




























































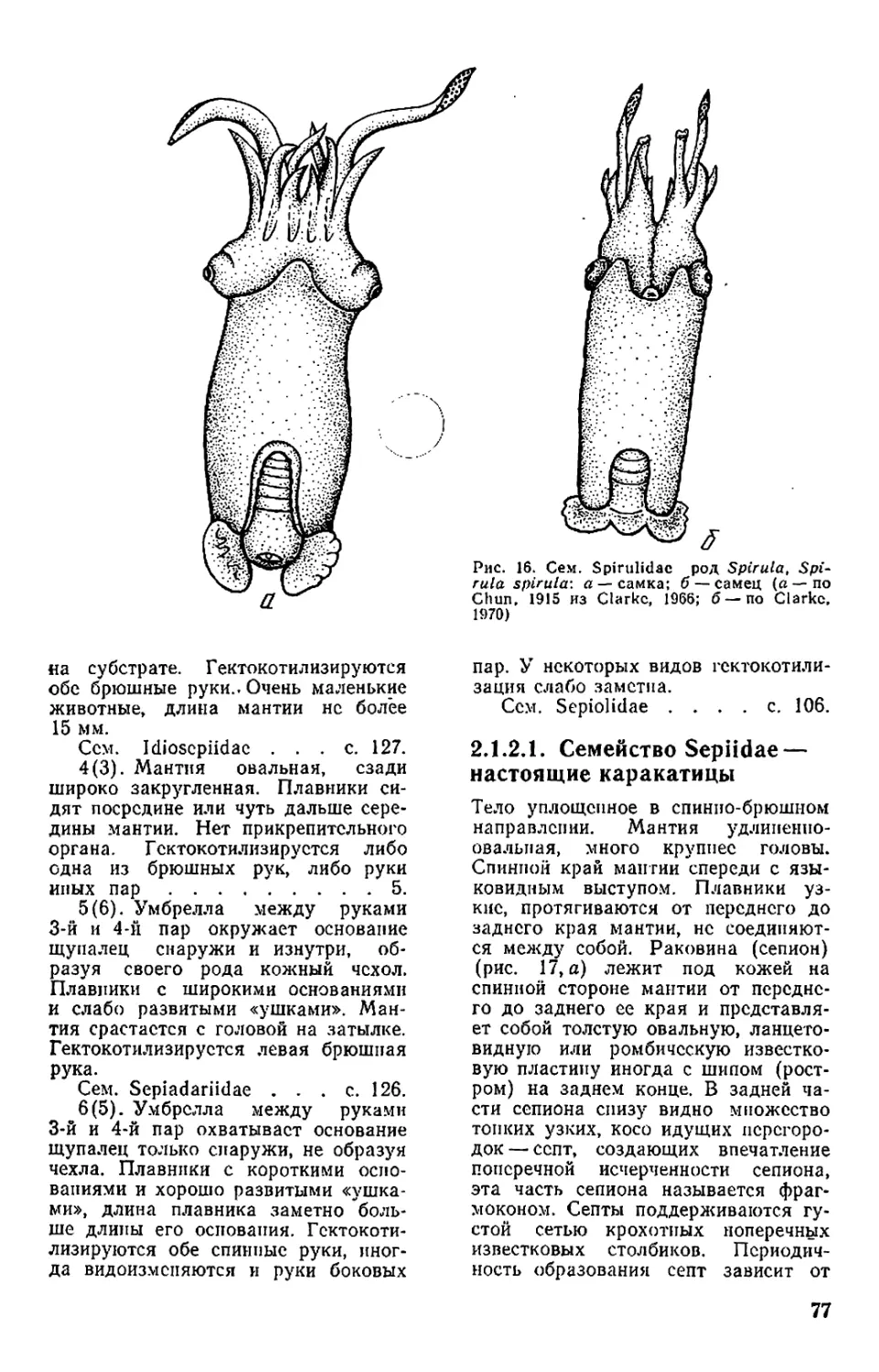

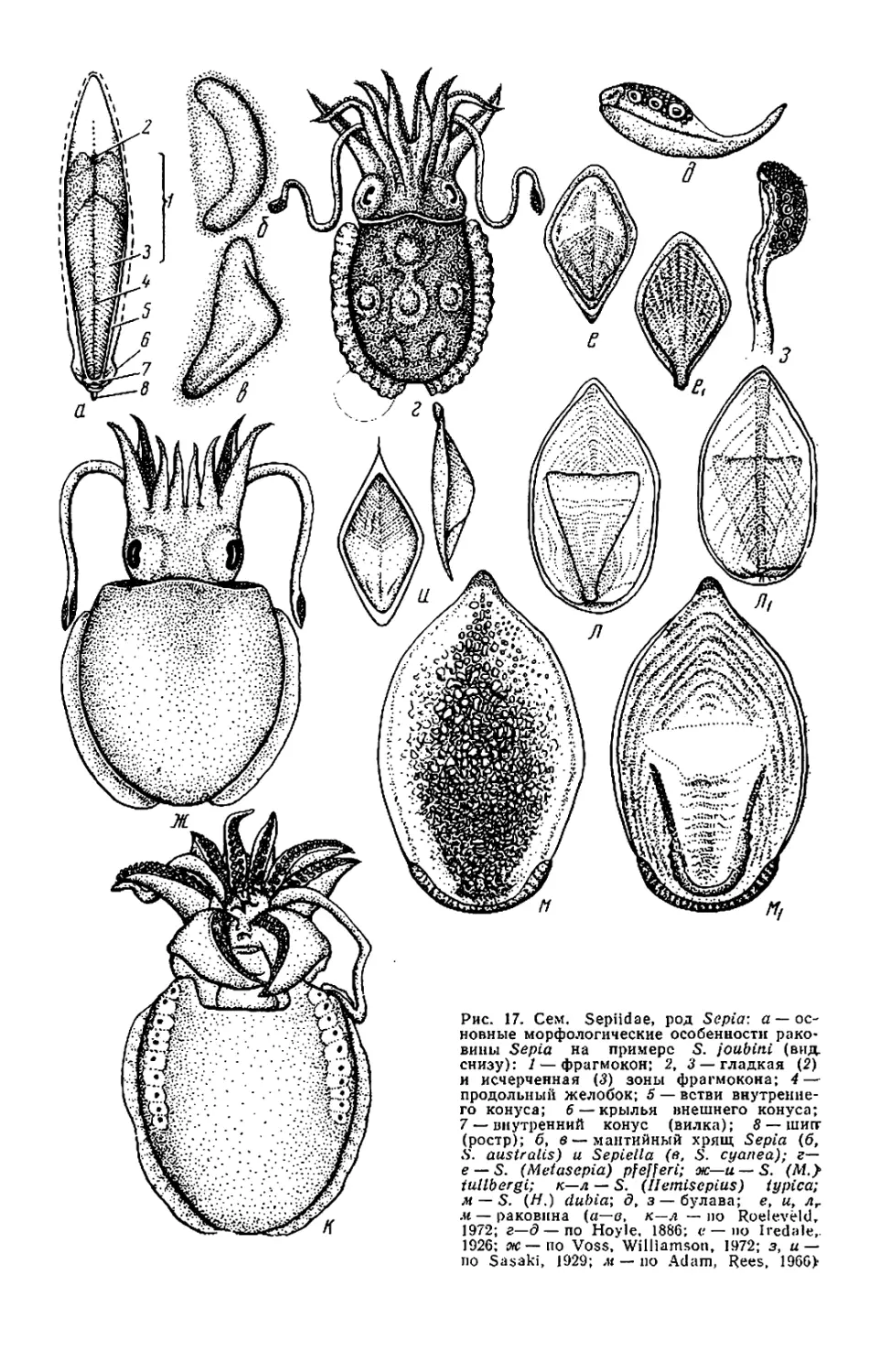


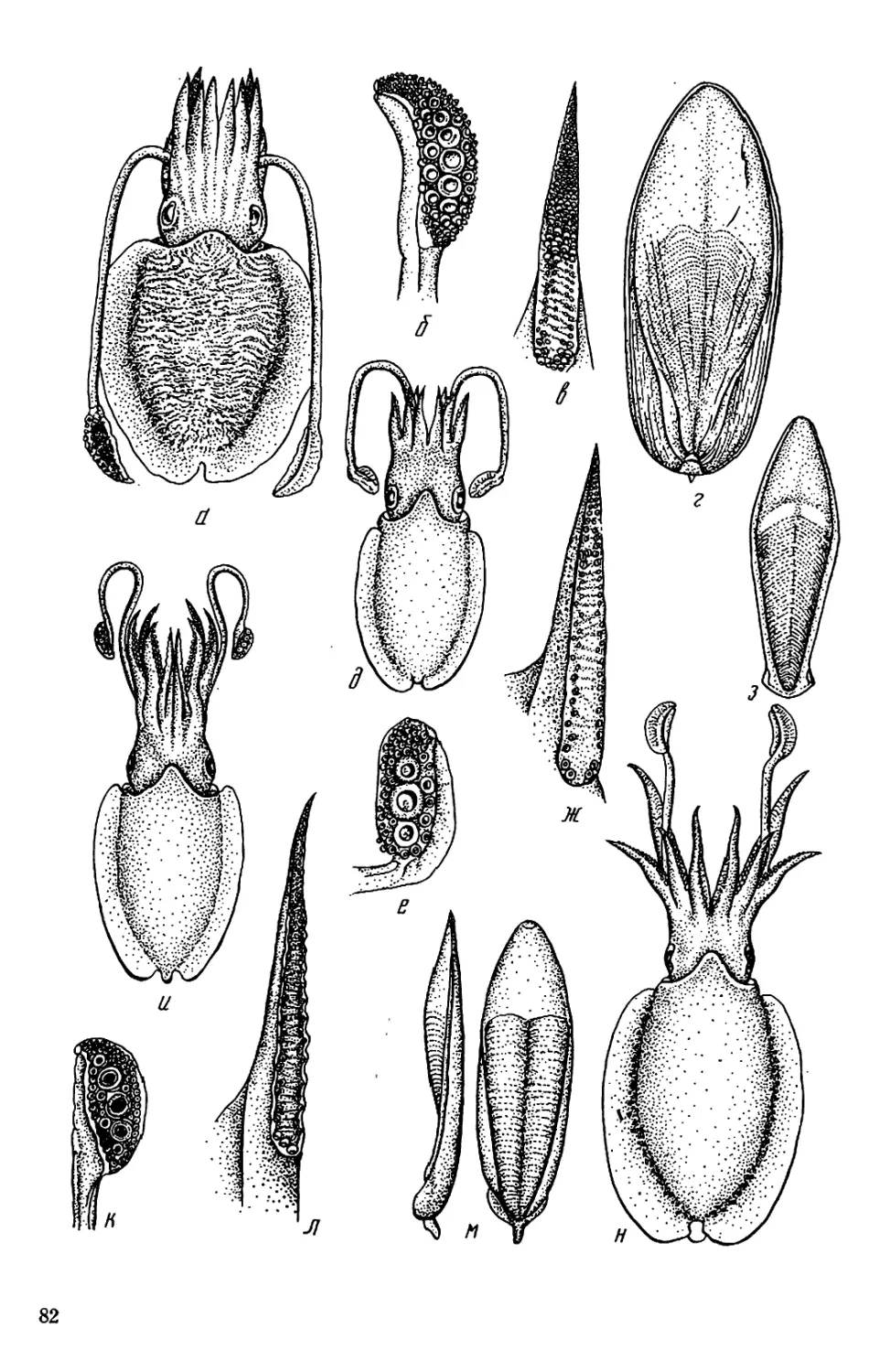
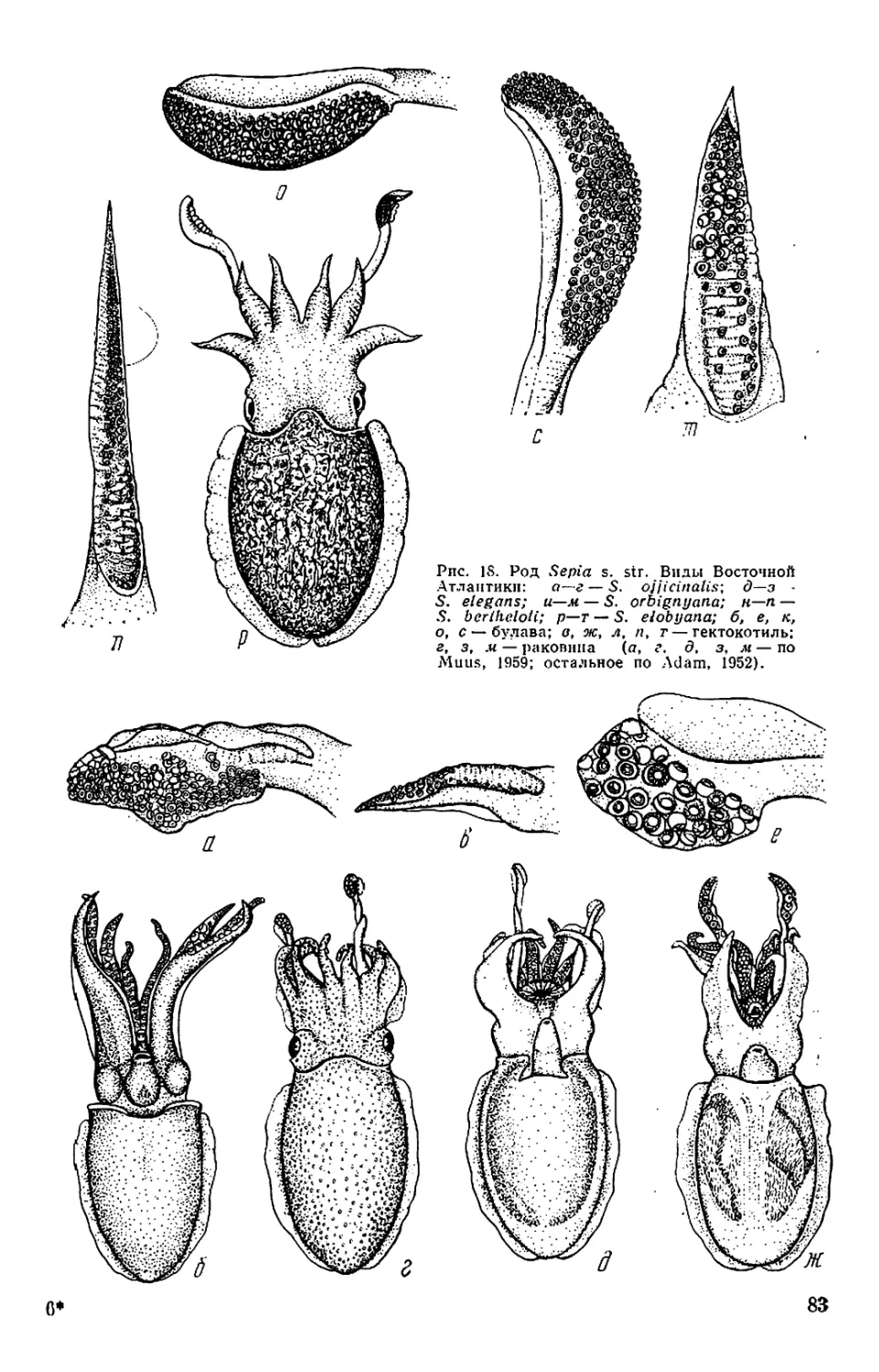
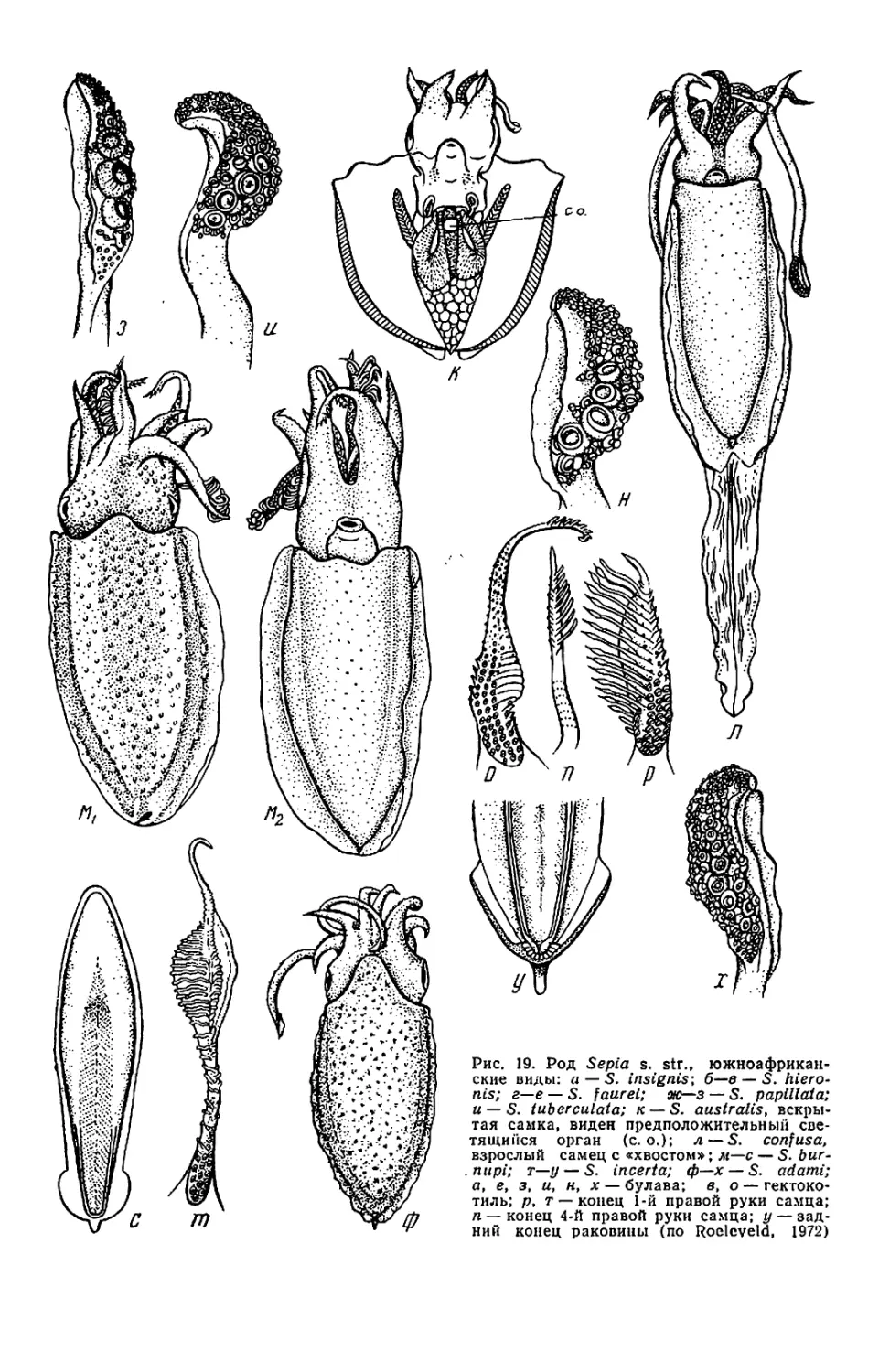




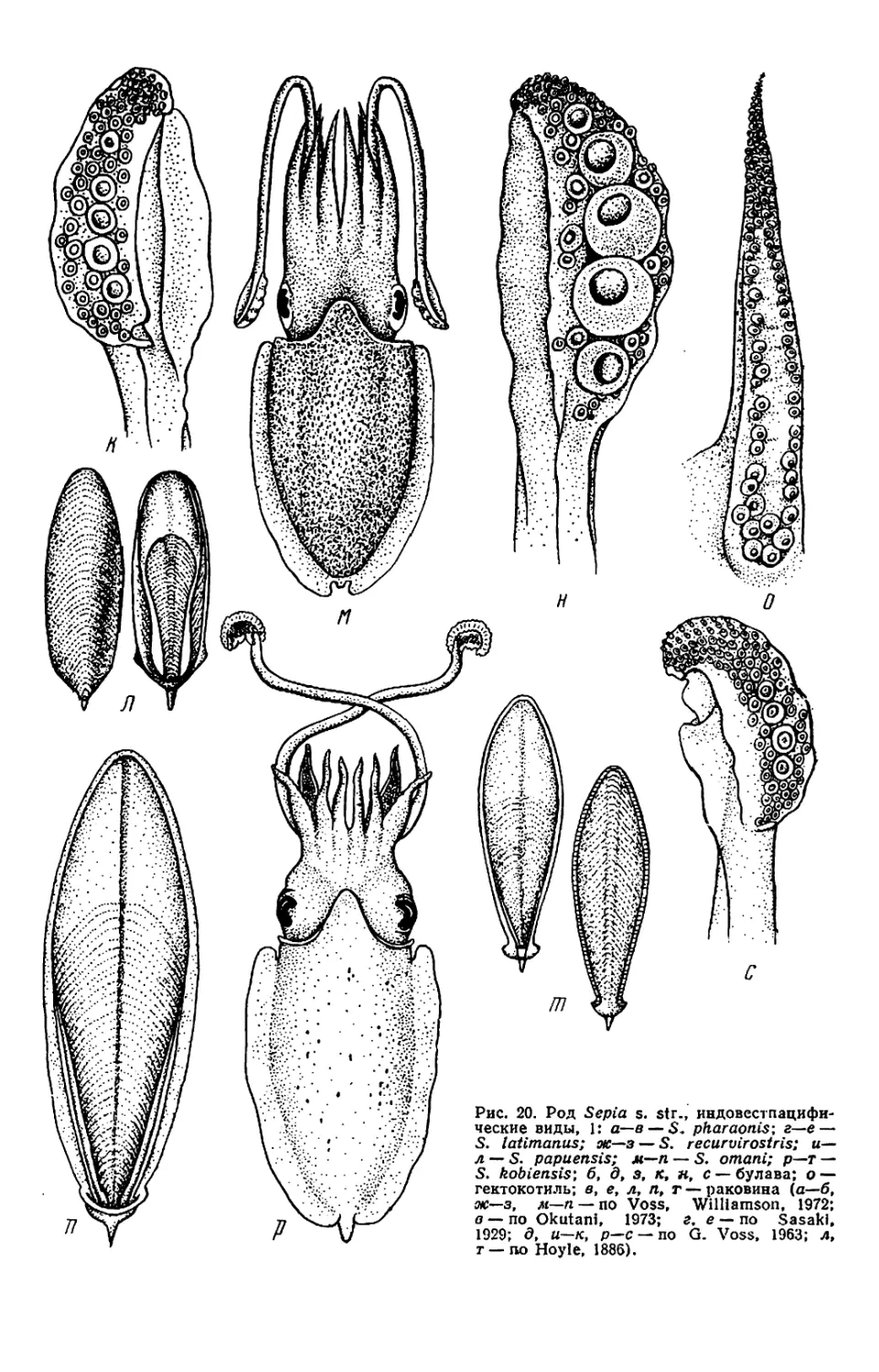


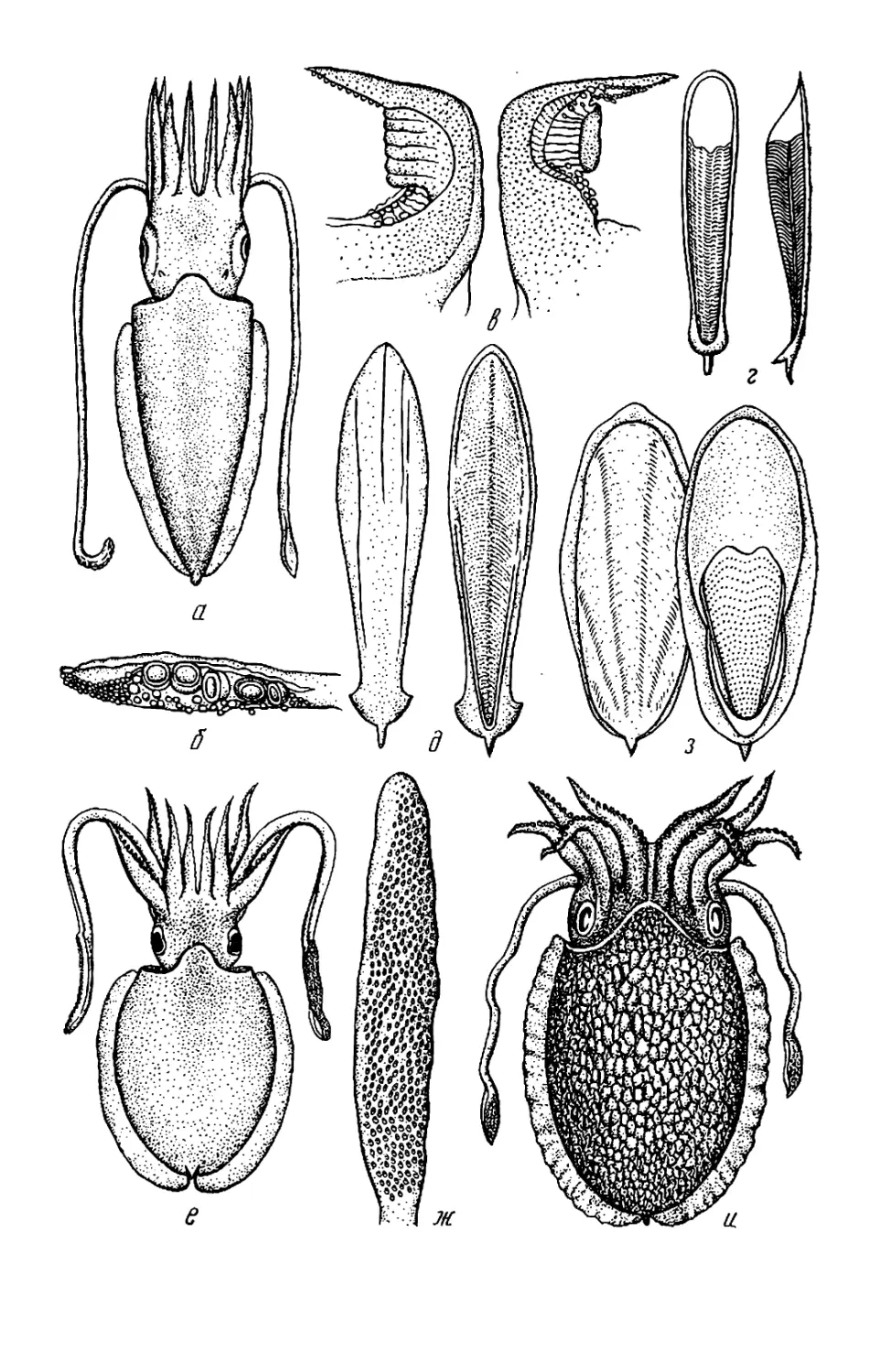



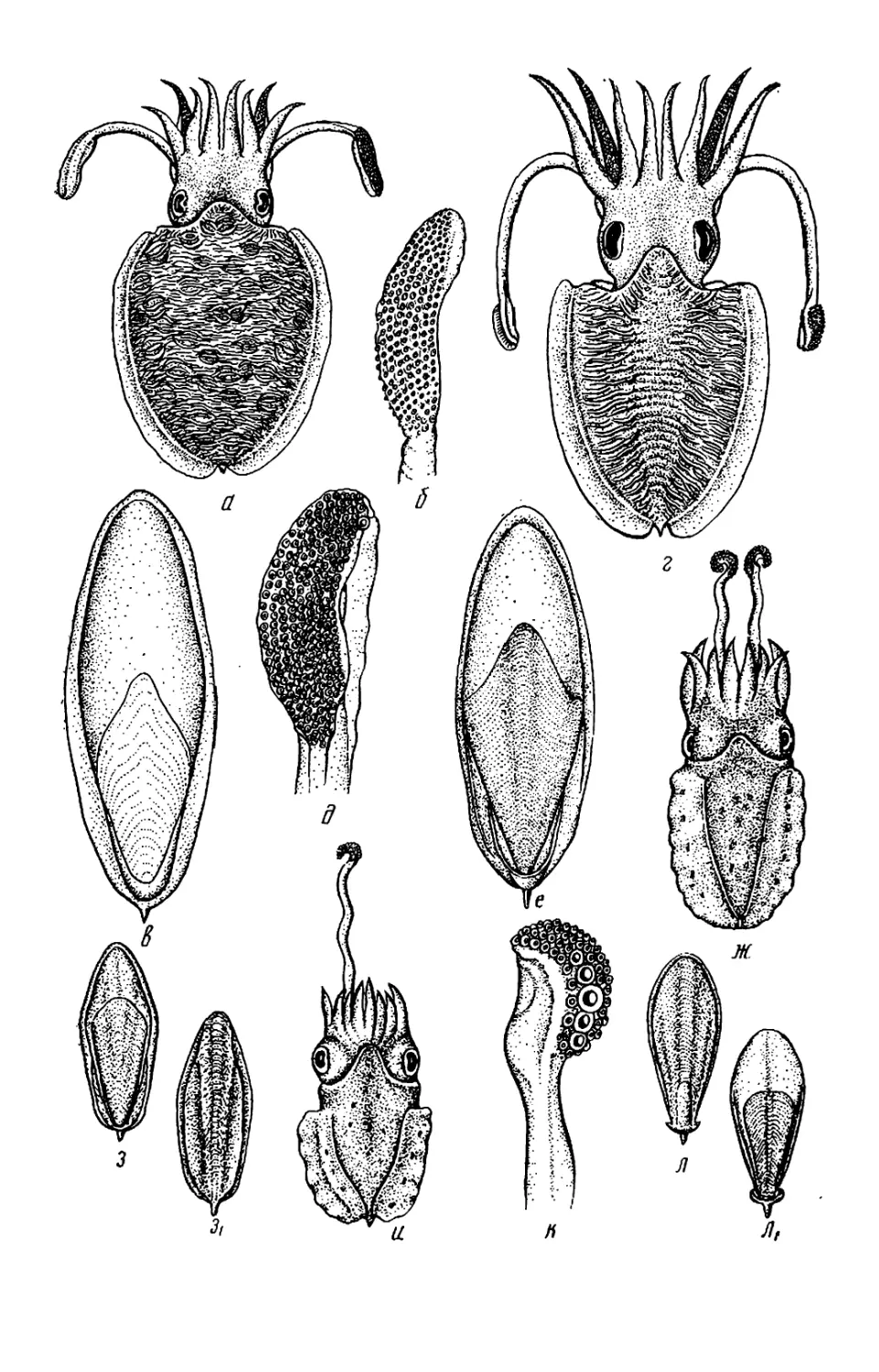


























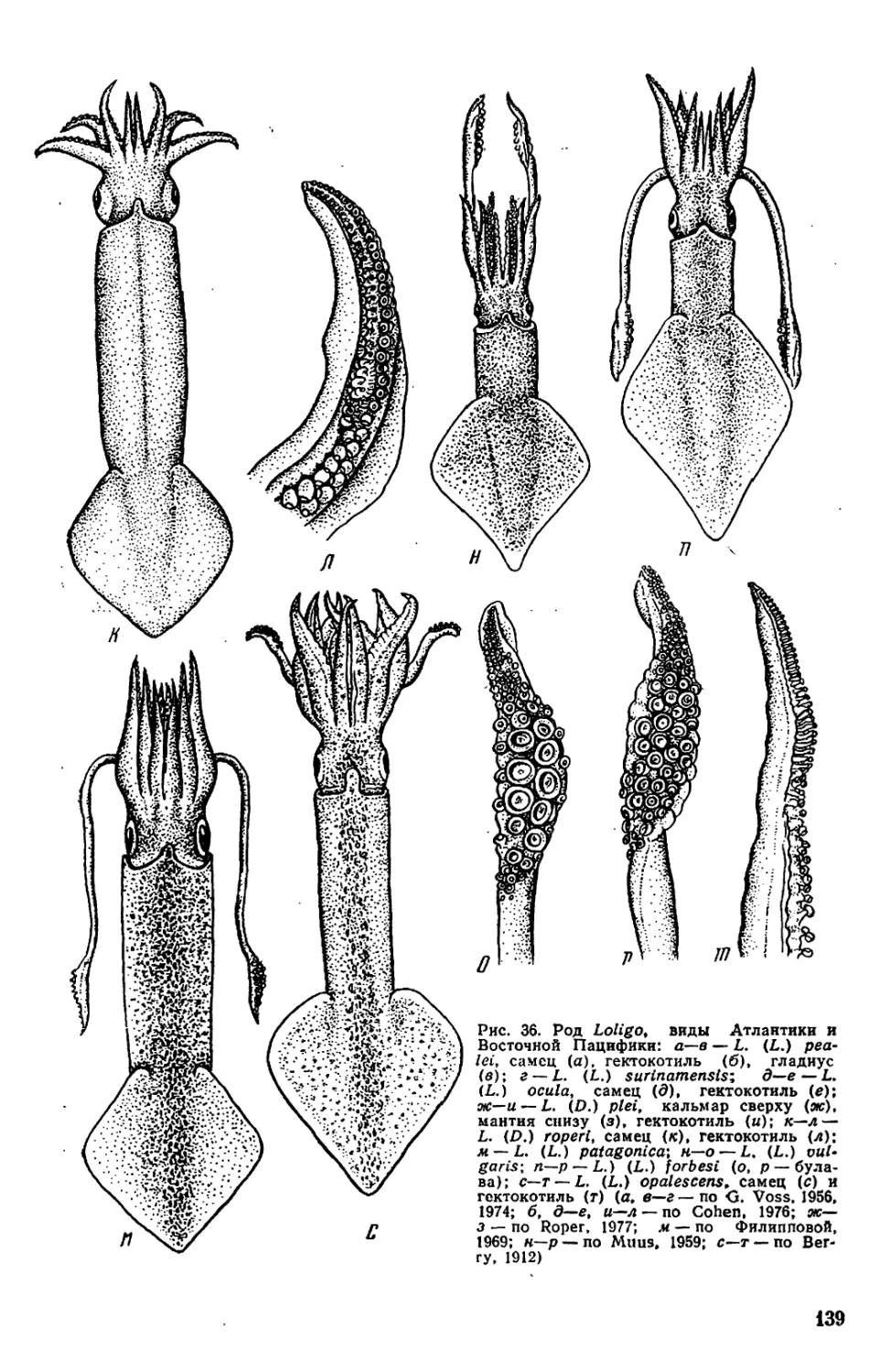


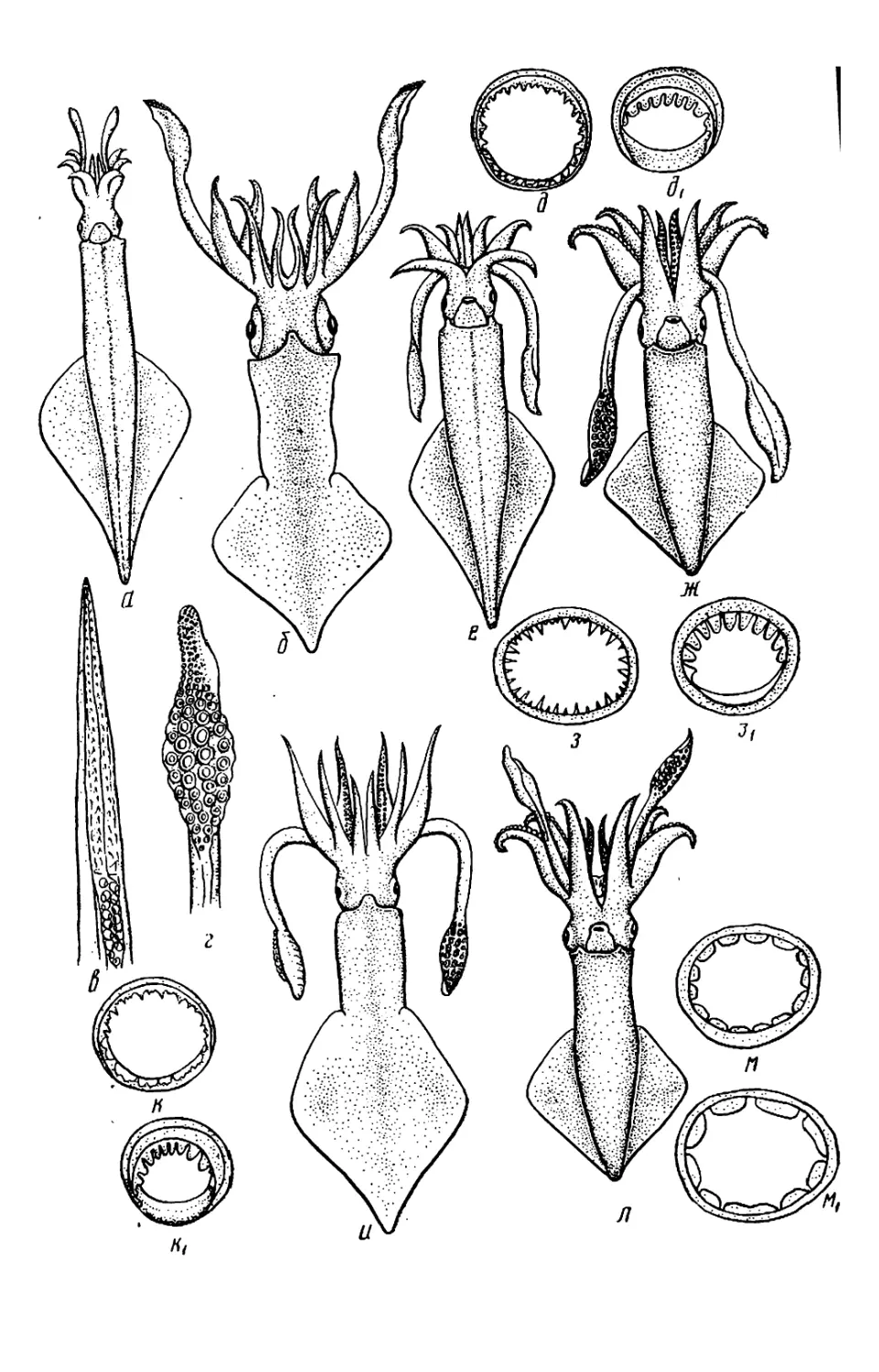
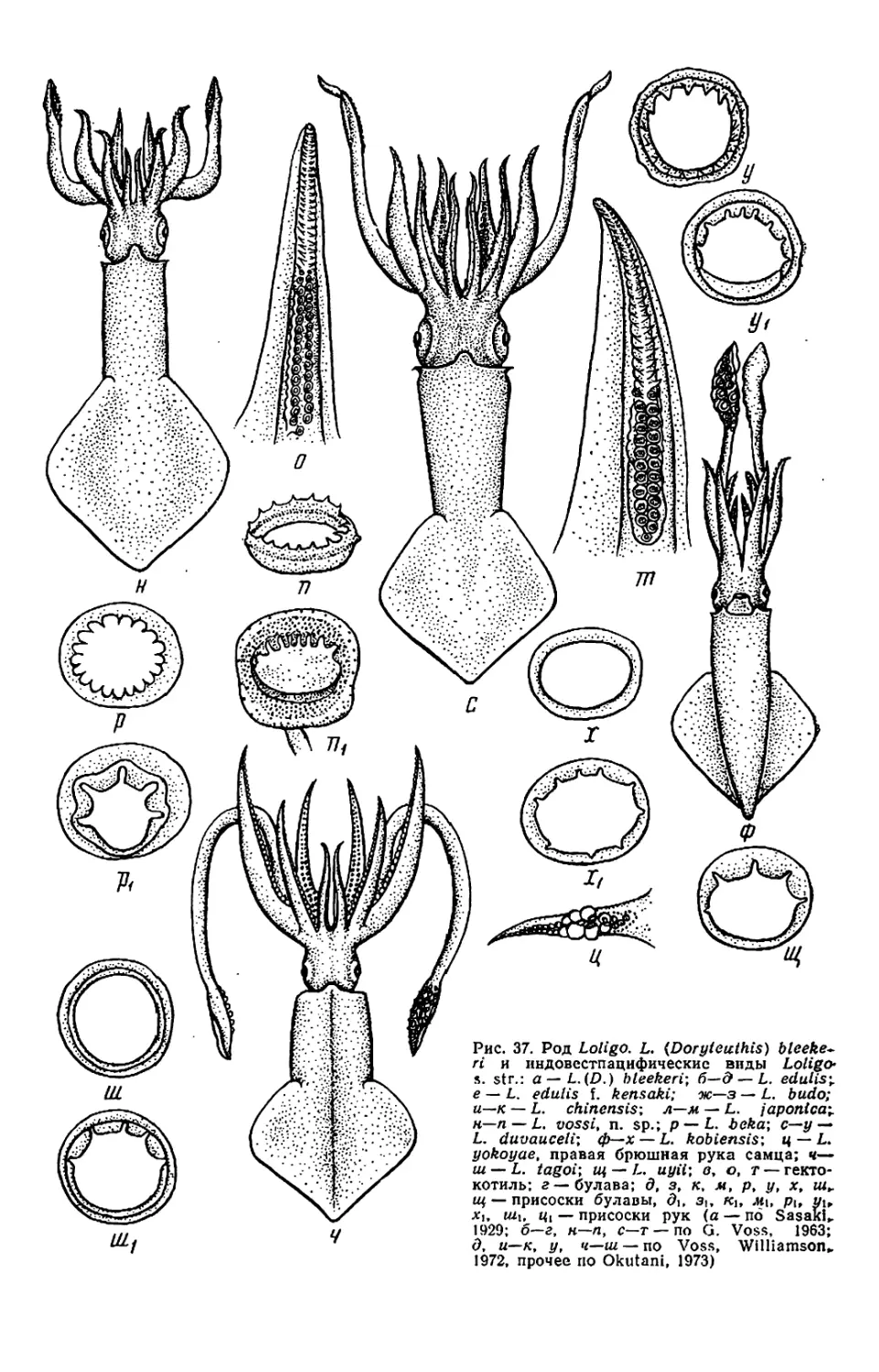







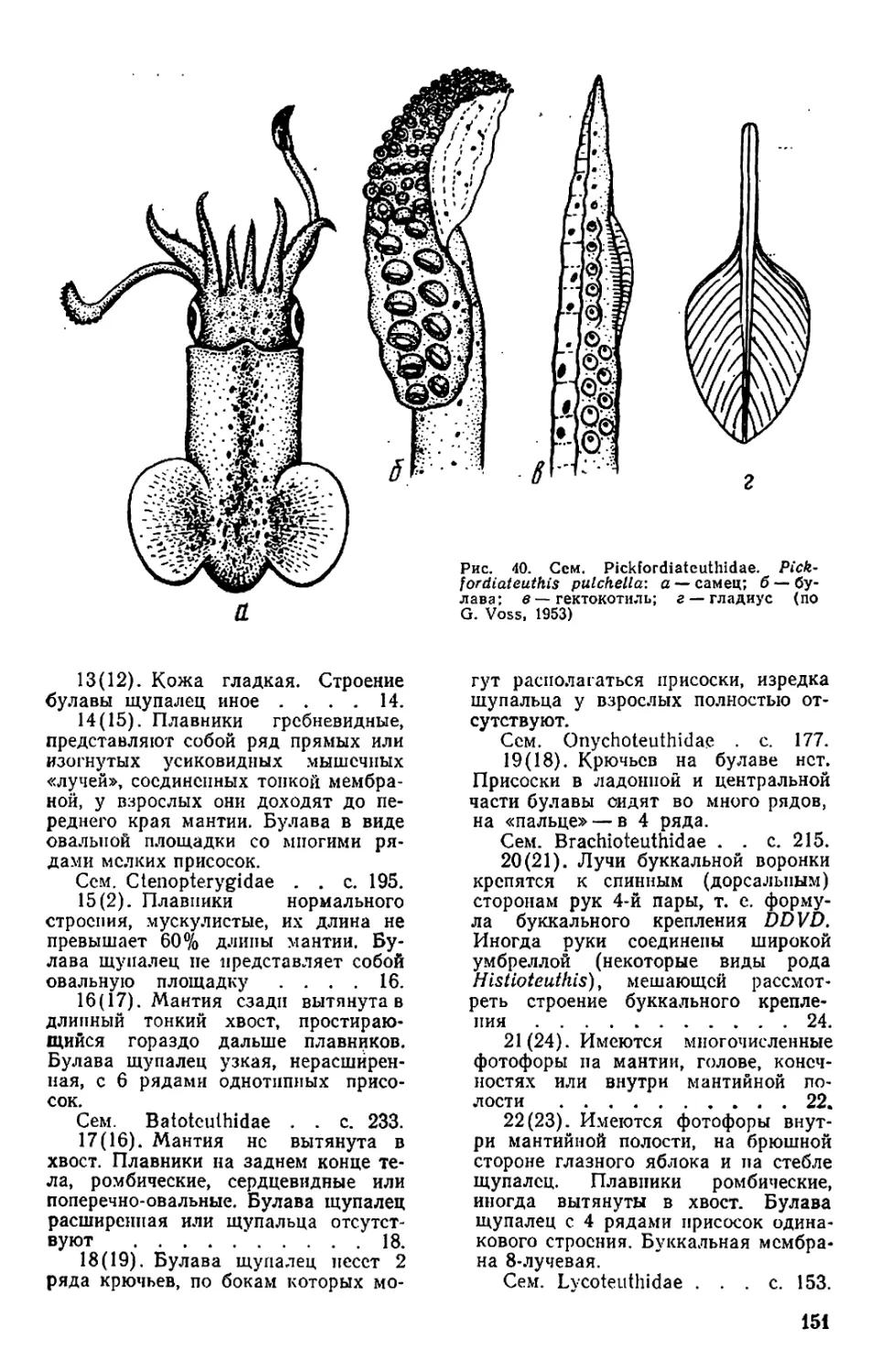



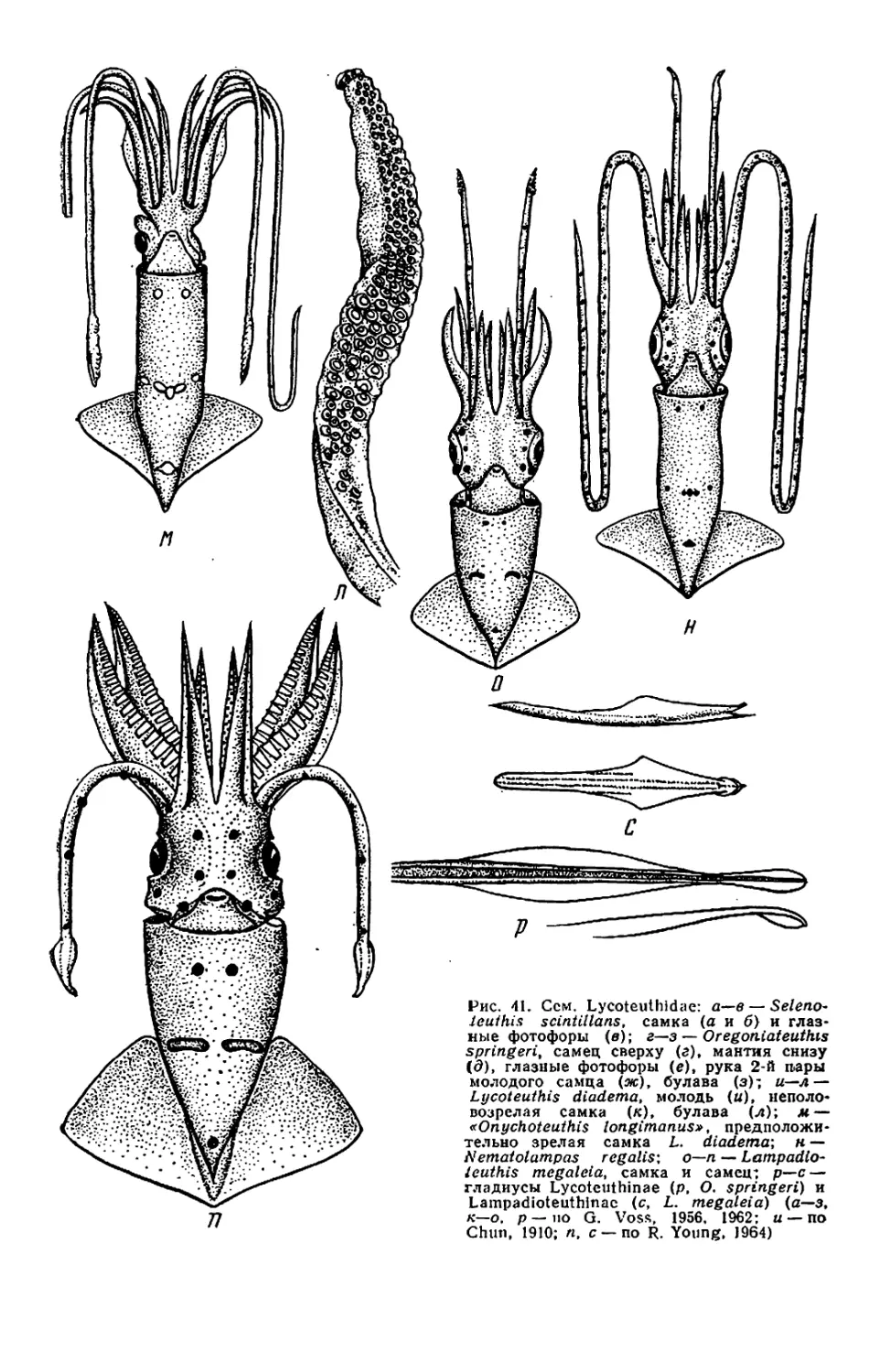




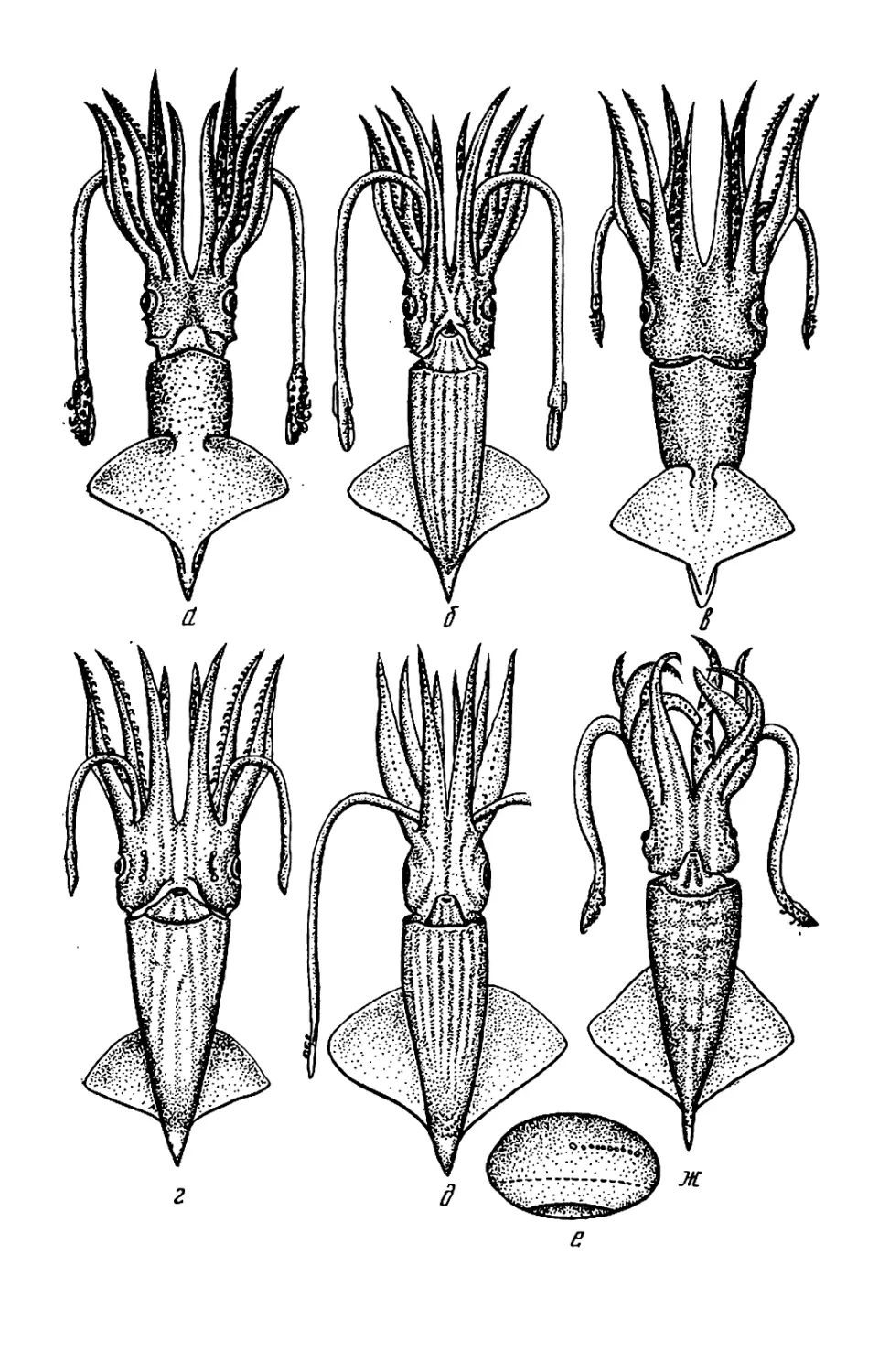
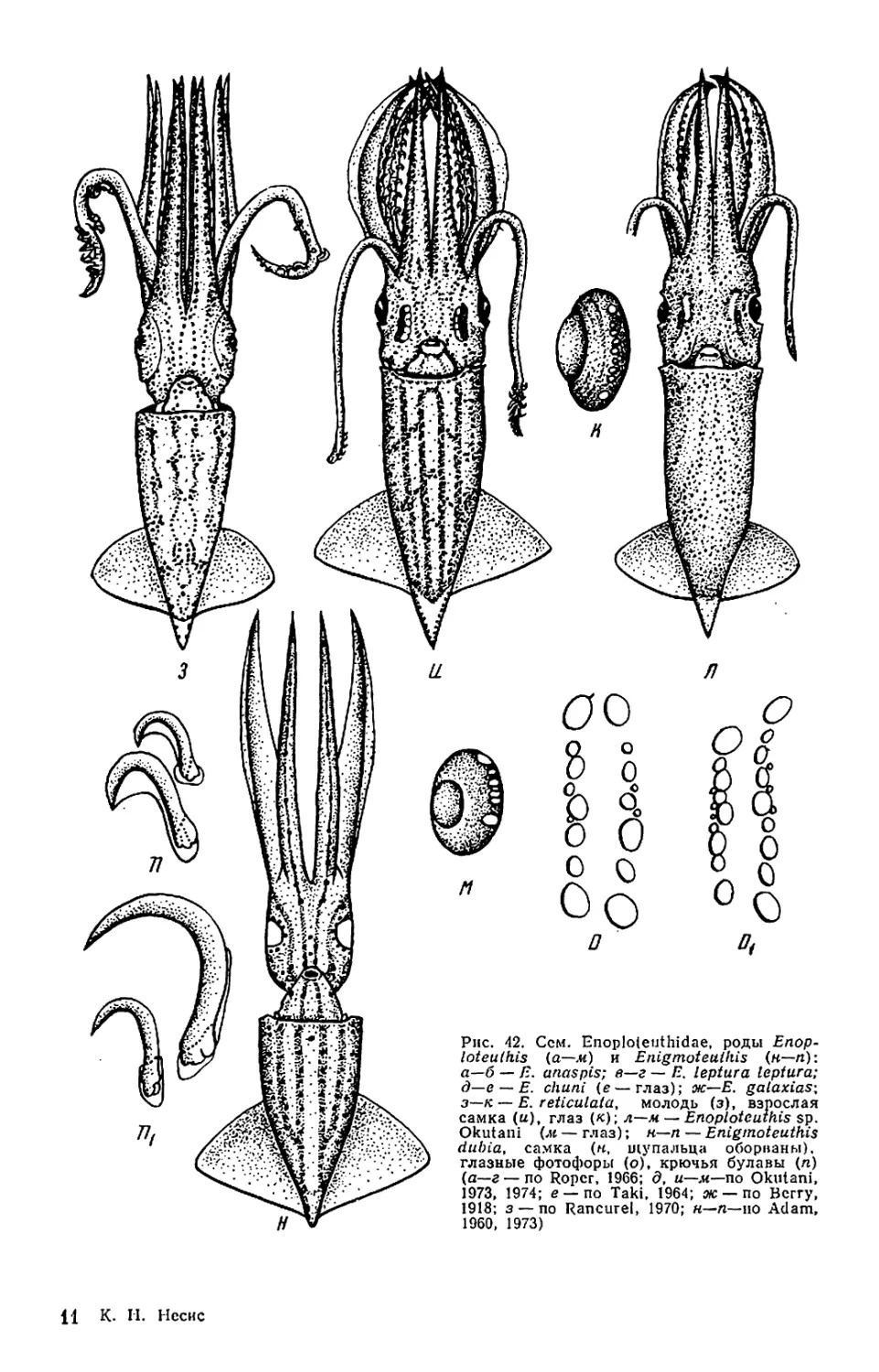


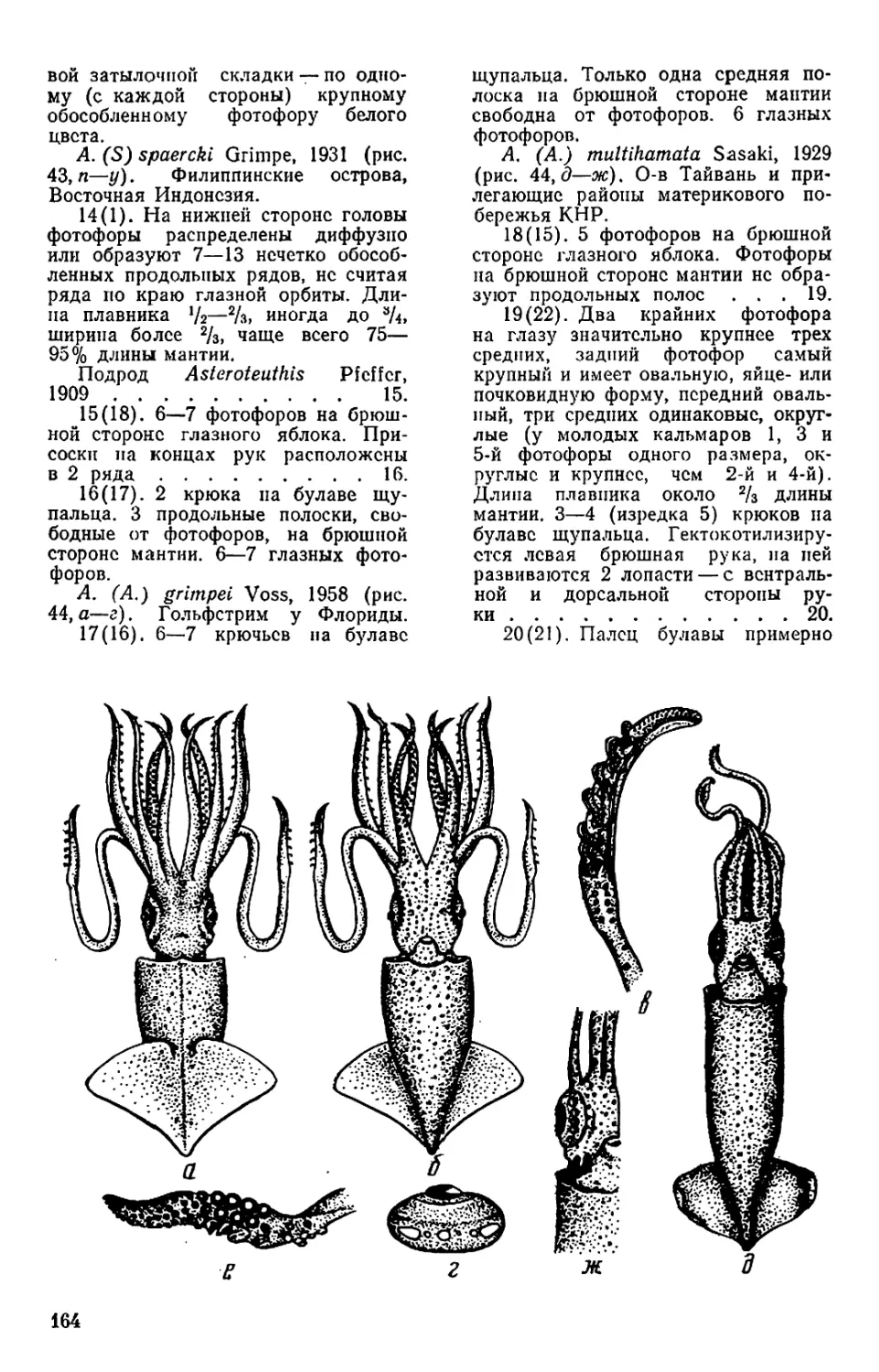
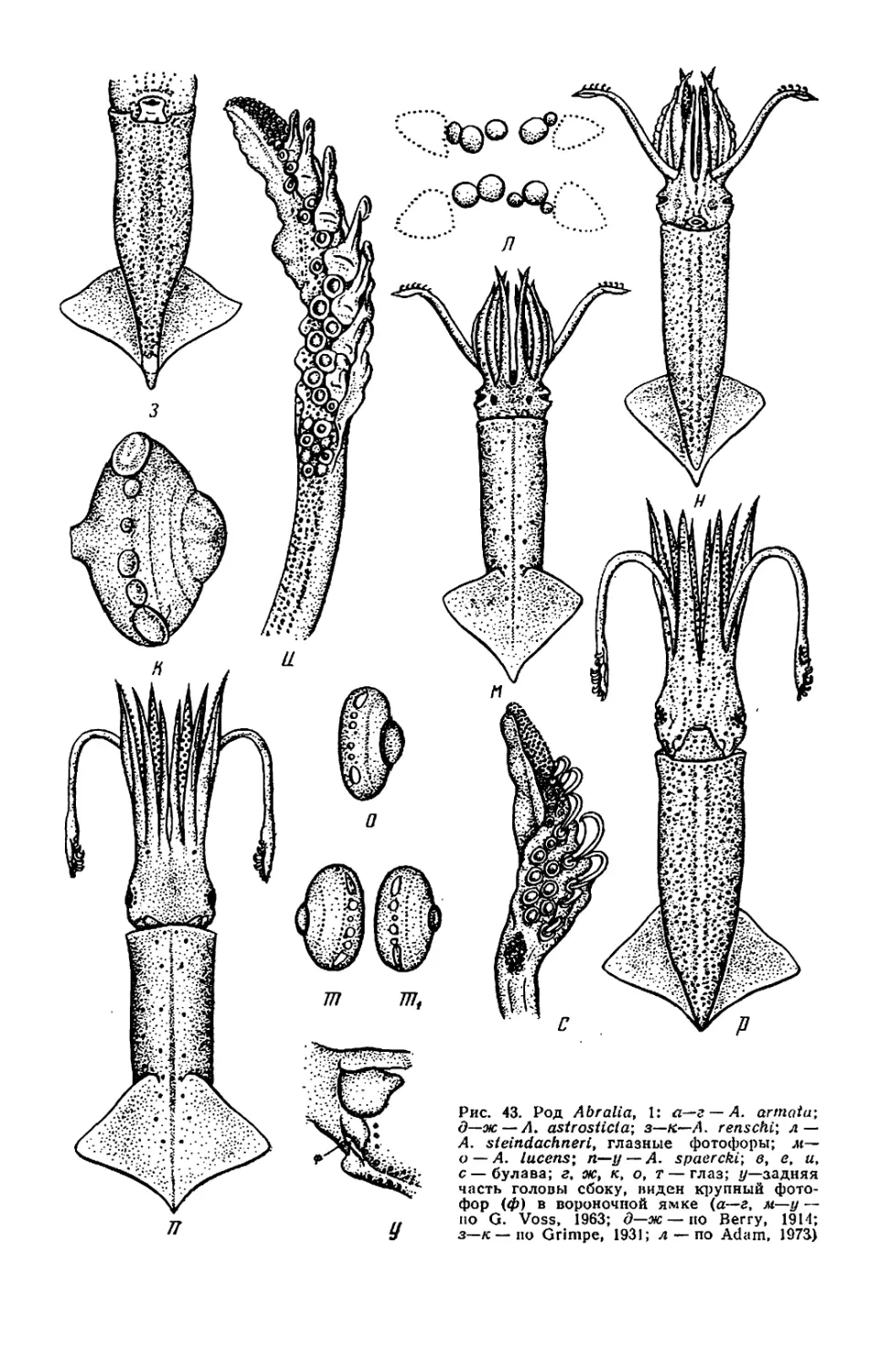
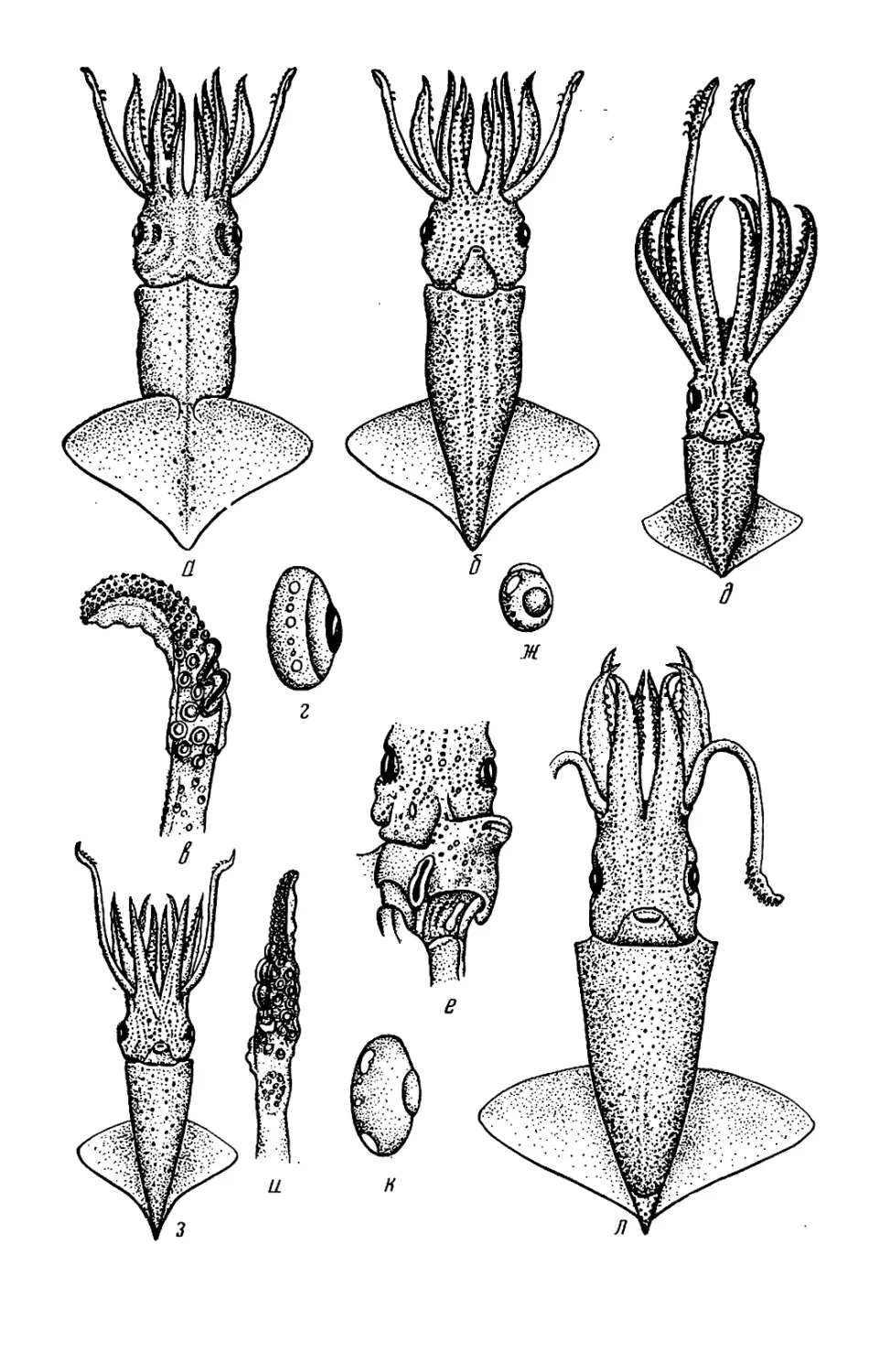
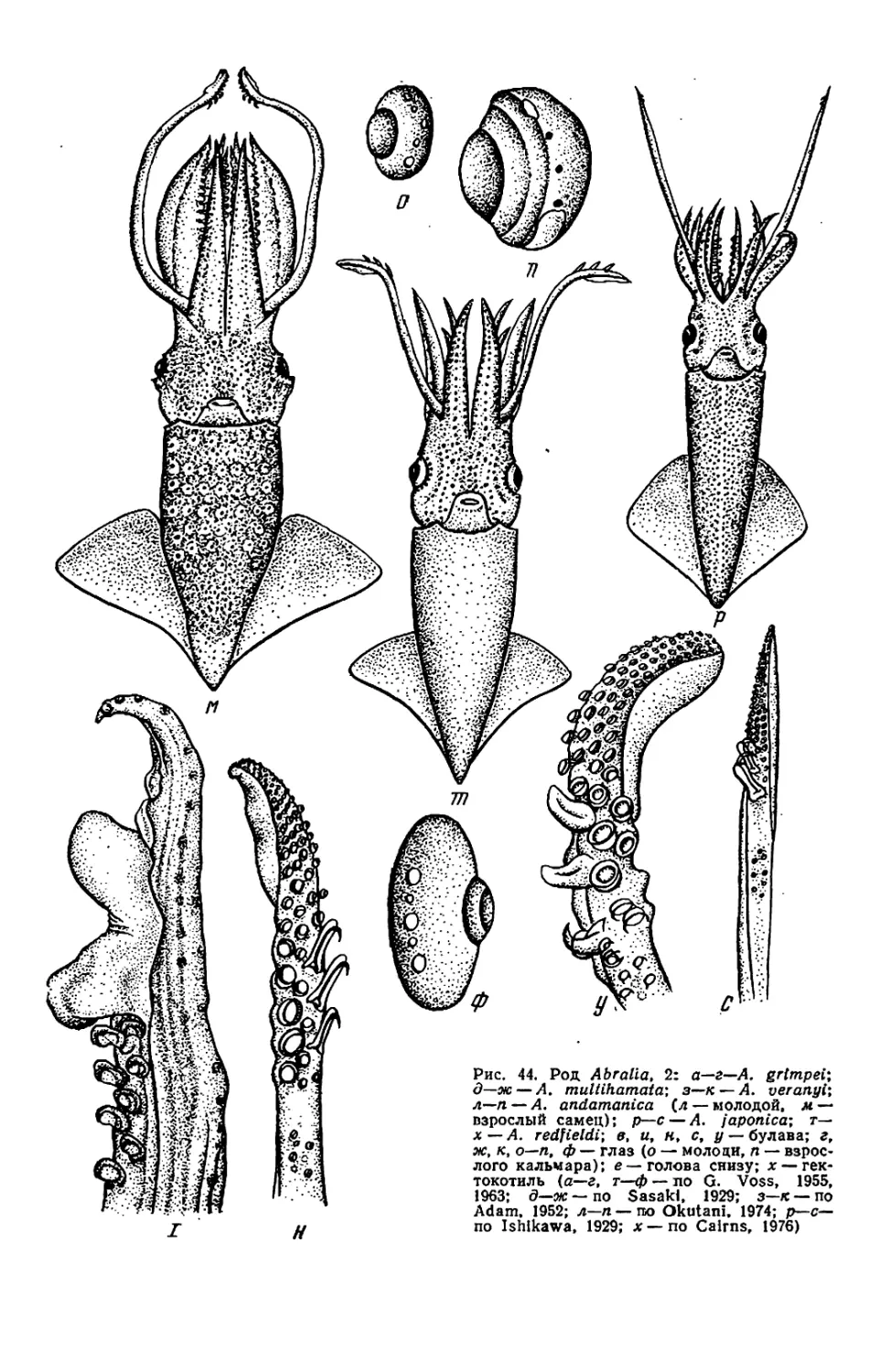


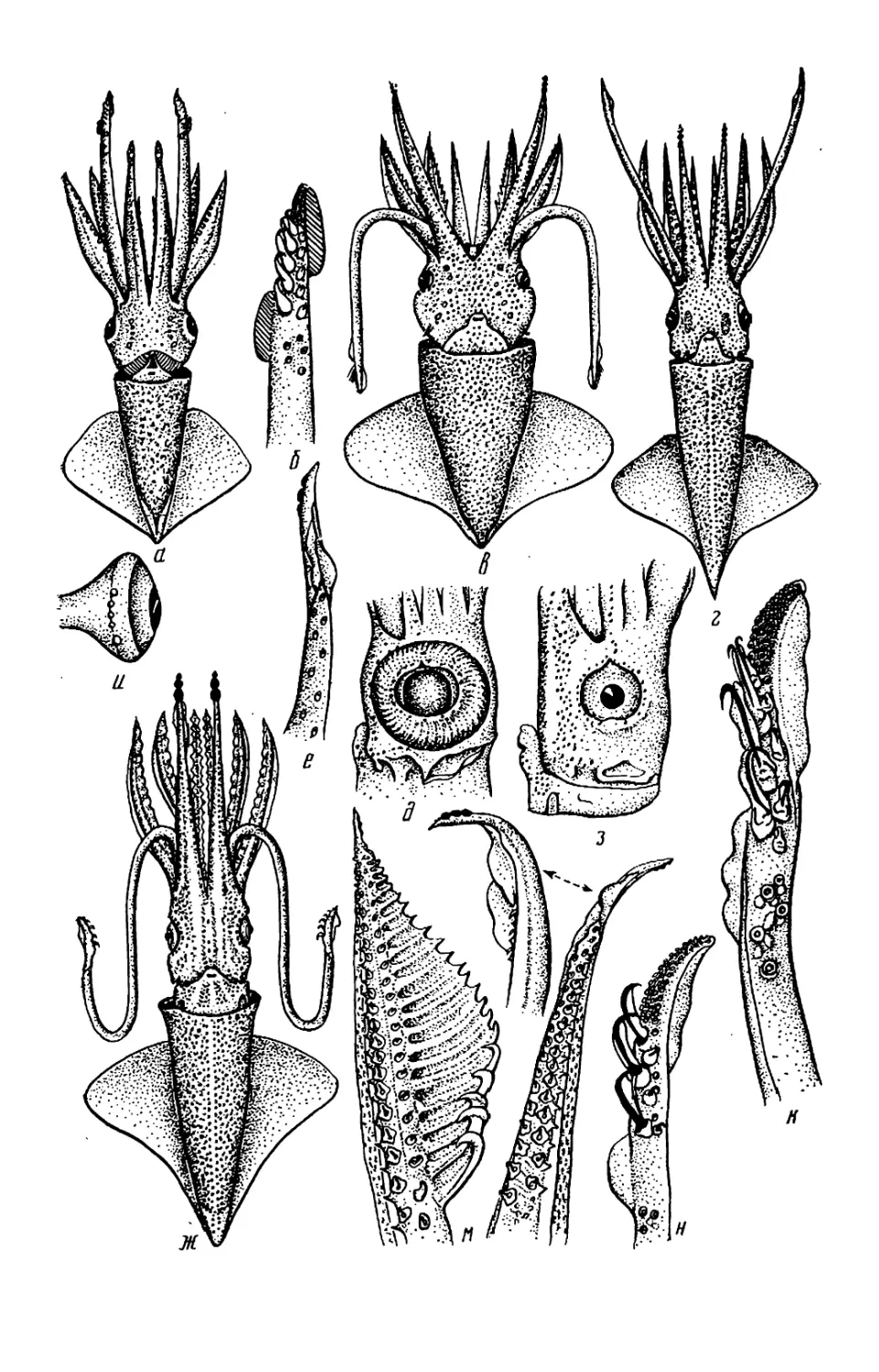






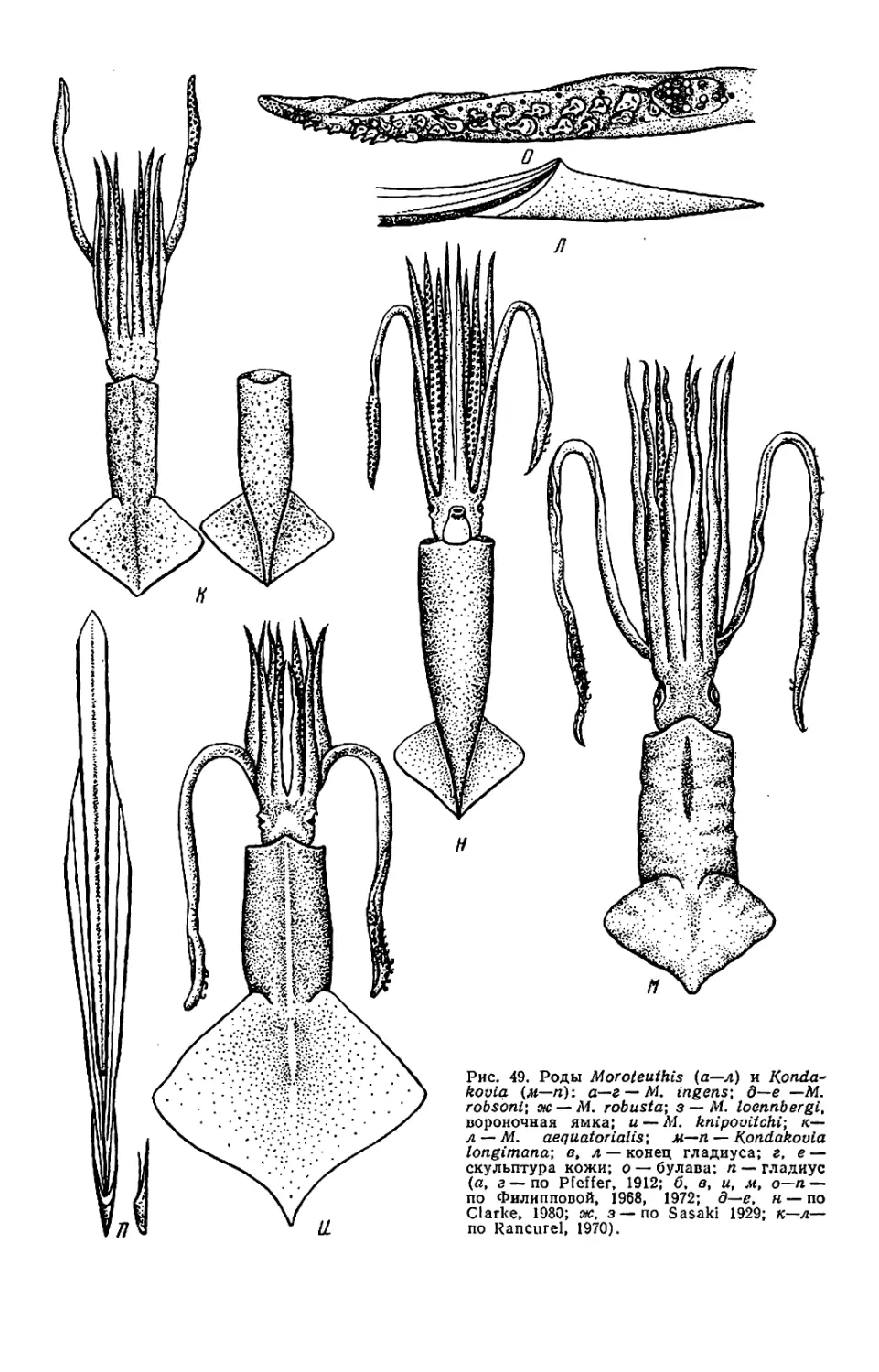


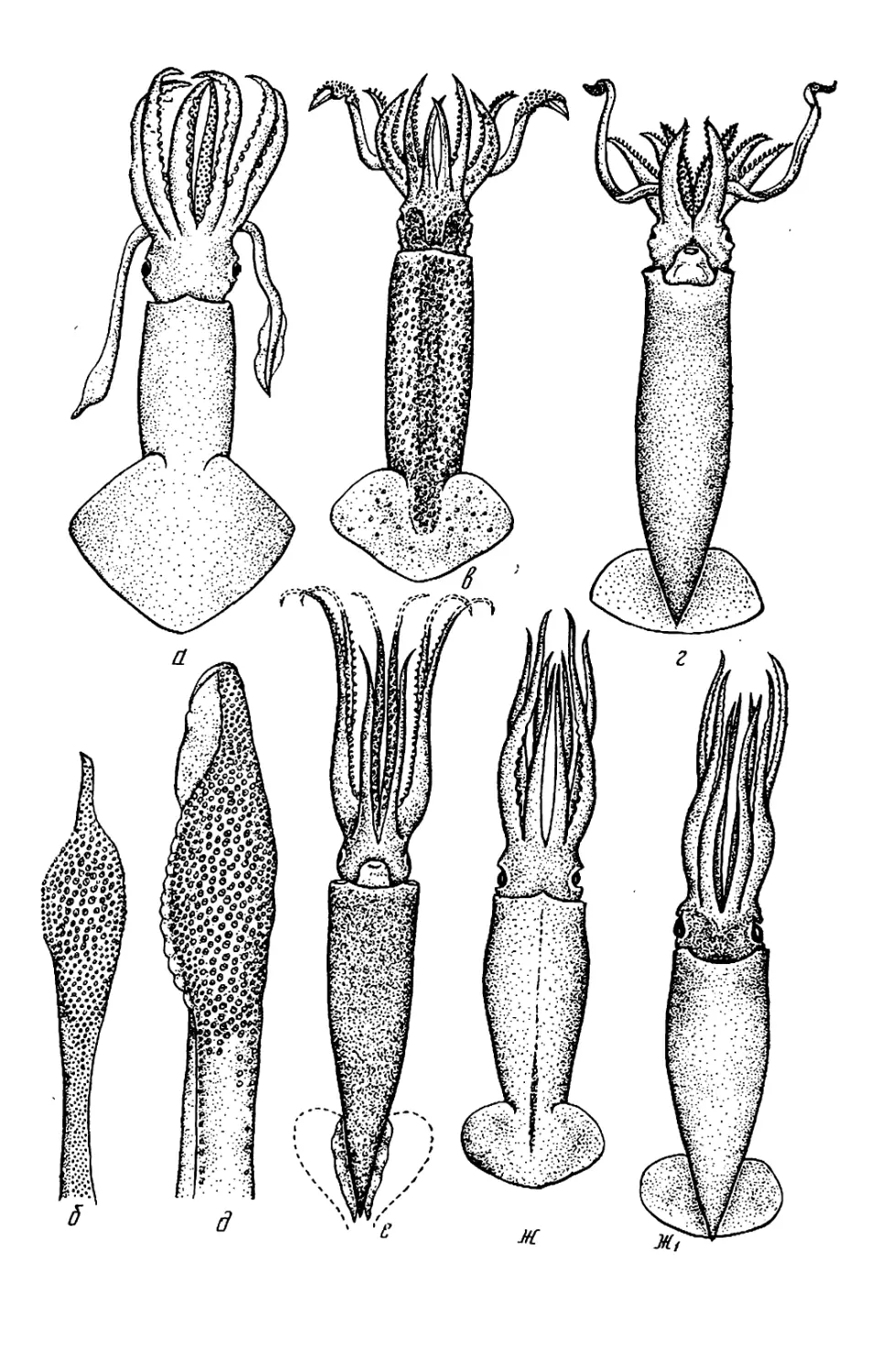
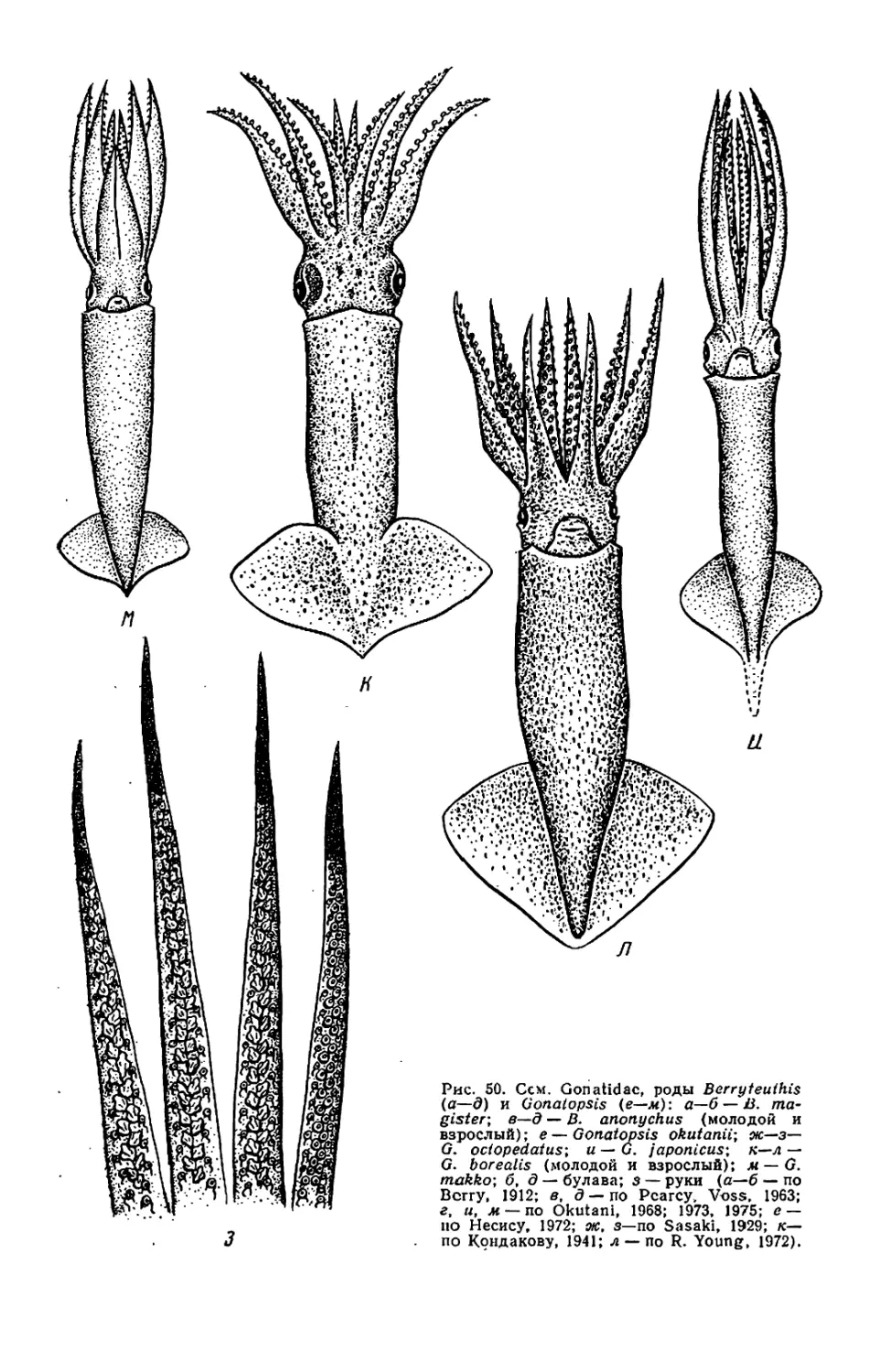


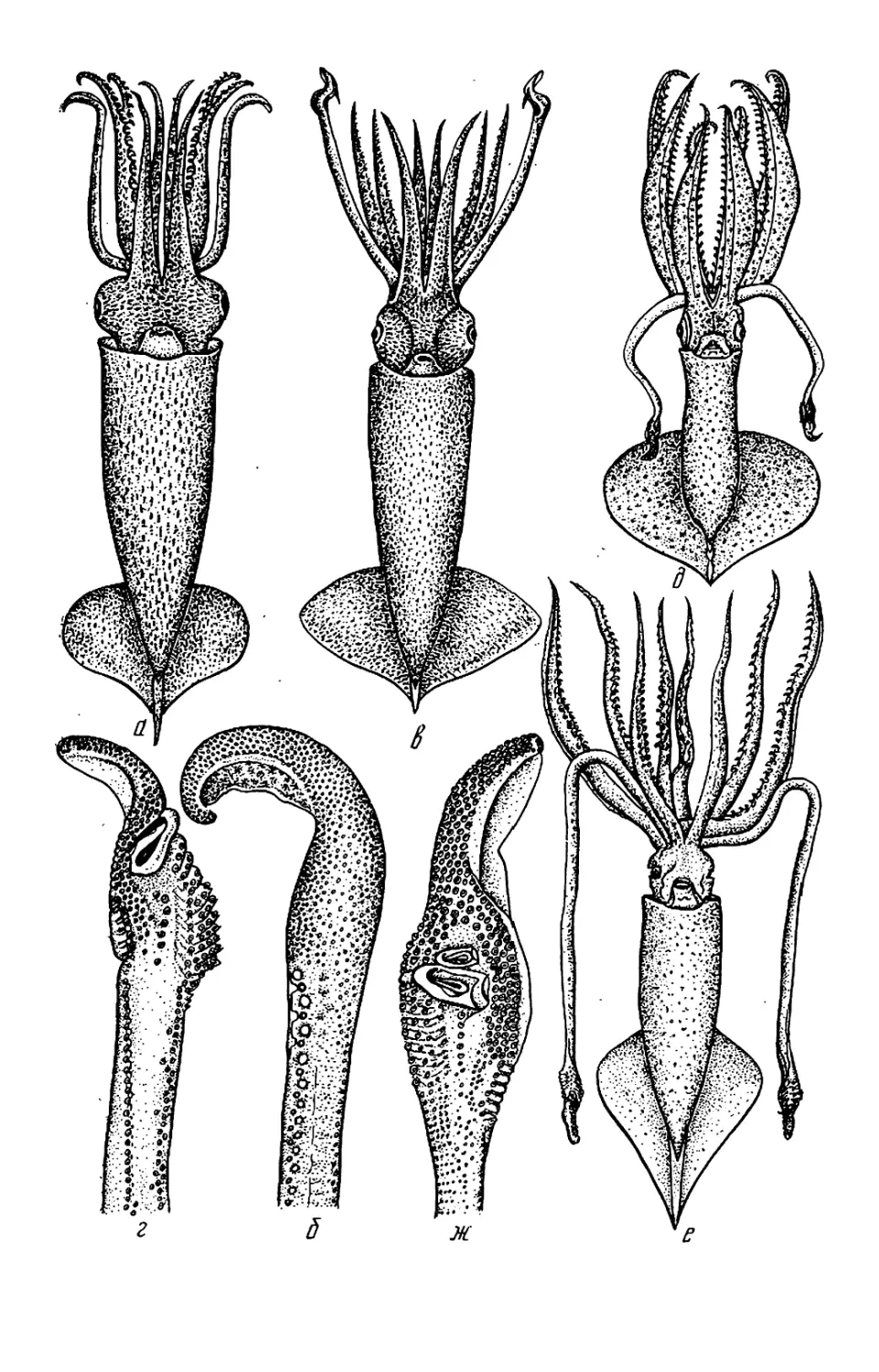
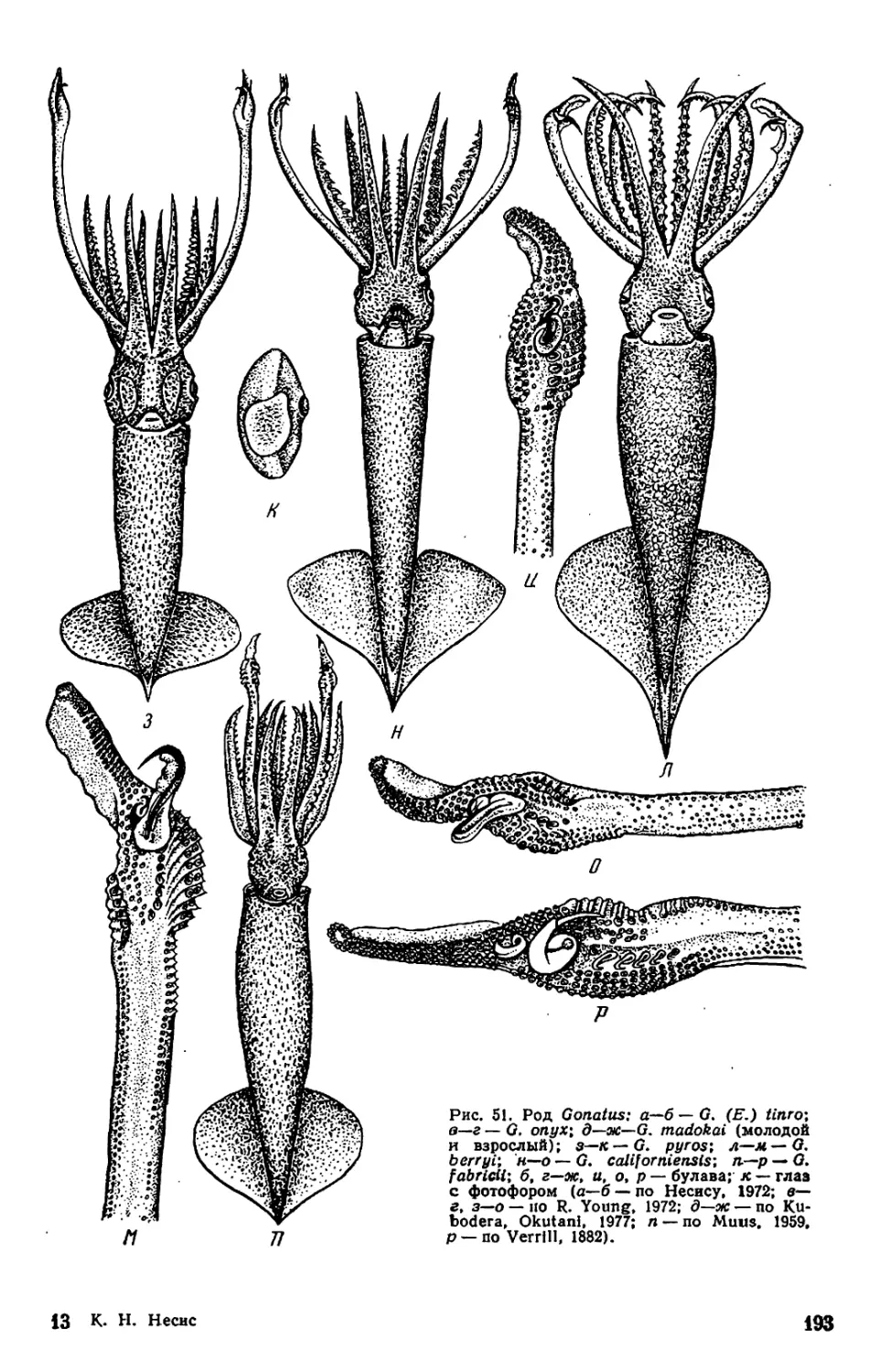


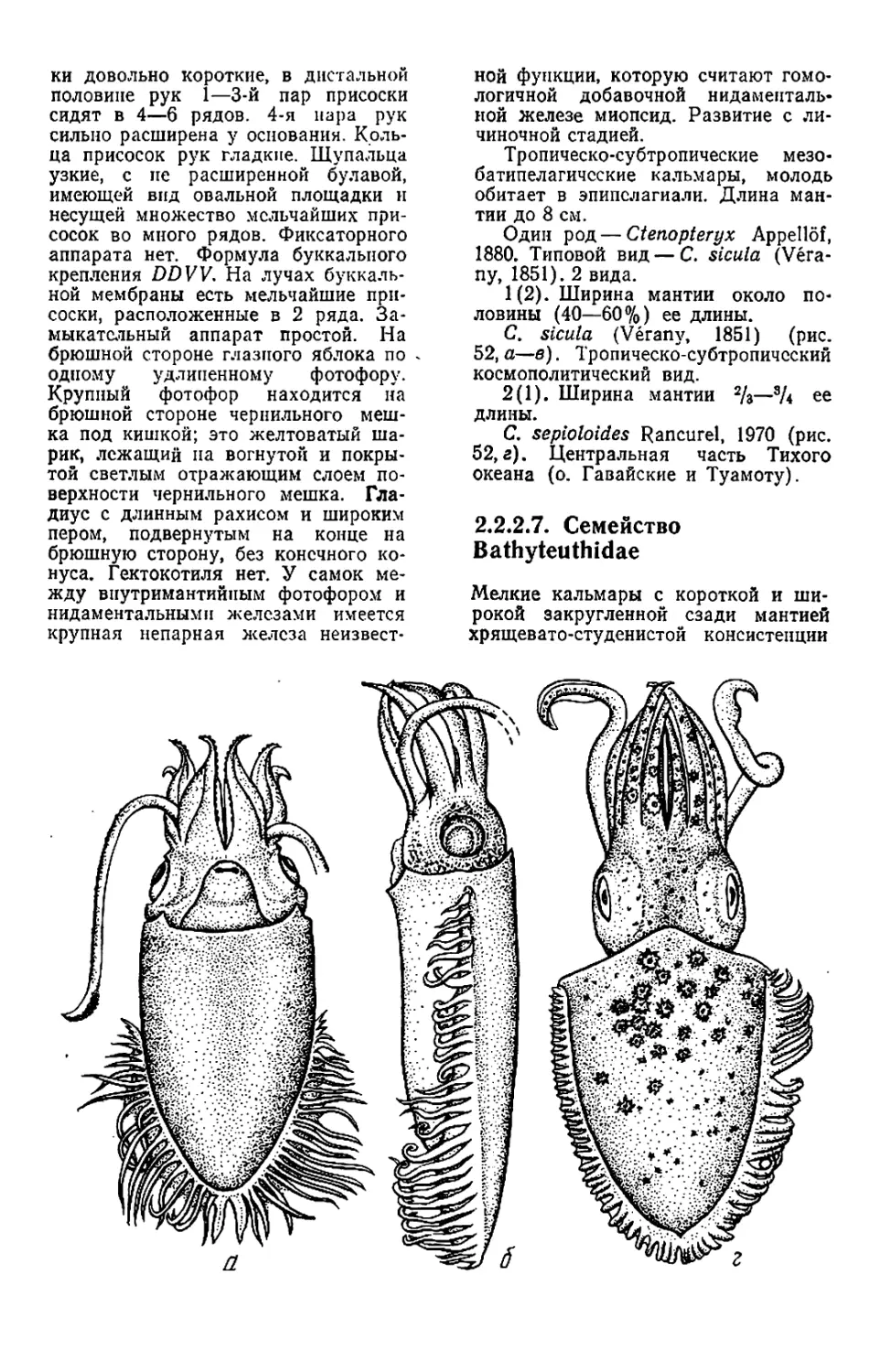
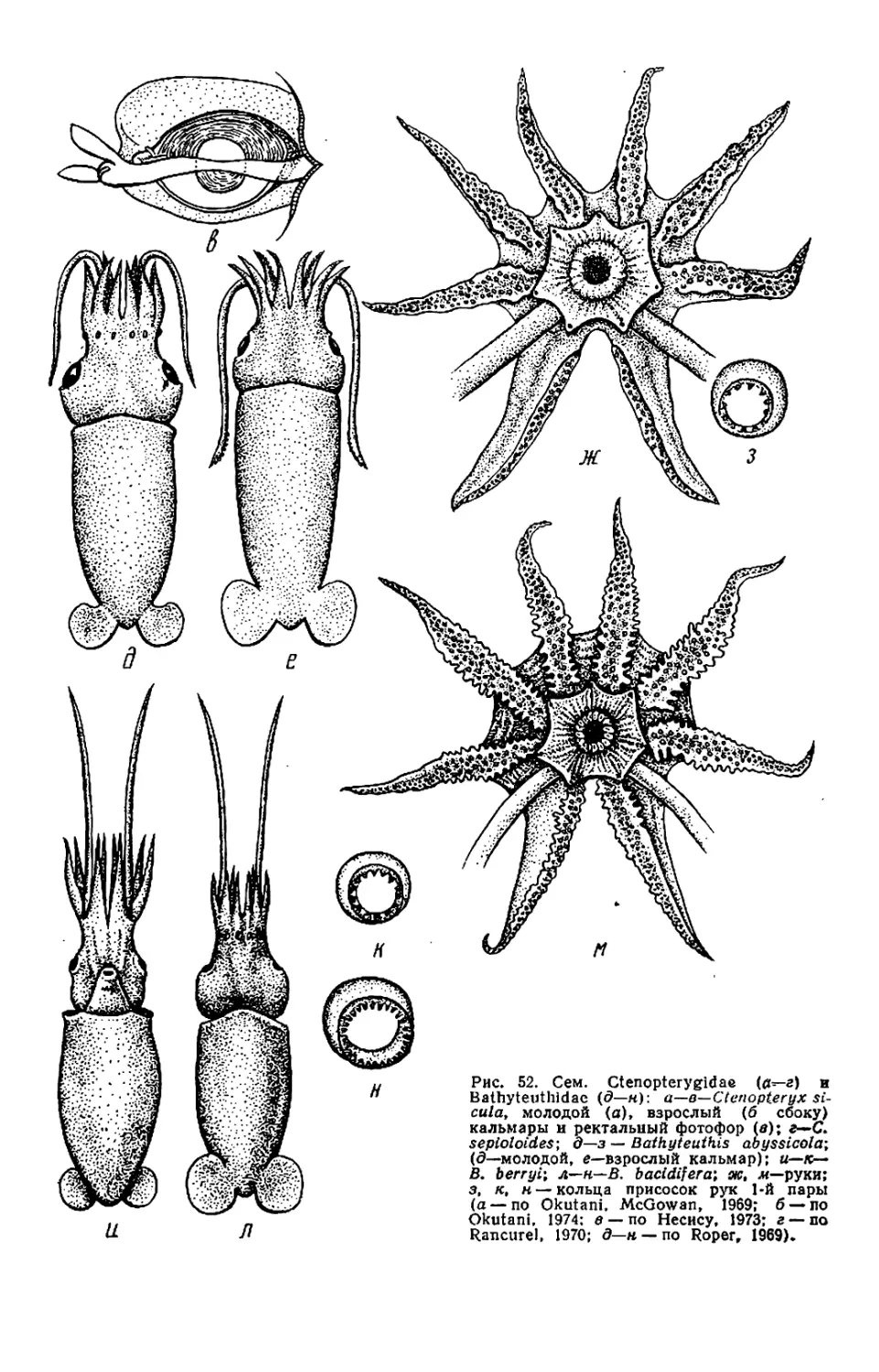



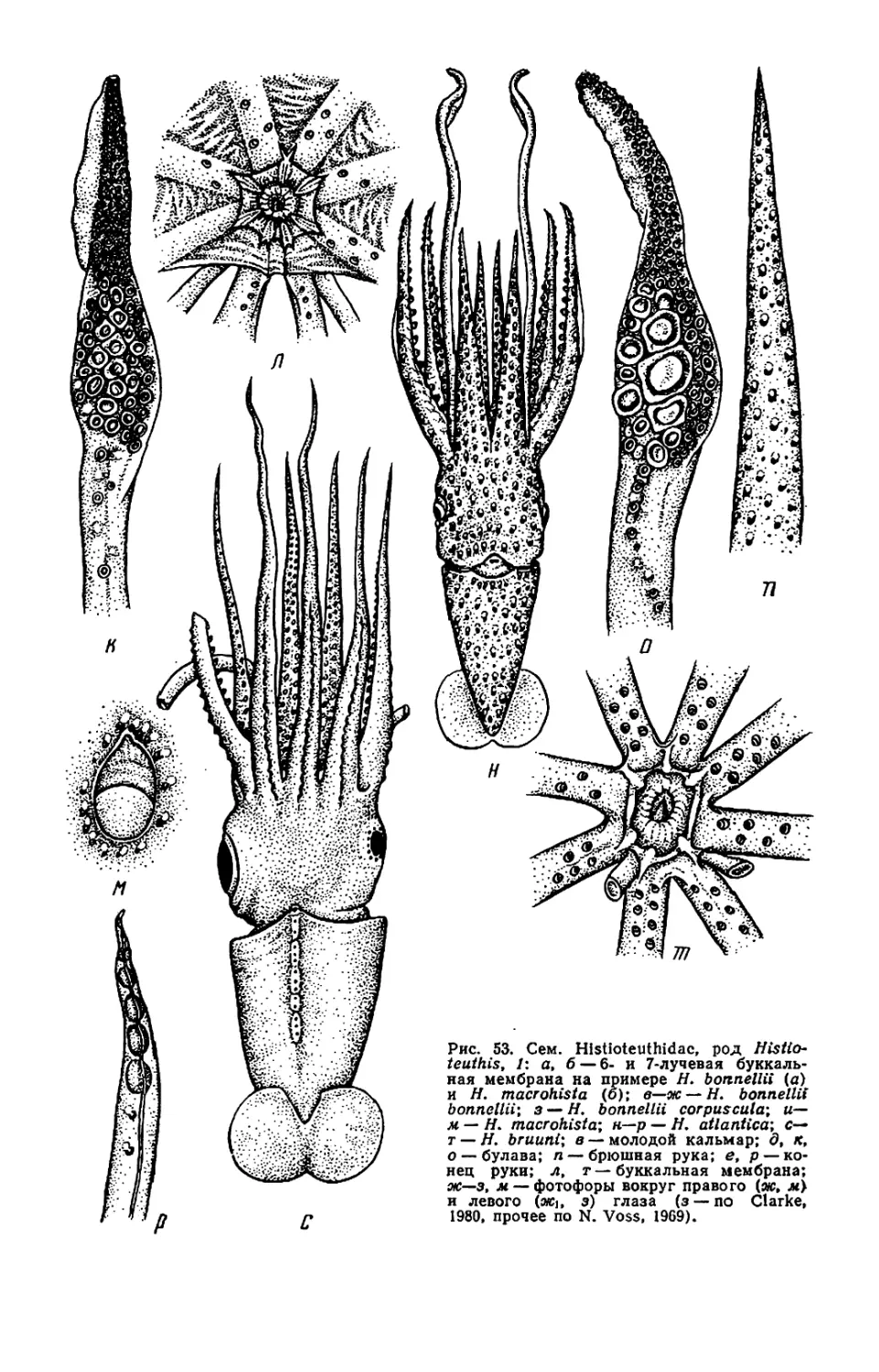


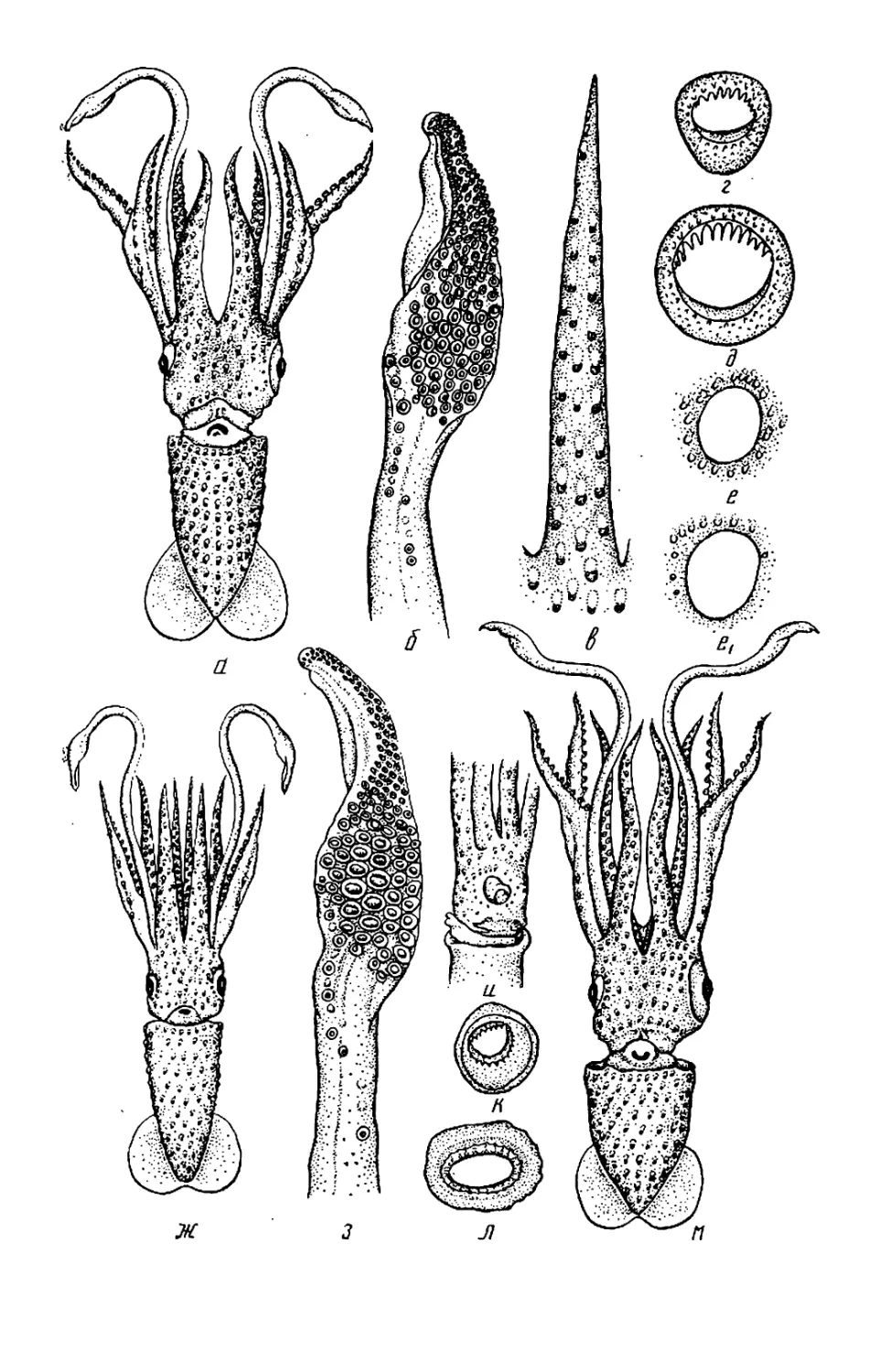
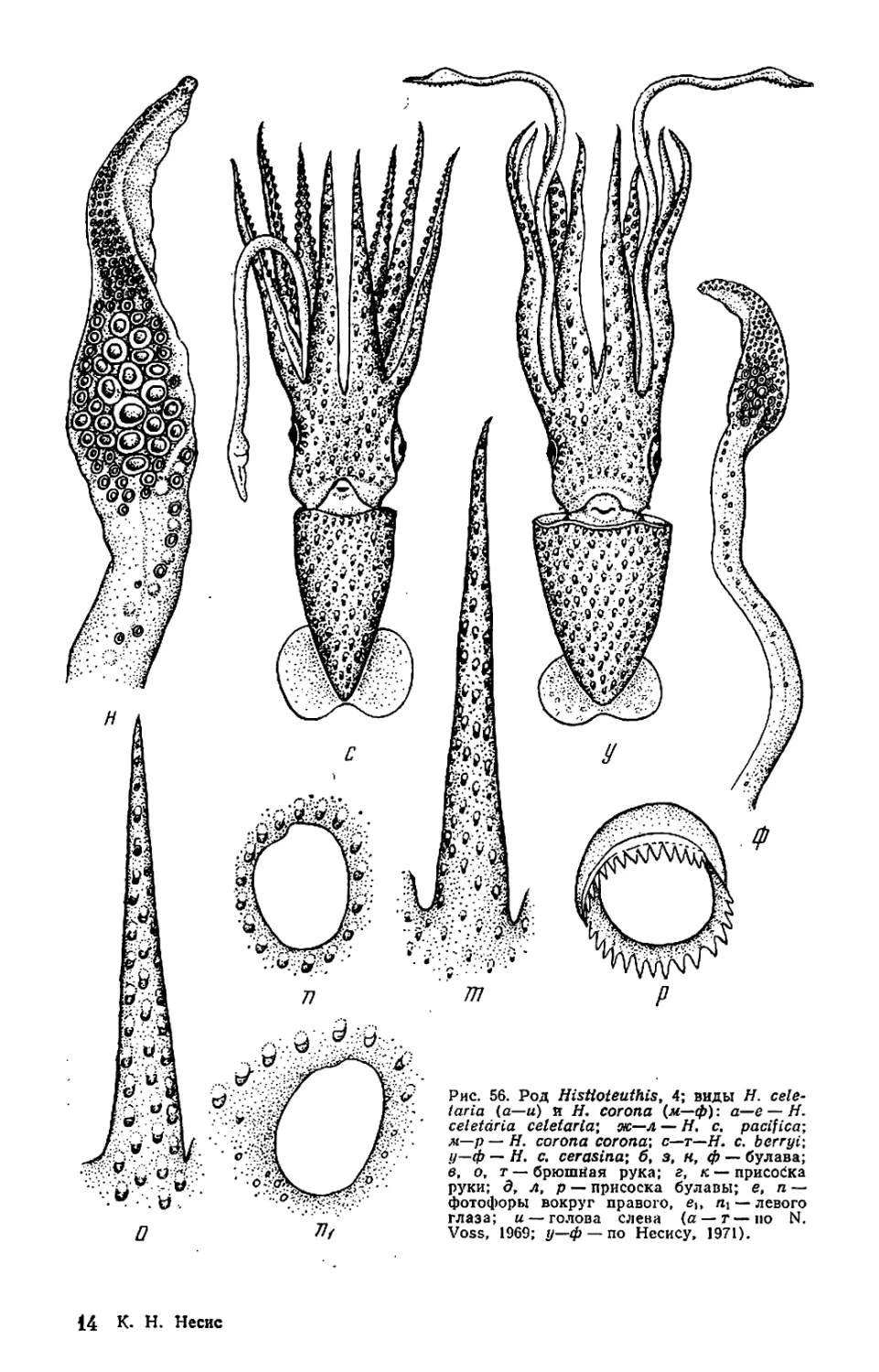

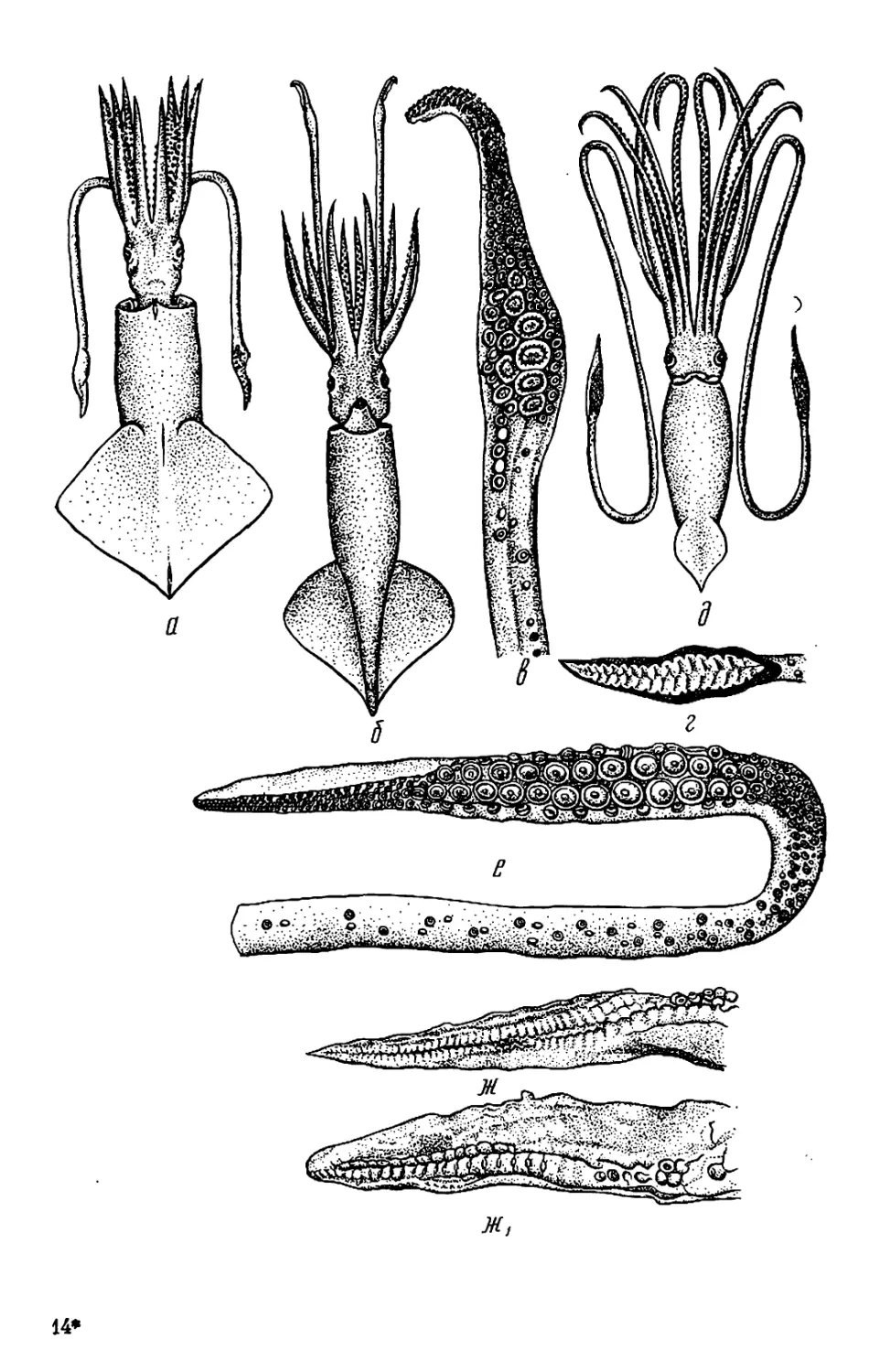
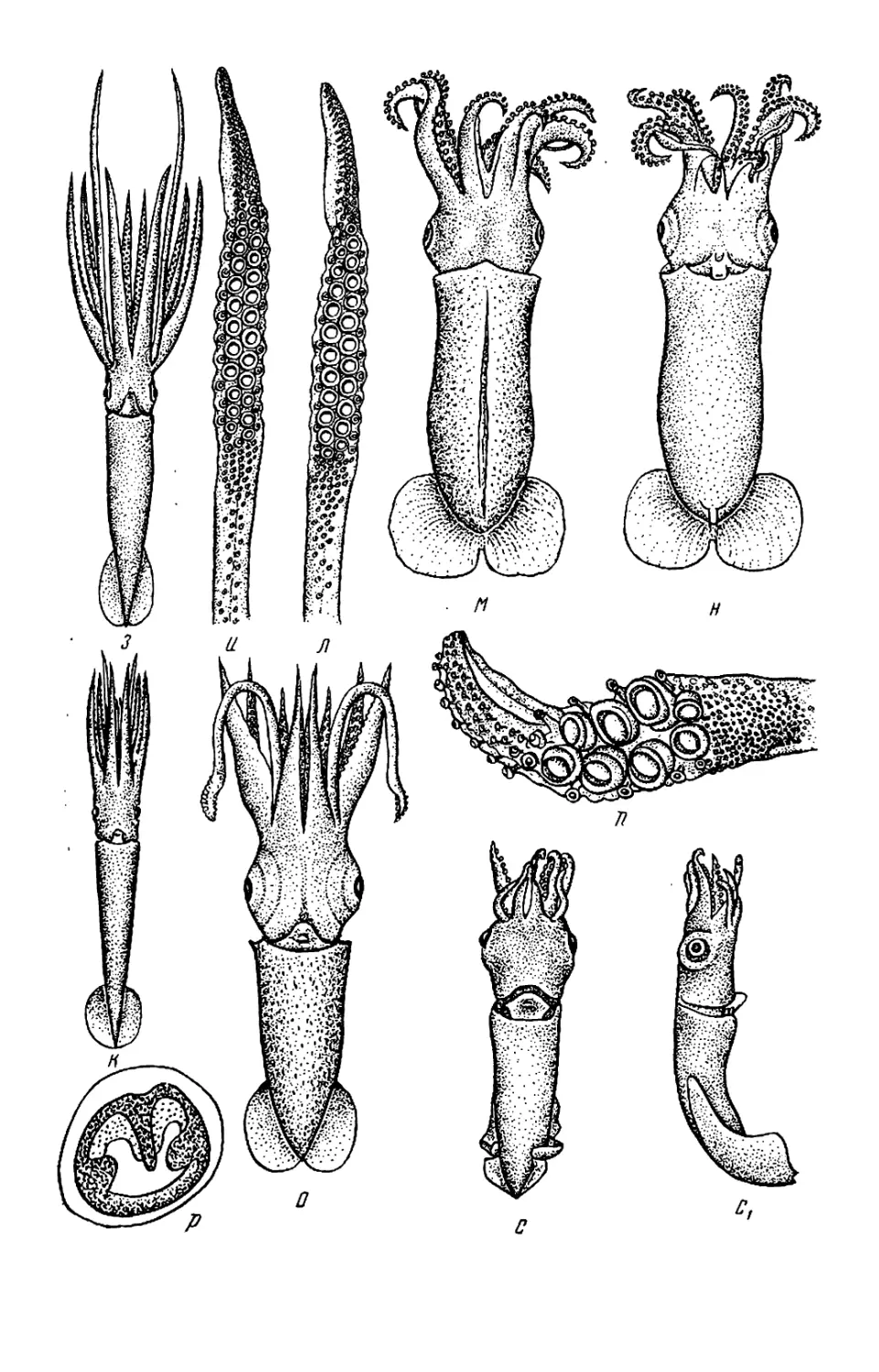
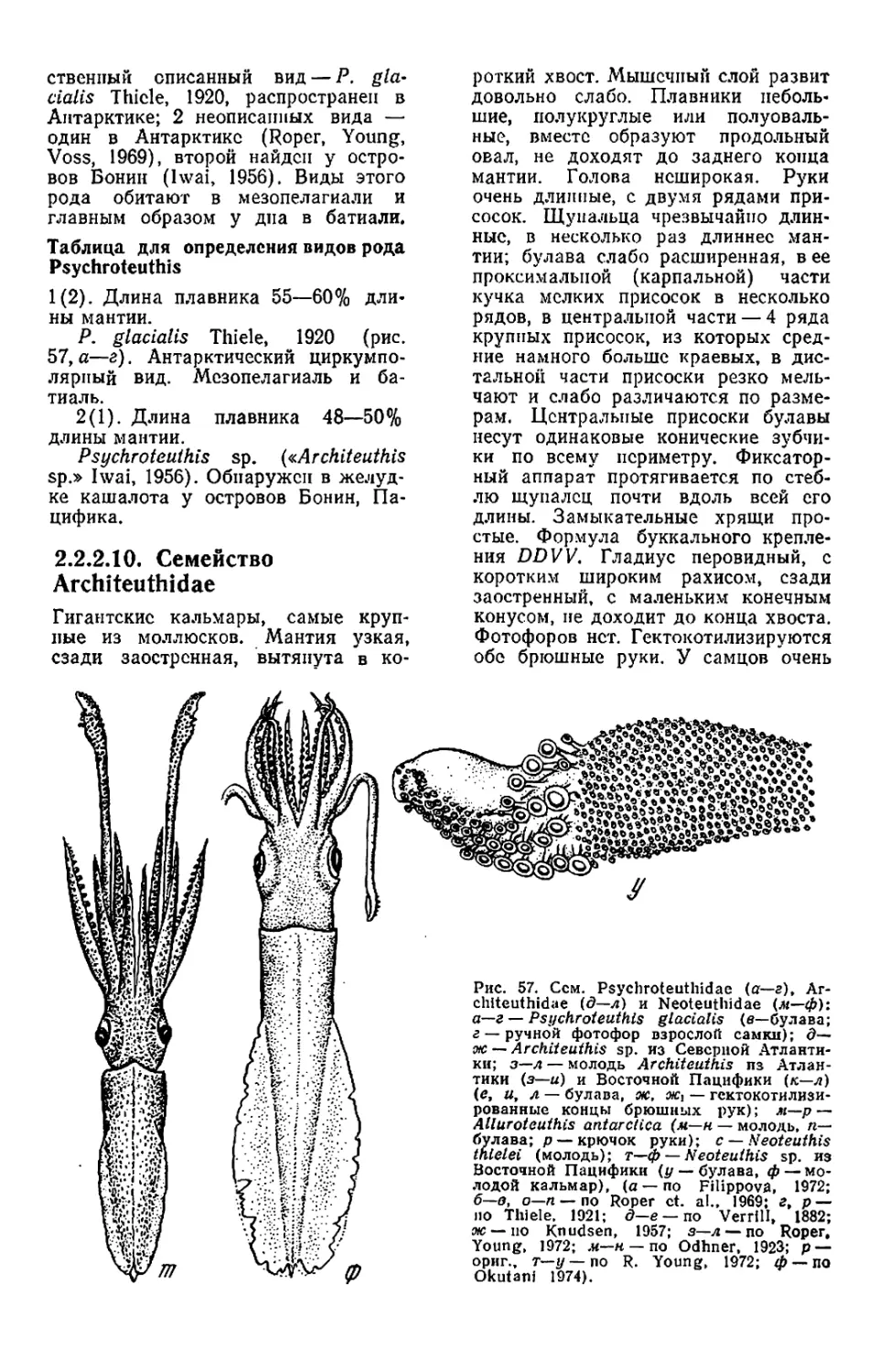



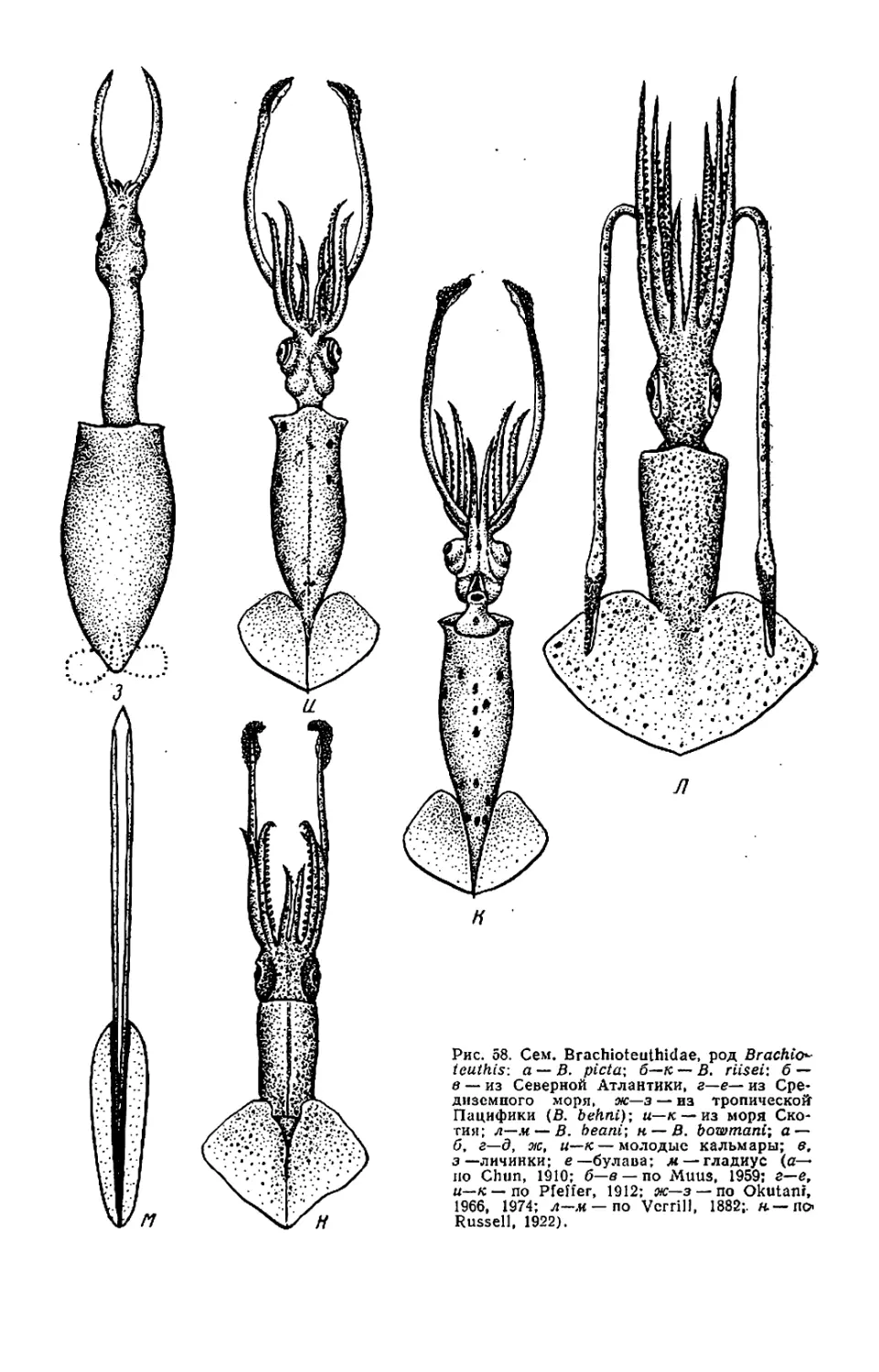



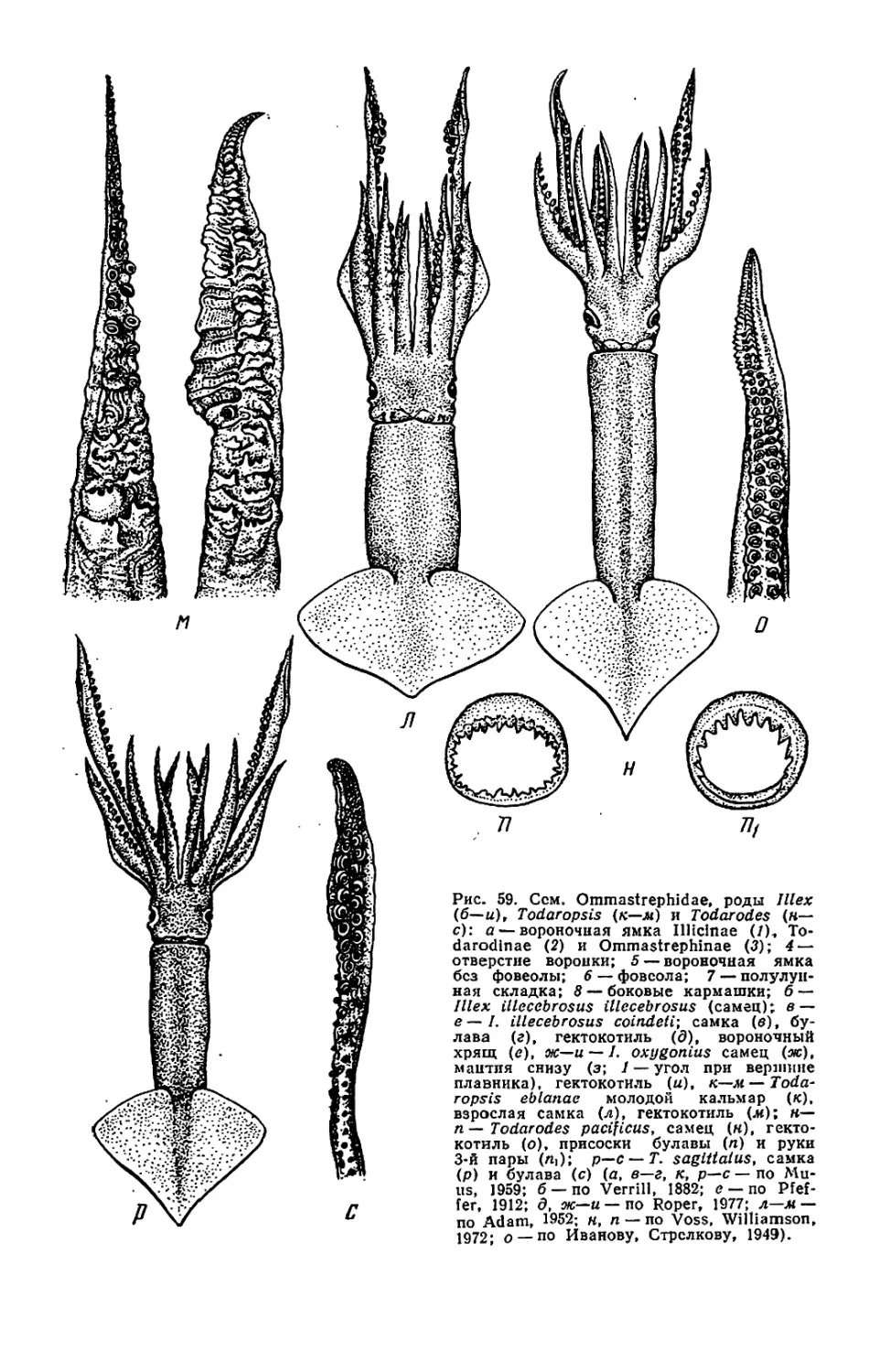







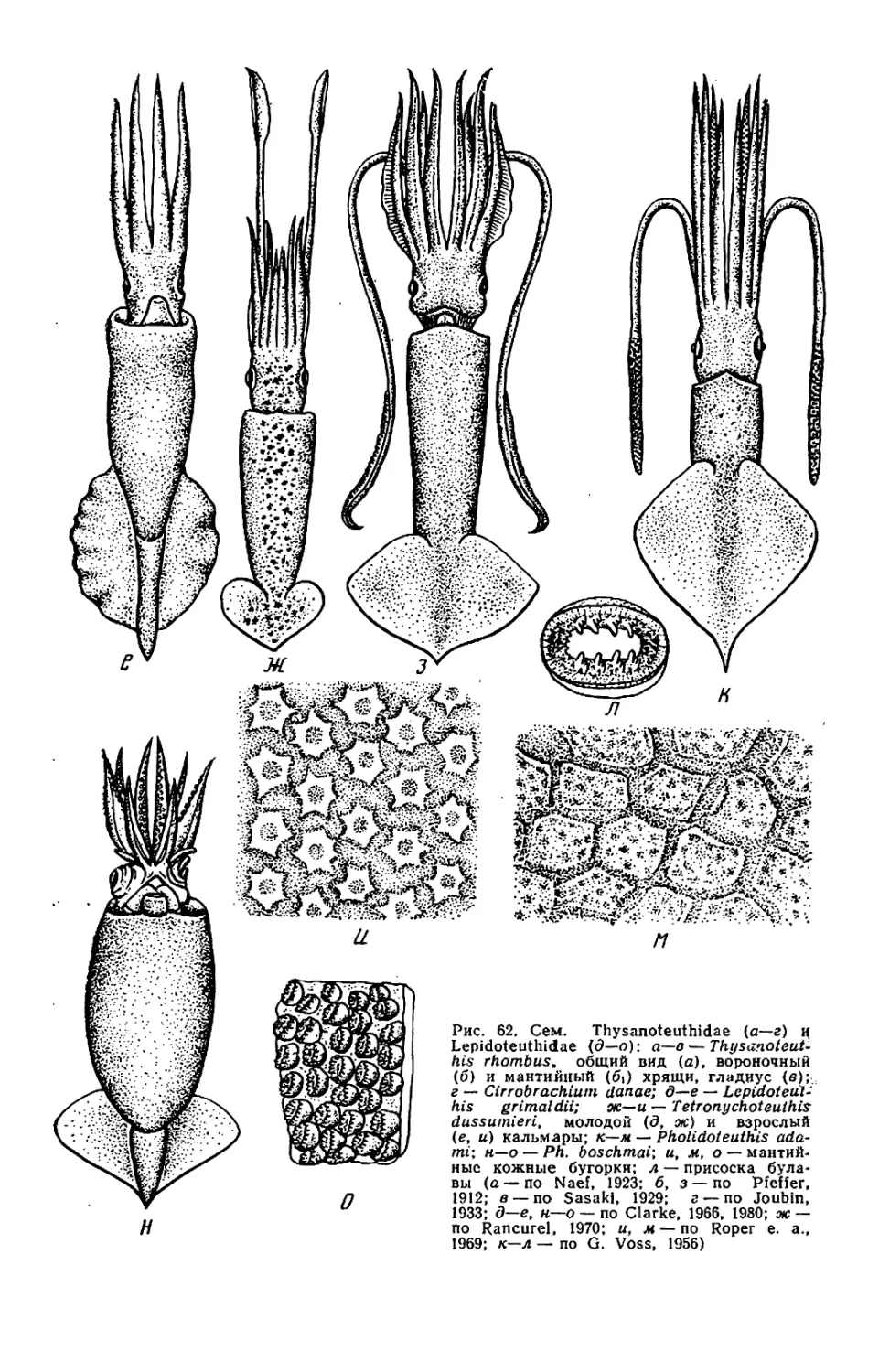



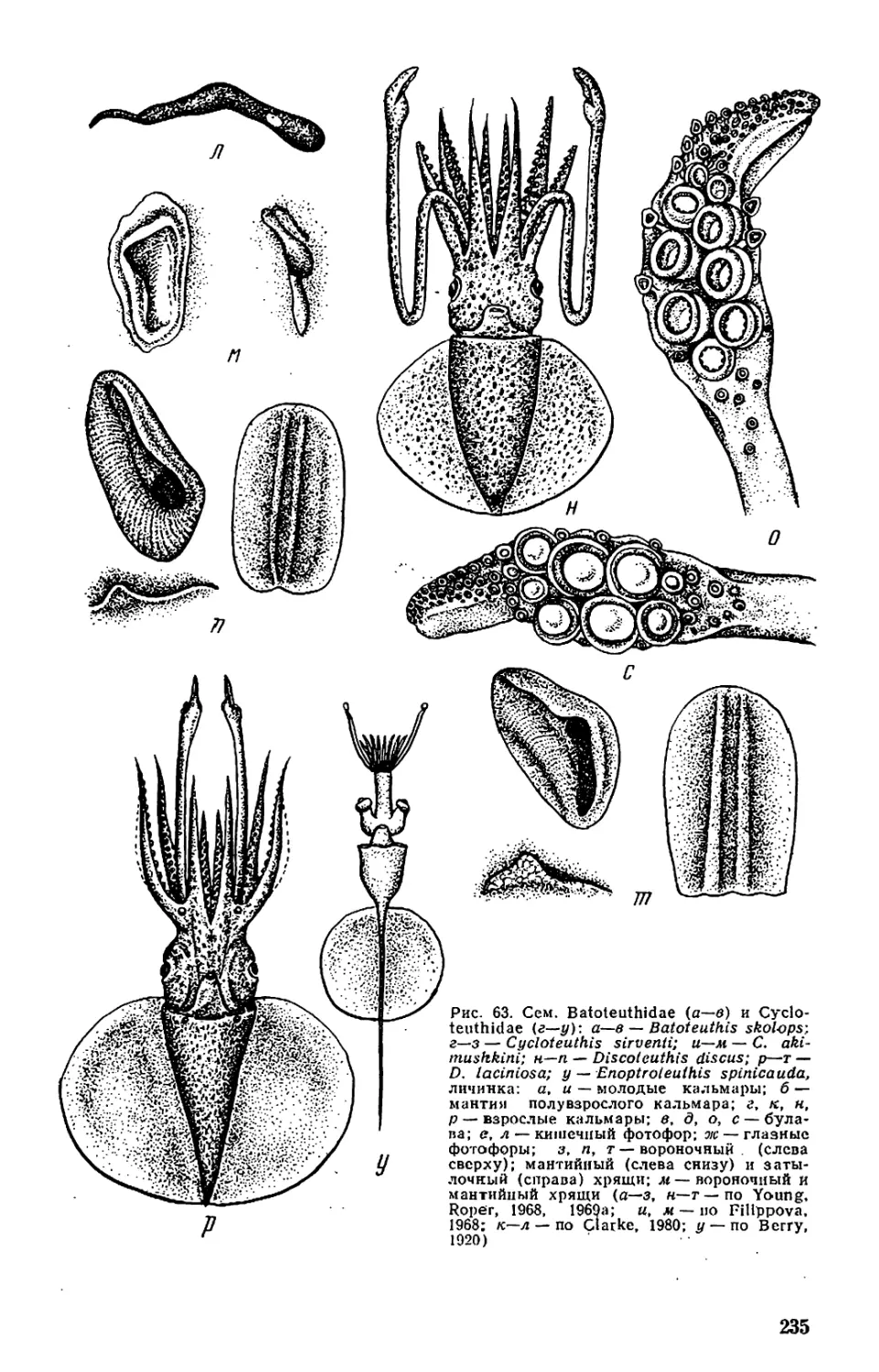


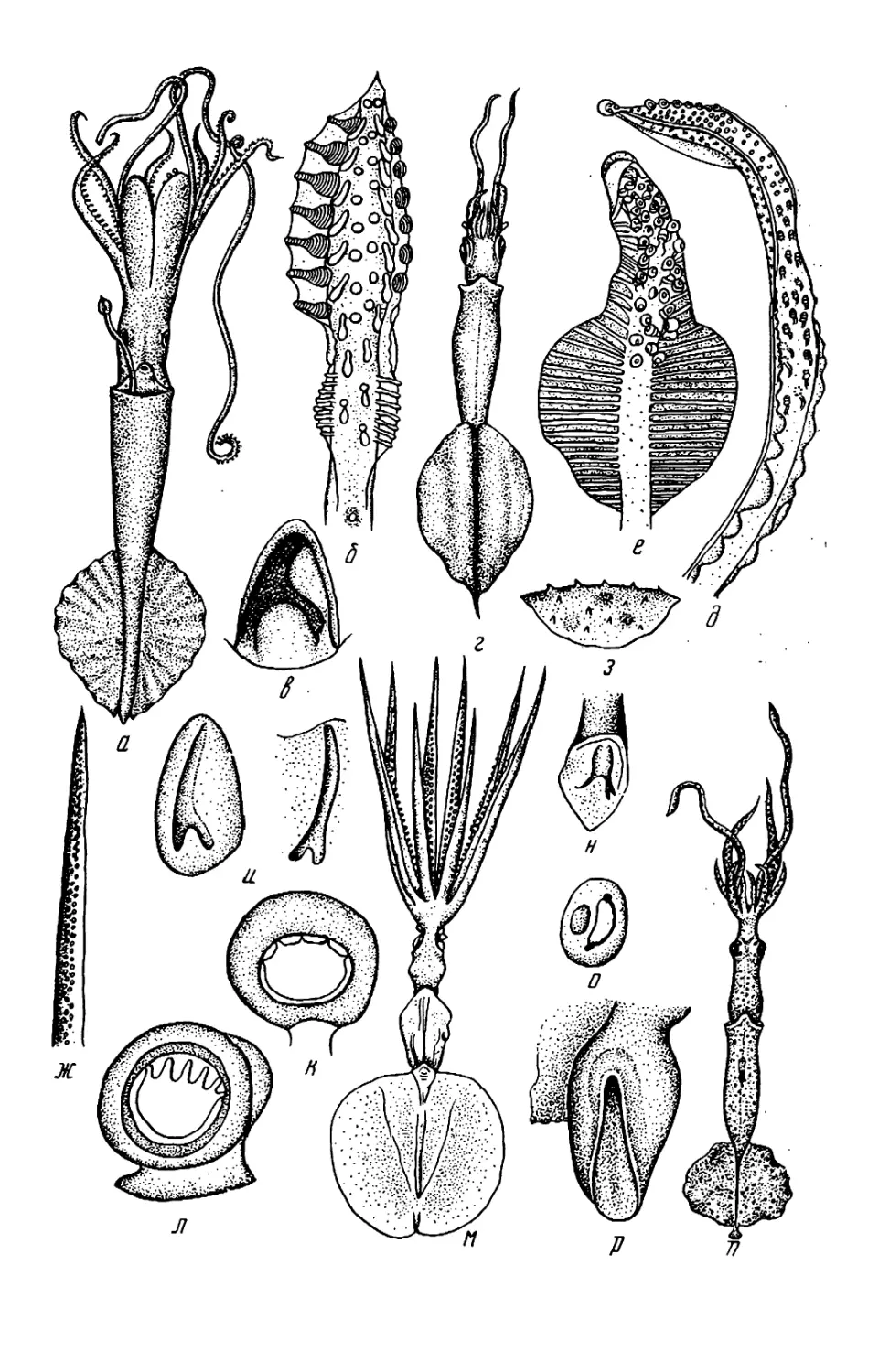






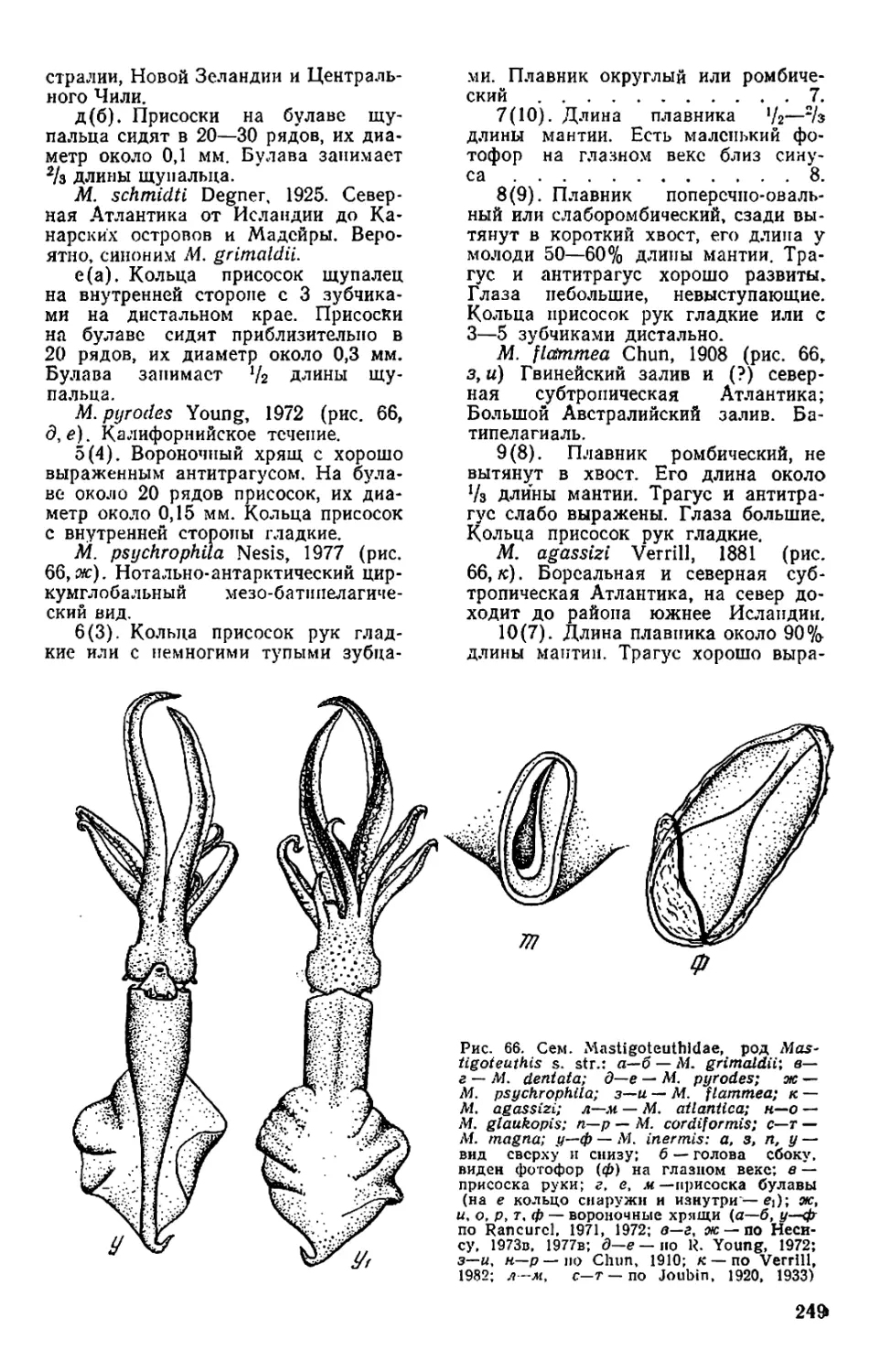
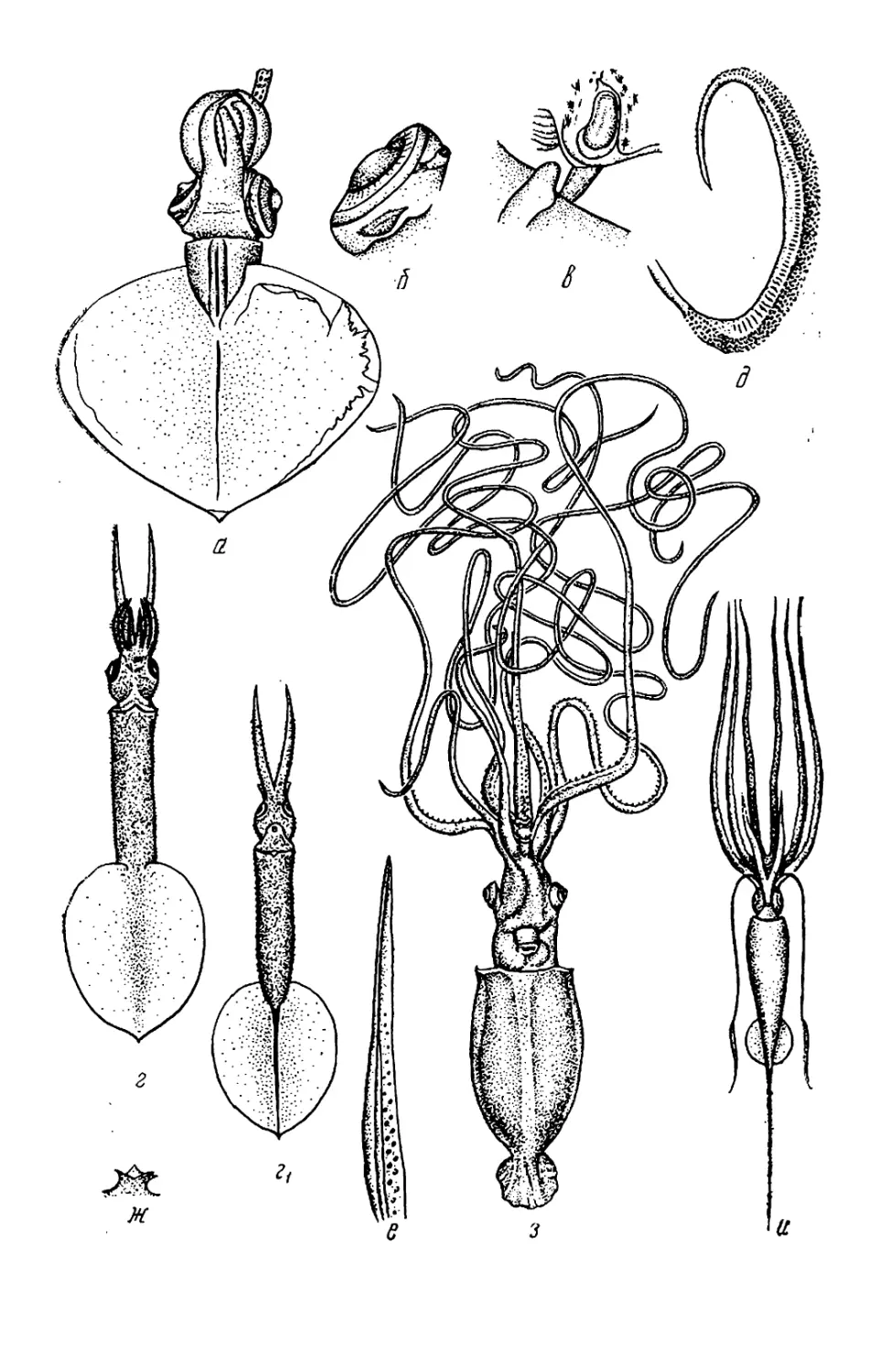


