























































































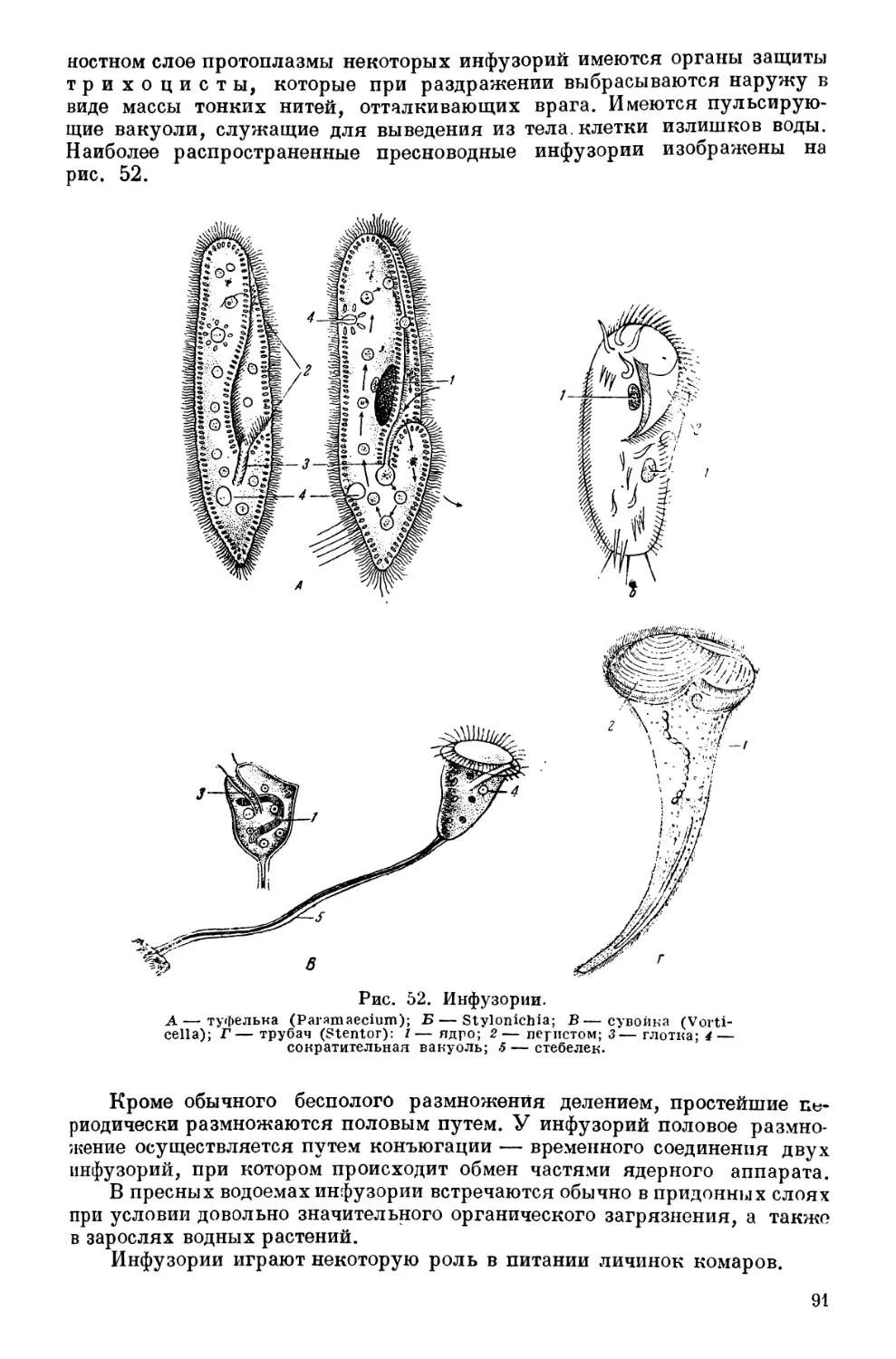








































































































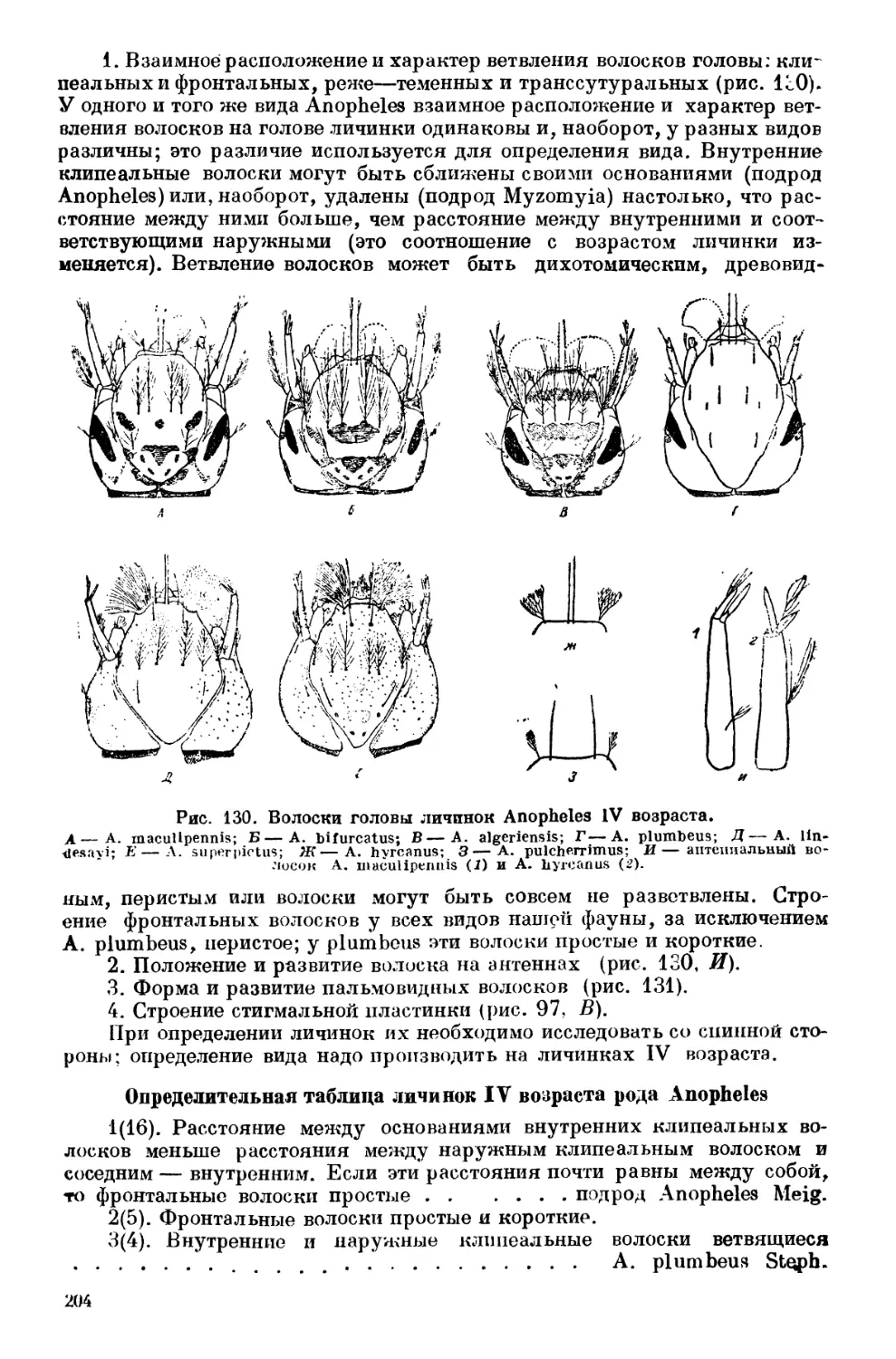

































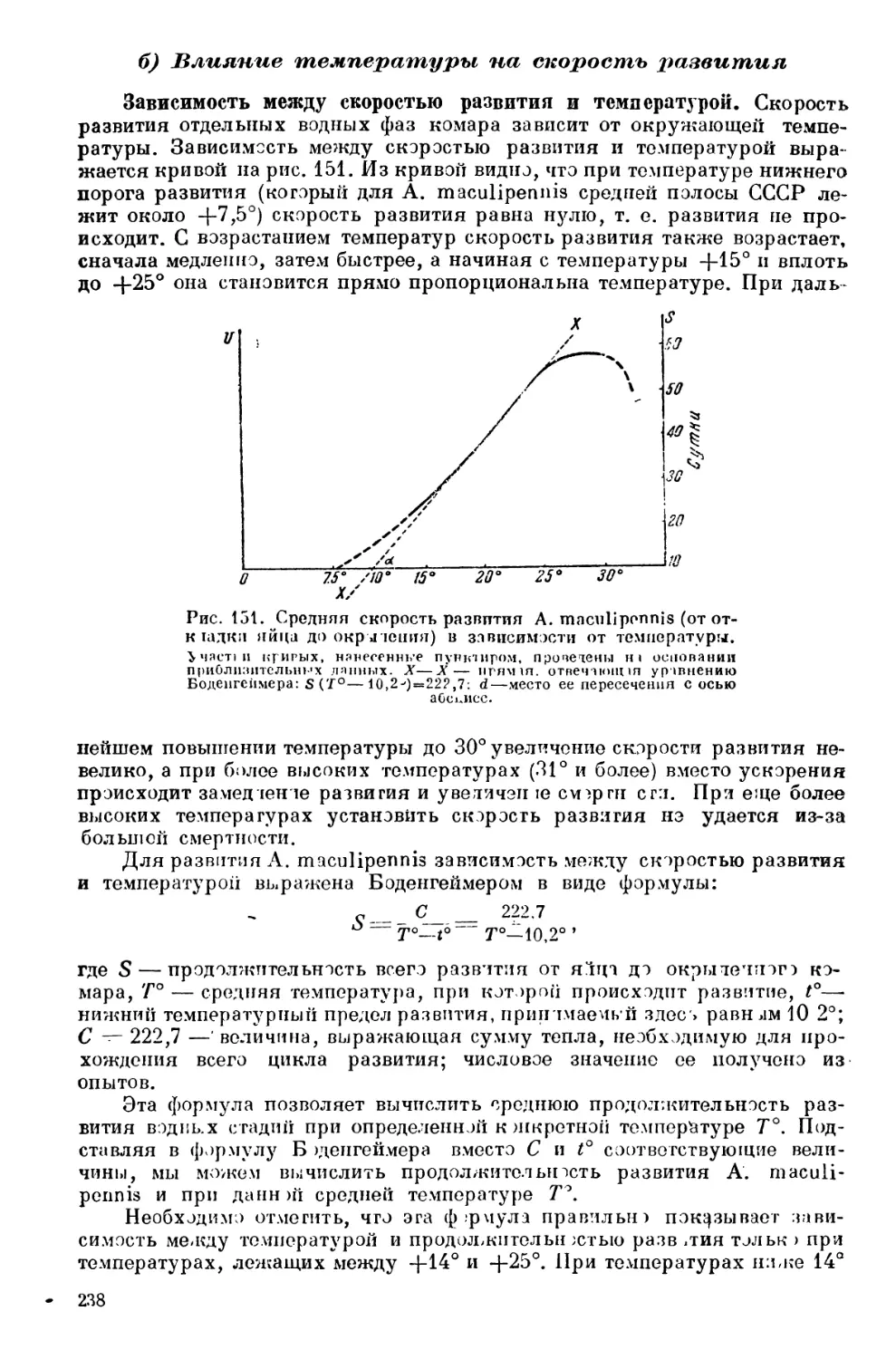























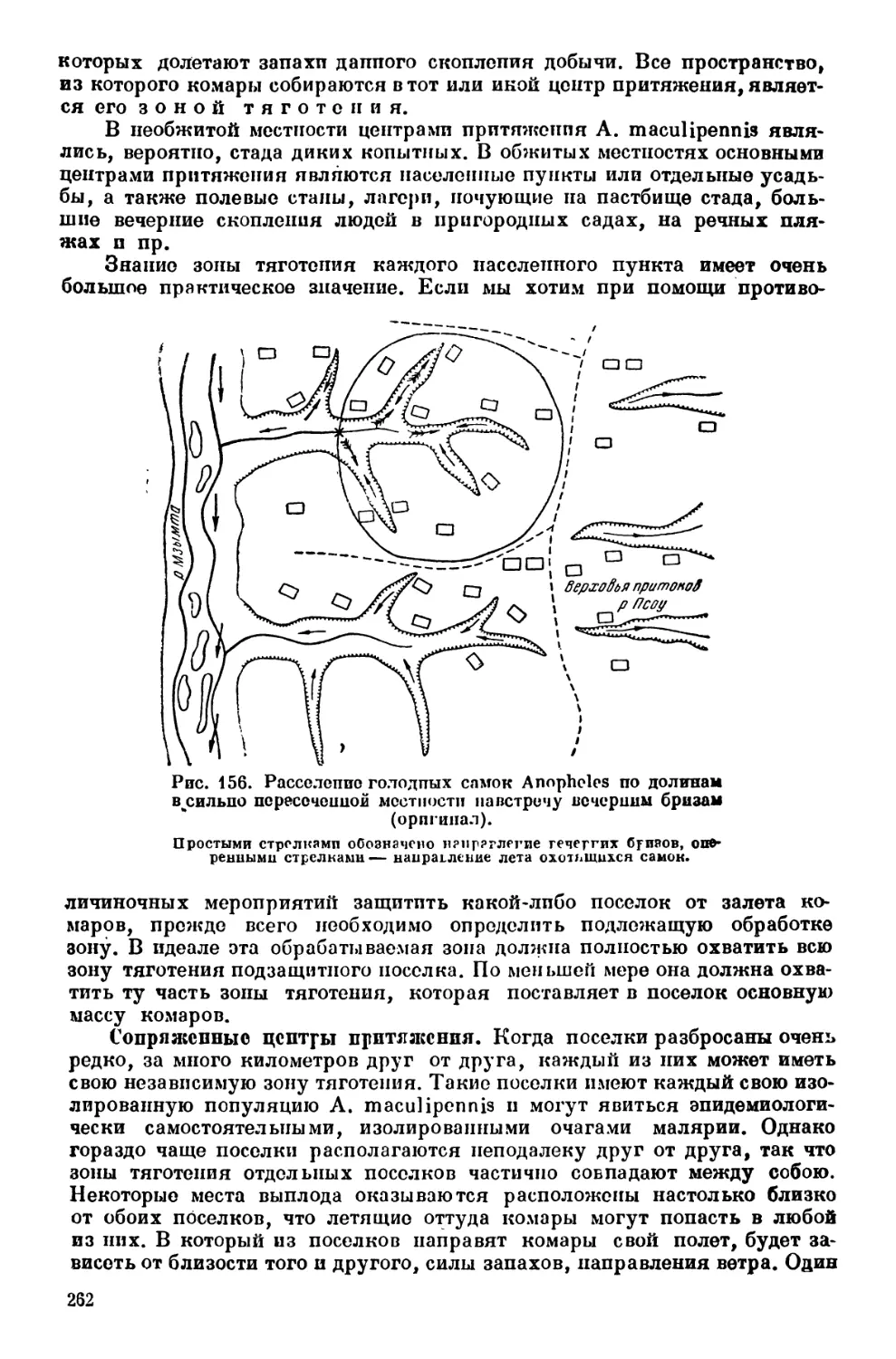











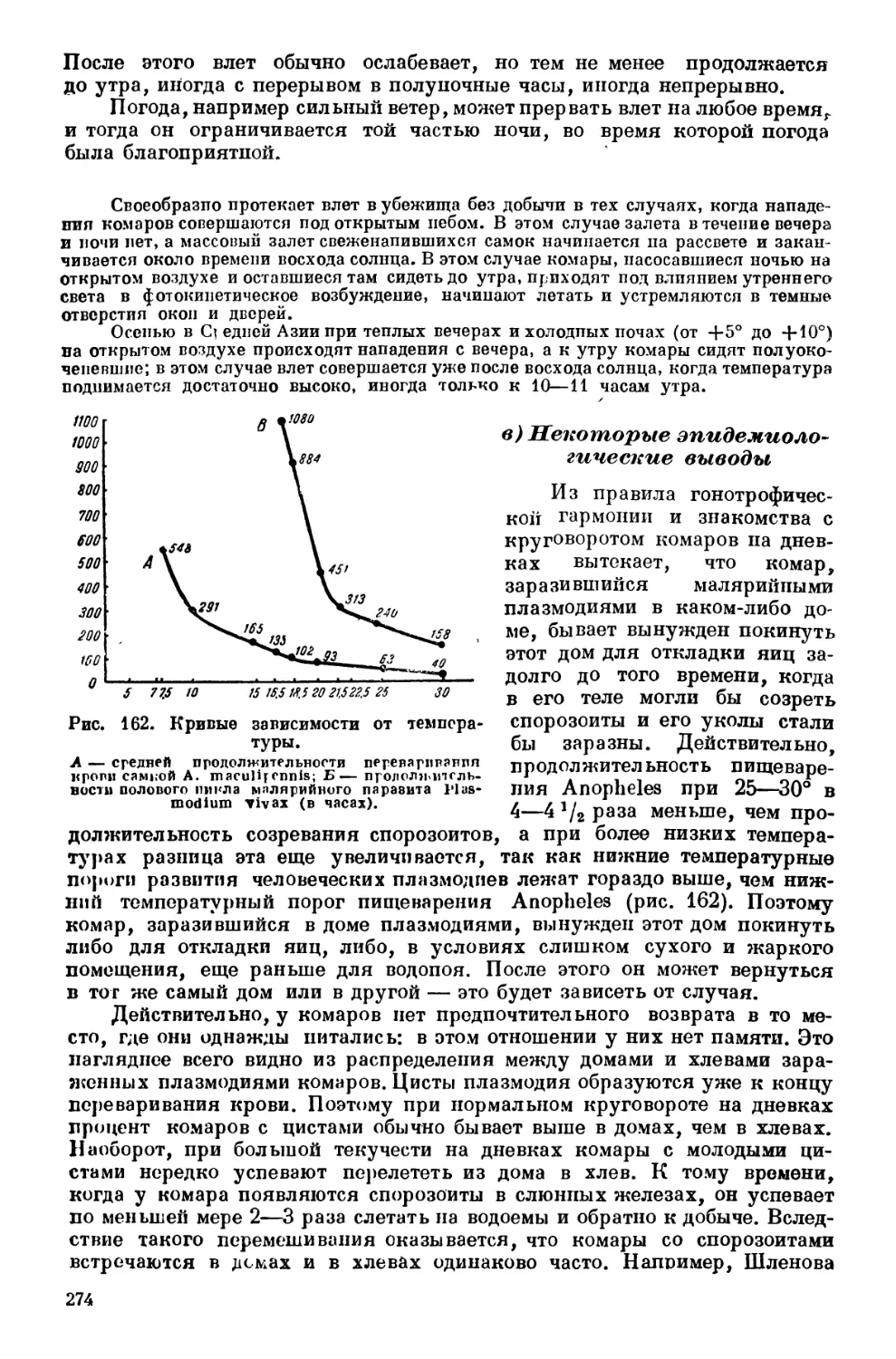

















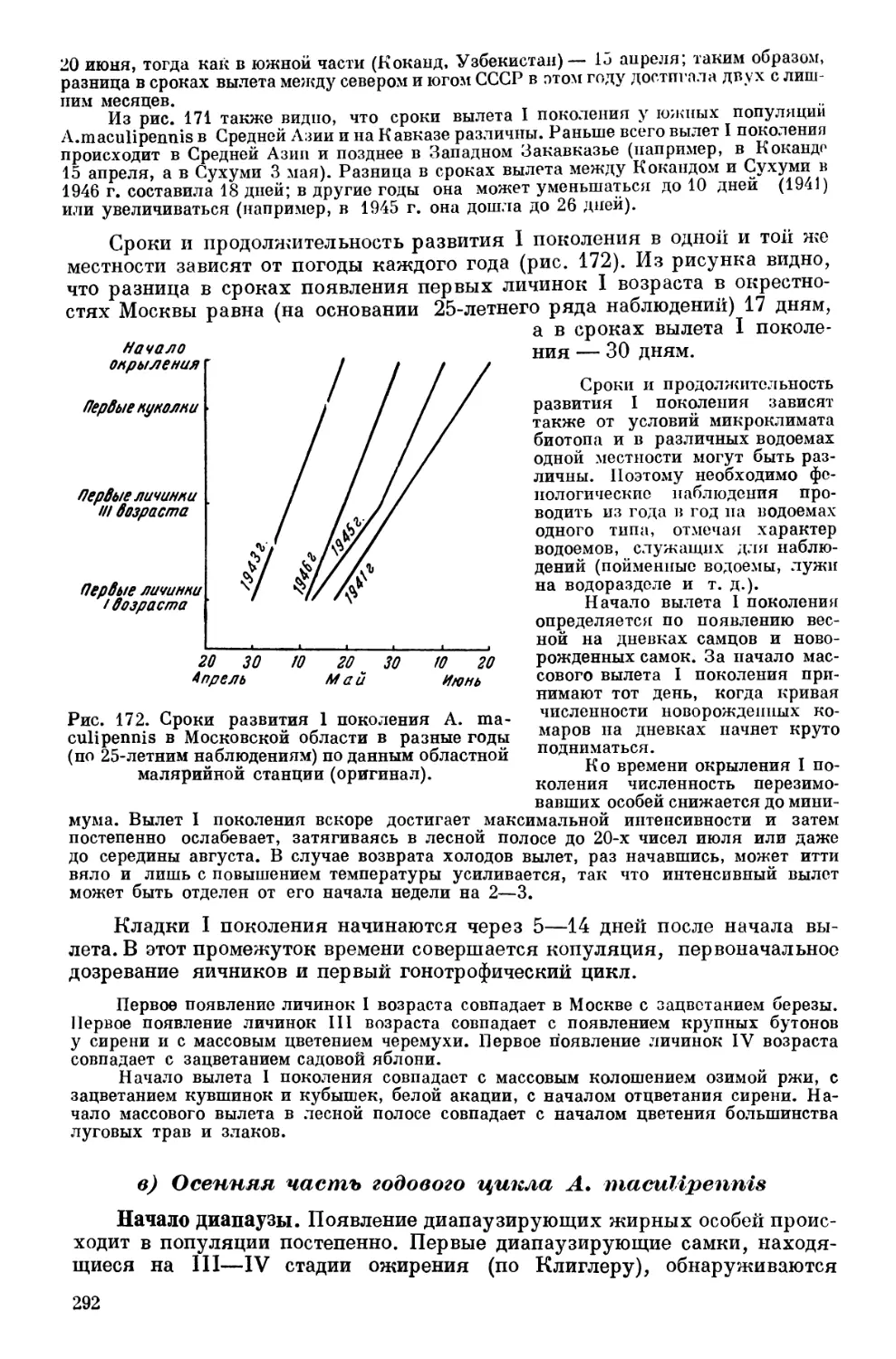



















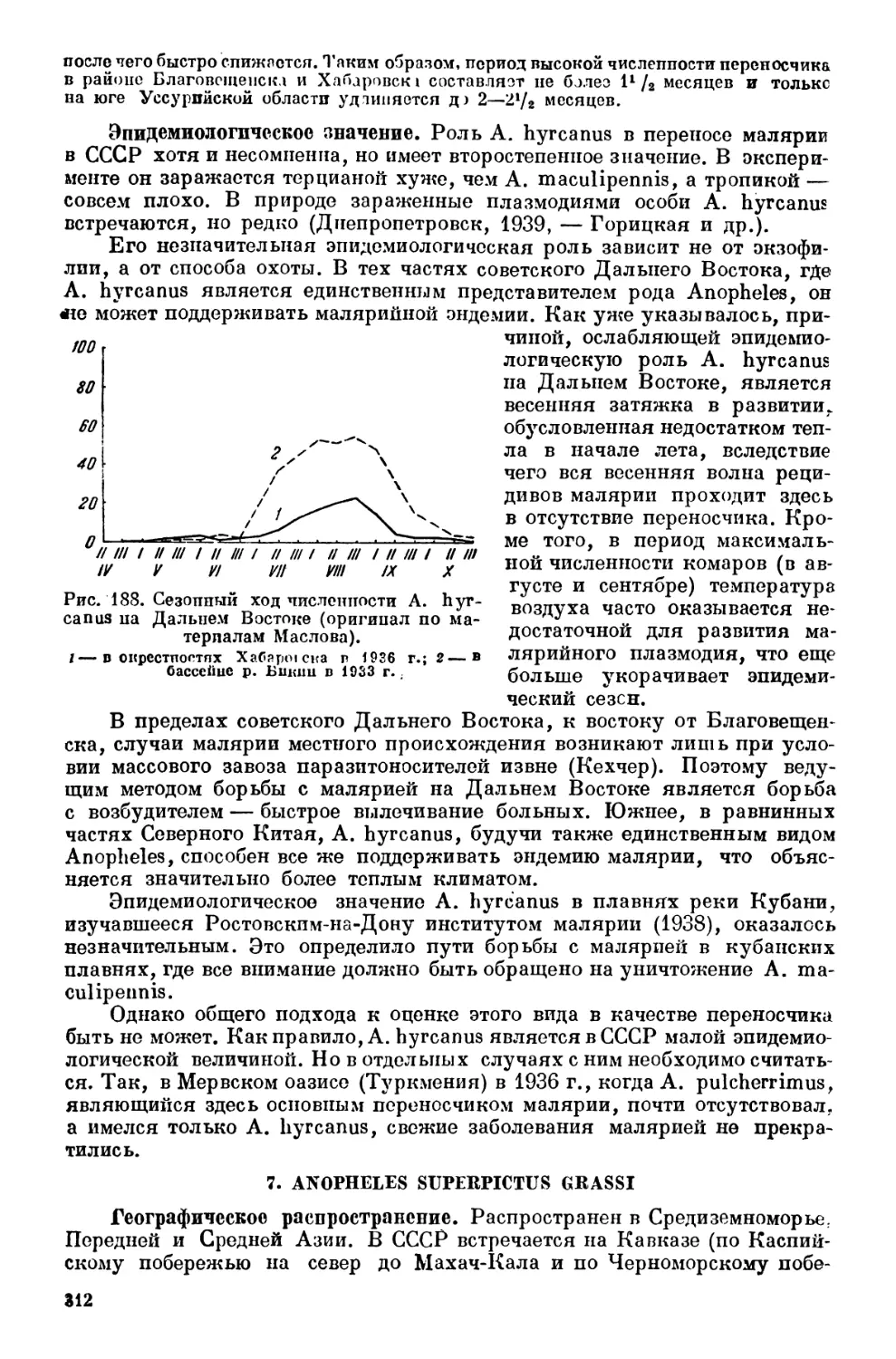
























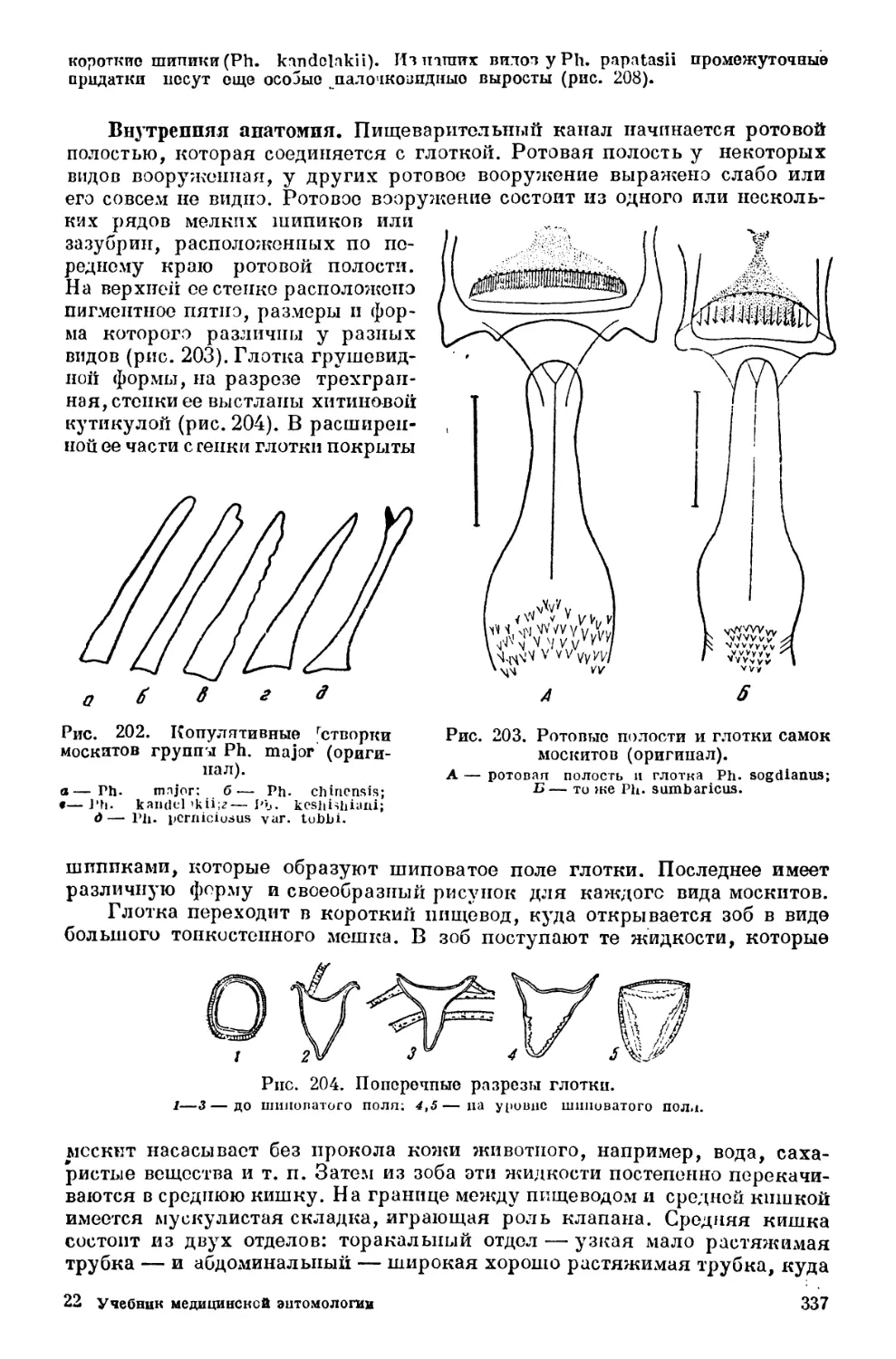







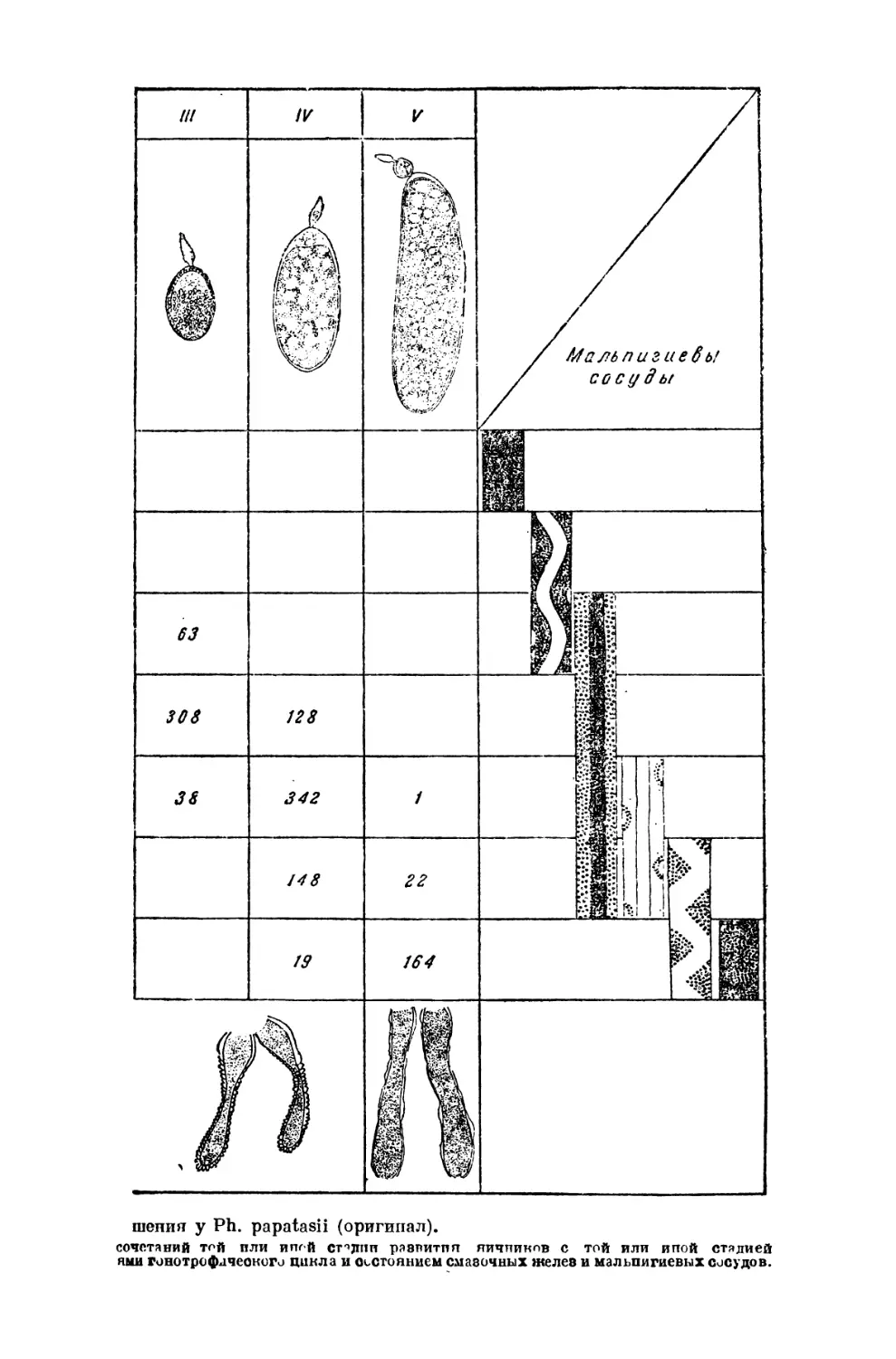



























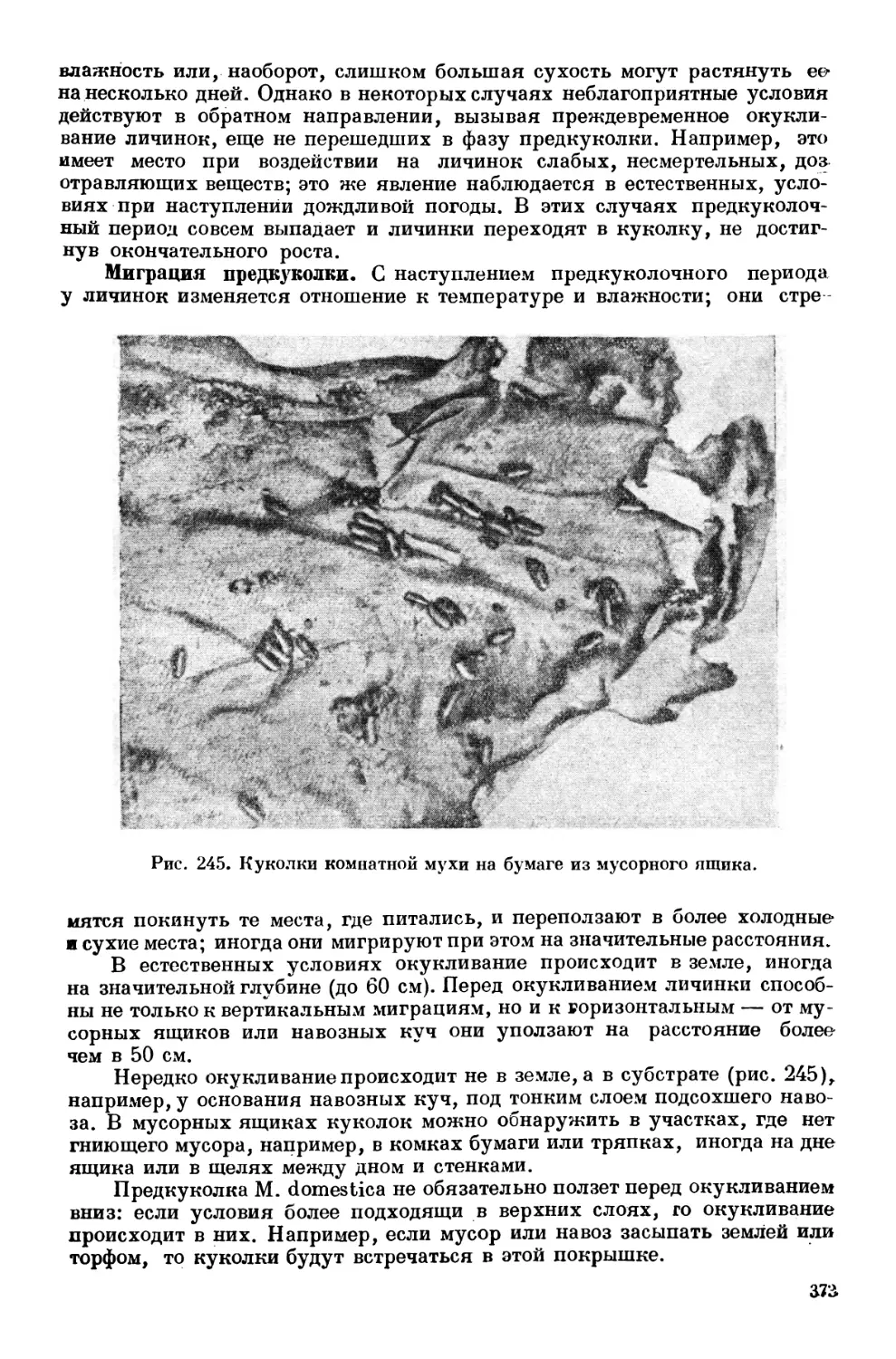






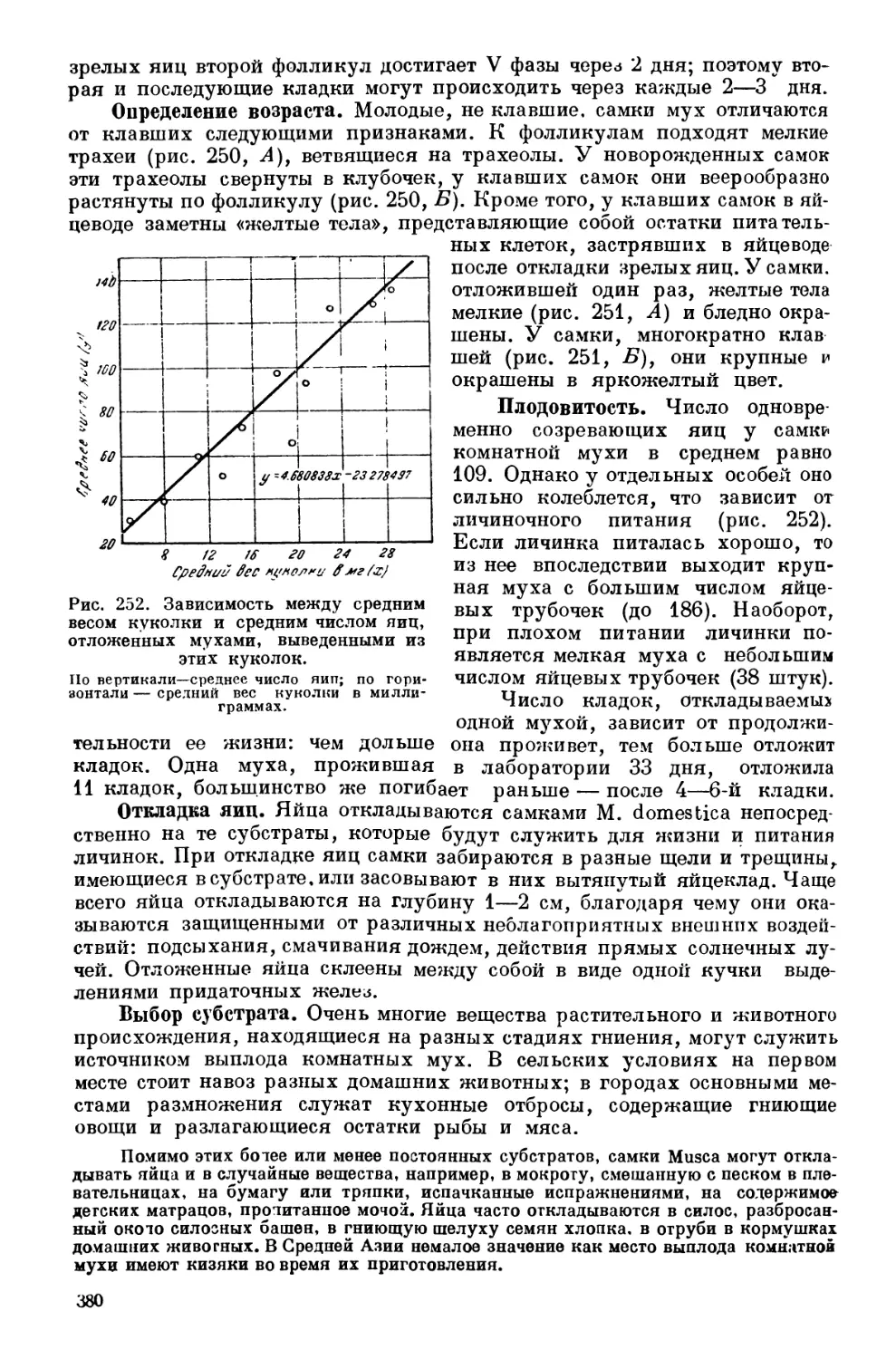





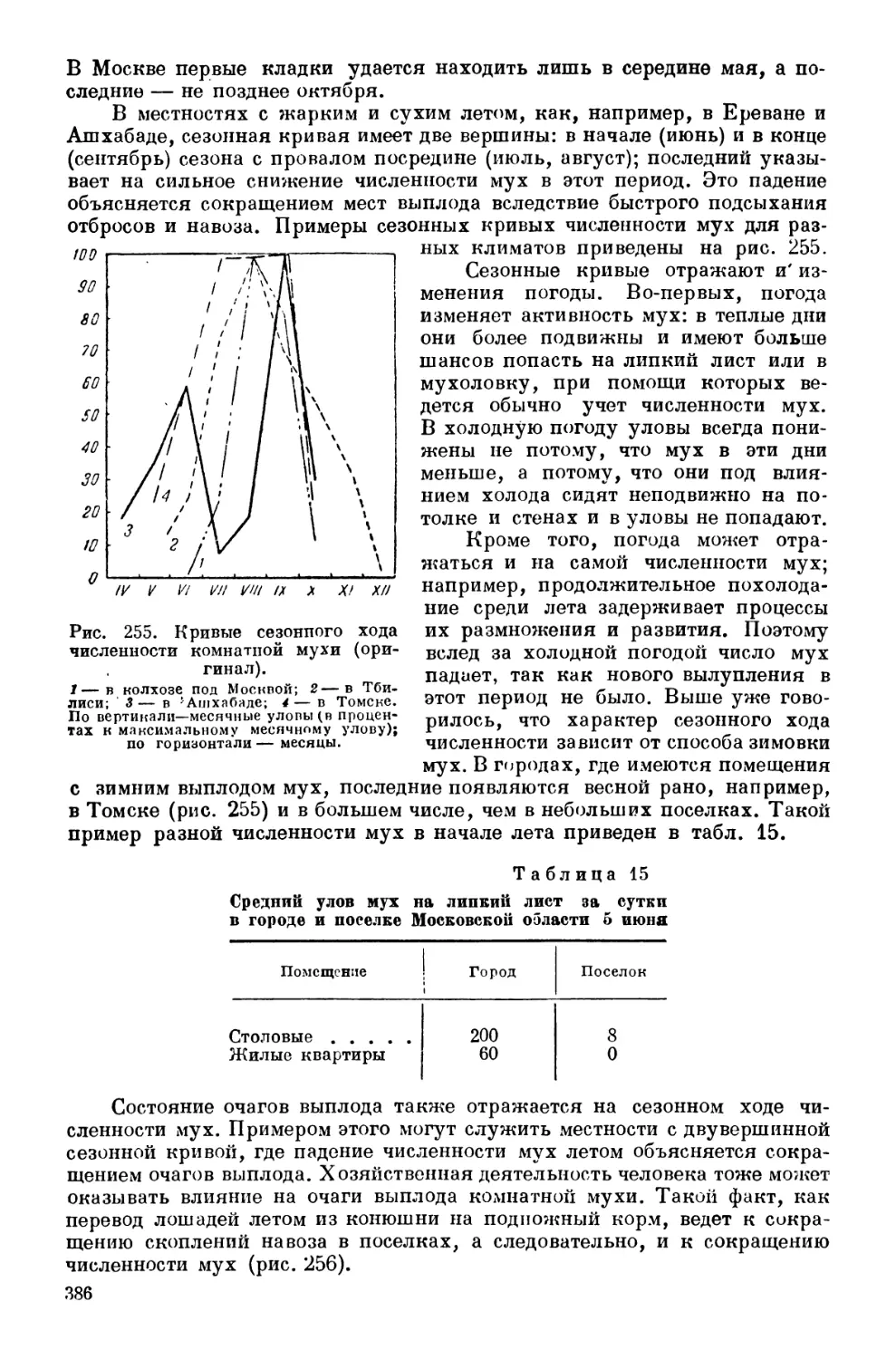












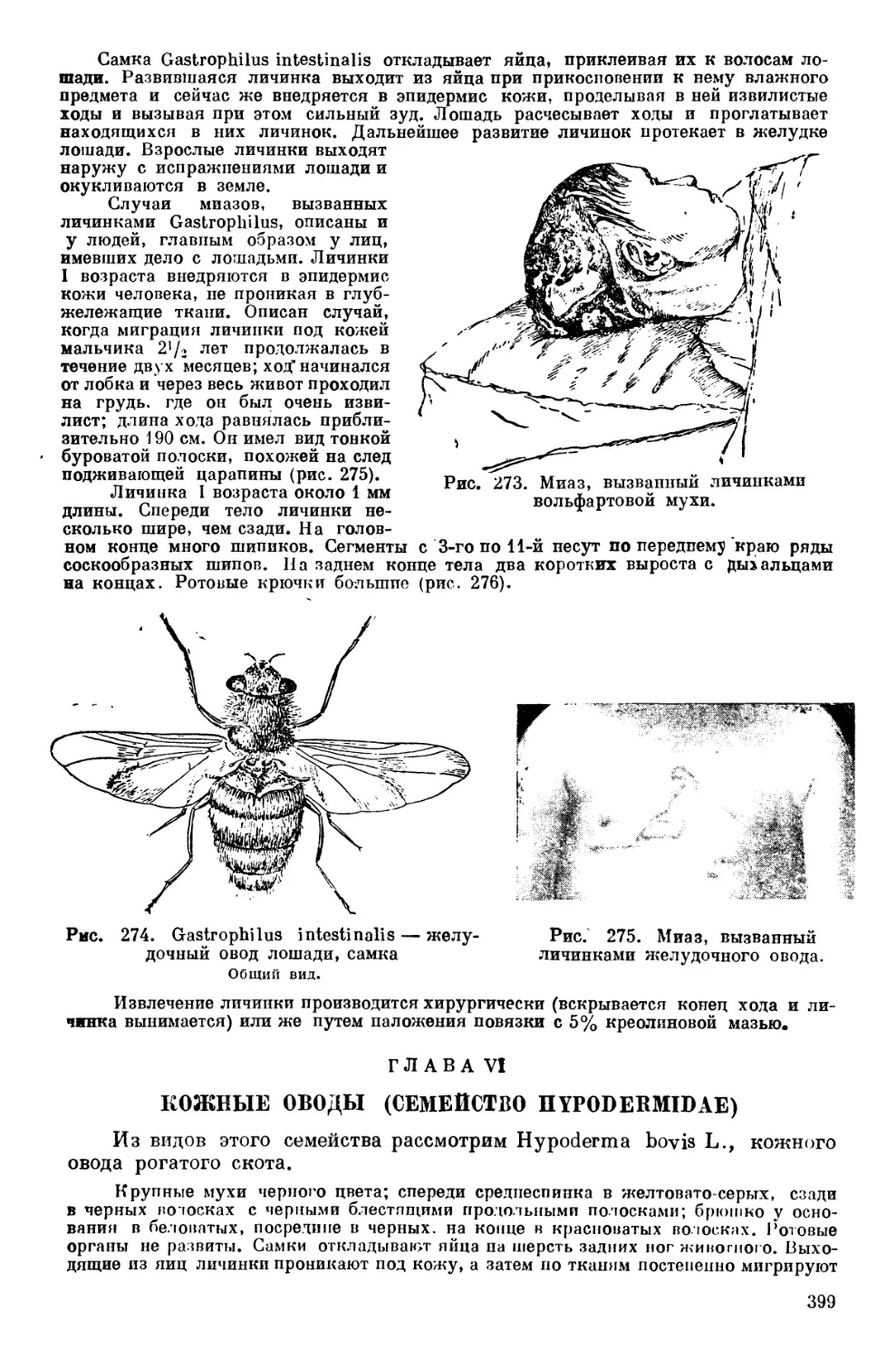






























































































Текст
УЧЕБНИК 4
МЕДИЦИНСКОЙ
ЭНТОМОЛОГИИ
В.Н БЕКЛЕМИШЕВА
МЕДГИЗ — 1949
УЧЕБНИК
МЕДИЦИНСКОЙ ЭНТОМОЛОГИИ
КОД РЕ.ДАКЦИ ЕЙ
Проф. В. Н. БЕКЛЕМИШЕВА
ЧАСТЬ I
МЕДИЦИНСКАЯ ЭНТОМОЛОГИЯ С ОСНОВАМИ
ОБЩЕЙ ЭНТОМОЛОГИИ И ГИДРОБИОЛОГИИ
Управлением
•средних медицинских учебных заведений
Министерства здравоохранения СССР
рекомендован дгч школ и отделений
помощников медицинских энтомологов
С ОС ТА В ИЛ И:
В. И. БЕКЛЕМИШЕВ». О. И. ВИНОГРАДСКАЯ,
В. II. ДЕРБЕНЕВА-УХОВА, А. В. ДОЛМАТОВА,
Э С. КУЗИНА. В. И. ОЛИФАН, М. В. ПОСПЕЛОВА-ШТРОМ
и Н. К. ШИПИЦИНА
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
МЕДИЦИНСКОЙ ЛИТЕРАТУРЫ
М Е Д Г И 3 • 1949 • МОСКВА
ПРЕДИСЛОВИЕ ч
Настоящий учебник предназначен прежде всего для учащихся школ
помощников медицинских энтомологов и охватывает три предмета учеб-
ного плана этих школ: 1) общую энтомологию, 2) гидробиологию с гидро-
фауной и гидрофлорой и 3) медицинскую энтомологию.
Первая часть, «Общая энтомология??, должна служить введением к
медицинской энтомологии; в ней приводятся элементарные зоологические
сведения о членистоногих и краткое изложение их строения и жизни.
Из классов членистоногих основное внимание уделено насекомым, из
насекомых — отряду двукрылых, так как этот отряд содержит наиболь-
шее число важнейших вредителей здоровья человека. Вторая ио значе-
нию группа членистоногих — клещи — в курсе общей энтомологии опи-
сывается лишь в общих чертах; более подробный очерк строения и жизни
клещей дан в посвященной им главе медицинской энтомологии-.
Вторая часть, «Гидробиология», представляет, в согласии с про-
граммен, курс маляриологичсской гидробиологии. Излагая в кратком
виде основные положения гидробиологической науки, она в то же время
дает учащемуся важнейшие предпосылки для изучения экологии личинок
комаров. Это касается главным образом главы IV — «Основные типы
материковых водоемов», в которой подчеркиваются и подробнее раз-
бираются типы апофелогеппых водоемов СССР. Таким образом, курс
гидробиологии отчасти является введением и к курсу противомалярий-
ной гидротехники, который проходится на пятом и шестом семестрах.
Третья часть, «Медицинская энтомология», в соответствии с местом,
которое этот предмет занимает в учебном плане (395 учебных часов), имеет
наибольший объем. Принимая во внимание профиль школы, готовящей
помощников энтомологов для малярийных станций, первое место мы уде-
лили малярийным комарам, биология которых освещена с надлежащей
полнотой. Из других членистоногих-вредителей здоровья более под-
робно рассматриваются мухи-переносчики кишечных заболеваний,
иксодовые клещи (включая Ixodidae и Argasidae) и москиты, борьба с
которыми зачастую входит в план работ малярийных станций. Биоло-
гия других кровососущих двукрылых, а также вшей, блох, клопов и пр.
изложена более кратко, однако в объеме, достаточном для того, чтобы
слушатель мог ориентироваться в этих группах и приступить к изучению
методов борьбы с ними.
Методы борьбы с насекомыми и клещами-вредителями здэрэвья
человека в настоящей книге не рассматриваются, — им нцсвящен от-
дельный курс: соответствующая вторая часть учебника должна вскоре
выйти в свег.
При изложении курса мы старались исходить из уровня знаний уча-
щихся, не вводить лишних терминов, переводить и объяснять все термины,
избежать которых было трудно и знание которых мы считали полезным.
Нам хотелось дать не только учебник, но и руководство, которым окон-
чивший мог бы пользоваться в дальнейшем в своей текущей работе:
«...школа - это только подготовительная ступень. Настоящая закалка ка-
дров получается па живой работе вне школы, на борьбе с трудностями,
на преодолении трудностей» (Сталин). Имея в виду указанную задачу, мы
включили в учебник ряд определительных таблиц, списаний, цифр, фено-
логических дат и других материалов, которые не предназначены для
запоминания, но необходимы на практике. Для выделения этих материа-
лов мы широко использовали мелкий шрифт.
Как уже указывалось, книга предназначена для учащихся школ
помощников медицинских энтомологов. Однако мы полагаем, что она будет
полезна также для энтомологов, врачей и гидротехников, работающих
в противомалярийной организации, а также для работников санитарных
станций и др. Ряд отделов книги (Анатомия и физиология комаров, сводка
по экологии всех видов Anopheles в СССР, сравнительное описание
биологии важнейших видов Aedes п Culex, биология комнатной мухи, систе-
матика и экология аргазид) печатается в таком объеме впервые. Курс
медицинской энтомологии, специально ориентированный на работников
малярийных станций и стоящий па уровне сегодняшнего дня, в нашей
литературе до сих пор вообще отсутствовал.
Между соавторами работа распределилась следующим образом:
В. Н. Беклемишев паппсал: «Общая энтомология»; «Введение в ме-
дицинскую энтомологию»; «Экология Anopheles maculipennis на имаги-
налыюй фазе»; «Географическое распространение A. maculipennis»; «Срав-
нение эпидемиологической роли отдельных видов Anopheles»; «Мошки
и мокрецы».
О. Н. Виноградская: «Наружное строение взрослого комара»; «Вну-
тренняя анатомия и физиология взрослого комара»; «Систематика комаров
(взрослые насекомые)».
В. П. Дербенева-Ухова: «Слепни»; «Высшие мухи».
А. В. Долматова: «Москиты».
О. С. Кузина: «Блохи»; «Вши»; «Клопы»; «Тараканы».
В. И. Олифан: «Гидробиология» (кроме главы IV).
М. В. Поспелова-Штром: «Клещи».
Н. К. Шипицина: «Морфология водных фаз комаров»; «Внутренняя
анатомия и физиология личинок и куколок комаров»; «Экология водных
фаз Anopheles maculipennis»; «Экология и распространение других ви-
дов Anopheles»; «Фенология A. maculipennis»; «Численность популя-
ции A. maculipennis и ее годовой ход»; «Некоторые важнейшие виды не-
малярийных комаров СССР»; «Систематика комаров (личинки)».
Совместно В. Н. Беклемишев и В. И. Олифан: «Гидробиология»
(глава IV): «Основные типы материковых водоемов».
ВВЕДЕНИЕ
РАЗВИТИЕ МЕДИЦИНСКОЙ ЭНТОМОЛОГИИ В НАШЕЙ СТРАНЕ
Медицинская энтомология как самостоятельная отрасль знания —
наука сравнительно молодая. Она всецело принадлежит XX веку. Уси-
ленное изучение насекомых и клещей, нападающих на человека, началось
главным образом после того, как стала выясняться их роль в переносе
заболеваний.
Первое обширное исследование по малярии и малярийным комарам
в России принадлежит В. В. Фавру (1903). Его книга содержит громадный
материал по распространению малярии и ее переносчиков на территории
России и до сих пор является необходимым источником при изучении
истории малярии в нашей стране. К этому же периоду относятся прекрас-
ные исследования по биологии малярийного комара Н. М. Кулагина
(1901), Г. А. Кожевникова (1901—1903), Н. В. Васильева (1912—1913),
И. А. Порчинского (1904).
Порчинский, талантливый наблюдатель, оставил после себя замеча-
тельные исследования по биологии также и других двукрылых—вредите-
лей здоровья: слепней, мух и оводов. Его работы и сейчас читаются с
захватывающим интересом.
Еще в 1889 г. начал свою деятельность по изучению блох проф.
Ю. Н. Вагнер, положивший начало изучения этой группы в нашей
стране. По вшам отличные работы оставил один из крупнейших русских
зоологов профессор Военно-медицинской академии Н. А. Холодковский.
Однако в тот период все эти исследования оставались разрозненными
и медицинской энтомологии как таковой еще не существовало. Она воз-
никла у нас только при советской власти.
После создания советской власти два человека первыми оценили все
значение для строительства здравоохранения в новом обществе паразито-
логии вообще и медицинской энтомологии в частности. Это были Е. И.
Марциновский, уже до революции работавший по малярии, и Е. Н. Пав-
ловский, ученик и преемник Н. А. Холодковского.
Основным делом жизни и незабываемой заслугой Е. И. Марциновского
является создание противомалярийной организации — разветвленной по
всему Советскому Союзу системы тропических институтов и малярийных
(тропических) станций с Всесоюзным институтом малярии и медицинской
паразитологии во главе. Этим самым был создан исследовательский п науч-
но-практический аппарат и для изучения паразитов и переносчиков, и
для широкой борьбы с ними. Таким разветвленным аппаратом для борь-
бы с малярией и другими паразитарными болезнями не располагает ни
одна другая страна в мире. В дальнейшем под руководствОхМ П. Г. Сер-
5
гнева противомалярийная организация, борьба с малярной, а также тео-
ретическая маляриология и паразитология получили още более широкий
размах. Наша противомалярийная организация является предметом за-
конной гордости советского здравоохранения, и мы должны направить
все усилия на то, чтобы непрерывно улучшать ее работу на благо нашей
великой родины.
Деятельность Е. И. Марциновского и создание противомалярийной
сети несомненно открыли дорогу для широкой работы в области меди-
цинской энтомологии. Но в научной организации этой работы гро-
мадную роль сыграл и Е. Н. Павловский. Работая над изучением раз-
нообразных паразитов и переносчиков, Павловский привлек к этой
работе целый ряд ученых и выработал таковых из своих учеников,
поручая им изучение отдельных групп. Таким образом, был создан
ряд книг и руководств, которые явились основой для морфологии, систе-
матики и зоогеографии важнейших в медицинском отношении членисто-
ногих в пределах СССР. Особенное значение имели сводки по комарам
(А. А. Штакельберг, А. С. Мончадский) и москитам (П. 11. Перфильев).
Много открытий было сделано Е. Н. Павловским и его сотрудниками в
результате непрерывно организуемых им паразитологических экспедиций.
В основу большинства этих экспедиций была положена идея всесторон-
него изучения природных очагов трансмиссивных заболеваний. Павлов-
ским был создан и перший на русском языке учебник медицинской пара-
зитологии, включающий и медицинскую энтомологию и выдержавший
ряд издании.
Преимущества советской пауки, вытекающие из огромных возмож-
ностей ее развития, создаваемых советской системой, особенно сказались
при изучении малярийных комаров. Дружная, планомерная работа гро-
мадной сети институтов и малярийных станций позволила нам быстро за-
нять в изучении биологии малярийных комаров одно из первых, если не пер-
вое место в рядах мировой пауки. Фауна малярийных комаров СССР изу-
чена лучше, чем в любой другой стране мира. Те знания по биологии личи-
нок Anopheles нашей страны, которыми мы располагаем, но только удовле-
творяют потребности противоличиночпых мероприятий, но и позволяют
нам при создании водохранилищ или каналов предвидеть степень их при-
годности для жизни личинок и заранее предпринимать защитные меры.
Такого рода прогноз (предвидение) анофелогенности будущих водохрани-
лищ систематически проводится только в СССР.
В области биологии взрослых комаров паша паука также во многом
опережает западную. Так, изучение возрастного состава комариной по-
пуляции, столь существенное при проверке успешности борьбы с взрос-
лыми комарами, стоит у нас гораздо выше, чем за границей (благодаря
главным образом работам Алмазовой, Половодовой и Детиповой). Изу-
чение фенологии комаров (т. о. календарных сроков наступления годового
цикла жизни) всюду за границей сводится к случайным и отрывочным на-
блюдениям, и только у нас представляет систему наблюдений, из года
в год ведущихся на громадной территории по единой программе. Естест-
венно, что в результате такой постановки работы получаются данные,
одинаково важные и для биологии комаров, и для практики борьбы с ко-
марами (Шипицина). В изучении физиологии комаров многое сделано
Виноградской и Ягужинской. Несомненно первое место заняла наша
наука и в вопросе о зоопрофплактике, об использовании отвлечения ко-
маров домашним скотом для защиты людей от малярии. Тщательное изу-
чение этого вопроса (Г. Е. Раевский и др.), а также и других причин,
влияющих на расселение комаров, позволило советским маляриологам
6
впервые разработать противомалярийные требования при планировке
населенных пунктов.
Большие успехи достигнуты нашей наукой и в области изучения ланд-
шафтной маляриологии, т. о. учения о приуроченности мест массового
выплода малярийных комаров, а следовательно, и очагов малярии к опре-
деленным ландшафтам, т. о. к определенным типам местности (Бекле-
мишев, Исаев, Прендель, Устинов, Шипицина и др.) Знания эти очень
полезны при планировании противомалярийных мероприятий и в особен-
ности при составлении генеральных планов оздоровления местности.
Фауна немалярийных комаров СССР начала изучаться с самого на-
чала работы противомалярийной сети, но более пристальное изучение
биологии этих комаров началось лишь после того, как в 30-х годах была
установлена их роль в распространении вирусных энцефалитов и туля-
ремии. В отношении переносчиков японского энцефалита особенно много
сделано П. А. Петрищевой, в отношении Aedes aegypti — Л. П. Калан-
дадзе и А. И. Таировой.
Целый ряд советских работ посвящен «гнусу», т. е. совокупности
кровососущих двукрылых, массами нападающих на человека и животных
в природе. В состав гнуса входят комары, слепни, мошки и мокрецы. На-
падения их иногда бывают настолько тягостны, что делают пребывание в
таких местах почти невыносимым. Над изучением гнуса особенно много
работали А. В. Гуцевич и А. С. Мончадский. Из числа насекомых, вхо-
дящих в состав гнуса, слепням (Tabanidae) посвящена монография Олсу-
фьева^ мошкам (Simuliidao) — Рубцова.
Мухи как переносчики кишечных заболеваний человека имеют гро-
маднее медицинское значение. Внимание к этой проблеме у нас впервые
привлек Е. И. Павловский. Первая лаборатория по изучению мух была
создана в Центральном институте малярии Е. С. Смирновым. В настоя-
щее время, главным образом благодаря работам этой лаборатории и в
особенности В. П. Дербеневой-Уховой, биология мух-переносчиков ки-
шечных заболеваний изучена в нашей стране лучше, чем где бы то ни
было. Однако задача избавления наших городов и сел от мух громадна, и
успешное ее выполнение требует дальнейшего тщательного изучения
этой проблемы.
Изучение москитов (Phlebotomus) и передаваемых ими заболеваний
также стоит у пас высоко. Благодаря работам Марциновского, Попова,
Щуренковой, Перфильева и др. фауна москитов СССР прекрасно изучена;
распространение москитов по различным убежищам, в различных ти-
пах ландшафта изучено в СССР, как нигде (Петрищева и др.). В изучении
биологии москитов большой шаг вперед представляют работы А. В. Дол-
матовой, в изучении их эпидемиологической роли — работы Н. И. Хо-
дукина и в особенности Н. II. Латышева.
Изучение аргазид, клещей-переносчиков возвратных тифов, началось
у нас давно [Бялыницкпп-Бируля (1900)]. Джунковский (1915) открыл
спирохет — возбудителей персидского возвратного тифа, передаваемых
плещем Alectorobius tholozani. Магницкий (1922) нашел эту болезнь у
нас в Средней Азии и Н. И. Латышев (1926) экспериментом на себе дока-
зал факт переноса этих спирохет клещом A. tholozani papillipes. С тех
пор на территории Средней Азии и Кавказа был открыт целый ряд спиро-
хето.юв, передаваемых клещами Ornithodorini. Биология и систематика
этих клещей в настоящее время изучены у нас во многих отно-
шениях лучше, чем где бы то ни было за границей, в особенности благо-
даря работам Павловского, Поспеловой-Штром, Москвина, Исаева, Кан-
делаки. Остроумовой и др.
7
Клещи семейства Ixodidae лишь недавно стали у нас изучаться
медицинскими энтомологами, а именно с тех пор, как в 1937 г. была об-
наружена первая на нашей территории болезнь, передаваемая клещами
этого семейства, — таежный энцефалит. Л. А. Зильбер первый высказал
предположение, что переносчиком этого заболевания является клещ;
В. С. Миронов (1938) показал, что этим клещом может быть только Ixo-
des persulcatus. Эти предположения были вслед за тем полностью под-
тверждены Павловским и Смородинцевым с сотрудниками, очень подробно
изучившими дальневосточные очаги таежного энцефалита. Следует заме-
тить, что до этих открытий опасные для человека и передаваемые клещами
вирусные заболевания вообще не были известны.
В последующие годы на территории СССР было открыто еще несколько
заболеваний человека, передаваемых иксодовыми клещами, и в настоящее
время изучение этой группы клещей занимает в работе медицинского
энтомолога крупное место.
Изучением блох у нас больше всего занимается противочухмная ор-
ганизация. Хорошо налаженная работа этой организации и усилия ряда
талантливых исследователей привели к тому, что блохи в настоящее время
являются одной из наилучше изученных групп членистоногих-перенос-
чиков в СССР, а СССР — одной из стран, наилучше изученных в отноше-
нии фауны блох. Очень хорошая сводная работа по экологии блох в свя-
зи с их эпидемиологическим значением (главным образом перенос чумы,
но также крысиного сыпного тифа, туляремии и пр.) дана И. Г. Поффом
(1941).
Итак, за цремя советской власти у нас создана медицинская энто-
мология, которой в царские времена, несмотря на усилия отдельных уче-
ных, у нас еще не было. Успешно развиваясь, советская медицинская
энтомология вышла на одно из первых мест, а по ряду разделов — на
первое место в мире.
Полное оздоровление нашей страны и освобождение ее от всех бо-
лезней, передаваемых членистоногими, требуют от нас дальнейшей боль
шой работы по изучению переносчиков и мер борьбы с ними.
Методам борьбы с членистоногими паразитами и переносчиками бо-
лезней человека посвящен II том настоящего учебника.
ЧАСТЬ 1
ОБЩАЯ ЭНТОМОЛОГИЯ
Г Л /X В А I
ПРЕДМЕТ И ЗНАЧЕНИЕ ЭНТОМОЛОГИИ
Предмет энтомологии — членистоногие. Энтомология—учение о чле-
нистоногих; членистоногие являются одним из подтипов животного цар-
ства. Таким образом, энтомология представляет собой раздел зоологии —
науки о животных, а сама зоология является одним из разделов биоло-
гии — науки о живых существах, т. е. животных и растениях.
Слово «энтомология» происходит от греческого слова «энтомон», что
буквально означает «насекомое», т. е. членистое животное, как бы покры-
тое насечками. Поэтому очень часто слово «энтомология» понимают как
науку только о насекомых, обозначая науку о паукообразных, как арахно-
логию С науку о ракообразных, как'карцинологию, и т. д. Однако пра-
вильнее обозначать словом «энтомология» науку о всех членистоногих
вообще, включая в это понятие ракообразных, многоножек, насекомых
п паукообразных.
Значение членистоногих. Членистоногие— одна из важнейших групп
животных. Они чрезвычайно разнообразны. По числу видов членисто-
ногие превосходят все остальные группы животных, взятые вместе. Число
особей членистоногих в природе также чрезвычайно велико; в море из
членистоногих особенно многочисленны ракообразные, на суше — насе-
комые и клещи. Достаточно сказать, что в лесной подстилке па площади
1 дм2 живет до 500 мелких клещей, до 100 мелких насекомых и, кроме
того, но нескольку пауков, многоножек и более крупных насекомых.
К тому же членистоногие очень активны, энергично питаются и размно-
жаются и в силу этого оказывают большое влияние па всю окружающую
природу. Некоторые из них питаются живыми частями зеленых растений,
другие — грибами п бактериями, третьи — разлагающимися расти-
тельными остатками, трупами животных, испражнениями, наконец, мно-
гие в качестве хищников пли паразитов питаются живыми животными.
Полезные членистоногие. Большое .значение в жизни природы имеют
насекомые, питающиеся нектаром и пыльцой цветов и при этом опыляю-
щие цветы. Оказывая громадное влияние на всю жизнь природы, члени-
стоногие имеют большое значение и для хозяйства человека. Многие из
них полезны: прямую пользу приносят съедобные членистоногие. Неко-
торые насекомые употребляются для изготовления лекарств, как, на-
пример, жуки-нарывники. Настоящими «домашними животными» стали
медоносная пчела и бабочки-шелкопряды. Косвенная польза, приносимая
членистоногими, не так наглядна, но в действительности она еще больше.
Арахнология — от греческого слова «арахнэ» — паук.
II
Без опыления цветов насекомыми невозможно было бы созревание се-
мян и плодов множества важнейших растений; ракообразные и в море,
и в озерах составляют очень важную часть пищи рыб; многие насекомые
и клещи почвы являются крошечными пахарями, обеспечивающими бла-
гоприятную структуру почвенного слоя, необходимую для поддержания
плодородия почвы. Огромную пользу приносят многие насекомые и пауко-
образные, истребляя вредных насекомых.
Вредные членистоногие. Вред, приносимый человеку членистоногими,
также колоссален. Прежде всего они вредят нам, повреждая полез-
ные растения. Насекомые и клещи уничтожают наши посевы, наши сады,
повреждают и уничтожают леса. Далее, они повреждают наши запасы,
одежду, мебель, жилища. В качестве паразитов и переносчиков болезней
животных насекомые и клещи поражают наших домашних животных и
причиняют громадный ущерб животноводству. В этом заключается их
ветеринарное значение. Многие паразитические ракообразные являются
вредителями рыб. И, наконец, нападая на самого человека, членистоно-
гие наносят ущерб его здоровью — в этом заключается их медицинское
значение.
Таким образом, и польза, и в особенности вред, приносимый члени-
стоногими, так великп и так близко затрагивают интересы человека, что
заставляют нас очень внимательно заняться изучением этих животных.
ГЛАВА II
ЭНТОМОЛОГИЯ КАК ЧАСТЬ БИОЛОГИИ. ЗООЛОГИЧЕСКИЕ
НАЗВАНИЯ
Биологические науки. Итак, энтомология является частью биологии,
науки о живых существах. Биология распадается на ряд отделов соот-
ветственно либо изучаемой области, либо точки зрения ее изучения. По
предмету биология прежде всего делится па зоологию (учение о животных)
и ботанику (учение о растениях) К
Зоология в свою очередь делится на протозоологию (учение о про-
стейших, Protozoa), гельминтологию (учение о гельминтах, червях), энто-
мологию (учение о членистоногих), ихтиологию (учение о рыбах), орни-
тологию (учение о птицах), териологию (учение о зверях) и т. д. По точке
зрения на предмет биология делится на целый ряд частных наук, из
которых важнейшие следующие: 1) морфология — учение о строении и раз-
витии живых существ; 2) физиология — учение о жизненных отправлениях
живых существ; 3) экология 1 2 — учение о взаимоотношениях организма
с окружающей средой, в которой он живет, т. е. о воздействии организма
на среду и среды на организм; 4) биогеография — учение о распростране-
нии различных организмов на земле; 5) биохронология — учение об исто-
рическом прошлом организмов и об их эволюционном происхождении;
6) систематика — учение о сходствах и различиях между всеми организ-
мами в том виде, как они сложились в результате процесса эволюции.
Прикладная энтомология. Прикладная биология изучает пользу и
вред, приносимые отдельными группами организмов, и разрабатывает
способы извлекать пользу и бороться с вредом, который они причиняют.
Прикладная биология опирается на все перечисленные частные науки;
1 Биология — от греческих слов: «bios» — жизнь и «logos» — слово, учение;
таким же образом зоология — от греческих Слов «zoon» — животное и «logos».
Ботаника — от греческого слова «bot» — растение, овощь.
2 Экология — от греческих слов «ecos» — дом и «logos» — слово, учение.
12
в частности, прикладная энтомология опирается на систематику членисто-
ногих, их экологию, морфологию, физиологию и т. д. Так, чтобы понять
вред, причиняемый нам малярийным комаром, и уметь его предотвратить,
мы должны прежде всего уметь отличать комаров от других насекомых и
малярийных комаров от немалярийных; этому учит нас систематика.
Далее, мы должны знать образ жизни взрослого комара и его личинки, ме-
ста, где они встречаются, пищу комаров, условия, при которых они на-
падают на человека, и т. д. Обо всем этом говорит экология комаров.
Но и систематика, и экология комаров опираются на знание строения и
развития комаров, которые составляют предмет морфологии. Далее, по-
нять образ жизни комара и все его взаимоотношения с внешним миром, о
которых говорит экология, невозможно и без знания законов его жизнен-
ных отправлений — дыхания, пищеварения, поведения, размножения
ит. д., о которых говорит физиология.
Основные понятия систематики. Вид. Прежде чем перейти к рассмо-
трению членистоногих, остановимся на некоторых практически наибо-
лее важных понятиях систвхматики.
Основное понятие систематики — вид. К одному виду принадлежат
все сходные между собой особи, различия между которыми не больше тех
различий, которые существуют между детьми одной пары родителей.
Если между особями какого-либо вида встречаются достаточно постоянные
отличия, мы разбиваем такой вид на несколько разновидностей. Раз-
новидности, которые встречаются но всюду, где водится данный вид, а
лишь на строго определенном участке занятой им территории, называются
географическими разновидностями, или подвидами.
Каждый вид животных и растений в какой-то мере отличается от всех
остальных видов и по своему строению, и по физиологическим отправле-
ниям, и по образу жизни; каждый вид распространен на определенной
части земного шара. Область распространения какого-либо вида назы-
вается его географическим ареалом.
Высшие единицы системы: род, семейство и т. д. Группа наиболее
близких между собой видов объединяется в род. Все виды одного рода
более сходны между собой, чем с любым видом другого рода. Наиболее
близкие между собой роды соединяются в трибы, трибы — в семейства,
семейства — в отряды, отряды — в классы, классы — в типы. Между
этими главными подразделениями иногда устанавливают и промежуточные:
подсемейства, подотряды, подклассы и т. д.
Диагноз вида. Каждый вид принято характеризовать кратким переч-
нем его отличительных признаков (видовой диагноз). Когда какой-либо
ученый открывает новый вид животных или растений, он должен опуб-
ликовать диагноз этого вида в печати и дать ему название, придерживаясь
устано вленны х пр а вил.
Правила зоологических обозначений (номенклатуры). Каждый ор-
ганизм обозначается двумя названиями: родовым и видовым. Оба назва-
ния даются на латинском языке. Родовое название всегда пишется с про-
писной буквы, видовое — всегда со строчной (видовые названия живот-
ных пишутся со строчной буквы даже в тех случаях, когда они образо-
ваны от имени собственного). Когда требуется особая точность, то после
видового названия пишут фамилию автора, который впервые описал дан-
ный вид, и год, когда он был описан. Так, обыкновенный городской комар
называется Culex pipiens L. 1758; здесь Culex — название рода, pipiens —
название вида, L. — сокращенная фамилия Linne — Линнея, впервые в
1758 г. описавшего данный вид. Если какой-нибудь вид имеет несколько
подвидов, приходится вместо двойного обозначения («бинарной», т. е.
13
двойной, номенклатуры) прибегать к тройному обозначению. Так, наибо-
лее распространенный в СССР подвид обыкновенного малярийного ко-
мара обозначается как Anopheles maculipennis messeae F'alleroni 1926;
здесь Anopheles —родовое название, maculipennis — видовое, messeae —
подвидовое, Falleroni — автор, описавший данный подвид, 1926 — год,
когда данный подвид был описан. Один из подвидов получает в качестве
подвидового названия видовое, например, A. mac. maculipennis. При со-
кращенном обозначении автор и год не пишутся или пишется только ав-
тор без года. При частом упоминании какого-либо вида родовое название
сокращают до заглавной буквы, видовое название сокращать в таких
случаях не принято; так, можно написать «А. maculipennis», но не сле-
дует писать «Anopheles macul.» или «A.macul.». В том случае, если указан
подвид, подвидовое название пишут обязательно полностью, видовое
и родовое могут быть сокращены, например, «А. mac. messeae».
Большие роды, содержащие много видов, разбивают на подроды.
Каждый подрод объединяет группу наиболее близких между собой видов
данного рода. Один из подродов обозначается тем же именем, как и весь
род в целсм, остальные подроды получают особые названия. Название
подрода пишется с прописной буквы и ставится в скобках после родового
названия. Так, например, виды рода Anopheles, водящиеся в СССР, от-
носятся к двум подродам: Anopheles sensu s trie to (т. e. в тесном смысле
слова) и Myzcmyia; представителем первого является A. (Anopheles) ma-
culipennis Meigen, представителем второго — A. (Myzomyia) superрictus
Grassi. Названия подродов разрешается писать сокращенно, например.
А. (М.) superpictus.
Название трибы образуется путем прибавления окончания-ini к корню
родового названия старейшего (т. е. наиболее давно. известного) рода
трибы, название подсемейства—путем прибавления окончания -inae, назва-
ние семейства — путем прибавления окончания -idae к корню старейшего
родового названия данного семейства или подсемейства. Названия вы-
шестоящих категорий системы (отрядов, классов) даются независимо от
родовых названий. Приведем перечень и примеры основных ступеней
системы.
Царство —например, животные, Zoa
Подцарство » многоклеточные, Metazoa
Тип » членистые, Articulata
Подтип » членистоногие, Arthropoda
Класс » насекомые, Insecta
Подкласс » крылатые, Pterygcgenea
Отряд » двукрылые, Diptera
Подотряд » прямошовные, Orthorhapha
Секция » длинноусые, Nematocera
Семейство » крсвососущие комары, Culicida<
Подсемейство » Culicinae
Триба » Anophelini
Род » Anopheles
Подрод » Anopheles (в тесном смысле)
Вид » A. (Anopheles) maculipennis
Подвид 1 » A. (A.) mac. messeae
1 Под’иды Anopheles maculipennis очень час^о обозшч; кг с я как ргсы; на-
пример, пгшут: «j ась messeae»; однако тькое cj.oi 05 noq c6j ei.re является непра-
вильным.
14
ГЛАВА III
ОСНОВНЫЕ ОСОБЕННОСТИ ЧЛЕНИСТОНОГИХ (ARTHROPODA)
1. ПОЛОЖЕНИЕ ЧЛЕНИСТОНОГИХ В СИСТЕМЕ ЖИВОТНЫХ
12
А
—17
-16
-18
VI
11-
Тип членистых. Членистоногие являются одним из подтипов членисты х
(Articulata) V Каждый из типов животного царства характеризуется
своим планом строения. Членистые—двусторонне симметричные животные^
тело которых слагается пз головной лопа-
сти, ряда члеников и анальной лопасти
(рис. 1). На границе между головной лопа-
стью и первым члеником тела располагается
ротовое отверстие (6), на анальной лопасти
(17) находится задний проход. Для всех
членистых характерно строение их нервного
аппарата. В головной лопасти (4) находится
надглоточный ганглий (мозг) (о), в каждом
из члеников тела (I —VII) лежит па брюшной
стороне пара нервных узлов (ганглиев) (10),
соединенных между собой поперечной пе-
ремычкой (комиссурой) (15) и снабжаю-
щих нервами (13) все органы своего члени-
ка. Каждая пара ганглиев продольными
перемычками (коннективами) (12) соединяет-
ся с соответствующими ганглиями как пре-
дыдущего, так и последующего членика
тела. Таким образом, все пары ганглиев ту-
ловища последовательно соединяются между
собой и образуют брюшную нервную це-
почку. Ганглии первого членика парой
окологлоточных коннективов соединяют-
ся с головным мозгом. Так образуется
окологлоточное нервное кольцо. Такой план
строения является общим для всех члени-
стых.
Подтипы типа членистых. К типу чле-
нистых относятся подтипы кольчатых червей
(Annelides), членистоногих (Arthropoda) и
некоторые другие. Примером кольчатых
червей могут служить всем известные дож-
девые черви (Lumbricidae). Кольчатые
черви представляют низшую группу чле-
нистых, более простого и менее совершен-
ного устройства; членистоногие являются
высшей группой, и многие из них достигают
высокой степени сложности и совершен-
ства.
Классы подтипа членистоногих. Гро-
мадный подтип членистоногих (Arthro-
poda) слагается в свою очередь из нескольких классов. Из них
важнейшими являются четыре: ракообразные (Crustacea), многоножки
Рис. 1. Схема строения
представителей типа членистых
Articulata (оригинал).
А — вид с брюшной стороны; Б —
вид сбоку: 1 — головные придатки
(усики); 2 — их нерв; 3 —глаза:
4 — головная л^насть; 5 - го-
ловной мозг; 6 — рог; 7 — перед-
няя шинка (глотка); s—ганглий
1-го членика тела (1 -и узел брюш-
ной нервной цепочки); 9— парные
конечности; ю — нервные узлы
брюшной нервной цепочки; 11 -—
средняя кишка; 12—продольные
перемычки (коннективы) брюшной
нервной цепочки; 13 — нервы;
14—сегментальные органы:
/•>—поперечные перемычки нерв-
ной цепочки; 1в •— задняя кишка;
17 — анальная лопасть; 18— ее
придатки; 19 — спинной кровенос-
1 В большинстве учебников ч пенисто то гиз рассматриваются как самостоятель-
ный тип Arthropoda; такая классификация неправильна, так как затушевывает
сходство и родство членистоногих с кольчатыми червями.
(Myriapoda), насекомые (Insecta) и паукообразные (Arachnoidea). Все
эти классы, в особенности ракообразные, насекомые и паукообразные,
встречаются в большом числе видов и в неисчислимом множестве особей.
В силу этого они играют в жизни природы первостепенную роль. Ракооб-
разные живут главным образом в воде; в настоящее время их известно
около 20 000 видов. Многоножки живут исключительно на суше; их из-
вестно 8 100 видов. Насекомые живут и на суше, и в воде и являются са-
мым большим из всех классов животных: число видов насекомых доходит
до 1 млн. Паукообразные заселяют главным образом сушу и по числу
видов уступают насекомым; их известно около 28 000 видов. Однако п<>
числу особей паукообразные, в частности,
клещи, в отдельных случаях даже прево-
сходят насекомых; роль их в жизни при-
роды колоссальна, хотя в целом они и
уступают в этом отношении насекомым.
Для здоровья человека основное значе-
ние имеют насекомые и паукообразные, в
частности, клещи.
Рис. 2. Строение кожных покровов.
Л —кольчатого червя; Б— членистоногого (схемы):!—кутикула червя; 2—наружный
эпителий кожи; 3 — слой кольцевых и 4 — слой продольных мышц; 5 — эпителий, выстила-
ющий полость тела червя; 6—эпикутикула; 7— экзокутикула; 8—эндокутикула; 9 — основ-
ная перепонка эпителия; 10—истинный волосок; 11—выделяющая его клетка; 12—ку-
тикулярные шипикв.
2. СТРОЕНИЕ КОЖНОГО ПОКРОВА КАК ОСНОВНАЯ ОСОБЕННОСТЬ
ЧЛЕНИСТОНОГИХ
Кожный покров кольчатых червей. Важнейшей особенностью члени-
стоногих, прежде всего отличающей их от кольчатых червей, является
устройство их кожного покрова. Кожный покров кольчатого червя (рис.
2, 4) состоит из одного слоя клеток, расположенных правильной пластин-
кой (как говорят, в виде однослойного эпителия). Клетки эпителия обра-
зуют на своей наружной поверхности плотный слой защитного вещества,
кутикулу (рис. 2, .4,1). Кутикула кольчатых червей настолько тонка, что
не мешает ни движениям, ни росту животного, тело остается мягким и
мало защищенным. Мускулатура тела образует сплошной мешок, под-
16
стилающий кожный эпителий, и в силу этого все тело может изгиоаться,
сокращаться и вытягиваться целиком или на любом участке.
Особенность кожного покрова членистоногих — толстая кутикула.
В отличие от этого кожный эпителий членистоногих покрыт толстой кути-
кулой, толщина которой обычно превосходит толщину самого эпителия,
иногда во много раз (рис. 2, Б), Такая кутикула представляет для жи-
вотного хорошую защиту, нередко настоящую броню. Но, с другой сто-
роны, толстая кутикула мало растяжима, а там, где она прочнее, она
теряет и гибкость, как соче-
тается прочность брони с гиб-
костью и подвижностью тела?
Это достигается следующим
путем. Броня, покрывающая
голо членистоногого, образо-
вана рядом утолщенных, же-
стких? твердых щитков, кото-
рые отделены друг от друга
участками тонкой и гибкой ку-
тикулы. Щитки носят название
склеритов, соединяющие их
участки тонкой кутикулы назы-
ваются соединительными пере-
понками. Соединительные пере-
понки, складываясь или рас-
правляясь, позволяют склери-
там передвигаться друг относи-
тельно друга.
Наружный скелет члени-
стоногих. Кожные мышцы, ко-
торые у кольчатых червей об-
разуют сплошной кожно-мы-
шечный мешок, у членисто-
ногих представлены отдельными
пучками, идущими от одного
склерита к другому. При со-
кращении этих мышц склериты
сближаются между собой и
этим путем осуществляют все
движения животного. Таким,
образом, склериты кутикулы
образуют не только броню, но
и наружный скелет членистоно-
гого. Наподобие наших костей,
они служат опорой для мышц,
но в отличие от нашего ске-
Рпс. 3. Наружный скелет и мускулатура
членистоногого. Схема поперечного разреза
через крылатый грудной сегмент насекомого.
1— тергит; 2-—продольные спинные мышцы; 3—кры-
лья (изображены частично); 4—дорзо-вентральные
(сииннобрюшные) мышцы; 5— плеврит; 6 — его
внутренний отросток; 7 — стерпит; 8 — его внут-
ренние ' отростнн; 9— промежуточный мускул;
10 и 11 — мышцы, идущие к основанию ноги;
12 и 13—мышцы тазика; 14—мускул — двигатель
бедра; 15— мускул —разгибатель голени; 16—мус-
кул — сгибатель голени; 17 и 18— мускулы —
сгибатели лапни;’!'?— мускул — разгибатель лапки;
20—сухожилие лапки.
лота, являющегося внутренним, скелет членистоногих наружный.
Обыкновенно в каждом членике тела бывает по меньшей мере два
щитка: спинной щиток (тергит, рис. 3, 1) и брюшной (стернит, рис. 3,7).
Иногда бывают и дополнительные щитки (рис. 3, 5), иногда, наоборот, тер-
гит и стернит могут сливаться между собой или со склеритами соседних сег-
ментов, образуя крупные щиты. У некоторых членистоногих, в виде исклю-
чения, склеритов совсем нет и вся кожа является более или менее гибкой.
Конечности. У примитивных членистоногих — многоножек (рис. 5, Б)
и многих раков (рис. 12, Г) — каждый членик тела несет пару придатков—
2 Учебник медицинской энтомологии
17
(рис. 3, 9—19).
Рис. 4. Зубец, об-
разованный вы-
ступом кожи и со-
держащий про-
должение поло-
сти тела. Разрез
через кожу насе-
комого.
конечностей. Конечности обычно бывают покрыты толстой кутикулой,
образующей отдельные трубчатые склериты, соединенные перепонками и
сочлененные между собой. Таким образом, каждая конечность разбита
на отдельные членики, откуда и происходит название членистоногих.
Конечности, состоящие из подвижно сочлененных члеников, снабжены
многочисленными мышцами: сгибателями, разгибателями и вращателями
Конечности образуют сложные рычаги, допускающие
быстрые, сложные и точные движения, и являются
замечательными органами плавания, беганья, хватания,
жевания, благодаря которым членистоногие оказы-
ваются в числе наиболее подвижных и активных жи-
вотных.
Придатки кожи. Чрезвычайно разнообразны соб-
ственные придатки самой кожи членистоногих
(рис. 2, Б). Их можно разделить на три группы:
1. Придатки, образованные поверхностью кутикулы
(ложные волоски, кутикулярные шипики, рис. 2, Б, 12);
кожный эпителий в них не проникает, они не имеют
подвижного сочленения с остальной кутикулой и обыч-
но очень малы. 2. Истинные волоски (рис. 2, Б9 10).
Каждый из них выделяется отдельной клеткой, отрос-
ток которой и заполняет собой волосок. Последний при
помощи тонкой хитиновой пленочки подвижно сочленен
с кожей. Если такого рода истинный волосок тонок,
он носит название волоска; более толстые волоски
называются щетинками. Волоски могут быть раз-
ветвленными (как, например, у личинок Anopheles),
вообще принимать разнообразные формы. Иногда у бабочек, комаров
и др. они бывают сплющены и принимают вид чешуек. 3. Зубцы, пред-
ставляющие собой вырост покровов, в который продолжается полость
тела (рис. 4).
3. РОСТ И ЛИНЬКА ЧЛЕНИСТОНОГИХ
Рост и кутикула. Под влиянием кожного покрова видоизменяется
и способ развития проста членистоногих. Толстая кутикула их и в особен-
ности все склериты нерастяжимы и, таким образом, препятствуют росту
животного. Молодое животное питается, количество живого вещества
в его теле увеличивается, но рост задержан; создается напряженное по-
ложение, которое разрешается наступающей линькой.
Линька. При линьке старая кутикула отстает от эпителия, в простран-
ство между ними изливается секрет имеющихся в коже линочных желез1,
под влиянием которого старая кутикула частично переваривается. После
этого она лопается, и животное вылезает из нее, как из футляра. Тело его,
выйдя на свободу, сразу настолько увеличивается в размерах, что уже
не смогло бы поместиться в старой шкурке. К этому времени наружный
эпителий уже успевает образовать новую кутикулу, пока еще мягкую и
бесцветную. Вскоре она достигает нормальной толщины, твердеет и тем-
неет. Таким образом, рост членистоногих по крайней мере у форм, сплошь
1 Железистыми называются те клетки организма, которые производят жидкие
вещества, необходимые организму, как, например, слюна, желудочный сок и т. п.
Вещества, приготовляемые и выделяемые железистыми клетками, называются секре-
тами. Железистые клетки могут размещаться поодиночке среди нежелезистых, как,
например, линочные железы насекомых, или могут быть собраны в целые органы —
многоклеточные железы, как, например, слюнные железы комара.
18
одетых в толстую броню, всецело приурочен к периодам линьки, в силу
чего является прерывистым, периодическим. У членистоногих с мягкими
покровами, вроде личинок мух, значительный рост происходит и между
линьками за счет растяжения кутикулы. Однако растяжимость кутикулы
и у них имеет пределы, и без периодических линек они тоже обойтись
не могут. У личинок комаров, имеющих твердую головную капсулу
(см. ниже) и тонкую кутикулу на остальном теле, рост тела происходит и
между линьками, а увеличение головы — исключительно в период линьки.
Возрастные изменения формы тела и его придатков — конечностей,
шипов, волосков и пр. — также совершаются исключительно во вре^ми
линек. В промежутке между линьками единственные возможные измене-
ния внешности членистоногого сводятся к пассивному растяжению мягких
участков кожи, соединительных перепонок. Поэтому смена стадий развития
членистоногих всегда совпадает с линьками; каждая линька является кон-
цом одной стадии, или возраста, и началом следующей.
4. РАСЧЛЕНЕНИЕ ТЕЛА ЧЛЕНИСТОНОГИХ
Отделы тела у многоножек. У многих кольчатых червей все членики
тела хорошо обособлены перетяжками и все более или менее одинаковы
между собой (рис. 5, . J. В отличие от этого у членистоногих такое одно-
образие резко нарушено. Их тело распадается на отделы, состоящие
каждый из нескольких члеников. Так, у многоножек (рис. 5, Б) имеется
голова и туловище. Голова многоножки состоит из предротовой, или го-
ловной, лопасти и четырех члеников, тергиты и стерниты которых более
или менее слились в одну головную капсулу или череп. Конечности головы
видоизменены либо в органы чувств (усики или антенны, принадлежащие
предротовой лопасти), либо в ротовые органы (одна пара верхних челю-
стей, или жвал, и две пары нижних челюстей). Вторая пара усиков, ле-
жащая впереди жвал, у зародыша многоножек появляется, но затем
исчезает и у взрослой многоножки отсутствует. Туловище многоножек
состоит из длинного ряда более или менее одинаковых члеников, несу-
щих по одной паре ног1.
Отделы тела } насекомых. У насекомых (рис. 5, В) имеется голова,
в состав которой вх дят те же членики и с такими же конечностями, как
в голове многоножек. Но в отличие от многоножек туловище насекомых
распадается на два отдела: грудь и брюшко. Грудь состоит из 3 члеников,
брюшко имеет не более 11 члеников. Членики груди сохраняют хорошо
развитые и служащие для передвижения ноги. Конечности брюшка боль-
шей частью совсем отсутствуют; если же они имеются, то не служат для
передвижения, а выполняют какие-нибудь другие функции: органов ося-
зания или половых придатков. Исключений очень мало; к числу исклю-
чений относятся гусеницы бабочек с их многочисленными парами нечлени-
стых брюшных ножек. Разделение туловища на грудь и брюшко — глав-
ное отличие насекомых от многоножек.
Отделы тела у ракообразных. Расчленение тела ракообразных
(рис. 5, Г) быв *ет весьма различно, но их головные конечности в основном
всегда похожи на головные конечности многоножек и насекомых: ракооб-
разные имеют две пары усиков, одну пару верхних челюстей (жвал) и две
пары нижних челюстей. Отличие заключается в том, что у ракообразных
1 У части мно'оно кек (огряд Diplopoda) членики тела сливаются попарно,
в силу чего создается впечатление, что каждый членик несет две пары пог; в дей-
ствительное ги каждая из них принадлежит отдельному сегменту, лишь утратившему
свою самостоятельность.
2*
19
vr///
15
15
Рис. 5.
Расчленение
тела коль-
чатого червя
и членисто-
ногих
(Lopadorhyn-
_ тремя парами
— задний проход: Б — мпого-
Л—мореной кольчатый червь
ehus): 1 — головная лопасть с
придатков; 2- . __Л __г__ ________
ножка Scutigerella (вид снизу): 1—IV—голо-
ва, образованная слиянием головной лопасти и 4 члеников; XVIII—членики туловища:
2—усики (остальные головные конечности плохо видны); 3—ноги; 4—церки (конечности
18-го членика туловища); В — насекомые. Тело жука, расчлененное па главные отделы (вид
сверху): 1— голова с усиками, глазами и ротовыми частями; 2— переднегрудь с первой парой
ног; 3 — среднегрудь со второй парой ног и первой парой крыльев (надкрыльями); 4—зад-
негрудь с третьей парой пог и второй парой крыльев; 5—брюшко; Г— ракообразное. Боки-
плав, G-ammarus (вид сбоку): I — VI — голова, образованная слиянием головной лопасти
и 5 члеников; VII—XIII — члеппкп груди; XIV—XIX—членики брюшка; 1 и 2—усики (пер-
вые и вторые); 3—верхние челюсти; 4*и 6 — две пары максилл (нижних челюстей); 5—че-
люстные ножки; 7 — 13— грудные ноги; 14— 19— брюшные конечности; 26— центральный
отдел нервного аппарата; Д—‘ паукообразные, лжескорпион, Chelifer (вид со спинной стороны):
i — хелицеры; 2— педипальпы; 3 — первая пара пог; 4 — четвертая пара ног; 5 — три щитка,
покрывающие передний отдел тела (просому), состоящий из головной лопасти и 6 члеников;
VIII, XVII и XVIII— тертиты остальных члеников, образующих задний отдел тела (метасому).
всегда хорошо развита вторая пара усиков, которая у многоножек и на-
секомых появляется на зародышевой стадии развития, но затем всегда
исчезает. Кроме того, ракообразные, как правило, дышат жабрами, мно-
гоножки и насекомые — трахеями (см. ниже).
Отделы тела у паукообразных. Паукообразные отличаются совершенно
иным наборОлМ конечностей (рис. 5, Д).
Передний отдел их тела образовался слияниехМ головной лопасти и
шести члеников и несет шесть пар конечностей, резко отличных от ко-
нечностей раков, многоножек и насекомых. А именно: первой пары уси-
ков у паукообразных совсем нет, вторая пара усиков, отсутствующая у
насекомых, представлена у паукообразных хелицерами, которые имеют
вид клешней или крючков и служат для хватания пищи; вместо жвал пау-
кообразные имеют ногощупальца (педипальпы); иногда педипальпы так-
же имеют вид клешней (например, у скорпионов), но чаще являются щу-
пальцами и входят в состав ротового аппарата. Четыре остальные пары
являются ходильными ногами.
Задний отдел тела паукообразных может состоять из различного числа
члеников, но не менее шести. Подобно брюшку насекомых, задний
отдел тела паукообразных либо вовсе лишен конечностей (например, у
бихорок, у клещей), либо несет конечности, не служащие для передвиже-
ния, а видоизмененные в половые придатки или в органы дыхания (напри-
мер, у скорпионов). Ввиду этого передний отдел тела паукообразных
иногда обозначают как головогрудь, задний — как брюшко, хотя такое
отождествление и не совсем правильно.
5. ОРГАНЫ ПИЩЕВАРЕНИЯ, ДЫХАНИЯ, ВЫДЕЛЕНИЯ
И КРОВООБРАЩЕНИЯ ЧЛЕНИСТОНОГИХ
Органы пищеварения. Кишечный канал всех членистоногих слагает-
ся из трех отделов: передней, средней и задней кишки. Передняя и зад-
няя кишка при развитии зародыша образуются впячивапием наружной
кожи (наружного зародышевого листка, пазываслмого также эктодермой).
В соответствии с этим они всегда бывают выстланы изнутри хитиновой
кутикулой. Средняя кишка при развитии образуется из внутреннего
зародышевого листка (энтодермы). Это собственно пищеварительный
отдел кишечника. Эпителий ее лишен хитиновой кутикулы.
Органы дыхания членистоногих. Многие членистоногие, живущие
во влажной среде, не имеют специальных дыхательных органов и дышат
просто через кожу. Более крупные ракообразные имеют разветвленные или
неразветвлепные жабры, сидящие на первых члениках некоторых пар
ног (рис. 6, А, 8). Тонкие стенки и большая поверхность жабер облегчают
газообмен между кровью животного и наружной водой.
У скорпионов и пауков имеются небольшие легочные мешки, откры-
вающиеся наружу дыхательными отверстиями на нижней стороне брюшка.
Одна из стенок легочного мешка образует множество складочек в виде
тонких листочков, служащих для увеличения дыхательной поверхности
легкого. У пауков бывает одна или две пары легочных мешков, у скор-
пионов — четыре пары.
У всех остальных наземных членистоногих органами дыхания явля-
ются трахеи (рис. 7, Б, 2- рис. 19). Трахеи представляют собой тонкие
трубочки, открывающиеся наружу дыхальцами (стигмами) и разветвляю-
щиеся внутри тела. Трахеи образованы врастанием наружной кожи и по-
тому также состоят из кожного эпителия и хитиновой кутикулы. Обычно
трахеи имеют очень характерный вид, так как стенка их представляет утол-
21
щение в виде пологой хитиновой спирали, придающее трахейным стволи-
кам как бы поперечную исчерченность. Это утолщение делает стенки тра-
хей более жесткими и мешает им спадаться и сплющиваться под влиянием
давления со стороны окружающих тканей. В то же время промежутки
между оборотами спирального утолщения остаются тонкими и легко до-
пускают прохождение газов из трахей в ткани и обратно. Лишены спи-
Рис. 6. Органы дыхания и кровообращения членистоногих.
А — речной рак (AstaciB): 1— сердце с тремя остиями; 2—головная аорта; 3—сяжко-
вая артерия; 4—задняя аорга; 5— нисходящш артерия; 6'—главная артерия брюшной сто-
роны; 7 — брюшной венозный сгвол; 8— капилляры жабер; .9— выносящие сосуды жабер,
несущие кровь к сердцу; Б — паук (Lycosa): 1, 3 и 4 — артерии ядовитой железы хелицер;
2-— глазная артерия; 5— передний слепой отросток кишки (остальные части кишки не
врисованы); 6 — аорта; 7— 8—задние артерии; 9— 10—боковые артерии сердца;
11 — легкое; 12— выносящий легочный сосуд; 13—16— артерии ног; 17— артерия пищевода;
18— артерия педипальп; 19— артерия хелицер; В— план кровообращения личинки стре-
козы: 1—сердце; 2—его входные отверстия, остии; 3— клапаны сердщ; 4—аорта
5 —жабры; стрелки показывают путь крови по полостям тела.
рального утолщения лишь самые тонкие концевые разветвления трахей —
трахеолы.
И жабры, и легкие служат только мостом, где кровь насыщается кис-
лородом и отдает углекислоту, кровь же разносит кислород по всему телу.
Наоборот, трахеи сами разветвляются по всему телу и непосредственно
доводят воздух до тканей.
22
Кровеносный аппарат членистоногих. В связи с этим кровеносный
аппарат (рис. 6) хорошо развит только у крупных ракообразных, имею-
щих жабры, у крупных паукообразных, имеющих легкие, и у многоно-
жек, имеющих слабо развитые трахеи. У насекомых и у всех мелких
форм из других классов, в том числе и у клещей, кровеносный аппарат
сильно упрощен: от него остается только сердце, лежащее у всех члени-
стоногих на спинном стороне тела,
над кишечником (в отличие от
позвоночных, у которых сердце
лежит на брюшной стороне).
У клещей сердце имеет вид неболь-
шого мешочка, у насекомых—длин-
ной трубки, тянущейся вдоль все-
го брюшка (рис. 6, В),
Рис. 7. Органы выделения членистоногих.
А— коксальная железа скорпиона: 1— концевой мешочек; 2— мочевой пузырен; 3— ар-
терия; 4— выводной канал; 5— лабиринт; 6— диафрагма; Б— клещ Haemaphysalis, вскры-
тый самец; вокруг заднего прохода вырезан кусочек хитина и органы пищеварения отведены
вперед: 1— мальпигиев сосуд; 2— пучок трахей; 3— слюнные железы; 4— отростки кишеч-
ника; 5— семяпровод(по Павловскому).
Выделительные органы. У ракообразных органами выделения явля-
ются сегментальные железы, открывающиеся наружу при основании ко-
нечностей. В отличие от кольчатых червей, имеющих сегментальные выде-
лительные органы во всех или во многих члениках тела (рис. 1), ракооб-
разные обычно имеют одну пару таких желез. Паукообразные также имеют
сегментальные железы, обозначаемые у них как коксальные железы
(рис. 7, А), так как они открываются на коксах (первых члениках ног).
У клещей имеется всего одна пара коксальных желез, открывающаяся
при основании первой пары ног.
Наряду с этим, у всех наземных членистоногих имеются выделитель-
ные органы другого типа, а именно мальпигиевы сосуды (рис. 7, Б, 1).
Это тенкие трубочки, слепые на одном конце, другим концом открываю-
щиеся в кишечник. У насекомых (рис. 18, 7) и многоножек они открывают-
ся в заднюю кишку, у паукообразных — в среднюю кишку. У большин-
ства насекомых мальпигиевы сосуды являются единственными выделитель-
ными органами. У некоторых клещей имеются и мальпигиевы сосуды, и
коксальные железы.
‘6. НЕРВНЫЙ АППАРАТ И ОРГАНЫ ЧУВСТВ ЧЛЕНИСТОНОГИХ
Нервный аппарат. Центральный отдел нервного аппарата всех чле-
нистоногих состоит из надглоточного ганглия, или мозга, окологлоточ-
ных коннективов и брюшной нервной цепочки (рис. 8). Мозг у больший-
23
ства членистоногих посылает нервы к глазам и усикам, у паукообразных —
к глазам и хелицерам, у всех членистоногих — к передней кишке. Все
остальное тело получает нервы от брюшной цепочки. Каждая пара ган-
глиев брюшной цепочки посылает нервы к органам своего членика: дви-
гательные нервы — к мышцам и железам и чувствительные нервы —к коже
п другим органам. У некоторых членистоногих брюшная нервная цепоч-
Рис. 8. Схемы строения центрального нервного аппарата
насекомых.
1—комарик, Tendipes plumosus; Б—слепень,’Tab anus bovinus;
В—мясная муха, Sarcophaga carnariaj Г—постельный клоп, Ci-
mex lectularis: 1—надглоточный ганглий; 2—основание усиков;
3— глазные ганглии; 4—сложные глаза; 5— окологлоточные кон-
нективы; 6 — один из грудных ганглиев; 7 — один из брюшных
ганглиев; 8 — периферические нервы.
ка бывает резко укорочена; благодаря укорочению коннективов, часть
ганглиев сближается между собой, соприкасается и сливается в один
сложный ганглий. Однако и внутри него ганглии отдельных члеников
сохраняют свою самостоятельность и каждый из них продолжает посылать
нервы к своему собственному членику, хотя сам нередко сдвинут из этого
членика куда-нибудь вперед, иногда на значительное расстояние. У мно-
гих форм (клопы, некоторые мухи и жуки) укорочение брюшной цепочки
доходит до того, что все ее ганглии сливаются в одну сплошную брюшную
нервную массу, или брюшной мозг (рис. 8, В и Г), а у клещей, благодаря
укорочению также и окологлоточных конпективов, весь центральный
отдел нервного аппарата сливается в сплошную массу, окружающую
пищевод.
Органы чувств. Раздражения, исходящие от внешнего мира, воспри-
нимаются членистоногими при помощи множества органов чувств. По функ-
ции органы чувств наземных членистоногих можно разделить следующим
образом: 1. Осязательные органы, воспринимающие механические раздра-
жения: прикосновение, давление и т. п. 2. Органы температурного чувства,
воспринимающие тепло и холод. 3. Органы, воспринимающие влажность
воздуха. 4. Органы, воспринимающие химические свойства предметов —
вкусовые и обонятельные. 5. Хордотональные органы, воспринимающие
колебания и сотрясения. 6. Слуховые органы, воспринимающие звуковые
колебания. 7. Зрительные органы.
Осязательные органы. Органы осязания бывают двоякого рода.
В местах с тонкой кутикулой имеются разветвленные чувствительные
24
клетки, прикасающиеся к коже многочисленными окончаниями и восприни-
мающие всякий прогиб ее при прикосновении (рис. 9,6); полученное раз-
дражение передается в ганглий по центростремительному отростку чувст-
вительной клетки (рис. 9, 7). В местах с толстой кутикулой органами ося-
зания служат волоски. Волосок бывает подвижно прикреплен к кутикуле
при помощи тонкой соединительной перепоночки. Каждый волосок имеет
свою клетку-образовательницу, которая проникает через всю толщу ку-
тикулы и продолжается внутрь волоска. Чувствительная клетка лежит
под кожей п ее воспринимающий отросток проходит рядом с клеткой-
образовательницей через тол-
щу кутикулы и заканчивает-
ся чувствительным оконча-
нием в месте причленения
волоска (рис. 9, 3); с другой
стороны от чувствительной
клетки отходит центростре-
мительный отросток, направ-
ляющийся к нервному узлу
(рис. 9 ,4). При малейшем
Рис. 9. Органы осязания насекомого (схема).
1 — хитиновая кутикула склерита; 2—чувствительная
клетка; 3 — ее чувствительный отросток с окончанием
при основании волоска; 4 — ес центральный отросток,
идущий к мозгу; 5—волосок; 6—чувствительная
клетка с многочисленными окончаниями на тонком
хитине сочленовной перепонки; 7 — ее центральный
отросток.
прикосновении к волоску он
наклоняется, надавливает на
воспринимающее окончание,
и животное воспринимает
раздражение. Таким образом,
несмотря на непроницаемую
броню, членистоногие сохра-
няют тонкое осязание.
Органы обоняния. Органы обоняния часто бывают также представ-
лены волосками, но в отличие от осязательных обонятельные волоски
покрыты чрезвычайно тонкой кутикулой, через которую легко проникают
пахучие вещества и воспринимаются заходящими в волосок окончаниями
чувствительных клеток (рис. 10, А и В). Часто обонятельный волосок
бывает укорочен и превращается в небольшой сосочек, едва выдающийся
над поверхностью кожи или даже скрытый в глубокой ямке кутикулы.
Обонятельные органы у насекомых особенно многочисленны на усиках,
у клещей — на последнем членике первой пары ног. У пксодовых кле-
щей здесь образуется сложный обонятельный орган (Галлеров орган)
(рис. 304, 12).
Органы вкуса. Органы вкуса также представляют собой видоизменен-
ные волоски и чаще всего имеют вид тонкостенных палочек или сосочков.
У насекомых они располагаются на ротовых придатках, но иногда и па
усиках, а у мух и бабочек — даже на лапках. У клещей они лежат на
ротовых придатках.
Органы, воспринимающие влажность, построены приблизительно
так же, как и органы химического чувства (обоняния и вкуса)'.
Хордотональные органы и органы слуха. Насекомым свойственны
хордотональные органы, воспринимающие сотрясения и вибрации
(рис. 10, Б). Когда вы неосторожно подходите к воде, личинки комара
воспринимают сотрясение почвы, передающееся воде, и тотчас ныряют:
они снабжены хордотональными органами.
Органы слуха воспринимают не вибрации предметов, а вибрации
воздуха, т. е. звуковые волны. Сложные органы слуха, построенные из
большого количества хордотональных органов, свойственны немногим
насекомым: кузнечикам, сверчкам, кобылкам, некоторым ночным бабоч-
25
кам. У некоторых членистоногих органами слуха служат осязательные
волсски, вибрирующие в тон определенному звуку; вибрации восприни-
маются чувствительной клеткой, находящейся у основания волоска. Та-
кие слуховые волоски имеются, например, на сяжках комаров. Но очень
многие членистоногие непосредственно звуковых волн не воспринимают
совсем и ощущают лишь вибрацию предметов, с которыми соприкасаются,
по примеру того, как мы ощущаем вибрацию, когда кладем руку на
спину говорящего человека.
Рис. 10. Обонятельные и хордотональные органы насекомых.
А — схема строения обонятельного органа: /—эпителий ножи; 2—обонятельные
клетки; 3— их чувствительные отростни; 4— их нервные (центральные) отростки;
5—чувствительный сосочек; Б— хордотональный орган насекомого (орган, вос-
принимающий сотрясения и колебания): 1—клетки эпителия кожи; 2—кутинула;
» — обкладочные клетки; 4—их кутикулярная выстилка; 5—чувствительная
клетка; 6— ее нервный отросток; 7— ее чувствительный отросток; 8— воспри-
нимающее окончание в виде утолщения на конце отростка; В—кончик .усика
(антенны) гусепипы Mamestra: 1— осязательная щетинка (конец обломан);
2—рудиментарный последний членик; 3—большие и 4—малые обонятельные
шипики.
Органы зрения. Глаза различных членистоногих устроены различно.
Большинство ракообразных и насекомых имеет сложные глаза (рис. 11).
Сложные глаза ракообразных и насекомых. Каждый сложный глаз
состоит из множества тесно сближенных маленьких глазков, называемых
омматидиями. Кутикула кожи образует над каждым омматидием прозрач-
ную, двояковыпуклую линзу. Под ней располагается светопреломляю-
щий «хрустальный» конус; еще глубже лежит световоспринимающий ап-
парат, состоящий из семи длинных клеток, совместно выделяющих одну
общую светочувствительную палочку, вокруг которой они и располага-
ются. От каждой из воспринимающих клеток идет к мозгу центростре-
мительное нервное волокно. Весь омматидий окутан пигментными клет-
ками. Благодаря этим клеткам, образующим вокруг омматидия непрозрач-
ный футляр, каждый омматидий представляет собой как бы узкую трубку.
Все омматидии, вместе образующие сложный глаз, представляют как
бы густой пучок узких зрительных трубочек, направленных в разные
стороны. В каждую трубку виден лишь крошечный участок находящегося
перед глазами предмета, но из множества таких участков слагается пол-
26
Рис. И. Схема построения изобра-
жения в сложном глазу насекомого.
1— хитиновая кутикула головы; 2—хру-
сталик отдельного омматидия (глаз-
ка); 3 — зрительные палочки, воспри-
нимающие свет; 4 — пигмент; 5 — зри-
тельный нерв; 6 — хрустальные конусы;
а, б, в, г, О, е— лучи, попадающие на
светочувствительные части глазков (ре-
тинулы) и дающие изображение отдель-
ных точек предмета; все остальные изо-
браженные на рисунке лучи— а-2, а3, at,
и т. д.—на ретинулы не попадают;
7—8 — изображение видимого предмета.
вовсе лишены глаз.
ное изображение. Ввиду того что поле зрения сложного глаза слагается,
как мозаика, из множества отдельных кусочков, зрение насекомых на-
зывают мозаичным.
Таким образом, насекомые, обладающие сложными глазами, разли-
чают форму и очертания предметов. Однако острота зрения насекомых,
вообще говоря, невелика, и сколько-
нибудь отчетливо они видят лишь на
близком расстоянии: все они близору-
ки. Все насекомые лучше замечают
движущиеся предметы, чем неподвиж-
ные. Многие насекомые различают цве-
та, например, пчела; у многих других
способность различать цвета отсутст-
вует или сомнительна.
Простые глаза насекомых. Кроме
пары сложных глаз, многие взрослые
насекомые имеют на лбу 1—3 простых
глазка (рис. 16, 6). Личинки также
большей частью имеют простые глазки,
иногда по нескольку с каждой стороны.
Некоторые насекомые даже и во взрос-
лом состоянии имеют только простые
глазки (вши, блохи). Зрение, дости-
гаемое такими глазками, большей
частью очень несовершенно, и во многих
случаях их обладатели способны вос-
принимать только разницу в яркости
света и направление световых лучей,
но не различают формы предметов.
Некоторые насекомые совсем лишены
глаз, как, например, личинки мух и
многие блохи. Клещи либо имеют
одну-две пары простых глазков, либо
«слепые» клещи и насекомые все же обычно отличают свет от тьмы; оче-
видно, они воспринимают свет какими-то чувствительными клетками кожи.
7. ПОЛОВОЙ АППАРАТ, РАЗМНОЖЕНИЕ И РАЗВИТИЕ ЧЛЕНИСТОНОГИХ
Половой аппарат. В огромном большинстве случаев членистоногие
раздельнополы. Основную часть мужского полового аппарата представ-
ляют семенники, в которых происходит образование мужских половых
клеток, живчиков (сперматозоидов). От семенников отходят семенные
протоки, открывающиеся наружу половым отверстием. Нередко имеются
мужские совокупительные органы, в разных группах устроенные весьма
различно. Женский половой аппарат состоит из яичников, в которых об-
разуются женские половые клетки, яйца. От яичников отходят яйце-
воды, также открывающиеся наружу половым отверстием. Иногда имеются
и женские совокупительные органы.
Размножение. Большинство членистоногих откладывает яйца, но
многие насекомые и паукообразные являются живородящими. У многих
ракообразных самки вынашивают отложенные яйца, приклеенные к
ножкам (речной рак) или заключенные в яйцевые мешки (циклоп), или
в выводковые камеры различного устройства (дафнии, мокрицы). Неко-
торые пауки (Lycosidae) и насекомые (например, тараканы) тоже носят
27
с собой своп окруженные яйцевыми капсулами яйца, иногда вплоть до
вылупления молоди. Во всех классах членистоногих мы сталкиваемся
и с более сложными формами проявления заботы о потомстве, например,
с постройкой гнезд и охраной находящихся в гнезде яиц и молоди. Наи-
более сложно проявление заботы о потомстве у некоторых насекомых.
Развитие членистоногих. Яйца членистоногих обычно сравнитель-
но крупные и переполнены запасными питательными веществами в виде
зерен желтка. В связи с этим из яйца обычно выходит сравнительно круп-
ная и хорошо развитая личинка. У насекомых и у большинства паукообраз-
ных новорожденная личинка имеет полное число члеников, свойственное
п взрослой форме. Наоборот, у многих ракообразных и многоножек только
что вышедшая из яйца личинка имеет немного члеников; например, ли-
чинка веслоногих раков имеет всего 3 членика (рис. 54, В). У такой ли-
чинки с каждой линькой прибавляется по одному членику до тех пор, пок<>
не образуется число члеников, свойственное взрослым особям данного вида.
Новые членики образуются на переднем крае анальной лопасти и, таким
образом, как бы вставляются один за другим между передним краем
анальной лопасти и самым задним из имеющихся члеников. При таком
способе .роста самый задний членик всегда является самым молодым.
Некоторые членистоногие (например, многие ракообразные) растут
и линяют в течение всей жизни. Но во многих группах членистоногих,
в том числе у громадного большинства насекомых и клещей, и рост, и
линька с достижением половой зрелости прекращаются. У таких форм
вся жизнь резко распадается на два периода: предимагинальный, в то-
чение которого членистоногое растет и линяет, но не размножается, и
имагпнальный, в течение которого членистоногое больше не растет и
не линяет, но размножается.
В том случае, если неполовозрелые стадии резко отличаются от взрос 7
лого животного своим строением и в особенности если они снабжены ка-
кими-либо органами, отсутствующими у взрослого, эти стадии получаю:
название личиночных; находящиеся на этой стадии молодые особи назы-
ваются личинками (larva). Взрослое животное в таких случаях называется
имаго (imago). Про развитие, в котором имеются личиночные стадии, го-
ворят, что оно сопровождается превращением (метаморфозом).
Названия личинка и имаго связаны со старинными представлениями
о метаморфозе насекомых. Слово «личинка» — уменьшительное от ли-
чина, что на древнерусском языке значит маска, по-латыни — larva.
В старину полагали, что гусеница — это как бы замаскированная бабоч-
ка, которая потом сбрасывает с себя маску (личиночную и кукол очную
шкурку) и является в своем истинном образе (слово «образ» — по-латыни
imago). Отсюда и возникло обозначение взрослого насекомого, или
клеща, словом «имаго».
ГЛАВА IV
КЛАССЫ ЧЛЕНИСТОНОГИХ
Класс I. Р а к о о б р а з н ы е (С г u s t а с о а) 1
Большей частью водные членистоногие, голова которых несет две
пары усиков, одну пару верхних челюстей, или жвал (мандибул), и две
нары нпжнпх челюстей (максилл). Дыхание в большинстве случаев осу-
ществляется при помощи жабер, являющихся придатками конечностей.
1 Некоторые важнейшие пресноводные ракообразные описаны во второй части
учебника («Гидробиология», стр. 92, 100).
28
Расчленение тела весьма разнообразное. Половое отверстие обычно распо-
ложено в задней части грудного отдела. Важнейшие отряды ракообразных:
А. Подкласс Entomostrac а, н изшие раки. Брюшко
лишено конечностей. Число члеников тела у различных представителей
различно.
Отряд 1-й. Phyllopoda, листопогие.
Подотряд Euphyllopoda, настоящие лпстоногие: Apus, щптень; Bran-
chipus (рис. 12, A); Limnadia.
Подотряд Cladocera, ветвистоусые (рис. 42,27, и 55): Daphnia, дафния;
Scapholebris (рис. 56, A); Moina; Bosmina.
Отряд 2-й. Ostracoda, ракушечные: Cypridina (рис. 12, Бу Noto-
dromas (рис. 56, Б). •
Отряд 3-й. Copepoda (рис. 54), веслоногие: Cyclops, циклоп; Diap-
tomus.
Б. Подкласс М alacost гас а, высшие раки. Каж-
дый членик брюшка несет по паре ножек. Тело всегда состоит из 19 чле-
ников: 13 члеников головогруди и 6 члеников брюшка.
Отряд 4-й. Amphipoda, бокоплавы: Gammarus (рис. 5, Г).
Отряд 5-й. Isopoda, равноногие, или мокрицы: Asellus, водяной ос-
лик (рис. 57, A); Porcellio, мокрицы (рис. 12, By
Отряд 6-й. Decapoda, десятиногие.
Подотряд Macrura, длиннохвостые: Astacus, речной рак (рис. 12, Гу
креветки.
Подотряд Brachiura, крабы; пресноводный краб Средней Азии и Кав-
каза, Potamon.
Медицинское значение из ракообразных у нас имеют только весло-
ногие рачки и пресноводные крабы (последние — на Дальнем Востоке):
и те, и другие являются промежуточными хозяевами некоторых глистов.
В частности, названные выше веслоногие являются первыми промежу-
точными хозяевами лентеца широкого (Diphyllobothrium latum), [в не-
которых местностях широко распространенного паразита человека.
Класс II. Многоножки (М у г i а р о d а). Наземные члени-
стоногие. Тело состоит из головы и туловища. Голова несет одну пару
усиков, одну пару жвал п одну или две пары нижних челюстей. Вторая
пара усиков появляется у зародыша, затем исчезает. Туловище слагается
из однообразных члеников, несущих более или менее одинаковые ножки.
Лишь немногие пары ног видоизменены в качестве половых придатков
пли ногочелюстей.
Важнейшие отряды:
Отряд Chilopoda, сороконожки. Хищные многоножки, с двумя парами
нижних челюстей. Первая пара туловищных ног превращена в мощные
ногочелюсти, снабженные ядовитыми железами. Последние три членика
тола бывают уменьшены/один из них несет половое отверстие; конечности
этих члеников превращены в половые придатки.
