анаэробные
бактерии кардинально изменили состав
земной атмосферы, которая с появлением
фотосинтеза превратилась из практически
лишенной молекулярного кислорода смеси
газов в атмосферу, содержащую 21%
кислорода. Появление кислорода в земной
атмосфере имело решающее значение для
дальнейшего развития жизни на Земле.
Можно думать, что после возникновения
в земной атмосфере кислорода какая-то
часть первичных анаэробных прокариот —
бактерий и архей — была вынуждена для
своего выживания либо искать
специальные экологические ниши, где сохранялись
анаэробные условия ввиду большой
токсичности для них кислорода, либо каким-то
образом адаптироваться к нему, развить
систему ферментов, способных
утилизировать кислород для более эффективного
окисления пищи. Таким образом могли
возникнуть гораздо более быстро растущие и
размножающиеся на Земле аэробные про-
кариотические организмы. Эти живые
организмы приобрели возможность более
полно окислять органические соединения, в
первую очередь углеводы. В частности,
глюкоза в отсутствие кислорода у анаэробных
бактерий и архей могла окисляться далеко
не полностью — например, до масляной и
молочной кислот или этилового спирта, тогда
как конечными продуктами полного
окисления являются Н20 и С02. Этот процесс,
называемый дыханием, приводил к
извлечению из молекулы сахара гораздо
большего количества энергии, аккумулируемой
в клетках в форме богатых энергией
фосфорных соединений (неорганических
полифосфатов, пирофосфата, АТФ и др.).
Как считают Альберте с соавторами,
важнейший этап жизни на Земле связан с
развитием у анаэробных прокариот в условиях
сильного обеднения среды обитания
пищевыми ресурсами (органическими
соединениями) способности к поеданию друг друга,
называемого фагоцитозом. По
предположению этих авторов, при фагоцитозе одних
организмов другими анаэробные
прокариоты могли вступить в симбиоз с поглощенными
ими аэробными клетками, образуя с ними
взаимовыгодную ассоциацию.
Фагоцитированные таким образом аэробные клетки могли
быть не утилизированы в качестве пищи,
а оставлены внутри клетки-хозяина для
более полного и эффективного окисления
органических веществ. Это, безусловно, могло
дать большие преимущества таким клеткам-
хозяевам в борьбе за существование.
Возникновение и развитие
гипотезы об эндосимбиотическом
происхождении
эукариотических клеток
Концепция симбиоза, т. е. образования
взаимовыгодной ассоциации двух и более
организмов, возникла в России во второй
половине XIX столетия. Еще в 1867 г. была
опубликована работа А. С. Фаминцина о том,
что лишайники являются взаимовыгодной
ассоциацией двух разных организмов:
грибов и водорослей. Само название этого
явления — симбиоз — было введено в
литературу немецким микологом и анатомом
растений Де Бари через несколько лет после
работы Фаминцина. Опираясь на
собственные исследования и работы своето
соотечественника А. Ф. Шимпера, показавшего
способность хлоропластов к саморепликации
в клетках растений, Фаминцин в 1907 г.
опубликовал книгу, в которой сделал
предположение о том, что хлоропласты могут
являться одноклеточными
водорослями-симбионтами растений. Эта идея была
поддержана и развита далее К. С. Мережковским
и Б. М. Козо-Полянским в 20-х гг. нашего
столетия. Причем они предполагали, что не
только хлоропласты, но и митохондрии
являются симбионтами, так как и те и
другие были способны к саморепликации в
клетках высших растений. (См. предисловие
Б. М. Медникова к работе Л. Маргелис.)
Однако об этой гипотезе, как весьма
экстравагантной, долгое время даже не
упоминали. О ней снова вспомнили только после
того, как в конце 50-х—начале 60-х гг.
биохимики получили данные о содержании в
хлоропластах растений и в митохондриях,
выделенных из разных организмов, ДНК,
отличной от ДНК ядер этих биологических
объектов и похожей на ДНК прокариот (в
частности, она имела кольцевое строение).
Кроме того, лабораторией Н. М. Сисакяна
и рядом других научных групп было уста-
28
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
новлено наличие в хлоропластах растений
рибосом прокариотического типа. Процесс
биосинтеза белка в хлоропластах и
митохондриях по ряду черт был весьма схож с
таковым у бактерий и существенно
отличался от биосинтеза белка, происходящего
на цитоплазматических рибосомах эукари-
отических клеток. Хлоропласты и
митохондрии оказались близки бактериальным
клеткам и по другим биохимическим признакам
(в частности, по наличию в их мембранах
специфически построенного фосфолипида —
кардиолипина, характерного только для
мембран бактерий и полностью
отсутствующего в плазматической мембране эукари-
от). Однако биохимики, а также
молекулярные биологи и цитологи получили довольно
большое число аргументов против этой
гипотезы. Главным из них была очень слабая
автономия жизнедеятельности, и в первую
очередь, процесса биосинтеза белка, в
хлоропластах и митохондриях. Оказалось, что
и в митохондриях, и в хлоропластах
синтезируется только сравнительно малая часть
необходимых для их формирования и
функционирования белков (в первую очередь,
ферментов).
Еще одним импульсом к пристальному
рассмотрению гипотезы об эндосимбиотичес-
ком происхождении эукариотических
организмов послужили работы и идеи
американской исследовательницы Л. Маргелис. Она,
в частности, показала очень большую
иммунологическую, т. е. структурную, близость
жгутиков и некоторых элементов цитоске-
лета эукариотических клеток
(отсутствующих у прокариот) со спирохетами —
бактериями, обладающими спиралевидной
формой клеток. Эти факты позволили
Маргелис предположить, что цитоскелет и
жгутики эукариотической клетки, возможно,
произошли от спирохетоподобных прокариот.
Сравнительно недавно было показано, что
АТФаза — один из универсально
распространенных во всех живых организмах АТФ-
утилизирующих ферментов, выделенная из
вакуолей дрожжей, по своей структуре
резко отличается от АТФаз других
субклеточных органелл этих низших эукариот и
практически идентична АТФазе некоторых
архей. На этом основании было выдвинуто
предположение, что предшественниками
вакуолей низших эукариот являлись
древние прокариоты, принадлежащие к архе-
ям и вступившие в симбиотические
отношения с хозяйской клеткой.
Совсем недавно в нашей лаборатории
было показано, что другой фермент — по-
лифосфатаза (утилизирующий
неорганические полифосфаты), — выделенный из
разных органелл дрожжей (ядер, митохондрий,
клеточной оболочки, цитозоля и вакуолей),
обладает совершенно разными
характеристиками и свойствами. Эти данные также
свидетельствуют в пользу того, что все или,
по крайней мере, многие внутриклеточные
структуры эукариотических клеток имеют
различное происхождение.
Решающие доказательства
эндосимбиотического
происхождения хлоропластов
и митохондрий
Самые существенные доказательства
происхождения важнейших органелл
эукариотических клеток — хлоропластов и
митохондрий из прокариот были получены в
последние годы при изучении структуры
одного из наиболее консервативных (в плане
возможности изменений в процессе
эволюции) компонентов живых клеток — рибосом-
ных РНК. Выдающийся американский
молекулярный биолог К. Воз с сотрудниками
показал уже в 80-х гг., что структура так
называемой 16S РНК из рибосом
хлоропластов растений совершенно не похожа на
соответствующую ей 18S РНК из
цитоплазматических рибосом растений и почти
идентична структуре аналогичной РНК из
некоторых цианобактерий.
В то же время эти исследователи
установили, что 16SPHK рибосом, полученных
из митохондрий разных эукариотических
организмов, не похожа по своей
структуре на соответствующую ей РНК рибосом из
цитозоля тех же самых биологических
объектов, но чрезвычайно схожа с 16SPHK
некоторых бактерий, в частности, бактерий
рода Paracoccus. Интересно, что и набор
дыхательных ферментов указанной
бактерии очень похож на «дыхательный ансамбль»
митохондрий животных.
Все эти данные достаточно ясно
свидетельствуют в пользу большой
вероятности эндосимбиотического происхождения
эукариотических клеток. Уместно также
сказать, что и по сей день явление эн-
досимбиоза чрезвычайно широко
распространено в природе. Так например,
анаэробная амеба Pelomyxa polustris, которая,
в отличие от других эукариот, не имеет
митохондрий, содержит, однако, когда-то
фагоцитированных ею аэробных бактерий-
симбионтов, выполняющих функцию
митохондрий. Кроме того, хорошо известно,
что в теле некоторых морских рыб и
других животных часто поселяются
светящиеся бактерии, которые не только
извлекают выгоду для себя (состоящую, в первую
очередь, в отсутствии необходимости
постоянно искать пищу), но и очень
полезны для организмов-хозяев. Они,
по-видимому, освещают им путь и помогают
ориентироваться среди «своих» и «чужих».
ПРОИСХОЖДЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
29
2
Ф
о.
со
Рис. 2. Предположительная
схема происхождения эукариот путем
многократного симбиоза аэробных
и анаэробных прокариот [из:
Альберте и др., 1986]
Предкоеый прокариотический организм
Существует и множество других
примеров взаимовыгодного эндосимбиоза
прокариот с различными эукариотическими
организмами.
О природе клетки-хозяина
Все вышесказанное достаточно
убедительно свидетельствует в пользу эндосим-
биотического происхождения эукариоти-
ческих клеток из гораздо меньших по
объему (на несколько порядков) клеток
прокариот. Однако самый главный и пока
весьма туманный вопрос этой проблемы
заключается в установлении природы
хозяйской клетки. Ведь клетки эукариот
обладают уже хорошо сформированным
ядром, имеющим оболочку, а
прокариоты такого ядра не имеют.
Вместе с тем Маргелис, например,
считает, что роль хозяйских клеток,
возможно, сыграли прокариоты — предшественники
современных бактерий, относящиеся к ми-
коплазмам — очень примитивно устроенным
прокариотам, практически лишенным
клеточных стенок.
Альберте с соавт. приводят следующую
предположительную схему происхождения
эукариот путем многократного симбиоза
аэробных и анаэробных, фотосинтезирующих
и нефотосинтезирующих прокариотических
организмов (рис. 2).
На этой гипотетической схеме все как
бы понятно, но все же остается неясным,
откуда же или, по крайней мере, когда
возникает клеточное ядро?
В последнее время появились и
широко обсуждаются работы японского
исследователя Т. Осимы. На основании
тщательного изучения структуры рибосомных
16SPHK многих про- и эукариот, в том
числе и многих представителей до сих пор
мало изученного надцарства архей, этот
автор приходит к выводу, что клеткой-
хозяином являлся один из предков
современных архей. Кстати, следует сказать,
что у представителей именно этого
надцарства (в отличие от бактерий) имеется
Рис. 3. Предположительная
схема происхождения эукариотичес-
ких клеток из хозяйской клетки
древних архей и бактериальных
предков митохондрий и хлоропла-
стов [из: Oshima, 1994]
Е. coli
Т. thermophilus ^
Т. tenax §
э
0)
о
D. mobilis о
0)
М. vanniellii
Митохондрии
Хлоропласты
Н. sapiens
P. polycephalum
m
с
1
Н. halobium §¦
Эукариоты
СП
0)
4
Ф
и
Археи
Н. morhuae sr
30
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
РАСТЕНИЯ ГРИБЫ
Покрытосеменные Голосеменные
Морковь
Горох
Фасоль
Табак
Лен
Лилия
Ascobolus
Neurospora
Папортники
Водный
папортник
(Azolla)
Бурые водоросли
Fucus
Красные
водоросли
Мхи,
печеночники
Зеленые
водоросли
Nitella
Acetabularia
Volvox
Gonium
uniamyaomonas
Циатомны
Членистоногие
Плодовая лягушка
Drosophila
Таракан
Клоп
Циклопы
Нематоды
Caenorhab-
ditis
Слизевики
водоросли
Dictyosteliui \
Цианобактерии
(синезеленые водоТюсли)
Пурпурные
оактерий"""
Saccharo-
myces
Хлоропласты
Митохондрии
Paracoccf°
Escherichicoli
Salmonella
Миксобактерии
i оамположительные бактери
им -tiixf|||5 С—
Бактерии
BaCIIIUS 5UMIH5
bar-
ЖИВОТНЫЕ
Позвоночные (хордовые)
Человек
Лягушка
Мышь
Курица
Перепелка
Тритон
Крыса
Корова
Сиг
Кит
Оболочники
Асцидии
Иглокожие
Морской еж
Морская звезда
Морской огурец
Моллюски
Кальмар
Улитка
Кишечнополостные
Гидра
Актиния
Губки
Амебы
Солнечники
ACTinospnaenum
Инфузории
vonlceiia
Paramecium
Динофлагеллаты
Архебактерии
Галобактерии
О
s
о.
со
со
0
2
X
У
о
о
X
5
J3
о о
О
S
Q.
2
О
Q.
С
Рис. 4. Общая предположительная схема эволюционного родства и происхождения основных типов живых
организмов [из: Альберте и др., 1986]
очень много общих с эукариотами
биохимических и молекулярно-биологических
особенностей.
На рис. 3 представлена схема
происхождения эукариотических клеток по Осиме. В
его последних работах приводятся доводы
в пользу того, что хозяйской клеткой при
возникновении эукариот является предок
архей, подобный одному из современных
представителей этих древних прокариот —
термоплазме. Каково будет окончательное
решение проблемы установления природы
клетки-хозяина, давшей начало эукарио-
тическим клеткам в процессе
множественного эндосимбиоза, покажут дальнейшие
исследования. Примером очень
осторожного отношения современных биологов к
этому вопросу является гипотетическая схема,
представленная Альбертсом и др. (рис. 4).
Заключение
На сегодняшний день наиболее
популярной и обоснованной гипотезой
происхождения эукариотических клеток является
достаточно обоснованное представление об их
образовании путем многократно
происходившего в процессе эволюции в течение
первых 1,5—2 млрд лет существования Земли
эндосимбиоза различных прокариот,
относящихся к бактериям и археям. Вопрос о
природе клетки-хозяина, в которую
включались те или иные прокариотические
предшественники эукариотических органелл,
пока является наиболее дискуссионным.
Литература
Альберте Б., Брей Д., Льюис Дж. и др.
Молекулярная биология клетки. Пер. с англ. М.: Мир,
1986, т. 1.
Клюйвер А., Ван Ниль К. Вклад микробов в
биологию. М.: Иностранная литература, 1959.
Кулаев И. С. Сорос, образоват. журн., 1996,
№2, с. 28.
Маргелис Л. Роль симбиоза в эволюции клетки.
Пер. с англ. М.: Мир, 1983.
Хахина Л. Н. В кн.: Андрей Сергеевич Фа-
минцын. Л.: Наука, 1981, с. 165.
Woese С. R.t Kandler О., Wheeliss M. L. Proc. Natl.
Acad. Sci. USA, 1990, vol. 87, p. 4576.
В. В. Малахов
ПРОИСХОЖДЕНИЕ ХОРДОВЫХ ЖИВОТНЫХ
Введение
Хордовые — один из крупнейших типов
животного царства, представители
которого освоили все среды обитания. В состав этого
типа входят три группы (подтипа)
организмов: оболочники (в том числе живущие на
дне морские сидячие организмы — асцидии),
бесчерепные (похожие на маленьких рыбок
морские существа — ланцетники),
позвоночные (хрящевые и костистые рыбы,
земноводные, пресмыкающиеся, птицы и
млекопитающие). Человек— тоже представитель
типа хордовых. Происхождение типа
хордовых— это важнейший этап в историческом
развитии животного мира, означающий
появление группы животных с уникальным
планом строения; именно он позволил в ходе
дальнейшей эволюции достичь максимальной
среди живых существ сложности строения и
поведения. Реконструкция этого отрезка
филогенеза означает в сущности реконструкцию
первых шагов на пути, приведшем к
человеку. Вот почему проблема происхождения
хордовых уже более полутора столетий
вызывает большой интерес биологов.
Происхождение хордовых относится к
тому периоду эволюции животного
царства, от которого сохранилось очень мало
палеонтологических остатков. Известно
лишь, что в среднем кембрии (около
550 млн лет назад) уже существовали
бесчерепные, родственные современным
ланцетникам. Более ранние этапы эволюции
хордовых приходится реконструировать на
основе изучения современных организмов,
преимущественно
сравнительно-анатомическими и эмбриологическими методами,
привлекая данные и из других областей
биологии.
Основные гипотезы
происхождения хордовых
Одна из первых концепций,
связывающих план строения хордовых с планами
строения групп беспозвоночных, не
принадлежащих к их числу, была разработана
выдающимся французским сравнительным
анатомом Э. Жоффруа Сент-Илером еще в
первой половине XIX в.
Он считал, что членистые животные
(кольчатые черви и членистоногие) могут
рассматриваться как перевернутые
позвоночные. «Взгляните на рака,
опрокинутого на спину, говорил он, — и вы увидите,
что различные системы его расположены
совершенно так же, как у высших
позвоночных животных». Именно эта концепция
была в центре публичного спора между
Жоффруа Сент-Илером и его
соотечественником Ж. Кювье, который состоялся в
Париже в 1830 г. Как известно, в том
знаменитом споре победил Кювье. Однако к идее
сравнения хордовых (и позвоночных) и
перевернутых кольчатых червей или
членистоногих зоологи возвращались неоднократно
на протяжении всего XIX в.
Эту точку зрения отстаивал выдающийся
зоолог и, как бы мы сейчас сказали,
«организатор науки» А. Дорн, построивший на
личные средства первую в мире и до сих
пор успешно работающую Неаполитанскую
зоологическую станцию. В начале XX в. идею
происхождения хордовых от
перевернувшихся на спинную сторону членистоногих
отстаивали английские биологи У. Гаскелл,
У. Паттен и другие.
Остроумные и хорошо объясняющие
план строения хордовых гипотезы их
происхождения от перевернутых аннелид или
членистоногих утратили свою популярность
прежде всего в связи с тем, что стали ясны
фундаментальные эмбриологические
различия между кольчатыми червями и
членистоногими, с одной стороны, и хордовыми —
с другой.
В начале нынешнего столетия были
разработаны гипотезы происхождения
хордовых от кишечнодышащих — обитающих в
толще грунта морских червеобразных
организмов. У этих животных (как и у
хордовых) имеются жаберные щели, а в передней
части тела — опорный вырост кишечника
(стомохорд), который сравнивали с хордой.
Однако кишечнодышащие резко
отличались от хордовых обратным направлением
тока крови, расположением сердца,
строением нервной системы и другими
важными особенностями организации. Делались
попытки выводить хордовых от иглокожих,
немертин, моллюсков и других групп
беспозвоночных, однако ни одна из них не
имела успеха.
Своеобразной реакцией на неудачи
зоологов решить проблему происхождения
хордовых традиционными способами явилась
разработка так называемых неотенических
гипотез, согласно которым хордовые
происходят от личинок асцидий. Взрослые
асцидии — прикрепленные морские
животные, фильтрующие воду через объемистый
жаберный мешок и улавливающие
взвешенные в воде мельчайшие органические ча-
32
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
стицы. Они не имеют ни хорды, ни нервной
трубки и совсем не похожи на хордовых
животных. Но, как давно известно (а
впервые это показал еще в конце прошлого
века выдающийся русский биолог А. О.
Ковалевский), личинки асцидий имеют и
хорду, и нервную трубку, утрачиваемые при
метаморфозе. На основании этого
некоторые биологи (Н. Берилл, М. Уитер, К. Бо-
ун и др.) предполагают, что хордовые
произошли от ставших половозрелыми личинок
асцидий. Правда, в этом случае остается
непонятным, откуда взялись сами асцидий.
Да и трудно представить, чтобы хорда и
нервная трубка возникли у живущих 1—2
дня и непитающихся личинок. Более
вероятно, что эти особенности — наследие сво-
бодноживущих предков асцидий, об
организации которых неотенические гипотезы
ничего не говорят. Ни одна из
рассмотренных гипотез в настоящее время не может
считаться истинной, но в каждой из них
содержатся отдельные элементы, которые
могут оказаться полезными при
реконструкции путей происхождения хордовых.
Первичноротые и вторичноротые
Все билатерально-симметричные
организмы делятся на две группы — первич-
норотых и вторичноротых и прежде чем
приступать к анализу проблемы
происхождения хордовых, следует выяснить, к
какой из этих двух групп они принадлежат.
Названия «первичноротые» (Protostomia) и
«вторичноротые» (Deuterostomia) указывают
на способ развития ротового отверстия в
эмбриогенезе. Так, у первичноротых
животных бластопор (возникающее в
эмбриональном развитии отверстие
первичного кишечника) частично или полностью
переходит в ротовое отверстие. У
вторичноротых (например, у иглокожих)
бластопор становится анальным отверстием, а
рот прорывается заново на переднем конце
личинки. В других случаях бластопор
закрывается, а рот и анус открываются
заново.
Помимо происхождения рта,
первичноротые и вторичноротые различаются и
другими эмбриологическими особенностями.
Во-первых, это характер дробления
оплодотворенного яйца: у первичноротых оно
спиральное, а у вторичноротых —
радиальное (рис. 1). Другое различие состоит
в способе закладки целома (вторичной
полости тела): у первичноротых стенки
вторичной полости тела происходят от двух
клеток, а у вторичноротых за счет
выпячивания карманов эмбрионального
кишечника (рис. 1). Третье важнейшее различие
между первично- и вторичноротыми —
Первичноротые
Вторичноротые
Спиральное
дробление
Радиальное
дробление
Мезодерма
Мезодерма
Закладка мезодермы
Первичный
мозг
Мезодерма
Личинка
первичноротых
Первичный
мозг
1 пара
целомов
II пара
целомов
III пара
целомов
Личинка
вторичноротых
Рис. 1. Особенности эмбрионального
развития первичноротых и
вторичноротых животных
ПРОИСХОЖДЕНИЕ ХОРДОВЫХ ЖИВОТНЫХ
33
Первичноротые
Вторичноротые
Кольчатые черви
Полухордовые
Иглокожие
Членистоногие
Моллюски
Круглые черви
Плоские черви
Хордовые
Погонофоры
Рис. 2. Представители основных групп первичноротых и вторичноротых животных
судьба зачатка первичного мозга: у
первичноротых он дает начало мозгу
взрослых форм, а у вторичноротых всегда
редуцируется, так что новый нервный
центр возникает в другом месте заново.
Вторичноротые — это вторичномозговые
животные.
Рот и анус хордовых животных
формируются без связи с бластопором. Хордовые
имеют типичное радиальное дробление
яйца, вторичная полость тела (целом)
формируется у них за счет выпячивания
карманов первичного кишечника, а в нервной
системе нет следов первичного мозга. Все
эти признаки указывают на принадлежность
хордовых к вторичноротым животным.
Помимо хордовых к ним относятся
погонофоры, полухордовые и иглокожие (рис. 2).
Первичноротые включают плоских,
круглых и кольчатых червей, моллюсков,
членистоногих (рис. 2).
Сравнение хордовых с другими
вторичноротыми
Хордовые — это вторичноротые
животные, развитие которых имеет ряд
особенностей. На рис. 3 представлены стадии
развития одной из групп вторичноротых —
погонофор и низших хордовых — асцидий.
Анимальный полюс яйца погонофор, на
котором располагается ядро яйцеклетки,
соответствует спинной стороне эмбриона,
а вегетативный, на котором
концентрируется желток, — брюшной. У низших
хордовых ситуация обратная: анимальный полюс
соответствует брюшной стороне, а
вегетативный — спинной. Бластопор (т. е. область
эмбриона, где впячивается энтодерма и
формируется первичный кишечник) у
погонофор соответствует брюшной стороне
личинки, а у низших хордовых — спинной.
Зачаток нервной системы, закладывающийся
по линии замыкания бластопора, у
погонофор становится брюшным нервным тяжем,
а у хордовых — спинной нервной трубкой
(рис. 3).
План строения низших хордовых также
существенно отличается от планов
строения других вторичноротых. Так, например,
у всех вторичноротых, кроме хордовых,
передние целомы диссимметричны: левый
передний целом больше правого, обычно
выполняющего функцию околосердечной
сумки. У хордовых же ситуация обратная:
правый передний целом больше левого.
34
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Целомы нехордовых вторичноротых
сообщаются с внешней средой отверстиями,
открывающимися на спинной стороне и
выполняющими осморегуляторные и половые
функции. У хордовых эти отверстия
располагаются на той стороне тела, которую мы
традиционно считаем брюшной.
Кровеносная система у хордовых и
прочих вторичноротых устроена по общему
плану и состоит из двух главных сосудов:
спинного и брюшного. У нехордовых
вторичноротых, как и всех других
беспозвоночных, кровь по спинному сосуду течет
вперед, а по брюшному назад. У хордовых
направление тока крови обратное: по
спинному сосуду кровь течет назад, а по
брюшному — вперед. Сердце у нехордовых
вторичноротых лежит на спинной стороне, а
у хордовых — на брюшной.
Суммируем эти отличия.
1. Бластопор хордовых соответствует не
брюшной, а спинной стороне.
2. Нервная пластинка хордовых,
закладывающаяся по месту замыкания бласто-
пора, превращается не в брюшной нервный
тяж, а в спинную нервную трубку.
3. Диссимметрия первой пары целомов
имеет обратный знак в сравнении с
другими вторичноротыми: правый передний
целом у хордовых больше левого.
4. Отверстия целомов у хордовых
открываются не на спинной, а на брюшной
стороне.
5. Кровь у хордовых по брюшной стороне
течет вперед, а по спинной — назад, и
сердце располагается не на спинной, а на
брюшной стороне.
Что же означают эти несоответствия?
Только одно: предки хордовых в своем
эволюционном развитии испытали инверсию
сторон тела, т. е. перевернулись и стали
передвигаться на морфологически спинной
стороне, которая стала функционировать как
физиологически брюшная. Морфологически
Погонофоры
Анимальный
полюс
i
О'О
Оболочники
Передне-задняя
ось
Анимальный
полюс
Передне-задняя
ось
Вегетативный
полюс
Вегетативный
полюс
Бластопор
Бластопор
Эмбрион на стадии
гаструляции
IV отдел тела
IV отдел тела (хвост)
Брюшной нервный тяж
Личинка
погонофор
Нервная трубка
Личинка
асцидий
Рис. 3. Особенности эмбрионального развития погонофор и низших хордовых
ПРОИСХОЖДЕНИЕ ХОРДОВЫХ ЖИВОТНЫХ
35
IV отдел тела
Околосердечная
Сердце сумка
—Щупальца
IV отдел тела
Погонофоры
Сердце
Околосердечная
сумка
Стомохорд
Кишечнодышащие
IV отдел тела
Щупальца
Сердце
Околосердечная
сумка
IV отдел тела
Околосердечная
сумка
Стомохорд
Крыложаберные
Хорда
IV отдел тела
Личинка асцидий
Щупальца
Хорда
Ланцетник
1
2
3
4
5
Рис. 4. Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник
изображены в перевернутом положении)
7— целомы первой пары; 2— целомы второй пары; 3— целомы третьей пары; 4 — кровеносная система; 5—
нервная система. Стрелки показывают направление тока крови
брюшная сторона предков хордовых стала
функционировать как физиологически
спинная.
Если мы примем этот вывод, все
перечисленные эыше несоответствия исчезнут:
бластопор хордовых станет соответствовать
их исходно брюшной стороне, нервная
трубка будет располагаться на морфологически
брюшной стороне, диссимметрия передних
целомов приобретет нормальный для
вторичноротых характер, отверстия целомов
окажутся на морфологически спинной
стороне, и направление тока крови, так же
как положение сердца, окажется типичным
для вторичноротых. На рис. 4
представители низших хордовых (личинка асцидий и
ланцетник) изображены перевернутыми,
чтобы продемонстрировать, что при таком
положении их системы органов
соответствуют таковым других вторичноротых.
36
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Переворот на спинную сторону мог стать
тем важнейшим эволюционным событием,
которое определило формирование
плана строения хордовых животных и его
коренное отличие от планов строения
прочих вторичноротых и других
беспозвоночных.
Пути эволюции вторичноротых
животных
Общий предок вторичноротых животных,
по-видимому, вел донный образ жизни,
ползая на исконной брюшной стороне с
помощью ресничной полоски, подостланной
вентральным нервным тяжем (рис. 5). Он
имел три пары целомических мешков и
щупальцевый аппарат, с помощью
которого собирал мелкие пищевые частицы. В
дальнейшем могло произойти обособление
самого заднего отдела тела, с помощью
которого предки вторичноротых временно
зарывались в верхние слои грунта. В
эволюции вторичноротых этому отделу
принадлежит очень важное место.
Предки иглокожих перешли к
сидячему образу жизни и стали собирать
взвешенные в воде пищевые частицы с
помощью щупалец. Радиальная симметрия
иглокожих — это наследие сидячего образа
жизни, сохраняемого среди современных
форм морскими лилиями, тогда как
морские звезды, морские ежи, офиуры и
голотурии способны медленно переползать с
места на место.
Полухордовые в качестве основного
органа движения стали использовать пред-
ротовой отдел тела: хобот кишечнодышащих
или головной щит крыложаберных.
Обычно полухордовых считают близкими
предками хордовых, о чем говорит и само
название этой группы. В действительности эта
группа не слишком близка к хордовым, в
частности потому, что имеет два нервных
тяжа, только один из которых может быть
гомологизирован с нервной трубкой
хордовых. У них имеется слепой вырост
кишечника — стомохорд; он сформировался как
опорная структура хобота и не имеет
прямого отношения к хорде.
Современные погонофоры — сидячие
организмы, обитающие внутри выделяемых
ими трубок. Во взрослом состоянии они
лишены кишечника и питаются за счет сим-
биотических хемосинтезирующих бактерий,
окисляющих сероводород или метан. Их
предки, вероятно, питались этими бактериями,
собирая их из придонного слоя воды с
помощью щупалец. Впоследствии они стали
культивировать бактерии в клетках
кишечника. Личинки одной из групп современных
погонофор — вестиментифер — на ранних
стадиях развития собирают бактерий с
поверхности грунта, а затем начинают их
культивировать внутри видоизмененного
кишечника, после чего рот и анус у них
редуцируются. Особенности эмбрионального
развития и строения нервной системы
позволяют рассматривать погонофор как
самую близкую к хордовым группу
вторичноротых.
Предки хордовых также
первоначально собирали пищевые частицы с помощью
околоротовых щупалец, которые до сих пор
сохранились у ланцетников. Одной из
особенностей эволюции хордовых явилось
использование самого заднего, IV отдела тела
как органа активного движения. Этот отдел
имеется у всех вторичноротых. Иглокожие
и крыложаберные используют его как
прикрепительный стебелек. У кишечнодышащих
этот отдел функционирует только у
молодых особей: с его помощью они
закапываются в толщу грунта. У погонофор этот
отдел активно функционирует всю жизнь: он
вооружен щетинками, торчит из заднего
конца трубки и, благодаря его движениям,
погонофоры заглубляют задний конец трубки
в толщу грунта по мере своего роста.
Хордовые — единственная группа
вторичноротых, использующая этот отдел как орган
активного плавания. Именно в связи с активной
двигательной функцией этого органа в нем
происходит полная сегментация мускулатуры
и развивается хорда (у личинок
оболочников хорда локализована именно в хвосте).
Активное использование хвостового
отдела как органа движения в какой-то мере
предопределило примат подвижности в
эволюции хордовых.
Другой важный момент эволюции
хордовых — это инверсия сторон тела, т.-е.
произошедший в филогенетическом
развитии хордовых переворот с
морфологически брюшной на морфологически спинную
сторону. Сам по себе этот переворот —
сравнительно-анатомический факт, следующий
из сопоставления плана строения хордовых
с таковыми других групп вторичноротых.
В то же время конкретные причины
этого переворота восстановить трудно, ведь
мы так мало знаем об условиях жизни в
морях того отдаленного геологического
периода и о конкретном облике предков
хордовых. Такой переворот мог совершиться
при переходе от улавливания с помощью
щупальцевого аппарата взвешенных в
толще воды пищевых частиц, как это
делают, например, современные
крыложаберные, к их сбору из поверхностного слоя
грунта, как это делают современные
ланцетники.
Заметим, что между личинками
оболочников, с одной стороны, и
бесчерепными и позвоночными, с другой, есть одно
важное различие. Если внимательно по-
ПРОИСХОЖДЕНИЕ ХОРДОВЫХ ЖИВОТНЫХ
37
смотреть на две нижние фигуры на рис. 4,
изображающие личинку асцидии и
ланцетника, то нетрудно заметить, что у
первых рот располагается на той же
стороне, что и нервная трубка (последняя даже
открывается в глотку особым отверстием),
т. е. на исходно брюшной стороне, а у
ланцетника (так же, как у позвоночных) — на
противоположной стороне тела.
Положение рта у личинок асцидий — первичное,
оно совпадает с тем, что имеет место у
других вторичноротых (см. рис. 4). Таким
образом, личинки асцидий по своей
организации более примитивны, чем
остальные хордовые, и отражают организацию
подвижных предков хордовых до их
переворота.
У бесчерепных и позвоночных рот
располагается на противоположной стороне,
гомологичной спинной стороне тела
прочих вторичноротых (см. рис. 4). Такое
положение рта не может быть первичным
и возникло уже после переворота.
Удивительно, но в индивидуальном развитии
бесчерепных сохранились следы
постепенного перемещения рта со спинной
стороны на брюшную по левой стороне тела. А
один из представителей бесчерепных
(Assymetron) так и застыл на стадии, когда
рот остался на левой стороне тела. Все это
свидетельствует о постепенном
перемещении рта со старой морфологически
брюшной стороны на новую физиологически
брюшную сторону.
Заключение
Современные представления о
происхождении хордовых объединяют
положения, существовавшие ранее в рамках
альтернативных гипотез: перевернутость
хордовых, их родство с вторичноротыми
и факт большей примитивности личинок
оболочников по сравнению с другими
хордовыми. В отличие от ранее
существовавших точек зрения хордовые
рассматриваются не как перевернутые кольчатые черви
и членистоногие, но как перевернутые вто-
ричноротые. Что же касается личинок
асцидий, то они не являются предками
остальных хордовых, как это принимается
в гипотезах неотенического происхождения
хордовых. Организация личинок асцидий
рекапитулирует (т. е. повторяет в сжатой,
видоизмененной форме) в индивидуальном
развитии определенный этап в
историческом развитии хордовых, а именно
строение предков хордовых до их переворота.
Дискуссия о происхождении хордовых не
закончена. Развитие биологической науки
приносит новые доводы в пользу той или
иной точки зрения. Иногда эти доводы при-
Иглокожие Крыложаберные Кишечнодышащие Погонофоры
Хордовые
Предок вторичноротых
Рис. 5. Филогенетические отношения между вторичноротыми животными
38
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
ходят с совсем неожиданной стороны. Так,
в 1993 г., в английском журнале «Nature»
появилась статья биологов Дж. Слэка,
П. Холланда и С. Грэма, анализирующая
многочисленные публикации, в которых
были выявлены зоны экспрессии одной
интересной группы генов (так называемой Ноя-
группы) в развитии различных
беспозвоночных и позвоночных животных. В этих
публикациях было показано, что у
беспозвоночных эти гены маркируют брюшную
сторону, а у позвоночных их экспрессия
наблюдается на спинной стороне. Этот факт
хорошо согласуется с идеей
перевернутости хордовых, принимаемой современной
сравнительной анатомией.
Литература
Жоффруа Сент-Илер Э. Избранные труды. М.:
Наука, 1970.
Иванова-Казас О. М. Очерки по филогении
низших хордовых. СПб.: Изд-во СПб. ун-та, 1995.
Малахов В. В. Журн. общ. биологии, 1977,
т. 38, № 4, с. 485.
Малахов В. В. Природа, 1982, №5, с. 12.
Slack J. M. W., Holland P. W. Я., Graham С. F.
Monthly Nature, 1993, vol. 1, № 2, p. 21.
Ю.П.Алтухов
ВИД И ВИДООБРАЗОВАНИЕ
С момента возникновения научного
интереса к проблеме биологического вида и
вплоть до наших дней вся эта область
исследования, равно как и проблема
эволюции в целом, представляет собой поле
непрекращающихся, подчас весьма
драматических дискуссий. Стремясь к
максимальной объективности, следует, по-видимому,
признать, что мы еще не располагаем
общепринятым определением вида и
завершенной теорией видообразования. Одновременно
с развитием цитологической,
биохимической и молекулярной генетики накапливается
все больше фактов, которые противоречат
наиболее широко распространенной
концепции, трактующей видообразование как
вероятностный процесс, протекающий на
популяционном уровне, и укрепляют
типологические представления о сальтационной
природе появления новых видов.
Попытаемся систематизировать в
сжатой форме существующие дискуссионные
моменты, уделив главное внимание самой
сути взглядов на вид и видообразование,
характерных для типологов и популяцио-
нистов.
* * *
Представление о виде как одном из
иерархических уровней организации
жизни, ее основной таксономической единице
не вызывает возражений у большинства
биологов. В логике виду соответствует
понятие, занимающее промежуточное
положение между более общим понятием «род»
и более частным понятием «индивидуум».
В этом смысле понятие «вид» может быть
весьма условным и не соответствовать
какой-либо объективно существующей
категории. Однако еще в Древней Греции
прослеживается стремление фиксировать
понятие «вид», придав ему онтологический
смысл. Согласно концепции идей Платона,
наблюдаемая изменчивость органического
мира — не более чем искажения неких
постоянных, идеальных сущностей
разнообразных форм жизни. Аристотель называл
видом «то в каждом, что оно, собственно,
и есть в первую сущность». Это
определение лежит в основе типологических
представлений о биологическом виде,
постулирующих существование истинных видовых
признаков.
Английский ботаник Дж. Рэй,
вероятно, был первым, кто дал определение
биологическому виду: «Видовое тождество быка
и коровы, мужчины и женщины вытекает
из того, что они происходят от одинаковых
родителей, часто от одной и той же
матери; у растений точно так же самый
верный признак принадлежности к одному виду
есть происхождение от одного и того же
растения. Формы, принадлежащие
различным видам, сохраняют неизменный
характер своего вида и никогда один вид не
возникает из семян другого, и наоборот»
[«Historia plantarum», 1686]. Очевидно,
принципиальный момент в таком понимании
природы видов — постулирование
репродуктивной изоляции между ними, хотя это
условие в явном виде и не присутствует в
определении Рэя.
К. Линнеем соответствующий подход был
положен в основание созданной им
системы природы, построенной на бинарной
номенклатуре, которая используется в
систематике и сегодня. Наиболее сходные
между собой виды (лат. species) Линней
объединял в роды (лат. genus) и каждому виду
давал двойное наименование.
Этот подход, заменивший прежние
многословные определения вида,
революционизировал биологию, открыв неизвестные
ранее возможности для систематизации
растительного и животного мира. Вместе
с тем в вопросе о происхождении видов
Линней, как и его предшественники,
стоял на библейских позициях, считая, что все
особи любого вида суть потомки одной
первоначально созданной пары и что после акта
творения на Земле не появлялся ни один
новый вид.
В конце жизни Линней уже не был столь
категоричен, допуская возможность
возникновения новых видов, например путем
гибридизации. Однако в биологии XVIII —
первой половины XIX вв. идея неизменности
видов все же была господствующей, чему
в немалой степени способствовал авторитет
Ж. Кювье. Он связал идею постоянства вида
с известным учением о геологических
переворотах, имевших место в различные
эпохи на Земле и приводивших к
исчезновению ранее существовавших фаун и флор,
которые заменялись новыми, никак
преемственно с ними не связанными. Для
животного и растительного мира каждой
геологической эпохи постулировался специальный
акт творения новых видовых форм.
Между тем уже с конца XVIII столетия
в биологии стала укрепляться
эволюционная идея, получившая наиболее
завершенное воплощение в материалистической те-
40
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
ории происхождения видов путем
естественного отбора, созданной Ч. Дарвином. С этого
времени типологическое представление о
биологическом виде как неизменной
таксономической единице постепенно
вытесняется популяционным принципом, приведшим
к возникновению так называемой
синтетической теории эволюции (СТЭ),
разделяемой сегодня многими учеными. Название этой
теории отражает то обстоятельство, что она
возникла в результате синтеза дарвиновой
концепции естественного отбора и
положений популяционной генетики, синтеза,
начатого в 1926 г. в известной работе
С. С. Четверикова и завершенного в 40-е гг.
текущего столетия. Главная черта СТЭ,
иногда называемой неодарвинизмом, в том,
что она представляет собой вероятностную
концепцию, ибо и теория естественного
отбора Дарвина, и наиболее успешные
модели популяционной генетики основываются
на неопределенной наследственной
изменчивости. Но такая изменчивость,
исследуемая как состояние или как процесс,
может быть выражена только в терминах
теории вероятностей. Неудивительно, что
свершившийся в свое время синтез был столь
органичным. Эта замена типологического
мышления популяционным, по мнению
выдающегося американского биолога
Э. Майра, «быть может, величайшая
идейная революция в биологии». Как отмечал
Майр [1968, с. 20], «способы мышления
популяционистов и типологов
диаметрально противоположны. Популяционист
подчеркивает неповторимость любого явления в
мире живого. Как среди людей нет двух
одинаковых индивидуумов, так нет их и
среди других видов животных и растений.
Все организмы и жизненные явления
обладают индивидуальными особенностями и
в совокупности могут быть описаны только
в статистических терминах. Индивидуумы
или любые другие единицы жизни
образуют популяции, для которых мы можем
определить арифметическое среднее и
статистически оценить изменчивость. Средние
цифры являются статистической
абстракцией, реальны только индивидуумы, из
которых состоит популяция. Конечные
выводы типолога и популяциониста прямо
противоположны друг другу. Для типолога
тип (eidos) реален, а изменчивость —
иллюзия, тогда как для популяциониста тип
(среднее) — это абстракция и реальна только
изменчивость. Трудно представить себе
взгляды на природу, которые различались
бы сильнее».
Действительно, эти различия четко
проявляются и в отношении к видовым
критериям и как следствие — к определениям вида
и к проблеме видообразования. Согласно
типологической концепции, как уже
указывалось, существуют истинные видовые
признаки, по которым особи одного и того
же вида должны быть тождественны друг
другу и в то же время отличаться от
особей других видов. Согласно популяционной
концепции, инвариантных видовых
признаков быть не может, ибо «разница между
видом и разновидностью не в сущности, но
в степени. Разновидность есть
зарождающийся вид, а вид — резко выраженная
разновидность». В редакции Майра [там же, с. 270]
этот дарвиновский тезис звучит так: «Все
признаки, которые используются для
разграничения видов, подвержены
географической изменчивости». Соответственно, с
позиций типологизма вид можно определить
как «совокупность особей, тождественных
по видовому признаку», тогда как с
позиций популяционизма виды определяют как
«группы действительно или потенциально
скрещивающихся популяций, репродуктивно
изолированных от других таких же групп»
[Майр, 1947, с. 334].
То же принципиальное различие
характерно и для трактовки механизмов
видообразования. Если не брать в расчет
недоступное рациональному объяснению
возникновение видов как творений
некоего высшего разума, а оставаться в
границах научного метода, то, с точки зрения
типолога, видообразование должно быть
процессом сальтационным (скачкообразным),
сопряженным с изменчивостью именно
видовых признаков, приводящей к
репродуктивной изоляции. С точки зрения
популяциониста, видообразование может быть
лишь градуальным (постепенным)
процессом, основанным на явлении
внутривидовой наследственной изменчивости,
вызывающей небольшие, но накапливающиеся
изменения фенотипа. В основе теории
градуальной эволюции при оценке ею
состояния среды, к которому должна
адаптироваться популяция, лежит идущий от геолога
Ч. Лайеля так называемый принцип акту-
ализма. В соответствии с этим принципом
медленные и незначительные естественные
изменения, подобные тем, которые
происходят в окружающей среде сегодня,
действовали и многие тысячелетия назад, шаг
за шагом вызывая дифференциацию
разновидностей (популяций), вплоть до их
полного видового обособления. В терминах
генетики популяций видообразование есть
длительный адаптивный процесс
постепенных замещений аллельных генов,
приводящий к темпоральной трансформации вида
как целого (анагенез, филетическая
эволюция), либо к расщеплению исходного вида
на дочерние виды вследствие
географической изоляции популяций (кладогенез,
«истинное видообразование»). Очень наглядно
процесс кладогенеза изображен на схеме
ВИД И ВИДООБРАЗОВАНИЕ
41
с
О.
а
Рис. 1. Схематическое изображение расщепления
исходного вида (а) на два (Ь и с) во времени [из:
Dobzhansky Th. Evolution, genetics and man. N.-Y.:
Wiley, 1955]
Переплетающиеся ветви— отдельные популяции
(рис. 1), предложенной в свое время
Ф. Г. Добжанским.
Таковы фундаментальные различия между
двумя концепциями вида,
пересекающимися лишь в одном пункте — в признании
репродуктивной изоляции в качестве
важнейшего видового критерия. Но и здесь сходство
только кажущееся: если в типологической
концепции вида репродуктивная изоляция
особей — первичное условие его
возникновения, то в популяционной она
рассматривается как побочный продукт далеко
зашедшей адаптивной дивергенции пространственно
изолированных популяций. Не случайно в
течение долгих лет в рамках неодарвинизма
была весьма популярна модель аллопатри-
ческого (географического) видообразования (см.
рис. 1) и нередко исключалась возможность
симпатрического видообразования, т. е.
видообразования без пространственного
разделения исходной популяции, как это,
например, бывает в случае полиплоидии. Но
поскольку такой путь видообразования все
же широко распространен в живой природе
(например, у растений), было предпринято
немало усилий, чтобы обосновать
качественное различие механизмов видообразования
у животных и растительных организмов.
Например, утверждалось, что, хотя у
животных-гермафродитов или
размножающихся партеногенетически (дождевых червей, тур-
беллярий, жуков-долгоносиков и др.)
полиплоидные ряды видов довольно обычны,
для их подавляющего большинства такое
скачкообразное видообразование исключается,
поскольку при этом, согласно правилу Г. Мел-
лера, нарушался бы хромосомный баланс в
определении пола и в потомстве полиплоида
появлялось бы слишком много генетически
несбалансированных и, стало быть, стериль-
1 2 3 4 5 6 7 8 9 10 11 12
б в
12 3 4 5
Л
12 3 4 5 6 7
г
1234 1234567
Рис. 2. Межвидовые и межродовые различия,
обнаруживаемые при электрофоретическом
исследовании множественных гемоглобинов рыб семейства
лососевых (Salmonidae) [из: Алтухов Ю. П., 1989]
а: 1—3 и 5—12— кета, Oncorhynchus keta\ 4 — кижуч,
О. kisutch (электрофорез в агаровом геле); б: 1—5 —
голец, Salvelinus leucomaenis; 6, 7— горбуша,
Oncorhynchus gorbuscha (электрофорез в
крахмальном геле); в: 1 — О. gorbuscha; 2— гибрид F^ 3—5 —
сима, Oncorhynchus masu (электрофорез в агаровом
геле); г: 1, 2 — 0. keta\ 3, 4 — 0. gorbuscha; д: 1, 2 —
Salvelinus leucomaenis; 3—7— О. keta
(электрофорез в крахмальном геле). Стрелка обозначает
стартовую позицию, «+» и «-»— анод и катод
соответственно. Хорошо видно, как при полном генотипи-
ческом тождестве особей внутри вида наблюдаются
качественные межвидовые различия. Более
детальную информацию о методе электрофоретического
разделения генетически контролируемых вариантов
белков [см.: Алтухов Ю.П. Генетика популяций и
сохранение биоразнообразия. Наст, том]
ных особей (если мать несет хромосомы
ХХХХ, а отец— XXYY, то среди потомков
будет много нежизнеспособных особей XXXY
и такой вид просто не сможет существовать).
Как указывал Майр [1968, с. 359]: «Даже если
будет показано существование полиплоидии
у некоторых из этих групп (у ракообразных,
чешуекрылых, рыб и др.— Ю. А.), все же
можно утверждать, что у животных это
явление исключительное в противоположность
тому, что имеет место у растений. У нас нет
доказательств в пользу того, что целые
группы видов или роды обязаны своим происхож-
42
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Время, с *-
Вид 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Примитивный
(без названия)
clnctipennis
hebes
potomaca
frontalis
versicolor
fairchildi
lucicrescens
pensylvanica
pyralomimus
aureolucens
caerulucens
tremulans
Рис. З. Характер световых вспышек у разных видов североамериканских светляков рода Photuris [из: Майр,
1968]
Высота и длина отметок показывают интенсивность и характер вспышек
дением полиплоидии, с чем мы постоянно
сталкиваемся в растительном царстве».
Именно поэтому в популяционной
концепции видовая категория оказывается
приложимой только к перекрестно
размножающимся обоеполым организмам.
Существование в природе однополых форм
создает для СТЭ непреодолимые трудности
в смысле определения их видового
ранга, поскольку такой критерий, как
нескрещиваемость, к ним неприложим.
Предлагалось даже использовать для таких форм
какой-либо нейтральный термин,
например «бином», сохранив термин «вид»
только для двуполых организмов.
Для типологической концепции
проблема видовой идентификации решается
однозначно, независимо от того, имеет ли дело
исследователь с одно- или с двуполыми
организмами. Для этого достаточно лишь
опереться на приведенное выше
типологическое определение вида и признать
существование диагностических признаков,
инвариантных (мономорфных) внутри вида, но
вместе с тем дифференцирующих виды в
пределах рода. Такой подход оказывается
плодотворным при распознавании по ряду
морфологических признаков «хороших
видов». Однако при изучении так называемых
видов-близнецов, когда не удается найти
заметных внешних (морфологических)
различий, задача видовой идентификации
кажется неразрешимой.
Тем не менее поиск межвидовых
различий на других структурных уровнях
организации, например на молекулярном или
клеточном, рано или поздно позволяет
отыскать такие признаки, которые дают
возможность успешно решить проблему
видовой идентификации. Хорошо известна,
например, видовая специфичность карио-
типов, составляющая основу кариосистема-
тики. То же характерно для ряда
мономорфных белковых систем как маркеров
соответствующих структурных генов, что
используется в биохимической
систематике (рис. 2). Нередко удивительные
примеры видовой специфичности и инвариантности
внутри вида демонстрируют многие
признаки, особенно связанные с репродуктивным
поведением (структура муравейников,
населенных различными видами; гнезд у
некоторых видов пчел; различия в пении и
сигнальных звуках и др.). Например,
изучение световых вспышек светлячков рода
Photuris Северной Америки привело к
открытию многих новых видов вместо двух-
трех описанных ранее (рис. 3).
Исследование белков лягушек, относимых ранее к
одному-единственному виду Ran a pipiens,
или подвидов чаек, объединяемых в
комплексе Larus argentatus, вскрыло
существование совокупности видов-близнецов.
Число таких примеров легко умножить.
Приведенные выше и многие другие
данные, полученные за последние 15—20
ВИД И ВИДООБРАЗОВАНИЕ
43
лет, в противоречии с распространенным
мнением некоторых ученых, не дают
оснований считать, что популяционная
(биологическая) концепция вида есть некая
«истина в последней инстанции», полностью
вытеснившая типологизм. Популяционная
концепция, бесспорно, внесла важный вклад
в науку, защищая представления о
реальности биологического вида как особого
уровня организации жизни, привлекла
внимание широкого круга исследователей к
явлению внутривидовой (географической)
наследственной изменчивости, обосновала
представление о политипическом виде,
подчеркнула значение и вскрыла
разнообразие форм репродуктивной изоляции и
т. д. Вместе с тем она усложнила работу
систематиков в той мере, в какой вместо
поиска истинных видовых признаков
стала уделять чрезмерное внимание явлению
внутривидовой изменчивости. По сути
общепринятая популяционная концепция
опирается лишь на два критерия вида:
морфологический и генетический. При таком
подходе морфологическим критерием вида
служит хиатус (пробел) в величине
какого-либо изменчивого внешнего признака,
и эта прерывистость обычно используется
как доказательство принадлежности
сравниваемых популяций к различным видам.
Однако морфологический критерий вида не
является универсальным, поскольку «для
характеристики подвида, вида и рода
используются совершенно одни и те же
признаки. То, что в одной группе служит
критерием рода или даже семейства, в другой
представляет собой лишь критерий подвида
или даже не имеет никакого
систематического значения» [Майр, 1947, с. 187].
Генетический критерий вида, лежащий и
в основе современной типологической
концепции, указывает на нескрещиваемость видов
при их совместном обитании в одном
биотопе. Этот критерий можно считать достаточ-
4л I
Зп 2л —'
III
2п 2п =*
I
2/7 2п
Рис. 4. Схема гибридогенного (сетчатого)
видообразования (до стадии аллотетраплоидии) у
позвоночных [из: Боркин, Даревский, 1980]
/—///— этапы последовательной гибридизации,
приводящие к повышению степени плоидности
гибридной формы. Синие кружки — бисексуальные виды,
зеленые кружки — однополые гибридные виды
но универсальным, поскольку он приложим
к любым группам организмов (кроме
однополых). Вместе с тем из-за разнообразия
изолирующих механизмов такая универсальность
имеет лишь чисто теоретический смысл и в
подавляющем большинстве случаев не
может быть использована на практике. Несмотря
на то обстоятельство, что биологическая
концепция вида получила широкое
признание среди ученых и вызвала к жизни
развитые статистические методы классификации,
например нумерическую таксономию, она не
внесла принципиальных перемен в
практическую работу систематиков, которая до сих
пор в основном базируется на
типологических представлениях о виде.
Более того, за последние годы взгляды
на вид и его эволюцию претерпели
существенные изменения. Наряду с градуализмом,
принимающим лишь модель географического
видообразования, все большее
распространение стали получать представления о
«квантовом» (сальтационном) видообразовании,
осуществляющемся симпатрически. Наиболее
распространенные механизмы такого
происхождения видов — полиплоидия и различные
типы хромосомных перестроек (перицентри-
ческие инверсии, реципрокные
транслокации и др.). Вопреки прежним
представлениям, за последние годы обнаружено немало
случаев полиплоидии у животных. Среди
соответствующих работ особое место
принадлежит исследованиям С. Оно, представившего
цитологические и биохимические
доказательства того, что тетраплоидизация геномов
играла важную интегрирующую роль в
эволюции позвоночных, включая, по-видимому, и
человека. Полиплоидные ряды видов (роды
и даже целые семейства) обнаружены у рыб,
амфибий и рептилий. Показано, что в
эволюции этих групп тесно связаны
межвидовая гибридизация, бесполое размножение
(партеногенез или гиногенез) и полиплоидия.
Такая эволюционная схема включает
следующие этапы.
1. При межвидовой гибридизации
образуется диплоидная форма, переходящая к
однополому размножению.
2. При скрещивании этих гибридных форм
с одним из родительских (или близких к
нему) бисексуальных видов образуется
однополая триплоидная форма,
размножающаяся гино- или партеногенетически.
3. Скрещивание триплоида с одним из
близких бисексуальных диплоидных видов
приводит к формированию тетраплоида,
способного возвратиться к бисексуальному
типу размножения (рис. 4).
Аллотетраплоидные виды могут
возникать и минуя стадию триплоидии, когда
гибридные формы производят
нередуцированные гаметы, и при скрещивании алло-
тетраплоидных самцов с несбалансирован-
44
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
ным хромосомным набором (Х^Х^) с
самками ХХХ2 образуется потомство Х1Х1Х2Х2
(самки) и X1XlX2Y2 (самцы); скрещивание
внутри этого потомства должно привести
к формированию стабильного аллотетрап-
лоидного вида.
Совокупность недавних открытий,
сделанных в биохимической генетике
популяций и сравнительной генетике вида,
заставляет признать, что процесс видообразования,
а тем более макроэволюции далеко не
всегда сводится к простой замене аллелей уже
существующих генных локусов и
репродуктивная изоляция не есть побочный продукт
длительного процесса дивергенции
популяций. Напротив, репродуктивная изоляция —
важнейшее первичное условие
возникновения нового вида, как то уже давно очевидно
из хорошо аргументированной роли
некоторых хромосомных аберраций, не
приводящих к внутривидовому полиморфизму, но
вместе с тем маркирующих уровень вида. При
этом принципиально важно следующее
обстоятельство: виды оказываются
гомозиготами по соответствующей мутации, сильно
снижающей фертильность ее носителей,
когда она находится в гетерозиготном
состоянии.
Возможность фиксации крупных
аберраций хромосом наиболее велика при подраз-
деленности вида на небольшие колонии, лишь
незначительно обменивающиеся друг с другом
генетическим материалом. Как показал
С. Райт, в такой системе популяций резко
возрастает вероятность того, что две
гетерозиготные по хромосомной мутации
особи встретятся в каком-либо из
периферических изолятов и дадут жизнеспособных
гомозиготных потомков, относящихся к
новому виду, который фактически за два
последовательных гиага окажется репродуктив-
но изолированным от предкового вида.
Обсуждая это обстоятельство в связи с
бесспорно доказанной ролью перицентричес-
ких инверсий в видообразовании у приматов
и обнаружением гомозиготности по
соответствующей мутации у потомства от инцестно-
го брака в современной человеческой
популяции, Фогель и Мотульски задаются вопросом
[1990, с. 15]: «Не происходит ли все
человечество от одной-единственной предковой
пары?» и высказывают соображение, что миф
об Адаме и Еве как предках человеческого
рода может иметь научную основу.
С учетом всех этих новых фактов
видообразование можно рассматривать как
следствие крупных реорганизаций генома,
непосредственно сопряженных с
репродуктивной изоляцией, а обычную
внутривидовую изменчивость (например,
полиморфизм), на которой базируется СТЭ,
трактовать лишь как адаптивную
«стратегию природы», поддерживающую
устойчивость и целостность вида в условиях
нормально колеблющейся среды. Такой взгляд
на эволюцию, предполагающий чередование
периодов видовой трансформации через
качественные реорганизации геномов с фазами
длительной стабильности видов, был
выдвинут в начале 70-х гг. в США и России.
Он основан на явлении генетического
мономорфизма видов и обнаружении роли
системной организации популяций в
длительном сохранении присущих им уровней
генетического полиморфизма. Эта модель
постулирует двойственность в
структурно-функциональной организации генетического
аппарата (т. е. указывает на существование
двух групп генов или признаков:
жизненно важных — видовых и адаптивно менее
значимых — популяционных) и
перекликается с известными эволюционными
представлениями генетиков Г. Де Фриза, Р. Гольд-
шмидта и цитофизиолога Б. П. Ушакова,
подчеркивавших качественное различие
между собственно процессом
видообразования и адаптивной внутривидовой
дивергенцией. Близкие взгляды на видообразование
развивает американский генетик X. Карсон.
По Карсону, у бисексуальных видов
следует различать две системы генетической
изменчивости: открытую и закрытую.
Первая представлена свободно сегрегирующи-
Отбора нет
Отбор Отбор
s
S
3"
о:
5.
С
О
с
л
\-
о
о
X
X
О)
с;
о
S
г
1
О Время
Рис. 5. Схема видообразования на основе
реорганизации «закрытой» генетической системы [из:
Carson. American Naturalist, 1975, vol. 109, p.83]
Популяция после фазы отбора (а) проходит стадии
расцвета (б) и краха {в). Пережившие катастрофу
особи-основатели (г) порождают новую популяцию,
снова стабилизируемую естественным отбором (д).
«Совместимые» с онтогенезом реорганизации
закрытой генетической системы вида возможны лишь
при ослаблении отбора, когда один или несколько
«дискордантных» индивидуумов могут выжить
ВИД И ВИДООБРАЗОВАНИЕ
45
Размер гаплоидного генома (пг)
0,5 1 5 10 50 100
Насекомые
Ракообразные
Паукообразные
Кольчатые черви
Сипункулиды
Эхиуриды
Головоногие
моллюски
Брюхоногие
моллюски
Двустворчатые
моллюски
Боконервные
Круглые черви
Плеченогие
Иглокожие
Полухордовые
Оболочники
Головохордовые
Хрящевые рыбы
Костные рыбы
Земноводные
Пресмыкающиеся
Птицы
Млекопитающие
Кишечнополостные
Губки
108 109 1010 1011
Размер гаплоидного генома (нуклеотидные пары)
Рис. 6. Изменчивость размеров генома у различных таксономических групп животных [из: Britten, Davidson.
Quart. Rev. Biol., 1971, vol. 46, p. 111]
За годы, прошедшие после публикации этой важной работы, получены новые данные о размерах генома многих
десятков видов, полностью подтверждающие обнаруженные закономерности
46
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Грубая оценка общего числа видов, описаных систематиками
[из: May. Science, 1988, vol. 241, p. 1441]
Беспозвоночные
Protozoa
Porifera
Coelenterata
Echinodermata
Nematoda
Annelida
Brachiopoda
Bryozoa
Entoproctra
Mollusca
Crustacea
Arachnida
bisecta:
Coleoptera
Diptera
Lepidoptera
Hymenoptera
Hemiptera
260 000
10 000
10 000
6 000
1 000 000 (?)
15 000
350
4 000
150
100 000
39 000
63 000
1 000 000 (?)
300 000
85 000
110 000
100 000
40 000
Позвоночные
Pisces
Amphibia
Reptilia
Aves
Mammalia:
Monotremata
Marsupialia
Insectivora
Dermoptera
Chiroptera
Primates
Edentata
Pholidota
Lagomorpha
Rodentia
Cetacea
Carnivora
Tubulidentata
Proboscidea
Hyracoidea
Sirenia
Perissodactyla
Artiodactyla
Pinnipedia
19 000
2 800
6 000
9 000
4 500
3
266
345
2
951
181
29
7
58
1702
76
231
1
2
11
4
16
187
33
что такого рода межвидовые различия не
могут быть результатом замещений аллель-
ных генов, а суть следствие крупных
геномных реорганизаций, связанных прежде
всего с тандемными дупликациями генов,
межвидовой гибридизацией и
полиплоидией. Более подробно затронутые вопросы
освещаются в монографиях Ю. П.
Алтухова, Л. 3. Кайданова, С. Оно, Л. П. Та-
таринова (см. литературу). Оригинальные
взгляды на видообразование через
системные мутации развивают также известные
российские генетики Л. И. Корочкин и
В. Н. Стегний.
* * *
Завершая статью, хотелось бы
подчеркнуть то, с чего мы ее начали: проблема
ми локусами, не влияющими существенно
на жизнеспособность (полиморфизм разных
типов, подвидовая, клинальная
изменчивость), вторая — внутренне
сбалансированными, коадаптированными блоками генов
(супергенов), настолько сильно
влияющими на приспособленность, что любая их
реорганизация отметается естественным отбором.
Такие блоки, защищенные от кроссингове-
ра инверсией или же просто сильными эпи-
статическими взаимодействиями генов,
варьируют между видами, но не внутри вида.
Видообразование осуществляется через
реорганизацию закрытой системы генома, и
новые виды берут свое начало от одной или
немногих особей-основателей (рис. 5).
На рис. 6 показано, как варьирует
размер генома у разных представителей
животного царства. Совершенно очевидно,
ВИД И ВИДООБРАЗОВАНИЕ
47
биологической эволюции все еще далека
от своего разрешения. Как нам
представляется, необходимым шагом на пути к
пониманию этого процесса является
осознание невозможности объективно разобраться
в центральной для всей эволюционной
теории проблеме вида только с позиций по-
пуляционизма. Необходимо ослабить накал
чисто теоретических споров, сосредоточить
усилия на экспериментальной разработке
вопроса. Ведь как это ни парадоксально,
до сих пор все еще нет даже надежной
оценки общего числа видов, слагающих
биосферу. На сегодняшний день более или
менее детально описано лишь около 1,5 млн
видов (см. таблицу), тогда как
предполагается, что всего их может быть больше
5 млн. Отсутствует и общепринятая
классификация типов видов в природе, о чем
свидетельствует многообразие
употребляемых на этот счет терминов (например,
«жорданон», «линнеон», «виды-двойники»,
«первичные виды», «вторичные виды»,
«зарождающиеся виды», «полувиды» и т. п.).
И тем не менее реальность
биологического вида как особого уровня организации
жизни сомнения у ученых не вызывает. Что
же касается нашей практической
деятельности, то она строится на признании
существования в природе многообразных и
генетически стабильных видовых форм
жизни. Более того, без обитания на Земле
множества разнообразных биологических
видов, имеющих не только хозяйственную,
но и, по-видимому, до сих пор до конца
неосознанную экологическую и
эстетическую ценность, само существование
человечества стало бы просто невозможным. Этот
вывод приобретает особое звучание в наши
дни, когда антропогенное воздействие на
живую природу достигло небывалого
размаха. Деградация среды обитания,
нерациональное использование биологических
ресурсов, насыщение биосферы мутагенами,
бурное развитие сельского хозяйства и
другие типы человеческой деятельности
приближают нас к экологической катастрофе.
Биосфера в настоящее время деградирует,
и средний темп вымирания видов превышает
все, что известно на этот счет из
палеонтологической летописи. За какие-нибудь
последние 100 лет человеческая
деятельность привела к тому, что исчезновение
угрожает или уже осуществилось для
более чем 25 тыс. видов животных и 15 тыс.
видов растений. По недавним оценкам
специалистов, к 2010—2015 гг. с лица Земли
может исчезнуть до 15—20% населяющих
ее видов.
В этой связи проблема сохранения
видового разнообразия жизни становится
поистине глобальной. С учетом современного
состояния наших знаний ее решение
возможно лишь на пути создания единой
стратегии охраны природы и рационального
использования природных ресурсов. В
теоретическом плане в ближайшие годы
следует ожидать значительного сближения
взглядов типологов и популяционистов на
ключевые аспекты проблемы вида и
видообразования.
Литература
Алтухов Ю. Генетические процессы в
популяциях. 2-е изд. М.: Наука, 1989.
Боркин Л. Я., Даревский И. С. Журн. общ.
биологии, 1980, т. 41, № 4, с. 485.
Воронцов Н. Н. Теория эволюции: Истоки,
постулаты и проблемы. М.: Знание, 1984.
Кайданов Л. 3. Генетика популяций. М.:
Высш. шк., 1996.
Майр Э. Зоологический вид и эволюция. Пер.
с англ. М.: Мир, 1968.
Майр Э. Систематика и происхождение видов.
Пер. с англ. М.: ИЛ, 1947.
Оно С. Генетические механизмы
прогрессивной эволюции. Пер. с англ. М.: Мир, 1973.
Стегний В. Н. Архитектоника генома,
системные мутации и эволюция. Новосибирск:
Новосиб. ун-т, 1993.
Татаринов Л. П. Очерки по теории
эволюции. М.: Наука, 1987.
Фогель Ф., Мотульски А. Генетика
человека. М.: Мир, 1990, т. 3.
Korochkin L. Evol. Biol., 1993, vol. 7, p. 153.
Е. К. Еськов
АКУСТИЧЕСКАЯ СВЯЗЬ У НАСЕКОМЫХ
Введение
Класс насекомых выделяется в
животном мире наибольшим разнообразием
жизненных форм. На долю представителей этого
класса приходится около 70% общего
видового состава животного мира. Обилию
видов насекомых соответствует
многообразие их адаптации к различным условиям
среды: температуре, освещенности,
влажности, газовому составу воздуха и т. п.
Насекомые не имеют себе равных по
разнообразию режимов питания и форм
взаимоотношений с другими животными,
растениями и микроорганизмами.
Эволюция человека проходила в тесном
взаимодействии с насекомыми. Вначале они
были только его врагами: сперва —
паразитами, а позже — вредителями продуктов
и построек. Со временем некоторым из них
было" найдено полезное применение. Так,
не менее 7 тыс. лет назад люди начали
добывать пчелиный мед и около 5 тыс. лет
назад освоили разведение тутового
шелкопряда для получения шелка. Современная
экологическая концепция отношения к
насекомым базируется на разработке
технологий, способствующих ограничению их
вредного воздействия на здоровье
человека, агроценозы, постройки, продукты и
другие сферы его интересов и
максимальном использовании их полезных качеств.
Успешное решение этих задач находится
в прямой зависимости от достижений в
изучении биологии насекомых. Например,
немаловажное значение для разработки
средств контроля за их состоянием и
управления их поведением имеет анализ
используемых ими способов пространственной
ориентации и осуществления коммуникаций.
В системе пространственной ориентации
и связи между насекомыми значительное
место занимает акустическая сигнализация.
Она широко используется в брачный
период, обеспечивая повышение вероятности
встречи особей противоположного пола и
снижая затраты энергии и времени на их
сближение. Это имеет особо важное значение для
видов, которые ведут сумеречный и(или)
ночной образ жизни. Призывные сигналы
обладают обычно выраженной видовой
специфичностью, чем обеспечивается усиление
репродуктивной изоляции близких видов.
Некоторые отличия в структуре сигналов
выполняют роль экологических, межрасовых
и популяционных барьеров (см.: Алтухов Ю. П.
Вид и видообразование. Наст. том).
У социальных насекомых (термитов,
муравьев, пчел, ос и шмелей)
разнообразная сигнализация используется для
оповещения членов своих семей об опасности.
Сигналы стимулируют различные защитные
реакции. Так, муравьи-листорезы,
оказавшиеся под обвалившимся муравейником,
издают акустические сигналы,
выполняющие роль призыва о помощи. Это
стимулирует муравьев, находящихся на
поверхности, раскапывать обвалившийся грунт и
спасать членов семьи, терпящих бедствие.
Посредством акустических сигналов могут
регулироваться внутригрупповые
иерархические отношения. Указанную функцию
выполняет пение у некоторых видов
сверчков. Поэтому в случае утраты слуха
сверчком, занимающим подчиненное положение,
он становится инициатором столкновений с
представителями своей группировки.
Акустическая мимикрия обнаружена у
тараканов, генерирующих звуки, которые не могут
использоваться самими этими насекомыми,
так как не воспринимаются ими. Но
сходство таких звуков со звуками грызунов, на
них охотящихся, способствует повышению
безопасности тараканов, которая
достигается в результате акустической
дезориентации грызунов.
Пчелы пользуются пульсирующими
звуковыми сигналами, чтобы указать членам
своей семьи расстояние до обнаруженного
источника корма. Информация о
расстоянии кодируется в количестве импульсов и
продолжительности периода их генерации.
Преодолеваемое пчелами расстояние от
источника корма до жилища оценивается
по энергетическим затратам крыловой
мускулатуры на полет. Столь совершенная
форма акустической связи уникальна и не имеет
аналогов даже у млекопитающих.
Генерация акустических сигналов
В царстве животных наибольшим
многообразием механизмов генерации
акустических сигналов характеризуются
насекомые и рыбы. При этом у рыб используются
преимущественно органы, основная
функция которых не имеет прямого отношения
к генерации звуков, например плавники,
позвоночник, плавательный пузырь и т. п.
В отличие от рыб, у насекомых генерация
акустических сигналов обусловливается
разнообразными специализированными
структурами, различающимися по
строению и локализации.
АКУСТИЧЕСКАЯ СВЯЗЬ У НАСЕКОМЫХ
49
Воздействие на опорный субстрат. К
простейшим способам генерации звуков
относится использование твердого опорного
субстрата. Чаще всего насекомые стучат или
скребут по субстрату. Ударным механизмом
пользуются для оповещения об опасности
термиты. Они стучат мандибулами и (или)
брюшком. Сходный механизм генерации
звуков обнаружен у садовых муравьев и
жуков-часовщиков, которые получили свое
название за ритмические постукивания по
древесине, напоминающие тиканье часов.
Личинки бумажных ос используют для
генерации звуков стенки ячеек сот. Звук
возникает в то время, когда личинка,
втягиваясь в ячейку, скребет о ее стенку
широко раскрытыми мандибулами. Этот
звуковой сигнал голодные личинки подают для
привлечения к себе взрослых ос.
Трепие специализированных участков
экзоскелета. Акустический аппарат
фрикционного (фрикция — трение) действия
обычно включает в себя два
специализированных экзокутикулярных образования. Одно
из них чаще всего представляет собой
ребристое или бороздчатое поле, другое —
заостренный вырост или край какого-либо
внешнего органа. Их трение порождает
импульсные вибрации экзоскелета.
Рис. 1. Звуковые аппараты
фрикционного типа
/ — местоположение (а) и
увеличенные фрагменты (б, в)
звукового аппарата муравья-
листореза; // — зазубренные
участки на 3—7 сегментах
брюшка личинки Epiophlebia
superstes; III — ребристое
поле (а) на переднегруди
термита и внутренний участок
ноги (б), трущийся об это
поле; IV— ребристое поле на
боковой поверхности
брюшка жука (а) и внутренняя
сторона задней ноги (б ); V—
заостренная жилка на крыле (а,
показана стрелкой) и
ребристое поле на задней ноге (б);
VI — генерация звука
саранчей; VII — трущиеся участки
(показаны стрелками) на
крыле и элитрах (а, б) жука; VIII —
трущиеся участки надкрылий
(а, б — показаны стрелками) у
сверчка (в — фрагмент
ребристого поля)
Временная структура сигнала
(количество импульсов, их длительность, частота
следования) зависит от скорости трения.
Спектр сигналов (частотно-амплитудная
характеристика) определяется резонансными
свойствами структур экзоскелета,
колебания которых возбуждаются в процессе
функционирования звуковых аппаратов.
Количество фрикционных аппаратов
зависит от их локализации на теле
насекомого. Обладание единственным
акустическим аппаратом наиболее вероятно в
случае его размещения на непарном органе или
на каком-то из двух парных органов. По
одному звуковому аппарату имеют полевые
сверчки. На одном из их надкрылий
находится ребристое поле, на другом —
скребок. Звук генерируется в периоды сведения
приподнятых надкрылий (рис. 1, VIII).
По одному, реже по два акустических
аппарата обнаружено на брюшных
сегментах примерно у половины изученных видов
муравьев. У многих из них заостренный
вырост локализуется на третьем дорсальном
сегменте брюшка, входящем в состав
стебелька (органа, обеспечивающего
подвижное сочленение брюшка с грудью), а
ребристое поле— на четвертом (рис. 1, I). У
некоторых видов муравьев сходный аппарат
50
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
имеется также на вентральной части брюшка.
Эти звуковые аппараты функционируют при
подъеме и опускании брюшка.
Парные звуковые аппараты имеют
многие виды насекомых, использующих в
качестве функционирующих органов ноги
(рис. 1, II—VI). На них, чаще всего на
голенях, локализуются ребристые поля или
заостренные выросты. У саранчовых
ребристые поля, находящиеся на третьей паре
голеней ног, трутся о заостренные края
крыльев (рис. 1, VI). У многих видов жуков и
некоторых личинок ребристые поля
расположены на брюшных или грудных отделах,
а заостренные выросты — на задних ногах
(рис. 1, II, IV). У некоторых термитов
ребристые поля локализуются по бокам
грудного отдела. По ним трутся заостренные
выросты, расположенные на передних ногах.
Некоторые жуки генерируют акустические
сигналы трением специализированных
участков крыльев о расположенные над ними
поверхности элитр (рис. 1, VII).
Вибрации мембран. У цикад звуковые
аппараты образуются склеротизированными
выпуклыми тимбальными мембранами
(жесткими хитиновыми пластинками),
прогибаемыми специализированными мощными
мышцами. Парные мембраны локализуются на
нижней стороне первого брюшного
сегмента. От внутренней полости мембраны
отделяются воздушными мешками.
Структура звуков, генерируемых
тимбальными мембранами, зависит от их
резонансных свойств и частоты прогибания. У многих
видов цикад мембрана имеет ребра
жесткости (см. рис. 5, TV у б). Прогибанию каждого из
них сопутствует генерация одного
затухающего звукового импульса. Частота
следования импульсов зависит от скорости сокращения
мышц. Частотное положение
энергетического максимума в спектре сигнала
определяется преимущественно частотой колебаний
мембраны. Некоторое влияние на спектр звука
оказывают резонансные свойства воздушных
полостей, примыкающих к мембранам.
У большинства цикад тимбальные мышцы
сокращаются поочередно. Этим
обеспечивается двукратное увеличение частоты
следования пакетов импульсов, генерируемых
насекомыми. Синхронное сокращение мышц,
обнаруженное у австралийских цикад,
способствует некоторому увеличению
интенсивности сигнала.
Использование летательных аппаратов.
Звуки, генерируемые летающими
насекомыми, нередко имеют для них важное
сигнальное значение. Широкие возможности
использования летательных аппаратов для
генерации специальных сигналов
обнаружены у представителей надсемейства
пчелиных. На медоносной пчеле доказано, что
механизм генерации звуков посредством
летательных аппаратов функционирует на
основе вибраций экзоскелета грудного
отдела, обеспечиваемых сокращением
летательных мышц. Положение энергетических
максимумов в спектре звуков зависит от
частоты сокращения мышц. Возбуждаемые
ими вибрации экзоскелета грудного
отдела передаются крыльям. Они усиливают
интенсивность колебаний, совпадающих с
собственными резонансными частотами. В тех
случаях, когда насекомое машет
крыльями, например в полете, основная частота
звуков определяется частотой взмахов.
Восприятие акустических
сигналов
У наземных насекомых в качестве
линий акустического канала связи
используется воздушная среда и (или) твердый
опорный субстрат. Соответственно этому
насекомые пользуются разными рецепторами.
Они различаются морфологически и
функционально.
Рецепторы звуковых
колебаний воздуха
Быстроадаптирующиеся трихоидные
(волосковые) сенсиллы. Рецепторы
низкочастотных акустических и электрических
колебаний, у которых в качестве
первичного преобразователя используются волоски,
впервые были обнаружены у пчел.
Волоски локализуются по бокам головы между
фасеточными глазами и затылочным швом.
Длина волосков, покрытых остевидными
шипами, равняется в среднем 640 мкм, их
диаметр у основания— около 10 мкм.
Волосок удерживается в равновесном
состоянии посредством сочленовной
мембраны. Она соединяет (сочленяет) основание
волоска со входом в кутикулярную полость.
Через нее к основанию волоска подходит
дендрит рецепторной клетки,
представляющей собой биполярный нейрон. Каждый
волосок иннервируется одним нейроном. Его
длина около 23, диаметр — 9 мкм (рис. 2).
Угол отклонения волоска, колеблющегося
под действием звука, зависит от
интенсивности и частоты звуковых колебаний. На
маятникообразные колебания волоска,
деформирующие окончание дендрита, нейрон
отвечает генерацией нервных импульсов
(потенциалов действия). Их количество и
частота следования возрастают с
увеличением амплитуды колебаний волоска.
Вместе с потенциалами действия генерируются
незатухающие электрические колебания,
совпадающие с частотой вибраций волоска
(рис. 3). Весьма вероятно, что отмеченные
электрические колебания, связанные с
амплитудой отклонений волоска, имеют
отношение к возбуждению нейрона и гене-
АКУСТИЧЕСКАЯ СВЯЗЬ У НАСЕКОМЫХ
51
12
13
11
13
10
12
Рис. 2. Микроструктура фонорецептора медоносной пчелы
а— поперечный срез через центральную часть кутикулярной капсулы; б— продольный срез: 1 — кутикула, 2—
проксимальная часть волоска, 3— шейка, 4— основание, 5— сочленовная мембрана, 6— вход в кутикулярную
капсулу, 7—дендрит, 8 — тело нейрона, 9 — ядро, 10 — сопутствующие клетки, 7 7 — полость капсулы, 12— внут-
риволосковый канал, 13— стенка капсулы
рации им потенциалов действия. При
прочих равных условиях под влиянием
увеличения продолжительности колебаний волоска
в 10 раз количество потенциалов действия,
генерируемых нейроном, возрастает в
среднем в б раз.
На прикосновение к волоску,
отклоняющему его от положения равновесия,
нейрон отвечает генерацией потенциалов
действия. Они генерируются только в то время,
когда происходит отклонение волоска от
положения равновесия и (или) возвращение
его в исходное положение (рис. 4).
Отклоненный волосок не генерирует потенциалов
действия. Этим быстроадаптирующиеся
сенсиллы отличаются от гравитационных три-
хоидных сенсилл у пчел и ветрочувствитель-
ных рецепторов у саранчи.
Тимпанальные органы.
Специализированные фонорецепторы — тимпанальные
органы — обнаружены у многих видов
насекомых, не имеющих эволюционного родства.
Различия в строении тимпанальных
органов соответствуют филогенетической
отдаленности насекомых. Функциональное и
морфологическое сходство этих органов
выражается в использовании в качестве
первичных преобразователей звуковых
колебаний так называемых тимпанальных
мембран (с этим и связано их название).
Последние представляют собой тонкие
участки кутикулы, находящиеся в натянутом
состоянии. Натяжение обеспечивается
участками трахей, примыкающих к мембранам,
или к утолщениям кутикулы, обрамляющим
мембраны.
Строение и локализация тимпанальных
органов являются таксономическими
признаками. У сверчков и кузнечиков
тимпанальные органы локализуются в голенях
передней пары ног (см. рис. 5, I—III). В месте
локализации слухового органа голень
расширяется и имеет по два щелевидных
отверстия (см. рис. 5, I, II, а). Они находятся
перед мембраной, которая плотно
прижимается к разветвлению ножной трахеи (см.
рис. 5, III). Вдоль нее размещаются
чувствительные клетки (см. рис. 5, III, в), которые
раздражаются вибрациями трахеи,
возникающими под действием акустических
колебаний тимпанальной мембраны.
У цикад, саранчовых и некоторых видов
бабочек тимпанальные органы
локализуются в брюшном или грудном отделах. Тимпа-
52
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
0,1 С
I I
Рис. 3. Электроответы (1) одной и той же трихоидной сенсиллы на звуки (2), различающиеся по частоте
нальные мембраны цикад, размещающиеся
в передней части брюшка, представляют
собой очень тонкие хитиновые пленки (см.
рис. 5, IV, V, а). С внешней стороны они
защищены толстыми складками экзоскелета и
волосками. С внутренней стороны к мембранам
примыкают стенки воздушных мешков. Ими
и специальными мышцами поддерживается
натяжение мембран. Их колебания
передаются чувствительным клеткам (сколопиди-
ям), локализующимся в хитиновых
капсулах (см. рис. 5, V, б, в). Они выступают по
бокам брюшного отдела цикады. Сходное
строение имеют слуховые органы
саранчовых и бабочек из семейства совок.
Чувствительность тимпанальных органов
зависит от акустических свойств мембран. Как
правило, максимум чувствительности
слуховых органов приходится на диапазон
интенсивных составляющих в спектрах акустических
сигналов, используемых насекомыми. Эта
универсальная для животных зависимость
является следствием сопряженной эволюции
механизмов генерации и восприятия
акустических сигналов. Совпадение интенсивных
составляющих в спектрах генерируемых
сигналов с диапазонами максимальной
чувствительности слуховых органов обеспечивает
надежную связь при минимальных затратах
энергии на сигнализацию.
Рецепторы колебаний
опорного субстрата
Специализированные органы,
воспринимающие колебания опорного субстрата, были
обнаружены Мае Индо в начале текущего
столетия. Они локализуются в верхней
части голеней, непосредственно под коленным
сочленением (субгенумом), и поэтому их
называют субгенуальными органами.
Чувствительные клетки (сколопидии) суб-
генуальных органов совмещают в себе
функции преобразователей и рецепторов
колебаний опорного субстрата. Сколопидии
прикрепляются в субгенуме в двух точках —
вверху и сбоку. Свободный край,
противоположный месту бокого прикрепления, под
действием силы тяжести веерообразно раз-
0,3 с
Рис. 4. Электроответы на однократное отклонение волоска (7) и его возвращение в исходное положение (2)
АКУСТИЧЕСКАЯ СВЯЗЬ У НАСЕКОМЫХ
53
/ // ///
а а г
а
а
б б
б
в в
г г
IV V VI в
б б б
а
д д
в г
а а
Рис. 5. Разновидности рецепторов акустических колебаний
/—/// — фонорецептор кузнечика: / — местоположение на ноге (а — щелевидные отверстия); // — поперечный
разрез (а — щелевидные отверстия, б — тимпанальные полости, в — трахейные полости, г — тимпанальные
мембраны); /// — продольный разрез (а — субгенуальный орган, б — промежуточный орган, в — слуховой
гребень, г — субгенуальный нерв; д — тимпанальные полости); IV, V— тимпанальный орган цикады: /У —
местоположение (а — тимпанальные мембраны, б —тимбальные мембраны); V— строение (а — тимпанальная
мембрана, б — слуховая капсула, в — сколопидии, г— полость воздушного мешка); VI — субгенуальный орган муравья
ворачивается (см. рис. 5, VI). Наличие
свободно подвешенных клеток способствует
усилению эффекта их деформации (растяжения
и сжатия) под действием сил инерции.
Несмотря на сходство в строении и
функционировании субгенуальных органов, они
у разных видов насекомых существенно
различаются по чувствительности к
колебаниям опорного субстрата. Самая низкая
виброчувствительность обнаружена у мух-
пчеловидок, самая высокая — у тараканов.
У первых максимум чувствительности
находится на частоте около 200 Гц, у вторых —
1400 Гц, а воспринимаемая минимальная
амплитуда колебаний на указанных
частотах равняется соответственно 6590и0,004 нм.
Возникновение и эволюция
акустической связи
Средствами акустической связи
достигается повышение адаптации к среде
обитания. Использованием акустической
сигнализации обеспечивается оперативность регуляции
комплексом внутривидовых отношений,
снижаются затраты энергии и времени на
реализацию биологически важного
взаимодействия. Изменчивость структуры акустических
сигналов связи имеет большое значение для
эволюции насекомых. Групповая
консолидация на основе сходства сигнальных средств
связи, наряду с их изолирующей функцией,
способствуют микроэволюционным
преобразованиям, приводящим к внутривидовой
дифференциации.
Использованию в качестве акустического
канала связи опорного субстрата, которое
встречается у самых разных насекомых,
очевидно, предшествовало возникновение
виброрецепции. Она, по-видимому,
зародилась на самых начальных фазах
формирования этого класса и совершенствовалась
как средство ориентации при адаптации к
наземному образу жизни. Естественный
отбор благоприятствовал повышению
надежности функционирования и чувствительности
виброрецепторов, так как этим достигалась
лучшая защита от врагов и хищников,
приближение которых часто порождает
усиление колебаний твердой опоры. Наличие
высокочувствительных виброрецепторов
послужило предпосылкой для возникновения
акустической связи без затрат на
образование специализированных звуковых
аппаратов.
Сигнализация, основанная на
использовании опорного субстрата для генерации и
передачи звуковых колебаний, обладает
высокой эволюционной пластичностью.
Однако объем информации, передаваемой
посредством воздействия на опорный
субстрат, имеет значительные ограничения по
54
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
принципам ее кодирования. Оно возможно
лишь по продолжительности воздействия на
опору, а также по количеству и частоте
следования вибрационных импульсов.
Отличия по спектральной структуре сигналов не
могут иметь информационного значения из-
за высокой изменчивости резонансных
свойств природных субстратов.
Значительные ограничения пределов
повышения надежности и экономичности
связи посредством опорного субстрата
побуждали развитие независимых от него
механизмов генерации и передачи
акустических сигналов. Конвергентно в разных
филогенетических ветвях класса насекомых
получили развитие специализированные
звуковые аппараты, функционирующие по
принципу трения или соударения экзоку-
тикулярных структур, расположенных на
теле насекомых и их локомоторных
органах. Для восприятия акустических
колебаний, распространяющихся по воздуху,
используются фонорецепторы, отличающиеся
по локализации, но сходные по принципу
функционирования. Минимизация
энергозатрат на связь и повышение ее надежности
достигается прогрессирующим сближением
диапазонов генерируемых и
воспринимаемых акустических колебаний.
Поскольку системы акустической
связи наследственно запрограммированы,
отбор благоприятствовал стабилизации
механизмов генерации и восприятия
акустических сигналов. Этим обеспечивалось
повышение надежности внутривидовых
акустических коммуникаций. Индивидуальные
различия по структуре сигналов снижают
эффективность связи, увеличивая
затраты времени и энергии на реализацию
акустического взаимодействия. Однако
возникновение групповых различий выступает и
в качестве изолирующего фактора,
ускоряющего внутривидовую дифференциацию.
Развитие акустической сигнализации в
направлении возрастающей специализации,
конкретные воплощения которой
отличаются в разных филогенетических ветвях
класса насекомых, обеспечило им высокую
надежность связи.
Литература
Еськов Е. К. Биофизика, 1975, т. 20, с. 646.
Еськов Е. К. Акустическая сигнализация
насекомых. М.: Знание, 1979.
Еськов Е. К. Акустическая сигнализация
общественных насекомых. М.: Наука, 1979.
Еськов Е. К. Этология медоносной пчелы. М.:
Колос, 1992.
Жантиев Р. Д. Биоакустика насекомых. М.:
Изд-во МГУ, 1981.
Морозов В. Занимательная биоакустика. М.:
Знание, 1987.
Попов А. В. Акустическое поведение и слух
насекомых. Л.: Наука, 1985.
А. К. Бродский
АЭРОДИНАМИКА ПОЛЕТА НАСЕКОМЫХ
Введение
Насекомые летают над поверхностью
земли не менее трети миллиарда лет. А наши
знания о том, как работает их крыловой
аппарат, до сих пор очень поверхностны.
Ключевым вопросом остается проблема
создания аэродинамических сил в машущем
полете. Выяснить природу этих сил
пытались еще первые исследователи полета
насекомых. Немецкий ученый Е. Хольст в
1943 г. первым экспериментально доказал,
что господствовавшие ранее представления
о полете насекомых как о «гребном» не
соответствуют действительному принципу
работы машущего крыла. На смену
упрощенным и неточным представлениям об
«отбрасывании» воздуха крылом пришел так
называемый квазистационарный подход, при
котором аэродинамические процессы
сводились к серии стационарных состояний,
поддающихся расчету методами классической
аэродинамики. Исследование полета
саранчи, проведенное в Лондонском противоса-
ранчовом центре в 1956 г., показало
высокое совпадение расчетных данных с реально
измеренными средними значениями
аэродинамических сил. Несмотря на
общепризнанность квазистационарного подхода,
никому больше не удалось добиться соответствия
расчетных значений сил реальным, что
заставляет предполагать наличие
дополнительных аэродинамических сил. Кроме того,
многие авторы отмечают нестационарный
характер создаваемых в полете сил и
необычное поведение воздушных струй за
летящим насекомым.
Физические основы полета
Локомоция в сплошных средах
(воздухе и воде) настолько отличается от
передвижения на границе раздела сред, что
природа сил, делающих такое движение
возможным, заслуживает особого внимания.
Сплошная среда представляет собой
континуум, в котором не на что опереться, не
от чего оттолкнуться. В вязкой жидкости,
каковой являются вода и воздух,
возможны две основные формы движения:
ламинарное, или слоистое, когда линии тока
параллельны друг другу, и турбулентное,
при котором происходит перемешивание
жидкости. В турбулентных течениях скорость
и давление в каждой точке пространства
непостоянны и характеризуются
нерегулярными высокочастотными пульсациями.
Всякое тело, движущееся в сплошной
среде, испытывает на своей поверхности
нормальные и касательные напряжения. Их
результирующая представляет собой силу
полного сопротивления среды движению
тела, причем вектор силы полного
сопротивления далеко не всегда совпадает с
направлением движения. Проекция полного
сопротивления тела на ось х в скоростной
системе координат (ось х направлена вдоль
скорости движения, ось z
перпендикулярна к ней в горизонтальной плоскости, а ось
у — в вертикальной) есть лобовое
сопротивление, проекция же на ось у — при
положительном ее значении есть подъемная
сила, при отрицательном — либо топящая
(в воде), либо отрицательная подъемная сила
(в воздухе), т. е. зависит от того, где
происходит движение. Лобовое сопротивление
складывается из сопротивления трения и
сопротивления давления, или
сопротивления формы (разности давлений впереди и
позади тела). Количественно лобовое
сопротивление и подъемная сила выражаются
формулами:
Q = pcxv2s/2,
Y = pcyv 2s/2,
где s — характерная площадь тела, v —
скорость движения, р — плотность среды,
схи с — безразмерные коэффициенты
соответственно лобового сопротивления и
подъемной силы. Они зависят от формы
движущегося тела, его ориентации в
пространстве, но прежде всего от
соотношения сил инерции (они проявляются в
давлении на поверхность тела) и трения,
определяемых вязкостью. Соотношение этих
сил дает безразмерную величину,
численное значение которой должно быть
одинаковым у подобных течений. Иными
словами, речь идет о масштабном критерии,
дающем возможность сравнивать движение
различных объектов, имеющих разные
характеристики. Этот критерий получил
название числа Рейнольдса (Re):
сила инерции pv2/l _ pvl
сила трения pv/l2 р '
где I — характерный размер, ц — вязкость
среды, р — плотность среды.
Диапазон Re биологических объектов
достаточно велик: от 10"6 (бактерии) до 107
(крупные китообразные). Этот ряд
начинают мелкие организмы, движение
которых целиком зависит от вязкости, и
заканчивают крупные плавающие животные,
56
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
на движение которых оказывают влияние
в основном силы инерции. Стоит
инфузории перестать двигать ресничками, как
она тут же останавливается. Рыбе же
достаточно один раз ударить хвостовым
плавником, чтобы сравнительно долго
скользить вперед. Когда такое мелкое
насекомое, как трипе, перестает двигать
крыльями, оно останавливается, птица же
при этом продолжает по инерции двигаться
вперед.
По мере увеличения роли
инерционных сил с ростом Re в движении
животных все большее значение приобретает
подъемная сила. При Re = 103 подъемная
сила в три раза и более превышает
лобовое сопротивление. Так, для крыла мухи
каллифоры отношение подъемной силы к
лобовому сопротивлению равно 3:1, а у
более крупных насекомых оно еще
больше. Там, где силы инерции преобладают
над силами трения, движение животных
основано главным образом на
использовании подъемной силы.
Строение крылового аппарата
Локомоторным центром насекомого
является грудь. Два ее сегмента — средне- и
заднегрудь, которые снабжены крыльями,
объединяют под общим названием «птеро-
торакс». В крыловых мышцах груди
осуществляется переход энергии из химической
формы в механическую. Таким образом,
птероторакс можно сравнить с двигателем
летательного аппарата. Усилие сокращения
мышц передается на крылья через скелет
и систему мелких пластинок в корне
крыла. Крылья совершают взмахи и
генерируют аэродинамические силы.
2
1
3
2
Рис. 1. Полетный механизм стрекозы (вверху) в
сравнении с мухой (внизу) [из: Бродский,
Львовский, 1990]
1 — мышцы-опускатели прямого действия; 2— мыш-
цы-подниматели: 3— мышцы-опускатели непрямого
действия. Стрелками показано направление
движения спинки и крыльев при взмахах
Крыло насекомого представляет собой
уплощенный вырост стенки грудного
отдела, покрытого двумя слоями кутикулы,
прочно соединенными друг с другом.
Дорсальная поверхность крыла постепенно
переходит в спинной отдел скелета,
вентральная — в боковую стенку сегмента. В области
сочленения находятся уплотненные
участки скелета — склериты; их взаимное
расположение между краем спинки и крылом
характеризуется строгой упорядоченностью,
здесь же сосредоточен ряд суставов.
Крылья насекомых, в отличие от
крыльев летающих позвоночных животных,
лишены собственной мускулатуры и
приводятся в движение сокращением мышц груди
(рис. 1). Морфофункциональную связь между
мышцами и крыльями осуществляют
скелет птероторакса и крыловые сочленения.
Усилие от сокращения мышц передается
сперва на спинную область сегмента — тер-
гум, а затем на основание крыльев.
Крыло насекомого представляет собой рычаг
первого рода, и для того, чтобы поднять
или опустить его, совсем не обязательно
прикладывать усилие к длинному плечу —
пластине крыла. Достаточно на небольшой
угол опустить или поднять короткое
плечо, надавив на него краем спинки (рис. 1).
Последняя уплощается или выгибается под
действием мышц, называемых мышцами
непрямого действия.
От этой схемы резко отличается
работа летательного аппарата стрекоз. У них
крыловые мышцы прикрепляются
непосредственно к основаниям крыльев (рис. 1).
Такие мышцы, называемые крыловыми
мышцами прямого действия, при сокращении
тянут крылья за основания вниз, опускйя
их на некоторый угол. У всех прочих
насекомых мускулы-опускатели относятся к
крыловым мышцам непрямого действия, так
как прикрепляются не к основаниям
крыльев, а к двум складкам спинки спереди и
сзади от крыла. Когда такие мышцы
сокращаются, спинка аркообразно выгибается,
приподнимая основания крыльев, вследствие
чего их лопасти опускаются. Мускулы-под-
ниматели опускают спинку, а с ней и
основания крыльев, что приводит к движению
крыльев вверх. У стрекоз мускулы-подни-
матели прикрепляются близко к друг
другу на спинке, а сама спинка маленькая и
не играет большой роли в движении
крыльев, однако крыловые мышцы прямого
действия развиты у них сильнее, чем у
других насекомых. Такая система движения
крыльев неспособна обеспечить их быстрые
взмахи, но обладает тем преимуществом,
что каждое из четырех крыльев работает
независимо, что позволяет стрекозам
совершать в воздухе различные сложные
маневры. Все прочие насекомые — мухи,
АЭРОДИНАМИКА ПОЛЕТА НАСЕКОМЫХ
57
перепончатокрылые, клопы — мало
уступают стрекозам в маневренности, которая
достигается взмахами крыльев правой и
левой стороны с подчас очень высокой
частотой. Кроме того, большинство насекомых
обладают способностью изменять наклон
плоскости взмаха по отношению к
продольной оси тела.
Подходы нестационарной
аэродинамики
Рассмотрим природу сил, создаваемых
при взмахах крыла насекомого. Крыло,
совершающее колебательные движения, то
ускоряется, то тормозится; в крайних
точках взмаха оно испытывает вращение
вокруг своей продольной оси. Такое движение
нестационарно, и для его описания
непригодны методы, разработанные в
классической аэродинамике для крыла или
пропеллера. Тем не менее существуют подходы,
цель которых состоит в приближении
существующих классических методов к
сложной картине движения крыльев насекомых
при взмахах. Два из них наиболее
популярны.
При описании машущего полета на базе
квазистационарного подхода допускают, что
крыло насекомого — тонкая пластинка,
обтекаемая потоком с постоянной скоростью
(силовые коэффициенты постоянны по
размаху и во времени), а аэродинамическое
взаимодействие между правым и левым
крылом отсутствует.
В центре современной теории крыла
находится постулат Чаплыгина—Жуковского:
задняя кромка крыла является линией, по
которой стекает поток с верхней и нижней
поверхности крыла. Как только крыло
начинает двигаться (рис. 2, а, 2), на его
задней кромке образуется вихрь (рис. 2, а, 2).
Этот вихрь быстро растет — до тех пор,
пока не прекратится движение жидкости
вокруг задней кромки крыла, т. е. пока она
не станет линией схода потока с верхней и
нижней поверхностей (рис. 2, а, 3). Как только
это произойдет, вихрь отрывается и
уносится с потоком. Отрыв разгонного вихря в
соответствии с теорией Томпсона
индуцирует определенную по величине
циркуляцию вокруг крыла, которую можно
представить так называемым присоединенным
вихрем (рис. 2, а, 2). Направление его вра-
/
/
/
Г
R
Рис. 2. Механизм создания полезных аэродинамических сил с позиции квазистационарного подхода
а — обтекание профиля крыла с образованием подъемной силы: 1-3— объяснения в тексте; б—
распределение давления по контуру профиля крыла (горизонтальная линия разделяет области повышенного и
пониженного давления); в— поляра Лилиенталя, которая показывает, как меняется сотношение подъемной силы (У) и
сопротивления крыла (О) при изменении угла атаки (а); г— траектория движения левого крыла насекомого,
показанная на фоне силуэта насекомого. Участки траектории, удовлетворяющие квазистационарному расчету
сил, отмечены двойной линией. Аэродинамическая результирующая сила (Я) раскладывается на подъемную
силу (У) и силу тяги (Т)
58
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
щения противоположно таковому разгонного
вихря. Наложение набегающего потока на
циркуляцию вокруг крыла создает хорошо
знакомое из классической аэродинамики
распределение давления по аэродинамическому
профилю (рис. 2, б), в этом случае
величина подъемной силы, приходящейся на
единицу размаха крыла, определяется из
теоремы Жуковского:
L = рг?Г,
где Г— циркуляция потока вокруг
профиля.
Зависимость коэффициентов подъемной
силы и лобового сопротивления от угла атаки
выражается посредством поляры Лилиен-
таля, которую можно представить как
кривую, описываемую вектором полной
аэродинамической силы R при изменении угла
атаки (рис. 2, в). В свою очередь, полная
аэродинамическая сила раскладывается на
вертикальный компонент (подъемная сила,
Y) и горизонтальный компонент
(сопротивление крыла, Q).
В соответствии с этим подходом
обтекание крыла, совершающего взмахи,
рассматривается как последовательность отдельных
стационарных ситуаций, когда
изменениями угла атаки и скорости набегающего
потока можно пренебречь. Моментов,
удовлетворяющих квазистационарному подходу, в
цикле взмаха два — один приходится на
большую часть нисходящей ветви
траектории, другой — на нижнюю треть
восходящей (рис. 2, г). При движении вниз крыло
создает подъемную силу и тягу, при махе
вверх — тягу и отрицательную подъемную
силу. Создает ли крыло какие-либо силы в
другие фазы взмаха, неизвестно, так как
процессы, происходящие в верхней и
нижней точках траектории, не поддаются
описанию с позиции квазистационарного
подхода.
Можно взглянуть на проблему
создания аэродинамических сил машущим
крылом и с другой стороны. В результате
взаимодействия движущихся крыльев с
потоком воздуха последний ускоряется и
отбрасывается вниз и назад. Импульс силы,
получаемый насекомыми, направлен
вперед и вверх. Оценка создаваемых сил по
импульсу потока воздуха,
отбрасываемому машущими крыльями, широко
применяется при изучении особого режима
машущего полета, когда насекомое как бы
«висит» на одном месте. Если
поступательный полет стараются понять, применяя
теорию крыла самолета, то для
зависающего полета пытаются применить теорию
пропеллера. Все теории пропеллера
сводятся, в конечном счете, к тому, как
образуется и отбрасывается струя воздуха. В
соответствии с данным подходом параметры
взаимодействия крыльев с потоком не
принимаются во внимание и
рассматриваются как черный ящик, на выходе которого
имеется поток, ускоренный работающими
крыльями.
Аэродинамические силы
генерируются благодаря тому, что над машущими
крыльями создается зона пониженного
давления, а под ними — зона повышенного
давления. Импульс силы, получаемый
насекомым, равен по величине и
противоположен по направлению моменту сил,
переданному машущими крыльями
окружающей среде. Как следствие
взаимодействия машущих крыльев с воздухом за
летящим насекомым остается
аэродинамический след, структура которого
содержит информацию о природе сил,
создаваемых в машущем полете.
Как образуются вихревые кольца?
Рассмотрим теперь работу машущего
крыла с позиции принципов, заложенных
в обоих подходах. Как уже упоминалось, как
только крыло начинает двигаться, у его
задней кромки образуется разгонный вихрь.
Этот вихрь через концевые вихри
смыкается с циркуляционным потоком вокруг
крыла, образуя кольцо (рис. 3, а). При
внезапной остановке движущегося крыла слой
приторможенного воздуха, наиболее
близкий к поверхности профиля (пограничный
слой), на верхней его дужке, движущийся
быстрее, нежели на нижней дужке,
обтекая заднюю кромку, сворачивается в вихрь,
который имеет противоположное по
сравнению с разгонным направление вращения
и такую же по абсолютной величине
интенсивность; он называется тормозным
вихрем. Таким образом, движущееся крыло
«несет» вихревое кольцо, которое
освобождается при его остановке.
Если мы теперь представим, что крыло
совершает колебания в плоскости,
перпендикулярной набегающему потоку, то
поочередно отделяющиеся от крыла разгонные и
тормозные вихри образуют цепочку сцепле-
ных друг с другом вихревых колец (рис. 3, б).
Кольцо, образовавшееся при махе вниз,
будет обладать импульсом, направленным
вниз и назад, и в то же время крыло из-за
перепада давления на его верхней и
нижней поверхности разовьет подъемную силу
и тягу. Аналогично, при махе вверх крыло
разовьет тягу и отрицательную подъемную
силу, а кольцо, образовавшееся в это
время, будет обладать импульсом,
направленным вверх и назад. Следовательно, в
возмущениях, которые производят в воздухе
машущие крылья, или, иначе, в
аэродинамическом следе, который они оставляют, в
как бы зашифрованной форме содержится
АЭРОДИНАМИКА ПОЛЕТА НАСЕКОМЫХ
59
Рис. 3. Вихри вокруг машущего крыла
а— вихревая система равномерно движущегося крыла состоит из циркуляционного потока вокруг самого
крыла, разгонного вихря и пары концевых вихрей, которые все вместе образуют вихревое кольцо; б—
колеблющееся крыло оставляет за собой цепочку сцепленных наклоненных вихревых колец; ее пересечение
продольной вертикальной плоскостью дает так называемую вихревую дорожку. Остальные объяснения в тексте
информация о характере взаимодействия
крыльев с воздушными потоками.
Структура следа — своего рода ключ к пониманию
природы сил, создаваемых машущими
крыльями.
Эволюция аэродинамики
полета насекомых
Работа крыла реального насекомого
отличается от рассмотренной схемы тем, что
только вершина крыла совершает
колебания относительно неподвижного основания.
Кроме того, само крыло в верхней и
нижней точках взмаха испытывает
вращательные колебания относительно своей
длинной оси. Тем не менее, когда удалось наконец
визуализировать (т. е. сделать видимым) след
летящего насекомого, то оказалось, что его
форма почти идентична форме следа,
который образуется за крылом, совершающим
колебания в плоскости, перпендикулярной
набегающему потоку (рис. 3, б). Впервые
трехмерную картину аэродинамического
следа за летящим насекомым удалось
получить для насекомого средних размеров с
относительно невысокой частотой крыловых
взмахов (30 Гц) — для бабочки-толстоголовки
(рис. 4). Какова же эта картина? Прежде
всего след представляет собой систему
попеременно наклоненных к оси вихревых
колец. Через отверстия колец проходит
толстая волнообразно изгибающаяся струя
воздуха. Если вертикальной продольной
плоскостью рассечь такой след, то мы
получим его плоское изображение (рис. 4, б),
так называемую вихревую дорожку: вокруг
центральной струи в шахматном порядке
располагаются вихри, вращающиеся
навстречу друг другу.
Изменения параметров взмаха крыльев,
таких как амплитуда колебания, частота,
наклон плоскости взмаха к продольной оси
насекомого и к направлению полета,
сопровождаются закономерным изменениями
формы аэродинамического следа.
Наиболее примитивной и, возможно,
исходной формой следа, если судить по
сравнительной простоте образования и
распространенности среди многих
примитивных насекомых, можно считать ту,
которая свойственна крылу, колеблющемуся
в плоскости, перпендикулярной
набегающему потоку (рис. 3, б). В этом случае
за телом образуется цепочка из
сцепленных вихревых колец, равнодействующая
импульсов которых определяет создание
аэродинамической силы, направленной
строго вперед (рис. 5, а). Последнее
обстоятельство вынуждает многих
примитивных насекомых летать с большим углом
возвышения, как бы приподнимая тело
над горизонталью и тем самым
направляя вихревую дорожку под углом к
горизонту для того, чтобы создать
подъемную силу. Образование сил в данном
случае как при махе вниз, так и при махе
вверх следует трактовать с позиции
квазистационарного действия крыла.
Следующий шаг в эволюции полета
насекомых состоял в том, что при махе вниз,
осуществляемом с большими значениями
60
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
угла атаки, чем при махе вверх, крылья
стали продуцировать вихревые кольца
большей интенсивности и, следовательно,
меньшего размера. Кольцо, сошедшее с
крыльев в конце маха вниз, имеет меньший
диаметр, вследствие чего ось следа
отклоняется вниз, а равнодействующая
импульсов колец направлена под углом вверх
(рис. 5, б), Образующаяся за крыльями
вихревая дорожка получила название косой,
а природа сил, создаваемых при взмахах
крыльев, здесь в принципе такая же, что
и в предыдущем случае.
У некоторых насекомых в полете за
крыльями образуется вихревой след, форма
которого аналогична той, которая
характерна для наиболее примитивных
насекомых (рис. 5, в). Есть, однако,
существенное отличие. Исследования показали, что
кольцо малого диаметра, образовавшееся
при махе вниз, во время подъема
крыльев расширяется. Как и в предыдущем
случае (рис. 5, б), в данном случае мах вниз
активнее, чем вверх, но из-за того, что
кольцо малого диаметра при подъеме
крыльев расширяется, след принимает вид
прямой вихревой дорожки. Расширяющееся
кольцо придает ускорение струе воздуха,
направленной косо вниз, что, по
предположению, компенсирует отрицательную
подъемную силу, создаваемую при махе
вверх. В итоге распределение сил в
цикле взмаха выглядит следующим образом:
подъемная сила создается при махе вниз,
а тяга — в течение всего цикла взмаха.
Следовательно, генерацию сил при махе
вверх можно объяснить с позиций
нестационарного действия крыла. Более того, при
развороте крыльев в верхней точке
взмаха они отталкивают ближайшее к телу
кольцо, а вместе с ним и всю цепочку назад, в
результате чего насекомое получает
небольшой толчок вперед. Образование сил в
верхней точке взмаха, стало быть, можно
объяснить действием механизма, близкого
к реактивному.
Роль последнего возрастает у
ширококрылых бабочек, которые в полете
отбрасывают дискретные вихревые кольца. У
этих насекомых по мере увеличения ско-
Рис. 4. Форма вихревого следа за летящим насекомым
а— вид сбоку; б— вид сверху. Стрелками показано направление потоков воздуха
АЭРОДИНАМИКА ПОЛЕТА НАСЕКОМЫХ
61
рости полета цепочка вихревых колец
сначала размыкается в верхней точке
взмаха (рис. 5, г), что достигается энергичным
хлопком крыльев над спинкой, а затем и
в нижней. В итоге, при наиболее
скоростном миграционном полете, а также при
взлете крылья бабочки отбрасывают
дискретные вихревые кольца: при хлопке
крыльев в верхней точке кольцо
отбрасывается назад, и бабочка получает
толчок вперед; в нижней точке взмаха
бабочка хлопает крыльями и отбрасывает
кольцо вниз, получая вследствие этого
толчок вверх. И, наконец, у насекомых с
высокой частотой взмаха крыльев
отбрасывание мелких дискретных колец
становится основным способом создания
полезных аэродинамических сил.
Заключение
Таким образом, объяснение природы
сил, создаваемых машущими крыльями,
нельзя свести исключительно к квазиста-
Рис. 5. Предполагаемое
изменение формы вихревого следа в
процессе эволюции крылатых
насекомых
а-г— объяснения в тексте. Я —
аэродинамическая
результирующая сила, возникающая как
реакция на моменты сил вихревых
колец, отбрасываемых при махе
вниз (d) и при махе вверх (и).
Толстая стрелка показывает
направление центральной струи
воздуха
62
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
ционарному действию крыла. У многих
насекомых при взмахе вверх, когда ранее
образовавшееся кольцо расширяется и
ускоряет струю воздуха назад, возникает
кратковременный импульс силы,
происхождение которого следует отнести на счет
нестационарного действия крыла.
Значение механизма, аналогичного
реактивному, когда насекомое отбрасывает назад
вихревые кольца, резко усиливается по
мере того, как непрерывная цепочка
колец разрывается. Существенную роль в
этом играют особые движения крыльев,
в частности их хлопок в верхней или
нижней точке взмаха. Немецкий исследователь
Нахтигаль рассмотрел несколько особых
движений крыльев, которые могут иметь
значение с точки зрения создания
аэродинамических сил способом, отличным от
квазистационарного. Эти движения
крыльев порождают различные, еще
недостаточно изученные, нестационарные
эффекты, роль которых в полете, несомненно,
возрастает по мере того, как
наблюдается рост частоты взмаха крыльев.
Литература
Бродский А.К. Механика полета насекомых
и эволюция их крылового аппарата. Л.: Изд-во
ЛГУ, 1988.
Бродский А. К., Львовский А. Л. Пауки,
насекомые. Л.: Лениздат, 1990.
Захваткин Ю. А. Курс общей энтомологии.
М.: Агропромиздат, 1986.
Шмидт-Ниельсен К. Размеры животных:
почему они так важны? М.: Мир, 1987.
Brodsky А. К. The Evolution of Insect Flight.
Oxford: Oxford University Press, 1994.
В. А. Ратнер
МОЛЕКУЛЯРНАЯ ЭВОЛЮЦИЯ
В последние годы теория эволюции
испытывает интенсивное изменение и
развитие в связи с появлением нового мощного
пласта экспериментальных данных и
теоретических методов. В ее поле зрения попали
кодирующие макромолекулы — ДНК, РНК,
а также белки, составляющие основу мо-
лекулярно-генетической организации клеток
и организмов. Теперь уже широко известна
роль ДНК и РНК как материальных
носителей наследственности, мРНК — как
переносчика генетической информации от генов к
белкам и белков — как исполнителей самых
разнообразных биологических функций (так,
ферменты катализируют биохимические
реакции, антитела осуществляют
иммунологическую защиту организма, другие
белки выполняют роль структурных, регулятор-
ных или транспортных элементов и т. д.).
Первые последовательности аминокислот
в белках были расшифрованы (секвенирова-
ны) в 50-е гг., нуклеотидов в ДНК и РНК —
в 60-е, но массовое секвенирование
началось только в 1977—1978 гг., став к
настоящему времени рутинной технологической
операцией. Результаты секвенирования с тех
пор накапливаются в компьютерных банках
данных. К весне 1993 г. крупнейшие
международные банки данных содержали
сведения о нуклеотидных последовательностях,
имеющих общую длину 130 млн
нуклеотидов (-110 тыс. генов и других фрагментов)
и аминокислотных последовательностях,
состоящих из 10 млн аминокислот (-30 тыс.
белков), причем объем этой информации
очень быстро возрастает.
Расшифровка нуклеотидных и
аминокислотных последовательностей создает новую
ситуацию, позволяющую изучать процесс
эволюции на молекулярном уровне
организации жизни. В классической генетике аллели
генов принято обозначать строчными и
прописными буквами или буквами с индексами:
В и Ь, А1 и А2, и т. п. Если в процессе
эволюции один аллель вытесняет другой
(например, В замещается Ь), то кажется, что
происходит как бы утрата предыдущего аллеля
(В) и распространение нового (Ь). В то же
время из молекулярной генетики известно,
что многие мутации состоят в замене лишь
одного или нескольких нуклеотидов.
Поэтому замещение аллеля обычно сводится к
замене и фиксации единственного нуклеоти-
да, а остальная, большая, часть гена остается
неизменной. Таким образом, замена В на Ъ
может быть эквивалентной, скажем,
замене TTCGAGGCTAG на TTCGCGGCTAG.
Подобные замещения приводят к
возникновению очень сходных последовательностей
аллельных генов (и соответственно белков).
Однако по мере накопления различий
сходство уменьшается. Чем раньше дивергировали
два гена, тем больше они различаются.
Таким образом, сходство
последовательностей макромолекул можно положить в
основу молекулярно-филогенетических
построений. Число пар различающихся
мономеров, находящихся в гомологичных
позициях сравниваемых молекул, служит мерой
эволюционной дивергенции.
В течение последних 30 лет было
разработано несколько десятков
компьютерных методов и пакетов программ для
построения филогенетических деревьев
макромолекул, что позволило
реконструировать сотни таких деревьев. Их анализ
дал важные и интересные результаты.
Например, оказалось, что многие
макромолекулы эволюционировали гораздо
медленнее, чем морфологические признаки, и
поэтому их филогенетический анализ
позволяет заглянуть в очень ранние периоды
эволюционной истории, отстоящие на 100—
1000 млн лет.
На основе полученных данных создана
и развивается теория молекулярной
эволюции. Здесь мы представим
филогенетические деревья — основной продукт этой
области науки — и покажем, к каким
любопытным выводам приводят их анализ и
интерпретация. Часть результатов, о
которых пойдет речь, получены автором и его
сотрудниками.
Разные макромолекулы
эволюционируют с разными скоростями. Наиболее
консервативными обычно являются гены и белки,
связанные с некоторыми базовыми, рано
возникшими генетическими процессами,
свойственными многим (или даже всем) формам
жизни. К ним принадлежат, например, гены
рибосомных РНК (рРНК— часть
механизма синтеза белка), некоторых гистонов —
белков, участвующих в компактизации ДНК
хромосом и др. Менее консервативны гены
и белки систем, которые встречаются у
меньшего, но все же достаточно большого
числа видов, образующих таксоны высокого
ранга (к таким молекулам относятся
глобины животных). Наконец, наиболее изменчивы
гены и белки РНК-содержащих вирусов,
которые стремительно изменяются в
борьбе с иммунной системой своих хозяев (это
молекулы вируса гриппа, ВИЧ, онкогенных
ретровирусов и др.).
64
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
Примером «глобального» дерева,
охватывающего все крупнейшие таксоны
биологического мира, является дерево 16S—18S-
рибосомных РНК, построенное американским
ученым К. Возом и сотр. (рис. 1). Наиболее
неожиданное свойство этого дерева —
наличие у него трех крупнейших ветвей,
объединяющих эубактерий (обычных бактерий),
архебактерии (особую экзотическую
группу бактерий, см.: Громов В. В.
Удивительный мир архей. Наст, том) и эукариот
(высших ядерных организмов). До этой работы
считалось, что бактерии составляют единое
надцарство, Prokariota, отделенное от
второго надцарства, Eukariota, существенной
дистанцией. На дереве Воза вершины всех
трех глобальных ветвей примерно равно
удалены друг от друга. Поэтому
архебактерии следует считать самостоятельным над-
царством. Таким образом, Воз ввел три
надцарства, или домена: Bacteria, Archaea,
Eucarya. Некоторые исследователи
предлагали для них термин империи (см.: Кула-
ев И. С. Происхождение эукариотических
клеток. Наст. том).
.Этот важный результат сразу привлек
внимание к изучению архебактерии,
причем вскоре было показано, что и по
многим другим свойствам они существенно
удалены как от эукариот, так и от эубак-
терий. Многие архебактерии существуют в
природе в весьма экзотических условиях:
при высокой температуре вблизи
подводных вулканов; в среде, насыщенной
метаном, соединениями серы и т. д. Время
разделения надцарств Bacteria и Archaea
оценивается примерно в 3,5 млрд лет.
Дерево Воза основано на данных о
фракциях ieS-подобных рРНК,
кодируемых не только ядерными генами, но и
генами клеточных органелл — митохондрий
и хлоропластов. Легко заметить, что
ядерная фракция рРНК кукурузы (Zea mays)
оказалась на ветви эукариот, как и
следовало ожидать, а фракции из
митохондрий и хлоропластов той же кукурузы — на
ветви эубактерий. Этот факт считается
самым веским аргументом в пользу «сим-
биотической» гипотезы эволюционного
возникновения эукариот, согласно которой
митохондрии происходят от симбиотичес-
ких предковых пурпурных бактерий, а
хлоропласты — от цианобактерий (синезе-
леных водорослей).
Белки глобины (гемоглобины крови,
миоглобины тканей) и их гены имеются не
у всех организмов. Они найдены только у
животных, хотя гомологичные им, но сильно
отличающиеся по первичной структуре
белки (леггемоглобины) есть и у растений;
у прокариот они не обнаружены. Поэтому
глобины образуют молекулярное надсемей-
ство, охватывающее лишь часть
филогенетического дерева жизни. Деревья эволюции
глобинов и их генов реконструировались
многими исследователями. На рис. 2
приведено одно такое дерево, построенное
А. А. Жарких в лаборатории генетики по-
ARCHAEBACTERIA
Halococcus morrhua
Halobacterium volcanii
Methanospirillum hungatei
Sulfolobus solfataricus
Thermoproteus tenax
Methanobacterium formicicum
Methanococcus vannielii
Homo sapiens
Xenopus laevis
Zea mays
proroc<
о Л'
^
*i°
^ ч<? & <?
$
f?
EUCARYA
CD
-4
3
о
3
о
s
g-
c'
3
s
en
CD
С
CD Г>
z. <a
СЛ
%.
О
Jeudomonastestoste,
Escherichia coli
roni
%.
О
¦о
5)*
ел
V
'<*
*»
Ь
'%
'Qf.
%
%
<4
4
%
*hh.
EUBACTERIA
Рис. 1.
с сотр.
<Глобальное» филогенетическое дерево надсемейства 16S—18S рибосомных РНК, построенное К. Возом
МОЛЕКУЛЯРНАЯ ЭВОЛЮЦИЯ
65
1
2
л
"5з
СО СО ф
'CD "с J?
.о а>
-О
2 Q.K СО О * -О
^^О-о^
32 3
JQ СО ОТ
от
осо^хососососо
CD СО .О JO О )5 О iP О
;g5S"o<<
2 О. К СО X * Л • .(ВТОишошошо ^ Ф^ .= ;J >.-* О
<i-c:^e<,P-g:trxirc;o^o^o^SQ.S S 2^со.
* _: Q.
СО
ii
СО >» i-CN
СО S S <° С0С0С0<О я
Q.^ OOOQOOO< DQ О О0Ш
/"s / \
а-цепи
р-цепи гемоглобинов
Миоглобины Эритрокруорины
хирономуса
с; i
о з 5
•
Леггемо-
глобины
Рис. 2. Филогенетическое дерево надсемейства глобинов, построенное А. А. Жарких для 40 аминокислотных
последовательностей белков
1 — фиксированные делеции; 2— инсерции. Цифры указывают на их локализацию внутри белка
пуляций Института цитологии и генетики
СО РАН. В целом оно неплохо соответствует
принятой систематике животных.
Из независимых палеонтологических
данных можно получить датировки времен
существования общих предков многих ныне
живущих биологических форм. Так, на
дереве глобинов были датированы б точек
ветвления. Поскольку методы
реконструкции деревьев позволяют определить длины
ребер дерева (числа фиксированных замен,
мутаций) на различных маршрутах, то зная
датировки, можно вычислить скорости
эволюции на разных этапах и в разных
ветвях дерева. Нами были получены
интересные оценки (рис. 3).
Во-первых, оказалось, что средняя
скорость эволюции молекулы этого белка была
не постоянна, а имела максимум
примерно 400—500 млн лет назад, в эпоху
выхода позвоночных животных из Мирового
океана на сушу (рис. 3, кривая 1).
Во-вторых, эта особенность выявилась еще
отчетливее для тех участков глобинов, которые
отвечают за образование их четвертичной
(мультимерной, агрегатной) структуры. Дело
в том, что миоглобины и гемоглобины
некоторых примитивных животных до сих пор
представляют собой протомеры (т. е. состоят
из одной субъединицы). Круглоротые
(миноги, миксины) имеют димерные
гемоглобины, а большинство других позвоночных —
тетрамерные (ос2р2), состоящие из 2 а- и 2
Р-субъединиц. Субъединицы соединяются при
помощи так называемых центров контакта,
которые обозначаются как ос1-Р1, ос1~Р2 и т. д.
Для современных гемоглобинов
аминокислоты, входящие в центры контакта,
известны. Поэтому можно вычислить скорости
эволюции непосредственно для этих
центров. Особенно отчетливо максимум
скорости выявляется для o^-Pg (рис. 3, кривая 2)
и регуляторного центра связывания дифос-
фоглицерата (ДФГ) (рис. 3, кривая 3). Итак,
при выходе позвоночных животных на сушу
их гемоглобин приобрел тетрамерную
структуру, в которой разные субъдиницы (а и
Р) связаны между собой центрами
контакта oc^Pg, и регуляторным центром ДФГ. В
Скорость фиксации замен х109 на позицию, в год
24
22
20
18
16
14
12
10
8
6
4
2
1
2
3
900
700
500 300
млн лет назад
100
Рис. 3. Адаптивное ускорение фиксации замен
аминокислот в гемоглобинах в эпоху выхода
позвоночных животных на сушу (400—500 млн лет назад)
Скорость эволюции: 7— в среднем по белку; 2— в
центре контакта оц-р^ 3— в регуляторном центре
связывания ДФГ
66
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
то время (или немного раньше) ген общего
предка гемоглобинов был дуплицирован, а
его копии в ходе дивергенции дали
начало двум родственным семействам — а и р.
И именно в это время отмечается
максимальная скорость эволюции центров контакта
и регуляции глобинов, которая затем в
течение сотен млн лет резко падала, подчас
до нуля, когда данные центры вообще не
изменялись. Иначе говоря, в этом случае
мы как бы наблюдаем отдаленный от нас
процесс формирования новых
функциональных структур молекул гемоглобина,
которые в дальнейшем сохраняются у всех
наземных форм позвоночных. Следует учесть,
что выход позвоночных на сушу и переход
к дыханию свободным кислородом воздуха
сопровождались резкой перестройкой всей
системы дыхания, в том числе, как мы
видим, структуры гемоглобинов.
Произошедшее тогда ускорение эволюции означает,
что указанные приобретения были
высокоадаптивными, т. е. обеспечивали
существенное преимущество их обладателям.
Пример наиболее быстрой и локальной
молекулярной эволюции дают гены,
кодирующие гемагглютинины НЗ вируса
гриппа (рис. 4), эволюционное дерево которых
1920 b
1930Ь
1940L
1950 k
1960L
19701-
i
19801- |
103/17
5/3
14/4
2
20/13
11/7
14/17
ш
s
3
Рис. 4. Филогенетическое дерево генов гемагглю-
тинина НЗ вируса гриппа типа А, построенное
Л. В. Омельянчуком и др.
«Висячие» вершины и ведущие к ним ребра
отражают эволюцию эпидемических штаммов (1, 2, 3,4, 5,
10). Индексы штаммов указывают на место и год
выделения. Внутренние вершины и ребра дерева
соответствуют неэпидемическим стадиям
циркуляции вируса (в «резервуарах»). У ребер указаны
числа синонимических/несинонимических замен нукле-
отидов
было построено Л. В. Омельянчуком с сотр.
Некоторые выпуклые участки белка кап-
сида этого вируса (антигенные
детерминанты) узнаются специфическими антителами
хозяина (человека) и иммунное
сопротивление хозяина препятствует размножению
вируса. Вирус гриппа имеет РНК-геном, в
котором мутации происходят с наибольшей
из известных частотой ~10~3—10'4 на
позицию за репликацию. Был изучен ряд
локальных эпидемий и пандемий гриппа,
случившихся в XX в. Первой —пандемия «испанки»
1918—1919 гг., затем — пандемия
«гонконгского» гриппа и др. В большинстве
случаев были собраны и сохранены в
коллекциях образцы эпидемических штаммов вируса.
После 1978 г. РНК этих штаммов были сек-
венированы и на основе полученных
данных построены филогенетические деревья.
На приведенном дереве (рис. 4) все
эпидемические варианты располагаются на
нижних тупиковых его ветвях (так
называемых висячих вершинах). Это означает,
что эпидемические штаммы, как правило,
не являются прямыми потомками других
эпидемических штаммов. Иначе говоря,
неэпидемический вирус, видимо,
циркулирует в каком-то «резервуаре» (локальной
популяции человека, где к вирусу есть
устойчивый иммунитет, или в популяции
животных, которые могут быть
промежуточными носителями вируса). Эпидемии
возникают тогда, когда появляется новый
вариант вируса, против которого нет
готовых антител, т. е. готового иммунитета
хозяина. Этот вариант как бы проникает в
человеческую популяцию и быстро в ней
распространяется, пока не будет
выработан адекватный иммунный ответ. Тогда
эпидемия затухает. Следующая эпидемия
возникает путем независимого проникновения
нового варианта вируса гриппа, не
связанного, как правило, с предыдущей эпидемией.
Но это не все. Поскольку все эпидемии
были датированы (см. рис. 4), оказалось
возможным определить скорости эволюции.
В эпидемических ветвях они оказались в 3—
5 раз выше, чем в неэпидемических,
причем особенно большое ускорение (в 36 раз)
было выявлено именно в позициях,
кодирующих антигенные детерминанты белка.
Иммунное сопротивление хозяина
размножению вируса является главным
селективным фактором, действующим на вирус.
Поэтому проникновение эпидемического
штамма, не встречающего иммунного
сопротивления хозяина, сопровождается
ускорением эволюции. Это размножение
высокоадаптивно для вируса и неадаптивно
для человека-хозяина. Напротив, после
выработки иммунного ответа спокойное
размножение вируса становится для него
и для хозяина нейтральным — в ожидании
МОЛЕКУЛЯРНАЯ ЭВОЛЮЦИЯ
67
Рис. 5- Филогенетическое дерево
штаммов вируса ВИЧ,
вызывающего заболевание СПИДом,
построенное В.-Х. Ли с соавт.
Числа у ребер— процент
дивергенции последовательностей. Справа
указаны регионы, где изолированы
штаммы
2,7
2.6
1,9
9,3
0,1
0,5
0,4
2,1
0,1
0,1
2,4
3,3
3,7
2,1
0,8
0,9
0,9
4,3
2,2
4,2
6,1
о'з ВН8/83
' PV22/83
Z** ВН10/83
JJ'~ BRU/83
НХВ/83
SF2/83
CDC/84
WMJ1/84
WMJ2/85
WMJg/85
RF/83
-^
v <
4
ELI/83
MAL/85
> CO
CO
3 4 5 6 7
Процент дивергенции
Z3/83
10 11
новых адаптивных мутаций и рекомбинаций.
В целом можно говорить о коэволюции
вируса и иммунного ответа системы хозяина.
Вирус стремится как бы мутационно
«выскользнуть» из-под готового иммунного
ответа хозяина, а хозяин стремится «догнать»
новые варианты вируса путем выработки
нового специфичного иммунного ответа (см.:
Агол В. И. Разнообразие вирусов. Наст. том).
Еще один пример — эволюция вируса
ВИЧ (вируса иммунодефицита человека,
английская аббревиатура — HIV),
вызывающего заболевание СПИД (синдром
приобретенного иммунодефицита, английская
аббревиатура — AIDS). Вирус ВИЧ
передается половым путем, а также нередко при
переливании крови, нестерильных
инъекциях и хирургических вмешательствах, при
родах — от матери к ребенку и в других
подобных ситуациях. Он был обнаружен в
начале 80-х гг. в крови больных,
погибавших от различных тяжелых заболеваний. Эти
заболевания имели одно общее свойство —
иммунная система больного была неспособна
противостоять как этому вирусу, так и
сопровождавшим его возбудителям других
заболеваний, от которых, собственно,
больные и погибали. Были обследованы
образцы из банков переливания крови, которые
созданы во многих странах для
медицинских целей. Оказалось, что многие образцы
содержали вирус ВИЧ.
Различные штаммы вируса ВИЧ,
выделенные в разных районах Африки, Вест-
Индии и США, были секвенированы и для
них были построены филогенетические
деревья. Одно из таких деревьев приведено
на рис. 5. Его анализ показывает, что
вирус ВИЧ существовал в Центральной
Африке (Заире) еще до 1960 г., а затем до
середины 70-х гг. был занесен в
Вест-Индию (Гаити), и до 1978 г.— в США. Иначе
говоря, его истоки лежат в Африке, где в
некоторых странах вирусоносители
составляют до половины населения, хотя
заболевание СПИДом проявляется далеко не у всех.
Видимо, размножение вируса все же
сдерживается их иммунной системой. Вирус ВИЧ
сходен с некоторыми вирусами обезьян,
поэтому многие ученые предполагают, что он
возник в результате изменчивости этих
вирусов и был занесен в популяцию человека
извне относительно недавно.
Интересно, что изменчивость вируса
ВИЧ (мутационная и рекомбинационная)
столь велика, что он успевает
измениться, стать еще более патогенным
непосредственно в ходе развития болезни, у
некоторых больных в течение 1,5—2 лет и даже
меньшего времени. Оценки изменчивости
показывают, что она превышает верхний
допустимый предел, в результате чего
иммунная система больного не успевает
«справиться» с выработкой новых
вариантов антител, и фактически распадается,
открывая путь процессу заболевания
СПИДом и другими болезнями. Именно в
этом, по-видимому, и состоит причина особой
патогенности вируса иммунодефицита.
Есть биологические проблемы, которые
всегда интересуют человека. Среди них
особое место занимает проблема происхождения
и дифференциации самого человечества. В
последние годы прогресс в этой области
связан в основном с филогенетическим
анализом макромолекул.
Американский ученый М. Гудмен с
соавт. построили филогенетические деревья
для ряда генов и белков приматов. На рис. 6
приведено одно из них. Из него следует,
что ближайшим филогенетическим
родственником человека является шимпанзе.
Исключительная близость этих двух
видов дала основание Дж. Дайамонду
назвать человека «третьим шимпанзе». По
оценкам этого автора, геномы человека и
68
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
V
NWM Rh Or G С H
Рис. 6. Филогенетическое дерево высших
приматов, построенное М. Гудменом с соавт. для некоди-
рующих последовательностей, смежных с глобино-
вым псевдогеном цгг|
Н — человек; С — шимпанзе; G — горилла; Or —
орангутан; Rh— макак резус; NWM— обезьяны Нового
Света. Цифры у ребер означают числа замен на 100
позиций нуклеотидов
шимпанзе отличаются примерно по 3,2Х107
одиночным заменам нуклеотидов из 3><109,
т. е. они различаются по каждому сотому
нуклеотиду, а остальные 99 из 100
нуклеотидов у них одинаковы. Время их
молекулярной дивергенции оценивается в 5—10 млн
лет. Эти факты являются очень веским
аргументом в пользу естественного
происхождения человека в процессе эволюции и
противоречат религиозным мифам о его
сотворении 5—10 тыс. лет тому назад.
Таким образом, молекулярно-филогене-
тический анализ позволяет приблизиться
к решению очень важных вопросов
эволюции. Иногда это единственная возможность
заглянуть далеко в глубь эволюционной
истории. При этом в преобладающем
большинстве случаев мы располагаем
последовательностями только ныне живущих видов.
«Ископаемых» последовательностей почти
нет. Правда, в последнее время удалось
выделить и секвенировать фрагменты ДНК из
замерзших ископаемых животных (мамонтов
и др.), а также мумифицированных тел
человека и животных (см.: Розанов А. Ю.
Современная палеонтология. Наст. том).
Особый интерес в этом плане представляет ДНК
мелких насекомых (мух, термитов, пчел,
жуков-долгоносиков), законсервированных
в природной смоле деревьев (янтаре). Ее
анализ вполне реален. Так, были
клонированы и секвенированы гены 18S рибо-
сомной РНК термита, пчелы и
долгоносика, найденных в янтаре. Полученные
данные использовались при построении
филогенетических деревьев этих молекул.
Возраст термитов и пчел — 25—40 млн лет,
а долгоносика— 120—135 млн лет.
В заключение следует отметить, что
молекулярный филогенетический анализ
становится сейчас одним из наиболее
эффективных методов таксономии. Например,
современная систематика приматов
опирается в значительной мере на молекулярные
данные. В систематике таксонов высшего
ранга, а также высших растений
решающую роль играют деревья рибосомных РНК.
Классификацию бактерий вообще
невозможно построить без молекулярных данных. Есть
основания полагать, что в будущем
молекулярный филогенетический анализ
приобретет еще большее значение.
Литература
Айала Ф. Введение в популяционную и
эволюционную генетику. Пер. с англ. М.: Мир, 1984.
Айала Ф., Кайгер Дж. Современная
генетика. Пер. с англ. М.: Мир, 1988, т. 3.
Кимура М. Молекулярная эволюция: теория
нейтральности. Пер. с англ. М.: Мир, 1985.
Ратнер В. А. Краткий очерк теории
молекулярной эволюции. Новосибирск: НГУ, 1992.
Ратнер В. А., Жарких А. А., Колчанов Н. А.
и др. Проблемы теории молекулярной эволюции.
Новосибирск: Наука, 1985.
Эволюция. Пер. с англ. М.: Мир, 1981.
Ratner V. A., Zharkikh A. A, Kolchanov N. А.
et al. Molecular Evolution. Berlin etc.: Springer-
Verlag, 1996.