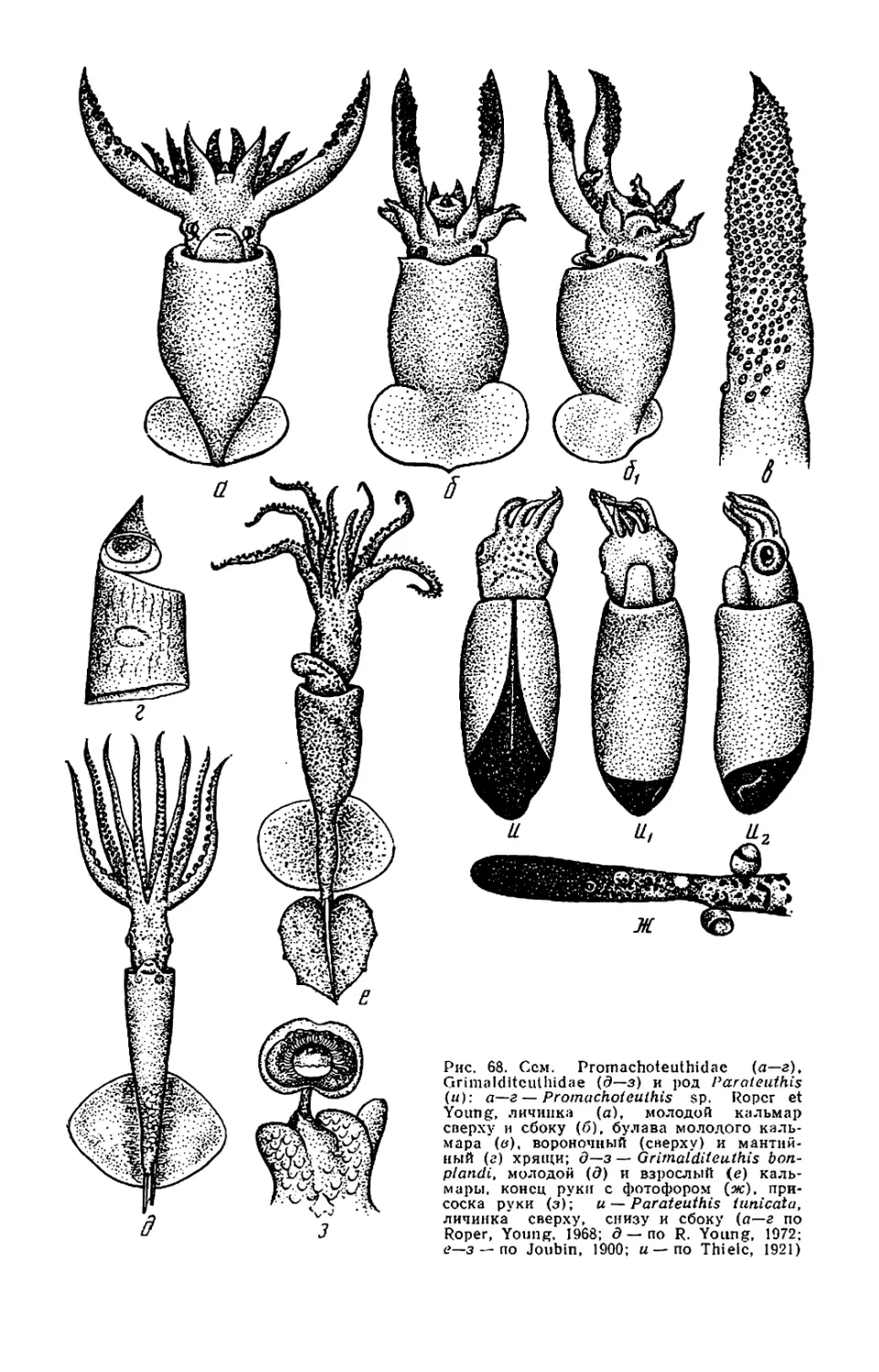





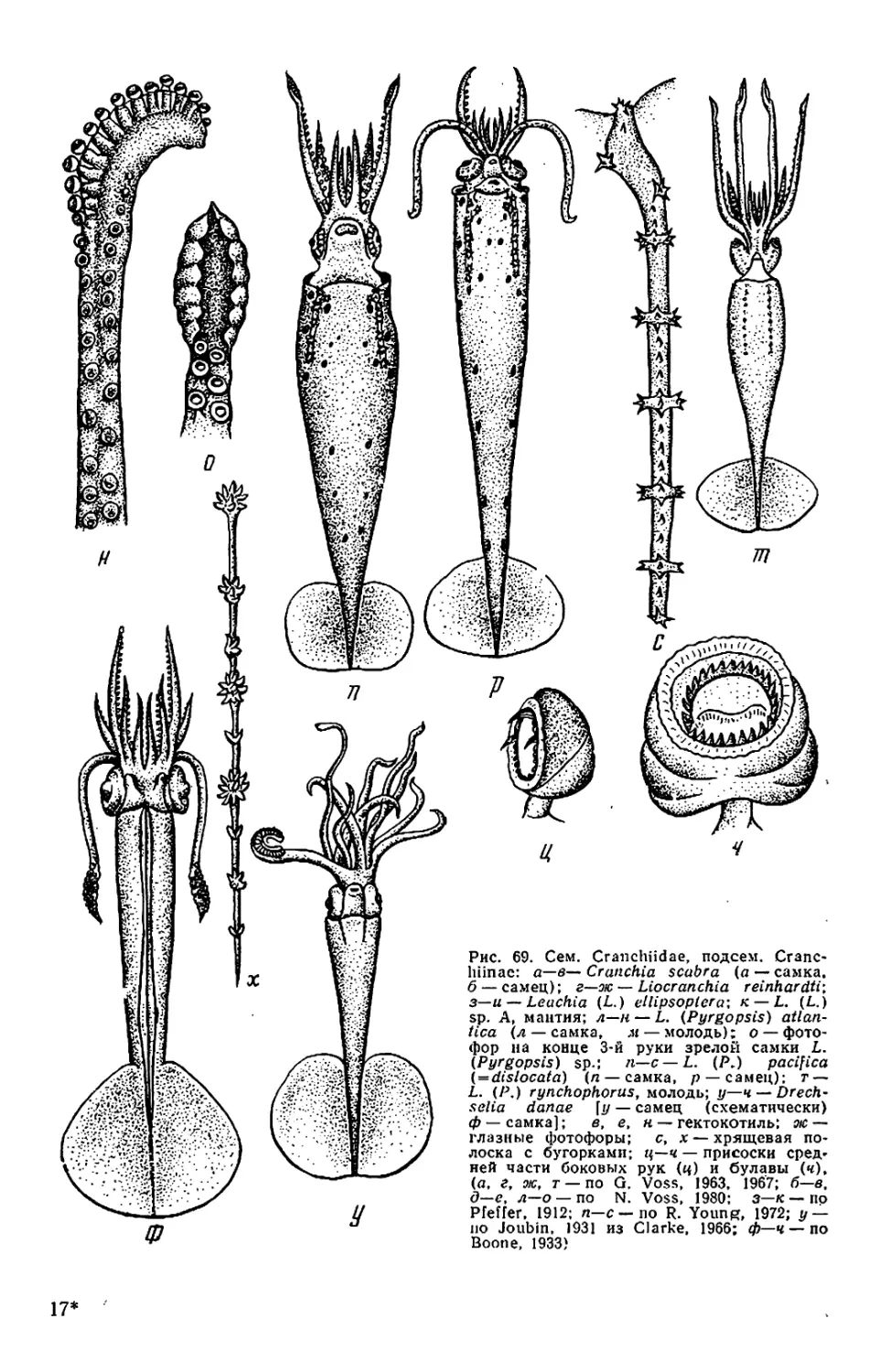







































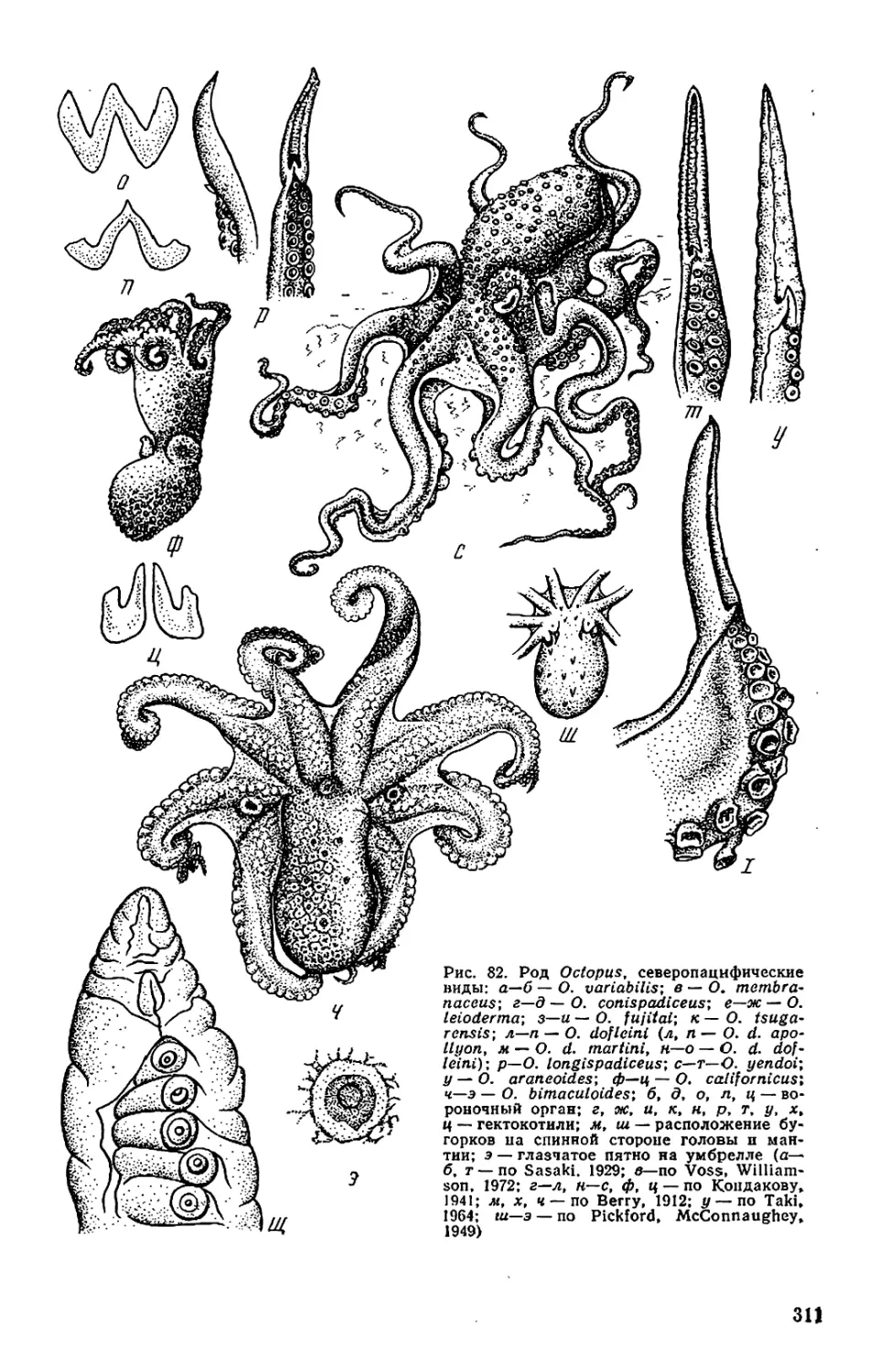



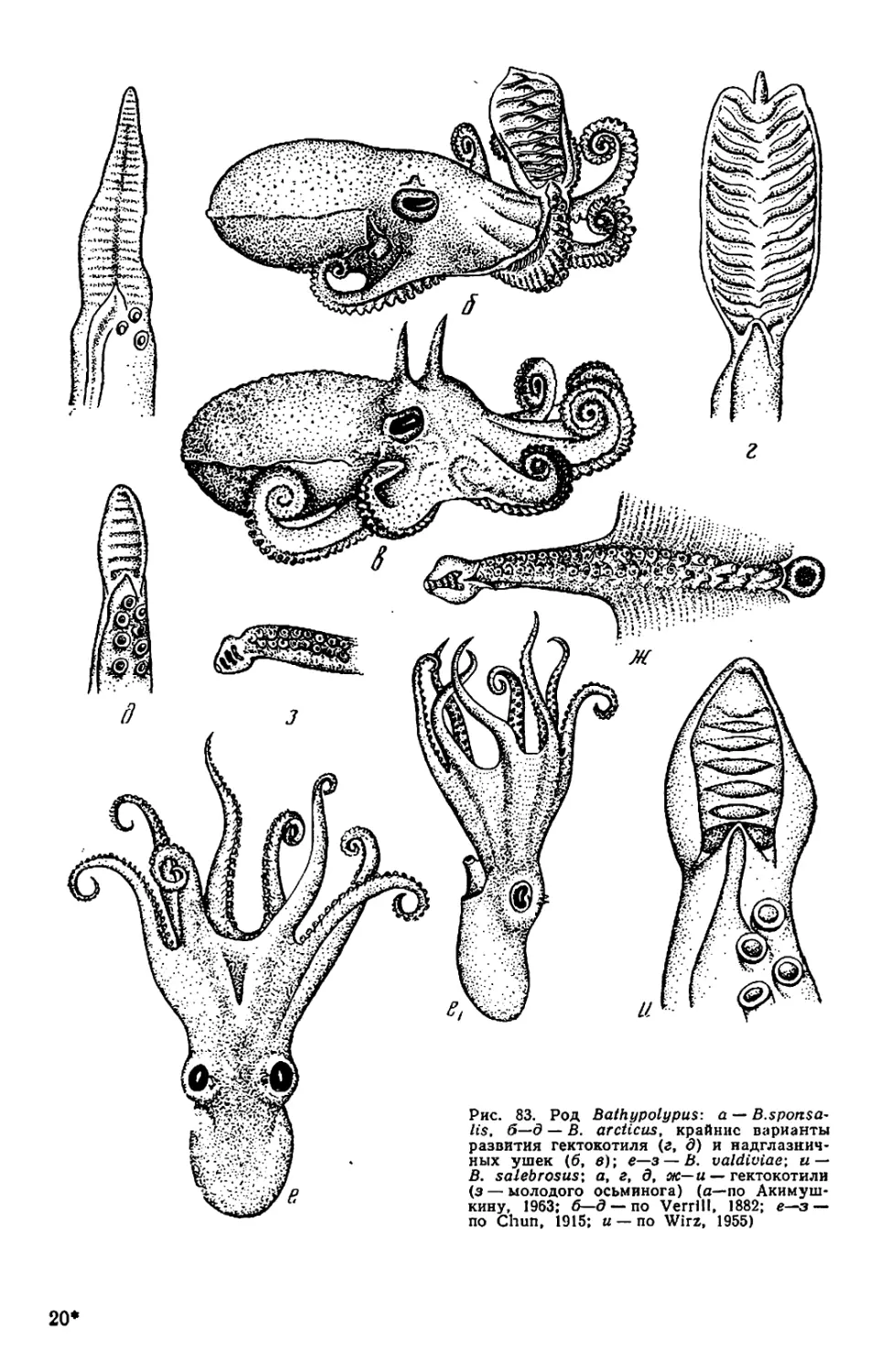


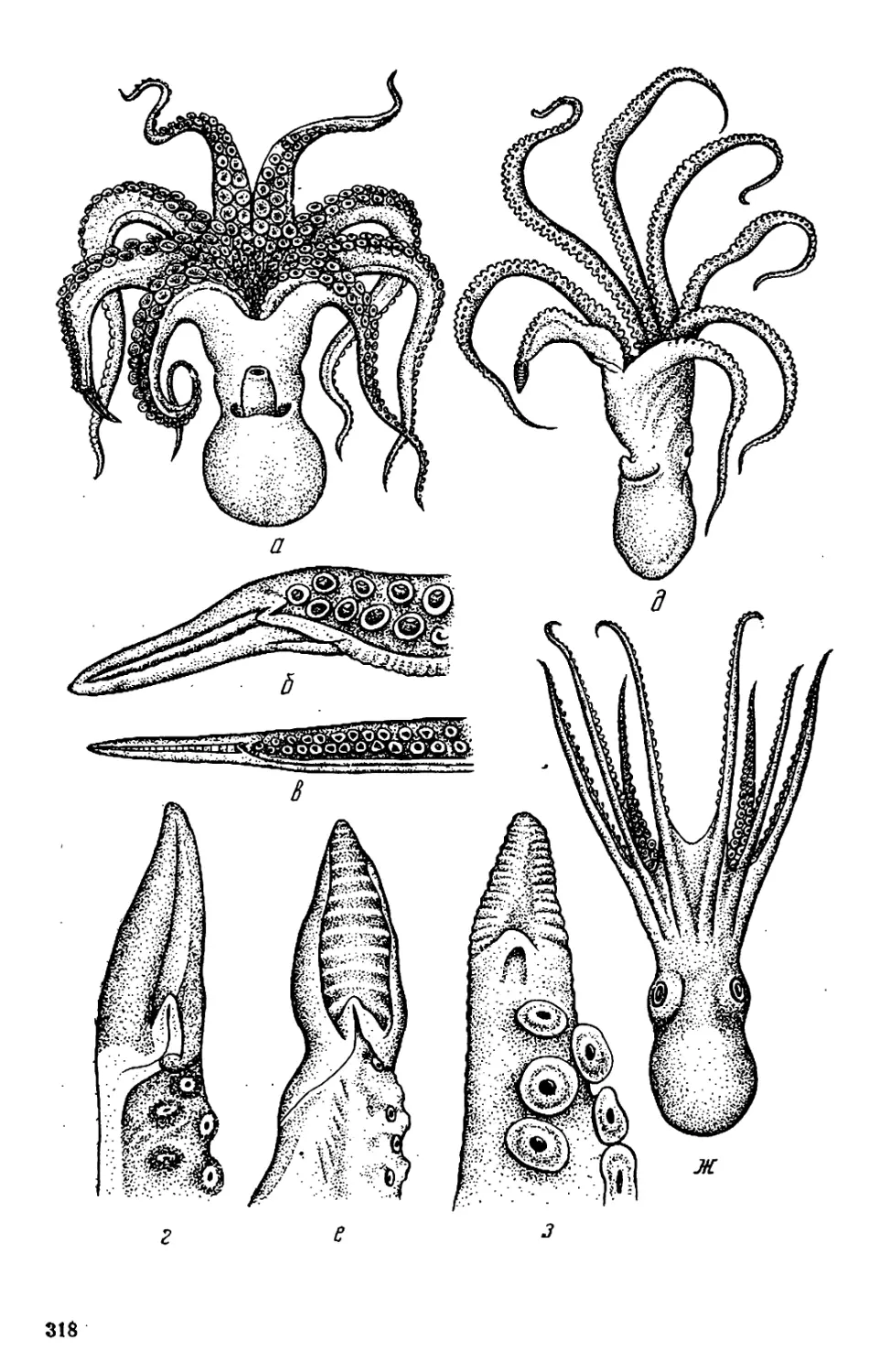




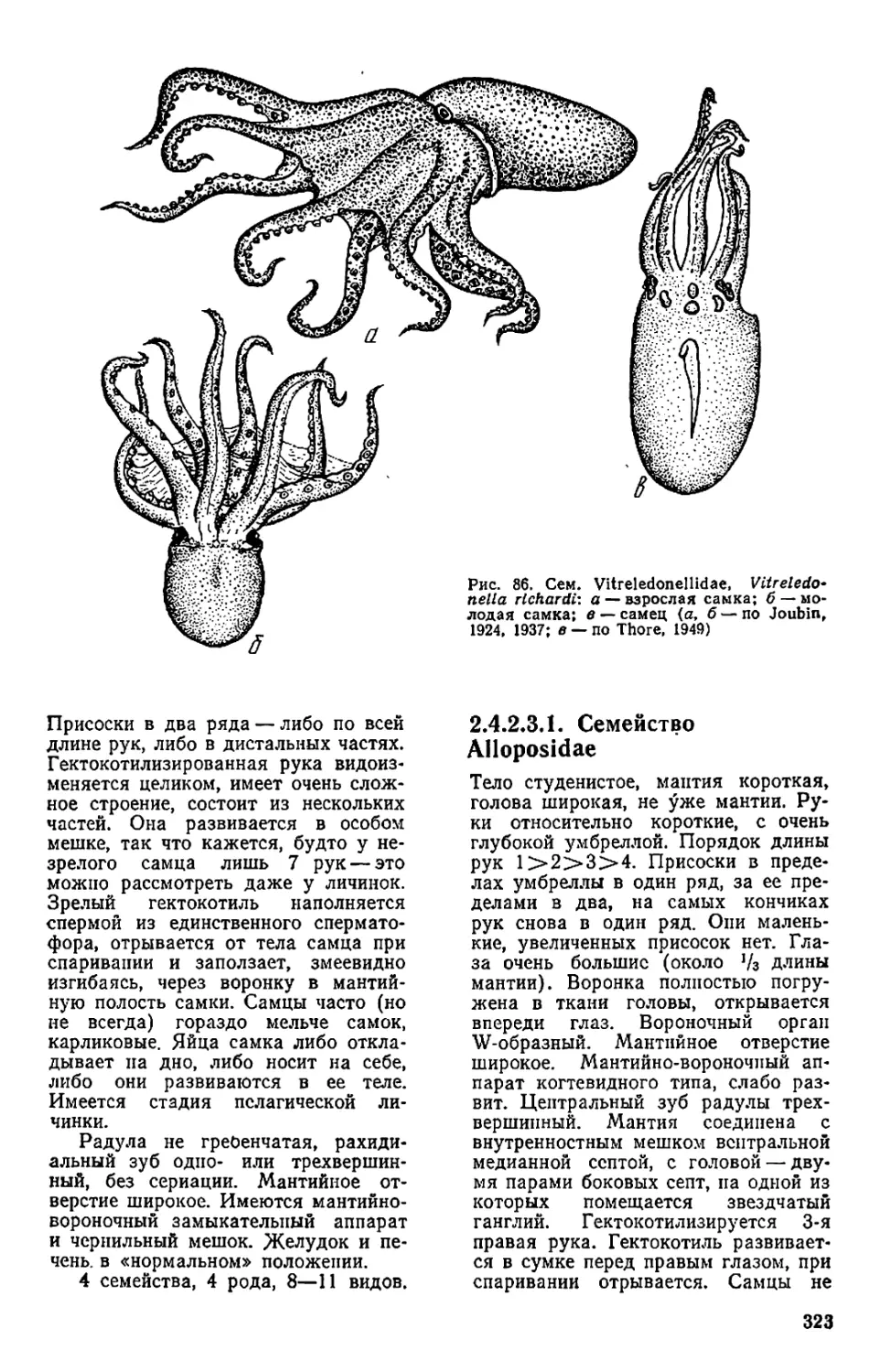








































Текст
К.Н.НЕСИС
Краткий определитель
головоногих
моллюсков
Мирового океана
Москва
•Легкая и пищевая промышленность-
1982
ББК 47.2
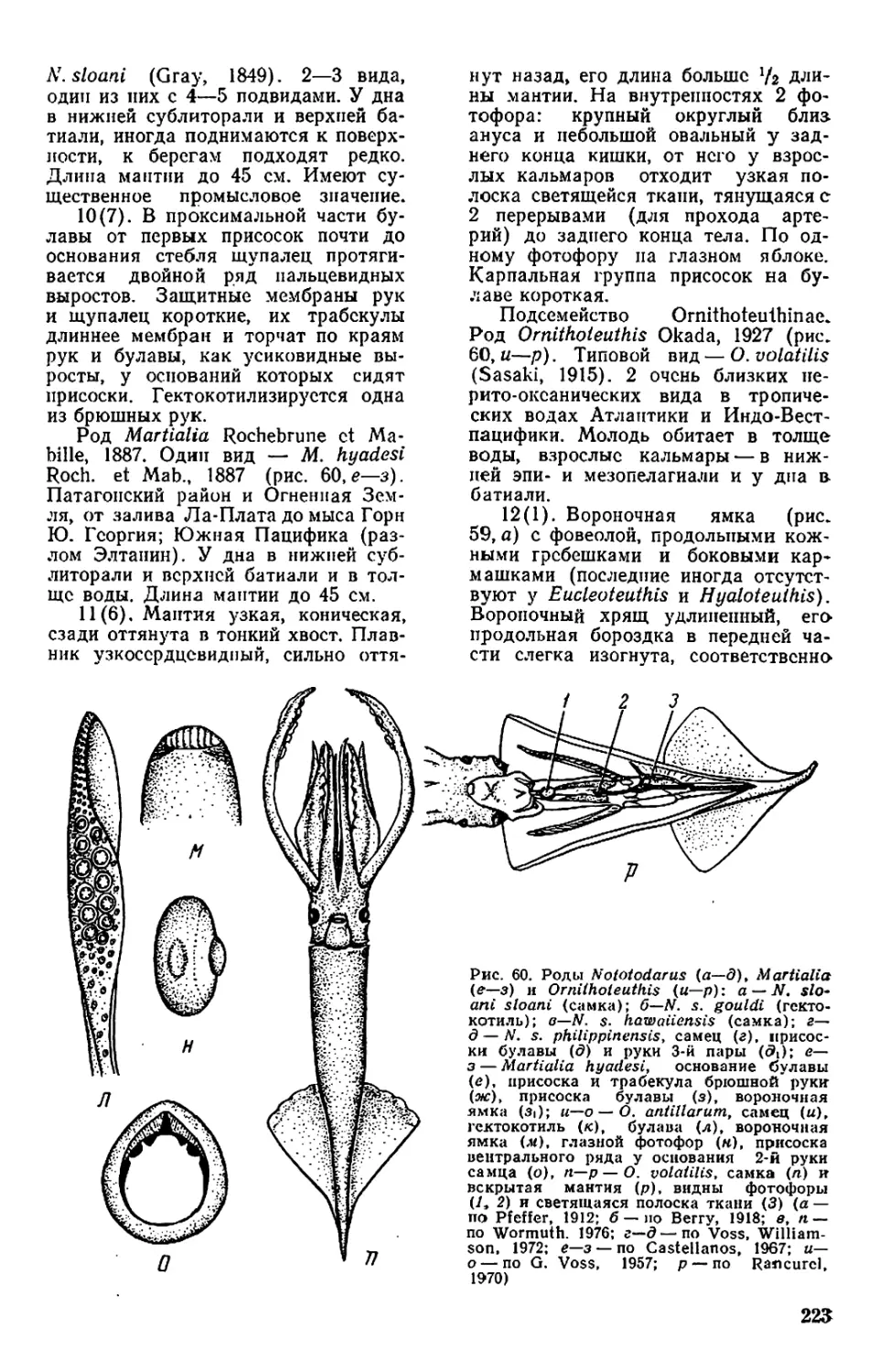
Н 55
УДК 594.582.1+639.27
К. н. Несис.
Краткий определитель головоногих моллюсков Мирового океа-
на.— М.: Легкая и пищевая пром-сть, 1982.—360 с.
Книга представляет собой первый в мировой литературе определитель
головоногих моллюсков Мирового океана. В нее включены все виды кальмаров
и каракатиц мировой фауны и большинство осьминогов, за исключением лишь
плохо изученных мелководных тропических осьминогов Индо-Пацификн, по
которым определение доводится до рода и подрода. Даны иллюстрированные
определительные таблицы отрядов, семейств, родов и видов головоногих
моллюсков. Для каждого вида приводятся сведения о распространении и
вертикальной зоне обитания. Общая часть определителя включает подробное
описание анатомии, морфологии и физиологии головоногих моллюсков.
Приведена библиография по строению, физиологии, таксономии и распростране-
нию головоногих моллюсков.
Предназначен для работников морских рыбохозяйственных учреждений и
промысловых разведок, морских биологов, зоологов, физиологов.
Ил. 88. Библ. — 470 назв.
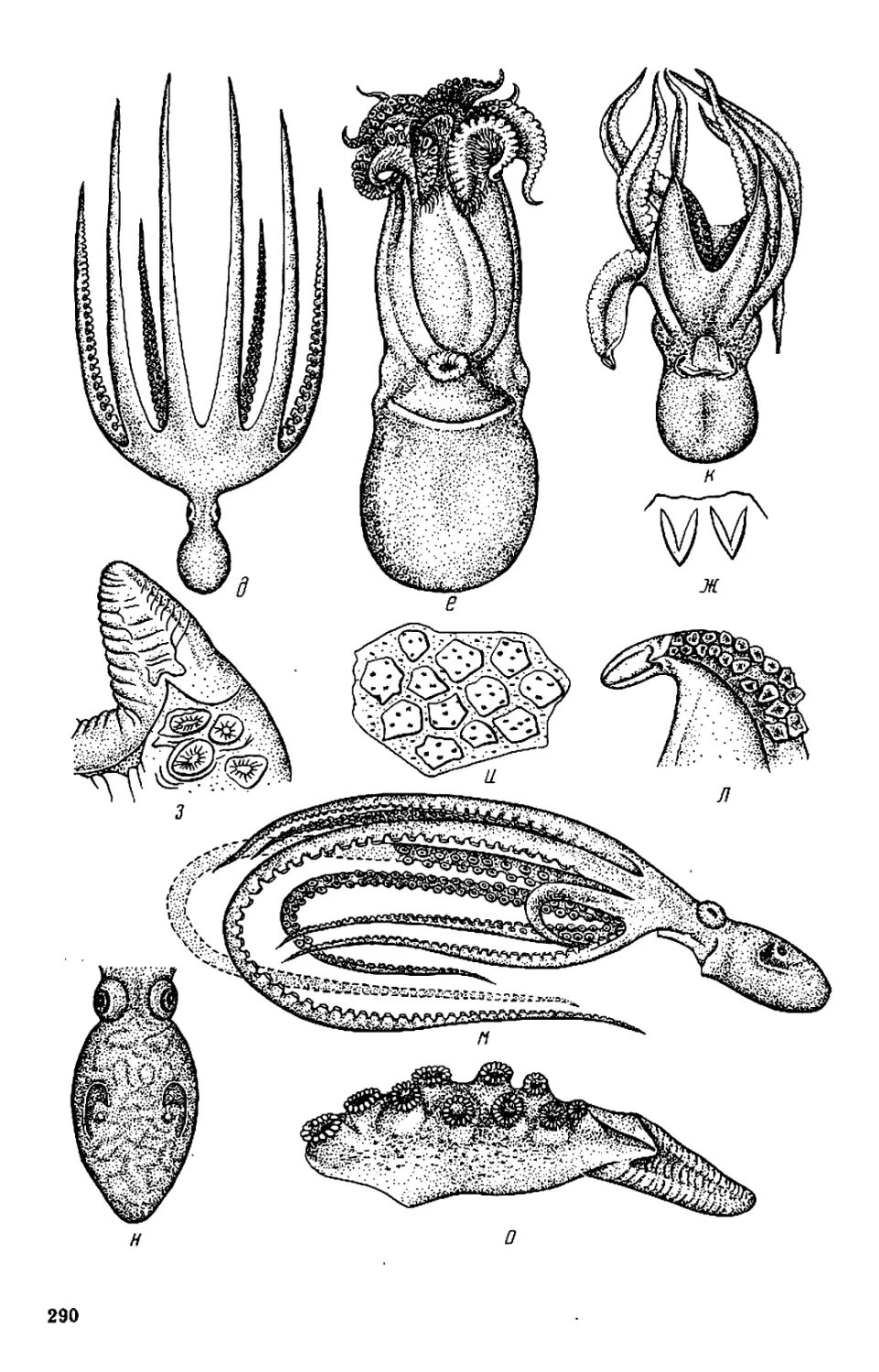
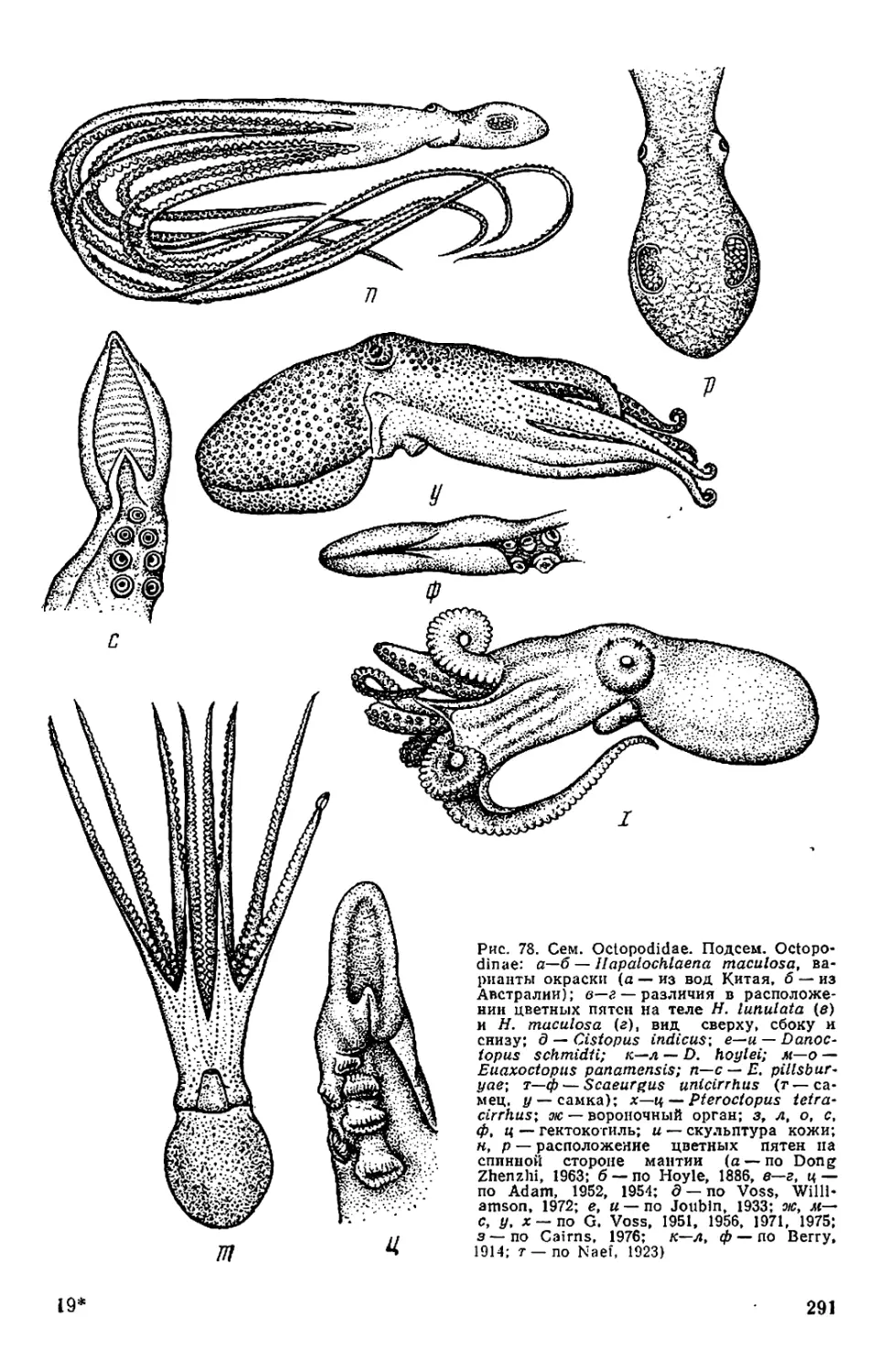
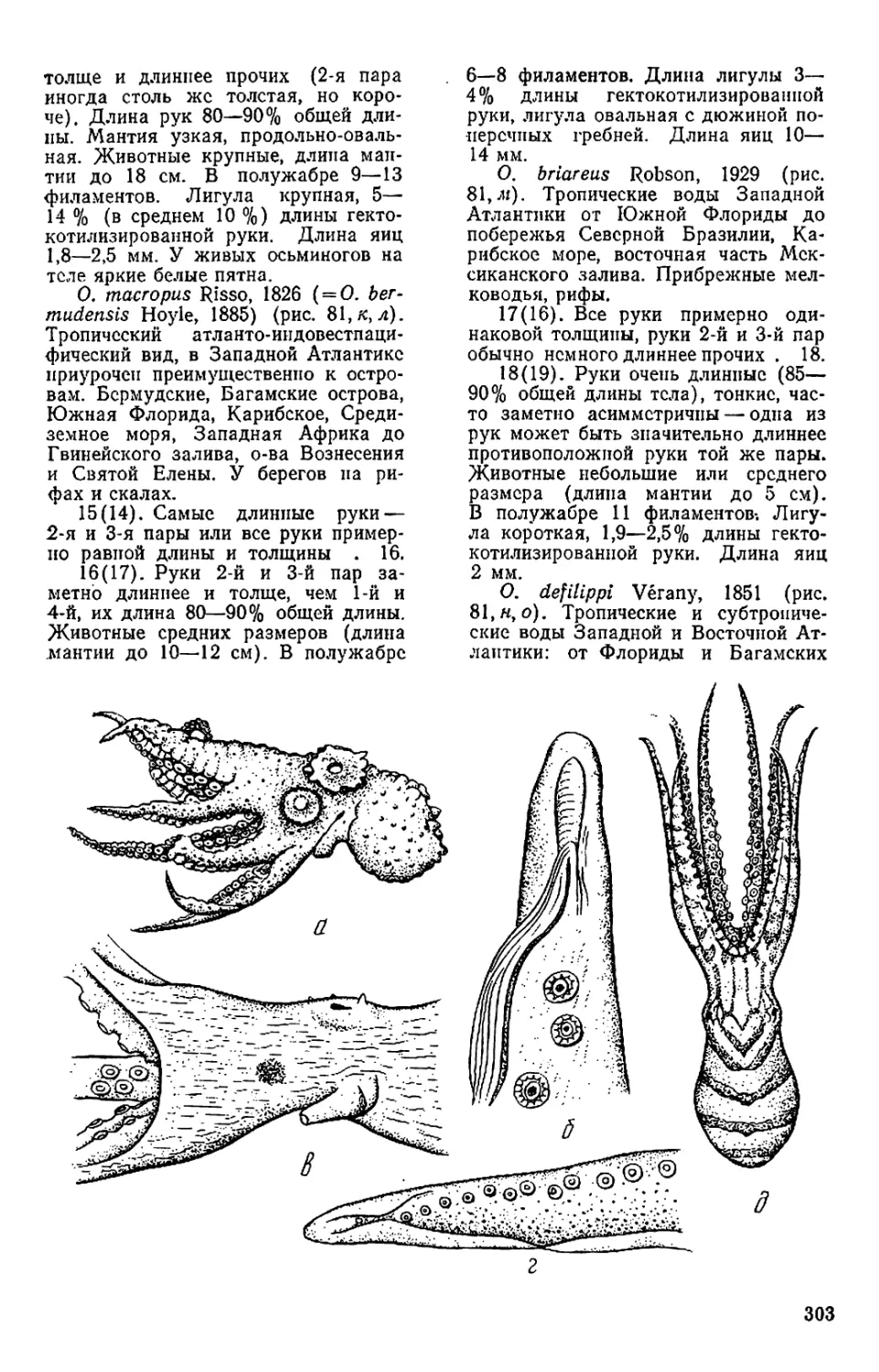
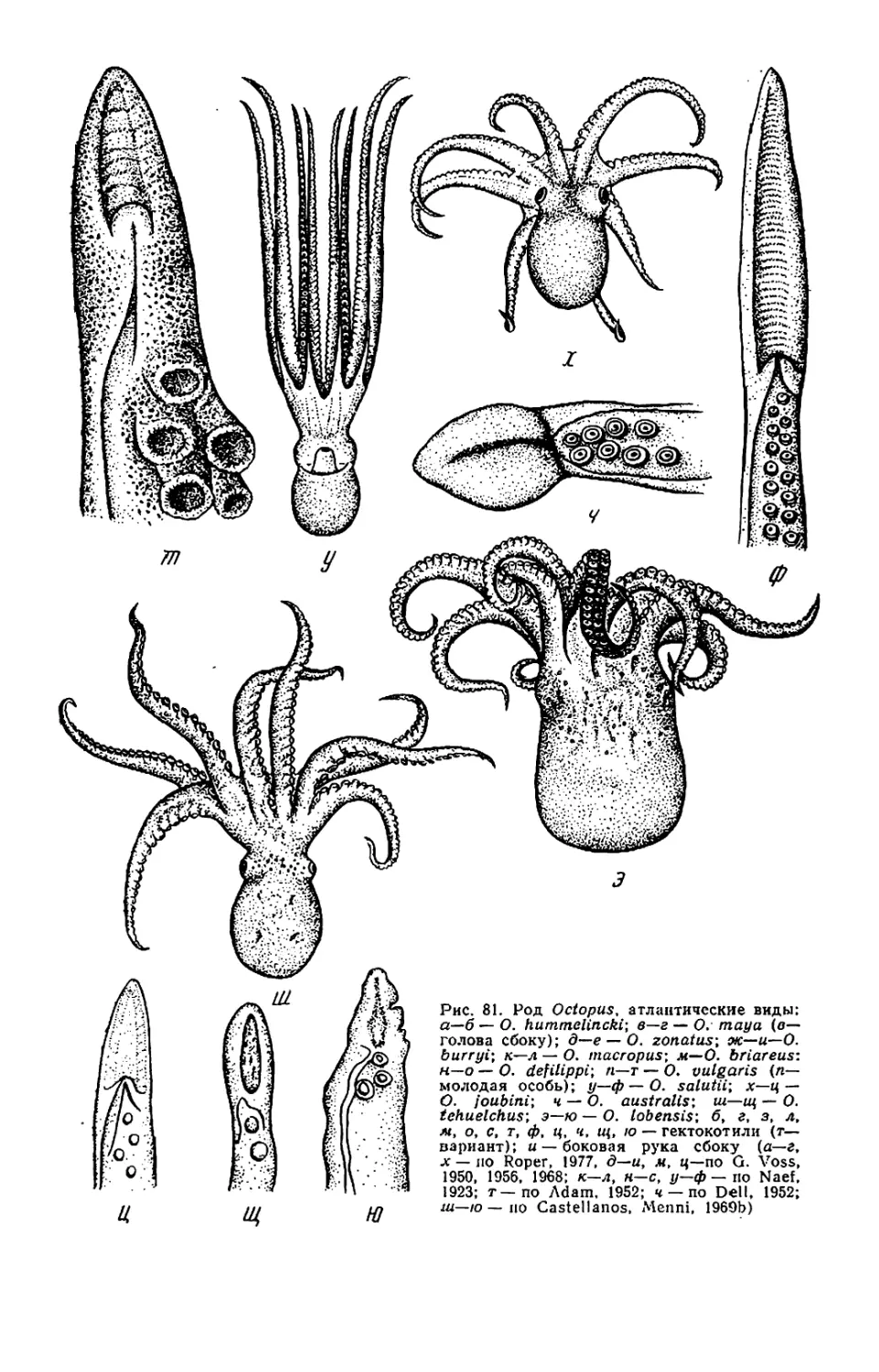
На форзаце: Octopus yendoi и Graneledone boreopacifica n. sp.
Рецензент P. H, Буруковский.
Иллюстрации к книге выполнены Н. А. ДУБОВКИНОЙ,
к форзацу —Н. Н. КОНДАКОВЫМ.
4003030000 IH?
II IH’’
04-1(01) Н',’
© Издательство «Легкая и пищевая
промышленность», 1982 г.
ПРЕДИСЛОВИЕ
За последние годы наблюдается непрерывный и быстрый рост ин-
тереса к головоногим моллюскам — наутилусам и спирулам,
кальмарам, каракатицам и осьминогам. Причины этого интереса
разнообразны. Головоногие моллюски — важный и чрезвычайно
перспективный объект промысла. Высокие пищевые качества, ко-
роткий жизненный цикл и необыкновенно быстрый рост (некоторые
тропические осьминоги достигают массы 3 кг за 8—10 мес) делают
их перспективными объектами марикультуры. Для нейрофизиоло-
гов кальмары — один из важнейших объектов для исследования
процессов проведения нервного импульса. Психологи интересуются
головоногими моллюсками как беспозвоночными с высокоразвитой
психикой («приматами моря»). Не меньший интерес представляют
они и для этологов. Новейшие открытия показали, что немногие из
морских животных могут соперничать с головоногими моллюсками
как источником важных и перспективных биологически активных
препаратов (противошоковых, обезболивающих, антипаразитарпых
веществ, мощных токсинов, цитостатиков и т. п.). Для палеонтоло-
гов изучение современных головоногих, в особенности наутилуса, —
ключ к познанию образа жизни и особенностей фоссилизации и за-
хоронения ископаемых паутилоидей, аммонитов, белемнитов.
Этот растущий поток исследований сталкивается с немаловаж-
ной трудностью — отсутствием определителей головоногих мировой
фауны. Определители локальных фаун обычно охватывают лишь
прибрежные виды, в последние же годы все большее значение при-
обретают виды открытого океана, а по ним определители практи-
чески отсутствуют. Предлагаемый вниманию читателя краткий оп-
ределитель головоногих моллюсков Мирового океана хотя бы отча-
сти заполняет этот пробел.. В него включены все без исключения
валидные роды и подроды современных головоногих моллюсков и
все виды наутилоидей, каракатиц, неритических и океанических
кальмаров и осьминогов, за исключением лишь некоторых австра-
лийских видов рода Sepia, описанных только по раковине, и дон-
ных осьминогов из родов Octopus и Benthoctopus. Эти роды не ре-
визованы, и составить полные определительные ключи для всех ви-
дов оказалось невозможным. Пришлось ограничиться составлени-
ем ключей для лучше изученных видов Benthoctopus п всех видов
Octopodinae Атлантического и северной части Тихого океанов.
3
Предлагаемый определитель краткий, так как он включает толь-
ко определительные таблицы, но не дает подробных описаний каж-
дого вида. Дать такие описания — значило бы ие только отложить
выпуск пособия на годы, но и сделать его чрезмерно объемистым
и малопригодным для использования в полевой обстановке.
Определительные таблицы предваряются сжатым очерком мор-
фологии и анатомии головоногих моллюсков с упором на описание
двужаберных (внутреннераковинных) и с предпочтительным опи-
санием тех признаков, которые имеют наибольшее значение при
определении видов. Обширная литература по частным вопросам
изучения современных головоногих моллюсков пе приводится,
в список литературы включены общие работы по головоногим мол-
люскам, определители по отдельным группам (отряды, семейства,
роды) и локальным фаунам (отмечены*), современные каталоги и
списки видов отдельных регионов, а также немногочисленные рабо-
ты, специально цитируемые в тексте.
Автор глубоко признателен своим друзьям, коллегам, помогав-
шим при сборе материала, передававшим свои сборы для опреде-
ления, читавшим в рукописи те или иные части книги и высказав-
шим свои замечания: В. Э. Беккеру, Г. М. Беляеву, Т. Н. Беляни-
ной, О. Д. Бородулиной, Р. Н. Буруковскому, А. Н. Вовку, Т. С. Ду-
бининой, Г В. Зуеву, Э. С. Кармовской, Н. И. Кашкину, Ч.М. Ниг-
матуллину, Н. В. Ларину, М. А. Пинчукову, Т. С. Рассу, Ю. И. Са-
зонову, Б. И. Федоряко, Ю. А. Филипповой, Ю. М. Фроермапу,
А. Г. Цокуру, Ю. Г Чиндоновой, В. М. Чувасову, Г. А. Шевцову,
Ю. Н. Щербачеву. Указатель составлен И. В. Никитиной и
Л. В. Перегудой.
ВВЕДЕНИЕ
Головоногие моллюски — наутилусы, каракатицы, кальмары, вам-
пироморфы и осьминоги — высший класс типа моллюсков. Ниже
приведены систематическое положение и подразделение класса
головоногих моллюсков.
Царство Animalia — животные
Подцарство Metazoa — многоклеточные
Надраздел Entcrozoa — энтерозои
Раздел Bilateralia — билатеральные
Надтип Trochozoa — трохофорные
Тип Mollusca — моллюски
Подтип Conchifera — раковинные
Класс Cephalopoda — головоногие
Класс головоногих моллюсков делится на 5 подклассов со мно-
жеством отрядов. В современной фауне представлено два подклас-
са: Nautiloidea с единственным, отрядом Nautilida, семейством Nau-
tilidae и родом Nautilus и Coleoidea, который делится на 4 отряда:
каракатиц Sepiida (подотряды спирул Spirulina и собственно ка-
ракатиц Sepiina), кальмаров Teuthida (подотряды неритических
кальмаров Myopsida и океанических кальмаров Oegopsida), вампи-
роморф Vampyromorpha (единственный вид Vampyroteuthis infer-
nalis) и осьминогов Octopoda (подотряды плавниковых осьминогов
Cirrata и собственно осьминогов Incirrata). Отряды каракатиц и
кальмаров иногда объединяют в группу Decapoda. В общей части
определителя будет дана характеристика только двужаберных го-
ловоногих. Характеристику четырехжаберных см. на с. 71.
Определительные таблицы составлены главным образом по.
внешним признакам, не требующим вскрытия животного, и, по воз-
можности, так, чтобы учесть признаки строения неполовозрелых;
и половозрелых особей обоего пола. К сожалению, это не всегда
удавалось, так как во многих группах каракатиц, кальмаров и ось-
миногов, особенно обитателей шельфа, виды уверенно различаются
лишь по взрослым самцам. Определение самок и неполовозрелых-
особей в этих группах крайне трудно, а иногда практически невоз-
можно, так что приходится ограничиваться лишь идентификацией
рода или подрода. Определение личинок и ранней молоди по при?
водимым в книге таблицам также практически невозможно; табл
лицы с 'описанием пелагических личинок й молоди головоногих мол-;
люсков публикуются в издаваемом Зоологическим инсти,тутом АЙ
СССР «Полевом определителе планктона».
5
ОБЩАЯ
ХАРАКТЕРИСТИКА
ГОЛОВОНОГИХ
моллюсков
Класс Cephalopoda — двусторонне-
симметричные моллюски с наружной
(Nautilus) или внутренней раковиной,
которая у настоящих каракатиц (Sc-
piidae) представлена известковой
пластинкой, у спирулы — закручен-
ной в «бараний рог» раковинкой, у
кальмаров — тонкой прозрачной рого-
вой пластинкой (гладиусом), у части
каракатиц и у осьминогов она ре-
дуцирована или отсутствует.
Туловище (рис. 1) одето кожно-
мускульным мешком, мантией, кото-
рая служит основным движителем
при быстром (реактивном) плавании.
Внутри мантии находятся органы
мантийного комплекса — жабры
(одна пара у двужаберных голово-
ногих и две у наутилуса) и органы
пищеварительной, кровеносной, по-
ловой и выделительной систем. Круп-
ная голова у моллюска обычно (за
исключением некоторых осьминогов)
хорошо отделена от туловища и не-
сет высокоразвитые (кроме наутилу-
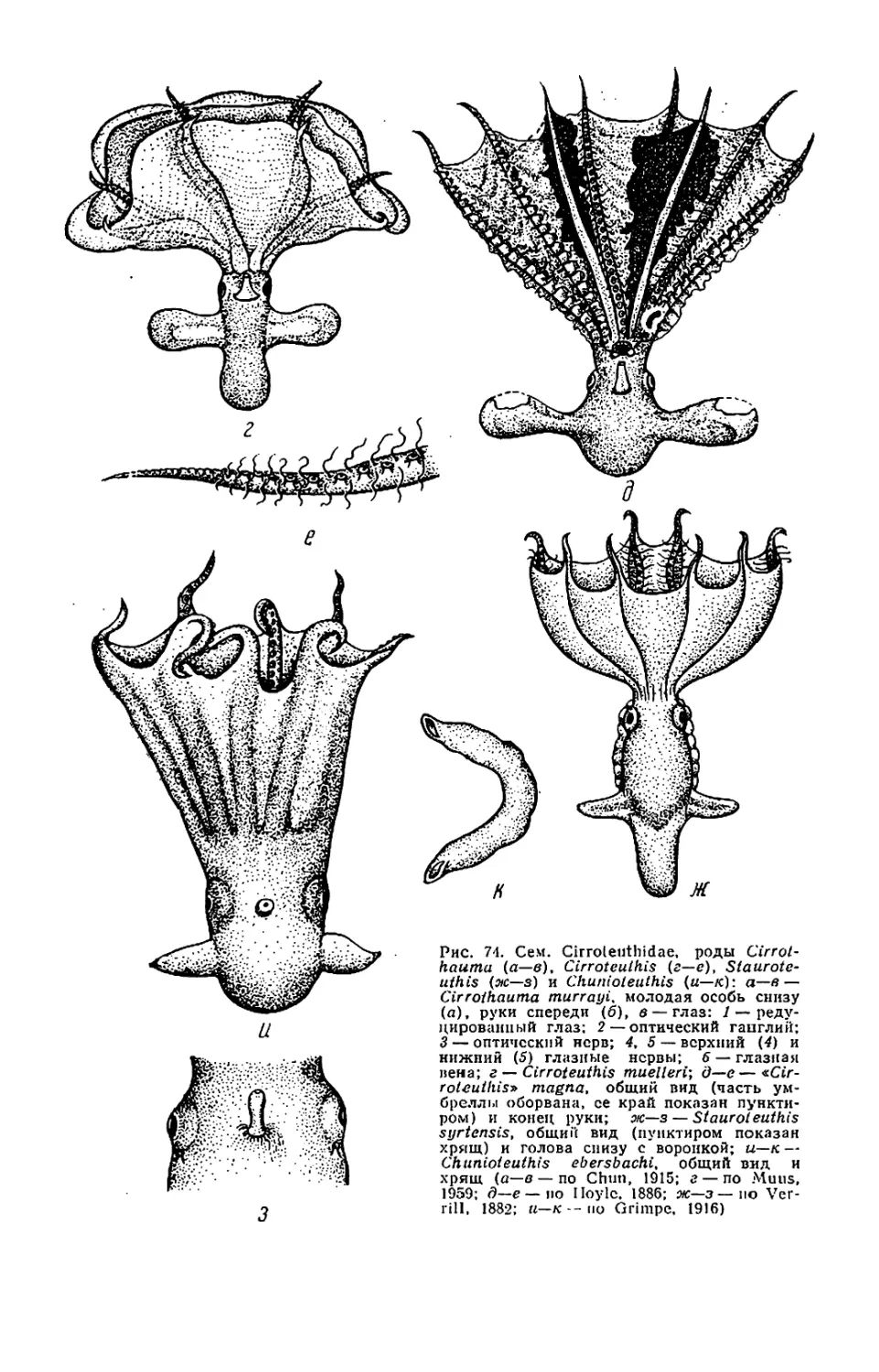
са и Cirrothaurna, см. рис. 74, в)
глаза, венец конечностей, окружаю-
щих ротовое отверстие, и воронку.
У двужаберных головоногих 8 или
10 пар конечностей: у кальмаров и
каракатиц всегда (по крайней мере,
на личиночной стадии) 8 рук и па-
ра щупалец, которые находятся ме-
жду брюшно-боковыми и брюшными
руками и резко отличаются от рук
по строению и вооружению; у вам-
пироморф 8 рук и пара тонких ните-
видных филаментов, которые поме-
щаются в особых сумках между
спинными и спинно-боковыми руками
(см. рис. 73); у осьминогов только
8 рук. Конечности двужаберных го-
ловоногих снабжены присосками, ко-
торые у кальмаров и каракатиц не-
сут роговые кольца с зубцами; у не-
которых океанических кальмаров
присоски превращаются в крючья.
В голове помещается крупный
6
высокоразвитый головной мозг, оде-
тый хрящевой капсулой. У кальма-
ров и каракатиц особенно развиты
оптические доли мозга, перерабаты-
вающие информацию, полученную
глазами. С оптическими железами
связан также орган внутренней сек-
реции (оптическая железа). На ниж-
ней стороне головы находится ворон-
ка — коническая, суженная впереди
трубка. Через нее выбрасывается во-
да из мантии при дыхании и реак-
тивном плавании, выводятся наружу
чернильная жидкость, моча, экскре-
менты, яйца и вещества, из которых
строится яйцевая капсула. Боковые
стороны воронки срастаются с ман-
тией или соединяются с ней посред-
ством замыкательных хрящей, дей-
ствующих по принципу застежки-
кнопки. Хрящи помещаются на пра-
вой и левой сторонах воронки и на
соответствующих им местах внутрен-
ней стороны мантии по бокам от
средней линии, недалеко от передне-
го края. Голова на затылке также
соединяется с мантией хрящом или
срастается с ней. Рот снабжен креп-
ким роговым клювом, похожим на
клюв попугая и состоящим из ниж-
ней и верхней мандибул; во рту по-
мещается радула (отсутствует у спи-
рул и плавниковых осьминогов).
На заднем конце или в средней
части мантии находится пара плав-
ников — основной пропульсивный ор-
ган при медленном плавании. У ось-
миногов Incirrata плавников нет, хо-
тя в эмбриональном развитии они
закладываются. Гладкая кожа со-
держит многочисленные пигментные
тельца — хроматофоры нескольких
типов, пассивно отражающие свет
«зеркальца» (иридоциты), и — у мно-
гих океанических кальмаров и неко-
торых осьминогов — органы свече-
ния, фотофоры.
К органам чувств двужаберных
головоногих относятся: глаза, слож-
но устроенные статоцисты — органы
р/1 нновесия и восприятия линейных и
угловых ускорений, помещающиеся
и задней части головной капсулы;
обонятельные папиллы, или ямки,
многочисленные хемо- и механорс-
центоры; особого рода внеглазпые
фоторецепторы, которые у каракатиц
и кальмаров помещаются на голов-
ном мозге (парольфакторные пузырь-
ки), у осьминогов — на внутренней
гюронс мантии на звездчатом ганг-
лии (эпистеллярные тельца), а у вам-
пироморф— в передней части ман-
тийной полости.
Органы дыхания — жабры. Ос-
новные органы кровеносной систе-
мы— сердце и пара дополнительных
жаберных сердец; кровеносная систе-
ма почти замкнута, основной дыха-
тельный пигмент — гемоцианин. Ор-
ганы выделения включают пару по-
чечных мешков и придатки жабер-
ных сердец; процессы выделения
происходят также в жабрах и при-
датках полой вены и пищеваритель-
ного протока.
Органы пищеварительной систе-
мы включают: две пары слюнных
желез, из которых задняя часто вы-
деляет токсины; пищевод; зоб (у не-
которых осьминогов и вампиро-
морф); желудок; цекум (слепой вы-
рост желудка); пищеварительную
железу, обычно называемую пече-
нью; пищеварительный проток с при-
датками («поджелудочная железа»)
и кишку, в которую впадает проток
чернильной железы.
Головоногие моллюски раздель-
пополы. Органы размножения вклю-
чают непарную гонаду (семенник,
яичник); у самок — парный или не-
парный яйцевод с придаточными же-
лезами — яйцеводной, придаточной
яйцеводной и пидаментальными
(имеются нс у всех головоногих);
у самцов — как правило, непарный
ссмепровод со сложной системой
придаточных желез, в которых обра-
зуются сперматофоры — особые фор-
мирования для хранения и передачи
спермы — и спсрматофорпый (нидхе-
мов) мешок. Для передачи спермы
самке обычно одна или несколько
рук самца трансформируются цели-
ком или только на конце; измененная
рука или ее часть называется гекто-
котилсм. Яйца откладываются либо
в воду, либо на дно. Как правило,
они окружены дополнительными
оболочками, образующими яйцевую
капсулу. Яйца богаты желтком,
дробление дискоидалыюе меробла-
стическое. Развитие может быть пе-
лагическим (полным или сокращен-
ным) или бентосным (прямым). При
пелагическом развитии из яйца вы-
лупляется личинка, при бентосном —
молодой моллюск.
Головоногие моллюски могут при-
надлежать к планктонной, нектонпой,
нектопслагической (придонно-планк-
тонной), нектобснтосной или бентос-
ной жизненным формам.
Класс насчитывает 600—650 со-
временных видов, распространенных
от Центрального Полярного бассейна
до Антарктиды и от литорали до
ультраабиссали, но только в полно-
соленых морях и оксанах.
Размеры и масса
Стандартный метод измерения голо-
воногих моллюсков — измерение дли-
ны мантии по спинной стороне.
У осьминогов измеряется расстояние
от заднего конца тела до линии, со-
единяющей центры глаз; у тех видов
отряда каракатиц, голова которых
срастается с мантией, — от заднего
конца тела до середины затылочной
ленты. В тексте определителя под
размером моллюсков всегда понима-
ется длина мантии. Иные измере-
ния — общая длина (до конца самой
длинной руки или до конца вытяну-
тых щупалец) — оговариваются.
Ниже перечислены самые крупные
и самые мелкие виды кальмаров, ка-
ракатиц и осьминогов. Установить их
точные размеры не всегда возможно:
сообщения о гигантских головоногих
зачастую оказываются рыбацкими
байками, а в числе самых маленьких
часто упоминают личинок и молодь,
ошибочно описанных под самостоя-
тельными видовыми названиями. На-
до к тому же учесть, что у многих
кальмаров и осьминогов самцы соз-
ревают гораздо раньше самок и мак-
симальные размеры их бывают раза
в два меньше размеров самок.
7
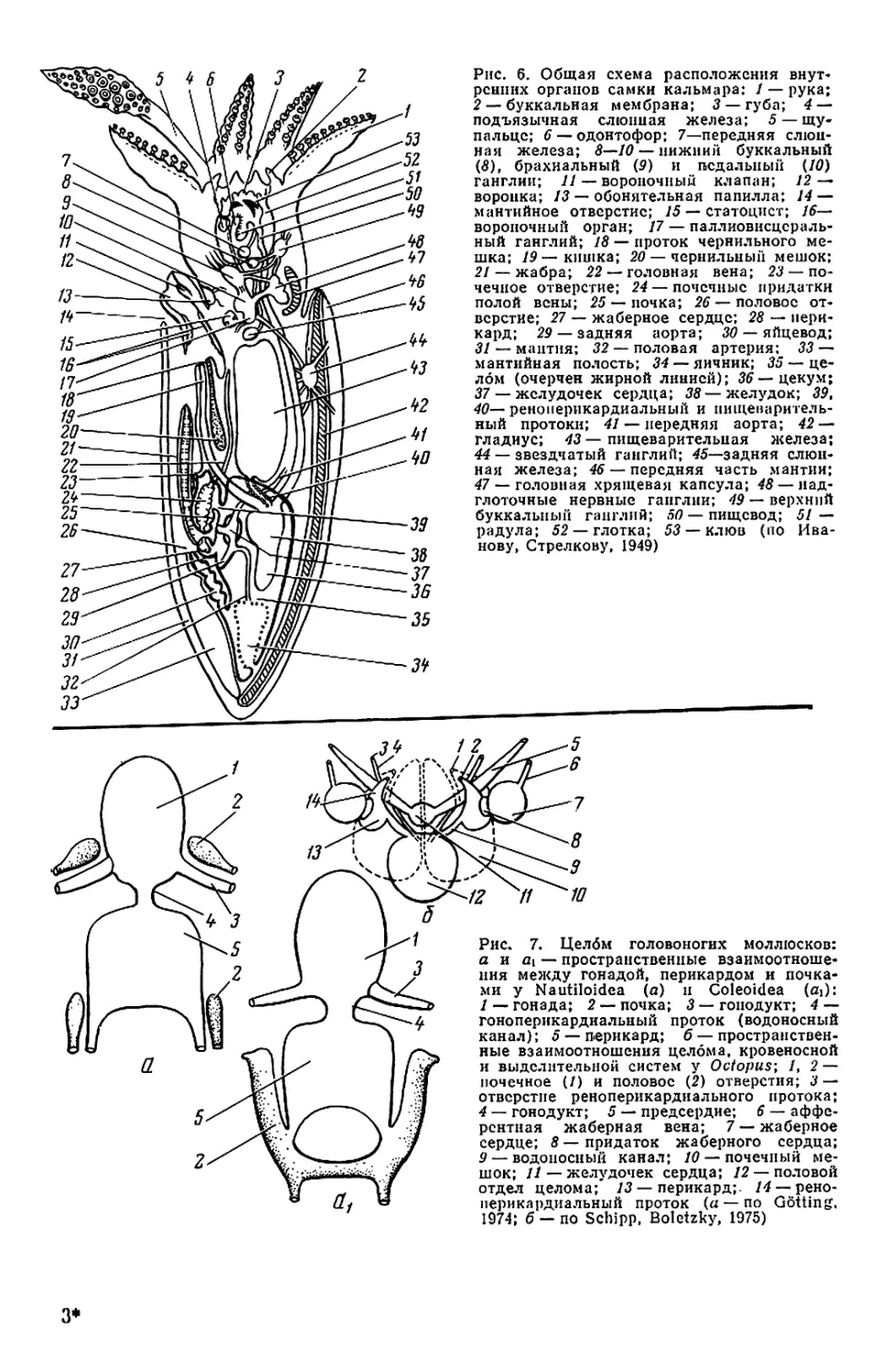
г
20
21
13
25
26
2
9
7
8
9
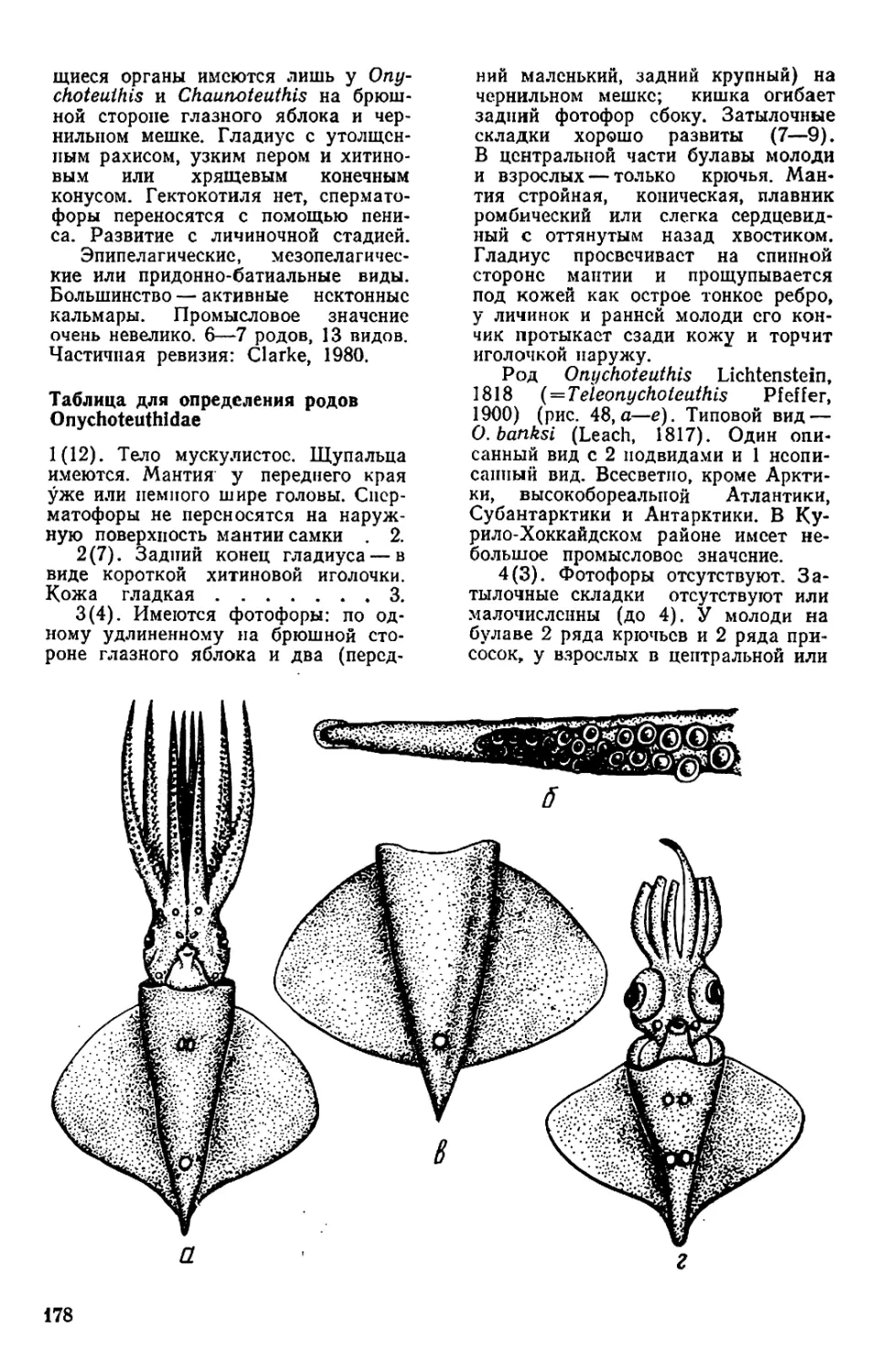
анатомия
кальмара
5
6
24
23
22
30 29 18
28
Б
7
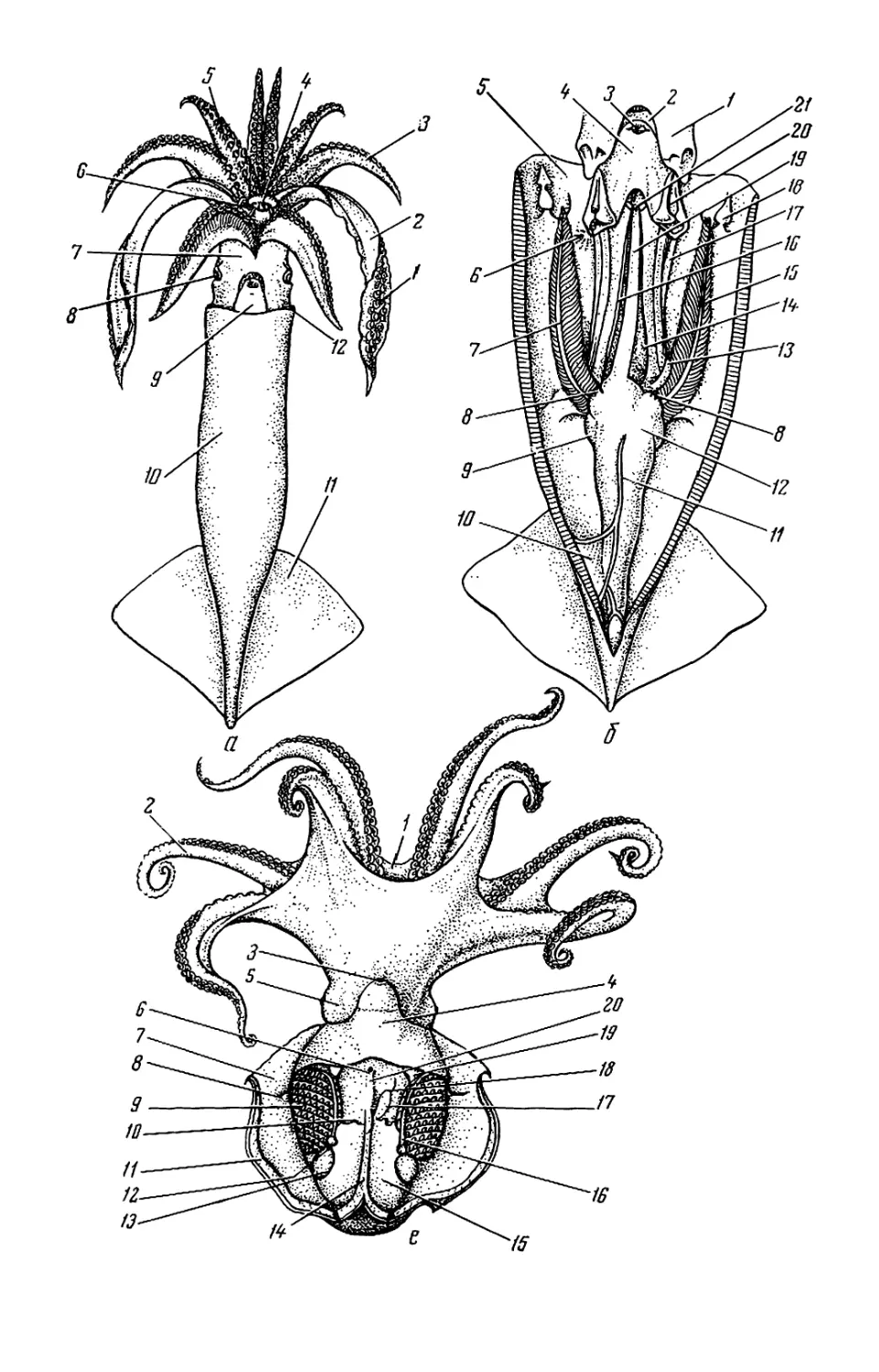
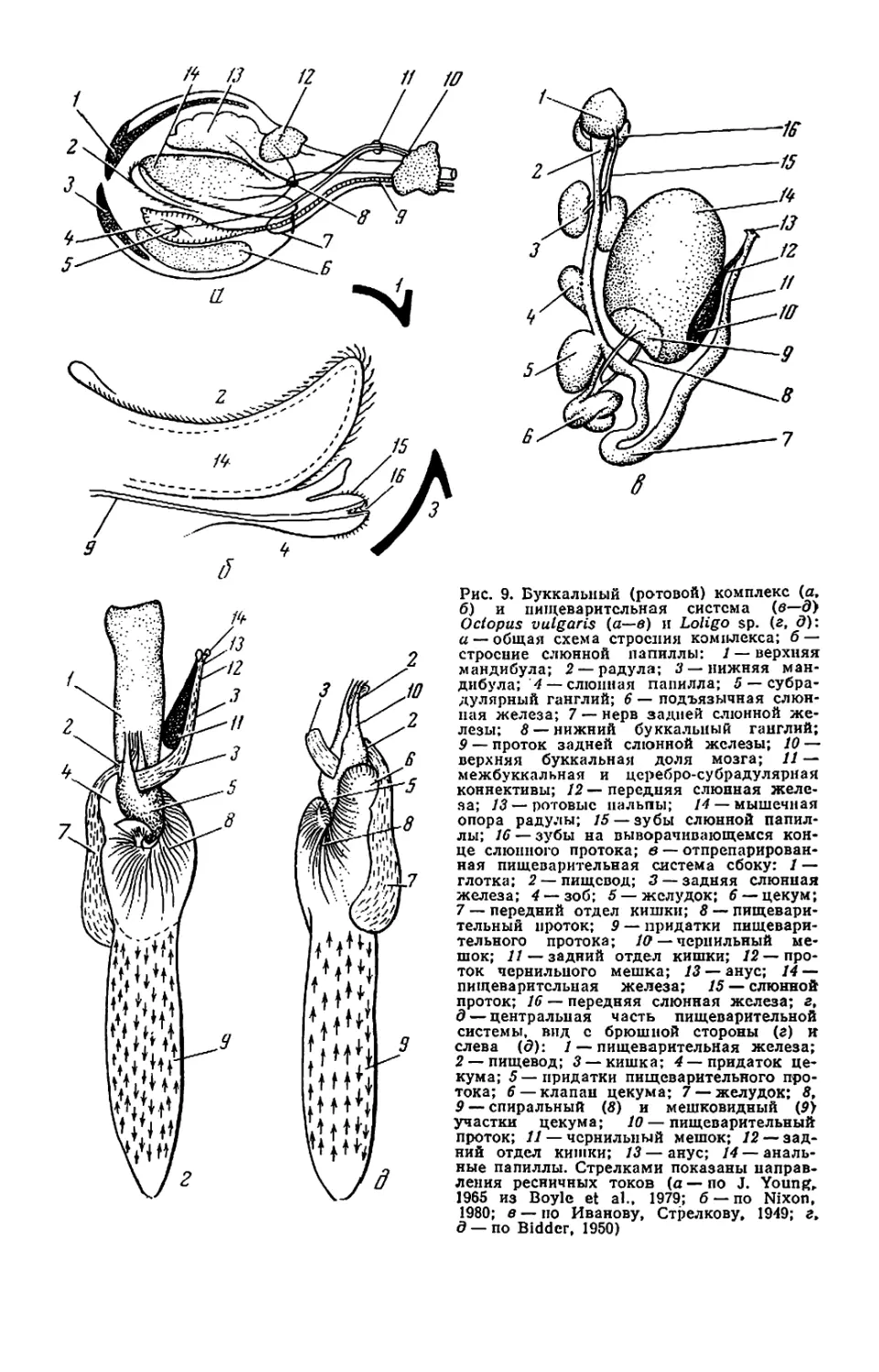
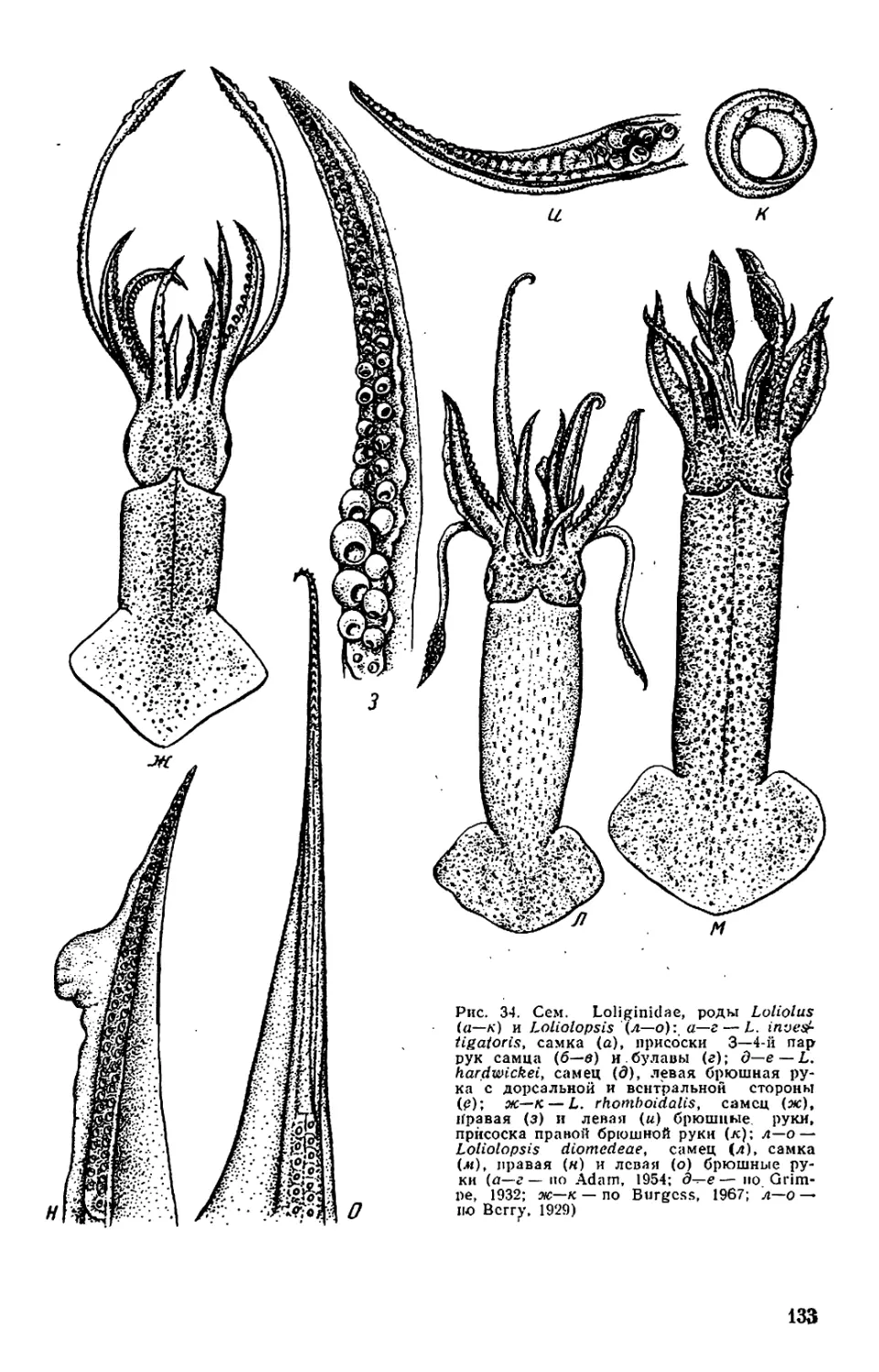
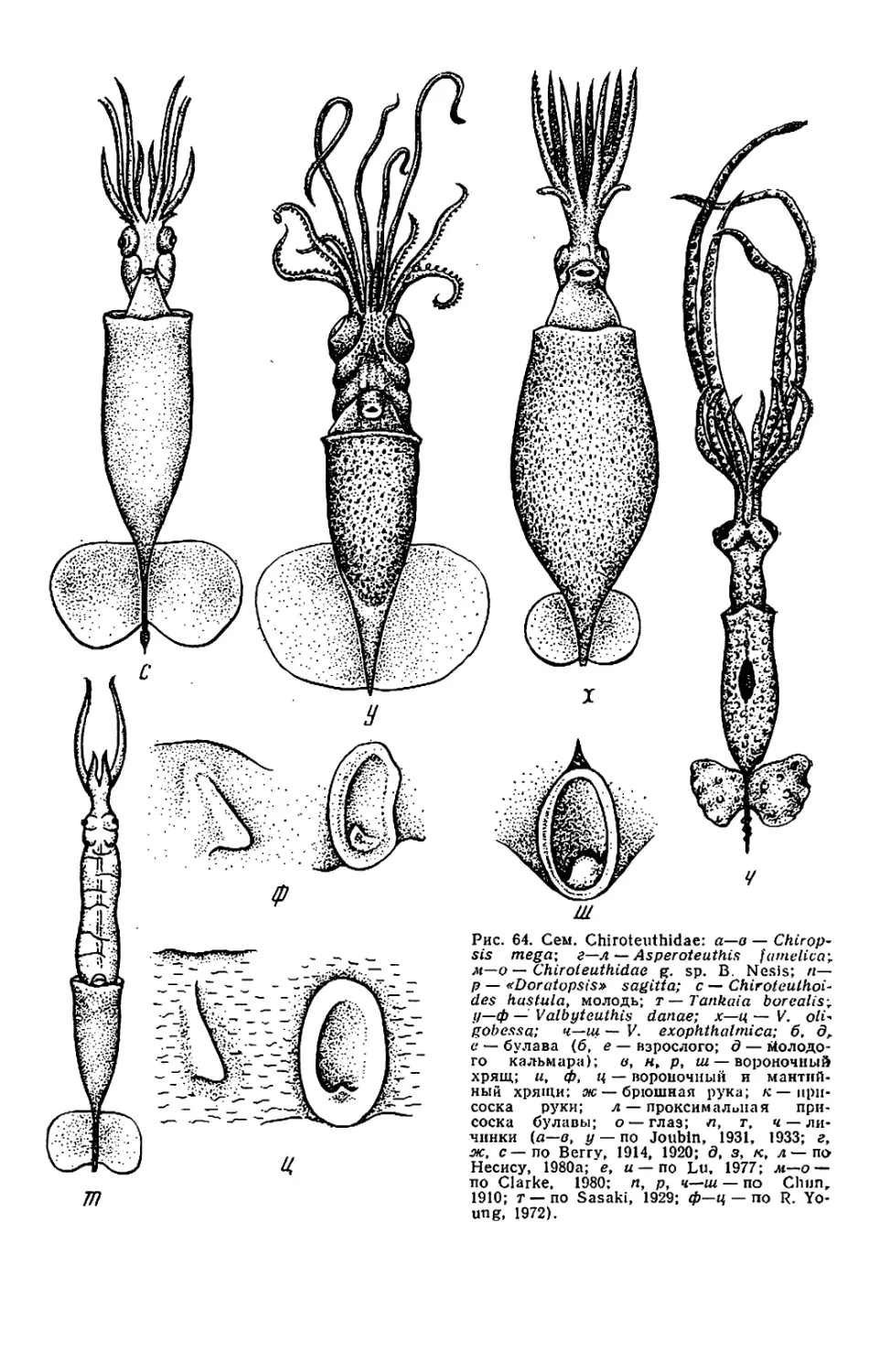
Рис. 1. Внешний вид и общая
мантийного комплекса органов _ г
(а—в), каракатицы (а) и осьминога (д—
ж): а—в—молодой самец Todarodes pad-
ficus-, а: 1 — булава щупальца; 2 — сте-
бель щупальца; 3 — рука; 4 — клюв; 5 —
присоски; 6 — буккальная (ротовая) во-
ронка; 7 — голова; 8 — глаз; 9 — воронка:
10— мантия; 11— плавник; 12— вход в
мантийную полость; б, в: / — голова; 2 —
вороночная ямка; 3 — вороночное отвер-
стие; 4 — воронка; 5 — мантия; 6 — звезд-
чатый ганглий; 7 — жабра; 8— почечное
отверстие; 9 — местоположение жаберного
сердца; 10 — брыжжейка; // — задняя аор-
та; 12 — внутренностная масса; 13 — пенис;
14 — чернильный мешок; 15 — жаберная
вена; 16 — головная вена; 17 — мускул-
втягнватель воронки; 18—мантийный хрящ;
19 — кишка; 20 — вороночный хрящ; 21 —
анус; 22—хрусталик глаза; 23— радужная
оболочка; 24 — глазной синус; 25 — мускул-
подниматсль воронки; 26 — вороночный
клапан; 27 — шейный мускул; 28 — пере-
понка. на которой подвешена жабра; 29—
мантийный компонент затылочного хряща;
30 — продольные шейные складки; г — сам-
ка Sepia officinalis'. 1 — щупальце; 2 — бук-
кальная воронка с ссмеприемником; 3 —
замыкательные хрящи; 4 — анус; 5 — по-
чечное отверстие; 6 — половое отверстие;
7 — жабра; 8 — внутренностная масса: 9 —
плавник; д, е — самец Octopus dofleini;
д: 1 — мантия: 2— надглазничный усик;
3 — глаз; 4 — голова; 5 — зрачок; 6 — рука;
7 —присоски; 8 — умбрелла; 9 — гектоко-
тиль: е: 1 — умбрелла; 2— рука; 3— во-
роночное отверстие: 4— воронка; 5 —глаз;
6 — анус; 7 — мантия; 8 — пленка, прикреп-
ляющая жабру к мантии; 9 — жабра; 10—
почечное отверстие: // — разрез стенки
мантии: 12 — жаберная железа: 13— жа-
берное сердце: 14 — мантийная септа: 15—
внутренностная масса; 16—жаберная ве-
на: 17 — пенис: 18 — половое отверстие;
19— мускул-втягпватель воронки; 20 —
кишка: ж—молодой Octopus vulgaris".
«конфликтная окраска» (а—в, д—е — по
Иванову, Стрелкову. 1949: г — по Kaestner,
1969; ж— по Packard, 1972)
Самый крупный кальмар — и са-
мое крупное беспозвоночное живот-
ное — гигантский кальмар Archi-
teuthis dux, общая длина со щупаль-
цами до 18 м, длина мантии до 5 м,
масса предположительно до 1 т.
Обычные размеры гигантских кальма-
ров: общая длина 6—13 м, длина
мантии 1,0—2,5 м, масса 50—300 кг.
•Самый маленький кальмар из Myop-
si da—Pickfordiateuthis pulchella имеет
длину мантии до 2,2 см, из Oegopsi-
<1а—Pterygioteuthis giardi — до 2,5 см.
Самые крупные каракатицы — за-
падноафриканская Sepia officinalis
hierredda и S. latimanus с о-вов Рю-
кю; длина мантии до 50 см, масса до
10—12 кг. Немного уступает им по
размерам австралийская S. арата,
длина мантии которой до 48 см. Са-
мые маленькие из настоящих кара-
катиц-сепиид — южноафриканская
5. typica: длина мантии самцов до
2 см, самок до 2,5 см. Еще мельче
тоже южноафриканские S. robsoni и
S. dubia (до 17 мм), по они извест-
ны лишь но единичным особям. Чрез-
вычайно мелки также каракатицы
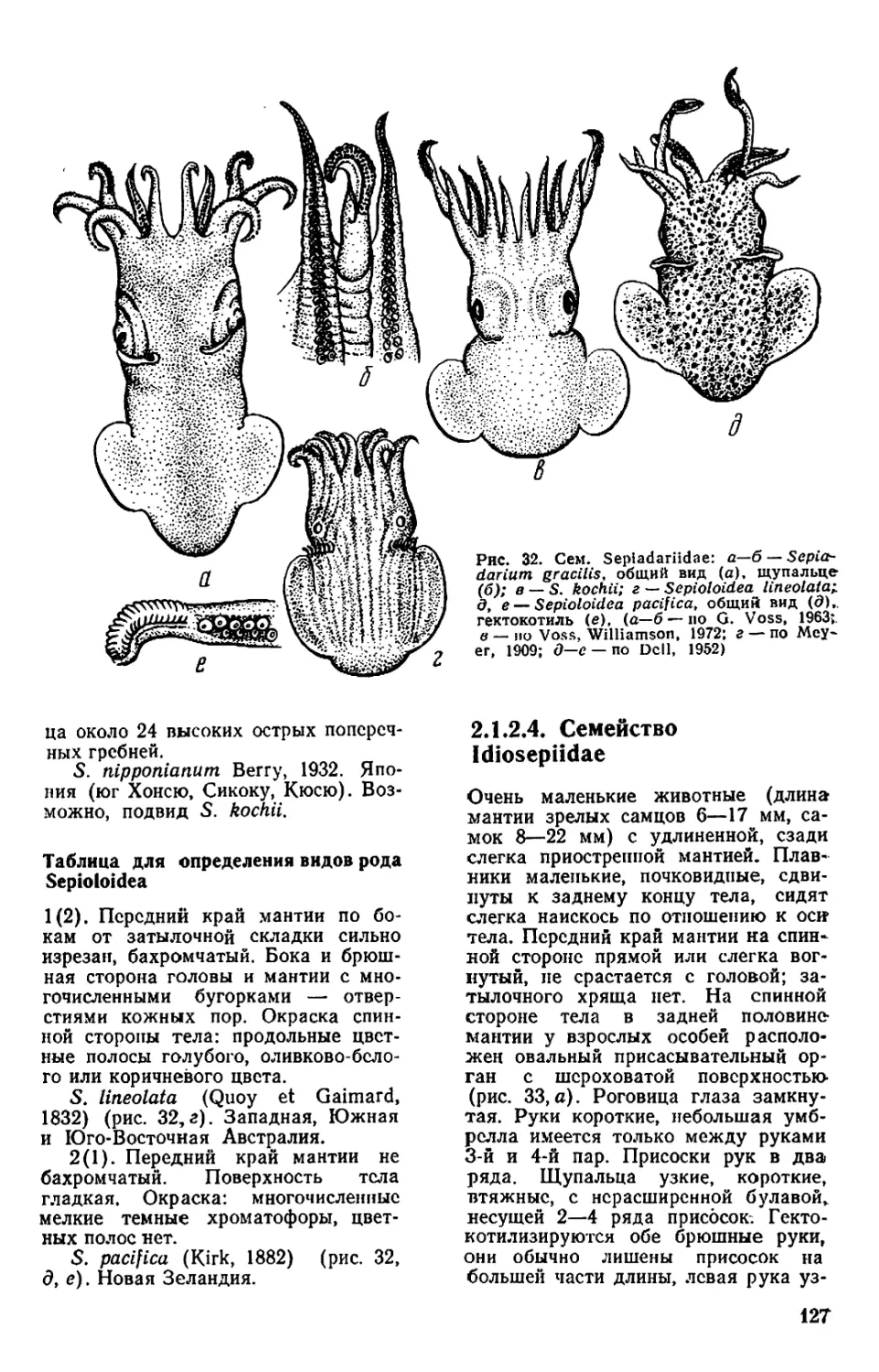
сем. Idiosepiidac (род Idiosepius):
длина мантии зрелых самцов 6—
17 мм, самок 8—22 мм, а также Ron-
deletiola minor из сспиолид (до
15 мм). В одном из рейсов «Витязя»
в море Хальмахсра (Индонезия) бы-
ла выловлена почти зрелая самка се-
пиолиды, относящаяся к новому ро-
ду и виду (в определитель не вклю-
чен). Длина ее мантии только
8,3 мм — это самый маленький из из-
вестных головоногих моллюсков.
Самый крупный из плавниковых
осьминогов — абиссальный Cirroteu-
this п. sp: его общая длина до 1,5 м.
Это самое крупное из абиссальных
беспозвоночных. Среди собственно
осьминогов рекордсмен — гигантский
тихоокеанский осьминог Octopus dof-
leini. По непроверенным данным; его
общая длина достигает 9,6 м, мас-
са — 270 кг. Достоверно измеренные
особи имели общую длину 4,5—5 м
и массу свыше 50 кг. К самым ма-
леньким осьминогам относятся кали-
форнийский О. micropyrsus (длина
мантии самца 10 мм, самки до 25 мм)
и красноморскпй О. nanus (длина
мантии зрелого самца до 13 мм, сам-
ки раза в два крупнее). Зрелые сам-
ки О. stictochrus с тихоокеанского
побережья Панамы имеют длину
мантии лишь 20 мм. Самки тропиче-
ского западноатлантического О. jou-
bini созревают при длине мантии 2—
2,5 см и общей длине около 10 см.
Масса этих осьминогов составляет
всего 10—15 г.
Единственный вид вампироморф—
Vampyroteuthis infernalis: длина ман-
тии до 13 см, общая длина до
37,5 см.
Внешнее строение
ГОЛОВА. У каракатиц и кальмаров
голова хорошо отделена от туловища
шеей (шейным перехватом). У ча-
сти сепиолид (под сем. Sepiolinae
и роды Sepiolina, Stoloteuthis, Irido-
ieuthis из подсем. Heteroteuthinae) и
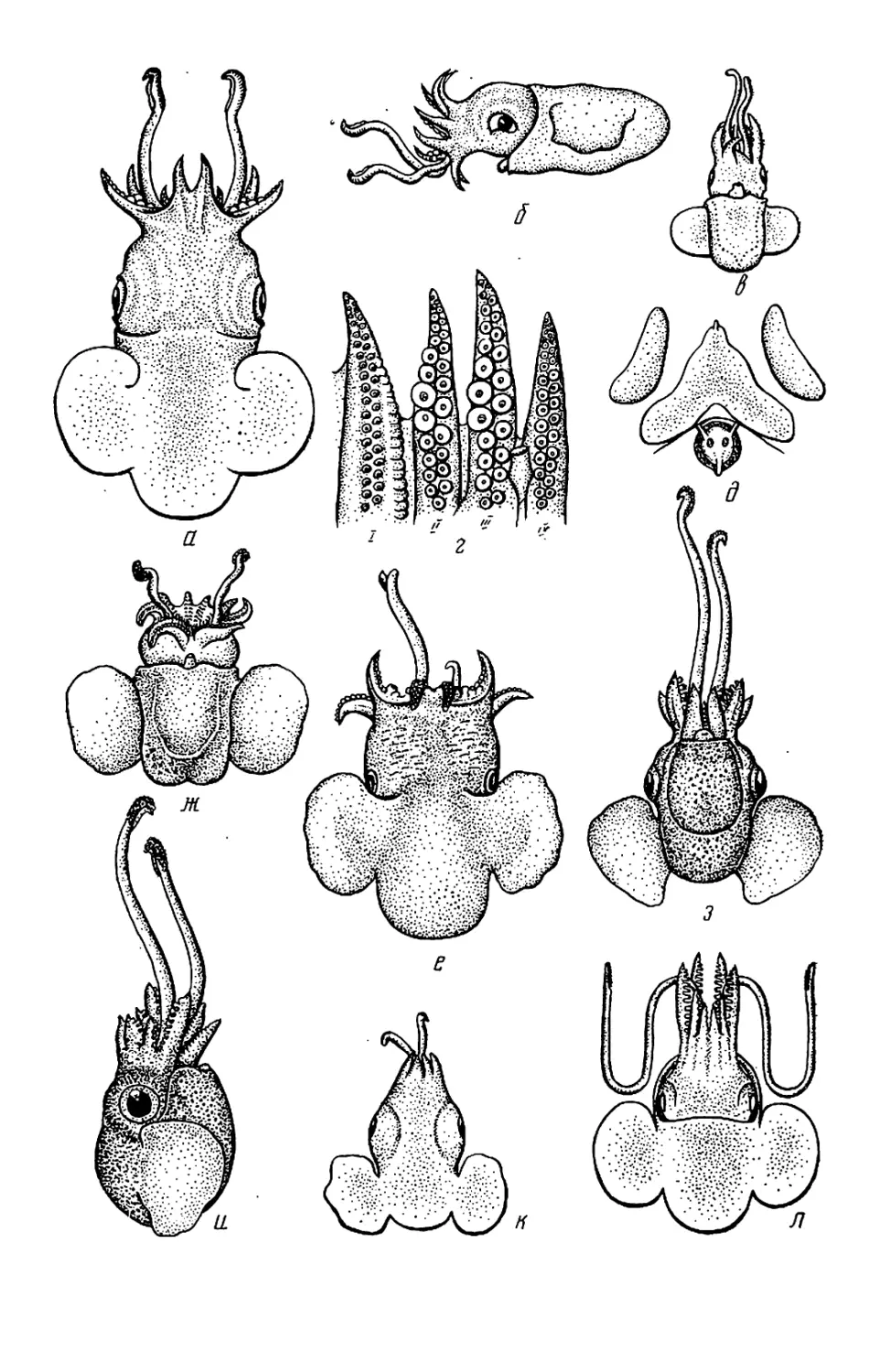
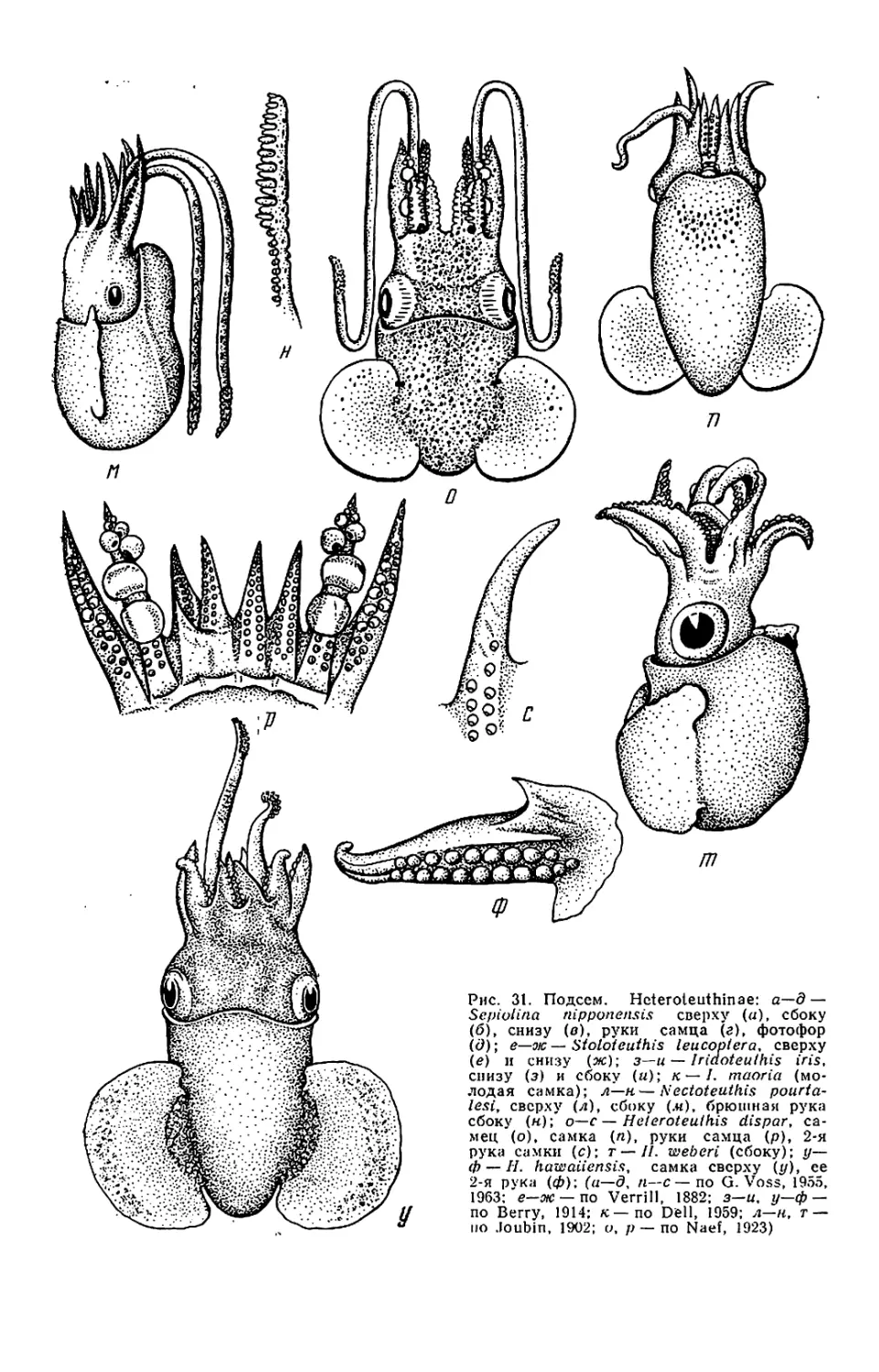
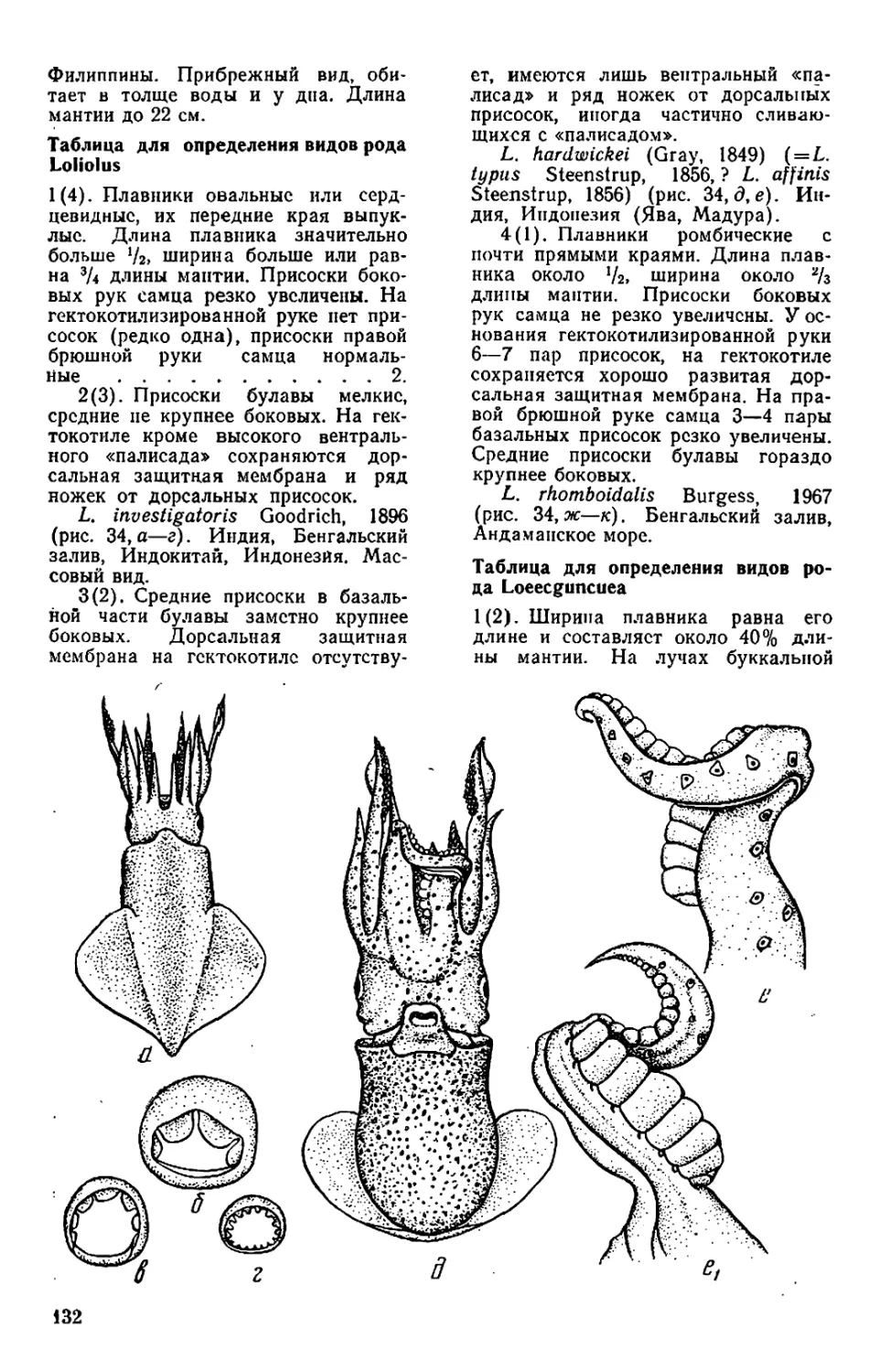
у Sepiadariidae (см. рис. 28—32)
мантия срастается с головой на спин-
ной стороне, соединяясь с пей узкой
или широкой затылочной кожной
лентой. У Idiosepiidae (см. рис. 33)
мантия не срастается с головой, по
затылочного хряща нет. Срастается
мантия с головой и у океанических
кальмаров семейств Promachoteuthi-
dae и Cranchiidae. У всех остальных
кальмаров и каракатиц мантия со-
единяется с головой посредством за-
тылочного хряща.
У вампироморф и осьминогов го-
лова плохо отделена от туловища.
10
Шейный перехват хорошо заметен у
некоторых Octopodidae, по у боль-
шинства пелагических и многих дон-
ных осьминогов голова плавно пере-
ходит в туловище. Обычно голова
мала по сравнению с туловищем, но
у кальмаров сем. Histioteuthidae (см.
рис. 53—56), напротив, она значи-
тельно крупнее. У плавниковых ось-
миногов сем. Opisthoteuthidac (род
Opisthoteuthis, см. рис. 76) голова с
туловищем представляет собой буго-
рок, лежащий сверху на растопы-
ренных и соединенных толстой пере-
понкой (умбреллой) руках.
Голова кальмаров и каракатиц
может быть шире, равна или уже
диаметра переднего края мантии.
В последнем случае она в принципе
может втягиваться в мантию. Это
свойственно некоторым океаническим
кальмарам преимущественно на ран-
них стадиях развития. Личинки ом-
мастрсфид Todarodes, Ornithoteuihis
и др. и некоторых кранхиид могут
втягивать в мантию и голову, и руки,
так что кальмар выглядит шариком.
Личинки ряда онихотсйтид также
втягивают голову в мантию. Единст-
венный вид кальмаров, сохраняю-
щий эту способность и во взрослом
состоянии, — Onychoteuthis banksi.
Он втягивает голову до середины
глаз и даже дальше. Это наблюда-
ется при резком раздражении, на-
пример при фиксации. У некоторых
каракатиц голова может частично
втягиваться под языкообразный выс-
туп спинной стороны мантии. Те ви-
ды, у которых голова срастается с
мантией, разумеется, втягивать ее пе
способны.
Ширина головы обусловлена
главным образом величиной глаз.
У многих, преимущественно глубоко-
водных кальмаров и осьминогов гла-
за столь велики, что занимают прак-
тически всю голову, почти смыкаясь
в ее середине. Напротив, у таких
(тоже глубоководных) пелагических
осьминогов, как Eledonella и Vitre-
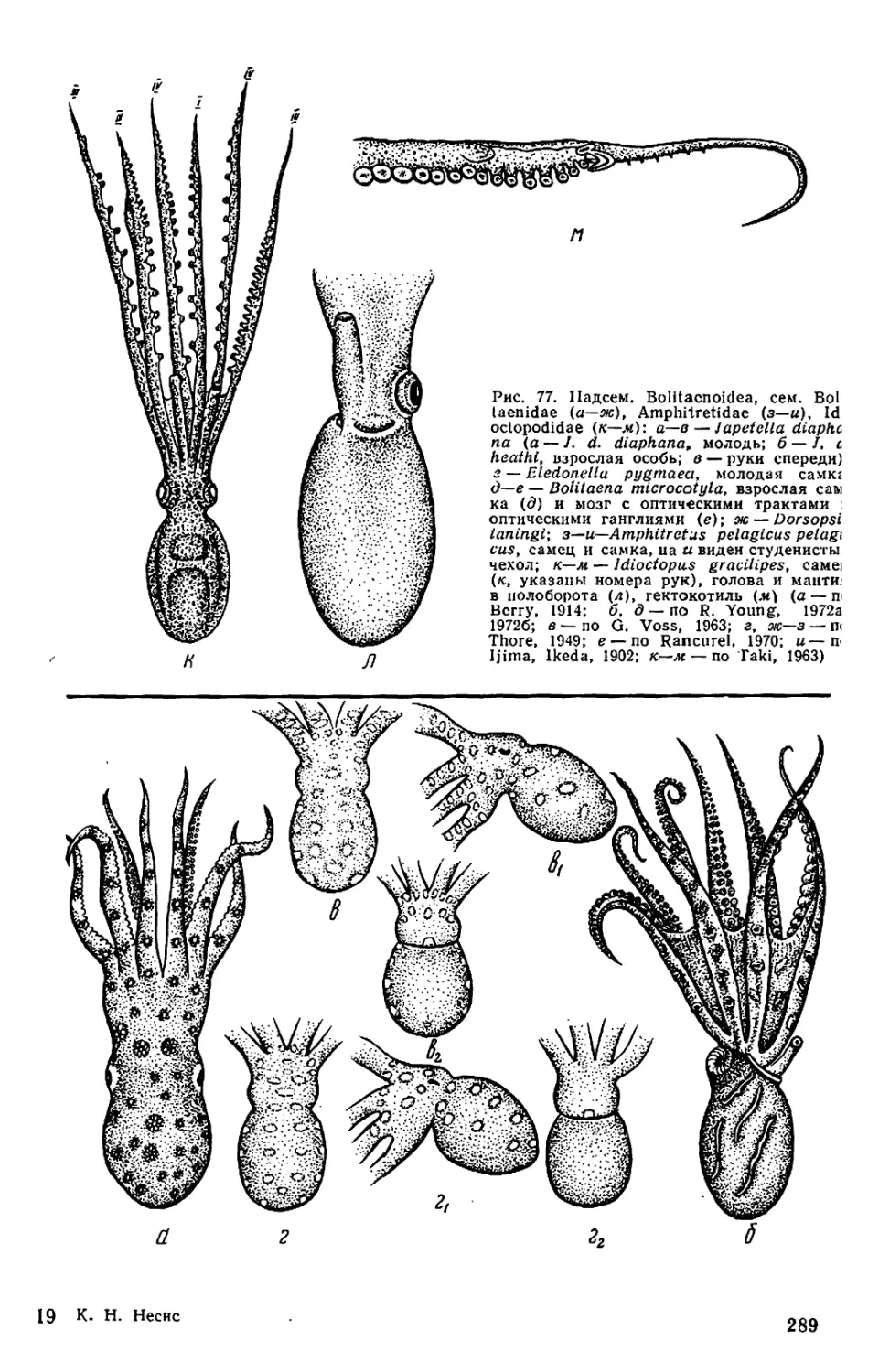
ledonella (см. рис. 77, г; 86), глаза
малы, по широко расставлены и со-
единяются с мозгом длинными нер-
вами— оптическими трактами. Поч-
ти у всех головоногих глаза распо-
ложены по бокам головы, но у Am-
phit г etas (Amphitretidae) и Dorsopsis
(Bolitaenidae) (см. рис. 77, ж—и) они
сближены и сидят па «макушке».
У Amphitretus, а также молоди неко-
торых океанических кальмаров глаза
имеют почти цилиндрическую форму
(«телескопические глаза»). Личинки
Cyclotcuthidae, личинки и молодь
большинства Cranchiidae (кроме Сгап-
chia и Liocranchia) имеют стебельча-
тые глаза, но с возрастом они стано-
вятся сидячими. Глазные стебельки
некоторых кранхиид, особенно Batho-
thauma lyromma, бывают чрезвычай-
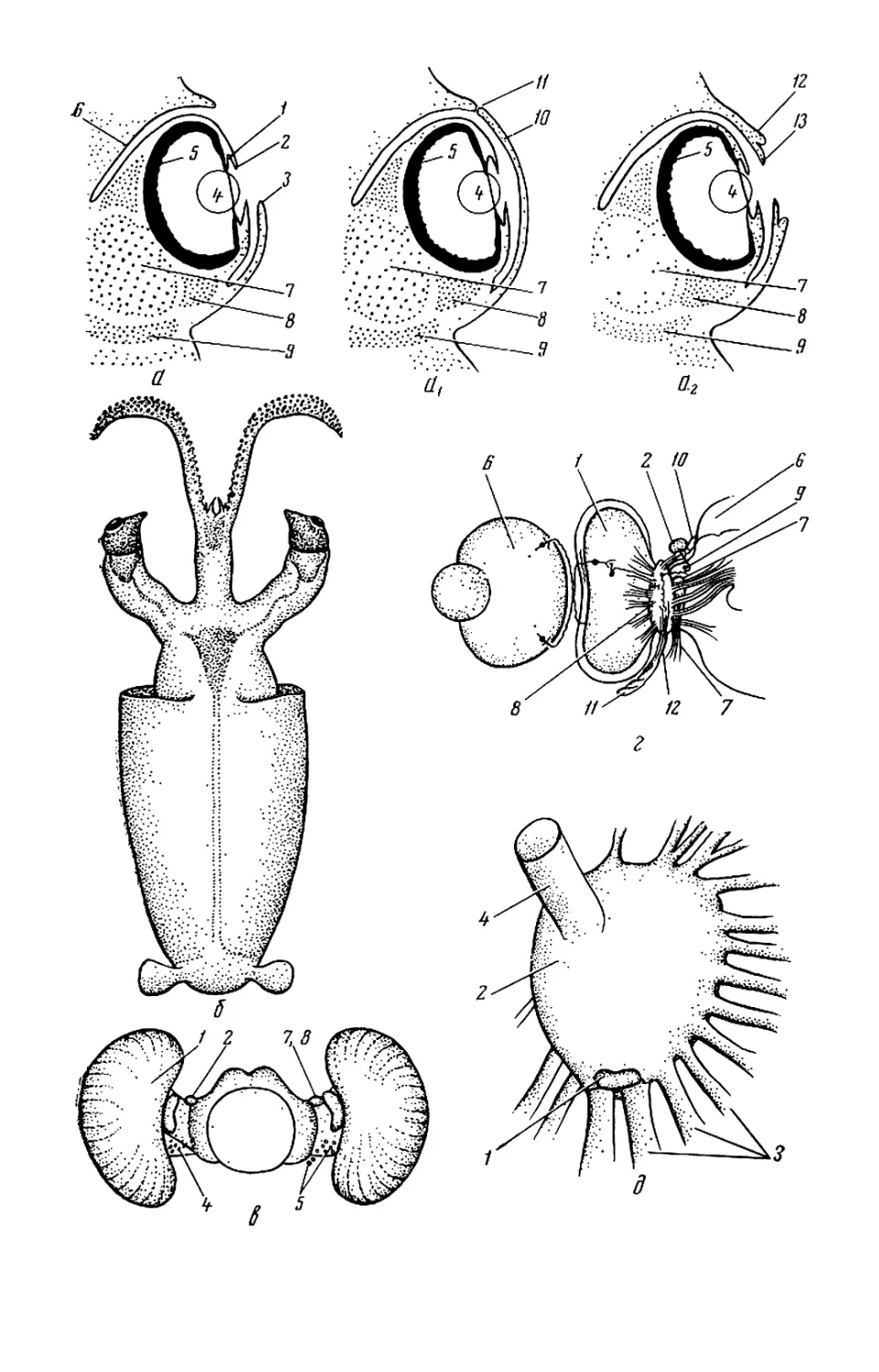
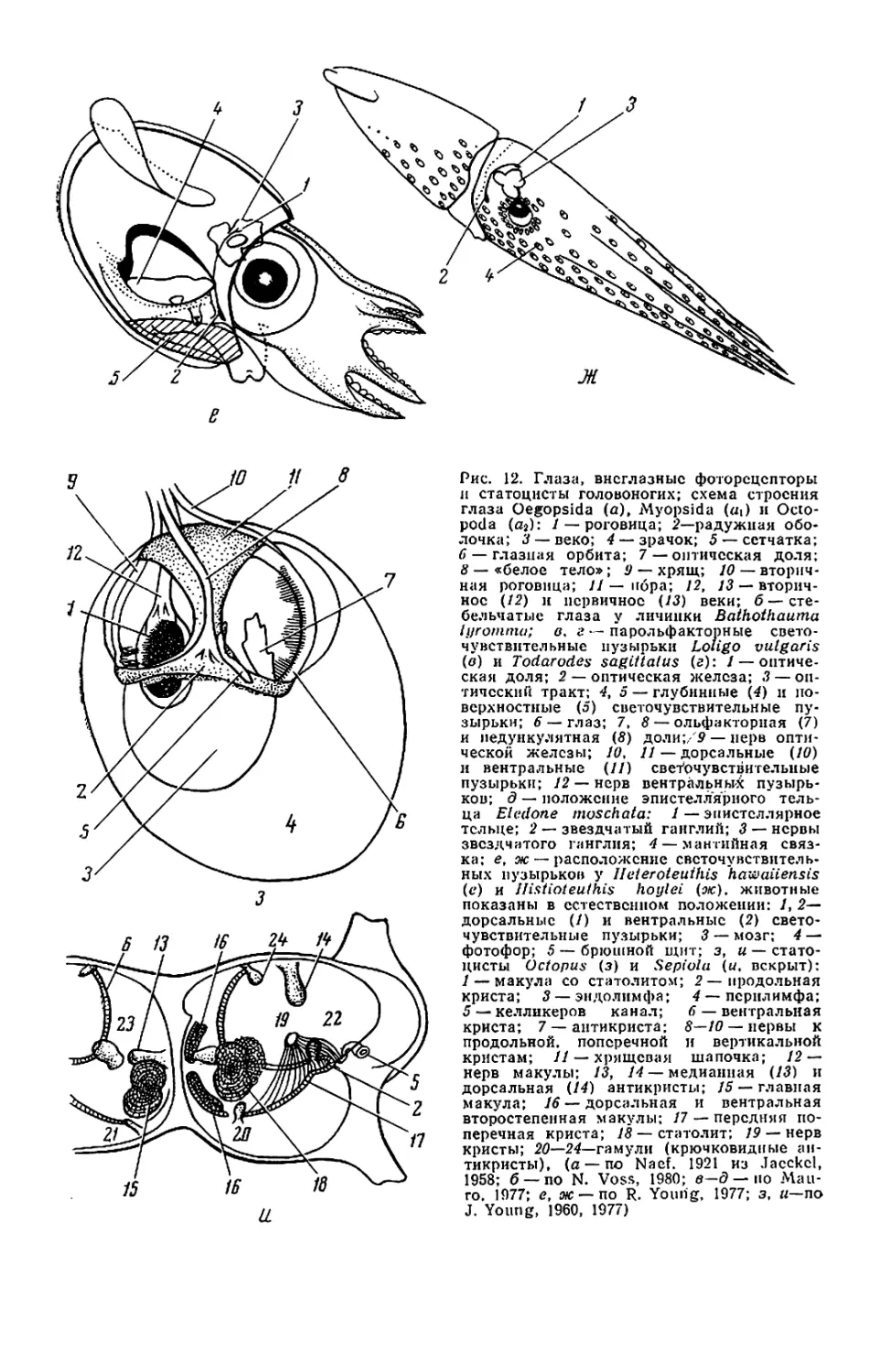
но длинными (рис. 12, б). Глаза личи-
нок и молоди кранхиид, а также ли-
чинок некоторых Cycloteuthidae
удлиненные, вытянутые в спинно-
брюшном направлении, иногда с изо-
гнутым выступом снизу в форме
туфли (Sandalops и др.). Обычная
же форма глаз округлая, почти ша-
рообразная. Как правило, оба глаза
имеют одинаковую форму и разме-
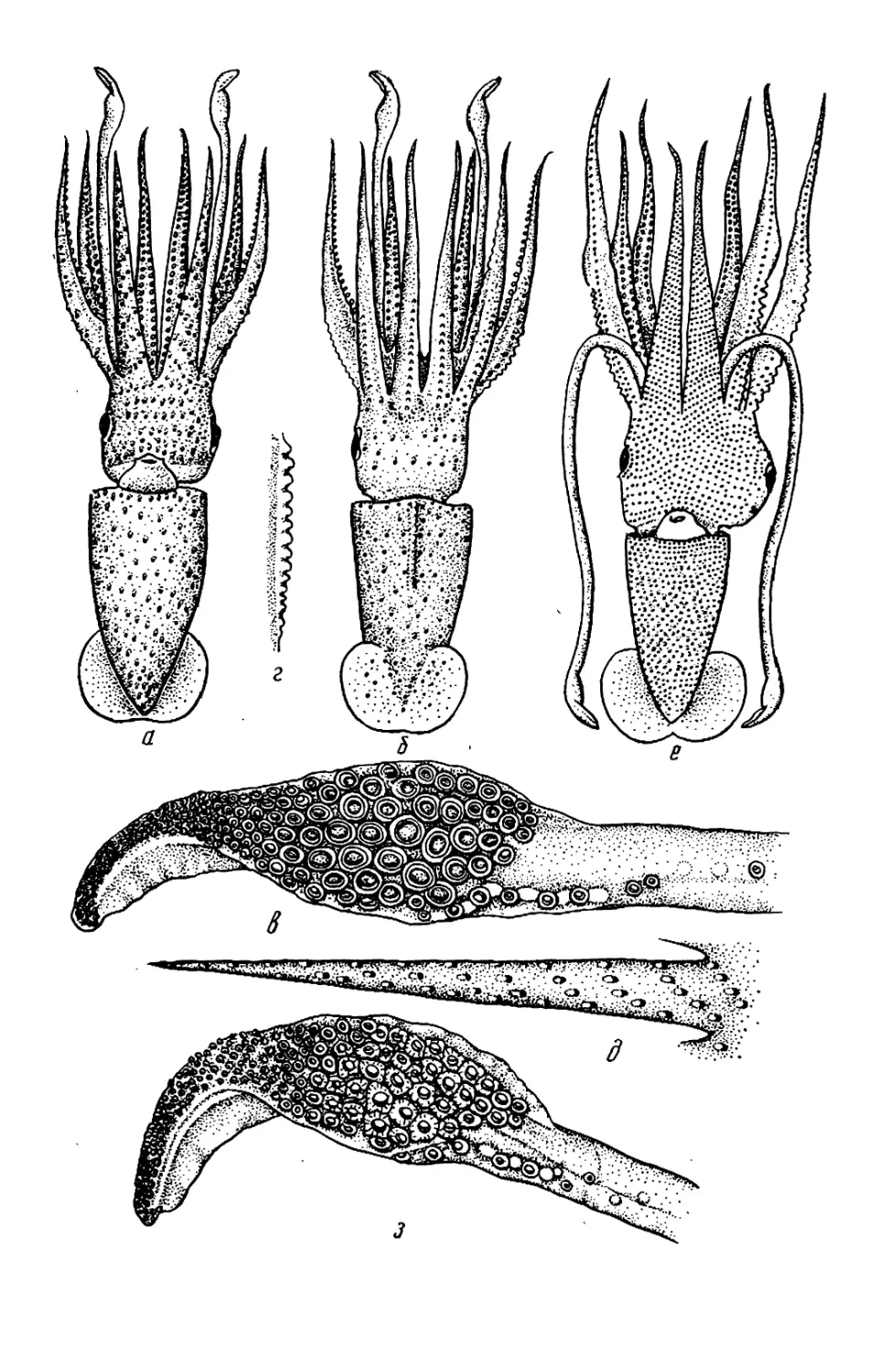
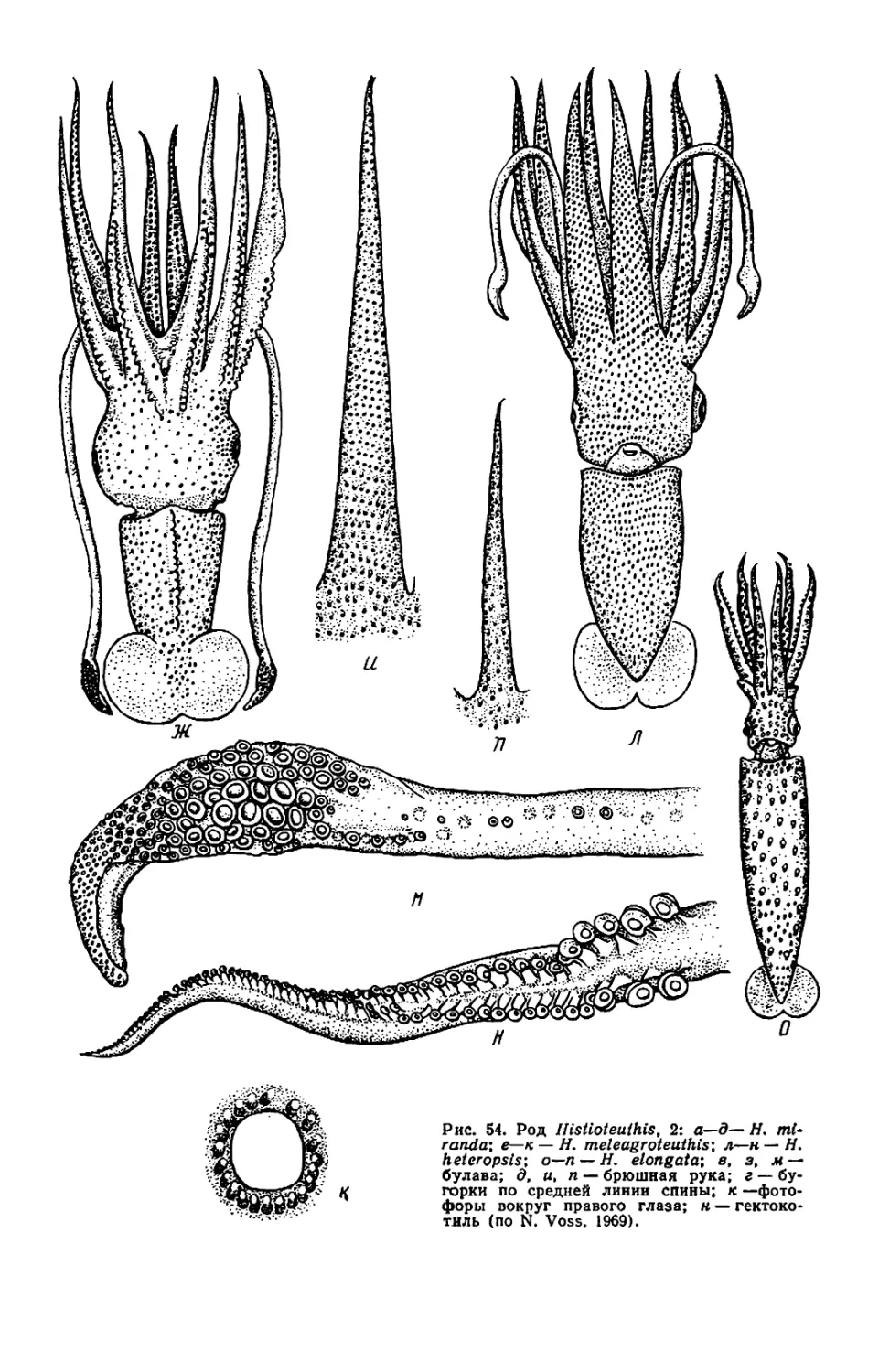
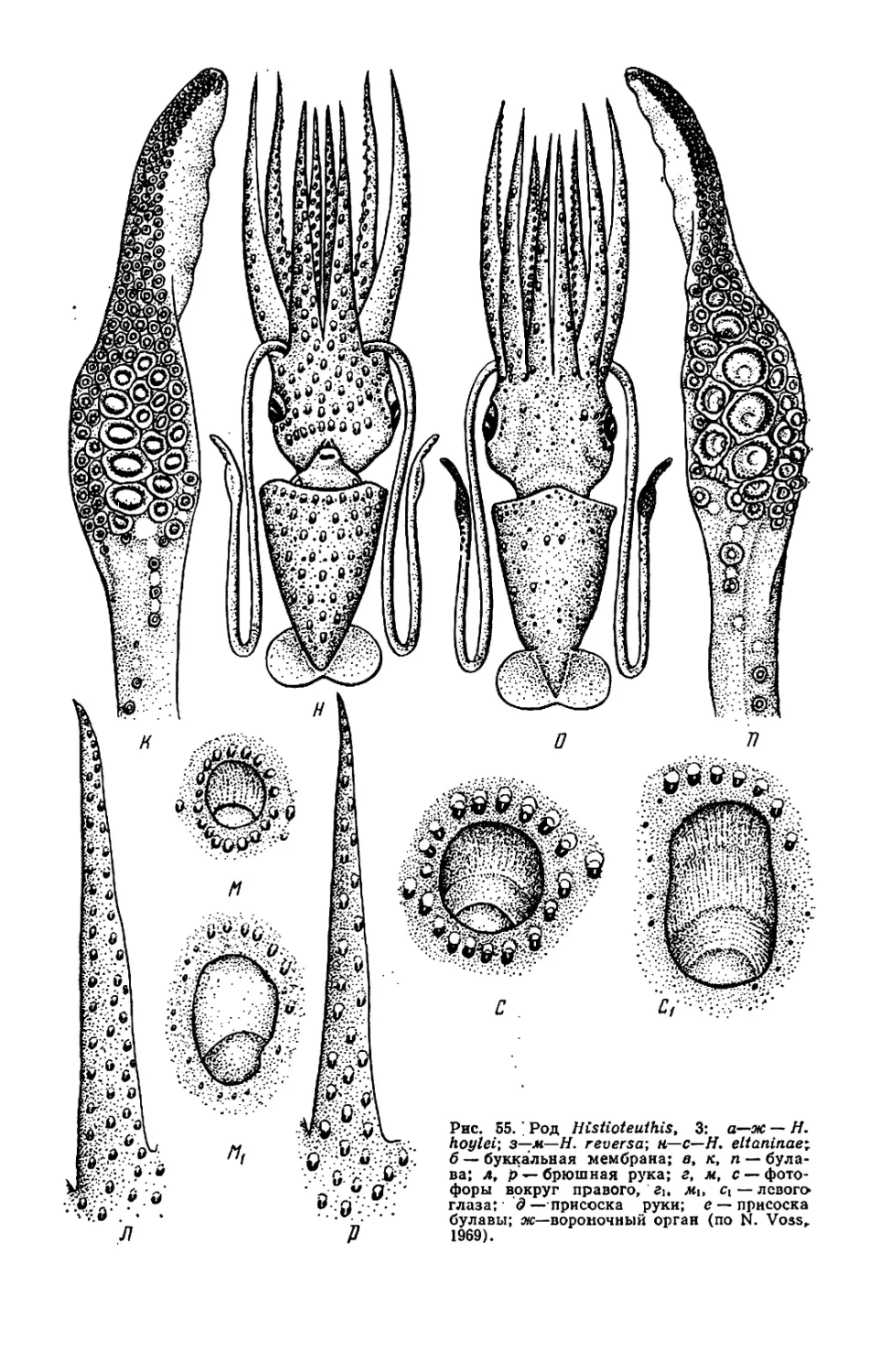
ры, но у Histioteuthis (см. рис. 53—
56) левый глаз значительно крупнее
правого.
Кроме глаз на голове находятся
обонятельные органы. У кальмаров,
каракатиц и вампироморф это па-
пиллы (обонятельная папилла), у
осьминогов — обонятельные ямки.
Расположены они по бокам головы
ближе к шее.
РУКИ И ЩУПАЛЬЦА. Руки
двужаберных головоногих в норме
имеют коническую форму и воору-
жены одним, двумя или четырьмя
(редко больше) рядами присосок.
Счет рук ведется от спинной к брюш-
ной паре: 1-я пара — спинная, 2-я —
спинно-брюшная, 3-я — брюшно-бо-
ковая, 4-я — брюшная. Длина рук
разных пар, как правило, разная, и
соотношение их длины, выражаемое
формулой рук (порядок длины рук
по мере уменьшения, например
4.3.2.1, т. е. 4>3>2>1), имеет
важное значение при определении,
особенно осьминогов. Правая и ле-
вая руки одной и той же пары обыч-
но имеют одинаковую длину, за ис-
ключением гектокотилизированной
руки, которая часто длиннее или ко-
роче противоположной. У некоторых
видов (из Scpiidac, Lycoteuthidae,
Chiroteuthidae, Mastigoteuthidac, Oc-
topodidae и др.) одна или несколько
пар рук могут быть резко удлинены
или вытянуты в длинный тонкий
хлыст; часто это свойственно лишь
зрелым самцам.
Руки кальмаров и каракатиц при-
крепляются к окружающей рот на-
ружной губе кожно-мускульной ото-
рочкой — буккальной (ротовой) во-
ронкой (рис. 2,а). Она имеет звезд-
чатую форму и состоит из 6, 7 или
8 лучей. Первоначальное их число —
8, по лучи, ведущие к рукам 1-й и
иногда 4-й пар, могут сливаться.
У ряда Loliginidae, Bathyteuthidae и
Ctcnopterygidae па лучах буккальной
воронки есть крохотные присоски.
Буккальная воронка крепится к ру-
кам особыми связками (буккальное
крепление), отходящими от вершин
лучей буккальной воронки. Если лу-
чи, ведущие к 1-й или 4-й паре рук,
слиты, то связка раздваивается и
имеет форму вилки. Связки буккаль-
ного крепления присоединяются к
дорсальной стороне рук 1-й и 2-й
пар, к вентральной — рук 3-й пары и
либо к дорсальной, либо к вентраль-
ной стороне рук 4-й пары. Послед-
11
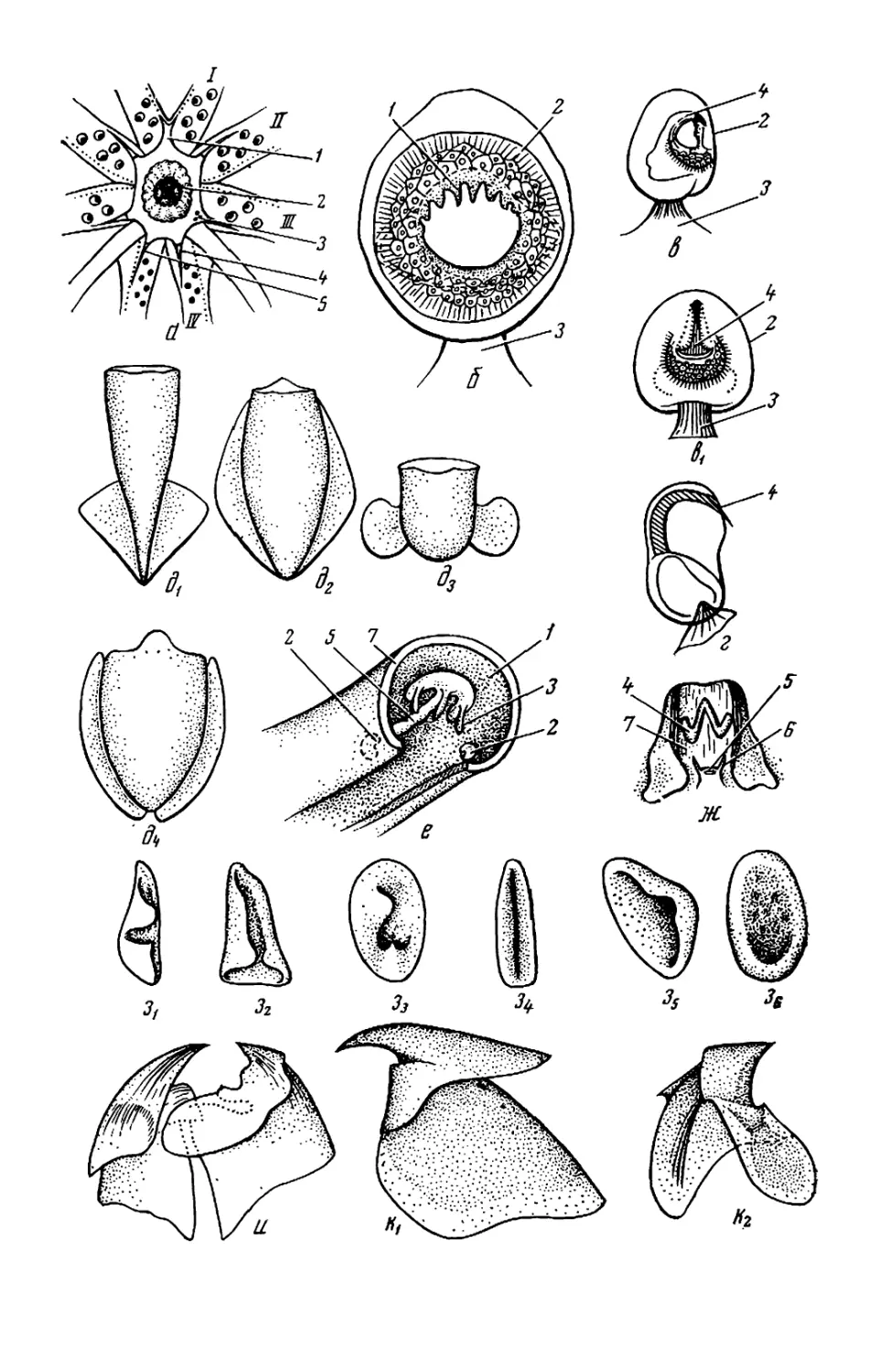
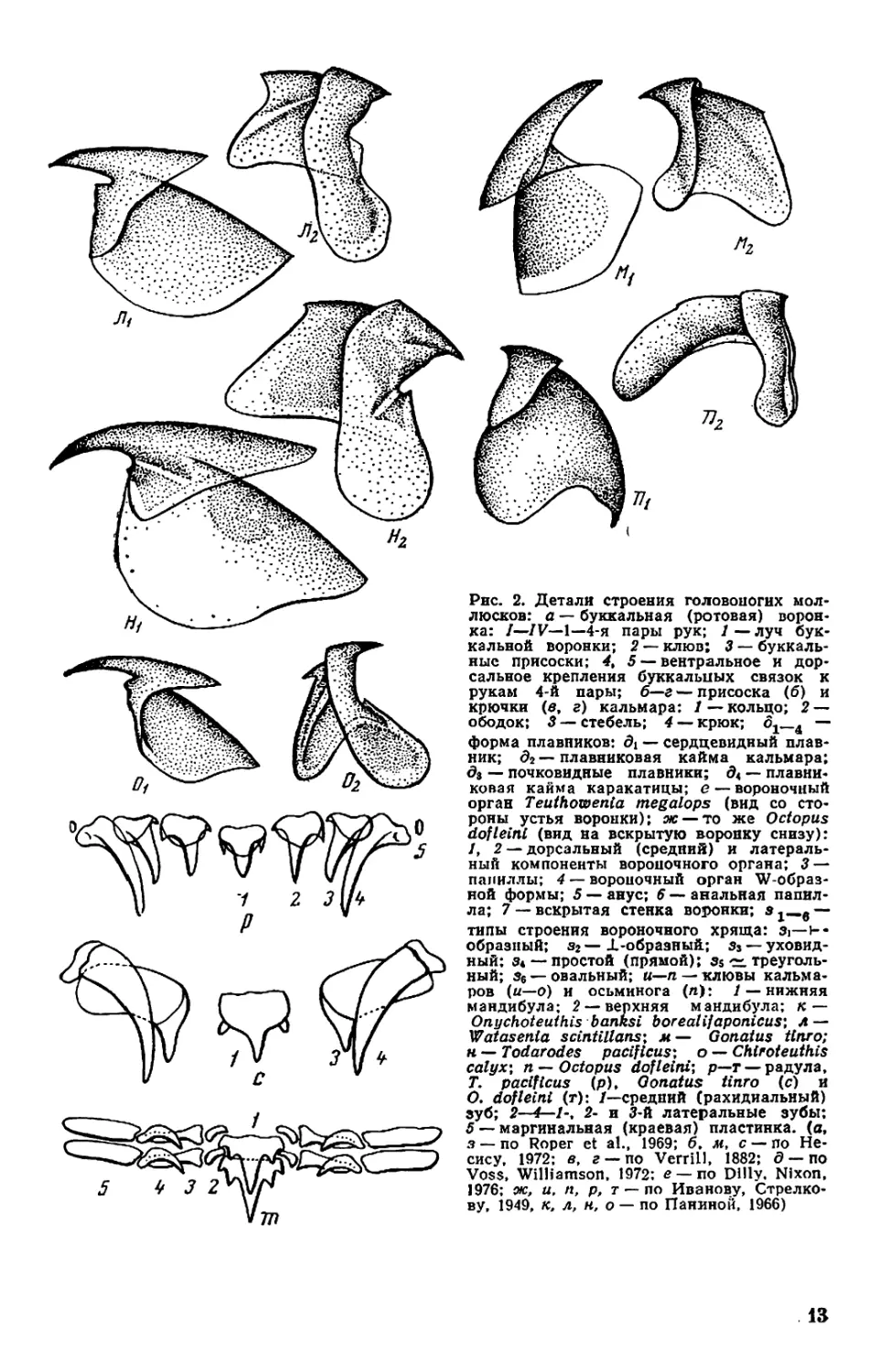
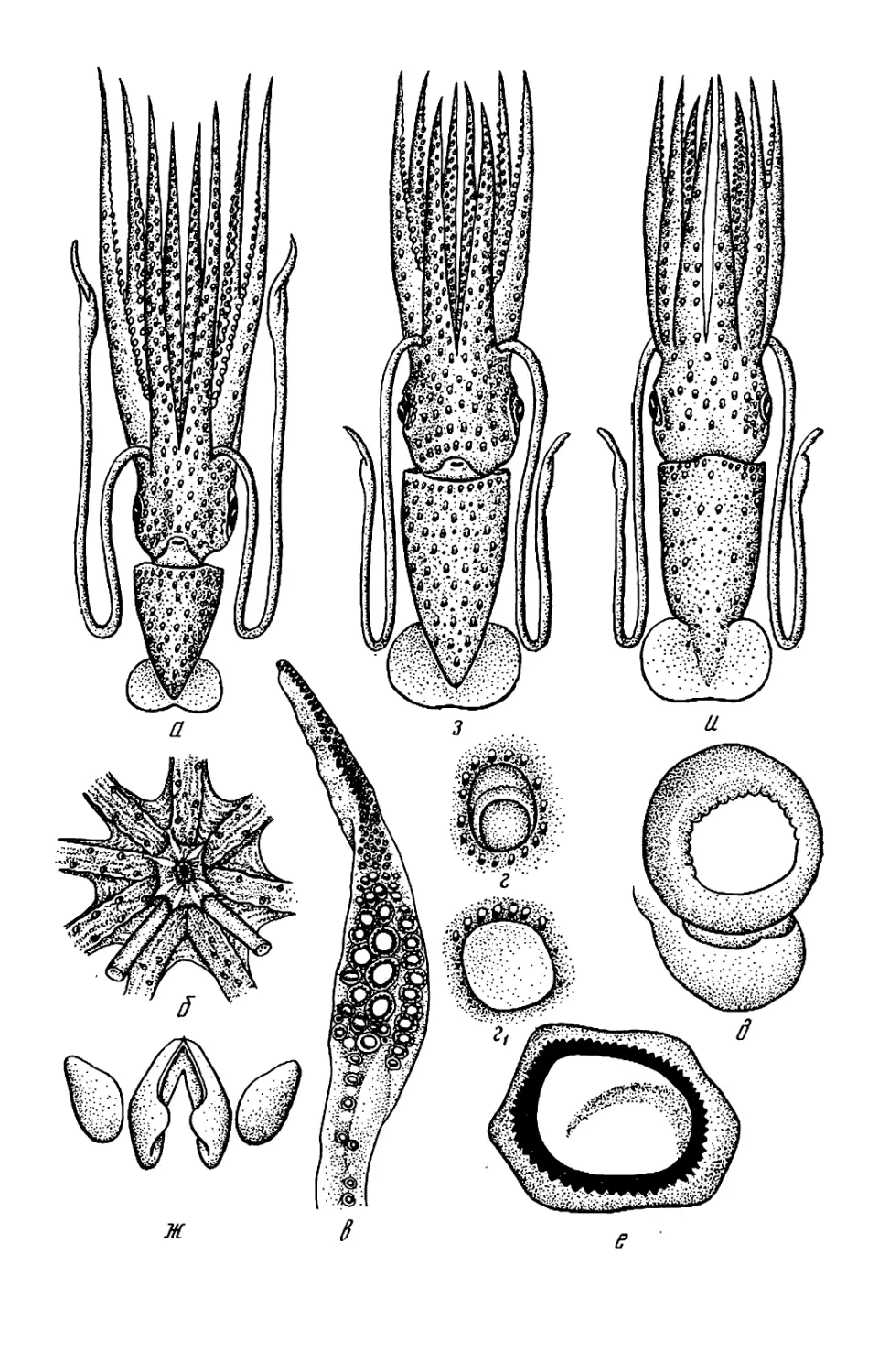
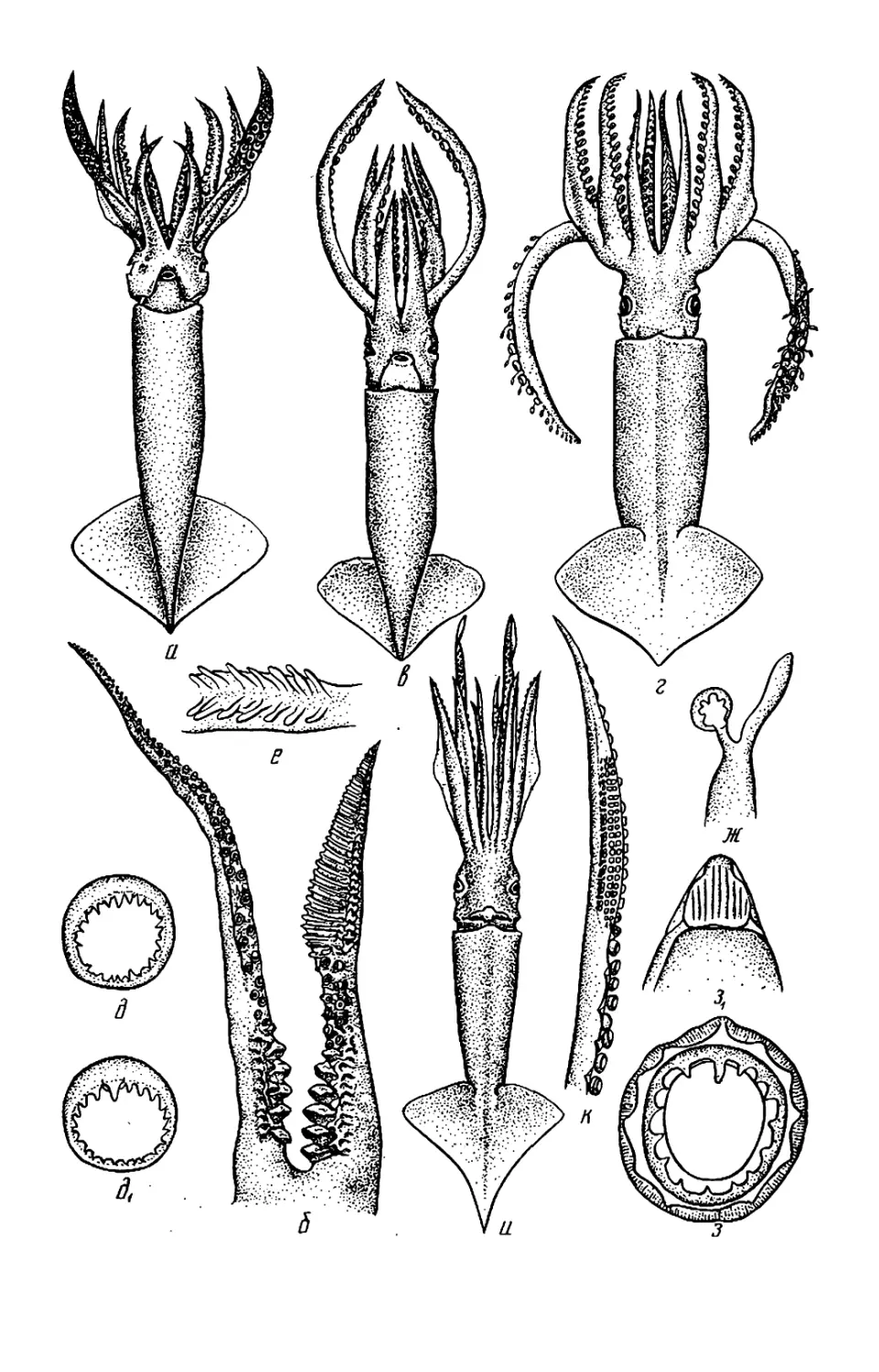
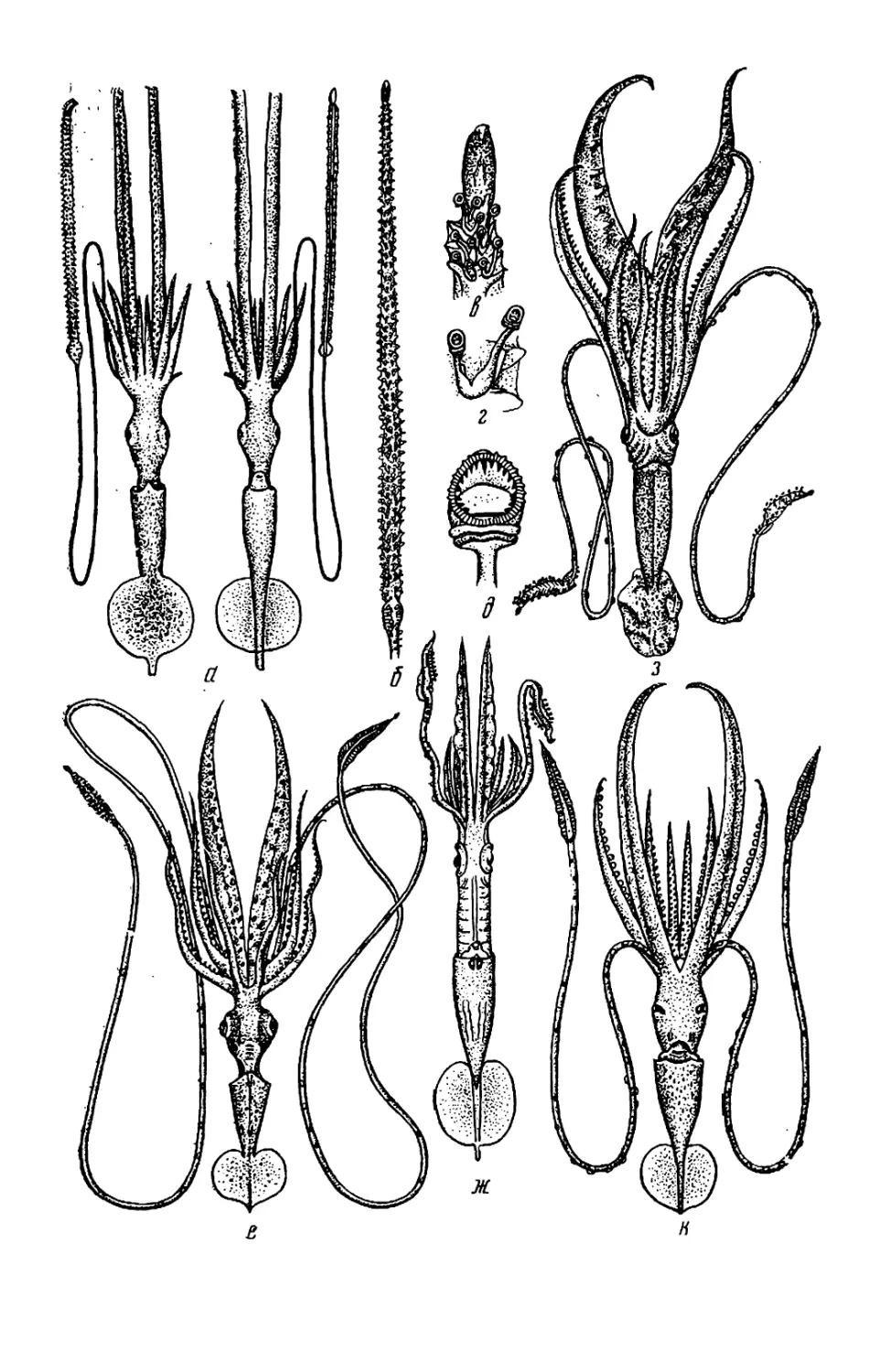
Рис. 2. Детали строения головоногих мол-
люсков: а — буккальная (ротовая) ворон-
ка: /—/V— 1—4-я пары рук; 1 — луч бук-
кальной воронки; 2 — клюв: 3 — буккаль-
ные присоски; < 5 — вентральное и дор-
сальное крепления буккальных связок к
рукам 4-й пары; б—г — присоска (б) и
крючки (в, г) кальмара: 1 — кольцо; 2 —
ободок; 3 — стебель; 4 — крюк; ^—4 —
форма плавников: 01 — сердцевидный плав-
ник; д2 — плавниковая кайма кальмара;
0з — почковидные плавники; 04 — плавни-
ковая кайма каракатицы; е — вороночный
орган Teuthowenia megalops (вид со сто-
роны устья воронки); ж — то же Octopus
dofleini (вид на вскрытую воронку снизу):
1, 2 — дорсальный (средний) и латераль-
ный компоненты вороночного органа; 3—
папиллы; 4 — вороночный орган W-образ-
ной формы; 5 — анус; 6 — анальная папил-
ла; 7 — вскрытая стенка воронки; з —
типы строения вороночного хряща: 3i—ь-
образный; з2—JL-образный; з3— уховид-
ный: з* — простой (прямой); з$ треуголь-
ный; з6 —овальный; «—п —клювы кальма-
ров (и—о) и осьминога (п): / — нижняя
мандибула; 2 — верхняя мандибула; к —
Onychoteuthis banksi boreal if aponicus* Л—
Watasenia scintillansx m— Gonatus ttnro;
н — Todarodes pacificusx о — Chiroteuthis
calyx\ n —Octopus dofleini; р—т — радула,
T. pacificus (p), Gonatus tinro (с) и
О. dofleini /—средний (рахидиальный)
зуб; 2—2- и 3-й латеральные зубы:
5 — маргинальная (краевая) пластинка, (а,
з — по Roper et al., 1969; б. м, с — по Не-
сису, 1972; в, г —по VerrilJ, 1882; 0 — по
Voss, Williamson, 1972: е— по Dilly. Nixon,
1976; ж, и, п, р, т — по Иванову, Стрелко-
ву, 1949, к, л, н, о — по Паниной, 1966)
13
нее имеет iiidkiiiic таксономическое
значение для кальмаров: у всех
продетакителей 8 семейств эгопенд
(Lycoteiillihhie, luioploteuthidae, Hi-
stiotciillildae, Psychroteuthidae, Ba-
tliyteutlildae, Neoicuthidae, Architeu-
thiduc, Oiiiiiiastrcphidae) связки кре-
пятся к рукам 4-й пары дорсалыю,
у всех представителей остальных 15
семейств эгопсид и у миопсид — вен-
трально. Эти отношения выражают-
ся формулой буккального крепления,
которая показывает, с какой сторо-
ны [дорсальной (D) или вентральной
(У)] крепятся связки буккальной
мембраны к рукам 1—4-й пар. Фор-
мула может быть либо DDVD (у упо-
мянутых 8 семейств), либо DDVV (у
остальных). Исключение составляет
род Enigmoteuthis (Enoploteuthidae):
ко всем рукам связки крепятся дор-
сально (формула DDDD). Основная
функция буккального крепления —
держать при плавании руки вместе,
конусом. У вампироморф и осьмино-
гов буккального крепления нет, и им
приходится затрачивать мускульные
усилия, чтобы руки при реактивном
плавании пе болтались и не тормо-
зили движения.
В разрезе руки кальмаров и ка-
ракатиц имеют обычно треугольную
форму: внутренняя поверхность уп-
лощена и несет присоски, наружная
заострена. Руки 1-й пары обычно не-
сут сверху небольшой наружный
гребень, 2-й и 3-й пар — острый тре-
угольный киль — плавательную мем-
брану. Он играет приблизительно ту
же роль, что и хвостовое оперение у
зенитных ракет: обеспечивает ста-
бильность горизонтального движе-
ния. Руки 4-й пары обычно уплоще-
ны и не несут наружного киля.
У малоподвижных глубоководных
кальмаров и каракатиц кили обычно
слабо развиты.
Несущая присоски внутренняя по-
верхность рук по бокам окаймлена
защитными мембранами — тонкими
пленками кожи, растянутыми на под-
порках — трабекулах, отходящих
от края руки обычно вперемежку с
присосками. Степень развития защит-
ных мембран сильно варьирует.
У Thysanoteuthidae (см. рис. 62, а) и
высших Ommastrephidae они очень
широки и служат для поимки мелкой
добычи в образованную мембранами
«корзинку». Вентральная защитная
мембрана боковых рук обычно шире
14
дорсальной, а у взрослых самок От-
mastrephes bartrami она вытянута в
громадную треугольную лопасть.
У некоторых видов Histioteuthis
обе защитные мембраны рук 1—3-й
пар сливаются, образуя широкую
внутреннюю перепонку (умбреллу),
как у осьминогов (см. ниже); к сере-
дине перепонки между руками 3-й
пары присоединяются перепонки, ве-
дущие к рукам 4-й пары, так что
щупальца остаются снаружи от ум-
бреллы и свободны (см. рис. 53, в).
Такая перепонка позволяет кальмару
двигаться по способу медузы, отча-
сти компенсируя слабость мантийных
мышц. У Н. hoylei аналогичная, но
короткая перепонка охватывает ос-
нования рук нс изнутри, а снаружи
(см. рис. 55, а, б).
Щупальца всегда находятся меж-
ду руками 3-й и 4-й пар, не соеди-
няются с буккальной воронкой, со-
стоят из эластичного стебля круглой
или четырехгранной формы и оваль-
ной расширенной булавы. У многих
видов каракатиц и океанических
кальмаров булава не расширена, но
всегда отличается от стебля по во-
оружению. Стебель щупалец карака-
тиц очень эластичен, и щупальца мо-
гут втягиваться в особые сумки или
чехлы (у Sepiadariidae); у кальмаров
щупальца могут слегка растягивать-
ся и сокращаться, но пе втягиваются
целиком. Если булава щупальца рас-
ширена, опа обычно несет киль на
спинной (противоположной присос-
кам) стороне и защитные мембраны
по бокам.
У Octopoteuthidae, некоторых Go-
natidae (Gonatopsis), Onychotcuthi-
dae (Chaunoteutnis) и Lepidoteuthidae
(Lepidoteuthis) щупальца имеют
только личинки, а у возрослых каль-
маров они отсутствуют или от них
остаются лишь рудименты стеблей
(Gonatopsis okutanii, Taningia da-
nae). Утрата щупалец, по-видимому,
происходит очень быстро — они от-
брасываются как хвост ящерицы.
Щупальца Mastigoteuthis и, по-види-
мому, Echinoteuthis таким же точно
образом отбрасываются (автотомиру-
ются) при поимке. Отбрасываются
щупальца и у зрелых самок многих
кальмаров — Gonatus, Histioteuthis,
Joubiniteuthis^ Leachia, Taonius, San-
dalops и др. Grimalditeuthis также
лишены щупалец и, поскольку их ли-
чинки не описаны, не известно, ког-
да это происходит. Утрата и автото-
мия щупалец присущи в основном
полупланктонным и планктонным
глубоководным и среднеглубинным
океаническим кальмарам, но не
свойственны каракатицам и нерити-
ческим кальмарам. Руки вампиро-
морф в сечении округлые, и па зна-
чительную часть длины соединены
перепонкой (умбреллой), гомологич-
ной защитным мембранам кальмаров
и каракатиц. У плавниковых осьми-
ногов умбрелла соединяет руки поч-
ти до кончиков, у Argonauta и Ocyt-
hoe она отсутствует, у большинства
осьминогов се глубина составляет от
’/4 до ’/2 длины наибольшей руки.
Пять секторов умбреллы имеют раз-
ную глубину, наибольшими могут
быть либо спинной, либо боковые
сектора, брюшной обычно наименее
глубок. Сектора обозначают буквами
А, В, С, D, Е, начиная со спинной
стороны. Глубину умбреллы измеря-
ют в растянутом виде по середине
секторов и выражают формулой,
аналогичной формуле длины рук.
Например, формула
>£ означает, что сектор между ру-
ками 1-й пары наиболее, а между
руками 4-й пары наименее развит.
У некоторых Cirroteuthidae сектора
умбреллы срастаются нс с боковыми
сторонами рук, как обычно, а непо-
средственно друг с другом, обходя
руки снаружи и присоединяясь к
спинной стороне рук дополнительной
перепонкой — вторичной умбреллой
(см. рис. 74). Это позволяет макси-
мально растопыривать руки: осьминог
становится похожим на раскрытый
зонтик. В большинстве случаев ум-
брелла кальмаров и осьминогов —
топкая пленка, но у Opisthoteuthidae
она утолщена, и соединенные ум-
бреллой руки напоминают блин или
лепешку (см. рис. 76).
Концы рук 1-й пары у самок пе-
лагических осьминогов tretnoctopus
и Argonauta сильно видоизменены.
У Tretnoctopus они превращены в
орудие защиты и отпугивания вра-
гов, у Argonauta — в орган сооруже-
ния раковины, в которой сидит сам-
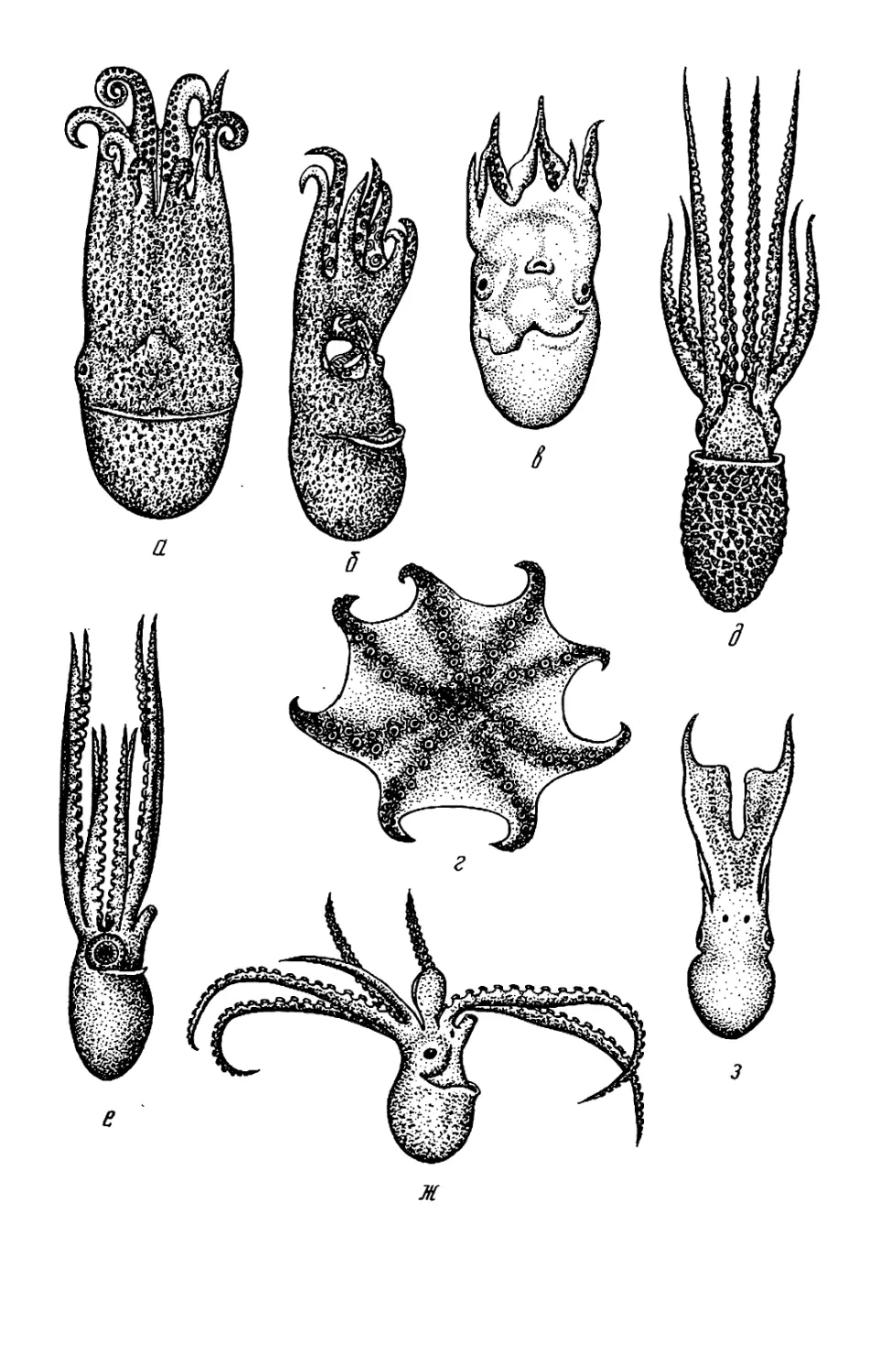
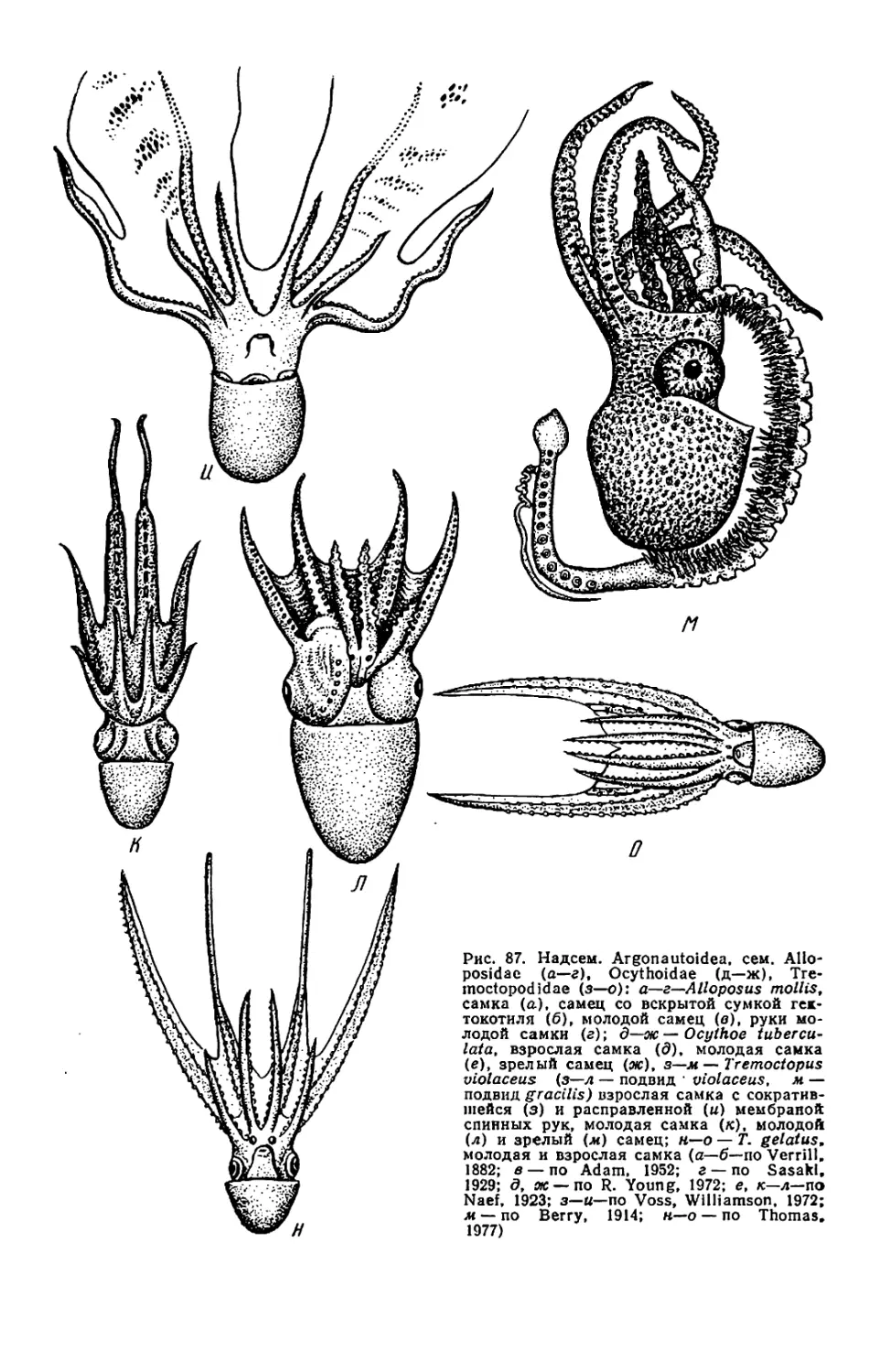
ка и вынашивает яйца (см. рис. 87—
ПРИСОСКИ. У кальмаров и ка-
ракатиц присоски имеют в норме
полушаровидную форму и сидят на
стебельке (рис. 2, б). Стебелек мо-
жет быть коротким или длинным,
тонким или толстым, иногда с выро-
стами посредине (присоски рук
Grimalditeuthis, см. рис. 68, з) или с
промежуточным расширением — ча-
шечкой (присоски щупалец Chiroteut-
his см. рис. 65, к).
Присоски рук кальмаров и кара-
катиц сидят обычно в 2, но иногда в
4 ряда. Последнее свойственно мно-
гим Sepiidae, Sepiolidae, а из каль-
маров— Gonatidae. У спирулы при-
соски сидят в 4 ряда в средних и
дистальных и в 5—8 рядов в базаль-
ных частях рук. У некоторых глубо-
ководных эгопсид присоски на руках
расположены в 4—6 рядов (Joubini-
teuthis) или в 2 ряда в базальных
и в 4—6 рядов в дистальных частях
рук (Ctenopteryx, Bathyteuthis).
Иногда умножение присосок захваты-
вает самые концы рук, это может быть
свойственно лишь самцам (многие-
Cranchiidac) или обоим полам (неко-
торые Sepiolidae, Abralia). В ряде
случаев присоски, особенно па руках
4-й пары, перестраиваются в один
ряд (Pyroteulhis), либо отсутствуют
(Pterygioteuthis) или имеются лишь
у основания руки (Valbyteuthis). По
вооружению руки 4-й пары нередко*
отличаются от рук других пар. Резко
отличается по вооружению также
гектокотилизированная рука. Самые
крупные присоски обычно располага-
ются посредине рук или ближе к их
основанию, к концу рук присоски
сильно мельчают.
Присоски булавы щупалец в
центральной части обычно располо-
жены в 4 ряда, но у многих океани-
ческих кальмаров, каракатиц и у спи-
рулы — в 6, 8 или более рядов.
У многих Sepiolidae и Mastigoteuthi-
dae булава сплошь усажена крохот-
ными присосочками в 20—30 или 50
продольных рядов. В центральной
части булавы (манус) обычно рас-
полагаются самые крупные присоски,,
на конце булавы (палец, или дакти*
лус) они мельчают. Число рядов при-
сосок на пальце часто больше, чем в
центральной части (например у II-
lex, Histioteuthis и др.). В ближай-
шей к стеблю (карпальной) части
булавы присоски тоже мелкие и час-
то расположены нерегулярно. В этой
же части булавы располагается фик-
саторный аппарат, состоящий из не-
скольких присосок с гладкими края-
ми и бугорков-кнопок.
Последние расположены так, что
15»
бугорки одного щупальца противо-
стоят присоскам другого. При быст-
ром плавании щупальца прижимают-
ся одно к другому и прочно скрепля-
ются. Фиксаторный аппарат есть
только у кальмаров, но нс у всех и
устроен очень разнообразно. Присос-
ки и бугорки могут располагаться
на округлой площадке, окруженной
кожным гребнем (например, у Ony-
choteuthis и Cranchiidae), протягива-
ться рядом вдоль края булавы
(Ommastrephidae) или вдоль стебля
щупальца (Gonatus, многие Cranchii-
dae). У Todarodes sagittatus (см. рис.
59, с) карпальная группа присосок
протягивается почти вдоль всего
стебля, а у близкого к нему вида
Martialia hyadesi (см. рис. 60, е) по
стеблю тянутся два ряда усиков-—
ножки редуцированных присосок.
У молоди некоторых кальмаров (на-
пример, кранхиид) личиночный фик-
саторный аппарат занимает весь сте-
бель щупальца; с возрастом он реду-
цируется. У личинок и молоди
Chiroteuthis вооружение булавы рез-
ко отличается от ее вооружения у
взрослых: «взрослая» (дефинитив-
ная) булава развивается ближе к
стеблю, чем личиночная, которая
впоследствии редуцируется или от-
брасывается.
Присоски кальмаров и каракатиц
вооружены роговыми кольцами. Край
кольца может быть гладким (на ру-
ках Onychoteuthidae, многих Cran-
chiidae, на щупальцах некоторых Lo-
liginidae) или нести зубцы. На ди-
стальном крае кольца зубцы выше и
острее, чем на боковых сторонах, а
на проксимальном крае кольца зуб-
цы могут вовсе отсутствовать или
редуцироваться до небольших за-
зубрин. Зубцы бывают заостренны-
ми — коническими или когтевидны-
ми — или плоскими, башневидными
(как на вершине шахматной ладьи).
Часто крупные зубцы чередуются с
мелкими (Loliginidae).
У некоторых океанических каль-
маров присоски превращаются в
крючья (рис. 2,в,г). Крючья имеют-
ся на руках большинства Gonatidae
(только на руках 1—3-й пар), всех
Enoploteuthidae (иногда лишь в сред-
них частях рук или только на руках
1—3-й пар) и Octopoteuthidae, а так-
же у Alluroteuthis и у гигантской
кранхииды Mesonychoteuthis hamilto-
ni, на булавах Gonatidae (только
16
подрод Gonatopsis s. str.), Enoploteut-
hidae (кроме Pterygioteuthis), Ony-
choteuthidae и некоторых кранхиид
(Belonella, Galiteuthis, Mesonycho-
teuthis). Крючья всегда развиваются
из присосок вследствие неравномер-
ного удлинения, изгибания и про-
дольного складывания дистального
края кольца или резкого удлинения
одного или двух зубцов (редко:
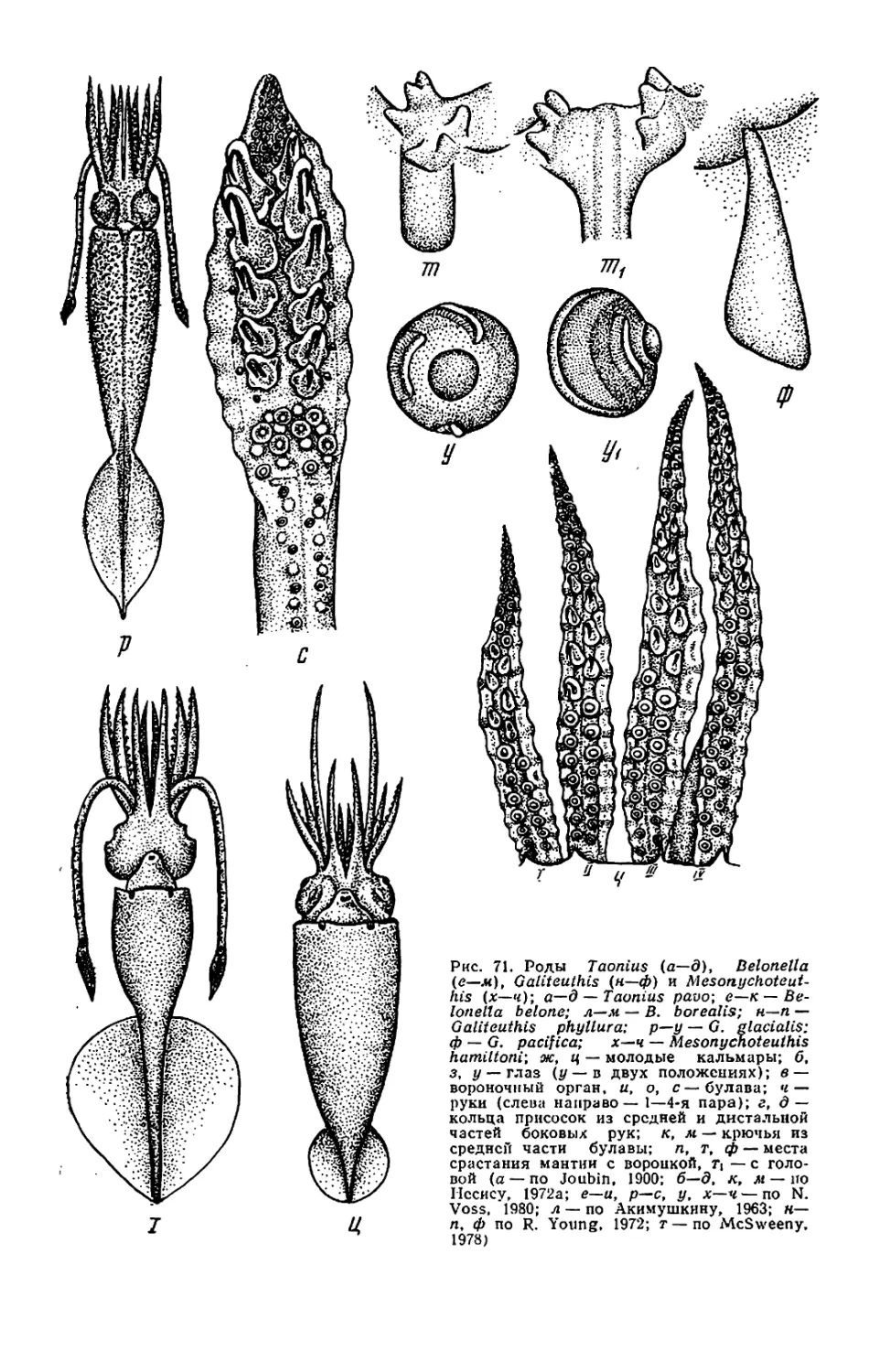
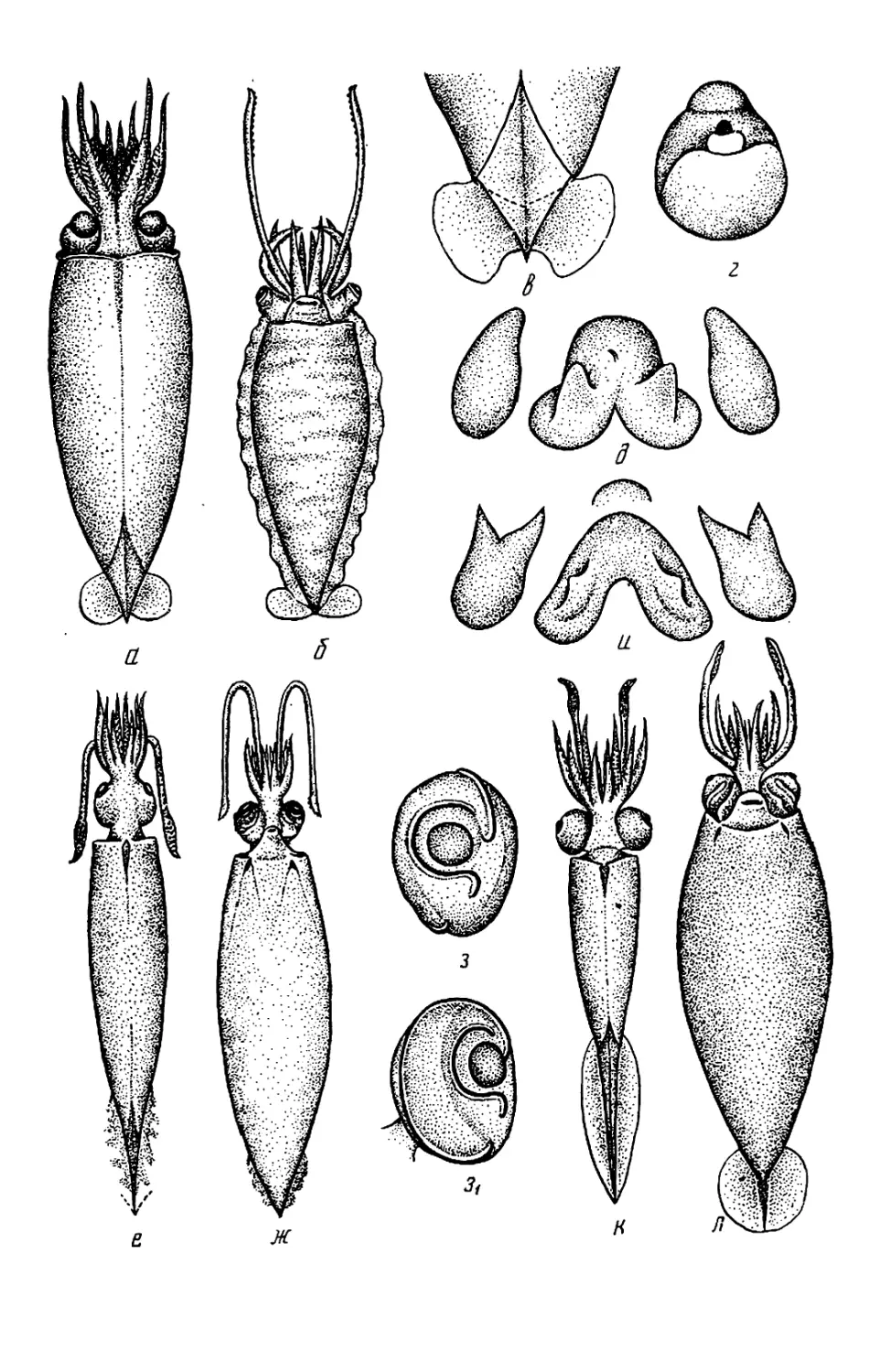
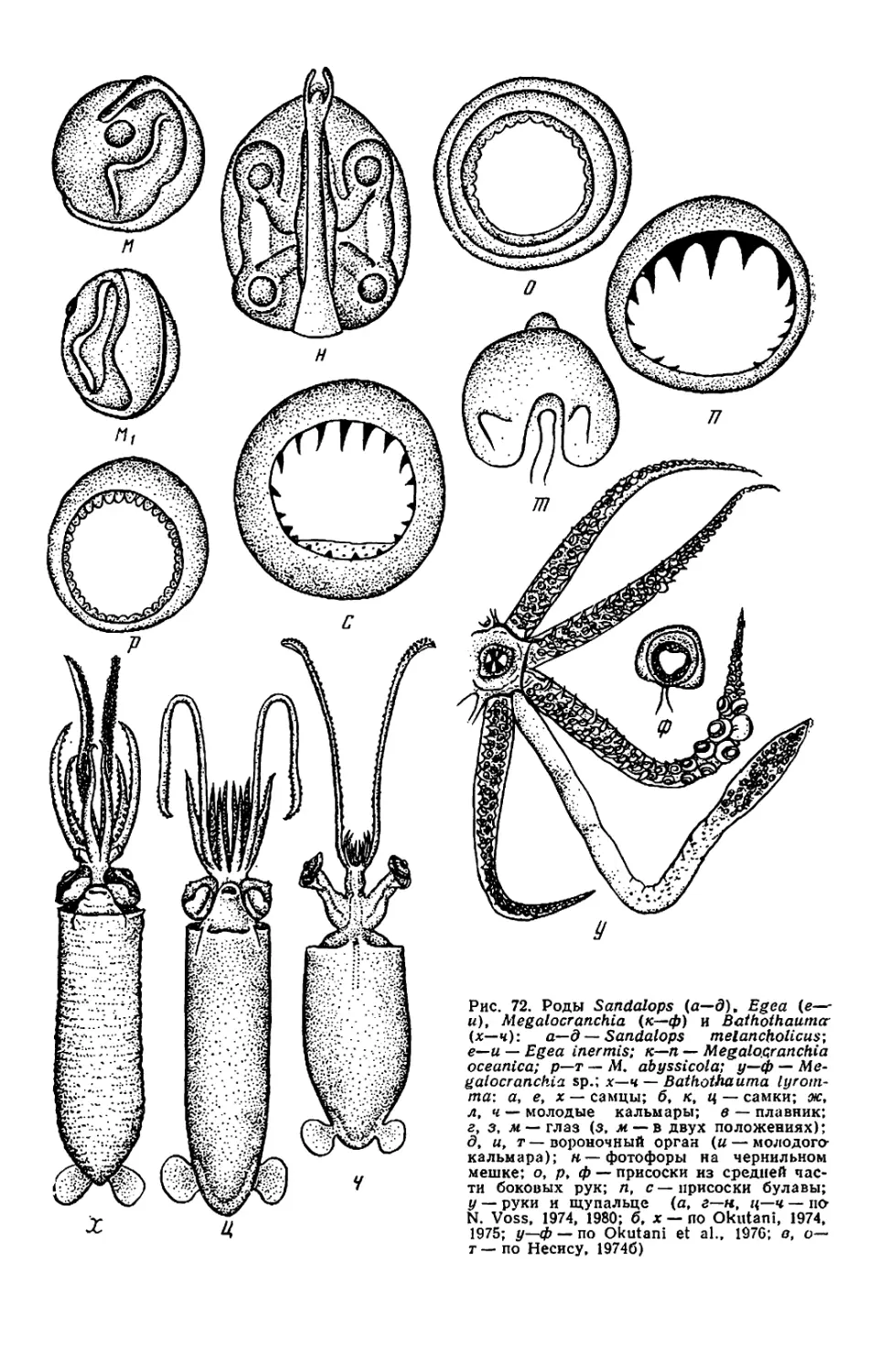
Belonella, см. рис. 71,к,и). Мяг-
кий край присоски превращается в
чехол, прикрывающий крюк так, что
наружу торчит лишь его кончик. Раз-
витие крючьев часто происходит на
поздней стадии онтогенеза. Например
у Berryteuthis magister крючья раз-
виваются на руках лишь при длине
мантии около 75 мм, у Galiteuthis
phyllura — на булаве при длине око-
ло 40 мм. Присоски, из которых в
будущем разовьются крючья, обыч-
но крупнее нормальных. Из-за посте-
пенного превращения присосок в
крючья число крючьев у молоди
меньше, чем у взрослых. Например,
у молоди Galiteuthis на булаве бы-
вает 5—7 пар крючьев, а у взрос-
лых— 13—14, у молоди Octopoteut-
his концы рук несут присоски, а у
взрослых крючья идут до самых кон-
чиков рук.
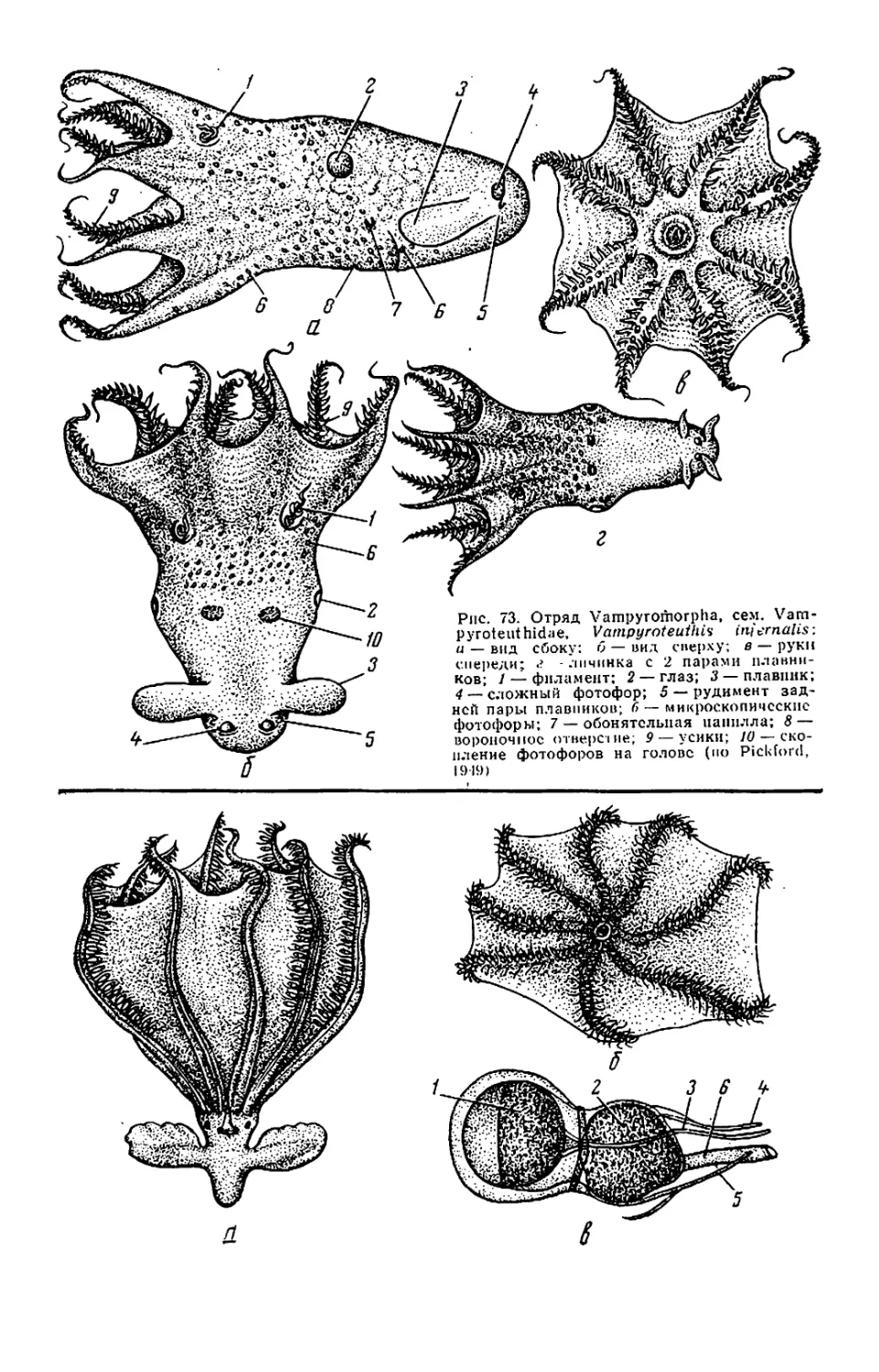
У вампироморф и осьминогов ру-
ки вооружены только присосками.
Присоски могут быть плоскими
(обычно), урноподобными или почти
шаровидными, по они всегда сидячие,
без стебельков и рогового кольца.
Внутренняя полость присоски выст-
лана нежной кожицей, которую ось-
миноги периодически сбрасывают
(«присоски линяют»). Присоски обыч-
но начинаются вблизи рта, но у вам-
пироморф— далеко отступя от пего
(рис. 73,а). Присоски вампироморф,
плавниковых осьминогов и многих
собственно осьминогов (Bolitaenidae,
Amphitretidae, Idioctopodidae, Vitre-
ledonellidae, а из Octopodidae— у
всех Eledoninae и некоторых Bathy-
polypodinae) — расположены в один
ряд, у остальных Octopodidae и у
Argonautoidea — в два ряда. Обычно
присоски быстро увеличиваются по
направлению от рта к дистальной
части руки, достигают максимума
приблизительно в конце первой трети
руки и зятем плавно уменьшаются к
концу. У самцов некоторых Octopus
вблизи края умбреллы несколько
присосок на всех или только боковых
руках резко увеличены: это опозна-
вательный признак взрослого самца.
Самец демонстрирует самке эти при-
соски, чтобы та на пего нс напала.
У самцов некоторых Opisthoteuthis и
Cirroteuthis резко увеличено несколь-
ко присосок на самом конце рук или
также в базальных частях, строение
этих присосок может сильно отли-
чаться от прочих. Присоски некото-
рых Cirroteuthidae, в том числе Cir-
rothautna (рис. 74,6), модифициро-
ваны— имеют всрстеновидпую фор-
му и не приспособлены к ловле до-
бычи. Их функция неясна (свечение?)
Вдоль ряда присосок па каждой
руке вампироморф и плавниковых
осьминогов протягивается справа и
слева по одному ряду усиков. Усики
(цирри) расположены вперемежку с
присосками и начинаются у вампиро-
морф ближе ко рту, чем первая при-
соска (см. рис. 73, в), у плавниковых
осьминогов — дальше. Они идут до
концов или почти до концов рук. На
концах рук вампироморф усики сли-
ваются в низкую мембрану. У вампи-
роморф и Opisthoteuthidae усики ко-
роткие, у некоторых Cirroteuthidae
(см. рис. 74, д) очень длинные. Их
функция тактильная: помогают на-
щупывать мелкую добычу (копепод
и др.).
МАНТИЯ. Ее форма очень разно-
образна. У каракатиц мантия обычно
уплощенная и овальная, у сепиолид
похожа на последний сустав боль-
шого пальца, у кальмаров в норме
веретеповидная, но бывает кониче-
ской, бокаловидной и даже совер-
шенно шарообразной, как у молоди
Cranchia и Liocranchia. Мантию ось-
миногов можно назвать яйцевидной
или мешковидной, а у Opisthoteuthis
она не более как бугорок.
Мантия нектонных кальмаров и
каракатиц имеет следующее строе-
ние; снаружи кожа, потом наруж-
ная подкожная оболочка (туника) с
многочисленными коллагеновыми во-
локнами и тонким слоем продольных
мышц, затем собственно мантия —
мышечный слой, тонкая внутренняя
туника (тоже с волокнами и немно-
гочисленными продольными мышца-
ми) и тонкая кожная выстилка ман-
тийной полости, в отличие от наруж-
ной кожи лишенная хроматофоров,
иридоцитов и пр. Собственно ман-
тия — основная съедобная часть
кальмаров и каракатиц — имеет тол-
щину до 2—3 см у активных нсктоп-
ных видов и около 0,5—1 см у ме-
нее подвижных. Она состоит из чере-
дующихся колец поперечных (коль-
цевых) и радиальных косоисчерчеп-
ных мышц. В состав мышечных
слоев входя г и эластичные коллаге-
новые волокна. Они направлены по-
переменно вправо и влево и как бы
навиваются спирально па мантию,
образуя конструкцию, хорошо рабо-
тающую па сжатие, типа «геодезиче-
ского купола». Попеременное сокра-
щение радиальных и кольцевых мышц
обеспечивает расширение и сокраще-
ние мантии, т. с. забор воды в ман-
тийную полость и выброс ее через
воронку, а эластичность коллагено-
вых волокон наружной и внутренней
туники и мышечного слоя — восста-
новление формы мантии после мы-
шечного сокращения.
Совсем иначе устроена стенка
мантии у полупланктонпых и планк-
тонных головоногих. У них мышцы
редуцированы и оводнены. В пре-
дельном случае — как у глубоковод-
ных осьминогов из Cirroteuthidae,
Bolitacnidae, Amphitretidae, Vitrcledo-
nellidae или у Tretnoctopus gelatus—
«мышечный» слой мантии представ-
ляет собой сильно оводненную бес-
клеточную основу, местами пронизан-
ную отдельными мышечными и кол-
лагеновыми волокнами. Такая ман-
тия почти прозрачна (сквозь тело
живого осьминога можно читать га-
зету) и по консистенции напоминает
колокол медузы. Толстая стенка ман-
тии океанических кальмаров из сем.
Octopoteuthidae, Histioteuthidae, Chi-
roteuthidae и др. заполнена вакуоля-
ми с раствором NH4CI. Он легче мор-
ской воды, так что кальмары в це-
лом имеют нейтральную плавучесть.
Такими же вакуолями заполнены
ткани их рук и головы, мускулисты
лишь плавники да стебли щупалец
(у Octopoteuthidae щупалец пет). Ра-
зумеется, такие животные для чело-
века несъедобны, они могут удов-
летворять вкус лишь кашалота.
Дальше зашли па этом пути кальма-
ры сем. Cranchiidae — «кальмары-ба-
тискафы»: у них раствор NH4CI хра-
нится в обширной полости тела — це-
ломе, мантия же стала плотной, ко-
жистой, однако нс мускулистой, так
как наличие в середине тела обшир-
ного резервуара с легкой жидкостью
(«поплавок батискафа») делает нс-
2 К. Н. Несис
17
возможными быстрые, частые и ин-
тенсивные сокращения мантии, с по-
мощью которых плавают нектонные
кальмары.
Нормальный цикл реактивного пла-
вания заключается в следующем: про-
дольные мышцы мантии расслабля-
ются, мантия становится более тон-
кой и выпуклой на брюшной стороне
(спинная сторона укреплена скелет-
ной пластинкой и не может менять
форму), щели между мантией и бо-
ковыми сторонами головы широко
раскрываются, устье воронки замы-
кается. Вода входит в мантийную
полость и заполняет ее. Затем мыш-
цы резко сокращаются, мантия утол-
щается, ее брюшная стенка поджи-
мается, мышцы-ретракторы головы и
воронки слегка поджимают голову,
щели по бокам головы замыкаются,
устье воронки широко раскрывает-
ся — вода с силой выбрасывается че-
рез узкое «сопло», давая животному
реактивный толчок назад. Такие
толчки следуют один за другим.
У полупланктонных и планктон-
ных головоногих со слабыми мыш-
цами сил хватает только на один-два
резких толчка — при необходимости
срочно уплыть от опасности. У моло-
дых кранхиид один толчок перемеща-
ет животное на 3—4 длины тела:
слишком мало, чтобы спастись от ак-
тивного хищника, во достаточно, что-
бы избежать хищпика-засадчика,
столь же безмускулыюго и малопо-
движного, как и сама жертва. Ман-
тийно-вороночный механизм служит
им главным образом для дыхания,
плавают они с помощью плавников,
а имеющие сильно развитую умбрел-
лу — и по способу медузы.
ПЛАВНИКИ. Всегда мускулистые,
даже у мсдузообразных головоногих.
Их форма разнообразна, можно вы-
делить 5 основных типов: плавнико-
вая кайма; ромбический, или серд-
цевидный; почковидный; округлый;
языковидный плавник (рис. 2, д).
Плавниковая кайма — узкая по-
лоска, протягивающаяся вдоль всей
мантии или (редко) лишь вдоль зад-
ней половины тела. Такие плавники
у настоящих каракатиц-сепиид, каль-
маров Sepioteuthis (Myopsida) и Neo-
teuthis (Oegopsida) (см. рис. 35, и—л\
57, с—т). Животные с такими плав-
никами плавают в горизонтальном
положении, довольно медлительны,
но очень маневрепны и поворотливы.
Ромбический, или сердцевидный,
плавник свойствен быстрым нектон-
ным или нектобентосным кальмарам
из сем. Loliginidae, Lycoteuthidae,
Enoploteuthidae, Gonatidae, Onycho-
teuthidae, Brachiotcuthidae, Psychro-
tcuthidae, Ommastrephidae, Thysano-
tcuthidac, Lepidoteuthidae. Самые
быстрые из них имеют ромбический
плавник. Кальмары с сердцевидным
плавником несколько уступают им в
скорости (меньше площадь при рав-
ной длине), но выигрывают в манев-
ренности. Обычно такой плавник за-
нимает заднюю часть мантии, его
длина от ‘/з До 2/з длины мантии, но
у Thysanoteuthidae он тянется вдоль
всей мантии. Ромбический плавник
имеют и некоторые полупланктонные
кальмары — Chaunoteuthis, Octopo-
teuthis, Cycloteuthidae. Они также
плавают в горизонтальном поло-
жении.
Почковидные плавники свойствен-
ны преимущественно мелким живот-
ным — Spirula, Sepiolidae, Sepiada-
riidae, Idiosepiidae, Pickfordiateuthi-
dae, Pyroteuthinae, Histioteuthidae,
Bathyteuthidae, Alluroteuthis, неко-
торые Cranchiidae. Из животных с
такими плавниками только полу-
планктонные Histioteuthidae и планк-
тонные Cranchiidae могут достигать
размеров, измеряемых дециметрами,
прочие — сантиметровых размеров.
Характерная особенность почковид-
ных плавников — выгнутый задний
край. Иногда такие плавники сидят
посредине мантии (Rossiinae, см. рис.
26) или даже сдвинуты вперед (не-
которые Sepiolinae), но прикрепля-
ются они, как правило, в задней ча-
сти мантии и иногда (Spirula, Bathy-
teuthis, см. рис. 16, 52, д—л) под уг-
лом к продольной оси тела. Живот-
ные с такими плавниками — скорее
«вертолеты», чем «самолеты». Для
многих из них обычное положение те-
ла — вниз головой (Spirula, Bathy-
teuthis), или же они могут зависать
в таком положении (Histioteuthis,
Cranchiidae). Сспиолиды используют
плавники для закапывания в песок:
воронкой размывают грунт, образуя
ямку, а плавниками и руками набра-
сывают грунт на спину; Pickfordia-
teuthidac и Idiosepiidae маневрируют
между стеблями морских трав.
Округлый (или эллиптический)
плавник свойствен малоподвижным
полуплаиктонпым и планктонным
18
кальмарам — Batoteuthidae, Chiro-
tcuthidae, IWastigoteuthidae (часть),
Grimalditeuthidae, Joubiniteuthidae,
Promachoteuthidae, многие Cranchii-
dac (Cranchiinae и часть Taoniinac).
Как правило, он сидит целиком или
задней своей частью на вытянутом
палочковидном гладиусе. Кальмары
с таким плавником могут держаться
в любой позе — и горизонтально, и
вертикально головой вниз, и наискось;
функция плавника — поддерживать
их в стабильном положении. Из всех
форм плавников эта наименее мус-
кулиста; округлый плавник, как пра-
вило, прозрачен. У Grimalditeuthis
(см. рис. 68, е) и некоторых Chiro-
teuthidae («Doratopsis» sagitta} за
обычным округлым плавником на
длинной игле гладиуса сидит второй,
листовидный или широколаицетовид-
ный и совсем прозрачный плавник.
Обычно он обламывается в молодом
возрасте, либо при опасности (на-
пример, при поимке). Известны лишь
единичные случаи поимки кальмаров
с двумя плавниками.
Языковидная или весловидная
форма плавника свойственна вампи-
роморфам и плавниковым осьмино-
гам. Языковидный плавник имеет
одинаковую ширину почти на всем
протяжении, весловидный сужен у
основания и расширен посредине, его
передний край выпуклый, задний
почти прямой. У Opisthoteuthis и
Cirroteuthidae плавники опираются
на края седловидного или подково-
образного скелетного хряща (см. рис.
75, л), обеспечивающего достаточную
жесткость конструкции движителя.
Обычная поза этих животных — го-
ловой вниз, горизонтальное положе-
ние они занимают при быстром пла-
вании (реакция бегства).
У гигантских кальмаров Architeu-
this, крупных Lepidoteuthis и Masti-
goteuthis форма плавника промежу-
точная между округлой и ромбиче-
ской. Наиболее разнообразна форма
плавника у крапхиид — от крохотных
лепестков у Helicocranchia = Asco-
cranchia до длинного узкого лан-
цета у Galiteuthis или Belonella кру-
га у Mesonychoteuthis или Leachia и
узкой каймы у Egea (см. рис. 69—
72). Плавник крапхиид в большинст-
ве случаев прикреплен к задне-боко-
вым сторонам ромбического расши-
рения задней части гладиуса — лап-
цсолы, но иногда его передняя часть
2*
прикреплена прямо к боковым сто-
ронам мантии (Тeuthowertia, Egea),
а у Bathothauma лапцеола редуциро-
вана, и плавнички крепятся к мантии
далеко один от другого.
Самый необычный плавник у Cte-
nopteryx — кальмара с «рыбьим плав-
ником». У взрослых Ctenopteryx
плавник — широкая пленчатая боко-
вая кайма, проходящая вдоль всей
мантии и растянутая па плотных со-
единительнотканных трабекулах,
очень похожих на мягкие лучи рыбь-
их плавников (см. рис. 52, а, б, г).
Плавники — пропульсивный ор-
ган при медленном плавании (при
быстром плавании они прижимаются
к телу или обвертываются вокруг
мантии). Волна, пробегающая по
плавнику, движет моллюска вперед.
Если плавник имеет форму длинной
каймы или ланцета, на нем умеща-
ется более одной волны (длина вол-
ны меньше длины плавника), если он
ромбический — половина или одна
волна. Кроме того, плавник может
создавать пропульсивную силу рез-
ким ударом сверху вниз. Для этого
лучше всего приспособлены ромби-
ческие и почковидные плавники, для
почковидных такой машущий способ
движения — единственный. Животное
взмахивает обоими плавниками син-
хронно. Осьминоги Cirrata могут,
взмахивать своими языковидными
или весловндными плавниками и син-
хронно, и попеременно; в последнем
случае плавники позволяют живот-
ному зависать на одном месте, подоб-
но вертолету. Каракатицы — сепии-
ды могут разворачиваться па одном
месте, направляя волну на одном
плавнике вперед, на другом — назад.
В онтогенезе форма плавника
кальмаров сильно меняется, тогда
как у каракатиц и плавниковых ось-
миногов эти изменения невелики.
Плавники новорожденного кальмара
крохотные, лепсстковидные, помеща-
ются на заднем конце тела и часто
нс соединяются между собой. По ме-
ре роста они удлиняются, перемеща-
ются вперед и принимают дефини-
тивную форму. У многих кальмаров
ко времени полового созревания вы-
растает длинный хвост, так что плав-
ник, первоначально занимавший зад-
нюю часть мантии, оказывается по-
средине тела (Alloteuthis, Uroteuthis^
Enoploteuthis, Ancistrocheirus), а си-
девший в середине тела — в перед-
19?
пей его половине (Octopoieuthis).
Если плавник заходит на хвост, то
его относительная длина в период
полового созревания быстро увели-
чивается (Alloteuthis, Gonatus tnado-
kai), если нс заходит — уменьшается
(Ancistrocheirus). У Mastigoteuthis и
некоторых других кальмаров с нача-
лом полового созревания прекраща-
ется рост плавника в длину, тогда
как ширина его продолжает увеличи-
ваться. Соответственно изменяется и
его форма.
ВОРОНКА. Это коническая труб-
ка, расширенная сзади и суженная
впереди. Она помещается на нижней
стороне головы в задней ее части
симметрично относительно продоль-
ной оси тела. У каракатиц и кальма-
ров воронка свободна по всей длине
и лежит в особом углублении, воро-
ночной. ямке, у осьминогов она час-
тично или почти полностью погру-
жена в ткани головы и часто лишь
копчик ее остается свободен. Перед-
няя часть вороночной ямки — перед
устьем воронки — у многих быстро-
плавающих кальмаров (Onychoteut-
hidae, Ommastrephidae) окаймлена
кожными гребнями, а у некоторых
оммастрефид отделена от остальной
части вороночной ямки кожной
складкой — фовеолой (см. рис. 59, а).
Мускулатура воронки очень со-
вершенна. Спинная сторона воронки
соединяется с нижней стороной го-
ловы парой мускулов-аддукторов
(уздечка), а задне-боковые стороны
присоединяются к гладиусу или к
спинной стороне мантийной полости
парой мощных и длинных мускулов-
ретракторов. Действием этих мышц
воронка может подтягиваться назад
й вверх и разворачиваться в стороны.
Слой кольцевых мышц в стенке са-
мой воронки обеспечивает сужение и
расширение ее диаметра. Передний
конец воронки очень подвижен и мо-
жет поворачиваться вбок и даже на-
зад, что обеспечивает возможность
разворота и заднего хода. Это на-
стоящее поворотное сопло.
У каракатиц, вампироморф и аб-
солютного большинства кальмаров
внутри воронки, па спинной стороне
перед устьем имеется небольшой
прямоугольный или треугольный кла-
гьац- Его функция — упрочнение
стенки воронки при повороте устья
ндзад для реактивного плавания го-
ловой, вперед. Клапан отсутствует у
осьминогов и некоторых малопо-
движных океанических кальмаров
(Valbyteuthis и почти все Cranchii-
dae, кроме Cranchia, Liocranchia, Ме-
golocranchia и Egea).
Дополнительная функция клапана
связана с выбрасыванием чернильной
жидкости: если чернила выбрасыва-
ются при поднятом клапане, они об-
разуют в воде удлиненную каплю,
отдаленно напоминающую самого
кальмара (псевдоморфа), если при
опущенном — обширное, висящее в
воде облако («дымовая завеса»).
На спинной же стороне воропки,
чуть отступя назад от клапана,
у всех двужаберных головоногих
располагается железистое образова-
ние— вороночный орган, или орган
Вериля (см. рис. 2,е,ж). У карака-
тиц и кальмаров он состоит из трех
частей в форме IAI, у осьминогов они
либо сливаются в W-образную ленту,
либо разделяются в середине — VV,
но иногда вороночный орган состоит
из 4 частей — III I. У некоторых ви-
дов Histioteuthidac и Cranchiidae на
ветвях средней части вороночного
органа развиваются выросты — ло-
пасти или папиллы, свисающие в
просвет воронки (см. рис. 55, ж\
70, к). Строение вороночного органа
имеет большое таксономическое зна-
чение, особенно у кранхиид и окто-
подид. Его функция слабо выяснена,
предполагают, что выделяемая воро-
ночным органом слизь помогает
прочищать воронку струей воды.
МАНТИЙНОЕ ОТВЕРСТИЕ И
ЗАМЫКАТЕЛЬНЫЙ АППАРАТ.
Мантийное отверстие — вход в ман-
тийную полость — расположено во-
круг шеи и представляет собой щель,
разделенную на три сегмента в мес-
тах (на затылке и на задне-боковых
сторонах воронки), где мантия сра-
стается или соединяется с головой.
У кальмаров и каракатиц в этих ме-
стах, как правило, располагаются
замыкательные хрящи — затылочный
и мантийно-вороночные. Затылочно-
го хряща нет у Sepiolinae, части He-
tero teuthinае (Sepiolina, Stoloteuthis,
hidoteuthis), Sepiadariidae, Idiosepii-
dac, Grimalditeuthidae и Promacho-
teuthidae. У двух видов Ommastrep-
hidae: Sthenoteuthis oualaniensis и
Eucleoteuthis luminosa (см. рис.
61, ле) — мантийный и вороночный
хрящи на послеличиночной стадии
срастаются на одной или обеих сто-
30=
ронах, но остаются вполне развиты-
ми и имеют типичную для семейства
форму. У Sepiadariutn, Grimalditeut-
his и Cranchiidae мантия срастается
с воронкой, мантийные и вороночные
хрящи отсутствуют.
У части каракатиц (Sepiolidae, Se-
piadariidae, Idiosepiidae) и всех ось-
миногов вентральная часть внутрен-
ностной массы соединяется с внут-
ренней поверхностью брюшной стен-
ки мантии недлинной продольной
мышечной септой, которая перегора-
живает мантийную полость и делит
ее на правую и левую половины. При
этом остается проход в передней и
задней частях полости. Септа может
подтягивать вентральную стенку
мантии к внутренностям, при ее со-
кращении на брюхе животного ста-
новится заметен неглубокий желобок.
У спирул, сепий, кальмаров и вампн-
роморф септа отсутствует, и мантий-
ная полость нс имеет продольной
брюшной перегородки.
Основной путь входа воды в
мантийную полость при «вдохе» и
заборе воды — вдоль боковых сторон
шеи, ближе к затылку. У быстропла-
вающих головоногих, особенно каль-
маров, в этих местах имеются осо-
бые направляющие гребни — про-
дольные и поперечные шейные склад-
ки. Продольные складки служат для
регулирования потока воды, попереч-
ные—для более плотного прилега-
ния края мантии к голове при вы-
бросе воды. У некоторых Onycho-
tcuthidae кроме них по обе стороны
затылка имеется серия тесно сбли-
женных продольных затылочных
складок {см. рис. 48,6).
У вампироморф и осьминогов
мантия срастается с головой на за-
тылке, по не срастается с воронкой.
Мантийное отверстие осьминогов
(кроме Argonautoidca) обычно в
большей или меньшей степени суже-
но. Иногда оно достигает уровня се-
редины или верхнего края глаза,
иногда занимает лишь нижнюю сто-
рону головы. Крайних пределов су-
жение мантийного отверстия дости-
гает у плавниковых осьминогов, Ani-
phitretidae и Idioctopodidae: у Opi-
stlioteuthidae и большинства Cirro-
tcuthidac мантийное отверстие реду-
цировано до узкой кольцевой щели
вокруг основания воронки, у Chunio-
huthis (см. рис. 74, и) полностью за-
растает — край мантии прирастает к
основанию воронки, у Idioctopodidae
и Amphitretidae мантия тоже сра-
стается с основанием воронки, но по
бокам головы остаются две щели
(круглые у Atnphiiretus и сарделько-
видные у Idioctopus) (см. рис.
77, з, л).
Затылочный хрящ — это удлинен-
ный прямоугольник или трапеция с
прямыми или слегка вогнутыми бо-
ковыми сторонами и, как правило,
тремя продольными гребнями — по
средней линии и по бокам. Его фор-
ма мало изменчива.
Значительно более изменчива
форма вороночного хряща, который
имеет чрезвычайно важное значение
для систематики, в первую очередь,
океанических кальмаров. Различают
следующие основные типы его строе-
ния: простой (прямой), _1_-образный,
Н -образный, треугольный, овальный,
уховидпый хрящ (см. рис. 2, з).
Имеются многочисленные варианты
основных типов. Простой вороноч-
ный хрящ — сильно удлиненный тре-
угольник с прямой, реже слегка изо-
гнутой или расширяющейся назад
бороздой, на мантии ему соответст-
вует низкий гребень. Такова (с не-
большими вариациями) форма замы-
кательных хрящей спирулы, сепио-
лид, неритических и большинства (14
из 23 семейств) океанических каль-
маров. У Sepiidae вороночный хрящ
удлиненно-овальный с глубокой бо-
роздой, мантийный — у Sepia оваль-
ный, у Sepiella — треугольный (см.
рис. 17, б—в). Характерный _1_-образ-
ный вороночный хрящ Ommastrepidae
имеет форму треугольника с двумя
перпендикулярными бороздами, у
места пересечения которых располо-
жены два крепких бугорка, мантий-
ный хрящ — два перпендикулярных
гребня, вертикальный перед местом
пересечения резко суживается. Похо-
жую, по повернутую на 90° форму
имеет 1- -образный хрящ Thysano-
teuthidae. Треугольный вороночный
хрящ с глубокой центральной ямкой
и отходящими от нес вперед и на-
зад бороздами свойствен C’ycloteut-
hidae. У высших Chiroteuthidae и не-
которых Mastigoteuthidae вороноч-
ный хрящ уховидпый — имеет форму
овала, внутрь которого выступают
два треугольных бугорка; более
крупный боковой (вентральный) —
трагус и более мелкий задний (ка-
удальный) — антитрагус, мантийный
21
хрящ имеет форму человеческого
носа (см. рис. 65—66). У низших
Chiroteuthidae вороночный хрящ мо-
жет быть простым, расширенным
сзади (кегле- или бутылковидным)
или овальным, с антитрагусом или
без него; у части Mastigoteulhidae он
овальный, с трагусом или без него,
у некоторых видов — бутылковидный
(с двумя трагусами и антитрагусом^.
Овальный вороночный хрящ без вы-
ступов свойствен также .Joubiniteut-
hidae и Promachoteuthidae (см. рис.
64, 66—68).
У наиболее специализированных
осьминогов надсем. Argonautoidea
также имеется замыкательный аппа-
рат. Он примитивен и в принципе
представляет собой крючковидную
структуру: задне-нижний свободный
край воронки сильно изогнут вперед
и зацепляется за передний край ман-
тии. Таково его строение у Alloposi-
dae. У Tremoctopodidae па мантии
развивается удлиненная ямка, в ко-
торую входит загнутый край ворон-
ки. У Ocythoidae край воронки завер-
нут и имеет вид улиткообразной
кнопки, входящей в соответствую-
щую ямку на мантии. Еще лучше
развит этот кнопковидный замыка-
тельный аппарат у Argonautidae.
РОТ, ЧЕЛЮСТИ И РАДУЛА.
Ротовое отверстие окружено наруж-
ной и внутренней губами. На наруж-
ной губе или па вентральной части
буккальной мембраны у самок мно-
гих видов кальмаров и каракатиц
помещаются ссмеприемники. Во рту
находится крепкий роговой клюв с
верхней и нижней половинами (ман-
дибулами). Верхняя мандибула клю-
ва короче нижней, так что нижняя
заходит за верхнюю. Обе мандибулы
состоят из боковых стенок (глоточной
пластины), похожих на сло-
женный «домиком» листок, и капю-
шона (фронтальной пластины), по-
крывающего переднюю часть боко-
вых стенок подобно крыше или тен-
ту над палаткой. Боковые стенки и
капюшон сливаются в передней час-
ти клюва, образуя рострум и режу-
щий край (плечи) мандибул. Верх-
ние части капюшона и боковых сте-
нок представляют собой гребни. Ка-
пюшон вентральной мандибулы
выступает за край боковых стенок,
образуя крылья. У всех двужабер-
пых головоногих боковые стенки
вентральной мандибулы более или
22
менее узкие и удлиненные, дорсаль-
ной — короткие и высокие, однако
форма, относительные размеры и
скульптура боковых стенок и капю-
шона, размер рострума, наличие или
отсутствие зубчика на режущем крае
и пр. сильно различаются. Строение
клюва в целом характерно для се-
мейства, а обычно — для рода и да-
же вида, что даст возможность оп-
ределять кальмаров и каракатиц по
их клювам (см. рис. 2, и—о). Манди-
булы осьминогов более однообразны
и труднее определимы. Клювы ось-
миногов легко отличаются от клювов
кальмаров и каракатиц коротким ту-
пым рострумом (см. рис. 2, п).
Клювы молодых головоногих по-
лупрозрачны. С возрастом они по-
степенно темнеют, начиная с крыль-
ев, и у взрослых особей почти
сплошь черные или бурые. Период
быстрого потемнения клювов совпа-
дает с половым созреванием.
В ротовой полости на мускулис-
том языкоподобном выступе — одон-
тофоре помещается радула (см.
рис. 2, р—т). Это длинная лента,
согнутая пополам в передней части
одонтофора и несущая зубы. Новые
зубы формируются в задней и ниж-
ней частях одонтофора, в верхней
передней его части находятся самые
старые, постепенно отпадающие зу-
бы. С ростом моллюска ширина ро-
товой полости увеличивается, соот-
ветственно и ширина самых молодых
рядов радулы больше, чем старых,
образовавшихся в молодом воз-
расте.
Обычно радула двужаберных го-
ловоногих состоит из 7 продольных
рядов зубов: средний (рахидиаль-
ный) зуб, 1-, 2-, и 3-й латеральные
(или латеральный, внутренний и
внешний маргинальные) зубы (см.
рис. 2, р). По краям от них поме-
щается по одной маргинальной
(краевой) пластинке. Пластинки
имеются далеко не у всех видов —
редуцированы или отсутствуют у
сспиолид и большинства эгопсид, но
сильно развиты у осьминогов. У спи-
рулы и плавниковых осьминогов ра-
дула отсутствует. У некоторых го-
натид (род Gonatus и подрод Gona-
topsis) радула состоит из 5 рядов
зубов (отсутствует 1-й латеральный
зуб) (см. рис. 2, с). У некоторых
Octopodidae (Thaurneledone, Vosse-
ledone) радула частично дегенериро-
вана: 1-й латеральный зуб еле заме-
тен, 3-й отсутствует. Отчасти дегснс-
рировапа радула также у Bentheledo-
пе, некоторых Graneledone и др.
Рахидиальный зуб у кальмаров,
каракатиц и вампироморф одно- или
трехзубый. У осьминогов он может
быть одно-, трех- или пятизубым
(иногда с 7—11 зубчиками), симмет-
ричным или асимметричным (см.
рис. 2, т). У ряда октоподид форма
рахидиалыюго зуба разная в сосед-
них поперечных рядах, но в точно-
сти повторяется в каждом третьем,
четвертом или пятом ряду (сериация
радулы). 1-й латеральный зуб у дву-
жаберпых головоногих однозубый с
широкой пяткой или двузубый (зу-
бец, который ближе к средней линии,
крупнее), 2-й и 3-й латеральные
(маргинальные) зубы у кальмаров и
каракатиц схожи: короткое основа-
ние и длинный саблевидный или кин-
жаловидный зубец; у большинства
осьминогов 2-й латеральный зуб, на-
против, более сходен с 1-м и лишь
3-й саблевидный. У иадсем. Bolitae-
noidea (Ctenoglossa, т. е. гребенчато-
зубые) рахидиальный, 1-й и 2-й лате-
ральные зубы гребенчатые; рахи-
диальный обычно семизубый (Idio-
ctopus имеет 9—И зубцов), латераль-
ные с 4—7 зубцами.
Радула кальмаров, каракатиц и
Argonautoidea служит главным об-
разом для захвата откусанных клю-
вом кусочков пищи и переноса их в
глотку (принцип действия снегоубо-
рочной машины). С этим и связано
сильное развитие 2-го и 3-го лате-
ральных зубов, удерживающих ку-
сочки пищи, и относительно слабое
развитие рахидиалыюго зуба, функ-
ция которого — пе допускать их про-
скальзывания вдоль радулярной
ленты.
У донных осьминогов функция
радулы несколько иная: с помощью
рахидиалыюго зуба они просверли-
вают раковины’ двустворчатых и
брюхоногих моллюсков и выцарапы-
вают кусочки мяса из конечностей
крабов Поэтому их рахидиальный
зуб очень велик и крепок. Этим обу-
1 В этом процессе участвуют мелкие
зубчики на конце слюнной папил-
лы — так называемая «вторая раду-
ла»; ферменты слюны размягчают
карбонат раковины.
словлено и сильное развитие марги-
нальных пластинок, служащих проч-
ной опорой поднятым в рабочее по-
ложение зубам. Латеральные же зу-
бы осьминогов, особенно 1-е, умень-
шены. Как действует гребневидная
радула Bolitaenoidea, неизвестно.
КОЖА И ХРОМАЮ ФОРЫ. Ко-
жа головоногих моллюсков тонкая,
но сложно устроенная. Верхний
слой — эпидермис образован одно-
слойным цилиндрическим эпителием
с многочисленными слизистыми клет-
ками. Выделяемая ими слизь делает
тело головоногих скользким и облег-
чает передвижение в воде. Ниже ле-
жит соединительнотканный слой, со-
держащий мышечные волокна, хрома-
тофоры и иридоциты, а под ним —
внешняя туника.
Поверхность кожи может быть
совершенно гладкой или шерохова-
той, бугорчатой, бородавчатой. Кожа
большинства кальмаров гладкая, но у
Lepidoteuthidae в соединительноткан-
ном слое развиваются чешуеподоб-
ные образования (см. рис. 62, и, м, о).
Они могут тесно соприкасаться по-
добно брусчатке мостовой (Tetrony-
choteuthis), или налегать друг на
друга, как чешуя рыб (Lepidoteut-
his— «чешуйчатый кальмар»). На
коже некоторых Chiroteuthidae (Аз-
peroteuthis), Mastigoteuthidae (Echi-
noteuthis) и Cranchiidae (см. рис. 64,
67, 69) имеются хрящевые бугорки
конической или звездчатой формы.
У Asperoteuthis, Echinoteuthis, С ran-
chia они покрывают почти все тело
(кроме плавников и щупалец), у Lio-
cranchia, Leachia и Drechselia идут
вдоль гиалиновых хрящевых полосок
на мантии, у некоторых Taoniinae
(Тeuthowenia, Galiteuthis, Mesonycho-
teuthis) сидят в местах срастания
мантии с воронкой или головой. Ко-
жа некоторых каракатиц (например,
Sepia tullbergi), кальмаров (Masli-
goteuthis cordiformis) и большинства
донных осьминогов покрыта бугорка-
ми, бородавочками и папиллами.
У осьминогов особенно крупные бу-
горки расположены над глазами:
надглазничные усики, или «ушки».
Папиллы некоторых, преимуществен-
но батиальных донных осьминогов
могут быть очень крупными- и сплошь
покрывать тело. Таковы, например,
звездчатые бородавки Graneledone,
некоторых Pareledone и Bathypolypus
(см. рис. 80, 83, 85). Многие осьми-
23
ноги и каракатицы могут поднимать
папиллы и бугорки или сглаживать
их. Это хороший способ камуфляжа.
На брюшной стороне мантии и рук
4-й пары некоторых каракатиц (5ё-
pia tuber culata, S. papillata) и спин-
ной стороне мантии Idioscpiidae име-
ются участки шероховатой кожи —
«кожные присоски» (см. рис. 19, д;
33). Каракатицы присасываются ими
к камням в полосе прибоя, а кро-
хотные Idiosepius— к листьям мор-
ских трав.
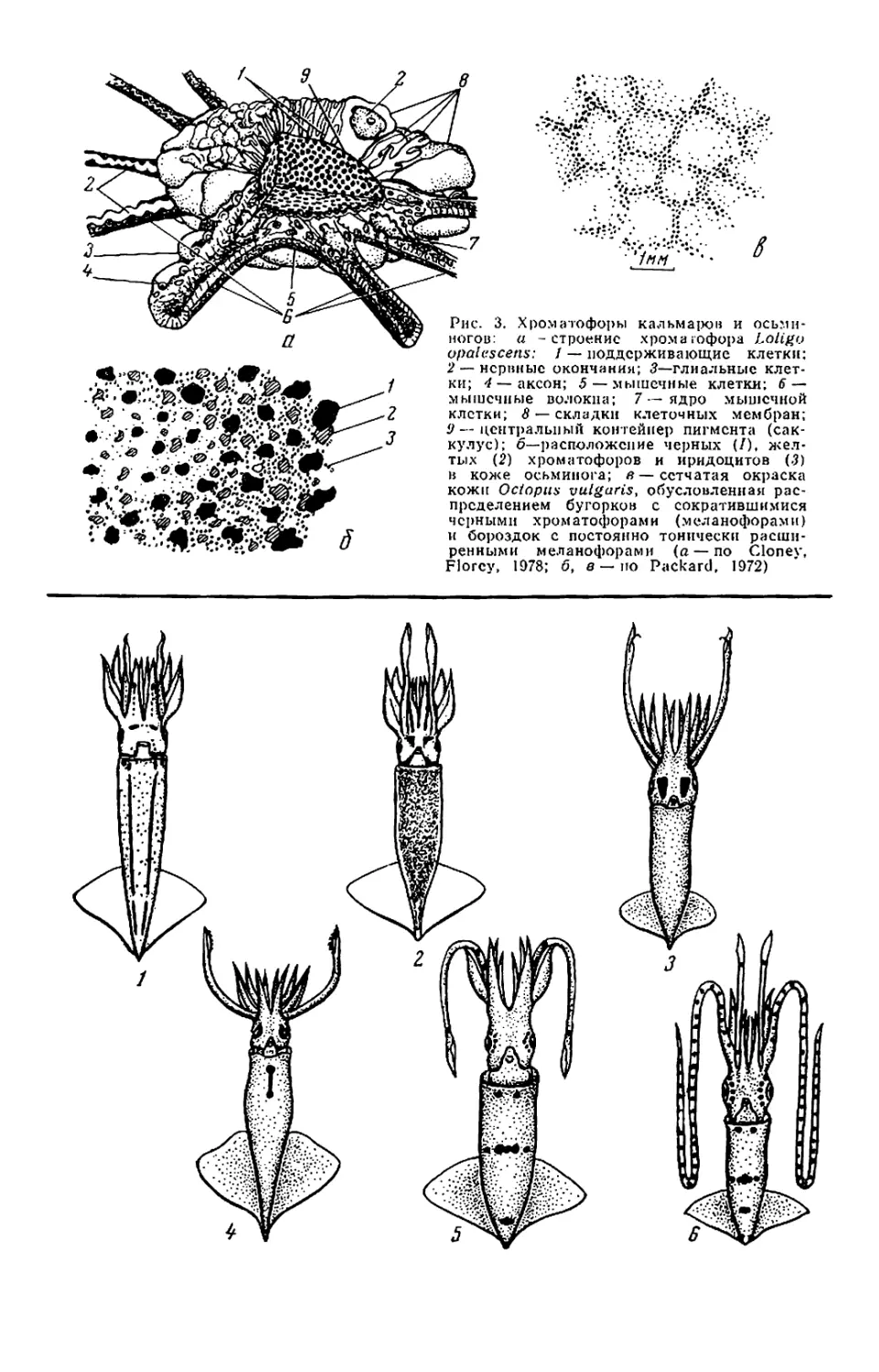
Хроматофоры — органы, содержа-
щие пигмент (рис. 3, а, б), лежат не-
сколькими слоями. Различают собст-
венно хроматофоры (красные, оран-
жевые и желтые) и меланофоры
(черные или бурые). В хроматофо-
рах имеются как минимум три сор-
та пигментов, относящихся к груп-
пам меланинов (эвмеланин, фсомела-
нины) и оммохромов, причем один и
тот же пигмент может давать раз-
ную окраску в зависимости от кон-
центрации окрашенных компонентов
(лигандов). Разные типы хромато-
форов распределены слоями незави-
симо друг от друга, меланофоры об-
разуют обычно верхний слой, самые
светлые хроматофоры — нижний.
Хроматофор имеет звездообразную
форму, состоит из центрального кон-
тейнера (саккулус), содержащего
пигмент, расходящихся от него по
радиусам мышечных клеток и сети
поддерживающих, нервных и глиаль-
ных клеток. Когда животное в спо-
койном состоянии, пигмент скон-
центрирован в середине саккулуса, и
хроматофор представляет собой
цветную точку («веснушку»). При
возбуждении хроматофорных нервов
мышечные волокна сокращаются и
растягивают пигмент ио всей клетке,
образуя цветное пятно в форме не-
правильного многоугольника. Хрома-
тофоры находятся под двойным
нервным контролем — местным и
центральным (от хроматофорной до-
ли мозга). Нервные волокна несколь-
ких соседних хроматофоров соедине-
ны, и такая группа реагирует на
нервные сигналы как единое целое
(рис. 3, в), образуя единицу окраски,
например крохотные глазчатые пят-
на, из которых складывается общий
фоп окраски большинства кальмаров
(темная точка окружена светлым ко-
лечком, которое в свою очередь ок-
ружено темным кольцом). Отдель-
24
ные единицы управляются разными
нейронами, поэтому при возбужде-
нии по телу моллюска могут пробе-
гать цветные «волны» или «обла-
ка» — тс или иные единицы то
«вспыхивают», то «гаснут». Местный
контроль хроматофоров выражает-
ся в возбуждении их под действием
местных раздражителей. Например,
если провести по телу кальмара
острой палочкой, то в этом месте тут
же вспыхнет темная линия.
Иридоциты — клетки, в которых
содержатся пластинчатые кристаллы,
хорошо отражающие и преломляю-
щие падающий свет. Кристаллы мо-
гут быть расположены параллельно
поверхности кожи или под разными
углами, так что могут работать как
микроскопические зеркальца и приз-
мы. Различают собственно иридоци-
ты (иридофоры), которые благодаря
разложению (дифференциальному
отражению) и последующей интер-
ференции световых лучей дают жел-
тую, зеленую и голубую окраску, и
лейкофоры, отражающие лучи с лю-
бой длиной волны и, следовательно,
дающие чисто-белый цвет. Благодаря
многообразным сочетаниям хромато-
форов и иридоцитов и быстроте нерв-
ного контроля головоногие моллюски
могут чрезвычайно быстро менять
окраску и камуфлироваться па лю-
бом фоне. Ие имея цветного зрения,
они прекрасно подгоняют свою ок-
раску под окраску субстрата, так
как хроматофоры создают общий
рисунок, а боковые лейкофоры дей-
ствуют как маленькие зеркальца, от-
ражая окраску фона. В отдельных
случаях иридоциты лежат не под
хроматофорами, а над ними. При
этом возникает интенсивная метал-
лически ирризирующая окраска чис-
тейшего синего тона, насыщенность
и контрастность которой моллюск
может изменять. Так устроены круп-
ные глазчатые пятна некоторых ось-
миногов и синие кольца самого кра-
сивого и самого опасного в мире
осьминога Hapalochlaena maculosa
(см. рис. 78, а, б). Глазчатые пятна
осьминогов располагаются на ум-
брелле между руками 2-й и 3-й пар
впереди глаз и имеют диаметр око-
ло 1—2 см (см. рис. 82,б,ч,э). Это
темное круглое пятно, окруженное
последовательно светлым, темным и
снова светлым кольцом (иногда по-
рядок тонов бывает обратным). При
опасности осьминог растопыривает
и подворачивает руки и показывает
нападающей рыбе два больших ши-
роко расставленных круглых «гла-
за» — этого достаточно, чтобы испу-
гать любого противника.
Богатые возможности изменения
окраски и структуры поверхности ко-
жи позволяют каракатицам и в осо-
бенности донным осьминогам Octopo-
didae принимать разнообразнейшие
облики, «надевать маски» — покрови-
тельственные, отпугивающие, устраша-
ющие, дезориентирующие врага, при-
влекающие самку и т. п. (см. рис.
1,ас). «Маски» обусловлены рисун-
ком светлых и темных пятен и по-
лос на теле и конечностях в сочета-
нии с изменениями текстуры поверх-
ности кожи, особыми позами и дви-
жениями — поднятие, опускание или
растопыривание рук, приподнимание
или втягивание головы, движения
глаз, изменение очертаний мантии,
учащение или замедление дыхания
и пр. Они резко различаются у ли-
чинок, молоди и взрослых животных.
Основные типы — «картины» обли-
ков — стандартны и сходны у раз-
ных видов и даже разных (но эко-
логически сходных) групп, как Sepia,
Sepioteuthis, Octopus', однако детали
«маски» не только видоспецифичны, но
и индивидуальны в такой же степе-
ни, как отпечатки пальцев у людей.
У пелагических кальмаров и осьми-
ногов репертуар обликов гораздо
беднее прежде всего потому, что нет
камуфлирующей и отпугивающей
«маски» (хотя некоторые лолигини-
ды могут, ложась на дно, принимать
дезориентирующую окраску). Еще
беднее репертуар обликов глубоко-
водных головоногих — одни из них
прозрачны, у других (вампиромор-
фы, некоторые глубоководные каль-
мары и осьминоги) хроматофоры ли-
шены мышечных элементов и цент-
ральной иннервации, так что окрас-
ка этих животных не меняется.
ОРГАНЫ СВЕЧЕНИЯ. Фотофо-
ры имеются у спирулы, вампиротей-
тиса, абсолютного большинства океа-
нических кальмаров: у всех Lycoteut-
hidae, Enoploleuthidae, Octopoteuthi-
dac, Ctenopterygidac, Bathyteuthidae,
Histioteuthidae, Cycloteuthidae, Gri-
malditeuthidae, Cranchiidae, по-види-
мому, всех Psychroteuthidae, у боль-
шинства родов Ommastrephidae, Chi-
roteuthidae, Mastigoteuthidac, некото-
рых Onychoteuthidae (2 рода), Gona-
tidae и Brachiotcuthidae (по 1 роду),
у 9 из 14 родов сепиолид, многих ло-
лигинид (из родов Loligo и Uroteut-
his), Bolitaenidae (с сомнением) у
отдельных видов сепиид (Sepia aus-_
tralis), осьминогов Octopus arakawai
( — 0. ornatus) и Treinoctopus vio-
laceus.
У головоногих моллюсков разли-
чают 2 типа свечения: симбиотичес-
кое (бактериальное) и собственное
(внутриклеточное). При симбиотиче-
ском свечении светятся бактерии ро-
да Photobacterium, живущие в желе-
зах, расположенных па чернильном
мешке по обе стороны кишки. Эти
органы имеются главным образом у
шельфовых донных головоногих —
сепиолид Sepiola, Rondeletiola„
Euprymna, Semirossia, лолигипид Lo-
ligo и Uroteuthis, а также у полу-
глубоководпых сепиолид подсем. Нс-
teroteuthinae (см. рис. 28, ф\ 29, л\.
31, д', 39, н). Предполагают, что спи-
рула также имеет бактериальное све-
чение, но твердых доказательств это-
го не получено. Бактерии могут све-
титься внутри железы. Слизь со взве-
шенными в ней бактериями может
также выбрасываться наружу через--
воронку, образуя светящееся облачко»
(«стрельба огнем»).
Собственное свечение обусловлена
реакцией окисления люциферина ато-
марным кислородом, катализируемой;
ферментом люциферазой. Химия это-
го процесса изучена у японского
кальмара-светлячка Watasenia scin-
tillans и оказалась весьма сход-
ной с классической формой люцифс-
рин-люциферазной реакции, свойст-
венной ракушковому рачку Cypridinai
и многим другим морским живот-
ным (кишечнополостным, гребневи-
кам, полихетам, десятиногим ракооб-
разным, рыбам). Люциферин ватасс-
нии имеет формулу C26H23O10N3S2-
Собственное свечение свойственно
преимущественно пелагическим голо-
воногим. Головоногие с этим типом
свечения распространены от поверх-
ности океана до абиссопслагиали.
При внутриклеточном свечении
свет испускают особые клетки — фо-
тоциты; светящееся вещество не вы-
ходит за пределы клетки. Таковы
фотофоры океанических кальмаров,
вампироморф и осьминогов.
Расположение фотофоров у голо-
воногих моллюсков чрезвычайно раз-
25
пообразпо (рис. 4). Их количество
может варьировать от одного (Spiril-
la) до многих сотен (Enoploteuthinae,
Ancistrochcirinae, Histioteuthis, Mas-
tigoteuthis) и тысяч (Ommastrephi-
nae). «Излюбленные» места располо-
жения фотофоров — там, где имеется
или легко может возникнуть черная
подложка — пигментная ширма, ко-
торая позволяет направлять луч све-
та: вентральная сторона мантии, го-
ловы, рук, глазного яблока, поверх-
ность чернильного мешка. Однако
фотофоры могут находиться также
на концах рук (Abraliopsis, Oulopo-
teuthis, Grimalditeuthis^ взрослые сам-
ки Psychroteuthis и ряда кранхиид)
и щупалец (Chiroteuthis), на спинной
стороне мантии (Sthenoteuthis, Vam-
pyroteuthis) , на заднем конце тела
(Spirula), стеблях щупалец (Lyco-
ieuthis), внутри мантийной полости
на внутренностях (Lycoteuthidae, Ру-
roteuthinae), на нервных стволах рук
(например, у Ommastrephinae), да-
же внутри конуса гладиуса (Руго-
teuthinae).
Общая особенность размещения
•фотофоров в том, что у животных,
плавающих в горизонтальном поло-
жении, они расположены, как пра-
вило, на вентральной стороне тела.
Это помогает светящимся головоно-
гим маскироваться и от хищников,
нападающих сверху, и от врагов, на-
падающих снизу (с помощью вен-
трального противосвсчсния, см, ни-
же) . Если фотофоры расположены
на заднем конце или спинной сторо-
не тела, значит, либо животное «не
•боится» врагов (Sth enote at his), либо
плавает вниз головой (Spirula, Bathy-
ieuthis, Vampyroteuthis, крапхииды).
Строение фотофоров, особенно у
океанических головоногих, столь же
разнообразно, как и их расположе-
ние. Простейший тип строения фото-
форов встречается у Ommastrephidae
и Vampyroteuthis. Фотофоры оммаст-
рефид — крохотные овальные или
•округлые желтоватые тельца, погру-
женные в соединительнотканный
слой кожи и рассеянные ио всему
телу, за исключением щупалец и
плавников. Каждое тельце — скопле-
ние фотоцитов, пронизанное крове-
носными сосудами и одетое оболоч-
кой из вакуолизированной ткани.
Тельца могут быть рассеяны пооди-
ночке или собраны в тесные группы,
например в спинном фотофорс Sthe-
26
noteuthis. Примерно так же устроены
и фотофоры Vampyroteuthis.
Наиболее сложный тип вырабо-
тался независимо у разных семейств
океанических кальмаров. В фотофо-
рах этого типа скопление фотогенной
ткани, пронизанное сосудами, под-
стилается толстым слоем отражаю-
щих свет структур (хитин и цито-
плазма), причем рефлектор часто
подразделен на 2—3 отдельных об-
разования. Он окружен пигментным
чехлом. Сверху лежит крупная лин-
за или мощный слой световодов
(светопроводящих волокон), еще вы-
ше могут лежать прозрачная оболоч-
ка, цветной фильтр или пигментная
ширма из мелапофоров. Детали
строения фотофоров чрезвычайно
разнообразны, у одного вида могут
быть фотофоры многих (до десятка)
типов. Фотофоры, снабженные лин-
зой (например, мантийные фотофо-
ры Enoploteuthidae или Histioteuthi-
dae), дают узко направленный луч
света; снабженные световодами, рас-
пределяющими свечение по всей по-
верхности органа (глазные фотофо-
ры Enoploteuthidae и Cranchiidae,
внутрнмантийпые фотофоры Lyco-
tcuthidae и др.), — разлитое свече-
ние. Очень варьирует и интенсив-
ность свечения. Например, у Wata-
senia scintillans свет мантийных фо-
тофоров еле заметен, а свечение фо-
тофоров на концах брюшных рук
подобно вспышке зажженной в тем-
ноте спички.
Фотофор неритических кальмаров
и сен иолид е бактериальным свече-
нием представляет собой железу,
соединенную протоком с мантийной
полостью/ Обычно она парная, ино-
гда обе железы сливаются в одну,
однако протоки остаются парными.
Имеются рефлектор, линза и обиль-
ная сеть кровеносных сосудов. Роль
пигментной ширмы играет поверх-
ность чернильного мешка, на кото-
рой лежит фотофор.
Наиболее просто устроены фото-
форы у поверхностных и самых глу-
боководных (бати-, абиссопелаги-
ческих) головоногих, наиболее слож-
ные и разнообразные фотофоры — у
среднеглубинных (мезопелагических)
видов. Наибольшее число видов све-
тящихся головоногих — также в ме-
зопелагиали.
Свечение у всех головоногих мол-
люсков, как и у большинства мор-
2 — нервные окончания; 3—глиальные клет-
ки; 4 — аксон; 5 — мышечные клетки; 6 —
мышечные волокна; 7 — ядро мышечной
клетки; 8 — складки клеточных мембран;
0 — центральный контейнер пигмента (сак-
кулус); б—расположение черных (/), жел-
тых (2) хроматофоров и нридоцитов (.3)
в коже осьминога; в— сетчатая окраска
кожи Octopus vulgaris, обусловленная рас-
пределением бугорков с сократившимися
черными хроматофорами (меланофорами)
и бороздок с постоянно тонически расши-
ренными меланофорами (а — по Cloney,
Florey, 1978; б, в — по Packard, 1972)
23
Рис. 4. Схема расположения фотофоров
(зачернены) на теле головоногих моллюс-
ков (из Herring, 1977): 1 — Eucleoteuthis
lutninosaz 2 — Otntnasirephes bartrami; 3 —-
Gonatus pyros', 4 — Onychoteuthis banksi
borealijaponicus\ 5 — Selenoteuthis scintitt-
ans; 6 — Nematolampas regalis', 7 — Masti-
goteuthis pyrodes; 8 — Octopoteuthis delet-
ron\ 9 — Chiroteuthis calyx', 10 — Ctenopte-
ryx sicula; 11 — Grimalditeuthis bonplandi’,
12— Histioteuthis bonnellii; 13—Bathyteuthis
abyssicola (juv.); 14 — Abraliopsis falco,
IS — A ncistrocheirus alessandrinii (j u v.);
16 — Pterygioteuthis gemmata; 17—Cranchia
scabra; 18 — Galiteuthis phyllura', 19 —
Megalocranchia oceanica*. 20 — Cycloteuthis
sir vend', 21 — Discot euthis spp. ( a — D.
laciniosa; 6 — D. discus); 22 — Vampyrote-
uthis infernatis', 23 — Loligo (Doryte'uthis)
sp.; 24 —Spirula spirula', 25 — Sepiolina
nipponensis
Рис. 5. Гладиус и хрящевой скелет: а—
в —гладиус кальмаров Gonatus tinro (а)
Todarodes pacificus (б) и Loligo sp. (в):
1 — ствол (рахис); 2 — перо; 3 — конечный
конус: г—головная капсула Octopus doj-
leini: 1 — углубление для глазного яблока;
2 — отверстие для сосуда; 3— заднее от-
верстие капсулы; 4— капсула для стато-
циста. (а — по Несису, 1972; б, г — по Ива-
нову, Стрелкову, 1949; в — no Roper, 1977)
ских животных, зслеповато-синее,
длина волны около 474 им. У неко-
торых видов, однако, цвет его может
меняться цветными фильтрами. Кро-
ме того, у некоторых мигрирующих
мезопелагнчсских кальмаров цвет
свечения зависит от температуры:
днем на глубине при низкой темпе-
ратуре свечение синеватое, ночью
при подъеме в верхние, хорошо про-
гретые слои воды оно становится
зеленоватым (Young, Mencher, 1980).
Цвет же самих фотофоров определя-
ется соотношением преломления и
отражения света в светоотражающих
и светораспрсделяющих структурах
и может быть самым разным — бе-
лым, желтым, красным, зеленым, ме-
таллически ирригирующим.
Бактерии светятся непрерывно.
Управление свечением фотофоров
симбиотического типа осуществляет-
ся по принципу шторы: тонкий слой
чернил из чернильного мешка, на ко-
тором лежит фотофор, закрывает
его и поглощает свет, затем моллюск
смывает его, и свечение возобновля-
ется. Интенсивность свечения регу-
лируется усилением и ослаблением
подачи крови к органу — усиленное
поступление кислорода стимулирует
свечение.
У животных с внутриклеточной
биолюминесценцией регуляция све-
чения может быть троякой. Отдель-
ные крупные фотофоры сложного
устройства иннервированы и предпо-
ложительно находятся под прямым
нервным контролем (например, Hi-
stioteuthidae). Большинство фотофо-
ров, однако, не иннервированы, по
богато снабжены кровеносными со-
судами, их свечение контролируется
усилением и ослаблением кровотока.
Этот способ регуляции свечения, од-
нако, довольно медленный. В тех
случаях, когда необходима быстрая
регуляция свечения, в ход идут хро-
матофорныс пигментные ширмы,
снабженные меланофорами, которые
могут почти мгновенно закрывать
фотофор и полностью поглощать
свет. Таковы, например, крупные
фотофоры на концах брюшных рук
Abraliopsis и Watasenia.
Фотофоры па концах рук Tanin-
gia и Cranchiidae снабжены особыми
лопастями, закрывающими светящую-
ся поверхность, подобно оконным
ставням (см. рис. 47, н; 69, о). Фото-
фор Spirula снабжен кольцевой
складкой кожи, закрывающей его
как зрачок глаза, а задние фотофо-
ры Vampyroteuthis — специальным
«веком». В итоге головоногие могут
светиться постоянно с периодическим
усилением и ослаблением или корот-
кими вспышками; в некоторых слу-
чаях наблюдаются вспышки на фоне
общего свечения. Вспышками светят
фотофоры, находящиеся под прямым
нервным контролем или снабженные
меланофорными ширмами; непрерыв-
ное свечение, которое то усиливает-
ся, то ослабевает, характерно для
сосудистой регуляции свечения.
О функции свечения у головоно-
гих моллюсков существует множест-
во гипотез, но фактические наблюде-
ния малочисленны. Функция свече-
ния понятна только в отношении
сепиолид, способных «стрелять ог-
нем», ослепляя и отпугивая напа-
дающего хищника. У океанических
кальмаров главной функцией свече-
ния фотофоров, расположенных на
брюшной стороне мантии, головы и
рук, вентральной стороне глаз и
внутренних органах, считают каму-
флирующее вентральное противосве-
чение. Нссветящийся кальмар, если
смотреть на него снизу днем или в
лунную ночь, должен выглядеть тем-
ным силуэтом на фоне светлого не-
ба, по если он будет тускло светить-
ся, то может стать незаметным.
Р. Янг и К. Ропер (R. Young,
1973, 1977; Young et al., 1980; Young,
Mencher, 1980; Young, Roper, 1976,
1977; Young, Roper, Wallers, 1979)
показали, что изученные ими виды
Enoploleuthidae, Octopoteuthis и Hi-
stioteuthis «включают» фотофоры,
если освещаются сверху, и «выклю-
чают» их в темноте; они способны
подгонять интенсивность и (до неко-
торой степени) цвет свечения фото-
форов под интенсивность и цвет па-
дающего света (солнечного и лун-
ного) в довольно широком диапазо-
не величин освещенности, при этом
совершенно «исчезая» для наблюда-
теля. Этот способ камуфляжа эффек-
тивен для глубин от 350—400 до
750—800 м, что совпадает с обычны-
ми глубинами дневного обитания
мезопелагических головоногих. Такой
механизм свечения должен быть
рассчитан на некоторых определен-
ных врагов, а именно, массовых ак-
тивных нектонных зрительных хищ-
ников (преимущественно, конечно,
30
рыб), охотящихся днем и обитаю-
щих в дневное время в нижней ме-
зопслагиалп и па границе мезо- и
батипслагиали. Большинство же
рыб — врагов головоногих моллюс-
ков — не отвечают этим требовани-
ям. Они либо охотятся ночью в верх-
них слоях воды, либо пассивно под-
стерегают добычу, либо ориентиру-
ются в охоте не па зрение, а на
другие органы чувств, либо, нако-
нец, настолько малочисленны, что их
давление па кальмаров не могло бы
стать эффективным фактором естест-
венного отбора. Кроме того, многие
кальмары имеют совершенно иное
расположение фотофоров, живут на
иных глубинах, а некоторые мезопе-
лагичсские виды с «подходящим»
расположением фотофоров (напри-
мер, Ancistrocheirus alessandrinii)
светятся по-иному. Очевидно, проти-
восвечепис — в любом случае не
единственная функция органов све-
чения океанических кальмаров.
Строгая видовая специфичность
распределения фотофоров по телу
(для различения некоторых видов
Enoploteuthidae и I listiotcuthidae
фотофоры служат единственным на-
дежным признаком) наводит на
мысль, что важная, а может быть, и
важнейшая функция свечения — опо-
знавание особей своего вида, парт-
неров по стае. Главная функция фо-
тофоров, развивающихся только к
моменту полового созревания или
незадолго до пего (спинной фотофор
Sthenoteuthis, фотофоры на концах
рук Psychroteuthis и некоторых
Cranchiidae), очевидно, — опознава-
ние потенциального полового парт-
нера и сигнализация готовности к
спариванию. Кроме того, фотофоры
могут быть использованы для ослеп-
ления врага внезапной яркой вспыш-
кой света, разбивки силуэта, прима-
нивания добычи (фотофоры па кон-
цах щупалец Chiroteuthis) и др. Ве-
роятно, одни и те же фотофоры
могут выполнять разные функции.
Так, если Heteroteuthis hawaiiensis
внезапно осветить в темном аква-
риуме, он выбрасывает светящееся
облако (ослепление врага), отпры-
гивает на некоторое расстояние в
сторону, несколько минут светится
постоянным светом (вентральное
противосвсчсние) и «гаснет» (Young,
Roper, 1976).
Предполагается, что люциферин
вырабатывается в пищеварительной
железе из пищи. Его источник у
головоногих с собственным свече-
нием, по-видимому, целентеразин —
C26H21O3N3, высвечивающийся при
окислении. Благодаря его присутст-
вию многие ткани кальмаров (орга-
ны пищеварительной системы, жаб-
ры, почки, кровь и т. п.) могут све-
титься при растирании их па воздухе.
Пищеварительная железа светится
даже у лишенной фотофоров молоди
пелагических осьминогов семейства
Bolitaenidac (R. Young ct al., 1979).
Внутреннее строение
ГЛАДИУС. Гладиус — скелетная
пластинка внутреннераковинных го-
ловоногих — лежит па спинной сто-
роне тела во средней линии под
слоем кожи или мантийных мышц.
У спирулы и сепиид это раковина, т. с.
известковое образование с системой
изолированных камер, разделенных
перегородками — септами, она слу-
жит для регуляции плавучести (по-
плавок). У примитивных сепиолид
(подсем. Rossiinae и Scpiolinae),
неритических и абсолютного боль-
шинства океанических кальмаров
(рис. 5, а—в) и у вампироморф это
собственно гладиус («гладиаторский
меч») — узкая псровидная, ланцето-
видная или игловидная роговая (но
с примесью хитина) пластинка, обес-
печивающая продольную жесткость
тела при плавании. У большинства
плавниковых осьминогов скелетная
пластинка — хрящевое образование
седловидной, пряжковидной или
подковообразной формы, поддержи-
вающее основания плавников и даю-
щее им упор при работе. У донных
осьминогов-октоподид гладиус реду-
цирован до двух тонких хрящевых
палочек, лежащих в толще мантий-
ных мышц по обе стороны от сред-
ней линии тела, а у высших сепиолид
(Heteroteuthinae), Sepiadariidae, Idio-
sepiidae и пелагических осьминогов
(включая Argonautoidea) и, возмож-
но, Promachoteuthidae он отсутствует.
31
Раковина спирулы — раскручен-
ная плоская спираль, свернутая эн-
догастрически (внутрь). У взрос-
лых она состоит приблизительно из
35 камер. Раковина занимает заднюю
часть тела и лежит так, что ее плос-
кость совпадает с сагиттальной
плоскостью. На заднем конце тела
раковина прикрыта лишь кожей
(см. рис. 16). При фиксации мантия
съеживается, и кожа часто проры-
вается, обнажая часть раковины.
Раковина настоящих карака-
тиц— сепиид —называется сспио-
ном. Ее строение очень характерно
для отдельных видов или групп ви-
дов (см. рис. 17, а). Это толстая из-
вестковая пластинка овальной или
языковидной формы, закрывающая
всю спинную часть тела животного
или, по крайней мере, ее середину и
покрытая сверху только кожей. Се-
пиоп состоит из множества тонких
пластин — септ, сложенных толстой
пачкой и наклоненных назад. Систе-
ма пластин носит название фрагмо-
кона. Септы соединены между собой
множеством «подпорных столбов» и
контрфорсов, хорошо заметных на
вентральной стороне сспиона и име-
ющих вид тонких полосок, извилис-
тых черточек. У каракатиц в тропи-
ческих водах новые септы формиру-
ются ежедневно, так что число чер-
точек приблизительно соответствует
возрасту животного. В умеренных
водах летом формируются 2—3 сеп-
ты в педелю, зимой рост приостанав-
ливается. Передняя часть сепиоиа
сплошная, снизу не исчерченная. Со-
отношение гладкой и исчерченной
зон фрагмокона характерно для мно-
гих видов, по может меняться по се-
зонам. Посредине нижней поверхно-
сти сепиопа обычно проходит про-
дольный желобок, иногда окаймлен-
ный двумя низкими валиками. Очер-
тания желобка и валиков отражают-
ся на форме передних полосок
исчерченной зоны фрагмокона. Спин-
ная сторона сспиона ровная, иногда
с продольным килем. Фрагмокон ото-
рочен с боков кантом, в передней
половине обычно роговым, в зад-
ней — обызвествленным. Передняя
часть фрагмокона обычно выпуклая,
задняя — вогнутая, поджатая, и зад-
ний край канта окружает ее, обра-
зуя внешний конус.
Между внутренней стороной
внешнего конуса и задним концом
фрагмокона у видов рода Sepia на-
ходится V- или U-образный узкий
наплыв, имеющий форму шнурка
или изогнутой стеночки — внутрен-
ний копус, или вилка. Ветви внутрен-
него конуса («зубцы вилки») протя-
гиваются вперед почти до переднего
конца исчерченной части. На самом
заднем конце сепиопа дорсальнее
внешнего конуса находится (не у
всех каракатиц) шип — рострум.
У некоторых видов Sepia раковина
слабо обызвествлена и в той или
иной степени редуцирована.
Гладиус кальмаров и сепиолид
состоит из прочного ствола (рахи-
са), обычно с продольными ребрами,
тонкого пера и (у многих видов
океанических кальмаров) конечного
конуса, образованного свернутыми
фунтиком краями задней части пера.
Форма гладиуса очень варьирует, по
строго постоянна в пределах рода.
В общем она определяется образом
жизни моллюска, поэтому иногда у
родов, далеко отстоящих друг от
друга в системе, гладиусы оказыва-
ются сходными, например у Loligo и
Histioleuthis, Rossia и Abralia, Sepio-
la и Brachioteuthis и т. п. У быстрых
нектонпых океанических кальмаров
с ромбическим плавником гладиус —
прямой, негибкий, тонкий стержень
с узким, смещенным в заднюю часть
пером и коротким конусом (рис.
5, б). У менее быстрых, но более по-
воротливых нектонпых кальмаров с
сердцевидным плавником перо сме-
щается в среднюю часть гладиуса,
конус удлиняется. Иногда, впрочем,
конус не удлиняется, по вместо пего
развивается хрящевая насадка на
заднем конце гладиуса, также назы-
ваемая рострумом (Ancistroteuthis,
Moroteuthis, Rondakovia, см. рис.
49, в, л, п) или оттянутая часть
плавника опирается не па конус гла-
диуса, а на плотный мускулистый
«хвост», являющийся продолжением
заднего конца тела (Gonatus). У по-
лупланктонпых кальмаров гладиус
обычно с широким пером, причем у
плавающих в горизонтальном поло-
жении обычно имеется длинный ко-
нус, а у плавающих в вертикальном
или косом положении конус часто
редуцирован и гладиус сзади закруг-
лен. Наконец, у планктонных каль-
маров гладиус — очень тонкий стер-
жень, без пера, лишь с узким ромби-
ческим расширением и длинным тон-
32
Ким конусом. Сам рахис обычно
жесткий, но бывает и гибким, при
этом кальмар может принимать ша-
рообразную форму (например Сгап-
chia); у Bathoihauma задняя часть
гладиуса редуцирована.
Гладиус неритических кальмаров
имеет короткий рахис и довольно
широкое перо с тонкими выгнутыми
или утолщенными прямыми краями
(рис. 5, в). Конус отсутствует, но у
Alloteuthis и Uroteuthis на конце гла-
диуса есть маленький ростр, подоб-
ный ростру у Onychoteuthidac. У сам-
цов и самок лолигинид форма гла-
диуса довольно сильно различается:
относительная ширина пера у самок
больше.
Гладиус Vatnpyroteulhis резко от-
личается от гладиусов кальмаров и
сепиолид, по форме напоминает гла-
диусы ископаемых Loligosepiidae. Он
чрезвычайно тонкий, прозрачный, с
очень широким стволом без каких
бы то ни было ребер и широким
пером с промежуточной перетяжкой.
Сзади гладиус подвернут внутрь, об-
разуя маленький чашевидный конус,
иногда с крохотным рудиментарным
ростру мом.
Из плавниковых осьминогов у
Cirroieuthis и Cirrothauma плавнико-
вый хрящ седловидный, расширен-
ный по бокам и суженный спереди, у
Grimpoteuthis, Chunioteuthis и Froeke-
nia — подковообразный, пряжко- или
дуговидный, у Stauroteuthis — V-об-
разиый, у Opisthoteuthis — дугооб-
разный или почти прямой.
Гладиус всех впутреннераковин-
ных головоногих свободно лежит
внутри замкнутого эпителиального
мешка, который в эмбриогенезе раз-
вивается как впячивание мантии.
Эпителий стенок раковинного мешка
и выделяет гладиус. Поэтому мышцы
и хрящи непосредственно к гладиусу
не прикрепляются: мускулы — ре-
тракторы воронки и головы и плав-
ники Cirrata крепятся к наружной
стороне раковинного мешка, но гла-
диус служит им прочной опорой.
ХРЯЩЕВОЙ СКЕЛЕТ. Основа
внутреннего хрящевого скелета — го-
ловная капсула («хрящевой череп»)
сложной формы, состоящая из трех
обособленных отделов, в которых
находятся мозг и статоцисты (рис.
5, г). В вогнутых боковых сторонах
головной капсулы помещаются гла-
за и оптические доли мозга, глаза
защищены также небольшими удли-
ненными хрящами, примыкающими к
головной капсуле спереди. Имеются
отверстия для прохода пищевода,
нервов и сосудов. Кроме того, у не-
которых< головоногих моллюсков
есть хрящевые образования в осно-
ваниях рук и вдоль основания плав-
ников (например, у сепиид и вампи^
роморф); напомню также о замыка-
тельных хрящах и хрящевых обра-
зованиях на коже некоторых океани-
ческих кальмаров.
ОРГАНЫ МАНТИЙНОЙ ПО-
ЛОСТИ (рис. 6). Целом. Вторич-
ная полость тела головоногих мол-
люсков гомологична целомическим
полостям кольчатых червей, но у
тех она связана с функцией движе-
ния (гидростатический скелет), а у
головоногих выполняет совершенно
иные функции. Целом головоногих
моллюсков разделен на два отдела:
перикардиальный и висцеральный,
или половой целом (рис. 7, а). У нау-
тилуса, кальмаров, каракатиц и вам-
пироморф целом обширен и оба его
отдела широко сообщаются друг с
другом гонопсрикардиальным про-
током. У наутилуса половой отдел
целома образует настоящую полость
тела: он включает гонаду, желудок,
часть кишечника и заходит в сифон.
Перикардиальный отдел целома ох-
ватывает только сердце и открывает-
ся в мантийную полость двумя от-
верстиями, независимыми от от-
верстий полового целома (половой
проток) и почек. Перикардиальные
железы лежат вне перикарда, а жа-
берных сердец нет. У кальмаров и
каракатиц обширный перикард со-
держит сердце, жаберные сердца и
перикардиальные железы и соединя-
ется с почечными мешками. Половой
отдел целома тоже велик и заключа-
ет гонаду и желудок. Начальные
части выводных половых путей при-
надлежат к целому, конечные же
имеют иное происхождение — обра-
зованы не мезодермой, как все части
целома, а эктодермой. Целом осьми-
ногов резко отличается от целома
других головоногих сильной редук-
цией (рис. 7,6). Сердце осьминогов
помещается вне целома. Перикард
мал и состоит лишь из сумок жабер-
ных сердец и коротких протоков,
соединяющих их с водоносными ка-
налами, через которые полость гона-
ды сообщается с почечными мешка-
3 К. Н. Несис
33
ми. У ряда осьминогов водоносные
каналы отсутствуют или имеются
лишь с одной (левой) стороны (Opist-
hoteuthis), но реноперикардиальные
каналы всегда имеются. Половой от-
дел целома охватывает лишь полость
гонады.
Основные функции целома дву-
жаберных головоногих моллюсков
состоят в защите сердца, участии в
процессах выделения и солевого об-
мена и в выведении половых продук-
тов. В ходе эволюции выделительная
функция целома особенно .усовер-
шенствовалась, а функция защиты
сердца постепенно утрачивалась.
Совершенно особую функцию вы-
полняет целом у кранхиид («кальма-
ров-батискафов»), у которых он очень
велик (до % объема мантийной по-
лости) и разделен вертикальной мем-
браной (мезентерием) пополам на
правую и левую половины. Мускул-
замыкатель воронки превратился в
тонкую горизонтальную мембрану,
которая протягивается через всю
мантийную полость, прирастая с бо-
ков к ее стенкам, а сверху — к ниж-
ней части целома. В задней части
мантийной полости, за жабрами,
мембрана поднимается к ее спинной
стороне и огибает целом сзади, раз-
деляя мантийную полость на верх-
нюю и нижнюю половины. В резуль-
тате передняя часть тела оказывает-
ся состоящей из 5 камер: две поло-
вины целома, две боковые дорсаль-
ные камеры между целомом и
стенкой мантии и вентральная каме-
ра, расположенная ниже горизон-
тальной мембраны. Целом замкнут
и содержит большое количество
раствора NH4C1 — «поплавок батис-
кафа». Боковые дорсальные камеры
соединяются с внешней средой от-
верстиями на боках шеи, открываю-
щимися при вдохе и замыкающимися
при выдохе; вентральная камера —
через воронку. Дорсальные камеры
соединены с вентральной двумя
округлыми отверстиями — спиракулу-
мами (дыхальцами), расположенны-
ми прямо над разнесенными в бока
жабрами.
Дыхательный ток воды у кранхи-
ид создается иначе, чем у других го-
ловоногих: согласованными движе-
ниями стенки полости тела и гори-
зонтальной мембраны. При вдохе пе-
редняя часть горизонтальной мем-
браны оттягивается книзу, и вода
34
входит по бокам шеи в боковые
дорсальные камеры. Одновременно
под давлением мембраны вода,
прошедшая через жабры при преды-
дущем вдохе, выжимается через
воронку наружу. Затем задняя часть
мембраны поджимается кверху, от-
верстия по бокам шеи замыкаются.
Волна перистальтического сокраще-
ния пробегает по мембране назад,
прогоняя воду через дыхательные
отверстия и жабры.
Кровеносная система (рис.
8, а). Состоит из сердца, жаберных
сердец, жаберных желез и сосудов.
Кровеносная система кальмаров и
каракатиц почти замкнутая, у ось-
миногов имеются пространства кро-
веносных синусов (головной, кишеч-
ный), где кровь перемещается между
органами. Кровь головоногих почти
бесцветная, слегка голубоватая. Пиг-
мент крови — гемоцианин. По своим
функциям он похож на гемоглобин,
но резко отличен от него по струк-
туре, вместо атома железа содержит
атом меди. Сродство гемоцианина к
кислороду значительно ниже, чем ге-
моглобина, поэтому и кислородная
емкость крови головоногих невелика
(3—5 % об. против 15—20 % об. у
млекопитающих и птиц), но артери-
альное давление высокое (систоличе-
ское давление в головной аорте ги-
гантского осьминога 45—70 см вод.
ст.) и обеспечивает интенсивную цир-
куляцию крови. Движение крови
обеспечивается биением сердца и обо-
их жаберных сердец. Частота биения
сердца Octopus vulgaris при темпера-
туре 22° С и в покое 40—50 ударов
в минуту. У наутилуса жаберных
сердец пет и перемещение крови
обеспечивается ритмическими сокра-
щениями почечных придатков и пе-
рикардиальных желез.
Сердце состоит из желудочка и
2 (у наутилуса 4) предсердий. Из
желудочка выходят две аорты; пе-
редняя (головная) и задняя (брюш-
ная, мантийная). Передняя аорта
снабжает кровью голову, конечности,
мозг, ротовой аппарат, воронку, пе-
реднюю часть мантии, желудок и пе-
чень, задняя — мантию, плавники,
гонаду, кишку, чернильный мешок.
Венозная кровь из передней части
тела собирается в головную вену,
которая переходит в пару полых вей.
Задняя часть головной и полые ве-
ны окружены почечными придатка-
Рис. б. Общая схема расположения внут-
ренних органов самки кальмара: / — рука;
2 — буккальная мембрана; 3 — губа; 4 —
подъязычная слюнная железа; 5 — щу-
пальце; 6— одонтофор; 7—передняя слюн-
ная железа; 8—10 — нижний буккальный
(5), брахиальный (Р) и педальный (10)
ганглии; 11 — вороночный клапан; 12 —
воронка; 13 — обонятельная папилла; 14 —
мантийное отверстие; 15 — статоцист; 16—
вороночный орган; 17 — паллиовисцсраль-
ный ганглий; 18 — проток чернильного ме-
шка; 19—кишка; 20 — чернильный мешок;
21 — жабра; 22 — головная вена; 23 — по-
чечное отверстие; 24 — почечные придатки
полой вены; 25 — почка; 26 — половое от-
верстие; 27 — жаберное сердце; 28 — пери-
кард; 29 — задняя аорта; 30 — яйцевод;
31 — мантия; 32 — половая артерия: 33 —
мантийная полость; 34 — яичник; 35 — це-
лом (очерчен жирной линией); 36— цекум;
37 — желудочек сердца; 38—желудок; 39,
40— ренонерикардиальный и пищеваритель-
ный протоки; 41 — передняя аорта; 42 —
гладиус; -/3 — пищеварительная железа;
44 — звездчатый ганглий; 45—задняя слюн-
ная железа; 46 — передняя часть мантии;
47 — головная хрящевая капсула; 48 — над-
глоточные нервные ганглии; 49 — верхний
буккальный ганглий; 50 — пищевод; 51 —
радула; 52 — глотка; 53 — клюв (по Ива-
нову, Стрелкову, 1949)
/4
7
Л?
5
6
fl
8
10
/J
7, Целбм головоногих моллюсков:
щ — пространственные взаимоотноше-
между гонадой, перикардом и почка-
Рис.
а и
ния _______ ____...... ______ - _________
ми у Nautiloidea (а) н Coleoidea (ai):
1 — гонада; 2 — почка; 3 — гоподукт; 4 —
гонопернкардиальный проток (водоносный
канал); 5 — перикард; б — пространствен-
ные взаимоотношения целома, кровеносной
и выделительной систем у Octopus-, 1, 2 —
почечное (/) и половое (2) отверстия; 3 —
отверстие реноперикардиального протока;
4 — гонодукт; 5 — предсердие; 6 — аффе-
рентная жаберная вена; 7 — жаберное
сердце; 8 — придаток жаберного сердца;
9 — водоносный канал; 10 — почечный ме-
шок; /i — желудочек сердца; /2 —половой
отдел целома; 13 — перикард;. 14 — рено-
перикардиальный проток (а— по Gotting,
1974; б — по Schipp, Boletzky, 1975)
3*
ми, относящимися к выделительной
системе, но неразрывно связанными
с кровеносной системой. Полые вены
впадают в жаберные сосуды, а те —
в жаберные сердца. В жаберные
сердца поступает венозная кровь и
из задней части тела — через брюш-
ные вены. Жаберные сердца — мус-
кулистые толстостенные пузырьки,
лежащие у основания жабр. Кроме
мышц в их стенках имеются железы.
Ритмически сокращаясь, жаберные
сердца проталкивают кровь в жаб-
ры. Обогащенная кислородом кровь
из жабр поступает в предсердия.
Кровеносные сосуды, особенно арте-
рии, имеют мускулистые стенки и
пульсируют, помогая трем сердцам
прокачивать кровь через капилля-
ры, особенно развитые в конечно-
стях и задней части мантии.
При плавании или иной работе
частота биения сердца осьминога
почти не меняется, но ударный объ-
ем сердца (объем крови, перекачива-
емый за одно сокращение) может
возрастать втрое. Частота сердечных
сокращений существенно возрастает
с ростом температуры.
Жаберная железа протягивается
вдоль жабры в том месте, где жаб-
ра прирастает к мантии. Эта желе-
за— место синтеза гемоцианина. Ге-
моцианин растворен в плазме крови.
Кровяные тельца у головоногих
представлены только лейкоцитами-
амебоцитами. Они синтезируются в
особом органе — «белом теле» (же-
леза Гензена), располагающемся в
голове вблизи глаза (см. рис. 12, а).
В этой же железе, а также в слюн-
ных и жаберной железах и жабрах
концентрируются фагоциты ретику-
ло-эндотслиальной системы.
Жабры. У наутилуса 2 пары
жабр, у всех прочих головоногих од-
на пара. Жабры лежат в мантийной
полости по обе стороны от внутрен-
ностей (рис. 8,6). Одна сторона
жабры прикрепляется к внутренней
стенке мантии топкой пленкой, про-
тивоположная сторона свободна.
Жабры кальмаров, каракатиц и вам-
пироморф удлиненно-конические, ле-
жат слегка под углом к продольной
оси тела — расходятся вперед. Жаб-
ры большинства осьминогов оваль-
ные, параллельные продольной оси
тела. У некоторых плавниковых ось-
миногов они напоминают половинку
разрезанного поперек апельсина и их
36
ось перпендикулярна продольной оси
тела.
По строению жабры головоногих
сходны с ктенидиями других мол-
люсков— двоякоперистые, состоят из
оси и попеременно отходящих от нее
вправо и влево лепестков — филамен-
тов, между которыми циркулирует
вода. Число филаментов у кальмаров
и каракатиц велико: от 20 до 40—80
с каждой стороны (в каждой полу-
жабре), и изменчиво; у донных ось-
миногов в каждой полужабре обыч-
но 6—13 филаментов, их число до-
вольно постоянно в пределах вида и
служит важным таксономическим
признаком. Жабры глубоководных
осьминогов уменьшены в размерах,
часто внутренняя полужабра редуци-
руется, число филаментов падает до
3—5.
Жаберные филаменты складчатые,
что значительно увеличивает дыха-
тельную поверхность. По обе сторо-
ны жаберных лепестков проходят
кровеносные сосуды. С той стороны,
где жабра прирастает к мантии, дви-
жется венозная кровь (афферентный
сосуд), с противоположной, свобод-
ной стороны — артериальная (эффе-
рентный сосуд). При дыхании жаб-
ры ритмически сокращаются, что так-
же облегчает кровоток.
Дыхательные движения те же,
что и при реактивном плавании:
вода входит в мантийную полость
при «вдохе», омывает жабры и вы-
брасывается через воронку при «вы-
дохе». Частота дыхательных движе-
ний сильно варьирует — у разных ви-
дов в зависимости от образа жизни,
у одного и того же — от состояния.
У нектонпых кальмаров, каракатиц и
шельфовых донных осьминогов она
составляет в покое 20—30 движений
в минуту, у холодноводных полуглу-
боководных видов ниже (до 10), у
пелагических видов значительно вы-
ше (у молодых тремоктопусов 65—
85, у аргонавтов при возбуждении
до 200, у наутилуса при 18—20° С
40—60 движений в минуту). При
плавании частота дыхательных дви-
жений у кальмаров составляет от
120—180 в минуту при медленном до
300—350 при быстром плавании. Та-
ким образом, чем быстрее кальмар
плывет, тем интенсивнее омываются
свежей водой его жабры.
Все головоногие, в особенности
кальмары и пелагические осьминоги,
Рис. 8. Кровеносная, выделительная и ды-
хательная системы головоногих; а — кро-
веносная система Todarodes pacificus с
брюшной стороны (артерии светлые, вены
темные): /, 2 —вены и артерии рук; 3 —
общая вена рук; 4, 5 — мозговая (4) и во-
роночная (5) вены; 6 — артерия пищевари-
тельной железы; 7 — головная вена; 8 —
почечные придатки полой вены; 9 — пред-
сердие; 10 — жаберная вена; 11 — желудо-
чек; /2 —задняя аорта; 13 — мантийная
артерия; 14 — контуры цекума; 15 — плав-
никовая артерия; 16 — контуры гонады;
17 — брюшная вена; 18 — половая артерия;
19 — половая вена; 20 — контуры желудка;
21 — жаберное сердце; 22 — придаток жа-
берного сердца; 23 — мантийная вена; 24—
полая вена; 25 — желудочная артерия;
26 — панкреатическая артерия; 27—жабер-
ная артерия; 28 — передняя аорта; 29—ки-
шечная вена; 30 — вена пищеварительной
железы; 31 — артерия переднего края ман-
тии; 32 — глоточная артерия; б — дыха-
тельная система Octopus vulgaris: /—звезд-
чатый ганглий; 2—мантийная вена; 3 —
генитально-мезентериальная вена; 4 —
жаберный нерв; 5— яйцевод; 6 — почка;
— полая вена; 8— жаберный ганглий;
) — жаберное сердце; 10—11 — афферент-
ный и эфферентный жаберный сосуд; 12—
жабра; 13 — жаберная артерия; 14 — жа-у
берная вена; 15 — жаберная железа; 16—
мантия; в — почечные придатки и сердце
Sepia officinalis: 1 — дорсальный почечный
мешок; 2—полая вена; 3 — генитально-
мезентериальная вена; 4 — правый вент-
ральный почечный мешок; 5 — передняя
аорта: 6 — правое предсердие; 7 — перед-
няя почечная артерия; 8 ^- половая арте-
рия; 9 — задняя аорта; 10 — левое пред-
сердие; 11 — перикардиальный отдел цело-
ма {а — по Иванову, Стрелкову, 1949; б —
по Taki, 1964; в—по Schipp, Boietzky, 1975), .
очень чувствительны к содержанию
кислорода в воде и при недостатке
его быстро слабеют и погибают. Для
О. vulgaris концентрация кислорода
2,5 мл/л пороговая, так как их ткани
полностью отбирают кислород из
крови и венозная кровь почти лише-
на кислорода.
Выделительная система.
Органы выделения у головоногих
весьма разнообразны и чрезвычайно
тесно связаны с органами кровенос-
ной и дыхательной систем (рис. 8, в).
Выделительная система включает по-
чечные мешки, перикардиальные же-
лезы (придатки жаберных сердец),
реноперикардиальный канал, почеч-
ные придатки полой вены и имею-
щиеся только у каракатиц придатки
пищеварительного протока (панкреа-
тические придатки). Почечные мешки
кальмаров и каракатиц обширны и
соединены друг с другом так, что
получается трсхлопастное образова-
ние из двух вентральных и одной
дорсальной части. Почечные мешки
осьминогов — пара полностью изоли-
рованных камер, соответствующих
вентральным мешкам кальмаров и
каракатиц. Почечные отверстия от-
крываются на маленьких сосочках в
мантийную полость впереди основа-
ний жабр.
Перикардиальные железы, или
придатки жаберных сердец, соседст-
вуют с жаберными сердцами и спле-
таются сосудами с ними. Комплекс
жаберное сердце — перикардиальная
железа — это орган ультрафильтра-
ции. Здесь под давлением крови рас-
творенные в плазме вещества, под-
лежащие удалению, переходят из
жаберного сердца в полость перикар-
диальной железы. У осьминогов они
затем поступают в рсноперикарди-
альный капал, где происходит обрат-
ное всасывание (реабсорбция) солей,
аминокислот, сахаров и других важ-
ных для организма мелких молекул,
прошедших сквозь фильтр. У кальма-
ров и каракатиц реабсорбция осуще-
ствляется в почечных придатках по-
лой вены и отчасти в жабрах. Почеч-
ные придатки полой вены и есть на-
стоящая почка, основной экскретор-
ный орган. Они участвуют также в
осморсгуляции. Дополнительный
секреторный oprair— перикардиаль-
ная железй. Придатки пищеваритель-
ного протока каракатиц кроме уча-
стия в пищеварении играют роль до-
38
полпительного органа осморегуляции
и формирования мочи. Значительную
роль в экскреции азотистых веществ
и осморегуляции играют жабры. Са-
ми почечные мешки — главным обра-
зом хранилища мочи (мочевой пу-
зырь), по они выполняют и секретор-
ную функцию. Периодические пуль-
сации почечных придатков полой ве-
ны передаются стенкам почечных
мешков, благодаря чему моча пере-
мешивается и растворяет гранулы
секретов, выделяемых железами
эпителия почечных мешков и при-
датков.
Головоногие — аммониотеличес-
кие животные, у них основной ком-
понент мочи — аммиак, точнее, ионы
аммония (Nl-lf) , в отличие от урико-
телических животных, основной ком-
понент мочи которых — мочевая кис-
лота. Молекулы NH3 диффундируют
из крови в первичную мочу, в кислой
среде которой идет реакция NH3+
Часть аммония выделя-
ется с мочой, часть — через жабры.
Выделение в воду значительных ко-
личеств аммиака — одна из причин
того, что головоногие моллюски пло-
хо выживают в замкнутом простран-
стве без смены воды: отравляют сами
себя аммиаком. У кальмаров, имею-
щих жидкостный механизм нейтраль-
ной плавучести, большая часть аммиа-
ка не выделяется во внешнюю среду,
а накапливается в форме NH4C1.
У крапхиид объем этой легкой жид-
кости так велик, что в «поплавке»
полости тела содержится до 40%
азота, выделенного в течение всей
жизни кальмара.
В почечных мешках донных ось-
миногов (Octopodidae) и каракатиц
(Sepiidae, Sepiolidae) обитают свое-
образные животные — дициемиды,
представители особого, нигде более,
кроме почек головоногих, не встреча-
ющегося класса Dicyemida (тип Ме-
sozoa). У шельфовых осьминогов они
так обычны, что осьминог без дицие-
мид — исключительная редкость. На-
против, в почках пелагических голо-
воногих они не встречаются. Счита-
лось, что дициемиды — типичные па-
разиты, питающиеся растворенной
органикой. Недавно, однако, была
высказана гипотеза (Lapan, 1975),
что это — полезные симбионты. Оби-
тая на границе между поверхностью
почечных придатков и мочой в почеч-
пых мешках, они подкисляют среду,
тем самым способствуя превращению
молекул аммиака в ионы аммония.
Пищеварительная систе-
м а. Пищеварительная система (рис.
9) состоит из буккального (ротово-
го) комплекса с клювом и радулой,
пищевода, слюнных желез, зоба, же-
лудка, пищеварительной железы (пе-
чени), придатков пищеварительного
протока (поджелудочной железы),
слепого отростка желудка (цекума)
кишки и ануса. Анатомически, но не
функционально с пищеварительным
трактом связан чернильный мешок.
Пищевод головоногих моллюс-
ков — тонкая трубка, соединяющая
ротовую полость с желудком и про-
ходящая в передней части через
мозг, в задней — через печень. По-
этому головоногие вынуждены рас-
кусывать пищу на довольно мелкие
куски — добыча, заглоченная цели-
ком, могла бы повредить жизненно
важные органы. Зоб имеется только
у некоторых осьминогов подотряда
Incirrata и служит местом хранения
размельченной пищи до поступления
ее в желудок.
Имеются 3 слюнные железы
(рис. 9, а): подъязычная, передняя
(парная) и задняя (парная или не-
парная). Подъязычная железа име-
ется у всех головоногих, задняя
отсутствует у спирулы и нау-
тилуса, а у плавниковых осьминогов
нет ни передней, ни задней железы.
Функция маленькой подъязычной же-
лезы не изучена. Передние слюнные
железы невелики, их секрет, по-види-
мому, содержит протеолитические
ферменты, задние — очень крупные у
донных осьминогов, у каракатиц по-
мельче, еще мельче у кальмаров. Их
функция двойная: это ядовитые же-
лезы и важный источник пищевари-
тельных ферментов, в первую оче-
редь протеолитических.
Слюна с ядовитым секретом вы-
деляется наружу через расположен-
ную под «языком» (одонтофором)
хоботковидную слюнную папиллу,
или субрадулярный орган (рис. 9,6).
Этот мускулистый «слюнной хобо-
ток» очень подвижен и может высо-
вываться изо рта. Слюна выпрыски-
вается под давлением: маленький
осьминог может «плюнуть» так, что
струйка слюны пролетает в воздухе
расстояние до 25 см. На конце слюн-
ной папиллы осьминога имеются мно-
гочисленные мелкие зубчики, участ-
вующие в процессе сверления рако-
вин моллюсков.
Яд осьминогов рода Octopus со-
держит 2 токсина: а- и р-цефалоток-
син. Оба токсина смертельны для
крабов, омаров, лангустов и других
ракообразных, служащих обычно пи-
щей осьминогам. Они погибают через
несколько минут после укуса. Для
человека токсин, как правило, не-
опасен, за исключением токсина На-
palochlaena maculosa, но укус неко-
торых мелких осьминогов (О. fitchi,
О. rubescens, О. joubini) иногда вы-
зывает довольно тяжелую местную
реакцию: укушенная рука, например,
может болеть 2—3 недели, хотя
обычно боль проходит через несколь-
ко часов. Яд действует только при
попадании в кровь, а па кожу, гла-
за и пр. влияния не оказывает.
Самый ядовитый головоногий мол-
люск — маленький австралийский
осьминог Hapalochlaena maculosa,
особенно самки, вынашивающие яй-
ца. Ядовитый секрет желез содержит
два токсина: макулотоксин и хапа-
лотоксин. Макулотоксин сходен с
тетродотоксином — ядом японской
собаки-рыбы, фугу. Укус взрослой
самки Я. maculosa может быть смер-
тельным. Яд парализует гладкую
мускулатуру, и человек задыхается.
Спасти его может только длительное
искусственное дыхание. На сердце яд
не действует.
Крупные осьминоги, в том числе
гигантский тихоокеанский О. dofleini,
не ядовиты.
Секрет задней слюнной железы
каракатиц также содержит цефало-
токсин и ядовит для ракообразных,
но для человека безвреден. Напро-
тив, у кальмаров этот секрет нс со-
держит токсинов. Единственный ядо-
витый кальмар — Onychoteuthis bank-
si banksi. Его укус напоминает ужа-
ление крупной осы; боль проходит в
течение 1 ч. Укус близкого к нему
О. banksi borealijaponicus не ядовит.
Желудок головоногих моллюсков
мускулистый и так же, как пищевод,
выстлан кутикулой. В него поступа-
ют раскусанная пища и пищевари-
тельные соки из ротовой полости,
«печени» и «поджелудочной железы»
(рис. 9, в—д). Мускулатура желудка
обеспечивает постоянное перемеши-
вание пищи и пищеварительных со-
ков. В желудке происходит первичное
39
переваривание пищи. Активность пи-
щеварительных ферментов у голово-
ногих очень высока, поэтому в по-
следующих отделах пищеваритель-
ного тракта оформленной пищи уже
не встречается. В связи с этим при
изучении питания голо.воногих мол-
люсков имеет смысл вскрывать толь-
ко желудок (у осьминогов еще и
зоб).
В желудке переваренная пища не
всасывается. Это происходит в це-
кумс, «печени» и «поджелудочной
железе». Из желудка смесь полупс-
реваренной пищи и пищеварительных
соков в виде мутной эмульсии посту-
пает в цекум (слепой отросток).
У каракатиц и большинства кальма-
ров и осьминогов это небольшой за-
крученный спиралью мешок. У лоли-
гинид он состоит из двух частей: ко-
роткой спиральной и длинной меш-
ковидной. У некоторых плавниковых
осьминогов, кроме цскума, есть еще
дополнительный мешковидный вы-
рост — «третий желудок». Обычно
желудок и цекум располагаются за
печенью, в средней части тела, у ло-
лигинид цекум протягивается почти
до заднего конца тела. У Amphitretus
и Vitreledonella желудок с цекумом
перемещаются в переднюю часть тела
и помещаются впереди «печени».
Внутренняя поверхность цекума вы-
стлана ресничным и слизистым эпи-
телием и в спиральной части несет
многочисленные лепестки. Лепестки
покрыты ресничками, в борозды ме-
жду ними поступает слизь, очищаю-
щая эмульсию от всех взвешенных
частиц. У живых кальмаров-лолиги-
нид по мешковидной части цекума
пробегают волны перистальтики. Ак-
тивное сокращение стенок то втяги-
вает эмульсию в цекум, то выталки-
вает се обратно в спиральную часть.
В цекуме завершается переваривание
пищи и начинается всасывание жи-
ров, аминокислот и углеводов.
Вблизи протока, соединяющего
желудок с цекумом и кишкой, в цс-
кум открывается пищеварительный
проток, соединяющий с цекумом пи-
щеварительную железу, в него же
открывается проток «поджелудочной
железы». Этот комплекс представля-
ет собой сложную систему с клапа-
нами и желобками, однозначно оп-
ределяющую направление передви-
жения жидкостей и эмульсий. Пище-
вая эмульсия поступает только из
40
желудка в цекум, а из пего — в пи-
щеварительную железу и кишку, при
этом крупные частицы из желудка в
цекум нс проходят. Сначала пища
переваривается в желудке, затем от-
крываются цекальный клапан, сфинк-
тер пищеварительной железы и пи-
щеварительный проток и в цекум
поступают одновременно пищевая,
эмульсия из желудка и ферменты из
«печени» и «поджелудочной желе-
зы». После этого клапан, сфинктер и
проток замыкаются и ферменты на-
чинают поступать не в цекум, а в
желудок, содержимое которого изо-
лируется от цскума. Время от време-
ни содержимое цекума поступает
через пищеварительный проток в «пе-
чень» и «поджелудочную железу»,
где всасывается; ферменты при этом
не выделяются. В конце периода пи-
щеварения остатки пищи начинают
подаваться в кишку.
«Печень», или пищеварительная
железа (железа средней кишки), —
объемистый овальный орган бурова-
того цвета, расположенный обычно
впереди желудка (кроме Amphitretus
и Vitreledonella, у которых печень
узко-веретеповидпая и лежит сзади
желудка). Название «печень» неточ-
но, так как многообразие функций
этого органа у головоногих значи-
тельно превосходит функции печени
позвоночных. Пищеварительная же-
леза головоногих производит и вы-
деляет пищеварительные ферменты,
в ней происходит всасывание амино-
кислот, она является местом хране-
ния запасных питательных веществ
(жира) и участвует в экскреции.
У осьминогов и каракатиц периоды
выделения пищеварительных фермен-
тов чередуются с периодами их син-
теза, так что пищеварительная желе-
за функционирует маятникообразпо,
у кальмаров оба процесса идут од-
новременно, что значительно ускоря-
ет пищеварение. Как орган всасыва-
ния переваренной пищи пищевари-
тельная железа играет основную
роль, поглощая 65—95% пищи. Об-
ширные ресурсы жира и каротинои-
дов накапливаются в железе в пе-
риод нагула и поддерживают суще-
ствование моллюска в период созре-
вания яиц и нереста, а у самок ось-
миногов— и в период «насижива-
ния» яиц, так как количество и ак-
тивность пищеварительных фермен-
тов в эти периоды резко сокращают-
8
Рис. 9. Буккальный (ротовой) комплекс (а,
б) и пищеварительная система (в—б>
Octopus vulgaris (а—в) и Loligo sp. (г, д):
а — общая схема строения комплекса; б —
строение слюнной папиллы: 1 — верхняя
мандибула; 2 — радула; 3 —нижняя ман-
дибула; 4— слюнная папилла; 5 — субра-
дулярный ганглий; 6 — подъязычная слюн-
ная железа; 7 — нерв задней слюнной же-
лезы; 8— нижний буккальный ганглий;
9 — проток задней слюнной железы; 10 —
верхняя буккальная доля мозга; 11 —
межбуккальная и церебро-субрадулярная
коннективы; 12 — передняя слюнная желе-
за; 13 — ротовые нальпы; 14 — мышечная
опора радулы; /5 —зубы слюнной папил-
лы; 16 — зубы на выворачивающемся кон-
це слюнного протока; в — отпрепарирован-
ная пищеварительная система сбоку: 1 —
глотка; 2 —пищевод; 3 — задняя слюнная
железа; 4 — зоб; 5 — желудок; 6 — цекум;
7 — передний отдел кишки; 8 — пищевари-
тельный проток; 9 — придатки пищевари-
тельного протока; /0 — чернильный ме-
шок; // — задний отдел кишки; 12 — про-
ток чернильного мешка; 13 — анус; 14 —
пищеварительная железа; 15 — слюнной
проток; 16 — передняя слюнная железа; г,
д — центральная часть пищеварительной
системы, вид с брюшной стороны (г) и
слева (б): / — пищеварительная железа;
2 — пищевод; 3 — кишка; 4 — придаток це-
кума; 5—придатки пищеварительного про-
тока; б —клапан цекума; 7 —желудок; 8,
9 — спиральный (5) и мешковидный (Р>
участки цекума; 10 — пищеварительный
проток; 11 — чернильный мешок; 12 — зад-
ний отдел кишки; 13 — анус; 14 — аналь-
ные папиллы. Стрелками показаны направ-
ления ресничных токов (а — по J. Young,
1965 из Boyle et al., 1979; б — по Nixon,
1980; в — по Иванову, Стрелкову, 1949; г»
д — по Bidder, 1950)
ся, а питание прекращается или
уменьшается в несколько раз. В «пе-
чени» синтезируется также люци-
ферин.
Придатки пищеварительного про-
тока обычно называют поджелудоч-
ной железой, что тоже неправильно,
так как это не секреторный орган.
Это небольшая разветвленная желе-
за светлого цвета, участвующая в
экскреции, осморегуляции, формиро-
вании мочи и всасывании углеводов
и аминокислот. У кальмаров и в осо-
бенности каракатиц это двойной ор-
ган, состоящий из внутреннего и на-
ружного эпителия. К пищеваритель-
ной системе относится только внут-
ренний эпителий, наружный же со-
ставляет часть выделительной си-
стемы. Только у осьминогов придат-
ки пищеварительного протока цели-
ком относятся к пищеварительной
системе как орган всасывания.
Кишка, как и цскум, выстлана
ресничным и слизистым эпителием и
перистальтически сокращается, по
всасывание в пей, если и происходит,
то в незначительной степени. Киш-
ка— основной поставщик слизи, по-
ступающей в желудок и обволакива-
ющей непереваренные остатки пищи,
которые, минуя цекум, попадают в
кишку. В кишку же поступают и сли-
зистые тяжи из цекума. Те и другие
выбрасываются через анус — щелс-
видное отверстие, обычно окружен-
ное двумя анальными папиллами, или
анальными лепестками — у кальма-
ров они похожи на заячьи уши.
У Neorossia анальных папилл нет.
Немного впереди ануса в кишку впа-
дает проток чернильного мешка.
Скорость пищеварения у голово-
ногих моллюсков зависит от образа
жизни, температуры воды, пола, воз-
раста и других факторов. Чем выше
температура, тем скорее переварива-
ется пища. Самцы осьминогов пере-
варивают пищу скорее, чем самки, а
молодь — быстрее взрослых особей.
Скорость пищеварения у кальмаров
значительно выше, чем у каракатиц
и осьминогов, у которых синтез и
секреция. пищеварительных фермен-
тов происходят попеременно. Дли-
тельность переваривания пищи у ло-
лигинид при температуре около 18° С
составляет 4—6 ч; иллекс, живущий
в более холодной воде (8—13°С),
переваривает пищу приблизительно
за 12 ч. Наблюдения над оммастре-
42
фидами тропических и умеренных вод
показывают, что пища, съеденная
ими вечером, к утру успевает полно-
стью перевариться. У Octopus vulga-
ris и О. суапеа переваривание пищи
длится в зависимости от температу-
ры 12—16 ч, у Sepia officinalis и Ele-
done cirrosa при 15—20° С оно зани-
мает 15—20 ч, а у Е. cirrosa при
10° С — до 30 ч (обзоры: Bidder,
1950; Bouchcr-Rodoni, Mangold,
1977). Таким образом, осьминоги и
каракатицы питаются в основном раз
в день, а при низкой температуре да-
же не ежедневно, тогда как кальма-
ры — дважды в день.
Чернильный мешок. Чрез-
вычайно характерен для головоногих
моллюсков. Он имеется у всех без
исключения кальмаров, почти у всех
каракатиц и большинства Incirrata,
отсутствует у наутилуса, вампиро-
морф, плавниковых осьминогов, Neo-
rossia и Bathypolypodin ае. Все виды,
лишенные чернильного мешка, оби-
тают исключительно или преимущест-
венно в глубинах океана. У некото-
рых глубоководных и полуглубоко-
водных форм чернильный мешок сла-
бо развит (Spirula, Bolitaenidae,
I diocto pus, Velodona), по есть и та-