Род Lithobius (рис. 13, А), длиною около 2 см, широко распространен.
Род Scolopendra, крупные (в тропиках до 26,5 см в длину) многоножки,
укус которых слегка ядовит, хотя для человека не опасен. Водйтся у нас
на юге.
Род Scutigera (рис. 13, Бу длинноногие, очень быстрые ночные жи-
вотные; встречаются у нас на юге в домах; полезны тем, что истребляют
комаров и мух.
Отряд Diplopoda, кивсяки, или тысяченожки (рис. 13, В). Всего одна
пара нижних челюстей; ногочелюстей не имеют. Членики тела слились
попарно; каждый двойной членик несет две пары ног. Половое отверстие—
29
Рис. 12. Ракообразные.
А — Branchipus (отряд листоногих, Phyllopoda): 1 — сохранившийся личиночный глаз;
2—антенны первые; 3—антенны вторые; 4—верхняя челюсть; 5—сложный глаз левой
стороны; 6— максиллярная железа (орган выделения); 7—сердце; 8—кишечник; 9—сово-
купительный орган; 10— грудные ножки; 11—брюшко; 12—вилка (придатки анальной
лопасти); Б—CypricHna (отряд ракушковых, Ostracoda); В—мокрица, Porcellio (наземный
рачок из отряда Isopoda, равноногих); Г—.речной рак, Astacus, самец (вид с брюшной сторо-
ны): 1— первые усики; 2— вторые усики; 3— ротовые конечности; 4— 8— грудные ноги;
9 — 14 — брюшные конечности; 15 — анальная лопасть с задним проходом.
в передней части туловища. Снабжены пахучими железами. Медицин-
ского значения не имеют.
Класс III. Насекомые (Insect а).
Тело распадается на голову, грудь и брюшко. Голова несет одну пару
усиков, одну пару жвал и две пары нижних челюстей. Вторая пара усиков
имеется только у зародыша, затем она исчезает. Грудь состоит из трех чле-
ников и несет три пары ног, а у большинства, кроме того, две пары крыльев.
Брюшко состоит из различного числа члеников, но не более И, обычно
их бывает меньше. Конечности брюшка или отсутствуют, или во всяком
случае служат не органами передвижения, а либо органами чувств, либо
половыми придатками. Исключение составляют лишь некоторые личинки.
Рис. 13. Многоножки.
А — Lithobius forficatus (итрял Chiluiuua): i— голова; 2— глаза;
3— полоска мягкой кожи, соединяющая членики; 4— нога XVI
членика* туловища; 5— задний проход; 6 — стигмы; 7 — мягкая ку-
тикула плевры; 8— нога 11 членика туловища; У— ногочелюсть
(нога I членика); 10— усик; < — Х.Х— тергиты члеников туло-
вища; Б—Sciitigera (отряд Chilopoua); В— кивсяк, Tachipodo-
julus (отряд Diplopoda).
Органами дыхания служат трахеи. Половое отверстие расположено в зад-
ней части брюшка. Насекомые делятся на два подкласса: Apterygogenea,
первичнобескрылые, и Pterygogenea, первичнэкрылатые.
Первичнобескрылые — мелкие формы, снабженные несколькими
парами видоизмененных брюшных конечностей; в некоторых отношениях
близки к многоножкам. Медицинского значения не имеют.
Первичнокрылатые, как правило, имеют крылья, хотя некоторые
формы их утеряли. Они имеют не более трех пар брюшных конечностей:
конечности 8-го и 9-го члеников, превращенные в половые придатки, и
конечности 11-го членика (церки), превращенные в осязательные придатки.
Подробнее о насекомых — см. главу V.
Класс IV. Паукообр азные (А г а с h п о i d е а).
Тело распадается на передний отдел (просома) и задний отдел
(опистосома). Передний отдел несет шесть пар конечностей: хели-
церы, педипальпы и четыре пары ног. Задний отдел имеет от 7 до 13 чле-
ников; конечности его видоизменены в половые придатки, легочные меш-
ки, паутинные бородавки или вовсе отсутствуют. Органы дыхания бывают
представлены легочными мешками и трахеями. Выделительными орга-
31
нами служат коксальные железы и мальпигиевы сосуды. Половое отвер-
стие всегда расположено в передней части опистосомы.
Важнейшие отряды паукообразных:
Отряд 1-й. Scorpionoidea, скорпионы (рис. 84). Крупные паукообраз-
ные с хорошо расчлененным телом. Хелицеры в виде коротких клешней,
пединальпы в виде длинных клешней. Просома покрыта спинным щитом,
несущим от трех до шести пар глаз. Опистосома несет пару гребневидных
органов (конечности 9-го членика тела) и четыре пары легочных мешков.
Последние четыре членика опистосомы и анальная лопасть образуют узкий
подвижный «хвост». Анальная лопасть заключает одну пару ядовитых
желез, протоки которых открываются на конце острого шипа, загнутого
к спинной стороне и служащего скорпиону оборонительным и наступатель-
ным оружием. Самки живородящи. Питаются насекомыми, пауками и т. п.
Распространены у нас на юге.
Рис. 14. Фаланга, или бихорка, Galcodes araneoides, самка,
готовая к нападению (класс наукообразных, итряд
Solifugae).
1— просома; 2— хелицеры; 3— педипальпъс 4—7— холильные ноги;
8 — опистосома (брюшке»).
Отряд 2-й. Solifugae, фаланги, или бихорки (рис. 14). Крупные пауко-
образные с хорошо расчлененным телом. Хелицеры в виде коротких, мощ-
ных клешней, пединальпы — в виде похожих на ноги щупалец. Просома
покрыта широким, коротким щитом, несущим одну пару глаз. Опистосома
мягкая, состоит из 10 члеников, лишена конечностей. Вихорки имеют хо-
рошо развитые трахеи. Хищники. Своими хелицерами могут наносить силь-
ные укусы, по ядовитых желоз не имеют. Самка кладет яйца. Разви-
тие без превращения. Распространены в жарких, сухих странах, у нас —
в Средней Азии, в Восточном Закавказье и в Прикаспийской низменности.
Отряд3-й. Aranoina, пауки (рис. 15). Средне!к величины, реже —круп-
ные паукообразные, с нерасчленепным телом. Опистосома соединяется с
просомой тонким стебельком. Хелицеры в виде острых крючков, снабжены
ядовитыми железами, служат для хватания добычи. Пединальпы в виде
щупиков, у самцов служат совокупительными органами. Просома покрыта
спинным щитом, несущим три-четыре пары глаз. Опистосома нерасчленен-
ная, у некоторых форм несет две пары легочных мешков, у большинства
задняя пара заменена двумя пучками трахей. Близ заднего конца
опистосомы имеется две-три пары паутинных бородавок, на которых
множеством мелких отверстий открываются мощные паутинные железы.
Самка кладет яйца, покрывая их коконом из паутины. Развитие без пре-
32
вращения. Самцы значительно меньше самок. Часть пауков охотится,
преследуя добычу (семейство Lycosidae) или подстерегая ее на цветах и ли-
стьях (семейство Thomisidae), на коре деревьев (семейство Attidae) и т. п.,
другие подстерегают добычу, сидя в трубках, сплетенных из паутины
(Agalenidae, Drassidae), третьи ловят добычу, сплетая из паутины не-
правильные тенета (Linyphiidae, Pholcidae и др.), тогда как Agriopidae
строят тенета в виде геометрически правильного колеса.
О ядовитых пауках см. стр. 153.
Рис. 15. Паук-крестовик Aranea diadema (отряд Araneina),
самка (вид со спинной стороны).
1—просома (гопоногру ib); 2— хелицеры; 3—педипальпы;
4—ходильные ноги; 5—опистосома (брюшко).
Отряд 4-й, Pseudoscorpiones, лжескорпионы (рис. 5, D). Маленькие
или средней величины плоские паукообразные. Просома покрыта спин-
ным щитом с одной парой глаз. Конечности ее напоминают конечности
просомы скорпионов. Опистосома состоит из 11—12 члеников и никаких
конечностей не несет. Дышат трахеями. Самка кладет яйца. Развитие
прямое. Живут под корой деревьев и в других укрытых местах, иногда
в домах, например, в книжных шкафах. Питаются мелкими насекомыми.
Медицинского значения не имеют.
Отряд 5-й, Opiliones, сенокосцы. Средней величины паукообраз-
ные. Тело округлое или яйцевидное. Хелицеры в виде коротких клешней.
Педипальпы в виде щупалец, ноги чрезвычайной длины. Просома покрыта
спинным щитом, с одной парой глаз; опистосома из 10 члеников, конеч-
ностей не несет. Дыхание трахейное. Самка кладет яйца, развитие без
превращения. Питаются насекомыми. Нередко встречаются в домах, где
могут поедать на стенах комаров и мух.
3 Учебник медицинской энтомологии 33
Отряд 6-й, Acarina, клещи. Очень мелкие, мелкие или средней ве-
личины паукообразные. Хелицеры и педипальпы превращены в ротовые
органы. Опистосома лишена конечностей, обычно без видимого расчле-
нения, не отграничена или нерезко отграничена от просомы. Глаз бывает
одна-две пары, чаще их вовсе нет. Дыхательные органы, если они имеются,
представлены трахеями, которые открываются наружу одной парой
дыхалец. Большинство клещей кладет яйца, немногие живородящи.
Развитие с метаморфозом. Выходящая из яйца личинка имеет только три
пары ног, четвертая пара появляется только после первой линьки.
Громадная группа, имеющая большое медицинское и хозяйственное
значение (см. часть 3, раздел И).
ГЛАВА V
НАРУЖНОЕ СТРОЕНИЕ НАСЕКОМЫХ
1. РАСЧЛЕНЕНИЕ ТЕЛА НАСЕКОМЫХ
Голова. Голова насекомых заключена в сплошную хитиновую коробку,
образующую череп, и подвижно сочленена с грудью. У некоторых
насекомых голова поставлена горизонтально, так что ротовые органы на-
Рис. 16. Голова таракана.
А—спереди; Б—сзади: 1—верхняя губа; 2—верхние челюсти; 3—налич-
ник; 4 — лоб; 6 — усики; 6' — глазки; 7 — глаза; 8— щека; 9— нгвы; 10 — темя:
11— затылок; 12— затылочное отверстие (место присоединения шеи к голове);
13—подподбородок; 14—подбородок; 15—язычки; 16— губные щупики;
17—основной членик правой нижней челюсти; 18 — ее стволик; 19— жевательные
лопасти; 20—нижнечелюстной щупик.
правлены вперед, как, например, у жука-плавунца (рис. 59, А), у других
она поставлена вертикально, так что ротовые органы направлены в вен-
тральную сторону или даже немного назад, как, например, у тараканов
(рис. 17). В голове насекомых для удобства описания различают следую-
щие части: на спинной стороне —затылок, темя, лоб и головной щит, по
бокам — щеки, на брюшной стороне —горло (рис. 16). Кроме того, у
большинства насекомых по бокам головы много места занимают сложные
глаза; иногда они настолько велики, что покрывают почти всю голову
и на спине сходятся между собой, как, например, у многих стрекози у
самцов слепней. У большинства личинок, а из взрослых насекомых, на-
34
пример, у блох и вшей, вместо сложных глаз по бока^м головы располо-
жены простые глазки. У многих других насекомых простые глазки встре-
чаются наряду со сложными глазами и в таком случае они располагают-
ся на лбу или на темени в числе 1—3 штук. Кроме того, голова несет уси-
ки и ротовые органы.
Грудь. Грудь насекомых состоит из трех члеников, которые обозна-
чаются как переднегрудь, среднегрудь и заднегрудь (рис. 5, В; рис. 17).
Каждый из члеников груди несет по одной паре ног, среднегрудь несет,
кроме того, передние крылья, заднегрудь—задние крылья. Таким об-
разом, в грудном отделе сосредоточены все органы передвижения насе-
комых. В связи с этим мускулатура груди развита особенно сильно, а
Рис. 17. Расчленение тела таракана Blatta orientalis.
А—самка: 1—перелнеспинка; 2—среднеспинка; 3—заднеспин-
ка; 4— надкрылья; 5—11—стерниты 2—8-го члеников брюшна;
12— тергит 10-го членика; 13— церки; 14—20—тергиты 7—1-го чле-
ников; Б— самец: 1 — надкрылья; 2—о — стерниты 2—9-го члеников
брюшка; 10—грифельки; 11— церки; 12—17 — тергиты 10—3-го
члеников брюшка.
это влечет за собой и более мощное развитие скелета груди.‘Кроме тер-
гита и стернита, имеющихся в каждом нормально развитом членике тела,
в члениках груди имеется множество дополнительных склеритов. У хо-
рошо летающих насекомых, например, у двукрылых, вся грудь одета
гибкой и прочной броней, состоящей из большого числа правильно располо-
женных пластинок (рис. 104).
Чем сильнее развиты придатки какого-либо членика груди, ноги и
крылья, тем сильнее развит и данный членик. Так, у двукрылых, имею-
щих только переднюю пару крыльев, сильнее всего развита несущая их
среднегрудь. У жуков и клопов, в полете которых основную роль играет
вторая пара крыльев, сильно развита заднегрудь; переднегрудь хорошо
развита у жуков и прямокрылых с хорошо развитыми передними ногами
и очень мала у стрекоз или дневных бабочек с маленькими передними но-
гами и т. д.
Брюшко. В противоположность груди, которая у взрослых насеко-
мых обычно бывает снабжена твердым скелетом и заполнена мускулатурой,
последний отдел тела, брюшко, содержит главным образом внутренности
и обладает сравнительно мягкими покровами и слабой мускулатурой.
Брюшко имеет различное число члеников—от 11 до 5. Скелет каждого
членика состоит из двух полуколец — тергита и стернита (рис. 17).
У примитивных насекомых (стрекозы, тараканы и многие другие) чле-
ников брюшка 11, и все они по размерам не очень сильно различаются
между собой. У высших насекомых число члеников бывает уменьшено
3* 35
и некоторые членики резко сужены; первый или второй членик иногда
образует узкий стебелек, при помощи которого брюшко как бы подвешено
к груди (мухи, осы).
2. ПРИДАТКИ ТЕЛА НАСЕКОМЫХ
Усики. Первая пара придатков головы — усики (сяжки, антенны).
В простейшем случае усики состоят из длинного ряда совершенно оди-
наковых коротких члеников (нитевидные усики). У других насекомых
членики усиков могут резко различаться между собой по форме и длине,
и тогда возникают разнообразные типы строения. Усики многих видов
насекомых несут огромное количество" чувствительных нервных оконча-
ний и являются органами обоняния и осязания.
Ротовые конечности. Следующие за усиками три пары головных
конечностей превращены у насекомых в органы принятия пищи, или ро-
товые органы. Сюда входят верхние челюсти (жвалы, мандибулы,
рис. 16, 2), нижние челюсти (максиллы, рис. 16, 17—20) и нижняя губа
(лабиум, рис. 16,13—10), образованная слиянием второй пары максилл.
Кроме того, у многих насекомых встречается верхняя губа (рис. 16, 1),
представляющая складку кожи над мандибулами, и гипофаринкс, представ-
ляющий непарный вырост стенки ротовой полости. Пища у разных на-
секомых весьма различна; столь же разнообразно и устройство их ротового
аппарата. В основном мы различаем грызущие ротовые органы у насе-
комых, питающихся твердой пищей, и различного типа сосущие ротовые
органы у насекомых, питающихся жидкой пищей.
Примером грызущих ротовых органов могут служить ротовые органы
тараканов (рис. 16). Верхняя губа имеет вид небольшой, сильно хитини-
зованной пластинки. Мандибулы — одночленистые, короткие, с за-
зубренной жевательной поверхностью; правая и левая мандибулы дви-
гаются навстречу друг другу как две половины щипцов. Каждая максилла
состоит из двух более мощных основных члеников, из которых второй не-
сет две жевательные пластинки и челюстный щупик. Последний подвижен
и состоит из четырех члеников, несущих множество вкусовых и осяза-
тельных органов. Нижняя губа непарная, но образована частичным слия-
нием двух максилл второй пары. Она состоит из двух основных члеников,
жевательных пластинок и пары губных щупалец. Гипофаринкс таракана
представляет небольшой непарный выступ нижней стенки ротовой по-
лости, наподобие языка.
При переходе на питание жидкой пищей ротовые органы насекомых
претерпевают различные видоизменения, превращаясь в сосущие (колю-
ще-сосущие или лижуще-сосущие) ротовые органы. С ротовыми аппара-
тами этих типов мы познакомимся ниже на ряде различных приме-
ров.
Ноги. Грудные ноги насекомых состоят из определенных частей,
носящих следующие названия: ляжка, или тазик (соха), вертлуг (tro-
chanter), Седро (femur), голень (tibia) и лапка (tarsus). Лапка обычно со-
стоит из нескольких члеников (1—5) и заканчивается коготками. Коготков
чаще всего бывает два. В зависимости от способа передвижения, ноги насе-
комых могут быть приспособлены как ходильные, бегательные, прыгатель-
ные, плавательные, роющие и т. п. Нередко встречаются хватательные ноги
(например, у богомолов, у стрекоз).
Брюшные конечности. Из брюшных конечностей у самок конечности
8-го и 9-го члеников брюшка нередко бывают приспособлены в качестве
яйцеклада (например, у кузнечиков, рис. 25, А и у наездников); у ос, пчел
36
и муравьёв яйцеклад видоизменен в жало, снабженное двумя ядовитыми
железами и являющееся оборонительным и наступательным оружием
(рис. 83). У самцов конечности девятой пары иногда бывают превращены
в хватательные органы, служащие для придержания самки при копуля-
ции (например, вальвы комаров). Конечности 11-го членика брюшка,
церки, являются осязательными органами и встречаются у низших на-
секомых (рис. 17, 11).
Крылья. Крылья насекомых представляют парные выросты или
складки кожи, расположенные на боках груди. Как правило, крыльев
бывает две пары: передние крылья расположены на среднегруд и и зад-
ние крылья — на заднегруди. Переднегрудь крыльев не несет. В отряде
двукрылых (Diptera) задняя пара крыльев редуцирована и превращена
в жужжальца, маленькие палочки с утолщением на конце. У многих
насекомых передние крылья утолщены и превратились в надкрылья,
служащие для защиты нежных задних крыльев, а также брюшка.Таковы
передние крылья многих прямокрылых, клопов и в особенности жуков.
Каждое крыло представляет плоскую складку кожи и, следовательно,
состоит из двух тонких пластинок, склеенных между собой и пере-
ходящих друг в друга на краях крыла. Между ними проходят трахеи и
нервы. В местах прохождения трахей хитин крыла утолщен и образует
так называемые жилки, представляющие опорный скелет крыла, при-
дающий ему необходимую прочность. Жилкование крыльев чрезвычайно
разнообразно и играет большую роль в систематике насекомых, в раз-
личении семейств, родов и видов. У примитивных насекомых жилок
очень много и они образуют густую сеть (стрекозы, поденки, прямокры-
лые, см. рис. 24 и 25). У высших насекомых число жилок, в особенности
поперечных, значительно сокращено (перепончатокрылые, двукрылые,
жуки и пр., см. рис. 30—32). Крылья встречаются только у первичнокры-
латых насекомых (Pterigogenea), но и среди них многие в процессе эво-
люции их утратили. В частности, паразитический образ жизни часто
ведет к утрате крыльев. Так, пухоеды и вши не имеют никаких следов
крыльев, у блох на стадии куколки встречаются зачатки передней пары
крыльев, но потом они бесследно исчезают. У постельного клопа сохра-
нились только рудименты крыльев, и т. д.
ГЛАВА VI
ВНУТРЕННЕЕ СТРОЕНИЕ НАСЕКОМЫХ
1. ПИЩЕВАРИТЕЛЬНЫЙ АППАРАТ
Пищеварительный аппарат насекомых, как и у всех членистоногих
состоит из трех отделов: передней, средней и задней кишки (рис. 18).
Передняя кишка. Передняя кишка начинается ротовой полостью,
за которой следует мышечная глотка, за глоткой — пищевод. Пищевод
иногда расширяется в зоб, как у прямокрылых (например, у таракана).
В других случаях зоб представляет мешковидный вырост пищевода, как
у двукрылых. Эпителий передней кишки выстлан хитиновой кутикулой
и нередко вооружен шипиками, как, например, в глотке комаров и мос-
китов. В ротовую полость открываются слюнные железы; у многих на-
секомых они очень сильно развиты и сложно устроены. Слюна насекомых
содержит ряд пищеварительных ферментов; при еде пища проглатывается,
будучи смешана со слюной, причем ферменты слюны играют большую
роль в пищеварении (например, у тараканов, у комнатной мухи).
37
Средняя кишка. Средняя кишка у многих насекомых также разделя-
ется на несколько подотделов. В передней части, близ места впадения
пищевода, она часто имеет несколько слепых выступов или отростков —
Рис. 18. Кишечник таракана
Blatta orientalis.
1—глотка; 2— пищевоч; 3—зоб;
4— жевательный желудок; 5— кар-
диальные придатки; 6 — средняя
кишка; 7 — мальпигиевы сосуды;
8 — тонкая кишка; 9 — толстая
кишка; 10—слепой вырост толстой
кишки; 11—12— прямая кишка;
13— резервуар слюнной железы;
14 —слюнная ?келеза; 15 — ее вывод-
ной проток; 16—общий выводной
проток обеих слюнных желез.
кардиальные отростки. Пищеварение у
насекомых полостное, т. е. пища перева-
ривается в полости кишечника отчасти при
помощи ферментов слюны, отчасти при
помощи ферментов, выделяемых стенкой
средней кишки. Всасывание переварен-
ной пищи также происходит в средней
кишке. Пища в средней кишке не лежит
свободно; она бывает обычно одета пе-
ритрофической оболочкой; у большин-
ства насекомых перитрофическая оболоч-
ка представляет трубочку с тонкими про-
зрачными стенками, легко проницаемыми
для пищеварительных соков и переварен
ной пищи. Перитрофическая оболочка вы-
деляется клетками средней кишки и слу-
жит для защиты стенок кишечника от
повреждения проходящей пищей. Обычно
она нарастает с переднего конца и посте-
пенно изнашивается на заднем конце.
Задняя кишка. Задняя кишка, как и
передняя, выстлана эпителивхМ, покрытым
хитиновой кутикулой. Последний вывод-
ной участок задней кишки обозначается
как прямая кишка; прямая кишка за-
канчивается задним проходом, открываю-
щимся на заднем конце брюшка. В перед-
нюю часть задней кишки впадают маль-
пигиевы сосуды — длинные, тонкие, сле-
пые на конце трубочки, играющие у на-
секомых роль выделительных органов,
наподобие почек у человека. Конечным
продуктом обмена азотистых веществ у
насекомых, как и у птиц, является мо-
чевая кислота, слабо растворимая в воде.
Она скопляется в клетках мальпигиевых
сосудов и из них в виде белой кашицы
поступает в просветы сосудов и далее в заднюю кишку и наружу.
Таким образом, задняя кишка служит для выведения из организма
не только непереваренных остатков пищи, испражнений, но и последних
продуктов обмена веществ, экскретов.
2. ДЫХАТЕЛЬНЫЙ АППАРАТ
Дыхальца. Все насекомые, за исключением нескольких мельчайших
форм первичнобескрылых, дышат трахеями. Трахейная система насеко-
мого открывается наружу одной или несколькими парами дыхалец (стигм).
У более примитивных насекомых имеется по паре дыхалец на 2-м и 3-м
члениках груди и на 8 первых члениках брюшка. У многих насекомых
число дыхалец уменьшается, у многих личинок двукрылых их остается
всего одна пара. Дыхальца обычно снабжены мускулатурой, при помощи
Рис. 19. Трахейная
система насекомо-
го: жук-шпанка
Lytta vesica tor ia
(семейство Meloi-
dae, нарывники),
личинка второго
возраста (сильно
увеличено.)
которой могут закрываться и открываться, а также различными, волос-
ками и щетинками, которые защищают их от проникновения пыли.
Трахеи» Трахейная система обычно состоит из двух продольных ство-
лов (рис. 19); стволы сообщаются с дыхальцами при помощи соединитель-
ных ветвей; от стволов отходит множество ветвей к мускулам, нервной
системе, половому аппарату, кишечнику и другим органам. Чем крупнее
насекомое, чем массивнее его органы, тем сильнее разветвлена трахейная
система, снабжающая воздухом эти органы. Большинство крупных на-
секомых производит дыхательные движения, в силу которых объем брюш-
ка то увеличивается, то уменьшается и в главных стволах трахейной си-
стемы происходит обмен воздуха. У многих хорошо
летающих насекомых (мухи, пчелы) трахейные стволы
бывают расширены в обширные воздухоносные мет-
ки, присутствие которых значительно облегчает вес
тела.
Дыхание водных насекомых. Большинство водных
насекомых и под водой продолжает дышать воздухом;
многие из них уносят с собой некоторое количество воз-
духа в виде серебристого слоя, окружающего брюшко
(многие водяные клопы, жуки водолюбы), или под
надкрыльями (жуки плавунцы); другие, живя в воде,
выставляют на поверхность дыхальца, нередко распо-
ложенные на специальных выростах—сифонах, как,
например, у многих водных личинок двукрылых, или
иного устройства дыхательные трубки, как клопы
«водяные скорпионы» (рис. 20, В). Некоторые водные
личинки насекомых имеют замкнутую трахейную си-
стему, заполненную, однако, газом; газообмен в таком,
случае совершается при помощи трахейных жабер —
расположенных на брюшке листочков или нитей, бога-
то снабженных петлями трахей. Такие трахейные
жабры имеют многие личинки поденок (рис. 20, В),
стрекоз и др.
Наконец, многие личинки имеют вместо этого
кровяные жабры: полые выросты стенки тела в виде
мешочков, покрытые очень тонкой кожей, через кото-
рую свободно совершается газообмен между кровью
личинки и окружающей водой; таковы жабры личи-
нок комаров и комариков тендипедид (рис. 20, Л).
3. ПОЛОСТЬ ТЕЛА И КРОВЕНОСНЫЙ АППАРАТ
Полость тела, гемолимфа, жировое тело. Все
пространство между стенкэй тела и отдельными ор-
ганами представляет собой полость тела насекомого. Полость тела
заполнена кровяной жидкостью, так называемой гемолимфой. Гемолимфа
содержит бесцветные амебоидноподвижные кровяные клетки. Среди этих
клеток имеются фагоциты, способные захватывать и переваривать про-
никших в организм насекомого микробов, или отмирающие, подлежа-
щие разрушению клетки тела. Некоторые кровяные клетки могут
накоплять в себе запасные вещества — жир, белок, гликоген 1 и, осе-
1 Гликоген, или животный крахмал, представляет собой нерастворимый углевод,
который образуется в клетках животных за счет сахара (глюкозы) их крови и по мере
надобности превращается опять в глюкозу.
39
дая, слагаются в «жировое тело». Лопасти жирового тела нередко за-
полняют собой значительную часть полости тела.
Сердце» Гемолимфа приводится в движение работой сердца (рис.6, В).
Сердце лежит в брюшке, на спинной его стороне, и представляет
у большинства насекомых трубку с мышечными стенками. В каждом чле-
нике тела оно имеет пару отверстий, снабженных клапанами. На перед-
нем своем конце сердце переходит в узкую трубку, аорту, которая тянется
Рис. 20. Дыхательные приспособления водных насекомых.
j — кровяные жабры личинки Tendipes; 1— жабры; 2— голова;
J — передние личиночные ножки; 4— задние личиночные ножки;
Ь —» трахейные жабры личинки поденки Heptageuia (зачатки кры-
льев и ноги левой стороны, жаберные листочки правой стороны об-
резаны); В—дыхательная трубка клопа — серого водяного скор-
пиона (Nepa cinerea). Клоп изображен с расправленными крыльями.
через всю грудь и заканчивается в голове, где и открывается прямо
в полость тела. Сердце прикрепляется к тергитам брюшка при помощи
нескольких пар крыловидных мышц.
Брюшная диафрагма» Многие насекомые, например, комары, снаб-
жены брюшной диафрагмой, которая представляет собой мускулистую
пленку, натянутую вдоль нижней стороны брюшка, над нервной цепоч-
кой (рис. 135). Диафрагма производит волнообразные движения, и, в то
время как сердце гонит кровь по спинной стороне тела сзади наперед,
диафрагма гонит ее по брюшной стороне спереди назад.
Кровообращение» Физиологический смысл кровообращения насе-
комых сводится к следующему.
В брюшке кровь насыщается пищевыми веществами, проникающими
в нее из кишечника, и при помощи мальпигиевых сосудов очищается от
продуктов обмена веществ; в голове и груди расположены основные ра-
бочие органы (мозг, органы чувств, жевательная мускулатура, мышцы
ног и крыльев). Сердце сокращаясь сзади наперед, засасывает через
отверстия (остии) кровь из полости брюшка и перегоняет ее в голову; от-
40
туда кровь возвращается в брюшко по промежуткам между органами
головы и груди, омывая те и другие, отдавая им питательные вещества и
вымывая из них продукты обмена.
Нервный аппарат и органы чувств насеко-
мых описаны выше (стр. 23—27).
4. ПОЛОВОЙ АППАРАТ НАСЕКОМЫХ
Мужской половой аппарат» Насекомые раздельнополы. Имеются
самцы иГсамки. Мужской половой аппарат состоит из пары семенников,
лежащих в брюшке, пары семяпроводов, открывающихся в непарный
семяизвергательный проток, и пары трубчатых мужских придаточных
желез. Обычно бывает и совокупительный орган, более или менее слож-
ного строения. Половое отверстие находится на 8-м членике брюшка.
Конечности этого членика у самцов многих насекомых превращены в
совокупительные придатки, служащие для придерживания самки во
время совокупления.
Женский половой аппарат. Женский половой аппарат состоит из
яичников, яйцеводов, влагалища и придаточных органов.
Яичники насекомых парные, лежат в брюшке. У многих насекомых
каждый яичник состоит из четырех яйцевых трубочек, у многих дру-
гих число их увеличивается и может доходить до нескольких сот (на-
пример, у комаров); в других случаях число яйцевых трубочек умень-
шается и, например, у мухи цеце или у куклородных мух, в каждом
яичнике имеется всего-навсего по одной единственной трубочке.
Каждая яйцевая трубка на одном конце заканчивается слепо, а дру-
гим концом открывается в яйцевод. На слепом конце трубки располагается
зона размножения; она образована зачаткдвыми клетками, которые де-
лятся и дают начало первичным яйцевым клеткам. Яйцевые клетки по
очереди, по мере возникновения, продвигаются вдоль яйцевой трубочки.
Там они и располагаются в один ряд по возрасту: самая старшая клетка
лежит ближе всего к выходу. Каждая яйцеклетка окружена мешочком
из эпителиальных клеток — фолликулом. Яйцевые клетки, и особенно
самая старшая из них, усиленно растут, накопляя в себе питательные
вещества в виде зерен желтка. Когда клетка достигает окончательной ве-
личины, в ее ядре происходят процессы созревания и она становится зре-
лым яйцом. К этому времени клетки фолликула выделяют вокруг нее
оболочку, называемую хорион; это будущая скорлупа яйца. С момен-
та образования хориона (скорлупы) яйцо готово к откладке. После
того как оно отложено, наступает очередь дозревания следующей яйце-
клетки и т. д.
Разные типы яйцевых трубок насекомых. В простейшем случае, на-
пример, у таракана (рис. 21, А), яйцевая клетка сама накопляет в себе
желток, правд а,через посредство окружающих ее клеток фолликулярного
эпителия. У многих других насекомых, например, у всех двукрылых,
каждый фолликул яйцевой трубочки содержит, помимо яйцевой клетки,
несколько питающих (рис. 21,Б). Они жадно вбирают в себя пищевые
вещества из гемолимфы и передают их яйцевой клетке; когда последняя
приближается к зрелости, питающие клетки, выполнившие свою зада-
чу, начинают уменьшаться и отмирают, превращаясь в крошечные ко-
мочки, после откладки яйца остающиеся в яйцевой трубочке.
У клопов в каждой яйцевой трубочке имеется одна питательная ка-
мера, расположенная тотчас после зоны размножения (рис. 21, В). Пита-
тельная камера втягивает в себя питательные вещества из гемолимфы и
41
б в
>абатывает их; от нее ко всем фолликулам идут тяжи, по которым
'ельные вещества направляются к растущим яйцеклеткам.
1йцевые трубки, составляющие яичник, могут лежать все независимо
от друга, лшпь^выводными концами соединяясь с яйцеводом (у боль-
шинства насекомых), или же все они окру-
жены общей оболочкой и слагаются в
компактные яичники, как мы видим у
комаров, москитов, мошек и слепней.
Вспомогательные органы женского
полового аппарата. Как бы ни были устрое-
ны яичники, каждый из них откры-
вается в один из двух парных яйцеводов.
Парные яйцеводы сливаются в непар-
ный яйцевод, который при помощи вла-
галища и полового отверстия откры-
вается наружу.
К числу вспомогательных органов
женского полового аппарата прежде всего
относятся смазывающие железы, парные
или непарные, открывающиеся во влага-
лище и служащие для смазывания и
склеивания яиц или для приклеивания
их к предметам. Далее, к вспомогатель-
ным органам относятся семяприемники,
служащие для хранения спермы самца,
полученной при совокуплении, и др.
ГЛАВА VII
РАЗВИТИЕ НАСЕКОМЫХ
F Яйца насекомых. Яйца насекомых
относительно крупной величины. Прото-
плазма яйца окружена оболочкой (хо-
рион), которая может быть мягкой и ко-
жистой или твердой и жесткой. У многих
насекомых хорион обладает очень сложной
структурой (например, у комаров). Как
правило, он бывает пронизан одним или
несколькими маленькими отверстиями
(микропиле, что значит «маленькие воро-
та»), через которые проникает спермато-
зоид, оплодотворяющий яйцеклетку.
Развитие. В оплодотворенном яйце
разыгрываются сложные процессы заро-
дышевого развития и формируется ма-
ленькая личинка (рис. 22). Личинка
выходит из яйца либо выталкивая кры-
шечку (в тех случаях, когда таковая
имеется), либо прорезая хорион «яйцевым
- особым шипом, расположенным на лбу молодой личинки, либо
я хорион каким-либо другим способом. Крышечкой снабжены,
р, яйца вшей и клопов, яйцевым зубом— личинки комаров. Тот
азвития, который протекает в яйце, носит название зародышевого,
тонального. Остальной период развития вплоть до достижения
Типы яйцевых трубок
насекомых
штический тип; обособлен-
ощлх клеток нет; Б— по-
тений тип: каждый яйце-
икул имеет свою группу
клеток; В — телотро-
тип: в каждой трубке
)дна общая группа питаю-
к, от которой идут питаю-
гры к каждой растущей
е: I—верхушечная камера,
размножения; 2 — концевая
— яйцевые фолликулы;
ощле клетки; 5 — молодые
и (ооциты); а—зрелые яйца;
щие шнуры; 8 — остатки
клеток; 9 — переход к
яйцеводу.
половой зрелости носит название постэмбрионального (т. е. послезаро-
дышевого). В течение постэмбрионального периода развития молодое
насекомое растет и несколько раз линяет. При этом изменяется не
только величина животного, но и его строение. В зависимости от сте-
пени изменений, которым подвергается развивающееся молодое насе-
Рис. 22. Три стадии зародышевого развития
малярийного комара.
Из них В — на последний день развития. 1 — по-
ловой зачаток: 2—головная лопасть; 3 — усики;
4 — верхние целости; 5 — нижние челюсти;
6 —нижняя губа; 7 — передняя кишка; 8 — задняя
кишка; 9 — средняя кишка, заполненная желт-
ком; 10— веера верх-« й губы; 11— головной
мозг; 12 — подглоточный ганглий; 13 — слюнные
железы; 14—жабры (по Ивановой-Казас).
комое, различают два основ-
ных типа развития: развитие
с полным и с неполным превра-
щением.
Развитие без превращения
и с неполным превращением.
У многих из первичнобескры-
лых насекомых молодые особи
так мало отличаются от взрос-
лых, что говорить о превраще-
нии вообще не приходится. У
первично крылатых насекомых
вполне развитые крылья и спо-
собность к полету появляются
только с достижением половой
зрелости. В низших отрядах
первичнокрылатых наличие раз-
витых крыльев и половых
придатков часто представляет
единственное крупное отличие
взрослого насекомого от непо-
ловозрелых стадий; так обстоит
дело у прямокрылых, у клопов
и пр. Поэтому неполовозрелые
стадии клопов не называют ли-
чинками,— их называют ним-
фами. В тех случаях, когда
взрослое насекомое и неполо-
возрелые стадии ведут резко
различный образ жизни, раз-
ница в строении между ними может быть довольно значительной, как,
например, у стрекоз, личинки которых живут в воде, тогда как взрослое
насекомое является воздушным хищником. Однако если переход личинки
во взрослое насекомое совершается непосредственно, без стадии ку-
колки, мы все-таки говорим о неполном превращении.
Вши утратили под влиянием паразитического образа жизни крылья
и ведут на всех стадиях один и тот же образ жизни. В связи с этим их
молодые особи ничем существенным, кроме недоразвития половых орга-
нов, не отличаются от взрослого насекомого и развитие их совершается
без превращения.
Полное превращение. Неполовозрелые стадии насекомых растут, но
ле размножаются; взрослое насекомое размножается, но не растет. В эво-
люции высших насекомых личинка все более и более приспособлялась
к тому, чтобы как можнэ больше есть и быстрее расти, взрослое насеко-
мое, — чтобы как можно успешнее размножаться, пользуясь полетом
расселяться и помещать свое потомство в наилучшие условия питания и
развития. В силу таких совершенно различных направлений эволюции,
различие в строении личинки и взрослого насекомого увеличивалось все
Дольше и больше. Крайней степени это различие достигает у мух. Взрос-
43
лая мясная муха обладает хорошими ногами, прекрасными крыльями,
мощной мускулатурой, быстрым полетом, хорошим зрением, необычай-
но тонким обонянием. Она способна издали почуять падаль и быстро под-
лететь к ней, чтобы отложить на нее свои яйца. Личинка мясной мухи
червеобразная, безногая, без глаз, без челюстей, без головной капсулы;
зато у нее тонкая, растяжимая кожа, могучее пищеварение и необычайно
быстрый рост; опа замечательно приспособлена к использованию и усвое-
нию мясной пищи, в которую погружена, благодаря энергичным поискам
своей матери.
Между червеобразной личинкой и высоко совершенным крылатым
насекомым постепенного перехода быть не может. Переход личинки мухи
во взрослое насекомое совершается скачком. И действительно, зрелая ли-
чинка линяет в куколку, которая остается внутри засохшей в боченочек
личиночной шкурки. Куколка мухи неподвижна и внутри нее бурно
Рис. 23. Забота о потомстве у одиночных ос: самка осы Sphex
тащит в гнездо парализованного ею кузнечика, предназначенного
в пищу ее личинке.
протекают процессы перестройки организации; почти все личиночные
органы, кроме нервной системы и зачатка половых органов, разрушаются,
растворяются. Одновременно происходит быстрый рост и развитие
органов взрослой мухи из зачатков, которые возникли еще у личинки,
но оставались в дремлющем состоянии и не развивались. В результате
по истечении нескольких дней из куколки вылетает взрослое насекомое,
по своему строению не имеющее почти ничего общего с личинкой, дав-
шей ему начало.
Полное превращение свойственно всем отрядам высших насекомых:
жукам, бабочкам, перепончатокрылым, двукрылым, блохам и пр. Ли-
чинки многих жуков (например, жужелиц, плавунцов) снабжены длин-
ными ногами и по внешнему виду напоминают некоторых низших первич-
нобескрылых насекомых; личинки бабочек и низших перепончатокрылых
(пилильщиков), так называемые гусеницы, имеют короткие грудные ножки
и, кроме того, несколько пар нечленистых брюшных ножек. Личинки
некоторых жуков, высших перепончатокрылых, низших двукрылых и
блох лишены ног, но имеют хорошо развитую головную капсулу. Наконец,
личинки высших двукрылых не имеют ни ног, ни головной капсулы.
Заботы о потомстве. Характернейшей чертой большинства взрослых
насекомых является забота о потомстве. В простейшем виде она выра-
жается в выборе места откладки яиц: мать помещает яйца в такое место,
44
где будущие личинки найдут подходящие для себя условия существова-
ния — пищу и убежище. В других случаях инстинкт заботы о потомстве
проявляется гораздо сложнее: мн >гие насекомые строят гнезда и накоп-
ляют запасы пищи, обеспечивающие жизнь и развитие их потомства. Та-
ковы среди жуков многие навозники и мертвоеды, среди перепончато-
крылых одиночные пчелы и осы (рис. 23). Но наибольшей сложности ин-
стинкты заботы о потомстве достигают у так называемых общественных
перепончатокрылых: пчел, ос, муравьев и у термитов, живущих большими
колониями (семьями). У интересующих нас насекомых — паразитов че-
ловека — забота о потомстве не идет дальше целесообразного выбора
места откладки яиц.
ГЛАВА VIII
ПОДКЛАССЫ И ОСНОВНЫЕ ОТРЯДЫ НАСЕКОМЫХ
Класс Insecta, насекомые
Подкласс I: Apterygogenea, первичнобескрылые. Примитивные насекомые,
всегда бескрылые, развиваются без превращения и имеют во взрослом виде остатки
брюшных но;кек обычно на нескольких члениках брюшка. Не имеют медицинского
значения.
Подкласс II: Pterygogenea, первичнокрылатые. Громадное большин-
ство представителей этого подкласса имеет крылья, но у отдельных пред-
ставителей и в целых отрядах крылья частично или полностью редуци-
рованы. Брюшных конечностей не более трех пар, а именно: конечности
8-го и 9-го сегментов, превращенные в половые придатки, и конечности
11-го сегмента брюшка — так называемые церки (хвостовые придатки).
Первичнокрылатые насекомые распадаются на две группы: Hemimetabola
(насекомые с неполным превращением) и Holometabola (насекомые с пол-
ным превращением). Каждая из этих групп содержит множество отрядов,
из которых остановимся лишь на некоторых.
Стрекозы. Отряд Odonata, стрекозы (рис. 24, Г, Е), Крупные, хорошо
летающие насекомые, с мощной грудью, стройным брюшком. Глаза боль-
шие, усики короткие. Крылья с густой сетью жилок, не сцеплены между
собой. Передние и задние крылья или одинаковой величины, или задние
немного шире. В покое крылья раскрыты в стороны или прикладываются
друг к другу в вертикальном положении, никогда не складываются гори-
зонтально или крышеобразно поверх брюшка. Ротовые органы жующие,
ноги хватательные. Охотятся налету за мелкими насекомыми, в том числе
закомарами. Личинки стрекоз живут в воде, также хищные (рис. 24, Д, Ж).
Нижняя губа личинки видоизменена в длинные щипцы, служащие
для схватывания добычи. Охотно поедают личинок малярийного комара.
Поден и. Отряд Ephemeroidea, поденки (рис. 24, В), Нежные на-
секомые, с короткими усиками, с четырьмя сетчатыми крыльями, из
которых задние много короче передних; в покое крылья прикладываются
друг к другу в вертикальной плоскости. На конце брюшка имеется две
или три хвостовые нити. Личинки живут в воде, снабжены хвостовыми
нитями и трахейными жабрами (рис. 24, А; 20,2>). Многие из них хищны и
могут поедать личинок комаров. Закончив развитие, личинка вылезает из
воды и линяет в крылатую, но все еще неполовозрелую форму (рис. 24,5),
которая через несколько минут линяет еще раз во взрослую поденку.
Последняя совсем не питается и после копуляции и откладки яиц умирает.
Поденки являются единственными насекомыми, линяющими в окрылен-
ном состоянии.
45
Отряд Plecoptera, веснянки (рис. 24, Я, Я), Нежные насекомые с удлиненным телом
равномерной ширины. Усики длинные. Крылья с менее густой сетью жилок, чем у стре-
коз и поденок, в покое складываются плоско вдоль спины. Задние крылья много шире
передних. И взрослые, и личинки снабжены двумя хвостовыми нитями. Личинки
(рис. 24, 3) живут в воде, главным образом в реках и ручьях. Взрослые пищи
не принимают.
Отряд Orthoptera, прямокрылые. В настоящее время прямокрылых
обычно разбивают на несколько самостоятельных отрядов, но мы
здесь для краткости рассматриваем их как одну группу. Большей
Рис. 24. Стрекозы, веснянки, поденки и водомерки.
А— личинка поденки; Б— крылатая неполовозрелая стадия поден-
ки. из которой линяет половозрелая поденка В; Г— стрекоза Libel-
lula; Д— зе личинка; Е— стрекоза Agrion (стрелка); Ж— личинка
стрекозы Calopteryx (лютка); 3—личинка веснянки Perla; И —
взрослая Perla с кладкой яиц; К—летящая веснянка Chloroperla;
Л — водомерка Gerris (отряд Rhynchota).
частью крупные, более или менее массивные насекомые с жующими рото-
выми органами. Крылья с многочисленными густыми жилками; передняя
пара у большинства превращена в кожистые надкрылья. Личшки, как
и взрослые, ведут наземный образ жизни. К прямокрылым, в старом ши-
роком смысле слова, относятся кузнечики (рис. 25, Л), кобылки (рис. 25,Z>),
сверчки, уховертки (рис. 25, Г), тараканы, богомолы (рис. 25, Н), термиты
и пр. Из всех этих насекомых некоторое медицинское значение имеют только
тараканы, так как многие из них являются домовыми сожителями челове-
ка. Уховерток народное мнение несправедливо обвиняет в том, что они за-
лезают в уши спящего человека. Кобылки (саранчевые) имеют очень боль-
шое значение как вредители посевов. Термиты замечательны тем, что
живут обществами — семьями, наподобие муравьев. Прогрызая ходы в
мертвой древесине, некоторые термиты могут наносить большой вред дере-
вянным постройкам, мебели и т. п; у нас водятся только в Средней Азии.
6
Отряд Thysanoptera, пузыреногие, или трипсы (рис.26, А). Очень мелкие насе-
комые, живущие на цветах; многие трипсы вредят растениям, некоторые из них иног-
да сосут кровь животных и человека (случайные кровососы).
Отряд Anoplura, вши и пухоеды. Мелкие эктопаразитические насе-
комые, без крыльев, без церков, с короткими усиками; молодые особи,
кроме размеров, мало отличаются от взрослых, развитие протекает с не-
полным превращением, фактически — почти без превращения.
Рис. 25. Прямокрылые.
А-— кузнечик Decticus, самка; Б— кобылка Locusta; В— богомол Mantis; Г— уховертка
Forficula.
Подотряд Mallophaga, пухоеды или власоеды (рис. 26, Б), Имеют грызу-
щие ротовые органы с сильными жвалами. Переднегрудь свободна. Парази-
тируют главным образом на птицах, но также и на млекопитающих. Жи-
вут в шерсти пли в оперении, там же приклеивая свои яйца. Питаются
роговыми образованиями, выделениями кожных желез, но нередко и
кровью. Паразитов человека среди пухоедов нет, но многие из наших
домашних и лабораторных животных имеют своих пухоедов.
Подртряд Siphunculata, вши. Имеют колюще-сосущие ротовые орга-
ны; членики груди слиты между собой. Сложных глаз нет, но имеется
пара простых глазков. Лапки одночленистые. Паразиты млекопитающих
питаются кровью, имеют большое медицинское значение (часть 3,
раздел «Е»).
47
Отряд Rhynchota, хоботные. Насекомые с неполным превращением
и колюще-сосущими ротовыми органами, образующими членистый хобо-
ток, внутри которого движутся колющие щетинки. Ни челюстных, ни
губных щупиков нет. Крыльев обычно две пары, но встречаются и бес-
крылые формы.
Подотряд Homoptera, цикады и тли. Голова поставлена более или мепее верти-
кально, почти неподвижна, хоботок отходит от ее нижней поверхности. Передние
крылья не обращены в надкрылья. Цикады иногда могут колоть человека хоботком.
Тли не имеют медицинского значения, но являются важными вредителями растений.
Подотряд Heteroptera, полужесткокрылые, или клопы. Голова по-
ставлена более или менее горизонтально, подвижна, хоботок отходит
от ее переднего конца. Передние крылья
частично утолщены, образуя надкрылья.
Большая группа. Многие семейства клопов
живут в воде. Представители их, будучи хищ-
ными, приносят пользу, истребляя личинок
Рис. 26. Трипсы, пухоеды и хоботные.
А— Thrips (сильно увеличено); В—собачий власоед Trichodectes canis (сильно увеличено);
В — южноамериканский кровососущий клоп Panstrongylus meg 1st us (семейство Triatomldae)
(увеличено в 3 раза).
комаров; таковы гладыш (Notonecta) (рис. 59, Т7), серый водяной скорпион
(Nepa) (рис. 20, В), плавт (Naucoris) (рис. 59, Д), гребляки (Gorixidae),
водомерки (Gr iris) (рис. 24, Л) и пр. Крупные водяные клопы (гладыш и др.)
могут очень больно колоть своим хоботком человека, главным образом
при самообороне. Наземные клопы питаются соками растений или напа-
дают на других насекомых; некоторые из них являются вредителями расте-
ний. Некоторые наземные клопы могут насасываться крови человека или
млекопитающих (случайное, или факультативное, кровососание). И, на-
конец, среди клопов есть и настоящие кровососы, питающиеся исключи-
тельно кровью. Таковы все представители семейства Cimicidae, к кото-
рому принадлежит и наш постельный клоп (Gimex lectularius, см. стр. 414),
п семейства Triatomidae (рис. 26, В); триатомиды — очень крупные окры-
ленные клопы, живут в тропиках и на территории СССР не встречаются.
В Южной Америке они являются переносчиками болезни Чагаса, тяже-
лого заболевания человека.
Holometabola, насекомые с полным превращением.
48
Отряд Coleopter а, жуки. Насекомые с полным превращением и жую-
щими частями рта; передние крылья превращены в жесткие надкрылья.
Громадный отряд. Упомянем лишь несколько важнейших семейств.
Семейство Dytiscidae, плавунцы (рис. 59, Л, В). Хорошо плавающие _
хищные водяные жуки; и сами они, и их личинки истребляют большое
количество личинок комаров. Крупные плавунцы и их личинки поедают
головастиков и мальков рыб.
Рис. 27. Жуки (отряд Goleoptera).
А — карапузик, Hister; Б— кожеед D^rmcstes lardarius; В—его куколка; Г—его личин-
ка; Д—Leptinus, жучок, паразитирующий в ппрсти грызунов.
Семейство Gyrinidae, вертячки (рис. 56, В). Быстро скользящие по
поверхности воды, металлически блестящие жучки, могут также и нырять.
Наряду с другой добычей, поедают и личинок комаров.
Семейство Staphylinidae, хищники. Отличаются длинным, узким
гелом и укороченными надкрыльями. Виды рода Paederus (рис. 81, 6)
ядовиты и при раздавливании на коже вызывают нарывы.
Рис. 28. Жуки (отряд Goleoptera).
А — мучной хрущ Tenebrio molitor; Б — его личинка и В —
куколка; Г — амбарный долгоносик.
Семейство Hysteridae, карапузики (рис. 27, Л). Подобно многим
хищникам, живут в разлагающихся органических веществах, падали,
навозе. Многие представители как этого семейства, так и предыдущего
полезны как активные истребители личинок мух.
Семейство Dermestidae, кожееды (рис. 27, 2>, В, Г). Питаются падалью,
животными остатками, но могут объедать живых птенцов и цыплят. Встре-
чаются на продовольственных запасах (ветчина и т. п.).
4 Учебник медицинской энтомологии
49
Семейство Hydrophilidae, водолюбы (рис. 59, Б). Живут в воде.
Личинки могут уничтожать личинок кохмаров.
Семейство Meloidae, нарывники (рис. 81). Кровь их содержит кан-
таридин и очень ядовита для людей и животных. Некоторые виды упо-
требляются для изготовления лекарств.
Семейство Tenebrionidae, чернотелки. К ним относятся мучной хрущ
Tenebrio molitor, вредящий продовольственным запасам (рис. 28).
Семейство Scarabaeidae, пластипчатоусые жуки. Сюда относится майский жук.
бронзовки, навозники и многие другие.
Рис. 29. Походный шелкопряд Cnethocampa (отряд Lepidoptera.
чешуекрылые или бабочки).
А— самка; Б— гусеница; В— кокон и выпутан из кокона куколка (в<с-
в естественную величину); Г — ядовитые волоски (сильно увеличено).
Отряд Neuroptera, сетчатокрылые. Насекомые с полным превращением, жующими
ротовыми органами и двумя парами более или менее одинаковых нежных крыльев с
густой сеткой жилок. Медицинского значения не имеют.
Отряд Trichoptera, ручейники. Насекомые с полным превращением.
Крылья с немногими поперечными жилками, покрыты длинными волос-
ками, ротовые органы недоразвиты; личинки живут в воде, строят домики
из различных частиц, склеенных паутиной (рис. 57, Б). Многие личинки
хищны и некоторые едят личинок комаров.
Отряд Lepidoptera, бабочки или чешуекрылые (рис. 29). Насекомые
с полным превращением. Крылья, как правило, покрыты чешуйками.
Ротовые органы большей частью сосущие, хоботок образован нижними
50
челюстями. Личинки бабочек, гусеницы червовидной формы; грудные
ноги гусениц короткие; кроме них, имеется еще несколько пар нечлени-
стых, мясистых брюшных ножек; чаще всего их бывает четыре пары на
3—6 члениках брюшка и одна пара — на последнем. Бабочки представ-
ляют очень большой отряд; многие из них являются важными вредителями
растений. Некоторые гусеницы имеют ядовитые волоски; попадая в глаза
пли вонзаясь в кожу человека, такие волоски вызывают очень сильное
раздражение. Личинки некоторых бабочек вредят продовольственным за-
пасам или шерстяным вещам (моль, Tinea pelionella).
Отряд Hymenop ter а, перепончатокрылые. Насекомые с полным пре-
вращением, жующими частями рта и четырьмя перепончатыми крыльями,
из которых задние значительно меньше передних. Крылья со сравнитель-
Рпс. 30. Перепончатокрылые.
А — самка наездника Ilunterellus hookcri, личинка которого паразитирует в нимфах
иксодовых клещей (сильно увеличено); ГБ — шершень, Vespa crabro (естественная
величина).
но немногими жилками. Самки бывают снабжены яйцекладом, или жалом,
представляющим видоизменение конечностей 8-го и 9-го членика брюшка.
К отряду перепончатокрылых под названием наездников (Ichneumo-
noidea) относится ряд семейств, личинки которых паразитируют в дру-
гих насекомых, а также в крупных клещах (рис. 30, А). Истребляя вре-
дителей, многие наездники приносят большую пользу. К жалоносным
перепончатокрылым относятся муравьи (семейство Formicidae), осы и
пчелы. Многие муравьи жала как такового не имеют, но снабжены ядови-
той железой и выпрыскивают ее содержимое на врага. Поселяясь в до-
мах и нападая на пищу человека, муравьи могут загрязнять ее и служить
переносчиками кишечных болезней. Общественные пчелы (Apidae), осы
(Vespidae) (рис. 30, Б) и муравьи по сложности и совершенству своих
инстинктов занимают первое место среди беспозвоночных животных и
с этой точки зрения представляют величайший общебиологический
интерес.
Отряд Aphaniptera, блохи. Насекомые с полным превращением, бес-
крылые, с колюще-сосущими ротовыми органами. Личинки без ног, но
с хорошо развитой головой. Все блохи — высоко приспособленные пара-
зиты млекопитающих или птиц (стр. 402).
Отряд Diptera, двукрылые. Насекомые с полным превращением, со-
сущими (сосуще-лижущими пли сосуще-колющими) ротовыми органами
п одной парой перепончатых крыльев. Имеются передние крылья; зад-
ние превращены в жужжальца. Все членики груди слиты между собой.
Личинка всегда безногая. Из всех отрядов насекомых двукрылые имеют
наибольшее медицинское значение. Усики (антенны) у примитивных дву-
крылых длинные и состоят из большого числа более или менее одинако-
4* 51
вых члеников. У высших двукрылых все членики, начиная с 4-го, сли-
ваются в одну палочку или щетинку, сидящую на утолщенном 3-м членике.
Сложные глаза обычно велики и у самцов мух часто крупнее, чем у самок
Простые глазки встречаются в числе двух или трех, но могут и отсутст-
вовать. Ротовые органы сосущие; челюсти (в особенности—мандибулы)
сохранились только в немногих семействах подотряда прямошовных,
где они используются в качестве колющих щетинок. Нижняя губа у
этих форм образует футляр хоботка, губные щупальцы образуют ло-
пастинки (лабеллы) на его конце. У многих мух, например, у жигалок,
кроме нижней губы, образующей хоботок, имеется только верхняя губа
Рис. 31. Низшие двукрылые (прямошовные, или Orthorhapha).
А— комар-долгоножка Tipula (семейство Tipulidас); Б— комарик
Tendipes (семейство Tendipedidae, толкунчики); В — Symphorom^ia,
муха из семейства Leptidac, родственного слепням; симфоромия
принадлежит к немногим членам этого семейства, самки котогь'Х яв-
ляются кровососами; Г— ктырь, хищная муха семейства Asilidae.
и гипофаринкс, тогда как верхние и нижние челюсти полностью отсут-
ствуют. Челюстные щупальцы сохраняются обычно и у тех мух, у ко-
торых нижние челюсти отсутствуют.
Лапки ног всегда пятичленистые. Кроме двух коготков, они могут
нести парные присоски (пульвиллы) или непарный придаток (эмподий);
благодаря присоскам мухи могут ползать по вертикально поставленным
гладким поверхностям, например, по стеклу.
Брюшко у низших двукрылых имеет 10 члеников, причем 10-й чле-
ник несет церки, являющиеся по своему происхождению придатками 11-го
членика. В других случаях два или несколько задних члеников брюшка
бывают недоразвиты или втянуты внутрь брюшка, и тогда видимое извне
52
число члеников брюшка значительно меньше. У некоторых паразитических
мух членики брюшка вообще неразличимы.
Личинки всех двукрылых безноги. Личинки низших двукрылых, на-
пример, комаров, имеют хорошо развитую голову с хитиновой головной
капсулой, глазами, усиками и жующими частями рта. У личинок высших
двукрылых головная капсула неразвита, ротовые органы редуцированы,
глаз нет, усики рудиментарны и в целом
голова не обособлена. Куколка у низших
двукрылых свободная, у высших двукрылых
окружена засохшей в виде боченочка, от-
ставшей, но не сброшенной последней личи-
ночной шкуркой. При вылуплении взрослого
насекомого оболочка куколки либо лопается
вдоль, образуя Т-образную щель (подот-
ряд прямошовных Orthorhapha), либо пе-
редний конец куколочной шкурки соскаки-
вает, как крышечка, отрываясь по кру-
говому шву (подотряд кругошовных
Gyclorhapha). Прямошовные, или низшие
двукрылые и кругошовные, или высшие
двукрылые представляют собой два подот-
ряда, на которые распадается весь отряд
Diptera. Orthorhapha делятся в свою оче-
редь на две секции: Nematocera (длинно-
усые, или комары) и Brachycera (корот-
Рис. 32. Высшие двукрылые (круглошовные, или Cyclorhapha).
А — самка Carnus haemapterus (ceMeiiCTBoAgromyzidae), кровососущая мушка, живущая в гнездах
птиц (сильно увеличена); Б — серая мясная муха Sarcophaga (семейство Larvivoridae); В — пти-
чья кровососка Ornithomyia avicularia (семейство Hippoboscidae), паразит шпц. главным
образом хищных и куриных.
коусые). Длинноусые имеют длинные усики, построенные из более или
менее одинаковых члеников. К длинноусым принадлежат семейства кро-
вососущих комаров (Culicidae), мошек (Simuliidae), мокрецов (Heleidae),
толкунчиков (Tendipedidae) (рис. 31, Б), долгоножек (Tipulidae)
(рис. 31, Л), бабочниц (Psychodidae) и ^многие другие. К прямошовным ко-
роткоусым (Orthorhapha Brachycera) принадлежит много семейств (рис. 31,
В и Г), из которых медицинское значение имеет семейство слепней (Та-
banidae).
53
Для всех кровососущих представителен подотряда прямошовных
двукрылых [семейств Culicidae (рис. 105 и 106), Simuliidae (рис. 215),
Heleidae (рис. 216), Tabanidae (рис. 217) и др.; Phlebotomus из семейства
Psychodidae (рис. 200)] характерно устройство ротового аппарата. Все они
имеют хорошо развитые мандибулы и максиллы в виде двух пар острых
щетинок или ножей, участвующих в проколе кожи. За пределами крово-
сосущих Orthorhapha хорошо развитых мандибул ни у кого из двукры-
лых нет. У остальных двукрылых, имеющих колюще-сосущий ротовой
аппарат, в его состав обычно входят только непарные части: нижняя
губа, верхняя губа, гипофаринкс.
Среди Cyclorliapha (рис. 32) наибольшее медицинское значение имеют
некоторые представители семейства Muscidae (стр. 358). Меньшее значе-
ние имеют семейства Piophilidae (стр. 358), а также Larvivoridae
(стр. 395), Gastrophilidae (стр. 398), Oestridae (стр. 400), Hippoboscidae
(стр. 401) и некоторые другие.
ЧАСТЬ 2
ГИДРОБИОЛОГИЯ
ГЛАВА 1
ВВЕДЕНИЕ
1. ОПРЕДЕЛЕНИЕ ГИДРОБИОЛОГИИ. ЕЕ ЗНАЧЕНИЕ.
МЕДИЦИНСКАЯ ГИДРОБИОЛОГИЯ
Определение гидробиологии. Гидробиология есть наука о взаимоот
ношениях между водными организмами и окружающей их средой.
Гидробиология понимает среду жизни широко, учитывая , при ее из-
учении не только физические и химические условия в воде, окружающей
организмы, но также и всех животных и растений, живущих в ней и ока-
зывающих воздействие друг на друга.
Слово «гидробиология» образовано из трех греческих слов: hydor —
вода, bios — жизнь и logos — наука (собственно — слово).
Гидробиология изучает не только жизнь водных организмов, но и
жизнь тех сообществ организмов (биоценозов), на которые распадается
все население водоема в связи с неоднородностью условий обитания в нем;
наконец, гидробиология изучает и водоемы как среду обитания организ-
мов.
Разделы гидробиологии. Развитие гидробиологии всегда было тесни
связано с практическими интересами человека. Потребности развития
рыбного хозяйства — рыболовства и рыбоводства — вызвали к жизни
рыбохозяйственную гидробиологию, изучающую условия существования
рыб и способы поднятия рыбной продуктивное^ («урожайности») водое-
мов.
Загрязнение рек фабричными сточными водами и отбросами жизни
городов вызвало развитие санитарной гидробиологии, изучающей пока-
затели степени загрязненности воды в реках и изыскивающей способы их
очищения.
Медицинская гидробиология. Тот факт, что многие водоемы являются
местом развития насекомых, переносчиков заболеваний человека и его
домашних животных, вызвал к жизни медицинскую гидробиологию.
Целый ряд кровососущих двукрылых откладывает яйца в воду и в
стадиях яйца, личинки и куколки живет в водной среде. Следовательно,
они являются объектами гидробиологии. В воде проходят свое развитие
все комары (Culicidae) и мошки (Simuliidae), часть мокрецов (Helei-
dae) и слепней (Tabanidae). Из всех этих форм первым мы назовем маля-
рийного комара - анофелес (Anopheles), далее — комара, обитающего
во влажных тропических и субтропических областях, переносчика
желтой лихорадки (A6des aegypti). В Японии, Китае, в Уссурийском крае
существуют очаги тяжелого заболевания — японского энцефалита; пе-
реносчиками его являются Culex bitaeniorhynchus, Culex tritaeniorhynchus,
57
A6des togoi, Aёdes japonicus, A6des esoensis. Причиной заболевании до-
машних животных являются мошки спмулпп (Simulium), которые массо-
выми укусами могут вызвать судороги и отеки у скота. Кроме названных,
еще целый ряд других двукрылых, развивающихся в воде, приносит вред
здоровью человека и его домашних животных.
Значение биологии водных фаз развития кровососущих двукрылых.
Для правильного понимания эпидемиологической роли различных ви-
дов кровососущих двукрылых, для рациональной организации мероприя-
тий по борьбе с ними важно знание биологии не только взрослых
двукрылых — переносчиков болезней, но также п ранних стадий их
развития.
Самые различные водоемы, начиная с больших водоемов, имеющих
заросшую водными растениями широкую прибрежную полосу, и кончая
лужицами в колеях дорог, могут быть местообитанием ранних стадий раз-
вития комаров. Кровососущие двукрылые откладывают яйца в водоемы
различного типа: одни комары заселяют пойменные водоемы, заводи рек,
береговую зону озер и других водоемов, имеющих чистые воды, заросших
погруженными водными растениями и хорошо освещаемых солнцем:
другие выбирают для откладки яиц водоемы умеренно загрязняемые, ли-
шенные всякой водной растительности, затененные. Мошки спмулии свое
развитие проходят на дне быстро текучих рек и ручьев п т. п.
Истребление кровососущих двукрылых на водных стадиях их раз-
вития всегда играло ведущую роль в системе мероприятий по борьбе с
переносчиками заболеваний. Для успешного проведения этих мероприя-
тий очень полезно знать требования, которые предъявляются к среде яй-
цами, личинками п куколками того пли иного вида комара — перенос-
чика заболевания. Умение различать благоприятные и неблагоприятные
для развития комара водоемы необходимо при оценке степени
опасности того п л п иного водоема в э п и д е м п о-
логическом отношении, что может значительно облегчить
работу при обработках водоемов и при проектировании оздоровительных
гидротехнических мероприятий. Ясное представление о тех условиях среды,
которые явно неблагоприятны пли гибельны для личинок комаров в
естественных водоемах, часто подсказывает новые методы борьбы — путем
искусственного создания в водоемах этих неблагоприятных условий или
же путехМ уничтожения условий благоприятных.
Основные задачи медацинской гидробиологии. Все сказанное при-
водит к выводу, что для успешной борьбы с комарами — переносчиками
болезней человека необходимо изучать водную среду, в которой они раз-
виваются, а также влияние ее на все стадии их развития. В частности,
должны быть изучаемы: 1) отношения яиц, личинок и куколок к различ-
ным условиЯхМ водной среды, 2) те сообщества, те биоценозы водоемов, в
состав которых входят стадии развития комаров; наконец, 3) те место-
обитания, или биотопы, к которым приурочены эти биоценозы, т. е. водое-
мы или участки водоемов.
Таким образом, основными объектами изучения медицинской гидро-
биологии являются: 1) водный организм; 2) сообщества водоемов; 3) сами
водоемы, т. е. как раз те объекты, которые находятся в центре внимания
гидробиологии.
Методика гидробиологии. Гидробиология в своих исследованиях поль-
зуется многими методами. При изучении животных и растений, населяю-
щих водоемы, она пользуется хметодпкой зоологии, ботаники п экспе-
риментальной физиологии, при изучении физико-химического режима
водоемов — методами физики и химии и т. д. Однако как самостоятельная
58
наука она имеет и свою собственную методику. К таковой надо отнести
методику сборов населения толщи воды и дна водоемов.
Для сборов микроскопически малых животных и растений, обитаю-
щих в толще воды, применяются «планктонные сетки». Сетки эти имеют
большей частью коническую форму и заканчиваются металлическим
стаканчиком (рис. 33, Л). Сетку шьют из очень тонкого шелкового «мель-
ничного газа», имеющего правильные ячеи; размер ячей бывает различен.
Для улавливания очень малых организмов применяют мельничный газ
№ 25 с размером ячей в 80 микронов (1 р. = 0,001 мм). Для лова более круп-
ных организмов пользуются другими но-
мерами газа (10 и 14)-—имеющими боль-
шие размеры ячей.
Планктонные сетки количественные
позволяют производить обловы определен-
ного объема воды и после подсчета вылов-
ленных организмов заключать о числен-
ностях различных видов в 1 м3 воды или
под 1 м2 поверхности водоема, а также о
соотношениях различных видов.
Рис. 33. Гидробиологические приборы:
А — планктонная сеть и К — дночерпатель.
Количественные данные можно еще получить путем фильтрования
через мелкоячеистую планктонную сеть определенного числа литров воды,
зачерпывая ее с поверхности водоема кружкой определенной емкости.
Из более глубоких слоев воду берут особым прибором, называемым ба-
тометром, который опускают в воду и закрывают на нужной глубине,
чтобы извлечь оттуда воду. Для изучения организмов, населяющих дно
водоема, пользуются дночерпателем (рис. 33, Б).
Ото тяжелая металлическая коробка с дном, раскрывающимся па две створки,
имеющие острые края. Дночерпатель опускают в открытом виде. Ои высекает на дне
опредетенную площадь грунта со всеми организмами, населяющими его, и закрывает-
ся при помощи особого механизма, после чего его поднимают па поверхность. Под-
считав чпсло пойманных этим прибором организмов и зная площадь дна, захватывае-
59
мую дночерпателем, можно определитьплотность донного населения. Более крупных
прибрежных животных, в том числе водных насекомых, улиток, обычно ловят с помо
щью сачков (надетых на палку мешков из грубой, крупноячеистой ткани).
Рис. 34. Изменение формы листа
лютика, развивающегося в водной и
воздушной среде.
А - часль листа в воздухе; Б—часть
листа иод водой.
2. ВОДА КАК СРЕДА ОБИТАНИЯ ОРГАНИЗМОВ
Организмы, живущие в водной среде, имеют ряд приспособлений к
водному образу жизни, которые отливают их от организмов, живущих
на суше и окруженных воздушной средой. Водная среда отличается от
воздушной большей плотностью, меньшим содержанием кислорода, нали-
чием растворенных в воде солей, кислот и т. п.
Сравнительно большая плотность воды позволяет многим организ-
мам, большей частью микроскопически малым, постоянно существовать
во взвешенном состоянии в толще воды.
Нервичноводные и вторичноводные организмы. Степень приспособ-
ленности к водному образу'жизни не у всех организмов, населяющих во-
доемы, одинакова. Организмы, отличающиеся очень большой приспособ-
ленностью к жизни в воде, обычно яв-
ляются такими видами, вся эволюция
которых протекала в водной среде.
Таковы инфузории, корненожки, губки
и кишечнополостные животные (при-
мером которых в пресной воде мо-
гут служить губка-бадяга и гидра),
ракообразные (речной рак, циклопы,
водяные блохи-дафнии), рыбы и многие
другие животные. Все эти организмы
можно назвать первичновод-
н ы м и. Такие водные организмы,
предки которых в отдаленные време-
на жили на суше, называются в т и-
р и ч н о в о д н ы м и. Несмотря на
свою приспособленность к водному
образу жизни, вторичноводные орга-
низмы имеют ряд особенностей, сви-
детельствующих о том, что их предки
жили в воздушной среде. К этим
вторичноводным организмам относятся
водные легочные моллюски (например,
прудовик), водные насекомые (водные жуки, клопы, личинки кома-
ров и других двукрылых и пр.), а также водные млекопитающие
(киты, тюлени, моржи). Несмотря на большую приспособленность к жизни
в водной среде, большая часть вторичноводных организмов сохранила спо-
соб дыхания, свойственный их далеким воздушным предкам: они дышат,
пользуясь не растворенным в воде кислородом, подобно всем первично-
водным организмам, а кислородом воздуха. Для того чтобы, живя в воде,
иметь возможность поглощать атмосферный кислород, вторичноводные
организмы имеют различные у разных видов приспособления. Живущие
в воде личинки и куколки кровососущих комаров дышат главным обра-
зом за счет кислорода воздуха. Это свидетельствует о том, что они
являются вторичноводными организмами.
Гетеротопные и земноводные организмы. То обстоятельство, что кро-
вососущие комары и мошки во взрослом состоянии живут в воздушной
среде, а на неполовозрелых стадиях развития—в водной среде, позво-
ляет отнести их к числу животных г е те р о т о п ны х, т. е. к тем ;ки-
60
вотпым, которые проводят разные стадии своего развития в различной сре-
де (слово «гетеротопный» — греческое, оно означает — живущий в не-
одинаковых местах). Некоторые другие водные’животные на одной и той
же стадии развития могут жить и в воздушной, и в водной среде. Такие
животные называются земноводными, или амфибийными; примером их
могут быть взрослые лягушки и тритоны, а также личинки комарика
Dixa и др.
Гидроморфозы. К земноводным организмам относятся также некоторые водные
растения, которые способны жить и на суше, и в воде, причем особи их, выросшие
в воде, отличаются строением от особей, выросших на суше (стр. 112). Здесь мы приведем
только один пример такого растения — водяной лютик. У водяных лютиков, расту-
щих в воде, листья рассечены на длинные узкие доли, у растущих па суше — листья
широкие, с вырезными лопастями пластинок. Можно поместить молодой воздушный
лист водяного лютика в воду таким образом, чтобы одна половина листа была погру-
жена в воду, а другая оставалась в воздухе. Через некоторое время погруженная часть
листа окажется глубоко рассеченной на узкие дольки, в то время как верхняя часть,
находящаяся в воздухе, сохранит, прежний вид широкой пластинки (рис. 34).
3. СВОЙСТВА ВОДЫ. ДЕЛАЮЩИЕ ЕЕ БЛАГОПРИЯТНОЙ СРЕДОЙ
ДЛЯ ЖИЗНИ
Вода играет исключительно большую роль в строении и жизни орга-
низмов и составляет от 60 до 95% веса их тела.
Благодаря каким же свойствам вода занимает такое большое место
в жизни организма? Целый ряд физических и химических свойств воды
ставит ее в это положение. Рассмотрим наиболее важные из них.
Температурные свойства воды. Высокая удельная теплоемкость и вы-
сокая скрытая удельная теплота плавления и испарения, способность к
расширению при замерзании — все эти свойства воды делают ее внешней
средой, весьма благоприятной для жизни.
Удельной теплоемкостью в физике называют то количество кало-
рий, которое должно быть затрачено для того, чтобы поднять температуру
1 г вещества на 1°. На нагревание 1 г воды на 1° расходуется 1 малая ка-
лория; при охлаждении 1 г воды на 1° выделяется 1 малая калория. По
сравнению с другими жидкостями это высокая удельная теплоемкость (для
хлороформа удельная теплоемкость—0,24, для спирта — 0,5—0,7).
Медленное нагревание водных масс в водоемах и медленное их охлажде-
ние, зависящие от значительной удельной теплоемкости воды, обуслов-
ливают сравнительное постоянство их температуры.
Скрытая теплота плавления воды также довольно высока: она равна
80 калориям. Это значит, что для переведения 1 г льда при 0° в жидкое
состояние надо затратить 80 калорий; столько же калорий выделяется при
превращении 1 г воды в лед. Благодаря тому, что при замерзании воды вы-
деляется большое количество тепла, в естественных водоемах не происхо-
дит одновременного замерзания всей массы воды. Если верхние слои воды
замерзли, то в дальнейшем, благодаря малой теплопроводности льда,
не будет происходить такого же быстрого охлаждения глубжележащих
слоев.
Теплопроводность воды по сравнению с теплопроводностью других
жидких веществ значительна. Это способствует сравнительно быстрому
выравниванию температуры в теле организмов.
Аномальная способность воды расширяться при замерзании создает
в период образования на водоемах ледяного покрова такие условия, ко-
торые препятствуют сплошному промерзанию водных масс до самого
дна.
61
Если бы вода при замерзании не становилась легче, образующийся в
поверхностных слоях водоемов лед опускался бы на дно, происходило бы
постепенное промерзание всего водоема и жизнь зимой на дне водоемов была
бы невозможна.
Благодаря расширению воды при замерзании на поверхности водое-
мов образуется покров из льда, более легкого, чем вода, и поэтому не по-
гружающегося, а представляющего сплошной мост, под которым вода не
замерзает.
Высокое поверхностное натяжение воды. Другим физическим свой-
ством воды, имеющим большое значение в жизни организмов, надо счи-
тать высокое поверхностное натяжение воды. Свободная поверхность вся-
кой жидкости, вследствие взаимного притяжения молекул, стремится со-
кратить свои размеры. Сила этого стремления к сокращению является мерой
поверхностного натяжения, которое у разных жидкостей различно. Исклю-
чительно высокое поверхностное натяжение имеет ртуть—43 дин см;
у этилового спирта оно равно 22 дип/см, у углекислоты — всего 1,3 дпн/см.
Поверхностное натяжение воды равно 75 дин/см. Такая большая ве-
личина поверхностного натяжения воды имеет большое значение в жизни
водных насекомых, в частности, в жизни личинок комаров.
Механизм удержания личинок кровососущих комаров в поверхностной
пленке воды. Как организмы вторичноводпые личинка и куколка комаров
дышат за счет атмосферного кислорода при помощи трахей; они поглощают
кислород из воздуха,выставляянаповерхности воды свои дыхальца (стигмы),
которые у личинки находятся на так называемой стигмальной пластинке,
на предпоследнем членике их тела (стр. 171). Личинки тяжелее воды и удер-
живаются на поверхности не с помощью активных движений, — их удер-
живает в поверхностной пленке воды сила ее поверхностного натяжения.
Каким образом действуют в этих случаях силы поверхностного натяжения
воды ? Если мы погрузим в воду предмет, который не смачивается водой, то
в местах соприкосновения его с водой поверхностная пленка воды, вслед-
ствие ее значительного натяжения, будет опускаться вокруг несмачиваемого
тела и, сжимаясь, как бы стремиться выталкивать его из воды. У личи-
нок комаров стигмальные пластинки не смачиваются водой и, благодаря
действию сил поверхностного натяжения, личинки удерживаются на по-
верхности водоема, несмотря на то, что они тяжеле воды. Благодаря силам
поверхностного натяжения, держатся на поверхности воды и яйца этих
комаров. Не смачиваются также лапки длинных и тонких ног водного
клопа-водомерки, который, находясь на поверхности воды, не тонет, а
скользит по ней, как на коньках (рис. 24, Л),
В практике борьбы с личинками кровососущих комаров широко при-
меняется заливка поверхности воды нефтью или керосином. Эти вещества,
обладая меньшим поверхностным натяжением, нежели вода, растекаются
по ее поверхности, смачивают стпгмальную пластинку личинок и прони-
кают в самые трахеи. Благодаря закупорке трахей нефтью, личинки по-
гибают от удушья или вследствие отравления некоторыми из веществ,
входящих в состав нефти.
Вода как растворитель. Пригодность воды для жизни обусловлена но
только ее физическими особенностями, но и химическими. Особенно боль-
шое значение в жизни организмов имеет способность воды растворять
неорганические вещества: соли, кислоты, щелочи. Без этой способности
невозможна была бы жизнь водных растений, поглощающих необходи-
мые питательные соли из воды, в .которой они растворены.
В дсстпллпрованной воде, лишенной солей, не могут жить и водные*
животные.
62
Неорганические вещества, растворенные в воде, находятся в ионизованном со-
стоянии. Вода обладает еще одним важным для жизни свойством. Известно, что соли»
кислоты и щелочи в растворенном состоянии распадаются на ионы— частички, несу-
щие электрические заряды: катионы с положительным зарядом и анионы с отрицатель-
ным зарядом. Между противоположно заряженными ионами происходит притяжение*
более пли менее сильное в зависимости от способности растворителя ослаблять элек-
тростатическую силу этого притяжения.
Благодаря тому, что вода отлпчается'очень значительной способностью ослаблять
эти притяжения, большая часть растворенных в ней неорганических веществ нахо-
дится в состоянии ионов.
ГЛАВА II
ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И ФАКТОРАМИ ВОДНОЙ СРЕДЫ
1. ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
О ролп отдельных условий водной среды. Рассматривая условия су-
ществования организмов в водной среде, мы можем выделить целый ряд
отдельных условий, или факторов, и изучить влияние каждого из них в
отдельности. Такими факторами являются: температура, освещение, со-
держание растворенного в воде кислорода, содержание солей, наличие
пищи и др. Среда обитания в различных водоемах и в различных биото-
пах одного и того же водоема бывает подчас весьма различна. Во-первых, от-
дельные факторы среды могут иметь различные значения: так, температура
может быть выше и ниже, пищи может быть больше и меньше и т. и. Во-
вторых, весьма различными оказываются и сочетания факторов; кроме
того, один и тот же фактор, например, высокая температура, может быть
благоприятным для одних организмов и неблагоприятным для дру-
гих.
Максимум, минимум и оптимум действия фактора. Каждый вид жи-
вотных и растений может существовать в определенных пределах темпе-
ратуры, света, кислорода и других условий внешней среды. Верхний пре-
дел каждого фактора, при котором организмы данного вида еще могут
существовать, называется максимумом этого фактора для данного вида, а
нижний его предел — его минимумом. При переходе интенсивности фактора
за эти пределы организм гибнет. Эти смертельные для данного вида зна-
чения фактора называются его летальными пределами. Между минимумом
и максимумом где-то находится та интенсивность фактора, при которой
данный вид чувствует себя лучше всего; это его оптимальная зона, опти-
мум фактора.
Для наглядности представим схему влияния какого-либо из факторов
среды на любой вид животного пли растения, например, действие темпе-
ратуры на личинку Anopheles maculipennis messeae (рис. 35). При темпе-
ратуре 7° личинки не развиваются, но и не гибнут, при температуре
—2° они уже гибнут. Поэтому температура 7° является нижним температур-
ным пределом развития, а —2Э—нижним летальным пределом для личинок
A. mac. messeae. Верхний летальный предел той же личинки ле-
жит при 37° и совпадает с верхним пределом ее развития. При темпера-
туре 25° жизнеспособность личинки выше всего, и эту температуру можно
считать оптимумом выживаемости личинки A. mac. messeae.
Для разных видов животных и растений длина отрезка шкалы между
минимумом и максимумом бывает различна; некоторые виды выносят боль-
шие колебания данного фактора, другие приспособлены к существованию
в очень узких его пределах.
63
Факторы абиотические и
можно разделить на две
верхний идеи ел t
Ч
Нижний предел
развития
верхний предел
развития
Нижний предел t
42*
409
38°
36е
34*
32*
30*
28*
26°
24*
20*
18°
Ю*
14°
12 °
//7°
8*
8*
~0
--4 е
-6°
~8'
--Ю •
—12 °
—14 9
Рис. 35. Схема, изображающая
влияние температуры на личинок
Anopheles maculipennis messeae.
биотические. Все факторы внешней среды
группы: 1) факторы абиотиче-
ские1, куда относятся воздействия на
организм неживой природы: движения
воды, температуры, света, кислорода, со-
лей и других физических и химических
условий существования, и 2) факторы био-
тические, куда относятся воздействия на
организм со стороны других организ-
мов, живущих с ним в одном биоценозе.
В этой последовательности мы и будем
рассматривать основные факторы водной
среды.
Понятие о биотопе и биоценозе. Все
факторы среды распределены по земной
поверхности (и на суше, и в воде) нерав-
номерно, и в связи с этим неравномерно
распределено население земли. Различия
в составе и в численности населения, свя-
занные с неоднородностью условий суще-
ствования, наблюдаются не только по гео-
графическим широтам (например, хоро-
шо известно различие в фауне и флоре по-
лярных, умеренных и тропических об-
ластей, связанное с различиями кли-
мата). Даже в одной и той же области
на рядом лежащих участках могут со-
здаваться различные условия среды, и это
всегда сопровождается образованием раз-
личного населения. Например, рядом
расположенные пруд и озеро всегда засе-
лены различными организмами. Мало то-
го, два участка одного и того же озера, даже одной и той же лужи, часто
представляют совершенно различные условия существования и содержат
различное население.
Всякий участок земной поверхности, отличающийся от окружаю-
щих его других участков по тем условиям существования, которые он
представляет для населяющих его организмов, является отдельным био-
топом (местообитанием). Совокупность организмов, населяющих биотоп,
называется его биоценозом.
2. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И ДВИЖЕНИЕМ ВОДЫ
Движение воды в текучих и стоячих водах. Только в небольших и не-
глубоких стоячих водоемах вода находится в спокойном состоянии.
В реках, ручьях, морях, озерах, даже в неглубоких прудах, но с большой
водной поверхностью, вода находится в движении — то более сильном,
то б *лое слабом. В реках и ручьях вода непрерывно движется вперед,
она непрестанно течет; поэтому все они называются текучими водами, или
водотоками. Их течение связано с наклонным положением их дна; чем
1 Абиотический — от греческих слов: а — без и bios — жизнь; абиотический —
неживой.
64
Рис. 36. Личинки
мошки Simulium на
камне на дне реки.
личинки направлен
более крутым является этот наклон, тем быстрее может оказаться их
течение.
В замкнутых стоячих водоемах (озера, пруды) движение водных масс
в основном зависит от воздействия дующих ветров. Если ветер долгое
время дует в одном и том же направлении, он сгоняет поверхностные слои
воды в ту сторону водоема, по направлению которой он дует. У этого бе-
рега уровень воды может на время повыситься, у противоположного—по-
низиться.
Эта разница уровней вызывает в более глубоких слоях воды вы-
равнивающее течение, идущее от подветренного берега, где уровень воды
выше, к надветренному. Кроме того, в стоячих во-
доемах ветры вызывают рябь или волнение.
Организмы, приспособленные к текучим водам.
Различные водные организмы к волнению и течению
относятся очень различно.
Существуют виды животных и растений, для
которых текучесть воды является необходимым
условием существования. В качестве примера на-
зовем обитателя быстро текучих рек и ручьев —ли-
чинку мошек симулий (Simulium). Личинки симу-
лий часто целыми колониями покрывают камни или
растения на дне рек. Чтобы не быть снесенными
водой, они прочно прикрепляются задним концом
тела к твердым предметам. Передний конец тела
всегда вниз по течению. Личинки симулии питаются, улавливая несомые
течением пищевые частицы, и в то же время приспособлены к дыханию
в текучей воде, всегда насыщенной кислородом (рис. 36). Если их
поместить в стоячую воду, они быстро погибают от недостатка кисло-
рода.
Личинки кровососущих комаров в водоемах со спокойной водой. Со-
вершенно иначе относятся к движению воды личинки кровососущих ко-
маров. Сколько-нибудь значительное движение воды для них, как пра-
вило, неблагоприятно. Они являются типичными обитателями спокойных
стоячих водоемов или медленно и спокойно текущих водотоков.
В больших стоячих водоемах заросли водных растений очень сильно
умеряют волнения, и часто среди растительности можно найти много ли-
чинок Anopheles даже в больших озерах. Это необходимо учитывать при
создании водохранилищ. Их береговая линия не должна быть сильно из-
резана, не должна иметь заливов, бухт и мелководий, чтобы волна не да-
вала возможности водным растениям завоевывать прибрежную зону.
Точно так же п в небольших речках и каналах густые заросли расте-
ний замедляют течение и могут у берегов на отдельных участках свести
его на-нет. Такие затишья охотно заселяются личинками Anopheles.
3. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И ТЕМПЕРАТУРОЙ
Температурные пределы жизни. Температурные условия среды играют
громадную роль в жизни организмов. Пределы температуры, в которых
возможно существование живых организмов, довольно широки: некото-
рые водные организмы остаются живы, проведя зиму в толще льда при
температуре ниже нуля Ч Так перезимовывают многие простейшие, не-
1 В соленых водоемах многие организмы живут при температуре ниже —2°.
б Учебник медицинской энтомологии 65
которые водные улитки и личинки насекомых, а также некоторые дру-
гие животные (С. А; Зернов). Личинки Anopheles зимовать в толще льда
неспособны.
Верхним температурным пределом для жизни можно считать темпера-
туру около 90°. Это температура горячих ключей, в которых живут не-
которые виды бактерий и водорослей, а также немногие животные: не-
которые простейшие живут при температуре до 51°, некоторые черви__
при температуре до 40°, некоторые водяные жуки — до 45°, а некоторые
улитки — до 53°.
Температурный режим водоемов. Основным источником тепла в водое-
мах является энергия солнечных лучей, которая, однако, поступает в них
в значительно ослабленном виде: часть ее поглощается атмосферой, часть
идет на испарение поверхностных слоев воды; значительное количество
солнечных лучей, падающих на поверхность водоема, отражается
от нее.
Количество солнечной энергии, получаемое водоемами, расположен-
ными в различных географических широтах, различно.
В зависимости от географического положения водоемов, их разделяют
на три группы: 1) водоемы тропические — расположены в тропиках и
субтропиках, температура воды в них никогда не бывает ниже 4°, 2) во-
доемы полярных областей и высокогорные озера, в которых температура
воды никогда не поднимается выше 4°, наконец, 3) водоемы умеренного поя-
са, отличающиеся тем, что зимой они, хотя бы ненадолго, покрываются
льдом, а летом, хотя бы ненадолго, прогреваются выше 4°.
Движение Земли вокруг Солнца и вокруг своей оси вызывает годич-
ную и суточную периодичность температурных изменений. В тропических
и полярных водоемах размах этих изменений невелик, но он очень широк
в большей части водоемов умеренного пояса.
Большие глубокие озера медленно нагреваются и медленно остывают.
Летом они не прогреваются до дна, зимой не остывают до дна. Летом для
больших озер характерно температурное расслоение: температура у по-
верхности озера теплее, чем у дна; зимой наблюдается обратное расслое-
ние: на поверхности озера замерзает, а теплее всего вода у дна. Весной
и осенью происходит полное перемешивание eo/ih озера. Таким обра-
зом, годовые изменения температуры в озере велики и сложны.
Наоборот, суточные колебания температуры озерной воды незначи-
тельны.
Температурный режим лужи прудов. Противоположную картину пред-
ставляют малые водоемы, например, лужи: солнечные лучи проникают в
них насквозь, а ветер полностью перемешивает их воду; поэтому сколько-
нибудь стойких различий в температуре воды у поверхности и у дна не
возникает.
Суточные колебания температуры велики. Если лужа лежит откры-
то и доступна прямым солнечным лучам, днем вода ее сильно про-
гревается и становится значительно теплее воздуха. За ночь она остывает,
но при ясной солнечной тихой погоде средняя суточная температура от-
крытой лужи все-таки выше средней суточной температуры воздуха. При
пасмурной, дождливой или ветреной погоде средние суточные температуры
лужи не отличаются от средних суточных температур воздуха. Но, так как
периоды солнечной и пасмурной погоды чередуются между собой, средняя
температура открытой лужи за более длительный срок, например, за ме-
сяц, всегда оказывается выше средней температуры воздуха за тот же пе-
риод; ясно, что это превышение создается за счет ясных дней и оказывается
тем больше, чем больше было таких ясных дней. Водоемы, средняя
66
температура которых выше средней температуры воздуха, называются
гипертер мическим и водоемам и.
Лужа, расположенная в тени, днем нагревается значительно меньше,
а ночью остывает почти настолько же, как и лужа, лежащая на откры-
том месте. Средняя суточная температура такой лужи более или менее
точно следует за средней температурой воздуха. Водоемы, средняя тем-
пература которых близка к средней температуре воздуха, называются
изотермическими1 (рис. 37).
Рис. 37. Суточный ход температуры.
А—в изотермической луже; Б—в гипертермической луже. --------------температура воздуха;
-------температура поверхности воды посреди лужи; —.— температура на глубине 30 см.
там же;-------------------------температура прибрежной зоны.
Летом многие лужи высыхают, а зимой могут промерзать до дна.
Пруды по своим размерам и по массе содержащейся в них воды зани-
мают промежуточное положение между озерами и лужами. Промежуточ-
ный характер носит и их температурный режим. Они согреваются и осты-
вают медленнее луж, но быстрее озер; температурное расслоение в них
выражено слабо; суточные колебания температуры меньше, чем в лужах,
но больше, чем в глубоких озерах. Маленькие мелководные заливчики
больших озер, если они хорошо отделены от озера, имеют такой же тем-
пературный режим, как пруды и мелкие озерки.
Влияние растительности на температурный режим водоемов; влияние
притока грунтовых вод. В мелких водоемах, заросших погруженными
водными растениями, как трехдольчатая ряска, роголистник, уруть,
температурный режим обычно своеобразен. Растительный покров на по-
верхности воды таких водоемов создает преграду для проникновения сол-
нечных лучей, и температура воды под этим покровом бывает значительно
ниже, чем в поверхностном слое. Так, например, в одном озере-пруде на
пойме реки Сыр-Дарьи, на поверхности, над густым ковром роголист-
1 Гинер-, изо- и гипотермический — слова, образованные греческими приста-
вками: hyper—выше, iso—равный, hypo — вине,и прилагательным термический,
т. е. имеющий отношение к теплоте. Таким образом, гипертермический буквально
значит: имеющий более высокую температуру (подразумевается в сравнении с воз-
духом).
5*
67
ника температура воды в летний день была 38°, а на глубине 30 см — всего
24°. Ночью эти резкие температурные различия сглаживаются.
Однако не только высшие водные растения, преграждая путь солнеч-
ным лучам, мешают им прогреваться до дна; почти такое же влияние ока-
зывает массовое развитие планктона.
Водоемы, питаемые грунтовыми водами, пробивающимися из-под
земли родниками, имеют особый температурный режим. Подземные
грунтовые воды обладают постоянной температурой, близкой к средней
Рис. 38. Суточный ход температуры в водоемах.
А — в водоемах, населенных личинками Anopheles maculipennis; Б — в
водоемах, населенных личинками Anopheles bifurcatus.
годовой температуре данной местности. В силу этого родники, обра-
зующиеся при выходе грунтовых вод па поверхность земли, летом имеют
очень постоянную температуру (рис. 38, 5), которая значительно ниже тем-
пературы воздуха. Зимой, наоборот, температура в них значительно выше
температуры наружного воздуха. В силу этого зимой родники обычно не
замерзают.
Температурный режим водоемов и личинки комаров. Выше мы видели,
что волнения и течения мешают существованию личинок в больших от-
крытых водоемах и в быстро текущих реках. Поэтому развитие личинок
комаров фактически происходит при температурном режиме, описанном
для луж, для небольших мелководных озер, прудов, копанок, заводей рек,
заливов озер. При этом водоемы, различающиеся по своему температурному
режиму (сильнее или слабее прогреваемые, имеющие большие пли мень-
шие суточные колебания температуры), заселяются личинками различ-
ных видов комаров (рис. 38).
68
4. ОТНОШЕНИЕ ВОДНЫХ ОРГАНИЗМОВ К СВЕТУ
Значение солнечного света для жизни. Источником света на земле яв-
ляется солнце. Лучи солнца, падая на воду, обусловливают и температур-
ный, и световой режим водоема. Значение солнечного света для жизни
заключается в том, что зеленые растения используют его для синтеза орга-
нического вещества, являющегося первоисточником пищи и заключенной
в ней энергии для всех остальных живых существ. Зеленые растения и на
суше, и в воде не могут существовать без света. Для всех животных свет как
таковой не является жизненно необходимым. Об этом говорит наличие
животных на очень больших глубинах океана, где свет совершенно от-
сутствует, а также в толще почвы п в абсолютно темных пещерах.
Однако поведение животных в отношении света, строение их органов
зрения, их окраска говорят о том, Что для многих из них свет является
важнейшим фактором внешней среды.
Световой режим водоемов. Световой режим в воде иной, чем на суше. Прежде
всего интенсивность света в водоемах значительно меньше. Солнечные лучи, падающие
на поверхность водоема, далеко не все проникают в воду, значительная часть их от-
ражается от поверхности и рассеивается.
Проникновение солнечных лучей в водоем зависит от угла их падения: косые лучи,
падающие в утренние и вечерние часы, в водоем не проникают, и ночь там на-
ступает раньше и длится дольше, чем на суше. Чем больше глубина, тем короче
день и длиннее ночь; чем мутнее вода, чем больше она содержит взвешенных частиц,
тем меньше ее прозрачность: лучи поглощаются сильнее и меньше проникают в глубину.
Не все лучи в одинаковой степени поглощаются водой: красные поглощаются больше,
синие — меньше. Поэтому очень чистая вода имеет синий цвет, менее чистая вода
имеет зеленый цвет.
С глубиной количество проникающего света падает очень резко, по в различного
типа водоемах в различной мере. Так, на глубине 1 м количество света составляет в
различных водоемах от 1 до 30% дневного света, а иа глубине 3—19 м — всего
лишь 0,01—1% дневного света. Для гидробиологических целей часто ограничиваются
измерением прозрачности воды при помощи простого прибора — белого диска, опу-
скаемого в воду. Глубина, на которой диск перестает быть видимым, является мерилом
прозрачности воды. Если говорят: «прозрачность данного озера равна 5 м», это значит,
что диск, опущенный в воду, перестал быть видимым на глубине 5 м.
Свет и водные организмы. От особенностей проникновения солнеч-
ного света в водоем зависит распределение в нем водорослей, которые со-
средоточены главным образом в поверхностной, лучше освещенной зоне.
Так же связано со световым режимом водоема и распределение высшей
водной растительности. Заросли водных растений занимают береговую
п прибрежную зону, — в глубинных областях озер они отсутствуют. Чем
больше прозрачность воды в озере, тем глубже опускается нижняя гра-
ница ассимиляции, а следовательно, и заросли растений. Стремясь к свету,
высшие водные растения чаще всего образуют длинные стебли, поднимаю-
щиеся к поверхности, вблизи которой и располагаются листья; некоторые
из них плавают по поверхности, другие поднимаются над поверхностью
воды, и, таким образом, и те, и другие получают больше света.
Распределение водных животных в водоемах часто также зависит
от света. Особенно это выражено у животных, обладающих фототак-
сисом. Фототаксисом называют ориентировку движений животных в
отношении направления световых лучей.
У сидячих животных и растений под влиянием световых лучей происходит ориен-
тировка не движений, а положения тела; такая ориентировка называется фототропиз-
мом. Суточные изменения в условиях освещения водоемов вызывают перемещения мно-
гих водных животных из поверхностных слоев в глубинные, и наоборот. Такие суточные
миграции совершают многие рачки: дафнии, моины и др.
69
Движение в направлении к свету называется положительным фото-
таксисом, а в направлении от света — отрицательным фототаксисом. По-
ложительный фототаксис играет большую роль в жизни водных насеко-
мых: когда нырнувший плавунец или личинка Anopheles возвращаются
на поверхность воды, они идут прямо на луч света и таким образом
быстро выбираются на свободную поверхность, минуя заросли водных
растений, преграждающие их путь.
5. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И РАСТВОРЕННЫМИ В ВОДЕ НЕОРГАНИЧЕСКИМИ ВЕЩЕСТВАМИ
Неорганические вещества в водоемах. Основным источником раство-
ренных неорганических веществ в водоеме являются те горные породы,
которые образуют его ложе, а также и те, по которым протекает питаю-
щая его вода. Поэтому в природе, как правило, водоемов, совсем лишен-
ных солей, нет. Наиболее бедны солями водоемы верховых болот —
сфагновые водоемы, питаемые атмосферными водами и имеющие ложе,
всецело образованное моховыми подушками, а не минеральным грун-
том.
Концентрация солей в водоемах колеблется в очень .широких преде-
лах — от 0,01 до 470 г в 1 л воды. Содержание солей в воде обозначается
различными способами, чаще всего в граммах на 1 л воды (так называемая
«соленость» воды).
Существует деление всех водоемов по солености на четыре группы:
1) водоемы пресные с соленостью от 0,01 до 0,3—0,5 г в 1 л воды; 2) водое-
мы солоноватые с соленостью от 0,5 до 16 г в 1 л воды (сюда относятся наши
внутренние ^моря-озера: Аральское и Каспийское, многие солоноватые
озера, приморские лиманы и др.; 3) океаны и моря с соленостью от 16 до
47 г на 1 л воды; 4) водоемы переосолоненные, плюющие концентрацию
солей от 47 до 370 г в 1 л воды и выше. Столь исключительно высокая со-
леность создается, например, в морских заливах, отделенных от моря в
результате постоянного испарения и при условии, если отсутствует приток
в них пресных вод. Бессточные озера, расположенные в сухих странах,
также могут иметь переосолоненпую воду (например, озера Эльтон и Бас-
кунчак). Такие озера встречаются в засушливых районах (в Прикаспий-
ских степях, в южной части Западной Сибири, в Казахстане и т. п.).
Роль солей в питании водных организмов. Соли, растворенные в воде
природных водоемов, имеют большое значение в жизни водных орга-
низмов.
Очень велика их роль в питании водных растений; растворенные соли
вместе с углекислотой являются тем материалом, из которого с помощью
световой энергии растения строят живое вещество своего тела. Особенно
большое значение для питания растений имеют соли азотной и фосфорной
кислот и калия, которые получили даже название‘«биогенных» элементов,
что значит — «дающих жизнь». Углекислота, являющаяся источником угле-
рода при синтезе органических веществ растениями, в свободном состоя-
нии обычно находится в воде естественных водоемов в небольшом коли-
честве, недостаточном для удовлетворения потребностей водных расте-
ний; основным ее резервом служат ее соли — карбонаты, растворенные в
воде в больших количествах [средние соли: СаСО3, MgCO3, и кислые соли:
Са (НСО3)2, Mg (НСО3)2]. Соли железа также очень важны для водных
растений, но избыток их действует губительно и на растения, и на многих
животных.
70
Ядовитое действие отдельных солей и уравновешенные растворы. Соли
калия, натрия, кальция и магния, каждая в отдельности, также ядовиты
для всех организмов; но, если растворы этих солей смешать в известной
пропорции, их вредное действие взаимно уравновешивается и смесь ока-
зывается неядовитой. Воды большинства природных водоемов представ-
ляют собой уравновешенные смеси, неядовитые для населяющих их
организмов.
Осмотическая роль солей. Помимо качественного состава растворенных
солей, большое значение имеет и общая их концентрация, от которой за-
висит осмотическое давление раствора. Осмотическое давление раствора
солей тем выше, чем больше их концентрация. У морских животных ос-
мотическое давление жидкостей тела высокое и близко к осмотическому
давлению морской воды, у пресноводных животных оно ниже, чем у мор-
ских, но значительно превышает осмотическое давление пресной воды.
Поэтому вода непрерывно насасывается в их тело через все полупроницае-
мые участки покровов (т. е. участки покровов, проницаемые для воды и
растворенных в ней газов, но не для растворенных солей).У водных жи-
вотных, дышащих всей поверхностью кожи, вся поверхность кожи про-
ницаема для воды; у водных животных, имеющих обособленные жабры,
проницаемым для воды является жаберный эпителий, что мы и видим, на-
пример, у личинок Anopheles младших возрастов.
У водных насекомых, дышащих исключительно атмосферным возду-
хом и лишенных кожно-жаберного дыхания, наружная поверхность тела
совершенно непроницаема для воды, как, например, у жуков-плавунцов
или у куколок комаров.
Благодаря тому, что жидкости тела пресноводных животных всегда
содержат больше солей, чем окружающая вода, через все проницаемые
для воды участки их покровов происходит непрерывное насасывание воды
в их тело; это насасывание должно было бы повести к разбавлению их
внутренней среды водой и к набуханию их тела. Однако этого не проис-
ходит благодаря процессу «осморегуляции», т. е. регуляции осмотиче-
ского давления. Этот процесс заключается в том, что почкп животного
(например, сегментальные органы кольчатых червей и раков, мальпигие-
вы сосуды личинок насекомых) непрерывно выводят из организма всю
лишнюю воду, проникающую в него через покровы или через жаберный
эпителий.
Организмы, живущие в солоноватых или соленых водоемах и имею-
щие осмотическое давление более низкое, чем во внешней среде, обладают
дыхательным эпителием, проницаемым не только для воды, но и для со-
лей. Вместе с водой соли поступают во внутреннюю среду, но избыток со-
лей выводится наружу почками. У рыб часть солей выводится через жабры.
Таким образом, концентрация солей во внутренней среде поддерживается
на постоянном уровне.
Солевыносливость и отношение к растворенным солям у различных
водных организмов весьма различны. Существуют строго пресноводные
организмы, неспособные жить в солоноватой воде; разноводные, или со-
левыносливые, организмы, приспосабливающиеся к жизни и в пресных,
и в солоноватых водоемах; чисто солоноводпые организмы, могущие жить
только в соленой воде. Личинки различных видов комаров представляют
примеры всех этих категорий, но среди наших видов Anopheles имеются
представители только двух из них: пресноводные (например, A. mac. mes-
seae) и солевыносливые (например, A. mac. atroparvus), хотя за пределами
СССР существуют и чисто солоноводные виды (например, индонезийский
A. sundaicus).
71
6. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И ОРГАНИЧЕСКИМИ ВЕЩЕСТВАМИ ВОДЫ
Органическое вещество встречается в воде в четырех основных
состояниях:
1) животные и растения, живущие в водоеме;
2) взвешенные в воде мертвые органические частицы, видимые про-
стым глазом пли под микроскопом;
3) органические вещества в виде коллоидов1;
4) растворенные органические вещества.
Органические вещества в водоемах. Каковы источники поступления
органических веществ в природные водоемы?
В самом водоеме ежедневно происходит отмирание организмов, на-
селяющих его дно и толщу воды; осенью, кроме того, отмирают заросли
водных растении. Остатки погибших водных животных и растении после
их разложения обогащают воду мельчайшими взвешенными частичками
(так называемый «органический детрит»), а также коллоидными и рас-
творенными органическими веществами. С другой стороны, органическое
вещество поступает в водоемы извне: из окружающей почвы, с болот пли
вместе со сточными водами, загрязненными продуктами жизни города;
в лесных водоемах большое значение как источник органического веще-
ства имеет листопад.
Благодаря жизнедеятельности бактерий, населяющих толщу воды
и дно водоемов, в них непрестанно происходят процессы распада сложных
органических соединений на более простые и в конечном итоге их минера-
лизация, т. е. распад на неорганические соединения и элементы. Как
указывалось выше, растворенные неорганические вещества потребля-
ются зелеными растениями, которые строят из них живое вещество своего
тела; таким образом, водоем вновь обогащается сложными органически-
ми веществами, заключенными в живых растительных организмах, которые
после отмирания и постепенного распада или после прохождения через
кишечники поглотивших их животных разлагаются на все более простые
и, наконец, на неорганические составные части.
Таким образом, органические вещества в водоемах находятся в по-
стоянном круговороте, благодаря жизнедеятельности бактерий, зеленых
растений и животных.
Зеленые растения за их синтезирующую органическое вещество дея-
тельность называются производителями живого вещества. Животных, за
то что они только потребляют органическое вещество, называют потре-
бителями. Бактерий за их способность разлагать сложные органические
вещества на более простые, вплоть до их минерализации, называют раз-
рушителями органического вещества. В водоемах разного типа кругово-
рот органического вещества бывает выражен различно,
Органических веществ в водоеме может быть больше или меньше (от
незначительных следов до 400 мг в 1 л воды). Больше всего органических
веществ в водоемах, загрязненных сточными водами.
Система сапробности водоемов. По степени органического загрязне-
ния (по их сапробности) все пресные водоемы разделяются на: 1) сильно
загрязненные, полисапробные водоемы, в которых протекают начальные
этапы разложения сложных органических соединений на более простые;
2) умеренно загрязненные, мезосапробные водоемы, где происходит окис-
1 То есть в виде взвешенных в воде мельчайших частиц, не видимых даже иод
микроскопом.
ленпе продуктов первых фаз разложения сложных органических веществ;
мезосапробпые водоемы с большим загрязнением называются а (альфа)-
мезосапробными, а с меньшим загрязнением—(бета)-мезосапробными.
Наконец, водоемы с очень малым содержанием органических веществ на-
зываются олпгосапробными; в них происходит полный распад и минера-
лизация органических веществ. Водоемы определенной степени сапроб-
пости характеризуются особым химизмом воды, количеством и видовым
составом бактерий, растений и
животных, населяющих толщу
воды и дно этих водоемов.
Для определения степени
сапробности какого-либо водо-
ема пользуются списками ви-
дов растений и животных, кото-
рые могут служить показателя-
ми степени загрязнения воды.
Нахождение в водах реки бак-
терий Sphacrotilus natans (рис. 39,M),
серобактерий Beggiatoa, водо-
росли Polytoma uvella, инфузорий
Paramaecium putrinum (рис. 39, Б),
червя-трубочника Tubifex tubifex по-
зволяет с уверенностью считать этот
Рис. 39. Полиса пробные организмы.
А— грибов- Sphaerotihis; Л — инфузория Param aecium puti;inum.
участок рек сильно загрязненным — полисапробным (рис. 39). Для водоема а-мезо-
сапробпого характерны: зеленая водоросль Stigeoclonium (рис. 40, А), ряд видов
синезелепых водорослей, главным образом виды Oscillator!а (рис. 40, Б), инфузория
трубач Stentor coeruleus, несколько видов коловраток, особенно живущих в при-
донных слоях Rotifer neptun ins (рис. 40, В), красные личинки мотыля Tendipes
plumosus (рис. 40, Г).
К водоемам ^-мезосапробным принадлежит большая часть наших прудов. Для
последних характерны сине-зеленые водоросли, главным образом Aphanizomenon flos
aquae (рис. 41, Л); при их массовом развитии диатомовые водоросли: Melosira varians
Diatoma, Navicula (рис. 41, Б) и др.; многие виды зеленых водорослей: протококко-
вые, конъюгаты (рис. 45) и др. Из числа микроскопических животных много разных
видов инфузорий, коловраток (рис. 41, В) и ракообразных. Впервые появляется выс-
шая растительность (роголистник) (рис. 61, В), несколько видов рыб (вьюн, карась,
линь, карп, уклея, верховна и др.).
Наконец, чистые воды— олигосапробные— характеризуются наличием следую-
щих видов: из водорослей — диатомовая водоросль Melosira italica, из коловраток —
Notholca longispina (рис. 42, Л), из рачков Daphnia longispina (рис. 42, Б), из рыб —
стерлядь, форель, гольян.
Кроме названных трех типов водоемов, характеризующихся различной степенью
загрязнения, выделяют еще четвертый тип — водоемы с совершенно чистой, богатой
кислородом и большей частью холодной водой.
73
Процессы самоочищения воды. Загрязненные органическими веществами воды
рек и других водоемов, благодаря деятельности населяющих их организмов, постепенно
очищаются; органические вещества распадаются на все более простые соединения и,
наконец, минерализуются. Говорят, что происходит «самоочищение» воды. Наибольшее
участие; в процессах «самоочищения» воды принимают бактерии; их роль особенно
велика в полисапробных зонах. В мезосапробпой зоне очень большую роль в процес-
сах самоочищения воды играют зеленые водоросли,
которые, благодаря процессам фотосинтеза, обогащают
воду кислородом, расходующимся на окисление раз-
лагающихся деятельностью бактерий органических
веществ.
Рис. 40. а-ме зоеа пробные «организмы.
А— водоросль Stigeoclonium; Б —водоросль Oscillatoria; В —
коловратка Rotifer neptunius; Г — мотыль (личинка комара
Tendipes plumosus).
Органические вещества как пища. Роль органических веществ в жиз-
ни водных организмов очень велика. Для многих животных органические
вещества в виде более крупных микроскопических частиц — раститель-
ного детрита — являются основной пищей; в питании других большую
роль играют наиболее мелкие фракции — коллоидно-распыленные орга-
нические вещества, состоящие из частиц, уже не видимых в микроскоп.
В питании личинок малярийного комара большое значение имеют и круп-
ные, и мелкие частицы органических веществ.
Влияние органического загрязнения на химические свойства воды. При
большом содержании в водоемах распадающихся органических веществ
вода их обычно становится бедна кислородом, который расходуется на
окисление продуктов распада; в таких водах способны жить лишь немно-
гие организмы.
74
Кроме дефицита кислорода, в водах, содержащих разлагающиеся
органические вещества, часто образуются ядовитые промежуточные ве-
щества, от действия которых многие организмы погибают.
Чувствительность к загрязнениям личинок кровососущих комаров.
Чувствительность личинок разных видов кровососущих комаров к со-
держанию органических веществ очень различна. Личинка обыкновен-
ного комара Culex pipiens переносит значительные загрязнения, личинка
малярийного комара Anopheles mac. messае очень чувствительна к орга-
ническому загрязнению: в водоемах з-мезосапробных она никогда не
встречается.
Рис. 41. ^-мезосапробные организмы.
А - сипезсленан водоросль Aphanizomenon flos aquae; Б — диато-
мовая водоросль T^ayicula; В—коловратка Anuraea aculeata.
Наименее чувствительна к содержанию органических веществ ли-
чинка A. plumbeus, обитающая в очень своеобразных водоемах — скоп-
лениях воды в дуплах деревьев. Дупляная вода содержит громадное ко-
личество органических веществ, но процессы их распада никогда не но-
сят там характера гниения.
Чувствительность личинки A. maculipennis к органическому загряз-
нению имеет большое практическое значение: водоемы, сопутствующие
жилью человека и очень сильно загрязненные продуктами его быта или
отбросами промышленности, как правило, не заселяются личинками.
7. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И РАСТВОРЕННЫМИ В ВОДЕ ГАЗАМИ
Состав газов, растворенных в воде. Из газов, растворенных в воде,
наибольшее значение для жизни организмов имеют кислород и угле-
кислота; азот значительно менее необходим для жизни; аммиак, сероводо-
75
род п метан присутствуют в воде очень редко, обычно только в водоемах
особого типа. Растворимость газов в воде сравнительно невелика, при-
чем кислород растворяется лучше азота, а углекислота — лучше кисло-
рода. Благодаря этому газы, растворенные в воде, находятся там в другом
соотношении, чем в атмосфере: в воде азота отно-
! сительно меньше, а кислорода и углекислоты от-
f носительно больше, чем в воздухе (табл. 1).
I.
Т а б л и п а 1
Содержание основных газов в воздухе и в воде
в кубических сантиметрах (при 0° и 760 мм давления)
1 л воздуха .... 210 780
1 л воды............ 10.26 18.32
Рис. 42. Олигосапробные организмы.
Л — ноловрать-а Nothulca 1 ongispinа; Б — ветвистоусый рачок Daphnia longispin ,.
А
Мы видим, что, несмотря на хорошую растворимость кислорода, кон
центрацпя его в воде значительно меньше, чем в атмосфере. Поэтому вода
является средой, менее благоприятной для дыхания, чем воздух.
Баланс газов в водоемах. Основным источником кислорода, азота и
углекислоты в воде является воздух: газы, входящие в состав атмосферы,
поглощаются поверхностными слоями воды различных водоемов. Однако
диффузия газов в воде происходит очень медленно; обогащение этими га-
зами происходит в основном не за счет их диффузии, а благодаря движе-
ниям водных масс, течениям, волнению, а также токам воды, вызываемым
различием температур в поверхностных и глубинных слоях.
В очень многих водоемах существует еще другой источник поступле-
ния кислорода: выделение его при фотосинтезе водных растений. В водое-
мах с массовым развитием водорослей пли с большими зарослями высших
растений количество выделяемого ими кислорода может быть очень вели-
ко, и вода становится перенасыщенной кислородом. Так, в некоторых озе-
рах в летние дни отмечено пересыщение воды кислородом до 364%.
76
Расход кислорода в водоемах вызывается дыханием водных животных
и растений, а также жизнедеятельностью бактерий, разлагающих мертвые
органические вещества. Процессы распада органического вещества проис-
ходят главным образом в придонных слоях воды и в грунте дна; в зави-
симости от большего или меньшего скопления гниющего органического
ила на дне водоема, происходит более или менее сильно выраженное обед-
нение воды кислородом.
Слоистость в распределении кислорода. Так создается слоистость (стратификация)
в распределении кислорода в водоемах. Поверхностные слои непрерывно обогащаются
кислородом как благодаря диффузии его из атмосферы, так и за счет кислорода, вы-
деляемого при ассимиляции растениями. Глубинные слои затрачивают кислород на
окисление органических веществ ила; чем ближе слой воды ко дну, тем он беднее кис-
лородом. Степень кислородного расслоения водоемов зависит от того, насколько бо-
гато население водоема, и в первую очередь от развития в нем зеленых водорослей.
В о юрах одиготрофных \ бедных питательными солями, как, например, в водоемах,
распо тоженных среди каменистых горных пород, взвешенных водорослей в водной тол-
ще мало. дно озера не имеет слоя органического ила, кислородное расслоение выражено
слабо. В озерах эвтрофных1 2 , богатых питательными солями, зеленые водоросли изо-
билуют. часто наблюдается «цветение» воды (вода окрашивается массой перепол-
няющих ее микроскопических водорослей). Отмирая, они образуют на дне озера мощ-
ный слой органического ила; и в этих водоемах очень резко выражено кислородное
расслоение.
Суточные изменения в содержании кислорода. В дневные часы ассимиляция зе-
леных организмов обогащает воду кислородом, который обычно пересыщает поверх-
ностные слои; глубинные слои беднее кислородом, так как здесь он расходуется на
окисление органических веществ. Ночью.когда продукция кислорода зелеными организ-
мами прекращается, происходит обеднение воды кислородом, особенно резко выражен-
ное в поверхностных слоях воды. Слоистость кислорода становится менее выра-
женной.
Летние и зимние заморы. В неглубоких водоемах с очень энергично протекающими
процессами разложения органических веществ летом в ночные часы кислород может
полностью исчезать, и тогда наступает гибель населяющих эти водоемы организмов;
наблюдаются так называемые летние заморы. В подобных водоемах происходят и зим-
ние заморы. Несмотря на низкую температуру, поглощение кислорода па окисление
органического ила все же происходит, и к концу зимы подо льдом вода полностью те-
ряет весь растворенный кислород; животные массами гибнут, в первую очередь рыбы.
В начале замора рыбы устремляются к прорубям пли к незамерзающим местам, где
нередко становятся добычей человека и животных. В водоемах, где зимует рыбка
амбузия (стр. 10)), явление замора нередко вызывает ее гибель.
, Углекислый газ. Как указывалось выше, углекислый газ растворяет-
ся в воде лучше, чем кислород и азот. Поглощаемая водой из атмосферы
углекислота связывается, образуя соли. Соли углекислоты называются
карбонатами. Обычно в водоемах больше всего карбонатов кальция, ко-
торые встречаются в виде монокарбонатовСаСО3 и бикарбонатов Са(НСО3)2.
Избыток углекислоты остается в свободнОхМ состоянии. Другим источни-
ком поступления углекислоты в водоем является выделение ее при дыха-
нии водных организмов и при разложении органических веществ бакте-
риями. Расходуется углекислота на процессы фотосинтеза водных растений.
При энергичнОхМ фотосинтезе свободной углекислоты, растворенной в воде,
оказывается недостаточно; после ее полного израсходования растения от-
нимают углекислоту от ее кислых солей — бикарбонатов, которые пере-
ходят при этом в средние соли — хмонокарбонаты.
Сероводород. Сероводород II2S образуется па дне очень загрязненных органиче-
скими веществами водоемов при распаде белков в условиях недостатка кислорода.
Этот процесс обычно наблюдается зимой на дне, покрытом мощным слоем гниющего
ила, при незначительном содержании, а часто и полном отсутствии кислорода в воде,
под толстым ледяным покровом. Наличие сероводорода в водоеме обычно сопровождает-
1 Олиготрофный — бедный пищей.
2 Эвтрофный —богатый пищей.
77
ся гибелью организмов, населяющих его. Только очень немногие организмы способны
существовать в присутствии сероводорода. Кроме того, сероводород встречается в
воде сернистых источников, где он бывает минеральною происхождения, так что вода
таких источников во всех остальных отношениях может быть вполне чистой.
Аммиак и метан. Аммиак NH3 в воде всегда является промежуточным продуктом
разложения белковых соединений в условиях дефицита кислорода, и наличие замет-
ных количеств аммиака говорит о сильном загрязнении воды.
Метан — болотный газ — СН4 образуется на дне водоемов, покрытом значитель-
ным слоем растительных остатков; он образуется за счет распада их клетчатки. В та-
ких водоемах пузырьки метана могут появляться па поверхности воды; при поднесении
к ним огня они загораются. Таково происхождение так называемых «блуждающих»,
«болотных» огней.
Роль кислорода и углекислого газа в жизни водных организмов.
Кислород и углекислый газ имеют очень большое значение в обмене ве-
ществ водных организмов (как всех организмов вообще): растворенный
кислород необходим для дыхания водных животных и растений, растворен-
ная углекислота — для питания погруженной водной растительности.
Различные животные неодинаково чувствительны к дефициту растворен-
ного кислорода: некоторые гибнут, когда содержание кислорода падает
до 4 см3 в 1 л, другие могут жить продолжительное время при полном отсут-
ствии кислорода, переключившись на иной тип обмена веществ (анаэроб-
ное — бескислородное — дыхание).
Личинки кровососущих комаров, как уже говорилось выше, дышат за
счет атмосферного кислорода; кроме того, они дышат и кислородом, рас-
творенным в воде, через кожу тела и особенно через тонкие покровы аналь-
ных жабер.
Углекислый газ при очень высоких концентрациях мог бы быть ядо-
вит для личинок Anopheles, но водоемов с такими его концентрациями очень
мало.
Колебания углекислоты в обычных пределах существенного зна-
чения для личинок не имеют.
То же можно сказать о роли сероводорода, аммиака и метана в био-
логии личинок Anopheles.
8. ВЗАИМООТНОШЕНИЯ МЕЖДУ ВОДНЫМИ ОРГАНИЗМАМИ
И АКТИВНОЙ РЕАКЦИЕЙ ВОДЫ
Мы уже говорили о том, что в воде естественных водоемов раство-
рены различные соли, кислоты и основания, а также о том, что мо-
лекулы этих веществ в растворе распадаются на ионы, т. е. частички,
несущие положительный или отрицательный электрический заряд.
Частички, несущие положительный заряд, называются катионами,
частички с отрицательным зарядом — анионами.
Например, поваренная соль — хлористый патрпй—NaC] распадается на катион
Na+ с положительным зарядом и анион С1~, несущий отрицательный заряд. Молекулы
соляной кислоты НС1 в растгоре распадаются па положительно заряженный ион
водорода Н + и на анион С1-. Щелочь КОН распадается на катион К+ и анион ОН-,
называемый гидроксильным ионом.
Концентрация водородных ионов. Из всех ионов наибольшей актив-
ностью обладают водородные и гидроксильные ионы. Можно сказать, что
все свойства кислот определяются водородными ионами, а все свойства
щелочей — гидроксильными ионами. В химически чистой (дестиллиро-
ванной) воде (Н2О) очень незначительная часть ее молекул (всего одна де-
сятимиллионная часть) также распадается на водородный и гидроксильный
ионы (Н.О—Л14 4" ОН-), причем в такой воде количество и водородных,
78
и гидроксильных ионов совершенно одинаково. Про такую воду говорят,
что она имеет нейтральную реакцию. Если в воде водородных ионов боль-
ше, чем гидроксильных, она имеет кислую активную реакцию^ при преоб-
ладании гидроксильных ионов над водородными вода имеет щелочную ак-
тивную реакцию.
Для обозначения степени кислотности или щелочности в жидкости
пользуются особым показателем pH, который представляет собой логарифм
концентрации водородных ионов, но без знака минус. Так, для химически
чистой воды pH равен 7,0, так как концентрация Н-ионов в такой воде
равна, как уже говорилось 0,0000001, а логарифм этого числа равен 7,00000.
В воде, богатой углекислыми солями, pH часто оказывается равным 8,0.
Это значит, что концентрация Н-ионов в этой воде меньше, чем в химически
чистой воде: она равна 0,00000001,а логарифм концентрации равен 8,00000.