А. Ю. Розанов
СОВРЕМЕННАЯ ПАЛЕОНТОЛОГИЯ
Палеонтология— это наука об
ископаемых организмах. Из-за специфики и
разнообразия изучаемых объектов палеонтология
использует множество различных методов. На
начальном этапе развития исследования в
палеонтологии в основном были направлены
на выяснение и описание разнообразия
органического мира прошлого, создание
систематик и выяснение филогенетических
взаимоотношений ископаемых групп. Позднее
четко обозначились вполне самостоятельные
направления: 1) таксономическое
(разнообразие, систематика, региональные
фауны и флоры); 2) морфофункциональное и
эволюционно-морфологическое; 3)
филогенетическое; 4) палеоэкологическое; 5)
микроструктурно-гистологическое; 6)
палеобиогеохимическое; 7) палеобиогеографическое;
8) биостратиграфическое и др. Наконец, в
самое последнее время обособились
палеонтология докембрия, молекулярная
палеонтология, бактериальная палеонтология,
биосферное направление и изучение
необычных (экстраординарных) биот. Такая
дифференциация палеонтологических исследований
определяется по крайней мере двумя
причинами. Во-первых, задачи палеонтологии
значительно обогащаются ее кооперацией с
другими областями биологических и
геологических наук, а во-вторых, расширяются с
развитием методической базы (внедрением
электронной микроскопии, томографии,
микроанализаторов, компьютерной техники).
Обособление новых направлений не
означает, однако, что результаты
традиционных исследований утрачивают свое
значение. Традиционные описания новых
таксонов и региональных фаун и флор всегда
были и будут оставаться фундаментом
палеонтологии, поскольку без пополнения
именно этих данных все остальные
исследования могут быстро потерять смысл,
лишившись основы. Остановимся на самых ярких
достижениях последних лет как в
традиционных, так и в новых направлениях.
Палеонтология докембрия. Сам термин
был введен сравнительно недавно (в
конце 60-х годов) Б.С. Соколовым. Это
направление связано с изучением органического
мира на интервале, равном примерно 7/8
длительности истории Земли. Развивается
оно в настоящее время очень бурно. Еще
не так давно считалось, что в докембрии
практически не было жизни, по крайней
мере макроскопически различимой.
Поэтому-то весь докембрий и носит название
«криптозой» (эра скрытой жизни). Огромное
количество новых находок от
макроскопических до молекулярных дало возможность
создать более или менее достоверную
общую картину эволюции органического мира
и биосферы докембрия. Еще недавно
считалось, что первые многоклеточные
организмы появились лишь в венде (~ 650—550
млн лет), а одноклеточные эукариоты —
приблизительно 1,2—1,3 млрд лет тому
назад. Теперь мы уже знаем о находках
низших грибов в породах, возраст которых
составляет около 2,2 млрд лет, и первых
многоклеточных животных и их следов в
породах, чей возраст оценивается 1,4 млрд лет.
Совершенно уникальные результаты
получены в результате анализа разнообразия и
пространственного распространения так
называемой вендо-эдиакарской фауны. Ее
специфика состоит в том, что все
представители этой биоты еще не имели скелета.
Сегодня уже описаны многие десятки
родов и видов, и обнаружены они на всех
континентах.
Бактериальная палеонтология. Это
направление сформировалось в последние
годы, хотя изучение ископаемых бактерий
в кремнях началось почти 50 лет тому
назад. Считалось, что кремни —
единственная порода, где в принципе можно
обнаружить сохранившиеся бактериальные
остатки. Однако развитие палеонтологии
докембрия и электронно-микроскопическое
изучение высокоуглеродистых пород и
фосфоритов выявили наличие фоссилизирован-
ных бактерий практически во всех
осадочных образованиях. Экспериментально на
современных цианобактериях было
показано, что процессы фоссилизации
(окаменения) могут происходить в считанные часы,
в связи с чем мы и находим в ископаемом
состоянии неразложившиеся бактерии
удивительно хорошей сохранности. Последствия
этих открытий пока трудно оценить. Но уже
сейчас ясно, что обнаружение, например,
бентосных цианобактериальных матов (ЦБМ)
в черных сланцах, многие из которых
считались показателями глубоководности (куда
лучи света практически не проникают),
заставляет пересмотреть устоявшиеся
палеогеографические построения — ЦБМ
должны были образовываться в фотической зоне
(в зоне проникновения света).
Повсеместное обнаружение бактерий в
осадочных породах дает основание думать,
что многие аспекты теории
осадкообразования должны быть пересмотрены,
поскольку сегодня уже трудно представить себе
70
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
процесс осаждения в эпиконтинентальных
бассейнах прошлого без участия бактерий.
Обнаружение же ЦБМ в докембрийских
породах ведет и к переоценке перспектив
нефтегазоносности древних толщ,
считавшихся обычно малоинтересными с точки
зрения накопления органического вещества.
Однако с учетом огромной продуктивности
современных ЦБМ такой взгляд
нуждается в пересмотре, поскольку нет оснований
полагать, что продуктивность древних ЦБМ
была ниже, чем современных. В
теоретическом плане данные бактериальной
палеонтологии тесно связаны с выяснением
огромного значения прокариотической
биосферы прошлого и ее трансформации в
эукариотическую.
Молекулярная палеонтология. Очень
серьезные результаты в последние годы были
получены в области изучения хемофосси-
лий, или биомаркеров. Это направление
чаще всего называют молекулярной
палеонтологией. Она изучает молекулярные
следы жизнедеятельности организмов прошлого,
выясняя по ним характер эволюции как
живых организмов, так и биосферы в
целом. Остатки органических молекул,
первичных или преобразованных, часто
устойчивы к внешним воздействиям и сохраняются
достаточно хорошо. Разнообразие
органических соединений (и, соответственно,
разнообразие хемофоссилий) возрастает с
эволюцией (усложнением) органического мира.
Кроме того, некоторые органические
соединения, сохраняющиеся в ископаемом
состоянии, специфичны для тех или иных
конкретных групп. Так, у бактерий для их
систематики используется жирнокислотныи
состав липидов, у растений — фенольные
соединения, алкалоиды и многое другое.
Очень интересно, что 7- и 8-метилгепта-
деканы характерны только для цианобак-
терий (синезеленые водоросли) и не
встречены у других бактерий, водорослей и
высших растений. Особенно интересны
результаты исследований докембрийских
хемофоссилий, поскольку они позволяют
наметить время возможного появления эука-
риот, затем многоклеточных и т. д.
В 1969 г. основоположник молекулярной
палеонтологии М. Кальвин, а позднее В. Шопф
с коллегами предложили первые сценарии
химической и биологической эволюции.
Современные данные позволяют внести
существенные коррективы. В основном они
связаны с постепенным «удревлением» находок,
свидетельствующих о появлении эукариот,
многоклеточных организмов и т. д. (рис. 1).
В последние 10 лет появились работы
по изучению ДНК вымерших форм. Эти
исследования открывают возможность
выявлять сходство и различия организмов на
основе комплекса молекулярных
характеристик. Можно получить количественную
оценку сходства сравниваемых форм,
вычислить генетические расстояния между
ними и даже оценить абсолютное
геологическое время расхождения групп.
Первые остатки ДНК вымершего
животного были выделены из шкуры квагги в 1984 г.
Исследование показало, что это животное
являлось подвидом современной зебры. Спустя
три года с помощью появившейся к тому
времени полимеразной цепной реакции
(ПЦР), позволяющей на основе матричного
синтеза размножать последовательности ДНК
и тем самым доводить их количество до
пригодного для биохимического анализа,
были получены многочисленные копии
участков генов из образца головного мозга
первобытного человека, погибшего,
по-видимому, около 7 тыс. лет тому назад.
Изучены последовательности ДНК,
выделенной из костей и мягких тканей
четырех видов новозеландских моа, вымерших
около тысячи лет назад. Оказалось, что эти
птицы гораздо древнее современных
нелетающих киви. Вероятно, киви попали на
острова значительно позже, чем вымершие
моа, жившие там уже 80 млн лет тому
назад. В то же время показано близкое
родство киви с австралийскими эму.
ДНК была успешно выделена из
ископаемых остатков мамонтов, датируемых от 10
до 50 тыс. лет. Выявлено, что виды
мамонтов отличались по ряду генетических
маркеров. Генетические дистанции мамонта от
каждого из родов современных слонов
примерно одинаковы, хотя морфологические
исследования показывают более близкое
родство мамонтов с индийскими слонами,
нежели с африканскими. Возможно, это
указывает на несоответствие темпов
эволюции на молекулярном и морфологическом
уровнях. Молекулярная палеонтология,
являющаяся частью палеогенетики, еще очень
молода, и ее основные открытия можно
ожидать в будущем. Как видит читатель,
исследования ДНК ископаемых остатков
организмов ограничены пока материалом очень
«молодого» возраста (тысячи, реже десятки
тысяч лет).
Биосферное направление.
Целенаправленные исследования в области эволюции
древних экосистем и биосферы в целом
являются в значительной мере типично
российским явлением, исторически связанным
с традицией, берущей начало от известных
работ В.И. Вернадского. Сегодня это
направление находится и в русле громадного
интереса человечества к современному
экологическому кризису, противостояние
которому немыслимо без учета предыстории
развития биосферы.
Каковы же основные результаты,
полученные в этом разделе палеонтологии?
СОВРЕМЕННАЯ ПАЛЕОНТОЛОГИЯ
71
Прежде всего на палеогеологическом
материале показано, что биосфера —
непрерывно развивающаяся авторегулируемая
система. Для ее развития характерна
цикличность разного масштаба. Было
выяснено, что только изменение разнообразия
является объективным критерием состояния
биосферы. На основе изучения динамики
разнообразия выявляются разной силы
кризисы в истории биосферы, а детальное
изучение их протекания показывает, что они
развиваются по сходному сценарию, то есть
все этапы кризиса (начало, главная фаза
и фаза выхода из кризиса) характеризуются
своими специфическими признаками
(симптомами), но всегда одними и теми же. Это
позволяет при выявлении естественных
трендов в эволюции биосферы более четко
оценить возможные последствия
антропогенных воздействий.
В предыдущие два десятилетия Д. Сеп-
коски (США) был создан банк данных по
морским организмам, позволивший
проанализировать динамику их разнообразия в
фанерозое (от кембрия — 540 млн лет
назад — до настоящего времени). Созданный
за последние годы в нашей стране банк
данных по наземным организмам дал
возможность сравнить динамику морского и
континентального разнообразия и показать
удивительную синхронность крупных
изменений, что наводит на мысль о
существовании глобальных причин, влияющих на
биосферу в целом. Биосферные
исследования потребовали разработки нестандартных
подходов и в палеобиогеографии.
Палеобиогеографическое направление.
На предыдущем этапе развития
палеобиогеографии самыми главными вопросами были
разработки принципов биогеографического
Данные
Возраст,
млрд лет
Интерпретация
.0,5
4,0
1,5
•2,0
2,5
т-3,0
i
i
Рис. 1. Хронологическая
последовательность главных событий в докемб-
рийской эволюции по данным
палеонтологии, петрографии и геохимии,
сравнительного изучения метаболизма и
биохимии современных организмов. По
В. Шопфу с дополнениями
Красным и зеленым цветом показаны
новые данные. В соответствии с этим
следует провести коррекцию и других
данных
Возраст Луны и метеоритов
3,5
4,0
4,5
Образование Земли
72
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
районирования и само районирование.
Мощным прорывом в то время было создание
СВ. Мейеном моделей происхождения флор,
заложивших основу реальной исторической
палеобиогеографии (рис. 2). Сегодня
активно ведутся исследования по созданию
моделей происхождения фаун (рис. 3). Теперь
уже достаточно уверенно можно
подтвердить высказанные ранее представления о
достаточно позднем (в девоне)
формировании бореальной области из-за перемещения
материков (континентальных плит),
расположенных ранее только в Южном
полушарии, в Северное. Можно также быть
уверенными в том, что со временем происходит
постепенная дифференциация (дробление)
биохорий (биогеографических единиц).
Таким образом, со временем идет нарастание
количества биохорий, и они в соответствии
с климатической зональностью
располагаются биполярно. Впервые сделана попытка
найти подходы к анализу внутренней
структуры биохорий на основе количественных
оценок.
Было показано, что фаунистические
комплексы проходят в своем развитии фазы
роста, расцвета и упадка. Фаза роста
характеризуется зарождением или
вселением в регион эврибионтных форм. Во время
фазы расцвета регистрируется массовое
Рис. 2. Эволюция фитохорий в позднем палеозое и
мезозое по СВ. Мейену
появление стенобионтных форм и
эндемиков. Механизм увеличения экологической
специализации можно представить как
дробление экологических ниш в результате
конкуренции за ресурсовое пространство.
Следующая фаза — фаза упадка связана с
неизбежным вымиранием неустойчивых
стенобионтных форм. Описанные фазы
аналогичны фазам экологических сукцессии.
Порядок чередования фаз развития фауни-
стического комплекса единообразен, так как
контролируется внутренними механизмами,
такими, как дробление ниш, конкуренция,
вымирание специализированных форм, а
время прохождения фаз может меняться в
зависимости от внешних воздействий.
Цикличная смена периодов космополитизма,
например: в раннем силуре, позднем
девоне, раннем карбоне, раннем триасе, и
провинциализма, например: в раннем ордовике,
раннем девоне, поздней перми,
принципиально прогнозируема, если учесть
упомянутые авторегуляторные циклы. Кроме
изоляционных механизмов, связанных с
палеогеографическими изменениями, кос-
мополитные фауны появляются всегда после
массовых вымираний, прежде всего в связи
с вымиранием специализированных форм.
Экстраординарные биоты. Как правило,
значительный элемент новизны в познании
древнего разнообразия связан с изучением
необычных экстраординарных захоронений
(лагерштаттов). Широко известно, например,
довольно давно открытое местонахождение
в среднекембрийских сланцах Бёрджес в
западной Канаде, откуда описаны многие
организмы, обычно не сохраняющиеся в
ископаемом состоянии. В последнее время А.Ю.
Иванцовым было открыто исключительной
важности местонахождение в
нижнекембрийских породах среднего течения р. Лены. Уже
сегодня оттуда известны губки, черви-палео-
сколициды, онихофоры, брадоириды и др.
Ценность этих находок, как и в случае с
Бёрджес, состоит в том, что мы имеем
возможность в таких местонахождениях изучать
остатки не только обычных раковинных
(скелетных) форм, но и бесскелетных
организмов или отпечатков мягких тканей скелетных
форм. Последний десяток лет интереснейшие
результаты были получены К. Мюллером
(ФРГ) и его коллегами по фосфатизирован-
ным мягким тканям верхнекембрийских ост-
ракод и других вымерших членистоногих. В
самое последнее время аналогичные
находки были сделаны в породах среднего
кембрия Сибири. Обычно это очень мелкие
(порядка нескольких миллиметров) организмы,
многие из которых ранее не описывались.
Удивительный результат получил И.С. Барсков,
обнаруживший фосфатизированные
кровеносные сосуды у верхнеюрского аммонита из
района Воробьевых гор в Москве.
СОВРЕМЕННАЯ ПАЛЕОНТОЛОГИЯ
73
Выше было сказано о возможности в силизации примыкает и проблема интерпре-
определенных условиях высочайшей скоро- тации загадочных ископаемых, особенно
сти фоссилизации. Сегодня мы уже пред- многочисленных в кембрии. Чрезвычайно
ставляем как условия этих захоронений, так интересна находка в кембрии Гренландии
и специфику пород, в которых подобное полного организма с покрывающими его тело
явление следует искать. К проблемам фос- склеритами и двумя образованиями, напо-
Рис. 3. Эволюция зоохорий в палеозое и триасе по Т.А. Грунт и А.Ю. Розанову
/— Южная Америка; //— Восточно-Европейская платформа; ///— Центральная и Южная Европа, Северная
Африка; IV— Северная Америка; V— Центральная и Южная Африка; VI— Индо-Пакистанский блок; VII—
Австрало-Антарктический блок; VIII— Южно-Китайская платформа и Индокитай; IX— Сибирская платформа, Алтае-
Саянская область, Казахстан
74
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
минающими раковины на концах. Понять,
что представляет собой такой организм, пока
невозможно, но достаточно ясно одно:
отдельные склериты такого организма
обычно описывались ранее как самостоятельные
таксоны родового и видового рангов.
Естественно, после находок в лагерштат-
тах происходят существенные изменения в
системе ряда групп организмов, а
количество реальных, признанных учеными
таксонов значительно сокращается. В связи с
изучением проблематичных организмов венда
и кембрия, естественно, с особой остротой
вновь встает вопрос о возможности
существования в прошлом многочисленных групп
(даже в ранге типов), быстро угасших еще
в кембрии или, точнее, в палеозое. Есть
исследователи, которые в этом архаическом
многообразии видят не многочисленные
вымершие таксоны высокого ранга, а
переходные формы между известными
типами. Но это не столь уж принципиальное
расхождение, поскольку определение
ранга того или иного таксона связано с
субъективными представлениями разных
специалистов.
Компьютерная томография.
Существенно новые возможности в изучении
ископаемого материала палеонтологи связывают
с компьютерной томографией,
исследованиями с помощью медицинского
томографа. Объект сканируется рентгеновскими
лучами с получением фронтальных срезов
через заданные интервалы. В результате
анализа полученных сечений моделируются
сечения в других плоскостях: сагиттальной
и фронтальной — и создается
компьютерная модель объекта. Метод позволяет
получить срезы необходимой области, не
повреждая при этом материала, выявить
невидимые снаружи структуры без допол-
36
нительной препаровки, которая приводит
подчас к уничтожению тонких деталей.
Совсем недавно, например, сотрудницей
Палеонтологического института РАН Т.А.
Тумановой совместно с американскими
коллегами на оборудовании Госпиталя при
Университете Пенсильвании (США) было
проведено томографическое исследование двух
черепов анкилозавров — панцирных
динозавров из коллекции
Палеонтологического института РАН (рис. 4). Сканирование
позволило воспроизвести внутренние
структуры бронированной головы анкилозавров:
носовые проходы, синусы, мозговую
полость. Неожиданный результат анализа —
обнаружение на одном из образцов,
возраст которого датируется около 80 млн лет,
патологического образования в области
нёба — остеомы. Вероятно, ее
возникновение связано с повреждением, полученным
животным при жизни, след от которого
имеется на крыше черепа.
Традиционные направления. Теперь
остановимся на некоторых результатах
традиционных направлений исследований в
палеонтологии. Пополнение сведений по
разнообразию организмов древних эпох
происходит обычно из трех источников. Это,
во-первых, новые поступления из
конкретных регионов, во-вторых, пополнение
информации за счет целенаправленных
сборов по конкретному временному отрезку,
в-третьих, целенаправленные сборы по
какой-либо группе организмов. Наиболее
мощный всплеск новой информации по
разнообразию прошлых эпох произошел в связи
с началом интенсивных палеонтологических
работ по Южной Америке, информация по
палеонтологии которой была ранее крайне
скудной. Мощный поток новой информации
продолжает поступать и из Китая.
Рис. 4. Последовательные томографические срезы черепа панцирного динозавра. Красной стрелкой
показана остеома, обнаруженная в области нёба
СОВРЕМЕННАЯ ПАЛЕОНТОЛОГИЯ
75
Описание все новых и новых
ископаемых организмов по некоторым интервалам
напрямую коррелирует с проведением
международных работ по совершенствованию
единой международной стратиграфической
шкалы. Уточнение границ, выбор страто-
типов стратиграфических подразделений,
который проводят международные рабочие
группы, приводят к интенсивному
изучению органического мира смежных
интервалов. Приведем лишь известный пример,
связанный с выбором границы кембрия и
докембрия (фанерозоя и криптозоя). В
начале работ (конец 60-х) в пограничных
отложениях кембрия и докембрия было
известно всего 15—20 видов, а уже через 10
лет были описаны сотни.
Третий источник — специальное
изучение какой-либо из групп организмов.
Например, за последние 15 лет произошел
настоящий информационный взрыв в области
изучения исторического развития класса птиц.
Только количество известных видов
ископаемых птиц стало в два раза больше, чем
было их установлено за предыдущие сто с
лишним лет изучения начиная с середины
XIX в. Были открыты новый инфракласс
птиц — энанциорнисы (Enantiornithes),
широко распространенные по всем
континентам на протяжении мелового периода, и
позднетриасовый (275 млн лет)
североамериканский протоавис (Protoavis), вероятный
предок всех настоящих веерохвостых птиц.
Значительно больше мы стали знать и о
других меловых и палеогеновых птицах.
Например, стало известно, что зубатые
нелетающие гесперорнисы позднего мела
обитали не только у побережий Северной
Америки, но и у берегов Европы и Азии.
Считавшиеся чисто южноамериканскими
кариамы (Cariamidae) и фороракосы
(Phorosrhacidae) были открыты в палеогене
Европы. Несмотря на открытие новых
археоптериксов (Archaeopteryx) — а теперь их
найдено семь экземпляров в верхней юре
(145 млн лет) южной Германии, — эту
известнейшую ископаемую птицу нельзя считать
прямым предком всех позднейших птиц.
Археоптериксы вместе с энанциорнисами
представляют боковую и тупиковую
(исчезнувшую в мезозое) ветвь в эволюции птиц
(рис. 5). Вопрос о непосредственных
предках птиц остается открытым, но едва ли ими
были динозавры, на чем настаивают
некоторые ученые.
Палеонтология дала огромный
материал для разработки теоретических аспектов
биологии, и прежде всего теории
эволюции. Необратимость эволюции, например,
не могла бы быть показана без наличия
палеонтологического материала. В последнее
время именно палеонтологический материал
дал возможность обосновывать одну из
важнейших теоретических концепций. На
разных группах позвоночных (кистеперые рыбы,
древние амфибии, парарептилии,
териодонты) было показано, что морфофункциональ-
ные переходы на более высокие
таксономические уровни происходили мозаично и
параллельно на базе исходной организации,
черты которой сохранялись долгое время у
представителей новой группы. В этой связи
можно говорить о существовании тетрапод-
ных признаков у палеозойских кистеперых
рыб—рипидистий, маммальных признаков у
териодонтов (рис. 6) и, наоборот, о
сохранении рыбных черт (чешуя, остатки
жаберного аппарата, связь позвоночника с
черепом и др.) древними амфибиями и
парарептилиями (работы Л.П. Татаринова,
Э.И. Воробьевой, О.А. Лебедева, М.Ф. Ивах-
ненко, Е.Н. Курочкина и др.).
В том же ключе можно рассматривать
и проблемы происхождения цветковых
растений. Внезапное появление в геологической
летописи и широкое распространение
покрытосеменных Ч. Дарвин назвал
отвратительной тайной, поскольку оно
совершенно не вязалось с его представлениями о
постепенности эволюции. Широкое
развитие палинологических исследований и
тщательное изучение местонахождений
макроскопических остатков флоры позволили в
корне изменить ситуацию. Палинология дала
целостную, хотя и не очень детальную
картину для всей Земли, так что не осталось
места для домыслов ни о географии, ни о
Рис. 5. Филогения класса птиц по Е.Н. Курочкину
76
ЭВОЛЮЦИЯ И СИСТЕМА ТИКА
о.
Q
о
аз
S
а
5
Q.
CD
I
X
а.
а>
СО
IV
///3
со
VI
VI
Eotheriodontia
V
IV
Рис. 6. Уровни появления
признаков млекопитающих в разных
группах вымерших
пресмыкающихся— териодонтов (зверозу-
бых) по Л.П. Татаринову
Кривые, пересекающие филети-
ческие линии, соответствуют
уровням (градам), на которых
приобретались характерные для
млекопитающих признаки; /—
звукопроводящий аппарат из трех
слуховых косточек, //— челюстное
сочленение между зубной и
чешуйчатой костями, ///— мягкие,
снабженные собственной
мускулатурой губы, IV— несколько
расширенные большие полушария
головного мозга, V— трехбугор-
чатые щечные зубы, VI— верхние
обонятельные раковины
временной привязке процесса
происхождения и распространения покрытосеменных.
Детальные исследования многих, в том
числе давно известных и бедных
остатками, местонахождений позволили получить
не только обычный материал по листьям
и древесинам, но и многочисленные цветы
и цветкоподобные органы непосредственных
предшественников покрытосеменных.
Оказалось, что многие группы голосеменных
растений, прежде всего гнетовые и бен-
неттиты, параллельно вырабатывали
признаки цветковых растений.
Таким образом, исследования
последнего десятилетия в палеонтологии дали
возможность коренным образом изменить наши
представления по двум основным позициям.
Во-первых, в результате интенсивного
изучения докембрия стало ясно, что
появление высокоорганизованных форм жизни
происходило гораздо раньше, чем мы обычно
себе представляли. И во-вторых, развитие
работ по изучению исторических
закономерностей эволюции биосферы (а это
возможно только на палеонтологическом
материале) ясно показало, что биосфера является
хорошо сбалансированной, циклически
развивающейся авторегулируемой системой,
которая пережила все внешние возмущения
за 4,5 млрд лет существования Земли.
Стало ясно, что только изучение естественных
трендов эволюции компонентов биосферы и
симптомов кризисов прошлого может дать
реальное представление об экологической
стратегии поведения человека в условиях
антропогенного кризиса.
Литература
Воробьева Э.И. Проблема происхождения
наземных позвоночных. М.: Наука, 1992.
Красилов В.А. Происхождение и ранняя
эволюция цветковых растений. М.: Наука, 1989.
Наймарк Т.Е., Пономаренко А.Г., Раутиан А.С.,
Розанов А.Ю. Экосистемные перестройки и
эволюция биосферы // Глобальные изменения прит
родной среды и климата / Под ред. Н.П. Лаве-
рова. М.: Наука, 1997.
Розанов А.Ю. Что произошло 600 миллионов
лет назад. М.: Наука, 1986.
Розанов А.Ю.у Заварзин Г.А. Бактериальная
палеонтология // Вестн. РАН. 1997. Т. 67, № 3.
Соколов Б.С.,Федонкин М.А. И еще 100
миллионов лет // Наука в СССР. 1983. № 5.
Современная палеонтология. М.: Недра, 1988.
Т. 1.
Современная палеонтология. М.: Недра, 1988.
Т. 2.
Kurochkin E.N. Synopsis of Mesozoic Birds and
Early Evolution of Class Aves // Archaeopteryx.
Munchen, 1995. Vol. 13.
Earth's Earliest Biosphere its Origin and
Evolution / Ed. W. Schopf. Princeton: Princeton
Univ. Press, 1983.
ГЕНЕТИКА
С. Г. Инге-Вечтомов
ЧТО ТАКОЕ ГЕН?
Введение
Ген — функционально неделимая
единица генетического материала; участок
молекулы ДНК (у некоторых вирусов —
РНК), кодирующий первичную структуру
молекулы белка, транспортной или рибо-
сомной РНК; в широком смысле
(обиходном) — любой наследственный задаток, или
наследственный фактор, передаваемый из
поколения в поколение и определяющий
наследуемые черты сходства между
предками и потомками.
Ген и генетическое кодирование
Термин «ген» ввел В. Иоганнсен в 1909 г.
в своем классическом труде «Элементы
точного учения о наследственности». Согласно
его определению, «свойства организма
обусловливаются особыми, при известных
обстоятельствах отделимыми друг от друга и в
силу этого до известной степени
самостоятельными единицами или элементами...,
которые мы называем генами» [Гайсинович,
1988].
Понятие «ген» вскоре стали связывать
с хромосомами ядра клетки. Согласно
хромосомной теории наследственности
Т. X. Моргана (Нобелевская премия 1933 г.),
гены — элементарные, или далее
неделимые единицы наследственности —
располагаются в хромосомах в линейной
последовательности.
Теория гена, сформулированная
Морганом в начале 20-х гг., утверждала, что ген
контролирует единичную, далее неделимую
функцию, которую определяют формально-
генетически на основании специальной
процедуры — функционального теста на алле-
лизм. Идея, на которой основан этот тест,
сводится к следующему. Если гибрид
между двумя мутантами имеет мутантный
фенотип, то соответствующие мутации относятся
к одному и тому же гену. Если же гибрид
между двумя мутантами имеет дикий или
нормальный фенотип, то соответствующие
мутации относятся к разным генам. Этот
критерий аллелизма используют и в
настоящее время.
Доказательство генетической роли и
расшифровка структуры ДНК
предоставили возможность простого объяснения
природы генов как участков ДНК с различной
последовательностью нуклеотидов,
кодирующих первичную структуру отдельных
молекул белков, а также рибонуклеиновых
кислот. В этих последовательностях есть
знаки начала и конца считывания гена.
Генетический код:
триплетен, поскольку каждому
аминокислотному остатку в полипептидной цепи
соответствуют три нуклеотида (основания)
в ДНК гена;
вчитывается с фиксированной
точки в пределах гена;
не имеет запятых, т. е. каких-либо
разделительных знаков между кодонами;
вырожден, или избыточен, так как
каждому кодируемому аминокислотному
остатку соответствует более одного трип-
летного кодона;
универсален, т. е. одинаков у всех
организмов. Это свойство кода описано позже
и нуждается в уточнении, поскольку
известны некоторые вариации кода у
отдельных видов. В настоящее время принято
говорить, что код квазиуниверсален.
Расшифровка всех 64 возможных
триплетов, составленных комбинированием
четырех нуклеотидов, правда, не в ДНК, а
в иРНК, переносящей информацию от ДНК
к месту синтеза белка на рибосомах, была
завершена в 1964—1965 гг.
Помимо генов, на которых в
результате транскрипции синтезируются
информационные РНК (иРНК), используемые для
последующей трансляции (синтеза белка на
рибосомах под контролем иРНК),
существуют гены, которые кодируют структуру
транспортных РНК (тРНК) и рибосомных
РНК (рРНК). Эти гены только
транскрибируются. В остальном структура гена
представлялась универсальной для всех живых
существ, так же как универсальным
считался и генетический код. Представления об
универсальном строении гена и
генетического материала в конце 60-х гг. лучше всего
характеризует высказывание,
приписываемое нобелевскому лауреату Ж. Моно: «То,
что справедливо для кишечной палочки,
справедливо и для слона».
Дальнейшее развитие теории гена
оказалось связанным именно с выявлением
различий между таксономически удаленными
друг от друга организмами. Это прежде
всего выяснение того, что оперонная
организация и регуляция действия генов
характерны только для бактерий, но отнюдь не
для эукариот. Бактериальный оперон (рис. 1),
как показали Ф. Жакоб и Ж. Моно,
объединяет несколько генов, регулируемых как
единица транскрипции, т. е. с них считы-
вается одна общая молекула иРНК, коди-
80
ГЕНЕТИКА
Ac
+
Р О
-Н-Т+-
Информационная Ў
РНК ЛЛЛЛЛЛАЛЛЛЛЛЛЛЛЛЛЛЛАЛЛАА
у NH2
*
АЛЛ
ч
Белки
р-галактоза
Рис. 1. Схема лактозного оперона Е. coli [по
Жакобу с изменениями; из: Гершкович, 1968]
Круг вверху— карта хромосомы Е. coli, на
которой показано положение лактозного (Lac)
оперона по отношению к другим маркерам.
Ниже— Lac-оперон в увеличенном виде: /' —
ген-регулятор, кодирующий белок репрессор;
о —оператор, который узнает репрессор, если
репрессор находится на операторе,
транскрипция оперона (синтез /.ас-иРНК) невозможна;
р-галактозид связывается с репрессором, что
приводит к изменению структуры последнего,
и он более не может связываться с
оператором; тем самым разрешается транскрипция;
р— промотор— место посадки РНК-полиме-
разы, которая и осуществляет транскрипцию;
z— структурный ген р-галактозидазы; у —
структурный ген р-галактозидпермеазы;
Неструктурный ген трансацетилазы. иРНК
лактозного оперона связывается с рибосомами
(заштрихованные кружки), на которых
осуществляется трансляция — синтез всех трех
белков, кодируемых Lac-опероном
Ас
рующая несколько белков (Нобелевская
премия 1965 г.). Гены эукариот не
объединяются в опероны и каждый из них, как
правило, регулируется отдельно, служит
матрицей для отдельной молекулы иРНК и
кодирует один полипептид.
Развитие методов генной инженерии и
прямого определения первичной
структуры молекул ДНК, т. е. последовательности
их нуклеотидов, позволило перейти от
косвенных, генетических критериев в
изучении генетического материала к прямой
физической характеристике генов, их
структуры и функции.
К концу 70-х гг. было обнаружено, что
ДНК генов эукариот часто значительно
длиннее, чем соответствующие им иРНК. Так было
открыто мозаичное или интрон-экзонное
строение генов эукариот. Как оказалось, в
ДНК гена и его первичном транскрипте
чередуются участки, представленные в
молекуле зрелой и?НК (экзоны), и участки, в
ней не представленные (интроны), которые
удаляются при созревании первичного
транскрипта (рис. 2). Этот процесс получил
название сплайсинга. Термин заимствован из
морского дела и означает сращивание канатов.
В этот же период еще два открытия
поколебали устоявшиеся было представления о
структуре и функции гена: обнаружение
перекрывающихся генов у некоторых вирусов
(рис. 3) и установление вариаций
генетического кода. Некоторые кодоны такого
квазиуниверсального кода имеют разное
значение не только у разных организмов, но и в
разных компартментах эукариотическои
клетки — в ядре и митохондриях.
В результате развития теории гена и
формирования области исследования,
которую Дж. Уотсон назвал «молекулярной
биологией гена», стало все труднее находить
вг молекулах ДНК границы того, что
обозначают привычные уже понятия «ген» или
«гены». Это связано прежде всего с тем,
что сигналы начала и конца таких
матричных процессов, как транскрипция (на ДНК)
и трансляция (на РНК), не совпадают как
по локализации, так и по сочетаниям
нуклеотидов. Наряду со структурными генами,
кодирующими первичную структуру белков
или РНК, существуют регуляторные
участки или регуляторные элементы, которые
не кодируют структуру биополимеров, но
необходимы для экспрессии структурных
генов, прежде всего их транскрипции. Нук-
леотидные последовательности этих
участков — операторы, промоторы (см. рис. 1) —
взаимодействуют с ДНК-зависимой РНК-
полимеразой, регуляторными белками —
репрессорами и активаторами
транскрипции. Регуляторные участки,
предшествующие структурным генам, обнаруживают и
в иРНК, если регуляция экспрессии гена
происходит при трансляции его иРНК.
Считать ли такие регуляторные участки
самостоятельными генами или частью
структурного гена, которому они предшествуют?
Противоречивость реальной ситуации
выражается, например, в том, что
рецессивные мутации, относимые на основании
ЧТО ТАКОЕ ГЕН?
81
Tag Sst Hinf Mbo II Pst I Mbo II Hae III
AUG
А В С D E F G
UAA
4,2
EcoRI EcoRI
a
2,4
1,8 9,5
Eco RI Eco RI Eco RI
С
G
Рис, 2. Физическая карта гена куриного овальбумина
а— чередование интронов (темные участки: A-G) и экзонов (светлые участки). Показаны сайты рестрикции —
места, расщепляемые различными специфическими эндонуклеазами— рестриктазами. Внизу— расстояния в
тысячах пар нуклеотидов между сайтами, узнаваемыми рестриктазой Eco RI. Показано положение кодонов:
инициатора— AUG и терминатора— UAA; б — результат гибридизации иРНК и ДНК гена: 1 — электронная
микрофотография; 2— графическая интерпретация той же картины. Более длинная молекула ДНК гибридизу-
ется с иРНК только отдельными участками (экзонами), представленными в обеих молекулах. Интроны,
присутствующие только в ДНК, но не в иРНК, образуют петли
классических критериев аллелизма
(например, функционального) к данному гену, могут
оказываться за пределами транслируемой и
даже транскрибируемой его части, тем не
менее нарушая экспрессию гена. Возможна
и обратная ситуация, когда мутантный ген,
потерявший участки инициации трансляции
или транскрипции в результате прямой
мутации, ревертирует благодаря
возникновению новых инициаторов, раздвигающих
прежние границы единицы экспрессируемой
генетической информации.
Экзотическими представлялись также
примеры существования самостоятельно
экспрессируемых генов в интронах.
Например, в одном из интронов митохондриаль-
ного гена box, кодирующего один из фер-
Рис. 3. Перекрывающиеся гены у бактериофагасрХ 174
ментов дыхательной цепи у дрожжей,
закодирован белок матураза, необходимый для
сплайсинга.
Наряду со структурными генами и ре-
гуляторными участками, у эукариот
обнаружены многочисленные повторяющиеся
последовательности, функции которых чаще
всего неизвестны. Они обычно
концентрируются в районах так называемого гетеро-
хроматина и характеризуются как сателлит-
ная, или эгоистическая ДНК, функция
которой в клетке состоит исключительно в
собственном воспроизведении. К этой же
категории «генов» в широком, обиходном
смысле слова относят и разнообразные
мигрирующие элементы, называемые также
множественными диспергированными
генами (мдг). Элементы, перемещающиеся по
геному, обнаружены у всех организмов,
исследованных на этот предмет: от бактерий
до человека. Их локализация на хромосомах
остается относительно постоянной в
стабильных условиях, при неизменном направлении
естественного или искусственного отбора. При
изменении направления отбора, как это было
показано для дрозофилы в работах Л. 3. Кай-
данова, В. А. Гвоздева и др., частота
перемещений мдг резко повышается. Кроме того,
эти мигрирующие элементы могут
захватывать «классические гены» и перемещать их
по геному. Таким образом, очевидно, что
яркая гипотеза «эгоистичной ДНК»,
предложенная для объяснения природы и функций мдг,
82
ГЕНЕТИКА
не исчерпывает возможных объяснений их
роли в геноме. Некоторые гипотезы
связывают эти генетические элементы с
эволюцией генетического материала, а также с
регуляцией экспрессии генов, поскольку
часто перемещающиеся элементы содержат
регуляторные последовательности нуклео-
тидов.
Таким образом, в понятиях
молекулярной биологии гена генетический материал
как совокупность всех материальных
носителей наследственной информации состоит
не только из генов, т. е. структурных
единиц этой информации, представление о
которых претерпело значительную эволюцию
со времен классической генетики.
Перемещение акцентов в разработке проблемы гена
на молекулярный уровень и применение
прямого секвенирования геномов породили
систему понятий, лишь косвенно связанных
с генетикой. Они нашли отражение,
например, в таких терминах, как:
открытые рамки считывания,
обозначающие участки ДНК, в которых
закодирована какая-то аминокислотная
последовательность, но в которых чередование
кодирующих триплетов не прерывается
триплетами-терминаторами. Такая рамка
(или фаза) считывания открыта на
значительном протяжении, однако что за белок
в ней закодирован, неизвестно;
нуклеотидные последовательности;
термин употребляется в тех случаях,
когда неясно, кодируют ли эти
последовательности что-нибудь;
консенсусы, или усредненные по
структуре последовательности регуляторных,
мигрирующих, теломерных,
автономно-реплицирующихся и других элементов генома.
Попытки установления точного
соответствия между «молекулярными» и
классическими генетическими понятиями вряд ли
будут успешными. Это связано с тем, что
организм или клетка представляют собой
многоуровневую систему, в которой
структуры и законы функционирования
низшего (молекулярного) уровня не
тождественны таковым на более высоких уровнях
организации.
Сравнительная молекулярная
биология и эволюция гена
Изучение молекулярной организации
генетического материала разных объектов
показало, что гены разных объектов и даже
гены одного объекта могут различаться по
структуре и характеру экспрессии. Как мы
уже знаем, гены могут быть организованы
в опероны, что, однако, не является
общим правилом. Ген может контролировать
одну или несколько ферментативных
реакций, содержать или не содержать интро-
ны. Наконец, гены могут перекрываться
своими кодирующими нуклеотидными
последовательностями, хотя, как правило, они
разделены межгенными интервалами.
Сопоставление этих структурных особенностей
генов, а также их функционирования у
таксономически удаленных организмов —
прокариот, эукариот и их вирусов,
позволяет выявить следующие основные
тенденции в эволюции генов.
1. Автономизацию генов эукариот по
сравнению с генами прокариот —
исчезновение оперонов, сопровождаемое потерей
способности эукариотических рибосом к
повторной инициации синтеза белка на одной
и той же матрице иРНК после его терми-
нации, даже если за кодоном-терминато-
ром находится кодон-инициатор.
По-видимому такая ситуация создает условия для
более тонкой, независимой регуляции
отдельных генов, а также открывает новые
пути эволюции генома за счет хромосомных
перестроек и транспозиций, т. е.
перемещений отдельных генов.
2. Олигомеризацию генов, которая
выражается в широком распространении
«сложных» генов (их также
называют-генами-кластерами) у эукариот. Тенденция к усложнению
генов ярко выражена в ходе прогрессивной
эволюции эукариот и часто приводит к
объединению нескольких генов, ранее
контролировавших отдельные ферментативные
реакции, в один, который теперь
контролирует несколько реакций, катализируемых
единым полипептидом. Так, например, три
(из тринадцати) реакции биосинтеза
пуринов контролирует один ген у
млекопитающих (человека) и птиц (голубя), два гена у
грибов (дрожжей, нейроспоры) и три гена- у
бактерий (кишечной палочки). У грибов
«собрались» вместе три реакции в биосинтезе
гистидина, пять реакций в биосинтнзе
ароматических аминокислот и т. д. Эта
тенденция получила название олигомеризации
генов по аналогии с олигомеризацией органов,
открытой в свое время В. А. Догелем.
3. Распространение мозаичной, или ин-
трон-экзонной, структуры генов у
эукариот, но не у эубактерий. У последних, за
очень редким исключением, интроны
отсутствуют. В то же время интроны
обнаружены у архебактерий. Это позволило
предположить, что архебактерий были общим
предком эукариот, сохранивших и
развивших мозаичную структуру генов, и
эубактерий, которые ее утратили. Тем самым,
согласно мнению У. Гилберта (Нобелевская
премия 1980 г.), эубактерий утратили и
возможности более гибкой эволюции генов
путем перетасовки экзонов за счет так
называемой негомологичной рекомбинации. В
то же время эукариоты с успехом
использовали этот путь эволюции, поскольку эк-
ЧТО ТАКОЕ ГЕН?
83
зоны обычно кодируют разные домены, т. е.
полуавтономные участки одной белковой
молекулы. На этом механизме скорее
всего и основана тенденция к олигомеризации
генов эукариот. Кроме того, механизмы
альтернативного сплайсинга открывают
новые возможности для поисков новых
функций в форме адаптивных
модификаций, когда вырезание интронов происходит
не всегда и зависит от внешних условий.
При сохранении этих условий
возникновение мутаций-генокопий, закрепляющих один
из вариантов сплайсинга и поддерживаемых
стабилизирующим отбором, способствует
дальнейшей адаптации организма.
4. Перекрывание генов (не путать с
перекрыванием кодонов, которого не
существует в генетическом коде), обнаруженное
у вирусов как про-, так и эукариотических
клеток связано со специализацией и
паразитизмом. Оно направлено на
максимальное использование небольших молекул ДНК
и РНК, повышающее их информационную
емкость. Перекрывание генов влечет за
собой их дальнейшую сопряженную эволюцию,
поскольку мутации одного гена в зоне
перекрывания будут мутациями и другого гена.
Строение гена вирусов в основном
отражает строение генетического материала
и клеточную организацию хозяина. Так,
например, обезьяний вирус SV40 в ДНК
своей минихромосомы имеет мозаичные
гены, не образующие оперонов.
Неспособность эукариотических рибосом к реиници-
ации трансляции отражается на экспрессии
генетической информации у РНК-содержа-
щих онкогенных вирусов (онкорнавирусов).
Их РНК одновременно служит
генетическим материалом и функционирует как иРНК.
Первичным продуктом трансляции у них
является единый полипептид —
полипротеин, который затем расщепляется протеа-
зами до отдельных белковых молекул,
необходимых для созревания вириона.
Сигналы, разделяющие отдельные гены у
этих вирусов, как бы вынесены на
полипептид. В то же время весь геном вируса
можно рассматривать как единый
ген-кластер. Отражение биологической
организации прокариот у бактериальных вирусов —
бактериофагов проявляется в существовании
оперонов.
Современные представления отводят
второстепенную роль в эволюции генов точ-
ковым мутациям — заменам отдельных пар
нуклеотидов, их вставкам и выпадениям.
Роль точковых мутаций сводится к
обеспечению умеренной дивергенции
гомологичных белков за счет фиксирования
ограниченного числа аминокислотных замен,
не нарушающих третичную структуру, а,
стало быть, и функциональную активность
белка. Это выражается в том, что
возникают целые семейства белков (а
следовательно и генов), обнаруживающих
структурную и функциональную гомологию у
разных объектов. Такие белки могут иметь
существенные различия (до 75%) по
аминокислотному составу, т. е. по первичной
структуре, но быть сходными по
третичной структуре. Удовлетворительное
объяснение подобных фактов дает концепция
нейтральной, или недарвиновской
эволюции, предложенная М. Кимурой, а также
Дж. Кингом и Т. Джуксом, согласно
которой большинство фиксирующихся
мутаций нейтральны по отношению к
естественному отбору. Такая эволюция сводится
к фиксации нейтральных мутаций в
результате дрейфа генов, т. е. протекает без
участия отбора.
Значительная изменчивость первичной
структуры белков и консервативность их
третичной структуры, т. е.
пространственной конфигурации белковой молекулы,
указывают на то, что новые белки и новые гены
не могут возникать только за счет замен
оснований в нуклеиновых кислотах и
аминокислотных остатков в белках. Новые гены
возникают благодаря локальным
удвоениям генетического материала —
дупликациям, их последующей дивергенции в
результате мутаций и естественного отбора, а
также в результате слияния и разделения
генов и их частей. Такой механизм, по
мнению С. Оно, является основой
прогрессивной эволюции.
О реальности этого процесса в геномах
эукариот свидетельствуют неактивные
копии нормальных генов, лишенные
интронов и несущие мутации — прежде всего
нонсенсы («бессмысленные» мутации) и
сдвиги рамки считывания,
препятствующие экспрессии генов. Эти так
называемые псевдогены гораздо быстрее, чем
функционирующие гены, накапливают мутации
и, возможно, служат материалом для
возникновения новых генов.
Образно говоря, новые структурные
гены возникают из старых генов путем
перекомбинирования их частей, кодирующих
отдельные домены. Действительно,
сравнение первичной структуры большого числа
генов довольно часто позволяет выявить
гомологию отдельных участков разных
генов. Это показывает, что эволюция
неоднократно использовала одни и те же блоки
генетической информации и служит яркой
иллюстрацией блочного или модульного
принципа биологической эволюции.
Эволюция генов тесно связана с
эволюцией систем регуляции и пространственной
организации генетического материала. Кроме
перехода от регуляции оперонов к
регуляции отдельных генов, в эволюции
возникали принципиально новые механизмы регу-
84
ГЕНЕТИКА
ляции, связанные с появлением
хроматина и возникновением эукариотической
организации клетки. Так, появились новые ре-
гуляторные элементы — энхансеры, или
усилители и сайленсеры, или глушители. Эти
нуклеотидные последовательности
действуют через изменение локальной укладки
хроматина, затрагивая (усиливая или
подавляя) экспрессию генов, находящихся на
значительном удалении от них — за сотни
и тысячи нуклеотидных пар.
Заключение
Теория гена, несмотря на все
сложности своего развития, демонстрирует пример
огромного практического значения хорошей
теории. Многочисленные системы
«ген—фермент», упомянутые ранее, стали основой
системы выявления генетической
(мутагенной, канцерогенной) опасности окружающей
среды, особенно ее антропогенных
факторов. Установление роли ДНК как
универсального носителя наследственной
информации и выяснение структуры и функций
многих конкретных генов разных объектов
легли в основу генной инженерии и
биотехнологии.
Литература
Гайсинович А. Е. Зарождение и развитие
генетики. М.: Наука, 1988.
Жаков Ф. Генетика бактериальной клетки.
Нобелевская лекция. 1965. Приложение XI //
В кн.: Гершкович И. Генетика. Пер. с англ. М.: Мир,
1968, с. 660.
Кимура М. Молекулярная эволюция: теория
нейтральности. Пер. с англ. М.: Мир, 1985.
Льюин Б. Гены. Пер. с англ. М.: Мир, 1987.
Молекулярная генетика. Пер. с англ. М.: ИЛ,
1963.
Оно С. Генетические основы прогрессивной
эволюции. Пер. с англ. М.: Мир, 1973.
Уотсон Дою. Молекулярная биология гена. Пер.
с англ. 1-е изд. М.: Мир, 1967.
Хесин Р. Б. Непостоянство генома. М.: Наука,
1984.
О. Н. Кунаева
ВЗАИМОДЕЙСТВИЕ ХЛОРОПЛАСТНОГО
И ЯДЕРНОГО ГЕНОМОВ
Введение
Главной особенностью растительной
клетки является наличие в ней зеленых хлоро-
пластов, в которых происходит фотосинтез —
образование органического вещества из С02
и воды за счет энергии света. Именно
благодаря этому процессу растению
принадлежит центральная роль в сохранении жизни
на нашей планете, т. к. в ходе фотосинтеза
синтезируется органическое вещество из
неорганического, пополняются запасы
кислорода в атмосфере и снижается
содержание С02, непрерывно поступающего в
результате дыхания живых организмов и
сжигания топлива в промышленности и на
транспорте (см.: Тихонов А. Н. Фотосинтез;
Климов В. В. Растение и биосфера. Наст. том).
Здесь мы рассмотрим, как устроен
хлоропласт, как эта клеточная органелла,
обладающая собственным генетическим
аппаратом и способная к делению, контролируется
ядерным геномом, как взаимодействуют
ядерный и хлоропластный геномы в регуляции
развития хлоропласта, почему хлоропласт
полуавтономен в клетке растений и какие
существуют гипотезы о его происхождении.
Структура и функция
хлоропласта
Хлоропласт представляет собою одну из
разновидностей специфичных для клеток
растений органелл — пластид. (Органелла-
ми называют клеточные структуры, своего
рода клеточные органы, выполняющие
специфические функции в жизни клетки.) В
клетках растений присутствует несколько
типов пластид с разными функциями: в
зеленой пластиде — хлоропласте — происходит
фотосинтез; в амилопластах синтезируется
и отлагается в запас крахмал; в
хромопластах накапливаются пигменты-каротиноиды.
Для пластид характерна оболочка,
состоящая из двух различных по свойствам
мембран. Пластиды имеют свой генетический
аппарат и белоксинтезирующую систему и
способны делиться внутри клетки.
Хлоропластом называют зеленую
пластиду, в которой происходит фотосинтез
(рис. 1, а). Как и все другие пластиды,
хлоропласт окружен оболочкой, состоящей из
наружной и внутренней мембран (рис. 1, б).
Он содержит сложно организованную
внутреннюю мембранную систему, в которую
входят мембранные «мешки» — тилакоиды,
уложенные в стопки, называемые
гранами. Тилакоиды гран соединены межгранными
мембранными каналами — ламеллами стро-
мы. В тилакоидах происходят световые
реакции фотосинтеза: улавливание квантов
света зеленым пигментом хлорофиллом,
фотоокисление воды, передача электронов
по электронтранспортной цепи с
образованием восстановленной формы никотинамид-
адениндинуклеотидфосфата (НАДФН) и
макроэргического соединения аденозинтри-
фосфата (АТФ). Все эти процессы
подробно рассмотрены в статье Тихонова. Здесь
следует лишь отметить, что образованные
в световых реакциях фотосинтеза АТФ и
НАДФН далее используются в ходе
построения органических соединений из С02 и
Н20. Эти реакции называют темновыми
реакциями фотосинтеза, потому что в них
кванты света непосредственно не участвуют.
Они протекают в строме хлоропласта. Стро-
мой называют внемембранное пространство
а
б
3
Рис. 1. Электронная микрофотография
хлоропласта, изолированного из семядолей тыквы (а), на
которой видны граны и ламеллы стромы (снимок И.М. Ку-
киной), и схема структуры хлоропласта (б)
7— тилакоиды граны; 2— ламеллы стромы; 3—
наружная мембрана оболочки хлоропласта; 4—
внутренняя мембрана оболочки хлоропласта; 5—ДНК
хлоропласта; 6— крахмальное зерно; 7— строма
хлоропласта; 8— полисомы (полирибосомы), синтезирующие
белок на мРНК
86
ГЕНЕТИКА
хлоропластов. В нем находятся ферменты,
ответственные за усвоение углекислоты, а
также генетический и белоксинтезирующий
аппарат хлоропластов.
Усвоение углекислоты в хлоропласте или,
как принято говорить, фиксация
углекислоты, т. е. включение ее углерода в состав
органических соединений, происходит в
сложном цикле реакций, обычно называемом по
имени американского ученого, внесшего
наибольший вклад в его открытие, циклом
Кальвина, а иногда циклом Кальвина—Бен-
сона. Ключевым ферментом этого цикла
является рибулозобисфосфаткарбоксилаза
(РБФК) — оксигеназа, которая
обеспечивает присоединение углекислоты к пяти-
углеродному соединению — сахару рибуло-
зобисфосфату. Образующийся при этом
короткоживущии шестиуглеродныи продукт
распадается с образованием двух трехугле-
родных молекул фосфоглицериновой кислоты:
сн2оро3'"
со;
со2 + с = о
I
неон
I
неон
сн2оро32-
I
+ Но0 -> 2 НСОН + 2Н+
I
сн2оро3'-
фосфоглицериновая
кислота (ФГК)
рибулозобисфосфат (РБФ)
Восстановление молекулы ФГК до фос-
фоглицеринового альдегида является
собственно восстановительной реакцией на пути
превращения углекислоты в молекулу
углевода.
со;
НСОН
I
СН2ОРО^
ФГК
НС = О
I
-» НСОН
глюкоза
сн2оро^-
фосфоглицериновый альдегид
Сложные превращения соединений в
цикле Кальвина обеспечивают регенерацию
молекул рибулозобисфосфата для
присоединения новой молекулы С02, а также
приводят к образованию стабильного продукта
фотосинтеза — шестиуглеродного углевода
глюкозы.
Генетический аппарат
хлоропласта
Хлоропласт имеет собственную ДНК, т. е.
собственный геном. В отличие от линейных
молекул ДНК в хромосомах ядра, хлоропла-
стная ДНК (хлДНК) представляет собою
замкнутую кольцевую двуспиральную молекулу.
Ее размеры варьируют у разных видов
растений в очень небольшом интервале, в
большинстве случаев от 130 до 160 тыс. пар
оснований. В настоящее время полностью
расшифрована нуклеотидная
последовательность хлДНК ряда видов, в том числе
табака и риса. При этом были выявлены общие
принципы организации хлоропластной ДНК
и сделан вывод о ее консервативности
(неизменности первичной структуры) в ходе
эволюции. ХлДНК содержит около 130 генов.
В ней представлено по 2 гена 4 типов рибо-
сомных РНК (рРНК), гены всех
транспортных РНК (около 30 видов), гены рибосомных
белков (около 20), гены субъединиц РНК-
полимеразы — фермента, осуществляющего
синтез РНК на хлДНК. Хлоропластный геном
кодирует около половины белков тилакоид-
ной мембраны, участвующих в
формировании комплексов электронтранспортной цепи.
Остальные белки тилакоидной мембраны
кодируются в ядре. ХлДНК содержит ген
большой субъединицы ключевого фермента
фотосинтеза РБФК.
По своей организации генетический
аппарат хлоропластов имеет много общего с
генетическим аппаратом бактерий. По про-
кариотическому типу организованы
промоторы, регулирующие начало транскрипции
(локализованы в области, отстоящей на 35-
10 пар нуклеотидов от точки начала
транскрипции), а также терминаторы,
определяющие ее окончание. Вместе с тем, в отличие
от прокариот, в ДНК хлоропластов
обнаружены интроны — нетранскрибируемые
области гена, не несущие информации о
структуре белка и характерные для генов эукариот.
Как известно, интроны вырезаются из
первичного транскрипта, а смысловые
участки сшиваются между собой (это называют
сплайсингом) в ходе созревания (процессинга)
РНК. Некоторые эукариотические черты
обнаружены и в промоторах отдельных хло-
ропластных генов.
Имея собственный генетический аппарат,
хлоропласт обладает и собственной белок-
синтезирующей системой, отличающейся от
белоксинтезирующей системы цитоплазмы,
в которой синтез белка идет на
информационных РНК (мРНК), синтезированных в ядре.
Цитоплазматические рибосомы принадлежат
к рибосомам эукариотического типа. Константа
их седиментации, отражающая скорость их
осаждения в растворе при ультацентрифу-
гировании, составляет 80 единиц Сведберга —
80S. Хлоропластные рибосомы мельче. Они
относятся к 70S типу, характерному для
прокариот. Вместе с тем по набору рибосомных
белков хлоропластные рибосомы
отличаются от прокариотических. Хлоропластный синтез
белка, подобно бактериальному, подавляется
антибиотиком — хлорамфениколом (левоми-
ВЗАИМОДЕЙСТВИЕ ХЛОРОПЛАСТНОГО И ЯДЕРНОГО ГЕНОМОВ
87
цитином), который не действует на синтез
белка на eOS-эукариотических рибосомах.
Синтез белка на eOS-рибосомах подавляется
другим ингибитором — циклогексимидом, не
влияющим на белковый синтез на 70Б-ри-
босомах бактерий и хлоропластов. Используя
поочередно два этих ингибитора, можно
установить, где в растительной клетке
происходит синтез того или иного белка: в
хлоропласте или в цитоплазме. Особенности
хлоропластного синтеза РНК и белка можно
исследовать в суспензии изолированных
хлоропластов. При этом легко убедиться, что в
хлоропласте синтез РНК и белка на свету не
нуждаются в поступлении макроэргических
соединений извне, т. к. эти процессы
используют АТФ, образованную в фотосинтетических
реакциях, протекающих в тилакоидных
мембранах. Поэтому синтез РНК и белка в хло-
ропластах резко активируется светом.
Итак, хлоропласт обладает собственным
геномом (совокупностью генов) и собственным
аппаратом реализации генетической
информации путем синтеза РНК и белка, причем
организация этих систем в хлоропласте
отличается от таковой у эукариот. Следует
заметить, что то же самое справедливо и для
других органелл клетки — митохондрий, но
митохондрии существуют во всех эукарио-
тических клетках, являясь их
энергетическим депо, тогда как хлоропласты присутствуют
только в клетках зеленых растений.
Хлоропласты размножаются в клетках
растений путем деления. Делению
хлоропласта предшествует удвоение
(редупликация) ДНК, однако размножение этих
органелл имеет свои пределы. Для каждого вида
характерно определенное число
хлоропластов в клетке, которое варьирует у разных
видов от нескольких единиц до величин,
превышающих сотню. Число хлоропластов
в клетке, а следовательно, их деление,
контролируется ядром. Например, ДНК-
полимераза, осуществляющая
редупликацию хлДНК, кодируется в ядре и
синтезируется на eOS-рибосомах цитоплазмы; после
своего образования она проникает в
хлоропласт, где обеспечивает синтез ДНК. В ядре
же кодируется, а в цитоплазме
синтезируется большое число других хлоропласт-
ных белков, что и определяет зависимость
хлоропласта от ядерного генома.
Двойное подчинение
хлоропласта собственному
и ядерному геномам
Хлоропласт, который обладает
собственной ДНК, собственным аппаратом синтеза
РНК и белка и способен к делению, тем не
менее «полуавтономен» в растительной
клетке, так как его рост, деление, развитие
тилакоидной системы и формирование
ферментативных комплексов темновых реакций
1
Рис. 2. Построение основных комплексов электронтранспортной цепи тилакоидов из белков, кодируемых
ядерным и хлоропластным геномами [из: Herrmann. In: Cell Organelles. Wien, N.Y.: Springer, 1992]
Зеленым цветом обозначены белки, кодируемые хлоропластной ДНК, желтым — ядерной ДНК. Лиловым
цветом помечены белки, кодируемые генами, геномная принадлежность которых не установлена.
1 — фотосистемаII; 2— b/f-комплекс; З— фотосистема!; 4— АТФ-синтазный комплекс
88
ГЕНЕТИКА
Субъединичная структура
РБФК
100 А
137 А
80S-
рибосомы
Предшественник Предшественник
МС РБФК 60-кДа белка
Хлоропласт
Рис. 3. Схема взаимодействия ядерного и хлоропластного геномов в процессе синтеза рибулозобисфосфат-
карбоксилазы (РБФК)
БС— большая субъединица РБФК; МС— малая субъединица РБФК, 60-кДа белок— шаперон. Отдельно
представлена структура РБФК из 8 больших и 8 малых субъединиц
фотосинтеза находятся под контролем не
только собственного, но и ядерного
генома. Это осуществляется таким образом, что
каждый функционально важный комплекс
хлоропластов состоит из белков, часть
которых кодируется и синтезируется в нем
самом, а часть кодируется в ядре,
синтезируется на 80Б-рибосомах цитоплазмы, и
лишь потом проникает в хлоропласт, где
включается вместе с образованными в нем
белками в построение функционально
активных комплексов. Например, для
формирования хлоропластных рибосом
используются синтезированные в хлоропластах
рибосомные РНК и часть рибосомных
белков, а другая часть белков, необходимых
для построения этих органоидов,
кодируется в ядре и синтезируется в цитоплазме.
Транспортные РНК, которые доставляют
аминокислоты в рибосому и находят им место
в растущей полипептидной цепи,
синтезируются в хлоропласте, но ферменты,
активирующие аминокислоту и помогающие
ей найти свою транспортную РНК и к ней
присоединиться (аминоацил-тРНК-синтета-
зы), кодируются в ядре и синтезируются
в цитоплазме. Точно так же около
половины белков, необходимых для построения че-
ВЗАИМОДЕЙСТВИЕ ХЛОРОПЛАСТИОГО И ЯДЕРНОГО ГЕНОМОВ
89
тырех основных комплексов электронтран-
спортной цепи тилакоидных мембран
кодируется и синтезируется в хлоропласте, а
остальные — кодируются в ядре и
синтезируются в цитоплазме, из которой они
проникают в хлоропласт, где, взаимодействуя
с хлоропластными белками, участвуют в
образовании этих комплексов (рис. 2). Так на
уровне формирования сложных пептидных
комплексов происходит интеграция работы
ядерного и хлоропластного геномов в
построении тилакоидной мембраны хлоропла-
стов, осуществляющей фотозависимые
реакции фотосинтеза. Формирование
ферментативного аппарата темновых реакций
фотосинтеза, в ходе которых происходит
ассимиляция (усвоение) С02, также
находится под контролем двух геномов, ядра и
хлоропласта. Это осуществляется за счет
того, что ключевой фермент цикла
Кальвина РБФК состоит из двух субъединиц
(полипептидов), из которых большая
субъединица (54 кДа) кодируется и синтезируется
в хлоропласте, а малая (14кДа)
кодируется в ядре и синтезируется в цитоплазме в
виде предшественника (рис.3). Затем она
проникает в хлоропласт и встречается с
Предшественники
хлоропластных
белков
синтезированной там большой
субъединицей. Для того чтобы эти два полипептида
образовали правильную конечную структуру
функционально активного фермента РБФК,
состоящего из 8 больших и 8 малых
субъединиц, необходим дополнительный 60-кДа
белок. Такие белки называют шаперонами
или шаперонинами от англ. слова chaperon,
которое обозначает провожатую, наставника
при молодой особе (см.: Наградова Я. К.
Внутриклеточная регуляция формирования на-
тивной пространственной структуры белков.
Том 8). 60-кДа шаперон обеспечивает
правильную укладку цепей двух субъединиц
РБФК. Этот белок также кодируется в ядре
и синтезируется в цитоплазме, проникая
затем в хлоропласт (рис. 3). Так ядро
контролирует формирование функционально
активной РБФК — ключевого фермента
фиксации углекислоты в процессе фотосинтеза.
Таким образом, формирование всех
важнейших структур хлоропласта
зависит от ядра и цитоплазмы. Поэтому идея
создания культуры изолированных хлоро-
пластов, где бы они самостоятельно
размножались на питательной среде,
невыполнима.
Двойной генетический контроль
формирования функционально-активных зеленых
хлоропластов эффектно демонстрируется в
опытах по слиянию протопластов
(представляющих собой покрытое клеточной
мембраной содержимое растительной клетки,
освобождающееся после разрушения
клеточной стенки специальными ферментами).
Для этого берут протопласты листьев, у
которых развитие зеленых хлоропластов
повреждено вследствие мутации в ядерной
или хлоропластной ДНК. У одного типа
листьев формирование зеленых
хлоропластов нарушено в результате мутации
ядерного гена, а генетический аппарат
пластиды не поврежден. У другого типа листьев
формирование хлоропластов подавлено в
результате мутации в хлоропластном геноме
при сохранении неизменным ядерного
генома. При слиянии таких протопластов в
гибридном протопласте присутствуют
неповрежденные хлоропластные геномы и
неповрежденный ядерный геном. Это
обеспечивает формирование полноценных зеленых
хлоропластов.
Рис. 4. Схема транспорта белков из цитоплазмы в
хлоропласт с помощью транзитного пептида [из:
Keegstra, von Heijne. In: Cell Organelles. Wlen, N.Y.:
Springer, 1992]
Транспорт белков в хлоропласты
Транспорт нуклеиновых кислот (ДНК
или РНК) через мембраны оболочки
хлоропластов не обнаружен, а вот транспорт
белков из цитоплазмы в хлоропласт
происходит очень активно. Без него было бы
невозможно построение внутренней
структуры хлоропласта. При этом одни белки,
синтезированные в цитоплазме для хло-
90 ГЕНЕТИКА
Хлоропласт
Этиопласт Хромопласт
1 мкм
Амилопласт ' ' Пропластида
Рис. 5. Схема развития хлоропластов, хромопластов и амилопластов из пропластид и взаимного превращения
пластид [из: Herrmann. In: Cell Organelles. Wien, N.Y.: Springer, 1992]
ропласта, встраиваются в мембраны хло-
ропластной оболочки, другие направляются
в строму, третьи встраиваются в тилако-
идные мембраны или, проходя через них,
оказываются во внутреннем постранстве
«тилакоидного мешка» (рис. 4). Как же
белки проникают через две мембраны
оболочки хлоропласта? И как они находят
свой адрес внутри хлоропласта? Как
выяснилось, в ядерных генах, кодирующих
хлоропластные белки, записана
информация не только об их структуре, но и
о локализации в хлоропласте. Адрес белка
содержится в специальной транзитной
аминокислотной последовательности,
которая располагается в начале
синтезируемой белковой цепи хлоропластного
белка (на N-конце) и состоит у белков стромы
примерно из 40, а у белков тилакоидов
более чем из 80 аминокислотных
остатков. Транзитная последовательность
находит рецептор на наружной мембране
хлоропласта и обеспечивает прохождение
через нее полипептида. Затем
специальный фермент стромы, специфическая
пептидаза, узнает транзитную
последовательность и отрезает ее от белка, гид-
ролизуя всего лишь одну пептидную связь.
Освобожденный от транзитного пептида
белок встраивается в соответствующие
ферментные комплексы в строме,
например, при формировании РБФК. Если же
белок предназначен для тилакоидной
мембраны, дополнительная сигнальная
последовательность помогает ему найти в ней свое
место и даже пройти через эту мембрану,
если его место находится во внутреннем
пространстве тилакоида. Затем
дополнительная сигнальная последовательность
отрезается от белка пептидазой тилакоида,
которая гидролизует одну пептидную связь,
соединяющую сигнальную
последовательность с основным белком. Так происходит
доставка хлоропластных белков к месту их
действия.
Развитие хлоропласта
из пропластиды
Хлоропласт развивается из
пропластиды — маленькой бесцветной органеллы
(несколько микрон в поперечнике),
окруженной двойной мембраной и содержащей
характерную для хлоропласта кольцевую
молекулу ДНК. Пропластиды не имеют
внутренней мембранной системы. Они плохо
изучены ввиду их крайне малых размеров. В
цитоплазме яйцеклетки содержится
несколько пропластид. Они делятся и передаются
от клетки к клетке в ходе развития
зародыша из оплодотворенной яйцеклетки. Этим
объясняется то обстоятельство, что
генетические признаки, связанные с ДНК
пластид, передаются только по материнской
ВЗАИМОДЕЙСТВИЕ ХЛОРОПЛАСТНОГО И ЯДЕРНОГО ГЕНОМОВ
91
линии. Такие признаки относят к так
называемой «цитоплазматической
наследственности».
В ходе развития хлоропласта из пропла-
стиды внутренняя мембрана ее оболочки
образует впячивания внутрь пластиды. Из них
развиваются мембраны тилакоидов, которые
образуют стопки — граны, и ламеллы стро-
мы (рис. 1, а, б). В темноте пропластиды дают
начало формированию предшественника
хлоропласта (этиопласта), который содержит
структуру, напоминающую кристаллическую
решетку (рис. 5). При освещении эта
структура разрушается и происходит
формирование характерной для хлоропласта
внутренней структуры, состоящей из тилакоидов гран
и ламелл стромы (рис. 5).
В клетках меристемы также
содержится несколько пропластид. При
формировании зеленого листа они делятся и
превращаются в хлоропласты. Например, в клетке
закончившего рост листа пшеницы
содержится около 150 хлоропластов.
В органах растений, запасающих
крахмал, например, в клубнях картофеля,
крахмальные зерна формируются и
накапливаются в пластидах, называемых амилоп-
ластами (рис. 5). Как выяснилось, амилоп-
ласты образуются из тех же пропластид и
содержат ту же ДНК, что и хлоропласты.
Они формируются в результате дифферен-
цировки пропластид по пути, отличному от
того, который приводит к образованию
хлоропластов (рис. 5). Известны случаи
превращения хлоропластов в амилопласты, и
наоборот. Например, часть амилопластов
превращается в хлоропласты при позеленении
клубней картофеля на свету. В ходе
созревания плодов томатов и ряда других
растений, а также в лепестках цветков и в
осенних красных листьях хлоропласты
превращаются в хромопласты — органеллы,
содержащие оранжевые пигменты-кароти-
ноиды. Такое превращение связано с
разрушением структуры тилакоидов гран и
приобретением органеллой совершенно иной
внутренней организации. Эту перестройку
пластид диктует ядро, и она
осуществляется с помощью кодируемых в ядре и
синтезируемых в цитоплазме особых белков.
Например, кодируемый в ядре 58-кДа
полипептид, образующий комплекс с кароти-
ноидами, составляет половину всего белка
мембранных структур хромопласта. Так на
основе одной и той же собственной ДНК в
результате ядерно-цитоплазматического
влияния пропластида может развиваться в
зеленый фотосинтезирующий хлоропласт,
белый, содержащий крахмал амилопласт или
оранжевый, заполненный каротиноидами
хромопласт. Между ними возможны
превращения, указанные на рис. 5. Это крайне
интересный пример различных путей диф-
ференцировки органелл на основе одной и
той же собственной ДНК, но под влиянием
ядерно-цитоплазматического «диктата».
Происхождение хлоропласта
Общепринятым в настоящее время
является представление об эндосимбиотичес-
ком происхождении хлоропластов в
клетках растений.
Хорошо известно, что лишайники
представляют собою форму сожительства
(симбиоза) гриба и водоросли, при котором
зеленые одноклеточные водоросли живут
внутри клеток гриба. Предполагают, что
подобным же образом несколько
миллиардов лет назад фотосинтезирующие циано-
бактерии (синезеленые водоросли) проникли
в эукариотические клетки и затем в ходе
эволюции потеряли свою автономность,
передав большое число важнейших генов
в ядерный геном. В результате
независимая бактериальная клетка превратилась в
полуавтономную органеллу, сохранившую
главную исходную функцию — способность
к фотосинтезу; при этом формирование
фотосинтетического аппарата оказалось под
двойным ядерно-хлоропластным контролем.
Под ядерный контроль перешло деление
хлоропластов и сам процесс реализации их
генетической информации, осуществляемой
по схеме ДНК -> РНК -> белок.
Убедительнейшие свидетельства прока-
риотического происхождения хлоропластов
получены при анализе нуклеотидных
последовательностей их ДНК. ДНК рибосомных
генов имеет высокую степень сродства
(гомологии) у хлоропластов и бактерий.
Сходные нуклеотидные последовательности
обнаружены у цианобактерий и хлоропластов
в генах АТФ-синтазного комплекса, а также
в генах аппарата транскрипции
(кодирующих субъединицы РНК-полимеразы) и
трансляции. Как уже упоминалось, регулятор-
ные элементы хлоропластных генов —
промоторы, определяющие начало
транскрипции, и их терминальные нуклеотидные
последовательности, определяющие ее
прекращение, организованы по бактериальному
типу. И хотя миллиард лет эволюции
внесли в хлоропласт массу изменений, его гены,
очевидно, сохранили большое сходство с
генами древнего предшественника
современных цианобактерий. Этот факт служит
неоспоримым доказательством происхождения
хлоропласта от прокариотического предка.
Заключение
Мы рассмотрели принципы организации
и развития хлоропласта в клетке растений
и показали, как скоординированная работа
двух геномов — ядерного и хлоропластно-
92
ГЕНЕТИКА
го — приводит к формированию сложнейшего
молекулярного аппарата,
осуществляющего процесс фотосинтеза. Высокое сходство
ДНК хлоропластов и цианобактерий
свидетельствует о происхождении хлоропласта от
древних предшественников современных
цианобактерий, проникших миллиарды лет
назад в эукариотическую клетку. Однако в
ходе последующей эволюции независимая
бактериальная клетка превратилась в
полуавтономную органеллу, находящуюся под
контролем ядра, которое определяет, по
какому пути она будет развиваться —
превратится в фотосинтезирующий хлоропласт,
станет местом накопления пигментов-каро-
тиноидов (хромопластом), или пойдет по пути
биосинтеза и отложения в запас крахмала
(станет амилопластом). Изучение
молекулярных механизмов дифференцировки трех
рассмотренных типов органелл и их взаимного
превращения — одна из увлекательных
задач современной молекулярной биологии
растений. Исследования в этом направлении
позволят лучше понять функционирование
систем сигнализации, обеспечивающих
скоординированную работу ядерного и пластид-
ного геномов.
Литература
Албертс Б., Брей Д., Льюис Дж. и др.
Молекулярная биология клетки. Т. 3. Пер. с англ. М.:
Мир, 1994.
Одинцова М. С. Организация и экспрессия
геномов хлоропластов и митохондрий (Итоги науки
и техники. Сер.: «Общие проблемы
физико-химической биологии»). М.: ВИНИТИ, 1987, т. 6, с. 5.
Юрина Н. П., Одинцова М. С. Молекулярная
биология, 1992, т. 26, №4, с. 757.
Cell Organelles. Wien; New York: Springer
Verlag, 1992.
Л. И. Корочкин
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ РАЗВИТИЯ КЛЕТОК
Введение
Живой организм состоит из миллиардов
самых разнообразных клеток. Их форма
может быть и очень простой и очень
причудливой: например, есть клетки,
напоминающие паука, снежинку, звездочку и другие
сложные образования (рис. 1). Этот факт был
установлен еще на заре развития
гистологии — науки о клетках и тканях, строящих
различные органы животных и растений.
Когда научились выявлять клетки с
помощью специфических красителей на тонких
(несколько микронов) срезах органов и
тканей, оказалось, что при использовании смеси
некоторых гистологических красителей можно
обнаружить множество клеток, которые
различаются не только по размерам и форме,
но и по сродству к этим красителям.
Например, при окраске слизистой желудка по
методике Доминичи-Кедровского видны
клетки оранжевого, фиолетового и
голубого цветов. Первые, называемые «обкладоч-
ными», вырабатывают соляную кислоту,
вторые, именуемые «главными», — синтезируют
пищеварительный фермент пепсин, а
третьи — «добавочные» — вырабатывают слизь.
В последующем были обнаружены и
химические «метки» разных клеточных типов: в
так называемых адренэргических клетках
нервной системы выявили тирозингидрокси-
лазу и катехоламины, а в холинэргических —
ацетилхолинэстеразу и холинацетилтрансфе-
разу — ферменты, ответственные за
образование и распад ацетилхолина.
В то же время известно, что генетическая
информация, содержащаяся в столь
различающихся по внешнему виду, функциям и
химической специфике клетках, одинакова.
Как же получается, что клетки,
содержащие одинаковую генетическую информацию,
приобретают столь различный «облик»?
Рис. 1. Многообразие клеточных форм в тканях животных
7 — эпителиальная клетка кишечника; 2— клетка гладкой мускулатуры; 3— фибробласт (клетка
соединительной ткани); 4— нервная клетка; 5— эритроцит птиц; 6— клетка поперечнополосатой мускулатуры
94
ГЕНЕТИКА
Две генетические «модели»,
объясняющие специфику
разных клеток
Начиная с 20—30-х гг. существуют две
«модели» объяснения описанного выше
феномена, сформулированные
американским ученым Т. Г. Морганом и немецким
исследователем Р. Гольдшмидтом. Эти
ученые были одними из создателей
современной генетики и экспериментальной
эмбриологии и потому хорошо понимали как
генетическую, так и эмбриологическую
проблематику.
Морган полагал, что, несмотря на
одинаковый набор генов во всех клетках
многоклеточного организма, в ходе
онтогенеза в клетках, расположенных в разных
частях развивающегося зародыша, и на
разных стадиях его развития функционируют
разные гены; поэтому клетки зародыша
приобретают сначала химическое, а затем
и морфологическое своеобразие. Активация
же разных генов в разных клетках
обусловлена различиями в химическом составе
цитоплазмы в разных частях зародыша,
сложившимися еще в ходе оогенеза.
Следовательно, процесс индивидуального
развития начинается не в момент дробления
или даже оплодотворения яйца, а в
период его созревания, оогенеза. Таким
образом, клеточная специализация является, по
Моргану, следствием дифференциальной
активности генов, или, как сейчас
принято говорить, дифференциальной
транскрипции (это называют регуляцией на уровне
транскрипции; рис. 2, а).
Гольдшмидт придерживался другой точки
зрения. Он предположил, что во всех
клетках одинаково работают все гены, но их
продукты попадают в разную клеточную
плазму (последнее перекликается с
гипотезой Моргана). В одной плазме способны
функционировать продукты одних генов, в
другой — других. В этом случае
специализация клеток будет осуществляться на
уровне дифференциального функционирования
генопродуктов, можно сказать — на
посттранскрипционном уровне, на уровне
дифференциальной трансляции, или на
уровне посттрансляционных модификаций, т. е.
в результате изменений, происходящих с
белком после его синтеза — трансляции
(рис. 2, б).
Развитие клетки
и функционирование генов.
Современные данные
Кто же прав — Морган или Гольдшмидт?
Возможность получения ответа на этот
вопрос появилась относительно недавно, начиная
Б
В
\AflEj
д
Е
Б В Г
Рис. 2. Две «модели» генетической регуляции
клеточной дифференцировки по Т. Моргану (а) и по
Р. Гольдшмидту (б)
1 — ядро; 2— цитоплазма; 3— хромосомы с генами
А, Б, В...; красным выделены функционально
активные гены, а—А, Б, В...— продукты генов, б—А, Б,
В... — продукты, функциональная активность которых
блокирована в цитоплазме. В каждом случае
показаны две клетки, дифференцирующиеся в разных
направлениях
с 60-х гг., когда в генетике и эмбриологии
стали использоваться методы молекулярной
биологии. Бельгийский эмбриолог Г. Дени
обнаружил, что на разных стадиях
эмбриогенеза и в разных участках эмбрионов
амфибий синтезируются различные фракции
РНК, подтвердив тем самым гипотезу
Моргана. Однако чуть позднее, в
исследованиях, проведенных в лаборатории Э.
Дэвидсона в США, были найдены «гольдшмидтовские»
варианты генетической регуляции развития:
в разных частях эмбриона синтезировались
одинаковые информационные РНК (мРНК),
которые, однако, по-разному
транслировались в разных клетках (см. рис. 3). Наконец,
оказалось, что в созревающей яйцеклетке
уже синтезируется глобиновая мРНК,
которая будет функционировать много позднее
в кровяных клетках. Проблема, таким
образом, осложнилась.
Автор статьи совместно с М. Б. Евгень-
евым и Б. А. Кузиным избрал для
проверки двух указанных моделей другой путь. Мы
воспользовались тем, что у самцов
знаменитой плодовой мушки дрозофилы в
репродуктивной системе есть орган, подобный
предстательной железе млекопитающих, —
это так называемая семявыносящая
луковица. Она представляет собой очень удоб-
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ РАЗВИТИЯ КЛЕТОК
95
ный для молекулярных исследований орган,
поскольку является своеобразным мешком,
стенка которого выстлана одним слоем
однородных и синхронно
дифференцирующихся эпителиальных клеток, вырабатывающих
белковый тканеспецифический секрет,
передаваемый в ходе оплодотворения в
половые пути самки. Дифференцировка этих
клеток была подробно изучена в нашей
111111 i 11111 11 11} 11 it
Структурный ген
а* Ь_
Конститутивный транскрипт
Ь а
с с
ss
Интегрирующие регуляторные ss
транскрипционные единицы
Набор транскриптов регуляторных единиц
е
b а с с d
Ь1 а1 с1 с1 с(1
7 2 3
Комплементарные комплексы транскриптов
структурных и регуляторных генов
Рис. 3. Схема регуляции генной экспрессии на
посттрансляционном уровне, согласно данным
Бриттена и Дэвидсона
В различных клетках и на разных стадиях развития
транскрибируются практически все структурные гены,
т. е. уникальные последовательности ДНК (как в
модели Р. Гольдшмидта). Однако в клетках,
специализирующихся в разных направлениях, активируются
разные умеренно повторяющиеся
последовательности ДНК; они и регулируют специфику трансляции
уникальных мРНК в каждой клетке. Продукты
умеренно повторяющихся последовательностей ДНК
несут участки (помечены красным), способные
опознавать комплементарные им участки транскриптов
структурных генов и образовывать соответствующий
специфический комплекс (транскрипты структурных
генов —транскрипты повторяющихся
последовательностей). В клетках, которые дифференцируются в
разных направлениях, существуют разные наборы
таких комплексов. Предполагается, что только те
продукты структурных генов подвергаются созреванию
с последующим транспортом зрелой мРНК в
цитоплазму, которые образуют комплекс с
соответствующим транскриптом повторяющихся
последовательностей. Остальные продукты структурных генов
уничтожаются специфическими ферментами нуклеазами
(на схеме перечеркнуты красным). Таким образом,
специфика дифференцировки клеток
обеспечивается процессами, происходящими на
посттранскрипционном уровне
лаборатории С. Асоновой. Основываясь на ее
результатах, мы выделяли на разных
стадиях развития клеток семявыносящих
луковиц мРНК, меченную радиоактивным
уридином, и гибридизовали ее с ДНК
гигантских, так называемых политенных
хромосом слюнных желез дрозофилы (в самих
семявыносящих луковицах хромосомы очень
маленькие и с ними неудобно работать).
Таким способом мы выявляли участки
хромосом, комплементарные мРНК,
выделенной на разных стадиях клеточной
дифференцировки, т. е. участки, функционально
активные на изучавшихся стадиях
развития клеток.
Оказалось, что когда клетка только
начинает специализироваться, в ней
работает большая часть генома, но тканеспе-
цифические гены еще неактивны. По мере
дифференцировки и роста клетки
количество активно работающих генов
уменьшается, но зато начинают функционировать
тканеспецифические гены,
транскрипционная активность которых постепенно
возрастает. Таким образом, получается, что, как
это часто бывает, правы оба — и Морган,
и Гольдшмидт. Однако разные «модели»
реализуются или на разных стадиях
развития, или в клетках разных типов.
Регуляторные гены
и развитие клетки
Тканеспецифические гены,
определяющие особенности клеточной
дифференцировки, испытывают давление системы
других генов, контролирующих их функции.
Эти гены можно разделить на
регуляторные, временные и «архитектурные».
Рассмотрим, как они работают, на примере
проявления в семявыносящей луковице
самцов Drosophila virilis (одного из
хорошо генетически изученных видов рода
Drosophila) гена, кодирующего
тканеспецифический изофермент (одну из форм
фермента) эстеразы. Этот изофермент,
попадая при оплодотворении в половые пути
самки, расщепляет в них цис-вакценила-
цетат с образованием феромона — пахучего
вещества, которое «сообщает» самцам, что
данная самка больше не нуждается в их
услугах.
На рис. 4 видно, что уровень
активности структурного гена этой эстеразы,
локализованного во 2-й хромосоме, зависит от
сигналов, поступающих от регуляторного
гена, разместившегося в половой Х-хромо-
соме. Последний определяет, сколько
молекул мРНК будет синтезировано в
клетке (как это предусмотрел в своей модели
Морган). Однако (как и предугадал в своей
модели Гольдшмидт), хотя мРНК
синтезируется и транспортируется в цитоплазму
96
ГЕНЕТИКА
Р Ядро
I г" «|
А л ^-—-*J
>.* г
Цитоплазма
Рис. 4. Генная система, контролирующая «работу»
тканеспецифического гена
1 — ядерная мембрана; 2— ядерные поры; 3 — тка-
неспецифический структурный ген; 4—
синтезированная им информационная РНК (мРНК); р— ген
регулятор активности, «включающий» тканеспецифичес-
кий ген и определяющий количество молекул мРНК,
которое он синтезирует в клетке; вр— временной
ген, продукт которого «разрешает» трансляцию мРНК
(синтез белка) тканеспецифического гена, а—
«архитектурный» ген, контролирующий прикрепление
белка к клеточной мембране; б—белок, продукт мРНК
клетки, синтез фермента не начинается,
пока не поступит сигнал от другого гена,
гена «времени», который расположен в 4-й
хромосоме и от которого зависит момент
начала синтеза молекул эстеразы на
матрицах мРНК.
Что же это за сигнал? По
предварительным данным, это появление одной из
транспортных РНК, несущих
аминокислоты к месту синтеза белка; она оказалась
особенно нужной для синтеза изофермен-
та рассматриваемой эстеразы. Однако
молекулы изофермента, которые
синтезируются в клетке, могут находиться там либо
в свободном, либо в связанном с белками
клеточных мембран состоянии. У разных
линий и видов мухи соотношение
«свободной» и связанной с мембранами фракций
может быть различным; оно определяется
специальным «архитектурным» геном,
отвечающим за синтез определенного белка,
встроенного в эти мембраны.
Итак, процесс генетической регуляции
клеточной дифференцировки очень сложен
и существует система генов, от которой
зависит синтез тканеспецифических
продуктов. Такие системы генов описаны и у
дрозофилы, и у млекопитающих
сотрудниками различных лабораторий как в нашей
стране, так и за рубежом, преимущественно
в США, Германии и Австралии.
Молекулярные данные
о генетической регуляции
специфических белков в клетке
Рядом со структурным геном существует
участок ДНК, который воспринимает
контролирующие сигналы (возможно,
связывающиеся с ДНК специфические
низкомолекулярные белки) и заставляет
структурный ген функционировать в
нужное время и в нужном месте с
определенной степенью активности. Утрата этого
участка влечет за собой нарушение функции
гена: он или не работает, или работает плохо
или необычно, что приводит к
нарушениям в развитии зародыша, а иногда и к его
гибели. Как же выявляют такие
специфические участки ДНК, выполняющие
командные функции при структурном гене?
Возможность их обнаружения появилась
благодаря соединению методов
молекулярной биологии и экспериментальной
эмбриологии, позволившему получать так
называемых трансгенных животных, т. е. животных,
в геном которых с помощью
микроманипуляций введены чужеродные гены.
Если, например, нужно узнать, от
какого участка ДНК зависит тканеспецифич-
ность экспрессии гена, кодирующего алко-
гольдегидрогеназу (фермент, расщепляющий
спирт) у дрозофилы, то с помощью
методов генной инженерии «изготавливают»
искусственные «конструкции», содержащие
интересующий нас структурный ген и
смежные участки ДНК, имеющие в разных
вариантах конструкции разную длину. Такие
конструкции соединяют с подвижным
генетическим элементом — Р-элементом,
способным перемещаться в геноме,
внедряться в его ДНК и закрепляться там, и затем
вводят в развивающиеся яйца дрозофилы,
принадлежащей к такой линии, у
представителей которой алкогольдегидрогеназа в
результате мутации утратила свою
активность. Трансгенных животных узнают,
таким образом, по появлению активности
алкогольдегидрогеназы. Среди них есть мухи,
у которых тканевая специфичность
экспрессии гена алкогольдегидрогеназы
восстанавливается полностью. В этом случае
смотрят, какой длины фрагмент регуляторной
ДНК попал в их геном, и заключают, что
он и определяет специфичность
активности структурного гена в определенном
наборе тканей и органов (рис. 5).
Гены и формообразование
Несмотря на успехи в изучении
молекулярной специализации клеток, долгое
время не удавалось перекинуть мостик между
молекулярными событиями, происходящи-
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ РАЗВИТИЯ КЛЕТОК
97
Рис. 5. Продукты тканеспецифического гена
Drosophila virilis в репродуктивных путях
трансгенных Drosophila melanogaster выявлены с помощью
иммуногистохимической реакции с антисывороткой
против тканеспецифического белка Drosophila virilis,
проведенной на гелевых блоках после электрофо-
ретического разделения белков семявыносящей
луковицы трансгенной мухи
5—самец; 10 и 12 — семявыносящие луковицы; 14 —
семявыносящие луковицы D. virilis. Остальные
пробы—либо другие органы, либо самки
ми в клетках во время индивидуального
развития, и процессами формообразования,
собственно и составляющими его суть. Было
лишь обнаружено соответствие
определенным молекулярным событиям
последовательных изменений формы зародыша и его
частей в разные периоды онтогенеза. Но
приводят ли именно эти молекулярные
события к определенным морфогенетическим
изменениям, существует ли между этими
Рис. 6. Сканирующая электронная микроскопия
эктопически развившегося у Drosophila melanogaster
глаза, по данным В. Геринга
а— эктопический глаз (помечен красным цветом) в
головной области; б— эктопический глаз в районе
крыла и на антенне; в— при большом увеличении;
г— эктопический глаз в районе крыла при большом
увеличении
двумя рядами событий каузальная
(причинная) связь или же процесс
формообразования зависит от неких специфических, так
называемых «морфогенетических полей»,
существование которых постулировалось
ранее, — этот вопрос дискутируется уже
достаточно давно.
И лишь в 1995 г. работы швейцарского
эмбриолога В. Геринга пролили некоторый
свет на эту сложную и запутанную
проблему. Он заинтересовался, в какой степени ген
«безглазия» (eyeless), мутация которого
обусловливает отсутствие глаз у дрозофилы, и
гомологичный ему ген «малые глаза» (small
eyes) отвечают за всю цепь
формообразовательных процессов, завершающихся
образованием глаза? И что получится , если этот
ген активировать в других частях
развивающегося зародыша, там, где глаза никогда
не возникают? Современные методы генной
инженерии позволяют это сделать.
Используя их, Геринг «сращивал»
нормальный аллель гена «безглазия» (eyeless)
с упоминавшимися регуляторными
фрагментами ДНК, смежными с разными
структурными генами, экспрессирующимися под
влиянием этих фрагментов в разных
частях зародыша, где глаз не бывает. Такие
необычные конструкции были введены в
ядра развивающихся яиц на самых ранних
стадиях развития дрозофилы с помощью
микроинъекции. Иногда (операция сложная
и не всегда удается) эти конструкции
«срабатывали», и тогда там, где удавалось
«вызвать к жизни» ген «безглазия»,
формировался глаз: на брюхе, на крыльях, на груди
и т. д. (рис. 6). Более того, если для
микроинъекций использовали конструкцию,
содержащую вместо гена «безглазия»
дрозофилы его гомолог, выделенный из клеток
лабораторных мышей, результат
получался тот же — развитие глаза на необычном
месте, там, где его не должно быть.
Иными словами, оказывается, что один-
единственный ген «безглазия» (а у мыши ген
«малых глаз») «запускает» каскад событий,
активируя множество определенных генных
систем, функционирование которых имеет,
в конечном счете, один итог —
формирование глаза. Это, пожалуй, первый случай,
когда продемонстрирована четкая связь
между последовательным включением в
функцию группы генов и обусловленным этим
включением морфогенетическим
процессом — образованием глаза. По-видимому,
молекулярно-генетический механизм этого
процесса в высшей степени консервативен
и един во всем животном мире, ибо ген
мыши способен произвести у дрозофилы тот
же эффект, что и ее собственный
гомологичный ген.
О существованиии единого
генетического консервативного механизма управления
98
ГЕНЕТИКА
формообразовательными процессами в
разных, филогенетически удаленных группах
животного мира свидетельствуют данные
недавних экспериментов. Так, итальянские
ученые доказали возможность установления
нервных связей между нервными клетками
насекомых и птиц в условиях культуры
тканей. В наших опытах, проведенных вместе с
С. Савельевым, получены данные о
переживании дифференцирующихся нервных
клеток дрозофилы при трансплантации их в
нервную трубку эмбрионов амфибий, а также
млекопитающих. Нейроны дрозофилы в
первом случае не только успешно
развиваются, но и устанавливают синаптические
контакты с клетками хозяина и оказывают
влияние на скорость развития хозяина и
дифференцировку его мозга, стимулируя
васкуляризацию (рост кровеносных сосудов)
и ветвление отростков нервных клеток, — один
из главных признаков их специфической
специализации.
Такое поведение нервных клеток
насекомых позволило добавлять их к
эмбриональной нервной ткани зародышей
человека, трансплантируемых в мозг пациентам,
страдающим болезнью Паркинсона
(постоянное дрожание различных частей тела) и
другими неврологическими расстройствами.
Стимулирующие факторы, выделяемые
тканью дрозофилы и, по-видимому,
консервативные, способствуют приживлению
трансплантата, быстрой дифференцировке
входящих в его состав клеток и тем самым
благоприятному исходу операции.
Так шаг за шагом расшифровываются
этапы генных влияний на процессы
клеточной дифференцировки и индивидуального
развития в целом.
Многие проблемы остаются
нерешенными, в частности, природа тех
биологически активных веществ, которые
играют роль в регуляции активности генов, хотя
в некоторых случаях она уже известна. Но
будем надеяться, что в ходе дальнейших
научных исследований уже в обозримом
будущем будут даны ответы если не на
все, то на многие интересующие нас
вопросы.
Литература
Альберте Б., Брей Д., Льюис Дж. и др.
Молекулярная биология клетки. Т. 3. Пер. с англ. М.:
Мир, 1994.
Аналитические аспекты дифференцирования.
М.: Наука, 1991.
Гилберт С. Биологическое развитие. Пер. с
англ. М.: Мир, 1994. Т. 2.
Зенгбуш Л. Молекулярная и клеточная
биология. В 3 т. Пер. с. англ. М.: Мир, 1982.
Корочкин Л. И. Взаимодействие генов в
развитии. М.: Наука, 1977.
Рэфф Р., Кофман Т. Эмбрионы, гены и
эволюция. Пер. с англ. М.: Мир, 1986.
И. Ф. Жимулев
ДЕЙСТВИЕ ГЕНОВ В РАННЕМ РАЗВИТИИ ДРОЗОФИЛЫ
В ходе развития организма формируются
многочисленные органы и ткани, совершенно
непохожие друг на друга. Они приспособлены
для выполнения определенных функций, и
каждая ткань поразительно отличается от
остальных. В исследовании дифференцировки
органов и тканей необходимо решить две
проблемы: каким образом ткани становятся
непохожими друг на друга и каким образом
дифференцированное состояние, характерное
для каждой клетки, наследуется в ряду
клеточных поколений.
В последние годы, благодаря
использованию в исследованиях недавно
разработанных методов молекулярной биологии и
клонирования ДНК, достигнуты огромные
успехи в понимании того, как развивается
организм и как происходит
дифференциация клеток.
Наиболее важные результаты
получены при изучении действия генов на самом
раннем этапе эмбрионального развития
дрозофилы. Трое генетиков, добившиеся
выдающихся успехов в этой области, Э. Лью-
Яйцо
б
exuperantia-
swallow
stauferr
Рис. 1. Распределение по длине яйца дрозофилы
матричной РНК, считанной с гена bicoid в
нормальной линии (черное пятно) (а), у мутантов exuperantia
(б), swallow (в) и staufen (г) (показаны штриховкой)
[Lawrence, 1992]
ис, К. Нюссляйн-Волхард и Э. Вишаус,
получили в 1995 г. Нобелевскую премию.
Еще в 50-е гг. сформировалось
представление о морфогенах как о веществах,
индуцирующих образование определенных
частей тела. Предполагали, что «эти
вещества диффундируют через ткань и их
распределение диктует тот или иной путь
развития клетки». Позднее теория морфогенов
получила значительное развитие. По
современным представлениям морфоген
выделяется из локального источника и во время
последующей диффузии в ткани
образуется градиент его концентрации. В каждой
группе клеток свой набор и концентрация
морфогенов, т. е. своя информация о
последующем развитии — то, что генетики
развития называют позиционной
информацией.
Лучше всего изучены градиенты
морфогенов, образующиеся в развивающемся
яйце дрозофилы.
Известно, что у дрозофил яйцо
созревает в особой камере — фолликуле. Эта
камера содержит ооцит — созревающее яйцо
и 15 огромных питающих клеток, функция
которых заключается в том, чтобы
синтезировать питательные вещества и
направлять их в ооцит. В этих клетках
функционируют так называемые «гены с материнским
эффектом», т.е. такие гены, которые
функционируют в питающих клетках ооцитов —
в организме матери еще до оплодотворения
яйца сперматозоидом и информация от
которых передается в ооцит.
Оказывается, что белки, кодируемые
генами, функционирующими в ходе
созревания яйца и транспортируемые туда из
питающих клеток, распределяются по оси
яйца, образуя градиент. На рис. 1
показано распределение в пределах яйца продукта
гена bicoid. Он занимает строго
определенный участок яйца. Чтобы он занял это
место, нужно, чтобы поработали и другие гены.
Если эти гены мутируют, продукт
распределяется неправильно.
Так, в нормальном яйце РНК гена bicoid
(Ъс) располагается в узко локальном
участке (рис. 1). Однако у ряда мутантов
распределение этой РНК в яйце сильно изменено:
в результате мутации гена exuperantia РНК
Ъс более или менее равномерно
распределена по всему яйцу с небольшим
градиентом от переднего полюса к заднему. У
мутантов swallow градиент этого морфогена
выражен сильнее, т. е. его распределение
ближе к нормальному: в передней части яйца
100
ГЕНЕТИКА
выявлено большое скопление РНК Ъс и
некоторое ее количество распределено в
остальной цитоплазме (рис. 1).
Совсем близкое к норме, но все еще
ненормальное, распределение продукта Ъс
обнаружено у мутантов staufen (рис. 1): у
них РНК Ъс вообще не переходит в заднюю
часть эмбриона.
Таким образом РНК считывается с гена
bicoid еще в питающих клетках ооцитов в
материнском организме и оттуда
поступает в яйцеклетки. Затем с помощью
продуктов других генов, в данном случае это гены
exuperantia, swallow и staufen, эта РНК
занимает определенное положение в
цитоплазме яйца, т. е. создается определенный
градиент в распределении этого морфоге-
на. В случае мутации любого из трех
перечисленных генов распределение РНК Ъс
изменяется, что приводит к серьезнейшим
нарушениям развития.
Известно, что в яйцо поступает РНК,
считанная с огромного числа генов.
Поскольку каждая из этих РНК еще и
распределяется по своим местам в яйце в результате
активности других генов, совершенно
очевидно, сколь велико число генов,
участвующих в формировании яйца.
В свою очередь, после занятия
правильного положения в яйце продукты таких
генов, как bicoid, вступают во
взаимодействие с другими генами, которые
активируются после оплодотворения и
образования зиготы (зиготические гены). Белки гена
bicoid связываются с контролирующими
районами зиготических генов и активируют их.
Понятно, что клетка, возникающая в
области локализации морфогена Ъс, будет
испытывать его влияние, и развитие
пойдет в одном направлении, а если клетка
расположена в задней части эмбриона, где
морфогена Ъс нет (рис. 1), она будет
развиваться в другом направлении. Таким
образом, набор определенных белков,
накопленных цитоплазмой к данной стадии
развития, способен активировать
определенный набор генов, благодаря чему либо
поддерживается данное дифференцированное
состояние, либо развитие продвигается
дальше.
Каким образом белковый продукт
одного гена может взаимодействовать с другим
геном? Надежно установлено, что
фактически любой ген состоит из трех элементов:
некодирующего и нетранскрибируемого
(регуляторного) участка, расположенного в
начале любого гена, а также кодирующих
фрагментов — экзонов, перемежаемых не-
кодирующими, но транскрибируемыми
участками — интронами. Оказалось, что регу-
hb
bed
GGAAGTCATAAAAACACATAATAATGATGTCGAAGGGATTAGG
gt
Кг
-1500 пн
Кг gt
-800
gt
Кг
gt Kr
Кг
Кг
bed
bed
bed
bed
hb
bed
hb
hb
bed
bed
GTTAATCCGTTTGCCATCAGCGAGATTATTAGTCAATTGCAGTTG
Kr
gt
Рис. 2. Часть регуляторного участка гена eve длиной 700 пар нуклеотидов (от 800 до 1500 пн) [Lawrence, 1992]
а — нить ДНК, выше и ниже которой соответствующими символами изображены участки посадки белков,
кодируемых генами Кг, gt, bed и hb\ б, в— последовательности нуклеотидов, связывающие данные белки
ДЕЙСТВИЕ ГЕНОВ В РАННЕМ РАЗВИТИИ ДРОЗОФИЛЫ
101
ляторные части генов содержат
специфические группы нуклеотидов (мотивы),
имеющие сродство к определенным
сочетаниям аминокислот (доменам) в молекулах
белков. Связывание различных белковых
факторов с соответствующими мотивами в
нити ДНК приводит к изменениям ее
пространственной организации и началу
транскрипции кодирующей части гена (если
белок является активатором) или блокированию
транскрипции (если белок является инак-
тиватором).
На рис. 2 изображен фрагмент (длиной
700 пар нуклеотидов) регуляторной части
гена eve (even-skipped), который
контролирует развитие правильной сегментации тела
дрозофилы. Видно, что мотивы
связывающих активирующие белки генов bed и hb,
часто перекрываются с нуклеотидными
последовательностями, на которые садятся
белки, подавляющие транскрипцию (гены
Кг и gt).
Эти данные свидетельствуют о том, что
расположение белков, синтезированных в
материнском организме, в определенной
части яйца (см. рис. 1) имеет
первостепенное значение для процесса активирования
генов в уже начавшем развитие эмбрионе.
Ясно, что ген eve будет функционировать
в той части эмбриона, в которой содержится
много материнских белков bed и hb, и не
будет функционировать в клетках,
содержащих избыток белков Кг и gt.
Одним из генов, осуществляющих
важнейшую функцию на ранних этапах
эмбрионального развития, является ВХ-С
(комплекс bithorax). Известно, что большинство
представителей животного мира (исключая
круглых червей) имеют сегментальное
строение (рис. 3), т. е. их тела состоят из серии
члеников или сегментов. У млекопитающих
сегментальное строение наблюдается на
самых ранних этапах эмбриогенеза и эти
сегменты носят название сомитов.
а б в
^У дрозофилы личинки имеют ярко
выраженные сегменты; у взрослых мух
сегментацию, особенно брюшка, легко
заметить даже невооруженным глазом (рис. 3).
Можно выделить всего 12 сегментов: один
головной, три грудных и восемь брюшных.
Каждый сегмент имеет уникальный набор
дифференцированных морфологических
структур. Например, мезоторакальный
сегмент несет пару крыльев и пару ног, мета-
торакальный — пару ног и пару гальтеров —
особых булавовидных образований,
помогающих удерживать равновесие в полете.
Характерный набор можно найти и на
сегментах личинки (рис. 3).
По мнению выдающегося
американского ученого Льюиса, мухи эволюционировали
из насекомых, имевших четыре крыла, а
насекомые, в свою очередь, произошли от
членистоногих, имевших множество ног. В
ходе эволюции мух у них должны были
сформироваться несколько групп генов: тех,
которые подавляют развитие ног на
брюшных сегментах многоножкоподобных
предков, и генов, подавляющих развитие
второй пары крыльев. Должна была также
появиться группа генов, формирующих
новые структуры: гальтеры и брюшные
сегменты.
Одним из генов, влияющих на эти
процессы, является ВХ-С. В ходе своих
экспериментов Льюис удалил ген ВХ-С с
помощью небольшой нехватки хромосомного
материала в том районе, где этот ген
локализован. Организм без этого гена
развивается до конца периода эмбриогенеза и
затем гибнет.
Результаты изучения погибшего
эмбриона оказались поразительными. Этот
организм имел очень характерную морфологию:
у него были только вторые торакальные
сегменты (или мезоторакальные, см. рис. 3).
Если бы этот организм вырос во взрослую
муху, то она бы имела 10 пар крыльев и
Рис. 3. Схема сегментального
строения личинки (а) и взрослой мухи
дрозофилы (в) [Lewis, 1978]
На обобщенной схеме (б) видно, что
как личинка, так и взрослая муха
имеют общий принцип сегментации. Они
имеют головной сегмент (HEAD), три
грудных сегмента (PRO, MS, MT), а
также 8 брюшных (от АВ1 до АВ8).
Каждый сегмент как у личинки, так и у
имаго имеет свой набор органов,
отличающий данный сегмент от остальных
102
ГЕНЕТИКА
а -120-100-80 -60 -60 -60 0 +20+40+60+80+100+120+140+160+180+200 тпн
Ubx+ abd-A+ Abd-B+
Рис. 4. Схема организации комплекса генов ВХ-С: Ubx, abd-A и Abd-B [Lawrence, 1992]
а— расстояние на карте ДНК от стартовой точки начала клонирования гена (0), т.е. масштаб длины ДНК в
тысячах пар нуклеотидов; б— затененные участки— регуляторные последовательности генов; Н— районы
локализации гомеодоменов; в— экзон-интронная структура. Кодирующие участки генов в виде черных
прямоугольников расположены в соответствующих участках генетической карты. Между ними расположены некоди-
рующие интроны. Экзоны соединены между собой зигзагообразными линиями. Только та РНК, которая считана
с экзонов, затем транслируется в молекулу белка
10 пар ног. Льюис сделал вывод о том, что
функция гена ВХ-С заключается в
инактивации генов, формирующих ноги и крылья
во всех последующих сегментах после
второго торакального и в формировании всех
структур на брюшных сегментах.
В дальнейших экспериментах оказалось,
что ВХ-С содержит три различных гена:
Ubx, abd-A и Abd-B (рис. 4). Каждый из них
контролирует формирование определенной
группы сегментов. Мутации этих генов
заставляют последующие сегменты развиваться
в предыдущие, и генетический порядок
мутантов примерно соответствует
пространственному порядку органов по оси тела
(рис. 5). Так, если все три гена удалены
{Ubx~, abd-A', Abd-B' на рис. 5), нормально
развиваются только первый торакальный (Т1)
и девятый брюшной (А9) сегменты,
контролируемые другими генами, остальные
сегменты (ТЗ и все последующие брюшные)
развиваются как более ранние Т2.
Если ген Ubx сохраняется, но
повреждаются abd-A и Abd-B, нормально
развиваются грудные сегменты, а брюшные
представлены только самым первым — А1 (рис. 5).
При повреждении Abd-B гена
нормально развиваются все грудные сегменты,
затем брюшные Al, A2 и A3, все
остальные представлены сегментом А4 (см. рис. 5).
В молекулярно-генетических
экспериментах выяснили, что все три гена комплекса
ВХ-С имеют гомологичные друг другу
участки, т. е. последовательности нуклеотидов в
них фактически одинаковы (сходство более
чем на 90%). Последовательность длиной 180
пар нуклеотидов, которая имела наибольшее
сходство, назвали гомеодоменом.
К настоящему времени найдены сотни
генов, обладающих гомеодоменом: у
человека, мышей, птиц, лягушек, червей, жуков.
Фактически все представители животного
мира, проходящие хотя бы на некоторых
этапах развития стадию
сегментированного зародыша, имеют гены, обладающие
Дикий тип
Ubx+
a abd-A+
Abd-B+
А1 А2 A3 А4 А5
Ubx"
б abd-A"
Abd-B-
A9
Ubx+
в abd-A"
Abd-B"
Ubx+
г abd-A+
Abd-B"
A1 A2 A3 A4 A4
Рис. 5. Схема нарушений дифференцировки в
результате мутаций генов Ubx, abd-A и Abd-B [Lawrence,
1992]
а— все три гена работают нормально, у эмбриона
нормально развиты грудные сегменты (Т1—ТЗ) и
брюшные (А1—А8); б—г— нарушения сегментации в
результате мутирования одного, двух или всех трех
генов
ДЕЙСТВИЕ ГЕНОВ В РАННЕМ РАЗВИТИИ ДРОЗОФИЛЫ
103
гомеодоменом. А у дрозофилы найдено около
100 генов, содержащих в своем составе го-
меодомен.
180 пар нуклеотидов гомеодомена
кодируют полипептид длиной 60 аминокислот. Эти
60 аминокислот скручены в 4 ос-спирали,
каждая из которых отделена от другой
наклоном оси вращения. Третья из этих
спиралей помещается в большую бороздку ДНК,
опознает последовательность нуклеотидов и
связывается с ними.
Такими свойствами структуры обладают
ДНК-связывающие белки-факторы
транскрипции. Процессы взаимодействия
нуклеотидов в ДНК и аминокислот
белка-фактора транскрипции организованы так, что
определенная последовательность
нуклеотидов связывается только с определенной
последовательностью аминокислот. Поэтому нук-
леотиды в гомеодомене и расположены в
такой консервативной последовательности у
представителей разных типов, классов, родов
и видов животных. Например, из 60
аминокислот в гомеодомене мухи дрозофилы и
лягушки ксенопуса 55 оказались одинаковыми.
Рассмотренные выше данные
несомненно свидетельствуют о том, что развитие —
это процесс последовательного включения
все более и более усложняющихся генных
систем. При этом продукты одних генов
находят специальные «посадочные
площадки» в регуляторных районах других генов,
садятся на них и включают эти гены в
активное функционирование. И так в ходе
всего развития — сплошное чередование
включений и выключений генов.
Литература
Гердон Дж. В кн.: Молекулы и клетки. Вып. 5.
Пер. с англ. М.: Мир, 1970, с. 19.
Хадорн Э. В кн.: Молекулы и клетки. Вып. 5.
Пер. с англ. М.: Мир, 1970, с. 54.
Корочкин Л. И. Взаимодействие генов в
развитии. М.: Наука, 1977.
Lawrence P.A. The making of a fly. The genetics
of animal design. Oxford: Blackwell Scientific
Publications, 1992.
Lawrence P. A, MotaraG. Cell, 1994, v. 78,
p. 181.
Lawrence P. A., StruhlG. Cell, 1996, v. 85,
p. 951.
Lewis E. B. Nature, 1978, v. 276, p. 565.
Martin С. Н. et al. Proc. Natl. Acad. Sci. U.S.A.,
1995, v. 92, p. 8398.
А. Ф. Смирнов, И. Ф. Жимулев
РЕГУЛЯЦИЯ ПОЛА
Введение
Пол — совокупность морфологических и
физиологических особенностей организма,
обеспечивающих половое размножение,
сущность которого сводится к оплодотворению,
т. е. слиянию мужских и женских половых
клеток (гамет) в зиготу, из которой
развивается новый организм. В изучении
полового размножения есть еще множество
нерешенных вопросов. В частности, до сих пор
обсуждаются преимущества и недостатки
полового размножения по сравнению с
бесполым, эволюционная связь этих способов
размножения. Предполагается, что
асексуальные системы менее приспособлены к
изменяющимся условиях внешней среды,
неспособны эффективно элиминировать
вредные мутации и относительно недолговечны.
Различают дифференциацию пола (фено-
типический пол), т. е. появление внешних
гениталиев, вторичных половых признаков
и первичное определение пола. Под
первичным определением пола понимают
появление гонады (репродуктивного органа
соматической природы) самки или самца —
яичника или тестиса. Считается, что
принципиальная схема этого процесса
консервативна. Существует конкретный
контролирующий сигнал, включающий некий ключевой
ген. Этот ген, в свою очередь, активирует
некое количество детерминант гонадогене-
за и, далее, факторов дифференцировки
половых признаков. Все компоненты этой
системы могут различаться у разных
животных. В табл. 1 представлены
принципиальные схемы определения пола у некоторых
представителей животного мира и
обозначены соответствующие гены.
Выделяют несколько типов определения
пола в зависимости от числа и состава
половых хромосом, например, у самца может быть
одна Х- и одна Y-хромосома, а у самки —
две Х-хромосомы. К такому типу относятся
человек, плодовая мушка дрозофила,
водяной клоп Ligaeus, по названию которого и
назван данный тип определения пола.
Еще один тип, названный по названию
другого водяного клопа Protenor и встречающийся
у некоторых бабочек и червей, связан с
наличием у самцов одной Х-хромосомы (набор
половых хромосом — ХО), а у самок — двух.
Особый тип хромосомного определения
пола найден у птиц, некоторых бабочек,
рыб, земноводных и цветковых растений. У
них гетерогаметным (т. е. с разными
половыми хромосомами) полом является
женский, и самки имеют набор половых хромо-
Таблица 1
Принципиальная схема определения пола у некоторых
представителей животного мира
Контролирующий
сигнал
i
Ключевой ген
Гены,
контролирующие гонадоге-
нез(полопреде-
ляющие)
i
Гены,
контролирующие половую
дифференцировку
Saccharomyces
cerevisiae
Транскрипция
НО гена
М4Г(а/а)
+/-
Caenorhabditis
elegans
Соотношение X хр
her(+l-)
xoll
sdcl, sdc2
herl
tra2, tra3
feml,fem2,
fem3, tral
+/-
Организм
Drosophila
melanogaster
омосом и аутосом
?*/(+/-)
sis-a, sis-b
da, Hz
fl(2)d
Sxl
tra, tra2
dsx, ix
+/-
Alligator
missippiensis
Внешняя
температура
TDF
эффекторные
молекулы,
гормоны
+/-
Mammalia
Y-хромосома
TDF-SRY
гормоны
+/-
Примечание. + и — означают включение или выключение соответствующего гена
РЕГУЛЯЦИЯ ПОЛА
105
сом ZW или Z0, в то время как самцы —
ZZ. Известны и некоторые другие типы
определения пола.
Совершенно очевидно, что в
разобранных выше случаях колоссальные и легко
фиксируемые даже на первый взгляд
различия в организации и физиологии особей
разных полов развиваются на фоне
полностью идентичных у самцов и самок
наборов генов, расположенных в большей
части хромосом, не связанных с половыми
различиями кариотипов — аутосомах.
Напротив, все половые различия
определяются сравнительно небольшим числом
генов, заключенных в половых хромосомах.
Основные принципы
детерминации пола
у млекопитающих
Классическими эмбриогенетическими
исследованиями установлены два правила
определения пола у млекопитающих.
Первое из них сформулировано в 60-х гг. А. Жо-
стом на основе экспериментов по удалению
зачатка будущих гонад (гонадного валика)
у ранних эмбрионов кролика: удаление
валиков до формирования гонады
приводило к развитию всех эмбрионов как самок.
Было высказано предположение о секре-
тировании гонадами самцов (тестисами)
эффектора (тестостерона), ответственного
за маскулинизацию плодов, и предсказано
наличие второго эффектора — антимюлле-
ровского гормона (MIS), непосредственно
контролирующего такие анатомические
преобразования. Результаты наблюдений
были сформулированы в виде правила:
«Специализация развивающихся гонад в
тестис или яичник определяет последующую
половую дифференциацию эмбриона».
Примерно до 1959 г. предполагалось, что
число Х-хромосом, которое, как известно,
равно двум у самок и одной у самцов,
является важнейшим фактором контроля пола
у млекопитающих. Однако обнаружение
индивидуумов с единственной Х-хромосомой,
развивающихся как самки, а также с
одной Y-хромосомой и множественными Х-хро-
мосомами, развивающихся как самцы,
заставило отказаться от таких представлений.
Сформулировано второе правило
определения пола у млекопитающих: «Y-хромосома
несет генетическую информацию,
требуемую для детерминации пола у самцов».
Комбинация приведенных выше двух правил
иногда называется принципом Жоста:
«Хромосомный пол, связанный с присутствием
или отсутствием Y-хромосомы,
определяет дифференциацию эмбриональной гонады,
которая, в свою очередь, контролирует фе-
нотипический пол организма». Подобный
механизм определения пола называют
генетическим и противопоставляют
основанному на контролирующей роли факторов
внешней среды или соотношению половых
хромосом и аутосом (см. табл. 1).
Биологической основой генетического
механизма определения пола является
бисексуальность эмбриональных гонад
млекопитающих. В таких прогонадах одновременно
присутствуют мюллеровы и вольфовы протоки —
зачатки половых путей соответственно самок
и самцов. Первичная детерминация пола
начинается с появления в прогонадах
специализированных клеточных линий — клеток
Сертоли. В последних синтезируется
предсказанный Жостом MIS, ответственный за
прямое или опосредованное ингибирование
развития мюллерова протока — зачатка будущих
фаллопиевых труб и матки (рис. 1).
Прогонада
Пронефрос
Мезонефрос
Мюллеров
проток
Метанефрос
Прогонада
Мезонефрос
Вольфов/
проток
Яичники
Урогенитальный синус
Тестисы
Эпидидимис
Яйцевод
Матка
Рога матки
Семенной пузырек
Рис. 1. Развитие гонад у млекопитающих
а— плод с недифференцированной прогонадой; б—
схема мочеполовой системы самцов и самок у
такого плода. Гонадный бугорок развивается вместе с
мезонефросом (почка). Прогонада морфологически
неразличима у XX- и XY-эмбрионов. Первые половые
различия между развивающимися гонадами
наблюдаются у человека через 6 недель после зачатия, у
мышей через 12,5 дней. Мюллеров проток прогона-
ды является предшественником матки, яйцеводов,
верхней части влагалища, вольфов проток —эпидидими-
са, семяпроводящих протоков, семенных пузырьков
106
ГЕНЕТИКА
Используя химерные клеточные линии
мышей, удалось показать, что для
нормального функционирования клеток Сертоли
необходим XY-кариотип. Установлено также,
что именно в этих клетках Y-хромосома де-
конденсирована и может быть транскрипци-
онно активна. Схематически роль Y-хромосомы
и сцепленного с ней гипотетического тести-
сопределяющего фактора (TDF) в развитии
недифференцированной бисексуальной гонады
в направлении тестисов и ингибировании
появления яичников представлена на рис. 2.
а
Оогонии
У-хромосома,
TDF
Клетки фолликулов
Прогонада Клетки Тэки Тестостерон
Эстроген
Недиффе-
б ренциро-
Формированная вание Дифферен-
гонада «гонадного циация
Догонадная (прогонада) пола» пола
Рис. 2. Схема определения пола у млекопитающих
а— определение тестисов из четырех клеточных
типов прогонады. Пунктирные стрелки обозначают пути
в направлении развития яичников; б— временная
диаграмма последовательных стадий определения
пола. 1 — догонадная. Отсутствие у эмбриона
половых органов; 2— прогонадная. Появление гонадного
валика и бисексуальной прогонады; 3—
формирование «гонадного пола» (первичное определение
пола). Синтез TDF. Появление клеток Сертоли,
окружающих половые клетки (семявыносящие канальцы);
4— дифференциация пола. Синтез MIS клетками
Сертоли. Появление клеток Лейдига, продуцирующих
андрогены. Маскулинизация внешних гениталиев
Тестисопределяющие гены
Возможность построения и
использования генетических карт животных, в
частности Y-хромосомы человека и мыши,
обеспечила установление гипотетического TDF.
На рис. 3 проиллюстрирована процедура
поиска этого гена на конкретном
интервале Y-хромосомы Homo sapiens. Его
идентификация состоялась также благодаря
анализу довольно редко встречающихся особей
с хромосомной инверсией пола — самцов XX
и самок XY. Подобного рода нарушения
описаны для 17 видов млекопитающих, и
согласно основному принципу определения
пола в этой систематической группе,
должны нести соответственно транслокацию или
делецию Y-хромосомного фрагмента с TDF.
У мышей описаны два типа самцов XX-
XXSxr и XXSxr'. Их появление
обусловлено дупликацией и переносом несколько
различающихся по размеру участков
короткого плеча Y-хромосомы на общий для обеих
гоносом (половых хромосом) так
называемый псевдоаутосомный (PAR) терминальный
район Х-хромосомы. Такие самцы
обладают тестисами, но их репродуктивная
система дефектна из-за аномалий
сперматогенеза. Важная особенность самцов XXSxr' —
отсутствие у них антигена гистосовмести-
мости (HY-антигена), первоначально
рассматривавшегося в качестве необходимого
атрибута гетерогаметного, т. е.
производящего гаметы с Х- или Y-хромосомой, пола
и наличие гипотетического TDF. Мужчины
с кариотипом XX, обладающие нормальными
тестисами, но стерильные, встречаются у
человека с частотой 1 : 20000.
В 1987 г. Д. Пэйдж и его коллеги,
исследуя мужчину XX, унаследовавшего 280-т.п.н.
фрагмент Y-хромосомспецифической ДНК,
и женщину XY с делецией (нехваткой),
захватывающей эту область в результате
обмена участками между хромосомами X и
22, казалось, обнаружили неуловимый TDF.
Им оказался присутствующий в Y-хромосоме
всех плацентарных млекопитающих
(Eutheria) и расположенный в области
размером 140 т.п.н. в 100 т.п.н. от границы псев-
доаутосомальной области ген ZFY. Гомолог
ZFY— ZFX обнаружен в Х-хромосоме,
причем он избегает характерной для генов,
в ней локализованных, инактивации. Оба эти
фактора кодируют белок, образующий
структуру так называемых цинковых пальцев
и обладающий ДНК-связывающей
активностью, который можно рассматривать как
фактор транскрипции.
Однако довольно быстро накопились
данные, не позволяющие отождествлять ZFY
и TDF. Так, у млекопитающих для
детерминации гонад не требуется присутствия
половых клеток, но это оказалось
необходимым для проявления активности одного
из двух мышиных гомологов ZFY — zfyl-тена.
Экспрессия этого гена выявлена в гонадном
бугорке TDF-мутантов Ytdyml, впоследствии
развивающихся как самки. Весомым
контраргументом против рассмотрения ZFY в
качестве TDF явилось его аутосомное
местоположение у сумчатых (подкласс Metatheria),
для которых, как и для Eutheria, характерно
участие Y-хромосомы в определении пола.
В последнее время появились данные,
отрицающие прямую зависимость между
наличием половых клеток и экспрессией гена
РЕГУЛЯЦИЯ ПОЛА
107
Yp
Псевдо-
аутосо-
мальная
область
1
1А1
(140т.п.н.)
1А2
(140 т.п.н.)
60 т.п.н.
SRY
35 т.п.н.
ZFY
Yq
4а
1959
1966
1986
1987
1989
1990
Рис. 3. Поиск тестисопределяющего фактора (TDF) на Y-хромосоме человека
1959—вся Y-хромосома рассматривается как полопределяющая (составляет 2-3% гаплоидного генома);
1966— гипотетический TDF связывают с коротким плечом;
1986— поиск сужается до 1 короткого плеча;
1987—дальнейшее ограничение поиска участком 1А1 и приграничных участков. Выявлен ген ZFY\
1989— обнаружены мужчины, не обладающие ZFY;
1990— идентифицирован вероятный TDF— ген SRY. Мутации, связанные с инверсией пола, захватывают лишь
небольшой консервативный мотив этого гена— HMG-бокс (80 аминокислотных остатков, 320 п.н.)
zfyl. Однако даже если это так,
ZFY^-подобные гены выполняют важную роль только
в последующей половой дифференциации,
а не при первичной детерминации пола.
Дальнейший детальный анализ
специфических последовательностей Y-хромосом у
особей с инверсией пола ограничил поиск
районом размером 35 т.п.н. и привел к
обнаружению гена, рассматриваемого как
истинный эквивалент классического TDF (рис. 3).
Такой ген получил название SRY (Sex
determining Region Y gene). Приведем
некоторые его характеристики. Предполагается,
что SRY расположен в полопределяющей
области и содержит консервативный домен
(HMG-бокс), кодирующий белок размером 80
аминокислотных остатков. Его активность
проявляется накануне периода
дифференциации прогонады в тестис— на 10,5-12-й
день эмбрионального развития у мыши и по
крайней мере на этой стадии не зависит от
присутствия половых клеток.
Специфические точковые мутации или делеции в HMG-
боксе этого гена у женщин XY приводят к
инверсии пола. Перенос фрагмента ДНК
длиной 14 т.п.н., содержащего этот ген с
фланкирующими участками, в оплодотворенную
яйцеклетку гомогаметной особи с помощью
микроинъекции (процедуры трансгеноза —
переноса генов) привел к появлению
«самца» с ХХ-кариотипом. Правда, у этого
животного отмечен дефектный сперматогенез.
Белок, кодируемый HMG-боксом SRY-
гена, специфически связывается с ДНК,
приводя к изгибанию ее молекулы. Такая
деформация структуры ДНК,
индуцируемая SRY-белком или родственными ему
молекулами (известно более 100 белков с
HMG-доменом), может механически
передаваться на расстояние и играть важную
роль в регуляции транскрипции,
репликации и рекомбинации. Имеется
экспериментальный материал, свидетельствующий в
пользу, скорее всего, опосредованного
положительного контроля со стороны
белкового продукта SRY-активности MLS-гена и
отрицательного в результате проявления
гена ароматазы, контролирующего
превращение мужских стероидных гормонов в
женские. На наличие такой специфической
активности гена SKY указывает и
внутриядерное расположение антител к его
продукту.
Только HMG-бокс является
консервативной частью гена SRY. Способность связываться
с ДНК, а также специфическая
локализация белкового продукта в ядре также
определяются этим доменом. За пределами этой
последовательности наблюдается
существенное различие между гомологами SRY даже
у близких видов. Человеческий SRY-тек имеет
небольшой размер, не содержит интронов
и кодирует белок размером 204
аминокислотных остатка. Его мышиный гомолог
содержит уже 395 аминокислот и в нем 58
остатков на N-конце, терминальном к HMG-
боксу, а не два, как у человека. Отмечена
значительная межвидовая дивергенция
аминокислотных остатков этого белка и
практическое отсутствие внутривидовой
изменчивости.
SRY— один из членов большого
семейства генов (их более 20), получившего
название SOX (SRY type HMG box). Для это-
108
ГЕНЕТИКА
го семейства характерна тканеспецифичес-
кая экспрессия в раннем эмбриогенезе. Soxl,
Sox2 и Х-хромосомный Sox3 активны при
развитии нервной системы. Sox4 работает как
активатор транскрипции в Т-лимфоцитах,
a Sox5 проявляет специфическую активность
во время сперматогенеза. Недавно изолирован
человеческий гомолог Sox9 гена мыши,
требующийся для нормального развития
скелета и ответственный за CD-синдром
(campomelic displasia).
TDF и другие гены, участвующие
в детерминации пола
В последнее время стал
конкретизироваться материал об участии других генов
по крайней мере в модификации эффекта
TDF. Замечено, что эмбрионы и гонады
будущих самцов растут быстрее эмбрионов
и гонад будущих самок и такие отличия
наблюдаются у человека, мышей и крыс еще
до появления первых клеток Сертоли и даже
гонадного бугорка. Высказано
предположение о том, что клетки прогонад должны
подвергнуться определенному числу
делений для дифференциации клеток Сертоли.
Контроль за опережающим развитием
мужских прогонад и эмбрионов может
осуществлять особый ген, отличный от TDF,
например гипотетический Gdy (growth and
development), расположенный в Sxr-учас-
тке Y-хромосомы мышей. У человека
описан ген WT1, вызывающий ряд
наследственных заболеваний (опухоль Вильмса, синдром
Дэнис-Драма), экспрессирующийся на 9-й
день эмбрионального развития и
контролирующий развитие недифференцированной,
бисексуальной гонады. Ранее упомянутый
ген Sox9, локализованный на хромосоме 11
мыши и имеющий гомолог на хромосоме 17
человека, помимо участия в возникновении
доминантного CD-синдрома — смертельного
нарушения развития костей, играет роль в
аутосомной инверсии пола (аутосомный локус
SRA1). Принимая во внимание раннюю
экспрессию этого гена (до 13-го дня
эмбрионального развития) в примитивной
зародышевой соединительной ткани (мезенхиме),
его считают модификатором влияния SKY
на экспрессию MIS, своеобразным
ограничителем рамок активности этого гена в
раннем эмбриогенезе.
Исследование более 100 XX
индивидуумов, являющихся фенотипическими
мужчинами, но не обнаруживающих
присутствия гена SKY, привело к предположению
о существовании рецессивного гена Z.
Продукт этого гена является отрицательным
регулятором развития тестисов: в норме он
функционирует у самок, а у самцов его
активность блокируется геном SKY.
Предполагается и существование аллеля Z\
нечувствительного к такому подавлению
(супрессии).
Недавно был выделен ген DAX1,
локализованный на участке Хр21, кодирующий
ядерный рецептор — молекулярный
комплекс на поверхности клетки, способный
распознавать химические группировки и
молекулы. При дупликации он, возможно,
вызывает зависимую от дозы реверсию
мужского пола к женскому (Ь^Я-мутация).
Предполагается, что DAX1 чувствителен
к гормонам и в увеличенной дозе
способен преодолевать сигнал SKY и сдвигать
развитие гонад в направлении яичника. Этот
ген рассматривается как реликт более
примитивной Х-хромосомной системы
детерминации пола. Кодируемый им рецептор
родствен стероидогенному фактору SF1,
экспрессия которого отмечена в
недифференцированной прогонаде у мышей. Мутации
по SFl-тену могут приводить к отсутствию
гонад у обоих полов. Предполагается и его
участие в контроле активности гена анти-
мюллеровского гормона (MIS) в клетках
Сертоли.
Наличием особого аутосомного гена
Tdalb, предположительно сцепленного с
хромосомами 2 или 4 и
взаимодействующего с TDF, объясняют появление овотестиса
или яичника у мышей с кариотипом XY.
Феномен наблюдается в том случае, когда
Y-хромосома Mus musculus domesticus (или
poschiavinus) — ydom попадает в геном с ауто-
сомами М. га. musculus. Обычно в таких
скрещиваниях используются самки линии C57BL/
CJ (B6). Степень выраженности этого
феномена зависит от типа УБОМ-хромосомы.
Интересно, что в присутствии трансгена,
содержащего аллель гена SKY от М. га. musculus, у
таких гибридов развиваются исключительно
тестисы. Для каждого из исследуемых
подвидов мышей обнаружены различия,
касающиеся размера продукта гена SKY и
тандема тринуклеотидов в его участке, кодирующем
богатый гистидиновыми и глютаминовыми
остатками С-концевой домен. Удалось
продемонстрировать, что ген SKY экспрессируется
нормально у особей с генотипом B6YD0M, хотя и
инактивируется несколько позднее. Экспрессия
же других генов, связанных с появлением
тестисов (MIS, Р450-ароматазы, 17(ос)-гид-
ролазы), задержана в овотестисах или
отсутствует в яичниках. Авторы считают, что
наблюдаемые эффекты — результат каких-либо
нарушений экспрессии генов, участвующих
в определении пола на более поздних
этапах, чем SKY, например гена Tdalb. У
мышей выявлены и другая аутосомная мутация —
Tas, расположенная в хромосоме 17, а
также ряд факторов со сходным эффектом,
приводящим к появлению XY-самок и
гермафродитов с овотестисами.
РЕГУЛЯЦИЯ ПОЛА
109
Таблица 2
Гены, предположительно участвующие в детерминации пола
у млекопитающих, помимо SRY и ZFY
Генетический фактор
вру
WT1
Sox9
Z
DAX1 (DSS)
Tdalb
Tas
MIST
Tfm(hAR)
Гены сперматогенеза
и спермиогенеза
SF1
Od
Участие в контроле детерминации пола
Контролирует скорость роста эмбрионов и прогонад
Влияет на развитие прогонады примерно на 9-й день эмбрионального развития. Связан с
наследственными заболеваниями — опухолью Вильмса, синдромом Дэнис-Драма
Содержит HMG-бокс. Ответствен за CD-синдром и аутосомную инверсию пола. Экспрессиру-
ется до 13 дня эмбрионального развития. Возможный модификатор влияния SRYua MIS ген
Отрицательный регулятор развития тестисов. В норме активен у самок, у самцов
заблокирован работой SRY. Существует мутация Z1, нечувствительная к эффекту воздействия TDF-SRY
Локализован в районе Хр21. В двойной дозе способен преодолевать эффект SR Y и сдвигать
развитие гонад в направлении яичника. Кодирует ядерный рецептор. Ответствен за дозозави-
симую реверсию пола
Предположительно связан с хромосомами 2 и 4 мыши. Приводит к появлению овотестисов у
B6Ydom гибридов
Аутосомная мутация на хромосоме 17 мышей, приводящая к появлению XY самок
Ген антимюллеровского гормона, секретируется клетками Сертоли в ответ на активность SRY. Инги-
бирует развитие мюллерова протока. Локализован на хромосоме 19 человека—р13.2-р13.3
Ген рецептора андрогенов. У человека локализован на Х-хромосоме (Xpl 1-12). Вызывает тес-
тикулярную феминизацию
Известен ряд факторов — Spy, SMSY, UBE1Y, TSPY, YRRM, проявляющих такую активность
Ген рецептора стероидных гидролаз. У человека предположительно локализован в участке
9q33. Необходим для развития недифференцированной прогонады и регуляции активности
MIS-гена, в клетках Сертоли
Гипотетический Х-хромосомный или аутосомный ген, ответственный за дифференциацию
прогонады в яичник и отрицательно регулируемый геном SRY
Клонирован и локализован в участке Xqll-
12 ген рецептора андрогенов человека,
ответственный за тестикулярную феминизацию
(мутация Т/га), известную для ряда
объектов. Изучение наследования XY-синдрома
реверсии пола у лошадей показало, что в
такой передаче участвуют по крайней мере
два гена: основной полопределяющий и ген-
модификатор или гены-модификаторы.
Надежные доказательства сцепления этих
генов не получены. У самцов американских
коккер-спаниелей с ХХ-кариотипом также
предполагается наличие аутосомного гена —
детерминатора инверсии пола. Описана
аутосомная мутация polled, приводящая к
появлению тестикулярных тканей у коз с
ХХ-кариотипом. Недавно сразу в двух
лабораториях путем молекулярной гибридизации
с пробами SRY- и ZFY-генов удалось
исключить возможность Y-хромосомных
транслокаций или химеризма для этого случая.
Сходная ситуация описана и применительно к
довольно частому (0,1-0,6%) появлению ин-
терсексов у свиней.
Следовательно, активность TDF-SRY
является недостаточным условием
появления семенников из потенциально
бисексуальных прогонад у млекопитающих. Для
появления клеток Сертоли, семявыносящих
канальцев, клеток Лейдига и т. д.
требуется его взаимодействие со многими аутосом-
ными и Х-хромосомными факторами, детали
которого еще только предстоит понять.
Некоторое представление о возможных
дополнительных генетических факторах
контроля первичной детерминации пола и
их взаимодействии дает табл. 2.
Y-хромосома и пол
Детерминация пола — сложный
многокомпонентный процесс, для понимания
которого нужно решить множество вопросов.
Один из них — вопрос о
биолого-генетической значимости Y-хромосомы. ДЛя этой го-
носомы характерны специфические черты,
резко отличающие ее от других хромосом
набора: 1) обедненность генами; 2) обога-
110
ГЕНЕТИКА
щенность повторяющимися блоками нукле-
отидов, и, в частности, многократно тан-
демно повторяющими (сателлитной ДНК);
3) большая доля гетерохроматиновых
участков; 4) наличие области гомологии с Х-хро-
мосомой — псевдоаутосомной области (PAR).
Y-хромосома, как правило, невелика —
составляет 2-3% гаплоидного генома. Тем
не менее кодирующей способности ее ДНК
у Homo sapiens достаточно по крайней
мере для нескольких тысяч генов.
Однако в человеческой Y-хромосоме
выявляется всего около 40 обогащенных ГЦ-па-
рами, так называемых ЦрГ-островков,
обычно фланкирующих большинство
генов. Реальный же список генетических
функций, связанных с этой хромосомой,
вдвое меньше. Ее фенотипическое
влияние у мышей ограничено весом тестисов,
уровнем тестостерона, серологического
HY-антигена, чувствительностью органов
к андрогенам и сексуальным поведением.
Обращает на себя внимание и тот факт,
что большая часть генов этой хромосомы
имеет Х-хромосомные аналоги. Это
касается и генов, непосредственно
вовлеченных в определение пола: ZFX/ZFY,
Sox3/SRY, UBE1X/UBE1Y, SMCX/SMCY.