кие обитатели глубин, у которых он
имеет вполне нормальное развитие.
Чернильный мешок лежит на
брюшной стороне внутренностной
массы с дорсальной стороны кишки.
У некоторых осьминогов он частично
погружен в ткани печени и плохо за-
метен снаружи. Состоит из черниль-
ной железы, лежащей на внутренней
стороне дорсальной стенки мешка,
самого мешка — резервуара для чер-
нильной жидкости, и длинного прото-
ка, снабженного двумя сфинктерами.
Чернила секретируются клетками же-
лезы в виде гранул пигмента мела-
нина. Гранулы взвешены в бесцвет-
ной жидкости. Готовая к выбросу
порция чернил накапливается в рас-
ширении протока между сфинктера-
ми. Из протока чернильная жидкость
поступает в кишку, затем через анус
и воронку выбрасывается наружу.
При опорожнении протока туда по-
ступает новая порция чернильной
жидкости. Цвет чернил головоногих
от коричневого до черного.
Основная функция чернильной
жидкости — дезориентация нападаю-
щего хищника (см. с. 20). Выбросив
чернила, моллюск бледнеет, резко
меняет траекторию движения и скры-
вается. Чернильная жидкость оказы-
вает раздражающее воздействие па
глаза и, как предполагают, временно
парализует органы обоняния, что за-
трудняет хищнику погоню за жерт-
вой. На глаза человека тоже дейст-
вует раздражающе (при попадании
чернил в глаза их надо немедленно
промыть пресной водой).
Пигмент головоногих моллюсков—
•один из наиболее стойких красите-
лей. Если чернила кальмара или ка-
ракатицы попали на одежду или па-
лубу судна и высохли, избавиться от
них исключительно трудно. У иско-
паемых головоногих чернила сохра-
няются в течение сотен миллионов
лет. Из чернильных мешков карака-
тиц издавна приготовляли высокока-
чественную коричневую краску — се-
пию (содержимое чернильного меш-
ка, высушенное и обработанное ед-
ким кали для удаления органики).
Какова функция чернил у обита-
телей глубин, можно лишь догады-
ваться. Напомним, что на черниль-
ном мешке многих кальмаров и кара-
катиц-сепиолид помещаются светя-
щиеся органы, для которых он слу-
жит пигментной ширмой. У Hetero-
teuthis чернила иногда выбрасывают-
ся вместе со светящейся жидкостью.
Могут ли они выбрасываться без нес
неизвестно.
Половая система (рис. 10).
Все головоногие моллюски раздель-
нополы. Пол определяется генетиче-
ски. У Argonautoidea самца можно
отличить от самки уже в момент вы-
лупления из яйца (у самок 8 рук,
у самцов 7). У других головоногих
вторичные половые признаки стано-
вятся заметны для невооруженного
глаза лишь у молоди, хотя и задолго
до начала созревания; при гистоло-
гическом исследовании определить
пол возможно даже у очень моло-
дых особей.
Женская половая система голо-
воногих состоит из яичника, яйцево-
дов, яйцеводных, нидаментальных и
добавочных (акцессорных) нидамен-
тальных желез (рис. 10, а—в). Яич-
ник всегда непарный, гроздевидного
строения, занимает заднюю часть
мантийной полости и имеет у каль-
маров коническую, у каракатиц и
осьминогов преимущественно полу-
шаровидиую форму. Зрелый яичник
очень велнк, часто отжимает вперед
все остальные органы мантийной по-
лости.
Яйцевод парный у эгопсид (кроме
Pyroteuthinae), вампироморф и Cirra-
ta, непарный — у Pterygioteuthis
(только правый), Pyroteuthis, всех
каракатиц, миопсид и плавниковых
осьминогов (только левый). У самок
Idiosepiidae имеются оба яйцевода,
но правый не функционирует. У каль-
маров постепенно созревающие яйца
перемещаются из яичника в яйцевод
(яйцеводы), где хранятся до нереста.
Перед самым нерестом объем яйце-
водов может значительно превосхо-
дить объем яичника. Поэтому у не-
зрелых кальмаров яйцеводы изогну-
ты и спирально закручены. У кара-
катиц и осьминогов и созревающие,
и зрелые яйца находятся до момента
нереста в половом целоме, яйцеводы
нс изогнуты.
Яйцеводные железы имеются у
всех головоногих, их число соответст-
вует числу яйцеводов. У кальмаров
и каракатиц они имеют сердцевид-
ную форму п насажены на яйцевод
близ его конца, кроме Enoploteuthi-
пае, у которых они чрезвычайно ве-
лики (не меньше самого яйцевода),
овальные и подразделены на две ча-
сти, большую и меньшую. У осьми-
ногов бубликовидпые яйцеводные
железы сидят посередине яйцевода
или ближе к проксимальному концу,
как кольцо па пальце (рис. 10,6).
Дистальнее них у некоторых видов
находится расширение яйцевода —
«влагалище». Яйцеводные железы
выделяют легкий и липкий секрет,
который у кальмаров и каракатиц
образует третичную оболочку яиц
(первичная оболочка — яйцевая мем-
брана, вторичная — хорион), а у ось-
миногов — конец стебелька и липкий
цемент, приклеивающий яйцо к суб-
страту. У Octopodinae внутри яйце-
водной железы находится семспри-
емник, где хранится сперма; в нем
же происходит оплодотворение.
У Eledone сперма проникает непо-
средственно в яичник. У кальмаров,
каракатиц и вампироморф опа хра-
нится в семеприсмниках вне половой
системы (см. ниже).
Нидамснтальные железы имеются
у кальмаров (кроме Enoploteuthinae)
и каракатиц. У зрелых самок они за-
нимают всю середину вентральной
стороны мантийной полости. Их фор-
ма у кальмаров языковидная, у ка-
43
е
Рис. 10. Половая система головоногих мол-
люсков: а—в — половая система самок
Sepia elegans (a), Loligo vulgaris (б) и
Octopus dofleini (в); а, б'. 1 — анус; 2 —
жабра; 3 — почечное отверстие; 4, 5 — от-
верстия придаточной нидаментальной (4)
и яйцеводной (5) желез; б, 7 — яйцеводная
(£) и придаточная нидаментальная (7) же-
лезы; 8 — отверстие нидаментальной желе-
зы; 9 — жаберная артерия; 10» 11 — жабер-
ная (10) и мантийная (11) вены; 12 — ос-
нование жабры; 13 — жаберное сердце;
14—придаток жаберного сердца; 15 — ни-
даментальная железа; 16 — яйцевод; 17 —
брюшная вена; 18 — брюшная артерия;
19 — мантийная артерия; 20 — начало яй-
цевода; 21 — яичник; 22 — цекум; в: 1 —
яйцевод; 2 — связка, на которой подвешен
Яичник; 3 — яичник; 4 — яйцеводная желе-
за; 5 — половое отверстие; г — яйцеводная
железа Octopus vulgaris: 1 — центральная
полость; 2 — семеприемник; 3, 4 — цент-
ральная (3) и периферическая (4) части
железы; 5 — яйцевод; д» е» ж — половая
система самца Loligo vulgaris reynaudi (д,
е) и Octopus dofleini martini (ж); д: 1 —
половое отверстие; 2 — пенис; 3 — отвер-
стие генитального мешка; 4 — оболочка
внутренностной массы; Б — проток гени-
тального мешка; 6 — придаточная железа
(простата); 7 — придаток (слепой мешок)
придаточной железы; 8 — генитальный ме-
шок; 9—10 — железы наружной и внутрен-
ней оболочки сперматофора; 11—12 — 1-я
и 2-я части слизистой (сперматофорной)
железы; 13 — аппендикулярная железа:
14 — железа эйякуляторного аппарата;
15 — сперматофорный мешок; 16 — слепой
вырост генитального мешка; 17 — семепро-
вод; 18 — дистальная часть сперматофорно-
го мешка (фундус); 19—ампула семепрово-
да; 20—целомостом (воронка семепровода);
21— семенник; 22 — половой отдел целома;
23—ресничный канал (канал Куна); 24— во-
ронка ресничного канала; 25—сперматофор-
ный проток; е—схема соединения сперма-
тофорного протока, придаточной железы и
желез оболочки сперматофора; 26 — желе-
зистая часть спсрматофорного протока;
27 — дорсальная складка, прочие обозна-
чения, как на д; ж: 1 — семенник; 2 —
семепровод; 3 — сперматофорный проток;
4» 5 — сперматофорная (4) и придаточная
(5) железы; 6 — пенис; 7 — слепой отро-
сток (дивертикул) пениса; 8 — спермато-
форпый мешок; з—к — сперматофор (з, и)
и семенное тельце (к) Octopus dofleini (з)
и Dosidicus gigas (и, к): / — задний конец
семенной нити; 2 — цементное тельце; 3 —
головка; щ, ич» и3 — то же, увеличено (а.
б — по Jaeckel, 1958; з, з — по Иванову,
Стрелкову, 1949; г — по Froesch, Marthy,
1975; д. е — по Badenhorst. 1974; ж—по
Mann ct al., 1970; и—к — по Несису, 1970)
ракатиц овальная. Они образуют
четвертичную оболочку яиц, т. е.
внешнюю яйцевую капсулу кальма-
ров, внутри которой в слизи, выде-
ляемой яйцеводными железами, рас-
полагаются яйца, а у каракатиц —
плотную наружную оболочку яиц.
Обычно секрет нидаментальпых же-
лез плотнее, чем яйцеводных, так что
кладка в целом тяжелее воды (у ви-
дов с донными кладками) или имеет
нейтральную плавучесть, секрет этих
желез также чрезвычайно липкий и
быстро застывает в воде. У лолиги-
нид он имеет удивительное свойство
подавлять синхронное биение ресни-
чек инфузорий и мелких многоклеточ-
ных с ресничным движением, что
спасает кладки от нападения множе-
ства мелких, но опасных врагов. Не
имеющие нидаментальпых желез
Enoploteuthinac выметывают, подоб-
но Vampyroteuthis, отдельные яйца,
окруженные только секретом яйце-
водных желез.
Небольшие добавочные, или при-
даточные (акцессорные), нидамен-
тальныс железы имеются у карака-
тиц и Myopsida. Они есть у обоих
полов, но у самцов рудиментарны.
У незрелых самок они белого цвета,
по мерс созревания меняют цвет от
желтого и оранжевого до красного
(у зрелых особей). Они состоят из
множества трубочек, в которых жи-
вут симбиотические бактерии — бе-
лые, желтые и красные, в каждой
трубочке особого цвета. Цвет бакте-
рий и красный цвет железы у зрелых
самок обусловлены выделением бак-
териями особого каротиноидного пиг-
мента — сепиаксаптпна. Бактерии
могут жить и вне тела кальмара, по
для синтеза сепиаксантипа им необ-
ходимы добавки особых веществ жи-
вотного происхождения. По-видимо-
му, его синтез контролируется секре-
торной деятельностью клеток доба-
вочной нидамептальной железы
(Bloodgood, 1977; Richard et al.,
1979). Функция добавочных нида-
ментальных желез загадочна.
При изучении процесса созрева-
ния самок головоногих следует раз-
личать периоды и фазы развития оо-
цитов, стадии развития гонады и ста-
дии зрелости репродуктивной систе-
мы в целом. Р. Н. Буруковский и др.
(Методические ..., 1977) выделяют
следующие периоды и фазы развития
ооцитов кальмаров Sthenoteuthis pte-
46
ropus, применимые в той или иной
степени ко всем головоногим:
1) оогонии; 2) премейотические-
ооциты; 3) период протоплазматиче-
ского роста (превителлогенез) с
3 фазами: превителлогенеза, первич-
ного фолликула, простого фолликула
(подфазы бокаловидного и оформ-
ленного фолликула); 4) интеркаляр-
ный период с 2 фазами: сложного
фолликула и гидратации; 5) период
трофоплазматического роста с 4 фа-
зами: вакуолизации, накопления
желтка, изгнания фолликула и зре-
лого ооцита. В ходе этого процесса
ооциты растут и окружаются клет-
ками фолликулярного эпителия.
В период превителлогенеза слабо
дифференцированные фолликулярные
клетки, клетки соединительнотканной
оболочки (теки) и капилляры охва-
тывают снаружи весь ооцит, а затем
начинают внедряться в его поверх-
ность. В результате появляются
складки, и ооцит при рассматрива-
нии снаружи становится сетчатым.
В период вителлогенеза (трофоплаз-
матического роста) фолликулярные
клетки синтезируют желток и постав-
ляют его в ооцит. В конце вителло-
генеза складки ооцита выворачива-
ются и изгоняются из цитоплазмы.
Одновременно корковый слой цито-
плазмы видоизменяется, образуя
первичную оболочку яйца, а фолли-
кулярные клетки специализируются
па формировании вторичной оболоч-
ки— хориона. Зрелый ооцит, ранее
мутный и беловатый, становится
гладким, прозрачным, обычно янтар-
ного цвета и выпадает в просвет яй-
цевода.
Для головоногих, кроме наутилу-
сов, характерна непрерывная асин-
хронность развития гонады. Яичник
кальмара — гроздь гнезд, представ-
ляющих собой как бы отдельные го-
надки, а каждое гнездо — гроздь
ооцитов, соединенных капиллярами
со своим кровеносным сосудиком.
Сеть сосудов образует нечто вроде
дерева. На любой стадии развития
гонады в каждом гнезде можно най-
ти ооциты на всех стадиях развития:
ближе к сосудам самые мелкие, чем
дальше от сосудов, тем крупнее.
В зависимости от того, на какой фа-
зе развития находится основная мас-
са ооцитов, выделяют 6 стадий раз-
вития гонады: ювенильная, незрелая,
подготовительная, созревшая, зрелая
гонада и стадия выбоя. Необходи-
мость одномоментного нсрсста при
непрерывной асинхронности развития
гонады повлекла за собой выработку
накопительного механизма. Такой на-
копитель у кальмаров — яйцевод
(яйцеводы). Весь процесс подготовки
к размножению делится на два пе-
риода: первый — развитие гопады,
яйцеводов и желез, второй — накоп-
ление зрелых яиц в яйцеводе (яйце-
водах). Выделяют 6 стадий зрелости
репродуктивной системы кальмаров
(Методические... 1977): I—III ста-
дии соответствуют незрелой гона-
де, IV — подготовительной стадии
и созревшей гонаде, V — зрелой го-
наде. Последняя в зависимости от
хода процесса накопления зрелых
яиц делится па три подстадии: Vi —
зрелые яйца немногочисленны (до
5—8% объема всех ооцитов), V2 —
зрелые яйца в яйцеводах составляют
10—60% объема ооцитов, V3 — яйце-
воды набиты зрелыми яйцами (60—
80% объема ооцитов). VI стадия —
выбой. Эта шкала применима для
всех кальмаров.
У осьминогов и каракатиц накоп-
ление яиц происходит непосредствен-
но в половом целоме, поэтому у них,
в отличие от кальмаров, самые зре-
лые яйца лежат в задней, а не в пе-
редней части мантии. К ним указан-
ная шкала не применима.
Мужская половая система состо-
ит из семенника, семепровода, спер-
матофорного органа, придаточной
железы (простаты), сперматофорпого
мешка (нидхемов оргап) и пениса
(рис. 10, д—ж). Почти у всех голо-
воногих мужские половые органы не-
парные и лежат в левой половине
мантии, только у Selenoteuthis, Ore-
goniateuthis, Histioteuthis hoylei пар-
ные. Семенник — округлый (у осьми-
ногов), удлипеппо-треугольиый или
сигарообразный (у кальмаров и ка-
ракатиц), белого или кремового цве-
та орган трубчатой структуры, ле-
жащий в задней части мантии, в зад-
ней половине полового целома. От
пего отходит топкий, очень длинный,
спирально завитой или закрученный
семспровод (или проксимальная
часть семепровода), переходящий в
сперматофорный оргап. Семепровод
не соединен непосредственно с семен-
ником, его начало — воронка, откры-
вающаяся в целом (целомостом).
Сперматофорный орган имеет
чрезвычайно сложное строение, все
его части плотно упакованы и окру-
жены оболочкой, образующей гени-
тальный мешок. У кальмаров ч(рис.
10, д—е) и каракатиц семепровод пе-
реходит в слизистую железу (ее на-
зывают также сперматофорной же-
лезой, или семенным пузырьком).
Она состоит из двух частей, следую-
щих одна за другой. Далее следуют
4 железы: железа эйякуляторного
аппарата, железы средней и наруж-
ной части оболочки сперматофора и
обширная придаточная железа (про-
стата) с придатком или слепым.меш-
ком. От места перехода железы
средней части в железу наружной
части оболочки отходит ресничный
канал (канал Куна), а от слепого*
мешка придаточной железы — тон-
кий трубчатый железистый вырост
(аппендикулярная железа). Затем
следует прямой сперматофорный про-
ток, или дистальная часть семепро-
вода (лежит уже за пределами ге-
нитального мешка). Генитальный
мешок имеет два длинных выроста.
Один из них идет параллельно спер-
матофорному протоку, но заканчива-
ется слепо, второй — параллельно пе-
нису и заканчивается у одних видов
отверстием, открывающимся в ман-
тийную полость, у других — тоже-
слепо.
Сперматофорный проток впадает в
обширный сперматофорный мешок,
(нидхемов орган), который перехо-
дит в пенис. У каракатиц и кальма-
ров, имеющих гектокотиль, пенис
довольно короткий и доходит только
до воронки, у кальмаров, лишенных
гектокотиля, особенно из семейств.
Onychotcuthidae, Gonatidae, Chiroleut-
hidae, Mastigoteuthidae, он очень дли-
нен и способен далеко высовываться
из мантийной полости. Очень длин-
ный он и у Architeuthis. Обычно пе-
нис — мускулистая цилиндрическая
трубка, но у Mastigoteuthis он крюч-
ковидно изогнут, а у Chiroteuthidae
заканчивается пиковидпой головкой.
У осьминогов (рис. 10, ж) сперма-
тофорный орган устроен значительно*
проще и состоит из длинной извили-
стой сперматофорной железы и объ-
емистой придаточной железы (про-
статы) с небольшим слепым мешком.
Ресничный канал отсутствует. Спер-
матофорный проток короткий, зато
сперматофорный мешок очень велик
(у Octopus dofleini около 1 м) и не
47
переходит в пенис, а пенис отходит
особо, невдалеке от места впадения
сперматофорного протока в мешок.
Пенис осьминогов невелик, но устро-
ен значительно сложнее, чем у нери-
тических кальмаров и каракатиц:
Т-образный, состоит из собственно
пениса (терминальный орган) и сле-
пого отростка — дивертикула, кото-
рый часто значительно длиннее пе-
ниса, иногда закручен, а у некоторых
видов, например О. australis, О. fon-
tanianus раздвоен.
Сперматогенез происходит по всей
длине трубочек семенника и их вы-
ростов. Сперматогонии возникают из
терминального эпителия и, проходя
стадии сперматид и сперматоцитов,
превращаются в сперматозоиды, име-
ющие узкую головку и чрезвычайно
длинный тонкий хвост-жгутик. Го-
ловка сперматозоида у кальмаров и
каракатиц ‘ веретеновидная; у одних
осьминогов головка или передняя
часть ее (акросома) штопоровидная,
у других — удлиненно-коническая.
Общая длина сперматозоида у Loli-
go pealei 0,05 мм, у Octopus dofleini
0,5 мм, у Eledone cirrosa 0,55 мм,
ширина головки 1—2 мкм. Харак-
терная черта сперматозоидов голово-
ногих моллюсков, отличающая их от
спермиев млекопитающих, — боль-
шое количество гликогена в головке
и способность использовать поступа-
ющие извне углеводы. Благодаря
этому сперматозоиды могут сохра-
нять жизнеспособность в течение
длительного времени (до 3,5 мес)
после проникновения в семеприемни-
ки самки. Жидкость, в которую по-
гружены сперматозоиды в спермато-
форе (см. ниже), подавляет их по-
движность и дыхание, а морская вода,
напротив, стимулирует их движе-
ние. В семеприемнике они вновь ста-
новятся неактивными.
Вполне оформленные спермато-
зоиды поступают в сперматофорный
орган, где формируются спермато-
форы, чрезвычайно характерные для
головоногих моллюсков. Они сложно
устроены, их форма весьма характер-
на- для отдельных видов и часто слу-
жит надежным критерием для раз-
личения даже очень близких видов
(в родах Illex, Octopus, Eledone
и др.).
Сперматофор кальмаров и кара-
катиц — слегка изогнутая трубка,
похожая на казацкую шашку с длин-
ным лезвием (рис. 10, и). Длина
сперматофора обычно измеряется не-
сколькими миллиметрами или санти-
метрами (вариации от 2—3 мм до
10—20 см). У осьминогов спермато-
форы похожи на длинные толстые и
постепенно суживающиеся упругие
нити (рис. 10, з). Их длина также
измеряется сантиметрами, но у
О. dofleini опа достигает 1,0—1,2 м.
Количество сперматофоров находит-
ся в обратной зависимости от их от-
носительных (по отношению к длине
мантии) размеров. Обычно их около
сотни или несколько сот, редко боль-
ше тысячи, но у осьминогов с круп-
ными спсрматофорами (О. dofleini,
Pteroctopus tetracirrhus) их всего
10—12, у Bathypoly pus до 7, а у
Benthoctopus ergasticus бывает лишь
один сперматофор, длина которого в
полтора с лишним раза превосходит
длину мантии. Особое положение за-
нимает Argonautoidea с единствен-
ным в жизни сперматофором и Cirra-
ta, у которых нет оформленных спер-
матофоров (см. ниже).
Сперматофор плавно сужается от
расширенного заднего конца, занято-
го семенным резервуаром (семенной
нитью), к головке. Между концом
семенного резервуара и задним кон-
цом сперматофора имеется свободное
пространство. С противоположной
стороны семенной резервуар перехо-
дит в цементное тельце. Между це-
ментным тельцем и головкой нахо-
дится эйякуляторный аппарат в виде
плотно сжатой спиральной пружины.
В расширенной головке спираль скру-
чена в комок. Сперматофор покрыт
двойной оболочкой, состоящей из на-
ружного и промежуточного (средне-
го) слоев. Семенной резервуар, це-
ментное тельце и эйякуляторный ап-
парат окружены еще третьим слоем—
внутренней оболочкой. Двуслойная
внешняя оболочка состоит в основ-
ном из мукополисахаридов, внутрен-
няя— из мукопротеинов. Кроме того,
резервуар, цементное тельце и пру-
жина эйякуляторного аппарата по-
крыты тонкой мембраной, отделяю-
щей эти части сперматофора одну от
другой. В передней части головки
имеется участок — «окошко», закры-
тое тонкой перепонкой. Вблизи нее от
головки отходит длинная нить. У ом-
мастрефид семенной резервуар вме-
сте со свободным дистальным про-
странством занимает около или не-
48
сколько более половины длины спер-
матофора, у лолигинид — 2/з—3?4-
В топких и гибких сперм атофорах
осьминогов семенной резервуар пред-
ставляет собой свернутый спиралью
канатик, его длина обычно равна
Уз—Уг длины сперматофора. Эйяку-
ляторный аппарат очень длинен.
В сперматофорс Loligo pealei со-
держится 7—10 млн. спермиев, Octo-
pus dofleini — 0,5—37 млрд.
Спсрматофор целиком формиру-
ется в сперматофорпом органе. Мас-
са сперматозоидов сначала одевает-
ся мембраной, затем добавляются
цементное тельце и эйякуляторный
аппарат, все это покрывается оболоч-
ками. В процессе формирования
сперматофор все время вращается
вокруг своей оси — именно поэтому
эйякуляторный аппарат и (у осьми-
ногов) семенная нить свернуты в спи-
раль. В задней части нидхемова ор-
гана имеются спиральные гребешки,
благодаря которым сперматофоры
тоже размещаются по спирали. В пе-
нисе они перестраиваются параллель-
но оси тела головками вперед.
Основная функция сперматофо-
ра — обеспечить передачу спермато-
зоидов от самца к самке без потерь.
Пространство между наружной и
внутренней оболочками и в заднем
конце сперматофора заполнено рас-
твором высокомолекулярных соеди-
нений, которые не могут выйти на-
ружу, тогда как для морской воды
оболочка проницаема. Осмотическое
давление внутри сперматофора вы-
ше, чем в морской воде. Как только
спсрматофор попадает в воду, она
начинает проникать внутрь оболочки,
сжимая спираль эйякуляторного ап-
парата. Если сперматофор попал па
тело самки, чувствительная нить раз-
рывает тонкую перепонку «окошка»
и проникновение воды внутрь обо-
лочки ускоряется. Упругая оболочка
сначала выдерживает давление воды
и сжимающейся пружины, по, нако-
нец, достигается предел прочности, и
сперматофор «взрывается». Пружина
вылетает через прорвавшуюся обо-
лочку (у лолигинид выворачивается
наизнанку), вытягивая за собой це-
ментное тельце и семенной резервуар.
Весь этот процесс—сперматофорная
реакция — длится у кальмаров около
1 с, у О. dofleini — 1,2 ч. Освобож-
денный от оболочки семенной резер-
вуар меняет форму и превращается
в семенное тельце, или семенной пу-
зырек (рис. 10, к). У оммастрефид и
лолигинид он похож на дубинку^, по
«булаве» которой размазано клейкое
содержимое цементного тельца. Це-
ментное тельце многих оммастрефид
оканчивается острием, подобным на-
конечнику шишака, этот «гарпун»
вонзается в кожу самки вблизи семс-
приемпика, а цемент прочно приклеи-
вает семенной пузырек. Пустая обо-
лочка сперматофора с вывернутой,
пружиной отпадает (самки некото-
рых кальмаров проглатывают ее).
У осьминогов сперматофорная'
реакция протекает внутри мантий-
ной полости самки и пе столь быст-
ро, как у кальмаров, семенной ре-
зервуар под давлением проникаю-
щей воды постепенно продвигается
вперед, и, лишь когда пружина сож-
мется до отказа (у О. dofleini — ког-
да объем содержимого сперматофо-
ра с проникающей внутрь оболочки
водой увеличится впятеро), происхо-
дит «взрыв». Семенной пузырек име-
ет яйцевидную форму. Пустая обо-
лочка сперматофора выбрасывается
из мантийной полости током дыха-
тельной воды.
Морская вода стимулирует по-
движность сперматофоров, и они за-
полняют семеприемники, вблизи ко-
торых самец размещает семенные
пузырьки. У каракатиц и лолигинид
семеприемник — подкововидное уг=-
лубление на ротовой мембране ниже
рта. У Шех и Berryteuthis спермато-
форы размещаются в мантийной по-
лости, образуя пучок на внутренней
стенке мантии, у других оммастре-
фид и у Gonatopsis — кольцом на ро-
товой мембране, у Enoploteuthidae
они помещаются на «затылке» самки,
у Chaunoteuthis — в специальных бо-
роздах в передней части наружной
стороны мантии по бокам брюшной
стороны, у Octopoteuthidae — на го-
лове и у оснований рук и т. п. У са-
мок Vampyroteuthis имеются 2 ссме-
приемпика на голове перед глазами.
У Cirrata нет оформленных спер-
матофоров, соответственно отсутст-
вуют или редуцированы некоторые
части сперматофорного комплекса (в
частности, слепой мешок простаты)
и дивертикул (слепой отросток) пе-
ниса, другие же части, такие, как
придаточные железы, напротив, силь-
но развиты. Роль сперматофоров иг-
рают семенные резервуары, закручен-
4 К. Н. Нссис
49
пые в спираль и окруженные слизис-
тым секретом.
У Argonautoidea в сперматофор-
1юй железе формируется единствен-
ный крупный спсрматофор. Прида-
точная железа ведет в обширный
слепой мешок, который соответствует
нидхемову органу. Тут снерматофор
«взрывается» и его содержимое за-
полняет нидхемов орган, а затем весь
слепой мешок каким-то образом пе-
реносится внутрь гектокотилизиро-
ванной руки. При спаривании рука
отрывается и заползает в мантийную
полость самки. Там она хранится в
течение всей ее жизни. Таким обра-
зом, нидхемов орган одновременно
выполняет функцию семенного пу-
зырька, а гектокотилизированпая ру-
ка — семеприемника. В зрелом гск-
токотиле сперма заполняет особую
полость (семенной резервуар) у ос-
нования или на конце руки. Внутри
него находятся также остатки обо-
лочек сперматофора. На конце гекто-
котиля расположен конический или
нитевидный, иногда очень длинный,
вырост. Его называют пенисом, хотя
он имеет совершенно иное происхож-
дение и его функция скорее всего
аналогична функции каламуса гекто-
котиля Octopodidae. Настоящего пе-
ниса у Argonautoidea нет.
В отличие от непрерывной асин-
хронности созревания ооцитов фор-
мирование спсрматофоров, по край-
ней мерс у некоторых кальмаров,
происходит не непрерывно. Размер
сперматофора определяется размера-
ми частей сперматофорного органа, а
они увеличиваются с ростом моллюс-
ка, хотя (у половозрелых особей) и.
медленнее, чем длина мантии. Поэто-
му у крупных кальмаров спермато-
форы абсолютно длиннее, а относи-
тельно короче, чем у мелких. Если
бы сперматофоры формировались не-
прерывно и накапливались в сперма-
тофорном мешке, как яйца в яйцево-
дах, то их размеры у одного самца
довольно заметно различались бы.
В действительности же размерный
ряд спсрматофоров у одного и того
же самца, как правило, узок — пре-
делы изменчивости около 10%. На
кальмарах Stenoteuthis pteropus было
показано (Залыгалин и др., 1977),
что вариационная кривая размеров
сперматофоров . одновершинна у не-
давно созревших самцов (стадия Vi)
и отчетливо двувершинпа па более
поздних подстадиях (V2—V3). Оче-
видно, самцы этого вида в онтогене-
зе производят две разные порции
сперматофоров, причем между пе-
риодами их формирования происхо-
дит перестройка сперматофорного
комплекса.
Созревание половых продуктов у
самцов и самок головоногих моллюс-
ков находится под контролем оптиче-
ской железы — железы внутренней
секреции, выделяющей гонадотроп-
ный гормон. Три внешних фактора
(температура, свет и питание) мо-
гут регулировать процесс созревания.
При высокой температуре головоно-
гие моллюски созревают скорее и при
меньших размерах, чем при пони-
женной. У видов, размножающихся
весной и летом в умеренных широ-
тах, короткий световой день (условия
зимнего освещения) стимулирует
половое созревание, но подавляет
откладку яиц; увеличение длины све-
тового дня, напротив, подавляет
созревание у неполовозрелых живот-
ных, ио стимулирует нерест у созрев-
ших. У берегов Западной Европы
каракатицы Sepia officinalis размно-
жаются весной и летом на мелко-
водьях. Молодь проводит зиму на
глубинах, где слабая освещенность
ускоряет созревание, но низкая тем-
пература не позволяет им созреть, а
короткий световой день — отложить
яйца преждевременно, до наступле-
ния весны. Весенняя миграция на
мелководья вызывает быстрое на-
ступление нереста. При этом вылу-
пившиеся из яиц молодые каракати-
цы успевают до отхода на глубины
захватить период максимальной тем-
пературы воды, что ускоряет их рост.
Есть и варианты: у самцов обык-
новенного осьминога Octopus vulgaris
и длинный световой день может ус-
корять созревание, вследствие чего
родившиеся весной самцы могут ус-
петь созреть до конца сезона размно-
жения. За несколько педель до не-
реста моллюски прекращают питать-
ся или резко сокращают рацион. По
принципу положительной обратной
связи голодание ускоряет дозревание
половых продуктов. Но в целом
внешние факторы играют скорее сиг-
нальную, чем управляющую роль, и
главные факторы созревания — внут-
ренние. Например, кальмар Gonatus
fabricii по всему ареалу от Северно-
го полюса до Ньюфаундленда начина -
50
ет размножаться приблизительно в
одно и то же время — в марте (от ян-
варя до середины или второй полови-
ны апреля), хотя световые, темпера-
турные и кормовые условия в высокой
Арктике и умеренных широтах в это
время чрезвычайно сильно различа-
ются. Большая часть головоногих
моллюсков тропических регионов со-
зревает в возрасте 6—12 мес и раз-
множается круглогодично, т. е. тоже
независимо от изменений температу-
ры и освещенности. Однако в уме-
ренных широтах и субтропиках не-
рест абсолютного большинства шель-
фовых и эпипелагических, как и мно-
гих мезопслагических видов голово-
ногих, приурочен к определенному се-
зону (обычно весна и лето), а про-
должительность жизненного цикла
около 1 года. Таким образом, их не-
рест четко скоррелирован с наступ-
лением определенных световых, тем-
пературных и кормовых условий —
естественных сигналов времени нача-
ла размножения.
Нервная система (рис. 11).
Нервная система головоногих мол-
люсков развита выше, чем у любых
других моллюсков, и по сложности
строения и совершенству организации
не уступает нервной системе рыб.
Центральная нервная система
(ЦНС). Головной мозг двужаберных
головоногих помещается в хрящевой
головной капсуле. По относительной
массе он равен или превосходит мозг
рыб, но уступает мозгу птиц и мле-
копитающих. Относительно самый
большой мозг у быстроплавающих
эпипелагических нектонных кальма-
ров подссм. Ommastrephinae и сем.
Thysanoteuthidae. Несколько меньше
мозг нектонных, но не столь стреми-
тельных Illicinae, Onychoteuthis, Go-
natidae, еще меньше он у нектобен-
тосных Loliginidae и Sepiidae, далее
следует мозг бентосных Octopodidae
и на последнем месте мозг планк-
тонных кальмаров Cranchiidae и ось-
миногов Bolitaenidae (Зуев, 1973,
1975). Таким образом, относитель-
ный размер мозга головоногих Свя-
зан с быстротой их движений, что в
первую очередь обусловлено необхо-
димостью переработки в кратчайший
срок большого количества зритель-
ной информации.
* ЦНС двужаберных головоногих
моллюсков (рис. 11,6—6) распола-
гается над и под пищеводом и состо-
ит из 4 основных отделов: надгло-
точного (супраэзофагеального); под-
глоточного (субэзофагеального); оп-
тических долей с оптическими трак-
тами и прилегающими к ним нервны-
ми узлами и железами; буккального
и брахиального ганглиев. Все отделы
соединены многочисленными комиссу-
рами. Над- и подглоточный отделы
делятся на доли, общее число кото-
рых у осьминога равно 64. Все отде-
лы состоят из симметричных правых
и левых половин, соединенных нерв-
ными коннективами. При их перерез-
ке каждая половина может работать
самостоятельно. Осьминога с изоли-
рованными половинами мозга мож-
но, например, научить брать правыми
руками шероховатый предмет и от-
вергать гладкий, а левыми — на-
оборот.
Буккальный ганглий иннервирует
буккальную массу, клюв, радулу,
слюнные железы, брахиальный —1
конечности. У осьминогов, особенно
плавниковых, и у вампироморф они
сближены и почти слиты с мозгом,
у каракатиц и кальмаров отделены
от пего и соединены с мозгом цереб-
ро-буккальной и церебро-брахиаль-
ной коннективами, а друг с другом —
буккалыю-брахиальной коннективой.
Это обусловлено постоянством рас-
стояния от ганглиев до органов, ко-
торые они иннервируют: буккальной
массы и места отхождения рук от
ротовой воронки. Кальмары и кара-
катицы питаются крупной добычей и
поедают се быстро, раскусывая на
довольно крупные куски. Для этого
необходимы значительные физиче-
ские усилия, поэтому клюв и ротовая
воронка должны быть выдвинуты
вперед, чтобы увеличить плечо рыча-
га клюва. Соответственно, буккаль-
ный и брахиальный ганглии отдаля-
ются от основного мозга. Одни ось-
миноги (Cirrata) питаются микро-
планктоном, другие (Octopodidae) —
крупной добычей, но поедают ее в
норе понемногу, маленькими кусоч-
ками. Больших мышечных усилий
при этом не требуется, плечо рычага
может быть коротким, буккальный и
брахиальный ганглии приближаются
к мозгу. Естественно, что при этом
мозг осьминогов более концентриро-
ванный, чем у кальмаров и. кара-
катиц.
Оптические доли — самая большая
часть мозга, У осьминогов обе доли
4*
51
г
3
занимают больше половины, а у
кальмаров — до объема мозга.
У пелагических головоногих, в жиз-
ни которых зрение играет основную
роль, они развиты сильнее, чем у
донных; сильнее всего развиты опти-
ческие доли у быстроплавающих нек-
тонных кальмаров. В оптических до-
лях в строго определенном порядке
разветвляются волокна оптического
нерва, здесь они вступают в контак-
ты с дендритами зрительных клеток.
В оптических долях осуществляются
кодирование зрительной информации,
ее классификация и передача ин-
формации в другие отделы мозга,
преимущественно в высшие мотор-
ные центры надглоточного отдела,
Рис. И. Нервная система Todarodes paci-
ficus (а, 6) и Octopus vulgaris (в—д): а—
общий вид нервной системы со спинной
стороны; 6 — мозг сбоку; 1 — нервы рук;
2 — интербрахиальная комиссура; 3 — рото-
вой комплекс; 4 — нерв луча ротовой мем-
браны; 5— ганглий ручного нерва; 6—
глазной нерв; 7, 8 — оптическая (7) и пал-
лиовицеральная (S) доли; 9 — кишка; 10,
15 — звездчатый (10) и мантийный (15)
ганглий; 11 — жабра; 12 — желудок; 13 —
желудочный ганглий; 14 — цекум; 16 —
жаберное сердце; 17 — гонада; 18 — ман-
тийный нерв; 19 — нерв гонады; 20 — при-
даток жаберного сердца; 21, 22, 24 —же-
лудочный (21), висцеральные (22) и обо-
нятельный (24) нервы; 23 — коннектива
звездчатых ганглиев; 25 — надглоточные
доли мозга; 26, 27— верхняя (26) и ниж-
няя (27) буккальные доли; 28 — буккаль-
ные нервы; 29, 30 — церебро-буккальная
(29) и церебро-брахиальная (30) коннекти-
вы; 31, 33, 35—вертикальная (31), брахи-
альная (33) и педальная (35) доли; 32 —
передние глазные нервы; 34 — брахио-пе-
дальная коннектива; 36 — вороночные нер-
вы; 37 — задний вороночный нерв; 38 —
статоцист; 39 — нерв пищеварительной же-
лезы; 40 — пищевод; 41 — нерв мускула-
втягивателя воронки; в, г — мозг сверху и
сбоку; 1,2— буккальная (/) и оптическая
(2) доли; 3 — глазные нервы; 4 — оптиче-
ская железа; 5 — оптический тракт (опти-
ческая доля удалена); 6—15 — верхнефрон-
тальная (6), пижнефронтальная (7), верти-
кальная (8), задняя базальная (9), задняя
хроматофорная (10), вазомоторная (11),
паллиовисцсральная (12), магноцеллюляр-
ная (13), педальная (14) и брахиальная
(15) доли; 16 — нервы рук; д —схема рас-
положения некоторых долей на продоль-
ном разрезе через мозг (передняя часть
тела — слева): I—10 — надглоточные доли:
1 — вертикальная; 2 — субвертикальная;
3—5 — дорсальная (3), средняя (4) и пе-
редняя (5) базальные; 6 — задняя бук-
кальная; 7 — субфронтальная; 8 — средняя
пижнефронтальная; 9 — верхняя буккаль-
ная; 10 — средняя верхнефронтальная; 11—
пищевод; 12, 13 — подглоточные доли: 12—
педальная; 13 — паллиовисцсральная. Ос-
тальные доли не показаны, (а, б — по Ива-
нову, Стрелкову. 1949: в—г —по Wells,
1966; д — по Sanders, 1975)
визуостатическйй центр, центр зри-
тельной памяти, магноцеллюлярпую
долю. Волокна, поступающие в опти-
ческие доли и выходящие из них,
сохраняют четкие и постоянные то-
пографические отношения друг с дру-
гом. Оптические доли управляют так-
же мышечным тонусом животного.
Оптические доли соединяются с
мозгом толстыми нервами — оптиче-
скими трактами. Обычно они корот-
кие, по у некоторых родов, в частно-
сти у Eledonella и Cirrothauma (см.
рис. 74, в; 77, е) длинные и топкие.
На оптических трактах справа и сле-
ва от мозга располагаются оптиче-
ские железы, важнейшие органы
внутренней секреции, регулирующие
половое созревание и развитие вто-
ричных половых признаков. С ними
же связаны субпсдункулятпые (рас-
положенные под оптическим трактом)
доли мозга, контролирующие дея-
тельность оптической железы (см.
ниже).
В подглоточном отделе мозга рас-
положены: центры управления топ-
кими движениями рук; передняя хро-
матофорная доля — центр регуляции
окраски головы и конечностей; лате-
ральная педальная доля — окуломо-
торный центр (регуляция движения
глаз); задняя латеральная педальная
доля — предположительно центр
контроля движений щупалец у каль-
маров и каракатиц; передняя педаль-
ная доля, управляющая движениями
рук и щупалец; задняя педальная
доля — центр управления плаванием
(регуляция работы воронки, ретрак-
торов головы и плавников); магно-
целлюлярпая доля, контролирующая
реактивное плавание и управляющая
работой звездчатого ганглия; паллио-
висцсральная доля, иннервирующая
мантию, плавники и чернильный ме-
шок; задняя хроматофорпая доля,
управляющая изменениями окраски
мантии, воронки, плавников; висце-
ральная доля, иннервирующая внут-
ренности и воропку; вазомоторная
доля, управляющая мускулатурой
стенок кровеносных сосудов; плавни-
ковая доля (отсутствует у InGirra-Ц).
В этом отделе мозга концентри-
руются мотонейроны, здесь ^хдепрла-
гаются промежуточные моторные
центры, активируемые сигналами с
периферии или из высших моторных
центров надглоточного отдела мозга.
Это — центры простых рефлексов;
53
в подглоточных долях имеется лишь
кратковременная память, например
о боли.
В вентральной части магноцел-
люлярпой доли у кальмаров, карака-
тиц и вампироморф располагается
пара гигантских клеток диаметром
до 750 мкм (большинство нервных
клеток подглоточных и высших мо-
торных центров имеют размер 5—
10 мкм, а центров сенсорной интегра-
ции и памяти — менее 5 мкм), от
которых отходят гигантские нервные
волокна, отдающие через посредство
паллиовисцсральной доли сигналы
нервам звездчатого ганглия. Магно-
целлюлярная доля связана с центра-
ми плавания подглоточного отдела,
ее работа регулируется сигналами,
поступающими от конечностей, глаз,
статоцистов, и находится под кон-
тролем высших моторных центров.
У Mastigoteuthis эта доля также по-
лучает сигналы от рук и щупалец,
однако характерных для этой доли
гигантских клеток нет: Mast Igot eat -
his — плохие пловцы.
Нижнюю часть надглоточного
(супраэзофагеального) отдела мозга
составляют высшие моторные цент-
ры — передняя, средняя, дорсальная
и латеральные базальные доли, а
также связанные с оптическим трак-
том педункулятная и ольфакторная
доли. Педункулятная доля — это
визуостатический центр; вместе с пе-
редними и средними базальными до-
лями она образует структуру, функ-
ционально сходную с мозжечком
позвоночных. Педункулятная доля
получает от статоцистов сигналы о
положении и движениях головы, а от
оптических долей — о положении и
движениях глаз и посылает сигналы
в базальные доли и окуломоторный
центр латеральной педальной доли.
Этот центр также получает сигналы
от статоцистов и управляет тонкими
движениями головы и глаз, необхо-
димыми для нацеливания на добычу
и слежения за ней. Передняя базаль-
ная доля управляет и центрами кон-
троля движений рук в передней пе-
дальной доле, а средняя базальная
доля — центрами контроля дыхатель-
ных и плавательных движений ворон-
ки и плавников в задней педальной
доле. Кроме того, базальные доли
управляют работой магноцеллюляр-
ной и паллиовисцеральпой долей, ла-
теральные базальные доли контроли-
54
руют хроматофорные центры, оль-
факторная доля — орган обоняния.
Верхнюю часть надглоточного от-
дела мозга составляют фронтальные,
субфронтальная, вертикальная и суб-
вертикальная доли — «молчащие
центры», высшие центры зрительной
и хемотактильной памяти и обуче-
ния. У осьминогов к ним присоединя-
ется буккальный ганглий, или бук-
кальная доля мозга. Согласно теории,
разработанной Дж. Янгом, каждая
из систем зрительной и тактильной
памяти и обучения включает две па-
ры центров: нижнюю и верхнюю.
Верхние центры тактильной памя-
ти— средняя и боковая нижнефрон-
тальпые доли, нижние — субфрон-
тальная и задняя буккальная, верх-
ние центры зрительной памяти —
средняя верхнефроптальная и верти-
кальная, нижние — боковая верхне-
фронталытая и субвертикальная до-
ли. Эти две системы (зрительная и
хемотактильная) полностью разделе-
ны на высших уровнях мозга — у го-
ловоногих моллюсков в буквальном
смысле слова «две памяти в одном
мозгу». В указанных центрах распо-
лагаются постулируемые Дж. Янгом
гипотетические элементарные едини-
цы памяти — мнемопы.
Кальмары и осьминоги — одновре-
менно и хищные, и робкие живот-
ные, их жизнь проходит в борьбе ме-
жду желанием напасть на незнако-
мый, но, возможно, съедобный объ-
ект и страхом получить отпор. Ос-
нова мнемона — классифицирующая
клетка, имеющая два аксона, идущих
к моторным центрам: один из них
инициирует нападение, другой — от-
ступление. Предположим, что при
встрече с незнакомым объектом каль-
мар или осьминог предпочтет на-
пасть на него. Если объект оказыва-
ется съедобным, коллатеральные
нервные волокна, исходящие из выс-
ших моторных центров, возбуждают
одну из многочисленных мелких нерв-
ных клеток, которые выделяют пере-
даточные вещества, подавляющие
прохождение нервных импульсов, а
именно ту, которая «запирает» аксон,
инициирующий отступление. Мнем ок
«запомнит», что сигнал, соответству-
ющий этому объекту, означает —
объект вкусен и безопасен. Если же,
напротив, моллюск получит отпор,
«запертым» окажется аксон, иници-
ирующий нападение, а аксон, иници-
ирующий отступление, будет рабо-
тать, и, если моллюск еще раз встре-
тится с этим же объектом, он пред-
почтет пе вступать в опасный кон-
такт. Функция верхних и нижних
центров зрительной и тактильной па-
мяти состоит в усилении сигналов
положительного («вкусно») и отри-
цательного («больно») подкрепления;
классифицирующие клетки системы
тактильной памяти располагаются в
задней буккальной и субфронтальной
долях, зрительной памяти — в опти-
ческих долях (J. Young, 1965; San-
ders, 1975).
Участие в системе кратковремен-
ной зрительной памяти — нс един-
ственная из функций вертикальной
доли (высшей доли мозга головоно-
гих). Эта доля участвует также в
системе долговременной памяти, вы-
полняя роль, аналогичную записыва-
ющему и считывающему устройству
хранилища памяти современных
ЭВМ. Именно поэтому на личиноч-
ной и послеличипочпой стадиях жиз-
ни головоногих вертикальная доля
растет быстрее остальных, и по мере
се роста улучшаются способности к
обучению. При удалении вертикаль-
ной доли способности осьминогов к
запоминанию и обучению резко ухуд-
шаются.
Мозг большинства двужаберных
головоногих построен по общей схе-
ме. Различия между осьминогами, с
одной стороны, кальмарами и карака-
тицами— с другой, частично отмече-
ны выше. У кальмаров и каракатиц
мозг менее концентрирован, отдель-
ные доли сильнее обособлены, опти-
ческие доли очень велики, вентраль-
ная магноцеллюлярная доля сильнее
развита, имеется система гигантских
аксонов, нижние фронтальные доли
слабее развиты, супрабрахиальная
комиссура отсутствует. У осьминогов
мозг сильнее концентрирован, опти-
ческие доли не так велики, вентраль-
ная магноцеллюлярная доля и систе-
ма гигантских нервных клеток и ак-
сонов отсутствуют, нижние фронталь-
ные доли (центры тактильной памя-
ти) сильно развиты (они увеличены
также у глубоководных кальмаров),
брахиальные ганглии соединены суп-
рабрахиальной комиссурой. Базаль-
ные доли у кальмаров, каракатиц и
пелагических осьминогов развиты
лучше, чем у донных осьминогов.
Мозг вампироморф сочетает черты,
свойственные осьминогам (компакт-
ность, хорошее развитие нижних
фронтальных долей) и десятируким
головоногим (наличие вентральной
магноцеллюлярной доли, отсутствие
супрабрахиальной комиссуры). Мозг
плавниковых осьминогов чрезвычай-
но'компактен, оптические доли малы
и слабо развиты, оптические железы
редуцированы, весь надглоточный от-
дел мозга значительно упрощен, в
нем доминируют центры тактиль-
ной системы, тем не менее имеются
некоторые элементы системы гигант-
ских волокон (J. Young, 1977).
Мозг головоногих моллюсков рас-
тет в течение всей жизни, но его
относительный размер по мере рос-
та уменьшается. За период жизни
осьминогов — от личинки массой 1 мг
до «гиганта» массой 10 кг — число
мозговых клеток увеличивается в
1000 раз. Основной прирост клеток
приходится на личиночный и ранний
послеличиночный периоды. Мозг зре-
лого осьминога содержит около
170 млн. нервных клеток, в том чис-
ле 130 млн. в оптических долях,
30 млн. в вертикальной доле, 6 млн. в
высших тактильных и 4 млн. в выс-
ших .моторных центрах, 2,5 млн. в
подглоточных долях (J. Young, 1963;
Wells, 1966).
Осьминоги и каракатицы (нерити-
ческие кальмары в этом отношении
слабо изучены) прекрасно обучают-
ся, некоторые задачи на обучение и
переучивание они решают столь же
успешно, как крысы. У них хорошая
память, причем различаются кратко-
временная, промежуточная и долго-
временная память. С другой стороны,
наутилусы, приповерхностные пела-
гические осьминоги и, по-видимому
(судя по строению надглоточных до-
лей мозга), глубоководные океаниче-
ские кальмары — «тупые» животные с
плохо развитыми способностями к
обучению.
Периферическая нервная система.
Все органы тела головоногих мол-
люсков богато иннервированы, и их
работа находится под контролем
ЦНС. Упомянем важнейшие части
периферической нервной системы
(рис. 11, а). От брахиального ганглия
отходят нервы рук, щупалец и фила-
ментов вампироморф. Нервы рук
соединяются впереди рта кольцевой
интербрахиальной комиссурой. Каж-
дый ручной нерв, проходящий вдоль
55
оси руки, состоит из пары нервных
стволов и многочисленных ганглиев,
кроме того, в руках проходят 4 внут-
римышечных нервных тяжа. Особый
нервный ганглий имеется у основа-
ния каждой присоски, в нем интег-
рируются сигналы от хеморецепторов
и мехапорсцепторов (рецепторов ося-
зания), располагающихся- па ободке
присоски. В целом число нервных
клеток в конечностях- очень велико,
у осьминога их 350. млн. — вдвое
больше, чем в ЦНС. Наличие боль-
шого числа нервных ганглиев дает
конечностям головоногих некоторую
степень самостоятельности, напри-
мер, отрезанная рука осьминога хМО-
жет передавать кусочек пищи от од-
ной присоски к другой в направле-
нии ко рту, а автотомирующаяся при
спаривании гектокотилизированная
рука Argonautoidea самостоятельно
'заползает внутрь мантийной полости
самки.
Глазные нервы управляют рабо-
той мышц, поворачивающих глаза.
Ольфакторный нерв иннервирует ор-
ган обоняния. От буккального ган-
тлия кроме нервов, идущих к губам,
радуле и слюнным железам, отходит
пара симпатических нервов, прохо-
дящих вдоль пищевода назад и со-
единяющихся с крупным желудоч-
ным ганглием. Он иннервирует основ-
ную часть пищеварительной системы
и управляет процессами пищеварения
и всасывания. От висцерального ган-
глия отходит пара крупных висце-
ральных нервов, иннервирующих
органы кровеносной, выделительной
систем, жабры, половые органы, ней-
росекреторную систему полой вены,
воронку и ее ретракторы и пр.
В частности, с висцеральными нерва-
ми связаны ганглии сердца и жабер-
ных сердец. У оснований жабр на
бранхиальном (жаберном) нерве так-
же находится небольшой ганглий.
Паллиалытый нерв, отходящий от
паллиовисцеральной, магноцеллюляр-
ной и плавниковой долей, иннерви-
рует самый крупный из лежащих вне
мозга ганглиев — звездчатый. Пара
звездчатых ганглиев расположена на
внутренней поверхности мантии в ее
передней части по бокам от средней
линии спины. От каждого ганглия
отходит свыше десятка нервов, со-
стоящих из одного гигантского (до
0,5—1 мм в диаметре) аксона. Чем
более удаленный от ганглия участок
56
мантии иннервирует нерв, тем тол-
ще аксон. Скорость проведения нерв-
ного импульса пропорциональна
квадратному корню из диаметра ак-
сона, самые толстые нервы проводят
импульс в 5 раз быстрее, чем самые
тонкие. Диаметры аксонов «подобра-
ны» так, что импульсы, исходящие
от ганглия одномоментно, прибывают
«на место» также одновременно. Мы-
шечные волокна, иннервируемые нер-
вами звездчатого ганглия, сокраща-
ются по принципу «все пли ничего»,
т. е. срабатывают с первого импуль-
са. Система центрального контроля
гигантских аксонов такова: гигант-
ские клетки 1-го порядка в вентраль-
ной магпоцеллюлярной доле — ги-
гантские клетки 2-го порядка в пал-
лиовисцеральной доле — гигантские
клетки 3-го порядка в звездчатом
ганглии. Синхронность срабатывания
мышц правой и левой половин тела
обеспечена наличием перекреста ги-
гантских аксонов 1-го порядка меж-
ду педальной и паллиовисцеральной
долями: правая доля управляет ле-
вым ганглием, и наоборот. В резуль-
тате сигналы, поступающие в магно-
целлюляркую долю от глаз, стато-
цистов, с кожи и сигнализирующие
об опасности, вызывают одновремен-
ное и мгновенное сокращение всей
мантийной мускулатуры и немедлен-
ное бегство кальмара (или каракати-
цы). Loligo vulgaris с длиной! мантии
20 см может развивать при прыжке
хвостом вперед скорость 2 м/с
(7,2 км/ч) всего за 0,08—0,12 с. Систе-
ма гигантских нервных волокон, от-
крытая проф. Дж. Янгом в начале
30-х годов, сыграла выдающуюся
роль в развитии нейрофизиологии:
гигантский аксон кальмара оказался
идеальным объектом для исследова-
ния основных механизмов нервного
возбуждения, проведения нервного
импульса, строения мембран, синап-
сов и т. п. Для этих направлений
гигантский аксон кальмара оказался
тем же, чем был нервно-мышечный
препарат лягушки для нейрофизио-
логии второй половины XIX в.
ОРГАНЫ ЧУВСТВ. Органы чувств
двужаберных головоногих —г глаза,
внеглазные фоторецепторы — свето-
чувствительные пузырьки (пароль-
факторные и эпистелляриые тельца),
статоцисты, обонятельные папиллы
(ямки), хемо- и механорецепторы
присосок, губы и др.
Глаз а. Глаза дву жаберных го-
ловоногих очень велики и устроены
столь лее совершенно, как глаза поз-
воночных. Они обычно помещаются
в углублениях хрящевой головной
капсулы и имеют роговицу, радужи-
ну со способным к сужению и рас-
ширению зрачком, подвижный хру-
сталик и сетчатку (рис. 12, а). Рого-
вица у эгопсидных кальмаров имеет
кольцевую форму, спереди широко
открыта, так что передняя камера
глаза свободно сообщается с мор-
ской водой. Снаружи от нее имеется
снабженная мышцами кольцевая
складка кожи — веко, в нижне-перед-
нем углу века есть узкая выемка —
глазной синус. У некоторых эгопсид,
в частности, у Onychoteuthidae, веко
может сжиматься, почти полностью
закрывая глаз. У миопсидпых каль-
маров роговица почти полностью
замкнута, обычно имеется лишь ма-
ленькое отверстие — «слезная -пора»,
соединяющая переднюю кал^еру гла-
за с внешней средой; века синуса
нет. У каракатиц и осьминогов-рого-
вица полностью замкнута, но имеет-
ся кольцевое веко, без синуса. У вам-
пироморф глаза эгопсидного типа.
Зрачок у кальмаров круглый,
у осьминогов обычно прямоугольный,
вытянутый горизонтально, у карака-
тиц имеет форму перевернутой бук-
вы М (латинское W) с низкими и
широко наклоненными боковыми па-
лочками. На ярком свету он сжима-
ется у осьминогов в тонкую щель,
у каракатиц так, что кажется, будто
у них два крохотных зрачка. Зрач-
ковая реакция зависит от знака сме-
ны освещенности: при резком увели-
чении освещенности зрачок каракати-
цы сжимается за 5 с, при резком сни-
жении он расширяется гораздо мед-
леннее— через 0,5—2 мин, а затем
снова сужается — это связано с пере-
распределением пигмента в клетках
сетчатки.
Хрусталик головоногих, как и у
рыб, почти шаровидный и подвешен
на мышцах ресничного тела. Его фо-
кусное расстояние в 2,5 раза больше
диаметра. По-видимому, преломляю-
щие свойства хрусталика снижаются
от центра к периферии, вследствие че-
го на всех участках сетчатки полу-
чается четкое изображение. Как пра-
вило, хрусталик прозрачен для всех
лучей видимого спектра, но хруста-
лики Onychoteuthis banksi и большо-
го глаза Histioteuthis желтоватые —
желтый пигмент отсекает синие луч»
спектра. Предполагается, что таким
образом эти кальмары могут «демас-
кировать» свою добычу — мезопсла-
гических животных с вентральной
биолюминесценцией.
К аккомодации хрусталик голо-
воногих способен лишь в ограничен-
ной степени. Аккомодация происхо-
дит путем перемещения хрусталика
вперед и назад, как у рыб, а не пу-
тем изменения формы, т. е. фокус-
ного расстояния хрусталика, как у
млекопитающих. Назад хрусталик
подтягивают мышцы ресничного те-
ла, а вперед он перемещается вслед-
ствие упругости сократившихся
мышц, окружающих глазное яблоко.
Однако значительная аккомодация
головоногим и не требуется: дальность
видения в воде нс так уж велика, а
ближе концов рук им незачем ви-
деть. К тому же клетки сетчатки
головоногих очень длинны, и некото-
рая доля аккомодации может осуще-
ствляться за счет перемещения в них
пигмента.
Сетчатка головоногих моллюсков-
необращенная: светочувствительные
элсмспты направлены к свету, нерв-
ные волокна отходят в противопо-
ложном направлении, поэтому в сет-
чатке нет слепого пятна. Светочувст-
вительные клетки представляют
собой рабдомы, расположенные го-
ризонтально и вертикально, каждый
рабдом состоит из 4 рабдомеров.
Длина рабдомов 0,15—0,20 мм. Чис-
ло светочувствительных клеток сет-
чатки чрезвычайно велико, у карака-
тиц, кальмаров и мелководных дон-
ных осьминогов оно доходит до 50—
100 тыс. на 1мм2, а у Bathyteuthis—
даже до 250 тыс. на1 мм2. У голово-
ногих, за исключением Bathyteuthis,
нет «желтого пятна» — участка наи-
более четкого видения па сетчатке,
но у каракатиц и осьминогов имеется
«полоса паилучшего зрения», вытяну-
тая вдоль «экватора» глаза — в ее-
пределах длина рабдомов и плот-
ность их распределения наибольшие.
У сепиид на краях этой полосы име-
ются еще две перпендикулярные к
пей полоски, они лежат против боко-
вых расширений зрачка. Сетчатка не-
которых глубоководных форм (Histio-
teuthis, Bathothauma, Japetella, Cir-
roteuthis, вероятно, Vampyroteuthis)
подразделена на два различающихся
57
В
5
4
4
J
/ J
Рис. 12. Глаза, внсглазныс фоторецепторы
и статоцисты головоногих; схема строения
глаза Oegopsida (a), Myopsida (uj и Octo-
poda (аг): / — роговица; 2—радужная обо-
лочка; 3— веко; 4 — зрачок; 5— сетчатка;
б — глазная орбита; 7 — оптическая доля;
8 — «белое тело»; 9 — хрящ; 10 — вторич-
ная роговица; //—пора; 12, /3 — вторич-
ное (12) и первичное (13) веки; б — сте-
бельчатые глаза у личинки Bathothauma
lyrornma; в, г — парольфакторные свето-
чувствительные пузырьки Loligo vulgaris
(в) и Todarodes sagittal us (г): / — оптиче-
ская доля; 2 — оптическая железа; 3 — оп-
тический тракт; 4, 5 — глубинные (4) и по-
верхностные (5) светочувствительные пу-
зырьки; 5 —глаз; 7, 8 — ольфакторная (7)
и педункулятная (3) доли;/9 —нерв опти-
ческой железы; 10, // — дорсальные (10)
и вентральные (11) светочувствительные
пузырьки; /2 —нерв вентральный пузырь-
ков; д — положение эпистеллярпого тель-
ца Eledone moschata: 1 — эпистсллярное
тельце; 2 — звездчатый ганглий; 3 — нервы
звездчатого ганглия; 4 — мантийная связ-
ка; е, ж— расположение светочувствитель-
ных пузырьков у Heieroteuihis hawaiiensis
(е) и Histioteulhis hoylei (ж). животные
показаны в естественном положении: 1, 2—
дорсальные (/) и вентральные (2) свето-
чувствительные пузырьки; 3 — мозг; 4 —
фотофор; 5 — брюшной щит; з, и — стато-
цисты Octopus (з) и Sepiola (и, вскрыт):
/ — макула со статолитом; 2 — продольная
криста; 3 — эндолимфа; 4 — перилимфа;
5 — келликеров канал; 6 — вентральная
криста; 7 — антикрнста; 8—10 — нервы к
продольной, поперечной и вертикальной
кристам; // — хрящевая шапочка; 12 —
нерв макулы; 13, /'/ — медианная (13) и
дорсальная (14) антикристы; /5 — главная
макула; 16 — дорсальная и вентральная
второстепенная макулы; 17 — передняя по-
перечная криста; 18 — статолит; 19 — нерв
кристы; 20—24—гамули (крючковидные ан-
тихристы), (а — по Naef. 1921 из Jacckcl,
1958; б —по N. Voss, 1980; в-д — по Ман-
го. 1977; е, ж — по R. Young, 1977; з, и—по
J. Young, 1960, 1977)
по строению участка, которым соот-
ветствует подразделение оптических
долей. Предполагается, что одни час-
ти сетчатки «смотрят» вперед и
вверх, другие — назад и вниз.
Головоногие имеют два светочув-
ствительных пигмента: родопсин и
ретинохром. Родопсин — основной
зрительный пигмент, содержится
в рабдомах и имеет спектр поглоще-
ния с максимумом у разных видов
на волне 470—500 нм, в том числе
у океанических кальмаров — 480 им.
Рстинохром содержится в сетчатке
вне рабдомов, максимум поглощения
сдвинут на 15—20 нм в сторону
длинных волн. Функция его состоит,
по-видимому, в том, что он постав-
ляет активный рстиналь (альдегид
витамина AJ, который в соединении
с белком опсином образует родопсин.
Свет вызывает фотоизомеризацию
ретиналя, пе разрывая его связи с
опсином, но меняя проницаемость
для ионов натрия мембраны свето-
чувствительной клетки, а это вызы-
вает нервный импульс и регистрацию
попавшего на клетку света. В свето-
чувствительных и опорных клетках
сетчатки содержатся гранулы пиг-
мента, на ярком свету мигрирующе-
го к дистальной части рабдома, па
слабом — к базальной части. Так
осуществляется адаптация к измене-
ниям освещенности. Кроме того, у
животных, долгое время находящих-
ся в полутьме, возрастает длина
рабдомов.
Большинство головоногих мол-
люсков имеют монокулярное зрение,
но их глаза так велики, что поле
зрения обоих глаз близко к 360° или
еще шире. Однако Octopus при вни-
мательном рассмотрении какого-ли-
бо предмета поднимают и сближают
глаза, так что он рассматривается
обоими глазами. Еще в большей сте-
пени бинокулярное зрение свойствен-
но каракатицам — сепиидам и сспио-
лидам. В норме у Sepia officinalis
перекрывание полей зрения правого
и левого глаза спереди составляет
10°, но, сводя глаза, они достигают
перекрытия полей зрения в 75° и сле-
дят за добычей обоими глазами.
При этом изображение добычи по-
падает на сетчатку в полосе паилуч-
шего зрепия обоих глаз.
Многочисленными опытами с вы-
работкой условных рефлексов, ана-
лизом элсктрорстинограмм и исполь-
60
зованием других методик показано,
что головоногие моллюски не разли-
чают цветов (цветно-слепые). Зато
они, как и другие животные, имею-
щие рабдомы, различают плоскость
поляризации света и могут ориенти-
роваться по поляризованному свету.
Острота зрения у них высока, они
способны воспринимать образы.
Сильно развитая мускулатура глаза
позволяет следить за перемещением
рассматриваемых объектов. Зрачки
осьминогов и каракатиц расширяют-
ся и сокращаются пе только при из-
менении освещенности, но и по же-
ланию животного: расширенный зра-
чок имеет сигнальное значение, в
частности, при ухаживании. Чувст-
вительность глаза максимальна в си-
не-зеленых лучах, проникающих в
море глубже других.
Имеется единственный слепой го-
ловоногий моллюск — абиссопелаги-
ческий осьминог Cirrothauma. Его
глаза — темные подкожные пятна,
лишенные хрусталика, ресничного
тела и радужины, с редуцированной
сетчаткой и отдельными нерегулярно
расположенными рабдомами (см.
рис. 74, в). Оптические доли и центры
зрительной памяти также редуциро-
ваны. В родах Cirroteuthis и Octopus
наряду с видами с нормально разви-
тыми глазами есть виды с глазами,
сильно уменьшенными в размерах.
Среди глубоководных кальмаров и
осьминогов есть виды с глазами не-
обычной формы: разноразмерными,
телескопическими, стебельчатыми.
Разноразмерные глаза у преиму-
щественно мезопелагических видов
Histioteuthis (см. рис. 53—56). Левый
глаз у них по диаметру в 1,5—2 раза
крупнее правого и часто отличается
по форме. Соответственно различа-
ются и размеры оптических долей.
Чувствительность и острота зрения
у крупного глаза выше, чем у ма-
ленького. Крупный глаз приспособ-
лен для зрения в верхних слоях во-
ды, маленький — в глубинах. Пред-
полагается, что в нормальном поло-
жении кальмара крупный глаз смот-
рит вверх и воспринимает свет, пада-
ющий сверху, а маленький — вниз
и несколько вперед и воспринимает
биолюминесцептное свечение (Битю-
кова, Зуев, 1976; R. Young, 1975а).
Стебельчатые глаза характерны
для личинок и молоди кранхиид
(рис. 12,6) и в небольшой степени
для личинок Cycloteuthidac. Струк-
турно стебельчатые глаза сходны с
телескопическими глазами Amphitre-
tus (у него, как и у другого полу-
глубоководного пелагического осьми-
нога Dorsopsis, глаза сближены и
расположены на «макушке», см. рис.
77, ж—и) и с удлиненными глазами
Bathyteuthis, личинок некоторых Chi-
rotcuthidae и др. Стебельчатый глаз
представляет собой как бы цилиндр,
вырезанный из центральной части
нормального полушаровидного гла-
за. Оптические свойства такого гла-
за нс лучше свойств обычного, его
преимущества не очень ясны. Сте-
бельчатые глаза присущи преиму-
щественно мезопелагическим видам и
мезопелагпческим личинкам батипе-
лагических видов. Предполагается
(R. Young, 1975b), что основная их
функция — камуфлирование глаз
кальмара от взгляда хищника, под-
карауливающего его днем снизу.
Кроме того, разнесение стебельчатых
глаз в стороны от узкой палочковид-
ной (камуфляж!) головы кальмара
облегчает установление точной ди-
станции до добычи — принцип стерео-
трубы (J. Young, 1970).
Нужно отметить, что хотя среди
глубоководных головоногих и име-
ются виды с редуцированными или
уменьшенными в размерах глазами,
по виды с крупными нормально раз-
витыми глазами встречаются на всех
глубинах от поверхности океана до
абиссали.'
Глаза наутилуса резко отличают-
ся от глаз двужаберных головоногих.
Они лишены хрусталика, имеют кро-
хотный (1—2 мм) зрачок, соединяю-
щий внутреннюю камеру глаза с
внешней средой (см. рис. 14). Зра-
чок сужается на ярком свету и рас-
ширяется в полумраке. Глаз наути-
луса — камера-обскура с регулиру-
емой диафрагмой (зрачком). Хотя
при ярком освещении наутилусы, ве-
роятно, и способны воспринимать
четкие образы, но, будучи строго
ночными животными, они ориентиру-
ются в основном не по зрению, а по
осязанию и обонянию.
Внеглазные фоторецеп-
торы (рис. 12, в—ж). Внеглазные
фоторецепторы, или светочувстви-
тельные пузырьки, — один из наибо-
лее загадочных органов головоногих
моллюсков. Первые из таких орга-
нов — эпистеллярные тельца осьми-
ногов— были открыты Дж. Янгом в
1929 г., парольфакторные пузырьки
кальмаров замечены С. Туре в
1939 г. и описаны Б. Бойкоттом и
Дж. Янгом в 1956 г. Предполага-
лось, что это — нейросекреторные об-
разования и лишь в середине — кон-
це 60-х годов выяснилось, что они
представляют собой тесно связанные
с нервной системой фоторецепторы,
своего рода «глаза в мозгу». Они
имеются у всех головоногих моллюс-
ков, кроме наутилуса (Mauro, 1977).
Светочувствительные пузырьки-
осьминогов — эпистеллярные тель-
ца— маленькие пузырьки, располо-
женные по одному на задней сторо-
не звездчатого ганглия (рис. 12, д).
Обычно они ярко-оранжевого или
желтого цвета, по у Octopus бесцвет-
ные. Лежат на внутренней, обращен-
ной к мантии стороне ганглия, и свет
к ним может проникать лишь через
толщу мантийной стенки или при
вдохе, когда мантия раздувается,
тогда звездчатый ганглий оказывает-
ся лежащим против входного отвер-
стия мантийной полости. Среди кле-
ток эпистеллярного тельца, заметны
многочисленные биполярные клет-
ки — фоторецепторы рабдомного ти-
па, содержащие родопсин. При осве-
щении тельца возникает нервный им-
пульс. Расположение и основные осо-
бенности их строения одинаковы у
всех осьминогов, включая Cirrata.
Светочувствительные пузырьки
кальмаров, каракатиц и спирулы рас-
положены в задней части головного
мозга в районе ольфакторной доли,
почему и называются парольфактор-
ными («около-ольфакторными») пу-
зырьками. Они построены по совер-
шенно иной, чем у осьминогов, схе-
ме и чрезвычайно разнообразны (рис.
12, в, г). У Loligo, Spirula и Sepia они
лежат на оптическом тракте преиму-
щественно на его вентральной стороне
вблизи педункулятной доли, с кото-
рой связаны нервами, и представля-
ют собой группу из нескольких (у
Loligo vulgaris 4—15 с каждой сто-
роны) мелких (в 4—5 раз мельче
эпистеллярных телец осьминогов)
пузырьков, часть из которых лежит
на поверхности тракта, а часть по-
гружена в глубь его тканей. Еще бо-
лее разнообразны их строение и рас-
положение у эгопсидных кальмаров.
Одни из них, как Onychoteuthidae,
Bathyteuthidae, Ctenopterygidae, Вга-
61
chioteuthidae, Ncoteuthidae, Chiroteut-
hidae, Grimalditeuthidae, Joubiniteut-
hidac, Cranchiidae, имеют только
один набор парольфакторпых телец
сзади и ниже оптического тракта, но
тельца эти могут быть небольшими и
просто устроенными (у Cranchiidae)
или крупными и сложными (Bathy-
teuthis, Ctenopteryx). Pyroteuthinae,
Histioteuthidae, Octopoteuthidae, Cyc-
Joteuthidae, Mastigoteuthidae имеют
две группы парольфакторных те-
лец— дорсальную и вентральную. У
оммастрефид и Ancistrocheirinae три
группы телец (дорсальная, централь-
ная и вентральная), а у Enoplothcut-
hinae даже 4 (дорсальная, две вен-
тральных и задняя). Строение па-
рольфакторных телец высоко специ-
фично для семейств (подсемейств) и
отличается деталями у разных родов
и видов одного рода. У полуглубо-
ководпых пелагических сепиолид
Het erot ent his парольфакторные тель-
ца построены не как у каракатиц, а
как у кальмаров: дорсальная груп-
па расположена выше оптического
тракта, вентральная — под глазом и
чуть позади него.
Совершенно по-иному расположе-
ны светочувствительные пузырьки у
Vampyroteuthis: группа пузырьков
лежит в передней части мантийной
полости, на се дорсальной стороне
под краем мантии и прикрыта про-
зрачным эпидермисом. Таким обра-
зом, этот орган расположен как раз
посредине между парольфакторными
пузырьками кальмаров и каракатиц
и эпистеллярными тельцами осьмино-
гов. В каждом органе около полусот-
ни пузырьков.
Как и у осьминогов, светочувстви-
тельные тельца десятируких голово-
ногих и вампироморф обычно жел-
тые или оранжевые и содержат све-
точувствительные клетки рабдомного
типа с родопсином, дающие импульс
при освещении. Дорсальные свето-
чувствительные пузырьки «смотрят»
вверх и немного назад, а у живот-
ных, нормальное положение которых
в воде хвостом кверху (Вathyteuthis)
или косое — ось тела наклонена под
углом 30—45° и хвост выше головы
(Heteroteuthis, Histioteuthis и пр.),—
прямо вверх («глаза на затылке»)
(рис. 12,е,ж). Над ними обычно
имеются прозрачные кожные «окош-
<и», особенно хорошо видные у
Snoploteuthidae. Таким образом, эти
52
органы могут воспринимать дневной
свет, падающий сверху. Вентральные
органы «смотрят» обычно на приле-
гающую к голове сторону воронки то-
же через «окошки». У Heteroteuthis
и Pyroteuthinae они могут видеть
свечение внутримантнйного фотофо-
ра (фотофоров), у Histioteuthis —
фотофоров переднего края мантии, а
у Enoploteuthinae и Ancistrocheiri-
nae — специальных фотофоров, рас-
положенных па стороне воронки,
прилегающей к голове (см. рис. 43, у)
Крапхииды могут принимать почти
любое положение, их светочувстви-
тельные пузырьки смотрят как вверх»
так и вниз. Однако непонятно, что
же они видят: никакого диоптричес-
кого аппарата светочувствительные
пузырьки пе имеют, и единственное,
на что они способны, это отличать
свет от темноты и, вероятно, как-то
оценивать интенсивность освещения.
Р. Янг (R. Young, 1972b, 1973, 1977,
1978; Young, Roper, Walters, 1979)
высказал предположение, что основ-
ная функция светочувствительных
пузырьков у мсзопелагических каль-
маров и сепиолид — подгонка интен-
сивности свечения собственных фото-
форов к интенсивности падающей
радиации (дневной свет), что необ-
ходимо для успешного камуфляжа
при создании вентрального противо-
свечения (см. с. 45). Интенсивность
свечения фотофоров изменяется до
тех пор, пока величина сигнала,,
поступающего в мозг от вентральных
светочувствительных пузырьков, вос-
принимающих собственную биолюми-
несценцию животного, пе сравняется
с величиной сигнала от дорсальных»
воспринимающих падающий сверху
свет. Эта гипотеза подтверждена
рядом экспериментов (см. выше), а
также тем, что крупные, разнообраз-
ные и сложно устроенные органы
свойственны в основном видам, оби-
тающим днем в нижней мезопела-
гиали, на глубинах от 400 до 700—
800 м, тогда как эпипелагические и
батипелагичсские виды имеют пре-
имущественно небольшие и однооб-
разно устроенные органы. Но такое
объяснение не относится к осьмино-
гам, каракатицам, лолигинидам, не
имеющим фотофоров оммастрсфи-
дам, спируле и Bathyteuthis, свето-
чувствительные тельца которых рас-
положены так, что свет своих фотофо-
ров на них пе попадает, и, разумеет-
ся, к Vampyroteuthis, который сво-
ими светочувствительными пузырь-
ками может видеть лишь то, что
находится внутри его мантийной по-
лости (R. Young, 1972b). Среди ось-
миногов самые крупные эпистелляр-
ные тельца имеют не мезопелагичес-
кие Amphitretus и батипелагические
Bolitaenidae и Vitreledonella, хотя у
них они и крупнее, чем у сублито-
ральных Octopus и эпипелагических
Argonauta, а абиссальные Cirroteut-
his и абиссопелагические Cirrothauma
(J. Young, 1977).
Несомненно, что функции свето-
чувствительных пузырьков головоно-
гих многообразны и, вероятно, раз-
ные у разных видов, но наиболее
вероятно, что основная их функ-
ция — давать животному представ-
ление об общем уровне освещенности
в окружающей его среде и долговре-
менных (например, сезонных) изме-
нениях освещенности. Такая инфор-
мация может «запускать» процессы
полового созревания при соответст-
вующих изменениях фотопериода,
инициировать сезонные миграции
нектонных и нсктобентосных видов
и т. п.
Среди других возможных функ-
ций этих органов можно назвать,
например, восприятие биолюминес-
ценции светящихся глубоководных
животных. Но в целом функции этих
органов столь же загадочны, как и
функции их аналога у рыб, амфибий
и рептилий — пинеального, или те-
менного органа, называемого иногда
«третьим глазом».
Статоцисты (рис. 12, з, и).
Пара статоцистов расположена в
затылочной части головной хряще-
вой капсулы. Статоцист—орган,
воспринимающий положение живот-
ного по отношению к силе тяжести
(орган равновесия), угловые и ли-
нейные ускорения. Статоцисты каль-
маров и каракатиц построены по об-
щей схеме, хотя и различаются в де-
талях, статоцисты осьминогов резко
от них отличаются.
Статоцист осьминога (рис. 12, з) —
округлый мешочек, заполненный
жидкостью (перилимфой) и заклю-
ченный в заднюю часть хрящевой
головной капсулы. Внутри находится
второй мешочек, заполненный эндо-
лимфой;, На его внутренней стенке
размещены группами рецепторные
волосковые клетки. Часть из них вос-
принимает сигналы о положении ось-
минога в поле тяготения (клетки ма-
кулы), часть — об ускорениях при
движении (клетки крист). Другие
клетки рассеяны в стенке статоциста.
Макула — овальное пятно па
стенке статоциста, к которому при-
креплен известковый (арагопито-
вый) статолит. Продольная ось ма-
кулы занимает строго вертикальное
положение. Статолит один и относи-
тельно велик — у Octopus vulgaris.
его масса в среднем 0,9 мг. Имеют-
ся 3 кристы, расположенные под.
прямым углом друг к другу: про-
дольная, поперечная и вертикальная.
Каждая криста разделена на 3:
участка, иннервируемых особыми
нервами. Рецепторные клетки каж-
дого участка образуют функциональ-
ную единицу — купулу (всего 9 ку-
пул). Купулы имеют форму ссгмен-
товидного паруса. Часть крист имеет
один ряд рецепторных клеток,,
часть—два ряда; одно- и двухряд-
ные секции чередуются. Сигналы об-
угловых и линейных ускорениях
воспринимаются по отдельности каж-
дой купулой и передаются в мозг
также особо, в целом создавая пол-
ную картину положения и перемеще-
ния тела осьминога. Рецепторные-
клетки макулы и крист укреплены
хрящевыми подпорками — выростами
стенки статоциста. На стороне, про-
тивоположной макуле, имеется хря-
щевой вырост стенки статоциста —
антикриста, защищающая вертикаль-
ную кристу от резких инерционных
движений эндолимфы, которые при
броске осьминога на добычу могли
бы повредить нежное образование.
Вблизи продольной кристы находится
кёлликеров канал — маленькая сле-
пая трубка, открывающаяся в эндо-
лимфатический мешок. Его функция
загадочна.
У кальмаров и каракатиц (рис.
12, и) статоцист врастает в заднюю
часть головной капсулы. Он имеет
приблизительно треугольную форму
и одностенный — заполнен только
эндолимфой. Имеются три макулы
и соответственно три статолита:
один более крупный (в основной ма-
куле), два помельче. Но и крупный
статолит мельче, чем статолит ось-
минога: у Todarodes pacificus он име-
ет массу около 0,05 мг. Есть те же
три взаимно перпендикулярные крис-
63
ты, но поперечная криста подразде-
лена на две. Вместо одной анти-
.кристы их 11—12: 4—5 крючковид-
.ных на концах крист и 7 прямых
палочковидных посередине. Они раз-
бивают поток жидкости при движе-
нии моллюска и тем повышают ско-
рость реакции рецепторов; возможно,
.антикристы и сами являются рецеп-
торами. Имеются 4 купулы. В целом
система крист, аптикрист и купул
.аналогична системе полукружных
каналов во внутреннем ухе позвоноч-
ных. П. Стивенс и Дж. Янг (Step-
hens, Young, 1978) полагают, что го-
ризонтальная криста, представляю-
щая собой почти замкнутый туннель,
регистрирует угловые ускорения при
рыскании (повороты вправо-влево),
продольная — крен на правый или
левый «борт», а .поперечная — диф-
ферент, причем поднимание головы
и хвоста воспринимается разными
участками этой кристы; она же, ви-
димо, регистрирует линейные уско-
рения-при движении головой и хвос-
том вперед.
У разных видов имеются вариа-
ции в строении статоциста, особенно
антикрист. Палочковидные антикри-
сты хорошо развиты у нектонных и
нектобентосных видов. У спирулы
•число антикрист уменьшено до 9,
у кранхиид палочковидных антикрист
вообще нет, а крючковидных лишь 3.
Кёлликеров канал у глубоководных
видов увеличен и расширен, предпо-
лагают, что он может воспринимать
вертикальные перем-ещения при су-
точных миграциях.
Строение статоциста Vatnpyroteu,-
this, как и строение светочувствитель-
ных пузырьков, промежуточное между
свойственным осьминогам и десяти-
руким головоногим: как у осьмино-
гов, есть внутренний и наружный
мешки с эндолимфой и перилимфой,
по криста разделена на 4 части, ан-
тикристы многочисленны, макул три.
Наружный мешок имеет обширный
выступ сзади, такой же выступ есть
у некоторых глубоководных кальма-
ров (Mastigoteuthidae, Cranchiidae).
У Cirroteuthis строение статоциста в
общем как у Incirrata, по антикрист
несколько.
Строение статоциста до некоторой
степени отражает образ жизни мол-
люсков: малоподвижные глубоковод-
ные виды не приспособлены к опре-
делению больших линейных и угло-
64
вых ускорений, с которыми они не
сталкиваются, зато они, возможно,
способны определять глубину, па ко-
торой находятся. Статоцист наутилу-
са — тонкостенный пузырек, запол-
ненный эндолимфой, статолитов нет,
есть лишь маленькие известковые
«песчинки».
Статолиты двужаберных голово-
ногих имеют характерную форму и,
несмотря па малый размер (1—2 мм),
достаточно богаты внешними призна-
ками, чтобы по ним можно было оп-
ределять, по крайней мере, семейст-
во, а иногда и вид. Неправильно-уд-
линенный статоцист состоит из дор-
сального и латерального куполов,
клюва и крыла. Куполы и клюв
жесткие, полупрозрачные, крыло
мягче, опаковое. Статолиты состоят
из мелких известковых кристалли-
ков — статокониев. Статолиты рас-
тут по мерс роста животного, но по
достижении определенного размера
их рост приостанавливается, так что
статолиты старых особей лишь не-
многим крупнее статолитов молодых.
На статолитах головоногих отмечены
многочисленные тонкие полоски, сре-
ди которых идентифицированы су-
точные, полумесячные и месячные
кольца роста. Таким образом, по ста-
толитам можно определять возраст
головоногих моллюсков.
Судя по строению статоцистов,
головоногие моллюски должны быть
полностью лишены слуха. Действи-
тельно, опыты с осьминогами пока-
зали, что они глухи. Однако карака-
тицы и кальмары слышат низкочас-
тотные звуки, соответствующие вос-
принимаемым слухом человека.
Каракатицы реагируют па звук с
частотой 180 Гц. Тихоокеанский
кальмар положительно реагирует па
чистый тон частотой 600 Гц, звук от-
рабатывающего задним ходом судна
(отсечены частоты ниже 150 Гц),
звуки падающего на поверхность мо-
ря дождя, звуки, создаваемые пи-
тающимся жслтохвостом, и т. п. Эк-
спериментально показано, что транс-
ляция таких звуков через подводный
гидрофон повышает «интерес» каль-
маров к блеснам вертикального каль-
мароловного яруса, и улов' увеличи-
вается на 30—60%, а в отдельных
случаях вдвое. Однако неясно, каким
органом кальмары и каракатицы
воспринимают звуки: играют ли тут
роль статолит и макула, реагирую-
щие на вибрацию с низкой частотой,
или работают механорецепторы при-
сосок, ощущающие колебания воды.
Хе мо- и механорецепт о-
р ы. Самый заметный и тем не менее
загадочный орган — ольфакторный
(обонятельный). У кальмаров, кара-
катиц и эпипелагических осьминогов
это — папилла, расположенная сбоку
головы между глазом и мантийным
отверстием, у донных осьминогов —
ямка, находящаяся приблизительно
там же. Очевидно, обонятельная па-
пилла (ямка) располагается па пути
входящей в мантийную полость во-
ды; по ее положению, строению и ин-
нервации можно предположить, что
она действительно воспринимает ка-
кие-то запахи. Однако четких экспе-
риментальных доказательств этого не
получено. Во всяком случае, это нс
запахи врага и пищи. Может быть,
она воспринимает половые феро-
моны?
Вкусовые рецепторы расположе-
ны главным образом на ободках при-
сосок и на губе. Это — разнообраз-
ные эпителиальные ресничные рецеп-
торные клетки. Количество их огром-
но: в ободках присосок до несколь-
ких сотен на 1 мм2. Головоногие
очень тонко различают вкусы, их
чувствительность к некоторым хими-
ческим веществам на 2—3 порядка
выше, чем у человека. Там же, в
ободках и воронках присосок, рас-
полагаются многочисленные и разно-
образные мсханорецепторы, в осо-
бенности осязательные рецепторы,
реагирующие на сжатие, растяжение
и изгибание. На одной руке среднего
по размерам осьминога располагает-
ся до V4 миллиона рецепторных кле-
ток. Тесное соседство вкусовых и
осязательных рецепторов и" единство
их представительства в мозгу поз-
воляют говорить о существовании у
головоногих особого хемотактиль-
ного — «вкусо-осязательного» чувст-
ва. Оно особенно развито у донных
обитателей, в частности у донных
осьминогов, и у глубоководных ви-
дов. У донных осьминогов хорошо
развиты хсмотактильная память, в
том числе способность находить до-
рогу домой на «вкусо-ощупь». Они
различают на ощупь виды моллюс-
ков и отличают живых моллюсков от
пустых раковин.
К1>оме того, у головоногих имеет-
ся множество внутренних и пару ж-
5 К. Н. Несис
пых пропрнорсцепторов в коже, мыш-
цах (особенно под звездчатым ганг-
лием), толще тканей присосок, кишке,
на поверхности жабр, внутренней
стороне роговицы и т. п. Рецепторы
в коже и мышцах позволяют ли-
шенному жесткого костяка животно-
му получать информацию о собст-
венной позе, об относительном поло-
жении частей тела, работе мускула-
туры. Функция многих рецепторов
нс выяснена.
Интересно, что головоногие неспо-
собны определять массу предметов
и отличать тяжелые предметы от
легких.
У наутилуса органами обоняния
служат ринофоры — пара небольших
конических бугорков с узкой порой,
расположенных под глазами, — а ор-
ганами осязания и вкуса — щупаль-
ца. Строение ринофоров сходно со
строением обонятельной ямки осьми-
ногов. Две пары коротких щупалец
спереди и сзади каждого глаза снаб-
жены механорецепторами, их зада-
ча— охранять глаз от механических
повреждений, давая сигнал опасно-
сти при соприкосновении с чужерод-
ным телом. Дистальные щупальца
несут на концах рецепторные клетки,
подобные клеткам вкусовых сосочков
языка. Вкусовые и осязательные ре-
цепторы особенно многочисленны на
концах длинных п^упалец, которые
волочатся по чхруцФу за плывущим
над дном наутилусом, как гайдроп
за аэростатом. С их помощью наути-
лус ищет добычу (преимущественно
падаль, спящих рыб и малоподвиж-
ных донных ракообразных). У наути-
лусов есть также осфрадий — парный
орган внутри мантийной полости, оп-
ределяющий химические свойства
поступающей в мантийную полость
воды.
ОРГАНЫ ВНУТРЕННЕЙ СЕК-
РЕЦИИ И НЕИРОСЕКРЕЦИИ.
Единственный известный в настоя-
щее время орган внутренней секреции
головоногих моллюсков — это опти-
ческая железа (у наутилуса отсутст-
вует). Оптические железы — малень-
кие округлые парные тельца желто-
ватого цвета, лежащие на оптиче-
ском тракте между оптическими
долями и ЦНС. Они состоят из сек-
реторных и опорных клеток. Гормон
(или гормоны) оптической железы
имеет гонадотропные свойства, сти-
мулирует деление зародышевых кле-
65
ток, вызывает синтез желтка в фол-
ликулярных клетках яичника, контро-
лирует конечные стадии развития
ооцитов и фолликулярных клеток,
секреторную активность добавочных
ни да ментальных желез, рост и раз-
витие яйцеводов, яйцеводных желез,
сперматофорного комплекса и пениса
(но не гсктокотиля), управляет фор-
мированием сперматофоров, контро-
лирует спаривание. У самок осьми-
ногов он также определяет резкое
сокращение или полное прекращение
питания перед нерестом, вызывает
рефлекс «насиживания» яиц и опре-
деляет неизбежную послеперсстовую
гибель. Если у выметавшей яйца сам-
ки осьминога удалить оптические
железы, опа немедленно бросает
кладку, через несколько дней вновь
начинает питаться, спаривается и
живет значительно дольше, чем нс
оперированные самки, которые поги-
бают вскоре после вылупления мо-
лоди, хотя больше уже не отклады-
вает яиц (Wodincky, 1977). Возмож-
но, такую же функцию этот гормон
выполняет и у всех двужаберпых го-
ловоногих с единственным в жизни
нерестом.
Активность оптической железы
контролируется ингибирующим нерв-
ным влиянием субпсдункулятной до-
ли мозга. Если у неполовозрелого
осьминога удалить эту часть мозга,
начинается выделение гонадотроп-
ного гормона, и осьминог созревает
значительно раньше, чем это имеет
место в природе. В свою очередь ин-
гибирующее влияние субпедункулят-
ной доли,, по крайней мере, отчасти,
регулируется зрением, в частности
через изменение длины светового
дня: ослепление осьминога или пере-
резка зрительных нервов также вы-
зывают преждевременное созревание.
В целом эта система аналогична ги-
поталамо-гипофизарной системе кон-
троля активности гонад у позвоноч-
ных. По-видимому, у головоногих,
как и у позвоночных, имеется и об-
ратная связь от гонады к оптической
железе (гормональная), хотя выде-
ление гонадой половых гормонов по-
ка не доказано. Гонадотропный гор-
мон оптической железы нсспецифичен
(одинаков у разных видов и родов
осьминогов) и, вероятно,- представ-
ляет собой стероидный гормон, хи-
мическая структура которого еще нс
установлена. Его синтез — не едипст-
66
венная функция оптической железы,
она участвует также в защите орга-
низма от чужеродных белков (анти-
генов).
Нейросекреторные клетки встре-
чаются в разных местах центральной
и периферической нервной системы
головоногих. Нейрогормоны секрети-
руются клетками нервной системы и
выделяются в кровь. Наиболее важ-
ные нейросекреторные органы распо-
ложены в венозных сосудах: перед-
ней части полой вены, в глазном
(фаринго-офтальмическая вена) и
буккальном синусах. Нейрогормон
полой вены выделяется паллиовисце-
ралыюй долей мозга и оказывает
длительное возбуждающее действие
на сердце головоногих; он функцио-
нально подобен эпинефрину позво-
ночных; секретируется при стрессо-
вых ситуациях и ускоряет работу
сердца. В фаринго-офтальмическую
вену выделяется нейрогормон суб-
педупкулятной доли мозга, его дей-
ствие па сердце аналогично (по не
идентично) действию нейрогормона
полой вены, главпая функция — ре-
гуляция давления крови в глазной
полости. Полоска ткани — продол-
жение субпсдункулятной доли — про-
ходит в переднюю камеру глаза и,
вероятно, осуществляет обратную
связь в системе регуляции внутри-
глазного. давления. Нейрогормон
буккального синуса выделяется ниж-
ним буккальным ганглием. Чрезвы-
чайно активные нейрогормоны голо-
воногих представляют большой ин-
терес для фармакологии и, возможно,
со временем войдут в арсенал ме-
дицины.
РЕГЕНЕРАЦИЯ И АВТОТО-
МИЯ. Головоногие моллюски обла-
дают высокоразвитой способностью
к регенерации поврежденных или
утраченных частей тела. Рапы па те-
ле заживают быстро и без следа.
Это одно из преимуществ использо-
вания головоногих как лабораторных
животных при изучении функции
ЦНС методом удаления отдельных
частей мозга. Утраченные (скажем,
откушенные хищником) руки и щу-
пальца восстанавливаются полностью
и довольно быстро. Например, у мо-
лодых Sepia officinalis при экспери-
ментальном удалении руки через, 5—
14 сут на месте рубца появляется
полушаровидная почка, которая рас-
тет, превращается в конус, удлиня-
ется, через 17—25 сут на ней появ-
ляются зачатки присосок, через ме-
сяц— хроматофоры, и па втором ме-
сяце после ампутации восстановив-
шаяся рука начинает функциониро-
вать. От обычной руки опа в первое
время отличается только мелкими
размерами присосок. Восстанавлива-
ется и гектокотилизироваппая рука.
Животные с регенерировавшими ко-
нечностями попадаются довольно
часто. Однако иногда, особенно у
осьминогов, регенерация протекает
аномально: восстанавливающаяся
конечность начинает разветвляться
или, напротив, срастается с соседней,
неповрежденной.
Полпая. регенерация наблюдается
только тогда, когда животные со-
держатся в хороших условиях. При
плохих условиях содержания на по-
врежденных участках кожи поселя-
ются бактерии и грибки, с которыми
ослабленное животное не в состоя-
нии справиться, быстро развивается
некротический дерматит, и моллюск
погибает. Это Sнеобходимо учиты-
вать при разведении.1 головоногих в
марикультуре. Начало полового со-
зревания влечет за собой ухудшение
регенерационной способности; ресур-
сы организма переключаются с со-
матического роста па генеративный
Автотомия — произвольное отбра-
сывание конечностей наблюдается
у ряда видов океанических кальма-
ров и пелагических осьминогов.
У зрелых самцов Argonautoidea за-
полненная спермой гсктокотилизиро-
ванная рука (см. рис. 87, ж; 88,6)
автотомируется при спаривании, от-
рываясь у основания, и передается
самке, после чего хранится в ее ман-
тийной полости. Щупальца Gonatop-
sis и, по-видимому, Lepidoteuthis ав-
тотомируются в конце личиночной
стадии, у самок Histioteuihis, Chau-
noteuthis, JoubiniteuMs и некоторых
Cranchiidae они автотомируются в
период полового созревания или во
время нереста. Mastigoteuthis отбра-
сывают щупальца при опасности, на-
пример в момент поимки: они отры-
ваются на определенном месте неда-
леко от основания. Регенерации ото-
рвавшихся конечностей пи у одной
из названных форм не происхо-
дит.
У самок Tremoctopus автотомия
части рук 1-й пары — средство де-
зориентации хищника. Руки 1-й па-
ры имеют широкую боковую отороч-
ку, украшенную продольными рядами
крупных цветных пятен (см. рис.
87,з—к). Автотомия происходит по
анатомически предопределенной ли-
нии, перпендикулярной оси руки. На
отрывающемся куске имеется одна
присоска и 1—3 цветных пятна. Ли-
шенный нервного контроля кусок ру-
ки мгновенно расширяется до разме-
ра носового платка, и внезапно вспы-
хивающие перед глазами хищника
яркие пятна па прозрачном куске ото-
рочки отвлекают его внимание и, воз-
можно, пугают его, позволяя осьми-
ножихе спастись бегством. Вероятно,
эти руки растут в длину постоянно,
но у взрослых самок Tremoctopus их
концы всегда оборваны и они неиз-
менно короче рук 2-й пары (у моло-
дых — наоборот)..
Донным осьминогам, за исключе-
нием Euaxoctopus, О. defilippi и др.,
автотомия рук несвойственна, но
при плохом содержании (в малень-
ких аквариумах с непроточной во-
дой) у них нередко наблюдается
автофагия, «самоедство»: объедание
собственных конечностей, как прави-
ло, заканчивающееся гибелью жи-
вотного.
5*
СИСТЕМАТИЧЕСКАЯ
ЧАСТЬ
Таблица для определения подклассов,
отрядов, подотрядов и надсемейств
класса головоногих моллюсков
(рис. 13).
1(2). Имеется прочная наружная ра-
ковина с перламутровым слоем, вы-
деляемая мантией моллюска. Руки
многочисленные — несколько десят-
ков, тонкие, без присосок, расположе-
ны двумя кругами. Голова и основа-
ния рук сверху прикрыты мясистым
щитом, закрывающим устье при втя-
гивании животного в раковину. Во-
ронка нс сросшаяся па брюшной
стороне. Две пары жабр.
Подкласс Nautiloidca, отряд Nau-
tilida. Одно семейство Nautilidae и
род Nautilus................с. 71.
2(1). Раковина внутренняя или
отсутствует. Если имеется наружная
раковина (самки Argonauta), то
очень хрупкая, без перламутрового
слоя, выделяемая не мантией, а кон-
цами спинной пары рук, животное
легко можно вынуть из раковины.
Число конечностей 8 или 10, они рас-
положены одним кругом и всегда во-
оружены присосками и (или) крючь-
ями. Голова и руки сверху нс при-
крыты щитом. Воронка срастается в
коническую трубку. Одна пара жабр.
Подкласс Coleoidea .... 3
3(10). 5 пар конечностей: 4 пары
рук и 1 пара щупалец, в редких слу-
чаях щупальца могут исчезать у
взрослых особей (у личинок всегда
имеются), но при этом животное име-
ет типичный для кальмара внешний
облик. Присоски сидят на стебель-
ках и имеют хитиновое кольцо обыч-
но с зубчатым краем, иногда кольцо
преобразуется в крюк. Руки обычно
не соединены умбреллой. Плавники
всегда имеются, разнообразной, но нс
весловидной формы. Есть буккаль-
ная воронка и буккальное крепление.
Как правило, имеется затылочный,
мантийные и вороночные хрящи;
иногда мантия срастается с головой
68
и (реже) с воронкой, в этих случаях
мантийная полость соединяется с
внешней средой тремя отверстиями—
па брюшной стороне тела и по бокам
головы..........................4.
4(7). Плавники овальные или поч-
ковидные, сидят по бокам тела или
близ заднего конца либо протягива-
ются вдоль всей мантии в виде бо-
ковой каймы, но не соединяются
между собой. Стебли щупалец втяж-
ные и могут втягиваться в специаль-
ные сумки у основания щупалец. Ру-
ки и щупальца вооружены только
присосками. Раковина в виде извест-
ковой пластины, известковой спива-
ли, тонкой хитиновой пластинки пе-
рсвидпой формы или вообще отсут-
ствует. Вороночные хрящи простые,
край мантии близ мантийных хря-
щей прямой без выступающих «угол-
ков». Яйцевод непарный, придаточ-
ные пидаментальпыс железы имеют-
ся. Все зубцы радулы одновершин-
ные, или радула отсутствует.
Отряд Sepiida................5.
5(6). Раковина в виде спирально
закрученного, круглого в сечении из-
весткового «рожка» лежит в задней
части мантии. Плавники маленькие,
округло-почковидные, сидят на зад-
нем конце тела, между ними — круп-
ный светящийся орган. Руки корот-
кие с широкой умбреллой, гсктоко-
тилизируются обе брюшные руки.
Радула отсутствует. Мезопелагичс-
скис животные.
Подотряд Spirulina. Одно семей-
ство — Spirulidae и род Spiru-
la .........................с. 73.
6(5). Раковина нс закручена спи-
рально и не сдвинута в заднюю часть
мантии, иногда отсутствует. Непар-
ного заднего фотофора пет. Плавни-
ки не па заднем конце тела. Радула
имеется. Обычно донные животные.
Подотряд Sepiina ... с. 76.
7(4). Плавники сидят в задней
половине тела или вдоль всей май-
Рис. 13. Некоторые признаки строения, ис-
пользуемые при определении головоногих
моллюсков, и основные измерения: а —
кальмары: 1 — присоски; 2 — буккальная
мембрана; 3 — вороночная ямка; 4 — эго-
псидный (незамкнутый) глаз; 5 — воронка;
6 — срастание мантии с воронкой; 7, 8 —
короночный (7) и мантийный (5) хрящи;
9 — мантия; 10 — фотофоры; 1! — плавник,
вогнутый сзади; /2 —хвост; 13— плавник,
выпуклый сзади; 14 — миопсидный (замк-
нутый) глаз; 15 — руки (I—IV лара); 16—
крючья; 17 — щупальце; 18 — карпальная
часть щупальца с фиксаторным аппара-
том; 19 — ладонь щупальца с присосками
и крючьями; 20 — палец щупальца; 21 —
стебель щупальца; основные измерения;
22 —длина мантии; 23 — длина головы;
24 — длина плавника; 25 — длина рук; б —
осьминоги: 1 — жаберные филаменты (в
мантийной полости); 2 —мантийное отвер-
стие; 3—воронка; 4 — глаз; 5 — глазчатое
пятно на умбрелле; 6 — умбрелла; 7 — ру-
ки (I—IV нары); 8—присоски; 9 — гскто-
котиль; основные измерения: 10 — общая
длина; // — длина мантии; 12 — длина го-
ловы; 13 — длина рук; 14 — глубина ум-
бреллы; в — гектокотиль кальмара: / —
измененная часть руки; 2 — пластинки;
3 — нормальные присоски; г—гектокотиль
осьминога: / — присоски; 2 —каламус; 3 —
лигула. Длина лигулы измеряется от пос-
ледней присоски, (а—в по Roper, 1977; г —
по Иванову, Стрелкову, 1949)
69
тип и, как правило, соединяются
вдоль средней линии или близ зад-
него конца. Стебли щупалец пе втяж-
ные. Многие виды имеют крючья на
булаве или (и) на руках. Раковина
никогда пе обызвествляется и пред-
ставляет собой тонкий хрящевой гла-
диус перовидной или стреловидной
формы (чрезвычайно редко гладиус
отсутствует). На переднем крас ман-
тии близ мантийных хрящей неболь-
шие выступы — «уголки», между ко-
торыми на брюшной стороне тела
край мантии слегка вырезан. Цент-
ральный зубец радулы трсхвершин-
ный (очень редко одновершинный).
Отряд Teuthida...............8.
8(9). Передняя камера глаза
замкнутая (миопсидпого типа). При-
соски рук всегда в 2, булавы — в 4
ряда, крючьев нет. Развит только ле-
вый яйцевод. Придаточные нида-
ментальныс железы имеются. Гскто-
котилизируются левая или обе брюш-
ные руки. Нет наружных фотофоров.
Подотряд Myopsida . . с. 128.
9(8). Передняя камера глаза от-
крытая (эгопсидный глаз). Вооруже-
ние рук и щупалец разнообразно,
у многих видов имеются крючья.
Развиты, как правило, оба яйцевода.
Придаточных нидаментальных желез
нет. Гсктокотилизация имеется или
отсутствует. Многие виды имеют фо-
тофоры на поверхности тела, головы,
конечностей или (и) па нижней сто-
роне глазного яблока.
Подотряд Cirrata ... с. 275.
14(13). Плавников пет. Гладиус
Присоски лишены хитиновых колец.
Руки обычно соединены более или
менее глубокой умбреллой. Плавни-
ков нет или они короткие, вссловид-
ной формы, сидят по бокам мантии.
Нет буккальной воронки и буккаль-
ного крепления. Мантия всегда сра-
стается с головой на затылке, но с во-
ронкой обычно нс срастается. Если
мантийные и вороночные хрящи есть,
то они простые, в форме бугорка и
ямки. В большинстве случаев ман-
тийная полость соединяется с внеш-
ней средой одним отверстием, нахо-
дящимся на брюшной стороне тела,
редко — двумя (по бокам голо-
вы) ...........................11.
11(12), Имеются особые щупаль-
цсвидпые придатки — длинные, тон-
кие, нитевидные, лишенные присосок
филаменты, способные втягиваться в
карманы, расположенные с паруж-
70
пой стороны умбреллы между рука-
ми 1-й и 2-й пар. Имеется одна пара
плавников (у молоди на определен-
ной стадии развития 2 пары, из ко-
торых задняя исчезает с окончанием
стадии малька) удлиненпо-овальпой
формы. Имеется большое количество
мелких простых и пара крупных,
сложно устроенных наружных фото-
форов. Присоски па руках располо-
жены в 1 ряд, параллельно им про-
ходят 2 ряда усиков. Проксимальные
части рук лишены присосок. Гладиус
имеет вид широкой, очень тонкой
пластинки. Глубоководные батипела-
гические животные.
Отряд Vampyromorpha. Одно се-
мейство— Vampyroteuthidae и род —
Varnpyroteuthis ".........с. 269.
12(11). Щупальцевидпых придат-
ков нет. Плавников одна пара или
они отсутствуют. Нет сложно устро-
енных наружных фотофоров, лишь у
немногих видов имеются простые фо-
тофоры. Присоски в 1—2 ряда, си-
дячие (изредка стебельчатые). Гла-
диус представляет собой хрящевую
опору плавников, либо редуцирован
до небольших хрящевых палочек, или
совсем отсутствует.
Отряд Octopoda1 .... 13.
13(14). Имеется одна пара весло-
видных плавников, поддерживаемых
U- или V-образным или седловидным
хрящом. Присоски па руках в один
ряд, параллельно им проходят два
ряда усиков. Мантийное отверстие
очень узкое — щель вокруг основа-
ния воронки, иногда зарастает. Ум-
брелла, как правило, очень глубокая,
иногда доходит до самых концов
рук. Нет чернильного мешка и раду-
лы. Развит только один левый яйце-
вод. Глубоководные придонные (ред-
ко пелагические) животные.
Подотряд Cirrata . . с. 275.
14(13). Плавников пет. Гладиус
редуцирован или отсутствует. При-
соски на руках в 1—2 ряда. Усиков
нет. Мантийное отверстие обычно
средних размеров или широкое, из-
редка редуцировано до двух отвер-
стий по бокам головы. Ум брел л а
обычно нс доходит до концов рук.
Радула всегда имеется, чернильный
1 Для определения семейств Octo-
poda рекомендуется пользоваться таб-
лицей па с. 274.
мешок имеется или отсутствует. Раз-
виты оба яйцевода.
Подотряд Incirrata .... 15.
15(16). Радула гребенчатая, ра-
хидиальный (центральный), 1-й и 2-й
латеральные (боковые) зубы много-
вершинные. Тело студенистой конси-
стенции, присоски в один ряд. Пела-
гические глубоководные животные.
Надсемейство Bolitaenoidea (Cte-
noglossa) ...................с. 286.
16(15). Радула нс гребенчатая,
рахидиальный (центральный), 1-й и
2-й латеральные (боковые) зубы с
1—3 вершинами. Тело обычно плот-
ное ...........................17.
17(18). Гектокотилизируется толь-
ко конец правой или левой руки 3-й
пары, зрелый гектокотиль нс отры-
вается от тела. Самцы нскарликовые.
Присоски рук в 1 или 2 ряда. Как
правило, донные формы (кроме пе-
лагического сем. Vitreledonellidae).
Надсемейство Octopodoidca (Не-
teroglossa)..................с. 287.
18(17). Гектокотилизируется це-
ликом правая рука 3-п пары; гскто-
котиль созревает в особом мешке и
сначала незаметен снаружи, так что
кажется, будто у самца лишь 7 рук,
при спаривании он отрывается от те-
ла. Самцы обычно намного мельче
самок, карликовые. Присоски всегда
в 2 ряда, по крайней мере на ди-
стальных частях рук. Пелагические
(преимущественно эпипелагические)
животные.
Надсемейство Argonautoidea с. 322.
1. Подкласс Nautiloidea
1.1. Отряд Nautilida
Раковина современных видов круп-
ная, до 20—25 см в диаметре, дву-
сторонне-симметричная, спирально
закрученная так, что снаружи заме-
тен лишь последний оборот; пупок
раковины открытый или закрытый
(рис. 14). Наружная поверхность ра-
ковины блестящая, фарфоровидная,
белого цвета, с изогнутыми радиаль-
ными рыжими полосками, внутрен-
няя — перламутровая, переливаю-
щаяся, очень красивая. На продоль-
ном распиле раковины видно, что она
состоит из обширной последней —
жилой — камеры, в которой поме-
щается тело моллюска, и многочис-
ленных предшествующих (нежилых)
камер, служащих гидростатическим
аппаратом. Перегородки этих камер
(септы) в середине пронизаны сплош-
ной трубкой (сифоном), проходящей
через все камеры от самой первой до
жилой. С помощью сифона наутилус
может откачивать жидкость из ка-
мер и тем самым компенсировать
увеличение веса раковины 'йо мере
роста. I
Голова наутилуса \ крупная, с
большими примитивно устроенными
глазами, лишенными хрусталика и
стекловидного тела, с маленьким
зрачком. Мантия коническая, кожи-
стая, слабо мускулистая, лишенная
хроматофоров. Руки тонкие, похо-
жие па усики, лишены присосок, рас=-
положены двумя кругами: наружный
круг из 38 ловчих рук и внутренний
из 24 (у самцов) или 48—52 (у са-
мок) ротовых рук. Основания спин-
ных рук наружного круга сливают-
ся с обширным толстым головным щи-
том (капюшоном), закрывающим
устье раковины, когда наутилус втя-
гивается внутрь. По 2 глазных щу-
пальца с каждой стороны. Иижняя
часть внутреннего круга рук самца
с правой или (чаще) левой стороны
преобразована в копулятивный ор-
ган (спадикс). Семеприемник самки
находится в особой сумке под рото-
вым конусом.
Воронка, нс сросшаяся в трубку,
ее края внизу просто закручены один
вокруг другого. Вороночного органа
пет. Клюв сильный. Радула состоит
из 11 продольных рядов зубов. Име-
ются 2 пары жабр, 4 предсердия,
4 почечных мешка. Чернильного меш-
ка и светящихся органов нет. Нерв-
ная система примитивного строения,
мозг состоит из 3 пар ганглиев.
Звездчатого ганглия и гигантских
нервных волокон нет. Имеется осф-
радий. Развиты оба яйцевода, одна-
ко функционирует только правый.
Имеются пидаментальные и прида-
точные нидаментальныс железы.
Только один (правый) ссмспровод.
Яйца крупные, донные, развитие без
метаморфоза.
Один род — Nautilus Linnc, 1758.
Типовой вид — N. pompilius Linne,
1758. 3—6 видов. Распространены в
Индомалайской зоогеографической
подобласти Индо-Вестпацифики й ме-
стами у Западной и Южной Авст-
ралии.
Наутилусы — хищные животные.
Живут близ дна, невдалеке от бере-
71
гов, но на глубинах от нескольких
десятков до 500—700 м, к берегам
подходят редко и только в холодный
период года. Активны по ночам. Пи-
таются малоподвижной донной до-
бычей, раками, креветками, рыбой,
падалью. Плавают медленно. Добы-
чу находят в основном по запаху.
Яйца очень крупные, длиной до 40 мм
(с оболочкой), откладываются пред-
положительно па мелководье на дно
поодиночке; самка откладывает лишь
десяток яиц с промежутками около
2 недель.
Пустые раковины наутилусов лег-
че воды и могут дрейфовать в тече-
ние длительного времени на поверх-
ности, уплывая за тысячи миль от
мест обитания наутилусов: вплоть до
Мадагаскара, Южной Африки, Япо-
нии, Тасмании, Новой Зеландии.
Наутилусы служат предметом
местного промысла на Филиппинах
и в Меланезии. Их ловят вершами и
ловушками, наживленными рыбой
или мясом. Мясо наутилусов съедоб-
но, но главную ценность представля-
ют раковины, используемые для из-
готовления сувениров и украшений.
Обзоры: Stenzel, 1964; Hamada et al.,
1980.
Таблица для определения видов
рода Nautilus
1(4). Раковина с закрытым пупком,
гладкая.......................2.
2(3). Цветные полосы на ракови-
не доходят до пупка и заходят бо-
лее или менее далеко на наружные
боковые стенки жилой камеры. Дли-
на раковины до 18 см.
N. pompUius L., 1758 (=ЛГ. alum-
nus Iredale, 1944) (рис. 15, а—г).
Никобарские, Андаманские острова,
моря Сулу, Сулавеси, Южно-Китай-
ское, Филиппинские и Молуккские
острова, Новая Гвинея с прилегаю-
щими островами, крайний север Ав-
стралии, Меланезия, Западная Поли-
незия до Фиджи и Самоа. На остро-
вах Палау (Южная Микронезия)
обитает особый вид или скорее под-
вид N. (pompUius) belauensis Saun-
ders, 1981, отличающийся большими
размерами (длина раковины 18—
23 см) и струйчатыми линиями на-
растания; окраска раковины та же,
что у /V. pompUius,
3(2). Цветные полосы заметны
лишь на периферии раковины, не до-
72
ходят до пупка и не заходят па на-
ружные боковые стенки жилой ка-
меры. Длина раковины до 20—25 см.
N. repertus Iredale, 1944 (рис.
15, ж). Южная и Юго-Западная
Австралия. Возможно, подвид N, рот-
pilius. Особи с такой окраской попа-
даются среди типичных AC pompUius.
4(1). Раковина с открытым пуп-
ком ...........................5.
5(6). Раковина гладкая, лишь с
микроскопическими линиями нара-
стания. Пупок довольно узкий, его
край без киля. Цветные полосы на ра-
ковине доходят до пупка и заходят
на наружные боковые стенки жилой
камеры. Длина раковины до 18 см.
N. macromphalus Sowcrby, 1849
(рис. 15,(9). Новая Каледония и
прилегающие острова.
6(5). Поверхность раковины мор-
щинистая с извилистыми радиальны-
ми складками. Пупок широкий, его
край с резко очерченным килем.
Цветные полосы на раковине замет-
ны лишь па периферии жилой каме-
ры, не заходят на ее наружные боко-
вые стенки и не доходят до пупка.
Длина раковины до 19—20 см.
N. scrobiculatus Solander, 1786
(=N. umbilicatus Lamarck, 1822) (рис.
15, з). Новая Гвинея, Новая Брита-
ния, Новая Ирландия, Молуккские и
Соломоновы острова, Торресов
пролив.
В таблицу не включен сомнитель-
ный вид N. stenomphalus Sowerby
1849 (рис. 15,г), самостоятельность
которого нуждается в подтверждении.
Он похож на N. pompUius, но имеет
пупок очень узкий, гораздо уже, чем
у N. macromphalus. Цветные полосы
не доходят до пупка. Обнаружен у
Новой Каледонии, в Торресовом про-
ливе и у Большого Барьерного рифа.
2. Подкласс Coleoidea
Раковина внутренняя, хитиновая, ре-
же обызвествленная, иногда отсутст-
вует. Мантия мускулистая, у глубоко-
водных форм мускулатура дегенери-
рует и замещается студенистой соеди-
нительной тканью. Хроматофоры хо-
рошо развиты, сложные. Часто име-
ются фотофоры. Глаза крупные,
сложные, высокоразвитые, с хруста-
ликом, радужной оболочкой, рогови-
цей и стекловидным телом. Статоцис-
ты также высокоразвитые. Осфрадии
отсутствуют. Конечностей 4 или
5 пар. Они мускулисты и вооружены
присосками, часто снабженными хи-
тиновыми кольцами. У многих (но
далеко не у всех) видов одна или
две руки самца преобразованы для
передачи самке спсрматофоров. Во-
ронка имеет вид мускулистой (кроме
глубоководных видов) конической
трубки. На се спинной стенке нахо-
дится вороночный орган. У быстро
движущихся форм имеется также
вороночный клапан. Мантия сраста-
ется с головой и воронкой или под-
вижно сочленена с ними системой
хрящей. Клюв сильный, хорошо раз-
витый. Радула состоит из 7 продоль-
ных рядов зубов (у одного рода из
5), у некоторых планктоноядных,
преимущественно глубоководных
форм радула редуцирована или со-
всем отсутствует. Имеются одна па-
ра жабр, 2 предсердия, 2 почечных
мешка, 2 жаберных сердца; черниль-
ный мешок имеется или отсутствует.
Нервная система чрезвычайно высо-
ко развита, имеется концентрирован-
ный головной мозг, состоящий из
нескольких ганглиев и долей. Звезд-
чатый ганглий хорошо развит. У
нектонных форм имеется система ги-
гантских нервных волокон, иннерви-
рующих мантийные мышцы. Гонада
непарная. Мужские половые прото-
ки, как правило, непарные (в редких
случаях парные), женские половые
протоки могут быть как парными,
так и непарными. Яйца либо мелкие,
либо крупные, в первом случае раз-
витие с метаморфозом, во втором —
прямое. Морские и океанические
хищники, бентосные, нектонпыс,
планктонные, нектобентосные или
бентопелагичсские животные. Рас-
пространены во всех полносолепых
морях и океанах от Арктики до Ан-
тарктики и от литорали до абиссали.
Движение осуществляется разны-
ми способами: с помощью выброса
воды через воронку при резком со-
кращении мантии; с помощью муску-
листых плавников; с помощью ум-
бреллы — перепонки, соединяющей
руки (движение по способу медузы);
некоторые осьминоги могут «ходить»
на кончиках рук. Основная пища:
у нектонных видов рыба, макро-
планктоп, головоногие, у бентосных—
подвижная придонная фауна, у
планктонных и полуплапктонных ви-
дов— зоопланктон. Добычу находят
при помощи глаз или хемотактиль-
пого чувства. Чаще всего стайные
животные. Большинство видов рас-
тут быстро, созревают в возрасте
1—2 лет и погибают после первога
нереста.
2.1. Отряд Sepiida
Характеристику см. па с. 68.
2.1.1. Подотряд Spirulina—
спирулы, сем. SpirUlidae
Животные маленькие, длина мантии
до 4,5 см. Тело всретеновидное, валь-
коватос, кожа гладкая, но мантия:
под кожей шагринированная. Ман-
тия плотная, мускулистая, не сраста-
ется с головой, ее передний край на
спине с одним, на брюшной стороне
с двумя языковидными выступами,
между которыми выступает устье
воронки. Замыкательный аппарат
простой. Голова может втягиваться
в мантию у молодых спирул цели-
ком, а у взрослых наполовину. Руки
короткие, соединены широкой умб-
реллой. Присоски рук мелкие, при-
мерно равных размеров, сидят в не-
сколько рядов (до 6 у основания
рук). Щупальца длинные, тонкие,
частично втяжные, булава слабо
расширенная, присоски булавы мел-
кие, одинаковых размеров, сидят в
несколько рядов (до 12). Глаза
большие, эгопсидного типа — с от-
крытой передней камерой. Воронка
короткая, прикрыта краем мантии.
Плавники маленькие, округлые, си-
дят па заднем конце мантии почти
под прямым углом к продольной оси
тела и пе соединяются между собой.
Между ними расположен крупный
фотофор, окруженный кольцевой
складкой кожи. Окраска тела крас-
но-коричневая.
Раковина белая, известковая, фар-
форовидная, многокамерная, круглая
в поперечном сечении, похожая на:
«рог изобилия» — раскрученная спи-
раль примерно в 2!/г оборота, завер-
нутая на брюшную сторону живот-
ного. Лежит в задней половине ман-
тии и просвечивает на спинной и
брюшной сторонах (у старых спирул
кожа над раковиной иногда разры-
вается). Начальная камера шаровид-
ная, остальные — бочонковидные. Ка-
меры разделены перегородками (сеп-
тами), пронзенными у брюшного
73
Рис. 14. Строение самки Nautilus macromp-
halus: а — внешний вид; б — поперечный
разрез: /, 2—внешние и внутренние руки;
3 — чехлы, в которые втягиваются руки;
4 — капюшон; 5 —глаз; 6 — воронка; 7 —
радула; 8, 9 — верхняя (8) и нижняя (9)
мандибулы; 10 — мантия; 11 — мантийная
полость; 12 — нервные ганглии; 13 — си-
фон; 14 — септа; 15 — камерная жидкость;
16 — задний конец мантии; 17 — яичник;
18 — сердце; 19 — пищеварительная желе-
за; 20 — анус; 21 — жабра; 22 — глазное
щупальце; 23 — пупок (по Ward et al.,
1980)
74
Рис. 15. Сем. Nautilidae, род Nautilus', а —
г — N. pompilius: а — плывущим наутилус;
б и б| — схема расположения рук самца
(б) и самки (б[); в — распил раковины
(видны септы); г—раковина; д — N. tnac-
romphalus; е — N. stenomphalus; ж — N.
repertus; з — N. scrobiculalus (а—б — по
м
Nacf, 1921; о—в, з — по Sowcrby in Reeve,
1861, ж — по Ircdale, 1944)
75
края трубкой (сифоном). Раковина
служит гидростатическим аппаратом
и выдерживает давление до 200 атм.
Радула отсутствует. Развит толь-
ко левый яйцевод. У самца гектоко-
тилизируются обе брюшные руки,
они гораздо длиннее прочих, не со-
единены умбреллой, лишены присо-
сок и глубоко видоизменены: правая
желобовидная, вогнутая снаружи, на
конце с ложковидным расширением,
острым концом и двумя пальцевид-
ными выростами; левая округлая,
па конце с двумя ложковидными и
одним пальцевидным выростами и не-
сколькими мягкими шипами. Семе-
приемник находится па ротовой мем-
бране самки. Яйца донные, развитие
длительное без метаморфоза.
Один род — Spirula Lamarck,
1801, один вид—S. spirula (Linne,
1758) (рис. 16). Тропический атлан-
то-индовестпацифичсский мезопела-
гический нсрито-океанический вид.
Спирулы распространены над скло-
ном, чаще всего встречаются в водах
Индонезии, Меланезии, Юго-Восточ-
ной Африки, между Канарскими
островами и Северо-Западной Афри-
кой, в Карибском море и Мексикан-
ckoiM заливе. Пустые раковины раз-
носятся течением чрезвычайно дале-
ко от мест обитания. Животные
стайные. Обитают днем на глубинах
500—1000 м, ночью преимуществен-
но на 100—300 м. Яйца откладыва-
ются на дне в батиали, молодь пер-
вое время живет у дна. Нормальное
положение спирул при плавании го-
ловой вниз. Продолжительность жиз-
ни 1—1,5 года. Обзоры: Clarke, 1966,
1970; Bruun, 1943, 1955.
2.1.2. Подотряд Sepiina —
каракатицы
Раковина в виде толстой известко-
вой пластины, хитинового «пера» цли
отсутствует. Раковина лежит в тол-
ще мышц спинной стороны мантии от
переднего до заднего конца, снару-
жи незаметна. Мантия не срастается
с воронкой, но иногда срастается с
головой па затылке. Тело овальное,
слегка уплощенное в' спиппо-брюш-
ном направлении. Голова не втяги-
вается в мантию. Плавники либо
вдоль всей боковой поверхности ман-
тии в виде узкой каймы, либо сидят
на боковых сторонах мантии, их фор-
ма в этом случае полукруглая или
почковидная; плавники не соединя-
ются между собой сзади. Продоль-
ная ось плавников примерно парал-
лельна продольной оси тела. Наруж-
ных светящихся органов нет, иногда
имеются светящиеся органы внутри
мантии на чернильном мешке. Глаза
миопсидпого типа — с замкнутой пе-
редней камерой. Руки обычно корот-
кие со слабо развитой умбреллой
или без нес. Присоски никогда не
превращаются в крючья. Часто при-
соски на руках и булаве неодинако-
вых размеров. Щупальца втяжные,
обычно короткие и мощные. Гсктоко-
тилизируются одна, две или несколь-
ко пар рук. Радула хорошо развита,
ее зубцы одновершинные. Развит ле-
вый яйцевод. Яйца донные, развитие
обычно длительное без метаморфоза
и чаще всего без пелагической ста-
дии. Преимущественно донные жи-
вотные, несколько родов вторично
перешли к пелагическому образу
жизни.
4 семейства, 19 родов, 150—180
видов.
Таблица для определения семейств
подотряда Sepiina
1(2). Раковина в виде толстой оваль-
ной или ромбической известковой
пластины. Тело массивное, голова
намного меньше мантии. Мантия не
срастается с головой на затылке, ее
передний край на спинной стороне с
языковидным выступом. Плавники
узкие, идут по бокам вдоль всей ее
длины.
Сем. Scpiidae .... с. 77.
2(1). Раковина рудиментарная
или отсутствует. Мантия небольшая,
голова часто ненамного меньше ман-
тии. Передний край мантии на спин-
ной стороне без языковидного высту-
па, иногда срастается с головой.
Плавники полукруглой или почко-
видной формы, сидят посредине ман-
тии или ближе к заднему концу . 3.
3(4). Мантия удлиненная, слегка
суженная и приостренная к заднему
концу. Плавники в задней половине
мантии. Передний край мантии не
срастается с головой. Па спинной
стороне мантии в се задней полови-
не имеется овальный шероховатый
участок — прикрепительный орган,
служащий для удержания моллюска
76
на субстрате. Гектокотилизируются
обе брюшные руки.. Очень маленькие
животные, длина мантии нс более
15 мм.
Сем. Idioscpiidac . . . с. 127.
4(3). Мантия овальная, сзади
широко закругленная. Плавники си-
дят посредине или чуть дальше сере-
дины мантии. Нет прикрепительного
органа. Гсктокотилизирустся либо
одна из брюшных рук, либо руки
иных пар.........................5.
5(6). Умбрелла между руками
3-й и 4-й пар окружает основание
щупалец снаружи и изнутри, об-
разуя своего рода кожный чехол.
Плавники с широкими основаниями
и слабо развитыми «ушками». Ман-
тия срастается с головой на затылке.
Гектокотилизирустся левая брюшная
рука.
Сем. Sepiadariidae . . . с. 126.
6(5). Умбрелла между руками
3-й и 4-й пар охватывает основание
щупалец только снаружи, не образуя
чехла. Плавники с короткими осно-
ваниями и хорошо развитыми «ушка-
ми», длина плавника заметно "боль-
ше длины его основания. Гсктокоти-
лизируются обе спинные руки, иног-
да видоизменяются и руки боковых
пар. У некоторых видов гсктокотили-
зация слабо заметна.
Сем. Sepiolidae .... с. 106.
2.1.2.1. Семейство Sepiidae —
настоящие каракатицы
Тело уплощенное в спинно-брюшном
направлении. Мантия удлиненно-
овальная, много крупнее головы.
Спинной край мантии спереди с язы-
ковидным выступом. Плавники уз-
кие, протягиваются от переднего до
заднего края мантии, нс соединяют-
ся между собой. Раковина (сепион)
(рис. 17, а) лежит под кожей на
спинной стороне мантии от передне-
го до заднего ее края и представля-
ет собой толстую овальную, ланцето-
видную или ромбическую известко-
вую пластину иногда с шипом (рост-
ром) на заднем конце. В задней ча-
сти сепиона снизу видно множество
топких узких, косо идущих перегоро-
док — септ, создающих впечатление
поперечной исчерченности сепиона,
эта часть сепиона называется фраг-
моконом. Септы поддерживаются гу-
стой сетью крохотных поперечных
известковых столбиков. Периодич-
ность образования септ зависит от
77
температуры, в тропиках они обра-
зуются ежедневно, так что но числу
поперечных полосок можно опреде-
лить возраст каракатицы; самая но-
вая септа — самая передняя. Нижняя
поверхность сепиона в задней поло-
вине вогнута и образует с боковыми
краями раковины «вилку» (внутрен-
ний конус). Раковина окаймлена бо-
ковым кантом, широким в задней и
узким в передней половине сепиона.
Глаза миопсидпого типа с замк-
нутой передней камерой. Руки с 2—
4 рядами присосок. Руки 4-й пары
значительно шире остальных, с мус-
кулистой плавательной мембраной.
Гсктокотилизнрустся левая брюшная
рука. Щупальца втяжные, булава с
4—6 или большим числом присосок
иногда резко неравных размеров.
Чернильный мешок хорошо развит.
Все зубцы радулы одновершинные.
Донные животные, обитатели
шельфа, иногда встречаются в верх-
ней батиали. Активны по ночам, днем
обычно сидят на дне, иногда забра-
сывают себя грунтом. 2 рода, 90—
ПО видов. Обзоры: Adam, Rees, 1965;
Adam, 1979; Roeleveld, 1972.
Таблица для определения родов и
подродов семейства Sepiidae
1(6). В задней части мантии на
спинной стороне нет крупной под-
кожной железы. Плавники сверху
без ярких цветных пятен. Внутрен-
ний конус раковины хорошо развит.
Мантийный хрящ овальный (рис.
17, б).
Род Sepia Linne, 1758. Типовой
вид — S. officinalis Linne, 1758 . 2.
2(3). Раковина толстая, полно-
стью обызвествленная, спереди за-
кругленная, иногда сзади вытянута
в шип, форма раковины языковид-
ная или ланцетовидная. Длина рако-
вины почти равна длине мантии.
Передний край мантии па спинной
стороне обычно с большим языко-
видным выступом. На брюшной сто-
роне мантии нет кожных пор.
Подрод Sepia s. str.(=Acanthose-
pion Rochebrunc, 1884; Ascarosepion
Rochebrune, 1884; Doratosepion Roche-
brunc, 1884; Lophosepion Rochebrune,
1884; Rhombosepion Rochebrunc,
1884; S pat hidose pion Rochebrune,
1884; Andreasepia Grimpe, 1922; Pa-
rasepia Naef, 1923; Platysepia Naef,
1923; Eusepia Naef, 1923; Amplisepia
78
Iredale, 1926; Arctosepia Iredale,
1926; Crumenasepia Iredale, 1926; De-
corisepia Iredale, 1926; Eiscisepia Ire-
dale, 1926; Glyptosepia Iredale, 1926;
Mesembrisepia Iredale, 1926; Ponderi-
sepia Iredale, 1926; Solitosepia Iredale,
1926; Tenuisepia Cotton, 1932; Blando-
sepia Iredale, 1940). 78—100 видов.
Крупные или средних размеров, реже
мелкие каракатицы. Распространены в
Восточной Атлантике от Северного
моря до Южной Африки и в Индо-
Вестнацификс от Красного моря до
Южной Австралии и Японских ост-
ровов, па восток до Маршалловых
островов и Фиджи. Раковины далеко
разносятся течениями. На берегах
Южного Приморья и Южных Ку-
рильских островов (СССР) иногда
попадаются раковины каракатиц, за-
несенные течениями от Японских ост-
ровов, а в Карибском море и Мекси-
канском заливе — от Африки.
3(2). Раковина приострена спере-
ди и расширена или заострена сзади,
короче мантии. Передний край ман-
тии на спинной стороне прямой или
слабо выгнутый без языковидного
выступа. Животные мелкие, длина
мантии 2—7 см...................4.
4(5). Раковина заметно короче
мантии, обычно -75—85% ее длины,
ромбическая, тупо заостренная спе-
реди и угловатая (реже округлен-
ная) на боках. Мантия необычно
толстая, плавники сидят заметно
выше средней линии боковой сторо-
ны. Присоски булавы резко нерав-
ных размеров расположены в 5—6
продольных рядов, присоски рук —
в 4 ряда.
Подрод Metasepia Hoyle, 1885
(рис. 17,г—и). Типовой вид — S.(M.)
pfefferi Hoyle, 1885. 2 вида в Индо-
малайской подобласти Ипдо-Вестпа-
цификп.
а (б). Раковина тупо заострена на
заднем конце, без шипа или с хити-
новым (не обызвествленным) шипом.
На булаве 3—4 средние присоски
громадные, занимающие всю середи-
ну булавы. Голова и мантия гладкие.
S. (Af.) pfefferi Hoyle, 1885 (рис.
17, г—е). Западная, Северная и Севе-
ро-Восточная Австралия, на юге до
Фримантла и островов Каприкорн.
Длина мантии до 6 см.
б(а). Раковина сзади оттянута в
острый шипик, на дорсальной сторо-
не которого сидит маленькая хитино-
вая пластиночка. На булаве 7—9 уве-
Sepiidae, род Sepia: a — ос-
Рис. 17. Сем. _____ a .
новные морфологические особенности рако-
вины Sepia на примере S. joubini (вид
снизу): 1 — фрагмокон; 2, 3 — гладкая (2)
и исчерченная (3) зоны фрагмокона; 4 —
продольный желобок; 5 — ветви внутренне-
го конуса; 6 — крылья внешнего конуса;
7 — внутренний конус (вилка); 8 — шигг
(ростр); б, в — мантийный хрящ Sepia (б,
S. australis) и Sepiella (в, S. суапеа); г—
e — S. (Metasepia) pfefferi; ж—u — S. (М.}
tullbergi; к—л — S. (Hetnisepius) typica;
м — S. (Я.) dubia; д, з —булава; е, и, лг
м — раковина (а—в, к—л — но Roeleveld,
1972; г-д — по Hoyle, 1886; с— no Iredale,.
1926; etc— по Voss, Williamson, 1972; з, и —
no Sasaki, 1929; л« — no Adam, Rees, 1966>
-лнченпых присосок; 4—5 присосок
второго (с дорсальной стороны) и
•3—4 третьего ряда. Голова, спинная
и боковая стороны мантии живых
каракатиц покрыты буграми, боро-
давками и треугольными выступами
кожи.
S. (М) tullbergi Appellof, 1886
(рис. 17,ж—и). От Южного Хонсю
до о-ва Тайвань и Гонконга (шельф).
Длина мантии до 7 см.
5(4). Раковина ненамного короче
мантии (85—90% ее длины), сужен-
ная спереди и сильно расширенная и
округленная сзади, очень топкая,
слабо кальцифицированная, без ши-
на. Мантия обычной толщины, плав-
ники обычного положения — посре-
дине боков. Присоски на булаве щу-
пальца одинаковых размеров. При-
соски рук в 2 ряда..
Подрод Hemisepius Steenstrup,
1875 (=Hemisepion Rochebrune,
1884) (рис. 17, к—л). Типовой вид —
5. (Н.) typica (Steenstrup, 1875).
2 вида. Южная Африка. Длина ман-
тии до 2—3 см.
а (б). На вентральной стороне
мантии в передней половине вдоль
основания плавников ряд приблизи-
тельно из 12 кожных пор. Раковина
с дорсальной стороны хитинизирова-
на, внутренний конус слабо выражен.
S. (Н.) typica (Steenstrup, 1875)
<рис. 17,к—л). Южная Африка, от
залива Салданья до Дурбана
(шельф).
б (а). Нет кожных пор на вен-
тральной стороне мантии. Раковина
•с дорсальной стороны обызвествлена,
внутренний конус хорошо выражен.
S. (Н,) dubia Adam et Rees, 1966
(рис. 17, м). Мыс Доброй Надежды.
6(1). В задней части мантии на
спинной стороне под кожей лежит
крупная железа, проток которой от-
крывается между плавниками в виде
.красного пятна. Вдоль основания
плавников протягивается ряд из
нескольких крупных ярких цветных
пятен. Раковина узко-языковидная,
•без шипа, внутренний конус редуци-
рован, внешний представляет собой
широкую хитинизироваппую (пе-
юбызвествлепную) оторочку вокруг
заднего конца раковины. Присоски
булавы одинаковых размеров, мел-
кие. Мантийный хрящ треугольный
(рис. 17, в).
Род Sepiella Gray, 1849 (=Dipht-
herosepion Rochebrune, 1884). Типо-
S0
вой вид — S. ornata (Rang, 1837).
6—7 видов. У Западной и Южной
Африки и в Индо-Вестпацифике. См.
таблицу для определения видов.
Таблица для определения видов рода
Sepia отдельных районов Мирового
океана
I. Восточная Атлантика (от Северно-
го моря до Анголы, включая Среди-
земное море)
1(8). Присоски на булаве заметно
неравных размеров. Раковина с ши-
пом (род Sepia, подрод Sepia
s. str).........................2.
2(3). Плавники начинаются непо-
средственно у переднего края ман-
тии и заметно заходят вперед него,
достигая уровня заднего края глаз;
в задней части мантии плавники
расширяются. Присоски рук в 4 ря-
да. На булаве 5 поперечных рядов
присосок, 5—7 присосок среднего
ряда заметно крупнее прочих, но
бросающейся в глаза разницы между
громадными средними и маленькими
остальными присосками нет. Шири-
на раковины у самцов 30—40 %,
у самок 33—42 % ее длины. Задняя
часть раковины расширена, края
внешнего конуса широкие, иногда
очертания раковины слабо вогнутые
с боков.
<S. (S.) officinalis Linne, 1756
(рис. 18, а—г). От Шетландских ост-
ровов, Южной Норвегии (заходы в
теплые годы) и южной части Север-
ного моря до мыса Доброй Надеж-
ды и вдоль юго-восточного побе-
режья Африки до Мозамбика. Весь
шельф, главным образом верхняя
сублитораль. 4 подвида: S. о. offici-
nalus L. (=5. filliouxi Lafont,
1868)—от Северного моря до мыса
Зеленый, длина мантии до 27 см;
S. о, mediterranea Ninni, 1884 —Сре-
диземное и Адриатическое моря,
длина мантии до 30 см; S. о. hierred-
da Rang, 1837 — побережье За-
падной Африки от мыса Кап-Блан
до Южной Анголы, длина мантии до
50 см (масса до 9—12 кг); 5. о. ver-
miculata Quoy et Gaimard, 1832—
Южная Африка от устья р. Оли-
фанте до Мозамбика (залив Дела-
гоа), раковины — до устья Замбези,
длина мантии до 22—24 см (редко
29 см). Важнейший промысловый
вид; добывается от Бискайского за-
лива до Западной Африки и в Сре-
диземноморском бассейне.
3(2). Плавники начинаются не-
много отступя от переднего края
мантии и не заходят за линию ее пе-
реднего обреза, пе расширяются в
задней части. Ширина раковины у
самцов 23—33%, у самок 26—35%
ее длины, задняя часть раковины не
расширена.......................4.
4(5). Присоски в базальных ча-
стях рук в 2 ряда. В центральной
части булавы 3 громадные присос-
ки, занимающие всю со середину.
Раковина сильно сужается к заднему
концу, шип очень /малёщ>кий — ско-
рее короткий известковой гребень,
чем шип. \ /'
S. (S.) elegans d’Ofbigny, 1826
(рис. 18, д—з). От Западной Шот-
ландии и Ла-Манша (раковины
встречаются в СевернОхМ море) до
Южной Анголы, все Средиземное
море. Внешний шельф и самая верх-
няя батиаль. Длина мантии до 9 см.
5(4). Присоски рук в 4 ряда.
Шип раковины хорошо развит, у
фиксированных каракатиц часто про-
тыкает мантию и торчит наружу . 6.
6(7). В центральной части була-
вы 5 очень крупных присосок, самая
большая — средняя. Раковина плав-
но сужается к переднему и заднему
концам, ее ширина у самцов около
33%, у самок 33—35% длины, спин-
ная сторона розоватого или бледно-
оранжевого цвета со слабо выра-
женным продольным желобком.
S. (S.) orbignyana Ferussac, 1826
(рис. 18, и—м). От Ирландского мо-
ря и Ла-Манша (раковины встреча-
ются в Северном море) до Южной
Анголы, все Средиземное море.
Шельф (преимущественно внешний)
и самая верхняя батиаль. Длина
мантии до 14 см. Второстепенный
промысловый вид.
7(6). Присоски плавно увеличи-
ваются от боковых краев булавы к
третьему (считая с дорсальной сто-
роны) ряду, резко увеличенных при-
сосок пет. Раковина плавно сужает-
ся в передней половине, в задней ее
края почти параллельны. Ширина
раковины самцов 23—30%, самок
26—33%длины, спинная сторона бе-
лого цвета с округленным продоль-
ным килем.
S. (S.) bertheloti d’Orbigny, 1838
(=S. mercatoris Adam, 1937) (рис.
18, н—n). От Канарских островов и
Западной Сахары до Анголы. Шельф.
Длина мантии до 18 см. Добывается
в незначительных количествах.
8(1). Присоски на булаве мелкие,
почти одинаковых размеров. Ракови-
на без шипа, сзади округленная. По-
верхность головы и мантии шерохо-
ватая, бугорчатая. На булаве 8 ко-
сых рядов присосок. Ширина рако-
вины 38—46% се длины, раковина
быстро сужается к переднему концу,
в задней половине ее боковые сто-
роны параллельны. Края внешнего
конуса очень широкие.
S. (S.) elobyana Adam, 1941 (рис.
18, р—т). От мыса Кап-Блан до Га-
бона. Длина мантии до 5 см. Очень
редкий вид.
II. Южная Африка от Нами-
бии до Южного Мозамбика
1(16). Присоски булавы приблизи-
тельно одинаковых размеров . . 2.
2(9). Мантия спереди на спинной
стороне выдается вперед треуголь-
ным выступом, прикрывая заднюю
часть головы. Присоски булавы в 8
продольных (косых) рядов . . 3.
3(6). Руки 1—2-й пар с 2 рядами
присосок. Раковина без шипа, при-
остренная спереди.............4.
4(5). Передний край маптии на
брюшной стороне с вырезкой у осно-
вания воронки. Раковина сильно за-
острена спереди и угловато-округлая
сзади, края внутреннего конуса уп-
лощены и прилегают к поверхности
фрагмокона.
S. (S.) insignis Smith, 1916 (рис.
19, а). От мыса Доброй Надежды до
Наталя. Внутренний шельф. Длина
мантии до 6 см.
5(4). Передний край мантии на
брюшной стороне слабо вогнут. Ра-
ковина угловато-округлая спереди и
плавно округленная сзади, края
внутреннего конуса вздуты и в виде
валика окаймляют заднебоковые
края фрагмокона.
S. (S.) hieronis (Robson, 1924)
(рис. 19, б—в). Известна у западно-
го побережья ЮАР от 29° ю. ш. до
мыса Доброй Надежды, у Наталя и
самой южной части Мозамбика.
Шельф (преимущественно внешний)
и верхняя батиаль, в основном- па
глубинах 200—450 м. Длина мантии
до 7 см. Каракатицы с западного и
восточного побережья Южной Афри-
ки заметно различаются по форме
раковины.
6 к. н. Несис
81
82
6*
83
Рис. 19. Род Sepia s. str., южноафрикан-
ские виды: а — S. insignis-, б—в — S. hiero-
nis; г—е — S. faurei; ж—з — S. papillata;
u — S. tuberculata; к — S. australis, вскры-
тая самка, виден предположительный све-
тящийся орган (с. о.); л — S. confusa,
взрослый самец с «хвостом»; м—с — S. Ьиг-
. nupi; т—у — S. incerta; ф—х — S. adami;
а, е, з, и, я, х — булава; в, о — гектоко-
тиль; р, т — конец 1-й правой руки самца;
п — конец 4-й правой руки самца; // — зад-
ний конец раковины (по Roeleveld, 1972)
6(3). Руки с 4 рядами присосок.
Раковина овальная с шипом . . 7.
7(8). Ротовая мембрана с не-
сколькими мелкими присосочками.
Раковина плавно округленная спере-
ди и суженная сзади.
S. (S.) zanzibarica Pfeffer, 1884.
См. с. 91.
8(7). Ротовая мембрана без при-
сосочек. Раковина суженная, слегка
приостренпая спереди и плавно окру-
гленная сзади.
3. (S.) acuminata Smith, 1916. На-
таль, Южный Мозамбик, Кения.
Внешний шельф и самая верхняя ба-
тиаль. Длина мантии до 11 см.
9(2). Мантия спереди на спинной
стороне плавно выгнута без выступа.
Присоски булавы не в 8 рядов. Ра-
ковина без шипа или с бугорком на
месте шипа.....................10.
10(11). Булава очень длинная, за-