Если в водоеме с кислой водой pH равен 4,0, это значит, что в нем отно-
сительно высокая концентрация водородных ионов — 0,0001 и логарифм
ее 4,0. Для измерения pH в водоемах обычно пользуются методом
двуцветных индикаторов, т. е. применяют расторы особых красящих
веществ, которые в кислой и щелочной водах имеют разную окраску. Так,
если к небольшому объему воды, имеющей кислую реакцию, прибавить
несколько капель индикатор а бромтимол синего, то он примет желтый цвет,
в щелочной воде он примет синюю окраску, а в нейтральной воде — зеле-
новатую.
Активная реакция воды естественных водоемов. В обычных пресных
водоемах реакция воды определяется главным образом углекислотой и
углекислыми солями кальция и магния и бывает нейтральной, слабо ки-
слой или слабо щелочной.
Ясно выраженная щелочная реакция бывает в особенности при на-
личии в воде большого количества карбонатов натрия, как, например,
в содовых озерах, которые встречаются в степной полосе Сибири и Север-
ного Казахстана.
Относительно очень кислая вода бывает в том случае, когда в рас-
творе имеется избыток кислот и в особенности нелетучих органических
кислот и серной кислоты, как в сфагновых болотах и в некоторых руднич-
ных водах.
Итак, в естественных водоемах реакция колеблется в довольно широ-
ких пределах. Очень значительное количество озер и рек, благодаря бо-
гатству их углекислыми солями, имеет щелочную реакцию (pH = 8—9).
Многие водоемы — озера, пруды и т. п. — имеют реакцию, близкую к
нейтральной. Наконец, существует целая группа водоемов с кислой реак-
цией. Это главным образом сфагновые озера и моховые болота с коричне-
вой, богатой гуминовыми веществами водой и своеобразным населением.
В этих водоемах pH может быть равен 3,5—5,5.
Колебания реакции воды. Реакция в одном и том же водоеме может
претерпевать значительные колебания в течение суток, а также в разные
времена года. Причина этих колебаний заключается в том, что на реак-
цию воды большое влияние оказывает жизнедеятельность водных орга-
низмов.
Особенно сильное воздействие на реакцию в водоемах оказывает
фотосинтетическая деятельность зеленых растений, благодаря которой
реакция воды становится более щелочной. Различие в интенсивности фо-
тосинтеза в различных частях водоема — например, в поверхностных, хо-
рошо освещаемых слоях воды и в глубинных, получающих ничтожное ко-
личество света — часто служит причиной различия реакции воды: у по-
верхности она является менее кислой, чем у дна, и даже щелочной. Среди
70
зарослей погруженных растений реакция может оказаться более щелоч-
ной, чем в свободной йоде.
Роль реакции воды в жизни водных организмов. Реакция воды (сте-
пень ее кислотности или щелочности) в свою очередь оказывает воздейст-
вие на жизнедеятельность водных организмов. Иногда сравнительно не-
большие изменения в реакции воды тормозят пли совсем останавливают
жизненные отправления организмов, главным образом различных микро-
организмов. Крупные животные и растения на небольшие изменения ак-
тивной реакции среды реагируют гораздо меньше. Разные виды животных
и растений могут иметь различные летальные (смертельные) и оптималь-
ные (наиболее благоприятные) пределы реакции среды. Этим иногда объяс-
няется тот факт, что водоемы, имеющие различную реакцию, имеют раз-
личное население. Так, воды моховых болот и других сфагновых водоемов
со сравнительно очень кислой водой населены отчасти такими организмами,
которые в водоемах с нейтральной или щелочной реакцией совсем не встре-
чаются.
Роль реакции воды в жизни личинок комаров. Личинки кровососущих
комаров обычно встречаются в водоемах, имеющих реакцию, близкую к
нейтральной или слабо щелочную и слабо кислую. В торфяных водоемах
с очень кислой реакцией (pH = 3,5—5,0) личинки Anopheles maculipen-
nis совсем не живут. Однако опыты, проведенные с личинками этих ко-
маров, показали, что отсутствие их в водоемах с умеренно кислой водой
обусловлено не вредным воздействием водородных ионов, а тем, что в кис-
лых сфагновых водах создаются какие-то иные условия (присутствие
вредных органических веществ, неполноценная пища), которые губи-
тельно отражаются на существовании этих личинок (Котляревская).
Лишь в немногих водоемах крайних типов (особенно кислых или осо-
бенно щелочных) реакция воды сама по себе препятствует благополучию
личинок комаров.
9. ВЗАИМООТНОШЕНИЯ ВОДНЫХ ОРГАНИЗМОВ
(БИОТИЧЕСКИЕ ФАКТОРЫ ВОДНОЙ СРЕДЫ)
Воздействия организмов друг на друга в водной среде. Процветание
того или иного вида животных и растений в любой среде зависит не только
от благоприятных условий температуры, света, кислорода и других физи-
ческих п химических факторов, которые, как говорилось выше, принято
называть абиотическими. Животные и растения появляются на свет, раз-
виваются, живут и умирают среди других животных и растений, от кото-
рых в большей пли меньшей степени зависит их существование. Сами они
также влияют тем пли иным путем на окружающие их организмы. Можно
сказать, что все члены биоценоза находятся между собой в более пли менее
тесной связи.
Всевозможные воздействия на жизнь организмов со стороны других
организмов называют, как уже указывалось выше, биотическими факто-
рами среды.
Пищевые взаимодействия. Наиболее важными являются пищевые взаи-
моотношения организмов. Питание водных растений, каки наземных, лишь
косвенно зависит от других организмов. Из растворенных в воде солей и
углекислоты растения с помощью световой энергии солнечных лучей
сами строят органическое вещество своего тела. Животные этой способ-
ностью не обладают, они существуют за счет поедания других организ-
мов, растительных и животных. Поэтому благополучие животных в любом
80
водоеме тесно связано с численностью организмов, служащих им пищей.
В питании личинок кровососущих комаров большую роль играют водные
организмы: бактерии, мельчайшие водоросли, простейшие, коловратки
и т. д.
При изучении питания водных животных можно установить более или
менее сложные пищевые цепи или ряды. Так, живущие па дно водоемов
личинки комаров толкунчиков-тендипедид питаются бактериями, диато-
мовыми и другими придонными водорослями; тендипедиды поедаются
молодью карпа, крупные хищные рыбы (щука, судак) поедают мальков
карпа. Получается цепь: диатомовые водоросли—^личинки тендипедид—>
молодь карпа—>щука. Во всех подобных цепях первым звеном являются
производители живого вещества — растения или их мертвые остатки. Для
характеристики пищевых цепей, в которых участвует личинка маля-
рийного комара, можно привести такой пример: водоросли —>коловратка—►
личинка Anopheles—>гамбузия.
Животные, питающиеся одним и тем же кормом, одинаковыми орга-
низмами, обычно находятся в состоянии взаимной конкуренции, которая
остро выражена в тех водоемах, где данный корм имеется в ограниченном
количестве. Для личинок комаров конкурентами являются многие дру-
гие животные, питающиеся тем же кормом: коловратки, рачки и др.
Далее, благополучие каждого вида организмов в большой мере за-
висит от наличия и численности нападающих на него и истребляющих его
врагов. В пищевых цепях организмы каждого последующего звена явля-
ются врагами по отношению организмов звена предыдущего. У личинок
комаров много врагов. Самым опасным их врагом являются рыбы, особен-
но те из них, которые питаются организмами, обитающими в толще воды
и на ее поверхности, а не у дна. Очень опасный враг для личинок Anophe-
les — небольшая рыбка гамбузия (Gambusia), которая иногда может в ко-
роткий срок полностью очистить водоем от этих личинок. Опасными вра-
гами для личинок Anopheles могут быть также водяные жуки, клопы
и их личинки, личинки стрекоз и ручейников, водные пауки и многие
Другие.
Влияние жизнедеятельности бактерий и растений на газовый режим
водоема и на другие условия жизни. Кроме пищевых связей, еще целый
ряд других взаимоотношений связывает между собой водные организмы.
Прежде всего одни организмы могут представлять субстрат, на котором
помещаются другие организмы; так, многие водоросли и мелкие животные
поселяются на листьях водных растений.
Во-вторых, одни организмы создают физические и химические усло-
вия, благоприятные или неблагоприятные для других организмов.
Как указывалось выше, круговорот органического вещества
п связанные с ним изменения в содержании кислорода и углекислоты за-
висят от жизнедеятельности бактерий. В меныпей степени на газовом ре-
жиме водоема сказывается дыхание животных. Степень освещения и темпе-
ратура в более глубоких слоях водоема часто снижены под влиянием мас-
сового развития взвешенных в воде малых организмов или погруженной
высшей водной растительности. Все эти влияния сказываются на громад-
ном количестве организмов; они благоприятны для одних и гибельны для
других. Так проявляются связи между организмами, изменяющими усло-
вия среды, и теми, кто живет в этих видоизмененных условиях.
Все сказанное заставляет признать, что существование водных орга-
низмов находится в тесной зависимости от целого ряда биотических факто-
ров. Это побуждает нас изучать не только физические и химические усло-
вия жизни в водоемах, но и организмы, населяющие их.
6 Учебник медицинской энтомологии
81
ГЛАВА III
ОСНОВНЫЕ ТИПЫ ВОДНЫХ БИОЦЕНОЗОВ И ХАРАКТЕРНЫЕ
ДЛЯ ПИХ ЖИВОТНЫЕ И РАСТЕНИЯ
1. ОСНОВНЫЕ ТИПЫ ПРЕСНОВОДНЫХ БИОЦЕНОЗОВ
Каждый водоем, большой или маленький, не представляет среды оби-
тания, однородной на всем своем протяжении; наоборот, каждый водоем
сложно расчленен. И прежде всего в каждом водоеме непременно присут-
ствует три различных типа биотопов: 1) водная масса, т. е. толща воды дан-
ного водоема, 2) дно водоема, 3) поверхностная пленка в месте соприкоснове-
ния воды с атмосферой. В соответствии с этим основным расчленением во-
доема разделяются и населяющие его биоценозы: толща воды занята планк-
тическими биоценозами, дно водоема — бентическими и поверхностная
пленка — нейстическими. Планктические биоценозы образованы орга-
низмами, или пассивно взвешенными в толще воды, или обладающими
слабыми активными движениями (планктоном), или активно плаваю-
щими в толще воды — сильными пловцами, составляющими нектон. Дон-
ные или бентические биоценозы образованы организмами, роющимися в
грунте водоема, прикрепленными ко дну иди ползающими по его поверхно-
сти. Все эти организмы обозначаются для краткости словом бентос. Ней-
стические биоценозы образованы организмами, активно или пассивно пла-
вающими на поверхности воды и очень часто поддерживаемыми там силами
поверхностного натяжения; все они обозначаются словом нейстон.
Наиболее подвижные организмы, по способу передвижения принад-
лежащие к нектону (например, очень многие рыбы), обычно бывают свя-
заны не с отдельными маленькими биоценозами, а с комплексами биоцено-
зов, занимающими целый водоем или значительную его часть.
Организмы, живым слоем обрастающие подводные предметы, обо-
значают словом перифитон; перифитон представляет часть бентоса.
Крупные водные растения могут быть бентические (прикрепляющие-
ся ко дну), планктические (плавающие в толще воды) и нейстические (пла-
вающие на поверхности воды). Но и те, и другие, и третьи обычно зани-
мают одни и те же участки водоема, и, располагаясь друг над другом или
в первхмежку, составляют единые биоценозы зарослей, которые мы и бу-
дем рассматривать отдельно (стр. 103).
2. БИОЦЕНОЗЫ ТОЛЩИ ВОДЫ (ПЛАНКТИЧЕСКИЕ БИОЦЕНОЗЫ)
а) Условия сугцествования, особенности
и состав планктона
В состав планктона входят организмы, принадлежащие к очень раз-
личным типам, классам и отрядам. Но, благодаря сходному образу жиз-
ни, они имеют общие черты, связанные с их «парением» в водной толще.
Какие же это приспособления?
Приспособления водных организмов к «парению». Всякое тело с удельным весом
бдлъшим, чем вес воды, при погруя ении в нее тонет, и тем быстрее, чем больше его ос-
таточный вес 1. Поэтому приспособления к «парению» в воде прежде всею направлены
на уменьшение остаточною веса. Организмы планктона обычно не имеют тяя елых по-
кровов, тело их готов или покрыто о епь легкими раковинами. Все другие части скелета
таки е тонки и ле! ки. Облегчение геса тела достигается такя е выделением слизи, обра-
зованием пузырьков воздуха внутри тела (газовые вакуоли), как, например, у корне-
ножки Difflugia hyJrostatica (рис. 43, А). Отложение запасов питательных веществ
1 Остаточный вес — это вес погруженного предмета минус вес вытесненной
нм воды (закон Архимеда).
82
в виде капелек жира, очень распространенное у организмов планктона, также
приводит к облегчению их веса (нащилер, у диатомей и у рачков-циклопов). Об-
легчение веса достигается также благодаря очень большому содержанию воды в
теле планктических организмов (95—96%).
Второе приспособление организмов планктона к «парению» в воде — форма тела,
замедляющая скорость его погружения; такое замедление достигается увеличением
Рис. 43. Различные способы приспособления организмов планктона к «парению»
в воде.
А—приспособление к «парению» путем образования воз-ушных вакуолей: корненожка
Difflugia hydrostatica; Б — плоская форма особей или колоний в виде дисков. Водоросли:
1—Pediastrum; 2—Coscinodiscus; 3—Asterionella; В—лентовидная форма колоний. Водоросли:
i — Fragillaria, 2—Desmidium; Г—«колючая», или «ежевидная», форма: 1—солнечник
Actinosphaerium; 2—яйцо коловратки Synchaeta.
1 — ядра; 2—сократительные вакуоли; 3—пищевые тела.
6*
83
поверхности тела по сравнению с его объемом, т. е. увеличением удельной поверхно-
сти. Известно, что удельная поверхность с уменьшением размеров тела увеличивается,
в связи с этим в планктоне преобладают микроскопически мелкие животные и расте-
ния, обладающие относительно большой удельной поверхностью. Очень распро-
странена у организмов планктона плоская форма тела в виде дисков, лент, пучков лент.
Например, водоросли Ре liastrum (рис. 43, Б}), Coscino.liscus (рис. 43, Б2) и Asterionella
(рис. 43, Б3) имеют форму дисков, водоросли Fragillaria (рис. 43, и Desmidium
(рис. 43, В2), имеют вид лент и т. п. Очень часто среди планктонных организмов встре-
чаются колючие формы с большим количеством выростов, шипов, игл, например, сол-
нечники (рис. 43, Z\), некоторые коловратки: Triartlira, Notholca (рис. 42, А) и др.
Основные группы планктона. По своим размерам планктические
организмы могут быть разделены на четыре группы: 1) очень малые,
имеющие размеры до 50 р, принадлежат к наннопланктону1; сюда относятся
бактерии, самые мелкие виды водорослей и др.; 2) организмы, имеющие ве-
личину от 50 pi до 0,5 мм, составляют микропланктон, к которому относятся
очень многие водорссли, рачки, коловратки; они большей частью с тру-
дом видны невооруженным глазом; 3) организмы, имеющие величину от
0,5 до 1 мм, составляют мезопланктон, сюда относятся многие ракообраз-
ные; 4) наконец, организмы, имеющие величину более 1 мм, относятся
к макропланктону.
Кроме этого деления по размерам, планктические организмы по си-
стематическому положению подразделяются еще на фитопланктон 1 2 —
растительные организмы — и зоопланктон 3 — животные организмы.
В состав наннопланктона входят организмы самых мелких размеров:
бактерии, водоросли и жгутиковые с длиной менее 50 р. В жизни водоемов
роль наннопланктона очень велика. Особенно большую роль играют бак-
терии. При изучении растворенных в воде органических веществ мы уже
говорили о том, что одной из основных движущих сил в круговороте орга-
нического вещества в водоемах является жизнедеятельность бактерий.
Кроме того, они служат пищей многим водным животным: их поедают и
простейшие, и коловратки, и ракообразные, и двустворчатые моллюски;
большую роль они играют и в питании личинок комаров, например, личинок
малярийного комара. Однако если поместить личинок анофелес в среду
с очень большим количеством бактерий, они там жить не будут. В такой
среде обычно накапливается много продуктов жизнедеятельности бакте-
рий, делающих среду непригодной для жизни личинок. Другие организмы
наннопланктона — водоросли, жгутиковые — также могут служить пи-
щей для личинок анофелес.
б) Обзор характерных представителей пресноводного
планктона
Фитопланктон. Основную массу фитопланктона образуют
первичноводные примитивные растения, называемые водорослями (Algae).
В пресных водах обитает много разных групп водорослей, представленных
многочисленными видами. Мы познакомимся лишь с самыми распространен-
ными из них.
1) Синезеленые водоросли (тип Cyanophyceae)
Очень примитивны по организации. Название свое получили за сине-
зеленую окраску. Большая часть синезеленых водорослей представляет
колонии клеток. У некоторых из них клетки соединяются в четкообраз-
1 От слова «панно» — карликовый.
2 От греческого слова phyton — растение.
3 От греческого слова zoon — животное.
84
ные, у других — в цилиндрические нити; часто нити соединяются
слизью в шары и т. д. Размножение происходит путем деления или путем
образования спор. Эти водоросли очень обычны в прудах, в озерах и в
Рис. 44. Сине зеленые водоросли.
А — Anabaena; Б — Gloeotrichia echinata.
медленно текучих реках. Летом массовое развитие сине-
зеленых придает синевато-зеленую окраску воде прудов
и озер; в таком случае говорят о «цветении» водоема сине-
зелеными водорослями. Это «цветение» очень часто вызы-
вается следующими синезелеными: Aphanizomenon flos
aquae, Microcystis, Anabaena, Gloeotrichia echinata и неко-
торыми другими (рис. 44). Синезеленые водоросли —бо-
гатая белками пища для водных животных,
поедаются личинками Anopheles.
Синезеленые водоросли входят в со-
став не только планктона, — целый ряд
видов входит в состав бентоса и перифи-
тона, некоторые виды живут и вне воды,
во влажных биотопах суши.
они часто
и
2) Зеленые водоросли (тип Chlorophyceae)
Зеленые водоросли названы так по
яркозеленой окраске, обусловленной при-
сутствием красящего вещества (пигмента)
хлорофилла. Хлорофилл, как и у высших
растений, не заполняет собой всю про-
топлазму клетки, а сосредоточен в хро-
матофорах («хромое» по-гречески — крас-
ка, «форо» — несу, «хроматофор» бук-
вально — носитель окраски). Хромато-
форы зеленых водорослей бывают разно-
образной формы; часто в них видны крах-
мальные зерна. У большинства зеленых
водорослей хорошо различимо клеточное
ядро (рис. 45). Размножаются зеленые
водоросли главным образом посредством
размножаются также и посредством спор. Некоторые зеленые водоросли
обладают половым размножением.
Рис.
45. Зеленые водоросли
(семейство зигнемовых).
А — Spirogyra; Б— Zygncma: 1 —
ядро; 2—хроматофор; 3—пире-
ноиды.
простого деления, но они
Одна из наиболее распространенных групп зеленых водорослей —
это сцеплянки (класс Conjugatae), которые отличаются особым способом
полового размножения, состоящим в слиянии содержимого двух клеток,
совершенно одинаково построенных и лишенных специальных органов
движения. В результате этого слияния образуется одна клетка «зигота»,
из которой развивается новая водоросль. Важнейшие семейства сцепля-
нок: зигнемовые (Zygnemaceae) (рис. 45) и
десмидиевые (Desmidiaceae) (рис. 46).
К семейству зигнемовых принадлежат неветвя-
щиеся нитчатые формы, например, спирогира (Spirogyra)
(рис. 45, А) и зигнема (Zygnema) (рис. 45, Б). Спиро-
гира, обычно обитатель стоячих и медленно текучих
водоемов, встречается часто большими скоплениями;
в пасмурные дни они находятся иногда на дне водо-
ема, в ясные дни — всплывают благодаря пузырькам
кислорода, образующимся при ассимиляции. Нить
спирогиры представляет ряд продолговатых клеток, в
которых вдоль стенок расположена спирально идущая
лента хроматофора. Середина клетки занята большой
Рис. 46. Десмидиевые водоросли.
4— Closterium: 1 — ядро; 2—пиреноиды; 3— вакуоль; Б— Cosmarium: В— Micrasterias.
полостью — вакуолью, в центре которой расположено подвешенное на протоплаз-
менных нитях крупное ядро (рис. 45, А).
Зигнема (Zygnema) имеет хроматофор в виде двух крупных звездообразных
пластинок, между которыми на тонком протоплазменном мостике расположено ядро
(рис. 45, Б).
Зеленые подушки, образуемые скоплениями нитей зигнемовых, —
излюбленное местообитание личинок малярийного комара. Личинки на-
ходят внутри этих скоплений «и стол, и дом», поедая отдельные нити во-
дорослей и прячась в их чаще от врагов.
К сцеплянкам относятся и десмидиевые водоросли (Desmidiaceae).
Это семейство в основном состоит из одноклеточных форм. Каждая особь
состоит из двух симметричных половинок, нередко разделенных перетяж-
кой. Десмидиевые населяют болота или прибрежную часть озер, воды,
богатые гуминовыми веществами, получаемыми при разложении расти-
тельных остатков. Некоторые широко распространенные формы десми-
диевых изображены на рис. 46.
Вольвоксов ы е (отряд Volvocales) также относятся к зеленым
водорослям. Для Volvocales характерно наличие жгутиков, при помощи
которых они плавают. Имеются как одноклеточные, так и колониальные
формы.
83*
Как пример одноклеточных форм, назовем хламидомонад (Ghlamydo-
monas) (рис. 47, Л), которые очень характерны для весенних луж, а также для мелких
прудов. Хламидомонады имеют яйцевидное или круглое тело с двумя жгутами, отхо
дящими от переднего конца, сзади них расположен красный глазок; к оболочке клетки
прилегает чашевидный зеленый хроматофор. При помощи жгутиков хламидомонады
быстро двигаются вводе, скопляясь главным образом иа ее поверхности (положитель-
ный фототаксис). Размножаются они делением.
Из колониальных вольвоксовых наиболее распространены: эвдорина (Eudorina)
(рис. 47, Б), Пандорина (Pandorina) (рис. 47, В) и вольвокс (Volvox). Шаровидные ко
Рис. 47. Вольвоксовые водоросли.
А—одиночная форма Chlamydomonas; Б—колониальная форма
Eudorina; В— то же Pandorina; Г— то же Volvox; дочерние шары
внутри материнского.
лонии всех этих водорослей бывают покрыты студенистой оболочкой; внутри шара —
небольшие зеленые клетки колоний: у эвдорины обычно 32 клетки, у пандорины —
16, у вольвокса— несколько тысяч клеток (до 50 000) в одной колонии. Наиболее
сложна колония вольвокса (рис. 47, Г). Клетки колоний расположены в один слой жгу-
тами наружу; движениями этих жгутов обусловлено движение всего шара в воде.
Все клетки колонии соединены между собой тонкими протоплазменными мостиками.
Делясь, некоторые из клеток колонии дают начало маленьким дочерним колониям,
которые проваливаются внутрь материнского шара. Подросши, они разрывают мате-
ринский шар, выходят «наружу и становятся самостоятельными колониями. Через
2—3 дня они уже становятся сами способны к беспотому размножению. Вольвокс об-
ладает и потовым размпо пением. В котонин развиваются круглые неподвижные лен-
ские и подвижные двужгутиковые мужские половые клетки. В результате оплодо-
творения образуется зигота, которая окружается защитной обо.точкой. В виде зиюты
вольвоксы переживают неблагоприятные периоды (высыхание, зимние холода).
87
Вольвоксы обитают в не очень загрязненных прудах и озерах. Они хо-
рошо поедаются личинками анофелеса.
Протококковые водоросли (отряд Protococcales)
(рис. 48) распространены очень широко. Они неподвижны, за исключением
периодов полового размножения, и лишены красного глазка, какой имеют
вольвоксовые. Среди них есть как одноклеточные, так и колониальные
формы. Наиболее важной формой этой группы является «водяная се-
точка» (Hydrodictyon) (рис. 48, Л). Колонии этой водоросли, имеющие
вид сеточки, часто достигают очень больших размеров, ячеи сеточки об-
разованы несколькими клетками, соединенными в пяти- и шестиуголь-
ники. Комья водяной сеточки охотно заселяются
личинками анофелеса. В петли водяной сеточки,
как в рыболовные сети, иногда попадают гамбу-
зии и гибнут, так что это растение не только
полезно для личинок анофелеса, но и гибельно
для их злейшего вага (Казанцев).
Рис. 48. Протококковые водоросли.
д — Hydrodictyon. крупная клетка с молодой колонией внутри; Б— Ulotrix:
1—нить; 2—макроспора; 3—зооспоры; В—Oedogonium: деление клетки.
К типу зеленых водорослей относятся и улотриховые водоросли (отряд Ulotriclia-
les, рис. 48, Б), Многие из них, подобно спирогире, являются нитчатыми формами, об-
разующими плавающие комья. Относящийся сюда род эдогоний (Oedogonium,
рис. 48, В) отличается наличием па многих клетках его нитчатых колоний у одного
из концов клетки— очень характерных колпачков, словно надетых друг на друга;
они образуются в связи с делением клеток. Эдогоний часто развивается на субстрате,
па погруженных в воду предметах, по образует таки е плавающие зеленые комья,
которые охотно заселяются личинками Anophe.es maculipennis.
3) Диатомовые водоросли (тип Diatomaceae)
Эти водоросли отличаются наличием тонкого кремневого панцыря, со-
стоящего из двух створок, которые покрывают одна другую, как крышка
коробку. Цвет диатомовых — золотистожелтый, так как к хлорофиллу
у них присоединены бурые пигменты, главным образом диатомин. Пан-
цырь диатомовых бывает покрыт тончайшим узором из точек и штри-
хов.
88
Помимо планктона, многочисленные диатомеи входят также в состав
бентоса и, в частности, перифитона.
Некоторые из числа распространенных диатомей изображены на рис. 49
(одиночные формы и колониальные формы). Наличие жировых вклю-
чений у диатомовых водорослей делает их очень калорийной пищей для
водных организмов, значительную роль играют они и в питании личинок
кровососущих комаров.
Рис. 49. Диатомовые водоросли.
А — одиночная форма
Pinnularia; Б— то же СугоЬеПа; В— то же Surirella; Г — колониаль-
ная форма Melosira; Д — то же Tabellaria.
4) Жгутиковые (Flagellata)
Громадный и разнообразный класс жгутиковых стоит на грани меж-
ду растениями и животными. Прежде всего жгутиковые, как правило,
активно подвижны. Многие из них способны поглощать растворенные го-
товые органические вещества или даже оформленную пищу (бактерий).
Однако большинство жгутиковых имеет хлорофилл, и при благоприятных
световых условиях они сами синтезируют живое вещество. К жгутико-
вым относятся:
Эвгленовые (отряд Euglenaceae), имеющие веретеновидную форму тела,
один единственный жгутик и красный глазок, расп» л он енный на переднем конпе тела.
Большая часть эвгленовых зеленого цвета, но некоторые из них бесцгетны и питают-
ся растворенными органическими веществами; характерным представителем является
эвглена (Euglena) (рис. 50,А).
Перидинеи (отряд Peridineae), отличающиеся тем, что оба их жгута рас-
положены в двух яелобках— продольном и поперечном; последний опоясывает все
тело. Цвет перидиней бурый. Наиболее типичный их представитель изображен на
рис. 50, Б.
Зоопланктон. Животные, составляющие зоопланктон, относятся к
очень различным группам — от простейших и до личинок рыб.
К простейшим (тип Prctczoa) зоопланктона пресных водоемов, если
не считать жгутиковых, стоящих на грани животных и растений, относятся
корненожки и инфузории.
89
5) Корненожки
Корненожки (класс Pseudopodiata х), обличаются тем, что двигаются, выпуская
отростки протоплазмы, называемые ложноножками, или псевдоподиями; эти же лож-
ноножки служат им для захватывания пищи. Гел о корнено ек пречставлнет собой бес-
форменный комок протоплазмы с ядром.
В пресноводном планктоне встреча-
ются голые корненожки, к которым при-
надлежат амебы (Amoebae) (рис. 51, Л) и
корненожки, покрытые раковинкой, как,
например, арцелла (Arcella) (рис. 51, В) и
дифлюгия (Difflugia) (рис. 51, В), домик
которой построен из песчинок. Хорошо
приспособлены к жизни в планктоне
солнечники (отряд Helioz.oa (рис. 43, Z\).
Их псевдоподии имеют вид тонких ни-
тей, расходящихся, во все стороны, как
лучи солнца, от шарообразного тела сол-
нечника; внутри каждой псевдоподии про-
ходит тонкая осевая нить; осевые нити
всех псевдоподий соединяются в центре
шара.
6) Инфузории
50.
— пире-
5 — со-
Рис.
А — Euglena viridis
ноиды; 3— жгутик, _
кратлтельная "’вакуоль;
Рис. 51. Корненожки.
А —голые: амеба (Amoeba): 1 — ядро; 2 — сократительная вакуоль; Б — покрытые
раковинкой: арцела (Arcella); В—дифлюгия (Difflugia).
Строение тела у инфузорий
(класс Ciliata) значительно сложнее,
чем у корненожек. Поверхность
тела инфузорий покрыта ресничками,
которые могут быт в. расположены
весьма различно (см. ниже). Реснич-
ки являются органами движения
инфузорий; при помощи ресничек
они направляют в рот пищу. Инфу-
зории имеют ротовое отверстие,
обычно ведущее в глотку. Заглочен
ная пища переваривается в пище
Жгутиковые.
>: 1 — ядро; 2
;; 4— глазок;
...___ Б — Ceratium hi-
rundinella.
варптельных вакуолях внутри протоплазмы. Непереваренные остатки
выбрасываются через особое отверстие на заднем конце тела. В поверх-
1 Иначе называемые Rhyzopoda, что в буквальном переводе и означает корне-
ножки.
90
постном слое протоплазмы некоторых инфузорий имеются органы защиты
трихоцисты, которые при раздражении выбрасываются наружу в
виде массы тонких нитей, отталкивающих врага. Имеются пульсирую-
щие вакуоли, служащие для выведения из тела, клетки излишков воды.
Наиболее распространенные пресноводные инфузории изображены на
рис. 52.
Рис. 52. Инфузории.
А—туфелька (Paramaecium); Б — Stylonichia; В—сувойка (Vorti-
cella); Г— трубач (Stentor): I— ядро; 2— перистом; 3— глотка; 4 —
сократительная вакуоль; 5 — стебелек.
Кроме обычного бесполого размножения делением, простейшие пе-
риодически размножаются половым путем. У инфузорий половое размно-
жение осуществляется путем конъюгации — временного соединения двух
инфузорий, при котором происходит обмен частями ядерного аппарата.
В пресных водоемах инфузории встречаются обычно в придонных слоях
при условии довольно значительного органического загрязнения, а также
в зарослях водных растений.
Инфузории играют некоторую роль в питании личинок комаров.
91
7) Коловратки (класс Rotatoria)
Коловратки относятся к типу низших червей (Scolecida). Это микро-
скопически малые животные (приблизительно тех же размеров, что и ин-
фузории). Обычно они в больших количествах населяют водную толщу
прудов, озер и пресных водоемов. Отличительной чертой этой группы яв-
ляется «коловращательный» орган на переднем конце тела, состоящий у
многих видов из двух кругов ресничек, движениями которых обусловле-
но плавание коловратки в толще воды. Тело коловраток одето покровами
большей или меньшей плотности; те из них, которые имеют твердые по-
кровы, носят название панцырных коловраток. Между кругами ресничек
расположено ротовое отверстие, куда поступают частички, увлекаемые
движением ресничек. Характерным признаком коловраток является также
жевательный желудок, внутри которого находится сложный орган из-
мельчения пищи. Размножаются коловратки в основном партеногенети-
чески \ т. е. девственные самки откладывают неоплодотворенные яйца,
из которых в свою очередь вылупляются опять партеногенетические сам-
ки. Летом такое девственное размножение может длиться на протяжении
многих поколений, однако при наступлении неблагоприятных условий
(зимние холода, ухудшение питания и пр.) самки откладывают яйца, из
которых развиваются самцы и самки, способные к оплодотворению. Сам-
цы коловраток очень сильно отличаются от самок: они значительно мень-
ше, у них не развит кишечник, так как они совсем не питаются, и очень
хорошо развиты органы чувств, органы движения и половые органы. Наи-
более обычны следующие коловратки.
Из панцырных — Brachionus. Это обычный обитатель прудов. Задний конец его
тела образует так называемую «ногу» в виде узкого стебелька; заканчивается «нога»
двумя пальцами, снабженными клейкими железками; при помощи пальцев Brachio-
nus прикрепляется к погруженным предметам (рис. 53, А). Передний конец панцыря
несет шипы. Широко распространен в озерах и прудах род Anuraea; ноги Anuraea не
имеет; ее панцырь снабжен шипами на переднем и заднем конце (рис. 53,Б). Для глубо-
ких озер характерна панцырная коловратка Notholca longispina (рис. 42, А)
с очень длинными и тонкими шипами панцыря.
Из беспанцырных коловраток мы назовем: 1) крупную, очень прозрачную Asplan-
chna (рис. 53 В); 2) мелкую Polyarthra (рис. 53, Г) с парными пучками плавательных
весловидных шипов по бокам тела, и Synchaeta (рис. 53, Д), Как пример коловраток,
обитающих в придонных слоях воды или на погруженных в воду частях растений,
назовем Rotifer (рис. 40,В), который, кроме плавания в воде с помощью коловраща-
тельного аппарата, может ползать по поверхности субстрата, попеременно прикреп-
ляясь то передним, то задним концом тела, как пиявка.
Коловраток находили в кишечниках личинок Anopheles, особенно личинок стар-
ших возрастов.
8) Ракообразные (класс Crustacea)
К ракообразным пресноводного планктона относятся представители
трех отрядов этого класса: 1) веслоногие (Copepoda); 2) подотряд ветви-
стоусых (Cladocera) отряда листоногих (Phyllopoda); 3) ракушковые
(Ostracoda). Веслоногие представлены в пресноводном планктоне глав-
ным образом циклопами (Cyclops) (рис. 54, Л) и диаптомами (Diaptomus)
(рис. 54, Б).
Тело веслоногих рачков расчленено на головогрудь и брюшко, которое оканчивает-
ся «вилкой», снабженной щетинками различной длины. Ножки головогруди расширены
в виде весел и служат для плавания. Жаберных придатков они не несут, дыхание со-
вершается всей кожей. На переднем конце головогруди расположено две пары усов;
усы первой пары наиболее длинные. В середине переднего края головогруди находится
1 От греческих слов «parthenos» — дева и «geneso» — рождаю.
%
В,
Рис. 53. Коловратки.
А — панцырная Brachionus; Б то же Anuraea cochlearis; В — бес-
ианцырная Asplanchna: 1 — общий вид; 2—жевательный аппарат;
Г—то же Polyarthra; Д—то же Synchaeta.
единственный простой непарный глаз Размножаются веслоногие половым путем. После
оплодотворения самка откладывает яйца, которые она носит на себе в мешках, при-
крепленных к брюшку; у циклопов таких мешков два — по обеим сторонам брюшка,
у диа томусов — один. Из яиц выходят личинки, совершенно непохожие на взрослых
веслоногих; они имеют всего три пары членистых придатков (антенны I, антенны 11
Рис. 54. Веслоногие рачки.
А—Cyclops: 1—глаз; 2— антенны I; 3—антенны II; 4—го-
ловогрудь; 5—брюшко; 6—вилка; 7 —яйцевые мешчи; Б—
Diaptomus: 1—общин вид самни с яйцевыми мешками; 2—1 ан-
тенна самца; 3 — II антенна самки; В — личинки веслоногих:
1, 2 — две стадии развития.
и мандибулы). Эти личинки называются науплиями (Nauplius рис. 54, В}). Путем на-
растания на заднем конце тела новых члеников науплии постепенно превращаются
во взрослых рачков-циклопов, или диаптомусов.
Многие циклопы ждвут на дне водоемов, диаптомусы — все в планк-
тоне. Циклопы, несмотря на свою малую величину, хищники и способны
поедать личинок Anopheles I возраста. Диаптомусы — фильтраторы и
питаются самыми мелкими частицами и нанноплактоном.
94
Ветвистоусые рачки (подотряд Cladocera) принадлежат
к отряду лпстоногих (Phyllopoda). Характерной осооенностью ветвисто-
усых рачков являются их очень сильно развитые усы (вторая пара антенн);
они состоят из двух ветвей и служат главным органом движения. Те-
ло сплющено с боков и, за исключением головы, покрыто тонкой ракови-
ной; оно состоит из головы, груди и брюшка; голова вытянута спереди
у большинства видов в вырост, напоминающий клюв. На голове располо-
Рис. 55. Ветвистоусые рачки.
А — Daphnia pulex—водяная блоха: 1— сложный глаз; 2— плава-
тельная антенна; 3—грудные конечности; 4— брюшко; 5 —
кишечник; 6 — сердце; 7 — выводковая камера с яйцами; Б —
Moina; В—Chydorus; Г— Bosmina; Д—Leptodora: 1—глаз;
2 — антенна II, 3 — грудные ножки; 4 — абдомен; 5 — выводковая
камера.
жен сложный подвижный глаз, у многих имеется еще маленький личиноч-
ный глазок; под клювом расположены короткие передние усы (антенны
первые), являющиеся органами чувств. Грудной отдел несет четыре-шесть
пар ножек, снабженных жаберными мешочками. Брюшко, как и у всех
низших раков, лишено ножек; оно заканчивается двумя коготками, у ос-
нования которых расположено анальное отверстие.
У самок под раковиной, на спинной стороне, находится выводковая
полость, куда откладываются яйца и где происходит их развитие. У боль-
95
шинства видов ветвистоусых наблюдается чередование партеногенетиче-
ского размножения с половым, как у коловраток. Осенью оплодотворен-
ные самки откладывают богатые желтком «покоящиеся», или зимние яйца,
толстая скорлупа которых образует так называемое «седлышко».
Наибодее распространенными являются следующие ветвистоусые рачки: дафния
(Daphnia), называемая часто «водяной блохой» (рис. 55, А), нередко заселяющая ле-
том временные до кдевые лужи моина (Moina, рис. 55, Б), хидорус (Ghydorus) имеет
меньшие размеры, чем дафния, раковинка хидоруса темно келтого цвета. Обычно он
живет в мелких заросших водными растениями водоемах; служит пищей личинкам
Anopheles (рис. 55, В).
Рис. 56. Организмы нейстона.
А — ветвисто усы it район Scapholebcris; В — ракушковый рачок Notodromas inonacha; В— во-
дяной жук Gryrimis (вертячка); Г — личинка комарика Dixa.
Наконец, назовем босмину (Bosmina) (рис. 55, Г) и характерного для
больших глубоких озер рачка-лептодору (Leptodora). Это очень прозрач-
ный крупный рачок (величиной до 10 мм). Раковина его мала и служит
лишь зародышевой камерой (рис. 55, Д). Громадное большинство ветви-
стоусых является фильтраторами, лептодора принадлежит к числу не-
многих хищных ветвистоусых.
Ракушковые ракообразные (отряд Ostracoda). Эти рачки в планктоне попадаются
случайно; они населяют придонные слои воды самых различных водоемов (реки, озе-
ра, пруды, лужи и т. д.). Тело их мало расчленено и целиком покрыто овальной дву-
створчатой раковинкой, из которой наружу высовываются их конечности: две пары
антенн и три пары ходильных ножек. Примером ракушковых рачков может быть
Gypridina (рис. 12, Б), Notodromas monacha (рис. 56, Б).
3. БИОЦЕНОЗЫ ПОВЕРХНОСТНОЙ ПЛЕНКИ ВОДЫ
(НЕЙСТПЧЕСКИЕ БИОЦЕНОЗЫ)
Как указывалось выше, организмы нейстона входят в состав
биоценоза поверхностной пленки воды; нейстонная пленка тесно связана
с планктоном того водоема, на поверхности которого она живет. В состав
нейстона входят некоторые виды бактерий, водорослей, жгутиковых, а
также немногие виды животных. Из жгутиковых эвглена кровавокрасная
(Euglena sanguinea) и хромулина (Chromulina) иногда развиваются в мас-
совом количестве, вызывая так называемое «цветение» поверхностной
пленки воды пруда. Ветвистоусый рачок скафолеберис (Scapholeberis)
(рис.56,4) обитает в поверхностной пленке воды небольших и неглубоких
водоемов, часто вместе с личинками Anopheles (A. maculipennis); он сколь-
96
зит по нижней поверхности пленки воды, спиной книзу и брюшными края-
ми раковины, покрытыми волосками и превращенными в полозья, при-
жимаясь к поверхностной пленке; задние концы его раковины несут по
одному горизонтальному короткому шипу. Такой же образ жизни ведет
часто встречающийся совместно со скафолеберисом и личинками Ano-
pheles небольшой ракушковый рачок — Notodromas monacha (рис. 56,77).
Обитателями поверхностной пленки воды являются некоторые водные
клопы, например, водомерка геррис (Gerris) (рис. 24, Л)—крупная, на
длинных тонких ногах, с длинными тонкими усиками; она сверху сколь-
зит по поверхностной пленке, не прорывая ее, благодаря тому, что ее
лапки не смачиваются водой. Быстро кружатся по поверхности воды
мелкие блестящие жучки-вертячки (Gyrinus) (рис. 56, В)3 обладающие
глазами, способными одновременно видеть под водой и над водой.
К нейстону можно отчасти отнести и личинку комарика диксы (Dixa)
(рис. 56, Z7), о которой говорилось выше, что она относится к земноводным
организмам; она располагается на поверхности воды, опираясь на вы-
ступающие из воды части растений таким образом, чтобы средняя часть
ее U-образно изогнутого тела была над водой, а передний и задний
концы — в поверхностной пленке. Среди кровососущих комаров личинка
Anopheles отличается своей приспособленностью к питанию за счет
нейстонной пленки.
4. БИОЦЕНОЗЫ ДНА (БЕНТИЧЕСКИЕ БИОЦЕНОЗЫ)
а) Условия существования, особенности
и жггзнедеятелъностъ бентоса
Жизненные формы бентоса. Дно водоема представляет иные условия
жизни для организмов, чем его водная масса. Прежде всего здесь имеется
твердый субстрат, и к жизни па субстрате или внутри него приспособле-
ны все организмы бентоса. Так, среди них много форм, имеющих тяжелые
раковины или панцири, благодаря которым они не могут быть снесены вол-
нами, например, моллюски; другие приспособились к жизни на дне путем
закапывания своего тела в грунт дна на большую или меньшую глубину
(например, малощетинковые черви); третьи являются прикрепленными
формами, как, например, гидры (рис. 64, Б, 7); четвертые, наконец,
ползают по дну, как, например, водяной ослик (Asellus aquaticus) (рис.
57, А) или личинки ручейников (рис. 57,77).
Состав бентоса определяется в основнОхМ характером грунта. Каме-
нистый грунт свойствен главным образом рекам и ручьям и прибойным
берегам озер. Организмы этого грунта, особенно в водоемах с текучей
водой, большей частью имеют уплощенное или снабженное присосками
тело или другие приспособления для удержани^^а^амйсх.
Песчаный грунт, бедный питательным^ веществами, обычйб населен
биоценозами, в основнОхМ состоящими идмё&ких животных и одноклеточ-
ных водорослей, живущих в толще песка; биоценозы песчанодо/грунта
объединяются под названием «псаммон».
Наиболее обильно заселены богатые органическими веществами или-
стые грунты. Обычными обитателями ила являются малощетинковые
черви-трубоч тики (рис. 57,7?) и личинки комаров-толкунчиков (рис. 57,Г)—
тендипедид (см. ниже).
Своей жизнедеятельностью трубочники и личинки тендипедид оказы-
вают большое влияние на донную жизнь, а этИхМ самым и на жизнь водое-
ма в целом. Личинки тендипедид строят в грунте ходы на глубине до 2 см
и покрывают их стенки затвердевающими выделениЯхМИ своих слюнных
7 Учебник медицинской энтомологии
97
г // ж
Рис. 57. Организмы бентоса.
А — водяной ослик Asellus aquaticus: 1 — первые антенны; 2 — вторьте антенны; 3 — хвататель-
ные конечности; 4 — грудные ходильные нсгП; 5—крышечка, закрыггющая жабры; 6 — брюшные
конечности (первая пара); 7 — выводковая камера; Б — личинка ручейника Phrygmea; В — червь-
трубочник Tubifex; Г — личинки комаров Tendipedidae; 1 и 2—личинки двух видов комара Тапу-
pus; 3 — мотыль; 4 — личинки мокреца Culicoides; Д — ложноконсная пиягка Hacmopis; Е—ры-
бья пиявна Pjscicola на голове карпа; Ж — личинка, стрекозы коромысла Aeschna.
желез. Благодаря этим ходам, грунт сильно разрыхляется, что способ-
ствует лучшему доступу кислорода в глубже лежащие слои ила. Еще боль-
ше благоприятствуют этому непрерывные колебательные движения личи-
нок в их ходах, а также то, что они, пропуская через свой кишечник глуб-
же лежащие частички ила, переносят их на поверхность грунта. Сходную
работу производят черви-трубочники. Они закапываются в ил головой
вниз, а задние концы их, поднимающиеся над поверхностью ила, произ-
водят волнообразные движения. В тех случаях, когда численность червей-
трубочников или личинок Tendipedidae очень велика, переработка идет
очень энергично. Так, в некоторых озерах на 1 м2 поверхности грунта
черви-трубочники выносят за год 3—12,5 кг сухого веса ила. Благодаря
описанной работе этих обитателей ила, значительно замедляется накопле-
ние органических веществ в водоемах, и мертвые остатки, отложившиеся
на дне, возвращаются в общий круговорот органического вещества.
б) Обзор мара камерным представителей, пресноводного
бентоса,
1) Малощетинковые черви (класс Oligochaeta)
Малощетинковые черви принадлежат к типу членистых (Articulata). подтипу коль-
чатых червей (Annelides). Живущие в почве дождевые черви — их близкие родствен-
ники. Из живущих в водоемах малощетинковых червей большое значение имеют тру-
бочники (семейство Tubificidae), которые в органических илах загрязненных водоемов
развиваются иногда в таком количестве, что придонный слой воды имеет краснова-
тый цвет.
2) Пиявки (класс Birudinea)
Пиявки также принадлежат к подтипу кольчатых червей (Annelides); они отли-
чаются сплющенным мелкокольчатым телом и двумя присосками, из которых одна рас-
положена па переднем, а другая па заднем конце тела; присоски служат для прикреп-
ления к субстрату; в центре передней присоски помещается рот. Некоторые пиявки
хищники; они поедают улиток, червей пт. п., другие пьют кровь позвоночных, являясь
периодически нападающими кровососами. Некоторые пиявки стали настоящими пара-
зитами, чаще всего паразитами рыб. На дне водоемов наиболее часто встречается круп-
ная темная ложноконская пиявка (Haemopis) (рис. 57, Д). На камнях держится неболь-
шая зеленовато-серая, с очень большими присосками рыбья пиявка (Piscicola)
(рис. 57, Е). Некоторые хищные пиявки, возможно, поедают личинок комаров.
3) Личинки комаров-толкунчиков (семейство Tendipedidae)
Голова их напоминает голову личинок комаров Culicidae; в отличие
от последних грудь личинок тендипедид не шире брюшка. Первый членик
груди несет пару передних «ложных» ножек; последний (9-й) членик брюш-
ка имеет на спинке два бугорка с длинными щетинками; позади них распо-
ложено четыре тонкостенных мешочка — анальные жабры; на конце тела—
пара задних «ложных» ножек. Некоторые виды, например, широко распро-
страненный «мотыль» Tendipes plumosus (рис. 57, Г3) окрашены в темно-
красный цвет благодаря пигменту гемоглобину, растворенному в его кро-
ви Ч Живет в органических илах. Взрослые толкунчики имеют недораз-
витые ротовые органы, не принимают пищи, откладывают яйца и уми-
рают.
1 Гемоглобин — красящее вещество красных кровяных шариков человека п
всех позвоночных. Он встречается у некоторых беспозвоночных, папример, у мотыля.
7* 99
4) Личинки ручейников (отряд Trichoptera)
Личинки ручейников (рис. 57,2?) имеют голову, вооруженную грызущи-
ми ротовыми частями, грудь с тремя парами довольно длинных ног и мягкое
брюшко. Живут они в домиках, которые строят себе из песчинок, кусочков
растений, мелких раковинок, склеиваемых выделениями слюнных желез
личинки. У некоторых ручейников домик прикреплен к камню, но боль-
шинство таскает свой домик повсюду с собой, высовывая из него при ходь-
бе голову и грудь с ногами п прячась в него при опасности. Встречаются
ручейники в различных водоемах, одни живут на дне чистых быстро те-
кущих вод, другие — в самых разнообразных стоячих водоемах. Личинки
ручейников являются хищниками; некоторые из них, живущие в зарослях
водных растений, поедают и личинок Anopheles.
Взрослые Trichoptera по внешности несколько напоминают ночных
бабочек.
5) Личинки стрекоз (отряд Odonata)
Личинки стрекоз также прожорливые хищники. Для ловли добычи
они пользуются нижней губой, видоизмененной в так называемую «маску».
Оба членика нижней губы сильно удлинены, образуя сложный рычаг, а же-
вательные пластинки образуют хватательные щипцы. В сложенном виде
маска располагается под головой. При виде добычи личинка выбрасы-
вает маску, захватывает добычу, подтягивая ее ко рту.
В стоячих водах наиболее распространены очень крупная личинка «коромысли»
(Aeschna) (рпс. 57, Ж), личинки стрекозы (LiLellula) <рис. 24. Д), а также стройные
личинки люточек («стрелок») (семейство Agrionidae).
6) Высшие раки (подкласс Malacostraca, класс Crustacea)
Сюда относятся: речной рак (Astacus) (рпс. 12. Г), распространенный главным
образом в Европейской части СССР, и пресноводные крабы, встречающиеся в пресных
водах Закавказья, Средней Азии (Potamon fluviatilis) и Дальнего Востока (Eriocheir
japonicus). И крабы, и раки относятся к отряду десятиногих (EecapoJa). Менее круп-
ными, но более широко распространенными представителями высших раков являются
бокоплавы (отряд Amphipola) (рис. 5. Г) и водяные ослики (род Asellus из отряда
равноногих Isopoda) (рис. 57, А).
7) Моллюски (тип Mollusca)
Мягкое тело этих животных заключено в раковину. Наиболее обыч-
ный моллюск беззубка (Anodonta) (рис. 58, А), принадлежащая к классу
пластинчатожаберных (Lamellibranchiata). Беззубка имеет двустворчатую
тонкую и хрупкую раковину. Обе половины раковины соединены на спин-
ном крае упругим тяжом. Класс брюхоногих или улиток (Gastro-
poda) представлен в пресных водах двумя отрядами. К отряду легочных
(Pulmonata) относятся: прудовик (Limnea) (рис. 58, В), катушка (Planer -
bis) (рис. 58, Б). К отряду переднежаберных относится живородка (Vi-
vipara) (рис. 58, Д) и битиния (Bythinia) (рис. 58, В); последняя является
одним из промежуточных хозяев сосальщиков Opistorchis, опасного пара-
зита человека. Раковина брюхоногих улиток спирально завернута либо
в одной плоскости, как у катушки, либо в виде башенки, как у прудовика,
битинии и живородки.
Прудовики часто поднимаются к поверхности водоема, где они пол-
зают, подвесившись к поверхностной пленке воды; они могут поедать яйца,
100
отложенные кровососущими комарами, а также оставляют на поверхности
воды слизь, к которой прилипают личинки Anopheles; после этого прудо-
вики поедают их.
Рис. 58. Моллюски.
А—беззубка Anodonta; Ъ— катушка Planorbis: В—прудовик
Limnea; Д—живородка Vivipara; Г—бптиния Bylhinia.
Рядом с раковинами живородки и^бптииип изображены их крышечки.
в) Бенпю-некпгические и нектические животные
Большая группа сравнительно крупных животных, только частично
связанных с грунтом дна или с другим субстратом водоема. Передвига-
ются они, активно плавая в толще воды. Этих животных следует отнести не
к бентосу, а к бенто-нектическим формам. Большая часть их принадле-
жит к водным насекомым, которые в жизни личинок комаров играют важ-
ную роль, являясь их врагами. Познакомимся с наиболее распространен-
ными из них.
Водяные жуки (отряд Goleoptera). Наиболее часто встречаются в во-
доемах следующие жуки.
1) Жуки-плавунцы (семейство Dytiscidae)
Хищные жуки. Усики нитевидные. Третий грудной сегмент снизу
вдается хмежду основанием третьей пары ног в виде треугольного клина,
задние ноги, плавательные, сплющены, имеют по краям густую бахромку
из волосков (рис. 59, А). Запас воздуха для дыхания берут, выставляя из
воды задний конец тела.
101
Рис. 59. Организмы бентонектона и нектона.
А — жук-плавунец Dyttacus; Б — водолюб Hydous; В — личинка плавунца Dytiscus; BL — вид
со спинной стороны; В2 — вид сбоку; — вид с брюшной стороны; Г — гладыш Nbtonecta;
Д — плавт Naucoris; Е — гребенчатый тритон Triturus cristatus; Ж — гамбузия Gambusia;
самка; Ж.— самец.
2) Жуки-водолюбы (семейство Hydrophilidae)
Водолюбы имеют характерные короткие усики с утолщениями на
концах. Эти жуки периодически выставляют над водой передний конец
тела для получения запаса воздуха (рис. 59, Б), Личинки жуков обоих
семейств имеют тело, расчлененное на голову и членистое тулови-
ще. Голова снабжена сильно развитыми ротовыми частями, среди ко-
торых выделяются мощные верхние челюсти — то узкие с острыми кон-
цами, то широкие зазубренные. Большинство относящихся сюда личинок—
хищники. Личинка плавунцов отличается подвижностью и прожорли-
востью (рис. 59, В).
3) Водяные клопы
Довольно распространены в пресных водоемах и водяные клопы (под-
отряд Hemiptera, отряд Rhynchota), которые вест, свой жизненный цикл
проводят в воде. Назовем здесь двух опасных хищников: гладыш
(Notonecta), энергично плавающий брюхом кверху, с обтекаемой формой
тела, с плавательными задними и хватательными остальными ногами
(рис. 59, Z7); плавт (Naucoris), шмеющий овальное, плоское тело;
плавает спиной кверху, как плавунцы (рис. 59, Д).
Из крупных подвижных обитателей дна, имеющих значение в жизни
личинок кровососущих комаров, мы назовем тритонов Triturus crista-
tus (рис. 59, Е) и Тг. vulgaris (класс Amphibia). И личинки, и взрослые
тритоны поедают личинок комаров. Типичные представители нектона,
рыбы являются опасными врагами личинок комаров, главным образом ры-
бы, питающиеся у поверхности воды. Особенно большую роль в истребле-
нии личинок Anopheles играет, как уже говорилось, небольшая рыбка
гамбузия (Gambusia affinis) (рис. 59, Ж).
г) Перифитон
Перифитон — это та часть бентоса, которая располагается не прямо
на грунте, а на погруженных в воду предметах: камнях, сваях, листьях
и стеблях погруженных в воду растений и т. п. В состав перифитона обыч-
но входят зеленые, с ине зеленые водоросли и диатомеи, инфузории (на-
пример, сувопкп), сидячие и ползающие коловратки, а также ресничные,
круглые и малощетинковые черви, мелкие личинки тендипедид и пр. Часто
среди организмов перифитона можно встретить ракообразных: циклопов,
ветвистоусых и ракушковых рачков.
В жизни личинок кровососущих комаров перифитон имеет очень боль-
шое значение. Организмы перифитона представляют обильный и пита-
тельный корм для личинок Anopheles и Aedes, которые своими ро-
товыми частями соскребывают его с субстрата. Личинки многих видов
A6des питаются почти исключительно перифитоном.
5. ВЫСШИЕ ВОДНЫЕ РАСТЕНИЯ
а) Предварительные замечания
Прибрежная область озер, пруды, медленно текущие реки и многие
другие водоемы обычно зарастают водными растениями. Заросли водных
растений являются особым типом биоценозов, как по составу населения
и по характеру взаимоотношений между его отдельными членами, так и
по характеру и степени своего воздействия на внешнюю неорганическую
среду.
103
б) Физиономические группы водныэс растении
и их характерные представители
Водные растения принято разделять на несколько групп по их «фи-
зиономии», т. е. по их внешнему облику и расчленению, а также по рас-
положению их ассимилирующих органов по отношению ко дну и к поверх-
ности водоема. К одной физиономической группе могут принадлежать
растения разных семейств, отрядов и даже классов, весьма различные
между собой по внутреннему строению и способам размножения, но сход-
ные по расчленению своего тела, по своему облику (физиономии) и по своей
роли в жизни водных биоценозов. Основных физиономических типов вод-
ных растений мы насчитываем три:
I. Свободноплавающие растения, называемые лемнидами.
II. Укореняющиеся растения.
III. Прикрепляющиеся растения.
Мы будем знакомиться с растениями по этим трем основным группам.
1 тип — лемниды. Л е м н н ды — это те водные растения, которые
свободно плавают в воде, не прикрепляясь ко дну или к каким-либо пред-
метам. Лемниды распадаются на две группы: 1) плавающие по поверхно-
сти воды *— нейстические лемниды, и 2) плавающие под поверхностью,
в толще воды — планктические лемниды.
Нейстические лемниды. К нейстическим лемнидам относятся прежде
всего широко распространенные в различных водоемах ряски: малая
ряска (Lemna minor) (рис. 60, Л) и многокоренник, или большая ряска (Spi-
rodela polyrrhiza) (рис. 60, Б). Тело рясок состоит из плотной зеленой
пластинки, представляющей собой стебель, выполняющий функцию листа.
У малой ряски размеры пластинки малы, а из середины нижней поверхно-
сти отходит только один корешок; у большой ряски (многокоренника)
пластинка значительно крупнее, и от ее нижней поверхности отходит целый
пучок корешков. Размножаются ряски вегетативным путем: от каждой
пластинки вырастают дочерние стебли-пластинки, которые первое время
плавают вместе с материнским растением, но скоро отделяются от него
и начинают сами производить новые пластинки. Это размножение идет
так энергично, что в короткое время ряска может затянуть сплошным
ковром поверхность пруда.
К нейстпческим лемнидам относится и лягушечник, или водокрас (Hydrocharis
morsus ranae) (рпс. (50, В), имеющий круглые, глубоко вырезанные у основания листья,
раскинутые па поверхности воды на длинных черешках, которые у основания
собраны в розетки. Цветет лягушечник белыми цветами. Но, кроме размножения по-
средством цветов, лягушечник имеет и вегетативное размножение; он образует в па-
зухах листьев побеги, на копнах которых развиваются новые розетки листьев. Пере-
зимовывает лягушечник в виде опускающихся осенью на дно водоема зимующих но-
чек. Для всех нейстических лемнид благоприятным биотопом являются закрытые о г
ветра, не имеющие течений и волнений спокойные уголки водоемов пли малые водоемы.
Планктические лемниды. К п л а н к т и ч е с к и м лемнидам
относятся: ряска трехдольчатая(Lemna trisulca) (рис. 60, Г) и пузырчатка
(Utricularia) (рис. 60, Д). Погруженные в воду тонкие стебли-пластинки
трехдольчатой ряски полупрозрачны и имеют форму остроконечных кре-
стиков; размножается’она, как и нейстическая ряска, вегетативным путем.
Стебли пузырчатки погружены в воду; корней она не имеет. Размножается пузыр
чатка вегетативным путем и посредством цветов. При цветении пузырчатка всплывает
на поверхность воды и выносит высоко в воздух свои яркожелтые цветы. Волосовид-
ные листья пузырчатки покрыты маленькими пузырьками, из-за которых это расте-
ние получило свое название. Пузырьки имеют верхнюю и нижнюю губы, отделенные
особым клапаном, который при самом незначительном давлении открывается, а после
прекращения давления сам собой закрывается. Это делает их ловушками для мелких
104
водных животных. Стенки пузырьков покрыты маленькими железками, выделяющими
ферменты, переваривающие попавших в ловушку животных. Попадают изредка в
эти ловушки-пузырьки и личинки Anopheles младших стадий.
Скопления зеленых нитчаток, плавающие погруженными под водной
поверхностью и образующие «водяную вату», также относят к планктиче-
ским лемнидам.
Рис. 60. Лемниды.
А — ряска малая Loma т. minor; Б — мчог жэроннкк Spirodela polyrrhiza;
В— лягушечник Hydrocharis mors as ranae. Итанктические лемниды; Г — ряска
трехдольчатая Lemna trisulca; Д — пузырчатка Utricularia.
II тип—укореняющиеся растения. Элодеиды* К первой группе расте-
ний, укореняющихся в грунте водоемов, относятся элодеиды. Все они
характеризуются погруженным целиком в воду стеблем, вдоль которого
расположены листья. К элодеидам относятся: элодея (Elodea canaden-
sis), УРУТЬ (Myriophyllum). роголистник (Ceratophyllum) и очень бо-
гатый видами род рдестов (Potamogeton).
Элодея (рис.61, А) имеет стебель, густо покрытый продолговатыми листьями, кото-
рые расположены мутовками по 3—4 листа. Элодея размножается исключительно
вегетативным путем, образуя боковые побеги. За свою способность к исключительно
105
л
Рис. 61. Элодеиды.
Е___рдест курчавый Potamogeton сг is pus; /К — рдест гребенчатый Potamogeton pectinatus.
Д — рдест блестящий Potamogeton lucena;
энергичному размножению элодея названа водяной чумой; в короткое время она
иногда образует такие густые заросли, что пользование водоемом в целях судоход
ства и пр. становится невозможным. Перезимовывает элодея зимующими почками.
Уруть (рис. 61, Б) имеет стебли, покрытые мутовками перисто рассеченных листьев.
Листья так нежны, что, будучи вынуты из воды, слипаются в кисточки. Размножается
уругь главным образом вегетативным путем: всякий обрывок растения легко пускает
корни и вырастает в целое растение. Цветы урутп малозаметны, собраны в верхушеч-
ный колос, внизу которого расположены пестичные цветы, а вверху — тычинковые.
Опыление цветков урути может происходить не только в воздухе, по и под водой:
эго указывает на высокую приспособленность этого вторичноводного организма к
годной среде.
Роготистник (рис. 61, В) имеет светлозотеные иго (ьчатые листья, разделенные на
две-чегыре дольки и сидящие мутовками на целиком погруженном в воду стебле. Неж-
ные в молодости, листья роголистника к старости становятся ломкими, жесткими, от-
чего роголистник получил свое название. Вегетативное размножение такое же, как и
у урути: любой обрывок растения дает новое растение. Цветы роголистнике!! мелкие,
незаметные, сидят по одному в пазухах листьев. Опыление, как и у цветов урутц. может
происходить и под водой.
Рдесты (Potamogeton) представлены в водоемах многими видами:
салятся они и у самого берега, и на довольно значительных глубинах.
В грунте рдесты образуют ветвящиеся подземные стебли (корневища),
в которых накапливаются запасные питательные вещества. Благодаря вет-
влению подземных стеблей, рдесты энергично размножаются, образуя
густые подводные заросли. Размножение происходит и посредством цве-
тов, которые малы в отдельности, но собраны в довольно крупные колоски,
поднимающиеся над поверхностью воды. Мы опишем здесь только пять
наиболее распространенных видов рдестов.
Рдест пронзспнолистный (Potamogeton perfoliatus) (рис. 61, Г) имеет продолго-
ватые или яйцевидные листья, при основании глубоко сердцевидные, охватывающие
стебель, отчего создается впечатление, что листья пронзены стеблем. Листья шерохо-
ватые, по краям с зубчиками. Цветоносные стебли небольшой длины. Корневища ко-
ленчато согнуты. Этот рдест встречается в разнообразных стоячих и текущих водое-
мах; предпочитает умеренно загрязненный черный ил. Не боится волнения.
Рдест блестящий (Potamogeton lucens) (рис. 61, Д) имеет крупные, блестящие
листья; верхние листья сближены, нижние отдалены друг от друга. Цветы собраны в
колосок на верхушке длинного цветоноса. Корневища очень толстые, глубоко зале-
гающие в грунте (до 0,5 м).
Рдест курчавый (Potamogeton crispus) (рис. 61, Е) назван так за волнистые края
листьев. Колосок несет очень редкие цветы. Корневище топкое, сильно ветвистое.
Рдест плавающий (Potamogeton natans) имеет плавающие на поверх-
ности воды листья с длинным черешком п широкой овальной пластинкой. Верхняя
поверхность плавающих листьев покрыта восковым налетом, отчего они не смачивают-
ся. Погруженные листья внизу стебля имеют цилиндрическую форму, а вверху—
ланцетовидную. Ко времени цветепия развивающиеся вначале погруженные листья
обычно отмирают, остаются одни плавающие листья; рдест приобретает чисто нимфеоид-
' лый облик. Длинные цветоносы его несут многоцветковые колосья. Корневище длпн-*
ное. сильно ветвистое, осенью имеет клубневидные утолщения.
Рдест гребенчатый (Potamogeton pectinatus) (рис. 61, Ж) имеет длинные листовые
влагалища, от верхушки которых отходят нитевидные листовые пластинки. Колоски
прерванные. Одно из наиболее излюбленных растений личинки Anopheles maculipen-
nis. Из всех рдестов выносит наиболее соленхю воду.
Валлиснерииды. Второй физиономический тпн укореняющихся растений составля-
ют валлиснерииды, имеющие лентовидные, погруженные листья, отходящие от корот-
кого стебля. Типичным представителем является валлиснерпя (Vallisneria spiralis)
(рис. 62, Л), обитающая в наших южпых водоемах. Ее стебель достигает всего 2 см.
Лентовидные листья до 80 см длины поднимаются от дна по направлению к поверхно-
сти водоема. Цветы собраны по три' в мелкие соцветия, расположенные в пазухах
листьев; мужские и женские цветы — на разных растениях.
Нимфеиды. Третья группа укореняющихся растений, нимфеиды, харак-
теризуется плавающими по поверхности листьями. К этой группе относятся
кувшинки (Nymphaea) (рис. 62, Б), кубышки (Nuphar) (рис. 62 В) п
плавающие рдесты в период цветения (Р. natans).
108
Кувшинки, или водяные лилии, характеризуются крупными, круг-
лыми, с глубоким вырезом у основания темнозелеными листьями, плава-
ющими на поверхности воды; листья сидят на длинных круглых черешках.
Верхняя поверхность листьев блестящая, покрыта восковым налетом, за-
ставляющим скатываться с них воду, и пронизана большим числом
устьиц для поглощения углекислого газа из воздуха. Цветы большие,
с многочисленными белыми лепестками, постепенно переходящими в ты-
чинки, образующие «золотое сердечко» цветка.
Кубышки имеют овальные плавающие листья с большим количеством
слизи для уменьшения трения, что предохраняет их от повреждения.
Крупные цветы имеют пять желтых лепестков, тычинки и очень характер-
ное рыльце с десятью лучами.
Линеиды. К четвертой группе укореняющихся растений принадлежат
линеиды, имеющие надводные удлиненные, узкие листья или стебли, за-
меняющие собой листья. Сюда относятся все осоки и многие злаки, а также
хвощи, сусак и некоторые другие.
Опишем несколько наиболее важных линеид.
Камыш (Scirpus lacustris) (рис. 62, Г) растет на сырых берегах и заходит довольно
глубоко в воду. Он имеет высокие цилиндрические стебли с небольшим числом узких
длинных листьев, покрытых восковым налетом. На верхушке стебля — соцветие
.(метелка) из носкотьких колосков, состоящих из простых цветков (с шестью длинными
щетинками, с тремя тычинками и пестиком, имеющим три рыльца). В грунте камыш
укрепляется толстым подземным стеблем-корневищем.
Тростник (Phragmites communis) (рис. 62. Д) расчет обычно мощными зарослями,
спускающимися с берега в воду па глубину до 3 м. Длинные, узкие, серо-зеленые
листья пропитаны кремнеземом, делающим пх жесткими, шелестящими при ветре.
Высота стебля до 3—4 м, но бывает и больше. Па верхушке стебля летом развивается
соцветие — фиолетовая, пушистая, раскидистая метелка. Быстрое разрастание за-
рослей тростника обусловлено способностью его к образованию отводков от корне-
вищ.
Рогоз (Thypha) (рис. 62, Е). У основания высокого стебля длинные и довольно
широкие листья; па верхушке стебля в период цветения развивается большой густой
бархатистый темнокоричневый колос, состоящий из олень мелких просто устроенных
цветков; в верхней части колоса тычинковые цветы, в нижней части — пестичные.
Между этими цветками — бесплодные цветки и темнокорпчпевые волоски, придающие
колосу его окраску п бархатистость. На семенах рогоза длинные летательные волоски,
помогающие пм разноситься по воздуху. В грунте рогоз укреплен толстыми подзем-
ными стеблями с многочисленными придаточными корнями.
Осоки (Сагех). К осокам относятся растения с невысокими стеблями и узкими ли-
нейными листьями. Их тычиночные и пестичные цветы собраны либо в колоски, либо
в небольшие метелки. Осоки не заходят глубоко в воду прудов и озер, оставаясь в пре-
делах заболоченных берегов. В болотах низового типа они являются господствующими
растениями. Осоки очень часто образуют большие кочки или целые сплошные заросли.
Хвощи (Equisetum). Принадлежат к числу низших сосудистых растений, размно-
жаются не семенами, а спорами. Споры хвощей созревают на «весенних» растениях
хвоща (очень мало похожих па «летние» хвощи), в маленьких мешечках-спорангиях,
расположенных в колоске на верхушке «весенних» хвощей. Летние вегетативные
побеги хвощей характеризуются мутовчатым ветвлением. Все растение очень жесткое,
так как клеточные оболочки его пропитаны кремнеземом.
Земноводная полевица (Agrostis stolonicens). Относится к злакам; она имеет узкие
линейные листья и цветы, собранные в мелкие колоски, которые образуют на верхушке
стебля метелку. По берегам неглубоких водоемов она появляется уже весной; в лужах
на лугах часто по всей поверхности раскинуты ее мелкие длинные листья. В зависимо-
сти от того, растет ли полевица на берегу или погружена в воду, она имеет различный
вид. В водоемах листья ее мельче; прибрежные растения имеют более широкие
листья. Это позволяет относить полевицу также к земноводным растениям, о которых
будет речь ниже.
Фолииды. В пятую физиономическую группу укореняющихся растений
объединены растения с широкими надводными листьями, называемые фо-
лиидами. К фолиидам относятся: череда, вахта, белокопытник и др.
109
Рис, G2. Валлиснерииды, нимфеиды.
4 валлпсиерия Vallisneria spiralis; Б — водяная лилия Kymphaea; В — кубышка Nuphar.
Рис. 62. (продолжение). Линеиды.
Г— камыщ Scirpus; Д — трэзгаик Phragmites; Е-ф рогоз Т^УрЦа,
Вахта (Menyanthes trifoliata). Называется иногда еще трифоль за
тройчатые широкие листья. Имеет стебель, заканчивающийся кистями
розовато-белых цветов. Сплетение ее стеблей и корней образует сплавину
по берегам зарастающих водоемов.
Рис. 63. Фолииды и амфиоииды.
А—"стрелолист Sagittaria; Б— частуха Alisma; В— водяная гречиха .Polygonum amphibium;
Г — водяной лютик Ranunculus aquaticus.
Амфибииды. Чрезвычайно’близок к фолиидам ряд прибрежных ра-
стений, также имеющих широкие надводные листья; но вследствие того,
что эти виды способны существовать погруженными в воду и в этом слу-
112
чае их листья имеют совершенно иную форму — рассечены на узкие мел-
кие дольки или же становятся плавающими по поверхности водоема, их
объединяют в особый физиономический тип — земноводных растений,
или амфибиид. Сюда относятся:
Стрелолист (Sagittaria ‘sagittifolia) (рис. 63, А). Вырастая на берегу, стрелолист
имеет листья, расчлененные на черешок и листовую пластинку, в виде наконечника
копья или стрелы. У растений, растущих в воде, листья двух типов: либо целиком
погруженные в воду, длинные, лентовидные, напоминающие листья валлиснерии, без
следа расчленения на черешок и пластинку, либо плавающие по поверхности, подобно
листьям^нимфеид; такие листья имеют длинный черешок, поднимающийся к поверх-
ности, и более или менее широкую пластинку с вырезкой у основания. Белые цветы
Рис. 64. Прикрепляющиеся растения.
А— сфагновый мох Sphagnum; Б—лучица (харован водоросль) Chara. 1 — пре-
сноводные полипы (Hydra) с длинными щупальцами, сидящие на харе и на ряске.
стрелолиста собраны по три в мутовки, образующие кисть на верхушке стебля. Пере-
зимовывает стрелолист с помощью зимних почек, опускающихся на длинных побегах
осенью в ил.
Частуха (Alisma) (рис. 63, Б). Погруженные в воду растения частухи имеют лен-
товидные листья. Гораздо чаще встречаются надводные листья с широкой овальной
листовой пластинкой и узким черешком. Небольшие белые цветы частухи расположены
на пирамидальной раскидистой метелке.
Земноводная гречиха (Polygonum amphibium) (рис. 63, В). Очень широко распро-
странена в стоячих водоемах, а также на их берегах. Воздушные листья ее узки, с за-
зубренными краями, морщинисты и покрыты волосками; у растущих в воде растений
листья широкие, гладкие, блестящие, с ровными краями, плавают на поверхности
воды, наподобие листьев нимфеид.
Водяной лютик (Ranunculus circinatus) (рис. 63, Г). Очень похож на наземные
лютики, но цветы у него не желтые, а белые. Плавающие на поверхности .листья лю-
тика имеют широкую лопастную пластинку, а погруженные — рассечены на узкие
дольки (рис. 34).
III тип. Прикрепляющиеся растения. Третьим крупным разделом
водных растений являются растения прикрепляющиеся. Сюда относятся
мхи и харовые водоросли. Некоторые виды мхов живут на дне довольно
глубоких водоемов, другие — в ручьях и реках, третьи — в водоемах
болотного типа и на суше. По своему расчленению, а часто и по роли
в жизни водоема они нередко приближаются к элодеидам. Большую, свое-
образную роль в жизни водных организмов играют сфагновые мхи.
8 Учебник медицинской энтомологии
113
Сфагновый мох (Sphagnum) (рис. 64, Л). Каждое растение имеет боль-
шое число листьев, сидящих на стебле; листья тонкие, состоят из одного
слоя клеток; часть этих клеток — пустые и представляют собой капил-
лярные пространства, жадно всасывающие воду, отчего подушки сфаг-
нума в болоте обычно пропитаны водой. Клеточные оболочки сфагнов
содержат сфагнол — вещество, близкое химически к фенолу. Наличие
сфагнола в водах сфагновых болот является, вероятно, одной из причин
их непригодности для жизни личинок малярийного комара и многих
других организмов.
Харовые водоросли, или лучицы (Charophyta) (рис. 64, Б). Сложной расчленен-
ностью своего тела и крупными размерами (до 50 см в длину) напоминают высшие вод-
ные растения. В их теле можно различить главные оси, напоминающие стебель, и бо-
ковые оси, мутовками сидящие на главной оси, напоминающие игольчатые листья.
Внизу находятся образования, укрепляющие растения в грунте и слегка напоминаю-
щие корни. Размножаются хары половым путем. Встречаются в стоячих водоемах,
богатых известью. Заросли хар, напоминая заросли мелкорассеченных элодеид, вроде
роголистника, представляют удобный субстрат для личинок Anopheles. Однако ча-
сто они до поверхности не доходят, и в таком случае бывают непригодны для поселения
этих личинок.
в) Взаимоотношения между водными растениями
и другими организмами
Влияние растений на химизм водоема. Мы уже неоднократно говорили
о том, что высшие водные растения оказывают очень большое влияние на
химизм водоема. Благодаря их фотосинтезу, происходит обогащение воды
кислородом и уменьшение содержания углекислоты; при уменьшении
содержания углекислоты вода становится менее кислой, нередко подще-
лачивается. Иногда при этом частично выпадает из раствора углекислый
кальций. Эти изменения химизма воды представляют способ воздействия
водных растений на другие организмы. Далее, водные растения играют
существенную роль в питании животных. Иногда они сами служат пищей,
в других случаях животные питаются покрывающим листья и стебли вод-
ных растений перифитоном.
Водные растения как субстрат для личинок Anopheles. Личинки ма-
лярийного комара используют погруженную водную растительность как
удобный субстрат для причала: они прикрепляются к краям тех частей
растения, которые соприкасаются с поверхностной пленкой воды.
Водные растения умеряют движения воды; волны и рябь, набегающие
с открытой поверхности озер и больших прудов, а также течения рек и
каналов разбиваются о погруженные части растений. Благодаря этому, в
зарослях растений создаются тихие уголки, благоприятные для личинок
Anopheles. Густые заросли растений, особенно погруженных (элодеиды,
планктические лемниды), могут служить убежищем от врагов для многих
водных животных, в том числе и для личинок малярийного комара.
Наконец, водные растения при сильном развитии оказывают влияние на
освещение и температуру воды (стр. 67, 69). Все сказанное позволяет
рассматривать заросли водных растений как своеобразный тип биоцено-
зов, отличный от других основных биоценозов водоемов, и говорить о
большом и многообразном значении водной растительности в жизни вод-
ных организмов вообще и в жизни личинок Anopheles в особенности.
6. СТАРЕНИЕ ВОДОЕМА И СВЯЗАННАЯ 0 ЭТИМ СМЕНА ЕГО БИОЦЕНОЗОВ
Юность водоема. Если мы будем вести наблюдения в течение ряда лет
над водоемом с момента его возникновения, мы убедимся, что с течением
времени его биоценозы изменяются всегда в определенном направлении.
Эти изменения носят название смены биоценозов, или сукцессии. Процесс
114
начинается с первого момента возникновения нового водоема. Вновь об-
разованный водоем не имеет на дне органических отложений, он беден пи-
тательными веществами и население его поэтому бедно. Особенно бедно жи-
вотное население, так как для питания животных необходимы готовые ор-
ганические соединения. Первые обитатели водоема — жгутиковые, а за-
тем некоторые водоросли (главным образом нитчатки) и инфузории. Это
период юности водоема. В этот период при наличии нитчаток водоем уже
может заселяться личинками Anopheles. Уже на второй год своего сущест-
вования водоем заселяется водными растениями. Чаще всего вначале по-
являются амфибииды — частуха, стрелолист; вслед за ними — некоторые
элодеиды — элодея, уруть ( в зависимости от того, какие растения имеют
возможность попасть в новый водоем и какие из попавших могут там най-
ти благоприятные условия для существования). С поселением первых
высших растений в водоеме создаются уже более благоприятные условия
для личинок Anopheles.
Зрелость водоема. В течение последующих лет происходит обогащение
водоема питательными солями и органическими веществами, и в связи с
этим его биоценозы делаются все более богатыми и разнообразными. Во-
доем становится более зрелым. Среди амфибиид и элодеид появляются но-
вые типы водных растений; заросли становятся густыми, структура их
осложняется, нередко они становятся многоярусными. В зарослях водных
растений мы можем различать три главных яруса: 1) надводный, образо-
ванный линеидами и фолиидами; 2) нейстический, образованный нейсти-
ческими лемнидами и плавающими листьями нимфеид; 3) погруженный, об-
разованный элодеидами, планктическими лемнидами и другими погружен-
ными растениями, а также подводными частями растений двух верхних
ярусов.
В распределении зарослей намечается зональность, которая впослед-
ствии становится очень ясно выраженной: от берега к центру водоема раз-
личные типы водных растений сменяются в определенном порядке, обра-
зуя ряд концентрических поясов, или зон. На берегах и в мелкой при-
брежной области водоема располагаются заросли тростника, камыша и
других крупных линеид, часто вместе с осоками и другими средними ли-
неидами; нередко под покровом надводного яруса имеются ярусы нейсти-
ческих и погруженных растений. Следующая зона — нимфеиды: кувшин-
ки и кубышки. За этими двумя зонами идет зона элодеид: роголистник,
уруть, элодея. Наконец, ближе к центру водоема расположена зона рде-
стов, среди которых есть виды, способные жить на больших глубинах.
Дно таких зарастающих водоемов покрыто значительным слоем черного
органического ила. В воде происходит интенсивный фотосинтез как погру-
женными высшими растениями, так и взвешенными в воде водорослями.
Это сильно сказывается на газовом режиме водоемов: днем вода насыщена
кислородом и не содержит углекислоты в поверхностных слоях, ночью
содержание кислорода снижается и появляется углекислота, что особенно
выражено в придонных слоях. В этот период зрелости водоема личинки
Anopheles заселяют его обычно в больших количествах, особенно зону
планктических лемнид и элодеид.
Старость водоема. С течениехМ времени происходит наступание с бе-
регов в воду тростника, камыша, рогоза и др.; площадь, занимаемая эло-
деидами, все уменьшается. Из года в год водоем все более засоряется мерт-
выми растительными остатками и постепенно мелеет; площадь свободной
воды становится с каждым годом все меньше и, наконец, совсем исчезает.
По всей площади поселяются рогоз, тростник, осока. Водоем превраща-
ется в болото низинного типа.
8*
115
Влияние выпаса скота. Нельзя пройти мимо того громадного влияния,
которое оказывает на прибрежную растительность водоемов выпас скота.
Пасущиеся домашние копытные, коровы, лошади и пр., выедают и вы-
таптывают не только прибрежную растительность, но и ту, которая на-
ходится в воде на глубине до 0,5 м. Различные растения в различной сте-
пени устойчивы по отношению к выпасу. Крупные линеиды, вроде трост-
ника и рогоза, уничтожаются в первую очередь; элодеиды и амфибииды
оказываются гораздо более устойчивыми. Поэтому при выпасе скота на
берегу глубокого водоема, имеющего зональное расположение раститель-
ности, ближайшая к берегу часть зоны линеид оказывается уничтоженной
и возникает «предтростниковая» зона, занятая мелкими элодеидами, стре-
лолистом, частухой, земноводной гречихой и т. п.; предтростниковая зона
защищена от волнения поднимающимися с большей глубины и потому
сохранившимися линеидами и, таким образом, часто представляет идеаль-
ные условия для выплода личинок Anopheles: заросшие элодеидами, хо-
рошо прогреваемые и защищенные от волнения мелководья.
Если водоем был весь неглубокий и сплошь заросший линеидами, скот
уничтожает их полностью, и они замещаются элодеидами и лемнидами, а
водоем весь превращается в сплошной очаг Anopheles.
Таким образом, выпас скота на берегу водоема, как правило, резко
улучшает условия заселения его личинками Anopheles. Однако при чрез-
мерном Ьыпасе водоем сильно загрязняется, иногда исчезает вся высшая
растительность, и в итоге создаются сапробные условия, не допускающие
жизни личинок Anopheles. Следовательно, умеренный выпас увеличивает
анофелогенную роль водоема, а чрезмерный может ее уменьшить
(В. Н. Беклемишев).
ГЛАВА IV
ОСНОВНЫЕ ТИПЫ МАТЕРИКОВЫХ ВОДОЕМОВ
1. ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Ежегодно с поверхности океанов и материков испаряется 379 000 км
воды (304 000 км3 с поверхности океанов и 75 000 км3 с поверхности ма-
териков). Вся эта вода выпадает обратно в виде атмосферных осадков —
дождя, снега и пр. Вода, выпавшая на поверхность материков в количестве
112 000 км3, частью испаряется, частью впитывается в землю, частью стекает
по поверхности земли. В конце концов, вся она либо через реки возвра-
щается в море (37 000 км3), либо, испаряясь, уходит в атмосферу. Вода,
впитавшаяся в землю, образует в толще ее грунтовые воды, пропитываю-
щие водопроницаемые породы: песок, гравий, трещиноватые известняки
и т. п. В понижениях рельефа — в долинах, оврагах, у подножья гор •—
грунтовые воды выступают на поверхность, образуя родники, или ключи
(рис. 65). Вода, стекающая по поверхности суши или скопляющаяся в ее
впадинах, образует поверхностные воды. Все поверхностные воды делятся
на стоячие воды, или водоемы, и текучие воды, или водотоки.
Разнообразие водоемов и водотоков, имеющихся на поверхности земли,
колоссально, и очень многие из них, от ничтожнейших луж и канав до
величайших рек и озер, могут при известных условиях служить местом
выплода кровососущих двукрылых. Поэтому нам необходимо рассмотреть
основные типы поверхностных вод, встречающихся в нашей стране, и
выяснить, какие из них и при каких условиях служат местом выплода
малярийных комаров и других кровососущих двукрылых.
116
2. ВОДОЕМЫ
Стоячие водоемы подразделяются в зависимости от: 1) степени их
постоянства и 2) глубины и величины водной массы. По степени постоян-
ства все стоячие водоемы делятся на постоянные и временные. Постоянными
называются все водоемы, непрерывно существующие в течение всего года
(и в течение нескольких лет подряд). Временными называются те водоемы,
которые существуют менее года. Постоянные водоемы, в зависимости от
глубины и величины водной массы, делятся на озера, пруды и водоемы
болотного типа.
Рис. 65. Схема расположения подземных вод (разрез через местность).
1У 2 — места выхода на поверхность (выклинивания) родников.
а) Озера
Определение. Озером называется скопление больших масс воды в глу-
бокой котловине. От других стоячих водоемов озеро отличается тем, что
хотя бы часть его настолько глубока, что недоступна зарастанию высшей
водной растительностью. Озера, имеющие исток, т. е. такие, из которых
вытекает какая-либо река или ручей, называются проточными, хотя тече-
ние в них обычно незаметно или почти незаметно. Из больших озер проточ-
ным является, например, Ладожское озеро, избыток вод которого выносится
Невой. Озера, не имеющие истока, называются бессточными. Если вся вода,
поступающая в озеро, уходит на испарение, озеро является действи-
тельно бессточным; таково, например, озеро Балхаш или Аральское море.
Такие озера всегда являются в той или иной мере солеными. Наряду с
этим, некоторые небольшие озера, не имеющие видимого истока, имеют
подземный сток, и такие озера обычно остаются пресными.
Происхождение озерных котловин. Котловина озера может быть раз-
личного происхождения. В некоторых случаях она образована опуска-
нием участка земной коры под влиянием горообразующих сил; такие озера
называются тектоническими (например, Байкал). В горах встречаются
плотинные озера, возникшие вследствие перегораживания глубокой
горной долины обвалом или оползанием ее склонов (например, озеро Рица
в Абхазии), или лавовым потокохм (озеро Севан в Армении). На севере СССР
широко распространены ледниковые озера, котловины которых созданы
работой ледника в ледниковый период. Некоторые озера представляют
морские заливы, отгороженные от моря наносами или оторвавшиеся после
отступания моря (остаточные озера). Громадными остаточными озерами яв-
ляются озера Ладожское и Онежское и величайшее из всех озер мира —•
Каспийское море.
Искусственные озера. Искусственные озера, или водохранилища, обыч-
но создаются путем искусственного преграждения плотиной течения реки
в глубокой долине. Водохранилища создаются для различных хозяйствен-
ных целей: для получения электрической энергии, улучшения судоход-
ства на реках, водоснабжения, питания оросительных систем и т. п. От
117
естественных проточных озер многие водохранилища отличаются зна-
чительными колебаниями уровня воды в связи с потреблением ее для хо-
зяйственных целей. Создание водохранилища, затопляющего участок реч-
ной долины, резко меняет условия выплода малярийных комаров. Целый
ряд анофелогенных водоемов оказывается погребенным под глубоким
слоем воды и перестает существовать. Зато новые места выплода — и часто
обширные — могут возникать на побережье водохранилища, а именно в за-
рослях растений и в скоплениях нитчаток в защищенных от прибоя мел-
ководьях. Такие условия создаются главным образом в различных за-
ливчиках и в мелководных верховьях водохранилища.
Личинки Anopheles в водохранилищах. Личинки Anopheles появляются
в водохранилищах нередко в первый же год их существования; в это
время высшая водная растительность на большей части побережья отсут-
ствует; она имеется лишь в тех местах верховий, где прежде существовав-
шие водоемы затоплены тонким слоем воды, позволяющим водным расте-
ниям подняться до поверхности. Личинки Anopheles поселяются прежде
всего в таких местах, но также и в скоплениях нитчаток и других планкти-
ческих лемнид, появляющихся вдоль защищенных берегов в первый же
год существования водохранилища. В последующие годы защищенные
побережья быстро зарастают амфибиидами и элодеидами и заселяются мас-
сами личинок Anopheles. В дальнейшем появление на побережье крупных
линеид и возникновение зонального распределения растительности могут
несколько снизить численность личинок Anopheles. Но более гибельной
является и для растительности, и для личинок неустойчивость уровня во-
дохранилища: сильное снижение уровня в летнее время ведет к обсыха-
нию берегов и гибели зоны элодеид вместе с личинками. Зимнее снижение
уровня на севере может повести к промерзанию дна и гибели растений, что
замедляет развитие зарослей летом и уменьшает площади личиночных
местообитаний.
Общая характеристика озер. В каждом озере различают две основные
области: прибрежную, более мелкую литоральную область, или литораль,
и глубоководную центральную, пелагическую, или пелагиаль. Нижней
границей литоральной области считается такая глубина, ниже которой
высшие водные растения не распространяются. Литоральная и пелаги-
ческая области во всех отношениях различаются между собой.
Пелагиаль. Пелагическая область озер отличается хорошо выражен-
ной слоистостью температуры, освещения и кислорода (стр. 66, 69 77). Для
населения пелагиали большое значение имеет отсутствие там твердого суб-
страта, вследствие отдаленности дна и отсутствия высших водных растений.
В силу этого пелагиаль населена планктоном, который по составу значитель-
но отличается от планктона литорали или от планктона малых водоемов.
Очень характерны для него: коловратка Notholca longispina (рис. 42, А)
и прозрачный ветвистоусый рачок Leptodoa (рис. 55, Д).
Благодаря волнению и отсутствию высших водных растений, пела-
гиаль непригодна для жизни личинок кровососущих комаров. Глубинные
придонные слои пелэгиали озер в летнее время отличаются пониженной
температурой, недостатком кислорода и света и мало заселены (рис. 66).
Для них характерна прозрачная хищная личинка комара коретры (Ghao-
borus, рис. 128). В илу пелагической области озер обитают личинки кома-
ров-толкунчиков (Tendipedidae) и черви-трубочники (Oligochaeta)
(рис. 57, В, Г).
Литораль. Литоральная область озер, благодаря своей мелководно-
сти, летом прогревается до дна, и обычно ее вода бывает до дна насыщена
кислородом. В защищенных от сильного волнения участках литорали
118
встречаются обширные заросли водных растений, которые в зрелых
озерах расположены зонами, поясами. Очень богаты и разнообразны
по видовому составу и другие типы биоценозов литорали; планктон,
бентос и др.
Богатое развитие мельчайших организмов в воде и в перифитоне на
стеблях и листьях водных растений создает благоприятные пищевые усло-
вия для многих водных животных. Присутствие в литоральной зоне хищ-
ных водных насекомых и их личинок делает борьбу за существование в
ней особенно напряженной. Для ли-
чинок Anopheles защищенные от вол-
нения участки литорали озер иногда
представляют благоприятные условия,
особенно зона элодеид и планктических
лемнид.
б) Пруды
Определение пруда и его харак-
теристика. В гидробиологии прудом
называют замкнутый водоем, на дне
которого, благодаря незначительной
глубине, на всем протяжении может
произрастать высшая водная расти-
Рис. 66. Схема температурного рас-
слоения озера летом.
1 — кривая температуры: 2 — кривая ки-
слорода; 3 — кривая свободной углекис-
лоты; 4 — кривая связанной углекис-
лоты.
тельность; следовательно, основное
отличие прудов от озер — это их мень-
шая глубина. В отличие от водоемов
болотного типа пруд настолько глубок,
что его наиболее глубокая часть мо-
жет зарастать только погруженной и
плавающей растительностью и недоступна для надводных растений.
В общежитии озерами часто называются такие водоемы, которые гидробио-
логия относит к прудам. Основные черты отличия прудов от озер следую-
щие: 1) отсутствие глубинных водных масс с их пониженной температурой
и ослабленным освещением; 2) отсутствие волнения; 3) отсутствие орга-
низмов, типичных для пелагиали озер; 4) значительно большая зависи-
мость температуры воды пруда от температуры воздуха: большая прогре-
ваемость до дна днем и охлаждение ночью или при снижении температуры
вэздуха; 5) большие суточные колебания в содержании кислорода; 6) бо-
гатое развитие жизни. Все перечисленное очень сближает условия жиз-
ни в прудах с условиями жизни в литоральной области озер. Основное
различие между ними заключается в возможности большого обогащения
воды пруда питательными солями и органическими веществами, благо-
даря малому объему его водной массы; наоборот, воды литоральной об-
ласти озер постоянно разбавляются водами пелагиали. Высокая концен-
трация питательных веществ в воде и в грунте пруда позволяет развиваться
в прудах гораздо более пышной водной растительности, а полное развитие
растительности делает пруды по сравнению с литоралью больших озер,
значительно более благоприятной средой для развития личинок Ano-
pheles.
Естественные пруды-озера. К естественным прудам-озерам относятся
неглубокие карстовые озера, очень многие из степных озер Западной Си-
бири и Северного Казахстана, большая часть так называемых поемных
озер в долинах рек (кроме самых крупных поемных водоемов, которые мо-
гут оказаться настоящими озерами).
119
В тех местностях, которые сложены из легко растворимых горных пород,
вроде гипса, грунтовые воды промывают себе подземные ходы, и создается
сложная система подземных пустот и пещер, по которым течет вода. Время
от времени земля, лежащая над такими пустотами, оседает и на поверхно-
сти образуются провальные ямы, или воронки. Если дно такой воронки
заполнится глиной, в ней может застаиваться вода, и при достаточно влаж-
ном климате образуется постоянный водоем. Такие водоемы, вначале ли-
шенные жизни, постепенно заселяются, зарастают и проходят все стадии
сукцессии: юность, зрелость и старость, в конце концов превращаясь
в небольшие болотца. В стадии юности и зрелости они представляют ти-
пичные естественные пруды; в стадии зрелости имеют хорошо выражен-
ные растительные пояса: прибрежный пояс линеид, пояс кувшинок и
пояс элодеид. В стадии юности и зрелости карстовые 1 водоемы могут
заселяться большим количеством личинок A. maculipennis. У нас карсто-
вые водоемы имеют малярийное значение в некоторых районах Пред-
уралья и в других местах.
Степные и лесостепные озера Западной Сибири. Лесостепная и степ-
ная полоса Западной Сибири, Зауралья, Северного Казахстана покрыта
неисчислимым множеством плоских котловинок, занятых озерами и бо-
лотцами всевозможных размеров. Большинство из них не имеет стока:
дожди вымывают из почвы воднорастворимые соли и стекающая вода сно-
сит их в озерные котловины; вода испаряется, соли остаются, и потому
бессточные степные озерки оказываются в различной мере осолоненными.
В очень слабо солоноватых озерках (до6°/00 солености) еще растут тростник
и гребенчатый рдест и могут жить личинки Anopheles; более соленые озера
лишены высшей растительности и личинок Anopheles, самые соленые не
имеют и нитчаток и бывают населены очень бедной и своеобразной фло-
рой и фауной из солелюбивых организмов (например, Artemia salina, рако-
образное из отряда Phyllopoda). Многие озерки во влажные годы перепол-
няются, и вода их, переливаясь через край, получает сток в соседние, ни-
жележащие озера. Такие периодически промываемые озерки имеют со-
вершенно пресную воду, богатую растительностью, и часто являются ме-
стом массового выплода малярийного комара. Малярийное значение их
еще усиливается благодаря тохму, что сельские населенные пункты очень
часто бывают приурочены к соседству подобных пресных озер. На се-
вере лесостепной полосы, где увлажнение является более постоянным,
многие пресные озерки уже закончили свое развитие и превратились в
моховые болота. В степной полосе, где осадков значительно меньше, мно-
гие озерки периодически высыхают и в высохшем виде представляют
занятые солончаком западины, которые во влажные годы превращаются
в небольшие соленые озерки-пруды.
Третий и наиболее широко распространенный тип естественных пру-
дов-озер представляют поемные озера, располагающиеся на поймах рек.
К ним мы вернемся несколько ниже, при рассмотрении жизни реки и ее
придаточных водоемов.
Искусственные пруды. Искусственные пруды создаются путем за-
пруживания небольших водотоков (ручьев и речек), путем создания
искусственной выемки (копаные пруды) и т. п.
Пруды на речках бывают самых разнообразных размеров — от не-
больших деревенских противопожарных запруд на ручейках и до боль-
1 Название это происходит от области Карст в Югославии, имя которой стало,
нарицательным для обозначения местности, сложенной из гипсов и богатой пещерами
провальными воронками и «карстовыми» водоемами.
120
ших мельничных прудов и заводски^ прудов Урала; последние своими раз-
мерами уже напоминают водохранилища.
Мельничные пруды. Мельничные пруды на равнинных речках по
своей многочисленности представляют один из важнейших типов ано-
фелогенных водоемов средней полосы Советского Союза. Если речка
имеет хорошо обособленную поймочку с боле€<или менее крутыми скло-
нами, с противомалярийной точки зрения наиболее выгодной является
такая плотина, гребень которой достигает наиболее крутой части склона
(рис. 67, 2). Возникший при таких условиях пруд будет иметь две части:
в нижней части он будет глубок, с крутыми берегами, со слабо развитой
растительностью; в верхней — дно пруда постепенно поднимается и глубина
становится все меньше; верховья обычно бывают покрыты обильной
водной растительностью; со временем они мелеют еще больше, отчасти
заиляемые отложением наносов, которые приносит речка, отчасти вслед
Поперечный разрез долины с нанесением уровня воды в приплотин-
ной части пруда: 1— пруд мал и для своей долины; 2— пруд впору
для своей долины; 3 — пруд велик для своей долины.
ствие накопления мертвых растительных остатков. Во всяком случае
заросшие мелководные верховья мельничных прудов обычно являются
местом мощного выплода малярийных комаров.
Если пруд велик для своей долины, то уровень воды в приплотинноп его
ча.сти станет выше наиболее крутой средней трети склонов; вода частично за-
топит верхнюю, пологую, их треть и здесь возникнут прибрежные мелко-
водья (рис. 67, 3); средняя частьпруда будет иметь крутые берега; верховья,
как и в других случаях, неизбежно окажутся мелководными. При этом
типе пруда анофелогенные участки возникнут и в приплотинной части, и
в верховьях. Наконец, если пруд мал для своей долины, т. е. плотина невы-
сока и дно долины оказывается затопленным лишь тонким слоем воды
(рис. 67,1), пруд оказывается мелководным на всем своем протяжении,
сплошь зарастает и сплошь или почти сплошь продуцирует малярийных
комаров. Очень старые пруды могут настолько заилиться, что становятся
мелководными на всем своем протяжении и сплошь зарастают анофелоген-
ной растительностью даже в том случае, если первоначально низовья их
были достаточно глубоки и неопасны в малярийном отношении.
Если долина реки имеет мягкие, пологие склоны, пруд, созданный в
такой долине, на всем своем протяжении неизбежно будет иметь хорошо
развитые прибрежные мелководья. В малярийном отношении такие пру-
ды всегда опасны.
Загрязненные пруды. Небольшие деревенские пруды, используемые
для водопоя скота, разведения уток и гусей, стирки белья и пр., обычно
подвергаются очень сильному загрязнению. Такие пруды отличаются
слабым развитием или почти полным отсутствием высшей водной расти-
121
тельности и обильным фитопланктоно^, в котором преобладают синезеле-
ные и протококковые, массовое развитие которых нередко вызывает
«цветение» воды. При несколько меньшем загрязнении наблюдается боль-
шое развитие нитчатых, включая и зигнемовых. В зоопланктоне преоб-
ладают коловратки, в особенности Brachionus. Дно покрыто толстым слоем
гниющего ила, населенного личинками тендипедид и червями-трубочни-
ками. Иногда подобные пруды затягиваются малой ряской. Описанные
условия неблагоприятны для личинок Anopheles, и очень загрязненные
пруды обычно неопасны в малярийном отношении.
Рис. 68. Тип приусадебного пруда (Туркменская ССР) (фото Г. Е. Раевского).
Хаузы. К числу копаных прудов относятся среднеазиатские хаузы,
рыбные пруды карповых хозяйств и пр. Хаузы (рис. 68) представляют собой
обычно четырехугольные небольшие копаные пруды; вода в них напускает-
ся по канавам из оросительных систем. Глубина хаузов нередко доходит
до 2 м; обычно они бывают обсажены деревьями. Личинки Anopheles, глав-
ным образом A. maculipennis sacharovi, населяют либо каемку наземных
трав, спускающихся своими концами в воду, либо иногда встречающие-
ся в хаузах заросли рдестов или других элодеид, или хлопья нитчаток.
Сильно загрязненные или заселенные гамбузией хаузы личинок не имеют.
Рыбные пруды. Те пруды рыбоводного хозяйства^ которые отводятся
под нерест (икрометание) рыбы (рис. 69), устраиваются обычно в откры-
той, хорошо прогреваемой солнцем местности. Они имеют незначитель-
ную глубину (от 15 до 50 см), малую площадь водного зеркала (300—500 ма)
и непроточны. Для них характерно наличие погруженной растительности,
служащей субстратом для развивающихся икринок рыбы. Все это создает
очень благоприятные условия для личинок малярийного комара. Очень
часто встречаются в них и личинки комара Culex.
Выростные пруды, служащие для выращивания мальков и сеголеток,
нагульные пруды, в которых рдстут рыбы-двухлетки и трехлетки (рис. 70),
бывают чаще всего слабо проточными или же периодически пополня-
122
ются водой. Вода этих прудов богата удобрительными веществами, что
способствует развитию планктона и бентоса. Высшая водная раститель-
ность в них тоже имеется, но в меньшем количестве, чем в прудах Пере-
Рис. 69. Общий вид нерестового пруда.
Рис. 70. Схематический план рыбоводных прудов полного
карпового хозяйства.
стовых. И эти пруды бывают заселены личинками Anopheles и Culex, но
в меньшем количестве, чем нерестовые, вследствие менее благоприятных
условий, а также вследствие выедания их молодью рыб.
123
в) Болота и заболоченности
Всякий избыточно увлажненный участок земной поверхности назы-
вается заболоченностью. Избыточно увлажненный участок, поверхность
которого покрыта слоем торфа (т. е. разложившихся и полуразложившихся
растительных остатков), называется болотом. Как болота, так и заболочен-
ности могут представлять собой просто поверхность, избыточно увлажнен-
ную, без видимых водоемов, или же покрытую множеством водоемов и ми-
кроводоемов, т. е. скоплений воды в углублениях микрорельефа: вмочежи-
нах между кочками осоки, в следах от копыт животных и т. п.
Несмотря на ничтожные размеры, такие микроводоемчики могут отли-
чаться значительным постоянством и не пересыхать, поскольку вся окру-
жающая почва пропитана водой. Наконец, на поверхности болота или забо-
лоченности могут быть значительные по площади водоемы болотного или
даже прудового типа. Водоемы болотного типа отличаются от прудов не-
значительной глубиной, позволяющей на всем их протяжении расти торча-
щим из воды растениям—линеидам, фолиидам или даже кустарнику.
Типы болот. В лесной полосе чаще всего встречаются болота моховые
и осоковые, и болотные водоемы чаще всего зарастают мхами или осоками.
На юге лесной полосы, кроме того, нередки черноольховые болота, порос-
шие лесом с расположенными под его пологом болотными водоемами. Юж-
нее, начиная с лесостепной полосы, очень распространены тростниковые
болота, водоемы которых зарастают тростниками (Phragmites communis)
и другими крупными линеидами. К этому типу болотных водоемов отно-
сятся плавни рек, впадающих в Черное, Азовское, Каспийское, Араль-
скоЪ моря и в озеро Балхаш.
Искусственные заболоченности, возникающие в результате несовер-
шенно проводимого сельскохозяйственного орошения, широко распростра-
нены во всех искусственно орошаемых районах. Рисовые поля (см. ниже,
стр. 125) являются искусственными заболоченностями,создаваемыми вполне
сознательно в хозяйственных целях. Многие другие типы искусственных
заболоченностей могут возникать вследствие плохого ведения водного
хозяйства.
Болота лесной полосы. Торфяные болота лесной полосы чаще всего
возникают либо в силу зарастания водоемов, либо вследствие оподзо-
ливания лесных почв и зарастания их сфагновыми мхами. Болота, возни-
кающие в силу зарастания водоемов, проходят в своем развитии две фазы:
фазу низинного болота и фазу верхового, или сфагнового, болота. При
заболачивании в связи с оподзоливанием лесных почв сразу образуется
сфагновое болото.
Низинное болото представляет собой непосредственный результат
зарастания пруда-озера. Оно бывает покрыто густой травянистой расти-
тельностью, осокой, хвощом, различными фолиидамй и пр. Многочислен-
ны и мхи, но сфагновых мхов мало или они отсутствуют. Вода таких болот
богата солями, кислородом, имеет слабо кислую, нейтральную или слабо
щелочную реакцию. Весной эти болота богаты водой, в их водоемах раз-
вивайся разнообразный планктон и появляются многочисленные водя-
ные насекомые и улитки. При наличии погруженной водной раститель-
ности встречаются и личинки Anopheles.
Сфагновые болота. Сфагновые болота могут возникать как дальней-
шая стадия развития низинных болот или путем самостоятельного забо-,
лачивания оподзоленных почв. Вода сфагновых болот очень бедна соля-
ми и очень богата органическими веществами, так называемыми гумино-
выми кислотами. Она обладает кислой, иногда очень кислой реакцией.
124
Высшая растительность в сфагновых водоемах почти отсутствует. Все эти
условия не благоприятствуют жизни большинства пресноводных живот-
ных, и все население сфагновых водоемов очень бедно и однообразно. Там
нет ни улиток, ни высших раков, ни червей-трубочников, ни пиявок, ни
рыб и т. д. Вместо всего этого имеется своеобразная флора и фауна
специально сфагновых форм: десмидиевых водорослей, корненожек, коло-
враток и пр. Личинки Anopheles там не живут, и естественные водоемы
сфагновых болот в малярийном отношении безопасны.
Торфяные карьеры. При многих способах разработки торфа в местах
его выемки образуются водоемы, называемые торфяными карьерами. Нэ
старых торфоразработках они занимают огромные площади. Карьеры
осоковых болот по своим физико-химическим условиям и населению мало
отличаются от обыкновенных озер-прудов, часто зарастают элодеидами
и дают приют многочисленным личинкам Anopheles. Карьеры эти очень
опасны в малярийном отношении. Карьеры верховых сфагновых болот
(см. выше) по своему химизму и населению напоминают воду естественных
водоемов сфагнового болота. Среди высших водных растений этих карь-
еров преобладает пузырчатка. В резко кислых сфагновых водах подоб-
ных карьеров личинки Anopheles отсутствуют. В водах, переходных
от сфагновых к осоковым, с умеренно кислой или слабо кислой реакцией,
личинки Anopheles встречаются в небольших количествах в зарослях
пузырчатки.
Тростниковые болота. Тростниковые болота плавней и речных долин
на юге и юго-востоке имеют воду, богатую органическими веществами, но
также и солями, жесткую и нередко слабо солоноватую. Реакция ее щелоч-
ная. Густая тень от высоких тростников, нередко поднимающихся на 4—э м
над водой, препятствует росту погруженной растительности и умеряет
температуру воды, которая приближается к температуре воздуха.
Иногда мертвые стебли тростника образуют над водой настил, за-
трудняющий комарам доступ к воде. Для личинок A. maculipennis густые
заросли тростника неблагоприятны и личинок этих там нет. Они заселяют
только опушки тростника и разреженные места, где свет проникает до по-
верхности воды и дает возможность появиться трехдольчатой ряске, рого-
листнику и другим благоприятствующим развитию личинки погруженным
растениям. Кроме того, среди большого тростникового массива нередко
встречаются озерины, или лиманы, открытые пространства, заросшие
погруженной растительностью, в частности, элодеидами. Это по существу
озерно-прудовые водоемы тростникового болота. Они нередко бывают густо
заселены личинками A. maculipennis (а в Средней Азии и A. pulcherrimus)
и дают колоссальный выплод Anopheles. Глубже всех в гуще тростников
из наших видов Anopheles встречаются личинки A. hyrcanus.
Тростниковые болота родникового питания нередко встречаются в
предгорьях Средней Азии и Кавказа и отличаются более прохладной и ме-
нее богатой органическими веществами водой. Для таких болот характерны
личинки: Anopheles algeriensis и отчасти A. bifurcatus и A. hyrcanus. Ли-
чинки A. maculipennis и здесь поселяются только на опушках и в разрежен-
ных местах.
Рисовые поля. Совершенно особым типом искусственных водоемов
болотного типа являются чеки рисовых полей (рис. 71). Рис — чрезвы-
чайно влаголюбивая культура. Поэтому на весь вегетационный период
обвалованные участки рисовых полей (чеки) заливаются водой и в период
с мая по октябрь представляют собой настоящие водоемы. Иногда рисовые
поля имеют постоянную проточность: из оросйтеля в них непрерывно по-
ступает вода со скоростью, обеспечивающей полную смену воды в течение
125
полутора-трех суток. В других случаях в чеки добавляется только то’коли-
чество воды, которое необходимо для покрытия убыли воды на испарение
и фильтрацию. На ярком солнце юга, при малой глубине (10—25 см), вода
рисовых полей хорошо прогревается (иногда до 42°). Обычная температура
воды летом 26—29°. По ночам температура воды в чеках, благодаря их
малой водной массе, понижается. Содержание кислорода в воде высокое,
благодаря множеству погруженных растений. Уменьшение содержания
кислорода ночью не бывает значительным, благодаря круглосуточному
поступлению воды из оросителя. Углекислый газ в летние дни отсутствует;
реакция почти нейтральная или слабо щелочная. На рисовых полях, кроме
риса, произрастает множество водных растений-сорняков. Наиболее обыч-
Рис. 71. Общий вид рисовых полей (до всхода риса). Самаркандская
область (фото В. Н. Беклемишева).
ны на рисовых полях следующие растения. Из линеид: камыш, тростник,
рогоз, куриное просо; из амфибиид: стрелолист, земноводная гречиха.
Встречаются и элодеиды: рдест пронзеннолистный, уруть, роголистник;
из планктических лемнид — пузырчатка. На поверхности часто наблюда-
ются скопления нитчаток и водяной сеточки, а на дне—заросли харовых
водорослей. Фитопланктон разнообразен по составу, но по численности бед-
нее фитопланктона прудов. Зоопланктон своим большим видовым разно-
образием напоминает планктон зарослей прудов-озер, но количественно он
беден. Небогат и бентос, состоящий из червей-трубочников и личинок тен-
дипедид. Иногда бывает много личинок стрекоз, поденок, плавунцов, во-
дяных клопов.
Благодаря мощному развитию мелко рассеченной растительности
и нитчатых водорослей, рисовые поля обычно очень заселены личинками
малярийных комаров Anopheles hyrcanus и A. maculipennis, а при сильном
прогревании A. superpictus и A. pulcherrimus, и часто являются опасными
очагами малярии. Хорошим способом борьбы с личинками Anopheles
на рисовых полях является периодическая их просушка: 7—10 дней
поливки, 4—5 дней перерыва в орошении, т. е. система прерывистого
орошения.
126
г) Временные водоемы
Водоемы, длительность существования которых не превышает не-
скольких месяцев и во всяком случае меньше года, носят название вре-
менных водоемов. Временные водоемы могут быть периодическими или
случайными. Периодические водоемы образуются на одних и тех же ме-
стах в определенное время года и в более или менее определенное время
исчезают. Случайные водоемы возникают нерегулярно и часто неожидан-
но в таких местах, где повторение их в последующие годы не является
обязательным.
К числу периодических водоемов относятся весенние лужи, остав-
шиеся после таяния снега; поемные лужи, остающиеся после паводка в
небольших углублениях речных пойм и в течение лета высыхающие; лужи,
образовавшиеся после летних ливней; родниковые заболоченности, воз-
никающие вследствие выступления грунтовых вод и исчезающие приумень-
шении притока последних в сухое время года, и т. п.
Временные водоемы случайного происхождения чаще всего возни-
кают под влиянием деятельности человека; таковы, например, лужи,
возникающие за счет перелива воды через валики оросительных каналов
или прорывы валиков; водоемы, образующиеся вследствие бессистемного
сброса поливных вод в низины, и т. п. Сюда же можно отнести лужи, воз-
никающие в необычных местах при необычайно больших разливах рек
или в необыкновенно дождливые годы, но водоемы этого типа занимают
промежуточные положения между периодическими и случайными.
Временные водоемы чрезвычайно различны по длительности своего
существования: некоторые лужи существуют всего несколько часов или
2—3 дня и пересыхают прежде, чем в них успеет развиться какая-либо
жизнь, другие существуют дольше, некоторые — по нескольку месяцев.
Комары могут развиться лишь в тех лужах, которые просуществуют до-
статочное для этого время, т. еФ даже при самых благоприятных для раз-
вития личинок температурах — не менее 10—12 дней. Развивающиеся во
временных лужах личинки A6des нередко в громадных количествах гиб-
нут при преждевременном высыхании луж, особенно после малоснежной
зимы, когда весенние лужи имеют меньше воды; и такие же личинки благо-
получно даютвыплод на другой год, когда после многоснежной зимы лужи
лучше заполнены водой и держатся дольше.
Временные водоемы весьма различны по своему температурному ре-
жиму и химизму, в зависимости от почвы и растительности места их рас-
положения.
Весенние лужи. Расположенные на открытых местах весенние лужи
хорошо прогреваются днем и сильно остывают ночью. Благодаря большой
длине весеннего дня, их средняя температура значительно выше сред-
ней температуры воздуха. Настоящей водной растительности они обычно
не имеют; на дне их большей частью — наземные травы и мхи, нередко
влаголюбивого характера. Иногда в таких лужах успевают развиться
нитчатые водоросли или заросли водяного лютика.
Животное население весенних луж очень характерно. В составе его
совершенно отсутствуют те чисто водные животные, которые не способны
переносить высыхание: рыбы, большинство улиток, личинки крупных
стрекоз и другие многолетние водные личинки насекомых, высшие раки
и т. п. Имеются крылатые водные насекомые (жуки, клопы), прилетаю-
щие сюда извне; быстро развивающиеся личинки насекомых (комары,
однолетние личинки поденок и стрекоз и т. п.); приходящие извне лягуш-
ки и жабы, их головастики и те из водных беспозвоночных, которые имеют
127
хорошо защищенные от высыхания покоящиеся стадии (яйца, цисты
ит. п.), как многие низшие раки (особенно характерны листоногие, напри-
мер, Apus), ресничные черви, коловратки и пр.
До того как весенние лужи пересохнут, в некоторых из них успевает
развиться первая генерация A. maculipennis. Самки большинства наших
видов комаров рода Aedes откладывают летом свои яйца во влажную поч-
ву на местах высохших весенних луж или по краям высыхающих луж.
Яйца ихпокоятся до весны, и когда после таяния снега лужи заполняются
водой, из яиц Addes вылупляются личинки, в неисчислимом множестве
заселяющие весенние лужи. Для некоторых видов Aedes весенние лужи
являются единственным местом выплода: их личинки вылупляются только
весной (например, Аё. communis). Другие виды ASdes имеют яйца, из
которых личинки вылупляются каждый раз, когда они оказываются за-
литыми водой (например, Аё. caspius). Эти комары, кроме весеннего, мо-
гут иметь и летний выплод — из временных луж, образовавшихся вслед-
ствие летних дождей или каких-либо других причин.
Летние дождевые лужи. Летние дождевые лужи отличаются от ве-
сенних более ровной температурой и отсутствием целого ряда типично
весенних организмов, вроде крупных листоногих раков и т. п. Они могут
служить местом выплода целого ряда комаров, в том числе Anopheles macu-
lipennis, а также нескольких видов A6des и Gulex.
Поемные лужи. Временные водоемы, образующиеся после половодья
на поймах наших больших равнинных рек, бывают расположены чаще
всего среди луговых трав; население их напоминает население весенних
луж с добавлением ряда речных обитателей — мальков рыб, некоторых
личинок насекомых, улиток и пр. В малярийном отношении эти лужи име-
ют большое значение, часто даже в том случае, если существуют в качестве
замкнутых водоемов всего несколько дней: в них нередко заканчивают
свое развитие личинки Anopheles, жившие перед тем среди луговой ра-
стительности в заливчиках полузатопленной поймы.
В предгорных местностях большое анофелогенное значение имеют
временные водоемы, возникающие при летних паводках горных рек, а так-
же временные водоемы грунтового питания, пересыхающие к осени вслед-
ствие уменьшения притока родниковых вод.
Временные водоемы при искусственном орошении. Временные водое-
мы, возникающие в связи с неисправностями или неправильной эксплоа-
тацией оросительных систем, также часто служат местом массового вы-
плода малярийных комаров (A. maculipennis, A. pulcherrimus, A. superpic-
tus). В распространении малярии водоемы данного типа играют громадную
роль. Борьба с ними требует тщательного надзора за исправностью, свое-
временным ремонтом и правильной эксплоатацией оросительных систем.
Дупляные водоемы. Своеобразный тип периодических водоемов пред-
ставляют собой скопления воды в дуплах деревьев. На юге нашей страны —
на Кавказе, в Крыму, в Средней Азии и на Украине — в дуплах дуба,
ясеня, клена, граба, каштана, ореха и других деревьев скопления дожде-
вой воды часто бывают заселены большим количеством личинок комаров.
Здесь живут личинки дупляного малярийного комара (Anopheles plum-
beus) и несколько видов комаров рода Аёйеэ. Особенно многочисленны
дупляные виды Aёdeз в лесах Уссурийского края. Основные особенности
дупляных водоемов — небольшой объем воды, слабое освещение, незна-
чительные суточные колебания температуры и главное— совершенно осо-
бые химические свойства воды. Окраска воды коричневая, вода очень бо-
гата органическими веществами: гуминовыми, ульминовыми кислотами
и дубильными веществами. Несмотря на большое количество органиче-
128
ских кислот, реакция воды не кислая, а щелочная, что объясняется избыт-
ком в воде солей калия, кальция, натрия, магния. Железо отсутствует,
так как образует с дубильными веществами нерастворимые соли. Несмотря
на исключительно большое содержание органических веществ, в дупляных
водоемах нет гниения: отсутствует аммиак, углекислый газ, нитриты. Ду-
пляная жидкость отличается большой кормностью, и число личинок ко-
маров, которые могут одновременно развиваться в одном дупле, очень ве-
лико. Существование дупляных водоемов всецело зависит от осадков, и
при длительной засухе они пересыхают. Яйца большинства дупляных ко-
маров хорошо переносят высыхание и при новом заполнении дупла водой
из них вылупляются личинки.
а. ВОДОТОКИ
а) Реки и ручьи
Течение.' Все водотоки, от ничтожных ручьев до больших рек, отли-
чаются наличием течения. Скорость течения тем больше, чем больше ук-
лон русла, чем меньше его шероховатость и чем больше количество воды,
протекающее по руслу в единицу времени, т. е. расход воды. При повы-
шении уровня воды в реке расходы также увеличиваются и скорость воз-
растает. Течение реки может быть спокойным или бурным; в первом случае
поверхность реки представляется зеркальной, во втором случае она дро-
жит, волнуется, покрыта водоворотами. Спокойным бывает медленное
течение при гладком русле. Бурное течение безусловно гибельно для по-
павших в него личинок Anopheles.
Питание реки. Река, текущая в богатой осадками местности, на всем
пути принимает притоки (рис. 72, А), и таким образом расходы ее посте-
пенно увеличиваются. Такова, например, Кама или Верхняя Волга. Про-
текая по засушливой местности, река может почти совсем не принимать
притоков, как, например, Волга ниже Саратова или Сыр-Дарья ниже впа-
дения реки Арыси. На таком участке расходы реки постепенно умень-
шаются, особенно если воду ее разбирают на орошение (рис. 72, Б). Многие»
горные реки Средней Азии и других засушливых стран, выйдя на равни-
ну, целиком разбираются на орошение (как, например, река Мургаб в Турк-
мении или большинство речек Ферганской долины) или сами теряются
в песках (как река Чу или Зеравшан).
И расход воды, и уровень большинства рек не остаются постоянными
в течение года. Большинство наших равнинных рек имеет весеннее поло-
водье — огромный разлив, происходящий за счет таяния зимнего снега п
поступления в реку талых вод (рис. 73). Летом эти реки питаются родни-
ковыми Xi дождевыми водами и в связи с дождевым питанием имеют не-
большие подъемы в период летних и осенних дождей. Горные реки, питаю-
щиеся талыми водами снежных вершин и ледников, имеют летний паводок
в период высокой температуры воздуха и усиленного таяния льда и снега
в высокогорье (рис. 74). В богатых осадками горах, например, на Север-
ном Кавказе, частые летние паводки речек обусловлены выпадением лив-
ней в горах. В горах, бедных летними осадками и не имеющих вечного сне-
га, речки многоводны до тех пор, пока продолжается таяние снегов, а
когда снег весь растает, переходят на грунтовое питание и мелеют; с этого
момента, благодаря устойчивому режиму, они становятся прекрасным ме-
стом выплода для личинок Anopheles superpictus и ряда других комаров.
Наиболее постоянным уровнем обладают реки, вытекающие из боль-
ших озер (например, Нева) или питаемые родниками речки предгорных
равнин («черные речки»).
9 Учебник медицинской энтомологии
129
Временные водотоки. Наряду с постоянными, широко распростране-
ны и временные водотоки — весенние или ливневые. Сами по себе они
малярийного значения обычно не имеют, но нередко оставляют после
себя стоячие водоемы, которые могут служить опасными местами выплода
малярийных комаров.
А Б
Рис. 72. Карты.
А — река с притоками; Б — низовья среднеазиатской реки, разбираемой на орошение.
Химизм реки. Благодаря наличию течения, вода реки непрерывно
перемешивается. Поэтому даже в больших реках температура воды и со-
держание растворенного кислорода от дна до поверхности обычно одни
и те же. Вода почти всегда насыщена кислородом. Содержание и состав
растворенных веществ речной воды зависят от содержания их в воде при-
токов, и потому на протяжении реки значительно изменяются. Реки лес-
130
ной полосы по сравнению с равнинными реками юга беднее солями и бога-
че органическими веществами. Горные реки также часто бедны солями;
реки, текущие по пустыне, значительно обогащаются ими.
Зарастание рек. Высшие водные растения поселяются только в медлен-
но текущих реках, т. е. главным образом в малых речках и в придаточных
Месяцы TJJiniVVVI^lwn:
Рис. 74. Кривая уровня горной реки, питаемой таянием
ледников (Терек у ст. Котляревская в 1934 г.).
водах больших рек. Среди этих растений есть и линеиды (например, тро-
стник) и элодеиды (различные рдесты, уруть и др.). Родниковые речки
Кавказа и Средней Азии густо зарастают различными фолиидами. Все эти
растения еще более замедляют течение и дают возможность поселяться боль-
шому количеству личинок анофелеса, так что заросшие, медленно текущие
речки во многих местностях (Украина, Предкавказье, южные республи-
ки) имеют большое значение в распространении малярии (рис. 75).
Рис. 75. Поперечный профиль сильно заросшей, небольшой степной
речки (Харьковская область).