ZNF ZNF
UBE1 UBEX1
Sox3 Sox3
про X, Y X
ZFY
UBE1Y
SRY
Y
Рис. 4. Схема эволюции генов Y-хромосомы в
связи с определением и дифференциацией пола
Гены Sox3 и UBE1 принадлежали к консервативной
части прогоносомы, ZNF— к более новой ее части.
На постепенно деградирующей Y-хромосоме аллель
Sox3 -> SRY приобретает тестисопределяющие
функции, а аллели UBE1 -» UBE1Y и ZA/F-» ZFY— роль в
сперматогенезе. Другие Х-хромосомные гены на
Y-хромосоме исчезли в результате мутаций, деле-
ций и т. д. Роль в сперматогенезе не панацея от
потери— ген UBE1Y утерян у приматов, возможно, за
счет выполнения его функций другими генами (SMCY,
YRRM и т.д.)
Как уже было сказано, в этой
хромосоме велика доля разнообразных повторов,
включая и многократные. Большинство
Y-хромосомных последовательностей
гомологичны ДНК Х-хромосомы или аутосом, и
лишь часть из них строго уникальна. Нередко
обнаруживается количественное
преобладание в этой хромосоме некоторых семейств
повторов. Наличие таких относительно
специфических для Y-хромосомы блоков нук-
леотидов рассматривают иногда как
специфический инструмент ее конденсации в
клеточном цикле или даже определения пола.
Наличие псевдоаутосомных областей,
обеспечивающих мейотическое спаривание
и рекомбинацию, обычно рассматривается
как необходимое условие фертильности.
Интересно, что размер участка мейотичес-
кого спаривания существенно длиннее PAR.
У человека имеются два псевдоаутосомных
района на вершине короткого (2,6*106 п.н.) и
длинного (0,4-Ю6 п.н.) плеч Х-хромосомы.
Однако только для первого из них
установлены постоянный обмен в мейозе, наличие
хиазм, влияние на фертильность.
Сравнительный анализ указывает на
существование различия в генетическом содержании
PAR у изученных видов (мышей, человека,
приматов).
Высказано предположение о
происхождении половых хромосом млекопитающих от
предковой аутосомы в результате
независимых циклов: добавление — рекомбинация —
деградация. PAR, по такой терминологии,
представляет собой лишь как бы реликт
такого последнего добавления. Далее
происходят деградация и потеря соответствующих
Y-хромосомных частей и инактивация
Х-хромосомы. Все гены, представленные в
Y-хромосоме, или имеют реальную
селективную ценность (SKY, ZFY, UBE1Y, YRRM,
TSPY1, RSP4Y), или находятся на пути
исчезновения (STS, KAL, AMEL, PBD,
псевдогены). Каждый Y-хромосомный ген, быстро
дивергирующий, амплифицирующийся (как
бы размножающийся) или склонный к
исчезновению, имеет своего гомолога в Х-хромо-
соме, более консервативного и активного у
обоих полов. Так, Sox3, предполагаемый
Х-хромосомный гомолог SRY, кодирующий
почти идентичные продукты у человека,
мыши и сумчатых, экспрессируется в нервной
системе обоих полов. SRY быстро диверги-
рует и активен только в гонадном бугорке.
Этот Y-хромосомный ген подвергается
амплификации у многих мышей и крыс. Схема
эволюции генов Y-хромосомы представлена
на рис. 4.
Таким образом Y-хромосома,
единственная в геноме млекопитающих, не работает
непосредственно на реализацию фенотипа.
Ее генетическая значимость связана с
преемственностью между поколениями, в час-
РЕГУЛЯЦИЯ ПОЛА
111
тности с контролем гаметогенеза,
первичной детерминацией пола (рис. 5). Жесткий
отбор действует только на немногие ее
гены, остальная ДНК селективно нейтральна.
Можно предположить связь между
спецификой детерминации и дифференциации
пола и репродуктивной межвидовой
изоляцией. Возможно, быстрая дивергенция этой
системы лежит в основе сформулированного
еще в 1922 г. английским ученым Д. Холдей-
ном постулата о стерильности или
отсутствии гетерогаметного пола при отдаленной
гибридизации. Существует, по крайней
мере, один их примеров вклада системы
определения пола в феномен
репродуктивной изоляции — феминизация самцов у
гибридов М. musculus и М. domesticus.
Разнообразие систем
определения пола
Первоначально Y-хромосома, очевидно,
контролировала «гаметный пол» — гамето-
генез — и не была связана с первичным
определением пола; эта функция приходит
к ней только у позвоночных. Ее влияние
особенно сильно у млекопитающих.
Предполагается, в частности, что такая
специфическая Y-хромосомная система
сформировалась специально для обеспечения
развития эмбрионов обоих полов в «море
материнских эстрогенов» — женских
половых гормонов. Прогонады у этих объектов
успевают сделать выбор в пользу тестисов
с помощью специального генетического
блокирующего механизма еще до воздействия
женских гормонов. Усиливается взаимосвязь
гаметного и соматического пола.
Направление дифференциации половых клеток на
определенном этапе зависит не от их
собственной хромосомной конституции, а от
окружения, созданного соматическими
клетками гонад. В критический пренатальный
(предродовой) период первичные половые
клетки в яичниках входят в мейоз в
отличие от таковых в тестисах, в которых мейоз
и вхождение в сперматогенез блокированы.
В дальнейшем важную роль опять играет
кариотип, а именно присутствие двух
Х-хромосом или Y-хромосомы.
Консерватизм гонадогенеза, наличие
«древних» генов детерминации не
отвергают, а скорее подразумевают радикальное
изменение полопределяющей системы даже
за время эволюции Eutheria. У
млекопитающих реально существуют варианты
определения пола без участия SRY-тена. или
Y-хромосомы. Хромосомная инверсия пола в
норме встречается у многих видов (Myopus
schisticolor, Acodon azazae, Microtus cambrerae
и др.) и происходит она за счет
генов-модификаторов Х-хромосомного, Y-хромосомного
или аутосомного происхождения. Описаны
даже фертильные кобылы с XY-кариотипом.
Для сумчатых характерна дифференциация
мошонки, молочных желез, направляющей
связки тестисов, влагалищного отростка еще
до появления гонад. Развитие же вольфова
и мюллерова протоков, как и у всех
плацентарных млекопитающих, находится под
гормональным контролем. Для кенгуру
отмечено практическое отсутствие X-Y-хромо-
сомного спаривания, рекомбинация может
быть ограничена только очень небольшой
областью гомологии. Y-хромосома бычьих не
содержит значительных блоков сателлитной
ДНК и гетерохроматина; тем не менее она
является наиболее вариабельным элементом
их кариотипа. ZW/ZZ-система определения
пола у птиц аналогична XX/XY-хромосом-
ному варианту млекопитающих. Гонадный пол
а
SRY-млекопитающие Птицы ZW (доза Z)
3
Рыбы
Ящерицы
3 2
1 Тестис
2
3 Температура
Змеи ZW Черепахи Крокодилы
б
ДНК
LHRH
ДНК
Гонада
Тестостерон
Эстрадиол
Рис- 5. Возможные пути генетического контроля
гонадогенеза у позвоночных
а— предполагается наличие некоторого сходного
многоэтапного механизма гонадогенеза и
разнообразие генетического контроля этого процесса в
разных систематических группах.
1 — этап гонадогенеза;
2 — положительный контроль;
3 — отрицательный контроль;
б— гипотетический механизм температурного
варианта определения пола. Этапы, взятые в рамки,
предполагают тем пературочувствительность. LH и LHRH —
лютеинизирующий гормон и его релизинг-фактор,
положительно контролирующий синтез LH.
Предполагаются полспецифические различия по ДНК
112
ГЕНЕТИКА
у птиц можно переделать с помощью
инъекции ингибитора ароматазы, что
невозможно у млекопитающих несмотря на сходство
метаболизма стероидов. Описаны виды рыб
с хорошо дифференцированными половыми
хромосомами, обладающие способностью
преодоления генетического механизма го-
надогенеза при помощи факторов внешней
среды.
Примеры разнообразных вариантов системы
определения пола можно продолжить. Какое-
то представление о принципиальных
возможностях вариации его определения дает рис. 5.
В настоящее время поняты лишь некоторые
узловые элементы такой системы. Даже
идентификацию предполагаемых TDF-SRY
подобных генов млекопитающих, переделку
гонадного пола при его переносе мышам нужно
рассматривать лишь как многообещающее
начало исследования такого истинного шедевра
природы, какими являются половой
диморфизм и сексуальная репродукция. Недавние
эксперименты по «заселению» стерильных
тестисов мышей при помощи переноса
стволовых клеток сперматогониев, —
единственных потенциально тотипотентных (способных
к дифференцировке по многим
направлениям) диплоидных клеток у взрослых
млекопитающих, — новый впечатляющий успех
исследований в этом направлении.
Основные принципы
детерминации пола у дрозофилы
У дрозофилы в Y-хромосоме генов
практически нет. К настоящему времени открыто
только 11 генов, влияющих на
формирование сперматозоидов, но не на
формирование половых признаков, характеризующих
взрослых самцов. Более того, особь ХО
(Y-хромосома утрачена) также является
самцом. Каким же образом формируются
самцы и самки у дрозофилы?
Балансовая теория определения пола у
дрозофилы. В 1921 г. американский генетик
у—> Дозовая компенсация
у.д ^ Q ./ > Половые признаки
Л,А ьх1 \~ в соматических тканях
х > Половые признаки
в зародышевых клетках
Рис. 6. Первичный сигнал, возникающий из
соотношения числа Х-хромосом и аутосом контролирует все
аспекты половой дифференцировки через действие
ключевого гена Sxl
У самок, имеющих отношение Х:А равное единице, Sxl
активен. У самцов, имеющих отношение Х:А равное
0,5, ген Sxl остается неактивным. Состояние
активности гена Sxl регулирует развитие трех процессов,
находящихся под его контролем: дозовой компенсации,
развития половых признаков в соматических и
зародышевых клетках [из: Nothiger, 1992]
К. Бриджес обнаружил нескольких самок,
имевших триплоидный набор хромосом, т. е.
ЗХ+ЗА— три набора Х-хромосом и три
набора аутосом. В результате скрещивания
этих самок с нормальными самцами (2A+XY)
в потомстве среди нормальных самок и
самцов были обнаружены особи с
промежуточным или необычным проявлением половых
признаков. Все потомство распалось на 8
классов в зависимости от соотношения
половых хромосом и аутосом:
1. ЗХ : ЗА — триплоидные самки.
2. 2Х : 2А — диплоидные самки.
3. (2X+Y) : 2А — самки.
В этих трех случаях отношение числа
Х-хромосом к числу аутосом составляет
единицу. Наличие мужской Y-хромосомы не
влияет на нормальное развитие самки.
4. Особи, имеющие хромосомную
конституцию XY : 2А, для которых отношение
числа Х-хромосом к числу аутосом равно
0,5, по фенотипу—нормальные самцы.
5 и 6. Особи 2Х : ЗА и (2X+Y) : ЗА, у
которых отношение числа Х-хромосом к
числу аутосом варьировало между 0,5 и 1.
Они имели смешанное проявление
мужских и женских половых признаков; их
Бриджес назвал интерсексами. У таких мух
полностью отсутствовали секторы тела,
детерминированные по полу, либо в ходе
развития до определенного момента
формировались органы, присущие одному полу,
а затем — органы другого пола.
7 и 8. И, наконец, если число наборов
аутосом увеличивалось до трех при
наличии одной Х-хромосомы (X : ЗА), развивался
«сверхсамец» — организм с
гипертрофированными признаками самца, однако
стерильный. Увеличение числа Х-хромосом при,
диплоидном наборе аутосом (ЗХ : 2А),
напротив, ведет к формированию
«сверхсамки» с ненормально развитыми яичниками и
другими нарушениями признаков пола.
Эти данные позволили Бриджесу
заключить, что пол у дрозофилы
определяется балансом числа половых хромосом и
наборов аутосом, т. е. Y-хромосома у
дрозофилы вообще не играет роли в
определении пола.
Другую роль выполняет Y-хромосома у
человека. Нормальные женщины и
мужчины имеют, как ясно из предыдущего,
набор XX и XY соответственно. Однако при
отсутствии Y-хромосомы формируется особь
женского пола при любом числе Х-хромосом.
Особь ХО развивается главным образом по
женскому типу, но имеет ряд отличий от
нормы, известных под названием «синдром
Тернера». Такие индивиды возникают с
частотой 1/5000 новорожденных.
При наличии трех или даже четырех
Х-хромосом, но в присутствии Y-хромосомы
формируется мужской тип тела, правда
РЕГУЛЯЦИЯ ПОЛА
113
тоже с отклонениями, известными под
названием «синдром Клайнфельтера» (1/500
новорожденных).
Балансовая теория оказалась
неприменимой не только к человеку, но и к
высшим растениям.
Таким образом, путей определения пола
оказалось довольно много, но большинство
из них определяется хромосомами и, в
конечном счете, генами.
Действие генов при определении пола у
дрозофилы. Каким образом соотношение
числа Х-хромосом и аутосом, согласно
балансовой теории Бриджеса, может влиять
на развитие пола? В последние годы у
дрозофилы открыты многочисленные гены,
влияющие на правильное формирование
пола, среди них такие как Sxl (Sex lethal),
da (daughterless), sis (sisterless), tra
(transformer), dsx (double sex) и др.
Эксперименты показывают, что
соотношения числа Х-хромосом и аутосом
влияют на работу гена Sxl на ранней стадии
эмбрионального развития. Этот ген
контролирует 3 различных направления диффе-
ренцировки: формирование половых
признаков в соматических клетках и в клетках
зародышевого пути, а также контроль до-
зовой компенсации (рис. 6). Каким образом
это происходит? По данным
швейцарского ученого Р. Нётигера, на начальных
этапах формирования пола у эмбрионов
действуют гены sis-a и sis-b, расположенные
в Х-хромосоме, и ген da, расположенный
в аутосоме. Белковые продукты этих генов
образуют комплексную молекулу. Продукт
гена da поступает в яйцеклетку из организма
матери, его количество всегда соответствует
двум дозам, так как он считывается с
генов, локализованных в двух материнских
аутосомах. Количество продуктов,
считанных с генов sis-a и sis-b, зависит от того,
сколько Х-хромосом у особи — две или одна.
Поэтому комплекс белков sis/da имеет
отношение составляющих его компонентов 1:2
у самцов или 2:2 у самок. Эти белковые
продукты поступают на регуляторную зону
ключевого гена, определяющего пол, — Sxl.
Эта зона содержит два участка,
стимулирующих транскрипцию РНК с данного гена:
ранний и поздний промоторы (рис. 7).
Комплексный белок sis/da может активировать
начало транскрипции с раннего промотора
(РЕ на рис. 7) только в том случае, если он
содержит 2 дозы sis. Это происходит в
самом раннем эмбриональном развитии, на
стадии бластодермы. Позднее транскрипция
может начаться и с позднего промотора как
у ХХ/АА, так и у Х/АА особей. Но
результаты включения транскрипции с каждого
из этих промоторов будут разными.
Ген Sxl содержит 8 участков,
кодирующих последовательность аминокислот (1 —
8 на рис. 8) — экзонов, — разделенных не-
кодирующими районами (рис. 8). У самцов
(X : А = 0,5) при активировании позднего
промотора (PL) считывается третий экзон
(рис. 8), в котором расположен кодон UGA,
после которого трансляция останавливается,
и белок получается усеченным. В отсутствие
нормального функционального белка гена
Sxl, ген tra, расположенный далее в
данном каскаде, также функционирует
неправильно, опять же давая короткую
нефункциональную молекулу белка (трансляцию
блокирует кодон UAG во втором экзоне —
рис. 8). Хотя белок другого гена — tra2
(рис. 8) — присутствует у обоих полов, он
не формируется до нормального состояния
в отсутствие функционального продукта гена
tra. Более того, в отсутствие нормальных
продуктов генов tra и tra2, формируется
особый dsxM (самцовый) dsx-белок,
считанный с определенного набора экзонов (рис. 8).
Такой белок репрессирует развитие
признаков женского пола. В результате
нарушений действия каскада этих генов развитие
направляется в сторону формирования
признаков самца.
У самок (X : А = 1) транскрипт гена Sxl
не содержит экзона номер 3 со стоп-кодо-
ном, в результате чего формируется пол-
а
Pl Pe Sxl Sxl°n Самка
б
1 Pl I 1 Ре |
| А >
Sx/<
Рис. 7. Модель механизма включения гена Sxl у
разных полов
У самок (а) и самцов (б), белки, кодируемые генами
из Х-хромосомы (прямоугольники), формируют
активные комплексы с белками, кодируемыми генами,
расположенными в аутосомах (кружки). Самки имеют в
два раза больше комплексных молекул, чем самцы.
Наличие большого числа этих комплексных молекул
позволяет им связываться с РЕ промоторным
элементом гена Sxl и активировать транскрипцию
(стрелка вправо — Sxlon). У самцов (б) число молекул
комплекса невелико и неспособно активировать РЕ
промотор. Ген Sxl не включается (Sxloff), что и приводит
в конечном счете к развитию половых признаков
самца. PL— поздний промотор [из: Belote, 1992]
114
ГЕНЕТИКА
ноценный белок Sxl, взаимодействующий с
геном tra, который, взаимодействуя с
белком гена tra2, регулирует образование
специфической для самок РНК dsxF (рис. 8).
Наличие специфического для самок
продукта dsxF способствует вовлечению в данный
каскад гена ix. Белки генов dsxF и ix инак-
тивируют многие гены, которые могли бы
привести к формированию самцов, и в
конечном итоге формируется самка.
По этой схеме формируются внешние
(соматические) половые признаки.
Компенсация дозы генов. Как уже было
отмечено выше, в генетическом смысле оба
пола как у дрозофилы, так и у
млекопитающих различаются только числом и
составом половых хромосом. При этом у самцов
гены, локализованные в Х-хромосоме,
представлены в одной дозе, а у самок — в двух.
Если бы Х-хромосомные гены
функционировали с одинаковой интенсивностью,
количество продуктов этих генов у самок было
бы вдвое больше, чем у самцов. Однако этого
не происходит. Существуют механизмы
компенсации дозы генов. Разумно априори
предположить, что для уравновешивания
интенсивности функционирования Х-хромосомных
генов можно заставить их функционировать
Х:А = 0,5
AUG UGA
Sxl
5 6 7
нефункциональный
Sxl
AUG UGA
tra
нефункциональный
tra
X:A= 1,0
AUG
UGA
5 6 7 8
AUG
UGA
AUG
UGA
dsx
подавление дифференцировки самки
Самец
AUG
UGA
j/^X^
подавление дифференцировки самца
Самка
Рис. 8. Каскад генных взаимодействий, приводящих к формированию соматических половых признаков самца
(а) и самки (б) [из: Belote, 1992]
В каскад вовлечены гены Sxl, tra, tra2, ix и dsx. Прямоугольники с цифрами обозначают кодирующие части
генов— экзоны.Зигзагами показаны интроны
РЕГУЛЯЦИЯ ПОЛА
115
Sxl~
msl~
Рис. 9. Морфология (а) и присутствие специфических белков (б) в Х-хромосомах у самцов и самок дрозофилы, а
также изменение этих свойств (в) у мутантов, влияющих на развитие пола [из: Henikoff, Meneely, 1993]
а — у самок все политенные хромосомы кариотипа, включая Х-хромосому, имеют одинаковую степень разрых-
ленности. У самцов Х-хромосома выглядит значительно более диффузной, чем остальные хромосомы у самцов
и все хромосомы у самок; б — белок, кодируемый геном mle, выявляется только в Х-хромосоме самца; в — у
мутантов, влияющих на развитие половых признаков, изменяется распределение белков mle. Так, у мутантов Sx/
белок mle появляется в Х-хромосомах самок, чего никогда не наблюдается в норме. Мутация гена msl
приводит к отключению дозовой компенсации в Х-хромосоме самца из-за отсутствия в ней белка mle
вдвое интенсивнее у самцов, или же инак-
тивировать одну из Х-хромосом самок.
Природа использовала оба механизма. У
дрозофилы дополнительно активируется
единственная Х-хромосома самца до
уровня двух Х-хромосом самки, а у
млекопитающих инактивируется одна Х-хромосома
и уровень экспрессии у самок
уменьшается до уровня единственной Х-хромосомы
самца.
Основные открытия в области дозовой
компенсации у дрозофилы были сделаны
в результате анализа политенных
хромосом слюнных желез личинок (см.: Жиму-
116
ГЕНЕТИКА
лев И. Ф. Молекулярно-генетическая
организация политенных хромосом. Наст. том).
У самцов (XY) единственная политенная
Х-хромосома должна была бы быть
наполовину тоньше двух спаренных аутосом или
двух спаренных Х-хромосом у самки.
Однако на цитологических препаратах она
тоньше только на 25% и выглядит
значительно более разрыхленной, чем
остальные хромосомы и Х-хромосомы самки
(рис. 9). Вообще известно, что разрыхлен-
ность структуры хромосом связана с более
активной транскрипцией в них.
Компактный материал транскрипционно неактивен,
декомпактизованный разрыхленный
материал обнаруживает высокую активность при
синтезе РНК.
Уже первые биохимические
эксперименты показали, что в единственной по-
литенной Х-хромосоме самца количество
негистоновых белков примерно в 1,5 раза
больше, чем могло бы быть в одной
Х-хромосоме самки. Разрыхленность
структуры и обогащенность негистоновыми
белками является структурной основой для
дозовой компенсации. Эксперименты
показали, что интенсивность транскрипции в
одной Х-хромосоме самца в два раза выше,
чем в одной Х-хромосоме самки.
Оказалось, что существует целый
механизм, контролирующий формирование
разрыхленной структуры единственной
Х-хромосомы самца. Выделены продукты 4 генов:
msl-1, msl-2, msl-3 и mle — MSL-белки,—
участвующие в этом процессе. Все четыре
MSL-белка образуют комплекс и связываются
с сотнями участков Х-хромосомы самца,
обеспечивая диффузность ее структуры.
Присутствие в комплексе каждого из MSL-
белков необходимо для нормального
функционирования всего комплекса.
В белке MSL-2 был обнаружен особый
участок, имеющий способность интенсивно
связываться с ДНК и называемый ринг-фин-
гером, MSL-3-белок, по-видимому имеет,
другой активный участок— «хромо-домен»,
который характерен для белков,
связывающихся с хроматином. Кроме MSL-белков в
этом процессе участвуют молекулы других
белков — гистонов Н4. Гистоны обычно
выполняют обратную функцию — в комплексе
с молекулой ДНК они образуют нуклеосо-
мы, в составе которых ДНК упаковывается
более плотно и становится более компакт-
Самка
Sxl включен
5'UTR
3'UTR
II
Самец
Sxl выключен
Рис. 10. Схема регуляции-дозовой компенсации у дрозофилы [из: Bashaw, Baker, 1996]
Детали см. в тексте
РЕГУЛЯЦИЯ ПОЛА
117
ной (см.: Ченцов Ю. С. Организация митоти-
ческой хромосомы. Наст. том). Гистоны Н4,
декомпактизирующие Х-хромосому,
отличаются от гистонов Н4, компактизирующих
ДНК, тем, что они модифицированы:
аминокислота лизин, находящаяся в 16-м
положении в молекуле Н4, у них ацетилиро-
вана, т. е. содержит ацетильный остаток.
Оказалось, что такие модифицированные,
или Н4Ас16, гистоны в хромосомах
локализуются в тех же самых районах, что и MSL-
белки.
Предполагают, что белки MSL
взаимодействуют с регуляторными элементами
хромосом, контролирующими транскрипцию
и структуру хроматина. Молекулярный
анализ гена mle показал, что он имеет
гомологию с известными ранее генами,
участвующими в расплетании двух цепей ДНК, что
необходимо для транскрипции.
Белки MSL обладают и другой
особенностью. Недавно удалось показать, что
антитела, выработанные на белки MSL
Drosophila melanogaster, прекрасно
связываются с Х-хромосомами самцов других
видов двукрылых насекомых. Это
свидетельствует о том, что все белки, участвующие
в дозовой компенсации,
высококонсервативны, а сам механизм дозовой компенсации
возник в эволюции очень давно.
Дозовая компенсация у дрозофилы
контролируется тем же геном Sxl, который
осуществляет и общий контроль за
развитием пола. Нормальный белок гена Sxl
предотвращает дозовую компенсацию у самок,
не позволяя белкам MSL расположиться на
их Х-хромосомах. В случае мутации гена
Sxl, белки MSL появляются в
Х-хромосомах самок (рис. 9), нарушая тем самым весь
процесс дозовой компенсации.
Полагают, что в первую очередь белок
гена Sxl взаимодействует с геном msl-2
следующим образом (рис. 10).
У самок, у которых отношение числа
Х-хромосом к числу аутосом составляет 1,
ген Sxl находится во «включенном»
состоянии (см. выше) и подавляет трансляцию
мРНК гена msl-2. Специфические районы
РНК гена msl-2, называемые UTR,
блокируются белком SXL (рис. 10, I). В итоге
продукт гена msl-2 не поступает в ядро. В
его отсутствие, другие белки MSL не
способны к ассоциации с Х-хромосомами, аце-
тилированные формы гистонов Н4Ас1б не
накапливаются, и Х-хромосома самки не
становится сверхактивной в транскрипции
(рис. 10, I).
У самцов, у которых отношение
Х-хромосом и аутосом составляет 0,5, ген Sxl
выключен (см. выше). В отсутствие белка Sxl
ген msl-2 экспрессируется полностью, и
полный набор белков MSL ассоциируется
с Х-хромосомой самца. В результате
происходит и ацетилирование гистонов Н4 и,
как следствие, — изменение структуры
хроматина и последующая гиперактивация
транскрипции.
Комбинация методов классической
генетики и молекулярной биологии, построение
генетических карт и использование приемов
генетического планирования позволяют не
только выделять отдельные гены, но и
исследовать механизмы молекулярно-генети-
ческого контроля некоторых сложных и
биологически важных признаков. Исследование
первичной детерминации пола — хороший
пример реальных возможностей
современной молекулярной генетики.
Литература
Жимулев И. Ф. Хромомерная организация
политенных хромосом. Новосибирск: Наука, 1994.
Кауфман 3. К. Эволюция и размножение пола.
Петрозаводск: Карел, науч. центр РАН, 1994.
Мак-Ларен Э. В сб.: Проблемы генетики и
теория эволюции. Новосибирск: Наука, 1991, с. 115.
Мак-Ларен Э. Онтогенез, 1993, т. 24, с. 5.
Максимовский Л. Ф. Сельскохозяйственная
биология, 1988, № 1, с. 10.
Нестерова Т. Б., Закиян С. М. Генетика, 1994,
т. 30, №3, с. 293.
Осипова Г. Р. Генетика, 1996, т. 32, №2,
с. 184.
Основы цитогенетики человека. М.: Медицина,
1969.
Попов Л. С, РекешА. И. Молекулярные
механизмы регуляции пола у млекопитающих и
проблемы фракционирования сперматозоидов. М.:
Изд-во ИАЭ, 1990.
Прокофьева-Бельговская А. А.
Гетерохроматические районы хромосом. М.: Наука, 1986.
Тихомирова М. М. Успехи соврем, генетики,
1985, т. 13, с. 173.
Bashaw G. J., Baker В. S. Curr. Opinion in
Genetics and Development, 1996, vol. 6, p. 496.
Belote J.M. Seminars in Devel. Biol., 1992,
vol.3, p. 319.
Bridges C.B. Science, 1921, vol.54, p. 252.
Henikoff S. Meneely P.M. Cell, 1993, vol.72,
p. 1.
Nbthiger R. Verh. Dtsch. Zool. Ges., 1992,
Bd. 85, №2, S. 177.
С. И. Малецкий
ГЕНЫ САМОНЕСОВМЕСТИМОСТИ ЦВЕТКОВЫХ РАСТЕНИЙ
Введение
Большинство возделываемых человеком
растений принадлежит к покрытосеменным,
так как семена у них возникают внутри плода,
а семенные покровы развиваются из стенок
семяпочки. Возникновение семенных покровов
сыграло выдающуюся роль в эволюции
растений: оно привело к возникновению цветков.
Цветок — видоизмененный побег,
приспособленный для образования мужских и
женских гамет, внутри которого протекает
интимный процесс слияния гамет двух типов
в зиготу, т. е. зарождение новых растений.
Существует огромное разнообразие цветков,
которое можно обнаружить даже в
пределах отдельной популяции: так, в
популяциях некоторых видов у одних растений
закладываются и формируются только
пестичные (женские) цветки, а у других —
только тычиночные (мужские). Такие
популяции называют раздельнополыми; к их числу
принадлежат популяции конопли, тополя,
облепихи. Доля растительных видов,
формирующих раздельнополые популяции, во
флоре умеренных широт невелика.
Подавляющая часть растений образует
совершенные (обоеполые) цветки, у которых есть и
тычинки и пестик. Однако и у них
перекрестное оплодотворение осуществляется вполне
успешно, не менее успешно, чем у
раздельнополых растений.
После распускания цветков пыльники
растрескиваются и из них высыпается
пыльца, легко переносимая ветром или
насекомыми. Если пыльцевое зерно попало на
рыльце собственного пестика, проросло,
сформировало пыльцевую трубку, трубка
достигла зародышевого мешка семяпочки и
спермии пыльцевой трубки слились с
яйцеклеткой и центральным ядром, происходит
самооплодотворение. Растения, у которых оно
преобладает, называют самоопылителями.
Среди известных самоопылителей —
пшеница, ячмень, томаты.
Большинство растений неспособны к
самооплодотворению. Пыльца, попав на рыльце
собственного цветка или на рыльце цветков
того же растения, прорастает, но вскоре
останавливает рост, и пыльцевые трубки
погибают, не достигнув зародышевого мешка.
Растения, неспособные к
самооплодотворению вследствие подавления роста
собственных пыльцевых трубок, называют
самонесовместимыми. Ясно, что самонесовместимые
растения окажутся бесплодными
(бессемянными), если на их цветки не попадет
пыльца с соседних (неродственных) растений.
Первое упоминание в литературе о
самонесовместимости можно найти в трудах
немецкого ботаника И. Кельрейтера,
опубликовавшего в 1765 г. сообщение о
самонесовместимости у коровяка Verbascum
phoeniceum. Распространение
самонесовместимости в мире растений и ее роль в
регуляции перекрестного размножения в
растительных популяциях были осознаны в
нашем веке после переоткрытия законов
Г. Менделя и рождения новой
биологической дисциплины — генетики.
Все цветковые растения с совершенными
цветками удобно разделить на две группы:
самосовместимых (СС), к которым относятся
самоопылители и некоторые перекрестно
размножающиеся растения, и
самонесовместимых (СН), включающих только перекре*-
стно размножающиеся растения. Чтобы
определить, как размножаются растения, есть
ли у растений конкретной популяции
признак самонесовместимости, надо провести
наблюдения по завязыванию семян при
контролируемых опылениях
(скрещиваниях) (см. табл. 1).
Ныне обнаружено более 10 тыс.
самонесовместимых растительных видов. Их
можно разделить по типу цветков.
Таблица 1
Способы обнаружения самонесовместимости у растений
Тип скрещивания
Размножение отдельного(ых) растения(й) под индивидуальным(и) изолятором(ами) или
на изолированном участке
Свободное переопыление двух или большего числа растений на изолированном участке
Искусственное самоопыление отдельных цветков на растениях
Образование семян
СН
Нет
Есть
Нет
СС
Есть
Есть
Есть
ГЕНЫ САМОНЕСОВМЕСТИМОСТИ ЦВЕТКОВЫХ РАСТЕНИЙ
119
Гетероморфная несовместимость
Самонесовместимые растения делят на две
большие группы, каждая из которых
содержит десятки семейств: растения с
гомоморфной несовместимостью и растения с гете-
роморфной несовместимостью. У растений с
гомоморфной несовместимостью нет различий
в строении цветков — в пределах вида или
популяции все цветки морфологически
идентичны. У растений с гетероморфной
несовместимостью в популяциях встречаются два-три
морфотипа цветков. Например, при дистилии
в популяциях представлены два типа
растений с разным строением цветков, при трис-
тилии — три типа. В популяциях дистильных
видов часть растений формирует цветки с
короткими столбиками пестиков и
длинными тычинками, а вторая часть — цветки с
длинными столбиками пестиков и
короткими тычинками (рис. 1).
Дистилия контролируется одним
супергеном (суперген — блок сцепленных генов,
передающийся потомству как единое целое):
генотип растений с короткостолбчатыми
цветками обозначают Ss, с длинностолбчатыми
цветками — ss. Между цветками одного
морфотипа самооплодотворение и перекрестное
оплодотворение не осуществляются (так
называемый иллегитимный вариант
опыления), но оно возможно между растениями
разных морфотипов (легитимный вариант
опыления).
В названия морфотипов (длинно- и корот-
костолбчатые цветки) включен лишь один
признак— длина столбиков пестиков, тогда
как реально гетеростилии сопутствует
изменение не отдельного признака, а некоторой
их совокупности, включающей: а) длину
столбиков; б) размер клеток сосочков рыльца (у
короткостолбчатых растений эти клетки
мелкие, а у длинностолбчатых крупные); в)
положение пыльника в венчике цветка
относительно рыльца пестика (у длинностолбчатых
тычинки располагаются ниже уровня
поверхности рыльца, у короткостолбчатых — выше
этого уровня); г) размер пыльцевых зерен
(крупные пыльцевые зерна у
короткостолбчатых цветков и мелкие пыльцевые зерна у
длинностолбчатых); д) форму и структуру эк-
зины (экзина — внешняя оболочка
пыльцевого зерна у цветковых растений); е)
молекулярные (белковые) механизмы распознавания и
ингибиции иллегитимных вариантов опыления.
Поэтому суперген S-s иногда записывают в
более развернутом виде с указанием трех
основных блоков генов, входящих в его
структуру. Генотип растений с короткостолбчатыми
цветками записывается GAP/gap, а генотип
растений с длинностолбчатыми цветками —
gap/gap (G — гинецей, А — андроцей, Р —
пыльца). Реализация иллегитимных
(несовместимых) вариантов опыления внутри
популяции основана на молекулярных
взаимодействиях (молекулярном распознавании) белков
поверхности рыльца и пыльцевого зерна.
Взаимодействие продуктов S-генов пыльцы и
рыльца пестика тормозит прорастание и рост
пыльцевых трубок: у самонесовместимых ге-
теростильных растений семена от самоопыления
не завязываются, несмотря на то что
цветки имеют нормально сформированные
андроцей и гинецей.
Гетероморфия обнаружена в 155 родах
24 семейств; 91 из этих родов
принадлежит к семействам мареновых (Rubiaceae),
бурачниковых (Boraginceae), свинчаткоцвет-
ковых (Plumbaginaceae) и примуловых
(Primulaceae). Среди сельскохозяйственных
видов к числу гетеростильных относится
гречиха (Fagopyrum esculentum).
Гомоморфная несовместимость
На примере гетероморфной
несовместимости можно очень наглядно
проиллюстрировать, каким образом осуществляется
регуляция перекрестного опыления и
оплодотворения в популяциях растений.
Большое число публикаций о гетероморфии
цветков объясняется также и тем
повышенным вниманием, которое уделяли ей
выдающиеся исследователи начиная с Ч.
Дарвина. А вот о другой аналогичной по сути
проблеме — гомоморфной
несовместимости — работ мало, хотя ее
распространенность в мире растений многократно
превышает распространенность гетероморфии.
Первая публикация на русском языке,
содержащая сведения об этом феномене,
появилась в 30-е гг., а последующие лишь в
60-е гг.
По характеру экспрессии различают два
типа гомоморфной несовместимости: гаме-
тофитный и спорофитный — в
зависимости от того, пыльца (гаметофит) или мате-
Ss ss
а б
Рис. 1. Схематическое строение цветков у
дистильных растений
а— короткостолбчатая форма с высокими
тычинками (генотип Ss); б —длинностолбчатая форма с
низкими тычинками (генотип ss)
120
ГЕНЕТИКА
Таблица 2
Список важнейших родов культурных растений, у которых описана
гомоморфная самонесовместимость
Семейство
Род
Однолокусная гаметофитная система несовместимости
Solanaceae (пасленовые)
Rosaceae (розоцветные)
Leguminosae (мотыльковые)
Scrophulariaceae (норичниковые)
Onagraceae (ослинниковые)
Liliaceae (лилейные)
Commelinaceae (коммелиновые)
Nicotiana, Petunia, Solarium, Lycopersicon, Physalis
Malus, Prunus, Pyrus, Cerasus
Trifolium, Medicago, Lotus, Prosopis
Antirrhinum, Nemesia
Oenothera
Lilium
Tradescantia
2-4-локусная гаметофитная система несовместимости
Chenopodiaceae (маревые)
Ranunculaceae (лютиковые)
Papaveraceae (макоцветные)
Gramineae (злаковые)
Beta
Ranunculus
Papaver
Secale, Festuca, Dactylis, Hordeum, Lolium, Phalaris
Спорофитная система несовместимости
Cruciferae (крестоцветные)
Compositae (сложноцветные)
Convolvulaceae (вьюнковые)
Corylaceae (лещинные)
Brassica, Raphanus, Eruca, Cardamine, Iberis
Crepis, Parthenium, Cosmos
Ipomoea
Corylus
ринское растение (спорофит) детерминирует
реакцию несовместимости между пыльцевым
зерном (пыльцевой трубкой) и поверхностью
рыльца столбика или другими тканями
пестика цветка. В табл. 2 приведены семейства
и роды, куда входят многие виды
культурных растений, обладающие гомоморфной
несовместимостью, и указано основное число
генов, детерминирующих этот признак.
Рассмотрим особенности экспрессии и
характер наследования реакции
несовместимости (реакции распознавания) между
пыльцевым зерном (пыльцевой трубкой) и
тканью пестика у растений с гомоморфной
несовместимостью.
Гаметофитный тип
несовместимости. Концепция гаметофитной
несовместимости у растений была первоначально выдвинута
Э. Истом и А. Мангельсдорфом в 1925 г. при
изучении генетического контроля
несовместимости на табаке Nicotiana sanderae. Были
сформулированы следующие принципы
функционирования системы несовместимости: а) в
пыльцевом зерне и ткани пестика активны
одни и те же факторы (гены),
контролирующие синтез распознающих друг друга
субстанций; б) экспрессия генов
несовместимости осуществляется независимо в гаплоидной
пыльцевой трубке и диплоидной ткани
пестика; в) результативное опыление
возможно, если аллели несовместимости в
пыльцевой трубке и ткани пестика различны; если
же они одинаковы, то рост пыльцевой трубки
ингибируется независимо от того, получено
ли пыльцевое зерно с цветка собственного
растения или любого другого, несущего тот
же самый аллель несовместимости; г) в
популяциях все растения гетерозиготны по
аллелям несовместимости; д) факторы (гены),
ответственные за самонесовместимость и внут-
рипопуляционную перекрестную
несовместимость, одни и те же; е) гены
несовместимости представлены сериями множественных
аллелей, и каждый аллель получает свой
порядковый номер по мере их открытия. Ло-
кус несовместимости (локус — место
локализации определенного гена в хромосоме) был
обозначен буквой S (от англ. self-
incompatibility), а его аллели — буквой S с
индексами: Sx, S2, S3 и т. д.
Исходя из .изложенных принципов
функционирования само- и внутрипопуляцион-
ГЕНЫ САМОНЕСОВМЕСТИМОСТИ ЦВЕТКОВЫХ РАСТЕНИЙ
121
нои несовместимости очевидно, что если
скрестить два растения, у которых все
четыре аллеля различны (например, SXS2 x
х S3S4), то в потомстве получится четыре
интерфертильные1, но при этом интрасте-
рильные2 группы; перекрестное
оплодотворение внутри каждой группы невозможно,
а вот скрещивания растений из разных групп
возможны. Так, в результате скрещивания
9 ЭД xCfS3S4 образуются группы: БД, SXS4,
S2S3, S2S4. Если же у скрещиваемых
растений есть один общий аллель (аллели —
различные состояния отдельного гена,
вызывающие фенотипические различия и
локализованные на гомологичных участках
гомологичных хромосом), то в потомстве
получаются две интрастерильные и
интерфертильные группы. Например,
скрещивание 9 S^jj *cf БД дает потомков S^ и S2S3,
так как пыльцевые трубки с аллелем Sx не
участвуют в образовании зигот по причине
несовместимости (рис. 2).
При гаметофитной однолокусной
несовместимости типа Nicotian а в популяциях
поддерживается огромное разнообразие аллелей,
неизвестное прежде для других признаков,
изучавшихся методами экспериментальной
генетики. В популяции энотеры (Oenothera
organensis) численностью всего 500 особей
описано 45 аллелей. Возможное число
аллелей у клеверов Trifolium pratense и Т. repens
близко к 400. Минимальное число S-аллелей
в популяции не может быть менее трех. Если
их всего три, эффективность перекрестного
опыления будет очень невысокой. Например,
при равной их частоте доля совместимых
вариантов переноса пыльцы не будет
превышать 1/3.
Однолокусная мультиаллельная
несовместимость гаметофитного типа
встречается не только у Nicotian a sanderae, но
и у других представителей семейства
пасленовых (Solan асеае). Кроме пасленовых
аналогичный тип несовместимости описан
и для видов семейства Rosaceae, к
которому относятся многие садовые растения
(яблоня, груша, слива, черешня).
Самонесовместимость плодовых деревьев,
размножаемых вегетативно, делает
необходимой закладку многосортовых садов, так
как высадка на большой площади деревьев
одного сорта (одного клона) исключает
возможность результативного перекрест-
1 Интерфертильность — совместимый
вариант опылений между растениями различных
S-генотипов, например, при переопылении
растений генотипов 9 SXS2 x С^ S3S4.
2 Интрастерильность — несовместимый
вариант опылений двух или большего числа
растений в популяции, имеющих идентичный
S-генотип, например, нескрещиваемость
растений одного типа — Sfi?
Генотип пыльцевого родителя S-jSg
S1 S2 S1 S3 S3 S4
Генотипы пестичных родителей
Рис. 2. Рост пыльцевых трубок при опылении трех
растений генотипов S^, S^ и S3S4 пыльцой
растения генотипа S^,, (гаметофитный тип
несовместимости)
ного опыления и делает такие сады
бесплодными.
Спорофитный тип несовместимости.
При спорофитном контроле реакция
несовместимости определяется не генотипом
пыльцевого зерна (гаметофита), а
генотипом самого растения. Это означает, что
S-аллели экспрессируются в диплоидных
тканях и затем их продукты попадают в
гаплоидные гаметофиты. Предполагается,
что S-аллели экспрессируются в тапеталь-
ных клетках3, выстилающих пыльник и
питающих микроспоры. Спорофитной
системе контроля само- и перекрестной
несовместимости, так же как и гаметофитной,
присущ множественный аллелизм.
Экспрессия S-аллелей в диплоидных клетках тапе-
тума создает условия для взаимодействия
продуктов активности двух аллелей:
между ними возникают отношения
доминирования (доминантность — проявление у ге-
терозиготы AXA2 только одного из аллелей,
например Ах) или кодоминирования (кодо-
минантность — проявление у гетерозиготы
AXA2 обоих аллелей). Ряд доминирования
принято записывать в виде неравенства Sj >
> S2 > S3 > S4. Доминирование возможно как
в пыльцевом зерне, так и в тканях
пестика. Если же аллели имеют независимое
проявление (кодоминирование), между ними
ставят знак равенства. Например, запись Sx =
= S2 означает, что аллели Sx и S2 имеют
независимый характер проявления, иными
словами в отдельном пыльцевом зерне (или
ткани пестика) представлены продукты обоих
аллелей (см. рис. 3).
3 Тапетум — выстилающий слой клеток в
пыльниках цветка, питающий пыльцевые
зерна в процессе их формирования.
122
ГЕНЕТИКА
Пыльца
Столбик пестика
Iberis
атага
Cosmos
bipenatus
Рис. 3. Межаллельные отношения в пыльце и
столбике Iberis amara и Cosmos bipenatus
Объяснения в тексте
Спорофитный тип несовместимости
характерен для растений семейства крестоцветных
(Cruciferae) и сложноцветных (Compositae).
Отсюда следует, что у таких важнейших
сельскохозяйственных растений, как
капуста, редис, репа, подсолнечник,
перекрестное опыление контролируется S-генами со
спорофитным типом действия.
Хотя сама реакция между пыльцевым
зерном и пестиком при гомоморфии
контролируется одним или несколькими генами,
у гомоморфных самонесовместимых
растений можно, как и у гетероморфных,
выделить комплексы признаков,
характеризующие эти системы контроля перекрестного
опыления (табл. 3).
Структура S-локуса и мутации
самофертильности. Рассмотрим структуру
S-локуса у пасленовых в связи с
феноменологией реакции несовместимости и
данными генетического анализа о строении
локуса несовместимости. При попадании
пыльцы на рыльце собственного цветка у
пасленовых происходит гидратация
пыльцевых зерен за счет выделений рыльца.
Гидратированные пыльцевые зерна
нормально прорастают, и пыльцевая трубка
растет в межклеточном пространстве
столбика. Вскоре после начала роста стенки
пыльцевой трубки начинают утончаться и
в ее кончике появляются многочисленные
частицы размером около 2 мкм. Она
разбухает и лопается, дальнейший ее рост
становится невозможным. Реакция
несовместимости в полости столбика у растений с
гаметофитным типом несовместимости, когда
происходит разрушение пыльцевой трубки,
напоминает картину вхождения пыльцевой
трубки в зародышевый мешок. Известно, что
клетки синергид4 выделяют вещество(а),
взаимодействие которого(ых) с пыльцевой
трубкой разрушает ее. Предполагается, что
в реакции несовместимости принимает
участие вещество-посредник из синергид или
аналогичный продукт синтезируется
непосредственно в столбике.
Из данных генетического анализа
следует, что субстрат несовместимости
синтезируется как в пыльцевой трубке, так и в ткани
столбика. Реакция несовместимости не
наступает до тех пор, пока в пыльцевой трубке
не произойдет второе митотическое
деление, приводящее к формированию клеток
спермиев.
Экспериментально установлено, что
S-локус имеет сложную структуру,
состоящую не менее чем из трех сегментов (рис. 4),
из которых один специфический и два
неспецифических. Один из неспецифических
сегментов активен в пыльце, а другой — в
ткани столбика. Показано, что продукты
S-генов — гликопротеиды (сложные
молекулы, составленные из молекул белка и
высокомолекулярных углеводов). Мономеры
4 Синергиды — два ядра зародышевого мешка
покрытосеменных растений, расположенные в
его микропиллярной части и участвующие в
двойном оплодотворении.
Таблица 3
Морфологические и физиологические особенности, отличающие спорофитную
и гаметофитную мультиаллельные системы несовместимости
Признак
Пыльцевые зерна
Дыхание
Жизнеспособность
Рост in vitro
Сосочки рыльца
Место ингибиции пыльцевых трубок
Место локализации каллозы в несовместимой пыльце
Тип несовместимости
гаметофитная
Двуядерные
Слабое
Длительная
Легкий
Влажные с «рваной кутикулой»
В столбике
Интина
спорофитная
Трехъядерные
Сильное
Короткая
Трудный
Сухие с целой кутикулой
На поверхности рыльца
Экзина
ГЕНЫ САМОНЕСОВМЕСТИМОСТИ ЦВЕТКОВЫХ РАСТЕНИЙ
123
Продукты S-аллелей
Три сегмента S-локуса
Место экспрессии
Пыльцевые трубки
(гаплоиды)
Ткани столбика цветка
(диплоиды)
В совместимых
вариантах нет
полной гомологии
мономеров пыльцевой
трубки и столбика
и потому димеры
(ингибиторы роста
пыльцевых трубок)
не образуются
Продукты Si- и Бг-аллелей образуют.
димерные молекулы гликопротеидов
в столбике
Эти димеры напоминают
по своей функции вещество-
посредник, синтезируемое в
синергидах, вызывающее
растворение апекса
пыльцевых трубок и
освобождение спермиев
Взаимодействие димеров (ингибиторов роста пыльцевых трубок) с поверхностью апекса
пыльцевых трубок ведет к прекращению их роста в ткани столбика
Рис. 4. Модель образования продуктов S-генов в ткани столбика цветка у гетерозиготы S^,,, приводящих к
разрушению пыльцевых трубок и нарушению самооплодотворения у пасленовых при гаметофитном контроле
несовместимости
S1 и S2— сегменты специфичности локуса несовместимости: РР (pollen part)— сегмент локуса, активный в
пыльцевой трубке; StP (style part)— сегмент локуса, активный в столбике пестика
гликопротеидов образуются как в столбике
цветка еще до начала роста пыльцевых
трубок, так и в кончике проросших пыльцевых
трубок. Эти мономеры (субъединицы, из
которых составлены белки) нефункциональны.
Если же в растущей пыльцевой трубке
встречаются идентичные мономеры из пыльцы и
столбика, то образуются функциональные
молекулы димеров (белков, составленных из
двух мономеров), которые крепятся к
поверхности ее апекса. Возникающие димеры
по своей природе должны быть идентичны
молекулам-посредникам из синергид,
вызывающих растворение наружной оболочки
пыльцевой трубки и излияние
содержимого трубки в зародышевый мешок при
оплодотворении.
Трехсегментная структура S-локуса
была впервые предложена Д. Льюисом в
1964 г. и подытожила данные многолетних
исследований о специфичности проявления
самофертильных радиомутантов. Само-
фертильными называют мутации S-генов,
приводящие к утрате реакции
несовместимости, так что у самофертильных
(самосовместимых) растений семена от
самоопыления завязываются столь же успешно,
как и от совместимого перекрестного
опыления. Независимо от того, в каком из
сегментов S-локуса произошла мутация,
носители таких мутаций становятся само-
фертильными — не наблюдается
торможения роста пыльцевых трубок в столбике
пестика. Пыльцевые трубки с мутацией в
пыльцевом сегменте ведут себя так, как
будто они несут другой (третий)
совместимый аллель. Однако в отличие от
растений — носителей новых аллелей —
растения с мутацией в пыльцевой части
S-локуса самосовместимы, т. е. способны
к самооплодотворению. Растения с
мутацией в пестичном сегменте S-локуса
также самофертильны, но пыльца таких
растений неотличима от пыльцы растений,
не затронутых мутацией.
Реакция несовместимости у
растений со спорофитным контролем.
Трехсегментная структура S-локуса, вероятно,
присуща и растениям со спорофитным
контролем несовместимости, у которых
реакция несовместимости протекает на
поверхности рыльца. Белки тапетума, вкрапленные
в тектум (слой, разделяющий интину и эк-
зину) пыльцевого зерна у крестоцветных,
прикрепляются к поверхности пыльцы с
помощью липопротеидов, образующих лип-
124
ГЕНЕТИКА
кий слой вокруг пыльцевого зерна. В
английской литературе этот слой получил
название «tryphine» («трифина»), а в
немецкой литературе — «pollenkitt» («полленкит-
та»). Белки трифина играют важную роль
в прикреплении (приклеивании) пыльцы к
рыльцу пестика, участвуя тем самым в
реакции распознавания между белками
пыльцевого зерна и тканью рыльца пестика и
препятствуя прорастанию собственной
пыльцы или пыльцы от растений с
идентичными S-генотипами.
Самонесовместимость и селекция
'растений. Самонесовместимость широко
используется в селекции, так как
садоводы давно заметили, что если высадить в
саду деревья только одного сорта (яблонь,
груш, слив), то такие сады оказываются
бесплодными. Каждый сорт — это один клон
(один генотип), так как размножение
сортов плодовых растений осуществляется
вегетативно, путем прививки. Чтобы сад был
плодоносящим, надо, чтобы в саду
одновременно росло два или большее число
сортов. Эта идея была использована в конце
40-х гг-. при получении гибридных сортов
овощных растений (различных видов
капусты). Для получения растений,
гомозиготных по генам несовместимости,
применялись приемы, позволяющие преодолеть
реакцию несовместимости и получить
потомство от самооплодотворения (опыление
незрелых цветков, температурная обработка
цветков). При одновременном выращивании
двух линий капусты, гомозиготных по
различным генам несовместимости,
образуется 100% гибридов.
Таким образом, самонесовместимость,
контролируя перекрестное опыление у
растений, препятствует гомозиготизации
генов и предотвращает негативные
последствия, возникающие в результате
родственных скрещиваний. Она сохраняет в
растительных популяциях высокий уровень
гетерозиготности генов, поддерживая на
определенном уровне эффект гетерозиса
(гибридную мощность) и позволяя растениям
лучше адаптироваться к разнообразным
условиям среды. Гены
самонесовместимости используются при получении гибридных
сортов. Информация о распространении
конкретных генов (аллелей)
самонесовместимости нужна при закладке садовых
плантаций.
Литература
Дарвин Ч. Собрание сочинений. М.; Л.:
Изд-во АН СССР, 1948, т. 7, с. 31.
Малецкий С. И. Генетика, 1969, т. 5, № 1,
с. 159.
Рыбин В. А. В кн.: Теоретические основы
селекции растений. М.; Л.: Гос. изд-во совхоз, и
колхоз, лит., 1935, т. 3, с. 463.
Суриков И. М. Успехи современной
генетики, М.: Наука* 1972, т. 4, с. 119.
Суриков И. М. Несовместимость и
эмбриональная стерильность растений. М.: Агропромиздат,
1991.
Л. И. Корочкин, И. Ф. Жимулев
ГЕНЫ И ПОВЕДЕНИЕ
Введение
Хорошо известно, что дети внешне
похожи на своих родителей вследствие
существования такого явления, как
наследственность. Суть его заключается в том, что
родители передают потомкам свои гены,
контролирующие различные признаки,
например, цвет глаз, волос, кожи, форму
носа и лица, рост, вес и др. В то же время
еще в IV в. христианский мыслитель Авгу-
100 а б
50
0
100 в
50
1234 51 2345
Рис. 1. Обучение избеганию электрического удара
в популяции нелинейных мышей (а), а также в
селекционированных лабораторных линиях Swiss (б),
DBA/2J (в) и СВА (г) по данным французских ученых
Видно, что в обычной популяции мышей велик
индивидуальный разброс по способности к обучению, а
в лабораторных линиях, прошедших процесс отбора,
мыши реагируют единообразно, обнаруживая либо
высокую, либо низкую способность к обучению.
Каждая кривая представляет собой поведение
отдельной мыши в течение пяти опытов избегания (ось
абсцисс) из 100 проб каждый. По оси ординат отмечен
процент избегания в каждом опыте
стин Блаженный заметил, что у рыбок не
только внешний вид, но и поведение
потомков напоминает родительское. Его
наблюдения можно было бы считать датой
рождения генетики поведения. Однако вопрос
о наследовании поведенческих признаков до
сих пор служит предметом дискуссий.
В 1923 г. вышла в свет известная
работа И. П. Павлова «Новые исследования по
условным рефлексам», в которой автор
высказал мнение, будто приобретенные в
течение жизни условные рефлексы могут
передаваться по наследству потомкам,
превращаясь со временем во врожденные,
безусловные рефлексы. Другой русский
ученый, Н. К. Кольцов, выступил с резкой
критикой представлений Павлова о
наследственной передаче благоприобретенных
признаков, в данном случае условных
рефлексов. Кольцов отметил, что Павлов
никогда не работал в области генетики и не
представляет себе всей сложности
генетических проблем. Павлов был вынужден
согласиться с критикой и отказался от
своего предположения. Это была, пожалуй,
первая серьезная полемика по вопросам
наследования поведенческих навыков. То,
что она возникла именно в России и в
начале нашего века, вполне объяснимо,
поскольку экспериментальные исследования
по генетике поведенческих признаков,
включая обучение, были начаты лишь на
рубеже XX в., а в числе первопроходцев были
российские ученые кольцовской школы.
В 1925 г. появилась статья М. П. Садов-
никовой-Кольцовой «Генетический анализ
психических особенностей крыс». Оказалось,
что в пределах одного и того же вида
разные особи обнаруживают различные
способности к обучению (рис. 1). Среди крыс,
мышей и других животных удается
выделить различные наследственные типы
психических способностей, очистить их путем
отбора в течение нескольких поколений и
затем перейти к установлению законов
наследования этих способностей.
Л. И. Корочкин с И. А. Шумской
провели тщательное исследование довольно
сложной поведенческой реакции — выработки пи-
щедобывательного двигательного условного
рефлекса. Крыса должна была в ответ на
звуковой и световой условные
раздражители прыгать на разные полочки и только
после успешного выполнения этих
упражнений получала пищевое подкрепление.
Оказалось, что лабораторные крысы
обнаруживают изменчивость по скорости выра-
126
ГЕНЕТИКА
ботки этого условного рефлекса: показатель
скорости варьирует от 3 сочетаний до 93.
Следовательно, налицо значительная
индивидуальная вариабельность животных по их
способности к обучению. Можно отбирать
быстро обучающихся крыс и скрещивать их
друг с другом и то же проделывать с
плохо обучающимися крысами. Через несколько
поколений (рис. 2) мы получили две линии
животных: представители одной обучались
хорошо, а другой — плохо. Это
свидетельствует о том, что способности к обучению
зависят в данном случае от многих генов.
Если бы этот признак определялся одним
геном, то разделение на две четкие
группы было бы выражено уже в первом
поколении, но на рис. 2 видно, что для
достижения данной цели понадобилось несколько
поколений.
В некоторых случаях установлены
физиологические основы генетических
различий в поведении. Так, Д. Скотт в 1966 г.
нашел большие различия между породами
собак по наследованию реакции лая.
Например, коккер-спаниели лают очень часто, а
африканские охотничьи собаки бейсенджи
почти не лают. Объясняются эти различия
разным генетически детерминированным
порогом реакции собак на внешние
раздражители. У спаниеля он очень низок, а у
бейсенджи высок. Гибриды первого
поколения близки по реакции лая к спаниелям,
следовательно, реакция эта является
доминантным признаком.
Наконец, ярким свидетельством
генетической детерминации форм поведения
являются наблюдения над осами НаЪгоЪгасоп,
проводившиеся сначала в 30-е гг., а затем
более основательно в 70-80-е. Известно, что
самки этих ос парализуют жалом личинок
Р > 0,95
Р > 0,99
36
32
28
24
20
16
12
8
4
0 1 23456789 10 11 12
Рис. 2. Скорость образования трехчленного пище-
добывательного условного рефлекса у крыс с
высокой (7) и низкой (2) способностью к обучению
По оси абсцисс отложены дни обучения, по оси
ординат— количество проб, необходимых для
освоения навыка
огневки мельничной, самцы же не
нападают на этих личинок, а ухаживают за
самками НаЪгоЪгасоп. У половых мозаиков, у
которых в ткани мозга участки клеток самцового
типа перемешаны с участками генетически
отличных клеток самок, поведение
нарушается: они пытаются ухаживать за
личинками огневки и жалить самок своего вида.
Норма реакции и поведение
Возникает вопрос, обрекают ли «плохие»
гены своего обладателя на тяжелую жизнь
с «испорченным» поведением или можно как-
то модифицировать, смягчить те невзгоды,
которые несут «плохие» гены.
Оказывается, можно. Ведь способности к обучению
формируются в ходе индивидуального
развития организма, которое осуществляется
не только в соответствии с сигналами
генов, но и на фоне воздействий внешней
среды, могущих как благоприятствовать
реализации наследственной информации, так
и блокировать ее на определенных этапах
становления организма. Американские ней-
робиологи провели любопытный эксперимент.
Мышат одного помета делили на две
группы: одну помещали в светлую просторную
клетку с множеством разнообразных
игрушек, а другую — в темную, тесную и
пустую. Авторы обнаружили различия в
развитии мозга испытуемых животных: у
первых толщина коры головного мозга была
больше, чем у контрольных мышей, а у
вторых — меньше. Соответственно
потомки одной и той же пары родителей в
зависимости от условий их содержания
обнаружили разные способности к обучению: те,
которые росли в особо благоприятных
условиях, оказались более способными
учениками. Однако толщина коры головного
мозга и уровень способностей к обучению
изменялись только в ограниченных
пределах, они не могли подняться выше и
опуститься ниже некоторой «планки», заданной
принадлежностью к той или иной линии,
породе, виду. Эти границы изменчивости
признака носят название нормы реакции. Их
существование принято связывать с тем,
что развитие различных признаков
определяется действием многих генов и
мутация каждого из них, если они
функционируют в мозге, отражается на характеристике
высшей нервной деятельности исследуемого
объекта (см.: Корочкин Л. И. Генетический
контроль развития клеток. Наст. том).
В связи с полученными данными
возникает вопрос о роли воспитания в развитии
различных поведенческих признаков. Иными
словами, «хорошие» наследственные
структурно-функциональные задатки мозга сами
по себе еще не обеспечивают правильной
их реализации в фенотипе — необходимы
ГЕНЫ И ПОВЕДЕНИЕ
127
соответствующие условия, чтобы эта
реализация осуществилась сполна. Точно так
же «плохие» наследственные структурно-
функциональные задатки нервной системы
еще не означают, что она будет безнадежно
«испорчена»: соответствующие
благоприятные условия могут ее «подправить» в
пределах нормы реакции. Но с помощью каких
механизмов гены контролируют поведение
и какова роль в этом процессов,
протекающих на молекулярном уровне?
Молекулярная генетика
и проблемы поведения
В 60-е гг., благодаря работам
шведского цитолога X. Хидена, возникло и
широко распространилось представление о так
S S
Рис. 3. Иллюстрация к концепции X. Хидена о роли
РНК в процессах запоминания
Автор предполагал, что молекулы памяти—
информационные РНК —синтезируют белок, ответственный
за ее хранение (его назвали S-100). На рисунке
представлены результаты опытов шведских ученых по
разделению методом микроэлектрофореза
водорастворимых белков нервной клетки из гиппокампа
контрольных крыс, пользующихся одной
предпочитаемой лапой (а), и крыс, натренированных
пользоваться непредпочитаемой лапой (б). Представлены
результаты количественной регистрации белка S-100,
обнаруженного в первом и втором случаях. Видно,
что в контроле обнаруживается одна фракция белка
(S), в опыте количество его возрастает и
обнаруживаются две фракции (S и S'). Хиден предполагал, что
данный белок локализован в ядре нейронов и
регулирует функции генов. В дальнейшем было
установлено, что в действительности он располагается в
цитоплазме глиальных клеток и, по всей
вероятности, не имеет отношения к процессам обучения
называемых молекулах памяти. Хиден
разработал изящный и тонкий метод,
позволяющий анализировать количество РНК и
даже ее качественный состав в отдельных
изолированных нервных клетках. В опытах
по обучению крыс (рис. 3) он и его
сотрудники обнаружили, что количество РНК в
тех нейронах, которые предположительно
имеют отношение к данной форме
обучения, увеличивается, а ее качественный
состав (содержание разных нуклеотидов)
изменяется. Исследователи пришли к
выводу, что в ходе обучения активируются
новые, ранее «молчавшие» гены, которые
синтезируют новую информационную РНК
(мРНК), являющуюся носителем памяти —
молекулами памяти. Благодаря их
образованию, т. е. функционированию в
соответствующих нейронах неких новых участков
ДНК, в мозгу животных запечатлеваются
определенные поведенческие навыки.
За этими работами последовало
множество публикаций, в которых описывались
опыты по влиянию торможения синтеза РНК
и белков в мозгу с помощью антибиотиков
актиномицина, пуромицина и циклогексимида
на процессы обучения разных видов
животных (крыс, мышей, рыб, птиц). В них было
показано, что такое вмешательство
препятствует овладению разными навыками, в
частности, мешает нахождению правильного
пути при обучении в лабиринте.
Результаты этих опытов как будто подтверждали
гипотезу Хидена. Дальше — больше: группа
американских исследователей выделила из
мозга обученных крыс РНК и инъецировала
ее в мозг необученных животных.
Получалось, будто подопытные животные
обучались быстрее контрольных. Наконец, стали
кормить необученных плоских червей —
планарий обученными и, регистрируя
скорость обучения, обнаружили ее
возрастание. Американский медик Д. Камерон
написал статью, в которой утверждал, что
кормление больных с потерей памяти
препаратами РНК способствует частичному ее
восстановлению.
Однако эта на первый взгляд
правдоподобная гипотеза не подтвердилась.
Методика определения качественного состава
РНК, разработанная Хиденом и в свое время
наделавшая много шума, оказалась
слишком грубой и не отражала реальной
ситуации, наблюдавшейся в клетке. Когда
появилась более точная методика, оказалось, что
качественных изменений РНК в нейронах
при обучении не происходит, могут иметь
место лишь изменения количественного
соотношения разных фракций мРНК, уже
существовавших в клетке. Шведский
биолог А. Эдсрем обнаружил, что торможение
синтеза РНК актиномицином в нейроне
рецептора натяжения рака никак не сказы-
128
ГЕНЕТИКА
вается на функции этой клетки. А недавно
проведенные опыты сотрудника
Института молекулярной биологии РАН Е.
Белявского показали, что при обучении улиток не
происходит включения новых генов, но
возрастает активность уже работающих зон
ДНК, что можно наблюдать не только при
обучении, но и при обычных
функциональных нагрузках. Иными словами,
наблюдаемые в процессе обучения количественные
изменения транскрипции в клетках мозга
отражают изменение функционального
состояния нейронов и не являются чем-то
специфическим для процессов обучения.
Что же касается влияния
антибиотиков на процессы обучения, то мы
исследовали их воздействие на морфологию
клеток мозга в месте их введения в самых
малых дозах, гораздо меньших, чем
использовали в опытах по обучению, и
показали, что даже эти малые дозы
вызывали патологические изменения в нервных
клетках, в том числе и их гибель.
Американцы нашли также, что антибиотики
могут блокировать процессы синаптической
передачи импульсов от нейрона к
нейрону, т. е. вызывать физиологические
нарушения в функционировании мозга
независимо от того, каково состояние работы
генетического аппарата клеток. Таким
образом, введение антибиотиков в мозг
препятствует обучению потому, что выводит
из строя целые функциональные
ансамбли нервных клеток в зоне введения
препарата. Регистрация скорости обучения в
опытах по введению РНК обученных
животных в мозг необученным оказалась
дефектной. В письме крупнейших
американских нейробиологов в журнал «Science» эти
данные были опровергнуты. В лаборатории
Корочкина в Новосибирске также
проводили эксперименты по проверке
подобного рода опытов с внутримозговымш
инъекциями РНК; результаты во всех случаях
были отрицательными. Что касается
кормления планарий, то оказалось: в теле
обученных животных образуются
стимуляторы нервных клеток, ускоряющие
синаптическую передачу импульсов от
нейрона к нейрону; именно это
обстоятельство и обусловило поразительный эффект
в экспериментах с каннибализмом.
Американский генетик Т. Тули
пытался обнаружить активацию генов в
процессе обучения дрозофилы, используя те же
антибиотики — актиномицин и пуромицин.
Его эксперименты отличались тем, что
были применены два способа обучения:
массированный и растянутый во времени. В
первом случае животных обучали
непрерывно, во втором делали перерыв после
опыта, а потом его возобновляли.
Оказалось, что в первом случае антибиотики,
подавлявшие синтез РНК и белка, не
влияли на ход обучения, во втором —
тормозили приобретение новых навыков.
Однако из его опытов не следует, что для
успешного обучения требуется активация
новых генов. Вполне возможно, что в
связи с возросшей функциональной нагрузкой
в нервных клетках начинают более
интенсивно транскрибироваться ранее уже
«работавшие» гены. Кроме того, им не был
проведен адекватный морфологический и
физиологический контроль, так что и в этом
случае нельзя говорить о вовлечении
генетического аппарата нейронов в
процессы обучения и памяти.
Приходится констатировать, что
генетические механизмы регуляции поведения
заложены, как и предполагал в начале века
испанский нейрогистолог, лауреат
Нобелевской премии С. Рамон-и-Кахал, в
структуре нейронных ансамблей, особенностями
формирования которых в онтогенезе и управляют
гены. Действительно, с морфофизиологичес-
кой точки зрения, тот или иной поведенческий
акт или процесс обучения обусловлен
формированием и замыканием новых нервных
связей. При этом, поскольку запоминание
часто происходит мгновенно, по всей
вероятности, физиологической основой этого
события является «пробой» не
функционировавших ранее синаптических контактов
между нейронами, включенными в
выполнение данной поведенческой реакции.
Синапсы эти являются предсуществующими: они
формируются в результате химического
взаимодействия дифференцирующихся в
онтогенезе нервных клеток и как бы ждут
своего часа, когда возникшие в ходе
жизни функциональные нагрузки «включат» их
в общую, активно работающую в
определенных условиях нервную сеть.
Раньше постулировали, что
образование новых нервных связей и
соответственно их включение в функционирование
нейронных ансамблей связаны с ростом новых
отростков, соединявших до того
изолированные друг от друга нейроны. Однако
скорость этого роста слишком мала
(миллиметры в сутки), чтобы объяснить быстрое
запоминание. Отмечаемые порой различия
в активности генетического аппарата или
отдельных генов между хорошо и плохо
обучающимися линиями животных являются
исходной характеристикой линий животных,
сложившейся в процессе дифференциров-
ки мозга в онтогенезе и обусловленной
генетически детерминированными
межлинейными различиями. В частности, в описанных
выше экспериментах по селекции хорошо
и плохо обучающихся крыс, выполненных
Корочкиным совместно с Шумской,
наблюдается именно такая ситуация. Можно
назвать и те генетически детерминированные
ГЕНЫ И ПОВЕДЕНИЕ
129
особенности мозга, от которых зависит
эффективность его функционирования:
1) масса ткани или количество
составляющих ее клеток, которое в свою очередь,
определяется соотношением темпов
размножения и гибели клеток в ходе онтогенеза;
2) способность нейронов к образованию
отростков и синаптических окончаний —
контактов между клетками: чем больше
отростков и синапсов образуется в ходе
онтогенеза и клеточной дифференцировки
и чем больше связей установится между
клетками, тем лучше;
3) способность образовавшихся синапсов
к функционированию;
4) региональные особенности
распределения клеток внутри органа, соотношение
нервных и вспомогательных, глиальных
клеток;
5) адаптивные и регулятивные
способности клеток;
6) способность определенных клеток
погибнуть в определенный момент
индивидуального развития, дабы обеспечить
более эффективное функционирование
оставшихся в живых нейронов (см.: Агол В. И.
30
431,0 ±6,0
20
10
CL
Ф
S
I
СО
*
Q.
ft
§30 г
о
I
I
§
487,0 ± 10,0
20
10
о
30
20
10
466,0 ± 11,0
о
ю
см
I
о
о
см
о
ю
со
I
о
о
со
о о
ю ю
^ ю
I I
о о
о о
^- ю
о
ю
CD
I
о
о
CD
Количество РНК, пг/нейрон
Рис. 4. Гистограмма распределения нейронов
вестибулярного ядра Дейтерса мозга крысы до и после
ряда аудиогенных эпилептических припадков
а— контрольные крысы, бив— крысы линии Кру-
шинского—Молодкиной, характеризующиеся
генетически детерминированной способностью реагировать
на звук эпилептическим припадком (до и после
припадков). Указано среднее количество РНК (в
пикограммах), приходящееся на один нейрон
Генетически запрограммированная смерть
клеток. Наст. том).
Молекулярные события являются
вторичными, что отчетливо прослеживается
в некоторых случаях генетически
детерминированной нейропатологии. К таким
случаям относится, например, генетически
детерминированная аудиогенная
(вызываемая прозваниванием) эпилепсия у крыс
линии Крушинского-Молодкиной,
селекционированных на кафедре высшей нервной
деятельности МГУ. В Новосибирском
Институте цитологии и генетики Сибирского
отделения АН СССР (ныне РАН) при
помощи современных методов молекулярной
цитологии исследовали нервные клетки в
тех отделах мозга, которые участвовали в
генезе аудиогенных эпилептических
припадков. Было установлено, что после
многократных индуцированных звуком
припадков количество РНК в соответствующих
нервных клетках снижалось, а после одно-
и двукратных припадков количество РНК
в этих клетках под влиянием повышенной
функциональной нагрузки повышалось
(рис. 4).
Сходные данные получены и при
помощи методов молекулярной биологии в
Институте биологии гена РАН в Москве, в
лаборатории А. П. Рыскова. Более того,
оказалось, что линия крыс, генетически
предрасположенная к аудиогенным
эпилептическим припадкам, исходно характеризуется
повышенным содержанием РНК в нейронах,
участвующих в генезе аудиогенного
эпилептического припадка, а снижение этого
количества путем подавления синтеза РНК
приводит к утрате подопытными животными
способности отвечать на воздействие
звука эпилептическим припадком.
Известны, однако, случаи, когда
молекулярные события запускают каскад
поведенческих реакций. Об этом свидетельствуют
изящные исследования регуляции
полового поведения у моллюска Aplysia,
выполненные группой Р. Шелера в США и в
некоторых других лабораториях. Они изучали
сложную поведенческую реакцию кладки яиц.
Эта кладка представляет собой длинный
шнур, содержащий более миллиона яиц. Как
только под действием сокращающихся мышц
протока гермафродитной половой железы
яйца начинают выталкиваться наружу,
улитка прекращает двигаться и питаться.
Она захватывает шнур яиц ртом и, двигая
головой вперед-назад, способствует его
выходу из протока железы, а затем
скручивает в моток. Специальная железа во рту
выделяет клейкое вещество, которое
прилипает к клубку яиц. В конце концов
животное энергичным движением головы
прикрепляет кладку к твердой основе, например
к поверхности камня.
130
ГЕНЕТИКА
Кислый
ГОЯ пептид
Предшественник
Пептиды
Пазушные
клетки
Брюшной
ганглий
L1, R1
L2, L3, L4, L6
R15
Проток
гермафродитной
железы
При отсутствии
пептида
В присутствии
пептида
Возбуждение
Торможение
Усиление
Сокращение
Рис. 5- Предшественник ГОЯ — белок, в состав которого входит несколько активных пептидов
В молекуле предшественника (вверху) имеются 10 точек (отмечены стрелками), в которых полипептидную цепь
могут разрезать ферменты эндопептидазы. При расщеплении по всем точкам должно образоваться 11
пептидов (второй рисунок сверху). Известно, что пазушные клетки выделяют четыре из них: а- и р-факторы, ГОЯ и
кислый пептид. Пептиды, выделенные цветом, функционируют в качестве медиаторов, влияя определенным
образом на активность конкретных нейронов брюшного ганглия; р-фактор возбуждает клетки L1 и R1; а-фактор
тормозит клетки L2, L3, L4, L6; ГОЯ усиливает пульсацию нейрона R15. ГОЯ выделяется также в кровоток и
действует как гормон, вызывая сокращение мускулатуры в протоке гермафродитной железы. Внизу
схематически показано, как действуют пептиды и ГОЯ на свои мишени
Были идентифицированы скопления
нейронов, регулирующих комплекс этих
поведенческих актов; затем удалось показать,
что нейроны активируются продуктами гена,
кодирующего гормон откладки яиц (ГОЯ),
причем сначала образуется
белок-предшественник, в состав которого входит
несколько активных фрагментов-пептидов (рис. 5).
Этот предшественник расщепляется
специальными ферментами, и продукты
расщепления влияют на функционирование нервных
клеток, управляющих кладкой яиц,
стимулируя одни клетки и тормозя другие. От
мозаики распределения активированных и
заторможенных нейронов и зависит успех
реакции в целом (см. рис. 5). Интересно, что
гомологичные пептиды недавно были
найдены у плодовой мушки дрозофилы, у
которой они тоже регулируют половое
поведение.
Упомянутые работы дают, пожалуй,
первый пример, показывающий каким образом
осуществляется связь гена и его продуктов
с точками приложения его влияния, в
данном случае с нейронными ансамблями,
управляющими совокупностью определенных
ГЕНЫ И ПОВЕДЕНИЕ
131
поведенческих актов. Известно множество
мутаций, так или иначе изменяющих
поведение. Например, у мышей описаны
«танцующие», «вращающиеся», агрессивные и
спокойные мутанты. Показана генетическая
детерминированность и форм полового
поведения. Генетик из Санкт-Петербурга Л. 3. Кай-
данов вывел мушек дрозофил, у которых
самцы могут характеризоваться очень
высоким или, наоборот, очень низким уровнем
половой активности, измеряемой по
количеству попыток ухаживать за самками и
временным интервалом между этими
попытками. У дрозофилы выделен ген, отвечающий
за гомосексуализм; предполагается наличие
сходного гена и у млекопитающих, у
которых подобные нарушения полового
поведения также известны. Функциональное
значение некоторых генов, ответственных за
поведение, идентифицировано, и начато
исследование механизмов их влияния на
функции нейронов и на само поведение.
Генетика поведения дрозофилы
Из генов, ответственных за поведение
дрозофилы, лучше всего изучены те,
которые контролируют такие простые
функции, как зрение, обоняние, способность к
обучению и брачное поведение.
Гены, контролирующие зрение
К настоящему времени изучено
несколько генов, контролирующих зрительные
функции:
Ген sevenless (sev). Мутанты по гену не
обнаруживают нормального фототаксиса на
ультрафиолетовый свет. Изменяется и
стереотип поведения: самки, у которых этот
ген поврежден, предпочитают откладывать
яйца на поверхности, окрашенные в
голубой цвет, а не в серый, желтый или
коричневый, которым обычно оказывается
предпочтение.
Ген optomotor-hlind (omb). Гомозиготы по
мутациям этого гена не реагируют на
вращающиеся вертикальные полосы;
особенно сильно действие мутаций проявляется
во время ходьбы, и несколько слабее — в
полете. У мутантов по гену отЪ
отсутствует гигантский тяж нейронов в оптической
доле головного мозга. У других двукрылых
насекомых, более крупных, чем
дрозофилы, эти тяжи необходимы для
обнаружения передвигающихся предметов.
Молекулярные размеры гена отЪ велики — около
100 тыс. пар нуклеотидов (т.п.н.). С этого гена
считывается около 20 видов различных
молекул мРНК.
Ген small-optic-lobes (sol). У мутантных
куколок дегенерирует около 50% клеток в
оптической доле головного мозга. В
результате у мух нарушаются система ориентировки
и правильность поведения при посадке после
полета. Ген sol клонирован, он
располагается в участке ДНК размером 14 т.п.н. С него
считываются два транскрипта длиной 5,8 и
5,2 т.п.н. Больший транскрипт кодирует
белок, состоящий из 1597 аминокислот. Этот
белок имеет способность связываться с ДНК,
т. е. может блокировать активность
регулируемых им генов.
Гены, контролирующие обоняние
Как на личиночной, так и на имагиналь-
ной стадии, дрозофилы могут различать
большое количество различных запахов.
Хирургические эксперименты показали,
что запахи улавливаются особыми
чувствительными органами, расположенными на
антенне. В этом органе есть поры, через
которые различные химические вещества
могут проникать и приближаться к нервным
окончаниям. От антенн идут нейроны к
головному мозгу. Нейронов очень много,
около тысячи, поэтому неудивительно, что
муха так легко различает многочисленные
запахи.
К настоящему времени обнаружены
шесть генов, контролирующих
чувствительность к запахам альдегидов и эфиров: olfA,
olfB, olfC, olfD, olfE, sbl. Ген olfE клонирован,
он имеет небольшие размеры, около 14 т.п.н.
С него считываются два транскрипта
длиной 5,4 и 1,7 т.п.н.
Гены, контролирующие способность
к обучению
Взрослые мухи дрозофилы обладают
способностью связывать ощущения запахов
с болевыми ощущениями от
электрического тока, т. е. у них можно вызывать
формирование условного рефлекса. К
настоящему времени открыты два гена, влияющие
на эффективность обучения. Это гены dunce
(due) и rutabaga (rut). Dunce («тупицы») —
мутанты, не способные обучаться.
Соответствующий ген кодирует фермент
циклическую фосфодиэстеразу. Он
замечателен тем, что имеет колоссальные
размеры — более 140 т.п.н. С него считывается
много различных типов мРНК. Его
функция осуществляется в клетках
определенной доли головного мозга (ганглия) мух —
в грибовидном теле. Если у древесных
муравьев хирургически разрушить это тело,
у них нарушаются основы их социального
поведения. Эти данные свидетельствуют о
роли грибовидного тела в осуществлении
сложных поведенческих реакций.
Интересно, что у молодых мух в грибовидном теле
быстро возрастает число новых нейронов,
что связывают с накоплением опыта в
процессе обучения. Ген rutabaga кодирует
фермент аденилатциклазу, индуцируемую
ионами кальция. Его находят также у
132
ГЕНЕТИКА
млекопитающих, у которых этот фермент
участвует в процессах осуществления
запоминания. Таким образом, по крайней мере
некоторые этапы обучения у насекомых и
млекопитающих имеют общие черты.
Гены, контролирующие
брачное поведение
К настоящему времени брачное
поведение мух, первые исследования которого
были проведены в 1956 г., описано во всех
деталях. И самцу и самке присущи довольно
сложные наборы движений, так
называемые брачные танцы. В это время
инициатива принадлежит самцу, он трогает
самку, бегает вокруг нее, преследует, исполняя
при этом «песню любви». Песня
представляет собой вибрацию крыльев (рис. 6, а)
продолжительностью 55 с, состоящую из
импульсов в 20-30 мс и межимпульсового
промежутка в 30-40 мс (рис. 6, б).
Очевидно, процесс ухаживания
довольно сложен и любые мутации, влияющие на
остроту зрения, обоняния, слуха, способность
выдерживать генетически закрепленную
периодичность «песни любви», приводят к
нарушению процесса ухаживания и его
эффективности, заключающейся в
скрещивании. Так, слепой самец не может увидеть
самку, в свою очередь, глухая самка не
может услышать «песню любви»,
исполняемую самцом. Оба пола, не различающие
запахов, все-таки могут вступить в
копуляцию, но у них на это уходит больше вре-
а
б
в
г
Д
Рис. 6. Положение самца и самки во время
ухаживания (а), акустические характеристики песни:
нормального самца дикого типа (wild-type) (б), мутантов
dissonance (e, г) и cacophony (д) (по Дж. Холлу)
мени, по крайней мере до начала
ухаживания. Эти нарушения полового поведения
ожидаемы, однако есть и более тонкие
механизмы. В результате исследований
американского ученого Дж. Холла обнаружены
мутации, по крайней мере, трех типов,
влияющие на исполнение «песни любви». В
результате мутации cacophony резко
увеличивается амплитуда колебаний во время
импульса, мутация dissonans изменяет песню
таким образом, что вместо правильного
чередования импульсов и межимпульсовых
промежутков наблюдается один мощный
продолжительный импульс (рис. 6, в—д).
Мутации гена clock (часы) изменяют
продолжительность одного цикла «песни любви».
Вместо общей продолжительности одного
цикла песни в 55 с некоторые мутации
уменьшают ее до 40 с, другие увеличивают до
80 с. Изменение акустической структуры
«песни любви» резко меняет и эффективность
ухаживания самца за самкой.
Кроме того, известен тел fruitless (fru) —
бесплодный. Мутации этого гена полностью
изменяют половое поведение самцов —
возникают сразу три нарушения: 1) они не
делают попыток ухаживать за самками,
2) ухаживают только за самцами —
такими же гомозиготами по этой мутации, 3)
стимулируют нормальных самцов ухаживать за
собой. Естественно, эти самцы потомства не
оставляют, это мухи-гомосексуалисты.
Мутанты fru выделяют специфическое
вещество с соответствующим запахом,
привлекающим других самцов. Ген fru активен
в небольшой антеннальной доле головного
мозга («грибовидном теле»), которая
получает информацию о запахах. Первые
данные свидетельствуют о наличии неболышэй
доли мозга, оценивающей информацию о
запахах, в которой происходит выбор, по
какому пути пойдет половое поведение —
по типу самца или самки. И этот выбор
контролируется геном fru, который
локализуется в третьей хромосоме. Данные
поистине удивительные: одна-единственная
мутация приводит к выделению только
одного нового вещества и самец, имеющий все
остальные признаки самца, становится
гомосексуалистом.
Гены, влияющие на биоритмы
У дрозофилы ген period (per)
контролирует правильное осуществление биоритмов.
Биоритмика у мух обнаружена при откладке
яиц, вылуплении эмбрионов и имаго из
куколок. Биоритмична также брачная
песня самцов. Выделены три группы мутаций:
perS укорачивают биоцикл с 24 до 19 ч,
мутации группы perL удлиняют его до 29 ч,
a perOl полностью нарушают ритмику
биологических процессов. Установлено, что ген
функционирует в клетках головного мозга,
ГЕНЫ И ПОВЕДЕНИЕ
133
он кодирует белок, состоящий из 1200
аминокислот. В середине данной белковой
молекулы находится 20-кратный повтор пары
аминокислот треонин-глицин. Именно этот
повтор влияет на продолжительность «песни
любви». Если его экспериментально удалить,
длина песни укорачивается с 55 до 40 с.
Таким образом, даже такая сложная
функция, как следование биоритмам или
«ощущение времени», контролируется только
одним геном, работающим в мозгу.
Генетика поведения человека
При изучении генетики поведения
человека используют главным образом метод
анализа идентичных близнецов, т. е. таких
близнецов, которые имеют одинаковые
наборы генов. Используют тесты — серии
вопросов и задач, в зависимости от числа
и качества решения которых выводится
количественная оценка ответа, такая же
условная, как школьные баллы. По этим
баллам и сравнивают двух близнецов. С их
помощью можно оценить такие различные
психические функции, как ум, внимание,
темперамент, фантазия, память и т. д.
При внутрипарных сравнениях
идентичных близнецов по различным признакам,
характеризующим интеллект, около 60%
пар обнаруживают разницу, не
превышающую 2,5 баллов, остальные 40%
различаются на величину от 2,5 до 15 баллов. При
аналогичных сопоставлениях неидентичных
близнецов 40% пар имеют разницу не
более 2,5 баллов, 20%— от 2,5 до 7,5,
остальные 40%— от 7,5 до 45 баллов. Такие
сопоставления ясно свидетельствуют о том,
что люди, имеющие одинаковые генотипы,
обладают очень близкими
интеллектуальными возможностями.
Были проведены сопоставления пар
идентичных и неидентичных близнецов по
признакам, характеризующим
предрасположенность к определенным сферам занятий,
таким, как наука, бизнес, религиозная или
артистическая деятельность. Результаты
показали, что у идентичных близнецов
коэффициенты корреляции между проявлением
одинаковых способностей значительно выше,
чем у неидентичных близнецов.
О возможности наследования
определенного уровня интеллектуальных
способностей свидетельствуют результаты
вычисления корреляций между коэффициентами
интеллектуальности (IQ) у людей,
состоящих в разной степени родства. Оказалось,
что самое низкое соответствие величин IQ
наблюдается у людей из различных
неродственных групп. Для них коэффициент
корреляции составляет 0-0,2. Корреляция между
отцом и приемным ребенком также
невелика— в пределах 0,15-0,35. Корреляции
между отцом и любым из родных детей
значительно выше — 0,2-0,8. Широкий размах
варьирования может свидетельствовать о
зависимости этой связи от расщепления
хромосом. У идентичных близнецов
коэффициенты корреляции еще выше — 0,35-
0,85.
Особый интерес представляют данные о
корреляциях в проявлении свойств
интеллекта у идентичных близнецов, особенно
таких, которые от рождения воспитывались
в разных семьях. В данном случае один и
тот же генотип реализуется в разных
условиях окружающей среды. У идентичных
близнецов, развивавшихся раздельно,
коэффициент корреляции варьировал в
пределах 0,6—0,85, у близнецов при
совместном воспитании значения были 0,75-0,95.
Различия в 0,10—0,15 можно целиком отнести
на счет окружающей среды. Видно, что это
влияние невелико и основной вклад в
формирование интеллектуальных
характеристик вносит генотип.
При анализе предрасположенности к
совершению преступлений был
использован семейный метод. Предположение, что
генетические факторы могут быть среди
причин криминального поведения, было
проверено на 14427 осужденных
приемных детях путем сравнения их с
биологическими и приемными родителями.
Никакой корреляции между приемными детьми
и родителями не было обнаружено. Однако
между детьми и их биологическими
родителями обнаружена достоверная
корреляция между частотой привлечения к
уголовной ответственности детей и числом
осуждений у их биологических отцов.
Таким образом, обстановка приемной семьи
мало повлияла на развитие наклонностей
усыновленных.
С середины прошлого века проводится
изучение предрасположенности к
алкоголизму. Уже в начале уходящего столения по
результатам анализа 4 тысяч алкоголиков,
которых на протяжении 35 лет лечили в одной
из клиник, было показано, что эта болезнь
имеет семейные корни. За более чем 80 лет,
прошедших после этой первой публикации,
сделанные в ней выводы получали все
новые и новые подтверждения. Однако
наличие семейных случаев не обязательно
свидетельствует о наследственном характере
недуга. И здесь близнецовый метод анализа
дал хорошие результаты. Наиболее
известна работа, проведенная во второй
половине этого века на 902 парах близнецов из
финской популяции. В этом исследовании
алкоголиков анализировали по трем
показателям: а) частоте выпивок, б) количеству
выпитого и в) потере контроля над собой при
опьянении. Только по последнему
признаку не было обнаружено генетической детер-
134
ГЕНЕТИКА
минированности. По первым двум
признакам коэффициенты корреляции составляли
0,39 и 0,40 соответственно. Аналогичные
данные были получены несколько позже
опять на финской, а также шведской и
американской группах близнецов.
Анализ семей с приемными детьми
показал, что если ни у биологических, ни у
приемных родителей алкоголизм не
отмечается, только около 5% усыновленных
становятся алкоголиками. Однако если у
биологических родителей отмечен
алкоголизм, а у приемных нет, уже 18% (в 3,5
раза больше) приемных детей становятся
алкоголиками.
Заключение
Рассмотренные выше результаты
экспериментов, выполненных на дрозофиле,
позволяют сделать два вывода: даже
сложные типы поведения, например процесс
ухаживания, можно разделить на серию
простых реакций. Огромную роль здесь
играют формирование зрительных и
осязательных образов, восприятие определенных
типов звуковых колебаний и их ритмики.
Твердо установлено, что за осуществление
всех этих более простых признаков
отвечают обычные гены, кодирующие белки и
ферменты. Исследование генетического
контроля наиболее сложных поведенческих
реакций у человека, таких, как мотивация
поступков, предрасположенность к
определенному типу поведения, выбор сферы
деятельности, еще только начинается. Тем не
менее не вызывает сомнения, что и здесь
роль наследственности является решающей.
Работы такого рода набирают силу,
однако генетика поведения таит в себе еще
много загадок, особенно в части,
касающейся молекулярно-генетических основ
запоминания (в частности, тех процессов,
которые происходят в синапсах), отношения
между молекулярными и морфогенетичес-
кими событиями, поведения нейронов при
различных формах патологии,
функциональной значимости вспомогательных глиальных
клеток и многого другого. Эти загадки ждут
решения, и тем, кто интересуется этими
проблемами, предоставляется обширное
поле деятельности.
Литература
Актуальные проблемы генетики поведения.
Л.: Наука, 1975.
Канаве И. И. Близнецы и генетика. Л.:
Наука, 1968.
Крушинский Л. В. В кн.: Актуальные
вопросы современной генетики. М.: Изд-во МГУ,
1965, с. 281.
Крушинский Л. В., Зорина 3. А., Полетаева
И. И., Романова Л. Г. Введение в этологию и
генетику поведения. М.: Изд-во МГУ, 1983.
Нейрогенетика и генетика поведения:
Успехи современной генетики. М.: Наука, 1991,
т. 17.
Полетаева И.И., Романова Л.Г. Генетические
аспекты поведения животных. (Итоги науки и
техники. Сер.: «Физиология человека и
животных». Т. 42). М.: ВИНИТИ, 1990.
В. П. Пузырев
ГЕНОМНЫЕ ИССЛЕДОВАНИЯ И БОЛЕЗНИ ЧЕЛОВЕКА
Введение
Объективность генетической
классификации болезней растет по мере успехов
молекулярной генетики и цитогенетики в
обнаружении дефектных генов,
вызывающих конкретные болезни. Это касается как
собственно наследственных (менделирую-
щих, или моногенных), так и мультифак-
ториальных (многофакторных) заболеваний
(МФЗ), в возникновении и развитии
которых существенны как генетические
факторы, так и факторы внешней среды.
Однако до сих пор доля болезней, зависящих
от известных конкретных «патологических»
генов, остается весьма небольшой. В
каталоге наследственных признаков человека за
1992 г. хромосомная локализация генов и их
первичные продукты указаны лишь для 322
из 5710 описанных признаков (т.е. всего в
5,6% случаев), а для 2,5 тыс. указана только
локализация. Возможное же число
наследственных болезней человека намного
больше и составляет не менее 50-100 тыс.
Выявить все такие болезни и хромосомные
синдромы невозможно, так как нарушения
структуры многих белков несовместимы с
жизнью и приводят к ранней
внутриутробной гибели плода; сейчас известно более
3000 из них.
Таким образом, наши знания о молеку-
лярно-генетических основах болезней
человека касаются пока относительно
небольшого их числа. В то же время в клинической
генетике известны примеры, показывающие
исключительное значение этих знаний для
диагностики, раннего выявления,
эффективного лечения и профилактики
конкретных наследственных болезней (например,
фенилкетонурии, муковисцидоза,
гипотиреоза, гиперхолестеринемии). Современные
достижения молекулярной генетики, а
точнее «методическая революция» 70-х гг.,
сделали возможным выделение
индивидуальных генов, химический анализ нуклеотидных
последовательностей ДНК, функциональное
изучение экспрессии генов и молекулярных
механизмов ее регуляции. Все это
создало предпосылки для формирования новых
направлений прикладной молекулярной
генетики — анатомии и патологической
анатомии генома человека, имеющих
непосредственное отношение к проблемам медицины.
Наиболее последовательно эти
направления развиваются в рамках проекта «Геном
человека». Проект действует во многих
странах, доказал свою жизнеспособность и
эффективность. Он пролагает путь в биологию
XXI в.
В этой статье рассказывается о целях
геномных исследований, технологии их
осуществления и некоторых приложениях,
которые они получили в медицине.
Груз наследственной патологии
в современных популяциях
человека
Понятие «генетического груза»
популяций человека введено американским
генетиком, лауреатом Нобелевской премии
Г. Меллером. Впоследствии он со своими
соотечественниками Н. Мортоном и Дж. Кроу
обосновали подход к оценке относительного
вклада в общий объем наследственной отя-
гощенности процессов мутаций
(мутационного груза) и расщепления
(сегрегационного груза), а также предложили
методический прием определения объема
генетического груза у человека. Однако
разработанная ими концепция, оказавшая
большое влияние на развитие популяци-
онной генетики человека, не получила
всеобщего признания, поскольку не всегда
дает реалистические оценки
генетического груза. Необходимо отметить, что
методологии точного измерения его объема до
сих пор нет. Но даже если современная
наука не может измерить генетический
груз, это не означает, что он не
существует. В настоящее время накапливается
информация о грузе наследственной
патологии среди населения, основанная на
результатах генетико-эпидемиологических
исследований и анализе медицинских
данных лечебно-диагностических учреждений.
Приведем некоторые литературные
данные, обобщающие эту информацию.
Среди лиц до 21 года у 10,6%
выявляются врожденные дефекты. Часто такие
дети и подростки рано умирают, другие
нуждаются в специальной медицинской
помощи и требуют больших затрат на их
содержание и социальную реабилитацию.
Ежегодно в мире рождаются свыше 5 млн
детей с тяжелыми врожденными
дефектами развития. Однако следует иметь в виду,
что некоторые наследственные аномалии
развития впервые проявляются в среднем,
зрелом и даже пожилом возрасте.
В основе моногенных (менделирующих)
болезней лежат мутации отдельных генов
(доминантные и рецессивные). Изменение
136
ГЕНЕТИКА
структуры и числа хромосом приводит к
хромосомным болезням. Однако во многих
случаях врожденные дефекты человека
возникают в результате одновременного
проявления комплекса разных мутаций.
Это — уже упоминавшиеся мультифакто-
риальные заболевания (МФЗ), или
полигенные болезни. Мультифакториальным
(полигенным) наследованием обусловлены многие
врожденные пороки развития. К МФЗ
относятся и такие широко распространенные
заболевания, как сахарный диабет,
гипертоническая болезнь, коронарная болезнь,
бронхиальная астма, шизофрения и др. Они
являются результатом сложного
взаимодействия множества генетических и средовых
факторов. Рис. 1 дает схематическое
представление об относительной роли
генетических факторов и факторов среды,
участвующих в возникновении и развитии МФЗ.
На схеме Г. Харриса область,
ограниченная внешней окружностью, это популяция
в целом, а площадь внутреннего круга —
те индивидуумы данной популяции,
которые наследственно предрасположены к МФЗ
определенного типа. Область, заключенная
между двумя радиусами большей
окружности, соответствует той (в данном случае
меньшей) части популяции, которая
подвергается воздействию факторов среды,
провоцирующих конкретное заболевание; ос-
а
в
Мультифакториальное
заболевание
Совокупность
генетических
факторов
б
G
Е
Средовые
факторы
Случайные
факторы
Главные гены
Полигены
(«генетический фон»)
тальная (большая) часть популяции не
подвергается воздействию этих факторов.
Заболевание развивается у небольшой части
популяции, т. е. у тех индивидуумов, у
которых генетическое предрасположение
сочетается с воздействием неблагоприятных
условий среды. На схеме Ф. Фогеля и А. Мо-
тульски (рис. 1) подчеркнута потенциальная
роль одного или нескольких «главных»
генов в развитии многих предположительно
полигенных признаков и МФЗ.
Предполагается, что достаточно малое число
главных генов может определять основной вклад
в генетическую этиологию МФЗ.
Совокупность всех других генов, против которых
действуют главные гены, образует
«генетический фон». Хорошо известно, что
генетический фон может модифицировать
экспрессию главных генов. Такая «игра
природы и судьбы», взаимодействие и
взаимообусловленность генетической конституции
и окружающего мира становятся
предметом пристального внимания исследователей.
Понять правила этой игры — одна из
важнейших задач экологической генетики.
Ю. П. Алтухов, обобщая литературные
данные за 30 лет, заключает, что в
европейском населении 15% человеческих
эмбрионов погибают на ранних стадиях
развития (спонтанные аборты), 3% составляют
мертворожденные, 2% приходится на нео-
натальную смертность, 3% — на смертность
до наступления репродуктивного возраста,
20% лиц не вступают в брак, 10% браков
бесплодны. Таким образом, не менее 50%
первичного генофонда не воспроизводится
в следующем поколении. Генетическая
компонента для всех этих явлений различна,
но в среднем составляет 20-30%. Следует
отметить, что генетическое
консультирование (одна из основных форм
профилактики наследственных болезней, наряду с
дородовой диагностикой и биохимическим
скринингом новорожденных), каким бы
квалифицированным оно не было, не в
состоянии устранить груз, а только
перераспределяет его. Можно дать совет не жениться
или не иметь детей, подобрать партнера,
который бы обеспечил здоровое потомство,
но и в этих случаях, констатируют
авторы, общество по-прежнему сохраняет
генетический груз.
Рис. 1. Общие представления об относительной
роли средовых и генетических факторов в развитии
мультифакторных заболеваний (МФЗ)
а — схема Г. Харриса [из: Харрис, 1973]; б: 1 —
собственно наследственная патология; 2— мультифак-
торные болезни; 3— ненаследственные болезни. По
абсциссе — средовые факторы (Е), по ординате —
генетические факторы (G); в— схема Ф.Фогеля и
А. Мотульски, с изменениями [из: Фогель, Мотульс-
ки, 1990]
Основные цели исследования
генома человека
Начальная цель геномных
исследований — создание серии описательных
диаграмм (карт) с высоким уровнем
разрешения для каждой хромосомы человека.
Картирование — это определение
положения и порядка генов и любых других
фрагментов ДНК на хромосомах. Следующий шаг
ГЕНОМНЫЕ ИССЛЕДОВАНИЯ И БОЛЕЗНИ ЧЕЛОВЕКА
137
после завершения картирования —
секвестрование, или определение
последовательности оснований во всех упорядоченных
(картированных) фрагментах ДНК.
Конечная цель исследования генома —
расшифровка его полной нуклеотидной
последовательности.
Картирование и секвенирование —
наиболее важные и трудоемкие части всех
геномных исследований, осуществляемых
в разных странах. Их результаты
должны привести к созданию подлинной
«молекулярной анатомии». Однако, как
заметил академик А. А. Баев, в итоге подобных
исследований мы получаем лишь
«мертвые символы нуклеотидных
последовательностей», и «нужно раскрыть их функцию».
И такая цель — провести
структурно-функциональный анализ генома человека в
отечественной программе «Геном человека»
сформулирована. Этот раздел программы,
вероятно, особенно важен для
клинической медицины, ставящей перед собой не
только задачи диагностики
наследственных болезней, но и их лечения — гено-
терапии.
Общее представление о физическом
объеме работы, которую предстоит
осуществить, иллюстрирует рис. 2. Если пос-
Геном человека: 200 томов
по 1000 страниц
Геном некоторых организмов
|| |||||| ||| Дрозофила 10 томов
D Дрожжи 1 том
We. coli (бактерия) 300 страниц
Рис. 2. Сравнительные размеры геномов человека
и некоторых организмов [из: Primer on Molecular
Genetics. Washington: US Department of Energy, 1992]
ледовательность секвенированной ДНК,
образованной 3 млрд п.н. и имеющей
совокупную длину 1,5 м, напечатать, то
получится 200 томов по 1000 страниц
каждый. Если бы последовательность генома
была опубликована, для ее прочтения
потребовалось бы не менее трети
человеческой жизни. Средние сроки осуществления
программы «Геном человека», которая
реализуется в США, европейских странах
и России— 15 лет. Автоматизация
методов и оптимизация технологии секвениро-
вания, мощная компьютеризация и
совершенствование специальных методов
информатики становятся главными
условиями эффективности геномных проектов.
Стратегия картирования
и секвенирования
Картирование в узком смысле —
определение положения гена или мутации в
хромосоме. Позднее этот термин получил
более широкое толкование и стал относиться
не только к гену, но к любому маркеру,
под которым подразумевают ген (мутацию),
участок ДНК с неопределенной функцией,
точку расщепления ДНК рестриктирующими
эндонуклеазами, т. е. любой наследуемый
признак, доступный идентификации тем или
иным способом. После установления
локализации любого из перечисленных
маркеров он используется для определения
относительного положения другого маркера.
Для определения положения маркера
(гена) в хромосоме используются
разнообразные методы: генетические, цитогене-
тические, молекулярно-биологические. В
зависимости от того, какие из них
применяются, различают два подхода к
картированию: генетический и физический.
Генетическое картирование —
определение положения фрагментов ДНК в
хромосоме с использованием генетических
методов, т. е. анализ сцепления и рекомбинации
генов на основе родословных. Карты,
полученные при использовании данного
метода, часто называют картами
генетического сцепления.
Маркеры, используемые в картировании,
должны быть полиморфными, т. е. должны
существовать альтернативные формы
признака, которые можно было бы легко отличить
и описать у членов семьи. В
последовательности ДНК варьирует (т. е. полиморфен) в
среднем один из 300—500 нуклеотидов. Вариации
ДНК в экзонах (кодирующих
последовательностях) могут приводить к видимым
изменениям — различию в цвете волос и глаз, группе
крови, особенностях строения тела,
деятельности физиологических систем, черт
характера, подверженности болезням и т. п.
Вариации же в интронах (некодирующих вставочных
последовательностях), а там появляется
большинство из них, имеют минимальный эффект
(или не имеют его) на внешние признаки и
функции организма. В то же время эти
изменения легко определяются на уровне ДНК
и могут использоваться как маркеры.
Маркеры этого типа включают, в частности: 1) ПДРФ
(полиморфизм длин рестрикционных
фрагментов) — изменение последовательности ДНК в
точках (местах, сайтах), которое «узнается»
рестрикционными ферментами
(эндонуклеазами) и 2) различия в числе тандемных
повторов — копий повторяющихся
последовательностей, которые приводят к легко измеримым
различиям по длине ДНК.
Карты генетического сцепления строят
на основе частот совместного наследования
138
ГЕНЕТИКА
пар маркеров. Два маркера, расположенные
на одной хромосоме вблизи друг от друга,
имеют тенденцию передаваться от родителей
к ребенку совместно. Во время нормальных
процессов формирования сперматозоидов и
яйцеклеток цепочка ДНК, локализованная
в тех или иных участках хромосомы или ее
гомолога, случайным образом разрывается,
а затем воссоединяется. Этот процесс,
называемый мейотической рекомбинацией,
может приводить к разделению двух
маркеров, первоначально расположенных на
одной хромосоме. Чем ближе расположены
маркеры друг к другу, тем «теснее» они
сцеплены, тем менее вероятно, что процесс
рекомбинации разделит их.
Вертикальные линии на рис. 3 изображают
пары 4-й хромосомы у каждого члена семьи.
Отец имеет два признака, которые в случае
передачи можно обнаружить у любого его
ребенка: известную короткую
последовательность ДНК, используемую в качестве
генетического маркера (М), и хорею Гентингто-
на (ХГ). То, что ребенок унаследовал только
один из этих признаков (М),
свидетельствует о рекомбинации генетического
материала в процессе сперматогенеза. Частота этого
события может быть использована для
определения расстояния между двумя
последовательностями ДНК на генетической карте.
Расстояние между маркерами
измеряют в сантиморганидах (сМ), названных так
в честь американского генетика Т.
Моргана. Когда частота рекомбинации между двумя
маркерами равна 1%, говорят, что они
находятся на расстоянии 1 сМ. Генетическое
расстояние в 1 сМ примерно равно
физической протяженности в 1 млн пар
оснований (м.п.н.). Уровень разрешения,
достигнутый на современной карте для большинства
регионов ДНК, составляет примерно 10 м.п.н.
Для медицины ценность генетических карт
состоит в том, что наследственная болезнь
Отец
Мать
М-
Маркер М
и ХГ хг4-
Дети
м-
м-
хг-
м-
хг-
Маркер М
иХГ
Только
маркер М*
Маркер М
иХГ
может быть локализована на них по данным
о наследовании маркера, который
присутствует у больных и отсутствует у здоровых,
причем даже в том случае, когда
молекулярные основы болезни или ответственный
за нее ген неизвестны. Сегодня уже
установлено точное хромосомное расположение
генов многих болезней человека.
Физическое картирование —
определение положения фрагментов ДНК в
хромосоме с помощью методов молекулярной и
клеточной биологии. Различные типы
физических карт различаются по уровню
разрешения. К низкоразрешающим относятся
хромосомные карты и карты кДНК1, к
высокоразрешающим — макрорестрикционная
карта и карта контиг2.
Хромосомные карты показывают
расположение генов или отдельных участков ДНК
на соответствующих хромосомах, а
расстояние между генами или фрагментами ДНК
указывают в парах оснований. Эти маркеры
могут быть физически связаны с
определенным сегментом хромосомы при гибридизации
in situ — методе, позволяющем пометить ДНК
специальной «видимой» (флуоресцентной или
радиоактивной) меткой. Локализация меченого
зонда устанавливается после того, как он
свяжется с комплементарной цепочкой ДНК
на интактной хромосоме. Точность
хромосомных карт была существенно увеличена
благодаря усовершенствованию метода
флуоресцентной гибридизации in situ и использования
для анализа хромосомы в интерфазной
стадии клеточного деления.
Карты кДНК показывают расположение
экспрессируемых участков ДНК (экзонов) по
отношению к регионам хромосом (бэндам).
На экспрессируемых участках ДНК транк-
рибируется (синтезируется) мРНК.
Используя в качестве матрицы молекулы мРНК,
в лабораторных условиях синтезируют их
копии, называемые сокращенно кДНК. Затем
каждый вид кДНК можно картировать в
геноме. Поскольку кДНК представляют эк-
спрессируемые участки генома, они
позволяют выявить те его части, которые
особенно значимы с медицинской точки зрения.
Для воссоздания оригинального
порядка фрагментов ДНК в геноме применяют
различные подходы. В некоторых из них
используют способность цепочек ДНК и (или)
РНК гибридизоваться — формировать двух-
цепочные участки, в которых цепочки
соединены водородными связями между
комплементарными основаниями. Для того чтобы
Рис. 3. Построение карты генетического сцепления
[из: Primer on Molecular Genetics. Washington: US
Department of Energy, 1992]
Звездочкой отмечен рекомбинант. Частота
рекомбинаций зависит от расстояния между маркером М
и геном хореи Гентингтона (ХГ)
1 кДНК — ДНК, комплементарная мРНК,
полученная путем обратной транскрипции;
используется как молекулярный зонд высокой
избирательности.
2 Контиги — группы клонов, охватывающие
перекрывающиеся участки генома.
ГЕНОМНЫЕ ИССЛЕДОВАНИЯ И БОЛЕЗНИ ЧЕЛОВЕКА
139
Хромосома
i i
Сверху
вниз
Макрорестрикционная
карта
(полная, но не детальная)
Карта контиг
(подробная, но не полная)
,-J II 1 III 1 IL1L-L,
Контиг
.-LLiiJ-,
1 ¦¦¦ 1 IIII,
, м i ш—ц
с±=ЬЬ
, II 1 1 1 1
сЬЬЬ
1111 1 Ш ,
,1111 1 III ,
2Му
Снизу
вверх
Поиск перекрывающихся фрагментов:
фингерпринтинг, картирование, сиквенс
Библиотека клонов
Рис. 4. Стратегия физического картирования [по: Primer on Molecular Genetics. Washington: US Department of
Energy, 1992]
определить, имеют ли фрагменты ДНК
общую последовательность и тем самым
перекрываются, используют метод фингерп-
ринтинга («отпечатков пальцев»).
На рис. 4 схематически представлена
стратегия картирования с высоким уровнем
разрешения. Стратегия картирования «снизу
вверх» заключается в разделении
хромосомы на небольшие участки, которые затем
клонируют и упорядочивают.
Упорядоченные фрагменты образуют протяженные
блоки ДНК (контиги). Получаемая в
результате библиотека клонов может включать от
10 т.п.н. до 1 м.п.н. Преимущество данного
подхода в доступности этих стабильных
клонов3 для исследований.
Сегодняшние технологические
достижения позволяют клонировать большие
участки ДНК с использованием искусственно
созданных хромосомных векторов, несущих
фрагменты ДНК человека размером до
1 м.п.н. Эти векторы используются в
клетках дрожжей в качестве «искусственной
хромосомы» (YAC)4.
Последняя стадия физического
картирования генома человека — определение всех
пар оснований для каждой хромосомы, т. е.
полной нуклеотидной последовательности ДНК.
Эта задача решается с помощью
современной технологии секвенирования, включающей
два основных этапа. Во-первых,
субклонирование фрагментов ДНК из космид5 или
библиотеки бактериофагов в специальные
векторы для секвенирования, несущие более
мелкие фрагменты ДНК по сравнению с
исходными. Во-вторых, дальнейшее
преобразование субклонированных фрагментов в
серию фрагментов, различающихся по длине
всего на 1 нуклеотид. Результаты
секвенирования объединяются затем в длинные
цепочки нуклеотидных последовательностей,
составляющих основу хромосомы.
Медицинские приложения
геномных исследований
Типы геномных карт представлены на
рис. 5, отражающем основные изложенные
выше подходы к картированию генома от
самого низкого уровня их разрешения до
самого высокого, т. е. до полного
выяснения нуклеотидной последовательности. Для
иллюстрации современного состояния
исследований по полному картированию генома
человека приведем генную карту его 1-й
хромосомы (рис. 6). В методическом плане
(т. е. по способу осуществления картирова-
Ген или
полиморфизм
Генетическая t
карта -Q
Ген или
полиморфизм
ч>
3 Клон — клеточная популяция, полученная
от одной клетки; в молекулярной генетике —
идентичные фрагменты ДНК.
4 YAC (сокр. от англ. Yeast Artificial
Chromosomes — искусственные хромосомы дрожжей) —
вектор, применяемый для клонирования очень
больших (до 400 т.п.н.) фрагментов ДНК.
5 Космида — искусственно созданный вектор
на основе генома фага лямбда, который
позволяет клонировать большие (до 45 т.п.н.) по
сравнению с плазмидами фрагменты ДНК.
Фрагменты
рестрикции I
nazi
Упорядоченная —
библиотека
Последовательность IIIIIIIIIIIIIIIIIIIIIIII1IIHHIIIIHIIIIIIIIIIIIIIIIIHHIII1IIIIIIIIIIIIIII1IIII1
оснований
Рис. 5. Типы карт генома [по: Primer on Molecular
Genetics. Washington: US Department of Energy, 1992]
140
ГЕНЕТИКА
р
ч
3
2
1
1
2
3
4
/ ^
36
34
32~
31 Е
22
21
13
12
21
23~
24 М
25
31 Н
32
„
42
i
¦
Рак груди (протоков)*
Недостаточность енолазы
Нейробластома*
?Гемолитическая анемия Rh-ноль
Эритробластоз плода
Эллиптоцитоз-1
Эритрокератодермия вариабельная
Гипофосфатазия, младенческая
Фукозидоз
Поздняя подкожная порфирия
Порфирия, гепатоэритропоэтическая
Недостаточность галактоэпимеразы
Ceroid lipofuscinos, нейронный-1, детский
Недостаточность С8, тип I и II
Лейкемия/лимфома, Т-клеточная
Недостаточность ацил-СоА-дегидрогеназы, средняя цепь
|_ Болезнь кленового сиропа, тип 2
Недостаточность миоаденилатдеаминазы
L Недостаточность АМФ-деаминазы эритроцитов
Гиперплазия надпочечников, тип II
L Гипотироидизм, незобный
Рис. 6, Патологическая
анатомия генома человека: гены
болезней, картированные на
первой хромосоме [из: Primer on
Molecular Genetics. Washington:
US Department of Energy, 1992]
Рамкой выделены аллельные
состояния; *—
новообразования, связанные со
специфической хромосомной перестройкой,
онкогеном или потерей гетеро-
зиготности опухолевыми
клетками; курсивом отмечены гены,
по которым наблюдается
несовместимость матери и плода
Миопатия из-за недостаточности ЦТФазы
Гипераммонемия из-за недостаточности ЦТФазы
Болезнь Гоше, 2 или более типов
]
Гемолитическая анемия в результате недостаточности ПК
Эллиптоцитоз-2
Пиропойкилоцитоз
Сфероцитоз, рецессивный
|_ Недостаточность антитромбина II
Болезнь Шарко-Мари-Туз, тип lb
Подверженность Vivax malaria
Синдром Криглера—Наджара
Немалиновая миопатия
Недостаточность фактора V
Lupus erythematosus, системный
Нейропения, иммунная
Катаракта опоясывающая, пулверулентная
Хронический грануломатоз в результате недостаточности
NCF-2
Гликогеноз VII
Недостаточность фактора XIIIB
Недостаточность CR1
Недостаточность фактора Н
Синдром Ушера, тип 2
Синдром ван дер Вуда
Недостаточность фумаразы
ния) полученные к настоящему времени
карты хромосом являются физическими,
хотя локализация отдельных генов
устанавливалась и генетическими методами, т. е. на
основе генетического сцепления. На этих
картах нанесены идентифицированные гены,
ответственные за наследственные
заболевания — «патологические» гены человека. Для
1-й хромосомы они перечислены справа от
ее изображения. Еще в 1980 г. В. Маккью-
сик провел интересную аналогию между
влиянием анатомии А. Везалия на
физиологию У. Гарвея и патологию Дж. Морганьи
и потенциальным влиянием «анатомии
генома человека» (понятие, введенное им
самим) на патологическую и функциональную
анатомию генома человека.
Информация о локализации гена
позволяет в настоящее время, используя ДНК-
технологии, осуществлять
ДНК-диагностику наследственных болезней. Ее основной
алгоритм состоит в следующем. ДНК,
выделенную из ткани любого органа, клеток крови
или клеток, культивируемых вне
организма, подвергают расщеплению рестрикцион-
ной эндонуклеазой. Среди множества обра-
ГЕНОМНЫЕ ИССЛЕДОВАНИЯ И БОЛЕЗНИ ЧЕЛОВЕКА
141
зовавшихся фрагментов ДНК (от нескольких
тысяч до десятков миллионов) необходимо
найти один или несколько, несущих
определенную нуклеотидную последовательность
и охарактеризовать эти фрагменты. Для этого
разделенные электрофорезом в геле
фрагменты «перепечатывают» на фильтр
(специальным образом обработанную бумагу,
нитроцеллюлозу или нейлон), фиксируют на нем,
и зафиксированные фрагменты ДНК
подвергают гибридизации с так называемым
«зондом» — олигонуклеотидом, меченным
радиоизотопом. Этот зонд, являющийся
ключевым элементом диагностической схемы,
представляет собой ранее клонированную с
помощью методов генетической инженерии
нуклеотидную последовательность гена,
подлежащего исследованию, или нуклеотидную
последовательность, синтезированную
искусственным путем. Благодаря тому, что нук-
леотидная последовательность зонда
специфически связывается с комплементарными
последовательностями фрагмента,
зафиксированного на фильтре, указанный фрагмент
выявляется путем экспозиции с рентгеновской
пленкой.
Таков принцип определения
специфических нуклеотидных последовательностей в
ДНК с помощью так называемой «блот-гиб-
ридизации», положенной в основу ДНК-
диагностики наследственных болезней и
выявления «генов-кандидатов» на роль
факторов, влияющих на возникновение МФЗ.
В последние годы появились новые
методические подходы к ДНК-диагностике
болезней человека, и в настоящее время уже
возможна молекулярная диагностика не
менее 300 наследственных болезней.
Список некоторых из них представлен в
таблице.
Благодаря геномным исследованиям, а
также совершенствованию
инструментальной диагностической медицинской техники,
появилась возможность обнаружения
«патологического» гена в пресимптоматическом
периоде, т. е. когда симптомов болезни еще
не наблюдается («болезнетворный» ген
проявит себя в более позднем возрасте), или
даже пренатальном (дородовом) периоде
(ДНК плода выделяют из ткани хорионичес-
кой оболочки плода, амниотической
жидкости, крови плода, полученных на разных
Наследственные болезни человека, для которых возможна ДНК-диагностика
Заболевание
Муковисцидоз
Мышечная дистрофия Дюшенна/Беккера
Фенилкетонурия
Гемофилия А
Гемофилия В
а-талассемия
р-талассемия
Серповидноклеточная анемия
Недостаточность 21-гидроксилазы (врожденная гиперплазия коры надпочечников)
Недостаточность ai-антитрипсина
Ретинобластома
Нейрофиброматоз Реклингаузена
Хорея Гентингтона
Атаксия Фридрейха
Миотоническая дистрофия
Спинальные амиотрофии (II, III)
Синдром Мартина—Белла
Семейная гиперхолестеринемия
Гепатолентикулярная дегенерация (болезнь Вильсона-
Поликистоз почек
-Коновалова)
Локализация гена
7q31,3^q32
хр21,2
12q24,l
xq28
xq27,l-xq27,2
16pter-pl3,3
11р15,5
11р15,5
6p21,3
14q32,l
13ql4,l-ql4,2
17dll,2
4pl6,3
9ql3-q21,l
19ql3,2^ql3,3
5ql2,2^ql3,3
xq27,3
19pl3,2-pl3,l
13ql4^q21
16pl3,31-pl3,12
142
ГЕНЕТИКА
стадиях беременности). Более того, возможна
преимплантационная диагностика, когда ряд
яйцеклеток матери оплодотворяются in vitro
(в пробирке), затем выращиваются зародыши
до стадии 8 клеток, от них берут 1-2 клетки
и проводят анализ на наличие
поврежденного гена. Зародыш, не содержащий
поврежденного гена, имплантируется в матку.
Геномные исследования и
идентификация генов, повреждение которых приводит
к заболеваниям, позволяют глубже понять
биохимические процессы, определяющие
интимные механизмы формирования
клинических проявлений болезни. Эти новые
знания служат основой для разработки
адекватных методов лечения заболеваний, в том
числе генотерапии — этиологической
(причинной) коррекции наследственных
болезней. Уже сейчас получено разрешение на
клинические испытания генотерапевтических
подходов к лечению семейной гиперхолес-
теринемии с помощью гена-рецептора ли-
попротеинов низкой плотности, разных форм
Р-талассемии с использованием рекомбинан-
тных ДНК, несущих гены тимидинкиназы,
дигидрофолатредуктазы и (3-цепи
гемоглобина, дефицита аденозиндезаминазы путем
введения активного гена
аденозиндезаминазы в Т-лимфоциты больных пациентов.
Результаты геномных исследований
находят широкое применение в судебной
медицине. Появление технологии геномной
диагностики (фингерпринтирования ДНК)
позволило решать важные проблемы
определения генетического разнообразия,
индивидуальности и родства людей (и других
организмов) с помощью анализа вариабельности
структуры ДНК. Приводится любопытный
пример использования возможностей моле-
кулярно-генетического анализа. В
Аргентине, в период диктатуры существовала
практика похищения детей репрессированных.
После восстановления демократии
организация «Grandmother» ведет поиск 200
пропавших детей и уже нашла 50 из них; их
родство устанавливается генетическими методами.
Возможности геномной дактилоскопии
использовались также для идентификации
останков Николая II и членов его семьи (см.:
Янковский Н. К. Молекулярная генетика и
судебная медицина. Наст. том).
Заключение
Впечатляющие успехи геномных
исследований, осуществляемых международной
и рядом национальных программ под
одинаковым названием «Геном человека»,
становятся достоянием не только
ученых-биологов, но и всего общества, озабоченного
моральными проблемами, возникающими
при использовании генетической
информации. Таким образом, у этих программ есть
«этическая компонента», которая
включает следующие положения: каждый член
общества и общество в целом должны ясно
понимать значение работ по картированию
и секвенированию генома человека;
постоянно контролировать социальные,
этические и правовые аспекты геномных
исследований; поощрять общественные дискуссии
по данным вопросам, в ходе которых
должна вырабатываться тактика действий,
гарантирующих, что генетическая
информация будет использована только во
благо отдельному лицу, членам его семьи и
обществу. В документах рабочей группы
американского проекта «Геном человека»
перечисляются некоторые, наиболее
важные сегодня, области «этической
компоненты» геномных исследований, на которые
следует обратить особое внимание:
использование генетической информации при
страховании и трудоустройстве
(предупреждение дискриминации носителей тех или иных
«особых» генов); ее использование в
процессах уголовного правосудия,
усыновления, проверки годности к военной службе;
проблема доступа к образованию с целью
получения той или иной специальности;
сохранение конфиденциальности
генетической, информации; совершенствование
программ здравоохранения по дородовой и
пресимптоматической диагностике
наследственных болезней; скрининг на выявление
носителей «больных» генов при отсутствии
методов лечения болезней, вызываемых
этими генами; генетическое образование
медицинского персонала, пациентов и
населения в целом; трактовка некоторых
вопросов истории генетики — евгенического
движения, генетики поведения.
В каждом обществе морально-этические
проблемы геномных исследований имеют свои
особенности. Однако приходится
констатировать, что разработка этих проблем
находится пока на самой начальной стадии.
Литература
Алтухов Ю. П. Генетические процессы в
популяциях. М.: Наука, 1989.
Баев А. А. Природа, 1995, №4, с. 64.
Дубинин Н. П. Некоторые проблемы
современной генетики. М.: Наука, 1994.
Фогель Ф., Мотульски А. Генетика человека.
В 3 т. Пер. с англ. М.: Мир, 1990.
Харрис Г. Основы биохимической генетики
человека. Пер. с англ. М.: Мир, 1973.
McKusick A. Mendelian Inheritance in Man.
Baltimor and London: Johns Hopkins University, 1992.
Primer on Molecular Genetics. Washington: US
Department of Energy, 1992.
Н. К. Янковский
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА И СУДЕБНАЯ МЕДИЦИНА
Введение
История изучения ДНК насчитывает
более века — с момента открытия в ядрах
клеток вещества, названного нуклеином.
Полвека назад было доказано, что
именно ДНК является носителем
наследственности, а затем установлена
принципиальная структура этой молекулы — двойная
спираль. В настоящее время молекулярная
генетика, т. е. изучение того, каким
образом хранится и реализуется записанная в
молекуле ДНК наследственная информация,
является одной из наиболее бурно
развивающихся областей науки вообще и
биологии, в частности. Для полной расшифровки
генетического текста человека
разработана и осуществляется крупнейшая за всю
историю биологии научная программа,
ежегодный объем финансирования которой
только в США составляет около 200 млн
долларов. В эту работу вовлечены сотни
лабораторий из разных стран. В России
исследования по этому направлению
выполняются в рамках собственной
государственной научно-технической программы «Геном
человека».
С появлением молекулярной генетики
методы генетических исследований
принципиально обогатились. От привычных по
школьным учебникам морфологических (цвет глаз,
цвет семян) и биохимических (группы крови
и варианты белков) признаков многие
генетики перешли к непосредственному изучению
собственно молекулы ДНК. Такой переход
связан с созданием хитроумных методов и
подходов, позволяющих работать с признаками,
выявляемыми при исследовании самой
молекулы ДНК. А это не так уж просто: ДНК
простенькой бактерии содержит миллионы пар
нуклеотидов, а геном человека — более трех
миллиардов пар. Вся ядерная ДНК в клетке
человека содержится в виде 23 пар молекул,
соответствующих хромосомам. Хромосомы
каждой пары называются гомологичными.
Линейные последовательности генов в них
одинаковы, размер соответствующих
гомологичным хромосомам молекул ДНК один и
тот же. Исключение составляют половые
хромосомы X и Y. Они сильно отличаются друг
от друга по размеру и содержанию генов.
Геном человека содержит от 50 до 100 тыс.
генов. Гомологичные гены (т. е. гены,
находящиеся в гомологичных хромосомах в
одних и тех же локусах) определяют один и
тот же признак (например, цвет глаз или
группу крови), но могут находиться в
разных аллельных состояниях, определяющих
различные фенотипические проявления
данного признака (глаза могут быть карими или
голубыми и т. п.). Аллельные гены
отличаются небольшими изменениями
последовательности нуклеотидов в ДНК, которые в
одних случаях приводят к изменению
кодируемого геном продукта и
соответствующему изменению фенотипа, а в других
случаях никакого заметного влияния на фенотип
организма не оказывают.
Уникальное сочетание аллельных
состояний всех пар генов и определяет
биологическую индивидуальность каждого человека.
Например, последовательности ДНК автора
и читателя (а также любых двух людей, кроме
монозиготных близнецов) почти одинаковы —
они различаются только одним нуклеотидом
из трехсот. Эти различия на молекулярном
уровне определяют то, чем мы друг от
друга отличаемся внешне, — цвет волос, глаз,
кожи, отчасти рост, вес. Выявление таких
замен, определение их значения для
развития того или иного признака организма в
норме и при патологии составляют предмет
фундаментальных исследований в этой области
науки. Полученные при этом результаты уже
имеют множество практических приложений.
Познакомимся с возможностями и принципами
современных методов молекулярно-генетичес-
кого анализа на конкретном примере,
возможно, известном читателю из сообщений
прессы.
Молекулярно-генетический
анализ ДНК из костных останков
последнего российского
императора Николая II
и членов его семьи
Назначение экспертизы и ее задачи
В 1991 г. в окрестностях Екатеринбурга
было обнаружено групповое захоронение
людей со следами насильственной смерти. По
некоторым данным, это могло быть
захоронение членов семьи и лиц из ближайшего
окружения последнего российского
императора, расстрелянных по приказу Уралсовета
осенью 1918 г. Было известно, что в
Екатеринбурге находилось 7 членов царской семьи:
император Николай Александрович,
императрица Александра Федоровна, их сын
царевич Алексей и четыре дочери — царевны
Ольга, Татьяна, Мария и Анастасия. После
расстрела трупы были обезображены, уби-
144
ГЕНЕТИКА
тых вывезли на грузовике и тайно
захоронили. Установление принадлежности
найденных останков членам царской семьи могло
пролить свет на тайну гибели императора и его
семьи.
По факту обнаружения останков
Генеральной прокуратурой и Бюро Главной
судебно-медицинской экспертизы РФ была
назначена комплексная экспертиза,
призванная ответить на следующие вопросы.
Какова половая принадлежность захороненных
лиц? Есть ли в группе останки
родственников и в какой степени родства были эти люди?
Соотносятся ли останки по каким-либо
признакам с ныне здравствующими
родственниками царской семьи?
Комиссия включала представителей
многих областей наук. В порядке
эксперимента в комиссию такого рода был впервые
включен специалист по молекулярной
генетике — П. Л. Иванов. Исследование
останков традиционными антропологическими
методами показало, что было захоронено
9 человек. Семь из них могли быть членами
царской семьи.
Определение пола индивидов
по анализу гена амелогенина
На первом этапе исследования
определяли пол людей, чьи останки находились в
Екатеринбургском захоронении. Из костных
останков 9 человек были получены
препараты ДНК. Хотя методы выделения ДНК из
различных тканей хорошо отработаны и
даже известен случай получения ДНК из
мумии фараона, задача была непростой.
Скелеты пролежали в земле более 70 лет
и сохранились не так хорошо, как в
саркофаге. Несмотря на это, из костей все же
удалось выделить ДНК, правда, в
количествах, измеряемых миллиардными долями
грамма (около 20 пг). Тем не менее этого
количества ДНК достаточно для анализа,
так как оно на порядок превышает
суммарный вес ДНК в одной клетке человека.
Определение половой принадлежности
по ДНК основано на том, что половые
хромосомы X и Y несут гомологичный ген,
который находится в них в разных аллель-
ных состояниях — ген в Y-хромосоме на 6
пар нуклеотидов длиннее, чем в Х-хромо-
соме. Этот ген кодирует компонент зубной
эмали амелогенин.
Для того чтобы выявить, содержится ли
в данном образце ДНК Y-хромосома (что
означает его принадлежность мужчине), из
всей геномной ДНК анализируют лишь
область амелогенинового гена, содержащую
участок, различный в Х- и Y-хромосомах.
Размер этой области составляет одну
тридцатимиллионную часть всего генома
человека. Для этого накапливают нужный
фрагмент в количествах, доступных для анализа,
при помощи полимеразной цепной реакции
(ПЦР). ПЦР позволяет получить большие
количества изучаемого фрагмента ДНК даже
ж
Рис. 1 - Схема полимеразной цепной реакции
а, б— исходную молекулу ДНК прогревают, чтобы разрушить водородные связи, соединяющие комплементарные
нити; в— реакционную смесь охлаждают в присутствии двух коротких^ 12-20 пар нуклеотидов) фрагментов ДНК,
один из которых комплементарен участку нити слева от изучаемого локуса, а второй комплементарен участку
другой нити справа от локуса. Эти фрагменты, называемые праймерами, связываются с соответствующими
участками ДНК; г— связавшиеся с нитью ДНК праймеры задают точку начала синтеза новой комплементарной
нити на матрице ДНК. Осуществляет этот синтез фермент ДНК-полимераза; д— раствор с полученными
копиями ДНК снова прогревают; е, ж— в следующем цикле вновь синтезированные нити ДНК используются в
качестве матрицы. Новая порция праймеров связывается с соответствующими участками и происходит новый цикл
синтеза. В живой клетке ДНК-полимераза осуществляет редупликацию ДНК при делении клетки, т. е. полностью
воспроизводит геномную ДНК. При проведении ПЦР синтезируется только небольшой фрагмент,
расположенный между праймерами. За 30 циклов количество синтезированных фрагментов составит около 1 млрд
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА И СУДЕБНАЯ МЕДИЦИНА
145
в том случае, если в распоряжении
исследователя имеется всего лишь одна
исходная молекула геномной ДНК.
Метод ПЦР лежит также в основе
идентификации личности по ее ДНК,
установления родства людей, выявления генов
наследственных болезней и т. д. Схема метода
ПЦР изложена в подписи к рис. 1.
Для определения размера полученных
ПЦР-фрагментов их разделяют в геле,
похожем на плотное желе. Через гель с
образцами ДНК пропускают постоянный
электрический ток. При этом, поскольку молекулы
ДНК несут отрицательный заряд, фрагменты
ДНК движутся к аноду. Чем меньше
фрагмент, тем быстрее он преодолевает поры в
геле и движется в нем и соответственно тем
дальше уходит от линии старта. Такой
процесс разделения фрагментов ДНК в геле
называется электрофорезом. При этом все
фрагменты ДНК из одной пробы движутся
по одной и той же «дорожке» на геле, а
фрагменты одного размера движутся с
одинаковой скоростью и после окрашивания
специальным красителем выглядят как тонкая
полоска в геле.
На рис. 2 показаны результаты
разделения фрагментов гена амелогенина после
проведения ПЦР на ДНК из костных останков.
На девяти дорожках справа — фрагменты,
полученные на ДНК из костных останков.
Слева — две дорожки с контрольными
образцами: фрагментами, синтезированными на
геномной ДНК, выделенной из крови
мужчины и женщины. У мужчин образуется два
фрагмента: длиной 112 пар нуклеотидов,
соответствующий Y-хромосоме, и 106 пар
нуклеотидов, соответствующий Х-хромосоме. У
женщин имеются две Х-хромосомы, каждая
из которых дает фрагмент 106 пар
нуклеотидов. В исследованных девяти образцах в 4
случаях видны две полосы, а в 5 случаях —
одна полоса. Следовательно, среди убитых
было 4 мужчин и 5 женщин. Именно столько
женщин было в царской семье в
Екатеринбурге. Но имеют ли эти образцы отношение
к членам царской семьи, остается неясным.
Тест позволяет установить только половую
принадлежность, но не индивидуальные
черты исследуемой ДНК.
Выявление семейной группы по анализу
аллельного состояния молекулярно-
генетических маркеров ДНК
На следующем этапе исследования
необходимо было ответить на вопрос: есть ли
в группе останки родственников и в какой
степени родства были эти люди? Для
установления генетического родства
используют локусы, которые имеют не два аллель-
ных состояния, как у гена амелогенина, а
больше — пять или десять. Локусы,
имеющие два или более аллельных состояния,
называются полиморфными маркерами. Если
проанализировать 5 локусов, каждый из
которых может быть в пяти аллельных
состояниях, то можно получить 55 = 3125
возможных вариантов сочетаний аллелей в
конкретной хромосоме, т. е. два
неродственных индивида, у которых совпадает аллель-
ное состояние всех пяти маркеров, будут
встречаться не так уж часто. А вот у
родственников аллельное состояние
полиморфных локусов-маркеров совпадает: чем
ближе родство, тем больше совпадений.
Удобно использовать такие аллели,
различия между которыми легко
тестировать, как, например, в приведенном выше
случае с геном амелогенина, по разной
длине ПЦР-фрагментов.
Для исследования ДНК из костных
останков был выбран локус ТН01 (его
название для понимания сути дела не
существенно). Локус ТН01 содержит многократно
повторяющуюся динуклеотидную
последовательность цитозин—аденин,
обозначаемую латинскими буквами СА. Такие
короткие повторы называются STR-повторами,
или STR-маркерами (от англ. short tandem
repeat). Существует 5 аллелей локуса,
содержащих от б до 10 копий повтора.
Аллели легко различимы по размеру
ПЦР-фрагментов, выявляемых в геле после
электрофореза.
Контроль
Старт (? 9
Образцы ДНК из костных останков
123456789
Y- 112 п. н.
Х-106 п. н.
Рис. 2. Определение половой
принадлежности по анализу ДНК из
костных останков
146
ГЕНЕТИКА
Старт
Число
повторяющихся
мономеров (СА)
Образцы ДНК из костных останков
1 23456789
10-
9-
8-
7-
6-
Рис. 3. Анализ аллельного
состояния локуса ТН01
Внизу указана семейная группа,
выявленная по анализу аллельного
состояния пяти полиморфных локусов
Выявлены
аллели
Семейная
группа
9 с/ с/ 9 9 9 9 с/ cf
9;10 8;10 7;10 8;10 7;8 8;10 8;8 6;9 6;6
На рис. 3 приведены результаты
тестирования аллельного состояния 9 образцов
ДНК из костных останков. Конечно, в
каждой дорожке мы видим только два из пяти
возможных фрагментов: ведь сколько бы
аллелей всего ни существовало, у
каждого человека в геноме присутствуют
только два аллеля по каждому локусу.