нимает почти половину щупальца.
Присоски булавы приблизительно в
24 продольных ряда. Мантия на вен-
тральной стороне спереди без выре-
за, плавно вогнута. Присоски рук в
4 ряда. Раковина имеет форму пра-
вильного овала, край внутреннего
конуса очень широкий сзади и резко
сужается впереди. На заднем конце
раковины тупой бугорок.
S. (S.) sitnoniana Thiele, 1920
(=S. natalensis Massy, 1925). От мы-
са Доброй Надежды до Наталя, ра-
ковины находят на севере до района
севернее устья, р. Олифанте. Шельф
преимущественно внутренний. Мно-
гочисленный вид. Длина мантии до
18,5 см.
И(10). Булава короткая, при-
соски булавы в 4—6 рядов. Мантия
на вентральной стороне спереди с вы-
резкой у основания воронки. При-
соски рук в 2 ряда. Раковина не
овальная, без бугорка на заднем кон-
це ............................12.
12(15). Концы рук 1-й пары от-
тянуты, пальцевидные, лишены при-
сосок. Раковина хитиновая или
обызвествленная................13.
13(14). На мантии и голове свер-
ху нет (или лишь немного) папилл.
Раковина спереди сильно заострена,
не обызвествлена (?).
S.(S.) robsoni (Massy, 1927).
V мыса Доброй Надежды. Верхняя
сублитораль. Длина мантии 2 см.
14(13). Спинная сторона мантии,
головы и основания рук густо по-
крыты мелкими кожными папиллами.
Раковина очень топкая, пе обызве-
ствлена, широко овальная, с при-
острепным передним и задним (?)
концами, передний конец фрагмокона
выпуклый.
3.(3.) faurei Roeleveld, 1972
(рис. 19,a—е). Южная Африка,
восточнее мыса Доброй Надежды.
Глубина 170 м. Длина мантии 2 см.
15(12). Концы рук 1-й пары нор-
мальные, присоски идут до концов
РУК.
Подрод Hetnisepius: S. (Н.) typica
(Stp.) и 3. (Н.) dubia Adam et Rees
(см. с. 80. и рис. 17, к—м).
16(1). Присоски булавы резко
неравных размеров.............17.
17(20). Поверхность мантии свер-
ху покрыта бугорками и бородавка-
ми (камуфляж). На брюшной сторо-
не мантии 2 крупных овальных пят-
на с морщинистой кожей, 2 полосы
такой же кожи тянутся вдоль вент-
ральной стороны брюшных рук (при-
способление для прикрепления к кам-
ням на прибое). Раковина без шипа,
иногда с маленьким тупым бугорком
сзади.........................18.
18(21). На булаве 3—4 резко уве-
личенные присоски, занимающие всю
середину булавы. Защитные мембра-
ны булавы не соединяются в кар-
пальной части. Концы рук 1—3-й пар
пе утончены. Вентральная сторона
раковины выпуклая, с продольным
желобком, толщина раковины 8—
12% ее длины.
S. (3.) papillata Quoy ct Gaimard,
1832 (рис. 19, ж—з), От залива Лю-
дериц-бей до Наталя. Внутренний
шельф. Многочисленный вид. Длина
мантии до 13,5 см.
19(18). На булаве 5 увеличенных
присосок, но размеры самых круп-
ных из них много меньше ширины
булавы и они не занимают всю ее
середину. Защитные мембраны бу-
лавы соединяются в карпальной ча-
сти. Концы рук 1—3-й пар утончены
и оттянуты. * Вентральная сторона
раковины плоская или вогнутая без
продольного желобка, толщина рако-
вины 4—9% ее длины.
3. (S.) tuberculata Lamarck, 1798
(рис. 19, и). Южная Африка от зали-
ва Салданья до Книсны. Раковины
встречаются до Мадагаскара. Лито-
раль и прибрежные мелководья. Дли-
на мантии до 8 см.
20(19). Поверхность мантии без
бугорков и бородавок. Нет пятен с
85
морщинистой кожей на брюшной сто-
роне мантии. Раковина с шипом . 21.
21 (24). Раковина овальная или
овально-ланцетовидная, ее ширина
29—46% длины. Края внешнего ко-
нуса имеют плавные очертания без
выступающих «ушек».. 5 центральных
присосок булавы значительно круп-
нее прочих....................22.
22(23). В средней части гектоко-
тилизированной руки 9—12 попереч-
ных рядов из 4 уменьшенных в раз-
мерах присосок. Раковина овальная,
внутренний конус хорошо развит и
срастается с внешним, края внешне-
го конуса широкие; ширина ракови-
ны 32—46% ее длины.
S. (S.) officinalis vermiculaia Quoy
et Gaimard, 1832 (см. с. 80). Весьма
многочисленный вид на мелководьях
(литораль, бухты, эстуарии и шельф)
Юж ной. Африки.
23(22). В средней части гектоко-
тилизированной руки 6—7 попереч-
ных рядов уменьшенных присосок,
причем вентральные пары присосок
перестраиваются в один продольный
ряд. Раковина овально-ланцетовид-
ная, суживается назад, ветви внут-
реннего конуса очень узкие, края
внешнего конуса узкие. Внутри ман-
тийной полости на чернильном мешке
непарный почковидный оргап неиз-
вестной функции, предположительно
фотофор (уникальное для сепиид об-
разование).
S. (S.) australis Quoyet Gaimard,
1832 (=S. capensis d’Orbigny, 1845)
(рис. 19, к). От устья р. Олифанте до
южной границы Наталя, банка
Агульяс. Отмечена (с сомнением) в
Красном море. Весь шельф и верх-
няя батиаль, преимущественно глу-
бины менее 250 м. Длина мантии до
6 см. Самый массовый вид южно-
африканских каракатиц.
24(23). Раковина ланцетовидная,
сильно суживается назад, се шири-
на 14—26% длины. Края внешнего
конуса образуют изгиб — «ушки» —
в задней части раковины ... 25.
25(32). Руки 2—3-й пар у самок
с утонченными и оттянутыми конца-
ми ...........................26.
26(29). Руки 1-й пары у самцов
нормальные с 4 рядами присосок.
У самок утончены и оттянуты дис-
тальные половины рук 2—3-й пар,
они несут 2 ряда присосок . . 27.
27(29). Плавники самца нормаль-
ные, не вытянуты в хвост. У самок
86
на конце рук 1-й пары защитные
мембраны сильно расширены и об-
кручены вокруг оральной поверхно-
сти руки. Задняя часть внутреннего
конуса раковины низкая, его ветви
прижаты к задне-боковым сторонам
фрагмокона.
S. (S.) joubini Massy, 1927. На-
таль. Шельф. Длина мантии до 5 см.
28(27). Плавники взрослых сам-
цов сзади срощепы и вытянуты в
длинный (иногда длиннее мантии)
хвост, у самок плавники нормаль-
ные. Защитные мембраны на концах
рук 1-й пары у самок не расширены.
Задняя часть внутреннего конуса ра-
ковины округленная, приподнятая,
без продольной борозды, образует
глубокую ямку над задним концом
фрагмокона.
S. (S.) confusa Smith, 1916 (рис.
19,л). От Занзибара до Наталя. Ра-
ковины встречаются па юге до райо-
на Порт-Элизабет. Отмечена на бан-
ке Сая-дс-Малья. Внешний шельф и
самая верхняя батиаль. Длина ман-
тии (без хвоста) до 15 см.
29(26). Руки 1-й пары у самцов
резко видоизменены. Руки 2—3-й пар
у самок утончены и оттянуты толь-
ко на концах и несут 4 ряда присо-
сок ...........................30.
30(31). Руки 1-й пары у самца
расширены и несут 2 ряда выростов
(длинные копьевидные на внутренней
и короткие конические на внешней
стороне). Присоски в дистальной по-
ловине рук 1-й пары в 2 ряда. Гек-
токотилизированы обе брюшные ру-
ки, левая длиннее правой, обе несут
на концах ряд пальцевидных вырос-
тов. Плавники самцов на заднем кон-
це расширены и налегают друг на
друга. Задняя часть внутреннего ко-
нуса раковины низкая. Полоски ис-
черченной зоны фрагмокона М-образ-
пыс, угловатые.
S. (S.) burnupi Hoyle, 1904 (=5.
exsignata Barnard, 1962) (рис.
19, м~с). Наталь. Внутренний шельф.
Длина мантии до 6 см.
31(30). Руки 1-й пары самца
сильно удлинены, хлыстовидные на
концах, несут присоски только в ба-
зальной части, в средней части обе
защитные мембраны расширены и со-
единены, образуя овальное расшире-
ние. Брюшные руки самца нс видо-
изменены, плавники нормальные.
Задняя часть внутреннего конуса ра-
ковины приподнята и разделена на-
двое глубокой продольной бороздой.
Полоски исчерченной зоны фрагмо-
кона А-образные, спереди округлен-
ные.
5. (3.) incerta Smith, 1916 (рис.
19, т—у). Восточное побережье Кап-
ской провинции и Наталь. Внутрен-
ний шельф. Длина мантии до 15 см.
32(25). Руки 2—3-й пар у самок
с не утонченными и пе оттянутыми
концами, несут 4 ряда присосок. Ра-
ковина, как у 5. joubini. Самцы не
описаны.
3. (S.) adami Roeleveld, 1972 (рис.
19,0—х). Наталь. Глубина 100 м.
Длина мантии до 6 см.
Примечание. В таблицу пе
включена известная только по рако-
винам 3. (S.) angulata Roeleveld,
1972. Раковина правильной овальной
формы, чуть суженная и угловатая
спереди и округленная сзади, очень
широкая (50—60% длины), упло-
щенная, без шипа, края внутреннего
конуса плоские, широкие, ” слиты с
внешним конусом, края последнего
очень широкие. Мыс Доброй Надеж-
ды—-мыс Игольный. Длина раковин
до 7,5 см.
III. Северная часть Индийского оке-
ана, воды Индонезии и Инд о китай-
ское о полуострова, Красное и Ара-
вийское моря, Персидский залив и
Восточная Африка до Мозамбика1
1(38). Раковина с шипом или хотя
бы маленьким бугорком на заднем
крае . . . *..................2.
2(21). Присоски булавы резко не-
равных размеров...............3.
3(14). Раковина широкая (30—
40% длины), овальная .... 4.
4(13). Присоски булавы посте-
пенно увеличиваются к среднему ря-
ду, в котором 5—7 присосок значи-
тельно крупнее прочих. Нет бросаю-
щейся в глаза разницы между гро-
мадными средними и маленькими
остальными присосками .... 5.
1 В таблицу нс включены: южноаф-
риканская 3. (3.) australis, отмечен-
ная (с сомнением) в Красном морс—
см. с. 86; доходящая до Занзибара
3. (S.) confusa (см. с. 86); сино-
янонские 3. (S.) lycidas и 3. (S.) es-
culenta, доходящие на юге до Филип-
пинских островов и Саравака — см.
с. 94, и недавно описанная 5.(3.) iva-
novi Khromov, 1982.
5(6). Защитные мембраны була-
вы в карпальной части идут парал-
лельно и закапчиваются на стебле
щупальца, пе соединяясь друг с
другом. Внутренний конус раковины
у взрослых особей прикрыт наплы-
вом темного цвета.
3. (S.) pharaonis Ehrenberg, 1831
( = S. torosa Ortmann, 1838, 5. rouxii
Ferussac et d’Orbigny, 1841,
3. formosana Berry, 1912, Crumena-
sepia hulliana Iredale, 1926, C. ursu-
lae Cotton, 1929, 3. tigris Sasaki,
1929) (рис. 20, a—e). От южной части
Суэцкого канала и Суэцкого залива
до Занзибара, Мадагаскара, Южно-
го Хонсю, Восточной Индонезии, За-
падной и Северо-Восточной Австра-
лии. Раковины попадаются на остро-
вах Каролинских, Лорд-Хау и Новая
Каледония. Внутренний шельф. Дли-
на маптии до 42 см, масса до 5 кг.
Важнейший промысловый вид кара-
катиц северной части Индийского
океана и Юго-Восточной Азии, до-
бывается от побережья Аравии до
КНР и о-ва Тайвань.
6(5). Защитные мембраны була-
вы срастаются в ее карпальной части
и отделяют несущую присоски по-
верхность от стебля. Внутренний ко-
нус нс закрыт темным наплывом . 7.
7.(8). Шип редуцирован до кро-
хотного тупого бугорка в задней ча-
сти раковины. Раковина овальная, ее
спинная сторона с сетчатой (ячеи-
стой) скульптурой.
3. (S.) bandensis Adam, 1939. Ява,
Восточная Индонезия, Филиппинс-
кие острова, Новая Гвинея, Маршал-
ловы острова, Прибрежные мелко-
водья. Длина маптии до 7 см.
8(7). Шип раковины хорошо раз-
вит, его длина 3—4% длины ракови-
ны. Спинная сторона раковины мор-
щинистая или гранулированная, без
сетчатой (ячеистой) скульптуры . 9.
9(12). Дорсальная сторона рук
1—3-й пар без бугорков. Нет «фун-
тика» на вентральной стороне задне-
го края раковины. Шип раковины в
сечении округлый...............10.
10(11). Передний край раковииы
плавно закруглен. Шип прямой.
5. (5J latimanus Quoy et Gaimard,
1832 (-S. hercules Pilsbry, 1894, Pon-
derisepia eclogaria Iredale, 1926, 5.
harmeri Robson, 1928) (рис. 20, г—e).
От Сингапурского пролива до Юж-
ного Кюсю, Северо-Западной и Се-
веро-Восточной Австралии, на во-
87
Рис. 20. Род Sepia s. str., нндовестпацифи-
ческие виды, 1: а—в — S. pharaonis', г—е —
S. latimanus; ж—з— S. recurvirosiris; u—
л — S. papuensis; м—п — S. omani; р—т —
S. kobiensis*, 6, dt 3, К, н, с —булава; о —
гектокотиль; в, е, л, п, г —раковина (а—б,
ж—з, м—п — по Voss, Williamson, 1972;
в — по Okutani, 1973; е, е — по Sasaki,
1929; д, и—к, р—с —по G. Voss, 1963; л,
т — по Hoyle, 1886).
-сток до Палау, Новой Каледонии и
Фиджи (указана с сомнением для
Мадагаскара и Южной Австралии).
Внутренний шельф, коралловые ри-
фы. Один из самых крупных видов
каракатиц, длина мантии до 50 см,
масса до 10 кг. Добывается в не-
больших количествах па островах
Рюкю, в КНР, на о-ве Тайвань и в
водах Индокитайского полуострова.
11(10). Передний край раковины
приостреп, округло-треугольный.
Шип загнут вверх.
S. (3.) recurvirostris Steenstrup,
1875 (=S. singaporensis Pfeffer, 1884)
(рис. 20,ас—з). Бирма, Западная Ин-
донезия, Индокитайский полуостров,
Южно- и Восточно-Китайское моря,
Филиппинские острова. Внешний
шельф. Длина мантии до 17 см. До-
бывается в небольшом количестве
в Южно-Китайском море и водах
Индокитайского полуострова.
12(9). Дорсальная сторона рук
1—3-й пар с разбросанными бугор-
ками, которые придают продольным
килям рук вид зубчатых гребней. Ра-
ковина спереди плавно закругленная.
Вентральный край внешнего конуса
загнут внутрь и образует «фунтик»
на заднем конце раковины. Шип пря-
мой с продольным. килем на вент-
ральной стороне.
3. (3.) papuensis Hoyle, 1885
(=3. galei Meyer, 1909, 3. prionota
Nqss, 1962, Solitosepia submestus Ire-
dale, 1926, 3. occidua Cotton, 1929?,
-3. genista Iredalc, 1954?, 5. lana Ire-
dale. 1954) (рис. 20, и—л). Филип-
пинские острова, Индонезия, Запад-
ная Австралия на юг до Фримантла,
Арафурское и Коралловое моря.
Шельф. Длина мантии до 7—11 см.
13(4). Вся центральная часть бу-
лавы занята 2 громадными и 2 не-
сколько меньшими присосками. За-
щитные мембраны булавы сближа-
ются в карпальной части по не со-
единяются. Передняя часть ракови-
ны сужена и закруглена.
3. (S.) prashadi Winckworth, 1936.
•От Красного моря и Оманского за-
лива до Мадагаскара, Маврикия и
Восточной Индии. Внешний шельф.
Довольно многочисленна в Красном
тг Аравийском морях. Местами добы-
вается. Длина мантии до 14 см.
14(3). Ширина раковины 15—30%
-ее длины................... . 15.
15(16). Раковина удлиненно-оваль-
ная, ее ширина 25—28% длины,
D0
плавно закругляющаяся к переднему
и заднему концу, место наибольшей
ширины — середина раковины. Була-
ва с 3—5 очень крупными присоска-
ми.
3. (3.) omani Adam ct Rees, 1966
(рис. 20,м—л). Известна в Оман-
ском заливе и Южно-Китайском мо-
ре у Гонконга. Глубины 50—200 м.
Длина мантии до 10 см.
16(15). Раковина узкая, ее шири-
на 15—25%, редко до 29 % длины,
ланцетовидная, приостренная к пе-
реднему и особенно заднему концу,
боковые стороны слегка угловатые,
место наибольшей ширины сдвинуто
в переднюю часть раковины. Присос-
ки булавы плавно увеличиваются к
среднему ряду, 5 центральных присо-
сок обычно самые крупные . . 17.
17(18). На гектокотилизированпой
руке самца 6—12 проксимальных по-
перечных рядов нормальных присо-
сок, затем 7—9 поперечных рядов или
пар резко уменьшенных присосок и йа
самом конце — нормальные, постепен-
но мельчающие присоски. Нет расши-
ренной боковой мембраны в средней
части руки, или она слабо развита и
представляет собой плавно изгибаю-
щуюся дугу.
3. (S.) kobiensis Hoyle, 1885 (=5.
andreanoides Hoyle, 1885) (рис.
20, p—т). По-видимому, полиморф-
ный вид или комплекс близких видов.
От Северного Хонсю (тихоокеанская
и япономорская сторона) и Желтого
моря до Филиппинских островов и
Тонкинского залива и в северной ча-
сти Индийского океана от Аравий-
ского моря и Персидского залива до
Бирмы (к этому виду, вероятно, от-
носятся каракатицы, описанные
Г. В. Зуевым (1971) из Аравийского
моря как «3. confusa (?)». .Преиму-
щественно внешний шельф. Дли-
на мантии до 7 см, редко до
9 см. Промысловый вид у Южной
Японии и КНР.
К этому виду близка S.(3.) kien-
sis Hoyle, 1885, обнаружена в море
Банда на глубине 250 м; се отличия
от 3. kobiensis неясны.
18(17). На гектокотилизировап-
ной руке самца 4—6 проксимальных
поперечных рядов нормальных при-
сосок, в средней части руки присос-
ки почти или полностью отсутствуют
и развивается широкая фестончатая
боковая мембрана, дистальная треть
занята мелкими присосками в 4 про-
дольных ряда...................19.
19(20). Раковина сильно вздута на
вентральной стороне, ее толщина
12%, ширина 16% длины. Присоски
на руках в 4 ряда.
5. (S.) elongata d’Orbigny, 1845
(рис. 21,а—г). Красное море. Длина
мантии до 10 см.
20(19). Раковина слабо вздута па
вентральной стороне, се толщина
7—9%, ширина 21—25% длины.
Присоски в дистальных частях рук,
особенно 1—2-й пар самца и 2—3-й
пар самки в 2 ряда.
S. (S.) irygonina (Rochebrune,
1884) (рис. 21, д). Красное и Ара-
вийское моря, Южная Индия. Шельф
и верхняя батиаль. Длина мантии
до 9 см.
21(2). Присоски булавы мелкие,
приблизительно одинаковых разме-
ров (иногда центральные слегка уве-
личены) .......................22.
22(35). Присоски булавы распо-
ложены в 8—14 косых рядов. Шири-
на раковины 30—50%, обычно 35—
45% ее длины...................23.
23(34). Раковина овальная, плав-
но закругляется к переднему и зад-
нему концу, края внешнего конуса
широкие .......................24.
24(27). Присоски булавы в 10—
14 косых рядов.................25.
25(26). Присоски булавы в 10—
12 рядов у самцов и в 13—14 у са-
мок. Булава длинная, защитные ме-
мбраны булавы не доходят до кар-
пальной части и не соединяются у
начала булавы. В средней части гек-
токотиля 4—6 поперечных рядов
очень мелких присосок, оба дорсаль-
ных ряда еле заметны или редуци-
рованы. Ширина раковины 30—37%
ее длины, у молоди — до 40%.
S. (S.) aculeata d’Orbigny, 1848
(рис. 21, е—з). От восточного побе-
режья Аравийского моря до Вос-
точной Индонезии, КНР и островов
Рюкю. Внутренний шельф. Длина
мантии до 23 см. Промысловый вид,
добывается у Индии, Шри-Ланки, в
южной части побережья КНР, у о-ва
Тайвань и др.
26(25). Присоски булавы в 10—
12 рядов у самцов и самок. Булава
короткая, защитные мембраны со-
единяются в ее карпальной части.
В средней части гектокотиля 7 по-
перечных рядов уменьшенных при-
сосок, присоски обоих вентральных
рядов немного меньше нормальных»
дорсальных — сильно уменьшены, но
нс редуцированы. Ширина раковины
42—48% ее длины.
S. (S.) elliptlca Hoyle, 1885
(=Acanthosepion ellipticum adjacens
Iredale, 1926) (рис. 21, и—л). От се-
верного побережья Аравийского мо-
ря до Южно-Китайского моря, Се-
верной и Восточной Австралии, на юг
до залива Эксмут и Нового Южного
Уэльса. Внутренний шельф. У Па-
кистана 1 экз. пойман на глубине
1000 м. Длина маптии до 11 (16?) см.
27(24). Присоски булавы в 8 ко-
сых рядов . . . -..............28.
28(29). Дорсальная защитная ме-
мбрана булавы значительно шире
вентральной, ее ширина в середине
булавы достигает половины ширины
несущей присоски части. Раковина
спереди сильно угловатая. Длина
шипа 4—9% длины раковины.
5. (S.) brevimana Steenstrup, 1875
( = S. winckworlhi Adam, 1939). От
Персидского залива и Аравийского
моря до Западной Индонезии и юж-
ной части КНР, на юг до банки Сая-
де-Малья. Шельф. Длина мантии до
11 см.
29(28). Обе защитные мембраны
булавы узкие. Раковина спереди
плавно округленная или очень слабо
угловатая. Длина шина 1,5—5,5%
длины раковины.................30.
30(31). Центральные присоски бу-
лавы довольно заметно крупнее бо-
ковых. Ширина раковины около 7з
ее длины. Основание шипа расшире-
но с боков.
5. (S.) zanzibarica Pfeffer, 1884.
У Восточной Африки (Кения, Танза-
ния, Мозамбик). Раковины находят
до Наталя и Мадагаскара. Отмече-
ны на банке Сая-де-Малья. Шельф.
Длина мантии до 25 см.
31(30). Присоски одинаковых или
почти одинаковых размеров. Шири-
на раковины около % се длины. Ос-
нование шипа не расширено с "бо-
ков ...........................32.
32(33). Ширина раковины 37—
40% ее длины. Длина шипа 3,0—
5,5% длины раковины.
5. (S.) thurstoni Adam et Rees,
1966. Бенгальский залив, Южная Ин-
дия, Шри-Ланка. Внутренний шельф.
Длина мантии до 11 см.
33(32). Ширина раковины 40—
44% ее длины. Шип крохотный, его
длина около 1,5% длины раковины.
S. (S.) savignyi Blainville, 1827.
91
Рис. 21. Род Sepia s. str., индовестпаци-
фические виды, 2: а—г — S. elongaia-, д —
S. trygonina; е—з — S. aculeata; и—л —
S. elliptica; м—п — <$. sulcata; б, ж, и —
булава; в, к, о —гектокотиль; а, д, з, л,
п — раковина (а—д — по Adam, 1959; а —
яс—no Voss, Williamson, 1972; з —по Oku-
tani, 1973; и—п — по Hoyle, 1886)
/Ж
Красное море, Аденский и Персид-
ский заливы. Шельф. Многочислен-
ный вид. Длина мантии до 19 см.
34(23). Раковина закругленная
на переднем и приостренная на зад-
нем конце, края внешнего конуса
узкие. Строение защитных мембран
булавы, как у S. brevimana (см.
с. 91). Присоски булавы в 6—8 ря-
дов. На гектокотилизировапной руке
2 продольных ряда присосок, разде-
ленных мясистым гребнем с борозд-
ками по бокам.
S. (S,) sulcata Hoyle, 1885 (рис.
21,.ад—л).. Море Банда. Глубина
250 м. Длина мантии 5 см.
35(22). Присоски булавы распо-
ложены в 5—8 продольных (косых)
рядов. Раковина ланцетовидная, рез-
ко сужающаяся к обоим концам, ее
ширина около V4 длины. Края
внешнего конуса очень узкие 36.
36(37). Мантия широкая (более
V2 длины). Присоски рук в 4, була-
вы в 6—8 рядов.
S. (S.) sewelli Adam et Rees, 1966.
Восточная Африка от мыса Гварда-
фуй до Занзибара. Шельф и зона пе-
региба к склону. Длина мантии до
3 см.
37(36). Ширина мантии около
% ее длины. Присоски в дистальной
части рук в 2 ряда, булавы в 5 рядов.
S. (S.) murrayi Adam et Rees, 1966.
Оманский залив. Внешний шельф.
Длина мантии до 4 см.
38(1). Раковина без шипа, сзади
закругленная. Присоски булавы при-
близительно одинаковых разме-
ров ..........................39.
39(40). На булаве 5—6 продоль-
ных (косых) рядов присосок. Рако-
вина узкая, ее ширина 15% длины,
на заднем конце опа приострена и
слегка изогнута вентрально. От зад-
него конца внутреннего конуса рас-
ходятся веером несколько гребней,
обызвествленных в центральной и
хитиновых в периферической части,
внешний конус окружает внутренний
в виде оторочки. На дорсальной сто-
роне заднего конца раковины — ма-
ленький хитиновый киль.
5. (S.) arabica Massy, 1916. Крас-
ное море, Персидский залив, Южная
Индия, Лаккадивские острова. Внеш-
ний шельф и зона перегиба к склону.
Длина мантии до 7 см.
40(39). На булаве 8 продольных
93
рядов присосок. Ширина раковины
35—50% се длины, она овальная, пе
приострепа на заднем конце. Нет
гребней и хитинового киля на зад-
нем конце раковины .... 41.
41(42). Раковина сильно вздута
горбом в середине вентральной сто-
роны, ее максимальная толщина 21 —
28% длины. В средней части гскто-
котиля уменьшены присоски только
двух дорсальных рядов.
S. (S.) gibba Ehrenberg, 1831
(=S. lejebrei d’Orbigny, 184Ь). Крас-
ное море. Коралловые рифы (по но-
чам укрывается среди игл ядовитого
длииноиглого черного морского ежа
диадемы). Длина мантии до 10 см.
42(41). Раковина не вздута гор-
бом на вентральной стороне, ее тол-
щина 14—19% длины. В средней ча-
сти гектокотиля все присоски силь-
но уменьшены.
S. (S.) dollfusi Adam, 1941. Крас-
ное море, южная часть Суэцкого ка-
нала. Длина мантии до 10,5 см.
IV. Японское, Желтое, Восточно- и
Южно-Китайское моря, воды Японии
1(34). Раковина овальная или
ланцетовидная, пе ромбическая, ее
длина почти равна длине мантии . 2.
2(13). Раковина овальная, плав-
но закругляющаяся к заднему кон-
цу ........................ ... 3.
3(6). Присоски булавы постепен-
но увеличиваются к среднему ряду,
в котором 5—7 присосок заметно
крупнее остальных...............4.
4(5). Защитные мембраны булавы
в карпальной части идут параллель-
но и заканчиваются па стебле щу-
пальца, не соединяясь друг с дру-
гом. Внутренний конус раковины у
взрослых особей прикрыт наплывом
темного цвета.
S. (S.) pharaonis Ehrenberg (см.
с. 87).
5(4). Защитные мембраны була-
вы срастаются в ее карпальной час-
ти и отделяют несущую присоски по-
верхность от стебля. Внутренний ко-
нус нс закрыт темным наплывом.
5. (S.) latimanus Quoy ct Gaimard
(см. с. 87).
6(3). Присоски булавы мелкие,
одинаковые, в 8—14 продольных
(косых) рядов ....... 7.
7(10). Задний край внутреннего
конуса вздутый, широкой дугой
окаймляет заднюю половину фраг-
мокона. Передняя часть раковины
плавно закругляется............8.
8(9). Присоски булавы в 8 рядов,
их количество 200—250. В средней
части гектокотиля 4 четверки (по-
перечных ряда) уменьшенных присо-
сок. Раковина сверху гладкая. Пе-
редний край фрагмокона представля-
ет собой направленный вперед угло-
ватый выступ. На спинной стороне
мантии взрослых особей характер-
ный рисунок; веретеновидные пятна
со светлой оторочкой и светлой по-
лоской посредине («женские губки»).
3. (S.)lycidas Gray, 1849 (=3. su-
baculeata Sasaki, 1914) (рис. 22, а—в).
От Южного Хонсю (тихоокеанская и
япономорская стороны) до Вьетнама
и Саравака. Внутренний и (чаще)
внешний шельф. Длина мантии до
38 см, масса до 5 кг. Добывается в
Японии, Южной Корее и КНР.
9(8). Присоски булавы в 10—12
рядов у самцов, в 13—14 у самок,
их количество более 500. В средней
части гектокотиля 4—6 пар крохот-
ных присосок двух вентральных ря-
дов. Раковина сверху с тремя округ-
ленными гребнями, лучеобразно рас-
ходящимися от заднего конца. Пе-
редний край фрагмокона представ-
ляет собой волнистую линию с двумя
направленными вперед выступами и
углублением между ними. Окраска
разнообразна, по пятен типа «жен-
ские губки» нет.
3. f3J aculeata d’Orbigny (см.
с. 91).
10(7). Задний край внутреннего
конуса тонкий, узкий. Передняя
часть раковины слегка приострена.
На спинной стороне мантии есть тем-
ные поперечные полоски, вдоль осно-
вания плавников — ряд из 6—7 низ-
ких треугольных выступов со светлой
каймой...................... 11.
И (12). Задний край внутреннего
конуса U-образных очертаний, ото-
гнут наружу и образует полукруг-
лую стеночку, окаймляющую заднюю
часть фрагмокона. Присоски булавы в
10—12 продольных рядов. В средней
части гектокотилизированной руки
4—6 поперечных рядов уменьшенных
присосок.
S. (S.) esculent a Hoyle, 1885 (=3.
hoylei Ortmann, 1888) (рис. 22, г—е).
От Центрального Хонсю (тихоокеан-
ская и япономорская стороны) и
Желтого моря до Вьетнама и Филип-
пинских островов. Преимущественно
94
внутренний шельф. Длина маптии до
18 см. Важнейший промысловый вид
в Японии, Южной Корее и КНР.
12(11). Задний край внутреннего
конуса V-образный, плотно прилегает
к задней части фрагмокона. Присос-
ки булавы в 8—10 продольных ря-
дов. В средней части гектокотилизи-
рованпой руки 8—10 поперечных ря-
дов уменьшенных присосок.
5 . (S J madokai Adam, 1939 (=S.
robsoni Sasaki, 1929, non Massy,
1927) (рис. 22, ж—з). У Южного
Хонсю, Сикоку, Кюсю, в Цусимском
проливе, Восточпо- и Южно-Китай-
ском морях. Шельф. Длина маптии
до 15 см. Второстепенный промысло-
вый вид в Южной Японии и КНР.
13(2). Раковина ланцетовидная,
приострсппая к переднему и резко
суженная к заднему концу . . 14.
14(15). Ширина раковины около
40 % длины. Присоски у оснований
и на концах рук в 2, в средних ча-
стях в 4 ряда. На булаве 5 средних
присосок заметно крупнее прочих.
S. (S.) carinata Sasaki, 1920 (рис.
22, и—л). Залив Сагами. Нижняя
сублитораль. Длина маптии до 3 см.
15(14). Ширина раковины 15—
30% длины.....................16.
16(21). Присоски булавы одина-
ковых размеров, расположены в
8 продольных (косых) рядов . 17.
17(20). У зрелых самцов руки 1-й
пары удлиненные, хлыстовидные.
Присоски в дистальпых частях рук *
1—3-й пар в 2 ряда. Ширина рако-
вины около ’/б ее длины . . . 18.
18(19). На спинной стороне ман-
тии множество (более 100) продоль-
ных удлиненных темных пятен. Пе-
редняя часть фрагмокона с двумя
низкими продольными гребнями по
бокам от средней линии раковины.
S. (S.) pardalis Sasaki, 1914
(рис. 22, At—w). У Центрального и
Южного Хонсю, Сикоку, Кюсю, Юж-
ной Кореи. Длина маптии до 23 см.
19(18). Нет темных пятен на
спинной стороне мантии. Руки 2—3-й
пар самца нормальные, у самки —
утонченные и оттянутые на концах
(но нс длиннее прочих рук). Перед-
няя часть фрагмокона с одним низ-
ким округлым гребнем по средней
линии раковины.
S. (S.) tenuipes Sasaki, 1929 (рис.
22, о—с). У Центрального и Южно-
го Хонсю, Сикоку. Длина мантии до
10,5 см.
20(17). Руки зрелых самцов при-
мерно одинаковой длины, концы рук
1—2-й пар утонченные, хлыстовид-
ные, с 2 рядами присосок, руки 3-й
пары с 4 рядами присосок. Ширина
раковины 25—30% ее длины.
S. (S.) appelloefi Wiilkcr, 1910
(рис. 23, а—я). У Южного Хонсю,
Сикоку, Кюсю, в Цусимском проливе.
Внешний шельф и самая верхняя ба-
тиаль. Длина маптии до 9 см.
21(16). Присоски булавы заметно-
неравных размеров..............22.
22(23). Шип раковины очень ко-
роткий, рудиментарный. Раковина
спереди плавно закругленная. Руки
1-й и 4-й пар у самца значительно-
длиннее прочих, их концы оттянуты и
лишены присосок (?). Концы рук 2—
3-й пар самца с 2 рядами присосок.
S. (SJ eroslrata Sasaki, 1929 (рис.
23, г—е). Залив Сагами. Длина ман-
тии 4,5 см.
23(22). Шип раковины хорошо-
развит. Руки 4-й пары самца не уд-
линены ........................24.
24(31). Руки взрослых самцов;
резко неравных размеров, 1-я или.
2-я пара гораздо длиннее прочих
(самки плохо различаются) . 25..
25(28). Удлинена 1-я пара рук
взрослых самцов, она в 2—3 раза
длиннее мантии, хлыстовидная, по-
утолщепа на конце. Присоски в ди-
стальных частях 1—2-й пар рук сам-
ца рудиментарные...............26.
26(27). Гектокотилизируются обе
брюшные руки самца. Фрагмокон с
резким продольным килем.
S. (S.) longipes Sasaki, 1914 (рис.
23, ж—з). Тихоокеанское и японо-
морское побережья Южного Хонсю.
Шельф. Длина мантии до 23,5 см.
27(26). Гектокотилизируется толь-
ко левая брюшная рука самца. Фраг-
мокон плавно выгнут по средней
линии.
5. (S.J lortgera Wulker, 1910 (рис.
23, и). У Южного Хонсю, Сикоку,
Кюсю, в Восточно-Китайском море.
Внешний шельф и верхняя ба-
тиаль. Длина мантии до 25 см. Вто-
ростепенный промысловый вид.
28(25). Удлинена 2-я пара рук
взрослых самцов. Фрагмокон с же-
лобком по средней линии ... . 29.
29(30). Руки 2-й пары взрослых
самцов в 3—6 раз длиннее маптии, к
концу постепенно утончаются и ста-
новятся нитевидными,
S. (S.) peterseni Appellof, 1886
95
Рис. 22. Род Sepia s. sir., японо-китай-
ские виды, 1: а—в — S. lycidas\ г—е—
S. esculenta; ж—з — S. madokai; и—л —
3. carinata; м—н — S. pardalis; o—c —
S. tenuipes; б, д, к, n— булава; p — 1—4-я
пары рук взрослой самки; в, в, з, л, н,
с — раковина (а, б, г- по Voss, William-
son, 1972; в, м—о, с —по Okutani, 1973;
д—по G. Voss, 1963; е — по Appellof, 1886;
ж—л, п—р — по Sasaki, 1929)
(рис. 23, к—лс). У Центрального и
Южного Хонсю, Сикоку, Кюсю, Юж-
ной Кореи. Внутренний шельф. Дли-
на мантии до 12 см.
30(29). Руки 2-й пары взрослых
самцов примерно такой же длины,
как мантия, почти цилиндрической
формы по всей длине, не нитевидные.
S. (S.) andreana Steen strup, 1875
( = ?S. sasakii Wakiya et Ishikawa,
1929) (рис. 23, н). Тихоокеанское по-
бережье Южного Хоккайдо, Север-
ного и Центрального Хонсю, Сангар-
ский пролив, у северной части КНР
(указания на находки у Гонконга
ошибочны). Шельф. Длина мантии
до 12 см. Второстепенный промысло-
вый вид у Северной Японии.
31(24). Руки самцов примерно
равной длины или 1-я пара чуть
длиннее прочих .............. 32.
32(33). Задний конец каждого
плавника оттянут назад и расширен
в прямоугольную утолщенную ло-
пасть.
S. (S.) tokioensis Ortinann, 1888
(=S. misakiensis Wulker, 1910) (рис.
23,o—n). От Сангарского пролива
до островов Кюсю и Осуми. Шельф.
Длина мантии до 9 см.
33(32). Задний конец плавника
нс заходит за задний конец мантии
и пе расширен в лопасть.
S. (S.) kobiensis Hoyle, 1885 (см.
с. 90).
34(1). Раковина ромбическая,
много короче мантии, заостренная
сзади.
S. (Metasepia) tullbergi Appellof,
1886 (см. с. 80).
V. Австралия 1
1(44). Раковина с шипом или хотя
бы маленьким бугорком на заднем
конце или заострена сзади . . 2.
2(43). Раковина овальная, лан-
1 Не включены описанные только по
раковине виды Sepia (S.) opipara
(Ircdale, 1926), S. glauerti (Cotton,
1929) ( = ? Solitosepia mestus Iredale,
non Gray), S. whitleyanum (Iredale,
1926) ( = ? S. elliptica Hoyle) и опи-
санная только по телу S. dannevigi
Berry, 1918.
7 К. Н. Несис
97
Рис. 23. Род Sepia s. str., японо-китай-
ские виды. 2: а—в — 5. appelloefr, г—е —
S. erostrata; ж—з — S. longipes; и — S. 1о-
rigera; к—м — S. peferseni; н — S. andreana;
o—n~S. iokioensis", б, д, л, п— булава;
в, е, з, и, м — раковина (а, в—е, з—и, о—
п — по Sasaki, 1929; б, дас, н — no Okutani,
1973; к—м — по ЛррсПбГ, 1886)
цетовидная или бисквитообразиая,
но нс ромбическая..............3.
3(42). Раковина с хорошо разви-
тым шипом (у взрослых S. арата
он покрыт выростом заднего края
внутреннего конуса) .... 4.
4(41). Раковина овальная с вы-
гнутыми или почти параллельными
боковыми сторонами, овалыю-ланце-
товидная или ланцетовидная, не
бисквитообразиая ............. 5.
5(28). Раковина овальная с вы-
гнутыми или почти параллельными
боковыми сторонами. Внутренний
конус хорошо развит, V- или U-об-
разный или полуовальный. Края
внешнего конуса широкие. Ширина
раковины 25—50% ее длины . . 6.
6(23). Присоски булавы резко
неравных размеров..............7.
7(22). Присоски булавы посте-
пенно увеличиваются к среднему ря-
ду, в котором 5—7 присосок значи-
тельно крупнее прочих. Нет бросаю-
щейся в глаза разницы между гро-
мадными средними и маленькими
остальными присосками ... 8.
8(9). Защитные мембраны була-
вы в карпальной части идут парал-
лельно и заканчиваются па стебле
щупалец, не соединяясь друг с дру-
гом. Внутренний конус раковины у
взрослых особей прикрыт наплывом
темного цвета.
S. (S.) pharaonis Ehrenberg (см.
с. 87).
9(8). Защитные мембраны булавы
сближаются или срастаются в ее
карпальной части и отделяют несу-
щую присоски поверхность от стебля.
Внутренний конус не закрыт темным
наплывом .......................10.
10(13). Задний край внешнего ко-
нуса подогнут на вентральную сто-
рону раковины, так что образуется
широкий воронковидный «фунтик».
Раковина с довольно широкими хи-
тинизированпыми каемками по бо-
кам в передней половине . . . 11.
11(12). Раковина спереди плавно
закругленная, ее ширина 33—
36(39) % длины. Полоски исчерчен-
ной зоны фрагмокона плавно дуго-
образно выгнуты. Голова и дорсаль-
ная сторона 1—3-й пар рук с разбро-
санными бугорками.
S. (S.) papuensis Hoyle (см. с. 90).
12(11). Раковина спереди приост-
рена, округло-треугольная, ее шири-
на 26—30%, у молоди — до 33%
7*
99
Рис. 24. Род Sepia s. str., австралийские
виды: a — S. plangon; б — S. tnesfus; в —
S. rozella; г-д — S. арата (молодая и
взрослая); е — S. irvingi*, ж — S. ostanes;
з—к — S. chirotrema; л—о — S. novaehollan-
diae; п—р — S. stniihi; с — S. rex; т—X —
S. cultrata; ц — 5. braggi; ч — S. rhoda;
tu — S. mira\ щ — S. baxteri' 3, м, ф — бу-
лава; и, н, у — гектокотиль; а—ж, к. О, р,
с, х—щ — раковина (а—г, jw, о, с, ч, щ
по Iredale, 1926; д — по Bell, Plant, 1977;
е— по Меуег, 1909; з—н — по Berry, 1018;
п—р, т—х — по Hoyle, 1886; ц — по Cotton,,
Godfrey, 1940; ш — по Cotton, 1931)
длины. Полоски исчерченной зоны
фрагмокона с узким треугольным
выступом, направленным вперед, и
боковыми «плечиками», напоминают
силуэт пера авторучки. Нет бугорков
на голове и руках.
S. (S.) plangon Gray, 1849 (рис.
24, а). Северо-Восточная и Восточная
Австралия (Квинсленд, Новый Юж-
ный Уэльс). Раковины встречаются
на о-ве Лорд-Хау и Северном остро-
ве Новой Зеландии. Многочислен-
ный вид. Длина мантии до 13,5 см.
13(10). Задний край внешнего
конуса не подогнут на вентральную
сторону раковины. Раковина без ши-
роких хитипизировапных каемок 14.
14(17). Ветви внутреннего конуса
раковины широкие, дуговидно изо-
гнутые, расходятся в задней части и
параллельны или слегка сходятся в
передней .....................15.
15(16). Вентральная сторона ра-
ковины уплощенная или с узкой не-
глубокой продольной бороздкой. Нет
гребней на дорсальной стороне. Ши-
рина раковины 37—45% (до 50%)
длины.
S. (S.) mestus Gray, 1849 (=As-
carosepion verreauxi Rochebrune,
1884, Solitosepia Uliana Iredale, 1926)
(рис. 24,6)., Северо:Восточная и
Восточная Австралия (Квинсленд,
Новый Южный Уэльс). Раковины
встречаются на о-ве Лорд-Хау. Про-
мышляется в небольших количествах.
Длина мантии до 14 см.
16(15). Вентральная сторона ра-
ковины с глубоким продольным же-
лобком, окаймленным широкими греб-
нями. Дорсальная сторона с тремя
невысокими продольными гребнями.
Ширина раковины 30—35% (у моло-
дых до 40%) длины. Внутренний
конус ярко-розового цвета.
5. (S.) rozella (Iredale, 1926)
( = Solitosepia rozella peregrina Ireda-
le, 1926) (рис. 24, в). Северо-Восточ-
ная и Восточная Австралия (Квин-
сленд, Новый Южный Уэльс) и за-
лив Шарк (?). Раковины встреча-
ются на о-ве Лорд-Хау. Длина
мантии до 15 см.
17(14). Внутренний конус рако-
виды узкий, V-образный с расходя-
щимися ветвями................18.
’ 18(19). Киль булавы примерно
Вдвое ’ длиннее несущей присоски
части ’ и продолжается па стебле
проксимальнее Карпальной части бу-
лавы. Внутренний конус состоит из
2 частей: узкой V-образной внутрен-
ней и более широкой наружной, сли-
той с внешним конусом и (у взрос-
лых каракатиц) продолжающейся
назад языковидным выступом. Шип
у взрослых особей полностью скрыт
этим выступом и незаметен. Ширина
раковины 35—45% длины. На голове
над глазом по 2—3 кожных «ушка».
S. (S.) арата Gray, 1849 ( = S. pal-
mata Owen, 1881, A mplisepia par у satis
Iredale, 1954) (рис. 24, a—д). Юго-
Западная, Южная и Восточная
Австралия (на север до мыса Се-
веро-Западного и Южного Квинслен-
да), Тасмания, о-ва Лорд-Хау и
Норфолк. Раковины известны на
Северном острове Новой Зеландии.
Длина мантии до 48 см. Массовый
вид внутреннего шельфа. Промышля-
ется в небольшом количестве у Юж-
ной и Восточной Австралии.
19(18). Киль булавы лишь немно-
го длиннее части, несущей присоски,
и заходит на стебель немного далее
карпальной части булавы. Нет язы-
ковидного продолжения внутреннего
конуса в задней части раковины.
Шип длинный, хорошо развит . 20.
20(21). Раковина правильно
овальная, боковые стороны равно-
мерно выпуклые. Ширина раковины
34—39% ее длины.
S. (S.) latimanus Q. ct G. (с. 87).
21(20). Боковые стороны ракови-
ны параллельны или слегка изгиба-
ются внутрь в начале задней трети
раковины. Ширина раковины 28—
35% ее длины. Раковина похожа на
раковину S. chirotrema (см. ниже),
но отличается слабо развитыми греб-
нями на дорсальной и вентральной
стороне и тем, что полоски исчер-
ченной зоны фрагмокона на перед-
нем крае имеют языковидный вы-
ступ.
S. (S.) irvingi Meyer, 1909 (рис.
24,6?). Западная Австралия. Длина
мантии до 18 см. Внешний шельф.
У Юго-Западной, Южной Австра-
лии и Тасмании встречается близкий
вид S. (S.) ostanes (Iredale, 1954)
(рис. 24, ж), известный лишь по ра-
ковине; у него полоски исчерченной
зоны фрагмокона спередц дуговидно
закруглены.
•22(7). Всю центральную часть
булавы занимают 3 громадные при-
соски. Конец гектокотилизированной
левой брюшной руки оттянут, сужен
и сжат с боков, несет микроскопи-
102
ческие присоски. Раковина с парал-
лельными или слегка изгибающими-
ся внутрь боковыми краями, на дор-
сальной стороне 3 хорошо заметных
округленных гребня, па вентраль-
ной — глубокая борозда, окаймлен-
ная двумя гребнями; полоски исчер-
ченной зоны фрагмокона спереди с
направленным вперед углом.
3. (S.) chirotrema Berry, 1918
(—Solitosepia hendryae Cotton, 1929)
(рис. 24,з—к). Юго-Западная и Юж-
ная Австралия, Большой Австралий-
ский залив. Внешний шельф и зона
перегиба от шельфа к склону. Дли-
на мантии до 20 см.
23(6). Присоски булавы прибли-
зительно одинаковых размеров . 24.
24(27). Присоски булавы в 8—
12(13) продольных (косых) ря-
дов ...........................25.
25(26). Видоизмененные присоски
в средней части гектокотиля распо-
ложены в 3 ряда и значительно
меньше обычных. Ширина раковины
25—35% ее длины, боковые края
параллельные, передний край узко-
закругленный, внутренний конус
V-образный, его ветви прижаты к
поверхности внешнего конуса.
S. (S.) novaehollandiae Hoyle, 1909
[==S. australis Ferussac, 1835, non
Quoy et Gaimard,.. 1832, S. hedleyi Ber-
ry, 1918, part, S. macandrewi (Iredale,
1926)] (рис. 24,л—о). Юго-Западная,
Южная и Юго-Восточная Австралия,
Тасмания, на север до мыса Северо-
Западного и Нового Южного Уэль-
са. Весь шельф и самая верхняя ба-
тиаль. Длина мантии до 16—18 см.
Многочисленный вид.
26(25). Видоизмененные присоски
в средней части гектокотиля распо-
ложены в 4 ряда, дорсальные — мно-
го меньше, вентральные — лишь не-
много меньше обычных. Ширина ра-
ковины 40—48% ее длины, боковые
края выпуклые, передний край ши-
роко-закругленный, ветви внутренне-
го конуса широкие, широко" расхо-
дятся (конус имеет полуовальную
форму), задний край внутреннего ко-
нуса приподнят и дуговидной сте-
ночкой окружает заднюю часть фраг-
мокона.
S. (5.) elliptica Hoyle (см. с. 91).
27(24). Присоски булавы прибли-
зительно в 20 поперечных рядов. Ра-
ковина овальная, ее ширина 35—40%
длины, внутренний конус полуоваль-
ный, его округленный задний край
образует дугообразный валик вокруг
задней части фрагмокона. Шип слег-
ка загнут вверх.
S. (о.) stniihi Hoyle, 1885 (=Acan-
thosepion pageorum I redale, 1954)
(рис. 24, n—p). Северная и Северо-
Западная Австралия, Тиморское,
Арафурское и Коралловое моря,
Большой Барьерный риф. Длина ман-
тии до 17 см.
28(5). Раковина овально-ромбиче-
ская, овально-ланцетовидная или
ланцетовидная. Внутренний конус
V-образный с очень узкими ветвя-
ми .............................29.
29(32). Ширина раковины 30—
40% се длины. Шип раковины с ки-
лем па вентральной, часто и на дор-
сальной стороне.................30.
30(31). Раковина овально:ромби-
ческая, плавно сужается назад и бо-
лее резко вперед. Место наибольшей
ширины приблизительно посередине
раковины. Дорсальная сторона рако-
вины с тремя невысокими, продоль-
ными ребрами, вентральная — с про-
дольным желобком. В средней части
булавы 4—5 очень крупных присо-
сок. В средней части гектокотилизи-
рованпой руки около 10 поперечных
рядов резкр уменьшенных в разме-
рах присосок.
S. (S.)rex (Iredale, 1926) ,(=? De-
.corisepia cottesloensis Cotton, 1929,
D.jaenschi Cotton, 1931) (рис. 24, c).
Юго-Западная, Южная, Восточная
Австралия, Тасмания, па север до за-
лива Эксмут и Южного Квинсленда.
Раковины отмечены на о-ве Лорд-
Хау. Шельф. Длина раковин до
12(15) см.
31(30). Раковина овально-ланце-
. товидная, треугольно-приостренная
спереди, место наибольшей ширины
сдвинуто в переднюю часть .рако-
вины. Дорсальная и вентральная ча-
сти раковины уплощены, с очень сла-
бо развитыми гребнями и без желоб-
ка. Присоски булавы в 5—6 продоль-
ных рядов, средние немного крупнее
боковых. В средней части гектокоти-
ля дорсальные присоски микроскопи-
ческие, по вентральные лишь немно-
го мельче нормальных.
S. (S.) cultrata Hoyle, 1885 [=S.
hedleyi Berry, 1918, part (раковина),
Glyptosepia gemellus Iredale, 1926,
G. macilenta Iredale, 1926] (рис.
24, r—x). Юго-Западная, Южная и
Восточная Австралия, па: север до
скал Хаутмен и Южного Квинслен-
103
да, Тасмания. Раковины встречают-
ся на о-ве Лорд-Хау. Внешний
шельф и верхняя батиаль, глубины
150—650 (820) м. Длина раковины
до 12 см. Обычный вид.
32(29). Ширина раковины 16—
30% длины. Киль в сечении округ-
лый ...........................33.
33(34). Раковина удлиненно-язы-
ковидная, сзади широко закруглен-
ная. Края внешнего конуса без ушко-
видного изгиба в задней части рако-
вины. Ширина раковины 25—28%
длины.
S. (S.) reesi Adam, 1979. О-в Рот-
нест близ Фримантла. Длина рако-
вины до 45 мм. Животное нс опи-
сано.
34(33). Раковина узко-ланцето-
видная, спереди закруглепно-приост-
рена, сзади вытянуто-заострена, как
отточенный карандаш. Место наи-
большей ширины — в передней тре-
ти длины. Ширина раковины 16—
23% длины. Края внешнего конуса
образуют изгиб — «ушки» в задней
части раковины.................35.
35(40). Вентральная поверхность
раковины с узкой продольной бо-
роздкой по всей длине. Толщина ра-
ковины 6—9% длины. Присоски бу-
лавы в 5 продольных рядов, 5—
6 средних присосок примерно вдвое
крупнее боковых................36.
36(39). Концы рук сайта оттяну-
ты и несут 2 ряда присосок. Длина
боковых рук самца Уз—Уг, иногда
руки 3-й пары до % длины мантии.
Брюшные руки самца не гектокоти-
лизированы. Нет обызвествленного
киля в задней части дорсальной
стороны раковины на границе с хи-
тиновой частью.................37.
37(38). Руки 3-й пары самца
сильно вытянуты — длина до у2—%
длины мантии, но не видоизменены.
Полоски исчерченной зоны фрагмо-
кона в средней части раковины М-об-
разно изогнуты.
S. (SJ braggi Verco, 1907
( = ? Arctosepia limata Iredale, 1926,
A. versuta Iredale, 1926, A. treba Ire-
dale, 1926) (рис. 24, ц). Юго-Запад-
ная, Южная и Восточная Австралия,
на север по крайней мере до Фри-
мантла и Южного Квинсленда, Тас-
мания. Шельф, преимущественно
внешний. Мелкий вид, длина мантии
до 5—6, редко 8 см.
38(37). Длина рук 3-й пары сам-
ца Уз—У2 длины мантии, они резко
видоизменены: в средней части руки
развиты толстые поперечные греб-
ни, проходящие попеременно от
внутреннего края одной защитной
мембраны до внешнего края проти-
воположной; па каждом гребне си-
дят по 2 присоски, дорсальная — по-
средине гребня, вентральная — почти
по краю мембраны. Полоски исчер-
ченной зоны фрагмокона слабо изог-
нуты.
S. (SJ cottoni Adam, 1979. Запад-
ная Австралия на север до Земли
Дампьера, Большой Австралийский
залив, Квинсленд (?). Весь шельф;
Массовый вид. Длина мантии до
6,5 см.
39(36). Руки 2—3-й пар самца
короткие — их длина около У4 дли-
ны маптии, значительно короче рук
1-й пары, руки 3-й пары нормаль-
ные» Руки 1—3-й пар с 4 рядами
присосок. Левая брюшная рука гек-
токотилизировапа, присоски средней
части руки резко уменьшены. На
дорсальной стороне раковины в ее
задней части обызвествленная часть
отделена от хитиновой резким изве-
стковым килем.
S. (S.) vercoi Adam, 1979. Запад-
ная Австралия. Шельф. Длина ман-
тии до 5 см.
40(35). Вентральная поверхность
раковины с продольным гребнем
вдоль фрагмокона, переходящим в
бороздку в передней части раковины.
Толщина раковины 11—12% се дли-
ны. Животное нс описано.
S.(S.)rhoda (Iredale, 1954) (рис.
24, ч). Западная Австралия.
41(4). Раковина бисквитообраз-
лая, удлиненно-языковидная, сужен-
ная на границе средней и задней
трети и расширяющаяся близ задне-
го конца. Наибольшая ширина рако-
вины (20% длины)—в первой трети
длины. Дорсальная сторона ракови-
ны гладкая кремового цвета. Внут-
ренний конус узкий. Шип прямой.
S. (S.) mira (Cotton, 1931) (рис.
24, ш). Северо-Восточная Австралия
(Квинсленд, Новый Южный Уэльс).
Длина раковины до 5,5 см. Живот-
ное не описано.
42(3). Раковина без шипа, на его
месте лишь маленький тупой буго-
рок. Раковина розовато-бурого цве-
та, удлиненно-овальная, спереди
слегка приостренная, ее ширина 41—
48% длины. Внутренний конус уз-
кий, полуовальный.
104
S. (S.) baxteri (Iredale, 1940)
(рис. 24, щ). О-в Лорд-Хау (восточ-
нее Австралии). Длина раковипы до
9 см. Животное не описано.
43(2). Раковина ромбическая, ту-
по заостренная спереди и угловатая
или округленно-угловатая на боках,
заостренная сзади, много короче ман-
тии. На булаве 3 громадные присос-
ки, занимающие всю ее середину.
S. (Metasepia) pfefferi Hoyle, 1885
(см. с. 78).
44(1). Раковина сзади округлен-
ная, без шипа и бугорка. Крупные
животные, длина раковипы до 48 см.
Sepia (S.) арата Gray, 1849 —
взрослые (см. с. 102).
Таблица для определения видов рода
Sepiella
1(4). Виды распространены в тропи-
ческих водах Восточной Атлантики и
юго-западной части Индийского окс-
ана. Присоски булавы в 10—14 ря-
дов ...........................2.
2(3). Ширина раковины 24—30%
ее длины (у самцов 24—27%, у са-
мок 27—30%).
w S. ornata (Rang, 1837) (рис.
25, а—б). Тропические воды Запад-
ной Африки от Мавритании до Анго-
лы. Шельф. Длина мантии до 10 см.
3(2). Ширина раковины 29—36%
ее длины (у самцов 29—33 %, у са-
мок 30—36%).
S. cyanea Robson, 1924 (см. рис.
17, а) Юго-Восточная Африка от
Дурбана до Порт-Элизабет, Мада-
гаскар. Внутренний шельф. Длина
мантии до 8 см.
4(1). Виды распространены в се-
верной и восточной частях Индийско-
го и западной части Тихого океа-
на ............................5.
5(6). Присоски булавы в 16—
32 ряда, обычно более чем в 20 ря-
дов. Ширина мантии около 30% се
длины.
S. japonica Sasaki, 1929 (=S.
maindroni Hoyle, 1886, non Rochebru-
ne, 1884) (рис. 25, в—d). От цент-
рального Хонсю до Гуанчжоу (КНР)
и Филиппинских островов, изредка
встречается в водах СССР (залив
Посьста). Внутренний шельф. В Япо-
нии, Южной Корее и КНР — важный
промысловый вид. Длина мантии до
20 см.
6(5). Присоски булавы в 8—
24 ряда, обычно в 8—16 рядов . 7.
106
7(8). Присоски булавы в 8—10
рядов. Раковина очень узкая, ее ши-
рина 20—25% длины.
S. ocellata Pfeffer, 1884 (рис.
25, е—ж). О. Ява. Длина мантии 5 см.
8(7). Присоски булавы в Ю и бо-
лее рядов. Ширина раковины более
25% се длины...................9.
9(10). Присоски булавы в 10—
12 рядов. Ширина раковины около
30 % ее длины.
S. weberi Adam, 1939 (рис. 25, з).
Восточная Индонезия (Сумба, Ти-
мор).
10(9). Присоски булавы в 12—24,
обычно в 13—20 рядов. Ширина ра-
ковины 33—43% се длины.
S. inermis (Ferussac et d’Orbigny,
1835) (рис. 25, и—л). От Персидско-
го залива и южной части Красного
моря до Восточной Индонезии и Тон-
кинского залива. Внутренний шельф.
Длина мантии до 10—12 см. Про-
мысловый вид, добывается в неболь-
ших количествах в Аравийском море,
у Южной Индии и в водах Индо-
китая.
Примечание. В таблицу не
включен сомнительный вид S. mel-
wardi Iredale, 1954 (рис. 25, л«) из
Северной Австралии, известный толь-
ко по раковине.
2.1.2.2. Семейство Sepiolidae
Небольшие животные (длина мантии
от 1 до 8 см) с короткой закруглен-
ной сзади мантией, передний край
которой на спинной стороне свободен
или срастается с головой, на брюшной
прикрывает основание воронки. Го-
лова широкая. Плавники почковид-
ные, спереди с хорошо развитыми
«ушками», максимальная длина
плавника заметно больше длины его
основания. Щупальца втяжные с хо-
рошо ограниченной булавой. Руки
довольно короткие, мощные, без за-
щитных мембран, с шарообразными
присосками. У самцов обычно при-
соски крупнее, чем у самок, и более
дифференцированы по размерам.
Гладиус хитиновый, рудиментарный
или отсутствует. Часто имеются све-
тящиеся органы на чернильном меш-
ке. Имеется продольная вентральная
мышечная септа в мантийной поло-
сти. Роговица глаза замкнутая с ма-
ленькой порой. Гектокотилизируются
одна или обе спинные руки. У самок
развит только левый яйцевод. Яйца
крупные, донные, развитие прямое
без метаморфоза.
Донные животные, обитатели
шельфа и склона, некоторые виды
живут в толще воды над склоном
или вблизи него.
3 подсемейства, 14 родов, 50—56
видов. Частичные обзоры: Joubin,
1902, Naef, 1921—1923; Sasaki, 1929;
G. Voss 1955, 1956, 1963.
Примечание. Большинство
видов сепиолид хорошо различаются
только по половозрелым самцам. Оп-
ределительные таблицы родов и ви-
дов сепиолид построены главным
образом по признакам строения зре-
лых самцов.
Таблица для определения родов
семейства Sepiolidae
1(18). Передний край мантии па
брюшной стороне пе вытянут вперед
и не прикрывает воронку. Руки 1-й
и 2-й пар нс соединены или соедине-
ны лйшь короткой умбреллой (пере-
понкой). Плавники умеренных раз-
меров: длина плавника много мень-
ше длины мантии. Донные живот-
ные, лишь некоторые виды в период
размножения поднимаются к поверх-
ности ..........................2.
2(11). Передний край мантии на
спинной стороне срастается с голо-
вой посредством затылочной кожной
лепты; затылочного хряща нет. Гск-
токотилизируется левая спинная ру-
ка. Гладиус рудиментарный или от-
сутствует.
Подсемейство Sepiolinae . . 3.
3(4). На гсктокотилизированной
руке самца резко видоизменяются
дистальные присоски: они превра-
щаются в мясистые папиллы и име-
ют щелевидиые устья с утолщенны-
ми «губами»; эти папиллы тесно
сближены, образуя своего рода «па-
лисад». В базальной части руки кро-
ме обычных присосок есть лишь 1—
2 небольшие папиллы. Присоски на
руках сидят в 4 ряда (у Е. phenax
в 2). 3-я пара рук самца нс загнута
внутрь. Присоски булавы очень мел-
кие, многочисленные, обычно более
чем в 10 рядов. Затылочная лепта
широкая, около половины ширины
головы. Имеется седловидный светя-
щийся орган в передней части чер-
нильного мешка.
Род Еиргутпа, Stecnstrup, 1887
(=Fidenas Gray 1849) (см. рис.
28, а—н). Типовой вид — Е, morsel
(Verrill, 1881), 7 видов в Индо-Всст-
иацифике — от Хоккайдо до Тасма-
нии, от Персидского залива и Юж-
ной Африки до Гавайских островов
и Полинезии. Шельф и верхняя ба-
тиаль. Некоторые виды имеют не-
большое промысловое значение.
4(3). Видоизменяется либо вся
гсктокотилизировапная рука, либо се
базальная часть, на которой разви-
вается специальный копуляторный
аппарат с разного рода выемками и
выростами. Присоски- на руках в 2
ряда (только на концах брюшных
рук иногда более чем в 2). 3-я пара
рук самца обычно сильно загнута
внутрь, ко рту.................5.
5(8). Имеется светящийся орган
на чернильном мешке. Затылочная
лента широкая, 7з—7г ширины го-
ловы ..........................6.
6(7). Светящийся орган располо-
жен на поверхности чернильного
мешка в его передней части и имеет
форму седла или двух «ушек». При-
соски на концах брюшных рук иног-
да сидят в 4—8 рядов. Присоски бу-
лавы обычно в 4—8 рядов.
Род Sepiola Leach, 1817 (см. рис.
30). Типовой вид — S. rondeleti Steen-
strup, 1856. 11—12 видов: 8—9 видов
у берегов Западной Европы от Нор-
вегии до Средиземного моря, у Севе-
ро-Западной Африки и (?) в Крас-
ном море, 3 в западной части Тихо-
го океана от Сахалина и Южных
Курильских до Филиппинских остро-
вов. Маленькие донные животные,
обитают в сублиторали и верхней
батиали, в период размножения
поднимаются к поверхности. Длина
мантии 1—3 см.
7(6). Светящийся орган лежит на
брюшной стороне чернильного меш-
ка и глубоко погружен в его
ткань. Присоски на всех руках в
2 ряда. Присоски булавы в 16 рядов.
Род Rondeletiola Naef, 1921. Один
вид — /?. minor (Naef, 1912) (рис.
29, е, к, л). Средиземное море и Ат-
лантика у побережий Европы и За-
падной Африки от Португалии до
Намибии. Мелкие животные (длина
мантии обычно не более 1,5 см).
Обитают на дне преимущественно в
нижней сублиторали, в период раз-
множения поднимаются к поверхно-
сти.
8(5). Нет светящихся органов на
чернильном мешке. Затылочная лен-
та узкая, обычно менее ’/з ширины
головы.........................9.
9(10). Гсктокотилизироваппая ру-
ка самца ложковидно расширена дис-
тальнее копуляторного аппарата.
Обе спинные руки самца сращены у
основания. Присоски булавы более
чем в 10 рядов.
Род Sepietta Naef, 1912 (=Sepl-
dium Levy, 1912) (рис. 29, a—dt
ж—и). Типовой вид S. oweniana
(d’Orbigny, 1839). 3—4 вида у бере-
гов Европы от Южной Норвегии до
Средиземного моря и у о-ва Мадей-
ра. У дна в сублиторали, иногда в
верхней батиали.
10(9). Гектокотилизированпая ру-
ка самца расширена в базальной
половине — в районе копуляторного
аппарата, дистальная часть руки
нормальная. Присоски булавы в 8—
10 рядов.
Род Inioteuthis Verrill 1881 (см.
рис. 28, о—ф). Типовой вид /.
japonica Verrill, 1881 [=/. inioteuihis
(Naef, 1912)]. 3 вида в Индо-Вест-
пацификс — от Японии до Южной
Африки. Сублитораль.
11(2). Передний край мантии па
спинной стороне не срастается с го-
ловой, затылочный хрящ развит. Гек-
токотилизируются левая или обе
спинные руки. Гладиус имеется.
Подсемейство Rossiinac . . 12.
12(13). Чернильный мешок реду-
цирован и не содержит чернил.
Анальные папиллы редуцированы до
крохотных бугорков. Нет светящего-
ся органа на чернильном мешке.
Присоски на руках в 2 ряда, в сред-
них частях боковых рук нет резко
увеличенных присосок. Гектокотили-
зируются обе спинные руки, но гек-
токотилизация слабо выражена. При-
соски булавы в .6—7 рядов.
Род Neorossia Boletzky, 1971.
Один вид — V. caroli (Joubin, 1902).
Восточная Атлантика от Юго-Запад-
ной Ирландии до Гвинейского зали-
ва, Средиземное морс, Северо-Запад-
ная Атлантика (южный склон Боль-
шой Ньюфаундлендской банки) и
Большой Австралийский залив. Ба-
тиаль (преимущественно средняя ба-
тиаль).
13(12). Чернильный мешок и
анальные папиллы развиты нормаль-
но ...........................14.
14(17). Нет светящихся органов
на чернильном мешке. Присоски рук
108
в 2 или 4 ряда. В средних частях
боковых рук нет резко увеличенных
присосок. Гсктокотилизируются обе
спинные руки................. . 15.
15(16). Булава щупалец расши-
рена, нс изогнута, присоски на була-
ве крупные или средних размеров, в
6-—12 рядов (изредка до 16 рядов,
в этом случае присоски в дисталь-
ных частях рук расположены в 4 ря-
да). Присоски рук в 2 или 4 ряда.
Присоски на гектокотилизироваппых
руках уменьшены (иногда очень сла-
бо), на измененных частях рук раз-
виваются вентральные защитные-
мембраны, отделенные от стебельков
присосок глубоким желобком. Нет
папилловидных желез по бокам пря-
мой кишки.
Род Rossia Owen, 1934 (=Epistij-
china Gistcl, 1848, Allorossia Gritnpe,.
1922) (см. рис. 26). Типовой вид — R.
palpebrosa Owen, 1834. 9—10 видов
в Арктике, бореальных водах Атлан-
тики и Пацифики и тропических во-
дах Западной Атлантики. Нижняя
сублитораль и батиаль. В водах
СССР 4—5 видов.
16(15). Булава щупалец узкая, не
расширенная, часто скрученная и
спиралевидно изогнутая, с микроско-
пическими присосками в 25—50 ря-
дов. Присоски рук в 2 ряда. В сред-
них частях гектокотилизированных
рук присоски резко увеличены, с
вентральной стороны развивается мя-
систая кожная складка. Пара папил-
ловидпых желез расположена по обе
стороны прямой кишки.
Род Austrorossia Berry, 1918 (см.
рис. 27,з—о). Типовой вид — A. aus-
tralis (Berry, 1918). 3—5 видов в
нижней сублиторали и верхней ба-
тиали (от 100—200 до 700—800 м)
тропических и субтропических вод
Западной и Восточной Атлантики и
Индо-Вестпацифики. Видовой состав
рода точно не установлен, различия
между видами сомнительные.
17(14). Имеется парный светя-
щийся орган в передней части чер-
нильного мешка. Присоски рук в
2 ряда, булавы в 5—8 рядов. В сред-
них частях боковых рук присоски
сильно увеличены в размерах. Гекто-
котилизируется только левая спинная
рука, в ее средней части развивает-
ся широкая наружная (вентральная)
мембрана, присоски сидят в 4 ряда.
Род Semirossia Steenstrup, 1881
(см. рис. 27, а—<w). Типовой вид —
3. tenera (Verrill, 1880). 3 вида у
восточного побережья Америки от
Новой Шотландии до Огненной Зем-
ли и у южной части Чили. Нижняя
сублитораль и верхняя батиаль.
18(1). Передний край мантии на
брюшной стороне вытянут в обшир-
ный выступ, прикрывающий снизу во-
ронку и иногда доходящий до уровня
середины глаз или далее; посредине
выступа имеется вырезка для устья
воронки. Руки первых трех пар соеди-
нены глубокой умбреллой (перепон-
кой). Присоски рук в 2 ряда, у сам-
цов некоторые присоски боковых рук
резко увеличены. Булава узкая, сла-
бо расширенная, с очень мелкими
присосками. Плавники большие, их
длина 60—100% длины мантии.
Имеется светящийся орган на брюш-
ной стороне чернильного мешка. Гек-
токотилизируются обычно обе спин-
ные руки. Гладиус отсутствует. Дон-
ные (главным образом батиальные)
или пелагические животные.
Подсемейство Heterotcuthi-
пае............................19.
19(24). Мантия срастается с го-
ловой на спинной стороне. Донные
или придонные животные, изредка
попадаются в пелагиали ... 20.
20(21). Тело слегка сжато в спин-
но-брюшном направлении. Длина
плавников 60—80% длины мантии.
У самца увеличены присоски 2-й и
3-й пар рук. Присоски булавы в 12
рядов. Затылочная лента узкая.
Род Sepiolina Naef, 1912. Один
вид — S. nipponensis Berry, 1911
(рис. 31, а—д). Южная Япония и
Филиппинские острова, Большой Ав-
стралийский залив. Верхняя батиаль.
21(22). Тело пе сжато в спинно-
брюшном направлении, «толстое».
Длина плавников обычно более 80%,
иногда более 100 % длины мантии.
У самца увеличены присоски 2-й или
3-й лар рук. Присоски булавы мел-
кие, от 8 приблизительно до 16 ря-
дов ...........................22.
22(23). Выступ на брюшной сто-
.роне мантии (брюшной щит) дохо-
дит только до уровня глаз. Места
срастания мантии с головой располо-
жены значительно дорсальнее уров-
ня середины глаза, ширина затылоч-
ной ленты 2/5—’/2 ширины головы.
Плавники прикрепляются в передней
половине мантии, их задние края не
доходят до конца тела. Глаза ма-
ленькие. Пара резко увеличенных
присосок находится в середине 2-й
руки самца.
Род Stoloteuthis Verrill, 1881.
Один вид — S. leucoptera (Verrill,
1878) (рис. 31, —ж). Северная Ат-
лантика: от залива Святого Лаврен-
тия до Флоридского пролива и в
Бискайском заливе. Нижняя субли-
тораль и верхняя батиаль. Близкая,
может быть, идентичная форма об-
наружена в верхней батиали у остро-
вов" . Кергелен, Принс-Эдуард и на
банке Дисковери.
23(22). Выступ на брюшной сто-
роне мантии (брюшной щит) захо-
дит за уровень переднего края глаза
и доходит до основания рук. Места
срастания мантии с головой находят-
ся па уровне середины глаза или
чуть выше, ширина затылочной леп-
ты почти равна ширине головы.
Плавники прикрепляются в середине
мантии, их задние края доходят до
заднего края мантии или заходят за
пего. Глаза очень крупные.
Род Iridoteuthis Naef, 1912 (рис.
31,з—к). Типовой вид — I.iris (Ber-
ry, 1909). 2 вида в Пацифике.
а (б). 3 пары увеличенных присо-
сок на 3-й руке самца. Плавники
сзади далеко заходят за задний
край мантии. I. iris (Berry, 1909)
(рис. 31,з—и). Гавайские острова.
Верхняя батиаль. Морс Серам (?).
Мезопслагиаль.
б (а). 3 пары увеличенных присо-
сок на 2-й руке самца (?). Плавни-
ки сзади доходят до заднего края
мантии или чуть заходят за пего 1.
1 Строение рук имевшегося в нашем
распоряжении самца /. maoria резко
отличается от указанного в перво-
описании. Концы рук 1—3-й пар
расширены и уплощены с боков; на
1-й парс присоски вентрального ряда
увеличены, а дорсального уменьшены
в размерах, на 2-й паре дистальная
часть руки занята крохотными при-
сосками, сидящими в один ряд на
очень длинных стебельках; в средней
части рук 3-й пары имеется несколь-
ко пар таких рудиментарных присо-
сок, причем стебельки присосок дор-
сального ряда много длиннее и тол-
ще, чем вентрального, дистальная
часть этой пары рук лишена присо-
сок. Ювенильный экземпляр из того
же трала по строению рук сходен с
.молодью из типовой серии.
109
I. maoria Dell, 1959 (рис. 31,л).
Северный остров Новой Зеландии,
пролив Кука, поднятие Чатем. Верх-
няя батиаль. Указание (Несис, 1974в,
с. 82) на находку у Кергелена оши-
бочно: в действительности оно отно-
сится к Stoloteuthis sp (см. выше).
24(19). Мантия нс срастается с
головой на спинной стороне. Брюш-
ной выступ доходит до уровня осно-
вания рук. Донные или пелагические
животные .......................25.
25(26). Присоски в дистальных
частях рук сидят на толстых (мно-
го толще самих присосок) длинных
ножках. Плавники прикреплены по-
средине тела и далеко заходят за
передний край мантии.
Род Necioteuthis Verrill, 1883 (см.
рис. 31, л—н). Один вид — N.pourta-
lesi Verrill, 1883 в батиали у Флори-
ды и Антильских островов.
26(25). Присоски сидят, как
обычно, на коротких тонких стебель-
ках. Плавники прикрепляются к зад-
ней половине мантии и не доходят
до ее переднего края. Пелагические
(эпи-мезопслагичсские) нерито-океа-
нические животные, взрослые встре-
чаются на дне в батиали.
Род Heteroteuthis Gray, 1849 (рис.
31,о—ф). Типовой вид — Н. dispar
(Riippell, 1845). 2 подрода: Hetero-
teuthis s. str. с двумя и Stephanoteut-
his Berry, 1909 [типовой вид — H.
(S.) haw aliens is Berry, 1909] c 2 по-
минальными видами. Тропики и суб-
тропики всех трех оксанов.
При определении видов рода He-
teroteuthis необходимо иметь в виду,
что строение рук (важнейший систе-
матический признак) у неполовозре-
лых особей резко отличается от зре-
лых самцов и (в отличие от прочих
сспиолид) зрелых самок.
Таблица для определения видов рода
Rossia
1 (4). Присоски в середине и дисталь-
ных частях рук сидят в 4 ряда. При-
соски булавы мелкие, одинаковые.
Тело плотное, кожа гладкая . . 2.
2(3). Присоски па булаве пример-
но в 10 рядов. Передний край плав-
ников не доходит до переднего края
мантии.
R. macrosoma (delle Chiaje, 1829)
(рис. 26,а). От Западной Норвегии
и Фарерских островов до Средизем-
ного, Адриатического морей и Севе-
ро-Западной Африки (Сенегал,
110
Азорские острова); отмечена также
в Гренландском море у берегов Грен-
ландии и Исландии. Преимуществен-
но верхняя батиаль.
3(2). Присоски на булаве пример-
но в 16 рядов. Передний край плав-
ников заходит за передний край
мантии.
R. brachyura Verrill, 1883 (рис.
26,6). Большие и Малые Антильские
острова.
4(1). Присоски на руках сидят в
два ряда; в средних частях рук они
иногда перестраиваются в 3—4 ряда,
но в дистальных частях — ни-
когда . ..................5.
5(6). Присоски булавы резко не-
равных размеров: в проксимальной
части булавы они значительно круп-
нее, чем в дистальной, самые круп-
ные присоски в дорсальных рядах.
В проксимальной части булавы при-
соски расположены в 4, в дисталь-
ной— в 6 рядов. Кожа гладкая, тело
мягкое, слегка студенистое.
R.moelleri Steenstrup, 1856 (рис.
26, в—д). Высокоарктический вид.
От Гренландии и Шпицбергена па
восток до моря Лаптевых, на запад
до- залива Амундсена, на юг до Се-
верного Лабрадора, в Норвежском,
Белом и южной части Баренцева мо-
ря отсутствует.
6(5). Присоски булавы примерно
одинаковых размеров, среди них нет
резко увеличенных .............7.
7(8). Спинная сторона головы и
мантии покрыта мелкими округлыми
папиллами, число и степень выра-
женности которых сильно варьиру-
ют; в типичном случае папиллы
крупные и редко расставленные, по
у молоди и некоторых локальных
форм (например, в Северо-Западной
Атлантике) папиллы мелкие и плохо
заметны. Присоски булавы в 7—10
рядов.
R.palpebrosa Owen, 1834 (рис.
26, е—и). Арктическо-бореальный
приатлантический вид. От Гренлан-
дии и Шпицбергена до Карского мо-
ря и Канадского Арктического архи-
пелага, на юг проникает в Западной
Атлантике до Южной Каролины, в
Восточной — до Шотландии и север-
ной части Северного моря. Обычен в
Баренцевом (кроме юго-восточной
части) и Карском морях, в Белом
море отсутствует (см. примечание на
с. 111).
8(7). Поверхность тела и головы
гладкая, лишена папилл ... 9.
9(10). Передний край плавников
заходит за передний край мантии.
Присоски булавы мелкие, в 8 или бо-
лее рядов. Присоски рук шарообраз-
ные с широкими отверстиями. Кон-
систенция тела довольно мягкая.
R. megaptera Verrill, 1881 (рис.
26, к). Северо-Западная Атлантика
от Дсвисова пролива до Новой
Англии.
10(9). Передний край плавников
не доходит до переднего края ман-
тии ..........................11.
11(16). Присоски рук шарообраз-
ные ..........................12.
12(15). Присоски рук с узкими
вертикально-щелевидными отверсти-
ями. Булава с 7—8 рядами присо-
сок ..........................13.
13(14). Консистенция тела плот-
ная. Затылочный хрящ быстро сужи-
вается назад. Присоски па гектокоти-
лизированных руках самца сильно
уменьшены в размерах. Присоски
булавы с широкими отверстиями.
В средних частях рук взрослых осо-
бей присоски обычно скучены в 3—
4 ряда.
R. pacifica Berry, 1911 (рис. 26, л,
л*)- Тихоокеанский бореальный вид.
От Берингова моря до Корейского
пролива и Калифорнии. Обычен в
Охотском и Японском морях и у
Курильских островов. В Японии до-
бывается.
14(13). Консистенция тела до-
вольно мягкая. Затылочный хрящ с
почти параллельными краями. При-
соски па гектокотилизировапных ру-
ках самца лишь немного меньше,
чем на боковых руках. Присоски бу-
лавы с узкими отверстиями. Присос-
ки па руках всегда в 2 ряда.
R. mollicella Sasaki, 1920 (рис.
26, я—о). Залив Петра Великого
[указанная в работе Н. Н. Кондако-
ва (1941) R. macrosoma принадлежит
к этому виду], тихоокеанское побе-
режье Хоккайдо и Хонсю.
15(12). Присоски рук с округлы-
ми отверстиями. Присоски булавы
с узкими отверстиями, расположены
в 10—12 рядов. Консистенция тела
довольно мягкая.
R.bullisi Voss, 1956 (рис. 26,л).
Мексиканский залив, Флоридский
пролив. Верхняя батиаль.
16(11). Присоски рук удлиненной
формы, бочонковидные, с узкими
вертикально-щелевидными отверстия-
ми. Присоски булавы с широкими
отверстиями, расположены в 10 ря-
дов. Консистенция тела довольно
мягкая.
R. tortugaensis Voss, 1956 (рис,
26, р). Найдена у Южной Флориды.
Примечание. В вид R. palpeb-
rosa Owen, 1834 (=/?. hyatti Verrill,,
1878) включена R. glaucopis Loven,
1846 (--R. sublaevis Verrill, 1878).
Пока неясно, разные это виды или
варистсты одного вида. R. palpebrosa
отличается от R. glaucopis тем, что у
нее присоски в проксимальных час-
тях рук перестраиваются в 4 ряда
(рис. 26, з—п), а бугорки на теле
значительно крупнее и заметнее, чем
у R. glaucopis, однако степень выра-
женности этих признаков очень из-
менчива. В Арктике R. palpebrosa
почти столь же обычна, как и
R. glaucopis, в южных частях ареала
встречаются только особи, сходные
с R. glaucopis.
Таблица для определения видов рода
Semirossia
1(4). Присоски булавы приблизитель-
но одинаковых размеров, дорсаль-
ные лишь немногим крупнее вен-
тральных. Увеличенные присоски в-
средних частях боковых рук самца
более или менее постепенно умень-
шаются по направлению к концам
рук . : . ......................2.
2(3). На гекто котил из иров ан ной
руке присоски расположены в прок-
симальной части (около трети) руки
в 2 ряда, в дистальной — в 4.
S. equalis (Voss, 1950) (рис. 27,а).
Западная Атлантика от Джорджии
до Флориды и Кубы, Мексиканский
залив, Карибское море.
3(2). На гектокотилизированной
руке присоски расположены в прок-
симальной части в 2 ряда, в сред-
ней — тесным зигзагом (кажется,,
будто в 4, но на самом деле в 2 ря-
да), па конце руки — в 4 ряда.
S.patagonica (Smith, 1881) (рис.
(рис. 27, б—г). Юг Южной Америки?
от о-ва Чилоэ (Чили) и залива Ане-
гада (Аргентина) до Огненной Зем-
ли и Фолклендских островов.
4(1). На булаве присоски двух
дорсальных рядов в 2—3 раза круп-
нее присосок двух вентральных ря-
дов. Крупные присоски в средней
части боковых рук самца резко сме-
няются мелкими присосками дисталь*
ИИ
шых частей рук. На гектокотилизиро-
ванной руке присоски сначала
(в проксимальной четверти или тре-
ти руки) сидят в 2 ряда, затем в 4,
а на самом конце руки — снова в 2
ряда.
S. tenera (Verrill, 1880) (рис.
27,д—ж). Западная Атлантика от
Новой Шотландии до Бразилии,
включая Мексиканский залив и Ка-
рибе кое море.
Таблица для определения видов рода
Austrorossia
1(2). Диаметр присосок 0,15—
Ю,25 мм.
A. antillensis (Voss, 1955) (рис.
27, з—и). Южная Флорида, Мекси-
канский залив, Куба.
2(1). Диаметр присосок в средней
•части булавы 0,07—0,15 мм, обычно
не более 0,1 мм.................3.
3(4) Присоски булавы располо-
жены в 24—30 рядов.
A. bipapillata (Sasaki, 1920), (рис.
27, к—м). От Южного Хонсю до
Филиппинских островов.
4(3). Присоски булавы располо-
жены в 30—50 нерегулярных ря-
дов ............................5.
5(6). Вид распространен у Аф-
рики.
A. mastigophora (Chun, 1915)
Г=? A. enigmatica (Robson, 1924)]
(рис. 27, я). Западная, Южная и
Восточная Африка, от Гвинеи и Со-
мали до мыса Доброй Надежды.
6(5). Вид распространен у Авст-
ралии.
A. australis (Berry, 1918) (рис.
27, о). Большой Австралийский залив
и Тасманово морс у Бассова проли-
ва. Возможно, сипопим предыдуще-
го вида.
Таблица для определения видов рода
Euprymna
1(2). Присоски па руках в 2 ряда.
У самца нет специально увеличенных
присосок. На гектокотилизированной
руке измененные присоски занимают
лишь самый копчик руки.
Е. phenax Voss, 1962 (рис. 28, а, б).
Филиппинские острова.
2(1). Присоски па руках в 4 ряда.
У самцов имеются резко увеличенные
присоски на руках. Измененные при-
соски занимают примерно половину
гектокотилизированной руки . . 3.
3(6). Увеличенные присоски на
.2—3-й и (если имеются) 4-й руках
самца располагаются только в вент-
ральном ряду ..................4.
4(5). Увеличенные присоски име-
ются только на 2-й и 3-й руках сам-
ца — по 2 в вентральных рядах.
E.tasmanica (Pfeffer, 1884). Вос-
точная и Южная Австралия и Тас-
мания.
5(4). Увеличенные присоски име-
ются в вентральных рядах 2-й, 3-й и
4-й пары рук самцов (приблизитель-
но по 10 шт.).
Е. mor sei (Verrill, 1881) (рис.
28, в—г). Япония, КНР, Филиппины,
Индонезия, Индия. Третьестепенный
промысловый вид.
6(3). По крайней мере, на 4-й па-
ре рук самца увеличенные присоски
располагаются и в дорсальном, и в
вентральном ряду ..............7.
7(8). Резко увеличенные присоски
располагаются в вентральном ряду
3-й (3—4 шт.), в дорсальном и вент-
ральном ряду 4-й пары рук самца.
В дорсальных рядах 2-й и 3-й пар
рук самца нет таких присосок.
Е. stenodactyla (Grant, 1883) (рис.
28, д—ж). Индо-Всстпацифика —
от Маскаренских островов до Квин-
сленда и Полинезии.
8(7). Резко увеличенные присоски
располагаются в дорсальном и вен-
тральном рядах 2-й и 4-й, иногда
также 3-й пары рук самца . . 9.
9(10). На 3-й паре рук самца нет
резко увеличенных присосок, они
есть на 1-й правой руке.
Е. albatrossae Voss, 1962 (рис.
28, з, и). Филиппинские острова.
10(9). На 3-й парс рук самца
есть резко увеличенные присоски; на
4-й правой руке нет резко увеличен-
ных присосок, краевые лишь немного
крупнее, чем присоски средних ря-
дов ..........................11.
11(12). 6—7 пар увеличенных
присосок па руках 4-й пары самца.
Е. berryi Sasaki, 1929 (рис. 28, к,
л). Япония, КНР, Тонкинский залив.
12(11). 3—4 пары увеличенных
присосок па руках 4-й пары самца.
E.scolopes Berry, 1913 (рис. 28, X
«). Гавайские острова.
Таблица для определения видов рода
Inioteuthis
1(2). Левая спинная рука самца в
базальной половине резко расшире-
на, с глубокой вырезкой на вентраль-
ной стороне ...................2.
112
8 К. Н. Несис
л
г/'С
и
о
77
р
Рис. 26. Сем. Sepiolidae, род Rossia: а —
R. macrosoma; б — R. b г achy ига; в—д —
R. moelleri, общий вид (в), булава (г),
гектокотилизированные руки (д), е—и —
R. palpebrosa, общий вид (е), булава (ас),
боковые руки R. palpebrosa (з) и R, glau-
copis (и); к—R. megaptera; л—м— R. ра-
ci]ica\ н—о — R. mollicella (м, о — затылоч-
ный хрящ); п — R. bullisi; р — R. tortu-
gaensis (а—е — по Muus, 1959; б, п, р — по
G. Voss, 1955, 1956; в, д — по Pfeffer, 1908;
г, ж—и, л, к —по Кондакову, 1937, 1941;
к —по Verrill, 1882; м, о —по Okutani,
1973)
114
Рис. 27. Роды Semirossia {а—ж} и Austro-
rossia (з—о): а — S. equalise б—г — S. ра-
tagonica, общий вид (б), боковая рука (в),
гектокотиль (г); д—ж— S. tenerat общий
вид (д), руки самца (е), булава (»с); з—
и —A. antillensts, общий вид (а), булава
(w): к—м — A. bipapillata, общий вид (к)>
булава (л), анус с анальными папиллами
и срединный компонент вороночного орга-
на (ле); н — A. masiigophora; о —A. aus-
- - •— 1955т
trails (а, з—м — no G. Voss» 1950, ________________,
1963: б—г —по Smith, 188Г из Joubin, 1902;
О—ж — по Verrill, 1882; н —по Chun, 1915;
о — но Berry, 1918)
в?
Рис. 28. Роды Еиргупъпа (а—н) и Inioieut-
his (о—ф): а—б — Е. phenax; в—г — Е. mor-
sei; д—ж — Е. stenodaciyla; з—и — Е. al*
batrossae; к—л — Е. berryi; м—н, — Е. s co-
lopes; о—fi — I. japonica; p—c — I. maculo-
sa; т—ф — I. capensis; a, e, 3, m, o, p —
самцы; в, д, т — самки; б, г, ж, и, л, л,
с — руки самцов; н, у -— гсктокотили; ф —
фотофор (а—б, е—и, р—ф — по G. Voss,
1962, 1963; в — по Арре! 16f, 1886; г, л, о—
л — по Sasaki, 1929; д — по Joubin, 1902;
к — по Voss, Williamson, .1972, м—н — по
Berry, 1914)
2(3). Левая спинная рука самца
намного короче правой. На дорсаль-
ной стороне расширенной базальной
части левой спинной руки имеется
несколько мелких присосок. В цен-
тральной части копуляторного аппа-
рата имеется крупная уковидпая ло- .
пасть без папилл.
I. japonica Verrill, 1881. [=/. inio-
teulhis (Naef, 1912)] (рис. 28, о, n).
Южная Япония, КНР.
3(2). Левая спинная рука самца
не укорочена. На дорсальной сторо-
не расширенной части этой руки ист
присосок. В центральной части копу-
ляторного аппарата две небольшие
языковидные папиллы.
1. maculosa Goodrich, 1896 (рис.
28, р, с). Северная часть Индийского
океана, Персидский и Бенгальский
заливы, Андаманское море, Индия,
Индонезия, Филиппины.
4(1). Левая спинная рука самца
очень слабо расширена в базальной
половине, без вырезки на вентральной
стороне. На дорсальной стороне ба-
зальной части руки несколько мел-
ких папилл. В центральной части
копуляторного аппарата с вентраль-
ной стороны две крупные треуголь-
ные папиллы, между ними одна ма-
ленькая.
L capensis Voss, 1962 (рис. 28, г—
ф). Южпая Африка, от залива Лю-
дериц-бей до залива Моссел-бей.
Таблица для определения видов рода
Sepietta
1(2). Присоски булавы крупные, при-
мерно в 12 рядов, присоски дорсаль-
ных рядов крупнее, чем вентральных.
Плавники округлые: место наиболь-
шей ширины плавника находится
посредине его длины. Левая спинная
рука самца слабо расширена ди-
стальнее копуляторного аппарата; в
этой части руки присоски дорсально-
го ряда крупнее, чем вентрального,
и плавно уменьшаются от копуля-
торного аппарата к концу руки. Рог
копуляторного аппарата короткий;
только 3 присоски находятся прокси-
мальнее копуляторного аппарата.
S. petersi. (Steenstrup, 1887)
(=S. obscura Naef, 1916) (рис.
29, а, ж). Средиземное море, Атлан-
тическое побережье Марокко (?).
Верхняя сублитораль.
2(1). Присоски булавы очень мел-
кие, более чем в 16 рядов, дорсаль-
ные не крупнее вентральных. Плав-
ники сердцевидные: их наружный
край угловатый, а место наибольшей
ширины сдвинуто к заднему краю.
Левая спинная рука самца сильно
расширена дистальнее копуляторно-
го аппарата; проксимальнее этого
аппарата находятся 4 присоски . 3.
3(4). На левой спипной руке сам-
ца в дорсальном ряду присосок ди-
стальнее копуляторного аппарата
находятся 2 крупные, затем 2—4
мелкие и снова 2 крупные присоски,
далее присоски равномерно умень-
шаются. Рог копуляторного аппара-
та длинный — более половины шири-
ны руки, закрученный. Животные
довольно крупные (общая длина
без щупалец до 7—9 см).
5. oweniana (d’Orbigny, 1839)
(рис. 29, 6,6, w). От Западной Норве-
гии и Фарерских островов до Среди-
земного моря, Мадейры и Маврита-
нии. Шельф (преимущественно внеш-
ний) и верхняя батиаль.
4(3). На левой спинной руке сам-
ца в дорсальном ряду присосок
дистальнее копуляторного аппарата
4 резко увеличенных присоски. Рог
копуляторного аппарата короткий,
значительно менее половины ширины
руки, слабо изогнутый. Животные
мелкие (общая длина без щупал ед
до 5 см).
S. neglecta Naef, 1916 (рис. 29, г,
д, з). От Южной Норвегии и Орк-
нейских островов до Средиземного
моря и у Атлантического побережья
Марокко. Верхняя сублитораль.
Неясно систематическое положе-
ние S. scandica (Steenstrup, 1887).
Одни авторы считают этот вид са-
мостоятельным, другие — лишь фор-
мой S. oweniana. S. ’scandica зарегист-
рирована в североевропейских водах
от Исландии и Западной Норвегии
до южной части Северного моря.
Таблица для определения видов рода
Sepiola
Побережья Европы, Северной Афри-
ки и Красное море.
1(14). Присоски на руках 4-й па-
ры в 2 ряда по всей длине ру-
ки ............................2.
2(5). Передний край мантии на
брюшной стороне с глубокой вырез-
кой против воронки и двумя полу-
круглыми выступами по бокам от
края вырезки. Присоски булавы мел-
кие, одинаковых размеров в дор-
118
Рис. 30. Род Sepiola, а—ф — свропейско-
североафриканские, х—щ — азиатские ви-
ды: а—в — S. pfefferi; г, о — 5. aurantiaca;
д, п — 5. ligulata; е, у ~ S. robusta; ж—
з, р — S. rondeleti; и, т — S, а[ finis; к, с —
S. intermedia', л,, л2, н — S. steenstrupiana;
Mi, м2, ф — S. atlantica: х—ц — S. birostra-
ta; ч—ш — S. trirostrala; щ — S. parva;
а, б, з — самки; г—ж, и—м, х, ч — самцы;
в, ц, ш— руки самца; н—ф, щ— гектоко-
тилнзированные руки; л2, л<2 — брюшная
рука (а, б, м2 — по Pfeffer, 1908; в — по
Grimpe, 1921; г—л, н—у — по Naef, 1923;
л<1, ф — по Muus, 1959; ' х—ц — по Sasaki,
1929; ш — по G. Voss, 1963; щ — по
Okutani, 1973)
сальных и вентральных рядах . 3.
3(4). Вырезка переднего края
мантии на брюшной стороне V-образ-
яая, ее глубина около Vs вентральной
длины мантии. Копуляторный аппа-
рат развит как обычно, только на
левой спинной руке самца.
S. pfefferi Grimpe, 1921 (рис.
30, а—в). Бореальный вид: от Фа-
рерских островов и Южной Норвегии
до Бретани (Франция). Шельф.
Возможно, пе самостоятельный вид,
а форма 5. aurantiaca,
4(3). Вырезка переднего края
мантии на брюшной стороне U-об-
разпая, ее глубина около JA вент-
ральной длины мантии. На правой
спинной руке самца имеется допол-
нительный копуляторный аппарат —
два небольших языковидных выроста
в базальной части руки.
S. aurantiaca Jatta, 1896 (рис.
30, г, о). От Южной Норвегии до
Средиземного моря. Внешний шельф
и верхняя батиаль.
5(1). Передний край мантии на
брюшной стороне волнообразно
изогнут, без глубокой вырезки . 6.
6(7). Левая спинная рука самца
резко ложковидно расширена, оба
ряда присосок дистальной части ру-
ки далеко расставлены. Копулятор-
ный аппарат состоит из двух круп-
ных языковидных лопастей (одна из
которых лежит в ложковидном рас-
ширении руки) и закрученного боко-
вого выступа. Передний край мантии,
на брюшной стороне сильно волно-
образно изогнут. Присоски булавы
мелкие, одинаковых размеров.
3. ligulata Naef, 1912 (рис.
30, д, п). Средиземное и Адриатиче-
ское моря. Глубины 200—250 м.
7(6). Левая спинная рука самца
нс расширена или слабо расширена.
Копуляторный аппарат состоит из
одной сложно устроенной изогнутой
лопасти. Передний край мантии на
брюшной стороне очень слабо изо-
гнут ......................а • 8.
8(9). Левая спинная рука самца
слегка расширена дистальнее копу-
ляторпого аппарата и резко искрив-
лена (закручена) на конце. Присоски
обоих рядов в дистальной части этой
руки примерно равных размеров.
Дорсальные присоски булавы круп-
нее вентральных.
5. robusta Naef, 1912 (рис.
30,е,у). Средиземное море. Внеш-
ний шельф.
9(8). Левая спинная рука самца
не расширена. Присоски обоих рядов
в дистальной части этой руки резко
неравных размеров..............10.
10(13). Левая спинная рука сам-
ца резко изогнута, начиная с сере-
дины. Дорсальные присоски булавы
несколько крупнее вентраль-
ных ............................И.
11(12). Все присоски дорсального
ряда в дистальной части левой спин-
ной руки самца намного крупнее со-
ответствующих присосок вентраль-
ного ряда.
S. rondeleti Steenslrup, 1856 (рис.
30, яс, з, р). Средиземное морс и лу-
зитанские воды, на юг до Западной
Сахары (?). Шельф и самая верхняя
батиаль.
12(11). Только 3—4 присоски
дорсального ряда в середине ди-
стальной части левой спинной руки
самца резко увеличены.
S. affinis Naef, 1912 (рис. 30, и, г).
Средиземное морс. Верхняя сублито-
раль.
13(10). Левая спинная рука сам-
ца слегка изогнута лишь на самом
конце. 2—3 присоски дорсального
ряда этой руки непосредственно за
копуляторпым аппаратом резко уве-
личены. Дорсальные и вентральные
присоски булавы равных размеров.
S. intermedia Naef, 1912 (рис.
30,к,с). Средиземное море. Шельф.
14(1). Присоски на концах рук
4-й пары более чем в 2 ряда. При-
соски дорсальных рядов на булаве
крупнее, чем вентральных . . 15.
15(16). На концах рук 4-й пары
3—4 ряда присосок. Левая спинная
рука самца пе изогнута и без особо
увеличенных присосок.
S. sleenstrupiana Levy, 1912 (=S.
ienera Naef, 1912) (рис. 30, л,«).
Средиземное море. Верхняя сублито-
раль. Красное морс (?).
16(15). На концах рук 4-й пары
4—8 рядов присосок. Левая спинная
рука самца сильно изогнута, начиная
с середины; в середине се дисталь-
ной части 3—4 присоски дорсального
ряда резко увеличены.
S. atlantica d’Orbigny, 1839 (рис.
30, Л1, ф). Бореально-лузитанский вид:
от Исландии, Фарерских островов и
Западной Норвегии до Марокко;
в Средиземном море отсутствует.
Шельф и зона перегиба к склону.
122
Побережья Восточной и Юго-Восточ-
ной Азии, западная часть Тихого оке-
ана и прилегающие моря
1(4). Копуляторный аппарат левой
спинной руки самца состоит из 2—
3 выростов, между ним и изменен-
ными присосками дистальной части
руки не более 1—2 увеличенных при-
сосок (в дорсальном ряду) . . 2.
2(3).. Левая спинная рука самца
с двумя рожковидными выростами
копуляторного аппарата. Присоски
дорсального и вентрального рядовка
правой спинной руке и руках 2-й па-
ры самца одинаковых размеров. При-
соски булавы мелкие, у основания их
4 ряда, в дистальной части булавы
до 16 рядов.
3. birostrata Sasaki, 1918 (рис.
30, х,ц). Южный Сахалин, Южные
Курильские острова, Приморский
край, Япония, КНДР и Южная Ко-
рея, северные районы КНР. Шельф.
3(2). Левая спинная рука самца
с двумя рожковидными и одним язы-
ковидным выростом копуляторного
аппарата. Присоски вентрального ря-
да на правой спинной руке и руках
2-й пары самца намного крупнее
присосок дорсального ряда. Присос-
ки булавы крупные, в 4 ряда, дор-
сальные присоски крупнее вентраль-
ных.
3. trirostrata Voss, 1962 (рис.
30,'/, ш). Филиппинские острова.
4(1). Копуляторный аппарат ле-
вой спинной руки самца состоит из
одного сосцевидного выроста, меж-
ду ним и измененными присосками
дистальной части руки несколько
крупных присосок в дорсальном
(4) и вентральном (2—3) ряду.
Присоски булавы мелкие, в 8 ря-
дов, дорсальные присоски не крупнее
вентральных.
3. parva Sasaki, 1914 (рис. 30, щ).
Южная Япония.
Таблица для определения видов рода
Heteroteuthis
1(4). У зрелых самцов глубина умб-
реллы между руками 1-й пары 1/2—
Уз длины руки. Руки 2-й пары нс
длиннее 1-й и 4-й, присоски на них
идут до концов.
Подрод Heteroteuthis s. str. . 2.
2(3). У зрелых самцов руки 1-й
и 2-й пар равны и значительно ко-
роче рук 3-й и 4-й пар. Увеличенные
присоски находятся па 3-й паре рук
самца: 2 громадные, в несколько раз
крупнее прочих, сидят под углом 90°
одна к другой, дистальнее от них
еще 3 помельче. Правые руки 1-й и
2-й пар с внутренней стороны на по-
ловину своей длины соединены мы-
шечной лентой. Концы рук 1-й и 2-й
пар у зрелых самок лишены присо-
сок, конец руки 2-й пары слегка
утолщен и снабжен килем на ораль-
ной стороне (в разрезе треугольный),
не отогнут наружу, не исчерчен по-
перечно. Передний край плавника
расположен на уровне середины
вентральной стороны мантии.
И. (И.) dispar (Ruppell, 1845)
(=Я. atlantis Voss, 1955) (рис.
31,о—с). Тропические и субтропиче-
ские воды Западной Атлантики от
Бермудских островов до залива Ла-
Плата, Северо-Восточная Атлантика
от Юго-Западной Ирландии до Гви-
неи, Карибское и Средиземное моря,
Мадейра, Канарские и Азорские ост-
рова. В Средиземном море — один из
самых массовых видов пелагиали,
встречается до Эгейского моря. Не-
рест у дна на склоне, личинки — в
мезо- и батипелагиали, взрослые осо-
би в нижней эпи- и мезопелагиали.
Популяции Западной и Восточной
Атлантики, по-видимому, изолиро-
ваны.
3(2). У зрелого самца руки при-
мерно равной длины, увеличенные
присоски находятся на 2-й руке, их 3,
и они примерно вдвое крупнее нор-
мальных. Передний край плавника
расположен немного дальше перед-
него края мантии. Самки неиз-
вестны.
Н. (И.) weberi Joubin, 1902 (рис.
31, т). Центральная Индонезия. Ба-
тиаль.
4(1). У зрелых самцов глубина
умбреллы между руками 1-й пары
составляет около 90% длины руки.
Нет мышечной связки между руками
1-й и 2-й пар. Руки 2-й нары равны
по длине рукам 3-й пары и значи-
тельно длиннее 1-й и 4-й пар. При-
соски на 1-й и 4-й парах рук крохот-
ные, на руках 3-й пары несколько
резко увеличенных присосок, концы
рук 2—3-й пар голые. У зрелых са-
мок руки 1-й и 2-й пар' короткие, их
концы лишены присосок и утолщены,
концы рук 2-й пары па оралыюй
стороне уплощены, поперечно исчер-
чены, отогнуты наружу.
123
Ум
н
Ж
77
/7
Z7
/о
'р
0’
70£
ж
fSSSjb:
№
&У с
<0 °-
ад
/77
ф
'Лё ч
1 IV
У
Рис. 31. Подсем. Heteroteuthinae: а—д —
Sepiolitia nipponensis сверху (а), сбоку
(б), снизу (в), руки самца (г), фотофор
(<?); е—ж—Sioloteuthis leucoptera, сверху
(е) и снизу (ж); з—и — Iriaoteuthis iris,
снизу (з) и сбоку (ы); к — /. maoria (мо-
лодая самка); л—н—N ectoteuthis pourta-
lesi, сверху (л), сбоку (.«), брюшная рука
сбоку (и); о—с — Heteroteuthis dispar, са-
мец (о), самка (л), руки самца (р), 2-я
рука самки (с); т — П. weberi (сбоку); у—
ф — Н. hawaiiensis, самка сверху (у), ее
2-я рука (ф); (а—д, п—с—по G. Voss, 1955,
1963: е—ж — по Verrill, 1882; з—и, у—ф —
по Berry, 1914; к—по Dell, 1959; л—н, т —
ио Joubin, 1902; о, р — по Naef, 1923)
Подрод Stephanoteuthis Berry,
1909. H. (S.) hawaiiensis (Berry, 1909)
( = ? 11. serventyi Allan, 1945) (рис.
31,i/, ф). Тропические и субтропиче-
ские воды Западной Пацифики: о-ва
Гавайские, Бонин, Рюкю, Индонезия,
Тасманово море (?). Большой Авст-
ралийский залив. Нижняя эпи- и ме-
зопелагиаль.
Н. (S.) dagamensis Robson, 1924.
Западная, Южная и Юго-Восточная
Африка. Батиаль. Соотношение этих
2 форм неясно.
2.1.2.3. Семейство
Sepiadariidae
Небольшие животные (длина мантии
2—4 см) с коротким туловищем и
узкими плавниками, сидящими в се-
редине или задней половине тела.
Плавники с широкими основаниями
и слабо выраженными «ушками».
Мантия на затылке срастается с го-
ловой. Руки соединены широкой ум-
бреллой, охватывающей основания
щупалец с обеих сторон, так что
стебли щупалец оказываются внутри
кожного чехла. Руки в проксималь-
ной части с двумя, в дистальной — с
двумя или четырьмя рядами присо-
сок. Гсктокотилйзируется левая
брюшная рука, дистальная часть ко-
торой лишена присосок. Гладиуса
нет. Светящиеся органы отсутству-
ют. Яйца крупные, донные, развитие
без метаморфоза. 2 рода, 7 видов.
Донные виды, обитатели шельфа
тропических и субтропических морей.
Таблица для определения родов се-
мейства Sepiadariidae
1(2). Мантия соединена с воронкой
широкой мышечной лентой, мантий-
ные и вороночные хрящи отсутству-
ют. Передний край маптии ровный.
Плавники сдвинуты в заднюю поло-
вину мантии.
Род Sepiadariurn Steenstrup, 1881
(рис; 32, а—в). Типовой вид — S. ko-
chii Steenstrup, 1881. 5 видов в Ин-
домалайской подобласти (на север
до Японии) и у Австралии.
2(1). Мантийно-вороночный за-
мыкательный аппарат обычного типа.
Передний край мантии ровный или
бахромчатый. Плавники сидят по-
средине тела.
126
Род Sepioloidea d’Orbigny, 1845
(рис. 32, г—е). Типовой вид — S. li~
neolata (Quoy et Gaimard, 1832).
2 вида у Австралии и Новой Зе-
ландии.
Таблица для определения видов рода
Sepiadariurn
1(2). Длина плавников 3Д—Vs Дли-
ны мантии. Ширина мантии немного
больше ее длины. 4-я пара рук пе
короче 2-й. Мантия и голова сверху
с маленькими круглыми красными
или коричневыми точками и черными
или темно-коричневыми пятнами.
S. auritum Robson, 1914. Северо-
Западная Австралия.
2(1). Длина плавников около по-
ловины длины маптии. 4-я пара рук
короче (реже равна) 2-й. Пигмент-
ные пятна светлые...............3.
3(4). Мантия узкая, ее ширина
составляет примерно % длины. При-
соски рук в 2 ряда, булавы в 6 ря-
дов.
S. gracilis Voss, 1962 (рис. 32, а,
б). Филиппинские острова.
4(3). Ширипа мантии примерно
равна се длине. Присоски рук, по
крайней мере местами, в 4 ряда, бу-
лавы в 6—8 рядов................5.
5(6). Присоски на булаве в 6 ря-
дов. Гектокотилизированная рука
самца дистально с серией конических
пластинок, без сплошной вертикаль-
ной мембраны.
S. austrinum Berry, 1921. Южная
Австралия.
6(5). Присоски па булаве в 8 ря-
дов. Гектокотилизированная рука
самца дистально с серией попереч-
ных весловидных пластинок, окай-
мленной вертикальной мембраной с
зубчатым краем..................7.
7(8). 1-я пара рук равна или
длиннее 2-й. В медианных частях
рук самки дистальнее середины при-
соски сидят редко, между ними есть
свободные пространства. На гектоко-
тилизированной руке самца 18—20
низких поперечных гребней.
S. kochii Steenstrup, 1881 (рис.
32, о). Северная часть Индийского
океана, Индонезия, Южно-Китайское
море.
8(7). 1-я пара рук короче 2-й.
Присоски на руках самки сидят
плотно, без свободных пространств.
На гектокотилизированной руке сам-
ца около 24 высоких острых попереч-
ных гребней.
S. nipponianum Berry, 1932. Япо-
ния (юг Хонсю, Сикоку, Кюсю). Воз-
можно, подвид S. kocfiii.
Таблица для определения видов рода
Sepioloidea
1(2). Передний край мантии по бо-
кам от затылочной складки сильно
изрезан, бахромчатый. Бока и брюш-
ная сторона головы и мантии с мно-
гочисленными бугорками — отвер-
стиями кожных пор. Окраска спин-
ной стороны тела: продольные цвет-
ные полосы голубого, оливково-бело-
го или коричневого цвета.
S. lineolata (Quoy et Gaimard,
1832) (рис. 32,а). Западная, Южная
и Юго-Восточная Австралия.
2(1). Передний край мантии не
бахромчатый. Поверхность тела
гладкая. Окраска: многочисленные
мелкие темные хроматофоры, цвет-
ных полос нет.
S. pacifica (Kirk, 1882) (рис. 32,
д, е). Новая Зеландия.
2.1.2.4. Семейство
Idiosepiidae
Очень маленькие животные (длина
мантии зрелых самцов б—17 мм, са-
мок 8—22 мм) с удлиненной, сзади
слегка приостренпой мантией. Плав-
ники маленькие, почковидные, сдви-
нуты к заднему концу тела, сидят
слегка наискось по отношению к ос»
тела. Передний край мантии на спин-
ной стороне прямой или слегка вог-
нутый, не срастается с головой; за-
тылочного хряща пет. На спинной
стороне тела в задней половине-
мантии у взрослых особей располо-
жен овальный присасывательный ор-
ган с шероховатой поверхностью
(рис. 33,а). Роговица глаза замкну-
тая. Руки короткие, небольшая умб-
релла имеется только между руками
3-й и 4-й пар. Присоски рук в два
ряда. Щупальца узкие, короткие,
втяжные, с нерасширенной булавой,
несущей 2—4 ряда присосок. Гекто-
котилизируются обе брюшные руки,
они обычно лишены присосок на
большей части длины, левая рука уз-
127
кая, с двумя лопастями на вершине:
правая широкая, уплощенная, окайм-
лена боковыми мембранами, иногда
имеет поперечные гребни и борозды.
Присоски всех рук самца увеличены.
Гладиуса нет. У самок развиты оба
яйцевода, но функционирует лишь
левый. Яйца довольно мелкие, дон-
ные, развитие с пелагической ста-
дией, но без метаморфоза.
Один род — Idiosepius Steenstrup,
1881 ( = ldiosepion Fischer, 1882, Mic-
.roteuthis Ortmann, 1888, Naefidium
Grimpe, 1920, Berryidiutn Grimpc,
1931). Типовой вид — I. pygmaeus
Steenstrup, 1881. 6 видов в Индо-
Вестпацификс.
Обитают у берегов в зарослях
морских трав и водорослей, при-
крепляются к субстрату спиной. Ли-
чинки/ молодь и «подростки» часто
встречаются в толще воды и у по-
верхности.
Таблица для определения видов рода
Idiosepius
1(2). Булава щупальца очень узкая
и длинная, с 2 рядами присосок, диа-
метр которых 0,25—0,3 мм. В базаль-
ной части брюшных рук самца по
4 присоски.
1. biserialis Voss, 1962 (рис.
33, е, ж). Южная Африка.
2(1). Булава щупальца с 4 ряда-
ми присосок......................3.
3(4). Булава щупальца расши-
ренная, присоски в середине булавы
заметно крупнее, чем у основания,
их диаметр около 0,33 мм.
I. macrocheir Voss, 1962 (рис.
-33,з—к). Южная Африка.
4(3). Булава щупальца пе расши-
ренная, присоски в середине булавы
нс крупнее, чем у основания, их диа-
метр около 0,12 мм...............5.
5(6). Брюшные руки самца несут
присоски по всей длине, 7—11 пар.
I. notoides Berry, 1921 (рис. 33, м).*
Южная и Восточная Австралия.
6(5). Брюшные руки самца несут
присоски только у основания. Пра-
вая брюшпая рука песет попереч-
ные гребни и борозды .... 7.
7(8). Правая брюшная рука сам-
ца намного короче и шире левой, на
обеих руках лишь по одной присоске
у основания.
I. picteti (Joubin, 1894) (рис. 33, л).
Восточная Индонезия.
8(7). Правая брюшная рука самца
не короче и лишь немного толще ле-
вой ..........................9.
9(10). У основания брюшных рук
самца по 4—7 присосок (изредка 3 на
правой руке).
I. paradoxus (Ortmann, 1888) (рис.
33, г, д). Южная часть Хоккайдо, Кю-
сю, Южная Корея.
10(9). У основания брюшных рук
самца по 1—3 присоски.
/. pygmaeus Steenstrup, 1881 (=/.
pygmaeus hebereri Grimpe, 1931) (рис.
33, a—в), Южно-Китайское море, Фи-
липпинские острова, Индонезия, Се-
верная и Северо-Восточная Австра-
лия.
2.2. Отряд Teuthida
Характеристику см. на с. 70.
2.2.1. Подотряд Myopsida
Передняя камера глаза целиком за-
крыта прозрачной роговицей, срос-
шейся с глазной орбитой, и связана
с внешней средой лишь крохотным-
отверстием — «слезной порой». Ман-
тия мускулистая. Плавники сидят в
задней половине мантии или протя-
гиваются вдоль большей части се
длины. Руки и щупальца только с
присосками, крючьев нет. Булава без
фиксаторного аппарата. Буккальное
крепление семилучевое, его формула
DDVV. Замыкательный аппарат про-
стой. Гладиус перовидпый, с корот-
ким узким рахисом, без. конечного
конуса. Фотофоры отсутствуют или
имеется пара на чернильном мешке,
свечение обусловлено развитием сим-
биотических светящихся бактерий.
Развит только левый яйцевод. Есть
парные нидаментальные и придаточ-
ные нидаментальные железы. Гекто-
котилизируются одна или обе брюш-
ные руки. Яйца крупные, донные, в
студенистых капсулах.
Обитатели шельфа тропических и
умеренных вод; некоторые виды мо-
гут встречаться и в верхней батиали;
2 семейства, 8 родов, 45—54 вида.
Обзоры: Зуев, Несис, 1971; Roper,
Young, Voss, 1969.
Таблица для определения семейств
Myopsida
1(2). Плавники доходят до зад-
него конца мантии и соединяются
128
9 К. П. Несис
между собой. Задний конец мантии
конический. Булава щупалец с 4 ря-
дами присосок. Длина мантии взрос-
лых кальмаров нс менее 3 см.
Семейство Loliginidae . . . 130.
2(1). Плавники почковидной фор-
мы, не доходят до заднего края ман-
тии и не соединяются между собой.
Задний конец мантии округленный.
Булава щупалец в проксимальной ча-
сти с 2, в дистальной с 4 рядами при-
сосок. Длина мантии взрослых каль-
маров 1,5—2 см (см. рис. 40).
Семейство Pickfordiatcuthidae
с. 148.
2.2.1 Л. Семейство
Loliginidae
Мантия мускулистая, удлиненная, ее
передний край на спинной стороне с
выступом. Плавники овальные, про-
долыго-ромбические или сердцевид-
ные, сидят в задней половине ман-
тии и доходят до ее заднего конца;
их длина часто превосходит половину
длины мантии, а иногда они доходят
до ее переднего края. Роговица гла-
за замкнутая. Руки с двумя рядами
присосок, роговые кольца которых
несут на дистальной стороне высо-
кие тупые (башневидные) зубцы,
проксимальная сторона чаще глад-
кая. Щупальца не втяжные, но их
стебли сократимы. Булава щупалец
расширенная, присоски в 4 ряда,
средние, как правило, крупнее боко-
вых. Кольца присосок несут зубцы
обычно по всему краю. Крючьев нет.
Фиксаторный аппарат не развит. За-
щитные и плавательные мембраны
на руках и булаве хорошо выраже-
ны. Буккальное крепление семилуче-
вое, его формула DDVV. Лучи бук-
кальной мембраны обычно с мелки-
ми присосками. Замыкательные хря-
щи простые. Гладиус по форме по-
хож на птичье перо с коротким ство-
лом. Иногда имеется пара фотофо-
ров на чернильном мешке. Гектоко-
тилизирустся левая брюшная рука,
редко обе. Развит только левый яй-
цевод. Сперматофоры переносятся
на^ротовую мембрану и (или) в ман-
тийную полость самки. Яйца донные,
в студенистых капсулах, развитие с
пелагической личиночной стадией.
7 родов, 44—53 вида. Обитают
на шельфе, лишь некоторые виды
иногда опускаются в верхнюю бати-
аль. Держатся у дна, но могут под-
130
ниматься в толщу воды и к поверх-
ности. Активные, подвижные некто-
бептосные кальмары. Многие виды,
особенно из рода Loligo, в меньшей
степени Sepioteuihis и Alloteuthis,
имеют важное промысловое значе-
ние. Частичные обзоры: Naef, 1921—
1923; Sasaki, 1929; Adam, 1939в,
1954; G. Voss, 1963; Cohen, 1976.
Таблица для определения родов
Loliginidae
1 (2). Плавники в виде овальной боко-
вой каймы, почти доходят до перед-
него края мантии. Тело овальное,
широкое.
Род Sepioteuthis Blainville, 1824
(= Chotidrosepia Ruppcll et Leuckart,
1828) (см. рис. 35, и—н). Типовой
вид — S. sepioidea (Blainville, 1823).
3—4 вида в тропических водах За-
падной Атлантики, Ипдо-Вестпаци-
фике и Австрало-Новозеландском
регионе. Обитают у берегов на ри-
фах и в зарослях морских трав на
открытом шельфе. Часто встречают-
ся близ поверхности. Длина мантии
до 40 см. Имеют промысловое зна-
чение.
2(1). Плавники поперечно-оваль-
ные, ромбические или сердцевидные,
далеко не доходят до переднего края
мантии. Тело коническое или цилинд-
рическое .......................3.
3(8). Маленькие кальмары (дли-
на мантии не более 10—12 см, обыч-
но до 5—6 см) с относительно ко-
роткой и широкой мантией и по-
перечно-овальным или сердцевидным
(реже ромбическим) плавником, ши-
рина которого равна или превосхо-
дит длину.......................4.
4(5). Плавник поперечно-оваль-
ный или эллиптический, сзади за-
кругленный, без оттянутого хвостика.
Род Lolliguncula Steenstrup, 1881
(см. рис. 35, а—з). Типовой вид —
L. brevis (Blainville, 1823), 5—6 ви-
дов: 5 видов в тропических водах За-
падной и Восточной Атлантики,
Красном море, Восточной Пацифике,
один (сомнительный) на Патагон-
ском шельфе. Прибрежные кальма-
ры, часто заходят в бухты и эстуа-
рии, по встречаются и на открытом
шельфе. Длина мантии обычно до
5—10 см. Два вида промысловые.
5(4). Плавник широко-сердцевид-
ный или ромбический, сзади заострен
или оттянут в короткий хвостик . 6.
6(7). Тело короткое, широкое.
Длина плавника равна или превосхо-
дит половину длины мантии, ширина
много больше длины. Гектокотилизи-
руется левая брюшная рука.
Род Loliolus Steenstrup, 1856 (см.
рис. 34, а—к). Типовой вид — L. iy-
pus Steenstrup, 1856. 3—4 вида в се-
верной части Индийского океана и
водах Индонезии. Мелкие (длина
мантии до 3—5 см) прибрежные
кальмары.
7(6). Тело короткое, но узкое и
довольно стройное. Длина плавника
у самок около Уз, ширина около 7г
длины мантии. Плавник широко-
сердцевидный, нс оттянут в хвостик.
Гектокотилизируются обе брюшные
руки, левая очень длинная (много
длиннее прочих рук), хлыстовидная,
без присосок, только с одним рядом
усиковидных выростов на конце, пра-
вая с широкой вентральной лопастью
в средней части.
Род Loliolopsis Berry, 1929. Один
вид — L. dipmedeae (Hoyle, 1904)
( = L. chiroctes Berry, 1929) (рис.
34, л—о). В тропических водах Вос-
точной Пацифики от Калифорнийско-
го залива до Перу. Длина мантии до
11 см. Прибрежные кальмары, час-
то встречаются у поверхности, в том
числе над довольно большими глу-
бинами. Имеют небольшое промысло-
вое значение.
8(3). Кальмары средних или
крупных размеров с узкой стройной
мантией и длинным продольно-ром-
бическим или узко-сердцевидным
плавником, длина которого обычно
превосходит ширину..............9.
9(10). Плавник продольно-ромби-
ческий, его передние края почти пря-
мые, задние прямые или слабо вогну-
тые. Нет длинного хвоста. Длина рук
7ч длины мантии. '
Род Loligo Lamarck, 1798 (см. рис.
36—38). Типовой вид — L. vulgaris
Lamarck, 1798. 2 подрода, 28—34 ви-
да в умеренных и тропических водах
Атлантического, Индийского и Ти-
хого океанов, на севере Атлантики
до Новой Шотландии, Фарерских
островов и Западной Норвегии, в Ти-
хом океане до Южных Курильских
островов и пролива Диксон-Энтранс,
на юге до Огненной Земли, банки
Бердвуд, Южной Африки и Тасма-
нии. Обитают близ берегов, на от-
крытом шельфе, местами в верхней
батиали. Держатся обычно у дна, но.
некоторые виды, особенно из подро-
да Doryteuthis, поднимаются к по-
верхности и подходят на свет. Длина
мантии до 50—75 см. Некоторые ви-
ды рода имеют очень большое про-
мысловое значение.
а (б). Боковые стороны гладиуса
дугообразно выгнуты и нс утолщены
(см. рис. 36, в). Ширина мантии в
3—6 раз меньше длины.
Подрод Loligo s. sir. 19—23 вида.
Ареал тот же, что и у рода в целом.
б (а). Боковые стороны гладиуса
прямые и утолщенные - (см. рис.
38,6). Ширина мантии в 4—9, обыч-
но в 6—8 раз меньше длины.
Подрод Doryteuthis Naef, 1912.
Типовой вид — D. plei (Blainville,
1823). 9—11 видов в тропических и
субтропических водах Западной Ат-
лантики и Индо-Вестпацифики, на
север до Хоккайдо и Южных Ку-
рильских островов.
10(9). Плавник сердцевидный или
продольно-ромбический, оттянутый
сзади в длинный (у самцов чрезвы-
чайно длинный, иногда >7г длины
мантии) тонкий хвост. Гладиус уз-
кий, почти не расширенный. Ширина
мантии в 6—15 раз меньше ее дли-
ны. Руки обычно ^7ч длины ман-
тии ...........................13.
11(12). Передние края плавника
выпуклые. Лучи буккальной мембра-
ны лишены присосок. На брюшной
стороне мантии самца нет продоль-
ного кожного гребня. Нет светящих-
ся органов на чернильном мешке.
Род Alloteuthis Naef in Wulker,
1921 (=Acruroieuthis Berry, 1920,
Acrololigo Grimpe, 1921) (см. рис.
39,a—к). Типовой вид — A. media
(Linne, 1758). Три вида в Восточной
Атлантике и Средиземном море, от
Южной Норвегии до Анголы. Массо-
вые верхнесублиторальные кальма-
ры, служащие объектами неболь-
шого промысла. Длина мантии до
20 см, чаще не более 10—15 см.
12(11). Передние края плавника
прямые. Лучи буккальной мембраны
с несколькими мелкими присосками.
На брюшной стороне мантии самца
вдоль ее средней линии проходит
кожный гребень. Два светящихся ор-
гана па чернильном мешке по обе
стороны кишки.
Род Uroteuthis Rehder, 1945. Один
вид—U. bartschi Rehder, 1945 (см.
рис. 39, л—н). Восточная Индонезия,
9*
131
Филиппины. Прибрежный вид, оби-
тает в толще воды и у дна. Длина
мантии до 22 см.
Таблица для определения видов рода
Loliolus
1(4). Плавники овальные или серд-
цевидные, их передние края выпук-
лые. Длина плавника значительно
больше Уг, ширина больше или рав-
на % длины маптии. Присоски боко-
вых рук самца резко увеличены. На
гектокотилизированной руке пет при-
сосок (редко одна), присоски правой
брюшной руки самца нормаль-
ные ............................2.
2(3). Присоски булавы мелкие,
средние не крупнее боковых. На гек-
токотиле кроме высокого вентраль-
ного «палисада» сохраняются дор-
сальная защитная мембрана и ряд
ножек от дорсальных присосок.
L. investigator is Goodrich, 1896
(рис. 34,а—г). Индия, Бенгальский
залив, Индокитай, Индонезия. Мас-
совый вид.
3(2). Средние присоски в базаль-
ной части булавы заметно крупнее
боковых. Дорсальная защитная
мембрана на гсктокотилс отсутству-
ет, имеются лишь вентральный «па-
лисад» и ряд ножек от дорсальных
присосок, иногда частично сливаю-
щихся с «палисадом».
L. hardwickei (Gray, 1849) ( = L.
typus Steenstrup, 1856, ? L. affinis
Steenstrup, 1856) (рис. 34, d, а). Ин-
дия, Индонезия (Ява, Мадура).
4(1). Плавники ромбические с
почти прямыми краями. Длина плав-
ника около У2, ширина около %
длины маптии. Присоски боковых
рук самца не резко увеличены. У ос-
нования гектокотилизированной руки
6—7 пар присосок, на гектокотиле
сохраняется хорошо развитая дор-
сальная защитная мембрана. На пра-
вой брюшной руке самца 3—4 пары
базальных присосок резко увеличены.
Средние присоски булавы гораздо
крупнее боковых.
L. rhomboidalis Burgess, 1967
(рис. 34,ж—к). Бенгальский залив,
Андаманское море.
Таблица для определения видов ро-
да Loeecguncuea
1(2). Ширина плавника равна его
длине и составляет около 40% дли-
ны мантии. На лучах буккальной
132
Рис. 34. Сем. Loliginidae, роды Loliolus
(а—к) и Loliolopsis (л—о): а—г— L. invest-
tigatoris, самка (а), присоски 3—4-й пар
рук самца (б—в) и.булавы (г); d—e — L.
hardwickei, самец (д), левая брюшная ру-
ка с дорсальной и вентральной стороны
(р); ж—к — L. rhomboidalis, самец (ж),
Правая (з) и левая (и) брюшные руки,
присоска правой брюшной руки (к); л—о —
Loliolopsis diomedeae, самец (л), самка
(м), правая (к) и левая (о) брюшные ру-
ки (а—г — по Adam, 1954; д—е— по. Grim*
не, 1932; ж—к — по Burgess, 1967; л—о
Q по Всггу, 1929)
133
л
%
ж.»
н
Рис. 35. Роды Lolliguncula (а—з) и Sepio-
teuthis (и—н): а—б — L. ellipsura, молодой
кальмар {а), кольцо присоски булавы (6);
в—г — L. mercatoris, самка (в) и гектоко-
тиль (г); <5—е — L. tydeus. самец (д) и
гектокотиль (в); ж — L, brevis, самка;
з — L. panamensis, самка; «—к — S. sepioi-
dea сверху и снизу; л—л — S. lessoniana,
самка (л) и гектокотиль (л<); н — S. ЫИ-
neata (а—б — по Hoyle, 1886; в—а — по
Adam, 1952; д—е — по Brakoniecki, -1980;
яс—no Verrill, 1882; з — по Berry, 1911;
и—к — по Roper, 1977; л — по Voss, Willi-
amson, 1972; м — по G. Voss, 1963; н — по
Dell. 1952)
л
мембраны по одной присоске. При-
соски боковых рук самца значитель-
но крупнее, чем прочих рук.
L. abulati Adam, 1955. Красное
морс.
2(1). Ширина плавника значи-
тельно больше его длины ... 3.
3(4). Длина плавника около 7з,
ширина менее половины длины ман-
тии. На лучах буккальной мембраны
по 2—3 присоски.
L. ellipsura (Hoyle, 1885) (рис.
35, а, б). Обнаружена на Патагон-
ском шельфе у Магелланова проли-
ва. Сомнительный вид.
4(3). Длина плавника 40—60%,
ширина больше половины длины
мантии...........................5.
5(6). Длина плавника менее по-
ловины (в среднем 40—45%), шири-
на в среднем 55—65% длины ман-
тии. Нет присосок на лучах буккаль-
ной мембраны. Присоски боковых
рук самца в вентральных рядах рез-
ко увеличены.
L, mercatoris Adam, 1941 (рис.
35, в, г). Западная Африка от Запад-
ной Сахары до Намибии.
6(5). Есть присоски на лучах бук-
кальной мембраны.................7.
7(10). Длина плавника около V2
длины мантии. Присоски рук несут
зубцы только по дистальному
краю........................... 8.
8(9). Гектокотилизированная ру-
ка значительно, в среднем на 7*, длин-
нее негектокотилизированной правой
брюшной руки, ее длина 45—60%
длины мантии.
L. tydeus Brakoniecki, 1980 (рис.
35, д, е). Тропические воды Восточ-
ной Пацифики от Южной Мексики
до Панамского залива, иногда в
сильно опресненных водах.
9(8). Гектокотилизированная рука
приблизительно той же длины, что
негектокотилизированная правая
брюшная рука.
L. brevis (Blainville, 1823) (рис.
35, ж). Тропические воды Западной
Атлантики от Чесапикского залива
до Ла-Йлаты, Мексиканский залив,
Карибское море. Имеет некоторое
промысловое значение.
10(7). Длина плавника около
60% длины мантии. Присоски рук
несут зубцы по всему периметру, хо-
тя па проксимальном краю они ред-
ки и плохо заметны. Гсктокотилизи-
рованная рука не длиннее правой
брюшной руки.
136
L. panamensis Berry, 1911 (рис.
35, з). Тропические воды Восточной
Пацифики от Калифорнийского за-
лива до Эквадора. Промышляется в
небольших количествах.
Таблица для определения видов рода
Sepioteuihis
1(6). Плавники равномерно сужают-
ся от места наибольшей ширины,
расположенного посредине или чуть
дальше середины тела, к переднему
и заднему концу мантии ... 2.
2(3). Буккальная мембрана лише-
на присосок. Наибольшая ширина
плавников посредине мантии. Макси-
мальное расстояние между противо-
положными краями плавников 60—
80%, в среднем 70—75% длины
мантии.
S. sepioidea (Blainville, 1823) (рис.
35, и, к). Тропические воды Западной
Атлантики от Бермудских островов
и Флориды до Южной Бразилии.
3(2). Буккальная мембрана с не-
сколькими (2—11, обычно 5—8 на
каждом луче) мелкими присоска-
ми ............................4.
4(5). Максимальное расстояние
между противоположными краями
плавников 50—75%, в среднем 60—
65% длины мантии. Место наиболь-
шей ширины плавников сдвинуто в
заднюю часть мантии, расстояние от
него до переднего края мантии 55—
72%, в среднем 60—65% ее длины.
S. lessoniana Lesson, 1830 (рис.
35, л, м). Индо-Вестпацифика — от
Красного моря и Южной Африки до
Хоккайдо, Гавайских островов и Се-
верной Австралии. Промысловый
вид.
5(4). Максимальное расстояние
между противоположными краями
плавников 65—85%, в среднем 70—
75% длины мантии. Место наиболь-
шей ширины плавников — посредине
мантии^
S. bilineata (Quoy et Gaimard,
1832) ( = S. australis Quoy et Gai-
mard, 1832) (рис. 35, я). Западная,
Южная и Юго-Восточная Австралия,
Новая Зеландия.
6(1). Плавники очень узкие в пе-
редней и широкие в задней половине
тела. Место наибольшей ширины
плавников находится па границе
средней и задней трети тела. Бук-
кальная мембрана лишена присосок.
5. loliginiformis (Riippell ct Leu-
ckart, 1828), Красное море. Сомни-
тельный вид.
Таблица для определения видов рода
Loligo
1 (26). Виды распространены в Ат-
лантическом оксане и его мо-
рях ............................2.
2(18). Виды распространены в за-
падной части Атлантического океа-
на .............................3.
3(12). Виды распространены в
Северо-Западной Атлантике: от Но-
вой Шотландии до северо-восточного
выступа Бразилии................4.
4(11). На всех лучах буккальной
воронки имеются мелкие присоски; об-
щее число присосок на всех лучах
больше 15, обычно 40—60. Более 26
поперечных рядов присосок на була-
ве .............................5.
5(10). Гладиус с выпуклыми и не
утолщенными краями. Отношение
максимальной ширины пера гладиуса
к максимальной ширине свободного
стебля (до начала пера) 2,4—3,7.
Левая брюшная рука самца видоиз-
меняется не до самого конца, при-
соски на конце гектокотиля одинако-
вых размеров в обоих рядах. Длина
плавника у кальмаров размером
свыше 5—6 см >-1/2 длины мантии.
Ширина мантии 7s—7з ее длины. На
брюшной стороне мантии обычно
нет длинных красных полосок . . 6.
6(9). Глаза некрупные: диаметр
глазного яблока 8-—18%, хрусталика
2—6% длины мантии .... 7.
7(8). Присоски боковых рук сам-
ца немного крупнее, чем рук 1-й и
4-й пар. Ножки присосок на видоиз-
мененном участке гсктокотилизиро-
ванной руки округлые или слабо уп-
лощенные, у части присосок доволь-
но узкие. Кольца присосок в середи-
не гектокотилизированной руки с
прямоугольными зубцами.- На коль-
цах крупных присосок булавы зубцы
обычно чередуются в таком порядке:
крупный — маленький — средний —
маленький — крупный. Длина плав-
ника у взрослых кальмаров в сред-
нем 55—60% длины мантии.
L. (L.) pealei LeSueur 1821 (рис.
36, а—в). Североамериканский, или
восточноамериканский длинноперый
кальмар. От Новой Шотландии (зал.
Фанди) до Юкатана и у южного по-
бережья Карибского моря. Встреча-
ется на шельфе и в верхней части
склона, к поверхности поднимается
редко. Длина мантии до 47 см, обыч-
но до 25—30 см. Имеет важнейшее
промысловое значение в Северо-За-
падной Атлантике.
8(7). Присоски боковых рук
самца резко увеличены и примерно
равны крупным присоскам булавы.
Ножки присосок на видоизмененном
участке гектокотилизированной руки
широкотреугольные, уплощенные.
Кольца присосок в середине гекто-
котилизированной руки с острыми
зубцами. На кольцах крупных при-
сосок булавы зубцы только двух раз-
меров, крупные и мелкие, чередуют-
ся без особого порядка. Длина плав-
ника у взрослых кальмаров в сред-
нем 50—55% длины мантии.
L. (L.) surinatnensis Voss, 1974
(рис. 36, г). От устья р. Ориноко до
устья р. Амазонки. Внутренний
шельф. Длина мантии до 12 см.
9(6). Глаза очень крупные: диа-
метр глазного яблока 15—21%, хру-
сталика 6—8 % длины мантии. При-
соски боковых рук самца не резко
увеличены. Ножки присосок на видо-
измененном участке гектокотилизиро-
ванпой руки широкие, треугольные.
Длина плавника около 7з длины
мантии.
L. (L.) ocula Cohen, 1976 (рис.
36, д, е). Карибское море и Большие
Антильские острова. Верхняя часть
склона (глубина 250—400 м). Дли-
на мантии до 13 см.
10(5). Гладиус с прямыми утол-
щенными краями. Отношение макси-
мальной ширины пера гладиуса к
максимальной ширине стебля 1,5—
2,4. Гектокотилизированная часть ру-
ки доходит до ее конца и занимает
74—7г длины руки. Диаметр присо-
сок дорсального ряда па конце руки
меньше половины диаметра присосок
вентрального ряда. Длина плавника
у кальмаров мелких и средних раз-
меров (до 10, иногда до 20 см)
меньше 7г длины маптии. Ширина
мантии обычно 7а—7б се длины.
У взрослого самца по середине
брюшной стороны мантии проходит
кожный гребень, по бокам брюшной
стороны — 6—8 извилистых продоль-
ных полосок красно-бурого^ цвета.
L. (D.) plei (Blainville, 1823) (рис.
36,ж—и). От мыса Хаттерас (изред-
ка доходит до мыса Код) и Бермуд-
ских островов до Северо-Восточной
137
- W-/
О
Loligo,
Рис. 36. Род Loligo,
Восточной Пацифики: _ _
lei, самец (а), гектокотиль (б),
(в); г — L. (L.) surinamensls*
-да:
Я
виды Атлантики и
a—fl — L. (L.) pea-
гладиус
____ . .. _________________ d—e — L.
(L.) ocula, самец (д), гектокотиль (e);
ж—u. — L, (D.) plei, кальмар сверху (ж),
мантия снизу (з), гектокотиль (ц); к—л —
L. {£>.) roperl, самец (к), гектокотиль (л):
м — L. (L.) patagonica', н—o — L. (L.) vul-
gar is \ n—p — L.) (L.) forbesi (о, p — була-
ва); c— t — L, (L.) opalescens, самец (с) и
гектокотиль (г) (a. в—а—по <j. Voss, 1956,
1974; б, д—е, и—л — по Cohen, 1976; ж—
з — по Roper, 1977; м — по Филипповой,
1969; н—р — по Muus, 1959; а—т — по Ber-
ry, 1912)
139
Бразилии. Многочислен в Мексикан-
ском заливе, Карибском море, у Ан-
тильских островов. Встречается и у
поверхности, и у дна, хорошо подхо-
дит па свет. Длина мантии до 50 см,
обычно до 30—35 см.
11(4). Нет мелких присосок па
двух вентральных лучах буккальной
воронки; общее число присосок на
остальных лучах пе более 6. Гладиус
с прямыми утолщенными краями.
Менее 25 поперечных рядов присосок
на булаве. Гектокотилизированная
часть руки доходит до ее конца и за-
нимает около % длины руки. При-
соски дорсального ряда на конце ру-
ки крошечные. Длина плавника Vs—
2/5 длины мантии, ширина мантии
Ve—‘Л ее длины.
L. (D.) roperi Cohen, 1976 (рис.
36, к, л). Флоридский пролив, Ан-
тильские острова, Карибское море.
У дна на шельфе, в верхней части
склона и у поверхности. Длина ман-
тии до 7 см.
12(3). Виды распространены у
Бразилии, Аргентины и Фолкленд-
ских островов...................13.
13(14). Длина плавника заметно
больше половины (в среднем 60%)
длины мантии, боковые углы плавни-
ка закругленные. На кольцах круп-
ных присосок булавы 15—25 (до 28)
зубцов одинакового размера.
L. (L.) brasiliensis Blainville, 1823
(=L. emmakina Gray, 1849). Брази-
лия и Северная Аргентина, па юг до
Росона (находки у Кубы сомнитель-
ны). Шельф. Длина мантии до 20 см,
обычно до 15 см. Имеет местное про-
мысловое значение.
14(13). Длина плавника нс более
V? длины мантии. На кольцах круп-
ных присосок булавы у взрослых
кальмаров 25 или более зуб-
цов ............................15.
15(16). Передние и задние края
плавника прямые, боковые углы при-
острены, самое широкое место плав-
ника— посередине его длины. У зре-
лых самцов нет продольного кожно-
го гребня посредине брюшной сторо-
ны мантии.
L. (L.) patagonied Smith, 1881
(рис. 36, м). Патагон'о-Фолклендский
шельф и склон от залива Ла-Плата до
банки Бёрдвуд. Преимущественно
вдали от берегов на глубинах 80—
800 м. Встречается также у Южного
Чили, к югу от 50° ю. ш. Длина ман-
тии до 24 см. Многочисленный, по-
140
тепциальпо промысловый вид.
16(15). Передние края плавника
слегка выпуклые, задние слегка во-
гнуты, боковые углы округленные,
самое широкое место плавника зна-
чительно впереди середины его дли-
ны. У зрелых самцов есть продоль-
ный кожный гребень посередине
брюшной стороны мантии.
L. (L.) gahi d’Orbigny, 1835 (см.
с. 146).
17(2). Виды распространены в
восточной половине Атлантического-
оксана и прилегающих морях . 18.
18(21). Виды распространены в
Северо-Восточной Атлантике, Север-
ном и Средиземном морях . . 19.
19(20). Присоски средних рядов
центральной части булавы в 3—4 ра-
за крупнее присосок краевых рядов.
Кольца крупных присосок булавы
гладкие или с нерегулярно располо-
женными мелкими зубчиками на ди-
стальном крас. Кольца мелких при-
сосочек па лучах ротовой мембраны
дистально с 7—10 длинными остры-
ми зубцами. Длина плавника у
взрослых около % длины мантии.
L. (L.) vulgaris Lamarck, 1798
(рис. 36, н, о). Обыкновенный, или
европейский длинноперый кальмар.
Распространен на шельфе от Фарер-
ских островов и Южной Норвегии до
Сенегала, в Северном и Средиземном
морях; особый подвид (см. с. 141)
у Южной Африки. Длина мантии до
45 см. Важный промысловый вид.
20(19). Присоски средних рядов
центральной части булавы немного
(примерно в 1,5 раза) крупнее присо-
сок краевых рядов. Кольца крупных
присосок булавы с зубцами по всему
периметру. Кольца мелких присосо-
чек па лучах ротовой мембраны
дистально с 12—15 тупыми зубчика-
ми. Длина плавника у взрослых
около 3/4 длины мантии.
L. (L.) forbesi Steenstrup, 1856
(рис. 36, п, р). Северный (североев-
ропейский) длинноперый кальмар.
На шельфе и в самой верхней части
склона от Средней Норвегии до Кап-
Блана, Канарских островов и Сре-
диземного моря, по южнее Бискай-
ского залива редок. Длина мантии до
75 см. Важный промысловый вид в
Северном море и районе Британских
островов.
21(18). Виды распространены в
Юго-Восточной Атлантике и у Юж-
ной Африки.....................22.
22(23). Присоски средних рядов
центральной части булавы и 3—4 ра-
за крупнее присосок краевых рядов,
их кольца гладкие или с нерегуляр-
но расположенными мелкими зуб-
чиками на дистальном крас. Кольца
присосок рук с 20—30 острыми кони-
ческими зубцами по всему перимет-
ру. Присоски боковых рук самца
значительно увеличены. Длина плав-
ника у взрослых около 2/з длины
мантии. Фотофоров нет.
L. (L.) vulgaris reynaudi d’Orbig-
ny, 1845 (см. с. 140). У Южной Аф-
рики от Анголы до Дурбана. Шельф.
Длина мантии до 45 см.
23(22). Присоски средних рядов
центральной части булавы ненамно-
го крупнее присосок краевых рядов,
их кольца несут острые зубцы по
всему периметру. Кольца присосок
рук с 5—12 острыми или тупыми
зубцами на дистальном крае . 24.
24(25). Присоски боковых рук
взрослых самцов сильно увели-
чены (крупнее присосок булавы), их
кольца с 9—12 низкими тупыми зуб-
цами на дистальном и боковых кра-
ях кольца. Кольца нс увеличенных
присосок рук с 5—8 низкими тупыми
зубцами на дистальном крае. При-
соски булавы с 15—20 зубцами. Дли-
на плавника у взрослых около V*
длины мантии. Есть парный фотофор
па чернильном мешке.
L. (L.) duvauceli d'Orbigny, 1839
(см. с. 145). До Южного Наталя.
25 (24) .Присоски боковых рук
самца нс увеличены. Кольца присо-
сок рук с 10—12 узкими конически-
ми зубцами на дистальном и несколь-
кими крохотными зубчиками на
проксимальном краях. * Присоски бу-
лавы с 23—24 зубцами. Длина плав-
ника у взрослых около % длины
мантии.
L. (Doryteuthis ?) sp. В. Robson,
1924. Наталь. Длина мантии 21 см.
Плохо описанный вид неясного си-
стематического положения.
26(1). Виды распространены в
Индийском и Тихом океанах . 27.
27(66). Виды распространены в
Индийском и западной части Тихого
океана и прилегающих морях, от
Красного моря и Южной Африки до
Японии, Австралии и Полинезии . 28.
28(29). Присоски булавы очень
мелкие, значительно мельче присо-
сок рук, почти одинаковых размеров.
В дистальной части булавы присоски
скучены и сидят более чем в 4 ряда.
Руки очень короткие, около !Д дли-
ны мантии. Длина плавника значи-
тельно больше l/z длины мантии.
Вдоль средней линии брюшной сто-
роны мантии самца проходит про-
дольный кожный гребень. 20—30
мелких присосочек па каждом луче
буккальной мембраны.
L. (D) bleekeri Keferstein, 1866
(рис. 37, я). Япония от Хоккайдо до
Кюсю, Южная Корея, северные
районы КНР. Шельф и самая верх-
няя батиаль. Во время нагула дохо-
дит до Южно-Курильского пролива.
Длина мантии до 35—40 см. Про-
мышляется в Японии в значительных
количествах.
29(28). Присоски булавы, как
правило, крупнее присосок рук; хо-
рошо различаются по размерам:
центральные крупнее боковых. В
дистальной части булавы присоски
сидят в 4 ряда. Руки, как правило,
больше ’Д. обычно около 7г длины
мантии. На каждом луче буккальной
мембраны обычно не более 10—13
присосок .....................30.
30(53). Боковые стороны гладиу-
са дугообразно выгнуты и не утол-
щены. Ширина мантии обычно в 3—
6 раз меньше длины . . . . 31.
31(52). Кольца крупных присосок
рук с зубцами.................32.
32(37). Зубцы на кольцах круп-
ных центральных присосок булавы
резко неравные: крупные зубцы чере-
дуются, особенно на дистальном
крае, с мелкими, малозаметными.
Кольца присосок рук дистально с
6—15 узкими высокими зубцами,
проксимально гладкие или слабо за-
зубренные. Присоски 3-й пары рук
лишь немного крупнее, чем на прочих
руках. Есть парный фотофор па чер-
нильном мешке.................33.
33(36). Кольца присосок рук ди-
стально с 6—12, чаще с 6—8 тупыми
зубцами. Наибольшая ширина гладиу-
са 14—17% его длины .... 34.
34(35). Длина булавы 1Д—!/з дли-
ны мантии. Диаметр наибольших
присосок булавы около 2% длины
мантии. Вдоль средней линии брюш-
ной стороны мантии самца проходит
продольный кожный гребень. Длина
гектокотилизироваппой части руки
1Д—% длины руки.
L. (L.) edulis Hoyle, 1885 (рис.
37,6—е). Ипдо-Вестпацифика от
Красного моря до Сангарского про-
141
У<
Рис. 37. Род Loligo. L. (Doryteuthis) bleeke~
ri и индовестпацифическис виды Loligo-
s. str.: a— L.(D.) bteekerv, 6—d— L. edulis\.
e — L. edulis f. kensaki; ж—з — L. budo;
u—k — L. chine ns is; л—м — L. japonica;.
н—n — L. vossi, n. sp.; p — L. beka; c—y —
L. duvauceli; ф—х — L. kobiensis; ц — L.
yokoyae, правая брюшная рука самца; ч—-
ш — L. tagoi; Щ — L. uyii; в, о, т — гекто-
котиль; г—булава; д, з, к, м, р, у, х, ш-
щ — присоски булавы, д\, зь «ь Ль Рь
Хи Mi, — присоски рук (а — по SasakL
1929; б—г, н—п, с—т — по G. Voss, 1963;
д, и—к, у, ч—ш — по Voss, Williamson^
1972, прочее по Okutani, 1973)
лива и Северной Австралии. Шельф.
Длина мантии до 20—40 см. Важ-
ный промысловый вид. В Северо-
Западной Пацифике представлен
двумя формами, таксономические от-
ношения которых неясны.
а. У основания гектокотилизиро-
в а иной руки самца 7—8 пар нормаль-
ных присосок. Нет присосочек на па-
пиллах гектокотиля. Длина мантии
до 20 см.
L. edulis f. edulis (=L. edulis f. na-
gasakiensis Sasaki, 1929) (рис.
37,6—d). От Красного моря до Сан-
гарского пролива.
б. У основания гектокотилизиро-
ванной руки самца 20—27 пар нор-
мальных присосок. Папиллы гекто-
котиля несут рудиментарные присо-
сочки. Длина мантии до 40 см.
L. edulis f. kensaki Wakiya et Ishi-
kawa, 1921 (=L. edulis f. edulis Sa-
saki, 1929, non Hoyle, 1885) (рис.
37, e). От Южной Японии до Синга-
пура.
35(34). Длина булавы %—V2
длины мантии. Диаметр наибольших
присосок булавы 3—3,5% длины
мантии. У основания гектокотилизи-
рованной руки самца 13—14 пар нор-
мальных присосок; нет присосок па
папиллах гектокотиля.
L. (L.) budo Wakiya et Ishikawa,
1921 (-L. edulis f. grandipes Sasaki,
1929) (рис. 37, ж, з). Южная Япония
(Внутреннее море, япономорское по-
бережье Хонсю, Кюсю). Длина ман-
тии до 26 см. Самостоятельность ви-
да вызывает сомнение, возможно,
это форма L. edulis.
36(33). Кольца присосок рук ди-
стально с 10—18 длинными острыми
коническими зубцами. На брюшной
стороне мантии самца нет продоль-
ного кожного гребня. Гектокотили-
зированпая часть руки занимает
Уз—% се длины.
L. (L.) chinetisis Gray, 1849 (=L.
itheridgei Berry, 1918, L.formosana
Sasaki, 1929, ? L. australis Gray, 1849)
(рис. 37, u, к). Ипдомалайский вид:
•от островов Рюкю и южной части
КНР до Северной, Западной и
Восточной Австралии и Тасмании.
Шельф. Длина мантии до 31 см.
Имеет существенное промысловое
значение.
37(32). Зубцы на кольцах цент-
ральных присосок булавы приблизи-
тельно одного размера, плавно
уменьшаются от дистального края к
144
проксимальному, либо кольца этих
присосок гладкие. Кольца присосок
рук с тупыми зубцами ... 38.
38(45). Кольца крупных централь-
ных присосок булавы с зубцами . 39.
39(44). Присоски средних рядов
в центральной части булавы резко
(в 2—3 раза) крупнее присосок
краевых рядов. Нет фотофора на
чернильном мешке..............40.
40(43) Кольца крупных присосок
рук с 7—13 зубцами. Гектокотилизи-
рованная часть руки занимает ]/2 ее
длины ........................41.
41(42). Крупные присоски булавы
с 20—25 низкими зубцами по всему
периметру. Кольца присосок рук не-
сут низкие широкие округленные зуб-
цы почти по всему периметру. При-
соски боковых (особенно 3-й) рук
самца резко увеличены, в 2—3 раза
крупнее присосок 1-й и 4-й рук и
крупнее центральных присосок була-
вы. Наибольшая ширина гладиуса
18—25% его длины.
L. (L,) japonica Stcenstrup in
Hoyle, 1885 (рис. 37, л, м). Япония
(от Хоккайдо до Кюсю), Желтое и
север Восточно-Китайского моря.
Прибрежные мелководья. Длина
мантии до 12 см. Промысловый вид.
42(41). Крупные присоски була-
вы примерно с 15 треугольными зуб-
цами, из которых 2—4 на дистальном
крае острые, когтсвидпыс, прочие
низки и мало заметны, иногда коль-
ца этих присосок лишь с немногими
треугольными зубцами на дисталь-
ном крае. Кольца присосок рук с вы-
сокими узкими тупыми зубцами
только на дистальном крас. Присос-
ки боковых рук самца гораздо мель-
че крупных присосок булавы.
L. (L.) vossi п. sp.' ( — Loligo sp.
A. Voss, 1963: 74—76, fig. 11 f—j)
(рис. 37, н—n). Филиппины, Шри-
Ланка, Аравийское море и Оманский
залив. Шельф. Длила мантии до
14 см.
43(40). Кольца присосок рук с
3—5 очень широкими низкими округ-
ленными зубцами. Кольца присосок
булавы примерно с 20 острыми трс-
1 Голотип: самец, длина мантии
136 мм, 7.IV. 1909 г, Порт-Лапгкан,
о-в Дум ар а и, Восточный Палаван,
Филиппины. Американский нацио-
нальный музей естественной истории
(U. S. National Museum), Вашинг-
тон.
угольными зубцами одинаковых раз-
меров. Присоски боковых рук самца
менее чем в 2 раза крупнее присосок
1-й и 4-й рук, по крупнее централь-
ных присосок булавы. Гектокотили-
зированная часть руки занимает 2/з
ее длины.
L. (L.) beka Sasaki, 1929 (рис.