Работа реки, размыв и отложение. Всякая река, достаточно быстро
текущая по земной поверхности, воздействует на свое ложе, производя
тройную работу: размыв, перенос и отложение. Размыв состоит в отрыве
частиц от дна и берегов водоема. Чем быстрее течение и рыхлее грунт, тем
сильнее размыв. Отмытые частицы река подхватывает и переносит вниз
по течению либо во взвешенном состоянии, либо влача по дну. Благодаря
наличию взвешенных частиц, речная вода почти всегда является мутной.
Чем больше скорость течения, тем крупнее и тяжелее те частицы, которые
9*
131
переносит река. Наконец, весь отмытый и подхваченный рекой материал
она где-то отлагает; а именно, как только течение реки на каком-нибудь
участке замедляется, оно становится не в силах нести дальше наиболее
тяжелые частицы, которые выпадают на дно, где и отлагаются. Наиболее
легкие частицы река может доносить до устья, до впадения в море или
озеро, где эти частицы, отлагаясь, образуют непрерывно растущую дель-
ту, вроде дельты Волги, Кубани, Аму-Дарьи, Дуная и пр.
Горные реки и их заселение личинками комаров. Горные реки проте-
кают в глубоких долинах, в которых чередуются участки с большими или
меньшими уклонами; в силу этого в речках чередуются участки
с ускоренным и замедленным течением. В местах с ускоренным течением
Рис. 76. Горная речка с галечным руслом, служащим местом выплода
Anopheles superpictus (фото В. II. Беклемишева).
преобладает размыв. Река здесь быстро течет по скалистому грунту, и
мест выплода комаров нет; они появляются здесь в тех случаях, когда реч-
ка наполовину пересыхает и в русле ее остаются выбоины, заполненные
стоячей водой. В местах с замедленным течением река отлагает валуны
или гальку, загромождает свое русло и течет множеством рукавов по ши-
рокому галечному ложу. Особенно характерна подобная картина для
горных рек после выхода их на равнину. В этих случаях прибрежные га-
лечники по краям протоков нередко оказываются полузатопленными, вода
едва сочится между галькой, и в промежутках между последними созда-
ются идеальные условия для личинок комаров, в частности, Anopheles
maculipennis maculipennis, A. superpictus, Culex mimeticus (рис. 76).
Вода таких галечных побережий прогревается днем на юге до 35° и
выше, иногда зарастает зелеными и синезелеными водорослями. Обилие
личинок Anopheles достигает 1 000 на 1 м2 и больше. Запрятанные среди
гальки личинки находятся в безопасности от нападения рыб. Главная
опасность для них — это паводки, и в тех речках, где бывают летние па-
водки, личинки отсутствуют. При отсутствии летних паводков даже га-
лечные побережья больших рек, например, Сыр-Дарьи при выходе ее из
Ферганских ворот, заселяются личинками. Галечные биотопы имеют у
нас на юге огромное значение в распространении малярии.
Участки равнинной реки. Равнинные реки очень часто имеют наи-
большие уклоны в своем верхнем течении, меньшие уклоны в среднем те-
132
чении и наименьшие в низовьях. В своих верховьях равнинная река часто
представляет ручей, текущий в овраге. Благодаря большим уклонам, те-
чение быстрое, река размывает свое ложе, углубляя долину. Весь отмытый
материал уносится вниз по течению; благодаря этому речка не «образует
наносов и не имеет придаточных вод. Такие речки, как правило, не за-
селяются личинками Anopheles, но, если речка пересыхает, личинки Ano-
pheles появляются в воде остающихся в ее ложе бочагов.
Если уклон меньше, отмытый материал сносится вниз по течению, но
отлагается тут же неподалеку в местах замедленного течения и русло
реки загромождается наносами. Скопление наносов заставляет речку от-
клоняться в сторону, делать излучины и расширять свою долину, подмы-
Рис. 77. Пойма реки с пологими излучинами.
Схема строения одного сегмента поймы: 1 — склон надпойменной
террасы; 2— размываемый рекой (верхний по течению) край сег-
мента; 3—нарастающий (нижний по течению) край сегмента; 4—бе-
реговой вал; 5— поемные озера промывного происхождения; 6—при-
террасное болото, образованное выклиниванием грунтовых вод
у подножья надпойменной террасы; 7 — притеррасная речка, выте-
кающая из притеррасного болота. Стрелки указывают направление
течения в реке в летнее время.
вая ее края. Во время половодья река заливает все свои наносы и отлагает
поверх них новый материал. В силу этого отлагаемые рекой наносы по-
степенно нарастают до той высоты, до которой поднимаются весенние па-
водки. Создается плоское дно долины, образованное речными наносами и
заливаемое рекой во время половодья; его и обозначают как пойму реки.
Река непрерывно перестраивает свою пойму, размывая ее в одних
местах и надстраивая в других. Поверхность поймы, в целом плоская и
соответствующая уровню половодья, покрыта, однако, гривами, борозди-
нами и множеством поемных водоемов.
Типы речных пойм равнинных рек. Существует три главных типа пой-
мы равнинных рек: 1) поймы с пологими излучинами; 2) поймы со срезыва-
нием излучин; 3) островные поймы.
1. Поймы с пологими излучинами свойственны рекам со сравнительно
значительными уклонами. Река полого извивается от одного берега к дру-
гому, оставляя то справа, то слева поемные сегменты, размывая каждый из
них в его верхней половине (рис. 77, 2) и надстраивая в нижней (рис. 77, 3).
Вдоль верхнего края сегмента большей частью расположен приречный
вал. Гривы и бороздины такой поймы образованы работой полых вод.
В бороздинах иногда имеются водоемы — поемные озерки или лужи скром-
ных размеров. Общая анофелогенная площадь таких пойм большей ча-
стью невелика.
2. Поймы со срезыванием излучин свойственны рекам с очень неболь-
шим уклоном долины. Пойма обычно широка, река сильно извивается, из-
133
лучины ее часто имеют вид петель (рис. 78). Время от времени река проры-
вает основание петли и спрямляет свое течение; при этом прежняя излу-
чина превращается в кольцевидный проток. Постепенно оба соединения его
с рекой забиваются наносами и проток превращается сначала в крючковид-
Рис. 78. Пойма реки, срезывающей свои
излучины.
1 — склоны надпойменной террасы; 2 — излучина реки;
3 — кольцевидный проток; 4 — крючковидный затон;
5 — подковообразное озеро.
ный затон, а затем в подковообразное озеро. Все эти образования — проток,
затон и озеро — являются на поймах такого рода староречьями, или ста-
рицами. Количество их бывает очень велико, выплод комаров с подобной
поймы колоссален. Этот тип поймы распространен наиболее широко.
Рис. 79. Пойма островного типа.
1 — склоны надпойменной террасы; 2 — размываемые верховья остро-
ва; 3 — нарастающее низовья острова; 4— заостровка; 5 — озера, пред-
ставляющее остатки зюстровкм; 6 — промывные озера.
3. Поймы островного типа (рис. 79). Речные наносы образуют острова,
которые растут, перестраиваются, иногда разрушаются рекой и исчезают,
иногда прирастают к берегу или другим островам и образуют островную
пойму. Протоки между островами (заостровки) часто остаются в качестве
проточных рукавов, соединенных двумя концами с рекой, частью отделя-
ются от реки наносами и превращаются в затоны или озерки, часто обла-
дают змеевидной формой. Наряду с характерными для них водоемами на
поймах второго и третьего типа, всегда имеются водоемы, характерные
для пойм первого типа, т. е. такие водоемы, котловины которых промы-
134
ты на поверхности поймы полыми водами. Общее количество водоемов на
поймах островного типа бывает колоссально (до 10% всей поверхности
поймы).
Островной поймой обладает, например, Волга на всем протяжении от
устья Камы до дельты.
Сукцессии поемных водоемов. Все поемные водоемы, независимо от их
происхождения, находятся под воздействием реки, ежегодно затопляю-
щей пойму. Полые воды, протекая по пойме, вымывают со дна ее водоемов
скопившийся там ил, заносят их песком, вместе с илом вымывают корне-
вища и зимующие почки растений, приносят новые растения.
Все эти воздействия реки уничтожают результаты собственного разви-
тия водоема, сводящиеся к зарастанию, заилению и постепенному превраще-
нию озера в болото; поэтому воздействие реки ведет к сохранению поемным
озером состояния длительной молодости. Чем ближе водоем к реке, тем
обычно сильнее воздействие на него полых вод.
Водоемы, расположенные в более древней части
долины, от которой река отошла далеко, под-
вергаются меньшему воздействию. Поэтому
ближайшие к реке поемные водоемы часто
имеют песчаное дно, растительность, состо-
ящую из одних амфибиид и элодеид, без пра-
вильного зонального распределения и обиль-
но заселены личинками Anopheles. Водоемы,
находящиеся под менее сильным воздейст-
вием реки, получили возможность дальше
подвинуться по пути своего развития и
обладают растительностью с хорошо выра-
женными зонами линеид, нимфеид и эло-
деид. Водоемы, находящиеся в отдаленных от
реки частях поймы и слабо затрагиваемые
водами половодья, прошли свое развитие до
конца и превратились в болота; в лесной
полосе это бывают болота осоково-моховые
Рис. 80. Личинки двукрылых,
живущие в родниках.
ИЛИ ольховые, В степной полосе ---тростни- A-Pertcoma; В — Liponeura.
ковые и т. д. Таким образом, поемные водоемы проходят те же стадии
развития, как и другие водоемы той же местности, но развитие это за-
держивается благодаря воздействию половодий. В силу этого громадное
большинство водоемов поймы чрезвычайно благоприятно для жизни ли-
чинок Anopheles, и речные долины являются главными местами выплода
малярийных комаров на большей части Европейской территории СССР.
б) Ключи, или родники
Определение. Ключом, или родником, называется естественный выход
на поверхность земли подземных грунтовых вод (рис. 65). Иногда ключ вы-
ходит на свет на склоне и дает начало ручью. В других случаях ключи бьют
из-под земли на дне небольшого углубления, образуя родниковый водоем
криницу с чистым песчаным или илистым дном, иногда заросшим растени-
ями. При переполнении этого водоема ключевая вода переливается через
край и при наличии уклона образует родниковый ручей; при отсутствии
уклона образуются большие скопления родниковой воды, ведущие к воз-
никновению родниковых болот.
Режим. Основной особенностью родниковых вод является их темпе-
ратура; она ниже температуры воздуха в теплое время года и выше ее
135
осенью и зимой. По этой причине даже при относительно суровой зимо
вода родников часто не покрывается льдом.
Другая особенность родников — бедность их воды кислородом. Обыч-
но содержание кислорода в родниковой воде не превышает 1 мг на 1 л воды.
Общее содержание и состав солей в воде зависят от тех горных пород, че-
рез которые пробиваются ключи под землей.
Организмы, населяющие родники. Своеобразный режим родников,
особенно их температурный и кислородный режим, определяет своеобраз-
ный состав их населения. В биоценоз ключей входят организмы, предпо-
читающие низкую температуру и неспособные переносить большие коле-
бания температуры. Типично родниковыми организмами являются ли-
чинки двукрылых: перикома (Pericoma), липоневра (Liponeura) (рис. 80)
и др.; на заболоченной почве, окружающей родники, часто ползает личин-
ка комара диксы (Dixa), селятся мокрицы, пауки, дождевые черви и др.
Из комаров наиболее обычным обитателем родниковых водоемов являются
личинки Anopheles bifurcatus, в известных условиях—A. algeriensis.
Личинки A. maculipennis и A. superpictus поселяются в родниковых водах
лишь после того, как вода в них хорошо прогрелась.
в) Искусственные водотоки
Искусственные водотоки — каналы и канавы — создаются для самых
разнообразных целей. Наибольшее значение в практической деятельности
маляриолога имеют каналы оросительных и осушительных систем, по-
дробнее рассматриваемые в курсе противомалярийной гидротехники.
ЧАСТЬ 3
МЕДИЦИНСКАЯ ЭНТОМОЛОГИЯ
РАЗДЕЛ А
ВВЕДЕНИЕ В МЕДИЦИНСКУЮ ЭНТОМОЛОГИЮ
Медицинская энтомология изучает членистоногих, вредящих здо-
ровью человека. Вредить здоровью человека членистоногие могут тремя спо-
собами: как паразиты человека, как переносчики болезней и как ядовитые
животные.
ГЛАВА I
ЧЛЕНИСТОНОГИЕ КАК ПАРАЗИТЫ ЧЕЛОВЕКА
Паразитами человека являются те членистоногие, которые нападают
на человека и питаются за счет его тела.
Паразитизм и хищничество. Паразит отличается от хищника прежде
всего тем, что хищник убивает свою добычу и съедает ее, паразит же пи-
тается за счет своего «хозяина», не убивая его, и может питаться за счет од-
ной и той же особи хозяина длительное время или повторно несколько раз.
Иногда паразит все-таки вызывает в конце концов гибель хозяина, но лишь
после длительного питания за его счет; так, личинки наездника (стр. 51)
в конце концов губят гусеницу бабочки, в которой паразитировали, но
лишь после того, как закончат за ее счет свое развитие. Все членисто-
ногие много меньше человека и не могут быть по отношению к нему хищ-
никами.
Таким образом, членистоногие, нападающие на человека и питаю-
щиеся за счет его тела, должны считаться паразитами человека.
Паразитизм членистоногих может принимать разнообразные формы.
Типичный паразит, например, человеческая вошь (Pediculus humanus,
стр. 408), тесно связан со своим хозяином, за счет которого он питается,
которому наносит вред и тело которого является для него постоянной
средой обитания.
Степень обязательности паразитизма. Вошь может жить только в ка-
честве паразита человека. Паразитический образ жизни для нее обязателен,
она является обязательным паразитом. Таким же обязательным парази-
том является, например, личинка вольфартовой мухи (стр. 397), живущая
в живых тканях млекопитающих и человека и никогда не живущая на па-
дали. Самка вольфартовой мухи откладывает своих личинок исключитель-
но на живых животных. С другой стороны, зеленая мясная муха Lucilia
sericata (стр. 397) откладывает свои яйца как на падаль, так и на живых
животных (например, овец), и личинки ее одинаково успешно развиваются
и в качестве паразитов, и в качестве мертвоедов. Паразитизм этих личи-
нок является факультативным, т. е. закономерно встречающимся, но не-
139
обязательным. Наконец, многие другие мясные мухи (например, синяя
мясная муха Calliphora), как правило, откладывают свои яйца или своих
личинок на падаль и лишь в виде редкой случайности на рану; однако, по-
пав на рану, личинки нормально развиваются; такой паразитизм является
случайным, а личинки Calliphora, живущие в ране человека, являются
случайными паразитами.
Источники пищи паразита. Различные членистоногие-паразиты, пи-
таясь за счет человека или животных, выбирают различные источники пи-
щи. Большинство из них питается кровью; таких паразитов мы называем
кровососами. Другие поедают живые ткани, как вышеупомянутая личин-
ка вольфартовой мухи. Третьи питаются за счет рогового слоя кожи и его
производных (пуха, шерсти), как пухоеды и многие мелкие клещи млеко-
питающих и птиц.
Место паразитирования. Паразиты, живущие и питающиеся на поверх-
ности тела хозяина, как, например, вошь, называются наружными пара-
зитами (эктопаразитами). Паразиты, живущие и питающиеся внутри тела
хозяина, называются внутренними паразитами (эндопаразитами). Эндо-
паразиты по месту своего паразитирования в свою очередь распадаются
на несколько категорий: 1) внутрикожные паразиты, живущие в толще ко-
жи (например, чесоточный зудень, стр. 429) или в ее железах (например,
угрица, стр. 428); 2) полостные паразиты, живущие в наружных полостях
тела — в носовой, ротовой полости, в носовых пазухах и т.п. (например,
личинки полостных оводов, стр. 400); 3) кишечные паразиты (напри-
мер, личинки желудочного овода, стр. 398); 4) тканевые паразиты, жи-
вущие в толще живых тканей (например, личинка вольфартовой мухи);
5) кровяные паразиты, живущие в крови; среди членистоногих паразитов
человека кровепаразитов нет, но они имеются среди червей, простейших
и пр. (например, малярийный паразит Plasmodium).
Некоторые паразиты проделывают внутри тела своего хозяина слож-
ные миграции (путешествия), как, например, личинка кожного овода
(стр. 399).
Способ доступа паразита к хозяину: постоянные паразиты. Степень
связи паразита с хозяином и характер этой связи зависят также и от спо-
соба доступа паразита к хозяину, как это видно из следующих примеров.
Вошь живет на теле хозяина. Она питается на нем, переваривает пи-
щу, приклеивает свои яйца. Покидает она тело хозяина лишь случайно
и долго оставаться во внешней среде не может. Как постоянный эктопа-
разит вошь все время имеет доступ к хозяину, а потому пьет кровь часто
и небольшими порциями. Она неспособна долго голодать и может оставаться
без пищи лишь в течение нескольких дней; живет вошь всего около 40—60
дней.
Вошь обладает присущими постоянному эктопаразиту приспособ-
лениями, помогающими ей удерживаться на теле хозяина: плоская форма,
цепкие лапки, могущие крепко зажимать волосок, за который она ухва-
тилась. В связи с постоянным риском быть раздавленной хозяином вошь
обладает значительной прочностью.
Домовые подстерегающие паразиты. Постельный клоп (стр. 414) живет
не на теле человека, а где-нибудь вблизи него: в постели, на стене, в пред-
метах обстановки. Некоторые виды того же семейства Cimicidae живут
в гнездах птиц и убежищах летучих мышей. Клоп регулярно нападает на
хозяина, но, насосавшись крови, тотчас же покидает его. Он перевари-
вает кровь и кладет яйца вне тела хозяина — в постели, на стене и т. п.
Ма хозяине он бывает только в момент принятия пищи, все остальное
время проводит в убежище, которое и является основной средой его обита-
140
ния. Паразиты, ведущие подобный образ жизни, обозначаются как до-
мовые, гнездовые или норовые паразиты. Лишь периодически нападая
на хозяина, домовые паразиты высасывают каждый раз значительно боль-
ше крови, чем постоянные эктопаразиты. Если хозяин покидает свое
убежище (дом, гнездо, нору), домовые паразиты способны очень долго,
месяцами и даже годами, переносить голодание. Все они отличаются
большой долговечностью.
Пастбищные подстерегающие паразиты. Пастбищный клещ, напри
мер, Ixodes persulcatus, также подстерегает добычу, но не тогда, когда
она возвращается к себе в убежище (в нору или гнездо), а когда
она случайно проходит мимо него на пастбище. В этих условиях шансы
на встречу с добычей значительно меньше. Поэтому пастбищный подсте-
регающий кровосос еще более прожорлив, чем домовый (норовой, гнездо-
вой). Кроме того, он обладает и громадной плодовитостью при почти
такой же способности выносить голодание.
Свободные кровососы. Комар также нападает на человека и насасы-
вается на нем крови. Без питания кровью он не может обойтись, так как
кровь необходима для созревания яиц в его теле1. Напившись крови,
комары улетают и многие из них во время переваривания пищи прячутся в
траве и кустах, вдали от добычи, а для откладки яиц все они улетают в поис -
ках водоемов, подходящих для жизни их личинок. Связь комара с хозяи-
ном — значительно слабее, чем у вши, для которой тело хозяина пред-
ставляет постоянное место обитания и единственный источник питания.
Для клопа постоянным местом пребывания служит убежище его хо-
зяина, для комара — главным образом внешний мир; правда, и комар
может задерживаться на некоторое время в доме человека или в норе
животного, но он не привязан к этим убежищам и быстро их поки-
дает. Вошь—постоянный эктопаразит человека, клоп — домовый (гнез-
довой, норовой) паразит, комар — свободный, активно нападающий
кровосос. По отношению к хозяину и клоп, и комар не постоянные,
а периодические паразиты. Свободные кровососы выпивают каждый
раз относительно больше крови, чем постоянные эктопаразиты, но
меньше, чем домовые или пастбищные подстерегающие паразиты, так
как при чрезмерной прожорливости они потеряли бы способность к
полету. Активно разыскивая добычу, а не поджидая ее, они не обладают
ни способностью длительно переносить голод, ни долговечностью под-
стерегающих паразитов.
Паразиты кругложизненные и возрастные. Клоп и вошь питаются кро-
вью на всех стадиях своего жизненного цикла: они являются кругложизнен-
ными паразитами; наоборот, комар питается кровью только на взрослой
стадии, тогда как личинка комара живет в воде и питается различными жи-
выми и мертвыми частицами, которые находит в воде. Со своей стороны,
вольфартова муха паразитирует на человеке или животных только в
личиночном возрасте, а взрослое насекомое кормится на цветах. И кома-
ры, и муха Вольфарта в отличие от клопов и вшей являются возрастными,
или фазовыми, паразитами, паразитирующими лишь в одном каком-либо
возрасте, или фазе, своей жизни — в личиночном, как вольфартова муха,
или в имагинальном, как комары.
Паразиты одноядные и многоядные. Человеческая вошь питается ис-
ключительно человеческой кровью и не живет ни на каком другом животном.
1 Лишь немногие виды кровососущих комаров способны отложить кладку яиц без
принятия крови, за счет запаса веществ, накопленных в теле во время личиночной
жизни.
141
Поэтому она является одноядным паразитом. Наоборот, «человеческая^
блоха нападает не только на человека, но и на многих других млекопи'
тающих (на свинью, собаку, на различных диких хищных и пр.): она яв-
ляется многоядным паразитом.
Способы попадания на хозяина. Членистоногие паразиты человека
могут попадать на него либо активно, либо пассивно. Активно нападает,
например, комар, слепень, разыскивающие добычу и настигающие ее бла-
годаря быстрому полету. Активно нападают и клещ или клоп, подстере-
гающие добычу и всползающие на нее, как только она окажется на доступ-
ном для них расстоянии. Вошь активно переползает с одного человека на
другого или переходит на соприкоснувшегося с ней человека из внешней
среды, в которую она случайно попала. Паразитические личинки мух по-
падают на хозяина пассивно. Взрослая муха отыскивает подходящего
хозяина и откладывает на него яйца или помещает на него своих личинок.
Степень связи с человеком. С точки зрения связи с человеком всех
нападающих на него членистоногих можно разделить на четыре группы:
1) паразиты тела и одежды; 2) сожители жилища; 3) сожители поселка;
4) паразиты из внешней природы. К числу паразитов тела и одежды отно-
сятся все постоянные эктопаразиты человека: вши, чесоточные зудни, уг-
рицы. К числу паразитов из числа сожителей жилища относятся клопы
(Cimex), домовые клещи (домовые Argasidae, стр. 433. Bdellonyssus,
стр. 433), человеческая блоха. К числу паразитов из числа сожителей по-
селка относятся комнатная муха и ряд других мух, а также москиты и ко-
мары в том случае, когда они выплаживаются внутри поселка. К числу
паразитов внешней природы относится большинство иксодовых клещей,
клещи-краснотелки, комары и москиты, нападающие на открытом воздухе
вне поселка, слепни, мошки, мокрецы и пр.
Блохи, москиты, мухи, нападающие на своего хозяина главным об-
разом внутри дома и выплаживающиеся как в доме, так и в поселке, за-
нимают промежуточное место между сожителями жилища и сожителями
поселка.
Комар Anopheles maculipennis, выплаживающийся во внешней при-
роде, но нападающий на человека в жилище или в поселке, занимает
промежуточное место между этими тремя группами.
Чем теснее данный паразит связан с человеком, тем тягостнее (при
прочих равных условиях) для последнего его присутствие. С другой сто-
роны, чем теснее связан он с человеком, тем уже поле его деятельности,
тем более он доступен воздействию наших мероприятий.
Вредоносность паразитов. Характерной чертой паразитизма является
вред, который паразит причиняет хозяину, и враждебная по отношению к
паразиту защита со стороны хозяина. Между хозяином и паразитом идет
борьба, взаимоотношения их антагонистичны. Тканевые эндопаразиты, на-
пример, личинки вольфартовой мухи, могут производить в теле человека
огромные разрушения: попав под веко, они могут целиком уничто-
жить глазное яблоко и т. п. Типичные эктопаразиты тоже вредоносны, хотя
и в меньшей степени. Так, вошь питается кровью человека; она впрыски-
вает ему свою слюну, которая вызывает покраснение кожи, образование
папулы, сильный зуд. При чрезмерном размножении вши становятся чрез-
вычайно тягостны для хозяина, вшивость становится настоящей болезнью.
Укусы вшей раздражают людей и животных (у многих млекопитающих
имеются свои собственные виды вшей) и потому они стремятся от них из-
бавиться. Они вылавливают или сбрасывают с себя паразитов и таким
путем защищают себя от чрезмерного их размножения. Опытами с жи-
вотными установлено, что наиболее поражены вшами бывают больные и
142
ослабленные особи, которые не могут достаточно энергично от них защи-
щаться.
Вред, причиняемый паразитом хозяину, и степень противодействия
(антагонизм) со стороны хозяина выражены различно. Так, многие пухоеды
(стр. 47), живущие на птицах и питающиеся их перьями, почти не прино-
сят вреда своему носителю, но при чрезмерном размножении они становят-
ся ему в тягость. Некоторые хищные клещики (семейство Gheyletidae), жи-
вущие в шерсти млекопитающих, уничтожают поселившихся там парази-
тических клещей и этим приносят хозяину несомненную пользу. Такое
сожительство приносит пользу и млекопитающему, и хейлетидам, и поэ-
тому взаимоотношения этих хейлетид с хозяином представляют уже не
борьбу, не антагонизм, а ^содружество, мутуализм (от латинского слова
mutuus — взаимный).
Симбиоз, мутуализм, паразитизм. Все формы тесного сожительства
двух организмов называются симбиозом (син — вместе, биос — жизнь, сим-
биоз — совместная жизнь)1. О симбиозе говорят в том случае, если хотя
бы для одного из сожителей совместная жизнь является существенно
важной. Если совместная жизнь выгодна для обоих сожителей, симбиоз
их является мутуалистическим сожительством, или мутуализмом. Если
совместная жизнь выгодна для одного из сожителей и безразлична для дру-
гого, сожительство является безразличным симбиозом. Если совместная
жизнь выгодна для одного из сожителей и вредна для другого, сожитель-
ство их представляет антагонистический симбиоз, т. е. паразитизм.
В природе между паразитизмом, безразличным симбиозом и мутуализ-
мом существует множество переходов. В отношении членистоногих —
сожителей человека и других теплокровных — мы можем признать, что
всякое членистоногое, питающееся за счет крови или тканей теплокров-
ного, является паразитом последнего.
Вредоносность и степень связи с человеком. Чем теснее связь какого-
либо паразитического членистоногого с человеком, тем скорее оно при
прочих равных условиях может стать тягостным и невыносимым.
Эндопаразитические членистоногие, главным образом паразитические
личинки мух, например, вольфартовой мухи, так раздражают чело-
века, даже в небольшом числе настолько опасны и неприятны, что
их следует немедленно удалять, как только они будут обнаружены.
Постоянные эктопаразиты, например, вши, также абсолютно нетер-
пимы. То же можно сказать о домовых паразитах (клопы, домовые клещи,
блохи), которые, размножаясь в больших количествах, делают дома почти
необитаемыми для человека. И хотя в малых количествах они могут быть
для жителей мало ощутимыми, полное искоренение таких паразитов в
жилищах безусловно обязательно.
Свободно нападающие кровососы (комары и слепни), если их немного,
ощутимы мало. В тех местах, где они встречаются массами, они становят-
ся настоящим бедствием. В мало обжитых местностях — в тундре, в тай-
ге, в плавнях — численность кровососущих двукрылых (гнуса) и интен-
сивность их нападения достигают таких пределов, что и для человека, и
для домашних животных жизнь там становится невыносимой. У нас в СССР
изучению гнуса особенно много труда отдал А. В. Гуцевич. Таким образом,
и свободно нападающие кровососы, как будто не тесно связанные с челове-
ком, при высокой численности приобретают большое значение и против
них приходится изыскивать меры защиты.
1 Во многих книгах слово «симбиоз» употребляется в более узком смысле, а именно
для обозначения мутуализма; такое словоупотребление неправильно.
143
ГЛАВА II
ЧЛЕНИСТОНОГИЕ КАК ПЕРЕНОСЧИКИ БОЛЕЗНЕЙ
Способы проникновения возбудителей болезней в организм человека.
Значение членистоногих как переносчиков болезней человека и животных
еще больше, чем значение их как паразитов.
Микробы — возбудители болезней человека попадают в организм по-
следнего различными путями. Очень многие из них способны некоторое
время просуществовать во внешней среде и в таком случае попадают в
организм человека прямо извне — через воздух, воду или пищу. Другие
попадают из одного организма в другой при непосредственном контакте
здорового человека с больным.
Очень многие микробы передаются переносчиками, которыми явля-
ются почти исключительно членистоногие.
Возбудители болезней человека, передаваемые членистоногими. Воз-
будители болезней человека, передаваемые членистоногими, чрезвычайно
разнообразны; они встречаются в следующих группах организмов: филь-
трующиеся вирусы, риккетсии, бартонеллы, бактерии, спирохеты, про-
стейшие и черви.
Фильтрующиеся вирусы. Фильтрующиеся вирусы — это настолько
мелкие существа, что их нельзя обнаружить даже с помощью микроскопа:
они проходят через фарфоровый фильтр, удерживающий всех бактерий
(откуда и происходит название «фильтрующиеся»). Все они паразитируют
внутри живых клеток животных и растений, вызывая у тех и у других раз-
нообразные заболевания. Некоторые вирусы, например, вирус гриппа,
передаются непосредственно от человека к человеку через воздух — с мель-
чайшими капельками, вылетающими изо рта, при кашле, чихании или даже
разговоре (капельная инфекция); очень многие вирусы передаются при
помощи членистоногих-переносчиков: комаров, москитов и клещей; так,
например, вирус желтой лихорадки передается комаром Aedes aegypti
(стр. 328), вирус лихорадки паппатачи — москитом Phlebotomus papatasii
(стр. 335), вирус таежного энцефалита — клещом Ixodes persulcatus
(стр. 459).
Риккетсии. Риккетсии (род Ricketsia) — паразитические организмы
близкие к бактериям. Они очень мелки, но все же хорошо видимы при
больших увеличениях микроскопа; способны размножаться обычно только
внутри клеток хозяина. Болезни, вызываемые риккетсиями, называются
риккетсиозами.
У человека риккетсии вызывают сыпной тиф (Ricketsia prowazeki)
и целый ряд сыпнотифозных лихорадок. Все эти болезни передаются
исключительно через членистоногих переносчиков (вошь, блохи, различ-
ные клещи).
Бактерии. Большинство бактерий — возбудителей болезней человека—
передается без помощи переносчиков. Однако в переносе бактерий, воз-
буждающих кишечные заболевания (брюшной тиф, паратифы, дизен-
терия, холера), насекомые-переносчики (главным образом мухи, см.
стр. 387) играют очень большую роль. С другой стороны, бактерии, сильно
размножающиеся в крови хозяина (чума, туляремия, сибирская язва),
могут распространяться кровососущими членистоногими. Перенос чумы
при помощи блох является основным способом распространения этой
болезни в природе.
Спирохеты. Спирохеты (класс Spironemacea) — подвижные микроорга-
низмы, имеющие вид нити, изогнутой штопором, нередко с жгутами на
144
концах. Многие из них паразитируют в самых разнообразных животных —
от червей и до человека. Болезни, вызываемые спирохетами, называются
спирохетозами. Наиболее распространенный трансмиссивный 1 спирохе-
тоз человека — возвратный тиф — передается вшами. Наряду с этим, во
всех теплых странах имеется множество клещевых спирохетозов, переда-
ваемых клещами семейства Argasidae. К числу нетрансмиссивных спиро-
хетозов, передаваемых непосредственным контактом здорового человека
с больным, относится сифилис.
Простейшие. Среди простейших (тип Protozoa), паразитирующих в
теле человека, надо различать паразитов кишечника и паразитов крови.
Кишечные простейшие (дизентерийная амеба, кишечные жгутиковые)
могут переноситься мухами с испражнений больного на пищу, но могут
передаваться и без помощи переносчика: через воду, грязные руки и т. п.
Все кровяные простейшие передаются почти исключительно при помощи
членистоногих переносчиков. Кровяные паразиты человека принадлежат
к двум классам: споровико^в (Sporozoa) и жгутиковых (Flagellata). К классу
споровиков, отряду кровяных споровиков (Haemosporidia), принадлежат
малярийные паразиты (Plasmodium). К классу жгутиконосцев (отряд
Protomonadina, семейство Trypanosomidae) принадлежат трипаносомы
(Trypanosoma, Schizotrypanum), вызывающие тяжелые болезни человека
(первая — в Африке я вторая — в Южной Америке), и лейшмании (Leish-
mania), возбудители лейшманиозов, передаваемых москитами (Phlebo-
tomus).
На территории наших южных республик имеется несколько форм
лейшманиозов человека.
Паразитические черви. Из червей при помощи уколов кровососущих
членистоногих передается несколько видов филярий (семейство Filari-
idae, класс Nematodes, круглые черви). Однако на территории СССР нет
ни одного вида филярий, паразитирующих в человеке. Для некоторых
других паразитических червей человека членистоногие служат промежу-
точными хозяевами, в которых поселяются неполовозрелые стадии пара-
зита, и человек заражается паразитом, поедая промежуточного хозяина.
Так, на Дальнем Востоке личинки сосальщика (Paragonimus) поселяются
в пресноводных крабах, и если человек или другое млекопитающее съе-
дает сырого краба, личинка проникает в легкие нового хозяина, где и раз-
вивается во взрослого сосальщика.
Связь между систематическим положением возбудителя и переносчика.
Определенные группы членистоногих в течение своей эволюции вступили
в связь с определенными группами возбудителей, переносчиками которых
они и стали. Табл. 2 показывает, для каких групп возбудителей являются
естественными 1 2 переносчиками представители той или иной группы крово-
сосущих членистоногих. В основном в таблицу включены только специфи-
ческие переносчики, в теле которых возбудитель питается и размножается.
Механические переносчики кровепаразитов (в теле которых размноже-
ние возбудителя не происходит) включены в таблицу только в тех слу-
чаях, когда они имеют несомненное значение в распространении данного
заболевания. Механические переносчики кишечных заболеваний челове-
ка не включены.
1 Значение этого слова — см. ниже, стр. 148.
2 Естественными переносчиками того или иного возбудителя являются те члени-
стоногие, которые являются переносчиками в естественной обстановке. Те членисто-
ногие, которые в условиях лабораторного эксперимента могут передавать данного воз-
будителя, но не связаны с ним в природе, в таблицу не включены.
Ю Учебник медицинской энтомологии
145
Таблица 2
Соотношение между систематическим положением переносчиков и возбудителей -
трансмиссивных заболеваний наземных позвоночных, включая человека
Возбудители Переносчики Виру- сы Рик- кет- сии Бак- терии Спиро- хеты Жгути- коносцы Споро- вики Филя- рии
Комары (Gulicidae) + + | М — — 1 4"+ 1 i 4-+
Мошки (Simuliidae) — — — — 1 4- +
Мокрецы (Heleidae) I м 1 — — — +
Москиты (Phlcbotcmus) + 1 — — -Н- 4- —
Слепни (Tabanidae) — — м — М — +
Мухи цеце (Glossina) — — — — ++ — —
Мухи-кровососки (Hippoboscidae) — ?_ — — 4- 4- —
л Блохи (Aphaniptera) — + + — 4-4- — —
Клопы (Hemiptera) I 1 — । ! — — — — —
Вши 4-
(Siphunculata)
Гамазоидные клещи (Gamasoidea) + + i + —1 1 ++ +
Иксодовые клеши (Ixodidae) ++ 1 1 1 ++ + + ++ —
Аргасовые клещи (Argasidae) ? — + ++ — ? —
Клещи-краснотелки (Trombidiidae) — + — — — — —
146
Крестик означает наличие данного сочетания переносчика и возбудителя; па-
пример, крестик, стоящий на пересечении вертикального столбца «споровики» и гори-
зонтального ряда «мошки», означает, что известен случай переноса споровика мошкой.
Два крестика означают наличие целого ряда случаев данного сочетания; другими ело
вами, два крестика означают, что данная группа переносчиков, например, комары,
передает целый ряд видов возбудителей из данной группы, например, споровиков;
буква М означает механический перенос возбудителя. Однако в таблицу включены лишь
те случаи, когда значение переноса в распространении данной болезни доказано.
Крестики или буква М жирные (Ц- или М) означают, что среди сочетаний данного
типа имеются болезни, поражающие человека.
Из этой таблицы видно, что особенно многочисленными и разносторон-
ними связями с возбудителями болезней обладают комары, москиты, гама-
зоидные и иксодовые клещи. Они являются переносчиками очень боль-
шого количества возбудителей. С другой стороны, мухи цеце и аргазиды
как бы специализировались на отдельных группах возбудителей: мухи
цеце являются переносчиками целого ряда жгутиконосцев (Flagellata) йз
рода трипаносома, аргазиды — переносчиками целого ряда спирохет.
Некоторые группы, наоборот, являются специфическими переносчиками
очень немногих возбудителей, как, например, слепни, или вовсе не явля-
ются переносчиками заболеваний, как, например, постельный клоп.
Способы переноса возбудителей членистоногими. Способы переноса
так же разнообразны, как и возбудители, переносимые членистоногими.
Комнатная муха непрерывно летает из уборной или помойки в столо-
вую или на кухню и обратно и таким образом переносит на лапках и на хо-
ботке с нечистот на пищу человека бактерий — возбудителей кишечных
болезней (брюшной тиф, дизентерия), цисты простейших, яйца глистов.
Частично она заглатывает нечистоты и загрязняет ими пищу человека, ис-
пражняясь и отрыгивая на нее бактерий. Таким образом, муха осущест-
вляет перенос болезней человека путем загрязнения (контаминации) пищи.
Садясь на раны, мухи могут и их загрязнять бактериями. В обоих случаях
муха является механическим переносчиком: возбудитель в ее теле не раз-
множается и не проделывает никаких стадий развития.
Такими же механическими переносчиками являются, например, слеп-
ни (Tabanidae) по отнешению к бактериям сибирской язвы. Если слепень
напьется на больном животном, кровь которого содержит множество си-
биреязвенных палочек, часть этих палочек останется на его хоботке и при
следующем сосании попадет в кожу нового хозяина (Олсуфьев). Так как
в данном случае возбудитель вводится в кожу путем укола, этот способ
переноса обозначается как прививка (инокуляция).
Совершенно иным путем переносит вошь возбудителя возвратного
тифа человека (Spirochaeta recurrentis). Спирохеты, попавшие вместе с
кровью больного в желудок вши, проникают оттуда в полость тела, где бла-
гополучно живут и размножаются. Но выйти из полости тела вши спи-
рохеты не могут и проникают в организм человека лишь в том случае, если
человек при почесывании раздавит вошь. Тогда спирохеты попадут на
поврежденную расчесами поверхность кожи или будут занесены руками
на слизистые оболочки — в рот, на конъюнктиву глаза и т. п. и оттуда
проникнут в организм.
Чумные палочки, возбудители чумы человека и грызунов, попав с
кровью больного в кишечник блохи, усиленно размножаются в нем и не-
редко закупоривают пищевод паразита; пробка бактерий препятствует
нормальному сосанию. Под влиянием усиленных сосательных движений,
которые производит блоха, при повторном сосании группы бактерий от-
рываются от комка и попадают врану. Происходит прививка (инокуляция)
при помощи отрыгиваемых из кишечника возбудителей.
10*
147
Наиболее совершенной формой переноса является прививка (ино-
куляция) при помощи зараженной слюны. Пример такого рода представ-
ляет малярийный паразит Plasmodium, который, попав из крови больного
в организм комара, проделывает в нем сложный цикл развития, размно-
жается, и образовавшиеся спорозоиты проникают в слюнные железы ко-
мара. При уколе они вводятся в рану вместе со слюной.
Кроме того, в ряде случаев членистоногие являются промежуточными
хозяевами паразитических червей человека (гельминтов, или глистов).
Так, маленький пресноводный рачок циклоп заражается, поедая микроско-
пических личинок ришты, которые проникают в полость его тела и там
остаются. Если человек случайно проглотит с водой такого рачка, он зара-
жается риштой (Федченко, Исаев). Животные, охотно поедающие своих
эктопаразитов, нередко заражаются таким путем не только глистами, но
и целым рядом других возбудителей.
* В дальнейшем мы остановимся на всех этих способах переноса.
Болезни нетрансмиссивные, трансмиссивные и со смешанным спосо-
бом переноса. Существует ряд болезней, которые могут передаваться от
человека к человеку или от животного к человеку и непосредственно, через
внешнюю среду, и при помощи переносчика. Так, брюшной тиф может пере-
носиться мухами, но возможны заражения через воду или через пищу, за-
грязненную без участия насекомых. Чуму переносят блохи, но в некото-
рых случаях она передается и через воздух при помощи мельчайших ка-
пелек слюны или мокроты больного. Туляремия может передаваться раз-
личными насекомыми и клещами, но также через воду или путем непосред-
ственного соприкосновения с больными грызунами, например, при снятии
шкурок с зараженные водяных крыс.
Наряду с этим, многие болезни передаются только через переносчика
и никаким другим способом перейти от одного теплокровного к другому
не могут. Типичным примером такого рода болезни служит малярия, ко-
торая в естественной обстановке может перейти от больного человека
к здоровому исключительно через посредство комара Anopheles.
Все болезни, передаваемые от одного теплокровного другому через
членистоногого переносчика, носят название трансмиссивных болезней
(Павловский) (от слова «трансмиссия», что означает — передача). Маля-
рия является примером чисто трансмиссивной болезни. Туляремия—
представитель болезней со смешанным типом распространения: вспышки
туляремии могут быть и трансмиссивного, и нетрансмиссивного происхо-
ждения. Таким образом, малярия является облигатно (обязательно) транс-
миссивной болезнью, так как никаким другим путем передаваться не мо-
жет; туляремия является факультативно-трансмиссивной болезнью.
Антропонозы и зоонозы. Среди трансмиссивных болезней некоторые
являются чисто человеческими. Таковы малярия, сыпной тиф, возвратный
тиф и др. В этих случаях переносчик передает возбудителя исключительно
от человека к человеку; никакие другие теплокровные в распространении
болезни роли не играют. Возбудитель оборачивается между двумя своими
хозяевами — человеком и комаром (в случае малярии), человеком и во-
шью (в случае сыпного и возвратного тифа). Болезни, свойственные толь-
ко человеку, называются антропонозами (от греческого слова antropos—
человек).
Многие другие трансмиссивные болезни человек получает от живот-
ных; такие болезни называются зоонозами х.
1 Зоонозами называются также и нетрансмиссивные болезни, получаемые чело-
веком от животных, например, сап, получаемый от лошадей, и ящур, получаемый от
коров.
148
Природные очаги трансмиссивных болезней. В дикой природе звери и
птицы подвержены множеству болезней, среди которых имеются трансмис-
сивные. Для человека некоторые из этих болезней не представляют угрозы,
так как ко многим из них он невосприимчив, например, к малярии птиц,
передаваемой комарами рода Culex, или к пироплазмозам млекопитающих,
передаваемым клещами. В других случаях человек к возбудителю хотя
и восприимчив, но переносчиком является какое-либо членистоногое,
совсем или почти не нападающее на человека (например, спирохетоз песча-
нок в Средней Азии, передаваемый клещом Alectorobius tartakovskii). Но
наряду с этим, в природе встречается целый ряд болезней, к которым чело-
век восприимчив и переносчик которых охотно нападает на него. В этих
случаях человек подвергается риску заражения и нередко заболевает.
Таких трансмиссивных болезней животных, которыми человек зара-
жается в дикой природе, существует очень много. Тайге свойствен клещевой
энцефалит—болезнь мелких млекопитающих и птиц, передаваемая кле-
щом Ixodes persulcatus. В ряде степных районов Сибири и Дальнего Восто-
ка существует клещевая сыпнотифозная лихорадка — болезнь грызунов,
передаваемая несколькими видами пастбищных клещей. Степной и
полупустынной зоне во многих местах свойственна чума — болезнь гры-
зунов, передаваемая блохами. Сухим пространствам Восточного Кавказа
и Средней Азии свойственны клещевые возвратные лихорадки — болезнь
различных диких млекопитающих, передаваемая клещами Ornitho-
dorini и т. д. г.
Одомашнение трансмиссивных зоонозов. Источником целого ряда забо-
леваний человека являются грызуны, поселившиеся в наших домах вместе
со своими эктопаразитами и норовыми паразитами, а также домашние жи-
вотные. Так, крысы принесли с собой в города чуму и крысиный сыпной тиф,
передаваемые блохами и клещиками Bdellonyssus, собаки (на юге) — внут-
ренностный лейшманиоз, свойственный их предкам шакалам (Латышев) и
передаваемый москитами и т. д. Охотно селятся в человеческих по-
стройках клещи Ornithodorini, переносчики спирохет клещевой возвратно-
тифозной лихорадки, поражающей и человека, и целый ряд мелких
млекопитающих из его окружения. Таким образом, целый ряд трансмис-
сивных болезней животных вместе с этими последними перешел в наши се-
ления и города.
Значение трансмиссивных болезней. Болезни, передаваемые членисто-
ногими, имеют для человечества колоссальное значение. Эпидемии чумы
или сыпного тифа неоднократно опустошали целые страны. Сыпной тиф
зимой и дизентерия летом в прежние века нередко истребляли ар-
мии. Передаваемые мухами кишечные заболевания — брюшной тиф и ди-
зентерия — во многих местах до сих пор являются бичом населения. Бес-
численные трансмиссивные болезни поражают людей во всех уголках зем-
ного шара. Однако по широте распространения, по массовому охвату на-
селения и по тяжести ущерба, наносимого человечеству, на первом месте
стоит малярия. Где влажно и зелено, — там есть малярия; другими сло-
вами, малярия там, где природа благоприятствует человеку (Торопов).
Сильнее всего она поражает лучшие теплые и плодородные места и часто
превращает их в очаги мучения и смерти.
Итак, в целом вред, наносимый человечеству трансмиссивными болез-
нями, колоссален, а следовательно, колоссален и вред, приносимый их
1 В СССР изучению природных очагов трансмиссивных болезней особенно много
внимания уделил акад. Е. Н. Павловский и его ученики и сотрудники. Е. Н. Павлов-
ским введен термин «природный очаг болезни».
149
переносчиками. Большой ущерб наносит малярия и ряд других трансмис-
сивных болезней, в частности, и нашей стране. Борьба с трансмиссивными
болезнями обычно успешнее всего осуществляется путем борьбы с их
переносчиками — кровососущими насекомыми и клещами. Очень часто
борьба с переносчиками является наилучшим, а иногда почти единствен-
ным способом профилактики (предупреждения) трансмиссивных болезней.
В силу этого борьба с переносчиками приобретает огромное значение.
Залогом успеха в этой борьбе является знание врага, знание члени-
стоногих переносчиков, их строения, развития и образа жизни. Эти знания
и дает нам медицинская энтомология.
ГЛАВА III
ЯДОВИТЫЕ ЧЛЕНИСТОНОГИЕ
Яды. Ядом является всякое вещество, которое, попадая во внутрен-
нюю среду организма, своим химическим действием вызывает в нем болез-
ненные изменения. В высоких концентрациях ядовитым может оказаться
почти любое вещество. Поэтому ядами в узком смысле слова называются
те вещества, которые оказывают ядовитое действие уже в небольших кон-
центрациях.
Действие ядов. Действие яда на организм может быть местным и об-
щим. Местное действие проявляется в месте проникновения яда в организм.
Оно может сказываться различным образом: в виде ожога (отмирание при-
шедших в соприкосновение с ядом живых тканей), воспаления и т. д. Об-
щее действие наступает после распространения яда по организму и может
в различной степени затрагивать разные органы: сердце, почки, нервную
систему и т. п. Если отравление было несмертельным, реакция организма
на повторное введение данного яда часто оказывается измененной; при по-
вторном введении одного и того же яда часто наблюдается либо повышен-
ная к нему чувствительность и гораздо более сильное его действие, чем
при первом введении, либо, наоборот, чувствительность к яду умень-
шается. В первом случае происходит сенсибилизация организма к данному
яду, во втором — иммунизация организма по отношению к данному яду.
Ядовитые животные. Многие животные и растения содержат или вы-
деляют вещества, ядовитые для других организмов. Много ядовитых видов
существует и среди членистоногих; среди них имеются такие, которые
в той или иной мере ядовиты для человека.
По своему значению для выделяющего их животного яды разделяются
на две основные группы: яды оборонительные и яды наступательные.
Значение оборонительных ядов. Оборонительные, или защитные, яды
очень часто содержатся в крови членистоногих, в других случаях они
являются продуктом выделения кожных желез, или, наконец, содержатся
в железистых волосках. Когда птица или зверь схватывает ядовитую гу-
сеницу, получает ожог и бросает добычу, для последней избавление часто
оказывается запоздалым, так как она уже смертельно повреждена клювом
или зубами. Но, ожегшись ядовитой гусеницей, животное очень скоро на-
учается избегать ей подобных, и таким путем данный вид гусениц извлекает
из своего ядовитого действия большую пользу. К тому же ядовитые насе-
комые большей частью ярко окрашены: эта окраска предостерегает хищни-
ка от нападения на данную добычу.
Ядовитые жуви. Из насекомых пассивнозащитные яды особенно рас-
пространены среди жуков и среди гусениц бабочек. Кровь многих жуков:
листоедов (Chrysomelidae), божьих коровок (Coccinellidae), нарывников
150
(Meloidae) (рис. 81) и пр. ядовита. Будучи схвачены врагом, они могут
выделять капельки крови в местах сочленений ног. Кровь жуков-нарыв-
ников содержит кантаридин; это вещество настолько ядовито, что 1 г чи-
стого кантаридина могли бы смертельно отравиться 20 000 человек.
В небольших количествах кантаридин, нанесенный на кожу человека,
вызывает нарывы; общее действие кантаридина в первую очередь сказы-
вается на почках.
Очень ядовит небольшой жук-хищник Paederus fuscus (рис. 81, 6)
(семейства Staphylinidae), живущий во влажных местах. Раздавленный на
коже, он также может вызвать нарыв (Павловский).
Рис. 81. Ядовитые жуки-нарывники (семейство
Meloidae) и хищники (семейство Staphylinidae).
1 — майна Ме1оё proscarab aeus, самец; 2 — то же, самка
(естественная величина); 3 — шпанка Lytta yesicatoria
(слегка увеличено); 4 — молодая личинка шпанки
(сильно увеличено); 5 — Mylabris (увеличено в два ра-
за); 6 — ядовитый хищник (семейство Staphylinidae)
Paederus (увеличено в четыре раза).
Рис. 82. Ядовитый
волосок гусеницы ба-
бочки Megalopyge в
продольном разрезе
(сильно увеличено).
1 — ядовитая железа;
2 — ее проток; 3 — хи-
тиновая стенка волоска.
Активно оборонительные яды, представляющие продукты кожных
желез, выделяют многие жуки.
Крупные жужелицы (Carabus) могут выпрыскивать едкие выделения
своих околоанальных желез на 20—30 см.
Ядовитые гусеницы. Очень многие мохнатые гусеницы, кроме простых
волосков, вооружены ядовитыми волосками, которые, попадая на кожу,
на слизистые оболочки или, еще хуже, в глаз человека, вызывают чрез-
вычайно сильное раздражение. Один из типов таких ядовитых волосков
изображен на рис. 82.
Отравления амбарными клещами. Из других членистоногих, защит-
ные яды которых могут иметь значение для человека, надо упомянуть мел-
ких клещей, поражающих продовольственные запасы (мучные клещи Ту-
roglyphidae и т. п., см. стр. 430). Пыль амбаров, содержащая шкурки, ис-
пражнения, волоски этих клещей, часто вызывает у рабочих высыпи
151
на коже, иногда приступы астмы и пр. Такие случаи, повидимому, воз-
никают большей частью при наличии сенсибилизации этих лиц по отно-
шению к ядовитым веществам клещей. Пища, содержащая большое ко-
личество клещей (сыр, мука и пр.), также может вызвать катарры кишеч-
ника у людей и животных.
Наступательные яды хищников. Основное назначение наступатель-
ных ядов хищников — быстро прекратить сопротивление добычи. Поэто-
му они впрыскиваются в тело противника и имеют сильное общее действие.
Обычно они впрыскиваются при помощи ротовых органов хищника; ис-
ключения составляют ядовитый шип скорпиона и жало перепончато-
крылых.
Рис. 83. Половой и ядоносный аппарат шмеля
Bombus.
1—ядовитая железа; 2—ее резервуар; 3—допол-
нительная железа; 4 — яичники; 5 — влагалище;
б — жало.
Жалящие перепончатокрылые. Жало перепончатокрылых, расположен-
ное на заднем конце брюшка, представляет собой самостоятельное ядови-
тое оружие и является видоизменением яйцеклада. Различные осы пользу-
ются жалом для убийства или парализации добычи (рис. 23), т. е. как чисто
наступательным оружием. Шмели и пчелы, питающиеся растительной пи-
щей (нектар, пыльца), пользуются жалом (рис. 83) как оружием активной
обороны. Пчелиный 1 яд по своей природе близок к некоторым змеиным
ядам. Укол вызывает местное действие (боль, покраснение, отек) и общее
действие, которое при множестве одновременных уколов может оказаться
очень сильным. Акад. Павловский считает, что 500 пчелиных уколов, полу-
ченных единовременно, могут убить человека. Уколы шершневой осы
(Vespa crabro) (рис. 30, Б) действуют еще сильнее. Пчеловоды под влия-
нием частых уколов приобретают значительную степень иммунитета про-
тив пчелиного яда. Известны случаи сенсибилизации людей к пчелиному
яду, проявляющиеся в очень сильных явлениях общего отравления после
единичного укола.
1 Здесь речь идет о домашней пчеле Apis mellifica.
152
Ядовитые многоножки. Из многоножек ядовитые железы свойственны
представителям отряда Chilopoda (рис. 13); они открываются на первой
паре ног, превращенной в ногочелюсти. Встречающиеся у нас на юге
крупные сколопендры (Scolopendra) могут прокусить кожу человека, но
укусы их не болезненнее укола пчелы.
Среди паукообразных имеют значение как ядовитые животные глав-
ным образом скорпионы и пауки.
Скорпионы и фаланги. Ядовитые железы скорпиона помещаются в
постанальной лопасти, заканчивающей его тело и несущей ядовитый шип
(рпс. 84). Загибая свое тонкое брюшко над головой, скорпион колет до-
Рис. 85. Ядовитый паук
каракурт batrodectes tre-
decimguttatus. Самка (уве-
личено) .
Рис. 84. Скорпион Buthus и его ядовитая железа.
1—скорпион, схвативший паука; 2—хвостовая лопасть
скорпиона, вскрытая сбоку: правая ядовитая железа сдви-
нута кверху; 3 — два последних членика тела и хвостовая
лопасть с ядовитым крючком; 4 — поперечный разрез через
хвостовую лопасть. Видны две ядовитые железы со складча-
тыми стенками и с мышечной обкладкой, служащей для вы-
прыскивания яда.
бычу, схваченную клешнями педипальп, и быстро убивает ее. Этим же
оружием он пользуется для самообороны, в том числе против человека.
Уколы скорпионов, живущих на территории СССР, для человека не опас-
ны, хотя очень болезненны. Немногие смертные случаи известны только
после уколов некоторых скорпионов тропических стран.
Бихорхи, или фаланги (отряд Solifugae), ядовитых желез не имеют.
Таким образом, несмотря на свою устрашающую внешность (рис. 14) и
мощные хелицеры, они неядовиты (Павловский).
Каракурт. Все представители отряда Агaneina (паутинные пауки) имеют
хелицеры, снабженные ядовитыми железами (рис. 6,2>), при помощи которых
они быстро убивают свою добычу. Однако лишь немногие пауки могут про-
кусить кожу человека, а из этих немногих не у всех укус очень болезнен
153
или опасен для человека. Из водящихся у нас пауков наиболее опасен кара-
курт (Latrodectes tredecimguttatus) (рис. 85), распространенный в су-
хих и жарких частях СССР. Это черный паук длиной около 1 см. Гнездо
его состоит из трубки — убежища и тенёт, представляющих сплетение
нитей, без особой правильности натянутых в траве. Гнезда чаще строятся
на невозделанной земле. Укус взрослой самки чрезвычайно ядовит. Она
убивает не только крупных насекомых, пауков и скорпионов, но и млеко-
питающих. Известны и случаи гибели человека от укуса каракурта, хотя
достоверных случаев немного. Действие укуса главным образом общее;
выражается оно в сильных болях по всему телу, дрожании, судорогах
и пр.
При ужалении перепончатокрылыми, уколах скорпионов и укусах
каракурта и других ядовитых членистоногих полезным противоядием
является перманганат калия, крепким раствором которого надо промы-
вать место укола.
Наступательные яды кровососущих членистоногих. Кровососущие
членистоногие при сосании вводят в тело человека или других позвоноч-
ных свою слюну, содержащую ряд более или менее ядовитых веществ. Яды
кровососущих оказывают обычно довольно сильное местное действие, вы-
зывая раздражение кожи хозяина, что может быть полезно для кровососа,
так как усиливает приток крови к месту укола. Однако обычно их яды име-
ют и общее действие, большей частью слабое, но при сенсибилизации хо-
зяина к уколам данного членистоногого резко усиливающееся. Особенно
ядовиты уколы мошек; ядовитым действием слюны объясняются также
параличи, иногда вызываемые присасыванием клещей. У людей, часто
подвергающихся уколам какого-либо кровососущего насекомого, нередко
вырабатывается по отношению к ним иммунитет, при котором и общая, и
местная реакция на уколы данного насекомого резко уменьшается. Имму-
нитет этот узко специфичен: так, иммунитет, выработанный против уколов
комаров рода A6des, нисколько не уменьшает реакцию кожи на уколы
комаров рода Anopheles.
РАЗДЕЛ Б
КОМАРЫ (СЕМЕЙСТВО CULICIDAE)
ГЛАВА I
ОБЩИЕ ЗАМЕЧАНИЯ
Гетерогенность. Все комары являются животными гетеротопными х:
яйца, личинки и куколки их живут в воде, взрослые комары — в воздухе.
Двойственное питание взрослой самки. Характернейшей чертой взрос-
лых комаров является двойственное питание: самцы питаются сладкими
соками, самки большинства видов — сладкими соками и кровью. При
этом самка комара, как и самец, может поддерживать свою жизнь, питаясь
одними сладкими жидкостями; но яйца в ее теле без принятия крови не со-
зревают, и, следовательно, кровяное питание необходимо для размножения.
В силу этого большинство видов комаров является обязательными кро-
вососами, ибо без кровяного питания вид не может существовать; однако
в отличие от вши или от постельного клопа, не принимающих никакой
пищи, кроме крови, комары не являются исключительными кровососами,
так как, кроме крови, они принимают и другую пищу.
Миграции, вызванные гетерогенностью комаров. Различия в образе
жизни между личинками и взрослыми особями заставляют самок комаров
совершать постоянные перелеты, и иногда на значительные расстояния.
С водоема, где она отродилась, самка летит разыскивать добычу, а позже
она должна вернуться на водоем, чтобы отложить туда яйца. В силу этого
самка комара должна обладать большой подвижностью. Кроме того, не-
обходимость покидать хозяина для откладки яиц не позволяет комарам стать
постоянными паразитами. Все они являются свободными, периодически
нападающими кровососами.
Степень прожорливости и причины, ее ограничивающие. Свободный
кровосос, раз отыскав добычу, заинтересован в том, чтобы взять от нее как
можно больше крови, так как неизвестно, удастся ли ему встретить добы-
чу во второй раз. Чем больше он выпьет крови, тем больше за ее счет смо-
жет он отложить яиц, оставить потомства. Однако, с другой стороны, ко-
личество крови, которое выпивает свободный кровосос, и количество яиц,
которые развиваются в его теле, ограничены известными пределами, ибо
он должен, как мы видели, сохранить свою подвижность. Из столкновения
этих двух противоречивых интересов вытекает основная особенность ко-
маров: комар выпивает ровно столько крови, сколько необходимо и до-
статочно для полного развития одной порции яиц. Если ему удалось вы-
пить полную порцию крови, к концу пищеварения в его яичниках оказы-
вается полная порция яиц, и, прежде чем пить кровь вторично, комар
должен слетать на водоем и отложить там эти яйца.
1 Значение этого слова см. стр. 60.
155
Гонотрофическая гармония и гонотрофический ритм. Точное соот-
ветствие между процессами переваривания крови и созревания яиц носит
название гонотрофической гармонии Ч Гонотрофическая гармония свой-
ственна всем кровососущим комарам и, невидимому, всем другим крово-
сосущим прямошовным двукрылым (москитам, мошкам, слепням и пр.).
Ограниченная прожорливость комара влечет за собой ограниченную
плодовитость. Самка комара выпивает количество крови, приблизитель-
но в полтора раза превосходящее вес ее тела, и за этот счет откладывает
в среднем около 250 яиц. Между тем образ жизни и личинки, и взрослого
комара вызывает очень большую смертность. Чтобы обеспечить существо-
вание вида, необходима гораздо большая плодовитость, которая и дости-
гается повторностью питаний и кладок, или, как говорят, повторностью
гонотрофических циклов: раз отложившая самка, если она осталась жи-
ва, вновь разыскивает добычу, вновь напивается, переваривает кровь и
откладывает яйца. Подобные циклы следуют один за другим до тех пор,
пока на одном из них самка не умрет. Таким образом, жизнь взрослой
самки комара представляет ряд повторяющихся гонотрофических циклов;
повторение этих циклов придает ее жизни ритмический характер. Это
выражают, говоря, что жизнь самки комара подчиняется гонотрофиче-
скому ритму.
Комары обладают полным превращением, причем, как выше упомянуто, их непо-
ловозрелые стадии живут в воде, а взрослые насекомые — па суше. Все'это обуслов-
ливает резкую разницу в строении и образе жизни, с одной стороны, неполовозрелых
стадий, а с другой стороны, взрослых комаров. Поэтому и анатомию, и физиологию,
и экологию личинок и взрослых комаров мы рассматриваем в отдельных главах. Кроме
того, особые главы посвящены биологии вида в целом; сюда относятся такие вопросы,
как сезонные явления в жизни комаров, движение их численности, географическое
распространение и эпидемиологическое значение комаров. При этом основное внима-
ние уделено малярийным комарам, образ жизни которых описан сравнительно подробно.
Из немалярийных комаров мы описываем лишь несколько видов, давая по одному-д в а
примера основных типов образа жизни встречающихся у нас комаров; при этом в каче-
стве примеров взяты по возможности виды, представляющие наибольшее практическое
значение как переносчики болезней или как широко распространенные кровососы.
ГЛАВА II
НАРУЖНОЕ СТРОЕНИЕ НЕПОЛОВОЗРЕЛЫХ ФАЗ КОМАРОВ
!• ЦИКЛ РАЗВИТИЯ КОМАРОВ
Из яиц, откладываемых самкой комара на поверхность воды, вылуп-
ляются маленькие личинки. От выхода из яйца и до окукления личинка
усиленно питается и растет; за это время личинка увеличивается в длину
более чем в 8 раз, а по объему — более чем в 500 раз. Такой усиленный рост
вызывает необходимость в периодических линьках, т. е. в сбрасывании
старых наружных покровов, оказывающихся тесными для растущей ли-
чинки, и в образовании новых, больших по величине. В течение своего
развития личинка линяет четыре раза, после четвертой линьки превра-
щаясь в куколку. Таким образом, личинка проходит четыре личиночных
возраста, отделенных друг от друга линьками. Новая кутикула после каж-
дой линьки бывает сначала мягкой и бесцветной, но постепенно затверде-
вает и темнеет.
Личинка Anopheles maculipennis, только что вылупившаяся из яйца,
имеет в длину несколько больше 1 мм, длина личинки II возраста 3—4 мм,
III возраста 5—6 мм и IV возраста 8—10 мм; с каждой линькой, помимо
1 То есть гармонии, или согласованности, процессов размножения и питания
(трофических).
156
увеличения размеров, усложняется и внутренняя организация личинки.
После завершения IV стадии личинка превращается в куколку. Куколка
не принимает пищи, а существует за счет запасов, накопленных в личи-
ночный период. Однако, в отличие от куколок большинства насекомых, ку-
колка комара подвижна, она может плавать. На стадии куколки проис-
ходит перестройка некоторых внутренних органов и возникают те органы
взрослого комара, которые отсутствуют у личинки (крылья, ноги, ротовые
части). После завершения развития взрослого насекомого комар освобож-
дается из куколочной шкурки.
2. ЯЙЦО
а) Способы откладки яиц
Способы откладки яиц комарами Anophelini и Culicini1 различны.
Яйца, отложенные самкой Anopheles, плавают на поверхности воды по-
одиночке или сцепившись по не-
скольку штук, не склеенные друг
с другом. Яйца A. superpictus мо-
гут развиваться и под водой, где
они прилипают к камням (Лисова).
У Culicini яйца откладыва-
ются или по отдельности, или
Рис. 87. Яйцо Anopheles ma-
culipennis.
1 — вид сверху; 2}— вид сб >ку.
А — поплавок; Б — микропиле.
Рис. 86. Яйца Anopheles |(вид
сверху).
А—A. maculipennis; Б— A. mac. sa-
charovi; В— A. bifurcatus; Г—A. al-
geriensis; Д—A. hyrcanus; Е —
A. pluinbeus; Ж — A. pulcherrimus; 3 —
A. superpictus.
склеиваются во время откладки в компактную массу, образуя плавающий
на воде плотик. Некоторые из видов комаров, откладывающих яйца по
отдельности, откладывают их на поверхность почвы в пониженных ме-
стах, подверженных затоплению водой при выпадении дождей (род A6des
и подрод Culicella рода Theobaldia); при втором способе откладки яйца
откладываются на воду (подрод Theobaldia, род Culex и род Mansonia).
1 О подразделении семейства Culicidae см. ниже, главу IV.
157
б) Наружная морфология яиц Culicinae
I. Яйца Anopheles
Длина яиц 0,6—0,8 мм. При рассматривании сверху яйца представ-
ляются удлиненно-овальной формы (рис. 86). Передний конец яйца не-
сколько расширен, задний — слегка сужен. Верхняя поверхность яйца
уплощена, нижняя — выпукла (рис. 87). Яйцо покрыто прочной скорлу-
пой, которая состоит из трех слоев (бесструктурная тонкая желточная обо-
лочка, эндохорион и экзохорион). Скорлупа образуется еще в яичнике, ее
выделяют клетки фолликулярного эпителия.
Рис. 88. Детали строения яйца Anopheles maculipennis.
А—продольный срез через передний конец яйца; Б—микропиле
яйца (вид сверху). 1—экзохорион; 2—эндохорион; 3—выросты
экзохориона, окружающие микропиле; 4—микропиле; 5—образо-
вание, закрывающее отверстие микропиле после оплодотворения
яйца; 6—желточная оболочка; В—поплавки яйца. Исчерченная
(7) и гладкая (8) межреберная перепонка (увеличено в 180 раз).
В свежеотложенных яйцах эндохорион студенистый и светлый, от-
чего яйца белого цвета; через несколько часов после откладки он темнеет
и затвердевает; для потемнения эндохориона необходимо соприкосновение
яиц с водой. Яйца, отложенные на сухую поверхность, не темнеют, а жел-
теют, засыхают и гибнут. Наружная оболочка — экзохорион, прозрачная
и эластичная. По бокам средней части яйца имеются образованные эк-
зохорионом два поплавка и оторочка, окаймляющая свободный край верх-
ней поверхности; они удерживают яйца в определенном положении, пло-
ской стороной кверху. У яиц всех видов Anopheles имеется либо одно из
этих образований, либо оба (рис. 86). Так, у A. algeriensis оторочка от-
сутствует, имеются только поплавки; у A. maculipennis sacharovi, A. plum-
beus, A. superpictus имеется только оторочка без поплавков; у остальных
подвидов A. maculipennis, у A. pulcherrimus, A. bifurcatus и A. hyrcanus
имеются оба эти образования.
Поплавки состоят из ребер и участков межреберной перепонки
(рис. 88). Экзохорион верхней поверхности яйца не гладкий, а образует
158
Рис. 89. Яйца Culicini (вид'сверху
и на поперечном разрезе через се-
редину).
А— Лёбеэ; Б— Theobaldia и Mansonia;
В— Culex; Г— Orthopodomyia. 1 —
венчик.
множество узких выростов в виде крошечных столбиков, вершины кото-
рых, соприкасаясь друг с другом, образуют мозаику верхней поверхности.
У A. bifurcatus, A. algeriensis и A. plumbeus верхняя поверхность яйца одно-
образно темная; у A. maculipennis есть рисунок из темных и светлых пя-
тен, который имеет значение для определения подвидов. Наличие рисунка
на верхней поверхности яйца связано с различной высотой столбиков эк-
зохориона: там, где они ниже и мельче,
яйцо кажется темнее, там, где они шире
и выше, яйцо кажется светлее.
На выпуклой поверхности расши-
ренного конца яйца находится микро-
пиле — отверстие, через которое в мо-
мент откладки происходит оплодотво-
рение яйца.
2. Яйца Culicini
Отдельное яйцо комара рода Culex
имеет удлиненную форму с закруг-
ленными концами; один конец — бо-
лее широкий и тупой, другой — не-
сколько суженный (рис. 89, В). На
переднем, расширенном конце каждого
яйца имеется блюдцеобразный венчик,
при помощи которого яйцо удержи-
вается на поверхности воды в верти-
кальном положении, расширенным кон-
цом книзу, чем и обусловливается форма лодочки с выпуклой нижней и
вогнутой верхней поверхностью (рис. 90). Так как отдельные яйца
Рис. 90. Кладка Culex pipiens.
в лодочке склеены друг с другом боковыми поверхностями и ориентиро-
ваны к поверхности воды вертикально, то при рассматривании сверху
лодочка имеет мелкоячеистое строение; в этом случае мы видим только
обращенные кверху концы яиц. Яйца Aedes имеют овальную или эллипсо-
идальную форму. На одном из концов яйца находится микропиле. Оболочка
яиц имеет сетчатый рисунок, образованный экзохорионом.
159
Подвиды Anopheles
Морфологические отличия* Эколого-биологиче
Группы подвидов и подвиды яйцо личинки |
воздушные камеры 1 рисунок на верхней поверхности
А. Группа maculipennis: A. maculipennis Mg. (typicus) Большие. Занимают около 60% длины яйца. Верхняя поверхность межреберной пере- понки морщинистая. У кавказской формы встречаются яйца с камерой меньших раз- меров (20—25% дли- ны яйца) и гладкой верхней поверхностью межреберной перепон- ки Две темные попе- речные полосы на уровне концов воз- душных камер; тем- ные пятна у каждого полюса. В промежут- ках между темными поперечными полоса- ми изредка одно-два темных пятна. Ос- тальная поверхность однородно снежпобе- лого или пепельносе- рого цвета Как в стоячих, так и в проточных водо- емах, иногда в лужах на галечниковых от- мелях, в руслах гор- ных речек I
Б. Группа messeae: a) A. mac. mes- seae Fall. I Хорошо развиты, занимая более 40% длины яйца. Верхняя поверхность меж ре- берной перепонки гру- бо исчерчена, мор- щиниста Поперечные темные полосы неотчетливо выделяются, являясь частью рисунка из темных пятен, встре- чающихся по всей по- верхности яйца В стоячих или мед- ленно текучих водо- емах , пресноводных заболоченностях, ри- совых полях
б) A. mac. me- lanoon Hack в) A. mac. sub- alpinus Hack. a. Le- wis 1 Камеры меньше. Межреберная пере- понка гладкая или с редкими поперечны- ми линиями Окраска однооб- разно темная, без ри- сунка. Отчетливые попе- речные темные поло- сы вблизи концов ка- мер и несколько тем- ных пятен в проме- жутке между попе- речными полосами. Общий фон яркобе- лый; рисунок кон- трастный В больших заболо- ! ченностях и в при- I брежной полосе боль- ших озер и прудов, на рисовых полях
В. Группа labranchiae-atro- parvus A. mac. atroparvus v. Thiel. Хорошо развиты, но меньше, чем у messeae. Верхняя поверх- ность межреберной перепонки гладкая или с немногими по- перечными линиями вблизи концов камер Рисунок образован темными клинообраз- ными пятнами, отхо- дящими от краев яй- ца и суживающимися к середине; иногда, сливаясь, пятна рас- полагаются попереч- ными полосами, но и в этом случае их рас- положение не связа- но с концами камер Солевыносливость большая, большая вы- носливость к азоти- стому загрязнению. В пресноводных и солоноватых стоячих водоемах морских побережий, плавней, степей
Возможно, является одной из форм изменчивости A. m. melanoon.
160
maculipennis Mg.
Таблица 3
с кие отличия 1
имаго Эпидемиологическое Распространение значение в пределах СССР
Копуляция на лету; зи- мовка в холодных или в теплых помещениях, в за- висимости от климата мест- ности; возможно питание кровью в зимнее время В Европейской части Советского Союза пере- носчик малярии там, где встречается в большом количестве. Основной и во многих местах един- ственный переносчик на Кавказе; поддерживает тяжелые эндемические очаги В Европейской части Со- ветского Союза найден во многих местах, по числен- но уступает A. m. messeae. Северная граница совпада- ет с границей вида; на юг идет дальше, чем А. ш. messeae (Закавказье, Север- ный Иран, Копет-Даг), на ; восток—не дальше Томска. На Кавказе преобладающая форма, а в горах — един- ственная
Копуляция на лету; зимовка в холодных поме- щениях; зимой крови нор- мально не пьют; имаги- нальная диапауза при на- рушенном пищеварении. Менее теплоустойчивая форма, чем другие подвиды Основной переносчик малярии на большей ча- сти территории Евро- пейской части Советско- го Союза и почти един- ственный в Сибири Северо-восточная форма. На севере естественной гра- ницей является южный край тайги, по с колони- зацией тайги вместе с че- ловеком продвигается даль- ше на север. На востока' встречается до Зейско-Бу рейнской низменности; на западе выходит за западные границы Советского Союза. Южная граница не везде изучена: доходит до север- ных склонов Киргизского хребта: в Предкавказье до берегов Терека и Кубани
Копуляция на лету. Зимуют в теплых поме- щениях, возможно питание кровью зимой Повидимому, важней- ший переносчик на влажных приморских низменностях Кавказа, в особенности в Ленко- рани Средиземноморская фор- ма, у пас распространенная главным образом на низ- менностях Кавказа (Абха- зия, Адлер, Дербент, Лен- корань, Колхида, Кахетия, Кабарда)
Копуляция в сидячем по- 1 ложении и на лету; легко : размножается в неволе; зи- i мовка в теплых помещени- ях; нормально зимнее пи- тание кровью; при имаги- нальной диапаузе пищева- рение нормальное Переносчик в Приазо- вье и везде, где встре- чается в большом числе По течению Днепра от устья до Орши, в Донбас- се, па Черноморском побе- режье, в Приазовской низ меппости и на Северном Кавказе: до Гудермеса и Адлера — на юг и до Эли- сты — на Восток
161
И Учебник медицинской энтомологии
Морфологические отличия Энолого-биологиче
Группы подвидов и яйцо
подвиды 1 воздушные камеры | рисунок на верхней поверхности личинки
Г. Группа sacharovi: A. mac. sacharovi Favre Отсутствуют или рудиментарны Одноцветно белое Солевынослив; ме- нее чувствителен к азотистому загрязне- нию
в) Отличия подвидов Anopheles maculipennis по яйцам
Среди видов Anopheles в СССР самый распространенный и самый
важный — A. maculipennis. A. maculipennis — не однородный вид; он
состоит из ряда подвидов. Подвиды отличаются между собой морфологи-
чески (главным образом по структуре оболочки яйца) и биологически,
благодаря чему они играют несколько различную роль и в эпидемиологии
малярии.
В СССР известно пять подвидов A. maculipennis: A. mac. maculipen-
nis Mg. (typicus) и его кавказская разновидность, A. mac. messeae Falle-
roni, A. mac. melanoon (subalpinus Hack, et Lewis), A. mac. atroparvus
v. Thiel и A. mac. sacharovi Favre.
Основным отличительным признаком подвидов A. maculipennis
является рисунок на верхней поверхности яйца (рис. 91) и характер верх-
ней поверхности межреберных перепонок, которая может быть гладкой,
блестящей или же с грубой поперечной или сетчатой исчерченностью
(рис. 88, В). Сводка отличий подвидов A. maculipennis дана в табл. 3.
3. ЛИЧИНКА
а) Наружное строение личинки
Тело личинки расчленяется на голову, грудь и брюшко (рис. 92)
Наружный покров тела образован кутикулой, непроницаемой для воды
(за исключением анальных жабер, проницаемых для воды и растворенных
в ней газов).
Кутикула несет многочисленные и разнообразные по строению во-
лоски и шипы. Волоски несут чувствительную (рис. 9) и двигательную
функцию. При помощи волосков личинка улавливает малейшие ко-
лебания в окружающей среде, реакцией на которые является быстрая пе-
ремена местоположения.
Для личинок Anopheles, живущих в поверхностном слое воды, харак-
терны укороченные или специально видоизмененные волоски спинной по-
верхности (например, пальмовидные, рис. 92, 3) и, наоборот, сильно раз-
162
Таблица 3 (продолжение)
сние отличим
имаго Эпидемиологическое значение Распространение в пределах СССР
Копуляция на лоту; зи- ( мовка в теплых и холод- 1 ных помещениях; в теплых 1 помещениях нормально ! зимнее питание кровью; 1 при имагинальной диапау- зе пищеварение нормаль- I ное. Наиболее теплолюби- вая и теплоустойчивая фор- ма Виновник тяжелых эндемий в некоторых мостах Средней Азии (Ферганская долина, Ташкентский оазис, до- лина Зеравшана) и на рагнинах Восточного Закавказья В Средней Азии един- ственная форма. На север идет до Аральского моря и Киргизского хребта, на Ка- спийском побережье Кавка- за к северу—до Махач-Кала. Сухие равнины восточного Закавказья. Не встречается в чрезмерно влажных мест- ностях (Черноморское по- бережье Кавказа, Ленко- рань)
витые боковые волоски; последние увеличивают трение поверхности тела
личинки о воду, и, следовательно, сопротивление тела падению под влия-
нием силы тяжести, и способствуют парению нырнувшей личинки в толще
воды. Для большинства волосков личинки Anopheles характерен перистый
тип ветвления. На теле личин.ки встречаются также темные, склероти-
зованные пластинки (склериты)н Их расположение обычно приурочено к
местам прикрепления мышц (спи ные пластинки брюшка у Anopheles, седло
9-го сегмента).
Голова. Форма головы личинки связана со способом питания. У ли-
чинок Anopheles и Dixa, питающихся с поверхности воды, общая форма
головы удлиненная в связи с продольным расположением мускулатуры
ротовых частей. У личинок Culicini голова сильнее развита в ширину, чем
в длину, что связано с диагональным расположением мышц, характерным
для питания планктоном и с субстрата. Голова обычно выпуклая со спин-
ной и уплощена с брюшной стороны. У хищных личинок форма головы
значительно видоизменена. Так, у Chaoborinae голова вытянута в длину
и у некоторых форм сужена, при этом основания антенн выдвинуты к пе-
реднему краю головы и сближены. У взрослых личинок относительные
размеры головы меньше, чем у ранних стадий. Покров головы толще, чем
на других частях тела. Головная капсула образована несколькими скле-
ритами: лобным (фронтальным), двумя щечными, горловым и затылочным
(воротничок) (рис. 93); спереди к лобному склериту примыкает наличник,
или клипеус.
Места прикрепления мышц, двигающих ротовые части и антенны,
к головной капсуле видны как темные пятна на поверхности головы, так
как эти участки кутикулы более уплотнены. Голова личинок несет до
20 пар волосков. В систематике, для различения видов имеют значение сле-
дующие волоски. На переднем крае фронтального склерита у Anopheles
имеется по паре наружных и внутренних клипеальных волосков; у осталь-
ных личинок Culicidae они обычно отсутствуют. Позади них помещается
пара слабее развитых задних клипеальных волосков. Следующий ряд
образуют три пары обычно мощно развитых фронтальных (постантенналь-
ных) волосков. В задней половине головы расположено три пары неболь-
11*
163
тих волосков: глазные, теменные и транссутуральные (по-латыни sutura —
шов, trans — по ту сторону; «транссутуральные» — находящиеся за швом).
По бокам головы расположены глаза. У личинок 1,11 и в начале III во-
зраста имеется одна пара простых глазков; это небольшие округлые пиг-
ментные пятна, которые остаются и у взрослого комара, прикрытые чешуй-
ками. У старших личинок (в конце III возраста и в IV возрасте) появляется
вторая пара — сложные глаза, являющиеся зачатками сложных глаз!
взрослого комара. Они полулунной формы и огибают глазки спереди и
со спинной стороны.
Рис. 93. Голова 'личинки Anopheles maculipenni
IV возраста (со спинной стороны).
А — фронтальный склерпт; Б — затылочный склерит
(«воротничок»): 1— антеннальный волосок; 2 и 3—
внутренний и наружный клипеальные волоски; 4 —
задний клипеальный волосок; 5, 6 и 7 — фронтальные
волоски; 8 —теменной волосок; 9—транссутуральный
волосок; 10—волосок на нижнечелюстном щупике.
Рис. 92. Личинка Anopheles
(вид сверху).
1—усик (антенна); 2— плечевые
лопасти; 3—пальмовидные волоски;
4— дыхальце (увеличение ХЮ).
Голова личинок несет придатки: усики (антенны) и ротовые органы
Антенны одночленистые и сравнительно мало подвижные (за исключением
Chaoborinae). Длина антенн у различных личинок комаров бывает различ-
на, от четверти длины головы до величины, в два раза ее превышающей, что
связано со способом питания. Наиболее длинные антенны бывают у личи-
нок, питающихся планктоном. У личинок, питающихся с поверхностной
пленки или обскребывающих подводные предметы, антенны короткие,
у хищных личинок Chaoborinae антенны сильно развиты и подвижны и слу-
жат для схватывания добычи. На вершине антенн имеются чувствительные
щетинки и другие органы чувств. На теле антенны расположен антенналь-
ный волосок, который может иметь у разных видов различное строение.
Строение ротовых частей. Строение ротовых частей личинок комаров
еще теснее, чем строение антенн, связано с типом питания. По способу пи-
тания личинки Culicidae делятся на фильтраторов, обскребы вате лей и
165
хищников. При первом способе питания различают питание с поверхност-
ной пленки, характерное для Anopheles и Dixinae, и питание изболев глу-
боких слоев воды взвешенными в воде частицаАми (планктонный тип пита-
ния), характерное для личинок Culex, Mansonia и некоторых Аёйез и Theo-
baldia (Мончадский). Питание обскребыванием, т. е. путем соскабливания
обрастаний из микроорганизмов с поверхности различных подводных пред-
метов, характерно для многих личинок A6des, Theobaldia и др.
У хищных личинок (подсемейство Chaoborinae), питающихся более крупной добы*
чей, все строение ротового аппарата приспособлено для захватывания и разжевывания
пиши и резко отличается от строения ротового аппарата фильтраторов и обскребыва-
телей.
Рис. 94. Голова личинки Anopheles IV возраста (с брюшной стороны).
1 — щетки; 2 — медиальная лопасть; 3 — подглоточник; 4 — мандибулы;
6 — максиллы; 6 — нижняя губа.
Каждая из групп волосков надглоточника изображена только на одной
какой-либо его стороне, т. е. либо справа, либо слева (оригинал).
Ротовой аппарат всех личинок комаров состоит из верхней губы с над-
глоточником, пары верхних и пары нижних челюстей, нижней губы и
подглоточника. Верхняя губа причленяется к преклипеусу (рис. 94,95) и
несет парные щетки, непарную срединную (медиальную) лопасть и над-
глоточник. У личинок Anopheles каждая щетка представляет пучок тон-
ких, гибких волосков, способных к быстрым ритмическим движениям. При
сокращении мускулов, управляющих щетками, волоски загибаются на
брюшную сторону, при их расслаблении они расправляются и выступают
впереди головы в виде желтого веера. Между щетками находится медиаль-
ная лопасть, свободный край которой несет ряд крупных, крепких, зазуб-
ренных шипов. Во время работы щеток эта лопасть пассивно следует за их
движением. Функция щеток, а у некоторых форм и медиальной лопасти —
166
вызывать в поверхностном слое воды токи и направлять их вместе с пище-
выми частицами ко рту. У личинок Anopheles медиальная лопасть с ее
зазубренными волосками служит органом добывания пищи при помощи
обскребы вания.
У личинок, питающихся планктоном (рис. 06, Л), волоски в каждой щетке рас-
полагаются двумя пучками: наружным, более мощным и длинным, направленным вбок,
и внутренним, более коротким, Направленным вперед и внутрь; щетки широко рас-
ставлены, медиальная лопасть имёёт вид широкого полукруга. У типичных обскребы-
вателей (рис. 96, Б) каждая Щётка состоит из одного пучка неоднородных волосков*
наружная часть пучка состоит из тонких волосков, а внутренняя — из S-образно
изогнутых, более коротких и толстых, зазубренных на концах волосков, служащих
для соскребывания пищи.
На брюшной стороне верхней губы (рис. 95, А) несколько позади ме-
диальной лопасти, находится выступ, свешивающийся над ротовым от-
верстием; это надглоточник. Он несет многочисленные волоски и шипы
различного строения. Шипы надглоточника располагаются рядами, об-
разуя целые гребенки. Пара мускулов приводит надглоточник в движение,
причем движения его совпадают со взмахами щеток. Основное назначение
надглоточника: 1) совместно со щетками верхней губы направлять ток воды
к ротовому отверстию; 2) препятствовать обратному выскальзыванию из
ротовой полости попавших в нее пищевых частиц (при помощи барьера
из волосков и шипов «гребенки»); 3) периодически очищать щетки от при-
ставших к ним частиц. Надглоточник образует переднюю границу рото-
вой полости; с боков она отграничена верхними челюстями.
Верхние челюсти, или мандибулы (рис. 95, Б),—массивные оваль-
ные пластинки; к ним прикрепляются мускулы, приводящие их в движе-
ние. Внутренний край челюсти несет хитинизованные зубы, шипы и во-
лоски, строение которых различно у разных родов и видов личинок,
в зависимости от способа их питания. У личинок Anopheles зубы хорошо
развиты, крепкие шипы несут на концах гребни или заострены.
У типичных обскребывателей шипы образуют по внутреннему краю мандибул
гребень. У планктоноядных форм вместо шипов имеется ряд длинных волосков. При
питании зубы мандибул совместно с выростами подглоточника производят раздроб-
ление крупных частиц. Шипы мандибул играют роль гребня, сквозь который проче-
сываются волоски щеток и счищаются приставшие к ним частицы; они служат также
для проталкивания крупных частиц в глотку. Пучки длинных волосков образуют сво-
его рода фильтр, препятствующий попавшим в ротовую полость пищевым частицам
уноситься обратным током воды; кроме того, они счищают с внутренней поверхности
нижних челюстей приставшие к ним частицы пищи.
Нижние челюсти (максиллы) (рис. 95, В) помещаются за мандибулами
и отграничивают ротовое отверстие с боков и частично с брюшной стороны.
Максилла состоит: из крупной пластинки четырехугольной (у Anopheles)
или круглой (у планктоноядных) формы, образующей тело максиллы, из
небольшого базального склерита и из одночленистого нижнечелюстного
щупика. Внутренняя створка (стенка) максиллы покрыта многочислен-
ными и разнообразными по величине и строению волосками, служащими
фильтром, задерживающим частицы, поступающие в ротовую полость.
У Anopheles щупик хорошо развит; верхний конец щупика несет чув-
ствительные выросты, отсутствующие у других личинок Culicidae; они
являются приспособлением к особенностям питания Anopheles.
У личинок-обскребывателей строение максилл и степень развития волосков на
них сходны с таковым у Anopheles, но щупик развит слабее. Максиллы у этих форм,
так же как и у Anopheles, слабо подвижны. У личинок с резко выраженным планктон-
ным типом питания максиллы конической формы, с длинным пучком волосков на вер-
шине, с длинными волосками вдоль внутреннего края и с очень слабо развитым щупи-
167
4
Рис. 96. Строение ротовых частей личинок Culicini, питающихся
планктоном (А) и обскребыванием (В).
1— верхняя губа; 2— мандибулы; .?—максиллы.
ком; они очень подвижны и служат вместе с верхней губой для привлечения пищи ко
рту. У хищных личинок (Megarrhinini, Chaoborinae) максиллы, в связи с малой ролью
их при питании, в той или иной степени редуцируются.
Нижняя губа (рис. 95, Г) расположена между основаниями максилл и
образует вместе с подглоточником брюшную стенку ротивэй полости. Эта
сложно устроенная, слегка подвижная треугольной формы трехслойная
пластинка несет по краям мощно развитые зубы; вместе с мандибулами
она участвует в схватывании и раздроблении пищи и проталкивании ее
в глотку. У основания нижней губы открывается непарное отверстие про-
токов слюнных желез. Кзади от нижней губы, несколько вглубь ротовой
полости, расположен подглоточник в виде хитинизованной, сильно бу-
гристой пластинки, образующей дно ротовой полости. Над ним скапли-
вается пищевой комок.
Описание механизма действия ротового аппарата при питании см.
на стр. 228.
Грудь. У личинок Culicidae, за исключением Dixinae, все три груд-
ных членика слиты вместе и образуют наиболее широкий отдел тела.
В груди находится шесть пар крупных имагинальных дисков (зачатки ног
и крыльев взрослого насекомого); размеры и строение дисков с раз-
витием личинки меняются. Три пары дисков на брюшной стороне впослед-
ствии дают начало трем парам ног, три пары на спинной стороне — зачат-
ки крыльев, жужжалец и дыхательных трубок куколки. Имагинальные
диски начинают интенсивно развиваться с III возраста; у личинок 1Увоз-
раста, в связи с их развитием, грудь сильно разбухает. В конце IV воз-
раста окончательно сформировываются дыхательные трубки куколки,
которые, просвечивая через кутикулу, являются признаком готовой к
окуклению личинки.
У личинок Anopheles по бокам переднего края спинной стороны гру-
ди помещаются плечевые лопасти (рис. 97, А) — полые прозрачные выросты
стенки тела. При помощи специального мускула они могут втягиваться
внутрь, а при его расслаблении и благодаря давлению гемолимфы и упру-
гости кутикулы, расправляться и выступать из полости. Они служат ли-
чинке Anopheles для прикрепления к поверхностной пленке воды. У осталь-
ных личинок комаров эти образования отсутствуют.
На груди имеются волоски, расположенные тремя поясами, соответ-
ственно трем слившимся членикам груди. Некоторые из них имеют зна-
чение в систематике (т. е. служат для различения родов и видов личинок
комаров).
Брюшко. Брюшко личинки состоит из девяти члеников. Первоначаль-
ное число члеников брюшка у личинок комаров равнялось десяти. Это пол-
ное число члеников брюшка, наряду с расчлененной грудью, в настоящее
время сохранили только личинки Dixinae. У остальных личинок Culici-
dae первоначальные 8-й и 9-й членики в процессе эволюции слились, об-
разовав сложный 8-й членик. Первые семь члеников сходны по величине.
Каждый членик брюшка несет волоски, из которых наиболее развиты
боковые (плевральные), способствующие парению личинок в толще воды.
На спинной стороне каждого членика брюшка, от 1-го до 7-го, личинка
Anopheles имеет парные пальмовидные или звездчатые волоски. На первом
членике они рудиментарны. У большинства личинок Anopheles из подрода
Myzomyia они хорошо развиты, начиная со 2-го членика, у остальных —
с 3-го. Каждый волосок состоит из стержня и многочисленных листовидных
пластинок (рис. 97, Б), в раскрытом состоянии расположенных в плоско-
сти, параллельной спинной поверхности брюшка. Назначение этих воло-
сков, подобно плечевым лопастями стигмальнойпластинке, прорывать по-
170
верхностную пленку и силами поверхностного натяжения удерживаться
на поверхности воды, тем самым удерживая личинку в горизонтальном по-
ложении. При погружении личинки вглубь лепестки волосков смыкаются,
захватывая между собой пузырек воздуха, при посредстве которого проис-
ходит разрыв поверхностной пленки и раскрывание лепестков при
подъеме.
Стигмальная пластинка. В строении 8-го и 9-го сегментов имеется ряд
особенностей, отличающих их от первых семи сегментов. На спинной сто-
Рис. 97. Органы прикрепления личинки Anopheles к поверх-
ностной пленке воды.
А— плечевые лопасти на переднем крае переднегруди; Б—паль-
мовидный волосок спинной стороны брюшка; В— стигмальная пла-
стинка: 1 — стигмы; 2 — стигмальные отростки; 3, 4 и 5 — задняя,
боковая и передние лопасти; 6' — центральная пластинка.
роне 8-го сегмента у личинок Anopheles и Dixinae на особой стигмальной
пластинке помещается пара дыхалец — стигм. Стигмальная пластинка
личинок Anopheles (и Culicini), подробно изученная А. С. Мончадским,
снабжена пятью лопастями: непарной передней, одной парой боковых и
одной парой задних (рис. 97, В) и поддерживается в горизонтальном поло-
жении при помощи специального образования — темной хитиновой дуги,
окружающей основание задних лопастей снизу, так, что их нижние поверх-
ности опираются на дугу. По бокам 8-го сегмента концы дуги переходят
в пигментированную треугольную хитиновую пластинку, задний край
которой несет хорошо развитые шипы, гомологичные гребню сифона ли-
171
чинок Culicini. Функция стигмальной пластинки рассматривается ниже
(стр. 208).
У личинок Culicini (и у некоторых родов подсемейства Chaoborinae}
от спинной поверхности 8-го сегмента отходит дыхательная трубка —
сифон, на вершине которой открываются дыхальца, окруженные стигмаль-
ной пластинкой (рис. 98). Длина и ширина сифона различны у разных ви-
Рис. 98. Личинка Culicini (Аёдев cine-
reus) (вид сверху). VIII сегмент пока-
зан повернутым па 90° для того, что-
бы были видны анальный сегмент и
щетка VIII сегмента.
1 — глаза; 2 — усики; 3 — щетки верхней
губы; 4 — голова; 5 — грудь; 6 — брюшко;
— гребень; 8 — сифон; 9 — стигма льная
пластинка; 10— пучок волосков; 11— хво-
стовые волоски (спинной стороны); 12 —
жабры; 13 — рулевые волоски (брюшной
стороны); 14—анальный сегмент; 15—
щетка.
Рис. 99. 8-й и 9-й членики брюшка ли-
чинки Anopheles I и IV возраста.
Схематизовано.
1 — чешуйки щетки у личинки I возраста;
2 — аппарат, поддерживающий стигмальную
пластинку (вид сбоку); 3 — седло; 4—5— хво-
стовые волоски; 6 — жабры.
дов. По бокам задней поверхности сифона, начиная от основания, имеется
два продольных ряда зубцов или шипов — гребни, строение которых раз-
лично у разных видов. На сифоне имеются также волоски, располагающиеся
большей частью на задне-боковой поверхности сифона в числе от одной до
восьми пар. Расположение и число этих волосков являются важнейшим
родовым признаком.
У личинок Culicini по бокам 8-го сегмента под основанием сифона
помещаются щетки, состоящие из чешуек, число и строение которых раз-
личны у разных видов. Щетка каждой стороны окружена пятью волосками.
172
У личинок Anopheles, начиная со II возраста, щетка отсутствует;
у личинок I возраста она представлена пятью-шестью длинными чешуйка-
ми.
Анальный сегмент. Последний (анальный) сегмент брющка несколь-
ко уже остальных и отогнут на брюшную сторону. На спинной стороне он
несет хитиновую пластинку — седло (рис. 99), которое у некоторых видов
обхватывает весь сегмент в виде кольца. Седло придает сегменту прочность,
необходимую при движении личинки. Анальный сегмент вместе с рулевыми
волосками, или плавником, расположенными на его брюшной стороне,
служит главным органом движения личинок. Плавник состоит из ряда силь-
но развитых ветвящихся волосков, основания которых сближены и располо-
жены в один зигзагообразный ряд. На спинной стороне последнего сегмен-
та, позади седла помещается две пары хвостовых волосков. Обе пары волос-
ков несут чувствительную и двигательную функции, а наружные у личи-
нок Anopheles крючкообразно изогнуты на концах и служат для прикрепле-
ния к стенкам сосуда, водяным растениям и другим предметам. Вокруг
анального отверстия расположены две пары жабер, длина и форма которых
различна у разных видов. Жабры представляют выросты, покрытые тон-
кой, проницаемой для воды кутикулой. Полости жабер сообщаются с по-
лостью тела личинки; они заполнены гемолимфой; в каждом из жаберных
мешочков разветвляется трахея.
б) Возрастные различия личинок Anopheles
В процессе роста, от выхода из яйца до окукления, линейные размеры
личинок A. maculipennis, как указывалось, увеличиваются в 8,3 раза,
при этом личинка проходит через четыре возраста, или стадии, разделенных
линьками. Рост личинки происходит как в периоды линек, так и в проме-
жутках между ними в течение всего периода личиночной фазы. На практике
для определения возраста личинок обращается внимание на соотношение
между шириной головы и размерами тела (длиной) (Каганова и Мончад-
ский). Определять возраст только по длине тела нельзя, так как по одному
этому признаку не всегда возможно отличить соседние стадии. Дело в том,
что грудь и брюшко личинки, покрытые тонким хитином, растут непрерыв-
но в течение отдельных личиночных фаз и почти не испытывают прироста
в моменты линек. Поэтому свежеслинявшие личинки IV возраста могут
иметь длину тела, одинаковую со взрослыми личинками III возраста. На-
оборот, части тела, покрытые твердыми хитиновыми покровами (голова,
стигмальная пластинка), увеличиваются только во время линек. В про-
межутках между линьками размеры головы не изменяются, так как твер-
дые хитиновые покровы головы не способны после затвердения растяги-
ваться. В силу таких особенностей роста головы ее ширина для каждой
личиночной фазы остается постоянной и подвержена лишь индивидуаль-
ной изменчивости, не связанной с ростом внутри фазы. Однако длина го-
ловы увеличивается и между линьками, вследствие роста в длину воротнич-
ка (Митрофанова). Воротничок образуется и растет путем утолщения ку-
тикулы шеи, примыкающей к голове. У новорожденной личинки I возраста
воротничок имеет вид узкой полоски, а у готовой к линьке личинки этого
возраста он достигает 70% длины головы (рис. 100). После линьки вновь
образовавшаяся головная капсула опять лишена воротничка, но к концу
стадии он вновь нарастает. Подобные изменения в росте воротничка проис-
ходят и у личинок III и IV возраста, но у них прирост воротничка по от-
ношению к длине головы значительно меньше, чем у младших возрастов.
У личинок Culicini роста воротничка не происходит.
173
Методика определения возрастного состава личинок Anopheles в полевых условиях
построена на изложенных закономерностях роста личинок. В основе ее лежит опре-
деление возраста по ширине головы; изменение размеров головы у личинок I—IV
возраста показано на рисЛЮО. Для определения возраста личинок необходимо знать
абсолютные размеры головы в каждом возрасте. При некотором навыке различия в
размерах головы каждого возраста легко улавливаются глазом. Определение возраста
личинок на практике производится следующим образом. Пробу с личинками выливают
в сосуд, из которого личинок вылавливают пипеткой по возрастам; при этом подсчиты-
вают число личинок каждого возраста. В случае присутствия в пробе личинок всех че-
тырех возрастов, что обычно и бывает на практике, прежде всего вылавливают самых
крупных личинок, с наибольшей шириной головы — личинок IV возраста, затем са-
мых мелких, с наименьшей шириной
головы—личинок 1 возраста; осталь-
ные личинки, независимо от разме-
ров их тела, на-глаз разбиваются на
две группы: одну с большей шириной
головы, чем у личинок I возраста
(II возраст), другую — с меньшей,
чем у личинок IV возраста (III воз-
раст). При небольшом навыке и
тренировке глаза этот способ дает
возможность быстро и безошибочно
определять возраст личинок в по-
левой обстановке.
Иногда требуется более
точное определение возраста
личинки—различить более ста-
рых и более юных личинок среди
личинок каждого отдельного воз-
раста или фазы. Для такого более
точного определения возраста у
личинок I—III фазы пользу-
ются измерением воротничка;
при определении возраста у
личинок IV фазы, у которых
прирост воротничка незначите-
лен, обращают внимание на со-
отношение между шириной го-
ловы и шириной груди: у взрос-
лой личинки IV фазы, в связи
с усиленным ростом имагиналь-
ных дисков, грудь разрастается
Рис. 100. Рост головы личинки и становится значительно шире
A. maculipennis. головы. Кроме того, для опре-
Слева — начало ф'зы (без воротничка), справа — ко- деления ТОЧНОГО возраста ЛИЧИ-
Н’Ц фазы (с ворогвичком) (но Ю. Г. Митре ф шивсй). цОк IV фазы МОЖНО ПОЛЬЗО-
ваться общей длиной тела, так
как за время IV фазы личинка вырастает в длину почти вдвое, и, таким
образом, личинки, находящиеся в начале IV фазы, много меньше, чем
личинки, готовые к окуклению. Однако надо помнить, что размеры ли-
чинки зависят от предшествующего питания и т. д.
Кроме изменений размеров тела, каждая из фаз отличима по строению и степени
разветвления волосков, по числу волосков в плавнике, по числу лепестков пальмовид-
ных волосков и т. п. Как правило, с каждой последующей стадией усиливается ветви-
стость волосков, увеличивается число шипов. Особенно резко отличаются, личинки
I возраста; у них полностью отсутствует плавник и аппарат, поддерживающий стиг-
мальную пластинку (рис. 99); все волоски на их теле простые, неразветвленные
(за исключением фронтальных) (рис. 101); пальмовидные волоски рудиментарны. Для
них характерно также наличие «яйцевого зуба» — твердого конусовидного образо-
174
Рис. 101. Личинка А.
bifurcatus, начало I воз-
раста.
1 — «яйцевой зуб».
вания, расположенного посредине близ заднего края фронтального склерита и служа-
щего личинке при вылуплении для разрыва оболочки яйца. У личинок I и II фазы
отсутствует волосок на нижнечелюстном щупике, который появляется только с
III фазы.
В оперативной практике для определения возраста изменениями в
степени развития волосков не пользуются, так как эти признаки требуют
применения микроскопа и потому не позволяют
иметь дело с массовым материалом.
Каждая линька является критическим пери-
одом в жизни личинки. К периодам линьки при-
урочена наибольшая смертность личинок. Уже за
несколько часов до линьки под кожей личинки
становятся видимы волоски следующей фазы,
обернутые вокруг тела под старой шкуркой; по
этому признаку можно судить о близости линьки.
Перед линькой из III возраста в IV появляются
сложные глаза, характерные для IV возраста.
Перед линькой IV возраста в куколку через
покровы груди просвечивают дыхательные труб-
ки куколки.
Только что слинявшие личинки отличаются
слабой пигментацией наружных покровов (осо-
бенно головы); тотчас после линьки голова белая,
полупрозрачная и по отношению к груди ка-
жется непропорционально большой.
Определение возраста личинок бывает необ-
ходимо для установления сроков противоличипоч-
ных мероприятий. Более точное определение в
пределах I возраста может характеризовать
эффективность опыления ядами: если спустя 1—2
дня после опыления водоема мы находим в нем
личинок I возраста без воротничков, то мы можем
заключить, что это свежевылупившиеся личинки,
вышедшие из яиц после того, как яд утонул,
и наличие их не говорит о плохом качество опыле-
ния; если же встретятся личинки с большим во-
ротничком, мы считаем, что они пережили опыление, что указывает на
недостаточную эффективность последнего.
4. КУКОЛКА
Куколка состоит из нерасчлененной широкой головогруди и узкого
брюшка из 9 сегментов, обычно подогнутого под головогрудь. Сквозь
прозрачные покровы головогруди просвечивают голова и грудь развиваю-
щегося комара и их придатки (ротовые части, глаза, конечности, крылья).
На спинной стороне головогруди расположена пара дыхательных трубочек,
суженные основания которых соединяются с трахейной системой. Нормаль-
но куколка держится у поверхности воды, выставляя стигмы в атмосферу.
Форма дыхательных трубок у Anophelini и Culicini различна и является
их различительным признаком. У Anophelini дыхательная трубка кониче-
ской формы, у Culicini — цилиндрической (рис. 102). На спинной стороне
первого членика брюшка находится пара древовидных волосков, служащих
для восприятия колебаний водной поверхности. Остальные членики брюш-
ка свободно подвижны. На брюшке имеются многочисленные волоски, рас-
положение и строение которых постоянны для отдельных видов. Система-
175
тическое значение имеет строение волосков, расположенных в задне-бо-
ковых углах 3—8-го сегмента. У Culicini они ветвисты, у Anophelini пред-
ставлены массивными шипами на 3—7-м сегменте и перистыми — на 8-м
(рис. 102). От заднего края 8-го сегмента отходит пара широких листовид-
ных пластинок — плавников. Короткий 9-й сегмент, расположенный с
брюшной стороны плавников, несет придатки, являющиеся зачатками на-
ружных половых органов будущего комара. По размерам и форме этих
придатков можно определить пол куколки (рис. 102, 4).
2 3
Рис. 102. Куколка комара.
1— общий вид сбоку; 2— дыхательные трубки при большом увеличении: А — Anopheles; К —
Culicini; 3 — последние сегменты брюшка куколок-; 4 — зачатки наружных гениталий самца (а)
ц самки (б); 5—плавники; 6— шипы у Anopheles; 7—ветвистые волоски у Culicini.
Куколки, в отличие от личинки, удельно легче воды, что обусловлено
наличием больших воздушных полостей в головогруди и первом членике
брюшка. Они удерживают куколку в определенном положении — всегда
вверх головогрудью. В развитии куколки различают три фазы (Чинаев):
в I фазе, т. е. тотчас после превращения из личинки в куколку, внутри ку-
колки намечаются только контуры футляров придатков головы, ног,
крыльев, гипопигия г; все тело прозрачно, шипы светлые. В III фазе, перед
вылуплением комара, все придатки хорошо сформированы и видны; ши-
пы и все тело куколки темнокоричневые. Куколки, находящиеся на
II фазе, имеют промежуточные признаки.
5. ВЫЛУПЛЕНИЕ КОМАРА
В последние часы перед вылуплением комара воздушные полости ку-
колки сильно увеличиваются, так что спинная поверхность головогруди
и брюшка распрямляется и выступает над водой. Усиление давления воз-
духа влечет разрыв кутикулы на спинке головогруди. При этом нарушается
связь дыхательных трубок с трахейной системой. Комар, освобождая из
куколочной шкурки голову, начинает заглатывать воздух, вследствие чего
его брюшко раздувается и, не вмещаясь в старом футляре, постепенно вы-
тягивается из него, при помощи также и активных движений. Комар вы-
ползает из чехла, который служит ему опорой как во время вылупления,
так и спустя некоторое время после вылупления. Только что вылупивший-
ся комар бесцветен, имеет мягкие наружные покровы и наполненное воз-
духом брюшко. Через несколько минут после вылупления крылья и ко-
нечности расправляются, кутикула приобретает нормальную твердость
и окраску и комар слетает с поверхности воды.
1 То есть половых придатков, см. стр, 186.
176
ГЛАВА III
НАРУЖНОЕ СТРОЕНИЕ ВЗРОСЛОГО КОМАРА
Взрослые комары — стройные двукрылые с вытянутым телом, ма-
ленькой головой, длинным тонким хоботком, большей частью с длинными
ногами. Крылья, покрытые вдоль жилок чешуйками (рис. 103), в покое
складываются горизонтально поверх брюшка, налегая одно на другое.
Тело хрупкое, механической прочностью не отличается.
Рис 103. Различные формы чешуек, покрывающих
тело окрыленных комаров.
4.9
Рис. 104. Наружное строение Anopheles bifurcatus.
1— лоб; 2—гортань; 3—затылок; 4—глаза; 5—щека; 6 — наличник;
7 — верхняя губа; 8 — фасетки глаз; 9 — усик; 10 — подглоточник; 11 — верхние
челюсти; 12 — нижние челюсти; 13 — нижнечелюстной щупик (/—V — его члени-
ки); 14 — нижняя губа; 15 — лабеллы; 16— шейный склерит; 17— переднегрудь;
18— переднеспинка; 19—проэпимера; 20—вентральная пластинка переднегру-
ди; 21—проэпистерна; 22—тазик; 23—вертлуг; 24— бедро; 25— передняя
голень; 26 — 1—5-й членики передних лапок; 27 — коготки; 28 — средне-
грудь; 29—среднеспинка; 30 —щиток; 31 — заднеспинка; 32—мезоэпистерна
верхний отдел); 33— мезоэпистерна (нижний отдел); 34— мезоэпимера; 35 — стиг-
ма среднегруди; 36—основание крыла (крыло удалено); 37 —средний тазик;
38 — среднегрудка; 39 — спинная пластинка заднегрудки; 40 — стигма задне-
груди; 41—метаэпистерна и метаэпимера; 42—жужжальце; 43 — задний тазик;
44— задний вертлуг; 45— заднее бедро; 46 — сегмент брюшка; 47—1-й—8-й тер-
киты брюшка; 48— 1—8-й стерниты брюшка; 49—церки; 50—стигмы брюшка.
12 Учебник медицинской энтомологии
177
t! <*
Рис. 105. Голова”'АпорЬе1е8.
А—наружный вид: 1— постфронтальный шов; 2—меж антен-
нальный шов; 3— щена; 4— нлипеус: 5—нижнсчслкcifop щупик;
б—ножны (тека) нижней губы; 7—галеа нижней челюсти; 8— верх-
няя губа; 9—л^белла; 10— язычок; 11— погглотсчнин;
12— верхняя челють* 13— бич усика; 14 — торус усика;
15— основание усика; Б— внутренний скелет: 1— тело тенто-
риума; 2—задняя ветвь тепториума; 3—стебелек нижней
челюсти; 4—протон слюнных желез; 5— щека; 6— шпора;
7 — слюнный насос; 8— нижнегубные ножны; 9 — нлипеальное
углубление; 10— гипофаринкс; 11—верхняя губа; 12— крыло
клипс уса; 13 — фулькральная нла< пинка; 14 — клипсус;
15 —фулькральный рожок; 16— передняя тенториальная ветвь;
17— дорсальная тенториальная гетвь; 18 — передняя глотка;
19 — задняя глотка.
1. ГОЛОВА
Голова круглая, спереди продолжается в хоботок; несет сложные гла-
за и усики (рис. 105). Поверхность головы перед глазами носит название
лба (по-латыни—frons). На лбу часто бывает хоходок из чешуек и волосков,
направленных вперед. К нижней части лба прикрепляется наличник (cly-
peus), обычно голый. Между глазами сверху и несколько сзади располо-
жено темя (vertex). Часть головы, обращенная к грудному отделу, —• это-
затылок (occiput). Темя и затылок покрыты чешуйками и волосками. Пла-
стинки перед глазами по бокам головы — щеки (genae).
Границами отдельных пластинок с годовиой капсуле являются швы. Самый за-
метный — это венечный шоз, идущий ог затылка, продо пкающийся по лбу и разветв-
ляющийся па две боковые ветви, не доходя до усиков. Другой шоз, межантеннальный,
начинается о г наличника и, продо пкаясь вверх, дает на лбу две боковые ветви; вместе
с боковыми ветвями венечного шва они ограничивают па лбу ромбическое пространство,
боковые углы которого касаются краев глаз.
41 2. СТРОЕНИЕ РОТОВЫХ ОРГАНОВ
Хоботок с брюшной стороны причленяется к перепончатому образо-
ванию, называемому рострумом, и со спинной стороны — к наличнику.
Строение хоботка. Хоботок образован полным набором ротовых ча-
стей: он состоит из верхней губы (labrum), двух верхних челюстей (man-
dibulae), подглоточника (hypopharynx), двух нижних челюстей (maxillae)
и нижней губы (labium). От основания нижних челюстей отходят нижне-
челюстные щупики, не входящие в состав хоботка и связанные с ним только
при основании (рис. 10G).
Нижняя губа примерно в четыре раза длиннее головы, от нижней перед-
ней части которой она протягивается вперед. На верхней, или дорсаль-
ной, стороне нижней губы имеется глубокая борозда, которая вмещает
все остальные части, расположенные следующим образом: в середине ле-
жит верхняя губа; верхние челюсти прилегают к вентральным, боковым
краям верхней губы; под ними, ближе к средней линии, лежит иодглоточ-
ник. Нижние челюсти тесно прилегают к боювым стенкам срединного
ребра подглоточника. В спокойном состоянии все эти части, как в футляр,
заключены в желобок нижней губы (рис. 10'6).
Для того чтобы рассмотреть ротозой аппарат, следует выделить части его, вло-
женные в нижнюю губу. Это достигается погружением го юзы комара в кипящую 20%
щелочь (NaOU или КОИ). После промывки в воде мо кно заключить препарат в гли-
церин или канадский бальзам.
Верхняя губа представляет собой узкую и длинную пластинку, края
которой отогнуты книзу, образуя трубку, открытую на конце, по которой
при сосании кровь поступает в глотку. Только в основании этой трубки
образуется щель, которая прикрывается подглуточником. На переднем
конце с вентральной стороны верхняя губа срезана в виде пиечэго пора.
У края этого среза с нижней стороны сидят два чувствительных бугорка,
повидимому, — органы вкуса.
Каждая верхняя челюсть представляет собой тонкую выпуклую поло-
ску хитина, передний конец которой расширен в плоское лезвие. Передняя
половина наружного края этого лезвия у сам ж рода Anopheles несет
ряд очень мелких зубчиков, число юторых колеблется от 30 до 50.
Подглоточник — длинная, тонкая пластинка, являющаяся выростом
вентральной стенки глотки. Вдоль его нижней стороны проходит ребро,
содержащее проток слюнных желез. Открывается этот проток наружу со-
всем близко от заостренного конца п эдгл тгэчнлка, так что кончик послед-
него напоминает кончик шприца (рис. 106, В).
12* 179
Каждая нижняя челюсть построена из двух частей: одна из них, узень-
кая полоска, лежащая в голове, носит название стебелька и прикрепляет-
ся к внутренней поверхности затылка; спереди к ней прикрепляется вто-
Рис. 106. Строение хоботка.
А —обппй план стр оения: 1 — нижняя губа; 2 — верхняя губа; 3 — нижние челюсти; / — верх-
ние челюсти; 5 — гипофаринкс; в — нижнечелюстные щупики; Б — разрез хоботка; 1 — нижняя
губа; 2 — внешний край нижней губы; 3 — верхняя губа; 4 — канал верхней губы; 5 — нерв верх-
ней губы; в — замыкание верхней губы; 7 — верхняя челюсть; 8 — нижняя челюсть; Я —проток
слюнных желез; 10— гипофаринкс; 11 — нерв; 12 — трахея; 13 — мускулы; В — окончания частей
ротового аппарата; 1 — верхняя губа; 2 — гипофаринкс; 3 — верхняя челюсть; 4 — нижняя че-
люсть; Г—]сги01ние нижней губы во время сосания крови — разрез через комара, вонзившего
хоботок в кожу (микрофотография).
рая, наружная часть нижней челюсти, носящая название галеа. На перед-
нем конце галеа заострена и несет 8—20 боковых зубцов, концами направ-
ленных назад. Нижнечелюстные щупики прикрепляются к основанию
галеа и отходят, таким образом, от основания хоботка. Обычно они со-
180
стоят из пяти члеников; могут быть или короткими (все самки Culicini,)
или длинными (самки и самцы Anophelini и самцы Culicini) (рис. 107).
Щупики часто покрыты чешуйками.
Нижняя губа образована длинным цилиндром (тека, или ножны),
несущим на конце пару лопастинок, называемых лабеллами, и между ни-
ми—непарный язычок (лингула) (рис. 105, А, 10). На дорсальной стороне
теки находится желобок; его верхние края соприкасаются и образуют
трубку, в которую помещены все остальные части ротового аппарата.
Рис. 107. Строение щупика самцов различных родов трибы Anophelini и
Culicini.
1—Anopheles maculipennis; 2—A6des caspius; 3—Theobaldia annulata;
4 — Culex pipiens.
Тело нижней губы пронизано косо расположенными тонкими трубочками, имею-
щими значение при сгибании и выпрямлении теки. Кроме того, она содержит мускулы,
при помощи которых изгибается при сосании, трахеи и нервы. Лабеллы несколько
длиннее язычка (рис. 105, А, 9). На внутренней стороне каждая лабелла имеет мно-
жество продольных складочек, представляющих некоторое сходство с псевдотра-
хеями мух. На концах лабелл расположена густая щетка из тонких волосков. Лабеллы
соответствуют губным щупальцам других насекомых (ср. рис. 16 —- ротовые органы
таракана).
3. РАБОТА РОТОВЫХ ОРГАНОВ
Нижняя губа в силу своей массивности не может проникать под кожу
и складывается в виде петли, только обоими своими концами продолжая
охватывать пучок колющих щетинок, вонзающийся в кожу (рис. 106, Г).
Нижняя губа направляет колющие стилеты, удерживает их во время со-
сания и возвращает на место после окончания акта питания. Железки вну-
тренней поверхности нижней губы выделяют вязкую смазывающую
жидкость, обеспечивающую совместное проникновение всех стилетов,
принимающих участие в проколе кожи.
181
Нижние челюсти играют активную роль в проколе кожи хозяина.
Работая попеременно и цепляясь зубчиками за ткани хозяина, они углуб-
ляются в кожу и облегчают вхождение в кожу всего остального пучка
колющих щетинок: верхней губы с надглоточником, верхними челюстями
и подглоточпиком. Так как нижние челюсти находятся в связи со щупика-
ми, то обычно при работе челюстей приходят в движение и щупики.
По верхней губе кровь из тканей хозяина проходит в глотку насеко-
мого. Назначение верхних челюстей состоит в том, чтобы своими расши-
ренными концами закрывать отверстие верхней губы при внедрении ро-
товых частей в кожу; во время сосания крови концы верхних челюстей
оттягиваются посредством соответствующих мышц, и входное отверстие
для крови открывается.
Подглоточник содержит проток слюнных желез, открывающийся около
его вершины.
4. ОРГАНЫ ЧУВСТВ ГОЛОВЫ
Усики (antennae) (рис. 105) прикреплены впереди глаз при помощи
довольно широкого первого членика, или так называемой основной плас
тинки. Второй членик основания усиков, размером больше всех остальных,
называется торус. Остальные 13 члеников усиков составляют бич (flagel-
lum). В торусе располагается Джонстонов орган, открывающийся изо-
гнутой щелью. Предполагают, что это орган слуха комаров. У самцов
торус и Джонстонов орган развиты сильнее, чему самок; и действительно,
самцы, невидимому, обладают более тонким слухом. Бичи усиков сам-
цов и самок настолько различны, что по их строению легче всего отли-
чать самцов от самок. У самок все членики бича почти одинаковые,
цилиндрические, п несут небольшое число коротких волосков. У самцов
членики бича усиков имеют коническую форму и волоски каждого чле-
ника многочисленнее и длиннее, чем у самок. Поэтому, если самца рассма-
тривать даже без увеличения, усики его кажутся пушистыми. Два послед-
них члепика усиков самца значительно длиннее остальных. Предполагают,
что усики самцов и самок служат для обоняния и восприятия движения
воздуха.
Сложные глаза занимают боковые поверхности головы, почти сопри-
касаясь на темени. Простые глазки отсутствуют.
5. ГРУДЬ И ЕЕ ПРИДАТКИ
Как п у всех двукрылых, три членика, составляющие грудь, развиты
очень неравномерно. Среднегрудь (mesothorax), несущая крылья и за-
ключающая в себе всю летательную мускулатуру, развита сильнее всего.
Наоборот, переднегрудь (prothorax) и заднегрудь (metathorax) развиты
слабо (рис. 104).
Переднегрудь. Наружный скелет переднегруди представлен тремя пластинками:
передпеспипкой (pronotum), нроэппмерой и проэпистерной, расположенными в перед
них частях бочков груди. Переднюю вентральную часть переднегруди занимают две
сросшиеся между собой пластинки неправильной формы. За счет переднегруди обра-
зуется и довольно длинная шея комара.
Среднегрудь. Среднегрудь—это наиболее развитая часть грудного
отдела. Тергит ее (спинная пластинка) распадается на три части: 1. Средне-
спинка (scutum), покрывающая собой большую часть спинки груди.
2. Щиток (scutellum), имеющий вид поперечно-выпуклой пластинки, соеди-
няющей места прикрепления крыльев. Щиток у комаров трибы Anopheli-
ni и Megarrbinini цельнокрайний, без выступов (рис. 108). У комаров три
182
Рис. 108. Щитки Anophelini (слева) и Culicini (справа).
Рис. 109. Расположение щетинок на бочках груди Theobaldia.
1— иронотальные; 2—проэнистернальные; 3—проэпимерные;
4— дыхальцевые; 5 — стерноплевральные; 6 — пред крыловые;
7 и 8— верхние и нижние мезэпимерные.
9
Рис. 110. Расположение щетинок на бочках груди Aedes
9^— задыхальцевые щетинки; прочие обозначения см. на
рис. 109. t .
183
бы Culicini щиток трехлопастной; задний его край имеет три выступае
У Anophelini волоски образуют на щитке равномерный ряд; у Culicini
группы волосков сосредоточены на каждой из трех выпуклостей щитка.
3. К среднегруди относится и лежащая за щитком заднеспинка (post-scute]
lorn, post-notum).
На бочках среднегруди мы, так же как и в переднегруди, различаем пластинки,
лежащие впереди, — мезоэпистерны, и пластинки, лежащие сзади, — мезоэпимеры.
У комаров мезоэпистерны имеют треугольную форму и разделяются каждая на два
отдела: верхний и нижний. Мезоэпимеры в нижней, вентральной части имеют по-
перечные двойные швы, отделяющие их
от вентральной пластинки (mesosternum).
На тонкой плевральной пере-
понке между проэпимерой и верхним
отделом мезоэпистерны лежит перед-
нее грудное дыхальце.
Рис. 112. Крыло Uranotaenia.
Рис. 111. Жилкование крыла Culex и
Anopheles.
Жилки: 1- костальная; 2—субкостальная;
3, 4 и 3— первая-третья радиальные; 6—зад-
няя радиальная Р4 + 5; 7 и 8—передняя и
задняя медиальные; 9 и 10— передняя и зад-
няя кубитальные; и —анальная; 12—ра-
дио-медйалъная или средняя поперечная;
13 — медно-кубитальная или задняя попе-
речная; J4 — плечевая.
Для того чтобы л} чше ориентировать-
ся в пластинках бочков груди, лучше всего
найти переднее дыхальце и, исходя от него,
искать расположенную впереди проэпимеру
и позади от него мезоэпистерну. Большое
значение в определении комаров имеют
щетинки, расположенные на отделах груди (рис. 109 и 110). Здесь мы различаем:
1. Проэпимерные щетинки, прикрепленные к заднему (обращенному к стигме) краю
проэпимеры. 2. Дыхальцевые, имеющиеся у комаров рода Theobald ia. лежащие не-
посредственно перед передним торакальным дыхальцем. Основания этих щетинок
следует искать в одной плоскости с самим дыхальцем, в то время как проэпимерные
прикрепляются на возвышении, образованном выпуклой проэпимерой. 3. Задыхаль-
цевые прикреплены к верхнему отделу мезоэпистерны. Следует заметить, что бочки
грудного отдела бывают обычно покрыты большим числом чешуек, среди которых еле
дует уметь различать возвышающиеся над ними щетинки (более крупные и толстые)
и волоски (более мелкие и тонкие).