Наличие двух полос указывает на гетерозиготное
состояние локуса ТН01, а одной полосы —
на гомозиготное. Например, в образце ДНК
на дорожке 7 видна одна полоса,
соответствующая гомозиготному состоянию
локуса ТН01, в котором присутствуют аллели с
числом повторов мономера (СА), равным 8.
Такой аллель называют ТН01(СА)8. На
дорожке 3 видны две полосы, которые
указывают на гетерозиготное состояние этого
локуса: там выявляются аллели ТН01 (СА)7
и ТН01 (СА)10.
Сравнение аллельного состояния
локуса ТН01 для любой пары индивидов
позволяет принять или отвергнуть гипотезу об
их генетическом родстве по вертикали
(родитель-ребенок). У ребенка размер одного
фрагмента совпадает с размером одного из
фрагментов матери, а размеры второго
фрагмента — с одним из фрагментов отца.
Если у двух индивидов не выявляются
одинаковые полосы (например, на рис. 3
индивиды 1 и 5 или 7 и 8), то их родство по
вертикали исключено, а если одинаковые
полосы присутствуют (т. е. присутствуют
одинаковые аллели локуса), то родство
возможно.
Например, из результатов анализа,
представленного на рис. 3, мы видим, что
индивид 3 (мужского пола по результатам
предыдущего ДНК-теста, см. рис. 2) несет
аллели (СА)7 и (СА)10 локуса ТН01 и что эти
аллели отсутствуют у индивидов 7, 8, 9.
Следовательно, индивид 3 не может быть
их отцом или сыном — у детей должен
присутствовать какой-либо из аллелей
родителя. В то же время индивид 3 может быть
либо отцом, либо сыном индивидов 1, 2,
4, 5 или 6. Попробуйте сами установить по
данным на рис. 3, родство между какими
индивидами возможно, а между какими
исключено.
Аналогичные картины распределения
аллелей были получены еще для четырех
полиморфных локусов. По данным
анализа, среди останков 9 скелетов была
выявлена семейная группа из 5 человек: отец,
мать и три дочери. Остальные 4 индивида
не обнаруживают родственных связей ни
между собой, ни с членами выявленной
семейной группы. Можно утверждать, что если
это семья царя, то отсутствуют останки
царевича Алексея и одной царевны.
Заметим, между прочим, что выявить
мужа и жену при сравнении только их
собственных образцов ДНК, конечно, нельзя.
Их родство в юридическом смысле
обычно подразумевает отсутствие генетического
родства. Супругов можно идентифицировать,
если в анализируемой группе есть их дети.
Идентификация родителей подтверждает,
в свою очередь, выявление братьев и
сестер в группе, потому что у них
обнаруживаются только комбинации родительских
аллелей и никаких других.
Пока генетические данные о семье,
выявленной в Екатеринбургском
захоронении, ничего не говорят о том,
действительно ли это семья царя Николая II и царицы
Александры. Ответ на этот вопрос можно
получить, сравнив образцы ДНК,
предположительно принадлежащие царю и царице,
с образцами ДНК их ныне живущих
родственников.
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА И СУДЕБНАЯ МЕДИЦИНА
147
Последовательности митохондриальной ДНК. Точками обозначены
нуклеотиды, совпадающие со стандартной последовательностью
Источник ДНК
Герцог Эдинбургский, образец крови
Скелет № 7, костные останки
Скелет № 4, костные останки
Скелет № 5, костные останки
Скелет № 6, костные останки
Скелет № 7, костные останки
Герцог Файф, образец крови
Графиня Шереметьева-Сфири, образец крови
Георгий Романов, костные останки
Стандартная последовательность
стссссттт
т с
т с
т с
т с
т с
. с . т т . . . .
. с т . т т . . .
. с т . т т . . .
. С Y . Т Т . . .
Результаты идентификации
Царица Александра
Царевна
Царевна
Царевна
Царь Николай
Анализ митохондриальной ДНК:
в Екатеринбургском захоронении
находятся останки императора
Николая II и его семьи
В последние годы накоплено множество
сведений о расположении различных
STR-маркеров в геноме человека. Созданы
международные компьютерные базы данных,
позволяющие выявлять одинаковые
комбинации маркеров не только у родителей и
детей, но и у отдаленных родственников.
Конечно, чем дальше родство, тем сложнее
его выявить, ведь индивид передает детям
50% маркеров, внукам — 25% и т. д. Но если
провести анализ по достаточно большому
количеству STR-маркеров, родство будет
установлено или исключено с большой
степенью вероятности. Однако на момент
проведения исследования останков из
Екатеринбургского захоронения было известно лишь
небольшое количество таких маркеров.
Поэтому для проверки гипотезы о
принадлежности останков Николаю II и его семье был
выбран другой метод, более трудоемкий —
определение последовательности нуклеотидов
митохондриальной ДНК. Дело в том, что при
оплодотворении цитоплазма сперматозоида,
содержащая митохондрии, не попадает
внутрь яйцеклетки. Зародыш получает
митохондрии только из клеток матери.
Последовательность нуклеотидов митохондриальной
ДНК идентична у лиц, восходящих к
общему предку по материнской линии. Как и в
ядерной ДНК, в митохондриальной есть
консервативные участки, одинаковые у всех
людей. Они чередуются с вариабельными
участками, нуклеотидная последовательность
которых часто изменяется в результате
мутаций. Один из таких вариабельных
участков, так называемая D-петля, и был выбран
для исследования. Как и в предыдущих
экспериментах, участок ДНК размножали
методом ПЦР, используя праймеры
(начальные участки синтеза молекул ДНК),
комплементарные к соседним с D-петлей
консервативным участкам. В качестве
объектов сравнения использовали ДНК,
выделенную из крови ныне живущих
родственников царя по женской линии (герцога Файфа
и графини Шереметьевой-Сфири) и царицы
(герцога Эдинбургского). Родословные
приведены на рис. 4 и 5. Данные анализа
показали, что по сравнению с наиболее часто
встречающейся в D-петле
последовательностью СТССССТТТ у родственника царицы
Александры выявлены две замены
нуклеотидов (они указаны в таблице). Те же
замены обнаружены в митохондриальной ДНК
всех женских скелетов семейной группы. У
обоих родственников царя выявлено
четыре замены нуклеотидов в стандартной
последовательности. Три из них полностью
совпадали с последовательностью
митохондриальной ДНК отца семейства из
захоронения, однако четвертая замена отличалась.
Принцесса Алиса -
дочь королевы Виктории
Николай II Царица
Александра
Принц Филипп
Герцог Эдинбургский
Рис. 4. Материнская генеалогическая ветвь
императрицы Александры Федоровны
Линии родственников, имеющих одинаковые
последовательности митохондриальной ДНК, выделены
красным цветом
148
ГЕНЕТИКА
Луиза Гессен-Кассель
Королева Дании
Николай II
Герцог Файф Графиня Ксения
Шереметьева-Сфири
Рис. 5. Материнская генеалогическая ветвь
императора Николая II
Линии родственников, имеющих одинаковые
последовательности митохондриальной ДНК, выделены
красным цветом
Возникло предположение, что это отличие
было вызвано мутацией. Хотя три
совпавшие замены однозначно указывают на
родство, все же для достижения полной
ясности была определена последовательность
митохондриальной ДНК из останков Георгия
Александровича Романова, брата Николая II.
Нуклеотидные последовательности D-петли
митохондриальной ДНК братьев оказались
полностью идентичными. Это позволило
сделать вывод, что обнаруженные в
Екатеринбургском захоронении останки действительно
принадлежат последнему российскому
императору Николаю II и его семье.
Результаты экспертизы, проведеннной П. Л.
Ивановым, позволили однозначно ответить на
каждый из поставленных перед экспертной
комиссией вопросов, причем два наиболее
важных вопроса — о родстве между убитыми
и их принадлежности царской фамилии —
были разрешены исключительно в
результате разработки и применения методов мо-
лекулярно-генетического анализа ДНК.
Проведенное исследование создает основу для
ДНК-тестирования лиц, считающих себя
потомками императора Николая II. Теперь
ответ на вопрос о генетическом родстве
может быть получен в тот же день, когда
будет проведен анализ. Однако никто из лиц,
объявлявших себя членами семьи
императора Николая II или их потомками, пока не
выразил желания пройти соответствующее
тестирование-
Заключение
Все методы, о которых рассказано в
этой статье, основаны на результатах
фундаментальных молекулярно-биологических
исследований. Об этом свидетельствует
присуждение Нобелевских премий в
соответствующих областях науки, начиная с
установления структуры ДНК (Дж. Уотсон
и Ф. Крик), исследования механизма
синтеза ДНК на ДНК-матрице (А. Корнберг),
разработки метода определения
последовательности нуклеотидов (Ф. Сэнгер) до
совсем недавнего открытия К. Мюллиса,
разработавшего схему полимеразной цепной
реакции. Упомянутые исследования не
носили прикладного характера, не имели
сиюминутной практической значимости, да
и целесообразность их проведения не была
очевидной. Однако сейчас основанные на
этих открытиях методики широко
применяются в криминалистической экспертизе,
медицине, сельском хозяйстве и
биотехнологической промышленности. Метод ПЦР-
анализа применим к любому геному, не
только к геному человека. Например, ДНК-
паспортизацию используют при продаже
элитных животных, чтобы исключить их
подмену. На основе ПЦР разработаны
методы высокоточной лабораторной
диагностики заболеваний, передающихся половым
путем (хламидиоза, уреаплазмоза, мико-
плазмоза, цитомегаловирусной инфекции
и СПИДа). Достаточно, чтобы в пробе,
полученной от больного или носителя,
чувствующего себя пока еще абсолютно
здоровым, присутствовал всего один
возбудитель заболевания, и он будет выявлен
при помощи ПЦР. Традиционные способы
диагностики в этих случаях
малоэффективны, а в первые несколько месяцев после
заражения ВИЧ вирус в организме
человека никаким иным методом, кроме ПЦР-
анализа, вообще выявить невозможно.
Что же касается исследований генома
человека, то в настоящее время имеется
техническая возможность
ДНК-паспортизации людей. Такой паспорт невозможно
потерять или подменить. Теоретически
возможна и всеобщая ДНК-паспортизация. Это
поможет избежать появления детей с
наследственными дефектами или проводить
профилактическое лечение для
предотвращения развития симптомов выявленных
наследственных заболеваний. Но в то же
время возможности тотальной
ДНК-паспортизации могут рассматриваться как
покушение на неприкосновенность личной
жизни. Принятие решения о применении
достижений науки, в том числе о
проведении ДНК-диагностики, например, при
подозрении на наличие ВИЧ-инфекции или
при ДНК-индивидуализации, т. е.
определении уникального сочетания молекулярно-
генетических маркеров в данном геноме,
лежит вне рамок самой науки — это право
общества и индивида. Однако пока ДНК-
индивидуализация чаще всего
проводится по предписанию суда. Основное поле
ее применения — установление
биологического отцовства ребенка или
установление идентичности ДНК
подозреваемого с ДНК, выделенной из биологических
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА И СУДЕБНАЯ МЕДИЦИНА
149
материалов с места преступления (капель
крови и спермы, волос, фрагментов
мягких тканей). Так, несколько лет назад в
США широко освещался уголовный
процесс по делу об убийстве Николь Симп-
сон и ее приятеля. Подозреваемый —
звезда американского футбола О. Дж. Симпсон.
Один из присяжных сказал в интервью,
что должен был принять решение о
виновности или невиновности Симпсона по
данным ДНК-идентификации следов
крови на месте преступления. Однако он не
понимает, почему, по мнению экспертов,
следы крови принадлежат именно
подозреваемому и никому другому. Поскольку
защита представила доказательства того,
что один из полицейских, участвовавших
в сборе вещественных доказательств, был
уличен в расистских высказываниях,
присяжные не приняли во внимание
результаты тестирования ДНК и, заподозрив
фальсификацию следов преступления,
оправдали О. Дж. Симпсона.
Исследования, о которых говорится в
этой статье, еще несколько лет назад были
известны только узким специалистам.
Однако теперь ДНК-индивидуализация и ДНК-
диагностика наследственных и
инфекционных заболеваний доступны каждому. Такого
рода анализы проводят в лабораториях
Москвы, Санкт-Петербурга, Новосибирска
и других крупных городов России.
Ю. П. Алтухов
ГЕНЕТИКА ПОПУЛЯЦИИ И СОХРАНЕНИЕ
БИОРАЗНООБРАЗИЯ
Введение
За последние 100 лет хозяйственная
деятельность человека привела к тому, что
с лица Земли исчезли (или близки к
исчезновению) до 25 тыс. видов высших
растений и более 1 тыс. видов позвоночных
животных. На грани вымирания находятся или
уже вымерли сотни уникальных пород
домашних животных. По имеющимся оценкам,
к 2010-2015 г. биосфера может утратить до
10-15% составляющих ее видов.
Чем же вызвано сокращение видового
разнообразия жизни? Только ли тем, что
возросло давление промысла или
изменились исторически сложившиеся условия
обитания? Но, может быть, дело еще и в
том, что мы просто нерационально
используем биологические ресурсы, нарушаем те
внутренние естественные механизмы,
которые позволяют видам животных и
растений устойчиво воспроизводиться в
последовательных поколениях, поддерживая
равновесие с окружающей средой?
Поиск ответов на эти вопросы ведется с
использованием разных подходов и методов.
Среди них важное значение имеет популя-
ционно-генетический подход. Причин тому две.
Во-первых, все биологические особенности
популяций суть производные их
наследственных особенностей, или, как говорят генетики,
их генофондов — совокупной наследственной
информации, передаваемой от родителей
потомкам. Во-вторых, в популяционной
генетике уже давно существует и
продолжает совершенствоваться количественная теория
тех естественных факторов, под давлением
которых облик популяций либо остается
неизменным, либо меняется в
последовательных поколениях, вызывая сопряженные
изменения биологически важных свойств.
Иными словами, популяционная генетика
использует математические модели.
Модели могут быть адекватными или
неадекватными природе, но они интересны в том
отношении, что определенным образом
позволяют спланировать исследование. Кроме
того, если мы наблюдаем соответствие между
природной ситуацией и моделью, то
получаем возможность количественной оценки
происходящих изменений и прогнозирования
их возможных последствий.
Цель этой статьи — представить факты
и вытекающие из них выводы, полученные
при генетическом исследовании природных
и сельскохозяйственных популяций. Мы
рассмотрим, во-первых, как нерациональная
хозяйственная деятельность разрушает по-
пуляционно-генетическую структуру видов
и, во-вторых, как принципы и методы
генетики популяций позволяют преодолевать
неблагоприятные явления и помогают
осуществлять разумное использование
биологических ресурсов, их длительное сохранение.
Основные принципы генетики
популяций
Популяция — это исторически
сложившаяся, самовоспроизводящаяся совокупность
организмов, принадлежащих одному и тому
же биологическому виду и обитающих в
определенной части его ареала.
Популяции животных и растений —
основной объект хозяйственной
деятельности человека. Чтобы обеспечить себя
продуктами питания, мы эксплуатируем поля,
засеянные теми или иными сортами
зерновых культур, стада различных пород
домашних животных, скопления рыб какого-
либо промыслового вида, обитающие в
разных водоемах. Все это — отдельные
популяции живых организмов. И если мы
хотим рационально использовать
биологические ресурсы суши и моря, то должны
понимать процессы, протекающие в
популяциях, а главное — сознавать, к каким
последствиям может привести то или иное
вмешательство в их жизнь.
Популяционная генетика зародилась в
начале нашего столетия, но долгое время
жизнь популяций можно было исследовать
лишь на тех, как правило, не имеющих
хозяйственной ценности организмах, у
которых есть какие-то внешние наследственные
признаки, позволяющие различать внутри
вида дискретные группировки. Например, на
надкрыльях божьих коровок имеются
пигментные узоры (пятна, точки) (рис. 1),
характерные для каждой особи, а популяцио-
нную принадлежность рыбок пецилий —
излюбленного объекта аквариумистов —
можно определить по рисунку на хвостах у
самцов. Большие серии популяционно-гене-
тических работ были выполнены также после
обнаружения индивидуальных различий в
строении хромосом (например, по наличию
или отсутствию инверсий).
Такого рода индивидуальная
прерывистая изменчивость носит название
полиморфизма (от греч. полиморфос — многообразный).
Английский генетик Э. Форд,
сформулировавший концепцию генетического
полиморфизма, определил это явление как «ноли-
ГЕНЕТИКА ПОПУЛЯЦИЙ И СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ
151
10
11
12
13
14
15
16
Рис. 1. Полиморфизм пигментного рисунка
надкрылий у божьей коровки Harmonia axyridis
Этот вид встречается в Сибири, Китае, на Корейском
п-ове и в Японии. Особи типа 1 преобладают в
Западной и Центральной Сибири, а далее на восток
популяция становится все более полиморфной,
частота различных фенотипов 2-8 увеличивается.
Особи типов 13-16 обнаружены только в популяциях
Дальнего Востока [из: Айала, Кайгер, 1988.
Воспроизведено с разрешения Ф. Айалы и Benjamin/
Cumings Publishing Company]
чие в одном u том же местообитании двух
или более дискретно отличающихся
внутривидовых форм и в таких количественных
соотношениях, что самая редкая из них не
может поддерживаться лишь давлением
повторяющихся мутаций» (см.: Алтухов, 1989).
Надо сказать, что и сама генетика как
наука обязана своим возникновением именно
явлению генетического полиморфизма:
Г. Мендель не смог бы открыть законы
наследственности, если бы семена
изучавшегося им гороха не варьировали по цвету и
форме.
Поскольку индивидуальные различия
между организмами контролируются аллель-
ными генами, можно дать несколько иное
определение полиморфизма, подразумевая
под ним наличие в популяции двух или
более аллелей одного локуса1,
встречающихся с ощутимой частотой. Обычно на
практике полиморфной считается популяция с
частотой гетерозигот по некоторому локу-
су, превышающей 1%.
Однако у многих видов полиморфизма
нет, они единообразны, мономорфны по
внешнему облику, — их гены скрыты от
наблюдателя, и, следовательно, генетический
анализ невозможен. Как преодолеть такого
рода затруднение? Развитие молекулярной
биологии позволяет теперь изучать
генетику любого вида, опираясь на скрытые
наследственные различия, «записанные» в
белковых структурах организма. Эти особенности
выявляются специальными процедурами:
либо иммунологическими тестами, либо при
электрофорезе белков. Аналогичный
полиморфизм обнаружен за последние 10—15 лет
при изучении у самых различных видов
первичной структуры ДНК2 — как ядерной, так
и неядерной (например, митохондриальной),
так что арсенал технических средств попу-
ляционной генетики необыкновенно
расширился. Однако высокая стоимость и большие
затраты времени на проведение
ДНК-тестов ограничивают их широкое применение.
Напротив, изучение белкового полиморфизма
требует сравнительно умеренных денежных
расходов на оборудование и реактивы, а
общепринятая методика позволяет работать
с массовым материалом (сотни образцов за
день) и используется во многих
лабораториях. Суть методики в следующем: кровь или
экстракты различных тканей от отдельных
растений или животных помещают в
специальные аппараты и в нейтральных
мелкопористых поддерживающих средах (гелях)
сортируют молекулы в электрическом поле
по заряду и размеру, детерминируемым
наследственностью. Оказалось, что особи
внутри популяции различаются составом
крови, а также некоторыми особенностями
строения определенных белков, и такие
отличия передаются неизменными от
родителей потомкам.
Достаточно посмотреть на рис. 2, где
видны различия между особями нерки,
Oncorhynchus nerka— одного из видов
тихоокеанских лососей — различия,
определяемые двухаллельным геном, кодирующим
синтез фермента фосфоглюкомутазы.
Множественные молекулярные формы одного и того
же фермента называют изоферментами, а
генетические варианты, приведенные на
рис. 2, получили название аллозимов. Расшиф-
1 Локус — местоположение гена на хромосоме. В данном контексте речь идет о генном локусе,
или просто об отдельном полиморфном гене. В современной генетической литературе слова «ген» и
«локус» нередко используются как синонимы.
2 Распространенное рабочее название этой изменчивости — «полиморфизм длин рестрикцион-
ных фрагментов ДНК»(ПДРФ). (См.: Янковский Н. К. Молекулярно-генетические методы в руках
детектива. Наст. том).
152
ГЕНЕТИКА
12 3 4 5
Рис. 2. Наследственный полиморфизм фермента
фосфоглюкомутазы из мышечной ткани
тихоокеанского лосося нерки, Oncorhynchus nerka
№2, 3 и 4 — гомозиготы АА и ВВ соответственно;
№ 1,5— гетерозиготы АВ. Электрофорез в
крахмальном геле. Стрелка указывает на стартовую позицию.
Направление миграции молекул в электрическом
поле — от катода к аноду
ровка схемы такова: один ген — одна полоса
на электрофореграмме; разными номерами
обозначены разные рыбы из одной и той же
популяции. У рыб № 2~3 выявляется один ген,
у № 4 — другой, у № 1 и 5 представлены
оба гена, это — гетерозиготы, т. е. типы,
промежуточные между двумя первыми,
гомозиготными генотипами. Таким образом, с
точки зрения генетики, популяция — смесь
генотипов. Многие биологические
особенности популяций и прежде всего численность
и продуктивность зависят от этой их
генетической структуры. Если соотношения числа
особей различных генотипов, а точнее —
частоты генов — в популяции неизменны в
поколениях, то популяция устойчива. Если
соотношения меняются, то говорят об
изменяющейся популяции или о так называемых
процессах микроэволюции, в самом общем
виде описываемых в теории популяционнои
генетики математическими выражениями,
приведенными на рис. 3. За время,
прошедшее с конца 60-х гг., когда метод
электрофореза белков стал широко использоваться
в популяционных исследованиях,
биохимическая наследственная изменчивость
обнаружена уже у более чем 2000 различных
видов — от микроорганизмов до человека.
Важно иметь представление о панмик-
тической и подразделенной популяциях. На
рис. 3, а схематически изображена панмик-
тическая популяция, в которой
принадлежащие ей особи свободно скрещиваются друг
с другом. Если соблюдаются определенные
условия (бесконечная численность,
отсутствие средовых воздействий и др.), то пан-
миктическая популяция, согласно
известному правилу Харди—Вайнберга, остается
равновесной, стабильной в поколениях, и
соотношение гомо- и гетерозиготных
генотипов в ней соответствует коэффициентам
разложения бинома Ньютона:
р2+ 2pq+ g2= l, (1)
где р и q — частоты соответствующих ал-
лельных генов.
Такие идеальные, бесконечные,
существующие вне среды популяции — лишь
абстракция, поскольку в природе
постоянно действуют факторы, нарушающие гено-
типическое равновесие. Например, может
действовать естественный отбор и
приводить к тому, что одни генотипы оставят
потомков больше, чем другие, и это
вызовет изменение генетической структуры
популяции. Может происходить миграция —
приток или отток генов, и популяция
также будет менять свою генетическую
характеристику. Наконец, может происходить так
называемый случайный генетический дрейф,
когда численность популяции резко падает
и она «ошибается» при воспроизводстве
генофонда следующего поколения. Но дрейф
генов имеет место не только в случае вне-
а
б
Рис. 3. Основные математические модели
популяционнои генетики
а — панмиктическая популяция неограниченной
численности, остающаяся в стабильном состоянии (Ар =
= 0) в отсутствие отбора (W), миграции (/г?) и
случайного дрейфа генов (N) — так называемое правило
Харди-Вайнберга, представленное формулой/, гдер
и q — частоты генов, случайным образом
комбинирующихся в гомозиготных (р2, q2) и гетерозиготных
(pq) генотипах; //, ///, IV— элементарные формулы для
отбора, миграции и случайного дрейфа генов как
основных факторов, определяющих динамику частот
генов в популяциях; б— подразделенная популяция,
представленная совокупностью субпопуляций,
испытывающих одновременное воздействие случайного
дрейфа и миграции генов, а также естественного
отбора; V— математическая функция С. Райта для
распределения частоты гена в такой популяции
ГЕНЕТИКА ПОПУЛЯЦИЙ И СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ
153
запного и резкого сокращения численности
популяции (так называемый «эффект
бутылочного горлышка»). Численность реальных
популяций ограничена, что создает условия
для постоянно текущего генетического
дрейфа и сопутствующего ему инбридинга
(близкородственного скрещивания).
Кроме того, и это очень важно,
генетически эффективная численность (Ne)
популяции всегда и существенно меньше ее
общей численности (Nt) уже по той простой
причине, что старшие и младшие
возрастные группы исключены из процесса
воспроизводства. У человека, например, если
обратиться к сельскому населению, величина
Ne составляет примерно треть от величины
Nf. У природных видов отношение Ne/Nt
может быть еще меньше, порядка 0,1-0,2.
Такая репродуктивная структура
способствует утрате генетического разнообразия.
Однако внутрипопуляционный инбридинг
может компенсироваться генными
миграциями, и в этом случае панмиктическая
популяция трансформируется в подразделенную,
т. е. в совокупность субпопуляций, связанных
миграционными взаимодействиями (рис. 3, б).
Когда центробежные (дрейф) и
центростремительные (миграция) процессы взаимно
уравновешиваются, распределение частот
генов в подразделенной популяции
становится стационарным, т. е. направленных
генетических изменений не происходит. Если же
одновременно действует стабилизирующий
отбор (гетерозиготы приспособлены лучше
гомозигот), то полиморфизм становится очень
устойчивым, генетический состав
подразделенной популяции оказывается неизменным на
протяжении сотен и тысяч поколений.
Напротив, направленный отбор (гетерозиготы
приспособлены хуже гомозигот) приводит к
нарушению устойчивости и утрате полиморфизма.
В разработку теории подразделенных
популяций решающий вклад внесен
американским генетиком С. Райтом. Такие
«популяции популяций» получили в
отечественной литературе название популяционных
систем, а за рубежом довольно широко
употребляется термин «метапопуляция».
Рассмотрим теперь, в какой мере
теоретические предсказания соответствуют
реальным ситуациям. Ведь хорошо известно, что
многие, если не все, биологические виды
представляют собой
сложноструктурированные популяционные системы.
Генетические процессы
в интактных популяционных
системах
Обратимся к изолированной популяции
непромыслового моллюска Littorina squalida,
обитающего в лагуне Буссе на Южном
Сахалине и имеющего рельефную субпопуля-
ционную структуру (рис. 4, а). У литторины
хорошо выражен полиморфизм окраски
раковины, трактуемый как проявление
двухаллельной системы с неполным
доминированием3 (рис. 4, б). Так же как и у
исключительно полно изученной садовой
улитки Cepaea nemoralis, различия генотипов
(фенотипов) выявляются и у ископаемых
форм литторины и таким образом имеется
возможность исследовать их распределения
в выборках из ныне живущих и древних
3 Неполное доминирование, или «эффект дозы
гена», когда гетерозигота Аа по фенотипу
промежуточна между гомозиготными формами АА
и аа.
7 2 3
Рис. 4. Местоположение лагуны Буссе на о-ве
Сахалин (а) и полиморфизм рисунка раковины в
сахалинских популяциях моллюска Littorina squalida (б)
1, 3— гомозиготы АА и ВВ соответственно; 2—
гетерозигота АВ
оА
j i i i i i i i i i
II 1 2 3 4 5 12 3
f0 f2000
Рис. 5. Генетическая стабильность популяционной
системы L squalida как целого (показано кружками)
в исследованном интервале поколений
t0— частоты генов (ордината) в выборках
ископаемого материала (п = 479 экз., q0 = 0,293 ± 015); f2000 —
частоты генов в выборках из современных
популяций (приближенно в двухтысячном поколении) (п =
1252 экз., q0 = 0,280 ±010). Вертикальные черточки —
удвоенные стандартные ошибки
154
ГЕНЕТИКА
популяций. В случае с рассматриваемой
популяцией литторины временной интервал,
на котором ее можно изучать, составляет
4-5 тыс. лет; это соответствует примерно
2-2,5 тыс. последовательных поколений. С
1969 по 1974 г. Б. А. Калабушкиным были
исследованы распределения морф в трех
выборках современного и в пяти выборках
голоценового материала. Представленные на
рис. 5 оценки генных частот показывают, что
если сравнивать отдельные современные
выборки с отдельными выборками
ископаемых улиток, то можно прийти к весьма
противоречивым выводам, обнаружив
проявления как генетического сходства, так и
различий во времени и в пространстве. Но
если опереться на ясные представления о
системной организации популяций и провести
сбор и анализ первичного материала по всем
элементам популяционной структуры, то
можно сделать лишь один вывод:
несмотря на изменчивость в частях, система как
целое устойчиво сохраняет генофонд,
унаследованный от прапопуляции.
Аналогичные результаты были
получены для различных видов животных и
растений, а также для человека.
То же самое обнаружено при
исследовании экспериментальных подразделенных
популяций Drosophila melanogaster и при
компьютерном моделировании генетических
процессов в популяционных системах: при
одинаковой численности панмиктической и
подразделенной популяций последняя
оказывается более устойчивой в смысле
сохранения генетического разнообразия. Так,
например, к 1000-му поколению
эксперимента из 10 модельных панмиктических
популяций численностью 500 особей
дая «вырождались» (т. е. становились
полностью гомозиготными) 8, тогда как ни одна
из подразделенных популяций той же
численности не утратила генетического
разнообразия. Если рассчитать «время жизни»
панмиктических и равных им по величине
подразделенных популяции, приняв за него
число поколений, соответствующее
потере 99% от исходного уровня гетерозигот-
ности, то для первого случая эта оценка
составит 2301 поколение, а для второго —
5341. Иными словами, всего лишь наличие
субпопуляционной структуры с
ограниченной интенсивностью генных миграций
(порядка 0,5—1%) замедляет темп убыли
генетической изменчивости так, как если бы
эффективная численность панмиктической
популяции удвоилась.
Этот вывод важен для природоохранной
биологии, ибо совершенно очевидна
несоизмеримость ресурсов (величина ареала,
количество пищи и т. п.), требуемых для
достижения одинакового практического
результата: в одном случае численность
«минимальной жизнеспособной популяции»
может быть, благодаря внутренней
фрагментации, равна всего сотням, тогда как в
другом — как минимум тысячам особей.
Для количественного описания структуры
генетической изменчивости в подразделенной
популяции американским генетиком М. Ней
развита теория, связывающая внутрипопу-
ляционную (Hs) и межпопуляционную (GST)
компоненты гетерозиготности (или, что то
же, — генного разнообразия) следующими
соотношениями:
GST=(HT-Hs)/HT,
#Г=1-ХЯ2>
#s = l//zX#s,
(2)
где р^ — частота г-го аллеля в
субпопуляции S, р. — средняя частота аллеля во всей
подразделенной популяции, состоящей из п
субпопуляций. Таким образом, Hs есть
средняя гетерозиготность субпопуляции, а НТ —
гетерозиготность всей подразделенной
популяции как если бы она представляла единое
панмиктическое сообщество. В современных
Таблица 1
Пространственная генетическая дифференциация на разных уровнях иерархии
популяционной системы тихоокеанского лосося нерки, Oncorhynchus nerka
Уровень иерархии
Регионы (Азия и Америка)
Субрегионы в рамках регионов
Реки в субрегионах
Показатель
вз
G2
Gl
Степень локальной генетической дифференциации
В единицах G^j
0,0254
0,0237
0,0249
В долях G$j4_t
34%
32%
34%
Примечание. GSTi_t = 0,074. Оценки локальной дифференциации основаны на собственных и
литературных данных о частотах генов аллозимных локусов в 45 выборках из природных популяций
нерки, размножающихся в бассейнах 20 рек Северной Америки и Северной Азии.
ГЕНЕТИКА ПОПУЛЯЦИЙ И СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ
155
работах как внутри-, так и межпопуляцион-
ная гетерозиготность оценивается
одновременно по многим полиморфным генам.
Величина GST, служащая мерой
локальной генетической дифференциации, есть
показатель разброса, дисперсии генных
частот на субпопуляционном уровне. Чем
сильнее генетические различия между
локальными группировками особей, тем
больше GST. Одновременно соответствующим
образом ведет себя и внутрипопуляционная
гетерозиготность Hs: она увеличивается,
когда GST уменьшается и, напротив,
уменьшается, если GST растет (поскольку Hs +
+ GST = 100% от Ят; см. формулу (2)).
Понятно, что если бы подразделенная
популяция устойчиво воспроизводила свой
генофонд в последовательных поколениях,
т. е. если бы процессы его дифференциации
и интеграции находились в равновесии, как
в рассматривавшемся случае с популяцией
литторины, соотношение внутри- и межпо-
пуляционной компонент генного
разнообразия также оставалось бы постоянным на
разных иерархических уровнях системы.
Такая авторегуляция действительно имеет
место, если условия воспроизводства попу-
ляционных генофондов не нарушены. Это
можно показать на примере
тихоокеанского лосося нерки, имеющего, как известно,
сложную субпопуляционную структуру,
представленную несколькими естественно
сложившимися уровнями иерархии (табл. 1). Та же
зависимость была описана и для других
видов, чья пространственная дифференциация
изучалась одновременно по многим генам.
Следовательно, в условиях естественного,
ненарушенного воспроизводства видовых
генофондов соотношение внутри- и меж-
популяционной компонент генного разно-
образия остается неизменным на разных
уровнях популяционной структуры вида.
Какими же могут быть последствия
антропогенных воздействий на эту
структуру? Рассмотрим соответствующие данные,
полученные для природных и
сельскохозяйственных популяций.
Генетическая динамика
природных популяционных
систем, испытывающих
антропогенные воздействия
Цикл многолетних работ, выполненных
на промысловых рыбах, включая лососевых,
в наибольшей мере отвечает интересующей
нас задаче — понять специфику
микроэволюционных процессов, протекающих в
популяциях при антропогенных
воздействиях. Рассмотрим эти данные.
Промысел. При изучении природных
популяций рыб (стад) обнаруживается их
ярко выраженная гетерогенность, диффе-
ренцированность на более мелкие,
генетически различающиеся субпопуляции.
Обнаружение системной организации
популяций имеет принципиальное
практическое значение. Очевидно, если мы хотим
осуществлять рациональный промысел, имея
дело с системой, то должны подходить к
ней как к целому, с учетом ее внутренней
структуры. Вместе с тем рыбаки обычно
игнорируют эту организацию стад,
вследствие чего происходит разрушение
подразделенных популяций.
Для иллюстрации сказанного обратимся
к результатам изучения последствий
промыслового воздействия на подразделенные
популяции нерки, размножающейся в озерах
Камчатки (рис. 6). Важная биологическая
особенность нерестовых популяций этого
вида — уникальная картина изменчивости
производителей по длине тела: самки
характеризуются одновершинным
распределением, тогда как для самцов
прослеживается четко выраженная двувершинность (рис. 7).
В период размножения в водоемах
обнаруживаются три легко распознаваемые
группы рыб: мелкие молодые самцы, старые
крупные самцы и самки, занимающие между
двумя группами самцов промежуточное
положение по признаку «длина тела» и по
возрасту. Интересно, что со временем в
исследуемой популяции в результате
промысла доля мелких половозрелых самцов
увеличивается. Такие, как правило,
трехлетние самцы (их местное название на
Камчатке — каюрки, канадцы и американцы
называют их джек), лишь с небольшой час-
Рис. 6. Брачная пара нерки (вверху— самец,
внизу— самка)
С помощью хвоста самка роет в гравии гнездо
глубиной 30-40 см, куда откладывает «3000 икринок.
Самец, охраняющий самку во время «строительных
работ», поливает затем икринки молоками. После
оплодотворения самка засыпает икринки гравием и
остается возле гнезда до самой смерти,
наступающей несколькими днями позже. Как и все
тихоокеанские лососи, нерка— моноциклический вид, т.е.
размножается лишь раз в жизни и погибает вскоре
после нереста
156
ГЕНЕТИКА
тотой встречаются в нативных, мало
облавливаемых стадах. Напротив, в популяциях,
испытывающих систематическое
промысловое воздействие, количество мелких, рано
созревающих самцов резко возрастает.
Ярким примером, иллюстрирующим это
правило, может служить стадо нерки
озера Дальнего (п-ов Камчатка), биология
которого изучалась с 30-х гг. Ф. В. Крогиус. Так,
если в 30-х гг. численность нерестовой
части дальнеозерского стада составляла
около 100 тыс. производителей, а доля каюрок
среди половозрелых самцов не превышала
0,2%, то в 60-70-е гг. численность
производителей сократилась до 2—5 тыс., а доля
каюрок увеличилась до 38%.
В чем же причина столь драматических
изменений? Исследования показали, что
главный фактор, вызывающий эти
изменения, — селективный морской промысел, из
поколения в поколение нарушающий
генетическую структуру стад нерки из-за
непропорционального изъятия жаберными
Число рыб, %
30 1971 Самки
Крупные
самцы
20
.. р. Мелкие
1 и самцы
30 1974
20
10
30 1978
20
10
35 55 75
Длина тела, см
Рис. 7. Изменчивость длины тела производителей
тихоокеанского лосося, нерки, в период нереста (оз.
Азабачье, п-ов Камчатка)
По оси абсцисс— длина тела, в см; по оси
ординат—частота соответствующих классов, в %. На
рисунках видно, что имеются три группы рыб, с
различной длиной тела: крупные самцы, средние по
величине самки и мелкие самцы. При этом если
распределение длины тела самок остается более или
менее однородным во времени, то распределение
длины тела самцов сильно варьирует за счет
резкого увеличения в отдельные годы количества мелких
самцов (сравните, например, 1971 и последующие
годы)
сетями крупных, более гомозиготных
старых самцов. Другие рыбы, идущие на
нерест и отличающиеся генетически от
крупных самцов, облавливаются промыслом или
равномерно (самки), или недолавливаются
(мелкие самцы), что и приводит к
резкому изменению исторически сложившейся
популяционно-генетической структуры стада.
Дело в том, что в нерестовых стадах
нерки существует весьма консервативная
система так называемых избирательных
скрещиваний. При формировании брачных пар
на нерестилищах самки отдают
предпочтение медленно растущим, более
гомозиготным крупным самцам и лишь в маловодные
годы и на мелководных нерестилищах, куда
крупные самцы не могут проникнуть,
репродуктивный успех сопутствует
быстрорастущим молодым, более гетерозиготным
самцам. Промысел нарушает естественную
систему воспроизводства и более
гетерозиготные (гетерозисные) мелкие самцы4 во все
большей мере передают свои гены
последующим поколениям. Доля крупных рыб в
стаде уменьшается, нарушается
равновесное соотношение полов, увеличивается
скорость полового созревания, сокращается
средняя продолжительность жизни и как
следствие возрастает темп смены
поколений. Одновременно падает численность
стада, так как мелкие самки имеют более
низкую плодовитость. Таким образом, в
условиях снижения воспроизводительной
способности стада даже постоянный по
интенсивности промысел, вполне
совместимый с изначальными продукционными
возможностями популяции, приводит к
сокращению ее численности в поколениях только
из-за непропорционального изъятия рыб
определенных генотипов.
Обнаруженные процессы свойственны не
только стадам тихоокеанских лососей, но и
другим видам рыб — объектам промысла. Во
всех до сих пор исследованных случаях
картина была однотипной— измельчание
промысловых стад, их омоложение, возрастание
доли рано созревающих мелких самцов.
Поскольку направление отбора оказывается
неизменным (в пользу гетерозигот), внутри-
популяционная компонента генного
разнообразия возрастает, тогда как межпопуля-
ционная — падает, приводя к снижению
локальной генетической дифференциации.
Более сложная картина наблюдается при
искусственном воспроизводстве рыбных
популяций.
Искусственное воспроизводство. Гене-
4 Обычно гетерозисные животные
характеризуются увеличенными размерами, однако у
нерки это не так, поскольку рыба
размножается только раз в жизни, и после нереста
производители погибают.
ГЕНЕТИКА ПОПУЛЯЦИЙ И СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ
157
Таблица 2
Генетическое разнообразие природных и искусственно поддерживаемых
популяций лососей
Вид,
регион
Salmo salar
Западная и Восточная
Атлантика
Salmo trutta
Франция
Испания
Hf
0,041
0,111
0,069
Природные
**S
0,038
29 популяций
0,050
(8 популяций)
0,027
(4 популяции)
GST
0,064
0,550
0,610
Искусственно поддерживаемые
Hj>
0,037
0,077
0,092
**s
0,030
"1А популяции'
0,072
(7 популяций)
0,083
(4 популяции)
GST
0,196
0,063
0,028
тические последствия искусственного
воспроизводства лучше всего рассмотреть на
примере лососей. Как уже подчеркивалось,
их стада — сложноструктурированные по-
пуляционные системы, состоящие из
множества дискретных субпопуляций, в
разное время заходящих на нерест в реки или
озера. Если мы воспроизводим такие
системы искусственно, на рыбоводных заводах,
то должны осуществлять сбор половых
продуктов на всем протяжении
нерестового хода, а не ограничиваться
использованием лишь части дифференцированного
генофонда. Чем рельефнее субпопуляционная
структура популяции, тем меньше шансов
воссоздать целое по его отдельной части.
К сожалению, это обстоятельство на
рыбоводных заводах нередко игнорируется и
как следствие нарушается соотношение
внутри- и межпопуляционной генетической
изменчивости.
Так, например, при сравнении
природных и искусственно поддерживаемых
популяций атлантических лососей — семги,
Salmo salar и кумжи, Salmo trutta —
обнаруживаются два противоположно
направленных процесса, связанных с
перераспределением внутри- и межгрупповой компонент
генного разнообразия (табл. 2). У семги,
воспроизводимой на рыбоводных заводах,
межпопуляционная генетическая
дифференциация существенно выше, а внутрипопу-
ляционный полиморфизм ниже, чем в
природных условиях. Прямо противоположная,
но еще более рельефная картина характерна
для испанских и французских стад кумжи.
Очевидно, в случае с семгой рыбоводная
деятельность приводит к нарастанию
инбридинга, чему способствует малая
численность производителей, используемых для
воспроизводства, тогда как в случае с
кумжей увеличение внутрипопуляционной ге-
терозиготности и стирание
межпопуляционной генетической дифференциации есть
следствие перемешивания генофондов
различных по происхождению маточных линий,
либо отбора в пользу гетерозигот. В
первом случае стада лососей страдают от
инбридинга, во втором — от аутбридинга, что
приводит к плохой выживаемости молодняка
и падению численности.
Итак, наблюдается довольно
однотипная картина изменений при антропогенных
воздействиях на подразделенные
популяции хозяйственно-ценных рыб.
Практически во всех изученных случаях имеют
место неблагоприятные генетические процессы,
т. е. такой тип воспроизводства видовых
генофондов, при котором нарушается эво-
люционно-сложившееся соотношение
внутри- и межпопуляционной компонент
генного разнообразия.
Эти процессы порождаются
игнорированием в хозяйственной деятельности
исторически сложившейся субпопуляционной
структуры. Даже рыбоводная практика,
преследующая, казалось бы, благородную
цель — искусственное воспроизводство
биологических ресурсов — может приводить к
нежелательным последствиям. Они связаны
с перераспределением генетического
разнообразия таким образом, что его внутрипо-
пуляционная компонента уменьшается, тогда
как межпопуляционная нарастает.
Ситуация — типичная для лососевых рыбоводных
заводов, использующих либо недостаточное
число производителей и тем самым
провоцирующих инбридинг, либо ведущих
бессознательный отбор в пользу гомозигот, что,
по сути, одно и то же. Этот процесс ина-
даптивен и может привести к необратимой
деградации популяций даже после
прекращения соответствующего воздействия.
Перераспределение разнообразия за счет
увеличения внутрипопуляционной гетерози-
готности обнаруживается при мониторинге
158
ГЕНЕТИКА
самовоспроизводящихся популяций —
объектов промышленного рыболовства, а также
при искусственном воспроизводстве
атлантических лососей на рыбоводных заводах и
при садковом выращивании. Этот процесс
адаптивен, однако конечный его
результат — тоже деградация популяций, ибо
плата за адаптацию оказывается
непомерно большой, например, происходит
замена проходных (мигрирующих)
высокопродуктивных популяций малопродуктивными, а
кроме того, повышается эмбриональная
смертность.
Таким образом, эволюционно
сложившееся соотношение внутри- и межпопуляци-
онной компонент генетического
разнообразия является оптимальным: как убыль
гетерозиготности, так и ее чрезмерное
нарастание неблагоприятны для
нормального функционирования популяции.
Этот факт позволил нам разработать
концепцию оптимального генного
разнообразия как важнейшего условия
благополучного существования популяций в
нормально колеблющейся природной среде. Зная
соотношение внутри- и межгрупповой
компонент наследственной изменчивости при
нормальном воспроизводстве, либо
непосредственно перед тем или иным
антропогенным воздействием, мы действительно
получаем уникальную возможность детально
изучить генетику любой популяционной
системы и разработать практические
рекомендации по ее рациональному
хозяйственному использованию.
Тот же подход оказывается
эффективным и в приложении к
сельскохозяйственным популяциям.
Генетическая динамика
сельскохозяйственных популяций
На сегодняшний день наиболее
надежная информация получена для сортов
ячменя, Hordeum vulgare, возделываемых в
Восточной Сибири, и для различных пород
кур, Gallus gallus.
За последние десятилетия было
выведено, районировано и внедрено в
производство около 60 новых сортов ячменя.
Учитывая, что практически все они
представляют собой популяции, а их
районирование проводится в различных агроэколо-
гических зонах, существенное значение
приобретает вопрос о направлении отбора
и его влиянии на генетическое
разнообразие таких популяций.
Анализ этих данных показывает, как
существенно изменилась гетерогенность
популяций: если раньше они были
представлены смесью различных генотипов, то в
настоящее время преобладают линейные
сорта. Уровень генетической изменчивости
существенно выше у местных, стародавних
сортов по сравнению с современными: у
первых доля разновидностей с 3-4 аллелями по
локусу HrdB достигает 30%, тогда как в
преобладающей части вторых обнаружены
лишь один, в лучшем случае, два аллеля
исследованных гордеиновых локусов. Ясно,
что за последние 60 лет в генотипическом
составе сортов ячменя, возделываемых в
Восточной Сибири, произошли значительные
сдвиги, связанные с уменьшением
наследственного разнообразия. Эти изменения
вызваны сложившейся селекционной
практикой — новые сорта являются потомством
одного или нескольких растений.
Та же тенденция к утрате
генетического разнообразия во времени отчетливо
прослеживается и при мониторинге
популяций кур. Среди факторов, вызвавших
снижение генетической изменчивости в
промышленном птицеводстве, следует
отметить резкое сокращение числа
используемых в коммерческих целях пород. В
состав теперешних промышленных кроссов
входят лишь 4-7 из более чем тысячи
пород, известных во всем мире. Что
касается России, то из 80 старых пород к
настоящему времени не сохранилось (или не
найдено) около 30, что соответствует
сокращению генетических ресурсов в плане
породного состава на 37,5% за последние
50 лет. Многие другие породы находятся на
грани исчезновения.
Исследование динамики генетической
изменчивости в птицеводстве и более
точная количественная ее оценка согласуются
с изложенными выше фактами. Были
использованы экспериментальные данные
(собственные и литературные) по
биохимическому полиморфизму 48 популяций кур
иностранного (средиземноморского и
азиатского) и российского происхождения, а
также диких предков домашних кур,
принадлежащих к подвиду G. g. gallus. Путем
усреднения частот аллелей у 47 изученных
пород реконструирована генетическая
структура гипотетической «прапопуляции».
Генетические «профили» прапопуляции
и некоторых отечественных пород кур
представлены на рис. 8. Мы видим, что одни
породы имеют уникальную структуру,
другие вследствие их синтетического
происхождения похожи на прапопуляцию. К первой
группе принадлежат пять популяций:
орловская, первомайская, русская белая,
ленинградская белая, московская; ко
второй — остальные девять популяций, чьи
генетические профили изображены на рис. 8.
Кроме того, обращает на себя внимание
следующий важный факт: близкие к
прапопуляции отечественные породы
характеризуются более высоким уровнем внутри-
популяционной гетерозиготности (Hs = 0,213)
ГЕНЕТИКА ПОПУЛЯЦИЙ И СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ
159
Юрловская голосистая
Рис. 8. Генетические «портреты» отечественных
пород кур и их «прапопуляции»
На радиусах отложены частоты аллельных генов,
кодирующих синтез белков крови и яиц: 1 — овальбу-
мин; 2-4 — глобулины; 5— трансферрин; 6—
альбумин; 7,8 — эстеразы. Интервал частот генов: 0— в
центре круга, 1 — на окружности. На линиях,
соединяющих отдельные породы с прапопуляцией,
приведены оценки их генетической удаленности от пред-
кового генофонда («генетические расстояния» по
М. Ней)
и низкой межпопуляционной изменчивостью
(GST = 0,0975) в сравнении с породами,
наиболее удаленными от прапопуляции {Hs =
= 0,183; GST= 0,2311).
Со статистической точки зрения так и
должно быть. Но существенно то, что
генетическая близость к прапопуляции
оказывается связанной с меньшей породной
специализацией: почти все девять
соответствующих пород имеют мясо-яичное
направление продуктивности, тогда как пять
удаленных более специализированы либо в
Рис. 9. Сорт подсолнечника «Енисей»
сторону яичного (русская белая,
московская), либо мясного (ленинградская белая)
направлений, а в основании наиболее
удаленной от прапопуляции орловской породы
находятся бойцовые куры.
Значительная межпопуляционная
генетическая дифференциация и вместе с тем
пониженный уровень гетерозиготности,
характерные для группы
специализированных пород, ясно указывают: селекция
сопровождалась потерей внутрипопуляцион-
ного генного разнообразия. Для группы
неспециализированных пород характерна
противоположная тенденция: рост
гетерозиготности и утрата породного своеобразия.
Очевидно, это все те же неблагоприятные
генетические процессы, порождаемые
человеческой деятельностью, которые, как
отмечалось выше, происходят и в
природных популяциях.
В свете этих выводов становятся
понятными те нежелательные явления, с
которыми постоянно сталкиваются работники
сельского хозяйства.
При создании новых сортов и пород
отбор ведется преимущественно по
признакам продуктивности. Однако зачастую
наряду с полезными признаками селекционеры,
сами того не ведая, отбирают и
закрепляют вредные, нежелательные. Пример тому —
сверхвысокомасличные сорта
подсолнечника, о которых в свое время немало
писали. На практике эти растения оказались
неустойчивыми к болезням, что
приводило к гибели урожая на корню. Создавая
новые сорта, вместе с генами высокой мае-
личности «соотобрали» гены
позднеспелости; получился подсолнечник, созревающий
под осень, в период дождей, когда
условия наиболее благоприятны для развития
корневой гнили. Высокая специализация
сорта приводит, как указано выше, к
потере внутрипопуляционного наследственного
разнообразия и соответственно к утрате
адаптивности. Избегать таких ошибок
позволяет разработанная в Институте общей
генетики им. Н. И. Вавилова РАН методика
селекции и семеноводства. Она дает
возможность сочетать направленный отбор по
признакам продуктивности со
стабилизирующим по признакам адаптивности, т. е.
получать популяции, одновременно
высокопродуктивные и устойчивые к
неблагоприятным воздействиям среды. По этой
методике, используемой с 1974 г., улучшен,
например, сорт подсолнечника «Енисей»; он
дает стабильный урожай, устойчив к
капризам погоды и заболеваниям,
приспособлен к возделыванию и уборке машинами.
Сейчас этот сорт занимает немалые площади
на полях страны и приносит хозяйствам
большую выгоду (рис. 9).
160
ГЕНЕТИКА
Заключение
Надеюсь, мне удалось показать, как
теоретические и практические работы в
области популяционной генетики
позволяют обосновать принципы оптимальной
эксплуатации природных и искусственных
популяций — принципы, которыми нельзя
пренебрегать, если мы хотим
пользоваться биологическими ресурсами нашей
планеты, не истощая их.
Действительно, проблема рациональной
промысловой эксплуатации и искусственного
воспроизводства биологических ресурсов
требует самого пристального внимания. Эво-
люционно сложившиеся уровни
внутривидового наследственного разнообразия
нарушаются не только в процессе промысла
(например, рыбного), но и в ходе селекции
и улучшения сельскохозяйственных
растений (ячмень) и животных (куры),
производимых с самыми благими намерениями, а
также при искусственном воспроизводстве
стад лососей на рыбоводных заводах. Во всех
случаях непропорциональное изъятие одних
генотипов и недоиспользование, либо
неравномерное воспроизводство других порождают
неблагоприятные процессы, приводящие к
снижению жизнеспособности популяций.
Механизм, лежащий в основе открытых
явлений, сопряжен не только с
уменьшением генетического разнообразия, но и с его
увеличением по отношению к исторически
сложившемуся оптимуму.
Есть основания утверждать, что во
многих случаях или, по крайней мере, в тех
из них, когда внутрипопуляционный
полиморфизм снижается, а меэюпопуляционная
пространственная дифференциация пара-
стает, пределы допустимых генетических
изменений уже превышены. Чтобы не
допустить дальнейшего развития
неблагоприятных процессов, нужно пересмотреть
стратегию взаимодействия Человека с Природой
таким образом, чтобы не разрушалась
системная организация популяций, а
соотношение внутри- и межпопуляционной
наследственной изменчивости удерживалось на
оптимальном уровне. Такой подход
предполагает: 1) сохранение генетического
разнообразия еще уцелевших популяционных
систем в процессе их промысла и
искусственного воспроизводства (неистощитель-
ное природопользование); 2) восстановление
тех систем, чья структура уже
нарушена; 3) создание новых систем популяций в
тех регионах, где существуют
необходимые естественноисторические и
экономические условия. Для сохранения
генетического разнообразия сельскохозяйственных
пород животных и сортов растений также
следует опираться на популяционно-гене-
тические принципы. Реализация всех этих
подходов будет способствовать не
экстенсивному росту и сопряженному с ним
разрушению биосферных генофондов, а
устойчивому существованию системы «Человек —
Биосфера» в неограниченно долгом ряду
поколений.
Литература
Айала Ф., Кайгер Дж. Современная генетика
Пер. с англ. М.: Мир, 1988, т. 3.
Алтухов Ю. П. Генетические процессы в
популяциях. М.: Наука, 1989.
Алтухов Ю. П. Генетика, 1995, т. 31, №10,
с. 1333.
Жизнеспособность популяций:
Природоохранные аспекты. Пер. с англ. М.: Мир, 1989.
Калабушкин Б. А. Журн. общ. биологии, 1976,
т. 37, №3, с. 369.
Корочкин JI. И., Серов О. Л., Пудовкин А. И. и
др. Генетика изоферментов. М.: Наука, 1977.
Крогиус Ф. В. Биология моря, 1979, т. 3, № 1,
с. 24.
Ли Ч. Введение в популяционную генетику.
Пер. с англ. М.: Мир, 1978.
Моисеева И., Банникова Л., Алтухов Ю.
Международный сельскохозяйственный журнал, 1993,
№5-6, с. 66.
Поморцев А. А., Калабушкин Б. А., Ладоги-
па М. П., Бланк М. Л. Генетика, 1994, т. 30, № 6,
с. 806.
КЛЕТОЧНАЯ БИОЛОГИЯ
Ю. /If. Васильев
ПОДВИЖНАЯ АРХИТЕКТУРА КЛЕТКИ
Введение
Наш организм есть федерация
огромного множества отдельных клеток. Однако мы
часто недооцениваем тот простой факт, что
каждая из этих клеток — сложный
индивидуум, обладающий собственными принципами
поведения. Если не понять эти принципы,
нельзя разобраться во взаимодействиях
клеток в организме. Изучать поведение
отдельных клеток лучше всего пользуясь
методом клеточных культур, т. е. выделяя
отдельные клетки из организма и помещая их
в сосуд с питательной средой. Если
наблюдать эти клетки под микроскопом и
фиксировать их поведение на кино- или
видеопленке, то легко убедиться в том, что каждая
клетка в такой культуре живет
самостоятельной сложной жизнью: прикрепляется ко
дну сосуда и ползает по этому дну
(подложке), меняя свою форму и направление
движения, выбрасывая и втягивая отростки.
Внутри клеток отдельные пузырьки-органел-
лы все время движутся. Долгое время
казалось, что разобраться в механизмах
этого сложного поведения клеток и их частей
почти невозможно.
Замечательное достижение последних
десятилетий — открытие и исследование
системы структур, ответственных за
подвижную архитектуру клетки, за ее
движения и форму. Этой системой в клетках эука-
риот оказался цитоскелет, состоящий из
белковых нитей, наполняющих цитоплазму
Рис. 1. Сеть актиновых микрофиламентов в
цитоплазме культивируемой клетки (фибробласта)
Микрофотография с трехмерной реплики цитоске-
лета под электронным микроскопом. Увеличение
х 45000. Уменьшение при печати в 1,6 раза.
Препарат Т. М. Свиткиной
(рис. 1). В настоящей статье
рассказывается том, как организован цитоскелет и
каковы основные типы его конструкций.
Полимеризация
и деполимеризация нитей —
основа динамики цитоскелета
Цитоскелет состоит из трех основных
типов нитей, образующих три системы:
микрофиламентов, микротрубочек и
промежуточных филаментов. Каждый тип нитей
состоит из одного—двух основных белков:
микрофиламенты — из актина,
микротрубочки — из тубулина, промежуточные фи-
ламенты — из специальных белков,
различных в разных тканях: кератинов — в
эпителиях; десмина — в мышцах; вимен-
тина — в тканях внутренней среды
(соединительной ткани, хряще, кости и др.); белков
нейрофиламентов — в нейронах.
Разумеется, белки цитоскелета, как и
любые белки клетки, закодированы в ДНК
и синтезируются на рибосомах. Клетка
может менять набор синтезируемых белков.
Однако конструкция цитоскелета может
быстро меняться даже без синтеза новых
молекул, за счет полимеризации и
деполимеризации нитей. Отдельные молекулы,
мономеры, растворенные в цитоплазме
клетки, способны соединяться, полимери-
зоваться в нити соответствующего типа.
Новые мономеры могут присоединяться к
концам нити, удлиняя ее. Полимеризация
обратима: мономеры могут отделяться от
концов нити, которая при этом
укорачивается и может исчезнуть совсем. В клетке
все время идет обмен между нитями и
раствором мономеров в цитоплазме (рис. 2). Во
многих клетках примерно половина
молекул актина и тубулина находится в виде
мономеров в цитоплазме и половина
входит в состав нитей микрофиламентов или
микротрубочек. Локальные условия
полимеризации могут часто меняться. Поэтому
одна и та же нить может то
укорачиваться, то удлиняться (см. рис. 2).
Клетка регулирует стабильность нитей
цитоскелета, присоединяя к ним
специальные белки, которые меняют скорость
полимеризации и деполимеризации. Поэтому
нить, состоящая из одного и того же
мономера, может иметь очень разную
продолжительность жизни. Например,
индивидуальные микротрубочки, входящие в
состав жгутика или реснички, обычно жи-
164
КЛЕТОЧНАЯ БИОЛОГИЯ
вут много часов и дней. Напротив, ка^к—
дая микротрубочка митотического веретена,
состоящая из того же тубулина, живет в
среднем лишь несколько минут.
Микротрубочки веретена все время растут и
распадаются, одни микротрубочки заменяются
другими. Между тем само веретено, т. е.
совокупность микротрубочек, идущих от
полюсов к хромосомам и экватору клетки,
сохраняется в течение всего митоза, лишь
постепенно меняя свою тонкую структуру.
Уже в середине митоза веретено состоит
из иных микротрубочек, чем в его начале.
Пример с веретеном иллюстрирует общий
принцип работы большинства цитоскелет-
ных систем, названный принципом
динамической нестабильности: отдельные нити
в системе могут появляться и исчезать в
результате
полимеризации—деполимеризации, и поэтому детальное строение
системы постоянно меняется, но, несмотря
на это, общий план организации системы
может сохраняться.
Рассмотрим теперь, как проявляется
динамическая нестабильность в работе
каждой из трех цитоскелетных систем.
Система микрофиламентов
Мономеры актина полимеризуются с
образованием микрофиламентов, диаметр
которых составляет около 6—7 нм (1 нм =
= 10"9 м). Микрофиламенты полярны: их концы
неодинаковы. Полимеризация микрофиламен-
та на одном конце, называемом
плюс-концом, идет легче, чем на другом, минус-конце.
Полимеризация и деполимеризация молекул
регулируются разными актинсвязывающими
белками. Некоторые из таких белков
присоединяются к одному концу нити,
блокируя на этом конце полимеризацию и
деполимеризацию, тогда рост и укорочение
микрофиламента идут лишь на другом
конце, не закрытом блокирующим белком.
Некоторые специальные белки соединяют
несколько мономеров в «зачаток» нити,
инициируя формирование нового
микрофиламента. В дальнейшем такие нити растут в
одну сторону, обычно в сторону
плюс-конца. Специальные белки могут присоединяться
к бокам нескольких микрофиламентов. При
этом одни белки связывают
микрофиламенты в сети, другие — в пучки.
Особую роль среди актинсвязывающих
белков играют миозины, так как они могут
двигаться по микрофиламенту. В настоящее
время известна структура свыше 80
вариантов молекул миозинов. У всех миозинов
молекула состоит из трех частей: головки,
шейки и хвоста. Головка способна
присоединяться к боку актинового
микрофиламента, и если снабжать эти головки
поставляющим химическую энергию веществом —
а
+
б
т2
Рис. 2. Динамичность нитей цитоскелета
а — микрофиламент, полимеризующийся из
мономеров (сиреневые кружки) на плюс-конце и деполиме-
ризующийся на мономеры на минус-конце, б—
динамическая нестабильность системы из трех
микротрубочек (отрезки прямых), полимеризующихся из
одного центра (кружок): О, Tv T2—
последовательные моменты времени. Верхняя микротрубочка
сильно укоротилась в интервале О—Т, и вновь выросла в
интервале Г7—Т2. Нижняя микротрубочка тоже
укоротилась (О—7"г), а затем выросла (Т^Т^. Здесь и на
рис. 3—6 все структуры изображены сугубо
схематически, без соблюдения точности деталей
АТФ, то головка движется вдоль
микрофиламента, от плюс- к минус-концу,
перескакивая с одного мономера на другой. Этот
процесс — основа очень многих движений
в клетке. Характер этих движений во многом
зависит от структуры того миозина,
который их осуществляет, от того, каковы у этой
молекулы головка и хвост. Например,
молекула обычного миозина из
поперечнополосатых мышц человека, так называемого
миозина II, имеет длинный хвост.
Переплетаясь хвостами, эти молекулы образуют
миозиновые филаменты с торчащими
наружу многими головками (рис. 3). В мышечной
клетке очень стабильные актиновые
микрофиламенты расположены параллельно
друг другу, на фиксированных
расстояниях друг от друга и от миозиновых фила-
ментов, помещающихся между ними.
Прикрепляясь к актиновым филаментам, головки
миозиновых нитей движутся вдоль этих
филаментов, и это скольжение — основа
всех мышечных движений (см. рис. 3). У
других миозинов, например у так
называемых миозинов I, хвосты очень короткие.
Поэтому такие миозины, в отличие от
миозинов И, переплетаться хвостами и
образовывать филаменты не могут. Вместо
этого молекулы некоторых миозинов I
могут поодиночке прикрепляться своими
короткими хвостами к мембранам разных орга-
нелл (например, митохондрий, лизосом и др.).
ПОДВИЖНАЯ АРХИТЕКТУРА КЛЕТКИ
165
а
б
Рис. 3. Взаимодействие актиновых микрофиламен-
тов (нити из синих кружков) с миозинами (красные
структуры)
а— схема движений в миофибрилле мышцы.
Молекулы миозина II соединены длинными хвостами в нить,
из которой наружу торчат в разные стороны головки.
Головки миозиновых молекул движутся по двум
параллельным актиновым нитям, вызывая скольжение
этих нитей в противоположных направлениях,
б—схема движения органеллы (зеленый круг) вдоль микро-
филамента при помощи миозина I. Молекулы
миозина I короткими хвостами прикреплены к
мембране органеллы, а концами головок—к актиновой нити
Если головка той же молекулы одновременно
прикрепится к актиновой нити, то она
сможет двигать органеллу вдоль этой нити (см.
рис. 3).
Комбинируя стандартные актиновые
микрофиламенты с различными миозинами
и другими актинсвязывающими белками,
клетка строит самые различные структуры,
отличающиеся по архитектуре и
подвижности. Мы уже упоминали об одной из таких
структур — миофибрилле, образующейся в
высоко специализированных мышечных
клетках. Так как в мышце все нити строго
параллельны друг другу, то их скольжение и
сокращение всей мышцы идут в одном
направлении и мышца может развить большое
напряжение. У большинства же клеток,
например в клетках соединительной ткани
(фибробластах), эпителия, лейкоцитах и
других, преобладающая часть микрофила-
ментов образует иную структуру — актино-
вый кортекс, располагающийся под
мембраной. Кортекс, подобно миофибрилле, может
сокращаться за счет взаимодействия
актиновых микрофиламентов с миозиновыми
молекулами. Однако, в отличие от миофиб-
риллы, в кортексе микрофиламенты
далеко не всегда параллельны друг другу и
часто образуют сложные сети. Поэтому сжатие
кортекса обычно идет в нескольких
направлениях. Кроме того, в кортексе, в отличие
от миофибриллы, микрофиламенты очень
динамичны; кортекс все время обновляется
и перестраивается путем полимеризации—
деполимеризации нитей. Если средняя
продолжительность жизни микрофиламента в
миофибрилле более 7 дней, то в кортексе
лейкоцита — всего лишь 5 с.
Основным и очень важным типом
перестроек кортекса являются псевдоподиаль-
ные реакции: выбрасывание, прикрепление
и сокращение псевдоподий. Рассмотрим
подробнее эти реакции. При выбрасывании
псевдоподии на поверхности клетки очень
быстро, в течение нескольких минут или
даже секунд, образуется вырост цитоплазмы.
Такой вырост может иметь разную форму,
например форму плоской пластинки (ламел-
лоподия), узкого цилиндра (филоподия) или
просто шаровидного пузыря. Внутреннее
строение всех типов псевдоподий просто: они
часто не содержат никаких структур, кроме
кортикальных микрофиламентов. При этом
в ламеллоподиях эти микрофиламенты
образуют густую уплощенную сеть, а в
пузырях — менее упорядоченный слой под
мембраной (рис. 4). Если микрошприцем
инъецировать в клетку раствор мономеров
а П Л
б
Рис. 4. Выросты поверхности клетки, образуемые
актиновыми микрофиламентами (синие линии)
а—два варианта псевдоподий: П— пузырь, в
котором под мембраной клетки имеется слой коротких
микрофиламентов, не имеющих упорядоченной
организации; Л— ламеллоподия — пластинчатый вырост,
в котором микрофиламенты соединены в
упорядоченную сеть молекулами специального актинсвязы-
вающего белка (изогнутые линии), б— стереоцилии
на поверхности двух соседних волосковых клеток
улитки внутреннего уха. Вертикальные параллельные
актиновые микрофиламенты в каждой стереоцилии
связаны друг с другом и с клеточной мембраной
молекулами миозинов и других актинсвязывающих
белков (горизонтальные красные линии). В одной
клетке разные ряды стереоцилии имеют разную
строго фиксированную длину. В соседних клетках
стереоцилии соответствующих рядов также имеют
разную фиксированную длину
166
КЛЕТОЧНАЯ БИОЛОГИЯ
актина, помеченных флуоресцирующей
краской, а затем наблюдать такую клетку
в флуоресцентном микроскопе, где краска
ярко светится, то можно видеть, что мик-
рофиламенты из меченых мономеров
появляются раньше всего именно в
псевдоподиях. Таким образом, псевдоподии являются
местом, где из мономеров полимеризуют-
ся микрофиламенты. Вероятно, под
мембраной в этих местах концентрируются
какие-то белки, вызывающие полимеризацию
новых микрофиламентов, но пока природу
этих белков мы еще точно не знаем.
Форма выпячивания может определяться
тем, с какими белками свяжутся вновь
возникшие микрофиламенты. Это
подтверждается недавними опытами американского
ученого Т. Штосселя. Он обнаружил, что
клетки одной из культивируемых
клеточных линий выпячивают на поверхности лишь
шаровидные пузыри, но не ламеллоподии.
Оказалось, что в геноме этих клеток
отсутствовал ген, кодирующий белок,
который связывает актиновые
микрофиламенты в сеть. Специальными методами генной
инженерии исследователи ввели в клетки
недостающий ген, и тогда клетки стали
делать не пузыри, а уплощенные
ламеллоподии. Таким образом, появление в ак-
тиновом кортексе одного дополнительного
белка направленно изменило архитектуру
псевдоподий.
Поверхность конца выброшенной
псевдоподии может прикрепиться к подложке,
по которой ползет клетка. При этом
образуется место прочного контакта, где
определенные белки мембраны наружным
концом молекулы соединяются с белками,
прикрепленными к подложке; внутренним
концом та же молекула соединяется, через
ряд промежуточных звеньев, с
актиновыми микрофиламентами псевдоподии.
Сразу после выбрасывания псевдоподия
содержит актин, а миозин II проникает в
псевдоподию (диффундирует) из внутренней
части клетки лишь несколько минут
спустя. Взаимодействие миозинов с актиновыми
нитями вызывает сокращение псевдоподии.
Это сокращение может иметь разные
последствия для клетки. Если псевдоподия не
прикреплена к подложке, она втягивается
и исчезает. Напротив, если псевдоподия,
выброшенная на одном из краев клетки,
успела прочно прикрепиться к подложке, то
ее сокращение смещает вперед все тело
клетки. Повторяя псевдоподиальные
реакции, клетка ползет по подложке. Если
много псевдоподий на разных краях клетки
выбрасываются и прикрепляются к подложке
одновременно, то они, стремясь сократиться,
натягиваются и растягивают клетку в
разные стороны, уплощая ее форму. Этот
процесс называют распластыванием.
Термин «псевдоподия» означает в
переводе — ложная ножка. Это действительно
ножка, которая двигает клетку вперед по
подложке. Вместе с тем это ножка особая:
в отличие от ноги человека псевдоподия
может вырасти заново из тела клетки,
образовать свои мышцы, сократиться и
исчезнуть за считанные минуты. Как мы
видели, преобразование псевдоподии является
результатом серии сложных молекулярных
реакций: полимеризации актиновых нитей,
присоединения к этим нитям других
белков, связывающих их в сети и
вызывающих их перемещение, а также связывания
нитей с белками мембраны.
Псевдоподии — лишь один из многих
типов отростков, содержащих актиновые
микрофиламенты. Расскажем еще об одном
виде весьма важных специализированных
отростков, а именно о так называемых сте-
реоцилиях (см. рис. 4). Эти отростки
располагаются пучками на верхней поверхности
волосковых клеток улитки внутреннего уха,
клеток, отвечающих за восприятие звуков.
Стереоцилии наполнены от основания до
верхушки параллельными друг другу
стабильными актиновыми микрофиламентами;
между ними и мембраной имеются
молекулы других белков, в том числе
специальных миозинов. В каждой клетке все
стереоцилии и их микрофиламенты имеют строго
определенные размеры; вариации такой
длины не превышают 5%. На поверхности
каждой клетки они располагаются подобно
трубам органа, рядами «по росту»: самые
короткие в переднем ряду, а самые
длинные — в заднем (см. рис. 4). При этом
длина и ширина стереоцилии самого длинного
ряда в разных волосковых клетках
закономерно меняются от клетки к клетке вдоль
поверхности улитки (см. рис. 4). Такая
точность размеров стереоцилии очень важна,
так как разные клетки улитки
активируются звуками разной частоты и
амплитуды: низкие звуки воспринимаются
клетками на одном конце улитки, высокие — на
другом. Полагают, что звуковая волна
вызывает колебания стереоцилии, притом на
различные частоты звука резонируют
стереоцилии разной длины и, следовательно,
активируются разные клетки.
Мы пока еще не знаем, как в
эмбриональном периоде создается замечательная
«музыкальная» архитектура стереоцилии; это
увлекательная проблема для исследователей.
Недавно было сделано еще одно
неожиданное открытие. Существует особая
наследственная глухота и слепота, так
называемый синдром Ашера; одно из его
проявлений — дегенерация волосковых клеток
уха. Большая международная группа
исследователей показала, что в основе болезни
лежит мутация, инактивирующая один из
ПОДВИЖНАЯ АРХИТЕКТУРА КЛЕТКИ
167
миозинов, так называемый миозин VIIА. Что
именно делает этот миозин в волосковых
клетках, как он участвует в реакциях сте-
реоцилии на звук, мы пока не знаем. Еще
менее понятно, почему тот же миозин
необходим для восприятия света клетками
сетчатки глаза, почему без одного этого белка
цитоскелета человек становится не только
глухим, но и слепым. Мы еще и еще раз
убеждаемся, как велико разнообразие
конструкций, которые клетка умеет строить на
основе одного основного элемента
цитоскелета — актиновых микрофиламентов.
Система микротрубочек
Микротрубочки представляют собой
цилиндры диаметром 25 нм с полостью
внутри. Их стенка образована мономерами
тубулина. Микротрубочки, подобно актиновым
микрофиламентам, полярны:
полимеризация из мономеров идет легче на
плюс-конце, чем на минус-конце. Система
микротрубочек, в отличие от актинового кортекса,
в большинстве клеток строго
централизована: в то время как в кортексе может
работать одновременно множество центров
полимеризации, из которых растут новые
гл
гл
гл
Рис. 5. Структуры, образуемые системой
микротрубочек и организующих их центров
а— параллельные микротрубочки ресничек,
растущие от базальных телец к поверхности клетки; б—
микротрубочки митотического веретена, растущие
навстречу друг другу из двух центров, расположенных
на полюсах веретена. Некоторые из микротрубочек
прикрепляются плюс-концами к особым участкам
(кинетохорам) хромосом. На схеме две хромосомы
в метафазе, т. е. до начала расхождения к полюсам
микрофиламенты, микротрубочки часто
имеют лишь 1—2 центра полимеризации на
клетку. Эти центры, организующие
микротрубочки (ЦОМТ), хорошо видны не
только под электронным, но и под световым
микроскопом. Практически все
микротрубочки в клетках растут из этих центров
плюс-концами к периферии, и поэтому
системы микротрубочек часто имеют вид звезд.
Наиболее распространенные варианты
ЦОМТ — центросомы, из которых растет
митотическое веретено и «звезды»
микротрубочек во многих клетках, а также ба-
зальные тельца, из которых растут
микротрубочки жгутиков и ресничек (рис. 5).
Замечательное свойство этих центров — их
способность репродуцироваться: новый центр
вырастает рядом со старым и затем
«материнский» и дочерний центры расходятся.
Долго искали в центрах ДНК, но не
нашли. Удвоение центров, видимо, имеет
совсем особый механизм, отличный от
удвоения ДНК, но природу его мы пока не знаем.
Как уже говорилось, микротрубочки
разных структур сильно различаются по
стабильности. Если инъицировать в клетки
раствор тубулина, меченного
флуоресцентной краской, то микротрубочки становятся
окрашенными, и в флуоресцентный
микроскоп можно непосредственно наблюдать, как
отдельные микротрубочки быстро растут от
центра к периферии, затем быстро
укорачиваются, иногда исчезая совсем, опять
растут и т. д. (см. рис. 2). Эта смена фаз
роста и укорочения — характерная черта
систем нестабильных микротрубочек. У
многих стабильных микротрубочек, например
в жгутиках, сохраняется постоянная
длина. Большую или меньшую стабильность
придают микротрубочкам особые белки,
связывающиеся с их наружной стенкой и
укрепляющие ее.
Некоторые растения образуют
специальные яды — вещества, которые избирательно
нарушают динамику микротрубочек в самых
разных типах клеток. Большая группа
таких веществ (колхицин, колцемид, винбла-
стин) деполимеризует нестабильные
микротрубочки. Точнее говоря, молекулы
этих веществ присоединяются к мономерам
тубулина и блокируют рост микротрубочек.
При этом их распад продолжается, и
через короткое время все микротрубочки
исчезают. В частности, у таких клеток в
митозе исчезает митотическое веретено и
хромосомы не могут разойтись к полюсам,
поэтому деление клеток не завершается.
Естественно, стабильные микротрубочки в
жгутиках мало чувствительны к действию
этих веществ.
Из коры тисса было выделено особое
вещество, таксол, действие которого на
микротрубочки противоположно действию
168
КЛЕТОЧНАЯ БИОЛОГИЯ
К
Рис. 6. Взаимодействия микротрубочек (цилиндры) с соответствующими моторными молекулами
Р— микротрубочки реснички. Молекулы динеина прикреплены хвостами (линии) и головками (черные кружки)
к соседним микротрубочкам. Перемещение головок вызывает сгибание обеих микротрубочек. К, Д —
движения органелл (большие круги) при помощи молекул кинезина (К) или динеина (Д) вдоль микротрубочки в
противоположных направлениях
веществ группы колхицина: молекулы так-
сола, связываясь с микротрубочками,
препятствуют их деполимеризации. Все
трубочки становятся стабильными и не могут
укорачиваться и исчезать. Во время
митоза обработанные таксолом клетки со
сверхустойчивыми микротрубочками не
могут разделиться, так же как
обработанные колхицином клетки, у которых нет
микротрубочек. Очевидно, для нормальной
работы веретена и, вероятно, многих
других систем микротрубочек существенно не
только присутствие трубочек, но и их
динамичность, постоянный рост и распад.
Вещества, действующие на микротрубочки
(винбластин, таксол и другие), широко
используются в клинике при химиотерапии
некоторых опухолей для остановки
деления опухолевых клеток.
Среди белков, прикрепленных к
микротрубочкам, очень важны моторные
молекулы — динеины и кинезины (рис. 6). Эти
молекулы одним концом прикрепляются
сбоку к микротрубочке и могут двигаться
по ней, если доставлять им энергию в виде
АТФ. При этом большинство вариантов
кинезина двигается по трубочке к ее плюс-
концу, а все динеины — к минус-концу.
Другим полюсом молекула динеина или
кинезина может прикрепиться к
мембранным органеллам или к другим
микротрубочкам. В результате эти молекулярные
моторы могут совершать много разных типов
движений. В жгутике и ресничке
молекулы динеина, прикрепляясь к двум
соседним микротрубочкам, заставляют их
скользить относительно друг друга. Так как своими
основаниями (минус-концами) микротрубочки
закреплены на базальных тельцах, то
скольжение соседних микротрубочек ведет к тому,
что жгутик изгибается (см. рис. 6).
Вызванное моторами взаимное скольжение
микротрубочек, отходящих от разных полюсов
митотического веретена, по-видимому,
приводит к расхождению этих полюсов в
противоположных направлениях.
Соединяясь с органеллами, микротрубочковые
моторы могут перемещать эти органеллы в
клетке. В зависимости от того, какой
мотор работает, направление движения
будет разным: кинезин «повезет» органеллу к
плюс-концу микротрубочки, т. е. к
периферии, динеин — к центру клетки. Иначе
говоря, органеллы могут «ездить» по клетке
по одним и тем же
рельсам-микротрубочкам туда и обратно, пользуясь разными
моторами.
Сейчас существуют замечательные
микроскопы, где контрастность изображения
сильно увеличивается с помощью
специальных компьютерных программ. Такие
приборы позволяют длительно наблюдать за
движениями отдельных органелл в живой
клетке. Эти наблюдения показывают, что
одна и та же органелла, активно двигаясь,
то идет к периферии клетки, то
возвращается обратно, нередко меняя
направление несколько раз в минуту. Что
определяет такую смену направлений? Одна из
возможностей, которая сейчас активно
проверяется, состоит в том, что на одной и той
же органелле сидят несколько разных
молекул-моторов, способных двигать органеллу
и к центру и от центра. Эти моторы могут
включаться и переключаться
специальными регуляторами, например ферментами,
присоединяющими к моторной группе
фосфатные группы и отщепляющими эти группы.
Кроме микротрубочковых моторов,
существуют, как мы уже говорили, еще мио-
ПОДВИЖНАЯ АРХИТЕКТУРА КЛЕТКИ
169
зиновые молекулы, способные перевозить
органеллы вдоль актиновых микрофиламен-
тов. Каково соотношение между этими двумя
видами транспорта в клетке? Недавно
предложено красивое сравнение: микротрубочки,
идущие на дальние расстояния, подобны
скоростным шоссе, по которым органеллы
быстро перемещаются между центром и
периферией клетки, тогда как микрофила-
менты — местные дороги. Доехав по
микротрубочке до нужной области клетки,
органелла может сменить мотор и пересесть
на местный микрофиламент, на котором
доедет точно до места назначения.
Напомним, что и сами рельсы, по которым
ездят органеллы — микротрубочки и микро-
филаменты — могут быстро менять свою
длину и положение в результате
полимеризации—деполимеризации. Таким образом,
распределение органелл в клетке в
каждый данный момент является
статистическим результатом сочетания их активных
перемещений по цитоскелетным путям и
динамики самих путей.
Промежуточные филаменты
Это третий основной компонент цито-
скелета, названный так потому, что его
нити по диаметру (8—10 нм) меньше, чем
микротрубочки, но больше, чем микрофи-
ламенты. Нити промежуточных филамен-
тов многочисленны в цитоплазме
большинства клеток; по-видимому, они растут из
многих центров, но вопрос еще
окончательно не решен. Промежуточные
филаменты— очень прочные структуры:
разными экстрагирующими солевыми
растворами можно удалить из клетки все ее
компоненты, а сеть промежуточных фи-
ламентов сохраняется, пока мы не
применим сверхсильные денатурирующие
агенты, например концентрированный
раствор мочевины. Другое отличие этих
филаментов от иных цитоскелетных нитей:
их мономеры легко полимеризуются, но
с большим трудом деполимеризуются,
поэтому в клетке свободных растворенных
мономеров почти нет. Впрочем, когда это
необходимо, клетка легко перестраивает
свою систему промежуточных филаментов.
Например, при митозе все филаменты
распадаются на фрагменты, по-видимому,
в результате того, что специальный
фермент присоединяет к их мономерам
фосфатные группы. После митоза
филаменты быстро востанавливаются.
Загадкой остается вопрос о том,
почему в разных тканях эти морфологически
сходные филаменты построены из разных
белков (см. выше). Особенно велико
внутриклеточное разнообразие белков
промежуточных филаментов эпителиальных
тканей — кератинов. Выделено уже более 30
кератинов, комбинирующихся по два типа
в каждой клетке. Разные наборы
кератинов имеются в различных типах эпители-
ев и даже в разных участках одного
эпителия. Например, в эпителии кожи,
покрывающем ладони и пятки человека,
обнаружен особый кератин (№ 9), которого
нет в эпителии других участков кожи или
каких-либо иных тканей. Неодинаковы по
белковому составу и промежуточные
филаменты (нейрофибриллы) разных типов
нервных клеток.
Вопрос о функциях всех этих филаментов
совершенно не ясен. Наиболее вероятная
гипотеза: промежуточные филаменты
укрепляют клетки и ткани механически,
делают их более прочными. Вспомним, что
кожа пятки и ладони испытывает разную
нагрузку и, возможно, молекулярные
различия кератинов делают филаменты
лучше приспособленными к разным нагрузкам.
Сильным аргументом в пользу
механической роли промежуточных филаментов
являются новые данные о том, что
основой некоторых наследственных кожных
болезней, при которых резко снижается
прочность кожного эпителия, являются мутации
генов определенных кератинов. В частности,
при мутациях упомянутого выше
кератина № 9, специфичного для пятки и ладони,
нарушается прочность кожи именно в этих
участках. Однако многие другие попытки
выяснить роль промежуточных филаментов
дали обескураживающие результаты.
Например, недавно группа исследователей
методами генной инженерии получила линию
мышей, у которых из генома был удален ген
виментина — белка, из которого сделаны
промежуточные филаменты в клетках
костей, хряща, соединительной ткани, крови
и костного мозга. Такие «безвиментиновые»
мыши развивались совершенно нормально
и разнообразные пробы на строение и
функции разных тканей не обнаружили у них
никаких дефектов. Зачем же в столь
многих клетках существуют многочисленные ви-
ментиновые филаменты? Вероятно, они
играют какую-то важную роль, но мы сегодня
еще не придумали эксперимент, который
выявил бы эту роль.
Заключение
Еще в начале века русский биолог
Н. К. Кольцов на основании большой серии
опытов по изучению свойств жгутиков
сперматозоидов пришел к выводу о том, что в
клетке есть скелетные структуры. Однако
термин «цитоскелет» получил широкое
распространение лишь в последние два
десятилетия. Первоначально предполагалось, что
цитоскелетные нити являются опорным
170
КЛЕТОЧНАЯ БИОЛОГИЯ
каркасом клетки, ее скелетом. Цитоскелет,
несомненно выполняет эту роль, но это
лишь одна из многих функций этих
структур в клетке.
Как мы видели, цитоскелет, особенно
актиновый, является двигательным
аппаратом клеток, а также аппаратом
движений органелл, в чем-то аналогичным
системе кровообращения многоклеточного
организма. Данные о стереоцилиях
иллюстрируют роль цитоскелета в восприятии
клеткой сигналов из внешнего мира. В
статье описаны эксперименты,
свидетельствующие о том, что цитоскелет, наряду с
клеточной мембраной, играет ключевую роль
в обобщении и запоминании результатов
реакций на внешние воздействия и в
определении поведения клетки. Эту функцию
можно сравнить с деятельностью мозга. Все
эти многообразные функции цитоскелет
выполняет, совершая реорганизации двух
типов: а) полимеризацию и
деполимеризацию цитоскелетных нитей и б) совершаемые
с помощью молекулярных моторов
движения одних нитей относительно других и
нитей относительно органелл. Эти
реорганизации, имеющие бесконечное число
вариантов, являются основой уникальной
подвижной архитектуры клетки, в которой
стабильность удивительно сочетается с
динамичностью.
Литература
Альберте А., Брей Д., Льюис Р. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ. М.:
Мир, 1994.
Чепцов Ю. С. Общая цитология. М.: Изд-во
МГУ, 1995.
StosselT.P. Science, 1993, vol.260, p. 1086.
WeilD. et al. Nature, 1995, vol.374, p. 60.
/О. М. Васильев
МЕХАНИЗМЫ ПЕРЕСТРОЙКИ ЦИТОСКЕЛЕТА
Введение
Изучение молекулярных механизмов
морфогенеза — одна из основных, может
быть, главная проблема биологии
ближайшего будущего. Мы уже много знаем о
генетических механизмах, определяющих
природу химических компонентов клетки и
особенно структуру белков. Вместе с тем
мы еще плохо понимаем, как особенности
этих химических компонентов определяют
сложнейшую структуру всего
многоклеточного организма. Мы не знаем, как
изменения генов и кодируемых ими белков
реализуются в изменениях формы и размещения
клеток, тканей и органов. Приведу только
один наглядный пример: мы хорошо
понимаем, как ДНК родителей определяет
первичную структуру любого белка их
ребенка, но совершенно ничего не знаем о том,
как эта ДНК определяет морфологические
черты его лица, т. е. форму подбородка, губ,
носа, ушей и т. д. А ведь о сходстве детей
с родителями мы обычно судим именно по
этим чертам.
Одним из наиболее перспективных
подходов к этой проблеме является
исследование механизмов изменений цитоскелета. В
предыдущей статье было показано, что
цитоскелет составляет основу подвижной
архитектуры клеток животных и растений.
Напомню, что он образован тремя
системами белковых нитей: микрофиламентами,
состоящими из белка актина,
микротрубочками, состоящими из белка тубулина, и
промежуточными филаментами,
состоящими из разных белков. Эти нити собираются
(полимеризуются) из соответствующих
белковых молекул и вновь разбираются на
отдельные молекулы. Благодаря такой
полимеризации—деполимеризации цитоскелет
непрерывно перестраивается, и эти
перестройки являются основой изменений
формы и движений клеток, лежащих в основе
образования клеток и органов.
В этой статье мы рассмотрим некоторые
простейшие цитоскелетные механизмы
таких движений и изменений формы.
Конкретнее, я попытаюсь объяснить, как клетка
(или часть ее) ползет по твердой
поверхности (так называемой подложке) и притом
ползет в определенном направлении,
проявляя зачастую поразительную способность
выбирать определенную ориентировку и в
одних случаях соблюдать эту
ориентировку, а в других — менять ее. На примере двух
разных клеточных типов — фибробластов
и нейронов будет показано, как «разумные»
движения отдельных клеток приводят к
образованию сложных многоклеточных
структур.
Фибробласты ползут к цели
Все клетки ползут, образуя на переднем
крае динамичные выросты — псевдоподии
разной формы. В псевдоподиях под
мембраной клетки полимеризуются актиновые
микрофиламенты, которые связываются с
миозином и другими белками (см.
предыдущую статью). Псевдоподии могут
прикрепляться к поверхности подложки и,
сокращаясь, тянут всю клетку вперед. Таков
основной механизм движения. Очевидно,
направление движения определяется тем,
на каком краю клетки будут образовываться,
прикрепляться и сокращаться псевдоподии.
Что же определяет места образования
псевдоподий? Для того чтобы лучше это
понять, рассмотрим движения одной из
клеток, чаще всего используемых в
экспериментах, клеток соединительной ткани —
фибробластов. Фибробласты, помещенные
в культуру и распластавшиеся на плоской
подложке, например, на дне чашки Петри
со средой, приобретают форму веера или
веретена. Они поляризованы, т. е.
образуют псевдоподии лишь на одном или двух
полюсах. Эти клетки могут ползти
направленно в сторону одного из активных полюсов.
Их боковые края неактивны.
Благодаря динамике цитоскелета фиб-
робласт может менять форму и
направление движений в ответ на изменения
окружающего внешнего мира: например, в ответ
на изменения питательной среды и
поверхности подложки. Ориентировка этих клеток
начинается с того, что клетка получает
направленный сигнал из внешнего мира.
Например, поместим возле одного из
краев клетки капилляр и будем из него
выпускать в среду раствор веществ, который,
попадая на поверхность клетки, вызывает
в этом месте поверхности образование
псевдоподий; в результате клетка начинает
направленно двигаться в сторону сигнала.
Это явление называется положительным
химиотаксисом. Веществами,
вызывающими такой химиотаксис у фибробластов,
являются некоторые специальные белки, так
называемые факторы роста. Химиотаксичес-
кие вещества связываются со специальными
белками-рецепторами в наружной
мембране клетки и активизируют их. Такая акти-
172
КЛЕТОЧНАЯ БИОЛОГИЯ
вация через какие-то еще неясные
промежуточные химические реакции вызывает
полимеризацию актина под
соответствующим местом мембраны и выпячивание
псевдоподии. Если концентрация активирующих
веществ с разных сторон клетки различна,
то на одном конце клетки будет
образовываться и прикрепляться к подложке
больше псевдоподий, чем на другом. Контакт с
другой клеткой может действовать
противоположно химиотаксису: если какой-то
участок активного края фибробласта
касается поверхности другой клетки, то
образование псевдоподий в этом месте края
немедленно прекращается; происходит
«контактное торможение» или «контактный
паралич» этого участка (рис. 1).
Механизмы такого паралича еще
неясны, но его биологический смысл очевиден:
благодаря параличу клетка не заползает на
другую клетку, но, коснувшись ее,
поворачивает туда, где есть свободная
поверхность подложки. Двигаясь, клетки
соблюдают взаимную вежливость. Третий внешний
фактор, меняющий распределение
псевдоподий, — различная адгезивность
(«липкость») разных участков поверхности
подложки. Например, посадим клетку не на
широкое плоское стекло, а на узкий
стеклянный цилиндр, диаметр которого (30 мкм)
лишь немногим больше диаметра самой
клетки. Тогда фибробласт начинает
выбрасывать псевдоподии во все стороны. Но лишь
те псевдоподии, которые выброшены вдоль,
а не поперек цилиндра, смогут коснуться
свободной поверхности стекла и
прикрепиться к ней; псевдоподии, выброшенные
поперек стекла, такой подложки не найдут,
и клетка втянет их обратно (рис. 2, 3).
Таким образом, во всех случаях под
влиянием внешних факторов у клетки возникает
первичная поляризация образования и
прикрепления псевдоподий. Однако такая
поляризация часто очень неустойчива. Чтобы
направленно двигаться, клетка должна
"запомнить и стабилизировать эффект внешних
факторов. Эта стабилизация выражается в
том, что клетка перестает выбрасывать
псевдоподии в тех направлениях, где их
прикрепление было менее удачно, и начинает
их выбрасывать более эффективно только
в наиболее удачных направлениях,
например, вдоль цилиндра или ближе к
источнику химиотаксического вещества (рис. 3, а).
Такая стабильная поляризация
достигается благодаря перестройкам архитектуры
двух цитоскелетных систем — актина и
микротрубочек. Первой ее стадией
является реорганизация актинового кортекса,
т. е. сети микрофиламентов,
расположенной под мембраной всего тела клетки. Как
мы знаем, прикрепленные псевдоподии
содержат актиновые микрофиламенты, со-
Рис. 1. Контактный паралич мышиного
фибробласта при встрече с другой клеткой
В кадрах микросъемки обведены контуры клеток.
Время между последовательными кадрами
составляет 20 мин. а — фибробласт, ползущий направо и
образующий широкие псевдоподии на правом
полюсе; б— правый активный полюс вступил в
контакт с краем другой клетки; б—г—образование
псевдоподий вдоль контакта почти прекратилось. Клетка,
сменив места образования псевдоподий,
вытягивается и движется параллельно краю другой клетки
единенные одним концом с подложкой
через мембрану, а другим концом с микро-
филаментами в кортексе тела клетки. Там,
где прикрепления к подложке прочны,
микрофиламенты, пытаясь сократиться, будут
тянуть в свою сторону весь кортекс. В тех
местах, где прикрепленных псевдоподий
больше, натяжение сильнее, и кортекс
будет вытягиваться вдоль этого
направления натяжений. При этом актиновые
микрофиламенты в кортексе будут,
натягиваясь, ориентироваться вдоль этого
направления. Каким-то неизвестным еще
образом ориентировка кортекса определя-
Рис. 2. Фибробласт, начавший вытягиваться на
цилиндрической подложке
Псевдоподии прикрепляются вдоль оси подложки, но
не поперек ее. (Сканирующий электронный
микроскоп)
МЕХАНИЗМЫ ПЕРЕСТРОЙКИ ЦИТОСКЕЛЕТА
173
ет места выбрасывания новых псевдоподий:
клетка перестает их выбрасывать на тех
боковых краях, которые параллельны
натяжениям. Эта роль натяжения
подтверждена прямыми опытами: если один из
участков поверхности клетки растянуть
механически микроиглой, то образование
псевдоподий в таком участке прекращается.
Стабилизация натяжением резко усиливает
небольшие количественные различия в
числе прикрепленных псевдоподий в
разных участках поверхности: происходит
качественное разделение поверхности на
стабильные и активные участки. У фиб-
робластов для стабилизации формы,
кроме актинового кортекса, необходима еще
и вторая цитоскелетная система,
микротрубочки. Если разрушить микротрубочки
колхицином или похожим веществом (см.
предыдущую статью), то фибробласт
теряет вытянутую форму, а его актиновые
микрофиламенты в кортексе теряют общую
ориентировку. Сосредоточить свои
псевдоподии на одном участке края такая
клетка не может, даже если имеется
ориентирующий внешний фактор, например,
цилиндрическая подложка или химиотак-
сический градиент (рис. 3, б). Клетка без
микротрубочек все время выбрасывает
псевдоподии по всему краю во всех
возможных направлениях. Разумеется, такая
клетка теряет способность направленно
двигаться и совершает лишь бессмысленный
«бег на месте».
Таким образом, у фибробластов
имеется две степени цитоскелетной стабилизации
10ц
7'
9'
19'
27'
8'
12'
10|х
68'
30'
12'
66'
18'
Рис. 3. Движения фибробласта, посаженного на узкую полоску стекла (параллельные линии)
Сверху вниз контуры одной клетки, сделанные по последовательным кадрам киносьемки; цифры рядом со
стрелками— временные интервалы между кадрами, а— контроль. Клетка постепенно вытягивается вдоль
полоски. Образование псевдоподий сосредоточивается на полюсах и прекращается на боковых краях клетки,
параллельных длине полоски (стабилизация боковых краев); б— клетка в среде с колцемидом, разрушившим
микротрубочки. Клетка несколько вытянулась, но полной стабилизации боковых краев не происходит:
псевдоподии образуются и вдоль и поперек длины полоски
174
КЛЕТОЧНАЯ БИОЛОГИЯ
формы и движений — актиновая и микро-
трубочковая. В отличие от фибробластов, у
некоторых слабовытянутых клеток,
например, у лейкоцитов, разрушение
микротрубочек почти не меняет форму и движения;
очевидно, у таких клеток «работает» одна
актиновая стабилизация. Лейкоцит меняет
ориентировку и направление движений
гораздо чаще, чем фибробласт. Можно
сравнить актиновую стабилизацию с
кратковременной памятью, которая запоминает
эффекты сигналов лишь на небольшое
время, а микротрубочковую стабилизацию фиб-
робласта — с долговременной памятью.
Остается нерешенным еще вопрос о механизмах
стабилизации: каким образом реорганизация
цитоскелета определяет места выбрасывания
псевдоподий? Вероятнее всего, актиновые
нити и микротрубочки транспортируют на
периферию, к определенным участкам края
какие-то органеллы, индуцирующие в этих
местах полимеризацию новых микрофиламен-
тов и, следовательно, выбрасывание
псевдоподий. Действительно, стабилизацию мест
образования псевдоподий можно нарушить
у фибробластов агентами, нарушающими
транспорт органелл, в особенности
инъекцией антител к кинезину — известному нам
«мотору», везущему органеллы к
микротрубочкам. Какие именно органеллы регулируют
образование псевдоподий, остается пока
неясным.
Движения фибробластов, как и движения
других клеток, удобнее изучать в культуре,
чем в организме, хотя бы потому, что
только в культуре и подложка (дно чашки
Петри), и среда прозрачны, и все детали
движений можно наблюдать под микроскопом.
Вместе с тем закономерности, открытые в
культуре, имеют общий характер и относятся
также к движениям клеток в организме.
Выберем только один пример — заживление
раны: кожа ранена каким-то орудием, из
поврежденных сосудов выходит кровь,
которая свертывается в ране. При свертывании
крови образуется сгусток фибриновых нитей,
и из разрушающихся клеток крови —
тромбоцитов выделяется тромбоцитарный фактор
роста. По всей вероятности, градиент этого
фактора в среде привлекает в рану фиброб-
ласты из окружающей соединительной
ткани. Нити фибрина, подобно цилиндрическим
подложкам в культуре, могут служить
«рельсами», направляющими дополнительно
движение фибробластов в рану. Наконец,
клетки, идущие в рану с разных сторон, благодаря
контактному параличу, ориентируются
упорядочение друг относительно друга. Фиброб-
ласты, направленно мигрировавшие в рану,
начинают там размножаться и делать кол-
лагеновые волокна. Образуется рубец, и рана
заживает. Заживление раны — лишь один из
вариантов процессов, при которых
направленные миграции фибробластов и близких к
ним клеток, например, костных клеток,
остеобластов, или хрящевых клеток, хонд-
робластов, восстанавливают поврежденную
структуру соответствующих тканей нашего
организма или меняют эту структуру в
ответ на действие внешних факторов.
Отросток нейрона ползет к цели
Рассмотрим теперь движения клеток,
которые составляют основу самой сложной
из существующих в природе организаций —
нашего мозга, нашей нервной системы.
Особая, наверное, главная черта этой
организации — система сложнейших
индивидуальных связей между клетками, по которым
через особые межклеточные контакты,
синапсы, сигнал передается от одной нервной
клетки (нейрона) к другому нейрону или
мышечной клетке. Для такого
направленного проведения сигналов нейроны в
процессе эмбрионального развития образуют
отростки, которые растут к клетке-мишени.
Иногда длина таких отростков может быть
очень большой: отростки нейронов коры
головного мозга, соединяющиеся с
двигательными нейронами спинного мозга, у
человека могут превышать 1 м. Рост отростков очень
точно направлен: они соединяются только
с нужными клетками-мишенями, т. е. с
определенной группой нейронов или
определенной мышцей. Важность такой точности
соединений для правильной работы мозга
очевидна: например, как бы мы двигали
рукой, если бы отростки двигательных
нейронов, заведующих движениями мизинца,
соединялись не с мышцами мизинца, а с
мышцами большого пальца или наоборот?
Как же осуществляется столь точно
ориентированный рост отростков нейронов?
Разберем кратко, как происходит такая
ориентировка у эмбриональных нервных
клеток, помещенных в культуру. Рост
отростков нервных клеток внешне совершенно
отличен от движений фибробластов: у
нейрона ползет по подложке лишь маленький
уплощенный фрагмент клетки — так
называемый конус роста, прикрепляющий на
переднем крае псевдоподии (рис. 4). Конус
роста очень похож на уменьшенную копию
безъядерного фибробласта. Задний конец
конуса роста соединен с телом клетки
цилиндрическим стволом, богатым
микротрубочками. Ни ствол, ни тело клетки
псевдоподий не образуют. Двигаясь вперед, конус
роста тянет за собой ствол, который при
этом удлиняется. Иногда сравнивают тело
нейрона с хозяином, который на
удлиняющемся поводке (стволе) держит бегущую
собачку (конус роста).
Направление движения конуса роста
определяется внешними сигналами, меня-
МЕХАНИЗМЫ ПЕРЕСТРОЙКИ ЦИТОСКЕЛЕТА
175
ющими образование и прикрепление
псевдоподий: а) градиентами концентрации
специальных белков, растворенных в среде (так
называемого фактора роста нервов) и б)
формой подложки: в частности, конус роста
хорошо ползет вдоль разных
цилиндрических поверхностей. Например, одним из
факторов стабилизации эффекта внешних
агентов является натяжение кортекса:
микроигла, натягивающая отросток в сторону,
может соответственно изменить
направление его роста. Для ориентировки отростка
необходима и система микротрубочек: при
разрушении этой системы рост отростка
прекращается, и сам отросток
сокращается. Таким образом, несмотря на внешние
различия, механизмы движений фибробла-
стов и роста нервных отростков сходны по
общим механизмам: они включают создание
внешними факторами неравномерности
прикрепления и стабилизацию этих различий
двумя цитоскелетными системами.
Особенность нейронов заключается в
чрезвычайно длительной и стойкой микро-
трубочковой стабилизации отростков, в
длительной «долговременной памяти».
Приобретя определенную ориентировку,
отростки сохраняют ее неопределенно долго,
часто до конца жизни организма. Именно
такая модификация цитоскелетного
механизма стабилизации отростков обеспечивает
правильную организацию нервной системы.
Ложные внутренние сигналы
активируют нерегулируемые
движения опухолевых клеток
Нормальная система регуляции
движений нормальных клеток может нарушаться
в результате мутаций, и эти нарушения
могут иметь самые серьезные последствия
для всего организма. Как мы видели,
движения нормальных клеток, таких как фиб-
робласты, вызываются и регулируются
внешними сигналами, например, факторами
роста, выделяемыми другими клетками,
такими как тромбоциты. Эти приходящие
извне молекулы активируют рецепторы
мембраны фибробласта и через ряд
промежуточных стадий реакций вызывают
образование псевдоподий. В некоторых клетках
могут произойти мутации генов,
кодирующих белки, участвующие в такой системе
проведения сигналов. В результате
синтезируются белки, постоянно активирующие
клетку, независимо от внешних сигналов.
Например, нормальный фибробласт не
синтезирует тромбоцитарный фактор роста;
ген, кодирующий этот белок, «молчит» в
нормальном фибробласте. Однако если этот
ген мутирует и активируется, то
фибробласт начинает синтезировать и выделять
такой фактор (рис. 5).
ТФР
ТФР
Рис. 4. Контуры отростка нейрона, растущего вправо
Справа— конус роста, слева — ствол отростка, тело
нейрона — на левом конце ствола вне рисунка.
Временной интервал между кадрами а и б— 5 мин. За
это время на активном краю конуса роста
вытянулось и сократилось несколько псевдоподий, тогда
как контур стабильного края ствола не изменился.
На стекле вне клетки— неподвижные точки; видно,
как сдвинулся вправо конус роста за время между
кадрами
Рис. 5. Нормальный фибробласт движется по
направлению к внешнему сигналу— градиенту тром-
боцитарного фактора роста (ТФР), выделяемого на
расстоянии тромбоцитами (вверху) и опухолевый
фибробласт, движущийся беспорядочно под
влиянием ТФР, выделяемого им самим (внизу). Это один из
вариантов стимуляции опухолевой клетки
собственными «внутренними» сигналами
176
КЛЕТОЧНАЯ БИОЛОГИЯ
Сев на рецепторы мембраны, этот белок
будет стимулировать ту же клетку,
которая его выработала: эта клетка начнет
двигаться независимо от внешних сигналов,
активируя саму себя.
Сейчас мы знаем, что опухолевые
клетки — это клетки с мутациями генов,
ответственных за проведение сигналов.
«Самостимуляция» клеточных движений в результате
некоторых из таких мутаций, по-видимому,
является основой самых опасных для
организма свойств некоторых опухолевых
клеток: их способности к инвазии и метастази-
рованию, т. е. способности выйти из той ткани,
где они возникли, и двигаться в другие ткани
и даже в другие органы, давая там начало
новым колониям, разрушающим нормальные
структуры организма.
Заключение
Подведем теперь некоторые итоги.
Выбрасывая псевдоподии в ответ на внешние
сигналы, клетка пробует и оценивает свое
окружение. Где можно лучше прикрепить
эти псевдоподии? Куда можно и нужно
ползти? Неравномерное образование,
прикрепление и натяжение псевдоподий в
разных участках поверхности клетки
вызывает перестройку всего цитоскелета, сперва
актинового, потом микротрубочкового.
Реорганизуя цитоскелет, клетки запоминают
результаты предыдущих проб, предыдущих
псевдоподиальных реакций и стабилизируют
направления будущих реакций.
Таким образом, клетка все время
исследует окружающий ее мир и меняет свое
поведение в зависимости от результатов
этого исследования. Один и тот же
механизм в разных его модификациях
обеспечивает и движения фибробластов при
заживлении раны, и движения отростков
нейронов при образовании нервной
системы, и, вероятно, множество других
процессов морфогенеза, приводящих к
формированию нашего многоклеточного организма.
При этом поведение каждой отдельной
клетки «социально разумно»: оно согласовано
с поведением окружающих ее клеток через
сигналы, поступающие от клетки к клетке
через жидкую среду или прямой клеточный
контакт. Каждый тип клеток реагирует на
сигналы по-своему. Эти различия
определяются химическими различиями белковых
молекул реагирующих систем, например,
рецепторов, цитоскелета и т. д.
Разумеется, эти различия определяются
особенностями кодирующих белки генов. Порча
механизмов оценки «социального» окружения
отдельных клеток приводит к тому, что эти
клетки перестают «разумно» реагировать на
внешние сигналы, и активируют сами себя.
Такая злокачественная трансформация клеток
может вызывать гибель всего организма. Мы
не знаем еще точной природы всех
нормальных и патологических вариантов этих
механизмов морфогенеза, но можно надеяться,
что мы уже начали понимать, где искать
единые принципы их функционирования.
Литература
Альберте А., Брей Д., Льюис Р. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ. М.:
Мир, 1994.
Чепцов Ю. С. Общая цитология. М.: Изд-во
МГУ, 1995.
Ю. С. Ченцов
ОРГАНИЗАЦИЯ МИТОТИЧЕСКОИ ХРОМОСОМЫ
Введение
При наблюдении за живыми клетками
или клетками после фиксации и окраски
внутри ядра выявляются зоны плотного
вещества, окрашиваемого разными
красителями, особенно основными. Благодаря
способности хорошо окрашиваться этот
компонент ядра получил название хроматин.
Способность хроматина воспринимать
основные красители указывает на его кислотные
свойства. Они определяются тем, что в
состав хроматина входит ДНК в комплексе с
белком. Такими же свойствами окрашива-
емости и содержанием ДНК обладают и
хромосомы, несущие ДНК тельца,
доступные наблюдению во время митотического
деления клеток.
Таким образом, хроматин интерфазных
неделящихся ядер представляет собой
хромосомы, которые теряют в это время
компактную форму, разрыхляются, деконден-
сируются. Степень деконденсации хромосом
может быть различной в ядрах разных
клеток. Как показано в многочисленных
работах, степень деконденсации хромосомного
материала — хроматина — в интерфазе
может отражать функциональную
нагрузку этой структуры. Чем более д^иффузен
хроматин интерфазного ядра, тем
активнее в нем синтетические процессы.
Снижение уровня синтеза ДНК и РНК в клетках
1
4
б 5
Рис. 1. Морфология митоти-
ческих хромосом
а — фазы митоза: 1 —
интерфаза; 2— профаза; 3— ме-
тафаза; 4—анафаза; 5, 6—
телофаза; б— одна из мета-
фазных хромосом: 1 —
сестринские хромосомы (хрома-
тиды); 2— плечи хромосом;
3— центромера; 4 —
вторичная перетяжка, ядрышковый
организатор; 5— теломеры;
6 — пучки микротрубочек;
в— хромосома! человека
при дифференциальной
окраске: в сегментах («бэн-
дах») локализованы
определенные гены
178
КЛЕТОЧНАЯ БИОЛОГИЯ
обычно сопровождается увеличением зон
конденсированного хроматина. Максимально
конденсирован хроматин во время митоти-
ческого деления клеток, когда он
обнаруживается в виде плотных телец —
хромосом. В этот период хромосомы не несут
никаких синтетических нагрузок, в них не
происходит включения предшественников
ДНК и РНК (рис. 1).
Исходя из этого, можно считать, что
хромосомы клеток могут находиться в двух
структурно-функциональных состояниях: в
рабочем, частично или полностью деконден-
сированном, когда с их участием в
интерфазном ядре происходят процессы
транскрипции и редупликации, и в неактивном —
в состоянии метаболического покоя при
максимальной их конденсации, когда они
выполняют функцию распределения и
переноса генетического материала в дочерние
клетки.
Общая морфология
митотических хромосом
- Морфологию митотических хромосом
лучше всего изучать в момент их наибольшей
конденсации — в метафазе и в начале
анафазы. Хромосомы животных и растений в этом
состоянии представляют собой
палочковидные структуры разной длины с довольно
постоянной толщиной, у большей части
хромосом удается легко найти зону первичной
перетяжки, которая делит хромосому на два
плеча (см. рис. 1). В области первичной
перетяжки находится центромера, где
расположен кинетохор; к нему подходят пучки
микротрубочек митотического веретена, идущие
в направлении к центриолям. Эти пучки
микротрубочек принимают участие в движении
хромосом к полюсам клетки при митозе.
Некоторые хромосомы имеют вторичную
перетяжку. Последняя обычно расположена вблизи
дистального конца хромосомы и отделяет
маленький участок, спутник. Вторичные
перетяжки называют, кроме того, ядрышковы-
ми организаторами, так как именно на этих
участках хромосом в интерфазе происходит
образование ядрышка. Здесь же
локализована ДНК, ответственная за синтез рРНК.
Плечи хромосом оканчиваются теломерами,
конечными участками. Теломерные концы
хромосом не способны соединяться с
другими хромосомами или их фрагментами.
Размеры хромосом у разных организмов
варьируют в широких пределах. Так, длина
хромосом может колебаться от 0,2 до 50 мкм.
Число хромосом у различных объектов также
значительно колеблется, но оно
характерно для каждого вида животных или
растений. Совокупность числа, величины и
морфологии хромосом называется кариотипом
данного вида. Кариотип — это как бы лицо
вида. Даже у близких видов хромосомные
наборы отличаются друг от друга или по
числу хромосом, или по величине хотя бы
одной или нескольких хромосом, или по
форме хромосом и по их структуре.
Около четверти века назад в практику
анализа хромосом стали широко входить
методы их дифференциального
окрашивания. Впервые метод был предложен
шведским ученым Т. Касперссоном. Он показал,
что при обработке препаратов митотических
хромосом флуорохромом акрихиниприта с
помощью флуоресцентного микроскопа
можно видеть поперечные светящиеся
полосы (бэнды), расположение которых
характерно для каждой хромосомы. Этот прием
цитологического анализа в сочетании с
генетическими наблюдениями уже в
настоящее время позволил составить
хромосомные карты человека, т. е. находить места
расположения генов на определенных
участках хромосом (см. рис. 1). Молекулярные
механизмы такой специфической окраски
до сих пор еще неясны, многие
исследователи связывают способность отдельных
участков хромосом к окрашиванию с их
химическими различиями. Большое число
наблюдений говорит о том, что
избирательное окрашивание связано с локализацией
так называемого гетерохроматина, ДНК
которого обогащена А—Т парами оснований.
Каково же пространственное
расположение отдельных деконденсированных
интерфазных хромосом в трехмерном объеме
клеточного ядра? Существует ли какой-либо
ц2
Рис. 2. Схема пространственного расположения
хромосом в интерфазном ядре
Т—теломеры, Ц—центромеры
ОРГАНИЗАЦИЯ МИТОТИЧЕСКОЙ ХРОМОСОМЫ
179
порядок в размещении хромосом в
интерфазном ядре или они хаотически разбросаны
внутри него? Было обнаружено, что в
объеме ядер хромосомы располагаются повторяя
ана-телофазную ориентацию. При этом
каждое плечо хромосомы занимает
определенную зону, объем которой не заходит в объем
соседних хромосом, хотя они примыкают друг
к другу. Каждая хромосома в нескольких
местах связана с ядерной оболочкой, как бы
фиксируясь на ней. Фиксированы на
ядерной оболочке и теломерные участки всех
хромосом, локализованных на одном из
полюсов интерфазного ядра. На
противоположном полюсе ядра располагаются центромер-
ные районы хромосом, также связанные с
ядерной оболочкой (рис. 2).
Суммируя общие представления о
формах организации хромосом, можно прийти
к заключению, что они могут находиться в
двух альтернативных состояниях, в двух
морфологических выражениях: 1)
максимально конденсированное, компактное,
метаболически неактивное транспортное
состояние, предназначенное для того, чтобы в
минимальном объеме без структурных
нарушений перенести во время клеточного
деления огромные по длине молекулы ДНК;
2) деконденсированное, при котором
линейная длина развернутых хромосом
увеличивается в десятки, а иногда и в сотни раз,
метаболически активное состояние,
связанное с синтезом ДНК и РНК (интерфаза).
Элементарные уровни
структуризации хромосомных
компонентов
Хроматин, основной компонент
клеточного ядра, достаточно легко можно
получить из выделенных интерфазных ядер и
из выделенных митотических хромосом. В
среднем в хроматине 40% приходится на
ДНК и около 60% на белки, среди
которых специфические ядерные белки-гистоны
составляют от 40 до 80% всех белков,
входящих в состав выделенного хроматина. В
структурном отношении хроматин
представляет собой нитчатые комплексные
молекулы дезоксирибонуклеопротеида (ДНП),
состоящие из ДНК, ассоциированной с ги-
стонами. Обнаружено, что в составе
хромосом длина индивидуальных линейных
молекул ДНК может достигать сотен
микрометров и даже нескольких сантиметров.
Было показано, что максимальная
молекулярная масса молекулы ДНК
дрозофилы равна 41 х 109, что соответствует
длине около 2 см.
Наблюдаются значительные различия по
составу и свойствам ДНК в различных
участках хромосом. Так, в области первичных
перетяжек располагается специфическая
центромерная ДНК с часто
повторяющимися короткими последовательностями нук-
леотидов, с которыми взаимодействуют
специальные белки кинетохоров — структур,
обеспечивающих связь микротрубочек
веретена с хромосомами. В теломерах также
располагаются особые участки ДНК,
предотвращающие укорачивание хромосом в
процессе репликации ДНК. В зонах
вторичных перетяжек располагаются
многочисленные участки ДНК, ответственные за
синтез рибосомных РНК. В основной массе тела
хромосом, в их плечах, располагается
основная часть ДНК, ответственная за
синтез многочисленных информационных РНК,
за работу структурных генов.
В клеточном ядре ведущая роль в
организации расположения ДНК, в ее компак-
тизации и регулировании функциональных
нагрузок принадлежит ядерным белкам. Как
уже указывалось, хроматин представляет
собой сложный комплекс ДНК с белками в
виде длинных неветвящихся нитей, или
фибрилл, хроматина. Белки в составе
хроматина очень разнообразны, но их можно
разделить на две группы: гистоны и негис-
тоновые белки. Взаимодействие гистонов с
ДНК происходит за счет солевых, или
ионных, связей и неспецифично в отношении
состава или последовательностей нуклеоти-
дов в молекуле ДНК. В отличие от
гистонов так называемые негистоновые белки
большей частью специфически
взаимодействуют с определенными
последовательностями молекул ДНК.
На ультратонких срезах интерфазных
ядер и митотических хромосом
обнаруживались хроматиновые фибриллы толщиной
25—30 нм. Такие же размеры имели
фибриллы в препаратах выделенного хроматина.
Крупным событием в изучении хроматина
было открытие нуклеосом — дискретных
частиц хроматина. Так, при осаждении на
подложку для электронной микроскопии
препаратов хроматина можно было видеть,
что нити хроматина представляют собой
нечто, напоминающее бусы на нитке:
небольшие, около 10 нм, глобулы, связанные
друг с другом отрезками ДНК длиной
около 20 нм (рис. 3).
Расшифровка молекулярной организации
одиночных нуклеосом показала, что эта
частица содержит 8 молекул гистонов,
образующих как бы сердцевину, по
поверхности которой располагается ДНК, образуя
1,75 оборота. Такие частицы связаны друг
с другом непрерывной молекулой ДНК.
Количество таких частиц-нуклеосом огромно.
Рассчитано, что на гаплоидное количество
ДНК человека приходится до 1,5 х 107
нуклеосом. При этом образуется хроматиновая
фибрилла, в которой ДНК первично ком-
180
КЛЕТОЧНАЯ БИОЛОГИЯ
0,1 MKM
I 1 МКМ .
Рис. 3. Начальные уровни компактизации хроматина
а — нуклеосомные фибриллы: «бусы на нити»; б— схема строения фибриллы толщиной 30 нм: 7 — соленоидная
укладка нуклеосомной фибриллы; 2—ее укладка в виде нуклеомеров («сверхбусин»); в — хромомеры —
петлевые домены из макронуклеуса инфузории [из: Сергеева и др.]
пактизована с плотностью упаковки,
равной б—7 (146 нуклеотидных пар ДНК
длиной в 68 нм уложены в глобулу диаметром
10 нм). Это так называемый первый, нук-
леосомный, уровень компактизации
хроматина, встречающийся как в интерфазных,
так и в митотических хромосомах. Однако
во многих электронно-микроскопических
исследованиях было найдено, что как в
митотических, так и в интерфазных ядрах
выявляются фибриллы хроматина
диаметром 25—30 нм. Было показано, что 25-на-
нометровая фибрилла хроматина может
обратимо менять свой диаметр, становясь
фибриллой толщиной 10 нм. Все это
говорило о том, что нуклеосомные цепочки
хроматина каким-то специфическим
образом уложены так, что возникает не
хаотическая агрегация нуклеосом, а
правильная нитчатая структура диаметром 25—30
нм. Относительно характера упаковки
нуклеосом в составе 25-нанометровой фибриллы
хроматина существуют, по крайней мере,
две точки зрения. Одна из них защищает
так называемый соленоидный тип укладки
нуклеосом. Согласно этой модели, нить
плотно упакованных нуклеосом диаметром 10 нм
образует в свою очередь спиральные
витки с шагом спирали около 10 нм. На один
виток такой суперспирали приходится 6—
7 нуклеосом.
Если исследовать хроматин в составе
ядер или в виде выделенных препаратов,
но при поддержании определенной
концентрации двухвалентных катионов (не ниже
1 мМ), можно наблюдать дискретность в
ОРГАНИЗАЦИЯ МИТОТИЧЕСКОЙ ХРОМОСОМЫ
181
составе 25-нанометровой фибриллы
хроматина: она состоит как бы из сближенных
глобул того же размера, из пуклеомеров.
В зарубежной литературе такие 25-нано-
метровые глобулы, или нуклеомеры,
получили название «сверхбусин» («супербид») (см.
рис. 3). В состав одного нуклеомера входит
отрезок ДНК, соответствующий 1600 парам
оснований или 8 нуклеосомам. Основная
25-нанометровая фибрилла хроматина
представляет собой линейное чередование нук-
леомеров вдоль компактизованной
молекулы ДНК. Нуклеомерный уровень укладки
хроматина обеспечивает сорокакратное
уплотнение ДНК, что важно не только для
достижения целей компактизации
гигантских молекул ДНК. Как нуклеосомный, так
и нуклеомерный (супербидный) уровни
компактизации ДНК хроматина осуществляются
за счет гистоновых белков, которые
участвуют не только в образовании нуклеосом,
но и в их кооперативном объединении в виде
фибрилл ДНП, где ДНК претерпевает
дополнительную сверхспирализацию. Все
остальные уровни компактизации связаны с
характером укладки 25-нанометровых
фибрилл в новые компактизационные уровди,
где ведущую роль играют негистоновые
белки.
Расшифровка принципа строения
элементарных хромосомных компонентов —
нуклеосом и 30-нанометровых фибрилл — еще мало
что дает для понимания основ трехмерной
организации хромосом как в интерфазе, так
и в митозе. Сорокакратное уплотнение ДНК,
которое достигается при сверхспиральном
характере ее компактизации, совершенно
недостаточно для получения реального (1 х 104)
уровня уплотнения ДНК. Следовательно,
должны существовать более высокие уровни
компактизации ДНК, которые, в конечном
счете, должны определять размеры и общие
характеристики хромосом. Такие высшие
уровни организации хроматина обнаружены
при искусственной его деконденсации,
когда было найдено, что поддержание их
связано с негистоновыми белками. В этом
случае специфические белки связываются с
особыми участками ДНК, образующей в
местах связывания большие петли, или
домены. Таким образом, следующие более
высокие уровни компактизации ДНК связаны не
с ее дополнительной спирализацией, а с
образованием поперечной петлистой
структуры, идущей вдоль интерфазной, или мито-
тической, хромосомы.
Признаки петлевой доменной организации
хроматина можно наблюдать с помощью
электронного микроскопа после помещения
ядер или хромосом в солевые растворы
низкой ионной силы в присутствии низких
концентраций двухвалентных катионов. При
этом в некоторых местах можно видеть, что
отдельные сгустки конденсированного
хроматина выявляют особую структуру. Это
розетковидные образования, состоящие из
многих петель 30-нанометровых фибрилл,
соединяющихся в общем плотном центре.
Средний размер таких плотных розеток
достигает 100—150 нм. Подобные розетки
фибрилл хроматина — хромомеры —
можно видеть в ядрах самых разнообразных
объектов: животных, растений, простейших.
Особенно демонстративно такие
хромомеры выявляются на тотальных препаратах
хроматина из макронуклеусов инфузории
Bursaria (см. рис. 3). В этом случае можно
видеть, что каждый хромомер состоит из
нескольких, содержащих нуклеосомы,
петель, которые связаны в одном центре.
Хромомеры связаны друг с другом
участками нуклеосомного хроматина, так что в
целом видна цепочка розетковидных
структур. Подобные розетковидные петлистые
структуры, хромомеры, можно видеть
также при разрыхлении митотических
хромосом как животных, так и растений.
Следовательно, хромосомные 30-нанометровые
фибриллы, состоящие из ДНК и гистонов,
упаковываются в виде петлистых
розетковидных структур, претерпевая еще
дополнительную компактизацию. Это третий
уровень структурной организации хроматина.
Размер отдельной петли совпадает с
размером средних репликонов (единица
репликации ДНК) и может соответствовать одному
или нескольким генам. На хромосому в
среднем приходится более 2000 таких
петельных доменов ДНК. В своих основаниях
петли ДНК связаны негистоновыми белками
ядерного матрикса, или остова, в состав
которых могут входить как ферменты
репликации ДНК, так и транскрипции. Такая пе-
тельно-доменная структура хроматина
обеспечивает не только компактизацию
хроматина, но и организует функциональные
единицы хромосом — репликоны и
транскрибируемые гены.
Высшие уровни структуризации
ДНК в составе митотических
хромосом
Возможно, главным в изучении
строения митотической хромосомы является
вопрос о том, каков порядок расположения,
упаковки гигантской (несколько
сантиметров!) молекулы ДНК в достаточно малом
объеме самой митотической хромосомы
(несколько кубических микрометров). Первые
элементарные уровни структуризации
приводят к укорочению фибриллярных
структур хроматина всего в 600—700 раз.
Дальнейшие этапы компактизации хроматина
еще окончательно не выяснены.
182
КЛЕТОЧНАЯ БИОЛОГИЯ
Как уже указывалось, одним из
промежуточных уровней компактизации
хроматина может быть хромомерный уровень.
Хромомеры хорошо выявляются в
интерфазных политенных хромосомах, в мейоти-
ческих хромосомах. Они видны в
электронном микроскопе при искусственной
деконденсации хроматина ядра и митоти-
ческих хромосом. В их строении
выявляется петлево-доменный принцип третьего
уровня компактизации хромосом.
При изучении ультраструктурных основ
строения митотических хромосом необходимо
учитывать хромонемный уровень
компактизации хроматина. Хромонему, или
нитчатую хроматиновую структуру, имеющую
среднюю толщину 0,1—0,2 мкм, удается
проследить в естественных условиях на
разных стадиях начальной конденсации
хромосом в профазе митоза и при
деконденсации хромосом в телофазе. Причем
такие хромонемы выявляются как в клетках
растений, так и животных (рис. 4).
Изучение профазных хромосом животных и
растений показывает, что процесс
конденсации хромосомного материала включает
промежуточный этап — образование
нитчатых хромонемных структур из фибрилл ДНП.
Хромонемные структуры подвергаются
последующей хромосомной структуризации.
В естественных условиях в составе ме-
тафазных хромосом хромонемные
элементы на ультратонких срезах не
выявляются. Но по мере деконденсации митотических
хромосом в поздней анафазе и ранней
телофазе снова можно видеть признаки хро-
монемной, нитчатой, организации хромосом.
В поздней анафазе, когда хромосомы
достигают противоположных полюсов клетки,
в их структуре снова выявляются хрома-
тиновые нитчатые образования толщиной
около 0,2 мкм. При этом вся структура
хромосом разрыхляется, что отражает
начало общей деконденсации митотических
хромосом. Эта начальная стадия деконденсации
связана не с разрыхлением фибрилл ДНП
внутри хромонем, а с расхождением,
обособлением участков хромонемы друг от
друга.
Обращает на себя внимание свойство
митотических хромосом обратимо изменять
свой объем при изменении ионного
окружения. Как уже указывалось, применение
гипотонических растворов приводит к
набуханию хромосом, но при возвращении их
в изотонические условия они вновь
приобретают исходную морфологию. Из этого
следует, что существует какой-то механизм,
стабилизирующий общую организацию
хромосомы. Было обнаружено, что хромосомы
не теряют морфологической целостности,
не распадаются даже при резком
набухании, вызванном удалением всех гистонов.
Рис, 4. Хромонема— хроматиновые нити толщиной
0,1—0,2 мкм на ультратонком срезе телофазных
хромосом клеток культуры ткани
Стрелками показаны сечения хромонемных нитей
Рис. 5, Латеральные петли ДНК (7) и осевые
компоненты (2) метафазной хромосомы после полного
удаления гистонов
ОРГАНИЗАЦИЯ МИТОТИЧЕСКОИ ХРОМОСОМЫ
183
10 нм
30 нм
300 нм
t
i
700 нм
t
Рис. 6. Схема некоторых уровней организации
хромосомы
Такие сильно набухшие, лишенные гистонов
хромосомы помещали на подложку и
рассматривали в электронный микроскоп.
Оказалось, что разрыхленные хромосомы
состоят из двух компонентов: из рыхлой сети
плотных фибрилл в центральных участках,
повторяющих контуры метафазных
хромосом (осевые компоненты), и
многочисленных длинных тонких петель, отходящих от
них в поперечном направлении (рис. 5).
Была показана белковая природа осевых
компонентов и присутствие ДНК в составе
петель. Средний размер боковых петель
составлял около 30 мкм. Если такие
препараты обработать ДНКазой, то можно
получить белковые остовы и анализировать
их состав. Оказалось, что в них присутствует
около 20 видов белков негистоновой
природы, сходных с белками интерфазного
ядерного матрикса. Исходя из этого, была
предложена модель структурной
организации митотических хромосом. В ее основе
лежит принцип поперечного расположения
петель ДНК вдоль белковой осевой
структуры. В принципе этот тип организации
митотическои хромосомы очень
напоминает хромосомы типа «ламповых щеток»,
встречающиеся в процессе мейоза. При
разных способах депротеинизации кроме
петель на периферии набухших хромосом
можно было выявить и розеткоподобные
структуры, состоящие из ДНК.
В последнее время получены данные,
говорящие о том, что осевые структуры
могут представлять собой артефакт,
получившийся в результате монтажа и
высушивания дегистонизированных хромосом на
подложке. На самом же деле в теле
хромосомы есть негистоновые белковые
связки (скрепки), сшивающие основания
боковых петель ДНК, но эти связки
разбросаны рыхло по объему хромосомы. Как бы то
ни было, принцип петлевой поперечной
укладки ДНК в теле хромосомы очень важен
для понимания ее ультраструктурной
организации.
Необходимо подчеркнуть, что на
поперечных и продольных сечениях
митотических хромосом, фиксированных в нативном
состоянии внутри клеток, никаких
центральных или осевых элементов не обнаружено.
Они выявляются только после удаления из
выделенных хромосом всего набора
гистонов, чему предшествует изоляция
хромосом в гипотонической среде.
На основании этих наблюдений
широкое распространение нашла схема,
объясняющая структуру митотическои хромосомы
(рис. 6). Согласно этой схеме, первым
уровнем компактизации ДНК является нуклео-
сомная фибрилла толщиной 10 нм, где
вокруг одной нуклеосомы оборачивается 146
нуклеотидных пар ДНК с коэффициентом
компактности, равным 6—7 (к.к. = б—7);
второй уровень — 30-нм фибрилла-соленоид
(к.к. = 40); третий уровень — петлевой
домен, в котором на петлю длиной в 0,2—
0,3 мкм приходится 60 тыс. нуклеотидных
пар (к.к. = 680). Далее отрезок примерно
с 18—20 петлевыми доменами образует
вокруг осевого элемента хромосомы один
виток диаметром 0,7—0,8 мкм (толщина хро-
матиды) с коэффициентом компактизации
12 х 104. Такой виток из петлевых доменов
может представлять собой минимального
размера бэнд, а набор из нескольких
витков — средний бэнд. Эта схема не учиты-
3
А
\
2
4_
Рис. 7. Схема различных уровней компактизации
хроматина [из: Ченцов, 1995]
1 — нуклеосомный; 2— нуклеомерный; 3— хромо-
мер, петлевой домен; 4—хромонема; 5—хрома-
тида
184
КЛЕТОЧНАЯ БИОЛОГИЯ
вает существования видимых в световом
микроскопе нитчатых, хромонемных
элементов.
По-видимому, процесс компактизации
ДНК, приводящий в конце концов к
построению плотного тела митотической
хромосомы, проходит через несколько
структурных уровней (рис. 7). Первый уровень —
нуклеосомный — обеспечивает
сверхскручивание ДНК по поверхности гистоновой
сердцевины. Второй — нуклеомерный
(сверхбусина), где идет объединение 8—10 нук-
леосом в виде глобулы. Так как все эти
уровни компактизации происходят на
огромных линейных молекулах ДНК, то ряд
сближенных нуклеомеров и образует 20—30-нм
фибриллу ДНП. Третий уровень — хромо-
мерный: на котором петли фибрилл ДНП,
объединенные скрепками из негистоновых
белков, образуют компактные тела (0,1—
0,2 мкм), которые при искусственной декон-
денсации дадут розетковидные структуры.
Расположение петлевых доменов, хромоме-
ров, может быть неравномерным: участки
тела митотической хромосомы,
обогащенные ими, могут соответствовать бэндам при
дифференциальной окраске хромосом.
Четвертый уровень — хромонемный:
сближенные в линейном порядке хромомеры
образуют толстые (0,1—0,2 мкм) нити, которые
можно уже наблюдать и в световом
микроскопе. Характер упаковки этой нити в теле
хроматиды еще недостаточно выяснен;
возможна спиральная укладка хромонемы, но
не исключено образование ею и еще
одного уровня петлистых структур. Конечно,
такая общая схема организации митотичес-
ких хромосом очень неполно отражает
особенности строения их специализированных
участков, таких как ядрышковый
организатор, теломеры и центромеры.
В заключение этого обзора можно прийти
к выводу, что при изучении
ультраструктуры хромосом исследователи
сталкиваются с парадоксальной ситуацией: чем ближе
мы подходим к высшим структурным
уровням организации митотических хромосом, тем
меньшей по объему и более низкой по
надежности становится информация об этой
важнейшей клеточной структуре. Наши
представления о структурной организации даже
элементарных компонентов ядра и о
структуре хромосом очень скудны и
противоречивы. Разрыв между успехами в
биохимическом изучении процессов биосинтеза ДНК
и РНК, с одной стороны, и чрезвычайно
медленным прогрессом в исследовании
тонкой организации клеточного ядра — с
другой, объясняется многими причинами. Одна
из основных причин заключается в том, что
современные методы не позволяют изучать
ядро и хромосомы в целостной
совокупности составляющих их элементов. Митотичес-
кая хромосома оказалась слишком мала для
детального анализа с помощью светового
микроскопа и слишком велика и плотна для
изучения в электронном микроскопе. Скорее
всего, расшифровка принципов организации
митотической хромосомы будет возможна при
использовании новейших методов
структурного анализа, таких как компьютерная
трехмерная реконструкция в сочетании с
методами конфокальной световой микроскопии.
Литература
Альберте Б., Брей Д., Льюис Дж. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ.
М.: Мир, 1994.
Восток К., Самнер Э. Хромосома эукариоти-
ческой клетки. Пер. с англ. М.: Мир, 1981.
Чепцов Ю. С. Общая цитология. М.: Изд-во
МГУ, 1995.
В. И. Агол
ГЕНЕТИЧЕСКИ ЗАПРОГРАММИРОВАННАЯ
СМЕРТЬ КЛЕТКИ
Организмы разных людей состоят из
более или менее одинакового количества
клеток. Различия есть, но они относительно
невелики. Если сравним количество клеток
в каком-нибудь органе у здоровых
взрослых людей, то также получим сходные
значения. Когда же, например, анализ крови
показывает изменение числа клеток, врач
начинает беспокоиться. Есть животные, у
которых число клеток не колеблется даже
в узких пределах. У очень мелкого,
длиной 1 мм, червя Caenorhabditis elegans ровно
945 клеток; из них нервных 302, ни
больше, ни меньше.
Как поддерживается такое постоянство?
Один из механизмов его поддержания
вполне очевиден. Он связан с регуляцией
клеточного деления. Клетка может разделиться
на две дочерние, либо остаться
неразделенной. Какая из этих возможностей
реализуется, зависит как от генетической
программы, так и от внешних сигналов, которые
клетка получает от своих соседей или из
окружающей среды. Но существует и
другой механизм, привлекший внимание
ученых лишь в последние годы. Оказывается,
наряду с программой, регулирующей
клеточное деление, есть особая генетическая
программа, реализация которой при
определенных условиях приводит клетку к
гибели. Гибнет клетка не от руки
какого-нибудь постороннего убийцы (хотя бывает и
так), она сама жертвует собой для блага
организма. Вот об этом самопожертвовании
клетки и пойдет речь.
Прежде всего о самом биологическом
явлении — программируемой клеточной
смерти. У упомянутого выше червя в
результате клеточных делений на самом деле
образуется не 945, а 1076 клеток, но 131
из них обязательно гибнет. При
формировании некоторых органов человека и
животных также первоначально возникает
намного больше клеток, чем потом
потребуется. Например, так бывает при
развитии нервной системы. Лишние клетки в
свое время мирно погибнут. Мирно —
значит, без воспаления. Клетка сморщивается
и постепенно распадается на части, обычно
поедаемые специальными клетками —
макрофагами. Но как узнать, какая клетка
лишняя, а какая нет? Какая из двух
клеток, рожденных действительно равными
(а генетически две сестринские клетки
идентичны), должна умереть, обеспечив
тем самым продолжение жизни своей
сестры?