37, р). От Южной Японии до о-ва
Хайнань и Тонкинского залива. Мел-
ководья. Длина мантии до 7 см.
Промысловый вид.
44(39). Присоски средних рядов в
центральной части булавы ненамно-
го (максимум в 1,5 раза) крупнее
присосок краевых рядов. Присоски
боковых (особенно 3-й) пар рук сам-
ца резко увеличены, у самцов из за-
падной части Индийского океана и
Красного моря они крупнее централь-
ных присосок булавы. Кольца присо-
сок рук несут 5—8 низких тупых
зубцов дистально, а резко увеличен-
ных присосок самца — до 9—13 та-
ких зубцов почти по всему перимет-
ру. Есть парный фотофор на чер-
нильном мешке.
L, (L.) duvauceli d‘Orbigny, 1839
(==L.indica Pfeffer, 1.884, L. galatheae
Steenstrup in Hoyle, 1885, L.oshimai
Sasaki, 1929) (рис. 37, с—у). Индо-
Вестпацифика, от Красного моря и
Южной Африки до южной части
КНР, о-ва Тайвань и Северной Авст-
ралии. Внутренний шельф, дно и
толща воды. Длина мантии до 18—
22 см, обычно до 15 см. Имеет неко-
торое промысловое значение.
45(38). 4—5 пар наибольших при-
сосок в центральной части булавы
имеют гладкие кольца; кольца про-
чих присосок булавы с зубцами.
Центральные присоски булавы очень
крупные, в 3—5 раз крупнее краевых.
Присоски боковых рук самца резко
увеличены . :.................46.
46(49). Кольца крупных присосок
боковых рук с 7—9 широкими тупы-
ми зубцами ...................47.
47(48). Присоски на правой
брюшной руке самца обычные, мел-
кие.
L. (L.) kobiensis Hoyle, 1885
( = ? L. sumatrensis d’Orbigny, 1839)
(рис. 37, фу x). Южная Япония, Юж-
ная Корея, КНР. По неподтвержден-
ным данным также Филиппины, Ин-
донезия и Мальдивские .острова.
Шельф. Длина мантии до 11 см. До-
бывается в небольших количествах.
48(47). В базальной части правой
10 К. Н. Несис
(не гектокотилизированной) брюш-
ной руки самца имеются 4 пары рез-
ко увеличенных присосок, самые
крупные из которых не мельче цен-
тральных присосок булавы.
L, (L.) yokoyae М. Ishikawa, 1925
(рис. 37, ц). Южная Япония.
49(46). Кольца присосок рук с
3—5 очень широкими тупыми зубца-
ми на дистальном крае ... 50.
50(51). Кольца присосок рук с
4—5 зубцами. На булаве 4 пары
крупных центральных присосок.
L. (L.) tagoi Sasaki, 1929 (=L. go-
toi Sasaki, 1929) (рис. 37, ч, ui). Юж-
ная Япония, КНР, Тонкинский залив.
Внутренний шельф. Длина мантии до
8 см. Возможно, синоним L. иуи (см.
ниже).
51(50). Кольца присосок рук с 3
зубцами. На булаве 5 пар крупных
центральных присосок.
L. (L.) uyii Wakiya et Ishikawa,
1921 (рис. 37,щ). Южная Япония,
возможно также КНР и Вьетнам.
Шельф. Длина мантии до 10 см. Вто-
ростепенный промысловый вид.
52(31). Присоски 3-й пары рук
самца резко увеличены, их кольца
гладкие.
L. (L.) aspera Ortmann, 1888. Юж-
ная Япония. Длина мантии зрелого
самца 5 см. Сомнительный вид, воз-
можно, относится к L. tagoi.
53(30). Гладиус узкий, его боко-
вые стороны прямые или почти пря-
мые, утолщенные. Ширина мантии в
4—9, обычно в 6—8 раз меньше дли-
ны ............................54.
54(59). Длина плавника у взрос-
лых значительно более V2 длины
мантии. Зубцы присосок булавы рез-
ко неодинаковые: крупные зубцы
числом 10—20, чередуются без осо-
бого порядка с мелкими н малоза-
метными; общее число зубцов 20—
40. Длина маптии взрослых кальма-
ров более 10—15 см .... 55.
55(58). Задний край плавника
вогнутый. Ширина плавника у
взрослых !/з—Чг длины мантии. Гск-
токотилизированная часть руки —
7з—% ее длины. Есть парный фото-
фор на чернильном мешке . . 56.
56(57). Присоски рук с 7—11 ши-
рокими зубцами дистально.
L. (D.) singhalensis Ortmann, 1891
(рис. 38, а—г). От Южного Китая и
о-ва Тайвань до Аравийского моря,
Сейшельских островов, Северной
Австралии и Соломонова моря.
145
Шельф. Встречается у дна и в тол-
ще воды до поверхности. Длина ман-
тии до 50 см, обычно до 20—25 см.
Важный промысловый вид.
57(56). Присоски рук с 18—24
зубцами по всему периметру, на ди-
стальной части кольца они выше и
шире, чем на проксимальной.
L. (D.) arabica (Ehrenberg, 1831)
(рис. 38, д—ж). Красное море. Дли-
на мантии до 27 см.
58(55). Задний край плавника
прямой. Ширина плавника у взрос-
лых менее Уз длины мантии. Гекто-
котилизированная часть руки зани-
мает У4 ее длины.
L. (D.) spectrum Pfeffer, 1884.
Маркизские острова. Длина мантии
до 28 см.
59(54). Длина плавника у взрос-
лых не более ’/2 длины мантии. Зуб-
цы присосок булавы без чередования
крупных и мелких, более или менее
одинаковые, их 10—20. Длина ман-
тии до 7—16 см.................60.
60(65). Присоски боковых рук с
зубцами ................. ... 61.
61(62). Длина плавника менее 2/б>
ширина около У4 длины мантии.
Присоски боковых рук самца резко
увеличены и намного крупнее при-
сосок булавы. Кольца присосок бо-
ковых рук самца и самки несут ши-
рокие зубцы по всему периметру.
7—8 пар нормальных присосок на
гектокотилизировапной руке.
L. (D.) pickfordae Adam, 1954
(рис. 38,з—к). Индонезия (о-в Фло-
рес). Длина мантии до 7 см.
62(61). Длина плавника равна или
более 40%, ширина 30—45% длины
мантии. Присоски боковых рук сам-
ца примерно равны по размерам
присоскам булавы. Кольца присосок
боковых рук с 7—9 широкими зуб-
цами на дистальном крае. Более
8 пар нормальных присосок на гек-
токотилизированной руке . . 63.
63(64). Кольца крупных присосок
булавы с 15—20 зубцами. Гектоко-
тилизируется только левая брюшная
рука, она несет 15—20 пар нормаль-
ных присосок, измененная часть за-
нимает 30—45% длины руки и несет
папиллы до самого конца. Имеются
2 фотофора на чернильном мешке.
L. (D.) sibogae Adam, 1954 (рис.
38, л—н). От о-ва Тайвань до Син-
гапура, Индонезии и западной части
Индийского оксана (банка Сая-де-
Малья). Шельф у дна и толща воды
146
до поверхности. Длина мантии до
16 см.
64(63). Кольца крупных присосок
булавы с 11—12 зубцами. Гектоко-
тилизируются обе брюшные руки.
Левая рука несет 9—10 пар нормаль-
ных присосок, измененная часть за-
нимает более V2 длины руки; правая
рука укорочена, присоски в ее сред-
ней части резко уменьшаются в раз-,
мерах, затем исчезают, остаются од-
ни ножки. Концы обеих брюшных
рук лишены присосок и папилл.
L. (D.) reesi Voss, 1963. (рис.
38, о—с). Филиппинские острова.
У дна и поверхности. Длина мантии
до 7 см.
65(60). Кольца присосок рук не-
ясно зазубрены, со слабо заметными
тупыми зубцами. Кольца крупных
присосок булавы с 3—5 узкими длин-
ными зубцами на дистальном крае,
несколькими широкими тупыми зуб-
цами ио бокам, проксимально глад-
кие. Длина плавника чуть меньше,
ширина чуть больше ’/2 длины ман-
тии.
L. (D.?) brevipinnis Pfeffer, 1884
( = L.pfefferi Hoyle, 1886). О-ва
Эллис (Тувалу, Тихий океан). Длина
мантии 5 см. Плохо описанный сом-
нительный вид.
66(27). Виды распространены в
восточной части Тихого океана, вдоль
берегов Северной и Южной Амери-
ки ...........................67.
67(68). Присоски рук с 9—12 ту-
пыми зубцами. Нет продольного кож-
ного гребня на брюшной стороне
мантии взрослых самцов.
L.(L.) opalescens Berry, 1911 (см.
рис. 36, с, т). Западноамерикаиский
длинноперый кальмар. От Британ-
ской Колумбии (Канада) до Мекси-
ки, возможно до Перу. У дна на
внутреннем шельфе и в толще воды
до поверхности. Длина мантии до
20 см. Важнейший промысловый вид
у Калифорнии.
68(67). Присоски рук с 6—7 ту-
пыми зубцами. Есть продольный кож-
ный гребень на брюшной стороне
мантии взрослых самцов.
L. (L.) gahi d'Orbigny, 1835. По-
бережья Перу и Чили. Шельф. Дли-
на мантии до 21 см. Промысловый
вид.
Таблица для определения видов рода
Alloteuthis
1 (2). Взрослые кальмары с корот-
ким (менее 1 см) хвостом. Длина
10*
плавника, включая хвост, менее по-
ловины длины мантии. Щуцальца
очень длинные, у живых кальмаров
они значительно длиннее головы и
маитии, вместе взятых, (при фиксации
стебли щупалец сокращаются). Бу-
лава широкая, диаметр наибольших
присосок 9—14% ширины головы,
пары средних присосок сидят почти
перпендикулярно оси булавы.
A. media (Linne, 1758) (рис. 39, а,
б). Лузитано-средиземноморский
вид: от южной части Северного мо-
ря до Средиземного моря и мыса
Кап-Блан.
2(1). Взрослые кальмары с длин-
ным (до 5—6 см у самцов, до 2 см
у самок) хвостом. Длина плавников,
включая хвост, равна или превосхо-
дит половину длины мантии. Щу-
пальца значительно короче головы и
мантии, вместе взятых. Присоски
булавы мелкие, диаметр наибольших
из них 6—8% ширины головы . 3.
3(4). Ширина плавника у взрос-
лых кальмаров более 25% длины
мантии. Длина наибольшей руки у
взрослых 20—25% длины мантии.
Булава узкая, пары средних присо-
сок сидят наискось к оси булавы
под углом примерно 45°. '
A.subulata (Lamarck, 1798) (рис.
39, в, г). Бореально-лузитанский вид,
от Шетландских островов и Южной
Норвегии до Кап-Блана., и Среди-
земного моря, в последнем редок.
4(3). Ширина плавника у( взрос-
лых кальмаров до 20% длины ман-
тии у самцов, до 30% у самок. Дли-
на наибольшей руки у взрослых 15—
.22% мантии. Булава широкая, пары
Средних присосок сидят перпендику-
лярно оси булавы.
A. africana Adam, 1950 (рис.
39, ж—к). Западная Африка от За-
падной Сахары до Анголы.
2.2.1.2. Семейство
Pickfordiateuthidae
Маленькие (длина мантии 1,5—2 см)
кальмары, внешне похожие на сепио-
лид, • с короткой. закругленной сзади
мантией и ’ почковидными .плавника-
ми, которые сидят по бокам спинной
стороны мантии у заднего конца те-
ла и пе соединяются между собой.
Присоски па руках в два ряда. Бу-
лава расширенная, без фиксаторно-
го аппарата, присоски в проксималь-
148
ной части булавы в 2, в дистальной
в 4 ряда. Роговица глаза замкнутая,
зрачок слегка: U-образный. Лучи
буккальной мембраны редуцированы
и не несут присосок, формула бук-
кального крепления DDVV. Замыка-
тельный аппарат простой. Гладиус с
коротким тонким стволом и очень
широким овальным пером. Фотофо-
ров нет. Гектокотилизируется левая
брюшная рука. Развит только левый
яйцевод. Семеприемник — на ротовой
мембране самки. Яйца крупные, дон-
ные. Окраска: яркие, крупные, крас-
ные, коричневые и желтые хромато-
форы на мантии, голове и конечнос-
тях. Плавники бесцветны, прозрачны.
Один род Pickfordiateuthis Voss,
1953 с единственным видом — Р.риГ-
chella Voss, 1953 (рис. 40). Обитает
на мелководьях тропической части
Западной Атлантики — у южного по-
бережья Флориды, в Карибском мо-
ре, па песке и в зарослях морских
трав. Обзор: G. Voss, 1953, 1956.
2.2.2. Подотряд
Oegopsida
Передняя камера глаза широко от-
крыта и непосредственно соприкаса-
ется с внешней средой. Глазное от-
верстие может закрываться лишь
кожными веками, веко имеет кольце-
вую форму с вырезом (синусом) в
нижнем переднем углу. Мантия ли-
бо мускулистая, либо полустудснис-
тая или топкая, почти лишенная
мускулатуры. Плавники разнообраз-
ной формы, могут сидеть близ зад-
него конца или впереди пего, иногда
протягиваются вдоль всей длины
мантии. Руки и щупальца могут
нести присоски и (или) крючья,
иногда щупальца автотомируются
или редуцируются в молодом воз-
расте. Булава часто имеет обособ-
ленный фиксаторный аппарат, состоя-
щий из присосок с гладкими кольца-
ми и соответствующих им бугорков-
кнопок, с помощью которых оба
щупальца могут прочно скрепляться
друг с другом (при этом бугорки-
кнопки одной булавы попадают в
присоски противоположной). Бук-
кальное крепление 6-, 7- или 8-луче-
вое, его формула DDVV или DDVD
(изредка DDDD). Замыкательный
аппарат простой или модифицирован-
Рис. 39. Роды Alloieuthis (а—к) и Uroteut'
his (л-«): а—б —A. media', в-г — А. subu-
lata\ д—е — сравнение пропорции тела (d)
и булавы (е) /1. media (т) и Л. subulata
(s)‘ ж—к—-A. africana (и — гектокотиль,
к,— булава); л—н—U. bartschi (я, «t
положение и строение светящегося орга-
на—с.о) (а, б — по Naef, 1921—1923; в, г—
по Mnus, 1959; д—е—по • Grimpe, .1925; ж—
к — по Adam. 1952; л—м — но G. Voss,
1963; я — по Haneda, 1963)
ный, иногда мантия срастается с во-
ронкой или головой на затылке, а у
сем. Cranchiidae--и с воронкой, и с
головой. Фотофоры имеются или от-
сутствуют, свечение биохимическое
без симбиотических бактерий. Фото-
форы часто сложно устроенные, раз-
ного строения на разных частях тела.
Развиты, как правило, оба яйцевода.
Нидаментальные железы имеются
(у Enoplotcuthinae отсутствуют).
Придаточные нидаментальные желе-
зы отсутствуют (у Ctenopterygidae
имеется непарное образование неяс-
ной функции, которое сопоставляют
с придаточной нидаментальной же-
лезой). Гектокотнль имеется или от-
сутствует, в последнем случае его
функцию обычно выполняет сильно
развитый пенис. Если гектокотнль
имеется, то он развивается па одной
или обеих руках брюшной пары, но
иногда дополнительно видоизменяют-
ся концы спинных или иных рук.
Яйца сравнительно мелкие, отклады-
ваются в студенистых капсулах в
толщу воды или на дно; виды, ли-
шенные нидаментальных желез, мечут
яйца в воду поодиночке.
Океанические кальмары, на шель-
фе и у берегов встречаются редко.
Обитают в толще воды или близ
дна — от поверхностных до глубинных
вод. 23 семейства, 77—81 род, 193—
226 видов. Обзоры: Chun, 1910;
Pfeffer, 1912; Clarke, 1966; Roper,
Young, Voss, 1969; Зуев, Несис, 1971.
Таблица для определения семейств
подотряда Oegopstda
*1(39). Мантия не срастается ни с
головой, ни с воронкой1 ... 2.
2(29). Мантийно-вороночный за-
мыкательный аппарат простой: ман-
тийный хрящ — длинный низкий гре-
бень, вороночный прямой или слабо
изогнутый, слегка расширенный сза-
ди .............................3.
3(8). На руках, по крайней мере
па первых трех парах, имеются
крючья, если их нет, то присоски на
руках в 4 ряда..................4.
1 У видов Eucleoteuthis luminosa и
Sthenoteuthis oualaniensis (сем. Om-
mastrephidae) мантийный и вороноч-
ный хрящи срастаются на одной
или обеих сторонах тела, при этом,
однако, сохраняют типичную для се-
мейства .L-образную форму.
150
4(5). Присоски и крючья на ру-
ках сидят в 4 ряда: на 1—3-й па-
рах рук по 2 ряда крючьев (средние
ряды) и 2 ряда присосок, а на 4-й
паре 4 ряда присосок, либо крючья
отсутствуют или слабо развиты, и
вооружение рук состоит из 4 рядов
присосок. Щупальца отсутствуют, а
если имеются, то булава вооружена
многими рядами мелких присосок,
среди которых могут быть один или
несколько крупных крючьев.
Сем. Gonatidae . . . . с. 187.
5(4). Присоски и крючья на ру-
ках сидят всегда в 2 ряда. Воору-
жение щупалец (если они имеются)
иное.............................6.
6(9). Щупальца имеются. Фото-
форы на концах рук, по крайней ме-
ре 1—3-й пар, отсутствуют. Формула
буккального крепления DDVD . 7.
7(8). Фотофоры имеются обычно
на поверхности мантии, вентральной
стороне глазного яблока, иногда
внутри мантии, на стебле щупальца
и др. На булаве, кроме присосок,
обычно 1—2 ряда крючьев, иногда
4 ряда присосок равных размеров.
Фиксаторного аппарата щупалец нет.
Сем. Enoploteuthidae . . с. 156.
8(7). Фотофоров нет. На булаве
4 ряда присосок резко неравных раз-
меров: средние гораздо крупнее крае-
вых. Фиксаторный аппарат щупалец
имеется.
Сем. Ncoteuthidae, род Alluroteut-
his (взрослые особи) (см. с. 214).
9(6). Щупальца имеются только
у личинок или ранней молоди, несут
на конце гроздь крупных присосок.
У взрослых кальмаров щупальца
полностью редуцируются. Имеются
фотофоры на концах 2-й или всех
пар рук. Формула буккального креп-
ления DDVV.
Сем. Octopoteuthidae . . с. 176.
10(3). Крючьев на руках не бы-
вает. Присоски па руках в 2 ря-
да ..............................И.
11(20). Лучи буккальной ворон-
ки крепятся к . брюшным (вентраль-
ным) сторонам рук 4-й пары, т. е.
формула буккального крепления
DDVV. Руки не соединены глубокой
умбреллой ......................12.
12(13). Мантия покрыта хряще-
выми бугорками или чешуйками. Бу-
лава щупалец с 4 рядами присосок,
или щупальца отсутствуют. Фотофо-
ров нет.
Сем. Lcpidoteuthidae . . с. .232.
Рис. 40. Сем. Pickfordiateuthidae. Pick-
fordiateuthis pulchella: a — самец; б — бу-
лава: в — гектокотиль; г — гладиус (по
G. Voss, 1953)
13(12). Кожа гладкая. Строение
булавы щупалец иное .... 14.
14(15). Плавники гребневидные,
представляют собой ряд прямых или
изогнутых усиковидных мышечных
«лучей», соединенных тонкой мембра-
ной, у взрослых они доходят до пе-
реднего края мантии. Булава в виде
овальной площадки со многими ря-
дами мелких присосок.
Сем. Ctenopterygidae . . с. 195.
15(2). Плавники нормального
строения, мускулистые, их длина не
превышает 60% длины мантии. Бу-
лава щупалец не представляет собой
овальную площадку .... 16.
16(17). Мантия сзади вытянута в
длинный тонкий хвост, простираю-
щийся гораздо дальше плавников.
Булава щупалец узкая, нерасширен-
ная, с 6 рядами однотипных присо-
сок.
Сем. Batotcuthidae . . с. 233.
17(16). Мантия нс вытянута в
хвост. Плавники на заднем конце те-
ла, ромбические, сердцевидные или
поперечно-овальные. Булава щупалец
расширенная или щупальца отсутст-
вуют ..............‘............18.
18(19). Булава щупалец несет 2
ряда крючьев, по бокам которых мо-
гут располагаться присоски, изредка
щупальца у взрослых полностью от-
сутствуют.
Сем. Onychoteuthidae . с. 177.
19(18). Крючьев на булаве нет.
Присоски в ладонной и центральной
части булавы сидят во много рядов,
на «пальце» — в 4 ряда.
Сем. Brachioteuthidae . . с. 215.
20(21). Лучи буккальной воронки
крепятся к спинным (дорсальным)
сторонам рук 4-й пары, т. е. форму-
ла буккального крепления DDVD,
Иногда руки соединены широкой
умбреллой (некоторые виды рода
Histioteuthis), мешающей рассмот-
реть строение буккального крепле-
ния .............................24.
21(24). Имеются многочисленные
фотофоры па мантии, голове, конеч-
ностях или внутри мантийной по-
лости ...........................22.
22(23). Имеются фотофоры внут-
ри мантийной полости, на брюшной
стороне глазного яблока и па стебле
щупалец. Плавники ромбические,
иногда вытянуты в хвост. Булава
щупалец с 4 рядами присосок одина-
кового строения. Буккальная мембра-
на 8-лучевая.
Сем. Lycoteuthidae . . . с. 153.
151
23(22). Многочисленные фотофо-
ры располагаются на мантии, голове
(в том числе вокруг века), на руках,
но их нет в мантийной полости, на
глазном яблоке и на стеблях щупа-
лец. Плавники почковидные, сзади с
вырезкой. Булава щупалец с несколь-
кими (4 или больше) рядами присо-
сок неравных размеров и неодина-
кового строения. Буккальная мем-
брана 6- или 7-лучевая.
Сем. Histioteuthidae . . . с. 198.
24(21). Фотофоры отсутствуют
или имеются лишь у основания или
на концах рук..................25.
25(26). Руки короткие, не длин-
нее или немногим длиннее головы,
присоски рук Мелкие и нерегулярно
расположенные — более чем в 2 ря-
да в середине и па концах рук. Бу-
лава не расширенная, более чем с
4 рядами мелких одинаковых при-
сосок. Плавники почковидные, сидят
на конце тела и нс соединяются
между собой. У молоди заметны фо-
тофоры у оснований первых 3 пар
рук. Есть мелкие присоски на бук-
кальной мембране.
Сем. Bathyteuthidae . . . с. 196.
26(25). Руки длинные. Присоски
крупные, всегда в . 2 ряда. Булава
расширенная, присоски разных раз-
меров. Плавники иные. Нет фотофо-
ров у оснований рук. Нет присосок
на буккальной мембране ... 27.
27(28). Плавники широкие, ром-
бической или сердцевидной формы.
Булава щупалец без кучки мелких
присосок в карпальной части, с 4 ря-
дами крупных присосок в централь-
ной и 5—7 рядами мелких — в дис-
тальной части. У взрослых самок
имеется крупный фотофор на концах
3-й пары рук.
Сем. Psychroteuthidae . . с. 210.
28(27). Плавники узкие, не ром-
бической и нс сердцевидной формы.
Карпальная, а иногда и центральная
часть булавы со множеством мелких
присосок, дистальная часть с 4 ря-
дами присосок. Фотофоры отсутст-
вуют ..........................29.
29(30). Щупальца не длиннее или
ненамного длиннее мантии. Плавники
доходят до заднего конца тела или
переходят за его конец, они обрат-
ио-сердцевидной формы с вырезкой
сзади или в виде боковой каймы. Не
более 4—5 пар крупных присосок в
центральной части булавы. Кальма-
ры небольших или средних размеров.
Сем. Neoteuthidae . . . с. 214.
30(29). Щупальца в несколько раз
длиннее мантии. Плавники ланцето-
видные, сидят на «хвостовом стебле»
и не доходят до заднего конца ман-
тии. Более 10 пар крупных присосок
в центральной части булавы. Каль-
мары достигают гигантских разме-
ров.
Сем. Architeuthidae . . . с. 213.
31(2). Мантийно-вороночный за-
мыкательный аппарат специализиро-
ванный. Мантийный хрящ не пред-
ставляет собой простого низкого
гребня.........................32.
32(35). Мантийный хрящ состоит
из двух гребней, смыкающихся под
прямым углом в виде ± или - ’ и
суженных в месте пересечения; во-
роночный хрящ — две смыкающихся
аналогично этому борозды с буграми
в месте пересечения. На руках всег-
да 2, на булаве 4 ряда присосок,
брюшные руки по длине и вооруже-
нию не отличаются резко от про-
чих ...........................33.
33(34). Мантийные и вороночные
хрящи 1-образные. Плавники по-
перечно-ромбические или сердцевид-
ные, их длина не превышает или не-
много превышает половину длины
мантии.
Сем. Ommastrephidae . . с. 218.
34(33). Мантийные и вороночные
хрящи -] -образные. Плавники про-
дольно-ромбические, доходят до пе-
реднего края мантии.
Сем. Thysanoteuthidae . . с. 229.
35(32). Вороночный хрящ ухо-
видный, овальный, треугольный, кег-
ле- или бутылковидный, мантийный—
носовидпый или бугорковидный. На
руках 1—3-й пар два или более, на
булавах четыре или более рядов при-
сосок .........................36.
36(41). На руках 1—3-й пар гря-
да присосок, 4-й пары 0—2 ряда.
Присоски крупные...............37.
.£7(38). Булава щупалец со мнр-
гими (до 20—40) рядами очень мел-
ких присосок примерно одинаковых
размеров и устройства, киля и фик-
саторного аппарата пет (щупальца
часто автотомируются близ основа-
ния, сохраняются лишь обрывки стеб-
лей). 4-я пара рук длиннее и толще
прочих, с 2 рядами присосок. Воро-
ночный хрящ овальный с 1—2 бугор-
ками, реже бутылковидный. На те-
ле и конечностях могут быть лишь
мелкие фотофоры в виде черных то-
152
чек, крупные фотофоры бывают лишь
на глазном веке или на нижней сто-
роне глазного яблока, иногда фото-
форов нет.
Сем. Mastigoteuthidae . . с. 243.
38(37). Булава щупалец с 4—6
рядами присосок разного размера.
Вороночный хрящ уховидный, оваль-
ный, ксглевидиый или треугольный.
Обычно имеются крупные фотофоры
либо на теле, либо на конечностях
(булава щупалец, руки 4-й пары),
часто еще и на кишке, у некоторых
видов фотофоров нет .... 39.
39(40). Вороночный хрящ тре-
угольный с глубокой бороздой, по-
средине или на заднем конце которой
находится глубокая ямка для бугор-
ковидного мантийного хряща. 4-я па-
ра рук не отличается от прочих по
длине, толщине и вооружению. Плав-
ник поперечпо-овальпый или округ-
лый, его длина более % длины ман-
тии, иногда равна ей. Булава щупа-
лец с килем и фиксаторным аппара-
том. Фотофоры всегда имеются, но
щупальца и руки 4-й пары без фото-
форов.
Сем. Cycloteuthidae ... с. 233.
40(39). Вороночный хрящ ухо-
видный или овальный с 1—2 бугор-
ками, либо удлиненно-треугольный
или ксглевидиый, мантийный — но-
совидный, реже бугорковидный. 4-я
пара рук длиннее и (или) шире про-
чих, а если незначительно отличается
от прочих по размерам, то несет при-
соски только в один ряд и только в
базалыюй части. Длина плавника
менее % длины мантии. Булава щу-
палец без фиксаторного аппарата и,
как правило, без киля. Фотофоры,
если имеются, располагаются вдоль
рук 4-й пары и (или) в дистальной
части булавы щупальца, могут быть
также на глазном яблоке и кишке.
Сем. Chiroteuthidac ... с. 236.
41(1). Мантия срастается с во-
ронкой и (или) с головой ... 42.
42(31). Мантия срастается и с
головой (на затылке), и с воронкой.
Гладиус имеется. Булава с 4 рядами
присосок или 2 рядами крючьев.
Сем. Cranchiidae .... с. 255.
43(42). Мантия срастается либо с
головой, либо с воронкой ... 44.
44(45). Мантия срастается с во-
ронкой, затылочный хрящ имеется.
Гладиус развит. Щупальца у взрос-
лых отсутствуют.
Сем. Grimalditcuthidae . . с. 254.
45(44). Мантия срастается с голо-
вой на затылке, затылочного хряща
нет. Мантийный хрящ овальный. Гла-
диус редуцирован. Щупальца имеют-
ся, булава с несколькими рядами мел-
ких присосок.
Сем. Promachoteuthidae . с. 254.
2.2.2.1. Семейство
Lycoteuthidae
Кальмары небольших, редко средних
размеров, крепкого сложения, с мус-
кулистой мантией конической формы.
Плавник широкий, ромбический,
иногда вытянут в хвост. Руки с 2,
щупальца с 4 рядами присосок. Од-
на—две пары рук у взрослых могут
быть чрезвычайно удлинены. Бук-
кальная мембрана 8-лучевая. Форму-
ла буккального крепления DDVD,
Замыкательный аппарат простой.
Гладиус узкий, с одним или двумя
расширениями, на конце ложковид-
пый, иногда с ростр умом. Имеются
многочисленные (от 23—24 до более
100) светящиеся органы разного
строения и цвета, располагающиеся
внутри мантийной полости, на брюш-
ной стороне глазного яблока, иногда
также на руках, концах рук, поверх-
ности мантии и па хвосте. У некото-
рых видов мужские половые органы
парные. Развитие с личиночной ста-
дией.
Тропическо-субтропические мезо-
пелагические и придонно-батиальные
кальмары. 2 подсемейства, 5 родов,
5 видов. Обзоры: G. Voss, 1962, R.
Young, 1964.
Таблица для определения родов и
видов Lycoteuthidae
1(8). На брюшной стороне глазного
яблока 5 светящихся органов, распо-
ложенных в один ряд. На стебле щу-
палец 2, в мантийной полости 8—10
светящихся органов. Гладиус (рис.
41, р) с перетяжкой на границе сред-
ней и задней трети длины, без рост-
рума. Мужские половые органы пар-
ные (изучено у Selenoteuthis и Ore-
goniateuthis), гектокотиля нет.
Подсемейство Lycoteuthinae . 2.
2(3). На заднем конце тела
имеется один крупный' круглый фо-
тофор. На концах рук 2-й и 3-й нар
у самцов по 1 шаровидному фото-
фору (у самок их нет).
15а
77
Рис. 41. Сем. Lycoteuthidae: а—в — Seleno-
ieuthis scintillans, самка (а и 6} и глаз-
ные фотофоры (в); г—з — Oregon, iateuthis
springeri, самец сверху (г), мантия снизу
(д), глазные фотофоры (е), рука 2-й пары
молодого самца (лс), булава (з); л —
Lycoteuthis diadema, молодь («), неполо-
возрелая самка (к), булава (л); м —
«Onychoteuthis longimanus», предположи-
тельно зрелая самка L. diadema; н —
N ematolampas regalis; о—п — Latnpadio-
Ieuthis megaleia, самка и самец: р—с —
гладиусы Lycoteuthinae (р, О. sprtngeri) и
Lampadioteutbinac (с, L. megaleia) (а—з,
к—о, р — по G. Voss, 1956, 1962: и — по
Chun, 1910; п, с —по R. Young, 1964)
Род Selenoteuthis Voss, 1958. Один
вид — S. scintillans Voss, 1958 (рис.
41, a—в). Тропические и субтропи-
ческие воды Западной Атлантики,
Карибское море, Мексиканский за-
лив, район Канарских островов. Ме-
зопелагиаль, по ночам поднимаются
и в эпипелагиаль. Длина мантии до
4,5 см.
3(2). Нет фотофора на заднем
конце тела. Концы рук без шаровид-
ных фотофоров..................4.
4(5). Задний конец тела вытянут
в длинный хвост. На спинной сторо-
не мантии и хвоста расположено
11 кожных фотофоров, еще 8 в тол-
ще тканей хвоста на дорсальной сто-
роне и 8 на голове. Задняя группа
внутримантийных фотофоров состоит
из трех обособленных органов. 2-я
пара рук сильно удлинена, ее конец
уплощен, расширен и лишен присо-
сок. 3-я пара рук нормальная. Име-
ется ряд фотофоров вдоль дорсаль-
ной стороны рук 2-й и 3-й пар по
всей их длине.
Род Oregoniateuthis Voss, 1956.
Один вид — О. springeri Voss, 1956
(см. рис. 41,г—з, р). Субтропические
воды Северо-Западной Атлантики и
Мексиканский залив. Батиаль.
5(4). Задний конец тела не вы-
тянут в хвост. Нет фотофоров на
спинной стороне мантии и хвоста.
Задний впутримантийный фотофор
состоит из трех слившихся вместе
фотофоров и имеет поперечно-вере-
теновидную форму. Удлинена либо
3-я, либо 2-я и 3-я пары рук . . 6.
6(7). Концы 2-й и 3-й пар рук у
взрослых (длина мантии больше 8—
9 см) удлинены, утончены и оттяну-
ты в нитевидный бич, лишенный при-
сосок; длина рук 2-й пары значи-
тельно больше, чем 3-й, и может в
2 с лишним раза превосходить длину
мантии; 3-я пара гораздо короче
мантии. У молоди концы рук нор-
мальные. Нет фотофоров па брюш-
ной стороне мантии у заднего конца.
Фотофоры на руках 2-й и 3-й пар тя-
нутся вдоль основания плавательной
мембраны (бокового киля): у взрос-
лых на руках 2-й пары почти по всей
длине, 3-й — лишь в дистальной по-
ловине.
Род Lycoteuthis Pfeffer, 1900
(=Thaumatolampas Chun, 1903, Ast-
henoteuthion Pfeffer, 1912, Leptodon-
toteuthis Robson, 1926). Один вид —
L. diadema (Chun, 1900) [=? Orego-
156
niateuthis longimanus (Steenstrup,
1857), ?O. lorigera (Steenstrup, 1875)]
(рис. 41, а—л). Тропические воды
Атлантики, Мексиканский залив и
циркумглобально в южносубтропиче-
ской и потальной зонах Мирового»
океана. Многочислен на Китовом
хребте, на склоне у Юго-Западной и
Южной Африки, у Северного остро-
ва Новой Зеландии. Батиаль, иногда
мезопслагиаль. Длина мантии до
18 см.
7(6). 3-я пара рук нитевидная,
много длиннее мантии, дистальная
часть руки лишена присосок. 2-я па-
ра рук нормальная. Несколько де-
сятков фотофоров расположено
вдоль 3-й пары рук, на 1-й и 2-й па-
ре по 1 фотофору недалеко от конца
руки. Пара фотофоров на брюшной
стороне мантии у заднего конца те-
ла в местах присоединения заднего
края плавников к мантии.
Род Nernatolampas Berry, 1913.
Один вид — N. regalis Berry, 1913
(рис. 41, н). Юго-западная часть Ти-
хого океана (о-ва Ксрмадек и пред-
положительно Северный остров Но-
вой Зеландии). Длина мантии 3 см.
10(1). На брюшной стороне глаз-
ного яблока 3 фотофора, еще 1 по-
зади края зрачка, все 4 далеко рас-
ставлены. По 1 фотофору у основа-
ния щупалец и по 4 на их стебле.
5 фотофоров внутри мантийной по-
лости: 2 анальных, 2 жаберных и
1 задний. Нет фотофоров на мантии,
голове и руках. Гладиус (рис, 41, с)
без перетяжки, расширен в задней
половине и несет маленький острый
рострум на дорсальной стороне над
ложковидпым концом гладиуса (уни-
кальный для кальмаров признак!).
Мужские половые органы непарные.
Гектокотилизируется правая брюш-
ная рука.
Подсемейство Lampadiotcuthinac.
Род Lampadioteuthis Berry, 1916.
Один вид — L. megaleia Berry, 1916
(рис. 41,о, п). Субтропические воды
Северной Атлантики и юго-западная
часть Тихого оксана. Длина мантии
до 3 см.
2.2.2.2. Семейство
Enoploteuthidae
Кальмары мелких, редко средних раз-
меров с мускулистой (изредка хря-
щевато-студенистой) мантией, обыч-
но конической формы, суживающейся
к хвосту. Плавники широкие, муску-
листые, ромбической или округлой
формы, сидят в задней половине или
в середине тела. Руки и щупальца
крепкие с сильно развитыми килями
и защитными мембранами. Руки не-
сут 2 ряда крючьев, присоски имеют-
ся лишь в базальных частях и на
концах рук; у некоторых видов на
руках 4-й пары нет крючьев. Булава
щупалец у молоди с 4 рядами при-
сосок, у взрослых кальмаров (кроме
Pterygioteuthis) на ней развиваются
один или два ряда крючьев, краевые
присоски в центральной части була-
вы могут исчезать. Замыкательный
аппарат простой. Буккальное крепле-
ние 8-лучевое, его формула DDVD
(у Enigmoteuthis DDDD), Имеются
многочисленные фотофоры, располо-
жение и строение которых строго
специфичны. Они могут располагать-
ся на брюшной (реже' на спинной)
стороне мантии, головы, рук, на стеб-
лях щупалец, на брюшной стороне
глазного яблока, внутри мантии и
даже внутри «фунтика» гладиуса.
Гладиус с коротким рахисом и рас-
ширенным пером, конечный конус
может быть хорошо развит или ре-
дуцирован. Гектокотилизируется од-
на из рук 4-й пары. Сперматофоры
переносятся па «затылок» самки.
Пелагические (от эпи- до батипе-
лагиали) океаничёские, реже нерито-
юкеанические кальмары, некоторые
виды встречаются у дна в батиали.
Размножение в пелагиали. Развитие
с личиночной стадией. Обитают jb
тропических и умеренных водах.
Многочисленны, но промыслового
значения почти не имеют. 3 подсе-
мейства, 8 родов, 33—37 видов.
Таблица для определения родов
семейства Enoploteuthidae
1(12). Фотофоры расположены на
поверхности мантии, головы и рук.
Внутри мантийной полости их нет
(у Abralia armata один). На брюш-
ной стороне глазного яблока один
ряд фотофоров или их нет совсем.
Задний конец мантии конический, не
вытянут в острый хвост, конечный
конус гладиуса слабо развит. Зад-
ний край плавников вогнутый или
прямой. Развиты оба яйцевода 2.
2(11). На нижней поверхности
мантии, головы и рук множество
(сотни) мелких фотофоров, различа-
ющихся по строению, но слабо раз-
нящихся по размерам. Есть фотофо-
ры на брюшной стороне глазного яб-
лока. Нет фотофоров на стебле щу-
палец. Нидаментальные железы
отсутствуют, их заменяют чрезвычайно
увеличенные и разделенные на 2 по-
ловины яйцеводные железы. Длина
мантии взрослых кальмаров до 13 см.
Подсемейство Enoploteuthinae . 3.
3(8). На брюшной стороне глаз-
ного яблока 5—8 фотофоров нерав-
ных размеров (2 крайних обычно
значительно крупнее средних), сидя-
щих поодаль друг от друга. Плавни-
ки доходят до заднего конца мантии.
В центральной части булавы щупа-
лец 1 ряд крючьев и 2 ряда присо-
сок или 2 ряда крючьев и несколько
мелких присосок между ними (у ли-
чинок и молоди — только присос-
ки) ............................4.
4(5). На концах брюшных рук
нет крупных шаровидных фотофоров
черного цвета (могут быть лишь
мелкие, меньше диаметра руки, свет-
лые). Крючья на булаве расположе-
ны в один ряд. На брюшной стороне
глазного яблока 5—8 фотофоров раз-
ных размеров и строения, обычно
задний отличается от прочих. Бук-
кальная мембрана розового цвета.
Гектокотилизируется левая (реже
правая) брюшная рука.
Род Abralia Gray, 1849 ( = Епор-
loion Pfeffer, 1912) (рис. 43—44). Ти-
повой вид — A. armata (Quoy et
Gaimard, 1831). 3 подрода: Abralia
s. str. Asteroteuthis Pfeffer, 1900 (ти-
повой вид A. (A.) veranyi (Ruppell,
1844), Stenabralia Grimpe, 1931, [ти-
повой вид — A. fS.) rertschi Grimpe,
1931]. 13—14 видов в тропических и
субтропических водах Атлантическо-
го, Индийского и западной части Ти-
хого океана. Чаще всего невдалеке
от склона. Мелкие (длина мантии до
5—7 см) активные кальмары.
5(4). На концах брюшных рук по
3 крупных фотофора, которые у фик-
сированных кальмаров выглядят как
черные шарики. На брюшной стороне
глазного яблока 5 круглых фотофо-
ров одинакового строения, оба край-
них немного крупнее средних, кото-
рые одинаковы. Буккальная мембра-
на темно-фиолетового цвета. Гекто-
котилизирустся правая брюшная
рука........................ . 6.
157
6(7). Крючья на булаве в два
ряда.
Род Abraliopsis Joubin, 1896 (см.
рис. 45, а—г). Типовой вид — A. pfef-
feri Joubin, 1896. 2 подрода: Abraliop-
sis s. str. и Mierabralia Pfeffer, 1900
[типовой вид — A. (M.) lineata (Go-
odrich, 1896). 8—9 видов и 2 подвида
в тропических, субтропических и от-
части умеренных водах Атлантическо-
го, Индийского и Тихого океанов.
Мелкие океанические мезо- и мезо-ба-
типелагические кальмары, по ночам
поднимаются в эпипелагиаль; местами
весьма обычны. Длина мантии до
6 см.
7(6). На булаве только 2 крюка,
в один ряд.
Род Watasenia С. Ishikawa, 1913.
Один вид W'. scintillans (Berry, 1911)
(см. рис. 45, у—ф), японский каль-
мар-светлячок. Северо-западная часть
Тихого океана от Южных Куриль-
ских островов до Южной Японии,
Восточно-Китайское, Японское и
южная часть Охотского моря. Эпи-
мезопелагический вид. Длина ман-
тии до 7 см. У япономорского побе-
режья Южного Хонсю в период не-
реста служит предметом небольшого
промысла.
8(3). На брюшной стороне глаз-
ного яблока ряд из 8—12 (редко 6—
7) фотофоров, из которых 2 крайних
крупные, прочие мелкие и сидят поч-
ти впритык друг к другу. Задний ко-
нец мантии у взрослых обычно вы-
тянут в мягкий конический хвостик,
заходящий за край плавников . 9.
9(10). На булаве 4 крюка, распо-
ложенных в один ряд. один из
крючьев значительно крупнее про-
чих, около каждого крюка — по две
присоски. Фотофоры на брюшной
стороне глазного яблока (кроме
краевых) неравных размеров: более
мелкие чередуются с более крупны-
ми. 6 рядов фотофоров на брюшной
стороне головы. Формула буккаль-
ного крепления DDDD, т. е. все кон-
нективы, в том числе и идущие к ру-
кам 3-й пары, крепятся к дорсаль-
ным сторонам рук. Плавник лишь
чуть-чуть не доходит до заднего кон-
ца мантии.
Род Enigmoteuthis Adam, 1973.
Один вид Е. dubia (Adam, 1960) (рис.
42, н—и), в прибрежной зоне Крас-
ного моря. Длина мантии до 4 см.
Обзор: Adam, 1973.
10(9). На булаве 2 ряда крючьев,
у взрослых центральная часть була-
вы лишена присосок. Фотофоры на
брюшной стороне глазного яблока,
кроме краевых, одинаковые, мелкие,,
часто соприкасающиеся. 61/г—Ю ря-
дов фотофоров на брюшной сторо-
не головы. Формула буккального
крепления обычная для семейства:
DDVD, т. е. коннективы, идущие к
рукам 3-й пары, крепятся к вен-
тральной стороне рук. Мантия у
взрослых обычно далеко заходит за
задний край плавников, образуя мяг-
кий хвостик.
Род Enoploteuthis d’Orbigny, 1848
(рис. 42, а—л(). Типовой вид — Е. 1ер-
tura (Leach, 1817). 5—6 видов и 2
подвида в тропических, субтропиче-
ских и местами в умеренных водах
Атлантического, Индийского и Тихо-
го океанов. Эпи-мезопелагические
океанические и нерито-океанические
кальмары. Длина маптии до 10—
13 см. Частичный обзор: Roper, 1966.
11(2). На нижней поверхности
мантии 22 крупных полушаровидных
фотофора, расположенных в строгом
порядке, кроме них крупные фото-
форы имеются на голове (вокруг
глаз, близ воронки, у основания 2-й
пары рук) и на стебле щупалец.
Множество мельчайших фотофоров
разбросано по телу, голове и рукам
4-й пары. Нет фотофоров внутри
мантии и на брюшной стороне глаз-
ного яблока. Задний конец мантии у
взрослых кальмаров оттянут в хвост,
заходящий на задний край плавни-
ков. Крючья на булаве в 2 ряда,
краевые присоски в центральной час-
ти булавы у взрослых отсутствуют.
Нидаментальные , железы имеются.
Длина мантии взрослых кальмаров
до 40 см.
Подсем. Ancistrocheirinae. Обзор:
Несис, 1978а.
Род Ancistrocheirus Gray, 1849
( = Thelidioteuthis Pfeffer, 1900) (см.
рис. 46, а—г). Типовой вид — А, 1е-
sueuri Ferussac et d’Orbigny, 1839.
2 вида, один из них сомнительный.
Эпи-мезопелагические и придонно-
батиальные кальмары тропических и
субтропических вод.
12(1). Фотофоры расположены
внутри мантийной полости (8—
10 шт. разного строения, размера и
цвета), в толще тканей стебля щу-
палец и на брюшной стороне глазно-
го яблока (12—15 шт. также разно-
го размера и строения, расположен-
158
пых несколькими группами). Задний
конец Мантии вытянут в острый
хвост (игловидный конечный конус
гладиуса). Плавники округлые (поч-
ковидные), их задний край выпук-
лый, они не доходят до конца тела.
Функционирует только один яйцевод.
Подсемейство Pytotcuthinae . 13.
13(14). Есть крючья па булаве
щупалец и па руках 4-й пары.
Крючья на булаве расположены в
1 ряд, на руках они идут в 2 ряда
по всей длине. На брюшной стороне
глазного яблока 12 фотофоров:
9 крупных (разного размера) и
3 мелких. В мантийной полости 10
фотофоров, в том числе на уровне
жабр 3 фотофора, расположенных
поперек продольной оси тела. На
стебле щупалец у молоди и взрос-
лых 6—7, иногда 8 фотофоров. Гек-
токотилизируется правая брюшная
рука. Развит только левый яйцевод.
Род Pyroteuthis Hoyle, 1904
( = Charybditeuthis Viviani, 1914)
(см. рис. 46, д—л). Типовой вид —
A margaritifera (Ruppell, 1844). 2 ви-
да в мезопелагиали тропических и
субтропических вод. Длина мантии
до 4—6 см.
14(13). Нет крючьев на булаве
щупалец. На руках 4-й пары крючьев
нет или не более 2. На руках 1—3-й
пар крючья имеются в небольшом
числе лишь в середине рук, в один
или два ряда. На брюшной стороне
глазного яблока 14—15 фотофоров:
10 крупных и 4—5 маленьких. В ман-
тийной полости 8 фотофоров, в том
числе на уровне жабр только 1, на
продольной оси тела. На стебле щу-
палец 4 фотофора. Гектокотилизи-
руется левая брюшная рука. Развит
только правый яйцевод.
Род Pterygioteuthis Fischer, 1896
(см. рис. 46,м—у). Типовой вид —
Р. giardi Fischer, 1896. 2 вида в эпи-
я мезопелагиали тропических и суб-
тропических вод. Длина мантии до
2,5—Зсм.
Таблица для определения видов рода
Enoploteuthis
1(6). Фотофоры на брюшной сторо-
не мантии расположены, по крайней
мере, в передней части мантии пря-
мыми продольными рядами. Между
рядами фотофоров пет соединяющих
их «перемычек» из обособленных ку-
чек фотофоров....................2.
2(3). На брюшной стороне мантии
4 продольных ряда фотоцюров, ru-
торые, как и свободная от фотофо-
ров полоска посредине брюшной сто-
роны, протягиваются до заднего кон-
ца тела. На брюшной стороне головы
6V2 продольных рядов фотофоров,
не считая идущих по краю глазной
орбиты. Непарный средний ряд фо-
тофоров па голове образуется из сли-
вающихся рядов, идущих по вен-
тральным краям рук 4-й пары, и кон-
чается на уровне передних краев
глаз. Первый и второй (считая от
средней линии) боковые ряды фото-
форов сливаются в передней (перед
основаниями рук 4-й пары) и зад-
ней (на уровне отверстия воронки)
частях головы. Ряд фотофоров на
руках 3-й пары доходит лишь до се-
редины руки. Щупальца сильные,
длиннее мантии.
Е. anaspis Roper, 1964 (рис.
42, а, б). Тропические и субтропиче-
ские воды Атлантики, Мексиканский
залив и Карибское море, и Индо-
Вестпацифика. Океанический вид.
3(2). На брюшной стороне ман-
тии в передней части тела 6—8 про-
дольных рядов фотофоров. Свобод-
ная полоска вдоль средней линии
брюшной стороны отсутствует или
протягивается не далее середины
тела. На брюшной стороне головы
7—10 продольных рядов фотофо-
ров ............................4.
4(5). На брюшной стороне ман-
тии 7 продольных рядов фотофоров,
3 средних протягиваются до конца
тела. На брюшной стороне головы
10 более или менее отчетливых про-
дольных рядов фотофоров, средняя
линия головы свободна от них. 6 ря-
дов фотофоров на воронке. Щупаль-
ца тонкие, слабые.
Е. leptura (Leach, 1817) (рис.
42, в, г). 2 подвида. Вид тропический,
атланто-индовестпацифичсский, нери-
то-океаничсский.
а (б). Непарный средний ряд фо-
тофоров на брюшной стороне ман-
тии ответвляется от правого или ле-
вого бокового ряда значительно даль-
ше переднего края мантии.
Е. leptura leptura (Leach, 1817).
Тропические воды Атлантики от
Бермудских островов и Мадейры до
Бразилии и Юго-Западной Африки.
б (а). Непарный средний ряд фо-
тофоров начинается, лишь чуть от-
ступя от переднего края мантии, и не
ответвляется от бокового ряда.
159
£
Сем. Enoploieuthidae, роды Епор-
(а~м) и Enigmoteiilltis (н—п):
Рис. 42.
loteuihis . , .
а—б — Е. anaspis; в—г — Е. leptura leptura;
д—е — Е. chuni (е — глаз); ж—Е. galaxias-,
з—к — Е. reticulata, молодь (з), взрослая
самка (и), глаз (к); л—м — Enoploteuihis sp.
Okutani (ле — глаз); н—п — Enigmoteuthis
dubia, самка (к, щупальца оборваны),
глазные фотофоры (о), крючья булавы (п)
(а—г —по Roper, 1966; д, и—м—по Okutani,
1973, 1974; е — по Taki, 1964; ж — по Berry,
1918; з — по Rancurel, 1970; н—п~ио Adam,
1960, 1973)
О О
р о
И К. II. Несис
E. leptura magnoceani n. ssp.1
[=K leptura d’Orbigny, 1839, part;
Atlas (Onychoteuthis), pl. 6; pl. 11 fig.
6—14; pl. 12 fig 10—24]. Тропиче-
ские воды Западной и Центральной
Пацифики и Австрало-Азиатские мо-
ря. Экваториальные воды.
5(4). На брюшной стороне ман-
тии в передней половине тела 6—8
продольных рядов фотофоров, в зад-
ней половине фотофоры распределя-
ются диффузно. На брюшной стороне
головы 7 продольных рядов фото-
форов, непарный средний ряд раз-
дваивается спереди и сзади и пере-
ходит на руки 4-й пары и края воро-
ночной ямки. 4 ряда фотофоров па
воронке. Щупальца крепкие, длин-
ные.
Е. chuni С. Ishikawa, 1914 (=£.
theragrae Taki, 1964) (рис. 42, д,е).
Тихоокеанское и япономорское побе-
режья Южной и Центральной Япо-
нии, о-ва Бонин, Изду-Осима, Кюсю,
Рюкю, Куросио до Гавайских остро-
вов. Нсрито-океанический вид.
6(1). Фотофоры на брюшной
стороне мантии расположены изви-
листыми продольными или косыми
рядами, соединенными «перемычка-
ми» из обособленных скоплений фо-
тофоров, так что образуется сетча-
тая структура..................7.
7(10). Фотофоры на брюшной
стороне мантии образуют извилис-
тые продольные ряды. 3 ряда фото-
форов па руках 4-й пары ... 8.
8(9). На брюшной стороне ман-
тии 4 извилистых продольных ряда
фотофоров, 2 средних соединены
многочисленными «перемычками» из
обособленных кучек фотофоров. На
брюшной стороне головы 5 продоль-
ных рядов фотофоров, не считая ря-
да по краю глазной орбиты, 3 сред-
них ряда представляют собой поло-
сы из многочисленных фотофоров,
лежащих бок о бок. 3 ряда фотофо-
ров на руках 4-й пары лежат на рав-
ных расстояниях друг от друга. Дли-
на рук 3/б—2/з длины мантии. Щу-
пальца намного длиннее рук. Хвосту
взрослых кальмаров длинный и
тонкий.
1 Голотип: самка, длина мантии
73 мм, Тихий океан (без точного
местонахождения). Национальный
музей естественной истории (Museum
national cTHistoire naturelle), Париж.
162
E. galaxias Berry, 1918 (рис.
42, xw).’Западная, Южная и Юго-Вос-
точная Австралия и Новая Зеландия.
Южносубтропический нерито-океапи-
ческий вид. Мезопелагиаль и ба-
тиаль.
9(8). На брюшной стороне ман-
тии 6 извилистых продольных рядов
фотофоров. Они соединены немного-
численными (3—4) «перемычками» из
обособленных скоплений фотофоров,
которые направлены поперек про-
дольной оси мантии в ее середине и
диагонально — на боках. На брюш-
ной стороне головы 6 продольных
рядов фотофоров, не считая ряда по
краю глазной орбиты, каждый ряд
состоит из одной цепочки фотофоров.
Средняя линия головы свободна от
фотофоров. 2 дорсальных ряда фо-
тофоров на руках 4-й пары тесно
сближены, крайний дорсальный ряд
тянется вдоль основания плаватель-
ной мембраны: Длина рук равна или
превосходит длину мантии. Щупаль-
ца не длиннее или лишь немного
длиннее рук. Хвост короткий.
Е. reticulata Rancurel, 1970 (=Е.
sp. № 1 Okutani, 1974) (рис. 42,
з—к), Индо-Всстпацифика от запад-
ной части Индийского океана до
района восточнее о-вов Маркизских
и Туамоту, экваториальные и цент-
ральные воды. Нижняя эпи- и мезо-
пелагиаль.
10(7). В передней части брюшной
стороны мантии 2 продольных и 4
косых, диагонально расходящихся
назад ряда фотофоров. В задней ча-
сти мантии фотофоры расположены
диффузно. 3 ряда фотофоров на ру-
ках 4-й пары. Щупальца тонкие,
слабые.
Enoploteuthis sp. Okutani, 1974
(р. 54, 56, fig. 12 с, d, f) (рис. 42,
л, At). Восточно-экваториальная Па-
цифика.
Таблица для определения подродов
и видов рода Abralia
1(14). На нижней стороне головы
5 четко обособленных продольных
рядов фотофоров, нс считая ряда по
краю глазной орбиты; средний ряд
раздваивается в задней половине го-
ловы ..........................2.
2(3). Длина плавника V2—2/з, ши-
рина больше 5/в длины мантии.
Имеется светящийся орган на чер-
нильном мешке. Фотофоры на брюш-
ной стороне мантии распределены
диффузно, без свободной средней
продольной полосы. 6 (редко 5 или
7) фотофоров на брюшной стороне
глазного яблока. 6 крючьев на була-
ве щупальца. Гектокотилизируется
правая брюшная рука.
Подрод Abralia s. str. А, (Л.) ar-
mata (Quoy et Gaimard, 1832) (рис.
43, a—г). Восточная Индонезия и
Филиппинские острова.
3(2). Длина плавника Vs—V2, ши-
рина Уз—8Д длины мантии. Нет све-
тящихся органов на чернильном меш-
ке. Фотофоры па брюшной стороне
мантии образуют 6—8 узких про-
дольных полос, из которых две сред-
ние четко обособлены от остальных,
продольная средняя полоса обычно
(не всегда) свободна от фотофоров.
Гектокотилизируется левая брюшная
рука.
Подрод Stenabralia Grimpe,
1931 .........................4.
4(7). 5 фотофоров на брюшной
стороне глазного яблока. 3 продоль-
ных ряда фотофоров на руках 4-й
пары. Вдоль средней линии брюшной
стороны мантии проходит узкая про-
дольная полоска, свободная от фо-
тофоров. На заднем конце тела по
бокам мантии вентрально — два
крупных фотофора. На концах рук
4-й пары по 3—б обособленных фо-
тофоров, более крупных, чем обыч-
ные. 4—5 крючьев на булаве щупаль-
ца. Присоски на концах рук — в 2
ряда.............................5.
5(6). Длина плавника равна или
немного меньше У2 длины мантии.
Продольные полоски фотофоров на
брюшной стороне мантии широкие,
четко обособленные, фотофоры в них
тесно сближены и слабо различаются
по размерам. 3 обособленных фото-
фора на концах рук 4-й пары.
A. (S) astrolineata Berry, 1914.
Восточная Австралия (Квинсленд) и
о-ва Керм а дек.
6(5). Длина плавника около Уз
длины мантии. Продольные полоски
фотофоров на брюшной стороне ман-
тии узкие, слабо обособленные, фо-
тофоры в них распределены диффуз-
но и резко различаются по размерам.
4—6 обособленных фотофоров на
концах рук 4-й пары.
Л. (S.) astrosticta Berry, 1909 (рис.
43, д—ж). Куросио, ^Маршалловы и
Гавайские острова. Пелагиаль и дно
в нижней сублиторали и верхней ба-
тиали.
7(4). 6—9 фотофоров па брюш-
ной стороне глазного яблока. 3—4
продольных ряда фотофоров на ру-
ках 4-й пары. 6—8, редко 5 крючьев
на булаве щупальца. Нет крупных
фотофоров па конце тела и концах
рук 4-й пары. Присоски на самых
кончиках рук расположены в 3—
4 ряда ...........................
8(11). 6—7, редко 8 фотофоров
на брюшной стороне глазного ябло-
ка: 5 крупных, 1—3. мелких. Длина
плавника более У3, ширина более У2
длины мантии...................9.
9(10). 6 фотофоров на брюшной
стороне глазного яблока: 2 крупных
по краям, между ними 3 крупных и
1 маленький. 3 ряда фотофоров на
руках 4-й пары.
A.(S) renschi Grimpe, 1931 (рис.
43,з—к). О-ва Мальдивские, Сумат-
ра и Ява.
' 10(9). 7—8 фотофоров на брюш-
ной стороне глазного яблока: 2 круп-
ных по краям, между ними 3 круп-
ных и 2—3 маленьких. 3 ряда фото-
форов на руках 4-й пары, но между
средним и вентральным рядом близ
основания руки — короткий неполный
дополнительный ряд из нескольких
фотофоров (только у взрослых).
A. (S.) steindachneri Weindl, 1912
(рис. 43, л) Красное, Аравийское мо-
ре, Восточная Африка, Маскаренский
хребет, Сейшельские острова.
11(8). 8—9 фотофоров на брюш-
ной стороне глазного яблока: 6 круп-
ных, 2—3 маленьких (изредка мел-
кие фотофоры отсутствуют). Длина
плавника У5—У4, ширина У4—Уз дли-
ны мантии....................: 12.
12(13). 3 продольных ряда фото-
форов на руках 4-й пары, между ни-
ми (у основания руки) до 2 обособ-
ленных дополнительных фотофоров.
Нет обособленных крупных фотофо-
ров в вороночной ямке.
A. (S.) lucens Voss, 1962 (рис.
43, —о). Филиппинские острова, По-
линезия (о-ва Лайн).
13(12). 4 продольных ряда фото-
форов на руках 4-й пары: 3 обыч-
ных и дополнительный ряд из 4—9
фотофоров между средним и вент-
ральным рядами; 1—2 дополнитель-
ных фотофора могут быть между
средним и дорсальным рядами у ос-
нования руки. В вороночной ямке по
бокам от воронки у основания пер-
11*
163
вой затылочной складки —- по одно-
му (с каждой стороны) крупному
обособленному фотофору белого
цвета.
A. (S) spaercki Grimpe, 1931 (рис.
43, п—у). Филиппинские острова,
Восточная Индонезия.
14(1). На нижней стороне головы
фотофоры распределены диффузио
или образуют 7—13 нечетко обособ-
ленных продольных рядов, нс считая
ряда по краю глазной орбиты. Дли-
на плавника ’/2—2/з, иногда до %,
ширина более %, чаще всего 75—
95% длины мантии.
Подрод Asteroteuthis Pfeffer,
1909 ........................ 15.
15(18). 6—7 фотофоров на брюш-
ной стороне глазного яблока. При-
соски па концах рук расположены
в 2 ряда......................16.
16(17). 2 крюка па булаве щу-
пальца. 3 продольные полоски, сво-
бодные от фотофоров, на брюшной
стороне мантии. 6—7 глазных фото-
форов.
A. (A,) grimpei Voss, 1958 (рис.
44,а—г). Гольфстрим у Флориды.
17(16). 6—7 крючьев на булаве
щупальца. Только одна средняя по-
лоска па брюшной стороне мантии
свободна от фотофоров. 6 глазных
фотофоров.
A, (A.) multihamata Sasaki, 1929
(рис. 44,0—лс). О-в Тайвань и при-
легающие районы материкового по-
бережья КНР.
18(15). 5 фотофоров на брюшной
стороне глазного яблока. Фотофоры
на брюшной стороне мантии нс обра-
зуют продольных полос ... 19.
19(22). Два крайних фотофора
на глазу значительно крупнее трех
средних, задний фотофор самый
крупный и имеет овальную, яйце- или
почковидную форму, передний оваль-
ный, три средних одинаковые, округ-
лые (у молодых кальмаров 1, 3 и
5-й фотофоры одного размера, ок-
руглые и крупнее, чем 2-й и 4-й).
Длина плавника около 2/з длины
мантии. 3—4 (изредка 5) крюков па
булаве щупальца. Гектокотилизиру-
стся левая брюшная рука, на пей
развиваются 2 лопасти — с вентраль-
ной и дорсальной стороны ру-
ки ...........................20.
20(21). Палец булавы примерно
164
3
равен по длине ладони. 3—4 ряда
крохотных присосочск на кончиках
рук 1— 3-й пар.
A. (A.) veranyi (Ruppell, 1844)
(=Enoploion eustictum Pfeffer, 1912)
(рис. 44,з—к). Тропические и суб-
тропические воды Западной и Вос-
точной Атлантики от средней части
Атлантического побережья США до
Мексиканского залива и от Среди-
земного моря и Мадейры до Ан-
голы. У дна в батиали и в толще во-
ды над склоном, иногда у поверхно-
сти. В открытом оксане вдали от
склона нс встречается. /Массовый
вид.
21(20). Палец булавы значитель-
но длиннее ладони. Присоски на кон-
чиках рук в 2 ряда.
A. (A.) andamanica Goodrich, 1896
(рис. 44, л—п). Тропические воды
Ипдопацифики от восточной части
Аравийского моря и Сейшельских ос-
тровов до Южной Японии, Гавайских
островов и Восточной Австралии.
По-видимому, отсутствует в запад-
ной части Аравийского моря. На вос-
токе немного не доходит до берегов
Центральной Америки. Встречается
в толще воды (эпи- и мезопслагиаль)
открытого океана и вблизи склона,
в том числе в батиали. Массовый
вид, распадается на несколько не-
четко обособленных форм, из кото-
рых описаны две.
а (б). Фотофоры на брюшной сто-
роне головы расположены продоль-
ными рядами, посредине брюшной
стороны мантии обычно заметна сво-
бодная от фотофоров продольная по-
лоска.
A. andamanica f. andamanica.
Преимущественно на склоне и над
ним.
б (а). Фотофоры па брюшной сто-
роне головы расположены диффуз-
но, посредине брюшной стороны ман-
тии обычно нет полоски, свободной
от фотофоров.
A. andamanica f. robsoni Grimpe,
1931. Преимущественно в открытом
океане.
22(19). Все глазные фотофоры
округлые, 1, 3 и 5-й крупнее, чем
2-й и 4-й. Присоски на концах рук —
в 2 ряда. Длина плавника около V2
длины мантии. 2—3 крюка на булаве
щупальца. Гектокотилизируется пра-
вая брюшная рука, на пей развива-
ется только 1 дорсальная мембрана,
имеющая форму двух или трех вы-
соких лопастей с общим основани-
ем .............................23.
23(24). 2 крюка на булаве щу-
пальца.
A. (A.) japonica М. Ishikawa,
1929 (рис. 44,р—с]. Южная часть
Японского моря, Куросио; близкая
форма в восточно-экваториальной
части Тихого океана. Вероятно, океа-
нический вид. Эпи- и мезопслагиаль.
24(23). У взрослых кальмаров
3 крюка па булаве щупальца.
A. (A.) redfieldi Voss, 1955 (рис.
44, т—х). Тропические воды Запад-
ной Атлантики от северной части
Саргассова моря до Северо-Восточ-
ной Бразилии, многочисленна у Ба-
гамских, Бермудских, Больших и
Малых Антильских островов, по из-
бегает материкового побережья
Америки. Обнаружена также у За-
падной Африки в районе Сенегала.
Эпи-мезопслагичсский нерито-океани-
чсскпй вид.
Примечание. В таблицу не
включен вид A. (Asteroteuthis) trigo-
nura Berry, 1913, описанный по
1 сильно поврежденному экземпля-
ру. Вид близок к A. andamanica, но
отличается тем, что фотофоры на
брюшной стороне мантии образуют
2 четкие и 4 плохо обособленные
продольные полосы. Распространен
у Гавайских островов в эпи- и мезо-
пелагиали, но не у поверхности.
Таблица для определения подродов
и видов рода Abraliopsis
1(4). Фотофоры на брюшной стороне
головы и мантии рассеяны диффузно
и пс образуют четких продольных ря-
дов. Левая брюшная рука самца не
видоизменена.
Подрод Abraliopsis s. str. . . 2.
2(3). На вентральной стороне
щупальца проксимальнее первого
крупного крюка имеется обособлен-
ная карпальная мембрана. Булава с
хорошо развитым килем. На булаве
4—5 пар длинных серповидно изог-
нутых крючьев. На руках 1—3-й пар
около 10 пар крючьев, идущих почти
до концов рук. Длина плавника 75—
80%, ширина равна или более 100%
длины мантии. Свободная от фото-
форов полоса посредине брюшпой
стороны мантии начинается значи-
тельно дальше ее переднего края
(иногда она слабо заметна), справа
168
и слева от нее — две широкие поло-
сы фотофоров, латерально ограни-
ченные плохо очерченным более, или
менее свободным от фотофоров про-
странством. Ряд фотофоров, идущий
по краю глазной орбиты, переходит
у глазного синуса в ряд, протягива-
ющийся по краю 3-й пары рук; меж-
ду рядом фотофоров, идущим по
краю орбиты, и тем, что идет под
брюшным краем глазного яблока,
еще минимум два ряда мелких фо-
тофоров.
A. (A.) hoylei (Pfeffer, 1884) (рис.
45, а—z?). 2 подвида: A. hoylei pfefferi
Joubin, 1896 (9—10, реже'8 крючьев
на булаве) — в тропических и суб-
тропических водах Атлантики, Мекси-
канском заливе, Средиземном море;
A. hoylei hoylei (как правило, 8
крючьев на булаве) — в тропических
и субтропических водах Индо-Вест-
пацифпки от Хоккайдо до Тасманова
моря и от Восточной Африки до Га-
вайских островов, отсутствует в Ара-
вийском море и Бенгальском заливе.
3(2). Булава щупальца без кар-
пальной мембраны и с очень слабо
развитым килем лишь в самой ди-
стальной части. На булаве 3, очень
редко 4 пары крючьев, они неболь-
шие (немного превосходят ширину
булавы), слабо изогнутые. На руках
1—3-й пар 7—9 пар крючьев и 5—10
пар присосок с зубчатыми кольцами.
Длина плавника 60—67%, ширина
75—90%, обычно 85% длины мантии.
Свободная от фотофоров полоса по-
средине брюшной стороны мантии
начинается от ее переднего края, ла-
теральные полосы не обособлены.
Ряды фотофоров, идущие по краю
глазной орбиты и по краю 3-й пары
рук, разделены свободным простран-
ством. Между рядами фотофоров,
идущими по краю орбиты и под
брюшным краем глазного яблока,
только один ряд фотофоров.
A. (A.) f.elis McGowan et Okutani,
1968 (рис. 45,г—e). Северо-Восточ-
ная Пацифика от штата Вашингтон
до Калифорнийского полуострова. На
запад доходит до района восточнее
Японских и Курильских островов.
4(1). Фотофоры на брюшной сто-
роне головы расположены 7—8 чет-
ко очерченными продольными ряда-
ми, между которыми фотофоров ли-
бо нет, либо они малочисленны и об-
разуют короткий дополнительный
ряд.
Подрод Micrabralia Pfeffer,
1900 ........................ 5.
5(6). Фотофоры на брюшной сто-
роне мантии распределены диффуз-
но. На брюшной стороне головы В
продольных рядов фотофоров: сред-
ний ряд состоит из двух параллель-
ных. Булава с обособленной удлинен-
ной карпальной мембраной и про-
дольным килем. Крючья очень длин-
ные, в 2—3 раза превосходят
ширину булавы. На левой брюшной
руке самца трабекулы защитной
мембраны удлинены и утолщены, но
пе соединены широкой мембраной.
А. (М.) gilchristi (Robson, 1924)
(=Enoploteuthis neozelanica Dell,
1959) (рис. 45,ле—к). Южные суб-
тропические воды Атлантического,
Индийского и юго-западной части Ти-
хого океанов.
6(5). Фотофоры на брюшной
стороне мантии сгруппированы в-
четко обособленные продольные ря-
ды. На брюшной стороне головы 7
продольных рядов фотофоров: сред-
ний ряд непарный, между ним и бо-
ковыми рядами иногда имеется по*
1 короткому вставочному ряду . 7.
7(10). Булава щупальца с килем и
обособленной карпальной мембраной.
Свободная от фотофоров полоса
посредине брюшной стороны мантии
не доходит до заднего конца тела.
Левая брюшная рука самца с расши-
ренной защитной мембраной оран-
жевого цвета. Нет дополнительных
вставочных рядов фотофоров между
основными рядами фотофоров на
нижней стороне головы ... 8.
8(9). Свободная от фотофоров
полоса посредине брюшной стороны
мантии кончается овальным расши-
рением на границе средней и задней
трети мантии. На булаве щупальца
4 (редко 3) пары маленьких крючь-
ев— их длина не более чем в 1,5 ра-
за превосходит ширину булавы.
А. (М.) affinis (Pfeffer, 1912)
(рис. 45, л—«). Тропические воды
Восточной Пацифики, от южной
части Калифорнийского полуострова
до северного побережья Чили, на за-
пад, по крайней мере, до 126° з. д.
9(8). Свободная от фотофоров по-
лоса посредине брюшной стороны
мантии сходит на конус в задней тре-
ти мантии, не расширяясь. На булаве
щупальца 3 (редко 4) пары крупных
—вдвое длиннее, чем ширина була-
вы,—серповидно изогнутых крючьев.
163
(а—т) и Wata-
pfefferi (на а
не изображе-
г—е — А. felis;
Рис. 45. Роды Abraliopsis
senia: а—б —A. hoylei
фотофоры, кроме глазных,
ны); в— A. hoylei hoylei; _ _ ... ___
ж~ к — Л. gilchristi-, л—н — A. al finis; о —
A. chuni n. sp.; п—с — А. falco; т — A. at-
laniica n. sp.; у—ф—Watasonia scintillans,
б, к, я —булава; д, з, р— голова сбоку;
е — гектокотиль; и — глаз; ж, с — брюшные
руки самца; ф — конец брюшной руки, (а—
б — по Naef, 1923; в — по Young, Roper,
1976; г-д, п—с — по R. Young, 1972; е — по
Okutani, McGowan, 1969; ж—к — по G.
Voss, 1967, л—н, у—ф — по Okutani, 1973,
1974; о, т — по Chun, 1910)
A. (M.) chuni n. sp.1 (=A. morisii
Chun, 1910, part: Taf. V, VIII fig. 3,
Taf. IX fig. 3—4) (рис. 45, о). Тро-
пические воды Индо-Вестпацифики,
от Восточной Африки до островов
Лайн. Преимущественно экваториаль-
ный вид.
10(7). Булава щупальца без обо-
собленной карпальной мембраны и
киля, есть лишь узкая вентральная
защитная мембрана на пальце була-
вы. Свободная от фотофоров полоса
посредине брюшной стороны мантии
проходит, постепенно сужаясь, до
заднего конца мантии . . . 11.
11(14). На брюшной стороне го-
ловы между средним (непарным) и
боковыми рядами фотофоров имеют-
ся отдельные дополнительные мел-
кие фотофоры, иногда образующие
короткий вставочный ряд; между
боковым и третьим от середины ря-
дом фотофоров такие мелкие фото-
форы разбросаны в беспорядке (у
молоди дополнительных фотофоров
обычно нет). Крючья на булаве ма-
ленькие, их длина не более чем в
1,5 раза превосходит ширину була-
вы ...........................12.
12(13). Левая брюшная рука сам-
ца нс видоизменена. Свободная от
фотофоров полоса посредине брюш-
ной стороны мантии обычно широ-
кая, четко очерченная.
А. (М.) lineata (Goodrich, 1896).
Тропические воды Ипдо-Вестпацифи-
ки, от Восточной Африки до Полине-
зии, в Северо-Западной Пацифике
доходит до Куросио, в юго-запад-
ной не южнее 15° ю. ш.
13(12). Левая брюшная рука сам-
ца с расширенной вентральной за-
1 Под названием Abraliopsis morisii
смешивали разные виды Abraliopsis.
Что такое настоящий A. morisi (Ve-
гапу, 1837), установить невозможно
ввиду неполноты первоописания.
К. Кун (Chun, 1910) соединял под
этим названием 3 вида, один из ко-
торых — A. lineata (со ст. 254), двум
другим даются новые видовые назва-
ния: A. chuni и A. atlantica. Голотип
A. chuni: самка, «Вальдивия», ст.
256, 01°49' с. ш., 45°29' в. д., лов
1134—0 м, длина мантии 38 мм. При-
родоведческий музей Университета
им. Гумбольдта (Museum fur Natur-
kunde an dor Humboldt-Universitat
zu Berlin), Берлин (ГДР).
172
щитной мембраной оранжевого цве-
та. Свободная от фотофоров полоса
посредине брюшной стороны мантии
обычно узкая, иногда плохо очерчен-
ная, в се передней части есть не-
сколько отдельных мелких фото фо-
ров.
А. (М.) falco Young, 1972 (рис.
45, п—с). Экваториальные воды
Восточной Пацифики, отсутствует в
Калифорнийском и Перуанском тече-
ниях; западная граница ареала не
установлена, вероятно, значительно
западнее 120° з. д.
14(11). Нет дополнительных мел-
ких фотофоров между средним (не-
парным) и боковыми рядами фото-
форов на брюшной стороне головы.
Свободная от фотофоров полоса
посредине брюшной стороны мантии
широкая, четко очерченная. Левая
брюшная рука самца с расширенной
вентральной защитной мембраной
оранжевого цвета (у A. sp. cf falco
этот признак не исследован) . 15.
15(16). Крючья на булаве малень-
кие, их длина не более чем в 1,5 ра-
за превосходит ширину булавы. Нет
дополнительных фотофоров между
боковым и третьим от середины ря-
дами фотофоров па брюшной сторо-
не головы.
А. (М.) atlantica n. sp.2 (=А. mo-
risii Chun, 1910, part: Taf. VI fig. 1;
Taf. VIII fig 1,5; Taf. X fig. 1). (рис.
45, т). Тропические и субтропические
воды Атлантики, Карибское море и
Мексиканский залив.
16(15). Крючья на булаве круп-
ные, их длина приблизительно вдвое
превосходит ширину булавы. Между
боковым и третьим от середины ря-
дами фотофоров на брюшной сторо-
не головы иногда бывает несколько
обособленных мелких фотофоров
впереди от вертикали переднего
края глаза.
А. (М.) sp. cf falco (=Abraliopsis
sp. A. Shevtsova, 1975, ^Abraliopsis
n. sp. Young, 1973) Северная Паци-
фика — Куросио, Г авайские остро-
ва (?).
2 Голотип: самец, «Вальдивия», ст.
54, 01°5Г с. ш., 00°31' в. д., лов
2000—0 м, длина мантии 20 мм. При-
родоведческий музей Университета
им. Гумбольдта, Берлин (ГДР).
Этот вид описан как Abraliopsis sp.:
Несис, 1974а, с. 53—54; Cairns 1976;
р. 245; Rocleveld, 1977, р. 123.
Таблица для определения видов рода
Ancistrocheirus
1(2). Крупные фотофоры на нижней
стороне мантии расположены чере-
дующимися поперечными группами
по 4 и 2, не образуют непарного
среднего продольного ряда и не за-
ходят на хвост. Формула их располо-
жения 4+2+4 + 2+4 + 2+4.
A. alessandrinii (Vcrany, 1851),
(=А. lesueuri auett non Fer. et d’Orb.)
(рис. 46, a—г). Тропическо-субтропи-
ческий космополитический вид. Ниж-
няя эпи- и мезопелагиаль, взрослые
кальмары обитают также у дна в
батиали.
2(1). Крупные фотофоры на ниж-
ней стороне мантии образуют кроме
пар и четверок, непарный средний
продольный ряд и заходят на хвост.
Формула их расположения 4+2+3+
+ 3+2+2+3 +1 +1 +1.
A. lesueuri Ferussac et d’Orbigny,
1839. Распространение и образ жиз-
ни неизвестны, вид сомнительный.
Таблица для определения видов рода
Pyroteuthis
1(2). Продольная мембрана гек-
токотиля длинная — около Va длины
руки, считая длину от первого крю-
ка; она начинается после 7—9-й па-
ры крючьев. На стебле щупальца
обычно пет дополнительного малень-
кого фотофора между первым (ба-
зальным) и вторым (по у особей из
вод Южной и Западной Австралии
он имеется).
' Р. margaritifera (Riippell, 1844)
(рис. 46, д—ж). Тропическо-субтро-
пический атланто-индовестпацифичес-
кий мсзопелагический (поднимается
в нижнюю эпипслагиаль) вид; в Вос-
точной Пацифике отсутствует, вос-
точная граница проходит в эквато-
риальных водах — в районе островов
Лайн, в южно-центральных — в рай-
оне 115° з. д. Вид распадается на
серию локальных форм, различаю-
щихся по расположению фотофоров
на стебле щупальца: у кальмаров из
экваториальных и западно-централь-
ных районов Тихого океана 7 (редко
6 или 8) фотофоров на щупальце,
нет дополнительного фотофора; у
особей из Куросио — 6, нет дополни-
тельного; у особей из Южной и Юго-
Западной Австралии — тоже 6, но
включая дополнительный.
2(1). Продольная мембрана гек-
токотиля короткая, менее длины
руки, она начинается после 5-й пары
крючьев. На стебле щупальца 7 фо-
тофоров, включая маленький допол-
нительный между первым (базаль-
ным) и вторым, ближе ко второму.
Р. addolux Young, 1972 (рис.
46, з—л). Восточная часть северо-
центральных вод Тихого океана от
Гаванских островов до Калифорнии
и в Калифорнийском течении. Мезо-
пелагиаль.
Таблица для определения видов рода
Pterygioteiithis
1(2). Крючья на руках 1—3-й пар
расположены только в середине вен-
трального ряда, их 2—8, чаще 3—7
шт. Руки 4-й пары с 2 рядами очень
мелких присосок. Гектокотилизиро-
ванная рука самца несет 2 непигмен-
тированных железистых поля при-
мерно равной длины, между которы-
ми находится гребневидная пластин-
ка с несколькими коническими
зубцами; на конце этой руки не-
сколько крохотных присосочек. 14
(редко 15) фотофоров на глазном
яблоке.
Р. gemmaia Chun, 1908 [=Р. tnic-
rolampas Berry, 1913. ?P. schneha-
geni (Pfeffer, 1912)] (рис. 46,p—y).
Преимущественно антиэкваториаль-
ный вид. Северные и южные цент-
ральные воды Атлантики и восточная
часть экваториальных вод (отсутст-
вует в западной части экваториаль-
ных вод, Карибском и Средиземном
морях и Мексиканском заливе); Ин-
дийский оксан (в южной части
центральных вод), Тихий океан (по
всей акватории северных централь-
ных вод и по южной периферии юж-
ных центральных). Калифорнийские
кальмары отличаются от атланти-
ческих и западнотихоокеанских по-
вышенным средним числом крючьев
на руках. Вид мсзопелагический, под-
нимается в нижнюю эпипелагиаль.
Длина мантии до 32 мм.
2(1). Крючья на руках 1—3-й пар
расположены в середине вентрально-
го и дорсального рядов, их 2—5 пар.
Руки 4-й пары у самок лишены при-
сосок. Гсктокотиль с 2 железистыми
полями, из которых проксимальное
ярко-оранжево-красного цвета и го-
раздо длиннее непигмептированного
дистального, между ними 2 изогпу-
173
Рис. 46. Роды Ancistrocheirus (а—г), Руго-
teuthis (б—л) и Pterygioteuthis (м—у): а—
г—Л. alessandrinii; д—ж—Pyr. margasitife-
га; з—л—Pyr. addolux; м—n—Pt. giardi; р—
у — Pi, gemmata. а — молодой кальмар; б,
д, р — самки (б — молодая)» з, м — самцы;
в — расположение фотофоров вокруг гла-
за; дас. и, п, у — глазные фотофоры; г, к—
булава; е, л, о, т •— гектокотиль (о — с
двух сторон); н — конечности самца; с —
брюшные руки самки (а, н, с—по Okutani,
1974; б, г —по Несису, 1978; в—по Degner^
1925; б— е — по Pfeffer, 1912; яс, jw, о, п».
т, у — по Chun, 1910; з—л, р—по R. Young,
1972)
тых крючковидных зубца. На правой
(не гектокотилизированной) брюш-
ной руке самца 1—2 крючка. 15 фо-
тофоров на глазном яблоке.
Р. giardi Fischer, 1896 (рис.
46, м—п). Тропическо-субтропический
космополитический мсзопелагический
(поднимается в эпипслагиаль) вид с 2
подвидами: Р. giardi giardi (на пра-
вой брюшной руке самца 1—2 до-
вольно крупных крючка), обитающий
в Атлантическом океане, и Р. giardi
hoylei (Pfeffer, 1912) (на правой
брюшной руке самца 1—2 крохотных
крючочка), распространенный в Ин-
дийском и Тихом океанах. Длина
мантии до 25 мм.
2.2.2.3. Семейство
Octopoteuthidae
(= Octopodoteuthidae, Veranyidae)
Кальмары средних и крупных разме-
ров. Тело студенистой консистенции.
Мантия коническая, сзади более или
менее заостренная. Плавники очень
большие, ромбические, протягивают-
ся почти вдоль всей длины мантии,
прирастают к ее спинной стороне и
срастаются друг с другом по сред-
ней линии спины, не сросшимся ос-
тается лишь небольшое U-образное
пространство близ переднего края
мантии. Голова широкая; руки ко-
роткие, толстые, их концы часто об-
ламываются. На руках два ряда ко-
ротких крючьев, покрытых капюшо-
ном из мягких тканей. Присоски,
если имеются, то лишь на самых
концах рук. Щупальца развиты толь-
ко у личинок или молодых кальма-
ров, они несут гроздь крупных при-
сосок на короткой булаве. У взрос-
лых кальмаров от них не остается
и следа. Замыкательный аппарат
простой. Формула буккального креп-
ления DDVV, Светящиеся органы
расположены на концах рук, иногда
также в толще тканей головы, рук и
мантии и внутри мантийной полости
близ ануса. Гладиус очень тонкий, с
широким пером, начинающимся пря-
мо. от переднего конца, и маленьким
конечным конусом. Гектокотиля нет,
сперматофоры передаются самке с
помощью длинного пениса. Развитие
с личиночной стадией.
Мезо-батипелагические и придон-
но-батиальные кальмары тропических
и умеренных вод. 2 рода, 7—10 ви-
дов.
176
Таблица для определения родов
Octopoteuthidae
1 (2). На концах 2-й пары рук имеет-
ся крупный сложно устроенный фо-
тофор, светящаяся поверхность ко-
торого может закрываться боковыми
кожными складками. Крупный округ-
лый непарный фотофор на чернильном
мешке. Нет фотофоров на концах
рук, по бокам от ануса и в задней
части мантии. Плавник у молоди
чрезвычайно широк, ширина его го-
раздо больше длины мантии, он до-
ходит до заднего конца тела, -но у
взрослых он далеко не доходит до
заднего конца мантии. Щупальца
редуцируются при длине мантии
40—45 мм, у взрослых кальмаров
сохраняются рудименты стеблей.
Длина мантии до 170 см.
Род Taningia Joubin, 1931 ( = С«-
cioteuthis auett.). Один вид—Т. danae
Joubin, 1931 ( = Cucioteuthis unguicu-
lata auett.) (рис. 47, м—н). Тропи-
ческо-субтропический (заходит в бо-
реальные и потальные воды) космо-
политический вид. Мсзопслагиаль и
у дна в батиали. Часто встречается
в пище кашалотов.
2(1). Концы всех рук несут по
одному удлиненному коническому
или веретеновидному фотофору без
боковых складок кожи. Нет непарно-
го фотофора на чернильном мешке.
Обычно имеются небольшие фотофо-
ры в толще тканей головы и задней
части мантии и внутри мантийной
полости близ ануса. Ширина плавни-
ка у молоди не превышает или лишь
немного превышает длину мантии.
Щупальца полностью редуцируются
в конце личиночной стадии (длина
мантии около 1—1,5 см). Длина
мантии до 30—50 см, обычно не бо-
лее 20 см.
Род Octopoteuthis Ruppell, 1844
(=Octopodoteuthis Krohn, 1845, Ve-
runia Krohn, 1847, Veranya auett.,
Octopodoteuthopsis Pfeffer, 1912) (рис.
47, a—л). Типовой вид—О. sicula
Ruppell, 1844. 6—9 видов. Тропические
и субтропические воды всех оксанов.
Мезо- и батйпелагиаль и батиаль.
Таблица для определения видов рода
Octopoteuthis
1(10). Имеется парный или непарный
фотофор в толще тканей задней
части мантии па брюшной стороне
тела (мантия над ним стекловидно
прозрачна). Есть парный фотофор
внутри мантийной полости по обе
стороны чернильного мешка: на дор-
сальной стороне мышц — втягивате-
лей воронки.....................2.
2(5). Один крупный фотофор в
толще тканей задней части мантии
на брюшной стороне .... 3.
3(4). Плавник протягивается
вдоль хвоста в виде тонкой отороч-
ки и доходит до заднего края ман-
тии. Есть фотофоры у основания рук
3-й и 4-й пар.
О. deletion Young, 1972 (рис.
47, а, б). Северо-Восточная Пацифи-
ка от Орегона до Калифорнийского
полуострова, найден также у север-
ного Перу и (?) у восточного побе-
режья Хонсю (у экземпляров из вод
Японии ширина плавника около
130% длины мантии против 70—95%
у взрослых О. deletion; возможно,
это — особый вид).
4(3). Плавник кончается немного
дальше хвостового фотофора и не
доходит до заднего конца мантии.
Octopoteuthis sp. A. Young, 1972
(рис. 47, в). Центральные воды Се-
веро-Восточной Атлантики.
5(2). Пара крупных фотофоров
находится в задней части мантии на
брюшной стороне по обе стороны от
средней линии тела..............6.
6(9). Хвостовые фотофоры нахо-
дятся на границе средней и задней
трети мантии. Нет фотофоров у ос-
нования рук 3-й и 4-й пар . . 7.
7(8). Плавник доходит до заднего
конца мантии.
O.danae Joubin, 1931. Северная
субтропическая Атлантика (Саргас-
сово морс, Сенегал).
8(7). Плавник доходит до задне-
го конца мантии, его задний край
расположен немного дальше уровня
хвостовых фотофоров, длина плавни-
ка около 3/4 длины мантии. Хвост
длинный, заостренный, уплощенный
сверху, с узкими боковыми оторочка-
ми.
О. megaptera (Verrill, 1885)
(рис. 47, а, яс). Тропическо-субтро-
пический космополитический вид.
Мезо- и батипелагиаль и батиаль, в
ночное время суток поднимается в
эпипелагиаль.
9(6). Хвостовые фотофоры лежат
на расстоянии ]/4—7S длины мантии
от заднего конца тела. Плавник до-
ходит до заднего конца мантии.
Есть фотофоры у основания' рук 3-й
и 4-й пар. Хвост короткий, у молоди
слабо выражен.
О. sicula (Riippell, 1844) (рис.
47, з). Т ропичсско-субтропический
атланто-индовестпацифический вид,
встречается в Средиземном море.
O.nielseni (Robson, 1948) (рис.
47,и). Тропический вид Восточной
Пацифики (от Мексики до северного
побережья Чили, па запад до Гавай-
ских островов).
Различия этих двух видов в на-
стоящее время не выяснены. Оба
обитают в мезо- и батипелагиали, по
ночам поднимаются в эпипелагиаль.
10(1). Нет хвостовых, головных и
внутримантийных фотофоров, имеют-
ся лишь удлиненные фотофоры па
концах рук. У молоди плавник дохо-
дит до заднего конца мантии, у
взрослых имеется короткий хвост.
Octopoteuthis rugosa Clarke, 1980
(=0. n. sp. Nesis, 1974a) (рис. 47, к,
л). Атланто-индовестпацифический
тропический вид: экваториальные
воды Атлантики, у Мавритании, Юж-
ной Африки, Юго-Западной Австра-
лии, в морях Сулу и Флорес. Мезо-
пелагиаль.
Примечание. В таблицу не
включен О. longiptera (Akimushkin,
1963) из южносубтропических вод
Атлантики (восточнее о-ва Мартин-
Вас), описанный крайне неполно.
2.2.2.4. Семейство
Onychoteuthidae
Кальмары от мелких до очень круп-
ных. Мантия удлиненно-коническая,
стройная. Тело обычно мускулистое,
редко водянистое. Плавник в задней
части мантии, ромбический, сердце-
видный или поперечно-овальный. Го-
лова неширокая, у некоторых видов
может втягиваться в мантию. Руки
с двумя рядами присосок, кольца ко-
торых обычно гладкие. Щупальца
крепкие, мощные (у Chaunoteuthis
имеются лишь па стадии личинки,
потом автотомируются). Булава с 2
рядами крючьев и 2 краевыми ряда-
ми присосок, последние у взрослых
кальмаров, как правило, исчезают
или сохраняются лишь как рудименты.
Фиксаторный аппарат щупалец
очень хорошо развит. Замыкатель-
ные хрящи простые. Формула бук-
кального крепления DDVV. Светя-
12 К. Н. Несис
177
щиеся органы имеются лишь у Опу-
choteuihis и Chaunoteuthis на брюш-
ной стороне глазного яблока и чер-
нильном мешке. Гладиус с утолщен-
ным рахисом, узким пером и хитино-
вым или хрящевым конечным
конусом. Гектокотиля нет, спермато-
форы переносятся с помощью пени-
са. Развитие с личиночной стадией.
Эпипелагические, мезопелагичес-
кие или придонно-батиальные виды.
Большинство — активные нсктонныс
кальмары. Промысловое значение
очень невелико. 6—7 родов, 13 видов.
Частичная ревизия: Clarke, 1980.
Таблица для определения родов
Onychoteuthidae
1(12). Тело мускулистое. Щупальца
имеются. Мантия у переднего края
уже или немного шире головы. Спер-
матофоры не переносятся на наруж-
ную поверхность мантии самки 2.
2(7). Задний конец гладиуса — в
виде короткой хитиновой иголочки.
Кожа гладкая....................3.
3(4). Имеются фотофоры: по од-
ному удлиненному на брюшной сто-
роне глазного яблока и два (перед-
ний маленький, задний крупный) на
чернильном мешке; кишка огибает
задний фотофор сбоку. Затылочные
складки хорошо развиты (7—9).
В центральной части булавы молоди
и взрослых — только крючья. Ман-
тия стройная, коническая, плавник
ромбический или слегка сердцевид-
ный с оттянутым назад хвостиком.
Гладиус просвечивает на спинной
стороне мантии и прощупывается
под кожей как острое тонкое ребро,
у личинок и ранней молоди его кон-
чик протыкает сзади кожу и торчит
иголочкой наружу.
Род Onychoteuthis Lichtenstein,
1818 ( = 7eleonychoteuthis Pfeffer,
1900) (рис. 48, a—e). Типовой вид —
O.banksi (Leach, 1817). Один опи-
санный вид с 2 подвидами и 1 неопи-
санный вид. Всесветно, кроме Аркти-
ки, высокобореальпой Атлантики,
Субантарктики и Антарктики. В Ку-
рило-Хоккайдском районе имеет не-
большое промысловое значение.
4(3). Фотофоры отсутствуют. За-
тылочные складки отсутствуют или
малочисленны (до 4). У молоди на
булаве 2 ряда крючьев и 2 ряда при-
сосок, у взрослых в центральной или
178
12*
е
з
Рис. 48. Сем. Onychoteuthidae, роды Ony-
choteuthis (а—<?), Onykia {ж—з), Ancistro-
teuthis (и) и Chaunoteuthis (к—м): а—в —
Onychoteuthis banks i banksi-, г—д — О.
banksi borealijaponicus; е — Onychoteuthis
sp. Rancurel; ж—з— Onykia carriboea, мо-
лодая и взрослая: и — Ancistroteuthis lich-
tensteini-, к—м — Chaunoteuthis mollis (сам-
ка); 6 — голова сверху, видны затылочные
складки; в, д, м — брюшные фотофоры
{а—б, и — по Pfeffer, 1912: в—д — по R.
Young, 1972; е, з — по Rancurel, 1970; ж —
по Chun, 1910; к—м — по Adam, 1972).
ж
Onychoteuthidae, роды Опу-
d
проксимальной части булавы, как
правило, сохраняется несколько от-
дельных мелких присосочек. Гладиус
у неповрежденных особей не просве-
чивает или просвечивает только в
задней части мантии между плавни-
ками. У личинок и молоди его зад-
ний конец нс протыкает кожу и не
торчит наружу...................5.
5(6). Плавник поперечно-оваль-
ный, его задний край слегка выпук-
лый или почти прямой. Длина плав-
ника около V2, ширина у взрослых
примерно равна длина мантии. Гла-
диус с толстым притупленным реб-
ром, у неповрежденных особей не
просвечивает. Спинная сторона у мо-
лоди темная, с синеватым отливом.
Затылочных складок нет.
Род Onykia LeSueur, 1821 ( = Oru/-
chia auett., Steensirupiola Pfeffer, 1884,
Teleoteuthis Verrill, 1885) (рис.
48, ж, з). Типовой и единственный
достоверно известный вид — О. саг-
riboea LeSueur, 1821 (рис. 48, ж, з).
Тропическо-субтропический космопо-
литический вид, молодь обитает у
поверхности (часто в гипонейстоне),
взрослые особи — в нижней эпи- и
верхней мсзопслагиали, к поверх-
ности не поднимаются. Многочислен-
ный вид. Длина мантии до 10 см.
Несколько номинальных видов описа-
но по молодым особям (см. Pfeffer,
1912), их таксономическое положение
неясно.
6(5). Плавник у ранней молоди
поперечно-овальный, у «подростков»
и взрослых ромбический, сзади угло-
ватый. Длина плавника 55—60%,
ширина 70—75% длины мантии. Гла-
диус с острым ребром в задней по-
ловине, просвечивает между плавни-
ками. Окраска светлая — ярко-жел-
тая у живых и коричневая у фикси-
рованных кальмаров. Имеются 3—4
короткие затылочные складки (у ран-
ней молоди плохо заметны).
Onykia (?) verrilli (Pfeffer, 1900).
По-видимому, нотальный циркумпо-
лярный (между 33 и 53° ю. ш., а воз-
можно, и южнее, во всех трех океа-
нах) эпи-мезопелагический (до
батипслагиали?) вид, молодь встреча-
ется по ночам у поверхности. Родо-
вая принадлежность неясна, вид со-
четает признаки Onychoteuthis, Ап-
cistroteuthis и Onykia. По-видимому,
это — особый род. Вероятно, каль-
мар, изображенный в работе
С. К. Клумова и В. Л. Юхова (1975,
182
с. 175, рис. 8) под названием Moro-
teuthis «широкий» — взрослая особь
этого вида. К нему же относятся «но-
тальная форма» Onychoteuthis banksi
(Несис, 1973 6, с. 222) и, предполо-
жительно, Moroteuthis sp. A. Clarke,
1980. Длина мантии наших молодых
экземпляров до 10 см.
7(2). Задний конец гладиуса —
длинный опаковый рострум хряще-
ватой консистенции в форме сильно
вытянутой трехгранной пирамиды со
скошенным основанием или (реже)
округлого сечения (конус). Фотофо-
ров нет. Кожа гладкая или морщи-
нистая. Гладиус не просвечивает на
спинной стороне мантии ... 8.
8(9). Затылочные складки хорошо
развиты (6—10). Гладиус очень уз-
кий, его перо почти не расширено в
середине. Плавник у взрослых каль-
маров (длина мантии более 10—
12 см) сердцевидный с оттянутым
хвостом. Кожа топкая, гладкая.
Род Ancistroteuthis Gray, 1849.
Один вид — A. lichtensteini (Ferussac
et d’Orbigny, 1839) (рис. 48,и). Тро-
пические и субтропические воды Ат-
лантики и Средиземное морс. Эпи-
мезопелагический и придонно-бати-
альный вид. Длина мантии.до ЗОсм.
В Средиземном море имеет третье-
степенное промысловое значение.
9(8). Затылочные складки отсут-
ствуют. Перо гладиуса в середине
значительно расширено. Кожа тол-
стая, морщинистая или бугорчатая,
реже гладкая...................10.
10(11). Булава щупалец у взрос-
лых кальмаров песет только крючья
(обычно не более 30). 10—20 присо-
сок па конце булавы. Руки не длин-
нее мантии. Длина плавника равна
или более V2 длины мантии. Длина
рострума гладиуса обычно 15—25%
длины мантии.
Род Moroteuthis Verrill, 1881
(=Moroteuthopsis Pfeffer, 1908) (рис.
49, a—л). Типовой вид — AL robusta
(Dall in Verrill, 1876). 6 видов в бо-
реальных, тропических, нотальных и
антарктических водах. Крупные и
гигантские (длина мантии до 2,5 м)
кальмары. У дна в батиали (редко —
в нижней сублиторали) и в толще
воды открытого океана, главным об-
разом над склоном и подводными
возвышенностями. Молодь пелагиче-
ская. Играют большую роль в пита-
нии кашалотов.
11(10). Булава щупальца у взрос-
лых кальмаров несет, кроме крючьев
(30—40), два ряда мелких краевых
присосок. Более 20 присосок на кон-
це булавы. Руки очень длинные, у
взрослых не меньше длины мантии.
Плавник слабый, короткий, широко-
сердцевидный, его длина около %,
ширина около % длины мантии. Дли-
на рострума гладиуса менее 10%
длины мантии.
Род Kondakovia Filippova, 1972,
Один вид — К. longimana Filippova,
1972 (рис. 49, л<—п). Антарктический
(по-видимому, циркумантарктиче-
ский) эпи-мезопслагический и при-
донно-батиальный вид. Длина ман-
тии до 80 см, возможно до 115 см.
12(10). Тело студенистое, дряб-
лое. Щупальца имеются только у ли-
чинок, а у взрослых самок (самцы
неизвестны) от них сохраняются
лишь рудименты стеблей. Ширина
мантии у переднего края намного
превосходит ширину головы. Плав-
ник ромбический, его длина больше
половины мантии, ширина лишь не-
много меньше длины мантии. Имеют-
ся фотофоры — по одному удлинен-
ному на брюшной стороне глазного
яблока и один на чернильном мешке.
Сперматофоры размещаются на на-
ружной поверхности мантии, по бо-
кам от средней линии в передней ча-
сти брюшной стороны — внедрены в
мантийные ткани в специальных про-
дольных разрезах, сделанных клю-
вом (?) самца.
Род Chaunoteuthis Appel lof, 1890.
Один вид — Ch. mollis Appellof, 1890
(см. рис. 48, к—л0. Тропическо-суб-
тропический, по-видимому, космопо-
литический мезопелагический вид.
Длина мантии до 17 см.
Таблица для определения видов и
подвидов рода Onychoteuthis
1(2). Плавник длинный, его длина
больше половины, обычно 55% дли-
ны мантии, форма плавника продоль-
но-ромбическая или сердцевидная,
задние края плавника, по крайней
мере в задней половине, вогну-
тые, и плавник оттянут назад в
небольшой хвостик; реже задние
края прямые, при этом ширина
плавника чуть больше его
длины.
О. banksi (Leach, 1817) (см. рис.
48, а—д). Теплые воды всех океанов,
на север до Северного и Берингова
морей, на юг до Тасмании. Преиму-
щественно эпипелагический вид. Дли-
на мантии до 30—37 см (такие круп-
ные особи известны только в Север-
ной Пацифике)...................2.
а (б). Задний фотофор па чер-
нильном мешке большой, округлый,
нс соединен с передним светящейся
тканью. На булаве 19—24, чаще 20—
21 крючок.
О. banksi banksi (Leach) (см. рис.
48,а—в). Теплые воды всех океанов,
в Атлантике на север до Северного
моря, в Тихом океане — до Южной
Японии, Гавайских островов и юж-
ной части Калифорнийского п-ва.
Молодь — по ночам у поверхности,
взрослые — глубоководные. Необыч-
ный размах колебаний ширины плав-
ника— от 55 до 85% длины мантии,
что при относительном постоянстве
длины плавника выражается в рез-
ких различиях его формы, заставля-
ет предполагать, что этот подвид —
смесь двух или более разных форм.
В тропических и субтропических во-
дах Атлантики и Пацифики узко-
хвостая и широкохвостая формы
встречаются в одних и тех же
местах.
б (а). Задний фотофор на черниль-
ном мешке маленький, очень узкий,
соединен с передним тонкой полос-
кой светящейся ткани. На була-
ве обычно 25—27 крючьев. Ши-
рина плавника 70—80% длины
мантии.
О. banksi borealijaponicus Okada,
1927 (см. рис. 48, г, д). Северная Па-
цифика, от Алеутских островов до
Японского моря, Южной Японии и
Калифорнийского полуострова (нет в
Охотском море). Эпипелагиаль. Про-
мысловый кальмар.
2(1). Плавник короткий и широ-
кий, его длина около 40%, ширина
70—85% длины мантии, форма — по-
перечно-ромбическая, задние края
прямые или слабо выпуклые, плав-
ник не оттянут в хвост. На булаве
8—11 пар крючьев. Фотофоры чер-
нильного мешка, как у О. banksi
banksi.
Onychoteuthis sp. Rancurel, 1970
(см. рис. 48, e). Южноцёнтральные
воды Индийского и западной части
Тихого океана, на восток распрост-
183
ранен до островов Туамоту. Фор-
мально не описанный вид. Длина
мантии до 7 см.
Таблица для определения видов рода
Moroteuthis
1(8). Кожа покрыта бугорками или
морщинистая....................2.
2(3). Плавник ромбический, не
оттянут в хвост, его длина равна
или немного больше !/2 длины ман-
тии, ширина больше длины. Кожа
покрыта плоскими удлиненными
«ли изогнутыми бугорками. Рост-
рум гладиуса в сечении треуголь-
ный.
М. ingens (Smith, 1881) (рис.
49,о—г). Нотальный, заходящий в
.Антарктику, циркумполярный вид.
На север проникает до северной ча-
сти Патагонского шельфа, Южного
Яили, островов Принс-Эдуард, Кро-
зе, Кергелен и южных островов Но-
вой Зеландии (о-ва Антиподов,
Кэмпбелл и др.). Чаще всего ловит-
ся в придонных слоях: нижняя субли-
тораль и батиаль. Длина мантии до
52 см.
3(2). Плавник сердцевидный, сза-
ди оттянут в хвост, его ширина
•обычно меньше длины .... 4.
4(5). Кожа покрыта плоскими
•бугорками неправильной формы.
Плавник взрослых особей очень
длинный, с сильно вытянутым хвос-
том, длина плавника около 2/з, ши-
рина около Уг длины мантии. Рост-
рум гладиуса в сечении треуголь-
ный.
№. robsoni Adam, 1962 (рис.
49, д—е). У дна и в пелагиали юж-
носубтропичсских и нотальных вод
(циркумглобально): Южная Атлан-
тика, Юго-Западная и Южная Афри-
ка, Южная Австралия, Новая Зелан-
дия, Перу, Чили. Длина мантии до
75 см.
5(4). Кожа покрыта продольными
•и косыми валиками (плоскими греб-
нями). Длина плавника около поло-
вины длины мантии..............6.
6(7}. Кожные валики длинные и
относительно широкие. Ширина
плавника заметно меньше его длины.
Рострум гладиуса в сечении округ-
лый. Вороночная ямка без попереч-
ной складки. Болес 30 крючьев на
булаве.
№. robusta (Dall in Verrill, 1876)
(рис. 49,ж). Тихоокеанский бореаль-
ный вид, распространен от Беринго-
ва моря и залива Аляска до Северо-
Восточной Японии и Калифорнии.
Придонные слои воды в нижней суб-
литорали и верхней батиали. Длина
мантии до 2,5 м.
7(6). Кожные валики короткие и
узкие, изгибаются и сливаются друг
с другом. Ширина плавника прибли-
зительно равна его длине или немно-
го больше ее. Рострум гладиуса в се-
чении узко-треугольный. В передней
части вороночной ямки имеется ду-
гообразная поперечная складка, вер-
шина которой соединена с вершиной
вороночной ямки продольным кож-
ным: гребнем. Не более 30 крючьев
на булаве. .
М. loennbergi Ishikawa et Wakiya,
1914 (рис. 49, з). Тихоокеанское по-
бережье Хонсю, на севере до Сан-
гарекого пролива, Куросио, Индий-
ский океан (банка Сая-де-Малья).
Эпи- и мезопслагиаль и батиаль. Дли-
на мантии до 35 см.
8(1). Кожа гладкая. Плавник ши-
рокий, ромбический, не оттянут или
очень слабо оттянут в хвост. Рост-
рум гладиуса в сечении треуголь-
ный .............................9.
9(10). Длина плавника 3/s длины
мантии. Длина наибольшей руки не-
много больше половины длины ман-
тии. Рострум гладиуса на конце
слегка изогнут на брюшную сторону,
его длина по вентральному (корот-
кому) ребру не более 10% длины
мантии. Все зубы радулы одновер-
шинные.
№. knipoviichi Filippova, 1972
(рис. 49, и). Антарктический (циркум-
антарктический) вид, на севере до
островов Крозе и Принс-Эдуард. Ме-
зопслагиаль и батиаль. Длина ман-
тии до 45 см.
10(9). Длина плавника около V2
длины мантии. Длина наибольшей
руки немного меньше длины мантии.
Рострум гладиуса прямой, его дли-
на по вентральному (короткому) реб-
ру 12—14% длины мантии. Рахиди-
альный зуб радулы трсхвершинный,
первые латеральные двухвершинные.
М. aequatorialis Thiele, 1920 (рис.
49, к, л). Вид описан из Экваториаль-
ной Атлантики. По-видимому, к не-
му же относятся кальмары из Мек-
сиканского залива и новокаледон-
ских вод, ошибочно определенные
как Ancistroteuihis lichtensteini (Voss,
1956; Rancurel, 1970), и Onykia japo-
186
nica Taki, 1964 из Южной Японии.
Предположительно тропический кос-
мополитический эпи-мсзопслагический
вид. Длина мантии до 40 см.
2.2.2.5. Семейство
Gonatidae
Кальмары средних размеров с узкой
мантией и большим ромбическим или
сердцевидным плавником. Консистен-
ция тела от мускулистой до полу-
студенистой. Руки с 4 рядами при-
сосок (у Gonatopsis octopedatus на
концах рук 8—12 рядов), на 1—3-й
парах рук два средних ряда присо-
сок у взрослых особей преобразуют-
ся в крючья (у Berryteuthis апопу-
chus крючья есть лишь у самок, да и
то в малом числе). Булава щупалец
длинная и широкая, несет много ря-
дов присосок, иногда сильно диффе-
ренцированных по размерам.
В центральной части булавы могут
развиваться 1—2 крупных и иногда
несколько мелких крючьев. Фикса-
торный аппарат щупалец хорошо
развит и дифференцирован. У рода
Gonatopsis щупальца имеются лишь
в молодом возрасте, у взрослых они
полностью утрачены. Развитие
крючьев па руках и булаве и утрата
щупалец происходят относительно
быстро при строго определенном раз-
мере кальмара. Замыкательный аппа-
рат простой. Формула буккального
крепления DDVV, Гладиус с узким
рахисом и фунтиковидным конечным
конусом, перо узкое с одним или
двумя расширениями. У видов с
сердцевидным плавником гладиус не
доходит до заднего конца плавника.
Светящиеся органы имеются лишь у
одного вида — на глазах. Гектокоти-
ля нет. Сперматофоры переносятся
либо в мантийную полость (Berry-
teuthis), либо на ротовую мембрану
самки (Gonatus, Gonatopsis). Разви-
тие с личиночной стадией.
Обитают в толще воды главным
образом в мезо- и батипслагиали, ре-
же в эпипелагиали или у дна в ба-
тиали. Распространены в умеренных
и холодных водах обоих полушарий,
богаче всего представлены в бореаль-
ной Пацифике. Играют важную роль
в пище морских позвоночных. Про-
мысловое значение очень невелико.
3 рода, 17—19 видов. Обзоры: Не-
сис, 1973, a; R. Young, 1972.
Таблица для определения родов
Gonatidae
1(4). Щупальца хорошо разви-
ты1 ............................2.
2(3). Щупальца вооружены толь-
ко присосками. Фиксаторный аппарат
состоит из присосок и бугорков-кно-
пок, без чередующихся поперечных
гребней и борозд. У молодых, а
иногда и у взрослых кальмаров
крючья на руках отсутствуют или
имеются в небольшом количестве.
Плавник ромбический или овальный,
не заходит за задний конец мантии.
Радула из 7 продольных рядов зу-
бов. Тело плотное, мускулистое.
Род Berryteuthis Naef, 1921 (рис.
50, а—д). Типовой вид — В. magister
(Berry, 1913). 2 вида в северной ча-
сти Тихого океана, один донный
сублиторально-батиальный, второй
пелагический. Имеют промысловое
значение.
3(2). Щупальца вооружены либо
только присосками, либо также од-
ним или несколькими крупными
крючками (если их несколько, то
один гораздо крупнее прочих). Фик-
саторный аппарат заканчивается в
ладонной части булавы серией из
нескольких чередующихся попереч-
ных гребней и глубоких борозд или
желобков. Крючья на руках хорошо
развиты, начиная с личиночной или
послеличиночной стадии. Плавник
сердцевидный, заходит за задний ко-
нец мантии, оттянут в хвостик с
хрящевым стержнем. Радула из 5
продольных рядов зубов. Тело и ко-
нечности часто слабо мускулистые,
водянистые.
Род Gonatus Gray, 1849 (=Lesto-
teuthis Vcrrill, 1880, Cheiloieuthis Ver-
rill, 1881) (рис. 51). Типовой вид —
G. fabricii (Lichtenstein, 1818). 2 под-
рода: Gonatus s. str. и Eogonatus
Nesis, 1972 (типовой вид—G. (E,)
tinro, Nesis, 1972). 10—11 видов в
арктических, бореальных и поталь-
ных водах Мирового океана, мезо- и
батипслагиаль. Многочисленные, по не
промысловые виды.
4(1). Щупальца имеются только у
личинок, взрослые кальмары их ли-
шены полностью.
Род Gonatopsis Sasaki, 1920 (рис.
1 У самок Gonatus на стадии выбоя
щупальца автотомируются.
187
к*
к
Мм
3
Рис. 50. Сем. Gonatidac, роды Berryteuihis
(а—д) и Gonatopsis (е—ле): а—б — J3. ma-
gister; в—д — В. anonychus (молодой и
взрослый); е — Gonatopsis okutanii; ж—з—
G. ociopedatus; G. japonicus; к—л —
G. borealis (молодой и взрослый); м — G.
tnakko; б, д — булава; з — руки (а—б — по
Всггу, 1912; в, д —по Рсагсу, Voss, 1963;
г, и, ле —по Oktitani, 1968; 1973, 1975; е —
по Несису, 1972; ж, з—по Sasaki, 1929; к—
по Кондакову, 1941; л — по R. Young, 1972).
-1W
в
50, е—м). Типовой вид — G. octopeda-
tus Sasaki, 1920. 2 подрода: Gonatop-
sis s. str, и Boreoteuthis Nesis, 1971
[типовой вид— G. (В.) borealis Sasa-
ki, 1923]. 5—6 видов в северной части
Тихого океана. От эпи- до батипела-
гиали, батиали и абиссали.
Таблица для определения видов рода
Berryteuthis
1(2). Плавник большой, ромбиче-
ский, его длина не менее 1/2 длины
мантии. Крючья на руках 1—3-й пар
хорошо развиты, появляются при
длине мантии около 75 мм. Булава
длинная, присоски в ее центральной
части заметно крупнее прочих.
В. magister (Berry, 1913) ( = Gona-
tus septemdentatus Sasaki, 1915) (рис.
50, а, б). Командорский кальмар. От
северной части Берингова моря до
Корейского пролива, Северо-Восточ-
ного Хонсю и Орегона. Почти все
Охотское море. У дна на глубинах от
30 до более 1200 м, чаще на глубине
100—600 м, молодь иногда поднима-
ется в толщу воды до поверхности.
Длина мантии до 35 см. Очень мно-
гочисленный, местами (возвышен-
ность Ямато, районы Курильских и
Командорских островов, западная
часть Берингова моря) промысло-
вый вид.
2(1). Плавник маленький, по-
перечно-овальный, его длина V4—Уз
длины мантии. Крючьев на руках у
самцов нет, у самок имеется не-
сколько штук в базальных частях
рук, появляются при длине мантии
между 30 и 60 мм. Булава короткая,
компактная, присоски на ней мелкие,
одинаковых размеров.
В. anonyenus (Pcarcy et Voss, 1963)
(рис. 50, в—д). Северо-Восточная Па-
цифика от залива Аляска и Алеут-
ских островов до южной части п-ва
Калифорния, на запад достигает при-
мерно 170° в. д. Эпипелагиаль, пре-
имущественно над склоном и «при-
брежной абиссалью», по ночам под-
нимается к поверхности. Длина ман-
тии до 15 см.
Таблица для определения подродов
и видов рода Gonatus
1(2). Нет крючьев па булаве. При-
соски булавы мелкие, одинаковых
размеров. Гребни и борозды ладон-
ной части фиксаторного аппарата
190
малочисленные (2—3), короткие.
Щупальца тонкие, довольно слабые.
Крючья на руках появляются при
длине мантии около 20 мм.
Подрод Eogonatus Nesis, 1972»
Один вид—G.(E.)tinro Nesis, 1972
(рис. 51, а, б). От Берингийского
склона до Охотского моря, Южных
Курильских островов и Британской
Колумбии. Мезо- и батипелагиаль,
также нижняя эпипелагиаль, взрос-
лые особи к поверхности не подни-
маются. Длина мантии до 9 см или
более.
2(1). На булаве щупальца один
или несколько крупных крючьев,,
центральный крюк очень большой и
острый, он развивается одновремен-
но с крючьями на руках или несколь-
ко раньше них (кроме G, berryi).
Дистальный и маленькие прокси-
мальные крючья (если имеются) раз-
виваются позже центрального. При-
соски булавы разных размеров; объ-
единены в несколько различающих-
ся по расположению групп. Гребни
и борозды ладонной части фиксатор-
ного аппарата многочисленные (4—
6), хорошо выражены. Щупальца
длинные, мощные.
Подрод Gonatus s. str. ... 3.
3(4). На булаве только 1 цен-
тральный крюк (изредка — только на
одной булаве — их бывает 2), появ-
ляется при длине мантии 15—22 мм.
На стебле щупальца между двумя
краевыми рядами присосок фикса-
торного аппарата нет или не более
10 мелких присосочек. Крючья на
руках появляются при длине мантии
25—26 мм. Ширина мантии 20—27%
ее длины, длина булавы 17—22%
длины мантии. 7—8 поперечных ря-
дов присосок и крючьев в базальной
половине рук 3-й пары, 8—9 по-
перечных рядов присосок в базаль-
ной половине рук 4-й пары. Менее
200 присосок на булаве.
G. (G.)onyx Young, 1972 (рис.
51, в, г). От Берингийского склона до
северо-восточной части Хонсю и се-
верной части п-ва Калифорния. От-
мечен в северной и центральной ча-
стях Японского моря. Личинки и
молодь встречаются и южнее: до зал.
Суруга и южной части п-ва Кали-
форния. Мезопелагиаль и нижняя
эпипелагиаль (ночью). Очень много-
численный вид. Длина мантии до
18 см.
4(3). На булаве кроме централь-
ного крюка имеются один дисталь-
ный крюк (меньше центрального) и,
как правило, несколько маленьких
проксимальных крючьев ... 5.
5(6). Тело и конечности очень
дряблые, почти студенистой конси-
стенции. Руки очень длинные, почти
равны или даже превосходят длину
мантии. Длина плавника 40—70%
длины мантии, 11—12 поперечных
рядов присосок и крючьев в базаль-
ной половине рук 3-й пары, 16 по-
перечных рядов присосок в базаль-
ной половине рук 4-й пары. Булава
длинная (около 20% длины мантии),
присоски на пальце булавы у молоди
заметно крупнее, чем па остальной
части булавы. Более 200 присосок па
булаве. Ширина мантии 23—27% ее
длины. Крючья па руках появляют-
ся при длине мантии около 25 мм,
центральный крюк на булаве при
длине мантии около 40 мм, дисталь-
ный — около 45 мм, проксималь-
ные — более 70 мм.
G. (G.) madokai Kubodera et Oku-
tani, 1977 (= Gonatus sp. A Nesis,
1973 a, Gonatidae type у Okutani,
1966. «Gonatus fabricii adolescent
stage» Sasaki, 1929) (рис. 51,(9—ж).
От северной части Берингова моря до
Южных Курильских островов и за-
лива Аляска, многочислен в Охот-
ском море. Эпи- и мсзопслагиаль.
Длина мантии до 33 см.
6(5). Тело и конечности не студе-
нистые. Длина рук 2/е—3А длины
мантии. Длина плавника 35—55%
длины мантии. Число поперечных ря-
дов присосок и крючьев в базаль-
ных половинах рук 3-й и 4-й пар со-
ответственно 6—13 и 8—17, обычно
6—11 и 8—15. Присоски на пальце
булавы у молоди нс крупнее осталь-
ных ........................... 7.
7(8). Вдоль вентрального края
стебля щупалец проходит двойной
ряд присосок. На брюшной стороне
глазного яблока имеется крупный
овальный фотофор беловатого цвета.
Ширина мантии 25—30% ее длины,
длина булавы 15—25% длины ман-
тии. Число поперечных рядов присо-
сок и крючьев в базальных полови-
нах рук 3-й и 4-й пар соответственно
7—9 и 8—10. Менее 200 присосок на
булаве. Крючья на руках появляют-
ся при длине мантии 17—21 мм,
центральный крюк на булаве при
длине 15—19 мм, дистальный — при
18—22, проксимальные—при 22—
27 мм.
6. (G.) pyros Young, 1972 (рис.
51,з—к). От центральной части Бе-
рингова моря до Восточного Хоиск>
и северной части п-ва Калифорния.
В толще воды (мезо- и нижняя эпи-
пелагиаль) и у дна в батиали. Дли-
на мантии до 16 см.
8(7). Вдоль вентрального края
стебля щупалец проходит один ряд
присосок. Фотофоров нет ... 9.
9(10). Булава очень длинная и
мощная, ее длина 25—37% длины
мантии. Ширина мантии 23—35% ее
длины. На вентральной стороне ла-
донной части булавы между краем,
булавы и центральным крюком 2 ря-
да крупных (ближе к краю) и 1—2*
ряда очень мелких присосок. Между
центральным и первым проксималь-
ным крюком на булаве обычно рас-
положены 1—2 крупные присоски.
Присоски вентрального ряда на паль-
це булавы мельче присосок других
рядов. Нет мелких присосок на стеб-
ле щупальца между двумя краевыми
рядами присосок фиксаторного аппа-
рата; вентральный ряд присосок
фиксаторного аппарата в 1,5—3 раза
короче дорсального. Число попереч-
ных рядов присосок и крючьев в ба-
зальных половинах рук 3-й и 4-й
пар соответственно 6—9 и 8—11.
Менее 200 присосок на булаве.
Крючья на руках появляются значи-
тельно раньше, чем на булаве, при
длине мантии 7—9 мм. Центральный,
дистальный и проксимальные крючья
на булаве появляются при длине
мантии соответственно 14—18, 20—
27 и 27—33 мм.
G. (G.) berryi Naef, 1923 (рис.
51, л, л<). От Бсрингийского склона дсх
южных Курильских островов (и Се-
веро-Восточного Хонсю?) и северной
части п-ва Калифорния. Мезо- и ба-
типелагиаль. Длина мантии до 19 см.
10(9). Булава короткая, ее длина
10—24% длины мантии (но у
G. steenstrupi до 32—36 %). Ширина
мантии 16—25% ее длины. На вен-
тральной стороне ладонной части бу-
лавы между краем булавы и цент-
ральным крюком либо 4 ряда одина-
ковых присосок, либо 3 ряда круп-
ных и 1 ряд мелких присосок. Прок-
симальные крючья на булаве (если
имеются) начинаются сразу после
центрального. Присоски на пальце
булавы более или менее одинакового
191
L4
о
р
/1
Рис. 51. Род Gonatus: а—б—G. (Е.) tinro;
в—г — G. опух\ д—ac—G. madokai (молодой
и взрослый); з—/с—(?. ругоз*, л—м—G,
berryi; н—о — G. californiensis; п—р —> G,
fabricii; б, г-ж, и, о, р — булава; к — глаз
с фотофором (а—б — по Несису, 1972; в—
г, з—о — ио R. Young, 1972; д—ж — по Ки-
bodera, Okutanl, 1977; л — по Мииз, 1959,
р — по Verrill, 1882).
77
13 К. Н. Несис
193
размера. На стебле щупальца между
краевыми рядами присосок фиксатор-
ного аппарата есть мелкие присосоч-
ки (иногда их много). Оба ряда при-
сосок фиксаторного аппарата при-
мерно равной длины .... 11.
11(14). Кальмары распростране-
ны в северной части Тихого океана.
9—11 поперечных рядов присосок и
крючьев в базальной половине рук
3-й пары, 10—14 поперечных рядов
присосок в базальной половине рук
4-й пары....................- 12.
12(13). Мантия очень узкая, ее
ширина 16—19% длины, довольно
мускулистая. Булава щупальца ко-
роткая—10—13(17)% длины ман-
тии. Проксимальные крючья на була-
ве обычно не развиты. Крючья на ру-
ках появляются при длине мантии
22—25 мм, центральный крюк на бу-
лаве — при длине около 50 мм, дис-
тальный — около 60 мм.
G. (G.) kamtschaticus (Middendorff,
1849) (=G. middendorffi Kubodera et
Okutani, 1981). Довольно редкий, по-
видимому, неритоокеанический вид,
известен в Беринговом и восточной
части Охотского моря, у Алеутских,
Командорских и Курильских остро-
вов, в северо-западной части Тихого
океана и в заливе Аляска. Длина
мантии до 30 см.
13(12). Ширина мантии 19—25%
ее длины, мантия слабо мускулистая.
Длина булавы щупальца 17—20%
длины мантии. Проксимальные
крючья у взрослых кальмаров обыч-
но имеются. Крючья на руках появ-
ляются при длине мантии 26—29 мм,
центральный, дистальный и прокси-
мальные крючья на булаве — соответ-
ственно при 17—23, 24—30 и 35—
40 мм.
G. (G.) californiensis Young, 1972
(рис. 51, н, о). У Калифорнии и п-ва
Калифорния, (?) Панамский залив.
Мезо-, реже батипелагиаль. Длина
мантии свыше 11 см.
14(11). Кальмары распространены
в Арктике, Северной Атлантике и во-
дах Южного океана. Ширина мантии
17—25% ее длины. 8—13 поперечных
рядов присосок и крючьев в базаль-
ной половине рук 3-й пары . . 15.
15;(18). Виды распространены в
северном; полушарии. 12—17 попереч-
ных рядов присосок в базальной по-
ловине рук 4-й пары .... 16.
16(17). Длина булавы 13—
17(20)% длины мантии. В базаль-
194
ных половинах рук 3-й и 4-й пар со-
ответственно 9—13 и 14—17 по-
перечных рядов присосок и крючьев.
Крючья на руках появляются при
длине мантии 30—35 мм, централь-
ный, дистальный и проксимальные
крючья на булаве соответственно при
длине 25—30, 30—35 и 35—40 мм.
G, (G.) fabricii (Lichtenstein, 1818)
(рис. 51, л, р). Вся глубоководная
часть Полярного бассейна, Норвеж-
ское, Гренландское и западная часть
Баренцева моря, Баффинов залив,
Девисов пролив, Ирмингерово море,
Лабрадор, северо-западная часть Ат-
лантики на юге до района южнее
мыса Код. Нижняя эпи- и мезопела-
гиаль, по ночам иногда поднимается
к поверхности. Встречается также у
дна на склоне. Личинки и молодь
живут в верхних слоях воды. Очень
многочисленный вид. Длина мантии
до 30—35 см. В прежние времена в
небольшом количестве добывался в-
Гренландии.
17(16). Длина булавы 20—36%,
обычно 23—32% длины мантии.
В базальных половинах рук 3-й и 4-й
пар соответственно 8—11 и 12—14
поперечных рядов присосок и крю-
чьев.
G. (G.) steenstrupi Kristensen, 1981.
Северо-Восточная Атлантика от Фа-
реро-Шетландского порога до Бис-
кайского залива. Мезопелагиаль.
Длина мантии до 13(19?) см.
18(15). Вид распространен в юж-
ном полушарии. Длина булавы 15—
24% длины мантии. 9—10 попереч-
ных рядов присосок в базальной по-
ловине рук 4-й пары. Крючья на ру-
ках появляются при длине мантии
около 25 мм, центральный, дисталь-
ный и проксимальные крючья на бу-
лаве — соответственно при длине
около 20—25, около 25—30 и бо-
лее 30 мм.
G (G.) antarcticus Lonnberg, 1898<
Нотальный, заходящий в Антаркти-
ку, циркумполярный вид, на севере
доходит до Южной Африки, проли-
ва Кука и Северного Перу. Личинки
и молодь в эпи- и мезо-, взрослые в
мезо- и батипслагиали. Длина ман-
тии до 35 см.
Примечание. По данным
Р. Янга (Young, 1972) в антарктиче-
ских водах обитает еще один, нс опи-
санный вид—Gonatus. Этот (?) вид
описан Имбсром (Imber, 1978) под
названном G. phoebetriae по сдинст-
венному клюву, извлеченному из же-
лудка дымчатого альбатроса с о-вр
Марион.
Таблица для определения видов рода
Gonatopsis
1(2). У взрослых кальмаров сохраня-
ются рудименты стеблей щупалец в
виде коротких обрывков, скрытых ме-
жду основаниями 3-й и 4-й пар рук.
Длина рук равна или превосходит
длину мантии. Мантия водянистая.
Радула из 5 продольных рядов зубов.
G. (Gonatopsis) okutanii Nesis
1972 (рис. 50, е). От Берингийского
склона до Южных Курильских остро-
вов и залива Аляска, Охотское море,
северная и центральная части Япон-
ского моря. Мезо- и батипелагиаль и
абиссаль. Длина мантии до 25
(?45) см.
2(1). Рудиментов щупалец нет. 3.
3(4). Конечная треть рук 1—3-й
пар занята очень мелкими присоска-
ми, сидящими в 6—12 продольных
рядов. Длина рук 80—85% длины
мантии, их концы утончены и изогну-
ты. Плавник маленький, почковид-
ный, его длина 20—30% длины ман-
тии. Мантия водянистая. Радула из
5 продольных рядов зубов. Щупаль-
ца исчезают и крючья на руках по-
являются при длине мантии между
30 и 60 мм.
G. (Gonatopsis) octopedatus Sasa-
ki, 1920 (см. рис. 50, ж, з). От Бе-
рипгийского склона до Японского мо-
ря, Северо-Восточного Хонсю и зали-
ва Аляска. Батипелагиаль и нижняя
батиаль. Длина мантии до 24 см.
4(3). Крючья идут почти до кон-
цов рук. Присоски на концах рук
в 4 продольных ряда. Длина рук
меньше или равна 70%, длина плав-
ника 30—55% длины мантии . . 5.
5(6). Плавник сердцевидный, сза-
ди оттянут в хвостик, его длина и
ширина 40—50% длины мантии.
Мантия узкая, се ширина около Vs
длины. Радула из 5 продольных ря-
дов зубов.
G. (Gonatopsis) japonicus Okiyama,
1969 (рис. 50, и). Южная часть Охот-
ского и центральная часть Японско-
го моря. Все известные особи пойма-
ны у поверхности. Длина мантии до
17^см.
6(5). Плавник ромбический, сза-
ди не оттянут в хвостик. Ширина
плавника больше его длины . . 7.
7(8). Плавник крупный, его дли-
на 37—45%, ширина 60—80% длины
мантии. Мантия широкая (ширина
около 25% длины), крепкая, муску-
листая. Руки короткие, их длина око-
ло V2 длины мантии. Радула из 7
продольных рядов зубов. Щупальца
исчезают при длине мантии 12—
15 мм. Крючья на руках появляются
при длине мантии 35—45 мм.
G. (Boreoteuthis) borealis Sasaki,
1923 (см. рис. 50, к, л). От Берипгий-
ского склона до северной части
Японского моря, Северо-Восточного
Хонсю и района южнее п-ва Кали-
форния. Охотское море. От эпи- до
батипслагиали, также у дна в батиа-
ли и абиссали. Молодые и взрослые
кальмары часто ловятся на свет у
поверхности в открытом океане. Дли-
на мантии до 29 см (изредка до
48 см). Массовый вид. Придавлива-
ется в небольших количествах при
ярусном лове кальмаров и сетевом
лове лососей.
8 (7). Плавник небольшой, его дли-
на около 30%, ширина .менее 50%
длины мантии. Мантия узкая (шири-
на около 20% длины), мягкая, водя-
нистая. Руки довольно длинные —
около % длины мантии.
G. (Gonatopsis) makko Okutani et
Nemoto, 1964 (рис. 50, л). От Берин-
гова до Японского моря и северо-
восточного побережья Хонсю, залив
Аляска. Мезопелагиаль и дно на глу-
бинах 650—1200 м. Самый крупный
вид гоиатид—длина мантии до 75 см.
В Беринговом море, у Алеутских
островов, в заливе Аляска и у Юж-
ных Курильских островов попадались
личинки Gonatopsis (Boreoteuthis)
sp., которых не удалось отнести ни к
одному из известных по взрослым
особям видов гонатид.
2.2.2.6. Семейство
Ctenopterygidae
Мелкие кальмарчики с широкой уп-
лощенной сверху мантией и ориги-
нальными «рыбьими» плавниками.
Плавник, короткий у молоди и почти
равный длине мантии у взрослых
особей, представляет собой серию
гибких мягких «лучей», особенно
длинных у заднего конца мантии,
соединенных на большей части дли-
ны тонкой кожной перепонкой. Голо-
ва широкая, но не шире мантии. Ру-
13*
195
ки довольно короткие, в дистальной
половине рук 1—3-й пар присоски
сидят в 4—6 рядов. 4-я пара рук
сильно расширена у основания. Коль-
ца присосок рук гладкие. Щупальца
узкие, с ие расширенной булавой,
имеющей вид овальной площадки и
несущей множество мельчайших при-
сосок во много рядов. Фиксаторного
аппарата нет. Формула буккального
крепления DDVV. На лучах буккаль-
ной мембраны есть мельчайшие при-
соски, расположенные в 2 ряда. За-
мыкательный аппарат простой. На
брюшной стороне глазного яблока по
одному удлиненному фотофору.
Крупный фотофор находится на
брюшной стороне чернильного меш-
ка под кишкой; это желтоватый ша-
рик, лежащий па вогнутой и покры-
той светлым отражающим слоем по-
верхности чернильного мешка. Гла-
диус с длинным рахисом и широким
пером, подвернутым на конце на
брюшную сторону, без конечного ко-
нуса. Гектокотиля нет. У самок ме-
жду впутримантийиым фотофором и
нидаментальнымн железами имеется
крупная непарная железа неизвест-
ной функции, которую считают гомо-
логичной добавочной нидаменталь-
ной железе миопсид. Развитие с ли-
чиночной стадией.
Тропическо-субтропические мезо-
батипелагичсские кальмары, молодь
обитает в эпипслагиали. Длина ман-
тии до 8 см.
Один род—Ctenopteryx Appellof,
1880. Типовой вид — С. sicula (Vera-
ny, 1851). 2 вида.
1(2). Ширина мантии около по-
ловины (40—60%) ее длины.
С. sicula (Verany, 1851) (рис.
52, а—в). Тропическо-субтропический
космополитический вид.
2(1). Ширина мантии 2/а—3Д ее
длины.
С. sepioloides Rancurel, 1970 (рис.
52, г). Центральная часть Тихого
океана (о. Гавайские и Туамоту).
2,2.2.7. Семейство
Bathyteuthidae
Мелкие кальмары с короткой и ши-
рокой закругленной сзади мантией
хрящевато-студенистой консистенции
к
Рис. 52. Сем. Ctenopterygidae и
Bathyteuthidae (д—к): а—в—Ctenopteryx si-
cula, молодой (а), взрослый (б сбоку)
кальмары и ректальный фотофор (а); г*С.
sepioloides; д—з — Bathyteuthis abyssicola\
(б—молодой, е—взрослый кальмар); и—/с—
В. berryii л—н—В. bacidifera; ж» м—руки;
з, к, н — кольца присосок рук 1-й пары
(а— по Okutani, McGowan, 1969; б —по
Okutani, 1974; в — по Несису, 1973; а —па
Rancurel, 1970; д—н — по Roper, 1969)»
и
л
и маленькими почковидными не со-
единяющимися плавниками на конце
тела. Голова очень широкая, глаза
выступающие, смотрят несколько
вперед [при естественном положении
кальмара (головой вниз) они смот-
рят вниз и вбок]. Руки короткие,
широкие у основания, их базальные
части соединены толстой перепонкой.
Руки 1—3-й пар с 2—4 рядами нере-
гулярно расположенных присосок,
руки 4-й пары с 2 рядами. Щупаль-
ца длинные, тонкие, булава не обо-
соблена, присоски на ней в 8—10 ря-
дов. Фиксаторного аппарата нет.
Присоски рук и булавы очень мел-
кие. Лучи буккальной мембраны с не-
многочисленными крохотными при-
сосками (имеются не у всех особей).
Формула буккального крепления
DDVD. Гладиус с длинным узким
рахисом и овальным пером, без ко-
нечного конуса. У основания рук 1—
3-й пар в толще подкожных тканей—
по одному фотофору, заметному
лишь у молодых кальмаров. Печень
с крупными жировыми включениями.
Гектокотиля нет. Развитие без личи-
ночной стадии. Окраска тела интен-
сивно красно-бурая. Длина мантии
до 7,5 см.
Батипелагические и мезо-батипс-
лагические кальмары, не совершаю-
щие заметных вертикальных переме-
щений.
Один $o& — Bathyteuthis Hoyle,
1885 (=Benthoteuthis Verrill, 1885,
IChunoteuthis Robson, 1921, Hndo-
teuthis Grimpe, 1922) (рис. 52, д—h).
3 вида. Обзор: Roper, 1969.
Таблица для определения видов рода
Bathyteuthis
1(4). Защитные мембраны рук хоро-
шо развиты, мясистые, их наружные
края прямые или слегка волнистые,
без торчащих свободных трабе-
кул ............................2.
2(3). Руки короткие, тупые. При-
соски рук немногочисленные — около
сотни на руках 1—3-й пар у взрос-
лых. Кольца присосок рук с 8—18
усеченными зубцами. Жабры корот-
кие, узкие.
В. abyssicola Hoyle, 1885 (=«Вгп-
thoteuthis megalops Verrill, 1855)
(рис. 52, д—з). Космополитический
вид, распространен в эвтрофных зо-
нах Мирового океана: в Атлантике
на севере до Ирландии, обитает глав-
ным образом в северной и восточной
частях океана (отсутствует в Кариб-
ском море и Мексиканском заливе),
в Индийском океане в основном у
Восточной Африки, в Тихом — пре-
имущественно в экваториальных (до
Гавайских островов) и юго-восточ-
ных районах, а также в Соломоно-
вом море. В тропиках вид относи-
тельно редок, в нотальных и антарк-
тических водах распространен цир-
кумглобально и чрезвычайно много-
числен. Обитает на глубинах 500—
3000 м (обычно 1000—2250 м), мо-
лодь изредка встречается в нижней
эпипелагиали.
3(2). Руки длинные, с оттянуты-
ми и утонченными концами. Присос-
ки рук весьма многочисленные —
250—300 па руках 1—3-й пар у
взрослых. Кольца присосок рук с
10-—14 треугольными зубцами. Жаб-
ры длинные, широкие.
В. berryi Roper, 1968 (рис. 52, и—
к). Калифорнийское течение, батипе-
лагиаль.
4(1). Защитные мембраны рук
слабо развиты, поддерживающие их
трабекулы не соединены между со-
бой и у основания рук торчат нару-
жу пальцевидными выступами. Руки
короткие, тупые. Присоски рук мно-
гочисленные — около полутора сотен
па руках 1—3-й пар у взрослых.
Кольца присосок рук с 18—34 усе-
ченными зубчиками. Жабры длинные,
широкие.
В. bacidifera Roper, 1968 (рис.
52, л—н). Индотихоокеапский тропи-
ческий (преимущественно экватори-
альный) мсзо-батипелагический вид.
2.2.2.8. Семейство
Histioteuthidae
Кальмары средних размеров и круп-
ные, с небольшой, обычно кониче-
ской мантией и большой головой.
Ткани студенисто-хрящеватой конси-
стенции. Плавники небольшие, округ-
лые, сзади соединяются, сидят на
заднем конце тела и иногда перехо-
дят за конец мантии. Глаза боль-
шие, асимметричные, левый обычно
намного больше правого. Руки длин-
ные, толстые у основания, с 2 ряда-
ми присосок. Щупальца очень длин-
ные, с небольшой булавой, несущей
4—8 рядов присосок обычно резко
неравных размеров, у зрелых самок
198
часто автотомируются. Фиксаторный
аппарат протягивается вдоль стебля
щупалец на расстояние в 2—3 раза
большее длины булавы. Буккальная
мембрана 6- или 7-лучевая (рис.
53, а, б). Формула буккального креп-
ления DDVD. Часто хорошо развита
внутренняя умбрелла — перепонка
между руками, оставляющая щупаль-
ца снаружи. Замыкательный аппарат
простой, вороночные хрящи сзади
слегка изогнуты и расширены. Вся
нижняя поверхность мантии, головы
и рук покрыта множеством сложно
устроенных фотофоров одного или
двух типов, светящих вперед; на
спинной стороне их мало, на плавни-
ках и щупальцах пет. Фотофоры рас-
положены на мантии и голове косы-
ми рядами, на руках — продольными..
Вокруг каждой глазной орбиты коль-
цо фотофоров, расположенных в
строгом и специфичном порядке.
Особые модифицированные фотофо-
ры имеются у некоторых видов на
концах рук. Гладиус тонкий, с корот-
ким рахисом и широким пером, без
конечного конуса. Гектокотилизиру-
ются обе спинные руки. Мужские по-
ловые органы непарные или (И. hoy-
lei) парные. Развитие без личиночной
стадии.
Батипелагические или мезо-бати-
пелагические, также придонно-бати-
альные кальмары. Иногда встречают-
ся в нижней эпипелагиали и нижней
сублиторали. Обитают всесветно,
кроме Арктики и Антарктики. Места-
ми многочисленны и служат пищей
кашалотам и другим хищникам.
Один род — Histioteuthis d’Orbig-
ny, 1841 (=Lolidona Risso, 1854, Cal-
liteuthis Verrill, 1880, Histiopsis Hoy-
Ze, 1885, Stigmatoteuthis Pfeffer, 1900,
Meleagroteuthis Pfeffer, 1900, Histio-
thauma Robson, 1948) (рис. 53—56).
Типовой вид —Я. bonnellii (Ferus-
sac, 1835). 13 видов и 8—11 подви-
дов. Обзоры: N. Voss, 1969; Несис,
1971.
Таблица для определения вида рода
Histioteuthis
1(4). Руки наполовину или на боль-
шую часть длины соединены внут-
ренней перепонкой. По одному круп-
ному длинному фотофору на концах
1—3-й пар рук...................2.
2(3). Буккальная мембрана шес-
тилучевая: связка от нее к рукам
4-й пары не раздвоена. Сегменты
внутренней умбреллы, отходящие к
рукам 3-й и 4-й пар, соединяются все
вместе вдоль одной линии. 17 фото-
форов вокруг правого глаза. Ветви
срединного компонента вороночного
органа гладкие.
Н. bonnellii (Ferussac, 1835) (=Я.
bonelliana auett. Я. ruppelli Verany
1846) (рис. 53, а, в—з). Атлантиче-
ский тропическо-субтропический (за-
ходящий в бореальные районы) вид,
распространенный также в юго-за-
падной части Индийского океана;
в Атлантике от Большой Ньюфаунд-
лендской банки и Исландии до южной
субтропической конвергенции; встре-
чается в Средиземном море, отсутст-
вует в Карибском море и Мексикан-
ском заливе. Преимущественно (но
не исключительно) нсрито-океаниче-
ский вид. Нижняя эпи- и мезопела-
гиаль (иногда батипелагиаль), у дна
в батиали (изредка в нижней субли-
торали). 2 подвида:
а. Нет крупных фотофоров сзади
и вентральнее левого глаза.
Я. b. bonnellii (Ferussac, 1835)
(рис. 53,в—ж). Северная Атлантика.
Длина мантии до 33 см;
б. 2—3 крупных округлых фото-
фора сзади и вентральнее левого
глаза.
Я. б. corpuscula Clarke, 1980 (рис.
53, з). Южная Атлантика и юго-за-
падная часть Индийского океана.
Длина мантии до 9 см.
3(2). Буккальная мембрана семи-
лучевая: связки от нее к рукам 4-й
пары полностью разобщены. Сегмен-
ты внутренней умбреллы, отходящие
к правым и левым рукам 3-й и 4-й
пар, соединяются попарно и объеди-
няются общей лентой. 16 фотофоров
вокруг правого глаза. Ветви средин-
ного компонента вороночного органа
с продольным гребнем.
Я. macrohista N. Voss. 1969 (рис.
53, б, и—м). Южпосубтропическо-но-
тальный циркумглобальный вид. Ме-
зо- и батипелагиаль. Длина мантии
до 7 см или более.
4(1). Глубина внутренней ум-
бреллы не более 30% длины наиболь-
шей руки. Фотофоры на концах рук
многочисленные, не удлиненные . 5.
5(6). Глубина внутренней умбрел-
лы 17—30% длины наибольшей ру-
ки. На концах рук 3-й пары — серия
из нескольких овальных черных фо-
тофоров, наибольший из которых в
199
Ж
&
*
о
е
Сем. Histioteuthidac, род Histio-
Рис. 53. ___ ____________, ____
teuihis, 1: а, 6 — 6- и 7-лучевая буккаль-
ная мембрана на примере Н. bonnellii (а)
и Н, macrohisia (6); в—ж — Н. bonnellii
bonnellii\ з — Н. bonnellii corpuscula; и—
м — Н. macrohista; н—р — Н. atlantica; с—
т — Н. bruuni; в — молодой кальмар; 6, к,
о —булава; п — брюшная рука; е, р —ко-
нец руки; л, г—буккальная мембрана;
яс—з, м — фотофоры вокруг правого (яс,
и левого (а»ь з) глаза (з —no Clarke,
1980, прочее по N. Voss, 1969).
2—3 раза крупнее обычных фотофо-
ров. 4 ряда фотофоров (не считая
мелких) на руках 4-й пары.
Н. atlantica (Hoyle, 1885) (=Я.
<cookiana Dell, 1951) (рис. 53, я—р).
Южносубтропическо-нотальпый цир-
кумглобальный вид. Мезо- и батипе-
VI аги аль, опускается также в абиссо-
пелагиаль. Длина мантии до 20 см.
6(5). Глубина внутренней умб-
реллы не более 15% длины наиболь-
шей руки. Фотофоры на концах рук
•обычного строения.............7.
7(14). Фотофоры на нижней сто-
роне мантии, головы и рук располо-
жены плотно: более 4 продольных
рядов фотофоров в базальной частй
рук 4-й пары...................8.
8(9). Буккальная мембрана шес-
тилучевая, асимметричная. Имеется
продольный ряд хрящевых бугорков
по средней линии спины в передней
половине мантии и вдоль рук I—
3-й пар в их базальных частях. Око-
ло 20 фотофоров вокруг правого
глаза.
Н. bruuni N. Voss, 1969 (рис.
53, с—г). Юго-Восточная Атлантика:
Китовый хребет, Намибия. Батипела-
гиаль. Длина мантии 8 см.
9(8). Буккальная мембрана семи-
лучевая, симметричная. Хрящевые
•бугорки на мантии и руках имеются
или отсутствуют...............10.
10(11). 5 продольных рядов фо-
тофоров в базальных частях рук 4-й
пары. На нижней стороне мантии 9—
10 косых (диагональных) рядов фо-
тофоров, в одном косом ряду, начи-
ная с бока, около 12 фотофоров.
16 фотофоров вокруг правого глаза.
Имеются хрящевые бугорки па спин-
ной стороне мантии и в базальных
частях 1—3-й пар рук.
Н. miranda (Berry, 1918) ( =
iiothauma oceani Robson, 1948) (рис.
54, a—д). Тропические и субтропиче-
ские воды Тихого океана от Филип-
пинских островов до Новой Гвинеи,
Южной Австралии и Новой Зелан-
дии на восток до островов Гавай-
ских, Лайн и Галапагосских;
Южная 'Африка, южная часть Ин-
дийского океана. По-видимому, нери-
то-океанический вид. Обитает пре-
имущественно в мезопелагиали и у
дна в батиали. Длина мантии до
24 см.
11(10). 8—9 продольных рядов
фотофоров в базальных частях рук
4-й пары. На нижней стороне мантии
более 20 косых (диагональных) ря-
дов фотофоров, в одном косом ряду
19—27 фотофоров. 19—21 фотофор
вокруг правого глаза .... 12.
12(13). Продольные ряды хряще-
вых бугорков на спинной стороне
мантии в ее передней части и в ба-
зальных частях рук 1—3-й пар име-
ются. На нижней стороне мантии
около 30 поперечных рядов фотофо-
ров, в одном косом (диагональном)
ряду, начиная с бока, 24—27 фото-
форов.
Н. meleagroteuthis (Chun, 1910)
(=Meleagroteuthis hoylei Pfeffer,
1908; At separata Sasaki, 1915) (рис.
54,e—к). Тропическо-субтропический
космополитический вид: в Атлантике
от 40° с. ш. до мыса Доброй Надеж-
ды, в Пацифике от Курильских ост-
ровов и Орегона (возможно, от Бе-
рингова моря) до Новой Зеландии.
Преимущественно нерито-океаниче-
ский вид. Нижняя эпи- и мезопела-
гцаль (иногда в батипелагиали) и у
дна в батиали. Длина мантии до
11 см.
13(12). Хрящевые бугорки на
мантии и руках отсутствуют. На
нижней стороне мантии около 20—
25 поперечных рядов фотофоров, в
одном косом ряду 19—22 фотофора.
Н. heteropsis (Berry, 1913) (рис.
54, л—я). Восточнотихоокеанский
тропическо-субтропический преиму-
щественно нерито-океанический вид:
от Калифорнии до Среднего Чили.
Мезопелагиаль. Длина мантии до
13 см.
14(7). Фотофоры на нижней сто-
роне мантии, головы и рук располо-
жены редко, в базальной части рук
4-й пары их 3—4 ряда, на нижней
стороне мантии в косом (диагональ-
ном) ряду, начиная с бока, 9—10
фотофоров .....................15.
15(16). Мантия очень удлиненная
и узкая, ее ширина !/ч—Уз длины.
Голова заметно ^же мантии. В зад-
ней части мантии на брюшной сторо-
не и в дистальных частях рук, кроме
обычных фотофоров, имеются своеоб-
разные крупные овальные черные фо-
тофоры без линзы. Левый глаз лишь
немного больше правого.
Н. elongata (N. Voss et G. Voss,
1962) (рис. 54, о, я). Субтропические
воды Северной Атлантики от Нью-
фаундленда до мыса Хаттерас и Ма-
дейры; Средиземное море. Мезопела-
гиаль. Длина мантии до 18 см.
202
16(15). Мантия коническая, ее
ширина %—% длины. Голова нс уже
или лишь немного уже мантии. Нет
черных фотофоров без линз. Левый
глаз значительно крупнее право-
го ............................17.
17(22) Кольца крупных присосок
рук с зубчатыми или зазубренными
краями (у взрослых Я. reversa этот
признак не всегда выражен). При-
соски ладонной части булавы резко
различаются по размерам: централь-
ные в 3—4 раза крупнее краевых.
Ветви срединного компонента воро-
ночного органа с продольным греб-
нем или лопастью...............18.
18(19). Руки очень длинные, у
взрослых в 2—2,5 раза длиннее ман-
тии. Имеется небольшая (до 14%
длины наибольшей руки) наружная
умбрелла у оснований рук. На ниж-
ней стороне мантии все фотофоры
однотипные, их размер постепенно
уменьшается от переднего к заднему
концу тела. 17 фотофоров вокруг
правого глаза. Ветви срединного
компонента вороночного органа со
свободно свисающей в просвет во-
ронки треугольной лопастью. Голова
очень крупная, ее длина V2—3А,
обычно около % длины мантии. Ко-
жа часто покрыта, особенно у взрос-
лых, мелкими мясистыми папиллами.
У самцов двойной набор половых ор-
ганов.
Я. hoylei (Goodrich, 1896) (=Я.
dofleini (Pfeffer, 1912), Stigmato-
teuthis chuni Pfeffer, 1912, S. arcturi
Robson, 1948) (рис. 55,a—ж). Ши-
роко распространенный тропическо-
субтропический, заходящий в боре-
альные и нотальпые районы, космо-
политический вид: Атлантика от
Гольфстрима и Гибралтара до под-
водной возвышенности Рио-Гранде,
Мексиканский залив, Карибское мо-
ре (редко); в Средиземном море от-
сутствует; в Индийском океане на
юге до Мозамбикского пролива, 'в
Пацифике от Алеутских островов до
40—45° ю. ш. От нижней эпи- до ба-
типелагиали, преимущественно в ме-
зопелагиали, также у дна в батиали.
Длина мантии до 21 см. По данным
Н. Восс (N. Voss, 1969), вид вклю-
чает два или более подвидов и воз-
можно, что Я. hoylei (типовое место-
нахождение — Андаманское море) и
Я. dofleini (типовое местонахожде-
ние — Южная Япония) — подвиды, а
пе синонимы.
19(18). Длина рук 100—150%
длины мантии. Наружная умбрелла
отсутствует. На нижней стороне ман-
тии крупные фотофоры перемежают-
ся с мелкими. 17 крупных и 1 ма-
ленький фотофор вокруг правого гла-
за. Ветви срединного компонента во-
роночного органа с продольным
гребнем, без лопасти. Длина головы
Vs—V2 длины мантии. Кожа гладкая.
Половые органы самцов нормаль-
ные ............................20.
20(21). В базальной части рук
4-й пары 4 ряда фотофоров: 3 ряда
крупных, 1 — мелких. Наружный
(дорсальный) ряд крупных фотофо-
ров состоит из 5—8 органов.
Я. reversa (Verrill, 1880) (=Calli-
teuthis meneghinii Pfeffer, 1912) (рис.
55, з—м). Атлантический тропичес-
ко-субтропический вид, заходящий в
бореальные и нотальные воды; на
севере распространен до Большой
Ньюфаундлендской банки и района
южнее Исландии, на юге приблизи-
тельно до 45° ю. ш., Средиземное
море; в Мексиканском заливе не об-
наружен, в Карибском море очень
редок. Обитает в мезопелагиали,
нижней эпипелагиали и у дна в ба-
тиали. Длина мантии до 13,5 см.
21(20). В базальной части рук
4-й пары только 3 ряда крупных фо-
тофоров. Наружный (дорсальный)
ряд состоит из 2—3 фотофоров.