Заднегрудь. Заднегрудь различима с бочков грудного отдела. Представлена мета-
эпистерной и метаэпимерой. В метаэпистерне лежит заднее торакальное дыхальце.
Края его хитинизованы слабее, чем края переднего дыхальца и поэтому оно труднее
различимо.
Придатки грудного отдела. Придатками грудного отдела являются:
одна пара крыльев, рудименты второй пары крыльев в виде жужжалец и
три пары ног.
Крылья имеют удлиненно-овальную форму. Они представляют собой
прозрачную перепонку, пронизанную сетью продольных и поперечных
жилок. Часть крыла, прилегающая к месту его прикрепления, носит на-
звание основания; противоположный, дистальный конец — это вершина
крыла. Край крыла, при распростертых крыльях направленный вперед,
носит название переднего края крыла, а противоположный ему — задне
го края крыла. Перепонка крыла покрйта мелкими волосками (микротри-
хиями); исключение составляет род (jranotaenia, у представителей кото-
рого перепонка крыла голая.
184
Жилки крь!ла (рис. 111 и 112). Костальная, или краевая, жилка огра-
ничивает крыло со всех сторон. Продольные жилки. 1. Субкостальная,
впадающая в передний край крыла около его вершины или ближе к осно-
ванию. 2. Четыре радиальные, впадающие близ вершины крыла. Сюда
относятся: первая радиальная, простирающаяся до основания крыла;
вторая и третья радиальные, расходящиеся недалеко от вершины крыла,,
образуя переднюю вилку крыла; четвертая радиальная называется р44_5
(двойное название связано с тем, что у других, более примитивных форм
она представлена двумя жилками). Она либо начинается от общего ствола
второй и третьей радиальной жилки (триба Culicini), либо самостоятельно
в центре крыла (триба Anophelini), перед пересечением поперечной радио-
медиальной жилкой. 3. Медиальные жилки (две) впадают в верхнюю часть
заднего края крыла. М | ;-2 и медиальная 3 образуют среднюю вилку крыла.
4. Две кубитальные жилки образуют заднюю вилку крыла, впадающую в
задний край крыла. 5. Анальная (одна) жилка, впадающая около середи-
ны заднего края крыла.
Поперечные жилки. 1. Плечевая, соединяющая костальную жилку
с субкостальной. 2. Радио-медиальная, соединяющая общий ствол второй
и третьей радиальной с общим стволом медиальных жилок. Эта жилка
построена у Anopheles из двух отрезков, называемых 2—3 и 3—4. 3. Ме-
диально-кубитальная, соединяющая общие стволы медиальных и куби-
тальных жилок.
В строении жилок крыла имеются особенности, характерные для три-
бы Anophelini и трибы Culicini. У Anophelini общий ствол передней вилки,
так же как и жилка р начинается самостоятельно в середине крыла, а не
от других продольных жилок; р^ 5 при этом пересекает поперечную радио-
медиальную. У Culicini общий ствол передней вилки начинается от пер-
вой радиальной, а радиальная р4 + 5 — от ствола передней вилки.
Крылья представителей подсемейства Culicinae покрыты чешуйками,
сконцентрированными на жилках. Скопление чешуек той или иной окрас-
ки образуют темные или светлые пятна, из которых слагается рисунок
крыла. На заднем крае крыла чешуйки составляют бахромку.
Жужжальца являются придатками заднегруди; их наружные покровы
богаты нервными окончаниями; предполагают, что эти придатки являются
органами чувств.
Ноги. Ноги (рис. 104) состоят из обычных частей: тазик (соха), верт
луг (trochanter), бедро (femur), голень (tibia) и лапка (tarsus). Лапка со-
стоит из 5 члеников; на последнем сидят два коготка. Под коготками могут
быть присоски (pulvilli), хорошо развитые у Culex и рудиментарные у Аё
des (рис. ИЗ). Коготки и пульвиллы служат для удержания комара на
вертикальных или опрокинутых поверхностях. Части ног, ближайшие к
месту прикрепления, носят название оснований, а части дистальные, наи-
более удаленные от места прикрепления, называются вершинами. На вну-
тренней стороне вершины задней голени расположен ряд плоских шипов,
образующих скребок (рис. 114).
6. БРЮШКО
Брюшко комаров построено из 10 члеников, из которых 9-й и 10-й
сильно изменены и используются как части наружного полового аппарата.
Каждый из 8 передних, нормально развитых члеников брюшка песет дор-
сальную пластинку (тергит) и вентральную (стернит). Тергиты и стерниты
брюшка соединены между собой несегментир о ванной эластичной перепон-
кой — плеврой. Растяжение брюшка и увеличение его объема при кро-
185.
вососании, созревании яиц и ожирении связаны с расправлением и рас-
тяжением этой плевры. При сокращении объема брюшка, например, при
откладке яиц, плевра образует складки. В плевре брюшка, начиная от
второго и кончая шестым сегментом, лежит шесть пар брюшных дыхалец.
По размеру они значительно меньше грудных и отличаются от них по строе-
нию (см. ниже, стр. 214).
Тергиты и стерниты двух последних члеников образуют пластинки,
окружающие анальное и половое отверстия. У самок на конце брюшка мож-
но видеть короткие придатки (церки). У самцов наружные придатки поло-
вого аппарата (так называемый гипопигий)
устроены значительно сложнее. Виды комаров
очень сильно различаются между собой строе-
нием своих гинонигиев, чем и пользуются при
•их определении.
1
Рис. 114. Соскребыватель-
ный аппарат Culex.
Рис. ИЗ. Пульвиллы (присоски) на концах лапок.
л — Aedes; Ь — Culex; В— Theobaldia: 1— коготки; 2--
эмподип; 3— пульвиллы.
Гипопигий самца. Для рассматривания гипопигиев необходимо изго-
товить постоянные препараты в канадском бальзаме. У самцов болыпиннства
двукрылых, в том числе и комаров, при выходе ттз куколок 9-й и 10-й сег-
менты поворачиваются на 180°, так что тергит оказывается на брюшной
стороне, а стсрнит наспинной, и в таком виде остаются на всю жизнь.
Гипопигий самца имеет следующее строение (рис. 115).
9-й тергит имеет вид неширокой пластинки, по бокам заднего края снабженной
выростами, называемыми лопастями 9-го тергпта. 9-й стерппт обычно имеет вид узкой
хитп новой полоски. Наиболее крупным отделом гипопигия являются боковые придатки
9-го сегмента, носящие название вальв. Каждая вальва состоит пз двух члеников.
Первый членик — массивный, второй, узкий, подвижно прикреплен к вершине пер
вого членика. Первый членик мо нет иметь с внутренней стороны выросты в виде бу-
горков— бородавок (триба Culicini); у Anophelini мы находим при его основании один
или несколько шипов. Среднюю часть гипопигия занимает собственно совокупительный
аппарат, называемый у комаров aedeagus. У Anopheles оы имеет вид трубки. обычно
снабженной па конце лисювпднымп придатками. Около aedeagus с топ и другой сто-
роны можно видеть базальные вентральные выросты первого членика вальв, носящие
название класпет. У Anophelini они разделены на лопасти. На вершинах этих ло-
пастей сидят шипы пли заменяющие их пластинки. Нея поверхность первого членика
вальв бывает покрыта чешуйками и длинными тонкими волосками. Дорсально по от-
ношению к aedeagus лежит анальный конус, несущий на конце заднепроходное отвер-
стие (Anophelini). У большинства Culicini можно различить утолщения около аналь-
ного отверстия, образованные 10-м перлитом, так называемые пар шрокгы.
Г Л А В А IV
СИСТЕМАТИКА КОМАРОВ
1. ОБЩИЕ ЗАМЕЧАНИЯ
Комары относятся к семейству Culicidae (комарообразных), которое
делится на 3 подсемейства: Culicinae, Dixinae и Chaoborinae. Подсемейство
Culicinae включает всех кровососущих комаров, Dixinae и Chaoborinae—
186
некровососущие формы и не имеют медицинского значения. Подсемейство
Culicinae делится на 3 трибы: Anophelini, Culicini и Megarrhinini. К трибе
Anophelini относится род Anopheles, представители которого являются
Рис. 115. Гинопигии Anopheles.
Л — вид с дорсальной стороны: 1— анальный ьонус; 2—2-й ч/нгик валы-t (класнер)
i — 1-й членик вальв (коксит); 4—конец эдеагуса; <5—класпеты: а—внутренняя лопасть;
<5 — наружная лопасть; 6 — шипы в основании 1-го члснина вальв; 7 — внутренний шип на
1'М членике вальв; Б— то же по удалении анального конуса; обозначения те же, что и в Д; В —
вид сбоку; обозначения те же, что и в А. кроме того, 8—9-й тершт; 9— 9-й стерпит.
переносчиками человеческих плазмодиев, возбудителей малярии; триба
Culicini объединяет ряд родов, некоторые представители которых также
имеют медицинское значение; комары трибы Megarrhinini крови не пьют
и не имеют медицинского значения. Признаки, отличающие подсемейства
трибы и роды Culicini, даны в определительных таблицах (см. ниже).
187
2. ОПРЕДЕЛЕНИЕ ПОДСЕМЕЙСТВ CULICIDAE ПО ВЗРОСЛЫМ КОМАРАМ
Определительная таблица
1(4). Усики с длинными^ щетинками. Крылья с многочисленными во-
лосками или чешуйками.
2(3). Хоботок довольно короткий...............................
............................подсемейство Chaoborinae (Corethrinae).
3(2). Хоботок длинный..................подсемейство Gulicinae.
4(1). Усики без длинных щетинок. Крылья с малочисленными шипи
ками или короткими волосками на жилках. Хоботок короткий ....
..............................................подсемейство Dixinae.
Представители подсемейства Gulicinae имеют удлиненный ротовой ап
парат, способный прокалывать кожу человека и животных и сосать кровь.
Этим признаком подсемейство Gulicinae — кровососущих комаров — от-
личается от двух других подсемейств, не имеющих длинного хоботка и
являющихся растительноядными. Виды подсемейства Gulicinae, распро-
страненные в СССР, принадлежат к 3 трибам и 8 родам.
* Б
Рис. 116. Брюшко Aedes и Anopheles
с дорсальной стороны.
А — ABdes punctor; Б—A. maculipen-
nis.
з. ОПРЕДЕЛЕНИЕ ТРИБ И РОДОВ ПОДСЕМЕЙСТВА CULICINAE,
РАСПРОСТРАНЕННЫХ В СССР, ПО ВЗРОСЛЫМ КОМАРАМ
(Из А. А. Штакельберга, с сокращениями и небольшими изменениями)
1(2). Щупальца самок длинные, равны по длине хоботку (рис. 117):
щиток, как правило, цельнокрайний (рис. 108). Брюшко без чешуек или
по меньшей мере стерниты на более или
менее значительном протяжении голые,
без чешуек (за исключением A. pulcher-
rimus Theo, у которого все брюшко по-
крыто чешуйками)...................................................
. триба Anophelini, род Anopheles Mg.
2(1). Щупальца самок короткие,
во много раз короче хоботка (рис. 117).
Как тергиты, так и стерниты брюшка
на всем протяжении покрыты чешуй-
ками (рис. 116, А).
3(4). Хоботок утолщенный, в вер-
шинной половине явственно утончаю-
щийся и изогнутый крючковидно книзу;
щиток цельнокрайний ...............
..............(триба Megarrhinini).
Крупные виды с пучками ярких
волосков на конце брюшка...........
род Megarrhinus R. — D
4(3). Хоботок тонкий, на всем про-
тяжении более или менее одинаковой
толщины или слегка утолщающийся к
концу, без резкого изгиба в вершин-
ной части; щиток трехлопастной (рис.
108).........триба Culicini.
5(6). Анальная жилка упирается в задний край крыла на уровне от-
ветвления общего ствола радиальной второй-4-третьей и радиальной чет-
вертой4~пятой (рис. 112). Перепонка крыла лишена коротких волосков
(микротрихий)..................................род Uranotaenia Агг.
18В
6(5). Анальная жилка упирается в задний край крыла на более или
менее значительном расстоянии за местом ответвления общего ствола
радиальной второй Ц- третьей и радиальной четвертой -|~ пятой (ближе к
Рис. 117. Головы самцов и самок Anophelini и Culicini.
/—самка Culicini; 2—самец Culicini; 3—самка Anophelini; 4 — самец Anophelini.
вершине крыла). Перепонка крыла покрыта короткими волосками (микро-
трихиями).
7(14). Пульвиллы под коготками на лапках отсутствуют или имеют-
ся в рудиментарном состоянии (у A6des).
8(9). Дыхальцевые щетинки имеются (рис. 109).................
..........................................род Theobaldia N. — L.
189
9(8). Дыхал ьцевыё щетинки отсутствуют.
10(13). Задыхал ьцевые щетинки отсутствуют; коготки самок без
зубчиков.
11(12). Первый членик передних лапок длиннее четырех следующих
члеников, вместе взятых; четвертый членик тех же лапок у обоих
полов сильно укороченный..................род Orthopodomyia Theo.
12(11). Первый членик передних лапок не длиннее четырех следующих
члеников, вместе взятых; четвертый членик тех же лапок у самок не уко-
роченный ...........род Mansonia Blanch (подрод CoquiHettidia Dyar).
13(10). Задыхальцевые щетинки имеются (рис. 110), коготки самок,
как правило, с зубчиками............................род A6des Mg.
14(7). Пульвиллы под коготками на лапках хорошо развиты
(рис. 113, Б).........................................род Culex L.
4. ОПРЕДЕЛЕНИЕ ВИДОВ РОДА ANOPHELES, ВСТРЕЧАЮЩИХСЯ В СССР,
ПО ВЗРОСЛЫМ КОМАРАМ
Род Anopheles. Щупальца у обоих полов по длине равны хоботку}
у самок равномерной толщины по всей длине, у самцов конечные членики
утолщены. Щиток цельнокрайний. Ноги длинные и тонкие. Радиальная
жилка р4+5 пересекает поперечную радио-медиальную и после этого
прерывается.
Определительная таблица подродов и видов рода Anopheles, встречающихся
в СССР, по взрослым насекомым
(По А. А. Штакельбергу с изменениями)
1(20). Костальная жилка по переднему краю крыла имеет не более
двух светлых пятен; сюда же относятся виды, имеющие только темные
пятна на крыльях, и виды, совсем не имеющие ни темных, ни светлых пя-
тен .........................................подрод Anopheles Meig.
2(15). Крылья с темными чешуйками, часто с темными пятнами, об-
разованными скоплениями чешуек еще более темного цвета, но без пятен
из белых чешуек. Сюда же относятся комары с крыльями, чешуйки на ко-
торых совсем не образуют скоплений (без пятен).
3(12). Крылья с более или менее одноцветными чешуйками (без пя-
тен); основание передней вилки крыла (радиальная вторая радиальная
третья) расположено явственно ближе к основанию крыла, чем основание
средней вилки (медиальная первая -р вторая и медиальная третья).
4(5). Пучок белых чешуек и волосков на лбу отсутствует (имеется
слабо развитой пучок желтоватых волосков). Среднеспинка одноцветная
(без продольной полосы), с довольно длинными темнобурыми волосками .
................................................A. algeriensis Theo.
5(4). Белый пучок на лбу имеется.
6(9). Преобладающая окраска черно-серая со свинцовым отливом;
на среднеспипке продольная серая полоса; пучок чешуек на переднем
крае среднеспинки сильно развит и чисто белого цвета.
7(8). Отрезок поперечной радио-медиальной жилки 3—4 довольно
значительно сдвинут к основанию крыла, так что непрерывной линии с
другим отрезком этой же поперечной жилки 2—3 не получается . . . .
.......................................A. plumbeus plumbous Steph.
8(7). Отрезок поперечной радио-медиальной жилки 3—4 лежит почти
на одной линии с жилкой 2—3. Концы их соприкасаются так, что образует-
ся непрерывная, чуть ломаная линия . . A. plumbeus barianensis James.
190
9(6). Преобладающая окраска желтобурая, бурая или сероватая^
пучок чешуек на переднем крае среднеспинки слабее развит и имеет желто-
ватый оттенок.
10(11). На среднеспинке ясно выраженная сероватая продольная
полоса.........................................A. bifurcatus Linn.
11(10). Среднеспинка без ясно выраженной сероватой полосы, желтая,
глянцевитая.......................A. marteri sogdianus Keschischian.
12(3). Крылья с темными пятнами; основания вилок передней и сред-
ней на крыльях расположены на одинаковом расстоянии от основания
крыла.
13(14). Общая окраска темнобурая; среднеспинка с широкой серой
продольной полосой, занимающей почти всю поверхность среднеспинки,
если смотреть на комара сверху. Темные пятна на крыльях выражены
явственно...................................A. maculipennis Meigen.
14(13). Общая окраска светложелтоватобурая; среднеспинка на всем
своем протяжении одноцветно светлобурая или желтая. Иногда можно раз-
личить полосу в отделе среднеспинки, прилегающехМ к щитку. Темные пят-
на на крыльях слабо выражены. ... A. maculipennis sacharovi Favre.
15(2). Крылья на переднем крае крыла по костальной жилке с од-
ним или двумя белыми пятнами, образованными скоплением белых че-
шуек *.
16(17). Костальная жилка с одним белым пятном у вершины; задние
бедра с явственным предвершинным широким белым кольцом...........
...............................................A. lindesayi Giles.
17(16). Костальная жилка с двумя светлыми пятнами; задние бедра
без предвершинного кольца.
18(19). В районе поперечных жилок имеется темнее полулунное коль-
цо.........................................A. hyrcanus sinensis Wied.
19(18). Темное полулунное кольцо в районе поперечных жилок отсут-
ствует.....................................A. hyrcanus hyrcanus Pall.
20(1). Костальная жилка по переднему краю крыла с четырьмя и бо-
лее светлыми пятнами....................подрод Myzomyia (Bl.) Chr.
21(22). По переднему, краю крыла промежутки между белыми пятнами
заполнены черными пятнами разнообразной формы и размера. Всего можно
различить шесть черных пятен. Два из них, в основании, очень маленькие,
следующие четыре, большие, расплывчатой формы. Брюшко сверху густо
покрыто чешуйками, образующими с боков по заднему краю торгитов тор-
чащие в сторону пучки. Последние членики задних лапок белые ....
............................................A. pulcherrimus Theo.
22(21). Темные пятна по костальной жилке крыла приблизительно все
одинаковые, удлиненной формы. Брюшко с боков без чешуек; последние
членики задних лапок темные...................A. superpictus Grassi.
5. ОПРЕДЕЛЕНИЕ ВИДОВ РОДА ANOPHELES ПО ГИПОПИГИЯМ САМЦОВ
(По А. А. Штакельбергу, с небольшими изменен! я ли)
1(14). Первый членик вальв при основании с внутренней стороны
по большей части с двумя (редко с одной или тремя) крепкими щетинками,
из которых по меньшей мере одна расположена на сосочковидном буг)р-
ке..................................... подрод Anopheles (Meig.) Chr.
1 При рассматривании этот о признака следует гостагить крыло гередпим краем
вверх на ребро, принимаются во внимание только белье пятна на косталы од л илке;
остальные, расположенные на других жилках крыла, в расчет не принимаются.
191
2(3). Первый членик вальв при основании с одной щетинкой .
„..................................................A. algeriensis Theo.
3(2). Первый членик вальв при основании с двумя-тремя щетинками.
4(5). Первый членик вальв при основании с тремя крепкими щетин-
ками, из которых две сближены своими основаниями и разветвлены .
...................................................A. bifurcatus Linn.
5(4). Первый членик вальв при основании с двумя простыми нераз-
ветвленными щетинками.
6(9). Шипы класпетт сближены, но не слиты.
7(8). Aedeagus короткий и широкий, без листовидных придатков .
...................................................A. plumbeus Steph.
8(7). Aedeagus длинный и узкий, на конце с листовидными придат-
ками..........................................A. maculipennis Meigen.
9(6). По крайней мере некоторые шипы класпетты слиты, образуя
подобие хитиновой пластинки.
10(13). Девятый тергит без выростов; пластинки класпетты длинные
и узкие.
11(12). Класпетты несут на вершине две пластинки, отходящие от
самостоятельных бугорков; aedeagus длинный ц узкий, несет на каждой
стороне 12—13 игловидных придатков и с каждой стороны по одному более
крупному ланцетовидному.............A. marteri sogdianus Keschischian.
12(11). На класпеттах по одной длинной и узкой пластинке ....
...................................................A. lindesayi Giles.
13(10). Девятый тергит с явственно развитыми длинными и узкими
выростами. Пластинка класпетт относительно короткая и широкая .
...................................................A. hyrcanus Pall.
14(1). Первый членик вальв при основании с четырьмя-шестью креп-
кими щетинками, сидящими непосредственно на теле членика, а не на со-
сочковидных его выростах................подрод Myzomyia (Blanch) Chr.
15(16). Aedeagus без листовидных придатков... A. pulcherrimus Theo.
16(15). Aedeagus с короткими листовидными придатками.............
..............................................[A. superpictus Grassi.
Примечание. Определения видов рода Anopheles без пятен на крыльях
следует проверять по строению гипопигиев самцов, так как окраска этих видов очень
изменчива. Например, весенние A. bifurcatus, вылупляющиеся из перезимовавших
личинок в Сухуми (Абхазия), своей темной окраской напоминают скорее A. plumbeus,
чем A. bifurcatus. A. algeriensis из Узбекской ССР имеют темноватую окраску в отли-
чие от более рыжих дагестанских особей. С другой стороны, A. mac. sacharovi в север-
ной части своего распространения близок ио окраске к другим подвидам A. maculi
pennis и может быть надежно определен только по строению яиц.
6. ОПИСАНИЕ ВИДОВ РОДА ANOPHELES (ВЗРОСЛЫЕ КОМАРЫ)
Подрод Anopheles
Признаки подрода. Костальная жилка по переднему краю крыла
имеет не более двух светлых пятен или пятен совсем нет, или есть темные пятна. Ги-
попигий: первый членик вальв при основании с одной-тремя крупными щетинками,
из которых хотя бы одна расположена на бугорке.
I группа видов подрода Anopheles — комары с темными пятнами на крыльях.
1. A. maculipennis Meig. (рис. 118). Самый широко распространенный в СССР
переносчик малярии. Общая окраска очень изменчива. Южные особи меньше ростом,
северные более крупные и темно окрашенные. На голове сверху между глазами име-
ется направленный вперед пучок длинных волосков и чешуек. На крыльях четыре-пять
темных пятен, расположенных в основаниях передней и средней вилок крыла, а также
в области радио-медиальной поперечной жилки и ближе к основанию крыла на общем
стволе радиальных жилок. Среднеспинка с широкой серой продольной полосой, в
передней половине которой обычно намечается две-три буроватые полоски. Брюшко
бурое, часто с более светлыми пятнами в боковых отделах тергитов; покрыто длин
192
яыми зочотистожелтыми или буроватыми волосками. Гипопигий самцов: в основании
первого членика вальв две крупные щетинки, из которых ближайшая к aedeagus си-
дит на большом бугорке, крайняя же — на едва заметном бугорке. Aedeagus длин-
ный и узкий, с листовидными придатками. Шипы па класпеттах сближены, но не сли-
ты. Этих шипов минимум три: вентральный, апикальный и дорсальный (иногда при
сутствуют еще промежуточные шипы). Ряд авторов обращал внимание па окончание
дорсального шипа; найдено, что у большинства особей atroparvus, labranchiae и sa-
charovi этот шип имеет заостренное окончание, у большинства особей messeae и typv
cus — тупое окончание. Этот признак по является удобным для определения каждой
особи, а поэтому в настоящее время пе применяется. У A. mac. messeae относительная
величина торакальных стигм больше, чем у A. mac. atroparvus.
Рис. 118. Anopheles maculipennis Meig.
Л — самка; Ь — гипопигий самца.
2. A. mac. sacharovi Favre (elutus Edwards) очень близок к основной форме,
от которой отличается более светлой окраской и расплывчатым рисунком пятен на
крыльях. У типичных экземпляров среднеспинка одноцветная, но в северных частях
области распространения этого вида встречаются особи с продольной полосой, как у
других подвидов. Иногда эта полоса начинается только с середины среднеспинки и
простирается до щитка.
11 группа видов подрода Anopheles— виды без пятен па крыльях.
1. A. algcriensis Theo. (рис. 119). Общая окраска тела светлокоричневая или
желтоватая (Дагестанская АССР), или более темноватая (Средняя Азия). Крылья без
пятен. Среднеспинка одноцветная, без продольной полосы, покрыта длинными темными
волосками. Задние лапки иногда сэ светлыми колечками. Гипопигий самцов отличается
от такового всех других Anopheles присутствием только одной хорошо развитой ще-
тинки в основании первого членика вальв, сидящей па большом плоском бугорке.
Aedeagus с листовидными придатками. Класпетты трехлопастные, широкие.
2. A. bifurcatus (Linn.) (рис. 119). Родниковый малярийный комар. Общая окраска
желтоватая (на юге) или светлошоколадная (на севере). Па лбу расположен направ-
13 Учебник медицинской энтомологии
193
Рпс. 119/ Л. algeriensis Theo, п Л. bifurcatus (L.).
А—гоп ни и среднсспинна Л. algeriensis (cipana) и A. bifurcatus
(ujijbJ; Б— гииииигнй A. algeriensis; В— гшюшьий A. bifurcatus.
Рис. 120. Anopheles plumbeus Steph.
A — самка Б — гипопигий самца.
13*
ленный вперед пучок белых чешуек и волоскоз. Среднеспинка бурая с беловато-серой
продольной полосой. Крылья без пятеп. Лапки одноцветно бурые. Гипопигий самцов:
при основании первого членика вальв три крупные щетинки, из которых две боковые
сближены своими основаниями и разветвлены. Внутренняя, расположенная ближе к
aedeagus, сидит на бугорке и не разветвлена.
3. A. plumbeus Steph, (--nigripes Staeger) (рис. 120). Дупляной малярийный комар.
Общая окраска черная; как чешуйкп, так и покровы тела черные. Голова черная, с
боков усаженная черными чешуйками; па лбу, между глазами, направленный вперед
пучок чисто белых чешуек. Усики самок черные, покрыты белым пушком при основа-
нии каждого членика. Грудной отдел чорнобурый, па переднем краю среднеспинки
белые чешуйкп. Среднеспинка с продольной пепельносерой полосой. Крылья без
пятен. Ноги черные или чернобурые, тазики и основания бедер светлые. Брюшко сверху
черное блестящее, снизу несколько светлее. Гипопигий самцов имеет в основании
первого членика вальв две крупные щетинки, сидящие па бугорке. Шипы на класпеттах
сближены, по не слиты. Aedeagus короткий и широкий, без листовидных придатков.
A. plumbeus barianensis James, описанный для Таджикистана, очень близок к
A. pl. plumbeus по всем признакам, за исключением строения радио-медиальной жилки,
оба отрезка которой образуют у A. pl. barianensis почти прямую линию. н-»
4. A. rnarteri sogdianus Keschischian (рис. 121) близок к средиземноморскому
A. marteri Sen. ct Prun.
Ш группа видов подрода Anopheles— с белыми пятнами по костальной жилке,
на переднем крае крыла.
1. A. li.idesayi Giles (рис. 122). Крылья в большей своей части покрыты темпыми
чешуйками. Ла переднем крае крыла, у его вершины, имеется крупное белое пят
по, распространяющееся па жилки — радиальную первую и вторую. Небольшие
участки белых чешуек имеются п на других жилках. Щупальцы и хоботок черные, с
прилегающими чешуйками. Ноги чернобурые; задние бедра перед вершиной с широким
чисто белым кольцом. Лапки темпобурые, с белыми колечками у вершины некоторых
члеников. Девятый тергит брюшка самца без отростка; первый членик вальв при оспо
ваппи с двумя крепкими щетинками. Па класпеттах длинная и узкая пластинка.
Aedeagus на вершине с листовидными придатками.
2. A. hyrcanus Pall. (рис. 123). Крылья в большей своей части покрыты темными
чешуйками; но переднему краю крыла па костальной жилке два больших белых пятна—
одно в начале дистальной трети, а другое перед вершиной крыла; первое из них
занимает жилки костальную, первую радиальную, второе — от костальной до второй
радпальпой. Хобогок и щупальцы темпобурые; последние у самок стремя белыми
колечками и белой вершиной. Усики самок темпобурые, покрытые белым пушком, у
основания члеников с розетками довольно длинных бурых волосков; первые 5—7
члеников усиков несут некоторое количество белых чешуек. Среднеспинка бурая, в
средней части разделенная темными потосками па две-четыре и более узкие серые
полосы. Лапки темнобурые, с белыми колечками у вершины первых трех или четырех
членикоз. Гипопигий: первый членик вальв при основании с двумя крепкими просты-
ми щетинками, сидящими на бугорках. Девятый тергит с длинными узкими выроста-
ми. На класпеттах широкая пластинка. Для A. hyrcanus описан целый ряд форм (см.
Штакельберг, 1937), которые являются в основном сезонными аберрациями.
Согласно ревизии этих форм Масловым (19 46), все ранее описанные в СССР разно
видностп не могут считаться таковыми и все относятся к A. hyrcanus hyrcanus. Ис
ключеппе составляет распространенный у нас в южной части Дальнего Востока
A. hyrcanus sinensis, являющийся устойчивой разновидностью. Отличие A. hyrcanus
hyrcanus Pall, or A. hyrcanus sinensis Wied., согласно описанию, данному Масловым,
это резкость рисунка крыла, равномерное распределение темных и светлых пятен,
резкое выделение более темных передних жилок у sinensis по сравнению с hyrcanus.
Кроме того, у A. hyrcanus sinensis в районе поперечных жилок имеется темное полулун-
ное пятно, отсутствующее у A. hyrcanus hyrcanus Pall.
Подрод Myzomyia (Blanch.) Chr.
Признаки подрода. Поперечные жилки и основания вилок крыла в
белых чешуйках; костальная жилка по переднему краю с четырьмя или более белыми
пятнами. Гипопигий самцов: в основании первого членика вальв четыре-шесть креп
ких щетинок, сидящих не на бугорках, а на самой поверхности членика.
1. A. pulcherrimus Theo. Белый малярийный комар (рис. 124). Общая окраска
белая с контрастным черным рисунком. Готова, грудь, брюшко покрыты в большей
части белыми, налегающими друг на друга чешуйками. Крылья по переднему краю с
шестью черными пятнами. Из них два, у основания крыла, очень маленькие, и четыре
крупные, с расплывчатыми о гертапиямп, распространяются на субкостальную и пер
вую радиальную жилку. Кроме пятен по переднему краю крыла, имеются скопления
темных чешуек и по всей поверхности крыла. Бахромка крыла состоит из чередующихся
196
Рис. 121. A. marteri sogdianus Kesh.
A — самка; Б —. гипопигий самца-
Рис. 122. Anopheles lindesayi Giles.
А — самка; Б — гипопигий самца.
между собой участков белых и черных чешуек. Готова с боков в светлобурых, сверху
в чисто белых чешуйках. Вершины всех членикоз щупикоз белые (широкие белые
кольца). У усиков каждый членик песет розетку длилных белых волосков. Брюш-
ко с ясно выраженными торчащими пучками чешуек по бокам тергитов. Лап-
ки с белыми колечками у вершины члеников; три последних членика задних лапок
сплошь белые. Гипопигий: первый членик вальв при основании с четырьмя-пятью креп-
кими щетинками, сидящими непосредстЕенно на поюрхвости этою членика. Класпет-
ты с длинной пластинкой; aedeagus простой, без листовидных придатков.
2. A. superpictus Grassi (рис. 125). Сильно вариирукщий в окраске вид. Общая
окраска серовато-желтоватая. Рисунок крыла иногда очень четкий (например, в Тад-
жикистане), иногда расплывчатый (Южная Туркмения). Крылья по переднему краю
с однообразным чередованием приблизительно одинаковых во форме темных и белых
удлиненных прямоугольничков. Скопления темных чешуек имеются и в других частях
крыла. Щупальцы темные, с резкими узкими белыми колечками и белой вершиной.
Среднеспинка, особенно в передней части, густо покрыта узкими беловатыми чешуй-
ками. Лапки одноцветно бурые или со слабо развитыми узкими белыми колечками.
Брюшко в довольно длинных волосках, без пучков чешуек по бокам. Гипопигий:
первый членик вальв, относительно короткий и толстый, при основании с пятью креп-
кими щетинками. Aedeagus па вершине с короткими листовидными придатками.
7. ОПРЕДЕЛЕНИЕ П ХАРАКТЕГИСТИКА ПОДСЕМЕЙСТВ СЕМЕЙСТВА
CULIC1BAE ПО ЛИЧИНКАМ
а) Таблица для определения подсемейств семейства
(Julicidae по личинкам
1(2). Грудпые сегменты не расширены, границы между ними ясно за-
метны. Брюшко из 10 явных сегментов. Стигмальпая пластинка состоит
из частей 8-го и 9-го брюшных сегментов и не образует одного целого...
................................................подсемейство Dixinae.
2(1). Грудь заметно шире брюшка; сегменты ее слиты в один комплекс,
границы между ними неясные. Брюшко из 9 явных сегментов. На 8-м
сегменте помещается либо слитая в одно целое стигмальная пластинка, либо
дыхательная трубка, либо, при их отсутствии, дыхательная система замк-
нута . ....................................(род Chaoborus)
199
3(4). Антенны видоизменены в органы схватывания добычи, с длинными
крепкими шипами на конце. В 7-м (иногда в 6-м и 7-м) сегменте брюшка
имеются трахейные пузыри...................подсемейство Chaoborinae.
4(3). Антенны не видоизменены в органы схватывания добычи. Тра-
хейные пузыри отсутствуют.....................подсемейство Culicinae.
Anophehnt С и Li ci ni
Anopheles
Aedes
Cut ex
Рис. 126. Главные отличительные признаки Anophelini и Culicini.
1 — нмилййки; 2 — анальные ж. бры; 3 — брюшко; 4—антенны; 3 — щетки; 3’— глаза;
1— хвостовые волосьчг, 8 — плечевые лопасти; Г — нижнечелюстные щупики; 10 — пальмовид-
ные волоски; 11 — х,ботон: 12 — 19—1-й и 8-й сегменты брюшка; 2J —- сг.фзп; 21 — дыхаль-
це; 22 — грудь; 23 — дыхательные трубки; 24 — поверхность воды.
б) Характеристика подсемейств семейства Culicidae
по личинкам
I. Подсемейство Dixinae. Личинки этого подсемейства наиболее
близки к Anopheles; они живут также на поверхности воды и имеют сходный способ
питания и дыхания (питаются организмами поверхностной пленки, дышат через стиг-
мы, расположенные на спинной поверхности 8-го сегмента). Однако между личинками
Dixinae и Anopheles имеются и значительные отличия (рис. 127). Так, личинки Di-
xinae, единственные из Culicidae, имеют полное число члеников тела: неслившиеся
грудные сегменты (в ширину не превышающие членики брюшка) и 10 брюшных чле-
ников (8-й и 9-й членики в отличие от таковых Anopheles у них не слились). Лопасти
стигмальной пластинки расположены у Dixinae на 8-м и 9 -м члениках брюшка, а не
образуют одну слившуюся пластинку, как это имеет место у Anopheles; края лопастей
оторочены волосками. Набрюшной стороне некоторых члеников брюшка имеются при-
способления для ползания по твердому субстрату: ложноножки и ползательные валики.
Личинки Dixinae передвигаются с одинаковым успехом как по поверхности воды, тан
и па твердом субстрате (листьях, камнях), тогда как к передвижению в толще воды они
не приспособлены ( у них отсутствует плавник).
2. Подсемейство Chaoborinae. У личинок подсемейства (Jhaa
borinae связь с поверхностью воды менее выражена, чем у личинок Dixinae и Culi
200
cinae, или полностью утрачена; они живут в толще воды или периодически поднимают
ся к поверхности для дыхания. Только у одного вида нашей фауны, Mochionyx mart г
nib функционирует сифон, у всех остальных представителей этого семейства имеет
место исключительно кожное дыхание; сифон рудиментарен или отсутствует.
Рис. 127. Личинка Paradixa.
1—голова; 2— антенна; 3—нижнечелюстные щупики: 4— лгетни
верхней губы; 5— мансиллы; 6— 1-й сегмент гпуди; 7— волоски
1-го сегмента; 8— 2-й и 3-й сегменты груди; 7—X сегменты брюшка;
9— стигмальная пластинка; 10— боковые лопасти стигмальной пла-
стинки; 11— ее задние лопасти; 12— центральная пластинка; 13—
жаоры; 14 — трахеи.
Рис. 128. Личинка Chaoborus.
1— голова; 2— глаза; 3— антенны; 4— верхняя губа; 5 — секи-
ровидный волосок; 6 — мандибулы; 7 — максиллы; 8 — надглоточ-
ник; 9— грудь; 10— рулевые волоски; 11—трахейные пузыри:
1—IX — сегменты брюшка.
Личинки Chaoborinae—хищники. Для схватывания добычи им служат усики»
очень подвижные, снабженные на вершине длинными крепкими щетинками, играющими
роль при схватывании добычи. Ротовые органы приспособлены для хватания и раз-
жевывания добычи. Личинки Chaoborinae являются активными пловцами; у них
хорошо развит плавник. Остальные волоски па теле слабо развиты, так как в против-
ном случае они оказывали бы большое сопротивление движению личинки при схваты-
вании добычи. В связи с активным характером двияения тело их приобретает более
обтекаемую форму (рис. 128). С переходом личинок Chaoborinae к жизни в толще воды
201
трахейная система у большинства видов теряет дыхательную функцию, становится
замкнутой и служит для уменьшения удельного веса личинок. Пузыревидные расши-
рения главных трахейных стволов, имеющиеся в груди и брюшке, позволяют телу ли-
чинок сохранять горизонтальное положение, удобное для схватывания добычи.
3. Подсемейство Culicinae, Личинки характеризуются наличием пары
функционирующих дыхалец, расположенных на спинной поверхности 8-го брюшного
членика или непосредственно, или на вершине дыхательной трубки. Широкая грудь
образована 3 слившимися члениками, а более узкое брюшко — 9 члениками. По
способу питания являются фильтраторами или обскребывателями; личинки Megar-
rhinus, в частности, М. christophi, встречающегося у пас на Дальнем Востоке, яв-
ляются хищниками. Органами схватывания добычи у Megarrhinini в отличие от
Chaoborinae являются не антенны, а верхняя губа, мандибулы и максиллы. Щетки
верхней губы у личинок этого подсемейства всегда хорошо развиты. Подсемейство
Culicinae включает 3 трибы: Anophelini. Culicini и Megarrhinini.
8. ОПРЕДЕЛЕНИЕ ТРИБ И РОДОВ ПОДСЕМЕЙСТВА CULICINAE
ПС'ЛИЧИНКАМ/
1(2). Стигмы расположены на спинной поверхности 8-го сегмента брю-
шка; окружены стигмальной пластинкой................................
...........................триба Anophelini (род Anopheles Meig.).
2(1). Стигмы, окруженные стигмальной пластинкой, помещаются на
вершине сифона, отходящего от спинной стороны 8-го сегмента брюшка
....................................................триба Culicini.
3(4). Сифон и клапаны стигмальной пластинки видоизменены в бу-
равящий аппарат (рис. 129, Д) 1 2..............род Mansonia Blanch.
4(3). Сифон и клапаны стигмальной пластинки развиты нормально
(т. е. не видоизменены в буравящий аппарат).
5(12). Сифон с одной парой пучков волосков на задней поверхности.
6(7). Пучки помещаются у основания сифона (рис. 129, В) . . . .
...........................................род Theobaldia Nev-Lem.
7(6). Пучки расположены около середины сифона или ближе к его
вершине.
8(9). По бокам 8-го сегмента брюшка имеется по хитиновой пластинке,
от заднего края которой отходят чешуйки щетки (рис. 129, Е). Зубцы гре-
бня расщеплены на концах.......................род Uranotaenia Arrib.
9(8). По бокам 8-го сегмента имеется только щетка из чешуек, распо-
ложенных в один ряд или пятном различной формы. Если хитиновые пла-
стинки имеются, то чешуйки щетки самостоятельны. Зубцы гребня (при
его наличии) обычно шиповпдно заострены на концах.
10(11). На спинной стороне 6—8-го брюшных сегментов имеются хи-
тиновые пластинки, развитие которых усиливается по направлению к
заднему концу. Гребень отсутствует (рис. 129, Г)....................
...........................................род Orlhopodomyia Theob.
11(10). Хитиновые пластинки на 6—8-м сегментах отсутствуют. Гре-
бень на сифоне имеется (рис. 129, А)................род A6des Meig.
12(5). Сифон с несколькими парами пучков волосков на задней и бо-
ковых его поверхностях (рис. 129, Б)...................род Culex L.
9. ОПРЕДЕЛЕНИЕ ВИДОВ РОДА ANOPHELES ПО ЛИЧИНКАМ
В систематике личинок рода Anopheles главными диагностическими
признаками являются:
1 В таблицу пе включена триба Megarrhinini, не имеющая у нас медицинского
значения.
2 Личинки Mansonia дышат воздухом, находящимся в стеблях водяных растений,
куда они погружают острый конец сифона, и не всплывают к поверхности для дыха-
ния (рис. 199).
202
Рис. 129. Спфопы личипок Culicini IV возраста:
A — ASdes (Аёаез) cinereus; Б — Culex pipiens; В — Theobaldia annulata; Г — Orthopodcmyia pulchrlpalpis (характерно
отсутствие гребня); 1 — задний конец сбоку; 9— 4 чешуйки щетки (9— крупные; 3— мелкие; * — вид соокул «и виап-
sonia rlchiardii; Ё — Uranotaenia unguiculata.
1. Взаимное расположение и характер ветвления волосков головы: кли-
пе а льны х и фронтальных, реже—теменных и транссутуральных (рис. 1с0).
У одного и того же вида Anopheles взаимное расположение и характер вет-
вления волосков на голове личинки одинаковы и, наоборот, у разных видов
различны; это различие используется для определения вида. Внутренние
клипе ал ьные волоски могут быть сближены своими основаниями (подрод
Anopheles) или, наоборот, удалены (подрод Myzomyia) настолько, что рас-
стояние между ними больше, чем расстояние между внутренними и соот-
ветствующими наружными (это соотношение с возрастом личинки из-
меняется). Ветвление волосков может быть дихотомическим, древовид-
Рис. 130. Волоски головы личинок Anopheles IV возраста.
А—A. maculipennis; Б—A. bifurcatus; В—A. algeriensis; Г—A. plumbeus; Д—А. Ип-
iiesayi; Е—A. superpictus; Ж—A. hyrcanus; 3—A. pulcherrimus; И— антеннальный во-
лоеок A. maculipennis (1) и A. hyrcanus (2).
ным, перистым или волоски могут быть совсем не разветвлены. Стро-
ение фронтальных волосков у всех видов нашей фауны, за исключением
A. plumbeus, перистое; у plumbeus эти волоски простые и короткие.
2. Положение и развитие волоска на антеннах (рис. 130, И).
3. Форма и развитие пальмовидных волосков (рис. 131).
4. Строение стигмальной пластинки (рис. 97, В).
При определении личинок их необходимо исследовать со спинной сто-
роны; определение вида надо производить на личинках IV возраста.
Определительная таблица личинок IV возраста рода Anopheles
1(16). Расстояние между основаниями внутренних клипеальных во-
лосков меньше расстояния между наружным клипе ал ьным волоскохм и
соседним — внутренним. Если эти расстояния почти равны между собой,
то фронтальные волоски простые................подрод Anopheles Meig.
2(5). Фронтальные волоски простые и короткие.
3(4). Внутренние и наружные клипеальные волоски ветвящиеся
.............................................. A. plumbeus Stqph.
204
4(3). Внутренние и наружные клипеальные волоски простые . . .
..........................................A. pl. barianensis James.
5(2). Фронтальные волоски длинные и перистые.
6(13). Наружные клипеальные волоски простые или слабо разветвлен-
ные на концах или около середины.
7(8). Внутренние и наружные клипеальные волоски, начиная от
середины или от второй трети от основания, несут тонкие короткие веточки
.............................................A. algeriensis Theob.
8(7). Внутренние и наружные клипеальные волоски не имеют тонких
коротких веточек.
9(10). Задние клипеальные волоски из двух-пяти ветвей. Лепестки
пальмовидных волосков брюшка без вытянутой концевой нити. Отростки
на стигмах отсутствуют............................A. bifurcatus L.
Рис. 131. Строение пальмовидных волосков IV членика брюшка.
А—Ano] holes lindesayi (часть волоска); Б— A. marteri.
10(9). Задние клипеальные волоски простые. Отростки на стигмах,
за редкими исключениями, имеются.
11(12). Пальмовидный волосок на заднегруди слабо развит, состоит
из 10—11 лепестков. Концы лепестков пальмовидных волосков брюшка
с вытянутой концевой нитью....................A. marteri Senev. Prun.
12(11). Пальмовидный волосок на заднегруди хорошо развит, из 15—22
лепестков. Концы лепестков пальмовидных волосков брюшка постепенно
заостряются и не имеют концевой нити (рис. 131, А),.................
...................................................A. lindesayi Giles.
13(6). Наружные клипеальные волоски древовидно ветвятся.
14(15). Волосок на теле антенны длинный, около половины ее дли
яы; отходит от середины верхней поверхности или лишь немного смещен
к основанию антенны. Внутренние клипеальные волоски простые пли
слабо разветвленные................................A. hyrcanus Pall,
15(14). Волосок на теле антенны очень короткий, не больше, чем
ее ширина, и отходит близ ее основания. Внутренние клипеальные волоски
явственно разветвлены на концах................A. maculipennis Meig.
16(1). Расстояние между основаниями внутренних клипеальных
волосков больше расстояния между наружным и соседним с ним внутрен
ним волоском; фронтальные волоски перистые . . подрод Myzomyia Blanch.
17(18). Наружные клипеальные волоски ветвистые, с 6—15 ветвями
..............................................A. pulcherrimus Theob.
18(17). Наружные клипеальные волоски простые....................
..............................................A. superpictus Grassi.
2()Г>
ГЛАВА V
ВНУТРЕННЯЯ АНАТОМИЯ И ФИЗИОЛОГИЯ личинок
и КУКОЛОК КОМАРОВ
1. ПИЩЕВАРИТЕЛЬНЫЙ АППАРАТ
Органы пищеварения личинок состоят из кишечника и пары слюнных
желез. Слюнные железы с парными придатками помещаются в груди, их
непарный проток открывается между основанием нижней губы и подгло-
точником.
Кишечник личинок представляет почти прямую трубку и состоит из
трех отделов: передней, средней и задней кишки (рис. 132). Передняя киш-
ка начинается короткой мускулистой глоткой, которая переходит в корот-
кий и тонкий пищевод; стенки ее образованы кольцевой и продольной мус-
кулатурой; изнутри она выстлана хитиновой кутикулой. Назначение
этого отдела — проведение пищи в среднюю кишку.
Рпс. 132. Схема внутренних органов личинки комара.
1—ппщевол; 2— мышечный сфинктер пищевода; 3— клетки, выделяющие
вещество перитрофической перепонки; 4 — кутикулярное кольцо; 5 — аорта;
6—септе; 7 — перигрофпческан перепонка; 8—тр»хен; 9— надглоточный ган-
глий; 19— подглоточный ганглий; 11— слюнная шелепа; 12— кардиальные при-
датки средней кишки; 13—брюшин нервно! цепочка; 14— задняя кишка;
- 15 — мальпигиевы сосуды.
В передней трети груди пищевод переходит в среднюю кишку. На
границе между передней и средней кишкой имеется впячивание передней
кишки в просвет средней, которое играет роль клапана, допускающего
прохождение пищи только в направлении от пищевода к средней кишке,
но не обратно. На протяжении клапана кольцевая мускулатура образует
сфинктер, т. е. замыкающую мышцу. Средняя кишка образует самый
длинней от/ет кишечника, простирающийся от середины груди до 6-го
членика брюшка. В передней части средней кишки имеется восемь «карди-
альных» выростов, образованных крупными клетками железистого эпи-
телия.
Секреторная функция средней кишки. Степки средней кишки выстла-
ны крупными эпителиальными клетками. Всасывание пищи и выделение
пищеварительных ферментов происходят только в средней кишке, свободной
от кутикулярной выстилки. Пищеварительные ферменты выделяются глав-
ным образом клетками заднего отдела средней кишки. Из пищеваритель-
ных ферментов у личинок комаров (Andes aegypli и Culex fatigans) най-
дены ферменты, переваривающие белки, жиры и некоторые углеводы.
Реакция содержимого средней кишки слабо щелочная, а в кардиальных
выростах — слабокислая.
Перитрофическая оболочка. К эпителию средней кишки изнутри при-
легает тонкая, прозрачная перитрофическая оболочка, проницаемая как
для ферментов, так и для продуктов пищеварения. Она имеет вид трубки,
открытой на обоих концах. Выделяется перитрофическая оболочка клет-
ками кардиальной (т. е. самой передней) части средней кишки. Мускула-
206
тура средней кишки развита очень слабо, поэтому перистальтика здесь
слаба. В 6-м сегменте брюшка средняя кишка переходит в заднюю кишку;
на их границе в последнюю впадает пять мальпигиевых сосудов. Задняя
кишка состоит из узкой, S-образно изогнутой тонкой кишки и короткой,
расширенной толстой кишки; последняя переходит в тонкую и короткую
прямую кишку, открывающуюся заднепроходным (анальным) отверстием.
Вся задняя кишка выстлана кутикулой и имеет сильную мускулатуру.
Механизм передвижения пищи по кишечнику. Передвижение пищи в
средней кишке происходит не под влиянием перистальтики, которая здесь
слаба, а под напором непрерывно поступающих из передней кишки пище-
вых масс. Скорость продвижения пищи по кишечнику и продолжительность
пребывания ее в нем зависят от интенсивности заглатывания пищи, а также
от размера заглатываемых частиц и их концентрации. Чем больше пищи
личинка заглатывает в единицу времени, тем быстрее пища проходит через
кишечник. При питании личинки в естественных водоемах прохождение
каждой порции пищи через кишечник продолжается 30—60 минут (при 20°).
Под действием пищеварительных ферментов частицы пищи переходят в
жидкое состояние; переваренная пища проникает (диффундирует) через
перитрофическую перепонку в узкое пространство между последней и стен-
кой кишечника. Здесь действием слабых волн сокращений, проходящих
по мускулатуре средней кишки сзади наперед, она перегоняется кпереди,
при этом постепенно всасываясь; главным местом всасывания углеводов
является задняя половина средней кишки, главным местом всасывания
жиров — передняя ее половина (Рахманова). Степень усвоения пищи в ки-
шечнике личинки, вообще очень невысокая, находится к тому же в за-
висимости от обилия пищи в окружающей среде. При изобилии пищи личин-
ка заглатывает ее непрерывно в большом количестве и содержимое кишеч-
ника быстро обновляется, так что большая часть пищи проходит через
кишечник в недопереваренном виде; в таких случаях нередко можно видеть
в задней кишке личинки живых простейших в неизмененном виде. И, наобо-
рот, чем меньше пищи в воде, тем дольше пища задерживается и перевари-
вается в кишечнике, тем, до некоторой степени, выше ее усвояемость.
Перестройка кишечника в стадии куколки. В стадии куколки кишечник
личинки заново перестраивается. Распад эпителия кишечника начинается
уже в последние часы перед окукливанием. Новый эпителий развивается
из особых зародышевых клеток, сохранившихся с ранних стадий развития
в стенках кишечника; усиленный рост и размножение этих клеток проис-
ходят с IV фазы. При метаморфозе перестройку претерпевают также слюн-
ные железы, мускулатура и кожа.
2. ДЫХАТГЛЬПКЙ АППАРАТ
Стигмальное и кожво-жаберпое дыхание. У личитток Culicinae суще-
ствует два способа дыхания: стигмальное и кожно-жаберное. Основное
значение для газообмена имеет стигмальное дыхание, т. е. дыхание атмо-
сферным воздухом через стигмы.
Личинки Сольп.лю часть времени проводят па поверхности воды, при-
чем стигмы их широко открыты в атмосферу. Воздух поступает через стигмы
в мощно развитую систему трахей, конечные разветвления которых —
тончайшие трахеолы — подходят к отдельным клеткам, доставляя кисло-
род. От стигм у Anopheles отходит два главных трахейных ствола, которые
идут по спинной стороне тела, по бокам от спинного кровеносного сосуда
и доходят до переднего отдела груди, откуда более тонкие ветви направляют-
ся в голову. Главные стволы связаны между собой поперечными ветвями.
207
От каждого из главных стволов в каждом сегменте отходят спинная, брюш-
ная и боковые ветви; последние, соединяясь между собой, образуют два бо-
лее тонких боковых трахейных ствола. В 8-м сегменте от главных стволов
отходит две трахеи в последний сегмент, где они дают четыре ветви, идущие
к жабрам. У личинок Anopheles (и Dixa) впереди стигм, а у Culicini впереди
основания сифона от главных стволов отходят многочисленные трахеолы,
образующие густое сплетение вблизи последней камеры сердца. Это род
трахейного легкого, проходя через которое гемолимфа перед поступлением
в сердце обогащается кислородом. Трахеи проницаемы для газов повсюду,
но всего более — в трахеолах, где стенки тонки. Кислород проводится от
стигм до трахеол и до тканей путем диффузии, благодаря разнице в его пар-
циальном давлении. Углекислота выделяется главным образом через жаб-
ры и кожные покровы. Трахеи, помимо дыхательной функции, служат
вместилищем для запасов воздуха и уменьшают удельный вес тела личинки.
При погружении в воду передняя лопасть стигмальной пластинки за-
гибается назад на 180° и прикрывает собой стигмы. В образовавшуюся
при этом полость захватывается воздух, служащий при подъеме для про-
бивания поверхностной пленки и установления контакта между воздухом
трахей и атмосферой. Замыкание клапанов не бывает полным, но вода не
проникает внутрь и не вытесняет воздух из полости в силу несмачиваемо-
сти стигмальной пластинки водой, что достигается .маслянистой смазкой,
выделяемой стигмальпыми железами, открывающимися около стигм.
Таким образом, назначение стигмальной пластинки состоит в следую-
щем: 1) защищать трахеи от проникновения в них воды во время погруже-
ния личинки в воду; 2) способствовать установлению контакта дыхатель-
ного аппарата с атмосферным воздухом при последующем подъеме; 3) удер-
живать личинку па поверхности воды. Строение стигмальной пластинки
у личинок различных родов и видов комаров очень различно и может слу-
жить для различения этих форм (М нчадский).
' Действие жидких ларвицидов. При действии па личинок нефтепро-
дуктов, налитых на поверхность воды, масло или керосин беспрепятствен-
но проникают через стигмы в трахеи (в силу малого поверхностного натя-
жения этих веществ) и вытесняют воздух. В таких случаях личинки по-
гибают от удушения; к этому присоединяется токсическое действие не-
которых составных частей нефти, которые, проникая через стенки тра-
хей, вызывают остановку деятельности сердца.
Кожпо-жаберное дыхание. Наряду с дыханием за счет атмосферного
воздуха, личинки Culicidae могут дышать и растворенным в воде кисло-
родом: так, лишенные доступа к воздуху, личинки A. maculipennis I—III
возраста гораздо дольше живут в воде, богатой кислородом, нежели в
воде, бедной кислородом. У личинок IV возраста способность к кожно-
жаберному дыханию менее выражена, они быстро гибнут в отсутствии ат-
мосферного воздуха, что связано с более толстой у них кутикулой жабер,
препятствующей деятельности этих последних. При низких температурах
(от 0 до + 10°) личинки младших возрастов могут долгое время оставаться
под водой, даже бедной кислородом, благодаря пониженному потреблению
последнего. Дыхание за счет растворенного кислорода совершается в ос-
новном при помощи анальных жабер и в небольшой степени — через кож-
ные покровы.
3. ОРГАНЫ ВЫДЕЛЕНИЯ
У личинок комаров имеется пять мальпигиевых сосудов; каждый из
них представляет трубку, один конец которой слепой, а другой открывает-
ся в заднюю кишку. Стенки сосудов состоят из одного слоя крупных
•208
железистых клеток с оольшим ядром: между клетками находится канал
для выведения экскретов. Сосуды лежат в полости тела и омываются
гемолимфой.
Экскреция у личинок. Конечным продуктом азотистого обмена у ко-
маров и других насекомых является мочевая кислота (так же, как у птиц).
Зерна мочевой кислоты образуются в клетках сосудов, причем у личинок
I—III возрастов они скапливаются здесь лишь в небольшом количестве.
Однако у личинок IV возраста происходит значительное накопление экс-
крета в клетках, которое продолжается в стадии куколки. Удаление этих
экскретов происходит лишь после первого питания самки кровью.
Отсутствие накопления мочевой кислоты в мальпигиевых сосудах ли-
чинок I—III возрастов объясняется постоянной циркуляцией воды, наса-
сываемой через жабры и выделяемой мальпигиевыми сосудами,
которые, таким образом, непрерывно промываются. У личинок IV возраста
кутикула анальных жабер уплотняется и они перестают функциониро-
вать; в связи с этим прекращается и циркуляция воды через мальпи-
гиевы сосуды, что влечет за собой накопление экскрета.
Подготовка перехода в наземному существованию. Редукция аналь-
ных жабер у этих личинок ведет к изоляции внутренней среды личинки от
окружающей воды; это является началом перестройки жизненных отпра-
влений для перехода к наземному образу жизни. Внутренняя среда ку-
колки уже полностью изолирована от окружающей воды: куколка не пи-
тается, дышит только за счет кислорода воздуха и переносит большую
концентрацию солей в воде, гибельную для личинок и яиц.
4. КРОВЕНОСНЫЙ АППАРАТ
Спинной кровеносный сосуд. Спинной сосуд личинок комаров распо-
ложен на спинной стороне, между главными трахейными стволами; он
проходит от 8-го сегмента брюшка в голову, где открывается. Часть труб-
ки, лежащая в брюшке, снабжена отверстиями с клапанами, через которые
кровь засасывается в сосуд, это — сердце; более узкая передняя часть,
находящаяся в груди и голове, лишенная боковых отверстий, называется
аортой. Волной сокращений кровь прогоняется по сердцу от заднего конца
сосуда к переднему — аорте, и из конца аорты изливается в голову.
Из брюшка в голову по спинному сосуду поступает кровь, насыщенная
пищевыми веществами и очищенная от экскретов. Под влиянием создан-
ной таким образом разности давлений кровь из головы стекает по поло-
стям тела в грудь, а оттуда возвращается в брюшко к стенкам сердца. Та-
ким образом, кровообращение возникает вследствие работы сердца, на-
сасывающего кровь из брюшной полости и перекачивающего его в полость
головы. Деятельность сердца регулируется нервной системой.
Функция кровеносного аппарата. Кровеносный аппарат служит для
доставки пищевых веществ к работающим и растущим органам и для уда-
ления из них продуктов обмена веществ (экскретов); кровь является от-
части также переносчиком кислорода, так как задний отдел сердца опле-
тен сетью трахеол, обогащающей гемолимфу кислородом.
5. ЖИРОВОЕ ТЕЛО
Жировое тело состоит из зеленовато-желтых, блестящих жировых
клеток, скопления которых могут образовать компактные жировые доли.
Жировое тело образует два слоя: наружный, прилегающий к кожному
покрову, неравномерно развитый по всей длине тела, и внутренний —
14 Учебник медицинской энтомологии
209
окружающий пищеварительный канал, постепенно исчезающий в послед-
них сегментах. В передней части груди имеются самые большие скопления
жира. Различают четыре степени ожирения личинок: 1) с незначительным
количеством жира в виде отдельных жировых клеток; 2) с жировым телом
в виде рыхлых скоплений жировых клеток, сосредоточенных преимуще-
ственно в груди; 3) с жировым телом в виде небольших долек; 4) с сильно
развитыми компактными жировыми долями. Степень ожирения личинок
A. maculipennis усиливается к осени (Литвинова). Осенью личинки послед-
него поколения достигают максимального ожирения и дают зимовочных
самок на II, иногда III стадии ожирения по Клиглеру (стр. 220).
6. ЦЕНТРАЛЬНЫЙ НЕРВНЫЙ АППАРАТ
Центральный нервный аппарат состоит из надглоточного и подглоточ-
ного ганглиев; от последнего отходит брюшная нервная цепочка, состоя-
щая из трех сближенных между собой грудных ганглиев и восьми брюш-
ных, из которых первый впоследствии, на стадии куколки, сближается с
грудными ганглиями.
7. ПОЛОВОЙ АППАРАТ
Зачатки половых желез находятся у личинок в 6-м сегменте брюшка,
по бокам кишечника. У личинок IV возраста они уже значительно развиты
и просвечивают сквозь покровы. По форме этих зачатков можно опреде-
лить пол личинки: зачатки семенников округлой формы, яичников —
удлиненной.
8. ИМАГИНАЛЬНЫЕ ДИСКИ
В течение личиночной жизни подготовляется образование органов
взрослого насекомого (имаго). Зачатки органов взрослого комара и прежде
всего ног и крыльев, так называемые имагинальные диски, закладываются
еще на зародышевой стадии (до выхода личинки из яйца); они образуются
из наружного слоя зародыша и имеют вид полых выростов. Усиленный рост
имагинальных дисков начинается в III возрасте личинки.
ГЛАВА VI
ВНУТРЕННЯЯ АНАТОМИЯ И ФИЗИОЛОГИЯ
ВЗРОСЛЫХ КОМАРОВ
1. КУТИКУЛА И ВНУТРЕННИЙ СКЕЛЕТ
Кутикула комаров почти непроницаема для воды и водных растворов;
только при растворении ее поверхностного слоя (эпикутикулы) возможно
проникновение в тело насекомого растворенных в воде веществ. Испарение
жидкости из тела комара почти целиком осуществляется через стигмы.
Эпикутикула плохо смачивается водой, в силу чего капли воды легко ска-
тываются с тела комара. Скатывание их еще облегчается чешуйками, во-
лосками и кутикулярными шипиками, густо одевающими тело комара.
Внутренний скелет комаров образован внутренними выростами частей
наружного скелета.
В голове внутренний скелет представлен двумя продольными пластинками, со-
ставляющими тенториум (рис. 105, Б); они служат для прикрепления мышц
ротовых частей. В грудном отделе внутренний скелет образуется: 1) тремя вилками,
отходящими внутрь тела от стернитов передне-, средне— и заднегруди, и 2) внутрен-
ними ребристыми выростами склеритов бочков этого отдела. Так же как и в голове,
внутренний скелет служит для укрепления тела и прикрепления мышц.
210
2. МУСКУЛАТУРА
В голове комаров имеются следующие мышцы: 1) мышцы-расши-
рители глотки, участвующие в насасывании крови; 2) мышцы, двигающие
ротовыми частями; 3) мышцы, связанные со слюнным протоком, накачиваю-
щие слюну в гипофаринкс. В груди находятся шейные мышцы, двигающие
головой, ножные мышцы и летательные мышцы.
Больше всего места в груди занимают летательные мышцы (рис. 133): 1) продоль -
ные, соединяющие переднюю часть среднеспинки с заднеспинкой; 2) дорсо-вептральные
мускулы, расположенные по бо-
кам от продольных. Дорсо-веиг-
ральные мышцы отходят от стер-
нита и вентральной части мезо-
эпистерны и поднимаются до боко-
вых частей среднеспинки. В том
же отделе прикрепляются муску-
лы ног, связанные со склеритами
заднегруди, среднегруди и перед-
негруди, и мышцы, соединяющие
боковые склериты грудного отдела
друг с другом (межсегменталь-
ные мышцы»
В брюшке в каждом сегмен-
те имеется по четыре мускула:
по два, соединяющих рядом ле-
жащие тергиты, и по два, соеди-
няющих рядом лежащие стерниты
(рис. 134). Эти мускулы способ-
ствуют изгибанию брюшка, на-
пример, при откладке самкой яиц.
Боковые плевральные перепонки
брюшка подостланы поперечными
мускульными тяжами, соединяю-
щими тергиты со стернитами. В брюшке лежит и брюшная диафрагма (рис. 135),
отделяющая нервную цепочку от кишечника. Брюшная диафрагма представляет
тонкую мускульную пластинку. Она прикрепляется к боковым передним краям
стернлтоз. Диафрагма сокращается от переднего края брюшка к заднему, совер-
шая вотнообразные движения. Движения диафрагмы имеют большое значение в
процессах дыхания и кровообращения комаров (см. ниже).
Рис* 133. Мускулатура грудного отдела самки
A. maculipennis.
1— продольная; 2— дорсо-ЕСнтральная.
3. ПИЩЕВАРИТЕЛЬНЫЙ АППАРАТJ
Передняя кишка. Передняя кишка выстлана хитином, и здесь не про-
исходит ни всасывания пищи, ни ее предварительной переработки. Пе-
редняя кишка начинается ротовой полостью (рис. 139, Z), находящейся в
трубке, образованной верхней губой; дальше в голове ротовая полость
переходит в глотку, имеющую два расширения: одно до головного нервного
ганглия и другое позади него (рис. 139, 6).
Передняя глотка снабжена мускулом-расширителем, прикрепляющимся одним
концом к верхней поверхности глотки, а другим к внутренней поверхности передней
части головы. Задняя глотка имеет три мускула-расширителя (дорсальный и два бо-
ковых). Каждый из этих мускулов прикрепляется одним концом к верхней поверхно-
сти задней глотки, а другим — к внутренней поверхности затылка. Дорсальный мус-
кул отходит от средней части верхней поверхности глотки, а боковые — с правой
и левой его стороны.
Слюнные железы. Придатками передней кишки являются: две слюн-
ные железы и три придатка пищевода. Слюнные железы лежат в передней,
вентральной, части грудного отдела (рис. 139). Каждая железа состоит
из трех долек (лопастей). Они построены из крупных железистых клеток.
Боковые, более длинные дольки служат для выделения секрета; средние,
более короткие являются вместилищем секрета. Протоки обеих долей
14* 211
ка.кдон железы соединяются в один общий проток. Протоки желез ооинх
сторон объединяются в непарный слюнный проток (рис. 139, 10), стенка
которого укреплена спиральным утолщением хитиновой кутикулы, в силу
чего представляется поперчно исчерченной наподобие трахейных стволов.
Непарный слюнный проток проходит в канал, лежащий в подглоточнике,
и открывается почти на самом его конце. Около места вхождения протока
в подглоточник в нем можно различить слюнный насос, снабженный
Рис. 134. Мускулатура брюшка
самки A. maculipennis:
А—тергитнап; Б—стернитная (оригинал).
Рис. 135. Брюшная диа-
фрагма самки A. ma-
culipennis (оригинал).
мускулом, назначение которого накачивать слюну в подглоточник
(рис. 139, 11).
Когда комар прокалывает кожу, слюна попадает через ранку в ткани
хозяина, вызывая расширение сосудов и усиление притока крови к месту
укола. Слюна A. maculipennis содержит ряд ферментов: антикоагулин,
задерживающий свертывание крови хозяина, гемолизин, разрушающий
заглоченные кровяные тельца, и агглютинин, агглютинирующий эти
последние, т. е. заставляющий их слипаться между собой.
Придатки пищевода. Придатками пищевода являются: вентральный,
непарный зоб (рис. 139, 14) и спинные придатки (рис. 139, 13) в виде двух
небольших мешочков, обычно заполненных пузырьками воздуха. Зоб
212
комаров — обширный, тонкостенный мешок, открывающийся в пищевод
вблизи преджелудка, в грудном отделе. Задний слепой конец зоба про-
стирается в брюшко, располагаясь вентральнее желудка. Его плоский
эпителий выстлан тонкой, очень растяжимой хитиновой кутикулой. Стенка
зоба оплетена сеточкой мышечных волокон.
При заполнении жидкостью зоб может очень сильно растягиваться, за-
полняя почти все брюшко. При впадении зоба в пищевод расположен коль-
цевой мускул-замыкатель (сфинктер). В тех случаях, когда комар прока-
лывает кожу человека или животных, вход в зоб закрывается и кровь по-
ступает в желудок. Когда же комар без прокола сосет свободные жидкости,
вход в зоб открывается, вход в желудок закрывается и жидкости поступают
в зоб.
В зобе жидкости не всасываются (в силу полной непроницаемости сте-
нок зоба), а постепенно, по мере надобности, волнообразными сокращени-
ями стенки зоба перекачиваются в желудок. Таким образом, зоб служит
вместилищем запасов воды и углеводной пищи. Находясь в сухом воздухе,
комары довольно быстро теряют воду путем испарения и легко могут по-
гибнуть от высыхания. Почувствовав жажду, комар напивается воды, ко-
торая при этом поступает в зоб, а оттуда небольшими порциями постепенно
перекачивается в желудок; в желудке вода быстро всасывается, и таким об-
разом концентрация гемолимфы поддерживается на нормальном уровне.
Иногда в зобе имеются пузырьки воздуха, заглоченные комаром; особенно
сильно наполнен зоб воздухом у только что вылупившихся комаров, что
помогает комару выталкиваться из куколки. В зобе можно обнаружить
дрожжевые грибки и бактерии, попадающие туда со сладкой пищей; быть
может, они вызывают брожение попадающих сюда углеводов.
Желудок. Средняя кишка комаров представляет собой желудок (рис.
139, 75—76), который является местом переваривания и всасывания пищи.
Желудок состоит из двух отделов: переднего узкого и заднего расширен-
ного. У самок, уже пивших кровь, это расширение особенно заметно: же-
лудок принимает грушевидную форму. У новорожденных, еще не пивших
крови самок, желудок имеет цилиндрическую форму с небольшим сужением
впереди. Самая передняя часть желудка представляет небольшое расши-
рение — преджелудок. Пищевод вдается в преджелудок небольшИлМ со-
сочком — пищеводным клапаном, который заключает в себе сфинктер
(замыкательную мышцу), препятствующий пище попадать обратно е пи-
щевод. Сфинктер этот также сокращается, когда комар сосет без прокола
свободные жидкости, и таким образом направляет их в зоб. Стенки
желудка состоят из эпителия, подстилающей его пограничной перепонки
и тонкого слоя мышц. Часть эпителиальных клеток является желе-
зистыми; они выделяют пищеварительные ферменты. В желудке Ano-
pheles доказано наличие ферментов, переваривающих жиры и белки
(Ягужинская).
Перитрофическая оболочка образуется у самок комаров каждый раз
после принятия крови. Она представляет прозрачный мешочек, образо-
ванный тонким слоем хитина, и является продуктом выделения клеток
передней части желудка. Перитрофическая оболочка одевает собой сгу-
сток крови; по окончании пищеварения обрывки ее выделяются наружу
через задний проход вместе с испражнениями, и при следующем принятии
крови перитрофическая оболочка образуется заново (Ягужинская, Ле-
витанская).
Мускулатура желудка пернстальтирует от переднего конца к заднему,
способствуя передвижению ферментов, а позже проталкиванию экскре-
ментов в заднюю кишку.
2 13 •
Утолщение мускулатуры при переходе желудка в тонкую кишку об-
разует задний сфинктер желудка. Непосредственно за сфинктером желудка
в тонкую кишку впадают органы выделения, мальпигиевы сосуды (см.
ниже).
Пищеварение. Полностью насосавшаяся самка имеет брюшко, заполненное
кровью от основания и почти до заднего его конца. Полный желудок растягивает брюш-
ко; растяжение брюшка достигается растяжением и расправлением боковых плевраль
ных перепонок. Внешняя картина процесса переваривания крови следующая: вначале
весь заполненный желудок равномерно мутнокрасного цвета. Потом происходит аг-
глютинация и оседание эритроцитов и кровь расслаивается на темнокрасный осадок
и светлую плазму. Вслед за этим начинается гемолиз, заключающийся в том, что про-
исходит разрушение эритроцитов, плазма окраппГвается гемоглобином и все брюшко
становится рубиновым, прозрачнокрасным. Одновременно с этим содержимое желудка
становится все более и более плотным в силу всасывания воды, входящей в состав
плазмы крови. Расщепление гемоглобина и вызванное им почернение кровяного комка
начинаются прежде всего в задне-вентральной его части, постепенно распространяясь
на всю поверхность комка. Во все время пищеварения задний сфинктер желудка
закрыт, и только тогда, когда все белки переварены и в желудке остается черная масса
гематина, открывается сфинктер желудка и экскременты выбрасываются в заднюю
кишку, а потом через анус наружу. Естественно, что во время переваривания крови в
желудке объем его неуклонно уменьшается.
Задняя кишка. Задняя кишка (рис. 139, 17) слагается из тонкой кишки
и расширения перед анальным отверстием, называемого прямой кишкой
(ректум). В тонкую кишку поступают экскременты из желудка и продукты
экскреции из мальпигиевых сосудов; путем перистальтики этого отдела
они перегоняются в прямую кишку, а затем выбрасываются наружу.
В прямой кишке помещается шесть ректальных желез (рис. 139, 18—20),
каждая из которых богато снабжена трахеями. Назначение этих желез не
выяснено не только для комаров, но и для других насекомых.
1. ТРАХЕЙНАЯ СИСТЕМА И ДЫХАТЕЛЬНЫЙ АППАРАТ
Трахейная система комаров открывается наружу при помощи двух
пар грудных дыхалец и шести пар брюшных. Брюшные дыхальца распо-
лагаются во 2—7-м члениках. Передние грудные дыхальца расположены
на боковой поверхности груди (рис. 104, 35). Они имеют удлиненно-оваль-
ную форму и расположены косо по отношению к грудному отделу.
Грудные дыхальца (рис. 136, А) обладают двумя сильно хитинизованными
выпуклыми губами, расположенными на хитиновой рамке-перитреме. Губами стигмы
снаружи и перитремой внутри ограничено так называемое преддверие. И губы, и пред-
дверие покрыты волосками, образующими цедильное приспособление, очищающее от
пыли воздух, поступающий в трахейную систему. Каждое из передних грудных дыха-
лец дает начало семи трахейным стволам (см. ниже). На живых комарах хорошо видно,
как закрываются и открываются грудные дыхальца. Закрывание их обусловлено со-
кращением мускула-замыкателя, прикрепляющегося одним концом к переднему краю
перитремы, а другим — к внутреннему выступу проэпимеры.
Задни е грудные дыхальца устроены так же, как и передние, расположены вер-
тикально в заднем отделе грудки; все составляющие их части слабее хитинизованы
(рис. 136, Б).
У различных комаров величина грудных дыхалец различна. Сухо-
устойчивые виды Anopheles (superpictus и pulcherrimus) имеют относитель-
но меньший размер грудных дыхалец, по сравнению с влаголюбивыми ви-
дами (например, A. algeriensis, A. mac. messeae, A. hyrcanus).
У сухоустойчивых видов малые размеры грудных дыхалец, наряду с
а ругими приспособлениями, ведут к сокращению поверхности испарения
и экономии воды в теле.
Брюшные дыхальца устроены сложнее грудных (рис. 136, В). В них можно раз-
личить перитремуи прилегающую к ней постстигма л ьную камеру, покрытую внутри
21ч
волосками. К постстигмальной камере подходит трахея. К основанию ноетстигмаль-
ной камеры примыкает клапан, снабженный мышцей-замыкателем и способный плотно
закрывать вход в трахею.
Клапан состоит из пластинки, приросшей к стенке камеры и снабженной рыча-
гом. Пучок мускульных волокон прикрепляется одним концом около вершины рычага,
а другим — к стенке постстигмальной камеры у места вхождения трахеи. При сокра-
Рис. 136. Дыхальца A. maculipennis.
А—переднее торакальное дыхальце: 1—губы; 2—волоски цедильного аппарата; и—мускул:
Б—заднее торакальное дыхтльце; обозначения те же, что и в переднем дыхальце; В— абдоми-
нальное дыхальце: 1—перитрема; 2—постстигмальная камера; 3—клапан; / — мускул-
замыкатель; Г— абдоминальные стигмы разных видов Anopheles и Agdes (оригинал): 1— A. sn-
perpictus; 3 — A. bifurcatus: 3 — Agdes cinereus.
щении мускула пластинка клапана втягивается в стенку камеры и закрывает отверстие
трахеи; при расслаблении мускула открывание стигмы происходит в силу упругости
камеры.
У различных видов комаров.наружное отверстие постстигмальной камеры может
быть резко суженным, например, у A. superp ictus, или широким (A. algeriensis) и даже
расширенным (Аёйез cinereus) (рис. 136, Г). Ширина наружного отверстия имеет здесь
215
то же значение, что и в торакальных стигмах: чем шире отверстие, тем сильнее проис-
ходит испарение с поверхности трахейной системы (Виноградская).
Наибольшее развитие трахей мы находим в груди (рис/137), где раз-
вита мускулатура, требующая для работы обильного притока кислорода.
От передних дыхалец отходят по семи трахейных стволов (в голову, к
хмышцам груди и ногам). Одна пара этих стволов соединяет передние дыхаль-
ца с задними. От заднего торакального дыхальца, кроме того, отходит мно-
жество более мелких трахей и по одному стволу с каждой стороны к брюш-
ку. Продольные стволы брюшка дают ответвления к брюшным дыхальцам.
В местах этих ответвлений от продольных стволов отходят также и тра-
хейные ветви к органам брюшка: кишечнику, половому аппарату, мус-
кулатуре, сердцу, жировому телу.
Дыхание. Смена воздуха в трахеях насекохмых происходит двояким пу-
тем: в большие стволы воздух входит и выходит через дыхальца, благодаря
сжатию и расширению трахей при движении насекомого; в тонких ветвях
обмен газов совершается путем диффузии. У спокойно сидящего комара
обмен воздуха в крупных трахеях совершается благодаря дыхательным
движениям брюшка, обусловленным сокращением диафрагмы. При полете
летательные мышцы, сокращаясь, ритмически сжимают лежащие между
ними мощные трахейные стволы груди, и чем быстрее работают мышцы,
тем сильнее проветривается трахейная система.
Дыхательный ритм. Дыхательные движения диафрагмы хорошо видны на жи-
вых комарах. По бокам тела, на краях стернитов, поочередно образуются впячивания,
причем волна сокращений пробегает вдоль брюшка от переднего его конца к заднему.
За несколькими (10—14) волнами сокращений следует перерыв в несколько секунд.
Во время остановки диафрагмы грудные дыхальца открываются, при возобнов-
лении дыхательных движений они закрываются. Чем выше окружающая температура,
тем быстрее дыхательные движения; число их достигает 50 в минуту (при 30°).
216
Рис. 138. Строение серд-
ца Anopheles maculipen-
nis (оригинал Л. В. Ягу-
жинской).
1 — тергиты брюшка;
2—сердце; 3—крыло.видные
мышцы; 4 — ОСТИИ.
крови осуществляется
5. ВЫДЕЛИТЕЛЬНЫЕ ОРГАНЫ
Выделительные органы взрослого комара по строению своему ничем
не отличаются от таковых личинки.
Работа мальпигиевых сосудов. К моменту вылупления комара из ку-
колки клетки мальпигиевых сосудов битком набиты зернами мочевой ки- _
слоты. Когда самка комара напивается крови, вода крови очень быстро
всасывается стенками желудка и попадает в полость тела комара. Отсюда
она выводится через мальпигиевы сосуды и по пути производит промывку
этих последних. Поэтому тотчас вслед за выде-
лением воды мальпигиевы сосуды выделяют и на-
копившуюся в них мочевую кислоту в виде белова-
той кашицы. У голодающих комаров освобож-
дение мальпигиевых сосудов приостанавливается,
накопление экскрета продолжается. Клетки опять
заполняются экскретом. Таким образом, маль-
пигиевы сосуды комаров подчиняются ритму
гонотрофических циклов так же, как и другие
органы (стр. 156).
У зимующих комаров, не принимающих пи-
щи, эпителий мальпигиевых сосудов бывает до
отказа набит зернами мочевой кислоты.
О процессах, происходящих в кишечнике и маль-
пигиевых сосудах комара на протяжении гонотрофического
цикла, можно судить также по количеству и цвету
пятен, выделяемых комаром через анальное отверстие.
Самка, свеженасосавшаяся крови, тотчас же, иногда
еще во время сосания, начинает выделять прозрачные
капельки. Это — вода крови, очень быстро всасываю-
щаяся из желудка в полость тела, .откуда она выво-
дится мальпигиевыми сосудами в заднюю кишку, а затем
наружу. Высыхая на поверхности стекла, она образует
«водянистые» пятна. Рассматривая их под микроскопом,
можно различить содержащиеся в них кристаллы солей.
Желтоватые пятна — это сплошная масса экскрета
(мочевой кислоты) из мальпигиевых сосудов; они начинают
выделяться через несколько часов после приема пищи и
продолжают выделяться до конца пищеварения.
Черные пятпа состоят из гематина — неперевари-
мого продукта расщепления гемоглобина крови; он вы-
деляется до тех пор, пока желудок не освободится от
гемоглобина.
Помимо мальпигиевых сосудов, выделительную функ-
цию несут и перикардиальные клетки (крупные клетки,
лежащие двумя цепочками по бокам сердца), а у зимую-
щей самки также и клетки жирового тела, накапливаю-
щие в себе продукты обмена веществ.
6. КРОВЕНОСНЫЙ АППАРАТ
У комаров, как и у других насекомых, замк-
нутой кровеносной системы нет. Кровь или гемо-
лимфа заключена в полости тела. Передвижение
благодаря работе сердца и брюшной диафрагмы.
Сердце. Сердце комаров (рис. 138) представляет собой тонкостенную
мускулистую трубку, не разделенную на камеры и лежащую под 1—7-м тер-
гитами брюшка. На границе груди сердце переходит в аорту; аорта уже
сердца и лишена мышц; на переднем своем конце она открывается в по-
лость головы. В каждом членике брюшка (от 1-го до 7-го) в сердце имеются
боковые отверстия (остии), снабженные клапанами. Кроме того, одна пара
217
l—нижняя губа; 2—•
(ОЛИ СЛЮННОЙ железы; - ....у--- w..«,»uv» ...««*«««., „ — и«кпиш vowuuma mcj*co; 1U--- ООЩИИ посте
слюнного Прогона; 12 — пищевод; /3 — парный придаток пищевода; 14— непарный придаток пищевода (зоб)- 15-
17 — задняя щипка; 18 п 29 — ректальные железы; 19 — задний проход; 21 — аорта; 22 — сердце- 23 — надгло
24 — ПОДГЛОТОЧНЫЙ ганГЛИП; 2 а—27 — t-ц, 2-Й И 3-Й грудные ганглии; 28—34 — 1 — 7-Й брюшные ганглии; 35— МЗЛышгисвы иииудш.
'Д~* 37—парный яйцевод; 38 — общий яйцевод; 39—придаточная железа; 40—сперматека; 41 проток сперматеки;
8-и тергцт; 43 — 1-й тергит; 44 спинная пластинка заднегруди; 45 и 48 — продольная мускулатура груди; 46 — щиток:
дорсо-вептральная мускулатура; 49—среднеспинка; 57— плечевой склерит; 57—затылок; 52—лоб; 53—переднеглоточное расшире-
ние; 54 — мускулы ротового аппарата; 55— клапан насоса слюнных желез.
Рис. 139. Внутреннее строение A. bifurcatus.
проток слюнных желез; 3 — верхняя губа; 4 — ротовая полость; 5 — усик; 6 — задняя глотка; 7 — боковые
; 8 • оредаяя доля слюнной железы; 9 — парные протоки слюнных желез; 10—общий пооток- 11—мускул
— пишаво.г- 13 — пяпиий ппггтта'гпь» ггтжгтг^тта. --------- б), 75—16 — желудок;
надглоточный ганглий;
мальпигиевы сосуды;
остии имеется на заднем конце сердца. Волны сокращения мускулатуры
проходят вдоль по сердечной трубке, прогоняя кровь из одного ее конца
в другой. Сердце комаров может сокращаться и гнать кровь как от зад-
него конца тела к переднему (прямая перистальтика), так п от переднего
конца к заднему (обратная перистальтика).
Кровообращение. При прямой перистальтике кровь из полости тела засасывается
в сердце через остии и, прогоняясь вдоль сердца вперед, в аорту, идет по аорте до
головы и там изливается опять в полость тела. Обратно из головы в брюшко кровь ча-
стично стекает по промежуткам между органами, орошая эти последние. Однако назад
кровь стекает медленно, и при длительной прямой перистальтике в переднем конце
тела скопляется слишком много крови. Тогда сердце начинает работать в обратном
направлении — волны сокращения проходят по сердцу спереди назад (обратная пери-
стальтика). При этом кровь засасывается из головы через аорту, проходит по сердцу
спереди назад и изливается через задние остии в полость брюшка (Ягужинская). Вен-
трально от сердца к его наружной поверхности прикрепляются в каждом сегменте
треугольные крыловидные мышцы. Эти мышцы с двух сторон срастаются между со-
бой под сердцем; они служат для прикрепления сердца, но не играют непосредствен-
ной роли в его растяжении. По бокам от сердца с каждой стороны лежит по одному ряду
крупных перикардиальных клеток. Под крыловидными мышцами лежит тонкая соеди
нительнотканная пленочка (спинная диафрагма).
Кровь. Кровь комаров — бесцветная жидкость; она принимает не-
большое участие в переносе кислорода; крупные и активно работающие
органы снабжены трахеями и получают кислород непосредственно из тра-
хей. Кровь является полостной жидкостью, циркулирующей во всех по-
лостях тела; ее функция — осуществлять химическую связь между отдель-
ными органами, тканями и клетками; она приносит им питательные веще-
ства и уносит из них продукты обмена.
7. ЖИРОВОЕ ТЕЛО
Жировое’тело имеет большое значение в жизни комаров, особенно
в период диапаузы, помогая им сохранять жизнеспособность в течение хо-
лодного периода года. На рис. 140 показан продольный разрез вполне жир-
Рис. 140. Медиальный разрез жирной самки и строение жировой ткани
A. maculipennis (оригинал).
1— жировые дольни в брюшке; 2— строение жировой ткани.
нои самки A. maculipennis. Жировые дольки развиваются главным обра-
зом в брюшке, а отчасти в грудном отделе и меньше в голове. В брюшке
они располагаются под тер гитами и над стер нитами, окружая желудок
219
и заднюю кишку. Каждая долька построена из крупных клеток, на пол-
ненных запасными веществами: капельками жира, гликогеном и зернами
белка. Иногда клетки жирового тела содержат также зерна пигмента и
экскретов. По аналогии с другими насекомыми можно думать, что и у ко-
маров клетки жирового тела образуются оседанием блуждающих клеток
гемолимфы, отлагающих в своей протоплазме запасные вещества.
Источники жира. Нередко комары вылупляются из куколки с доволь-
но хорошо развитым жировым телом. При кормлении самки комара са-
харистой пищей жировое тело ее значительно увеличивается, она жиреет,
в особенности если содержится при прохладной температуре. При голода-
вещества расходуются и жировое тело уменьшается. При
кровяном питании нормальная летняя самка не жиреет,
так как усвоенная пища тратится, помимо поддержания
жизни, на развитие яичников.
Осеннее ожирение. Осенью личинки A. maculipennis
в водоемах значительно жирнее, чем летом, и дают начало
более жирным самкам (Литвинова). У всех комаров, зи-
мующих в состоянии диапаузы (A. maculipennis, A. hyr-
canus, Culex pipiens и др.), диапаузирующие самки осенью
сильно жиреют, частью за счет углеводного, частью за
счет кровяного питания. Огромное количество накоплен-
ных запасных веществ при низких температурах зимних
убежищ тратится медленно и постепенно, что позволяет
комарам пережить зиму (см. ниже, стр. 284).
При максимальном развитии жирового тела по
внешности самки можно подумать, что брюшко ее за-
полнено зрелыми яйцами. Но все же отличия совершенно
отчетливы. Жирная самка имеет сероватое, просвечи-
вающее брюшко, у самок же со зрелыми яйцами брюшко
желтовато-белое и не просвечивает.
Степени ожирения. Клиглер и Мер (1930) предло-
жили различать по внешнему виду следующие степени
ожирения диапаузирующих самок A. mac. sacharovi
(рис. 141).
Стад ия 1. Брюшко слегка изогнуто кверху и плевры
не растянуты, образуют по бокам брюшка слегка выгнутые бо-
розды. На поперечном разрезе через брюшко тергиты п стер ни гы
тесно сближены.
Стадия 2. Плевры слегка растянуты, бороздок не об-
разуют, брюшко еще изогнуто.
3. Заполнение брюшка жиром придает ему цилиндрическую форму,
брюшко выпрямляется.
Стадия 4. Вентральная сторона брюшка становится слегка выпуклой.
Описание степени ожирения, обнаруживаемой при вскрытии комаров, дается
Беклемишевым и Митрофановой, устанавливающими четыре степени: 1) полное от-
сутствие капель жира; 2) жировое тело, развитое в пределах летней нормы, т. е. от-
дельные клетки по поверхности кишечника и кольцо вокруг сперматеки, по обособ-
ленных долек нет; 3) небольшие или умеренные доли жирового тела (полужирные
самки); 4) брюшко раздуто от крупных долей жирового тела (вполне жирпые самки).
По-настояшему жиреющими можно считать только две последние категории.
нии запасные
4
Рис. 141. Степе-
ни ожирения.
1 — брюшко изо-
гнуто, плевры не-
видимы; неожи-
ревшая самка;
2-— показались
плевры, брюшко
изогнуто (начало
ожирения); з —
б рюш но прямое,
цилиндрическое;
4 — брюшко г
профиль выпук-
лое (жирная сам-
ка).
Стадия
8. нервный аппарат
Центральный нервный аппарат (рис. 139), как и у всех членистоногих
состоит из надглоточных ганглиев (головного мозга) и брюшной нервной
цепочки. Надглоточный ганглий окологлоточными коннективами соеди-
220
няется с подглоточным, который образован у насекомых слиянием трех
первых ганглиев брюшной цепочки и расположен также в голове. У ко-
маров окологлоточное кольцо узкое и как бы сжимает глотку, которая
в силу этого перетянута на две части — переднюю и заднюю.
Надглоточный ганглий иннервирует органы чувств головы—глаза и
усики, а также верхнюю губу и оказывает регулирующее влияние на ра-
боту ганглиев брюшной цепочки. Подглоточный ганглий управляет рото-
выми органами (кроме верхней губы). Все три грудных ганглия тесно сбли-
жены между собой и к ним присоединен первый брюшной ганглий. Сле-
дующие пять брюшных ганглиев, 2—6-й, лежат каждый в своем членике
и соединены друг с другом длинными коннективамп. В 7-м членике лежит
сложный ганглий, образованный слиянием
ганглиев 7-го и всех последующих члеников
брюшка (см. рис. 124, Б).
Грудные ганглии управляют движением
ног, крыльев и жужжалец. Брюшные ган-
глии управляют каждый своим сегментом,
и от них отходит всего по одной паре нер-
вов. Последний брюшной ганглий комаров,
кроме последних члеников брюшка, посы-
лает нервы еще и к задней кишке и половому
аппарату. Передняя кишка у всех насеко-
мых обслуживается симпатической нерв-
ной системой, связанной с надглоточным
ганглием.
0. ПОЛОВОЙ АППАРАТ
Мужской половой аппарат. Самец имеет
Рис. 142. Половой аппарат сам’
ки комара.
1—яичники; 2—парный яйце-
вод; 3 — ампулы непарного яйце-
вода; 4— влагалище; 5— спер-
матека; 6—придаточная железа.
два семенника, от задних концов которых
отходит пара семявыносящих протоков, кото-
рые сливаются в непарный семяизвергатель-
ный проток. Имеются также придаточные
железы. Семяизвергательный проток откры-
вается наружу на конце наружного совоку-
пительного органа (aedeagus) (см. выше, стр. 186). Живчики (сперматозо-
иды) комаров длинные, нитевидные, подвижные.
Женский половой аппарат. Половой аппарат самки состоит из двух
яичников, двух яйцеводов, общего яйцевода, сперматеки и придаточной
железы (рис. 142). У Culicini, кроме перечисленных частей полового ап-
парата самки Anopheles, имеется совокупительная сумка, и вместо одной
сперматеки — их три.
Каждый яичник построен из большого числа коротких яйцевых тру-
бочек. Число их колеблется от нескольких десятков до 200 с лишним, в сред-
нем около 150. Яйцевые трубочки располагаются вокруг центрального яй-
цевода, в который они и открываются. Внутренний яйцевод идет вдоль
всего яичника; спереди он кончается слепо, задний его конец переходит
в парный яйцевод, лежащий уже вне яичника. Снаружи яичник одет со-
единительнотканной оболочкой. На переднем его конце эта оболочка про-
должается в концевую нить, которая прикрепляется к стенке брюшка в
4-м сегменте. Яйцеводы, как внутри яичника, так и по выходе из него,
снабжены сильной мускулатурой. Имеются мышечные волокна и между
яйцевыми трубками. Вся эта мускулатура служит для выведения зрелых
яиц из яичника. Яичники обильно снабжены нервами и трахеями.
221
Каждая яйцевая трубочка состоит из концевой камеры и двух фол-
ликулов. В концевой камере находится зона размножения; старший фол-
ликул лежит ближе к яйцеводу, младший — ближе к зоне размножения.
После созревания старшего фолликула и удаления его яйца второй фол-
ликул становится старшим, а за ним образуется третий, который и стано-
вится теперь младшим. Таким образом, в каждой яйцевой трубочке
последовательно образуются и развиваются один за другим несколько
фолликулов. Фазы развития фолликулов описаны Кристоферсом, а затем
подробнее Мером и исцользуются при изучении биологии комаров
(рис. 143).
Фазы развития яйцевого фолликула комаров по Кристоф ер су-Меру
Развитие яйцевого фолликула. Фаза I. Фолликул состоит из малень-
ких групп недиференцированных клеток......................фаза Nc.
Происходит обособление фолликулярных клеток от центральных.
Фолликулярные клетки слагаются в правильный кубический эпителий
фаза N Мера......................................................
Фолликул становится овальным, ядро яйцевой клетки становится
отличимым от ядер фолликулярных клеток...............фаза I Мера.
Фаза II. В протоплазме яйцевой клетки появляется вокруг ядра
венец из мелких желточных зерен...................фаза I—II Мера.
Появляются более крупные и более многочисленные желточные зерна
....................................................фаза II Мера.
Яйцо увеличивается по сравнению с желточными клетками до половины
размеров фолликула
Фаза III. Яйцо последовательно занимает 2/3, 3;4 фолликула.
Ядро его больше не видимо, будучи закрыто массой желтка.
Фаза IV. Фолликул удлиняется. Питательные клетки становятся
все менее отчетливо видимыми.
Фаза V. Появляются поплавки, и становится заметна структура
хориона. Яйцо откладывается.
Желтые тела. Хорион образуется только около яйцевой клетки, пи-
тательные клетки остаются вне его, в проксимальном, т. е. расположен-
ном ближе к зоне размножения, конце фолликула (рис. 143). После от-
кладки яиц остатки питательных клеток лежат в небольших расширениях
яйцевых трубок, впереди от новых развивающихся фолликулов, в виде
прозрачных или желтоватых капель. Эти желтые тела при последую-
щих кладках выталкиваются прочь, но расширения яйцевых трубочек
остаются и по числу их можно различать возраст самок (см. ниже).
Созревание яичников идет параллельно пищеварению: с уменьшением
объема крови в желудке увеличивается объем яичников. Увеличиваясь
в объеме, яичники оттесняют желудок вперед и вентрально, так что к кон-
цу пищеварения все брюшко оказывается заполненным беловато-желто-
ватой массой зрелых яиц.
Трахейная система яичников; клубочки трахеол. Каждый яичник по-
лучает два трахейных ствола, которые сильно ветвятся внутри яичника.
Каждая самая тонкая веточка делится на пучок тончайших трахеол
и в этом месте бывает смотана в клубочек. Множество таких клубочков
лежит среди яйцевых трубок. Концы трахеол выходят из клубочка и
оплетают соседние яйцевые трубочки вплоть до концевых камер
(рис. 144, А, 1).
При развитии яичника созревающие фолликулы вырастают во много
раз. По мере их роста прикрепленные к ним трахеолы разматываются, и
is моменту полной зрелости яичника вся его возросшая масса оказывается
пронизана густой сетью трахеол. Таким образом, трахейные клубочки
222 #
Рис. 143. Созревание яиц
A. maculipennis messeae. Вид отдельных яйцевых
трубок.
У
в фа£еР1— ПФМераК7вЛтоВпоФй— в В«Т>Й— в фа8е N°: г~ “в₽вый фолликул
Мера, второй_в Фазы Na* 4_прппмй «ьАппп первый фолликул в начале фазы II Кристоферса-
пер^йфолликул в кон^ ^азы в КОжЦе Ча8? П’ второ/- в фазе Nfcepa;
фазы, второй—в фазе I-JI- 7 — пеп^мГЛазе 1; первый фолликул в конце IV
У-•желтое тело»;/- питательта?клеХ-^ Ф^8Ы’ ВТ°Р°Й~ в фазе II;
22 — нона V» ПОПЛ?ВКИ; ^—Фолликулярный эпителий;
ла вона размножения; 13 — яйцевая клетка.
Рис. 144. Определение возраста самок.
А — по окончаниям трахей в яичниках: 1 — самки, не откладывав-
шие яйца (трахеолы свернуты в клубочки); 2 — самки, откладывавшие
яйца (клубочки развернуты) (по Детиновой); Б—по расширениям в яй-
цевых трубочках! 1 — яичники и яйцеводы у новорожденной самки;
2—то же после первой кладки; 3—то же после трех кладок;
В — по структуре ампул (оригинал В. П. Половодовой).
представляют приспособление, обеспечивающее непрерывное снабжение
воздухом быстро растущего яичника.
После откладки яиц трахеолы не могут обратно смотаться в клубочки
и в спавшемся яичнике отложившей самки образуют густую сеть волни-
стых нитей (рис. 144, А, 2). Правда, иногда остаются отдельные неразмо-
танные клубочки в связи с тем, что отдельные яйцевые трубки могли не
участвовать в развитии яичников. Тем не менее отличить яичники ни
разу не клавшей самки от яичников самки, уже однажды отложившей,
по строению яичников очень легко (Детинова) х.
Гонотрофический цикл. Мы видели уже, что в связи с периодическим
принятием пищи вся жизнь взрослой самки комара распадается на ряд
последовательных гопотрофических циклов. Каждый гонотрофический
цикл можно разделить на три большие фазы: 1) поиски добычи и напа-
дение; 2) переваривание крови и созревание яиц; 3) поиски водоема и от-
кладка яиц. После откладки яиц самка снова оказывается голодной
особью, ищущей добычу, т. е. вступает в первую фазу следующего цик-
ла (Беклемишев).
Селла (1920) дал более дробное разделение гонотрофического цикла
Anopheles на 7 фаз (рис. 145), легко различимых у большинства видов по
внешнему виду брюшка самки, благодаря просвечивающим через покровы
желудку и яичникам.
К 1 стадии по Селла относятся самки голодные, с пустым желудком и
неразвитыми яичниками, ко II стадии по Селла — самки, только что
напившиеся крови, к III, IV и V стадии — самки, находящиеся на про-
межуточных фазах пищеварения и созревания яичников. К VI стадии
относятся самки с почти переваренной кровью и яичниками на последней
стадии развития. На VII стадии крови в кишечнике уже нет, она вся пе-
реварилась и непереваренные остатки выведены наружу, яичники до-
стигли полного развития.
Созревание яичников у комаров совершается под воздействием гормона, выделен-
ного железой внутренней секреции, corpora allata. Corpora allata у самок A. maculi
pennis расположены в виде двух долек по бокам переднею конца аорты, в задней ча-
сти шеи (рис. 146). Поступление гормона в кровь происходит в первые часы после
принятия самкой крови и служит непосредственным толчком к развитию яичников
(Детинова).
Непарный яйцевод, ампулы. Непарный яйцевод выстлан железистым
эпителием и имеет хорошо развитый мышечный слой. У самок рода Ano-
pheles в местах впадения парных яйцеводов непарный яйцевод образует
расширения, называемые ампулами. У неклавшей самки эпителий непар-
ного яйцевода не содержит секрета. Во время созревания яичников он
заполняется секретом, отчего объем эпителиальных клеток увеличивается.
Особенно заметны эти изменения на ампулах, которые при этом увеличи-
ваются в размерах и становятся менее прозрачными. При каждой откладке
яиц часть секрета расходуется, и ампулы чуть-чуть уменьшаются; но при
каткдом созревании яичников они опять несколько увеличиваются, хотя
и не так сильно, как при первом созревании. Изменения ампул яйцевода
служат одним из критериев для определения возраста самки (Мер, Поло-
водова) (см. ниже, стр. 283).
1 Для изучения трахейной системы яичников последние выделяют обычным спо-
собом. ведя вскрытие не в физиологическохм растворе, а в чистой воде. Не покрывая
покровным стеклышком, яичники подсушивают; трахеолы, при жизни частично за-
полненные жидкостью, заполняются воздухом и становятся хорошо видимыми. Под-
сушенные яичники рассматриваются в сухом виде при среднем увеличении микро-
скопа.
15 Учебник медицинской энтомологии
22
Семяприемник. Семяприемник (сперматека) у Anopheles шаровидной
формы; наружный слой его стенки клеточный, а внутренний хитиновый,
пронизанный рядом маленьких круглых отверстий. Тонким стебельком
сперматека соединяется с влагалищем, т. е. выводной частью непарного
яйцевода.
В сперматеке в течение всей жизни самки сохраняются жизнеспособ-
ные сперматозоиды. Одна копуляция может обеспечить оплодотворение
яиц во всех кладках данной самки. Даже во вермя зимы у зимующих
Рис. 145. Стадии переваривания крови и развития яичников
(вид на Срюшко самки сбоку).
1— без кропи, яичники не развиты; 2— бри ш ко полно кропи, бо-
лее или менее красное; яичники не раз> иты; 3— много крови, еще
красноватой; сзади свободны 2—2.5 стернита и 4—5 тергитов; 4—
кровь черная, свободны от нее 2 5—3 стернита и 5—6 тергитов;
5—кровь черная, свободны 2.5—3.5 стернита и 6.5—7 5 тергита; 6—
кровь черная только на брюшной стороне, под конец стадии остается
в ничтожном количестве; 7 — кровь совершенно переварена, яйца
зрелые.
Рис. 146. Расположение согрета allata
поотно1иению кдругпм органам самки
A. maculipennis (п перечный срез
через шею комара).
1— доли corpora allata; 2— аорта; 3—
стенки пищевода; 4—стеши тела;
5 — мышцы; 6 — жировые доли.
самок сохраняются жизнеспособные сперматозоиды, и весной эти самки
откладывают оплодотворенные яйца без дополнительной копуляции.
Выходя из яичника, яйцо проталкивается в парный яйцевод, потом
в непарный общий и во влагалище. Сюда же открывается проток спер-
матеки, и здесь происходит оплодотворение проходящего яйца. Сперма-
тозоиды проникают в яйцо через микропиле, отверстия, находящиеся на
концах яйца.
ГЛАВА VII
ЭКОЛОГИЯ ВОДНЫХ ФАЗ ANOPHELES MACULIPENNIS
[ 1. УСЛОВИЯ СУЩЕСТВОВАНИЯ яиц
Как уже упоминалось, самки A. maculipennis откладывают свои
яйца на поверхность воды, где они и плавают, поддерживаемые силами
поверхностного натяжения, вплоть до момента вылупления личинки.
Причины гибели лиц в природе. В природных условиях яйца кома-
ров могут попадать в условия, неблагоприятные для их существования.
Одной из причин, вызывающей нередко их гибель, является высыхание.
226
Яйца, оказавшиеся после откладки вне воды, не темнеют и гибнут. Яйца,
попавшие под воду, чернеют, но также не развиваются, в отличие от
яиц A. superpictus. Яйца подвергаются высыханию, будучи выброшен
ными волной на берег или при колебаниях уровня водоема, когда они
могут оказаться вне воды на обнажившейся прибрежной растительности.
Япца A. maculipennis плохо переносят даже кратковременное высыха-
ние. Особенно чувствительны к высыханию свежеотложенные яйца;
после сформирования личинки устойчивость к высыханию повышается.
Так, свежеотложепные яйца A. maculipennis не выдерживают и 15-ми-
нутного пребывания на сухом субстрате при 24—26° (и дефиците влаж-
ности 10—13 мм). В этих же условиях часть яиц со сформировавшейся
личинкой сохраняла жизнеспособность и после 29-часового пребывания
на сухом субстрате. При более высокой температуре продолжительность
высыхания, которую может пережить яйцо, еще меньше. На влажном
грунте яйца сохраняют жизнеспособность дольше.При температуре около 0J
на влажной земле яйца остаются жизнеспособными неделями.
В природе яйца Anopheles подвергаются также замерзанию. Так,
весенние ночные заморозки, сопровождающиеся образованием поверх-
ностной корочки льда в мелких водоемах, днем хорошо прогреваемых,
ведут к значительной гибели отложенных яиц. В экспериментах замора-
живание яиц при —4—8° уже через 24 часа губит значительный процент
яиц. Однако часть яиц A. maculipennis messeae в Омске выживала послз
их охлаждения до —18° (Нецкий).
Отношение яиц к солености воды. Яйца A. maculipennis, отклады-
ваемые в очень соленую воду (например, 3%), изменяют форму и поги-
бают в результате чрезмерного осмотического давления; в воде меныпэй
солености (2—1%) происходит также частичная их гибель. Однако и в
нормальных условиях среды яйца A. maculipennis, как правило, не дают
полной «всхожести»; обычно она бывает не выше 95%.
Яйца Anopheles могут поедаться врагами (улитками, личинками
IV возраста того же вида комара).
Влияние температуры. Продолжительность развития яиц зависит <т
температур»]. При 10—12° оно продолжается 8 суток, при 16—19'—
5 суток, при 20—25° — 3 суток, при 25°—2 суток и при 27—29° —
46 часов. Яйца A. maculipennis одной кладки при одинаковых условиях
развиваются с неодинаковой скоростью, и вылупление личинок из яиц
происходит неодновременно; вылупление небольшого процента личинок
иногда затягивается до 5 и даже до 18 дней.
2. ПРИСПОСОБЛЕНИЕ ЛИЧИНКИ* К (ЖИЗНИ В ПОВЕРХНОСТНОЙ ПЛЕНКЕ
ВОДЫ. POehb СУБСТРАТА
Прикреплеспе личинки к поверхности воды. Личинки Anopheles
Гдлывую часть времени проводят на поверхности воды в г )ризонтальнэм
положении, прикрепившись к поверхностной пленке стигмальной пластин-
кой, пальмовидными волосками и плечевыми лопастями. Эти образования
позволяют личинке удерживаться у поверхности без каких-либо
усилий с ее стороны: силы поверхностного натяжения поддерживают
личинку, несмотря на то, что опа удельно тяжелее воды.
Ес и мы будем искусственно снижать го' ерхиостпое натяжение голы ниже олре-
долетшо о пге’ета (например, прибавляя к re.i слабый мыльный распор), личинки
теряют сго’он ость прикрепляться к г o’epxi ости; в такой годе их органы крику вяле-
ния теряют clojcti о песмачивсемозти. Однако в естественных ано^ело: енных l од осмат
15*
227
поверхностное натяжение всегда значительно выше того предела, который допускает
прикрепление личинок Anopheles к поверхности воды.
Личинка обычно держится на поверхности около какого-либо пред-
мета, прикрепившись к нему крючками хвостовых волосков.
Такое положение часто достигается пассивным путем: личинка, оказавшаяся
вблизи берега или какого-либо соприкасающегося с поверхностью воды предмета,
притягивается к нему силами поверхностного натяжения: при этом задний конец ли
чинки со стигмальпой пластинкой притягивается сильнее переднего. С другой стороны,
личинка, находящаяся на поверхности воды без прикрепления хвостовыми крючками
к какому-либо предмету, приходит в возбуждение и начинает перебегать по поверхно
стиводы и продолжает это до тех пор, пока не найдет точку опоры. Прикрепление ли-
чинки задним концом тела к предмету резко снижает частоту ее самопроизвольных дви-
жений.
Значение субстрата для существования личинки. Таким образом,
личинке необходимы предметы (субстрат) для прикрепления, точка при
чала. Субстратом является берег водоема или растительность, достигаю-
щая поверхности, или различные плавающие на поверхности предметы.
Поэтому личинки встречаются в водоемах среди доходящей до поверх-
ности растительности, плавающего растительного мусора или бревен,
или просто у берега. Роль берега как субстрата особенно резко выражена
в биотопах A. superpictus и A. mac. typicus, «микроводоемах» галечного
русла горных речек (т. е. заполненных водой небольших промежутках
между гальками), часто в отсутствие нитчаток и высшей растительности
(рис. 76).
Наличие субстрата, дающего причальную линию, — одно из условий
массового развития личинок.
3. ПИТАНИЕ ЛИЧИНКИ ANOPHELES
а) Способы питания
Способы принятия пищи. Личинки комаров Culicidae пользуются
разнообразными способами добывания пищи: активной безвыборной филь-
трацией, обскребыйанием субстрата, заглатыванием крупной добычи.
Многие личинки комаров узко специализованы на одном спо-
собе питания, в то время как личинки Anopheles пользуются своим
ротовым аппаратом разнообразно: 1) при помощи быстрых, со скоростью
до 200 взмахов в минуту, ритмических движений щеток верхней губы
спереди назад они способны вызывать в поверхностном слое мощные
токи воды по направлению корту, с которыми притекают частицы пищи;
это основной способ питания личинки Anopheles, называемый фильтра-
цией 2) они способны соскребывать пищу с субстрата; 3) разгрызать
нитчатые водоросли и некоторых планктонных животных.
При работе щеток личинка находится у поверхности воды в горизон-
тальном положении, прикрепляясь к поверхностной пленке служащими
для этой цели органами (стр. 227); при этом голова личинки поворачи-
вается на 180°, ртом кверху. Большую часть времени (97%) личинка
фильтрует, прекращая фильтрацию только в периоды линек. Личинка
является безвыборным фильтратором, привлекающим в рот все взвешен-
ные в воде частицы, независимо от их питательной ценности. На этом осно-
вано применение пылевидных инсектицидов — ядовитых порошков,
распыляемых на поверхности воды (парижская зелень и др.) с целью
уничтожения личинок.
1 Термин «фильтрация» мы применяем в смысле способа добывания пищи: филь-
труют воду (в смысле отделения частиц от воды) не щетки, а челюстной аппарат.
228
6) Механизм действия ротового аппарата
Основная роль верхней губы с ее щетками — направлять ток воды
с пищевыми частицами ко рту, в промежуток между мандибулами. Сюда,
на фильтр челюстного аппарата, собирается весь притекающий мате-
риал. Вылов частиц производится волосками мандибул и максилл, ниж-
ней губой и подглоточником. Мелкие пищевые частицы задерживаются
густым покровом тонких волосков максилл, откуда счищаются волос-
ками мандибул и направляются в рот. Более крупные частицы задержи-
ваются выступами терки нижней губы и зубами мандибул, раздробля-
Рис. 147. Приносящие токи, создаваемые фильтрующей личинкой
A. maculipennis в поверхностном слое воды (вид сверху).
А — личинка IV возраста; Б— личинка I возраста; обе изображены
в одном масштабе.
ются ими и движениями выступов нижней губы, подглоточника и шипов
мандибул проталкиваются в глотку. В ротовой полости частицы скап-
ливается в пищевой комок, который, как только он достигнет известной
величины, тотчас заглатывается. Вода при этом заглатывается в неболь-
шом количестве. Не все частицы, притекающие ко рту, улавливаются;
значительная часть их не попадает в рот, а уплывает прочь — вылов
частиц несовершенен
Эффективность фильтрации. Щетки верхней губы обладают большой
сферой действия, собирая пищу с большой площади поверхностной пленки.
Токи, возникающие в поверхностной пленке вокруг фильтрующей ли-
чинки, сходятся к щеткам с различных сторон, охватывающих около
250° дуги окружности (рис. 147). Наиболее сильные токи возникают
впереди щеток. Наибольшее расстояние, с которого щетки могут привле-
кать пищевые частицы ко рту у личинок IV возраста, равно 20 мм, чти
в 2,6 раза превышает длину их тела. Впереди фильтрующей личинки
I возраста токи заметны только на расстоянии до 2,5 мм. Площадь по-
верхностной пленки, облавливаемая щетками, зависит от размеров щеток
и степени развития их мускулатуры; эти величины меняются с возрастом.
Поэтому старшие возрйеты личинок имеют более ^мощный облавливающий
аппарат, чем младшие.
229
Вся площадь, с которой пища привлекается к личинкам IV возраста,
составляет около 8 см2, а к личинкам I возраста — 20 мм2 (при 20°), т. е.
личинки IV возраста облавливают площадь, в 40 раз большую, чем ли-
чинки I возраста. В 1 час личинка IV возраста облавливает при этих
условиях 108 см2 водной площади, а личинка I возраста — 14,4 см .
Фильтрацияп уничтожение личинок пылевидными ядами. Размер площади поверх
постной пленки, облавливаемой личинками, является исходной ьеличввой для вычи-
сления дозировок пылевидных ларвицидов (ядов). Для того чтобы дозировка яда была
смертельна для всех личинок всех возрастов, необходимо, чтобы при расчете ее при-
нимался во внимание размер площади, облавливаемой личинками младшего возраста.
Весовая дозировка парижской зелени, высчитанная на основании размера площади,
облавливаемой личинкой I возраста, и теоретически достаточная, чтобы отравить
всех наличных личинок, во много раз меньше применяемых на практике: она равняется
26 г парижской зелени па 1 га водной площади и в условиях лабораторных опытов
оказывается эффективной в отношении личинок как I возраста, так и IV возраста.
Однако несовершенная техника распыления порошкообразных ядов заставляет на
ппактике увеличивать дозировки минимум до 300 г парижской зелени па 1 га.
Рис. 148. Направление входящих и выходящих токов воды,
возникающих при фильтрации (оригинал). г
Д - вхздящ ie. В — ВЫХЭДЯ1Ц1С токи воды; Б — токи, отбрасываемые
разгибательным движением щзток.
Большое количество плавающих на поверхности воды частиц (пы-
ли и т. п.) оказывает хмеханическое сопротивление работе щеток и снижает
эффективность фильтрации. Поэтому примешивание к яду слишком боль-
ших количеств пыли или других безразличных примесей нецелесообраз-
но, так как уменьшение площади облова уменьшает шансы на захват яда,
замедляет гибель личинок.
Фильтрующая личинка улавливает как плавающие, так и взвешен-
ные в воде частицы. В отношении взвешенных частиц интенсивность филь-
трации измеряется объемом профильтровываемой за сутки воды. Личинка
A. maculipennis IV возраста профильтровывает за сутки около 2 000 мм3
воды*
Вода, отработанная ротовым аппаратом личинки, пьтбрасыгается пргерх средней
части максилл двумя струями, которье гроходят в несколько Со.т се глубоких слоях,
не мешая движению приносящих нжов Кроме этих струй, при фильтрации личинки
возникают еще две мощные струп, отбрасываемые раз] ибательпым движением щеток;
они гонят воду, не успевшую пройти через фильтр челюстною аппарата, и являются
показателями непроизводительной работы щеток (рис. 148).
ej Размеры чатиц, вылавливаемых личинкой Anopheles
Личинки используют в пищу не только микроскопические, но час-
тично и ультрамикрсскспическпе частицы, настолько мелкие, что они
не видимы в микрссксп. С другой стороны, наибольший размер заглаты-
ваемых личинкой частиц соответствует диаметру глотки. Поэтому чем
старше и больше личинка, тем больше размер частиц, которые она может
проглотить. Так, максимальный размер частиц, заглатываемых личин
230
ками 1 возраста, равен 341 (=0,034 мм), а IV возраста — 165р. (=0,105 мм).
Частицы порошкообразного яда, чтобы быть доступными для заглатывания
любой личинке I возраста, не должны превышать 20л Парижская зелень
содержит много частиц в 10—20,1 и поэтому легко заглатывается личинками
всех возрастов.
г) Состав пищи
Личинки отфильтровывают и используют в пищу все то, что ветре
чается в поверхностном слое воды: ультрамикроскопические частицы,
взвесь органического детрита (т. е. мертвые растительные и животные
остатки), бактерии, простейших (инфузорий, жгутиконосцев и др.),
мелкие водоросли, коловраток, споры, пыльцу и пр. Количество отдель
ных видов пищи зависит от состава населения водоема и от размеров
личинки. У младших личинок основную роль в питании играют детрит,
бактерии и мелкие формы простейших, тогда как у старших добавляется
более крупная пища — коловратки, очень мелкие рачки и нитчатки. Выс
ших растений личинки Anopheles не поедают. Большую роль в питании
личинок играет обскребывание перифитона (т. е. обрастаний из вэдорос
лей, бактерий, простейших и мелких многоклеточных на подводных
предметах); перифитон является гораздо более концентрированной пищей,
нежели взвешенный в воде планктон. Личинки Anopheles могут поедать и
себе подобных. Однако активно на добычу они не бросаются, а схватывают
и едят только то, что само попадает им в рот при работе щеток; в этих
случаях они поедают обычно ослабленных личинок младших возрастов
или трупы погибших.
Для нормального роста личинок необходима пища, содержащая вита-
мины. При голодании, которое легко наступает при воспитании личинок
в лаборатории, наблюдается большая смертность, замедленное развитие н
отставание роста. Смертность при голодании больше поражает женских
личинок, чем мужских, поэтому из сосудов с голодающими личинками
выплаживается больше самцов. Из голодных личинок развиваются мелкие
комары; мелкие самки имеют меньшее число яйцевых трубочек в яичниках
и поэтому менее плодовиты, чем нормальные. Яичники у новорожденных
самок, полученных из голодавших личинок, оказываются в среднем на
более ранних стадиях развития, чем у самок, полученных из нормально
питавшихся личинок.
4. ДВИЖЕНИЕ ЛИЧИНОК- ДВИГАТЕЛЬНЫЕ РЕАКЦИИ.
ДВИЖЕНИЯ КУКОЛОК
Реакции бегства и нырка. Всякое резкое внешнее раздражение ^прп
косновение, колебание водной поверхности и т. д.) приводит находящую-
ся у поверхности воды личинку в движение, вызывая перебежку по по-
верхности или нырок. При движении по поверхности „личинка Ano-
pheles, изгибаясь, производит небольшие, короткие взмахи плавником
анального сегмента, перебегая на расстояние нескольких сантиметров.
При плавании личинки двигаются задним концом тела вперед. При ны-
рянии личинка движениями плавника сначала опускается вниз, затем
некоторое время остается неподвижной, отдыхая на дне или в толще воды
(«парит»), после чего вновь поднимается к поверхности. Личинка обладает
небольшой мышечной силой и, поднимаясь после нырка к поверхности,
быстро устает; устав, она перестает двигаться и начинает медленно опу
скаться, парашютируя благодаря громадному сопротивлению своих пе-
ристых волосков; отдохнув, она опять начинает подниматься. Таким обра-
зом, поднимаясь со сколько-нибудь значительной глубины, личинка
231
действует по правилу: два шага вперед, шаг назад. В итоге подъем совер-
шается медленно. Поднимается личинка хвостом вперед — кверху. Глу-
бина нырка у личинки A. maculipennis IV возраста —до 45 см. Максималь-
ная глубина, с которой может происходить подъем личинок IV возраста
в условиях эксперимента (в узком сосуде), — 2,37 м. Личинки младших
возрастов ныряют менее охотно и менее глубоко, чем личинки IV возраста.
Вообще чем слабее личинка, тем меньше глубина, на которую она ныряет.
Ни одна личинка при нырке не опускается на глубину, большую той,
с которой она может активно подняться.
Самопроизвольные движения личинок: реакции на свет и температуру.
Помимо движений, возникающих под влиянием внешних раздражений,
Рис. 149. Распределение личинок A. maculipennis
между затененной и освещенной солнцем половинами
кристаллизатора (фотография).
у личинок наблюдаются самопроизвольные (спонтанные) движения. Ча-
стота самопроизвольных перебежек зависит от того, уташла ли личинка
точку опоры, а также от освещения и температуры. В полной темноте
личинки малоподвижны. На свету число самопроизвольных перебежек
увеличивается, и тем больше, чем ярче освещение. А
Явление возрастания подвижности с усилением освещения есть проявление
фотокинеза, т. е. возбуждение под влиянием света. При длительном пребывании на
свету одйбй-хиды-наступает привыкание, личинка становится менее чувствительной
и подвижность снижается.
Фотокпнез влияет па распределение личинок внутри небольшого водоема или
сосуда. Если на освещенный сосуд с личинками падает тень, то все личинки через не
которое время соберутся в тени (рис. 14_£). В этом случае тень действует," как ловушка:
личинки, подвижные на свету, случайно забегают в тень, где становятся менее подвиж-
ными, вследствие чего и остаются на затененной части сосуда. Таким-Образом, переход
личинок в тень вызывается не отрицательным фототаксисом (бегством от света),
а является результатом фо i окипетического возбуждения.
Особенности реакций у нырнувшей личинки. Наряду с фотокинети
ческой реакцией, личинки обладают положительным фототаксисом, кото
рый заставляет их двигаться по направлению источника света. Положи-
232
тельный фототаксис больше выражен у нырнувших личинок, нежели
у находящихся на поверхности; нырнувшей личинке он помогает при ос
возвращении на поверхность: ориентируясь на просвет среди раститель
яости, личинка прямым путем возвращается на свободный, ничем не за
громожденный участок поверхности. Резкое изменение освещения, напри-
мер внезапно упавшая на нее тень, заставляет личинку IV возраста
нырять.
На подвижность личинок оказывает влияние также температура.
При низких температурах (около 5°) личинки A. mac. messeae неподвиж-
ны и даже не реагируют на прикосновение; эти температуры вызывают
полное прекращение движений (зона торможения холодом). При не
сколько более высоких температурах (около 4-10°) появляются редкие
самопроизвольные перебежки. При более высокой температуре частота
перебежек увеличивается, достигая максимума около 4"^^° (зона воз-
буждения умеренно низкими температурами). При более высоких тем-
пературах подвижность личинок вновь снижается и между 4”25 и 4-30а
становится минимальной; это зона минимального
возбуждения при оптимальной температуре
(рис. 150). Температура 25—30° для личинок
A. maculipennis является оптимальной; усло-
вия питания и роста у них при этои~темпера-
туре оказываются наиболее благоприятными.
Дальнейшее повышение температуры вызывает
новое нарастание подвижности, которая около
37° достигает второго максимума; это зона
возбуждения высокими температурами. При
дальнейшем повышении температуры подвиж-
ность резко снижается. При температурах
около 40° личинки становятся неподвижными,
они лишь судорожно извиваются; это зона
теплового окоченения. Увеличение подвиж-
ности личинок под влиянием неблагопри*ят-
Рис. 150. Кривая числа
спонтанных перебежек, про
изводимых личинками А.П1 а
culipennis messeae, в
зависимости от темпера
туры.
ных температур (термокинез) влияет на распределение личинок в
сосуде или в небольшом водоеме. Личинки собираются в той части
водоема или сосуда (при прочих равных условиях), где температура мень
ше их раздражает, т. е. при температуре, более близкой к оптимуму
(25—30°); такое распределение является следствием частых беспорядоч-
ных перебежек в зонах слишком высоких или слишком низких темпера-
тур и прекращением таких перебежек при попадании в зону оптимальных
температур. Таким образом, температура 25—30° является температурой.