Ситуации бывают разные. У Caenorhabditis
elegans судьба клетки определена
абсолютно жестко, и поменять эту судьбу у
здоровой особи практически невозможно; есть
клетки (те самые 131), у которых, можно
сказать, «на роду написано» покончить жизнь
самоубийством. В других случаях задача
решается не столь однозначно. Например,
при развитии организма млекопитающих те
нервные клетки, которые не успевают
установить контакты — синапсы — между
собой или с иннервируемыми органами, и есть
лишние. Их существование не имеет
смысла. Для клеток других тканей сигнал к
включению программы смерти может быть иным,
но биологический смысл самопожертвования
можно найти всегда.
Самопожертвование осуществляется по
определенному ритуалу, при участии ряда
факторов, многие из которых еще не
известны науке. Схематически сценарий этого
процесса можно разбить на несколько
основных этапов. На первом этапе клетка
получает «послание» о том, что она должна
пожертвовать своей жизнью для благополучия
организма. Это известие приходит из
окружающей среды — либо от соседних клеток,
либо от межклеточных веществ, твердых или
жидких. Чтобы воспринять такое «послание»,
клетки имеют специальные органы чувств,
называемые рецепторами (от лат. recipere —
получать). Рецепторы представляют собой
белковую молекулу, обычно состоящую из
трех частей: внеклеточной,
внутриклеточной и промежуточной, пронизывающей
клеточную мембрану. Наружная часть рецептора
способна узнавать молекулы строго
определенного строения, которые могут либо
свободно плавать во внеклеточной жидкости,
либо быть фиксированными на
поверхности других клеток или межклеточных
волокон. Эти сигнальные молекулы и рецепторы
подходят друг к другу, как ключ к замку.
Информация передается через различные
рецепторы или сочетание рецепторов.
Информацией может являться и отсутствие
специфического вещества в окружающей клетку
среде.
В результате контакта сигнальных
молекул с наружной частью белка-рецептора
этот рецептор претерпевает структурные
изменения: некоторые атомы меняют
положение относительно других атомов.
Структурная перестройка захватывает и
внутриклеточную часть молекулы рецептора. Она
может либо обладать определенной
ферментативной активностью сама, либо быть тесно
186
КЛЕТОЧНАЯ БИОЛОГИЯ
связанной с некоторыми клеточными
ферментами. Изменение структуры рецептора
сказывается на работе этих ферментов,
поэтому в результате контакта рецептора
с внеклеточным веществом внутри клетки
происходят разные биохимические
изменения. Часто речь идет об изменении
концентрации ионов кальция, а также некоторых
относительно мелких фосфорсодержащих
органических соединений, относящихся к
классу нуклеотидов. Активные соединения
появляются и в результате гидролиза
определенных липидов клеточной мембраны.
В свою очередь, все это ведет к
присоединению или отсоединению остатков
фосфата от молекул белковых регуляторов,
способных влиять на генетический аппарат
клетки. Фосфорилирование и дефосфори-
лирование, а также некоторые другие
биохимические модификации меняют активность
этих регуляторов.
Во втором действии драмы
внутриклеточные регуляторы-посланники, получив
важные инструкции, вносят поправки в
работу отдельных генов. Работа эта, как
известно, заключается в образовании РНК,
а затем и белков. Таким образом, в
результате срабатывания генетической
программы, первоначально запущенной сигналом с
рецептора, изменяется набор
внутриклеточных РНК и белков. В конечном счете,
появляются или активируются ферменты,
способные разрушать клеточные белки и
нуклеиновые кислоты; их называют проте-
азами (протеин — белок) и нуклеазами. В
заключительном акте клетка теряет свою
целостность и становится пищей для
макрофагов. Морфологические и
биохимические изменения в клетках-самоубийцах
весьма схожи, даже если это клетки разных
органов и разных организмов. Хроматин —
главный внутриядерный компонент,
содержащий ДНК и белки, — уплотняется, а само
ядро в конечном счете распадается на
мелкие фрагменты (рис. 1). Вся клетка также
дробится на отдельные фрагменты (рис. 2).
Этот комплекс изменений, характерный для
программируемой гибели клеток, часто
обозначают термином апоптоз, что в
переводе с греческого означает «опадание
листьев».
Существуют и несколько иные
варианты сценария программируемой смерти клетки.
Например, сигналы, исходящие от
рецепторов, могут активировать гидролитические
ферменты без промежуточного изменения
работы генетического аппарата. Важную роль
при этом могут играть митохондрии,
высвобождая особые активаторы.
Программа, принимающая крайне
ответственное и иногда непоправимое решение —
жить или не жить, должна быть
предельно осмотрительной, поэтому клетка стара-
Рис. 1. Клетки, в которых ядерный хроматин
окрашен красителем, флуоресцирующим при освещении
ультрафиолетом
а — клетки в состоянии апоптоза. В апоптозных
клетках хроматин уплотнен и фрагментирован; б—
контрольные клетки
Рис. 2. Электронные микрофотографии контрольной
клетки (а) и клетки в состоянии апоптоза (б)
Апоптозная клетка раздроблена на отдельные части,
многие из которых содержат фрагменты
уплотненного хроматина (на фото — темноокрашенное
вещество)
ГЕНЕТИЧЕСКИ ЗАПРОГРАММИРОВАННАЯ СМЕРТЬ КЛЕТКИ
187
ется сделать все, чтобы не ошибиться.
Перед вынесением окончательного приговора
(активирования разрушающих клетку
ферментов) сигналы, получаемые извне,
подвергаются всестороннему анализу. Этот
анализ начинается уже на уровне первичных
сигналов, получаемых клеткой. Так, в ткани,
в которой идет интенсивное клеточное
деление, обычно находятся разнообразные
белковые вещества, называемые факторами
роста. Эти факторы стимулируют деление.
Их отсутствие является либо указанием на
то, что потребность в «молодых» клетках
уже удовлетворена, либо серьезным
сигналом неблагополучия. Разные факторы
роста взаимодействуют с разными
рецепторами и клетка часто ориентируется на
информацию, получаемую одновременно от
нескольких рецепторов.
Анализ этой информации внутри
клетки происходит при участии многих белков.
В последнее время открыты
белки-регуляторы как способствующие, так и
препятствующие развитию апоптоза. Исход
процесса часто зависит от относительной
концентрации этих регуляторов. Подобно
присяжным заседателям, некоторые из них
обычно «голосуют» за смертный приговор,
а другие — за помилование. В ряде
случаев решение принимается простым
большинством голосов. Например, это может
происходить следующим образом (рис. 3). Есть
такой белок Вах, молекулы которого
имеют сродство друг к другу и образуют ди-
меры. Повышенное содержание белка Вах
способствует развитию апоптоза. Другой
белок — Вс1-2 также умеет формировать
димеры, но он может также присоединиться
и к белку Вах, образуя при этом Вах/Вс1-2
гетеродимеры. В результате происходит
нейтрализация апоптозной активности белка
Вах. Таким образом, при прочих равных
условиях преобладание белка Вах будет
Вах/Вах Bax/Bcl-2 Bcl-2/Bcl-2
^Z7
Апоптоз Предотвращение апоптоза
Рис. 3. Схема регуляции апоптоза при помощи
взаимодействующих между собой белков с
разнонаправленными активностями
Комплексы, состоящие из двух молекул белка Вах
(гомодимеры), способствуют развитию апоптоза.
Однако при наличии достаточного количества
молекул белка Вс1-2 происходит образование Bax/Bcl-2
гетеродимеров, которые лишены способности
вызывать апоптоз
способствовать гибели клетки (при наличии
соответствующего сигнала), а при
преобладании белка Вс1-2, наоборот, клетка с
большей вероятностью будет защищена от
гибели.
Интересно, что некоторые из генов,
контролирующих апоптозную реакцию у
людей, являются очень древними.
Например, ген человека, направляющий синтез
белка Вс1-2, имеет структурное сходство
с геном Caenorhabditis elegans, в котором
закодирован антиапоптозный белок Ced-9.
При нарушении функции белка Ced-9
гибнет не 131 клетка, как в норме, а
большее их число. В специальных опытах было
показано, что в определенных пределах
белки Вс1-2 и Ced-9 взаимозаменяемы. Есть
и другие примеры родственных связей
между участниками систем,
контролирующих апоптоз у людей и червей. Так,
некоторые из протеаз человека,
принимающих участие в финале апоптозной
драмы, — родственники другого белка
(Ced-3) Caenorhabditis elegans.
Но в целом эта система у
млекопитающих все же значительно сложнее и
включает большее число компонентов. Аппарат
наших клеток, несмотря на их
микроскопические размеры, огромен, в том числе
и та его часть, которая занимается апоп-
тозом. Но это излишество кажущееся. В
частности, система, контролирующая
апоптоз, контролирует и клеточное деление.
Иными словами, некоторые белки
одновременно регулируют и апоптоз, и деление
клетки. Таким образом, системы регуляции
клеточного деления и клеточной смерти
оказываются тесно переплетенными
между собой. Это обстоятельство имеет очень
важные биологические последствия. Одно
из них заключается в том, что апоптоз —
мощное и важнейшее средство
естественной профилактики раковых и других
злокачественных новообразований. Есть
специальные гены, которые так и называются —
антионкогены. Среди них важное место
занимает ген, кодирующий белок с
маловыразительным названием р53 (название
характеризует массу этого белка в кило-
дальтонах). Белок р53 внимательно «следит»
за работой генов, способных вызвать
несвоевременное деление клетки, и вообще за
неполадками в ДНК — генетическом
материале клетки. В случае необходимости р53
сдвигает равновесие в пользу апоптоза и
потенциально опасная клетка гибнет. Если
же мутирует сам р53, то система
регуляции клеточного деления остается без
должного присмотра.
Все это не просто абстрактные
рассуждения: в клетках злокачественных
новообразований человека нередко
обнаруживают мутации в белке р53, нарушающие его
188
КЛЕТОЧНАЯ БИОЛОГИЯ
работоспособность. Без присмотра
нарушается дисциплина, и вместо того, чтобы
погибнуть, клетка начинает бесконтрольно
делиться. Возникает опухоль. Если же р53
нормален, система программируемой
клеточной смерти резко снижает частоту
раковых заболеваний. Сходным образом эта
система следит и за некоторыми другими
неполадками в работе генетического
аппарата. Значительно легче своевременно
устранить потенциально опасную клетку, чем
потом бороться с ее многочисленным
непослушным потомством.
Многие противораковые лекарства
обладают лечебным действием именно
потому, что способны вызывать апоптоз в
раковых клетках. Эти лекарства часто
воздействуют на генетический аппарат
делящихся клеток, вызывая нарушения в его
работе. А как мы уже знаем, клеточные
белки, в частности р53, реагируют на
такие нарушения включением апоптознои
системы. В то же время при лечении
противораковыми препаратами нередко
возникает устойчивость к этим препаратам —
одна из главных причин недостаточной
эффективности химиотерапии
злокачественных опухолей. Эта устойчивость имеет в
разных случаях разную природу, но одна
из причин — нарушение работы
апоптознои системы. Клетка как бы
отказывается жертвовать собой для общего блага,
несмотря на полученные инструкции.
Кончается такой «эгоизм» скверно. Лучевая
терапия раковых заболеваний также
основана на способности облучения вызывать
апоптоз (при участии р53) прежде всего
в активно делящихся клетках.
Нарушение физиологического
равновесия между делением и гибелью клеток
лежит в основе и некоторых других —
неопухолевых — заболеваний. В частности,
есть основания считать, что при СПИДе
(синдроме приобретенного иммунодефицита)
уменьшение содержания в крови
определенного класса лейкоцитов, играющих важную
роль в иммунитете, обусловлено их
апоптознои гибелью. В основе некоторых
других дегенеративных заболеваний также,
возможно, лежит нарушение функции
апоптознои системы.
Большую роль играет апоптоз и в защите
организма от возбудителей инфекционных
заболеваний, в частности, от вирусов.
Многие вирусы вызывают такие глубокие
нарушения обмена веществ зараженной
клетки, что она воспринимает эти нарушения
как сигнал к экстренному включению
программы гибели. Биологический смысл такой
реакции вполне понятен. Смерть
зараженной клетки еще до того, как в ней
образуется новое поколение вирусных частиц,
предотвратит распространение инфекции по
организму. Однако у некоторых вирусов
выработались специальные приспособления,
направленные на подавление апоптоза в
заражаемых клетках. Тут могут
использоваться разные средства. В одних случаях в
вирусном генетическом материале
закодированы вещества, которые и по строению,
и по функции очень похожи на клеточные
антиапоптозные белки-регуляторы (такие,
например, как уже упоминавшийся Вс1-2
или белки, подавляющие активность про-
теаз). В других случаях вирус
стимулирует синтез клеткой ее собственных анти-апоп-
тозных белков. Так или иначе, создаются
предпосылки для беспрепятственного
размножения вируса.
Подведем некоторые итоги. В
генетическом аппарате каждой клетки
многоклеточного организма имеется специальная
программа, которая при определенных
обстоятельствах может привести клетку к
гибели. При нормальном развитии эта
программа направлена на удаление избыточно
образовавшихся клеток, а также клеток,
переставших выполнять свои функции.
Другая важная роль клеточной гибели —
удаление клеток с серьезными
нарушениями структуры или функции генетического
аппарата. В частности, апоптоз — один из
основных механизмов самопрофилактики
онкологических заболеваний.
Система программируемой клеточной
смерти — существенный фактор
иммунитета, поскольку гибель зараженной клетки
может предотвратить распространение
инфекции по организму. Нарушения системы
программируемой гибели клетки приводят
к серьезным болезням. Ослабление
способности к апоптозу может вызвать развитие
злокачественных опухолей. Избыточный
апоптоз тоже вызывает заболевания,
например, дегенеративные повреждения
нервной системы.
Воздействие на программу клеточной
гибели — перспективное направление
лекарственного лечения. Так, одна из
важных задач противораковой терапии —
стимуляция апоптознои системы. В других
случаях задача врача, наоборот, состоит в
том, чтобы предотвратить вредное для
организма самоубийство клетки.
Таким образом, смертоносный механизм
апоптоза не только необходим, но, в
конечном итоге, очень полезен. Без системы
программируемой клеточной гибели мы с
вами не смогли бы появиться на свет
такими, какими мы рождаемся. И поддержание
порядка в наших организмах в течение
дальнейшей жизни в значительной степени
обеспечивается именно способностью наших
клеток к программируемой смерти.
Ю- М. Васильев
НОРМАЛЬНЫЕ И ОПУХОЛЕВЫЕ КЛЕТКИ
Изучение опухолей — основа
анализа нормального
поведения клеток
Всякий знает, что злокачественные
опухоли (раки, саркомы) и близкие к ним
по природе разрастания кроветворных
клеток (лейкозы) — тяжелейшие заболевания
человека и что предотвращение и
лечение этих заболеваний — важнейшая
задача медицины. Вместе с тем выяснение
природы опухолевых заболеваний — одна
из центральных проблем современной
клеточной биологии. Дело в том, что
основой этих заболеваний являются появление
и размножение в организме особых
генетически измененных (мутантных) клеток.
Исследования показали, что каждая
опухоль возникает обычно в результате
появления в какой-либо ткани одной-един-
ственной измененной клетки, затем
опухоль растет только за счет
размножения потомков этой клетки. Как говорят
биологи, каждая опухоль есть клон, т. е.
колония, выросшая из одной мутантной
клетки. Речь идет о мутациях, меняющих
такие свойства клеток, о которых
раньше мы знали очень мало, — мутациях,
изменяющих «социальное» поведение клеток.
Что это означает? Каждая клетка в
нашем и любом многоклеточном организме
является, по определению, членом
сложнейшего сообщества. Еще более 100 лет
назад знаменитый немецкий патолог Р. Вир-
хов очень точно назвал наш организм
«государством клеток». И в государстве, и в
организме поведение индивидуума (клетки
или человека) разумно и нормально
тогда, когда он адекватно реагирует на
социальные сигналы — сигналы от других
членов сообщества. Человек, неадекватно
реагирующий на социальные сигналы,
часто становится преступником. Клетка,
неадекватно реагирующая на социальные
сигналы, может дать начало опухоли.
В биологии создание и изучение
организмов с различными дефектами
определенной функции часто становятся основным
способом анализа нормального механизма этой
функции. Например, выясняя природу
мутаций, нарушающих разные этапы
свертывания крови у человека, исследователи
сумели расшифровать «устройство» сложнейшей
системы белков, контролирующих этот
процесс. Аналогичным образом, анализ свойств
опухолевых клеток, выяснение сути их
генетических дефектов оказался наиболее
эффективным способом расшифровки
молекулярных механизмов нормальных реакций
клеток на социальные сигналы. Рассмотрим
некоторые результаты этих исследований.
Онкогены и антионкогены
Прежде всего отметим, что вызвать
превращение нормальных клеток в
опухолевые (начальную опухолевую
трансформацию) или увеличение степени
ненормальности этих клеток (последующие
трансформации) способны мутации
многих генов, кодирующих множество
белков. Обычно различают две большие
группы таких генов: онкогены и антионкогены
(рис. 1). Онкогенами называют гены,
мутации которых вызывают трансформацию
потому, что в клетке их продукт — он-
кобелок — появляется в увеличенном
количестве или приобретает измененную
структуру; иными словами, присутствие
онкобелка в клетке необходимо для
появления и поддержания трансформации.
Как известно, у всех клеток с двойным
(диплоидным) набором хромосом имеется
по два гена каждого вида. Для
трансформации достаточна мутация даже одного
гена из пары нормальных генов —
предшественников онкогена (такие
нормальные гены называют протоонкогенами). Т. е.
мутации, превращающие протоонкоген в
онкоген, доминантны.
Антионкогены обладают совсем
иными, во многом противоположными
свойствами. Для появления или усиления
трансформации необходима инактивация
обоих членов каждой пары антионкогенов,
имеющихся в клетке. Инактивация
только одного из генов пары к трансформации
не приводит. Таким образом, мутации,
инактивирующие антионкогены,
рецессивны.
К настоящему времени в геноме разных
видов млекопитающих и птиц выявлено
несколько десятков типов протоонкогенов
и более дюжины типов антионкогенов.
Вероятно, в ближайшем будущем число
известных генов, принадлежащим к этим двум
группам, существенно увеличится.
Развитие опухоли не всегда бывает результатом
единственной мутации: внутри разросшегося
клона с поломкой одного гена могут
появиться клетки с поломкой другого гена, затем
третьего и т. д. Раковые опухоли человека
обычно результат целой серии поломок
разных генов.
190
КЛЕТОЧНАЯ БИОЛОГИЯ
а б
АО+ АО+
по по
Прото- Прото- Белок Белок
онкобелок онкобелок
АО+ АО-
ПО О
Прото- Онкобелок Белок
онкобелок
АО- АО-
Рис. 1. Схема изменений двух типов генов,
участвующих в опухолевых трансформациях
а— изменения протоонкогенов (ПО). Изменения
одного из генов пары ПО, превращение его в
онкоген (О), кодирующий патологический онкобелок,
достаточно для трансформации; б — изменения
антионкогенов (АО+). Инактивация одного из этих генов
(АО-), например, утрата части гена или всего гена, не
вызывает изменений клетки, так как второй ген пары
продолжает нормально функционировать. Только
потеря функции обоих генов пары ведет к
трансформации
Итак, в клетке имеется сложная
система генов, изменения которых вызывают
или тормозят опухолевую трансформацию.
Каковы нормальные функции этих генов?
Что касается протоонкогенов, то впервые
они были выявлены по их способности,
мутируя, превращаться в онкогены и
вызывать трансформацию, однако абсурдно
было бы полагать, что до этого
изменения, к счастью редкого, протоонкогены
ничего не делают в нормальной клетке.
Ниже мы рассмотрим данные,
показывающие, что продукты протоонкогенов и
антионкогенов являются компонентами
системы реакций клетки на внешние
сигналы. Однако сначала попробуем
разобраться в том, о каких сигналах и каких реакциях
идет речь.
Клетки выделяют молекулы,
регулирующие размножение
других клеток
Наиболее изученный вариант
социальной регуляции — индукция размножения
клеток гуморальными факторами, т. е.
молекулами, выделяемыми другими
клетками в жидкую среду, окружающую данную
клетку (кровь или тканевую жидкость в
организме, питательную среду в культуре).
Среди таких факторов всем известны
гормоны — молекулы, синтезируемые
клетками специальных эндокринных желез и
разносимые кровью по всему организму.
Попадая с кровью в чувствительную ткань,
гормон, наряду с другими факторами,
может стимулировать размножение клеток этой
ткани. Например, женские половые
гормоны, выделяемые клетками яичника, могут
активировать размножение клеток таких
органов, как матка или молочная железа.
Разные гормоны, выделяемые гипофизом,
могут избирательно стимулировать клетки
определенных эндокринных желез,
расположенных вдали от гипофиза. Например,
тиреотропный гормон гипофиза стимулирует
клетки щитовидной железы, адренокорти-
котропный гормон гипофиза — клетки коры
надпочечников и т. д.
Наряду с давно открытыми гормонами,
за последние два десятилетия обнаружена
другая большая группа молекул —
регуляторов размножения клеток — так
называемые факторы роста. Показано, например,
что определенные клетки крови —
тромбоциты, — разрушаясь, выделяют белок,
стимулирующий размножение фибробластов
соединительной ткани. Этот белок был
назван тромбоцитарным фактором роста (ТФР).
ТФР, по-видимому, играет важную роль в
механизме заживления ран. Когда кровь из
разорванных кровеносных сосудов
изливается в рану, то содержащиеся в ней
тромбоциты разрушаются. Выделенный ТФР
может активировать миграцию и
размножение окружающих рану фибробластов,
приводящих, в конце концов, к появлению рубца
на месте раны. ТФР, выделившийся из
тромбоцитов, всегда содержится в сыворотке
крови, которую добавляют к среде
клеточных культур для стимуляции размножения
фибробластов и других клеток.
Когда мы говорим, что гормон или
фактор роста стимулирует размножение,
вхождение клетки в митотический цикл, то речь
идет о глубокой многосторонней и
многофазной перестройке всей деятельности
клетки. Напомним, что обычно различают
следующие фазы цикла, завершающегося
делением (рис. 2). Состояние клетки до
получения стимула к делению называется
фазой G0, она может длиться относитель-
НОРМАЛЬНЫЕ И ОПУХОЛЕВЫЕ КЛЕТКИ
191
Сигнал
Gt S G2 Митоз
Блок размножения
Повреждение Накопление
ДНК р 53
Апоптоз
Гибель
Рис. 2, Упрощенная схема стадий митотического
цикла (вверху) и системы, охраняющей геном:
повреждение ДНК лучами или другими агентами
невыясненным еще путем приводит к накоплению
белка р53, включающего реакции, блокирующие митоти-
ческий цикл на границе фаз G1 и S, а позднее
включающего реакции, ведущие к
программированной гибели клетки (апоптозу)
но долго. После получения стимула клетка
вступает в фазу Gv когда идет подготовка
к синтезу ДНК. В следующей фазе — S —
идет синтез ДНК, геном клетки
удваивается. В короткой фазе G2 после окончания
синтеза ДНК идет подготовка к делению, а
затем сам митоз (фаза М). Внешние для
клетки факторы, стимулирующие деление,
регулируют в основном переход G0-Gr
При опухолевых трансформациях
клетка приобретает способность входить из G0 в
Gj и проходить весь цикл даже без внешней
или по крайней мере при ослабленной
внешней стимуляции. Пролиферацию опухолевых
клеток отличает от нормы именно эта
независимость от стимулирующих факторов,
нерегулируемость размножения, а не его
скорость: суммарная продолжительность фаз
Gj + S + G2+M после трансформации
может не уменьшаться по сравнению с
нормой. Даже клетки самых злокачественных
опухолей делятся не чаще, чем
стимулированные нормальные клетки. Например,
клетки рака молочной железы делятся даже
реже, чем клетки нормальной молочной
железы в начале беременности, в период
максимальной гормональной стимуляции. Беда
в том, что опухолевые клетки не требуют
такой стимуляции для своего размножения
и потому могут размножаться непрерывно.
Клетки вызывают смерть
своих собратьев
В ответ на внешний сигнал клетки
могут не только делиться, но и гибнуть. Эту
реакцию называют программированной
гибелью, или апоптозом. Непрограммирован-
ная смерть клетки (некроз) возникает в
результате повреждения клетки внешним
воздействием: механического разрыва,
перегрева, переохлаждения и т. д. Напротив,
при апоптозе внешний агент сам по себе
безвреден для клетки, но включает цепь
биохимических изменений, приводящих ее
к гибели. Одно из таких конечных
изменений — активация специального фермента,
ДНКазы, разрезающего ДНК на куски и,
делающего клетку недееспособной.
Примеры апоптоза многочисленны. Например,
известно, что при метаморфозе у лягушек,
т. е. при превращении хвостатого
головастика в бесхвостого лягушонка, происходит
исчезновение (резорбция) хвоста. Такая
резорбция вызывается гормоном, который
во время метаморфоза начинает усиленно
вырабатывать щитовидная железа
головастика. Недавно было показано, что при этом
в клетках хвоста включается программа
апоптоза, которая и приводит к резорбции.
Апоптоз лимфоцитов в культуре
вызывают некоторые гормоны коры надпочечников
(стероиды). В некоторых случаях сигналом
к апоптозу является, по-видимому, не
гормон, а его исчезновение, выключение
гормонального сигнала. Например,
прекращение лактации (вскармливания молоком
детенышей) вызывает быструю гибель
большей части клеток молочных желез у самки
мыши (и, по всей вероятности, у других
млекопитающих). Окончание вскармливания
быстро вызывает у мыши падение уровня
гормона гипофиза (пролактина) и других
гормонов, активировавших молочную
железу. Это резкое понижение
гормонального сигнала, по-видимому, приводит к
массовому и быстрому апоптозу клеток.
Опухолевые трансформации делают
клетку менее чувствительной или совсем
нечувствительной к сигналам, вызывающим
апоптоз. Например, прекращение
кормления мыши-самки приводит к апоптозу
нормальных клеток ее молочной железы, но
не вызывает гибели клеток
доброкачественных и злокачественных опухолей этого
органа, если такие опухоли у мыши имеются.
Очень интересные результаты дал
генетический анализ одного из вариантов
лимфомы человека (доброкачественной
опухоли из лимфоидных клеток). Оказалось, что
для клеток этой лимфомы характерна
мутация неизвестного до того протоонкогена,
названного онкогеном Ъс1-2. Анализ
показал, что мутация, его активирующая,
тормозит апоптоз лимфоидных клеток, который
все время происходит в норме,
уравновешивая размножение этих клеток. После
трансформации клетки продолжают
размножаться, подобно нормальным, но перестают
гибнуть, и поэтому число мутантных кле-
192
КЛЕТОЧНАЯ БИОЛОГИЯ
а М
ФР
ФР
б М
в М
Рис. 3. Упрощенная схема передачи сигалов,
индуцирующих размножение
а — передача сигнала в нормальной клетке.
Молекула фактора роста (ФР) прикрепляется к наружной
части молекулы специфического рецептора (Р),
пересекающей клеточную мембрану (М). В результате
такого связывания активируется внутренная часть
молекулы, что условно обозначено красной точкой.
Активация передается затем по цепи специальных
промежуточных белков (ПБГ ПБ2 ПБП) на
факторы транскрипции (ФТ), которые, активируясь,
связываются с определенными участками ДНК и
запускают синтезы, необходимые для подготовки к делению;
б, в — передача «ложных сигналов» в опухолевых
клетках: б— мутация в протоонкогене, кодирующем
белок-рецептор, делает этот белок неспособным
связываться с фактором роста; патологический
рецептор активирован постоянно и без такого связывания
(красная точка) и передает сигналы по цепи внутрь
клетки; в— мутация гена одного из промежуточных
белков делает этот белок постоянно
активированным и вызывают активацию следующих
компонентов цепи
ток непрерывно увеличивается, возникают
опухолевые узлы. Открытие Ъс1-2 стало
началом изучения группы родственных ему
генов, регулирующих апоптоз.
Как клетка чувствует
и проводит внешние сигналы
Мы уже говорили о реакциях клеток на
сигнальные молекулы (гормоны или факторы
роста). Как они осуществляются? Как
молекула, находящаяся вне клетки, вызывает
перестройку всех процессов внутри
клетки, приводящую в конце концов к ее
делению или гибели? Лучше всего в этом
плане изучены реакции на ростовые
факторы, активирующие размножение (рис. 3).
Первой стадией таких реакций является
связывание молекулы-сигнала с
соответствующей ей молекулой-рецептором. Для этого
сигнал и рецептор должны специфично
соответствовать друг другу (подходить друг
к другу, как ключ к замку). Клетка
каждого типа имеет специфичный для нее набор
рецепторов, определяющий круг сигналов,
на который она реагирует. Например, как
мы уже видели, молекулы тиреотропного
гормона гипофиза циркулируют в крови и
соприкасаются со всеми тканями, но
связываются лишь с клетками щитовидной
железы, поскольку только у них есть
рецепторы для этих гормонов. Все
рецепторы — белки. Рецепторы для большинства
сигнальных молекул находятся на
поверхности клетки. Такие рецепторные
молекулы «протыкают» клеточную мембрану
насквозь: наружная их часть торчит на ее
наружной поверхности, средняя —
проходит через нее, а внутренняя —
находится в цитоплазме под мембраной. Сигнальная
молекула, находящаяся снаружи клетки,
связывается с наружной частью
рецептора, торчащей из клетки, и обратимо
меняет форму всей его молекулы: наружной,
средней и внутренней ее частей.
Изменение формы приводит белок-рецептор в
особое активированное состояние. В этом
состоянии внутренняя часть рецепторной
молекулы становится способной вызывать
в цитоплазме цепь химических реакций,
проводящих сигнал. Таким образом,
начальные стадии процесса можно описать
так:
сигнал + рецептор —> активированный
комплекс; сигнал — рецептор —> начало
реакций в цитоплазме.
Главную роль в осуществлении этих
реакций играет большая группа
специальных ферментов — протеинкиназ,
присоединяющих фосфатные группы к
аминокислотным группам различных белков (рис. 4).
Различают киназы, присоединяющие
фосфат к аминокислоте тирозину (тирозин-
киназы) или к аминокислотам треонину и
серину (серин-треонин-киназы). У многих
рецепторов внутренняя цитоплазматическая
)
Рис. 4. Схема одного из механизмов активации
белков в цепях передачи сигналов
Специфические ферменты— киназы присоединяют
фосфатный остаток к белку, активируя его. Другие
ферменты — фосфатазы — отщепляют фосфат,
прекращая активацию
НОРМАЛЬНЫЕ И ОПУХОЛЕВЫЕ КЛЕТКИ
193
часть молекулы является тирозин-киназои,
которая активируется после присоединения
молекулы-сигнала к наружной части
рецептора. Присоединение фосфата к
определенному белку-ферменту в цитоплазме
активирует этот белок, он активирует следующий
белок и т. д. Таким образом создается цепь
активации специальных белков-ферментов,
чаще всего киназ. При активации
размножения в конце такой цепи находятся
белки, расположенные в ядре. В некоторых
случаях фосфорилирование белков
заставляет их перейти из цитоплазмы в ядро.
Многие из них, так называемые факторы
транскрипции, при активации
связываются с определенным участком ДНК,
стимулируя синтез РНК на определенных генах.
Эти РНК выходят в цитоплазму и служат
матрицами для синтеза новых белков,
необходимых для прохождения митотичес-
кого цикла или, наоборот, для развития
апоптоза. Итак, цепь проведения сигнала
начинается со связывания сигнальной
молекулы с рецептором, проходит через серию
активаций промежуточных белков в
цитоплазме и завершается активацией
определенных участков генома. Все этапы такого
проведения сигнала обратимы. Реакция
прекращается, как только сигнальная молекула
отделяется от рецептора, например, если
резко уменьшается ее концентрация во
внешней среде. Обратимы и промежуточные
этапы цепи реакций. Например, фосфатные
группы, присоединенные киназой к белку,
могут быть отсоединены ферментом
противоположного действия — фосфатазой. Сейчас
интенсивно изучается роль разных фосфатаз
в прерывании цепей проведения сигналов,
но конкретно о таких прерывателях мы
знаем еще мало. Сигнал может также
прекратиться в результате быстрого распада
самого сигнального белка. Расшифровать
все эти сложные цепи реакций удалось
благодаря изучению их нарушений в разных
типах опухолевых клеток. Теперь мы
можем ответить на вопрос о том, что
делают в нормальной клетке белки —
продукты протоонкогенов. Все они — компоненты
цепей проведения сигналов: сигнальные
молекулы, молекулы рецепторов, киназы и
другие белки цитоплазмы, активируемые
сигналом, наконец, транскрипционные
факторы в ядре. Когда протоонкоген
мутирует в онкоген, его белок-продукт меняет свои
свойства так, что он остается
активированным все время независимо от того,
действует ли на начало цепи сигнальная
молекула из внешней среды. Так, в
некоторых опухолях вместо получаемых извне
сигнальных молекул клетка сама
начинает продуцировать такие молекулы (см.:
Васильев Ю. М. Механизмы перестройки ци-
тоскелета. Наст. том). В других опухолевых
клетках мутировали гены, кодирующие
рецептор одного из факторов роста. Такой
мутантный белок-рецептор остается
активным все время даже без связывания
сигнальной молекулы. Сходным образом,
мутации генов промежуточных белков цепи и
факторов транскрипции могут вызвать
постоянную активацию размножения без
внешнего сигнала, т. е. изменения,
типичные для опухолевых клеток.
Отметим, что о нормальных сигналах
системы проведения сигналов мы знаем по
существу пока очень немного. Например, мы
не знаем, почему один сигнал включает
деление клеток, а другой — их гибель,
апоптоз. Чем различаются соответствующие
цепи?
Система внутренней
безопасности клетки
Важнейший результат недавних
исследований биологии опухолевых
трансформаций — открытие и изучение особой
функциональной системы клетки: системы,
охраняющей постоянство генома (см. рис. 2).
Началось с открытия факта, который не
казался особенно важным: в некоторых
опухолях был обнаружен неизвестный белок с
молекулярным весом 53000; белок был
назван соответственно р53 (р от англ. protein —
белок). Оказалось, что р53 — мутантный
продукт соответствующего гена р53. Далее
было показано, что нормальный белок р53
в клетке появляется в больших количествах
после воздействия агентов, повреждающих
ДНК, например после облучения
ультрафиолетовыми или рентгеновскими лучами,
которые вызывают разрывы цепей ДНК и
другие повреждения этих цепей.
Было показано, что накопление
продуктов нормального гена р53 останавливает
прохождение клеткой митотического
цикла на границе фаз Gx и S. В результате
поврежденная клетка не может удвоить
поврежденную ДНК и передать
испорченный геном своим клеткам-потомкам при
делении. У клетки есть специальные
ферменты, ремонтирующие испорченную ДНК.
Если через какое-то время повреждение
исправлено, то вызванный р53 блок
деления снимается, и клетка может вновь
размножаться. Однако если повреждение ДНК
долго не исправляется, то р53
«приговаривает» поврежденную клетку к смерти:
включает программу апоптоза.
Таким образом, из клеточного
сообщества путем блокирования размножения или
апоптоза исключаются клетки с поврежденной
ДНК; тем самым предотвращается
передача потомкам клеток с испорченным геномом
и потому резко уменьшается вероятность
194
КЛЕТОЧНАЯ БИОЛОГИЯ
накопления мутантных клеток. Поэтому р53
называют «сторожем клеточного генома». Как
уже говорилось, во многих, особенно
злокачественных, опухолях имеется дефектный
или инактивированный ген р53. В таких
опухолях геном становится непостоянным: клетки
с изменениями ДНК, возникшие по каким-
либо причинам, перестают выбраковываться
из опухолевой ткани и в ней появляется много
клеток с различными мутациями. В
частности, накопление мутаций протоонкогенов и
антионкогенов способствует отбору клеток с
повышенной степенью трансформации, т. е.
повышению злокачественности опухоли.
Таким образом, мутации р53 сами по себе не
нарушают проведения сигналов, но
способствуют накоплению клеток с такими
нарушениями. Сейчас идет чрезвычайно
интенсивное исследование механизмов действия
р53 и связанных с ним белков, образующих
«систему внутриклеточной безопасности» и
охраняющих клетку от порчи генома.
Заключение
Как мы видели, развитие опухолей —
результат одной или многих мутаций в
генах, кодирующих белки, которые
контролируют размножение и гибель клеток. При
этом для появления патологического
белка, активирующего размножение,
достаточно доминантной мутации одного из двух
протоонкогенов, кодирующих этот белок.
Напротив, для прекращения синтеза
нормального белка, ограничивающего
размножение клеток или вызывающего их гибель,
необходима инактивация (рецессивная
мутация) обоих кодирующих этот белок
антионкогенов (см. рис. 1). Таким образом,
успехи изучения опухолей привели к открытию
двух сложнейших функциональных систем,
контролирующих социальное поведение
нормальных клеток, систем проведения сигналов
и системы внутриклеточной безопасности. Мы
еще очень многого не знаем об этих
системах, но уже догадываемся, что и где
искать.
Механизмы поведения каждой клетки не
проще механизмов поведения целого
организма, состоящего из таких клеток.
Замечательно, что эти механизмы познаваемы.
Литература
Альберте А., Брей Д., Льюис Р. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ. М.:
Мир, 1994.
Чепцов Ю. С. Общая цитология. М.: Изд-во
МГУ, 1995.
Б. В. Громов
ЭНДОЦИТОБИОНТЫ КЛЕТОК ЖИВОТНЫХ
Введение
Под эндоцитобиозом понимают развитие
клеток определенных микроорганизмов
внутри клеток других организмов. К
настоящему времени эндоцитобионты
обнаружены только в клетках эукариот,
эндоцитобионты прокариот — бактерий и архей (см.:
Громов Б. В. Удивительный мир архей. Наст,
том) пока неизвестны и, возможно, не
существуют. Эндоцитобионты есть среди
эукариот, например, одноклеточные
водоросли являются симбионтами большого числа
простейших, пресноводной гидры, а в
морской среде населяют клетки многих
кораллов и фораминифер. Есть они и среди
грибов. Однако чаще всего это представители
прокариот. Иногда эндоцитобионты приносят
хозяину более или менее значительный
вред, т. е. являются паразитами. В других
случаях они полезны хозяину, иногда он
без них просто не может существовать, т. е.
находится с ними в симбиотических
отношениях.
Интересно, что внутриклеточные
паразиты обитают в клетках самых
разнообразных животных, а также и человека, тогда
как внутриклеточные симбионты
встречаются только у беспозвоночных, а у
позвоночных пока не обнаружены. В большинстве
случаев эндоцитобионты в результате их
адаптации к существованию в чужой
клетке уже неспособны жить вне ее, т. е.
являются или облигатными (обязательными)
паразитами или симбионтами.
Паразитические эндоцитобионты
Известны бактерии, паразитирующие в
амебах, в клетках членистоногих и пауков,
причем заражение некоторыми из этих
бактерий приводит к заболеванию и гибели
хозяев. Среди возбудитей опасных болезней
млекопитающих, в том числе человека, есть
риккетсии и хламидии. Риккетсии
циркулируют в природе, развиваясь в организме
членистоногих (клещей, вшей) и
млекопитающих. Членистоногие получают их при
питании на зараженных млекопитающих, а
потом передают незараженным особям. У
млекопитающих риккетсии вызывают
заболевания, так называемые риккетсиозы, к
которым относится такая опасная болезнь,
как сыпной тиф. Хламидии — возбудители
венерических болезней и трахомы (болезни
глаз, приводящей к слепоте) у человека, а
также пситтакоза птиц. Вдыхание
человеком воздуха, содержащего частички птичьего
помета с хламидиями, вызывает воспаление
дыхательных путей. Хламидийная инфекция
у сумчатых медведей коала приводит к
бесплодию, что способствует вымиранию этих
млекопитающих. Как было недавно
установлено, хламидии могут развиваться в
клетках амеб, что облегчает распространение
хламидии в природе.
Они потеряли способность к синтезу
многих необходимых соединений, которые
забирают у хозяина. В частности, они
неспособны образовывать универсальный
клеточный энергоноситель АТФ (аденозин-
трифосфат), но в их мембране
функционирует эффективная транспортная
система, переправляющая АТФ из клетки
хозяина в бактерию. У хламидии
наблюдается смена стадий жизненного цикла:
сравнительно крупные вегетативные
клетки, находясь в вакуолях в клетке
хозяина, растут и делятся обычным образом,
но затем превращаются в более мелкие
и плотные «элементарные тельца» (рис. 1).
В вегетативной стадии бактерии
неустойчивы к воздействию внешней среды и вне
клетки хозяина сразу погибают; в этой
стадии хламидии не могут заражать новые
клетки. «Элементарные тельца»
неспособны к делению, но являются
инфекционными; после гибели и разрушения
зараженной клетки они сохраняются во
внешней среде и обеспечивают заражение
Рис. 1. Клетки хламидии (Chlamydia psittaci) в
вакуоли зараженной клетки
Светлые— вегетативные клетки, плотные темные —
«элементарные тельца». Ультратонкий срез,
электронный микроскоп. Масштабная линия = 1 мкм [из:
Matsumotoetal., 1991]
196
КЛЕТОЧНАЯ БИОЛОГИЯ
новых клеток и новых хозяев. В отличие
от риккетсий хламидии не нуждаются в
членистоногих переносчиках.
Симбионты инфузорий
Бактерий обнаруживали в клетках
разных простейших, но наиболее изучены из
них симбионты инфузорий. Однако говорить
в данном случае о симбиозе можно
только условно, поскольку польза этих
бактерий для животного не очевидна, а иногда
они приносят хозяину несомненный вред.
Бактерии, заселяющие ядра инфузории
туфельки, были изучены и описаны еще в
1890 г. учеником И. И. Мечникова доктором
В. В. Хавкиным. Он назвал их голоспорами —
Holospora. Голоспоры в ядрах проходят цикл
развития, включающий вегетативные клетки
(это — обычные бактерии,
размножающиеся бинарным делением), и длинные
плотные клетки, не способные к делению, но
выходящие в среду и заражающие
инфузорий, еще не имеющих симбионтов (рис. 2).
Подобный цикл развития ни у каких
других бактерий пока не обнаружен.
Существует много бактерий, способных
развиваться в цитоплазме инфузории
туфельки. Бактерии рода Caedobacter, что в
переводе означает «бактерия-убийца»,
придают инфузории-хозяину свойства
киллера. Если встретятся инфузория с такими
симбионтами и инфузория, их лишенная, то
последняя либо будет заражена, либо убита.
Это зависит от состояния клеток бактерий,
вышедших в среду и поглощенных незара-
женной инфузорией. Если это обычные
вегетативные клетки бактерии, инфузория
будет заражена, но если в этих бактериях
активировались гены находящегося в них
вирусного генома, она погибнет. В последнем
Рис. 2. Клетки голоспоры (Holospora obtusa) —
симбионта ядер инфузории туфельки (Paramecium
caudatum)
а — тотальный препарат вегетативной клетки и
споры; электронный микроскоп, увеличение х 14 000; б—
ультратонкий срез зараженного ядра; электронный
микроскоп, увеличение х 8 800 [из: Громов и др., 1976]
Рис. 3. Светопреломляющее тело в клетке
Caedobacter
а— срез клетки, электронный микроскоп,
увеличение х 60 000; б— развернутая белковая лента,
тотальный препарат; электронный микроскоп,
увеличение х 40 000 [из: Beale era/., 1969]
ЭНДОЦИТОБИОНТЫ КЛЕТОК ЖИВОТНЫХ
197
случае бактерии перестают делиться, в них
обнаруживаются вирусные частицы и
крупные преломляющие свет «11»-тела (рис. 3).
Эти «Ё»-тела образованы скрученной
белковой лентой, которая в вакуолях
инфузории, захватившей бактерий, может
раскручиваться, разрушая окружающую вакуоль
мембрану. Видимо, «11»-тело — одно из
возможных орудий убийства лишенных
симбионтов туфелек. Туфельки, содержащие
симбионтов, к «11»-телам нечувствительны.
Очевидно, в популяции инфузорий в
результате описанного эффекта должно
возрастать число особей с симбионтами. Это
может способствовать процветанию бактерий,
но едва ли особенно полезно для
инфузорий. Приведенный пример
свидетельствует о том, что эндоцитобиоз способен
влиять на характер взаимоотношений между
организмами.
Эндоцитобионтами инфузорий могут быть
и метанобразующие археи. Подобный
симбиоз наблюдается у инфузорий, обитающих
в отсутствие молекулярного кислорода —
в кишечнике животных, в илах и т. п. В
таких условиях кислородное дыхание
невозможно, и энергетический обмен приводит
к освобождению молекулярного водорода. У
таких животных отсутствуют митохондрии,
но в их клетках имеются специальные орга-
неллы — гидрогеносомы, ответственные за
образование водорода. Археи, которые могут
присутствовать в клетках таких инфузорий
в качестве симбионтов, окисляют водород
углекислотой, также накапливающейся в
процессе жизнедеятельности инфузории. В
результате образуется метан. От такого
сожительства археи получают очевидную
выгоду, а для инфузории польза
заключается в удалении конечного продукта
энергетического метаболизма.
Эндоцитобиоз в глубинах океана
Бактерии обладают огромными
биосинтетическими возможностями, в том числе
способностями к утилизации «необычных»
субстратов и к синтезу разнообразных
компонентов клетки за счет простых
соединений. При этом они могут использовать
самые разнообразные источники энергии.
Таких возможностей часто лишены эука-
риоты, и поэтому взаимодействие с
определенными прокариотами в симбиотических
системах может оказаться для них крайне
полезным. Иногда симбиотические
ассоциации бывают приурочены к определенным
экологическим условиям. Так, в морской
среде широко распространен симбиоз
различных беспозвоночных с бактериями,
способными к синтезу органических веществ за
счет энергии окисления молекулярным
кислородом соединений серы, прежде всего
сероводорода. Соединения восстановленной
серы содержатся в воде глубоководных
холодных или горячих источников, они
также образуются в результате деятельности
микроорганизмов при отсутствии
молекулярного кислорода. В среде, содержащей
кислород, происходит микробиологическое
окисление этих соединений. Окисляющие серу
бактерии могут населять клетки моллюсков,
губок, нематод, погонофор и других
беспозвоночных, они снабжают хозяина
органическими веществами, что-то получая от
него взамен, и находя в его клетках
место для жизни и защиту от
неблагоприятных влияний окружающей среды. Иногда,
например, у погонофор синтезированные
бактериями вещества составляют
единственный источник пищи животного. Так, в
различных районах Мирового океана на
глубинах 1500 — 6000м вблизи выходов горячих
вод обитают своеобразные бескишечные
беспозвоночные — вестиментиферы (их
относят к типу погонофор). Самые крупные
вестиментиферы из рода Riftia достигают
длины 1,5 м при диаметре около 4 см.
Питание животного обеспечивается
процессами, проходящими в трофосоме — особом
органе, включающем клетки-бактериоциты,
содержащие плотно упакованные клетки
бактерий, концентрация которых
достигает 3,7 х 109 на 1 г ткани. В воде источника
содержится сероводород, который и
окисляют находящиеся в бактериоцитах
бактерии. Трофосома имеет развитую систему
сосудов. Через кровь осуществляется
снабжение бактерий сероводородом,
кислородом, необходимым для окисления
сероводорода, и углекислотой, из углерода которой
бактерии синтезируют органическое
вещество, служащее единственным источником
питания как бактерий, так и животного. В
местах выхода метана обитают
представители другой группы погонофор, в
трофосоме которых поселяются метанокисляющие
бактерии (рис. 4).
Эндоцитобионты насекомых
У 10-20% насекомых, относящихся к
различным систематическим группам, в
клетках можно обнаружить палочковидные
бактерии из рода вольбахия (Wolbachia). У
представителей одного вида вольбахии,
видимо, могут передаваться от одного
насекомого другому. Иногда они не оказывают
на насекомое заметного влияния, но в
других случаях заражение приводит к
определенным последствиям. Особенно часто
вольбахии с выгодой для себя вмешиваются
в процессы размножения насекомых,
например долгоносиков, божьих коровок,
паразитических перепончатокрылых.
Потомству насекомых эти бактерии передаются
198
КЛЕТОЧНАЯ БИОЛОГИЯ
с яйцами, т. е. по материнской линии.
Потомство зараженных самок содержит воль-
бахий и жизнеспособно независимо от того,
был ли заражен самец. Потомство незара-
женных самок и зараженных самцов воль-
бахий не содержит, но нежизнеспособно.
Не содержащее вольбахий потомство
появляется лишь тогда, когда оба родителя
их не содержат. Если количество
зараженных особей в популяции увеличивается, то
для бактерий это выгодно. Кроме того,
зараженные насекомые способны к
партеногенезу, при котором самки без
оплодотворения рождают самок, содержащих
вольбахий. Это тоже приводит к накоплению в
популяции зараженных насекомых. Выгода
для бактерий здесь очевидна, у насекомых
же так или иначе происходит нарушение
нормальных процессов размножения, что
едва ли может способствовать их
процветанию. В научной литературе содержатся
Рис. 4. Схема строения маленькой погонофоры
(Siboglinum fiordicum), погруженной в морской
осадок [из: Southward etal., 1986]
1 — органическая трубка, окружающая тело
животного; 2— щупальце; 3— задняя часть тела животного,
наполненная бактериями-симбионтами; 4— опис-
тосома — копающий орган животного, лишенный
симбионтов и выступающий из трубки
также сообщения о вызванных вольбахи-
ями заболеваниях насекомых и других
членистоногих.
Однако во многих случаях эндоцитоби-
онты полезны или даже совершенно
необходимы насекомому. Это случаи истинного
симбиоза.
В некоторых группах насекомых
симбионты обычны у форм, питающихся
древесиной, соком растений или кровью. К ним
относятся многие представители
равнокрылых (тли, цикады, белокрылки, червецы,
листоблошки), некоторые жуки, клопы. У
таракановых симбионты присутствуют всегда
и у всех видов независимо от характера
питания. Видимо, такие насекомые не
получают некоторых необходимых им
соединений с пищей, и эндобионты снабжают их
этими соединениями.
У тараканов бактерии, относимые к роду
Blattabacterium, населяют клетки-бактерио-
циты, расположенные в жировом теле на
брюшке животного (рис. 5). Они
проникают в яйца в процессе их формирования и
таким образом передаются потомству. Если
таракана лишить симбионтов, обработав его
антибиотиками, он становится бледным,
мало активным и в конце концов
дегенерирует.
Описано более 50 различных
симбионтов цикад. У цикад бактерии населяют клетки
бактериоциты, находящиеся в довольно
сложно устроенных органах-бактериомах.
Интенсивность размножения бактерий
контролируется гормонами хозяина, а
специальные мигрирующие клетки насекомого
обеспечивают перенос бактерий в
формирующееся яйцо, где они образуют
характерные округлые скопления. Без
симбионтов насекомые не могут нормально
развиваться.
Симбионты насекомых, как правило,
неспособны существовать вне клеток
хозяина, поэтому до недавнего времени не было
возможности исследовать особенности их
физиологии и механизмы взаимодействия
с организмом хозяина. Успехи,
достигнутые в разработке методов молекулярной
биологии в течение последних лет,
позволяют получать и исследовать не только
клетки симбионтов, но и их нуклеиновые
кислоты, отдельные гены и группы генов,
включенные и сохраняемые в геноме клеток
кишечной палочки. Таким образом,
можно выяснить последовательности оснований
в ДНК этих генов и их возможную
функциональную роль. К настоящему времени
молекулярно-генетическими методами
исследованы симбионты тлей.
Тли питаются соком растений, который
они сосут при помощи стилета —
трубочки, внедряемой в ткани флоэмы растения.
Тли приносят растениям не только непос-
ЭНДОЦИТОБИОНТЫ КЛЕТОК ЖИВОТНЫХ
199
редственныи вред, но и служат
переносчиками их многочисленных вирусов. Эти
насекомые проходят сложный жизненный
цикл, в котором сменяются периоды
полового и бесполого размножения. Одна
самка за месяц может оставить 50-60
потомков. Внутри тела большинства видов тлей
имеется специальный двулопастной
орган— бактериом, состоящий из 60-90
полиплоидных клеток бактериоцитов. В
вакуолях этих клеток находятся округлые
или овальные бактерии, относимые к роду
Buchnera. Размножение бактерий находится
под строгим контролем со стороны
хозяина, и их число в бактериоме
определяется его размерами. Бактерии насекомое
получает от матери, вне организма
хозяина они существовать не могут, так же
как не могут передаваться от одного
насекомого другому. Если бактерий убить,
обработав насекомое антибиотиками, оно
перестает размножаться. Результаты
сравнительного исследования
последовательностей оснований в рибосомных генах
симбионтов свидетельствуют о том, что
эволюция этих бактерий шла очень
быстрыми темпами. Симбиоз бактерий и тлей
начал формироваться 200-250 млн лет
назад. Несколько неожиданной оказалась
очевидная родственная связь симбионтов
с кишечной палочкой, обитающей в
кишечнике человека и животных.
Предполагают, что предками эндобионтов были
бактерии, близкие к кишечной палочке и
развивавшиеся в кишечнике древних тлей.
В растительном соке, которым
питаются тли, содержится много углеводов, но
мало аминокислот. Между тем, тли, как и
другие животные, должны получать с
пищей около 10 различных аминокислот,
которые они сами синтезировать не могут.
Бактерии-симбионты способны синтезировать
все аминокислоты, и они снабжают ими
хозяина. Установлено, что в число этих
аминокислот входят триптофан, метионин,
лейцин, изолейцин, валин и фенилаланин.
Активность генов бактерии, определяющих
их синтез, регулируется таким образом, что
бактерия синтезирует аминокислот
гораздо больше, чем это ей необходимо, и их
избыток достается насекомому.
получили признания, были осмеяны и почти
забыты. В 60-е гг. нашего столетия
американская исследовательница Л. Маргелис
возродила и развила идею симбиотического
происхождения эукариотическои клетки, исходя
уже из данных, полученных современными
молекулярно-генетическими и
цитологическими методами. К настоящему времени в
подобных представлениях еще остается много
неясного и спорного, однако то, что
митохондрии и хлоропласты произошли от про-
кариот-эндобионтов, сомнений, кажется, уже
не вызывает, т. е. идея симбиогенеза себя
оправдала.
Заключение
Эндоцитобиоз способствует увеличению
биоразнообразия, так как в результате
тесного взаимодействия двух организмов
образуется, по существу, новый организм с
новыми свойствами. С другой стороны,
внутриклеточные симбионты или паразиты
нередко приобретают свойства, для свобод-
ноживущих форм не характерные. Иногда
в результате симбиоза жизнь животного
становится возможной в условиях,
непригодных для его самостоятельного
существования (например, в случае погонофор). В
большинстве случаев эндоцитобионты уже
Эндоцитобиоз и эволюция клетки
В конце прошлого — начале нашего
столетия русские биологи Ф. С. Фаминцин,
К. С. Мережковский, В. М. Козо-Полянский и
ряд других выдвинули гипотезу о симбиоти-
ческом происхождении сложной
эукариотическои клетки. В соответствии с этой
гипотезой, клеточные органеллы в действительности
являются более или менее измененными
самостоятельными организмами. Эти идеи не
Рис. 5. Симбиотические бактерии таракана (Blatta-
bactehum) в клетках жирового тела насекомого
Ультратонкий срез, электронный микроскоп,
увеличение х 40 000 [из: Громов, Мамкаева, 1980].
200
КЛЕТОЧНАЯ БИОЛОГИЯ
не могут существовать самостоятельно, и
в этом отношении они напоминают
клеточные органеллы. Некоторые ученые
считают, что их и следует рассматривать в
качестве клеточных органелл, тем более, что,
как было сказано, митохондрии и хлороп-
ласты скорее всего действительно возникли
в результате эндоцитобиоза. Однако
между органе л лами и эндобионтами есть, по всей
видимости, одна весьма существенная
разница. Развитие и функционирование
клеточных органелл контролируются в том
числе и генами, находящимися в ядерных
хромосомах (возможно, в процессе
эволюции происходил переход некоторых генов из
генома симбионта в геном хозяина), тогда
как для эндобионтов такого явления пока
отмечено не было.
Литература
Громов Б. В. Успехи микробиологии, 1978,
т. 13, с. 50.
Маргелис Л. Роль симбиоза в эволюции клетки.
Пер. с англ. М.: Мир, 1983.
Ваитапп P., Baumann L., Lai C-Y. et al. Ann.
Rev. Microbiol., 1995, vol.49, p. 55.
/О. Af. Васильев
КЛЕТКИ И ПОСТРОЕНИЕ ТКАНИ
Введение
Поведение клетки в организме
определяется средой, непосредственно ее
окружающей. Такая микросреда состоит из
компонентов трех типов: а) соседних
клеток; б) внеклеточных поверхностей, с
которыми контактирует клетка, например,
поверхностей коллагеновых волокон и
других волокон, выделяемых клеткой (так
называемого внеклеточного матрикса);
в) жидкой среды: тканевой жидкости,
крови и лимфы. В упрощенных условиях
культуры клетки также окружены трехкомпо-
нентной микросредой: жидким компонентом
здесь является питательная среда, а среди
внеклеточных поверхностей, наряду с
волокнами матрикса, имеется внутренняя
поверхность культурального сосуда —
подложка культуры. Все поведение
нормальной клетки определяется сигналами,
получаемыми из микросреды.
Механизмы регуляции деления и гибели
клеток молекулами жидкого компонента
микросреды — факторами роста и
гормонами — описаны в другой статье (см.:
Васильев Ю. М. Нормальные и опухолевые
клетки. Наст. том). Здесь мы рассмотрим
реакции клетки на контакт с двумя
другими компонентами микросреды —
внеклеточным матриксом и поверхностью других
клеток. Сложность этих взаимодействий в
том, что они двусторонни: при контакте
между клетками меняются обе клетки; при
контакте с матриксом меняется не
только клетка, но и матрикс. В результате
множества таких взаимодействий группы
клеток «обустраивают» ту территорию, на
которой они живут, формируют ткани и
органы. Эта деятельность клеток аналогична
деятельности людей, обустраивающих свою
локальную территорию: дом и участок
вокруг дома.
Изучению механизмов контактных
реакций, ведущих к построению тканей (мор-
фогенетических реакций), очень помогает
сравнение поведения нормальных и
опухолевых клеток. Как мы знаем, у опухолевых
клеток реакции на внешние сигналы
нарушаются в результате мутаций
определенных генов: протоонкогенов и антионкогенов.
Рассмотрим нарушения способности к мор-
фогенетическим реакциям, возникающие в
результате таких мутаций.
Клетки обустраивают
территорию культуры
Простейший вариант обустройства своей
территории группой клеток —
организация многоклеточной структуры, подобной
ткани из изолированных клеток эпителия
Рис. 1. Топография поверхности нормальных и трансформированных фибробластов в культуре
Сканирующие электронные микрофотографии Ю. А. Ровенского: а— нормальный мышиный фибробласт
в жидкой среде. Сферическая клетка с многочисленными пузырчатыми выпячиваниями на поверхности;
б — нормальный мышиный фибробласт, прикрепившийся к плоскому дну сосуда. Клетка распласталась на
подложке, ее поверхность стала гладкой. Впереди— уплощенная ламелла с рядом плоских псевдоподий;
сзади—узкое тело клетки; s—трансформированный мышиный фибробласт, прикрепившийся к плоскому
дну сосуда. Клетка плохо распласталась на подложке, и ее поверхность не стала совершенно плоской. На
этой поверхности видны многочисленные короткие ворсинки, а на концах отростков— фестончатые
псевдоподии
202
КЛЕТОЧНАЯ БИОЛОГИЯ
или соединительной ткани (фибробластов),
высаженных в культуру, где клетки
прикрепляются ко дну (подложке) сосуда,
заполненного жидкой питательной средой.
Одна из важных реакций, возникающих
при такой организации, — выработка и
выделение специальных белков
внеклеточного матрикса. Эти белки прикрепляются
ко дну сосуда, а также полимеризуются
вокруг клетки в особые нити, например,
в коллагеновые волокна. Фибробласты в
культуре, как и в организме, образуют
рыхлую сеть таких волокон, окружающих
клетки со всех сторон. Эпителиоциты
образуют более густую решетку из особых
коллагеновых волокон — так называемую
базальную мембрану между клеткой и
подложкой.
Клетка, коснувшаяся подложки с
сорбированными молекулами белков
матрикса или волокон матрикса, сделанных ею
самой или другими клетками, начинает тут
же реагировать на этот контакт, меняя
форму и перестраивая цитоскелет (рис. 1).
Мы уже рассматривали механизмы таких
реакций в другой статье (см.: Васильев Ю. М.
Механизмы перестройки цитоскелета. Наст,
том). Повторим коротко суть того, что было
сказано о происходящих при этом
процессах. Центральной реакцией здесь
является образование подвижных отростков —
псевдоподий, наполненных внутри
актиновыми филаментами. Поверхность
псевдоподий образует затем специальные
структуры — фокальные адгезии, где скопления
белков-рецепторов прикрепляют мембрану
этой клетки к белкам матрикса и через них
к подложке. С внутренней стороны
мембраны молекулы тех же рецепторов
соединяются с актиновыми филаментами
цитоскелета. Сокращения натянутых и
прикрепленных к матриксу филаментов могут
вызвать подтягивание всей клетки к
фокальной адгезии. Многократное повторение
Рис. 2. Схема взаимоотношений между клетками
(светло-коричневый цвет) и волокнами матрикса
(синий цвет) в нормальной соединительной (вверху) и
эпителиальной ткани (внизу)
этих реакций — основа движений клетки
по подложке.
Клетки реагируют также на контакт с
поверхностью другой клетки. Этот сигнал
вызывает две реакции: остановку
образования псевдоподий в направлении другой
клетки (контактное торможение движений)
и образование адгезионных структур
второго типа (межклеточных адгезии).
Таким образом, комплекс морфогенети-
ческих реакций в культуре включает
секрецию белков матрикса, образование и
сокращение псевдоподий, образование адгезии
между мембраной псевдоподий и матрик-
сом и мембранами соседних клеток, а
также контактное торможение движений. В
результате многократного повторения этих
реакций клетки организуются в
многоклеточные системы, очень похожие на те
ткани, которые те же клетки (эпителиоциты
и фибробласты) строят в организме.
Эпителиоциты соединяются друг с другом в
пласт из клеток, плотно спаянных адгези-
ями и сидящих на базальной мембране, что
очень похоже на однослойный эпителий в
организме.
Фибробласты приобретают в культуре
вытянутую форму, образование которой
зависит от цитоскелетной системы
микротрубочек. Межклеточные адгезии у
фибробластов, в отличие от эпителиоцитов,
непрочны, они легко рвутся при движениях
и контактирующие клетки отделяются друг
от друга. Поэтому фибробласты в
культуре образуют систему из параллельно
расположенных удлиненных клеток, между
которыми располагается густая сеть волокон
матрикса. В организме соединительные
ткани и родственные им хрящевые и
костные ткани построены по тому же типу —
из клеток, отделенных друг от друга
межклеточным матриксом разного состава
(рис. 2).
Контактные реакции регулируют
размножение и гибель клеток
Реакции с матриксом и подложкой не
только приводят к построению тканей, но
и участвуют в регуляции
внутриклеточных процессов, и прежде всего
размножения клеток (рис. 3). Нагляднее всего это
видно из опытов, демонстрирующих
зависимость размножения нормальных
фибробластов от их прикрепления ко дну
культуры и волокнам матрикса на этом дне
(подложке). При выращивании обычной
культуры взвесь (суспензия) отделенных
друг от друга клеток добавляется в
питательную среду культурального сосуда,
например в чашку Петри. Под влиянием
силы тяжести клетки взвеси опускаются
на дно сосуда, прикрепляются к нему и
КЛЕТКИ И ПОСТРОЕНИЕ ТКАНИ
203
лишь затем начинают размножаться, т. е.
вступают в митотический цикл (см.:
Васильев Ю М. Нормальные и опухолевые
клетки. Наст. том). Прикрепление клеток
к подложке можно предотвратить, если
добавить в среду вязкое вещество (агар
или метилцеллюлозу), мешающее клеткам
оседать на дно. Клетки остаются
взвешенными в таком «студне» и не приступают к
размножению. Достаточно, однако,
поместить в вязкую среду очень маленькие
стеклянные палочки, чтобы клетки, не
осевшие на дно, но прикрепившиеся к
поверхностям этих висящих палочек,
немедленно вступили в митотический цикл и
образовали через несколько дней колонии
клеток вокруг каждой палочки. Очевидно,
митотический цикл здесь индуцируется
прикреплением клетки к подложке или
сходной с ней поверхности палочки.
В отличие от контактов с подложкой,
контакты с другими клетками,
по-видимому, приводят к торможению
размножения (рис. 3). Действительно, когда в
результате размножения клеток культура
становится густой и на дне сосуда не
остается незанятых клетками мест, они
перестают делиться, т. е. выходят из мито-
тического цикла в так называемую фазу
покоя GQ. Если «нанести рану» (удалить
бритвой со дна сосуда небольшой участок
густой культуры), то контактное
торможение на ее краях исчезает и клетки
начинают мигрировать на освободившуюся
подложку. Вскоре мигрировавшие клетки
приступают к активному размножению и
делятся до тех пор, пока не зарастет вся
рана. Эти опыты свидетельствуют, что
размножение зависит от локальной
плотности населения культуры. Хотя они не дают
строгого доказательства того, что
именно прямые контакты клетка—клетка
тормозят размножение, такая возможность
представляется очень вероятной.
Контроль размножения соседними
клетками и подложкой в культуре аналогичен
процессам, происходящим в организме.
Приведем только один пример. Представьте
себе царапину, нарушившую целостность
эпителия кожи. Клетки эпителия по краю
царапины начинают мигрировать на
поверхность свободного матрикса и
размножаются до тех пор, пока эта поверхность будет
полностью закрыта.
Так регулируется локальное
соотношение клеток и территорий, на которых они
находятся. Нормальная клетка
размножается только тогда, когда у нее есть
свободная «площадь для жилья». Благодаря
этим регуляциям поддерживается и
восстанавливается после повреждений
правильная тканевая структура (происходит
заживление ран).
Нарушения морфогенетических
реакций при опухолевых
трансформациях
Опыты с культурами показали, что при
опухолевых трансформациях эпителиоци-
тов и фибробластов резко нарушаются все
морфогенетические реакции на контакты
с другими клетками и подложкой.
Трансформированные клетки производят меньше
матрикса, хуже прикрепляются к нему,
подложке и другим клеткам.
Нарушаются также контактные регуляции
размножения. Например, клетки, взвешенные в
вязкой среде, продолжают размножаться
без прикрепления к подложке. Эти
клетки размножаются и в густой культуре
независимо от числа соседей. В
результате тканевые системы эпителия и
соединительной ткани в культуре и организме
становятся хуже организованными, а в
предельном случае вообще не
образуются, а распадаются на отдельные клетки
и группы клеток. В организме
отделившиеся от ткани клетки начинают двигаться
на соседние территории и размножаться
там. Например, трансформированные
клетки эпителия, отделившись от пласта,
начинают мигрировать через дефектную
базальную мембрану в окружающую
соединительную ткань, а затем проникать
в просвет сосудов, откуда они током лимфы
а в
б г
Рис. 3. Регуляция размножения клеток в культуре
Жидкая среда синяя, стенки и дно сосуда черные.
У неразмножающихся клеток ядра не окрашены, у
клеток в митотическом цикле красные, а, б —
нормальные клетки, а — зависимость размножения
клетки от подложки: вступила в цикл лишь клетка,
прикрепившаяся к поверхности дна сосуда
(подложке), но не клетки, взвешенные в жидкой среде;
б— зависимость размножения от плотности
населения культуры. Среди клеток, прикрепленных к
подложке, размножаются лишь те, которые
граничат со свободной поверхностью этой подложки, но
не клетки, окруженные со всех сторон соседями; в,
г— опухолевые клетки размножаются без
подложки в жидкой среде (е) и на подложке независимо
от числа соседей (г)
204
КЛЕТОЧНАЯ БИОЛОГИЯ
или крови могут переноситься в другие
органы. Такие крайние изменения
способности к морфогенетическим реакциям и
построению тканей обусловливают
наиболее опасные свойства опухолевых клеток —
способность к врастанию в нормальные
ткани (инвазия) и к образованию колоний
в других органах (метастазирование).
Отметим, что способность к инвазии и ме-
тастазированию свойственна лишь
клеткам опухолей, наиболее резко измененным
генетически. Врачи называют такие
опухоли злокачественными (рис. 4). Клетки с
менее измененными морфогенетическими
реакциями образуют доброкачественные
опухоли, в которых имеются различные
нарушения организации тканевых
структур, но нет инвазии и метастазирования.
Анализ генома показал, что клетки
злокачественных опухолей человека имеют
целую серию мутаций разных онкогенов
и антионкогенов, тогда как клетки
доброкачественных опухолей обычно имеют
меньшее число подобных мутаций. Таким
образом, степень нарушения морфоге-
нетических реакций и соответственно
развитие опухоли определяются числом и
степенью изменений генома опухолевых
клеток, одной мутации для развития рака
обычно недостаточно.
Механизм регуляции
контактных реакций
Ранее мы рассмотрели молекулярные
механизмы действия специальных
сигнальных молекул, находящихся в жидкой среде
и регулирующих размножение клеток (см.:
Васильев Ю. М. Нормальные и опухолевые
клетки. Наст. том). Напомним, что такие
молекулы связываются со специальными
Рис. 4. Два опухолевых клона среди клеток
нормального эпителия. Нормальные клетки
светло-коричневые. Клетки клона, образующего
доброкачественную опухоль —полип (темно-коричневый цвет),
дают избыточный рост и образуют выпячивание над
поверхностью пласта, но не проникают в
соединительную ткань под эпителием. Напротив, клетки
злокачественного (ракового) клона (красный цвет)
проникают под базальную мембрану в
соединительную ткань
рецепторами в наружной мембране
клетки и активируют эти рецепторы, а те, в
свою очередь, активируют цепь
промежуточных белков в цитоплазме, что в
конце концов приводит к активации синтеза
ядерных белков, необходимых для
удвоения генома (ДНК) и подготовки деления.
Онкогены опухолевых клеток —
мутировавшие гены, которые кодируют белки
участвующие на разных этапах в
проведении сигналов. При опухолевых
трансформациях онкогены, вызывающие
нарушения реакций на гуморальные факторы,
индуцируют, как мы видели, и
нарушения морфогенетических реакций. Отсюда
следует, что многие этапы обеих групп
реакций на сигналы контролируют одни и
те же белки, в частности специальные
ферменты, присоединяющие фосфат к
белку (киназы). Конечными этапами цепей
активации морфогенетических реакций
становятся, вероятно, какие-то белки
цитоскелета, например, белки,
связывающиеся с актиновыми филаментами (см.:
Васильев Ю. М. Подвижная архитектура
клетки. Наст, том) и вызывающие
изменения сборки, сокращения псевдоподий и
образование контактных адгезии.
Действительно, у некоторых таких белков
присоединение фосфата резко повышено в
трансформированных клетках, однако пока
еще неясно, каково значение изменений
каждого конкретного белка в
нарушениях морфогенеза. Другой нерешенный
вопрос: какие рецепторы определяют
контактную регуляцию размножения нормальных
клеток? Вероятное предположение состоит
в том, что активация и торможение
размножения зависят от тех же белков,
которые прикрепляют клетку к подложке и
другим клеткам, т. е. от
белков-рецепторов мембраны адгезионных структур,
соединяющихся с белками подложки или
соседних клеток. Оказалось, что
адгезионные структуры с подложкой (так
называемые фокальные контакты, рис. 5)
содержат не только белки, необходимые для
механического соединения белков матрикса
через мембрану с цитоскелетом, но и
несколько десятков видов других белков,
функции которых неясны. Среди этих
«непонятных» белков имеются различные про-
тоонкобелки, участвующие в проведении
сигналов, в том числе ферменты-киназы.
Зачем они здесь? Естественно
предположить, что фокальные адгезии не только
механические «склейки», но и «органы
осязания» клетки, передающие сигнал от
соприкосновения с другой поверхностью с
наружной мембраны внутрь клетки и
вызывающие активацию размножения.
Возможно также, что рецепторы адгезии
клетка—клетка, наоборот, индуцируют сигна-
КЛЕТКИ И ПОСТРОЕНИЕ ТКАНИ
205
Индукция
размножения Актин
л к л
Индукция
псевдоподий __ л
Мембрана
Матрикс
Рис. 5. Упрощенная схема фокального контакта
клетки с матриксом
Молекулы-рецепторы белков матрикса (Р)
наружной частью прикрепляются к волокнам матрикса, а
на внутренней стороне мембраны те же рецепторы
через посредство специальных белков — линкеров
(Л) соединяются с концами актиновых филаментов
цитоскелета (зеленый цвет), которые могут
натягивать контакт. В фокальных контактах содержатся
также специальные регуляторные белки (киназы-К),
которые могут присоединять фосфат к другим
белкам, меняя состояние и прочность контакта.
Красным пунктиром обозначены гипотетические цепи
проведения сигналов от фокальных контактов.
Через ряд промежуточных белков (красные круги)
такие цепи могут активировать размножение клеток и
вызывать образование новых псевдоподий на
поверхности клетки
лы, тормозящие размножение. Все эти
гипотезы сейчас активно проверяются в
разных лабораториях мира.
Взаимодействие системы, проводящей сигналы от
мембраны, с цитоскелетом определяет ту
«разумную» организацию движений и
изменений формы клеток, тот «разум
цитоскелета», о котором говорилось в другой
статье этого тома (см.: Васильев Ю. М.
Механизмы перестройки цитоскелета. Наст,
том).
Заключение
Исследования онкогенов и
антионкогенов показали, что в клетке имеется
сложнейшая система восприятия
окружающего мира и многообразных реакций на этот
мир. Мы описали реакции на отдельные
сигналы (гормоны, контакты), но ведь на деле
клетка как-то обобщает разные сигналы и
реагирует на весь комплекс таких
сигналов комплексом разных реакций
(движениями, делением). Поведение клетки не
проще, чем поведение человека, каждая
реакция которого включает множество
сигналов и сложных ответных действий.
Подобно этому, при нанесении раны
клеточной культуре клетки оценивают целый
комплекс факторов (появление свободной
подложки, исчезновение контакта с
другими клетками, состав жидкой среды) и
дают в ответ комплекс реакций: движение
в рану, включающее реорганизации
цитоскелета и мембраны, и почти
одновременное вхождение в митотический цикл,
включающее синтезы множества белков и РНК,
удвоение ДНК, движения хромосом и т. д.
Мы уже знаем молекулярные механизмы
многих этапов этих реакций. Мы еще не
знаем, однако, как все эти реакции
интегрируются в клетке.
Как уже говорилось, биологическая
суть опухолевой болезни — разрастание
клонов клеток, у которых имеются
мутации «генов социального поведения». Такие
мутации в опухолях могут возникать
последовательно, пока не появятся
злокачественные клетки с целой серией
мутировавших онкогенов и антионкогенов. Эти
клетки реагируют не на внешние
сигналы, а лишь на свои внутренние ложные
сигналы, вызываемые мутантными
белками. Сейчас врач может спасти больного от
таких «антисоциальных» клеток, удалив
весь клон хирургически или убив их
рентгеновскими лучами и специальными
веществами, которые губят делящиеся
клетки. Данные методы при надлежащем
искусстве врача высокоэффективны и
излечивают очень многих. Однако все эти
способы лечения не очень избирательны:
нередко они не истребляют все опухолевые
клетки или, наоборот, убивают не
только опухолевые, но и нормальные
делящиеся клетки (например, клетки
костного мозга) и потому могут погубить организм
вместе с опухолью.
Чем больше мы узнаем о
молекулярных механизмах регуляции клеток, тем
больше надежда на то, что все же будут
разработаны лекарства, убивающие
избирательно только опухолевые клетки или,
еще лучше, исправляющие поведение этих
клеток. Можно представить себе два
подхода к созданию таких «исправляющих»
лекарств:
с помощью генетической инженерии, т. е.
введением в геном опухолевой клетки
недостающих антионкогенов или, наоборот,
разрушением онкогена. Генетики называют
такое избирательное разрушение одного
гена «нокаутом»;
посредством инактивации
патологической функции онкобелков, например
избирательным подавлением их киназной
активности, т. е. способности вызывать ложные
сигналы путем присоединения фосфата к
другим белкам.
Сейчас многие лаборатории и
фармакологические фирмы ведут активную работу
в этом направлении. Однако пока ни
одного эффективного лекарства на основе этих
подходов еще не создано.
Тем временем не надо забывать, что если
не все, то многие мутации, приводящие к
206
КЛЕТОЧНАЯ БИОЛОГИЯ
опухолевой трансформации, можно не
только пытаться исправить, но и эффективно
предотвращать. Для этого необходимо
предотвратить контакты нормальных клеток
организма с мутагенными агентами:
излучениями и химическими веществами, которые
повреждают ДНК. Рассмотрение действия
таких агентов — задача отдельной статьи,
но следует напомнить о самом
распространенном мутагене, губящем ежегодно огромное
число людей, — о табачном дыме. По
подсчетам ученых-эпидемиологов, курение
вызывает во многих странах до трети всех
злокачественных опухолей. За многие годы
в клетках легких у курильщика возникает
целая серия мутаций онкогенов и
антионкогенов. Обычно нужно накопить около десятка
таких мутаций, чтобы измененная клетка
дала начало злокачественному клону. Если
человек перестает курить на несколько лет
раньше, чем появились последние мутации,
то рак не возникнет. Попытаемся сохранять
наши клетки нормальными, способными
правильно строить ткани и правильно вести себя
в нашем организме.
Литература
Альберте А., Брей Д., Льюис Р. и др.
Молекулярная биология клетки: В 3 т. Пер. с англ. М.:
Мир, 1994.
Чепцов Ю. Общая цитология. М.: Изд-во МГУ,
1995.
ИММУНОЛОГИЯ
Г. И. Абелев
ВРОЖДЕННЫЙ ИММУНИТЕТ. ВОСПАЛЕНИЕ
Введение
Воспаление — реакция организма на
микроорганизмы и корпускулярные
продукты тканевого распада, основной механизм
естественного (врожденного) иммунитета,
а также начальный и заключительный этап
иммунитета приобретенного. Оно
выполняет повседневную работу по очистке
внутренней среды организма от
микроорганизмов. При местной инфекции или остром
повреждении тканей воспалительная
реакция сопровождается покраснением,
сопряженным с локальным повышением
температуры, набуханием (отеком) и болью.
Как и всякая защитная реакция,
воспаление должно сочетать способность
распознавать чужеродную для организма частицу
с действенным способом ее обезвреживания
и удаления из организма.
Феноменология
Рассмотрим классический пример —
воспаление, вызванное занозой, загрязненной
бактериями и прошедшей под кожу. В
течение первых часов поврежденное место
краснеет, в нем повышается температура, оно
распухает и становится чувствительным к
прикосновению — налицо покраснение,
нагревание, отек и боль. Затем, в первые дни
после возникновения этих признаков,
образуется нарыв —вокруг занозы происходит
нагноение, после чего нарыв прорывается,
и заноза вместе с накопившимся гноем
выбрасывается из организма. Что стоит за этой
картиной?
Заноза проникает в ткань, разрушая ее
и разрывая мелкие сосуды. Происходят
кровоизлияние и свертывание крови в
поврежденном участке. Капилляры кровеносной
системы в этой зоне расширяются,
увеличивая приток крови в район
формирующегося воспаления. Внешне это выглядит как
покраснение и местное повышение
температуры. Далее сквозь стенки капилляров
в данное место просачивается плазма
крови, возникает отек, выглядящий как
припухлость. В норме стенки кровеносных
сосудов, включая капиллярную сеть,
непроницаемы для компонентов крови — как
плазмы, так и форменных элементов —
эритроцитов и лейкоцитов. Повышенная
проницаемость для плазмы крови —
следствие специфического изменения стенки
сосудов— образования «щелей» между
плотно прилегающими друг к другу
клетками эндотелия1. В этом же участке
наблюдается торможение движения
эритроцитов и лейкоцитов (клеток белой крови),
поскольку они начинают как бы липнуть к
стенкам капилляров, перекатываясь по ним
и образуя «пробки» в непрерывном потоке
клеток, проходящих по сосудам. Два типа
лейкоцитов — моноциты и нейтрофилы —
начинают активно проникать из крови в
окружающую ткань между клетками
эндотелия в очаг формирующегося воспаления.
Этот процесс, называемый диапедезом,
приводит к накоплению в районе
воспаления моноцитов и нейтрофилов — клеток,
предназначенных для фагоцитоза —
поглощения и разрушения посторонних частиц —
бактерий и обломков ткани. Моноциты и
нейтрофилы активно передвигаются к
источнику воспаления. Такое целенаправленное
активное движение к очагу воспаления
носит название хемотаксиса. В очаге
воспаления моноциты превращаются в
макрофагов — активно фагоцитирующие клетки
с «липкой» поверхностью, подвижные, как
бы ощупывающие все, что находится в
ближайшем окружении. Нейтрофилы
также приходят в очаг воспаления, где их
фагоцитирующая активность возрастает.
Фагоцитирующие клетки накапливаются
в очаге воспаления, они активно поглощают
и разрушают (внутриклеточно) бактерии и
обломки клеток, выделяют ферменты,
разрушающие межклеточное вещество
соединительной ткани (см.: Ширинский В. П.
Межклеточные взаимодействия и
контактное ингибирование. Том 8). Процессы,
ведущие к фагоцитозу и заканчивающиеся
им, составляют суть нагноения. Кожный
покров, окружающий очаг воспаления
(нарыв), истончается и прорывается: заноза,
обломки клеток и накопившиеся фагоциты
выбрасываются из организма. Пораженный
участок ткани постепенно
восстанавливается.
Таким образом, в воспалении
участвуют два компонента — сосудистый,
определяющий динамику выхода плазмы и клеток
в очаг воспаления, и собственно клеточный,
связанный с накоплением и
функционированием фагоцитирующих клеток.
1 Эндотелий — клеточный пласт,
образующий стенку кровеносного капилляра. Клетки
эндотелия плотно смыкаются друг с другом,
создавая непроницаемую для жидкости и
непроходимую для клеток границу.
210
ИММУНОЛОГИЯ
Механизмы воспаления
Каков же механизм описанных событий?
Активация трех главных систем,
участвующих в воспалении, определяет состав и
динамику действующих факторов. Это
система образования кининов, система
комплемента и система активированных
фагоцитирующих клеток.
Образование кининов
Заноза (кусочек древесины) —
отрицательно заряженная чужеродная для
организма поверхность, содержащая
высокополимерные молекулы целлюлозы. На ней
адсорбируется фрагмент так называемого
фактора Хагемана (ФХ) — одного из
начальных компонентов системы свертывания
крови (см.: Зубаиров Д. М. Механизмы
свертывания крови и фибринолиза. Том 8). Этот
белок, предшественник специфической
протеазы2, присутствует в крови и спонтанно
диссоциирует на два фрагмента — ФХа и
ФХЬ, составляющих, однако, лишь
незначительную долю от недиссоциированной
формы. ФХа имеет сродство к
отрицательно заряженным поверхностям, поэтому
адсорбируется и стабилизируется на
поверхности занозы. Он обладает протеолитической
активностью, его субстратом является сам
ФХ и другой белок — прекалликреин —
также предшественник протеазы (рис. 1). Под
действием ФХа от прекалликреина
отщепляется фрагмент, превращающийся в про-
теазу калликреин.
Калликреин резко усиливает
образование ФХа из ФХ, и одновременно
действует на новый субстрат, так называемый
высокомолекулярный белок (ВМБ). Этот белок
под действием калликреина отщепляет
короткий пептид — брадикинин, состоящий
из 9 остатков аминокислот, и является
одним из главных медиаторов воспаления'.
Брадикинин действует на эндотелий
сосудов, вызывая «размыкание» краев клеток
сосудистого эндотелия и тем самым открывая
путь плазме крови в очаг воспаления.
Таким образом, данная система
обнаруживает чужеродное тело по его
отрицательно заряженной поверхности. Поверхности же
собственных клеток устроены так, что они
не адсорбируют ФХа, не стабилизируют его
и не индуцируют тем самым дальнейшую цепь
событий. Другая особенность системы
образования кининов — ряд каскадных усилений
начальной реакции. Каскадное усиление
определяется ферментативными звеньями
реакции, резко усиливающими эффект
первичных взаимодействий. В рассмотренной
цепи реакции таких каскадных усилений три.
Первое — при адсорбции ФХа на
поверхности и образовании положительной обратной
связи ФХа —> ФХ -> ФХа+ФХЬ; второе —
при образовании калликреина, энзиматически
действующего на ФХ с образованием ФХа,
который, в свою очередь, вновь ведет к
образованию калликреина; и третье — при
ферментативном действии калликреина на
предшественник брадикинина. Таким образом,
«точечная» начальная реакция на
чужеродной поверхности порождает
макроскопические, видимые простым глазом
физиологические изменения в формирующемся очаге
воспаления. Активированный ФХ индуцирует
и систему свертывания крови, что
способствует изоляции очага воспаления (рис. 1).
Следует указать, что брадикинин
является наиболее характерным, но не
единственным кинином, образующимся при
воспалении.
Комплемент
Вторая, более тонкая система
врожденного иммунитета — система комплемента (С).
Она включает одиннадцать белков крови,
представленных в основном неактивными
предшественниками протеаз. Активация
системы комплемента в естественном (т. е.
врожденном) иммунитете начинается с его
третьего компонента (СЗ). СЗ спонтанно
диссоциирует на СЗа и СЗЬ, образуя
следовые количества этих фрагментов. СЗЬ
ковалентно связывается с поверхностью
бактериальной клетки, стабилизируется там
и присоединяет к себе белок В 3,
способствуя превращению его во фрагмент ВЬ
(рис. 2). Образующийся при этом фермента-
тивно активный комплекс СЗЬВЬ оказыва-
3 Белок В — компонент сыворотки крови, не
относящийся собственно к системе комплемента.
ФХ
Поверхность (-)
ФХа
(следы) фХа
(Система
свертывания
крови
Прекалликреин
Положительная обратная
связь
ВМБ
(кининоген)
Калликреин
Брадикинин
2 Протеазы — ферменты, расщепляющие
белки. Протеолиз — ферментативное
расщепление белка.
Рис. 1. Генерация брадикинина
ФХ — фактор Хагемана; ФХа — ферментативно
активный фрагмент ФХ; поверхность ^ —
отрицательно заряженная поверхность; ВМБ—
высокомолекулярный белок (кининоген); —> —ферментативная
реакция
ВРОЖДЕННЫЙ ИММУНИТЕТ. ВОСПАЛЕНИЕ
211
ет действие на исходный СЗ и следующий
компонент комплемента С5, который он
расщепляет на С5а и С5Ь. Таким образом,
на мембране бактериальной клетки
формируется стабильный ферментативно
активный комплекс, обладающий двойной энзи-
матической активностью — он генерирует
новые молекулы СЗа/СЗЬ и С5а/С5Ь.
Компоненты СЗЬ и С5Ь фиксируются на
мембране, а СЗа и С5а остаются в среде.
Последние представляют собой полипептиды,
состоящие соответственно из 77 и 74
аминокислотных остатков, и являются
сильнейшими медиаторами воспаления (см. ниже).
Компонент С5Ь образует на мембране
новые центры ферментативной активности,
предназначенные для активации особого
комплекса, атакующего мембрану. Этот
комплекс состоит из нескольких
компонентов, последовательно активирующих друг
друга и совместно фиксирующихся на
клеточной мембране (С6-С9). Конечный
компонент системы комплемента (С9) входит в
комплекс, атакующий мембрану, и
становится начальным звеном полимеризации.
Присоединяя к себе 10-15 таких же, как и
он сам, молекул, он погружается в
мембрану, полимеризуется в кольцо и
образует поры, «продырявливающие» оболочку
клетки, что ведет к ее гибели.
Таким образом, система комплемента
распознает чужеродную клетку и
запускает цепную реакцию стимуляции биологически
активных белков, что приводит в конечном
итоге к гибели бактериальной клетки.
Кроме того, компонент СЗЬ, фиксированный на
поверхности бактериальных тел, резко
усиливает их фагоцитоз. Последнее
обусловлено присутствием на мембране
фагоцитирующих клеток рецепторов к СЗЬ, благодаря
чему существенно повышается сродство
фагоцитов к бактериям (опсонизация). Это
один из самых важных феноменов в
антибактериальном иммунитете.
Иная судьба у растворимых факторов —
СЗа и особенно С5а. Эти биологически
активные пептиды обладают рядом свойств,
важных для развития воспаления: прямым
действием на проницаемость сосудов, хе-
мотаксической активностью и самое
главное способностью активировать тучные
клетки (рис. 2). Тучные клетки
синтезируют и хранят большие запасы мощного
медиатора воспаления — биологически
активного амина гистамина. Они рассеяны
повсеместно в соединительной ткани и осо-
C3R
Фагоцитоз
СЗ СЗа
С5 С5а
Гистамин
Воспаление
С6С7С8
Плазма
С9
%^
Рис. 2. Путь активации комплемента при воспалении
СЗ-С9— компоненты системы комплемента; СЗЬ, С5Ь — фрагменты СЗ и С5, связанные с мембраной
бактериальной клетки (размеры бактериальной клетки увеличены в несколько тысяч раз); СЗа и С5а— пептиды,
отщепившиеся соответственно от СЗ и С5; С6-С8— компоненты комплекса, атакующего мембраны; С9— конечный
компонент комплекса, полимеризующийся в мембране и образующий поры; ВЬ — фрагмент белка В, связанный
с мембраной; — каскадно-усиливающиеся компоненты реакции; МФ— макрофаг; C3R— рецептор к
СЗЬ-компоненту комплемента; /С— капилляр; Э— эндотелиальная выстилка капилляра; Н и М— диапедез
нейтрофила и моноцита
212
ИММУНОЛОГИЯ
бенно вдоль кровеносных сосудов. Тучные
клетки несут на своей поверхности
рецепторы к СЗа и С5а; при присоединении
пептидов СЗа и С5а эти клетки секретируют
гистамин в окружающую среду. Роль гис-
тамина в воспалении многогранна. Он
быстро и заметно влияет на сосудистую
капиллярную сеть. Эндотелий капилляров под его
воздействием выделяет сосудорасширяющие
вещества, и проток крови через очаг
воспаления существенно возрастает, что
приводит к нагреву и покраснению. Между
клетками эндотелия образуются «щели», плазма
выходит из капилляров в зону воспаления,
свертывается и тем самым предотвращает
распространение инфекции из очага. По
градиенту концентрации гистамина фагоциты
«поднимаются» к источнику воспаления.
Таким образом, гистамин действует подобно
брадикинину, но более активно и быстро,
благодаря чему и является медиатором
острой фазы воспаления.
Возвращаясь к комплементу, следует еще
раз обратить внимание на широту его
действия (он токсичен для микроорганизмов,
усиливает фагоцитоз, генерирует
медиаторы воспаления) и на каскадный механизм
усиления его активности. При этом опять
возникает вопрос, каким образом начальный
компонент комплемента СЗЬ отличает
«чужую» поверхность от «своей», как
ограничивается каскадно-разрастающиися процесс
и почему он не захватывает весь организм?
Компонент СЗЬ может фиксироваться на
биологических мембранах «своих» или
бактериальных клеток, но мембраны собственных
клеток защищены от его действия
специальными белками, которые препятствуют
образованию комплекса СЗЬВЬ и делают СЗЬ
IL-8
Int у
Хемотаксис
N0
•N--6 Индукция белка
острой фазы
Повышение г°,
активация лимфоцитов
А фнп ^ Индукция
^пи «белка прилипания»
СЮ" Н90р 102
Кислородный
взрыв
Рис. 3. Активированный макрофаг: медиаторы
воспаления
МФ — макрофаг; ФНО — фактор некроза опухолей;
lnt-у— у-интерферон, секретируемый лимфоцитом
(Л); IL-1, 6, 8 — интерлейкины 1, 6 и 8; А— аутости-
муляция МФ ФНО; N0— окись азота; СЮ"— гипо-
хлорит; 102— синглетный кислород; кислородный
взрыв — продукция активных форм кислорода;
внутри макрофага— гранулы, содержащие бактерии и
их фрагменты
чувствительным к протеазе сыворотки
крови, разрушающей его. Эта же протеаза
разрушает СЗЬ, находящийся в несвязанной
форме, и тем самым препятствует
распространению «каскада» по организму. Такова
система комплемента. Очевидно, она играет
центральную роль в формировании
воспаления и непосредственно связана с его
важнейшим компонентом — фагоцитозом.
Фагоцитоз
Фагоцитоз был открыт и понят как
важнейший элемент воспаления и
естественного иммунитета И.И.Мечниковым в 1882г.
Громадная роль фагоцитоза не только во
врожденном, но и в приобретенном
иммунитете (см.: Абелев Г. И. Основы
приобретенного иммунитета. Наст, том) становится все
более очевидной благодаря работам двух
последних десятилетий. Как отмечалось выше,
фагоцитоз начинается с диапедеза
фагоцитов и их накопления в очаге воспаления.
Главную роль в этом процессе играют
моноциты и нейтрофилы. Моноциты, придя в
очаг воспаления, превращаются в макрофаги,
тканевые фагоцитирующие клетки.
Фагоциты, взаимодействуя с бактериями или их
продуктами, активируются, их мембрана
становится «липкой», в цитоплазме
накапливаются гранулы, наполненные мощными
протеазами. Возрастают поглощение
кислорода и генерация активных форм кислорода
(кислородный взрыв), включая синглетный
кислород, перекись водорода и гипохлорит,
а также окись азота (см.: Владимиров Ю. А.
Свободные радикалы в биологических
системах. Том 8) (рис. 3). В дополнение к
перечисленным признакам активации
макрофаги начинают выделять в среду мощные
медиаторы воспаления, среди которых
особой активностью отличаются так
называемый фактор некроза опухолей (ФНО) и
интерлейкины 1, 6 и 8 (IL-1, IL-6, IL-8)
(см.: Фрейдлин И. С. Клеточные механизмы
иммунной защиты организма. Цитокины. Наст,
том). Все они являются биологически
активными пептидами, существенно
дополняющими и расширяющими действие медиаторов
воспаления, рассмотренных выше.
Какова же их роль в воспалении? ФНО —
небольшой «многопрофильный» белок,
синтезируемый и секретируемый макрофагами.
Он активирует макрофаги, а также
индуцирует синтез и экспрессию на мембране
клеток сосудистого эндотелия особых
белков, благодаря которым поверхность
эндотелия становится «липкой» для моноцитов и
нейтрофилов. ФНО и IL-1 при их
совместном действии на эндотелий сосудов
индуцируют экспрессию рецепторов, имеющих
высокое сродство к мембранам моноцитов и
нейтрофилов, что приводит к остановке этих
клеток у стенок капилляров и диапедезу.
ВРОЖДЕННЫЙ ИММУНИТЕТ. ВОСПАЛЕНИЕ
213
IL-8 наиболее эффективен в создании
градиента для хемотаксиса
фагоцитирующих клеток. Именно IL-8, наряду с
другими медиаторами воспаления, стимулирует
нейтрофилы к диапедезу и к миграции по
его градиенту в очаг воспаления.
Фагоциты имеют рецепторы к хемотаксическим
медиаторам, которые «чувствуют»
разницу в концентрации медиатора на стороне,
обращенной к его источнику, и на
противоположной стороне, и направляют свое
движение по оси, вдоль которой различия
максимальны.
IL-1 и IL-6 обладают также и
активностью, выходящей за пределы очага
воспаления. Так, IL-1 и IL-6, воздействуя на
гипоталамус, вызывают повышение температуры
тела (озноб), a IL-6 еще индуцирует в клетках
печени синтез белка острой фазы,
способствующего фагоцитозу бактерий.
Особую роль в развитии воспаления
играет у-интерферон (Int-y), вырабатываемый
лимфоцитами, активируемыми в очаге
воспаления.
Int-y— также «многоцелевой» медиатор
воспаления. Одна из главных его функций —
активация макрофагов и нейтрофилов. Как
уже упоминалось, в этих клетках резко
усиливается синтез протеаз, которые
накапливаются в специальных цитоплазматичес-
ких органеллах — лизосомах. В них
происходит кислородный взрыв — они производят
активные формы кислорода и окись азота,
высокотоксичные для микроорганизмов.
Поверхность фагоцитов становится «липкой»,
поскольку количество различных
рецепторов на ней, включая рецепторы к СЗЬ,
резко увеличивается, как увеличивается и
«ощупывающая» подвижность цитоплазмы этих
клеток. Когда такая клетка встречается с
бактерией, она «прилипает» к поверхности
фагоцита, обволакивается его
псевдоподиями4 и оказывается внутри клетки, где и
разрушается протеазами.
Так заканчивается цикл «воспаление-
фагоцитоз». Мы видим, насколько мощный
и многогранный защитный механизм
действует в этом случае. Однако слабость его
в том, что он срабатывает одинаково при
встрече с различными «врагами».
Естественно, что многочисленные популяции
микроорганизмов вырабатывают обходные пути
для проникновения в организм. Одни
микробы начинают синтезировать белки,
функционально сходные с теми, которые не
дают СЗЬ фиксироваться на собственных
клетках организма, другие «одеваются» в
капсулу, резистентную к токсическому
действию комплемента. Есть микробы
синтезирующие специфические протеазы,
«срезающие» хеморецепторы моноцитов и
нейтрофилов, в результате чего клетки теряют
направление движения в очаг воспаления.
Наконец, имеются и такие
микроорганизмы (например, бактерии, вызывающие
туберкулез или проказу), которые после
захвата их макрофагом, поселяются в нем,
используя его в качестве безопасного
«дома», где они развиваются и
существуют. В таких ситуациях естественный
иммунитет бессилен, и в дело вступают уже
более изощренные и гибкие механизмы
иммунитета приобретенного (см.: Абелев Г. И.
Основы приобретенного иммунитета. Наст,
том).
Литература
Пол У. Иммунология. Пер. с англ. М.: Мир,
1987, т. 1.
Ройт А. Основы иммунологии. Пер. с англ. М.:
Мир, 1991.
Улъянкина Т.И. Зарождение иммунологии. М.:
Наука, 1995.
Хэм А., Кормак Д. Гистология. В 5 т. Пер. с англ.
М.: Мир, 1983, т. 1.
Janeway Ch., Travers P. Immunobiology.
London, N.Y.: Curr. Biol. LtD, Garland Publ. Inc.,
1994.
4 Псевдоподии — выросты цитоплазмы,
непрерывно образующиеся и исчезающие, как бы
ощупывающие микроокружение клетки.
Г. И. Абеле в
ОСНОВЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА
Введение
Иммунитет — защита организма от
инфекции или, в более широком смысле, —
реакция организма на чужеродные
макромолекулы, микроорганизмы и клетки. Защита
осуществляется с помощью двух систем —
неспецифического (врожденного,
естественного) и специфического (приобретенного)
иммунитета. Эти две системы могут
рассматриваться и как две стадии единого процесса
защиты организма. Неспецифический
иммунитет выступает как первая линия защиты
и как заключительная ее стадия, а система
приобретенного иммунитета выполняет
промежуточные функции специфического
распознавания и запоминания болезнетворного
агента (или чужеродного вещества) и
подключения мощных средств врожденного
иммунитета на заключительном этапе процесса.
Система врожденного иммунитета
действует на основе воспаления и фагоцитоза (см.:
Абелев Г. И. Врожденный иммунитет.
Воспаление. Наст, том) Эта система реагирует только
на корпускулярные агенты (микроорганизмы,
занозы) и на токсические вещества,
разрушающие клетки и ткани, вернее, на
корпускулярные продукты этого разрушения.
Вторая и наиболее сложная система —
приобретенный иммунитет — основана на
специфических функциях лимфоцитов —
клеток крови, распознающих чужеродные
макромолекулы и реагирующих на них либо
непосредственно, либо выработкой
защитных белковых молекул.
Феноменология
Рассмотрим элементарную реакцию
специфического иммунитета на примере двух
внешне совершенно непохожих моделей —
выведении чужеродного белка и
отторжении чужеродной ткани. Если в кровь
животного, например, кролика, ввести
непосредственно, минуя тканевые барьеры, белок
крови животного другого вида (например,
альбумин1 лошади) в смеси с собственным
гомологичным белком (в данном случае
альбумином), то в первые дни после
введения оба белка будут вести себя одинаково,
т. е. выводиться с постоянным и довольно
длительным периодом «полужизни»2 (рис. 1).
Чужеродный белок в нашем случае не
токсичен и молекулярно-дисперсен. Он не вы-
1 Альбумин — основной белковый компонент
жидкой части крови.
2 Каждый белок, находящийся в клетке или
крови, имеет свой постоянный период
полураспада, или «полужизни», равный времени, за
которое данное количество белка уменьшается вдвое.
С, %
100
50
25
12,5
САК
САЛ
САК
САЛ
С, %
100
50
САК
САЛ
САЧ
САК
САЛ
6 8
С, %
100
До рождения
САЛ сп
50
25
12,5
-4-2 0
10
2
25
12,5
12 0
Дни
САК
САЛ в
САЧ
САЧ
4 6
Дни
2
8
4 6
Взрослые
САК
САЛ
10 12
САЧ
Рис. 1. Кривые выведения
собственного и чужеродного белков из крови
кроликов
САК— сывороточный альбумин
кролика; САП— сывороточный альбумин
лошади; САЧ—сывороточный
альбумин человека; а — первичный ответ;
б— повторный ответ; в— индукция
толерантности
ОСНОВЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА
215
зывает ни воспаления, ни фагоцитоза, т. е.
беспрепятственно минует первую линию
защиты и непосредственно встречается со
второй. Примерно на 7-й день кривые,
отражающие динамику выведения
собственного и чужеродного белка, расходятся —
первая продолжает «двигаться» по своей
линии «полужизни», а вторая сильно
отклоняется — чужеродный белок начинает
выделяться из кровотока с резко возросшей
скоростью (рис. 1, а). Организм,
следовательно, отличает «свое» от «не своего»; это —
первая особенность реакции
специфического иммунитета. Отличие запоминается —
повторное введение лошадиного белка
приводит к сокращению латентного периода и
ускоренной реакции (наклон кривой
выведения увеличивается). Это так называемая
иммунологическая память — вторая
характерная черта реакции специфического
иммунитета. Память специфична
(запоминается контакт лишь с лошадиным альбумином,
но не с каким-либо третьим белком) и это
третья особенность реакции
приобретенного иммунитета (рис. 1, б). Иммунный ответ на
чужеродную макромолекулу можно
избирательно подавить, если ввести ее в
развивающийся организм внутриутробно или в
первые часы после рождения. Способность
отличать введенный чужеродный белок от
собственного у такого животного
утрачивается после рождения (рис. 1, в). Это явление,
носящее название толерантности
(терпимости), составляет четвертую особенность
элементарной реакции приобретенного, или
специфического, иммунитета.
Перечисленные признаки реакции
иммунитета неразделимы, вне зависимости от
того, в какой системе она разыгрывается.
Например, отторжение кожи или органов
у генетически разнородных животных и
человека, внешне совсем непохожее на
выведение чужеродного белка, подчиняется
тем же закономерностям. Так, если взять
мышей какой-либо генетически однородной
линии3, то особи этой линии будут
генетически идентичны, например, все будут
иметь белую окраску. Если таким мышам
пересадить лоскут кожи (трансплантат)
другой инбредной линии, скажем черной,
а в качестве контроля — лоскут кожи от
генетически идентичной линии, то
сначала приживутся оба трансплантата, но на
12-14-й день черный лоскут будет окружен
валом лейкоцитов4, затем его
кровоснабжение начнет ухудшаться и через 3—4 дня он
3 Так называемые чистые (инбредные)
линии мышей, все особи которых генетически
идентичны, как однояйцевые близнецы,
получают путем длительного брато-сестринского
скрещивания — инбридинга.
4 Лейкоциты — белые клетки крови.
будет отторгнут, в отличие от контрольного,
генетически идентичного трансплантата.
Очевидно, мы вновь встретились здесь со
способностью организма отличать «свое» от
«не своего». Эта особенность запоминается:
вторичная пересадка черной кожи на белую
мышь ведет к отторжению трансплантата
в более короткие сроки и более
интенсивно из-за иммунологической памяти.
Запоминание специфично — лоскут кожи от
мыши «коричневой» линии, пересаженный
при повторной трансплантации «черного»
трансплантата, отторгается по типу
первичного, а не повторного ответа. И,
наконец, толерантность: при пересадке тканей
она выявляется еще лучше, чем в системе
с чужеродным белком. Введение
животному живых клеток крови генетически
чужеродной линии в процессе внутриутробного
развития делает его на всю жизнь
невосприимчивым к пересадке тканей и органов
линии донора крови. Именно на этой
модели толерантность и была впервые
обнаружена в 1953 г.
Таким образом, в организме человека и
позвоночных животных (рыб, земноводных,
пресмыкающихся, птиц, млекопитающих)
имеется система иммунитета, способная
отличать «свое» от «не своего», запоминать
встречу с «не своим», причем с высокой
специфичностью, отторгать «не свое» и
отвечать иммунологической ареактивностью
(толерантностью) на контакт с чужеродным
веществом, предварительно введенным в
процессе раннего развития. Эта система, лежит
в основе приобретенного, или
специфического, иммунитета.
Приобретенный иммунитет широко
используется для вакцинации, т. е. введения
ослабленных или убитых микроорганизмов,
или выделенных из них макромолекул,
вызывающих иммунологическую реакцию на эти
микроорганизмы. Вакцинация является
основным способом предупреждения таких
страшных заболеваний, как оспа,
туберкулез, полиомиелит, сибирская язва и многих
других. Приобретенный иммунитет
составляет основное препятствие для пересадок
органов (сердца, почек, печени и тканей) от
одного человека другому. Для преодоления
этого барьера несовместимости пользуются
препаратами, подавляющими иммунную
систему.
Вещество, способное вызывать реакцию
приобретенного иммунитета, носит
название антигена. Антигеном может быть не
всякое вещество. Оно должно быть
чужеродным, макромолекулярным (с
молекулярным весом более 10000-12000) и иметь
устойчивую химическую структуру. К
типичным антигенам относятся белки и
полисахариды. В первом примере (см. рис. 1)
антигеном является сам чужеродный белок —
216
ИММУНОЛОГИЯ
Акт. ц.
Акт. ц.
Акт. ц.
S-S
S-S
S-S
S-S
Н Н
Антитело
S-S
а р
РТК
м
Рис- 2. Принципиальная схема строения антител и рецепторов Т-клеток (РТК)
Н и L— тяжелая и легкая цепи антител соответственно; VL и VH— вариабельные районы цепей антител; Va и Vp—
вариабельные районы а- и р-цепи РТК; Акт. ц. — активный центр антител и РТК; прямоугольники — повторяющиеся
сходные звенья (домены) в структуре полипептидных цепей; М — клеточная мембрана. Другие пояснения в тексте
альбумин крови, во втором — особые
белки, присутствующие на мембранах
пересаженных клеток, так называемые антигены
тканевой совместимости.
Антитела и антиген-
распознающие рецепторы
лимфоцитов
Что же происходит в точке перелома
кривых выведения и отторжения? В
первом случае, когда свободный молекулярно-
дисперсный антиген циркулирует в крови,
в ответ на него появляются защитные
белки — антитела, которые специфически
распознают его, образуют с ним комплекс,
обезвреживают, если антиген токсичен (как,
например, бактериальный токсин, змеиный
яд или болезнетворный вирус), и
способствуют его ускоренному выведению из
организма.
При отторжении трансплантата главная
роль принадлежит особой разновидности
лимфоцитов — Т-лимфоцитам-киллерам5,
«убийцам». Эти лимфоциты несут на своей наруж-
5 Лимфоциты, созревающие в тимусе (ви-
лочковой железе), в отличие от В-лимфоцитов —
предшественников антителобразующих клеток,
развивающихся в костном мозге.
ной мембране антителоподобные рецепторы —
рецепторы Т-клеток (РТК). РТК специфически
распознают антиген, находящийся на
мембране чужеродных клеток, и обеспечивают
прикрепление киллера к клетке-мишени, что
необходимо для осуществления им своей
смертоносной функции. Киллеры, прикрепившись
к мембране клетки-мишени, выделяют
особый белок, «продырявливающий» эту
мембрану, в результате чего клетка гибнет.
После этого они открепляются от мишени и
переходят на другую клетку, и так
несколько раз. При повторной встрече организма с
антигеном образуется больше антител и
киллеров, они появляются в более короткие
сроки, чем при первой встрече. В этом и
проявляется иммунологическая память, на
которой основан эффект вакцинации.
Как устроены антитела и рецепторы
лимфоцитов? Очевидно, их структура должна
быть необычной, так как они строго
специфически распознают громадное множество
различных антигенов — любой чужеродный
белок, полисахарид или синтетическую
молекулу, вообще в природе не
встречающуюся. При этом антитела и Т-лимфоциты, как
правило, не реагируют с белками и
клетками собственного организма. Как природа
решает эту задачу?
Принцип строения антитела и РТК
представлен на рис. 2. Антитело состоит из двух
ОСНОВЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА
217
идентичных пар полипептидных цепей: Н
(от англ. heavy — тяжелый) с
молекулярным весом примерно 50000 и L (от англ.
light — легкий), имеющих молекулярный вес
около 25000. Цепи соединены друг с
другом ковалентными (дисульфидными) связями.
Рецептор лимфоцитов, в отличие от
антител, — гетеродимер, т. е. состоит из пары
различных цепей — а и (3 — с
молекулярным весом около 50000, соединенных между
собой одной дисульфидной связью. Н-, L-,
а- и р-цепи имеют сходный план строения
и относятся к одному большому
суперсемейству белков-иммуноглобулинов. В основе их
структуры лежит сходное повторяющееся
звено (домен), состоящее примерно из 110
аминокислот, свернутых в глобулы,
напоминающие друг друга (см. рис. 2, где эти
звенья обозначены как прямоугольники).
Каждый домен в молекуле антитела или РТК
выполняет свою биологическую функцию,
а наиболее важную из них —
распознавание и связывание с антигеном —
осуществляют концевые домены. Пара концевых
доменов (один из Н-цепи, другой из L-цепи,
а в РТК — один из ос-цепи, другой из Р-цепи)
образуют активный центр — уникальную по
структуре полость, распознающую в
молекуле антигена небольшие дискретные
участки, состоящие из 4-8 аминокислот. Эти
Семейство
а Семейство l/'-генов С-генов
б
(1-95) (96-100) (101-110)
Рис. 3. Упрощенная схема семейства генов антител
(а) и сборки гена, контролирующего вариабельный
район антител (б)
а — Принципиальная схема участка хромосомы,
включающего семейство вариабельных (V) и
константных (С) генов. Численность l/'-генов в отдельных
семействах варьирует в зависимости от типа цепей и
вида животных от 50 до 1000, а С-генов от 1 до 9. б—
Сборка У-гена из фрагментов. V— фрагмент гена,
контролирующий первые 95 аминокислот У-района;
D— короткий фрагмент, контролирующий от 2 до 10
аминокислот , начиная с 96. (Число D— до 20, в
зависимости от типа цепи и вида животных); J—
короткий концевой фрагмент 1/-гена, контролирующий
от 5 до 15 аминокислот. Районы ДНК, разделяющие
V-D и D-J, вырезаются и утрачиваются при сборке
гена. Транскрибируется (т. е. переписывается в
информационную РНК) лишь собранный VDJ ген.
Участок от VDJ до С-гена вырезается и утрачивается при
синтезе информационной РНК
участки антигена подходят к структуре
активного центра, «как ключ к замку»,
образуя прочные нековалентные связи
антигена с антителом. Разные антитела (и РТК
также) отличаются друг от друга
структурой активных центров. На химическом
уровне различия активных центров определяются
первичной структурой концевых доменов
Н- и L-, а- и Р-цепей, т. е.
последовательностью аминокислот в них. Участки Н и L
цепей (равно как а и (5), образующие
«стенки» активного центра, носят название
вариабельных (или V) районов. Остальная часть
этих молекул постоянна и носит название
константного (С) района. Вариабельных
районов среди молекул антител тысячи,
константных — единицы. Главная функция
вариабельных районов — образование
активных центров антител и РТК. Разнообразие
активных центров очень велико, их не
менее 107-108, что вполне достаточно для
более или менее специфического
распознавания любого произвольно взятого антигена.
Таким образом, структура антител и РТК,
а, точнее, их активных центров
обеспечивает узнавание любого чужеродного
белка или полисахарида, попавшего в
организм.
Генетический контроль
синтеза антител
Как же осуществляется контроль синтеза
полипептидных цепей, один из районов
которых постоянно варьирует при переходе от
одного антитела (или РТК) к другому
(вариабельный, V-район), а остальные строго
постоянны (константные, С-районы). Эта
задача решается благодаря уникальному
генетическому механизму — независимому
генетическому контролю V- и С-районов,
составляющих одну полипептидную цепь, и
сборке гена, контролирующего
вариабельный район, из фрагментов в процессе
созревания лимфоцита.
Гены, контролирующие структуру одной
полипептидной цепи антитела или РТК,
объединены в семейства, расположенные в
одном участке хромосомы. При этом гены,
соответствующие V-районам, «разорваны» на
2 или 3 фрагмента — основной V и один или
два дополнительных — D и J6. V'-фрагментов
в геноме недифференцированных лимфоцитов
много— от 50 до 1000 для разных цепей и
все они отличаются друг от друга по своей
структуре. D представлены несколькими
десятками копий, a J— единичными,
причем каждая из копий имеет уникальную
6 D — от англ. diversity (разнообразие), J —
от англ. joining (соединяющий), V— фрагмент
гена, контролирующего первые 95 аминокислот
в V-доменах, D— от 2 до 10 и J— от 5 до 15.
218
ИММУНОЛОГИЯ
структуру. В процессе созревания лимфоцита
фрагменты V-гена объединяются на
случайной основе в V'DJ-ген (рис. 3), причем
число возможных сочетаний равно
произведению чисел V-, D- и J-генов. Для различных
цепей антител или РТК число вариантов
составляет от тысячи до десятков тысяч.
Кроме того, в процессе сборки возможны
ошибки и химические модификации
объединяющихся фрагментов, что резко
увеличивает число возможных комбинаций.
Дальнейшее разнообразие активных центров
возникает за счет объединения
вариабельных районов пары цепей, составляющих этот
центр. Таким образом, благодаря сборке
V-гена из фрагментов и образованию
активных центров из разных цепей, создается 107-
108 уникальных по своей структуре
участков молекулы антитела (или РТК), среди
которых всегда найдутся способные
взаимодействовать с любым произвольно взятым
антигеном. Все это разнообразие
составляет репертуар антител или РТК. Следует
подчеркнуть, что этот репертуар возникает до
встречи с антигеном и независимо от него,
при этом большая часть репертуара
антител не потребуется в течение всей жизни
особи.
Клеточные механизмы
иммунитета
Итак, в организме предсуществуют
антитела и РТК к любому произвольно
взятому антигену. Эти антитела и РТК
расположены на поверхности лимфоцитов, образуя
там антиген-распознающие рецепторы.
Чрезвычайно важно, что один лимфоцит может
синтезировать антитела (или РТК) только
одной специфичности, не отличающиеся
друг от друга по структуре активного
центра. Это формулируется как принцип «один
лимфоцит — одно антитело».
Каким же образом антиген, попадая в
организм, вызывает усиленный синтез
именно тех антител, которые специфично
реагируют только с ним? Ответ на этот вопрос
дала теория селекции клонов австралийского
исследователя, Нобелевского лауреата
Ф. М. Бернета (1899-1985). Согласно этой
теории, сформулированной в 1957 г. и
полностью подтвержденной последующими
экспериментами, одна клетка синтезирует лишь
один тип антител, которые локализуются на
ее поверхности. Репертуар антител
формируется до и независимо от встречи с
антигеном. Встреча антигена с предсуществую-
щим антителом приводит к активированию
этой клетки. Активированный лимфоцит
вступает в деление и дифференцировку. В
результате из одной клетки возникает 500-1000
генетически идентичных клеток (клон),
синтезирующих один и тот же тип антител,
способных специфически распознавать антиген
и соединяться с ним. В ходе дальнейшей диф-
ференцировки лимфоцит превращается в
клетку, не только синтезирующую данное
антитело, но и секретирующую его в
окружающую среду.
Таким образом, суть иммунного ответа
заключается в селекции нужных клонов и
их стимуляции к делению. Динамика
первичного и повторного ответов, согласно
теории Бернета — это отражение динамики
размножения клонов клеток,
продуцирующих антитела к данному антигену.
Толерантность — утрата клона клеток вследствие
их контакта с антигеном в процессе
созревания лимфоцита.
В основе образования
лимфоцитов-киллеров лежит тот же принцип: селекция
антигеном Т-лимфоцита, несущего на своей
поверхности РТК нужной специфичности, и
стимуляция его деления и дифференцировки.
В результате образуется клон однотипных
киллеров, несущих на своей поверхности
большое количество РТК,
взаимодействующих с антигеном, входящим в состав
чужеродной клетки, и способных убивать эти
клетки.
И здесь мы встречаемся с проблемами,
выходящими за пределы клонально-селек-
ционной теории иммунитета. Первая из них:
как РТК узнают антиген? Дело в том, что
киллер, в отличие от антитела, ничего не
может сделать с растворимым антигеном —
ни обезвредить его, ни удалить из организма.
Но лимфоцит-киллер очень эффективно
убивает клетки, содержащие на
поверхности чужеродный антиген. Для этого
существует специальный механизм, так
называемое «распознавание в контексте». Он
заключается в том, что РТК не узнают
соответствующий им антиген, если он
находится в свободном виде, но строго
специфически реагируют с ним, если он
находится в комплексе с антигеном тканевой
совместимости. Эти антигены всегда
присутствуют на поверхности любых клеток
организма и обладают способностью комплекси-
роваться с чужеродными белками, вернее,
с их фрагментами. Таким образом,
антигены тканевой совместимости образуют
«контекст», и только в нем РТК распознают
чужеродный антиген, активируя лимфоцит
и стимулируя его к делению и дифферен-
цировке в полноценный киллер.
Вторая проблема, выходящая за
пределы клонально-селекционного принципа,
связана с ролью лимфоцитов-помощников.
Детальное изучение реакций иммунитета
показало, что для образования клона
клеток, продуцирующих антитела, или клона
киллеров необходимо участие специальных
лимфоцитов-помощников. Сами по себе они
не способны ни вырабатывать антитела, ни
ОСНОВЫ ПРИОБРЕТЕННОГО ИММУНИТЕТА
219
убивать клетки-мишени. Но, распознавая
чужеродный антиген, они реагируют на него
выработкой ростовых и дифференцировоч-
ных факторов, необходимых для
размножения и созревания антителообразующих и
киллерных лимфоцитов. В этой связи
интересно вспомнить о вирусе СПИДа,
вызывающем сильнейшее поражение иммунной
системы (синдром приобретенного
иммунодефицита — СПИД). Этот вирус поражает
именно лимфоциты-помощники, делая
иммунную систему неспособной ни к
выработке антител, ни к образованию киллеров.
И, наконец, еще одна очень важная
проблема: как вырабатывается толерантность к
антигенам собственного организма? В
полном соответствии с теорией Бернета было
показано, что если незрелый лимфоцит,
несущий антительный рецептор или РТК к
собственным антигенам, встречается с
таким антигеном, то он инактивируется или
погибает. Таким образом, организм лишается
клонов лимфоцитов, способных реагировать
с собственными антигенами, не ослабляя
своего ответа на антигены чужеродные.
Важно отметить, что при некоторых
заболеваниях сохраняются «запрещенные» клоны,
отвечающие антителами или киллерами на
антигены собственных клеток. В этом
случае возникают тяжелейшие заболевания,
такие, например, как красная волчанка, при
которой поражаются собственные ткани
организма.
Эффекторные механизмы
иммунитета
Как антитела или лимфоциты-киллеры
удаляют из организма чужеродные вещества
или клетки? Очевидно, в случае киллеров
РТК выполняют лишь функцию
«наводчика» — они распознают соответствующие
мишени и прикрепляют к ним
клетку-убийцу. Так распознаются клетки, зараженные
вирусом. Сам по себе РТК не опасен для
клетки-мишени, но «идущие» за ним Т-клетки
обладают огромным разрушительным
потенциалом. В случае антител мы встречаемся
со сходной ситуацией. Сами по себе
антитела безвредны для клеток, несущих
антиген, и не обладают физиологической
активностью, ведущей к разрушению антигена.
Правда, они подавляют биологическую
активность опасных антигенов, таких, как
дифтерийный или столбнячный токсины, или
змеиный яд. Они также способны
блокировать вирус, находящийся в крови (но не в
клетках). Но это лишь особые случаи
действия антител. При встрече с обычными
антигенами (нетоксичными),
циркулирующими или входящими в состав клеточной стенки
микроорганизма, к антителам
подключается система комплемента (см.: Абелев Г. И.
Врожденный иммунитет. Воспаление. Наст,
том), резко усиливающая эффект действия
антител. Во-первых, происходит каскадное
усиление реакции, почти каждый
последующий этап реакции активации
комплемента является ферментативным, при котором
продуктов реакции образуется несравнимо
больше, чем исходных реагирующих веществ.
Во-вторых, на поверхности бактерии
фиксируются компоненты комплемента, резко
усиливающие фагоцитоз этих клеток, т. е.
сродство их к фагоцитирующим клеткам
организма. В-третьих, при ферментативном
расщеплении белков системы комплемента
образуются фрагменты, обладающие
мощной воспалительной активностью. И,
наконец, при включении в комплекс антиген-
антитело последнего компонента комплемента
этот комплекс приобретает способность
«продырявливать» клеточную мембрану и тем
самым убивать чужеродные клетки.
Однако комплемент активируется любым
комплексом антиген-антитело, вредным или
безвредным для организма. В связи с этим
воспалительная реакция, возникающая на
безвредные антигены, регулярно
попадающие в организм, может вести к
аллергическим, т. е. извращенным, реакциям
иммунитета, тяжело переносимым человеком
или животным. Аллергия развивается при
повторном, как правило, многократном
попадании антигена в организм, например, при
повторном введении антитоксических
сывороток, или у мукомолов на белки муки, или
при многократной инъекции
фармацевтических препаратов, в частности, некоторых
антибиотиков. Правда, есть аллергические
реакции, не требующие для своего
развития комплемента. Борьба с аллергическими
болезнями состоит в подавлении либо
самой реакции иммунитета, либо в
нейтрализации образующихся при аллергии
веществ, вызывающих воспаление.
Итак, мы рассмотрели сложную и
удивительно целесообразно устроенную
систему защитных реакций организма. Одной из
важнейших проблем современной биологии
является вопрос о том, как и из чего она могла
возникнуть в процессе эволюции. Подходы к
этой проблеме пока только намечаются.
Литература
Ройт А. Основы иммунологии. Пер. с англ. М.:
Мир, 1991.
Альберте Б., Брей Д., Льюис Дою. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ.
М.: Мир, 1994, гл. 18.
Галактионов В.Г. Иммунология. М.: Изд-во
МГУ, 1998.
Г. И. Абелев
МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
Введение
При введении в организм животных и
человека чужеродных макромолекулярных
веществ — белков или полисахаридов
(антигенов) — в крови появляются защитные
белки — антитела (см.: Абелев Г. И.
Основы приобретенного иммунитета. Наст, том),
для которых характерна необыкновенная,
уникальная специфичность. Каждое антитело
узнает только свой антиген, вернее, одну
его детерминантную группу. Детерминан-
тная группа состоит из нескольких
аминокислот, обычно из б~8, образующих
пространственную структуру, характерную для
данного белка. В одном белке, состоящем из
нескольких сотен аминокислот, имеется
несколько — 5—15 разных детерминант, поэтому
к одному белку образуется целое семейство
различных по своей специфичности антител.
Даже к одной детерминанте образуется
целый спектр антител, отличающихся по
структуре, степени специфичности и
прочности связывания с ней. То же относится и
к полисахаридным антигенам, детерминан-
тные группы которых образуются 3—б
остатками моносахаридов.
Таким образом, при введении антигена
возникает семейство антител к разным
детерминантам этого антигена; есть и различия
между антителами, направленными к
одной и той же детерминанте. В крови
иммунизированных животных появляется
богатое и уникальное по составу семейство
антител, обеспечивающее их практически
абсолютную специфичность в
распознавании данного антигена.
Антитела давно и широко
используются для нейтрализации бактериальных
токсинов (дифтерийного, столбнячного, буту-
линического), змеиных ядов (кобры, гадюки),
вирусов, попавших в кровь (особенно
эффективно — вируса кори), и для
идентификации индивидуальных белков (и других
антигенов), находящихся в клетке или в
сложнейших тканевых экстрактах.
Однако иногда требуются не
многокомпонентные смеси антител, возникающие в
крови в ответ на введение антигена, а
отдельные, элементарные составляющие этой
смеси, направленные лишь к одной
детерминанте антигена и имеющие одни и те же
характеристики. Такие антитела бывают
нужны как для изучения их собственной
природы, так и для практического
использования, например, для доставки в
опухоли токсических веществ.
Как получить однородные
антитела?
Очевидно, путем иммунизации, т. е.
введения животному индивидуального антигена
или только одной его детерминантной
группы1, этого сделать, как правило,
невозможно. Дело в том, что в организме в
процессе созревания антителообразующих клеток
(АОК) образуется громадное количество
генетически однородных семейств клеток —
клонов, каждый из которых
специализируется на синтезе только одного варианта
антител. Таких клонов много больше, чем
требуется антител для распознавания
любого, случайно взятого антигена. Антиген,
попадая в организм, стимулирует
размножение тех клонов, которые продуцируют
антитела к его детерминантам. Таких
клонов сотни, и в этом причина большого
разнообразия антител, индуцируемых одним
антигеном.
Казалось бы, выход прост: надо
вырастить отдельные клоны антителообразующих
клеток в пробирке — в культуре тканей, и
они будут продуцировать моноклональные
антитела, т. е. антитела одной строго
определенной специфичности, являющиеся
продуктом одного клона. Но и это оказалось
невозможным — нормальные клетки
смертны, вскоре после высаживания в
культуру они погибают. Дело не доходит до
образования клонов АОК. Добавление в культуру
факторов роста несколько продлевает их
жизнь, но тоже не решает проблемы.
Решение проблемы
с помощью опухолей
Оказалось, что проблему можно решить,
используя злокачественные опухоли. Уже
давно известны опухоли человека — плаз-
моцитомы, вырабатывающие и секретиру-
ющие в кровь иммуноглобулины2, по
структуре своей неотличимые от антител. Причем,
каждое такое «антитело» слегка отличалось
от другого, вырабатываемого другой плаз-
моцитомой. Образовалась как бы коллекция
случайных антител к неизвестным
антигенам. Когда накопились сотни таких
«антител» и когда они были испытаны с сотнями
1 Это может быть простая химическая группа,
например, состоящая из арсаниловой кислоты,
ковалентно присоединенной к белку.
2 Иммуноглобулины — белки, имеющие план
строения, свойственный антителам.
МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
221
наугад взятых антигенов, оказалось, что
в этой коллекции обнаружились
специфически реагирующие пары
«антиген-антитело».
Почему возникла идея о возможности
использования опухолей для получения
моноклональных антител? Тому ряд
причин, и все они коренятся в самой природе
опухолевой клетки. Во-первых, она всегда,
или почти всегда, сохраняет свойства и
функции клетки, из которой произошла.
Плазмоцитома происходит из «юных»
плазматических клеток, которые как раз и
синтезируют антитела. Это свойство
сохраняется и у опухолей, возникших из них.
Очень важной особенностью опухолей
является то, что они возникают из одной
генетически измененной (мутантной)
клетки. Поэтому опухоль зарождается и
развивается как клоп, в нашем случае — как клон
иммуноглобулин-образующих клеток.
Причем они продуцируют строго однородный по
всем свойствам моноклональпый
иммуноглобулин.
Нормальные плазматические клетки (или
их предшественники — лимфоциты) —
смертны, их срок жизни — несколько дней.
Опухоль, и в этом ее принципиальное
отличие от нормальных предшественников, —
бессмертна. Ее можно культивировать в
пробирке или пересаживать от одного
животного другому неограниченное число раз
и в течение неограниченного времени.
В отличие от нормальной ткани опухоль
автономна, организм «хозяина» не
способен (за очень редкими исключениями)
остановить неограниченный рост
злокачественного опухолевого клона.
Плазмоцитомы возникают не только
спонтанно, т. е. непредсказуемо, как бы
случайно, но их можно довольно легко
индуцировать у мышей и крыс и получить, таким
образом, неограниченно растущий,
перевиваемый клон клеток, продуцирующих
иммуноглобулины, иногда обладающие
специфичностью антител, причем, антител
моноклональных. Вполне естественно было
желание иммунологов научиться получать
плазмоцитомы, продуцирующие антитела
заданной специфичности. Для этого мышей
вначале интенсивно иммунизировали, а
затем индуцировали у них плазмоцитомы,
чтобы получить опухоли тех клонов клеток,
которые производили антитела к антигенам,
использованным для иммунизации. Но тщетно.
Это практически не удавалось. Слишком
редки были совпадения. Тогда попробовали
индуцировать опухоли антителообразующих
клеток опухолеродными вирусами.
Результаты были лучше. Это, хотя и редко, но
удавалось. Однако создать простой и
универсальный метод получения
моноклональных антител на этом пути не удалось.
Успех пришел, как всегда,
неожиданно и не специально, а как побочный
продукт исследования, имевшего иные цели.
В начале 70-х гг. молодой немецкий
иммунолог, Г. Кёлер, получивший стипендию
для работы в знаменитом Базельском
институте иммунологии, заинтересовался
вопросом о генетической изменчивости
антител. В то время возникло
предположение, что антитела мутируют
(генетически изменяются) с большей частотой, чем
другие белки. Для исследования этого
вопроса надо было изолировать клон АОК,
продуцирующий антитела определенной
специфичности, получить из него
стабильную клеточную линию, поддерживаемую
в пробирке (в культуре), и проследить, с
какой частотой появятся там генетически
измененные варианты. Для реализации
такого проекта Кёлер поехал в Англию,
в лабораторию Ц. Милыптейна, изучавшего
клоны плазмоцитом, и они вместе
разработали совершенно оригинальный подход
к этой проблеме. Они решили получить
гибрид нормальной АОК и опухолевой
клетки. В случае успеха такой гибрид
унаследовал бы от нормальной клетки
способность к синтезу антител, а от
опухолевой — бессмертие и способность к
неограниченному и бесконтрольному росту.
Это им удалось осуществить.
Гибридомы
Методы гибридизации соматических (т. е.
не половых) клеток к тому времени были
хорошо известны и широко применялись для
разных целей. При этом использовался
вирус, способствующий слиянию клеток3.
Разнородные клетки образовывали двуядерные
гибриды, которые сохраняли способность к
клеточному делению. В процессе
клеточного деления хромосомы обоих ядер
перемешивались и образовывали общее ядро.
Таким образом, возникал истинный гибрид,
потомок двух соматических клеток, или
гибридома. Гибридому можно было
получить и между нормальной АОК и
опухолевой, плазмоцитомной клеткой. Плазмоцитома
была взята потому, что она ближе всего
соответствовала АОК по типу дифферен-
цировки. Весь ее синтетический аппарат был
настроен на синтез иммуноглобулинов.
Проблема была в том, как отделить заданную
гибридому от присутствующих в системе
отдельных неслившихся клеток и от гибридов
иного состава или иной специфичности, чем
требуемые.
3 Впоследствии симпластобразующий вирус
был заменен простым органическим веществом —
полиэтиленгликолем, вызывающим тот же
эффект — слияние оболочек клеток.
222
ИММУНОЛОГИЯ
Для достижения этой цели авторы
разработали специальную схему,
использующую отбор клеток в селективной среде.
Прежде всего был получен особый мутант
мышиной плазмоцитомы, рост которого
можно было контролировать составом
питательной среды.
Для получения такого мутанта
использовались особенности синтеза нуклеиновых
кислот (ДНК и РНК). Известно, что
имеются два пути синтеза предшественников
нуклеиновых кислот — основной и
резервный. Основной путь — это путь
новообразования нуклеотидов — звеньев, входящих
в состав нуклеиновых кислот. Этот путь
включает несколько этапов и блокируется
противоопухолевым препаратом — амино-
птерином (А). Однако клетки не гибнут от
этого препарата, поскольку обладают
резервным путем — способностью
синтезировать нуклеотиды и нуклеиновые кислоты,
реутилизируя продукты распада ранее
Смесь антигенов
Культура плазмоцитомы
Клетки селезенки (АОК) Клетки плазмоцитомы (ПЛ)
+ ПЭГ
Слияние
АОК; АОКхАОК; АОКхПл ; ПЛхПЛ;
ПЛ;1
Селективная ГАТ-среда
\
АОКхПл
АОКхПл
Гибридомы (анти-А, анти-В, анти-С, ...)
Клонирование и отбор
нужных продуцентов (анти-А)
Повторное клонирование и отбор
анти-А продуцентов
Гибридома, продуцирующая анти-А
(хранение при -70°С, размножение
в культуре или In vivo)
Рис. 1. Схема получения гибридом (объяснение в
тексте)
А,В,С,...— многокомпонентная смесь антигенов,
использованная для иммунизации; АОК— антителооб-
разующие клетки селезенки; ПЛ— клетки
плазмоцитомы, не растущие в селективной ГАТ-среде; ПЭГ —
полиэтиленгликоль; ГАТ—среда, содержащая гипок-
сантин, аминоптерин, тимидин; анти-А, анти-В,
анти-С— моноклональные антитела соответственно к
А, В, С-антигенам
синтезированных нуклеиновых кислот —
гипоксантина (Г) и тимидина (Т).
Добавление Г и Т в питательную среду,
содержащую А, снимает токсический эффект
последнего.
Для селекции гибридом надо было
получить мутант плазмоцитомы, не способный
пользоваться резервным путем, и,
следовательно, погибающий в среде, содержащей
Г, Т и А (ГАТ-среда). Такой мутант
получили путем добавления в среду токсичных
аналогов Г и Т. Все клетки, способные
усваивать Г и Т, включали их токсические аналоги
и погибали. Выживали лишь те редкие
мутанты, которые были неспособны усваивать
Г и Т, т. е. были лишены резервного пути.
Из потомства этих клеток дополнительно
отбирались еще и такие мутанты, которые
утратили способность к синтезу собственных
иммуноглобулинов. Теперь все было готово
для получения гибридом, т. е. гибридов
нормальных АОК и плазмоцитомных клеток
(рис. 1).
Мыши интенсивно иммунизировались
определенным антигеном — белком,
бактериальной клеткой или клеткой животного
происхождения. Когда в их крови
появлялись антитела, у них брались селезенка и
лимфатические узлы — места скопления
АОК, — и из них готовилась взвесь клеток.
К ней добавлялись в большом избытке
клетки мутантной плазмоцитомы и полиэтилен-
гликоль (ПЭГ). После короткой инкубации,
требующейся для слияния клеток, они
отмывались от ПЭГ и помещались в ГАТ-среду.
Теперь в системе находились: гибриды АОК
и АОК, плазмоцитомы и плазмоцитомы,
АОК и плазмоцитомы, а также
оставшиеся свободными АОК и клетки
плазмоцитомы. Из них нужно было отобрать только
гибриды АОК и плазмоцитомы. После
недолгого (несколько дней) культивирования
одиночные АОК, а также гибриды АОК и
АОК погибали, так как нормальные
клетки смертны и быстро погибают в культуре.
Плазмоцитомные клетки и их гибриды сами
собой погибали, так как А блокировал
основной путь синтеза предшественников
нуклеиновых кислот, а Г и Т их не спасали.
Выживали, следовательно, только
гибриды АОК и плазматических клеток, так как
бессмертие они унаследовали от
плазмоцитомы, а резервный путь от нормальной
клетки. Такие гибриды, гибридомы, сохраняли
способность синтезировать и секретировать
антитела.