Я. eltaninae N. Voss, 1969, (рис.
55, я—с), Нотальный циркумглобаль-
ный, лишь изредка заходящий южнее
антарктической конвергенции, мезо-
батипелагический вид. Длина мантии
до 7 см и более.
22(17). Кольца крупных присосок
рук гладкие (зазубренными могут
быть только кольца самых мелких
присосок в дистальных частях рук).
Присоски ладонной части булавы
различаются по размерам не более
чем втрое. Ветви срединного компо-
нента вороночного органа глад-
кие ............................23.
23(24). Присоски средней части
булавы слабо различаются по раз-
мерам: центральные не более чем в
1,5 раза больше краевых. Кольца
присосок крайнего вентрального ря-
да в ладонной части булавы значи-
тельно расширены. На концах рук
самая дистальная группа фотофоров
наружного (дорсального) ряда отде-
лена от предыдущих фотофоров «пу-
стым» участком, как будто 1—2 фо-
203
3
Рис. 54. Род Ilistioteuthis, 2: а—д— Н. ml*
randa} е—к — Н. meleagroteuthis} л—н — Н.
heteropsls\ о—п — Н. elongata*» з, з, м —
булава; д, и, п — брюшная рука; г — бу-
горки по средней линии спины; к —фото-
форы вокруг правого глаза; « — гектоко-
тиль (по N. Voss, 1969).
ж g
Рис. 55. ’ Род Histioteuthis, 3: а—ж — Н.
hoylei\ з—м—Н. reversa-, н—с—Н. eltaninae*r
б — буккальная мембрана; в, к, п — була-
ва; л, р’—брюшная рука; г, м, с —фото-
форы вокруг правого, 2j, Mi, — левого
глаза; д — присоска руки; е — присоска
булавы; ж—вороночный орган (по N. Voss,.
1969).
ж
14 к. Н. Несис
тофора «выпали». Мантия удлинен-
ная, цилиндроконическая, ее перед-
няя часть с почти параллельными
краями.
Н, celetaria (G. Voss, 1960) (рис.
56, а—е). Тропический мезопелагичс-
ский (или мезо-батипелагический) и
придонно-батиальный вид. Длина
мантии до 28 см. 2 подвида:
а. На брюшной стороне головы
имеются 2 свободные от фотофоров
площадки между глазом и воронкой.
Кольца крупных присосок ладонной
части булавы несут зубцы только на
дистальном крае.
Н. с, celetaria (G. Voss, 1960)
(рис. 56, а—е). Атлантика от Бер-
мудских островов и Мадейры до
30° ю. ш.
б. Нет свободных от фотофоров
площадок на нижней стороне головы.
Кольца крупных присосок ладонной
части булавы несут зубцы по всему
периметру.
Н. с. pacifica (G. Voss, 1962)
( = Calliteuihis japonica Massy, 1916,
?non Pfeffer, 1912) (рис. 56"ж—л).
Ипдо-Вестпацифика от Южных
Японских островов до района южнее
Мадагаскара, Северной Австралии и
Гавайских островов.
24(23). Центральные .присоски
средней части булавы в 2—3 раза
больше краевых. Кольца присосов
крайнего вентрального ряда в ладон-
ной части булавы не расширены.
Наружный (дорсальный) ряд фото-
форов на руках тянется до конца
рук без перерывов. Мантия кониче-
ская или бокаловидная.
Я. corona (N. Voss et G. Voss,
1962) (рис. 56, м—ф). Тропическо-
субтропический, заходящий в поталь-
ные воды, мезо-батипелагический
(поднимающийся в нижнюю эпипе-
лагиаль) и придонно-батиальный вид.
Длина мантии до 16 см. 4 подвида:
а. 4 ряда фотофоров в базальной
части рук 4-й пары. Кольца крупных
присосок в ладонной части булавы с
30 зубцами. Длина плавника 40—
45% длины мантии.
Н. с. berryi N. Voss, 1969 (рис.
56, с—т). Калифорнийское течение.
Гавайские острова.
б. 3 ряда фотофоров в базальной
части рук 4-й пары. Кольца крупных
присосок в ладонной части булавы с
20—25 зубцами. Длина плавника
45—55% длины мантии.
Н. с. inermis (Taki, 1964). Южная
210
Япония (Тихоокеанское побережье),
в. 3 ряда фотофоров в базальной
части рук 4-й пары. Кольца крупных
присосок в ладонной части булавы с
33—35 зубцами. Длина плавника
35—50% длины мантии.
Я. с. corona (N. Voss ct G. Voss,
1962) (рис. 56, м—p). Атлантика от
мыса Хаттерас и Гибралтара до Юж-
ного тропика, Индийский океан
(у Восточной Африки).
г. 3 ряда фотофоров в базальной
части рук 4-й пары. Кольца крупных
присосок в ладонной части булавы с
50—60 зубцами. Длина плавника
30—40% длины мантии.
Н, с. cerasina Nesis, 1971 (рис.
56, у, ф). Восточная экваториальная
часть Тихого океана от экватора до
Южного тропика, на запад до остро-
вов Лайн.
Кальмар с признаками, промежу-
точными между последними подвида-
ми (возможно, самостоятельный под-
вид), обнаружен в нотальных водах
па склоне Новозеландского плато ме-
жду островами Баунти и Антиподов.
2.2.2.Э. Семейство
Psychroteuthidae
Кальмары средних и крупных разме-
ров. Мантия цилиндрическая в пе-
редней и коническая в задней поло-
вине, мускулистая. Плавник ромби-
ческий с оттянутым концом. Голова
маленькая. Руки с двумя рядами
присосок. Булава щупалец расши-
ренная, с 4 рядами резко неравных
присосок в проксимальной и 5—7 ря-
дами мелких присосок в дистальной
части. Фиксаторный аппарат протя-
гивается вдоль стебля. Замыкатель-
ные хрящи простые, вороночный
слегка изогнут, не расширен.. Форму-
ла буккального крепления DDVV.
Гладиус перовидный, с коротким ра-
хисом и длинным, сначала резко рас-
ширяющимся, потом постепенно су-
жающимся пером, на конце ложко-
видный, не доходит до конца хвоста.
У зрелых самок на концах рук 3-й
пары по одному крупному сложно
устроенному фотофору, состоящему
из расположенных зигзагом светлых
поперечных пластинок и закрываю-
щих их черных лопастей — ширм.
Длина мантии до 44 см.
Один род — Psychroteuthis Thiele,
1920 (рис. 57, а—г). Типовой и един-
14*
с
ственпый описанный вид — Р. gla-
cialis Thiele, 1920, распространен в
Антарктике; 2 неописанных вида —
один в Антарктике (Roper, Young,
Voss, 1969), второй найден у остро-
вов Бонин (Iwai, 1956). Виды этого
рода обитают в мезопелагиали и
главным образом у дна в батиали.
Таблица для определения видов рода
Psychroteuthis
1(2). Длина плавника 55—60% дли-
ны мантии.
Р. glacialis Thiele, 1920 (рис.
57, а—г). Антарктический циркумпо-
лярный вид. Мезопелагиаль и ба-
тиаль.
2(1). Длина плавника 48—50%
длины мантии.
Psychroteuthis sp. (Architeuthis
sp.» Iwai, 1956). Обнаружен в желуд-
ке кашалота у островов Бонин, Па-
цифика.
2.2.2.10. Семейство
Architeuthidae
Гигантские кальмары, самые круп-
ные из моллюсков. Мантия узкая,
сзади заостренная, вытянута в ко-
роткий хвост. Мышечный слой развит
довольно слабо. Плавники неболь-
шие, полукруглые или полуоваль-
ные, вместе образуют продольный
овал, не доходят до заднего конца
мантии. Голова неширокая. Руки
очень длинные, с двумя рядами при-
сосок. Щупальца чрезвычайно длин-
ные, в несколько раз длиннее ман-
тии; булава слабо расширенная, в ее
проксимальной (карпальной) части
кучка мелких присосок в несколько
рядов, в центральной части — 4 ряда
крупных присосок, из которых сред-
ние намного больше краевых, в дис-
тальной части присоски резко мель-
чают и слабо различаются по разме-
рам. Центральные присоски булавы
несут одинаковые конические зубчи-
ки по всему периметру. Фиксатор-
ный аппарат протягивается по стеб-
лю щупалец почти вдоль всей его
длины. Замыкательные хрящи про-
стые. Формула буккального крепле-
ния DDVV. Гладиус перовидный, с
коротким широким рахисом, сзади
заостренный, с маленьким конечным
конусом, не доходит до конца хвоста.
Фотофоров нет. Гектокотилизируются
обе брюшные руки. У самцов очень
Рис. 57. Сем. Psychroteuthidae (а—г), Ar-
chiteuthidae (д—л) и Neoteuthidae (м—ф):
а—г — Psychroteuthis glacialis (в—булава;
г — ручной фотофор взрослой самки); д—
ж — Architeuthis sp. из Северной Атланти-
ки; з—л — молодь Architeuthis из Атлан-
тики (з—и) и Восточной Пацнфики (к—л)
(е, и, л — булава, ж, — тсктокотилизи-
рованные концы брюшных рук); м—р —
Alluroieuthis antarctica (м—н — молодь, /г—
булава; р — крючок руки); с — Neoteuthis
thlelei (молодь); т—ф — Neoteuthis sp. из
Восточной Пацнфики (у — булава, ф — мо-
лодой кальмар), (а — по Filippova, 1972;
б—в, о—п — по Roper ct. al., 1969; г, р —
по Thiele, 1921; д—е — по Verrill, 1882;
ас —no Knudsen, 1957; з—л — по Roper,
Young, 1972; м—н — по Odhner, 1923; р —
ориг., т— у — по R. Young, 1972; ф — по
Okutani 1974).
длинный пенис. Длина мантии до
5 м, чаще 1—2 м.
Один род — Architeuthis Steen-
strup, 1856 ( = Architeuthus auctt., Me-
gcdoteuthis Kent, 1874, Dinoteuthis
More, 1875, Mouchezis Vela in, 1877,
Megateuthis Hilgendorf, 1880, Plecto-
ieuthis Owen, 1881, Steenstrupia Kirk,
1882, Dubioteuthis Joubin, 1899)
(рис. 57, d—л). Типовой вид — A. dux
Steenstrup, 1875. Описано 15 номи-
нальных видов, но их различия не
установлены, и число реально суще-
ствующих видов неизвестно, предпо-
ложительно от 1 до 3. Род Archi-
teuthis антитропический, встречается
в Северной Атлантике и прилегаю-
щих морях от Южного Лабрадора
до Мексиканского залива и от Север’-
ной Норвегии до Азорских островов
(в Средиземном море отсутствует);
в Северной Пацифике от Берингова
до Японского моря, Южных Япон-
ских островов и Калифорнии; в по-
тальных и южносубтропических во-
дах Южного океана; изредка —
в тропических водах Атлантики и ат-
лантическом секторе Антарктики.
Молодь обитает в толще воды (эпи-
пелагиаль, возможно, верхняя мезо-
пелагиаль), взрослые особи — у дна
в батиали и над подводными возвы-
шенностями. Большинство известных
до настоящего времени архитейтисов
были либо подобраны издыхающими
или дохлыми на берегах или поверх-
ности воды, либо извлечены из же-
лудков кашалотов.
2.2.2.11. Семейство
Neoteuthidae
( = AI1uroteuthidae)
Небольшие и средних размеров каль-
мары с конической мантией и боль-
шой головой. Плавники без передних
«ушек», почковидные обратно-сердце-
видные (узкие спереди и широкие
сзади, с вырезкой на заднем крае)
или в виде боковой каймы. Руки с 2
рядами присосок. У взрослых АНи-
roieuthis присоски на руках 1—3-й
пар превращаются в крючья. Присос-
ки на руках 4-й пары значительно
мельче, чем на 1—3-й паре. Булава
щупалец несет в проксимальной ча-
стй множество мелких присосок бо-
лее чем в 10 рядов, в центральной
части булавы 4 ряда крупных при-
.сосок равных или разных размеров,
на дистальном конце 3—4 ряда по-
214
степенно мельчающих присосок. Фик-
саторный аппарат щупалец имеется
или отсутствует. Замыкательные хря-
щи простые. Формула буккального
крепления DDVD. Фотофоров пет.
Гладиус с коротким рахисом и ши-
роким пером. Развитие с личиночной
стадией. 2 рода, 3 вида.
Таблица для определения родов и
видов Neoteuthidae
1(2). Плавники почковидные, немно-
го заходят за задний конец мантии,
их длина и ширина около половины
длины мантии. Щупальца тонкие,
слабые, у молоди короче рук, у
взрослых особей немного длиннее.
В центральной части булавы 4 пары
присосок двух средних рядов резко
увеличены, во много раз крупнее
краевых присосок. Зона мелких при-
сосок в проксимальной части булавы
занимает примерно 7з длины бу-
лавы. Фиксаторный аппарат из при-
сосок и бугорков-кнопок имеется.
У взрослых особей присоски на ру-
ках 1—3-й пар превращаются в
крючья в результате сильного раз-
растания среднего зубца в дисталь-
ной части кольца.
Род Alluroteuthis Odhner, 1923.
Один вид — A. antarctica Odhner,
1923 (рис. 57, м—р). Антарктический
циркумполярный батипелагический
вид. Длина мантии более 7 см.
2(1). Плавники узко-обратно-серд-
цевидныс или в виде узкой боковой
каймы, ширина плавника гораздо
меньше его длины. Щупальца длин-
ные, крепкие. Присоски средних ря-
дов центральной части булавы не
крупнее краевых. Зона мелких прок-
симальных присосок занимает подав-
ляющую часть длины булавы. Фикса-
торного аппарата и крючьев на ру-
ках нет.
Род Neoteuthis Naef, 1921 (рис.
57, с—ф). Типовой и единственный
описанный вид — /V. thielei Naef, 1921.
2 вида..........................3.
3(4). Плавник постепенно расши-
ряется к заднему концу и заходит за
задний конец мантии, наибольшая
ширина плавника — у заднего края.
N. thielei Naef, 1921 (рис. 57,с).
Тропические и субтропические воды
Атлантики, Карибское море. Мезо-ба-
типелагический вид. Длина мантии
до 17 см.
4(3). Плавник в виде узкой боко-
вой каймы, не заходит за задний
край мантии, наибольшая ширина
плавника в его середине.
Neoieuthis sp. Young, 1972 (рис.
57, т—ф). Восточная часть Пацифи-
ки от п-ва Калифорния до экватора
и у Гавайских островов. Мезо- и ба-
типелагиаль.
Кальмары этого рода были добы-
ты также в нотальных водах Тихого
океана восточнее Новой Зеландии
(Young, 1972).
Примечание. К. Ропер,
Р. Янг и Г. Восс (Roper, Young, Voss,
1969) включают в семейство Neoteut-
thidae антарктический вид Parateuthis
tunicata Thiele, 1920, считая его пред-
положительно личинкой Л. antarctica.
По нашим данным, Р. tunicata — са-
мостоятельный вид, не относящийся
к семейству Ncoteuthidae (см. с. 254).
2.2.2.12. Семейство
Brachioteuthidae
Небольшие кальмары нектонного
внешнего облика, но с тонкой, слабо-
мускулистой мантией, узкой, строй-
ной, сзади заостренной. Голова ма-
ленькая, узкая, у личинок — на длин-
ной шее. Плавник ромбический или
сердцевидный. Руки с двумя рядами
присосок. Булава щупалец в прокси-
мальной части со множеством рядов
мелких присосок, в дистальной — с
4—6 рядами сначала крупных, затем
постепенно мельчающих присосок.
Фиксаторный аппарат вдоль стебля
щупальца. Замыкательные хрящи про-
стые. Формула буккального крепле-
ния DDVV. Гладиус с очень узким
рахисом, расширяющимся лишь в
задней трети, и хорошо развитым уз-
ким фунтиковидным конусом. Фото-
форы отсутствуют или имеются на
вентральной стороне глазного ябло-
ка. Развитие с личиночной стадией.
Длина мантии до 14 см.
Молодые кальмары обитают в
эпи- и мезопелагиали, взрослые пре-
имущественно в мезо- и батпислагиа-
ли, но иногда поднимаются в эпипе-
лагиаль.
Один род — Brachioteuihis Verrill,
1881 ( = Г г achelot euthis Steenstrup,
1882, Verrilliola Pfeffer, 1884, Ento-
mopsis Rochebrunc, 1884) (рис. 58).
Типовой вид — В, beani Verrill, 1881.
4 вида, из них один не описан, еще
два вида сомнительные.
Таблица для определения видов рода
Brachioteuthis
1 (2). Мантия очень узкая, слегка
расширяется у переднего края и рез-
ко сужается перед плавником. Плав-
ник неправильно-продольно-ромбиче-
ский, его ширина и длина составляют
примерно 7г длины мантии, отноше-
ние ширины к длине 0,9—1,1.
Имеется удлиненный колбасовидный
фотофор на вентральной стороне
глазного яблока. Окраска светло-
пурпурно-коричневая или шоколад-
ная. Кожа пе имеет сетчато-волокни-
стой структуры.
В. picta Chun, 1910 (рис. 58, а).
Экваториальные и южные субтропи-
ческие воды и южная перифериче-
ская зона Атлантического, Индий-
ского и Тихого океанов, на юге до
40—45° го. ш. Мезо- и батипелагиаль,
молодь в эпипслагиали. Длина ман-
тии до 8 см.
2(1). Мантия цилиндрической
формы, без резкого сужения перед
плавником. Плавник поперечно-ром-
бический или широкосердцевидный,
не оттянут или слабо оттянут назад,
отношение ширины к длине плавника
обычно 1,2—1,4..................3.
3(4). Длина плавника 35—50%,
ширина 45—60% длины мантии.
Мантия почти бесцветна, с мелкими
редкими хроматофорами. Глазные
фотофоры имеются или отсутствуют.
В. riisei (Steenstrup, 1882) (рис.
58,б—к). По существующим пред-
ставлениям, космополитический вид,
распространенный главным образом в
продуктивных водах (кроме бореаль-
ной Пацифики). В Атлантике рас-
пространен от южной части Норвеж-
ского моря и Исландии до крайнего
юга, имеется в Средиземном морс,
но отсутствует в тропических водах
Западной Атлантики, Мексиканском
заливе и Карибском морс; в Индий-
ском оксане обитает почти повсеме-
стно, кроме Аравийского моря и се-
верной части Бенгальского залива;
в Тихом океане — к югу от северной
границы центральных вод; в Южном
оксане циркумглобально. Многочис-
лен в Северной Атлантике, Субан-
тарктике и Антарктике, редок, в тро-
пических водах. Встречается в эпи-
и мезопелагиали, взрослые особи —
также в батипелагиали. Длина ман-
тии до 8 см (у антарктических каль-
215
к
I
Рис. 58. Сем. Brachioteuthidae, род Brachio-
teuthis: а — В. picta\ б—к — В. nisei: б —
в — из Северной Атлантики, г—е— из Сре-
диземного моря, ж—з — из тропической
Пацифики (В. behni); и—к— из моря Ско-
тин; л—.и — В. beani; н — В. bowmani; а —
б, г—д, ж, и—к — молодые кальмары; в,
з—личинки; е— булава; лс —гладиус (а—
по Chun, 1910; б—в — по Muus, 1959; г—е,
и—к —по Pfeffer, 1912; ж—з — по Okutani,
1966, 1974; л—я — по Verrill, 1882;. я—по
Russell, 1922).
маров до 14 см). Вид сборный. Сре-
ди личинок и молоди достаточно чет-
ко выделяются 2 типа:
а. Шея умеренной длины. Глаза
направлены вперед и вбок под ост-
рым углом к продольной оси тела.
B.riisei (Steenstrup, 1882) [=И.
nytnpha (Pfeffer, 1884), В, velaini
(Rochebrune, 1884)] (рис. 58, б—е).
Северная Атлантика, Средиземное
море, умеренные воды южных частей
Атлантического, Индийского и Тихо-
го оксанов, тропические воды восточ-
ной части Тихого океана. Взрослые
кальмары этой формы имеют глазные
фотофоры.
б. Шея у личинок чрезвычайно
длинная. Глаза направлены под пря-
мым углом к продольной оси тела.
Фотофоров нет.
В. behni (Steenstrup, 1882) [=В.
gracilis (Pfeffer, 1884),? В. clouei
(Rochebrune, 1884)] (рис. 58, жу з).
Экваториальные воды Атлантики, эк-
ваториальные и центральные воды
Индо-Пацифики.
Нотально-аптарктические «5. rii-
seb>, по-видимому, также представля-
ют собой особый вид (рис. 58, ц, к).
Глазных фотофоров у них нет, кожа
взрослых особей (длина мантии бо-
лее 6 см) под эпидермисом имеет сет-
чато-волокнистую структуру, обус-
ловленную переплетением соедини-
тельнотканных волокон (заметно
лишь у очень хорошо сохранившихся
экземпляров).
4(3). Длина плавника 50—55%,
ширина 60—67% длины мантии. Ок-
раска интенсивно-пурпурно-коричне-
вая. Фотофоров нет (?).
В. beani Verrill, 1881 (рис. 58, л, м)
(Северо-Западная Атлантика у вос-
точного побережья США) и В. bow-
mani Russell, 1909 (рис. 58. н) (Севе-
ро-Восточная Атлантика у Фареро-
Исландского порога). Длина мантий
до 6 см. Оба вида ловились у дна в
батиали. Их систематическое поло-
жение неясно.
2.2.2.13. Семейство
Ommastrephidae
Быстрые и сильные нектонные каль-
мары преимущественно средних и
крупных размеров. Мантия мускули-
стая, цилиндрической или конической
формы, плавник конечный, попереч-
но-ромбический или сердцевидный.
218
Голова крупная. Руки мощные, с хо-
рошо выраженными защитными мем-
бранами и плавательными килями,
несут два ряда присосок с зубчаты-
ми кольцами. Щупальца с расширен-
ной булавой, несущей 4 ряда присо-
сок (у Шех па конце булавы — 8 ря-
дов); присоски средних рядов в цен-
тральной части булавы сильно уве-
личены. Фиксаторный аппарат корот-
кий. Вороночная ямка (рис. 59, а)
треугольная, ее передняя часть у
многих родов отделена дугообразной
поперечной складкой кожи (фовеола)
и иногда несет продольные кожные
гребешки; за фовеолой, ближе к
хвосту, по обе стороны вороночной
ямки может быть еще несколько ма-
леньких дугообразных кожных кар-
машков. Вороночные хрящи с двумя
глубокими бороздами, пересекающи-
мися в виде перевернутой буквы
Т(_1_), мантийные состоят из двух пе-
ресекающихся гребней: поперечного
и продольного, соответствующих
форме вороночного хряща. Формула
буккального крепления DDVD.
У многих видов имеются светя-
щиеся органы — тысячи крохотных
желтых зернышек неправильной фор-
мы, разбросанных в толще мышц
маптии, головы и рук, иногда кон-
центрирующихся в округлые или
овальные пятна или продольные по-
лосы; у ряда видов есть фотофоры
на глазном яблоке и внутри мантии.
Гладиус узкий, без пера, сужа-
ющийся назад и слегка расширяю-
щийся у заднего конца, с небольшим
конечным конусом. Гектокотилизи-
руются одна или обе брюшные руки.
Сперматофоры переносятся на рото-
вую мембрану, реже — в мантийную
полость самки. Яйца довольно мел-
кие, кладка донная или пелагическая.
Личинка — рипхотсйтис — имеет хо-
боток из сросшихся вместе щупалец,
в конце стадии личинки хоботок рас-
щепляется, начиная с основания, и
щупальца обособляются.
Оммастрефиды обитают у поверх-
ности, в толще воды или у дна в
средней и нижней сублиторали и в
батиали. Большинство видов широко
эврибатны и совершают суточные
вертикальные миграции, поднимаясь
ночью к поверхности. Распростране-
ны от высокобореальных (изредка
даже арктических) до потальных
вод. Большинство видов мигрируют
на далекие расстояния (до лесколь-
ких тысяч миль) для нагула и нере-
ста. Хищники питаются мелкой ры-
бой, макропланктонпыми ракообраз-
ными и мелкими кальмарами.
Самое важное в промысловом от-
ношении семейство головоногих мол-
люсков. В умеренных широтах омма-
стрефиды доминируют в мировом
промысле кальмаров, перспективен их
промысел в некоторых районах тро-
пиков.
4 подсемейства, 11 родов, 17—18
видов, 10—11 подвидов. Обзоры:
Зуев и др., 1975; Wormuth, 1976;
Нигматуллин; 1979.
Таблица для определения
подсемейств и родов Ommastrephidae
1(12). Вороночная ямка (рис. 59, а)
без фовеолы или с фовеолой, но без
боковых кармашков. Вороночный
хрящ короткий, с почти прямой про-
дольной бороздой. Фиксаторный ап-
парат щупалец слабо развит, без
бугорков-кнопок. Кольца крупных
присосок средней части булавы без
крупных зубцов, выделяющихся сво-
ими размерами...................2.
2(5). Вороночная ямка простая,
без фовеолы. Карпальная группа
присосок на булаве короткая. Светя-
щиеся органы отсутствуют.
Подсемейство Illicinae . . 3.
3(4). На конце булавы 8 попереч-
ных рядов мелких присосок. Шири-
на мантии в 4—6 раз меньше длины.
Плавник сердцевидный или узко-ром-
бический, его ширина 45—60% дли-
ны мантии. Гсктокотилизируется од-
на из брюшных рук.
Род Illex Steenstrup, 1880 (рис.
59,6—и). Типовой вид — I. illecebro-
sus (LeSueur, 1821). 2 вида и 3 под-
вида в Атлантическом океане от Юж-
ной Гренландии и Исландии до Фол-
клендских островов и Намибии, в
Карибском и Средиземном морях. Оби-
тают у дна в средней и нижней суб-
литорали и верхней батиали, в уме-
ренных широтах во время нагула
иногда поднимаются к поверхности
и подходят к берегам. Нерест проис-
ходит па дне. Молодь обитает в пе-
лагиали, иногда вдали от берегов.
Род. имеет важное промысловое зна-
чение, главным образом в Северо-За-
падной, Юго-Западной и Восточно-
Центральной Атлантике и Средизем-
ном море. Обзор: Roper, Lu, Mangold,
1969.
4(3). На конце булавы 4 попереч-
ных ряда присосок. Ширина мантии
втрое меньше длины. Плавник широ-
коромбический, его ширина 75—85%
длины мантии. Гектокотилизируются
обе брюшные руки.
Род Todaropsis Girard, 1890. Один
вид — Т. eblanae (Ball, 1841) (рис.
59, к—м). Восточная Атлантика от
Шетландских островов до мыса Доб-
рой Надежды, Индийский океан —
банка Агульяс, Маскарепский хребет,
Тиморское море. У дна в нижней суб-
литорали и верхней батиали, к по-
верхности не поднимается и к бере-
гам не подходит. Длина мантии до
27 см. Имеет небольшое промысло-
вое значение в Северо-Восточной Ат-
лантике.
5(2). Вороночная ямка (рис.
59, а) с фовеолой, как правило, с
продольными кожными гребешками
(не всегда заметны у Ornithoteuthis),
без боковых кармашков. Нет мелких
подкожных фотофоров ... 6.
6(11). Мантия цилиндрическая.
Плавник ромбический, слегка оттянут
назад, его длина меньше или равна
У2 длины мантии. Светящиеся орга-
ны отсутствуют. Карпальная группа
присосок на булаве растянута и за-
нимает значительную часть стебля.
Подсемейство Todarodinae . 7.
7(10). На стебле щупальца про-
ксимальнее первых присосок нет
пальцевидных выростов. По краям
рук и булавы нет усиковидных выро-
стов, выступающих за пределы за-
щитных мембран ...... 8.
8(9). Гсктокотилизируется одна
из брюшных рук (правая, редко ле-
вая). Виды обитают в умеренных и
холодных водах, отсутствуют в тро-
пических.
Род Todarodes Steenstrup, 1880
[ = Omtnastrephes или Omtnalostrephes
auett., non d’Orbigny; Sagitiatus Ris-
so, 1954] (рис. 59, н—с). Типовой
вид — Т. sagittatus (Lamarck, 1799).
3 вида, встречающиеся как у дна,
так и в пелагиали. Имеют большое
промысловое значение.
9(8). Гектокотилизируются обе
брюшные руки, одна — в проксималь-
ной и дистальной, другая только в
проксимальной части. Виды распро-
странены в тропических и субтропи-
ческих водах Ипдо-Вестпацифики и
Австрало-Новозеландском районе.
Род Nototodarus Pfeffer, 1912
(рис. 60, а—д). Типовой вид —
219
5
77
й
в
fe
Рис. 59. Сем. Ommastrephidae, роды Шех
(б—и), Todaropsis {к—м} и Todarodes («—
с): а — вороночная ямка Illiclnae (/), То-
darodinae (2) и Ommastrephinae (5); 4 —
отверстие воронки; 5 — вороночная ямка
без фовеолы; 6 — фовеола; 7 — полулун-
ная складка; 8 — боковые кармашки; б —
Шех illecebrosus illecebrosus (самец); в —
e—I. illecebrosus coindeti', самка (в), бу-
лава (а), гектокотиль (д), вороночный
хрящ (а), ж—u — I. oxygonius самец (ж),
мантия снизу (з; 1 — угол при вершине
плавника), гектокотиль (и), к—ле — Toda-
ropsis eblanae молодой кальмар (к),
взрослая самка (л), гектокотиль (.и); к—
п — Todarodes pacificus, самец («), гекто-
котиль (о), присоски булавы (л) и руки
3-й пары (/21); р—с — Т. sagittalus, самка
(р) и булава (с) (а, в—г, к, р—с — по Mu-
ns, 1959; б — по Verrill, 1882; е — по Pfef-
fer, 1912; д, ж— и — по Roper, 1977; л—м —
по Adam, 1952; н, л — по Voss, Williamson,
1972; о —по Иванову, Стрелкову, 1949).
N.sloani (Gray, 1849). 2—3 вида,
один из них с 4—5 подвидами. У дна
в нижней сублиторали и верхней ба-
тиали, иногда поднимаются к поверх-
ности, к берегам подходят редко.
Длина мантии до 45 см. Имеют су-
щественное промысловое значение.
10(7). В проксимальной части бу-
лавы от первых присосок почти до
основания стебля щупалец протяги-
вается двойной ряд пальцевидных
выростов. Защитные мембраны рук
и щупалец короткие, их трабекулы
длиннее мембран и торчат по краям
рук и булавы, как усиковидные вы-
росты, у оснований которых сидят
присоски. Гсктокотилизируется одна
из брюшных рук.
Род Martialia Rochebrune ct Ма-
bille, 1887. Один вид — Af. hyadesi
Roch. et Mab., 1887 (рис. 60, e—з).
Патагонский район и Огненная Зем-
ля, от залива Ла-Плата до мыса Горн
Ю. Георгия; Южная Пацифика (раз-
лом Элтапин). У дна в нижней суб-
литорали и верхней батиали и в тол-
ще воды. Длина мантии до 45 см.
11 (6). Мантия узкая, коническая,
сзади оттянута в тонкий хвост. Плав-
ник узкосердцевидный, сильно оття-
нут назад, его длина больше у2 Д;ш’
ны мантии. На внутренностях 2 фо-
тофора: крупный округлый блиа
ануса и небольшой овальный у зад-
него конца кишки, от него у взрос-
лых кальмаров отходит узкая по-
лоска светящейся ткани, тянущаяся с
2 перерывами (для прохода арте-
рий) до заднего конца тела. По од-
ному фотофору па глазном яблоке.
Карпальная группа присосок на бу-
лаве короткая.
Подсемейство Ornithoteuthinae.
Род Ornithoieuthis Okada, 1927 (рис.
60, и—р). Типовой вид—O.volatilis
(Sasaki, 1915). 2 очень близких пе-
рито-оксанических вида в тропиче-
ских водах Атлантики и Индо-Вест-
пацифики. Молодь обитает в толще
воды, взрослые кальмары — в ниж-
ней эпи- и мезопелагиали и у дна а
батиали.
12(1). Вороночная ямка (рис.
59, а) с фовеолой, продольными кож-
ными гребешками и боковыми кар*
машками (последние иногда отсутст-
вуют у Eucleoteuthis и Hyaloteuihis).
Вороночный хрящ удлиненный, его
продольная бороздка в передней ча-
сти слегка изогнута, соответственно
/ 2 3^
р
Рис. 60. Роды Nototodarus (а—3), Martialia
(е—з) и Ornithoieuthis (и—р): a — N. slo-
ani sloani (самка); б—N. s. gouldi (гекто-
котиль); в—N. s. hawaiiensis (самка); г—
д — N. s. philippinensis, самец (г), присос-
ки булавы (б) и руки 3-й пары (6t); е—
з — Martialia hyadesi, основание булавы
(<?), присоска и трабекула брюшной руки
[ж), присоска булавы (з), вороночная
ямка (3|); и—о — О. aniillarum, самец (и),
гсктокотиль (к), булава (л), вороночная
ямка (ж), глазной фотофор («), присоска
вентрального ряда у основания 2-й руки
самца (о), п—р — О. volaiilis, самка (л) и
вскрытая мантия (р), видны фотофоры
U* 2) и светящаяся полоска ткани (3) (а —
па Pfeffer, 1912; б — по Berry, 1918; в, fi-
ne Wormuth. 1976; г—б— по Voss, William-
son, 1972; е—з — по Castellanos, 1967; и—
о — по G. Voss, 1957; р — по Rancurcl,
1970)
223
слегка искривлена и передняя часть
мантийного хряща. Фиксаторный ап-
парат с одной или несколькими бу-
горками-кнопками, перемежающими-
ся с присосками с гладкими кольца-
ми. Кольца крупных присосок була-
вы либо с 1 крупным зубцом в дис-
тальной части, либо с 4 крупными
зубцами, расположенными крест-на-
крест как бы по углам квадрата.
Есть мелкие подкожные фотофоры
на голове, «мантии, плавниках и ру-
ках, часто есть также глазные, внут-
римантийные и крупные поверхност-
ные фотофоры.
Подсемейство Ommastrephinae. 13.
13(16). Нет крупных фотофоров
на глазном яблоке, кишке и поверх-
ности тела. Мантия цилиндрическая,
плавник ромбический с не оттянутым
или слегка оттянутым концом. Коль-
ца крупных присосок булавы с 4
крупными зубцами крест-накрест. 14.
14(15). Концы всех рук у взрос-
лых кальмаров утончены и плетевид-
но вытянуты, несут множество (до
100—200 пар) мелких присосок. Коп-
цы трабекул защитных мембран вы-
ступают за края мембран в виде уз-
ких треугольных или усиковидных
выростов. Вентральная защитная
мембрана 3-й пары рук узкая, не пре-
вышает ширину руки. В карпальной
части булавы проксимальнее первого
бугорка-кнопки нет присосок с зуб-
чатыми кольцами. Плавник ромбиче-
ский, его конец не оттянут.
Род Dosidicus Steenstrup, 1857
(рис. 61,а—г). Один вид — D. gigas
(d’Orbigny, 1835) — псруано-чилий-
ский гигантский кальмар. Восточная
часть Тихого океана от п-ва Кали-
форния и Калифорнийского залива
до Центрального Чили, в отдельные
годы на севере доходит до залива
Монтерей (Калифорния), на юге —
до острова Чилоэ. На западе в эква-
ториальном районе распространен до
125—140° з. д. Обитает у поверхно-
сти и в толще воды (эпи- и мезопе-
лагиаль) открытого оксана, на нагу-
ле иногда массами подходит к бере-
гам. Длина мантии до 120 см. Имеет
небольшое промысловое значение, но
перспективы промысла у Перу и Чи-
ли очень велики.
15(14). Концы рук не оттянуты,
па руках около 35 пар присосок.
Рис. 61. Подсем. Ommastrephinae: а—а —
Dosidicus gigas, взрослая (а) и молодая
(б) самки, гектокотиль (в), основание (а)
и кончик (2|) боковой руки; д-ж — От-
mastrephes bartratni, сверху, снизу, булава
(ж), з—и — Sthenoteuthis pteropus, самка
(ф — фотофор) и основание булавы (и);
/с—м — S. oualaniensis, самка, присоски
булавы (л) и руки 3-й пары (Л1), сраста-
ние вороночного (слева) и мантийного
хряща (м); н—о — Eucteoteuthis I и mi nos а
(на о черным показаны фотофоры); л —
Hyaloieuthis pelagica (а, г—д, м — по
Pfeffer, 1912; б—в, л — по Wormuth, 1976;
е—ж, н—о —по R. Young, 1972; з—и — по
Adam, 1952; к—л — по Voss, Williamson,
1972)
15 К. Н. Нссис
225
Концы трабекул защитных мембран
не выступают за их края. Вентраль-
ная защитная мембрана 3-й пары рук
очень широкая, у взрослых самок
вытянута в большую треугольную
лопасть. В карпальной части булавы
проксимальнее первого бугорка-кноп-
ки 4—7 присосок с зубчатыми коль-
цами. Конец плавника слегка оття-
нут назад. Вдоль брюшной стороны
мантии почти от переднего края и до
уровня начала плавника тянется ши-
рокая серебристая продольная по-
лоска.
Род Ommastrephes d’Orbigny, 1839
( = ? Cycria Leach in Gray, 1849, Lo-
litnnites Risso, 1854).
Один вид — О. bartrami (LeSueur,
1821) (рис. 61,(5—ж) с тремя фор-
мально не описанными подвидами.
Распространение. бисубтропичсское
(антиэкваториальное): североатлан-
тический подвид — от Большой Нью-
фаундлендской банки и Северного
моря до п-ва Флориды, юга Сар-
гассова моря и Мавритании, северо-
тихоокеанский подвид — от Южных
Курильских островов (изредка — Ко-
мандорских и Западных Алеутских
островов) и юга Британской Колум-
бии до островов Тайвань, Бонин и
Западной Мексики, южный подвид —
от Бразилии и района мористее
Фолклендских островов до Анголы
и Южной Африки и далее на восток
между 20—25 и 35—40° ю. ш. до
Большого Австралийского залива
(135° в. д.) и в юго-западной части
Тихого океана от Новой Каледонии и
Тасмании до 150° з. д., возможно и
восточнее. Длина мантии до 76 см
(североатлантический подвид). Мно-
гочисленный эпи-мезопелагический
вид, добывается в больших количест-
вах в Курило-Хоккайдском районе.
16(13). Есть фотофоры на брюш-
ной стороне глазного яблока (по 1)
и на кишке (1 или 2), обычно также
имеются крупные поверхностные фо-
тофоры на брюшной или спинной
стороне мантии. В карпальной части
булавы% проксимальнее первого бу-
горка-кнопки не более 2 присосок с
зубчатыми кольцами. Нет серебри-
стой полоски на брюхе . . . 17.
17(18). Два фотофора па кишке—
у ануса и близ заднего конца. Нет
светящихся пятен и полос на брюш-
ной стороне мантии, головы и рук
4-пары. У взрослых обычно имеется
крупный овальный фотофор на спи-
226
не в передней части мантии («на за-
гривке»). Кольца крупных присосок
булавы с 4 крупными зубцами крест-
накрест. Мантия цилиндрическая,
плавник ромбический, его конец не
оттянут. Вентральная защитная мем-
брана на 3-й паре рук примерно рав-
на ширине руки.
Род Sihenoteuthis Verrill, 1880
(=Symplectoteuthis Pfeffer, 1900)
(рис. 61,з—ле). Типовой вид — S. pte-
ropus (Steenstrup, 1855). 2 вида в
эпипелагиали тропической Атлантики
п Индо-Пацифики.
18(17). Один фотофор на кишке,
на границе ее средней и задней тре-
ти. Имеются светящиеся пятна и
(или) полосы на брюшной стороне
мантии, головы и брюшных рук, пет
спинного фотофора. Кольца крупных
присосок ‘ булавы с одним крупным
зубцом на дистальном крае. Защит-
ные мембраны 3-й пары рук уз-
кие ...........................19.
19(20). Вдоль брюшной стороны
мантии по обе стороны от середины
протягиваются 2 длинные полоски
светящейся ткани, разделенные про-
светом на уровне наибольшей шири-
ны плавника; по 2 овальных светя-
щихся пятна у переднего края ман-
тии по бокам брюшной стороны, по
1 у основания и в середине брюш-
ных рук. Мантия коническая, сужа-
ется назад, плавник сердцевидный, с
оттянутым концом. Мантийный и во-
роночный хрящи у молоди и взрос-
лых срастаются.
Род Eucleoteuthis Berry 1916.
Один вид:— Е. luminosa (Sasaki,
1915) (рис. 61, я, о). Бипериферичс-
ский вид, распространён в северной
части Тихого океана полосой от Юж-
ных Курильских островов и Японии
(25—45° с. ш.) до Калифорнии и За-
падной Мексики (15—40° с. ш.), в
южных частях Атлантического, Ин-
дийского и юго-западной части Ти-
хого океана (в Атлантическом океане
между 10 и 35° ю. ш., в Индийском
и Тихом океанах между 15 и 35°
ю. ш.), на восток проникает до 80°
з. д. Эпипелагиаль. Длина мантии до
22 см.
20(19). На брюшной стороне ман-
тии в строгом порядке располагают-
ся 19 округлых светящихся пятен,
образующих сближенные или раздви-
нутые пары; по 1 пятну у основания,
в середине и близ концов брюшных
рук. Мантия цилиндрическая, плавник
ромбический, с прямыми задними
краями. Мантийный и вороночный
хрящи не срастаются.
Род Hyaloteuthis Gray 1849. Один
вид — Н. pelagica (Bose, 1802) (рис.
61, л). Океанический бицентральный
вид: Атлантический, Тихий и, веро-
ятно, Индийский океаны, па востоке
Тихого океана встречается и в эква-
ториальных водах. Эпи- и верхняя мс-
зопелагиаль, • иногда батипелагиаль.
Длина мантии до 9 см.
Таблица для определения видов рода
Шек
1(6). Плавник широкий с более или
менее тупым хвостом; ширина плав-
ника в 1,3—1,5 раза больше его дли-
ны; угол, образованный задними
краями плавника, прямой или тупой
(80—100° и более). Ширина головы
не больше ширины мантии у перед-
него края. Длина наибольшей руки у
самцов 50—90%, у самок 45—60%
длины мантии.
I. illecebrosus (LeSucur 1821) (см.
рис. 59,6—е). Три аллопатрических
подвида.........................2.
2(3). Плавник сердцевидный, его
ширина в 1,3—1,4 раза больше дли-
ны, угол при вершине плавника пря-
мой (80—100°). Относительная дли-
на рук у самцов и самок одинакова,
наибольшая рука — 45—55% длины
мантии. Присоски на боковых руках
самцов не увеличены, их диаметр
1,5—1,8% длины мантии. Длина гек-
токотиля 15—25% гектокотилизиро-
ваннон руки:
I. i. illecebrosus (LeSueur, 1821)
(см. рис. 59, б). Северо-Западная Ат-
лантика. Северная граница в период
нагула — Лабрадор, в период нерес-
та — Новая Шотландия, южная — у
Восточной Флориды, восточная — на
Срединно-Атлантическом хребте. Из-
редка встречается у Южной Грен-
ландии, Исландии, на банке Поркью-
пайн, в Северном море. Во время на-
гула на севере ареала обитает у по-
верхности, часто подходит к берегам,
в других районах держится у дна на
глубинах до 700 м (обычно 100—
500 эд). Длина мантии до 31 см.
В больших количествах добывается у
Ньюфаундленда, Новой Шотландии,
на Джорджсс-банке.
3(2). Плавник ромбический, его
ширина в 1,4—1,5 раза больше дли-
ны, угол при вершине обычно ту-
пой— 90—100° и больше. Длина наи-
большей руки у самцов 55—90%,
у самок 48—57% длины мантии. При-
соски боковых рук самца заметно
увеличены, их диаметр 3—3,5% дли-
ны мантии. Длина гсктокотиля не ме-
нее 30% гектокотилизированной ру-
ки .............................4.
4(5). Длина гсктокотиля Vs—2/з
гектокотилизированной руки.
I. i. argentinus (Castellanos, 1960).
Юго-Западная Атлантика от Рио-де-
Жанейро до Фолклендских островов.
У дна в нижней сублиторали и ба-
тиали (до 1350 м), иногда у поверх-
ности, молодь — у берегов. Длина
мантии до 38 "см. Добывается в
больших количествах на Патагонском
шельфе и у Уругвая.
5(4). Длина гектокотиля около ]/з
гектокотилизированной руки.
I. i. coindeti (Verany, 1837) (см.
рис. 59, в—в). Восточная Атлантика
от Южной Англии до Намибии,
Средиземное море, тропическая За-
падная Атлантика, Мексиканский за-
лив п Карибское морс — от мыса
Хаттерас до Венесуэлы; отмечен с
сомнением в КраснОхМ морс. Обитает
у дна на глубинах от 40—50 до
500—600 м и в толще воды. Длина
мантии до 26 см. Добывается в не-
больших количествах в Средиземном
морс и у Северо-Западной Африки.
6(1). Плавник узкий, оттянут в
длинный острый хвост. Ширина
плавника в 1,1 раза больше его дли-
ны; угол при вершине плавника
(между его задними краями) острый
(50—70°, редко до 80°). Голова ши-
ре мантии у переднего края. Руки
короткие, длина наибольшей руки у
самцов 45—55 %, у самок 35—45 %
длины мантии. Длина гектокотиля
около 1/з длины гектокотилпзирован-
ной руки.
/. oxygomtis Roper, Lu, Mangold,
1969 (см. рис. 59, at—и). Тропические
и субтропические воды Западной
Атлантики от залива Делавэр до
Кубы. Отмечен в Гвинейском заливе.
У дна на глубинах 50—600 м. Длина
мантии до 24 см.
Таблица для определения видов рода
Todarodes
1(2). Плавник ромбический, нс вытя-
нут в хвост, его длина около 40%
длины мантии. Булава короткая и
узкая с 10—12 мелкими присосками
15*
227
в проксимальной части и 8—9 пара-
ми крупных присосок в средних ря-
дах центральной части.
Т. pacificus Steenstrup, 1880 (упо-
треблявшееся ранее название От-
mastrephes sloani pacificus ошибоч-
ное) (см. рис. 59, н—п). Северо-за-
падная часть Тихого океана, Охот-
ское, Японское, Желтое (южная
часть), Восточно-Китайское моря и
северная часть Южно-Китайского
моря Северная граница в период
нагула — Восточная Камчатка (Ко-
мандорские острова?), центральная
часть Охотского моря и северная
часть Татарского пролива, южная
граница — близ Гонконга. Эпипелаги-
аль над шельфом и склоном, а в про-
дуктивных водах — и над большими
глубинами, иногда опускается ко дну
(для откорма и откладки яиц). Длина
мантии до 35 см. Важнейший объект
промысла в Японии, СССР, КНДР и
Южной Корее.
2(1). Плавник сердцевидный, сза-
ди вытянут в хвост, длина плавника
около половины длины мантии. Бу-
лава длинная и широкая, в средних
рядах центральной части булавы
12—15 пар крупных присосок . 3.
3(4). Несущая присоски часть
щупальца составляет 3А—5/б общей
длины щупальца, в проксимальной
части булавы 10—12 пар мелких при-
сосок, в центральной части 14—15
пар крупных присосок.
Т. sagittaius (Lamarck, 1799) (см.
рис. 59, р, с). Северо-Восточная Ат-
лантика и прилегающие воды. На се-
вер во время нагула заходит до
Баренцева моря (изредка до Карско-
го и Белого морей) и Исландии, на
юг — до Гвинеи и Канарских остро-
вов, на запад — до Бермудских ост-
ровов и района к югу от Новой
Шотландии (банки Кельвин), обы-
чен в Средиземном море. В откры-
том оксане и у берегов, у поверх-
ности и у дна на глубинах до 2500 м,
«о чаще всего над склоном и у дна
на склоне. Длина мантии до 60—
65 (75) см, обычно до 30—40 см.
В отдельные годы добывается в зна-
чительных количествах в Северной
Европе и средиземноморских странах.
4(3). Несущая присоски часть
щупальца составляет 2/3—3/4 его об-
щей длины. В проксимальной части
булавы 2—5 пар мелких присосок, в
центральной части 12—14 пар круп-
ных присосок.
228
Т. angolensis Adam, 1962 ( = Т. fi-
lippovae Adam, 1975). Потальный цир-
кумглобальный вид, в районах хо-
лодных течений заходит на север —
у Африки до Анголы, у Новой Зе-
ландии— до бухты Каипара, у Аме-
рики до Южного Перу. Молодь оби-
тает в эпипелагиали и, по-видимому,
также в мезопелагиали, взрослые —
у дна на склоне. Встречается как у
берегов, так и в открытом океане.
Длина мантии до 50 см.
Таблица для определения видов рода
Nototodarus
1(2). Поверхность тела гладкая.
Мантия стройная, ее ширина в сред-
нем Vo—’Д длины. Плавник относи-
тельно узкий, его ширина менее 70%
длины мантии. Угол, образованный
задними краями плавника, близок к
прямому (90—110°).
N. sloani (Gray, 1849) (рис. 60, а—
д). 4 подвида. V. sloani sloani (Gray,
1849) (рис. 60, а) обитает у Новой
Зеландии, островов Окленд, Кэмп-
белл, Чатем, длина мантии до 40—
45 см. N. sloani gouldi (/McCoy, 1888)
(рис. 60, б) распространен у Юго-
Западной, Южной и Юго-Восточной
Австралии, Тасмании и (?) в восточ-
ной части Индийского океана, длина
мантии до 33 см. N. sloani philippinen-
sis Voss, 1962 (рис. 60, в—д) рас-
пространен в Южно-Китайском
море, на юге Восточно-Китайского
моря и у Филиппин, длина мантии
до 18 см. N. sloani hawaiiensis (Ber-
ry, 1912) обитает у островов Гавай-
ских и Мидуэй, длина мантии до
16 см. Кальмары держатся у дна в
нижней сублиторали и верхней
батиали, но в период нагула подни-
маются к поверхности и иногда под-
ходят к берегам. Имеют большое
промысловое значение в новозеланд-
ских и временами тасманийских во-
дах и незначительное — в других
районах. Особый, не описанный под-
вид распространен в северной и за-
падной частях Индийского океана —
от Андаманского до Аравийского
моря, у Восточной Африки и на
Маскаренском хребте, длина мантии
до 29 см. Эти кальмары похожи на
N. s. philippinensis, но отличаются бо-
лее длинными, чем у большинства
N, sloani, руками (длина наибольшей
руки 7г—длины мантии) и более
широкой ГОЛОВОЙ (^4—Уз длины ман-
тии).
2(1). Поверхность мантии и голо-
вы на ощупь шершавые» кожа покры-
та мозаикой мельчайших бугорков и
чешуек. Ширина мантии около Уз ее
длины. Ширина плавника 70—90%
длины мантии. Угол, образованный
задними краями плавника, тупой
(120—130°). Длина наибольшей руки
У2—3А длины мантии.
N, nipponicus jOkutani et Uemura,
1973. Обнаружен у юго-восточного
побережья Хонсю и в Восточно-Ки-
тайском морс, у дна на глубинах
100—250 м. Длина мантии до 27 см.
В небольшом количестве придавлива-
ется к Todarodes pacificus.
Примечание. В таблицу не
включен плохо описанный вид
N. insignis (Gould, 1846) с островов
Фиджи (длина мантии 25 см). По-ви-
димому, это одна из форм N. sloanL
Таблица для определения видов рода
Ornithoteufhis
1(2). В средней части вентральной
стороны гектокотилизированной руки
самца находится сотовидная кожная
скульптура из 4—5 продольных ря-
дов ямок, около 20—25 ямок в ряду.
Ямки двух рядов, расположенных
ближе к оральной (несущей присос-
ки) поверхности руки, значительно
крупнее прочих, а ямки одного-двух
рядов, расположенных ближе к або-
ральной (противоположной) поверх-
ности, более мелкие. Распространен в
Атлантике.
O.antillarum Adam, 1957 (рис.
60, и—о). Амфиатлантический тропи-
ческий вид. Западная Атлантика от
района мористее Джорджес-банки
до Уругвая, Мексиканский залив,
Карибское море; Восточная Атланти-
ка от островов Зеленого Мыса до
Анголы. У дна в батиали и в толще
воды (нижняя эпи- и мезопслагиаль)
над склоном, молодь может встре-
чаться в толще воды над большими
глубинами экваториальной зоны.
Длина мантии до 20 см.
2(1). В средней части вентральной
стороны гектокотилизированной руки
самца гребенчато-сотовидная кожная
скульптура из 2—21/2 продольных ря-
дов небольших ямок, около 10—15 в
ряду, от ближайшего к оральной
стороне руки ряда ямок к оральной
поверхности отходят поперечные
кожные гребни (по 2 от каждой ям-
ки), сливающиеся или разветвляю-
щиеся. Распространен в Иидо^Вест-
пацифике.
О. volatilis (Sasaki, 1915) (см*
рис. 60, п, р). Индо-всстпацифический
тропический вид: от Аравийского
моря до Юго-Восточной Африки, от
Южного Хонсю до Кораллового мо-
ря, па восток доходит до островов
Лайн. Образ жизни, как у предыду-
щего вида. Длина мантии до 22 см.
Таблица для определения видов рода
Sthenoteutliis
1(2). Мантийный и вороночный хря-
щи срастаются между собой на од-
ной или обеих сторонах (на послели-
чпночной стадии). 6—8 пар присо-
сок на гектокотилизированной руке.
Распространен в Тихом и Индийском
оксанах.
S. oualaniensis (Lesson, 1830) (см.
рис. 61, к—м). Индопацифический
узко-тропический вид: от Красного
моря до Мозамбикского пролива и
Наталя (в- теплые годы — до банки
Агульяс), от Южной Японии до
Квинсленда и от юга п-ва Калифор-
ния до Северного Перу. Длина ман-
тии до 35 см. Массовый, потенциаль-
но промысловый эпипелагический
вид. Добывается на островах Рюкю
и Тайвань.
2(1). Мантийный и вороночный
хрящи не срастаются (кроме очень
редких случаев). 11—13 пар присо-
сок па гектокотилизированной руке.
Распространен в Атлантике.
S.pteropus (Steenstrup, 1855) (см.
рис. 61,з, и). Атлантический узкотро-
пический вид: от Флориды* (летом
единично до Джорджес-банки) и
Мексиканского залива до Уругвая и
от Мадейры и Канарских островов
до района южнее Намибии; в Среди-
земное море не заходит. Длина ман-
тии до 60—65 см. Массовый, особен-
но в продуктивных зонах, потен-
циально промысловый эпипелагичес*
кий вид.
2.2.2.14. Семейство
Thysanoteuthidae
Крупные нектонные кальмары с мощ-
ной мускулистой мантией и очень
длинным * продольно-ромбическим
плавником, протягивающимся вдоль
всего тела. Руки короткие, с 2 ряда-
229
ми присосок и сильно развитыми за-
щитными мембранами, растянутыми
па длинных усиковидных трабекулах.
Щупальца с 4 рядами присосок. Фик-
саторный аппарат развит. Вороноч-
ный хрящ с ч -образной бороздой,
мантийный.— с гребнями соответст-
вующей формы. Затылочный хрящ с
двумя буграми, входящими в ямки
па переднем крас мантии. Формула
буккального крепления DDVV. Фото-
форов нет (?). Гладиус напоминает
длинный наконечник копья на корот-
ком древке. Гсктокотилизируется
левая брюшная рука. Кладка пела-
гическая, имеет вид длинного (до
1—2 м) толстого цилиндра, или
«чулка» из прозрачной, полужидкой,
но прочной слизи: в наружном слое
«чулка» спиралью по всей его длине
в один ряд расположены яйца розо-
во-фиолетового цвета. Развитие с ли-
чиночной стадией.
Один род — Thysanoteuthis Tro-
schel, 1857 с единственным видом
Т. rhombus Troschel, 1857 (рис.
62, а—в) — тропическо-субтропичес-
кий космополитический эпипелаги-
ческий вид. Обитает в открытом
океане, к берегам подходит редко,
Имеет небольшое промысловое зна?
челне в Японии. Длина мантии до
100 см.
К семейству Thysanoteuthidae при-
числяют также род Cirrobrachium
Hoyle, 1904 с 2 видами, известными
лишь по молодым особям. Типовой
вид рода — С. filiferum Hoyle, 1904 —
личинка Т. rhombus. Систематическое
положение второго вида С. danae
Joubin, 1933 (он известен по единст-
венному экземпляру с длиной мантии
1,5 см из Центральной Атлантики)
неясно, отличить его от молоди Г.
rhombus можно так:
а (б). Защитные мембраны на ру-
ках 1-й нары очень слабо развиты,
ширина защитных мембран па 2—
3-й парах рук увеличивается к кон-
цу руки, достигая максимума в
дистальной четверти рук.
С. danae Joubin, 1933 (рис. 62, а),
б (а). Защитные мембраны на ру-
ках 1-й пары широкие. Ширина за-
щитных мембран на руках 1—3-й пар
максимальна в середине руки и плав-
но уменьшается к концу. Т. rhombus
Troschel = С. filiferum Hoyle.
280
/7
Н
62. Сем. Thysanoteuthidae (а—г) ц
ж
Рис. ___ _____ ___________________
Lepidoteuthidae (б—о): а—в — Thysanoteut-
his rhombus, общий вид (а), вороночный
(б) и мантийный (61) хрящи, гладиус (в);
г — Cirrobrachiutn clanae; д—е — Lepidoteul-
his grimaldii; ж—u — Tetronychoteuthis
dussutnieri, молодой (д, ж) и взрослый
(е, и) кальмары; к—м — Pholidoteuthis ada-
mi; н—о — Ph. boschmai; и, я, о — мантий-
ные кожные бугорки; л — присоска була-
вы (а —по Naef, 1923; б, з — по Pfeffer,
1912; в —по Sasaki, 1929; г — по Joubin,
1933; д—е, н—о — по Clarke, 1966, 1980; ж —
по Rancurel, 1970; и, м — по Roper е. а.,
1969; к—л — по G. Voss, 1956)
2.2.2.15. Семейство
Lepidoteuthidae
( = Pholidoteuthidae)
Чешуйчатые кальмары — крупные
животные со стройной цилиндричес-
кой мантией, поверхность которой
сверху и снизу, за исключением
участка между плавниками, покрыта
плоскими хрящевидными соедини-
тельнотканными пластинками, похо-
жими на ганоидную чешую прими-
тивных рыб (многоперов, панцир-
ных щук), а в молодости — зубча-
тыми бородавками, напоминающими
паксиллы морских звезд. Плавник
округлый, продольно-овальный, ром-
бический или сердцевидный, сзади
вытянут в короткий или длинный
хвост/ Руки длинные, с двумя ряда-
ми присосок. Щупальца длинные,
тонкие, с 4 рядами присосок; у Lepi-
doteuthis во взрослом состоянии от-
сутствуют. Замыкательные хрящи
простые. Формула буккального креп-
ления DDVV. Гладиус с узким пе-
ром, имеющим обычно перетяжку в
задней трети длины, и более или ме-
нее длинным конечным конусом. Фо-
тофоры не описаны (возможно, име-
ются на глазах у некоторых видов).
Развитие с личиночной стадией.
Придонно-батиальные и мезо-ба-
типелагические кальмары. Довольно
активные глубинные хищники. Игра-
ют заметную роль в питании каша-
лотов, дельфинов, акул. 3 рода, 4
вида.
Таблица для определения родов и
видов семейства Lepidoteuthidae
1(2). Чешуйки крупные, округлые
или неправильно прямоугольные, в
передней части мантии слегка нале-
гают друг на друга подобно чешуе
рыб. Щупальца есть только у личи-
нок и молоди, у взрослых пол-
ностью редуцируются. Плавник у
молоди округлый, у взрослых кальма-
ров удлиненно-овальный, не доходит
до заднего конца мантии, вытянуто-
го в длинный хвост.
Род Lepidoteuthis Joubin, 1895.
Один вид — L.grimaldli Joubin, 1895
(рис. 62, д—е). Тропические и суб-
тропические воды Атлантики (захо-
дит в нотальную зону), южная часть
Индийского и западная часть Тихого
232
океанов (от Хонсю до Новой Кале-
донии, на восток до Гавайских ост-
ровов). Длина мантии до 1 м.
2(1). Чешуйки мелкие, много-
угольные, округлые или зубчатые, у
взрослых соприкасающиеся, но не
налегающие друг па друга, у молоди
не соприкасающиеся. Щупальца хо-
рошо развиты. Плавник ромбический
или сердцевидный, доходит до зад-
него конца мантии ..............3.
3(4). Чешуйки расположены пра-
вильными косыми рядами, сближены,
но не соприкасаются; у молоди они
похожи на звездчатые хрящевые бо-
родавки с 6—9 остриями, у взрослых
округлые. Плавник у молоди почко-
видный, у взрослых ромбический,
нс вытянут или почти нс вытянут в
хвост. Присоски булавы мелкие,
округлые.
Род Тetronychoteuthis Pfeffer,
1900. Один вид — Т. dussumieri (d‘Or-
bigny, 1839) ( = Т. massyae Pfeffer,
1912) (рис. 62, яс—u). Тропические,
субтропические и нотальныс районы
Атлантического, Индийского и Тихого
океанов, бореальные районы Атланти-
ки. Длина мантии 72 см.
4(3). Чешуйки расположены непра-
вильными рядами, они четырех-,
пяти- или шестиугольные, плоские,
без зубцов, с приподнятыми краями,
тесно соприкасающиеся (напомина-
ют блинчатый лед). Плавник ромби-
ческий, вытянут в хвост. Присоски
булавы удлиненно-овальные, как бы
сплющенные с боков. Длина мантии
до 78 см.
Род Pholidoteuthis Adam, 1950
(рис. 62, к—о). Типовой вид —
Р. boschmal Adam, 1950. 2 вида , 5,
5(6). Длина и ширина плавника
у взрослых около половины длины
мантии. Гладиус с перетяжкой в зад-
ней трети.
Р- boschmai Adam, 1950 (рис.
62,«, о). Описан из моря Флорес
(Индонезия). Предположительно
обитает у Австралии и Южной Аф-
рики и на хр. Наска.
6(5). Длина плавника у взрослых
70—75%, ширина 60—70% дли-
ны мантии. Гладиус со слабо
расширенным пером, без пере-
тяжки.
P.adami Voss, 1956 (рис. 62, к—
лс). Тропические и субтропические
воды Атлантики, заходит и в умерен-
ные воды. У дна в батиали, чаще на
глубине 500—2000 м.
2.2.2.16. Семейство
Batoteuthidae
Небольшие кальмары с узкой длин-
ной мантией и .маленьким поперечно-
овальным или сердцевидным плавни-
ком, вытянутым сзади в длинный
тонкий заостренный хвост. Голова
очень маленькая. Руки с двумя ря-
дами присосок. Щупальца без обо-
собленной булавы, несут почти по
всей длине 6 рядов присосок. Замы-
кательный аппарат простой. Форму-
ла буккального крепления DuVV.
Гладиус палочковидный, без пера, с
длинным фунтиковидным конусом.
Внутренности расположены в самой
передней части мантийной полости,
лишь гонада сдвинута в се задний
конец. Развитие с личиночной стади-
ей. По-видимому, малоподвижные
батипелагические кальмары.
Один род — Batoteuthis Young et
Roper, 1968. Единственный вид —
В. skolops Young et Roper, 1968 (рис.
63, a—в). Обнаружен в южной части
Тихого океана, море Скотия и у
Фолклендских островов. Предполо-
жительно нотальный, заходящий в
Антарктику, мезо-батипслагический
вид. Длина мантии до 27 см. Описа-
ние: Young, Roper, 1968.
2.2.2.17. Семейство
Cycloteuthidae
Кальмары средних размеров с широ-
коконической мантией полустуде-
нистой консистенции и большим
округлым плавником, длина которого
превышает 2/з длины мантии. Руки с
2 рядами присосок. Щупальца длин-
ные, булава компактная, расширен-
ная, с 4 рядами присосок и слабо
развитым фиксаторным аппаратом.
Вороночный хрящ треугольный или
неправильно-овальный, с глубокой
ямкой для носовидного или бугорко-
видного мантийного хряща. Форму-
ла буккального крепления DDVV.
Фотофоры имеются, они могут поме-
щаться на глазах, поверхности ман-
тии и головы, чернильном мешке и
др. Гладиус с коротким или длинным
стволом и расширенным пером, сза-
ди закругленным или вытянутым в
более или менее длинный фунтико-
видный конус. Гектокотиля нет. Раз-
витие с личиночной стадией. Длина
мантии до 60 см.
Кальмары обитают в нижней эпи-
и мезопелагиали, иногда у дна в
батиали, распространены в тропиках
и субтропиках всех оксанов.
2 рода, 4—5 видов. Вероятно, к
этому же семейству, по неясно, к
какому роду, относится личинка, опи-
санная из Саргассова моря под наз-
ванием Enoptroteuthis spinicauda
Berry, 1920 (рис. 63,у). Обзор се-
мейства: Young, Roper, 1969 a; Filip-
pova, 1968.
Таблица для определения родов и
видов Cycloteuthidae
1(4). Мантия сзади вытянута в хвост.
Длина плавника 60-—80% длины
мантии. Присоски средних и крайних
рядов центральной части булавы
равной величины. Имеются фотофор
на чернильном мешке и серия из
15—30 мелких фотофоров разных
размеров и формы на радужной обо-
лочке глаза вокруг зрачка. На по-
верхности тела нет фотофоров. Гла-
диус узкий, тонкий, с длинным ко-
нечным конусом.
Род Cycloteuthis Joubin, 1919 (рис.
63, а—м). Типовой вид — С. sirventi
Joubin, 1919 ................2.
2(3). Мантия цилиндроконичес-
кая, плавно сужается от начала плав-
ника к хвосту, голова шире мантии.
Мантийные хрящи располагаются
близ переднего края мантии. Плав-
ник поперечно-овальный или ромби-
ческий, его ширина значительно
больше длины. У взрослых длина
плавника около 80%, ширина около
90% длины мантии. Нет продольного
мышечного гребня на спинной сторо-
не мантии.
С. sirventi Joubin, 1919 (рис.
63,2—з). Тропические и субтропи-
ческие воды Атлантики и (?) Индо-
Вестпацифика.
3(2). Мантия воронковидная, рез-
ко сужается перед началом плавни-
ка. Голова уже мантии и может втя-
гиваться внутрь нее. Мантийные хря-
щи находятся на значительном рас-
стоянии от переднего края мантии
Плавник округлый, его ширина при-
близительно равна длине и составля-
ет около 50% длины мантии. Вдоль
спинной стороны мантии проходит
продольный двойной мышечный гре-
бень.
С. akimushkini Filippova, 1968
(рис. 63, и—лг). Тропические и суб-
23:
к
а
р
Рис. 63. Сем. Batoteuthidae (а—в) и Cyclo-
teuthidae (г—у): а~в — Batoteuthis skolops;
г—з — Cycloteuthis sirventi; и—м — С. aki-
tnushkini; н—п — Discoteuthis discus; p—г —
D. iaciniosa; у — Enoptroleuihis spinicauda,
личинка: а, и — молодые кальмары; 6 —
мантия полувзрослого кальмара; г, к, н,
р — взрослые кальмары; в, д, о, с — була-
ва; а, л — кишечный фотофор; ж — глазные
фотофоры; з, п, т—вороночный, (слева
сверху); мантийный (слева снизу) и заты-
лочный (справа) хрящи; м — вороночный и
мантийный хрящи (а—з, н—т — по Young,
Roper, 1968, 1969а; и, м — ио Filippova,
1968; к—л — по Clarke, 1980; у — по Berry,
1920)
235
тропические воды Индийского и за-
падной части Тихого оксана, субтро-
пические воды Южной Атлантики.
4(1). Мантия не вытянута в хвост.
Плавник у взрослых очень большой,
его длина почти равна или даже
несколько превосходит длину мантии.
В центральной части булавы 4 пары
присосок средних рядов резко уве-
личены, а присоски крайних рядов
очень малы. Нет фотофоров на чер-
нильном мешке. Имеются один или
несколько фотофоров на поверхности
тела. Перо гладиуса широкое, силь-
но утолщенное, конечный конус ко-
роткий или отсутствует.
Род Discoteuthis Young et Roper,
1969 (рис. 63, н—т). Типовой вид —
D. discus Young et Roper, 1969.
5(6). У заднего конца тела на
нижней стороне мантии имеется не-
парный фотофор. Несколько фото-
форов вокруг зрачка. Нет фотофо-
ров у переднего края мантии и на
голове. Крупные присоски булавы
шаровидные. Передний край мантии
ровный. Вороночный хрящ с одной
бороздой, отходящей вперед от глу-
бокой ямки. Гладиус сзади закруг-
лен.
D, discus Young et Roper, 1969
(рис. 63, н—п). Тропические и суб-
тропические воды Атлантического,
вероятно, также Индийского и Тихо-
го океанов.
6(5). Два фотофора располагают-
ся у переднего края мантии с брюш-
ной стороны вблизи мантийных хря-
щей, еще два — на перепонке, соеди-
няющей основания рук 3-й и 4-й пар.
Нет непарного хвостового и (?) глаз-
ных фотофоров. Крупные присоски
булавы уплощенные. Передний край
мантии на брюшной стороне и боках
с несколькими (4 или более) высту-
пающими вперед короткими пальце-
видными кожными выступами (толь-
ко у молоди). Вороночный хрящ с
двумя бороздами, расходящимися
под тупым углом от центральной ям-
ки. Гладиус с коротким конечным ко-
нусом.
D.laciniosa Young et Roper, 1969
(рис. 63,p—г). Тропические и суб-
тропические воды Атлантики, Индий-
ского и Тихого океанов.
В Саргассовом море была добыта
личинка Discoteuthis, не принадлежа-
щая ни к одному из двух видов. Опа
близка к D. laciniosa, но от однораз-
мерных личинок этого вида отлича-
236
ется плавником без передних и зад-
них «ушек» и очень крупными гла-
зами.
2.2.2.18. Семейство
Chiroteuthidae
( = Valbyteuthidae)
Кальмары небольших и средних раз-
меров (длина мантии до 20—40 см)
со студенистыми тканями. Мантия
коническая или мешковидная, сзади
оттянута в тонкий и длинный хвост,
сохраняющийся только у личинок и
молоди. Плавник небольшой, округ-
лый, продольно- или поперечно-
овальный. Голова длинная, обычно
узкая. Руки 4-й пары по строению
или вооружению обычно резко отли-
чаются от остальных; либо они зна-
чительно толще и, особенно у моло-
ди, длиннее других рук, либо несут
очень мелкие присоски, которые час-
то расположены в один ряд или толь-
ко в базальной части. Щупальца
обычно очень длинные с тонким
стеблем и, как правило, расширенной
булавой, несущей 4 (редко 6) ряда
присосок. Строение булавы у личи-
нок и взрослых кальмаров некоторых
родов существенно разное, дефини-
тивные присоски развиваются в про-
ксимальной части булавы, после чего
личиночная булава редуцируется.
Булава с хорошо развитыми защит-
ными мембранами, иногда с килем.
Вороночный хрящ уховидный, оваль-
ный, удлиненно-треугольный или бу-
тылковидный, чаще с двумя бугорка-
ми на вентральной (трагус) и задней
стороне (антитрагус), иногда только
с антитрагусом или вовсе без бугор-
ков. Мантийный хрящ носовидный.
Формула буккального крепления
DDVV. Фотофоры могут распола-
гаться па глазном яблоке, черниль-
ном мешке, руках 4-й пары и на
конце булавы; у ряда форм отсутст-
вуют. Гладиус очень тонкий, палоч*
ковидпый, с длинным конечным ко-
нусом, заходит далеко за задний ко-
нец плавника; у личинок и молоди
некоторых видов па конусе может
сидеть второй, дополнительный плав-
ник или несколько маленьких плав-
ничков. У взрослых конец гладиуса
обламывается приблизительно на
уровне заднего конца плавника или
чуть дальше него. Гектокотиля нет.
Пенис длинный, с пиковидной голов-
кой. Сперматофоры переносятся в
мантийную полость самки, иногда в
яичник. Развитие обычно со слож-
ным метаморфозом: личинки кроме
упомянутого дополнительного плав-
ника имеют часто очень длинную
шею и вытянутое рыло (пространст-
во между ртом и глазами), так что
длина тела личинки (от рта до кон-
ца гладиуса) может превышать дли-
ну тела кальмаров более поздних
возрастов.
Батипелагические или мезо-батн-
пелагические кальмары; иногда встре-
чаются у дна в батиали.
7—9 родов, 16—18 видов, 2 под-
вида. Некоторые роды и виды опи-
саны только по личинкам и их си-
стематическое положение сомнитель-
но. Обзор: Несис, 1980а.
Таблица для определения родов
семейства Chiroteuthidae
1(4). Руки 4-й пары намного шире,
обычно также длиннее прочих. Есть
фотофоры на конце булавы и па
глазах. Вороночный хрящ уховидный,
с трагусом и антитрагусом. Булава
щупалец с 4, руки 4-й пары с 2 ря-
дами присосок. Личинки с длинной
шеей п обычно хорошо развитым
рылом ...........................2.
2(3). Булава щупалец длинная, в
ее ладонной части несколько десят-
ков поперечных рядов присосок. Ряд
фотофоров по всей длине рук 4-й па-
ры. Один фотофор па конце булавы.
Руки 4-й пары гораздо длиннее про-
чих. Нет или 2 фотофора на чер-
нильном мешке.
Род Chiroteuthis d’Orbigny, 1839
( — Chirothauma Chun, 1910, Bigelowia
Macdonald et Clench, 1934) (см. рис.
65). Типовой вид — Ch. veranyi (Fe-
russac, 1835). Личинки известны под
«родовым» названием Doratopsis Ro-
chebrune, 1884 ( = Leptoteuthis Verrill.
1884, Chiridioteuthis Pfeffer, 1912, To-
roteuthis Tomlin, 1931, Diaphanoteuthis
Tomlin, 1931). 5 видов и 2 подвида
обитают во всех океанах от Субарк-
тики до субануарктики. Мезо- и ба-
типелагиаль и у дна в батиали.
3(2). Булава щупалец короткая,
в ее ладонной части не более 10 по-
перечных рядов присосок. Руки 4-й
пары гораздо шире, по не длиннее
прочих и лишены фотофоров (?). На
конце булавы пара небольших фото-
форов бок о бок. Один фотофор па
чернильном мешке.
Род Chiropsis Joubin, 1933. Один
вид — Ch. mega Joubin, 1933 (рис.
64, а—б), обитает в Саргассовом мо-
ре в батипелагиали.
4(1). Руки 4-й пары не шире про-
чих и нс несут фотофоров. Вороноч-
ный хрящ без трагуса, часто и без
антитрагуса, удлиненный, кегле- или
бутылковидный, овальный, но не ухо-
видный. Булава щупалец с 4—6 или
более, руки 4-й пары с 0—2 рядами
присосок ......................5.
5(8). Имеется фотофор на конце
булавы и (?) на глазном яблоке.
Проксимальная часть булавы у
взрослых с расширенной базальной
мембраной, присоски начинаются
дистальнее начала мембраны. На ру-
ках 4-й пары присоски идут до кон-
цов ...........................6.
6(7). Кожа покрыта крохотными
коническими хрящевыми бугорками.
Вороночный хрящ с узкой глубокой
бороздой, изогнутой в виде бумеран-
га и вилкообразно расщепленной на
заднем (каудальном) конце, настоя-
щих трагуса и антитрагуса нет. Нет
ректального фотофора. Присоски
булавы в 4 ряда, их кольца с 3—5
низкими коническими зубцами без
выделяющегося своими размерами
центрального зубца. Присоски рук с
3—4 низкими округленными зубцами.
Плавник продольно-овальный. Ли-
чинки с короткими шеей и рылом и
короткими руками 3-й пары.
Род Asperoteuthis Nesis, 1980. Ти-
повой и единственный вид — A. fame-
Пса (Berry, 1909) ( — Chiroteuthis fa-
melica Berry, 1909, Ch. acanthoderma
Lu, 1977) (рис. 64,г—л). Тихоокеан-
ский североцснтралытый мезо-бати-
пслагический вид.
7(6). Кожа гладкая. Вороночный
хрящ овальный, с зубовидным анти-
трагусом. Есть ректальный фотофор.
Присоски булавы у личинок в 4, у
взрослых в 6 рядов. Кольца присо-
сок рук с 8—9 длинными тупыми
зубцами. Плавник округлый. Личин-
ки с очень длинными шеей и рылом.
Chiroteuthidae п. g. В Nesis (рис.
64, м—о). Один неописанный вид —
«Chiroteuthis п. sp.» Nesis 1974а
(^Mastigoteuthis? A Clarke, 1980).
По-видимому, атлантический южно-
субтропнческо-нотальпый (на юг до
Южной Георгии) мезо-батипелагиче-
ский вид.
8(5). Фотофоров нет. Базальная
мембрана булавы нс расширена . 9.
237
л
Рис. 64. Сем. Chiroteuthidae: а—а — С her ap-
sis mega; г—л — Asperoteuthis janielica'r
м—о —Chiroteuthidae g. sp. В. Nesis; n—
p — «Doratopsis» sagitta; c — Chiroleulhoi-
des hastula, молодь; т— Tankaia borealis,,
у—ф — Valbyteuthis danae; х—ц — V. oh-
gobessa; ч—ш — V. exophthaltnica; 6, d,
e — булава {б, e — взрослого; d — Молодо-
го кальмара); а, я, p, ш — вороночный
хрящ; и, ф, ц — вороночный и мантий-
ный хрящи: ж — брюшная рука; к—при-
соска руки; л — проксимальная при-
соска булавы; о —глаз; л, г, ч — ли-
чинки (а—в, у — по Joubin, 1931, 1933; г,
ж, с—по Berry, 1914, 1920; д, з, к, л—по
Несису, 1980а;’е, « — по Lu, 1977; м—о —
по Clarke, 1980; л, р, ч—iu — по Chunr
1910; т — по Sasaki, 1929; ф—ц — по R. Yo-
ung, 1972).
9(14). Присоски на руках 4-й па-
ры идут до конца рук в два ряда,
либо сначала в 2, а потом в 1 зиг-
загообразный ряд. Вороночный хрящ
без трагуса и аититрагуса . . 10.
10(11). Присоски па булаве в 4
ряда. Вороночный хрящ бутылковид-
ный. Плавник поперечно-овальный,
за ним на гладиусе сидит крупный
второй плавник ланцетовидной фор-
мы, обычно обламывающийся. Оль-
факторная папилла сдвинута назад,
в район воронки. Мантия личинок
гладкая, руки 3-й пары развиты нор-
мально.
«Dor atop sis» sagitta Chun, 1908
(рис. 64, л, р). Известны только ли-
чинки, распространенные в тропиче-
ских водах Атлантического, Индий-
ского и Тихого океанов. Системати-
ческое положение этого кальмара но
установлено, возможно, особый род.
11(10). Присоски на булаве в 6
или более рядов. Вороночный хрящ
простой (кеглевидный), овальный или
треугольно-овальный. Руки 3-й пары
у личинок недоразвиты, значительно
короче прочих. Плавник поперечно-
овальный ...................... 12.
12(13). Вороночный хрящ простой,
сзади слегка расширенный (кеглевид-
пыи). Личинки с короткой шеей.
Род Chiroteuihoides Berry, 1920.
Типовой вид — Ch. hastula Berry,
1920. 2 вида, один из них нс описан
я известен лишь по личинке.
а (б). Плавник личинки овальный,
в форме оо, руки 4-й пары длинные.
Ch. hastula Berry, 1920 (рис. 64, с).
Тропические воды Атлантики, Кариб-
ское море. Мезо- и батипелагиаль.
б (а). Плавник личинки сердце-
видный. руки 4-й пары короткие.
Chiroteuthoides sp. Nesis. Эквато-
риальные воды Западной Пацифики.
Известны только личинки.
13(12). Вороночный хрящ оваль-
ный или треугольно-овальный. Ли-
чинки с длинной шеей.
Род Tankaia Sasaki, 1929. Извест-
ны только личинки: Т. borealis Sasa-
ki. 1929 (рис. 64,г) из бореальной
Пацифики у Хоккайдо и Tankaia sp.
Nesis из моря Флорес, Индонезия.
14(9). Присоски па руках 4-й па-
ры располагаются в один ряд только
в базальной части рук, дистальная
•часть лишена их. Вороночный хрящ
обычно (кроме Valbyteuthis levimana
я У. sp.) с антитрагусом . . . 15.
15(16). Округлый плавник цели-
ком сидит па вытянутой цилиндриче-
ской задней части мантии. Передняя
часть мантии узко-коническая. Воро-
ночный хрящ удлиненно-треугольный,
антитрагус представляет собой по-
перечную складку с небольшим вы-
ступом. Вороночный клапан имеется.
На руках 4-й пары только по одной
присоске у основания руки. Щупаль-
ца автотомируются.
Chiroteuthidae n. g. A Nesis,
1975 а. Один нс описанный вид, Ка-
рибскос море, батипелагиаль.
16(15). Мантия резко сужается
на уровне передней части или сере-
дины плавника, так что, но крайней
мерс, часть его сидит на узком кону-
се гладиуса. Плавник поперечно-
овальный. Передняя часть мантии
бокаловидная или мешковидная. Во-
роночный хрящ овальный с хорошо
развитым зубовидным антитрагусом
или удлиненно-треугольный с глубо-
кой изогнутой выемкой и без анти-
трагуса. Вороночного клапана нет.
На руках 4-й пары более одной при-
соски. Щупальца не автотомируются.
Личинки с длинной шеей.
Род Valbyteuthis Joubin, 1931
(рис. 64, у—ш). Типовой вид— V.
danae Joubin, 1913. 4—5 видов в ба-
типелагиали (реже в мезо-батипсла-
гиали) тропических и умеренных вод.
Из личинок, относимых к «роду»
Planktoteuthis Pfeffer, 1912, по край-
ней мерс, часть принадлежит к Valby-
teuthis.
Дополнительная таблица для
определения Chiroteuthidae
преимущественно молодых или
поврежденных экземпляров
1(10). Вороночный хрящ без трагуса
и аититрагуса ................. 2.
2(7). Вороночный хрящ простой,
удлиненный, или слегка расширенный
сзади (ксглевидный, бутылковидный)
или раздвоенный на конце. Присос-
ки на руках 4-й пары идут до конца
руки............................3.
3(4). Вороночный хрящ с узкой
глубокой бороздой, изогнутой в ви-
де бумеранга и вилкообразно рас-
цепленной на заднем конце. Плавник
продольно-овальный. Кожа покрыта
крохотными коническими бугорками.
Личинки с короткой шеей, руки 3-й
пары короткие. У взрослых есть фо-
тофор на конце булавы.
Asperoteuthis famelica (рнс. 64,
г—л).
240
4(3). Вороночный хрящ кегле-
или бутылковидный, нераздвоенный.
Плавник поперечно-овальный или
сердцевидный. Кожа гладкая. Фото-
форы неизвестны.................5.
5(6). Присоски на брюшных руках
в два ряда до конца руки. Личинки
с длинной шеей и хорошо развитыми
руками 3-й пары. Вороночный хрящ
прямой, сзади заметно расширенный
(бутылковидный).
«Doratopsis» sagitta (рис. 64, nt р).
6(5). Присоски на брюшных ру-
ках идут в два ряда в базальной час-
ти и в один зигзагообразный ряд в
дистальной части. Личинки с короткой
шеей и недоразвитыми руками 3-й
пары. Вороночный хрящ сзади слабо
расширен (кеглевидный) и слегка
изогнут.
Chiroteuthoides spp. (рис. 64, с).
7(2). Вороночный хрящ овальный,
треугольно-овальный или удлиненно-
треугольный ....................8.
8(9). Присоски на брюшных ру-
ках в два ряда до конца руки. На
булаве 6 или более рядов присосок.
Личинки с очень длинной шеей и
практически без рыла.
Tankaia spp. (рис. 64, т).
9(8). Присоски па брюшных ру-
ках расположены в 1 ряд, не далее
середины руки. На булаве 4 ряда
присосок. Личинки с недлинной шеей
и рылом примерно такой же длины,
как шея.
Valbyteuthis — часть видов: V. 1е-
vimana («Doratopsis» lippula), V. sp.
См. таблицу для определения видов
Valbyteuthis.
10(1). Вороночный хрящ с тра-
гусом и антитрагусом или только ан-
титрагусом ....................Il-
li (16). Вороночный хрящ без
трагуса, удлиненно-треугольный или
овальный.......................12.
12(15). Присоски имеются только
в базальной части или у основания
рук 4-й пары, в 1 ряд. Внутриман-
тийных фотофоров нет .... 13.
13(14). Вороночный хрящ оваль-
ный. Вороночного клапана нет.
Valbyteuthis — большинство ви-
дов: V. oligobessa, V. danae, «Doratop-
sis> exophthalmica (рис. 64, у—ш).
См. таблицу для определения видов
Valbyteuthis.
14(13). Вороночный хрящ удли-
ненно-треугольный, с глубокой изо-
гнутой выемкой. Вороночный клапан
имеется.
Chiroteuthidae n. g. А.
15(12). Присоски в два ряда по
всей длине рук 4-й нары. Имеется
внутримантийный фотофор на чер-
нильном мешке.
Chiroteuthidae n. g. В (рис.
64, м—о).
16(11). Вороночный хрящ ухо-
видный, с трагусом и антитрагу-
сом ...........................17.
17(18). На чернильном мешке ли-
бо два фотофора, либо ни одного.
Имеются фотофоры на руках 4-й
пары.
Chiroteuthis spp. (см. рис. 65).
18(17). На чернильном мешке
один непарный фотофор. Нет (?) фо-
тофоров на руках 4-й пары.
Chiropsis mega (рис. 64,а—в).
Таблица для определения видов рода
Valbyteuthis
1 (8). Вороночпый хрящ овальный
Булава щупалец не расширенная ил*
слабо расширенная, со слабо разви
тыми защитными мембранами, бе:
заметного киля...................2
2(7). Вороночный хрящ с разви
тым антитрагусом. Ширина плавни
ка' значительно больше его длины
Булава щупалец без киля ... 2
3(4). На руках 4-й пары лиш
2—3 (4) присоски. Эти руки у взрос
лых не длиннее прочих рук (у моле
ди значительно длиннее и толще)
Антитрагус вороночного хряща елг
бо развит — маленький бугоро!
Длина плавника 7ч—7б длины мав
тии. Булава слабо расширенна}
Присоски рук с 25—35 узкими высс
кими тупыми зубцами.
V. oligobessa Young, 1972 (ри<
64, х, ц). Экваториальные воды Тг
хого океана от Индонезии до Южно
Калифорнии и п-ва Калифорния. Бг
типелагиаль.
4(3). На руках 4-й пары десятс
или более присосок. Эта пара ру
самая длинная. Вороночный хрящ
хорошо развитым зубовидным анп
трагусом ..........................
5(6). Булава щупалец компак
пая, слегка расширенная. На рука
4-й пары 12—15 присосок, тянущи
ся почти до середины руки. Присо
ки рук с 7—10 низкими широкик
зубцами па дистальном крае. Длит
плавника равна или несколько пр
восходит половину длины манти
V. danae Joubin, 1931 (pi
16 К- Н. Несис
2
64, г/, ф). Батипелагиаль тропической
восточной’Панифики от Калифорнии
до Северного Чили.
, 6(5). Булава щупалец длинная,
как бы растянутая, нерасширенная;
«Doratopsis (Planktoteuihis)» exo-
phthalmica Chun, 1908 (рис. 64, ч, ш).
Известны только личинки, встре-
чающиеся в северных субтропических
водах Атлантического океана и в
Мексиканском заливе.
7(2). Вороночный хрящ без анти-
трагуса. Плавник округлый, его дли-
на и ширина около 7з длины мантии.
Булава щупалец с низким килем.
Valbyteuthls sp. ( = Chiroteuthis sp.
Rancurel, 1970). He описанный вид,
возможно, широко распространенный
в южносубтропических водах Атлан-
тического, Индийского и Тихого окса-
нов (см. Roper, Young, 1967).
8(1). Вороночный хрящ удлинен-
ный, с глубокой выемкой, без анти-
трагуса. Булава щупалец компакт-
ная, расширенная, с хорошо выра-
женными защитными мембранами и
килем. Руки 4-й пары много длиннее
прочих, несут 6—11 присосок. При-
соски рук с низкими широкими зуб-
цами. Длина плавника около % дли-
ны мантии.
V, levimana (Lonnberg, 1896). Ат-
лантический тропическо-субтропиче-
ский мезо-бати пелагический вид. «Do-
ratopsis (Planktoteuthis)» lippula
Chun, 1908 — личинка, ио-видимому,
этого вида. Похожие на «D». lippula
личинки попадались в Пацифике у
Гавайских островов и Новой Гвинеи.
Таблица для определения видов рода
Chiroteuthis
1(2). Нет фотофоров на черниль-
ном мешке. Фотофоры на вентраль-
ной стороне глазного яблока распо-
ложены двумя параллельными полос-
ками. Булава очень длинная и узкая,
одинаковой ширины на всем протя-
жении, ножки присо'сок булавы с
крыловидной боковой оторочкой.
Мантия узко-коническая в передней и
цилиндрическая в задней части, за-
ходит за задний конец округлого
плавника.
Ch. capensis Voss, 1967 ( = ? Bige-
lowia atlantica Macdonald et Clench,
1934) (рис. 65, а—д). Атлантический
тропический мезо-батипелагический
вид.
2(1). Два фотофора на черпиль-
242
пом мешке. Булава расширена в
средней, а иногда и в базальной час-
ти. Мантия широкая..............3.
3(6). Фотофоры на вентральной
стороне глазного яблока расположе-
ны двумя параллельными полосками,
между концами полосок спереди на-
ходится один, сзади два округлых
фотофора. Мантия коническая, у
взрослых заходит чуть дальше зад-
него края плавника. Ножки присосок
булавы с промежуточным расшире-
нием в виде темноокрашенпой чашеч-
ки с ребристой стенкой, из середины
чашечки поднимается цилиндриче-
ская ножка собственно присоски . 4.
4(5). Кольца присосок рук па ди-
стальном крае с длинными острыми
зубцами. Диаметр присосок в сере-
дине рук 3-й пары не более чем
вдвое превосходит диаметр присосок
рук 4-й пары
Ch. ver any i (Ferussac, 1835) (рис.
65, e—и). 2 подвида.
а (б). Плавник округлый. Фотофо-
ры на чернильном мешке округлые
или яйцевидной формы. Защитные
мембраны булавы с небольшим суже-
нием (перетяжкой) в середине или
на границе базальной и средней тре-
ти булавы. Ch. veranyi veranyi (Ferus-
sac, 1835) (рис. 65, e, ж). Восточная
Атлантика от Бискайского залива до
Анголы Средиземное море, Азор-
ские, Канарские острова, Мадейра,
нотальныс и южносубтропические
воды Индийского и Тихого оксанов
от района южнее Африки до Перу и
Чили; в восточной части Тихого окса-
на на севере доходит до эквато-
риального района. В Северной Па-
цифике отсутствует. У дна в батиа-
ли и в мезо- и верхней батипелагиа-
ли над склоном. Личинка — Dora-
topsis vermicularis (Riippell, 1844).
б (а). Плавник .продольно-оваль-
ный. Фотофоры на чернильном меш-
ке удлиненно-овальные, усеченные на
концах. Защитные мембраны булавы
без сужения (перетяжки).
Ch. veranyi lacertosa Verrill, 1881
(рис. 65, з, и). Северо-Западная Ат-
лантика от Новой Шотландии до зал.
Делавэр, Мексиканский зал., М. Aib
тильские о-ва. Личинка предположи-
тельно Doratopsis diaphana (Verrill,
1884).
5(4). Кольца присосок рук с дис-
тальной стороны с тупыми усеченны-
ми зубцами или гладкие, лишь со
слабо заметными насечками. Присос-
ки в середине рук 3-й нары крупные,
их диаметр более чем в 3 раза нре-
восходит диаметр присосок рук 4-й
пары.
Ch. calyx Young, 1972 (рис. 65,
к.—я). Северо-тихоокеанский бореаль-
ный вид, распространен от южной
части Берингова моря и залива Аляс-
ка до Северо-Восточного Хонсю и
п-ва Калифорния, преимущественно
в мезопслагиали близ склона. Ли-
чинка — Doratopsis cf. ver mi cut ar is
Berry, 1963.
6(3). Фотофоры на вентральной
стороне глазного яблока округлые,
многочисленные, расположены двумя
или тремя продольными рядами . 7.
7(8). Фотофоры па вентральной
стороне глазного яблока красновато-
золотистые, расположены тремя про-
дольными рядами, тесно сближены
(кроме краевых). Ножки присосок бу-
лавы удлиненно-конические, иногда с
крыловидной боковой оторочкой. За-
щитные мембраны булавы без рас-
ширения в базальной части. Мантия
цилиндрическая в передней части,
продолжается за задний конец плав-
ника и образует на заднем конце ве-
ретенообразное расширение, окайм-
ленное узкой мембраной. Кольца
присосок с дистальной стороны с при-
тупленными зубцами, проксимально
гладкие.
Ch. picteti Joubin, 1894 ( = Ch. mac-
rosoma Goodrich, 1896, Ch. imperator
Chun, 1908) (рис. 65, p—т). Тропиче-
ские воды Индо-Всстпацифики от
Аденского залива до Южного Хонсю,
Индонезии и Гавайских островов.
Мсзопелагиаль и батиаль. Личинка—
Doratopsis pellucida (Goodrich, 1896).
8(7). Фотофоры на вентральной
стороне глазного яблока серебристые,
неравных размеров (крупные и мел-
кие), расположены двумя продольны-
ми рядами, на неодинаковых рас-
стояниях друг от друга. Ножки при-
сосок булавы с небольшим проме-
жуточным расширением в виде тсм-
ноокрашенной чашечки или пленки.
Защитные мембраны булавы резко
расширены в базальной трети: в
этом месте они кожистые, трабекулы
незаметны, так что кажется, будто
на булаву насажен овальный плав-
ник. В остальной части булавы
мембраны пленчатые, с хорошо за-
метными трабекулами. Мантия без
веретенообразного расширения за
задним краем плавника. Кольца при-
сосок рук дистально с тесно сбли-
женными округлыми или приострен-
пыми зубцами, проксимально гладкие.
Ch. joubini Voss, 1967 ( = Ch. la-
certosa Joubin, 1933, non Verrill)
(рис. 65, у, ф). Экваториальные во-
ды Атлантики (Joubin, 1933); восточ-
ная часть Саргассова моря, Индоне-
зия (море Банда) — наши данные;
Юго-Восточная Африка (Clarke,
1980). Описание составлено по на-
шим экземплярам: в первоописании
(Joubin, 1933) указывается, что фо-
тофор па чернильном мешке непар-
ный, а ножки присосок без проме-
жуточного расширения, кеглсвид-
ные.
2.2.2.19. Семейство
Mastigoteuthidae
Кальмары средних размеров (длина
мантии до 15—30 см) с хрящевато-
студенистыми тканями. Передняя
часть мантии цилиндрическая, зад-
няя— коническая. Плавник длинный,
округлый, овальный, ромбический
или сердцевидный, его длина обыч-
но не меньше половины длины ман-
тии. Руки 4-й пары, как правило,
значительно длиннее прочих. При-
соски на руках в 2 ряда. Щупальца
длинные, иногда в несколько раз
длиннее мантии, с тонким стеблем и
нерасширенной или почти нерасши-
ренной булавой, несущей множество
мельчайших присосок во много ря-
дов. Булава без киля и фиксаторно-
го аппарата, присоски занимают ’/2—
3А се длины. При поимке кальмара
щупальца часто автотомируются.
Формула буккального крепления
DDVV. Вороночный хрящ уховидный
или овальный, с одним или двумя
бугорками (трагусом и антитрагу-
сом), реже с тремя бугорками или
без них; мантийный хрящ носовид-
ный или в виде прямого угла. У мно-
гих видов имеются фотофоры, обыч-
но они рассеяны по поверхности
мантии, на нижней стороне головы и
брюшных рук, иногда па плавнике и
нижнем веке у глазного синуса.
Гладиус узкий, с длинным конечным
конусом, обычно заходит за задний
конец плавника в виде тонкой хво-
стовой нити. Гсктокотиля нет. Раз-
витие без личиночной стадии. Бати-
пелагичсские и мезо-батипелагиче-
16*
243
Рис. G5. Род Chiroteuthis: а—д — Ch. сареп-
sis; е—ж — Ch, veranyi veranyi, з—u—Ch. ve-
ranyi lacertosa; к—n—Ch. calyx; р—т—Ch.
picteti*, у—ф—Ch. joubini: a, e, з, к, p—
взрослые; ж, л — молодь (послеличиночвая
стадия); б, и, м, ф — булава; в —конце-
вой фотофор булавы; г, н, с — присоски
булавы; д — кольцо присоски булавы; о —
вороночный хрящ; п, т, у — глазные фото-
форы (я—d —по G. Voss, 1967; е — по
Pfeffer, 1912; ж—з, у—ф — по Joubin, 1924,
1931, 1933; и — по Verrill, 1882; к—п — по
R. Young, 1972: р—т — по Chun, 1910)
ские кальмары, некоторые виды при-
донные. 2 рода, 14—18 видов. Обзор:
Несис, 1977.
Таблица для определения родов
семейства Mastigoteuthidae
1(2). Присоски на руках 4-й пары
идут до концов рук. Булава щупаль-
ца не расширена, круглая в сечении,
присоски сидят почти по всему ее
периметру (кроме М. laiipinna). На
коже пет звездчатых бородавочек.
Плавник ромбический, округлый или
сердцевидный, хвостовая нить отсут-
ствует или очень короткая. Длина
плавника обычно значительно более
72 длины мантии (кроме взрослых
самок некоторых видов). Передний
край мантии на спинной стороне пря-
мой или со слабым угловатым вы-
ступом.
Род Mastigoteuthis Verrill, 1881
( = Chiroteuthopsis Pfeffer, 1900, Idio-
teuthis Sasaki, 1929) (рис. 66, 67, a—
в). Типовой вид — M agassizi Verrill,
1881. 2 подрода, 12—16 видов во
всех океанах, кроме Северного Ледо-
витого.
2(1). Присоски па руках 4-й па-
ры сидят лишь в базальной части
рук и не переходят за их середину.
Булава щупалец слегка расширена,
ее ладонная поверхность уплощена,
присоски сидят только на ладонной
поверхности. Кожа головы, мантии и
отчасти плавников покрыта крохот-
ными хрящевыми бугорками или
звездчатыми бородавочками. Плав-
ник продольно-овальный или широ-
ко-сердцевидный, его длина немного
более 7г длины мантии, сзади он вы-
тянут в длинную (иногда длиннее
самого плавника) хвостовую нить.
Передний край мантии на спинной
стороне с большим языковидным
выступом. Вороночный хрящ с тра-
гусом и небольшим антитрагусом.
Глазные фотофоры отсутствуют.
Род Echinoteuthis Joubin, 1933
{рис. 67, г—ж). Типовой вид — Е. da-
nae Joubin, 1933. 2 вида, один из них
не описан.
* Таблица для определения подродов
и видов рода Mastigoteuthis
1(22). Нет фотофоров на вентраль-
ной стороне глазного яблока. Плав-
ник не доходит до переднего края
246
мантии, его ширина значительно
меньше длины мантии. Фотофоры
разбросаны по поверхности тела или
отсутствуют.
Подрод Mastigoteuthis s. str.
(рис. 66) ...................2.
2(11). По всей поверхности тела,
особенно на брюшной стороне, раз-
бросано множество мелких черных
фотофоров, окруженных светлым ко-
лечком. Имеется маленький (менее
1 мм) фотофор на глазном веке из-
нутри близ синуса (не описан у М.
ialismani). Вороночный хрящ с тра-
гусом (у М. agassizi слабо выражен),
аптитрагус имеется или отсутству-
ет .............................3.
3(6). Кольца присосок рук на дис-
тальной стороне с 10—20 острыми
зубцами, проксимально с низкими
уплощенными зубцами или гладкие.
Плавник сердцевидный или ромбиче-
ский, сзади вытянут в небольшую
хвостовую нить, его длина у молоди
около 67%, у взрослых самцов 55—
60%, взрослых самок 40—50% дли-
ны мантии. Глаза крупные, выступа-
ющие ...........................4.
4(5). Аптитрагус отсутствует или
очень слабо выражен, вороночный
хрящ сзади как бы подрезан.
Группа видов М. grimaldii (рис.
66, а—е). 4 поминальных батинела-
гических вида, распространенных в
тропических и субтропических водах
всех оксанов. Различаются только по
строению и расположению присосок
па булаве (надо учесть, что щупаль-
ца, как правило, автотомируются при
поимке кальмаров). Самостоятель-
ность видов вызывает сомнение.
а(е). Кольца присосок щупалец с
внутренней стороны гладкие' . . б.
б(д). Присоски на булаве распо-
ложены в 30—40 рядов \ . . . в.
в (г). Булава занимает 2/з—3А
длины щупальца. Диаметр присосок
булавы 0,2—0,4 мм.
М grimaldii (Joubin, 1895) (рис.
66, а, б). Тропические и субтропиче-
ские воды Атлантики, Мексиканский
залив.
г (в). Булава занимает около 7г
длины щупальца. Диаметр присосок
булавы 0,15—0,25 мм.
А4. dentata Hoyle, 1904 (рис. 66,
ву г). Тропические и субтропические
воды Ипдо-Пацифики (кроме Кали-
форнийского течения).. Па север до-
ходит до Курильских островов, на
юг до Южной Африки, Южной Ав-
стралии, Новой Зеландии и Централь-
ного Чили.
д(б). Присоски на булаве щу-
пальца сидят в 20—30 рядов, их диа-
метр около 0,1 мм. Булава занимает
% длины щупальца.
М. schmidti Degner, 1925. Север-
ная Атлантика от Исландии до Ка-
нарских островов и Мадейры. Веро-
ятно, синоним Л4. gritnaldii.
е(а). Кольца присосок щупалец
на внутренней стороне с 3 зубчика-
ми на дистальном крае. Присоски
на булаве сидят приблизительно в
20 рядов, их диаметр около 0,3 мм.
Булава занимает 7г длины щу-
пальца.
М. pyrodes Young, 1972 (рис. 66,
д, е). Калифорнийское течение.
5(4). Вороночный хрящ с хорошо
выраженным антитрагусом. На була-
ве около 20 рядов присосок, их диа-
метр около 0,15 мм. Кольца присосок
с внутренней стороны гладкие.
М. psychrophila Nesis, 1977 (рис.
66,ж). Нотально-антарктический цир-
кумглобальный мезо-батипелагиче-
ский вид.
6(3). Кольца присосок рук глад-
кие или с немногими тупыми зубца-
ми. Плавник округлый или ромбиче-
ский ...........................7.
7(10). Длина плавника !/г—2/з
длины мантии. Есть маленький фо-
тофор на глазном веке близ сину-
са .............................8.
8(9). Плавник поперечно-оваль-
ный или слаборомбический, сзади вы-
тянут в короткий хвост, его длина у
молоди 50—60% длины мантии. Тра-
гус и антитрагус хорошо развиты.
Глаза небольшие, невыступающие.
Кольца присосок рук гладкие или с
3—5 зубчиками дистально.
М. flammea Chun, 1908 (рис. 66,
з, и) Гвинейский залив и (?) север-
ная субтропическая Атлантика;
Большой Австралийский залив. Ба-
типелагиаль.
9(8). Плавник ромбический, не
вытянут в хвост. Его длина около
Уз длины мантии. Трагус и антитра-
гус слабо выражены. Глаза большие.
Кольца присосок рук гладкие.
Af. agassizi Verrill, 1881 (рис.
66, к). Бореальная и северная суб-
тропическая Атлантика, на север до-
ходит до района южнее Исландии.
10(7). Длина плавника около 90%
длины мантии. Трагус хорошо выра-
Рис. 66. Сем. Mastigoteuthidae, род Mas-
tigoteuthis s. str.: a—б — M. grimaldii; в—
г — Al. dentata; д—е — M. pyrodes; ж —
Al. psychrophila; з—и — M. flammea; к —
Al. agassizi; л—м — M. ailantica; н—о —
M. glaukopis; n—p — Af. cordiformis; c—т —
Af. magna; у—ф — M. inermis: а» з, n, у —
вид сверху и снизу; б — голова сбоку,
виден фотофор (ф) на глазном веке; в —
присоска руки; г, е, .и—присоска булавы
(на е кольцо снаружи и изнутри —еО; ж,
и, о, р,т,ф — вороночные хрящи (а—б, у—ф
по Rancurci, 1971, 1972; в—г, ж — по Неси-
су, 1973в, 1977в; д—е — по R. Young, 1972;
з—и, н—р—ио Chun, 1910; к — по Verrill,
1982; я—м, с—т — по Joubin, 1920, 1933)
249i
жен, антитрагуса нет. Кольца присо-
сок рук гладкие.
М talismani (Fischer et Joubin,
1906). Тропическая и северная суб-
тропическая Атлантика — район ост-
ровов Азорских и Зеленого Мыса.
11(2). Поверхность тела лишена
фотофоров, иногда имеется фотофор
на глазном веке близ синуса . 12.
12(17). Кольца присосок рук с
зубцами. Вороночный хрящ с трагу-
сом (у взрослых М. glaukopis слабо
выражен)......................13.
13(16). Есть крупный (1—4 мм),
хорошо заметный светлый фотофор
изнутри па глазном веке близ сину-
са. Кожа гладкая..............14.
14(15). Присоски рук с 10—15
острыми зубцами па дистальном
крае, проксимальные края гладкие.
Присоски булавы крупные (0,3—
0,5 мм), их кольца несут 2—3 длин-
ных притупленных зубца на боко-
вых сторонах. Трагус хорошо выра-
жен, аптитрагус слабо заметен.
М. atlaniica Joubin, 1933 ( = ?М.
iselini .Macdonald et Clench, 1934)
(рис. 66, л, л). Тропическая и север-
ная субтропическая Атлантика (на
север до Бискайского залива) и Ин-
до-Вестпацифика. Батипелагиаль.
15(14). Присоски рук с 5—10 низ-
кими притупленными зубцами па дис-
тальном крас, проксимально с упло-
щенными зубцами или гладкие. При-
соски булавы мелкие (0,06—0,1 мм),
с гладкими кольцами. Трагус слабо
развит или отсутствует, аптитрагус
не выражен.
М. glaukopis Chun, 1908 (рис.
66, н, о). Западная часть Индийско-
го океана и море Флорес. Батипела-
гиаль.
16(13). Фотофоры отсутствуют.
Кожа головы, мантии и оснований
рук покрыта мелкими кожными бу-
горками. Присоски рук с низкими
округленными или слабо приострен-
пыми зубцами. Присоски булавы с
острыми зубцами по всему перимет-
ру. Вороночный хрящ с трагусом и
слабо выраженным антитрагусом.
Длина плавника 70—80% длины
мантии.
М. cordiformis Chun, 1908 (рис.
66, п, р). Южная Япония, Филиппин-
ские острова, Индонезия. Обитает
преимущественно у дна в батиали.
17(12). Кольца присосок рук
гладкие. Фотофоры отсутствуют. Во-
роночный хрящ без трагуса и анти-
трагуса .......................18.
Л
/7
Рис. 67. Сем. Mastigoteuthidac, роды Ato-
tigoteuihis (Mastigopsis) (а—в) и Echino-
leuthis (г-ж)’ сем. Joubiniteuthidae (з—н):
а—в — Mastigoteuthis hjorti, общий вид
(а), глаз (б), вороночный и мантийный
хрящи, (в); г—ж — Echinoteuthis danae,
молодь сверху и снизу (г). булана (б),
брюшная рука (е). кожный бугорок (лс);
з—н — Joubiniieuthis poriieri, молодой (а)
и взрослый (и) кальмары, вороночный и
мантийный хрящи (к), руки 3-й (л) и 4-й
Gw) пар, булава («) (а—в — по Rancurcl,
1973; г—з — по Joubin, 1924, 1933: и—н — по
Young, Roper, 19б9в)
251
18(21). Вороночный хрящ бутыл-
ко- или ксглевидный: суженный спе-
реди, расширенный и округленный
сзади. Булава щупалец округлая в
сечении, с зачаточными защитными
мембранами, присоски микроскопи-
ческие, с гладкими кольцами. Длина
плавника около % длины ман-
тии ..........................19.
19(20). Ширина плавника больше
или равна его длине и составляет
70—80% длины мантии.
М. magna Joubin, 1913 (рис. 66,
с, г). Тропическая и северная субтро-
пическая Атлантика, Индийский оксан
и Тасманово море. Батипелагиаль и
батиаль, по ночам поднимаются в ме-
зопелагиаль.
20(19). Ширина плавника меньше
его длины и составляет около ’А
длины мантии.
М inermis Rancurel, 1972 (рис.
66, у, ф). Экваториальная Атланти-
ка, Гвинейский залив. Батипелагиаль.
21(18). Вороночный хрящ оваль-
ный или яйцевидный. Проксимальная
часть булавы слегка расширена и
уплощена, с хорошо развитыми за-
щитными мембранами. Присоски
проксимальной части булавы не
мельче присосок рук, их кольца на
дистальном крае с зубцами. Длина
плавника около 5/в длины мантии.
М. latipinna (Sasaki, 1916). От
Южной Японии до Северо-Западной
Австралии. У дна в батиали.
22(1). В передней и задне-меди-
анной части брюшной стороны глаз-
ного яблока 2 крупных овальных
фотофора ярко-белого цвета: перед-
ний располагается на уровне участ-
ка между основаниями 2-й и 3-й пар
рук, задний — на уровне переднего
конца воронки. Нет фотофоров на
глазном веке и поверхности тела.
Плавник очень большой, почти дохо-
дит до переднего края мантии, его
ширина заметно больше длины ман-
тии. Вороночный хрящ овальный,
сзади слегка расширенный, без тра-
гуса и антитрагуса. Присоски рук на
дистальном крае с 10—15 длинными
притупленными зубцами. Присоски
булавы микроскопические.
Подрод Mastigopsis Grimpe, 1922.
Один вид — М. hjorli Chun, 1913
(рис. 67,а—в). Тропическая и север-
ная субтропическая Атлантика, Ин-
дийский океан, Тасманово морс,
(?) Восточно-Экваториальная Пацн-
фика. Мезо- и батипелагиаль. Моло-
252
дой кальмар, относящийся к этому
подроду (особый вид?), пойман в мо-
ре Сулу.
Таблица для определения видов рода
Echinoteuthis
1(2). Передняя часть мантии до на-
чала плавника узкая, цилиндриче-
ская. Кожные хрящевые бугорки
звездчатые.
Е. danae Joubin, 1933 (рис. 67,
г—ж). Тропическая и северная суб-
тропическая Атлантика. Предполо-
жительно мезо- и батипелагиаль.
2(1). Передняя часть мантии до
начала плавника — коническая (бо-
каловидная). Кожные хрящевые бу-
горки округлые.
Echinoteuthis sp. Nesis (=Masii-
goteuthis grimaldii Allan, 1945, non
Joubin). Южная часть Тасманова
моря и Большой Австралийский за-
лив. Мезо- и батипелагиаль.
2.2.2.20. Семейство
Joubiniteuthidae
Кальмары средних размеров с узкой
цилиндрической мантией хрящевато-
студенистой консистенции, вытяну-
той сзади в очень длинную (больше
длины мантии) тонкую хвостовую
нить, у взрослых она обычно обла-
мывается. Плавник маленький,
овальный или округлый. Голова уз-
кая, на длинной шее. Глаза неболь-
шие. Руки первых трех пар исключи-
тельно длинные, тонкие, особенно на
концах, гибкие, несут очень мелкие
присоски, расположенные в 6 рядов.
Руки 4-й пары значительно короче
остальных, с 4 рядами присосок.
Щупальца у молодых кальмаров
очень тонкие и слабые, короче рук
1—3-й пар, с нитевидным стеблем и
длинной, сжатой с боков булавой,
несущей 8—12 рядов мельчай-
ших присосок. Защитные мембраны
булавы развиты только на ее дис-
тальном конце, фиксирующего аппа-
рата нет. У взрослых особей щупаль-
ца полностью редуцируются. Воро-
ночный хрящ овальный, без трагуса
и антитрагуса, мантийный — в виде
удлиненного бугорка. Формула бук-
кального крепления DDVV. Фотофо-
ры отсутствуют. Гладиус игловид-
ный, не расширенный посредине, с
длинным фунтиковидным конусом.
Гектокотиль отсутствует. Окраска
коричнево-фиолетовая.
Один род Joubiniteuthis Berry,
1920 (= Valdetnaria Joubin, 1931).
Единственный вид — J. portieri (Jou-
bin, 1912) (= Valdetnaria danae Jou-
bin, 1931) (рис. 67,3—«), обитаю-
щий в мезо- и батипелагиали тропи-
ческой и субтропической Атлантики,
включая Карибское море, и в север-
ной части Тихого океана (Японские,
Гавайские острова). Обзор семейст-
ва: Young, Roper, 1969b.
2.2.2.21. Семейство
Promachoteuthidae
Мантия широкая, толстая, полусту-
депистая, срастается с головой на за-
тылке. Голова маленькая; глаза
очень маленькие, сильно редуцирова-
ны в размерах. Руки с 2 рядами при-
сосок. Булава щупалец не расшире-
на, без защитных мембран и фикси-
рующего аппарата, с многочисленными
(6—20 рядов) мелкими присосками.
Плавник широко-сердцевидный или
поперечно-овальный. Вороночный
хрящ овальный, без трагуса и антн-
трагуса, мантийный хрящ бугорко-
видный. Формула буккального креп-
ления DDVV, Фотофоров нет. Гла-
диус отсутствует или редуцирован.
Один род — Promachoteuihis
Hoyle, 1885 (рис. 68, а—г) с двумя
видами (один из них нс описанный),
известными лишь по очень молодым
особям, добытым в бати- и абиссо-
пелагиали. Различия молоди этих ви-
дов таковы:
1(2). Длина рук примерно равна
длине мантии. Длина плавника око-
ло 3/ч длины мантии, ширина в пол-
тора раза превосходит се.
Р. megaptera Hoyle, 1885. Япон-
ский глубоководный желоб южнее
Хонсю.
2(1). Длина рук около J/4 длины
мантии. Длина плавника меньше Уг,
ширина — около 2/з длины мантии.
Р. sp. Roper et Young, 1968. Юго-
восточная часть Тихого океана за-
паднее Среднего Чили и юго-залад-
нее Магелланова пролива, (?) Юж-
ная Атлантика. Обзор семейства:
Roper, Young, 19(58.
254
2.2.2.22. Семейство
Grimalditeuthidae
Кальмары средних размеров со сту-
денистой узкоконической мантией,
вытянутой сзади в длинный хвост.
Плавник поперечно-овальный или
округлый, его длина около Уг длины
мантии, считая до конца плавника.
За ним па длинной хвостовой нити
находится второй плавник пример-
но того же размера, но сердцевидной
формы. Обычно конец гладиуса со
вторым плавником обламывается.
Голова узкая и длинная, срастается
с воронкой, но нс срастается с голо-
вой на затылке. Руки длинные, тон-
кие, с 2 рядами присосок, ножки ко-
торых поднимаются из толстых бу-
горков с тремя тупыми вершинами.
На концах всех рук у взрослых каль-
маров по одному удлиненному тем-
ному фотофору. Щупальца полно-
стью утрачиваются, даже у молодых
сохраняются лишь короткие руди-
менты стеблей. Формула буккально-
го крепления DDVV. Гладиус очень
узкий, с длинным конечным кону-
сом. Гектокотиля нет.
Один род — Grimalditeuthis Jou-
bin, 1898 с единственным видом — G.
bonplandi (Verany, 1837) (рис. 68,
д—з). Распространен в мезо- и бати-
пелагиали тропической и субтропи-
ческой Атлантики и северной Паци-
фики (Гавайские острова, Калифор-
ния). Обзор: R. Young, 1972.
2.2.2.23. Семейство
incertae sedis
Род Parateuthis Thiele, 1921. Един-
ственный вид — Р. tunicaia Thiele,
1921 (рис. 68, и). Известен только по
личинкам с длиной мантии 5—8 мм,
добытым в Антарктике с глубин
2000—3425 м. Мантия веретеновид-
ная, мускулистая в передней и тон-
кая, пленчатая, полупрозрачная в
задней части, где просвечивают за-
пасы внутреннего желтка. Плавники
крохотные, лепестковидные. Мантия
не срастается с головой и воронкой,
но замыкательные хрящи плохо раз-
виты (мантийный и вороночные —
почти по выражены). Голова узкая,
с большими глазами; руки короткие,
с 2 рядами присосок. Гладиус отсут-
ствует или сильно редуцирован.
Строение булавы и буккального креп-
ления не известно.
Личинки нс принадлежат ни к
одному из известных семейств эгоп-
сид.
2.2.2.24. Семейство
Cranchiidae
Малоподвижные планктонные и по-
луиланктонные-полупектонные каль-
мары от мелких до гигантских.
Мантия срастается с воронкой и го-
ловой на затылке, она топкая, плен-
чатой, кожистой или полустудеии-
стой консистенции. Поверхность ман-
тии гладкая или несет маленькие
хрящевые бугорки, обычно распола-
гающиеся в определенных местах, ре-
же (СгапМа) по всему телу. Плавни-
ки очень разнообразной формы: лопа-
то-, ночко-, яйце-, ланцетовидные, от
маленьких до очень длинных, по их
длина не превышает 60% длины ман-
тии. Голова короткая и узкая, глаза
очень большие, у взрослых особей,
как правило, шаровидные и сидячие,
у личинок и молоди обычно стебель-
чатые, уплощенные и трубчатой фор-
мы. Руки с 2 рядами присосок (не-
сколько рядов на концах рук у сам-
цов некоторых видов), у Mesonycho-
teuihis часть присосок в середине рук
превращается в крючья. Имеются за-
щитные мембраны и слабо развитые
кили. Щупальца длинные со слабо
расширенной булавой, несущей 4 ря-
да присосок с зубчатыми кольцами,
иногда присоски средних рядов пре-
вращаются в крючья. Фиксаторный
аппарат хорошо развит, обособлен
или протягивается вдоль стебля.
У взрослых кальмаров щупальца
иногда автотомируются. Буккальное
крепление семилучевое, его формула
DDVV. Гладиус тонкий, палочковид-
ный, сзади с ромбическим расшире-
нисм — ланцеолой, задний конец ко-
торой может быть вытянут в длин-
ный острый конус. В последнем
случае мантия вытянута в хвост.
Задний конец мантии может не до-
ходить до заднего конца гладиуса.
Плавники либо целиком, либо своей
задней частью прикрепляются к зад-
не-боковым сторонам ланцеолы.
У Bathothauma ланцеола редуциру-
ется и плавники прикрепляются не-
посредственно к мантии. Всегда име-
ются фотофоры на глазном яблоке
(несколько округлых или 1—2 удли-
ненных), иногда имеются фотофоры
на концах рук (у взрослых самок}
и печени. Тело у молоди обычно по-
лупрозрачное. или совершенно про-
зрачное, слабо пигментированное, ок-
раска взрослых особей обычно пур-
пурная, коричневая или желто-
ватая.
Характерна обширная замкнутая
полость тела (целом), содержащая
значительное количество раствора
NH4C1. Плотность этого раствора ни-
же плотности морской воды, так что
целом служит поплавком, поддержи-
вающим кальмара в состоянии ней-
тральной плавучести («кальмары-ба-
тискафы»). Мантийная полость разде-
лена горизонтальной перегородкой
па верхнюю (включающую целом) и
нижнюю части, соединяющиеся меж-
ду собой круглыми отверстиями —
дыхальцами, расположенными над
жабрами; дыхательный ток воды
проходит вдоль верхней камеры, по
обеим сторонам целома назад, затем
через дыхальца мимо жабр вперед к
голове и через воронку выходит на-
ружу. Нидамснтальные железы пар-
ные (каждая железа подразделена
на две) или непарные. Гсктокотнли-
зируются концы одной руки или не-
скольких рук. Сперматофоры пере-
носятся па поверхность тела, головы
или рук или в мантийную полость,
самки (к половым отверстиям или
прямо в яйцевод). Развитие с личи-
ночной стадией. У личинок очень ко-
роткие руки и длинные щупальца*
крохотные плавнички, часто стебель-
чатые глаза удлиненной формы, за-
остренные книзу (с рострумом), при-
соски тянутся вдоль всего или поч-
ти всего стебля щупалец.
Преимущественно мезопслагнче-
скис или мсзо-батипелагические каль-
мары, молодь некоторых видов оби-
тает в эпипслагиали.
2 подсемейства, 15 родов, 32—38
видов. Обзор: N. Voss, 1980.
Синонимия семейства чрезвычай-
но запутана, так как одни роды и
виды описаны по личинкам, другие
же — по взрослым особям. В связи
с этим иногда разные возрастные
стадии одного и того же вида отно-
сили к разным родам и иод одним
видовым названием объединяли
сходные стадии разных кальмаров.
Определительные таблицы составле-
ны по признакам взрослых или полу-
взрослых особей.
255
Таблица для определения
подсемейств и родов Cranchiidae
1(8). На брюшной стороне мантии
имеются 2 или 4 гиалиновые полос-
ки, отходящие от мест срастания
мантии с воронкой и несущие хря-
щевые бугорки с одной или несколь-
кими острыми вершинками. Глазные
фотофоры многочисленные, округ-
лые, охватывают глазное яблоко
кольцом или двумя дугами. Плавни-
ки округлые или полукруглые, не вы-
тянуты в длинный хвост. Боковые
стороны воронки у взрослых при-
растают к брюшной поверхности го-
ловы. Гсктокотилизируется одна из
брюшных рук самца.
Подсемейство Cranchiinae . . 2.
2(3). Вся поверхность мантии и
верхняя сторона плавников покрыты
хрящевыми бугорками, вершины ко-
торых несут 3—5 острых шипиков.
Плавники маленькие, округлые, вме-
сте напоминают восьмерку. Мантия
шаро- или бочонковидная. На глаз-
ном яблоке 14 фотофоров: 6 вокруг
зрачка, 7 дугой по нижней стороне
яблока и 1 между ними. У взрослых
самок имеется крупный фотофор на
концах всех рук. Глаза у личинок си-
дячие. Вороночный клапан имеется.
Род Cranchia Leach, 1817. Один
вид —С. scabra Leach, 1817 (рис.
69, а—в). Тропическо-субтропический
космополитический мезо-батипелаги-
ческий вид. Молодь обитает в эпи- и
мезопелагиали. Длина мантии до 15
(20?) см,
3(2), Хрящевые бугорки и шипи-
ки имеются только на гиалиновых по-
лосках и иногда вдоль средней линии
спины...........................4.
4(5). В передней части брюшной
стороны мантии 2 пары гиалиновых
полосок, V-образно расходящихся от
мест срастания мантии с воронкой.
У L. reinhardti полоска хрящевых ши-
пиков проходит ,и вдоль средней ли-
нии спины. Число глазных фотофоров
от 4 до 14. Глаза у личинок сидячие.
Мантия у личинок и молоди шаро-
или бочонковидная, у взрослых ши-
роковсрстеновидная. Плавники у мо-
лоди вместе напоминают восьмерку,
у взрослых — правильный круг или
овал. Вороночный клапан имеется.
Род Liocranchia Pfeffer, 1884
( = Fusocranchia Joubin, 1920) (рис.
69, г—ж). Типовой вид — L. rein-
hardti (Steenstrup, 1856). 2—3 вида
256
в мезо- и батипслагиали (молодь в
эпи- и мезопелагиали) тропических и
субтропических вод. Длина мантии
до 20—25 см.
5(4). В передней части брюшной
стороны мантии 2 гиалиновые полос-
ки, параллельные продольной оси
тела. Нет хрящевых бугорков на
спине. Число глазных фотофоров от
5 до 21: внешняя и внутренняя дуги
или кольца вокруг зрачка. Мантия
удлиненная, цилиндрическая или
узковсрстсновидная. Глаза у личинок
стебельчатые. Плавники округлые
или поперечно-овальные. Нет воро-
ночного клапана................6.
6(7). Длина плавника 20—30%
длины мантии. Концы рук не оття-
нуты, руки 3-й пары менее чем с сот-
ней присосок. Кольца присосок рук
с несколькими зубцами па дисталь-
ном крае. Крупный фотофор может
быть только на концах 3-й пары рук
взрослых самок.
Род Leachia LeSueur, 1821 (=Pe-
ra this Rathke, 1835, Dyctidiopsis
Rochebrune, 1884) (рис. 69, з—т). Ти-
повой вид — L. су dura LeSueur, 1821.
2 подрода: Leachia s. str, с 2—3 ви-
дами и Pyrgopsis Rochebrune, 1884.
(типовой вид — Р. rynchophorus Ro-
chebrune, 1884) с 4—5 видами в мезо-
и батипелагиали (молодь — и в эпи-
пелагиали) преимущественно тропи-
ческих и субтропических вод. Длина
мантии до 15—20 см.
7(6). Длина плавника 33—40%
длины мантии. Концы рук оттянуты,
руки 3-й пары у взрослых более чем
с двумя сотнями присосок. Кольца
присосок в средней части 3-й пары
рук с тремя крупными когтевидпы-
ми зубцами на дистальном крае, они
равной длины или средний крупнее
боковых. Крупный фотофор на кон-
цах всех пар рук у взрослых самок.
Род Drechselia Joubin, 1931. Один
вид — D. danae Joubin, 1931 (рис.
69, у—ч). В мезо-батипелагиали тро-
пической части Восточной Пацифики
(Панамский залив). Длина мантии
до 17 см.
8(1). Нет гиалиновых полосок с
хрящевыми бугорками. Отдельные
хрящевые бугорки могут иметься
только в местах срастания мантии с
воронкой и из1&дка в местах ее
срастания с головой. 1—2 фотофора
на брюшной стороне глазного ябло-
ка, обычно полулунной или серповид-
ной формы, охватывающих глазное
яблоко полукольцом; изредка 3 фо-
тофора (2 длинных и 1 округлый).
Плавники разнообразной формы, ко-
роткие или длинные, но не округлые,
иногда мантия вытянута в длинный
хвост. Боковые стороны воронки не
прирастают к голове. Глаза у личи-
нок стебельчатые. Гектокотилизиру-
ются концы 1-й и (или) 2-й, иногда
3-й или трех первых рук.
Подсемейство Taoniinae . . 9.
9(22). Мантия взрослых кальма-
ров сзади вытянута в длинный
заостренный хвост, конец гладиуса
игловидный или узкоконический.
Плавник длинный, ланцетовидный,
яйцевидный или продольноовальный.
Вороночный клапан имеется или от-
сутствует .....................10.
10(15). На булаве щупалец не-
сколько присосок средних рядов пре-
вращаются в крючья. Хвост длин-
ный, игловидный, плавник ланцето-
видный или листовидный. Нет фото-
форов на концах рук. Нет вороноч-
ного клапана ..................11.
11(14) Нет крючьев на руках.
Плавник ланцетовидный, его ширина
гораздо меньше длины. Взрослые
кальмары средних размеров — дли-
на мантии до 0,6 м.............12.
12(13). Крючья на булаве однозу-
бые, с расширенным основанием и
редуцированным до небольшой щели
просветом, покрыты кожным капю-
шоном. Краевые присоски централь-
ной части булавы редуцированы или
исчезают. Кольца присосок рук
гладкие или (реже) с низкими округ-
лыми зубцами. Обычно (но нс всегда)
имеются хрящевые бугорки в местах
срастания мантии и воронки.
Род Galiteuthis Joubin, 1898
( = Taonidium Pfeffer. 1900, Cry st allo-
teuthis Chun, 1906, IZygaenopsis
Rochebrunc, 1884, ? Zygocranchia Hoy-
le, 1909, ^Euzygaena Chun, 1910,
^Phasmatoteuthion Pfeffer, 1912) (cm.
рис. 71, н—ф). Типовой вид—G. ar-
mata Joubin, 1898. 5 видов в батипе-
л а гнали и батиали (молодь в эпи- и
мезопелагиали) всех океанов, кроме
Северного Ледовитого. Длина ман-
тии до 60 см.
13(12). Крючья на булаве возника-
ют в результате сильного разраста-
ния одного или двух зубцов кольца
присосок, основание таких крючьев не
расширено, просвет кольца не реду-
цирован, крюк не покрыт кожным
капюшоном. Кроме главных крючьс-
подобных зубцов на кольце часто
имеются еще маленькие боковые
зубчики. Краевые присоски централь-
ной части булавы хорошо развиты.
Кольца присосок рук с низкими зуб-
цами (у молоди на дистальном крае,
у взрослых по всему периметру).
Хрящевых бугорков в местах сра-
стания мантии и воронки нет.
Род Belonella Lane, 1957 ( — То-
леита Chun, 1906, Taonius auett., non
LeSueur) (см. рис. 71,6?—м). Типовой
вид—В. belone (Chun, 1906). 3 вида
в батипелагиали и батиали (молодь
в эпи- и мезопелагиали) всех окса-
нов, кроме Северного Ледовитого.
Длина мантии до 54 (60?) см.
14(11). В средних частях рук по
нескольку пар крючьев. Крючья на
булаве щупалец однозубые, покры-
тые капюшоном, как у Galiteuthis.
В местах срастания мантии с ворон-
кой по одному 4—6-вершинному бу-
горку; один конический бугорок на
переднем крае мантии в середине
спинной стороны (у взрослых все эти
бугорки почти незаметны). Плавник
овальный у молоди, широколанцето-
видный у взрослых. Взрослые каль-
мары гигантских размеров, длина
мантии 200—225 см.
Род Mesonychoteuthis Robson,
1925. Один вид, М. hamiltoni Robson
1925 (см. рис. 71, х—ч), антаркти-
ческий (молодь встречается и в но-
талыюй зоне до субтропической кон-
вергенции) циркумполярный бати-
пелагический вид. Обычный компо-
нент пищи кашалотов.
15(10). Нет крючьев па булаве.
Хвост игловидный или узкоконичес-
кий, плавник ланцетовидный, про-
дольно-овальный или яйцевидный.
У взрослых самок обычно имеются
фотофоры на концах рук . . 16.
16(19). Вороночный клапан име-
ется. Срединный компонент вороноч-
ного органа с двумя треугольными
лопастями, без медианной папиллы.
Есть крупные светящиеся органы на
концах 1-й, 2-й или 3-й пары рук
взрослых самок. Кольца крупных
присосок в центральных частях рук
с низкими округленными зубцами. 17.
17(18). На печени 2 крупных ган-
телевидных фотофора с парой круг-
лых линз каждый. Плавник ланцето-
видный (у молоди яйцевидный), его
передние края не переходят на бо-
ковые стороны мантии, прикрепля-
ясь у места наибольшей ширины
17 КН. Несис
257
и
п
ж
и
т
г
у
1
§
о
&
Ml
4
&
*1
$
[А
Й
[1
«3
1
н
5®fW
йнда;
эд
л
£
<Р
л
р
И
[а
И
Z77
5*1
С
У
X
[г/ — самец
в, е, н — гектокотиль: ж —
с, х — хрящевая по-
Ф
У
Рис. 69. Сем. Cranchiidae, подсем. Сгапс-
liiinae: а—в— Cranchia scabra (а — самка.
б — самец); г—ж — Liocranchia reinhardti;
з—и —Leachia (L.) cUipsoptera-, к — L. (L.)
sp. А, мантия; л—н — L. (Pyrgopsis) atlan-
tica (л —самка, м — молодь); о — фото-
фор на конце 3-й руки зрелой самки L.
(Pyrgopsis) sp.; n—c — L. (Р.) pacifica
(=dislocata) (л —самка, p — самец); г —
L. (P.) rynchophorus, молодь; у—ч — Drech-
selia danae [i/ — самец (схематически)
ф — самка];
глазные фотофоры;
лоска с бугорками; ц—ч — присоски сред-
ней части боковых рук (ц) и булавы (ч),
(а, г, ж, т — по G. Voss, 1963, 1967; б—в,
д—е, л—о —по N. Voss, 1980; з—к — пр
Pfeffer, 1912; л—с—no R. Young, 1972; у —
no Joubin, 1931 из Clarke, 1966; ф—ч — по
Boone, 1933)
17*
ланцсолы или чуть впереди. У взрос-
лых особей на концах рук крупные
присоски с мелкими округлыми зуб-
цами резко сменяются мелкими при-
сосками, несущими на дистальном
крае кольца несколько острых клы-
ковидных зубцов. В дистальной час-
ти рук 3-й (в меньшей степени 2-й)
пары несколько резко увеличенных
присосок. Крупный удлиненный фо-
тофор па концах 3-й или первых
трех пар рук взрослых самок.
Род Megalocranchia Pfeffer, 1912
(= Corynomma Chun, 1906, Ascoteut-
his Berry, 1920, Carynoteuthis G. Voss,
1960, Phasmatopsis auett., non Rochc-
brune) (см. рис. 72, к—ф). 2—3 мезо-
батипелагичсских вида в тропических
и субтропических водах всех океанов.
Типовой вид — М. maxima (Pfeffer,
1884). Длина мантии до 81 см.
18(17). Нет фотофоров на печени.
Плавник удлиненно-ланцетовидный,
его передние края у взрослых пере-
ходят на боковые стороны мантии
впереди места наибольшей ширины
ланцсолы. На концах рук нет присо-
сок с клыковидными зубцами. Нет
резко увеличенных присосок на ру-
ках. Крупные удлиненные фотофоры
на концах всех пар рук взрослых
самок.
Род Egea Joubin, 1933, Один
вид — E.inermis Joubin, 1933 (=Phas-
matopsis lucifer G. Voss 1963, ? Teat-
howenia elongata Sasaki, 1929) (cm.
рис. 72,e—и). Тропические и субтро-
пические воды Атлантики и Индо-
Вестпацифнкн. От эпи- до батипела-
гиали. Длина мантии до 42 см.
19(16). Нет вороночного клапана.
Срединный компонент вороночного
органа с медианной папиллой. Нет
фотофоров па руках и печени. Коль-
ца крупных присосок рук гладкие
или неясно зазубренные . . 20.
20(21). Мантия у взрослых сту-
денистой (желеобразной) консистен-
ции, щупальца автотомируются. Ру-
ки короткие, слабые, в их дисталь-
ных частях нет заметно увеличенных
присосок. Глаза смотрят несколько
вперед. Воронка огромная, закрыва-
ет снизу всю голову. Пет хрящевых
бугорков в местах срастания мантии
с* воронкой. Срединный компонент
вороночного органа с конической
медианной папиллой и двумя низки-
ми треугольными лопастями на боко-
вых ветвях. Передние края плавни-
ков не переходят на боковые сторо-
260
ны мантии. Задний конец гладиуса
игловидный.
Род Taonius Steenstrup, 1861
(=Desmoteuthis Verrill, 1881,?РЛа$-
matopsis Rochcbrune, 1884) (см. рис.
71, a—д). Один вид — Т. pavo (Le-
Sueur, 1821). Широко распространен-
ный, но редкий тропическо-субтро-
пический вид, заходит в бореальные
и потальные воды. Предположитель-
но мезопелагиаль. Длина мантии до
40—45 см.
21(20). Мантия кожистой кон-
систенции, плотная. Руки крепкие,
в дистальных частях боковых пар
рук несколько сильно увеличенных
присосок. Щупальца не автотомиру-
ются. Глаза смотрят вбок. Воронка
обычных размеров. Есть один или
несколько хрящевых бугорков в
местах срастания мантии с воронкой.
Срединный компонент вороночного
органа с тремя коническими или
пальцевидными папиллами. Передние
края плавников у поздней молоди и
взрослых прикрепляются к боковым
сторонам мантии впереди места наи-
большей ширины лаицеолы. Задний
конец гладиуса узкоконический, не
игловидный. У взрослых самок име-
ется крупный фотофор на концах
всех пар рук.
Род Teuthowenia Chun, 1906
( = Desmoteuthis Verrill, 1881, part,
Megalocranchia Pfeffer, 1884, part,
Verrilliteuthis Berry, 1916, Anomalo-
cranchia Robson, 1924) (см. рис.
70, к—т). Типовой вид — Т. megalops
(Prosch, 1849). Два вида. Бореаль-
ные и северные субтропические воды
Атлантики, южные субтропические и
мотальные районы Атлантического,
Индийского и юго-западной части
Тихого океанов. Мезо- и батипсла-
гиаль, молодь в эпипелагиали. Длнпа
мантии до 35 см.
22(9). Мантия не вытянута в
длинный хвост, конец гладиуса ши-
рококонический, или ланцеола реду-
цирована и не определяет форму
заднего конца тела. Плавники ко-
роткие, полукруглые, почковидные,
лопатковндпые или языковидные,
прикрепляются к задне-боковым сто-
ронам ланцсолы или прямо к мантии.
Нет фотофоров на концах рук у
самок. Вороночного клапана нет . 23.
23(28). Ланцеола имеется. Плав-
ники прикрепляются к задне-боковым
сторонам ланцеолы. Мантия сзади
тупо закругленная или приостренная.
Рыло и глазные стебельки у личинок
короткие или умеренной длины . 24.
24(27). Плавники полукруглые
или почковидные, ширина каждого
плавника заметно меньше длины.
Задний конец гладиуса заходит за
конец мантии. Лаицеола большая,
ромбическая. 2 глазных фотофора.
Воронка доходит до основания рук
4-й пары ......................25.
25(26). Плавники полукруглые
или полуэллипсовидные, вместе обра-
зуют почти правильный круг. Они
заходят за задний конец гладиуса и
разделяются узкой щелью или сое-
диняются друг с другом. Руки уме-
ренной длины. В дистальных частях
боковых рук несколько увеличенных
присосок. Кольца крупных присосок
рук с низкими коническими или баш-
невидными зубцами. Срединный ком-
понент вороночного органа с 3 кони-
ческими или пальцевидными папил-
лами. Щупальца не автотомируются.
Иногда имеются маленькие бугорки в
местах срастания мантии с воронкой.
Глаза личинок на длинных стебельках.
Род Ligurlella Issel, 1908 («=Vos-
SGteuthis Nesis, 1974) (см. рис.
70, e—и). Типовой вид — L. podophtal-
та Issel, 1908. 2—4 мсзопелагичсских
вида в тропиках, субтропиках и по-
тальных районах Атлантического,
Индийского и западной части Тихо-
го океана. Длина мантии до 24 см.
26(25). Плавники почковидные,
не заходят за задний конец гладиуса
и не сближены друг с другом. Руки
очень короткие, без увеличенных при-
сосок. Кольца присосок рук гладкие
или слабо зазубренные. Срединный
компонент вороночного органа с не-
большой медианной папиллой и дву-
мя широкими треугольными лопастя-
ми на боковых ветвях. Щупальца у
взрослых автотомируются. Нет бу-
горков в местах срастания мантии с
воронкой. Глаза личинок на корот-
ких стебельках.
Род Sandalops Chun, 1906 ( = Ura-
noteuthis Lu et Clarke, 1974). Один
вид — S. melancholicus Chun, 1906
( = Uranoteuthis bilucifer Lu et Clarke,
1974) (см. рис. 72, a—д). Тропические
и субтропические районы Атлантики
(главным образом в центральных
водах) и Индо-Всстпацифики. Мезо-
и батипелагиаль, личинки и молодь
иногда встречаются в нижней эпи-
пелагиали. Длина мантии до 11 см.
27(24). Плавники лопатко- или
языковидные, ширина каждого плав
ника много больше длины. Задний
конец мантии подогнут вниз и мо
жет чуть заходить за задний конец
гладиуса или чуть не доходить дс
пего. Ланцсола маленькая, коротка?
и узкая. Срединный компонент воро
ночного органа с тремя коническим?
папиллами. Один крупный овальны?
или треугольный фотофор на брюш
ной стороне глазного яблока. Ворон
ка громадная, прикрывает снизу век
голову и заходит за основания руг
4-й пары.
Род Helicocranchia Massy, 1901
( = ? Hensenioteuthis Pfeffer, 1900, As
cocranchia G. Voss, 1962) (см. рис
70, а—д). Типовой вид H. pfeffer
Massy, 1907. 3 вида в эпипелагиал?
(молодь) и мезопелагиали тропиче
ских и субтропических вод. Длин*
мантии до 8 см.
28(23). Ланцеола редуцирована
Задний конец мантии широко округ
леи, лопатковидные плавники при
крепляются прямо к мантии вперед?
ее заднего конца и довольно далеке
друг от друга. Мантия цилиндричес
кая, пленчатой консистенции. Глаз;
у взрослых особей сидячие, па ниж
ней стороне глазного яблока одш
крупный полулунный фотофор. Сре
динный компонент вороночного орга
на без медианной папиллы, с двум?
коническими папиллами на боковые
ветвях. Личинки с чрезвычайно длин
пыми глазными стебельками и длин
ным тонким рылом.
Род Bathothauma Chun, 190(
( = Leucocranchia Joubin, 1912). Одш
вид — В. lyromma Chun, 1906 (=Leu
cocranchia pfefferi Joubin, 1912) (cm
рис. 72, x—ч). Тропическо-субтропи
ческий космополитический мезо-ба
типслагический вид (взрослые особ?
обитают в батнпелагнали). Длин*
мантии до 20 см.
Таблица для определения видов род;
Liocranchia
1(2). Вдоль средней линии спинь
над гладиусом проходит ряд хряще
вых бугорков, достигающий начал;
плавника. 14 фотофоров на глазно*
яблоке: 6 кольцом вокруг зрачка, '
дугой по вентральной стороне глаз
ного яблока, 1 между ними, У поло
возрелых самок имеется крупны!
фотофор па концах рук 3-й пары
26
Концы 3-й пары рук зрелых самцов
утончены н песут 4—8 рядов очень
мелких присосок.
L. reinhardti (Steenstrup, 1856)
(=L. intermedia Robson, 1924, Fusoc-
ranchia alpha Joubin, 1920) (см. рис.
69, г—ж). Тропическо-субтропический
космополитический мезо-батипелаги-
ческий вид. Обычен и многочислен.
2(1). Нет хрящевых бугорков
вдоль средней линии спины. 4 фото-
фора дугой па глазном яблоке.
Строение половозрелых особей неиз-
вестно.
L. valdiviae Chun, 1906. Индопаци-
фический тропический батипелагичес-
кий вид, личинки и молодь в эпи- и
мезопелагиали.
Примечание. В таблицу не
включена L gardineri Robson, 1921,
описанная по личинке из западной
части Индийского океана.
Таблица* для определения видов рода
Leachia
1(6). У зрелых самок пет фотофора
па концах рук 3-й пары. Глаза си-
дячие при длине мантии более 4 см.
Подрод Leachia s. str ... 2.
2(5). Длина гиалиновых полосок
с хрящевыми бугорками составляет
‘А—’/з длины мантии .... 3.
3(4). 6 глазных фотофоров при
длине мантии 7—9 см: 5 во внешнем
ряду (при длине мантии 4—6 см их
4), 1 вблизи зрачка.
L, (L.) ellipsoptera Adams et. Reeve,
1845 (—L. cyclurd auett., part) (cm.
рис. 69, з, и). Тропическая и северная
субтропическая Атлантика.
4(3). 8 глазных ‘ фотофоров при
длине мантии 6—7 см: 5 во внешнем
ряду и 3 вблизи зрачка.
L. (L.) eyelwra LeSueur, 1821
[=£. eschscholtzi (Rathke, 1835),
£. guttata (Grant, 1835)]. Тропические
и южносубтропические воды Индо-
Вестпацифики.
5(2). Длина гиалиновых полосок
с хрящевыми бугорками составляет
2А—3А длины мантии. 8 глазных фо-
тофоров при длине мантии 8,5—
10 см.
L. (LA sp.A Nesis (=L. eschscholt-
zi Pfeffer, 1912: 655—656, Taf. 47 fig.
♦ Таблица предварительная, так как
полный, цикл развития описан пока
лишь для одного вида — L. pacifica.
262
12—14, non Rathke) (см. рис. 69, к).
Юго-Западная Атлантика (восточнее
Северной Аргентины) и (?) новозе-
ландские воды. По-видимому, южно-
периферический вид.
6(1). У зрелых самок есть круп-
ный удлиненный фотофор па концах
рук 3-й пары (см. рис. 69, о). У личи-
нок и молоди при длине мантии ме-
нее 5—10 см стебельчатые глаза.
Подрод Pyrgopsis Rochebrune,
1884 ...................... 7.
7(14). Фотофоры на 3-й паре рук
самки развиваются при длине мантии
4—12 см или более .... 8.
8(13). Длина гиалиновых полосок
с хрящевыми бугорками до ’А длины
мантии. Ширина плавника превосхо-
дит его длину в 1,2—1,6 раза . 9.
9(10). Плавник с выпуклым пе-
редним и почти прямым задним кра-
ем, место наибольшей ширины плав-
ника находится заметно дальше се-
редины его длины. Полоска хряще-
вых бугорков прямая, се длина около
1/7 длины мантии.
L, (Р.) atlantica (Degncr, 1925)
(=? Pyrgopsis sp. Adam, 1960,
? P. lemur Mercer, 1966, non Berry)
(см. рис. 69, л—н). Тропическая и се-
верная субтропическая Атлантика,
на север доходит до района южнее
Большой Ньюфаундлендской банки и
Бискайского залива.
10(9). Плавник округлый или по-
перечно-овальный, место наибольшей
ширины плавника находится пример-
но посредине его длины . . . И.
11(12). Плавник поперечно-оваль-
ный, его ширина в 1,3—1,6 раза,
обычно в 1,35—1,5 рйза больше дли-
ны. Полоска хрящевых бугорков у
крупных особей явственно изогнута
в передней части: второй от края
мантии бугорок смещен и находится
вентральнее остальных (это заметно
при длине мантии 6—7 см, но у каль-
маров из Калифорнийского течения
иногда уже при 2 см). Длина полос-
ки ’/«—V» длины маптии.
L.(P.) pacifica (Issel, 1908)
( = L. dislocata Young, 1972, ? Pyrgop-
sis schnehageni Pfeffcr, 1884, ? P. le-
mur Berry, 1920) (см. рис. 69, n—c).
Тропическо-субтропический космопо-
литический вид.
12(11). Плавник почти округлый,
его ширина в 1,2—1,3 раза больше
длины. Полоска хрящевых бугорков
прямая, се длина около 75 длины
маптии.
L. (P.) sp. В Nesis ( = ? P, lemur
Voss, 1958, non Berry). Тропические
воды Атлантики.
13(8). Длина гиалиновых полосок
с хрящевыми бугорками ’/<—V2 Дли-
ны мантии. Плавник широкооваль-
ный, его ширина в 1,6—1,9 раза
больше длины, место наибольшей
ширины плавника немного дальше
середины его длины.
L. (Р-) rynchophorus Rochebrune,
1884 (=Р. pacifica Massy, 1916; Rob-
son, 1924; Allan, 1945; G. Voss, 1967,
non Issel) (см. рис. 69, г). Южносуб-
тропический вид, распространен от
Южной Африки до Новой Зеландии.
14(7). Фотофоры на концах 3-м
пары рук развиты при длине мантии
3,5—4 см.
L. (Р.) sp. С Nesis. Тропические
воды Индийского океана.
Таблица для определения видов рода
Helicocranchia
1(2). Плавники лопатковидные с уз-
ким основанием и расширенной ди-
стальной частью. Основания плавни-
ков соединены узкой пленкой ди-
стальнее конца гладиуса. Поверх-
ность мантии гладкая. Присоски в
средней части рук 3-й пары у самок
не увеличены. Кольца присосок рук
с дистальной стороны с нескольки-
ми низкими округлыми зубцами,
проксимально гладкие. Окраска свет-
лая с редкими крупными овальными
хроматофорами оранжевого или тем-
но-коричневого цвета, которые на
боках и вентральной стороне мантии
образуют поперечные ряды. Мелких
черпых хроматофоров нет.
H.pfefferi Massy, 1907 (=Н. bee-
bei Robson, 1948) (рис. 70, а, б). Тро-
пическо-субтропический космополити-
ческий мезопелагический вид (молодь
встречается в эпипелагиали и даже у
поверхности, взрослые — в мезо- и
батипслагиали).
2(1). Плавники языковидные, оди-
наковой ширины у основания и в се-
редине переднего края, сидят на
ланцеоле гладиуса, задние края
обоих плавников просто срастаются.
Присоски в средней части рук 3-й
пары у самок с гладкими кольцами
и значительно крупнее, чем на дру-
гих руках. Присоски других рук с
мелко зазубренными кольцами. Ок-
раска красно-бурая..............3.
3(4). Мантия толстая, кожистая,
мускулистая. Поверхность мантии
(особенно в передней части) и во-
ронки покрыта многочисленными
маленькими острыми кожными па-
пиллами.
H.papillata (Voss, 1960) (=? Неп-
senioteuthis joubini Pfeffer, 1912,
Teuthowenla megalops Joubin, 1933,
non Prosch) (рис. 70, в, г). Централь-
ные воды Северной Атлантики. Ме-
зо- и батипелагиаль.
4(3). Мантия тонкая, полустудс-
нистой консистенции, гладкая или
почти гладкая. Окраска однотонно
красно-бурая или кирпичная с немно-
гочисленными светлыми овальными
пятнами и множеством очень мелких
почти черных точек, разбросанных
по всей поверхности мантии.
Н. joubini (Voss, 1962) (рис. 70, б).
Тропические и субтропические воды
Атлантики, юго-западная часть Тихо-
го оксана. Мезо- и батипелагиаль.
Таблица для определения видов
рода Teuthowenla
1(2), В местах срастания мантии с
воронкой по одному крохотному
хрящевому бугорку.
Т. megalops (Prosch, 1849) [=Des-
moleuthis hyperborea (Steenstrup,
1856), D.tenera Verrill, 1881, D.thori
Degner, 1925, Verrilliteuthis megalops
auett, part, Megalocranchia megalops
auett, Desmoteuthis megalops auett.].
Североатлантический бореально-суб-
тропический вид; от Юго-Западной
Гренландии, Датского пролива, Фа-
реро-Исландского и Фареро-Шет-
лаидского порогов до Саргассова мо-
ря, островов Антильских, Канарских
и Зеленого Мыса. Единично встреча-
ется в Карибском море и Гвинейском
заливе (стерильная зона выселения),
отсутствует в Норвежском, Северном
и Средиземном морях. Обычный
и многочисленный кальмар.
2(1). В местах срастания мантии
с воронкой по 2—5 конических хря-
щевых бугорков (у ранней молоди
по одному).
Т. richardsoni (Dell, 1959)
(=? Desmoteuthis pellucida Chun,
1910,? Anomalocranchia impennis Rob-
son, 1924, Megalocranchia. megalops
australis G. Voss, 1967) (рис. 70, о—
г). Нотально-южпосубтропический
вид: в южных частях Атлантическо-
го, Индийского и юго-западной час-
ти Тихого океана до Новой Зелан-
дии и островов Кермадек.
263
Таблица для определения видов рода
Liguriella
1(2). Кольца крупных присосок бу-
лавы с 5—7 длинными острыми зуб-
цами дистально. В местах срастания
мантии с воронкой по 1—2 малень-
ких конических бугорка.
L. podophlalma Issel, 1908 (=Vos-
soteuthis pellucida Nesis, 1974, non
Chun 1910, «Bathothauma lyromma»
Allan, 1940, non Chun, 1906) (рис.
70, e—u)t Южносубтропическо-но-
тальный мезопелагичсский вид, обна-
ружен в южных частях Атлантиче-
ского и Тихого океанов, в Тасмано-
вом море и у Новой Зеландии.
2(1). Кольца крупных присосок
булавы с приблизительно 15—25 ост-
рыми коническими зубцами по всему
периметру (более высокими на дис-
тальном крае)...................3.
3(4). Около 25 невысоких кони-
ческих зубцов на крупных присосках
булавы.
L. pardus (Berry, 1916). Южносуб-
тропический, возможно мезопелаги-
ческий вид, найден у островов Кср-
мадек и (?) у Юго-Восточной Ав-
стралии, Тасмании и в Новогвиней-
ском море.
4(3). Около 15 высоких острых
зубцов на крупных присосках була-
вы. Нет бугорков в местах сраста-
ния мантии с воронкой.
Liguriella sp. Nesis ( = <tTeuthowe-
nia megalops» Morales et Guerra,
1977 non Prosch). Северо-Западная
Африка мористее мысов Зеленый,
Кап-Блан и Тимирис. Мезопелагиаль.
Систематическое положение вида не-
ясно.
Личинки неидентифицированного
вида Liguriella найдены (Несис,
1974 б) в северных центральных во-
дах Западной Пацифики и Филип-
пинском море.
Таблица для определения видов рода
Belonella
1(4). Защитные мембраны рук уз-
кие, их ширина примерно равна
диаметру присосок рук .... 2.
2(3). На самых крупных присос-
ках центральной части булавы два
264
3
и
ВФ$
to
й‘И
ЬУ
А)
gl.j
О
М
S':
л
л
ж
н
71
р
7П
Подсем. Taoniinae, роды Helicoc-
Рис. 70. __.____ __________, ____
rartchia (а—д), Liguriella (е—и) и Teutho-
wenia (к—т): а—б — Helico cranchia pfeffe-
ri; в—г — Н. papillata; д — Н. joubini- е—
и — Liguriella podophtalma; к—н — Teutho-
wenia megalops; о—т — T. richardsoni: а„
ж, л, о — молодые кальмары; б, и, м —
глаз (б. м — в двух положениях); г — го-
лова снизу, видны кожные бугорки; з, р —
места срастания маптии с воронкой; н —
вороночный орган; п — булава; с, т — при-
соски булавы и средней части боковых
рук (а — по Okutaiii, 1974; б—в, е—к, м—
w — по N. Voss, 1980; г—д, о—т — по’
G. Voss. 1960, 1962, 1967; г — по VcrrilL.
1882)
Рис. 71. Роды Taonius (а—д), Belonella