которую личинки предпочитают.
Передвижение личинок п/тем ползания. Кроме движений по поверх
ности воды и под водой, личинки Anopheles всех возрастов способны пол-
зать по влажной поверхности при помощи ^движений брюшка и головы
(Никольский). Ползающие личинки, в отличие от находящихся в водег,-имеют
резко выраженный отрицательный фототаксис (т. е. уползают От света).
Движения куколок. Перемещается куколка при помощи взмахов брюш-
ка, которое сгибается в спинно-брюшном направлении (тогда как у личи-
нок оно сгибается в стороны). При каждом распрямлении брюшка перед-
ний конец тела движется книзу, при сгибании — кверху, поэтому движения
куколки не бывают прямолинейны и горизонтальны. При движении она
всегда отрывается от поверхности, перебежек без отрыва от поверх-
ности у нее не бывает. После прекращения активных движений куколка
пассивно, но быстро вертикально поднимается к поверхности в силу того,
233.
что она удельно легче воды. При нырке куколка может удержаться под во-
дой лишь в том случае, если зацепится за подводные предметы. Макси-
мальная7 глубина нырка, наблюдавшаяся у куколки, равняется 40 см.
Движется куколка, как и личинка, в ответ на сотрясение, реагируя по
сравнению с личинкой на более слабые раздражения. Этим обусловлена
большая «пугливость» куколок. Прикосновение к какому-нибудь твер-
дому предмету, так же как и у личинок, ослабляет чувствительность ку-
колки к внешним раздражениям и делает ее менее подвижной. Реакции
на Лвет у куколки менее выражены, чем у личинки. Способность куколок
комаров к передвижению отличает их от куколок большинства насекомых.
х
г». ВЛИЯНИЕ ВЕТРА, ВОЛНЕНИЯ, ТЕЧЕНИЯ И КОЛЕБАНИЙ УРОВНЯ ВОДЬ\
ВЛИЯНИЕ ГЛУБИНЫ ВОДОЕМА
Спокойная поверхность водоема как условие существования личинок.
Необходимым условием Нормального существования личинок Anopheles
является наличие спокойной поверхпосш водоема, хорошо защищенной
от ветра и волнобоя. Уже мелкая рябь заставляет личинку повторно ны-
рять, мешает питанию, вызывает утомление; волнение, крохме того, по-
вреждает личинок и может их выброгДуь на Дерет. Поэтому большие откры-
тые водоемы, подверженные волнению, непригодны для развития личинок.
Мы находим личинок Anopheles только в'небэлыпнх водоемах, защищен-
ных от ветра берегами или проросших растительностью, в которых ветер
не может вызвать значительной ряби. Таким образом, для того, чтобы
вновь сооружаемые искусственные водоемы не заселялись личинками
Anophles, необходимо обеспечивать прибойность их берегов.
Влияние течения. Личинки Anopheles заселяют стоячие или слабо
проточные водоехмы. При слабом точении личинка прикрепляется хвосто-
выми крючочками к подводным'предметам и таким образом удерживается
па месте; при временпохм отрывании от субстрата она может сноситься
течением. С возможностью такого переноса личинок спокойной текучей
водой следует считаться, так как этим путем возможен занос личинок
внутрь подзащитной зоны в промежутках между обработками водоемов.
В качестве меры борьбы с заносом личинок по оросительпыхМ каналам
рекомендуют устанавливать на каналах поперечные пловучие заграждения,
удерживающие выше себя полосу нефти; личинки, приносимые течением
в полосу нефти, погибают. При больших скоростях течение становится бур-
ным, поверхность воды волнуется и дрожит, личинки не могут противо
стоять течению и при сносе их гибнут.
Скорость течения на стрежне капала, при которой личинки уже
не могут жить у берегов, равна для умеренно заросших каналов 0,2—0,4 хм
в секунду. Эта норма меняется в зависимости от устройства каналов
и от степени их зарастания растительностью. Поэтому при строительстве
оросительных и о.сушительщдх,капалов маляриологи должны требовать
создания их с максимальными уклонами и скоростями течения, техниче-
ски возможными и допустимыми в каждом конкретном случае.
Значение колебаний уровня воды. Опасны для жизни личинок также
резкие колебания уровня воды в водоемах. При опускании уровня воды
личинки, населяющие зону прибрежной растительности, оказываются
вместе с растительностью вне воды, обсыхают и гибнут. Паводки рек,
при которых происходит поднятие уровня и увеличение скорости тече-
ния, гибельны для личинок. Особенно губительны для личинок паводки
м торных речках, внутрирусловые и пойменные водоемы которых явля-
ются у нас в Советском' Союзе местами выплода A. mac. typicus и A. su-
234
perpictus. Гибель личинок в этих случаях вызывается удушением и махани*
ческим повреждением их во время сноса бурным течением.
Так как всякое резкое изменение водного режима в анофелогенных
водоемах в неблагоприятную для личинок сторону (увеличение скорости
течения, колебания уровня воды) вызывает итгибель, то искусственным
изменением водного режима пользуются как мерой борьбы. Так, искус-
ственные кратковременные колебания уровня водохранилищ как мера
борьбы с личинками Anopheles применяются в США. Этот метод борьбы
может иметь значение и в СССР в связи со строительством водохранилищ.
Для борьбы с личинками Anopheles в ручьях, речках и каналах применяют
искусственные попуски воды, чем достигается промывка каналов и гибель
личинок при сносе их бурным течением и осаждении па берегах. С этой
целью на речках устраиваются небольшие плотины, накапливающие воду,
которая затем периодически спускается. У нас в СССР такие опыты про-
мывки каналов с противомалярийной целью производились па Украине.
Перемежающийся водоток. Для борьбы с личинками в ороситель-
ных системах в местностях с искусственным орошением применяется ме-
тод перемежающегося водотока, при котором вода пускается по каждой
канаве лишь в течение определенного времени, после чего канавы остав-
ляют на несколько дней без воды; в этих случаях личинки погибают от вы-
сыхания. Путем периодических просушек предотвращается выплод ко-
маров и на рисовых полях (прерывистое орошение риса, см. стр. 126). Про-
должительность просушки при проведении этих мероприятий зависит
главным образом от метеорологических условий, определяющих ско-
рость высыхания почвы, па которой остаются личинки. Наблюдения по-
казывают, что при высокой влажности воздуха и облачном небе личинки
могут в течение недели оставаться живыми на влажной почве. В сухом
климате Таджикистана при просушке рисового поля полная гибель личи-
нок A. hyrcanus наблюдалась в густом рисе через двое суток, в разрежен-
ном — через сутки; личинки, укрывшиеся под нитчатками, жили до трех
суток (Петрищева).
Неблагоприятные условия в глубоких водоемах. Личинки встречаются
в глубоких водоемах только при наличии растительности, образующей
у поверхности «второе дно» Причиной отсутствия личинок в глубоких
водоемах является не глубина как таковая, а сопутствующие ей обстоя-
тельства: отсутствие субстрата из растительности, в котором личинки нуж-
даются для прикрепления и для защиты от врагов. Кроме того, глубокие
водоемы обычно обширны и подвержены волнению, и это также' делает
их непригодными для жизни личинок. К тому же в глубоких водоемах
для личинок недоступно дополнительное питание за счет обскребывания
подводных предметов.
В искусственных водоемах — таких, как колодцы, цистерны, копан-
ки, личйнки живут при больших глубинах, чем это имеет место в не-
заросших естественных водоемах, так как поверхность их невелика,
а крутые берега защищают ее от волнения; кроме того, в них меньше вра
гов и доступнее питание путем обскребывания.
G. ВЛИЯНИЕ СВЕТА £
Лучи солнца, падая на водоем с личинками Anopheles, действуют
на них различно:
1) нагревая воду, определяют температурный режим водоема;
2) вызывая фотосинтез зеленых растений, оказывают влияние на хв
мизм водоема; , ,
235
3) свет оказывает непосредственное влияние также и на поведение и
жизнедеятельность личинок.
Личинки A. maculipennis теплолюбивы и поэтому населяют хорошо
освещенные и прогреваемые> солндам_водо^ны. Только на юге при конти-
нентальном климате в жаркие летние месяцы температура воды в откры-
тых водоемах становится неблагоприятной для личинок этого вида, и они
развиваются в слегка затененных месхах,- Изменения химизма воды в во-
доемах, происходящие под воздействиелМ света на зеленые растения, также
чрезвычайно благоприятны для личинок Anopheles. В силу воздействия
лучей солнца на температуру и химизм воды личинки A. maculipennis
и бывают обычно приурочены к хорошо освещенным водоемам/
Однако свет, необходимый личинкам некоторых видов в естественных водоемах.
оказывается для них ненужным в условиях лаборатории, когда искусственно обеспе-
чиваются благоприятные условия температуры и химизма. Опыты показывают, что
личинки A. maculipennis, воспитываемые в абсолютной темноте, но в чистой воде и в
тепле, развиваются нормально.
Как мы уже видели, свет оказывает на личинок и прямое, резко возбуждающее
действие (фотокинез), в силу чего личинки при ярком свете собираются в затененной
части сосуда или микроводоема. Но на распределении личинок вдоль берега большого
водоема, измеряемого метрами, этот фактор не сказывается: если часть берега затенена
деревом, переселения личинок в тепь не происходит, и это место обычно оказывается
беднее личинками, а не богаче.
Интенсивность освещения и цвет фона влияют на окраску личинок. Личинки и
куколки, воспитанные на темном фоне или в темноте, становятся темнее. Отложение
в жировом теле личинки зеленого пигмента хлорофилла, получаемого личинкой с пи-
щей растительного происхождения, часто придает личинке окраску окружающих ее
водорослей. Приспособительная окраска к фону делает личинок менее заметными для
врагов.
Распределение личинок пр водоемам в зависимости от освещения.
Таким образом, на севере, т. е. на большей части Европейской терри-
тории Советского Союза и в Сибири, личинки A. maculipennis встречаются
преимущественно в хорошо освещенных водоемах. В водоемах, располо-
женных под пологом леса, по берегам прудов, затененных густыми де-
ревьями, в зарослях густых и высоких злаков личинки этого вида не
встречаются. Наоборот, на юге (в Средней Азии, на Кавказе и даже в
Нижнем Поволжье) личинки A. maculipennis в разгар лета населяют
более затененные места; однако слишком большого затенения в водоемах
они избегают и на юге.
Практические выводы. Из факта отсутствия личинок в затененных во-
доемах сделан практический вывод — искусственное затенение водоемов
(речек, каналов, небольших прудов) насаженными по их берегам деревья-
ми используется как мера борьбы с личинками. На севере эта мера может
быть особенно эффективна, на юге она может дать эффект, если тень до-
статочна густа. Искусственное затенение достигается также при помощи
искусственного перекрытия канав или копанок соломенными шитами
и т. п.
7. ВЛИЯНИЕ ТЕМПЕРАТУРЫ
а) Верхн/ий и нижний температурный предел жимм
Нормальная жизнедеятельность личинок A. maculipennis в средней
полосе СССР протекает при температурах между 4-10° и +35°. Опти-
мальной температурой является 25—30°. Температуры ниже 10° и выше
35° угнетают развитие. Ниже 4-7,5° начинается те^мпература холодного
окоченения, выше Зр° — температура теплового окоченения. Окоче-
нение ст холода обратимо, и после нескольких часов окоченения отогретые
личинки возвращаются к нормальному состоянию.
236
Влияние низких температур. Однако длительное действие умеренно
низких температур (от 0° до 4-10°) влечет вымирание личинок A. ma-
culipennis в водоемах поздней осенью. Некоторые виды комаров
зимуют в личиночном состоянии (A. bifurcatus, A. algoriensis, A. pul-
chcrrimus) или развиваются ранней весной (подрод Ochlerotatus),
личинки этих видов переносят умеренно низкие температуры без вреда
для себя.
Что касается действия на личинок температур ниже 0°, то при мед
ленном замораживании в течение нескольких часов даже при —10° часть
личинок при последующем оттаивании оживает.
Однако быстрое замораживание при той же температуре ведет к
гибели всех, личинок. Гибнут личинки и куколки и при продолжитель
ном пребывании во льду. Вероятно, личинки остаются живыми тогда,
когда жидкое® их тела остаются в переохлажденном состоянии и не за-
мерзают, и гибнут в тех случаях, когда происходит фактическое замер
зание тканевых жидкостей.
Верхний температурный предел жизни. Верхним температурным пре
делом жизни, температурой, при которой наступает мгновенная смерть,
для A. jaiaculipennis средней полосы СССР является температура 4-45°.
При кратковременном воздействии несколько более низких температур
(нЪ выше 39°) и последующем переносе личинок в оптимальные температуры
состояние окоченения может прекратиться и личинкй остаются живы;
при более продолжительной! действий этих температур наступает смерть.
При еще более низких температурах (+36°, +37°) личинки могут жить
и развиваться,'нб' смертность среди них значительно выше,чем при опти-
мальных температурах.
Относительность температурных границ. Указанные выше темпера-
турные пределы развития и жизни, характерные для личинки A. ma-
culipennis средней полосы СССР, не являются постоянными. Существует
индивидуальная изменчивость у личинок в пределах одной популяции,
в силу которой у одних личинок нижний температурный порог активности
лежит несколько выше, а у других — ниже. Кроме того, воспитывая личи-
нок при повышенных или пониженных температурах, можно значительно
сдвинуть в ту или другую сторону пороги активности (индивидуальное
привыкание, адаптация). Наконец, личинки одного вида из разных мест-
ностей или принадлежащие разным видам и подвидам ведут себя по отно-
шению к температуре различно. На юге все цифры, характеризующие от-
ношения к температуре личинок A. maculipennis, сдвинуты кверху. Так,
в Средней Азии личинки A.mac.sacharovi встречаются в водоемах с дневными
температурами до 38—40°, где нормально развиваются (однако и здесь —
при условии некоторого охлаждения воды ночью). На юге перегрев воды
с целью прекращения выплода A. maculipennis был впервые успешно
применен на рисовых полях Самаркандского района (Исаев). В этом случае
перегрев воды в отдельных клетках (чеках) рисового поля достигался путем
расположения чеков цепочкой вместо непосредственного питания каждого
чека холодной водой, поступающей прямо от оросителя.
Знание предельных и оптимальных температур развития личинок
дает возможность установить календарные сроки начала и конца раз-
вития личинок в водоемах и его наибольшей интенсивности. Весной, с
момента, когда вода в анофелогенных водоемах прогревается до 10°, водо-
емы следует держать под наблюдением. Осенью, когда температура воды
снизится до 10э и, следовательно, выплод комаров прекратится, обра-
ботка водоемов становится совершенно бесполезной.
237
б) Влияние температуры на скорость развития
Зависимость между скоростью развития и температурой. Скорость
развития отдельных водных фаз комара зависит от окружающей темпе-
ратуры. Зависимость между скоростью развития и температурой выра-
жается кривой на рис. 151. Из кривой видно, что при температуре нижнего
порога развития (который для A. maculipennis средней полосы СССР ле-
жит около +7,5°) скорость развития равна нулю, т. е. развития не про-
исходит. С возрастанием температур скорость развития также возрастает,
сначала медленно, затем быстрее, а начиная с температуры -J-150 и вплоть
до 4-25° она становится прямо пропорциональна температуре. При даль
Рис. 151. Средняя скорость развития A. maculipennis (от от-
к тадкп яйца до окр ячеиия) в зависимости от температуры.
Учаепп кгирых, накесеннь’е пунктиром, проверены m основании
приблизительных данных. X—X— пгямш. отвечаютш уравнению
Боденгеймера: S (Т°—10,2-')=22?,7: d—место ее пересечения с осью
абсцисс.
пейшем повышении температуры до 30° увеличение скорости развития не-
велико, а при более высоких температурах (31° и более) вместо ускорения
происходит замедление развития и увеличен ie см^ргн сги. При еще более
высоких температурах установить скорость развития не удается из-за
большой смертности.
Для развития A. maculipennis зависимость между скоростью развития
и температурой выражена Боденгеймером в виде формулы:
о _ С_ _ 222.7
*0 /J1Q ^6 у»°__ |Q 90 »
где S — продолжительность всего развития от яйца до окрылечпогз ко-
мара, Т° — средняя температура, при которой происходит развитие, t°—•
нижний температурный предел развития, принимаемый злее » равн ям 10 2°;
С — 222,7 —' величина, выражающая сумму тепла, необходимую для про-
хождения всего цикла развития; числовое значение ее получено из
опытов.
Эта формула позволяет вычислить среднюю продолжительность раз-
вития водных стадий при определенной к яшретной температуре Т°. Под-
ставляя в формулу Б )денгеймера вместо С и t° соответствующие вели-
чины, мы можем вычислить продолжительность развития A. maculi-
pennis и при данн >й средней температуре Г °.
Необходимо отметить, что эта ф ормула правильна показывает зави-
симость между температурой и продолжительностью разв ,тия тольк ) при
температурах, лежащих между -|-14о и 4-25°. При температурах ниже 14°
238
и выше 25° эта формула не применима; при температурах ниже 4-14*
продолжительность развития, вычисленная по формуле, будет больше,
чем в действительности; при температурах выше 4-25° она будет значи-
тельно меньше. Формула Боденгеймера находит применение в практиче-
ской работе для приблизительного вычисления срока очередной обра- -
ботки водоемов.
Влияние температуры на скорость развития A. maculipennis изуча-
лось в природных условиях и в эксперименте рядом авторов (Мартини,,
Киселевой и др.). Полученные ими данные сведены в табл. 3.
Таблица 3
Продолжительность развития A. maculipennis в сутках
Температура Продол ж 1тел1 н >сть развития ! 1 Все развитие
яйца I воз- раст п воз- раст III воз- раст IV воз- раст Ку- колки
16—19° 5 6 4 6'/а ЗЭ>/,
20—22° 3 2’/4 2 2*/» 43А 3 18
24- 27° 2 2 1*А 2*/г ^4 2>/2 14«/2
Из таблицы видно, что минимальная средняя продолжительность раз-
вития, наблюдаемая у A. maculipennis при 24—30°, составляет 14 дней.
Это — средняя величина; отдельные особи могут развиваться или несколь-
ко быстрее (11—12 дней), или медленнее; при расчетах сроков проведения
противоличиночных мероприятий с этим обстоятельством необходимо
считаться. Срок развития самцов летом на один день короче срока разви-
тия самок. На скорость развития могут сильно влиять другие факторы,
например, количество пищи. При недостатке пищи развитие может затяги-
ваться вдвое и больше по сравнению с развитшш нормально питающихся
особ» й при Тей Ж } Т Win 'P ITVp о
Влияние колебаний темиераачгы на скорость развития. В одной и
той же местности личинки живут в водоемах с различным температурным
режимом, в связи с этим и скорости их развития бывают различны. В при-
роде развитие личинок происходит в условиях больших колебаний тем-
пературы, особенно весной и осенью, которые не вредят личинкам, но от-
ражаются на продолжительности развития, в одних случаях ускоряя раз-
витие, в других замедляя его. Так, при постоянной температуре -j-10°
развитие личинок A. maculipennis почти не происходит, однако при той
же средней температуре и колебаниях от 4-5 до 4-15° личинки развиваются
во время теплых периодов; это имеет место в природе ранней весной.
в) Температурный режим личиночных биотопов
Как мы видели, оптимальными температурами для существования
личинок A. maculipennis в средней полосе СССР являются 25—30°; од-
нако средние температуры в<злуха в летнее время здесь этих величин до-
стигают редко. Поэтому личинки A. maculipennis в сродней полосе СССР
находят наилучн.ие температд] нь е условия в небольших водоемах, откры-
тых для солнечных лучей, со средней температурой виды, всегда превышаю-
239
щей таковую воздуха, т. е. в водоемах гипертермичных (рис. 37, А): в откры-
тых лужах, в прудах и озерках среди зарослей погруженной растительно-
сти, в небольших медленно текущих и хорошо прогреваемых речках и кана-
вах, во всевозможных микроводоемах: ямах от копыт, колеях от колес,
открыто лежащих на избыточно увлажненной почве. Среди зарослей по-
груженной растительности (роголистника, рдестов, урути, нитчаток),
достигающих поверхности воды, температурные условия особенно благо-
приятны для личинок, так как температура поверхностного слоя воды
среди зарослей растительности всегда выше температуры воды вне зарослей
или под слоем растительности (стр. 67).
В силу неблагоприятных температурных условий личинки A. macu-
lipennis в средней полосе СССР редко встречаются в затененных водоемах,
средняя температура которых равна температуре воздуха (водоемах изо-
термичных) (рис. 37, А) и обычно совершенно не встречаются в водоемах,
питающихся холодными грунтовыми водами, со средней температурой
воды более низкой, чем температура воздуха (гипотермичных) (рис. 38,ZJ).
Микробиотопы личинок. Каждый отдельный водоем в целом — не
однородная среда жизни для личинок Anopheles; не однороден он и в тем-
пературном отношении. Маленькая, ведущая полусндячий образ жизни
личинка тесно связана с условиями микробиотопа, т. е. с микроместооби-
таниями, и только они влияют на нее непосредственно и определяют дейст-
вительные условия ее существования. В отдельных участках водоема,
в зависимости от его расчленения (наличия заливов и мелководий), от ха-
рактера растительности (затеняющей или способствующей прогреванию
поверхности воды) и наличия или отсутствия притока грунтовых вод, тем-
пературные условия могут быть весьма различны. Поэтому и температур-
ные условия существования личинок Anopheles в разных микробиотопах
одного водоема могут оказаться различными. С температурным расчлене-
нием водоемов необходимо считаться, когда мы обследуем водоемы в поис-
ках личинок, а также при определении сроков обработки водоемов. Так,
может оказаться, что личинками A. maculipennis будут заселены лишь
наиболее благоприятные в температурном отношении участки водоема,
в то время как в другой части водоема они будут отсутствовать или там
будут встречаться личинки других видов Anopheles, в соответствии с тем-
пературными требованиями последних. При определении сроков обработки
водоема необходимо принимать во внимание температурные условия
наиболее благоприятных для развития личинок участков водоема, где раз-
витие протекает наиболее быстро.
Весенние местообитания личинок. Весной, когда крупные водоемы
(пруды и озерки) еще не прогрелись, личинки A. maculipennis заселяют
главным образом временные водоемы — лужи (образующиеся от таяния
снега или от разливов рек), средняя температура которых значительно
выше средней температуры воздуха. В дневные часы температура в лужах
поднимается настолько, что обеспечивает быстрый рост личинок, несмотря
на то, что по ночам эти же лужи могут сильно остывать и даже подерги-
ваться ледком.
Перемещение центров обилия личинок в летние месяцы. К моменту
окончания метаморфоза большинством личинок I поколения весенние лужи
обычно в значительной мере высыхают; в средней полосе СССР это
обычно происходит в начале июня — конце мая, на юге — на полмесяца-
месяц раньше. Но параллельно с высыханием луж идет прогревание более
крупных, постоянных водоемов — прудов и озерков. В них развиваются
и поднимаются к поверхности элодеиды (стр. 247), среди которых создаются
благоприятные условия температуры, химизма и питания для личинок,
240
з связи с чем в эти водоемы самки Anopheles начинают откладывать свои
яйца, и здесь развиваются личинки летних поколений. Таким образом,
центр личиночного обилия в течение сезона перемещается из одного типа
биотопов в другой. Это перемещение бывает связано, однако, не только
с изменениями температурных условий в водоемах в течение сезона, но
и с сезонными изменениями других факторов (химизма, растительности
и пр.).
Как указывалось, на юге личинки A. maculipennis живут при более
высоких температурах, чем личинки средней полосы СССР; температур-
ный оптимум их лежит около 30°. Тем но менее во многих местах Средней
Азии и Закавказья в жаркие летние месяцы температурный оптимум этого
вида оказывается превышенным и открытые, сильно прогреваемые водое-
мы становятся неблагоприятными для жизни личинок. В связи с этим
в июле и августе личинки A. maculipennis чаще встречаются в умэрепно
затененных водоемах.
8. ВЛИЯНИЕ ХИМИЧЕСКИХ ФАКТОРОВ СРЕДЫ НА ЛИЧИНКУ И КУКОЛКУ
В воде личиночных биотопов всегда содержится большое количество
различных неорганических и органических веществ в виде грубой взвеси
(сестона), в виде коллоидных (ультрамикроскопических) частиц или в виде
настоящих растворов; имеются также растворенные газы.
а) Влияние растворенных солей
Солевой состав водоемов. Солевой состав воды, количественное соот-
ношение отдельных солей, играет большую роль в жизни живых организ-
мов, населяющих водоемы. Качественные различия солевого состава водое-
ма могут влиять на распределение отдельных видов Anopheles и подвидов
A. maculipennis по водоемам. Большие количества солей железа ядовиты
для многих животных, в том числе и для личинок A. maculipennis, поэто -
му в водоемах, богатых железом, эти личинки обычно отсутствуют.
Действие солевых смесей. Присутствие в воде значительных количеств
извести (кальция) заметно улучшает условия существования личинок A. ma -
culipennis. Из подвидов A. maculipennis лучше других выносят низкие кон-
центрации солей кальция личинки A. mac. atroparvus. Наоборот, A. super-
pictus чрезвычайно чувствителен к недостатку кальция.
Соли магния всеми подвидами A. maculipennis и A. superpictus переносятся лишь
в присутствии солей кальция. Не только абсолютное количество солей кальция и же-
леза, но и соотношение их концентрации с концентрацией других солей влияет на при-
годность воды для существования личинок. Растворы отдельных солей (NaCl, КС1,
MgS04 и СаС12) менее благоприятны для личинок Anopheles, чем их смеси или чем мор-
ская вода, представляющая сложную смесь всех этих и ряда других солей. Смеси,
содержащие все четыре соли, оказываются физиологически более уравновешенными
и более благоприятными для личинок, чем более простые смеси.
Если помещать личинок в разбавленную морскую воду, наиболее вы-
носливыми к повышению концентрации оказываются личинки A. mac. sacha-
rovi, и получается следующий ряд, в котором, каждая последующая форма
менее вынослива, чем предыдущая, кроме messeae и typicus, которые ведут
себя приблизительно одинаково: sacharovi> superpictus, atroparvus >
subalpinus > messeae, typicus.
В сильно разбавленной морской воде все подвиды A. maculipennis живут лучше,
чем в дестиллированной воде. В дестиллированной воде (с хлебными крошками) лучше
всего выживал typicus и subalpinus, хуже — atroparvus, еще хуже — sacharovi
и хуже всех — superpictus. A. mac. sacharovi в присутствии солей кальция и магния
более других подвидов чувствителен к недостатку солей натрия.
16 Учебник медицинской энтомологии
241
Осмотическое давление водной среды. Влияние общей концентрации
солей на личинок сказывается через осмотическое давление раствора. Вну-
тренняя среда пресноводных животных, в том числе и личинок комаров,
обладает большей концентрацией солей, чем окружающая вода. Благодаря
втому окружающая вода непрерывно насасывается в тело личинки
через жабры. Разбухания тела личинки, несмотря на всасывание воды,
не происходит, так как избыток воды непрерывно удаляется из тела ли-
чинки мальпигиевыми сосудами; таким образом, разница осмотических
давлений между наружной и внутренней средой активно поддерживается
работой мальпигиевых сосудов. Способность поддерживать эту разницу
у личинок ограничена, и при повышении осмотического давления наруж-
ной среды выше известных пределов личинки погибают.
Предельные концентрации солей, выдерживаемые отдельными видами
Anopheles и подвидами A. maculipennis, различны. В условиях экспе-
римента личинок A. mac. messeae не удавалось воспитывать при солености
выше Э'/оо1-
Наоборот, личинки A. mac. atroparvus живут и при более высокой
концентрации солей — до 14/ )0. В этих условиях их внутренняя среда имеет
меньшее осмотическое давление, чем наружная, что характерно для соло-
новатоводпых животных. Определение концентрации солей в природных
местообитаниях личинок показало, что верхний предел солености для
A. mac. atroparvus лежит около 13%0; для messeae он лежит около 5—6э/00
(данные Половодовой для Западной Сибири).
Наилучшей средой для развития messeae является слабо солоноватая
вода (1—2°/о<>), а для atroparvus — вода около 3—10°/оо солености. Макси-
мум солености, выносимый A. mac. maculipennis, a sacharovi —
20°/1О.
Куколки A. maculipennis выдерживают без вреда большие концентра-
ции солей (например, 24,8°/00), так как они не имеют полупроницаемых
покровов и в осмотическом отношении изолированы от внешней среды.
Личинки IV возраста, благодаря прекращению физиологической дея-
тельности жабер, подобно куколкам, обладают большей устойчивостью
к действию высокой концентрации солей, нежели личинки младших воз-
растов.
Знание пределов солености, переносимых отдельными видами и под-
видами Anopheles, используется для борьбы с малярией. Так, путем осс-
лопсния приморских лагун, дававших выплод A. mac. sacharovi, был
оздоровлен г. Дураццо в Албании. У нас в СССР па Черноморском побе-
режье применение осол-нения водоемов с противомалярийной целью
также может иметь значение.
Действие' минеральных вгвесей па личинок. Минеральные вещества
встречаются в водоемах не только в растворимом виде, нои в виде взвешен-
ных глинистых частиц. Вода, содержащая в избытке взвешенные мине-
ральные частицы, оказывает неблагоприятное действие на личинок Ano-
pheles.
Благордаря безвыборной фильтрации, личинка заполняет кишечник
неудобоваримым материалом, что мешает нормальн )му усвоению пищи.
В силу этого рисовые поля, залитые мутной речной вод<й, не азселяются
личинками Anopheles до тех пор, пока вода в них не осветлится
(У литче ва).
1 Стегепь общей солепости годы часто выражается в граммах соли па 1 000 г
Ноды п осозиачаеюя S0/^ (pro mille, т. е. ва тысячу).
242
б) Влияние органическим веществ воды
Значение продуктов распада органических веществ. Органические
вещества встречаются в естественных водах в большом количестве в виде
взвешенных в воде частиц (сестона), коллоидных и истинных растворов.
Органические вещества разлагаются в воде главным образохм при уча-
стии бактерий. Исходные органические вещества большей частью без-
вредны для личинок и пригодны в пищу; конечные продукты распада —
соли, СО2,Н2О, N2 — также безвредны. Но промежуточные продукты рас-
пада очень часто вредны и ядовиты для личинок. Загрязнение воды про-
межуточными продуктами распада органических веществ оказывает, как
правило, чрезвычайно неблагоприятное действие на личинок Anopheles.
Поэтому личинки A. maculipennis обычно отсутствуют в сильно загряз-
ненных водоемах и встречаются в водоемах, вовсе не загрязненных или
очень слабо загрязненных.
Показателем степени загрязненности водовхмов является сапробность
волы, определяемая наличием показательных организмов (стр. 72—75).
Наиболее благоприятны для личинок воды олигэсапробные и р-мезосап-
робные. Личинки A. maculipennis избегают водоемов, загрязненных как
богатыми азотом веществами (например, навозом), так и растительными
остатками (древесиной, опилками, щепками, корой). Вода водоемов, за-
грязненных большим количеством древесины, обычно бывает бедна кисло-
родом, так как на поверхности погруженной древесины развивается бо-
гатая грибковая и бактериальная флора. Но если в таком загрязненном
водоеме имеется погруженная зеленая растительность, личинки находят
в нем благоприятные условия существования. Положительное действие
растений в загрязненном водоеме сводится к следующему: 1) в процессе
фотосинтеза выделяется много кислорода, что ведет к более быстрому раз-
ложению промежуточных продуктов распада органических веществ;
2) водяные растения могут использовать в качестве источника азота некото-
рые ядовитые для личинок азотистые продукты распада.
Однако при очень большой загрязненности воды никакая раститель-
ность не может сделать воду пригодной для личинок; мало того, в черес-
чур загрязненной воде не могут развиваться и сами зеленые растения.
В сфагновых болотах, вода которых чрезвычайно богата органическими
веществами и в то же время бедна солями и имеет кислую реакцию, личинки
A. maculipennis обычно также не встречаются.
В водоемах с чистым минеральным дном и бедной органическими ве-
ществами водой личинки A. maculipennis встречаются и при отсутствии
растительности (например, на галечниковых отмелях). Органическое за-
грязнение воды —причина отсутствия личинок A. maculipennis во многих
типах водоемов: в различных внутригородских и внутрипоселкэвых во-
доемах— прудах, лужах, канавах, загрязненных отбросами, в ямах для
мочки льна и кснзпли (пока в них производится мочка), в речках и ручьях,
загрязненных сточными водами, в лужах на территориях лесопильных
заводов, загрязненных древесными остатками, в деревянных бочках с во-
дой и т. д.
Искусственное загрязнение водоемов как метод борьбы. Искусствен-
ное загрязнение воды как биологический метод борьбы с личинками
Anopheles практикуется кое-где в небольшом масштабе. Этот метод приме-
ним только к хозяйственно не нужным водоемам, а из хозяйственно нуж-
ных— к копанкам, служащим для полива сельскохозяйственных культур,
п к рисовым полям. Навознсе удобрение копанок и рисовых полей делает
их временно непригодными для заселения личинками Anopheles, но ко-
16*
243
личество навоза должно быть для этого большим. Для загрязнения не-
больших стоячих водоемов можно пользоваться, кроме навоза, свежей
травой, листьями, ветками.
в) Значение степени насыщения воды кислородом
Биологическое значение степени насыщения воды кислородом» Как
относятся личинки A. maculipennis к концентрации кислорода в воде?
Как мы знаем, личинки A. maculipennis, особенно младших возрастов, мо-
гут долгое время дышать за счет растворенного в воде кислорода. Следо-
вательно, концентрация кислорода в воде для них не безразлична. Однако
в условиях эксперимента личинки могут жить в воде, очень бедной кисло-
родом, и это объясняется тем, что основным способом дыхания их является
дыхание при помощи стигм атмосферным воздухом.
В природе тем не менее личинки A. maculipennis встречаются в водое-
мах или частях водоемов, богатых кислородом, вода которых днем имеет
свыше 100% кислорода, а ночью — не меньше 60—80%. Приуроченность
личинок A. maculipennis к водоемам, богатым кислородом, объясняют глав-
ным образом косвенным значением кислорода: высокая концентрация ки-
слорода препятствует накоплению ядовитых для личинок промежуточ-
ных продуктов распада органических веществ.
Таким образом, растворенный кислород является условием и показа-
телем благоприятного для личинок A. maculipennis химизма водоема.
г) Действие активной реакции воды на личинок?*
В условиях эксперимента личинки A. maculipennis развиваются в довольно ши-
роких пределах активной реакции воды — от слабокислой до значительно щелочной;
оптимальные условия создаются при очень слабокислой реакции (pH—6,8). Резко ки-
слые и резко щелочные реакции вредны для личинок. В природе личинки A. maculi-
pennis встречаются обычно в водах нейтральных или слабо щелочных днем и очень сла-
бокислых — ночью. При очень слабокислых дневных реакциях личинки встречаются
редко и лишь в исключительных случаях — при сколько-нибудь значительной кислот-
ности воды. При сопоставлении наблюдений в природе и экспериментальных данных
приходят к выводу, что умеренно кислая реакция, сама по себе безвредная для личинок,
в природных водоемах действует на них косвенно, через посредство других организ-
мов: так, например, водоросли или бактерии сами очень чувствительны к различиям
в реакции воды и играют важную роль в жизни личинок.
9. РОЛЬ ВОДНОЙ РАСТИТЕЛЬНОСТИ В ЖИЗНИ ЛИЧИНОК]
а) Т*олъ растительности
Личинки A. maculipennis живут в водоемах чаще всего среди зарослей
растительности. В водоемах, лишенных растительности, они встречаются
скорее в виде исключения. Это правило, однако, распространяется не на все
виды личинок Anopheles в равной степени. Так, личинки A. plumbeus
живут в воде, всегда лишенной растительности. Наоборот, личинки
A. pulcherrimus, как правило, живут среди растительности. Личинки
A. superpictus, A. bifurcatus встречаются и в заросших, и в незаросших
водоемах. Роль растительности в жизни личинок A. maculipennis хорошо
изучена, и дальнейшее будет относиться к этому виду.
Водная растительность как субстрат для личинок. Значение раститель-
ности для личинок A. maculipennis разнообразно: растения создают ли-
чинке субстрат, они являются источником пищи и улучшают химический
состав воды. Растительность, достигающая поверхности воды, создает
наиболее удобный субстрат для личинок. Роль растительности как суб-
244
страта распадается на ряд функций: причальная, волнорезная, защитная
(убежище от врагов), посадочная для кладущей самки; в глубоких водое-
мах растительность играет роль второго дна, а в текучих — ослабляет
силу течения.
Листья и стебли растений, соприкасаясь с поверхностью воды, обра-
зуют линию причала. Чем гуще и мельче разветвлена растительность
около поверхности воды, тем больше длина причальной линии, тем боль-
ше личинок может разместиться на единице площади, следовательно, тем
лучший субстрат для прикрепления личинок она представляет. Доходя-
щие до поверхности заросли водяных растений убивают рябь, вызываемую
ветром, и создают условия спокойной водной поверхности, необходимые
для нормального существования личинок. Эта волнорезная функция ра-
стительности проявляется тем в большей степени, чем больше и гуще
разветвляются растения у поверхности воды.
Растительность служит также убежищем от врагов, в частности,
от рыб, являющихся опаснейшими врагами личинок. Наилучшую защиту
личинки находят среди растительности, образующей густую сеть, т. е.
ветвящейся около поверхности. Очень важна роль растительности как
второго дна. Глубокие водоемы, лишенные растительности, не заселяются
личинками A. maculipennis. Но в комьях нитчаток или в густых зарослях
рдестов, роголистника, урути, образующих под самой поверхностью во-
ды густые ковры, как бы «второе дно», личинки могут существовать над
большими глубинами. В зарослях этих растений личинки находят себе
пищу и скрываются от врагов.
Растительность как источник пищи. Растительность служит пищей
для личинок Anopheles. Многие водоросли идут непосредственно в пищу
личинок, высшие растения являются источником перифитона, т. е. пред-
ставляют пастбище. Перифитон доступен для личинок лишь в том случае,
если погруженные растения достигают поверхности воды. Чем больше пло-
щадь погруженных растений, достигающих поверхности, тем больше
количество доступного для личинок перифитона.
Химическая роль погруженных растений. Химическая роль погру-
женных растений связана с разложением ими углекислоты (фотосинте-
зом). Интенсивность фотосинтеза погруженных растений и воздействие
его на химизм водоема тем сильнее, чем больше поверхность погружен-
ных зеленых частей растений — листьев и стеблей.
Кроме рассмотренных значений растений в жизни личинок, стру-
ктура растительности влияет также на освещение, на температурный режим
водоемов и т. д.
Из всего вышесказанного вытекает, что не всякая растительность оди-
наково благоприятна для личинок Anopheles. Растительность тем благо-
приятнее для личинок Anopheles, чем больше площадь погруженных зеле-
ных органов (т. е. чем мельче рассечены листья), соприкасающихся с по-
верхностью воды. Таким образом, степень благоприятности растений для
личинок зависит не от видового состава растительных сообществ, а от фи-
зиономии растительности, т. е. от общего облика — характера расчлене-
ния листьев и от расположения листьев и стеблей по отношению к по-
верхности воды.
Водные растения, сходные по строению вегетативных органов, объеди-
няются в группы, или физиономические типы (рис. 152). Различают не-
сколько физиономических типов водной растительности, отличающихся
по степени пригодности их для существования личинок Anopheles. Фи-
зиономическая классификация водных растений в применении к эко-
логии личинок Anopheles (Беклемишев) чрезвычайно упрощает понима-
245
ние роли отдельных групп водной растительности в экологии личинок
и тем самым облегчает определение анофелогенного значения водных ра-
стительных сообществ и, следовательно, водоемов. Поэтому при обследо-
вании водоемов и оценке их зараженности личинками Anopheles мы обра-
щаем прежде всего внимание па характер водной растительности и по со-
ставу растительных форм уже можем определить, могут ли в них быть
личинки Anopheles и в каком примерно количество.
При обследовании водоемов на зараженность личинками необходимо
всегда учитывать условия микробиотопа. Личинки Anopheles относительно
малоподвижны и бывают распределены в водоемах очень неравномерно,
Рис. 152. Схемы тпппчпых представителей важнейших фи-
зиономических групп водной растительности.
а—ллан|ст11'1С<*|.'пе лемпнлы (пролетят птель— Lemna trisuloa);
б— нейстические лемниды (Lemna minoi); «—ji.3««этилы (Isoetes);
г—ялодеп u»i (Elodea сапэнгпяь); 0— нимфеиды (N хm । iiaea); е —
линеиды (Phragmites coin пить); эк-— яемны ольье (S; gilUria
Lavlulia); а—филмиды (McniantLts ti w
в зависимости от топкого расчленения среды обитаппя. Поэтому, чтобы их
находить в водоемах, в первую очередь нужно брать пробы именно в той
растительности, в которой личинки встречаются в наибольшем количестве.
Рассмотрим роль главнейших физиономических типов водной расти-
тельности в жизни личинок A. maculipennis (ср. стр. 104—114).
б) Ролъ основных физиономгшеских типов
ра ыпипгелъиоелш
1. Лсмппды—свсбсдпо плавен щпе, ncjKcpcnricmneca растения
Планктические лемппдьт. В этой группе различают две подгруппы:
плапктических п псйстпчсских лемппд. К плапктпчсским лемнидам от-
носятся нитчатые водсрссли, ряска трехлопастная (Lemna trisulca),пузыр-
чатка (Ulricularia) и др. Эти растения при небольших обилиях обычно
плавают в толще воды, п в жизни личинок Anopheles почти не играют
роли. Попри увеличении обилия они всплывают на поверхность, созда
вая при этом исключительно благоприятные для существования личинок
условия химизма и питания и большую причальную линию. Идеальная
среда создается в комьях нитчаток, обилие личинок в которых достигает
иногда громадных размеров — до 2 746 личинок на 1 м2 (пойма реки
Урала у Магнитогорска летом 1931 г.). В комке водорослей личинки могут
встречаться даже в водоемах явно неподходящего химизма (например,
246
в водоемах, содержащих сероводород), так как внутри комка водорослей
личинки фактически живут в условиях химически обособленного от окру-
жающей воды микробиотопа. При чрезмерном обилии планктических
водорослей и Lemna trisulca, всплывая, они покрывают поверхность воды
сплошным толстым слоем, частично выступающим из воды. В этих слу-
чаях поверхность воды становится недоступной для личинок. Таким обра-
зом, при чрезмерном размножении планктические лемниды становятся
непригодными для .жизни личинок.
Пейстические лемниды. К Теистическим лемнидам относятся расте-
ния, свободно плавающие па поверхности воды, как, например, ряска
малая (Lemna minor), лягушатник (Hydrocharis morsus ranae), сальвиния
(Salvinia natans), водяной орех, или чилим (Trapa natans) и др. При ма-
лых обилиях их химическое влияние па воду ничтожно, при больших оби-
лиях вредно для личинок: по выделяя сами кислорода в воду и создавая
затенение, они затрудняют доступ света и атмосферного кислорода в тол-
щу воды, препятствуют фотосинтезу погруженных растений, вследствие
чего в водоеме создается кислородный дефицит п, следовательно, небла-
гоприятные для личинок условия. При больших обилиях этих растений
сходит на-пет и причальная линия, так как промежутки свободной воды
уменьшаются. При сплошном покрытии поверхности водоема ряской ма-
лой существование личипок Anopheles в нем становится невозможным.
Таким образом, эта группа растений неблагоприятна для жизни личинок
Anopheles. Однако при умеренном покрытии поверхности воды этими рас-
тениями личинки Anopheles среди них встречаются, хотя и в небольшом
числе.
2. Укореняющиеся растения
Элодеиды. Элодеиды — укореняющиеся погруженные растения с
длинными стеблями и отходящими по всей длине их листьями. Сюда от-
носятся элодея (Elodca canadensis), роголистник (Ceratophyllum), уруть
(Myriophyllum), многочисленные виды рдестов, водяной лютик и др.
Стебли этих растений, поднимающиеся с глубины 2—3 м (иногда с глуби-
ны 5—6 м) и лишь вблизи поверхности начинающие сильно ветвиться,
образуют густые кроны в виде сплошных подводных ковров. Пока эло-
деиды не достигли поверхности воды, они имеют для личинок умеренное
значение, влияя лишь па химизм воды. Но, достигнув поверхности, они
дают большую причальную линию и создают для личипок Anopheles
наилучшие условия химизма п питания, что делает элодеиды, наряду
с планктическими лемпидами, наиболее благоприятными для личинок
растениями. Обилие личинок A. maculipennis в зарослях элодеид достигает
750 личинок па 1 м2 (пойма реки Урала у Магнитогорска, 1931)
и больше. Особенно благоприятны мелкорассеченные элодеиды: роголист-
ник, уруть, рдест гребенчатый; несколько менее благоприятны крупно-
листные рдесты: Potamogeton lucens и Р. perfolialus. С достижением по-
верхности рост элодеид прекращается (над поверхностью воды они выстав-
ляют только цветы и плоды), поэтому личинки всегда находят среди
элодеид достаточно места для прикрепления па поверхности воды.
Только при сильном понижении уровня водоема элодеиды могут выступать
из воды; в этом случае их отмирание и загнивание ухудшают химизм воды
и ведут личинок к гибели.
Пимфеиды. Нимфеиды —укореняющиеся растения с плавающими
на поверхности воды листьями. Сюда относятся белые й желтые кувшинки
(Nymphaea и Nuphary, ужовник (Limnanthemum nymphaeoides), рдест
плавающий (Potamogeton natans) и др.
247
По своему влиянию на личинок нимфеиды сходны с нейстическими
лемнидами. Нередко кувшинки почти сплошь покрывают поверхность,
воды листьями; в этих случаях существование личинок становится невоз-
можным. Обилие личинок A. maculipennis среди нимфеид в поемных во-
доемах реки Урала у Магнитогорска в 1931 г. составляло от 0 до 2 личи-
нок на 1 м2; при наличии погруженных растений в промежутках между
нимфеидами оно повышалось. В водоемах Средней Азии нимфеиды отсут-
ствуют.
Линеиды. Линеиды — укореняющиеся растения, получившие свое
название от линейной формы листьев, выступающих над поверхностью
воды. Сюда относятся злаки и осоки (камыш — Scirpus, тростники —
Phragmites, рогозы — Thypha, рис, ситняг—Heleocharis), а также хво-
щи, сусак — Butomus, аир — Acorus и др. Большинство этих растений
имеет мощные’ многолетние корневища; стебли их и листья ежегодно
отмирают осенью, а весной появляются новые молодые побеги. Благодаря
сильному вегетативному размножению, образуют густые прибрежные
заросли, обычно не идущие на глубину, большую 2 м.
Значение линеид для личинок Anopheles меняется в зависимости от*
стадии их вегетации, густоты насаждения и величины растений. Веснойг
побеги линеид, не достигшие поверхности воды? не имеют значения для
жизни личинок. Только когда они достигнут поверхности воды, создаются
условия для существования личинок — возникает причальная линия.
Редкая и низкая поросль их мало затеняет водоем, не препятствует фото-
синтезу погруженных растений и, следовательно, не оказывает вредного
влияния на химизм воды. На юге умеренное затенение поверхности воды
может благоприятно отразиться на температурном режиме водоема для
тенелюбивых видов: A. hyrcanus, A. bifurcatus, A. algeriensis, а в раз-
гар лета и для A. maculipennis. Некоторые из линеид никогда не дости-
гают большой высоты, и поэтому заросли их всегда умеренно благоприят-
ны для личинок A. maculipennis. Таковы ситняг—Heleocharis, боль-
шинство культивируемых у нас сортов риса. Однако при сильном развитии
зарослей крупных линеид, достигающих большой высоты (гигантолинеиды),
таких, как тростник, камыш, рогоз, создаются условия, исключающие
существование в них личинок A. maculipennis, а зачастую даже тенелю-
бивых видов. Под тенью этих зарослей на севере может создаться небла-
гоприятный температурный режим; кроме того, сильное затенение пре-
пятствует развитию погруженной растительности, что вместе с ежегодным
засорением воды остатками отмирающих стеблей и листьев создает не-
благоприятный для личинок химизм водоема. Таким образом, заболочен-
ности, заросшие гигантолинеидами, непригодны для развития личинок
A. maculipennis, личинки этого вида могут развиваться только на опуш-
ках таких зарослей или среди «окон», заросших погруженной раститель-
ностью.
Фолииды. Фолииды — укореняющиеся растения с широкими над-
водными листьями. Сюда относятся: сабельник (Comarum palustre), вах-
та (Menyanthcs trifoliata), белокопытник (Calla palustris) и др. Значение-
фолиид для экологии личинок A. maculipennis аналогично значению
линеид.
Амфибииды. Амфибииды — укореняющиеся земноводные полиморфные
растения, форма листьев которых может изменяться в зависимости от сте-
пени их затопления водой. Так как эти растения могут встречаться в не-
скольких модификациях —- с погруженными, плавающими или надвод-
ными листьями, то их нельзя причислить к определенному физиономи
ческому типу. Таковы стрелолист (Sagittaria), ежеголовка (Sparganium),
248
земноводная гречиха (Polygonum amphibium), земноводная полевица
(Agrostis stolonicens) и др. Наиболее благоприятны для жизни личинок
Anopheles те амфибииды, которые имеют наибольшую поверхность по-
груженных листьев. Так, например, заросли мелколистного злака поле-
вицы земноводной (Agrostis stolonicens), появляющейся ранней весной
во временных водоемах и в периферической зоне постоянных водоемов,
создают чрезвычайно благоприятные условия для жизни личинок A. ma-
culipennis; обилие личинок в зарослях этого злака достигает 2 855
на 1 м2 (Половодова).
3. Сезонные перемещения центров обилия личинок
В течение сезона анофелогенная роль переходит от одного типа расти-
тельного сообщества к другому. Сезонная смена состояний (аспектов) расти-
тельности ведет к перемещению центра обилия личинок из одного типа сооб-
щества в другие. Так, в средней полосе СССР центр личиночного обилия
весной находится в зарослях амфибиид и невысоких линеид (Heleocharis,
Glyceria, осок) в прибрежной зоне озерков и во временных лужах. В этих
биотопах в это время создается наиболее благоприятный температурный
режим. Летом, когда временные водоемы пересыхают и частично пересы-
хает и зоца амфибиид в постоянных водоемах, центр обилия личинок пе-
реходит к элодеидам, которые к этому времени поднимаются к поверх-
ности воды; температурный режим постоянных водоемов к этому времени
становится также благоприятным для личинок. Осенью личинки засе-
ляют как заросли элодеид, так и заросли планктических лемнид, которые
развиваются в это время в больших количествах. Наибольшее значение
для развития комариной популяции имеет летний состав растительности;
в поемных водоемах это будут элодеиды.
В Средней Азии и на Кавказе места выплода личинок A. maculipen-
nis часто возникают в местах прорывов каналов оросительной сети; та-
кие места обычно содержат земноводную флору: пальчатник (Cynodon
dactylon), куриное просо (Echinochloa crus galli), водяные лютики, гре-
чихи, солянки и пр., которые при заливании водой сразу создают благо-
приятные для личинок условия существования.
4. Выводы для практики
Уничтожение водной растительности, благоприятной для существо-
вания личинок (элодеид, планктических лемнид), является хорошим мето-
дом борьбы с личинками A. maculipennis.
В канавах осушительной и оросительно!! сети механическое уничто-
жение водной растительности применяется и должно применяться как
мероприятие, необходимое для правильной их эксплоатации, так как
зарастание канав уменьшает скорость течения в них и, следовательно,
препятствует их работе. Вполне применимо уничтожение водной расти-
тельности и в небольших стоячих водоемах: декоративных бассейнах,
хаузах 1 и т. п. В больших водоемах (водохранилищах, озерах, прудах)
механическое уничтожение зарослей элодеид и планктических лемнид
в настоящее время не является широко распространенным мероприятием,
так как обходится дорого. Это мероприятие может иметь будущее лишь
при условии механизации этой работы и хозяйственном использовании
1 Хаузы — небольшие копаные пруды на оросительных системах в Средней Азии
(рис. 68).
249
добываемых водных растений (может быть, в качестве корма для домашней
птицы, свиней). Что касается нейстических лемнид и крупных линеид,
то к уничтожению их надо подходить с большой осторожностью, так
как эти растения при сильном развитии сами предотвращают возмож-
ность развития личинок в водоеме. В оросительных и осушительных ка-
налах тростники безусловно подлежат уничтожению, так как их заросли
любой густоты мешают работе каналов. В остальных случаях тростники
следует истреблять там, где они слабо развиты и в них выплаживаются
личинки, и не трогать густых зарослей, не содержащих личинок. Опыты
показывают, что расчистка таких густых зарослей гигаптолинеид приво-
дит к заселению водоемов светолюбивыми растениями: нитчатками и т. д.
и, следовательно, личинками Anopheles. Часто тростниковые массивы за-
селяются личинками лишь по периферии — опушке, па 2—3 м от края,
куда проникает свет. В этих случаях следует опылять парижской зе-
ленью заселенную личипкамп зону, по не расчищать массив.
Биологическим методом борьбы с личинками Anopheles при помощи
воздействия па растительность является обсадка деревьями речек и ка-
налов, отчасти п стоячих водоемов, с целью создания густой тени, препят-
ствующей развитию водных растений и снижающей анэфелогенную про-
дукцию водоема (стр. 23J).
в) Влияние сукцессионного возраста водоемов
на анофелъную продукцгью
Производя оценку водоемов с точки зрения их апэфологенности, следует
иметь в виду, что каждый водое^м в течение своего существования не остается
неизменным, а на протяжении ряда лот претерпевает изменения, проходя
через стадии юности, зрелости п старости, и чтЬ все эти стадии развития
неравноценны по своим гидрохимическим и биотическим свойствахМ для
существования личинок Anopheles. Такие изменения в водоеме являются
результатом смены биоценозов или сукцессии. Места выпледа A. macu-
lipennis приурочены к водоемам юным и зрелым, т. е. находящимся на
первых стадиях сукцессии. Поэтому мы находим их на тех участках мест-
ности, где непрерывно возникают новые водоемы, например, в речных
долинах. Много новых водоемов создает п сам человек. Наоборот, в тех
случаях, где новых водоемов больше не образуется, а старые уже успели
завершить цикл своего развития (например, ва древних террасах речных
долин), апэфологеппых очагов мы по встречаем, а если они имеются, то
большей частью созданы рукой человека (ср. также стр. 114—116).
10. ВРАГИ ЛПЧППОК
В естественных условиях враги личинок Anopheles многочисленны.
Существует много животных, которые способны поедать личинок. Унич-
тожение личинок хищниками,повидимому, одна из главных причин есте-
ственной смертности личинок в водоемах. Враги личинок имеются среди
различных представителей животного царства, таковы: гидры, ресничные
$ерви7турбеллярии, пиявки (рис. 57, D), улитки-прудовики (Limnaea
palustris и L. ovata) (рис. 58, В), которые часто ползают снизу по поверх-
ности воды, бокоплавы (Ganimaridae) (рис. 5, Г), циклопы (рис. 54, А),
водные пауки (Argyroneta aqualica). Многочисленны враги личинок среди
водных насекомых: личинки стрекоз, водяные клопы-гладыши (Noto-
necta, рио. 59, Г), водяные скорпионы [Nepa (рис. 20, В), Ranatra] и пр.,
водомерки; водяные ткуки-плавунцы (Dyliscidae) (рис. 59, А), принадле-
250
жат к. числуособенно опасных врагов. В неволе один гладыш может съесть
»до 22 личинок A. maculipennis в сутки, а некоторые виды плавунцов —
до 26 личинок в сутки. Поедают личинок также личинки многих ручей-
ников. (рис. 57, Б) и хищные личинки комаров Chaoborinae и Ceratopogon.
Важнейшими врагами личипок являются рыбы. Личинки могут существо-
вать в одном водоеме с рыбами только в присутствии растительности
или помещаясь в полузатопленном галечнике, где они становятся недо-
ступными для рыб. При отсутствии возможности укрыться от рыб личинки
выедаются ими нацело. Особенно опасны рыбы, питающиеся с поверхно-
сти воды, а также мальки. Рыбы — единственные из врагов, используе-
мые в качестве биологической меры борьбы.
Гамбузия. Наибольшую известность в качестве особо опасных хищ-
ников, уничтожающих личинок Anopheles, получили в Европе и в СССР
гамбузии — рыбки, происходящие из Северной Америки (рис. 59, Ж).
Гамбузии относятся к отряду зубастых карпов: Cyprinodontiformes,
к семейству Pceciliidao. У нас в СССР имеется два подвида: Gambusia
affinis affinis и G. affinis holbroochi. Эти рыбки были завезены в СССР
в 1925 г. Николаем Павловичем-Ру хадзо. Гамбузия имеет перед нашими
рыбами ряд преимуществ.
В европейской фауне почти пет рыб, приспособленных к малым во-
доемам, так как эти водоемы не могут существовать непрерывно — они
промерзают на севере или высыхают (в Средиземноморье). Но во влажном
и теплом климато юга США такие водоемы и рыбы многочисленны. При-
способлением к жизни в малых водоемах у гамбузии являются: 1) неболь-
шие размеры: самки — 6,8 см, самцы — 3,5 см; 2) живорождение, воз-
никшее в результате неблагоприятных условий для развития эмбриона
во внешней среде: неустойчивый водный режим, большая доступность
врагам; 3) быстрота размножения (для развития- эмбрионов- при благо-
приятной температуре требуется месяц); 4) большая плодовитость (дает
4—7 пометов в сезон* при числе мальков в каждом помете в среднОхМ 100,
максимально 400); 5) вынесли весть в отношении всех факторов среды: а) мо-
гут жить в сильно загрязненных водоемах (но в присутствии раститель-
ности)^) выдерживают концентрацию солей до ЗО^/оо, в) способны длитель-
ное время голодать.
Следующие особенности биологии гамбузии делакЗт применение ее
в качестве врага личипок особенно эффективным: 1. Гамбузия — хищник,
питающийся с поверхности воды добычей как раз нужного для нас раз-
мера и преимущественно в прибрежной зоне, в местах концентрации ли-
чинок Anopheles. При этом она чрезвычайно прожорлива: в аквариуме
за одни сутки съедает до 145 личинок (Калапдадзо и Таирова). 2. Быст-
рота размножения гамбузий ведет к перенаселению ими водоемов, а непре-
рывное размножение — к постоянному наличию мальков, что чрезвычайно
повышает их эффективность в водоеме. Мальки гамбузий лучше взрослых
проникают в заросли растительности и выедают личинок Anopheles.
Однако при чрезмерной густоте растительности гамбузии проникать в за-
росли и выедать всех личинок не могут. В этом случае рекомендуют унич-
тожение растительности или применение гамбузий совместно с карпом;
последний выедает растительность, и личинки становятся доступнее для
гамбузий. 3. Имеет значение* и то обстоятельство, что гамбузия — чуж-
дый элемент в нашей фауне. Искусственное введение нового хищника
резко меняет условия биоценозов, вызывая иногда почти полное уничто-
жение животного населения водоема.
Для Кавказа и Средней Азии применение5гамбузии является важней-
шим методом борьбы с малярией. В ряде мест, например, на приморских
251
низменностях Черноморского побережья (в Абхазии, Аджарии), гамбузия
дала огромный оздоровительный эффект.
Однако применение гамбузии ограничено теплым климатом. Опти-
мальной температурой для ее жизнедеятельности является 20—30°, при
температурах ниже 15° она не размножается, а при 7—8° идет на зимовку.
Зимует на дне водоемов, в илу под камнями, в водорослях. В водоемах,
покрывающихся льдом, может перезимовывать при условии достаточного
количества кислорода.
Северной границей применения гамбузии в СССР является на Северном
Кавказе линия Кизляр—Армавир; на Украине гамбузия зимует только
на крайнем юге.
При использовании гамбузии для уничтожения личинок необходимо
помнить, что: 1) гамбузия непригодна для использования в рыбохозяйст-
венных прудах, так как выедает мальков; 2) гамбузия не выносит сильного
течения и поэтому применение ее в речках и каналах с бурным течением
z дает меньший эффект' 3) водная растительность в водоемах не должна
быть слишком густа, в силу чего необходима периодическая очистка от ра-
стительности 1.
Кроме указанных животных, врагами личинок является еще водоплавающая
птица: утки, дикие и домашние,,гуси. Однако для очистки водоема от личинок необхо-
димо значительное перенаселение его птицей/В этом случае птицы, помимо выедания
личинок, выедают также водную растительность и загрязняют водоем, делая его не-
пригодным для развития личинок.
11. ЛИЧИНОЧНАЯ СМЕРТНОСТЬ У ANOPHELES MACULIPENNIS
Размеры личиночной смертности. Размеры личиночной смертности
устанавливаются путем количественного учета личинок и куколок в водое-
мах в течение сезона. Учет показывает, что личиночная смертность в сред-
нем за-сбзон по* всем растительным сообществам равна 93—90%. В уло-
вах, как правило, преобладают личинки младших возрастов; чем личинки
старше,' тем их меньше. До стадии куколки доживает 7,5—10% особей
от числа проходивших первую личиночную фазу. Однако личиночная
смертность не всегда и не везде одинакова. Так, весной во время разви-
тия I поколения смертность личинок ниже, чем летом и осенью. Как по-
казывают наблюдения, весной 1930 г. в водоемах Урала у Магнитогорска
до IV возраста доживало почти 100% личинок, летом смертность повыша-
лась и осенью она была наиболее высокой.
Причины смертности. Причина высокой смертности осенью — резкое
' изменение физико-химических условий в водоемах в неблагоприятную
для личинок сторону (отмирание и загнивание растительности и пониже-
ние температуры). В условиях неволи голод, наряду с загниванием воды,
является главной причиной личиночной смертности. Летом главной при-
чиной смертности личинок в природных условиях является, повидимому,
поедание их врагами. Шансы на поедание врагами увеличиваются при
более густой популяции, так как при большом числе личинок на единице
площади шансы на встречу их с хищниками резко повышаются.
г Факт меньшей смертности среди первых весенних личинок объясняется
тем, что весной в водоемах плотность населения, в том числе и врагов,
меньше, вследствие чего враги причиняют меньше вреда; кроме того, в это
время меньше и конкуренция из-за пищи. Борьба за существование про-
текает более интенсивно в благоприятных для личинок биотопах, где боль-
1 Инструкция по применению гамбузии напечатана в Сборнике организационно
методических материалов по малярии и гельминтозам за 1943 г.
252
ше плотность личиночного населения и больше конкуренция, поэтому смерт-
ность личинок в густо заселенных биотопах всегда выше, чем в менее
заселенных (так, например, в элодеидах она больше, чем в нимфеидах и
линеидах).
12. АНОФЕЛЬНАЯ ПРОДУКЦИЯ ВОДОЕМОВ
Каковы размеры анофельной продукции водоемов, или, иными сло-
вами, какое число комаров окрыляется на единице поверхности водоема
в единицу времени, например, на 1 м2 в сутки?
Определение анофельной продукции водоемов. Определение анофельной про-
дукции водоема возможно двумя путями. Первый путь — улавливание окрыляющихся
комаров при помощи опущенного на водоем садка. Второй путь — вычисление
продукции из обилия куколок и продолжительности фазы куколок. Средняя за сезон
анофельпая продукция разных типов сообществ в поемных водоемах Магнитогорска
определялась Беклемишевым и Половодовой при помощи второго из указанных спо-
собов. Для этого число куколок, полученное при количественном учете, делилось на
продолжительность фазы куколки при средней температуре развития куколки, и та-
ким образом определялась суточная продукция.
В результате такого определения было установлено, что абсолютная
величина анофельной продукции в среднем за сезон для всех растительных
сообществ долины реки Урала у Магнитогорска составляла в 1931 г.
один комар на 1 м2 в сутки. За сезон продолжительностью в 100 дней это
составляет 100 комаров на 1 м2, или 1 млн. комаров на 1 га содержащих ли-
чинок сообществ. В более южных местах, например, на Кавказе или в Сред-
ней Азии, и сезон дольше, и скорость развития больше, в результате чего
число комаров, вылетающих в год с 1 га заселенной личинками водной
площади, может достигать нескольких миллионов. <
Средняя анофельная продукция в различных растительных сообще-
ствах различна: так, элодеиды с планктическими лемнидами дают (по на-
блюдениям в Магнитогорске) около 2 комаров на 1 м2 в сутки, тогда как
нимфеиды и гигантолинеиды — всего около 0,1 комара на 1 м2 в сутки,
т. е. в 20 раз меньше. Следовательно, тип растительности является важ-
нейшим индикатором при установлении анофелогенного значения водоема.
ГЛАВА VIII
ЭКОЛОГИЯ ANOPHELES MACULIPENNIS—ВЗРОСЛОГО КОМАРА
1. ПОВЕДЕНИЕ КОМАРА И ДЕЙСТВИЕ НА НЕГО ВНЕШНИХ ФАКТОРОВ
а) Посадка, геотаксис, движения
Днем взрослые A. maculipennis обычно столь же мало подвижны,
как и личинки. Они сидят на стенке, на потолке или на каком-нибудь пред-
мете и перелетают с места на место только под влиянием внешних раз-
дражений.
Сидят они под углом к субстрату, тело вытянуто прямолинейно, зад-
ние ноги обычно свободны и иногда по очереди раскачиваются. Поза си-
дящего Anopheles очень характерна и позволяет сразу отличить его от ко-
маров из трибы Culicini (рис. 153). Последние гораздо более горбатые,
и у сидящего брюшко направлено косо к стене. Впрочем, на зимовке
самки Anopheles часто прижимаются брюшком к стене (рис. 153, В), и
неопытный глаз может принять такую зимовочную самку Anopheles за
самку Culex. На стене все комары всегда сидят головой кверху. В этом
253
проявляется отрицательный геотаксис 1 комара. Отрицательный геотаксис
проявляется также в стремлении комара занять наиболее высокие места в
доступном ему пространстве. В силу этого A. maculipennis па дневках
располагается на потолке или в верхних частях стен, если этому не.
препятствует действие других факторов (свет, температура и пр.).
Вспугнутый Anopheles обычно склонен лететь кверху, тогда как
вспугнутый Culex скорее бросается книзу.
Полет Anopheles отличается от равномерного полета других комаров
порывистым, прыгающим характером, отчего издаваемая им при полете
песенка приобретает определенный ритм. Поэтому Anopheles можно отли-
чить от AeJes или Culex и на лету, и даже в темноте. Полет A. superpictus
еще более порывист, чем полет A. maculipennis, и скачки, которые он де-
лает в воздухе, значительно больше.
Рис. 153. Посадка комаров.
А—Anopheles maculipennis, но| мольная посадка; Г— то же, полуяи-
мовочная посадка; В—то же. зимоночная посадка; Г—Culex pi-
pitns, нормальная посадка.
Скорость полета самки A. maculipennis, преследующей добычу, на-
пример, едущий экипаж, достигает 3 км в час (Яценко), т. е. 83 см в се-
кунду.
Anopheles могут и ползать,например, когда им приходится пробираться
в узкие отверстия. Они проползают в щели окна при влете и вылете из по-
мещения или забираются в узкие пространства, прячась от света,.жары и
сухости. Поэтому искать комаров в хворостяной или тростниковой крыше
бывает трудно, а при механической защите домов сетками или людей по-
логами надо тщательно следить, чтобы не оставалось щелей.
б) Действие света
Фотокипсз. Свет оказывает огромное влияние па поведение взрослого
комара. Прежде всего взрослому комару, так же как и личинке, свой-
ственно явление фотокинеза: яркий свет приводит комара в возбуждение.
1 J еотакспс — буквально «| асгсгон опте го отов ению к земле», т. е. припп-
тие насекомым о.ц еделенно) о i олои ения i oo rou опию к направлению силы тля ее»и.
Отрицательный igoiukchc—расшлол енне 0 о..о^ой н]о<ив направления силы тяже-
сти, т. е. кверху.
254
Чем ярче свет, тем чаще под его влиянием взлетают сидящие комары. Если
помещение освещено неравномерно, комары в светлой его части взлетают
с места чаще, чем в темной, и в силу этого через некоторое время соби-
раются в темной части, где и успокаиваются. Поэтому на дневках A. ma-
culipennis собираются в наиболее за тененных Местах помещения. Полной
темноты они, однако, избегают, и в абсолютно темных частях помещения
их не бывает. Причиной этому является положительный фототаксис кочень
слабому свету (см. ниже).
Реакция бегства. Если комар подвергается какому-либо резкому раз-
дражению — механическому (например, прикосновением), химическому
(например, дымом), тепловому, световому, он улетает, обращается в бег-
ство. При реакции бегства появляется положительный фототаксис, кото-
рый мы наблюдали при нырке у личинки. Вспугнутый комар всегда бро-
сается к свету. На этом основан метод выкуривания комаров в комаро-
ловку (способ Ферми): все окна помещения завешивают, а в единственное
незавешенное окно вставляют ловушку; вспугнутые дымом комары устрем-
ляются на свет и попадают в ловушку. Фототаксисом вспугнутых
комаров пользуются при переселении комаров из одного садка в другой,
затемняя садск, в котором сидят комары, оставляя открытый вход в
освещенное, новое помещение и вспугивая комаров постукиванием или
встряхиванием. *
При одномоментном раздражении реакция бегства длится недолго, и
с ее прекращением исчезает притягательная сила светлого пятна. Поэтому
хотя вспугнутые в местах дневок комары сначала бросаются к свету, обычно
они тотчас «приходят в себя», поворачивают полет к стене или потолку и,
полетав немного, садятся. Но при длительном действии раздражителя ко-
мары долгое время находятся в состоянии «бегства» и все это время стре-
мятся к свету. Так, например, ведет себя выкуриваемый комар, длитель-
ное время раздражаемый дымом.
Комары, находящиеся под влиянием голода, жажды, полового инстин-
кта, приходят в возбуждение и потому ведут себя почти так же, как и вспуг-
нутые комары. Голодный или жаждущий комар также стремится к свету,
но в отличие от вспугнутого комара — только к слабому свету. Поэтому
из темного помещения голодные комары вылетают в более светлое, вечером
вылетают из убежища наружу. Но яркий свет их не привлекает, и при
ярком свете они остаются неподвижно сидеть в затененных местах. Однако
голодные комары занимают в помещении более светлые места по сравнению
с сытыми комарами.
Комар видит и отдельные предметы. Он видит приближающуюся к не-
му руку и реагирует бегством. Он не только чует, но и видит добычу,
правда, только вблизи и главным образом движущуюся добычу. Самка
пользуется зрением и при поисках водоема для откладки яиц; она видит
темные пятна окоп, когда утром прячется в убежище.
в) Денстсне тсл.нс'ратуры и ела ок н сети.
Дневные убеэкигца
Нижний порог активности зимовочной самки A. maculipennis лежит
очень низко: еще при температуре—1,08° комары слегка шевелят но-
гами в ответ на прикосновение, ползать могут при 4-2°, летать— при +5°.
Летом самки A. mac. messeae летают и нападают, начиная с температуры
-J-7P, A. mac. sacharcvi — начиная с температуры -j-10°.
О нижнем температурном пороге жизни комаров см. ниже, стр. 285.
255
Верхний порог активности для A. mac. messeae лежит между 25° и 30°:
при 25° они активно нападают, при 30° не нападают ни при каких условиях
влажности и освещения. Верхний порог жизни для A. mac. messeae
лежит около 35°.
Летние самки A. mac. messeae при возможности выбора собираются
в местах, имеющих 19—21°, и при этой же температуре среди них на-
блюдается наименьшая смертность. Идущие на зимовку самки выбирают
гораздо более низкие температуры.
Водный баланс. A. maculipennis принадлежат к числу насекомых,
непрерывно испаряющих большое количество воды. Испарение происходит
тем скорее, чем больше дефицит влаги в атмосфере, чем выше температура,
чем больше скорость ветра. Если A. maculipennis поместить в иссушаю-
щие условия (высокая температура, низкая влажность, ветер), он быстро
погибает от потери воды, теряя к моменту гибели 30—40% от перво-
начального веса тела.
В условиях опыта, при возможности выбора и температурах ниже 25°, северные
A. maculipennis собирались при относительных влажностях 70—96%; при температу-
рах 26—30° они выбирают наиболее влажные места. На дневках, если влажность выше
65—70%, северные A. mac. messeae располагаются внутри помещения, совершенно
не считаясь с влажностью; если же в помещении сухо (влажность ниже 65%), комары
придерживаются наиболее влажных мест. Таким образом, они начинают избегать су-
хого воздуха только при влажности пиже 65%. Для более суховыносливого A. mac.
.sacharovi этот порог пиже: он считается с влажностью лишь в том случае, если она
ниже 40%, и только при этих условиях собирается в наиболее влажных частях помеще-
ния. Однако воздуха, насыщенного водяными парами (относительная влажность 100%),
A. maculipennis избегает и, будучи помещен в такую среду, обращается в бегство,
а если остается в ней, то гибнет.
Итак, A. maculipennis хорошо себя чувствует в довольно широких
пределах влажности; он ищет не наибольшей влажности, а такой, при
которой он может дожить хотя бы до вечера, не умерев от жажды.
Распределение в дневных убежищах. Войдя в хлев или дом, в кото-
ром находится большая дневка A. maculipennis, мы находим, что они рас-
полагаются в помещении большей частью неравномерно и не всегда оди-
наково. Отчего зависит расположение комаров в дневном убежище?
Прежде всего отрицательный геотаксис заставляет комаров держаться
при прочих равных условиях в верхней части помещения. Если в поме-
щение врывается яркий свет, комары в силу фотокинеза располагаются
в наиболее затененных местах; в пасмурные дни, когда яркого света
нигде нет, они располагаются независимо от освещения. Если температура
близка к оптимуму (20—25°), мелкие различия температуры отдельных
углов помещения не оказывают влияния на расположение комаров, но
если в помещении слишком холодно, комары жмутся в тех местах, где
хотя бы на 1—2° теплее, чем в других. Если в помещении слишком жарко,
они теснятся к самым прохладным местам. Если в летнюю жару крыша на-
калилась и от нее идет жар, комары перекочевывают вниз и сидят против
своего обыкновения на нижней части стен. Если в помещении влажно,
комары располагаются независимо от степени влажности; если в поме-
щении слишком сухо, они жмутся в тех местах, где влажность хотя бы
на 2—3% выше, чем в других местах, и избегают малейшего сквозняка.
Таким образом, распределение комаров внутри помещения зависит от всех
метеорологических факторов, но сильнее всего сказываются те из них,
которые больше всего уклоняются от предпочитаемого комарами оптимума.
Понимание причин, обусловливающих распределение комаров внутри
убежища, очень помогает при обследовании дневок.
256
Суточный ход метеорологических факторов. Как изесстпо, в ясные и тцхие дпп
температура воздуха с утра и до 2—3 часов дня непрерывно попытается, относительная
влажность падае г, а с 2—3 часов дня и до восхода солнца температура падает, влаж-
ность все время пли часть времени поднимается. Эти колебания температуры и влаж-
ности в ясные летние дни могут достигать значительного размаха, особенно при кон-
тинентальном климате. Близ поверхности земли они бывают еще бо iee резкими, чем
на высоте 2 м, где обычно производятся метеоротопические наблюдения. В пасмурные,
дождливые пли ветреные дни суточный ход температуры и влажности менее выражен,
по дневной подъем температуры, а иногда и дневное падение влажности более илп
менее сохраняются.
Рис. 154. Дпевпые убежища A. mac. mes-
seae в дупле осокоря на пойме Волги про-
тив г. Куйбышева.
Обычный суточный ход метеорологических факторов делает дневные
часы непригодными для активности A. maculipennis и для пребывания его
на открытом во щухе.Ол изжарится
на солнце и высохнет под влиянием
высокого дефицита влажности и
ветра. Ему необходимо укрыться
в убежище с благоприятным мик-
роклиматом, т. е. не испытываю-
щем резких суточных колебаний
температуры и влажности, защи-
щенном от прямого солнца и
ветра. Таким убежищем может
явиться либо растительность, либо
естественные углубления в зем-
ле—пещеры,трещины скал и т.п.,
либо норы животных и постройки
человека.
Типы дневных убежищ. Уже в
густых зарослях травы около по-
верхности земли создается зате-
ненное от солнца и защищенное от
ветра пространство, в котором тем-
пература днем ниже и влажность
выше, чем на открытом воздухе.
Под прикрытием деревьев или
кустов густая трава создает еще
более защищенные условия. Вну-
три обросшего травой куста та-
мариска (в Туркмении, в июль-
ский день) температура равнялась
35° и относительная влажность
35%, в то время как на наруж-
ном воздухе было 46° тепла и 15% влажности; в кусте находилась
дневка A. pulcherrimus и A. hyrcanus (Митрофанова). Таким образом,
заросли часто служат дневными убежищами комаров, в том числе и
A. maculipennis. В Средней Азии дневки его находили, например, в лю-
церне (Правиков).
Встречаются дневки A. maculipennis и в дуплах деревьев (рис. 154),
в особенности обращенных на север или на восток или находящихся
в тени (Шлёнова). Норы, пещерки, колодцы, мосты и тому подобные места
также нередко служат местами дневного убежища, если они расположены
поблизости от мест выплода или добычи.
Наконец, важнейшими дневными убежищами нашего вида являются
постройки человека. Чем больше помещение изолировано от внешней
среды (толстые стены, хорошая крыша, отсутствие дыр, закрытые окна
17 Учебник медицинской энтомологии
253
и двери, наличие потолка), тем меньше отражаются внутри него колеба-
пия температуры и влажности внешней среды. Летом такое помещение
обычно хорошо защищает комаров от дневной жары и сухости и представ-
ляет для A. maculipennis хорошее дневное убежище. Наоборот, навесы,
балконы, палатки, шалаши, легкие хлева с плетеными стенками и мно-
гочисленными дырами, войлочные юрты — все эти помещения представ-
ляют небольшую защиту и в крайнем случае по своим условиям могут почти
не отличаться от наружного воздуха. Однако и в этом случае они могут
служить убежищем для комара. Действительно, открытый навес среди го-
лой степи все же защищает сидящих под ним комаров от прямых лучей
солнца и дает им возможность сидеть на высоте 2 м от раскаленной земли,
т. е. на такой высоте, на которой температура много ниже и влажность
выше, чем непосредственно над поверхностью почвы. И дневки A. macu-
lipennis в подобного рода убежищах встречаются очень часто, тем более
что они обычно гораздо доступнее для залета комаров, чем хорошо защи-
щенные убежища. Никогда дневки комаров не бывают лишь в тех убе-
жищах, в которых днем условия хуже, чем на открытом воздухе, напри-
мер, летом в сильно натопленных помещениях.
В холодные ночи, особенно весной или осенью, когда температура
наружного воздуха ниже предпочитаемой комарами, хорошо защищен-
ные помещения бывают теплее наружного воздуха и в силу этого сильна
привлекают к себе комаров. Сточки зрения распространения малярии ми-
кроклимат убежищ особенно важен на севере, при температурах, при-
ближающихся к нижнему порогу развития плазмодия в комаре
В этих условиях иногда бывает, что развитие плазмодиев невозможно или
идет очень медленно при температуре наружного воздуха и неплохо идет
в хлевах, средняя температура которых на 2—3° выше наружной.
Распределение комаров между постройками и природными убежи-
щами. Возникает вопрос: как распределяются комары A. maculipennis
между постройками человека и природными убежищами (растительность,
поры, пещеры и т. п.), какой процент особей сидит в постройках и какой
процент— в природных убежищах? Прежде всего распределение зрелых,
охотящихся самок очень отличается от распределения самц)в и молодых
самок. Первые сильно привлекаются добычей, вторые — нет. Поэтому боль-
шинство молодых самок и самцов очень часто остается в природных убе-
жищах; они встречаются лишь в постройках, очень близко расположенных
от мест выплода. Зрелые, охотящиеся самки чаще всего к шцентрирукися
в поселках, многие из них проникают в помещения — дома и хлева — с до-
бычей, пьют там кровь й остаются переваривать ее в том же помещении;
другие напиваются на двэре и частью влетают переваривать кровь в поме-
щения, а частью рассеиваются в растительн юти и других н вружных
убежищах. Поэтому в непосредственной близости поселка всегда имеется
некоторое количество переваривающих самок во внешней среде. Количе-
ство их зависит от климата местности и наличных убежищ. При теплом,
влажном, мягком климате и богатой растител ьн юти в последней будет
больше комаров; при резком континентальном климате, сильных ветрах,
открытой стопной местности большая часть переваривающих сам ок A. ma-
culipennis держится в помещениях. Наконец, в тех случаях, к >гда вдали
от поселков ночуют стада или группы людей, охотящиеся самки юрмятся
ва их счет и здесь же, во внешней среде, находят себе дневные убе-
жища.
В итоге распределение взрослых A. maculipennis между постройками
и природными убежищами в различных случаях бывает весьма различно,
но причины того или иного распределения вполне понятны. Распределе-
258
ние это имеет большое значение с точки зрения борьбы с комарами: чем
больше процент комаров, сидящих в легко доступных для нас помещениях,
тем легче и успешнее может проводиться противоимагинальная борьба.
2. ОКРЫЛЕНИЕ П ПЕРВЫЕ ДПП ПМАГИПАЛЬПОИ ЖИЗНИ
Окрыление. Вылупление A. maculipennis из куколок происходит во
все часы дня, но главным образом утром и вечером. Во время вылупле-
ния комар беспомощно торчит из куколки и в это время легко может стать
жертвой врагов или потонуть от порыва ветра.
Числа самцов и самок при вылуплении приблизительно одинаковы
или слегка преобладают самки. В лаборатории при недоедании вылупляет-
ся больше самцов, т. е. женские личинки больше страдают от голода.
Признаки новорожденной самки. В лаборатории новорожденную сам-
ку можно накормить кровью уже часов через 5 после окрыления, по в при-
роде потребность в крови возникает у нее только через один или несколь-
ко дней. За это время она успевает копулировать и происходит первона-
чальное доразвитие яичников.
Новорожденную самку можно отличить по целому ряду признаков.
Чешуйчатый покров тела и крыльев еще цел, что придает комару яркую
контрастную окраску, бахрома крыльев цела. Желудок имеет почти ци-
линдрическую форму, стенки его прозрачны, он содержит зеленоватую
массу (меконий), представляющую остатки сброшенного при метаморфозе
эпителия кишечника личинки. Жировое тело нередко окрашено в зелено-
ватый цвет. Яичники маленькие, прозрачные; старшие фолликулы нахо-
дятся на стадиях N, 1 или I—11. Трахейные клубочки яичников пераз-
мотаны, ампулы яйцеводов прозрачны и малы. Сперматека не содержит
спермы.
Первоначальное дозревание самки. В течение первых дней жизни исче-
зает зеленый цвет жирового тела, меконий выделяется через анальное от-
верстие наружу, старшие фолликулы яичников доразвиваются до II ста-
дии Кристоферса. Копуляция уже произошла и сперматека заполнена спер-
мой. Доразвитие яичников до II стадии происходит отчасти за счет запас-
ных веществ жирового тела самки, но в эти дни она может и принимать
сладкую пищу, например, сок цветов; для менее упитанных самок такое
дополнительное питание обязательно.
До тех пор, пока старшие фолликулы яичников находятся па ранних
стадиях развития, самка еще не подчиняется закону гонотрофичоскэй гар-
монии, и если ее напоить кровью, ее яичники только доразвиваются до
11 фазы. Лишьс того момента, когда яичники доразвилисьдо 11 фазы, один
прием крови становится достаточным для того, чтобы они развились
до конца, и именно с этого времени у самки просыпается инстинкт жажды
крови, и она начинает искать добычу: она становится зрелой и вступает
в период активной гонотрофической жизни. Период первоначального до-
зревания при 20° длится около 10 дней; при 25° срок сокращается до 4 дней.
В этот же период происходит и копуляция.
Жизнь самцов. Самцы держатся днем в тех же убежищах, где и пеопло-
дотворенные самки. В помещениях с добычей процент самцов невелик,
так как здесь они разбавлены большим количеством кровососущих самок.
При этом чем дальше отстоят такие помещения от мест выплода, тем меньше
в них самцов. Относительно много самцов бывает в уборных. Самцы
A. mac. atroparvus, в отличие от самцов других подвидов, всюду следуют
за самками, встречаясь также и вдали от мест вь плода. На дневках они со-
ставляют иногда 40% от общего числа особей. Так же ведут себя и самцы
17*
259
A. superpictus, тогда как самцы A. pulcherrimus и других наших видов
Anopheles в помещениях вообще почти не попадаются.
Копуляция. Большинство подвидов A. maculipennis копулирует
на открытом воздухе, на лету. В тихие вечера самцы образуют рои и тол-
кутся в воздухе, издавая жужжание; единичные самки влетают в рой;
образуются парочки, которые улетают из роя. Длится копуляция несколь-
ко секунд. Самцы A. mac. sacharovi иногда роятся в помещении, где в
таком случае и происходит копуляция. A. mac. atroparvus и A. superpi-
ctus, как правило, копулируют в помещении, хотя могут роиться и копу-
лировать также и на открытом воздухе.
В неволе A. mac. sacharovi может копулировать в садке объемом с компату,
A. superpictus легко копулирует в садке объемом 1 м3, Л. mac. atroparvus даже в садочке
в 1 дм3. Потому последний вид особенно удобен для разведения в неволе
8. ПОИСКИ ДОБЫЧИ И НАПАДЕНИЕ
а) Поиски добычи
После оплодотворения и первоначального дозревания яичников самка
Anopheles переходит к нормальной гонотрофическоп жизни. Начинается
ее первый гопотрофический цикл, его первая фаза — поиски добычи и на-
падение. Жажда крови приводит комара в возбуждение; у него появляется
положительный фототаксис к слабому свету, и с наступлением сумерек
комар вылетает на свет и покидает убежище в поисках добычи.
Способ охоты. A. maculipennis, подобно A. superpictus, принадлежит
к числу тех комаров, которые совершают сравнительно дальние полеты
по направлению к более крупным скоплениям добычи. Эти виды комаров
специализованы на питании за счет крупных стадных животных.
Вы можете провести ночь вдали от жилья, среди обширных водое-
мов, в которых происходит выплод многочисленных A. maculipennis
или A. superpictus, п почти не подвергнуться нападению этих видов кома-
ров (хотя на вас будут нападать другие виды). И в то же время в 2 км
A. maculipennis будет кишеть в деревне и в ее ближайших окрестностях.
Вы подвергнетесь их нападению в домах деревни, па улице, а также на
протяжении 100—200 м за пределами деревни, по вдали от деревни —
меньше всего. Самки A. maculipennis, летящие с водоемов в поисках до-
бычи, не замечают одинокого путника, но издалека чуют запах крупного
скопления добычи и туда направляют свой полет.
Дневные поиски комаров в природе в такой местности, где нет ночую-
щих в поле людей и животных, также но обнаруживают зрелых самок
A. maculipennis — все они сосредоточены в поселках или вокруг посел-
ков. Но если летом вдали от селений живут группы людей или стада жи-
вотных, окружающая их растительность может оказаться обильно засе-
ленной зрелыми самками A. maculipennis.
Роль обоняния. Нахождение добычи голодной самкой A. maculipen-
nis распадается на два этапа: отыскание крупного скопления добычи иногда
на большом расстоянии (до 3 км, а может быть, и больше) и отыскание
самой добычи, когда комар находится от нее уже близко. На первом этапе
комар, очевидно, руководится преимущественно обонянием и, следова-
тельно, ищет добычу, идя против ветра, приносящего запахи.
Значение ветра. В этом отношении большее значение имеют местные
ветры — бризы, возникающие под влиянием неравномерного нагревания
и остывания поверхности земли х. Так, Мартини наблюдал домик с затя-
1 Па берегу моря дует дневной брпз с моря тта сушу и почкой брпз с суп:и па
море; такие же бризы дуют па берегах больших | ек. оэср цт. п. В горах дневной брпз
дует вверх по склонам* ночной бриз — вниз но долинам.
260
нутыми сеткой окнами, из которых одно было обращено к морю, другое —
в противоположную сторону. Вечером, когда самки Anopheles начинали
биться снаружи в оконную сетку, стараясь проникнуть в дом, еще дул
дневной бриз с моря и все они бились исключительно в окно, обращенное
прочь от моря. В определенный момент ветер менялся — начинал дуть
ночной, береговой бриз, и тотчас комары у обвеваемого им теперь окна ис-
чезали и появлялись у противоположного окна, обращенного к морю;
все это повторялось ежедневно с точностью механизма. Очевидно, ветер,
проходящий через дом, выносит оттуда запахи, и комары, ищущие добычу,
проникают в дом, идя навстречу ветру, руководствуясь запахом. В от-
личие от этого самцы и сытые самки, которые ищут только убежища,
влетают в дом независимо от направления ветра, так как они летят йена
запах, а на темные пятна окон и дверей, руководствуясь зрением.
Рис. 155. Проникновение голодных самок Anopheles в дом,
стоящий над поймой реки, совершающееся против вечернего
бриза (оригинал).
Простыми стрелками обозначено напрапление вечернего бриза, опе-
ренными стрелками — направление лета комаров. Точки около до-
мика— комары, стремящиеся проникнуть в окно, расположенное
на подветренной стороне дома.
Таким же образом в Европейской части СССР комары поднимаются
с речных пойм в города и деревни, лежащие на материковом берегу; подни-
маются они навстречу дующему оттуда на пойму ночному бризу (рис. 155).
В горах ночью холодный воздух стекает по всем долинам боковых прито-
ков и оврагов в долину главной реки, образуя систему ночных горных бри-
зов. Если по этим долинам и оврагам расположено какое-либо жилье,
запах его примешивается к стекающему воздуху, и комары из долины реки
поднимаются навстречу бризу и, как по путеводной нити, находят жилье
и добычу. Навстречу горным бризам комары могут подниматься на сотни
метров вверх по долинам или склонам, но при этом они не преодолевают
ни малейшего перевальчика, если дома не стоят па самом гребне: дейст-
вительно, они не могут ничего узнать о добыче, находящейся по ту сто-
рону водораздела, так как оттуда воздух стекает уже в ином направлении
(рис. 156). Вообще при сколько-нибудь выраженном рельефе обычно су-
ществуют бризы, ночью, в период активности комаров, дующие из повы-
шенных мест, где чаще располагаются поселки, в пониженные места, где
располагаются водоемы, и, такихм образом, приносящие комараАм запахи
добычи. На совсем ровных местах, где небольшие водоемы могут быть
разбросаны со всех сторон, правильных бризов обычно не бывает, и в таких
случаях в тихую погоду слабые местные воздушные токи постоянно ме-
няют свое направление, относя запахи жилья то в одну, то в другую сто-
рону, и комары получают возможность попасть в поселок со всех сторон.
Центры притяжения. Каждое место, где проводят ночь люди и жи-
вотные, является ц ентром притяжения для охотящихся самок
A. maculipennis, а также A. superpictus и A. pulcherrimus. Сюда устрем-
ляются комары из всех мест выплода и из убежищ голодных самок, до
261
которых долетают запахи даппого скоплепия добычи. Все пространство!
из которого комары собираются в тот или иной центр притяжения, являет-
ся его зоной тяготения.
В необжитой местности центрами притяжения A. maculipennis явля-
лись, вероятно, стада диких копытных. В обжитых местностях основными
центрами притяжения являются населенные пункты или отдельные усадь-
бы, а также полевые станы, лагери, ночующие па пастбище стада, боль-
шие вечерние скоплепия людей в пригородных садах, на речных пля-
жах и пр.
Знание зоны тяготения каждого насоленного пункта имеет очень
большое практическое значение. Если мы хотим при помощи противо-
Рис. 156. Расселение голодных самок Anopheles по долинам
всильпо пересечен пои местности навстречу вечерним бризам
(оригинал).
Простыми стрелками обозначено напраглегие гечергих бризов, опе-
ренными стрелками— наираьление лета охотящихся самок.
личиночных мероприятий защитить какой-либо поселок от залета ко-
маров, прежде всего необходимо определить подлежащую обработке
зону. В идеале эта обрабатываемая зона должна полностью охватить всю
зону тяготения подзащитного поселка. По меньшей мере она должна охва-
тить ту часть зоны тяготения, которая поставляет в поселок основную
массу комаров.
Сопряженные центры притяжения. Когда поселки разбросаны очень
редко, за много километров друг от друга, каждый из них может иметь
свою независимую зону тяготения. Такие поселки имеют каждый свою изо-
лированную популяцию A. maculipennis и могут явиться эпидемиологи-
чески самостоятельными, изолированными очагами малярии. Однако
гораздо чаще поселки располагаются неподалеку друг от друга, так что
зоны тяготения отдельных поселков частично совпадают между собою.
Некоторые места выплода оказываются расположены настолько близко
от обоих поселков, что летящие оттуда комары могут попасть в любой
из них. В который из поселков направят комары свой полет, будет за-
висеть от близости того и другого, силы запахов, направления ветра. Один
262
а тот же комар, кладущий яйца в данном водоеме, может питаться то в од-
ном, то в другом поселке. Такие центры притяжения, зоны тяготения ко-
торых частично совпадают, называются сопряженными центрами (рис. 157).
Между сопряженными центрами все время происходит обмен комарами.
Комар, напившийся крови в деревне, являющейся одним из взаимно
сопряженных центров, и заразившийся там малярией, другой раз может
пить кровь в другом из них (в соседней деревне) и заразить там человека.
В силу этого два сопряженных центра не являются взаимно независимыми
Рис. 1^7. Поле тяготения четырех соттряжеппых пептров притяжения, расположен-
ных над краем Волжской наймы.
Нанесены границы одно-, двух- и трехкилометровых зон вокруг всех населенных пунктов»
а также водоемы (в пределах трехкилометровой зоны вокруг центрального поселка, учебного
хозяйства).
очагами малярии. Обмен комарами обусловливает и обмен малярийными
паразитами. Поэтому знание сопряженности центров необходимо и для по-
нимания местной эпидемиологической обстановки, и для правильного про
ведения таких противомалярийных мероприятий, как общественная хи-
миопрофилактика или борьба со взрослыми комарами.
Раздражители, руководящие нападением. Теперь посмотрим, чем ру-
ководствуется самка A. maculipennis при отыскании добычи па близком
расстоянии. Комары, запертые в комнате с человеком, сравнительно легко
находят его и напиваются крови. Если их запереть в той же комнате с кро-
ликом, многие из них не смогут его найти и погибнут от голода. А между
тем в тесном садке все они находят кролика и напиваются на нем. Чем
крупнее животное, тем легче находят его комары. И действительно, чем
крупнее животное, тем легче на него натолкнуться при случайном блуж-
263
дапип. В то же время, чем крупнее животное, тем легче ого может увидать
комар своими близорукими глазами; чем крупнее животное, тем больше
пахнущая поверхность его тела, тем легче его найти по запаху. И, на-
конец, чем больше животное, тем больше оно выделяет и тепла.
Зрением комары при поисках пищи на близком расстоянии несомненна
руководствуются; это доказано для A. hyrcanus и для AeJes aegypti, но
весьма вероятно и для других комаров.
Запах тела п еще больше запах выдыхаемого животными воздуха
очень сильно привлекают комаров и на близком расстоянии. В экспери-
менте самок A. maculipennis сильно привлекает углекислота, которая для
человека не имеет никакого запаха. A. hyrcanus сильно привлекается за-
пахом свежего навоза.
Особенно большое значение при нападении па добычу имеет тепловое
чувство. Голодный комар нападает на каждый теплый предмет, папример,
на горячий чайник, завернутый в материю, и пытается его колоть. Он не
сосет руку человека, если температура ее поверхности меньше 4-28%
и охотно нападает на нее, если температура ее поверхности 4"32э или
больше. Человек, разогретый физической работой, или температурящий
больной сильнее привлекает комара, чем неразогретый и пелихорадящий
человек.
б) Пищевые предпочтения и внешние факторы,
влияющие на выбор добычи
Пищевые предпочтения. Комары, как и все свободно нападающие кро
вососы, редко бывают узко специализовапы на каком-либо одном виде
добычи, однако известная степень специализации у них имеется. Так,
Culex pipiens pipiens охотно нападает на птиц. Наоборот, A. maculipennis
питается в основном за счет крупных млекопитающих и па птиц почти
по нападает. Что касается подвидов A. maculipennis, то большинство "за-
падных исследователей принимает, что A. mac. maculipennis, A. mac.
messeae, A. mac. subalpinus и A. mac. atroparvus предпочитают копытных
человеку, тогда как A. mac. sacharovi и A. mac. labranchiae нападают оди-
наково часто и на тех, и на других. В силу этого многие считают два по-
следних подвида более опасными переносчиками малярии по сравнению
со всеми остальными. Однако все подвиды A. maculipennis при известных
обстоятельствах очень охотно нападают на человека, и каждый из них
может создавать тяжелые очаги малярии.
На выбор добычи самками A. maculipennis очень большое влияние
оказывает внешняя обстановка.
Значение относительной численности разных видов добычи. Прежде
всего имеет значение относительная численность каждого вида добычи.
Наибольший процент комариных желудков с кровью человека мы встре-
чаем в тех местах, где меньше всего других животных. Так, в полевых
бригадах села Хамидиэ (Кабарда), где имелось большое количество лютей
и незначительное количество домашних животных (лошади), желудки 80%
самок A. mac. maculipennis содержали кровь человека. И в то же время
в с. Ново-Ивановке, где на один двор приходилось в среднем 4,2 головы
скота, кровь человека была обнаружена в желудках только 4% комаров
того же подвида, а 96% желудков содержали кровь животных (Раевский).
Из многих подобных примеров совершенно очевидно, что увеличение ко
личества скота, т. е. уменьшение относительной численности человека,
влечет за собой уменьшение процента самок A. maculipennis, питающихся
за счет человека. То же правило относится и к другим животным, т. е.
чем больше, например, коров по сравнению с лошадьми, тем больше про
264
цент комаров, питающихся на коровах, и тем меньше процент комаров^
питающихся на лошадях. В силу этого правила увеличение количества
скота, независимо от его размещения (см. ниже), уже само по себе ослабляет
связь между комаром и человеком и способствует снижению малярии.
Доступность добычи. Кроме численности отдельных видов добычи,
степень использования каждого вида комарами очень зависит и от доступ-
ности каждого из них для комаров. В этом отношении прежде всего имеют
значение механические препятствия для нападения. Если люди все спят
в хороших, плотных домах, с засеченными окнами, а животные стоят под
навесом, доступность людей резко падает и процент комаров, пьющих
Таблица 5
Процент самов A. mac. maculipen-
nis, содержащих в жэлудках кровь
человека, в селениях с различным
количеством животных (Кабарда,
1935; по Г. Е. Ра вскому)
Число голов скота в сред- нем hi один двор % комаров
С кров’ ю человека с кротгю Ж (BOTH яХ
4,5 3 8 96 2
0.9 23,0 77,0
Рис. 158. Планы домэв — нерациональные
(1, 2) п рациональный (3) для малярийном
местности.;
кровь человека, снижается. С этой точки зрения механичэская профилак-
тика: заделка всех щелей, защита окон домов металлической сеткой, две-
рей — тамбурами, защита спящих пологами, имеет величайшее значение
и при тщательном проведении может очень сильно снизить заболеваемость
малярией. Ряд особенностей в устройстве жилого дома может помочь
защитить его от залета комаров. План дома должен представлять простой
прямоугольник, без выступов, входящих углов (рис. 158) и т. п., так как
всякий входящий угол создает дополнительную защиту от ветра и таким
образом облегчает проникновение комаров в дом. Открытых балконов,
крылечек, портиков в сильно малярийной местности следует избегать;
наоборот, полностью затянутые сеткой террасы, тянущиеся вдоль всего
фасада, весьма целесообразны. Если в данном месте имеются бризы, вход-
ные двери должны располагаться со стороны, обвеваемой вечерним бри-
зом. Открываться они должны наружу, что затрудняет залет комаров вме-
сте с входящим в дом человеком. Наоборот, окна должны открываться
внутрь дома, что позволяет их открывать и закрывать, не вынимая окон-
ной сетки. Комнаты не должны быть чрезмерно высоки, так как это
затрудняет истребление залетевших комаров, и должны иметь как можно
меньше темных и мало доступных закоулков.
Доступность разных видов добычи для комаров очень зависит от того,
где находятся люди и животные в момент нападений комаров, а потому
очень важно знать время и условия активности последних.
На открытом воздухе благоприятные условия для нападения воз-
никают обычно вечером, незадолго до захода солнца, и длятся до утра.
В теплые тихие ночи чаще всего наблюдается два максимума активности:
вечером — после захода солнца и утром — перед восходом, и минимум
активности после полуночи. Сильное падение температуры (до 7—10°)
во второй половине ночи может полностью прервать активность комаров
265
на открытом воздухе. Такое же действие может произвести в любой час
сильный ветер или дождь. И наоборот, утренняя активность иногда затя-
гивается летом часов до 7 утра, что наблюдала, например, Шлёнова на
волжских пристанях.
Дневные нападения на открытом воздухе часто наблюдаются ранней
весной, когда по ночам еще слишком холодно, а днем достаточно влажно
и не слишком жарко. Лишенные возможности летать ночью и подгоняемые
голодом, комары преодолевают тормозящее влияние света и нападают
среди дня.
В общем же, как правило, для A. maculipennis па открытом воздухе
доступна та добыча, которая находится там в вечерние, ночные и отчасти
ранние утренние часы.
Нападение в помещениях. Еще чаще, чем на открытом воздухе, A. ma-
culipennis нападает в помещениях. Мы уже рассмотрели вопрос о влиянии
доступности помещений. Большую роль играет также их микроклимат.
Нередко комары с вечера залетают в дом пли хлев и, благодаря более вы-
сокой температуре этих помещений, могут всю ночь нападать там на
людей и животных, в то время как на дворе холод тормозит их актив-
ность. В летние дни комары иногда нападают в прохладе и полумраке
комнат, когда па открытом воздухе дневные нападения отсутствуют.
С другой стороны, в ряде случаев сухость и жара, бывающие в некоторых
помещениях, резко снижают активность комаров. Сухие, светлые, хорошо
проветриваемые комнаты, несомненно, несколько защищают своих обита-
телей от нападений A. maculipennis, хотя защита такого рода очень от-
носительная.
В каких же случаях нападения происходят в помещении и в каких
под открытым небом? В основном это зависит от места вечернего и ночного
пребывания добычи: комары следуют за добычей. Кроме того, имеет значе-
ние климат и погода: слишком суровые условия — чрезмерно холодные
ночи или, наоборот, жаркие, сухие ночи мешают нападениям] на откры-
том воздухе.
Влияние условий ночевки на доступность людей п животных. В тех 'случаях,
когда вся добыча ночует в совершенно одипакозых условиях, например, в такой де-
ревне, где п люди, и скот спят во дворах под открытым небом, условия нападения на
тех и на других создаются одинаковые. Но если люди спят в домах, а скот — на от-
крытом воздухе, то условия нападения па тех п других становятся сразу неравными,
и, в зависимости от по.’ оды п типа домов, в более уязвимом положении могут оказаться
то люди, то животные’ и теплую и влажную воль, при хмало доступных для залета
домах, нападения будут происходить главным образом на открытом воздухе — на
скот; если же ночь холодна пли ветрена, то нападения будут происходить главным
образом в домах — на людей.
В тех случаях, когда челочек почует в доме, а скот в хлевах, различия в микро-
климате тех и других, в степени доступности, в запахе также создают для человека
и животных совершенно различные шансы подвергнуться укусу, причем преимущество
будет то на одной стороне, то на другой, в зависимости от обстановки и вида
Anopheles.
Роль размещения людей и животных. В громадной мере связь комаров
с человеком или домашними животными определяется топографическим
размещением тех и других. Мы видели, что голодные самки А/ maculi-
pennis привлекаются издалека скоплениями скота и людей. К ка-
кому-либо поселку комары подлетают со стороны окружающих водоемов
и прежде всего нападают на его окраину; если на окраине поселка они
находят добычу, они насасываются крови и дальше не летят. Чем ближе
центр притяжения к личиночным очагам, том больше, при прочих равных
условиях, привлекает он к себо комаров. И чем ближе отдельные дома
266
или хлева находятся к окра ине поселка, обращенной к личипочпым оча-
гам, тем больше они подвергаются залету и нападению комаров (рис. 159).
В поселке, изображенном на рис. 159, были только люди. Но пред-
ставим себе, что со стороны риса впереди домов стояли бы хлева, —
в этом случае большинство комаров задержалось бы в этих последних
и питалось бы преимущественно за счет их обитателей; люди в поселке
почти перестали бы подвергаться нападениям. Пример такого рода при-
водят Раевский и Бейлин: в Прималкинском совхозе, в Кабарде, большие
хлева были расположены между главной усадьбой совхоза и прудом, слу-
жащим местом выплода комаров; пока в хлевах стояли свиньи, они удер
4 150
Рис. 159. Распределение комаров в одном из поселков
Каратальского рисового совхоза (оригинал). Рисунок изобра-
жает схематический план поселка.
Кривая А— средняя численность комаров (щи болипой их числен-
ности) на один дом в поперечных рядах ломов по ме|е улэле! ия от
рисового поля, в начале августа; кривая Б—то же, в конце августа,
при малой численности комаров.
живали йольшипство комаров и в дома проникало только 4% от общего
количества последних; после того как свиньи были выведены в отдаленные
лагери, комары мимо пустых хлевов полетели в усадьбу, и в домах ока-
залось 75% их общего количества.
Хлева как заслон от комаров. Описанное здесь влияние размещения
добычи на распределение комаров и их связь с человеком имеет величай-
шее практическое значение: па нем основана система зоопрофилактнкв
малярии. Принцип ее заключается в создании заслона из хлевов между
человеческим жильем и личиночными очагами и в недопущении, таким
образом, комаров до человека, в силу чего становится невозможным
и распространение малярии.
Опыт зоопрофилактики был поставлеп перед войной Центральным институтом
малярии в ряде точек. Рассмотрим в качестго примера с. Шитхала Кабардинской
АССР. Деревня расположена в зоне выклинивания грунтовых вод па предюрной рав-
нине северного склона Большою Кавказскою хребта, средп мпоючисленных мест
выплода A. maculipennis. Деревня построена в одну улицу, хлева расположены непо-
средственно около домов. В 1938 г. на двор было в среднем 1,8» а в 1940 г., в силу от-
гона скота в горы, всего 0,93 головы рогатого скота. В 1939 г. хлева были отнесены от
267
дом оз, а в 1941 г. сверх того прослежено, чтобы по возможности все хлева были запиты
животными. Несмотря на уменьшение количества скота, создание .защитней линии
из хлевов вызвало резкое отвлечение комароз от человека. Если в 1938 г. в среднем за
лето 37% всех комаров поселка сид зли па дневках в домах и 21% всех комаров
поселка имели в желудке кровь человека, в 19 40 г. в жилых помещениях сидело
всего 5% всех комаров и лишь 3 5% всех комаров имели в желудке кровь чело-
века. Таким образом, проведение зоопрофплактики даже при недостаточном количе-
Рис. 160. Тпповой проект противомалярийной планировки с учетом зоо-
профилактики д тя колхозного селения па 84 двора (оригинал М. И. На-
эаретова и С. М. Зелеитух).:
С севера и северо-востока поселок за щи щеп общественными хлевами, с востока —
хлевами индивидуального пользовании. Стрелками показаны направления лета
комаров.
ствескота в поселке снизило процент комаров, питающихся за счет человека, в
6 раз. При этом и все оснэвлыэ показатели малярийной заболеваемости снизились
примерно в 4 раза.
При проведении зоопрофилактики следует соблюдать целый ряд
правил. При защите колхозов (рис. 160) или совхозов в качестве первой
линии защиты надо использовать обобществленный скот — фермы, кол-
268
хозпые дворы и т. п., в качестве второй линии защиты — хлева инди-
видуального пользования. Последние должны находиться от домов на рас-
стоянии не ближе 25—35 м. Отверстия хлевов не должны обвеваться ве-
черними бризами, а должны находиться с подветренной стороны здания
(см. выше, стр. 231).
Важное преимущество зоопрофилактпки заключается в том, что
раз созданная защитная линия действует дальше автоматически, до тех
нор, пока в хлевах стоит скот, не требуя особой противомалярийной
эксплоатации, расходов или забот.
Специально перестраивать селения для создания зообарьера было
бы дорого, но при всяком новом строительстве животноводческих ферм
или целых селений зоопрофилактика не стоит ни копейки: расположить
пх разумно стоит ничуть не дороже, чем расположить нелепо. Поэтому
Совет народных комиссаров СССР постановлением № G29 от 29/V 1944 г.
обязал все строящие организации при создании новых селений или вос-
становлении селений, разрушенных войной, соблюдать правила зоопрофи-
яактики.
Иногда против зоопрофилактики возражают, указывая, что усиленное
нападение комаров может оказаться вредным для скота. Однако укусы
Anopheles очень мало отражаются на скоте по сравнению с укусами слеп-
ней, клещей, мошек или даже Acdes; с другой стороны, обычно и без
зоопрофилактики 90% самок A. maculipennis питаются на скоте; при-
бавка еще 9% будет для скота незаметна, а отвлечение этих 9% комаров
от человека во много раз снизило бы заболеваемость малярией.
Таким образом, противомалярийная организация должна следить
за тем, чтобы при всяком новом строительстве в сельских местностях поме-
щения для скота были как можно лучше использованы в целях защиты
населения от малярии.
в) Нападение на добычу
Найдя добычу, самка Anopheles садится на нее, прокалывает кожу
и напивается. Акт сосания продолжается при 20° в среднем 2 минуты, ми-
нимум 30—40 секунд. Хоботок погружается в кожу на глубину до 4 мм.
Комар выпивает количество крови, несколько превосходящее перво-
начальный вес его тела. Напившийся комар очень быстро вытаскивает
свой хоботок и слетает с добычи.
Если самку потревожить, пока она выпила очень мало крови, она сле-
тает и тотчас нападает вновь. Вследствие этого один комар может в одну
ночь заразить малярией двух или трех человек. Если комар успел выпить
значительное количество крови и особенно если с момента прерванного
сосания прошло некоторое время, он не возобновляет нападения до тех
пор, пока не переварит свою неполную порцию крови.
Самка Anopheles легко колет через топкую ткань одежды; шерсть
животных (например, коровы) также не служит помехой для сосания.
Но, например, у длинношерстных кроликов для нападения доступны
только мордочка и уши.
4. ПИЩЕВАРЕНИЕ И СОЗРЕВАНИЕ ЯИЦ
а) Вторая фаза гонотрофическсго цикла
В тот момепт, когда напившийся комар слетает с добычи, начинается
новая фаза гопотрофического цикла. Состояние и поведение самки резко
меняются. Из состояния активности она переходит в состояние относи-
тельного покоя.
269
Поведение свеженасосавшейея самки. Если самка A. maculipennis
напилась где-либо в помещении, слетая с добычи, опа направляется отяже-
левшим полетом к ближайшей стене и, поднявшись вверх вдоль стены,
садится на нее; если ничто ее не потревожит, она остается здесьдо утра.
Утром, раздражаемая светом, она перебирается в более темное ме-
сто. Если комар напился под открытым небом, слетев с добычи, он тоже
садится, где попало, на первый ближайший предмет, и часто остается
здесь до утра. С рассветом он приходит в фотокипетическое возбуждение,
начинает летать, находит убежище и залетает туда. В тех местах, где
добыча ночует во дворе, этот утренний залет свеженасосавшихся самок
носит массовый характер.
Гонотрофическая гармония. Процессы пищеварения и созревания
яичников рассмотрены выше, в главе IV.
При температурах, близких к оптимуму, летние самки A. maculipen-
nis строго подчиняются правилу гонотрофической гармонии. Оптимум
для подмосковных A. mac. messeae лежит при 20—25°. В лабораторных
условиях чем больше уклоняется температура кверху или книзу от опти-
мума, тем больше наблюдается случаев дисгармонии (т. е. неполное раз-
витие яичников при переваривании полной порции крови), а при низких
температурах (+10°) наблюдаются и случаи полного торможения яич-
ников.
Таблица 6
Распределение самок A. mac. messeae по фа-
вам развития, которых достиг и их яичники
после однократного перевариваю я крови при
разных т мп ратлрах. Число комаров выр аж < но
в процентах от общего числа бл» гополучно за-
кончивших пищеварение при каждой данной тем-
пературе (по М. Ф. Шлёцовсй)
Ф <3*J Р 8 *ИТИЯ яичнлков Температура I—II III-1V • V
10° 2.64 12,5 61 1
15° 20.0 10,0 7J 0
2Э° 4,1 0,0 95.9
25° 0,0 4,4 95,6
3J° 4,4 15,6 80,0
В природе летом случаи дисгармонии встречаются еще реже, чем
при тех же температурах в лаборатории.
Продожительвость пищеварения. Продолжительнссть переваривания
полном порции крови зависит от температуры (табл. 7).
Т аблица 7
Средняя продолжительность (в часах) псрсгарггапгя почтой порппи крови самкой
А» mac. messeae при относите/ь ой важности 70 - М«% и разных температурах
(по М. Ф. Шлсиово* )
Температура 5° 7,5° 10° 12.5° ! 5° 20° 250 3(Р ЗЬ°
Средняя продолжитель- ность ыпцеварсичя в часах 00 543 231 247 1 I 165 S3 59 41 43
270
Влажность воздуха влияет на продолжительность переваривания
лишь в той мере, в какой она влияет на собственную температуру тела
комара: при температурах -J-200 и выше низкая влажность довольно силь-
но замедляет пищеварение, при -f-10° слегка ускоряет его.
Индивидуальная изменчивость продолжительности пищеварения '
очень значительна: при одних и тех же условиях одни самки заканчивают
пищеварение много раньше большинства, другие — много позже.
Плодовитость. Больше всего одноразовая плодовитость самки A. ma-
culipennis, т. е. число одновременно созревающих в ее теле яиц, зависит
от ее размеров: мелкие самки имеют яичники с меньшим числом яйцевых
трубочек, нежели крупные. Поэтому мелкие самки, получившиеся из го-
лодавших личинок, менее плодовиты, чем крупные самки, получившиеся
из хорошо питавшихся личинок. Отражаются на плодовитости и сезонные
колебания размеров комаров: первые, весенние, комары, наиболее круп-
ные, отличаются и наибольшей плодовитостью; в течение лета и размеры
вылупляющихся комаров, и число яиц в кладках падают (табл. 8).
Таблица 8
Годовой ход плодовитости самок A. mac. meseeae (по Т. С. Д типовой)
Месяц Срегн'е чи- сло я рев lo- in х яиц Число вскрыть X самок Применения
Май 130,8±6,2 53 Перезимовавши^ самки
Июнь 185,1±8,6 95 Перелиновавшие и молодые самки
Июль 244 4±5 3 162 Самки начала 1 по»'ол иия
Аьгуст ......... 159,0±8,4 65 Самки более позднего от- рожденчя
Ноябрь 131,5±7 0 28 Зимовочные самки
Декабрь 131 ,5±5 Д 101 То же
Март 13Э,0±5,4 41 »> »
Между подвидами A. maculipennis также существуют раличия в пло-
довитости. В одной и той же местности более крупный messeae отклады-
вает больше яиц, чем более мелкий atroparvus (Калита), что имеет значение
в борьбе за существование между обоими подвидами.
Поведение в благоприятных убежищах. Свеженапившаяся кровью сам-
ка A. maculipennis отличается от голодной или жаждущей самки малой
подвижностью и почти полным отсутствием положительного фототаксиса.
При благоприятных условиях дневки эти свойства сохраняются комаром
до конца пищеварения. Напившийся комар может днями неподвижно оста-
ваться на одном и том же месте. Перелетает он с места на место только
под влиянием внешних раздражений: вспугивание, дым, яркий свет, не-
благоприятная температура и влажность ит. п. Как размещаются комары
под влиянием этих факторов внутри убежища, мы уже видели (стр. 256).
Подвижность и положительный фототаксис возвращаются только с окон-
чанием пищеварения к самкам со зрелыми яйцами.
Поведение в слишком с\хих помещениях. Иначе обстоит дело в поме-
щениях, неблагоприятных для дневки: слишком сухих, жарких и вет-
реных. При влажности около 60—70% и температуре до 20—25° количе-
ства влаги, выпитой с кровью, вполне хватаетдо конца пищеварения и ко-
мары не испытывают жажды; в связи с этим они и остаются неподвижными
на дневках вплоть до конца пищеварения. Наоборот, в условиях боль-
271
шего дефицита влажности имеющегося в теле запаса воды самке нахватает;
задолго до окончания пищеварения количество свободной воды в теле
настолько уменьшается, что дальнейшее уменьшение могло бы уже
угрожать ее жизни. Одновременно с этим у комара появляется возбужде-
ние, как при голоде, и точно так же появляется положительный фото-
таксис к сумеречному свету. Если комары находят воду в этом же поме-
щении, они могут здесь же напиться и успокаиваются. Если же они воды
не напьются, то с наступлением вечера вылетают в окно.
Тот же фототаксис заставляет покидать помещение и других неудов-
летворенных самок — голодных (почему-либо не напившихся крови), а
также со зрелыми яйцами и неоплодотворенных, юных, находящихся
под влиянием полового инстинкта. Таким образом, каждый вечер часть
комаров, находящихся на дневке, стремится ее покинуть; с одной стороны,
мы имеем нормальный вылет самок со зрелыми яйцами, голодных и нуж-
дающихся в копуляции, с другой стороны, — преждевременных! вылет
самок, находящихся на промежуточных стадиях пищеварения, под влия-
нием жажды.
б) Биологические типы убежищ; круговорот комаров
на дневках
Состав комариного населения убежищ. Как уже говорилось (стр. 257),
дневные убежища комаров в экологическом отношении весьма разно-
образны: они могут бытьприродными и искусственными по происхождению,
защищенными от колебаний температуры и влажности или незащищенны-
ми, легко или трудно доступными для залета, могут содержать или не со-
держать добычу. Наряду с этим, убежища можно разделить и по биологи-
ческому составу их комариного населения: они могут содержать либо
дневки с самцами и юными самками, либо дневки с переваривающими
самками.
Первый тип дневок располагается обычно в необитаемых помещениях,
легко доступных для залета, расположенных вблизи мест выплода.
Наряду с преобладающим количеством самцов и юных самок, в них мо-
жет быть примесь самок со зрелыми яйцами, которые перебрались сюда
из каких-нибудь убежищ, расположенных поодаль от водоемов, а если
неподалеку есть добыча, то бывает и примесь самок с кровью.
Находящиеся на дневке переваривающие самки располагаются в по-
мещениях, либо содержащих добычу, либо расположенных вблизи до-
бычи, и в последнем случае легко доступных для залета. В обоих случаях
дневки этого типа характеризуются преобладание^м самок с кровью па
разных стадиях переваривания; наряду с ними, всегда встречается из-
вестный процент голодных самок, из которых большинство — не юные,
а самки, нуждающиеся в питании кровью, которым почему-либо не уда-
лось удовлетворить свой голод. Всегда имеется и некоторое количество
самок со зрелыми япцами. Самцов на таких дневках бывает немного, и
тем меньше, чем дальше помещение отстоит от мест выплода; исключение
составляют самцы A. mac. atroparvus.
Круговорот комаров на дневках. Ни на дневках I типа, пи на дпевках
II типа состав комаров не является постоянным. В первом случае почти
все комары ежедневно покидают убежище и возвращаются в него лишь
частично, заменяясь новыми. Поэтому убежища с самцами и юными сам-
ками обозначают как однодневные убежища. В убежищах с пере-
варивающими самками также более или менее значительная часть комаров
каждый вечер покидает убежище, по разница заключается в том, что часть
272
вызываю-
Рис. 161. Распределение
самок A. maculipennis по
стадиям Селла на дневке
с малой текучестью, при
двухдневной продолжи-
тельности пищеварения.
но сборам, произведен-
ным в 6 часов утра.
убежище на 3-й день,
какой бы час мы ни
зта меньше, ив силу этого оборот комаров совершается медленнее. Однако
в этом отношении убежища с переваривающими самками не все одинаковы.
Степень текучести комариного населения оказывается в них весьма
различной. В одних случаях мы встречаем нормальный круго-
ворот, при котором в убежище влетают голодные самки (если в убё-
жище имеется добыча) или свеженапившиеся (если в убежище добычи нет),
а вылетают из него главным образом самки со зрелыми яйцами, закон-
чившие здесь пищеварение. Нормальный круговорот имеет место в убе-
жищах с благоприятным для комаров дневнььм микроклиматом, нс*
вызывающим у них чрезмерной потери воды.
Наряду с этим типом круговорота, нередко встречается другой тип,
сличающийся повышенной текучестью комариного населения, доходя-
щей иногда до почти полной ежедневной смены всех комаров. Причиной
такой текучести является неблагоприятный для комаров микроклимат
убежища с большим дефицитом влажности в дневные часы,
щим большую потерю воды комарами, следствием
которой является жажда и необходимость прежде-
временно покинуть убежище в поисках воды.
Состав комаров на дневках по стадиям пи-
щеварения. Легче всего объяснить состав ко-
маров в тех убежищах, где температура дер-
жится около 25—30° и пищеварение, таким обра-
зом, длится двое суток (рис. 161). Если притти в
такое помещение на восходе, то комары, напив-
шиеся сегодня ночью, все будут на 2-п фазе Селла.
В полуденные часы они будут на 3-й фазе, к ве-
черу—на 4-й фазе, на следующее утро большинство
будет на 5-й фазе, к полудню второго дня — на
6-й фазе, к вечеру — на 7-й фазе; с закатом солнца
все комары улетят. Первые фазы пищеварения у
всех протекают очень дружно, но на второй
день намечается значительное расхождение: одни
отстают, другие опережают, немногие из них не
заканчивают пищеварения к вечеру и остаются в
Тем не менее картина получается отчетливая; в
произвели обследование, мы легко различаем две группы комаров: на-
пившихся сегодня и напившихся накануне, и численность обеих групп
приблизительно одинакова.
При более низких температурах продолжительность пищеварения
больше, соответственно увеличивается и срок пребывания комаров в убе-
жище, п картина становится более запутанной.
Часы вылета комаров. Вылет A. maculipennis из помещений совер-
шается в основном вечером. Обычно он начинается незадолго до захода
солнца, причем в пасмурную погоду раньше, чем в ясную. Больше всего
комары устремляются в окна и двери, обращенные к закату. Самцы начи-
нают вылетать немного ранее самок. С окончание^м сумерек и наступлением
темноты вылет прекращается и ночью не происходит. На севере при свет-
лых ночах он затягивается иногда до полуночи; на юге, а осенью и на се-
вере, массовый вылет длится каких-нибудь полчаса. На утренней заре
вылет возобновляется, но в ничтожной степени.
Ввиду того что вылет приурочен к заходу солнца, время его наступ-
ления изменяется в течение всего сезона соответственно времени заката.
Часы влета комаров в помещение. Влет начинается несколько позже
вылета и достигает максимума через х/2—2 1/2 часа после максимума вылета.
18 Учебник медицинской энтомологии 27.»
После этого влет обычно ослабевает, но тем не менее продолжается
до утра, иногда с перерывом в полуночные часы, иногда непрерывно.
Погода, например сильный ветер, может прервать влет на любое времяг
и тогда он ограничивается той частью ночи, во время которой погода
была благоприятной.