Моноклональные антитела
Следующий этап после получения
гибридом — клонирование и отбор нужных
клонов. Выжившие в ГАТ клетки, рассева-
лись в специальные пластиковые планше-
МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
223
ты, содержащие обычно 96 лунок,
емкостью, примерно, по 0,2 см3. В каждую
лунку помещалось в среднем по 10 гибридом-
ных клеток, которые культивировались в
присутствии «кормящих» клеток, не
имеющих отношения к гибридомам, но
способствующих их росту. После нескольких
дней культивирования содержимое
каждой лунки проверялось на присутствие
антител нужной специфичности. При этом
использовались микрометоды для
определения антител к соответствующему
антигену. Клетки из лунок, содержащих
нужные антитела, клонировались, т. е.
повторно рассевались по таким же лункам,
но уже из расчета 1 клетка на лунку, вновь
культивировались и проверялись на
присутствие нужных антител. Процедура
повторялась 1-2 раза. Таким образом
отбирались клоны, продуцирующие монокло-
нальные антитела. Полученные клоны
можно заморозить при —70°С и хранить до
тех пор, пока они не потребуются. Их
можно культивировать, накапливая
антитела в культуральной среде, а можно
привить мышам (так как гибридомы — это
опухолевые клетки), где они будут
расти и накапливать колоссальные
количества моноклональных антител. От одной
мыши можно получить антител не
меньше, чем от кролика. Эти антитела
настолько однородны физико-химически, что
могут рассматриваться как чистые
химические реактивы.
Применение моноклональных
антител
Обычные поликлональные антитела
(иммунные сыворотки) давно и широко
применяются для определения биологически
активных веществ — белков крови и
других биологических жидкостей, гормонов,
ростовых факторов, клеточных рецепторов,
медиаторов воспаления и иммунитета,
бактериальных и вирусных антигенов,
различных ядов и т. п. Моноклональные антитела
в силу своей высочайшей специфичности,
стандартности и технологичности получения
успешно вытесняют и заменяют иммунные
сыворотки.
Метод получения гибридом создает
уникальные возможности для их
использования в аналитических целях: они могут
применяться как «иммунологический микроскоп»
с чрезвычайно высоким, если не
абсолютным разрешением. Так, например, если
нужно сравнить две клеточные линии,
различающиеся по одному или немногим
антигенам, то применение гибридом
позволяет выявить такие различия. Проиммуни-
зировав мышей одной из линий и получив
сотни гибридом, продуцирующих антитела
к антигенам этой линии, можно найти одну
или две с антителами только к данной
линии. Размножив такую гибридому в пробирке
или вырастив ее на мышах, можно
получить огромное количество антител к
уникальному антигену (или детерминантной
группе), затерянному среди других
компонентов клетки, подобно иголке в стоге сена.
Это будет продукт одного клона. В крови
иммунизированного животного среди
множества других антител он никак не
проявится в силу чисто количественных отношений.
Благодаря же гибридомам его можно не
только обнаружить, но и вывести в линию
и получить любое количество
соответствующих антител.
С помощью гибридом можно обнаружить
антигены, характерные для опухолей
определенных тканей, получить к ним
антитела и использовать их для диагностики и
типирования опухолей. Такие
моноклональные антитела нашли самое широкое
применение в онкологической клинике.
Наконец, во всем мире ведутся активные
исследования по использованию
моноклональных антител в качестве
специфических переносчиков токсических веществ в
опухолевые клетки.
Пока же с помощью моноклональных
антител доставляются в опухоль и в ее
метастазы радиоактивные вещества,
позволяющие обнаружить локализацию
небольших узелков опухоли по накоплению в них
радиоактивности.
Гибридомы сыграли и продолжают
играть огромную роль в фундаментальной и
прикладной иммунологии. Они созданы на
основе клонально-селекционнои теории
иммунитета и явились самым ярким и
окончательным доказательством этой теории.
Гибридомы сделали реальностью предпола-
Рис. 2. Флуоресцентное окрашивание клетки
соединительной ткани (фибробласта) моноклональным
антителом к тубулину— белку микротрубочек,
образующих скелет клетки
224
ИММУНОЛОГИЯ
Рис. 3. Последовательные срезы через желудок
(жел) и пищевод (пищ) мыши, окрашенные двумя
моноклональными антителами
7 — первое моноклональное антитело реагирует с
эпителием пищевода и слабее— с эпителием
желудка; 2— второе моноклональное антитело
реагирует только с эпителием желудка
гаемые клоны антителообразующих клеток
и позволили даже обнаружить их
существование в организме до введения
соответствующего антигена.
Гибридомы революционизировали
иммунологическую промышленность и создали в
ней совершенно новые области. Благодаря
гибридомам возникли новые методы
диагностики многих заболеваний и открылись
совершенно новые пути для изучения
злокачественных опухолей. И хотя гибридомы
скорее относятся к гениальным
изобретениям, а не к открытиям, работы по их
изучению были отмечены в 1984 г.
Нобелевской премией — высшей научной наградой,
присуждаемой за выдающиеся открытия.
Если бы Кёлер и Милыытейн
запатентовали свой метод, они вскоре стали бы
миллиардерами, так как все, кто
использует гибридомы в коммерческих целях,
должны были бы платить за право пользоваться
патентом. Авторы гибридом, несомненно,
понимали это, но в интересах развития
науки не пошли на такой шаг. Метод
гибридом беспрепятственно вошел во все сферы
иммунологии, и сами авторы всемерно
способствовали этому, предоставляя свою
клеточную линию плазмоцитомы для
использования в научных целях всем желающим.
Первые гибридомы в нашей стране,
полученные в 1979—1980 гг., также были созданы
на основе клеток, ведущих происхождение
из лаборатории этих ученых, и с их
любезного разрешения.
Литература
Кеннет Р. Г.,Мак-Керн Т. Дж. и Бехтол К. Б.
Моноклональные антитела. Гибридомы: новый
уровень биологического анализа. Пер. с англ. М.:
Медицина, 1983.
Ройт А. Основы иммунологии. Пер. с англ. М.:
Мир, 1991.
В. Г. Галактионов
генетический контроль взаимодействия
иммунокомпетентных клеток
Введение
Современный ландшафт
иммунологических знаний очень разнообразен. Сегодня мы
видим то, что было скрыто еще 10—15 лет
тому назад.
Защитная реакция организма от
патогена или любого другого чужеродного
биологического материала осуществляется
двумя системами иммунитета. В тех случаях,
когда организм инфицируется бактериями,
основная нагрузка падает на так
называемую В-систему иммунитета. В состав этой
системы входят костный мозг — основной
источник клеток, продуцирующих
специфические антитела и получивших название
В-лимфоцитов (от англ. bone marrow —
костный мозг), собственно В-лимфоциты и
набор различных классов антител.
Конечным результатом работы В-системы
является накопление специфических антител,
которые нейтрализуют бактерии или их
токсины. Если же организм столкнулся с
вирусной инфекцией, то в работу вступает
Т-система иммунитета. Составляющие
элементы Т-системы: тимус (вилочковая
железа) — лимфоэпителиальный орган,
расположенный за грудиной, различные
субпопуляции тимусзависимых клеток —
Т-лимфоцитов, антиген-распознающие
рецепторы, находящиеся на поверхности этих
клеток (Т-клеточные рецепторы — ТКР), а
также группа регуляторных молекул (ци-
токинов). Одна из клеточных субпопуляций
этой системы — Т-киллеры (цитотоксичес-
кие Т-лимфоциты) — является основным
«действующим лицом» антивирусного
иммунитета. Т-киллеры контактируют с
инфицированными вирусом клетками и
разрушают их. Попавшие в кровоток вирусные
частицы нейтрализуются антителами.
Изучая тонкие механизмы этих
процессов, исследователи поняли, что ни В-клет-
ки, ни Т-киллеры не в состоянии развить
максимально эффективную реакцию
самостоятельно. Для наиболее полного ее
проявления необходима помощь со стороны
других клеток. Эту вспомогательную
функцию выполняют как фагоцитирующие
клетки (макрофаги, дендритные клетки), так и
Т-хелперы (от англ. helper — помощник).
Именно через процесс взаимодействия
различных типов иммунокомпетентных клеток
формируется наиболее выраженный
иммунный ответ.
Параллельно и независимо от проблем
взаимодействия клеток, ответственных за
иммунный ответ, проводились
исследования по генетической организации главного
комплекса гистосовместимости (ГКГ). Этот
комплекс генов был открыт в связи с
пересадками (трансплантацией) тканей от одной
особи другой особи того же вида. Отсюда и
название комплекса. До конца 60-х-нача-
ла 70-х годов была известна лишь одна
функция этого комплекса — контроль
синтеза антигенов, вызывающих иммунную
реакцию отторжения пересаженной ткани.
Позднее при изучении генетического
контроля силы иммунного ответа и особенно
при анализе механизмов взаимодействия
генетически отличающихся клеток был
выявлен довольно широкий спектр
биологической активности комплекса. Под
контролем ГКГ проходят такие
иммунологические процессы, как регуляция силы
гуморального (В-клеточного) и клеточного
(Т-клеточного) иммунного ответа,
обеспечение иммуногенности проникшего в
организм антигена, селекция специфических
Т-клеток в тимусе.
И наконец, изучение участия генов ГКГ
во взаимодействии иммунокомпетентных
клеток привело к пониманию одного из
существенных явлений иммунитета —
распознавания антигена Т- и В-клетками.
Главный комплекс
гистосовместимости
ГКГ у мышей представляет собой
группу тесно сцепленных генов,
локализованных на 17-й хромосоме. У человека этот
комплекс располагается на б-й хромосоме.
Гены Н-2К, H-2D и H-2L ответственны за
синтез молекул I класса. Гены Ар, Aa, Е~ и
Еа обеспечивают образование
полипептидных цепей, которые формируют молекулы
II класса. Известны два вида таких
молекул: АрАа и ЕрЕа (рис. 1).
Молекулы I класса являются гликопро-
теидами с молекулярной массой 44 кДа,
состоящими из двух полипептидных цепей. С
основной, тяжелой ос-цепью этого белка не-
ковалентно связан Р2-микроглобулин (Р2-М),
имеющий близкую гомологию с
иммуноглобулинами. Молекулы II класса построены из
двух полипептидов ((3- и ос-цепей ).
Молекулы I и II класса, а также р2-мик-
роглобулин по характеру строения и
высокой гомологии по последовательности
аминокислот с иммуноглобулинами входят в состав
единого суперсемейства. Этот факт сам по
себе примечателен. Он указывает на то, что
226
ИММУНОЛОГИЯ
Гены
Н-2К
АР Аа Е$ Еа
H-2D H-2L
Белки-антигены
а
р2-м
Р а р а
р2-м
I класс
II класс
I класс
Рис. 1. Организация главного комплекса гистосовместимости (Н-2 комплекса) у мышей
Н-2К, H-2D, H-2L— гены, контролирующие молекулы I класса. А„ Аа, Ept Ea— гены, контролирующие молекулы
II класса, а, р— полипептидные цепи молекул I и II классов. Р2-М— р2-микроглобулин
роль ГКГ состоит не только в маркировке
индивидуальности, но, вероятно, и в
реакциях иммунологической природы, о чем
будет сказано ниже.
Взаимодействие генетически
идентичных (сингенных) клеток
Проблема межклеточных взаимодействий
возникла в биологии достаточно давно
(вероятно, около 100 лет тому назад, вскоре
после определения клетки как единицы
жизни). Исследователям, занимающимся
изучением вопросов клеточной биологии,
хорошо известно взаимное влияние
различных типов клеток в таких процессах, как
дифференцировка, эмбриональное развитие,
регуляция роста тканей и др. Однако в
иммунологии изучение вопросов клеточного
взаимодействия как одного из механизмов
осуществления иммунологических функций
до середины 60-х годов не проводилось.
Казалось, что клетки Т- и В-систем
работают автономно: клетки Т-систем — для
реализации клеточного типа ответа,
клетки В-систем — гуморального.
В 1963 г. появилась публикация Г. Клэ-
мэна с соавт., в которой было показано, что
для полноценной продукции антител
необходима кооперация по крайней мере двух
типов клеток — Т- и В-лимфоцитов.
Исследователи провели простые, но при этом
очень показательные опыты. Летально
облученным мышам, лишенным собственных
иммунокомпетентных клеток, вводили либо
только клетки тимуса (источника Т-лимфо-
цитов), либо клетки костного мозга
(источника В-клеток), либо смесь этих клеток. В
первых двух случаях регистрировали лишь
следы антител после введения антигена. В
третьем варианте опытов величина ответа
была значительно выше суммы ответов двух
предыдущих групп. Налицо был явный си-
нергический эффект. Именно синергизм при
кооперации двух типов клеток вызвал у
иммунологов особый интерес. Однако в
работе был существенный недостаток.
Опыты проводили на животных (in vivo),
поэтому не было возможности учесть всех
участников клеточного взаимодействия.
Позднее, в 1968 г., Д. Мозир и Л. Коп-
лсон провели эксперименты с культурой
клеток. Получив очищенные от
посторонних клеток популяции Т-, В-лимфоцитов
и макрофагов (МФ), они стали
комбинировать различные типы клеток in vitro. Из
всех возможных вариантов: Т-лимфоциты
плюс В-лимфоциты, Т-лимфоциты плюс
макрофаги, В-лимфоциты плюс
макрофаги и, наконец, Т-лимфоциты плюс
В-лимфоциты плюс макрофаги — продукция
антител наблюдалась только при сочетании
трех типов клеток. Стало очевидным, что
полноценное развитие гуморального
иммунного ответа требует участия не двух, а по
крайней мере трех типов клеток. Функция
каждого клеточного типа в антителопро-
дукции строго предопределена.
Макрофаги и другие фагоцитирующие клетки
поглощают, перерабатывают и экспрессируют
антиген в иммуногенной, доступной для Т-
и В-лимфоцитов форме. Т-клетки, а
точнее — одна из субпопуляций этих клеток,
так называемые Т-хелперы (помощники),
после распознавания антигена начинают
продукцировать цитокины (см.: Фрейд-
лип И. С. Клеточные механизмы иммунной
защиты организма. Наст, том), помогающие
В-клеткам. Получив специфический стимул
от антигена и неспецифический — от Т-кле-
ток, они приступают к выработке антител
(рис. 2).
При формировании клеточного типа
иммунного ответа также необходима
кооперация различных типов клеток.
Предшественник клеточного ответа, например
Т-киллер, после распознавания антигена
испытывает на себе действие Т-хелперов
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ВЗАИМОДЕЙСТВИЯ ИММУНОКОМПЕТЕНТНЫХ КЛЕТОК
227
ПК
Цитокин
Антиген
Антитела
Цитокин
МФ
ПК
Рис. 2. Взаимодействие иммунокомпетентных клеток
МФ — макрофаг (или другие фагоцитирующие клетки, способные перерабатывать антиген); Т —Т-клетки
(Т-хелперы); В— В-клетки; ПК— плазматические клетки
и вспомогательных клеток (фагоцитирующих
мононуклеаров). Только после
взаимодействия предшественника с этими клетками
появляется возможность его развития до
зрелой эффекторной клетки.
Сам процесс созревания эффекторов
осуществляется под влиянием
биологически активных молекул — цитокинов,
которые синтезируются клетками,
вступившими в кооперативные отношения.
Взаимодействие генетически
отличающихся клеток
С середины 60-х гг. начинает
формироваться направление, задача которого —
выяснение генетических механизмов,
регулирующих силу иммунного ответа. Были
обнаружены гены иммунного
реагирования —Ir-гены (Ir — сокр. от англ. immune
response), которые находятся в ГКГ.
Сразу после локализации в хромосоме 7г-генов
возник вопрос о типе клеток, в которых
проявляется их активность. Создание
комбинированных клеточных культуральных
систем, составленных из клеток животных
с разной способностью к иммунному ответу
на те или иные антигены (несингенные, или
аллогенные клетки), позволило ответить на
заданный вопрос. Эта же клеточная
система принесла пользу в решении проблемы
распознавания антигенов Т-лимфоцитами.
Дело в том, что до середины 80-х гг.
природа антиген-распознающих рецепторов
Т-клеток (ТКР) оставалась неизвестной. И
только совместные усилия клеточных и
молекулярных иммунологов привели к
пониманию характера распознавания
антигенов тимусзависимыми клетками. Первые
опыты, заставившие думать об особом
характере ТКР, отличном от антигенрас-
познающих рецепторов В-клеток
(поверхностных иммуноглобулинов — slg), были
проведены на культуре клеток.
Так, например, в опытах in vitro была
предпринята попытка оценить реакцию
Т-клеток (конкретно Т-хелперов) на
чужеродный антиген, который был
ассоциирован с макрофагами. Выяснилось, что при
генетической идентичности между
макрофагом и Т-хелперами последние
развивают сильную ответную реакцию. Ситуация
менялась, если Т-клетки генетически
отличались от макрофагов. Во всех случаях,
когда различия касались генов,
контролирующих молекулы II класса, Т-хелперы
оставались ареактивными. Подобная
инертность казалась странной. Действительно,
если чувствительные к определенному
антигену Т-хелперы распознают только тот
антиген, к которому они были
сенсибилизированы, им все равно с каким
макрофагом взаимодействовать. Однако факты
свидетельствовали об обратном. Оставалось
предположить, что Т-хелперы распознают
не только чужеродный антиген на
поверхности макрофагов, но и свои собственные
молекулы II класса, локализованные на тех
же макрофагах (рис. 3).
Если Т-хелперы распознают комплекс
чужеродного антигена с молекулами II класса
ГКГ, то генерация цитотоксических Т-лим-
фоцитов (Т-киллеров) связана с
ограничениями по молекулам I класса. Такой вывод
был получен после проведения серии
исследований по индукции Т-киллеров под
контролем ГКГ. Специфически
сенсибилизированные к определенному вирусу Т-киллеры
228
ИММУНОЛОГИЯ
ТКР
ТКР
Антиген I класса
Антиген II класса
Рис. 3. Распознавание
антигена, ассоциированного с
продуктами главного комплекса
гистосовместимости, Т-клеточ-
ными антиген-распознающими
рецепторами
ТКР— Т-клеточный рецептор;
АПК— антиген-представляю-
щая клетка
способны разрушать в условиях in vitro
генетически идентичные (сингенные) клетки-
мишени, зараженные тем же вирусом. Смена
одного вируса на другой отменяла реакцию
лизиса, что указывало на специфичность
анализируемых Т-киллеров. При этом
специфичность Т-киллеров была ограничена
молекулами I класса ГКГ (Н-2К или H-2D).
Реакция на специфический вирус
развивалась только при идентичности по
молекулам I класса ГКГ между примированными
Т-клетками и клетками-мишенями,
зараженными соответствующим вирусом (рис. 3).
Итак, Т-киллеры, так же как и Т-хел-
перы, распознают не собственно
чужеродный антиген, но его комплекс с продуктами
ГКГ: Т-помощники —комплекс с
молекулами II класса, Т-киллеры — с молекулами
I класса. Схематично генерация и
последующее функциональное проявление двух
типов клеток выглядят следующим образом.
Проникший в организм чужеродный
антиген (например, вирус) захватывается и
перерабатывается фагоцитирующей клеткой. В
результате внутриклеточной переработки
фрагменты антигена выходят на клеточную
поверхность, где и образуют комплекс с
молекулами I или II класса. Включение в
распознаваемый комплекс молекул ГКГ и
является тем генетическим ограничением,
которое не позволяет развиваться ответу в
тех случаях, когда в комплекс включены
молекулы иного генотипа.
Клеточная феноменология распознавания
комплексного антигена Т-хелперами и
Т-киллерами указывает лишь на двойной
характер такого распознавания
(распознавания «своего» и «чужого»), но ничего не
говорит о природе собственно антиген-рас-
познающих структур Т-клеток.
Только в середине 80-х гг. была,
наконец, понята молекулярная природа Т-кле-
точных рецепторов. Они представляют
собой гликопротеиды клеточной поверхности,
построенные из двух полипептидных
цепей — а и р. Каждая цепь включает два
домена — вариабельный (V) и константный
(С). Антиген-распознающий центр, так же
как и у антител, формируется при
взаимодействии V-доменов каждой из цепей.
Сходство с антителами касается и
организации генов, контролирующих синтез
рецепторов Т-клеток. Между двумя типами
рецепторов имеется гомология по
последовательности аминокислот в полипептидах.
Предполагается, что Т-клеточные
рецепторы были эволюционными
предшественниками антител. Черты сходства между
этими структурами позволили объединить их,
наряду с другими близкими белками, в
единое суперсемейство.
Тимус как место обучения
незрелых Т-клеток
В середине б 0-х гг. была установлена
ключевая роль тимуса в формировании пула
зрелых Т-клеток. Позднее стало очевидным, что
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ВЗАИМОДЕЙСТВИЯ ИММУНОКОМПЕТЕНТНЫХ КЛЕТОК
229
именно в тимусе происходят процессы диф-
ференцировки Т-клеток на субпопуляции
(Т-киллеров, Т-хелперов, Т-супрессоров). И
наконец, в середине 80-х гг. иммунологи
поняли, каким образом в тимусе Т-клетки
приобретают свое главное свойство — способность
к двойному распознаванию.
Путь созревания и «обучения»
предшественников Т-клеток в тимусе выглядит
следующим образом (рис. 4). Незрелые
клетки (пре-Т-лимфоциты), лишенные как
маркеров дифференцировки, так и Т-клеточ-
ных, антиген-распознающих рецепторов
(ТКР), мигрируют из костного мозга в суб-
капсулярную зону органа. Здесь
осуществляются первые шаги внутритимусной
дифференцировки лимфоцитов. В результате на
поверхности клеток начинается
одновременная экспрессия двух маркеров
дифференцировки (CD4 — маркера Т-хелперов и
CD8 — маркера Т-киллеров). Однако у ти-
моцитов субкапсулярной зоны еще
отсутствуют ТКР. Их умеренная экспрессия
начинается позднее, после миграции клеток
Рис. 4. Внутритимусная дифференцировка Т-клеток
CD4— маркер Т-хелперов; CD8— маркер Т-киллеров; ТКР— Т-клеточный (антигенраспознающий) рецептор
230
ИММУНОЛОГИЯ
в кору тимуса. С момента появления
антигенных рецепторов начинается основной этап
обучения. Из множества пришедших в кору
клеток успех в дальнейшем развитии
сопутствует только тем, чьи рецепторы
способны взаимодействовать с молекулами ГКГ,
обильно представленными на стромальных
клеточных элементах. Те клетки,
рецепторы которых в силу специфической конфор-
мации активного центра неспособны
взаимодействовать с антигенами ГКГ, погибают.
Селекция различных клонов тимоцитов —
основное событие, разворачивающееся в
одном из центральных органов иммунитета.
Этот отбор клеток по способности
распознавать свои собственные молекулы
является определяющим условием дальнейшего
внутритимусного развития Т-лимфоцитов. В
тех случаях, когда распознаются молекулы
I класса, лимфоциты «выбирают» путь
развития в сторону Т-киллеров (маркер CD8).
Если же распознавание связано с
молекулами II класса, провзаимодействовавшие
тимоциты трансформируются в Т-хелперы
(маркер CD4).
Предполагается, что узнавание
«своего» осуществляется не всем антиген-рас-
познающим центром ТКР, а лишь его
частью. Оставшаяся часть центра будет
взаимодействовать с возможными в
будущем чужеродными антигенами. Именно
тогда принцип двойного распознавания
«своего» и «чужого» найдет свое реальное
проявление.
В свете этих данных находит
разрешение иммунологическая загадка прошлого.
Долгое время оставалось непонятным —
почему в тимусе происходит гибель более
90% поступающих из костного мозга
ранних предшественников Т-клеток. Известный
австралийский иммунолог Ф. Вернет
предполагал, что в тимусе происходит гибель
тех лимфоцитов, которые способны к
аутоиммунной агрессии, иными словами, тимус
выступал в качестве заслона
нежелательным или даже «вредным» клонам. Сегодня
в данной проблеме имеется определенноя
ясность. Основная причина столь массовой
гибели связана с жесткостью селекционных
процессов — положительным отбором только
тех клеток, которые способны реагировать
со своими собственными молекулами ГКГ.
Все остальные, не прошедшие контроля на
специфичность, погибают.
Заключение
Изучение генетического контроля
взаимодействия иммунокомпетентных клеток
привело к пониманию наиболее тонких
механизмов работы иммунной системы и, в
первую очередь, — к выяснению природы
антиген-распознающих рецепторов
Т-клеток. Весь путь познания в данном случае
можно разбить на ряд этапов.
1. Определение тимуса как
центрального органа иммунитета, где закладываются
основы клеточного типа реагирования.
2. Формирование представлений о двух
системах специфической защиты — Т- и
В-системах иммунитета.
3. Установление взаимодействия
различных типов клеток при развитии иммунного
ответа.
4. Открытие в системе взаимодействия
генетически различающихся клеток
способности Т-клеточных рецепторов к двойному
распознаванию.
5. Выяснение молекулярной природы
Т-клеточного антиген-распознающего
рецептора.
6. Установление селекционирующей роли
тимуса — места формирования
специфических Т-клеток, способных к двойному
распознаванию — собственных антигенов гис-
тосовместимости и комплексированных с
ними чужеродных антигенов.
Литература
Галактионов В. Г. Графические модели в
иммунологии. М.: Медицина, 1986.
Иммунология. В 3 т. Пер. с англ. М.: Мир, 1987.
Janeway С. A., Travers P. Immunology. London
etc.: Curr. Biol. Ltd, 1994.
И. С. Фрейдлин
КЛЕТОЧНЫЕ МЕХАНИЗМЫ ИММУННОЙ
ЗАЩИТЫ ОРГАНИЗМА
Введение
На определенном этапе эволюции в
многоклеточном организме появились клетки,
призванные защищать его от
микробов-паразитов. Постепенно сформировалась особая
система органов и клеток, обеспечивающих
соответствующую защиту (иммунитет)
организма, которая получила название
иммунной. Клетки, входящие в состав иммунной
системы, были названы иммунокомпетент-
ными (см.: Галактионов В. Г. Генетический
контроль взаимодействия иммунокомпетен-
тных клеток. Наст. том).
Что такое цитокины?
Иммунокомпетентные клетки
происходят от одной родоначальной стволовой клетки
костного мозга, которая порождает разные
линии клеток — гранулоциты, моноциты,
Т-лимфоциты, В-лимфоциты и др. (рис.1).
Т-лимфоциты для окончательного
созревания перемещаются из костного мозга в
тимус, а оттуда расселяются в селезенку и
лимфоузлы, проходя через кровоток и лим-
фоток. В-лимфоциты, так же как
гранулоциты и моноциты, созревают в костном
мозге, откуда зрелые клетки выходят в
кровяное русло. В-лимфоциты также могут
покидать кровяное русло, оседая в
селезенке и лимфоузлах, и превращаться в
плазматические клетки. Моноциты,
выходя из кровеносных сосудов, попадают в
разные органы и ткани, где
превращаются в тканевые макрофаги. Гранулоциты
проходят свой жизненный цикл внутри
сосудов, циркулируя в крови, а покидают
сосудистое русло только в случае особой
необходимости, т. е. в случае
непосредственной угрозы вторжения и распространения
по организму микробов-паразитов. Такая
угроза возникает, например, при ранении,
когда нарушается целостность защитного
барьера кожи или слизистой оболочки. Рана
служит входными воротами для микробов-
паразитов. Проникновение микробов через
входные ворота, их размножение в тканях
организма служат сигналом к мобилизации
защитных клеток на борьбу с микробами.
Каким образом защитные клетки,
циркулирующие в крови или осевшие в
органах и тканях иммунной системы,
отдаленных от входных ворот инфекции, получают
и воспринимают сигнал об опасности
микробной агрессии? Как обеспечивается
строгая последовательность включения
отдельных типов клеток в борьбу с инфекцией?
Откуда узнает иммунная система, какие
именно защитные механизмы следует
использовать при данной инфекции?
Чтобы найти ответы на эти вопросы,
следует обратиться к семейству молекул,
получивших название цитокинов (от греч.
«цитос» — клетка и «кинетос» —
движущийся). Название отражает основное
назначение этих молекул, которые являются
переносчиками, передатчиками сигналов от
клетки к клетке. В геноме клетки имеются
специальные гены, ответственные за
синтез определенных цитокинов. До поры до
времени эти гены молчат, ничем не
проявляя своего присутствия. Однако как
только клетка получает сигнал о внедрении в
организм микробов-паразитов, гены
цитокинов переходят в активное состояние. С них
считывается информация о структуре
соответствующих белков, затем осуществля-
Единая стволовая
клетка
Миело-моноцитарный
предшественник
Миелобласт Монобласт
Лимфоцитарный
предшественник
Монобласт пре-В-
лимфоцит
ПромиелоцитПромоноцит j?]??™* D 3Ре^ый
к тимоцит В-лимфоцит
Гранулоцит Моноцит Т-хелпер
Макрофаг
Цитотоксический
Т-лимфоцит
Плазматическая
клетка
Рис. 1. Схема происхождения иммунокомпетентных
клеток из единой стволовой клетки костного мозга.
Гранулоциты и моноциты имеют раннего общего
предшественника, который может дать начало мие-
лопоэзу (продукции гранулоцитов) или моноцитопо-
эзу (продукции моноцитов, макрофагов). Из общего
раннего предшественника происходят Т-лимфоциты
и В-лимфоциты
232
ИММУНОЛОГИЯ
ется синтез этих белков, и готовые
молекулы цитокинов начинают выделяться (сек-
ретироваться) клеткой в окружающую среду.
Для восприятия и распознавания
различных сигналов клетки несут на своей
поверхности специальные сложно устроенные
молекулы-рецепторы. Для каждого цитокина
существует свой особый рецептор, к
которому молекула цитокина подходит как ключ
к замку. Как только ключ-цитокин входит
в скважину предназначенного для него
замка-рецептора, с поверхности клетки к ее
ядру передается соответствующий сигнал,
включающий определенные гены:
информация передана, воспринята и реализуется
(рис. 2).
Цитокины, являясь единицами
своеобразного межклеточного языка, позволяют
клеткам общаться, взаимодействовать, объединяя
свои усилия в борьбе с
микробами-паразитами. На внедрение и размножение микробов
организм отвечает мобилизацией защитных
клеток и продукцией защитных молекул —
иммунным ответом. Чтобы иммунный ответ
состоялся, оказался достаточно
эффективным, выполнил свои защитные функции и
был своевременно выключен за
ненадобностью, необходимы четкие межклеточные
взаимодействия, которые обеспечиваются
цитокинами. Рассмотрим, как
складываются такие межклеточные взаимодействия.
Роль макрофагов
Макрофагов называют
клетками-мусорщиками из-за их способности захватывать,
поглощать, убивать и переваривать все, с
чем они соприкасаются. Одни макрофаги
ведут оседлый образ жизни, находясь в
селезенке, в одном из лимфоузлов, в
печени, в легких или даже в костях. Другие —
блуждают, ползая по серозным оболочкам
и передвигаясь внутри рыхлой
соединительной ткани. Все они происходят от
моноцитов крови, а те, в свою очередь, от промо-
ноцитов костного мозга, возникающих в
результате постепенного созревания более
ранних клеток-предшественников. У
макрофагов есть с ними обратная связь: зрелый
макрофаг может продуцировать
специальные цитокины — ростовые факторы,
которые с кровью поступают в костный мозг и
усиливают процессы деления и созревания
клеток-предшественников. При инфекции,
когда многие макрофаги погибают в
борьбе с микробами, им на смену приходят
новые молодые макрофаги, ускоренно
созревающие в костном мозге (рис. 3).
На поверхности макрофагов находятся
разные молекулы, часть из них может
служить рецепторами других молекул.
Внедрившиеся в организм микробы-паразиты
вырабатывают и выделяют свои молекулы —
токсины. Соответствующие рецепторы на
поверхности макрофагов распознают такие
молекулы и макрофаги направляются к очагу
инфекции, к месту максимальной
концентрации токсинов. В очаге инфекции
начинается главная работа «мусорщиков»: захват
(фагоцитоз) микробов, попытка убить их и
переварить во внутриклеточных вакуолях,
т. е. макрофаги стремятся очиститься сами
и способствовать очищению всего
организма от микробов. Действие микробных
токсинов на соответствующие рецепторы
служит сигналом к активации многих генов в
геноме клеток макрофагов. При этом
активируется продукция цитокинов. Работая в
очаге инфекции, макрофаги синтезируют и
выделяют молекулы интерлейкина-1 (ИЛ-1),
Бактерия
ИЛ-1
ФНО
ИЛ-1
Рис. 2, Продукция и рецепция цитокинов
На поверхности бактерии-паразита имеется сигнальная молекула (ЛПС), которая распознается специальным
рецептором на поверхности макрофага. От рецептора идет сигнал активации гена, ответственного за синтез
интерлейкина-1 (ИЛ-1), начинается синтез этой молекулы и ее секреция. Другой макрофаг несет рецептор для
ИЛ-1, через который идет сигнал к его гену, ответственному за синтез фактора некроза опухолей (ФНО), и
запускаются синтез и секреция этого цитокина
КЛЕТОЧНЫЕ МЕХАНИЗМЫ ИММУННОЙ ЗАЩИТЫ ОРГАНИЗМА
233
которые с током крови попадают в мозг и
действуют на центр терморегуляции. В
результате у больного повышается
температура тела, т. е. вступает в действие один
из механизмов защиты, поскольку
большинство микробов-паразитов медленнее
размножаются при повышенной температуре, а
защитные клетки при такой температуре
более активны. Те же молекулы ИЛ-1
действуют через свои рецепторы на
лимфоциты, передавая им сигнал активации. Это
важно в тех случаях, когда одним
макрофагам не справиться с инфекцией, и
возникает необходимость развернутого
включения иммунного ответа. ИЛ-1 способен
запускать каскад продукции других цито-
кинов, получивших названия интерлейки-
нов-2, 3, 4, 5, 6, 7 и т.д. (ИЛ-2, 3, 4, 5,
б, 7 и т. д.), которые находят
соответствующие рецепторы на Т-лимфоцитах, В-лим-
фоцитах и других клетках, передавая им
сигналы, активирующие отдельные
функции (рис. 3).
Среди продуктов, нарабатываемых
макрофагами в очаге инфекции, есть особые
молекулы, получившие название «фактор
некроза опухолей» (ФНО). Название
связано с цитотоксичностью этих молекул, т. е.
с их способностью убивать клетки-мишени,
в том числе опухолевые клетки.
Рецепторы для этого фактора обнаружены на
поверхности всех ядерных клеток организма.
ФНО имеет непосредственное отношение к
мобилизации клеток в очаг инфекции.
Попадая в кровяное русло, эти молекулы
действуют через свои рецепторы на
эндотелиальные клетки внутренней оболочки
сосудов, активируя гены, ответственные за
синтез специальных «липких» молекул,
обеспечивающих прилипание к сосудистой
стенке циркулирующих в крови гранулоцитов,
моноцитов и лимфоцитов. Прилипание к
эндотелию для них — первый шаг к выходу
из сосуда и мобилизации в очаг инфекции.
Таким образом, ФНО помогает макрофагам,
способствуя увеличению их количества в
очаге инфекции. У самих макрофагов
также имеются рецепторы для ФНО, через
которые он может в свою очередь
активировать макрофаги, стимулируя включение
генов, нужных для организации ответа на
заражение (рис. 3).
При таком разнообразии продуцируемых
макрофагами молекул, по-видимому, было
бы уместнее назвать их
«клетками-диспетчерами» или «сигнальными клетками», чем
клетками-мусорщиками, так как продукты
активности макрофагов запускают и
контролируют иммунный ответ и работу других
участвующих в нем клеток.
Роль Т-лимфоцитов
Хотя иммунный ответ запускает
макрофаг, только лимфоциты имеют
специальные рецепторы для распознавания
чужеродных молекул, получивших название
антигенов, и только лимфоциты
обеспечивают иммунный ответ. От поверхности
Т-лимфоцитов к ядру идут одновременно два
Стволовые
клетки
ФНО
КСФ
Эндотелиальная
клетка
Рецепторы
Мозг- ИЛ-1
центр терморегуляции
ФНО
ФНО
Опухолевая
клетка
ИЛ-1
ИЛ-6
ИЛ-2,3,4,5,6,7
Рис. 3. Макрофаг— продуцент цитокинов
Макрофаг продуцирует и секретирует колониестимулирующий фактор (КСФ), интерлейкины (ИЛ-1 и ИЛ-6),
фактор некроза опухолей (ФНО). ИЛ-1 и ИЛ-6 вызывают лихорадку и активируют Т-лимфоциты, которые
индуцируют синтез и секрецию следующей серии интерлейкинов (ИЛ-2, 3, 4, 5 и т. д.); последние активируют Т- и
В-лимфоциты. ФНО разрушает опухолевые клетки, оказывает влияние на эндотелиальные клетки и на сами
макрофаги
234
ИММУНОЛОГИЯ
ИЛ-4
ИЛ-2 ИЛ-5
ИЛ-1 Антиген
ил-6 Антиген
ИЛ-
тор
иген-
шающий
ептор
1 2 сЗ 3 Антитела
Рис. 4. Взаимодействие Т-лимфоцитов и В-лимфоцитов
Получив два сигнала активации: от макрофага (интерлейкин-1, ИЛ-1) и в результате распознания антигена,
Т-лимфоциты начинают синтезировать и секретировать свой цитокин — интерлейкин-2 (ИЛ-2), который
распознается их же рецепторами и служит сигналом, вызывающим продукцию следующей серии интерлеикинов (ИЛ-4,
ИЛ-5, ИЛ-6). Под влиянием этих интерлеикинов и второго сигнала активации, обусловленного распознанием
антигена, В-лимфоциты превращаются в антитела-продуценты. 1 — геном клетки, 2—активированные гены, 3 —
рибосомы
сигнала активации: от антиген-распознаю-
щего рецептора и от рецептора, связавшего
ИЛ-1. Под действием этого двойного
сигнала в геноме Т-лимфоцитов
активируются гены как самого ИЛ-2, так и гены
рецепторов, специфичных для ИЛ-2. После
этого продукт Т-лимфоцитов ИЛ-2 начинает
воздействовать на клетки, в которых он и
был синтезирован. В них активируется
процесс деления, и за счет этого усиливаются
функции всей популяции Т-лимфоцитов,
участвующих в специфическом иммунном
ответе на данный антиген (рис. 4).
Характер иммунного ответа зависит от
присутствия определенных цитокинов в
микроокружении Т-лимфоцитов в момент
распознавания антигена и активации. Если в
данный момент в окружающей среде преобладает
ИЛ-4, клетки Т-лимфоцитов превращаются
в активированных Т-хелперов (помощников)
и начинают синтезировать тот же ИЛ-4, а
также ИЛ-5, б, 7, 10. Эти интерлейкины
активируют через соответствующие
рецепторы деление В-лимфоцитов, их созревание
в плазматические клетки и синтез
специфических для того или иного антигена антител —
иммуноглобулинов. Вот почему в данном
случае Т-лимфоциты выступают в роли
Т-хелперов, т. е. помощников В-лимфоцитов в их
основном деле — наработке запаса защитных
молекул-антител (рис. 4).
Нередко в момент контакта с антигеном
в окружении Т-лимфоцитов преобладает
другой цитокин — у-интерферон. Молекулы
интерферона (от англ. interfere — мешать,
препятствовать) принято дополнительно
обозначать буквами греческого алфавита (а, Р, у)
в зависимости от того, какие клетки их
продуцируют (лейкоциты, фибробласты,
лимфоциты). Если превалирует у-интерферон, то
активация идет по такому пути:
Т-лимфоциты начинают продуцировать еще большие
количества у-интерферона, а также
вырабатывать молекулы ФНО и других цитокинов,
участвующих в клеточном иммунном ответе
(иммунном воспалении). В последнем случае
Т-лимфоциты выступают в качестве
помощников макрофагов, так как их продукт
(у-интерферон) призван активировать
функции макрофагов в борьбе с
микробами-паразитами, у-интерферон не сам борется с
микробами, а повышает антимикробную
активность макрофагов. В клеточном
иммунном ответе основную роль играют
активированные макрофаги и Т-лимфоциты. Среди
Т-лимфоцитов существует разновидность ци-
тотоксических Т-клеток, которых называют
еще Т-киллерами (убийцами) за способность
убивать другие клетки, в том числе
клетки, зараженные вирусами и микробами.
Но и этим не исчерпываются возможные
функции Т-лимфоцита. Т-лимфоциты держат
весь иммунный ответ под контролем, не
допуская чрезмерной активации отдельных
иммунокомпетентных клеток, что чревато
осложнениями. Инструментами такого
контроля служат цитокины, способные не только
активировать (усиливать), но и подавлять
(ингибировать) функции других клеток. Так,
ИЛ-4, активатор В-лимфоцитов,
ответственных за синтез антител, является
ингибитором клеточного иммунного ответа,
подавляет синтез у-интерферона. у-интерферон, со
своей стороны, являясь активатором
макрофагов, стимулирующим продукцию всех
воспалительных цитокинов (ИЛ-1, б и ФНО),
в то же время способен ингибировать
продукцию ИЛ-4 и зависящие от этого цито-
кина процессы биосинтеза антител.
Некоторые продукты Т-лимфоцитов, например
ИЛ-10, выполняют сугубо ингибиторные
функции. Так, ИЛ-10 способен затормозить
КЛЕТОЧНЫЕ МЕХАНИЗМЫ ИММУННОЙ ЗАЩИТЫ ОРГАНИЗМА
235
синтез всех воспалительных цитокинов, в том
числе ИЛ-1, ФНО, у-интерферона и даже
свой собственный синтез. В связи с этим в
противоположность воспалительным цитоки-
нам он получил название
противовоспалительного.
Между Т-лимфоцитами и макрофагами
существует двусторонняя связь, в
динамике развития иммунного ответа протекают ци-
токиновые «диалоги». Первые получают от
макрофагов сигнал активации в виде
молекулы ИЛ-1, для восприятия которого они
имеют на поверхности соответствующие
рецепторы. От них идет сигнал активации
генов Т-лимфоцитов, отвечающих за синтез
ИЛ-2 и у-интерферона. Рецепторы
Т-лимфоцитов распознают ИЛ-2, а после того, как
последний «садится» на рецептор, от него
поступает сигнал, вызывающий дальнейшую
активацию синтезов в клетках
Т-лимфоцитов и начало деления клетки. Что касается
у-интерферона, то эти молекулы
направляются в виде ответного «послания» макрофагу,
на поверхности которого эту молекулу ждут
соответствующие рецепторы, у-интерферон
не зря называют макрофаг-активирующим
фактором. Связавшись со своим рецептором
на внешней поверхности клетки макрофага,
он посылает к ядру этой клетки сигналы
активации для нескольких десятков генов,
в том числе для гена, ответственного за
синтез ИЛ-1. В результате Т-лимфоциты
получают от активированного макрофага
новую порцию активирующих их молекул
ИЛ-1 (рис. 5).
Как организм защищается
от бактерий
Место проникновения бактерий в организм
называют «входными воротами» инфекции.
Уже здесь на борьбу с бактериями
поднимаются фагоцитирующие клетки — гранулоциты
и макрофаги. Первый сигнал к мобилизации
эти клетки получают от самих
бактерий-агрессоров в виде их токсинов. Одновременно
с фагоцитозом бактерий макрофаги
начинают синтезировать и выделять
воспалительные цитокины— ИЛ-1, ФНО и др. Под
влиянием цитокинов усиливается прилипание
циркулирующих лейкоцитов к эндотелию
сосудов, их выход из сосудов и мобилизация
в очаг инфекции. Те же цитокины
усиливают антибактериальную активность фагоцитов.
Если фагоцитирующие клетки не
справляются с очищением очага инфекции от
бактерий, ИЛ-1 выполняет роль межклеточного
сигнала, вовлекающего в процесс активации
Т-лимфоциты и включающего механизмы
специфического иммунного ответа.
Активированные Т-лимфоциты
пополняют ресурсы воспалительных цитокинов,
синтезируя у-интерферон, активирующий
макрофаги. Существенную помощь
фагоцитирующим клеткам в борьбе с бактериями
оказывают продукты В-лимфоцитов —
специфические антитела-иммуноглобулины.
Взаимодействуя с антигенами бактерий,
антитела как бы подготавливают бактерии
в пищу фагоцитам, делают их более
«удобоваримыми». Кроме того, специфические
антитела против бактериальных токсинов
нейтрализуют последние самостоятельно:
токсин, связавшийся со своими
специфическими антителами, утрачивает токсичность
и больше не представляет опасности для
организма. Особая разновидность
образующихся при специфическом иммунном
ответе антител, способных выходить на
поверхность слизистых оболочек, обеспечивает их
местную защиту, не допускает внедрения
бактерий через эти входные ворота, мешает
бактериям прикрепиться к эпителиальным
клеткам.
Как организм защищается
от вирусов
Специфические антитела на
поверхности слизистых оболочек защищают организм
и от внедрения вирусов. Встречаясь с
вирусом в крови или межклеточных
пространствах, специфические антитела способны
обезвредить его. Однако особенность виру-
ИЛ-2
ИЛ-1
ИЛ-2р
ИЛ-2
Гамма-интерферон
Рис. 5. Взаимодействие Т-лимфоцита и макрофага (цитокиновый диалог)
Для лимфоцита сигналом активации служит молекула интерлейкина-1 (ИЛ-1), которую синтезирует и секрети-
рует макрофаг. Активированный Т-лимфоцит начинает продуцировать интерлейкин-2 (ИЛ-2) для собственной
дальнейшей активации и у-интерферон — активатор макрофагов. Условные обозначения те же, что и на рис. 4
236
ИММУНОЛОГИЯ
са как паразита состоит в том, что он
предпочитает внутриклеточный паразитизм, т. е.
жизнь и размножение исключительно
внутри клеток хозяина и за счет клеток
хозяина. В таких условиях бороться против
вируса-паразита можно двумя путями: или
атаковать и убивать зараженные
вирусами клетки вместе с вирусами, или каким-
то образом воспрепятствовать
внутриклеточному размножению вирусов, если не
удалось помешать их внедрению. Первый
способ используют цитотоксические
клетки-киллеры разных типов, защищающие
организм от вирусов. Распознав на
поверхности зараженной клетки чужеродные
антигены, клетки-киллеры впрыскивают в
такую клетку-мишень содержимое своих
цитоплазматических гранул с ФНО, проте-
олитическими и липолитическими
ферментами и другими молекулами,
повреждающими клетку-мишень. Результатом атаки
киллера, как правило, является гибель
клетки-мишени вместе с
внутриклеточными паразитами. Правда, гибель и
разрушение собственных клеток организма
небезразличны для его жизнедеятельности. При
некоторых вирусных инфекциях такого рода
защитные реакции приносят больше
вреда, чем пользы.
Другой механизм защиты от вирусов —
молекулярный. Ответственные за
противовирусную защиту интерфероны
синтезируются клетками-продуцентами в ответ на
заражение вирусом. Клетка-продуцент секретирует
молекулы интерферона, которые соединяются
с соответствующими рецепторами на
поверхности клеток, зараженных вирусом. Как в
любом другом случае, взаимодействие цито-
кина (в данном случае интерферона) со
своим специфическим рецептором влечет за
собой передачу внутриклеточного сигнала к ядру
клетки, что приводит к активации генов,
ответственных за синтез белков,
препятствующих самовоспроизведению вируса в этой
клетке. Таким образом, интерферон
блокирует биосинтез вирусных частиц в
зараженной клетке. Это позволяет использовать
препараты интерферона в качестве лечебных при
вирусных инфекциях.
Клеточные и молекулярные механизмы
защиты от вирусов и бактерий работают
согласованно, приходя на помощь друг другу.
Молекулы интерферонов, кроме
противовирусного действия, оказывают влияние на
функции защитных клеток, в частности,
у-интерферон, как отмечалось, является
активатором макрофагов.
Активированные у~интеРФеРоном
макрофаги могут пополнить армию
клеток-киллеров, но только при участии
специфических противовирусных антител, которые
образуют своеобразные мостики между
макрофагами и зараженными клетками-
мишенями. Специфический ответ на
вирусные антигены неизбежно вовлекает
популяцию Т-клеток-помощников, начинающих
в ответ на активацию усиленно
синтезировать и секретировать ИЛ-2. А этот цитокин
известен своей способностью резко
активизировать клетки-киллеры.
Заключение
Клетки, связанные друг с другом
посредством цитокинов, образуют своеобразную
сеть, которая служит для многоканальной
передачи сигналов от клетки к клетке,
обеспечивает восприятие этих сигналов и
соответствующий ответ. Информация от клетки
к клетке передается в виде молекулы ци-
токина. Восприятие такой информации
гарантируется наличием на клеточной
поверхности соответствующего рецептора, от
которого сигнал передается к ядру клетки
и реализуется за счет включения
определенных генов.
Литература
Кетлинский С. А., Симбирцев А. С,
Воробьев А. А. Эндогенные иммуномодуляторы. СПб.:
Гиппократ, 1992.
Маянский А. Я., Маянский Д. Н. Очерки о
нейтрофиле и макрофаге. Новосибирск: Наука,
1989.
Ройт А. Основы иммунологии. Пер. с англ. М.:
Мир, 1991.
Фрейдлин И.С. Система мононуклеарных
фагоцитов. М.: Медицина, 1984.
И. С. Фрейдлин
ЗАЩИТА ОТ ВИРУСОВ С ПОМОЩЬЮ КЛЕТОК-УБИЙЦ
Эта вирусная инфекция известна всем
с детства: при простуде на губах или
крыльях носа появляются болезненные
пузырьки, потом пузырьки лопаются и
превращаются в болячку, покрытую корочкой; если
ее не трогать, она сама постепенно
отваливается. Но в следующий раз при
очередной простуде пузырьки снова высыпают на
том же месте. Это значит, что вирус
(вирус простого герпеса) остается в
организме в скрытом виде. Способность вируса
длительно сохраняться в организме называется
персистенцией. Такая вирусная инфекция,
при которой возбудитель пребывает в
организме длительное время в скрытой форме
и лишь иногда проявляет свое присутствие,
называется латентной. Попробуем
проанализировать причины такого поведения
вируса и возможную роль иммунной системы
в защите от него.
Защита от вирусов для иммунной
системы — более сложная задача по сравнению
с защитой от бактерий. Вирусы являются
облигатными (обязательными)
внутриклеточными паразитами. Проникнув внутрь
клетки организма-хозяина, вирус безнаказанно
распоряжается там: в клетке начинается
репликация вирусной нуклеиновой кислоты, идет
синтез вирусных белков, происходит
сборка новых вирионов. Все эти события
происходят внутри клетки, за счет клетки и под
прикрытием клеточной оболочки, через ко-
6
4
Рис. 1. Действие клетки-убийцы — цитотоксическо-
го лимфоцита на зараженную вирусом клетку-мишень
1— вирусный антиген; 2— антиген главного
комплекса гистосовместимости 1-го класса (ГКГ1); 3 —
антигенраспознающий рецептор цитотоксического
Т-лимфоцита (ЦТЛ); 4 — адгезионная молекула ЦТЛ;
5— поляризация гранул ЦТЛ, содержащих цитоток-
сины и цитокины; б— секреция цитотоксинов и ци-
токинов ЦТЛ; 7— мембраноассоциированная
форма ФНО; 8— рецептор для ФНО на клетке-мишени
(М)
торую в клетку не могут проникнуть какие-
либо защитные молекулы (например,
антитела). А результатом становятся
повреждение клетки, выход нового поколения вирусов,
которые тут же проникают в соседние
клетки, и все события вирусной инфекции
воспроизводятся вновь.
Иммунной системе ничего не остается,
как только убить клетку, зараженную
вирусом. Для этого в организме существуют
специализированные клетки-киллеры
(убийцы) (см.: Абелев Г. И. Основы приобретенного
иммунитета. Наст. том). Главная их задача —
распознать клетки, зараженные вирусом,
чтобы случайно не убить здоровую
нормальную клетку. Одна разновидность убивающих
клеток получила название «естественные
киллеры», которое подчеркивает присущую
им от природы (естественную) готовность
убивать клетки, зараженные любыми
вирусами. После первой встречи организма с
каким-либо вирусом в организме
накапливаются киллеры со специфическим
сродством к клеткам, зараженным именно
данным конкретным вирусом. Такие киллеры
из числа Т-лимфоцитов получили название
цитотоксических Т-лимфоцитов (ЦТЛ).
Кроме того, клетки-убийцы используют для
распознавания клеток-мишеней,
зараженных вирусами, образовавшиеся в
организме специфические антитела против
вирусных антигенов. В таком случае говорят об
антителозависимой клеточной цитотоксич-
ности.
Когда попавший в организм вирус
размножается в клетках эпителия кожи или
слизистых оболочек, он вызывает
интенсивный и эффективный иммунный ответ. В
эпителиальных клетках антигены вируса
(образовавшиеся при распаде его белков короткие
пептиды) объединяются с собственными
антигенами организма, так называемыми
антигенами главного комплекса
гистосовместимости первого класса— ГКГ1. Такие
комплексы размещаются (презентируются)
на поверхности зараженных вирусами
эпителиальных клеток для того, чтобы их
опознали специальные рецепторы
специализированных ЦТЛ (рис. 1). Прежде всего
клетка-убийца, в роли которой выступает
ЦТЛ, с помощью специальных липких
(адгезионных) молекул очень прочно
прикрепляется к клетке-мишени. Содержащиеся в
цитоплазме ЦТЛ гранулы поляризуются, т. е.
все концентрируются непосредственно в
точке контакта ЦТЛ с клеткой-мишенью.
Именно в этой точке ЦТЛ начинает секре-
238
ИММУНОЛОГИЯ
цию повреждающих молекул (см. рис. 1).
Первыми выбрасываются так называемые
перфорины, которые полимеризуются и
формируют поры (перфорации) в мембране
клетки-мишени. Через эти поры в клетку
устремляется вода, что может привести к
ее осмотическому шоку (рис. 2, а). Кроме того,
формирование пор облегчает последующее
проникновение в клетку-мишень других
продуктов ЦТ Л: грензимов и цитокинов. Грен-
зимы — это протеолитические ферменты,
которые участвуют в разрушении
компонентов клетки-мишени. Интересно, что сами ЦТЛ
относительно устойчивы к действию секре-
тируемых ими разрушающих молекул типа
перфоринов и грензимов. Благодаря
поляризации выброса таких молекул в точке
контакта, обусловленного распознаванием
специфического вирусного антигена, ЦТЛ
избирательно убивают только зараженные
данным вирусом клетки, не затрагивая
соседние незараженные клетки.
Особую роль в реализации цитотокси-
ческих эффектов ЦТЛ играют
продуцируемые и секретируемые ими цитокины:
у-интерферон (у-ИФН) и фактор некроза
опухолей (ФНО). Цитокины действуют на
специфические рецепторы, находящиеся на
мембранах клеток-мишеней. у-ИФН, как и
другие интерфероны, может индуцировать
в клетке-мишени синтез белков,
препятствующих репликации и транскрипции
нуклеиновых кислот вируса. Название
«интерферон» и происходит от способности этой
молекулы интерферировать с вирусной
инфекцией, т. е. мешать репродукции вирусов.
Но у-ИФН обладает еще дополнительной
способностью усиливать экспрессию молекул
ГКГ1 на мембране клетки-мишени. В
результате у зараженной вирусом клетки-мишени
резко возрастает шанс быть распознанной
и убитой соответствующим по
специфичности киллером-ЦТЛ.
Цитокины, как правило, секретируют-
ся клетками-киллерами, но иногда эти
молекулы остаются связанными с мембраной
синтезировавшей их клетки. При этом они
сохраняют способность соединяться со
специфическими для них рецепторами на
мембранах клеток-мишеней (см. рис. 1).
Связывание ФНО со своим
специфическим рецептором на клетке-мишени служит
для нее сигналом включения различных, до
того «молчавших» генов. Среди этих генов
индуцируется и ген апоптоза (рис. 2, б).
Апоптоз — это генетически
запрограммированная активная гибель клетки,
начинающаяся с фрагментации ядра (см.: Агол В. И.
Генетически запрограммированная смерть
клеток. Наст. том). В данном случае
зараженная вирусом клетка-мишень отвечает
апоптозом (самоубийством) на сигнал,
поступивший от рецептора ФНО.
Избирательность действия обеспечивается за счет
первоначального связывания ЦТЛ с
зараженной клеткой-мишенью через специфический
антигенраспознающий рецептор. Только
после этого мембраноассоциированная форма
молекулы ФНО на поверхности ЦТЛ
«нащупывает» свой рецептор на поверхности
клетки-мишени и связывается с ним.
Все описанные события ведут к гибели
зараженных вирусом простого герпеса
эпителиальных клеток, вместе с которыми
гибнут и вирусы. Обломки погибших клеток
захватываются и перевариваются стекающимися
к месту инфекции лейкоцитами.
Воспалительная реакция заканчивается очищением «поля
боя» от трупов погибших клеток и
восстановлением (регенерацией) исходной структуры
ткани: болячка отваливается. Однако
небольшая часть выживших вирусов укрывается
внутри нервных клеток — чувствительных
нейронов, в которые они проникают по
отросткам (аксонам) и распространяются вплоть
до тройничного нервного ганглия, где
«отсиживаются», ничем себя не проявляя, до
следующего благоприятного случая. Нейроны
существенно отличаются от эпителиальных
клеток по многим свойствам. В нейронах
вирусы простого герпеса не находят
подходящих условий для размножения. Вирусная
инфекция в этих клетках протекает как ла-
а б
* 4
5
I \
3 7
Рис. 2. Гибель клетки-мишени, зараженной вирусом,
путем лизиса (некроза) (а) и апоптоза (б)
1— секреция повреждающих молекул: перфорина,
грензимов; 2— образование пор в мембране
клетки-мишени; 3— гибель клетки путем лизиса
(некроза); 4— мембраноассоциированная форма ФНО; 5—
рецептор ФНО на мембране клетки-мишени; б—ядро
клетки-мишени (М); 7— гибель клетки путем
апоптоза
ЗАЩИТА ОТ ВИРУСОВ С ПОМОЩЬЮ КЛЕТОК-УБИЙЦ
239
тентная. Вирусная нуклеиновая кислота
реплицируется очень медленно, вирусных
белков синтезируется мало и поэтому
вирусных пептидов не хватает для презентации
на мембране клеток. Кроме того, нейроны
несут на своих мембранах очень мало
молекул ГКГ1, необходимых для презентации
вирусных антигенов клеткам ЦТЛ. При
таком низком уровне экспрессии антигенов
ГКГ1 резко снижен риск того, что клетка
будет атакована ЦТЛ. Это создает
определенные преимущества для нейронов,
которые, как известно, не могут регенерировать.
Вместе с тем нейроны представляют собой
безопасное убежище для неактивных (пер-
систирующих) вирусов. Персистирующий
вирус простого герпеса может быть
реактивирован под влиянием каких-либо
неблагоприятных воздействий на организм:
переохлаждения, переутомления, голодания,
заражения другой инфекцией. В этом
случае вирус проделывает обратный путь по
аксону чувствительного нейрона и вновь
проникает в эпителиальные клетки, на
страже которых стоят, как мы уже знаем,
клетки-киллеры, убивающие избирательно
зараженные вирусами эпителиальные клетки
ради того, чтобы защитить остальные клетки
нашего организма от вирусов.
Литература
Кетлинский С. А., Симбирцев А. С,
Воробьев А. А. Эндогенные иммуномодуляторы. СПб.:
Гиппократ, 1992.
Ройт А. Основы иммунологии. Пер. с англ. М.:
Мир, 1991.
Фрейдлин И. С. Сорос, образоват. журн., 1996,
№ 7, с. 19.
Abbas А.у Lichtman A.f Pober J. Cellular and
Molecular Immunology. N.Y.: Saunders Co., 1991.
Janeway Ch. A., Travers P. Immunobiology. L.:
Curr. Biol. Ltd., 1994.
ФИЗИОЛОГИЯ ЧЕЛОВЕКА
И ЖИВОТНЫХ
М. Г. Пшенникова
СТРЕСС И АДАПТАЦИЯ
Стресс есть жизнь и жизнь есть стресс.
Г. Селье
Введение
В настоящее время трудно найти
человека, который не слышал бы о стрессе.
Популярно мнение, что стресс — это
явление, присущее главным образом
сегодняшней непростой жизни и, кроме того,
явление вредное, с которым надо
бороться. И то, и другое отчасти верно, но лишь
отчасти и в очень небольшой степени.
Термин «стресс» ввел канадский
физиолог Г. Селье, который в 1936 г. впервые
охарактеризовал это явление как «общую
неспецифическую нейрогормональную
реакцию организма на любое предъявленное ему
требование». Селье заимствовал этот
термин из техники, где он означает
«напряжение, нажим, давление».
В общем виде стресс можно определить
как состояние организма, всегда
возникающее при действии на него различных
факторов, или стрессоров, будь то так
называемые физические стрессоры (резкие
изменения температуры, влажности,
атмосферного давления, физическая нагрузка и
т. п.) или психические (эмоциональные)
стрессоры, которые обычно называют
стрессорными ситуациями (опасность,
горе, радость, конфликт, цейтнот и др.). В
настоящее время люди часто попадают в
стрессорные ситуации, чем и объясняется
особый интерес к стрессу и желание
исследователей разобраться в его природе и
механизмах.
В ответ на стресс человек или
животное стремятся либо избавиться от действия
стрессора, либо, если избавление
невозможно, приспособиться, адаптироваться к
этим новым требованиям среды. При
первичном действии стрессора возникает так
называемая срочная адаптация, которая
весьма несовершенна и не обеспечивает
организму эффективное устойчивое
приспособление. Устойчивая долговременная
адаптация, характеризующаяся высокой
резистентностью к стрессору, формируется
лишь в результате его повторных
воздействий.
Феномены стресса и адаптации,
интуитивно предсказанные и в той или иной
форме описанные многими поколениями
естествоиспытателей и врачей, включая
Гиппократа, Р. Декарта, К. Бернара, У. Кен-
нона, Г. Селье, И. М. Сеченова и А. А.
Ухтомского, в настоящее время, благодаря
успехам биологии и медицины, получают
новое содержание.
Феномен стресса
Стресс как биологическая категория —
сложное явление, присущее всему
живому, от простейших до Homo sapiens. Он
неотделим от самого явления жизни;
организмы находятся в сложном динамическом
равновесии с окружающей средой и жизнь
без взаимодействия с ней невозможна.
Непременным условием нормальной
жизнедеятельности организма и сохранения самой
жизни является поддержание постоянства
внутренней среды, или гомеостаз. Этот
термин был предложен Кенноном в 1929 г., но
сама идея была впервые высказана в XIX в.
Бернаром, который утверждал, что
«постоянство внутренней среды —
непременное условие свободной и независимой
жизни». Гомеостаз означает поддержание
основных физических и химических
параметров организма (температуры,
артериального давления, концентрации глюкозы,
количества форменных элементов и
парциального давления кислорода в крови и
т. п.) на постоянном уровне с минимально
допустимыми отклонениями. Стресс
представляет собой состояние нарушенного
гомеостаза, а стрессоры — это факторы,
вызывающие нарушение гомеостаза. При
этом стрессоры могут быть как внешними
(факторы окружающей среды), так и
внутренними (формирующимися в организме).
Последние могут возникать в результате
влияния внешних стрессоров, но
нарушение гомеостаза (например, возникновение
очага воспаления, боли и др.) вызывают
самостоятельно.
В ответ на действие стрессоров в
организме возникают адаптивные процессы,
направленные на восстановление гомеостаза
и сохранение нормальной
жизнедеятельности. Эти процессы, т. е. реакцию на стресс,
Селье назвал общим адаптационным
синдромом. Таким образом, сам стресс и стресс-
реакция по своей сути — процессы
необходимые и «полезные», а представление о
«вредности» стресса проистекает из тех
патологических явлений, которые возникают
при чрезмерно сильных и (или)
длительных воздействиях стрессора а также при
нарушениях работы систем,
осуществляющих стресс-реакцию и формирование
адаптации.
244
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
В основе явлений стресса и
стресс-реакции, которые иногда объединяют под
общим термином «стресс», лежит особое
свойство живой материи — раздражимость,
т. е. способность ощущать воздействия
внешних факторов и реагировать на них.
Раздражимость — это ключ к пониманию
стресса и адаптации.
Содержание стресс-реакции организма
зависит от сложности организации живого
существа. Одноклеточные организмы
реализуют свою стресс-реакцию с помощью
таких изменений в цитоплазме, которые
позволяют избежать опасности и сохранить
жизнь, например, посредством хемо- и
фототаксиса, а также цистообразования (как
у амебы). У млекопитающих, в том числе
и у человека, реакция на стресс очень
сложна. Она проявляется в комплексе
изменений основных функциональных систем
организма. Изменяется поведение — возникают
возбуждение, настороженность, обострение
внимания и познавательных способностей,
подавление пищевого и сексуального
поведения, что в целом отражает
«ориентировочную реакцию» и «боевую готовность».
Мобилизуется функция систем кровобраще-
ния и дыхания, ответственных за
доставку кислорода и питательных веществ в
органы и ткани, изменяется активность иммунной
системы.
Структура стресс-реакции при
воздействии физических стрессоров, механизм
действия которых исследован лучше, чем
эмоциональных, представлена на рис. 1.
Стресс через высшие регуляторные
центры активирует регуляторную
стресс-систему, которая объединяет определенные
отделы нервной и эндокринной систем и
«неспецифически» активируется в ответ на
любой стрессор, а также функциональную
систему, объединяющую органы и ткани,
«специфически» ответственные за
приспособление к конкретному стрессору.
Например, при физической нагрузке
функциональная система объединяет
соответствующие моторные центры в головном и
спинном мозге и управляемые ими
скелетные мышцы, выполняющие
необходимую мышечную работу. Активируются
также органы дыхания и кровообращения,
обеспечивающие увеличенный приток
кислорода и питательных веществ к
работающим мышцам и центрам управления. При
этом стресс-система не только
обеспечивает более полную мобилизацию
функциональной системы, но и координирует
адаптационные процессы. Иначе говоря,
она осуществляет «настройку» органов и
тканей, вовлеченных в адаптацию, на
деятельность в новых условиях. При этом
влияние стресс-системы в случае
достаточно сильного стрессорного воздействия
Рис- 1. Схема адаптивной стресс-реакции в ответ
на действие физических стрессоров
Пояснения в тексте
может оказаться избыточным и приводить
к побочным неблагоприятным эффектам, в
частности — к стрессорным повреждениям.
Стресс-система
В соответствии с данными новейших
исследований стресс-система состоит из
центрального звена и двух
периферических ветвей, которые осуществляют связь
центрального звена со всем организмом
(рис. 2). Центральное звено находится в
головном мозге — гипоталамусе и других
отделах ствола мозга. Гипоталамус
выполняет функции «дозорного» центральной
нервной системы, он получает информацию
о появлении стрессора и «запускает»
работу стресс-системы.
Центральное звено стресс-системы
объединяет три основных группы нейронов:
1) нейроны, вырабатывающие кортикотро-
пин-рилизинг-гормон1 (КРГ), который
стимулирует секрецию адренокортикотропно-
го гормона (АКТГ) в гипофизе и тем самым
активирует гипоталамо-гипофизарно-адре-
наловую систему (ось); 2) нейроны,
вырабатывающие гормон аргинин-вазопрессин,
обладающий рядом регуляторных функций
и в том числе усиливающий влияние КРГ
на секрецию АКТГ в гипофизе; 3) группы
нейронов в гипоталамусе и других отделах
ствола мозга (синем пятне), синтезирующих
катехоламины, в основном норадреналин.
Периферические ветви стресс-системы
представлены: 1) гипоталамо-гипофизарно-
1 Т. е. гормон, способствующий
освобождению кортикотропина.
СТРЕСС И АДАПТАЦИЯ
245
адреналовой осью, активируемой КРГ и
АКТГ; ее конечным продуктом являются
глюкокортикоиды, выделяемые корой
надпочечников; 2) симпато-адреналовой
системой, в которую входят симпатическая
нервная система (иннервирующая все органы
и ткани) и мозговой слой надпочечников;
конечным продуктом этой системы являются
катехоламины — норадреналин и адреналин.
Таким образом, основным результатом
активации стресс-системы является
увеличенный выброс в кровь глюкокортикоидов
Эмоциональные
стрессоры
Адаптивное
поведение
ГАМК
КРГ НА
ОП
Физические
стрессоры
Гипофиз
НГГУ"
Гипоталамус
и синее пятно
Глюкокортикоиды
Адреналин
Норадреналин
Активация функции
и метаболизма
органов и тканей,
ОТВЕТСТВЕННЫХ
ЗА АДАПТАЦИЮ
Рис. 2. Схематическое изображение центрального
и периферических звеньев стресс-системы и
взаимодействия компонентов этих звеньев между собой
и со стресс-лимитирующими системами
Обозначены области гипоталамуса и других
отделов ствола мозга, нейроны которых продуцируют
многие гормоны: КРГ —кортикотропин-рилизинг-гор-
мон, стимулирующий секрецию АКТГ (адренокорти-
котропного гормона гипофиза); АВ— аргинин-вазоп-
рессин; ГАМК—у-аминомасляная кислота;
НА—норадреналин; ОП— опиоидные пептиды; НП«У»-
нейропептид «Y», вырабатываемый симпатическими
нейронами; SP— субстанция «Р», вырабатываемая
клетками гипоталамуса и миндалевидного тела;
ГнРГ — гонадотропин-рилизинг-гормон — стимулятор
секреции половых гормонов; РГГР— рилизинг-гор-
мон гормона роста; ТРГ— гормон, стимулирующий
секрецию тиреоидных гормонов (гормонов
щитовидной железы), п.v. — центр блуждающего нерва.
Красным цветом обозначены компоненты
стресс-системы и стрелки, означающие активирующее влияние,
синим— компоненты стресс-лимитирующих систем
и стрелки, означающие тормозное, угнетающее
влияние. Зелеными стрелками отмечены адаптивные
реакции
и катехоламинов — главных
стресс-гормонов, которые способствуют мобилизации
функций органов и тканей, ответственных
за адаптацию. Это ключевое действие
стресс-системы реализуется путем
взаимодействия гормонов с рецепторами —
специальными воспринимающими структурами
наружной мембраны клеток органов и
тканей (рис. 3).
Рецепторы избирательно реагируют на
определенные гормоны. В результате этого
контакта активируются связанные с
рецепторами ферменты и образуются так
называемые вторичные мессенджеры (посредники) —
циклический аденозинмонофосфат (цАМФ),
инозитолтрифосфат (ИФ3), диацилглицерол
(ДАГ) и др. Они служат для передачи
внешнего гормонального сигнала внутрь клетки.
Мессенджеры «запускают» внутриклеточные
процессы, приводящие к повышению
концентрации Са2+ — универсального регулятора
внутриклеточных процессов (первичного мес-
сенджера) и активности протеинкиназ,
находящихся в цитоплазме. Эти ферменты
активируют специфическое функционирование
клетки, процессы ее энергообеспечения и
генетический аппарат, что является основой
формирования устойчивой адаптации при
повторных действиях стрессора.
Как выяснилось в последние 3-5 лет,
помимо прямых продуктов
стресс-системы, в стресс-реакцию вовлечены также
вещества, вызывающие или опосредующие
эффекты стресс-системы: ангиотензин II,
цитокины (например, интерлейкины), та-
хикины (нейропептид «Y», субстанция «Р»)
и др. В частности, цитокины опосредуют
воздействие стресс-системы на систему
иммунитета. Нейропептид «Y», содержащийся
наряду с катехоламинами в
симпатических нервных волокнах и надпочечниках,
усиливает сосудосуживающее действие
катехоламинов. Вместе с тем
нейропептид «Y», выделяющийся из отростков
НА-нейронов в гипоталамусе, оказывает
угнетающее действие на высвобождение
норадреналина, но потенциирующее — на
КРГ-нейроны. КРГ, открытый в 1955 г.
М. Сафраном и А. Шэлли в качестве
«фактора высвобождения АКТГ из гипофиза»,
является, кроме того, самостоятельным
нейрогормоном и участвует в изменении
функций внутренних органов и иммунной
системы при стрессе.
По концепции Селье, стресс-система
активируется в ответ на любой стрессор
неспецифически, т. е. продуцирует
одинаковый набор гормонов и медиаторов
независимо от вида стрессора. Однако недавно
появились данные, позволяющие полагать,
что в ответ на физические стрессоры
происходит преимущественная активация
КРГ-нейронов, а в ответ на психологичес-
246
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
СТРЕСС
Активация стресс-системы
Высвобождение стресс-гормонов
Глюкокортикоиды Са^+
ФИ
ДАГ
Иф3 АТФ
MX ЦАМФ
пк-с Са2+ ^ цАМФ"пк
с? Са-КМ-ПК
белки
мРНК
Рис. 3. Схема регуляции функционирования клетки и ее генетического аппарата (на примере клетки
сердечной мышцы) с помощью универсального активатора внутриклеточных процессов— ионов кальция (Са2+)
и потенциации этой регуляции стресс-гормонами через посредство вторичных мессенджеров (ДАГ, цАМФ,
ИФ3)
Приходящий к клетке импульс возбуждения (обозначен черной стрелкой) вызывает деполяризацию
клеточной мембраны (М), что приводит к открытию потенциал-зависимых Са2+-каналов, входу внеклеточного Са2+ в
клетку, выделению Са2+ из саркоплазматического ретикулума (СПР) и митохондрий (MX) и повышению
концентрации этого иона в клетке. При соединении Са2+ со своим внутриклеточным рецептором кальмодули-
ном (КМ) происходит активация КМзависимой протеинкиназы (Са-КМ-ПК), которая запускает
внутриклеточные процессы, приводящие к мобилизации функции клетки. Одновременно Са2+ участвует в активации
генетического аппарата клетки. Гормоны, воздействуя на соответствующие рецепторы в мембране, активируют
эти процессы через посредство вторичных мессенджеров, образующихся в клетке с помощью ферментов,
сопряженных с рецепторами через специальный G-белок. Воздействие на оц-адренорецепторы активирует
сопряженный с ним фермент фосфолипазу С, с ее помощью из фосфолипида мембраны фосфатидилинози-
тола (ФИ) образуются вторичные мессенджеры — диацилглицерол (ДАГ) и инозитолтрифосфат (ИФ3). ДАГ
активирует протеинкиназу С (ПК-С), ИФ3 стимулирует высвобождение Са2+ из СПР. Воздействие на р- и
а2-адренорецепторы и вазопрессиновый рецептор (V2) приводит к активации аденилаткиназы, сопряженной
с этими рецепторами, и образованию вторичного мессенджера циклического аденозинмонофосфата (цАМФ)
из АТФ. цАМФ активирует соответствующую протеинкиназу (цАМФ-ПК), которая вызывает активацию
внутриклеточных процессов. Глюкокортикоиды, проникая в клетку, взаимодействуют с внутриклеточными
рецепторами стероидных гормонов (СТ-рецепторами) и активируют генетический аппарат клетки. Это выражается в
экспрессии генов регуляторных и структурных белков, образовании соответствующих информационных РНК
(мРНК), синтезе указанных белков и росте клеточных структур, ответственных за адаптацию, что приводит к
формированию структурного следа адаптации. Тонкие стрелки указывают последовательность процессов,
широкие стрелки означают активирующее влияние
СТРЕСС И АДАПТАЦИЯ
247
кие стрессоры — преимущественная
активация НА-нейронов и катехоламинового
звена.
Стресс-лимитирующие системы
Подобная активация стресс-системы и
системы, специфически ответственной за
адаптацию, позволяет восстановить гомео-
стаз (т. е. адаптироваться к данному
стрессору) и завершить стресс-реакцию при
условии, если сила и продолжительность
действия стрессора умеренны. Чрезмерное
стрессорное воздействие приводит к
нарушению функций и повреждениям органов и
тканей. Повреждающие эффекты стресс-
реакции возникают как «издержки»
активации стресс-системы в ответ на сильный
стресс и связаны с избыточным выбросом
стресс-гормонов (на рис. 1 — «побочные
эффекты стресс-реакции»).
Умеренный выброс катехоламинов из
надпочечников и из окончаний
симпатических нервов активирует функционирование
и энергетический метаболизм клеток, но
чрезмерное действие катехоламинов может
вызывать повреждение клеток и
нарушение функции органов. Вазопрессин и ка-
техоламины повышают артериальное
давление, что необходимо как адаптивная
реакция, однако сосудосуживающее
действие этих гормонов при их избытке
может оказаться чрезмерным и вызвать
нарушение мозгового кровообращения и
инсульт. Стрессорные язвы органов
желудочно-кишечного тракта также связаны с
сосудосуживающим эффектом
катехоламинов и возникающей вследствие этого
ишемией слизистой оболочки этих органов.
Избыточная секреция КРГ и глюкокортикоидов
подавляет секрецию половых гормонов и
гормона роста, функцию щитовидной
железы. Недавно установлено, что вместе с
увеличенной секрецией цитокинов избыток
КРГ вызывает угнетение системы
иммунитета, что возможно является причиной
возникновения раковых заболеваний при
тяжелых или затяжных стрессах.
Тяжелые повреждения у людей
возникают главным образом при эмоциональных
стрессах. Известны случаи так называемой
«внезапной сердечной смерти», наступившей
после получения трагических или очень
радостных известий. Согласно последним
данным, виновником такой смерти,
сопряженной с тяжелой аритмией — нарушением ритма
сердечных сокращений, является избыток
стресс-гормонов, главным образом
катехоламинов. Конкретный механизм
катехоламинового повреждения клеток связан с резким
повышением концентрации Са2+ в них, а
также с чрезмерной активацией свободно-
радикального окисления, продукты которого
оказывают повреждающее действие.
Сходное детергентное действие вызывает и
избыток свободных жирных кислот,
образующихся в органах при активации распада
липидов под влиянием катехоламинов.
Подобным же образом нарушается функция
сердца, сосудов, почек и других органов при
хроническом действии даже умеренных по
интенсивности психосоциальных стрессоров.
Эффективность адаптивной реакции на
стресс и вероятность возникновения стрес-
сорных повреждений и болезней в
значительной мере определяются, помимо
интенсивности стрессора, состоянием стресс-системы:
ее базальной (исходной) активностью и
реактивностью, т. е. степенью активации при
стрессе. Они обусловлены генетически, но
могут меняться в процессе индивидуальной
жизни. Хронически увеличенная базальная
активность стресс-системы и (или)
избыточная ее активация при стрессе
сопровождаются повышением артериального давления,
нарушением функции органов пищеварения,
подавлением иммунитета. Сниженная
базальная активность стресс-системы и (или)
неполноценная ее активация при стрессе также
неблагоприятны: они снижают способность
организма адаптироваться к окружающей
среде, решать жизненно важные задачи, что
может приводить к развитию депрессивных
состояний.
Нежелательные побочные эффекты
чрезмерной активации стресс-системы делают
целесообразным существование в
организме определенных естественных
ограничителей. Существуют два основных механизма,
регулирующих активность и реактивность
стресс-системы: механизм саморегуляции и
механизм внешней регуляции (рис. 2).
Механизм саморегуляции реализуется за
счет влияния друг на друга компонентов
самой системы. Так, по механизму
отрицательной обратной связи гормоны,
вырабатываемые в системе, ограничивают свою
собственную продукцию. Между КРГ-ней-
ронами и НА-нейронами существуют
нервные связи, приводящие к
взаимоактивации этих нейронов, так что выработка КРГ
активирует НА-нейроны и секрецию ими
НА, и наоборот. Кроме того, глюкокорти-
коиды ограничивают активность норадрена-
линового звена стресс-системы, угнетая
синтез, высвобождение и обратный захват
норадреналина в симпатических нейронах.
Механизм внешней регуляции
осуществляется модуляторными регуляторными
системами, не входящими в стресс-систему,
но тесно с ней связанными. Это так
называемые стресс-лимитирующие системы,
которые способны ограничивать активность
стресс-системы и чрезмерную
стресс-реакцию на центральном и периферическом
уровнях регуляции.
248
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
К центральным структурам стресс-лими-
тирующих систем относятся ГАМК-эргичес-
кая система (система нейронов,
продуцирующих ГАМК — у-аминомасляную кислоту,
обладающую тормозным действием на
нейроны головного и спинного мозга) и опиои-
дэргическая система, объединяющая нейроны
в гипоталамусе и секреторные клетки в
гипофизе, которые вырабатывают опиоидные
пептиды (ОП), также обладающие
тормозным действием. Важно отметить, что
существуют прямые связи нейронов
стресс-системы с ГАМК-нейронами и ОП-нейронами.
Выделяемые нейронами стресс-системы
гормоны активируют ГАМК-нейроны и ОП-ней-
роны, они секретируют ГАМК и опиоидные
пептиды, которые, в свою очередь,
ограничивают активность стресс-системы. Схема
(рис. 4) демонстрирует это положение на
примере ГАМК-системы.
Доказано, что в нейронах гипоталамуса
стресс-гормоны локализуются вместе с опи-
оидными пептидами (вазопрессином и динор-
фином). Показано также, что в гипофизе
АКТГ и опиоидные пептиды (р-эндорфины)
образуются из общего предшественника (про-
пиомеланокортина) и выделяются при стрессе
в эквимолярных количествах. В регуляции
стресс-системы участвуют субстанция «Р» —
пептид, образующийся в гипоталамусе и
миндалевидном теле и оказывающий тормозное
действие на КРГ-нейроны, и нейропептид «Y»,
обладающий угнетающим действием на
НА-нейроны, но стимулирующий
КРГ-нейроны в гипоталамусе (см. рис. 2).
Тормозное действие ГАМК и опиоидных
пептидов на катехоламиновое звено стресс-
системы осуществляется не только в
центральной нервной системе, но и на периферии:
рецепторы для ГАМК и опиоидных пептидов
локализованы на аксонах симпатических
нейронов, иннервирующих органы и ткани, и в
самих органах; это ограничивает
высвобождение катехоламинов и их действие.
Доказано совместное присутствие стресс-гормонов
и опиоидных пептидов в хромаффинных
пузырьках (катехоламинов и энкефалинов) и в
коре надпочечников (глюкокортикоидов и
энкефалинов). Поэтому выброс стресс-гормонов
может сопровождаться выходом таких же
количеств опиоидных пептидов.
Действие стресс-системы в органах и
тканях ограничивают системы локальной
регуляции — локальные или
периферические стресс-лимитирующие системы,
продуцирующие простагландины, аденозин,
опиоидные пептиды и другие вещества в самих
органах и периферических нейроэндокрин-
ных структурах. Они угнетают
высвобождение катехоламинов из нервных окончаний и
надпочечников а также их действие; тем
самым они ограничивают чрезмерную стресс-
реакцию и ее повреждающее воздействие
на органы и ткани. Важную роль в
ограничении активации свободно-радикального
окисления и повреждений при стрессе
играют антиоксидантные системы в органах и
тканях, включающие в себя
антиоксидантные ферменты (каталазу, супероксиддисму-
тазу, глутатионпероксидазу и др.) и
естественные антиоксиданты (а-токоферол,
витамин А, аскорбиновую кислоту и др.).
Таким образом, как базальная активность
стресс-системы, так и ее активация при
стрессе зависят от активности стресс-лимитиру-
ющих систем. Нарушение регуляции стресс-
системы приводит не только к нарушению
реакции организма на стресс, но и к
возникновению психических и эндокринных
заболеваний, к болезням системы кровообращения,
иммунной системы, обмена веществ или к
предрасположенности к этим заболеваниям.
В качестве примера можно привести данные
о том, что у линейных крыс с
наследственной гипертонией наблюдается врожденно
повышенная активность симпатического звена
стресс-системы, сочетающаяся с врожденным
уменьшением активности ГАМК-нейронов в
гипоталамусе, иннервирующих центры
симпатической регуляции сердечно-сосудистой
системы, и со сниженным содержанием
опиоидных пептидов в тканях.
Рис. 4. Схема взаимодействия стресс-системы и
ГАМК-эргической системы
Активация ГАМК-системы при стрессе приводит к
ограничению стресс-реакции и стрессорных
повреждений
Адаптация
Приспособление организма к
окружающей среде (в частности к периодическому
действию стрессоров) в процессе индивиду-
СТРЕСС И АДАПТАЦИЯ
249
альной жизни называется фенотипической
адаптацией в отличие от генотипической
адаптации, сформировавшейся в процессе
эволюции и определяемой генотипом
организма.
Фенотипическая адаптация — это
процесс, в результате которого организм
приобретает ранее отсутствовавшую
устойчивость к определенному фактору (или
факторам) среды и таким образом получает
возможность жить в условиях, ранее
несовместимых с жизнью, решать задачи,
ранее неразрешимые. Она базируется на
генотипе, т. е. генетической программе
организма, которая обеспечивает не заранее
готовую адаптацию, а возможности для
формирования адаптации в соответствии с
конкретными требованиями среды.
Результаты такой адаптации не передаются по
наследству, поскольку потомство может
столкнуться с совсем другими условиями
жизни и должно приспосабливаться к ним.
В основе формирования адаптации лежит
переход от срочной адаптации,
возникающей в ответ на первое действие
стрессора, к долговременной адаптации,
формирующейся при повторных действиях
стрессора и обеспечивающей устойчивость
к длительно действующему стрессору.
Срочная адаптация — немедленный
ответ организма на стрессор,
реализующийся в виде первой стресс-реакции,
подробно рассмотренной выше. Она обеспечивает
либо поведенческую реакцию избегания,
избавления от действия неблагоприятного
стрессора, либо мобилизацию
функциональных систем, ответственных за
приспособление к этому фактору. Селье называл этот
этап адаптации фазой тревоги.
Очевидными проявлениями срочной адаптации
являются, например, увеличение
теплопродукции в ответ на холод, а теплоотдачи — в
ответ на тепло, учащение дыхания и
увеличение минутного объема сердца в ответ
на недостаток кислорода в воздухе и т. п.
Важнейшая черта этого начального этапа
адаптации состоит в том, что деятельность
организма протекает при почти полной
мобилизации функционального и
энергетического резервов. Однако даже это не всегда
может обеспечить необходимый
адаптационный эффект. Например, бег
нетренированного человека происходит при близких
к максимальным величинах работы сердца,
легких, при максимальной мобилизации
энергетических ресурсов в скелетных
мышцах, сердце, печени. При этом бег не
может быть ни достаточно быстрым, ни
достаточно длительным, очень скоро наступает
истощение. Тренированный же человек
может бежать быстрее и дольше, на
выполнение той же мышечной работы ему
потребуется существенно меньшая мобилизация
систем кровообращения, дыхания и
меньшие энергетические затраты по сравнению
с нетренированным.
При действии эмоциональных стрессоров
именно на этом срочном этапе адаптации
могут возникать стрессорные повреждения.
Это объясняется тем, что эффективность
существующих физиологических механизмов
может оказаться недостаточной для нагрузки,
связанной с действием данного стрессора,
и организм вынужден работать «на износ».
Для того чтобы эта срочная, но
несовершенная адаптивная реакция сменилась более
совершенной долговременной адаптацией,
необходимы время и повторные действия
стрессора. В результате в органах и тканях
функциональной системы, вовлеченной в
адаптивную реакцию, формируется так
называемый структурный след адаптации —
материальная база повышения
устойчивости организма к действию стрессора, основа
тренированности.
Ключевое звено формирования
структурного следа и, следовательно,
устойчивой адаптации — активация синтеза
нуклеиновых кислот и белков, в результате чего
происходит увеличенная наработка регуля-
торных и структурных белков в клетках
органов и тканей, ответственных за
адаптацию к данному фактору. При этом
активируется синтез белков тех клеточных
структур, которые испытывают повышенную
функциональную нагрузку при действии
стрессора. Например, при адаптации к
недостатку кислорода в результате
повторных сеансов гипоксии в органах дыхания
активируется синтез структурных белков,
что приводит к гипертрофии (увеличению
массы) нейронов дыхательного центра,
дыхательной мускулатуры (диафрагмы и
межреберных мышц) и самих легких, в
которых увеличивается количество альвеол.
В результате возрастает функциональная
мощность аппарата внешнего дыхания,
увеличивается дыхательная поверхность
легких и коэффициент утилизации
кислорода, в результате чего возрастает
экономичность функции дыхания. В системе
кроветворения активация синтеза нуклеиновых
кислот и белков в костном мозге приводит
к увеличенной продукции эритроцитов, что
обеспечивает увеличение кислородной
емкости крови. В целом это обеспечивает
достаточное поступление кислорода к мозгу,
сердцу, другим органам и тканям,
несмотря на гипоксию, т. е. недостаток кислорода
в крови. Наконец, в клетках самих органов
и тканей активируется синтез белков
митохондрий, что постепенно приводит к росту
числа митохондрий в клетках и
активности в них ферментов синтеза АТФ. В итоге
способность клеток утилизировать кислород
из крови и образовывать АТФ возрастает,
250
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
несмотря на недостаток кислорода. В
результате этих процессов организм приобретает
устойчивость к недостатку кислорода и
возможность сохранения жизненной
активности в условиях гипоксии. Такие
особенности жизнедеятельности свойственны жителям
высокогорья.
В процессе адаптации к различным
стрессорам повышаются функциональная
мощность и эффективность как самой
стресс-системы, так и стресс-лимитирую-
щих систем. Это обусловлено
формированием структурного следа в клетках этих
систем. В исследованиях последних лет
показано, что при повторном действии
стрессоров активируется экспрессия генов,
кодирующих синтез ферментов,
ответственных за наработку катехоламинов в
гипоталамусе и надпочечниках а также
синтез КРГ в гипоталамусе. Показано также,
что при этом происходит увеличение
мощности системы синтеза ГАМК, опиоидных
пептидов, простагландинов, антиоксидантов.
Эти процессы оптимизируют регуляцию
стресс-системы и предупреждают развитие
чрезмерных стресс-реакций в
адаптированном организме.
Изучение механизма перехода срочной
адаптации в устойчивую, выявление
стимула, запускающего активацию экспрессии
генов в структурах, ответственных за
адаптацию, является одной из ключевых
проблем биологии и медицины. По-видимому,
основные процессы формирования
структурной (материальной) базы устойчивой
адаптации происходят в клетках органов и
тканей, ответственных за адаптацию. В них под
влиянием катехоламинов, глюкокортикои-
дов и других стресс-гормонов запускается
цепь событий, в результате которой в
цитоплазме активируются протеинкиназы.
Наряду с потенцированием функций
клетки, протеинкиназы участвуют в
механизме активации синтеза нуклеиновых кислот
и белков. Именно это лежит в основе
наработки белков клеточных структур,
которые обеспечивают увеличение мощности и
повышение эффективности работы клеток
и органов, вовлеченных в адаптацию к
данному стрессору. Установлено, что данный
процесс может реализоваться лишь при
повторном неповреждающем воздействии
стрессора, т. е. при повторном
кратковременном увеличении функции клеток, и
требует определенного времени.
Особенностью устойчивой адаптации
является наличие так называемых
перекрестных защитных эффектов. Оказалось, что
адаптация к одному стрессору повышает
устойчивость организма не только к
действию этого стрессора, т. е. обладает не
только прямым защитным эффектом, но
может также повышать устойчивость и к
действию других стрессоров. Например, у
животных, адаптированных к гипоксии в
барокамере, имитирующей условия
высокогорья, повышена резистентность к
физическим нагрузкам, повреждающему
действию эмоциональных стрессоров, в том
числе повышена устойчивость иммунной
системы к угнетающему действию стрессов.
В основе такого перекрестного защитного
эффекта адаптации лежит, как и в основе
прямого защитного эффекта, наличие
структурного следа адаптации, который
повышает функциональные резервы
органов и систем. Поэтому, чем больше
органов и систем вовлечены в процесс
адаптации к данному стрессору, т. е. чем богаче
структурный след, тем большим числом
перекрестных защитных эффектов обладает
данная адаптация.
Наличие перекрестных защитных
эффектов открывает широкие перспективы
использования адаптации к факторам среды для
повышения устойчивости здорового
человека к стрессорным ситуациям и болезням,
а также для лечения некоторых
заболеваний. Достижения в этой области уже есть,
и они составили основу появления нового
раздела медицинской науки —
адаптационной медицины. На базе этих достижений уже
успешно внедрено в клинику
использование адаптации к гипоксии в барокамере в
качестве метода профилактики и лечения
нарушений функции иммунной системы
(аллергических заболеваний), неврозов,
гипертонической болезни и др. Однако многие
стороны механизма адаптации к различным
стрессорам и формирования защитных
эффектов остаются неясными и требуют
дальнейших исследований.
Важный аспект проблемы адаптации
составляют явления деадаптации и
реадаптации. Деадаптация — обратное развитие
процесса долговременной устойчивой
адаптации, т. е. частичная или полная потеря
прямых и перекрестных защитных
эффектов в результате прекращения действия
стрессора, к которому адаптировался
организм. Процесс деадаптации выражается в
постепенном исчезновении структурного
следа, составляющего основу данной
адаптации.
Деадаптация является по существу
процессом, обратным процессу формирования
адаптации, и механизм ее состоит в том,
что непосредственно после прекращения
действия стрессора резко снижается
нагрузка на любой из органов функциональной
системы, ответственной за адаптацию к
данному стрессору. В результате в этом
органе уменьшается интенсивность
синтеза РНК и белков, возникает отставание
синтеза белков от их распада и как
следствие уменьшается масса структур орга-
СТРЕСС И АДАПТАЦИЯ
251
на, увеличенная в процессе адаптации. Так,
после прекращения физических нагрузок у
тренированного к ним животного или
человека происходит уменьшение массы
скелетных мышц и сердца, т. е. исчезновение
гипертрофии, уменьшение в них количества
митохондрий. После выхода из условий
гипоксии (возвращения из высокогорья на
равнину) исчезает гипертрофия правого
желудочка сердца и легких, уменьшаются
количество эритроцитов и содержание
гемоглобина в крови. При этом процесс
исчезновения гипертрофии (уже в течение
первых дней) начинается прежде всего со
снижения скорости синтеза белков в
сердечной мышце; существенное уменьшение
массы сердца происходит за 1-2 недели, а
полное восстановление — за 3—4 недели.
Деадаптация сопровождается утратой
устойчивости к гипоксии, приобретенной в
процессе долговременной адаптации, и
возвращением ее к уровню, свойственному
неадаптированному организму. Таким
образом, прекращение действия стрессора
постепенно приводит к утрате (как бы за
ненадобностью) наработанных в процессе
адаптации клеточных структур. В последние
годы выяснилось, что в механизме разборки
клеточных структур и деградации
структурного следа при деадаптации наряду с
активацией лизосомальных ферментов про-
теолиза (распада белков) важную роль
играет активация свободно-радикального
окисления. Однако конкретный механизм,
который запускает процесс разборки
структур и возвращения к исходной, доадапта-
ционной норме, пока неясен.
Процесс исчезновения структурного
следа адаптации и самой адаптации с
возвратом к состоянию, которое было до действия
стрессора, т. е. к условной норме,
называется физиологической деадаптацией. Ее
следует отличать от деадаптации,
вызванной специальными условиями, например
гипокинезией или невесомостью, при
которых в нормальном здоровом организме
может начаться процесс, приводящий к
необычному уменьшению массы мышечных
клеток и снижению их работоспособности.
Такая деадаптация является очевидной
предпосылкой многих заболеваний и
называется патологической деадаптацией. Она
развивается на основе тех же механизмов,
что и физиологическая деаптация. Однако
в отличие от нее при патологической
деадаптации происходит снижение
функциональных возможностей органов и систем уже
ниже уровня условной нормы.
Реадаптация — явление восстановления
утраченного при деадаптации
структурного следа адаптации с помощью повторных
воздействий данного фактора, т. е. с
помощью повторной адаптации. Наиболее
наглядно это явление наблюдается при
возобновлении тренировок у спортсменов, длительное
время лишенных физических нагрузок, а
также у жителей высокогорья, длительное
время проживавших на равнине и вновь
переселившихся в горы. Поскольку при
адаптации возникает определенное напряжение
синтетического аппарата клеток органов и
систем, ответственных за адаптацию,
повторы цикла «деадаптация-реадаптация»
могут привести к «изнашиванию» этого
аппарата, потере способности адекватного
обновления клеточных структур и, как
следствие, к возникновению заболеваний. Такая
ситуация особенно характерна для людей
пожилого возраста, у которых происходят
естественные изменения в
функционировании генетического аппарата клеток.
Поэтому поддержание структурного следа
необходимой в будущем адаптации путем
сравнительно небольших периодических
нагрузок на ответственную за адаптацию
функциональную систему представляет
гораздо более экономный и менее
истощающий вариант процесса, чем повторение
циклов «деадаптация-реадаптация».
Литература
Меерсон Ф.З., Пшенникова М.Г. Адаптация к
стрессорным ситуациям и физическим
нагрузкам. М.: Медицина, 1988.
Меерсон Ф.З., Пшенникова М.Г. Стресс-лими-
тирующие системы организма и новые принципы
профилактической кардиологии. М.: НПО «Со-
юзмединформ», 1989.
Селье Г. Стресс без дистресса. Пер. с англ. М.:
Мир, 1979.
Физиология адаптационных процессов. М.:
Наука, 1986.
В. И. Кулинский
ДВЕ СТРАТЕГИИ ВЫЖИВАНИЯ ОРГАНИЗМА
Введение
Живые организмы нередко попадают в
трудные, а иногда и в экстремальные
условия. Это связано с нарушениями
основных параметров среды обитания
(температуры, содержания 02, химического состава,
уровня облучения и др.) и патологическими
изменениями в самом организме (болью,
травмами, заболеваниями). В результате
возникает опасность для здоровья и даже для самой
жизни особи. Важнейшая задача организма
в этих условиях — выстоять, а при
длительном их сохранении — адаптироваться к ним.
Эта задача решается регуляторными
системами организма, реализующими различные
стратегии адаптации.
Еще в 1878 г. К. Бернар указал на одно
важное различие между низко- и
высокоорганизованными животными. Первые на
неблагоприятные условия реагируют анабиозом
(состоянием скрытой жизни), например,
лягушка при охлаждении впадает в окоченение.
Вторые сохраняют возможность свободной и
независимой жизни благодаря поддержанию
постоянства внутренней среды организма.
У. Кеннон в 1929 г. назвал такое постоянство
гомеостазом и установил роль симпато-адре-
наловой системы и ее гормонов — катехола-
минов в реакциях млекопитающих на самые
разные экстремальные воздействия. Г. Селье
в 1936 г. ввел понятие «стресс» («состояние
напряжения»), обозначающее стандартную
реакцию организма на любое неблагоприятное
воздействие, и открыл, что важное
значение в ней играют гормоны коры
надпочечников — глюкокортикостероиды.
В настоящее время в экологической
физиологии признается существование двух
качественно различающихся стратегий
приспособления к неблагоприятным условиям —
путем повышения резистентности
(сопротивляемости) и посредством увеличения
толерантности (переносимости, выносливости).
Более простые организмы и зимнеспящие
млекопитающие реализуют толерантную
стратегию. Для млекопитающих в общем и
целом характерна резистентная стратегия
(стресс). Однако теперь уже ясно, что
большинство млекопитающих могут с успехом
реализовывать обе стратегии.
Стратегия резистентности (стресс)
Стратегия резистентности характерна
для реакций при отрицательных эмоциях
(гневе, ярости, страхе), тяжелой
мышечной работе, остром охлаждении, при
возникновении боли и травмах. Обычно эта
стратегия реализуется как стресс-реакция
с характерным для нее активным
противодействием внешней среде,
максимизацией функций основных физиологических
систем (головного мозга,
сердечно-сосудистой, дыхательной, мышечной и
гормональной систем). Такая реакция приводит
к сохранению гомеостаза и даже к
состоянию гиперкомпенсации. В результате
увеличивается резистентность организма, а
при длительных или повторяющихся
воздействиях возникает устойчивая
адаптация (см.: Пшенникова М. Г. Стресс и
адаптация. Наст. том).
Метаболическая основа данной
стратегии — преобладание катаболизма
(распада) эндогенных питательных веществ над
анаболизмом (синтезом). Распадаются
биополимеры: гликоген печени — до глюкозы,
гликоген мышц — до молочной кислоты,
жир — до жирных кислот и глицерина,
белки — до аминокислот. Значительно
возрастает потребление 02 митохондриями,
клетками, органами и организмом в целом
(калоригенный эффект), увеличиваются
теплопродукция и синтез АТФ.
Происходит мобилизация всех энергетических
ресурсов и их расходование на
компенсаторные нужды организма.
Ведущими гормонами стратегии
резистентности являются катехоламины и
глюкокортикостероиды (глюкокортикоиды).
Первые через Рх, (32 и о^-адренорецепторы1
участвуют в запуске всех, вторые —
большинства описанных реакций; кроме того,
эти гормоны стимулируют глюконеогенез —
образование углеводов из органических
кислот (метаболитов глюкозы, глицерина
и аминокислот). При этом катехоламины
активируют основные ферменты
катаболизма, а глюкокортикостероиды индуцируют
их (увеличивают синтез). В стресс-реакции
участвуют и другие гормоны, многие из
которых активируются катехоламинами:
глюкагон, паратирин (паратиреоидный
гормон), ангиотензин II, в ряде ситуаций —
вазопрессин, альдостерон, лейкотриены,
а также аминокислоты (глутаминовая и ас-
парагиновая).
1 Адренорецепторы — белковые молекулы на
поверхности клеток, узнающие молекулы адре-
нергических гормонов — адреналина и норадре-
налина.
ДВЕ СТРА ТЕГИИ ВЫЖИВАНИЯ ОРГАНИЗМА
253
Главное достоинство стратегии
резистентности в том, что она позволяет
организму в трудных и даже экстремальных
условиях выжить, успешно выполнять свои
функции и, более того, решать
сверхважные задачи, добиваясь выдающихся
результатов. Однако эта стратегия предельно
неэкономична, расточительна: за
резистентность, адаптацию организму
приходится платить дорогую цену — возникают так
называемые болезни адаптации. К
последним относят такие наиболее
распространенные в развитых странах заболевания,
как атеросклероз, гипертония, ишемичес-
кая болезнь сердца, язва желудка и
двенадцатиперстной кишки, депрессия.
Ф. 3. Меерсон выдвинул представление о
стресс-лимитирующих системах как
тормозных механизмах, ограничивающих
интенсивность или длительность
стресс-реакции.
Стратегия толерантности
Многие живые существа способны
использовать качественно отличающуюся от
стресса стратегию приспособления —
стратегию толерантности, известную также под
названиями «торпидность» (оцепенение),
«гибернация2», «гипометаболизм», «гипобиоз»
(снижение интенсивности жизненных
процессов). Для нее характерны «подчинение»,
«уступка» условиям внешней среды,
минимизация функций основных
физиологических систем (головного мозга,
сердечно-сосудистой, дыхательной и мышечной систем)
и уменьшение выработки энергии. Это
неизбежно приводит к определенным
нарушениям гомеостаза, которые, однако, не
настолько чрезмерны, чтобы привести к
смерти организма. Стратегия
толерантности направлена не на поддержание
гомеостаза, а на сохранение жизни в крайне
тяжелых условиях.
Эта стратегия наиболее типична для эво-
люционно более древних холоднокровных
животных, а также для низших
позвоночных с их достаточно низким потреблением
02, небольшим количеством митохондрий,
отсутствием бурого жира и мышечной
дрожи, необходимых для усиленной
теплопродукции, и слабой термоизоляцией.
Периодические спячка или оцепенение встречаются,
примерно, у двухсот видов млекопитающих
(суслика, хомяка, летучей мыши, медведя
и др.) при охлаждении или недостатке пищи
или воды.
Главное достоинство данной страте-
2 Гибернация — состояние минимального
клеточного метаболизма, достаточное лишь для
поддержания структуры, но не функции
клеток.
гии — возможность выжить в тяжелых и
крайне опасных условиях. Но снижение
интенсивности жизненных процессов
неизбежно ограничивает функциональные
возможности организма, делает невозможным
решение особо важных задач.
Неблагоприятные последствия этой стратегии пока
неизвестны.
Стратегия толерантности,
сочетающаяся со снижением потребления 02 и
температуры тела, гипометаболизмом,
реализуется посредством ослабления действия
катехоламинов на ах- и р-адренорецепторы
и усиления их действия на ос2-адрено-
рецепторы, а также эффектов, связанных
с действием серотонина, аденозина и аце-
тилхолина. Указанные гормоны и
синтетические антагонисты их рецепторов
значительно повышают устойчивость
организма к облучению, ишемии головного
мозга, гипоксии и перегреванию. Это
хорошо согласуется с давно известным резким
увеличением толерантности организма к
различным экстремальным факторам при
спячке, гипотермии, гипобиозе.
Выбор стратегии
Принципиальные различия между
описанными стратегиями очевидны. Выбор
организмом той или другой из них определяется
биологической целесообразностью, зависящей,
очевидно, от характера действующего
фактора, его силы и длительности, наличия
эндогенных и доступности экзогенных
ресурсов питания и кислорода, наследственности,
приобретенного опыта и функционального
состояния организма.
Стратегия толерантности, очевидно,
используется прежде всего при
чрезмерном действии повреждающих факторов;
при этом толерантная стратегия может
быть как первичной, так и вторичной, т. е.
использоваться тогда, когда первичная
стратегия резистентности оказалась
неэффективной. Иными словами, она может
быть как «другим выходом» из тяжелой
ситуации, так и второй фазой реакции на
действительно экстремальные факторы.
Стратегия толерантности может заменяться
на резистентную: чрезмерное снижение
температуры тела прерывает зимнюю
спячку — опасность для жизни пробуждает
животное. Это доказывает, что стратегия
толерантности — не результат истощения
защитных сил организма, питательных
веществ или стрессовых гормонов, а
оптимальный путь приспособления в
определенных условиях.
Наличие у млекопитающих не только
стрессовой стратегии резистентности, но
и альтернативной стратегии
толерантности значительно расширяет диапазон воз-
254
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
можных защитных реакций. Кроме того, это
позволяет разрабатывать качественно
различные протекторы рецепторного действия,
характеризующиеся высокой
эффективностью и низкой токсичностью.
Литература
Кулинский В.И. Вопр. медицинской химии,
1994, т. 40, №6. с. 14.
Кулинский В.И.9 Ольховский И.А. Успехи
соврем, биологии, 1992, т. 112, с. 697.
Тепперман Дж., ТепперманХ. Физиология
обмена веществ и эндокринной системы. Пер. с
англ. М.: Мир, 1989.
Физиология адаптационных процессов. М.:
Наука, 1986.
Хочачка П., Сомеро Дж. Биохимическая
адаптация. Пер.с англ. М.: Мир, 1988.
Экологическая физиология животных. В 3 ч.
Л.: Наука, 1979-1982.
О. Г. Газенко, А. И. Григорьев, А. Д. Егоров
АДАПТАЦИЯ ЧЕЛОВЕКА К НЕВЕСОМОСТИ
Введение
Запуск первого искусственного спутника
Земли возвестил о начале практического
освоения космического пространства и
создал надежные предпосылки для
космических полетов человека. Непосредственное
проникновение человека в космос — полет
12 апреля 1961г. Ю.А.Гагарина на
корабле «Восток» — явился одним из самых
значительных событий в истории человечества.
Оно стало возможным пртому, что уже
были накоплены знания об окружающем
Землю пространстве и ожидаемых реакциях
человека в космическом полете. В течение
последующих 35 лет были достигнуты
впечатляющие результаты: в космосе (по
состоянию на 31.12.96 г.) побывало около 400
человек, а продолжительность
пилотируемых полетов возросла со 108 мин до 438
суток.
В развитии медико-биологических
исследований в космических полетах можно
выделить четыре основных этапа.
Первый этап —биологическая
индикация космических полетов. На этом этапе
(50-е и начало 60-х гг.) во время полетов
ракет и первых спутников Земли
проводились исследования с использованием
широкого круга биологических объектов и
подготовка человека к полету в космическое
пространство. Полученные результаты
явились достаточным основанием для решения
вопроса о возможности кратковременного
полета человека.
Второй этап — кратковременный
полет человека в космос.
Медико-биологические исследования, проведенные в 60-е и
начале 70-х гг. во время кратковременных
пилотируемых полетов космических
кораблей, свидетельствовали о возможности
безопасного пребывания человека в
условиях невесомости длительностью до 2 — 3
недель и его активной деятельности вне
корабля, в том числе и на поверхности
Луны. При этом в послеполетном периоде
у космонавтов были выявлены заметные
изменения, которые оказались весьма
существенными после полетов длительностью
14-18 суток. Последнее обстоятельство
стимулировало разработку и создание
системы соответствующих профилактических
мероприятий и углубленных медицинских
обследований с целью обеспечения надежной
безопасности пребывания человека в
космосе при увеличении продолжительности
полетов.
Третий этап — исследования во время
длительных полетов пилотируемых
станций. Медико-физиологические исследования,
проведенные в 1971-1996 гг. во время
пилотируемых полетов длительностью от 1 до
14,5 месяцев на российских орбитальных
станциях «Салют» и «Мир» и трех
экспедиций длительностью 28, 56 и 84 суток на
американской станции «Скайлэб»,
показали, что как мужчины, так и женщины
достаточно хорошо приспосабливаются и могут
эффективно работать в условиях
невесомости длительностью до 438 суток.
Важным дополнением к программам
медицинских исследований в пилотируемых
полетах были биологические эксперименты
в космосе в период 1973-1996 гг.,
проводившиеся с целью углубленного изучения
механизмов влияния условий космического
полета, прежде всего невесомости, на процессы
жизнедеятельности организмов различного
уровня организации. К настоящему
времени исследования проведены на 10
биоспутниках, длительность полета которых
составляла от 5 до 22 суток.
Достигнутая длительность пилотируемых
космических полетов и перспективы ее
дальнейшего увеличения определяются
способностью человека адаптироваться к
невесомости и реадаптироваться к силе тяжести после
возвращения на Землю. На основании
обобщения накопленного опыта складывается
впечатление, что все практически здоровые
люди могут с меньшей или большей
выраженностью реакций адаптироваться к
условиям космических полетов длительностью, по
крайней мере, до года. Важно отметить, что
повторные полеты, как правило, переносятся
легче. Физиологические изменения в
организме человека во время и после космических
полетов проявляются в ряде закономерно
развивающихся симптомокомплексов (табл. 1).
При оценке изменений физиологических
функций необходимо учитывать то
обстоятельство, что на всем протяжении
длительных полетов экипажами используется
бортовой комплекс средств профилактики
неблагоприятного влияния невесомости.
Основной принцип проводимых в
космическом полете профилактических
мероприятий — как это ни парадоксально звучит —
противодействие адаптации к невесомости.
Это достигается созданием осевой
нагрузки на тело, вызывающей определенную
степень деформации опорно-двигательного
аппарата и стимуляцию в нем
соответствующих механорецепторов; физическими уп-
256
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
ражнениями, направленными на
поддержание наиболее важных систем, сохранение
работоспособности организма, стимуляцию
некоторых групп рецепторов и сохранение
двигательных навыков. Важное значение
имеет имитация в невесомости с помощью
физических средств эффекта
гидростатического давления крови, задержка
жидкости в организме и воспрепятствование ее
накоплению в нижних конечностях.
Реализация в длительных полетах указанного
комплекса профилактических воздействий
купирует или в значительной степени
ослабляет возникающие под влиянием
невесомости сдвиги со стороны различных
систем организма (табл. 2).
Рассмотрим основные результаты
исследований реакций организма человека в
пилотируемых космических полетах, а также
некоторые данные, полученные при
изучении влияния невесомости на структурно-
функциональные показатели организма
экспериментальных животных.
Изменения физиологических
функций в космических полетах
Действие невесомости характеризуется
устранением взаимного давления частей тела
друг на друга и тонического напряжения
мышц, удерживающих туловище в
условиях гравитации, а также снятием
гидростатического давления крови, в результате в
основных системах организма возникают
специфические симптомокомплексы (табл. 1).
Следствием этого является уменьшение
афферентной (чувствительной) импульсации с
проприоцепторов тела, что нарушает
взаимодействие афферентных систем,
возникают сенсорные конфликты, являющиеся
основной причиной возникновения космической
болезни движения. В результате снижается
активность гипоталамуса и гипофиза,
ослабляется активизирующее влияние
ретикулярной формации на кору головного мозга.
У человека, находящегося в
вертикальном положении в условиях Земли, стенки вен
Таблица 1
Изменения со стороны основных систем организма, развивающиеся
в длительных космических полетах
Сенсорные системы (начальный период полета):
нарушение взаимодействия сенсорных систем и развитие сенсорных конфликтов;
снижение статической и повышение динамической возбудимости вестибулярных каналов;
развитие космической болезни движения (в 50—60% случаев).
Двигательная система:
изменение чувствительности проприоцептивных и сенсорных входов;
субатрофия/атрофия и снижение массы и силы антигравитационных мышц;
снижение эффективности двигательного регулирования.
Костная система:
снижение минеральной плотности костной ткани в нижней части тела (нижние конечности,
поясничные позвонки и их дорсальные элементы);
увеличение минерализации в костях черепа и рук;
установление отрицательного кальциевого баланса.
Сердечно-сосудистая система:
более частые сердечные аритмии у некоторых космонавтов при нагрузках;
снижение устойчивости к ортостатическим и физическим нагрузкам;
некоторые знаки венозного застоя во внутренних органах.
Эндокринная система:
уменьшение в плазме крови уровня АКТГ и увеличение кортизола;
снижение концентрации АДГ в плазме крови и моче и активация ренин-ангиотензин-альдостероновой
системы.
Водно-солевой баланс:
гипогидратация организма — уменьшение объема плазмы и межклеточной жидкости;
установление отрицательного баланса ряда ионов.
Метаболизм:
отрицательный азотистый баланс и превалирование процессов катаболизма;
уменьшение аминокислотного пула крови;
прогрессирующее замедление утилизации глюкозы при сахарной нагрузке.
Система крови:
развитие функциональной эритроцитопении;
морфологические и метаболические изменения в эритроцитах;
уменьшение активности Т-клеточной системы иммунитета.
АДАПТАЦИЯ ЧЕЛОВЕКА К НЕВЕСОМОСТИ
257
Таблица 2
Средства профилактики неблагоприятного влияния невесомости
1. Постоянное ношение специальных костюмов «Пингвин», создающих нагрузку по продольной оси тела, для
имитации веса, воспроизведения определенной степени деформации и стимуляции механорецепторов в
опорно-двигательном аппарате.
2. Физические тренировки на тредбане с системой притяга и велоэргометре, направленные на поддержание
тренированности наиболее важных систем, сохранение земных двигательных навыков и работоспособности
организма и стимуляцию некоторых групп рецепторов.
3. Механические средства (ОДНТ, бедренные окклюзионные манжеты), воспроизводящие в невесомости
эффект гидростатического давления крови, направленные на уменьшение перераспределения крови и на
стимуляцию нервно-рефлекторных механизмов, регулирующих кровообращение.
4. Средства задержания жидкости в организме (отрицательное давление на нижнюю часть тела и
водно-солевые добавки).
5. Средства, препятствующие депонированию крови в нижних конечностях (эластические и противоперегрузоч-
ные костюмы) и способствующие повышению переносимости перегрузок и ортостатической устойчивости.
6. Электр о стимуляция некоторых мышц для поддержания силовых характеристик, статической и
динамической выносливости мышц.
нижних конечностей, растянутые скопившейся
в них кровью, имеют более высокое
напряжение, чем стенки вен шеи и головы,
находящиеся в спавшемся состоянии. Снятие
гидростатического давления приводит к тому,
что этот существующий в условиях Земли
градиент эластических сил стенок вен,
расположенных ниже и выше сердца,
способствует перемещению крови из вен нижних
конечностей в сторону головы, вследствие чего
увеличивается переход жидкости в интерсти-
циальное пространство сосудистых областей,
расположенных выше сердца.
Снятие весовой нагрузки (устранение
опоры) и уменьшение мышечных усилий при
выполнении статической и динамической
работы являются основной причиной
снижения активности, а также развития де-
тренированности мышц, участвующих в
поддержании позы.
Важно отметить, что результаты
исследований, полученные как в пилотируемых
орбитальных полетах, так и в экспериментах
на животных по программе «Бион»,
показали, что в состоянии микрогравитации
отсутствуют типичные признаки стресса. Вместе с
тем неспецифические приспособительные
реакции отчетливо проявляются у
космонавтов при реадаптации к условиям Земли
после завершения космических полетов.
Вестибулярная1 и другие
сенсорные системы
Ослабление афферентной импульсации,
поступающей в ЦНС с вестибулярного ап-
1 Вестибулярный аппарат — нейродинамичес-
кая система ориентации тела в пространстве.
парата, не согласуется с активностью
других сенсорных систем. Механизмы
нейрофизиологической адаптации в условиях
невесомости, направленные на устранение
возникшего рассогласования, реализуются
посредством изменения поведения (резкого
снижения скорости и амплитуды
движений головы, а также временного отказа
или частичного ограничения приема пищи
и воды). Эти реакции, однако, не
достаточны для устранения симптомов
космической болезни движения. Полезный для
организма приспособительный эффект
может быть достигнут лишь за счет
включения механизмов долговременной
адаптации посредством формирования
соответствующей функциональной системы и
компенсации развивающихся сенсорных
конфликтов; организму требуется
определенное время для приобретения
индивидуального опыта, поиска и выбора
оптимальной формы реагирования.
Двигательная система
В начальной фазе микрогравитации
обнаружены существенные изменения точности
прилагаемых мышечных усилий и
скорости движений. При длительном пребывании
в условиях невесомости снижаются силовые
показатели мышц голени, уменьшаются
объем и плотность мышц спины. Эти
сдвиги зависят главным образом от
перестройки биомеханической структуры движений
и двигательного стереотипа, изменения
афферентного входа, развития детрениро-
ванности и атрофических процессов позной
мускулатуры вследствие пониженной
нагрузки и изменения активности ряда гор-
258
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
монов (кортизола, инсулина, тироксина и
гормона роста).
В послеполетном периоде выявлено
повышение чувствительности сенсорных
опорных и мышечных входов, субатрофия,
атония, уменьшение массы, снижение силовых
и скоростных характеристик
антигравитационных мышц, нарушение межконечност-
ной синергии (согласования) и системы
двигательного регулирования. Степень
выраженности этих изменений мало
зависела от длительности полета и в основном
определялась видом, интенсивностью и
объемом выполненных во время полета
физических тренировок.
Костная система
Изменения костной системы после
длительных космических полетов связаны с
перераспределением минеральных веществ
таким образом, что минерализация в костях
черепа и рук повышается, а в поясничном
отделе позвоночника и костях нижней
половины тела, особенно в костях таза —
снижается. Общая масса скелета не претерпевает
заметных изменений. Снижение плотности
костной ткани объясняется в настоящее время
уменьшением депонирования кальция и
фосфора, а также нарушением вещества кол-
лагеновой матрицы в костях, несущих
весовую нагрузку. Эти изменения могут быть
связаны с уменьшением нагрузки на скелет
и модификацией поля сил деформации, что
нарушает процессы саморегуляции и
питания костной ткани.
Важная роль в поддержании гомеоста-
за костной ткани принадлежит
продуцируемому лимфоцитами остеокласт-активиру-
ющему фактору. Данный фактор в сочетании
с интерлейкином-1 и некоторыми другими
факторами участвует в механизме
поддержания равновесия между функциями
остеобластов и остеокластов2. Пребывание в
условиях невесомости, сопровождающееся
сдвигами в системе иммунитета с
изменением содержания и активности популяций
ряда иммунокомпетентных клеток, может
приводить к изменению продукции ос-
теокласт-активирующего фактора и как
следствие способствовать потере кальция
клетками костной ткани. В результате
увеличивается содержание ионов кальция и
фосфора в крови и их выведение из
организма. В конечном счете уменьшается
минеральная насыщенность и развиваются
деструктивные процессы в костной ткани.
Однако пока даже в самых длительных
полетах потеря кальция у человека, а,
следовательно, и изменение прочности
скелета не имели угрожающего характера.
2 Клетки костной ткани.
Сердечно-сосудистая система
Основными факторами, определяющими
изменения кровообращения и водно-солевого
баланса в условиях невесомости, являются
перераспределение жидкости в организме
из-за падения гидростатического давления
крови, а также уменьшение роли
мышечной системы в гемодинамике из-за
развивающейся детренированности. Перемещение
жидкости в верхнюю часть тела вызывает
включение депрессорных рефлексов с
артериальных механорецепторов этой области.
Сократительная способность миокарда,
средние величины ударного и минутного
объема сердца и артериального давления не
изменяются. Изменения соотношений
длительности основных фаз сердечного цикла
левого желудочка указывают на увеличение
систолической работы сердца и его
присасывающей функции. Ультразвуковые
исследования обнаруживают признаки венозного
застоя (возрастание размера ряда
внутренних органов и увеличение поперечного
сечения магистральных сосудов брюшной
полости). В ближайшем послеполетном периоде
наблюдается существенное снижение
физической и ортостатической устойчивости.
Долговременные адаптивные реакции
проявляются в активации некоторых нейро-
гуморальных механизмов, поддерживающих
циркуляторный гомеостаз, в частности, в
увеличении активности ренин-ангиотензин-
-альдостероновой системы. В результате
устанавливается новый уровень
функционирования системы кровообращения и
взаимосвязанных с ней систем. Этот уровень
характеризуется перераспределением жидкости с
увеличением ее содержания в верхней
части тела, снижением роли мышечного
фактора в кровообращении, а также
уменьшением активности симпато-адреналовой системы
и некоторых прессорных гормонов (кате-
холаминов, АКТГ, АДГ). На этом фоне
дозированные тесты с приложением
отрицательного давления к нижней части тела (ОДНТ)
и с нагрузкой на велоэргометре
сопровождаются более значительным, по сравнению
с условиями на Земле, перемещением крови
из сердечно-легочной системы в нижние
конечности и изменениями ударного объема
сердца и частоты сердечных сокращений.
Описанные реакции, наблюдавшиеся у ряда
космонавтов в полетах, свидетельствуют о
развитии в условиях микрогравитации
ортостатической и физической
детренированности.
Водно-солевой обмен
Состояние водно-солевого баланса
характеризовалось в отдельные периоды
длительных полетов снижением содержания в
АДАПТАЦИЯ ЧЕЛОВЕКА К НЕВЕСОМОСТИ
259
крови и увеличением выведения с мочой ряда
ионов (натрия, кальция и фосфора),
отрицательным балансом воды (на протяжении
первых б суток полета), некоторым
снижением положительного баланса калия и
магния. В длительном 438-суточном полете при
помощи радиоизотопной методики
обнаружено прогрессирующее снижение объема
внеклеточной жидкости на 4~5-е сутки
полета. Это снижение составило 9—10% при
примерно одинаковом уменьшении
сосудистой и внеклеточной фракций, а к концу
полета — 18% при неизменном уровне
объема внутриклеточной жидкости.
В основе изменений водно-солевого
баланса лежит включение срочных
приспособительных механизмов (нейрональных,
гуморальных и прямых гидравлических),
которые реагируют на перемещение крови
в верхнюю часть тела и увеличение ее внут-
рисосудистого и центрального объема и
способствуют удалению «кажущегося»
избытка жидкости из организма, уменьшению
объема плазмы крови и увеличению
выведения ряда электролитов почками. В ответ
на уменьшение объема плазмы крови
активируется ренин-ангиотензин-альдостеро-
новая система, что способствует развитию
устойчивого отрицательного баланса калия.
Он также поддерживается из-за
уменьшения калийдепонирующих возможностей
мышц в связи с развитием в длительном
полете их детренированности и потерей
мышечной массы.
Система крови
Основные изменения крови проявлялись
в развитии синдрома функциональной эрит-
роцитопении (снижении числа эритроцитов
в единице объема крови), сопряженного с
преобразованием структуры части
эритроцитов и снижением содержания
гемоглобина. Сниженное количество эритроцитов в
единице объема крови сохранялось в
течение 2-3 недель от момента старта и после
60 дней полета начинало
восстанавливаться вследствие снижения концентрации
гемоглобина в крови, которое стимулирует
процесс образования эритроцитов (эритро-
поэза). К концу 438-суточного полета
показатель гематокрита (отношение объема
форменных элементов к объему плазмы) мог
даже возрастать из-за снижения объема
плазмы.
Развитие функциональной эритроцито-
пении под влиянием невесомости может быть
связано с торможением эритропоэза.
Снижение стимуляции эритропоэза в
начальной стадии космического полета, скорее
всего, вызывается уменьшением объема
плазмы крови и должно рассматриваться
как приспособительная реакция,
направленная на сохранение нормального
соотношения эритроцитов и гемоглобина в единице
объема крови. Дополнительным фактором
может служить снижение уровня
метаболизма и кислородного запроса ряда тканей
из-за уменьшения функциональной нагрузки
на мышечную и некоторые другие
системы организма. Можно ожидать, что в
условиях невесомости (вследствие
уменьшения афферентного входа с гравирецепторов)
снижается активность заднего отдела
гипоталамуса, который может стимулировать
образование эритроцитов путем усиления
продукции эритропоэтина (стимулятора
эритропоэза) и посредством изменения
деятельности эндокринных желез, в
частности, гипофиза. Развивающиеся в костной
ткани деструктивные процессы, в свою
очередь, могут быть причиной нарушения
метаболизма эритроцитов и костного
мозга, а также торможения эритропоэза.
Заключение
Общие итоги медицинских исследований,
выполненных в кратковременных и
длительных космических полетах, свидетельствуют,
что человек достаточно хорошо
приспосабливается и эффективно работает в
условиях невесомости длительностью до 14,5
месяцев. Наблюдавшиеся при этом изменения ряда
функций носили приспособительный
характер, соответствовали воздействующим
факторам и были обратимыми. Реакции в
послеполетном периоде в значительной степени
определялись полнотой выполнения
профилактических мероприятий, а также
зависели от индивидуальных особенностей членов
экипажей и в некоторой мере от
длительности полетов. В целом, со стороны основных
физиологических систем, на которых, в
первую очередь, могло сказаться
неблагоприятное влияние факторов космического полета,
не выявлено существенных изменений,
которые могли бы препятствовать
дальнейшему планомерному увеличению
продолжительности космических полетов.
Адаптация человека к невесомости
сопровождается развитием срочных и
долговременных приспособительных реакций и
удовлетворительно достигается за счет
физиологической и, в определенной
степени, структурной перестройки организма.
Особенность развития адаптационных
процессов при длительном пребывании в
невесомости состоит в том, что в этих
условиях, наряду с увеличением нагрузки на ряд
функциональных систем, снижается нагрузка
на опорные структуры организма. Это
сопровождается частичной утратой свойств и
качеств, приобретенных человеком в процессе
индивидуального развития и жизни в
условиях воздействия земной силы тяжести.
260
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
Выявленные в космических полетах
изменения основных функций организма не
сказываются существенным образом на
работоспособности и самочувствии
космонавтов и их деятельности, связанной с
реализацией программы полета, включая работы
вне космического летательного аппарата.
Исключение составляет лишь начальный
период адаптации к условиям
космического полета, во время которого у
большинства космонавтов ухудшается самочувствие
в связи с появлением симптомов болезни
движения, ощущения переполнения
головы кровью, заложенности носа и т. д.
Развивающиеся другие сдвиги в
организме человека (уменьшение объема
нижних конечностей, атрофия или субатрофия
антигравитационных мышц, увеличение
размеров ряда внутренних органов,
изменение показателей водно-солевого
баланса и т. д.) субъективно не проявляются и
не сказываются на самочувствии
космонавтов, что свидетельствует о
достаточности механизмов адаптации организма
человека к новой среде жизнедеятельности.
После -острого периода адаптации в
системе «организм-среда» устанавливается
равновесие, которое обеспечивает
адекватный уровень работоспособности,
необходимый для конкретной деятельности в
условиях космического полета.
При возвращении космонавтов на
Землю в ближайшем периоде реадаптации
возникают гравитационно-обусловленные
острые реакции организма со снижением его
работоспособности.
В предпринятых до сих пор полетах
практически не возникало серьезных
психологических проблем. Это достигалось
высокой мотивацией космонавтов,
эффективностью психологического отбора,
сбалансированным режимом труда и отдыха,
специальными мерами по психологической
поддержке экипажа, созданием
надлежащего комфорта и уюта на станции,
рациональным построением режима труда и
отдыха, адекватным по продолжительности
сном и полноценным питанием.
Рассмотренные в настоящей работе
результаты физиологических исследований не
исчерпывают всех проблем, связанных с
пребыванием человека в длительном
космическом полете. Вполне понятно, что
возможность выполнения пилотируемых полетов
неразрывно связана с созданием,
непрерывным совершенствованием и модернизацией
систем медицинского обеспечения
безопасности, направленных на сохранение
здоровья и работоспособности экипажей.
Литература
Атъков О. Ю., Бедненко В. С. Гипокинезия,
невесомость, клинические и физиологические
аспекты. М.: Наука, 1989.
Физиологические проблемы невесомости. М.:
Медицина, 1990, с. 15.
Н- И. Волков
ФИЗИЧЕСКАЯ РАБОТОСПОСОБНОСТЬ ЧЕЛОВЕКА
Введение
Изучение основ физической
работоспособности человека традиционно проводится
при обследовании спортсменов. Один из
основателей современной биохимии и
физиологии спорта лауреат Нобелевской премии
А. В. Хил л в своей книге «Мышечные
движения человека», изданной в 1927 г., писал:
«Почему мы исследуем спортсменов? На этот
вопрос можно дать два ответа: во-первых,
процессы, происходящие при выполнении
физических упражнений, просты и
измеримы, упражнения выполняются с большой
напряженностью и, как правило, до предела
человеческих возможностей — этого нет в
трудовых процессах; во-вторых, сами
спортсмены, будучи здоровыми и в состоянии
динамического равновесия, могут быть
объектом изучения и без всякой опаски для
здоровья повторять вновь и вновь с
совершенной точностью заданные упражнения».
Действительно, чтобы достичь высоких
результатов и побеждать в борьбе со столь
же одаренными и работоспособными
соперниками, спортсмену необходимо овладеть
всем арсеналом сложных двигательных
навыков и достичь в ходе многолетней
подготовки наивысшего уровня развития тех
качеств, от которых в наибольшей мере
зависит успех в избранном виде
упражнений. Вместе с тем следует подчеркнуть, что
все основные закономерности
работоспособности, установленные в результате
исследований на спортсменах, в полной мере
приложимы и к трудовой деятельности
нетренированных людей.
Факторы, лимитирующие
физическую работоспособность
человека
Физическая работоспособность
человека определяется как общими для всех
видов двигательной активности факторами, так
и специальными факторами, действие
которых проявляется только в условиях
конкретного вида деятельности. Основными
факторами общего значения являются:
1) биоэнергетические (аэробные и
анаэробные) возможности человека;
2) нейромышечные функции (сила и
техника выполнения упражнения);
3) психологические факторы (мотивация
и тактика ведения спортивного состязания).
Мышечная сила и биоэнергетические
возможности составляют группу факторов
потенций, а техника, тактика и
психологическая подготовка объединяются в
группу факторов производительности. Последние
играют важную роль в том, чтобы в
конкретных условиях избранного вида
деятельности силовые и биоэнергетические потенции
проявились в полной мере.
Сила и мощность
В проявлениях мышечной силы и
мощности1 определяющее значение имеют
структурная организация и
ферментативные свойства сократительных белков мышц.
Величина усилия, развиваемого мышцей
в процессе сокращения, пропорциональна
числу поперечных соединений (мостиков)
между актиновыми и миозиновыми
нитями в миофибриллах. Потенциально
возможное число этих соединений, а,
следовательно, и максимальная мышечная сила зависит
от длины саркомеров миофибрилл,
определяющей степень возможного контакта
между актиновыми и миозиновыми
нитями. Максимальная сила мышцы,
отнесенная к площади ее поперечного сечения,
достигает 7 кг/см2.
Другой фундаментальной
характеристикой мышцы является скорость укорочения
при перемещении определенного груза. Чем
больше груз, тем меньше скорость
сокращения, это так называемая зависимость
сила—скорость.
Произведение этих двух величин есть
мощность. Скорость сокращения, а также
и мощность мышцы, зависят от АТФазной
активности миозина, которая
определяется количеством актомиозиновых связей,
образующихся в единицу времени. В интакт-
ной мышце человека развиваемая мощность
пропорциональна величине суммарной
АТФазной активности, т. е. общей
скорости расщепления АТФ (рис. 1).
Мышечные волокна разделяются по
скорости сокращения на два основных
класса — «быстрые» и «медленные» (рис. 2).
Первые предназначены для выполнения
быстрых, но кратковременных усилий, вторые —
для относительно медленных, но
длительных усилий. Медленные мышцы (ST-тип) в
связи с высоким содержанием в них
митохондрий и интенсивным аэробным (с
участием кислорода) обменом имеют красный
1 В теории и практике спорта эти
физические качества обычно объединяются в понятии
скоростно-силовой подготовленности спортсмена.
262
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
цвет, а быстрые мышцы (FT-тип) с низким
содержанием митохондрий и
преимущественно анаэробным (без участия кислорода)
обменом имеют белый цвет.
Более высокая скорость сокращения
быстрых, белых волокон обусловлена
повышенной АТФазной активностью миозина по
сравнению с его активностью в медленных,
красных волокнах. Соответственно
максимальная величина относительной
мощности быстрых волокон составляет около 155 Вт
на 1 кг веса мышц, а медленных волокон —
40 Вт на 1 кг веса мышц.
Быстро и медленно сокращающиеся
волокна различаются также по порогу
раздражения, что определяет
неодновременное их включение в работу. Работа
умеренной интенсивности, сопряженная с невысокой
частотой возбуждений, приходящих к
мышцам по нервам, выполняется в основном
медленными двигательными волокнами. С
ростом интенсивности работы, когда частота
возбуждений превышает пороговое
значение для быстрых двигательных волокон,
происходит вовлечение в нее быстро
сокращающихся мышечных волокон.
В скелетных мышцах человека быстро и
медленно сокращающиеся волокна находятся
в разных соотношениях. Чем больше процент
быстро сокращающихся мышечных волокон
в составе скелетной мышцы, тем выше ее
скоростно-силовые характеристики. Изменения
этой пропорции соответственно
сказываются на функциональных свойствах мышцы.
Поэтому бегуны-спринтеры, в икроножной
мышце которых содержание быстро
сокращающихся белых волокон достигает 60%,
заметно превосходят бегунов на длинные
дистанции, у которых только 35% белых волокон,
по значениям максимальной мощности (120 Вт
на 1 кг веса мышц против 85 Вт). Величина
же максимальной силы для быстрых и
медленных волокон примерно одинакова. Так как
в большинстве скелетных мышц красные и
белые волокна находятся в определенных
пропорциях, сократительные свойства этих мышц
будут относиться к той области на рис. 2,
которая заключена между экстремальными
значениями для красных и белых волокон.
Биоэнергетические способности
человека
Самыми важными факторами,
определяющими физическую работоспособность
человека, являются его биоэнергетические
возможности (см.: Скулачев В. П. Эволюция
биологических механизмов запасания
энергии. Том 8). Образование энергии в мышцах
может осуществляться аэробным или
анаэробным путем. В анаэробных условиях
синтез АТФ происходит при ферментативном
распаде углеводов — гликолизе, конечным
100
§
if
н
о
о
|50
о:
со
х
л
§
1
0 0,01 0,1 1,0
АТФазная активность миозина S
кол-во актина, усл. ед.
Рис. 1. Зависимость максимальной мощности,
развиваемой мышцей, от величины суммарной
АТФазной активности миофибрилл
продуктом которого является молочная
кислота (лактат). В аэробных условиях
источником энергии служит окисление пищевых
веществ, главным образом углеводов и
1,0
0,8
5:
со
§0,6
о
о:
со
i
с;
0
?0,4
о
о
i
I-
О
0,2
0 12 3 4 5 6
Максимальная скорость сокращения, /0/с
Рис. 2. Зависимость между относительной силой
(максимальная сила принята за 1) и максимальной
скоростью укорочения (в единицах длины мышцы за
1 с) для мышц, состоящих из медленных (левая
кривая) и быстрых (правая кривая) волокон
ФИЗИЧЕСКАЯ РАБОТОСПОСОБНОСТЬ ЧЕЛОВЕКА
263
Таблица 1
Критерии мощности, емкости и эффективности для аэробного
и анаэробного биоэнергетических процессов
Метаболический источник
Максимальная мощность, на 1 кг
веса, кДж/мин
Максимальное время удержания, с
Максимальная емкость, кДж/кг
Метаболическая эффективность, %
Алактатный анаэробный
процесс (АТФ+КрФ*)
3770
6
630
80
Анаэробный
гликолиз
2500
60
1050
44
Аэробный процесс
(окисление углеводов и жиров)**
1250
600
оо
60
* КрФ — креатинфосфат.