(е—.и), Galiteuthis (н—ф) и Mesonychoteut-
his (х—ч); а—О — Taonius pavo; е-к — Ве-
lonelta belone; л—м — В. borealis; н—п —
Galiteuthis phyllura: р—у — G. glacialis:
ф — G. pacifica; х—ч — Mesonychoteuthis
hamiltoni- ж, ц — молодые кальмары; б,
з, у — глаз {у— в двух положениях); в —
вороночный орган, и, о, с—булава; ч —
руки (слева направо — 1—4-я пара); г, д —
кольца присосок из средней и дистальной
частей боковых рук; к, м — крючья из
средней части булавы; п, т, ф — места
срастания мантии с воронкой, г, — с голо-
вой (а — по Joubin, 1900; б—д, к, м — по
Несису, 1972а; е—и, р—с, у, л—ч —по N.
Voss, 1980; л — по Акимушкину, 1963; к—
л, ф по R. Young, 1972; т — по McSweeny,
1978)
(редко один) крупных крючковидных
зубца, по бокам от них нет мелких
зубчиков.
В. belone (Chun, 1906) ( = В. paci-
fica pacifica Nesis, 1972, Galiteuihis
triluminosa Lu et Clarke, 1974, Taonius
pavo auctt., non LeSucur) (рис. 71,
e—к). Тропические и субтропические
воды Атлантики, тропические воды
Индийского и Тихого океанов; в Ти-
хом океане от острова Кюсю до ост-
ровов Тонга. Батипелагиаль и бати-
аль, молодь обитает преимуществен-
но в мсзопелагиали.
3(2). На самых крупных присос-
ках центральной части булавы 1—2
крупных крючковидных зубца, по бо-
кам от них с каждой стороны коль-
ца по 2—3 маленьких зубчика.
В. borealis Nesis, 1972 ( = Taonius
pavo Akimushkin 1963, non LcSueur,
B. pacifica borealis Nesis, 1972a) (cm.
рис. 71, л, м). Тихоокеанский боре-
альный вид: от Берингова моря до
Восточного Хонсю и Южной Кали-
форнии, также в Охотском море. Ме-
зо- и батипелагиаль, батиаль и абис-
саль.
4(1). Защитные мембраны рук
широкие, примерно равны ширине
руки. На самых крупных присосках
центральной части булавы один зу-
бец намного крупнее остальных, кро-
ме него имеются 1—5 маленьких зуб-
чиков.
Belonella sp. Nesis, 1974 а. Ноталь-
но-антарктический батипелагический
и батиальный вид, найден в море
Скотия и предположительно в индо-
океанском секторе Антарктики и у
островов Антиподов.
Таблица для определения
видов рода Galiteuthis
1(6). В местах срастания мантии с
воронкой, а иногда и с головой име-
ются хрящевые бугорки ... 2.
2(3). По 2—4 маленьких кониче-
ских одновершинных бугорка в мес-
тах срастания мантии и воронки
(у взрослых их нелегко заметить, но
легко нащупать пальцем). Нет бугор-
ков па спинной стороне мантии. При-
соски рук с гладкими кольцами. По-
верхность мантии гладкая. Перед-
ний глазной фотофор узкий, палоч-
ковидный.
G. armata Joubin, 1898 [=? Zygo-
cranchia zygaena (Verany, 1851),
Taonidium pfefferi Russell, 1909, G.
suhmi Chun, 1910, non Hoyle, 1885].
Тропическая и северная субтропиче-
ская Атлантика, от Бискайского до
Гвинейского залива и Флоридского
пролива, Средиземное море. Мезо-
батипелагический и придонно-бати-
альный вид.
G. phyllura Berry, 1911 (=Crystal-
loteuthis beringiana Sasaki, 1920)
(рис. 71, н—n). Бореальный тихооке-
анский мезо-батипслагический и при-
донно-батиальный вид, распростра-
ненный от Берингова моря до Север-
ной Японии и п-ва Калифорния и в
Охотском море. Различия между ат-
лантическим и тихоокеанским вида-
ми очень невелики и нуждаются в
подтверждении на большом мате-
риале.
3(2). Бугорки в местах срастания
мантии с воронкой многовершинные,
крупные . 4.
4(5). В местах срастания мантии
и воронки по одному бугорку с не-
сколькими (у взрослых до 5—6)
вершинами. Нет бугорков на спинной
стороне мантии. Присоски рук с глад-
кими кольцами. Поверхность мантии
гладкая.
G. suhmi (Hoyle, 1885) ( = Taoni-
dium suhmi Hoyle, 1885, non 1912).
Южносубтропическо-нотальный цир-
кумглобальный мезо-батипелагичс-
ский вид.
5(4). В местах срастания мантии
и воронки по два сближенных трех-
вершинных хрящевых бугорка, обра-
зующих короткий гребень. В хместе
срастания мантии с головой два ма-
леньких трсхвершинных бугорка, раз-
несенных по обе стороны переднего
конца гладиуса. Кольца присосок рук
дистально с низкими округленными
зубцами. Поверхность мантии, осо-
бенно в передней части, шероховатая,
«шагреневая» из-за множества мик-
роскопических кожных шипиков.
G. glacialis (Chun, 1906) ( = Теи-
thowenia antarctica Chun, 1910, Crys-
tal tote ut his glacialis Chun 1906, G. as-
pera Filippova, 1972, ? Taonidium
suhmi Hoyle, 1912, non 1885) (рис.
71,p—у). Антарктический циркумпо-
лярный мезо-батипслагический вид,
поднимающийся и в нижнюю эпипе-
лагиаль. Один из самых обычных и
многочисленных кальмаров Антарк-
тики. Севернее антарктической кон-
вергенции встречается лишь в мес-
тах выноса больших масс антаркти-
ческих промежуточных вод.
268
6(1). Нет хрящевых бугорков в
местах срастания мантии с воронкой
и головой. Кольца присосок рук
гладкие. Поверхность мантии глад-
кая. Передний глазной фотофор
округлый или овальный.
G. pacifica (Robson, 1948) ( = Тао-
nidium pacificum Robson, 1948) (рис.
71, ф). Тихоокеанский тропический
вид, распространен от Филиппинско-
го моря и Южной Калифорнии до
Новой Каледонии и Северного Чили.
Мсзо-батипслагический вид.
Таблица для определения видов рода
Megalocranchia
1(4). Кольца крупных присосок бу-
лавы с 6—12 сближенными крупны-
ми зубцами на дистальном крас
кольца, проксимальный край с мел-
кими зубчиками..................2.
2(3). Кольца крупных присосок
булавы с 10—12 крупными зубцами
на дистальном крас, проксимально с
мелкими зубчиками. Крупные присос-
ки 3-й пары с тесно сближенными
правильными башнсвидными зуб-
цами.
М. abyssicola (Goodrich 1896)
(= ? Megalocranchia maxima Pfeffer,
1884, Corynomma speculator Chun,
1910.. part. ? Helicocranchia fisheri
Berry, 1909, M. maxima Sasaki, 1929)
(рис’ 72, p—т). Индотихоокеанский
тропическо-субтропический мсзо-ба-
типелагический (часто поднимается в
нижнюю энинелагиаль) вид.
3(2). Кольца крупных присосок
булавы с 6—8 крупными зубцами на
дистальном крае, проксимально с
мелкими зубчиками. Крупные при-
соски рук 3-й пары с неправильными
низкими округлыми зубцами.
М. oceanica (Voss, 1960) ( = Cory-
nomma speculator Chun 1910, part,
? Teuthowenla (Ascoteuthis) corona
Berry 1920, M. abyssicola Joubin, 1924,
non Goodrich, 1896, Carynoteuthis
oceanica G. Voss, 1960, ? Phasmatop-
sls cymoctypus Clarke, 1962, non Ro-
chebrune, 1884) (рис. 72, к—n). Ат-
лантический тропическо-субтропиче-
ский (преимущественно бицептраль-
ный) мсзо-батипелагнческий вид, ли-
чинки и молодь встречаются также
в эпипелагнали.
4(1). Кольца крупных присосок
булавы с 15—18 зубцами по всему
периметру, из них 2—3 дистальных
зубца очень широкие и далеко от-
ставлены друг от друга, прочие бо-
лее узкие и теснее сближены. Круп-
ные присоски 3-й пары рук с десят-
ком башневидных зубцов.
Megalocranchia sp. Nesis (рис.
72, у, ф). Тихий океан северо-восточ-
нее Хонсю и в Куросио. Системати-
ческое положение вида неясно.
2.3. Отряд Vampyromorpha
Животные средних размеров (общая
длина до 37,5 см, длина мантии до
11—13 см). Ткани студенистой кон-
систенции. Ширококоническая ман-
тия сращена на затылке с широкой
головой так, что шейного перехвата
пет. Два весловидных плавника на-
ходятся в задней части мантии.
Мантийное отверстие широкое, дохо-
дит до уровня середины глаз. Глаза
эгопсидного типа, крупные, без си-
нуса. Восемь рук. Руки недлинные,
соединенные глубокой умбреллой,
несут один ряд присосок, начинаю-
щихся далеко от основания рук. При-
соски без хитиновых колец, сидя-
чие в средних и стебельчатые в дис-
тальных частях рук. По обе стороны
от присосок тянутся два ряда корот-
ких усиков (по паре между кажды-
ми двумя присосками). Концы рук
тонкие, оттянутые, лишены присосок;
усики начинаются проксимальнее
первой присоски и тянутся до кон-
цов рук, у самых концов они сраста-
ются в мембрану. На спинной сто-
роне тела между основаниями 1-й и
2-й пар рук пара очень длинных,
тонких, нитевидных чувствительных
филаментов, которые могут полно-
стью втягиваться в специальные кар-
маны. Это — органы осязания, но они
гомологичны щупальцам кальмаров.
Имеются статоцисты, обонятельные
папиллы и парольфакторные пузырь-
ки. Два крупных сложных фотофора
фасеточного строения находятся в
задней части мантии по бокам, сзади
плавников. Множество мелких мик-
роскопических фотоФоров разбросано
ио поверхности (преимущественно
брюшной) мантии, головы, на плав-
никах, вдоль рук; на спинной сторо-
не головы за глазами они образуют
два овальных скопления.
Буккальной мембраны нет. Ворон-
ка почти целиком погружена в тка-
ни нижней стороны головы. Вороноч-
ный клапан имеется. Вороночный ор-
ган имеет вид двух небольших вали-
266
£
Ж
К
и
''ij
Рис. 72. Роды Sandalops (а—д), Egea (е—
«), Megalocranchia (к—ф) и Bathothaunur
(х—ч): а—д — Sandalops melancholicus;
е—и — Egea inermis; к—п — Megalo.Qranchia
oceanica; р—т — M. abyssicola: у—ф -Me-
galocranchia sp.; х—ч — Bathothauma lyrotn-
ma: a, e, x — самцы; 6, к, ц — самки; гмс,
л, ч — молодые кальмары; в — плавник:
г, з, ле—глаз (з, м — в двух положениях);
д, и, т—вороночный орган (и — молодого-
кальмара); «—фотофоры на чернильном
мешке; о, р, ф — присоски из средней час-
ти боковых рук; п, с—присоски булавы;
у — руки и щупальце (а, г—к, ц—ч — по
N. Voss, 1974, 1980; б, х — по Okutani, 1974,
1975; у—ф — по Okutani et al., 1976; в, о—
т — по Несису, 19746)
ков. Имеется примитивный мантийно-
вороночный замыкательный аппарат.
Гладиус тонкий, прозрачный, очень
широкий, сзади постепенно сужаю-
щийся. Медианной септы в мантий-
ной полости нет. Радула хорошо раз-
вита, все зубцы одновершинные.
Чернильный мешок отсутствует. Зоб
и задние слюнные железы маленькие.
Слепой мешок желудка (цскум) нс
закручен спирально/ Жабры хорошо
развиты. Почечные мешки полностью
разделенные. Звездчатый ганглий
рудиментарный, гигантских воло-
кон нет.
Самцы и самки одинаковых раз-
меров. Гектокотнлизацин пет. Пенис
обособлен от нидхсмова мешка. Раз-
виты оба яйцевода, их выводные от-
верстия расширены и напоминают по
•форме присоски. Нет нидаменталь-
ных и придаточных нидаментальных
желез. Два семеприсмннка находятся
на голове самки перед глазами. Яй-
ца крупные, диаметром 3—4 мм, вы-
метываются в воду поодиночке. Ли-
чинки внешне похожи на взрослых
•особей, ио проходят длительный
путь развития со сменой плавников:
личиночные плавники сидят у самого
конца тела, за сложными фотофора-
ми, по мере роста они укорачивают-
ся, а впереди фотофоров появляются
•окончательные плавники, так что на
определенной стадии развития моло-
дые животные имеют две пары плав-
ников. Затем личиночные плавники
редуцируются, а дефинитивные до-
стигают окончательных размеров.
Хроматофоры примитивные, не-
способны расширяться и сокращать-
ся. Окраска тела, головы и наруж-
ной стороны рук темно-фиолетовая
или темно-пурпурная, внутренняя
сторона рук бархатисто-черная.
Одно семейство Vampyroteuthidae,
•один род Vampyroteuthis Chun, 1903
(=Melanoteuthis Joubin, 1912, Hyme-
noieuthis Berry, 1916, Watasella Sa-
saki, 1920, Retroteuthis Joubin, 1929,
Hansenoteuthis Joubin, 1929, Dana-
teuthis Joubin, 1929). Один вид— V. in-
femails Chun, 1903 (рис. 73). Тропиче-
ско-субтропический батипелагический,
опускающийся и в абиссопелагиаль
вид. Обитает главным образом на
глубинах 700—1500 м, но в местах с
неглубоким залеганием главного тер-
моклина молодь встречается на глу-
бинах 300—500 м. Не стайные жи-
вотные, не совершающие заметных
272
вертикальных перемещений. Плава-
ют, вероятно, вниз головой. Обзор:
Pickford, 1946, 1949, 1959.
2.4. Отряд Octopoda —
осьминоги
Животные от мелких до очень круп-
ных (Octopus dofleini— общая дли-
на до 3—5 м). Тело может быть
плотным, мускулистым у прибреж-
ных видов или мягким, студенистым,
даже желеобразным у глубоковод-
ных видов. Мантия обычно мешко-
видная, на затылке широко сраста-
ется с головой. Мантийное отверстие
широкое, узкое или редуцированное.
Мантийно-вороночный замыкательный
аппарат слабо развит или отсутству-
ет, иногда мантия срастается с во-
ронкой. Поверхность мантии глад-
кая или с разного рода бугорками,
бородавочками, усиками (часто над
глазами). Плавники имеются (одна
пара) или отсутствуют. Глаза обыч-
но крупные, с замкнутой роговицей,
с веками или без них, в редких слу-
чаях уменьшены в размерах, теле-
скопические или редуцированные.
Всегда восемь рук. Присоски в один
или два ряда, обычно уплощенные,
редко шаро- или веретен овидные,
всегда без хитиновых колец. Усики
по бокам присосок (вероятно, орга-
ны осязания) имеются или отсутст-
вуют. Руки часто соединены перепон-
кой — умбреллой; у форм с глубокой
умбреллой она играет роль движите-
ля (медузоидпый способ движения).
У многих видов умбрелла слабо раз-
вита или отсутствует. Чувствитель-
ных филаментов нет. Статоцисты хо-
рошо развиты. Обонятельных папилл
нет, есть обонятельные ямки (редко
гребешки). Светящиеся органы име-
ются у очень немногих видов и
представляют собой простые скопле-
ния фотогенной ткани. Хроматофоры
сложного строения, способны расши-
ряться и сокращаться, но у некото-
рых, преимущественно глубоковод-
ных, форм простого строения. Ворон-
ка свободна или срастается с голо-
вой. Вороночный клапан имеется.
Вороночный клапан хорошо развит,
цельный или подразделенный на 2—
4 части. Буккальной мембраны нет.
Внутренностный мешок не соеди-
нен с мантией на спинной стороне,
но обычно соединен с ней па брюш-
ной стороне медианной мантийной
септой и мускулом — мантийным ад-
дуктором. Гладиус представлен либо
хрящевой поддержкой плавников,
либо редуцирован до пары крохот-
ных палочек, либо отсутствует.
Радула, зоб, задние слюнные же-
лезы и чернильный мешок обычно
хорошо развиты, различного строе-
ния, но иногда могут отсутствовать.
Желудок обычно мускулистый, сле-
пой мешок желудка (цекум) хорошо
развит, обычно нс закручен спираль-
но. Печень крупная, как правило,
впереди желудка. Две половины каж-
дой жабры параллельны и сраста-
ются на обеих сторонах или напоми-
нают по форме дольки апельсина.
Кровеносная система незамкнутая.
Перикард редуцирован. Почечные
мешки разделены. Мозг очень сильно
развит, сконцентрирован, с хорошо
обособленными долями. Оптические
доли обычно невелики. Звездчатый
ганглий имеется, с ним связаны эпи-
стеллярные тельца — дополнительные
органы фоторецепции. Системы ги-
гантских нервных волокон нет.
Гектокотиль обычно имеется, ви-
доизменяется обычно одна из рук
3-й пары. Гектокотнлизация захваты-
вает либо конец руки, либо всю руку
целиком, в последнем случае опа
развивается в особом подкожном
мешке и отрывается при спаривании
от тела самца. У самцов иногда име-
ются увеличенные присоски на неко-
торых руках. У самок нет семепри-
емпиков, нидаментальных и прида-
точных нидаментальных желез.
Яйцеводы парные или непарные
(только левый). Сперматофоры вво-
дятся в мантийную полость самки,
спермин проникают в яйцеводы или
даже в яичник. Яйца от очень ма-
леньких до очень крупных, удли-
ненные и снабженные стебельком.
У донных форм они откладываются
па дно, самки некоторых осьминогов
охраняют яйца и ухаживают за ни-
ми вплоть до вылупления всех личи-
нок. Яйца пелагических осьминогов
либо развиваются внутри тела самки
(яйцеживорождение), либо самка
носит их на себе, или они отклады-
ваются в воду, сплетенные стебель-
ками в кучки. Инкубационный пе-
риод обычно длительный. Развитие
прямое или со стадией пелагической
личинки. Личинки похожи на взрос-
лых особей. Два подотряда, 11 се-
мейств, около 40 родов, примерно
200 видов.
Донные, придонные или пелаги-
ческие животные, обитают от лито-
рали и эпипелагиали до абиссали и
абиссопелагиали. Распространены от
Арктики до Антарктики. Как прави-
ло, это одиночные животные, актив-
ные в сумерках или ночью. Планк-
тофаги, бентофаги или хищники.
Темп роста быстрый, большая часть
видов достигает половой зрелости в
возрасте одного года или нескольких
месяцев. Самки большинства, а мо-
жет быть, и всех видов гибнут после
первого нереста. Некоторые виды се-
мейства Octopodidae имеют большое
промысловое значение. Обзор: Rob-
son, 1929, 1932.
Таблица для определения семейств
отряда Octopoda
1 (4). Плавники имеются. Усики на
руках есть. Мантийное отверстие
очень узкое или редуцированное.
Подотряд Cirrata.............2.
2(3). Животные имеют форму ко-
локольчика, мантия удлиненная, хо-
рошо развита. Умбрелла тонкая, го-
раздо тоньше рук, изредка отсутст-
вует. Плавники посредине или у зад-
него конца мантии. Воронка направ-
лена вперед. Придонно-пелагические
(бентопелагичсские) или пелагические
виды.
Сем. Cirroteuthidae ... с. 276.
3(2). Животные имеют форму
толстой лепешки, мантия очень уко-
рочена и представляет собой бугор,
лежащий на растопыренных руках.
Умбрелла толстая, руки погружены
в нес и выступают сверху лишь как
валики. Воронка направлена назад.
Плавники сидят «на макушко за
глазами. Донные (наддоппые) виды.
Сем. Opisthoteuthidae. Один род—
Opisthoteuthis Verrill, 1883 с. 281.
4(1). Плавников и усиков на ру-
ках пет. Присоски в один или два
ряда. Мантийное отверстие узкое или
чаще широкое.
Подотряд Incirrata .... 5.
5(14). Тело студенистое или по-
лустуденистое. Присоски на руках в
один ряд (или в один ряд в преде-
лах умбреллы и в два в дистальных
частях рук). Мезо- и батипелагиче-
ские, редко придонные виды . . 6.
6(11). Мантийное отверстие ши-
рокое. Воронка не срастается с ман-
18 К. Н. Несис
273
тией ............................7.
7(10). Присоски на руках в один
ряд. Диаметр глаза не более */< дли-
ны мантии. Ширина головы обычно
в 1,5—2,5 раза меньше длины ман-
тии. Гектокотилизации пет или опа
захватывает только кончик руки сам-
ца, гектокотиль нс отрывается от те-
ла при спаривании. Рахидиальный
(центральный) зуб радулы многовер-
шинный. Тело обычно папу прозрач-
ное. Океанические пелагические ви-
ды ..............................8.
8(9). Руки не длиннее туловища.
Диаметр присосок 3—7% длины
мантии. Глаза округлые или оваль-
ные. Печень в нормальном положе-
нии— впереди желудка, овальная.
Радула гребенчатого строения. Ман-
тия 'полупрозрачная, пигментирован-
ная.
Сем. Bolitaenidae ... с. 286.
9(8). Руки у взрослых в 2—3 ра-
за длиннее туловища. Проксималь-
ные присоски мелкие, далеко рас-
ставленные, дистальные очень круп-
ные и сближенные, у взрослых осо-
бей их диаметр 10—17% длины ман-
тии. Глаза узкие, почти прямоуголь-
ные. Печень длинная, топкая, ’заост-
ренная сзади, лежит позади желуд-
ка. Радула не гребенчатая. 1-й и 2-й
латеральные зубцы одновершинные.
Мантия почти прозрачная и бес-
цветная.
Сем. Vitreledonellidac. Один род. и
вид — Vitreledonella richardi Joubin.
1918.............................с. 322.
10(7). Присоски па руках в один
ряд в пределах умбреллы, в два ря-
да за се пределами и снова в один
ряд па самых копчиках. Глаза огром-
ные, их диаметр Уз—% длины .ман-
тии, ширина головы равна или пре-
восходит длину мантии. 3-я правая
рука самца гектокотилизируется пол-
ностью, развивается в особом мешке
под глазом и отрывается от тела
при спаривании. Радула не гребенча-
тая, рахидиальный зуб трехвершин-
иый. Ткани студенистые, но непро-
зрачные. Нерито-океанические жи-
вотные, обитают у дна в батиали и
в толще воды над склоном (.молодь
может быть встречена и вдали от
склона).
Сем. Alloposidae. Один род и
вид — Alloposus mollis Verrill,
1880 •..........................с. 323.
• 11(6). Мантия срастается с во-
ронкой, мантийное отверстие редуци-
ровано до двух щелей, расположен-
ных по бокам головы. Руки в 1,5—
3 раза длиннее туловища. Воронка
очень длинная. Печень лежит позади
желудка. Радула гребенчатая . 12.
12(13). Глаза телескопические,
сближены основаниями и смотрят
вверх. Умбрелла хорошо развита.
Воронка не прирастает снизу к голо-
ве. Тело полупрозрачное, почти бес-
цветное, покрыто студенистым чехлом.
Сем. Amphitretidae. Один род и
вид — Amphitretus pelagicus Hoyle,
1885 ......................с. 287.
13(12). Глаза нормального строе-
ния. Умбрелла неглубокая, тонкая.
Воронка снизу прирастает к голове,
свободен лишь ее передний конец. Те-
ло полупрозрачное, по со многими
подкожными хроматофорами. Нет
студенистого чехла.
Сем. Idioctopodidae. Один род и
вид — Idioctopus gracilipes Taki,
1962 ......................с. 287.
14(5). Тело мускулистое, па
ощупь плотное или мягкое, по не сту-
денистое и не полупрозрачное. При-
соски либо в один, либо в два ряда
по всей длине руки. Донные или эпи-
пелагические виды...............15.
15(16). Донные животные. Сам-
цы и самки слабо различаются по
размерам. Гектокотилизируется
только конец 3-й руки, гектокотили-
зированная рука развивается нор-
мально и при спаривании не отрыва-
ется. Яйца обычно откладываются
на дно. Мантийно-вороночные замы-
кательные хрящи отсутствуют. При-
соски в 1 или 2 ряда.
Сем. Octopodidae ... с. 292.
16(15), Эпипелагические живот-
ные. Самцы карликовые, во много
раз мельче самок. Гектокотилизиру-
ется вся 3-я рука самца, гектокотиль
развивается в особом мешке, так что
кажется, будто у неполовозрелого
самца лишь’7 рук (это заметно да-
же у личинок). При спаривании гек-
токотиль отрывается от тела и про-
никает в мантийную полость самки.
Самки живородящие либо носят яй-
ца на себе (на руках или в ракови-
не). Мантийно-вороночные замыка-
тельные хрящи имеются. Присоски в
2 ряда..........................17.
17(18). Самка выделяет тонкую
ребристую наружную раковину, в ко-
торой сидит сама и куда она от-
кладывает яйца. На концах рук 1-й
пары самки широкая, тонкая, очень
274
эластичная кожная лопасть, секрети-
рующая раковину; у живого осьми-
нога она покрывает всю раковину
снаружи. Умбрелла очень слабо раз-
вита" 3-я пара рук немного короче
прочих. Задний конец мантии самки
слегка загнут вверх. Кожа гладкая.
Кожных пор на голове пет. У самца
гектокотилизуется 3-я левая рука, все
прочие руки примерно равной длины.
Сем. Argonautidae. Один род —
Argonauta Linne, 1758 . . с. 327.
18(17). Раковины нет. Руки 1-й
пары на концах без лопасти. Задний
конец мантии самки нс изогнут вверх.
Гектокотилизируется 3-я правая ру-
ка .......................... 19.
19(20). Руки 1-й и 2-й пар соеди-
нены широкой умбреллой. Концы рук
1-й нары у взрослых самок автото-
мируются, несут один ряд присосок,
эти руки часто значительно короче
рук 2-й пары. Руки 1-й и 2-й пар у
самцов и самок гораздо длиннее, чем
3-й и 4-й. Мантия снизу гладкая. На
голове 2 пары кожных пор — у осно-
ваний рук 1-й и 4-й пар. Яйца вы-
нашиваются на руках 1-й пары.
Семейство Tremoctopodidae. Один
род — Tremoctopus delle Chiaje,
1830 .......................с. 326.
20(19). Умбрелла отсутствует. Ру-
ки 1-й и 4-й пар у самцов и самок
значительно длиннее, чем 2-й и 3-й.
Концы рук не автотомируются. При-
соски в 2 ряда до концов рук. Ман-
тия взрослых самок снизу с попереч-
ными гребнями, образующими сетча-
тую структуру. На голове 1 пара
кожных пор — у основания 4-й пары
рук. Живородящие животные.
Сем. Ocythoidae. Один род и
вид-т-Ос//Мо<? tuberculata Rafincsque,
1814........................с. 326.
2.4.1. Подотряд Cirrata
Животные небольшие пли средних
размеров, редко крупные. Ткани сту-
денистые, желеобразные. Мантия
мешковидная, широкоовальпая. В се-
редине или у заднего конца тела па-
ра весловидных или языковидных
плавников, поддерживаемых седло-
видным, U* либо . У:образпым хря-
щом (видоизмененный гладиус). Ру-
ки с одним рядом плоских, шаровид-
ных, редко веретеновидных присосок,
сидячих или стебельчатых. Умбрел-
ла,-как правило, очень глубокая, час-
то достигает концов рук и вместе с
плавниками служит основным дви-
жителем. Сектора умбреллы либо
соединяются как обычно, прямо с ру-
ками, либо срастаются между собой,
присоединяясь к рукам дополни-
тельной перепонкой — вторичной ум-
бреллой. Вороночное отверстие очень
узкое, редуцировано до кольцевой
щели вокруг основания воронки,
иногда вовсе зарастает. Мантийно-
вороночный движитель используется
лишь при бегстве от опасности. Ман-
тийно-вороночного замыкательного
аппарата нет. Воронка нс срастается
с головой. Глаза хорошо развиты,
иногда редуцированы. Хроматофоры
примитивного строения. В окраске
преобладают фиолетовые, пурпур-
ные, бурые, шоколадные тона, неко-
торые виды полупрозрачны. Фото-
форов нет.
Медианная мантийная септа
обычно имеется. Радула, зоб, чер-
нильный мешок и задние, «а иногда и
передние слюнные железы .отсутству-
ют. Желудок мускулистый, слепой
мешок не закручен спирально. Жаб-
ры маленькие, с небольшим числом
филаментов.
Гсктокотиль отсутствует, или гек-
токотилизируются концы рук 1-й па-
ры, у самцов некоторых видов не-
сколько присосок на всех руках рез-
ко увеличены. Развит только левый
яйцевод. Яйца крупные, в кожистой
оболочке, откладываются поодиночке
па дно. Развитие прямое, без псляги*
ческой стадии.
Глубоководные придонные или
бсптопслагические животные, немно-
гие виды постоянно или время от
времени обитают в пелагиали.
Систематика подотряда находит-
ся в хаотическом состоянии. Боль-
шинство исследователей выделяют
3 семейства: Cirroteuthidae, Stauro-
teuthidae и Opistholeuthidac. Само-
стоятельность последнего из них не-
сомненна, что же касается первых
двух, то одни и те же роды разные
исследователи относят то к одному,
то к другому семейству. «Крайние»
роды различаются столь же сильно,
как .обычно различаются роды раз-
ных семейств, но разделение проме-
жуточных форм весьма условно, по-
этому я отношу всех Cirrata, кроме
Opisihoteuthis. к семейству Cirroteuthi-
dae. Однако, вероятно, когда будет
исследована анатомия ныне не изу-
18*
276
ченнЫХ ВИДОВ, роды Cirroteuthis и
Cirrothauma, с одной стороны, Gritn-
poteuthis и Chunioteuthis, с другой,
будут отнесены к разным семействам.
7—8 родов, 27—32 вида.
Большинство видов Cirrata опи-
сано по единичным, как правило,
сильно поврежденным экземплярам,
а иногда просто по обрывкам. Мно-
гие виды определялись с сомнением.
Размах индивидуальной, возрастной
и половой изменчивости для абсо-
лютного большинства видов нс из-
вестен. Поэтому определительными
таблицами для Cirrata необходимо
пользоваться с большой осторожно-
стью, учитывая, что некоторые при-
водимые в таблицах признаки для
различения видов могут в действи-
тельности оказаться не имеющими
таксономического значения.
2.4.1.1. Семейство
Cirroteuthidae
(= Stauroteuthidae)
Животные средних размеров и круп-
ные (общая длина до 1,3—1,5 м).
Консистенция тканей студенистая,
желеобразная. Мантия . коническая
или мешковидная, обычно плохо обо-
соблена от головы. По виду живот-
ные напоминают колокольчик. Ман-
тийное отверстие имеет вид кольце-
вой щели или зарастает. Воронка
направлена вперед. Плавниковый
хрящ седловидный (без направлен-
ных вперед «рогов»), U- или V-об-
разный, пряжковидный или полу-
круглый (с направленными вперед
«рогами»). Умбрелла, как правило,
имеется, доходит с одной или с обе-
их сторон руки почти до концов или
до самых концов рук. Опа тонкая,
пленчатая, гораздо тоньше рук. Иног-
да соседние сектора умбреллы сра-
стаются друг с другом и присоеди-
няются к руке дополнительной (вто-
ричной) умбреллой. Глаза хорошо
развиты или редуцированы.
Продольная ось жабры либо па-
раллельна, либо перпендикулярна
оси тела; в первом случае жаберные
филаменты лежат, как обычно, один
за другим, во втором они лежат в
продольной плоскости тела, так что
жабра напоминает половинку разре-
занного поперек апельсина. Зоба нет,
но иногда имеется особый вырост пи-
щевода («третий желудок»). Гекто-
276
котиль отсутствует или (редко) из-
меняются концы рук 1-й пары.
Придонные (бснтопелагичсские)
батиальные (главным образом нижне-
батиальные) и абиссальные виды;
один род бати-абиссопелагический.
6—7 родов, 19—24 вида.
Таблица для определения родов
Cirroteuthidae
1 (2). Глаза редуцированы, просвечи-
вают через кожу как маленькие тем-
ные шарики. Руки в 3—4 раза длин-
нее тела. Вторичной умбреллы нет.
Присоски в средних и дистальных
частях рук модифицированы, бочон-
ковидной формы, с крохотными от-
верстиями. Плавники крупные, пре-
восходят ширину тела, сидят посре-
дине туловища. Плавниковый хрящ,
мантийное отверстие и жабры, каку
Cirroteuthis (см. ниже).
Род Cirrothauma Chun, 1911. Один
вид — С. murrayi Chun, 1911 (рис.
74, а—в). Обитает в бати- и абиссо-
пелагиали (1500—3600 м) северной
части субтропической Атлантики и
желобов Пуэрто-Рико и Кайман.
Осьминоги этого рода (вид не опре-
делен) добыты в абиссопелагиали
(2900—4100 м) моря Скотия и Юго-
Восточной Пацифики у Южного Чи-
ли и (?) в Центральной Арктике у
поверхности. Длина мантии до 22 см,
общая длина до 1 м.
2(1). Глаза развиты . . . . 3.
3(4). Умбрелла отсутствует. Руки
в 5 раз длиннее тела. Плавниковый
хрящ полукруглый.
Род Froekenia Hoyle, 1904. Один
вид—F. clara Hoyle, 1904. Обитает
в батиали Панамского залива. Сом-
нительная форма.
4(3). Умбрелла имеется . . 5.
5(12). Мантийное отверстие в ви-
де узкой кольцевой щели вокруг ос-
нования воронки.................6.
6(9). Сектора основной умбреллы
срастаются друг с другом снаружи
от рук и соединяются с дорсальной
стороны рук посредством вторичной
умбреллы. Умбрелла симметрична:
ее протяженность с дорсальной и
вентральной стороны рук- одинако-
ва .............................7.
7(8). Умбрелла доходит до самых
или почти до самых концов рук. Уси-
ки доходят до концов рук. Плавни-
ковый хрящ седловидный, без на-
правленных вперед «рогов». Про-
дольная ось жабры параллельна
продольной оси тела.
Род Cirroteuthis Eschricht, 1838
(рис. 74,г—е). Типовой вид С. tnuel-
leri Eschricht, 1838. 3 вида и еще 3
неясного систематического положе-
ния, относимые к этому роду услов-
но (см. ниже). Обитают у дна в абис-
сали, в Арктике поднимаются в ба-
тиаль и (редко) эпипелагиаль. Са-
мые крупные из плавниковых осьми-
ногов и абиссальных беспозвоночных
(общая длина до 1,2—1,5 м).
8(7). Умбрелла не доходит до
концов рук примерно на треть их
длины. Усики кончаются перед краем
умбреллы. Плавниковый хрящ V-об-
разный с направленными вперед
«рогами». Продольная ось жабры
перпендикулярна продольной оси
тела.
Род Stauroteuthis Verrill, 1879.
Один вид — S. syrtensis Verrill, 1879
(рис. 74, жу з). В средней и нижней
батиали Северо-Западной Атлантики.
Общая длина около 52 см.
9(6). Вторичной умбреллы нет.
Основная умбрелла несимметрична:
с дорсальной стороны рук она при-
крепляется к спинной части руки и
доходит, постепенно утончаясь, поч-
ти до конца, с вентральной — при-
крепляется к боковой части рук и
далеко не доходит до конца (часто
лишь до середины руки). В месте
присоединения умбреллы к боковой
стороне руки с 'вентральной стороны
обычно имеется полое вздутие округ-
лой или овальной формы с плотными
стенками.......................10.
10(11). Плавниковый хрящ сед-
ловидный, без направленных вперед
«рогов». Усики доходят только до
места прикрепления умбреллы к
вентральной стороне рук.
3 вида неясного систематического
положения, каждый описан лишь
по единственному экземпляру. Их
условно относят к роду Cirroteuthis
(см. таблицу для определения ви-
дов).
11(10). Плавниковый хрящ полу-
круглый, подковообразный, U-образ-
ный или пряжковндпый, с направлен-
ными вперед «рогами» (см. рис.
75, л). Усики обычно доходят до кон-
цов рук. Продольная ось жабры пер-
пендикулярна продольной оси тела.
Род Grimpoteuthis Robson, 1932
(см. рис. 75). Типовой вид G. umbel-
lata (Fischer, 1883). 11—12 видов в
ба гнали (обычно средней и нижней)
и абиссали всех океанов, кроме Се-
верного Ледовитого. Общая длина
до 58 см (возможно, до 1—1,2 м).
12(5). Мантийное отверстие пол-
ностью зарастает — край мантии при-
растает к основанию воронки. Вто-
ричная умбрелла имеется в прокси-
мальных частях рук. Умбрелла сим-
метрична. Она не доходит до концов
рук, усики доходят только до края
умбреллы. Нет вздутий в местах
прикрепления умбреллы к вен-
тральной стороне рук. Плавниковый
хрящ широко V- или U-образный.
Род Chunioteuthis Grimpe, 1916
(рис. 74,к). Типовой вид Ch. ebers-
bachi Grinipe, 1916. 2 вида. Общая
длина до 40 см.
а (б). В середине каждого секто-
ра умбреллы имеется небольшой за-
остренный бугорок. Руки в 3,5—4 ра-
за длиннее тела. Плавниковый хрящ
широко V-образный. Ch. ebersbachi
Grimpe, 1916 (рис. 74, и, к). Нижняя
батиаль южного склона Большой
Ньюфаундлендской банки и (?) се-
веро-восточного побережья США.
б (а). Нет бугорков на умбрелле.
Руки в 5 раз длиннее тела. Плавни-
ковый хрящ широко U-образный,
слабо изогнутый. Ch. gilchristi (Rob-
son, 1924). Абиссаль у Южной Аф-
рики.
Таблица для определения видов,
принадлежащих и условно относимых
к роду Cirroteuthis
1 (6). Умбрелла симметрична. Вто-
ричная умбрелла имеется ... 2.
2(5). Глаза маленькие, их диа-
метр около 10% длины мантии. Дли-
на рук V2—% общей длины. Живот-
ные некрупные, общая длина 20—
30 см ..........................3.
3(4). Длина рук около 2/3 общей
длины. Присоски конические.
С. muelleri Eschricht, 1838 (рис.
74, <?). Центральный Полярный бас-
сейн, Скандская впадина, Баффинов
залив. У дна в батиали и абиссали и
в толще воды (мезо- и батипелаги-
аль), изредка у поверхности (?).
4(3). Длина рук чуть более V2
общей длины. Присоски в средних
частях рук высокие, бочонковидныс,
с крохотными устьями, в самых про-
ксимальных и дистальных частях ко-
нические.
Cirroteuthis п. sp. Nesis. Плато
277
5
нижпий (5) глазные нервы; 6 — глазная
вена; г — Cirroteuthis muelleri; д—е—*Cir-
roteuihis» magna, общий вид (часть ум-
бреллы оборвана, се край показан пункти-
ром) и конец руки; ж—з— Stauroteuihis
syrtensis, общий вид (пунктиром показан
хрящ) и голова снизу с воронкой; и—к —
Chunioteuthis ebersbachi, общий вид и
хрящ (а—в — по Chun, 1915; г — по Muns,
1959; д—е — по Hoyle. 1886; ж—з— по Ver-
3
rill, 1882; и—к--по Grimpe, 1916)
Натуралиста у Юго-Западной Ав-
стралии. Верхняя абиссаль.
5(2к Глаза крупные, их диаметр
до 40% длины мантии. Длина рук
2/з—4/s общей длины. Общая длина
до 1,3—1,5 м.
Cirroteuthis sp. Roper et Brundage,
1972. Северная Атлантика от Канар-
ских островов до Дакара и Кариб-
ского моря. Абиссаль, глубины 2500—
5200 м, в апвеллинге у мыса Кап-
Блан поднимается на глубину 1300—
2000 м (Несис, Головань, 1975).
5(1). Умбрелла несимметричная.
Вторичной умбреллы пет ... 7.
7(8). Руки 1-й пары вдвое толще
прочих. Усики сидят но бокам от
каждой присоски, а не между двумя
соседними присосками, их длина не
превосходит диаметр наибольшей
присоски. В базальных частях рук
есть несколько увеличенных присо-
сок, диаметр которых достигает 8—
9% длины мантии. Есть полое взду-
тие в местах прикрепления края умб-
реллы к вентральной стороне рук.
«С.» massyae (Grimpe, 1920). Бис-
кайский залив юго-западнее Ирлан-
дии. Нижняя батиаль. Для этого ви-
да предложен особый подрод Cirro-
teuthopsis Grimpe, 1920.
8(7). Все руки примерно одина-
ковой длины и толщины. Усики си-
дят, как обычно, между соседними
присосками, их длина в 10—15 раз
больше диаметра присосок. Специ-
ально увеличенных присосок пет. Нет
вздутий в местах прикрепления ум-
бреллы к рукам..................9.
9(10). Диаметр присосок около
2—2,5% длины мантии.
«С.» hoylei Robson, 1932. Запад-
нее Центрального Чили. Абиссаль.
10(9). Диаметр присосок около
4,5% длины мантии.
magna Hoyle, 1885 (рис.
74, дуе). Южный океан между ост-
ровами Принс-Эдуард и Крозе и к
северо-востоку от острова Буве.
Нижняя батиаль и абиссаль. Общая
длина до 115 см.
Таблица для определения видов рода
Grimpoteuthis
1 (6). Ширина плавников значитель-
но превосходит ширину тела (шири-
на плавников измеряется по их длин-
ной оси, т. е. по оси, перпендикуляр-
ной оси тела, длина — по короткой
оси, параллельной оси тела). Плав-
280
никовый хрящ подковообразный, за-
кругленный ....................2.
2(3). Ширина плавников в 2—3
раза больше ширины тела и пример-
но равна длине тела. Тело в 6—7 раз
короче рук. Глаза маленькие.
G. megaptera (Verrill, 1885).
Абиссаль тропической и северной
субтропической Атлантики.
3(2). Ширина плавников не более
чем в 1,5—2 раза больше ширины
тела. Тело в 2,5—4 раза короче рук.
Диаметр присосок 3—4% длины ман-
тии ...........................4.
4(5). Усики на руках начинаются
после 3—7-й присоски. Глаза не-
большие.
G. pacifica (Hoyle, 1885). Абис-
саль Кораллового моря и Южного
океана к югу от Тасмании.
5(4). Усики на руках начинаются
после 2-й присоски. Глаза крупные.
6. wuelkeri (Grimpe, 1920) (рис.
75, а). Субтропическая Северо-Вос-
точная Атлантика. Абиссаль.
6(1). Ширина плавников равна
или меньше ширины тела ... 7.
7(14). Ширина плавников при-
близительно равна ширине тела . 8.
8(13). Тело в 2—3 раза короче
рук. Наибольшая ширина тела — на
уровне глаз....................9.
9(12). Присоски мелкие, их диа-
метр 1,5—6% длины мантии . 10.
10(11). Плавниковый хрящ пряж-
ковидный: его прямая или слабо за-
кругленная задняя часть переходит в
направленные вперед «рога» не плав-
но, а с резким углом. Диаметр гла-
за около % длины мантии.
G. meangensis (Hoyle, 1885) (рис.
75,6). Индийский океан, Индонезия,
о-ва Ксрмадск, (?) Куба. Батиаль.
11(10). Плавниковый хрящ сзади
плавно закругленный или почти пря-
мой, нет уголков в местах перехода
задней части хряща в «рога». Глаза
маленькие, их диаметр 7s—7е длины
мантии.
G. utnbcllata (Fischer, 1883) (рис.
75, в). Тропическая и северная суб-
тропическая Атлантика, Карибское
море, (?) Юго-Западная Индия.
Нижняя батиаль и абиссаль (1100—
5400 м).
12(9). Диаметр наибольшей при-
соски (примерно 19-я) 8—9% длины
мантии.
G. glacialis (Robson, 1930) (рис.
75, г). Антарктический полуостров.
Верхняя батиаль.
13(8). Тело в 1,5 раза короче рук.
Глаза небольшие. Наибольшая шири-
на тела — за глазами. Присоски мел-
кие.
Q. mawsoni (Berry, 1917) (рис.
75, д). Антарктида (Земля Адели).
Верхняя батиаль.
14(7). Ширина плавников значи-
тельно меньше ширины тела . 15.
15(18). Тело короткое, в 4—6 раз
короче рук, сзади широко закруглен-
ное. Ширина тела гораздо больше
его длины. Ширина плавников в 3—
4 раза меньше межглазничного рас-
стояния. Плавниковый хрящ дугооб-
разный. Глаза крупные . . . 16.
16(17). Диаметр максимальных
присосок (6—7-я) у молоди 8—9%
длины мантии, у взрослых значи-
тельно меньше. Усики идут до кон-
цов рук, их длина превосходит диа-
метр наибольших присосок. Гектоко-
тилизируются концы спинных рук
самца: несколько присосок близ кон-
цов рук резко увеличены, шаровид-
ные.
G. albatrossi (Sasaki, 1920) (рис.
75, е—и). Севсротихоокеанский бо-
реальный вид, распространен от Бе-
рингова до Охотского моря и Юж-
ной Калифорнии. Глубина 136—
3400 м.
17(16). Диаметр максимальных
присосок (4—7-я) около 15—20%
длины мантии. Усики пе доходят до
концов рук, их длина меньше диамет-
ра присосок в средних частях рук (не
самых крупных). Гектокотиль неиз-
вестен.
G. gritnaldii (Joubin, 1903) [=?(?.
caudani Joubin, 1896)] (рис. 75, к).
Северо-Восточная Атлантика: район
Азорских островов, Бискайский за-
лив. Нижняя батиаль.
18(15). Тело довольно длинное, в
1,5—2 раза короче рук. Ширина тела
равна или меньше его длины. Шири-
на плавников не более чем в 2 раза
меньше меж глазничного расстояния.
Плавниковый хрящ подковообразный
(не описан у G. plena). Глаза ма-
ленькие .....................19.
19(20). Ширина тела примерно
равна длине. Диаметр присосок 4—
4,5% длины мантии.
G. plena (Verrill, 1885). Северо-
Западная Атлантика, восточнее Че-
сапикского залива. Глубина около
2000 м.
20(19) Ширина тела примерно %
его длины. Диаметр присосок около
3% длины мантии.
G. hippocrepium (Hoyle, 1904),
Тропическая зона Восточной Паци-
фики. Абиссаль.
Не определенные до вида Grimpo-
teuthis найдены в Антарктике и Суб-
антарктике, у Южной Африки, в се-
верной части Индийского океана, ин-
донезийских водах и в тропической?
зоне восточной части Тихого океана
в батиали и абиссали. Среди них бы-
ли особи размером до 1—1,2 м (об-
щая длина).
2.4.1.2. Семейство
Opisthoteuthidae
Животные мелкие и средних разме-
ров. Тело уплощенное, мантия реду-
цирована до степени бугра, лежаще-
го сверху на растопыренных и со-
единенных толстой умбреллой руках.
Животное напоминает толстую ле-
пешку. Ткани полустуденистые. Ум-
брелла простирается почти до копцов-
рук. Вторичной умбреллы пет. Плав-
ники короткие, сидят на вершине
мантии, па «макушке». Плавниковый
хрящ дугообразный или почти пря-
мой. Воронка очень короткая, на-
правлена назад. Мантийное отвер-
стие— узкая кольцевая щель. Глаза*
хорошо развиты. Продольная осы
жабры перпендикулярна оси тела.
У зрелых самцов одних видов уве-
личены присоски базальных частей
рук, у других особо увеличенные
присоски располагаются также (или
только) на концах рук некоторых
или всех пар. Иногда базальные уве-
личенные присоски бочонковидной,,
дистальные — шаровидной формы.
Один род—Opisthoteuthis Verrill,
1883. Типовой вид — О. agassizi Ver-
ril, 1883. 8 видов. Обитают в батиа-
ли (125—2250 м) тропических и бо-
реальных вод, держатся на дне и
над дном, в толщу воды поднимают-
ся редко. Диаметр тела с руками до
60 см. Надежное определение видов
возможно лишь по зрелым самцам,
однако для половины видов они нс
описаны.
Таблица для определения видов рода
Opisthoteuthis
1(8). Виды встречаются в Атланти-
ческом и северной части Тихого
океана .........................2.
281
К
и
Рис. 75. Род Grimpoteuthis: a — G. wttelke-
ri, схема расположения умбреллы (4-я ле-
вая, 4-, 3-, 2-, 1-я правая, 1-я левая рука):
б — G. meangensis, хрящ сверху и спереди;
в — G. umbellata; г — G. glacialis; д — G.
mawsoni', e—u — G. albatrossi, взрослый са-
мец (е) молодой экземпляр (дас), руки (з)
и гектокотиль («) самца; к — G. gritnaldti-,
л — Grimpoteuthis sp., схема положения
хряща (х) в теле (а — по Grimpe, 1920:
б — по Hoyle, 1886; в, к—по Joubin, 1900.
1920; г —по Robson, 1930; д, ж — по Berry,
1912, 1917; е— по Sasaki, 1929; з—и — по
Кондакову, 1941; л — по Muus, 1959)
Q
Рис. 76. Сем. Opisthoteuthidae, род Opis-
thoteuthis: а—в — О. agassizi, общий вид
(а), хрящ (б), гектокотилизированиая рука
(в); г—ж — О. californiana, дистальная и
проксимальная часть гектокотилизирован-
ной руки (а), схема расположения присо-
сок на левой и правой спинных руках
самца (д), вскрытая мантия с жабрами
(е), хрящ (ж); з—к — О. depressa, общий
вид (з), руки самца (и), хрящ (к); л—м—
О. medusoides, молодой экземпляр сверху
и сбоку, руки; н—о—О. exiensa, общий
вид самки и руки (а—б— no G. Voss,
1956; в — по Morales, 1959; г-ж, к—по
Taki. 1963: з—и — по Sasaki, 1929; л—о —
по Chun, 1915)
2(3). У зрелых самцов увеличены
4—5 базальных присосок (обычно с
6—7-й по 9—11-ю) и 2—3 дисталь-
ные присоски (обычно с 23—24-й по
25—26-ю) па всех руках. Ширина
плавников более 7г межглазпичного
расстояния, они языковидные, с за-
кругленными концами. Вид распро-
странен в Атлантике.
О. agassizi Vcrrill, 1883 (рис. 76,
а—в). От Джорджес-банки до Ка-
рибского моря и от Фарерской банки
до Намибии; западная часть Среди-
земного моря. Глубина 500—2250 м.
3(2). У зрелых самцов увеличен-
ные дистальные присоски отсутству-
ют или имеются лишь на концах 1-й
пары рук. Ширина плавников мень-
ше V2 межглазничного расстояния.
Виды распространены в Тихом океа-
не ............................4.
4(5). У зрелых самцов увеличены
несколько базальных присосок (обыч-
но с 3—4-й по И—12-ю) на всех ру-
ках и 5—8 (обычно 6—7) дисталь-
ных присосок на 1-й паре рук. Плав-
ники конические с приострсппым
концом, их ширина в 1,5—2 раза
больше длины.
О. caUforniana Berry, 1949 (рис.
76, а—ж). Северотихоокеанский бо-
реальный вид: от Берингова моря до
Центрального Хонсю и Калифорнии.
Глубины 125—1100 м, обычно более
300 м.
5(4). У самцов увеличены только
базальные присоски (10—15 шт.).
Они так крупны, что кажутся распо-
ложенными в 2—3 ряда вместо од-
ного . '.......................6.
6(7). Плавники короткие и широ-
кие, их ширина в 1,5—2 раза больше
длины, концы плавников закруг-
ленные или притупленные. Окраска
верхней стороны тела пятнистая,
пятна расположены продольными
рядами на верхней стороне тела и
вдоль рук. На каждой руке самца
около 15 увеличенных присосок.
О. depresses Ijima et Ikeda, 1895
(рис. 76,з—/с). Тихоокеанское побе-
режье Центрального и Южного Хон-
сю. Глубина 130—1100 м.
7(6). Плавники языковидные, их
ширина в 5 раз больше длины, кон-
цы плавников заострены. Окраска
однотонная пурпурно-фиолетовая,
без пятен. На каждой руке самца
около 10 увеличенных присосок.
О. japonica Taki, 1962. Тихоокеан-
ское побережье Южного Хонсю. Глу-
бина 150 м.
8(1). Виды встречаются в Индий-
ском океане и у Южной Австра-
лии ...........................9.
9(10). Тело полупрозрачное, сла-
бопигментированное. Руки соедине-
ны умбреллой почти до самых кон-
цов. 19—20-я присоски на руках 4-й
пары слегка увеличены.
О. medusoides Thiele, 1915 (рис.
76, л, л). Восточная Африка. Глуби-
на 400 м.
10(9). Тело интенсивно окрашено.
Концы рук не соединены умбрел-
лой .........................11.
11(12). На верхней стороне тела
нет глазчатых пигментных пятен.
Глубина умбреллы в промежутках
между руками примерно равна поло-
вине длины рук.
О. extensa Thiele, 1915 (рис. 76,
н—о). У южного берега Суматры.
Глубина 770 м.
12(11). На верхней стороне тела
разбросаны маленькие глазчатые пиг-
ментные пятна. Глубина умбреллы в
промежутках между руками около
2/з длины рук.................13.
13(14). Присоски проксимальных
частей рук (5—7-я) крупнее про-
чих. Плавники сидят близ заднего
края глаз. В окраске доминируют
бурые тона. У самцов увеличивают-
ся 3—4 присоски в дистальных час-
тях 3—4-й пар рук близ края ум-
бреллы.
О. pluto Berry, 1918. Большой Ав-
стралийский залив. Глубины 275—
1100 м.
14(13). Все присоски одинаково
мелкие. Плавники сидят посредине
тела. В окраске доминируют серые
тона.
О. persephone Berry, 1918. Боль-
шой Австралийский залив и Бассов
пролив. Глубины 275—550 м.
Не идентифицированные (новые?)
виды Opisthoteuthis обнаружены в
абиссали у Перу и в батиали у о-ва
Кергелен и в Калифорнийском за-
ливе.
2.4.2. Подотряд Incirrata
Типичные осьминоги. Плавников и
усиков на руках нет. Умбрелла не
доходит до концов рук. Вторичной
умбреллы не бывает. Присоски всег-
да сидячие, уплощенные, реже ша-
ро- или бочонковидные. Глаза разви-
285
ты. Вороночное отверстие разного
размера, но никогда не зарастает.
Гладиус сильно редуцирован или от-
сутствует. Радула всегда имеется,
хотя у некоторых глубоководных ви-
дов редуцирована. Продольная ось
жабры параллельна осн тела. Гекто-
котиль, как правило, имеется — из-
меняется одна из рук 3-й пары. Раз-
виты оба яйцевода.
3 надсемейства, 9 семейств, 33—
35 родов, 165—180 видов. Распро-
странены всесветно в полносоленых
морях и океанах, на всех глубинах,
кроме ультраабиссали и абиссопсла-
гиали.
2.4.2.1. Надсемейство
Bolltaenoidea
(Ctenoglossa)
Глубоководные осьминоги со студе-
нистым, обычно полупрозрачным те-
лом. Присоски в один ряд. Радула
гребенчатого строения — рахидиаль-
ный и оба латеральных зуба много-
вершинные.
3 семейства, 6 родов, 6—7 видов
в тропических и субтропических
(редко бореальных) районах. Обзо-
ры: There, 1949; Taki, 1963.
2.4.2.1.1. Семейство
Bolitaenidae
Тело студенистое, пигментированное.
Руки короткие, короче мантии,
третья пара длиннее прочих. Умбрел-
ла средних размеров. Присоски в
один ряд. Мантийное отверстие ши-
рокое. Желудок позади печени. Ра-
дула гребенчатая. Чернильный ме-
шок имеется, но небольшого размера.
4 рода, 4—5 видов. Мезо- и батипе-
лагиаль.
Таблица для определения родов и
видов Bolitaenidae
1(6). Глаза нормальных размеров,
расположены по бокам головы . 2.
2(5). Присоски сближены, рас-
стояние между соседними присоска-
ми не более чем вдвое превосходит
диаметр присоски...............3.
3(4). Глаза крупные (диаметр
превышает 18% длины мантии). Оп-
тические нервы короткие, оптические
ганглии приближены к мозгу (это
хорошо заметно из-за полупрозрач-
ности тканей). Присоски крупные, их
диаметр составляет в среднем 6%
длины мантии, расстояние между со-
седними присосками меньше диамет-
ра присоски. Максимальная ширина
умбреллы '/<—Уз длины наибольшей
руки. Гектокотиля нет, у самцов
лишь несколько увеличены присоски
в средней части 3-й правой .руки.
В окраске преобладают коричневые
тона.
Род Japetella Hoyle, 1885 (=СЛи-
nella Sasaki, 1920, Bolitaenella Grim-
pe, 1922). Один вид — /, diaphana
Hoyle, 1885 ( = ? Octopus brevipes
d’Orbigny, 1838) (рис. 77, a—e).
Тропическо-субтропический космопо-
литический батипслагический вид, мо-
лодь встречается также в эпи- и ме-
зопелагиали. В северной части Тихо-
го океана проникает в бореальные во-
ды — па севере до залива Аляска, об-
разует особую форму.
/. diaphana heathi (Berry, 1911)
(рис. 77, б). Встречается в водах
СССР восточнее Курильских остро-
вов. /. diaphana — один из самых
обычных глубоководных пелагиче-
ских осьминогов. Заметных суточных
вертикальных миграций не соверша-
ет. Яйца мелкие, соединены по не-
скольку штук стебельками. Личинки
одеты липкой студенистой оболочкой,
облегчающей парение в воде. Длина
мантии до 10 см.
В водах Калифорнии обнаружена
особая форма (самостоятельный
вид?) Japetella sp. Young, 1972. отли-
чающаяся от /. diaphana отсутствием
яркого ирризирующего . серебристого
слоя, одевающего глаза с внутренней
стороны и печень.
4(3). Глаза маленькие, . диаметр
глаза у взрослых особей (длина ман-
тии более 6 см) 9—12%, у молоди
менее 17% длины мантии. Оптиче-
ские нервы длинные, оптические
ганглии сидят на большом расстоя-
нии от мозга. Присоски мелкие (в
среднем 4% длины мантии), расстоя-
ние между соседними присосками
равно или больше диаметра присос-
ки. Максимальная ширина умбреллы
Уз—Уз длины наибольшей руки. Гек-
токотилизируется 3-я левая рука, на
3-й правой руке самца дистальнее
середины сидят 2—3 резко увеличен-
ные горшковидные присоски. В ок-
раске преобладают красные тона.
Род Eledonella Verrill, 1884. Один
вид — Е. pygmaea Verrill, 1884 (рис.
77,г). Тропическо-субтропический, не
заходящий в бореальные воды, бати-
пелагический вид, молодь встречает-
ся также в нижней эпи- и мезопела-
гиали. Более редка, чем /. diaphana.
Яйца мелкие, сцепленные стебелька-
ми. Длина мантии до 20 см.
5(2). Присоски широко расстав-
лены в пределах умбреллы и сбли-
жены за се пределами. Глаза круп-
ные. Оптические нервы длинные, оп-
тические ганглии удалены от мозга.
Максимальная ширина умбреллы
’/г—% длины наибольшей руки. Ок-
раска тела светло-пурпурная, ум-
брелла изнутри темно-пурпурная.
Род Bolitaena Steenstrup, 1859.
Один вид — В, microcotyla Steenstrup
in Hoyle, 1886 (рис. 77, d, e). Редкий
тропический космополитический ме-
зо-батипелагический вид. Яйца круп-
ные, самка посит их на руках в об-
разованной умбреллой сумке до вы-
лупления личинок и во время инку-
бации не питается. Длина маптии до
4 см.
6(1). Глаза очень крупные (око-
ло ‘/4 длины маптии), сидят па спин-
ной стороне, тесно сближены и смот-
рят вверх. Оптические нервы корот-
кие. Присоски тесно сближены. Ши-
рина умбреллы около Уз длины
наибольшей руки.
Род Dorsopsis Thore, 1949. Один
вид — D. taningi Thore, 1949 (рис.
77, ж). Пойман у Дакара в батипс-
лагиали. .
2.4.2Л .2. Семейство
Amphitretidae
Тело студенистое, полупрозрачное,
почти бесцветное, заключено в сту-
денистый чехол. Руки длинные, с од-
ним рядом присосок, умбрелла очень
глубокая. Мантия срастается с во-
ронкой, мантийное отверстие пред-
ставлено двумя отверстиями, распо-
ложенными по бокам головы. Ворон-
ка очень длинная. Глаза расположе-
ны на спинной стороне головы,
телескопические, смотрят вверх, их
основания сближены. Радула гребен-
чатая. Желудок лежит впереди пе-
чени, яичник — над печенью. Чер-
нильный .мешок развит. Гектокотили-
зируется 3-я правая рука, лигула уз-
кая и длинная, уплощенная, каламус
очень короткий. Яйца крупные. Длин-
на мантии до 9 см.
Один род — Amphitretus Hoyler
1885. Один вид — A. pelagic us Hoyler
1885 с 2 подвидами: 4. pelagicus pe-
lagicus Hoyle (рис. 77, з, и) — тропи-
ческий ипдотихооксапскнй (в Тихом
океане экваториально-западноцент-
ральный) и A. pelagicus thielei Rob-
son, 1930 — потальио-южпосубтропи-
ческий атлантический. Атлантический
подвид отличается от индотихоокеан-
ского узкой, вытянутой в длину ман-
тией (у А. р. pelagicus она округлая
или широко-овальная). Вид мезо-
батипелагичсский, молодь встречает-
ся и в эпинслагиали.
2.4.2.1.З. Семейство
Idioctopodidae
Тело удлиненно-овальное с хороша
выраженным шейным перехватом.
Ткани студенистые, полупрозрачные.
Руки тонкие, длинные, сжатые с бо-
ков, присоски в один ряд. Умбрелла»
неглубокая. Воронка срастается с
мантией и (па большей части длины)
с нижней поверхностью головы. Ман-
тийное отверстие — две узкие щели»
по бокам головы, доходящие до
уровня нижнего края глаз. Радула
гребенчатая. Зоб и слюнные железы
имеются. Чернильный мешок руди-
ментарный. Желудок впереди пече-
ни. Гсктокотилизируется 3-я правая
рука, лигула длинная, узкая, круглая
в сечении, каламус имеет вид тре-
угольного выроста.
Один род — I dioctopus Taki, 1962;
один вид — I. gracilipes Taki, 1962
(рис. 77,/с—м). Найден у тихоокеан-
ского побережья Южпой Японии у
дна в верхней батиали. Длина ман-
тии 5 см, общая длина 20 см. Обзоре
Taki, 1963.
2Л.2.2. Надсемейство
Octopodoidea
(Heteroglossa)
Дойные или пелагические осьминоги.
Присоски на руках в один или два
ряда. Радула не гребенчатого строе-
ния, рахидиальный зуб многовер-
шинный, средний зубец самый круп-
ный; первый и второй маргинальные
зубы одно- или двухвершинные.
Форма рахидиалыгого зуба в сосед-
287
и
3
*
Рис. 77. IТадсен. Bolitaonoidea, сем. Bol
taenidae (а—ж), Amphitretidae (з—и). Id
octopodidae {к—м): а—в — Japetella diaphc
na (a — J. d. diaphana, молодь; 6 — J, c
heathi, взрослая особь; в —руки спереди)
2 — Eledonella pygmaea, молодая самкг
д—е — ВоШаепа microcotyla, взрослая сам
ка (д) и мозг с оптическими трактами :
оптическими ганглиями (е); ж — Dorsopsi
taningi; з—u—Amphitretus pelagicus pelagi
cus, самец и самка, иа и виден студенисты
чехол; к—м — Idioctopus gracUipes, саме1
(/с, указаны номера рук), голова и маити:
в полоборота (л), гектокотиль (.и) (а — п<
Всггу, 1914; б, д — по R. Young, 1972а
19726; в — по G. Voss, 1963; г, ж—з — п<
Thore, 1949; е — по Rancurel, 1970; и— п*
Ijima, Ikeda, 1902; к—м. — по Taki, 1963)
19 К- Н. Несис
289
H О
290
&&&££&
Octopodidae. Подсем. Octopo-
и
Рис. 78. Сем. Octopodidae. Подсем. Octopo-
dinae: а—б — Hapalochlaena maculosa, ва-
рианты окраски (а — из вод Китая, б — из
Австралии); в—г — различия в расположе-
нии цветных пятен на теле Н. lutiulata {в)
и Н. maculosa (г), вид сверху, сбоку и
снизу; д — Cistopus indicus; e-u — Danoc-
topus schmidti; — D. hoylei; m—o —
Euaxoctopus panamensis; n—c — E. pillsbur*
yae; т—ф — Scaeurgus unicirrhus (т — са-
мец, у — самка); x—ц — Pteroctopus tetra-
cirrhus; etc — вороночный орган; з, л, о, с,
ф, ц — гектокогиль; и — скульптура кожи;
н, р — расположение цветных пятен па
спинной стороне мантии (а—-по Dong
Zhenzhi, 1963; б — по Hoyle, 1886, в—г, ц —
по Adam, 1952, 1954; д — по Voss, Willi-
amson, 1972; е, и — по Joubin, 1933; ж, м—
с, у, х —по G. Voss, 1951, 1956, 1971, 1975;
з—по Cairns, 1976; к—л, ф — по Berry,
1914; т — по Naef, 1923)
19*
291
пих поперечных рядах зубов разли-
чается, но повторяется через каждые
3—4 ряда (сериация). Гектокотиль
всегда имеется — видоизменяется 3-я
левая или правая рука. Гсктокотили-
зация захватывает лишь конец руки,
она не развивается в особом мешке
и не отделяется от тела при спари-
вании. Самцы не карликовые. 2 семей-
ства, 23—25 родов, свыше 150 видов.
2.4.2.2.1. Семейство
Octopodidae
Тело плотное, мускулистое или дряб-
лое, слабомускулистое, ио не студе-
нистое и нс полупрозрачное. Руки
мускулистые, обычно в несколько
раз длиннее туловища, с 1—2 ряда-
ми присосок. Умбрелла короткая или
средних размеров, обычно не превос-
ходит половину длины наибольшей
рукн. Мантийное отверстие не реду-
цировано. Радула не гребенчатая,
характерна сериация рахидиального
зуба. Печень впереди желудка и го-
нады. Гсктокотилизирустся правая
или левая рука 3-й нары, у самцов,
кроме того, часто увеличиваются
присоски в середине некоторых рук,
иногда видоизменяются концы всех
рук. Гектокотиль с лигулой и, как
правило, каламусом. Яйца донные,
охраняются самкой, реже она вына-
шивает их на руках. Развитие пря-
мое или со стадией пелагической
личинки.
Донные животные, распростране-
ны .от Арктики до Антарктики и от
литорали до абиссали. Некоторые
виды — промысловые.
3 подсемейства, 22—24 рода,
150—160 видов.
Систематика семейства как па ви-
довом, так и на родовом уровне раз-
работана очень слабо, многие роды,
считающиеся самостоятельными, пло-
хо отличаются от остальных. Большая
половина видов семейства (около 90)
относится к роду Octopus, чрезвы-
чайно гетерогенному и крайне нуж-
дающемуся в ревизии.
В таблицу для определения ро-
дов не включены плохо описанные и
сомнительные роды Laetmoteuthis
Berry, 1913 и Haptochlaena Grimpe,
1922; сомнительный род Pinnocto-
pus cTOrbigny, 1845 провизорно вклю-
чен в род Octopus. Ввиду не-
разработанности системы рода Octo-
292
pus таблицы для определения всех
видов составить не удалось, по при-
водятся таблицы для определения
всех атлантических и северотихооке-
апских видов.
Таблица для определения
подсемейств и родов семейства
Octopodidae
1(32). Чернильный мешок имеется.
Иногда он уменьшен в размерах и
глубоко погружен в печень, но чер-
нильная жидкость всегда выделяет-
ся .............................2.
2(21). Руки с двумя рядами при-
сосок.
Подсемейство Octopodin ас . 3.
3(16). Гектокотилизируется пра-
вая 3-я рука....................4.
4(13). Тело крепкое, более или
менее мускулистое. Вороночный ор-
ган W- или VV-образный. Мантий-
ное отверстие нс сужено или слабо
сужспо..........................5.
5(10). Гектокотиль хорошо раз-
вит, с более или менее длинной диф-
ференцированной лигулой и каламу-
сом. Мантийное отверстие широ-
кое ............................6.
6(7). В окраске не бывает ярких
цветных колец, разбросанных по те-
лу и рукам, пет глазчатых пятен на
мантии, могут быть лишь круглые
глазчатые пятна па умбреллс перед
глазами, а также различного рода
полосы, пятна, разводы и т. д., реже
окраска одноцветная.
Род Octopus Lamarck, 1798
(=Polypus Schneider, 1784, ? Eledo-
nenta Rochebrune, 1884, Macrotritopus
Grimpe, 1922) (см. рис. 81—82). Ти-
повой вид — О. vulgaris Lamarck,
1798. В род Octopus мы включаем
Pinnoctopus d’Orbigny, 1845, Tritaxe-
opus Owen, 1881, Amphioctopus Fi-
scher, 1882, Enteroctopus Rochebrune
et Mabille, 1887, Paroctopus Naef, 1923
( = Pseudoctopus Grimpe, 1923), Mac-
roctopus Robson, 1929, Robsonella
Adam, 1938 t—Joubinia Robson, 1929,
ргаеосс) и Callistoctopus Taki, 1964.
Эти «роды» не имеют однозначной
характеристики, которая позволила
бы отличить все их виды от других
видов Octopus. Род Octopus в широ-
ком понимании включает около 90
видов, распространенных в сублито-
рали и верхней батиали всех окса-
нов, кроме Арктики и Антарктики.
Северная граница ареала проходит в
районе пролива Лонг-Айленд, в юж-
ной части Северного моря и у Бе-
рингова пролива, южная граница —
у мыса Горн и островов Фолкленд-
ских, Крозе, Кергелен, Окленд и
Кэмпбелл. Осьминоги от очень мел-
ких до очень крупных. Общая длина
самого крупного вида — О. dofleini—
до 3—5 м. У одних видов мелкие яй-
ца и развитие со стадией пелагиче-
ской личинки, у других — крупные
яйца и прямое развитие. Многие ви-
ды (О. vulgaris, О. dofleini, О. may а
и др.) имеют важное промысловое
значение. В Японии налажено искус-
ственное выращивание осьминогов.
В водах СССР на Дальнем Восто-
ке обитает около 10 видов (Конда-
ков, 1941; Акимушкин, 1963).
7(6). На мантии и руках имеются
яркие голубые кольца или есть глаз-
чатые пятна на спинной стороне ман-
тии ...........................8.
8(9). На спинной стороне мантии
посредине между головой и задним
концом тела по обе стороны от сред-
ней линии имеется пара крупных
глазчатых пятен, окруженных с
внешней стороны неполным (С-образ-
ным) неокрашенным гребнем. Руки
очень длинные и тонкие, их длина
окаю 90% общей длины, концы рук
часто обламываются. Вороночный
орган W-образный (?). В полужабре
11—13 филаментов. Умбрелла очень
короткая, ее глубина 5—6% длины
наибольшей руки.
Род Euaxoctopus Voss, 1971, вид
Е. pillsburyae Voss, 1975 (рис. 78,
п—с). См. тезу 17(18).
9(8). На мантии, голове и руках
имеются многочисленные ярко-голу-
бые кольчатые пятна или также из-
вилистые полосы, ирризирующис при
возбуждении осьминога. Глазчатых
пятен на мантии нет. Руки короткие
(окаю 2/з общей длины), приблизи-
тельно одинаковые, не утонченные,
не обламываются. В полужабре 6—
7 филаментов. Умбрелла глубокая,
25—45% длины наибольшей руки.
Род Hapalochlaena Robson, 1929
(рис. 78,а—г). Типовой вид — H.lu-
nulata (Quoy et Gaimard, 1832).
2 плохо разграниченных вида рас-
пространены в Индо-Вестпацифике.
Верхняя сублитораль, иногда лито-
раль. Длина мантии до 3—6 см.
а (б). На голове и мантии имеют-
ся только голубые кольца. Черниль-
ный мешок заметно редуцирован.
Самка откладывает яйца на дно.
Длина яиц 3,5 мм.
Н. lunulata (Quoy et Gaimard,
1832) (рис. 78, в). Филиппинские ост-
рова, Индонезия, Меланезия, Север-
ная и Западная Австралия.
б (а). На голове, мантии и руках,
кроме (или вместо) колец, обычно
имеются извилистые голубые полос-
ки. Чернильный мешок хорошо раз-
вит. Самка вынашивает яйца на ру-
ках. Длина яиц 7—8 мм.
Н. maculosa (Hoyle, 1883) (=Oc-
topus pictus Brock, 1882, non Blain-
ville, O. fasciata Hoyle, 1886) (рис.
78, а, б, г). От Южного Хонсю до
Тасмании, на запад до Мальдивских
островов. Самый опасный вид голо-
воногих моллюсков. В секрете зад-
них слюнных желез содержатся 2
мощных токсина: макулотоксип (схо-
ден с тетродотоксином) и хапало-
токсин. Укус взрослой самки может
быть смертелен. В Австралии отме-
чено несколько смертных случаев.
Человек погибает от паралича дыха-
тельной мускулатуры.
10(5). Гектокотиль очень корот-
кий с маленькой недифференцирован-
ной лигулой, без каламуса . . И.
11(12). У оснований каждой ру-
ки имеется кожная пора, открыва-
ющаяся с одной стороны на внутрен-
ней поверхности умбреллы (на уров-
не 3-й пары присосок), с другой —
на наружной поверхности головы
(они плохо заметны у фиксирован-
ных особей). Руки с утонченными
концами, 1-я пара самая длинная,
в 5—7 раз длиннее туловища. При-
соски на 1-й и 2-й парах рук самца
резко увеличены. Ширина умбреллы
7в—Ув длины наибольшей руки. Го-
лова заметно уже мантии. Мантий-
ное отверстие широкое. Воронка
длинная, вороночный орган W-образ-
ный.
Род Cistopus Gray, 1840. Один
вид — С. indicus (d’Orbigny, 1840)
(рис. 78, <5). Прибрежный тропиче-
ский индомалайский вид; распростра-
нен от Западной Индии до Южного
Китая и Индонезии. Длина мантии
до 11 см. Имеет небольшое промыс-
ловое значение.
12(11). Кожных пор нет. Руки
примерно такой же длины, как ту-
ловище, голова не уже мантии; ман-
тия узкая, удлиненная, так что ось-
миног по форме тела похож на каль-
мара. Присоски у самцов не увели-
19а К. Н. Несис
293
чены. Ширина умбрсллы около 7з
длины наибольшей руки. Мантийное
отверстие довольно узкое. Воронка
короткая, почти целиком погружена
в ткани головы, вороночный орган
W-образный.
Род Macrochlaena Robson, 1929.
Один вид — М. winckworthi (Robson,
1929). На мелководьях у Южной
Индии.
13(4). Тело слабомускулистое,
мягкое на ощупь, с подкожным слоем
пол у студен истой ткани. Голова и
мантия широкие. Руки короткие, в
1,5—3 раза длиннее тела. Вороноч-
ный орган VV-образный . . . 14.
14(15). Мантийное отверстие уз-
кое. Воронка почти целиком погру-
жена в ткани головы. Руки в 1,5—
2,5 раза длиннее мантии. Ширина
умбреллы Vs—Va длины наибольшей
руки. Глаза обычных размеров. Ко-
жа покрыта низкими широкими бу-
горками, как бы шагреневая (это за-
метно главным образом на боках и
задней стороне маптии и только под
увеличением). Чернильный мешок
заметно редуцирован.
Род Danoctopus Joubin, 1933
(=Berrya Hoyle, 1939, Hapaloctopus
Taki, 1962) (рис. 78,e—л).. Типовой
вид — D. schmidti Joubin, 1933. 2—
3 вида в верхней батиали (реже
нижней сублиторали) тропической
Западной Атлантике и Ипдо-Вестпа-
цифнки. Их различия невелики и не-
достаточно четко установлены.
а (б). Длина гектокотиля 4—8%
длины гектокотилизированной руки.
D. schmidti Joubin, 1933 (рис.
78, е—и). Карибское море, Флорид-
ский пролив, Багамские острова.
Глубины 300—1200 м.
б (а). Длина гектокотиля обычно
10—13% длины гектокотилизирован-
ной руки.......................в.
в (г). Глубина умбреллы Vs—%
длины наибольшей руки
D. hoylei (Berry, 1909) (рис.
78,/с, л). Иидо-Всстпацифика от Пер-
сидского залива до Южной Японии
и Гавайских островов. Глубины 50—
850 м, обычно более 200 м.
г (в). Глубина умбреллы 2/5—V2
длины наибольшей руки.
D. keralensis (Oommen, 1966)
[=? D. аппае (Oommen, 1980)]. Ара-
вийское море у Юго-Западной Ин-
дии. Глубины 200—400 м. Возможно,
синоним предыдущего вида.
15(14). Мантийное отверстие ши-
рокое. Воронка свободна на Vs своей
длины. Руки в 3 раза длиннее ман-
тии. Ширина умбреллы около Vs
длины наибольшей руки. Глаза очень
большие, почти соприкасающиеся на
«темени». Кожа гладкая, не шагре-
невая. Чернильный мешок хорошо
развит. -
Род Sasakinella Taki, 1964 *. Один
вид — 5. eurycephala Taki, 1964. Тихоь
океапское побережье Южного ХонсЮ.
16(3). Гектокотилизирустся левая
3-я рука.......................17.
17(18). Тело удлинеппо-яйцевид-
ной формы. Руки очень длинные и
тонкие, их длина 80—85% общей
длины, концы рук легко обламыва-
ются. Глубина умбреллы лишь око-
ло 9% длины наибольшей руки. Над-
глазничных усиков нет. На спинной
стороне маптии посредине между
головой и задним концом тела по обе
стороны от средней линии пара яр-
ких пятой крюковидпой формы, окру-
женных светлой полоской. Вороноч-
ный орган VV-образный. Гектокоти-
лизированпая рука в 3—6 раз короче
противоположной руки. Длина мап-
тии до 3 см.
Род Euaxoctopus Voss, 1971. Вид—
Е. panamensis Voss, 1971 (рис. 78,
м—с).
Род Euaxoctopus (типовой вид —
£. panamensis Voss, 1971) включает
2 вида, у которых гектокотилизиру-
ются разные руки 3-й пары — един-
ственный случай такого рода у ось-
миногов.
а (б). Гектокотилизирустся 3-я ле-
вая рука. Вороночный орган W-об-
разный. В полужабрс 1J—13 фила-
ментов. Глазчатые пятна па спиппой
стороне маптии крюковидной формы.
Е. panamensis Voss, 1971 (=?Гге-
moctopus scalenus Hoyle, 1904) (рис.
78,м—о). Панамский залив Тихого
океана. Верхняя сублитораль.
б (а). Гсктокотилизируется 3-я
правая рука. Вороночный орган
W-образный (?). В полужабрс 7 фи-
ламентов. Глазчатые пятна на ман-
тии С-образной формы.
♦ Самцы Sasakinella не описаны.
Предположение, что у них гектокоти-
лизирована 3-я правая рука — теза
3(16), я сделал на основании общего
сходства этого рода с Danoctopus,
для которого характерен этот при-
знак.
294
E. pillsburyae Voss, 1975 (рис.
78, л—с). Южная часть Карибского
моря и побережье Гвианы. Верхняя
сублитораль. См. тезу 8(9).
18(17). Тело обычной для осьми-
ногов широко-овальной формы. Ру-
ки умеренной длины и толщины. Ум-
брелла глубокая. Надглазничные
усики имеются. Нет глазчатых пятен
на мантии. Гектокотилизированная
рука не укорочена. Длина мантии
до 12—16 см...................19.
19(20). Тело на ощупь плотное.
Кожа сплошь покрыта мелкими ок-
руглыми плосковершиниыми папил-
лами. Над каждым глазом по одно-
му усику. По бокам мантии обычно
проходит низкий продольный кож-
ный гребень или складка. Ширина
умбреллы 20—30%, редко до 40%
длины наибольшей руки. Концы рук
не обламываются. Вороночный орган
W-образный, редко VV-образный.
Лигула гектокотиля длинная (8—
10% длины руки) с закрученными
внутрь краями, каламус тоже длин-
ный.
Род Scaeurgus Troschcl, 1857.
Один вид — S. unicirrhus (delle
Chiaje, 1830) (=S. patagiatus Berry,
1913) (рис. 78, г—ф). Тропическо-
субтропический атл а нто-ин довестпа-
цифичсский нижнссублнторальпо-
верхпебатиальпый вид.
20(19). Тело на ощупь мягкое,
дряблое. Кожа почти гладкая или с
редкими бородавочками. Над каж-
дым глазом по два усика или бугор-
ка. Ширина умбреллы 30—40% дли-
ны наибольшей руки. Боковой кож-
ной складки пет. Концы рук часто
обламываются. Вороночный орган
VV- или W-образный. Лигула и ка-
ламус короткие, длина гектокотиля
3—7% длины руки.
Род Pteroctopus Fischer, 1882
(рис. 78, х, ц). Типовой вид —Р. tet-
racirrhus (delle Chiaje, 1830). 2 вида
распространены в Атлантическом
океане и Охотском море.
а (б). Руки в 3—4 раза длиннее
мантии. Вороночный орган VV-об-
разный. Диаметр присосок 4—6%
длины маптии.
Р, tetracirrhus (delle Chiaje, 1830)
(рис. 78, х, ц). Атлантический тропи-
ческо-субтропический нижнесублито-
рально-верхнебатиальный вид: от
Южной Флориды, Мексиканского за-
лива и Карибского моря до Уругвая
и от Средиземного моря до Габона.
Обычно на глубинах 200—500 м.
б (а). Руки в 2,3 раза длиннее
мантии. Вороночный орган W-образ-
ный. Диаметр присосок 11% длины
мантии.
Р. witjazi Akimushkin, 1963. Най-
ден па глубине 100 м у юго-западно-
го побережья Камчатки. Вид сомни-
тельный.
21(2). Руки с одним рядом при-
сосок.
Подсемейство Eledoninae . . 22.
22(29). Тело обычно без бугорков
со звездчатыми основаниями (про-
стые бугорки часто имеются). Воро-
ночный орган в форме W или VV .23.
23(24). Гектокотиль очень корот-
кий, с почти неразвитыми лигулой и
каламусом. Концы всех остальных
рук самца с видоизмененными при-
сосками, преобразованными в па-
пиллы или узкие пластинки. Обита-
тели бореальных, тропических, суб-
тропических вод.
Род Eledone Leach, 1817 (=Мо-
schites Schneider, 1784, Ozoena Rafi-
nesque, 1814, Epistrophea Gistcl, 1848,
Hoylea Rochebrune, 1886, Acantheledo-
ne Fort, 1941) (см. рис. 80, a—л).
Типовой вид — E. moschata (Lamarck,
1798). 6 видов в Атлантическом океа-
не и прилегающих морях от Север-
ной Бразилии до Центральной Ар-
гентины и от Лофотенских островов
до Южной Африки, также в Крас-
ном море (?). Обычные обитатели суб-
литорали (преимущественно нижней)
и верхней батиали. Е. moschata и Е.
cirrosa имеют значительное промыс-
ловое значение в Южной Европе.
24(23). Гектокотиль с хорошо
развитыми лигулой и каламусом
(у Pareledone carlgreni каламуса
нет). Концы остальных рук самца не
изменены. Обитатели Антарктики и
умеренных вод Южного полуша-
рия ..........................25.
25(28). Животные небольших и
средних размеров (длина маптии до
10 см, общая длина до 35 см, обыч-
но меньше) Голова шире, равна или
немного уже мантии. Зоб имеется.
Чернильный мешок хорошо развит.
Кожа гладкая или бугорчатая . 26.
26 (27). Радула дсгенерировавша я;
1-й латеральный зуб едва заметен.
2-й без зубца, 3-й отсутствует. Воро-
ночный орган VV-образный. Кожа
покрыта коническими бугорками.
Над глазами по 2 усика.
19а*
295
Род Vosseledone Palacio, 1978.
Один вид—V. charrua Palacio, 1978
(см. рис. 79, а, б). От Южной Брази-
лии до Северной Аргентины, преиму-
щественно на глубинах 100—800 м.
27(26). Радула нормальная. Во-
роночный орган VV- или W-образ-
ный.
Род Pareledone Robson, 1932 (см.
рис. 80, м—с). Типовой вид—Р. char-
coti (Joubin, 1905). 7—8 видов: 6—
7 распространены у берегов Антарк-
тиды и островов дуги Скотин в суб-
литорали и батиали (2 вида прони-
кают на север вдоль Патагонского
шельфа до района Рио-де-Жанейро),
1 — у Южной Африки. В Антаркти-
ке эти осьминоги обычны и много-
численны.
28(25). Крупные животные, длина
мантии до 20—25 см, общая длина
до 70—75 см. Голова намного уже
мантии. Зоб отсутствует. Чернильный
мешок слабо развит. Кожа лишена
бугорков.
Род Megaleledone Taki, 1961. Один
вид — М. senoi Taki, 1961. Восточная
Антарктида. Сублитораль и батиаль.
Самый большой осьминог Антаркти-
ки. Яйца этого вида — самые круп-
ные из всех двужаберных головоно-
гих моллюсков: длина яйца без сте-
белька 34 мм.
29(20). Тело покрыто бугорками
со звездчатыми основаниями. Воро-
ночный орган двойной VV или чет-
верной НИ. Надглазничные усики
имеются .......................30.
30(31). Вороночный орган VV-об-
разный. Руки за пределами умбрел-
лы с широкими оторочками на вен-
тральной стороне. По одному усику
над каждым глазом. Лигула и кала-
мус хорошо развиты.
Род Velodona Chun, 1915. Один
вид—V. togata Chun, 1915 (рис.
79, в, г). Найден у Кении на глуби-
не 750 м. У Наталя на глубине 400—
450 м поймана V. togata чат. сареп-
bis Robson, 1924, отличающаяся бо-
лее плотным расположением бугор-
ков й наличием усиков на теле.
31 (30). Вороночный орган ПП-
образный. Руки без широких оторо-
чек. По два усика над каждым гла-
зом. Лигула короткая, слабодиффе-
ренцированпая.
Род Тetracheledone Voss, 1955.
Один вид — Т. spinicirris Voss, 1955
(рис. 79, д—ж). Распространен в верх-
ней батиали тропической Западной
296
Атлантики и прилегающих морях, от
Северной Каролины до Карибского
моря.
32(1). Чернильный мешок отсут-
ствует. Преимущественно глубоко-
водные виды (батиаль, абиссаль), но
в высоких широтах выходят на
шельф.
Подсемейство Bathypolypodi-
пае............................33.
33(40). Присоски рук в л-ва -ря-
да ................... . ./ . . 3 4>
34(37). Вороночный орган W- или
VV-образный. Задние слюнные желе-
зы не редуцированы.............35.
35(38). Присоски рук мелкие, не
более 15%, чаще 3—8% длины ман-
тии (кроме высокоарктического
Benthoctopus sibiricus см. рис. 84,
а, б). Каламус гектокотиля корот-
кий, тонкий....................36.
36(37). Лигула короткая, узкая, ее
длина 4—15%, обычно 5—7% длины
гектокотилизированной руки. Руки
обычно в 2,5—6 раз длиннее мантии.
Кожа гладкая, надглазничных усиков
нет. Зоб имеется.
Род Benthoctopus Grimpe, 1921
(r=Atlantoctopus Grimpe, 1921) (см.
рис. 84). Типовой вид—В. pisedtorum
(Verrill, 1879). 15—23 вида (в том
числе несколько неописанных), рас-
пространенных в батиали и у под-
ножья материкового склона всех
океанов от Арктики до Субантаркти-
ки. В высоких широтах обитают и па
шельфе, в тропиках обычно на глу-
бине более 500 м. Длина мантии до
10—13 см. Длина яиц до 28 мм.
37(36). Лигула длинная, кониче-
ская, ложковидная или округлая,
часто с резкими поперечными гребня-
ми, ее длина обычно 13—35%, иног-
да до 40% длины гектокотилизиро-
ванной руки. Руки обычно в 1,5—
3 раза длиннее тела. Кожа, как пра-
вило, покрыта бугорками и борода-
вочками либо по всему телу, либо на
голове и у основания рук. На голове
над глазами часто имеются кониче-
ские усики (рожки). Вороночный ор-
ган, как правило, VV-образный. Зоб
редуцирован или отсутствует.
Род Bathypolypus Grimpe, -1921
(рис. 83). Типовой вид — В. arcticus
(Prosch, 1849). 4—5 видов в нижней
сублиторали и батиали Арктики, бо-
реальных районов Атлантики и Па-
цифики, в тропических водах Атлан-
тики, Средиземном море и у Южной
Африки. Длина маптии до 10 см,
обычно до 4—5 см.
38(35). Присоски очень крупные,
их диаметр у самца до 22 % длины
мантии. Каламус гектокотиля очень
длинный и толстый, почти равный
по длине лигуле, которая отогнута
наружу под прямым углом. Гектоко-
тилизированная рука вдвое короче
противоположной 3-й левой руки.
Руки в 4—5 раз длиннее мантии.
Кожа с мелкими бугорками. Воро-
ночный орган W-образный.
Род Grimpella Robson, 1928. Один
вид — G. thaumastocheir Robson,
1928 (рис. . 79,з). У Южной Австра-
лии на мелководье.
39(34). Вороночный орган из 4
частей, ПП-образный. Руки в 2—3
раза длиннее мантии. Кожа гладкая.
Зоб имеется. Задние слюнные желе-
зы редуцированы.
Род Teretoctopus Robson, 1929.
Типовой вид — Т, indicus Robson,
1929. 2 вида в батиали северной час-
ти Индийского оксана и прилегаю-
щих морей.
а (б). Длина рук около 70% об-
щей длины. Диаметр присосок 4%
длины мантии. Глубина умбреллы
35—40% длины наибольшей руки.
Длина лигулы около 6% длины гек-
токотилизированной руки.
Г. indicus Robson, 1929. Найден в
батиали Аравийского моря.
б(а). Длина рук 75—80% общей
длины Диаметр присосок 5—7%
длины мантии. Глубина умбреллы
30—35% длины наибольшей руки.
Длина лигулы 12—16% длины гек-
токотилизированной руки.
Т, alcocki Robson, 1932. Аравий-
ское море, Бенгальский залив, Анда-
манское море. Верхняя и нижняя ба-
тиаль.
40(31). Присоски рук в один
ряд............................41.
41(44). Руки в 2—5 раз длиннее
маптии. Ширина умбреллы меньше
»/2 длины наибольшей руки. Зоб сла-
бо развит или отсутствует. Латераль-
ные зубы радулы имеются (у Gra-
neledone antarctica иногда отсутству-
ет 1-й латеральный зуб) ... 42.
42(43). Спинная сторона мантии
и головы, часто также рук 1—3-й
пар покрыта коническими или слож-
ными звездчатыми папиллами или
бородавками. Вороночный орган VV-
образный. В полужабре 5—8 фила-
ментов.
298
Род Graneledone Joubin, 1918 (см.
рис. 85). Типовой вид — G. verrucosa
(Verrill, 1881). 5—6 видов в нижней
батиали и абиссали северных частей
Атлантического Тихого океанов,
Антарктики у'’ тропических районов
Тихого океана. \
43(42). Тело гладкое. Вороноч-
ный орган V-образный (?). В полу-
жабре 4—6 филаментов. Глаза не-
большие.
Род Bentheledone Robson, 1932
Spnc. 79, и). Типовой вид — В. rotun-
а (Hoyle, 1885). 2 вида в Юж-
ном океане (индоокеанский сектор)
и не определенные до вида формы в
абиссали Восточной Пацифики.
а (б). Руки вдвое длиннее мантии.
Диаметр присосок составляет 5%
длины мантии. Окраска темно-пур-
пурная.
В. rotunda (Hoyle, 1885) (рис.
79, и). Найдена в абиссали Австрало-
Антарктической котловины.
б (а). Руки втрое длиннее маптии.
Диаметр присосок 8% длины мантии.
Окраска серовато-белая.
В. albida (Berry, 1917). Найдена
в абиссали севернее Земли Уилкса,
Антарктида.
44(41). Руки в 1,3—1,7 раза длин-
нее мантии. Ширина умбреллы 3/5—
2/з длины наибольшей руки. Тело
покрыто мелкими бугорками или гра-
нулами. Вороночный орган двух-
или четырехчастный. 4—6 филамен-
тов в полужабре. 1-й и 2-й латераль-
ные зубы' радулы сильно редуциро-
ваны или отсутствуют. Глаза очень
крупные.
Род Thaumeledone Robson, 1930
(рис. 79, к, л). Типовой вид — Г. bre-
vis (Hoyle, 1885). 2 вида в батиали
Южной 'Атлантики.
а (б). Вороночный орган ПП-об-
разный. Три надглазничных усика.
Т. brevis (Hoyle, 1885) (рис. 79, к).
Юго-Западная Атлантика у Монте-
видео. Нижняя батиаль.
б (а). Вороночный орган W-об-
разный. Надглазничных усиков нет.
Т, gunt eri Robson, 1930 (рис.
79, л). В районе Южной Георгии.
Верхняя батиаль.
Таблица для определения видов рода
Eledone
1(6). Все тело густо покрыто мелки-
ми папиллами, между ними разбро-
саны многочисленные более крупные.
Надглазничные усики или бугорки
имеются. Руки примерно одинаковой
длины. На спинной стороне тела нет
крупных темных пятен, могут быть
лишь темные разводы. Осьминоги не
имеют мускусного запаха ... 2.
2(5). По бокам тела проходит
низкая периферийная кожная склад-
ка или гребень. Присоски у самцов
не увеличены, их диаметр 9—13%
длины маптии. На концах всех рук
самца, кроме гектокотилизированной,
по два ряда коротких папилл (рис.
80, г)..........................3.
3(4). Вороночный орган W-образ-
ный. Над глазом одна усиковидная
папилла. Окраска желто-бурая или
светло-красная. Внутренняя стенка
передней части сперматофора покры-
та мелкими шипиками.
Е. cirrosa (Lamarck, 1798) (рис.
80, а—г). Бореально-лузитано-среди-
земном орский вид: от Исландии и
Лофотенских островов (Норвегия)
до Касабланки (Марокко) и Среди-
земного моря, на восток до Мрамор-
ного моря. Сублитораль и верхняя
батиаль до глубины 500—800 м,
обычно на глубинах 50—200 м. Име-
ет промысловое значение.
4(3). Вороночный орган VV-об-
разный. Над глазом 3—4 усиковид-
ные папиллы. Окраска пурпурно-
красная с темными разводами. Спер-
матофоры слабодифференцированы,
без шипиков.
Е. massyае Voss, 1964. Тропиче-
ская и южносубтропическая Юго-За-
падная Атлантика — у побережий
Бразилии, Уругвая и Аргентины от
о-ва Тринидад до Росона. Верхняя
сублитораль.
5(2). Нет периферийной кожной
складки. Присоски в проксимальных
частях рук самцов резко увеличены,
их диаметр больше 20% длины ман-
тии. На самых кончиках всех рук
самца, кроме гектокотилизированной,
имеется щетка из усиковидных па-
пилл, расположенных в 3—4 ряда
(рис. 80, е). Сперматофоры как у
Е. cirrosa.
Е. thysanophora Voss, 1962 (рис.
80, д, е). Прибрежные воды Южной
Африки.
6(1). Тело гладкое или с редки-
ми мелкими папиллами на спинной
стороне. Периферийной кожной
складки нет. Присоски в базальных
частях рук 1-й и 2-й пар самца рез-
ко увеличены, их диаметр 11—22%
длины мантии....................7.
7(10). Руки примерно равной дли-
ны .............................8.
8(9). Кожа гладкая, лишь со сле-
дами морщинистости. Окраска темно-
коричневая, почти черпая.
Е. nigra (Hoyle, 1910). Намибия
(залив Людериц-бей). Литораль и
верхняя сублитораль.
9(8). Спинная поверхность ман-
тии, головы и умбреллы с редкими
мелкими папиллами, над глазами
имеются крупные «рожки». Тело жел-
товатое, серо-бурое или серо-желтое
с крупными резко очерченными тем-
ными пятнами. Чернильная жидкость
издает сильный мускусный запах, те-
ло даже фиксированных осьминогов
пахнет мускусом. На концах всех рук
самца, кроме гектокотилизированной,
2 ряда коротких, уплощенных папилл
(рис. 80, и). Сперматофоры без ши-
пиков.
Е. moschata (Lamarck, 1798) (рис.
80,ж—и). Средиземное море, Кадис-
ский залив. Сублитораль и самая
верхняя батиаль, обычно верхняя
сублитораль. Имеет небольшое про-
мысловое значение.
10(7). 1-я пара рук на V3—Ч2
длиннее 2-й, а 2-я заметно длиннее
3-й и 4-й пар. Тело гладкое, надглаз-
ничных усиков нет. Окраска серова-
то-лиловая или красновато-бурая без
крупных темных пятен. На концах
всех рук самца, кроме гектокотили-
зированной, 2 ряда уплощенных пла-
стинок (рис. 80,л). Сперматофоры
без шипиков.
Е. caparti Adam, 1950 (рис. 80, к,
л). Тропические воды Западной Аф-
рики от Мавритании (?) и Сьерра-
Леоне до Анголы. Сублитораль.
Таблица для определения видов рода
Pareledone
1(2). Верхняя сторона туловища, го-
ловы и рук покрыта сближенными
или изолированными бугорками и бо-
родавочками, над глазом обычно
имеется по 1—2 более крупных бу-
горка. Вороночный орган VV-образ-
ный.
Р. charcoti (Joubin, 1905) [=Р.
aurorae (Berry, 1917)] (рис. 80, м, н).
Антарктика (циркумполярно), дуга
Скотия, Фолклендские острова, Па-
тагонский шельф, на север распрост-
ранена до района Рио-де-Жанейро.
299
п н
В Антарктике от литорали до батиа-
ли, возможно до абиссали, у Фолк-
лендских островов и Патагонии в
нижней сублиторали и верхней ба-
тиали. Самый, обычный осьминог Ан-
тарктики.
2(1). Верхняя сторона туловища,
головы и рук гладкая или с неболь-
шими разбросанными бугорками . 3.
3(6). Кожа гладкая. Вороночный
орган W-образный. Периферийная
складка кожи на туловище обычно
отсутствует ...................4.
4(5). Диаметр присосок 7—11%
длины маптии, длина рук около 70%
общей длины животного. В каждой
полужабре 6—7 филаментов.
Р. adelieana (Berry, 1917). Вос-
точная Антарктида. Сублитораль и
верхняя батиаль.
5(4). Диаметр присосок 4—6%
длины мантии. Длина рук около 60%
общей длины. В каждой полужабре
8 филаментов.
Р. utnitakae Taki, 1961 (рис. 80, о).
Море Космонавтов (Антарктика).
Батиаль.
6(3). Кожа покрыта бугорками и
бородавочками ....... 1.
7(8). Диаметр присосок 14—15%
длины мантии. Длина рук около
общей длины. Вороночный орган
VV-образный. Периферийная складка
кожи отсутствует. Кожа с тесно сбли-
женными мелкими гранулами.
Р. carlgreni Thore, 1945. У мыса
Доброй Надежды.
8(7). Диаметр присосок 5—11%
длины мантии. Длина рук около %,
общей длины. Кожа с редкими гра-
нулами или папиллами. Виды рас-
пространены в Антарктике и на Па-
тагонском шельфе................9.
9(12). Вороночный орган VV-об-
разный ........................10.
10(11). Кожа с редкими мелкими
гранулами, почти гладкая. Перифе-
рийная складка кожи отсутствует..
В полужабре 9—11 филаментов.
Р. turqueti (Joubin, 1905) (рис.
80, и). Антарктика (циркумполярно)^
дуга Скотия, Южная Георгия, Пата-
гонский шельф, на севере до района
Рио-де-Жанейро. От верхней субли-
торали до верхней батиали.
11(10). Кожа с рассеянными, на
хорошо выраженными папиллами.
Периферийная складка кожи па теле-
развита. В иолужабре 8—10 фила-
ментов.
Р. harrisoni (Berry, 1917). Циркум-
полярно в Антарктике. От верхней
Рис. 80. Роды Eledone (а—л) и Pareledone
{м—с)-. а—г—Eledone cirrosa (а — самец,
б —самка); д—е — Е. thysanophora’, ж—и—
Е. moschata; к—л — Е. caparti’ м—н —Pa-
reledone charcoti; о — Р. umitakae; п — Р.
turqueti-, р—с — Р. polymorpha; в, д, з, к,
о, с — гектокотиль; г, е, и, л — конец не
гектокотилизированной руки самца (а, в—
г, ж—и — по Naef, 1923; б — по Muns, 1959;
д—е — по G. Voss. 1962b; к—л — по Adam,
1952; м—н, л — по Joubin, 1905; о — по Ta-
ki, 1961; р—с — по Robson, 1930)
30t
сублиторали до верхней батйали.
12(9). Вороночный--орган W-об-
разный. Периферийная складка кожи
•имеется. Кожа с редкими, хорошо
выраженными папиллами. В полу-
жабре 7—9(10) филаментов.
Р. polytnorpha (Robson, 1930)
(рис. 80, р\ с). Южная Георгия, Ю.
Шетландские и 10. Сандвичевы о-ва.
Сублитораль и самая верхняя ба-
тиаль.
Примечание. В таблицу не
включен вид В. aniarctica (Thiele,
1920), описанный (крайне неполно),
•из моря Дейвиса. Возможно, сино-
ним Р. harrisoni.
Таблица для определения видов
подсемейства Octopodinae
Атлантического океана
1(34). Гектокотилизируется 3-я пра-
вая рука . . ..................2.
2(33). Тело крепкое, более или
.менее мускулистое. Мантийное от-
верстие нс сужено..............3.
3(4). На спинной стороне мантии
по бокам от средней линии тела име-
ется по одному крупному глазчатому
пятну полукруглой или С-образной
формы. Руки очень длинные (80—
В4% общей длины), часто обламыва-
ются на концах. В полужабре 7 фи-
ламентов. Длина лигулы 7—10%
длины гектокотилизированной руки.
Вороночный орган W-образиый (?).
Длина мантии до 2,5 см.
Euaxoctopus pillsburyae Voss, 1975
(см. с. 294—295).
4(3). Нет глазчатых пятен на
спинной стороне мантии (род Octo-
pus) ..........................5.
5(26). Виды распространены в тро-
пических и субтропических водах
Атлантики, на юг до Южной Брази-
лии и мыса Доброй Надежды * . 6.
6(9). С наружной стороны ум-
бреллы между руками 2-й и 3-й пар
впереди глаза расположено по одно-
му округлому глазчатому пятну . 7.
7(8). Вокруг центра глазчатого
пятна имеется голубое кольцо. Жи-
вотные мелкие (длина мантии до
7 см, чаще до 3—4 см). В полужабре
6—7 филаментов. Длина лигулы 3—
5% длины гектокотилизированной ру-
ки. Длина яиц 1,6—1,8 мм.
* У Южной Африки может встре-
титься О. fontanianusi см. тезу
29(30).
302
О. hutnmelincki Adam, 1936 (рис.
81, а, б). Тропические воды Западной
Атлантики от Багамских островов и
Южной Флориды до Баия (Брази-
лия), Карибскос морс. Преимущест-
венно близ островов, на рифах, в
верхней сублиторали.
8(7). Нет голубого кольца вокруг
центра глазчатого пятна. Животные
крупные, длина мантии до 20 см.
В полужабрс 9—10 филаментов.
Длина лигулы 1,4—1,9% длины гек-
токотилизировапиой руки. Длина яиц
около 17 мм.
О. тауа Voss et Solis, 1966 (рис.
81, в, г). Прибрежные мелководья
залива Кампече и п-ва Юкатан.
Важный промысловый вид.
9(7). Нет глазчатого пятна на ум-
брелле.........................10.
10(11). На мантии, голове и ру-
ках проходят широкие поперечные
полосы темно-коричневого или жел-
товато-коричневого цвета на светло-
желтоватом фоне. Животные мелкие
(длина мантии до 3 см). В полужаб-
ре 6—7 филаментов. Длина лигулы
6—9% длины гектокотилизированной
руки. Длина яиц около 6 мм.
О. zonatus Voss, 1968 (рис. 81,6,
е). Южная часть Карибского моря.
Внутренний шельф.
11(10). Нет поперечных полос ко-
ричневого цвета на мантии и голо-
ве .................. , ... 12.
12(13). Мантия, голова и руки по-
крыты тесно сближенными мелкими
остр о-округленным и папиллами. По
дорсальной стороне каждой руки
вдоль всей се длины проходит тем-
но-пурпурная или бурая полоска.
Длина мантии до 7 см.. В полужабре
8—10 филаментов. Длина лигулы 4—
6% длины гектокотилизированной
руки. Длина яиц 2—2,5 мм.
О. burryi Voss, 1950 (=О. Vincenti
Pickford, 1955) (рис. 81, от—и). Тро-
пические воды Западной и Восточной
Атлантики: от Джорджии до Мекси-
канского залива, Карибского моря и
Северной Бразилии, Западная Афри-
ка от Сенегала и островов Зелено-
го Мыса до Анголы. Внутренний (ре-
же внешний) шельф, на песчаном и
илистом дне.
13(12). Мантия, голова и руки
без папилл или с отдельными раз-
бросанными папиллами главным об-
разом между глазами. Нет темной
продольной полосы на руках . 14.
14(15). Руки 1-й пары заметно
толще и длиннее прочих (2-я пара
иногда столь же толстая, но коро-
че), Длина рук 80—90% общей дли-
ны. Мантия узкая, продольно-оваль-
ная. Животные крупные, длина ман-
тии до 18 см. В полужабре 9—13
филаментов. Лигула крупная, 5—
14 % (в среднем 10 %) длины гекто-
котилизированной руки. Длина яиц
1,8—2,5 мм. У живых осьминогов на
теле яркие белые пятна.
О. macropus Risso, 1826 ( = 0. ber-
mudensis Hoyle, 1885) (рис. 81, к, л).
Тропический атланто-индовестпаци-
фический вид, в Западной Атлантике
приурочен преимущественно к остро-
вам. Бермудские, Багамские острова,
Южная Флорида, Карибское, Среди-
земное моря, Западная Африка до
Гвинейского залива, о-ва Вознесения
и Святой Елены. У берегов па ри-
фах и скалах.
15(14). Самые длинные руки —
2-я и 3-я пары или все руки пример-
но равной длины и толщины . 16.
16(17). Руки 2-й и 3-й пар за-
метно длиннее и толще, чем 1-й и
4-й, их длина 80—90% общей длины.
Животные средних размеров (длина
мантии до 10—12 см). В полу жабре
6—8 филаментов. Длина лигулы 3—
4% длины гектокотилизированной
руки, лигула овальная с дюжиной по-
перечных гребней. Длина яиц 10—
14 мм.
О. briareus Robson, 1929 (рис.
81,л). Тропические воды Западной
Атлантики от Южной Флориды до
побережья Северной Бразилии, Ка-
рибское море, восточная часть Мек-
сиканского залива. Прибрежные мел-
ководья, рифы.
17(16). Все руки примерно оди-
наковой толщины, руки 2-й и 3-й пар
обычно немного длиннее прочих . 18.
18(19). Руки очень длинные (85—
90% общей длины тела), тонкие, час-
то заметно асимметричны — одна из
рук может быть значительно длиннее
противоположной руки той же пары.
Животные небольшие или среднего
размера (длина мантии до 5 см).
В полужабре 11 филаментов-. Лигу-
ла короткая, 1,9—2,5% длины гекто-
котилизированной руки. Длина яиц
2 мм.
О. defilippi Verany, 1851 (рис.
81, к, о). Тропические и субтропиче-
ские воды Западной и Восточной Ат-
лантики: от Флориды и Багамских
303
©
©
£
ф
Рис. 81. Род Octopus, атлантические виды:
а—б — О. hummelincki; в—г — О. тауа Се-
голова сбоку); д—е —О. zonatus; ж—и—О.
burryi; к—л — О, tnacropus; м—0. briareus:
н—о — О. defilippi; п—т — О. vulgaris (п—
молодая особь); у—ф — О. salutii; х—ц —
О. j'oubini; ч — О. australis; ш—щ — О.
tehuelchus; э—ю — О. lobensis; б, г, з, л,
м, о, с, т, ф, ц, ч, щ, ю — гектокотили (т—
вариант); u — боковая рука сбоку (а—г,
х — по Roper, 1977, д—и, м, ц—по G. Voss,
1950, 1956, 1968; к—л, н—с, у—ф—ио Naef,
1923; т—по Adam, 1952; ч — по Dell, 1952;
ш—ю — по Castellanos, Menni, 1969b)
островов до южной части Карибско-
го моря и от Средиземного моря до
Анголы, по-видимому, Индийский и
юго-западная часть Тихого океана.
Весь шельф. Личинка — макротрито-
пус (Macrotritopus spp.).
19(18). Длина рук не более 85%
общей длины, их концы не утончены.
Руки симметричны...............20.
20(25). Длина рук 70—85% об-
щей длины. Длина маптии взрослых
животных больше 3 см. В полужабре
7—11, обычно 8—10 филаментов . 21.
21(24). Лигула короткая, се дли-
на 2,5—3% гектокотилизированной
руки...........................22.
22(23). Лигула с хорошо разви-
тым каламусом (!/г длины лигулы)
и продольной бороздой. Глубина ум-
бреллы 20—25% длины наибольшей
руки. Диаметр наибольших присосок
у самца в среднем 15—16 % длины
мантии. Кожа покрыта небольшими
бугорками и бородавочками, на го-
лове и в средней части спины имеет-
ся несколько «рожков». Длина ман-
тии до 20(30) см. Длина яиц 1,8—
2,5 мм.
О. vulgaris Lamarck, 1798 [=0. ru-
gosus (Bose, 1792) (рис. 81, п—т).
Тропическо-субтропический космопо-
литический вид. В Атлантике от Лонг-
Айленда до Южной Бразилии и от
южной части Северного моря до мы-
са Доброй Надежды; многочислен в
Мексиканском заливе, Карибском и
Средиземном морях, встречается так-
же у океанических островов. Весь
шельф, иногда верхняя часть скло-
на, но обычно на прибрежных мел-
ководьях. Важнейший промысловый
вид.
23(22). Лигула очень короткая,
без каламуса, с поперечной бороздой.
Глубина умбреллы Уз длины наи-
большей руки. Диаметр наибольших
присосок па руках самца 22% дли-
ны мантии. Кожа гладкая. Длина
мантии 5 см.
О. schulizei (Hoyle, 1910). Нами-
бия (залив Людериц-бей).
24(21). Лигула очень длинная, ее
длина 12—16% длины гектокотили-
зированной руки. Умбрелла далеко
протягивается вдоль вентральных
краев рук. Средняя глубина умбрел-
лы 24% длины наибольшей руки.
Кожа гладкая, с небольшими бугор-
ками, над глазами по одному «рож-
ку». Длина мантии до 12 см. Длина
яиц 5 мм.
306
О. salutii Vcrany, 1851 (рис.
81,1/, ф). Бискайский залив и Среди-
земное море. Верхняя батиаль и
нижняя сублитораль.
25(20). Длина рук 65—70% об-
щей длины. Животные мелкие (длина
маптии до 5 см, обычно до 2—3 см).
В полужабре 5—6 (реже 7) фила-
ментов. Длина лигулы 4—7% длины
гектокотилизированной руки. Длина
яиц 6—8 мм.
О. joubini Robson, 1929 (=О. mer-
catoris Adam, 1937) (рис. 81, x, ц).
Тропические воды Западной Атланти-
ки, Мексиканский залив, Карибское
море — от Джорджии до Венесуэлы
и Гвианского побережья. Прибреж-
ные мелководья и внутренний шельф.
26(5). Виды распространены у бе-
регов Уругвая, Аргентины, Фолкленд-
ских островов и Огненной Зем-
ли ♦..........................27.
27(28). Присоски в средних час-
тях всех рук самцов и самок увели-
чены, их диаметр 16—20% длины
мантии. Животные крупные, длина
мантии до 8—19 см. Длина рук 72—
83% общей длины. Длина лигулы
5—9% длины гектокотилизированной
руки. Умбрелла далеко протягивает-
ся вдоль вентральной стороны рук.
Кожа гладкая.
О. megalocyathus Gould, 1846
Ь=О. patagonicus Lonnberg, 1898,
. brucei (Hoyle, 1912)]. Магелланов
пролив, Огненная Земля, банка
Бердвуд, Фолклендские острова.
28(27). Увеличенные присоски
имеются лишь в средних частях рук
2-й и 3-й пар. Диаметр присосок у
самок 7—12% длины мантии. Живот-
ные мелкие, длина мантии 2—4 см.
Длина рук 60—80% общей длины.
Умбрелла обычного строения . 29.
29(30). Кожа шероховатая, на
спинной стороне маптии, головы и
рук много мелких бугорков и боро-
давочек, над глазами по одному
«рожку». Лигула со вздутыми боко-
выми сторонами и очень узкой, глу-
бокой, почти замкнутой бороздой,
* В таблицу не включены О. hyadesi
Rochebrune et Mabille, 1887, О. pen-
therinus Rochebrune et Mabille, 1887
и О. membranaceus (Rochebrune et
Mabille, 1887, non Quoy et Gaimard,
1832), описанные из района мыса
Горн по молодым экземплярам (дли-
на мантий 12—20 мм) и крайне не-
полно.
длина лигулы 5—9% длины гектоко-
тилизированной руки. В средних ча-
стях боковых рук самца 1—2 пары
резко увеличенных присосок, их диа-
метр 13—17% длины мантии. Длина
рук 70—80% общей длины. В полу-
жабре 8—11 филаментов. Длина яиц
3,5 мм.
О. fontanianus d’Orbigny, 1835
(=Robsonella fontaniana, Joubinia
fontaniana auctt). Южная часть Юж-
ной Америки от Северного Перу и
залива Голфо-Нуэво (Аргентина) до
мыса Горн. Крайне близкие формы
распространены у Наталя (Южная
Африка) — О. fontanianus var. afri-
canus (Robson, 1929) и в Австрало-
Новозеландском районе у Южной и
Юго-Восточной Австралии, Новой Зе-
ландии, островов Норфолк, Стьюарт,
Кэмпбелл, Окленд, Антиподов, Ча-
тем (О. australis lloyle, 1885) (рис.
81, ч). Возможно, это локальные
формы одного циркумглобалыюго но-
талыю-южносубтропического вида.
Вид распространен от литорали до
верхней батиали.
30(29). Кожа гладкая, на голове,
в передней части мантии и у осно-
вания рук редкие крошечные буго-
рочки; надглазничных усиков нет.
Резко увеличенных присосок нет.
Длина рук в среднем 65—70% общей
длины. Края лигулы не вздуты, бо-
розда не резко сужена . . . . 31.
31(32). Тело овальное, голова уз-
кая, шейный перехват обычно хоро-
шо заметен. Кожа гладкая, блестя-
щая. 6—7-я пары присосок на боко-
вых руках самца и самки немного
увеличены. Лигула с глубокой и до-
вольно узкой бороздой. Длина яиц
10—15 мм.
О. tehuelchus d’Orbigny, 1835 (рис.
81, ш, щ). От Южной Бразилии до
залива Голфо-Нуэво (Аргентина).
Отмечен у мыса Горн (?). Литораль
и прибрежные мелководья.
32(31). Тело округлое, голова ши-
рокая, шейный перехват нс выражен.
Кожа шероховатая, опаковая. Уве-
личенных присосок нет. Лигула с ши-
рокой мелкой бороздой.
О. lobensis Castellanos et Mcnni,
1969 (рис. 81,э,ю). Заливы Сан-Ма-
тиас и Голфо-Нуэво (Аргентина),
прибрежные мелководья.
33(2). Тело слабомускулистос,
мягкое па ощупь, с подкожным сло-
ем оводненной полустуденистой тка-
ни. Мантийное отверстие сужено.
Воронка почти целиком погружена в
ткани головы. Голова и мантия ши-
рокие, руки короткие, 60—65% об-
щей длины. Глубина умбреллы — Уз
длины наибольшей руки. Вороночный
орган VV-образный. Кожа покрыта
низкими широкими бугорками, как бы
шагреневая, в центре каждого бу-
горка— светлое пятнышко, над гла-
зами— по паре «рожек». Окраска
оранжевая. В полужабре 9—10 фи-
ламентов. Длина мантии до 4 см.
Danoctopus schmidti Joubin, 1933
(см. с. 294).
34(1). Гектокотилизируется 3-я
левая рука.....................35.
35(36). Спинная сторона мантии,
головы и оснований рук густо покры-
та крупными округлыми папиллами,
иногда выстраивающимися в про-
дольные ряды. Над каждым глазом
по одному длинному «рожку». По бо-
кам мантии между спипной и брюш-
ной сторонами у фиксированных осо-
бей обычно имеется кожная складка.
Тело на ощупь плотное, концы рук
не обламываются. Глубина умбреллы
Уб—Уз, иногда до 2/6 длины наиболь-
шей руки. Диаметр присосок 6—10%
длины мантии. Длина лигулы 8—
10% длины гектокотилизированной
руки. Вороночный орган, как прави-
ло, W-образный. В полужабре 12—
14 филаментов. Цвет оранжевый или
орапжсво-красный. Длина яиц 2 мм.
Длина мантии до 12 см.
Scaeurgus unicirrhus (delle Chiaje,
1830) (см. с. 295). В Атлантике от
Джорджии до Бразилии и от Бис-
кайского залива до Анголы, в Мек-
сиканском заливе, Карибском и Сре-
диземном морях. Главным образом
па глубинах 100—400 м.
36(35). Кожа спинной стороны
покрыта микроскопическими тесно-
сближенными пупырышками, почти
гладкая. Над каждым глазом по па-
ре «рожков». Боковой кожной склад-
ки нет. Концы рук часто обламыва-
ются. Кожа на ощупь мягкая, жела-
типообразной консистенции (особен-
но у живых особей). Глубина ум-
бреллы Уз—2/б длины наибольшей
руки. Диаметр присосок 4—6% дли-
ны мантии. Длина лигулы 3—7%
длины гектокотилизированной руки..
Вороночный орган W-образный.
В полужабре 8—10 филаментов.
Цвет желтовато- или красно-корич-
невый. Длина яиц 7—8 мм. Длина
мантии до 16 см.
307
Pteroctopus tetracirrhus (delle
‘Chiaje, 1830) (cm. c. 295).
Таблица для определения видов
подсемейства Octopodinae
Северной Пацифики (от Берингова
пролива до Корейского полуострова,
Центрального Хонсю и
Калифорнийского полуострова)
1(30). Виды распространены в севе-
ро-западной части Тихого океана,
Беринговом, Охотском и Японском
морях, па юг до полуостровов Ко-
рейского, Ното и Босо .... 2.
2(3). Руки резко неравной длины:
1-я пара гораздо длиннее и толще
прочих (вдвое длиннее рук 3—4-й
пар), ее длина около 80% общей
.длины. Умбрелла очень короткая
(6—10% длины, наибольшей руки).
Вороночный орган VV-образный. Гек-
токотиль крупный (И—23% длины
гектокотилизированной руки). Дли-
на яиц 8 мм.
Octopus variabilis (Sasaki, 1929)
[=0. minor variabilis (Sasaki, 1920),
'O. macropus auett. non Risso] (рис.
«82,'а, б). Япония от Хоккайдо до
Рюкю, Корея, КНР. Шельф, обычно
на илах. Дина мантии до 10 см.
3(2). Руки слабо различаются по
.длине и толщине, иногда 1-я пара
.длиннее, но не толще прочих. Длина
наибольшей руки обычно менее 80%
•общей длины. Глубина умбреллы
Ve—Уз, обычно 7s—V4 длины наиболь-
шей руки.........................4.
4(5). Имеются крупные (до 7 мм)
глазчатые пятна на умбрелле перед
глазами, между руками 2-й и 3-й
пар — по одному справа и слева.
Поперек головы кирпично-красная
полоса. Тело покрыто гранулами, по
’бокам обычно полоска из нескольких
удлиненных бугорков, над глазом
пара крупных папилл. Гектокотиль
короткий, длина лигулы 5—7% дли-
ны гектокотилизированной руки. Дли-
на яиц 10—13 мм.
О. membranaceus Quoy et Gaimard,
1832 ( = 0. fangsiao d’Orbigny, 1840,
O. areotatus de Haan in Ferussac et
d'Orbigny, 1840, O. ocellatus Gray,
1849, 0. pulcher Brock, 1887, O. brocki
Ortmann, 1888) (рис. 82, в). Индо-
вестпацифический тропическо-суб-
тропический вид, на севере доходит
до Хоккайдо. Прибрежные мелко-
водья. Длина мантии до 8 см. Второ-
308
степенный промысловый вид в Юго-
Восточной Азии.
5(4). Нет глазчатых пятен па ум-
брелле, красной поперечной полосы
на голове и удлиненных бугорков по
бокам .........................6.
6(27). Тело гладкое или покрыто
папилл ами, гранулами, бугорками,
усиками, по не звездчатыми бородав-
ками ..........................7.
7(12). Тело гладкое, иногда лишь
с малозаметными морщинками или
крохотными бородавочками па голо-
ве; надглазничные усики имеются . 8.
8(9). Вороночный орган VV-об-
разный. Консистенция тела плотная,
мясистая. Периферической складки
кожи нет. Умбрелла глубокая, 30—
40% длины наибольшей руки. Гекто-
котиль длинный (13—21% гектоко-
тилизированной руки), открытый.
Диаметр наибольших присосок 8—
15% длины мантии. Длина яиц 22—
28 мм.
О. conispadiceus (Sasaki, 1917)
(рис. 82, г, д). Низкоборсальный вид,
распространенный от Татарского
пролива и Южных Курильских остро-
вов до Корейского пролива в Внут-
реннею моря Японии. Сублитораль,
преимущественно песчаные грунты
верхней сублиторали («песчаный
осьминог» Приморья). Длина мантии
до 25 см, общая длина до 1,5 м.
Важный промысловый вид, добыва-
ется в Северной Японии, КНДР,
Южной Корсе и па Дальнем Востоке
СССР.
9(8). Вороночный орган W-образ-
ный. Консистенция тела дряблая, во-
дянистая. Периферическая складка
кожи обычно имеется.«Диаметр наи-
больших присосок 7—9% длины ман-
тии ...............: .... 10.
10(11). 1-я пара рук самая длинная.
Глубина умбреллы около Уа длины
наибольшей руки. Гектокотиль длин-
ный (7—15% длины гектокотилизи-
рованной руки). Над каждым гла-
зом по одному коническому бугорку.
O.leioderma (Berry, 1911) (рис.
82, е, ж). Высокобореальный вид: от
Берингова пролива до Курильских
островов и Северной Калифорнии, в
Японском море не обнаружен. Суб-
литораль и батиаль (у берегов Ва-
шингтона, Орегона и Калифорнии
только в нижней сублиторали и ба-
тиали). Длина мантии до 6 см.
11(10). Руки приблизительно рав-
ной длины. Глубина умбреллы около
]/4 длины наибольшей руки. Гектоко-
тиль короткий, около 4% длины гек-
токотилизированной руки. Над каж-
дым глазом по 3 небольших бугорка,
средний крупнее других и уплощен с
боков. Длина яиц 9 мм.
О. ochotensis (Sasaki, 1920). Охот-
ское море. Нижняя субли1. ораль.
Длина мантии до 5 см.
12(7). На теле имеются бородавки
(не звездчатые), папиллы или уси-
ки ............................13.
13(22). Надглазничные усики име-
ются — конические «рожки» или уп-
лощенные с боков «ушки» . . 14.
14(17). На голове и верхней сто-
роне маптии разбросаны отдельные
немногочисленные бородавочки, па-
пилл и усиков нет..............15.
15(16). Окраска спинной стороны
тела мраморная (темные разводы па
розово-буром фоне). Длина рук око-
ло 80% общей длины. Диаметр наи-
больших присосок 9—12% длины
мантии. Длина лигулы 14% длины
гектокотилизированной руки.
O.fujitai (Sasaki, 1929) (рис.
82, з, и). Южное побережье Японско-
го моря. По Н. Н. Кондакову (1941)
и И. И. Акимушкину (1963), также
Олюторский залив и Охотское морс.
Нижняя сублитораль. Длина мантии
до 5 см.
16(15). Окраска спинной стороны
однотонная, красно-бурая, между
глазами несколько темных попереч-
ных полос или пятен. Длина рук око-
ло 70% общей длины. У самца на
руках 1-й пары несколько резко уве-
личенных присосок близ края ум-
бреллы, их диаметр 15—16% длины
мантии. Длина лигулы 9% длины
гектокотилизированной руки.
О. tsugarensis (Sasaki, 1920) (рис.
82, к). Сангарский пролив (глубина
356 м), Охотское морс. Длина ман-
тии до 4 см.
17(14). Голова и верхняя сторона
мантии покрыты многочисленными
низкими гранулами и бородавками,
среди которых выделяются отдель-
ные более крупные усики или «рож-
ки» ...........................18.
18(19). Над каждым глазом по
одному уплощенному округленному
«ушку», кроме того, могут быть и
конические папиллы. Гектокотиль
очень длинный (8—25 %, обычно 10—
20 % длины гектокотилизированной
руки), лигула узкая, у свежепойман-
ных особей почти закрытая (трубча-
тая). Диаметр наибольших присосок
10—16% длины мантии. На голове
и мантии имеется несколько коничес-
ких «рожков». Длина яиц 6—8 мм.
Размеры очень крупные.
О. dofleini (Wiilker, 1910) [ = О*
gilbertianus (Berry, 1912), О. apollyon
(Berry, 1913), Paroctopus asper Aki-
mushkin, 1963, O. punctatus auett., non
Blainville, O. hongkongensis auett., non
Hoyle, ? O. madokai (Berry, 1921)]
(рис. 82, л—n). Тихоокеанский бо-
реальный вид, обитает в сублиторали
и верхней батиали. Самый крупный
из современных осьминогов: длина
мантии до 60 см и более, общая дли-
на до 3—5 м. Массовый вид, важ-
ный промысловый объект, добывает-
ся у Северной Японии, в КНДР,
Южной Корее, на Дальнем Востоке
СССР, на Алеутских островах, у Бри-
танской Колумбии, Вашингтона и
Орегона. Объект интереса аквалан-
гистов и подводных охотников. Дер-
жится преимущественно в скалах и
среди камней. 3 подвида: O.dofleini
dofleini (Wiilker) [ = ? О. madokai
(Berry)] (рис. 82, я, о) — Южные Ку-
рильские острова, Южный Сахалин,
Приморье, Корейский полуостров,
северные берега Желтого моря, Хок-
кайдо, Хонсю, Сикоку;
О. d. apollyon (Berry) [=0. gUber-
tianus (Berry), P. asper Akimushkin,
O. punctatus * auett., non Blainville]
(рис. 82, л, n)—от Берингова моря
до Курильских островов, северной
части Японского моря и залива
Аляска; О. d. martini Pickford, 1964
(рис. 82, м) — от Британской Колум-
бии до Южной Калифорнии.
19(18). Надглазничные усики не
уплощенные — конические или ци-
линдрические. Длина лигулы до 10%
длины гектокотилизированной ру-
ки ............................20.
20(21). 1-я пара рук — самая
длинная. На руках 1—2-й пар у сам-
ца есть несколько резко увеличенных
присосок, их диаметр 15—24% длины
маптии. Лигула длинная, около 10%
длины гектокотилизированной руки.
Конических папилл («рожков») на
спинной стороне мантии пет.
O,longispadiceus (Sasaki, 1917)
(рис. 82, р). Охотское, Японское мо-
ря, Тихоокеанское побережье Ку-
рильских островов, Хоккайдо, Хонсю.
Глубины порядка 100—800 м. Длина
маптии до 8 см.
21(20). 1-я пара рук обычно са-
309
310
И
Рис. 82. Род Octopus, северопацифические
виды: а—б — О. variabilis; в — О, membra-
naceus; г—д—О. conispadiceus; е—ж — О.
leioderma; з—и — О. fujitai; к — О. tsuga-
rensis; л—п — О. dofleini (л, п — О. d. аро-
Uy on, м — О. d. martini, н—о — О. d. dof-
leini); p—O. longispadiceus; c—t—O. yendoi;
у — О. araneoides; ф—ц — О. californicus;
ч—э — О. bimaculoides; б, д, о, л, ц — во-
роночный орган; г, ж, и, к, н, р, т, у, х,
ц — гектокотили; м, ш — расположение бу-
горков на спинной стороне головы п ман-
тии; э — глазчатое пятно на умбрелле (о—
б, г —по Sasaki. 1929; в—по Voss, William-
son, 1972; г—л, н—с, ф, ц — по Кондакову»
1941; м, х, ч — по Berry, 1912; у — по Taki,
1964; ш—э — по Pickford, McConnaughey,
1949)
311
мая короткая. Диаметр наибольших
присосок на руках 1—2-й пар у
самца 12—16% длины мантии. Лигу-
ла короткая, 2—5% длины гектоко-
тилизированпой руки. На спинной
стороне мантии несколько крупных
конических папилл. Яйца мелкие
(1,8—2,5 мм).
О. vulgaris Lamarck, 1798 [ = 0. ru-
gosus (Bose, 1792)1 (см. с. 306). В
Северо-Западной Пацификс доходит
на север до Сангарского пролива, но
обычно не встречается севернее полу-
островов Ното и Босо. Самый частый
осьминог Центральной и Южной Япо-
нии. Важнейший промысловый вид,
объект искусственного выращивания.
Длина мантии в Японии до 12 см.
22(13). Надглазничные усики от-
сутствуют. Голова и верхняя сторона
мантии покрыты редкими мелкими
бородавочками и пустулами . 23.
23(26). Гектокотилизирустся пра-
вая рука 3-й пары. Длина лигулы
6—12% длины гсктокотилизирован-
ной руки .....................24.
24(25). Воронка почти целиком
погружена в ткани нижней стороны
головы. Длина рук 70—82 %, обычно
75% общей длины. Диаметр наиболь-
ших присосок у самок 9—12%, у
самцов 13—16% длины мантии. Дли-
на лигулы 6—7% длины гсктокоти-
лизированной руки. Длина яиц 17 мм.
O.yendoi (Sasaki, 1920) (рис.
82, с, т). Охотское, Японское моря и
Тихоокеанское побережье Северной
Японии. Нижняя сублитораль и са-
мая верхняя батиаль. Длина мантии
до 6 см.
25(24). Воронка свободна на по-
ловину своей длины. Длина рук 65—
70 % общей длины. Диаметр наи-
больших присосок у самок 5—8 %, у
самцов 9—10 % длины мантии. Дли-
на лигулы 9—12% длины гсктокоти-
лизированпой руки. Длина яиц 13 мм.
О. arancoides (Taki, 1964) (рис.
82,у). Северо-Восточный Хонсю (Ку-
сиро). Мелководье. Второстепенный
промысловый вид. Длина мантии до
9—10 см.
26(23). Гсктокотилизируется ле-
вая рука 3-й пары. Длина лигулы 5%
длины гектокотилизированной руки.
Pteroctopus witjazi Akimushkin,
1963 (см. с. 295).
27(6). Голова, мантия и основа-
ния рук густо покрыты крупными
звездчатыми или дольчатыми боро-
давками ......................28.
312
28(29). Вороночный орган W-об-
разный. Длина рук 60—65% общей
длины. Умбрелла глубокая, 30—33%
длины наибольшей руки. Спинная
сторона тела серо-коричневатого
цвета.
О. spinosus (Sasaki, 1920). Сап-
гарский пролив, Хоккайдо. Нижняя
сублитораль и верхняя батиаль. Дли-
на мантии до 2,5 см.
29(28). Вороночный орган VV-об-
разный. Длина рук 70—75% общей
длины. Глубина умбреллы 22—30%
длины наибольшей руки. Спинная
сторона тела от темно-оранжевой до
кирпично-красной.
О. californicus (Berry, 1911) (рис.
82, ф—ц). См. тезу 37(38).
30(1). Виды распространены в
Северо-Восточной Пацифике от зали-
ва Аляска до Калифорнийского по-
луострова .....................31.
31(56). Длина рук 2/3—7/8 общей
длины. Глубина умбреллы 15—30%
длины наибольшей руки. Присоски в
2 ряда по всей длине руки, начиная
с 3-й ......................32.
32(35). Имеются крупные (до
20 мм) глазчатые пятна на умбреллс
перед глазами, между руками 2-й и
3-й пар — по одному справа и слева
(рис. 82,а). Гектокотиль крохотный,
длина лигулы 2—3% длины гектоко-
тилизировапной руки. Голова и ман-
тия покрыты мелкими гранулами и
коническими папиллами ... 33.
33(34). Диаметр наибольших при-
сосок у самца 16—26%, в среднем
19%, у самки 9—18%, в среднем
13% длины мантии. Длина рук 75—
80% общей длины. Длина яиц 2—
4 мм.
О. bimaculatus Vcrrill, 1883. Юж-
ная Калифорния, п-ов Калифорния,
Калифорнийский залив. Литораль и
верхняя сублитораль, преимуществен-
но на скалах. Длина мантии до 10—
18 см. Третьестепенный промысло-
вый вид.
34(33). Диаметр наибольших при-
сосок у самца 10—15%, в среднем
13% длины мантии, у самки 7—13%,
в среднем 10%. Длина рук обычно
80—90% общей длины. Яйца круп-
ные, длина 10—17 мм.
О. bitnaculoides Pickford et McCon-
naughey, 1949 (рис. 82, ч—э). Юж-
ная Калифорния и п-ов Калифорния.
Преимущественно на илистой лито-
рали. Массовый вид. Длина мантии
до 7 см.
35(32). Нет глазчатых пятен на
умбрелле ......................36.
36(55), Концы рук нормальные,
присоски па концах рук не видоиз-
менены ........................37.
37(38), Поверхность головы, ман-
тии и основания рук густо покрыта
крупными звездчатыми бородавками.
Окраска красная или оранжевая [см.
тезу 29(28)]. Вороночный орган
W-образный. Гектокотиль крупный
(длина лигулы 14—17% длины гек-
токотилизировапной руки). Длина
яиц 12—15 мм.
О. californicus (Berry, 1911) (рис.
82, ф—ц). Калифорния, нижняя суб-
литораль и верхняя батиаль. Япон-
ское море (залив Петра Великого и
Татарский пролив), верхняя субли-
тораль. Длина мантии до 12 см.
38(37). Поверхность тела гладкая
или покрыта гранулами, папиллами,
усиками, но не звездчатыми бородав-
ками. Вороночный орган не VV-об-
разный ........................39.
39(40). Руки чрезвычайно длин-
ные (около 85% общей длины), очень
хрупкие, их концы часто обламыва-
ются. Умбрелла короткая (14—22%
длины наибольшей руки). Гектоко-
тиль крохотный (2—2,5% длины
гектокотилизированной руки). По те-
лу разбросаны отдельные папиллы
трех размеров, самые крупные —
сложные; папиллы заходят и па
внутреннюю сторону умбреллы.
O.alecto (Berry, 1953). Калифор-
нийский залив. Мелкий вид (длина
мантии 2—3 см).
40(39). Длина рук 2/з—Vs общей
длины. Глубина умбреллы Vs—7з
длины наибольшей руки . . . 41.
41(42). 1-я пара рук самая длин-
ная (около 70% общей длины). Гек-
токотиль длинный (длина лигулы
7—15% длины гектокотилизирован-
ной рукну. Кожа гладкая, иногда с
отдельными плохо заметными мор-
щинками и бугорочками на голове.
Над каждым глазом по одному кони-
ческому бугорку. Периферийная
складка кожи обычно имеется. Резко
увеличенных присосок пет.
О. leiodeftna (Berrv, 1911) (см.
с. 308).
42(41). Руки приблизительно рав-
ной длины или 1-я пара короче дру-
гих. Периферийная складка кожи
отсутствует ...................43.
43(44). Длина рук % общей дли-
ны. Вороночный орган IAI-образный
Умбрелла протягивается вдоль вен-
тральных сторон рук 1—3-й пар до
их концов в виде широкой парусо-
видной мембраны. Голова, мантия,
основания рук и внутренняя сторона
умбреллы покрыты тесно сближенны-
ми папиллами. Жабры очень круп-
ные: в полужабре 15—17 филамен-
тов.
О. veligero Berry, 1953. П-ов Ка-
лифорния. Глубина 90 м. Длина ман-
тии 5—7 см.
44(43). Длина рук 70—80% об-
щей длины. Вороночный орган W-об-
разный. Нет широкой мембраны
вдоль вентральных сторон рук. Жаб-
ры обычных размеров .... 45.
45(46). Осьминоги гигантских
размеров. Гектокотиль очень длин-
ный, узкий, почти трубчатый, длина
лигулы 8—25%, обычно 10—20%
длины гектокотилизированной руки.
Диаметр наибольших присосок 10—
16% длины мантии. Над каждым
глазом по одному уплощенному с бо-
ков округленному «ушку», кроме
того, могут быть конические «рожки».
Тело покрыто морщинами, складками
и папиллами, на голове и мантии
несколько конических папилл. Длина
яиц 6—8 мм.
О. dofleini (Wiilker, 1910), подви-
ды О. d. apollyon (Berry, 1913) и
О. d. martini Pickford, 1964 (см. с.
309).
46(45). Осьминоги мелкие или
средних размеров (общая длина дс
30 см). Длина лигулы до 10—12%
длины гектокотилизированной руки
Уплощенных надглазничных «ушекя
не бывает — лишь конические «рож-
ки» ............................47
47(50). Кожа гладкая, лишь
иногда с малозаметными папиллам*
или бородавочками на голове . 48
48(49). Одна из присосок 4-й па-
ры на 1—3-й руках самца резко уве-
личена (у самок несколько увеличе
ны присоски 5—6-й пар). Лигула от-
крытая, коническая, ее длина 6—7%
длины гектокотилизированной рукн
Каламус очень длинный — V2 длинь
лигулы. Длина яиц 10—12 см.
О. micro руг sus Berry, 1953. Юж
ная Калифорния. Литораль и верхняя
сублитораль. Один из самых мелки:
видов осьминогов (длина мантии зре
лого самца 10 мм, максимальный раз
мер 25 мм).
49(48). Резко увеличена одна и:
присосок 5-й или 6-й пары на 1—3-i
20 К. II. Несис
31!
руках самца и самки. Лигула узкая,
удлиненная, се длина 9—12 °/о длины
гектокотилизированной руки, Кала-
мус маленький.
О. fitchi Berry, 1953. Калифорний-
ский залив. Многочислен на литора-
ли и на мелководьях под камнями.
Длина маптии взрослых самцов око-
ло 2 см. Укус болезнен, но не опа-1
сен.
50(47). Тело покрыто сетью мел-
ких низких бородавочек, соединенных
узкими гребешками-морщинками, или
гранулированное (низкие папиллы,
разделенные узкими бороздками); на
голове (в том числе над глазами) и
на спине могут быть небольшие
«рожки».......................51.
51(54). Одна-две присоски на 1—
3-й руках самца резко увеличены.
Гектокотиль длинный (7—11% дли-
ны гектокотилизированной руки).
Внутренняя сторона умбреллы глад-
кая ..........................52.
52(53). Глаза крупные, выступа-
ющие. 2-я пара рук обычно длиннее
прочих. На лигуле 20—24 поперечных
складки. Яйца мелкие, 2—4 мм.
О. rubescens Berry, 1953. От юж-
ной Аляски до п-ова Калифорния,
Калифорнийский залив. . Многочис-
ленный мелкий (длина мантии до
5—10 см) сублиторальный вид.
53(52). Глаза очень маленькие.
Руки равной длины. На лигуле около
10 поперечных складок. Яйца круп-
ные (длина 9—10 мм).
O.digueti Perrier et Rochebrune,
1894. Калифорнийский залив, па юг
доходит до Масатлана. На мелко-
водьях, часто в пустых раковинах
двустворок. Длина мантии до 6 см.
54(51). Резко увеличенных присо-
сок нет, хотя в целом присоски боко-
вых рук крупнее, чем 1-й и 4-й пар.
Гектокотиль маленький, длина лигу-
лы 1,5% длины гектокотилизирован-
ной руки. Внутренняя поверхность
умбреллы и оснований рук покрыта
папиллами.
О. hubbsorum Berry, 1953. Кали-
форнийский залив. Длина мантии до
5—7 см.
55(56). На концах рук самца
(кроме гектокотилизированной) вмес-
то присосок щеточка мелких пальце-
видных папилл в 4 ряда. Резко уве-
личенных присосок нет. Длина лигу-
лы 10—11% длины гёктокотилизи-
рованной руки. Тело гладкое, не-
сколько папилл над глазами. На те-
314
ле и голове темные пятна, на руках
пятна и полоски.
О. penicillifer Berry, 1954. Крайний
юг п-ва Калифорния. Верхняя субли-
тораль. Длина мантии до 3—4 см.
56(31). Длина рук 62% общей
длины. Глубина умбреллы 7з длины
наибольшей руки. Первые 5—6 при-
сосок на руках расположены в один
ряд.
Octopus (?) pricei (Berry, 1913).
Калифорния. Систематическое поло-
жение неясно. Длина мантии 2 см.
Примечание. В таблицу не
включен вид О. oshimai (Sasaki,
1929), описанный из вод Тайваня.
Н. Н. Кондаков (1941) и И. И. Аки-
мушкин (1963) указывают его для
Южного Приморья и юго-западной
части Охотского моря; определение
нуждается в подтверждении.
Таблица для определения видов рода
Bathypolypus
1(2). Гектокотиль узкий, конический,
его длина около 14% гектокотилизи-
рованной руки. Лигула с многочис-
ленными нерезко выраженными по-
перечными бороздками, каламус ко-
роткий. Все тело покрыто тесно сбли-
женными бугорками. Надглазничных
усиков нет. Присоски крупные (8—
12% длины мантии). 9—10 филамен-
тов в полужабре.
В. salebrosus (Sasaki, 1920) (рис.
83, и). Тихоокеанский призиатский
бореальный вид: Охотское море, Ти-
хоокеанское побережье Курильских
островов, Хоккайдо и Северо-Восточ-
ного Хонсю. Батиаль. Длина мантии
до 4—5 см.
2(1). Г ектокотиль значительно
шире руки, ложковидный или округ-
лый, иногда с боковыми лопастями.
Лигула с несколькими (от 4 до 16)
резкими поперечными гребнями.
Скульптура разнообразная, от силь-
но бугорчатой до почти гладкой ко-
жи. Надглазничные усики обычно
имеются, часто представляют собой
высокие конические «рожки». При-
соски мелкие, от 2,5 до 7—8% дли-
ны мантии, 6—9 филаментов в полу-
жабре ..........................3.
3(4). Гектокотиль с параллельны-
ми краями или расширяющийся ди-
стально, без боковых лопастей. Ли-
гула с 8—16 поперечными гребнями.
Каламус короткий. У молодых сам-
цов лигула дистально сходит па ко-
20*
нус, у взрослых на ее вершине сре-
динный бугорок и два выступа по
бокам его. Тело более или менее
плотно покрыто звездчатыми бугор-
ками, иногда слабо заметными, в от-
дельных случаях почти гладкое. Ко-
нические надглазничные «рожки»
почти всегда хорошо выражены.
В. arcticus (Prosch, 1849) [—5. ba-
irdii (Verrill, 1873), В. lentus (Ver-
rill, 1880), B.obesus (Verrill, 1880),
B. faeroensis (Russel, 1909)] (рис.
83,6—д). Атланто-арктический
арктическо-бореальный вид. Приат-
лантическая Арктика на восток до
прол. Вилькицкого, па запад до за-
лива Франклин, возможно циркумпо-
лярно; Норвежское, Гренландское,
Баффиново моря; Северная Атланти-
ка, на юге до Флоридского пролива,
Юго-Западной Испании, северней
части Северного * моря и пролива
Каттегат. Обычен в Баренцевом и
Карском морях, отсутствует в Белом
и Печорском. Длина мантии до 7 см.
Сублитораль (преимущественно ниж-
няя) и батиаль, на юге ареала толь-
ко в батиали. Сильно изменчивый
вид, представлен двумя формами,
таксономический статус которых не
выяснен, возможно, разные виды.
а (б). Голова уже туловища, ее
ширина 60—90% длины мантии. Дли-
на лигулы не больше 20% длины гск-
токотилизированной руки. 8—10 фи-
ламентов в полужабре.
B.arcticus f. arcticus (Prosch).
Преимущественно арктические рай-
оны.
б (а). Голова равна или шире ту-
ловища, ее ширина не меньше длины
маптии. Длина лигулы У5—У3, иногда
до % длины гектокотилизированной
руки. 6—7 филаментов в полужабре.
В. arcticus f. proschi Muus, 1962.
Преимущественно бореальные райо-
ны.
4(3). Гектокотиль округлый или с
параллельными краями в базальной
половине и сходящий на конус в
дистальной. Боковые лопасти обычно
хороню выражены и могут целиком
скрывать' внутреннюю поверхность
лигулы. Лигула с 4—7 поперечными
гребнями, Каламус длинный (J/3—2/б
длины лигулы). Длина гектокотиля
12—18% длины гектокотилизирован-
ной руки. 6—7 филаментов в полу-
жабре .........................5.
5(6). Гектокотиль округлый, поч-
ти шаровидный, у взрослых с хоро-
316
шо выраженными боковыми лопастя-
ми. Надглазничные усики острые, ко-
нические. Кожа на голове с немного-
численными бугорками. Глубина ум-
бреллы Уз—% длины наибольшей
руки. Лигула с 4—5 поперечными
гребнями.
B.valdiviae (Chun et Thiele, 1915)
( = B. grimpei Robson, 1924) (рис.
83, e—з). Южная, Юго-Западная
(Намибия) и Юго-Восточная Афри-
ка, банка Агульяс и прилегающие
районы. Батиаль. Длина мантии до
4—5 см.
6(5). Гектокотиль с параллельны-
ми или слегка выпуклыми краями в
базальной половине, в дистальной
сходит на конус. Кожа на голове
вокруг глаз с многочисленными мел-
кими округлыми бугорками с 1—4
низкими округлыми вершинами. Над-
глазничные усики отсутствуют или
слабо заметны. Глубина умбреллы
Уе—Уз длины наибольшей руки. Ли-
гула с 6—7 поперечными гребнями.
В. sponsalis (Р. et Н. Fischer, 1892)
(рис. 83, а), Западная часть Среди-
земного моря и Северо-Восточная
Атлантика от Бискайского залива до
мысов Кап-Блан и Зеленый. Батиаль,
преимущественно средняя и нижняя.
Длина мантии до 10 см.
Таблица для определения видов рода
Benthoctopus
Виды рода Benthoctopus плохо изу-
чены, не ревизованы, поэтому состав-
ление общего ключа невозможно.
Ниже приводятся таблицы для опре-
деления арктобореальных и поталь-
ных видов и список тропических ви-
дов рода.
Арктика и бореальные области
1(4). У взрослых самцов несколько
присосок в средних частях рук резко
увеличены, их диаметр значительно
больше 10% длины мантии. Присос-
ки самок не увеличены, их диаметр
8—9 % длины мантии. Вороночный
орган в форме W................2.
2(3). Диаметр наибольших при-
сосок самца (8—12-я пара) около У4
длины мантии, диаметр неувеличен-
ных присосок около У6 длины мантии.
Длина лигулы около 12 % длины гек-
токотилизированной руки. Глубина
наибольшего сектора умбреллы око-
ло 30% длины наибольшей руки.
В, sibiricus Loyning, 1930 (рис.
84, а, б). Восточная Арктика — моря
Лаптевых, Восточно-Сибирское, Чу-
котское, Бофорта (?). Средняя и
нижняя сублитораль и, вероятно,
верхняя батиаль. Самый мелковод-
ный и холодноводный вид рода.
3(2). Диаметр наибольших присо-
сок у самца (14—17-я пара) около
15% длины мантии. Длина лигулы
7,5% длины гектокотилизированной
руки. Глубина умбреллы ’/4 длины
наибольшей руки.
В. abruptus (Sasaki, 1920) (рис.
84, в). Южное побережье Хонсю.
Нижняя батиаль.
4(1). Резко увеличенных присосок
нет. Диаметр присосок у самцов и
самок не более 10% мантии . . 5.
5(8). Вороночный орган в форме
VV. Диаметр присосок около 6%
длины маптии...................6.
6(7). Лигула без поперечных
гребней, ее длина 5% длины гекто-
котилизированной руки. Глубина ум-
бреллы около l/s длины наибольшей
руки.
В. fuscus Taki, 1964 (рис. 84,г).
Восточное побережье Хонсю. Ба-
тиаль.
7(6). Лигула с 8—10 хорошо вы-
раженными поперечными гребнями,
ее длина 7% длины гсктокотилизи-
рованпой руки. Глубина умбреллы
около !/4 длины наибольшей руки.
В. ergasticus (Р. et. Н. Fischer,
1892) (рис. 84, д, е). Северо-Восточ-
ная Атлантика от Юго-Западной Ир-
ландии до Сенегала. Средняя и ниж-
няя батиаль, глубины 450—1400 м.
8(5). Вороночный орган в фор-
ме W ..........................9.
9(12). В полужабре 6—10 фила-
ментов. Глубина умбреллы ’Д—Уз
длины наибольшей руки . . . 10.
10(11). Лигула с десятком по-
перечных гребней.
В. piscatorum (Verrill, 1879) (рис.
84, ж, з). Северная Атлантика и
Приатлантическая Арктика от Нью-
фаундленда, Датского пролива и За-
падного Шпицбергена до района за-
лива Делавэр и Юго-Западной Ир-
ландии. Глубины 80—2500 м, на се-
вере ареала обычно менее 750 м, на
юге, как правило, более 900 м.
11(10). Лигула без поперечных
гребней или с неясно выраженными
гребнями.
В. profundorutn Robson, 1932
Г=В. januarii (Sasaki, 1929), non
Hoyle]. От Берингова моря до остро-
ва Кюсю и залива Аляска, вероятно,
также западное побережье Канады
и США до Калифорнии. Охотское
море. Глубины 150—3400 м. Обыч-
ный и многочисленный вид.
12(9). В полужабре 11—13 фила-
ментов .......................13.
13(14V Глубина умбоеллы ’А
длины наибольшей руки. Мантийное
отверстие очень широкое.
В. violescens Taki, 1964. Восточное
побережье Хонсю.
14(13). Глубина умбреллы У4
длины наибольшей руки. Мантийное
отверстие довольно узкое.
В. hokkaidensis (Berry, 1921)
( = Polypus glaber Sasaki, 1929, non
Wiilker) (рис. 84, и, к). От мыса Ha-
варин до Южного Хонсю и Орегона.
Охотское море. Верхняя батиаль,
глубины 130—1000 м. Довольно час-
тый вид.
Нс идентифицированный вид Ben-
thoctopus зарегистрирован в бухте
Провидения (Чукотка).
Мотальная область
1 (6). Вороночный орган W-образ-
ный. Кожа гладкая..............2.
2(5). Диаметр присосок 7—12%
длины мантии. Юго-Западная Атлан-
тика ..........................3.
3(4). Диаметр присосок около
12% длины мантии. Сектор умбрел-
лы А самый или один из самых ко-
ротких.
В. eureka (Robson, 1929). От Се-
верной Аргентины до Фолклендских
островов. Сублитораль.
4(3). Диаметр присосок 7—8%
длины мантии.
Вид или несколько видов, близких
к В. januarii. Фолклендские острова,
банка Бердвуд и прилегающие райо-
ны. Глубины 80—450 м.
5(2). Диаметр присосок 3—4%
длины мантии. Сектора умбреллы
приблизительно равной глубины.
В. berryi Robson, 1924. Юго-запад-
нее Кейптауна. Глубина 2200 м.
6(1). Вороночный орган W-об-
разный.........................7.
7(8). Диаметр присосок 9—13%
длины мантии. Глубина умбреллы
7з длины наибольшей руки. Сектор
умбреллы А самый или один из са-
мых глубоких. Кожа шероховатая.
Юго-Западная Атлантика.
В. magellanicus Robson, 1930. Па-
тагония, Фолклендские острова, Ог-
ненная Земля. Нижняя сублитораль.
8(7). Диаметр присосок 6—9%
317
Ж?
318
thieleii n—p — В. fanuarU; б—г, e, s, u, m,
o, p — гектокотиля; к —вороночный орган
(а—б — по Н. Н. Кондакову; в, а — по Sa-
saki. 1929; г — по Taki, 1964; О—е, з — по
Massy, 1909; ж — по Verrill, 1882; к — по
Кондакову, 1941; л—м, л—р—по Hoyle,
1886; я— по Chun, 1915; о — по Thiele,
1921)
310
длины мантии. Сектор умбреллы А
укорочен. Кожа гладкая. Южная
часть Индийского оксана ... 9.
9(10). Глубина умбреллы Vs—%
длины наибольшей руки. Длина гек-
токотиля 7—9% длины гектокотили-
зированной руки. Диаметр присосок
6—8% длины мантии.
В. levis (Hoyle, 1885) (рис. 84,
л, ж). О. Херд. Нижняя сублитораль
и верхняя батиаль.
10(9). Глубина умбреллы Дли-
ны наибольшей руки. Длина гектоко-
тиля 13% длины гектокотилизирован-
ной руки. Диаметр присосок 9%
длины мантии.
В. thielei Robson, 1932 (рис.
84, н, о). О. Кергелен. Нижняя суб-
литораль и (реже) верхняя батиаль.
Обычен и многочислен па Кергелен-
ском плато.
Тропическая зона Мирового океана.
1. Западная Атлантика
а (б). Вороночный орган W-образ-
ный. Руки в 4—6 раз длиннее ман-
тии. Гектокотилизированная рука
приблизительно вдвое короче про-
тивоположной (3-й левой) руки, дли-
на лигулы 6—9% длины гектокоти-
лизированной руки.
В, januarii (Hoyle, 1885) (рис.
84, п, р). От Мексиканского залива до
Юго-Восточной Бразилии. Верхняя
батиаль.
б (а). Вороночный орган W-об-
разный. Руки в 2,5—4 раза длиннее
мантии. Гектокотилизированная ру-
ка на Vs—V4 короче противополож-
ной руки, длина лигулы 3—5% дли-
ны гектокотилизированной руки.
В. oregonae Toll, 1981. Южная
часть Карибского моря, батиаль.
2. Восточная Атлантика. В. ergas-
ticus (Р. et Н. Fischer, 1892) (см.
с. 317). От Юго-Западной Ирландии
до Сенегала. Глубины 450—1400 м.
Отсутствует в Средиземном море. От
внешне похожего Bathypolypis spon-
satis (см. с. 316) отличается строе-
нием гектокотиля.
3. Индо-Вестпацифика. Комплекс
из 3—4 неописанных видов, близких
к В. januarii и В. profundorum. Обна-
ружены в Андаманском море и индо-
незийских морях. Глубины 500—
1900 м.
4. Восточная Пацифика. В. sp. aff.
januarii Nesis, 1973. От о-ва Кокос
320
до Северного Перу. Глубины 570—
1850 м.
Сомнительные виды: В. lothei
(Chun, 1913) (=? В. ergasticus):
у Канарских островов на глубине