Своеобразно протекает влет в убежища без добычи в тех случаях, когда нападе-
ния комаров совершаются под открытым небом. В этом случае залета в течение вечера
и ночи пет, а массовый залет свеженапившихся самок начинается па рассвете и закан-
чивается около времени восхода солнца. В этом случае комары, насосавшиеся ночью на
открытом воздухе и оставшиеся там сидеть до утра, приходят под влиянием утреннего
света в фот ©кинетическое возбуждение, начинают летать и устремляются в темные
отверстия окон и дверей.
Осенью в С| едней Азии при теплых вечерах и холодпых почах (от +5° до 4-10°)
на открытом воздухе происходят нападения с вечера, а к утру комары сидят полуоко-
ченевшпе; в этом случае влет совершается уже после восхода солнца, когда температура
поднимается достаточно высоко, иногда только к 10—11 часам утра.
Рис. 162. Кривые зависимости от темпера-
туры.
А — средней продолжительности перенарпранпл
кропи самкой A. maculifcnnis; Б— продолжитель-
ности полового никла малярийного паразита Plas-
modium vivax (в часах).
должительность созревания спорозоитов
турах разница эта еще увеличивается,
в) Некоторые эпидемиоло-
гические выводы
Из правила гонотрофичес-
кой гармонии и знакомства с
круговоротом комаров па днев-
ках вытекает, что комар,
заразившийся малярийными
плазмодиями в каком-либо до-
ме, бывает вынужден покинуть
этот дом для откладки яиц за-
долго до того времени, когда
в его теле могли бы созреть
спорозоиты и его уколы стали
бы заразны. Действительно,
продолжительность пищеваре-
ния Anopheles при 25—30° в
4—4 */2 раза меньше, чем про-
, а при более низких темпера-
так как нижние температурные
пороги развития человеческих плазмодиев лежат гораздо выше, чем ниж-
ний температурный порог пищеварения Anopheles (рис. 162). Поэтому
комар, заразившийся в доме плазмодиями, вынужден этот дом покинуть
либо для откладки яиц, либо, в условиях слишком сухого и жаркого
помещения, еще раньше для водопоя. После этого он может вернуться
в тот же самый дОхМ или в другой — это будет зависеть от случая.
Действительно, у комаров пет предпочтительного возврата в то ме-
сто, где они однажды питались: в этом отношении у них нет памяти. Это
нагляднее всего видно из распределения между домами и хлевами зара-
женных плазмодиями комаров. Цисты плазмодия образуются уже к концу
переваривания крови. Поэтому при нормальном круговороте на дневках
процент комаров с цистами обычно бывает выше в домах, чем в хлевах.
Наоборот, при большой текучести на дневках комары с молодыми ци-
стами нередко успевают перелететь из дома в хлев. К тому времени,
когда у комара появляются спорозоиты в слюнных железах, он успевает
по меньшей мере 2—3 раза слетать па водоемы и обратно к добыче. Вслед-
ствие такого перемешивания оказывается, что комары со спорозоитами
встречаются в демах и в хлевах одинаково часто. Наппимер, Шленова
274
ъ поселках около Мацесты нашла в 1937 г. встречаемость спорозоитов
у A. mac. maculipennis равной 0,8%, как у комаров, пойманных в домах,
так и у комаров, пойманных в коровниках.
Процент желудков с человеческой и коровьей кровью у комаров
со спорозоитами бывает такой же, как и у незараженных комаров. Дру-
гими словами, зараженные комары, однажды уже пившие человеческую
кровь, имеют не большую склонность нападать на человека, чем все
остальные комары.
Итак, комар, заразившийся в доме малярией, обязательно улетает
оттуда, прежде чем сможет в свою очередь кого-нибудь в этом доме зара-
зить, и имеет столько же шансов вторично залететь в этот дом, как и вся-
кий другой комар той же популяции. Такими образом, больной малярией,
живущий с вами в одной комнате, представляет для вас не большую опас-
ность, чем для соседа через улицу. В летнее время внутридомовых очагов
малярии не бывает. Какие изменения порой вносит в это положение на
ступление осенней диапаузы, — мы увидим ниже (сгр. 287).
С другой стороны, все рассмотренные в этой главе данные полно
стью оправдывают нижеследующее рассуждение Рональда Росса. Если
в данной местности 9/10 комаров питаются на скоте, то здесь, по сравнению
с такой местностью, в которой все комары питаются только на людях, шан-
сы на заражение комаров снижаются в 10 раз. Далее, при следующих
сосаниях из заразившихся комаров опять ’/10 будут питаться на скоте,
и шансы на заражение человека, уже сниженные благодаря меньшему
проценту заразившихся комаров, снижаются еще в 10 раз благодаря
уменьшению процента зара?кающих укусов, падающих на человека. Дру-
гими словами, с уменьшением доли комаров, кусающих человека, в 10 раз,
шансы на перенос малярии от человека к человеку уменьшаются в 102,
т. е. в 100 раз. При этих рассуждениях Росс допускал отсутствие пред-
почтительного возврата комаров к прежнему месту питания и к прежней
добыче, и, как мы видим, факты вполне подтверждают это допущение.
5. ОТКЛАДКА ЯИЦ
Последний период гонотрофического цикла — поиски водоема и от-
кладка яиц. Яйца окончательно созревают чаще всего к моменту заверше-
ния пищеварения, но еще до опорожнения кишечника, т. е. па 6-й фазе
Селла, однако вылетают из убежища главным образом самки па 7-й фазе
Селла, т. е. уже освободившиеся от непереваренных остатков крови. Но
и они не всегда откладывают яйца в первую ночь. Многие из них к утру
с неотложенными яйцами влетают обратно. Многие перебираются в убе-
жища, расположенные поближе к водоемам. Все же часть самок откла-
дывает, вероятно, в первую ночь, а большинство — не позже двух-трех
суток.
Рассеяние кладущих самок. Если все водоемы, подходящие для от-
кладки яиц, расположены в непосредственной близости от мест питания,
самки откладывают туда яйца и никуда дальше им лететь не приходится.
Но если ближайшие водоемы мало подходят для развития личинок, гро-
мадное большинство самок летит мимо и откладывает яйца в более отда-
ленные, но более благоприятные водоемы. Если, наконец, вокруг убе
жища имеется неограниченное множество водоемов, как, например, на тор-
форазработках, в плавнях и т. п., распределение кладок получается очень
характерное: в ближайшие водоемы откладывается очень много яиц, но
часть самок пролетает мимо вполне благоприятных ближайших водоемов
и откладывает яйна в более удаленных. Чем дальше отстоят водоемы от
18*
275
убежища с переваривающими самками, тем меньше в них попадает яиц,
тем меньше будет там личиночное обилие. Последние кладки оказываются
отложены примерно в 3 км от центра, а когда кладущих самок очень мно-
го, — то и дальше (рис. 163). Тацим образом, наблюдается рассеивание
кладущих самок по водоемам, объясняющееся, повидимому, тем, что часть
самок случайно не замечает ближайших водоемов и пролетает мимо.
Ореховских торфоразработок личинками A. maculipennis.
Концентрические зоны вокруг обоих поселков проведены через каждые 0,25 км.
кроме периферических зон вокруг поселка 46-го, которые проведены через 0,5 и
1 км. Цифры обозначают число личинок на 0,20 м* в среднем для всех водоемов
данного сектора каждой зоны за весь май 1936 г. А — Озерепкий поселок, Б —
поселок №46.
Рассеяние кладущих самок имеет для вида большое значение. Мы
видим, что охотящиеся самки A. maculipennis обладают способностью со-
бираться с расстояния 3—5 км к месту питания. Если бы все они, не рассеи-
ваясь, отложили яйца тут же, в непосредственной близости от места пита-
ния, ближайшие водоемы оказались бы страшно перенаселенными личин-
ками, что вызвало бы огромную личиночную смертность п резкое сокраще-
ние анофельной продукции, а большие площади более отдаленных водое-
мов оказались бы неиспользованными. Рассеяние кладущих самок обес-
печивает более равномерное использование водоемов комарами.
В то же время рассеяние кладущих самок A. maculipennis чрезвы-
чайно осложняет борьбу с этим видом. Если бы все самки откладывали
яйца только в ближайшие водоемы, достаточно было бы уничтожить ли-
чинок в узкой полосе вокруг населенных пунктов и не было бы надоб-
ности расширять защитную зону до 3 км и больше, тем более что ничто
так не затрудняет и не осложняет борьбу с личинками A. maculipennis,
как большие размеры, которые приходится придавать защитной зоне
если мы хотим добиться действительного успеха.
Выбор места кладки. Самки комаров не откладывают яйца куда по
пало; как п другие насекомые, они обладают инстинктом выбора места
276
кладки. В силу этого*в тех водоемах, которые непригодны для развития
личинок (слишком холодные, слишком теплые, слишком соленые, слиш-
ком грязные и т. п.), яйца, как правило, не откладываются. Каким обра-
зом самка находит благоприятные для личинок водоемы?
Когда самка издалека летит к водоемам, ею, очевидно, руководит обо-
няние и чувство влажности; как мы видели, комары очень тонко различают
степень влажности воздуха. Руководствуясь чувством влажности, кома])
находит крупные водоемы или заболоченные участки с многими мелкими
водоемами и, повидимому, не замечает маленьких вместилищ, вроде бочки
с водой и т. п. В таких вместилищах кладки A. maculipennis встречаются
чрезвычайно редко.
Приблизившись к водоему, самка совершает «предоткладочную пля-
ску» — она летает низко над землей, время от времени круто ныряя вниз
и прикасаясь лапками к земле. При этом ее привлекает влажность, привле-
кают темные и блестящие поверхности. Если при одном из опусканий она
прикоснется лапками к воде, она останавливается над этим местом в воз-
духе, дрожа крыльями, и на лету начинает ронять в воду яйца. Часто опа
садится на воду или на растения и продолжает класть сидя. Иногда самка
садится раньше, чем начинает класть и предварительно пробует воду хо-
ботком. Прикосновение к воде, хотя бы лапками на лету, является непре-
менным условием начала кладки. Движение воды, низкая или высокая
температура (например, 10°, 35°), чрезмерная соленость воды, гнилост-
ный запах отпугивают самку, и кладка в такие воды не происходит. Таким
образом, при откладке самка руководствуется всеми своими чувствами:
и зрением, и обонянием, и вкусом, и чувством влажности и пр.
Когда самок очень много, некоторое количество кладок попадает
и в неблагоприятные водоемы, в которых личинки жить не могут. При-
чиной таких ошибок являются, очевидно, различные случайности, пости-
гающие ту или иную самку из множества кладущих.
Продолжительность процесса откладки. Вся откладка у сидячей сам-
ки занимает 20—30 минут, срок этот, естественно, меняется в зависимости
от температуры и от численности яиц. При 20—25° яйца появляются из
полового отверстия кладущей самки через каждые 6—8 секунд. Нор-
мальное число яиц в кладке 120—350, максимальное — около 450.
В лаборатории для того, чтобы вызвать откладку яиц, проще всего
отрезать у самки крылья и посадить ее на поверхность воды в чашке или
блюдце. Откладка начинается иногда уже через полминуты и обыч-
но не позже, чем через 2—3 минуты (при 20—25°).
6. ДАЛЬНОСТЬ ПОЛЕТА
Вопрос о дальности полета комаров довольно сложен, слова «даль-
ность полета» употребляются в нескольких смыслах. Можно различать
дальность полета физиологическую (максимальное расстояние, которое
комар мог бы покрыть по своим силам) и экологическую (расстояния,
которые он фактически покрывает по своим потребностям). G другой сто-
роны, следует различать дальность одноразового полета и дальность
расселения путем повторных перелетов.
Физиологическая дальность полета. A. maculipennis — неплохой ле-
тун. Он может летать быстро и далеко. За один перелет он может, по-
видимому, покрыть до 3 км, а может быть, и больше. Во всяком слу-
чае он может перелетать через наиболее широкие из наших рек.
Экологическая дальность полета. Экологическая дальность однократ-
ного перелета в основном зависит от топографии местности. Если места
кормежки и места выплода находятся совсем близко друг от друга, на-
пример, в 50 м, то большинство комаров и будет летать на это расстояние
как при поисках добычи, так и при откладке яиц, и лишь меньшая часть
их будет лететь дальше и рассеиваться по окружающей местности. Если,
наоборот, места кормежки и места выплода разделены расстоянием в
1—2—3 км, комары и при полете на водоем, и при обратном полете будут
Рис. 164. Схема проникновения самок А.
superpictus в г. Бьявконово из мест вы-
плода по р. Буонамико.
Пунктиром намечена т] ехиглометрс гая зона
вокруг г. Бьянкопоьо п шм.руг дер. Казиньяна
(подробности см. в тексте).
вынуждены покрывать это рас-
стояние.
Если рядом с водоемом, на-
пример, с небольшим рисовым по-
лем, начинается поселок, то боль-
шинство комаров оседает на его
окраине, и в глубине поселка, на
расстоянии 0,5 км от риса, мы мо-
жем не найти ни одного комара;
наоборот, в том случае, когда
поселок отодвинут от водоема на
0,5 км, несмотря па это расстоя-
ние, в его первых домах мы на-
ходим множество комаров. В пер-
вом случае 0,5 км, заполненных
домами, оказались непреодолимым
препятствием, или, лучше сказать,
отвлечением, для летящих с во-
доема комаров; во втором случае
0,5 км пустого поля преодолева-
ются комарами с легкостью.
Рассеяние комаров путем по-
вторных перелетов. В каком-либо
изолированном очаге самка А.
maculipennis может всю жизнь
летать из одного и того же по-
селка на одни и те же водоемы и
обратно. Но там, где имеются со-
пряженные центры притяжения,
судьба ее может сложиться со-
всем иначе. Действительно, отро-
дившись в каком-либо водоеме,
она может полететь в поселок,
расположенный в 2 км на восток. Оттуда для откладки яиц она может
вернуться в свой родной водоем, но если дальше на восток в 1—2 км
от поселка имеется другой водоем, самка может полететь и туда. После
кладки, вместо того, чтобы возвратиться к месту первого питания, она
может полететь в другой поселок, расположенный хотя бы к северу или
к югу от того водоема, где опа отложила яйца. Таким образом, в мест-
ности, богатой как водоемами, так и местами питания, комар может с
каждым гонотрсфичсским циклом перелетать все в новые места.
Пример. Городок Бьянкоиово в Италии расположен на берегу моря (рис. 164).
Вблизи пего протекает несколько маленьких речек, служащих местом выплода
A. superpictus. В радиусе 2,5—3,5 км все водоемы опылялись парижской зеленью, и в
начале лета комаров в городе не было. Однако в августе на контрольных дневках вдруг
появилось много самок A. superpictus. Раздумывая о том, откуда появились эти ко-
мары, руководители борьбы с комарами пришли к следующему предположению: за
пределами защитной зоны протекала река Буонамико, служившая обильным местом
выплода комаров, которые оттуда летели в ближайшую деревню Казиньяна. Напившись
278
гам крови, часть самок возвращалась для откладки яиц па Буонамико, другие летели
откладывать их на ручьях защитной зоны. В последнем случае вышедшие из яиц ли-
чинки при следующем опылении водоемов погибали, а самки, отложив яйца, частью
возвращались в прежнюю деревню или в большинстве случаев летели к более мощному
центру привлечения — в г. Бьянконово. Таким образом, в три приема они преодоле-
вали расстояние в 7 км. Исходя из этих рассуждений, па следующий год реку Буопа-
мико включили в зону обработки, и в результате этого залет комаров в г. Бьянконово
больше не наблюдался.
Приведенный пример очень поучителен: во-первых, он иллюстрирует
явление расселения малярийных комаров в несколько приемов; во-вто-
рых, он показывает, что населенный пункт, расположенный между оча-
гом выплода и интересующим нас поселком, может служить для послед-
него не только заслоном от залета комаров, но и, наоборот, этапом, об-
легчающим их расселение.
Роль промежуточных населенных пунктов. Такую роль промежуточ-
ный населенный пункт приобретает в тех случаях, в которых, как в опи-
санном примере, между обоими местами кормежки существуют водоемы,
являющиеся возможным местом кладки (хотя бы они и не были сами по себе
мощными очагами выплода).
Если между двумя поселками водоемов нет, а один из них лежит
ближе к местам выплода и заслоняет от них другой, в таком случае пер-
вый поселок задерживает у себя комаров и препятствует их доступу во
второй.
Роль случая. Случайность играет большую роль в расселении комаров.
В силу случая комар, летящий с водоема, может пропустить ближайшее
место питания и проникнуть в более удаленные, или комар, летящий класть
яйца, может пропустить ближайшие водоемы и отложить свою кладку
где-нибудь дальше. От целого ряда случайных обстоятельств может зави-
сеть, в какую сторону полетит кладущая самка, когда водоемы имеются
с нескольких сторон от места ее дневки, пли к какому из сопряженных
центров притяжения полетит голодная самка. Поэтому чем больше ко-
маров вылупляется из какого-нибудь водоема, тем больше вероятность,
что некоторые из них в силу разных случайностей разлетятся от него
на дальнее расстояние. И действительно, мы видим, что небольшой водоем
заполняет умеренным количеством комаров только ближайшие хлева,
тогда как большой анофелогенный водоем переполняет ближайшие хлева
колоссальным множеством комаров, и в то же время значительные коли-
чества комаров разлетаются из него и на 3 км, и даже на 5 км. Поэтому
при защите от комаров какого-либо населенного пункта мы вправе пре-
небречь небольшими анофелогенными водоемами, расположенными дальше
3 км от подзащитного объекта, но часто бываем вынуждены обрабатывать
крупные водоемы, лежащие от него на расстоянии 4 км и даже 5 км. Точно
так же, когда выплод комаров невелик, они проникают только на окраины
населенных пунктов; когда в этом же месте выплод увеличивается, ко-
мары в громадном количестве переполняют окраины населенного пункта
и в то же время проникают глубоко внутрь его.
Не иначе обстоит дело и с кладками: если в населенном пункте кладу-
щих самок много, они заваливают яйцами ближайшие водоемы и некото-
рые из них рассеиваются по более отдаленным водоемам на 3 км и дальше.
Если же число кладущих самок резко сократится (например, вследствие
истребления их противоимагинальной борьбой), немногие появляющиеся
кладки оказываются все приурочены к ближайшим водоемам.
Наряду с элементом случайности, важнейшую роль в расселении ко-
маров играет топография местности. Из топографических факторов основ-
ным является взаимное расположение личиночных очагов и мест пита-
279
ния, о чем мы уже говорили. Кроме того, приходится считаться «: рель
ефом местности и ветрами, в особенности с бризами (см. выше, стр. 260 *.
В связи с этим зоны тяготения комаров к населенным пунктам в гори-
стых местностях могут представлять весьма определенные очертания,
охватывая отрезки долины, расположенные ниже соответствующих насе-
ленных пунктов (рис. 156).
Лес вряд ли служит препятствием для расселения A. maculipenni>.
Пассивный занос ветром. Обычно охотящиеся самки летят прети г
слабого ветра. Однако большую роль играет и пассивный занос комаров
по ветру. В ряде случаев наблюдается резкое увеличение численности
A. maculipennis в каких-либо селениях после сильных ночных ветров,,
дувших со стороны крупных мест выплода.
Завоз транспортом. В том случае, когда поезда пли пароходы останав-
ливаются вечером в местах, богатых комарами, последние могут проникать
туда в большом числе. Залет комаров, с одной стороны, представляет он а с
ность заражения малярией для пассажиров и, с другой стороны, может
повлечь за собой завоз их в другие города, иногда очепь отдаленные.
В Астрахань и Сталинград самки A. mac. messeae регулярно зг)во-
зятся дачными пароходами (наблюдения Покровского и Поликарповой;
Блахова и Купцовой). В ряде случаев наблюдается завоз комаров самоле-
тами. В тропиках, в связи с опасностью завоза самолетами опасных пере-
носчиков болезней из числа комаров, на больших воздушных путях
существует служба дезинсекции самолетов и защиты аэродромов от w
никновения туда комаров.
7. ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
Календарный и физиологический возраст комаров. В неволе при бла
гоприятных условиях жизни отдельные самки A. maculipennis летом могут
прожить больше 2 месяцев. Высокая температура, даже если она недо-
статочна, чтобы повредить комару, укорачивает его жизнь, так как при
высокой температуре скорее протекают все жизненные процессы. Ввиду
того что и в природе комары живут при различных температурах и таким
образом проходят свой жизненный путь с различными скоростями, мы
различаем календарный возраст комара, определяемый числом прожитых
дней, от физиологического возраста, определяемого числом проделанных
гонотрофичоскпх циклов. Самки, прожившие активной половой жизнью
неделю при 25—30° или месяц при 15°, проделают одинаковое число го-
нотрофических циклов и ока;кутся в одном физиологическом возрасте.
Продолжительность жизни в неволе при питании кровью. Изучая
в неволе физиологическую продолжительность жизни самок A. maculi-
pennis, мы находим, что у отдельных особей она оказывается весьма раз-
личной. Даже при самых благоприятных условиях содержания часть
особей погибает после первого гонотрофического цикла, другие особи —•
после 2 циклов, после 3 циклов и т. д., самые последние особи — после
17—18 циклов (рис. 165). Процент особей, умирающих на каждом цикле,
по отношению к числу особей, благополучно проделавших предыдущий
цикл, остается почти постоянным: если для первых четырех циклов эта
величина в среднем равняется 20%, то к 13—16-му циклу она возрастает
лишь до 27%. Численность одновозрастной комариной популяции при
питании кровью непрерывно сокращается, и в начале, при самых первых
гонотрофических циклах, сокращение это идет наиболее быстро (если
выражать его в абсолютных цифрах).
280
Продолжительность жизни в природе. В природе продолжительность
жизни комаров легче всего наблюдать в какой-либо местности, прослежи-
вая вымирание наличного состава взрослых комаров с того момента, когда
по каким-нибудь причинам прекратился выплод из водоемов.
Число
кладок 1 2 3 4 5 5 7 3 9 10 11 12 13 14 15 16 17
Рис. 165. Кривая вымирания одновозрастной популяции самок^ А.
mac. atroparvus при кровяном питании и вполне благоприятных
.условиях содержания в неволе.
Столбики и цифры под ними обозначают число комаров, доживших до
того числа кладок, которое обозначено соответствующими цифрами ниж-
него ряда.
Пример. В 1932 г. в К аратальско.м рисовом совхозе производилось, опыление
парижской зеленью, прошедшее в общем удачно. Однако, вследствие запоздания оче-
комаров, который
начался вылет
Рис. 166. Ход численности самок A. ma-
culipennis в двух поселках Караталь-
ского рисового совхоза: нарастание
численности дневок при прорыве в авиа-
опылении и последующее естественное
вымирание популяции.
редного опыления, с 1/VIII на одном из участков
длился 13 дней и был прекращен очеред-
ным опылением (рис. 166). На двух посел-
ках, получавших комаров с этого поля,
численность их непрерывно возрастала до
последнего дня вылета. После прекраще-
ния вылета со следующего же дня нача-
лось крутое падепие числа комаров. По
мере того как число комаров уменьшалось,
кривая вымирания приобретала более по-
логий характер. Последние комары на
этих поселках были пойманы 5/IX, через
23 дня после прекращения вылета.
Сходный характер носит выми-
рание комариной популяции и в дру-
гих случаях удачных противоличи-
ночных работ, при сплошном пере-
сыхании мест выплода и т. п. Суще-
ственно во всех этих случаях крутое
падение численности взрослых кома-
ров, наступающее тотчас по прекра-
щении выплода. Оно ,указывает, что в природе, как в лаборатории,
смертность равномерно распределена по всем гонотрофическим циклам.
Благодаря этому мы и можем проверять степень успеха наших проти-
воличиночных мероприятия по немедленному падению численности
взрослых комаров.
Причины смертности. Отчего зависит смертность среди взрослых са-
мок A. maculipennis? Она имеет ряд причин: 1) смерть от истощения при
откладке яиц; 2) неблагоприятные условия микроклимата дневок; 3) не-
достаточно быстрое нахождение отложившей самкой добычи и смерть от
голода; 4) рассеяние комаров — залет в поисках мест кладок или мест пи-
тания по таким направлениям, где нужных объектов не оказывается;
5) враги и паразиты.
В убежищах врагами комаров являются пауки, многоножки (Scutigera,
рис. 13, Б), ящерицы-гекконы, хищные клопы (Reduviidae), осы, муравьи.
На водоеме кладущую самку могут схватить клопы-водомерки (Gerris
и др.), бегающие пауки. В воздухе комаров ловят стрекозы, ласточки, лету-
чие мыши, хищные мухи (рис. 31, Г) и пр. Собственная добыча также губит
комаров: люди убивают некоторое количество, собаки катаются по земле,
пытаясь их раздавить, птицы и мелкие грызуны стараются их съесть; безо-
паснее всего, вероятно, копытные.
Какая из всех перечисленных причин смертности губит больше всего
комаров? Можно думать, что в различных местностях размеры этих опас-
ностей бывают различны. Условия Каратальского совхоза (см. рис. 166),
где максимальная продолжительность жизни A. maculipennis ока-
залась 23—25 дней, по ряду признаков весьма суровы: сухой климат,
сильные ветры, редкость населенных пунктов, допускающая гибель ко-
маров во время рассеяния. В местах, более благоприятных для жизни
комаров, вымирание одновозрастной группы самок затянулось бы на бо-
лее продолжительное время.
Значение возрастного состава комариной популяции. Продолжитель
ность жизни комаров и возрастной состав комариной популяции имеют
большое значение. Действительно, при каждом сосании самка имеет из-
вестные шансы заразиться плазмодиями. Чем больше она проделала го-
потрофических циклов, тем большее число раз она подвергалась шансам
заразиться. Поэтому среди старых самок зараженные особи со спорозои-
тами Plasmodium встречаются чаще, чем среди молодых. С другой стороны,
и умирают самки на всех гонотрофических циклах; поэтому чем старше
какая-либо группа самок, тем она малочисленнее. Чем больше процент
старых особей в комариной популяции, тем опаснее может оказаться эта
популяция в эпидемиологическом отношении.
Когда мы истребляем взрослых комаров на дневках, первая цель,
которую мы себе ставим, — настолько увеличить их смертность, чтобы
как можно меньше комаров доживало до того физиологического возраста
(3—4 гонотрофических цикла), когда у них могут оказаться спорозоиты
в слюнных железах, когда они смогут стать действительными переносчи-
ками. Поэтому иуспехпротивопмагиналыюй борьбы мы измеряем сниже-
нием возрастного состава комариной популяции.
Методы определения возраста комаров. Для того чтобы определить
возраст комара, мы пользуемся возрастными изменениями, которые на-
ступают с течением времени в его организме. Эти изменения касаются,
во-первых, чешуйчатого покрова и, во-вторых, женского полового аппа-
рата комара.
На оценке целости чешуйчатого покрова крыльев основан метод
Перри. Перри различает четыре степени потертости крыльев.
1. Весь чешуйчатый покров, включая и заднюю бахромку крыла, цел.
2. Бахрома слегка повреждёпа, чешуйки на верхней поверхности
крыла целы.
282
3. Бахрома сильно повреждена, верхняя поверхность потерта, но
пятна на крыльях отчетливы.
4. От бахромы — только остатки, верхняя поверхность сильно по-
терта, пятна на крыльях почти незаметны.
Преимущество этого метода состоит в том, что он применим также и
к диапаузирующим самкам, недостаток — в том, что он не позволяет
выделить ни разу не клавших самок. Кроме того, этот метод требует боль-
шой осторожности при ловле комаров и дальнейшем с ними обращении,
так как в противном случае чешуйки крыльев легко стираются и комар
может показаться старее, чем
он есть на самом деле.
Метод Мера основан на из-
менении размеров1 ампул яйце-
водов (стр. 225).
Размеры ампул увеличи-
ваются при каждом созревании
яичников.
Так, у A. mac. atroparvus сред-
ние размеры ампул следующие:
до 1-го гопотрофического цикла —
0,010 мм2,после 1-Го цикла-0,021 мм2,
после 2-го цикла — 0,028 мм2, после
3-го цикла — 0,031 мм2; у новорож-
денной самки A. mac. messeae —
0,013 мм2 и после 1-го цикла —
0,029 мм2, у новорожденной самки
A. mac. sacharovi — 0,006 мм2 и
после 1-го цикла — 0,015 мм2 (По-
ловодова, Солодовникова).
Однако индивидуальная
лзменчивость размеров ампул
велика, а рост их с последу-
ющими циклами постепенно за-
медляется и кривые изменчи-
Рис. 167. Кривая измепчпвости размеров ам-
пул самок A. mac. messeae, собранных в хле-
вах с 12 июля по 8 августа 1934 г. в Молотов-
ской области. Нормальный возрастной состав
популяции A. maculipennis в летнее время на
дневках с добычей.
(По горизонтальной осн — размеры ампул, по вер-
тикальной оси — число самок.)
вости размеров ампул после-
довательных возрастных классов все более и более заходят одна на
другую. Поэтому метод Мера не позволяет точно решить, сколько цик-
лов проделала каждая самка в отдельности. Но он позволяет прибли-
зительно судить о количественном соотношении особей, проделавших
различное число гонотрофических циклов. Для этого, вскрыв боль
шое количество самок и измерив их ампулы, строят кривую изменчи-
вости ампул (рис. 167). Если комары собраны в убежищах с перевари-
вающими самками, кривая имеет асимметричный вид. Вершина ее обычно
приходится на самок, находящихся в конце первого и начале второго
гонотрофического цикла. Слева кривая падает круто ввиду малочислен-
ности в таких убежищах самых юных самок. Справа кривая идет более
полого, образуя небольшой хвост за счет старших возрастов.
В настоящее время чаще всего пользуются методом изучения возраст-
ного состава популяции по качественным признакам. Среди вскрытых
самок ни разу не клавшие легко отличаются по присутствию в яичниках
трахейных клубочков и полному отсутствию желтых тел. Самки, отло-
жившие яйца один раз, узнаются по размотанным трахеям яичников,
1 Размеры ампул условно выражаются в виде произведения их длины на ширину,
у. е. в долях квадратных миллиметров.
283
по слабо пигментированным яичникам и по наличию в каждой из яйцевых
трубочек не более 1 желтого тела. Совсем старые самки распознаются
по сильно пигментированным яичникам с крупными, ярко окрашенными
желтыми телами (обычно оранжевого и красного цвета). Число проделан-
ных гонотрофических циклов определяется по числу расширений (желтых
тел) на яйцевых трубочках (Половодова, Детинова).
Прослеживая количественные соотношения самок разного возраста,
мы можем следить за жизнью комариной популяции, за появлением и
судьбой последовательных поколений комаров и за эффективностью на-
ших истребительных мероприятий.
8. ЗИМОВКА
а) Общие замечания
Приспособление A. maculipennis к зимовке выражается в появлении
в конце лета самок, резко отличных по своему физиологическому строю
от нормальных летних самок. Эти «зимовочные» самки одни переживают
зиму, тогда как личинки, самцы и нормальные летние самки с наступле-
нием холодов вымирают. Основная особенность зимовочных самок — диа-
пауза \ выражающаяся у них в торможении развития яичников.' Они
больше но подчиняются правилу гонотрофической гармонии; если даже
они и пьют кровь, яичники их но развиваются. Наряду с этим, в организме
самки появляется ряд других особенностей, помогающих ей переносить
зиму; важнейшей из этих особенностей является ожирение.
к Отдельные подвиды A. maculipennis очень различны в отношении
биологи изпмовки. A. mac. mossoae приспособлен к перенесению только
суровой зимы, A. mac. atroparvus и A. mac. subalpinus, наоборот, способны
зимовать только в условиях мягкой зимы; наконец, A. mac. maculipennis
и A. mac. sacharovi одинаково успешно переносят как мягкую, так и суро
вую зиму.
б) Зимовка /1. mac. wesseae
Происхождение зимовочных самок. Начиная с определенного' момента
(•озона, вновь окрыляющиеся самки A. mac. messeae оказываются в со-
стоянии диапаузы. В Московской области первые зимовочные особи по-
являются в конце июля, в Сольвычегодске—в начале июля. Постепенно
число их увеличивается, и к середине сентября в Московской области
нежирные летние самки встречаются уже единично. Все они — старые
особи, отродившиеся еще в июле, продолжающие класть яйца и посте-
пенно вымирающие.
Источники ожирения. Осенью личинки имеют жировое тело, несколь-
ко более развитое, чем это бывает летом; поэтому и вылетающие осенью
диапаузирующие самки несколько жирнее летних самок. Диапаузирую-
щие самки питаются в основном сладкими растительными соками, отчасти
и кровью, и постепенно достигают полного ожирения. Яичники их при
этом не развиваются и остаются на самых ранних фазах I и I—II. Раз ожи-
рев, самки A. mac. messeae до конца зимовки крови больше не пьют.
Предзимовочное поведение. С наступлением диапаузы и ожирения
меняется и поведение самок. Вместо помещений с добычей они начинают
выбирать прохладные и пустые убежища, часто долго задерживаются
1 Диапауза есть временная остановка в морфологических процессах развития
насекомого, возникающая под влиянием раздражителей, подействовавших на одной
из предыдущих стадий развития.
284
где-нибудь в природе. При этом во многих местностях наблюдаются пред-
зимовочные миграции: зимовочные самки покидают речные поймы и дру-
гие влажные, низменные места, где воздух в осенние ночи насыщен вла-
гой, и переселяются на верхние террасы речных долин и водоразделы,
иногда за несколько километров от своих летних мест пребывания
(рис. 168).
Типы зимних убежищ. С наступлением холодов все зимовочные самки
постепенно собираются в зимние убежища. Убежища эти разнообразны;
часть особей зазимовывает в природе: в пещерах, в дуплах, в прикорневых
ямах, норках и т. п. В человеческих
постройках зимовки также помещают-
ся в различных местах. При более теп-
лых зимах зимовки бывают на черда-
ках домов и хлевов. Чем суровее зи-
ма, тем более защищенные места вы-
бираются для зимовки: в Европейской
части Союза и в Сибири это чаще всего
погреба, овощехранилища, подполья
крестьянских изб, хлева. Однако в
условиях суровой континентальной зи-
мы зимовки в хлеву не ведут к зимнему
кровососанию, так как при господству-
ющих в хлевах Урала или Сибири от-
рицательных температурах, и актив-
.50 X
у я ///
Г/// IX
Рис. 168. Предзимовочное рассеяние
самок A. mac. messeae из долины Се-
верной Двины (Красноборский рай-
он Архангельской области). Сезон-
ный ход численности по трем зонам.
кость комаров, и расход жира незна-
чительны.
Условия для зимовки. Благопри-
ятными условиями для зимовки А.
mac. messeae являются ровная и низкая
температура, умеренная влажность, от-
/ — деревни, расположенные непосред-
ственно над поймой Северной Двиньг
2—деревни, расположенные в 1,5—2
им от поймы; 3 — деревни, расположен-
ные в 4—5 км от поймы. Вне поймы ано-
фелогеиных водоемов практически нет и
осеннее нарастание численности в отда-
ленных деревнях обусловлено исклю
чительпо переселением из I зоны.
сутствие сквозняка, полумрак. Слиш-
ком высокая температура (выше -4-5°) вызывает повышенный расход жира
и смерть от голода. СлишкохМ низкая температура (ниже —5°) вызывает
гибель части особей от холода. Слишком высокая влажность (100%)
вызывает вымирание всех комаров, слишком низкая (меньше 60%)
тоже опасна.
Однако много зимовок бывает и при температурах и влажностях,
уклоняющихся от оптимума, и в таких случаях наблюдается большая
смертность среди зимующих комаров; иногда они вымирают все. Но часть
комаров доживает до весны и при не совсем благоприятных условиях.
В частности, на севере, в помещениях, температура которых не отличается
от наружной и спускается временами до —30° и ниже, часть комаров все
же благополучно перезимовывает.
Поведение на зимовке. При отрицательных температурах зимующие
комары находятся в состоянии окоченения (полная неподвижность с —2°);
при температуре 4-5° и выше жирные самки ведут себя приблизительно,
как сытые летние самки: неподвижно сидят в полутемных местах и взле-
тают только при раздражении. Но на теплой зимовке комар, истративший
свой жир, начинает вести себя, как голодная самка: у него появляется
положительный фототаксис к слабому свету. Под его влиянием комар
может полететь на окно и там гибнет от холода пли вмерзает в лед и по-
гибает. Но если зимовка находилась в подполье, комар может вылететь
в избу п там напиться на людях. Такие нападения вылетевших с зимовки
комаров во второй половине зимы — не редкость, особенно на севере
283
и в Сибири, но долго прожить в условиях отапливаемой избы комар не
может.
Прекращение зимовочного состояния. Диапауза представляет собой
как бы особую фазу, вставленную в жизненный цикл комара, на протяже-
нии которой деятельность яичников заторможена. В течение зимы она
постепенно изживается. Самки выходят из диапаузы не все сразу, а одни
раньше, другие позже. В Московской области уже в начале ноября по-
являются отдельные самки, способные класть яйца после одного сосания
крови, т. е. вышедшие из диапаузы. К началу декабря опи составляют
большинство, в январе активными в половом отношении стали уже почти
все. Таким образом, в условиях долгой, холодной зимы диапауза заканчи-
вается еще в начале зимовки. Однако это раннее прекращение диапаузы
не вредит сохранению вида: самки к этому времени находятся в прохлад-
ных помещениях, в которых, как правило, им па всю зиму хватает соб-
ственных жировых запасов; в силу этого они крови не пьют и отсутствие
диапаузы не может вызвать у них несвоевременного созревания яцц. Диа-
пауза нужна была, чтобы войти в зимовку, чтобы не растратить осенью
силы на откладку яиц, из которых все равно не могли бы развиться
комары.
При холодных убежищах самки тратят свой жир только весной,
когда на дворе уже тепло и когда нагревается и убежище. Яичники к это-
му времени давно способны к активности и за счет усвоения жира дораз-
виваются до II фазы Кристоферса. Это обеспечивает самке возможность
доведения яиц до полной зрелости за счет одного принятия крови. Истра-
тившие жир самки приобретают положительный фототаксис, в первый
теплый день покидают зимнее убежище и начинают искать добычу — зи-
мовка закончилась.
На юге (Киев, Ростов), в связи с более теплой и затяжной осенью,
диапауза A. mac. messeae затягивается месяца на 11/2—2 дольше, чем
в таежной полосе.
в) Зимовка A. mac. atroparvus
Условия зимовки. В отличие от A. mac. messeae A. mac. atroparvus
приспособлен к зимовке в сравнительно теплых помещениях и к жизни
в странах с мягкой зимой. В Приазовье зимовки его встречаются в хлевах
и неотапливаемых, полу жилых помещениях, где температура зимой колеб-
лется от +3° до -|-9о и только очень ненадолго спускается ниже 0°. При
этих температурах жир тратится довольно быстро, и комар вынужден в те-
чение зимы неоднократно пить кровь; поэтому зимовки atroparvus всегда
находятся вблизи добычи. Диапауза у A. mac. atroparvus еще более дли-
тельна и глубока, нежели у messeae. Начинаясь в Приазовье у части осо-
бен уже в середине августа, она полностью прекращается только в конце
февраля, на месяц позже,чем у A. mac. messeae в тех же местах.
Осенняя фиксация на дневках. Осенью, в то время когда messeae
вместо обычных дневок идет в зимовочные убежища, atroparvus попреж-
нему остаются в домах и хлевах, несмотря на начавшуюся и у Них диа-
паузу. Они продолжают пить кровь, но яичники их не развиваются, а
за счет крови развивается жировое тело. Самка теряет необходимость уле-
тать из убежища для откладывания яиц, и если микроклимат благопри-
ятен, она может оставаться в одном и том же убежище длительное время,
повторно питаясь там кровью. После каждого сосания самка жиреет,
а затем постепенно тратит жир и, истратив его, опять пьет кровь. Однако
пока тепло, слабый круговорот па дневках все же идет. С наступлением
286
холодов самки окончательно остаются в тех помещениях, где их застанет
зима, и продолжают там также периодически сосать кровь.
Выход из зимовки. Прекращение зимовки у atroparvus совершается
весьма незаметно. С приближением весны комары начинают чаще питаться.
В то же время подходит к концу и диапауза. С ео окончанием следующий
же прием крови ведет к развитию яичников, и тогда вскоре, если будет
благоприятствовать погода, комары вылетают из убежища для откладки
яиц. Место зимовки постепенно превращается в обычное убежище с
добычей.
Осенние внутридомовые очаги. Особенности зимовки A. m. atro-
parvus имеют определенное эпидемиологическое значение. Уже тот факт,
что самки atroparvus осенью жиреют за счет повторного питания кровью,
значительно удлиняет сезон возможного переноса ими малярии, как пока-
зала Половодова в Ростове-на-Дону. Но еще большее значение имеет фи-
ксация самок в определенных убежищах и прекращение круговорота на
дневках, связанного с размножением. Этим создается возможность воз-
никновения осенних внутридомовых очагов малярии. Действительно,
если в каком-либо доме имеется обильная дневка atroparvus и живет ма-
лярик-гаметоноситель, заражающиеся в этом доме комары летом имеют
весьма мало шансов вернуться туда же со спорозоитами (см. выше, стр. 274),
наоборот, большинство их рассеется по хлевам и не сможет никого зара-
зить; если же эти комары и попадают опять в дом, то необязательно в тот,
где они заразились.
Другое дело осенью, когда самки atroparvus перестают улетать для
откладки яиц, а температура еще достаточйо высока; в силу этого они
вынуждены довольно часто пить кровь. Если среди жителей имеется гаме-
тоноситель, кормящиеся на нем комары заражаются, а так как они никуда
не улетают, процент зараженных комаров в таких домах непрерывно воз-
растает и имеющиеся там здоровые люди в громадном большинстве под-
вергаются заражению. Возникает осенний внутридомовой очаг малярии.
В наших условиях такое положение длится до начала отопительного
сезона. Когда помещение начинают отапливать, зимовочные самки A. mac.
atroparvus либо гибнут, либо спасаются бегствОхМ и очаг па этот год ликви-
дируется. В тех же странах, где дома, благодаря мягкой зиме, осенью поч-
ти не отапливаются, как, например, в Голландии, зимовка в человече-
ском жилье сохраняется дольше. В этих условиях комары продолжают
заражаться в комнатах до начала ноября и число зараженных спорозоита-
ми особей достигает чудовищных размеров, доходя до 20, иногда до 30%
(в летнее время даже встречаемость спорозоитов в 2—3% считается уже
чрезвычайно высокой и наблюдается только при тяжелых эпидемических
вспышках).
► В тех случаях, когда имеются подобные очаги, своевременное истреб-
ление зимующих вблизи человека комаров является совершенно обяза-
тельным.
г) Зимовки других подвидов
A. mac. subalpinus, подобно A. mac. atroparvus, зимует в присут-
ствии добычи и зимой пьет кровь.
Зимовка A. mac. maculipennis. A. mac. maculipennis в отношении*
способа зимовки представляет значительную гибкость. На севере (Кали-
нинская, Киевская, Харьковская области, Алтай) он зимует в холодных
убежищах вдали от добычи, подобно A. mac. messeae. Так же он ведет
себя и на юге в горах. Наоборот, в тех местах, где зимы слишком теплы,
287
чтобы можно было провести их без пищи, A. mac. maculipennis зимует
в помещениях с добычей и питается кровью в течение всей зимы (Абха-
зия, Грузия, Плоскостной Дагестан).
Зимовка A. mac. sacharovi. Точно так же ведет себя и A. mac. sacha
rovi: в тех местах, где зима холоднее, он зимует в холодных помещениях
и крови зимой не пьет (например, в предгорьях около Ташкента); там, где
зимы теплее, он зимует вблизи добычи и пьет кровь всю зиму (например,
на низменностях Азербайджана).
Итак, на низменностях юга (Закавказье, Средняя Азия) все имеющи-
еся там подвиды A. maculipennis могут пить кровь в течение зимы, и при
ходится считаться с возможностью зимнего контакта всех их с человеком
и осенне-зимних внутридомовых заражений малярией. Осуществление
этой возможности в значительной мере зависит от жилищных и бытовых
условий каждой местности. На севере такая возможность имеется толькс
в отношении A. mac. atroparvus.
ГЛАВА IX
ЭКОЛОГИЯ A. MACULIPENNIS КАК ВИДА В ЦЕЛОМ
1. ФЕНОЛОГИЯ
а) Служба фенологии на малярийных станциях
На огромной территории СССР, при разнообразии ее климатов, сроки
наступления сезонных явлений у Anopheles различны. Для установления
рациональных сроков проведения противомалярийных мероприятий тре
буется точное знание сезонной биологии: средних дат наступления основ
ных сезонных явлений в жизни комариных популяций и пределов уклоне
ний от этих средних дат, а также знание сезонного хода численности
Anopheles. Для этого необходимы многолетние ряды фенологических на-
блюдений, проведенных по единой программе и методике \ необходима хо-
рошо налаженная служба фенологии на малярийных станциях. Энтомо-
логи-маляриологи должны собирать материалы по фенологии Anophe-
les, проверять и обрабатывать их, передавать в республиканские и област-
ные малярийные станции для составления сводок и их опубликования
и в то же время отсылать копии наблюдений в энтомологический отдел
Центрального института малярии для составления общей сводки по СССР.
Результаты собственных наблюдений должны храниться на самой маля-
рийной станции, образуя ее фенологический архив. Совершенно необхо-
димо обеспечить полную научность фенологических наблюдений и сравни-
мость получаемых фенологических дат.
б) Весенняя часть годового цикла A. maculipennis
Вылет с зимовок. У подвидов, зимующих в присутствии добычи (напри-
мер, A. mac. atroparvus), вылет с зимовок происходит для откладки яиц,
у подвидов, зимующих в холодных помещениях без добычи (например,
A. mac. messeae), вылет с зимовок совершается в поисках добычи.
Для комаров, зимующих в присутствии добычи, весенние фенологиче-
ские наблюдения начинаются с установления времени окончания диапаузы
и появления на зимовках (дневках) самок с созревающими яичниками
1 Программа наблюдений по фенологии была напечатана в журнале «Медицин-
ская паразитология», в. 3. 1935, и в сборнике организационно-методических материале!
ио малярии и гельминтозам, 1943 п 1947; дополнительные методические указания
даны в статье Шиппцпной в журнале «Медицинская паразитология», в. I, 1947,
288
(Ill—V фазы Кристофорса). Сроки прекращения диапаузы и начала раз-
вития яичников у подвидов A. maculipennis в разных климатических
условиях их ареалов различны.
Раньше всего самки с развивающимися яичниками в СССР появляются
в Западном Закавказье, где климат теплый и мягкий.
В Сухуми диапауза у части комаров прекращается в феврале (средняя темпера-
тура -(-6,3°), полностью - в марте (средняя температура -р-9,5°). Массовое созревание
яичников начинается с середины марта или с апреля. Первые кладки имеют место
в начале — середине марта (Устинов), реже — в конце февраля (Рухадзе, 1923). Мас-
совая откладка яиц происходит к 20 марта или в апреле.
В Араратской долине (Центральное Закавказье), с ее более континентальным
климатом, выход самок из диапаузы происходит несколько позднее, чем в Западном
Закавказье. В феврале гоноактивные самки составляют здесь только 4%, остальная
часть популяции находится в диапаузе. Массовое развитие яиц начинается с сере-
дины—конца марта (средняя температура 6,6°), прекращение диапаузы у всех перези-
мовавших особей происходит в апреле (Чубкова).
В Восточной Грузии и Азербайджане развитие яичников у самок начинается
приблизительно в то же сроки, что и в Араратской долине, причем у A. m. sacharovi
несколько позже, чему A. m. maculipennis. Первые кладки яиц в водоемах Централь-
ного и Восточного Закавказья появляются вначале, чаще во второй половине марта
(Лемер).
В Средней Азии появление половоактивпых особей A. m. sacharovi начинается
с февраля—марта при средней температуре воздуха 4,4° и убежищ 6,8°. Первый гоно-
трофический цикл длится 10—15 дней (Чинаев). Первые кладки наблюдаются в пер-
вой декаде марта (Ташкент), во второй-третьей декаде марта (Самарканд, Коканд) и в
первой половине апреля (Н. Ургенч) при среднедекадной температуре воздуха 9,8°
и воды 12,7°.
Среди приазовских A. mac. atroparvus первых самок с яйцами па IV—V фазе
Кристофорса находят уже в середине февраля, когда еще бывает холодно; массовое
появление их отмечается только в третьей декаде марта. Первые кладки имеют место
в конце марта или в начале апреля.
Таким образом, наиболее ранний срок созревания яиц у A. macu-
lipennis на юге СССР — начало февраля; в это время средняя температура
воздуха в большей части Закавказья, Средней Азии и Приазовья не пре-
вышает 4° и откладка яиц в водоемы не всегда возможна. Те яйца, которые
все же откладываются в феврале, дают в марте выплод личинок, однако
процент окрыляющихся из них комаров снижен гибелью личинок от хо-
лода (наблюдения Дагестанского института малярии).
Вылет A. maculipennis из холодных убежищ разного типа происходит
неодновременно и продолжается нередко около 1—2 месяцев. Растяну-
тость вылета связана с неодновременным прогреванием разных помещений.
Из медленно прогреваемых убежищ: каменных подвалов, овощехранилищ,
ям и т. п., вылет совершается значительно позже, чем из легко прогре-
ваемых надземных псстрсек: подполий, хлевов и т. п.
Встречаются и различия, связанные с местными климатами различных элементов
ландшафта, на которых расположены места зимовок (рис. 169); так как на возвышен-
ности теплее, чем в долине реки или в лесу, то и вылет комаров с зимовок начинается
раньше всего из помещений, расположенных открыто и па возвышенных участках рель-
ефа. Колебания погоды также отражаются па интенсивности вылета, как это видно
из рис. 170.
Начавшийся с первым потеплением вылет комаров может прекратиться при
временном возврате холодов и возобновляется после нового повышения температуры.
При дружной весне вылет бывает менее растянут, чем при затяжной.
Сроки вылета в различных местностях различны (рис. 171). Из ри-
сунка видно, что в северной части СССР массовый вылет A. mac. maculi-
pennis и messeae начинается обычно не ранее 15 мая, а в южной 10—20
марта; таким образом, разница в сроках вылета между севером и югом СССР
достигает 2—2х/2 месяцев.
19 Учебник медицинской энтомологии
289
В одной и той же местности различия, зависящие от погоды, могут
быть значительными. Так, под Москвой в 1934 г. массовый вылет начался
18 апреля, а в 1946 г. — 10 мая; разница в сроках составляла 23 дня.
20
4 6 8 10 12 14 16 18 20 22 24 26 28 80 2 4 6 8 10 12 14 16 18
А прель Май
Рис. 169. Вылет комаров с зимовок из помещений, расположенных
на различных элементах ландшафта. Ижевск, Удмуртская АССР,
1937 (оригинал Поздеева).
1—подполье, расположенное на горе; 2— подполье, расположенное на
пойме; 3— ааирошенное овощехранилище и лесу.
К моменту вылета первых самок среднесуточные температуры воздуха
к началу массового вылета достигают 4-4°,
4-8° (Гуцевич). Помимо средне-
суточных температур, оказыва-
ют влияние и суточные максиму-
мы, а также характер подъема
сезонной температурной кри-
вой; поэтому в местностях с
континентальным климатом вы-
лет начинается при более низ-
ких среднесуточных темпера-
турах (4,5—6°), а в местностях
с менее континентальным кли-
матом вылет происходит при
более высоких средних темпе-
ратурах (6—7,5°).
Вылет с зимовок происходит
во все светлые часы суток, достигая
максимума к заходу солнца (Логи-
новский).
I Начало массового вылета с зи-
мовок в лесной полосе СССР совпа-
дает с началом распускания почек
листьев черемухи, березы и сморо-
дины, с началом цьеюния калуж-
ницы.
едва поднимаются над нулем
210л
200-
190
180
170
160
150
140
Ч 130'
Й 120'
^110
100'
90'
80
70
60
50
40
30
20
10
21
20
49
48
г/7
16
15
14
'13
12
11
10
9
8.
100
95
90
85
80
75
70
65
60
55
50
45
40
35
30
25
20
15
10
I
4
8
1
1 2 345 6 7 8 9 1011121314151617181920
Маи
Дни fa/лета Anopheles maculipennis
Рис. 170. Ход вылета A. mac. messeae из под-
полья, в зависимости от погоды. Дер. Заюрчим
Молотовскоп области, май 1941 г.
1— числи вь'летак ших комаров; 2—среднее}-
Точьан температура; 3—относительная влажность.
Время, протекающее от начала вылета с зимовок п до первых кладок,
зависит от продолжительности пищеварения и созревания яиц у кома-
ров, определяемых температурой убежищ. Среднесуточные температуры
хлевов в лесной полосе СССР в это время года бывают на 2—4° выше
наружных, т. е. достигают 9—12°. При такой температуре первые кладки
происходят только через 7—15 и более дней после начала вылета.
Весь период активной жизни перезимовавшего поколения продол-
жается в средней полосе СССР до 2—3 месяцев.
290
Так, в 1947 г. первый вылет единичных самок A. maculipennis с зимовок в окрест-
ностях Москвы произошел в конце первой декады апреля, тогда как последние пе-
резимовавшие самки встречались до 9 июля.
Перезимовавшие комары могут весной заражаться плазмодиями
малярии. Попадая в благоприятные условия микроклимата убежищ, часть
из них может дожить до момента созревания спорозоитов и фактически
оказаться переносчиками малярии.
К концу массового вылета с зимовок число комаров на дпевках дости-
гает максимума, который сменяется быстрым, а затем замедляющимся
уменьшенивхМ численности вследствие вымирания. В соответствии с этим
находится и распределение кладок во времени. Поэтому следует отмечать
начало кладок, их массовое появление и конец откладки яиц перезимовав-
шими самками.
начало окрыления
Первые личинки
Начало откладки
яиц
Начало массового
вылета с зимовок
—. 1 1 1 1 1 1 1 1 1—1 — »—«—с
20 30 10 20 30 Ю 20 30 10 20
Март Апрель май Июнь
Рис. 171. Сроки выплода I весеннего поколения A. maculi-
pennis в СССР в 1946 г. (оригинал).
Развитие весеннего поколения. Первые кладки происходят обычно
при среднесуточных температурах воздуха 7—9°; температура воды в мел-
ких открытых лужах, куда откладываются весной яйца, бывает в это
время в средней полосе градуса на 4 выше — около 11—13°.
Развитие первых яиц первой кладки продолжается большей частью 4—8 дпей и
происходит при среднесуточной температуре воздуха 7—11° и температуре воды
И—17°. На Черноморском побережье Кавказа, где весна более ранняя, но затяж-
ная, развитие первых яиц приходится на март и длится несколько дольше — 8—20 дней.
В Европейской части СССР все развитие первых особей 1 поколения в водоемах
продолжается свыше месяца, колеблясь от 32 до 49 дпей, протекая при средней тем-
пературе воздуха 10—14° и закапчиваясь при 13,5—16°. Развитие большей части
особей протекает при ботее высоких температурах воздуха и воды (14—20°) и продол-
жается от 20 до 28 дней.
При ботее крутом ходе весны в Средней Азии развитие A. mac. sacharovi идет
при ботее высоких температурах и про екает быстрее. Так, в Узбекистане развитие
I покотения длится окото 30—40 дней, протекает при средней температуре воздуха
10—16,5° и заканчивается при 19,8° (Чинаев).
Паибоюе растянутые сроки развития наблюдаются в СССР па Черноморском
побережье Кавказа. В Сухуми развитие первых особей I поколения длится около
50 дней (1938 и 1939 — Устиноз). из которых 20 дней приходятся па развитие яиц, и
проге <ает при средне декадных температурах вощуха от 4° до 14,4°, развитие боль-
шинства особей 1 поколения проходит при И—16,1° и продолжается около месяца
(Батуми 1945—1946, Чихладзе).
Сроки выплода A. maculipennis I весеннего поколения в СССР в 1946 г. показаны
на рис. 171, из ко* орого видно, что в северной части Евро зейской части СССР (Великий
Устюг) и в Восточной Сибири (Чита) вылет 1 поколения A. maculipennis начался только
19*
291
20 июня, тогда как в южной части (Коканд, Узбекистан)— 15 апреля; таким образом,
разница в сроках вылета между севером и югом СССР в этом году достигала двух с лиш-
ним месяцев.
Из рис. 171 также видно, что сроки вылета I поколения у южных популяции
A.maculipennis в Средней Азии и на Кавказе различны. Раньше всего вылет I поколения
происходит в Средней Азии и позднее в Западном Закавказье (например, в Коканде
15 апреля, а в Сухуми 3 мая). Разница в сроках вылета между Кокандом и Сухуми в
1946 г. составила 18 дней; в другие годы она может уменьшаться до 10 дней (1941)
или увеличиваться (например, в 1945 г. она дошла до 26 дней).
Рис. 172. Сроки развития 1 поколения А. та-
culipennis в Московской области в разные годы
(по 25-летним наблюдениям) по данным областной
малярийной станции (оригинал).
Сроки и продолжительность развития I поколения в одной и той же
местности зависят от погоды каждого года (рис. 172). Из рисунка видно,
что разница в сроках появления первых личинок I возраста в окрестно-
стях Москвы равна (на основании 25-летнего ряда наблюдений) 17 дням,
а в сроках вылета I поколе-
ния — 30 дням.
Сроки и продолжительность
развития I поколения зависят
также от условий микроклимата
биотопа и в различных водоемах
одной местности могут быть раз-
личны. Поэтому необходимо фе-
нологические наблюдения про-
водить из года в год па водоемах
одного типа, отмечая характер
водоемов, служащих для наблю-
дений (пойменные водоемы, лужи
на водоразделе и т. д.).
Начало вылета 1 поколения
определяется по появлению вес-
ной на дневках самцов и ново-
рожденных самок. За начало мас-
сового вылета I поколения при-
нимают тот день, когда кривая
численности новорожденных ко-
маров па дневках начнет круто
подниматься.
Ко времени окрыления I по-
коления численность перезимо-
вавших особей снижается до мини-
мума. Вылет I поколения вскоре достигает максимальной интенсивности и затем
постепенно ослабевает, затягиваясь в лесной полосе до 20-х чисел июля или даже
до середины августа. В случае возврата холодов вылет, раз начавшись, может итти
вяло и лишь с повышением температуры усиливается, так что интенсивный вылет
может быть отделен от его начала недели на 2—3.
Кладки I поколения начинаются через 5—14 дней после начала вы-
лета. В этот промежуток времени совершается копуляция, первоначальное
дозревание яичников и первый гонотрофический цикл.
Первое появление личинок I возраста совпадает в Москве с зацветанием березы.
Первое появление личинок III возраста совпадает с появлением крупных бутонов
у сирени и с массовым цветением черемухи. Первое появление личинок IV возраста
совпадает с зацветанием садовой яблони.
Начало вылета I поколения совпадает с массовым колошением озимой ржи, с
зацветанием кувшинок и кубышек, белой акации, с началом отцветания сирени. На-
чало массового вылета в лесной полосе совпадает с началом цветения большинства
луговых трав и злаков.
в) Осенняя часпгъ годового цикла A. maculipennis
Начало диапаузы. Появление диапаузирующих жирных особей проис-
ходит в популяции постепенно. Первые диапаузирующие самки, находя-
щиеся на III—IV стадии ожирения (по Клиглеру), обнаруживаются
292
в лесной п лесостепной полосе СССР с момента вылета комаров последней
генерации, т. е. обычно со второй декады июля. Массовое ожирение и
диссоциация имеют место с начала — середины августа и совпадают с
массовым вылетом комаров последней генерации. К концу августа около
половины популяции находится уже в состоянии диапаузы, имеет развитое
жировое тело и не пьет крови; во второй декаде сентября в диапаузе
находится не менее 70—80% популяции и только немногие старые летние
самки продолжают пить кровь и развивать яичники.
*
кп а
Рис. 173. Ход численности личи-
нок A. maculipennis в контроль-
ном водоеме и численности диапа-
узирующих самок. Дер. Заюрчим
Молотовской области, 1941.
1 — среднедекадная температура воз-
духа; 2 — среднедекаднан температу-
ра воды; 3— обилие личинок на 1 м3;
4 — процент диапаузирующих самок
на контрольных дневках.
На севере большая часть самок messeae, вылупляющихся в состоянии диапаузы,
в августе не летит в места питания (хлева, дома) или зимние убежища, а скопляется
во временных «предзимовочных» убежищах без добычи вблизи мест выплода (забро-
шенные постройки ит. п.), где и концентрируется
с тем, чтобы при наступлении более холодных
температур в начале сентября переселиться в бо-
лее защищенные зимние убежища. Поэтому для
правильного установления процента диапаузи-
рующих самок в популяции необходимо прини-
мать во внимание численность жирных самок
как в местах питания (в домах и хлевах), таки в
предзимовочных убежищах (заброшенных хле-
вах, банях и т. п.).
Вследствие резкого уменьшения числа
гоноактивных самок к концу августа,
число кладок быстро сокращается, что
влечет снижение численности водных фаз
уже к концу августа — началу сентября
(рис. 173). Последние кладки яиц встре-
чаются в лесной зоне в конце первой или
во второй декаде сентября, однако самки
с развивающимися яичниками находятся
на дневках в небольшом числе вплоть до
конца сентября — начала октября.
Со снижением температуры воды в
сентябре до 10° развитие личинок прекра-
щается и они постепенно вымирают.
С конца сентября — начала октября, когда
дневные температуры не превышают -|-6о,
сосание крови и развитие яичников у
комаров прекращаются.
Уход на зимовки. Параллельно с ожи-
рением начинается уход диапаузирующих самок в зимние убежища. Так,
в лесной полосе в конце июля—начале августа зимующие самки уже
встречаются в подпольях, в то время как основная масса комаров нахо-
дится в летних и в предзимовочных убежищах. Массовый уход на зимовки
и связанное с этим уменьшение численности комаров на дневках проис-
ходят во второй-третьей декаде августа или в первой декаде сентября при
среднесуточной температуре наружного воздуха 15—12°. В дальнейшем
залет на зимовки и перемещение комаров происходят вплоть до конца
октября. Ход залета комаров на зимовки и изменение температуры воз-
духа в этот период показаны на рис. 174.
Ход ожирения и гонотрофической диссоциации у южных популяций
A. maculipennis (sacharovi и typicus) в Средней Азии и на Кавказе
различен.
В Средней Азии, с ее континентальным климатом, с большими суточ-
ными колебаниями температуры, ночные похолодания (вплоть до замороз-
293
Рис. 174. Залет комаров A. maculipennis ua зимовки в подполье. Борисоглебск, 1944 (оригинал Марчуковой).
/— число комаров, залетевших в комароловки; 2— абсолютный минимум температуры воздуха; 3— среднесуточная температура
наружного воздуха..
ков) местами начинаются уже в начале сентября; в связи с этим диапауза
у A. mac. sacharovi наступает здесь рано.
Первые вполне жирные самки с неразвитыми яичниками появляются с третьей
декады августа; массовое ожирение и диссоциация имеют место со второ'! декады
сентября.
К концу сентября диапаузирующие и не пьющие крови особи составляют
73—90% о г общего количества комаров. Полный охват нолуляции диапаузой проис-
ходит в октябре (Самарканд, Ташауз).
Последние личинки 1 во траста в Ташкенте обнаруживались 10 октября (1936),
личинки IV во траста — в конце октября— начале ноября (при средней температуре
воздуха 12,6°), куколки—12 ноября (при температуре 10,9°). Массовый залет па зи-
мовку происходит в конце сентября — начале октября (при средней температуре
14,2—8,7°) и заканчивается к 20 ноября.
В Закавказье, особенно в западной его части, климат мягче; на Чер-
номорском побережье Кавказа температура воздуха в осенне-зимние
месяцы, как правило, не опускается ниже нуля. В связи с более теплой
осенью диапауза, а также прекращение выплода наступают здесь на
1х/а—2 месяца позднее, чем в Средней Азии.
Первые полностью ожиревшие самки с неразвитыми яичниками появляются
и здесь в августе; однако сроки массового о кирепия и диапаузы в различных мостах
Закавказья различны и зависят от сроков наступления низких ночных температур.
Раньше всего эти явления наступают в Азербайджане (например, в Хачмасе, Ди-
вичи).— в конце сентября — начале октября (Лемер, 1947), позднее — в Центральном
(Араратская долина) и Западном Закавказье.
В Сухуми количество диапаузирующих самок A. mac. maculipennis резко возра-
стает лишь в октябре (53%); в ноябре оно достигает 88—90%. С декабря диапауза
охватывает всю популяцию и длится только 2 месяца — декабрь и январь. Однако
единичные самки с развивающимися яичниками встречаются даже и в этот период.
Блиюк к описанному ход гонотрофической диссоциации и ожирения у A. mac.
typicus в Араратской долине (Чубкова).
Последние яйца и личинки I во фаста обнаруживались в Сухуми вплоть до
16 ноября (Устинов) при среднедекадпой температуре воздуха 12,3°, личинки IV воз-
раста — до 7 декабря при среднедекадпой температуре воздуха 7° и температуре
воды 11,5°. Выплод комаров может продолжаться здесь вплоть до декабря (средняя
температура ноября 12,4°). Массовый уход на зимовку в Сухуми происходит с декабря.
г) Число поколений
Под числом поколений понимают то максимальное число поколений,
которое может появиться в данной местности в течение сезона развития
комара.
Для комаров характерно взаимное налегание поколений, при кото-
ром различить отдельные генерации путем полевых наблюдений нередко
бывает трудно. Однако путем фенологических наблюдений, наблюдений
за сезонным ходом численности самок и личинок разных возрастов и
появлением самцов удается обычно установить сроки появления I и II по-
коления.
В северной части ареала вида, где имеется всего только два поколе-
ния, сроки развития последних хорошо различимы. Дальнейшее расчле-
нение отдельных поколений в полевых условиях становится обычно трудно
осуществимым, ввиду установления сплошного потока размножения и
одновременного наличия в водоемах кладок и личинок нескольких поко-
лений.
Существует несколько способов определения максимального числа поколений.
Число поколений можно установить путем воспитания комаров в инсектарии
при темпер «туре наружной среды, причем после первых но кладок каждою поколе-
ния все взрослые ком фы каждым раз удаляются по шестью. Этот способ применим
только к подвидам, могущим размножаться в неволе.
295
Этим путем Пикуль с сотрудниками в Махач-Кала (Дагестан), поль-
зуясь колоссальным инсектарием, установил 5—6 поколений A. mac.
maculipennis.
Второй способ основан па одновременном проведении наблюдений над феноло-
гией, численностью и возрастным составом комариной популяции в течение сезона,
причем продолжительность всех фаз жизненного цикла отдельной генерации прове-
ряется путем учета се зависимости от температуры. 11ри этом для определения продол-
жительности водных фаз пользуются формулой Боденгеймера и данными по измерению
температуры водоема; для определения промежутков времени, протекающих от окры-
ления до первой кладки, принимают во внимание темпера уры воздуха.
Вылет и начало кладок I поколения определяются легко. На севере начало вы-
лета II и III поколения также определяется по подъему численности особей и одно-
временному увеличению процента новорожденных. Непосредственным наблюдением
устанавливается и конец размножения комаров осенью.
Определения по этому способу устанавливают для всей таежной зоны
2 поколения (Карело-Финская АССР и Ленинград — Шуб; Восточная
Сибирь— Сибирякова и др.). Для южной части лесной зоны установлено
2—3 поколения (Москва), для лесостепной зоны (Магнитогорск, Куй-
бышев, Харьков и пр.) возможны 3—4 поколения, для степной полосы
(Ростов, Днепропетровск) — 4—5 поколений, для Средней Азии (Ташкент)
и низменной части Закавказья (Сухуми, низовье Куры)—6—7 поколений,
2. ЧИСЛЕННОСТЬ ПОПУЛЯЦИИ И ЕЕ ГОДОВОЙ ХОД
Численность популяции A. maculipennis имеет большое эпидемиоло-
гическое значение;шансы на перенос малярии, при прочих равных условиях
прямо пропорциональны численности переносчика (Росс). Поэтому сниже-
ние численности комаров до возможного предела — основная задача про-
тивокомариных мероприятий. Путем учета численности популяции мы
оцениваем эффективность истребительных работ.
Что такое численнссть популяции. Под численностью комариной по-
пуляции в какой-либо местности понимают общее число комаров, нахо-
дящихся на ее территории как в постройках, так и вне их. Однако опре-
деление абсолютных размеров комариной популяции практически неосу-
ществимо. Поэтому устанавливают относительную численность комаров,
пропорциональную общей численности популяции. Так, об изменениях
общего числа эндофильных 1 комаров судят по изменению числа комаров
на дневках в домах, хлевах и других постройках. Однако нужно помнить,
что изменение числа комаров на дневках может означать не только изме-
нение абсолютной численности популяции, но и перераспределение ко-
маров между убежищами разных типов (в связи с изменениями погоды,
перераспределением добычи, наступлением диапаузы). Поэтому в слу-
чаях резких изменений численности необходимо рассмотреть причины,
их вызывающие, чтобы отличить кажущиеся изменения численности от
д ейс т ви тел ьны х.
Для учета относительной численности экзофильных 2 видов (например,
A. hyrcanus, A. bifurcatus) пользуются числом комаров, прилетающих
на добычу (на человека, теленка и т. п.) в единицу времени.
Численность комариной популяции зависит от числа ежедневно окры-
ляющихся особей-, определяемого площадью и продуктивностью личи-
ночных очагов и температурой, и от пмагинальной смертности. Оелабле-
1 Эндофильными называются те ком 1ры, которые в качестве дневного убежища
часто избирают постройки человека (дома, хлева и т. п.).
2 Экзофильннми наняв ются комары, которые на дневках в помещениях не оста-
ются или ост потея в ничтожном проценie, даже и в том случае, если они залетели туда
б поисках добычи»
296
ние или прекращение вылета, вызываемое пересыханием очагов, истреб-
лением личинок или понижением температуры, вызывает немедленное па-
дение численности популяции.
Годовой (сезонный) ход численности популяции. В СССР, где имеется
смена времен года (в отличие от влажных тропиков), численность популя-
ции в течение года изменяется; эти изменения в основном определяются
климатом. Поэтому в местностях с разными климатами сезонный ход чи-
сленности может оказаться различным. Этот климатически обусловленный
сезонный ход численности может видоизменяться под влиянием сезонных
изменений площади и характера личиночных биотопов. В силу этого мест-
ности, обладающие одинаковым климатом, но отличающиеся системой
стока, могут давать различные виды сезонного хода численности комаров.
Влияют на сезонный ход численности также различия в погоде отдельных
лет, различия в размерах паводков рек. Но при всем разнообразии типов
сезонного хода численности в разных местностях и в разные годы общая
схема хода кривой будет следующая: в течение зймы выплода комаров
не происходит, но идет постепенное вымирание и уменьшение популяции,
поэтому число вылетающих с зимовок комаров всегда во много раз ниже
числа комаров, пошедших на зимовку.
Типы годового хода численности популяции. В пределах этой схемы
сезонного хода численности A. maculipennis различают три главных, кли-
матически обусловленных типа: 1) северный, 2) южный и 3) тип A. mac.
sacharovi на северной границе его распространения.
Нормальный северный тип хода численности. На севере (в Европей-
ской части СССР, в Сибири, Предкавказье, Средней Европе) численность
популяции A. maculipennis после вылета I весеннего поколения непрерыв-
но увеличивается вплоть до конца сезона, когда размножение прекра-
щается. С прекращением выплода начинается зимнее падение численности,
продолжающееся до момента вылета I весеннего поколения; в период, пред-
шествующий вылету I поколения, имеется глубокий годовой минимум
численности.
Таким образом, для северного типа сезонного хода численности ха-
рактерно наличие одного годового максимума, приходящегося на конец
периода размножения, и одного годового минимума в период, предше-
ствующий вылету I поколения.
Календарный срок наступления годового максимума численности со-
впадает со сроком осеннего прекращения выплода (если только он не ви-
доизменяется под влиянием местных особенностей мест выплода), который
в свою очередь зависит от температуры. Поэтому чем дальше на юг, тем
позже наступает годовой максимум популяции.
В тех местностях, где имеется чистая популяция A. mac. messeae,
самки которого зимуют в холодных помещениях без добычи, непрерывного
осеннего увеличения числа комаров на дневках не бывает, а, по мере ухо-
да самок на зимовкуЛв местах летних дневок комары исчезают (рис. 175).
Осеннее падение численности комаров на дневках начинается и прогрес-
сирует вместе с осенним ожирением комаров; оно начинается уже с конца
июля в лесной полосе и не позже середины — конца августа в степной
полосе и продолжается в сентябре.
В популяциях A. mac. atroparvus, у которого места летних и зимних
убежищ совпадают (главным образом хлева), наблюдается рост числен-
ности комаров на дневках вплоть до прекращения выплода, т. е. до сен-
тября— октября (рис. 176). С окончанием выплода кривая численности
в силу начавшейся зимней смертности идет вниз. Таким образом, различия
в сезонном ходе численности между A. mac. messeae и A. mac. atroparvus
297
касаются не действительного хода численности популяции, который у этих
подвидов сходен, а лишь хода обилия комаров на дневках с добы-
чей; последний отражает действительный ход численности популяции
у A. mac. atroparvus и не полностью отражает его у A. mac.j messeae.
Рис. 175. Сезонный ход численности A. ппс.
messeie (средняя по торфоразработками Мос-
ковской области за 1942 г. пи иятидневкам).
Рис. 176. Сезонный ход числен-
ности A. mac. atroparvus в
окрестностях Амстердама.
1—сроднее количество комаров в
коровниках; 2—обилие личинок в
водоемах; 3— первое и последнее об-
наружение самцов.
наиболее жарких частях Средней
Месяцы
Рис. 177. Численность A. mac. sacharovi па
контрольных дневках (зимовках) Кокандского
района в 1942 г.
Южный тип хода численности. В
Азии и Закавказья годовой ход численности A. maculipennis отличается
от рассмотренного северного типа наличием двух минимумов — весен-
него и летнего — и двух максимумов — ранне-летнего и осеннего (рис. 177).
Мы видим увеличение чис-
ленности дневок в феврале и
марте, связанное с вылетом с
зимовок (как у messeae), вы-
мирание перезимовавших самок
в апреле и последующий рост
популяции. Однако рост этот
идет только до конца мая; в
июне, с наступлением жаркой
погоды (среднемесячная темпе-
ратура 25—26°), превышающей
температурный оптимум этого
вида, численность A. mac. sa-
charovi начинает резко падать,
снижаясь в июле (наиболее жар-
ком месяце, со средней темпе-
ратурой 28—29°) до летнего
минимума. В августе, с пони-
жением температуры, начи-
который продолжается вплоть до
Этот осенний подъем но достигает
как значительная часть выплажи-
нается новый подъем численности,
октября (средняя температура 13°).
величины весеннего максимума, так
ваютцихся в это время комаров уходит в продзимовочные и зимовочные
убежища, но содержащие добычи, и не учитывается при обе лед о в шии
.контрольных помещений. Поэтому и уменьшение численности A. mac.
298
sacharovi на дневках в осенне-зимние месяцы, показанное на кривой
рис. 177, не отражает действительного соотношения численности вида
в это время года.
Такой же характер сезонного хода численности наблюдается у
A. mac. sacharovi в Ташаузе (Туркмения — Правиков), у A. mac. macu-
lipennis в Араратской долине (Чубкова) и в Кара-Кала (Туркмения —
Мельникова).
Сезонный ход численности A. mac. sacharovi близ северной гра-
ницы его распространения. С другой стороны, сезонный ход численности
A. mac. sacharovi на северной гра-
нице его ареала дает явление весен-
ней затяжки в росте популяции,
связанное с недостатком тепла вес-
ной и в начале лета (рис. 178). Мы
видим, что численность здесь дер-
жится вплоть до середины лета на
очень низких цифрах, и только после
наступления высоких летних темпе-
ратур начинается бурный рост по-
пуляции.
3. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕ-
НИЕ A. MACULIPENNIS
И ЕГО ПОДВИДОВ
A. maculipennis распространен
очень широко. Область его распро-
странения (рис. 179) почти сплош-
ным кольцом охватывает умеренные
широты северного полушария. В со-
став ее входят Европа, Северная
Африка, Передняя, Средняя и Север-
ная Азия, часть Аляски, Канада,
западные штаты США и Мексикан-
ское нагорье.
Границы ареала. На севере есте-
ственной границей A. maculipennis
служит южный край тайги; действи-
тельно, в условиях сплошных лесов,
Рис. 178. Сезонный ход численности А.
mac. sacharovi в Кзыл-Орле и его окрест-
ностях (оригинал Маслова).
I— в 1941 г.; 2— в 1942 г.; 3— в 1943 г. На
оси ординат — среднее число комаров на
дневку за один о.‘лов.
при прохладном лете, отсутствуют
типы водоемов, пригодные для развития личинки, и в нетронутой тайге
этот вид отсутствует. Под влиянием заселения и раскорчевки тайги усло-
вия меняются, и в историческое время A. maculipennis, вместе с человеком
передвигаясь на север, занял значительную часть таежной зоны.
Здесь он местами уже подошел к абсолютной климатической границе
своего распространения, к той линии, севернее которой летнего тепла уже
недостаточно для развития двух поколений. Эта граница приблизительно
совпадает с изолинией 85 дней в году с температурой выше
Южная граница A. maculipennis в Северной Африке и Передней Азии,
несомненно, обусловлена чрезмерно высокими температурами и недоста-
точной влажностью летних месяцев. В силу .этих причин A. maculipennis
отсутствует и в ряде районов Средней Азии, например, в долинах Тед-
жена и Мургаба и вдоль среднего течения Аму-Дарьи. В Северной Аррике
он живет только к северу от Сахары.
299
Наименее понятной является восточная граница распространения
A. maculipennis в Азии. До Тихого океана он здесь нигде не доходит.
По Амуру его можно встретить только до Зейско-Буреинской низменности,
немного восточнее Благовещенска, и на северо-западе Маньчжурии.
В различных частях своего громадного ареала A. maculipennis подвер-
гается воздействию весьма различных климатических условий. Столь вы-
сокая приспособляемость к различным условиям отчасти зависит от того,
что в пределах вида A. maculipennis существуют подвиды, приспособлен-
ные к существованию в различных климатах.
Распространение A. mac. messeae. Из всех подвидов A. maculipennis
A. mac. messeae распространен наиболее широко. Он заселяет Северную
Азию, Северную и Среднюю Европу. Северная и восточная его границы со-
впадают с границами вида. На западе он доходит до Атлантического побе-
режья Франции и Англии. Южная его граница проходит на большом
Рис. 179. Ареал Anopheles maculipennis в целом.
Границы ареала нанесены пунктиром. Южная граница в Азии плохо прослежена.
протяжении по территории Китая и Монголии; в Казахстане и Северной
Киргизии messeae доходит на юг до северного склона Киргизского хребта.
Дальше его граница идет через центральный Казахстан на низовья Эмбы.
В нлжнем течении Сыр-Дарьи его, повидимому, нет. В Предкавказье
A. mac. messeae в изобилии встречается до Терека и Кубани. К югу от
этих рек он встречается редко и нигде не доходит до Главного хребта. От-
сутствует он в южной половине Балканского полуострова, в южной
половине Италии и в Испании.
Итак, в основном A. mac. messeae — форма северная, из всех подви-
дов A. maculipennis наилучше выносящая недостаток тепла и хуже всех
других переносящая летнюю жару и в особенности теплые зимы. Дей-
ствительно, A. mac. messeae неспособен зимовать в теплом помещении и
питаться кровью во время диапаузы.
В качестве личиночных биотопов A. mac. messeae придерживается
преимущественно стоячих, пресных водоемов низменностей.
Распространение A. mac. melanoon(=subalpinus). A. mac. melanoon
(=subalpinus) встречается на южных полуостровах Европы, всюду исклю-
чительно на низменности. У нас он встречается на влажных низменно-
стях Кавказа (в Абхазии, Кахетии, Кабарде, Дагестане), но всюду мало-
числен. В громадном количестве и местами чистыми популяциями населяет
он приморские низменности южного побережья Каспия: Ленкорань и иран-
ские провинции Гилян и Мазандаран. Личинки этого подвида живут глав-
ным образом в пресных, стоячих водоемах низменности.
300
Распространение A. mac. maculipennis. A. mac. maculipennis засе-
ляет Францию, Скандинавию, всю Среднюю Европу, Италию, Балканы,
Малую Азию, Кавказ, северный Иран, Европейскую часть СССР и Запад-
ную Сибирь. Через Иран проникает в южную Туркмению (Копет-Даг).
На востоке он неизвестен дальше Томска. A. mac. maculipennis в основном
форма горная. В горах Кавказа и Ирана он является единственной фор-
мой A. maculipennis. В Западной Европе, на Урале и в Сибири он также
приурочен к горам. В Восточной Европе обильнее всего на более возвышен-
ных местах: Валдайская, Приволжская, Волыно-Подольская возвышен-
ности. В Западно-Сибирской низменности крайне редок. Личинки живут
в пресных водах, предпочитая слабо текучую воду и каменистый субстрат.
Рис. 180. Места нахождения A. mac. sacharovi в СССР.
Распространение A. mac. atroparvus. A. mac. atroparvus обладает
сравнительно узким ареалом. Это форма, связанная с умеренным, атлан-
тическим климатом. Он заселяет Испанию, Северную Италию, северную
часть Балканского полуострова, Францию, Южную Англию, Южную
Швецию, всю Среднюю Европу. У нас он найден по Днестру, почти по все-
му течению Днепра, от устья до Орши, в Донбассе, на северном побережье
Черного моря, в Приазовье и дальше по Северному Кавказу до Адлера и
Гудермеса на юге и до Астрахани на востоке. A. mac. atroparvus бывает
многочислен только в местностях с солоноватыми водоемами и вне их
встречается лишь в качестве небольшой примеси к другим подвидам. Но
в Северной и Центральной Испании, где он является почти единственным
подвидом, он многочислен и в местностях с чисто пресными водами. Пови-
димому, он не столько солелюбив, сколько солевынослив.
Летние жары A. mac. atroparvus выносит хуже, чем A. mac. maculi-
pennis или A. mac. subalpinus, и потому не заходит в Южную Европу и
Закавказье. С другой стороны, он не выносит суровых зим, так как зимует
обязательно в активном состоянии, вблизи добычи. Этим обусловлена его
граница в СССР, совпадающая с изотермой января — 7°.
Распространение A. mac. sacharovi. A. mac. sacharovi населяет Сред-
нюю Азию, Иран, Турцию, Месопотамию, Сирию, Палестину, Балкан-
ский полуостров и заходит на Кавказ, в Италию, Корсику и Сардинию.
В Средней Азии по Аму-Дарье и Сыр-Дарье доходит до Аральского моря
(рис. 180). Он заходит в Китайский Туркестан (Кашгар), но его нет в
301
Семиречье. На Каспийском побережье Кавказа идет к северу до Махач-
кала; он населяет широкие речные долины Восточного Закавказья, но
отсутствует в горах и на влажной низменности Ленкорани. Нет его и
на Черноморском побережье Кавказа.
A. mac. sacharovi — наиболее теплолюбивая и тепловыпосливая из
наших форм A. maculipennis. В то же время он способен зимовать и в пас-
сивном состоянии и не боится суровых зим. Это форма жаркого конти-
нентального климата.
ГЛАВА X
ЭКОЛОГИЯ И РАСПРОСТРАНЕНИЕ ДРУГИХ ВИДОВ ANOPHELES
1. ANOPHELES BIFURCATUS (L.)
Географическое распространение. Широко распространенный вид, в ос-
новном европейский и западноазиатский. Встречается в Европе, Европей-
ской части СССР, Западной Сибири (до Томска), на Кавказе, в Средней
Азии, в Передней Азии (Северный Иран, Турция, Сирия, Палестина)
и в Северной Африке. Северная граница этого вида лежит несколько юж-
нее границы A. maculipennis. Южная граница его обусловлена средней
годовой температурой около 20°. Поэтому в Северной Африке он встре-
чается только в горах.
Биология личинок. Характерной чертой биологии личинок этого вида
является их холодолюбивость и приуроченность к жизни в узких темпе-
ратурных границах. Личинки A. bifurcatus живут в родниковых водоемах,
питаемых грунтовыми водами. Температура таких водоемов летом значи-
тельно ниже температуры воздуха и обычно ниже 20°. Оптимальной
температурой для существования личинок A. bifurcatus является 14—16°.
Температурные пределы развития: нижний 7—8°, верхний 21°. При
температуре выше 21° наступает торможение развития и высокая смерт-
ность. Продолжительность развития при оптимальной температуре, по
наблюдениям Мельниковой в Туркмении, 18—20 дней, по наблюдениям
Маркович в Москве—32—33 дня.
Личинки A. bifurcatus хорошо выносят затенение и встречаются
обычно в затененных (тростником, камышом, деревьями), мало прогревае-
мых водоемах, как при наличии, так и при отсутствии погруженной расти-
тельности. Они хорошо переносят загрязнение воды органическими ве-
несгвами растительного происхождения, а также большое количество
гуминовых веществ; азотистое загрязнение воды для них вредно. Для мест
ьып юда A. bifurcatus характерен низкий процент насыщения воды кислоро-
дом (часто около 50% в дневные часы).
В большом числе A. bfiurcatus встречается главным образом в горных
и предгорных местностях. Выплод его приурочен к местам выклинивания
грунтовых вод, родников, по склонам оврагов, по краю пойм рек, у под-
ножья гор, в зоне конусов выноса, где образуются заболоченности грун-
тового питания и замедленно текущие, заросшие родниковые речки. Ли-
чинки A. bifurcatus выплаживаются также в колодцах и ямах с грунтовой
водой,в Палестине развиваются в цистернах с питьевой водой. Этот вид
зимует в состоянии личиночной диапаузы (личинки III и IV возраста)
в тех же водоемах, где развивается летом. Зимующие личинки малопо-
движны и находятся или на поверхности воды, или, чаще, на дне. Ко вре-
мени прогревания воды весной диапауза заканчивается и личинки окукли-
ваются. На крайнем юге своей области распространения, в Палестине,
302
A. bifurcatus не имеет диапаузы, а продолжает размножаться и разви-
ваться круглый год.
Биология окрылеавого комара. A. bifurcatus — влаголюбивый вид.
Дневки его встречаются во влажных и прохладных местах, главным обра-
зом в растительности.
По способу охоты A. bifurcatus резко отличается от A. maculipennis:
он преимущественно охотится, подстерегая добычу вблизи мест выплода.
В поисках добычи не разлетается далеко от водоема; рассеяние от мест вы-
плода значительно меньше, чем у A. maculipennis. В постройках человека
встречается главным образом тогда, когда они расположены вблизи мост
выплода. При таком способе охоты и малой подвижности A. bifurcatus
обычно слабо связан с человеком, в силу чего и роль его в переносе малярии
невелика. Но там, где A. bifurcatus выплаживается вблизи жилищ чело-
века, он может оказаться переносчиком (например, на окраинах Баку,
где он выплаживался в колодцах «овданах» — Ахундов; в предгорьях
Восточного Тянь-Шаня и в предгорьях Кавказского хребта — на Черно-
морском побережье, где он очень многочислен).
Активность. Максимум активности у A. bifurcatus наблюдается в ве-
черние, ночные и ранние утренние часы при температуре 10—26°, чаще
при температуре 19—13°, и при относительной влажности 51—95%;
среди густой растительности он нападает и днем. A. bifurcatus охотнее
нападает на добычу на открытом воздухе. Различий в выборе пищи меж-
ду A. bifurcatus и A. maculipennis нет, и в отношении человека оба вида
ведут себя при укусах па открытом воздухе одинаково. Резко выраженное
у A. bifurcatus предпочтение к залету в хлева, а не в дома связано, неви-
димому, не с выбором добычи, а с более сильной реакцией на запах хлева,
чем у A. maculipennis.
Экзофилыюсть. Комары, напившиеся крови в домах и хлевах, поки-
дают их на заре, вылетая в природные убежища, где происходит перевари-
вание крови и созревание яичников. Таким образом, A. bifurcatus от-
носится к экзофильпым видам, т. е. к комарам, как правило, перевари-
вающим кровь не на дневках — мостах питания, а в открытой природе, тогда
как A. maculipennis является эндофилом — комаром,’ переваривающим
кровь в тех же помещениях, где насосался.
Экзофилия сама по себе не является признаком, снижающим эпиде-
миологическое значение вида. Известны многие виды Anopheles, например,
A. minimus и A. maculatus па юго-востоке Азии, которые также являются
экзофилами, но питаются в жилище человека и поэтому опасны как пере-
носчики малярии.
Все экзофильные виды комаров в отличие от эндофильных сохраняют
в сытом состоянии положительный фототаксис к сумеречному свету; по-
следний является для них стимулом к вылету из помещения в ранние
утренние часы. Так как насосавшиеся крови комары не задерживаются
в помещениях, то среди A. bifurcatus на дневках отсутствуют самки на
поздних стадиях пищеварения (4—7-я стадия Селла).
Особенности пищеварения. Т.ереварпвание крови и созревание яиц при 14—16°
закапчиваются в 5—б Д1 ей (Мельникова). Внешняя картина переваривания крови
У A. bifurcatus несколько отличается от картины, описанной Селла для A. maculipen-
nis: изменение тега крови более замедлено и неравномерно, на 3-й и 4-й стадии Селла
кровь остается еше яркокрасной, чернеет только задний край кровяною комка. Не-
равномерность ноюмнения крови, невидимому, связана с неравномерным распределе-
нием железистых клеток в эпителии желудка (Маркович).
Роение самцов происходит на открытом воздухе в вечерние часы.
Разновидности. В СССР имеется две разновидности A. bifurcatus:
автогенная и неавтогенная (Маркович).
303
У новорожденных самок автогенной разновидности развитие первой
порции яиц происходит без питания кровью, за счет запасных веществ,
накопившихся в теле личинки. На повторных гонотрофических циклах
способность к автогенному развитию яиц у этих комаров исчезает: вторая
и дальнейшие порции яиц развиваются только за счет питания кровью,
по правилу гонотрофической гармонии.
У неавтогенной разновидности развитие яиц (как первой порции, так
п последующих) происходит только при условии питания кровью. Авто-
генная разновидность A. bifurcatus широко распространена в СССР (Се-
верный Кавказ, Восточная Грузия, Московская область, Алма-Атинская
область, Туркмения), неавтогенная известна из Абхазии (Сухуми) и дру-
гих мест Черноморского побережья Кавказа.
Морфологические различия между обеими разновидностями незна-
чительны.
Джер-Курган, /940г.
1940г.
Сухуми. /937г
Первые
весенние ш/ары
Москва, 1946г
Первые
весенние ну нал ни
1 10 20 301 10 20 301 10 20 301 10 20 301 10 20 30
Февраль Март Апрель Май Июнь
Рис. 181. Сроки выплода I весеннего поколения A. bifurcatus.
Оригинал по материалам Чинаева, Устинова и Маркович.
Самки A. bifurcatus осенью не жиреют, в течение летнего сезона
они встречаются лишь с небольшим количеством жира; с наступлением
холодов комары вымирают.
Фенология. Окончагие дичпаузы и появление первых куколок весной njonc-
ходят в Узбекистане в начале февраля — марта, а в средней полосе Европейской
части СССР •— со второй декады мая.
Выплод комаров I поколения начинается в Узбекистане (по данным Чинаева)
со второй декады февраля, массовое появление их происходит во второй половине
марта, т. е. примерно па месяц раньше, чем у A. mac. sacharovi, и на 2 месяца
раньше, чем у Л. superpictus и A. pulcherrimus.
В средней полосе Европейской части СССР (Московская область данные
Маркович) выплод I поколения приходится на середи iy — конец мая (рис. 181).
Личинки I возраста II (летнего) поколения появляются в Узбекистане во
второй поло-ине марта, а в Московской области — в июле. Раз и из их продол-
жается в Узбекистане в среднем 20—25 дней, в Mockoi ской области — до 42 дней.
Выптод II генерации в окрестностях Москвы происходит с июля до конца ав-
густа. Со второй декады июля по август происходят массовые кладки комаров
II поколения и появление личинок I возраста, в августе и сентябре появляются
личинки III и IV возрастов, которые и переживают зиму.
Таким образом, в условиях лесной полосы Европейской части СССР имеется
одна летняя генерация и одна зимующая. Окрыленные комары A. bifurcatus
встречаются в Узбекистане в течение 5 месяцев.
Сезонный ход численности. На юге интенсивное размножение
A. bifurcatus происходит весной и осенью; первый максимум численности
обычен в мае — июне; в жаркие летние месяцы численность падает и
вновь нарастает лишь в сентябре — ноябре (рис. 182).
Такой ход численности объясняется тем, что весной и осенью, когда
температура воды в родниках низкая, площади выплода A. bifurcatus
велики; летом выплод происходит только в наиболее прохладных местах,
304
в небольшом количестве сохранившихся водоемов, так как значительная
часть родниковых вод в разгар лета пересыхает или перегревается. Рез-
кое снижение численности этого вида комара в разгар лета, т. е. в период,
благоприятный для переноса малярии, снижает его роль как переносчика.
Рис. 182. Сезонный ход численности A. bifurcatus
в колхозе Кармир-Дрошак Ад герского района в
1938 г. (подекадно).
По вертикали — среднее число комаров па одну дневку.
10~2Ь 20’30 1~10 10’20 20’30 1~10 10’20 20~301’10 10-20 20-30
VI VII VIII IX
Рис. 183. Сезонный ход численности A. bifurcatus в Московской j j
области в 1943 г. (оригинал Маркович).
В горных местностях, где много осадков и снега и где родники летом
менее прогреваются, снижение численности в разгар лета менее выражено
(Северный Кавказ, Тянь-Шань). В средней полосе СССР, где родни-
ковые водоемы летом не прогреваются выше 20° и площади выплода
в течение лета резко не изменяются, сезонный ход численности A. bifur-
catus дает один максимум (июльско-августовский, рис. 183).
20 Учебник меД'.гц шской энтомологии
305
Эпидемиологическое значение. В лаборатории A. bifurcatus хорошо
заражается всеми тремя обычными видами человеческих плазмодиев.
Однако в Европейской части Союза и в Сибири он малочислен и эпидемио-
логического значения не имеет. В предгорьях Кавказа и гор Средней Азии,
где он местами многочислен и выплаживается вблизи населенных пунк-
тов, он кое-где может играть роль переносчика малярии.
2. ANOPHELES ALGERIENSIS THEOBALD
Географическое распространение. A. algeriensis Theob. распространен в Среди-
земноморье (включая Алжир) и по Атлантическому побережью Западной Европы.
В СССР встречается на Кавказе и в Средней Азии. На Кавказе идет по Каспийскому
побережью к северу до рекиСулака, в западной части Кавказа — до Майкопа, в Сред-
ней Азии — до Алма-Аты. В горах отсутствует. Северная граница этого вида в Со-
ветском Союзе 43° с. ш., в Англии до 53° с. ш.
Личинки этого вида холод любивы, живут в больших тростникево-камышевых
болотах грунтового питания, температура воды в которых в летнее время не превы-
шает 18°, т. е. значительно ниже, чем температура воздуха и открытых водоемов, и
более постоянна. Продолжительность развития при температуре 10° — 23 дня (Ени-
колопов) .
Вода биотопов A. algeriensis обычно содержит большое количество органических
остатков растительного происхождения и гуминовые вещество. Личинки этого вида
тенелюбивее всех других вид в Anopheles нашей фауны, но в самых густых зарослях
гигантолинеид все же не встречаются. Зимуют в состоянии личиночной диапаузы
(IV возраст) в н?замерзающих водоемах (ключах).
A. algeriensis требует для своего существ ования длительного теплого периода:
не менее 190 дней в году с температурой, равной 10° или выше. Такие условия A. al-
geriensis находит в СССР на Кавказе и в Средней Азии. Только в условиях климата
этих местностей возможно развитие двух поколений этого вида, медленно развиваю-
щегося в воде с низк >й температурой.
Биология имаго. Anopbeks algeriensis чрезвычайно влаголюбив. Благодаря круп-
ным отверстиям стигм на груди и брюшке, этот комар, попадая в сухой воздух, теряет
путем испарения очень много в )ды и быстро гибнет (Виноградская). Поэтому он
всегда живет в условиях высок й влажности (чаще в камышевых зарослях). В по-
исках добычи далеко от мест выплода не улетает. Питается главным образом на ди-
ких животных (кабанах, птицах, изобилующих в зарослях); с человек )м связан мало;
в постройки человек 1 залетает лишь тогда, к )гда они расп л )жены в щп срсдстванп й
близости от мест выплода. Нападает A. algeriensis при условии большой влажно-
сти в >здуха (80—83%) и в пределах температур 8—19° (Еник л п. в).
Эпидемиологическое значение. В силу указанных ос бшн стей биологии
A. algeriensis (питание на диких животных, б >льшая влаголюбив сть), а также срав-
нительной рщкости, этот вид у нас в СССР не является переносчиком малярии и
борьба с ним не ведется. В усл виях лаб 'ратории A. alg riensis способен заражаться
Pl. maliriae и в очшь сл )бой степ ши Pl. fdciparum и Pl. vivix.
3 и 4. ANOPHELES L1NDESAVI GILES и A. MARTERI SENEVET
Распространение. Ареал р icnp >странения A. lind -sayi прныз р ш шы Индии,
Индокитая, Филиппин, Ффм^зы, Яп )нии, Китая. В СССР найден в Таджикистане
(центральная и северная части) и па Западгь м Памир , в Ванче (Кешишьяи).
Ареал A. mart ri: горные районы Средиз< мш морья (Северная Африка, Испания,
Греция, Сирия), а также Св рный Иран. В СССР сн представлен подвидом A. mart,
sogdianus (Таджикистан — Кешишьяи).
Места выплода. В Таджикистане личинки A. lindesayi встречаются на высоте
300—1 900 м, a A. marteri — на высоте 900—1 500 м.
Лет )м личинки об шх вид в встречаются в холодных, бурных потоках, текущих
с крутых склонов гор. Русло поток )в каменистое, лишенное растительности. Темпе-
ратура воды не выше 11—15°. Вода чистая, прозрачная, мягкая, слабо минерали-
зованная, относительно б шатая кальцием (Глаголева).
Личинки выплаживаются в излучинах потока, в мелких прибрежных скопле-
ниях вады, хорошо затененных прибрежными скалами и растительностью. Течение
в этих местах замедленное, но все же заметное. На большем течении селятся A. marteri,
в более защищенных местах — A. lindesayi.
Зимовка. A. lindesayi зимует в стадии личинки. Зимующие личинки сносятся
осенними паводками в расположенные ниже по течению реки прибрежные мелкие за-
306
болоченпости, где и переживают зиму. В Греции Л. marteri зимует во взрослом со
стоянии. Комары A. marteri и A. lindesayi очень влаголюбивы. Оба вида живут в ди-
кой природе, вдали от человека. Эпидемиологического значения в СССР не имеют.
В условиях лаборатории A. lindesayi japonicus способен заражаться плазмодиями
малярии. Коидзуми считает этого комара переносчиком терцианыи квартаны на Фор-
мозе, а Фенг — в горной части Северного Китая.
5. ANOPHELES PLUMBEUS THEOBALD
Места выплода. Это единственный в Европе вид Anopheles, выплажи-
вающийся в дуплах деревьев (бука, ясеня, граба и т. п.). Он заселяет
лиственные леса влажного и умеренно теплого климата, в условиях кото-
рого дупляные водоемы могут существовать более или менее длительное
время. В дуплах создается своеобразный биотоп: полное отсутствие вол-
нобоя, небольшая глубина, темнота, своеобразный химизм. Вода дупел бо-
гата органическими веществами, частью питательными для личинок; ко-
личество их может составлять до 8 303 мг на 1 л (Рейнгард и Забудько),
вода приобретает темный, коричневый цвет. Количество личинок в одном
дупле может быть очень велико. Здесь же встречаются личинки других
комаров: Аё. geniculatus, Аё. pulchritarsis, Orthopodomyia pulchripalpis.
Температура воды в дуплах постоянна и по сравнению с летними тем-
пературами воздуха и открытых водоемов низка (не выше 20°), но выше,
чем в родниковых водоемах. В воде, температура которой значительно
превосходит 20°, личинки неспособны нормально развиваться. Развитие
личинок A. plumbeus идет медленно: так, при 20° продолжительность раз-
вития равна 25 дням, т. е. больше, чем у A. maculipennis (22 дня). При
температуре 8—12° развитие идет, по чрезвычайно медленно. У личинок
A. plumbeus резко выражен отрицательный фототаксис, им свойственно
прятаться в темноту.
Водный режим в дуплах непостоянен: дупло то заполняется дожде-
вой водой, то пересыхает. В полупересохших дуплах личинки могут жить
во влажном илу на дне дупла в течение нескольких недель, яйца — не-
сколько месяцев. В период высыхания дупел самки A. plumbeus отклады-
вают яйца в колодцы, в выбоины скалистого русла ручьев, в цистерны и
другие искусственные вместилища с твердыми стенками, но не кладут
яиц в водоемы с землистыми берегами.
Зимуют личинки (всех стадий), а там, где дупла промерзают, зимуют
яйца. При недолгом замерзании воды в дуплах часть личинок выживает.
Длительное промерзание дупел мешает распространению этого вида на
север.
Географическое распространение. В мягком климате Западной Европы
A. plumbeus встречается в Англии, Южной Швеции, Германии. В более
континентальном климате СССР северная граница его проходит значи-
тельноюжнее: через Киев, Полтаву, Харьков, Ростов-на-Дону, Краснодар,
Пятигорск и, далее, по Тереку. На Кавказе A. plumbeus встречается
всюду в горах. Через влажные районы Ленкорани и Северного Ирана
проникает в облесенные горные ущелья южной Туркмении (Копет-Даг).
В облесенных горах южного Таджикистана живет североиндийский подвид
этого вида — A. pl. barianensis (Кешишьян).
Биология окрыленного комара. A. plumbeus крайне влаголюбив, он
плохо переносит сухость воздуха. Поэтому он выбирает влажные убе-
жища: дупла деревьев, углубления в скалах и на склонах оврагов, це-
ментированные колодцы и ямы, густую растительность. При поисках убе-
жищ его привлекает те вое отверстие входа в убежище и большая влаж-
ность воздуха. A. plumbeus нападает на добычу как на открытом воздухе,
20*
307
Рис. 184. Сезопный ход численности A. plumbeus
в Псырцха (Новый Афон) в 1938 г.
1—число комаров на один облов; 2—количество
осадков в миллиметрах.
так и в помещениях. Напившись крови в домах и хлевах, комары по-
кидают их на заре, вылетая в открытую природу, где и заканчивают пище-
варение. Таким образом, A. plumbeus является экзофильным комаром.
Подобно A. maculipennis, A. plumbeus способен концентрироваться
в местах скопления крупной добычи: дневки его в природе обычно бывают
приурочены к периферии населенных пунктов или к посещаемым людьми
и скотом лесным дорогам. В глубине леса, вдали от животных и человека,
взрослых самок A. plumbeus значительно меньше (Шленова).
Роение самцов совершается на открытом воздухе незадолго до наступ
ления сумерек, но может происходить и днем в тени деревьев. Описано
роение самцов и копуляция в лаборатории, в садке (размер садка
50 X 50 X 50 см). Это указывает па возможность разведения этого вида в
лаборатории, подобно A. superpictus и A. m. atroparvus.
Фенология. Окрыление
комаров из перезимовавших
личинок на Новом Афоне
(Черноморское побережье
Кавказа) началось в конце
апреля — начале мая 1937 г.;
вылет комаров I летней гене-
рации произошел в начале
июня (Шлёлова).
Существует тесная зави-
симость между ходом чис-
ленности окрыленного кома-
ра A. plumbeus и количест-
вом осадков за период, пред-
шествовавший окрылению.
Чем больше осадков выпа-
дает в летние месяцы, тем
больше воды в дуплах, тем
больше выплаживается там
комаров (рис. 184). Поэтому
только в дождливое лето
бывает много A. plumbeus.
Эпидемиологическое значение. В специфических условиях курортов
Черноморского побережья Кавказа, где скота, отвлекающего комаров
от человека, мало, а людей, гуляющих по вечерам в парках, много, A. plum-
beus может являться переносчиком малярии. Это подтверждается: 1) наход-
ками зараженных малярийными плазмодиями комаров в природе и 2) дан-
ными реакции преципитации, показывающими большой процент самок
(свыше 10%), питающихся кровью человека. Однако многочислен этот вид
только в дождливые годы, и только в такие годы он может играть сущест-
венную роль в переносе малярии (Шлёнэва).
Борьба с личиьками. Борьба с личинками этого комара сводится к плом-
бировке дупел цементом (Павловский и Месс) или засыпке их пескам и
землей; работы эти удобнее проводить зимой, когда малярийная станция
свободна от других противоличиночных работ.
6. ANOPHELES HYRCANUS PALL.
Географическое распространение. A. hyrcanus обладает чрезвычайно
широким ареалом распространения. Он заселяет Азию (на север ди 50°
с. ш.) и Южную Европу. Центр обилия этого вида находится в Китае,
308
Индокитае и па Зондских островах. В СССР он встречается па Дальнем
Востоке (в Уссурийском крае п по всему течению Амура), на Алтае
(в окрестностях озера Зайсап), в устьях рек Урала и Волги, на юге Укра-
ины, в Молдавии, на Кавказе и в Средней Азии.
Экология личинок. Личинки A. hyrcanus живут в более узких темпера-
турных пределах, чем A. maculipennis. Температурный оптимум личинок
и имаго около 25—30°, т. о. совпадает с оптимумом личинок A. maculi-
pennis, но температурные амплитуды обоих видов различны: A. hyrcanus
не переносит таких низких и высоких температур, при которых
может существовать A. maculipennis. Так, нижний температурный пре-
дел развития у A. hyrcanus значительно выше, чем у A. maculipen-
nis (около 12°), а верхний — ниже (около 35°). Это ведет к тому,
что па Украине, в Приамурье и в Приморье, т. е. вблизи северной
границы своего распространения, A. hyrcanus заселяет наиболее прогре-
ваемые, освещенные солнцем водоемы, заросли погруженных растений
(элодеиды, нитчатки), избегая тени и стремясь максимально исполь-
зовать солнечное тепло. Так как температурные требования у A. hyr-
canus выше, чем у A. maculipennis, то весной на Дальнем Востоке он раз-
вивается медленнее и появляется в значительном количестве позднее
(только в июле). Эта весенняя затяжка в развитии ослабляет его эпидемио-
логическую роль па Дальнем Востоке. Вся весенняя волна рецидивов
малярии проходит здесь в отсутствие переносчика.
Наоборот, в более теплом климате Средней Азии и Кавказа, в условиях
температурного оптимума вида, A. hyrcanus только ранней весной заселяет
открытые водоемы, летом же встречается в сильно затененных водоемах,
не слишком прогреваемых (изотермичных), в зарослях линеид. Заселяет
болота не грунтового, а поверхностного питания: поймы рек, дельты, плав-
ни, рисовые поля (те их участки, где вода летом не слишкохМ прогревается).
В Средней Азии A. hyrcanus не дает весенней затяжки в развитии.
В субтропиках (Северная Индия, север Индокитая) A. hyrcanus
развивается в прохладное время года, в тропиках встречается только
при мягком океаническом климате (Ява, Суматра).
Биология окрыленных комаров. A. hyrcanus влаголюбивый вид, но
более устойчивый к сухости, чем A. bifurcatus и A. algeriensis. Дневки его
встречаются в относительно влажных и прохладных местах. В теплом и
влажном климате юго-восточной Азии A. hyrcanus находит благоприятные
условия микроклимата в разнообразных убежищах: в постройках человека,
в растительности. В жаркохм и сухохм климате Средней Азии встречается
па дневках почти исключительно среди растительности, микроклимати-
ческие условия в которой наиболее удовлетворяют его требованиям.
Убежища. В растительности A. hyrcanus обычно располагаются на
нижней поверхности листьев, невысоко над землей. Здесь, вследствие
испарения воды с поверхности листьев, воздух бывает несколько влажнее
и прохладнее, чем снаружи. Так, например, в Мервском оазисе (Туркме-
ния) внутри кочки больших злаков, где находилась дневка A. hyrcanus,
температура днем равнялась 35° и относительная влажность 35%, тогда
как снаружи куста температура была 44°, а влажность 15%. Характер
распределения A. hyrcanus и A. pulcherrimus в растительности показывает
прямо противоположное отношение этих двух видов к влажности и тем-
пературе воздуха: A. hyrcanus держится в наиболее прохладных и влаж-
ных участках куста, a A. pulcherrimus, наоборот, в более прогреваемых
п сухих (Митрофанова) (рис. 185 и 186).
В поведении A. hyrcanus зрение играет большую роль. Так, днем
A. hyrcanus наиболее охотно нападает на добычу темного цвета, а вече-
309
ром — на добычу белого цвета. Люди, одетые в черную одежду, вечером
подвергаются менее интенсивному нападению, чем люди в белых одеждах.
Ночью A. hyrcanus стремится залетать в освещенные окна. Этот комар
не боится нападать при ярком дневном свете (если температура позво-
ляет это).
Способ охоты» По способу охоты A. hyrcanus сходен с комарами рода
Аёйев. Он питается за счет равномерно распределенной по местности
и большей частью мелкой добычи (мелкие млекопитающие, птицы и т. п.)
и в гораздо меньшей мере привлегается отдаленными скоплениями круп-
ных стадных млекопитающих, хотя и активно нападает на них при слу-
чайной встрече.
Рис. 186. Встречаемость A. hyrca-
nus и A. pulcherrimus на дневках в
растительности в зависимости от
влажности.
Рис. 185. Встречаемость A. hyrca-
nus и A. pulcherrimus на дневках
в растительности в зависимости
от температуры.
В силу этого A. hyrcanus заселяет местность более или менее равно-
мерно, встречаясь п в поселках, и в растительности. Но часть популяции
находящаяся в поселке, незначительна по сравнению с топ, которая живет
в растительности. Больше всего A. hyrcanus нападает около водоемов,
в тростниках. В растительности можно найти самок этого вида на различ-
ных стадиях гонотрофического цикла, а также самцов. Вышеописанный
способ охоты за добычей снижает эпидемиологическую роль A. hyrcanus.
A. hyrcanus ведет себя, как экзофил ытый вид: если он и залетает в по-
мещения человека и животных для питания кровью, то, напившись,
под утро улетает. Дневки A. hyrcanus в домах находят в тех местностях,
где его численность очень велика; здесь, среди большого числа залетаю-
щих комаров, шансы на то, что часть из них задержится в помещении,
повышаются.
Зшмовка. Зимуют A. hyrcanus в окрыленном состоянии (оплодотворен-
ные, диапаузирующие. жирные самки). Никогда не зимуют в постройках,
а только в природе, среди растительности: в тростниках, в кустарнике и т. п.
Условия зимовки в различных местах ареала различны. На Дальнем
Востоке комары переносят суровую зиму, когда температура может опу-
скаться до —40°, в полной пассивности. В Туркмении и в Дагестане, где
зима мягкая, A. hyrcanus, находясь в диапаузе, зимуют в активном со-
стоянии и в периоды потепления пьют кровь (Правиков, Ениколопов).
310
Фенология. Питание кровью у A. hyrcanus во Владивостоке (1941—1945) начи-
нается с конца второй декады апреля, в Хабаровске и Благовещенске — с конца треть-
ей декады апреля при среднедекадной температуре 3—4°; массовое кровососание и
начало развития яичников наблюдаются со второй декады мая при среднедекадной
температуре 9,8° (Хабаровск, 1936) 1.
В Узбекистане массовое кровососание отмечается с 12 февраля— 10 марта (Тер-
мез, Ташкент, 1938—1944), а начало развития яичников — с середины или с конца
февраля.
Сроки выплода I весенней генерации A. hyrcanus в СССР показаны на рис. 187.
Из рисунка видно, что вылет I генерации в южной части Уссурийского края проис-
ходит примерно на 1—2 декады раньше, чем в районе Благовещенска и Хабаровска,
но на месяц позже, чем в Ташкенте, и на 2 месяца позже, чем в Термезе.
Развитие I генерации в Узбекистане происходит при средней температуре воды
И—18,7° и продолжается 30—40 дней; вылет самцов происходит при средней темпе-
ратуре 16°.
Лаоало окрыления
бла?овещенск/934г.
Владивосток, /943г.
Термез,/939г Таи/кент,/939г Хабаровск
Первые личинки
//возраста
/936г
Первые личинки
/возраста
Главки
Массовый в/ лег
с зимовок ю 20 зо w 2Q 3Q 20 1(J 20 3Q
Февраль Март Апрель Май Июнь
Рис. 187. Сроки выплода I весеннего поколения A. hyrcanus
в СССР (оригинал по материалам Маслова и Чинаева).
A. hyrcanus имеет в Благовещенске и Хабаровске две полных и одну неполную
(частичный выплод) генерации и в южной части Уссурийского края три полных гене-
рации. Число генераций для Средней Азии не установлено.
Массовое ожирение A. hyrcanus на Дальнем Востоке начинается с третьей де-
кады августа (Благовещенск, 1937) — первой декады сентября (Уссурийский край).
В Средней Азии ожирение происходит позднее, чем па Дальнем Востоке и чем у
A. mac. sacharovi. Так. в Ташкенте ожирение A. hyrcanus закапчивается только к концу
октября — началу ноября (в те же сроки, что и у A. superpictus), тогда как у A. mac.
sacharovi уже к началу сентября жирные самки составляют большинство популяции
(Ходукин, наблюдения 1925—1926 гг.). В Байрам-Али (Туркмения) Правиков кон-
статировал появление первых диаи аудирующих жирных самок (IV стадии Клиглера)
в 1936 1. с 23 сентября, а массовое ожирение — в октябре.
Массовый уход на зимовку происходит в Хабаровске с первой декады сентября
(1936), на кие Уссурийского края — со второй декады сентября (1933), в Средней
Азии—с октября.
Кровососание и развитие яичников у летней популяции A. hyrcanus заканчива-
ются в районе Благовещенска к концу сентября, а в Уссурийском крае и в Средней
Азии — в первой декаде октября; последние личинки, куколки и самцы исчезают
к 20 октября.
Период нормальной половой активности A. hyrcanus в Средней Азии 6—7 меся-
цев, в Уссурийском крае — около 5 месяцев.
Сезонный ход численности. Ход численности A. hyrcanus на Дальнем Востоке
показан на рис. 188. В районе Хабаровска численность A. hyrcanus увеличивается со
второй декады июля (с началом вылета II генерации), достигает максимума к третьей
декаде августа, после че! о резко снижается. В бассейне реки Бикин (Уссурийский край)
численность нарастает с первой декады июля (с вылетом II 1енерапии), достигает ма-
ксимума уже к концу июля и держится на высоком уровне до второй декады сентября,
1 Данные по фенологии A. hyrcanus взяты из следующих источников.- для Узбе-
кистана — из сводки Чинаева (1944), по Дальнему Востоку — из материалов Маслова
и малярийной станции Приморского края (энтомологи Кондратьев и Нельзина).
311
после чего быстро снижается. Таким образом, период высокой численности переносчика
в районе Благовещенска и Хабаровск! составляет не бэлеэ 11/2 месяцев и только
на юге Уссурийской области удлиняется д> 2—21/2 месяцев.
Эпидемиологическое значение. Роль A. hyrcanus в переносе малярии
в СССР хотя и несомненна, но имеет второстепенное значение. В экспери-
менте он заражается терцианой хуже, чем A. maculipennis, а тропикой —
совсем плохо. В природе зараженные плазмодиями особи A. hyrcanus
встречаются, но редко (Днепропетровск, 1939, — Горицкая и др.).
Его незначительная эпидемиологическая роль зависит не от экзофи-
лии, а от способа охоты. В тех частях советского Дальнего Востока, где
A. hyrcanus является единственным представителем рода Anopheles, он
«не может поддерживать малярийной эндемии. Как уже указывалось, при-
Рис. 188. Сезонный ход численности A. hyr-
canus па Дальнем Востоке (оригинал по ма-
териалам Маслова).
I—D окрестностях Хабартска н 1936 г.; 2—в
бассейне р. Бишш о 1933 г. .
чиной, ослабляющей эпидемио-
логическую роль A. hyrcanus
па Дальнем Востоке, является
весенняя затяжка в развитии,,
обусловленная недостатком теп-
ла в начале лета, вследствие
чего вся весенняя волна реци-
дивов малярии проходит здесь
в отсутствие переносчика. Кро-
ме того, в период максималь-
ной численности комаров (в ав-
густе и сентябре) температура
воздуха часто оказывается не-
достаточной для развития ма-
лярийного плазмодия, что еще
больше укорачивает эпидеми-
ческий сезен.
В пределах советского Дальнего Востока, к востоку от Благовещен-
ска, случаи малярии местного происхождения возникают лишь при усло-
вии массового завоза паразитоносителей извне (Кехчер). Поэтому веду-
щим методом борьбы с малярией на Дальнем Востоке является борьба
с возбудителем — быстрое вылечивание больных. Южнее, в равнинных
частях Северного Китая, A. hyrcanus, будучи также единственным видом
Anopheles, способен все же поддерживать эндемию малярии, что объяс-
няется значительно более теплым климатом.
Эпидемиологическое значение A. hyrcanus в плавнях реки Кубани,
изучавшееся Ростовским-на-Дону институтом малярии (1938), оказалось
незначительным. Это определило пути борьбы с малярией в кубанских
плавнях, где все внимание должно быть обращено на уничтожение A. ma-
culipennis.
Однако общего подхода к оценке этого вида в качестве переносчика
быть не может. Как правило, A. hyrcanus является в СССР малой эпидемио-
логической величиной. Но в отдельных случаях с ним необходимо считать-
ся. Так, в Мер веком оазисе (Туркмения) в 1936 г., когда A. pulcherrimus,
являющийся здесь основным переносчиком малярии, почти отсутствовал,
а имелся только A. hyrcanus, свежие заболевания малярией не прекра-
тились.
7. ANOPHELES SUPERPICTUS GRASSI
Географическое распространение. Распространен в Средиземноморье,
Передней и Средней Азии. В СССР встречается на Кавказе (по Каспий-
скому побережью на север до Махач-Кала и по Черноморскому побе-
212
режью до Сочи) и в Средней Азии (па север до Киргизского хребта), пре-
имущественно в предгорных и горных районах. Далеко вглубь равнины
он идет только по течению Аму-Дарьи, достигая Хорезма.
Экология личинок. Личинки A. superpictus теплолюбивы. Оптималь-
ные среднесуточные температуры лежат около 4-30°, дневные максимумы
температуры воды в биотопах A. superpictus обычно много выше:
+35—38°.
Средняя продолжительность развития при среднесуточной темпера-
туре воды 30° составляет 11 дней, 25°—14 дней (Улитчева) и 19,6° —
24 дня (Гасанов).
Другой характерной чертой личиночной экологии A. superpictus
является потребность в большом содержании растворенного кальция.
Эта потребность привязывает его к хорошо прогретым грунтовым водам
(которые, как правило, богаты кальцием).
Благоприятные условия температуры и химизма личинки A. super-
pictus находят в мелководных водоемах, образующихся в руслах горных
рек. Характерные биотопы A. superpictus — галечниковые побережья
горных рек, т. е. места отложения грубых наносов, среди которых река
течет многочисленными рукавами; здесь, в местах замедленного течения,
хорошо прогреваемых солнцем, обычно среди нитчаток (присутствие
которых, однако, необязательно), личинки A. superpictus встречаются
в большом количестве.
В биотопах A. superpictus никогда не встречаются элодеиды или гу-
стая надводная растительность: каменистое ложе галечниковых русел
препятствует развитию высшей растительности.
Личинки A. superpictus — реофилы, т. е. они предпочитают проточ-
ную воду, но они могут жить и в стоячей воде, если вода удовлетворяет их
условиям температуры и химизма (рисовые поля, орошаемые родпиково;
речной водой, копанки с грунтовым питанием).
Эта приуроченность мест выплода A. superpictus к текучим водам и
обусловливает распространение этого вида в условиях средиземномор-
ского климата, для которого характерно сухое, жаркое лето и мягкая,
обильная осадками зима. Обилие летних осадков делает выплод A. super-
pictus невозможным, так как личинки его погибли бы во время паводков,
горных рек. Отчасти по этой же причине A. superpictus предпочитают речки
не ледникового, а родникового питания, имеющие более устойчивый
режим уровня.
В СССР наиболее благоприятные климатические условия для разви-
тия A. superpictus существуют в Средней Азии и в восточной части Кав-
каза, где лето жаркое и сухое. На северном склоне Кавказа A. superpictus
изредка встречается только в пределах Дагестана; его распространению
здесь препятствует дождливое лето, а не недостаток тепла. На Кавказском
побережье Черного моря он редок из-за большого количества выпадающих
в летнее время осадков.
Места выплода A. superpictus встречаются в СССР до высоты 2 800 м
над уровнем моря (Западный Памир).
Экология окрыленного комара. A. superpictus теплолюбивее и сухо-
устойчивее всех видов подрода Anopheles, распространенных в СССР.
Дневки его встречаются в теплых и сухих местах: так, например, его мож-
но найти в туркменских войлочных юртах, где температура летом подни-
мается до 35°, а влажность спускается до 15% и где A. maculipennis но
мог бы существовать. Правда, на дневках с большим дефицитом влаги
наблюдается большая текучесть комаров, вплоть до ежедневной полной
смены всего состава; в этих случаям стимулом для вылета является жажда,
313
возникающая в результате большой потери воды путем испарения (Бек-
лемишев, Митрофанова и Детинова).
Эпидемиологическое значение теплолюбия A. superpictus заключается
в том, что на дневках он выбирает более теплые места, нежели A. macu-
lipennis, в результате чего скорее переваривает, чаще сосет кровь, в нем
быстрее развиваются спорозоиты и он оказывается более интенсивно функ-
ционирующим переносчиком.
Большая сухоустойчивость A. superpictus обусловлена наличием у него
приспособлений для уменьшения потери воды через испарение (малень-
кие грудные стигмы и небольшие выходные отверстия брюшных стигм).
Большой влажности воздуха A. superpictus избегает (Виноградская).
Этот вид очень подвижен и активен. Нападает A. superpictus при
значительно большей жаре и сухости, нежели A. maculipennis.
Подобно A. maculipennis, он охотитсй за крупной стадной добычей,
привлекается скоплениями этой добычи в поселки, где и концентрируется.
Вне поселков встречается только тогда, когда скопления добычи имеют-
ся в открытой природе. Нападает на добычу как на открытом воздухе, так
я в помещениях. Таким образом, A. superpictus тесно связан с человеком
и в силу этого контакта с человеком является опасным переносчиком
малярии.
A. superpictus — эндофильный вид; напившись крови в помещении,
он там же остается переваривать пищу; напившись крови на дворе, он
обыкновенно залетает переваривать кровь в помещение.
A. superpictus еще больше, чем A. maculipennis, избегает яркого
света и на дневках прячется в темные места.
Самцы A. superpictus, подобно самцам A. mac. atroparvus, следуют
за самками в места их питания — дома и хлева, где встречаются в боль-
шом количестве (до 40% от общего числа комаров). Это связано с тем, что
копуляция у этого вида может происходить не только на открытом воз-
духе, но п в помещении. Вейлу этого A. superpictus хорошо размножается
в лаборатории.