** При среднем значении максимального потребления 02 = 60 мл/кг • мин.
жиров, в реакциях цикла Кребса,
протекающих в митохондриях.
В соответствии с этим принято выделять
три обобщенных функциональных свойства,
определяющих физическую
работоспособность человека:
— алактатную анаэробную способность,
связанную с процессами анаэробного
расщепления АТФ и креатинфосфата в
работающих мышцах;
— гликолитическую анаэробную
способность, отражающую возможность усиления
при работе гликолитического процесса;
— аэробную способность, связанную с
возможностью выполнения работы за счет
усиления аэробных процессов в тканях при
одновременном увеличении доставки
кислорода к работающим мышцам и
утилизации его.
Метаболическая производительность
каждого из отмеченных выше источников энергии
может быть охарактеризована при помощи
количественных критериев трех типов:
— критерия мощности, отражающего
скорость образования энергии в
метаболическом процессе;
— критерия емкости, отражающего
размеры доступных для использования
субстратных фондов или общий объем
метаболических превращений в работающих мышцах за
время работы;
— критерия эффективности,
определяющего степень использования энергии для
выполнения специфической мышечной
работы.
Аэробные и анаэробные
биоэнергетические процессы заметно различаются по
значениям мощности, емкости и
эффективности (табл. 1). Наибольшая скорость
энергопродукции, соответствующая максимальной
мощности алактатного анаэробного
процесса, достигается при очень кратковременной
работе. Выход энергии в гликолитическом
анаэробном процессе происходит медленнее,
но он более продолжителен. Максимальная
мощность аэробного процесса наименьшая и
достигается при работе, длительность
которой составляет не менее 2—3 мин. Величины
максимальной мощности аэробного,
гликолитического и алактатного процессов
примерно соотносятся как 1:2:3.
Мощность гликолитического и
алактатного анаэробных процессов быстро
понижается с увеличением длительности работы. Это
связано с относительно небольшими
значениями их энергетической емкости. Аэробный
процесс по данному критерию во много раз
превосходит алактатный и гликолитический
анаэробные процессы. Субстратные фонды
для процессов окисления в митохондриях
скелетных мышц включают не только
внутримышечные запасы углеводов и жиров, но
также глюкозу, жирные кислоты и
глицерин крови, запасы гликогена в печени и
резервные жиры различных тканей
организма. Это позволяет поддерживать работу
весьма продолжительное время.
Заметно отличаются и показатели
эффективности для аэробных и анаэробных
биоэнергетических процессов. Наибольшая
эффективность преобразования энергии характерна для
алактатного анаэробного процесса,
наименьшая — для анаэробного гликолиза, а в
аэробном процессе метаболическая эффективность
промежуточная (табл. 1).
Приведенные в табл. 1 данные
получены путем непосредственных определений
биоэнергетических критериев у
высококвалифицированных спортсменов. У людей,
ведущих умеренно активный образ жизни,
мощность и емкость биоэнергетических
процессов заметно ниже.
Выносливость
Выносливость обычно определяют как
способность работать без утомления и
противостоять утомлению, когда оно возника-
264
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
ет в процессе выполнения работы. В
соответствии с наличием трех путей
производства энергии выделяют и три составляющих
выносливости — алактатную, гликолитичес-
кую и аэробную. Общее проявление
выносливости, оцениваемое по времени работы до
отказа (В), в этом случае может быть
представлено суммой таких времен для
каждого из упомянутых выше процессов — алак-
татного, гликолитического и аэробного:
В = В +В +В .
ал гл аэ
Каждое слагаемое представляет собой
отношение соответствующих емкости (Е, кКал)
и мощности (М, кКал/мин), помноженное на
относительную эффективность (Э, %),
В = (Е/М)х(Э, %).
Биоэнергетические критерии
выносливости могут быть установлены на основе
точных эргометрических измерений
выполняемой механической работы или путем
прямых физиологических и биохимических
измерений соответствующих
биоэнергетических функций. Для этого применяются
стандартизованные лабораторные и
специальные («полевые») тесты, ориентированные
на избирательную оценку каждого
отдельного компонента выносливости. Некоторые
из наиболее информативных показателей,
используемых в качестве
биоэнергетических критериев аэробного и анаэробных
компонентов физической работоспособности,
приведены в табл. 2.
Возрастные изменения
физической работоспособности
Аэробные и анаэробные возможности
возрастают по мере физиологического
созревания организма человека. Взросление
сопровождается увеличением общей мета-
болизирующей массы тела, количества и
активности ключевых ферментов
аэробного и анаэробного обмена в скелетных
мышцах, запасов энергетических веществ в
тканях. Параллельно совершенствуется работа
системы кровообращения, ответственной за
доставку к мышцам кислорода и
питательных веществ, а также устранение продуктов
распада.
Все эти показатели обычно достигают
наибольшего развития к 20-25 годам, в пору
полной физиологической зрелости
человека. В этом возрасте спортсмены достигают
своих лучших результатов в тех видах
спорта, где требуется значительное
проявление выносливости. После 40 лет
показатели физической работоспособности
человека постепенно понижаются и к 60 годам
становятся примерно вдвое меньше, чем в
20-летнем возрасте.
В процессе возрастного развития
наблюдаются определенные различия в
динамике отдельных биоэнергетических
показателей. Так, максимальная анаэробная мощность
у мужчин быстро увеличивается, достигая
своего максимума к 20 годам, сохраняется
на этом уровне примерно до 30 лет, а
затем начинает снижаться. У женщин этот
показатель характеризуется более быстрым
приростом в юном возрасте (максимум
достигается к 18 годам) и более выраженным
снижением в старшем возрасте. Показатель
мощности аэробного процесса —
максимальное потребление 02 — у мужчин достигает
своих наибольших значений к 25 годам,
удерживается на этом уровне до 40 лет и
затем понижается. У женщин наивысшие
значения максимума потребления 02
отмечаются к 20 годам и начинают снижаться
после 35 лет.
Способность к накоплению максимальных
количеств молочной кислоты в тканях (ана-
Таблица 2
Биоэнергетические критерии аэробного и анаэробных
компонентов выносливости
Критерии
Мощность
Емкость
Эффективность
Биоэнергетические показатели
Аэробные
Максимальное
потребление О2, критическая
мощность
Время удержания
максимального потребления С>2,
максимальный С^-приход
Кислородный эквивалент
работы, порог
анаэробного обмена
Гликолитические анаэробные
Максимум прироста лактата в
крови, максимум
«избыточного» выделения СС>2
Максимум накопления
молочной кислоты, общий С^-Долг,
наибольший сдвиг рН
Лактатный эквивалент работы,
отношение прироста рН на
единицу прироста работы
Алактатные анаэробные
Скорость распада КрФ,
максимальная анаэробная мощность
Размеры алактатного С^-Долга,
максимум расхода КрФ
Скорость оплаты алактатного
02-долга, отношение скорости
распада КрФ на единицу
прироста работы
ФИЗИЧЕСКАЯ РАБОТОСПОСОБНОСТЬ ЧЕЛОВЕКА
265
эробная гликолитическая мощность) как у
мужчин, так и у женщин достигает своих
наибольших значений к 22 годам и быстро
утрачивается в более зрелом возрасте.
Для показателей емкости и
эффективности биоэнергетических процессов
характерен замедленный темп развития.
Наивысших значений эти показатели достигают к
25-30 годам, и при систематической
тренировке могут удерживаться вблизи
максимально высокого уровня вплоть до 40-
45 лет. Темпы их снижения в пожилом и
старческом возрасте более выражены у
женщин.
Эти особенности возрастной динамики
показателей физической
работоспособности человека должны учитываться при
разработке программ по физическому
воспитанию подрастающего поколения и при
проведении занятий по оздоровительной
физической культуре с лицами разного
возраста и пола.
Влияние тренировки на уровень
физической работоспособности
Тренировка, имеющая целью повысить
максимальные показатели силы,
мощности или выносливости, должна быть
целенаправленной. На основе установленной
зависимости между силой и скоростью
сокращения мышц можно определить
основные требования для упражнений,
применяемых с целью развития скоростно-силовых
качеств человека. Так, при развитии
собственно силовых возможностей (повышения
максимальной силы мышц) величина
преодолеваемого сопротивления должна
составлять от 70 до 100% индивидуального
максимума силы. При этом в клетках возрастает
90
80
I70
|б0
840
Q.
Езо
С20
10
0 11 13 15 17 19 21
Возраст, годы
Рис. 3. Динамика прироста показателей различных
биоэнергетических функций в процессе
многолетней тренировки юных пловцов
МАМ— максимальная аэробная мощность; Ехс.
С02—выделение С0; V0 max— максимальное
потребление кислорода; max HLa— максимальное
выделение лактата; ПАНО — порог анаэробного обмена
содержание сократительных белков,
особенно актина, что обеспечивает увеличение
общей массы мышц.
При развитии способности выполнять
упражнение с максимальной скоростью
величина преодолеваемого сопротивления
должна составлять от 20 до 40%
индивидуального максимума силы, а при
совершенствовании способности к комплексному
проявлению силы и скорости сокращения,
т. е. мощности, оптимальные величины
внешнего сопротивления должны быть равны 40—
70% индивидуального максимума силы.
Биоэнергетические факторы
спортивной работоспособности также могут быть
заметно улучшены в процессе
тренировки. Это наглядно иллюстрирует график
(рис. 3), где представлена динамика
прироста показателей различных
биоэнергетических функций в процессе
многолетней тренировки юных пловцов. Наиболее
значительно и быстро возрастает
показатель максимального потребления
кислорода, достигающий 180-190% исходного
значения. Менее выраженные сдвиги, не
превышающие 50% исходного значения,
характерны для показателей «избыточного»
выделения С02 и максимальной
анаэробной мощности. Наименьшие темпы прироста
и наиболее длительные сроки развития
адаптации к тренировке отмечены для
показателей максимума накопления
молочной кислоты в крови и порога
анаэробного обмена.
Представленная картина изменений
биоэнергетических показателей в ходе
тренировки отражает общую закономерность
развития долговременной адаптации: ее
начальные этапы всегда осуществляются за
счет улучшения показателей мощности,
затем — показателей биоэнергетической
емкости и на заключительном этапе — за счет
улучшения биоэнергетической
эффективности. После прекращения тренировки
развитие процесса деадаптации происходит в
обратном порядке: прежде всего снижаются
показатели биоэнергетической
эффективности, затем — биоэнергетической емкости и
в последнюю очередь — показатели
мощности биоэнергетических процессов (см.: Пшен-
никова М. Г. Стресс и адаптация. Наст. том).
При направленной тренировке
наибольшие темпы развития и более длительное
поддержание максимально высокого уровня
характерны для биоэнергетических
показателей, определяющих выносливость к
длительной работе, т. е. аэробную способность.
Значительно медленнее поддаются
воздействию тренировки и недолго
удерживаются на максимально высоком уровне
биохимические и физиологические показатели,
составляющие основу скоростной
выносливости и скоростно-силовых качеств человека.
266
ФИЗИОЛОГИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ
1. Максимум потребления
02, мл/кг в мин
2. Максимальная
анаэробная мощность, кпм/мин
20
40 60
80
1
4Sx103 0
3. Скорость
образования молочной
кислоты, г/кг в мин
12 3 4
Рис. 4. Показатели мощности аэробного и анаэробных процессов у спортсменов различных специализаций
Номера соответствуют разным видам спорта
Выносливость, так же как и
физическая работоспособность в целом, зависит от
соотношения в уровне развития аэробной
и анаэробных способностей, которое
складывается под влиянием наследственности,
условий жизни и специализированной
тренировки. Так, самые высокие показатели
максимальной аэробной мощности
отмечаются у бегунов на длинные дистанции,
лыжников-гонщиков, конькобежцев и вело-
сипедистов-шоссейников (рис. 4).
Наибольшие значения алактатной анаэробной
мощности демонстрируют бегуны на короткие
дистанции, хоккеисты и велогонщики-тре-
ковики. Самыми большими величинами гли-
колитической анаэробной мощности обладают
велогонщики-трековики, бегуны на средние
дистанции, хоккеисты и ватерполисты. В то
1. Время удержания
максимума потребления
мин
8 10 0
2. Размеры алактат-
ного 02-долга, мл/кг
20
40
60
3. Максимум
молочной кислоты, мг%
0 50 100 150 200 250 300
Рис. 5. Показатели энергетической емкости аэробного и анаэробных процессов у спортсменов различных
специализаций
ФИЗИЧЕСКАЯ РАБОТОСПОСОБНОСТЬ ЧЕЛОВЕКА
267
Таблица 3
Влияние биоэнергетических факторов на уровень работоспособности
спортсменов, специализирующихся в разных видах упражнений
% общей дисперсии выборки, обусловленной влиянием изучаемого фактора)
Критерий
Аэробная мощность
Аэробная емкость
Аэробная эффективность
Гликолитическая анаэробная мощность
Гликолитическая анаэробная емкость
Алактатная анаэробная мощность
Алактатная анаэробная емкость
Метаболическая масса тела
Бег на
короткие
дистанции, №=84
37,0
?
?
9,7
12,9
17,9
7,8
?
Бег на
длинные
дистанции, №=56
41,0
17,0
7,7
6,2
14,8
3,6
?
3,6
Лыжные
гонки,
№=42
27,5
39,0
12,0
4,6
11,7
4,4
?
2,4
Плавание,
№=112
51,1
6,06
6,8
5,7
6,3
9,03
?
?
Скоростной
бег на
коньках, №=66
7,0
5,6
35,7
12,5
21,0
9,0
5,7
?
Баскетбол,
№=31
8,5
6,6
14,6
?
33,0
6,2
?
10,0
же время, как это видно из рис. 5,
наибольшую аэробную емкость имеют велоси-
педисты-шоссейники, лыжники-гонщики,
бегуны на средние и длинные дистанции.
Самые высокие величины алактатной
анаэробной емкости демонстрируют бегуны на
короткие дистанции, баскетболисты и борцы.
Наибольшие величины гликолитической
анаэробной емкости отмечаются у бегунов
на средние дистанции, велогонщиков-тре-
ковиков и хоккеистов.
Приведенные в табл. 3 данные
характеризуют относительное влияние отдельных
биоэнергетических факторов на общий
уровень физической работоспособности
спортсменов, специализирующихся в разных
видах упражнений. Как следует из приводимых
в таблице данных, в каждом виде спорта
существуют свои «ведущие»
биоэнергетические факторы, которые оказывают
определяющее влияние на уровень спортивных
достижений. Так, результаты в плавании,
беге на длинные дистанции и в лыжных
гонках зависят главным образом от аэробной
мощности, аэробной емкости и
гликолитической анаэробной емкости; в скоростном беге
на коньках — от аэробной эффективности и
гликолитической анаэробной емкости, в
плавании — от аэробной мощности и
алактатной анаэробной способности, в баскетболе —
от гликолитической анаэробной емкости и
аэробной эффективности. Таким образом,
каждому виду спорта присущ специальный
комплекс биоэнергетических факторов,
оказывающих основное влияние на уровень
физической работоспособности.
Литература
Биохимия. М.: Физкультура и спорт, 1986.
Astrand P. О., Rodahl К. Textbook of Work
Physiology (3rd ed.). New York: McGraw-Hill,
1986.
Wilmore J. H., Costill D. L. Physiology of sport
and exercise. Champaign: Human Kinetics, 1994.
ФИЗИОЛОГИЯ
И БИОХИМИЯ РАСТЕНИЙ
О. Н. Кулаева
СТРЕССОВЫЕ БЕЛКИ
Введение
Одно из общих свойств клеток всех
типов живых организмов состоит в том, что
в ответ на увеличение температуры они
включают синтез специфического набора
белков, называемых белками теплового шока
(БТШ), которые помогают клетке выжить
в условиях температурного стресса и
вернуться после его прекращения к
нормальной жизни. Сходство аминокислотной
последовательности (гомология) некоторых из
БТШ у самых различных организмов (от
бактерий до человека) говорит об их
высокой консервативности в эволюции,
характерной для жизненно важных белков. Все
это сделало их изучение одной из
важнейших областей биологических исследований,
в которой можно выделить три основных
направления:
1) изучение индукции БТШ при
тепловом шоке (ТШ), которая служит хорошей
моделью молекулярных механизмов
включения генов, регуляции их активности и
перестройки белоксинтезирующей системы;
2) выяснение молекулярных основ
устойчивости (резистентности) клеток к
стрессу, обусловленной БТШ;
3) изучение особой функции БТШ,
которая состоит в том, что они влияют на кон-
формацию других белков, препятствуя их
агрегации (см.: Hazpadoea H. А.
Внутриклеточная регуляция формирования нативной
пространственной структуры белков. Том 8).
В этой статье мы рассмотрим роль БТШ
в устойчивости клеток к тепловому шоку,
уделив особое внимание БТШ растений.
Будет показано, что у растений БТШ
участвуют в сложном комплексе программ,
вызываемых (индуцируемых) стрессом и
необходимых для выживания клеток в
стрессовых условиях.
Открытие БТШ
История открытия БТШ началась в 1962 г.
с работ итальянского генетика Ф. Ритоссы
на политенных хромосомах слюнных желез
личинок дрозофилы. Политенные
хромосомы можно изучать под световым
микроскопом. В них различимы отдельные «диски»,
соответствующие генам, и видно, как при
активации генов плотная структура диска
переходит в «пуф» — вздутие, образующееся
за счет разрыхления структуры
хроматина в активном участке и накопления
синтезирующихся молекул РНК. Ритосса
обнаружил, что повышение температуры с 20
до 37°С приводит к образованию пуфов там,
где они не появлялись при нормальной
температуре. Так были открыты гены теплового
шока. Правда, позднее было обнаружено,
что их можно активировать рядом
воздействий и при нормальной температуре.
Кодируемые этими генами белки были
идентифицированы только в 1974 г. Они получили
название БТШ. Вскоре выяснилось, что
синтез БТШ в ответ на увеличение
температуры характерен для клеток всех живых
организмов: бактерий, грибов, низших и
высших растений, животных и человека.
У всех организмов БТШ представлены
большим набором полипептидов и их
принято именовать в соответствии с
молекулярной массой, выраженной в килодальто-
нах (кДа). Например, БТШ с молекулярной
массой 70 кДа называют БТШ70. О важной
роли БТШ в жизни клеток говорит их
высокая консервативность в эволюции.
Например, аминокислотные последовательности
БТШ70 очень сходны у насекомых, птиц,
млекопитающих, грибов и растений.
Отдельные участки в БТШ70 сохраняют свыше
90% гомологии у бактерий и человека.
У растений БТШ были обнаружены в
1980 г. БТШ растений, как и БТШ других
организмов, весьма разнообразны. Они
образуют группы высокомолекулярных (110-
60 кДа) и низкомолекулярных (35-15 кДа)
белков. При двумерном электрофорезе
обнаруживается, что БТШ с одинаковым
молекулярным весом представлены
несколькими полипептидами. Их существование
обусловлено, с одной стороны, тем, что
каждое семейство БТШ кодируется, как
правило, несколькими генами (семейством
близких генов), различающимися главным
образом по регуляции их активности, а с
другой — модификациями, происходящими
после завершения синтеза БТШ. К числу
таких модификаций относится, например,
обратимое фосфорилирование.
Отличительными чертами растений являются наличие
множественых форм низкомолекулярных
БТШ и высокая интенсивность их синтеза
при ТШ.
Переключение клеток на синтез
БТШ при тепловом шоке
Синтез БТШ — стрессовая программа,
включаемая тепловым шоком (рис. 1). Он
происходит при подъеме температуры на
8-10°С выше нормальной. Например, у
272
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
ТШ
ТШ
Клеточная
стенка
Рис. 1. Схема индукции синтеза белков теплового шока (БТШ) в клетках
ТШ—тепловой шок, ТФ — трансфактор генов БТШ, РЭ— регуляторный элемент генов БТШ, включающий их
экспрессию при ТШ. Зеленые волнистые линии— мРНК БТШ. Зеленые спирали— БТШ. Красными стрелками
обозначена система ответа клетки на ТШ вплоть до поступления БТШ во все структуры клетки, включая ядро,
митохондрии, хлоропласты и цитоплазматические гранулы ТШ. Синей волнистой линией обозначена дошоко-
вая мРНК (в дошоковых полисомах и гранулах ТШ)
широко применяемого в генетических и
физиологических исследованиях растения
арабидопсис (резушка Таля — Arabidopsis
thaliana), имеющего малый размер
генома, синтез БТШ происходит при
температуре 28-37°С, в листьях ячменя
наибольшая скорость синтеза БТШ достигается при
40°С, а в листьях риса — при 45°С.
Необходимо заметить, что индукция синтеза
отдельных представителей БТШ может
происходить при разных температурах
нагрева и его продолжительности. Поэтому
различают ранние и поздние БТШ.
Переключение нормальной жизни клетки
на стрессовую программу — событие
чрезвычайное, которое осуществляется
одновременно на многих уровнях регуляции. Оно
включает в себя репрограммирование
генома — тормозится экспрессия генов,
активность которых характерна для жизни клетки
в нормальных условиях, и активируются
гены теплового шока. В результате в
клетках растений, как и в клетках дрозофилы,
через 5 мин от начала стресса появляются
мРНК, кодирующие БТШ. Тепловой шок
вызывает не только репрограммирование
генома и, следовательно, изменение состава
вновь синтезируемых мРНК, но и
репрограммирование рибосом — распад полисом,
синтезирующих белки, типичные для
нормальных условий обитания, и
формирование полисом, синтезирующих БТШ (рис. 1).
Стремительное включение синтеза БТШ не
только на транскрипционном (синтез РНК
на ДНК), но и на трансляционном (синтез
белка на мРНК) уровне достигается в
результате многих событий. ТШ вызывает
изменения в мРНК, синтезированных в
клетке до шока и модифицирует белковые
факторы трансляции и рибосомные белки.
Кроме того, мРНК БТШ имеют отличия от
мРНК обычных белков. Все это приводит к
ослаблению, а затем и прекращению
синтеза обычных белков в клетках при ТШ и
переключению аппарата белкового
синтеза на синтез БТШ. В результате в клетках
уже через 15 мин после начала теплового
воздействия обнаруживаются БТШ; их
синтез постепенно нарастает, достигая
максимума за 2-4 ч ТШ, а затем начинает
ослабевать. У дрозофилы синтез БТШ происходит
на фоне прекращения синтеза белков,
типичных для нормальной температуры.
Однако у растений это выражено не так резко.
СТРЕССОВЫЕ БЕЛКИ
273
У них синтез разных белков проявляет
различную устойчивость к нагреву.
На рис. 2 представлена флуорограмма
радиоактивных белков, выделенных из
семядолей тыквы и их хлоропластов. Синтез
белков в семядолях происходил в течение
1,5 ч в присутствии (358)-метионина.
Включение (358)-метионина в белки во время их
синтеза позволяет получать меченые
белковые молекулы. После выделения белки
фракционировали при помощи
электрофореза в полиакриламидном геле. Рис. 2
показывает, что в условиях ТШ ослабевал синтез
ряда белков, характерных для нормальной
температуры, хотя синтез большой и
малой субъединиц (БС и МС) рибулозобисфос-
фаткарбоксилазы (РБФК) — ключевого
фермента фотосинтеза — продолжался и при
40°С. ТШ усиливал синтез высоко- и
низкомолекулярных БТШ. Набор БТШ
хлоропластов существенно отличался от набора
БТШ семядолей.
На рис. 3 видно, что синтез различных
БТШ хлоропластов происходит при разной
температуре. Синтез высокомолекулярных
БТШ активировался в диапазоне 34-42°С,
ослабевал при 44°С и резко снижался при
46°С. Индукция синтеза низкомолекулярных
БТШ была особенно сильной при 40-42°С.
Существенное угнетение синтеза БС РБФК
происходило только при 44°С. Необходимо
подчеркнуть, что почти все из
обнаруженных БТШ хлоропластов кодируются в ядре
и синтезируются в цитоплазме, откуда
потом транспортируются в хлоропласт, где и
выполняют защитную функцию во время ТШ.
Включение генов БТШ при высокой
температуре определяется регуляторны-
кДа
кДа
БС
-98
-94
-82
-74
-72
-64
94,0
67,0
43,0
-94
-85
-74
«-60
БС
«-36
30,0
-26
-24
«-20
«-18
«-26
«-24
20,1
МС
14,4
МС
28° 40°
Хлоропласты
28° 40°
Семядоли
28° 34°
36° 40°
42° 44°
46°
Рис. 2. Флуорограмма радиоактивных белков
семядолей тыквы и их хлоропластов
Синтез белка происходил в семядолях в течение
1,5 ч при нормальной (28°С) и шоковой (40°С)
температуре в присутствии (358)-метионина. Затем
белки извлекали из семядолей и выделенных из них
хлоропластов и разделяли электрофорезом в
полиакриламидном геле. На рисунке представлена
флуорограмма радиоактивных белков. Цифрами
отмечены молекулярные массы БТШ в килодальтонах.
БС и МС — большая и малая субъединицы рибуло-
зобисфосфаткарбоксилазы
Рис. 3. Температурная зависимость синтеза белков
хлоропластов семядолей тыквы
Условия получения флуорограммы меченых белков
те же, что и на рис. 2. Синтез белка в присутствии
(358)-метионина происходил в семядолях при
температурах, указанных на рисунке. Слева указано
положение на электрофореграмме стандартных
белков и их молекулярных масс, которые позволяют
судить о молекулярных массах меченых хлоропластных
белков. Цифры справа означают молекулярные
массы БТШ в килодальтонах. БС и МС— большая и
малая субъединицы РБФК [из: Kusnetsov, Mikulovich,
Kukina etal. FEBS Lett. 1993, vol. 321, № 2/3, p. 189]
274
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
ми элементами генов БТШ (РЭ БТШ), т. е.
специфическими нуклеотидными
последовательностями ДНК в промоторной (регу-
ляторной) зоне этих генов. РЭ генов
разных БТШ сходны и обладают высокой
гомологией у всех эукариот. Например, в
промоторе гена БТШ70 кукурузы
обнаружены два таких РЭ, близких РЭ БТШ70
дрозофилы. Это говорит об эволюционном
консерватизме не только структуры БТШ,
но и механизма включения их синтеза.
РЭ ТШ включают гены БТШ после
взаимодействия со специфическими регулятор-
ными белками — факторами транскрипции
или трансфакторами этих генов. Эти
трансфакторы присутствуют в цитоплазме при
нормальных условиях (см. рис. 1). Тепловой
шок вызывает их модификацию, например
увеличение фосфорилирования, после чего
они приобретают способность
взаимодействовать с РЭ генов БТШ и тем самым
активировать эти гены. К настоящему времени
выявлены некоторые различия в механизмах
активации тепловым шоком трансфакторов
генов БТШ эволюционно далеких
организмов. Однако важнее подчеркнуть
принципиальную общность таких механизмов у всех
эукариот. Лучше всего об этом говорит
следующий факт. Введение в геном
растительной клетки генетической конструкции,
содержащей чужеродный репортерный ген (от
«репортер» — дающий информацию),
находящийся под контролем РЭ гена БТШ 7 0
дрозофилы вызывало экспрессию этого гена в
клетках растения при тепловом шоке.
Следовательно, трансфактор генов БТШ,
присутствующий в цитоплазме растительной
клетки, узнавал в условиях теплового шока РЭ
гена БТШ70 дрозофилы и включал
экспрессию репортерного гена.
После окончания теплового шока
синтез БТШ прекращается и возобновляется
синтез белков, характерных для клетки в
нормальных температурных условиях. При
этом при нормальной температуре мРНК
БТШ быстро разрушаются, тогда как сами
белки могут сохраняться существенно
дольше, обеспечивая, по-видимому, повышение
устойчивости клеток к нагреву. Длительное
пребывание клеток в условиях ТШ
обычно также приводит к ослаблению и
прекращению синтеза БТШ. В этом случае
включаются механизмы регуляции синтеза БТШ
по принципу обратной связи.
Доказательства защитной
роли БТШ
Переключение клеток живых
организмов в условиях ТШ на синтез БТШ
вызывает естественное предположение, что эти
белки защищают клетки при стрессе.
Однако нужны прямые доказательства их
защитной функции. Общность структуры
БТШ и молекулярных механизмов
включения их синтеза у всех представителей
живого мира позволяет, при поиске таких
доказательств, привлекать материалы,
полученные на разных организмах.
Представление о защитной роли БТШ
подтверждают следующие данные:
1) выключение синтеза белка
специфическими ингибиторами во время теплового
шока, когда происходит синтез БТШ,
приводит к гибели клеток;
2) клетки можно закаливать, повышая
их термоустойчивость путем
предварительного кратковременного воздействия
повышенных температур. Оказалось, что
условия такой закалки совпадают с условиями
индукции синтеза БТШ. Это хорошо
показано на культуре клеток растений;
3) синтез БТШ индуцируется не
только ТШ, но и рядом других
неблагоприятных воздействий. У животных таких
дополнительных индукторов синтеза БТШ очень
много, у растений к ним относятся,
например, соли кадмия и арсенит. Оказалось, что
синтез БТШ, вызванный солями Cd или
другими нетемпературными воздействиями,
повышает устойчивость клеток к нагреву;
4) мутации (изменения в структуре гена),
повреждающие синтез БТШ, приводят к
потере устойчивости клеток к нагреву. Было
показано, что термочувствительность
дрожжей, возникшая в результате мутации гена
БТШ 104, может быть исправлена с
помощью генной инженерии путем введения в
геном дрожжей гена БТШ101 арабидопси-
са. Это не только прямое доказательство
ответственности БТШ 104 за
терморезистентность клеток, но и убедительное
свидетельство общности ее механизмов у
дрожжей и растений;
5) убедительное доказательство
защитной роли БТШ при нагреве получено на
клетках соединительной ткани крыс — фиб-
робластах. В клетки вводили антитела к
БТШ70. Это приводило к гибели клеток при
тепловом шоке из-за того, что антитела,
взаимодействуя с БТШ70, мешали
осуществлению их функции в условиях шока,
препятствуя, в частности, проникновению
БТШ70 в ядро, существенному для
проявления их защитного действия. При
нормальной температуре антитела к БТШ70 не
вызывали гибели клеток.
Совокупность представленных аргументов
убедительно подтверждает предположение
о защитной роли БТШ при тепловом шоке.
Однако как это происходит, пока не ясно.
Для выяснения молекулярного механизма
защитных систем клеток при стрессе
необходимы исследования конкретной функции
каждого БТШ.
СТРЕССОВЫЕ БЕЛКИ
275
Функции БТШ и гомологичных
им белков, синтезирующихся
при нормальной температуре
Как выяснилось, большинство БТШ
имеют родственные белки, которые
синтезируются при нормальной температуре
(постоянно или на определенных стадиях
развития организма). Например, у
дрожжей семейство БТШ70 кодируется
девятью генами, из которых шесть
усиливают экспрессию при тепловом шоке, два
снижают, а один не реагирует на
тепловой шок. У дрозофилы один из БТШ70
индуцируется тепловым шоком, а семь других
экспрессируются при нормальной
температуре. В последнее десятилетие
достигнут прогресс в понимании того, что
делают эти белки. БТШ70 присоединяется к
другим белкам, вызывая их
разворачивание и препятствуя их агрегации, которая
помешала бы белку приобрести нативную
конформацию, необходимую для его
функциональной активности. Взаимодействие
БТШ70 с другими белками АТФ/АДФ
зависимо. Замена связанной с БТШ70 АТФ
на АДФ происходит за счет АТФазной
активности БТШ70. Разворачивание ряда
белков с помощью БТШ70 необходимо для
их проникновения через мембрану хлоро-
пластов, митохондрий и эндоплазматичес-
кого ретикулума. Предполагают, что
БТШ70 совершает АТФ-зависимый цикл
работы по протаскиванию белков через
мембрану, в основе которого лежит
изменение конформации БТ1П70 в
зависимости от связи с АТФ или АДФ.
Агрегация белков резко усиливается при
повышении температуры. Активация
синтеза БТШ70 в этих условиях должна
защищать другие белки от необратимого
повреждения. Поскольку БТШ70 влияет на
конформационное состояние этих белков,
его относят к группе шаперонов (от англ.
chaperon — провожатая, наставница при
молодой особе). (См.: Наградова Н. А.
Внутриклеточная регуляция формирования
нативной пространственной структуры
белков. Том 8.)
БТШ70 присутствует во всех «отсеках»
клетки. На рис. 1 представлена схема
ответа клетки на ТШ синтезом БТШ и их
последующим распространением по всей
клетке. БТШ70 обнаруживается в ядре
и ядрышке, где он накапливается при
тепловом шоке, и в цитоплазме в связи с
элементами цитоскелета, в связи с
клеточной мембраной, в митохондриях, хло-
ропластах и эндоплазматическом ретику-
луме. БТШ70 способствует прохождению
синтезированных в цитоплазме
предшественников хлоропластных белков через
мембрану хлоропластов, играя тем самым
важную роль в биогенезе этих органелл.
Такую же роль он играет и в биогенезе
митохондрий.
Другая группа белков теплового шока —
БТШ60 — тоже относится к шаперонам (их
еще называют шаперонинами). Эти белки
обеспечивают правильную сборку
четвертичной структуры клеточных белков.
Такую функцию БТШ60 выполняют у
бактерий, а также в митохондриях и хлоро-
пластах эукариотических клеток. Они
образуют два наложенных друг на друга
кольца, в каждом из которых
присутствуют по семь субъединиц БТШ60. Этот
сложный молекулярный комплекс направляет
упаковку клеточных белков при
образовании четвертичной структуры; например,
он участвует в формировании ключевого
фермента фотосинтеза РБФК, который
строится из восьми малых и восьми
больших полипептидов (субъединиц) и
локализуется в строме хлоропластов.
К группе шаперонов относят и БТШ90.
Этот белок играет крайне важную роль в
образовании комплекса стероидных
гормонов с их рецептором. Вместе с БТШ70 и
другими белками БТШ90 контролирует
функциональную активность рецепторов
стероидных гормонов в клетке.
Установлено, что БТ1П90 образует комплексы и
с другими белками, например, с
некоторыми протеинкиназами, контролируя их
активность. Как известно, протеинкиназы
фосфорилируют самые различные
клеточные белки и благодаря этому регулируют
их активность. БТШ90 обнаружен в
клетках растений, однако сведений о его
функции у растений пока нет. Таким образом,
БТШ60, 70 и 90 образуют комплексы с
другими белками клетки, изменяя их
структуру и функциональное состояние.
Предполагают, что во время теплового шока
они защищают клеточные белки от
повреждения, хотя многое в этом направлении
еще предстоит выяснить.
К БТШ относится также убиквитин —
белок, присоединение которого к N-концу
полипептида превращает этот полипептид в
мишень для разрушения протеазами. При
помощи убиквитина происходит выбраковка
поврежденных, недостроенных и
функционально неактивных полипептидов.
Ассоциированный с убиквитином белок разрушается
в особых мультикомпонентных комплексах,
называемых протеасомами. Тепловой шок
приводит к появлению в клетках
недостроенных и поврежденных белков, для
удаления которых нужен убиквитин. Интересно, что
появление таких поврежденных и
недостроенных белков служит сигналом к синтезу БТШ
даже при нормальной температуре, а
факторы, стабилизирующие структуру белков,
тормозят синтез БТШ.
276
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
Рис. 4. Ультраструктурные изменения клеток меристемы корня тыквы под действием теплового шока (40°С;
1,5 ч)
а— контрольная клетка при нормальной температуре 28°С. Видны ядро, ядрышко, митохондрии, каналы эндо-
плазматического ретикулума и небольшие вакуоли; б— ядро и ядрышко клетки при тепловом шоке. Видны
электронноплотные образования; в— эндоплазматический ретикулум при тепловом шоке; г— цитоплазмати-
ческая гранула теплового шока; д— локализация низкомолекулярных БТШ (17кДа) в цитоплазматической
грануле (ЦГ) теплового шока в клетках листа Arabidopsis thaliana. БТШ выявляли с помощью комплекса,
состоящего из антител к БТШ 17, протеина А и коллоидного золота. Видные под электронным микроскопом черные
точки коллоидного золота соответствуют локализации БТШ17. Протеин А— белок массой 42кДа из
золотистого стафилококка, распознающий иммуноглобулины, к которым относятся антитела к БТШ17. Я— ядро; Яд —
ядрышко; М— митохондрия; ЭНД— эндоплазматический ретикулум; ЦГ— цитоплазматические гранулы; X —
хлоропласт
В число БТШ входят, как уже
отмечалось, низкомолекулярные белки. У
дрожжей такие БТШ кодирует всего один ген,
а у растений— больше 30. Молекулярная
масса этих БТШ лежит в диапазоне от 16
до 40 кДа. Они локализуются главным
образом в цитоплазматических гранулах
теплового шока (рис. 4), которые появляются
в цитоплазме во время ТШ и исчезают после
его окончания. В гранулах теплового шока
обнаруживается также мРНК, синтез
которых происходил в нормальных
температурных условиях. Поэтому полагают, что
цитоплазматические гранулы теплового
шока предохраняют «дошоковые» мРНК, что
дает возможность вновь использовать эти
мРНК для синтеза белка после окончания
шока.
На рис. 4 показаны изменения в
ультраструктуре растительных клеток при дей-
СТРЕССОВЫЕ БЕЛКИ
277
ствии теплового шока. Видно появление
электронноплотных образований в ядре за
счет конденсации хроматина и скопления
нуклеопротеидных частиц, содержащих
предшественники мРНК — премРНК. В
этих частицах обнаруживается БТШ70,
который, по-видимому, защищает хроматин
и синтезированные до ТШ премРНК для
«дошоковых» белков от необратимых
повреждений при высокой температуре. Эн-
доплазматический ретикулум приобретает
характерный вид ватрушки. В
цитоплазме видны скопления электронноплотных
частиц — цитоплазматических гранул
теплового шока. Специальный метод с
использованием антител к БТШ 17 кДа,
протеина А (белка из золотистого стафилококка,
взаимодействующего с иммуноглобулинами
и, следовательно, с антителами к БТШ)
и коллоидного золота позволяет
обнаружить скопление низкомолекулярных БТШ
в цитоплазматических гранулах (рис. 4).
Важно, что такие же структурные
изменения обнаружены нами в клетках
растений при других стрессах: недостатке воды
и действии высоких концентраций солей.
Низкомолекулярные БТШ у растений
обнаруживаются не только в
цитоплазматических гранулах теплового шока, но и
в других клеточных структурах, в том
числе в хлоропластах. Предполагают, что они
защищают от теплового шока тилакоидные
мембраны, где осуществляются процессы
трансформации световой энергии в
химическую. Низкомолекулярные БТШ
вариабельны по своей первичной структуре, но
у всех представителей этого семейства есть
участки, гомологичные по аминокислотной
последовательности ос-кристаллину — белку
хрусталика глаза. Значимость этого
наблюдения и вопрос о функции
низкомолекулярных БТШ требуют дальнейшего
выяснения. Следует упомянуть, что многие из
них синтезируются на определенных
стадиях развития растений и имеют органную
специфичность. В период мейоза в
нормальных температурных условиях у лилии,
кукурузы, а также у дрозофилы и дрожжей
осуществляется синтез одного из
низкомолекулярных БТШ. Возможная регулятор-
ная роль БТШ в онтогенезе организмов
согласуется с тем, что у дрозофилы гормон
линьки — экдизон — инициирует синтез
БТШ. БТШ синтезируются в
эмбриогенезе растений, обеспечивая устойчивость
зародышей к ТШ в первые часы набухания
семян.
Долгое время одним из аргументов
против участия БТШ в термотолерантности
клеток служило отсутствие их синтеза при
ТШ в пыльце. Однако впоследствии у
некоторых растений (но не у всех) был
обнаружен конститутивный (неиндуцированный)
синтез БТШ во время формирования
пыльцы. Следовательно, предсинтезированные
БТШ могли обеспечить термостабильность
пыльцы при ТШ, хотя нельзя исключить
наличие у пыльцы и других механизмов
устойчивости к нагреву. Важно подчеркнуть,
что синтез БТШ, происходящий у
растений в жаркие часы при выращивании в
полевых условиях, по-видимому,
обеспечивает нормальную функцию растений при
смене температур.
Мы рассмотрели защиту клеток всех
живых организмов от теплового шока при
помощи БТШ. Это не исключает других
защитных механизмов, действующих на
уровне синтеза химических протекторов,
изменения свойств мембран и ионного го-
меостаза клеток. Не следует также
забывать, что, помимо клеточных механизмов
защиты от теплового шока, в которых БТШ
играют важнейшую роль, существуют
другие, организменные уровни, связанные с
морфологическими изменениями растений
или усилением транспирации (испарения
воды через устьица), снижающей
температуру листьев.
Другие стрессовые
белки растений
Репрограммирование генома при
стрессе, приводящее к синтезу специфических
стрессовых белков и тем самым
помогающее клетке пережить экстремальные
воздействия, является общим принципом
ответа клеток на многие стрессы. Например,
кислородное голодание (анаэробиоз),
которое чаще всего испытывают корни и
прорастающие семена при затоплении
почвы, вызывает синтез специфических
стрессовых белков анаэробиоза. К ним
принадлежат ферменты гликолиза, которые
позволяют обеспечить клетки энергией при
недостатке кислорода. Крайне активно
синтезируются несколько форм алкоголь-
дегидрогеназы. Синтез белков
анаэробиоза включается на уровне активации их
генов через регуляторный элемент
анаэробного шока, который гомологичен у
разных генов ответа на анаэробиоз и у
различных растений. Эта генетическая
программа позволяет растениям пережить
непродолжительный недостаток
кислорода. Так, клубни картофеля гибнут при
анаэробиозе через 48 ч, проростки
кукурузы — через 3 дня, риса — через 7 дней.
Но есть среди растений рекордсмены,
например растение-амфибия Acorus calamus,
клубни которого сохраняют
жизнеспособность в течение двух месяцев
анаэробиоза. Такие растения представляют особый
интерес для генных инженеров как
потенциальные доноры генов устойчивости при
278
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
получении стресс-резистентных
трансгенных растений.
Обезвоживание клеток растений
вызывает синтез большого набора белков с
невыясненной пока функцией. Их индуктором
является фитогормон абсцизовая кислота
(АБК). Эти белки синтезируются при
нормальном водоснабжении растений в
зародышах на стадии позднего эмбриогенеза,
когда происходит естественное
обезвоживание созревающих семян; синтезу
предшествует накопление в семенах АБК,
которая его индуцирует. Поэтому их называют
белками позднего эмбриогенеза или
белками ответа на АБК. В листьях при
нормальном водоснабжении они отсутствуют, но
недостаток воды вызывает накопление в
листьях АБК, которая индуцирует синтез
белков обезвоживания. Эти белки и
кодирующие их гены подвергаются пристальному
изучению с тем, чтобы понять механизм
устойчивости клеток к обезвоживанию.
Особое внимание уделяется таким
рекордсменам засухоустойчивости, как, например,
Craterostigma plantageneum, которая
способна высыхать до воздушно-сухого состояния,
а после предоставления воды быстро
зеленеть и возвращаться к нормальному
функционированию.
Растения испытывают недостаток воды не
только при засухе, но также при засолении
почвы и холоде. В настоящее время
обнаружено, что при каждом из этих стрессов
синтезируются как общие, так и специальные
для каждого из них белки. Важную роль в
реакциях на такие стрессирующие
воздействия играют низкомолекулярные осмопро-
текторы — вещества, которые,
накапливаясь в клетках в высоких концентрациях,
препятствуют их обезвоживанию и выполняют
защитную функцию по отношению к
внутриклеточным структурам, включая
мембраны и различные белковые комплексы. Осмо-
протекторную функцию выполняют сахароза,
пролин, полиамины, олигосахариды,
полиспирты, например маннит и сорбит, и
многие другие соединения. Их накопление
коррелирует с засухоустойчивостью. Например,
у Mesembryanthemum crystallinum —
растения, обитающего в пустынях Намибии и
обладающего устойчивостью к засухе,
засолению и холоду одновременно, — концентрация
осмопротекторов в цитоплазме и хлороплас-
тах достигает при стрессе 700 мМ, а в
специализированных эпидермальных клетках
превышает 1 М.
Среди белков, чей синтез у арабидоп-
сиса активируется при засолении,
обнаружен аквапорин — белок, образующий
каналы в мембране для прохождения в клетку
воды. Он гомологичен у растений и
животных. Введение мРНК аквапорина растений
в ооциты лягушки Xenopus повышает их
проницаемость для воды. Следовательно, у
растений и животных существуют общие
молекулярные механизмы поступления воды
в клетки. Это не противоречит факту
существования осморегуляторной функции
вакуоли, в мембране которой (тонопласте)
также обнаружен аквапорин. Все
сказанное свидетельствует об общности ответа
клеток растений и других организмов на
различные стрессы.
Заключение
Стрессовые белки синтезируются в
растениях в ответ на различные
экстремальные воздействия: анаэробиоз,
окислительный стресс, повышенные и пониженные
температуры, обезвоживание, высокие
концентрации соли, при действии
тяжелых металлов, вредителей, а также при
раневых эффектах и ультрафиолетовой
радиации. Каждый из перечисленных стрессов
вызывает синтез специфических для него
белков. Вместе с тем есть белки, которые
индуцируются при самых разнообразных
стрессах. Например, БТШ70 синтезируется
при поранении растений, недостатке воды,
засолении, действии АБК. Арсенит и соли
Cd индуцируют у растений полный спектр
БТШ, действуя не через РЭ БТШ, а
через другие регуляторные элементы генов
БТШ. Очевидно, способность этих генов
отвечать на различные неблагоприятные
воздействия и обеспечивать защитные
реакции клеток возникла в ходе эволюции.
Поэтому можно предположить, что наряду
со специфическими механизмами
устойчивости клеток к неблагоприятным
факторам, существуют и универсальные
механизмы. Они могут иметь много общего
у различных организмов. Выше мы
показали, что такая общность структуры,
функции и регуляции синтеза БТШ присуща
всем изученным в этом отношении
организмам. Восстановление
термотолерантности у мутантов дрожжей под действием
гена БТШ растений доказывает единство
механизмов устойчивости их клеток к
тепловому шоку.
Обнаружение общих осмопротекторов
при обезвоживании клеток и действии на
них засоления у бактерий, дрожжей и
растений также свидетельствует о
существовании сходных путей защиты клеток
разных организмов при стрессах. Открытие у
растений белка аквапорина говорит об
общности молекулярных механизмов
снабжения клеток водой.
Ни в коем случае не забывая о
специфике структуры и функции клеток
растений и растительного организма в целом,
следует подчеркнуть, что системы стресс-
резистентности растений на клеточном и
СТРЕССОВЫЕ БЕЛКИ
279
молекулярном уровнях имеют общие черты
с подобными системами других организмов.
Это делает возможным использование
подходов, разработанных на нерастительных
организмах, для изучения
стресс-резистентности растений и поиска в их геноме
генов, гомологичных тем, которые
участвуют в программах устойчивости видов, не
принадлежащих к растениям, к
неблагоприятным внешним факторам. Наряду с
изучением генов и белков, определяющих
особую толерантность представителей дикой
флоры, исследования в рассмотренном
направлении должны способствовать
получению высокорезистенных трансгенных
растений, т. е. таких, в геном которых введен
чужеродный ген, придающий им новые
свойства. На этом пути предстоят большие
трудности, хотя бы потому, что
стресс-резистентность, как правило, является полигенным
признаком. Однако работа в этом
направлении обещает принести успех, масштабы
которого сейчас невозможно предсказать.
Литература
Войников В. К., Иванова Г. Г., Рудиковский А. В.
Физиология растений, 1984, т. 31, № 5, с. 970.
Кулаева О. Я., Микулович Т. П., Веселова Т. В.
и др. Докл. АН СССР, 1988, т. 300, № 5. с. 1277.
Кулаева О. Я., Микулович Т. П., Хохлова В. А.
Стрессовые белки растений. В кн.: Современные
проблемы биохимии. М.: Наука, 1991, с. 174.
Лозовская Е. Р., Левин А. В., Евгеньев М. Б.
Генетика, 1982, т. 18, №2, с. 1749.
Bohnert Я., Nelson D., Jensen R. G. Plant. Cell,
1995, vol. 7, p. 1099.
Georgopoulos C, Welch W. J. Ann. Rev. Cell Biol.,
1993, vol.9, p. 901.
GlickB.S. Cell, 1995, vol.80, p. 11.
Nover L., Hellmund D., Neumann D. et al. Biol.
Zbl., 1984, Bd. 103, S. 357.
А. Н. Тихонов
ФОТОСИНТЕЗ
Введение
История изучения фотосинтеза ведет
свое начало с августа 1771 г., когда
английский теолог, философ и
натуралист-любитель Дж. Пристли (1733-1804) обнаружил,
что растения могут «исправлять» свойства
воздуха, меняющего свой состав в
результате горения или жизнедеятельности
животных. Пристли показал, что в присутствии
растений воздух снова становится
пригодным для горения и поддержания жизни
животных. В ходе дальнейших
исследований голландского врача и
естествоиспытателя Я. Ингенгауза, швейцарских
естествоиспытателей Ж. Сенебье и Н. Т. Соссюра,
французского агрохимика Ж. Б. Буссенго и
других ученых было установлено, что при
освещении растения выделяют кислород и
поглощают из воздуха углекислый газ. Из
углекислого газа и воды растения
синтезируют органические вещества. Этот процесс
был назван фотосинтезом. Немецкий врач
и физик Ю. Р. Майер, открывший закон
сохранения энергии, в 1845 г. высказал
предположение, что растения превращают
энергию солнечного света в энергию
химических соединений, образующихся при
фотосинтезе. По его словам,
«распространяющиеся в пространстве солнечные лучи
«захватываются» и сохраняются для
использования в дальнейшем по мере
надобности». Впоследствии русским ученым К. А.
Тимирязевым было убедительно доказано, что
важнейшую роль в использовании
растениями энергии солнечного света играют
молекулы хлорофилла, входящего в состав
зеленых листьев.
Образующиеся при фотосинтезе
углеводы (сахара) используются как источник
энергии и строительный материал для
синтеза различных органических соединений
у растений и животных. У высших
растений процессы фотосинтеза протекают в хло-
ропластах — специализированных энерго-
преобразующих органеллах растительной
клетки. Схематическое изображение
хлоропласта показано на рис. 1. Под двойной
оболочкой хлоропласта, состоящей из
наружной и внутренней мембран, находятся
замкнутые мембранные структуры,
называемые тилакоидами. Мембраны тилакои-
дов состоят из двух слоев молекул липи-
дов (жироподобных веществ), в которые
включены макромолекулярные
фотосинтетические белковые комплексы. В хлоро-
пластах высших растений тилакоиды
группируются в граны, которые представляют
собой стопки сплюснутых и тесно
прижатых друг к другу тилакоидов, имеющих
Внешняя
мембрана
Грана
Тилакоид
граны
Внутренняя
мембрана
Межмембранное
пространство
Строма
Межгранный
тилакоид
Рис. 1. Строение хлоропласта
ФОТОСИНТЕЗ
281
форму дисков. Продолжением отдельных
тилакоидов гран являются выступающие из
них межгранные тилакоиды. Пространство
между оболочкой хлоропласта и тилакои-
дами называется стромой. В строме
содержатся хлоропластные молекулы РНК, ДНК,
рибосомы, крахмальные зерна, а также
многочисленные ферменты, включая те,
которые обеспечивают усвоение С02
растениями.
Световые и темновые стадии
фотосинтеза
Фотосинтез представляет собой ряд
фотофизических и биохимических процессов,
в результате которых растения за счет
энергии солнечного света синтезируют
углеводы (сахара). Многочисленные стадии
фотосинтеза принято разделять на две большие
группы — световые и темновые.
Световыми стадиями фотосинтеза принято
называть совокупность процессов, в
результате которых за счет энергии света
синтезируются молекулы аденозинтрифосфата
(АТФ, лат. АТР) и происходит образование
восстановленного никотинамидадениндинук-
леотидфосфата (НАДФН, лат. NADPH) —
соединения, обладающего высоким
восстановительным потенциалом. Молекулы АТФ
выполняют роль универсального источника
энергии в клетке. Энергия макроэргических
(т. е. богатых энергией) фосфатных связей
молекулы АТФ, как известно, используется в
большинстве биохимических процессов,
потребляющих энергию. Световые процессы
фотосинтеза протекают в тилакоидах,
мембраны которых содержат основные
компоненты фотосинтетического аппарата растений —
светособирающие пигмент-белковые и
электрон-транспортные комплексы, а также АТФ-
синтазный комплекс, который катализирует
образование АТФ из аденозиндифосфата
(АДФ, лат. ADP) и неорганического
фосфата (P.) (ADP + Р. -> АТР 4- Н20). Таким
образом, в результате световых стадий
фотосинтеза энергия света, поглощаемого
растениями, запасается в форме
макроэргических химических связей молекул АТФ и
сильного восстановителя НАДФН, которые
используются для синтеза углеводов в так
называемых темновых стадиях фотосинтеза.
Темновыми стадиями фотосинтеза
обычно называют совокупность биохимических
реакций, в результате которых
происходят усвоение растениями атмосферной
углекислоты (С02) и образование углеводов.
Цикл «темновых» биохимических
превращений, приводящих к синтезу
органических соединений из С02 и воды, по имени
ученого, внесшего решающий вклад в
исследование этих процессов, называется
циклом Кальвина. В отличие от электрон-
транспортного и АТФ-синтазного
комплексов, которые находятся в тилакоидной
мембране, ферменты, катализирующие
«темновые» реакции фотосинтеза,
локализованы в строме.
В результате превращений ряда
органических соединений в цикле Кальвина из
трех молекул С02 и воды в хлоропластах
образуется молекула глицеральдегид-3-
фосфата, имеющего химическую формулу
СНО-СНОН-СН2ОР02". При этом на одну
молекулу С02, включающуюся в глицераль-
дегид-3-фосфат, расходуются три
молекулы АТФ и две молекулы НАДФН. Для
синтеза органических соединений в цикле
Кальвина используется энергия,
выделяющаяся в ходе реакции гидролиза
макроэргических фосфатных связей молекул АТФ
(реакция АТР + Н20 -> ADP + Р.), а также
сильный восстановительный потенциал
молекул НАДФН. Основная часть
образовавшихся в хлоропласте молекул глицераль-
дегид-3-фосфата поступает в цитоплазму
растительной клетки, где превращается во
фруктозо-б-фосфат и глюкозо-6-фосфат,
которые в ходе дальнейших превращений
образуют сахарозу. Из оставшихся в
хлоропласте молекул глицеральдегид-3-фосфата
синтезируется крахмал.
Преобразование энергии
в фотосинтетических
реакционных центрах
Фотосинтетические энергопреобразую-
щие комплексы растений, водорослей и
фотосинтезирующих бактерий хорошо
изучены. Установлены химический состав и
пространственное строение энергопреоб-
разующих белковых комплексов, выяснена
последовательность процессов
трансформации энергии. Несмотря на различия в
составе и молекулярном строении
фотосинтетического аппарата, существуют общие
закономерности процессов трансформации
энергии в фотореакционных центрах всех
фотосинтезирующих организмов. В
фотосинтетических системах как растительного,
так и бактериального происхождения
единым структурно-функциональным звеном
является фотосистема, включающая све-
тособирающую антенну, фотохимический
реакционный центр и связанные с ним
молекулы — переносчики электрона.
Рассмотрим сначала общие принципы
превращения энергии солнечного света,
характерные для всех фотосинтетических
систем, а затем более детально
проиллюстрируем их на примере
функционирования фотореакционных центров и цепи
электронного транспорта хлоропластов высших
растений.
282
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
Светособирающая антенна.
Поглощение света, миграция
энергии к реакционному центру
Самый первый элементарный акт
фотосинтеза — поглощение света молекулами
хлорофилла или вспомогательных пигментов,
входящих в состав специального пигмент-
белкового комплекса, называемого светосо-
бирающей антенной. Светособирающая
антенна представляет собой макромолекулярный
комплекс, предназначенный для
эффективного улавливания света. В хлоропластах
антенный комплекс содержит большое
число (до нескольких сотен) молекул хлорофилла
и некоторое количество вспомогательных
пигментов (каротиноидов), прочно
связанных с белком. На ярком солнечном свету
отдельная молекула хлорофилла поглощает
кванты света сравнительно редко, в
среднем не чаще, чем 10 раз в секунду. Однако
на один фотореакционный центр
приходится большое количество молекул
хлорофилла (200—400), что обеспечивает достаточно
частое срабатывание реакционного центра
даже при относительно слабой интенсивности
света, падающего на лист в условиях
затенения растения. Т.,е. ансамбль пигментов,
поглощающих свет, действительно
выполняет роль антенны, которая за счет своих
достаточно больших размеров эффективно
улавливает солнечный свет и направляет его
энергию к реакционному центру.
Тенелюбивые растения имеют, как правило, больший
размер светособирающей антенны по
сравнению с растениями, произрастающими в
условиях высокой освещенности.
У растений основными светособирающи-
ми пигментами служат молекулы хлорофилла а
и хлорофилла Ь, поглощающие видимый свет
л Светособирающие пигменты
Свет
е- е-
> >
Донор электрона Р Акцептор электрона
Фотореакционный центр
Рис. 2. Схема миграции энергии возбуждения по
пигментной матрице светособирающей антенны к
фотореакционному центру Р (димер хлорофилла)
Молекула D — первичный донор электрона для
окисленного центра Р+; молекула А— первичный
акцептор, принимающий электрон от возбужденного
фотореакционного центра Р*
с длиной волны Х< 700-730 нм.
Изолированные молекулы хлорофилла поглощают свет
лишь в двух сравнительно узких полосах
солнечного спектра— при \~ 660-680 нм
(красный свет) и X ~ 430-450 нм
(ультрафиолетовый свет), что, разумеется,
ограничивает эффективность использования всего
спектра солнечного света, падающего на зеленый
лист. Однако спектральный состав света,
поглощаемого светособирающей антенной, в
действительности значительно шире.
Объясняется это тем, что спектр поглощения
агрегированных форм хлорофилла, входящих
в состав светособирающей антенны,
сдвигается в сторону больших длин волн. Кроме
этого, наряду с хлорофиллом, в светособи-
рающую антенну входят вспомогательные
пигменты, увеличивающие эффективность
собирания света за счет его поглощения в тех
областях спектра, в которых сравнительно
слабо поглощают свет молекулы
хлорофилла. У растений вспомогательными
пигментами являются каротиноиды, поглощающие свет
в области X ~ 450-480 нм; в клетках фотосин-
тезирующих водорослей это красные и
синие пигменты — фикоэритрины у красных
водорослей (к « 495—565 нм) и фикоцианины у
синезеленых водорослей (Х~ 550-615 нм).
Поглощение кванта света молекулой
хлорофилла (Chi) или вспомогательного
пигмента приводит к ее возбуждению
(переходу электрона на более высокий
энергетический уровень):
СЫ + hv -> Chi*.
Энергия возбужденной молекулы
хлорофилла Chi* передается соседним пигментам,
которые, в свою очередь, могут передать
ее другим молекулам светособирающей
антенны:
СЫ* + Chi -> СЫ + Chi*.
Благодаря этому энергия возбуждения
может мигрировать по пигментной
матрице до тех пор, пока возбуждение не
попадет на фотореакционный центр Р
(схематическое изображение этого процесса
показано на рис. 2).
СЫ* + Р -> СЫ + Р*.
Характерные времена жизни
возбужденных состояний молекул хлорофилла и
других пигментов очень малы, т~ 10"10-10"9с.
Поэтому существует определенная
вероятность того, что на пути к реакционному
центру Р энергия короткоживущих
возбужденных состояний пигментов может
потеряться — диссипировать в тепло или
выделиться в виде кванта света (явление
флуоресценции). В действительности,
однако, эффективность миграции энергии к
фотосинтетическому реакционному центру
очень велика. В том случае, когда реакци-
ФОТОСИНТЕЗ
283
онный центр находится в активном
состоянии, вероятность потери энергии
составляет, как правило, не более 10-15%.
Такая высокая эффективность использования
энергии солнечного света обусловлена тем,
что светособирающая антенна
представляет собой высокоупорядоченную структуру,
обеспечивающую очень хорошее
взаимодействие пигментов. Благодаря этому
достигается высокая скорость переноса энергии
возбуждения от молекул, поглощающих
свет, к фотореакционному центру. Среднее
время «перескока» энергии возбуждения от
одного пигмента к другому, как правило,
составляет т« 10~12-10~и с. Общее время
миграции возбуждения к реакционному
центру обычно не превышает 10"10— 10"9с.
Фотохимический
реакционный центр. Перенос
электрона, стабилизация
разделенных зарядов
Современным представлениям о
строении реакционного центра и механизмах
первичных стадий фотосинтеза
предшествовали работы А. А. Красновского, открывшего,
что в присутствии доноров и акцепторов
электрона возбужденные светом молекулы
хлорофилла способны обратимо
восстанавливаться (принимать электрон) и окисляться
(отдавать электрон). Впоследствии у
растений, водорослей и фотосинтезирующих
бактерий Б. Коком, Г. Т. Виттом и Л. Дюйзен-
сом, работавшими в США, Германии и
Голландии, соответственно были
обнаружены особые пигменты хлорофилловой
природы, названные реакционными центрами,
которые окисляются при действии света и
являются, по сути дела, первичными
донорами электрона при фотосинтезе.
Фотохимический реакционный центр Р
представляет собой особую пару (димер)
молекул хлорофилла, которые выполняют
роль ловушки энергии возбуждения,
блуждающего по пигментной матрице светосо-
бирающей антенны (см. рис. 2). Подобно тому
как жидкость стекает со стенок широкой
воронки к ее узкому горлышку, к
реакционному центру направляется энергия
света, поглощаемого всеми пигментами све-
тособирающей антенны. Возбуждение
реакционного центра инициирует цепь даль-
S
1_
Q.
О)
X
СО
DP*AB
DP+A-B
АЕ
hv
DPAB
Свет
DP+AB-
D+PAB-
Акцептор
е~
Донор
Рис. 3, Схема энергетических уровней фотосистемы при различных состояниях реакционного центра (Р),
первичного донора электрона (D), первичного (А) и вторичного (В) акцепторов электрона
Внизу—схема разделения зарядов в мембране, инициированного фотовозбуждением реакционного центра
284
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
нейших превращений энергии света при
фотосинтезе.
Последовательность процессов,
происходящих после возбуждения реакционного
центра Р, и диаграмма соответствующих
изменений энергии фотосистемы
схематически изображены на рис. 3. Наряду с ди-
мером хлорофилла Р в фотосинтетический
комплекс входят молекулы первичного и
вторичного акцепторов электрона, которые
мы условно обозначим символами А и В,
а также первичный донор электрона —
молекула D. Возбужденный реакционный
центр Р* обладает низким сродством к
электрону и поэтому он с легкостью отдает его
находящемуся рядом с ним первичному
акцептору электрона А:
D(P*A)B -> D(P+A")B.
Таким образом, в результате очень
быстрого (т« 10"12с) переноса электрона от Р*
к А реализуется второй принципиально
важный этап преобразования солнечной
энергии при фотосинтезе — разделение
зарядов в реакционном центре. При этом
образуются сильный восстановитель А"
(донор электрона) и сильный окислитель Р+
(акцептор электрона).
Молекулы Р+ и А" расположены в
мембране асимметрично: в хлоропластах
реакционный центр Р+ находится ближе к
поверхности мембраны, обращенной внутрь
тилакоида, а акцептор А" расположен
ближе к внешней стороне. Поэтому в
результате фотоиндуцированного разделения
зарядов на мембране возникает разность
электрических потенциалов Аф = ф.п - cpout >
>0. Индуцированное светом разделение
зарядов в реакционном центре подобно
генерации разности электрических потенциалов
в обычном фотоэлементе. Следует, однако,
подчеркнуть, что в отличие от всех
известных и широко используемых в технике
фотопреобразователей энергии
эффективность работы фотосинтетических
реакционных центров является очень высокой. КПД
разделения зарядов в активных
фотосинтетических реакционных центрах, как
правило, превышает 90-95%.
Стабилизация разделенных зарядов
обеспечивается главным образом за счет
вторичных процессов электронного
транспорта, следующих за переносом электрона от
Р* к А. От восстановленного первичного
акцептора А" электрон очень быстро
(характерное время этого процесса обычно не
превосходит 10"10-10~9с) уходит на
вторичный акцептор электрона В:
D(P+A")B -> D(P+A)B".
При этом происходит не только
удаление электрона от положительно
заряженного реакционного центра Р+, но и
заметно снижается энергия всей системы (рис. 3).
Следовательно для переноса электрона в
обратном направлении (переход В" —> А) ему
потребуется преодолеть достаточно
высокий энергетический барьер АЕ « 0,3-0,4 эВ,
где АЕ — разность энергетических уровней
для двух состояний системы, при которых
электрон находится, соответственно, на
переносчике А или В. По этой причине для
возвращения электрона назад, от
восстановленной молекулы В" к окисленной
молекуле А, потребовалось бы гораздо
больше времени, чем для прямого перехода
А" —> В. Иными словами, в прямом
направлении электрон переносится гораздо
быстрее, чем в обратном. Поэтому после
переноса электрона на вторичный акцептор В
существенно уменьшается вероятность его
возвращения назад к рекомбинации с
положительно заряженной «дыркой» Р+.
Вторым фактором, способствующим
стабилизации разделенных зарядов, служит
быстрая нейтрализация окисленного
фотореакционного центра Р+ за счет электрона,
поступающего к Р+ от донора электрона D:
D (Р+А)В- -> D+(PA)B".
Получив электрон от молекулы
донора D и вернувшись в свое исходное
восстановленное состояние Р, реакционный
центр не может принять электрон от
восстановленных акцепторов и он готов к
повторному срабатыванию — отдать
электрон находящемуся рядом с ним
окисленному первичному акцептору А. Такова
последовательность событий, происходящих
в фотореакционных центрах всех фотосин-
тезирующих систем.
Цепь электронного транспорта
в хлоропластах
В хлоропластах высших растений
имеются две фотосистемы — фотосистема 1
(ФС1) и фотосистема 2 (ФС2),
различающиеся по составу белков, пигментов и
оптическим свойствам. Светособирающая антенна
ФС1 поглощает свет с длиной волны Л, < 700—
730 нм, а ФС2 — свет сА,< 680-700 нм.
Индуцированное светом окисление
реакционных центров ФС1 и ФС2 сопровождается их
обесцвечиванием, которое
характеризуется изменениями их спектров поглощения при
Х~ 700 и 680 нм. В соответствии с их
оптическими характеристиками реакционные
центры ФС1 и ФС2 получили название Р700
И Р680 •
Две фотосистемы связаны между
собой посредством цепи электронных
переносчиков (рис. 4). ФС2 является
источником электронов для ФС1. Инициируемое
светом разделение зарядов в
фотореакционных центрах Р700 и Рб80 обеспечивает
ФОТОСИНТЕЗ
285
перенос электрона от воды, разлагаемой
в ФС2, к конечному акцептору
электрона — молекуле НАДФ+. Цепь электронного
транспорта (ЦЭТ), соединяющая две
фотосистемы, в качестве переносчиков
электрона включает в себя молекулы пласто-
хинона, отдельный электронтранспортный
белковый комплекс (так называемый
b/f-комплекс) и водорастворимый белок
пластоцианин (Рс). Схема,
иллюстрирующая взаимное расположение электрон-
транспортных комплексов в тилакоидной
мембране и путь переноса электрона от
воды к НАДФ+, приведена на рис. 4.
В ФС2 от возбужденного центра Р*80
электрон переносится сначала на
первичный акцептор феофетин (Phe), а затем на
молекулу пластохинона QA, прочно
связанную с одним из белков ФС2:
Y(P*80Phe)QAQB -> Y(P+80 Phe")QAQB ->
^Y(P-80Phe)QAQB.
Затем электрон переносится на вторую
молекулу пластохинона QB, а Р?80 получает
электрон от первичного донора электрона Y:
Y(P^80Phe)QA QB^ Y+(P680Phe)QAQB.
Молекула пластохинона, химическая
формула которой и ее расположение в бис-
лойной липидной мембране показаны на
рис. 5, способна принять два электрона. После
двукратного срабатывания реакционного
центра ФС2 молекула пластохинона QB
получит два электрона:
QR + 2e-
->q;
Q
Отрицательно заряженная молекула
~2 обладает высоким сродством к ионам
водорода, которые она захватывает из
стромального пространства. После прото-
нирования восстановленного
пластохинона Q"2(Q-2+ 2Н + -> QH2) образуется
электрически нейтральная форма этой
молекулы QH2, которая называется плас-
тохинолом (рис. 5). Пластохинол выполняет
роль подвижного переносчика двух
электронов и двух протонов: покинув ФС2,
молекула QH2 может легко перемещаться
внутри тилакоидной мембраны,
обеспечивая связь ФС2 с другими
электрон-транспортными комплексами.
Окисленный реакционный центр ФС2
Pg80 обладает исключительно высоким
сродством к электрону, т. е. является очень
сильным окислителем. Благодаря этому в ФС2
происходит разложение воды —
химически устойчивого соединения. Входящий в
состав ФС2 водорасщепляющий комплекс
(ВРК) содержит в своем активном центре
NADP+-
-> NADPH
2Н+
2е-
Бислойная
липидная
мембрана
НоО
2Н+
2Н+ + 1/202
Фотосистема 2 > b/f-комплекс
Рс
Фотосистема 1
Рис. 4. Расположение электрон-транспортных комплексов (ФС1, ФС2 и b/f-комплекса) и их
взаимодействие в тилакоидной мембране
В состав фотосистемы 2 (ФС2) входят: фотореакционный центр Р680 первичный акцептор— феофетин (Phe),
вторичные акцепторы— молекулы пластохинона (QA и QB) и водорасщепляющий комплекс. Рс— пластоцианин.
Р700— энергетическая ловушка и реакционный центр фотосистемы 1 (ФС1). В состав (ФС1) входят также:
первичный акцептор электрона — молекула хлорофилла a (Aq), вторичные акцепторы — молекула филлохинона (А^
и три переносчика белковой природы (ферредоксина), у которых в активном центре находятся атомы железа и
серы (Fe-S). В переносе электрона от акцепторов ФС1 к НАДФ+ участвуют растворенный в строме белок
ферредоксин (Fd) и связанный с мембраной специальный электрон-транспортный комплекс— ферредоксин-
НАДФ-редуктаза (FNR), функционирующие на внешней стороне тилакоидной мембраны. При восстановлении
одной молекулы НАДФ+ до НАДФН на нее переносятся два электрона и один ион водорода, который
захватывается из стромы
286
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
группу ионов марганца (Мп2+), которые
служат донорами электрона для Р+80. Отдавая
электроны окисленному реакционному
центру Pg80, ионы марганца становятся
«накопителями» положительных зарядов,
непосредственно участвующих в реакции
окисления воды. В результате
последовательного четырехкратного срабатывания
реакционного центра Рб80 в Mn-содержащем
активном центре ВРК накапливаются четыре
сильных окислительных эквивалента (или
четыре «дырки») в форме окисленных ионов
марганца (Мп4+), которые, взаимодействуя
с двумя молекулами воды, катализируют
реакцию разложения воды:
2Мп4+ + 2Н20 -> 2Мп2+ + 4Н+ + 02.
Таким образом, после последовательной
передачи четырех электронов от ВРК к Р?80
происходит синхронное разложение сразу
двух молекул воды, сопровождающееся
выделением одной молекулы кислорода и
четырех ионов водорода, которые
попадают во внутритилакоидное пространство
хлоропласта.
Образовавшаяся при функционировании
ФС2 молекула пластохинола QH2
диффундирует внутри липидного бислоя тилакоид-
ной мембраны к b/f-комплексу (рис. 4 и 5).
При столкновении с b/f-комплексом
молекула QH2 связывается с ним, а затем
передает ему два электрона. При этом на
каждую молекулу пластохинола,
окисляемую b/f-комплексом, внутрь тилакоида
выделяются два иона водорода. В свою
очередь, b/f-комплекс служит донором
электрона для пластоцианина (Рс) — сравнитель-
Н
О | О |
^Y4- -2Н ^Х4^
О О
Н
2Н+
\
((
1
2Н+
Рис. 5. Окислительно-восстановительные
превращения пластохинола и предполагаемая схема его
расположения в мембране
но небольшого водорастворимого белка, у
которого в состав активного центра входит
ион меди (реакции восстановления и
окисления пластоцианина сопровождаются
изменениями валентности иона меди Си2+ +
+ е" <-> Си+). Пластоцианин выполняет роль
связующего звена между b/f-комплексом и
ФС1. Молекула пластоцианина быстро
перемещается внутри тилакоида, обеспечивая
перенос электрона от b/f-комплекса к ФС1.
От восстановленного пластоцианина
электрон поступает непосредственно к
окисленным реакционным центрам ФС1 — Р+00 (см.
рис. 4). Таким образом, в результате
совместного действия ФС1 и ФС2 два электрона
от молекулы воды, разлагаемой в ФС2, через
цепь электронного транспорта
переносятся, в конечном итоге, на молекулу НАДФ+,
обеспечивая образование сильного
восстановителя НАДФН.
Зачем хлоропластам нужны две
фотосистемы? Известно, что фотосинтезирую-
щие бактерии, которые используют в
качестве донора электрона для восстановления
окисленных реакционных центров
различные органические и неорганические
соединения (например, H2S), успешно
функционируют с одной фотосистемой. Ответ на
вопрос о происхождении двух фотосистем
заключается в том, что энергии одного
кванта видимого света недостаточно для
того, чтобы обеспечить разложение воды и
эффективное прохождение электроном всего
пути по цепи молекул-переносчиков от воды
к НАДФ+. Приблизительно 3 млрд лет
назад на Земле появились цианобактерии,
которые приобрели способность использовать
воду в качестве источника электронов для
восстановления углекислоты. В настоящее
время считается, что ФС1 ведет свое
происхождение от зеленых бактерий, а ФС2 —
от пурпурных бактерий. После того, как в
ходе эволюционного процесса ФС2
«включилась» в единую цепь переноса электрона
вместе с ФС1, стало возможным решить
энергетическую проблему — преодолеть
довольно большую разницу в
окислительно-восстановительных потенциалах пар
кислород / вода и НАДФ+ / НАДФН.
Возникновение фотосинтезирующих
организмов, способных окислять воду, явилось
одним из важнейших этапов развития
живой природы на Земле (см.: Климов В. В.
Растения и биосфера. Наст. том). Во-первых,
водоросли и зеленые растения, «научившись»
окислять воду, овладели неисчерпаемым
источником электронов для восстановления
НАДФ+. Во-вторых, разлагая воду, они
наполнили атмосферу Земли молекулярным
кислородом, создав, таким образом,
условия для бурного эволюционного развития
организмов, энергетика которых связана с
аэробным дыханием.
ФОТОСИНТЕЗ
287
Сопряжение процессов
электронного транспорта
с переносом протонов и синтезом
АТФ в хлоропластах
Перенос электрона по ЦЭТ, как
правило, сопровождается понижением энергии.
Этот процесс можно уподобить
самопроизвольному движению тела по наклонной
плоскости. Уменьшение уровня энергии
электрона в ходе его движения вдоль ЦЭТ не
означает, однако, что перенос электрона
всегда является энергетически бесполезным
процессом. В нормальных условиях
функционирования хлоропластов большая часть
энергии, выделяющейся в ходе
электронного транспорта, не пропадает
бесполезно, а используется для работы
специального энергопреобразующего комплекса,
называемого АТФ-синтазой. Этот комплекс
катализирует энергетически невыгодный
процесс образования АТФ из АДФ и
неорганического фосфата Pt. (реакция ADP + Р. —>
—> АТР + Н20). В этой связи принято
говорить, что энергодонорные процессы
электронного транспорта сопряжены с
энергоакцепторными процессами синтеза АТФ.
Важнейшую роль в обеспечении
энергетического сопряжения в мембранах тила-
коидов, как и во всех остальных энерго-
преобразующих органеллах (митохондриях,
хроматофорах фотосинтезирующих
бактерий), играют процессы протонного
транспорта. Синтез АТФ тесно связан с
переносом через АТФ-синтазу трех протонов из
тилакоидов (3H+J в строму (3H+ut):
ADP + P. + ЗН+ -> ATP + Н90 + ЗН+ ,.
г гп 2 out
Этот процесс становится возможным
потому, что вследствие асимметричного
расположения переносчиков в мембране
функционирование ЦЭТ хлоропластов
приводит к накоплению избыточного
количества протонов внутри тилакоида: ионы
водорода поглощаются снаружи на стадиях
восстановления НАДФ+ и образования пла-
стохинола и выделяются внутри
тилакоидов на стадиях разложения воды и
окисления пластохинола (рис. 4). Освещение
хлоропластов приводит к существенному (в
100-1000 раз) увеличению концентрации
ионов водорода внутри тилакоидов.
Итак, мы рассмотрели цепь событий, в
ходе которых энергия солнечного света
запасается в форме энергии высокоэнерге-
тичных химических соединений — АТФ и
НАДФ. Эти продукты световой стадии
фотосинтеза используются в темновых стадиях
для образования органических соединений
(углеводов) из углекислого газа и воды.
Основные этапы преобразования энергии,
приводящие к образованию АТФ и НАДФН,
включают следующие процессы: 1)
поглощение энергии света пигментами светособи-
рающей антенны; 2) перенос энергии
возбуждения к фотореакционному центру;
3)окисление фотореакционного центра и
стабилизация разделенных зарядов; 4)
перенос электрона по цепи электронного
транспорта, образование НАДФН; 5)
трансмембранный перенос ионов водорода; 6) синтез
АТФ.
Литература
Альберте Б., Брей Д., Льюис Дж. и др.
Молекулярная биология клетки. В 3 т. Пер. с англ.
М.: Мир, 1994, т. 1.
Кукушкин А. К.у Тихонов А. Я., Лекции по
биофизике фотосинтеза растений. М.: Изд-во
МГУ, 1988.
Николе Д. Д. Биоэнергетика. Введение в хе-
миосмотическую теорию. Пер. с англ. М.: Мир,
1985.
Скулачев В. П. Энергетика биологических
мембран. М.: Наука, 1989.
О. Н. Кунаева
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЖИЗНИ РАСТЕНИЙ
Введение
Жизнь каждого растения на всех ее
этапах регулируется гормонами, которые
называют фитогормонами.
В настоящее время активно исследуются
следующие из них: ауксины, цитокинины,
гиббереллины, абсцизовая кислота и
этилен (рис. 1). Кроме того, в последнее
время в растениях открыты также стероидные
гормоны, салициловая и жасминовая
кислоты, регуляторные пептиды и олигосаха-
риды, которые тоже можно отнести к фи-
тогормонам. Очевидно, со временем список
фитогормонов будет увеличиваться, и это
расширит наши представления о том, как
гормональная система регулирует онтогенез
растений и как она участвует в ответе
растений на различные внешние воздействия.
Все фитогормоны объединяют некоторые
общие свойства: они синтезируются в
самом растении и являются
высокоэффективными регуляторами физиологических
программ. Их действие проявляется при крайне
низких концентрациях (10~5-10~12 М) ввиду
исключительно высокой чувствительности
к ним растительных клеток. С помощью
фитогормонов клетки и ткани одних типов
регулируют физиологические процессы в
клетках и тканях других типов.
К числу физиологических программ,
регулируемых фитогормонами, относится
развитие и созревание семян, их
прорастание, рост и морфогенез растений,
переход растений к цветению, плодоношение,
старение, опадение листьев, покой
клубней, почек, семян и многое другое.
В последние годы наука достигла
больших успехов в понимании того, как
гормоны регулируют жизнь растений, вызывая
переключение генетических программ,
определяющих последовательность этапов
развития, а также ответ растений на
внешние воздействия.
Все, что известно сегодня о
механизме действия фитогормонов, крайне важно
для того, чтобы понять, как происходят рост
и развитие растений, т. е. как реализуется
их онтогенетическая программа, а также как
растение реагирует на стрессы в течение
своей жизни. Эти знания крайне важны для
решения практических задач сельского
хозяйства и биотехнологии. Однако многие
вопросы о действии фитогормонов
остаются еще невыясненными и представляют
собою увлекательные задачи для будущих
исследований.
Учение о гормонах растений ведет свое
начало с 1880 г., когда была опубликована
работа Ч. Дарвина и его сына, доказавших,
что росточки канареечной травы, изгибаясь
в сторону света, воспринимают его своей
верхушкой, а реагируют на него изгибом
ниже расположенной части. Они сделали
вывод о существовании в растении
сигнальной системы, которая воспринимает сигнал
в одном участке и передает его в другой
участок, где и осуществляется ответ на
принятый сигнал. Дальнейшее развитие эти идеи
получили в 20-30-е гг. нашего столетия, когда
Ф. Вент в Америке и Н. Г. Холодный в
нашей стране независимо и одновременно
разработали основные положения теории
гормональной регуляции роста растений,
которая во всем мире называется теорией
Вента—Холодного.
Ауксин
Химическая структура первого
открытого гормона растений — ауксина — была
установлена в 1934 г. Он представляет
собою Р-индолилуксусную кислоту или
сокращенно ИУК (рис. 1). Этот фитогормон
активирует рост клеток, вызывает образование
корней и регулирует многие другие
процессы. Его способность вызывать корнеоб-
разование широко используется для
укоренения черенков.
Ауксин образуется в верхушке стебля
растения и движется в расположенные ниже
части, достигая корневой системы. На своем
пути вниз по растению ауксин участвует в
регуляции деления и роста клеток. Одновре-
j
-снгсоон
,сн2он
н
р- Индол илуксусная
кислота
г/ н
HN-CH2-CH=C4
т EN>
сн.
Цитокинин (зеатин)
ООН
Гибберелловая кислота Абсцизовая кислота
N20=0142
Этилен
Рис. 1. Структурные формулы основных
фитогормонов
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЖИЗНИ РАСТЕНИЙ
289
Рис. 2. Схема совместного действия ауксина и ки-
нетина на рост и процессы дифференцировки у
каллуса сердцевины стебля табака
Концентрация ауксина во всех колбах— 2 мг/л.
Содержание цитокинина: а— 0; б— 0,02 мг/л; в— 0,5-
1 мг/л; г— 5мг/л
менно он задерживает рост боковых почек,
расположенных в пазухах листьев. Если
срезать верхушку стебля, поступление
ауксинов к пазушным почкам прекратится и
начнется их рост. При этом получится растение
с боковыми побегами. Ауксины необходимы
для клеточных делений, однако это их
свойство, как правило, проявляется в присутствии
другого гормона растений — цитокинина*
Цитокинины
Цитокинины были открыты Ф. Скугом с
сотрудниками в США в 1955 г. Скуг изучал
рост изолированных тканей растений в
стерильных условиях на питательной среде. Он
помещал на агаровую среду, содержащую
сахарозу, витамины, аминокислоты,
минеральные элементы, а также ауксин, кусочки
изолированной сердцевины стебля табака и
хотел вызвать переход его клеток к
делению с образованием каллуса — бесформенной
массы недифференцированных клеток.
Однако это ему не удавалось — клетки не
делились. Ф. Скуг и К. Миллер вызвали
деление клеток с помощью открытого ими
Рис. 3. Влияние цитокинина и абсцизовой кислоты
на изолированные семядоли тыквы
Слева направо: 1 — контрольные семядоли на воде;
2— на растворе синтетического цитокинина — (3-бен-
зиламинопурина (5 Ю_6М); 3— на растворе
абсцизовой кислоты (10~5М)
кинетина — вещества, образующегося при
деградации ДНК. Кинетин является
аналогом широко распространенного в
растениях цитокинина транс-зеатина. (В последнее
время он обнаружен в животных тканях.)
Благодаря открытию кинетина основные
свойства цитокининов были изучены за 10
лет до выделения из растений зеатина.
Как показали Скуг и Миллер, для
деления клеток необходимы два фитогормона:
ауксин и цитокинин. В их опытах было
также установлено, что корнеобразование у
каллуса вызывает ауксин, а дифференци-
ровку побегов цитокинин. На рис. 2
показано, что в отсутствие кинетина у отрезков
сердцевины стеблей табака клеточное
деление не происходит (а), добавление
кинетина в питательную среду вызывает деление
клеток и бурный рост каллуса (б),
увеличение концентрации кинетина приводит к
образованию стеблей (в), но дальнейшее ее
повышение подавляет рост каллуса (г).
Так, в опытах Скуга и Миллера были
разработаны основы современной культуры
изолированных клеток и тканей растений
и найдены пути регенерации целых
растений из недифференцированных клеток
каллуса.
Зеатин (см. рис. 1), был выделен
значительно позже, в 1964 г., Д. Литамом в
Австралии. Чтобы выделить 1 мг вещества и
установить его структуру, было
переработано 60 кг семян кукурузы в стадии
молочной спелости, когда семена особенно
богаты цитокининами. Этот пример
показывает, насколько малы концентрации
природных гормонов в клетках растений.
Именно в крайне низких концентрациях гормоны
и осуществляют свое регуляторное
действие. Если действовать фитогормоном на
растительные ткани или органы, которые
сами его не синтезируют и поэтому дают
на него четкий ответ, легко выяснить, что
уловить действие гормона удается уже при
концентрации 10"7—10"8М, а иногда даже
10"10М. Пропорциональное увеличение
реакции растений на гормон обычно
обнаруживается при изменениях его
концентрации от 10~8 до Ю-5 М. На этом основаны
биотесты определения фитогормонов.
Кроме установленной в опытах Скуга
способности вызывать деление клеток и
индуцировать дифференцировку побегов,
цитокинины активируют рост листьев и
семядолей двудольных растений (рис. 3),
стимулируют формирование хлоропластов,
задерживают старение листьев. Если
раствором цитокинина опрыснуть одну половину
листа, это задержит ее пожелтение и
старение, тогда как другая половина
пожелтеет и отомрет (рис. 4). Цитокинины
вызывают приток питательных веществ к месту
их нанесения.
290
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
В целом растении цитокинины
синтезируются в растущих кончиках корней, а
затем по сосудам ксилемы поступают вместе
с пасокой (раствором, который корни
нагнетают в побеги) в надземные органы, где
и принимают участие в регуляции
различных физиологических процессов.
Снабжение побегов цитокининами из
корней было открыто автором этой статьи в
1962 г. Если срезать растение и надеть на
перерезанный стебель стерильный приемник
для сбора пасоки (рис. 4, б), можно
обнаружить, что корень продолжает нагнетать воду
и растворенные в ней минеральные
вещества, поглощенные из почвы, а также
органические вещества, которые
синтезируются в корнях. Как выяснил автор, корневая
система посылает в побеги и цитокинины,
т. е. гормональный сигнал, с помощью
которого корни контролируют процессы,
протекающие в надземных органах.
Гиббереллины
Открытие гиббереллинов берет свое
начало из другой области исследований —
науки о болезнях растений —
фитопатологии. Болезнь риса, вызываемая грибком
Gibberella fujikuroi, сопровождается резким
вытягиванием стебля. Исследование этой
болезни в 20-е гг. нашего века в Японии
привело к выделению вещества, с помощью
которого грибок усиливает рост стебля
зараженного растения. Структура этого
вещества, названного гиббереллином, была
установлена в 1955 г. английским ученым
Б. Кроссом (см. рис. 1). Было доказано, что
гиббереллин представляет собою гормон ра-
6
Цитокинин
Рис. 4. Задержка старения половины листа
махорки в результате ее опрыскивания раствором кинети-
на (20 мг/л) (а) и обнаружение в пасоке веществ ци-
токининового типа (б)
стений, который активирует рост стебля и
вызывает цветение. С его помощью можно
вызвать пробуждение из состояния покоя
клубней и семян растений. Паразитический
грибок, синтезируя гиббереллин в больших
количествах, использует его в своих целях
для поражения больного растения.
Как выяснилось, очень многие
паразиты растений как грибного, так и
бактериального происхождения используют
различные фитогормоны, которые они активно
синтезируют, для «химической атаки» на
растение-хозяина. Такие грибы
культивируют в промышленных условиях для
выделения и получения в чистом виде фитогор-
монов, в частности, гиббереллина.
Гиббереллины представлены в растениях
большим семейством соединений. Они играют
крайне важную роль в переходе растений
к цветению, или, как принято говорить, в
индукции цветения у растений, называемых
длиннодневными, потому что весной в
условиях короткого дня они не зацветают даже
при наступлении теплых дней. Эти
растения умеют «определять» длину дня и
«отсчитывать» количество длинных дней, после
которого переходят к цветению. Это
приспособление выработалось эволюционно как
защита цветков и развивающихся плодов от
весенних заморозков. Теплые дни могут
смениться холодными, а удлинение дня с
переходом от зимы к лету абсолютно
стабильный фактор. Поэтому отбор в
эволюции растений, которые зацветают только
по прошествии определенного числа
длинных дней, обеспечил северным растениям
стабильное воспроизведение потомства.
Гипотеза о гормональной регуляции
цветения растений была выдвинута М. X. Чай-
лахяном еще в 1937 г. на основании
большого экспериментального материала.
Гипотетический в то время гормон
цветения был назван им флоригеном. Как
показали позднее А. Ланг (в США) и Чайлахян
(в нашей стране), переход длиннодневных
растений к цветению вызывает гиббереллин.
Именно его синтез в листьях растений
задерживается в условиях короткого дня и
осуществляется в условиях длинного дня.
Образовавшийся в листьях гиббереллин
транспортируется в точку роста и
вызывает рост стебля и формирование цветков. В
условиях короткого дня такие растения
остаются в форме розетки, но если на их
точку роста капать раствором
гиббереллина, начинается рост стебля, а затем
формирование цветочных зачатков, и растение
зацветает.
Химическая природа индуктора
цветения короткодневных растений — антезина
остается до сих пор не установленной.
Поскольку гиббереллин активирует рост
стебля, с его помощью удается получать ги-
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЖИЗНИ РАСТЕНИЙ
291
гантские растения. Способность гибберелли-
на пробуждать клубни из состояния покоя
находит применение на юге для получения
двух урожаев картофеля в год. Гиббереллин
применяют также для увеличения урожая
бескосточковых сортов винограда.
Абсцизовая кислота
К числу гормонов растений относится
абсцизовая кислота (АБК), выделенная в
1963 г. одновременно и независимо
английским ученым Ф. Уорингом из листьев
березы и явора и американскими
специалистами Ф. Эддикотом, Б. Милборроу и К. Окумой
из молодых коробочек хлопчатника. Ее
формула была установлена в 1965 г.
английским ученым Дж. Корнфордом и Окумой
(рис. 1).
АБК подавляет ростовые реакции
растений, вызванные ауксином, цитокинином
и гиббереллином, она тормозит дифферен-
цировку хлоропластов, активируемую
цитокинином. На рис. 3 показано угнетение
роста и позеленения изолированных
семядолей тыквы под действием абсцизовой
кислоты. Накопление АБК в созревающих
семенах, почках деревьев осенью и в клубнях
растений обеспечивает состояние покоя их
клеток в течение зимнего периода.
Снижение содержания АБК служит условием их
пробуждения и роста с наступлением
весны.
Фитогормоны регулируют не только рост
и процессы дифференцировки новых
органов у растений, но и многие другие
физиологические процессы, не имеющие
отношения к росту. Например, с помощью АБК
можно закрыть отверстия устьиц растений —
специальных структур, через которые лист
растений испаряет воду, поглощает С02 при
фотосинтезе или выделяет С02 при
дыхании, а также выделяет или поглощает 02.
В регуляции движения устьиц, как и во
многих других физиологических процессах, у
растений четко прослеживается антагонизм
двух гормонов: АБК закрывает устьица при
недостатке воды и тем самым сохраняет ее
резерв в растении, а поступающий из
корней с пасокой цитокинин открывает их при
восстановлении водоснабжения. Открытые
устьица обеспечивают поступление С02
в листья, необходимое для фотосинтеза,
а также поддерживают испарение воды
(транспирацию), обеспечивающее ее
передвижение вверх по растению и
поступление с ней из корней в побеги элементов
минерального питания и различных
органических веществ, синтезируемых в
корнях, включая цитокинины.
АБК не только ингибирует процессы,
вызываемые другими гормонами, но и
играет важную роль в индукции ряда
физиологических программ, например, синтеза
запасных белков семян, формирования
клубней, дифференцировки зимующих почек
ряски. Как и все другие гормоны, она
представляет собою регулятор физиологических
программ у растений, который ингибирует
одни и активирует другие
физиологические процессы.
АБК играет крайне важную роль в
ответе растений на стрессовое воздействие —
обезвоживание, засоление, действие низких
температур. В ответ на перечисленные
воздействия в растениях начинается усиленный
синтез АБК, которая, в свою очередь,
резко изменяет экспрессию генетических
программ в клетках: подавляет синтез мРНК и
соответствующих им белков, характерных для
нормальных условий, и индуцирует работу
генов и, следовательно, синтез
специфических белков, называемых белками ответа на
АБК. Их довольно много и их свойства
сейчас усиленно изучаются. По-видимому, они
защищают клетку от гибели в условиях
глубокого обезвоживания. Интересно, что эти
белки при нормальном развитии растений
синтезируются в период позднего
эмбриогенеза, когда происходит естественное
обезвоживание семян. Синтез этих белков в
позднем эмбриогенезе индуцируется АБК,
которая накапливается в семенах перед
началом их обезвоживания. В листьях они в
нормальных условиях не обнаруживаются.
Засуха вызывает накопление в листьях АБК,
и она индуцирует синтез белков позднего
эмбриогенеза, необходимых для выживания
клеток листа во время засухи. Исследование
таких белков и кодирующих их генов крайне
важно для создания устойчивых к засухе
сортов сельскохозяйственных растений. На
помощь здесь может прийти генная
инженерия, которая позволяет трансформировать
растения нужными генами, т. е. вводить
соответствующие гены в их ДНК и создавать
новый генотип. С этой точки зрения
большой интерес представляет поиск в природе
растений-рекордсменов по устойчивости к
неблагоприятным условиям, в частности, к
засухе. Один из таких рекордсменов —
Craterostigma plantagineum. Ее листья
высыхают при глубоком обезвоживании до
воздушно-сухого состояния, но через несколько
часов после предоставления им воды снова
восстанавливают нормальное
функционирование. Обезвоживание листьев индуцирует
синтез в них АБК, а она, в свою очередь,
индуцирует синтез de novo ряда белков.
Этилен
К гормонам растений относится также
этилен. Это его свойство было открыто в
1901 г. в Санкт-Петербургском университете
Д. Н. Нелюбовым. Он показал, что светиль-
292
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
ный газ (в который входит этилен)
оказывает тройное воздействие на проростки
гороха: подавляет рост стебля, вызывает его
утолщение и изменяет ориентацию стебля
в пространстве. Это открытие хотя и было
опубликовано, но долго оставалось не
оцененным научной общественностью. Этилен
как гормон растений, ускоряющий
созревание плодов, снова был «открыт» в 20-х гг.
В последующем было показано, что
растения синтезируют этилен, и был
установлен путь его биосинтеза. Этилен, как и все
другие гормоны растений, обладает
широким спектром регуляторного действия. Он
вызывает укорочение и утолщение стебля,
изменяя направление роста клеток. Это
свойство этилена крайне важно для
борьбы с полеганием хлебов. В районах с
плодородной почвой и влажным климатом
стебель не выдерживает тяжести колоса и
ложится на землю. Хлеба «полегают». Это
наносит огромный ущерб сельскому
хозяйству. Для борьбы с этим явлением существует
три подхода: 1) обработка растений этилен-
продуцентами, в частности этефоном,
который легко разлагается с образованием
этилена, вызывающего укорочение и
утолщение стебля; 2) обработка растений
ретардантами, подавляющими биосинтез гиб-
береллина — в результате тоже получаются
короткостебельные, устойчивые к
полеганию растения; 3) создание сортов
низкорослых растений с прочным и толстым
стеблем. Последний путь предпочтителен, так
как он избавляет сельское хозяйство от
внесения химических соединений,
ухудшающих экологические условия. Получение
низкорослых сортов, в частности, связано
с изменением гормональных характеристик
растений, обеспечивающих формирование
короткого, прочного стебля.
Этилен вызывает опадение листьев и
плодов растений. Обработка
этилен-продуцентами помогает удалить листья
хлопчатника для последующего машинного сбора
созревших коробочек. Этилен вызывает
старение листьев и ускоряет созревание
плодов. Этим объясняется тот факт, что
завернутые плоды помидоров или бананов быстрее
дозревают, чем находящиеся на воздухе, так
как выделяемый ими этилен сохраняется в
атмосфере хранения плодов и ускоряет их
созревание. Выращивание томатов в
условиях короткого лета основано на сборе
зеленых плодов и их последующей обработке
этилен-продуцентами. При транспортировке
плодов на далекие расстояния встает
другая задача — задержать быстрое
дозревание плодов. Здесь также на помощь
растениеводам приходит наука о гормонах
растений. Ученые проследили путь
биосинтеза этилена и деградации его прямого
предшественника, 1-аминоциклопропан-1-кар-
боновой кислоты (АЦК). В США получен
трансформант (генетически измененная
линия) томатов со сниженным биосинтезом
этилена, что продлевает хранение томатов
в свежем виде на целый месяц. Другой путь
продления срока хранения плодов, а также
сохранности в свежем виде срезанных
цветов, например, гвоздики, основан на
получении соединений, которые блокируют
рецептор этилена и не дают проявляться его
действию.
Этилен принимает участие в ответе
растений на различные патогены
грибного, бактериального и вирусного
происхождения. Растение воспринимает сигнал
патогена и включает в ответ синтез этилена,
а тот, в свою очередь, запускает сложную
программу химической защиты растений,
в которую, в частности, входит синтез
фитоалексинов, играющих роль
растительного противоядия против паразитов.
Интересно, что в систему защиты
растений входит также салициловая кислота
(производным которой является всем
известный аспирин). По многим признакам ее
можно рассматривать в качестве еще
одного гормона растений, участвующего не
только в антипатогенных реакциях, но и в
регуляции онтогенетических программ.
Возвращаясь к этилену, следует добавить, что
он играет важную роль при затоплении
растений, индуцируя развитие в стебле
аэренхимы — специальной ткани,
обеспечивающей поступление кислорода из надземных
частей растения в находящиеся под водой
корни, — и тем самым спасая их от гибели
в результате кислородного голодания.
Фитогормоны в культуре
изолированных клеток
и тканей растений
Особо следует подчеркнуть, что все
теоретические исследования, проводимые на
стерильной культуре клеток и тканей
растений, а также их практическое
применение в биотехнологии, основаны на
использовании фитогормонов, которые представляют
собою главные химические средства
воздействия на растительные культуры. Так,
размножение клеток in vitro (т. е. вне растения)
основано на индукции клеточных делений
ауксином и цитокинином (иногда лишь
синтетическим аналогом ауксинов). Это
позволяет культивировать в заводских условиях
клетки-продуценты ценных лекарственных
препаратов.
Регенерация целых растений основана на
индукции образования побегов под действием
цитокининов с их последующим укоренением
под действием ауксинов. (Существует еще
путь образования эмбриоидов — эмбрионо-
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЖИЗНИ РАСТЕНИЙ
293
подобных структур в суспензионной
культуре клеток.)
Регенерация растений особенно ваткна
при получении трансформированных
(генетически измененных) растений. Эта
техника позволяет выяснить — как, где, в
какое время в растении происходит экспрессия
гена, интересующего исследователя.
Кроме того, это путь получения растений с
новым генотипом, содержащим чужеродные
и притом, как принято говорить,
хозяйственно ценные гены, например, гены
устойчивости к вирусам, к грибным
патогенам, к листогрызущим насекомым, даже к
колорадскому жуку.
С помощью цитокининов
осуществляется техника микроклонального размножения
растений. На питательную среду
помещают изолированную стеблевую точку роста
растения, которая дает под действием
цитокининов образование не одного, а
нескольких побегов. При этом растения
размножаются без изменения генотипа и, кроме того,
обеспечивается получение безвирусного
материала. Этот метод применяется также
для сохранения и размножения исчезающих
видов растений.
Паразитические бактерии
растений — природные
генные инженеры
Крайне интересно, что гормоны растений
используют природные генные инженеры —
паразитические бактерии Agrobacterium
tumefaciens и Agrobacterium risogenes. В
клетках A. tumefaciens обнаружены
мини-кольцевые ДНК, называемые плазмидами (а
именно, Ti-плазмиды), которые содержат гены,
отвечающие за синтез ауксина (1-й и 2-й ген)
и за синтез цитокинина (4-й ген). При
инфицировании растений часть плазмидной ДНК
(так называемая Т-ДНК) встраивается в
растительную ДНК. В результате зараженные
клетки содержат в геноме бактериальные гены
синтеза ауксина и цитокинина и еще гены,
обеспечивающие синтез специфических
питательных веществ для бактерий. В клетках
начинается усиленный синтез ауксина и
цитокинина — двух необходимых индукторов
клеточных делений. В результате этого клетки
переходят к делению и образуется опухоль.
Таким образом, бактерия поступает так же,
как исследователь, который для
размножения клеток растений в культуре добавляет
туда ауксин и цитокинин. Методами генной
инженерии можно повредить в Ti-плазмиде
гены синтеза ауксина; тогда в зараженных
клетках будут работать только гены
биосинтеза цитокинина, и вместо образования
опухоли возникнет пучок
дифференцирующихся побегов. Если же повредить ген биосинтеза
цитокинина, то в трансформированных
клетках будут работать гены биосинтеза ауксина
и вместо опухоли получится «борода» корней.
Другая бактерия, A. risogenes,
трансформирует растение по-иному, образуя огромную
массу корней. Ее Ri-плазмида привносит в ДНК
зараженных клеток гены, повышающие
чувствительность клеток к ауксинам в 10000 раз.
Таким образом, оба типа бактерий
трансформируют растения, модифицируя их
гормональные свойства.
Последние достижения
в изучении механизма действия
фитогормонов
В последние годы в центре внимания
науки о гормональной регуляции жизни
растений стоит вопрос о механизме действия
фитогормонов: как воспринимается
гормональный сигнал клеткой и как он
передается на генетический аппарат, вызывая
включение, подавление или переключение
генетических программ? В связи с этим
особый интерес вызывают рецепторы
фитогормонов в клетках растений, которые
специфически узнают гормональный сигнал
и образуют гормон-рецепторный комплекс,
который в свою очередь определяет цепь
событий в клетке, необходимых для ее
высокоспецифичного ответа на фитогормон.
Чем глубже идут исследования рецепторов
гормонов растений, тем больше общего
обнаруживается в их свойствах с
рецепторами гормонов животных, к настоящему
времени хорошо изученных. Роль
рецептора в восприятии и передаче
гормонального сигнала может быть представлена
следующим образом:
Г + Р <=^ ГР —» система трансформации и
передачи сигналов в клетке —> индукция
физиологических программ.
Для гормонов животных установлены
рецепторы двух основных типов.
1. Рецепторы не проникающих в клетку
гормонов, расположенные в мембране. Они
узнают молекулу гормона на внешней
поверхности мембраны и в результате
взаимодействия с ним изменяют свое конфор-
мационное состояние (пространственную
структуру); это изменение передается
через специальные белки (G-белки)
ферментам, участвующим в синтезе вторичных
посредников (ферментам-эффекторам).
Посредники же участвуют в усилении
(размножении) сигнала и его передаче к
различным компонентам клетки, в частности,
ферментам, называемым протеинкиназами,
которые фосфорилируют клеточные белки
и в результате изменяют их свойства. Так
происходит изменение функциональной
активности ферментных, регуляторных и
294
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
Влияние зеатинсвязывающего белка с молекулярной массой 67 кДа
из цитозоля листьев ячменя в комплексе с зеатином (10~8М) на синтез
РНК in vitro в системе, содержащей ядра, выделенные из листьев ячменя
(данные представлены в % от синтеза РНК в контроле)
67-кДа белок
l + 1 +
Зеатин
. + +11
РНК-полимераза I
100
99
97
330
РНК-полимераза II
100
101
98
502
структурных белков клетки. Волна этих
изменений передается на генетический
аппарат, и происходит включение
(выключение) характерных для действия гормона
программ.
2. Рецепторы второго типа (к ним
относятся рецепторы стероидных гормонов)
взаимодействуют с гормонами в цитоплазме или
ядре; образуемый ими гормон-рецепторный
комплекс непосредственно участвует в
регуляции генетических программ, индуцируя
или репрессируя экспрессию генов (рис. 5).
Большие успехи генной инженерии
позволили с помощью генетических зондов
обнаружить в геноме растений гены,
ответственные за все основные звенья каскада
размножения и передачи гормонального
сигнала по первому типу. Локализованные
в мембране гормон-связывающие белки —
вероятные кандидаты на роль рецепторов
первого типа — обнаружены для ауксина,
этилена, гиббереллина. Из мембран коле-
оптилей кукурузы выделен ауксинсвязыва-
ющий белок (АСБ) — димер из двух
субъединиц с молекулярным весом 22 кДа каждая.
(Колеоптиль — бесцветный колпачок,
покрывающий росток при появлении всходов
злаков. Потом растущий лист прорывает
колеоптиль и он отмирает.) Для АСБ колеоптилей
кукурузы установлена аминокислотная
последовательность и найден участок
связывания ауксина. Доказано, что он расположен
на внешней стороне мембраны.
Взаимодействие ауксина с рецептором вызывает
гиперполяризацию мембраны, выделение
ионов Н+ из клетки, приводящее к
разрыхлению структуры клеточной стенки,
которое необходимо для увеличения размеров
клетки в ходе ее роста. Накопленные
данные делают вероятным предположение, что
АСБ колеоптилей кукурузы
функционирует по типу мембранных рецепторов
гормонов животных с использованием вторичных
посредников.
Есть также сведения о быстром репрог-
раммировании под действием ауксина
генетического аппарата растительных клеток.
Осуществляется ли это с помощью
сигналов, приходящих с мембранного
рецептора или через рассмотренные выше
рецепторы второго типа, — пока не ясно.
В последнее время выделен ген
рецептора этилена и установлена его нуклеотидная
последовательность. Ее анализ показал, что
рецептор представляет собой особого типа
протеинкиназу, в которой сигнал
вызывает фосфорилирование гистидинового остатка
с последующей передачей фосфатной группы
на остаток аспарагиновой кислоты.
По-видимому, так включается цепь процессов,
опосредующих воздействие этилена на
растения.
В нашей лаборатории выделен рецептор
цитокинина из цитоплазмы закончивших рост
листьев ячменя, обладающих высокой
чувствительностью к цитокинину. (Цитокинин,
как уже отмечалось, задерживает
старение срезанных листьев и активирует в них
синтез РНК.) Выделенный белок с
молекулярной массой 67 кДа, обладал всеми
признаками рецептора цитокинина. Он
высокоспецифично и обратимо связывал меченый
природный цитокинин — зеатин. Зеатин-
белковый комплекс активировал синтез РНК-
in vitro. Эти опыты проводились в системе,
содержащей хроматин из клеток листьев
ГР
М
Ц
синтез белка
конечный эффект
Рис. 5, Схема прямой регуляции экспрессии
генома гормон-рецепторным комплексом
Г—гормон; Р—рецептор; М —мембрана; Я —ядро;
Ц— цитоплазма
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЖИЗНИ РАСТЕНИЙ
295
ячменя (хроматин — комплекс ДНК и
белков, входящих в состав хромосом) и
связанную с ним РНК-полимеразу I,
осуществляющую синтез РНК. Цитокинин-белковый
комплекс проникал в изолированные из
листьев ядра и активировал в них синтез
РНК как РНК-полимеразой I,
ответственной за синтез рибосомных РНК, так и РНК-
полимеразой II, ответственной за синтез
информационных РНК (см. таблицу).
Таким образом, цитокинин-рецепторный
комплекс позволил промоделировать in vitro
типичный эффект цитокинина — активацию
в листьях синтеза РНК. Насколько нам
известно, это первые данные о выделении
функционально активного рецептора цито-
кининов из растений.
Несмотря на большие успехи последних
лет в изучении рецепции и передачи
гормональных сигналов у растений, многое
остается еще не выясненым. Исследования в этом
направлении необходимы как для разработки
теоретических основ регуляции жизни
растений, так и для практического
применения научных результатов в генной
инженерии, биотехнологии и сельском хозяйстве.
Литература
КулаеваО. Н. Физиология растений, 1962,
т. 9, с. 229.
Кулаева О. Я. Цитокинины, их структура и
функции. М.: Наука, 1973.
Кулаева О. Я. Гормональная регуляция
физиологических процессов у растений на уровне
синтеза РНК и белка. М.: Наука, 1982.
Кулаева О. Я. Физиология растений, 1995,
т. 42, с. 661.
Полевой В. В. Фитогормоны. Л.: Изд-во ЛГУ,
1982.
Уоринг Ф., Филлипс И. Рост растений и диф-
ференцировка. Пер. с англ. М.: Мир, 1984.
Чайлахяп М. X. Регуляция цветения высших
растений. М.: Наука, 1988.
В. А. Опритов
ЭЛЕКТРИЧЕСКИЕ СИГНАЛЫ У ВЫСШИХ РАСТЕНИЙ
Введение
Впервые мысль о том, что растения
обладают раздражимостью, т. е. способны
быстро реагировать на внешние воздействия
и передавать сигнал об этом воздействии от
одного органа к другому, высказал Ч. Дарвин
(1875 г.). Его внимание привлекли так
называемые насекомоядные растения
(росянка, венерина мухоловка, альдрованда и др.),
которые с помощью специальных
приспособлений захватывают мелких насекомых и
используют их в пищу. Ловчие органы этих
растений очень быстро механически
реагируют на прикосновение насекомого. В этом
Дарвин увидел большое сходство с
поведением животных. Однако он не знал, что
лежит в основе такой высокой
чувствительности. В 1887 г. Бердон-Сандерсон показал,
что быстрое движение венериной мухоловки
сопровождается распространением в ее
лопастях электрических импульсов, очень
напоминающих потенциалы действия в нерве.
Решающее значение в изучении
процессов раздражимости и возбудимости у
растений имели исследования выдающегося
индийского ученого Д. Боса, работавшего
в начале XX в. Бос проводил свои опыты
на мимозе стыдливой, которая, так же как
и насекомоядные растения, проявляет
способность к быстрым движениям в ответ на
механическое раздражение. Используя
весьма чувствительную экспериментальную
технику, Бос установил, что раздражение
листа мимозы вызывает возникновение в
черешке электрических импульсов,
которые, распространяясь до листовых
подушечек, приводят к их сокращению и
опадению листа. Электрические импульсы в
черешке мимозы оказались очень
похожими на те, что возникают в ответ на
раздражение у животных. Проводя
многочисленные эксперименты с мимозой, Бос все
больше убеждался в сходстве восприятия
и передачи раздражения у животных и
растений. Этому сходству Бос придавал очень
большое значение, справедливо видя в нем
убедительное подтверждение мысли о том,
что растения и животные при всем
кажущемся различии в их образе жизни, не
отличаются принципиально по своим
реакциям на внешние воздействия.
В опытах Боса были получены данные,
показывающие, что весьма быстрые
электрические ответные реакции на внешние
раздражители можно наблюдать и у некоторых
обычных высших растений, не обладающих
быстрыми двигательными реакциями.
Однако решающее значение в подтверждении
этого положения сыграли работы
отечественного ученого И. И. Гунара, работавшего в 40-
60-е гг. в Московской сельскохозяйственной
академии им. К. А. Тимирязева. Им и его
сотрудниками было убедительно показано, что
способность растений отвечать на внешние
раздражители генерацией электрических
импульсов, распространяющихся по нему и очень
напоминающих потенциалы действия в нервах,
свойственна всем высшим растениям.
Возникновение электрических
сигналов у растений
Если подвести к стеблю растения два
электрода, соединить их с усилителем и
самописцем, а затем на лист растения
нанести раздражение, то спустя короткое
время прибор зарегистрирует
электрический импульс (рис. 1). Он распространяется
от листа к корням и является сигналом о
внешнем воздействии. Аналогичный
электрический импульс, но распространяющийся
от корней к листьям, можно наблюдать,
если, например, подействовать на корни
0,1 н. КС1. Возникает он при действии
раздражителя и на другие органы растения
(стебли, усики и т. д.).
а
б
мВ
40
30
20
10
0 12 3 4
мин
32р
Рис-1. Потенциалы действия, возникающие при
раздражении листа растения
а— ПД, который распространяется по проводящим
пучкам, достигает корней и вызывает временное
изменение поглощения веществ (например, меченого
фосфата в форме КН232Р04); б— ПД,
распространяющийся от места раздражения на листе к корням
ЭЛЕКТРИЧЕСКИЕ СИГНАЛЫ У ВЫСШИХ РАСТЕНИЙ
297
Набор раздражителей, вызывающих
появление электрического сигнала,
весьма разнообразен. Это может быть
изменение температуры, механическое
воздействие, облучение участка растения светом
различного спектрального состава и т. д. При
этом было бы неверно думать, что
растения обладают меньшей способностью
реагировать на внешний стимул, чем
животные. Наоборот, в ряде случаев клетки
растений способны отвечать генерацией
электрических сигналов на чрезвычайно слабые
воздействия. Например, отрезок волоса весом
всего 0,000822 мг при соприкосновении с
щупальцем росянки вызывает ответную
биоэлектрическую реакцию и заметное
движение щупальца. Понижение температуры
от 23°С всего на 1-2°С вызывает генерацию
распространяющихся электрических
сигналов в стебле тыквы.
Таким образом, возникновение
электрических сигналов у растений может быть
обусловлено не только действием сильных,
повреждающих факторов, но и весьма
слабыми изменениями окружающей среды,
которые постоянно происходят в
естественных условиях.
Типы электрических сигналов
у растений
Удалось выявить, по крайней мере, три
типа электрических сигналов, возникающих
у растений в ответ на внешние воздействия.
Первый тип — это потенциалы действия
(ПД). Такое название данный тип
электрических сигналов у растений получил
благодаря тому, что по ряду признаков и
механизму возникновения он соответствует ПД,
генерируемым в нервах животных. На рис. 2
представлены записи ПД стебля тыквы и
[юмВ a J25mB б
1 мин Гмс
J-IOmB e I500 мкВ
i—i
л ' 1 С
1 МИН
Рис. 2. Типы импульсной электрической активности
у высших растений
а — потенциал действия, в — вариабельный
потенциал, г— микроритмы. Для сравнения приведена
запись потенциала действия аксона кальмара (б)
аксона кальмара. Видно, что внешне они
очень сходны между собой и состоят из двух
ветвей — восходящей (или фазы
деполяризации, во время которой происходит
уменьшение потенциала возбудимой мембраны)
и нисходящей (или фазы реполяризации,
в ходе которой мембранный потенциал
восстанавливается до исходного уровня).
Амплитуда обоих ПД составляет несколько
десятков милливольт. Их внешнее
различие состоит в том, что в нервном волокне
процессы деполяризации-реполяризации
происходят значительно быстрее, что
связано с особенностями строения возбудимых
мембран. Поэтому в аксоне кальмара общая
длительность ПД составляет всего несколько
миллисекунд, в то время как длительность
ПД в стебле тыквы достигает нескольких
секунд и даже нескольких десятков секунд.
Второй тип электрических сигналов у
высших растений — это так называемые
вариабельные потенциалы (ВП), которые
возникают при действии весьма сильных
раздражителей (ожог, механическое
повреждение ткани и др.). Как видно из рис. 2,
они лишь частично напоминают ПД. Как и
у ПД, у них четко выражена фаза
деполяризации. Однако фаза реполяризации очень
растянута. Таким образом, ВП имеют
природу, несколько отличную от природы ПД.
Наконец, рядом исследователей у
высших растений с помощью специальной
чувствительной техники были зарегистрированы
микроритмы (рис. 2), имеющие очень
небольшую амплитуду (обычно несколько
микровольт) и носящие весьма нерегулярный
характер. Природа микроритмов пока
остается неясной.
Из всех типов электрических сигналов
у растений особое внимание уделяется ПД,
поскольку его генерация и
распространение представляют собой один из
универсальных способов передачи информации о
внешнем воздействии в живой природе.
Пути распространения
ПД в растении
Возникнув в той или иной части
растения, ПД, передающие известие о внешнем
раздражении, распространяются по
растению обычно со скоростью несколько см/с
(или см/мин). Как известно, у животных
проводниками ПД являются нервные волокна.
Их возникновение в ходе эволюции было
большим шагом вперед в развитии этих
организмов. У растений ту же функцию
выполняют проводящие пучки.
Д. Бос одним из первых
экспериментально доказал причастность проводящих
пучков высших растений к распространению ПД.
Для этого он использовал разработанный им
метод электродного зондирования. Его суть
298
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
состояла в том, что в ткани растения с
помощью микровинтов погружался
металлический микроэлектрод, соединенный с
измерительной установкой. Таким образом
можно отводить электрические сигналы от
разных зон стебля или черешка. На
основании этих опытов Бос пришел к выводу,
что распространение ПД происходит только
в проводящих пучках. Существенно, что
электрические импульсы
распространяются не по крупным сосудам, а по мелким
клеткам пучка (мелким клеткам флоэмы и
протоксилемы). Это свидетельствует о том,
что каналы передвижения веществ и
распространения электрических импульсов в
проводящих пучках пространственно
разделены. Следовательно, хотя у растений и
отсутствуют специальные образования
(наподобие нервов), приспособленные только
для проведения ПД, в проводящих пучках
все же имеются особые ткани,
выполняющие данную функцию.
В месте раздражения ПД примерно
одинаковой амплитуды возникают не только в
указанных выше мелких клетках пучка, но
и в клетках окружающей его основной
паренхимы (рис. 3). Однако на некотором
расстоянии от этого места ПД регистрируются
только в проводящих пучках. Таким
образом, ПД генерируют как пучковые, так и
внепучковые клетки, но проводить его
могут только первые. Причина этих различий
лежит в особенностях межклеточных связей.
У мелких пучковых клеток такие связи (в
частности, с помощью специальных пор-
Время, мин
0 12 3 0 12 3
плазмодесм) выражены гораздо лучше, что
и обеспечивает их лучшую способность
проводить ПД. Плазмодесмы представляют
собою отверстия в клеточной стенке, через
которые сообщается «содержимое» двух
соседних клеток. Эти отверстия обеспечивают
связь цитоплазмы клеток данной ткани в
единый симпласт, что, очевидно, важно для
распространения электрических сигналов.
Механизмы генерации
и распространения
ПД у растений
При генерации ПД в растении, так же
как и в нерве, возникают ионные потоки
(рис. 4, а). Вначале под влиянием
внешнего раздражителя увеличивается проница-
Время
10
20
00
5
-50
-80
ПД
UI
Е-120
-160
-200
Са2+
Внутри
Мембрана
Снаружи
СГ К+
-50
-100
-150
0
-50
-100
-150
1 2 3
-50
-100
-150
0 12 3
-50
-100
-150 2
Рис. 3. ПД в стебле проростка тыквы при его
раздражении охлаждением
а— в месте раздражения; б— на расстоянии 50мм
от этого места; 1 — ПД в клетках, окружающих пучки;
2— ПД в мелких пучковых клетках
Внутри
Мембрана
Снаружи
Рис. 4. Схема ионных потоков, лежащих в основе
генерации ПД у высших растений (объяснение в
тексте) (а) и схема распространения ПД в возбудимой
мембране (б)
При возбуждении между невозбужденными
участками мембраны и возбужденным участком протекают
местные токи, которые вызывают реполяризацию
возбужденного участка. В то же время соседние с
возбужденным участки деполяризуются. Когда
величина деполяризации впереди фронта
возбуждения достигает порогового уровня, здесь возникает
ПД. Этот механизм обеспечивает его
распространение. Сзади фронта возбуждения ПД не возникает,
так как там имеется состояние невозбудимости (реф-
рактерности)
ЭЛЕКТРИЧЕСКИЕ СИГНАЛЫ У ВЫСШИХ РАСТЕНИЙ
299
емость мембраны для ионов кальция в
результате открывания кальциевых каналов.
Ионы кальция входят внутрь проводящих
ПД клеток. Войдя внутрь возбудимых
клеток, они активируют хлорные каналы, что
приводит к их открыванию. Это
сопровождается возникновением направленного
наружу потока ионов хлора. Поток
отрицательно заряженных ионов хлора наружу
приводит к деполяризации мембраны,
поскольку ее внешняя сторона заряжена
положительно, а внутренняя —
отрицательно. Возникает восходящая ветвь ПД.
Деполяризация мембраны способствует
открыванию калиевых каналов и
возникновению направленного наружу потока ионов
калия. Этот поток будет оказывать на
мембранный потенциал реполяризующее
действие, т. е. приводить к восстановлению его
исходного значения.
Данный механизм очень напоминает то,
что происходит при генерации ПД в нерве,
только вместо ионов натрия в качестве
деполяризующего иона у высших растений
выступают ионы хлора. Это сходство
свидетельствует об общности механизмов
генерации ПД в живой природе.
Механизм распространения ПД у
растений также подобен таковому у животных
и носит электрический характер.
Деполяризация участка ткани в месте генерации ПД
приводит к возникновению круговых
местных токов, протекающих между
деполяризованным возбужденным участком ткани и
соседними участками, где мембранный
потенциал клеток сохраняет нормальный
уровень. Эти токи деполяризуют соседние с
возбужденным участком области, что
приводит к возникновению в них ПД и, таким
образом, к его распространению от
исходного места (рис. 4, б). Яркое подтверждение
такого механизма получено в опытах с
изменением электропроводности окружающей
среды. Если вокруг участка проводящего
пучка растения поместить раствор
вазелинового масла (непроводящая среда,
препятствующая возникновению круговых токов),
то, дойдя до этого места, ПД дальше не
распространяется.
Роль ПД у высших растений
Полученные в настоящее время
результаты позволяют утверждать, что у высших
растений распространяющиеся ПД
выполняют вполне определенную
функциональную роль. Они служат самым быстрым
сигналом об изменениях в среде обитания.
Однако у растений нет центральной нервной
системы — этой диспетчерской, откуда
управляющие сигналы после поступления туда
информации о внешнем раздражителе
направляются к различным органам. У
растений ПД сам несет в себе возможность
непосредственно влиять на функции
органов и тканей, по которым он
распространяется. Это связано прежде всего с тем,
что при прохождении ПД по данному
участку ткани, а также в месте, до которого
он дошел, сильно меняются ионный состав
и особенно содержание ионов калия и хлора,
которые, как мы видели, выходят из
возбудимых клеток при генерации импульса. В
результате их концентрации в окружающих
проводящий пучок тканях могут увеличиться.
Меняется соотношение и других ионов, хотя
и в меньшей степени. В то же время
известно, что уровень обменных процессов в
ткани сильно зависит от ионного состава.
Поэтому ПД в состоянии оказывать
влияние на органы или ткани, по которым они
распространяются или которых они
достигают.
Возникновение ПД в ответ на действие
внешнего раздражителя неспецифично, т. е.
самые разные воздействия вызывают, как
правило, однотипную электрическую
реакцию. Кроме того, у растений в ответ на
действие раздражителя обычно
генерируются одиночные импульсы (в отличие от
животных, у которых возникают ритмически
повторяющиеся ПД). Исходя из этого, можно
заключить, что у высших растений
распространяющиеся ПД, по-видимому, не
имеют специфической информационной
нагрузки, а являются скорее сигналом о каком-то
внешнем воздействии. Сам по себе ПД как
сигнал неспецифичен, но в тканях и
органах, наряду с общими неспецифическими
явлениями, он вызывает изменение ряда
специфических процессов, свойственных
данному органу (например, в листьях —
изменение фотосинтеза, в корнях —
усиление поглощения веществ и т. д.).
Сигнальная роль ПД проявляется
прежде всего в ряде естественных процессов.
Например, при попадании пыльцы на рыльце
пестика в нем возникают многочисленные
электрические импульсы,
распространяющиеся по направлению к завязи. Это
запускает цикл преобразований, подготавливающих
завязь к восприятию пыльцы и
оплодотворению. ПД возникают и в усиках вьющихся
растений при соприкосновении с
механической опорой, по-видимому, способствуя их
лучшей ориентации в пространстве.
При умеренных изменениях в состоянии
окружающей среды также могут возникать
ПД, причем иногда они генерируются в ответ
на очень слабые воздействия (например, при
перепаде температур всего 1-2°С).
Генерация ПД растением в этом случае имеет
определенное значение, состоящее в
своеобразном «предупреждении» его органов и
тканей о вполне вероятных существенных
изменениях во внешних условиях. Напри-
300
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
мер, незначительный перепад температур
с сторону охлаждения сам по себе может
быть и незначим для растения, однако он
может свидетельствовать о возможном
предстоящем заметном понижении
температуры окружающей среды.
«Предупреждение» посредством
генерации ПД приводит к временному повышению
устойчивости органов и тканей растения к
неблагоприятным воздействиям. Это
временное повышение устойчивости носит, по-
видимому, неспецифический характер (т. е.
проявляется по отношению к разным
воздействиям) и может рассматриваться как
своеобразная преадаптация. Оно служит как
бы подготовкой к глубокой адаптации, если
вслед за «предупреждением»
действительно наступит усиление данного внешнего
фактора (например, вслед за
незначительным понижением температуры — резкое
похолодание).
Таким образом, можно считать, что при
действии слабых и умеренных
раздражителей мы имеем дело с ролью ПД,
заключающейся в опережающем отражении
действительности. Отсюда можно сделать вывод,
что растениям свойственна элементарная
недифференцированная чувствительность.
Постулируя у высших растений наличие
определенной чувствительности, было бы
неверным утверждать, что эта
чувствительность соответствует эмоциональным
восприятиям животных (например, существует
ошибочное мнение, что растения узнают
повредившего их человека и на его
появление реагируют усиленной генерацией
электрических импульсов). Это именно
элементарная чувствительность, но она,
очевидно, играет существенную роль во
взаимоотношениях растения со средой.
Наконец, при действии сильных
раздражителей ПД выполняют роль первичной
экстренной сигнальной связи, позволяющей
растению оперативно начать перестройку
своих жизненных функций в
экстраординарных условиях.
Итак, по современным представлениям
сигнальной функции ПД принадлежит
вполне определенная роль в осуществлении
быстрых взаимодействий высших растений
с окружающей средой.
Литература
Бос Д. Ч. Избранные произведения по
раздражимости растений. В 2 т. М.: Наука, 1964.
Гунар И. И., Синюхин А. М. Докл. АН СССР,
1962, т. 142, №4, с. 954.
Дарвин Ч. Насекомоядные растения. М.; JL:
Изд-во АН СССР, 1948. (Соч.; т. 7), с. 305.
Опритов В. А., Пятыгин С. С, Ретивин В. Г.
Биоэлектрогенез у высших растений. М.: Наука,
1991.
В. А. Пасешниченко
БИОЛОГИЧЕСКИ АКТИВНЫЕ СОЕДИНЕНИЯ
РАСТИТЕЛЬНОГО ПРОИСХОЖДЕНИЯ
Введение
Одна из примечательных особенностей
растений — многообразие путей обмена
веществ, в ходе которого синтезируется
огромное число специфических соединений,
называемых вторичными метаболитами. К ним
относятся алкалоиды, терпеноиды, гликозиды,
флавоноиды и ряд других. Известны многие
тысячи представителей этих классов
соединений, и каждый год их число
стремительно увеличивается, благодаря
совершенствованию методов извлечения, разделения и
анализа химической структуры веществ.
Многие соединения растительного
происхождения обладают высокой биологической
активностью. Их биосинтез, по-видимому, связан
с «биохимической» адаптацией растений к
условиям существования. Главную роль в этом
явлении играют вещества с аллелопатичес-
кими (воздействующими на соседние
растения), инсектицидными (губительными для
насекомых), противогрибковыми и
антибактериальными свойствами, а также токсины
и яды, действующие на животных и рыб.
Многие из фенольных соединений, тер-
пеноидов и особенно алкалоидов могут
воздействовать на организм человека и
животных. На этом базируется медицинское
применение растительных препаратов и
созданных на их основе лекарств. Природные
вещества растительного происхождения служат
и еще долго будут служить моделями для
химического синтеза полезных соединений.
Известный пример подобной модели —
салициловая кислота, выделенная из коры ивы.
На ее основе было создано такое
популярное и действенное лекарство, как аспирин
(ацетилсалициловая кислота).
Несмотря на колоссальные успехи
химии, большое число лекарственных
препаратов получают непосредственно из растений
или в результате последующих
превращений выделенных из растений веществ,
например, стероидов. Это объясняется
сложностью структуры многих растительных
соединений. Некоторые из таких
соединений представлены в таблице.
Число биологически активных вторичных
метаболитов растений настолько велико, что
их классификация невозможна без
выяснения того, как они образуются. На рисунке
представлена схема путей биосинтеза
некоторых важнейших классов растительных
метаболитов, используемых в медицине,
пищевой и парфюмерной промышленности.
со2
н2о
Фотосинтез
Целлюлоза
Пектины
Крахмал
Сахара
Шикимовая кислота
Мевалоновая
кислота
Ментол
Стерины
Таксол
Каучук
Каротин
Пировиноградная
кислота
Ацетил-КоА
Малонил-КоА
Аминокислоты
Резерпин
Кодеин
Морфин
Атропин
Никотин
Винкристин
Винбластин
Схема путей биосинтеза некоторых классов
вторичных метаболитов
Названия классов вторичных метаболитов обведены
красной рамкой
Алкалоиды
Алкалоидами называют природные
соединения, содержащие один или более атомов
азота, включенных обычно в состав
гетероциклических колец, построенных в основном
из атомов углерода. Алкалоиды очень
разнообразны по структуре и происхождению.
В соответствии с химическим строением
входящих в них гетероциклов их делят на
основные группы, а именно на производные
пирролидина и пиперидина, хинолина, изо-
хинолина, индола, имидазола, пурина (все
эти гетероциклы содержат один или более
атомов азота), пептидные алкалоиды и
алкалоиды терпенового (или изопреноидного)
происхождения, в том числе дитерпеновые
и стероидные.
Отдельные представители каждой из этих
групп обладают самой разной
фармакологической активностью и находят применение в
современной медицине. Например, индольные
алкалоиды винбластин и винкристин,
выделенные из барвинка, в настоящее время
рассматриваются как одни из наиболее
эффективных средств борьбы с раком. Находят
медицинское применение и другие индоль-
ные алкалоиды, в частности резерпин и ай-
302
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
Некоторые лекарственные средства,
Соединения
Гормоны коры надпочечников,
половые гормоны
Сердечные гликозиды
Опийные алкалоиды (морфин, кодеин)
Резерпин
Алкалоиды барвинка (винкристин,
винбластин)
Алкалоиды белладонны
Хинидин, хинин
Пилокарпин
Кокаин
ос-тубокурарин (кураре)
Таксол
Растительный источник
Dioscorea spp.
получаемые из растений
Digitalis spp., Strophanthus kombe
Papaver somniferum
Rauvolfia serpentina
Catharanthus roseus
Atropa bella-donna
Cinchona spp.
Pilocarpus spp.
Erythroxylon coca
Strychnos spp.
Taxus spp.
Сфера применения
Оральные контрацептивы,
анаболики, кортикостероиды
Карденолиды
Обезболивающие
Гипотензивные (понижающие
давление), психотропные
Противораковые
Сосудорасширяющие
Антималярийные, антиаритмические
Антиспазматическое
Местное анестезирующее
Релаксирующее скелетные мышцы
Противораковое
малин из корней растения раувольфии
змеиной {Rauvolfia serpentina). Лечебные свойства
резерпина отражены в таблице. Аймалин же
используют в качестве эффективного
антиаритмического средства. Тетрациклический
алкалоид пахикарпин из бобового растения со-
форы толстоплодной (Sophora pachycarpa)
повышает тонус и усиливает сокращение матки.
Его применяют при стимуляции родовой
деятельности и для улучшения функции мышц
при миопатии (дистрофии мышц).
В лесах горных склонов Крыма,
Кавказа, Карпат растет одно из самых ядовитых
растений, которое в народе называют «сонная
одурь», а в науке оно известно как красавка,
или Atropa bella-donna. К настоящему
времени из листьев и корней красавки
выделили семь различных алкалоидов, главные
из которых — гиосциамин и скополамин. Из
красавки готовят медицинские препараты,
используемые для снятия спазм гладкой
мускулатуры. Самые известные из них — бе-
салол, беллалгин, беллатаминал, солутан.
Другим ядовитым растением, содержащим
ценные алкалоиды, является аконит, или
борец (Aconitum). Существует предание, что
знаменитый хан Тимур был отравлен ядом
аконита, соком которого была пропитана его
тюбетейка. В Китае и Древней Греции из
аконита получали яд для стрел. Токсичность
аконита обусловливают присутствующие в нем
ядовитые алкалоиды — аконитины. Мнение
древних о том, что каждый яд является и
лекарством (все зависит только от дозы)
подтверждается на примере аконита. Так, в
тибетской медицине аконит считался королем
медицины и его использовали для лечения
столь тяжелых болезней, как сибирская язва,
воспаление легких и др. В настоящее время
аконит широко используется только в
гомеопатии, для которой растения до сих пор
играют основную роль в качестве источника
лекарственных средств.
Терпеноиды, или изопреноиды
Терпеноиды, или изопреноиды,
представляют класс природных соединений, самый
многочисленный по числу веществ с
установленным строением. Молекулы терпеноидов
построены из нескольких разветвленных
пятиуглеродных (изопреновых) единиц
НзС
н„с
^ с—сн=сн,.
В зависимости от числа таких единиц все
терпеноиды делятся на монотерпены (С10),
сесквитерпены (С15), дитерпены (С20), сес-
терпены (С25), тритерпены (С30), тетратер-
пены (С40) и политерпены (С45 и больше).
Молекулы стеринов и стероидов состоят
обычно из 24-29 углеродных атомов, но
поскольку они образуются в растениях из
тритерпеноида — циклоартенола, их
относят к тритерпенам. Каждый из
перечисленных классов терпеноидов представлен очень
большим разнообразием структур, главным
образом циклических. Особенно велико их
число у сескви- и дитерпеноидов.
Большинство терпеноидов имеет
полициклическое строение. Среди такого рода
соединений одну из самых важных в
фармакологическом отношении групп образуют
сердечные гликозиды, или карденолиды. Их
молекулы построены из стероидной части
БИОЛОГИЧЕСКИ АКТИВНЫЕ СОЕДИНЕНИЯ РАСТИТЕЛЬНОГО ПРОИСХОЖДЕНИЯ
303
(агликона) и нескольких сахарных остатков,
как, например, у дигитоксина из растения
наперстянки. Продуценты карденолидов —
строфант и наперстянка — наиболее
значимые для медицины растения. К настоящему
времени из наперстянки выделено более 50
карденолидов, отличающихся друг от
друга строением агликона или сахарной части,
в состав которой, кроме глюкозы, входят
сахара весьма необычного строения,
например, дигитоксоза. Лиана Strophanthus kombe
произрастает во влажных тропических
лесах Африки. Местное население с давних
времен использовало сок этого растения в
качестве яда для стрел. Впоследствии из семян
строфанта выделили природный гликозид —
строфантин, который широко вошел в
медицинскую практику и является
незаменимым помощником врачей скорой помощи —
он действует на сердце уже через 1-3
минуты после внутривенного вливания.
Строение строфантина настолько сложно, что
получить его синтетическим путем пока не
удается. В нашей стране строфант не
растет, однако в результате целенаправленных
поисков были найдены растения со строфан-
топодобным действием (горицвет весенний,
желтушник и др.).
Изучение механизма действия
карденолидов на сердечную мышцу показало, что
они влияют на систолическое сокращение
миокарда, воздействуя на транспорт ионов
калия и натрия, осуществляемый
мембранной АТФазой.
Сердечные гликозиды являются
действующим началом еще одного широко
известного лекарственного растения — ландыша
майского (Convallaria majalis). Его
лекарственные свойства известны очень давно, но не
утратили своего значения и до сих пор. В
России начало широкому применению
препаратов ландыша, послуживших основой
известных «ландышевых капель», положил
знаменитый врач С. П. Боткин. Уже в наше
время из ландыша был выделен ряд
карденолидов, причем некоторые из них,
например, конваллотоксин, по активности
превосходят другие сердечные гликозиды (в
частности, дигитоксин и цимарин).