1365 м; В. pseudonymus Grimpe,
1922): у Азорских островов на глу-
бине 1600 м. Родовая принадлеж-
ность неясна.
Таблица для определения видов рода
Graneledone
1(2). Мантия и голова сверху по-
крыты немногочисленными, очень
крупными бородавками, состоящими
из нескольких конических шипиков
на общем основании; между боро-
давками разбросаны отдельные кони-
ческие шипики. Верхняя сторона рук
гладкая. Над каждым глазом по од-
ной очень крупной сложной бородав-
ке. Вороночный орган VV-образный.
В полужабре 7 филаментов. 2-й ла-
теральный зуб радулы маленький,
с двумя зубчиками, с широким осно-
ванием.
G. macrotyla Voss, 1976 (рис.
85, а—я). Море Скотия. Глубина
1650—2050 м.
2(1). Мантия, голова, часто и
верхняя сторона рук покрыты мно-
гочисленными небольшими папилла-
ми, состоящими из нескольких остро-
конечных шипиков...............3.
3(4). Папиллы разбросаны по
спинной стороне мантии и головы и
почти не заходят на верхнюю сторо-
ну умбреллы и рук. Они расположе-
ны редко: 13—15 папилл по линии
поперек спинной стороны от одного
бока до другого в средней части ман-
тии. Папиллы звездчатые, в одной
папилле до 7—8 небольших шипиков.
Над каждым глазом по 3—5 увели-
ченных папилл. Длина лигулы 3,5—
7% длины гектокотилизированной ру-
ки. В полужабре 7—8 филаментов.
G. verrucosa (Verrill, 1881)
(=? G. verrucosa var. media Joubin,
1918) (рис. 85, г, д). Северо-Запад-
ная Атлантика от района южнее Ис-
ландии до района северо-восточнее
мыса Хаттерас. Глубины 850—2300 м.
4(3). Папиллы покрывают верх-
нюю сторону мантии, головы, ум-
бреллы и, по крайней мере, базаль-
ных частей рук 1—3-й пар. Они рас-
положены густо: 25—30 папилл по
линии поперек спинной стороны в
средней части мантии. Папиллы не
звездчатые, в одной папилле до 12—
14 шипиков. 2-й латеральный зуб
радулы крупный, однозубый, с узким
•основанием .....................5.
5(6). Вороночный орган состоит
из двух удлиненно-овальных вздутий,
•слегка расщепленных на передних
концах. Лигула с небольшим каламу-
сом и неглубоким продольным же-
лобком с 12—13 поперечными склад-
ками. Длина лигулы около 3% дли-
ны гектокотилизированной руки, ка-
ламуса — 40—50% длины лигулы.
В полужабре 6 филаментов. Диаметр
присосок 6—8% длины мантии. По
2—3 увеличенных папиллы над каж-
дым глазом. Рахидиальный зуб ра-
дулы однозубый, симметричный.
G. antarctica Voss, 1976 (рис.
35, е—и). Море Росса. Абиссаль. По-
хожая форма обитает у островов
Кергелен и Херд в верхней батиали.
6(5). Вороночный орган VV-об-
разный. Лигула с крупным каламу-
сом и глубоким продольным желоб-
ком без заметных поперечных скла-
док. Нет резко увеличенных папилл
над глазами. Рахидиальный зуб ра-
дулы с тремя зубчиками, слабо асим-
метричный .......................7.
7(8). Папиллы на голове и верх-
ней стороне умбреллы тесно сбли-
жены, почти соприкасаются, их диа-
метр 1—2 мм. Длина лигулы 3,3%
длины гектокотилизированной руки,
каламуса 55% длины лигулы. Диа-
метр присосок 7% длины мантии.
5—6 филаментов в полужабре.
G. challenged (Berry, 1916). У ост-
ровов Кермадек. Глубина 1150 м.
8(7). Папиллы на голове и верх-
ней стороне умбреллы расположены
редко, как и на верхней стороне ман-
тии, их диаметр 2—4 мм. Длина ли-
гулы 4—5% длины гектокотилизиро-
ванной руки, каламуса — 70% длины
лигулы. Диаметр присосок 5—6%
длины мантии. 7—8 филаментов в
полужабре.
G. boreopacifica Nesis, n. sp.1
(рис. 85,к,л). Охотское море и Ти-
хий океап северо-восточнее Хонсю
1 Голотип: зрелый самец, длина ман-
тии ПО мм. Охотское море,
50°04' с. ш., 15Г35' в. д., глубина
1350 м, 1.VIII 1977 г., сбор Г. В. Ав-
деева. Хранится в Зоологическом
институте АН СССР, Ленинград.
322
(берег Санрику), (?) Орегон и Ка-
лифорния. Глубины 1165—1500 м.
Не идентифицированный вид Gra-
neledone обитает в тропических во-
дах Восточной Пацифики, глубины
1865—2500 м.
2Л.2.2.2. Семейство
Vitreledonellidae
Глубоководные пелагические осьми-
ноги со стекловидно прозрачным и
почти бесцветным телом желеобраз-
ной консистенции. Руки средней
длины, у молоди равны, у взрослых
в 2—3 раза длиннее мантии. Присос-
ки в один ряд, в пределах умбреллы
очень широко расставлены, за ее пре-
делами сближены и резко увеличены:
диаметр крупных дистальных присо-
сок у взрослых до 10—17% длины
мантии. На концах рук присоски
мелкие. Глубина умбреллы у взрос-
лых %—% длины наибольшей руки.
Глаза небольшие, узкие, почти пря-
моугольные, их диаметр около 20%
длины мантии. Они смотрят вбок.
Оптические нервы очень длинные,
оптические ганглии далеко отстоят
от мозга. Мантийное отверстие очень
широкое. Радула с многовершинным
рахидиальным и одновершинными
1-м и 2-м латеральными зубами
Печень длинная, равна или превос-
ходит ширину головы, узкая, сзади
заостренная. Желудок впереди пече-
ни, яичник над ней. Задние слюнные
железы слиты в одну. Чернильный
мешок развит. Гектокотилизируется
3-я левая рука, она короче правой,
лигула короткая. Предположительно
живородящие формы, вынашивающие
яйца до рождения личинок.
Один род — Vitreledonella Joubin,
1918, один вид—V. richardi Joubin,
1918 (рис. 86). Тропическо-субтропи-
ческий космополитический батипела-
гический вид, молодь живет в эпи- и
мезопелагиали. Длина мантии до
11 см, общая длина до 45 см. Об-
зор: Thore, 1949.
2.4.2.3. Надсемейство
Argon autoidea
Пелагические или придонно-пелагиче-
ские осьминоги с мускулистой, реже
полустуденистой консистенцией тка-
ней. Глаза очень хорошо развиты.
Присоски в два ряда — либо по всей
длине рук, либо в дистальных частях.
Гектокотилизированная рука видоиз-
меняется целиком, имеет очень слож-
ное строение, состоит из нескольких
частей. Она развивается в особом
мешке, так что кажется, будто у не-
зрелого самца лишь 7 рук — это
можно рассмотреть даже у личинок.
Зрелый гектокотиль наполняется
спермой из единственного спермато-
фора, отрывается от тела самца при
спаривании и заползает, змеевидно
изгибаясь, через воронку в мантий-
ную полость самки. Самцы часто (но
не всегда) гораздо мельче самок,
карликовые. Яйца самка либо откла-
дывает па дно, либо носит на себе,
либо они развиваются в ее теле.
Имеется стадия пелагической ли-
чинки.
Радула не гребенчатая, рахиди-
альный зуб одно- или трехвершин-
ный, без сериации. Мантийное от-
верстие широкое. Имеются мантийно-
вороночный замыкательный аппарат
и чернильный мешок. Желудок и пе-
чень. в «нормальном» положении.
4 семейства, 4 рода, 8—11 видов.
2.4.2.З.1. Семейство
Alloposidae
Тело студенистое, мантия короткая,
голова широкая, не уже мантии. Ру-
ки относительно короткие, с очень
глубокой умбреллой. Порядок длины
рук 1>2>3>4. Присоски в преде-
лах умбреллы в один ряд, за ее пре-
делами в два, на самых кончиках
рук снова в один ряд. Они малень-
кие, увеличенных присосок нет. Гла-
за очень большие (около 7з длины
мантии). Воронка полностью погру-
жена в ткани головы, открывается
впереди глаз. Вороночный орган
W-образный. Мантийное отверстие
широкое. Мантийно-вороночный ап-
парат когтевидного типа, слабо раз-
вит. Центральный зуб радулы трех-
вершипный. Мантия соединена с
внутренностным мешком вентральной
медианной ссптой, с головой — дву-
мя парами боковых септ, па одной из
которых помещается звездчатый
ганглий. Гектокотилизируется 3-я
правая рука. Гектокотиль развивает-
ся в сумке перед правым глазом, при
спаривании отрывается. Самцы не
323
ж
Рис. 87. Надсем. Argonautoidea. сем. АПо-
posidae (а—г), Ocythoidae (д—ж), Тге-
moctopodidae (з—о): а—г—Alloposus mollis,
самка (а), самец со вскрытой сумкой гек-
токотиля (б), молодой самец (о), руки мо-
лодой самки (г); д—ж — Ocythoe tuber си-
lata, взрослая самка (д), молодая самка
(е), зрелый самец (ж), з—м — Tremoctopus
violaceus (з—л — подвид ' violaceus, м —
подвид gracilis) взрослая самка с сократив-
шейся (з) и расправленной (и) мембраной
спинных рук, молодая самка (к), молодой
(л) и зрелый (м) самец; н—о — Т. gelatus,
молодая и взрослая самка (а—б—по Verrill,
1882; в — по Adam, 1952; г — по Sasaki,
1929; д, ж —по R. Young, 1972; е, к—л—по
Naef, 1923; з—u—по Voss, Williamson, 1972;
м — по Berry, 1914; н—о — по Thomas,
1977)
карликовые. Яйца средних размеров,
вероятно, донные.
Один род — Alloposus Verrill, 1880
(=? Haliphron Sleenstrup, 1859, Л/Zo-
posina Grimpe, 1922, Heptapus Joubin,
1929). Один вид — Д. mollis Verrill,
1880 (рис. 87, а—г). Широко распро-
страненный (от тропиков до север-
ных и южных умеренных вод) кос-
мополитический вид. Личинки и мо-
лодь обитают в пелагиали (от эпи-
до батипелагиали), главным образом
(но не исключительно) над склоном
и подводными возвышенностями,
взрослые особи у дна в батиали.
Встречается в водах СССР (северо-
западная часть Тихого океана). Дли-
на мантии до 40 см, общая длина до
2 м. Обзор: Thore, 1949.
2.4.2.3.2. Семейство
Ocytholdae
Самки крупные, с плотным телом.
Руки длинные, 1-я и 4-я пары зна-
чительно длиннее 2-й и 3-й. Присос-
ки мелкие, в 2 ряда. Умбрелла не
развита, оторочек на руках нет. На
нижней части головы у основания
рук 4-й пары — одна пара кожных
пор. Глаза крупные. Воронка очень
длинная, вороночный орган 1Л1-об-
разный. Замыкательные хрящи слож-
но устроенные, кнопкообразные. На
брюшной стороне мантии взрослых
самок — сетчатая скульптура из пе-
ресекающихся кожных гребней и бу-
горков в местах пересечения. Внутри
мантийной полости па спинной сто-
роне — гидростатич еский орган.
Самцы мелкие. Гектокотилизиру-
ется 3-я правая рука. Гектокотиль
с очень длинным тонким «пенисом»,
присоски в два ряда. Яйца развива-
ются в яйцеводах самки, наружу вы-
ходят личинки.
Один род — Осythoe Rafinesque,
1814 (рис. 87, д—ж) (=Parasira
Stccnstrup, 1861). Один вид — О. tu-
ber culata Rafinesque, 1814 (рис.
87, д, ж). Бицснтральный (бисубтро-
пический) космополитический припо-
верхностный вид. Длина мантии са-
мок до 30 см, самцов до 3 см. Сам-
цы часто сидят в пустых оболочках
бочопочников или сальп. Обзор: Ro-
per, Sweeney, 1976.
326
2.4.2.3.3. Семейство
Tremoctopodidae
Самки крупные, общая длина до 2 м.
Тело обычно плотное. Присоски мел-
кие, в два ряда. Руки первых двух
пар соединены умбреллой, продол-
жающейся в широкую оторочку по
обеим сторонам рук 1-й пары и дор-
сальной стороне рук 2-й пары. Вдоль
оторочек продольными рядами рас-
полагаются крупные глазчатые пят-
на. На 3-й и 4-й парах рук оторочки
нет, умбрелла слабо развита. Руки
1-й и 2-й пар значительно длиннее,
чем 3-й и 4-й. У молодых самок ру-
ки 1-й пары очень длинные, присоски
на их концах расположены в один
ряд и далеко расставлены, поперек
руки проходят бороздки. При напа-
дении хищника кусок этой руки, не-
сущий одну присоску, 1—3 глазча-
тых пятна и (иногда) светящиеся
железы, отрывается по бороздке и
резко расширяется, дезориентируя и
отпугивая нападающего. Вследствие
этого у взрослых самок концы 1-й
пары рук всегда оборваны.
Кожа гладкая. На голове у осно-
ваний 1-й и 4-й пар рук имеются
2 пары кожных пор. Воронка сред-
них размеров. Вороночный орган
ПП-образный. Замыкательный аппа-
рат округлой или треугольной фор-
мы. По краям оторочки рук 1-й па-
ры иногда расположены светящиеся
органы.
Яйца мелкие, самка носит их на
базальных частях спинных рук. Мо-
лодые самки и самцы обычно носят
на этих руках кусочки жгучих щупа-
лец физалий — дополнительное сред-
ство защиты от врагов. Самцы очень
маленькие — длина мантии до 15 мм,
общая длина до 4 см. Гектокотилизи-
руется 3-я правая рука. В базальной
и средней частях гектокотиля два
ряда мелких присосок, кроме них в
базальной части оторочка из мелких
усиковидных папилл. Дистальная
часть преобразована в семенной ме-
шок с «пенисом». У незрелых самцов
он развивается в мешке между во-
ронкой и глазом.
Один род — Tremoctopus de lie
Chiaje, 1830 (=Philonexis d’Orbigny,
1835) (рис. 87,з—о). Типовой вид —
Т. violaceus delle Chiaje, 1830. Два
вида, 2 подвида, распространенных в
тропиках и субтропиках всех океа-
нов. Обзор: Thomas, 1977.
Таблица для определения видов рода
Tremoctopus
1(2), Тело плотное, мускулистое.
Взрослые самки сверху темно-сине-
пурпурного цвета, снизу светло-золо-
тистого, самцы и молодь светлые, с
мелкими темными точками. В полу-
жабре 13—16 филаментов у самки и
9—11 у самца.
Т, violaceus delle Chiaje, 1830
(рис. 87, з—ж). Тропические и суб-
тропические воды всех океанов от
Г олъфстрима, Бискайского залива,
Южных Курильских островов и Ка-
лифорнии до Уругвая, Южной Афри-
ки, Новой Зеландии и Чили. Эпипе-
лагиаль, по ночам поднимаются к по-
верхности. Длина мантии самок до
0,3—0,5 м. 2 подвида.
а (б). У взрослых самок глазча-
тые пятна расположены в один ряд
только на вентральной (наружной)
стороне оторочки рук 1-й пары. На
гектокотиле 15—19 пар присосок в
средней части и 22—23 пары в прок-
симальной.
Т. violaceus violaceus d. Ch. (рис.
87,з—л). Атлантический океан, Ка-
рибское и Средиземное моря.
б (а). У взрослых самок глазча-
тые пятна расположены в два ряда
или сдвоенным рядом на вентраль-
ной (наружной) и в один ряд на дор-
сальной (внутренней) стороне ото-
рочки рук 1-й пары. На гектокотиле
19—22 пары присосок в средней и
27—29 пар в проксимальной части.
Т. violaceus gracilis (Souleyet,
1852) (рис. 87, ле). Индийский и Ти-
хий океаны.
2(1). Тело студенистое, настолько
прозрачное, что через тело жи-
вой самки можно читать газету. Ок-
раска самок светло-оранжево-крас-
ная. Глаза очень крупные. Черниль-
ный мешок уменьшен в размерах.
В полужабре 8—11 филаментов у
самки, 7—8 у самца.
Т. gelatus Thomas, 1977 (рис. 87,
я, о). Атлантический океан у Южной
Флориды, Индийский океан южнее
Мадагаскара, Пацифика севернее и
северо-восточнее Гавайских остро-
вов. Возможно, бисубтропический
вид. Предположительно мезо- или
батипелагический вид, самки извест-
ные лишь по выбросам на берегах и
находкам в желудках алепизавров,
самец выловлен на поверхности. Дли-
на мантии самок до 33 см.
2.4.2.3.4. Семейство
Argonautidae
Самки строят тонкую, сжатую с бо-
ков, известковую раковину с одной
камерой, имеющую плоский киль„
окаймленный двумя рядами бугор-
ков; боковые стороны раковины с
радиальными ребрами; центр рако-
вины вдавлен или с отогнутыми на-
ружу острыми «рожками». Руки 1-й
пары самые длинные, на концах не-
сут широкую парусовидную отороч-
ку (лопасть), которая выделяет в
формует раковину. Самка сидит в»
раковине головой вперед, руки 1-й
пары обычно выставлены наружу, их
лопасти расширены и покрывают всю-
раковину снаружи, остальные руки
внутри раковины, удерживают ее на
теле самки. Умбрелла очень слабо-
развита. Присоски мелкие, в 2 ряда.
На голове нет кожных пор. Воронка
длинная, вороночный орган IAI-об-
разный. Замыкательные хрящи кноп-
ковидные. Кожа гладкая. Задний ко-
нец мантии обычно слегка отогнут
кверху.
Самцы крохотные. Гектокотилизи-
руется 3-я левая рука. Гектокотиль с
длинным тонким «пенисом» и много-
численными присосками в 2 ряда
(рис. 88,6). Он довольно долго хра-
нится в мантийной полости самки.
Яйца очень мелкие, самка вынашивает
их в раковине до вылупления личи-
нок. Нерест порционный.
Один род — Argonauta Linne,.
1758 (=*Argonautina Steinmann, 1925r
Argonautella Steinmann, 1925) (рис..
88). Типовой вид — A. argo Ьшпёг
1758. 4 достоверных и 2—3 сомни-
тельных вида в эпипелагиали (но не
только у поверхности!) тропических
и субтропических вод. Общая длина
самок до 45 см и более, самцов до*
2 см; длина мантии самок до 10 смг
самцов до 1,1 см.
Таблица для определения видов рода
Argonauta по раковине
1(4). Раковина крупная (диаметр до
20—30 см), уплощенная с боков, с
узким килем (его ширина до 15%
диаметра раковины). Ребра много-
численные, сближенные, часто раз-
дваивающиеся. Встречаются преиму-
щественно вблизи берегов. Диаметр*
раковины до 25—30 см .... 2.
2(3). Киль очень узкий, его ши-
327
рипа редко больше 6 % диаметра ра-
ковины. Ребра гладкие, на крупных
раковинах их больше 50. Каждое
ребро заканчивается на киле острым
бугорком.
A. ar go Linne, 1758 (рис. 88, а—в).
Тропическо-субтропический космопо-
литический вид, на севере достигает
мыса Код, Португалии, Средиземного
моря, Хонсю и Калифорнии, на
юге — Южной Америки, Новой Зе-
ландии и Перу. Молодь этого вида
очень похожа на молодь A. hians.
3(2). Ширина киля 10—15 % Диа-
метра раковины. Ребра представляют
собой цепочки отдельных бугорков
или узелков. На крупных раковинах
их 30—40.
A. nodosa Solander, 1786 (рис.
88, г). Тропическая Индо-Вестпаци-
фика от Красного моря и Южной
Японии до Южной Африки, Тасма-
нии и Новой Зеландии, на восток до
Полинезии и (?) Чили.
4(1). Раковина мелкая, диаметр
до 10 см, не уплощенная с боков.
Киль широкий (10—15% диаметра
раковины). Ребра немногочисленные.
Океанические виды...............5.
5(6). Поверхность раковины
обычно гладкая. Центр спирали не
вдавлен. «Рожки» в центре раковины
имеются или отсутствуют. Нс более
35 ребер, они редкие, расставленные.
Бугорки на киле бурого цвета, обычно
их 15—20. Диаметр раковины до
10 см.
A. hians Solander, 1786 (рис.
88, д). Тропический космополитиче-
ский вид. Не заходит в Средиземное
море, в остальном ареал сходен с
ареалом A. argo.
6(5). Поверхность раковины
обычно тонко гранулирована, т. с.
покрыта очень мелкими бугорочками.
Центр спирали слегка вдавлен.
«Рожки» отсутствуют. Ребра частые,
сближенные, до 40—45. Бугорки на
киле белого цвета, их до 20—33.
Диаметр раковины до 7 см.
A. boettgeri Maltzan, 1881 (рис.
88,е,ж), Тропический индо-вестпаци-
фический вид, распространен от за-
падной части Индийского оксана до
Гавайских островов и от Японии до
Новой Зеландии.
Сомнительные виды Argonauta\
A. cornuta Conrad, 1854 (рис.
88,з) — близка к A. hians, ио киль
более узкий, ребра более тесные (до
45), бугорков до 30, киль вогнутый,
несет мелкие бугорки (между круп-
ными по краям киля), поверхность
гранулирована, «рожки» сильно раз-
Ж
U
3
Рис. 88. Сем. Argonautidae, род Argonautaz
а—в — Л. argo, самка в раковине (а), са-
мец с нерасправленным и расправленным
гектокотилем (б), раковина (в); г, б —ра-
ковины A. nodosa (г) и A. hians (б); е—
ж — A. boettgeri, самка и раковина; з—и—
раковины A. cornuta (сзади) и A. nouryi
(а—по Voss, Williamson, 1972; б — по
Naef, 1923; в—д — по Sovcrby in Reeve,
1861; с—ж — по Berry, 1914; з—и— по Keen,
1968)
21 К. Н. Несис
32»
виты (последнее отличает Д. cornuta
от A. boettgeri). Восточная часть Ти-
хого океана от Калифорнийского за-
лива до Перу.
Д. nouryi Lorois, 1852 (рис. 87, и)—
раковина удлиненная, вытянутая,
овальных очертаний, края устья поч-
ти прямые (при взгляде сбоку),
«рожек» нет, ребра низкие, многочис-
ленные, киль узкий, вогнутый, бугор-
ки на нем слабо заметны, поверх-
ность раковины топко гранулирова-
на. Восточная часть Тихого океана
от Калифорнийского залива до Пе-
ру, па запад до Маркизских островов.
Кроме того, восточнопацифиче-
ских A. argo часто относят к особо-
му виду Д. pacifica Dall, 1869, хотя
их отличия от типичных Д. argo
точно не установлены.
Таблица для определения видов
Argonauta по строению тела самок
1(2). Руки 4-й пары значительно
(в 1,5—2 раза) Длиннее рук 3-й и на
10—20 % длиннее рук 2-й пары.
Д. argo Linne (рис. 88, а).
2(1). Руки 4-й пары короче рук
2-й, нс длиннее рук 3-й пары . 3.
3(4). Руки 4-й пары составляют
около 70—90% длины рук 2-й и
примерно равны рукам 3-й пары.
Д. nodosa Solander.
4(3). Руки 4-й пары значительно
короче рук 2—3-й пар, их длина у
крупных самок обычно около 60%
длины рук 2-й пары (у мелких самок
руки 4-й пары относительно длиннее).
A. hians Solander и Д. boettgeri
Maltzan (рис. 88, е).
СПИСОК ЛИТЕРАТУРЫ
Общая часть
* Акимушкин И. И. Головоногие мол-
люски морей СССР. М.; Л.: Изд-
во АН СССР, 1963. — 236 с.
Битюкова Ю. Е., Зуев Г. В. Асим-
метрия глаз кальмаров семейства
Histioteuthidae в связи с их эко-
логией. — Биология моря, Киев,
1976, вып. 38, с. 63—67.
Буруковский Р. Н.> Вовк А, Н, Неко-
торые вопросы оогенеза у северо-
американского кальмара (Loligo
pealei Les.) банки Джорджсс. —
Арх. анат. гист., эмбриол., 1974,
т. 66, № 5, с. 44—50.
Залыгалин В. П., Зуев Г. В., Нигма-
туллин Ч. М. Особенности проду-
цирования сперматофоров и пло-
довитость самцов кальмаров Sthe-
noteuihis pteropus (St.). — В сб.:
Тез. докл. Весе. конф, по пром,
бсспозв. М., 1977.— с. 37—38.
* Зуев Г. В, Функциональные основы
внешнего строения головоногих
моллюсков. — Киев: Наукова дум-
ка, 1966. — 140 с.
Зуев Г. В, Особенности строения
мантии головоногих моллюсков.—
Биология моря, Киев, 1969. вып.
16, с. 102—110.
Зуев Г. В. Аллометрический рост
мозга головоногих моллюсков. —
Биология моря, Киев, 1973, вып.
31, с. 135—152.
Зуев Г. В. Рост и размеры мозга в
онтогенезе океанических кальма-
ров.— Экология, 1975, № 4,
с. 70—73.
* Зуев Г. В., Несис К Н. Кальмары
(биология и промысел). М.: Пи-
щевая пром-сть, 1971. — 360 с.
Иванов А. В. Класс Cephalopoda —
головоногие. — В кн.: Большой
практикум по зоологии бсспозв.,
т. 2. М., 1946, с. 194—218.
Иванов А. В. Промысловые водные
беспозвоночные. М.: Советская
наука, 1955. — 356 с.
♦ Иванов А. В., Стрелков Д. Д. Про-
мысловые беспозвоночные дальне-
восточных морей. Описание стро-
ения и атлас анатомии. Владивос-
ток: ТИНРО, 1949. —104 с.
♦ Кондаков Н. Н. Класс головоногих
моллюсков (Cephalopoda). —
В кп.: Руководство по зоологии,
т. 2. М.; Л.: 1940, с. 548—609.
Методические основы разработки
шкал зрелости репродуктивной
системы кальмаров на примере
Sthenoteuthis pteropus/[Р. Н. Бу-
руковский и др.]—Зоол. журн.,
1977. т. 52, вып. 12, с. 1781—1791.
Моллюски. Авторефераты докладов,
сб. 2—6. Л.: Наука, 1965, 101 с.;
1968, 96 с.; 1971, 160 с.; 1975,
244 с.; 1979, 264 с.
* Несис К. И. Экологическая класси-
фикация (жизненные формы) го-
330
ловопогих моллюсков. — Итоги
науки и техники, серия Зоология
беспозвоночных, т. 2, М.: ВИНИТИ,
1973, с. 8—59.
Несис К. Н. Эволюция жизненных
форм современных головоногих
моллюсков. — Труды Института
океанологии АН СССР, 19756,
т. 101, с. 124—142.
Несис К. Н. Вертикальное распро-
странение пелагических головоно-
гих моллюсков. — Журнал общей
биологии, 19776, т. 38, № 4,
с. 547—558.
Несис К. Н. Эволюционная история
нектона. — Журнал общей биоло-
гии, 19786, т. 39, № 1, с. 53—65.
Орлов О. Ю., Бызов А. Л. Зрение го-
ловоногих моллюсков. — Приро-
да, 1962, № 3, с. 115—118.
★Проссер JL (ред.). Сравнительная
физиология животных. В 3-х т.
М.: Мир, 1977—1978.— 1837 с.
Статоцисты головоногих моллюсков/
[Я. А. Винников и др.]. — В кн.:
Рецептор гравитации. Пробл. кос-
мич. биол. Л.: Наука, 1971. Т.
12. —523 с.
Aldrich М. M.t Barber V. С., Emer-
son С. J. Scanning electron micro-
scopical studies of some cephalo-
pod radulae. — Can. J. Zool., 1971,
v. 49, № 12, p. 1589—1594.
Altman J. S.t Nixon M. Use of the
beak and radula by Octopus vul-
garis in feeding. —J. Zool., 1970,
v. 161, № 1, p. 25—38.
• AppelldJ A. Die Schalen von Sepia,
Spirula und Nautilus. — Kgl. Sven-
ska VctAkad. Handl., 1893, Bd. 25,
№ 7, S. 1—106.
Arnold J, M., Young R. E., King M. V.
Ultrastructure of a cephalopod
photophore, I—II. — Biol. Bull.,
1974, v. 147, № 4, p. 507—534.
Atkinson B. G. Squid nidamental
gland extract: isolation of a fac-
tor inhibiting ciliary activity. — J.
Exptl. Zool., 1973, v. 184, № 3,
p. 335—340.
♦ Badenhorst J, H. The morphology
and histology of the male genital
tract of the squid Loligo reynaudi
(d’Orbigny). — Ann. Univ. Stellen-
bosch, 1974, v. 49A, № 1, 36 p.
Bandel K., Boletzky S. v. A compara-
tive study of the structure, deve-
lopment and morphological rela-
tionships of chambered cephalopod
shells. — Vciiger, 1979, v. 21, №3,
p. 313-354.
Barber V. C. A neurosecretory tissue
in octopus. — Nature, 1967, v. 213,
№ 5080, p. 1042—1043.
Barber V. C. The structure of mollusc
statocysts, with particular referen-
ce to cephalopods. — Symp. Zool.
Soc. Lnd, № 23, 1968, p. 37—62.
Baumann F., Mauro A., Milecchia R,
et al. The extra-ocular light recep-
tors of the squids Todarodes and
fllex. — Brain Res., 1970, v. 21,
№ 2, p. 275—279.
★Bidder A. M. The digestive mecha-
nism of the European squids Loli-
go vulgaris, Alloteuthis media, and
Alloteuthis subulata. — Quart. J.
Microscop. Sci., 1950, scr. 3, v. 91,
№ 1, p. 1—43.
★Bidder A, M. Feeding and digestion
in cephalopods. — In: Physiology
of Mollusca, v. 2, NY—Lnd, 1966,
p. 97—124.
Bidder A. M. New names for old: the
cephalopod <mid-gud gland». —
J. Zool., 1976, v. 180, p. 441—443.
Blank R. Neurosekretorische Komple-
xe bei Loligo. — Zool. Jb., Anat.,
1978, Bd. 99, № 4, S. 505—526.
Bloodgood R. A. The squid accessory
nidamental gland: ultrastructure
and association with bacteria. —
Tissue a. Cell, 1977, v. 9, № 2,
p. 197—208.
♦ Boletzky S. v. The larvae of Cepha-
lopoda: a review. — Thalassia Ju-
gosl.; 1976, v. 10, p. 45—76.
Borer К. T., Lane С. E. Oxygen requi-
rements of Octopus briareus Rob-
son at different temperatures and
oxygen concentrations. — J. exp.
mar. Biol. Ecol., 1971, v. 7, № 3,
p. 263—269.
Bottazzi F, Ricerche sulla ghiandola
salivare posteriore dei Cefalopo-
di. — Pubbl. Staz. Zool. Napoli,
1916, v. 1, p. 59—146.
Bottke W. The fine structure of the
ovarian follicle of Alloteuthis su-
bulata Lam. — Cell Tissue Res.,
1974, v. 150, № 4, p. 463—479.
Boucaud-Camou E., Boucher-Rodoni R.,
Mangold K. Digestive absorption
in Octopus vulgaris. — J. Zool.,
1976, v. 179, № 2, p. 261—271.
Boucaud-Camou E., Yim M. Fine
structure and function of the diges-
tive cell of Sepia officinalis. —
Ibid., 1980, v. 191, № 1, p. 89—105.
Boucher-Rodoni R. Vitesse de la dige-
stion d'Octopus cyanea. — iVlar.
21
331
Biol., 1973, 18, № 3, p. 237—
242.
Boucher-Rodoni R. Vitesse de la dige-
stion chez les Cephalopodes Eleao-
ne cirrosa (Lamarck) ct Illex ille-
cebrosus (Lesucur). — Cah. Biol.
Mar., 1975, T. 16, № 2, p. 159—
175.
Boucher-Rodoni R., Mangold K. Expe-
rimental study of digestion in Oc-
topus vulgaris. — J. Zool., 1977,
v. 183, № 4, p. 505—515.
Boycott В. B. The chromatophore sy-
stem of cephalopods. — Proc. Linn.
Soc. London, 1953, v. 164, № 2,
p. 235—240.
Boycott В. B. The functioning of the
statocysts of Octopus vulgaris. —
Proc. R. Soc. London, 1960, v. 152B,
№ 946, p. 78—87
Boycott В. B. The functional organi-
zation of the brain of the cuttle-
fish, Sepia officinalis. — Ibid.,
1961, v. 153B, № 953, p. 503—534.
Boyle P. R., Mangold K.f Froesch D.
The organization of beak move-
ments in Octopus. — Malacologia,
1979, v. 18, p. 423—430.
Branden C. van den, Gillis M.f Ri-
chard A. Carotenoid producing
bacteria in the accessory nidamen-
tal glands of Sepia officinalis L. —
Comp. Biochem. Physiol., 1980,
v. 66B, № 2, p. 331—334.
Brocco S. L., Cloney R. A. Reflector
cells in the skin of Octopus doflei-
ni. — Cell Tissue Res., 1980, v. 205,
№ 2, p. 167—186.
Budelmann B.-U. The function of the
equilibrium receptor systems of
cephalopods. — Proc. Neurootol.
Equilibriometric Soc., 1979, v. 6,
. p. 15—63.
♦ Bullock T. H., Hor ridge G. A. Struc-
ture and function in the nervous
systems of invertebrates. 2 v. San
Francisco, 1965, 1719 p.
Caviello L., Zanetti L. a- and 0-ccpha-
lotoxin: two paralyzing protein
from posterior salivary glands of
Octopus vulgaris. — Comp. Bio-
chem. Physiol., 1977, v. 57C, № 2,
p. 169—174.
♦ Clarke M. R. The identification of
cephalopod «beaks» and the rela-
tionship between beak size and to-
tal body weight. — Bull. Brit. Mus.
(Nat. Hist.), Zool., 1962, v. 8,
№ 10, p. 419—480.
Clarke M. R. The cephalopod stato-
lith — an introduction to its
form. — J. mar. biol. Assoc.
U. K., 1978, v. 58, № 3, p. 701—712.
Clarke M. R., Denton E. J., Gilpin-
Brown J. B. On the use of ammo-
nium for buovancy in squids. —
Ibid., 1979, v. 59, № 2, p. 259—276.
Cloney R. A., Florey E. Ultrastructu-
re of cephalopod chromatophore
organs. — Z. Zellforsch., 1968, Bd.
89, S. 250—280.
Collewijn H. Oculomotor reactions in
the cuttlefish, Sepia officinalis. —
J. Exptl. Biol., 1970, v. 52, № 2,
p. 369—384.
Cousteau J.-Y., Diole P. Octopus and
squid, the soft intelligence. Lon-
don, 1973, 304 p.
Cowden R. R. Cytological and cyto-
chemical studies of oocyte develop-
ment and development of the folli-
cular epithelium in the squid, Loll*
go brevis. — Acta Embryol. Mor-
phol. Experim., 1968, v. 10, № 2,
p. 160—173.
Cowden R. R. Some cytological and
cytochemical observations on the
leucopoietic organs, the «white bo-
dies», of Octopus vulgaris. — J. In-
vertebr. Pathol., 1972, v. 19, № 1,
p. 113—119.
* Denton E. J., Gilpin-Brown J. B.
Floatation mechanisms in modern
and fossil cephalopods. — Adv. mar.
Biol, 1973, v. 11, p. 197—268.
Denton E. J., Land M. F. Mechanism
of reflexion in silvery layers of
fish and cephalopods. — Proc. R.
Soc. London, 1971, v. 178B, № 1050,
p. 43—61.
Dijkgraaf S. Vcrsuche fiber Schall-
wahrnehmung bei Tintenfischen. —
Naturwiss, 1963a, Bd. 50, № 2,
S. 50.
Dijkgraaf S. Nystagmus and related
phenomena in Sepia officinalis. —
Expcricntia, 1963b, v. 19, X? 1,
p. 29—30.
Dilly P. N., Nixon M. The cells that
secrete the beaks in octopus and
squids. — Cell. Tissue Res, 1976a,
v. 167, № 2, p. 229—241.
Dilly P N., Nixon M. The dermal tu-
bercles of Cranchia scabra: surface
structure and development. — J. Zo-
ol., 1976b, v. 179, № 3, p. 291—295.
Dilly P. N,, Nixon M., Packard A.
Forces exerted by Octopus vulga-
ris. — Pubbl. Staz. Zool. Napoli,
1964, v. 34, № 1, p. 86—97.
Drew G. A. Sexual activities of the
squid, Loligo pealeii (Les.), I—II.—
332
J. Morphol., 1911, v. 22, № 2,
p. 327—359; 1919, v. 32, p. 379—
435.
Emery D. G. The histology and fine
structure of the olfactory organ of
the squid Lolliguncula brevis Blain-
ville.— Tissue a. Cell, 1975a, v. 7,
№ 2, p. 357—367.
Emery D. G. Ciliated sensory cells and
associated neurons in the lip of
Octopus joubini Robson. — Cell
Tissue Res., 1975b, v. 157, № 3,
p. 331—340.
Feral J.-P. La regeneration des bras
de la seiche Sepia officinalis L.
I—II. — Cah. Biol. Mar., 1978,
T. 19, № 3, p. 355-361; 1979, T. 20,
№ 1, p. 29—42.
★Fields W. G. The structure, develop-
ment, food relations, reproduction,
and life history of the squid Loligo
opalescens Berry. — Fish Bull., Ca-
lif., 1965, № 131. 108 p.
* Flor kin M., Scheer В. T. (ed.) Che-
mical Zoology, v. 7. Mollusca.
NY—Lnd, 1972, 567 p.
Fort G. Le spermatophore des Cepha-
lopodes, I—II. — Bull. Biol. France
Belg., 1937, T. 71, p. 353—373;
1941, T. 75, p. 249—256.
Franzen Aa. Spermiogenesis and sper-
matozoa of the Cephalopoda. —
Ark. Zool., 1967, v. 19, № 4, p.
323—334.
Freeman S. E., Turner R. J. Maculo-
toxin, a potent toxin secreted by
Octopus maculosus Hoyle. — Toxi-
col. Appl. Pharmacol., 1970, v. 16,
№ 3, p. 681—690.
Froesch D. Antigen—induced secretion
in the optic gland of Octopus vul-
garis. — Proc. R. Soc. London,
1979, v. 205B, № 1160, p. 379—
384.
Froesch D., Mangold K. On the struc-
ture and function of a neurohemal
organ in the eye cavity of Eledone
cirrosa. — Brain Res., 1976, v. Ill,
№ 2, p. 287—293.
Froesch D., Marthy H.-J. The structu-
re and function of the ovi ducal
gland in octopods. — Proc. R. Soc.
London, 1975, v. 188B, № 1090,
p. 95—101.
Froesch D., Messenger J. B. On leuco-
phores and the chromatic unit of
Octopus vulgaris.—J. Zool., 1978,
y. 186, № 2, p. 163—173.
Ghiretti F. Cephalotoxin: the crab pa-
ralyzing agent of the posterior sa-
livary glands of cephalopods. —
Nature, 1959, 183, № 4669,
p. 1192—1193.
Ghiretti-Magaldi A., Ghiretti F., Salva-
to B. The evolution of haemocya-
nin. — Svmp. Zool. Soc. London,
N<> 38. 1977, p. 513—523.
Girsch S. J., Herring P. J., McCap-
ra F. Structure and preliminary
biochemical characterization of the
bioluminescent system of Ommast-
rephes pteropus (Stecnstrup). — J.
mar. biol. Assoc. U. K., 1976, v. 56,
№ 3, p. 707—722.
Giuditta A., Prozzo N. Postembryonic
growth of the optic lobe of Octopus
vulgaris Lam. — J. Comp. Neurol.,
1974, v. 157, № 2, p. 109—116.
Goto T.f Inoue S., Rakoi H. ct al.
Squid bioluminescence, I—IV.—
Tetrahedron Letters, 1976, № 26,
p. 2321—2324; 1976, № 34, p. 2971—
2974; Chemistry Letters, 1975, № 2,
p. 141—144; 1977, N° 3, p. 259—
262.
* Gotting K.-J. Malakozoologie. Grun-
driss der Weichtierkunde. Stuttgart,
1974. 317 S.
Graupneri H., Fischer J. Das Tinten-
driiscnepithel von Sepia vor, wah-
rend und nach dcr Pigmentbil-
dung. — Z. Zcllforsch., 1934, Bd. 21,
№ 3, S. 329—341.
Graziadei P. P. C., Gagne H. T. Sen-
sory innervation in the rim of the
octopus suckcr. — J. Morphol., 1976,
v. 150, Nb 3, p. 639—679.
Hanson D., Mann T., Martin A. W.
Mechanism of the spermatophorie
reaction in the giant octopus of the
North Pacific, Octopus dofleini mar-
tini. — J. Exptl. Biol., 1973, v. 58,
№ 3, p. 711—723.
Hara T., Hara R. Rctinochrome and
rhodopsin in the extraocular photo*
receptors of the squid, Todarodes. —
J. Gen. Physiol., 1980, v. 75, № 1,
p. 1—19.
Hara T., Hara R., Takeuchi J. Vision
in octopus and squid. — Nature,
1967, v. 214, № 5088, p. 572—573,
Harrison F. M., Martin A. IT. Excreti-
on in the cephalopod. Octopus do-
fleini.— J. Exptl. Biol., 1965, v. 42,
№ 1, p. 71—98.
♦ Herring P, J. Luminescence in cepha-
lopods. — Symp. Zool. Soc. Lon-
don, № 38, 1977, p. 127—159.
Hobbs M. J., Young J. Z. A cephalo-
pod cerebellum. — Brain Res., 1973,
v. 55, № 2, p. 424—430.
Hoffmann H.-I. Tintenfische — ihre
333
Preparation und mikroskopische
Untersuchung, I—III. — Mikrokos-
mos, 1970, Bd. 59, №№ 5—7, S.
142—147, 171—177, 201—205.
Hotta H. Identification of squid and
cuttie fish in the adjacent waters
of Japan, using the characteristics
of beaks. — Bull. Seikai Reg. Fish.
Res. Labor., 1973, № 43, p. 133—
147.
Hubbard S. J. Hearing and the octo-
pus statocyst. — J. Exptl. Biol.
I960., v. 37, № 4, p. 845-853.
* Isgrove A. Eledone. — Liverpool Mar.
Biol. Comm. Moms, v. 18, 1909.
105 p.
Ishikawa №. On the statocyst of Ame-
rican cephalopod genera. — J. Mor-
phol. Phvsiol. Philadelphia, 1929,
v. 48, № 2, p. 563—584.
Iverson I. L. K., Pinkas L. A pictorial
guide to beaks of certain Eastern
Pacific cephalopods, — Fish Bull.,
Calif., 1971, № 152, p. 83—105.
*Jaeckel S. G. A. Cephalopoden.—
Tierwelt der Nord- und Ostsee,
Lief. 37, Teil IXb3, 1958, S. 479—
723.
Jaeckel S. H. Kopffusser (Tintenfis-
che). —Neue Brehm-Biicherei,
№ 190. Wittenberg, 1957. 81 S.
Jander R., Daumer K., Waterman T. H.
Polarized light orientation by two
Hawaiian decapod cephalopods. —
Z. vergl. Physiol., 1963, Bd. 46,
№ 4, S. 383—394.
* Kaestner A. Lehrbuch der speziellen
Zoologie, Bd. 1, Teil. 1. 3. Aufl.
Jena, 1969, 898 S. (Cephalopoda:
S. 424—451).
Kristensen T. K. Periodical growth
rings in cephalopod statoliths. —
Dana, 1980, № 1, p. 39—51.
* Lane F. W. Kingdom of the octopus.
The life history of Cephalopoda.
London, 1957. 287 p. 2nd ed., NY,
1960, 300 p.
Lange №. №. On the regeneration and
finer structure of the arms of the
cephalopods. — J. Exptl. Zool.,
1920, v. 31, p. 1—57.
Lapan E.A. Studies on the chemistry
of the octopus renal system and an
observation of. the symbiotic rela-
tionship of the dicyemid Mcso-
zoa. — Comp. Biochem. Physiol.,
1975, v. 52Л, № 4, p. 651—658.
Leishman G.t Cohn D. H., Nedl-
son К. H. Bacterial origin of lu-
minescence in marine animals. —
Science, 1980, 208, № 4449, p.
1271—1273.
* MacGinitie G. E., MacGinitie N. Na-
tural history of marine animals.
2nd ed. NY, 1968. 523 p.
Maginnis L. A., Wells №. J. The oxy-
gen consumption of Octopus cya-
nea. — J. Exptl. Biol., 1969, v. 51,
№ 3, p. 607—613.
* Mangold-Wirz K. Biologie des Сё-
phalopodes benthiques et nectoniqu-
es de la mer Catalane. — Vie Mi-
lieu, suppl. № 13, 1963. 285 p.
Mangold K., Boucher-Rodoni R. Role
du jeune dans ^induction de la
maturation genitale chez les femel-
les d‘Eledone cirrosa.— C. R. Acad.
Sci. Paris, 1973, T. 276D, № 13,
p. 2007—2010.
Mangold K., Fioronl P. Morphologic
et biometrie des mandibules de
quelqucs Cephalopodes mediterra-
neens.— Vie Milieu, 1966, T. 17,
№ ЗА, p. 1139—1196.
* Mangola-Wirz K., Fioroni P. Die
Sonderstellung der Cephalopoden.
— Zool. Jb., Syst., 1970, Bd. 97,
№ 4, S. 522—631.
Mangold K., Froesch D. A reconside-
ration of factors associated with
sexual maturation in Cephalo-
pods.— Symp. Zool. Soc. London,
№ 38, 1977, p. 541—555.
Mangold K., Froesch D., Boucher-Ro-
doni R., Rowe V. L. Factors affect-
ing sexual maturation in cephalo-
pods.— Pubbl. Staz. Zool. Napoli,
1975, v. 39, suppl. 1, p. 259—266.
Mangold K., Portmann A. Dimensions
et croissance relatives des Octopo-
dides mediterraneens. — Vic Milieu,
suppl. № 17, 1964, p. 213—233.
Mann T., Martin A. W., Thiersch J. B.
Male reproductive tract, spermato-
phores, and spermatophoric reaction
in the giant octopus of the North
Pacific, Octopus dofleini martini. —
Proc. R. Soc. London, 1970,
v. 175B, № 1038, p. 31—61.
Marchand W. Der mannliche Lcitungs»
apparat der Dibranchiaten. — Z.
wiss. Zool., 1907, Bd. 86, № 3, S.
311—415.
Marchand W. Ober die Spermatopho-
ren. — Zoologica, Stuttgart, 1913,
Bd. 26, № 1—3, S. 171—200.
Marthy H.-J., Hauser R., Scholl A.
Natural tranquilliser in cephalopod
eggs. — Nature, 1976, v. 261,
№ 5560, p. 496—497.
* Martin A. W. Physiology of the ex-
£34
cretory organs of cephalopods. —
Fortschr. Zool., 1975, Bd. 23, № 2—
3, S. 112—123.
Martin A. W., Thiersch J. B.t
Dott H, M. et aL Spermatozoa of
the giant octopus of the North Pa-
cific, Octopus dofleini martini. —
Proc. R. Soc. London, 1970,
v. 175B, № 1038, p. 63—68.
Martin R. The giant nerve fibre sys-
tem of cephalopods. Recent structu-
ral findings. — Symp. Zool. Soc.
London, № 38, 1977, p. 261—275.
Matus A. I. Fine structure of the pos-
terior salivary gland of Eleaone
cirrosa and Octopus vulgaris. —
Z. Zellforsch., 1971, Bd. 122, № 1,
S. 111—121.
* Mauro A. Extra—ocular photorece-
ptors in cephalopods. — Symp. Zo-
ol. Soc. London, № 38, 1977, p.
287—308.
McCapra F. M., Hart R. The origin of
marine bioluminescence. — Nature,
1980, v. 286, № 5774, p. 660-661.
Messenger J. B. Parolfactory vesicles
as photoreceptors in deep-sea
squid. — Nature, 1967, v. 213,
№ 5078, p. 836—838.
Messenger Л B. Reflecting elements
in cephalopod skin and their impor-
tance for camouflage. — J. Zool.,
1974, v. 174, № 3, p. 387—395.
Messenger J. B. Evidence that octopus
is colour blind. — J. Exptl. Biol.
1977, v. 70, № 1. p. 49—55.
Messenger J. B. The nervous system
of Loligo, IV. The peduncle and
olfactory lobes. — Philos. Trans. R.
Soc. London, 1979a, v. 258B,
№ 1008, p. 275—309.
Messenger J. B. The eyes and skin of
octopus: compensating for sensory
deficiencies. — Endeavour, 1979b,
N. S., v. 3, № 3, p. 92—98.
Messenger J. B., Muzii E. O.t Nardi G.,
Steinberg H. Haemocyanin synthe-
sis and the branchial gland of oc-
topus. — Nature, 1974, v. 250,
№ 5462, p. 154—155.
Meyer W. T. Die Anatomie von О pi-
sthoteuthis depressa (Ijima und
Ikeda). — Z. wiss. Zool., 1906, Bd.
85, № 2, S. 183—269.
* Meyer W. T, Tintenfische. — Monogr.
einheim. Tiere, Bd. 6. Leipzig,
1913. 148 S.
Mochizuki A. An antiseptic effect of
cuttlefish ink. — Bull. Jap. Soc.
Sci. Fish., 1979, v. 45, Ns 11,
p. 1401—1403.
Morales E. Contribution al estudio de
la morfologia, estructura у anato-
mia microscdpica de la regi6n vis-
ceral de Pteroctopus tetracirrhus
D. Ch.— Invest. Pesq., 1973,
T. 37, № 2, p. 353—518.
Morishita T. Studies on the protein
digestive enzymes of octopus,
Octopus vulgaris Cuvier. — Bull.
Fac. Fish. Mie Univ., 1978, № 5,
p. 179—282.
* Morton J. E. Molluscs. 4th ed. Lon-
don, 1967, 244 p.
Muntz W. R. A., Johnson M. S. Rho-
dopsin of oceanic decapods. — Vi-
sion Res., 1978, v. 18, № 5, p. 601—
602.
Nishioka R. S., Yasumasu L, Packard
A. et. al. Nature of vesicles asso-
ciated with the nervous system of
cephalopods. — Z. Zellforsch., 1966,
Bd. 75, № 1, S. 301—316.
Nixon M. Growth of the beak and ra-
dula of Octopus vulgaris. — J.
Zool., 1969, v. 159, № 3, p. 363—
379.
Nixon M. Hole-boring in shells by Oc-
topus vulgaris Cuvier in the Me-
diterranean.— Malacologia, 1979,
v. 18, № 1—2, p. 431—443.
Nixon M. The salivary papilla of Octo-
pus as an accessory radula for
drilling shells. — J. Zool., 1980,
v. 190, № 1, p. 53—57.
* Nixon M., Dilly P. N. Sucker surfa-
ces and prey capture. — Symp.
Zool. Soc. London, № 38, 1977,
p. 447—511.
* Nixon M., Messenger J. B. (ed.).
The biology of cephalopods. Symp.
Zool. Soc. London, № 38, 1977,
615 p.
O'Dor R. K., Wells M. J. Reproduction
versus somatic growth: hormonal
control in Octopus vulgaris. — J.
Exptl. Biol., 1978, v. 77, № 1,
p. 15—31.
Packard A. Operational convergence
between cephalopods and fish: an
exercise in functional anatomy. —
Arch. Zool. Ital., 1966, v. 51, № 2,
p. 523—542.
Packard A. Visual acuity and eye
growth in Octopus vulgaris (La-
marck). — Monit. Zool. Ital., 1969,
N.S., v. 3, № 1, p. 19—32.
* Packard A. Cephalopods and fish:
the limits of convergence. — Biol.
Revs, 1972, v, 47, № 2, p. 241—
307.
Packard A., Albergoni V. Relative
335
growth, nucleic acid content and
cell numbers of the brain in Octo-
pus vulgaris (Lamarck). — J. Exptl.
Biol., 1970, v. 52, p. 539—552.
Packard A., Hochberg F. G. Skin pat-
terning in Octopus and other ge-
nera. — Symp. Zool. Soc. London,
№ 38, 1977, p. 191—231.
Packard A., Sanders G. D. Body pat-
tern of Octopus vulgaris and ma-
turation of the response to distur-
bance.— Anim. Behav., 1971, v. 19,
№ 4, p. 780—790.
Person P. Cartilaginous dermal sca-
les in cephalopods. — Science, 1969,
v. 164, № 3886, p. 1404—1405.
Peters W. Occurrence of chitin in MoL
lusca. — Comp. Biochem. Physiol.,
1972, v. 41B, № 3, p. 541—550.
Philpott D. E., Person P. The biology
of cartilage, 2. Invertebrate carti-
lages: squid head cartilage. — J.
Morphol., 1970, v. 131, № 4,
p. 417—430.
Potts W. T W. Todd M. Kidney func-
tion in the octopus. — Comp. Bio-
chem. Physiol., 1965, v. 16. № 4,
p. 4/9—489.
* Purchon R. D. The biology of the
Mollusca. 2nd ed. Oxford e. a.,
1977. 560 p.
Richard A. Analyse du cycle sexuel
chcz les Cephalopodes: mise en
evidence cxperimentale d’un
rythme conditionne par les varia-
tions des facteurs externes et in-
ternes. — Bull. Soc. Zool. France,
1970, T. 95, № 3, p. 461—469.
Richard A., Branden C. van den Dec-
leir IF. The cycle of activity in the
accessory nidamental gland from
cephalopods. — In: 13th Europ. Mar.
Biol. Symp., Oxford e:a., 1979,
p. 173—180.
Richard A„ Lemaire J. Determination
ct differcnciation sexuelle chez la
seiche Sepia officinalis L. — Pubbl.
Staz. Zool. Napoli, 1975, T. 39,
suppl. 1, p. 574—594.
Robson G. C. On seriation and asym-
metry of the cephalopod radula. —
J. Linn. Soc. London, 1925, v. 36,
№241, p. 99—108.
Rowe V. L., Mangold K. The effect of
starvation on sexual maturation in
Illex illecebrosus (LeSueur). — J.
exp. mar. Biol. Ecol., 1975, v. 17,
№ 2, p. 157—164.
♦ Sanders G. D. The cephalopods. —
In; Invertebrate learning, v. 3,
NY-Lnd, 1975, p. 1—101.
Savage I. V. E., Howden M. E. H.
Hapalotoxin, a second lethal toxin
from the octopus Hapalochlaena
maculosa. — Toxicon, 1977, v. 15,
№ 5, p. 463—466.
* Schipp R.f Boletzky S. v. Morpholo-
gy and function of the excretory
organs in dibranchiate cephalo-
pods.— Fortschr. Zool., 1975, Bd.
23. № 2—3, S. 89—111.
Schipp R., Boletzky S. v. The pancrea-
tic appendages of dibranchiate
cephalopods, I. — Zoomorphologie,
1976, v. 86, № 1, p. 81—98.
Schipp R„ Mollenhauger S., Boletz-
ky S. v. Electron microscopical
and histochemical studies of diffe-
rentiation and function of the
cephalopod gill (Sepia officinalis
L.). —Ibid., 1979, v. 93, № 3,
p. 193—207.
Schipp R., Pfeiffer K. Vergleichende
cytologische und histochemische
Untersuchungen an der Mittel-
darmdriise dibranchiatcr Cephalo-
poden. — Zool. Jb., Anat., 1980, Bd.
104, № 3, S. 317—343.
Sheumack D. D., Howden M. E. H.,
Spence /., Quinn R. J. Maculoto-
xin: a neurotoxin from the venom
glands of the octopus Hapalochlae-
na maculosa identified as tetrodo-
toxin. — Science, 1978, v. 199,
№ 4325, p. 188—189.
Solem A., Roper C. F. E. Structures
of recent cephalopod radulae —
Veliger, 1975, v. 18, № 2, p. 127—
133.
Stephens P. R., Young J. Z. The sta-
tocyst of Vampyroteuhis inferna-
lis. — J, Zool., 1976, v. 180, № 4,
p. 565—588.
Stephens P. R., Young J. Z. Semicir-
cular canals in squids. — Nature,
1978, v. 271, № 5644, p. 444—445.
Stuart A. E. The reticulo-endoihelial
apparatus of Eledone cirrosa. — Vie
Milieu, 1967, T. 18, № 1A, p. 175—
187.
Sutherland N. S. Shape discrimination
in rat, octopus, and goldfish: a
comparative study. — J. Comp.
Physiol. Psychol., 1969, v. 67,
p. 160—176.
Sutherland S. K., Lane W. R. Toxin
and mode of envenomation of the
common ringed or blue-banded oc-
topus.—Med. J. Australia, 1969,
v. 1, № 18, p. 893—898.
Takahashi N., Yahata T. Histological
studies of the maturation of the
336
ovary in the squid, Todarodes pa-
cificus. — Bull. Fac. Fish. Hokkai-
do Univ., 1973, v. 24, № 2, p. 63—
68.
Taki I. Observations on autophagy in
octopus. — Annotat. Zool. Jap.,
1936, v. 15, № 3, p. 352—354.
♦ Thore S. Beitragc zur Kcnntnis der
vergleichenden Anatomic des zent-
ralcn Ncrvcnsystcms der dibran-
chiaten Cephalopoden. — Pubbl.
Staz. Zool. Napoli, 1939, v. 17,
№ 3, p. 313—506.
*Tompsett D. H., Sepia. — Liverpool
Mar. Biol. Comm. Mems, v. 32,
1939, 184 p.
Ward D. V. Locomotory function of
the squid mantle. — J. Zool., 1972,
v. 167, № 4, p. 487—499.
Ward D. V., Wainwright S. A. Loco-
motory aspects of squid mantle
structure. — Ibid., 1972, v. 167,
№ 4, p. 437—449.
* Wells M. J. Brain and behaviour in
cephalopods. London, 1962. 171 p.
* Wells M. J. Learning by marine in-
vertebrates. — Adv. mar. Biol.,
1965, v. 3, p. 1—62.
* Wells M. J. Cephalopod sense or-
gans. The brain and behaviour of
cephalopods. — In: Physiology of
Mollusca, v. 2. NY-Lnd, 1966,
p. 523—590.
Wells M. J. Hormonal control of re-
production in cephalopods. — In:
Perspectives in experimental bio-
logy, v. 1. Oxford e. a., 1976a,
p. 157—166.
* Wells M. J. Proprioception and lear-
ning.— In: Structure and function
of proprioceptors in the invertebra-
tes. London, 1976b, p. 567—604,
* Wells M. J. Octopus. Physiology and
behaviour of an advanced inverte-
brate. London, 1978. 424 p.
Wells M. J. The heartbeat of Octopus
vulgaris. — J. Exptl. Biol., 1979,
v. 78, p. 87—104.
Wells M. J. Nervous control of the
heartbeat in octopus. — Ibid., 1980,
v. 85, p. 111—128.
Wells M, J., Wells J. Pituitary analo-
gue in the octopus. — Nature, 1969,
v. 222, № 5190, p. 293—294.
* Wilbur К. M., Yonge С. M. (ed.).
Physiology of Mollusca. NY—Lnd,
v. 1, 1964, 473 p.; v. 2, 1966. 645 p.
* Williams L. The anatomy of the com-
mon squid, Loligo pealeii LeSueur.
Leiden, 1909. 92 p.
Wirz K. Etude biometrique du systeme
nerveux des Cephalopodes. — Bull.
Biol. France Belg., 1959, T. 93,
№ 1, p. 78—117.
Wodinsky J. Mechanism of hole boring;
in Octopus vulgaris. — J. Gen.
Physiol., 1973, v. 88, № 2, p. 179—
183.
Wodinsky J. Hormonal inhibition of
feeding and death in octopus: cont-
rol by optic gland secretion. —
Science, 1977, v. 198, № 4320,.
p. 948—951.
Young J, Z. The statocysts of Octopus;
vulgaris. — Proc. R. Soc. London,.
1960. v. 152 B, № 946, p. 3—29.
* Young J. Z. Learning and discrimi-
nation in the octopus. — Biol. Revs,.
1961, v. 36, № 1, p. 32—96.
Young J. Z. The number and sizes of
nerve cells in octopus. — Proc.
Zool. Soc. London, 1963, v. 140,
№ 2, p. 229—254.
♦ Young J. Z. A model of the brain.
Oxford, 1964. 348 p.
Young J. Z. The central nervous sy-
stem of Nautilus. — Philos. Trans.
R. Soc. London, 1965, v. 249B,
№ 754, p. 1—25.
Young J. Z. The memory system of the
brain. Oxford, 1966. 148 p.
Young J. Z. The stalked eye of Batho-
thauma. — J. Zool., 1970, v. 162,
№ 4, p. 437—447.
* Young J. Z. The anatomy of the
nervous system of Octopus vulga-
ris. Oxford, 1971. 690 p.
Young J. Z. The organization of a>
cephalopod ganglion. — Philos^
Trans. R. Soc. London, 1972,
v. 263B, № 854, p. 409—429.
* Young J. Z. The central nervous sy-
stem of Loligo. I—III, V. — Ibid.,
1974, v. 267B, № 885, p. 263—302;
1976, v. 274B, № 930, p. 101—167;
1977, v. 276B, № 948, p. 351—39&;
1979, v. 285B, № 1009, p. 311—
354.
Young J. Z. The «cerebellum» and the
control of eye movements in cepha-
lopods. — Nature, 1976, v. 264,
№ 5586, p. 572—574.
♦ Young J. Z. Brain, behaviour and
evolution of cephalopods. — Symp,
Zool. Soc. London, № 38, 1977,
p. 377—434.
Young R. E. Function of extra-ocular
photoreceptors in bathypelagic
cephalopods. — Deep-Sea Res.,
1972, v. 19, № 9, p. 651—660.
Young jR. E. Information feedback
from photophores and ventral co-
337
untershading in mid-water squid. —
Pacif. Sci., 1973, v. 27, № 1,
p. 1—7.
Young R. E. Function of the dimor-
phic eyes in the midwater squid
nistioteuthis dofleini. Ibid., 1975a,
v. 29, № 2, p. 211—218.
Young R. E. Transitory eye shapes
and the vertical distribution of two
midwater squids. — Ibid., 1975b,
v. 29, № 3, p. 243—255.
Young R. E. Leachia pactfica: spaw-
ning habitat and function of the
brachial photophores. — Ibid.,
1975c, v. 29, № 1, p. 19—25.
* Young R. E. Ventral bioluminescent
countershading in midwater cepha-
lopods. — Symp. Zool. Soc. Lon-
don, № 38, 1977, p. 161—190.
* Young R. E. Vertical distribution and
photosensitive vesicles of pelagic
cephalopods from Hawaiian wa-
ters. — Fish. Bull. U. S., 1978, v. 76,
№3, p. 583—615.
Young R. E., Kampa E. M., May-
nard S. D. et al. Counterillumina-
tion and the uppermost limits of
midwater animals. — Deep-Sea Res.,
1980, v. 27, № 9A, p. 671—691.
Young R. E., Mencher F. M. Biolumi-
nescence in mesopelagic squid: diel
color change during counterillu-
mination.— Science, 1980, v. 208,
№ 4449, p. 1286—1288:
Young R. E., Roper C. F. E. Biolumi-
nescent countershading in midwa-
ter animals: evidence from living
squid. — Ibid., 1976, v. 191, № 4231,
p. 1046—1048.
Young R. E.j Roper C. F. E. Intersity
regulation of bioluminescence du-
ring countershading in living mid-
water animals. — Fish. Bull. U. S.,
1977, v. 75, № 2, p. 239—252.
Young R. E., Roper C. F. E., Mangold
K. et al. Luminescence from non-
biolumincscent tissues in oceanic
cephalopods. — Mar. Biol., 1979,
v. 53, № 1, p. 69—77.
Young R. E., Roper C. F. E., Wal-
ters J. F. Eyes and extraocular
photoreceptors in midwater cepha-
lopods and fishes: their roles in de-
tecting downwelling light for
counterillumination. — Ibid., 1979,
v. 51, № 4, p. 371—380.
Систематическая часть
* Зуев Г В. Головоногие моллюски
северо-западной части Индийско-
338
го оксана. Киев: Наукова думка,
1971. —223 с.
Зуев Г. В., Несис К. Н., Нигматул-
лин Ч. М. Система и эволюция
родов Omtnastrephes и Symplecto-
ieuthis. — Зоологический журнал,
1975, т. 54, вып. 10, с. 1468—
1479.
Зуев Г. В., Несис К. Н., Нигматул-
лин Ч. М. Распространение родов
Ommastrephes d’Orbigny, 1833,
Sthenoteuthis Verrill, 1880 и To-
darodes Steenstrup, 1880 в Атлан-
тическом океане. — Бюллетень
МОИП, отдел биологии, 1976,
т. 81, № 4, с. 53—63.
Клумов С. К-, Юхов В. Л, Mesony-
choteuthis harniltoni Robson, 1925
и его значение в питании кашало-
та антарктических вод. — В сб.:
Антарктика, 1975, вып. 14, с. 159—
189.
Кондаков Н. Н. Головоногие моллюс-
ки (Cephalopoda) Карского мо-
ря. — Труды Арктического инсти-
тута, 1937, вып. 50, с. 61—67.
* Кондаков Н. II. Головоногие мол-
люски (Cephalopoda) дальневос-
точных морей СССР. — Исследо-
вания дальневосточных морей,
1941, вып. 1, с. 216—255.
Кондаков Н. Н., Москалёв Л. И.,
Несис К. Н. Осьминог Bcnthocto-
pus sibiricus Loyning — эндемик
восточной Арктики. — В сб.: Эко-
логические исследования шельфа.
М.: Институт океанологии АН
СССР, 1981, с. 42—56.
Несис К. Н. Биология перуано-чилий-
ского гигантского кальмара Dosi-
dicus gigas. — Океанология, 1970,
т. 10, вып. 1, с. 140—152.
Несис К. Н. Новая форма кальмара
рода Histioteuthis из восточной
части Тихого океана. — Зоологи-
ческий журнал, 1971, т. 50,
вып. 10, с. 1463—1471.
Несис К. Н. Пересмотр родов каль-
маров Taonius и Belonella.—Зо-
ологический журнал, 1972а, т. 51,
вып. 3, с. 341—350.
Несис К. Н. Два новых вида каль-
маров семейства Gonatidae из се-
верной Пацифики. — Зоологиче-
ский журнал, 19726, т. 51, вып. 9,
с. 1300—1307.
* Несис К. Н. Система, филогения и
эволюция кальмаров семейства
Gonatidae. — Зоологический жур-
нал. 1973а, т. 52, вып. 11,
с. 1626—1638.
Несис К. Н. Типы ареалов головоно-
гих моллюсков северной Паци-
фики. — Труды Института океа-
нологии АН СССР, 19736, т. 91,
с. 213—239.
Несис К. Н. Головоногие моллюски
восточно-экваториалыюй и юго-
восточной частей Тихого океа-
на. — Труды Института океаноло-
гии АН СССР, 1973в, т. 94,
с. 188—242.
Несис К. Н. Океанические головоно-
гие моллюски юго-западной Ат-
лантики.— Труды Института оке-
анологии АН СССР. 1974а, т. 98,
с. 51-75.
Несис К, Н. Пересмотр родов каль-
маров Согупотта, Megalocran-
cfiia, Sandalops и Liguriella. —
Труды Института океанологии
АН СССР, 19746, т. 96, с. 5—22,
♦ Несис К. Н. Система современных
головоногих моллюсков. — Бюлле-
тень МОИП, отдел биологии,
1974в, т. 79, № 5, с. 81—93.
Несис К. Н. Гигантские кальмары. —
Природа, 1974, № 6, с. 55—60,
* Несис К. Н. Головоногие моллюски
Американского Средиземного мо-
ря.— Труды Института океаноло-
гии АН СССР, 1975а, т. 100,
с. 259—288.
Несис К. Н. Географические группи-
ровки головоногих моллюсков пе-
лагиали тропической западной
Пацифики. — Труды . Института
океанологии АН СССР, 1977а,
т. 107, с. 7—14.
Несис К. Н, Mastigoteuthls psych-
rophila sp. п. из Южного океа-
на. — Зоологический журнал,
1977в, т. 56, вып. 6, с. 835—841.
Несис К. II, Биология Argonauta bo-
ettgeri и A. hians в западной Па-
цифике и морях Малайского ар-
хипелага.—Зоологический журнал,
1977, т. 56, вып. 7, с. 1004—1014.
Несис К. II. Подсемейство Ancistro-
cheirinae (Enoploteuthidae). —
Зоологический журнал, 1978а,
т. 57, вып. 3, с. 446—449.
Несис К. Н. Краткий очерк зоогеог-
рафии пелагиали Австрало-Ново-
зеландского региона (по голово-
ногим моллюскам).— Труды Ин-
ститута океанологии АН СССР,
1979а, т. 106, с. 125—139.
Несис К. Н. Кальмары сем. Ommast-
rephidae Австрало-Новозеландско-
го региона. — Труды Института
океанологии АН СССР, 19796,
т. 106, с. 140—146.
Несис К. Н. Систематическое поло-
жение Chiroieuthis famelica Ber-
ry. — Бюллетень МОИП, отдел
биологии, 1980а, т. 85, № 4,
с. 59—66.
Несис К. Н. Сепииды и лолигиниды:
сравнительный обзор распростра-
нения и эволюции неритических
головоногих моллюсков. — Зооло-
гический журнал, 19806, т. 59,
вып. 5, с. 677—688.
Несис К. Н., Головань Г А. Гигант-
ский глубоководный осьминог. —
Природа, 1975, № 5, с. 112—113.
Несис К. Н., Нигматуллин Ч. М.
Придонные кальмары Патагоно-
Фолклепдского района. — Труды
АтлаптНИРО, 1972, вып. 42,
с. 170—175.
Несис К. Н., Нигматуллин Ч. М. Рас-
пространение и биология родов
Ornithoteuthis Okada, 1927 и Нуа-
loteuthis Gray, 1849. — Бюллетень
МОИП. отдел биологии, 1979,
т. 84, № 1, с. 50—63.
Несис К. Н., Никитина И. В. Макро-
тритопус — планктонная личинка
донного осьминога Octopus defilip-
pi: идентификация и распростра-
нение. — Зоологический журнал,
1981, т. 60, вып. 6, с. 835—847.
Несис К. Н„ Шевцов Г. А. Неритиче-
ские кальмары сем. Loliginidae в
водах Советского Дальнего Вос-
тока. — Биология моря, Владиво-
сток, 1977а, № 3, с. 70—71.
Несис К. Н., Шевцов Г А. Первые
сведения об абиссальных голово-
ногих моллюсках Охотского мо-
ря. — Биология моря, Владивос-
ток, 19776, № 5, с. 76—77.
Нигматуллин Ч. М. Биология и про-
мысел каракатиц шельфа северо-
западной Африки. — Труды Ат-
лантНИРО, 1972, вып. 42, с. 57—
85.
Нигматуллин Ч. М. Основные этапы
эволюции кальмаров семейства
Ommastrephidac. — В сб.: Вопро-
сы эволюционной морфологии жи-
вотных. Казань: Изд-во КазГУ,
1979, с. 210—219.
Новые данные о распространении
кальмаров семейства Ommastrep-
hidae в Мировом океане/[10. В.
Корзун, К. п. Несис, Ч. М. Ниг-
матуллин и др.]. — Океанология,
1979, т. 19, вып. 4, с. 729—733.
339
Филиппова Ю. Л. О фауне кальма-
ров Южной Атлантики. — Зооло-
гический журнал, 1969, т. 48, вып.
1, с. 51—63.
* Филиппова Ю. А. Распространение
и биология кальмаров. — Итоги
науки и техники, серия Зоология
беспозвоночных, т. 2, М.:
ВИНИТИ, 1973, с. 60—101.
Филиппова Ю. А., /Охов В. Л. Видо-
вой состав и распределение голо-
воногих моллюсков в мезо- и ба-
типелагиали антарктических вод.
— В сб.: Антарктика, 1979, вып.
18, с. 175—187.
Шевцов Г Л. К фауне головоногих
моллюсков юго-восточной и цен-
тральной частей Тихого океана.—
В сб.: Тропическая зона Мирово-
го океана и связанные с ней гло-
бальные процессы. М.: Наука,
1973, с. 225—230.
Шевцова С. П. К распределению и
биологии кальмаров Abraliopsis
из вод Куросио. — В сб.: Ком-
плексные исследования в Миро-
вом океане. М.: Институт океано-
логии АН СССР, 1975, с. 351—
352.
Юхов В. Л. Находки гигантских
кальмаров. — Природа, 1974, № 6,
с. 60—63.
Adam U7. The Cephalopoda in the In-
dian Museum, Calcutta/—Rec. In-
dian Mus., 1939a, v. 41, № 1, p.
61—110.
Adam W. Cephalopoda, I. Le genre
Sepioteuthis Blainville, 1824. — Si-
boga-Exped., Monogr. 55a, Livr.
134, 1939b, p. 1—34.
Adam UZ. Cephalopoda, II. Revision
du especes Indo—malaises du gen-
re Sepia Linne, 1758. Revision du
genre Sepiella (Gray) Steenstrup. —
Ibid., Monogr. 55b, Livr. 135,
1939c, p. 35—122.
Adam W. Un Ccphalopode nouveau:
Pholidoteuthis boschmai gen. et sp.
nov. — Verh. Akad. Wet. Amster-
dam. 1950, v. 53, № 10, p. 1592—
1598.
* Adam W. Cephalopodes. — Res. Sci.
Exped. Oceanogr. Beige eaux co-
heres afric. Atlant, sud (1948—
49), v. 3, № 3, 1952. 142 p.
Adam W. Cephalopoda, III. —Sibo-
ga-Exped. Monogr. 55c, Livr. 144,
1954, p. 123—198.
* Adam UZ Les Cephalopodes de la
mcr Rouge. — Mission R. Ph. Doll-
fus en Egypte (dec. 1927 — mars
1929), Res. Sci., pt. 3, № 28, 1959,
p. 125—193.
Adam W. Cephalopoda from the Gulf
of Aqaba. — Bull. Sea Fish. Res,
Stat Haifa, 1960a, № 26, p. 1—27.
Adam W. Les Cephalopodes de Flnsti-
tut francais d‘Afrique noire, II.—
Bull. IFAN, 1960b, v. 22, Ns 2A,
p. 465—511.
Adam W Notes sur les Cephalopodes,
24. Contribution a la connaissance
de Thectocotyle chez les Ommast-
rephidae. — Bull. Inst. roy. Sci. na-
tur. Belg., 1960c, v. 36, № 19, 10 p.
Adam HZ. Cephalopodes de FArchirel
du Cap-Vert, de F Angola et du Mo-
zambique. — Mem. Junta Invest.
Ultramar, ser. 2, 1962, № 33, p.
7—64.
Adam W. Cephalopoda from the Medi-
terranean Sea. — Bull. Sea Fish.
Res. Stat. Haifa, 1967, № 45, p.
65—78.
Adam W. Notes sur les Cephalopodes,
25. Contribution a la connaissance
de Chaunoteuthis mollis Appellof,
1891. — Bull. Inst. roy. Sci. natur.
Belg., 1972, v. 48, № 12. 7 p.
Adam UZ. Cephalopoda from the Red
Sea. — Contr. Knowl. Red Sea,
1973, Ne 47, p. 9—47.
Adam F. The Scpiidae (Cephalopoda,
Decapoda) in the collection of the
Western Australian Museum.—
Rec. W. Austral. Mus., 1979, v. 7,
№ 2, p. 109—212.
* Adam UZ., Rees W. J. A review of
the cephalopod family Sepiidae.—
Sci. Repts John Murray Exped.
1933—34, v. 11, № 1, 1966. 165 p.
Aldred R. 6. Structure, growth and
distribution of the squid Batho-
thauma lyromma Chun. — J. mar.
biol. Assoc. U. K., 1974, v. 54, № 4,
p. 995—1006.
Aldred R. G., Nixon M., Young J. Z.
The blind octopus, Cirrothauma. —
Nature, 1978, v. 275, № 5680, p.
547—549.
Allan J. Planktonic cephalopod larvae
from the Eastern Australian coast.
— Rec. Austral Mus., 1945, v. 21,
p. 317—350.
* Allan J. Australian shells. Melbour-
ne, 1959. 487 p.
Anderson M. E. Notes on the cephalo-
pods of Monterey Bay, California,
with new records for the area.—
Veliger, 1978, v. 21, № 2, p. 255—
262.
340
Berry S. S. A note on the genus Lo~
lliguncula.— Proc. Acad. Nat. Sei.
Philadelphia, 1911, v. 63, p. 100—
105.
* Berry S. S. A review of the cephalo-
pods of western North America.
— Bull. U. S. Bureau Fish., 1912,
v. 30, p. 267—336.
* Berry S. S. The Cephalopoda of the
Hawaiian waters. — Ibid., 1914
v. 32, p. 255—362.
Berry S. S. Cephalopoda of the Ker-
madec Islands. — Proc. Acad. Nat.
Sci. Philadelphia, 1916, v. 68, № 1,
p. 49—66.
Berry S. S. Report on the Cephalopo-
da obtained by the F. I. S. «En-
deavour» in the Great Australian
Bight and other Southern Austra-
lian localities. — Biol. Res. Fish.
Experiments «Endeavour» 1909—
14, v. 4, № 5, 1918, p. 201—298.
Berry S. S. Preliminary diagnoses of
new cephalopods from the Western
Atlantic. — Proc. U. S. Nat. Mus,
1920, v. 58, № 5335, p. 293—300.
Berry S. S. Loliolopsis chiroctes, a
new genus and species of squid
from the Gulf of California.—
Trans. San Diego Soc. Nat. Hist.,
1929, v. 5, № 18, p. 263—282.
Berry S. S. Cephalopods of the gene-
ra Sepioloidea, Sepiadarium, and
Idiosepius. — Philippine J. Sci.,
1932, v. 47, № 1, p. 39—55.
Berry S. S. The flapjack devilfish,
Opisthoteuthis, in California. — Ca-
lif. Fish a. Game, 1952, v. 38, № 2,
p. 183—188.
Berry S. S. Preliminary diagnoses of
six West American species of Octo-
pus.— Leafl. Malacol, 1953, v. 1,
№ 10, p. 51—58.
Berry S, S. The male flap jack devil-
fish.—Calif. Fish a. Game, 1955,
v. 41, № 3, p. 219—224.
Boletzky S. v. Neorossia n. g. pro
Rossia (Allorossia) caroli Joubin,
1902, with remarks on generic sta-
tus of Semirossia Steenstrup,
1887, —Bull. Alar. Sci, 1971, v. 21,
№ 4, p. 964—969.
Brakoniecki T. F. Lolliguncula tydeus,
a new species of squid from the
Pacific coast of Central America. —
Ibid. 1980, v. 30, № 2, p. 424—
430.
Bruun A. F. The biology of Spirilla
spirula (L.). — Dana-Rept, 1943,
№ 24. 46 p.
* Bruun Л. F. Cephalopoda. — Zool.
Iceland, v. 4, pt. 64, 1945. 15 p.
Bruun A. F. New light on the biology
of Spirula, a mesopelagic cephalo-
pod. — In: Essays in the natural
sciences in honor of Captain Allan
Hancock. Los Angeles, 1955, p.
61—72.
Burgess L. A. A study of the morpho-
logy and biology of Octopus hum-
melincki Adam, 1936. — Bull. Mar.
Sci, 1966, v. 16, № 4, p. 762—
813.
Burgess L. A. Loliolus rhomboidalis, a
new species of loliginid squid from
the Indian Ocean. — Ibid, 1967,
v. 17, № 2, p. 319—329.
Cairns S. D. Cephalopods collected in
the Straits of Florida by the R/V
«Gerda». — Ibid, 1976, v. 26, № 2,
p. 233—272.
Castellanos Z. J. A. de. Contribution al
conocimento biologico del calamar
argentine, I Ilex illecebrosus argen-
tine. — Bol. Inst Biol, marina,
1964, № 8. 37 p.
Castellanos Z. J. A. de. Rehabilitation
del genero Martialia Roch. et Mab,
1887. — Neotropica, 1967, v. 13,
№ 42, p. 121—124.
Castellanos Z. /. A. de, Cazzaniga N.
Aclaraciones acerca de los Loligi-
nid ae del Atlantico sudoccidental.
— Ibid, 1979, v. 25, № 73, p. 59—
68.
Castellanos Z. J. A. de, Menni R. C.
Los Cefaldpodos de la expedition
«Walther Herwig». — Notas Com.
Invest. Cient. Prov. Buenos Aires,
1968, v. 6, № 2, p. 1—31.
* Castellanos Z. Л A. de, Menni R. C.
Nota preliminar sobre distribution
de los Cefalopodos del Atlantico
sudoccidental, — An Soc. cient. Ar-
gentina, 1969a, v. 188, № 5—6,
p. 205—226.
Castellanos Z. J. A. de, Menni R. C.
Sobre dos pulpos costeros de la
Argentina. — Neotropica, 1969b,
v. 15, № 47, p. 89—94.
* Chun K. Die Cephalopoden. I. Oego-
sida. II. Myopsida, Octopoda.—
Wiss. Erg. Dtsch. Tiefsee-Exped.
«Valdivia» 1898— 1899, Bd. 18,
1910, 1915. 552 S.+Atlas.
Chun C, Cephalopoda. — Rep. Sci. Res.
«М. Sars» North Atlant. Deep-Sea
Exped, 1910, v. 3, № 1, 1913. 21 p.
Clarke M. R. Young stages of Lepido-
teuthis grimaldii. — Proc. Malacol.
341
Soc. London, 1964, v. 36, Ks 2,
p. 69—78.
* Clarke M. R. A review of the sys-
tematics and ecology of oceanic
sqiuds — Adv. mar. Biol., 1966,
v. 4, p. 91—300.
Clarke M. R. к deep-sea squid, Tanin-
gia danae Joubin, 1931. — Symp.
Zool. Soc. London, № 19, 1967,
p. 127—143.
Clarke M. R. Cephalopoda collected
on the SOND cruise. — J. mar.
biol. Assoc. U. K., 1969, v. 49, № 4,
p. 961-976.
Clarke M. R. Growth and development
of Spirula spirula. — Ibid., 1970,
v. 50, № 1, p. 53—64.
* Clarke M. R. Cephalopoda in the
diet of sperm whales of the Sou-
thern hemisphere and their bearing
on sperm whale biology. — Disco-
very Repts, 1980, v. 37. 324 p.
Clarke M. R.t Lu С. C. Vertical distri-
bution of cephalopods at 30° N,
23° W in the North Atlantic.— J.
mar. biol. Assoc. U. K., 1974, v. 54,
№ 4, p. 969—984.
Clarke M. R., Lu С. C. Vertical dis-
tribution cephalopods at 18° N,
25° W in the North Atlantic.—
Ibid., 1975, v. 55, № 1, p. 165—
182.
Clarke M. R., Maul G. E. A descripti-
on of the «scaled» squid Lepidote-
uthis grimaldii Joubin, 1895. —
Proc. Zool. Soc. London, 1962,
v. 139, № 1, p. 97—118.
Cohen A. C. The systematics and dis-
tribution of Loligo in the Western
North Atlantic, with description of
two new species. — Malacologia,
1976, v. 15, № 2, p. 299—367.
* Cotton В. C., Godfrey F. K. The mol-
luscs of South Australia. Adelaide,
1940, v. 2, p. 317—600.
Degner E. Cephalopoda. — Rep. Da-
nish Occanogr. Exped. 1908—10
Mediterranean, v. 2, С 1, 1925.
94 S.
* Dell R. K. The recent Cephalopoda
of New Zealand. — Bull. Dominion
Mus., 1952, v. 16. 157 p.
Dell R. K. Some additional New Zea-
land cephalopods from Cook
Strait. — Zool. Pubis Victoria Univ.
Wellington, 1959a, Ke 25. 12 p.
Dell R. л. Cephalopoda. — Rep. B. A.
N.Z.A.R. Expcd. 1929—1931, v.
8B, № 4, 1959b, p. 89-106.
Dell R. A. A specimen of the giant
squid Architeuthis from New Zea-
land.— Rec. Dominion Mus., 1970,
v. 7, № 4, p. 25—36.
Dilly P. N., Nixon M. Growth and de-
velopment of Taonius megalops,
and some phases of its life cycle. —
J. Zool., 1976, v. 179, № 1, p. 19—
83.
Dilly A N., Nixon M.t Young J. Z.
Mastigoteuthis, the whip-lash squ-
id. — Ibid., 1977, v. 181, № 4, p.
527—559.
♦ Dong Zhenzhi. A preliminary taxono-
mic study of the Cephalopoda
from the Chinese waters. — Stud.
Mar. Sinica, 1963, № 4, p. 125—
162.
Dong Zhenzhi On three new species
of the genus Octopus from the Chi-
nese waters. — Ibid., 1976, № 11,
p. 211—215.
*Ferussac A. de, d'Orbigny A. Histo-
ire naturelie generate et particu-
liere des Cephalopodcs acetabuli-
feres vivants et fossiles. Paris,
1835—1848, 2T. 361 p.+Atlas.
Filippova J. A. A new species of the
genus Cycloteuthis. — Malacol.
Rev., 1968, v. 1, p. 119-124.
Filippova J. A, New data on the
squids from the Scotia Sea (An-
tarctic).— Malacologia, 1972, v. 11,
№ 2, p. 391—406.
Garcia Cabrera R. C. Especes du gen-
re Sepia du Sahara cspagnoi. —
Rapp. Cons. int. Expl. Mer, 1970,
№ 159, p. 132—139.
Goodrich E. C. Report on a collection
of Cephalopoda from the Calcutta
Museum. — Trans. Linn. Soc. Lon-
don, Zool., 1896, v. 7, № 1. 24 p.
* Grimpe G. Zur Kenntnis der Cepha-
lopodenfauna der Nordscc. — Wiss.
Meeresuntcrs. Helgoland, 1925,
N. F., Bd. 16, II. 1. 122 S.
* Grimpe G. Die Cephalopoden des
arktischen Gebietes. — Fauna Arcti-
ca, Bd. 6, II. 5, 1933, S. 489—514.
* Hamada T., Obata L, Okutani T.
(ed.). Nautilus macromphalus in
captivity. Tokyo, 1980, 80 p.
* Hoyle W.E. Report on the Cephalo-
poda. — Rcpt. Sci. Res. Challen-
ger, v. 16, pt. 44, 1886, 246 p.,
Atlas.
Hoyle W. E. Report on the Cephalopo-
da.— Repts dredg. operat. «Albat-
ross». Bull. Mus. Comp. Zool. Har-
vard, 1904, v. 43, № I. 71 p.
* Hoyle W. E. A list of generic names
342
of dibranchiate Cephalopoda with
their type species. — Senckenbergi-
ana, 1910, Bd. 32, S. 407—413.
Imber M. J. The squid families Cran-
chiidae and Gonatidae in the New
Zealand area. — N. Z. J. Zool.,
1978, v. 5, № 3, p. 445—484.
Iredale T. The cuttle-fish «bones» of
the Sydney beaches. —Austral. Zo-
ol., 1926, v. 4, p. 186—196.
Iredale T. Cuttle-fish «bones» again. —
Ibid., 1954, v. 12, № 1, p. 63-82.
Iwai E, Description of unidentified
species of dibranchiate cephalo-
pods, I—II.— Sci. Repts whales
Res. Inst., 1956, № 11, p. 139—
161.
Joubin L. Contribution a I’etude des
Cephalopodes de 1’Atl antique
Nord. — Res. Camp. Sci. Monaco,
fasc. 9, 1895. 63 p.
Joubin L. Cephalopodes provenant
des campagnes de la «Princesse —
Alice» (1891—1897).— Ibid., fasc.
17, 1900. 135 p.
Joubin L. Revision des Sepiolides.—
Mem. Soc. Zool. France, 1902,
T. 15, № 1, p. 80—145.
Joubin L. Contribution a I’etude des
campagnes de la «Princesse —
Alice» (1898—1910). — Res. Camp.
Sci. Monaco, fasc. 54, 1920. 95 p.
Joubin L. Contribution a I’etude des
Cephalopodes de I’Atlantique Nord,
4. — Ibid., fasc. 67, 1924. 113 p.
Joubin L. Notes preliminaires sur les
Cephalopodes des croisieres du
«Dana» (1921—1922), 3, 4. —Ann.
Inst. Oceanogr., N. S., 1931, T. 10,
№ 7, p. 167—211; 1933, T. 13,
№ 1, p. 1—49.
Joubin L. Les Octopodes de la croisie-
re du «Dana» 1921—22. — Dana-
Rept, № 11, 1937. 49 p.
Kubodera T., Okutani T, Description
of a new species of gonatid squid,
Gonatus madokai n. sp., from the
Northwest Pacific........ — Venus,
1977, v. 36, № 3, p. 123—151.
Kubodera T., Okutani T. Gonatus mid-
dendorffi, a new species of gonatid
squid from the northern North Pa-
cific, with notes on morphological
changes with growth and distribu-
tion in immature stages. — Bull.
Nat. Sci. Museum, Tokyo, 1981,
v. A7, № 1, p. 7—26.
Lonnberg E. On the cephalopods col-
lected by the Swedish Expedition
to Tierra del Fucgo 1895—96. —
Svenska Exped. Magellanslandeme,
Bd. 2, № 4, 1898, S. 49—64.
Lu С. C. A new species of squid, Chi-
roteuthis acanthoderma, from the
Southwest Pacific. — Steenstrupia-
na, 1977, v. 4, № 16, p. 179—188.
Lu С. C., Clarke M R. Vertical dis-
tribution of cephalopods at 40° N,
53° N and 60° N at 20° W in the
North Atlantic. — J. mar. biol. As-
soc. U. K., 1975a, v. 55, № 1,
p. 143—163.
Lu C. C.t Clarke M. R. Vertical distri-
bution of cephalopods at 11° N,
20° W in the North Atlantic.—
Ibid., 1975b, v. 55, № 2, p. 369—
389.
Lu С. C., Roper C. F. E. Cephalopods
from the Deepwater Dumpsite 106*
(Western Atlantic): vertical distri-
bution and seasonal abundance.—
Smithson. Contribs Zool., 1979,
№ 288, 36 p.
Massy A. L. The Cephalopoda Dibran-
chiata of the coasts of Ireland. —
Fisheries, Ireland, Sci. Invest, for
1907, 1909, № 1, 39 p.
Massy A. L., Mollusca, 2. — Cephalo-
poda. — Brit. Antarctic («Terra
Nova») Exped. 1910, Nat. Hist.
Rept, Zool., v. 2, № 7, 1916a,
p. 141—176.
Massy A. L. The Cephalopoda of the
Indian Museum. — Rec. Indian
Mus., 1916b, v. 12, № 5, p. 185—
247.
Massy A. L. The Cephalopoda of the
Irish coast. — Proc. R. Irish Acad.,
1928, v. 38B, № 2, p. 25—37.
McSweeny E. S. Description of the
juvenile form of the Antarctic
squid Mesonychoteuihis hamiltont
Robson. — Malacologia, 1970, v. 10,
№ 2, p. 323—332.
McSweeny E. S. Systematics and
morphology of the Antarctic cran-
chiid squid Galiteuthis glacialis:
(Chun). — Antarctic Res. Series,
1978, № 27, 39 p.
Mercer M. C. First record of Pyrgop-
sis lemur from the Northwest At-
lantic.— J. Fish. Res. Bd. Canada,
1966, v. 23, № 8, p. 1257—1258.
♦ Mercer M. C. A synopsis of the re-
cent Cephalopoda of Canada. —
Proc. Symp. Mollusca, v. 1, 1968,
p. 265—276.
Morales E., Guerra A. Teuthoidea:
Oegopsida del NW de Africa. —
Invest. Pesq., 1977, v. 41, № 2,
p. 295—322.
343
Muus В. J. Development and distribu-
tion of a North Atlantic pelagic
squid, family Cranchiidae. — Medd.
Danmarks Fisk.- og Havunders.,
N.S., 1956, v. 1, № 15. 15 p.
* Mu us B. J. Skallus, sotaender, blaek-
sprutter. — Danmarks Fauna, №65,
Kobenhavn, 1959. 239 p.
Muus B. J. Cephalopoda. The Godthaab-
Expedition 1928. — Mcddr. Gron-
land, 1962, v. 81, № 5. 23 p.
* Naef A. Die Cephalopoden. — Fauna
und Flora des Golfes von Ncapel,
Monogr. 35. Bd. 1, 1921—1923,
863 S.; Bd. 2, 1928, 357 S.
Natsukari У. Description and new re-
cords of Loligo sibogae Adam, 1954
from Formosa. — Venus, 1976,
v. 35, № 1, p. 15—23.
Natsukari Y., Okutani T. Identity of
Loligo chinensis Gray, 1849, re-
description of the type specimen
and taxonomic review. — Ibid.,
1975, v. 34, № 3—4, p. 85—91.
•Nishimura S. Notes on the occurrence
and biology of the oceanic squid,
Thysanoteuthis rhombus Troschel,
in Japan. — Pubis Seto Mar. Biol.
Lab., 1966, v. 14, p. 327—349.
Nishimura S. A preliminary list of the
pelagic Cephalopoda from the
Japan Sea.— Ibid., 1968a, v. 16,
№ 1, p. 71—83.
Nishimura S. Glimpse of the biology
of Argonauta argo Linnaeus in the
Japanese waters. — Ibid., 1968b,
v. 16, № 1, p. 61—70.
Odhner N. H. Die Cephalopoden. —
Further Zool. Res. Swedish Antarc-
tic Exped. 1901—1903, v. 1, № 4,
1923. 7 p.
Okiyama M. A new species of Gona-
topsis from the Japan Sea, with
the record of a specimen referable
to Gonatopsis sp. Okutani, 1967. —
Pubis Seto Mar. Biol. Lab., 1969,
v. 17, № 1, p. 19—32.
Okutani T. Studies on early life histo-
ry of decapodan Mollusca, I—IV.
Bull. Tokai Reg. Fish. Res. Lab.,
1965, № 41, p. 23—31; 1966, №45,
p. 61—79; 1968, № 55, p. 9—57;
1969, № 58, p. 83—96.
Okutani T. Preliminary catalogue of
decapodan Mollusca from Japane-
se waters. — Ibid., 1967, № 50,
p. 1—16.
'•* Okutani T. Guide and keys to squid
in Japan. — Ibid., 1973, № 74,
p. 83—111; 1975, № 83, p. 41—
44.
”344
Okutani T. Epipelagic decapod cepha-
lopods collected by micronekton
tows during the Eastropac Expe-
ditions, 1967—1968 (Systematic
part). —Ibid., 1974, № 80, p. 29—
118.
♦ Okutani T. Useful and latent cuttle-
fish and squids of the World. 3rd
ed. Tokyo, 1980. 67 p.
Okutani T., Hasegawa M. A note on
decapod cephalopods (squids) traw-
led from the Southwest Atlantic
during May 1977 through March
1978.— Bull. Tokai Reg. Fish. Res.
Lab., 1979, № 99, p. 9—21.
Okutani T., McGowan J. A. Systema-
tics, distribution, and abundance
of the epiplanktonic squid larvae
of the California Current, April,
1954—March, 1957.— Bull. Scripps
Instn Oceanogr., 1969, v. 14, 90 p.
Okutani T., Satake У. Squids in the
diet of 38 sperm whales caught in
the Pacific waters off Northeastern
Honshu, Japan, February 1977.—
Bull. Tokai Reg. Fish. Res. Lab.,
1978, № 93, p. 13—27.
Okutani T., Satake Y., Ohsumi S>, Ka-
wakami T. Squids eaten by sperm
whales caught off Joban District,
Japan, during January—February,
1976. —Ibid., 1976, №'87, p. 67—
113.
Okutani T. Uemura K. A new species
of the genus Nototodarus from
Japan. — Venus, 1973, v. 32, №2,
p. 39—47.
Palacio F. J. Vosseledone charrua: a
new Patagonian cephalopod (Octo-
podidae) with notes on related ge-
nera.— Bull. Mar. Sci., 1978, v. 28,
№ 2, p. 282—296.
Pearcy W. G.f Voss G. L. A new spe-
cies of gonatid squid from the
Northeast Pacific. — Proc. Biol.
Soc. Wash., 1963, v. 76, p. 105—
112.
Per eyr a T. New records and obser-
vations on the flapjack devilfish,
Opisthoteuthis calif orniana Ber-
ry.— Pacif. Sci., 1965, v. 19, № 4,
p. 427—441.
Рёгег-Gandaras G., Guerra A. Estu-
dio sobre algunas especies del ge-
nero Bathypolypus halladas en las
costas de Galicia. — Invest. Pesq.,
1978, v. 42, № 1, p. 189—211.
* Pfeffer G. Die Cephalopoden der
Plankton-Expedition. — Erg. Plank-
ton-Exped. Humboldt-Stiftung, Bd.
2, F.a., 1912. 815 S. 4-Atlas.
Pickford G. E. Le poulpe americain:
a study of the littoral Octopoda of
the Western Atlantic. — Trans.
Connecticut Acad. Arts Sci. 1945,
v. 36, p. 701—812.
* Pickfora G. E. Vampyroteuthis infer-
nalis Chun, an archaic dibranchiate
cephalopod, I—II. — Dana-Rept,
1946, № 29, 45 p.; 1949, № 32,
132 p.
Pickford G. E. Vampyromorpha. —
Galathea-Rept., v. 1, 1959, p. 243—
253.
Pickford G. E. Octopus dofleini (Wiil-
ker), the giant octopus of the
North Pacific. — Bull. Bingham
Oceanogr. Coll., 1964, v. 19, № 1.
70 p.
Pickford G. E. Cistopus indicus (Or-
bigny): a common Indo-Malayan
species of octopus. — J. mar. biol.
Assoc. India, 1974 (1975), v. 16,
№ 1, p. 43—48.
Pickford G. E., McConnaughey В. H.
The Octopus bimaculatus problem:
a study in sibling species. — Bull.
Bingham Oceanogr. Coll., 1949,
v. 12, № 4. 66 p.
♦ Powell A. W. B. Antarctic and Sub-
antarctic Mollusca. — Rec. Auck-
land Inst. Mus., 1960, v. 5, № 3—
4, p. 117—193.
Rancurel P. Les contenus stomacaux
d’Alepisaurus ferox dans le sud-
ouest Pacifique (Cephalopodes). —
Cah. ORSTOiM, ser. Oceanogr.,
1970, v. 8, № 4, p. 3—87.
Rancurel P. Mastigoteuthis grimaldii
(Joubin, 1895), Chiroteuthidae. pcu
connus de 1’Atlantique tropical. —
Ibid., 1971, v. 9, № 2, p. 125—145.
Rancurel P. Mastigoteuthis inermis,
espece nouvelle de Chiroteuthidae
du Golfe de Guinee. — Bull. Soc.
Zool. France, 1972, v.- 97, № 1,
p. 25—34.
Rancurel P. Mastigoteuthis hjorti Chun,
1913. Description des trois echanti-
llions provenant du Golfe de Gui-
nea. — Cah. ORSTOM, ser. Ocea-
nogr., 1973, v. 11, № 1, p. 27—32.
Rees wl J. The Macrotritopus prob-
lem. — Bull. Brit. Mus.. (Nat.
Hist.), 'Zool., 1954, v. 2, № 4,
p. 67—100.
Rees W. J. Notes on the European
species of Eledone with special
reference to eggs and larvae. —
Ibid.', 1956, v. 3, № 6, p. 283—293.
* Rees W. J., Maul G. E. The Cephalo-
poda of Madeira^ Records and di-
stribution. — Ibid., 1956, v. 3, № 6,
p. 259—281.
Robson G. C. On the Cephalopoda
obtained in South African waters
by Dr. J. F. D. Gilchrist in 1920—
21. — Proc, Zool. Soc. London,
1924, v. 39, № 1, p. 589—686.
Robson G. C. Cephalopodes des mers
d’Indochine. — Note Serv. Ocea-
nogr. PSches Indochine, 1928,
№ 10. 53 p.
* Robson G. C. A monograph of the
recent Cephalopoda based on the
collections in the British Museum
(Natural History). London, pt. 1,
1929, 236 p.; pt. 2, 1932, 359 p.
Robson G. C. On Architeuthis clarkei,
a new species of giant squid, with
observations on the genus. — Proc.
Zool. Soc. London, 1933, № 3,
p. 681—697.
Robson G. C. The Cephalopoda Deca-
poda of the «Arcturus» oceanogra-
phic expedition 1925. — Zoologica,
NY, 1948, v. 33, pt. 3, № 7,
p. 115—132.
* Roeleveld M. A. A review of the Se-
piidae of Southern Africa. — Ann.
S. Afr. Mus., 1972, v. 59, Ns 10,
p. 193—313.
Roeleveld M, A. A revision of Massy’s
checklist of «South African» Cep-
halopoda. — Ibid., 1975, v. 66, № 11,
p. 233—255.
Roeleveld M. A. Cephalopoda from the
tropical Eastern Atlantic Ocean.—
Galathea-Rept., v. 14, 1977, p. 123—
132.
* Roper C. F. E. A study of the genus
Enoploteuthis in the Atlantic Oce-
an with a rcdescription of the type
species, E, leptura (Leach, ,1817).—
Dana-Rept., № 66, 1966. 46 p.
* Roper C. F. E. Systematics and zoo-
geography of the worldwide bathy-
pelagic squid Bathyteuthis. — Bull.
U.S. Nat. Mus., 1969, v. 291,
210 p.
Roper C. F. E., Brundage N. L. Jr. Cir-
rate octopods with associated deep-
sea organisms: new biological da-
ta based on deep benthic photo-
graphs. — Smithson. Contribs. Zool.,
1972, № 121. 46 p.
Roper C. Ft E.t Lu С. C., Mangold K.
A new species of Illex from the
Western Atlantic and distributional
aspects of other Illex species. —
Proc. Biol. Soc. Wash., 1969, v. 82,
p. 295—322.
Roper C. F. E., Sweeney M. J. The pe-
22 К. H. Несис
345
lagic octopod Ocythoe tuberculata
Rafinesque, 1814. — Bull. Amer.
Malacol. Union for 1975, 1976,
p. 21—28.
♦ Roper C. F. E.t Young R. A re-
view of the Valbyteuthidae and an
evaluation of its relationship with
the Chiroteuthidae — Proc. U. S.
Nat. Mus., 1967, v. 123, № 3612.
9 p.
♦ Roper C. F. E.f Young R. E. The Pro-
machoteuthidae, I: A reevaluation
of its systematic position based on
a new material from the Antarctic
and adjacent waters. — Antarctic
Res. Series, 1968, v. 11, p. 203—
214.
Roper C. F. E., Young R. E, First re-
cord of juvenile giant squid, Ar-
chiteuthis.— Proc. Biol. Soc. Wash.,
1972, v. 85, № 16, p. 205—222.
♦ Roper C. F, E., Young R. E. Ver-
tical distribution of pelagic cepha-
lopods.— Smithson. Contribs Zool.,
1975, № 209. 51 p.
* Roper C. F. E., Young R. E., Voss
G. L. An illustrated key to the fa-
milies of the order Teuthoidea. —
Ibid., 1969, № 13. 32 p.
Ruby G., Knudsen J. Cephalopoda
from the Eastern Mediterranean. —
Israel J. Zool., 1972, v. 21, p. 83—
. 97.
Rudolph H. Die Sepiolinen der Ad-
ria.— Zool. Anz., 1932, Bd. 101,
S. 112—120.
Sasaki M. Report on cephalopods col-
lected during 1906 by the U. S. Bu-
reau of Fisheries steamer «Albat-
ross» in the Northwestern Paci-
fic.— Proc. U. S. Nat. Mus., 1921.
v. 57, № 2310, p. 163—203.
♦ Sasaki M. A monograph of the di-
branchiate cephalopods of the Ja-
panese and adjacent waters. —
J. Fac. Agric. Hokkaido Imp. Univ.
1929, suppl. 20. 357 p.
Silas E. G. Cephalopoda of the West
coast of India collected during the
cruises of the R. V. «Varuna», with
a cataloquc of the species known
from the Indian Ocean. — Proc.
Symp. Mollusca, v. 1, 1968, p. 277—
359.
Steenstrup J. The cephalopod papers
of Japetus Steenstrup. Transl. into
Engl. Copenhagen, 1962. 330 p.
* Stenzel H. B. Living Nautilus. —
Treatise on Invertebrate Paleonto-
logy» Р*- K, Mollusca. v. 3. 1964,
p. 59—93.
Stephen A. C, The species of Archite-
uthis inhabiting the North Atlan-
tic. — Proc. R. Soc. Edinburgh,
1961, v. 68B, № 2, p. 147—161.
Taki L On two new eledonid octopods
from the Antarctic Sea.— J. Fac.
Fish. Anim, Husb. Hiroshima
Univ., 1961, v. 3, № 2, p. 297—
316.
Taki I. On four newly known species
of Octopoda from Japan. — Ibid.,
1963, v. 5, № 1, p. 57—93.
Taki I. On eleven new species of the
Cephalopoda from Japan, including
two new genera of Octopodinae.—
Ibid., 1964, v. 5, № 2, p. 277—343.
Thiele J. Die Cephalopoden der Deut-
schen Siidpolar-Expcdition 1901—
1903. — Dtsch. Siidpolar-Exped.
1901—1903, Bd. 16, H. 4, 1921,'
S. 431—466.
* Thiele J. Handbuch dcr systemati-
schen Wcichtierkunde. Bd. 3, Jena,
1934, S. 779—1022 (Cephalopoda:
S. 948—995).
* Thomas R. F. Systematics, distribu-
tion, and biology of cephalopods
of the genus Tremoctopus.— Bull.
Mar. Sci., 1977, v. 27, № 3,
p. 3o3—392.
* Thore S. Investigations on the «Da-
na» Octopoda, 1. Bolitaenidae,
Amphitretidae, Vitreledonellidae,
and Alloposidae. — Dana-Rept.,
№ 33, 1949, 85 p.
Thore S. Cephalopoda. — Repts Lunds
Univ. Chile Exped. 1948—49, №33,
Lunds Univ. Aarsskr., N. F. Avd.
2, 1959, Bd. 55, № 1. 19 p.
Toll R. B. Benthoctopus oregonae, a
new species of octopod from the
Southern Caribbean, with a rede-
scription of Benthoctopus januarii
(Hoyle, 1885). — Bull. mar. Sci.,
1981, v. 31, № 1, p. 83—95.
* Verriil A. E. Report on the cepha-
lopods of the northeastern coast of
America. — Rep. U. S. Comm. Fish.,
1882, v. 7, p. 211-455.
Verrill A. E. Supplementary report on
the «Blake» cephalopods. — Bull.
Mus. Comp. Zool. Harvard, 1883,.
v. 11, № 5, p. 105—115.
Voss G. L. A new family, genus and
species of myopsid squid from the
Florida Keys. — Bull. Mar. Sci,.
Gulf Caribb., 1953, v. 2, № 4,
p. 602—609.
Voss G. L. The Cephalopoda obtained
by the Harvard—Havana Expediti-
on off the coast of Cuba in 1938—
346
39. — Ibid., 1955, v. 5, № 2, p. 81—
115.
♦ Voss G, L. A review of the cepha-
lopods of the Gulf of Mexico.—
Ibid., 1956, v. 6, № 2, p. 85—178.
Voss G. L. Observations on Ornitho-
ieuthis antiliar um Adam, 1957, an
ommastrephid squid from the West
Indies. — Ibid., 1957, v. 7, № 4,
p. 370—378.
Voss G. L. The cephalopods collected
by the R/V «Albatross» during the
West Indian cruise of 1954. — Ibid.,
1958, v. 8, № 4, p. 369—389.
Voss G. L. Bermudan cephalopods. —
Fieldiana, Zool., 1960, v. 39, № 40,
p. 419—446.
♦ Voss G. L. A monograph of the Ce-
phalopoda of the North Atlantic. I.
The family Lycotcuthidae. — Bull,
mar. Sci. Gulf Caribb., 1962a. v. 12,
Ns 2. p. 264—305.
Voss G. L. South African cephalo-
pods.— Trans. R. Soc. S. Afr.,
1962b, v. 36, № 4, p. 245—272.
* Voss G. L. Cephalopods of the Phi-
lippine Islands. — Bull. U. S. Nat.
Mus., 1963, № 234. 180 p.
Voss G. L. Some bathypelagic cephalo-
pods from South African wa-
ters. — Ann. S. Afr. Mus., 1967,
v. 50, № 5, p. 61—88.
Voss G. L. Octopods from the R/V
«Pillsbury» Southwestern Caribbe-
an cruise, 1966, with a description
of a new species, Octopus zona-
tus. — Bull. Mar. Sci., 1968, v. 18,
№ 3, p. 645—659.
Voss G. L. Cephalopods collected by
the RA’ «John Elliott Pillsbury» in
the Gulf of Panama in 1967.—
Ibid., 1971, v. 21, № 1, p. 1—34.
Voss G. L. Loligo surinamensis, a new
species of loliginid squid from
Northeastern South America. —
Zool. Mededel., 1974, v. 48, Ka 6,
p. 43—o3.
Voss G. L. Euaxoctopus pillsburyae,
new species from the Southern Ca-
ribbean and Surinam. — Bull. Mar.
Sci., 1975, v. 25, № 3, p. 346—352.
Voss G. L. Two new species of octo-
pods of the genus Graneledone
from the Southern Ocean. — Proc.
Biol Soc. Wash., 1976, v. 88, № 42,
p. 447—458.
Voss G. L., Opresko L., Thomas R.
The potentially commercial species
of octopus and squid of Florida,
the Gulf of Mexico and the Carib-
bean Sea. — Univ. Miami Sea
Grant Field Guide Series, № 2,
1973. 33 p.
♦ Voss G. L., Williamson G, R. Cepha-
lopods of Hong Kong. Hong Kong,
1972, 138 p.
♦ Voss N. A. A monograph of the Cep-
halopoda of the North Atlantic. The
family Histioteuthidae. — Bull.
Mar.’ Sci., 1969, v. 19, № 4,
p. 713—867.
Voss N. A. Studies on the cephalopod
family Cranchiidae. A redescripti-
on of Egea inermis Joubin, 1933,—
Ibid., 1974, v. 24, № 4, p. 939—956.
* Voss V. A. A generic revision of the
family Cranchiidae. — Ibid., 1980,
v. 30, № 2, p. 365—412.
Wirz K. Bathypoly pus sponsalis (P. et
H. Fischer), espece commune dans
la partie ouest de la Mediterra-
nee.—Vie Milieu, 1955, T. 6, №1,
p. 129—147.
* Wirz K. Cephalopodcs. — Faune mari-
ne des Pyrenees—Orientates, fasc.
1. Paris, 1958. 59 p.
* Wormuth J. H. The biogeography and
numerical taxonomy of the oegop-
sid squid family Ommastrephidae
in the Pacific Ocean. — Bull.
Scripps Instn Oceanogr., 1976,
v. 2§. 90 p.
Yamamoto K., Okutani T. Studte* On
early life history of decapodan
Mollusca, V. — Bull. Tokai Reg.
Fish. Res. Labor., 1975, № 83,
p. 45—96.
Young R. E. A note on three speci-
mens of the squid Lampadioteuthis
megaleia Berry, 1916 from the At-
lantic Ocean, with the description
of the male. — Bull. Mar. Sci. Gulf
Caribb., 1964, v. 14, p. 444—452.
* Young R. E. The systematics and
areal distribution of pelagic cepha-
lopods from the seas off Southern
California. — Smithson. Contribs
Zool., 1972, № 97. 159 p.
Young R. E., Roper C. F. E. The Ba-
toteuthidae, a new family of squid
from Antarctic waters. — Antarctic
Res. Series, 1968, v. 11, p. 185—202.
* Young R. E., Roper C. F. E. A mo-
nograph of the Cephalopoda of the
North Atlantic. The family Cyclo-
teuthidac. — Smithson. Contribs
Zool., 1969a, № 5. 24 p.
♦ Young R. E., Roper C. F. E. A mo-
nograph of the Cephalopoda of the
North Atlantic. The family Joubinir
teuthidae. — Ibid.,. 1969b, № l*x
10 p.
22*
Указатель
латинских
названий1
Abralia 15. 32, 157, 162*, 165, 167
Abralia subg., Abralia 157, 163*
Abraliopsis 27, 30, 153*, 168*, 171. 172
Abraliopsis subg., Abraliopsis 158, 168*
abruptus, Benthoctopus 317*, 319
abulati, Lolliguncula 136*
abyssicola, Bathy tenth is 29, 197, 198*
abyssicola, Megalocranchia 269*, 271
Acantheleaone 295
acanthoderma, Chiroteuthis 237
Acanthosepion 78
Acrololigo 131
Acruroteuthis 131
aculeata, Sepia (Sepia) 91*, 93, 94*
acuminata, Sepia (Sepia) 84*
adami, Pholidoteulhis 231, 232*
adami. Sepia (Sepia) 85, 87*
Bddolux, Pyroteuthis 173*, 175
adelieana, Pareledone 301*
adjacens, Acarthosepion elliptlcum 91
aequatorialis, Moroteuthis 185, 186*
affinis, Abraliopsis (Micrabralia) 169*, 171
affinis, Loliolus 132
affinis, Sepiola 121, 122*
africana, Alloteuthis 148*, 149
africanus, Octopus fontanianus var. 307
agassizi, Mastigoteuthis 246, 249*
agassizi, Opisthoteuthis 281. '284*, 285
akimushkini, Cycloteuthis. 233*, 235
albatrossae, Euprymrfll 114*, 117
albatrossi, Grimpoteuthis 281*, 283
albida, Bentheledone 298*
alcocki, Teretoctopus 298*
alecto, Octopus 313*
alessandrinii, Ancistrocheirus 29, 31, 173*,
175
Alloposldae 22, 275, 323*, 325
Alloposina 326
Attoposus 275, 325, 326*
Allorossia 108
Alloteuthis 19, 20, 33, 130, 131, 146*, 149
Alluroteuthidae 214
Alluroteuthis 16, 18, 150, 213, 214*
alpha, Fusocranchia 262
alumnus, Nautilus 72
Amphioctopus 290
Amphitretidae 11. 16, 17, 21, 275, 287*, 289
Amphitrctus 11, 21, 40, 61, 63, 275, 287*, 289
Amplisepia 78
anaspis, Enoploteuthis 159*, 181
Ancistrocheirinae 27, 62, 158*
Ancistrocheirus 19, 20, 158, 173*, 175
Ancistroteuthis 32, 181, 182*
andamanica, Abralia (Asteroteuthis) 167,
168*
andamanica, Abralia (Asteroteuthis) ahda*
manica f. 168*
andreana, Sepia (Sepia) 97*, 99
andreanoides, Sepia 90
Andreasepia. 78
T‘ Синодимы напечатаны курсивом, страни-
цы, на которых помещен рисунок, набраны
полужирным шрифтом, описания отмече-
ны звездочкой *.
angolensis, Todarodes 228*
angulata, Sepia (Sepia) 87*
annae, Danoctopus 294
Anomalocranchia 260
anonychus, Berryteuthis 187, 189, 190*
antarctica, Alluroteuthis 213, 214*, 215
aiitarctica, Graneledone 298, 321, 322*
antarctica, Pareledone 302
antarctica, Teuthowenia 268
antarcticus, Gonatus (Gonatus) 194*
antillarum, Ornithoteuthis 223, 229*
antillensis, Austrorossia 114*, 115
apama, Sepia (Sepia) 10, 101, 102*, 106*
apollyon. Octopus dofleini 309*, 311, 313*
appelloefi, Sepia (Sepia) 95*, 99
arabica, Loligo (Doryteuthis) 146*, 147
arabica, Sepia (Sepia) 93*
arakawai, Octopus 26
araneoides, Octopus 311, 312*
Architeuthidae 14, 152, 213*
Architeuthis 19, 47, 213, 214*
Architeuthus 214
arcticus. Bathypolypus 296, 315, 316*
arcticus, Bathypolypus arcticus f. 316*
Arctosepia 78
arcturi, Stigmatoteuthis 203
areolatus, Octopus 308
argentinus, Illex illecebrosus 227*
argo, Argonauta 327, 329*, 330*
Argonauta 15, 36, 63, 68, 276, 327*, 329
Argonautella 327
Argonautidae 22, 276, 327*, 329
Argonautina 327
Argonautoidea 16, 21—23 31, 43, 48, 50,
56, 67, 71, 322*. 325
armata, Abralia (Abralia) 157, 163*, 185
armata, Galiteuthis 257, 268*
Ascarosepion 78
Ascocranchia 19, 261
Ascot euthis 260
asper, Paroclopus 309
aspera, Galiteuthis 268
aspera, Loligo (Loligo) 145*
Asperoteuthis 23, 237», 239
Asteroteuthis subg., Abralia 157, 164*
AsthenOleuthion 156
astrolineata, Abralia (Stenabralla) 163*
astrosticta, Abralia (Stenabralla) 163*, 165
atlantica, Abraliopsis (Micrabralia) 171, 172*
atlaniica, Bigelowia 242
atlantica, Histioteuthis 201, 202*
atlantica, Leachia (Pyrgopsis) 259, 262*
atlantica, Mastigoteuthis 249, 251*
aurantiaca, Sepiola 121, 122*
atlantis, Heteroteuthis 123
Atlantoctopus 296
aurantiaca; Sepiola 121, 122*
auritum, Sepiaaarlum 126*
aurorae, Pareledone 299
atistralis, Austrorossia 108, 114, 115
australis, Loligo 144
australis, Megalocranchia megalops 263
australis, Octopus 48, 305, 307
australis, Sepia (Sepia) 26, 79, 85, 86*, 87,
J03
australis, Sepioteuthis 136
348
austrinum, Sepiadarium 126*
Austrorossia 108, 114*, 115
bacidifera, Bathyteuthis 197, 198*
bairdii, Bathypolypus 316
bandensis. Sepia (Sepia) 87*
banksi, Onychoteuthis 11, 57, 178, 181, 182,
183*
banksi, Onychoteuthis banksi 39, 181, 183*
bartraml, Ommastrephes 14, 29, 225, 226*
bartschi, Uroteuthis 131*, 149
Bathothauma 19, 33, 57, 255, 261*, 271
Bathypolypodinac 16, 42, 296*, 297
Bathypolypus 23, 48, 296, 314*, 315
Bathyteuthidae 11, 14, 18, 26, 61, 152, 196*
197
Bathyteuthis 15, 18, 27, 57, 61, 62, 197, 198*
Batoteuthidae 151, 233*, 235
Batoteuthis 19, 233*, 235
baxtcri, Sepia (Sepia) 101, 106*
beani, Brachioteuthis 215, 217, 218*
beebel, Helicocranchia 263
behni, Brachioteuthis 217, 218*
beka, Loligo (Loligo) 143, 145*
belauensis, Nautilus (pompilius) 72*
belone, Bclonella 257, 287, 268*
Belonella 16, 19, 257, 264*, 267, 268
Bentheledone 23, 297, 298*
Benthoctopus 3, 296, 316*, 317, 319
Benthoteuthis 198
beringiana, Crystalloteuthis 268
bcrmudensis, Octopus 303
Berrya 294
berryi, Bathyteuthis 197, 198*
berryi, Benthoctopus 317*
berryi, Euprymna 114*, 117
berryi» Gonatus (Gonatus) 190, 191*, 193
berryi^ Histioteuthis corona 209, 210*
Berryidiunt 128
Berrytcuthis 49, 187, 189, 190*
bertheloti. Sepia (Sepia) 81*, 83
Bigelowia 237, 242
bilineata, Sepiotcuthis 135, 136*
bilucifer, Uranoteuthis 261
bimaculatus, Octopus 312*
bimaculoides, Octopus 311, 312*
bipapillata, Austrorossia 114*, 115
birostrata, Sepiola 121, 123*
biserialis, Idiosepius 128*, 129
Blandosepia 78
bleekeri, Loligo (Doryteuthis) 141 ♦, 143
boettgeri, Argonauta 329*, 330*
Bolitaena 287*, 289
Bolitaenella 286
Bolitaenidae 11, 16, 17, 26, 31, 42, 51, 63, 275,
286», 289
Bolitacnoidea 23, 71, 286*
bonelliana, Histioteuthis 199
bonnellii, Histioteuthis 29, 199*, 201
bonncllii. Histioteuthis bonnellii 199*, 201
bonplandi, Grimalditcuthis 29, 253, 254*
borealijaponicus, Onychoteuthis banksi 13,
29, 39, 181. 183*
borealis, Belonella 267, 268*
borealis, Belonella pacifica 268
borealis, Gonatopsis (Boreoteuthis) 189,
190, 195*
borealis, Tankaia 239, 240*
boreopaciflca, Granelcdone 321, 322*
Boreoteuthis subg., Gonatopsis 190, 195*
boschmai, Pholidoteuthis 231, 232*
bowman!, Brachioteuthis 217, 218*
Brachioteuthidae 18, 26, 61, 151, 215*. 217
Brachioteuthis 32, 215*, 217,
brachyura, Rossia 110*. 113
braggi, Sepia (Sepia) 101, 104*
brasiiiensis, Loligo (Loligo) 140*
brevimana, Sepia (Sepia) 91*, 93
brevipes, Octopus 286
brevipinnis, Loligo (Doryteuthis?) 146*
brevis, Lolliguncula 130, 135, 136*
brevis, Thaurnelcdone 297, 298*
briareus, Octopus 303*, 305
brocki, Octopus 308
brucei, Octopus 306
bruuni, Histioteuthis 201, 202*
budo, Loligo (Loligo) 143, 144*
bullisi, Rossia 111*, ИЗ
burnupi, Sepia (Sepia) 85, 86*
burryi, Octopus 302*, 305
californiana, Opisthoteuthis 284*, 285
californicus, Octopus 311, 312*, 313*
californicnsis, Gonatus (Gonatus) 193, 194*
Callistoctopus 290
Calliteuthis 199
calyx, Chiroteuthis 13, 29, 243*, 245
caparti, Eledone 299*, 301
capensis, Chiroteuthis 242*, 245
capensis, Inioteuthis 117, 118*
capensis, Sepia 86
capensis, Velodona togata var. 296*
carinata, Sepia (Sepia) 95*. 97
carlgreni,- Pareledone 295, 301*«
caroli, Neorossla 108*
carriboea, Onykia 181, 182*
Carynoteuthis 260
caudani, Grimpoteuthis 281
celctaria, Histioteuthis 209, 210*
celetaria, Histioteuthis celetaria 209, 210*
cctasina, Histioteuthis corona 209, 210*
challcngeri, Graneledonc 322*
charcoti, Pareledone 296; 299*, 301
charrua, Vosselcdonc 296*, 297
Charybaiteuthis 159
Chaunotcuthis 14, 18, 49, 67, 177. 178, 181,
183*
Cheiloteulhis 137
chinensls, Loligo (Loligo) 143, 144*
Chiridioteuthis *237
chirocies, Loliolopsis 131
Chiropsis 237*, 239
Chiroteuthidae 11, 17, 19, 21-23, 26, 47, 61,
62, 153, 236* 237, 239
Chiroteuthidae n. g. A 240*, 241*
< В 237*, 239. 241*
Chiroteuthis 15, 16, 27, 31, 237, 241*. 242*.
245
Chiroteutholdes 239, 240*, 241*
Chiroteuthopsis 246
Chirothauma 237
chirotrema, Sepia (Sepia) 101, 102, 103*
Chondrosepia 130
Chunella 286
chuni, Abraliopsis (Micrabralia) 171, 172*
chuni, Enoploteuthis 161, 162*
chuni, Stigrnatoteuthis 203
Chunioteuthis 21, 83, 277, 278*, 279
Chunoteulhis 198
Cirrata 5, 19, 33, 43, 48, 49, 51 61, 70, 273,
275*
Cirrobrachium 230*, 231
cirrosa, Eledone 42, 48, 295, 299*, 301
Cirroteuthidae 15, 17, 19, 21, 273, 275, 276*,
279
Cirroteuthis 10 17, 33, 60, 63, 64, 273,
275*, 279, 280
Cirroteuthopsis subg., Cirroteuthis 280
Cirrothauma 6, 17, 33, 53 60, 63, 277*, 279
Cistopus 291*, 293
clara, Froekenia 277*
clouei, Brachioteuthis 218
coindeti, Illex illcccbrosus 221, 227*
Coleoidca, 5, 35, 68, 72*
confusa, Sepia (Sepia) 85, 86*, 87, 90
conispadiceus, Octopus 308*, 311
cookiana, Histioteuthis 202
cordiformis, Mastigoteuthis 23, 249, 251*
cornuta, Argonauta 329*, 330
corona, Histioteuthis 209, 210*
corona, Histioteuthis corona 209, 210*
corona, Teuthowenia (Ascoleuihis) 269
corpuscula, Histioteuthis bonnellii, 199* 201
Corynomtna 260
cottesloensis, Decorisepia 103
cotton!, Sepia (Sepia) 104*
349
Cranchia 11, 17, 20, 23, 33, 255, 256*, 259
Cranchiidae 10, 11, 15-21, 23. 26, 30, 31,
34, 51, 62, 64, 67, 150, 153, 255*, 256, 259
Cranchiinae 19, 256*, 259
Crunienasepia 78
Crysialloteuthis 257
Ctenoglossa 23, 71, 286
Ctenoptcrygidae 11, 26, 61, 150, 195*,
197
Ctenopteryx 15, 19, 62, 196*, 197
Cucioieutnis 176
cultrata, Sepia (Sepia) 101, 103*
cyanea, Octopus 42
cyanea, Sepiella 79, 106*
Cycloteuthidae 11, 18, 21, 26, 61, 62, 153,
233*» 235
Cycloteuthis 233*, 235
cyclura, Leachia (Leachia) 256, 262*
Cycria 226
cymoctypus, Phasmatopsis 269
dagamensis, Heteroteuthis (Stephanoteut-
his) 126*
danae, Cirrobrachium 230*, 231
danae, Drechsclia 256*. 259
danae, Echinoteuthis 246, 251, 252*
danae, Octopoteuthis 177*, 179
danae, Taningia 14 .176*, 179
danae, Valbyteuthis 239, 240, 241*
danae, Valdemar la 254
Danateuthis 272
dannevigi, Sepia (Sepia) 97
Danoctopus 293, 294*
Decapoda 5
Decorisepia 78
defilippi, Octopus 67, 303*, 305
deletron, Octopoteuthis 29, 177*, 179
dentata, Mastigoteuthis 246*, 249
depressa, Opisthoteuthis 284*, 285
Desmoteuthcs 260
diadema, Lycoteuthis 155, 156*
diaphana, Doratopsis 242
diaphana, Japetella 286*, 287, 289
diaphana, Japetella diaphana 289
Diaphanoteuthis 237
digueti, Octopus 314*
Dinoteuthis 214
diomedeae, Loliolopsis 131*, 133
Diphtherosepion 80
Dlscoteuthis 29, 235, 236*
discus, Discoteuthis 29, 235, 236*
dislocata, Leachia 259, 262
dlspar, Heterotcuthis (Heteroteuthis) 110,
123*. 125
dofleini, Histioteuthis 203
dofleini, Octopus 9, 10, 13, 29, 39, 45, 47—
49, 272, 291, 309*, 311, 313*
dofleini, Octopus dofleini 309*. 311
dollfusi. Sepia (Sepia} 94*
Doratopsis 237
Doratosepion 78
Dorsopsis 11, 61, 287*, 289
Doryteuthis subg., Loligo 29, 131*, 141, 147
Dosldicus 225*
Drechselia 23, 256*, 259
dubia, Enigmoteuthis 158*, 101
dubia, Sepia (Hemisepius) 10, 79, 80*, 84
Dubioteuthis 214
dussumieri, Tetronychoteuthis 231, 232*
duvauceli, Loligo (Loligo) 141*, 143, 145*
dux, Architcuthis 10, 214*
Dyctidiopsis 256
ebersbachi, Chunioteuthis 278*, 279
eblanae, Todaropsis 219*, 221
Echinoteuthis 14, 23, 246. 251, 252*
eclogaria, Ponderisepia 87
edulis, Loligo (Loligo) 141*. 143, 144
cdulis, Loligo (Loligo) edulis f. 144*
Egea 19, 20, 260*, 271
Eledone 43, 48, 295*, 298*, 301
Eledonella 11, 53, 286*, 289
Eledonenta 290
Eledoninae 16, 295*, 297
elegans, Sepia (Sepia) 45. 81*, 83
ellipsoptera, Leachia (Leachia) 259, 262*
elllpsura, Lolliguncula 135, 136*
elliptica, Sepia (Sepia) 91*, 93, 97, 103*
ellipticum, Acanthosepion 91
elobyana, Sepia (Sepia) 81*, 83
elongate, Histioteuthis 202*, 205
elongata. Sepia (Sepia) 91*, 93
elongata, Teuthowenia 260
eltaninae, Histioteuthis 203*, 207
emtnakina, Loligo 140
enigmatica, Austrorossia 114
Enigmoteuthis 14, 158*, 161
Enoploion 157
Enoploteuthidae 14, 16, 18, 26, 27, 30, 31, 49,
62, 150, 156*, 161
Enoploteuthinae 27, 43, 46. 62, 150 157*
Enoploteuthis 19. 158, 159*, 161, 162
Enoptroteuthis 233, 235
Enteroctopus 290
Entomopsis 215
Eogonatus subg., Gonatus 187, 190*, 193
Epistrophea 295
Epistychina 108
cqualis, Semirossia 111*, 115
ergastlcus, Benthoctopus 48, 317*, 319, 320*
erostrata. Sepia (Sepia) 95*, 99
eschscholtzi, Leachia 262
esculenta, Sepia (Sepia) 87, 94*. 97
ether idgei, Loligo 144
Euaxoctopus 67, 291*, 293, 294*
Eucleoteuthis 223, 225, 226*
Euprymna 26. 107, 114*, 117
eureka, Benthoctopus 317*
eurycephala, SasaKinella 294*
Eusepia 78
eustictum, Enoploion 168
Euzygaena 257
exophthalmica, «Doratopsis (Planktoteut-
his)» 241, 242
exophthalmica, Valbyteuthis 239, 241*, 242*
exsignata, Sepia 86
extensa, Opisthoteuthis 284*, 285
fabricii, Gonatus (Gonatus) 50, 187, 193,
194*
faeroensis. Bathypolypus 316
falco, Abraliopsis (Micrabralia) 29,
172*
famclica, Asperoteuthis 237*. 239, 240*
fatnelica, Chlroteuthis 237
fangsiao, Octopus 308
fasciata, Octopus 291
faurei, Sepia (Sepia) 84*, 85
felis* Abraliopsis (Abraliopsis) 169*, 171
Fidenas 107
filiferum, Cirrobrachium 230
filippovae, Todarodes 228
filliouxi, Sepia 80
Fiscisepia 78
fisher i, Helicocranchia 269
fitchi, Octopus 39, 314*
flammca, Mastigoteuthis 249*
fontaniana, Jouoinia 307
fontaniana, Robsonella 307
fontanianus. Octopus 48, 302, 307*
forbesi, Loligo (Loligo) 139, 140*
formosana, Loligo 144
formosana, Sepia 87
Frockenia 33, 277*
fujitai, Octopus 309*, 311
fuscus, Benthoctopus 317*, 319
Fusocranchia 256
gahi, Loligo (Loligo) 140*, 146*
galatheae, Loligo 145
galaxias, Enoploteuthis 161, 162*
galei. Sepia 90
Galitcuthis 16, 19. 23, 257, 267, 268*
gardineri, Liocranchia 262
gelatus, Tremoctopus 17, 325, 327*
gemellus, Glyptosepia 103
350
gemmata, Pterygioteuthis 29, 173*, 175
genista, Solitosepia 90
giardi, Pterygiotcuthis 10, 159, 175, 176*
giardi, Pterygioteuthis giardi 176*
•gibba, Sepia (Sepia) 94*
gigas, Dosidicus 45, 225*
gilbertianus, Octopus 309
gilchristi, Abraliopsis (Micrabralia') 169*,
171
gilchristi, Chuniotcuthis 278*
glaber, Polypus 317
glacialis, Crystalloteuthis 268
glacialis, Galiteuthis 267, 268*
glacialis, Grimpoteuthis 280*, 283
glacialis, Psychroteuthis 213*
glaucopis, Rossia 111*, 113
glauerti, Sepia (Sepia) 97
glaukopis, Mastigoteuthis 249, 251*
Glyptosepia 78
Gonatidae 14—16, 18, 26, 51, 150, 187*
189. 191
Gonatopsis 14, 49, 67, 187, 189, 195*
Gonatopsis subg., Gonatopsis 16, 22, 190,
195*
Gonatus 14, 16, 22, 32, 187, 190* 191. 193,
194
Gonatus subg., Gonatus 187, 190*
gctoi, Loligo 145
gouldi, Nototodarus sloani 223, 228*
gracilipes, Idioctopus 275, 287*, 289
gracilis, Brachioteuthis 218
gracilis, Sepiadarium 126*, 127
gracilis, Tremoctopus violaceus 325, 327*
grandipes, Loligo (Loligo) edulis f. 144
Graneledone 23, 298, 320*, 321, 322
grimaldii, Grimpoteuthis 281*, 283
grimaldii, Lepidoteuthis 231, 232*
grimaldii, Mastigoteuthis 246*, 249, 252
Grimalditeuthidae 19, 20, 26, 62, 153 253,
254*
Grimalditeuthis 14, 15, 19, 21. 27, 253, 254*
grimpei, Abralia (Asteroteuthis) 164*, 167
grlmpei, Bathypolypus 316
Grimpella 297, 298*
Grimpoteuthis 33, 277 278, 280*, 281, 283
gunteri, Thaumeledone 297, 298*
guttata, Leachia 262
Haliphron 326
hamiltoni, Mesonychoteuthis 16, 257*, 267
Hansenoteuthis 272
Hapalochlaena 291*, 293
Hapaloctopus 294
Haptochlaena 290
hardwickei, Loliolus 132*, 133
harmeri, Sepia 87
harrisoni, Pareledone 301*, 302
hastula, Chiroteuthoidcs 239, 240*
hawaiiensis, Heteroteuthis (Stephanoteut-
his) 31, 59 110, 125, 126*
hawaiiensis, Nototodarus sloani 223. 228*
heathi, Japetella diaphana 286*, 289
hebereri, Idiosepius pygmaeus 128
hedleyi, Sepia 103
Helicocranchia 19, 261, 263*. 265
Hemisepion 80
Hemisepius subg., Sepia 79, 80*. 84
hendryae, Solitosepia 103
Hensenioteuthis 261
Heptapus 326
hercules, Sepia 87
Heteroglossa 71, 287
heteropsis, Histioteuthls 202*, 205
Heteroteuthinac 10, 20, 26, 31, 109* 125
Heteroteuthis 43, 62, 110, 123*, 125
Heteroteuthis subg., Heteroteuthis 123*
hians, Argonauta 329*, 330*
hieronis. Sepia (Sepia) 81*. 85
hierredda. Sepia (Sepia) officinalis 10, 80*
hippocrcpium. Grimpoteuthis 281*
Hlsfiiopsis 199
Histloteuthidae 10, 14, , 18, 20, 26, 27, 30.
31, 62. 152, 198*, 201
Histioteuthis 11. 14, 15, 18, 27, 30, 32, 47.
57, 60, 62, 67, 151, 199*, 201, 205, 207, 209
Histiothauma 199
hjorti, Mastigoteuthis (Mastigopsis) 251,
252*
hokkaidensls, Benthoctopus 317*, 319
hongkongensis, Octopus 309
Hoy lea 295
hoylei, Abraliopsis (Abraliopsis) 169*, 171
hoylel, Abraliopsis (Abraliopsis) hoylei
169*, 171
hoylci, «Cirroteuthis» 280*
hoylei, Danoctopus 293, 294*
hoylei, Histioteuthis 14, 59, 199, 203*, 207
hoylei, Meleagroteuthis 202
hoylei, Pterygioteuthis giardi 176*
hoylei. Sepia 94
hubbsorum Octopus 314*
hulliana, Crutnenasepia 87
hummelincki, Octopus 302*, 305
hyadesi, Martialia 16, 223*
hyadesi, Octopus 306
Hyaloteuthis 223, 225, 227*
hyatti, Rossia 111
Hymenoteuthis 272
hyperborea, Desmoteuthis 263
Idioctopodidae 16, 21, 275. 287*. 298
Idioctopus 21, 23, 42, 275, 287*, 289
Idiosepiidae 10» 18 20, 21, 24, 31, 43, 77,
127*, 129
Idiosepion 128
Idiosepius 10, 24, 128*, 129
Idioteuthis 246
illecebrosus, Illex 219, 221. 227*
lllecebrosus, Illex illecebrosus 221, 227*
Illex 15, 48, 49, 218, 219, 221, 227*
Illicinae 51, 219*, 221
inipennis, Anomalocranchia 263
imperator, Chiroteuthis 243
incerta, Sepia (Sepia) 85, 87*
Incirrata 5. 6. 39, 42, 53, 64, 71, 274, 285*
indica, Loligo 145
indicus, Cistopus 291*, 293
indicus, Teretoctopus 298*
Indoteuthis 198
inermis, Egea 260*, 271
inermls, Histioteuthis corona 210*
inermis, Mastigoteuthis 249, 252*
inermls. Sepiefla 105, 106*
hifernalis, Vampyroteuthis 5, 10, 29, 272*,
ingens, Moroteuthis 185, 186*
Inioteuthis 108, 114*. 117
irdoteuthis, Inioteuthis 108, 118
insignls, Nototodarus 229
insignis. Sepia (Sepia) 81*, 85
intermedia, Liocranchia 262
intermedia, Sepiola 121, 122*
investigatoris, Loliolus 132*. 133
Iridoteuthis 10, 20, 109*, 125
iris, Iridoteuthis 109*, 125
Irvingi, Sepia (Sepia) 101, 102*
iselini, Mastigoteuthis 251
ivanovi, Sepia (Sepia) 87
jaenschi, Decorisepia 103
januarii, Benthoctopus 317, 319, 320*
Japetella 57. 286*, 289
Japonica. Abralia (Asteroteuthis) 167, 168*
japonica, Calliteuthis 210
japonica, Inioteuthis 108, 117, 118*
japonica, Loligo (Loligo) 143, 144*
japonica, Onykia 186
japonica, Opisthoteuthis 284*
japonica, Sepiella 105, 106*
japonicus, Gonatopsis (Gonatopsis) 189,
195*
joubini, Chiroteuthis 243*, 245
joubini, Helicocranchia 263*, 265
joubini, Hensenioteuthis 263
Joubini, Octopus 10, 39, 305, 306*
351
Joubini, Sepia (Sepia) 79, 86*, 87
Joubinia 290
Joublniteuthidae 19, 22, 62, 251, 252*
Joubiniteuthis 14, 15, 67, 2Б1, 254*
kamtschaticus, Gonatus (Gonatus) 194*
kensaki Loligo (Loligo) edulis f. 143, 144*
keralensis, Danoctopus 294*
kiensis, Sepia (Sepia) 90*
knipovitchi, Moroteuthis 185, 186*
kobiensis, Loligo (Loligo) 143, 145*
kobiensis, Sepia (Sepia) 89, 90*, 97*
kochii, Sepiadarlum 126*, 127
Kondakovia 32, 183*, 185
laccrtosa, Chiroteuthis veranyl 242*, 243,
245
laclniosa, Discoteuthis 29, 235, 236*
Laetmoteuthis 290
Lampadiotcuthinae 155, 156*
Lampadiotcuthis 155, 156*
lana, Solitosepia 90
latimanus, Sepia (Sepia) 10, 87*, 89, 94*,
102*
latipinna, Mastigoteuthis 246, 252*
Leachia 14, 19, 23, 256, 259, 262* 263
Leachia subg., Leachia 256, 259, 262*
lefebrei, Sepia 94
leioderma, Octopus 308*. 311, 313*
lemur, Pyrgopsis 262, 263
lentus, Bathijpolypu$ 316
Lepidoteuthidae 14, 18, 23, 150, 231, 232*
Lepidoteuthis 14, 19. 23, 67, 231, 232*
Leptodontoteuthis 156
Leptoteuthis 237
leptura, Enoploteuthis 158, 159*, 161, 162
leptura, Enoploteuthis leptura 159*
lessonlana, Sepioteuthis 135, 136*
Lestoteuthis 187
lesueurl, Ancistrocheirus 158, 173*
Leucocranchia 261
leucoptera, Stoloteuthls 109*, 125
levimana, Valbyteuthis 240, 241*, 242*
levis, Benthoctopus 319, 320*
lichtcnstelni, Ancistroteuthls 181, 182*, /85
ligulata, Sepiola 121, 122*
Liguriella 261, 264*, 265
Uliana, Solitosepia 102
limata, Arctosepia 104
lineata, Abraliopsis (Micrabralia) 158, 172*
lineolata, Sepioloidea 126, 127*
Liocranchia 11, 17, 20, 23, 256, 259, 261*
lippula, «Doralopsis (Planktoteuthis)» 241,
lobensis, Octopus 305, 307*
loennbergi, Moroteuthis 185, 186*
Lolidona 199
Loliginidae II, 16, 18, 51, 130*, 133
loliginiformis, Sepioteuthis 137*
Loligo 26, 29, 32, 41, 61, 130, 131, 137* 139,
143, 144, 147
Loligo subg., Loligo 131*
Loligosepiidae 33
Lolimnites 226
Loliolopsis 131*, 133
Loliolus 131. 132*, 133
Lolliguncula 130, 132*, 135
longlmana, Kondakovia 183*, 185
longimanus, Onychoteuthis 155
longimanus, Oregoniateuthis J56
longipes, Sepia (Sepia) 95*. 99
longiptcra, Octopoteuthis 177
longispadiceus. Octopus 309*, 311
Lophosepion 78
lorigera, Oregoniateuthis 156
lorigera, Sepia (Sepia) 95*, 99
lothei, Benthoctopus 320
lucens, Abralia (Stenabralia) 163*, 165
lucifer, Phasmatopsis 260
luminosa, Eucleoteuthis 20, 29, 150, 225.
226*
lunulata, Hapalochlaena 291*, 293
lycidas, Sepia (Sepia) 87, 94*, 97
352
Lycotcuthidae II, 14, 18, 26, 27, 151, 153*
155
Lycoteuthinae 153*, 155
Lycoteuthis 27, 155, 156*
lyromma, Bathothauma 11, 59, 261*, 264,
271
macandrewi, Sepia 103
macilenta, Glyptosepia 103
macrocheir, Idiosepius 128*, 129
Macrochlaena 294*
Macr octopus 290
macrohista, Histioteuthis 199*, 201
macromphalus, Nautilus 72*. 73, 75
macropus, Octopus 303*, 305, 308
macrosoma, Chiroteuthis 243
macrosoma, Rossia 110*, 111, 113
Macrotritopus 290, 306
macrotyla, Graneledone 320*, 321
maculosa, Hapalochlaena 24, 39. 291*, 298
maculosa, Inioteuthis 117, 118*
madokai, Gonatus (Gonatus) 20, 191*, 193
madokai, Octopus 309
madokai, Sepia (Sepia) 95*, 97
magellanicus, Benthoctopus 317*
magister, Berryteuthis 16, 187, 189, 190*
magna, «Cirroteuthis» 279, 280*
magna, Mastigoteuthis 249, 252*
magnoceani, Enoploteuthis leptura 162*
maindroni, Sepiella 106
makko, Gonatopsis (Gonatopsis) 189, 195*
maoria, Iridoteuthis 109, 110*. 125
margaritifera, Pyroteuthis 159, 173*, 175
Martialia 223*
martini, Octopus dofleini 45, 309*, 311,313*
niassyae, «Cirroteuthis» 280*
massyae, Eledone 299*
massyae, T etronychoteuthis 232
mastigophora, Austrorossia 114*, 115
Mastigopsis subg., Mastigoteuthis 251, 252*
Mastigoteuthidae 11, 15, 19. 21—23, 26, 47,
62, 64, 153, 243*. 249, 251
Mastigoteuthis 14, 19, 20, 27, 47, 54, 67, 237,
246*, 249, 251
Mastigoteuthis subg., Mastigoteuthis 246*,
249
mawsoni, Grimpoteuthis 281*, 283
maxima, Megalocranchia 260, 269
maya, Octopus 291, 302*, 305
meangensis, Grimpoteuthis 280*, 283
media, Alloteuthis 131, 148*, 149
media, Graneledone verrucosa var. 320
mediterranea, Sepia (Sepia) officinalis 80*
medusoides, Opisthoteuthts 284*. 285
mega, Chiropsis 237*, 239, 241*
megaleia, Lampadloteuthis 155, 156*
Megaleledone 296*
Mcgalocranchia 20, 260, 269*, 271
megalocyathus, Octopus 306*
megalops, Benthoteuthis 198
megalops, Desmoteuthis 263
megalops, Megalocranchia 263
mega lops, Teuthowenia 13, 260, 263*, 264
265
megalops, Verrilliteuthls 263
Megaloteuthis 214
megaptera, Grimpoteuthis 280*
megaptera, Octopoteuthis 177*, 179
megaptera, Promachoteuthls 254*
megaptera, Rossia Hl*, 113
Megateuthis 214
melancholicus, Sandalops 261*, 271
Melanoteuthis 272
Meleagroteuthis 199
meleagroteuthis, Histioteuthis 202*, 205
melwardi, Sepiella 105, 106
membranaceus, Octopus, 306, 308*, 311
meneghinii, Calliteuthis 203
mercatoris, Lolliguncula 135, 136*
mercatoris, Octopus 306
mercatoris, Sepia 81
Mesembrisepia 78
Mesonychoteuthls 16, 19, 23, 255, 257*. 267
mestus, Sepia (Sepia) 101, 102*
mestus, Solitosepia 97
Metasepia subg., Sepia 78*, 79
Micrabralia subg., Abraliopsis 158, 169*
microcotyla, Bolitaena 287*, 289
microlam pas, Pterygioteuthis 173
niicropyrsus, Octopus 10, 313*
Microteuthis 128
middendorffl, Gonatus (Gonatus) 194
minor, Rondeletiola 10, 107*. 119
minor. Octopus 308
mlra, Sepia (Sepia) 101, 104*
miranda, Histioteuthis 202*, 205
misakiensis, Sepia 97
mocllerl, Rossia 110*. 113
mollicella, Rossia 111*, 113
mollis, Alloposus 275, 325, 326*
mollis, Chaunoteuthis 181, 183*
mor is it, Abraliopsis 172
Moroteuthis 32, 182, 185, 186*
Moroteuthopsis 182
morsel, Euprymna 107, 114*, 117
moschata, Eledone 59, 295, 299*, 301
Moschites 295
Mouchezis 244
muelleri, Cirroteuthis 278*, 279
multihamata, Abralia (Asteroteuthis) 164*,
167
murrayi, Cirrothauma 277*, 279
murrayi, Sepia (Sepia) 93*
Myopsida 5, 10, 18, 46, 59, 70, 128*
Naefidium 128
nagasahiensis, Loligo (Loligo) edulis f. 144
nanus, Octopus 10
natalensis, Sepia 84
Nautilida 5, 68, 71*
Nautilidae 5, 68, 71*, 73, 75
Nautiloidea 5, 35, 68, 71*
Nautilus 5, 6, 36, 68, 71*, 73, 75
Nectoteuthis 110*, 125
neglecta, Sepietta 118*, 119
Nematolampas 155, 156*
Neorossia 42, 108*
Neoteuthidae 14, 62. 150, 152, 213, 214*, 215
Neoteuthis 18, 213, 214*, 215
neozelanica, Enoploteuthis 169
nielseni, Octopoteuthis 177*, 179
nigra, Eledone 299*
nipponensis, Sepiolina 29, 109*, 125
nlpponianum. Sepiadarium 127*
nipponicus, Nototodarus 229*
nodosa, Argonauta 329*. 330*
notoldcs, Idiosepius 128*, 129
Nototodarus 219, 223, 228*
nouryi, Argonauta 329, 330*
novaehollandiae, Sepia (Sepia) 101, 103*
nympha, Brachioteuthis 218
obesus, Bathypolypus 316
obscura, Sepietta 118, 119
occidua, Solitosepia 90
oceani, Histiothauma 202
oceanica, Carynoteuthis 269
oceanica, Corynomma 269
oceanica, Mcgalocranchia 29, 269*. 271
occllata, Sepiclla 105, 106*
ocellatus, Octopus 308
ochotensis, Octopus 309*
octopedatus, Gonatopsis (Gonatopsis) 187,
189, 190, 195*
Octopoda 5, 59, 70, 272*, 274
Octopodidae 10, 11, 16, 22, 26, 38, 50, 51,
274, 275, 292*, 293
Octopodinae 3, 43, 292*, 293, 302*
Octopodoidea 71, 287*
Octopodoteuthidae 176
Octopodoteuthis 176
Octopodoteuthopsis 176
Octopotcuthidae 14, 16, 17, 26, 49, 62, 150,
176*, 179
Octopoteuthis 16, 18. 20, 27, 30, 176*, 177,
179
Octopus 3, 16, 26, 35, 39, 48, 59, 60, 61, 63r
292*, 305, 311
ocula, Loligo (Loligo) 137*, 139
Ocythoe 15, 276, 325, 326*
Ocytholdae 22, 276, 325, 326*
Oegopsida 5, 10, 18, 59, 70, 148*
officinalis, Sepia (Sepia) 9, 37, 42, 50, 60,
66, 78, 80*. 83
officinalis, Sepia (Sepia) officinalis 80*
okutanii, Gonatopsis (Gonatopsis) 14, 189,.
oligobessa, Valbyteuthis 239, 241*
omani. Sepia (Sepia) 89, 90*
Ommastrephes 219, 225, 226*
Ommastrephidae 14, 16, 18, 20, 21, 26, 27,-
152, 218*, 219, 221
Ommastrephinae 27, 51, 221, 225*
Ommatostrephes 219
Onychia 182
Onychoteuthidae 14, 16, 18, 20, 21, 26, 33r
47, 57, 61, 151, 177*, 181
Onychoteuthis 16, 51, 162, 178, 181, 182, 183*
Onykia 181, 182*
onyx, Gonatus (Gonatus) 190*, 193
opalescens, Loligo (Loligo) 25, 139, 146*
opipara, Sepia (Sepia) 97
Opisthoteuthidae 10, 15, 21, 274, 276,
281*, 285
Opisthoteuthis 10, 19, 33, 34 , 274, 276,
281*. 284 , 285
orbignyana, Sepia (Sepia) 81*, 83
oregonae, Benthoctopus 320*
Oregoniateuthis 47, 153, 155, 156*
ornata, Sepiella 80, 105, 106*
ornatus, Octopus 26
Ornlthoteuthinae 223*
Ornithoteuthis 11, 219, 223, 229*
oshimai, Loligo 145 ,
oshimal, Octopus 314
ostanes, Sepia (Sepia) 101, 102*
oualaniensis, Stnenoteuthis 20, 150, 225, 229*
oweniana, Sepietta 108, 118*, 119
oxygonius, Illex 221, 227*
Ozoena 295
pacifica, Argonauta 330
pacifica, Belonella pacifica 268
pacifica, Galiteuthis 207, 269*
pacifica, Grimpoteuthis 280*
pacifica, Histioteuthis celetaria 209, 210*
pacifica, Leachia (Pyrgopsis) 259, 262*, 263
pacifica, Rossia 111*, 113
pacifica, Sepioloidea 127*
pacificum, Taonidium 269
pacificus, Ommastrephes sloani 228
pacificus, Todarodcs 9, 13, 29, 37, 53, 63,
221, 228*, 229
pQgeorum, Acanthosepion 103
paimata, Sepia 102
palpebrosa, Rossia 108, 110*, 111*, 113
panamensis, Euaxoctopus 293, 294*
panamensis, Lolliguncula 135, 136*
papillate, Helicocranchia 263*, 205
papillata, Sepia (Sepia) 24, 84*. 85
papuensis, Sepia (Sepia) 89, 90*, 99*
paradoxus, Idiosepius 128*, 129
Parasepia 78
Parasira 326
Parateuthis 253, 254*
pardaiis, Sepia (Sepia) 95*, 97
pardus, Liguriella 264*
Pareledone 23, 296, 299*, 301
Paroctopus 290
parva, Sepiola 121, 123*
par у sat is, Amplisepia 102
patagiatus, Scaeurgus 295
patagonica, Loligo (Loligo) 139, 140*
patagonica, Semirossia Ill*, 115
patagonicus, Octopus 306
pavo, Taonius 260*, 267, 268
pcalei, Loligo (Loligo) 48, 49, 137*, 139
pelagica, Hyaloteuthis 225, 227*
pelagicus, Amphitretus 275, 287*, 289
353
pelagicus, Amphitretus pelagicus 287*
tpellucida, Desmoteuthis 263
pellucida, Doratopsis 243
pellucida, Vossoteuthis 264
penicillifer, Octopus 314*
ipentherinus. Octopus 306
.peregrina, Solitosepia rozella 102
Perothis 256
ipersephone. Opisthoteuthis 284*
peterseni, Sepia (Sepia) 95*, 99
pctersi, Scpietta 118, 119
pfefferi, Abraliopsis (Abraliopsis) hovlcl
158, 169*, 171
pfefferi, Helicocranchia 261, 263*, 265
pfefferi, Leucocranchia 261
pfefferi, Loligo 146
pfefferi, Sepia (Mctasepia) 78*, 79, 106*
pfefferi, Sepiola 121, 122*
pfefferi, Taonidium 268
ipharaonis, Sepia (Sepia) 87*, 89, 94*, 99*
Phasmatopsis 260
Phasmatoteuthion 257
phenax, Euprymna 107, 114*, 117
philippinensts, Nototodarus sloani 223, 228*
Philonexis 326
.phoebetriae, Gonatus (Gonatus) 194
Pholidoteuthidae 232
Pholidoteuthis 231*, 232*
phyllura, Galiteuthis 16, 29, 267, 268*
ipickfordae, Loligo (Doryteuthis) 146*, 147
Pickfordiateuthidae 18, 130, 148*, 151
Pickfordiateuthis 148*, 151
picta, Brachioteuthis 215*. 217
picteti, Chiroteuthis 243*, 245
ipicteti, Idiosepius 128*, 129
pictus, Octopus 291
Pillsburyae, Euaxoctopus 291*, 293, 294*;
Pinnoctopus 290
ipiscatorum. Benthoctopus 296, 317*, 319
plangon, Sepia (Sepia) 101, 102*
Planktoteuthis 240
Plat у sepia 78
Plectoteuthis 214
jplei, Loligo (Doryteuthis) 131, 137*, 139
plena, Grimpoteuthis 281*
pluto, Opisthoteuthis 284*
podophtalma, Liguriella 261, 264*, 265
polymopha, Pareledone 301, 302*
Polypus 290
pompilius, Nautilus 71, 72*, 75
Ponaerisepia 78
portieri, Joubinitcuthis 251, 254*
pourtalesi, Nectoteuthis 110*, 125
prashadi, Sepia (Sepia) 90*
pricei, Octopus (?) 314*
prionota, Sepia 90
profundorum, Benthoctopus 317*, 320
Promachoteuthidae 10, 19, 20, 22, 31, 153,
253, 254*
Promachoteuthis 253, 254*
proschi, Bathypolypus arcticus f. 316*
Pseudoctopus 290
pseudonymus, Benthoctopus 320
psychrophila, Mastigoteuthis 249*
Psychroteuthidae 14, 18, 26, 152, 210*, 213
Psychroteuthls 27, 31, 210*, 213
Pteroctopus 293, 295*
pteropus, Sthenotcuthis 46, 50, 225, 229*
Pterygioteuthis 15, 16, 43, 159, 173*, 175
pulchella, Pickfordiateuthis 10, 148*, 151
pulcher, Octopus 308
.punctatus, Octopus 309
pygmaea, Eledonclla 287*, 289
pygmaeus, Idiosepius 128*, 129
Pyrgopsis subg., Leachia 256, 259, 262*, 263
pyrodes, Mastigoteuthis 29, 249*
pyros, Gonatus (Gonatus) 29, 191*, 193
Pyroteuthinae 18, 27, 43, 62, 159*
Pyroteuthis 15, 43, 159, 173*, 175
recurvirostris, Sepia (Sepia) 89, 90*
redfieldi, Abralia (Asteroteuthis) 167, 168*
354
reesi, Loligo (Doryteuthis) 146*, 147
reesi, Sepia (Sepia) 104*
regalis, Nematolampas 29, 155, 156*
reinhardti, Liocrancnia 256, 259, 262*
rcnschi, Abralia (Stenabralla) 157, 163*, 165
repertus, Nautilus 72*, 75
reticulata, Enoploteuthis 161, 162*
Retroteuthis 272
reversa, Histioteuthis 203*. 207
rex, Sepia (Sepia) 101, 103*
rcynaudi, Loligo (Loligo) vulgaris 45, 141*
rhoda, Sepia (Sepia) 101, 104*
rhomboidalis, Loliolus 132*, 133
Rhombosepion 78
rhombus, Thysanoteuthis 230*, 231
richardi, Vitreledonelia 275, 322*, 323
rlchardsoni, Teuthowcnia 263, 265
riisei, Brachioteuthis 215*, 217, 218*
Robsonella 290
robsoni, Abrala (Asteroteuthis) andama-
nica f. 168*
robsoni, Moroteuthis 185, 186*
robsoni. Sepia (Sepia) 10, 84*, 95
robusta, Moroteuthis 182, 185, 186*
robusta, Sepiola 121, 122*
rondeleti, Sepiola 107, 121, 122*
Rondeletiola 26, 107*, 119
roperi, Loligo (Dorvleuthis) 139, 140*
Rossia 32, 108, 110*, 113
Rossiinae 18, 31, 108*
rotunda, Bentheledone 297, 298*
rouxii, Sepia 87
rozella, Sepia (Sepia) 101, 102*
rubescens, Octopus 39, 314*
rugosa, Octopoteuthis 177*, 179
rugosus, Octopus 306. 312
ruppelli, Histioteuthis 199
rynchophorus, Leachia (Pyrgopus) 256, 259,
263*
sagitta, «Doratopsis» 19, 239, 240*, 241*
Sagittatus 219
sagittatus, Todarodes 16, 59, 219, 221, 228*
salebrosus, Bathypolypus 314*, 315
salutil, Octopus 305, 306*
Sandalops 11, 14, 261*, 271
sasakii. Sepia 97
Sasakinella 294*
savignyi. Sepia (Sepia) 91*
scabra, Cranchia 29, 256*, 259
Scaeurgus 293, 295*
scalenus, Tremoctopus 294
scandica, Sepietta 118*
schmidt!, Danoctopus 293, 294*, 307*
schmidti, Mastigoteuthis 249*
schnehageni, Pterygioteuthis 173
schnehageni, Pyrgopsis 262
schultzei, Octopus 306*
scintlllans, Selcnoteuthis 29, 155, 156*
scintillans, Watasenia 13, 26, 27, 158*, 171
scolopes, Euprymna 114*, 117
scrobiculatus, Nautilus 72*, 75
Selenoteuthis 47, 153, 155, 156*
Semirossia 26, 108, Hl*. 115
senoi, Megaleledone 296*
separata, Meleagroteuthis 202
Sepia 3, 21, 26, 32, 61, 78, 79, 80*
Sepia subg., Sepia 78*, 80, 83, 85, 89, 97,
99, 101
Sepiadariidae 10, 14, 18, 20, 21, 31, 77, 126*,
127
Sepiadarium 21, 126*, 127
Sepidium 108
Sepiella 21, 79, 80, 105, 106*
Sepietta 108, 118*, 119
Sepiida 5, 68*, 73
Scpiidae 6, 11, 15, 21, 38, 51, 76, 77*, 78. 79
Scpiina 5, 68, 76*
sepioidea, Sepioteuthis 130, 135, 136*
Sepiola 26, 32, 59, J07, 118*, 121
Sepiolidae 15, 18, 21, 38, 77, 106*, 107, 113
Sepiolina 10, 20, 109*, 125
Sepiolinae 10, 18, 20, 31, 107*
Sepioloidea 126, 127*
sepioloides, Ctenopteryx 196*. 197
Sepioteuthis 18, 26, 130, 135, 136*
septemdentatus, Gonatus 190
serventyi, Heteroteuthis 126
sewelli, Sepia (Sepia) 93*
sibiricus, Benthoctopus 296, 316*, 319
sibogae Loligo (Doryteuthis) 146*. 147
sicula, Ctenopteryx 29, 196*, 197
sicula, Octopoteuthis 176, 177*, 179
simoniana, Sepia (Sepia) 84*
singaporensis, Sepia 90
•slnghalensis, Loligo (Doryteuthis) 145*, 147
sirventi, Cycloteuthis 29, 233*, 235
skolops, Batoteuthis 233*, 235
sloani, Nototodarus 223, 228*, 229
sloani, Nototodarus sloani 223, 228*
smith!, Sepia (Sepia) 101, 103*
Solitosepia 78
spaercki, Abralla (Stenabralia) 164*, 165
Spathidosepion 78
spectrum, Loligo (Doryteuthis) 146*
speculator, Corynomma 269
spinicauda, Enoptroteuthis 233, 235
spinicirrls, Теtracheledone 296*, 297
spinosus, Octopus 312*
Spirula 18, 26, 27, 30, 32, 42, 61, 68, 76*, 77
spirula, Spirula 29, 76*, 77
Spirulidae 68, 73*, 77
Spirulina 5, 68, 73*
sponsalls, Bathypolypus 315, 316*
springer!, Oregoniateuthis 155, 156*
Stauroteuthidae 276, 277
Stauroteuthis 33, 278*, 279
steenstrup!, Gonatus (Gonatus) 191, 194*
Steenstrupia 214
steenstrupiana, Scpiola 121, 122*
Steenstrupiola 182
steindachneri, Abralla (Stenabralia) 163*,
165
Stenabralia subg., Abralia 157, 163*
stenodactyla, Euprymna 114*, 117
stenomphalus, Nautilus 72*, 75
Stephanoteuthis subg., Heteroteuthis 110,
126*
Sthenoteuthis 27, 31, 225, 226, 229*
stictochrus. Octopus 10
Stigmatoteuthis 199
Stoloteuthis 10, 20, 109*, 110, 125
subaculeata, Sepia 94
sublaevis, Rossia 111
submestus, Solitosepia 90
subulata, Alloteuthis 148*, 149
suhmi, Galiteuthis 268*
suhmi, Taonidium 268
sulcata, Sepia (Sepia) 93*
sumatrensis, Loligo 145
surinamensis, Loligo (Loligo) 137*, 139
Symplectoteuthis 226
syrtensis, Stauroteuthis 278*, 279
tagoi, Loligo (Loligo) 143, 145*
talismani, Mastigoteuthis 246, 251*
taningi, Dorsopsis 287*, 289
Taningia 30, 176*, 179
Tankaia 239, 240*, 241*
Taonidium 257
Taoniinae 19, 23, 257*, 265
Taonius 14, 257, 260*. 267
tasmanica, Euprymna 114*
tehuelchus, Octopus 305, 307*
Teleonychoteuthis 178
Teleoteuthis 182
tenera, Desmoteuthis 263
tenera, Semirossia 109, 114*, 115
ten era, Sepiola 122
tenuipes, Sepia (Sepia) 95*, 97
Tenuisepia 78
Teretoctopus 298*
Tetracheledone 296*, 297
tetracirrhus, Pteroctopus 48, 293, 295* 308*
Tetronychoteuthis 23, 231, 232*
Teuthida 5, 70*, 128
Teuthowcnia 19, 23, 260, 263*. 265
thaumastocheir, Grimpella 297, 298*
Thaumatolampas 156
Thaumeledone 22, 297, 298*
Thelidioteuthis 158
theragrae, Enoploteuthis 162
thlelei, Amphitretus pelagicus 287*
thielei, Benthoctopus 319, 320*
thiclei, Neoteuthis 213, 214*
thori, Desmoteuhtis 263
thurstonL Sepia (Sepia) 91*
thysanopnora, Eledone 299*. 301
Thysanoteuthidac 14, 18, 21, 51, 152, 229*,
230, 231
Thysanoteuthis 230*, 231
tigris, Sepia 87
tinro, Gonatus (Eogonatus) 13, 29, 187,
190*, 193
Todarodes 11, 219, 221, 227*
Todarodinae 219*, 221
Todaropsis 219*. 221
togata, Vclodona 296*, 297
tokioensis, Sepia (Sepia) 97*, 99
iorosa. Sepia 87
Toroteuthis 237
tortugaensls, Rossia 111*, 113
Toxeuma 257
Tracheloieuthis 215
treba, Arctosepia 104
Tremoctopodidae 22, 276, 325, 326*
Tremoctopus 15, 36, 67, 276, 325, 326*
trigonura, Abralia (Asteroteuthis) 168*
triluminosa, Galiteuthis 268
trirostraia, Scpiola 121, 123*
Tritaxeopus 290
trygonina. Sepia (Sepia) 91*, 93
tsugarensis, Octopus 309*, 311
tuberculata, Ocythoe 276, 325, 326*
tubcrculata, Sepia (Sepia) 24, 84*. 85
tullbergi, Sepia (Metasepia) 23, 79, 80*, 97*
tunicata, Parateuthis 215, 253, 254*
turqueti, Pareledone 301*
tydeus, Lolliguncula 135, 136*
typica, Sepia (Hemisepius) 10, 79, 80*, 84
fypus, Loliolus 131
umbellata, Grimpoteuthis 278, 280*. 283
umbilicatus, Nautilus 72
umitakae, Pareledone 301*
unguiculata, Cucioteuthis 176
unicirrhus, Scaeurgus 293, 295*, 307*
Uranoteuthis 261
Uroteuthis 19, 26, 33, 131*. 149
ursulae, Crumenasepia 87
uyii, Loligo (Loligo) 143, 145*
Valbyteuthidae 236
Valbyteuthis 15, 20, 239, 240, 241*. 242
Valaemaria 254
valdiviae, Bathypolypus 315, 316*
valdiviae, Liocranchia 262*
Vnmpyromorpha 5, 43, 70, 269*, 273
Vampyroteutnidae 70, 272* 273
Vampyroteuthis 5, 26, 27. 30,33, 46, 49, 57,
62—64, 70, 272*, 273
variabilis, Octopus 308*, 311
variabilis, Octopus minor 308
velaini, Brachioteuthis 218
veligero, Octopus 313*
Velodona 42, 296*. 297
Verania 176
Ver an ya 176
veranyi, Abralia (Asteroteuthis) 157, 187,
168*
veranyi, Chiroteuthis 237, 242*, 245
veranyi, Chiroteuthis veranyi 242*, 245
Veranyidae 176
vercoi, Sepia (Sepia) 104*
vermicularis, Doratopsis 242, 243
vermiculata, Sepia (Sepia) officinalis 80*»
355
verreauxi, Ascarosepion 102
vcrrilli, Onykia (?) 182*
Verrilliola 215
Verrllliteuthis 260
verrucosa, Graneledone 298, 320*, 321
versuta, Arctosepia 104
vincenti, Octopus 302
violaceus, Trcmoctopus 26, 325, 326, 327*
vlolaceus. Trcmoctopus violaceus 325, 327*
violesccns, Benthoctopus 317*
Vitreledonella 11, 40, 63, 275, 322», 323
Vitreledonellldae 16, 17, 71, 275, 322*, 323
volatills, Ornlthoteuthis 223, 229*
Vosseledone 22, 296*, 297
vossl, Loligo (Loligo) 143, 144*
Vossoteuthis 26!
vulgaris, Loligo (Loligo) 45, 56, 59, 61,
131, 139, 140*
vulgaris. Octopus 9, 25, 34, 37, 38, 41, 42,
45, 50, 53, 63, 290, 291, 305, 306*, 312*
Watasella 272
Watasenla 30, 158*. 171
weberi, Heteroteuthis (Heteroteuthis) 123*,
125
weberi, Seplella 105, 106*
whitleyanum, Sepia (Sepia) 97
wlnckworthl, Macrochlaena 294*
winckworifu, Sepia 91
witjazi, Pteroctopus 295*, 312*
wuelkeri, Grimpoteuthis 280*, 283
yendoi. Octopus 311, 312*
yokoyae, Loligo (Loligo) 143, 145*
zanzibarica, Sepia (Sepia) 84*, 91*
zonatus, Octopus 302*, 305
zugaena, Zygocranchia 268
Zygaenopsis 257
Zygocranchia 257
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ 3 2.2.2.3. Семейство Octopo-
ВВЕДЕНИЕ 5 tcuthidae 176
ОБЩАЯ ХАРАКТЕРИСТИКА 2.2.2.4. Семейство Onycho-
ГОЛОВОНОГИХ teuthidae 177
МОЛЛЮСКОВ 6 2.2.2.Б. Семейство tidae Gona- 187
РАЗМЕРЫ И МАССА 7 2.2.2.6. Семейство Cteno-
ВНЕШНЕЕ СТРОЕНИЕ 10 pterygidae 2.2.2.7. Семейство 195
ВНУТРЕННЕЕ СТРОЕНИЕ 31 Bathy-
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ 68 teuthidae 196
1. Подкласс Nautiloidea 71 2.2J.8. Семейство teuthidae Histio- 198
1.1, Отряд Nautilida 2. Подкласс Coleoidea 71 72 2.2.2.Э. Семейство roteuthidae Psych- 210
2.1. Отряд Sepiida 2.1.1. Подотряд Spirulina 73 73 2.2.2.10. Семейство tcuthidae Archi- 213
2.1.1.1. Семейство Spirulidae 2.1.2. Подотряд Sepiina 76 2.2.2.11. Семейство teuthidae Neo- 214
2.1.2.1. Семейство Sepii- dae 2.1.2.2. Семейство Sepioli- dae 2.1.2.3. Семейство Sepiada- riidae 77 L VtLA LUI VI14^ 2.2.2.12. Семейство Bra-
11 106 chioteuthidae 2.2.2.13. Семейство Om- 215
AW 126 mastrephidae 2.2.2.14. Семейство Thysa- 218
2.1.2.4. Семейство Idiose- noteuthidae 229
piidae 2.2. Отряд Teuthida 127 128 2.2.2.15, Семейство doteuthidae Lepi- 232
2.2.1. Подотряд Myopsida 2.2.1.1. Семейство Loligi- nidae 128 2.2.2.16. Семейство teuthidae Bato- 233
130 2.2^2.17. Семейство Cyclo-
2.2.1.2. Семейство Pickfor- teuthidae 233
diateuthidae 148 2.2.2.18. Семейство Chiro-
2.2.2. Подотряд Oegopsida 148 tcuthidae 236
2.2.2.1. Семейство Lyco- 2.2.2.19. Семейство Masti-
teuthidae 153 goteuthidae 243
2.2.2.2. Семейство Enoplo- 2.2.2.20. Семейство Joubi-
teuthidae 156 niteuthidae 252
357
2.2.2.21. Семейство Pro-
machoteuthidae 254
2.2.2.22. Семейство Gri-
malditeuthidae 254
2.2.2.23. Семейство incer-
tae sedis 254
2.2.2.24. Семейство Cran-
chiidae 255
2.3. Отряд Vampyromor-
pha 269
2.4. Отряд Octopoda 272
2.4.1. Подотряд Cirrata 275
2.4.1.1. Семейство Cirro-
tcuthidac 276
2.4.1.2. Семейство Opistho-
teuthidae 281
2.4.2. Подотряд Incirrata 285
2.4.2.1. Надсемейство Bo-
litaenoidea (Ctenoglossa) 286
2.4.2.1.1. Семейство Boli-
tacnidae 286
2.4.2.1.2. Семейство Am-
phitretidae 287
2.4.2.1.3. Семейство Idioc-
topodidae 287
2.4.2.2. Надсемейство Oc-
topodoidca (Heteroglossa) 287
2.4.2.2.1 Семейство Octo-
podidae 292
2.4.2.2.2. Семейство Vitre-
ledonellidae 322
2.4.2.3. Надсемейство Ar-
gonautoidea 322
2.4.2.З.1. Семейство Allo-
posidae 323
2.4.2.3.2, Семейство Ocyt-
hoidae 326
2.4.2.3.3. Семейство Tre-
moctopodidae 326
2.4.2.3.4. Семейство Argo-
nautidae 327
СПИСОК ЛИТЕРАТУРЫ 330
Обшая часть 330
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ 338
УКАЗАТЕЛЬ ЛАТИНСКИХ
НАЗВАНИЙ 348
КИР НАЗИМОВИЧ НЕСИС
КРАТКИЙ ОПРЕДЕЛИТЕЛЬ
ГОЛОВОНОГИХ
моллюсков
МИРОВОГО ОКЕАНА
Редактор
Е. П. Яковлева
Художник
Ю. П. Тропиков
Художественный редактор
Е. К. Селихова
Технический редактор
Т. С. Пронченкова
Корректоры
Т. А. Лашкина, . И. Гурычсва
ИБ № 1504
«Сдано в набор 27.01.82. Подписано в печать
27.08.82. Т-11999. Формат 60х90’/«б. Бумага
мелованная. Литературная гарнитура. Вы-
сокая печать. Объем 22,5 п. л. Усл. п. л.
22,5. Усл. кр. отт. 22,75. Уч.-изд. л. 32,64.
Тираж 3850 экз. Заказ 979. Цена 3 р. 70 к.
Издательство «Легкая и пищевая промыш-
ленность», 113035, Москва, М-35, 1-й Када-
шевский пер., 12.
Владимирская типография «Союзполиграф-
прома» при Государственном комитете
СССР по делам издательств, полиграфии
и книжной торговли
600000, г. Владимир, Октябрьский прос-
пект, д, 7