Стероидные и терпеновые
гликозиды
Другой важной для медицины группой
терпеноидов являются стероидные и тритер-
пеновые гликозиды, или сапонины.
Последнее название связано со способностью этих
соединений к пенообразованию (от англ.
soap — мыло), вызванной их высокой
поверхностной активностью. По биологическим
свойствам сапонины сильно отличаются от
карденолидов, хотя их объединяют общие
пути биосинтеза агликоновой (стероидной или
тритерпеновой) части, а также
принадлежность обеих групп к гликозидам. Подобно
карденолидам, сапонины состоят из агликона
и сахарной части, в состав которой входят,
однако, самые обычные сахара, например,
глюкоза, рамноза, ксилоза, арабиноза и др.
Примером стероидных гликозидов, агли-
кон которых построен из 27 углеродных
атомов, могут служить широко
распространенные среди растений рода Dioscorea гликозиды
диосгенина. Подобно многим другим
стероидным гликозидам, они обладают
гемолитической активностью (способностью вызывать
лизис эритроцитов), противогрибковым,
антимикробным и иммуномодулирующим
действием. Однако основное значение этих
соединений для медицины состоит в том, что
они служат исходным сырьем для синтеза
многих гормональных и противозачаточных
лекарственных средств. Химический синтез
стероидного ядра с определенными, очень
важными для сохранения биологической
активности особенностями стереохимической
(пространственной) конфигурации
чрезвычайно дорог и нецелесообразен. Поэтому
удобнее и дешевле использовать уже готовое
стероидное ядро, присутствующее в агли-
коне стероидных гликозидов растений. Для
этой цели служит в основном агликон —
диосгенин, получаемый в результате
кислотного гидролиза (расщепления)
гликозидов, накапливающихся в корневищах
различных видов лиан из рода Dioscorea,
произрастающих в Мексике и Центральной
Америке. В настоящее время все
стероидные лекарственные препараты, а их более
50 (это главным образом аналоги кортико-
стероидов с противовоспалительным
действием и контрацептивы — аналоги половых
гормонов), получают из диосгенина, годовая
потребность в котором составляет 1000 т.
Превращение диосгенина в кортикосте-
роиды и аналоги половых гормонов
осуществляется в несколько стадий, включающих
химическую обработку и модификацию
получаемых продуктов под действием
микроорганизмов.
Тритерпеновые гликозиды отличаются
от стероидных структурой агликона,
построенного из 30 атомов углерода. Они
являются действующим началом таких
известных в медицине биостимуляторов, как
женьшень, аралия и др. Настойки женьшеня
обладают тонизирующим, стимулирующим
и адаптогенным действием, оказывающим
благотворный эффект при гипотонии,
нервных заболеваниях и депрессии.
В качестве примера сравнительно
недавно открытого растительного соединения с
высокой биологической активностью можно
назвать полициклический дитерпеновый
спирт форбол, содержащийся в молочае.
Форбол (в виде эфиров) вызывает рост опу-
304
ФИЗИОЛОГИЯ И БИОХИМИЯ РАСТЕНИЙ
холей, воздействуя на активность
фермента протеинкиназы С, поэтому его широко
применяют в исследованиях по энзимологии.
Флавоноиды
Флавоноиды, молекула которых содержит
два ароматических кольца, представляют
собой самую многочисленную группу феноль-
ных соединений. Они накапливаются во многих
лекарственных растениях: в корнях солодки
Glyzyrrhiza glabra, траве пустырника Leonurus
cardiaca, цветках бессмертника Helichriyzum
aren avium и т. д. и отличаются широким
спектром фармакологического действия. Эти
вещества обладают желчегонным,
бактерицидным, спазмолитическим, кардиотоническим
действием. В медицине широко
используется свойство многих флавоноидов уменьшать
проницаемость и ломкость капилляров,
особенно в сочетании с аскорбиновой кислотой
(Р-витаминный эффект). У флавоноидов
выявлено также противораковое,
радиозащитное и противолучевое действие; они могут
связывать и выводить из организма
радионуклиды. Отсутствие токсических свойств и
избирательность действия на организм
человека увеличивают ценность флавоноидных
соединений и открывают большое будущее
для создания на их основе новых
лекарственных препаратов.
Культивирование клеток и органов
лекарственных растений
Исчезновение природных ресурсов диос-
кореи, женьшеня и ряда других ценных для
медицины растений сильно стимулировало
развитие метода культуры клеток и органов
растений, например, корней с целью
получения биологически активных вторичных
метаболитов. Этот метод незаменим и в тех
случаях, когда требуется получить вещества
из редких и исчезающих видов растений, или
же из растений, не произрастающих в
наших климатических условиях, например,
раувольфии или барвинка. Метод
культивирования клеток и тканей растений на
питательных средах позволяет также избежать
неблагоприятных воздействий окружающей
среды, что неизбежно в условиях
агрикультуры. Извлечение полезных соединений из
клеточной массы в этом случае нередко
значительно упрощается ввиду отсутствия
окрашенных примесей (пигментов). Вместе с тем
следует отметить, что биосинтез вторичных
метаболитов — это свойство
дифференцированных (специализированных) органов и
тканей, поэтому получение культур клеток
лекарственных растений, накапливающих
биологически активные вещества, — задача
сложная. Для ее решения применяют
методы химического мутагенеза и генетической
трансформации клеток с целью создания
новых штаммов — сверхпродуцентов. Хороших
результатов достигают и с помощью
изменения условий культивирования, меняя,
например, состав питательной среды. В нашей
стране были получены культуры клеток и корней
женьшеня, раувольфии, диоскореи и других
растений. Эти культуры являются основой
биотехнологического способа производства
природных биологически активных соединений,
которому принадлежит большое будущее.
Можно предположить, что в XXI в. с
помощью биотехнологии будут получены многие
незаменимые пока в медицине растительные
лекарственные препараты.
Заключение
К настоящему времени химически
обследовано от 5 до 15% всех обитающих на
Земле видов растений, а их
насчитывается не менее 250 тыс. Часто такие
обследования проводились с целью обнаружения
веществ только одного типа строения,
обладающих нужной биологической
активностью. Однако в современном мире прогресс
цивилизации сопровождается быстрым
уничтожением флоры. Например, исчезают с лица
Земли тропические леса Амазонии, и
вместе с ними человечество невозвратно
теряет многие соединения, свойственные
только произрастающим в них растениям,
которые могли бы принести большую пользу
медицине будущего. Этих невосполнимых
утрат можно избежать только путем
расширения фронта исследований в области
фитохимии, биохимии растений и
биотехнологии, что, несомненно, принесет пользу
медицине, сельскому хозяйству и
фундаментальным наукам. Поражающие
воображение биосинтетические возможности
растений еще далеко на раскрыты и ждут
своих энтузиастов — исследователей,
которые сумеют поставить их во всей полноте
на службу человеку.
Литература
Биология культивируемых клеток и
биотехнология растений. М.: Наука, 1991.
Гудвин Т., Мерсер Э. Введение в биохимию
растений. Пер. с англ. М.: Мир, 1986, т. 2.
Запрометов М. Н. Фенольные соединения.
Распространение, метаболизм и функция в
растениях. М.: Наука, 1993.
Ловкова М. Я., Рабинович А. М., Пономарева С. М.
и др. Почему растения лечат? М.: Наука, 1989.
Лукнер М. Вторичный метаболизм у
микроорганизмов, растений и животных. Пер. с англ.
М.: Мир, 1979.
Пасешниченко В. А. Биосинтез и
биологическая активность растительных терпеноидов и
стероидов (Итоги науки и техники. Сер.:
«Биологическая химия». Т. 25). М.: ВИНИТИ, 1987.
экология
Б. В. Громов
ЦИАНОБАКТЕРИИ В БИОСФЕРЕ
Любые организмы, живущие на Земле,
занимают определенное и уникальное
место в составе биоценозов; все они
незаменимы и заслуживают тщательного изучения.
Однако роль некоторых групп в эволюции
и существовании биосферы
представляется особенно значительной. Такой группой,
по данным современной науки,
несомненно, являются цианобактерии.
Раньше их называли синезелеными
водорослями. Это название широко
используется и в настоящее время, особенно в
ботанической литературе, где они
рассматриваются как таксон высокого ранга —
отдел или тип в системе низших растении.
Впрочем, еще в прошлом веке и в начале
нашего столетия некоторые ученые
обращали внимание на несомненное сходство
синезеленых водорослей и бактерий. Одним
из значительнейших достижений биологии
второй половины XX столетия является
установление того факта, что все живые
организмы, имеющие клеточное строение,
могут быть разделены на две четко
различающиеся группы (надцарства) в соответствии
с особенностями организации их клеток:
прокариот, имеющих примитивные
безъядерные клетки, и эукариот, клетки которых
Рис. 1. Морфология и строение некоторых цианобактерии
а— почкующаяся клетка Chamaesiphon sp. Фазово-контрастный микроскоп, увеличение хЮОО; б— клетки
Chaemosiphon sp.,y верхней клетки виден слизистый прикрепительный стебелек. Фазово-контрастный
микроскоп, увеличение х 1000; б —дробящаяся клетка Myxosarcina. Фазово-контрастный микроскоп, увеличение х 1000;
г— планктонная азотфиксирующая цианобактерия АпаЬаепа karakumica. В клетках видны скопления газовых
везикул (светлые точки), стрелкой обозначены гетероцисты, слева виден проросток, выходящий из остатка
оболочки акинеты. Фазово-контрастный микроскоп, увеличение хЮОО; д— фотография ультратонкого среза
клетки A. karakumica. Видны скопления газовых везикул и мембраны тилакоидов. Стрелкой показан плотный
слой клеточной стенки. Электронный микроскоп, увеличение х40 000
308
ЭКОЛОГИЯ
устроены более сложно. Прокариотическая
клетка лишена органелл, т. е. хлоропластов
и митохондрий, в ней нет окруженного
оболочкой ядра и вообще дифференцированных
внутриклеточных мембранных систем и
вакуолей, нет цитоскелета и митотического
аппарата, а ее хромосома устроена проще
хромосом эукариот. К прокариотам относят
бактерий и архей (см.: Громов Б. В.
Удивительный мир архей. Наст. том).
Синезеленые водоросли по характеру
своей клеточной организации вполне
соответствуют грамотрицательным бактериям и
представляют их самостоятельную
эволюционную ветвь; для них характерны
высокая морфологическая сложность и
способность к осуществлению фотосинтеза с
выделением молекулярного кислорода.
Таким образом, термин «цианобактерии» вполне
оправдан. Хотя цианобактерии, с точки
зрения формальной систематики, не могут
рассматриваться в качестве таксона
высокого ранга, в эволюции жизни на Земле они
сыграли особую роль; большое значение они
имеют и в функционировании современной
биосферы. Описано более 1500 видов сине-
зеленых водорослей, среди которых есть
формы одноклеточные, размножающиеся
делением, почкованием (рис. 1, а) или
дроблением клетки на ряд дочерних клеток
(рис. 1,в), а также формы колониальные и
нитчатые (рис. 1, г). Нити последних могут
быть простыми или ветвящимися. Размеры
клеток значительно варьируют: их диаметр
у одних видов может составлять доли
микрометра, а у других — десятки
микрометров. Колонии цианобактерии, или дерновин-
ки, образованные нитчатыми формами,
могут иметь макроскопические размеры.
Отдельные клетки или нити у некоторых
цианобактерии способны ползать по
плотному субстрату.
Клетка цианобактерии — типичная
прокариотическая клетка. Она окружена
клеточной стенкой грамотрицательного типа,
т. е. между двумя мембранами такой
стенки — внутренней (плазмалеммой) и
внешней — находится слой плотного полимера
(рис. 1,6). Клеточные органеллы и ядро
отсутствуют, нити ДНК расположены
ближе к центру клетки в участках,
выглядящих светлыми на срезах клеток (рис. 2).
Фотосинтетический аппарат представлен
сдвоенными мембранами (рис. 2), их
называют тилакоидами. Количество тилакоидов
и характер их расположения в клетке
различны у разных видов. На поверхности
тилакоидов расположены фикобилисомы —
структуры, улавливающие фотоны и
передающие их реакционным центрам
фотосинтетического аппарата. Клетка содержит
также полиэдральные тела, или карбокси-
сомы (рис. 2), образованные молекулами
ключевого фермента фотосинтеза рибуло-
зодифосфаткарбоксилазы. В клетках
цианобактерии, живущих в планктоне, обычно
присутствуют газовые везикулы. На
поперечном срезе они выглядят как
шестигранники (рис. 1,6), а на продольном имеют вид
ромба. Газовые везикулы образуют более
или менее крупные скопления (рис. 1,6); в
световом микроскопе они выглядят как
светлые сверкающие зерна (рис. 1, г). Они
содержат газ и придают клеткам лучшую
плавучесть. Следует отметить, что газовые
везикулы обнаруживаются только у
прокариот.
Некоторые цианобактерии способны к
клеточной дифференцировке — образованию
специализированных клеток. Это могут быть,
в частности, акинеты, представляющие
собой крупные покоящиеся клетки с
утолщенной оболочкой. Их образуют многие
планктонные нитчатые цианобактерии. В
Рис. 2. Фотография
ультратонкого среза клетки Nostoc sp.
Видны тилакоиды — мембраны
фотосинтетического аппарата с
находящимися на их поверхности
фикобилисомами. Электронный
микроскоп, увеличение х 20 000
ЦИАНОБАКТЕРИИ В БИОСФЕРЕ
309
условиях умеренного климата некоторые
планктонные цианобактерии при
похолодании образуют акинеты, которые опускаются
на дно водоемов, где перезимовывают.
Весной они прорастают, и молодые
снабженные газовыми везикулами клетки всплывают
к поверхности водоема, к свету. Впрочем,
акинеты используются не только для
перезимовки, но и вообще для выживания
организма в различных неблагоприятных
условиях.
Цианобактерии — фототрофные
прокариоты, использующие для своей
жизнедеятельности энергию света. Они
осуществляют так называемый оксигенный фотосинтез,
т. е. синтезируют органическое вещество из
углекислого газа и воды, освобождая при этом
молекулярный кислород. Это единственные
прокариоты, способные к оксигенному
фотосинтезу. Цианобактерии содержат
хлорофилл а растительного типа и
водорастворимые фикобилиновые пигменты: голубые —
фикоцианины и красные — фикоэритрины.
Эти пигменты находятся в фикобилисомах.
Большинство цианобактерии — облигат-
ные фототрофы, неспособные к жизни в
темноте за счет органического субстрата,
хотя есть и исключения. Среди
цианобактерии много форм, способных к азотфиксации.
Под азотфиксацией понимают возможность
использования организмом молекулярного
азота, который содержится в огромном
количестве в атмосфере. Почему-то никто из
эукариот не обладает способностью к
азотфиксации, это привилегия некоторых
прокариот. Азотфиксация связана с
активностью ферментного комплекса нитрогеназы. В
ходе этого процесса происходит
восстановление азота до аммиака, а затем
образуются азотсодержащие органические
молекулы. Данный процесс носит сугубо
восстановительный характер, поскольку в
присутствии молекулярного кислорода нит-
рогеназа инактивируется. Между тем
цианобактерии живут в присутствии
молекулярного кислорода, более того, они его образуют.
Поэтому формы цианобактерии, способные
к азотфиксации, должны как-то защитить
свой фермент от кислорода. Некоторые виды
решают эту проблему, осуществляя
фиксацию азота при отсутствии фотосинтеза, в
ночное время, или посредством
чередования фотосинтеза и азотфиксации,
подчиняющегося определенному эндогенному ритму.
Так обстоит дело, например, у морской
планктонной цианобактерии Trichodesmium, массы
которой населяют теплые моря и океаны и
фиксация азота которой имеет огромное
значение для жизни океана. У других
цианобактерии азотфиксация происходит в
специализированных клетках — гетероцистах.
Гетероцисты обладают дополнительными
слоями оболочки, защищающими их от про-
Рис. 3. Угнетение роста зеленых водорослей на
поверхности среды в чашке Петри гербицидом,
образованным цианобактерией (гербицидом
пропитана бумажка, положенная на поверхность среды)
никновения кислорода извне. Фотосинтез, в
ходе которого выделяется кислород, в них
не идет, но активно осуществляется
процесс азотфиксации. Образующиеся соединения
связанного азота передаются соседним
вегетативным клеткам через микроплазмодес-
мы, а от них поступает органический
субстрат, необходимый для фиксации азота.
Гетероцисты обычно неспособны к росту и
делению и со временем отмирают, их
единственная функция — фиксация азота. Таким
образом, цианобактерии дают единственный
пример прокариотического
многоклеточного организма, так как у них происходит
разделение функций между разными
клетками. Это еще одна уникальная особенность
цианобактерии.
Различные виды цианобактерии обладают
разнообразными адаптационными
механизмами, определяющими успешное их
развитие в тех или иных условиях окружающей
среды. Одни формы, Scytonema например,
образуют пигмент, концентрирующийся на
поверхности клетки и эффективно
защищающий ее от ультрафиолетовых лучей, что
определяет возможность развития этой
цианобактерии при прямом солнечном
освещении. Одни цианобактерии синтезируют си-
дерофоры — вещества, связывающие ионы
железа и делающие их доступными для
бактерии. Другие, развивающиеся в
прибрежной зоне водоемов, синтезируют сурфак-
танты — поверхностно-активные соединения.
При волнении вода у берега становится
мутной из-за взвешенных частиц песка и
детрита. В присутствии сурфактанта
частички слипаются и оседают на дно, вода
310
экология
становится прозрачной и цианобактерии
получают достаточно света. Многие
цианобактерии синтезируют биологически
активные вещества. Это могут быть антибиотики
с гербицидной активностью, препятствующие
росту других цианобактерии, водорослей и
высших растений (рис. 3). Значение
способности к синтезу таких веществ при
конкуренции в природе очевидно. Было бы
крайне желательно организовать производство
таких гербицидов промышленностью,
поскольку они экологически безопасны и не
токсичны для человека и животных. Реже
цианобактерии образуют антибиотики,
активные против грибков и гетеротрофных
бактерий (рис. 4). Приспособительное
значение таких антибиотиков менее очевидно, но
следует отметить, что существуют грибки
и бактерии, паразитирующие на цианобак-
териях.
К сожалению, довольно многочисленные
цианобактерии образуют токсины,
губительные для животных и человека. Это
представители родов Microcystis, Anabaena,
Nodularia, Nostoc, Aphanizomenon, Oscillatoria
и др. Способность к образованию яда — не
видовой признак, а свойство штамма или
клона, т. е. здесь наблюдаются
внутривидовые различия. Яды образуют главным
образом планктонные формы. Токсины
бывают двух типов — нейротоксины и гепа-
тотоксины. Первые представляют собой
алкалоиды, действующие на нервную систему.
Цианобактерии, образующие нейротоксины,
встречаются относительно редко. Гепатоток-
сины — это циклические гепто- или пен-
тапептиды (т. е. короткие цепи белковой
природы, составленные из пяти или
шести аминокислот), содержащие необычные
аминокислоты. К настоящему времени
описано множество разновидностей таких пеп-
Рис. 4. Угнетение грибка и зеленых водорослей на
поверхности среды вокруг бумажки, пропитанной
антибиотиком, образованным цианобактерией
тидов, различающихся по структуре и
степени токсичности. Гепатотоксин, попавший
в организм животного, вызывает
разрушение печени, и через несколько часов
наступает смерть. Низкие дозы этого яда
вызывают развитие рака. Случаи гибели
людей от гепатотоксинов цианобактерии в
литературе не описаны, но
представляется несомненным, что некоторые люди,
умершие от рака печени, были убиты циа-
нобактериями. Токсины находятся в
клетках цианобактерии и только после их
разрушения выходят в воду. Они весьма
устойчивы и не разрушаются при
хлорировании воды; сохраняются они и в сухих
клетках. Если вы вынуждены иметь дело с
водой, содержащей взвесь цианобактерии,
лучше с такой водой не контактировать;
особенно следует избегать ее попадания в
глаза. Для питья необходимо использовать
только отфильтрованную и тщательно
прокипяченную воду. При кипячении яды
разрушаются.
Среди цианобактерии есть формы,
способные к участию в симбиотических
ассоциациях. Около 8% лишайников содержат
в качестве фикобионта цианобактерии,
которые могут иметь различное
систематическое положение. Цианобактерии
поставляют грибу, входящему в состав лишайника,
продукты фотосинтеза и, если это азотфик-
сирующие формы, — соединения связанного
азота, так что польза ассоциации для
гриба понятна. Какую пользу извлекают из
симбиоза с ним цианобактерии, непонятно.
Азотфиксирующие представители родов
Nostoc и Anabaena могут быть симбионтами
некоторых мхов, имеющих на листьях
наполненные слизью полости, куда
проникают и где поселяются цианобактерии.
Известен единственный случай симбиоза
цианобактерии с папоротником — это
ассоциация тропического папоротника Azolla с
цианобактерией Anabaena, которая
поселяется в специальных полостях на листьях
папоротника и за счет азотфиксации может
обеспечить азотом как свое развитие, так
и рост папоротника. Цианобактерии из
родов Nostoc или Anabaena образуют клубеньки
на корнях некоторых цикадовых, причем
только у поверхности земли, что дает
возможность этим цианобактериям
осуществлять фотосинтез. Nostoc образует
клубеньки на листьях или стеблях некоторых видов
Gunnera — покрытосеменного растения,
обитающего в Южном полушарии. В
данном случае развитие цианобактерии
происходит внутри клеток растения-хозяина. В
морской среде известны случаи симбиоза
цианобактерии с губками или асцидиями.
Цианобактерии живут внутри клеток
некоторых водорослей и жгутиконосцев. В данном
случае симбиоз может быть облигатным, т. е.
ЦИАНОБАКТЕРИИ В БИОСФЕРЕ
311
обязательным, поскольку участвующие в
нем цианобактерии уже не могут жить
самостоятельно. Таких цианобактерии
называют цианеллами. Они представляют собой
нечто среднее между цианобактерией и
клеточной органеллой. В эволюции
биосферы способность цианобактерии к симбиозу
имела чрезвычайно большое значение.
Как принято считать, прокариоты, в том
числе и цианобактерии, появились на Земле
в самом начале существования биосферы.
Никто не знает, от кого произошли
бактерии, поэтому появляются домыслы, что они
занесены на Землю разумными
существами для эксперимента, или что они
разносятся кометами и т. п. Но большинство
ученых считают, что бактерии произошли от
более примитивных организмов, природа
которых пока неизвестна. Большое
значение для формирования наших
представлений об эволюции жизни на Земле имели
успехи палеомикробиологии, развитие
которой началось в 50-е гг. нашего столетия.
Давно было известно, что в осадочных
породах можно обнаружить окаменевшие
остатки древних растений и животных,
однако клетки прокариот даже если они там
и присутствуют, достоверно
идентифицированы быть не могут. Для известняков,
песчаников, каменного угля и т. п. это
действительно так. Но оказалось, что
мумифицированные клетки прокариот можно
обнаружить в теле осадочной кремнистой
породы. Для этого нужно изготовить очень
тонкую и потому прозрачную пластинку
кремня, что не представляет особых
затруднений. При микроскопировании таких
пластинок можно обнаружить колонии и
нити, совершенно идентичные колониям и
нитям современных цианобактерии.
Остатки цианобактерии обнаружены в кремнях,
взятых на разных континентах и имеющих
разный возраст. Наиболее древние из
таких кремней, найденные в Австралии,
имели возраст около 3500 млн лет,
больший, чем у прочих известных осадочных
пород. В этих кремнях присутствуют 7
различных довольно сложных по своей
морфологии типов (видов) прокариот, сходных
с цианобактериями. Ископаемые
цианобактерии обнаружены и в кремнях, найденных
на Урале, а также в других районах
нашей страны. Присутствие цианобактерии в
древнейших отложениях, удивительное само
по себе, означает, что в то время в
земной атмосфере уже присутствовал кислород.
Существование цианобактерии в докембрии
подтверждается и другого типа ископаемыми.
Это строматолиты. В наше время
строматолиты образуются в результате
жизнедеятельности сообществ нитчатых
цианобактерии, развивающихся в толстых пленках —
матах — в условиях солоноводиых
мелководий, например на побережье Австралии.
На пленке постепенно нарастает слой
кальцита, нити бактерий отмирают или
переползают наверх. Постепенно образуется
слоистая структура — строматолит.
Строматолиты имеют характерное строение и
форму, кальцит, из которого они состоят,
иногда замещается кремнем. Подобные
строматолиты обнаруживают и в докембрийс-
ких отложениях. То, что цианобактерии
образовывали кислород уже в самые
начальные периоды развития жизни на Земле,
представляется спорным. Не подлежит,
однако, сомнению, что именно деятельность
цианобактерии обеспечила постепенное
формирование богатой кислородом
атмосферы, в условиях которой стало
возможным возникновение растений и животных.
Считается, что прокариоты появились на
Земле 3-4 млрд лет назад, тогда как эука-
риоты, по разным оценкам, только 2,5-
1,5 млрд лет назад. Приблизительно к
этому времени (1,2—1,4 млрд лет тому назад)
за счет деятельности цианобактерии
содержание кислорода в атмосфере достигло
примерно 0,2% (в современной атмосфере —
21%). В дальнейшем кислород
накапливался за счет активности не только
цианобактерии, но и появившихся к этому времени
растений; около 440 млн лет назад в
нижнем силуре в атмосфере было уже около
2% кислорода, что определило вспышку
эволюции аэробных эукариот. Роль
цианобактерии, однако, не исчерпывается их
влиянием на состав атмосферы; они
явились еще предшественниками хлоропластов
растений. Данные молекулярной биологии
с несомненностью свидетельствуют о том,
что при возникновении эукариотической
клетки большое значение имело
превращение симбиотических прокариот в клеточные
органеллы. Получено много доказательств
того, что хлоропласты растительных
клеток образовались из симбионтов — древних
цианобактерии. Таким образом, в процессе
эволюции биосферы цианобактерии не
только создали условия, необходимые для
возникновения и жизни эукариот, —
кислородную атмосферу, но также явились предками
хлоропластов растений, а без растений не
возникли бы и животные и человек.
Поэтому утверждение об исключительной роли
цианобактерии в эволюции жизни
представляется вполне оправданным.
Вероятно, каждый человек когда-либо
обращал внимание на слизистые
темно-зеленые, иногда почти черные налеты или
корочки на поверхности влажной земли,
камней и т. п., так же как и на появление
синезеленой или бурой взвеси в озерной воде.
Но мало кто знает, что такие налеты и
взвеси представляют собой скопления
цианобактерии. Цианобактерии в современной
312
ЭКОЛОГИЯ
биосфере распространены повсеместно. Они
отсутствуют только в очень кислых средах —
на моховых болотах и в кислых сернистых
источниках, хотя некоторые формы могут
жить даже при рН 4. Конечно, их
присутствие, как правило, нельзя определить
невооруженным глазом. Цианобактерии
являются пионерами при освоении минерального
-субстрата, например лавы после
извержения вулкана, или поверхности скал.
Осуществляя фотосинтез, они обогащают
субстрат органическим веществом, а за счет
азотфиксации накапливают соединения
связанного азота, необходимого для роста
растений. Некоторые редкие формы могут жить
в пещерах при ничтожном освещении, иногда
вокруг их нитей откладывается кальцит и
тогда образуются своеобразные живые
сталактиты. Цианобактерии участвуют в
почвообразовательном процессе и составляют
обязательный компонент сообщества
почвенных микроорганизмов. Особенно много
цианобактерии бывает в почвах рисовников.
Азотфиксирующие виды обогащают почву
азотом и делают ее более плодородной.
Любые пресные или солоноводные
водоемы населены цианобактериями. Они
находятся на поверхности донных отложений,
на поверхности растений и в планктоне.
Цианобактерии служат пищей для водных
животных и таким образом вносят свой вклад
в продуктивность водоемов.
Азотфиксирующие виды обогащают воду связанным
азотом. При создании определенных условий,
однако, происходит массовое развитие
цианобактерии — цветение водоема.
Накапливается огромная биомасса цианобактерии,
которые быстро отмирают и начинают гнить.
Из воды исчезает кислород, появляются
продукты гниения, и среда становится
непригодной для жизни водных организмов,
в том числе рыб.
По данным мировой статистики,
примерно в 40-50% случаев цветения происходит
развитие токсигенных цианобактерии и в
воде присутствуют токсины (см. выше). Такая
вода опасна для животных. Еще в 1878 г. в
журнале «Nature» была опубликована
статья, в которой сообщалось о гибели
лошадей, свиней, собак в результате
потребления воды из водоема, в котором наблюдалось
массовое развитие синезеленых водорослей.
Подобные случаи наблюдались время от
времени и в последующие годы, однако
сейчас развитие токсигенных
цианобактерии приобретает глобальный характер, что
связано с усилением антропогенного
загрязнения водоемов. Токсичные цветения озер
объявлены национальной проблемой в
Англии, Финляндии, Норвегии; здесь
созданы специальные центры для их изучения
и контроля. Мы изучали развитие
токсигенных цианобактерии в ряде озер Карелии и
Невской Губе. Один миллиграмм биомассы
цианобактерии из красивейшего
карельского озера оказался смертельным для
белой мыши при парэнтеральном введении. По
нашим наблюдениям, очень много
цианобактерии в воде Ладожского озера и их
количество постоянно увеличивается. Пока
нет данных о развитии здесь токсигенных
форм, однако такая возможность реальна.
Цветение Ладожского озера,
обусловленное размножением токсигенных
цианобактерии, сильно затруднило бы снабжение
Санкт-Петербурга питьевой водой. Опасность
массового развития токсигенных
цианобактерии здесь тоже вполне реальна.
Среди пресноводных цианобактерии есть
формы термофильные, т. е. растущие при
высоких температурах. Верхний
температурный предел для них +74°С. Термофильные
цианобактерии обитают в горячих
источниках, которые имеются на всех
континентах, в особенности в районах
вулканической активности. У нас много горячих
источников на Дальнем Востоке, и в них
происходит обильное развитие
цианобактерии, образующих мощные темно-зеленые
налеты. Среди термофильных
цианобактерии есть одноклеточные и нитчатые.
Цианобактерии населяют также моря и
океаны. Они развиваются в планктоне, особенно
в наннопланктоне, т. е. среди самых мелких
форм. Мелкие одноклеточные, окрашенные
в красноватый цвет и способные к
эффективному использованию зеленого света
цианобактерии встречаются на значительных
глубинах. Их активность очень важна для
обеспечения продуктивности океана. В
тропических районах нередко наблюдается
массовое развитие азотфиксирующей
нитчатой цианобактерии Trichodesmium. В
условиях северных морей происходит
массовое развитие другой нитчатой
цианобактерии— Nodularia spumigena. Последняя
также фиксирует азот, но, кроме того, она
образует гепатотоксин, поэтому ее
массовое развитие в Балтийском море
вызывает большое беспокойство в странах Балтии.
В условиях морской среды цианобактерии
развиваются также на дне, на
поверхности различных предметов и организмов.
Существуют эндолитические виды,
пробуравливающие отверстия в известняках или в
раковинах моллюсков и живущие в
образующихся норках.
Цианобактерии характеризуются
исключительно высокой приспособляемостью, в том
числе и к крайне неблагоприятным условиям
существования. В засушливых районах
цианобактерии, прежде всего виды Nostoc, при
наличии минимума влаги образуют налеты
на поверхности почвы, которые после
подсыхания сохраняются длительное время в
виде хрупких корочек и вновь оживают при
ЦИАНОБАКТЕРИИ В БИОСФЕРЕ
313
увлажнении. Но особенно интересны
цианобактерии, развивающиеся в так называемых
криптоэндолитических сообществах (от греч.
«криптос» —скрытый). Криптоэндолитичес-
кие сообщества обнаружены как в холодных
сухих пустынях Антарктиды, так и в
горячих сухих пустынях, например в пустыне
Негев в Израиле. В этом случае развитие
организмов происходит в порах пористой
породы или в полостях, образовавшихся в
плотной породе в результате процессов
выветривания. Поверхность камня покрыта
тонкой минеральной пленкой, образовавшейся
в результате физико-химических процессов.
Полости, в которых оказываются
микроорганизмы, имеют микроскопические
размеры, они полностью или почти полностью
изолированы от окружающей среды.
Микроорганизмы оказываются защищенными от
внешних неблагоприятных факторов, но
вместе с тем они изолированы и от
источников пищи. Энергетически такие
сообщества зависят от солнечного света,
круговорот элементов в них может быть почти
замкнут, а обмен с окружающей средой
минимален. Жизнь здесь едва теплится. В
состав криптоэндолитических сообществ
входят лишайники и цианобактерии.
Итак, цианобактерии — важнейший
элемент биосферы. Они были связаны с ней уже
на самых начальных этапах ее развития и
оказали решающее влияние на эволюцию
земной атмосферы и живых организмов; они
обнаруживаются и там, где жизнь
кончается. В современной биосфере
цианобактерии распространены повсеместно. Развиваясь
в составе наземных, пресноводных или
морских сообществ, они увеличивают их
продуктивность. Вместе с тем массовое
развитие цианобактерии может представлять
угрозу для жизни животных и человека,
поскольку некоторые формы способны к
образованию сильных ядов, а постоянно
возрастающее антропогенное загрязнение
природы способствует массовому развитию
таких форм.
Литература
Голлербах М. М., Косинская Е. К.,
Полянский В. И. Определитель пресноводных
водорослей СССР. Вып. 2: Синезеленые водоросли. М.:
Советская наука, 1953.
Громов Б. В. Ультраструктура синезеленых
водорослей. Л.: Наука, 1976.
Гусев М. В., Никитина К. А. Цианобактерии.
М.: Наука, 1979.
Ю. Т. Дьяков
ГРИБЫ И ИХ ЗНАЧЕНИЕ В ЖИЗНИ ПРИРОДЫ
И ЧЕЛОВЕКА
Место грибов в системе
организмов
К. Линней, создавая свою знаменитую
классификацию, поместил грибы вместе с
некоторыми другими организмами в особую
категорию, названную им «хаосом». В самом
деле, грибы ведут прикрепленный образ
жизни, как растения, их клетки покрыты
оболочками, как у растений, но в состав
оболочек входит хитин, который есть у
животных, но отсутствует у растений (табл. 1).
В табл. 1 указана еще одна особенность
грибов, отличающая их как от растений, так
и от животных, — осмотрофный способ
питания. В отличие от других эукариотных
организмов грибы питаются, всасывая
питательные вещества из окружающей среды.
Поэтому грибы можно определить как
гетеротрофные эукариотные организмы,
питающиеся осмотрофно. На этом основании
американский ботаник Р. X. Уиттейкер в 1969 г.
предложил систему живой природы из пяти
царств (рис. 1), основанную, с одной
стороны, на строении клетки (прокариоты, эука-
риоты), а, с другой, — на способах
получения энергии и способах питания,
разделивших эукариотные организмы на царства
Plantae (фототрофы), Animalia (зоотрофы)
и Fungi (осмотрофы). Три важнейших
способа питания, возникшие еще у
прокариот, определили морфологическую и
физиологическую эволюцию биоты вплоть до
высших эукариотных организмов.
Однако трофическая общность вовсе не
свидетельствует о филогенетической
общности среди осмотрофных организмов. Так,
большая и очень важная группа грибов —
оомицеты — представляет собой
потерявшие хлорофилл гетероконтные водоросли
и, по определению английского альголога
Ф. Фрича, должна быть отнесена к
водорослям. Миксомицеты, конечно, животные,
близкие к амебам, к тому же, кроме ос-
мотрофного питания, они могут питаться
зоотрофно, поглощая целиком клетки
микроорганизмов, однако они тоже
традиционно относятся к грибам. Рассмотренные
примеры подчеркивают сложность
определения царства грибов.
Осмотрофный способ питания сказался
на морфологии и физиологии грибов.
Таблица 1
Признаки, характерные для растений, животных и грибов
Признак
Геном
Размер (х10 )кДа
Процент повторов
Цитология
Рост клеток
Цитокинез
Клеточная стенка
Центральная вакуоль
Метаболизм
Конечный продукт азотного обмена
Запасные углеводы
Структурные углеводы
Синтез меланина
Физиология
Тип питания
Способ извлечения энергии
Растения
1000-2000
Около 30
Растяжением, изодиаметри1
Сопряжен с митозом
Есть
В фазе метаболической активности
Аспарагин, глютамин
Крахмал
Целлюлоза, гемицеллюлоза, пектин
В мертвых клетках
Фототрофный
Автотрофный
Животные
100-1000
Около 40
ческий
Нет
Мочевина
Гликоген
Хитин
Грибы
10-30
Около 10
Апикальный
Не сопряжен
Есть
При старении
Мочевина
Гликоген, трегалоза
Хитин, глюкан
В живых клетках
Зоотрофный
Осмотрофный
Гетеротрофный
ГРИБЫ И ИХ ЗНА ЧЕНИЕ В ЖИЗНИ ПРИРОДЫ И ЧЕЛОВЕКА
315
Таблица 2
Биологическое разнообразие в различных группах организмов
[из: Bull et ah Ann. Rev. Microbiol., 1992, vol.46, p. 219]
Группа организмов
Вирусы
Бактерии
Грибы
Водоросли
Мохообразные
Голосеменные
Покрытосеменные
Простейшие
Нематоды
Насекомые
Рыбы
Птицы
Млекопитающие
Число видов, тыс.
известное
5
4,7
69
40
17
0,75
250
30,8
15
800
19
9,2
4,17
расчетное
130
40
1500
60
25
270
100
500
8000
21
Количество известных видов, %
4
12
5
67
68
93
31
3
10
90
Ок. 100
Ок. 100
1. Тело большинства грибов (или таллом)
представлено мицелием, или грибницей,
состоящей из сильно разветвленных нитей
(гиф). Такое строение позволяет грибу
максимально оккупировать субстрат для
извлечения из него питательных веществ. У грибов
Растения
Грибы
Животные
Простейшие
эукариоты
(протисты)
Прокариоты
Рис. 1. Эволюционная схема биоты Р. Уиттейкера из
пяти царств
Растения, грибы и животные— три эволюционные
линии, возникшие из протистов на основе фототроф-
ного, осмотрофного и зоотрофного питания [из:
Science, 1969, vol. 163, p. 150]
нет специальных структур,
приспособленных для питания, они всасывают
питательные вещества всем телом.
2. Осмотрофный способ питания
заставляет все вегетативное тело гриба
максимально погружаться в субстрат, но при этом
ему становится трудно распространяться и
занимать новые субстраты. Поэтому споры,
которыми грибы размножаются,
выносятся над субстратом с помощью специальных
выростов мицелия. То, что обычно
называют грибами, представляет собой лишь
органы размножения, несущие внутри или
на поверхности споры (рис. 2).
3. Тело грибов не может иметь очень
большие размеры, ибо поступление
питательных веществ с помощью экзоосмоса в
клетки, находящиеся в глубине таллома,
затруднительно. Может быть, поэтому грибы
не достигли столь высокой и сложной
организации, как высшие растения и животные.
4. В качестве источников энергии
грибы используют сложные органические
соединения, которые вследствие большой
молекулярной массы не могут проходить
через клеточные покровы. Поэтому грибы
выделяют в окружающую среду ферменты,
которые разрушают высокомолекулярные
полимеры до отдельных звеньев —
мономеров, способных проходить в клетку. Такие
ферменты называют гидролазами или де-
полимеразами. Грибы — источники высоко-
316
экология
Рис. 2. Мицелии и органы спороношения грибов
1 — клубочки мицелия эндомикоризного гриба Ceratobasidium в клетках корня орхидеи; 2— эндофитный
(внутритканевой) мицелий паразита картофеля Phytophtora infestans и высовывающиеся через устица
спорангиеносцы с зооспорангиями на концах; 3— плесневый гриб Мисог. Мицелий развивается в субстрате (почве),
спорангиеносцы со спорангиоспорами поднимаются над субстратом; 4— культивируемый шампиньон Agaricus
bisporus: зрелое плодовое тело (после разрыва частного покрывала освобождается пластинчатый гименофор
с базидиоспорами)
активных деполимераз. Таким образом,
пищеварительный сок, который у
животных выделяется в просвет кишечника, у
грибов выделяется наружу, непосредственно
в субстрат.
5. Грибы должны создавать в клетках
высокое тургорное давление для того, чтобы
вода с растворенными в ней питательными
веществами поступала из субстрата в
мицелий. Гигантское давление, создаваемое
грибами, можно проиллюстрировать
частыми случаями разрыва асфальтового
покрытия растущими плодовыми телами
шампиньонов.
Вопрос о масштабах биологического
разнообразия в царстве грибов весьма далек от
своего решения. В последних справочниках
приведены сведения о 65 тыс. видов
описанных грибов. Однако известный шведский
миколог Е. М. Фриз более 150 лет назад
высказал мнение, что грибы являются самой
большой группой в orbis vegetabilia и
занимают в ней такое же положение, как
насекомые в orbis animalia. Сейчас расчеты
показывают его глубокую правоту (табл. 2). По
современным представлениям, грибы, как
упоминалось выше, выделяются в
отдельное царство.
Такое гигантское видовое разнообразие
указывает на то, что грибы —
процветающая и широко распространенная в
природе группа, которая не могла не наложить
глубокий отпечаток на всю эволюцию био-
ты, функционирование современных
биоценозов, включая агроценозы, и даже нд
развитие цивилизации.
Роль грибов в эволюции
биосферы
Осмотрофный способ питания ставит
грибы в определенный разряд организмов
в пищевой цепи превращения веществ и
энергии. Наряду с бактериями, грибы —
редуценты, разлагающие сложные
органические соединения до более простых,
причем по способам питания их разделяют на
сапротрофов, т. е. мертвоядов, которые
усваивают сложные органические
соединения из мертвых субстратов, и паразитов,
усваивающих питательные вещества из
живых организмов. И сапротрофы и
паразиты связаны в своем питании в основном
с растительными тканями. Связь грибов с
растениями, по-видимому, сложилась очень
давно, вероятно, на самых ранних этапах
их эволюции. Самые примитивные грибы —
хитридиомицеты и оомицеты
паразитируют на примитивных растениях —
водорослях. Некоторые микологи считают, что грибы
ГРИБЫ И ИХ ЗНА ЧЕНИЕ В ЖИЗНИ ПРИРОДЫ И ЧЕЛОВЕКА
317
вышли на сушу под покровом вышедших на
сушу растений, как их паразиты и
симбионты. Практически нет грибов, живущих в
симбиозе с животными, но огромное
число видов грибов находится в постоянных
симбиотических связях с растениями.
Ферментативный аппарат грибов —
гидролитические ферменты, которые выделяются в
окружающую среду, — настроены на
разложение углеводов — строительного
материала и запасных питательных веществ
растений. Поэтому не только
паразитические грибы избрали объектами нападения в
основном растения, но и сапротрофы
питаются «трупами» растений. Большая группа
грибов (копрофилы) питается навозом
животных (но опять-таки травоядных),
содержащим непереваренные растительные
остатки. Ну а в разложении мертвой древесины
участвуют почти одни грибы.
Вместе с тем водные оомицеты часто
поражают икру и рыбную молодь, нанося
большой вред рыбному хозяйству. Есть виды,
питающиеся почвенными беспозвоночными
(нематодами, амебами). Некоторые из них
преобразуют отдельные гифы в ловчие
кольца — липкие капканы для нематод; эти
кольца сжимаются при контакте с жертвой,
а затем высасывают ее содержимое с
помощью выделяемых ферментов (рис. 3).
Большая группа грибов паразитирует на
насекомых, часто вызывая у них эпизоотии.
Рис. 3. Мицелиальные «ловчие кольца» хищного
гриба Arthrobotrys, захватившие нематоду
Многие грибы (микофилы) паразитируют на
других грибах (плодовых телах шляпочных
грибов, мицелии микромицетов). Наконец,
известны грибы, специализированные на
питании белком кератином, из которого
построены покровы млекопитающих (кожа,
волосы, ногти). Они вызывают дерматоми-
козы, от которых страдают как дикие и
домашние животные, так и люди. Но
среди общей массы грибов все эти группы
немногочисленны.
Все больше данных накапливается о том,
что важнейшие арогенезы (прогрессивные
эволюционные преобразования) биоты
связаны с эндосимбиозами. В результате
объединения геномов двух и более прокариот-
ных организмов возникла эукариотная
клетка, а ее симбионты превратились в орга-
неллы, такие, как центриоли и цитоскелет,
митохондрии, хлоропласты и др. В
дальнейшем симбиозы между эукариотными
организмами из разных царств приводили к
объединению их свойств и созданию
принципиально новых организмов, способных
осваивать новые ниши, недоступные
каждому симбионту в отдельности. Самый яркий
пример — лишайник, получивший в
результате объединения гриба с водорослью
своеобразную морфологию, физиологию и
образ жизни. Есть гипотезы, что огромная и
интересная группа, главным образом
морских водорослей багрянок, представляет
собой морские лишайники, возникшие в
результате симбиоза древних аскомицетопо-
добных грибов и синезеленых водорослей.
Более того, накапливается все больше
данных, что выход растений на сушу и
возникновение наземных сосудистых растений
были бы невозможны без симбиоза с
грибами. Во всяком случае, самые ранние ри-
нофиты из силура и нижнего девона
имели в стелющихся стеблях грибной мицелий.
Полагают, что мицелий симбиотических
оомицетов дал начало сосудам.
Возникновение более поздней эктотроф-
ной микоризы привело к расцвету
древесных растений в карбоне. Заражение клеток
грибами, как симбиотрофными, так и
паразитическими, вызвало эволюцию защитных
механизмов растений и привело к
формированию лигнина — трехмерного
разветвленного полимера ароматических спиртов,
затрудняющего проникновение грибов в клетку
растений. В свою очередь, у грибов базидио-
мицетов возникли ферментные системы,
разлагающие лигнин: специфические лак-
казы, пероксидазы и др. Продукты
разложения лигнина — гуминовые соединения
стали основой почвенного гумуса. Эктотрофная
микориза дала возможность в меловом
периоде распространиться древесным породам
из тропиков в умеренные зоны с
неустойчивым климатом и бедными почвами.
318
ЭКОЛОГИЯ
Находящийся в зараженных клетках
растений мицелий выделяет биологически
активные вещества, обладающие
гормональной активностью и вызывающие приток
питательных веществ к зараженным тканям
и их разрастание. Такие болезни, как рак
картофеля, кила капусты, пузырчатая
головня кукурузы вызваны грибами.
Роль грибов в функционировании
современных экосистем
Грибы-симбиотрофы. Около 80% видов
современных растений имеют на корнях
микоризу. Микоризные грибы, во-первых,
увеличивают всасывающую поверхность
корней; во-вторых, производят многие
биологически активные вещества, используемые
растениями; в-третьих, переводят трудно-
усваиваемые соединения фосфора почвы в
растворимую форму, доступную растениям;
в-четвертых, защищают корни от
заражения потенциальными паразитами,
обитающими в почве; и наконец, в-пятых, нити
мицелия микоризных грибов, выходящие из
корней разных растений в почву, сливаются
и по нему мигрируют от одного растения к
другому метаболиты, включая питательные
вещества, источники энергии, гормоны,
токсины, а возможно, и генетическая
информация. Таким образом, микориза не
только расширяет экологические ниши растений
(например, вересковые, благодаря микоризе,
растут на бедных песчаных, тундровых,
горных почвах), но и интегрирует
популяции и даже разновидовые сообщества в
единый гигантский организм с
последствиями, которые пока трудно поддаются
анализу.
Недавно в злаках, затем в осоках и
других растениях нашли грибы, названные
эндофитами. Они являются
родственниками хорошо известной спорыньи, живут в
надземных частях растений и, образуя
токсичные алкалоиды, защищают хозяев от
травоядных животных, включая насекомых,
слизней и жвачных. Эндофиты усиливают
рост вегетативных органов зараженных
растений, но подавляют продукцию семян,
тем самым изменяя соотношение
вегетативного и семенного способов размножения.
Грибы-почвообразователи. Они имеют
огромное значение для существования среды
обитания 80% наземных организмов,
включая почти все высшие растения (табл. 3).
Протяженность грибных гиф в почве
превышает протяженность корней высших
растений, причем, как в зоне корней —
ризосфере, так и в зоне гиф грибов — гифос-
фере вследствие выделения метаболитов
создается специфическая среда,
благоприятная для одних групп микроорганизмов и
невыносимая для других.
Грибы (включая лишайниковые) вместе
с актиномицетами не только создали почву,
но и продолжают активно участвовать в
почвообразовательном процессе. Отмершие
растения сразу же заселяются грибами,
которые, передавая их, как по конвейеру,
от одних видов другим, осуществляют
превращение растительной биомассы в почву.
Грибы-паразиты растений. Им нет
числа, и они чрезвычайно разнообразны;
могут быть узкоспециализированными,
поражающими только близкородственные виды
растений или, наоборот, абсолютно
неразборчивыми; среди них есть виды,
приспособленные к заражению только всходов или,
наоборот, стареющих растений; способные
извлекать энергию лишь из живых,
активно функционирующих клеток, или же из
клеток, которые они предварительно
убили своими токсинами. Все эти паразиты в
природных фитоценозах выполняют важные
экологические функции. В 50-х гг. известный
зоолог В. Н. Беклемишев написал статью
«Паразиты как члены биоценозов», в
которой определил две важные функции
паразитов: увеличение видового разнообразия,
а следовательно, и устойчивости биоценоза
и защита сложившегося биоценоза от
вторжения новых сочленов. Можно привести
множество примеров того, как фитопатоген-
ные грибы выполняют эти функции. И если
Таблица 3
Воздушно-сухая биомасса (в кг/га)
организмов в лесной почве
[из: Satcell, 1970]
Актиномицеты
Прочие бактерии
Грибы
Простейшие
Нематоды
Дождевые черви
Энхитреиды
Моллюски
Клещи
Коллемболы
Двукрылые
Прочие членистоногие
Общая микрофлора
Общая микрофауна
Общая биомасса
0,2
7
454
1
2
12
4
5
1
2
3
6
461
36
496
ГРИБЫ И ИХ ЗНА ЧЕНИЕ В ЖИЗНИ ПРИРОДЫ И ЧЕЛОВЕКА
319
грибные болезни растений — страшный бич
сельского хозяйства во всех странах, то это
лишь оттого, что люди в погоне за
высокими прибылями разрушили эволюционно
сложившиеся связи между растениями в фи-
тоценозах, заменив сложные фитоценозы,
состоящие из многих видов, простыми и
следовательно, неустойчивыми, в которых
доминирует один вид.
Грибы-ксилотрофы. Древние связи
грибов с древесными растениями привели к
возникновению у грибов уникального
комплекса ферментов, разрушающих древесные
полимеры: целлюлозу и лигнин. Не будь
грибов в лесу, деревья до вершин были бы
покрыты мертвыми ветками, т. е. грибы —
важнейшие санитары леса. Но поскольку из
мертвой древесины делают дома, фонарные
столбы, железнодорожные шпалы и т. д.,
дереворазрушающие грибы, помимо пользы,
приносят большой экономический ущерб.
Роль грибов в развитии
цивилизации
Быстрое накопление биомассы и синтез
различных биологически активных веществ
сделали грибы важными продуктами
питания и объектами микробиологической
промышленности.
Одноклеточные грибы были первыми
микроорганизмами, использованными
человеком для улучшения питательных
свойств растительной и животной пищи.
Дрожжи с незапамятных времен дали
человечеству два важнейших продукта, без
которых развитие цивилизации было бы
немыслимо: хлеб и вино. С грибами
связаны две революции в медицине нового
времени. Первая — открытие пенициллина.
Этот первый, нашедший клиническое
применение, антибиотик спас от смерти
больше людей, чем все остальные лекарства
вместе взятые. С его открытием стало
возможным лечение таких болезней,
считавшихся ранее летальными, как перитонит
и сепсис. И хотя затем было выделено
огромное число антибиотиков из прокариот,
главным образом актиномицетов, грибные
антибиотики из группы бета-лактамов — пе-
нициллины и цефалоспорины — остаются
вне конкуренции. Общая выручка от
продажи продуктов биотехнологии в 1990 г.
составила 16 млрд долларов. Из них свыше
8 млрд было получено от продажи пени-
циллинов и цефалоспоринов. Таким
образом, продукты двух грибов составили
половину всего мирового биотехнологического
рынка. Кроме того, в мире ежегодно
производятся миллионы тонн дрожжей —
продуцентов органических кислот, ферментов,
фитогормонов и других продуктов.
Вторая фармакологическая революция
произошла недавно. Всем известны опыты
по трансплантации органов человека,
методы пересадки которых были
разработаны когда-то на животных. Но несмотря на
то, что технически проблема трансплантации
давно решена, практически она не
находила широкого применения вследствие
иммунной несовместимости пересаженных
органов и их отторжения. И только после
открытия грибных антибиотиков из группы
циклоспоринов, которые оказались
высокоактивными иммунодепрессантами, эти
операции стали обычным клиническим приемом.
Наконец, грибоводство — важная отрасль
сельского хозяйства. Это сложное
производство, требующее стерильности и четкого
воспроизведения технологических режимов.
Грибы — традиционная пища человека,
важный источник пополнения белками его
рациона. Однако в последние годы собирать
грибы стало небезопасно даже людям,
умеющим отличать съедобные виды от
ядовитых. Дело в том, что грибы как осмотрофы
активно поглощают из почвы и воздуха
вредные вещества. Поэтому во многих
развитых странах дикорастущие грибы вообще
не собирают. Продолжающееся ухудшение
экологической обстановки свидетельствует
о том, что в недалеком будущем это про-
Производство грибов в мире в 1980 г.
Таблица 4
Вид
Agaricus bisporus
Lentinus edodes
Flammulina velutipes
Pleurotes spp.
Tremmela, Auricularia
Stropharia rugoso-annulata
Гриб
Шампиньон
Спитаке
Зимний опенок
Вешенка
Серебряное ухо
Кольцевик
Производство, тыс. т
870
170
60
32
10
2
%
71,9
14,0
5,0
2,7
0,8
0,2
320
ЭКОЛОГИЯ
изойдет и в России. И единственным
грибным продуктом станут грибы,
культивируемые в искусственных условиях (табл. 4).
Как видно, чемпионом среди них
остается шампиньон, растущий в богатой
перегноем почве; его выращивают на специально
приготовленных компостах (рис. 4).
Производство шампиньона очень дорогое, он никогда
не будет дешевым продуктом. Другие
широко используемые грибы — это ксилотрофы,
растущие в природе на мертвой древесине.
Их производство дешевле, чем шампиньона;
кроме того, при их выращивании решается
еще одна важная практическая задача —
утилизация отходов сельского хозяйства, пищевой
и деревообрабатывающей промышленности.
Для выращивания этих грибов используют
солому, опилки, лузгу семечек, хлопковые
очесы и другие отходы. Оставшийся после
сбора плодовых тел субстрат, во-первых,
обогащен грибным мицелием, и, во-вторых, в
нем частично или полностью произошло
разложение стойких полимеров — целлюлозы и
лигнина, поэтому его можно использовать в
качестве удобрений, добавок в корм скоту и
для других целей.
Но грибы играли не только
положительную роль в истории человечества. Уже
говорилось о болезнях полезных растений,
разрушении древесных изделий и других вредных
воздействиях. Когда болезни растений
становились массовыми, это сказывалось на судьбе
целых народов. Далеко не единственный
пример — фитофтороз картофеля в Западной
Европе в середине прошлого века. В то время
в Ирландии из 4 млн населения около 1 млн
человек питались исключительно
картофелем, а для 2 млн картофель составлял 70%
пищевого рациона. Два неурожайных года
свели в могилу более миллиона ирландцев,
а полтора миллиона жителей страны
эмигрировали в Америку. Хорошо известно, что
остров Цейлон (Шри Ланка) — один из
центров мирового производства чая, а Южная
Америка — кофе. Но так было не всегда. В
прошлом веке холмы острова были
засажены кофейными деревьями, однако эпидемия
ржавчины привела к их гибели. Опустевшие
плантации были засажены чаем, а кофе
укрылось от ржавчины за Атлантическим
океаном. Паразитические грибы приводили и к
изменениям ландшафтов, как природных, так
и созданных человеком. Восточные штаты
США почти полностью лишились каштановых
лесов, уничтоженных грибом Criphonectria
parasitica, вызывающим раковые поражения
коры. В некоторых западноевропейских
странах погибли от болезни все ильмы.
Нельзя не отметить еще одно чрезвычайно
важное воздействие грибов на человека.
Заселяя органы растений, особенно во
влажную погоду, грибы выделяют в них
продукты жизнедеятельности, которые нередко
Рис. 4. Плантация шампиньона
оказываются токсичными для людей и
сельскохозяйственных животных. В средние века
массовая гибель людей происходила в основном
от двух болезней — чумы и эрготизма —
смертность от которых превышала смертность от
всех остальных вместе взятых. Эрготизм —
это отравление алкалоидами спорыньи,
попавшими в муку из зерен ржи, зараженных
Рис. 5. Токсиногенные грибы
а— склероции спорыньи в колосе ржи; б— колос
пшеницы, пораженный фузариозом (виден розовый
налет пигментированного мицелия)
ГРИБЫ И ИХ ЗНА ЧЕНИЕ В ЖИЗНИ ПРИРОДЫ И ЧЕЛОВЕКА
321
склероциями (рис. 5, а). Алкалоиды спорыньи
вызывают сокращения мышц. Высокие их дозы
приводят к мучительной смерти, низкие —
к сильным болям, умственным расстройствам,
агрессивному поведению. Интересно, что
начало многих войн в средневековой Европе
совпадало с массовым отравлением
спорыньей. Сейчас в связи с нарушением
агротехники, повышением влажности климата в
основных зернопроизводящих зонах России
огромную угрозу приобрел фузариоз колоса
зерновых, возбудитель которого также очень
опасный токсиногенный гриб (рис. 5, б).
Грибы — модели биологических
исследований
Грибы как объекты исследований
обеспечили поступательное движение науки во
многих направлениях, но прежде всего в
генетике.
Это вызвано тем, что грибы, так же как
и бактерии, можно легко культивировать на
питательных средах со стандартным составом;
опыты с ними воспроизводимы и к ним
применимы почти все генетические приемы
работы с бактериями, такие как выделение
единичных мутантов или рекомбинантов из
неограниченной популяции на селективных
средах. Геном грибов почти такой же
маленький, как и бактериальный. Но в отличие от
бактерий, грибы — эукариоты, у них есть
хромосомы, митоз. Данные, полученные на
грибах, можно с большой долей уверенности
экстраполировать и на других эукариотов,
включая человека. Поэтому например,
молекулярной генетикой дрожжей занимается
большая лаборатория в Кардиологическом
центре в Москве. Выдающийся американский и
немецкий ученый, нобелевский лауреат
М. Дельбрюк ввел в генетику в качестве
объекта исследований бактериофаг, а вслед за ним
и мукоровый гриб Phycomyces, у которого спо-
рангиеносцы чрезвычайно чувствительно
реагируют на свет, изгибаясь в его
направлении. Этот гриб служит простой моделью для
изучения сенсорных ответов на внешние
раздражители. Другой известный немецкий
генетик, К. Эссер, был избран почетным членом
Международного геронтологического общества
за работы по молекулярной генетике гриба
Podospora, живущего в навозе. Мицелий
этого гриба после нескольких дней роста
начинает стареть — гифы утончаются, вакуоли-
зируются, на их концах образуются вздутия,
рост прекращается. Эссер установил
причины старения и обнаружил, что в одном из
митоходриальных генов участок, вырезаемый
перед трансляцией (интрон), покидает
митохондрию, замыкается в кольцо,
реплицируется и вызывает описанные перерождения.
С грибами как объектами связано много
выдающихся открытий. Американские
ученые Дж. У. Бидл и Э. Л. Тейтум впервые
получили и исследовали биохимические мутанты
сумчатого гриба Neurospora и на основании
своих работ выдвинули важнейшее
положение молекулярной генетики: один ген — один
фермент. Их работа, заложившая основы
нового направления— биохимической
генетики, была удостоена Нобелевской премии. На
сумчатых грибах американский генетик
К. Линдгрен разработал блестящий метод
генетического анализа — тетрадный анализ,
который дает не статистически близкое к
расчетному, а абсолютное менделевское
расщепление. Использование тетрадного анализа
показало, что при внутригенных рекомбинациях
часто наблюдаются отклонения от менделев-
ского расщепления, названные конверсией
генов. Анализ этого явления привел
английского генетика Р. Холлидея к открытию
молекулярных механизмов рекомбинации,
которые оказались едиными для всех
организмов — от бактериофагов до
человека. Клеточный миксомицет Dictyostelium тоже
гриб и более интересной, чем он, модели
клеточной интеграции пока не найдено. В
богатой среде (в лаборатории на
бактериальном газоне) этот организм существует в форме
амебообразных клеток, размножающихся
делением. Но при истощении среды клетки
начинают синхронно собираться в центры
инициации и, наползая друг на друга,
формировать вертикальную колонку (споранги-
еносец) с расширением на конце
(спорангием). «Амебы» в спорангии покрываются
оболочками, превращаются в споры, которые
прорастают на богатом субстрате и снова
преобразуются в «амеб».
Из сказанного ясно, что изучение
грибов имеет большое теоретическое и
практическое значение, которое обычно
недооценивают. Автор хотел бы привлечь внимание
преподавателей и учащихся к этой важной
и интересной группе организмов.
Литература
Гарибова Л. В. В царстве грибов. М.: Лес. пром-
сть, 1982.
Горленко М. В., Гарибова Л. В., Сидорова И. И.
и др. Все о грибах. М.: Лес. пром-сть, 1985.
Дараков О. Грибной огород — и здоровье и
доход. М., 1995.
Дудка И. А., Вассер С. П. Грибы: Справочник
миколога и грибника. Киев: Наук, думка, 1987.
Дьяков Ю. Т. О болезнях растений. М.: Аг-
ропромиздат, 1985.
Жуков А. М., Миловидова Л. С. Грибы — друзья
и враги леса. Новосибирск: Наука, 1980.
Каратыгин И. В. Ботан. журн., 1994, т. 79,
№ 2, с. 13.
Мазин В. В., Шашкова Л. С. Грибы, растения,
люди. М.: Агропромиздат, 1986.
Мюллер Э., Леффлер В. Микология. М.: Мир, 1995.
Хуксворт Д. Л. Микология и фитопатология,
1992, т. 26, вып. 2, с. 152.
В. В. Климов
РАСТЕНИЯ И БИОСФЕРА
Введение
Основным и практически неиссякаемым
источником энергии на поверхности Земли
является энергия солнечного излучения.
Полный поток солнечного излучения
(измеренный за пределами земной атмосферы)
близок к 1400 Вт/м2. Значительная часть этой
энергии приходится на область видимого и
ближнего инфракрасного излучения (0,3-
1,0 мкм) — фотосинтетически активную
радиацию, эффективно поглощаемую
пигментами, участвующими в фотосинтезе растений
и фотосинтезирующих бактерий.
Фотосинтез растений заключается в
преобразовании и запасании солнечной энергии.
В результате этого процесса из простых
веществ — углекислоты и воды —
синтезируются углеводы и выделяется
молекулярный кислород. При этом за счет энергии
поглощенного света происходит окисление воды
с образованием молекулярного кислорода,
а электроны, которые отнимаются от воды,
используются на восстановление С02 до
первичных Сахаров (см.: Тихонов А. Н. Фотосинтез.
Наст. том). В общем виде процесс
фотосинтеза можно представить в виде
несложного уравнения (см. рисунок).
Несмотря на кажущуюся простоту
фотосинтеза, на Земле нет другого
процесса, который смог бы так же сильно
преобразовать нашу планету.
Фотосинтез
Фотоокисление воды
| Светт
Н20 + С02 ->02 + jjj(C6H1206) (+114 ккал/моль)
I Растения А
Ассимиляция (восстановление) С02
Дыхание
-^
Уравнение фотосинтеза, осуществляемого кислород-
выделяющими фотосинтезирующими организмами
Запасание энергии
Как следует из приведенного уравнения
(см. рисунок), на каждый
ассимилированный в процессе фотосинтеза моль
углекислоты запасается 114 ккал энергии. В чем же
состоит достоинство запасания солнечной
энергии растениями по сравнению с
неорганизованной («нефотосинтезирующей»)
системой?
Любое вещество, поглощая квант
солнечной энергии, переходит в возбужденное
состояние, что можно рассматривать как
преобразование энергии электромагнитного
излучения и ее запасание. Однако энергия
возбуждения электронов вещества,
поглотившего свет, очень быстро (за 10"13~10"п сек)
растрачивается в тепло или же вновь
излучается в пространство (в случае сложных
органических молекул типа хлорофилла этот
процесс происходит за 10"9-10~8с);
следовательно, в виде возбужденных состояний
энергия света может запасаться лишь на
незначительные доли секунды. В
результате же фотосинтеза энергия поглощенного
кванта света (или, лучше сказать, часть этой
энергии) запасается надолго: от минут и часов
до сотен и даже миллионов лет (как это
имело место, например, при образовании
горючих ископаемых — нефти, природного
газа, каменного угля, торфа — в
результате разложения наземных и морских
растений или животных). Но этим, конечно, не
исчерпывается специфика фотосинтеза в
использовании солнечной энергии.
Еще одно преимущество фотосинтеза
состоит в том, что запасание солнечной
энергии происходит в очень удобной форме —
молекулярной, в виде богатых энергией
органических соединений, которые в любой
необходимый момент могут быть
использованы растениями или «съевшими» их нефо-
тосинтезирующими (гетеротрофными)
организмами для покрытия своих энергетических
потребностей и синтеза собственных
высокомолекулярных соединений. Масштабы
фотосинтетического преобразования и
запасания солнечной энергии огромны: каждый
год за счет фотосинтеза на Земле
образуется около 200 млрд т биомассы, что
эквивалентно энергии, равной 3-Ю21 Дж, или
7,2-Ю20 кал. При этом необходимо иметь в
виду, что фотосинтез — единственный
биологический процесс, протекающий с
запасанием (увеличением) свободной энергии.
Остальные процессы как в растениях, так
и у животных осуществляются за счет
химической энергии, накапливаемой в
фотосинтезирующих организмах в результате
преобразования поглощенного солнечного
света. Следовательно, практически вся
живая материя на Земле представляет собой
прямой или отдаленный результат
фотосинтетической деятельности растений, которые
являются посредниками между неиссякае-
РАСТЕНИЯ И БИОСФЕРА
323
мым источником энергии — Солнцем — и всем
живым миром нашей планеты. Именно
поэтому мы говорим о фотоавтотрофии
биосферы Земли и о фотоавтотрофии
человечества.
Население Земли ежегодно потребляет
около 1 млрд т продуктов питания (если
считать численность населения равной 5 млрд
человек), что соответствует 151018Дж.
Следовательно, человечество потребляет в виде
органических веществ лишь около 0,5% всей
энергии, запасаемой в результате
фотосинтеза. Общее потребление энергии в
мировом масштабе составляет 3-41020Дж в год,
т. е. около 10% всей энергии, запасаемой за
то же время благодаря фотосинтезу.
Разведанные запасы ископаемого топлива
(нефти, газа, угля, торфа) по накопленной в
них энергии соответствуют продукции
фотосинтетической деятельности на Земле
приблизительно за 100 лет. Примерно
такое же количество энергии
аккумулировано во всей биомассе, находящейся в
настоящее время на нашей планете.
Ассимиляция двуокиси углерода
Ежегодная ассимиляция углекислого газа
на Земле в результате фотосинтеза
составляет около 280 млрд т, что эквивалентно
7,81010т углерода, и это связывание
углерода компенсируется выделением практически
такого же количества С02 в результате
дыхания нефотосинтезирующих организмов.
Количество С02, вовлекаемого в цикл
фотосинтез-дыхание, составляет около 10%
массы углекислого газа в атмосфере,
которая в 1980 г. была эквивалентна 7,110й т
углерода. В то же время до I860 г.
атмосфера содержала лишь б,1-Ю11 т углерода в виде
С02. Произошедшее за 120 лет 15%-ное
увеличение количества С02 в атмосфере
связывают прежде всего с дополнительным
количеством С02, образующимся вследствие
интенсивного сжигания ископаемого
топлива, которое в настоящее время
эквивалентно 5109т углерода в год и увеличивается в
среднем на 4,3% в год.
Необходимо отметить, что
значительно большая часть углерода содержится в
виде карбонатов в осадочных породах —
5,51016т, а в живых (это в основном леса)
и отмерших организмах— 3,51012т. В
Мировом океане содержится в 60 раз больше
углерода, чем в атмосфере (3,5Ю13т), что
связано с очень высокой растворимостью С02
в воде с образованием Н2СОэ.
Следовательно, можно было бы предположить, что
незначительное дополнительное
поступление С02 в атмосферу за счет сжигания
ископаемого топлива (которое составляет
менее одного процента в год от
содержания углекислого газа в атмосфере) не
должно приводить к заметному увеличению
содержания в ней С02. Однако обмен
углерода с атмосферой осуществляется
достаточно быстро (за 6-7 лет) лишь в верхних
слоях океана, содержащих только 1,5%
углерода, растворенного в воде; для
установления такого равновесия с глубинными
слоями океана требуется несколько
тысячелетий.
Вследствие этого сжигание
ископаемого топлива в промышленном масштабе
привело к увеличению содержания С02 в
атмосфере с 0,027% (в доиндустриальную
эпоху) до 0,034% в настоящее время.
Расчеты показывают, что к 2035 г. содержание
углекислого газа в атмосфере удвоится, т. е.
будет составлять около 0,06%. Основным
последствием этого будет, как
предполагают, глобальное потепление климата,
обусловленное так называемым парниковым
эффектом, связанным с тем, что
углекислый газ «прозрачен» для основной части
солнечного света, но задерживает
(поглощает) тепловое (инфракрасное) излучение
от нагретой солнцем поверхности Земли.
Увеличение концентрации С02 в
атмосфере в 2 раза может привести к повышению
температуры поверхности Земли на 2~3°С,
причем оно будет минимальным в
тропической зоне и максимальным в высоких
широтах (8-11°С). Такое увеличение
температуры может вызвать таяние льдов,
особенно в Антарктиде, и как следствие
повышение уровня моря на 5 м с
затоплением значительной части суши. Поэтому
возможность глобального потепления
климата становится сейчас проблемой всего
человечества. Согласно Международной
конвенции, принятой в 1992 г., развитые
индустриальные страны будут проводить
политику ограничения промышленного
выброса С02 в атмосферу, а также защиты и
увеличения поглотителей и резервуаров С02
(в основном растительного и почвенного
покрова Земли). Обсуждается даже
предположение, что страны с повышенными
выбросами С02 должны платить
компенсацию странам, где потребление С02
превышает его продукцию. Согласно оценкам,
проведенным российскими учеными, Россию,
наряду с северными территориями Канады,
можно отнести к странам с увеличенным
потреблением С02, что связано главным
образом с «отставанием» минерализации
органического вещества от
фотосинтетической ассимиляции С02 в условиях
переувлажненных почв и невысоких температур в
северных областях нашей страны. Интересно
также отметить мнение академика А. Л.
Яншина о том, что для России,
значительная часть территории которой
расположена в зоне вечной мерзлоты, повышение
концентрации С02 и связанное с ним потеп-
324
ЭКОЛОГИЯ
ление климата выгодны. При этом следует
также учитывать, что двукратное
повышение содержания С02 в атмосфере
приведет к 60%-му повышению скорости
фотосинтеза на Земле.
Выделение молекулярного
кислорода
Более 2 млрд лет назад в процессе
эволюции растения приобрели способность к
фотосинтетическому окислению воды, в
результате которого две молекулы воды
теряют четыре электрона и четыре
протона, что приводит к образованию молекулы
02 (см. рисунок). Это вызвало поистине
революционные преобразования на Земле, три
из которых следует отметить особо.
1. Фотосинтезирующиё организмы, а
через их посредство и вся живая природа
получили доступ к практически
неиссякаемому источнику «мобильных» электронов
(участвующих во всех биоэнергетических
процессах) — воде, что естественно
привело к резкому возрастанию масштабов
фотосинтеза и поступления энергии в
биосферу. Ранее в качестве источника
электронов для фотохимических реакций могли
использоваться лишь восстановленные
вещества типа H2S, представленные на
Земле в довольно ограниченном количестве.
2. Продукт фотосинтетического
окисления воды — молекулярный кислород,
содержание которого в атмосфере возросло в
результате фотосинтеза практически от нуля
в древней атмосфере до 21% в настоящее
время, вызвал значительные изменения в
живой природе. Появление в атмосфере
свободного 02 привело к массовой гибели
существовавших тогда организмов, ибо
такой сильный окислитель, как кислород,
оказался для них ядом, нарушающим
протекание биоэнергетических процессов. В то
же время в результате длительной
эволюции на фоне возрастающей концентрации
02 в атмосфере появились новые аэробные
организмы. Они включили 02 в свой
метаболизм в качестве конечного акцептора
электронов в дыхательной цепи, что
позволило за счет более эффективного
«сжигания» органики значительно повысить
эффективность биоэнергетики. Так, например,
выход макроэргического эквивалента —
аденозинтрифосфата (АТФ) — на 1
молекулу исходной гексозы составляет 2
молекулы при молочнокислом брожении и 36-
38 молекул (т. е. в 18—19 раз больше) при
аэробном дыхании.
Фотосинтетическое образование 02,
сопровождающееся поглощением С02, в сочетании
с появившейся возможностью аэробного
дыхания, когда происходит обратный процесс —
потребление 02 и выделение С02 (см. рисунок),
привело к образованию замкнутых циклов 02
и С02, имеющих глобальные масштабы.
Совместное осуществление этих циклов
позволяет восполнять содержание как С02, так и
02, а их участие в едином цикле
фотосинтез-дыхание обеспечивает постоянное
поступление энергии в биосферу.
В результате фотосинтеза в атмосферу
ежегодно поступает около 2-Ю11 т
молекулярного кислорода. Если учесть, что в
настоящее время в атмосфере содержится
1,21015т 02, то для полного обновления
запаса кислорода в атмосфере под действием
фотосинтеза потребуется около шести
тысяч лет. Необходимо отметить, что на
окислительное дыхание гетеротрофных
организмов и на окисление мертвого органического
вещества из атмосферы ежегодно
потребляется примерно такое же количество 02,
которое выделяется при фотосинтезе.
Разница между фотосинтетическим выделением
02 и его потреблением в дыхательных
процессах составляет около 108 т кислорода (или
около 0,04% общего количества 02,
ежегодно выделяющегося при фотосинтезе).
Существенная часть этого кислорода
расходуется на окисление горных пород, а
также газов (Н2, H2S, CO, S02),
попадающих в атмосферу из глубоких слоев Земли.
Значительное количество
атмосферного кислорода потребляется в результате
сжигания топлива— около 1,ЗЮ10т в год.
Это примерно в 130 раз больше, чем
указанная выше разница между
фотосинтетическим выделением 02 и его потреблением
в дыхательных процессах, что приводит к
понижению содержания 02 в атмосфере.
Необходимо, однако, отметить, что даже
при сжигании всех разведанных запасов
ископаемого топлива содержание 02 в
атмосфере уменьшится лишь на доли процента.
В качестве источника 02 на Земле
рассматривается также фотодиссоциация
молекул Н20 на атомы Н и О в верхних слоях
атмосферы под действием
ультрафиолетового света с длиной волны 175-200 нм. При
этом более легкие атомы Н с большей
вероятностью покидают атмосферу,
преодолевая силу тяготения Земли, чем атомы О,
которые, объединяясь, образуют
молекулярный кислород. Считается, однако, что в
результате такого процесса может
образоваться лишь незначительная часть 02,
находящегося сейчас в атмосфере Земли. Это
во многом связано с так называемым
эффектом Юри, который состоит в том, что
образующийся 02 даже если он присутствует
в небольших количествах (0,2%),
эффективно поглощает ультрафиолетовую радиацию,
наиболее активную в процессе
фотодиссоциации паров воды; в результате этот
процесс подавляется.
РАСТЕНИЯ И БИОСФЕРА
325
3. Появление кислородвыделяющих
организмов привело к тому, что практически все
процессы на поверхности Земли приобрели
биогеохимический характер. Именно в
данный период значительного повышения
количества 02 в атмосфере произошло
окисление соединений железа, серы, марганца,
причем на это было израсходовано более
95% кислорода, выделенного фотосинтези-
рующими организмами за всю историю
биосферы. До появления кислородвыделяющих
фотосинтезирующих организмов атмосфера
Земли была разреженной. Она состояла
главным образом из С02 и аммиака. По мере
возрастания концентрации 02 в атмосфере
увеличивалось, благодаря окислению
аммиака, и содержание молекулярного азота. Сам
процесс фотосинтеза, сопровождающийся
образованием и запасанием органического
вещества, привел к значительному (более чем
в 100 раз) уменьшению количества С02 в
атмосфере.
В итоге первичная разреженная
атмосфера превратилась во вторичную плотную
азотно-кислородную оболочку Земли. Теперь
солнечная и другая космическая радиация,
прежде чем достичь поверхности, должна
была проникать через мощный слой
атмосферы. Кроме того, под действием
фотохимических процессов в верхних слоях
атмосферы двухатомный молекулярный кислород
превращается в трехатомную молекулу
озона, имеющего интенсивное поглощение в
области жесткого ультрафиолета (0,2-
0,3 мкм). В результате образовавшийся
озоновый экран стал задерживать наиболее
опасную для живых организмов
ультрафиолетовую радиацию.
Таким образом, фотосинтез способствовал
защите живой материи от гибельного
действия космической радиации.
В настоящее время фотосинтез лежит
в основе биологического круговорота энергии
и веществ на Земле, от масштабов
которого зависит жизнь на нашей планете, ее
разнообразие и возможности.
Исследование фотосинтеза привлекает
внимание не только современных биологов,
но и физиков, химиков, математиков.
Познание молекулярных механизмов
фотосинтеза будет иметь большое значение для
обеспечения человечества экологически
чистой энергией за счет практически
неиссякаемого источника — солнечного
излучения (например, на основе фоторазложения
воды на молекулярный водород и кислород),
для повышения фотосинтетической
продуктивности растений, лежащей в основе
обеспечения человечества пищей, для
использования принципов фотопреобразования
световой энергии при фотосинтезе в фото-
биотехнологических и фотобиотехнических
системах, для обеспечения длительных
космических экспедиций органикой и
молекулярным кислородом, для решения проблем
экологической безопасности отдельных
регионов, а также для сохранения и
развития биосферы в целом.
Литература
Будыко М. И., Ронов А. Б., Яншин А. Л.
История атмосферы. Л.: Гидрометеоиздат, 1985.
Климов В. В. Сорос, образоват. журн., 1996,
№10, сб.
Мак-Ивен М., Филлипс Л. Химия атмосферы.
Пер. с англ. М.: Мир, 1978.
Фотосинтез. В 2 т. Пер. с англ. М.: Мир, 1987.
Холл Д., Рао К. Фотосинтез. Пер. с англ. М.:
Мир, 1983.
Шувалов В. А. Первичное преобразование
световой энергии при фотосинтезе. М.: Наука,
1990.
С. Г. Инге-Вечтомов
ЭКОЛОГИЧЕСКАЯ ГЕНЕТИКА: НОВЫЙ ВЗГЛЯД
Введение
Словосочетания, включающие слова
экология, экологический, встречаются
сегодня очень часто. Это отражает
объективное стремление человека осознать свое
место в мире живых существ и вообще в
мире как единой системе взаимозависимых
факторов, формирующих среду обитания.
Задача настоящей публикации — рассказать
читателям о содержании сравнительно
молодой области биологии — экологической
генетике, приобретающей все большее
практическое значение.
Несколько определений
Что же такое экологическая генетика?
Определяя содержание этой области знания,
Э. Б. Форд в 60-е гг. впервые сформулировал
понятие об экологической генетике как о
генетике популяций в природных условиях.
В последующем некоторые стали понимать
ее более широко, как область науки,
перекрывающую почти всю современную генетику.
Нам представляется, что, определяя
предмет экологической генетики, необходимо
исходить из исторической логики развития
биологии, а также содержания и
методологических возможностей обеих наук—
экологии и генетики, давших название этой
новой пограничной дисциплине.
Общепринято определение экологии как
науки об отношениях организмов с
окружающей средой, а также с другими
организмами, составляющими часть этой среды.
Определение генетики как физиологии
наследственности и изменчивости, данное
еще в 1906 г. У. Бэтсоном, сохраняет свое
значение до сих пор.
Отталкиваясь от этих общих
представлений, попытаемся дать новое определение
экологической генетики.
Экологическая генетика это область
знания, исследующая взаимовлияние
генетических процессов и экологических
отношений. При этом как раздел генетики она
опирается на мощную методологию генетического
анализа и одновременно использует весь
методический арсенал экологии.
Структура экологической генетики
упрощенно представлена в таблице.
Обсуждение этой таблицы поможет нам
рассмотреть структуру и задачи экологической
генетики более конкретно.
Генетические аспекты
Генетические подходы в экологической
генетике базируются на двуединстве
методологии генетического анализа,
оперирующего понятиями «наследственность» и
«изменчивость». Наследственность — это
свойство сходства родственных организмов,
их способности передавать определенные
признаки из поколения в поколение. При этом
в качестве признаков могут фигурировать:
наличие или отсутствие органов, их
число, размер, цвет; способность осуществлять
те или иные биохимические реакции;
свойства нервной системы, тип поведения и т. д.
Генетический анализ вскрывает гены,
контролирующие все это разнообразие
признаков, изучает их наследование и
локализацию в геноме.
Кроме того, генетический анализ
вскрывает причины изменчивости, т. е. отвечает
не только на вопрос: «Почему организмы
имеют сходство между собой?», но и на
вопрос: «Почему организмы отличаются друг
от друга?» Особое внимание при этом
уделяется причинам, механизмам и
последствиям мутационной изменчивости, т. е.
наследуемых изменений генетического материала.
Как известно, мутации могут быть
спонтанными — возникающими, казалось бы, без
Таблица 1
Общая структура экологической генетики
Генетические аспекты
Генетический контроль признаков
(наследственность)
Влияние различных факторов на
генетические процессы (изменчивость)
Экологические аспекты (типы экологических отношений)
Синэкология
Эколого-генетические модели
Биологические факторы
изменчивости (мутагенеза)
Аутэкология
Генетика устойчивости к
факторам среды
Генетическая токсикология
ЭКОЛОГИЧЕСКАЯ ГЕНЕТИКА: НОВЫЙ ВЗГЛЯД
327
какой-либо внешней причины, и
индуцированными — возникающими под
действием различных внешних агентов:
физических, химических и т. д.
Мутационный процесс обычно
связывают с так называемыми генетическими
процессами. К ним обычно относят:
'репликацию, т. е. воспроизведение генетического
материала; рекомбинацию, т. е. различные
способы пересортировки генов и их частей,
происходящие при смене поколений, и
репарацию, т. е. процессы, поддерживающие
нативную структуру генетического
материала, постоянно повреждаемого под
влиянием как внутренних факторов —
физиологических, метаболических, так и внешних
факторов — температуры, излучений,
химических воздействий. Спонтанные или
индуцированные нарушения этих
процессов, лежащих в основе наследственной
передачи генетического материала,
являются причиной наследственной изменчивости —
мутационного процесса.
К числу генетических процессов
относят также транскрипцию — синтез РНК на
матрице ДНК и трансляцию — синтез
белков на матрицах инфомационных РНК. Это
уже процессы, лежащие в основе
реализации генетической информации, в основе
действия генов. Их нарушения в
значительной степени ответственны за модификаци-
онную изменчивость.
Механизмы наследования признаков, а
также механизмы наследственной
изменчивости изучены довольно подробно, чего
нельзя сказать о механизмах модификаций.
Это приходится учитывать в работе, тем
более, что адаптивные модификации играют
важную роль в экологических отношениях.
Следует оговориться, что в генетике
часто употребляют также понятие
«генетические процессы в популяциях», т. е.
процессы, влияющие на частоты генов
(точнее их аллелей) в популяциях организмов
и обусловливающие микроэволюционные
преобразования (см.: Алтухов Ю. П.
Генетика популяций и сохранение
биоразнообразия. Наст. том).
Типы экологических отношений
Экологические отношения делят на син-
экологические, т. е. отношения между
организмами, и аутэкологические, т. е. отношения
организмов с окружающей средой. При всей
полезности такого деления следует признать
его относительность, поскольку многие
факторы окружающей среды имеют
биотическое происхождение. Тем не менее мы
будем пользоваться таким подразделением,
поскольку оно позволяет формализовать
ситуацию, что необходимо на первых
порах знакомства с новой областью знания.
Синэкология исследует как отношения
между организмами одного вида, так и
отношения между организмами разных видов,
объединяемых в экосистемы. Чаще всего эти
отношения основываются на
взаимозависимости разных видов, составляющих
различные звенья пищевых цепей . Знание
структуры пищевых цепей в природе необходимо
для прогнозирования последствий любых
воздействий на экосистемы.
Хорошо известен пример того, к чему
привело игнорирование структуры
пищевых цепей при попытке уничтожить
комаров на озере Клир Лэйк в США. После
использования для этой цели инсектицида
ДДТ (4,4-дихлор-дифенилтрихлорэтана) его
концентрация в воде составила 0,02 части
на 106, в планктоне— 10 на 106, в рыбах,
питающихся планктоном— 903 на 106, в
хищных рыбах— 2690 на 106, а в
рыбоядных птицах— 2134 на 106. Таким образом,
концентрация ДДТ по мере продвижения
«вверх» по пищевой цепи увеличилась в
100 000 раз, что привело к сокращению
численности птиц на озере Клир Лэйк.
Главная опасность ДДТ заключается в том, что
его токсичность сочетается со
стабильностью, характерной для всех хлорорганических
соединений, к которым он относится.
Аутэкология рассматривает отношения
живых существ с факторами окружающей
среды, преимущественно абиотического
происхождения. При этом подобные
абиотические факторы могут быть
естественными, с которыми живые организмы
сталкивались неоднократно в ходе эволюции. Это —
температурные воздействия, земное
тяготение, различные виды излучений: от
излучений в видимой части спектра до
рентгеновских, многие химические агенты и т. д.
В ходе эволюции живые существа
выработали адаптивные реакции на такого рода
воздействия, в результате чего возникала
устойчивость (в определенных пределах)
организмов к действию повреждающих
факторов. Многим химическим агентам живые
существа противостоят путем включения их
в собственный метаболизм или в пищевые
цепи экосистем.
Сложнее обстоит дело с новыми, как
правило антропогенными, факторами
внешней среды, которые никогда не встречались
в природе в ходе биологической эволюции.
Так, например, многие инсектициды —
хлорированные углеводороды, никогда не
существовали в природе. Они не
трансформируются в пищевых цепях и потому
неразложимы биологическим путем, что не
учитывалось при их применении. К ним
относятся полихлорбифенилы, в частности
пестициды: 2,4-дихлорфеноксиуксусная
кислота (2,4-Д) или 2,4,5-трихлорфеноксиуксус-
ная кислота (2,4,5-Т) — эффективные дефо-
328
ЭКОЛОГИЯ
лианты. Притчей во языцех становятся в
последнее время диоксины, также
относящиеся к полихлорированным бифенилам и
представляющие собой самые активные яды,
известные в настоящее время. Один из них
входил в состав печально известного
«оранжевого реактива», применявшегося армией
США во Вьетнаме в качестве боевого
дефолианта. Диоксины образуются также при
сжигании мусора на заводах по
уничтожению городских отходов.
В списке наиболее значимых факторов
загрязнения среды (из 19 наименований)
первые 5 мест занимают пестициды,
тяжелые металлы, диоксид углерода, диоксид
серы и другие продукты ее окисления,
взвеси, разливы нефти, сточные воды
промышленных предприятий.
При этом радиоактивные отходы,
например, обладающие несомненной
генетической активностью, как загрязнители стоят
только на двенадцатом месте. Однако надо
подчеркнуть, что непосредственное
влияние загрязнений на комфорт и здоровье
человека может быть несопоставимо с
отдаленными последствиями тех или иных
изменений окружающей среды.
Эколого-генетические модели
Применение методов генетического
анализа связано с выявлением элементарных
признаков. Это же справедливо и для
генетического анализа последствий
экологических воздействий, которые обычно весьма
сложны. Поэтому необходима разработка
специальных эколого-генетических моделей.
Большую помощь при построении таких
моделей оказывает знание пищевых цепей,
особенно, если в них организмы одной
экосистемы выступают как продуценты и
потребители каких-либо метаболитов. В
подобные взаимоотношения вступает,
например, почвенная бактерия Agrobacterium
tumefaciens, размножающаяся вблизи корней
крестоцветных растений. Агробактерии столь
тесно взаимодейстуют с корешками высшего
растения, что передают ему часть своей
плазмидной ДНК (часть так называемой
Ti-плазмиды), которая встраивается в его
хромосомы. Это приводит к образованию
растительных опухолей, которые
начинают интенсивно синтезировать некоторые
аналоги аминокислот — опины (производные
лизина, гистидина, орнитина или
аргинина). Образовавшиеся соединения, в свою
очередь, служат дополнительным источником
азота для агробактерии и тем самым
стимулируют их размножение. Такое
взаимоотношение бактерий и растения
получило название генетической колонизации
(рис. 1). Стоит отметить, что способность
Ti-плазмиды трансформировать клетки
растений легла в основу ряда методов генной
инженерии растений. Здесь важно
подчеркнуть, что биосинтез опинов происходит
частично под контролем генов растения, но
завершается под контролем генов
агробактерии, передаваемых в клетки растения.
Генетический контроль этих процессов, а
следовательно, и взаимосвязи агробактерия-
растение довольно подробно исследованы
Ti-плазмида
Т-ДНК
Хромосома
Опины
Корончат.
галл-
Agrobacterium
tumefaciens
A. tumefaciens
Хромосомная ДНК
Т-ДНК
Трансформированная
растительная клетка
Рис. 1. Генетическая колонизация высшего растения бактерией A. tumefaciens
7 — A. tumefaciens существует в ризосфере (корневой сфере) растения; 2— в клетках бактерии наряду с ее
хромосомой существует Ti-плазмида; З— Ti-плазмида проникает в клетку растения и часть ее— Т-ДНК —
встраивается в геном растения; 4— это приводит к образованию опухоли— корончатого галла и к синтезу
опинов
ЭКОЛОГИЧЕСКАЯ ГЕНЕТИКА: НОВЫЙ ВЗГЛЯД
329
и представляют собой хорошую эколого-ге-
нетическую модель. Интенсивно
разрабатываются и другие эколого-генетические
модели, включающие других почвенных
бактерий, в частности азотфиксирующих.
Практические выгоды от разработки этого
направления очевидны: мы получаем
возможность управления процессом азотфик-
сации и симбиотическими отношениями
бактерий и высших растений.
Еще один пример элементарной эколо-
го-генетической модели дают
взаимоотношения насекомых и высших растений. В этих
системах насекомые-вредители сельского
хозяйства и сельскохозяйственные
растения связаны как потребители и
продуценты стеринов. Стерины, которые насекомые
не могут синтезировать самостоятельно,
являются для них незаменимыми
метаболитами, и они получают их с растительной
пищей.
Так или иначе, выявление генов,
отвечающих за элементарные экологические
отношения, позволяет использовать
генетический контроль для регулирования этих
отношений и тем самым выбирать
оптимальную стратегию «сдерживания» вредителей
сельского хозяйства вместо того, чтобы
вести с ними тотальную войну на
уничтожение. Как мы показали, такая война
часто оборачивается против самого человека.
Генетика устойчивости
к факторам среды
Изучение генетического контроля
устойчивости модельных объектов, особенно
сельскохозяйственных растений, животных и
человека, к неблагоприятным внешним
факторам имеет большое значение для
селекции, медицины и поддержания
оптимальной среды обитания человека.
Прежде всего остановимся на так
называемых молекулярных болезнях
человека. Известны, в частности,
наследственные аномалии репликации и репарации
ДНК. У человека, например, существуют
различные формы пигментной ксеродермы.
Эта болезнь возникает вследствии
рецессивного аутосомного дефекта репарации,
в частности дефекта ДНК-полимеразы,
принимающей участие в репарации.
Больные пигментной ксеродермой проявляют
повышенную чувствительность к
солнечному свету, который вызывает у них рак
кожи. Дефекты систем репарации выявлены
и при других наследственных
заболеваниях: анемии Фанкони, синдроме Луи Бар
и др. При радиотерапии таких больных
наблюдаются осложнения, часто со
смертельным исходом.
В человеческих популяциях
обнаруживается генетическая гетерогенность по
многим признакам, в том числе по
чувствительности к факторам окружающей среды,
устойчивости к стрессирующим агентам и
производственным вредностям. Так, у
людей с низкой активностью глюкозо-6-фос-
фатдегидрогеназы происходит гемолиз при
действии сульфаниламидов. Другие лица с
генетическими дефектами реагируют
повышением внутриглазного давления на
прием глюкокортикоидов. Некоторые мутант-
ные формы гемоглобина чувствительны к
окислителям и их применение приводит к
гемолизу. Люди с повышенной активностью
арилгидрокарбонгидроксилазы чаще
заболевают раком легких при контакте с
полициклическими углеводородами. Последние
под действием этого фермента
превращаются в эпоксиды, обладающие высоким
уровнем канцерогенности.
Такие факты необходимо учитывать при
планировании медицинских мероприятий,
приеме людей на работу и в деятельности
по профориентации. Очевидно, эти же
факты могут быть причиной и ряда социально-
экономических проблем. Возникают, в
частности, такие вопросы: Можно ли допускать
дискриминацию при приеме на вредные
производства по генетическим
противопоказаниям? Каковы должны быть условия
страхования людей, имеющих
дополнительные факторы риска в виде генетической
предрасположенности к тем или иным
заболеваниям? Простых ответов на эти и
подобные им вопросы пока нет, но не
ставить их нельзя.
Генетическая токсикология
Следует отметить, что загрязнение
окружающей среды опасно не только для ныне
живущих, но часто и для грядущих
поколений, поскольку многие загрязнители му-
тагенны (или, что почти то же самое,
генетически активны). Выявление и устранение
генетически активных факторов из среды
обитания человека — задача генетической
токсикологии. Генетическая токсикология
представляет собой наиболее активно
развивающийся раздел экологической
генетики. Это объясняется ее огромным
прикладным значением.
Парадоксально, но факт, что открытие
индуцированного мутационного процесса
потребовало значительных усилий от
исследователей: вспомним, что Г. Дж. Мёллер
получил Нобелевскую премию за свое
открытие мутагенного действия рентгеновских
лучей (1927 г.). Теперь же мутагены
обнаруживаются повсеместно.
Многие продукты производственной
деятельности человека, появляющиеся как
результат технического прогресса, обладают
генетической активностью. При этом мы
330
экология
говорим не только об отходах производства.
Это могут быть лекарства, консерванты,
пищевые добавки и красители, косметика,
инсектициды и пестициды, не говоря уже
о дыме сигарет и излучениях,
сопровождающих эксплуатацию атомных станций, а
также производство и особенно испытание
оружия массового уничтожения — ядерного,
химического, и т. д.
Что же такое генетически активные
факторы? В генетической токсикологии
принято говорить не только о мутагенах,
но и более широко — о генетически
активных факторах. Не всегда удается
определить непосредственно мутагенный эффект
того или иного воздействия, но можно
показать его влияние на кроссинговер, т. е.
на рекомбинацию генов или на индукцию
репаративного синтеза ДНК,
сопровождающего многие повреждения генетического
материала.
Таким образом, мутагенез, рекомбиногенез
и индукция репаративного синтеза ДНК —
это показатели генотоксичности или
генетической активности вызвавшего их
фактора.
Генетически активные факторы
делятся на физические, химические и
биологические. Физические: температура,
ионизирующая радиация, ультрафиолетовый свет,
по-видимому, высокочастотное
электромагнитное излучение, ультразвук и т. д.
Химические генетически активные факторы
гораздо труднее поддаются перечислению
и классификации. Достаточно сказать, что
к ним относятся любые вещества, прямо
или косвенно нарушающие структуру и
воспроизведение молекул ДНК. Так
выхлопные газы автотранспорта и выбросы в
атмосферу производственных предприятий
содержат алкилирующие соединения (их
еще называют радиомиметиками),
органические соединения ртути, полициклические
углеводороды и пр., обладающие
генетической активностью. Многие химические
соединения сами по себе не проявляют
генетической активности, но их легко активируют
взаимодействия с другими соединениями.
Например, распространенные соли азотной
кислоты легко превращаются в нитриты
(соли азотистой кислоты) — мутагены, де-
заминирующие основания ДНК. В кислой
среде желудка млекопитающих нитриты и
аминосоединения дают нитрозосоединения —
супермутагены, нарушающие репликацию
ДНК. Многие вещества — так называемые
промутагены активируются в организме
млекопитающих при действии цитохрома
Р450. Этот фермент, синтезируемый в
печени, относится к классу неспецифических
монооксигеназ и предназначен для
инактивации чужеродных соединений,
попадающих в организм. Тем не менее некоторые
промутагены, так же как и потенциальные
канцерогены — вещества, вызывающие рак,
он, напротив, активирует.
Необходимо отметить высокий уровень
корреляции между мутагенным и
канцерогенным эффектами многих факторов,
прежде всего физических и химических.
Биологические же факторы, о которых будет
сказано ниже, в этом отношении
исследованы хуже всего.
С некоторых пор (у нас — с 1979 г.), все
новые химические соединения (а всего их
более 4,5 млн) проходят проверку на
генетическую активность. Это работа
своеобразной службы генетической безопасности,
использующей богатый арсенал различных
тест-систем для выявления генетической
активности. Такие системы позволяют
учитывать мутации и рекомбинации генов,
потери и другие аберрации хромосом,
нарушения делений ядра, индукцию репарации
ДНК и т. д. При этом используются
различные объекты: бактерии, дрожжи и другие
низшие грибы, плодовая мушка
дрозофила, растения, культуры клеток животных
и человека и т. д.
Наибольший интерес представляет,
конечно, генетическая активность
исследуемых агентов при их воздействии на
человека. Поскольку прямое исследование
такого действия невозможно,
приходится ограничиваться результатами,
получаемыми на перечисленных модельных
объектах. Эти результаты в значительной
степени справедливы и для человека в силу
биологической универсальности свойств
генетического материала (ДНК). Тем не
менее экстраполяция получаемых
результатов на человека всегда представляет
некоторые сложности, так как наряду с
принципом биологической
универсальности, следует учитывать и специфику
объектов, имеющих свои особенности
реагирования на мутагены.
В качестве примера расскажем только
об одной тест-системе, получившей
широкое распространение при первичном
выявлении генетической активности. Это
система, разработанная в 60-е гг. XX в.
американским исследователем Б. Эймсом,
который длительное время изучал
мутации в генах, контролирующих биосинтез
гистидина у Salmonella typhimurium.
Работа Эймса — прекрасный пример того, как
первоначально чисто теоретическое
исследование, направленное на выяснение
структуры и функции гена, приобрело сугубо
практическое значение. Имея в своем
распоряжении подробно охарактеризованные
мутанты сальмонеллы, нуждающиеся в
гистидине, и зная молекулярную природу
соответствующих мутационных изменений,
среди которых были замены, вставки и
ЭКОЛОГИЧЕСКАЯ ГЕНЕТИКА: НОВЫЙ ВЗГЛЯД
331
выпадения пар оснований, а также более
крупные перестройки генетического
материала, Эймс предложил изучать реверсии
гистидиновых мутантов, т. е.
восстановление у них в результате воздействия
различных мутагенов и способности
синтезировать гистидин, и следовательно расти на
среде без гистидина.
Этот тест очень прост: достаточно
засеять среду без гистидина мутантом
сальмонеллы, нуждающимся в гистидине (который
естественно не растет на такой среде), и
нанести в центр используемой для этого
чашки Петри испытуемое химическое
соединение; через 2-3-е суток можно увидеть
появление колоний мутантов (в данном случае
ревертантов) вокруг пятна нанесенного
вещества, если оно обладает генетической
активностью (рис. 2). Это пример так
называемого спот-теста (от англ. spot — пятно).
В настоящее время тест Эймса
усовершенствован: наряду с хорошо изученными
мутациями потребности в гистидине в геном
сальмонеллы вводят делецию по одному из
генов репарации, т. е. инактивируют этот
процесс, тем самым повышая
чувствительность бактерии к мутагенам. Вводят также
мутацию, блокирующую синтез липополиса-
харидной капсулы для повышения
проницаемости клеток, а также плазмиды,
повышающие чувствительность клеток к агентам,
усиливающим рекомбинацию. Наконец,
испытуемое вещество стали наносить вместе
с экстрактом мышиной или крысиной
печени, содержащим цитохром Р450 для
активации промутагенов. Тест-системы для
выявления генетической активности могут
совершенствоваться и в дальнейшем, причем
в значительной степени генетическими
методами. С помощью теста Эймса впервые
были показаны мутагенные эффекты
сигаретного пепла, некоторых пищевых
консервантов, красителей для волос и т. д.
Биологические факторы
мутагенеза
Особый интерес представляют
биологические генетически-активные факторы,
поскольку их существование указывает на
генетическую активность синэкологических
отношений. В конце 30-х гг. С. М. Гершен-
зон установил мутагенный эффект ДНК и
вирусов. Позже было выяснено, что
вирусы оспы, кори, ветряной оспы, гриппа,
гепатита и др. могут также вызывать
хромосомные аберрации в соматических клетках.
Стрептолизин-О-токсин гемолитического
стрептококка повышает частоту мутаций в
культуре эмбриональных фибробластов
человека. В контроле частота хромосомных
аберраций составляет 4,0±0,5%, а при
действии токсина — 24,3±0,6%. Ю. Я. Керкис
показал мутагенный эффект
иммунологического стресса при пересадке и отторжении
вследствие тканевой несовместимости
кожного лоскута у мышей. Мощным мутагеном
биологического происхождения оказался
афлатоксин — продукт жизнедеятельности
плесневого гриба Aspergillus flavus.
б
Рис. 2. Схема выявления мутагенной активности химического соединения— нитрозогуанидина в спот-тесте с
использованием системы Б. Эймса
а— контрольная чашка Петри, содержащая среду без гистидина и засеянная культурой мутанта S. typhimurium
His-, не способного синтезировать гистидин. Вырастают только редкие спонтанные ревертанты His~-> His+. В
центр помещен кружок фильтровальной бумаги, смоченный растворителем, но не содержащий мутагена; б—
такая же чашка, в центр которой помещен кружок фильтровальной бумаги, смоченный раствором мутагена.
Вокруг него повляется кольцо индуцированных мутантов-ревертантов His+
332
ЭКОЛОГИЯ
C57BL/6 СВА
s
X
о
о
о
§ ю
7 2 3 4
Рис. 3. Влияние феромонального стресса на
частоту хромосомных аберраций у мышей
Гистограмма показывает частоту аберраций в
сперматогенезе: 1 — без дополнительного воздействия;
2— в результате помещения мышей 30-дневного
возраста на чистую подстилку; 3,4— в результате
помещения мышей того же возраста на подстилку
после пребывания на ней самцов 3-4-месячного
возраста из генетических линий C57BL6 и СВА
соответственно
Под руководством М. Е. Лобашева еще в
60-е гг. на кафедре генетики и селекции
Ленинградского университета были
начаты эксперименты, доказавшие, что нервная
система принимает участие в контроле
частоты хромосомных аберраций в
соматических клетках (роговице глаза) у мышей.
Развивая это направление, ученики Лобашева
(Р. И. Цапыгина, С. Н. Новиков, Е. В. Даев
и др.) показали мутагенный эффект
феромонального стресса у мышей. При этом
важно, что речь идет уже о мутациях не в
соматических, а в генеративных клетках —
при сперматогенезе. Схема проведенного
эксперимента представлена на рис. 3.
Известно, что запах во
взаимоотношениях мышей выполняет функции
своеобразного языка. Феромоны — летучие вещества,
содержащиеся в моче этих животных,
играют роль сигналов, вызывающих реакцию
подчинения, агрессии и т. д. Пользуясь
этими сигналами, старые самцы держат в
подчинении самок и молодых самцов. Оказалось,
что запах взрослого самца при однократном
воздействии повышает частоту
цитологических нарушений в сперматогенезе у молодых
самцов, увеличивает частоту аномальных
сперматозоидов и доминантных летальных
мутаций, выявляемых после их спаривания
с самками, не подвергавшимися воздействию.
Ю. Я. Керкис, Д. К. Беляев и П. М. Бородин
(Институт цитологии и генетики
Сибирского отделения РАН, Новосибирск) показали,
что стресс у мышей, вызываемый
некоторыми физическими воздействиями, повышает
у них частоту кроссинговера.
Таким образом, синэкологические
отношения, т. е. отношения между
организмами, могут быть источником наследственной
изменчивости и тем самым служить
дополнительным фактором эволюции. Эти
исследования в последние годы развиваются, к
сожалению, недостаточно активно.
Заключение
В этой статье мы попытались в общих
чертах рассказать о содержании и
некоторых основных разделах сравнительно
молодой науки — экологической генетики. Было
показано, что она решает как
фундаментальные, так и прикладные проблемы. При
этом данная область науки, еще вчера
представлявшая чисто теоретический интерес,
сегодня позволяет решать сугубо
конкретные, практические задачи, связанные -с
селекцией, медициной и сохранением
оптимальной среды обитания человека.
Литература
Дажо Р. Основы экологии. М.: Прогресс, 1975.
Жученко А. А. Экологическая генетика.
Кишинев: Штийница, 1980.
Инге-Вечтомов С. Г. Генетика с основами
селекции. М.: Высшая школа, 1989.
Кайданов Л. 3. Генетика популяций. М.:
Высшая школа, 1996.
Киселев А. В., Худолей В. В. Отравленные
города. М.: Greenpeace, 1995.
Пирузян Э. С, Андрианов В. М. Плазмиды
агробактерий и генетическая инженерия
растений. М.: Наука, 1989.
Риклефс Р. Основы общей экологии. М.: Мир,
1979.
Тихомирова М. М. Генетический анализ. Л.:
Изд-во ЛГУ, 1990.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
предметный указатель
А
Абсцизовая кислота 288, 291
Агробактерии 328
Агроценоз 316
Адаптационная медицина 250
Адаптация 243, 248, 250, 252, 253
долговременная 243
срочная 243
Адгезия
межклеточная 202
фокальная 202
Аденозиндифосфат (АДФ, лат. ADP) 281,
275
Аденозинтрифосфат (АТФ, лат. АТР)
19, 85, 87, 168, 195, 262, 275, 281,
324
Адреналин 245
Адреналовая ось 245
АДФ, см. Аденозиндифосфат
Азотфиксация 309, 310, 312
Акинеты 308, 309
Аксоны 238
Активирование генов 101
Актин 163, 166
Акустическая сигнализация 48
Алкалоиды 301
Алкоголизм 133
Аллели 63, 121
Аллелизм 79
критерий 79
тест 79
Аллергия 219
Аллозимы 151
Альбумин 214
о^-адренорецепторы 252, 253
Амелогениновый ген 144, 145
Амилопласты 90, 91
Аминокислотные последовательности 63
Аминокислоты 63
Аминоптерин 222
Анабиоз 252
Анаболизм 252
Анагенез 40
Анатомия генома человека 135, 140
Анаэробиоз 277
Ангиотензин II 245
Андроцей 119
Анкилозавры 74
Аномалии развития 135
Антибиотики 127, 128, 310
Антигены 215, 216, 218, 220, 228, 233,
234
гистосовместимости 106
тканевой совместимости 218
Антимюллеровский гормон 105, 108
Антиоксидантные ферменты 248
Антиоксиданты 248, 250
Антионкоген 187, 189, 204
Антионкогены 227
Антитела 216, 217, 218, 220, 225, 226, 237
моноклональные 220, 221, 222, 223
поликлональные 223
Антителообразующие клетки 220
Анус 32
Апоптоз 186, 187, 191, 238
Арабидопсис 272, 274, 278
Аргинин-вазопрессин 244, 245
Ароматаза 112
Архебактерии 17, 64, 82
Археи 17, 26, 195, 197, 308
галофильные 19
метанобразующие 19
Археоптериксы 75
Асцидия 31
АТФ, см. Аденозинтрифосфат
АТФаза 28
АТФ-синтаза 287
Ауксины 288, 289, 293, 294
Аутосома 104
Аутэкология 326, 327
Аэробное дыхание 324
Аэробные организмы 27, 324
Аэродинамика 57
Аэродинамические силы 55
Аэродинамический след 58
Б
Бактерии 17, 26, 79, 81, 195, 225, 235,
236, 307, 308
автотрофные 9
гетеротрофные 9
Бактериофаги 9, 22
Безусловные рефлексы 125
Белки 24, 63, 79, 99, 187
теплового шока (БТШ) 271, 272
Белок-рецептор 193, 202
Беспозвоночные 38, 195
Бесчерепные 31
Р2-микроглобулин 225
Билатерии 15
вторичноротые 15
первичноротые 15
Бинарная номенклатура 7
Биогеографическое районирование 71, 72
Биоразнообразие 150
Биоритмы 132
Биосинтез белка 28
Биосфера 47, 70, 71, 76, 307, 308, 311,
312, 324
Биоты 72, 316
Биоценоз 318
Биохории 72
336
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Близнецы
идентичные 133
неидентичные 133
В-лимфоциты 225, 226, 231, 233, 234
Б лот-гибридизация 141
Болезни 25
моногенные 135
полигенные 136
Болезнь Паркинсона 98
Брадикинин 210, 212
Брачное поведение 132
Бэнд 138
В
Вакуоль 8
Вазопрессин 247, 248
Вента-Холодного теория 288
Вестибулярный аппарат 257
Виброрецепторы 53
Виброрецепция 53
Виброчувствительность 53
Вид 7, 39, 40
Видовое разнообразие 47
Видообразование 39, 40, 46, 47
аллопатрическое 41
симпатрическое 41
Вилочковая железа, см. Тимус
Виментин 163, 169
Вирионы 21, 237
Вирус
гриппа 63
иммунодефицита человека 63, 67
простого герпеса 237, 238
Вирусные инфекции 225, 237, 238
Вирусология 9
Вирусы 9, 21, 66, 82, 235, 236, 237,
238
ВИЧ, см. Вирус иммунодефицита человека
Водно-солевой баланс 256
Возбудимость растений 296
Воспаление 209, 210, 214
Воспитание 126
Врожденные дефекты 135
Врожденные пороки развития 136
Вторичноротые 32
Вымирание видов 47
Выносливость 263, 264, 265
Г
Газовые везикулы 308, 309
Галофилы 19
Гаметогенез 111
Гаметофит 121
Гаметофитный тип несовместимости 119,
120
Гаметы 104
у-аминомасляная кислота 248
Гемоглобины 65
Ген 63, 64, 79, 137
Генеалогические связи 7
Генетика поведения 125, 142
Генетика
популяций, см. Популяционная генетика
экологическая 326
Генетическая токсикология 326, 329
Генетически-активные факторы 330, 331
Генетические процессы в популяциях 327
Генетический груз 135
Генетический дрейф 152
Генетический код 79
Генетический критерий вида 43
Генетический полиморфизм 150, 151
Генетический фон 136
Генетическое разнообразие 155, 160
Генетическое консультирование 136
Генная инженерия 274, 277, 293, 294
Геном человека 136, 137, 142, 143, 147
«Геном человека», программа 135, 137,
143
Геном 25, 43, 45, 46, 200
хлоропластный 88, 89, 91
ядерный 88, 89, 91
Геномные исследования 135, 137, 142
Генотерапия 137, 142
Генотип 119, 121, 124
Генофонд 136, 150, 155
Гены 99, 100, 104, 125, 136, 217, 225,
231, 236
иммунного реагирования 227
с материнским эффектом 99
Геохронология 7
Гепатотоксины 310, 312
Гербициды 309
Гесперорнисы 75
Гетеростилии 119
Гетеротрофные организмы 322, 324
Гетерохроматин 81
Гетероцисты 309
Гиббереллин 288, 290
Гибридизация
in situ 138
межвидовая 43
Гибридома 221, 222
Гинецей 119
Гипертермофилы 18
Гипоксантин 222
Гипоталамо-гипофизарно-адреналовая
система 244
Гистамин 211, 212
Гистоны 63
Гиф 315, 321
Главный комплекс гистосовместимости
225, 237
Гликозиды 301, 303
сердечные 302, 303
стероидные 303
тритерпеновые 303
Гликолиз 262, 263
Гликопротеиды 225, 228
Глобины 63, 64, 66
Глюкокортикоиды 245, 246, 247, 252
Гомеодомен 102, 103
Гомеостаз 243, 247, 252
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
337
Гомология 271
Гонадный бугорок 105, 106
Гонадогенез 104, 111
Гонадотропин-рилизинг-гормон 245
Гонады 105
Гоносомы 106
Гормоны 190, 252
Грамотрицательные бактерии 308
Гранулоциты 231, 233, 235
Граны 85, 91, 280, 281*
Грензимы 238
Грибница 315
Грибы 13, 314
Д
Двигательная система 256
Деадаптация 250, 251
патологическая 251
физиологическая 251
Деполяризация мембраны 297
Десмин 163
Детерминантная группа 220
Детерминация пола 105, 108, 112
Дефосфорилирование 186
Диагностика 135, 142
Диапедез 209, 211, 212
Динеин 168
Динорфин 248
Диоксины 328
Диски 271
Дистилия 119
Дифференциация клеток 99
ДНК 63, 70, 79, 90, 99, 127, 137, 138,
140, 143, 144, 145, 146, 147, 177,
330
митохондриальная 147, 148
ядерная 147
ДНКаза 191
ДНК-диагностика 140, 141
ДНК-зависимая ДНК-полимераза 23, 24
ДНК-зависимая РНК-полимераза 23
ДНК-полимераза 144
Докембрий 69
Домены 17, 26, 64, 83, 101, 180, 181
Доминантность 121
Доядерные организмы 8, 9
Дрейф генов 83
Дрозофила 94, 96, 97, 99, 101, 104, 112,
128
Дупликации 83
Дыхание 27
Е
Естественный отбор 83, 152
направленный 153
стабилизирующий 153
Естественный тренд 76
Jit
Животные 8, 13
многоклеточные 14
настоящие многоклеточные 14
радиально-симметричные, или
двуслойные 15
двустороннее-симметричные, или
трехслойные 15
примитивные многоклеточные 14
простейшие, или одноклеточные 14
3
Заболевания 25, 135
Зародышевые листки 15
мезодерма 15
эктодерма 15
энтодерма 15
Зародышевый мешок 118
Зеатин 294
Зиготы 100, 104
Злокачественная опухоль 189
Змеиный яд 219
Зонд 141
Зрение 131
И
Иглокожие 33
Изменчивость 326
Изоляция репродуктивная 43, 44
Изопреноиды 302
Изоферменты 96, 151
Иллегитимный вариант опыления 119
Иммунитет 209, 214, 215, 218, 225, 231
врожденный 214
неспецифический 214
приобретенный 214, 215
Иммунный ответ 225, 227, 232, 233
Иммуноглобулины 220, 225, 227, 234,
235
Иммунокомпетентные клетки 225
Иммунологическая память 215
Инактивация генов 102
Инбридинг 153
Инвазия 204
Инверсия пола 106, 107
Инсектициды 327, 330
Интеллект 133
Интерлейкины 212, 232, 234
Интерферон 234, 236, 238
Интерфертильность 121
Интрастерильность 121
Интроны 80, 86, 100, 137, 321
Инфекция 231, 232, 235, 236
Информационная РНК (иРНК, мРНК) 63,
79, 85, 86, 94, 95, 96, 127, 138, 272,
274, 276
Ионы
калия 299
кальция 299
хлора 299
Ископаемое топливо 323
Искусственная хромосома 139
338
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
К
Калликреин 210
Каналы
калиевые 299
кальциевые 299
хлорные 299
Канцерогены 329
Карбоксисома 308
Карденолиды 302, 303
Кардиолипин 28
Кариамы 75
Каротиноидные пигменты 19
Карта
контиг 138
макрорестрикционная 138
хромосомная 138
Картирование 136, 137, 138, 139
Катаболизм 252
Катехоламины 244, 245, 247, 253
кДНК 138
Кембрий 71
Кератин 169
Киназа 192, 193, 204
Кинезин 168
Кинины 210
Кислород 309, 311
Кишечнодышащие 31
Кладистика 7
Кладогенез 40
Клетка 8, 24, 165, 171, 185, 186, 201,
202, 204, 235
Клетки
прокариот 26
Сертоли 105, 108
эукариот 26
Клетки-киллеры 238
Клетки-убийцы 237
Клеточная дифференцировка 94
Клеточная специализация 94
Клеточные органеллы, см. Органеллы
Клон 190, 218, 221
Кодирующие макромолекулы 63
Ко доминантность 121
Кодон 80
Кодон-инициатор 82
Кодон-терминатор 82
Коллагеновое волокно 201, 202
Кольчатые 33
Коммуникация 48
Компенсация дозы генов 114
Комплемент 210, 211, 212, 219
Консенсусы 82
Концепция вида
биологическая, см. популяционная
популяционная 40, 41, 42, 43
типологическая 40, 41, 42, 43
Кора головного мозга 126
Кортикотропин-релизинг-гормон 244
Космиды 139
Космос 255
Костная система 256
Костный мозг 225
Коэволюция 67
Коэффициент интеллектуальности (IQ) 133
Кремни 69
Кренархеоты 17, 18
Криптоэндолические сообщества 313
Кровь 256, 257
Круглоротые 65
Крыло насекомого 56
Крысы 125
Культивирование клеток 304
Л
Ламеллы стромы 85, 91
Леггемоглобины 64
Легитимный вариант опыления 119
Лейкоз 189
Лейкоциты 209, 238
Летательные аппараты 50
Лимфоциты 214, 216, 217, 218, 233, 233
Липиды 21
Лишайники 313
Лобовое сопротивление 55
М
Макрофаг 185
Макрофаги 209, 212, 225, 226, 231, 232,
233, 235
Матка 105
Матрикс 202
Мейотическая рекомбинация 138
Мембраны 17, 85, 308
Метаморфоз 191
Метан 19
Метапопуляция, см. Популяционная
система 153
Метастазирование 204
Микориза 317, 318
Микробы 231, 232, 233
Микроритмы 297
Микротрубочка 163, 167, 171, 178
Микрофиламент 163, 164, 171
Миоглобины 65
Миозин 164, 165, 261, 262
Митоз 178
Митотический цикл 191, 203
Митохондрия 8, 26, 64, 87, 272
Мицелий 315, 316
Млекопитающие 105
Множественные диспергированные гены
(мдг) 81
Мозг 33, 126
Молекулярная генетика 143
Молекулярная эволюция 63
Моноклональные антитела 220, 221, 222, 223
Моноклональный иммуноглобулин 221
Мономорфизм 44
Моноциты 209, 211, 212, 231, 233
Морфогенез 171, 176
Морфогенетическая реакция 201
Морфогены 99
Морфотип 119
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
339
мРНК, см. Информационная РНК
Мультифакториальные заболевания
(МФЗ) 136
Мутагены 329, 331
Мутанты 79, 102
Мутации 63, 79, 102, 135, 137, 189, 192
нейтральные 83
спонтанные 326
индуцированные 327
Мутационный груз 135
Мышечные волокна 261
Мыши 106
Мышцы 164, 261
Н
НАДФ, см. Никотинамидадениндинуклео-
тидфосфат
Надцарство 29
Насекомоядные растения 296
Насекомые 49, 56, 68, 197, 329
Наследственная изменчивость 327
Наследственность 125, 326
Наследственные болезни 135
Невесомость 255, 256
Нейронные ансамбли 128
Нейроны 50, 127, 171, 174, 238, 239
Нейропептид «Y» 245, 248
Нейротоксины 310
Нейрофиламент 163
Нейтрофилы 209, 211, 212
Некроз 191
Нервная трубка 32
Несовместимость
гетероморфная 119
гомоморфная 119
Нефтегазоносность 70
Никотинамидадениндинуклеотидфосфат,
(НАДФ, НАДФН, лат. NADPH) 85,
281, 285
Норадреналин 244, 245
Норма реакции 126
Нуклеаза 186
Нуклеиновые кислоты 21
Нуклеомера 181
Нуклеосома 180, 181
Нуклеотидные последовательности 63,
82
Нуклеотиды 63, 147
О
Обезвоживание 278
Оболочники 31
Обоняние 131
Обучение 128
Общий адаптационный синдром 243
Оксигенный фотосинтез 309
Онкобелок 189, 190
Онкоген 189
Онтогенез 128
Ооциты 99
Оператор 80
Оперон 82
Опиоидные пептиды 245, 250
Опсонизация 211
Опухолевая клетка 175, 176, 189, 201
Опухолевая трансформация 190, 206
Опухоль 188, 205
злокачественная 189
Органеллы 26, 85, 90, 91
Осадочные породы 311
Осмотрофный способ питания 314, 315
Остеобласты 257
Остеокласты 257
Открытые рамки считывания 82
П
Палеонтология 69
бактериальная 69
докембрия 69
молекулярная 70
Палеобиогеография 72
Память 128
Пандемии 66
Паразиты 195, 316
Парарептилии 75
Парниковый эффект 323
ПДРФ (полиморфизм длин рестрикцион-
ных фрагментов) 137
Первичное определение пола 104
Первичноротые 32
Пёрсистенция вируса 237
Персистирующий вирус 239
Перфорины 238
Пестик 118, 119
Пестициды 328, 330
Пигментная ксеродерма 329
Питающие клетки 99
Пищевые цепи 327, 328
Плазмалемма 26
Плазмодесмы 298
Плазмоцитомы 221
Планктон 312
Пластиды 85, 91
Пластохинон 285
Пластоцианин 285, 286
Плацентарные млекопитающие 106
Поведение 125, 126
Погонофоры 33
Подъемная сила 58
Позвоночные животные 65, 66
Позиционная информация 99
Покрытосеменные 118
Пол 104, 109
детерминация 105, 108
дифференциация 110
инверсия 107
определение 111
первичное определение 104
фенотипический 104
Полет насекомых 55
Полимеразная цепная реакция (ПЦР) 70,
144, 145, 147, 148
Полиморфизм 44, 46, 150, 151
340
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Полипептид 225, 228
Полиплоидия 41, 42
Полисомы 272
Политенные хромосомы 271
Полиэтиленгликоль 221
Половое размножение 104
Половые хромосомы 104
Полухордовые 33
Поляра Лилиенталя 57
Популяционизм 40, 47
Популяционная генетика 150
Популяционная система 153
Популяция 135
панмиктическая 152
подразделенная 152
Постулат Чаплыгина-Жуковского 57
Потенциалы действия 296, 297
Правило Харди-Вайнберга 152
Праймер 147
ПремРНК 277
Приматы 68
Принцип актуализма 40
Проводящие пучки 297, 298
Прогенот 17, 26
Прогона ды 105
Прокариотическая клетка 308
Прокариоты 8, 9, 10, 17, 26, 82, 195,
307, 311
анаэробные 26
аэробные 26
Пролиферация 191
Промежуточный филамент 169, 171
Промоноциты 232
Промотор 80, 86, 274
Промутагены 330, 331
Проприорецепторы 256
Простагландины 248, 250
Протеаза 186
Протеасомы 275
Протеинкиназы 245, 250
Протоавис 75
Протобионты 26
Протоонкобелок 204
Протоонкоген 189
Процессинг 86
Псевдогены 83
Псевдоподия 165, 166, 171, 201
Психические функции 133
Птероторакс 56
Пуф 271
ПЦР, см. Полимеразная цепная реакция
Пыльники 118, 119
Пыльца 118, 119
Пыльцевая трубка 118, 120
Р-элемент 96
Р
Работа 264
Раздражимость 244
Раздражимость растений 296
Рак 310
Раковые заболевания 187, 188
Растения 8, 11, 118, 301, 304
высшие 12
низшие 11
семенные 13
споровые 12
Растительная клетка 85
Реадаптация 250, 251
Реакции фотосинтеза
световые 281
темновые 85, 89, 281
Ревертаза 24
Регуляторные гены 95
Регуляторные части генов 100, 101
Регуляторные элементы генов 273, 274
Редупликация хлДНК 87
Рекомбинация 327
генов 137
Репарация 327
Репликация 21, 23, 327
Репликон 181
Репортерный ген 274
Репродуктивная изоляция 39, 40, 41, 43,
44, 48
Рецептор 185, 192, 204
Рибонуклеиновые кислоты (РНК) 17, 21,
63, 79, 86, 87, 100, 127, 178, 186
рибосомные (рРНК) 17, 28, 79
транспортные (тРНК) 17, 79, 88
Рибосомы 17, 22, 28
Рибулозобисфосфат 86
Рибулозобисфосфаткарбоксилаза (РБФК)
86, 88, 273
Риккетсии 195
Риккетсиозы 195
Рйпидистии 75
РНК, см. Рибонуклеиновые кислоты
РНК-зависимая РНК-полимераза 23, 24
РНК-полимераза 86
Род 7
Рот 32
Рыбы 48
С
Сайленсеры 84
Самонесовместимые растения 118
Самооплодотворение 118
Самоопылители 118
Самосовместимые растения 118, 123
Сантиморганиды (сМ) 138
Сапонины 303
Сапротрофы 316
Сателлитная, или эгоистическая ДНК 81
Световые стадии фотосинтеза 281
Светособирающая антенна 282, 283
Сегрегационный груз 135
Секвенирование 63, 137
Сенсорные системы 256
Сера 20
Сердечно-сосудистая система 256
Силур 311
Симбиогенез 8, 199
Симбиоз 27, 196, 310, 311
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
341
Симбиозы 317
Симбионты 195, 196, 310, 317
Симпато-адреналовая система 245
Симпласт 298
Синапсы 128
Синдром
Клайнфельтера 113
Тернера 112
приобретенного иммунодефицита 219
Синезеленые водоросли 307
Синергиды 122
Синтез белков 271, 274
Синтез РНК 193
Синтетическая теория эволюции (СТЭ)
40
Синэкология 326, 327
Систематика 7
Сколопидии 52
Скорость эволюции 65
Современный экологический кризис 70
Сочленовная мембрана 50
Сперматогенез 106
СПИД, см. синдром приобретенного
иммунодефицита
Сплайсинг 80, 83, 86
Спорофитный тип несовместимости 121,
122
Спортсмен 261
Способность к обучению 126
Стереоцилия 166
Стратегия адаптации 252, 253
Стресс 243, 244
Стрессорные ситуации 243
Стрессоры 243, 244, 250
STR-повтор 145, 147
Стресс-реакции 244, 245, 252, 253
Стресс-система 244, 245, 247
Строма 85, 91, 280, 281
Строматолиты 10, 311
Структурный след адаптации 249, 250
СТЭ, см. Синтетическая теория эволюции
Субгенуальные органы 52
Субстанция «Р» 248
Судебная медицина 142
Сурфактанты 309
Сцепление генов 137, 140
Т
Ti-плазмиды 293
Таксономическая категория 7
Таксономия 7
Таксоны 7
Таллом 315
Тапетум 121
Тахикины 245
Теломера 178, 184
Темновые стадии фотосинтеза 281
Теорема Жуковского 58
Тепловой шок 271, 272, 276, 277
Тергум 56
Териодонты 75, 76
Термоплазмы 20
Терморегуляция 233
Термофильные цианобактерии 312
Терпеноиды 301, 302
Тест Эймса 331
Тестис 104, 105, 106
Тестисопределяющий фактор 106
Тестостерон 105, 106
Тест-системы 330
Тилакоиды 85, 280, 281, 308
Тимбальные мембраны 50
Тимидин 222
Тимоциты 230
Тимпанальные мембраны 51
Тимпанальные органы 51
Тимус 225, 226, 228, 229
Типологизм 40, 43
Тканеспецифическая экспрессия генов 96
Ткань 201, 202
Т-киллеры 225, 226, 227, 228
Т-клетки 229
Т-лимфоциты 225, 226, 227, 231, 233, 234
Токсины 232, 310, 312
Толерантность 215, 218, 219
Транскрипция 24, 94, 106, 327
Трансляция 83, 94, 327
Трансплантат 215, 216
Трансплантация 225
Транспозиции 82
Трансфактор 272, 274
Тренировка 265
Триплет 79
Тромбоцит 175, 190
Тромбоцитарный фактор роста (ТФР)
174, 190
Т-супрессоры 229
Тубулин 163
Тургорное давление 316
Тучные клетки 211, 212
Т-хелперы 225, 226, 227, 234
У
Убиквитин 275
Условные рефлексы 125
Ф
Фаги 21
Фагоцитирующие клетки 210
Фагоцитоз 27, 209, 212, 214, 215
Фаза (развития фаунистического
комплекса)
роста 72
расцвета 72
упадка 72
Фазы митоза 177
Фактор некроза опухолей 212, 233
Фактор роста 187, 190
Фактор Хагемана 210
Факторы транскрипции 103, 193
Фаллопиевы трубы 105
Фанерозой 71
Феромоны 95, 332
342
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Фибробласт 163, 171, 172, 173, 174, 201,
202
Физиологические функции 255
Физическая работоспособность человека 261
Физическое картирование 139
Фикобилиновые пигменты 309
Фикобилисомы 308
Фикобионт 310
Фикоцианины 309
Фикоэритрины 309
Филамент 202
Филетическая эволюция 40
Филогенетическая приемственность 7
Филогенетические деревья 67
Филогенетический анализ 63
Фингерпринтинг 139
Фитогормоны 288, 290, 292
Фитофтороз картофеля 320
Флавоноиды 301, 304
Фонорецепторы 51
Фороракосы 75
Формообразование 96
Фоссилизация 69, 73
Фосфатаза 193
Фосфоглицериновая кислота 86
Фосфорилирование 186, 274
Фотическая зона 69
Фотоавтотрофия 323
Фотосинтез 9, 26, 85, 86, 89, 92, 275,
280, 281, 308, 309, 310, 322
Фотосистема 281
Фотохимический реакционный центр 281,
283
X
Хемосинтез 9, 26
Хемотаксис 209
Хемофоссилии 70
Хламидии 195
Хлоропласт 85, 86, 87, 88, 91, 92, 280,
284
Хлоропластная ДНК (хлДНК) 86, 87
Хлоропласты 26, 64, 89, 90, 272, 273,
308, 311
Хлорофилл 280
Хорда 32
Хордовые 31
Хс^рея Гентингтона 138
Хроматида 183
Хроматин 177, 186, 271
Хромомер 181
Хромонема 182
Хромопласты 90
Хромосома 177, 178, 181
Хромосомная карта 138
Хромосомные аберрации 331
Хромосомные болезни 136
Хромосомные перестройки 82
Хромосомы 79, 136
человека 143, 144
Х-хромосома 105
человека 144, 145
Y-хромосома 104, 105, 111
человека 144, 145
ц
цАМФ, см. Циклический аденозинмоно-
фосфат
Царская семья 145
Царство 8
Цветки 119
длинностолбчатые 119
короткостолбчатые 119
Целом 32
Центромера 178, 184
Цианобактериальные маты (ЦВМ) 69
Цианобактерии 27, 307, 308, 309, 310,
311, 312, 313
Цианобионты 9, 10
Цикл Кальвина 86, 89, 281
Цикл Кребса 263
Циклический аденозинмонофосфат
(цАМФ) 245
Цитокинины 288, 289, 293, 294
Цитокины 225, 227, 231, 232, 234, 236,
237, 238, 245
Цитоплазма 8
Цитоплазматическая мембрана 26
Цитоплазматическая наследственность 91
Цитоплазматические гранулы 272
Цитоскелет 28, 163, 169, 171, 202
Цитотоксические Т-лимфоциты 237
Цитохром Р450 331
ч
Частота гена 152
Человек 31, 67, 133, 135, 255, 264, 265
Членистоногие 33, 37
ш
Шаперон 88, 275
Шаперонин 89
Шимпанзе 67
э
Эволюционная теория 47
Эволюция 31, 59, 63, 101, 307
генов 82
Эвриархеоты 17, 18
Экзина 119
Экзоны 80, 100, 137, 138
Экзоскелет 49
Экологическая генетика 136, 326
Экологические отношения 326
Экология 326
Эколого-генетические модели 328, 329
Экосистемы 327
Экспрессия гена 95, 96
Экстремофилы 17
Электрический импульс 296, 300
Электронтранспортная цепь 85, 86, 87
Электрофорез 141, 271
Эмбриогенез 101
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
343
Эмбрион 100, 102
Энанциорнисы 75
Эндоплазматический ретикулум 277
Эндосимбиоз 8, 26, 317
Эндоцитобиоз 195
Эндоцитобионты 195
Энкефалины 248
Энхансеры 84
Эпидемии 66
Эпителий 239
Эпителиоцит 202
Эстераза 95
Эстрогены 106
Этилен 288, 291
Этиопласт 91
Эубактерии 64, 82
Эукариотическая клетка 311
Эукариоты 8, 10, 11, 17, 26, 64, 82, 87,
307, 311
Эффект Юри 324
Я
Ядерные организмы 8, 11
Ядро 8, 26, 178, 179
Ядрышковый организатор 178
Яды 310
Яичник 104
Яйцеклетка 118
СОДЕРЖАНИЕ
Эволюция и систематика 5
И. А. Михайлова,0. Б. Бондаренко. Система органического мира 7
Б. В. Громов. Удивительный мир архей 17
В. И. Агол. Разнообразие вирусов 21
И. С. Кулаге. Происхождение эукариотических клеток 26
В. В. Малахов. Происхождение хордовых животных 31
Ю. П. Алтухов. Вид и видообразование 39
Е. К. Еськов. Акустическая связь у насекомых 48
A. К. Бродский. Аэродинамика полета насекомых 55
B. А. Ратнер. Молекулярная эволюция 63
А. Ю. Розанов. Современная палеонтология 69
Генетика 77
C. Г. Инге-Вечтомов. Что такое ген? 79
О. Н. Кулаева. Взаимодействие хлоропластного и ядерного геномов 85
Л. И. Корочкин. Генетический контроль развития клеток 93
И.Ф.Жимулев. Действие генов в раннем развитии дрозофилы 99
A. Ф. Смирнов, И. Ф. Жимулев. Регуляция пола 104
С. И. Малецкий. Гены самонесовместимости цветковых растений 118
Л. И. Корочкин, И. Ф. Жимулев. Гены и поведение 125
B. П. Пузырев. Геномные исследования и болезни человека 135
Н. К. Янковский. Молекулярная генетика и судебная медицина 143
Ю. П. Алтухов. Генетика популяций и сохранение биоразнообразия 150
Клеточная биология 161
Ю. М. Васильев. Подвижная архитектура клетки 163
Ю. М. Васильев. Механизмы перестройки цитоскелета 171
Ю. С. Ченцов. Организация митотической хромосомы 177
В. И. Агол. Генетически запрограммированная смерть клетки 185
Ю. М. Васильев. Нормальные и опухолевые клетки 189
Б. В. Громов. Эндоцитобионты клеток животных 195
Ю. М. Васильев. Клетки и построение ткани 201
Иммунология 207
Г. И. Абелев. Врожденный иммунитет. Воспаление 209
Г. И. Абелев. Основы приобретенного иммунитета 214
Г. И. Абелев. Моноклональные антитела 220
В. Г. Галактионов. Генетический контроль взаимодействия иммунокомпетентных клеток. 225
И. С. Фрейдлин. Клеточные механизмы иммунной защиты организма 231
И. С. Фрейдлин. Защита от вирусов с помощью клеток-убийц 237
Физиология человека и животных 241
М. Г. Пшенникова. Стресс и адаптация 243
В. И. Кулинский. Две стратегии выживания организма 252
О. Г. Газенко, А. И. Григорьев, А. Д. Егоров. Адаптация человека к невесомости 255
Н. И. Волков. Физическая работоспособность человека 261
Физиология и биохимия растений 269
О. Я. Кулаева. Стрессовые белки 271
A. Н. Тихонов. Фотосинтез 280
О. Н. Кулаева. Гормональная регуляция жизни растений 288
B. А. Опритов. Электрические сигналы у высших растений 296
В. А. Пасетниченко. Биологически активные соединения растительного происхождения. 301
Экология 305
Б. В. Громов. Цианобактерии в биосфере 307
Ю. Т. Дьяков. Грибы и их значение в жизни природы и человека 314
B. В. Климов. Растения и биосфера 322
C. Г. Инге-Вечтомов. Экологическая генетика: новый взгляд 326
Предметный указатель 333