Переваривание крови и развитие яичников в условиях высокой тем-
пературы дневок летом протекают очень быстро (в течение около двух
суток).
Со второй декады августа и в сентябре появляются первые жиреющие
и диапаузпрующие (с заторможенными яичниками) самки, продолжаю-
щие пить кровь, которая идет на образование жира. Таким самкам нет
надобности вылетать из помещений для откладки яиц, и они повторно пьют
кровь в одном п том же помещении. Эти особенности осенней биологии
A. superpictus имеют эпидемиологическое значение: осеннее ожирение са-
мок за счет питания кровью удлиняет сезон возможного переноса ими ма-
лярии, а фиксация их в убежищах и прекращение круговорота на дневках,
связанные с прекращением размножения, создают возможность возник-
новения осенних внутридомовых очагов малярии (при наличии гамето-
носителя).
У жирных самок, пьющих кровь, -желудок не может сильно растяги-
ваться, и поэтому они сосут кровь чаще, чем летом. В связи с этим морфо-
логическая картина пищеварения у жиреющих A. superpictus несколько
иная, чем описанная Селла для A. maculipennis (рис. 189) (Виноградская).
Зимовка. Зимуют оплодотворенные самки в состоянии имагинальной
диапаузы обязательно вблизи добычи, обычно в хлевах, реже в домах, в
надворных постройках, в дикой природе (например, в пещерах). Всю зиму
они пьют кровь, прерывая питание лишь в холодное время. Диапауза
у A. superpictus длится дольше, чем у A. maculipennis в той же местности;
314
весной в Узбекистане она кончается на месяц позже, чем у A. mac.
sacharovi (Исаев).
Фенология. Развитие яичников у A. superpictus начинается в Узбекистане,
с конца февраля или в марте при среднедекадной температуре убен ищ 10,5° и длится
в среднем около 10 дней. Первая порция яиц созревает к середине марта — началу
апреля. Первые вылеты с зимовок в северной части Узбекистана (Ленинске Н.Ургенче)
происходят между 7 и 15 апреля, а в южной части (Термез)— на 3 недели раньше.
Массовый вылет с зимовок в Узбекистане происходит обычно в первой-третьей декадах
апреля при среднедекадной температуре воздуха 14°.
Рис. 189. Схема переваривания крови при развитии жирового слоя у
A. superpictus осенью.
Стадии Селла 2—6; 2— красная кровь в 1—6-м сегментах; 3— красноватая
кровь в 1—5-м сегментах; 4—темнокрасная кровь в 1—4-м сегментах; 5 —
коричневая кровь во 2—4-м сегментах; 6 — черная кр« вь в 3—4-м сегментах.
Даты развития I генерации к Узбекистане и на Кавказе показаны на рис. 190.
Из рисунка видно, что вылет I генерации A. superpictus в северном Узбекистане (Ленин-
ске) происходит во второй половине мая, т. е. примерно на Р/а— 2 декады позднее,
чем в южном (Термез), и на 3—4 недели позже, чем у A. maculipennis.
Развитие личинок и куколок в водоемах происходит при средней температуре
воды от 16° до 21°, вылет I генерации — при среднедекадной температуре 22,6°
Рис. 190. Сроки выплода I весеннего поколения A. superpictus в Средней
Азии и на Кавказе (оригинал по материалам Чинаева и Гасанова).
Развитие водных стадий I весенней генерации длится 20—30 дней.
Первые перезимова! шие самки с гонотро(]ич( сксй гармоние й появляются
в Узбекистане в апреле и мае при среднесуточной температуре 21°. Массовое ожиро-
ние самок происходит в октябре, т. е. на 1—Р/2 месяца позже, чем у A. mac. sacbarovi.
Период нормальной половой активности у A. superpictus составляет 4—4>/2 месяца.
Последние личинки, куколки и самцы исчезают в конце октября —начале ноября.
В Дагестане (Ирганай) A. superpictus имеет три генерации (Гасанов).
Сезонный ход численности. На севере Средней Азии и на Кавказе,
т. е. у северной границы распространения A. superpictus, наблюдается
весенняя затяжка роста популяции; в мае и июне вид малочислен, что
315
связано с недостатком тепла в эти месяцы, а также с промыванпем мест
выплода — русел речек — весенними паводками. В связи с поздним окон-
чанием диапаузы и весенней затяжкой в развитии этого теплолюбивого
вида, годовой минимум чис-
ленности популяции наблю-
дается здесь в мае, т. е. почте
па месяц позднее, чем у A. ma-
culipennis в тех же местах.
В таких местностях A. super-
pictus появляется в значитель-
ном количестве лишь с июля,
с наступлением высоких летних
температур, являясь, таким
образом, позднелетним видом.
Максимум численности A. su-
perpictus наблюдается в ноябре,
когда комары залетают из
природных убежищ в более
теплые хлева с добычей, где и
концентрируются.
Таким образом, годовой
Рис. 191. Численность комаров Anopheles па
контрольных дневках (зимовках) Кокандско-
го района в 1942—1943 гг.
1— A. superpictus; 2— Л. pulcherrimus в 1942 г.;
3—A. pulcherrimus в 1943 г.
ход численности A. superpictus
на севере Средней Азии и па Кавказе имеет один поздне-весенний мини-
мум и один осенне-зимний максимум (рис. 191). На крайнем юге Средней
Азии (например, в Кара-Кала) весенняя затяжка в развитии A. superpic-
tus менее выражена; увеличение численности начинается уже со второй
половины мая (рис. 192).
Рис. 192. Сезонный ход численности A. superpictus в
Кара-Кала (Туркмения) (по материалам 1930—1931 гг.
П. А. Петрищевой).
Эпидемиологическое значение. По эпидемиологическому значению
A. superpictus занимает у нас второе место после A. maculipennis. Он яв
ляется важнейшим переносчиком малярии в предгорных и горных районах
Восточного Закавказья (в Восточной Грузии, Армении, Азербайджане)
и Средней Азии. Там, где A. superpictus — поздний вид, он продолжает
перенос возбудителя малярии, начатый A. maculipennis. В тех местностях,
где он развивается без весенней затяжки, он является интенсивным пе-
реносчиком в течение всего сезона. A. superpictus отличается большой
восприимчивостью к заражению плазмодиями малярии, в том числе Plas-
modium falciparum.
316
Важное эпидемиологическое значение имеет сезоппоо распределение
численности вида: максимум численности A. superpictus совпадает с жар-
ким временем года (июль—август — сентябрь) и с максимумом тропиче-
ской малярии. Вследствие большой восприимчивости к заражению Pl. fal-
ciparum, A. superpictus становится опасным переносчиком тропической
малярии. Проявление в сентябре гонотрофической диссоциации с частыми
повторными кровососаниями усиливает роль A. superpictus в переносе
малярии.
8. ANOPHELES PULCHERRIMUS THEOBALD
Географическое распространение. Распространен в оазисах среди
пустынь Средней Азии, Месопотамии, Афганистана, Ирана, северо-западной
части Индии. В СССР встречается в Средней Азии (долины рек Сыр-
Дарьи, Аму-Дарьи, Зеравшана, Мургаба, Теджена и пр.) и в Восточном
Закавказье (Кура-Аракспнская низменность Азербайджана). Распростра-
нен только на равнине; выше 1 000 м не идет.
Экология личинки. Этот вид является резко выраженным термофи-
лом. Оптимум развития личинок лежит при среднесуточной температуре,
превышающей 30°; температура биотопов личинок этого вида па 1—2°
выше, чем у A. superpictus (Петрищева), при средних суточных максиму-
мах в 35° и более. Водоемы с дневной температурой 25° для развития ли-
чинок уже менее благоприятны.
Типичными местами выплода являются крупные стоячие постоянные
водоемы (наиболее благоприятны — поемные), где личинки заселяют
заросли элодеид. В затененные водоемы не идут в силу термофил ытости.
Выплаживаются также во временных водоемах, в разливах ороситель-
ных каналов и т. п.; на рисовых полях — преимущественно при непро-
точной системе орошения, когда в застойной воде, особенно последнего
чека, создается высокая температура.
Личинки A. pulcherrimus более выносливы к азотистому загрязне-
нию воды, чем личинки A. maculipennis.
Зимуют личинки III возраста в стоячих, постоянных, непромерзае-
мых водоемах (в сбросах — «заурах», карьерах и ямах с грунтовой
водой, в поемных озерах). Борьба с зимующими личинками A. pulcher-
rimus путем заселения этих водоемов гамбузией очень эффективна.
Водоемы, где зимуют личинки A. pulcherrimus, обычно удобны и для зимов-
ки гамбузии.
Экология имаго. A. pulcherrimus — наиболее теплолюбивая и сухо-
устойчивая форма по сравнению с другими видами Anopheles, встречающи-
мися в СССР. В сухом воздухе он теряет путем испарения очень мало во-
ды, что связано с малыми размерами грудных и брюшных дыхалец
(Виноградская). В силу большой теплолюбивости и сухоустойчивости A. pul-
cherrimus часто остается на дневках в убежищах с высокой температурой
воздуха (сараи, навесы, пещеры, щели в каменных стенах, заросли рас-
тительности).
A. pulcherrimus меньше всех других видов боится света, встречается
па дневках в более освещенных местах, нежели A. maculipennis и A. su-
perpictus. Это единственный из наших Anopheles, который не безусловно
избегает прямого солнечного света и иногда располагается па дневках
на освещенных солнцем местах. Белый плотный покров из чешуек, покры-
вающих его тело, является приспособлением для отражения солнечных
лучей и уменьшения нагрева тела комара. Нападает A. pulcherrimus
в пределах температур 12—33,5°, наиболее активно при 19—28°. В'теп-
лые летние ночи он активен от захода до восхода солнца; в холодные
317
весенние и осенние ночи при температурах, лежащих ниже порога актив-
ности (12°), A. pulcherrimus не летает; в этих случаях он нападает и ле-
тает на водоемы для откладки яиц вечером и утром, нередко после вос-
хода солнца, когда температура повышается.
Низкая влажность (30 и даже 10%) не ограничивает его активность
(Крохина, Чинаев), нападения при 30—35% относительной влажности
обычны. По способу охоты за добычей A. pulcherrimus несколько отличает-
ся от A. maculipennis и A. superpictus: этот комар может нападать в
поселках, его привлекают скопления крупной добычи, но более многочи-
слен он в дикой природе, где питается на животных. При нападениях в
поселке A. pulcherrimus остается на дневках в местах питания (хлевах,
домах) до конца переваривания крови. Таким образом, A. pulcherrimus
является в значительной степени эндофильным видом.
Рис. 193. Сроки выплода I весеннего поколения A. pulcherrimus
в Узбекистане (оригинал по материалам Чинаева).
Однако A. pulcherrimus, хорошо перенося микроклимат хлевов и убе-
жищ в природе, меньше стремится в жилье человека, чем A. mac. sa-
charovi. Благодаря этому в Средней Азии A. pulcherrimus менее связан
с человеком, чем A. mac. sacharovi, который, в силу меньшей сухоустой-
чивости, нападает в большинстве случаев в жилых помещениях, где усло-
вия микроклимата для него благоприятнее (влажность выше), чем в хле-
вах или на открытом воздухе. При вскрытиях, проведенных в Сыр-Дарв-
инской, A. pulcherrimus оказался заражен малярийными плазмодиями
вчетверо меньше, чем A. mac. sacharovi (Баклемпшев и Митрофанова).
Исследования видового состава крови из кишечников комаров пока-
зывают, что у A. pulcherrimus кровь скота встречается много чаще, чем
у A. mac. sacharovi (Ходукин).
В поисках добычи A. pulcherrimus разлетается на большие расстояния
от мест выплода (4—5 км).
Фенология. Длительность периода времени с момента появления первых окры-
ленных комаров A. pulcherrimus до момента исчезновения последних в условиях
Узбекистана составляет З1^—5 месяцев, в то время как A. superpictus и A.m. sacharovi
встречаются здесь на дневках в более или менее значительных количествах в течение
7—10 месяцев.
Весной развитие личинок в водоемах идет медленно, и поэтому вылет комаров
I поколения происходит сравнительно поздно (рис. 193) в северной части Узбекистана
(в Н. Ургенче и Ленинске) — не ранее третьей декады мая — первой декады июня,
на юге, в Термезе, — на Р/а—2 месяца раньше.
Появление окрыленных комаров I генерации у этого наиболее теплолюбивого
из видов Anopheles нашей фауны отмечается на севере Узбекистана на 1—2 месяца
позже, чем вылет I поколения A. mac. sacharovi, и на г12—Р/г месяца позже, чем
A. superpictus.
318
^Кладки и личинки 1 возраста II поколения появляются на севере Узбекистана
(в Н Ургенче) 20 июня (1938), а на юге (в Термезе) — 17 апреля (1940).
Цикл развития II поколения в водоемах продолжается в среднем 20—25 дней.
Выплод II генерации происходит на севере Узбекистана (Н. Ургенч, 1938) в первой
декаде июля, а на юге (в Термезе) в первой декаде мая (1940—1941).
Сезонный ход численности» На севере Средней Азии наблюдается ве-
сенняя затяжка в росте численности этого вида, связанная с недостатком
тепла для его развития. Здесь он появляется в значительном количестве
лишь с июля, достигает максимальной численности в августе и первой по-
ловине сентября, после чего численность его быстро снижается, и к сере-
дине октября этот вид на дневках исчезает, вследствие вымирания имаго
(рис. 191).
На юге Средней Азии, например, в бассейне реки Мургаба, A. pul-
cherrimus появляется значительно раньше, чем в северной Туркмении.
Здесь численность его повышается уже со второй половины апреля, ма-
ксимум наблюдается в конце июля и в августе, падение — в середине сен-
тября. В годы с теплой затяжной осенью в Мервском оазисе A. pulcher-
rimus встречается на дневках до второй декады ноября (Правиков).
Для этого вида характерны большие колебания численности от года
к году, иногда приводящие почти к полному временному его исчезнове-
нию в той или иной местности. Эти колебания приписываются способу зи-
мовки этого вида — мало совершенному, делающему его уязвимым в хо-
лодные зимы, когда промерзание водоемов губит зимующих личинок.
Эпидемиологическое значение. В некоторых местах Средней Азии
A. pulcherrimus безусловно является важнейшим переносчиком малярии,
главным образом во второй половине лета и в начале осени. Так, например,
в Мервском оазисе (Туркмения) во многие годы 1 это единственный эффек-
тивный переносчик малярии (если не принимать во внимание A. hyrcanus),
и заболеваемость бывает очень высока. Естественная зараженность A. pul-
cherrimus плазмодиями малярии может быть значительна; так, по данным
экспедиции Центрального института малярии, в 1933 г. в Сыр-Дарьин-
ской она достигала 1%, что при большом количестве комаров составляло
достаточно высокую цифру.
ГЛАВ А XI
СРАВНЕНИЕ ЭПИДЕМИОЛОГИЧЕСКОГО ЗНАЧЕНИЯ ОТДЕЛЬНЫХ
ВИДОВ ANOPHELES СССР
Если мы желаем добиться снижения малярийной заболеваемости
при помощи борьбы против переносчика, прежде всего мы должны уста-
новить, какие виды и подвиды Anopheles имеются в данном очаге и какова
роль каждого из них в поддержании очага. После этого мы должны на-
править свои мероприятия только против тех видов переносчика, которые
играют существенную роль в переносе малярии и в поддержании очага.
И, наконец, план борьбы, который мы составим и будем проводить, дол-
жен соответствовать особенностям биологии тех видов переносчика, с кото-
рыми мы должны бороться.
Отчего же зависит роль каждого из видов Anopheles в передаче маля-
рии, или, как говорят, его эпидемиологическая роль? В этом отношении
имеет значение ряд условий.
Значение заражаемости плазмодиями. 1. Заражаемость ко-
маров плазмодиями. Почти все изученные в этом отношении
виды Anopheles способны заражаться человеческими плазмодиями и по-
1 В другие годы там же вместо A. pulcherrimus преобладает A. superpictus.
319
еле этого заражать своим уколом здорового человека. Однако различные
виды Anopheles заражаются различными видами Plasmodium не одина-
ково легко. Из наших видов «хорошими» переносчиками являются
A. maculipennis, A. bifurcatus, A. plumbeus, A. superpictus; при благо-
приятных обстоятельствах они заражаются и возбудителем трехдневной
малярии (Plasmodium vivax), и возбудителем тропической малярии (Р1.
falciparum) в очень высоком проценте случаев. С другой стороны, A. hyr-
canus, A. pulcherrimus и A. algeriensis, при прочих равных условиях,
заражаются в меньшем проценте; A. lindesayi и A. marteri как переносчики
изучены недостаточно.
Различия в степени заражаемости между «хорошо» и «плохо» зара-
жающимися видами большей частью невелики: если на каком-либо больном
трехдневной малярией заражается 80—90% особей A. maculipennis
или A. superpictus, то A. pulcherrimus или A. hyrcanus заражается 30—40%
особей; другими словами, «хорошие» переносчики заражаются раза в 2—3
лучше «плохих». Это различие, конечно, довольно существенно, по оно
не является решающим, так как другие факторы, определяющие эпиде-
миологическую роль видов Anopheles, колеблются в гораздо более широ-
ких пределах. Но иногда различия в заражаемости бывают очень значи-
тельны: так, тропикой A. hyrcanus заражается раз в 30 хуже, чем Л. su-
perpictus; такая низкая заражаемость резко снижает роль A. hyrcanus
в переносе тропики.
Значепие числепЕОСТи. 2. Ч и с л е и п о с т ь. Шансы на перенос ма-
лярии каким-либо видом комаров, при прочих равных условиях, прямо
пропорциональны числу комаров данного вида в данном очаге. Поэтому
только многочисленные виды Anopheles могут быть опасными переносчи-
ками. А численность каждого вида в том или ином очаге больше всего за-
висит от наличия и размеров благоприятных для пего мест выплода. По-
этому во многих местах многочисленными могут быть виды, приспособлен-
ные к развитию в широко распространенных типах водоемов, как, напри-
мер, A. maculipennis, A. pulcherrimus, A. hyrcanus, A. superpictus.
Наоборот, виды, приспособленные к более редким типам водоемов, оста-
ются всюду малочисленными, как A. marteri, A. lin lesayi, или много-
численны в немногих местах, как A. algeriensis, A. bifurcatus и A. plum-
beus.
Зкачгние сезонности. 3. Сезонность. При прочих равных
условиях наиболее опасными переносчиками являются те виды Anophe-
les, которые многочисленны в течение всего жаркого сезона, когда воз-
можна передача малярии; таков, например, A. maculipennis в центре
своего ареала. Однако многие виды бывают ми нечисленны только на про-
тяжении части сезона; например, A. superpictus на севере Средней Азии
многочисленны только в конце лета. В тех местах, где он является един-
ственным переносчиком, отсутствие переноса в начале лета резко снижает
малярийнссть местности и A. superpictus оказывается плохим перенос-
чиком. В тех местах, где, кроме нег), имеется A. mac. sacharovi, присут-
ствующий все лето, добавление осенью A. superpictus резко усиливает пе-
ренос малярии в осенние месяцы и отягощает малярийную обстановку
(Исаев). A. bifurcatus, на юге многочисленный только осенью и весной,
когда температура не благоприятствует переносу малярии, в силу одного
этого во многих местах теряет эпидемиологические значение.
Значение степени связи с человеком. 4. Связь с человеком.
Из всех условий, определяющих эпидемиологическую роль отдельных
видов Anopheles, наибольшее значение имеет степень связи с человеком.
Степень связи какого-нибудь вида комара с человеком измеряется про-
320
центом особей, пьющих кровь человека, по отношению ко всему числу пью-
щих кровь особей данного вида. Связь комара с человеком в свою очередь
зависит от нескольких факторов. Прежде всего — пищевые предпочтения.
Существуют комары, обладающие хорошо выраженным предпочтением
к крови человека, предпочитающие человека любой другой добыче; таков,
например, самый страшный из переносчиков малярии, африканский A. gam-
biae. Наряду с этим, некоторые виды Culex предпочитают кровь птиц, не-
которые виды Anopheles — кровь копытных.
Большинство наших видов Anopheles хотя, может быть, и предпочи-
тает кровь тех или иных млекопитающих, но предпочтения эти не настоль-
ко сильны, чтсбы их легко было доказать, и, повидимому, не играют ре-
шающей роли в переносе малярии. Так, A. mac. messeae, повидимому,
предпочитающий кровь копытных, тем не менее является одним из наших
важнейших переносчиков малярии. Очевидно, степень его предпочтения
к скоту не настолько велика, чтобы при всех обстоятельствах слишком
сильно ослабить его связь с человеком.
Второй фактор, определяющий степень связи с человеком, — спо-
соб охоты. В этом отношении наши Anopheles разбиваются на три группы.
Способы охоты, а) Охотники за крупными стадными животными, с да-
лекого расстояния собирающиеся вокруг скоплений добычи — поселков,
пасущихся стад и пр. Таковы A. maculipennis, A. superpictus, в известной
мере A. pulcherrimus и, может быть, A. plumbeus. Такой способ охоты резко
увеличивает связь комара с человеком и повышает эпидемиологическую
роль этих видов.
б) Охотники за рассеянными, большей частью мелкими животными.
По этому способу охотятся многие виды A6des (см. ниже, стр. 323),
а из видов Anopheles,—повидимому, A. hyrcanus. Такие комары сравни-
тельно редко имеют возможность повторно питаться на человеке, а потому
обычно не могут явиться эффективными переносчиками малярии; зато
они часто становятся переносчиками зоонозов с мелких млекопитающих
и птиц на человека (перенос японского энцефалита, туляремии и пр.).
в) Подстерегающие хищники, остающиеся близ места выплода и
подстерегающие здесь приближение добычи. К этой категории относятся
A. bifurcatus, A. algeriensis. Подобные виды оказываются тесно связан-
ными с человеком лишь в том случае, если вылупляются в непосредственной
близости от жилья. Это происходит, например, в тех случаях, когда A. bi-
furcatus поселяется в городах, как это имело место в Палестине или на
окраинах Баку (стр. с02). Вылупляясь здесь в тесном соседстве с человеком,
A. bifurcatus, как раз в силу свсей относительно малой подвижности, ока-
зывается особенно тесно связанным с человеком и становится опасным пе-
реносчиком малярии.
Помимо пищевых предпочтений и способа охоты, на связь комара с че-
ловеком влияет множество внешних обстоятельств: быт населения, чи-
сленность скота, его расположение, характер помещений и пр. Влия-
ние этих обстоятельств уже рассмотрено нами выше (стр. 266—269).
В итоге всех этих сложных взаимоотношений складывается эпиде-
миологическая роль каждого вида Anopheles в каждом очаге. Опыт пока-
зывает, что, в зависимости от обстановки, один и тот же вид Anopheles
может в одной местности оказаться опаснейшим переносчиком и не играть
никакой роли в другой местности. Так, A. mac. messea# при длинном и
жарком лете и умеренном количестве скота обычно создает тяжелые очаги
малярии, но при коротком и прохладном лете (все же допускающем зара-
жение комаров) и большом количестве скота даже порядочное количество
комаров этого вида может совсем пе создать малярии. С дру-
21 Учебник, медицинской энгомолсгии
321
гой стороны, Л. bifurcatus, обычпо почти но участвующий в перенос© маля-
рии, в известных случаях становится, как мы видели, опасным перенос-
чиком.
Разделение видов Anopheles СССР по их эпидемиологической роли.
И все же, несмотря на это разнообразие, обобщая весь наш опыт, мы можем
разделить виды Anopheles СССР по степени их участия в переносе малярии
на три группы:
1. Основные переносчики: A. maculipennis, A. superpictus, A. pul-
cherrimus.
2. Второстепенные переносчики: A. hyrcanus, A. bifurcatus, A. plum-
beus.
3. Виды, фактически пе являющиеся па нашей территории переносчи-
ками малярии: A. algeriensis, A. lindesayi, A. marteri.
Первые три вида почти всюду опасны, и если в данном очаге борьба
с переносчиком признана необходимой, она в первую очередь должна быть
направлена па этих опаснейших переносчиков. При этом мы безусловно
вправе пренебречь видами третьей категории и не тратить сил и средств
на их уничтожение. Что касается трех второстепенных переносчиков (ка-
тегория 2), необходимость борьбы с ними должна устанавливаться в каж-
дом случае в отдельности, в зависимости от обстановки.
ГЛАВА XII
ПЕКСТС1ЫЕ ВАЖНЕЙШЕЕ ГИДЫ НЕМАЛЯРПЙНЫХ КОМАРОВ
СССР
а) Род Aedes
Род A6des широко распространен, встречаясь от Крайнего Севера
до тропиков; в странах с влажным климатом он богаче видами; большин-
ство видов Aedes является назойливыми кровососами.
Способ откладки яиц. Самки Aedes откладывают яйца всегд а по одному,
по краям пересыхающего водоема, периодически заполняемого водой.
Развитие зародыша в яйце происходит только во влажной среде: на сы-
рой почве или под водой. До сформирования личинок яйца неустойчивы к
высыханию. Зрелые яйца сохраняют жизнеспособность в течение длитель-
ного времени (у Аё. dorsalis по крайней мере до Злет), переживая небла-
гоприятные условия (высыхание,.низкие температуры, длительное пребы-
вание под водой).
В пашем климате все виды Aedes зимуют на стадии яйца.
Биология личинок. Местами выплода большинства наших АёЛез яв-
ляются небольшие временные водоемы в лесу, на опушке, па лугу: канавы,
ямы, лужи, заболоченности, образующиеся после таяния снега, выпадения
осадков, разлива воды из (росительпой сети или родников и т. д. Дно та-
ких водоемов обычно покрыто затопленной травой, опавшей листвой, на
которых развивается богатый перифитон. Ряд видов выплаживается в дуп-
лах деревьев< (Аё. aegypti — в Закавказье, Аё. geniculatus — в Европей-
ской части СССР и в Закавказье, Аё. gallcisi, Аё. alectorovi и Аё. niveus—
на Дальнем Востоке, Аё. pulchritarsis — па Украине, в Крыму, па Кав-
казе и в Сродней Азии).
Своеобразны личиночные биотопы Аё. togoi — каменные ванны (т. е.
углубления) на скалистом морском побережье Приморского края. Неко-
торые виды A6des могут выплаживаться в различных искусственных во-
доемах (бочках, цистернах, консервных банках и т. п.); таковы Аё. aegyp-
ti, Аё. japonicus, изредка Аё. togoi и Аё. galloisi. ’
322
Большее количество разлагающихся органических веществ при от-
сутствии зеленых растений и прямого солнечного освещения но вредит
личинкам. Личинки Аё. vexans встречаются иногда почти в навозной
жиже. Многие виды Andes выносят значительную засоленность (напри-
мер, Аё. dorsalis, Аё. caspius, Аё. togoi и др.).
Личинки AёJes находятся в водоеме обычно па дне, где питаются,
обскребывая перифитон, и лишь периодически поднимаются на поверх-
ность воды для дыхания; связь с поверхностной пленкой у них меньше
выражена, чем у личинок Anopheles и Culex. Обилие личинок в водоеме
может быть очень велико (до 4 000 в 1 л). Преждевременное высыхание
водоема нередко губит огромную массу личинок.
Приспособлением, повышающим шансы на выживание потомства,
является неодновременное вылупление личинок из яиц одной кладки;
иногда оно растягивается на несколько суток и даже месяцев. Если пре-
ждевременное высыхание водоема погубит личинок, оставшиеся яйца
при следующем затоплении их водой дают начало новой порции личинок.
Биология имаго. Самки большинства видов Aedes — активные охот-
ники за мелкой, диффузно рассеянной добычей, главным образом за мел-
кими млекопитающими и птицами, хотя при случае они жадно нападают па
человека и особенно па крупных копытных.
От мест выплода многие комары (например, Аё. dorsalis, Аё.
caspius и др.) рассеиваются в поисках добычи иногда па большие расстоя-
ния (до 35 км по долинам рок), равномерно заселяя доступное простран-
ство. Такие виды, как Ac. aegypti, Аё. Logoi, наоборот, малоподвижны и
разлетаются недалеко gt мест выплода (обычно не далее 50—100 м).
Большинство видов нападает главным образом па открытом воздухе
вблизи мест выплода: в лесу, по опушкам леса, на лугу в зарослях травя-
нистой растительности; в закрытые помещения — жилища и хлева — они
залетают в заметных количествах лишь при большей численности их в
природе. Иску сети ппьй источник света способствует привлечению
комаров в помещение.
Характерной является способность комаров Aedes развивать хотя бы
часть яиц за счет принятия неполной порции крови, обеспечивающая
размножение особей и продолжение вида даже в случае прерванного про-
весе саппя. Таким образом генотрофическая гармония доведена у Aedes
до крайнего предела (Детгпша, Кзлаьдадзе и Таирова).
Самки пьют кровь и откладывают яйца многократно. Средняя поло-
вая продукция самок разных видов Aedes от 38 до 160 яиц. Меньшая по
сравнению с Anopheles и Culex плодовитость компенсируется значитель-
ней продолжительностью имагипалыгой жизни и большой повторностью
кладок. Так, отдельные особи весенних видов доживают в природе до осе-
ни, т. е. живут более 3 месяцев. Ac les, как и другие комары, наиболее
активны вечером (в период сумерек), в первые часы ночи и в ранние утрен-
ние часы. В тени лесов и внутри помещений Аё les нападают и днем. При
пасмурной и теплой погоде комары активны большую часть суток, кроме
наиболее прохладных ночных часов. Главным фактором, стимулирующим
их активность, является ослабление освещения. Нападения большин-
ства северных видов (Аё. cxcrucians, Аё. cincrcus, Аё. vexans) происходят
в лесной полосе в пределах температур от 7—11° до 28° и при влажности
воздуха 50—100% (Шленова).
Большинство видов A6dcs очень влаголюбиво. Дневки их находятся
в растительности: в траве, на лугах и по берегам водоемов (например,
у Аё. caspius, Аё. dorsalis, Аё. excrucians), в кустарнике, в дуплах деревьев,
в ямах, в расщелинах скал, в пещерах.
21»
323
Сезонный ход численности и число генераций. Максимальная числен'
ность большинства видов A6des в средней полосе наблюдается задолго до
середины лета (обычно в июне); к осени количество их сильно уменьшается.
Лишь отдельные, более теплолюбивые виды появляются в массе во вто-
рой половине лета, таковы Аё. togoi и Аё. aegypti.
Разные виды A6des дают за сезон от одной до нескольких генераций.
Большинство наших видов имеет только одну генерацию (например, Аё.
communis, Аё. annulipes, Аё. flavescens и др.); личинки появляются весной
или ранним летом, но взрослые комары могут жить несколько месяцев,
вплоть до осени. Некоторые другие виды Аёс1ёэ могут иметь в год две ге-
нерации (например, Аё. dorsalis, Аё. caspius, Аё. cinereus в средней полосе
СССР). В теплом климате отдельные виды имеют по нескольку генераций
(например, Аё. togoi и Аё. aegypti).
Однако в силу биологических особенностей яиц Аёдеэ (способность
противостоять длительному высыханию, разновременность вылупления
личинок), генерации их ясно не разграничены. Появление личинок отдель-
ными порциями создает впечатление множества поколений и затрудняет
их учет.
1. Aedes (Ochlerotatus) communis Deg.
Широко распространен в лесной и лесостепной зонах СССР, Северной Европы,
США, Канады. По образу жизни — типичный предста вит ель подрода, один из наибо-
лее ранних весенних видов.
В лесной полосе СССР личинки вылупляются и з перезимовавших яиц в конце
марта или в начале апреля, когда с таянием снегов яйца заливаются водой. Отдельные
яйца могут давать еще более поздний выплод— па втор ую весну. Личинки развиваются
в лужах и канавах в хвойных и лиственных лесах и по лесным опушкам. Окукле-
ние происходит в конце апреля — в мае; вылет имаг о начинается в начале мая. Раз-
витие личинок и куколок происходит при средней температуре воды 11—12° и про-
должается 42 дня (1947г. — Шленова). Максимум численности в окрестностях Москвы
наступает в конце июня; с конца первой декады июля их число быстро снижается;
последние комары были найдены в 1947 г. 31 июля (Ш ленова). Имеют одну генерацию.
Самки Аё. communis — назойливые кровососы. Их укусы, как и укусы большин-
ства Аёйея, вызывают сильное раздражение.
2. Aedes (О.) caspius dorsalis Meigen.
Широко распространен в Северной и Средней Е вропе, найден в Китае и Монго-
лии, Северной Америке. В СССР встречается в Европейской части, в Крыму, на Север-
ном Кавказе, Среднем Урале и в Сибири, вплоть до восточной оконечности. Харак-
терен для лесной и лесостепной зоны.
Выплаживается в открытых, временных лужах, образующихся после таяния
снега, выпадения осадков, разливов рек, прорывов оросительной сети, а также в ми-
кроводоемах в местах повышенного стояния грунтовых вод (следы колес, копыт скота,
следы человека и т. д.). Выносят значительную засоленность. Вылупление личинок
происходит под слоем воды пе менее 0,6 см и не более 1 м; они не отрождаются при слу-
чайном смачивании водой или незначительном затоплении. При температуре ниже 9°
вылупления личинок не происходит (Хелевип).
При 22—26° развитие зародыша продолжается 5—6 дней, развитие личинок и
куколок при 15° — 28 дней, при 20° — 15 дней, при 25° — И дней и при 30° —
7х/а Дпей (Хелевин).
Аё. с. dorsalis — один из ранних комаров. В лесной полосе личинки его начи-
нают отрождаться из перезимовавших яиц в середине апреля; вылет окрыленных ко-
маров происходит в середине мая; период активности и размножения — с середины
мая до конца сентября. Может давать повторный выплод в течение лета, главным обра-
вом за счет развития личинок в дождевых лужах.
Всюду является злостным кровососом. В США — переносчик энцефаломиэлита
лошадей.
3. Aedes (Aedes) cinereus Meig.
Распространен в Сегерной и Центральной Европе и в Северной Америке.
В СССР встречается на большей части Европейской территории Союза, на Среднем
Урале, Северном Кавказе, в Сибири и на Дальнем Востоке.
324
Личинки живут во временных водоемах, единичные личинки изредка встречаются
в водоемах грунтового происхождения, вместе с Anopheles bifurcatus (Шленова).
Взрослые комары чрезвычайно влаголюбивы. Местами дневок являются густая
травянистая растительность (осоки и др.) и кустарники, в зарослях которых комары
глубоко прячутся. В дневные часы нападают только среди растительности, в нижнем
ее ярусе, где влажность наиболее высока и температура ниже. В вечерние и ночные
часы залетают иногда для питания в хлева, но на длительное время здесь не остаются.
Первые личинки в окрестностях Москвы в 1947 г. были обнаружены 23 апреля,
вылет окрыленных комаров I генерации произошел 20 мая. В течение лета личинки по-
являлись неоднократно во временных водоемах, образовавшихся после дождей, да-
вая вылет имаго в середине июня, в начале августа и сентября. Таким образом, этот
вид полициклическии. Развитие личинок в водоемах в летние месяцы (июнь—август)
происходило при средней температуре 15,5—19,5° и продолжалось 19—11 дней. Мак-
симальное количество комаров этого видав 1947 г. под Москвой наблюдалось во вто-
рой половине июня; последние самки были найдены 7 октября (Шленова).
В условиях опыта доказана возможность передачи вируса энцефаломиэлита ло-
шадей укусами экспериментально зараженных комаров Аё. cinereus (Муратова).
Неоднократно встречались в природе самки этого вида, зараженные туляремией
(В. М. Попов и др.).
4. Aedes (Aedes) esoensis Jam.
Распространен на Дальнем Востоке: на Сахалине, в Японии; в СССР встречает-
ся в безлесных и, реже, в таежных районах Приморского края.
Аё. esoensis — ранне-летний вид. Личинки встречаются па юге Приморья с
конца апреля, начало окрыления — с конца второй декады мая — начала июня.
Развитие I генерации протекает при температуре 11—14,5° (Чагин). Наибольшая чи-
сленность вида наблюдается в период вылета I генерации, т. е. с конца мая по первую
декаду июля (рис. 196); с августа численность резко снижается; в дальнейшем, до конца
325
сентября, встречается лишь неоочыиое число особей. Таким образом, Ac. esdensis
в Приморском крае имеет одну генерацию. В годы с небольшим количеством осадков
он малочислен, в дождливый сезон численность его довольно высока.
Личинки выплакиваются в мелких естественных и искусственных, хорошо про-
греваемых водоемах: в лужах на лугах и среди кустарников, в канавах и ямах с доста-
тошо развитой водной растительностью. Оптимальная температура для развития
водных стадий 18—22°, верхний порог развития — 32°. Средняя продолжительность
развития при 15—16° — 32,5 дня, при 19—20° — 23,5 дня, при 24—25° — 18,5 дня
(Чагин). Личинки чувствительны к загрязнению воды загнивающими органическими
веществами. Дневки комаров — в растительности па лугах. Агрессивен но отношению
к человеку; нападает вблизи мест дневок, однако способен и к активному лоту на боль-
шое расстояние (Чагин). В жилище залетает в небольшом числе. Актинон до конца сен-
тября. В условиях лаборатории самки живут 50—76 дней. Вместе с Culex tnL.eniorhyn-
Chus является переносчиком японского энцефалита в безлесных районах Приморья.
Заражения происходят во время пребывания людей на сенокосных угодиях в период
сенокошения (Иетрищеза).
5. Aedes (Finlaya) japonicus Theob.
Распространен в Японии, Корее, Китае; в пределах СССР встречается в лесной
зопе Приморского края. У пас это синантропный 1 вид, живущий на территории на-
селенных пунктов или вблизи них и выплаживающийся вблизи жилья.Типичными
местами выплода являются водоемы искусственного нр< исхо кдения: кадки, цистерны,
баки с дождевой водой, неглубокие заброшенные колодцы, реже — дождевые лужи-
иногда прикорневые дупла (Петрищева). Личинки избегают яркого света и встреча,
ются в затененных водоемах; крайне пугливы, будучи вспугнуты, скрываются надолго
па дне в илу. Вылупление личинок из яиц происходит большей частью дружно
(в 2—3 дня) и редко затягивается до 8 дней (Чагин). Оптимальной температурой для
развития личинок является 20—26°; верхний ворог развития 30—32и. При 9—11° ли-
чинки старших возрастов и куколки еще развиваются, но медленно. Средняя продол-
жительность развития водных стадий при 24—26° равна 18 дням, ирн 17—18° —
287г дням (Чагин).
Окрыленные комары встречаются на дневках в траве, дуплах, ямах, па внутрен-
них затененных стенках бочек и цистерн. В жилье (и хлевах) встречаются редко. Охот-
но нападают на человека, еще чаще — па скот и на кур. Нападают больше вне помеще-
ний, почти в любые часы суток (Петрищева).
В лаборатории Ac. japonicus может копулировать в больших садках, по с трудом.
Поэтому хуже, чем Аё. togoi, разводится в неволе (Чагин). Личинки встречаются в при-
роде с середины мая, взрослые комары — с конца мая до конца сентября — начала
октября. Наибольшая численность комаров наблюдается в июле и августе (рис. 196).
При благоприятных условиях (достаточно дождей и тепла) в Приморье дает до четырех
генераций (Чагин).
Аё. japonicus является одним из опасных переносчиков японского энцефалита
в Приморье; он — основной переносчик в лесной зоне (Петрищева).;
6. Aedes (Finlaya) togoi Theob.
Распространен в Японии, Корсе, Маньчжурии, в СССР — па побережье п остро-
вах Японского моря.
Местами выплода являются небольшие годоемы па скалистом побережье: запол-
пепнье голой трещины скал, углубления поюрхл ости камней и т. л.; года в пих по
происхождению — дождевая, родниковая пли морская; го концентрации солей — от
пресной до горькосоленой, нередко со. оное морской годы. Дно водоема, покрытое
илом п выброшенными морем органическими остатками, не;едко обрастает водоросля-
ми, которые, наряду с детритом, служат нищей личинкам.
Личинки встречаются преимущественно в открытых, хорошо прогреваемых во-
доемах. в меньшем числе — в затененных водоемах. В конце сезона, с пересыханием
водоемов на побережье, комары переселяются ближе к поселкам п личппки попадаются
в бочках, деревянных колодах для водопоя лошадей, металлических чанах и т. и.
Самка при откладке приклеивает яйца к камню, погружая их наполовину в воду.
Число созревающих яиц зависит от количества выпитой крови и достигает 160.
Развитие личинок возможно при температуре от 5° до 35—37°, оптимум при
24—27°. Продолжительность развития при 25° равна 29—5P/a суткам; колебание
температуры ускоряет развитие (Чагин).
1 От греческих слов: <?syn> — вместо и eantropos»—человек; гид, живущий вме-
сте с человеком, тесно связанный с человеком.
326
Дневными убежищами взрослых комаров являются пещеры, трещиныскал, заросли.
Копуляция монет происходить и в садко (Чагин, Лисова). В вожсках добычи самки
разлетаются о г мест выплода обычно не далее 0,5 км. Питаются они па морских
птицах (чайках, бакланах) и больше всего их бывает вблизи гнездовий этих птиц.
Однако при случае они нападают также па человека и млекопитающих. Развитие яиц в
условиях лаборатории заканчивается при 15° через 9—И суток, при 25°—через
G—8 суток, при 3J° — через 5—7 суток.
Рио. 195. A5des (Finlaya) togoi Thoob., самка.
В южных районах Приморья личинки Аё. to£oi починают отрождаться пз пере-
зимовавших яиц в начало апреля; при средней температуре 5—7° куколки появляются
в первой декаде мая, вылет 1 генерации начинается со второй половины мая. На юге
Приморья имеет не менее трех генерации. Вылет комаров 11 генерации начинается с
конца июня, 111 генерации — с конца июля. Сезонный максимум численности — в конце
июля и в августе; с первой половины сентября обилие их падает, и к середине октября
имаго исчезают (рис. 196). Развитие личипок в водоемах заканчивается во второй по-
ловине октября, когда средняя температура падает до 7—9°.
Аё. lo^oi является переносчиком японского энцефалита. Морские птицы служат
па побережье резервуаром вируса. Случаи заболевания по побере кью и па островах
Японского моря связаны только с этим видом (Пстрпшова, Чагин). Особенно опасен
для групп населения, работа которых связана с пребыванием в вечернее и водное
время на скалистых берегах (рыбаки, портозые рабочие).
327
7. Aedes (Stegomyia) aegypti (L).
Распространен от 43° с. ш. до 40° ю. ш. во влажных тропиках и субтропиках.
В СССР живет на Черноморском побережье Кавказа (к северу — до Сухуми, во всей
Западной Грузии) и в окрестностях Баку; в Тбилиси встречается спорадически, при
завозе комара по железной дороге; перезимовывать там он не может.
Типичный домашний вид, у нас не встречающийся вдали от человека.
Местами выплода являются разнообразные скопления воды (преимущественно
дождевой). В природе выплаживается в дуплах деревьев и других эпифитиых 1 водое-
мах. В населенных пунктах личинки развиваются в различных искусственных водое-
мах как вокруг, так и внутри жилищ: в бочках с дождевой водой, цистернах, ваннах,
кувшинах, вазах для цветов, жестяных банках, затопленных подвалах и т. п.
Самки откладывают яйца только в водоемы с твердыми стенками — на стенки во-
доема несколько выше поверхности воды или на воду. Яйца сохраняют жизнеспособ-
ность при высыхании длительное время. Особенно стойки зимующие яйца (в течение
японского энцефалита
Рис. 196. Кривая численности переносчиков японского энцефалита
в Приморском крае в 1941 г.
А — окрыленные комары; Б — личинки.
17 месяцев), но вылупление личинок из таких яиц сильно задерживается (на 100—
119 дней) (Каландадзеи Таирова). Яйца Аё. aegypti не выносят длительного замора-
живания (до 148 часов), вследствие чего они не могут перезимовывать там, где имеет
место продолжительное замерзание водоемов (например, в Тбилиси).
Личинки переносят значительное загрязнение воды органическими веществами г
но большая соленость им вредит; затененные места благоприятнее для развития, чем
освещенные.
Нормальное развитие личинок и куколок возможно при температуре от 18° до
35—37°. Нижний порог развития личинок 15° (Каландадзе и Таирова). При 15—16е
развитие идет вяло и много личинок гибнет. При 10° личинки неподвижны и при тем-
пературе ниже 7° быстро гибнут. Оптимум около 26—30°. При этой температуре идо-
статочном питании развитие от яйца до имаго продолжается 9х/2 дней: яйцо 2 дня.
личинка 6 дней и куколка 36 часов. Температура 38—40° дает большую смертность,
при температуре свыше 40° личинки существовать не могут.
Таким образом, личинки Аё. aegypti приспособлены к жизни в узких температур-
ных пределах (15—37°), что характерно для обитателей тропического леса, выходцем
из которого этот комар является. Личинки этого вида дольше выдерживают пребыва-
ние на сухом (5—100 минут) и влажном (12—108 часов при 18—22°) субстрате, чем
личинки Anopheles и Culex pipiens, и могут развиваться и окукливаться даже в жид-
кой грязи.
Личинки и куколки Аё. aegypti (подобно A. plumbeus) обладают отрицательным
фототаксисом.
Взрослый комар живет вблизи человека и является назойливым домашним пара-
зитом. Это подстерегающий хищник, который остается все время в помещениях, где
спят люди. Он летает недалеко (обычно не далее чем за 100—300 м) и часто всю жизнь
проводит в том месте, где выплодился, перелетая в другие помещения лишь в случае
1 От греческих слов «эпи» — на, «фитоп» — растение; эпифитный — находящийся
на растениях.
328
отсутствия поблизости подходящих мест для откладки яиц. Копуляция происходит в
помещениях. Нападает как днем, так и попью. Охотно сосет кровь человека, домашних
животных, а также холоднокровных и птиц. За счет одного приема крови развивается
40—120 яиц (Шахов). Оптимальные условия для взрослого комара: температура 25—
35° и очень большая влажность воздуха; при температурах ниже 17° комар становится
вялым, при 7° может находиться в окоченении долгое время, а при 6° гибнет; при 0е
комары погибают в течение суток. Температура выше 37° также гибельна для них.
Средняя продолжительность жизни самок свыше месяца, но при благоприятных усло-
виях отдельные особи могут жить в
лаборатории до 5 месяцев.
В тропиках размножается весь
год. В Закавказье зимует в стадии
яйца; личинки зимовать здесь не
могут, так как зимой температура
воды даже в самой теплой зоне па-
дает до 2—4°. Личинки перезимо-
вывают только там, где температура
водоемов не понижается на длитель-
ное время ниже 4—5°.
В Сухуми личинки появляются
с конца апреля — начала мая, има-
го — с мая. Осенью личинки исче-
зают к концу октября, куколки в
ноябре и имаго в конце декабря.
С октября самки перестают нападать
(Рухадзе).
Развитие водных фаз весной и
осенью длится 19—31 день, а летом
10—14 дней (Каландадзе и Таирова).
Максимум численности комаров — в
августе и первой половине сентября
(Шахов). За сезон Аё. aegypti мо-
жет дать в южной части Черномор-
ского побережья Кавказа 3—4 поко-
ления (Гуцевич).
Эпидемиологическое значение.
Аё. aegypti является переносчиком
желтой лихорадки — опасной бо-
лезни человека, широко распростра-
ненной в Центральной и Южной
Америке, в Западной и Центральной
Африке и ввозившейся раньше время
от времени в портовые города Запад-
ной Европы и США, где она вызы-
,™ЭПИДеМИИ С ВЫСОКОЙ Смерт' Рис. 197. Aedes (Stegomyia) aegypti (Ь.), самка,
н остью.
Возбудитель болезни относится
к фильтрующимся вирусам. Комар становится заразным через 9—14 дней после
заражения от человека и остается носителе^м инфекции до конца жизни. Одно время
считали, что желтая лихорадка ликвидирована в городах Южной Америки.
Однако оказалось, что она переносится не только городским Аё. aegypti, но
многими комарами, населяющими леса как в Африке, так и в Южной Америке, при-
чем носителями вируса в дикой природе являются обезьяны и некоторые другие ди-
кие млекопитающие. Таким образом, о полной ликвидации желтой лихорадки в
этих странах пока не может быть и речи.
Аё. aegypti является также переносчиком лихорадки денге, болезпи с низкой ле-
тальностью, но широко распространенной в тропиках и субтропиках. Возбудитель ее—
также фильтрующийся вирус. Денге встречается на побережье Индийского океана
и восточной части Средиземного моря, а потому следует считаться с возможностью
завоза этого заболевания в порты Черноморского побережья Кавказа; такая возмож-
ность заставляет особенно внимательно относиться к истреблению Аё. aegypti.
t6) Род Culeoc
Большинство видов рода Culex распространено преимущественно
в теплом климате (в тропиках и субтропиках), в северную зону СССР
(и Европы) проникают лишь 3—4 вида.
329
Рис. 198. Culex pipiens L., самка.
Биология водных фаз. Самки Culex откладывают яйца на поверх-
ность воды склеенными в компактную массу («лодочку») (рис. 90).
Местами выплода Culex являются главным образом постоянные,
длительно существующие водоемы (прибрежные заболоченности рек, озер,
прудов, рисовые поля), а также различные искусственные водоемы:
ямы, канавы, карьеры, цистерны, бочки с дождевой водой.
Личинки и окрыленные комары Culex теплолюбивы. Оптимум разви-
тия личинок большинства видов пашей фауны лежит при температурах
не ниже 20—25°, при низких
температурах 10—15° развитие
их идет чрезвычайно медленно.
Вылупление личинок из яиц
одной кладки происходит более
или менее одновременно, в раз-
гар лета через 1—2 дня. Ли-
чинки Culex живут у поверх-
ности воды, прикрепляясь вер-
шиной сифона к поверхностной
пленке, остальное тело их ви-
сит вниз под углом к поверх-
ности.
Питаются планктоном и
детритом, находящимися в слое
воды под поверхностной плен-
кой (чистые фильтраторы).
Биология имаго. По типу
гонотрофических взаимоотно-
шений Culex сходны с Anophe-
les: только прием полной пор-
ции крови обеспечивает разви-
тие до конца полной порции
яиц. Подобно Anopheles п
Aedes, самки Culex пьют кровь
и откладывают яйца много-
кратно. Среднее число яиц в
кладке у самки Culex значи-
тельно больше, чем у Aedes.
Зимуют оплодотворенные, жир-
ные, дпапаузирующие самки,
у одних видов — в защищен-
ных помещениях (погребах, подпольях), у других — в природе (в расти-
тельном сухостое, дерне, пещерах и норах).
В лесной полосе СССР наблюдается весенняя затяжка в сезонном ходе
численности Culex в связи с недостатком тепла; максимум численности
наблюдается лишь в конце лета.
Отдельные виды Culex дают в течение сезона от двух до нескольких
генераций.
1. Culex pipiens L.
Распространен очень широко большая часть Палеарктики (за исключением Арк-
тики), Америки и Африки. Ь СССР встречается всюду, кроме Крайнею Севера.
В Европейской части СССР этот вид представлен двумя подвидами: С. pipiens
pipiens L. и С. pipiens mo’.eslus F.. которые различаются главным образом биологи-
чески; морфологические отличия их но всегда отчетливы А.
1 См. статью Федорова, Медицинская паразитология,2В- 2, 1916.
330
G. pi pions pi pions (L.)
Более распространен в сельских местностях, чем в городах. Самки откладывают
«лодочку» из 150—300 яиц. Личинки развиваются в бочках с дождевой голой, канавах,
цистернах, прудах, обычно в довольно чистой воде, но могут приспосабливаться к
водам, значительно загрязненным и солоноватым.
Оптимум развития личинок при 25—26°, температуры выше 27° угнетают их раз-
витие. Продолжительность развития i одних стадий при 25° около 13 дней.
Комары предпочитают крозь птиц и па человека нападают редко. Однако в поис-
ках убежищ и добычи (домашних птиц) часто следуют за человеком в поселки.
Более влаголюбивы, чем A. maculipennis, и встречаются па дневках и зимовках
в более влажных местах. Колуляция происходит на открытом воздухе.
Максимум численности в средней полосе СССР в конце августа — начало сентя-
бря; в сентябре самки уходят па зимовку. Зимуют го влажных прохладных }бен ишах,
погребах, пещерах, всегда в отсутствие добычи. Взятые с зимовки комары упорно от-
казываются пить крозь.
С. pipiens molestus F.
Менее широко распространен, чем С. р. pipiens (найден во многих странах Евро-
пы, в Северной Америке, в Египте), чаще— на кие; па севере исключительно в ю-
родах. В СССР обнаружен в Ленинграде, Москве, Днепропетровске, Киеве.
Резко выраженный домашний вид.
Яйца откладывает в виде маленьких кучек («кусочков лодочки»), число
яиц в кладке мепыие, чем у С. р. pipiens, — от 50—70 до 110.
Личинки могут развиваться в оюпь сильно загрязненной воде. Они хорошо вы-
носят сильное затенение и развиваются даже в полной темно о (в подземных гал-
лереях, погребах и колодцах). Для своего развития требует более высокой темпера-
туры воды, чем С. р. pipiens.
Самки питаются не только па птицах, во нападают охотно на человека, обычно
внутри жилья пно гыо.
Могут копулировать в узком пространстве, без роепия. Поэтому легко размно-
жаются и разводятся в лаборатории.
Оплодотворенные самки свою первую кладку могут откладывать без предвари-
тельного питания кровью (авто енное созревание яиц), пагютобве A. bifurcatus.
Самки не способны к диапаузе и предзимовочпому ожирению; они продолжают
питаться крозыо и откладывать яйца в течение всего юда, независимо от сезона.
Перезимовывают в активном состоянии вблизи человека. Па севере зимними ме-
стами выплода служат водоемы теплых подвальных помещений с температурой
-f-16—18°, где отмечаются и случаи массовых зпмиих нападений С. р. moiestus на
человека.
С. pipiens является переносчиком плазмодиев птичьей малярии. В Японии, а
также п у нас в Приморье (Петрищева) С. pipiens v. pallcns— переносчик японского
энцефалита; однако в Приморье этот вид, невидимому, редок (Чагин).
2. Culex tritaeniorhynchus Giles
Распространен в Африке, Палестине, Месопотамии, Иране, Ипдии, Китае, Япо-
нии. В СССР встречается в теплом и влажном климато: в Восточном Закавказье (Лен-
корань) п в южной части Уссурийского края.
Типичными местообитаниями (стациями) С. triteeniorliincbus являются безлес-
ные районы с преобладанием луговой растительности. Местами выплода являются
рисовые поля, прибрежные заболоченности рек, ручьев и пресноводных п солоноватых
озер, временно затопляемые низины, чате на скошенных лугах, а также различные
искусственные водоемы: ямы, каланы, карьеры и т. д.
Личинки развиваются при температурах 9—30°, оптимальной является темпе-
ратура 20—25° (Чагин). Продолжительность развития водных стадий при 24—25°
равна 22л/г Дням, а при 19—20,5°—34’/з дням ( в лабораторных условиях).
Комары встречаются на дневках среди растительности и в жилищах. В Приморье
этот вид тяготеет к жилью челоюка, где питается и переваривает кровь. В поисках
добычи разлетается от мест выплода на рассто.лнпя до 2—6 км (Петрищева).
Активно нападает в пределах 9—29°, максимум активности между 15° и 21°
(Чагин).
Продолжительность пищеварения прп 17—20° 9 дней, при 25° — 5 дней и
при 30°—4’/. дня (Чагип). Гояотрофическая диссоциация наблюдается с пергой поло-
вины сентября. Зимуют самки в густом растительном сухостое, в дерне, трещинах
почвы, реже—в порах грызуноз (Петрищева).
В Приморском крае С. tritaeniorhynchus является поздпе-летпим видом. Лёт
перезимовавших начинается с середины июня (Чагин), численность их крайне мала.
331
Первые личинки— в начале июля. Вылет I генерации начинается с конца июля—на-
чала августа. Генераций бывает три. Наиболее многочисленна II генерация, ее
окрыление заканчивается в начале сентября. Окрыление малочисленной III генерации
заканчивается в первой декаде октября. Максимум численности — с конца первой
декады августа до конца первой декады сентября (рис. 116).
С. tritaeniorhynchus один из основных переносчиков японского энцефалита в Япо-
нии и в Приморском крае (Петрищева). Сезонный ход заболеваемости вполне совпа-
дает с ходом численности этого вида.
в) JPod Mansonia
Рис. 199. Личинка Manso-
nia richiardii (вид сбоку).
Род распространен преимущественно в тропиках и субтропиках. В СССР встре-
чается только один вид— М. richiardii Ficalbi, который найден в Европейской части
Союза (на север — до широты Ленинграда), на Среднем Урале, на Кавказе, в Средней
Азии, на юге Сибири; широко распространен в Европе,
за исключением северной ее части, найден в Пале-
стине .
Личинки этого рода резко отличаются от личи-
нок других комаров способом дыхания. Сифон их
укорочен и несет на вершине сильно хитинизованные
лопасти стигмальной пластинки, видоизмененные в бу-
равящий аппарат (рис. 123, Д’), при помощи которого
личинки прорезают стебли водных растений и прони-
кают отверстиями стигм в воздухоносные полости ра-
стений, используя воздух последних для дыхания
(рис. 199). Личинки ведут придонный, малоподвижный
образ жизни, прикрепляясь к растениям вблизи дна.
В связи с особенностями дыхания и экологии способ-
ность к плаванию развита слабо; чаще всего они пол-
зают при участии плавника и волосков брюшка и гру-
ди. Удельный вес личинок больше воды. Для личинок
этого вида, живущих на дне в затененных водоемах,
характерны слабо развитые глаза, молочнобелый цвет
тела. Личинки живут в постоянных водоемах, богатых
водной растительностью (макрофитами). Прикрепляются
к стеблям ежеголовки, аира, манника, рогоза, рдеста
плавающего, водяного лютика. При обычных способах
вылова личинок комаров из водоемов при помощи сачка
личинки, как правило, не попадают в уловы. Лучший
способ добывания их пз водоемов — вырывать стебли
растений со дна и помещать их в стеклянные банки;
искать личинок следует на стеблях растений. Зимуют
личинки старших возрастов; окукливание их происходит весной.
Куколки имеют видоизмененные дыхательные трубки, вбуравливающиеся в стеб-
ли; подобно личинкам, опи дышат воздухом воздухоносных полостей. Перед вылупле-
нием имаго куколки всплывают на поверхность. Благодаря придонному образу жизни,
личинки не поддаются действию обычных ларвицидов (парижская зелень, нефть).
Окрыленные комары являются назойливыми кровососами. В СССР М. richiardii
может иметь эпидемиологическое значение в качестве переносчика туляремии.
г) Комары — переносчики болезней
Комары подсемейства Gulicinae являются переносчиками многих
болезней человека и животных.
1. Комары рода Anopheles являются переносчиками человеческих
плазмодиев — возбудителей малярии.
2. Некоторые виды родов Culex и Aedes имеют значение в переносе
вируса, вызывающего японский (летне-осенний) энцефалит.
Японский энцефалит — эпидемическое заболевание с острым
течением и высокой летальностью (до 60%). Болезнь эта распростра-
нена в Японии, Северном Китае, Маньчжурии, Корее и в Приморском крае
СССР, где впервые описана экспедицией П. Г. Сергиева в 1938 г.
332
В Приморье установлены следующие виды переносчиков: Culex tri-
taeniorhynchus, Aedes japonicus, Аё. togoi, Аё. esoSnsis, C. bitaeniorhyn-
chus и C. pipiens v. pallens (Шубладзе, Петрищева). Два последних вида
играют меньшую роль в переносе заболевания на человека, так как С. bi-
taeniorhynchus редко нападает на людей (но, вероятно, поддерживает
круговорот вируса в природе среди животных), а С. pipiens в Приморье
сравнительно редок.
Комары считаются основным резервуаром вируса японского энце-
фалита в природе (Петрищева, Смородинцев). Установлена естественная
зараженность комаров в условиях дикой природы, где комары связаны
только с птицами и дикими животными. Вирус сохраняется в комарах дли-
тельное время. Люди и животные, заражаясь от комаров, сохраняют вирус
недолго, так как они либо гибнут, либо выздоравливают. Развитие вируса
в комаре возможно лишь при высокой температуре: 21—27° и выше. По-
этому распространение японского энцефалита у нас ограничивается южной
частью Приморья. Передача вируса комарами возможна тахмсконца июля.
В июле, августе и сентябре отмечается максимальная численность перенос-
чиков. Эпидемические вспышки японского энцефалита возникают в конце
августа — в сентябре. Заболевает главным образом сельское население.
В городах Приморья эпидемий не бывает в отличие от Японии, где болезнь
распространена и в городах.
Основные типы очагов японского энцефалита в Южном Приморье
приурочены к местам обитания переносчиков и свойственны различным
ландшафтам (Петрищева), а именно:
1) Безлесные районы с преобладанием луговой растительности, исполь-
зуемой для сенокошения, с наличием заболоченных участков, мелководных
озер и рек. Переносчики: Аё. е8оёп81з, С. tritaenicrhynchus и С. bitaenio-
rhynchus.
2) Обжитые лесные районы на юго-востоке Приморья (Сучанская до-
лина). Переносчики: Аё. japonicus, С. tritaeniorhynchus.
3) Скалистое побережье и острова Японского моря. Единственный
переносчик Аё. togoi.
3. «Лошадиный» энцефа ломи элит. А. П. Муратова (1939) показала,
чтов СССР имеются виды рода A6des, способные переносить вирус энцефа-
ломиелита лошадей (Аё. cincreus, возможно—Аё. dorsalis). Это заболевание,
распространенное в Америке, в некоторых частях Центральной Европы,
в СССР, Индии и Японии, может поражать и других животных, а также
и человека. Клещи также играют роль в поддержании очагов этого за-
болевания.
Целый ряд заболеваний человека и животных, вызываемых фильтрую-
щимися вирусами и передаваемых комарами, не встречается на терри-
тории СССР (желтая лихорадка, депге и др.).
4. Туляремия. Комары родов A6des, Culex, Mansonia и Anopheles, на-
ряду с другими кровососущими насекомыми и клещами, могут иметь зна-
чение как механические переносчики туляремии. Исследования Олсу-
фьева, а также Романовой и Даниловой показали, что комары Аё. vexans,
Аё. caspius, Аё. lutescens, Аё. cinereus, Mansonia richiardii, Culex modestus,
Anopheles hyrcanus и A. maculipennis после кормления на больной водяной
крысе могут сохранять в себе некоторое время жизнеспособных туляремий-
ных микробов (в условиях опыта Аё. vexans — свыше месяца, М. ricliiar-
dii — до 16 дней) и передавать затехМ уколами инфекцию другим живот-
ным. Способность передачи инфекции уколами сохранялась у комаров до
27 суток после их кормления на больном животном. Зараженные комары
выделяют Bacterium tularense с испражнениями до 9—10 суток. Раздав-
333
ливапие инфицированных комаров на неповрежденной коже животных
(морских свинок) вызывало у последних заболевание туляремией. Уста-
новлена естественная зараженность возбудителем туляремии комаров Ао.
cinereus и Аё. excrucians в Томской области (Карпов и Попов). Комары
могут передавать эту инфекцию как среди грызунов, так п от грызунов че-
ловеку.
б. Фпляриидозы. Комары также являются переносчиками целого ряда
фпляриозов, т. е. болезней, вызываемых паразитированием круглых чер-
вей из отряда нитчаток (Ьilariata). На территории СССР филяриидэзов,
поражающих человека, не существует, по в жарких странах они широко
распространены.
РАЗДЕЛ В
ДРУГИЕ СЕМЕЙСТВА КРОВОСОСУЩИХ НИЗШИХ
ДВУКРЫЛЫХ (Orthorhapha)
Кроме комаров (семейство Culicidae' в состав пизших двукрылых
(подотряд Orlhcrhaplia отряда Diptera) входит множество семейств, среди
которых — несколько семейств, состоящих преимущественно из крово-
сосущих форм. Таковы мошки (Simuliidao), мокрецы (Heleidae), слепни
(Tabanidae) и из семейства бабочниц (Psychodidae) — обширный род
москитов (Pblebotomus). Наибольшее медицинское значение из всех
этих групп имеют москиты, являющиеся переносчиками целого ряда
заболеваний человека, и па этой группе мы остановимся в первую очередь.
В заключение кратко коснемся остальных упомянутых семейств.
ГЛАВ А I
МОСКИТЫ (ГОД rnitBCTCMUS, СЕМЕЙСТГО rSYCHOBIDAE)
1. СТРОЕНИЕ
Псд названием <москиты» у нас известны мелкие кровососущие дву-
крылые, принадлежащие к роду Phlebotcmus.
Род Piilebclcinus относится к семейству бабочниц, Psychodidae. Ос-
тальные роды этого семи ства, как правило, не являются кровососущими,
по представители рода Phlebotcmus все питаются кровью.
Наружное стрдгие. Москиты — мелкие насекомые, длиной от 1,3
до 3,5 мм. Голова, тело и крылья густо покрыты желтоватыми или темно-
ватыми волосками. Голева москита — небольшая, несет пару больших
сложных глаз чо] него цвета; кпереди голова продолжается в большой на-
личник. Усики 16-члениковые,первый членик короткий, цилиндрический,
второй — шаровидный, остальные членики — узкие и удлиненные. Хо-
боток довольно длинный, колющий, состоит из тех же частей, как и хоботок
комаров. Нижнечелюстные щупики пятичленистые, 1-й членик наиболее
короткий, 5-й самый длинный. Крылья широкие, остроконечные, волоса-
тые, без чешуек, в псксе приподняты (рис. 200). Ноги довольно длинные
и тонкие, однако короче и крепче, чем у комаров, покрыты волосками,
чешуйками и щетинками. Лапка состоит из пяти члеников и закапчивается
парой коготков.
Брюшко состоит из 10 члеников, причем последние два видоизменены
и образуют наружные части полового аппарата. У самок они состоят из
пары верхних и пары нижних пластинок, покрытых мелкими волоскахми
и шипиками. У самцов 9-й и 10-й членики превращены в наружные поло-
335
вые члены (гипопигий), устроены довольно сложно и разнообразно и
•имеют значение для систематики. Гипопигий самца (рис. 201) состоит
из следующих частей.
Рис. 200. Общий вид москита (Phlebotomus),
самец.
1. Верхние щипцы двучле-
нистые: первый, более мощный,—
проксимальный членик и меньшей
величины — дистальный. Верхние
щипцы, как и все части наруж-
ного полового аппарата самца,
густо покрыты волосками. Кроме
того, у некоторых видов имеются
еще особые пучки волосков, рас-
положенные непосредственно на
основном членике верхних щип-
цов; иногда они сидят па специ-
альных выступах или буграх и
носят название «кисточки». Кис-
точка находится на основании
членика, с внутренней его сто-
роны. Второй членик верхних
щипцов песет 4—5 длинных ши-
пов, расположение которых раз-
лично у разных видов москитов.
Форма шипов бывает тоже раз-
лична: они могут быть прямые
или изогнутые, тупые пли заост-
ренные. Шипы, находящиеся на
вершине членика, называются
апикальными, смещенные кзади—
субапикальными и расположенные
в середине членика носят на-
звание медиальных и субмеди-
ал ьпых; шипы, сидящие у осно-
вания членика, называются ба-
зальными.
2. Нижние щипцы — длин-
ные, одночлениковые придатки.
У большинства видов они устрое-
ны более или менее одинаково,
только у Ph. papatasii они несут
шипы и называются вооружен-
ными. У других наших видов
на нижних щипцах шипов пет.
3. Копулятивные створки —
плотные хитиновые отростки. Фор-
ма их в большинстве случаев
треугольпая, только у группы
Ph. major они более удлиненные.
Конец копулятивных створок мо-
жет быть закруглен, раздвоен,
иметь шип или вилку, особенно у
группы Ph. major (рис. 202).
4. Срединные пластинки со-
стоят из непарного придатка и
двух пластинок. Представляют
собой сильно измененный аналь-
ный сегмент брюшка.
5. Промежуточные придат-
ки — парные выросты, являю-
щиеся ветвью нижних щипцов и
москита.
з — про-
створки;
Рис. 201. Наружные гениталии самца
1 — нижние щипцы; 2 — верхние щипцы;
межуточные придатки; 4— копулятивные
5 — срединные пластинки.
расположенные по бокам копуля-
тивных створок. Промежуточные
придатки состоят из двух частей:
проксимальной и дистальной. Ди-
стальная часть обычно покрыта. волосками, тогда как на проксимальной части
их пет. У некоторых видов с дорсальной стороны промежуточных придатков, на
границе перехода проксимальной части в дистальную имеются бугорки, несущие
336
короткие шипики (Ph. kandelakii). Из innnrx видов у Ph. papatasii промежуточные
придатки несут еще осоэыо палочковидные выросты (рис. 208).
Внутренняя анатомия. Пищеварительный канал начинается ротовой
полостью, которая соединяется с глоткой. Ротовая полость у некоторых
видов вооруженная, у других ротовое вооружение выражено слабо или
его совсем не видно. Ротовое вооружение состоит из одного или несколь-
ких рядов мелких шипиков или
зазубрин, расположенных по пе-
реднему краю ротовой полости.
На верхней ее стенке расположено
пигментное пятно, размеры п фор-
ма которого различны у разных
видов (рис. 203). Глотка грушевид-
ной формы, на разрезе трехгран-
ная, стенки ее выстланы хитиновой
кутикулой (рис. 204). В расширен-
ной ее части сгепки глотки покрыты
Рис. 203. Ротовые полости и глотки самок
москитов (оригинал).
А — ротовая полость и глотка Ph. sogdianus;
Б — то же Ph. sumbaricus.
Рис. 202. Копулятивные гстворки
москитов группы Ph. major (ориги-
нал).
а—Ph. major: б— Ph. chincnsis;
9—Ph. kandelikii;z—РЪ. keshhhiani;
d— Ph. pcrniciusus var. tubbi.
шиппками, которые образуют шиповатое поле глотки. Последнее имеет
различную форму и своеобразный рисунок для каждого вида москитов.
Глотка переходит в короткий пищевод, куда открывается зоб в виде
большого тонкостенного мешка. В зоб поступают те жидкости, которые
Рнс. 204. Поперечпые разрезы глотки.
1—3—до шиповатого поля; 4,5—па уровне шиповатого поля.
москит насасывает без прокола кожи животного, например, вода, саха-
ристые вещества и т. п. Затем из зоба эти жидкости постепенно перекачи-
ваются в среднюю кишку. На границе между пищеводом и средней кишкой
имеется мускулистая складка, играющая роль клапана. Средняя кишка
состоит из двух отделов: торакальный отдел — узкая мало растяжимая
трубка — и абдоминальный — широкая хорошо растяжимая трубка, куда
22 Учебник медицинской энтомологии 337
Рис. 205. Впутреппге полпрьто
органы самца флоботомус (схема).
I— сгменлиь»:; 2— мгщкл (> ь’;
3—семянкой м< ш )чет.;4—сомяпзгс! га-
т«.1Ы1 ыи канал: б — пологой iiawc;
б — копулятивные нити.
поступает насасываемая пасекомым кровь. При поступлении в желудок
кровь обволакивается перитрофической перепонкой.
За желудком идет короткая тонкая кишка, которая переходит в тол-
стую кишку, расширяющуюся в ректальный мешок с двумя наискось ле-
жащими ректальными железами. Потом идет прямая кишка и анальное
отверстие.
На границе средней и задней кишки в эту последнюю впадает 4 маль-
пигиевых сосуда, которые перед впадением соединяются попарно. Работа
мальпигиевых сосудов у самок москита
подчиняется готютрэфичэск >му ритму, как
и у самок комаров (стр. 217); у голодной
самки опи сильно заполнены экскретом,
после принятия крови происходит выде-
ление секрета в проток сосуда и оттуда
в заднюю кишку (рис. 212). Между срод-
ной и топкой кишкой имеется клапан,
состоящий из мускулистой складки.
Слюнные железы — парное пузыре-
видные органы, расположенные по бокам
переднегруди. Протоки их, сливаясь в
один общий, открываются в подглотэчпик.
Мужские половые органы. Половые
органы самцов состоят из парных семен-
ников овальной формы (рис. 205); ко-
роткие семяпроводы впадают в большой
семенной мешок грушевидной формы, от-
куда выходит узкий семяизвергательный
канал. Последний открывается в поло-
вой насос, хитиновое образование, кото-
рое обеспечивает поступление спермато-
зоидов в копулятивный орган. Половой*
насос состоит из двух хитиновых юлою-
лов, соединенных хитиновым стержнем,
п по краям — мышечными пучками. Се-
мяизвергательный канал впадает в зад-
нюю, выводную часть насоса, в так на-
зываемую капсулу. Из последней выхо-
дят копулятивные нити, представляющие
собой тонкие и длинные хитин )вые трубки.
Они могут высовываться наружу через отверстие, находящееся на концах
описанных выше копулятивных створок. При копуляции нити служат
дтя введения спермы в сперматеки самки.
Женские половые органы. Половые органы самки состоят из двух
овальных яичников, из которых выходят короткие яйцеводы; последние
сливаются в непарный яйцевод. Яичники по строению сходны с яичника-
ми комаров, но содержат меньшее число яйцевых трубок (40—50). В вы-
водную часть непарного яйцевода открывается пара сильно развитых при-
даточных желез. Степень заполнения этих желез секретом зависит от фи-
зиологического состояния самки (рис. 212): у ни разу но клавшей самки в
железах секрета пет, он начинает вырабатываться с принятием пищи и раз-
витием яиц; при наличии в яичниках зрелых яиц придаточные желозы дости-
гают максимального заполнения секретом. Поело откладки яиц в железах
можно обнаружить только следы оставшегося секрета, основная же мас-
са его израсходовалась при откладке яиц. Семяприемников (сперматек)
338
всегда бывает два. Сперматека самки москита состоит из хитиновой ка-
псулы и выводного протока.
Строэтпте кшсулы р1зпоэ5р1зпо. Она бывает сегметгирозапшй и посегмоптиро-
ваппой. В первой стучав кшсула созготг из р пли шо°о числа сегментов, причем по-
следний сегмент имеет угзуб тонио, в котором помещ ierca го точка сперматеки, окружен-
ии группой желошстых клеток в виде шапки. Несегментирозанпые сперматеки де-
лятся па трубчатые и к incyтарные (рис. 236). Выводные прогони бывают различны по
длине и форме одни с гладкими стенками, кор )ткие, в конце сливающиеся вместе, дру-
гие ис юрченпые, дтинные, открывающиеся в вагину отдельно. На просветленных пре-
паратах сперматеку нужно искать в оэласти 7—8-го сегментов брюшка.
Особенности ротовой полости, глотки и сперматеки являются систе-
матическими признаками для определения
Отличие от комаров во внутреннем
строении. Внутренние органы мсскитэв
по своему устройству очень близки к вну-
тренним органам комаров. Главные отли-
чия следующие:
1. Из трех придатков пищевода кома-
ров москиты имеют только один зоб;
парные придатки отсутствуют. 2. Слюн-
ные железы состоят каждая из одной
единственной дольки, тогда как v кома-
ров они состоят из трех долек. 3. М юкиты
имеют не 5, а только 4 мальпигиевых сосу-
да. 4. Смазочная железа полового аппара-
та у комаров непарная и слабо развита;
у москитов опа парная и обо доли ее
сильно развиты. 5. Трахейная система
москита, как и у всех мелких насеко-
мых, развита сравнительно слабо, в част-
ности, отсутствуют клубочки трахеол в
яичниках.
самок москитов.
2. РАЗВИТИЕ
Только что отд о конные яйца светлые,
но вскоре они темнеют и приобретают
корпчневато-сорую окраску. Длина яйца
от 0,35 до 0,38 мм. Форма удлипенн)-
овалыгая, с выпуклой спинн >й и несколь-
ко вогнутой брюпшш поверхностью. На
Рпс. 20G. Формы семяприемников
(сперматек) самок москитов (ори-
гинал).
/—3—сперм «тени, сегментированные
без шейки; •/ и 5— сясрматгки. сег-
ментиповинные с шейкой; в — спер-
М ‘1<*кц с неявно выраженной сегмен-
тацией; 7 и 8—несегм шти зозлшые
сперматеки (схема).
поверхности яйцевой оболочки имеется сетчатый рисунок (рис. 207, Л),
который, видимо, различен у разных видов москитов (Шевченко, 1920).
Личинка червеобразной формы, покрыта волосками. Число члеников
туловища 13, из них 3 грудных и 10 брюшных. Длина только что вышед-
шей из яйца личинки равна 0,5—0,8 мм. Тел) личинки светложелтого
цвета, голова тсмнскоричпевая. На заднем конце тела помещаются удлинен-
ные хвостовые щетинки; у личинки I возраста их 2, у личинок всех осталь-
ных трсх возрастов их 4 (рис. 207, Б, В), Голова личинки почти округ-
лой фермы. На голове личинки 1 возраста имеется яйце вэй зуб, который
предназначен дня вскрытия оболочки яйца и после первой линьки исчезает.
Усики трехчле пиковые, первые два их членика маленькие, цилиндриче-
ские, третий шаровидный, с коротким волсск )м на верхушке. Ротов нт ап-
парат грызущего типа. Нижнечелюстной щупдк однэчдениегый. Между
22*
339
9-м и 10-м члеником брюшка помещается пара стигм, а начиная со
II возраста можно различать и переднюю пару стигм, лежащую между
2-м и 3-м члеником груди.
Личинка четыре раза линяет и после четвертой линьки превращается
в куколку. Куколка имеет около 3 мм в длину; она желтоватого цвета,
булавовидной формы (рис. 207, Г), Последние членики брюшка куколки
одеты шкуркой личинки IV возраста, благодаря которой куколка при-
крепляется к субстрату. Куколка неподвижна.
Рис. 207. Предимагинальпые стадии москитов.
А—яйцо москита; Б—личинка москита I стадии; В—личинка москита IV/стадии;
Г—куколка москита/"
' 8. СИСТЕМАТИКА
Водящиеся в СССР виды москитов делят па четыре группы по типу
наружного полового аппарата самца и по форме сперматеки самки. Эта
классификация не является совершенной, так как некоторые виды мос-
китов не укладываются в эти группы (самки Ph. chinensis п Ph. clydei), но
она является наиболее удобной и помогает разобраться в наших москитах.
Первая группа — Ph. papatasi. Последний членик верхних щипцов удлиненный,
почти цилиндрический, с 5 короткими шинами, из коюрых 3 лежат па гершпие членика
и но 2 по бокам. Нижние щипцы вооруженные (самцы). Сперматека сегментированная
(8—10 сегментов), без шейки. Сюда относится единственный вид Ph. papatasii Scop,
(рис. 208).
Вторая группа — Ph. sergenti. Последний членик верхних щпппов укорочен,
песет 4 длинных шипа. При основании первого членика имеется кисточка (3)« Спер-
матека сегментированная, без шейки (Q). Сюда относя гея Ph. se.genti, Ph. sergenti
var. mongolonsis, Ph. sergenti vac. alexaadri n Pn. caucasicus (рис. 209).
Третья группа — Ph. major. Москиты этой группы характеризуются удлиненны-
ми члениками верхних щипцов, длинными шипами вт орот о членика, из которых 3 рас-
положены в средней части членика на хорошо развитых бугорках, а 2— па ксице. Ко-
пулятивные створки длинные (3). Сперматека самки сегментированная, с шейкой,
(см. рис. 206. 4 и 5). Особняком стоит самка Ph. chinensis, сперматека которой имеет
исчерченные стопки капсулы, без ясного подразделения па сегменты и лишена длин-
ной шейки. Самцы отдельных видов этой группы различаются между собой строением
копулятивных створок и промежуточных придатков. У нас наиболее часто встречаются
340
следующие виды: Ph. major, Ph. kandelakii, Ph. perniciosus var. tobbi, Ph. perfili-
cvi. Ph. perfilievi var. transcaucasicus, Ph. wenyoni, Ph. keshishiani Ph. chincnsig
и Ph. smirnovi (рис. 210). ’
Четвертая группа — Ph. minutus. Второй членик верхних щипцов самца имеет
форму вытянутого треугольника, несколько изогнуто! о подлине, и несет 4 длинных кон-
цевых шипа. Ротовая полость вооруженная. Сперматека самки песегмептированная
Рис. 208. Ph. papatasii.
Гипопигий самца, глотка и сперматека.
Рис. 209. Ph. sergenti.
Гипопигий самца, глотка и сперматека.
Рис. 210. Ph. chinensis.
Гипопигий самца, глотка п сперматека.
Рис. 211. Ph. minutus.
Гипопигий самца, глотка и сперматека.
(исключение представляет Ph. clydei Sinton, у которого сперматека сегментированная»
по самец вполне подходит под общую характеристику группы). Сюда относятся следую-
щие виды, встречающиеся у нас: Ph. sogdianus, Ph. graekovi, Ph. squamipleuris,
Ph. minutus var. arpaklensis, Ph. sequens, Ph. sumbaricus и др. (рис. 211).
4. ЭКОЛОГИЯ
Биотопы. В отличие от комаров москиты не имеют связи с водой.
Их личинки встречаются во влажном, но не мокром субстрате. Яйца и ли-
чинки младших возрастов довольно требовательны к влажности, но стар-
шие личинки и куколки выживают в более сухой среде. Москиты живут
как в дикой природе, вдали от человеческих поселений, где человек
для них не может быть источником пищи, так и вблизи человека, в селе-
ниях и городах. Когда человек впервые поселяется в необжитой местно-
сти, некоторые из живших там видов москитов приспособляются к новой
обстановке и массами размножаются во вновь созданных человеком ме-
341
стообитанйях (Петрищева). Москиты откладывают яйца в темные и влаж-
ные места, богатые органическими веществами.
Температурные требования личи/ок. Креме влажности и пищи, для
развития личинок и их последующего превращения необходима довольно
высокая температура. В условиях эксперимента при 28° развитие яйца
Ph. papatasii длится 7 дней, личиночное развитие —28—35 дней. Колебания
температуры удлиняют срок развития до 2 месяцев и больше. Первый ли-
чиночный возраст длится в среднем 5—6 дней, второй — 7—8 дней, третий
8—9 дней и четвертый — сколи 10 дней. Куколка развивается в точение
10—12 дней. Оптимальная температура в лабораторных условиях 26—30°.
При температуре ниже 18° развитие прекращается. Продолжительность
развития предпмагинальных стадий отдельных видов москитов при одних
и тех же температурах различна (Шевченко, Балашева, Лисова, Кама-
лов и др.) (табл. 9).
Таблица 9
Продолжительность (в днях) предиуагппалыюго развития
некоторых видов москитов в :ависи ости от температуры
20-28° 22—28° |2’«° ( «О.1С- б< It IH I 2 1-23°) 2 5-30°
Ph. papatasii 41—59 44-58 69 47—56
Ph. sergenti 37—44 53
Ph. caucasicus .... 34-46 58
Ph. chinensis 49—45 58 42—52
В большинстве случаев москиты зимуют в стадии личинки IV возраста,
но личинки довольно чувствительны к холоду. Наблюдения Соколова и
Тарвит и др. показали, что личинкп Ph. papatasii погибают после 41/,
дней при 0° и в 2—3 дня при —2°, через 2*/2 часа при —6° и через 30 ми-
нут при —10°. Наиболее выносливы к холоду личинки Ph. chinensis.
По наблюдениям в Китае личинки этого вида длительнее время (более
месяца) выживают при 0°. Яйца москитов, попавшие на сухой субстрат,
не развиваются, по сохраняют свою всхожесть в течение 25 дней пребыва-
ния на сухом субстрате при умеренной влажности окружающего воздуха
(Петрищева). По наблюдениям Исаева, яйца после месячного пребывания
в воде сохраняют жизнеспособность. Из яиц Ph. papatasii и Ph. caucasicus,
выдержанных в течение часа на солнце, личинки не развиваются (Мирзоян).
Биология личиьок; разведегие мосььтов. Личинки питаются органи-
ческими веществами как животного, так и растительного происхождения.
В природных условиях личинок разыскать очень трудно, и почти все,
что нам известно о личинках москитов, является результатом наблюде-
ния в лаборатории. Методика разведения москитов в лаборатории заклю-
чается в следующем.
Самок со зрелыми яйцами помещают в пробирки, стаканы или банки, па дпо кото-
рых помещается ватная подстилка и сверху фильтровальная бумага. На сухую бумагу
самка яйца не откладывает. Видимо, влажная поверхность является раздражителем,
вызывающим кладку яиц. Через несколько часов, а иногда через 1—2 дня самка от-
342
кладывает яйца. Число яиц, откладываемых одной самкой, значительно колеблется —
от нескольких до 90—100. Позле этого в пробирку насыпается субстрат — помет
кроликов, коз или листья с деревьев. Предварительно субстрат измельчается и просу-
шивается при 60" в точе! и? не менее I — 17а пасов. После эюго он слабо увлажняется
и насыпается в пробирку пли стаканы па отложенные япца* Этоi субстрат является
средой об in алия п нишей для личинок. Он должен все время поддерживаться в увлаж-
ненном соло шии, для чего необходимо ежедневно в пробирки вливать несколько ка-
пель вохы на фильтровальную бумагу. Вместо это; о о юнь удобен метод, предложенный
А. 11 Щуронкозоя: в химические пробирки вливается 3—4 см* воды, затем вставляется
ватный тампон, ко:орый плотно подюняется к поверхности воды; на тампон кладут кру-
жо тки фильтровальной бумаги, и пускают самку для откладки яиц. Здесь исключается
необходимость ежедневно: о увлажнения пробирок.
Окукление происходит на поверхности субстрата. Вылет взрослых
москитов из одной и той же кладки происходит неравномерно, интервал
может быть в несколько дней, от 10—20 и больше. Процесс откладки яиц
в лабораторных условиях сильно истощает самку, и в большинство слу-
чаев опа при первой кладке яиц гибнет. В природе происходит не так: там
многие самки остаются после откладки живыми и дают иногда по нескольку
кладок, проходя ряд повторных гонотрофических циклов. Очевидно, мы
еще не умеем создавать в лаборатории достаточно хорошие условия для
кладущей самки.
Методы обнаружения личипок. Поискам мост выплода москитов
в природе уделяется большое внимание.
Псрсопачальпым методом отыскания личинок служил просмотр подозреваемого
субстрата год лупой. Таким способом огысказь личинок довольно трудно, так как
они очень малы, мало подвижны и, kj оме тою, к пх телу, густо покрытому волосками,
прилипают часзицы земли и маскируют их. Второй способ—испытание подозревае-
мых субстратов па выплод москитов. Субстрат приносят в лабораторию и покрывают
садком или колпаком; вынлодившнеся москиты задерживаются в садке. Третий ме-
тод— изолируют отдельные места, накрывая их опрокинутыми ящиками, обклеенными
внутри липкой бума: ой. Вылетающие москиты застревают иа липких стейках ящиков.
В сараях, курятниках, хлевах устраиваются ловушки па влет и вылет (Буракова и
Мирзоян). Кроме того, существует метод промывки подозреваемою субстрата на ситах
с ячеями последовательно уменьшающеюся диаметра. Последний промыв погружается
в насыщенный раствор нозаренпой соли, и имеющиеся там личинки всплывают на по-
верхность. Петрищева и Взюмская несколько видоизменили этот метод: субстрат по-
мещался в тазы, заливался водой и размешивался. После каждою размешивания жид-
кость отстаивалась 5—10 минут, и всплывший мусор осторожно выбрасывался. Так
повторялось несколько раз. Тщательно очищенный субстрат процеживался через
мелкоячеистое сито п заливался насыщенным раствором поваренной соли. За сезон им
удалось обнаружить 162 личинки из 6 т промытою субстрата.
Места выплода. Основными местами выплода москитов в Севастополе
являются подпольные пространства, что обнаружено и Долматовой
в Ленинабаде. Рядом советских авторов личинки были найдены в сле-
дующих местах: 1) в естественных биотопах: поры черепах, туркестанской
крысы, пластинчатозубой крысы-незокии, большой песчанки, тонкопа-
лого суслика, дикобраза; дупла яблони и орешника, пещеры разного рода
и т. д.; 2) в хозяйственных биотопах: свинарники, норы домовых грызунов,
мусорные ящики, скотные дворы, растительный субстрат в огороде, под
полом в жилой комнате, в подстилке собачьей будки и т. п.
Фегология. В Крыму москиты появляются в конце мая и исчезают
в начале октября. В Средней Азии они появляются в апреле и исчезают
в октябре. За этот период замечается два максимума численности
москитов — один в июне, другой в конце августа, очевидно, отвечающие
двум поколениям. Иногда эти сроки смещаются в зависимости от темпера-
туры. В Семиречье Ph. chinensis дает один единственный максимум чи-
сленности и, повидимому, одну генерацию в год. Зимуют москиты в стадии
343
Стадии X. Я и U и и К О В Стадии ч. п ищебарения / и
. 1
/ 344 30
2 г.; 54 93
3 vL 1 ик 50 545
4 Lb 57
5
6
7 Ч 1 1
Женские придаю о ч н ь/е <ж е л е з 6/ 0 d с/ 1 в й ш
Рис. 212. Гопотрофичоские взаимоотио-
Цпфры таблицы указывают относительную частоту, с которой встречается копире из
переваривания крови. Кроме того, наглядно показано соответствие между сгади-
шепия у Ph. papatasii (оригипал).
сочетаний той пли ипой стадии развитии яичников с той или ипой стадией
ями гоно трофического цикла и состоянием смазочных желез и мальпигиевых сосудов.
личинки IV возраста. В южной Туркмении Петрищева наблюдала лёт
москитов из нор круглый год.
Характер паразитизма москитов. Подобно комарам, москиты обла-
дают двойственным питанием, хотя самки москитов прибегают к углевод-
ному питанию реже, чем самки комаров. Для созревания яиц кровяное
питание необходимо. Таким образом, москиты, как и комары, являются
обязательными, но не исключительными кровососами.
Москиты, как и комары, являются периодически нападающими, сво-
бодными хищниками. Обычно они отличаются гораздо меньшей степенью
гетеротопности, так как откладывают яйца не в воду, а в почву, и притом
часто в тех же убежищах, где прячутся взрослые особи: в порах, в пеще-
рах, трещинах скал, в хлевах, уборных, домах или в их ближайшем со-
седстве. Эти же места часто служат и убежищами для хозяев москитов —
разных млекопитающих, птиц и рептилий. Таким образом, москиты часто
не имеют надобности далеко летать за добычей: они находят и добычу,
и места для откладки яиц в непосредственной близости друг от друга.
Москиты летают значительно хуже комаров и па меньшие расстояния.
Наибольшая дальность полета москитов в полупустынном ландшафте—
1,5 км (Латышев). В условиях города они обычно летают не более чем
на десятки метров: так, Петрищева в Севастополе путем выпуска окрашен-
ных москитов 1 установила наибольшую дальность полета в 25 м. Пра-
виков этим же методом в Ашхабаде нашел, что москиты, выплодившиеся
в данной усадьбе, как правило, в течение своей жизни не вылетают за ее
пределы. Обычно москиты передвигаются короткими перелетами, скач-
ками; в дом через открытые окна большая часть москитов проникает в
припрыжку по подоконнику и сторонам окна, меньшая часть — прямым
полетом. Эта их особенность облегчает борьбу с пими посредством стой-
ких контактных ядов, наносимых па стоны1 2 3. В то же время москиты го-
раздо лучше бегают и прыгают, чем комары, что позволяет им свободно
и ловко двигаться в узких порах и щелях и избегатьпападений со стороны
хозяина поры. Этому способствует и их малая величина.
Однако настоящими поровыми паразитами они не стали: способность
к полету избавляет их от необходимости сидеть в поре, покинутой хозяи-
ном, и дожидаться там какой-нибудь добычи. Голодный москит покидает,
как комар, свое убежище и улетает искать добычу. Благодаря этому у мо-
скитов в их эволюции не выработалась пн долговечность, ни способ-
ность выносить голодание, ни большая прожорливость, свойственные
настоящим поровым паразитам, вроде клопов или клещей аргазид.
В основном они сохранили жизненную схему свободного кровососа, свой-
ственную другим кровососущим низшим двукрылым, хотя и с некоторыми
особенностями.
Гопотрофическая гармония, повторность гопотрофических циклов.
Подобно комарам, самки москитов всецело подчиняются закону гонотро-
фической гармонии и проделывают в течение своей жизни по нескольку
гопотрофических циклов (Долматова). Все физиологические отправле-
ния самки подчиняются гонотрофическому ритму (рис. 212). У москитов,
как и у комаров, известный процент самок умирает па каждом гонотро-
фическом цикле, но некоторые особи проделывают большое число циклов.
В летней популяции москитов встречается 20—40% самок на повторных
гопотрофических циклах и 60—80% самок на первом гопотрофическом
1 Окраску москитов производят по методу Мошкозского. Москитов помещают в
большую банку. Стенки банки густо обсыпают тонким порошком анилиновой краски,
зернышки которой застревают между волосками москита.
3 Методы борьбы с москитами описаны в курсе борьбы с насекомыми.
346
цикле (еще не клавших). Это указывает па несколько большую смерт-
ность самок москитов, чем самок Anopheles maculipennis, все же не пре-
пятствующую им служить переносчиками заболевании (см. ниже).
Определение возраста. Отличить клавших самок москитов от ни разу
не клавших, молодых, можно двумя способами: по структуре яични-
ков и по состоянию смазочных желез.
Первый способ основан на том, что после откладки яйцевые трубоч-
ки в той части, где было зрелое яйцо, некоторое время остаются растя-
нутыми, что придает яичнику сетчатый вид (рис. 213). Правда, через не-
которое время стенки яйцевой трубочки снова сокращаются и определение
уже невозможно. Как правило, на следующем за откладкой гонотрофи-
ческом цикле растяжение трубочек бывает
хорошо заметно на I и II стадиях пище-
варения и изредка — на III стадии.
Второй способ основан па том, что у
новорожденных самок, еще по пивших
крови, в смазочных железах совершенно
пет секрета; он появляется там лишь
после принятия крови и с началом раз-
вития яичников. К моменту созревания
яиц смазочные железы бывают перепол-
нены секретом. При откладке яиц секрет
используется, но часть его остается в же-
лезах. Наличие остаточного секрета в
Рпс. 213. Яйцевые трубочки сам-
ки москита (оригинал).
А — оТ ни разу нр |.лан1псп самки;
Б—от све>кеитЛ('ии1Р1ьеп с;»м« н; ди-
стальн ’н часть т| уиьи сильно растя-
нута; В—дистальная часть 1рубкн
начала соь.ращачься.
смазочных железах у самок с пустыми
желудками и неразвитыми яичниками и
служит признаком того, что самка уже
отложила яйца.
Активность, типы убежищ. Как и
комары, москиты — сумеречные насеко-
мые. Активный лет их и нападение па
добычу совершаются перед
заходом солнца и в первые часы после захода; днем они нуждаются в убе-
жище и прячутся в укромные, затемненные моста, не проявляя активности.
Некоторые убежища удовлетворяют всем потребностям самки, предостав-
ляя ей и защиту от неблагоприятных метеорологических условий (жары,
сухости, ветра, холода, сырости, дождя), и добычу, п благоприятные ме-
ста для откладки яиц. Таковы, например, поры грызунов, а также некото-
рые постройки человека, например, хлева. Другие помещения служат им
только местом питания или местом питания и дневным убежищем, или
только местом откладки и т. д. Из тех убежищ, которые удовлетворяют
всем потребностям москита, они, казалось бы, могли бы п совсем не выле-
тать. Однако Латышев в населенном пункте, расположенном в полупусты-
не с массой нор грызунов, наблюдал, что в вечерние часы москиты выле-
тали из пор и летели в поселок. У москитов, как и у комаров, существует
положительный фототаксис к сумеречному свету. Не все москиты поки-
дают вечером ноуы. Вылетают главным образом те из них, у которых име-
ются внутренние раздражители — голод, жажда и половой инстинкт
(Долматова). На рис. 214 указан состав москитов, вылетающих из нор,
и мы видим, что среди вылетающих москитов большинство либо голодны
и летят в поисках пищи, либо ото самки со зрелыми яйцами, летящие
в поисках мест откладки яиц; па промежуточных стадиях гонотрофического
цикла вылетают лишь немногие москиты для утоления жажды. Из не-
обитаемых пор (рис. 214, слева) вылетает, естественно, голодных моски-
тов больше, чем из нор, имеющих хозяина (рис. 214, справа). Вылетая
347
из нор, москиты влетают в такие же норы или, как сказано выше, летят в
близлежащие человеческие поселения. В помещениях человеческих посе-
лений, каки в колониях пор, происходит такой же круговорот москитов
на дпевках. Круговорот москитов на дневках, как и повторность гонотро-
фических циклов, играет большую роль в распространении ими болезнен.
Сборы п хранеппе москитов. Для изучения фауны москитов в тол или дру-
гой местности^ сборы москитов нужно произгодить не только в жилых помеще-
Рис. 214. Биологический состав москитов,
вылетающих из нор. Распределение по ста-
ниях и иадгорных постройках, по и
вдали от жилья: в норах, пещерах и
тому подобных местах. В комнатах
москитов можно ловить рано утром или
вечером, так как днем они или выле-
тают из помещения, или забираются
под висящую одежду и в щели стен.
Взатененных помещениях москитов мож-
но обнаружить и днем. Москиты хорошо
видны на побеленных стенах, па сером
фоне они слабо заметны. Москитов, как
и комаров, можно собирать длинными
химическими пробирками. Если москп-
тов нужно сохранить живыми, то про-
бирку с ними следует держать в прох-
ладном и темпом месте. Если москитов
дням пищеварения самок.
На ординате — числа москитов (в процентгх: на
абсциссе—стадии пищеварении; 1—москиты,
вылетающие из обитаемых нор; 2— москиты, вы-
летающие из необитаемых нор.
сохранить живыми не нужно, на дно
пробирки кладут кусочек ваты, смолен-
ной эфиром; в такую пробирку можно со-
брать до 20 москитов. Хранить москитов
можно в 75° спирту или в виде сухого
материала — в пробирках или коробках. Применяется еще вылов москитов на лип-
кие листы бумаги, которые смазывают касторовым маслом. С липучек москитов сни-
мают кисточкой, смоченной 96° спиртом, а для храпения переносят в 75° спирт. Вы-
лавливать москитов можно и ловушками, как комаров.
5. ЭПИДЕМИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
Уколы москитов чувствительны и могут вызвать сильную местную
реакцию и нестерпимый зуд, особенно после некоторой сенсибилизации
(стр. 150) человека к их укусам. У постоянных жителей местностей, богатых
москитами, вырабатывается известный иммунитет к их укусам. Но главное
значение москитов для человека заключается в том, что они являются пере-
носчиками ряда заболеваний — лихорадки паппатачи и лейшманиозов. Эти
заболевания встречаются только там, где есть москиты.
Лихорадка паппатачи. Возбудителем этого заболевания является
фильтрующийся вирус.
Вирус в периферической крови больного циркулирует короткое гремя — одни
сутки перед заболеванием и в течение первых суток болезни, т. е. всего двсе суток.Толь-
ко в это время самка москита и может заразиться, напившись крови больней о. Человек
заражается через укус зараженной самки москита. Инкубационный период колеблется
от 2х/2 до 6—7 дней. Болезнь начинается быстрым подъемом температуры до 39—4Г
и держится 3—5 дней.
Заболевания лихорадкой паппатачи встречаются только в период
лёта москитов. Обычно наблюдается две волны заболеваемости, которым
предшествуют две волны вылета москитов. Кривая лёта москитов подни-
мается на несколько дней раньше, чем кривая заболеваемости лихорадкой,
причем интервал этот равен продолжительности инкубации. Поэтому
учет сезонного хода численности москитов в эндемичных местностях имеет
большое значение. Москит, насосавшийся крови больного, может заражать
здоровых людей только через 6—8 дней после кормления па больном.
Очевидно, этот срок необходим для того, чтобы вирус проник в слюнные
железы москита.
348
Работами советских авторов в 1936 п 1937 гг. был установлен факт
трапсовариалыюй передачи вируса лихорадки паппатачи дочернему и
даже внучатному поколению москитов (М ликовский, Петрищева и др.).
Москит, происходящий от зараженной лихорадкой паппатачи матери, мо-
жет заражать человека при. первом же сосании крови.
В пределах СССР лихорадка паппатачи встречается в некоторых мест-
ностях Крыма, Молдавии и Кавказа и в республиках Средней Азии.
Основным переносчиком лихорадки паппатачи является Ph. рара-
tasii, а в некоторых местах в переносе, может быть, участвуют и другие
виды москитов.
Кожный лейшманиоз. Возбудителем кожного лейшманиоза является
Leishmania tropica (класс Flagellata, отряд Protcmonadina, семейство
Trypanosomidae). Кожный лейшманиоз распространен в Африке но бе-
регам Сред иземного моря, на Балканском полуострове, в Передней и
Средней Азии. В пределах СССР имеется в среднеазиатских республи-
ках и Восточном Закавказье; наиболее поражена Туркменская ССР. Для
СССР известны два типа кожного лейшманиоза: поздно изъязвляющийся
и остро некротлзующийся (К лкэвпикэв).
Поздно изъязвлятсптийся лейшманиоз (ашхабадская язва, кокапдка, годовик)
встречается преимущественно в городах. Инкубационный верьод обычно 2—6 месяцев,
нередко 1—2 года. Первоначально появляется небольшая папула, которая через
3—6 месяцев изъязвляется. Язва держится год и больп е. Первичное заболевание воз-
можно па протяжении всего года. Никаких друтих гозгогочвых-хозяев, кроме че-
ловека, пока неизвестно. Лица, перенесшие этот тип кожною лейшманиоза, могут
заболеть вторым типом.
Остро некротизующийся лейшмаппоз (пендинская язва, мургабская язва)
встречается преимущественно в сельских поселках, па окраинах городов и в пустын-
ных местностях. Инкубационный период 1—4 педели. Появляется папула в виде фу-
рункула, ко орый через 1—3 педели изъязвляется. Язва держится 3—6 месяцев.
Свежие забо ювапия возможны только в летне-осенние месяцы. Хранителем паразита
являются дикие грызуны пустыни: толстая песчанка, тонкопалый суслик и др. (Ла-
тышев).
Переносчиком кожного лейшманиоза у нас в СССР являются моски-
ты группы Ph. sergenti и Ph. papatasii. Лсйшманпи в организме москита
проходят определенный цикл развития, и заразными становятся москиты,
выдержанные не менее 7—8 дпей после инфицирующего питания (Ходу-
кин, Софиев и др.).
Висцеральный лейшмагиоз. Возбудителем висцерального лейшмани-
оза является L. donovani. Морфологически она неотличима от L. tropica.
В СССР висцеральный лейшманиоз распространен в среднеазиатских
республиках и Восточном Закавказье. За пределами СССР распростра-
нен в Африке, Средиземноморье, Индии и Китае. Болезнь очень тяжелая,
летальность ее (т. о. процент умирающих от этой болезни к общему числу
заболевших) очень велика. Лейшмании поражают внутренние органы че-
ловека: селезенку, лимфатические железы, костный мозг, печень; их
можно также найти в клетках кожи и в крови.
Ипкубтцчотпый период длятся от пемпогих педель до песколькпх (3—9) месяцев.
Болезнь сопровождается неправильной и упорной лихорадкой; селезенка и печень по-
степенно увеличиваются, ипо.’да до громадных размер ов. Болезнь идет быстро или
перохощт в хроническую форму (длится 1—3 года). Наибольшее число свежих забо-
леваний приходится па начало весны и на осень. В Средней Азии и Закавказье больше
всею бо leioT дети, в Индии главным образом поражается юношеский возраст. Кроме
человека, сходное забозевание бывает и у собак. Возбудителем его является L. canis.
В 1946 г. Латышев в Средней Азии обнаружил лейшманиоз шакалов в естественных ус-
ловиях. Возможно, что лейшманиоз шакалов, собак и человека тождественны.
349
Переносчиками висцерального лейшманиоза являются москиты труп-
пы Ph. major. У пас в СССР обвиняются Ph. chinensis и Ph. kandelaki,
в Индии — Ph. argentipes, в Китае — Ph. chinensis и в Средиземноморье—
Ph. perniciosus.
6. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Москиты распространены во всех частях света, главным образом в тро-
пической, субтропической и прилегающих к ним зонах. Это объясняется
тем, что москиты теплолюбивы: они не выносят холодной зимы и нуждаются
в жарком лете. Они имеются в Средиземноморье, Африке, Америке,
Индии, Индокитае и Китае. В пределах СССР они встречаются во всех
среднеазиатских республиках, включая Киргизию и южную часть Ка-
захстана, на Кавказе, на юге Украины, в Молдавии и Крыму. Северная
граница их распространения лежит между 45° и 50° с. ш. Во влажной части
Черноморского побережья Кавказа (Сочи-Батуми) москитов нет.
ГЛАВА II
МСШКП (СЕМЕЙСТВО SIMULHDAE)
Мелкие двукрылые, плотным телосложением напоминающие скорое
мух, нежели комаров. У наиболее крупных видов тело по длинное A мм,
нередко около 2 мм. Усики короткие и толстые, по состоят из 22 более или
менее одинаковых члеников. Крылья относительно велики. Самцы легко
отличимы по громадным глазам, сливающимся между собой по средней
линии.
Личинки живут в текучих ведах. Голова напоминает голову личинок
комаров, по усики трехчленистые. Верхняя губа разделена на два при-
датка, несущие мощные веера щетинок, служащие для уловления добычи.-
На переднегруди имеется попарная <ножка». Брюшко к концу вздуто, и
последний (девятый) членик песет присоску, вооруженную множеством
шипиков, при помощи которой личинка присасывается к подводным пред-
метам — камням, кустам, корням и т. п. Опа может ползать, как пиявоч-
ка, и при этом прядет питьпаутины,при помощи которой может вернуться
па свей камень или пень, если ее случайно оторвет течением. Личинки
всегда живут в текучей воде, насыщенной кислородом, и в стоячей воде
быстро задыхаются. Вырссшая личинка прядет домик, приклеенный к пред-
мету, на котором она жила: домик имеет вид колпачка с отверстием в сто-
рону течения. Куколка снабжена длинными трахейными жабрами. Взрос^
лая менка поднимается на погерхшеть реки в пузырьке воздуха;
пузырек лопается, и спа вылетает сухая из воды.
Взрослые мешки имеют двойственное питание, подобно комарам. Сам-
цы роятся в воздухе, копуляция происходит на лету. Мошки обладают
исключительно дневным зрением и нападают на добычу только днем, на
открытом воздухе. Ни в сумерки, ни в помещении они нападать не спо-
собны. Массовые нападения мошек причиняют людям и живот-
ным большие страдания, а для последних бывают иногда гибельны. Слюна
мошек имеет резко выраженное ядовитое действие, и сенспбилизованный
предшествующими укусами скот нередко гибнет от их укусов. Особенно
известны в этом отношении некоторые местности Румынии и Венгрии,
где массовый падеж скота от укусов симулий не представляет редкости.
У человека отравление укусами мошек встречается очень редко, но массо-
вые их нападения и для людей крайне тягостны.
350
б
Охотящиеся самки мошек разлетаются иногда па несколько кило-
метров от реки, где живут их личинки. Самка полностью подчиняется закону
гонотрофической гармонии: один прием крови необходим и достаточен для
созревания одной порции яиц (Рубцов). Самка может откладывать в те-
чение жизни несколько кладок. Для откладки яиц самка спускается под
воду и приклеивает целые кладки к подводным предметам.
Симулий существует довольно много видов. В соответствии с этим
места выплода довольно разнообразны и встречаются в самых различных
водотоках: от больших рек, вроде Волги, до маленьких ручейков и от гор-
Рис. 216. Мокрецы (семейство Ilclcnlae).
А—Culicuides pulicarius. гзгоелпя самка со
спины; Ь—личинка л В— кхкплка одни го
из видов мокрецов, р; si пвактпхся в i од« ;
Г—личинка п Д—к\колка одн< го из гидов
мокрецов, ррз! иг; ЮЩ1 хсн на суше (в лесной
^подстилке) (сильно увеличены).
ных потоков до медленно текущих
речек. В стоячих водах они, одна-
ко, никогда по встречаются. Мно-
гие виды имеют у пас два поколе-
ния в год. Зимуют личинки. Симу-
лин встречаются всюду, где име-
ются постоянные (неперссыхаю-
щие) водотоки.
В Африке мошки являются пе-
реносчиками одного из филариидо-
зов человека.
ГЛАВА III
МОКРЕЦЫ
(СЕМЕИСТ1О HELEIDAE)
Крошечные комарики, некото-
рые из них имеют в длину менее
1 мм, так что являются самыми
мелкими из всех кровососущих дву-
крылых. По телосложению строй-
нее, уже и значительно нежнее си-
мулий. Ноги у них не такие длин-
ные, каку комаров. Усики состоят
из 13 или 14 члеников; колющий
хоботок относительно короткий.
Личинки некоторых видов
мокрецов живут в стоячих водах,
других видов — во влажных ме-
стах па суше: в лесной подстилке,
в дуплах, под корон, даже в отбро-
сах. Водные личинки узкие и длин-
ные, плавают, змеевидно извива-
ясь (см. также рис. 57, Г, 4), охотно поселяются в плавающих скопле-
ниях нитчаток. Некоторые виды живут в солоноватой воде (в соленых
озерах, в заливах Аральского моря). Наземные личинки значительно
короче и, подобно личинкам тендипедид (стр. 98—99), имеют по паре
ножек на 1-м членике груди и 9-м членике брюшка.
Самки нападают на людей п животных и днем во влажных ме-
стах, и в сумерки, и под открытым небом, и в помещениях. Невидимому,
они подчиняются закону гонотрофической гармонии. В тех местах, где
мокрецы встречаются в больших количествах, нападения их крайне тя-
гостны (например, в лесах Уссурийского края по Мончадскому). Распро-
странены повсеместно.
352
Мокрецы — большая груйпа, с разнообразным образом жизни, не-
достаточно изученная. Некоторые виды мокрецов нападают и на беспо-
звоночных: червей, насекомых. Представители рода Culicoides иногда напа-
дают на насосавшихся крови комаров (Anopheles, Culex) и высасывают
кровь из их брюшка, вызывая гибель комара.
Некоторые виды мокрецов являются переносчиками филариидозов
человека (Африка, Южная Америка).
ГЛАВА IV
СЛЕПНИ (СЕМЕЙСТВО TABANIDAE)
; 1. СТРОЕНИЕ'И СИСТЕМАТИКА СЛЕПНЕЙ
К семейству Tabanidae — слепней — относятся* крупные и сред-
ней величины мухи с большой, вытянутой в поперечном направлении го-
А В
Рис. 217. Слепни.
Рис. 218. Голова Tabanns
сбоку.
1— глаза; 2— усики:
3 — хоботок.
А—ТчЬапаз—слепень (общяй вид со спинной стороны); Б—. Chrysops.
златоглазик (общий вид со спинной стороны); В— Chrysozona, дождевка (общин
вид со спинной стороны)'
ловой (рис. 217 и 218). Бблыпую часть головы
занимают крупные фасеточные глаза, у живых
мух красиво окрашенные в зеленый, желтый, крас-
ный и другие цвета с металлическим отливом.
Усики состоят из 6—7 члеников, из которых 3-й
сильно увеличен, а все дальше лежащие, начиная
с 4-го, значительно меньше по величине и ча-
стично сливаются между собой. Ротовые органы,
в виде хоботка, приспособлены как для проколов
кожи и сосания крови из сосудов, так и для под-
лизывания свободных жидкостей, например, ра-
стительных соков или воды. В состав хоботка вхо-
дят те же части, как и у других кровососущих
Orthorhapha, в том числе одна пара максилл и
одна пара мандибул. Мощное развитие ротовых
органов придает им режуще-колющий характер.
Крылья у некоторых форм прозрачные, у дру-
гих — с темными перевязями и пятнами,
у третьих — с темным сетчатым рисунком. По-
следний членик лапки несет 3 пульвиллы (присоски). Брюшко широкое,
сдавленное сверху вниз, состоит из 7 члеников. Крепкие щетинки на
теле отсутствуют.
23 Учебник медицинской энтомологии 35 3
Определительная таблица главных родов семейства Tabanidae
1 (2). Задние голени на конце со шпорами (крупными щетинками); 2-й членик
усиков, вытянутый в длину; крылья с темными по херечными полосами-----------
'................................................Ghrysops Meig. — златоглазик.
2 (1). Задние голени без шпор; 2-й членик усиков короткий.
3 (4). Конец усика веретеновидный; состоит из 4 члеников; крылья с сетчатым ри-
сунком ......................................~...Ghrysozona Meig. — дождевка.
4 (3). Конец усика сдавлен с боков, сверху с треугольным выступом; состоит из
.“> члеников; крылья прозр ачные-----------abanus L. — слепень.
2. БИОЛОГИЯ И ЭКОЛОГИЯ 'СЛЕПНЕЙ
Яйца. Самки слепней откладывают яйца кучкой; у большинства видов
они приклеиваются к нижней стороне листьев или стеблям растений на
берегу водоемов над самой водой
Рис. 219. Кладка слеп-
ня, приклеенная к стеб-
лю травы.
(рис. 219). Кладки чаще всего
встречаются на местах, хорошо
освещаемых солнцем, вблизи
пастбищ, водопоев животных
и т. д. Некоторые виды отклады-
вают яйца на камни по берегам
потоков. Только что отложенные
яйца белые, но постепенно тем-
неют. Через несколько дней
после откладки (от 3 до 8 дней)
из яиц вылупляются личинки
I возраста. С растения они пада-
ют в воду,где и продолжают раз-
виваться. Самки немногих ви-
дов слепней откладывают яйца
на растения вдали от водоемов
и их личинки развиваются не в
воде, а в почве. Личинки слеп-
ней ведут полухйщный образ
жизни: они питаются как рас-
тительными остатками, так и
разными мелкими организмами
(мелкими моллюсками, насеко-
Рис. 220. мыми, червями).
Личинка Личинки. Личинки слепней
слепня. (рис. 220) имеют веретенообраз-
1 — передний конец, ную форму с заостренными пе-
редними и задними концами.
Их тело состоит из головы и 12 члеников. Новорожденная личинка Taba-
nus имеет в длину 2,5—2,7 мм, взрослая достигает 25—30 мм. Голова ли-
чинки (рис. 221) вытянута в трубку; на ее переднем конце расположены
ротовые органы: крючкообразная верхняя губа, пара острых верхних челю-
стей, пара более широких нижних челюстей, которые несут щупики, и
раздвоенная на конце нижняя губа. По бокам головы находятся усики,
глаза и особые площадки, усаженные щетинками, при помощи которых
личинка сцепляется с жертвой. Задний членик тела личинки вытянут в
узкую дыхательную трубку, на конце которой расположены дыхальца.
Личинки слепней встречаются в водоемах следующих типов: по бе-
регам рек и ручьев, в запущенных канавах, в копанках, карьерах, в по-
стоянных и временных болотах. В водоемах личинки слепней не живут
354
в открытой воде, — они могут встречаться под водой в прибрежном илу
или песке или в поверхностных слоях водной растительности. Нередко
их можно встретить на берегу в старых пнях, кочках и т. д. В течение
жизни личинка линяет 6 раз, т. е. проходит 7 возрастов. Полувзрослая ли-
чинка зимует, а весной вновь продолжает свое развитие. Зимовка проис-
ходит в тех же местах, где и питание.
Рис. 221. Голова личинки слепня (вид сбоку).
1 — ] j сп жэйном состоянии; ?Б—при нападении: 2—верхняя губа; ? — верхние челюсти;
3—нижние челюсти; 4— нижняя губа; 5 — площадка со щетинками; 6—* глаз.
Личинки слепней очень плохо переносят сухость, но весьма устойчивы
к другим неблагоприятным факторам внешней среды. Так, например,
в полувзрослом или взрослом состоянии они могут
длительно существовать при температурах от 0 до 8° и
довольно хорошо переносят понижение температуры
до—3°; они чувствуют себя нормально при высоких
температурах до 30° и начинают проявлять беспокой-
ство лишь при дальнейшем повышении температуры.
Личинка VII возраста выбирается из водоема, заби-
рается в почву или моховой покров и здесь превра-
щается в куколку (рис. 222). Окукливание происхо-
дит иногда довольно далеко от воды. Куколка раз-
вивается, в зависимости от температуры, в срок от
6 дней до 3 недель. В средней полосе Советского Союза
вылупление из куколок взрослых слепней начинается
в конце мая — начале июня.
Экология взрослых слепней. Самцы слепней пита-
ются сладкими растительными соками и встречаются
главным образом среди растительности. Самки также
пьют сладкие жидкости, но для откладки яиц им необ-
ходима кровь. Добычей им служит главным образом
рогатый скот, но они могут нападать и на мелких мле-
копитающих, и на человека. Нападение происходит на
открытом воздухе; особенно многочисленны слепни
на пастбищах. Они проявляют особую активность
в жаркие солнечные дни. Исключение составляют
представители рода Chrysozona — дождевки, которые
Рис. 222. Куколка
слепня (вид сбо-
ку).
1 — передний конец.
охотно нападают в пасмурную погоду перед дождем.
За одно кровососание самки крупных видов могут выпить до 200 мг
крови.
В местностях, где слепни очень обильны, их массовые нападения
сильно досаждают и людям, и домашним животным, заставляя иногда
переходить на ночные работы в поле и на ночной выпас скота.
23*
У слепней, как у всех кровососущих низших двукрылых, существует
гонотрофическая гармония — одного приема крови самке хватает для раз-
вития одной порции яиц (Олсуфьев). Созревание яиц длится 5—10 дней.
Число одновременно созревающих яиц у разных видов колеблется от 400
до 1 000. В течение жизни самки могут отложить не одну кладку.
Слепни — влаголюбивые насекомые и в те периоды, когда они не на-
падают на животных, держатся в затененных и влажных местах. Во время
пребывания на пастбищах они время от времени летят к водоему,
на мгновение опускаются в воду, затем взлетают, унося на нижней поверх-
ности тела каплю воды, которую и всасывают хоботком.
Слепни встречаются на всем земном шаре, но особенно обильны при
наличии следующих условий: 1) водоемы, подходящие для развития личи-
нок; 2) лесная, кустарниковая или тростниковая растительность, в кото-
рой взрослые самки скрываются от слишком большой сухости воздуха;
3) скопления крупных млекопитающих, служащих для них добычей.
Эпидемиологическое значение. Эпидемиологическое значение слепней
заключается в том, что они могут механически переносить целый ряд ин-
фекционных заболеваний, в том числе сибирскую язву и туляремию. Уку-
сив больное животное, а иногда труп погибшего от болезни животного,
они захватывают вместе с кровью и возбудителей болезни. Нападая затем
на здоровое животное или человека, они передают им в кровь и заразное
начало. Споры сибирской язвы сохраняются в хоботке слепня до 5 дней;
в хоботке одной самки их находили до 600. В таежной полосе около 80%
случаев сибирской язвы передается слепнями (Олсуфьев). Дождевка
и златоглазик были найдены в природе, зараженные туляремией (Сомов
и Данилова). В некоторых местностях в период лёта слепней наблюда-
ются эпидемические вспышки этого заболевания.
В Африке слепни-златоглазики являются переносчиками одного из
филариидозов человека.
Мероприятия. Основная мера борьбы со слепнями — очистка и осуш-
ка водоемов, где происходит их развитие.
Для предупреждения распространения болезней домашних живот-
ных, передаваемых через слепней, следует изолировать больных живот-
ных от переносчика, что вполне доступно, так как слепни в помещениях
не нападают.
РАЗДЕЛ Г
ВЫСШИЕ ДВУКРЫЛЫЕ, ИЛИ КРУГЛОШОВНЫЕ
(подотряд CyclorhapKa)
Г Л А В’А I
ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
К подотряду Cydorhapha -—круглошовных— относятся высш не дву-
крылые, или мухи. У круглошовных превращение в куколку происходит
внутри последней личиночной шкурки, которая образует чехол, носящий
название puparium — пупарий. При выходе имаго пупарий открывается
круглой щелью таким образом, что его передний конец отходит в виде
крышечки. От формы щели подотряд и получил свое название.
Многие представители круглошовных приносят большой вред здо-
ровью человека. Мухи являются переносчиками ряда заболеваний. За очень
немногими исключениями, перенос осуществляется чисто механическим
путем — никакого цикла развития большинство микроорганизмов в теле
мухи не проходит. Питаясь зараженными веществами, муха захватывает
вместе с пищей и возбудителей болезни, после чего, перелетая на продук-
ты питания или на посуду, оставляет здесь микробов, которые впоследст-
вии могут быть заглочены человеком. Наряду с бактериями, мухи переносят
таким же образом цисты кишечных простейших и яйца глистов. Кровосо-
сущие формы могут переносить инфекции $т больного животного в кровь
здорового животного или человека.
Паразитизм личинок мух. Личинки некоторых видов мух могут парази-
тировать в тканях или полостях животного или человека, нанося иногда
значительный ущерб здоровью. Заражение какой-либо части тела живот-
ных личинками мух носит название миаза. Миазы могут быть тканевые
и полостные. Среди мух одни виды являются возбудителями случайных
миазов, часто представляющих лишь зачаточный пли даже мнимый па-
разитизм. Примеры такого рода бывают в тех случаях, когда человек
проглатывает пищу, зараженную яйцами или личинками мух; личинки
не перевариваются кишечными соками человека, но неспособны и нормаль-
но там развиваться. Некоторое время они обитают в кишечнике и вызывают
кишечный (полостной) миаз, но затем обычно выводятся наружу. Иногда
самки тех видов мух, которые обычно размножаются в различных гниющих
веществах, откладывают яйца на незащищенную рану человека; вылупив-
шиеся из яйца личинки развиваются в ране и вызывают наружный (тка-
невой) миаз. Возбудители таких миазов бывают случайными или фа-
357
культативными (необязательными) паразитами. Личинки других
видов мух развиваются исключительно в животных тканях и являются
обязательными паразитами — возбудителями миазов. Обязательные па-
разиты из числа личинок мух бывают как тканевые, так и полостные.
ГЛАВА II
СЕМЕЙСТВО PIOPHILIDAE
Piophila easel L. — сырная муха.
Взрослые мухи мелкие (4—5 мм) (рис. 223), блестяще черные; передние ноги со
светлыми тазиками и коленями, средние и задние ноги на большом протяжении светлые.
Встречаются на всем земном шаре.
Яйца (от 40 до 80 штук) откладывают на соленую и копченую рыбу, сыр, ветчину,
свиное сало. В этих и других белковых веществах животного происхождения и проте-
кает развитие личинок. Личинка (рис. 224) проходит три возраста. В III возрасте она
достигает 9 мм длины и отличается своеобразным способом передвижения, за который
и получила название «прыгун»: она отгибает
назад головной конец и цепляется ротовыми
крючками за задний конец брюшка, затем вне-
запно выпрямляется и в результате этого пе-
репрыгивает на значительное расстояние.
Куколки встречаются в остатках зара-
женных продуктов или в щелях пола про-
дуктовых складов.
Рис. 223. Piophila casei — сырная Рис. 224. Личинка сырной мухи (общий
муха (общий вид со спинной сто- вид сбоку).
роны). 1 — головной конец.
Личинки сырной мухи вызывают кишечный миаз. Это происходит в тех случаях,
когда они заглатываются вместе с зараженными продуктами. Личинки очень устойчивы
к пищеварительным сокам человека. Попадая в кишечник в большом числе, они могут
вызвать изъязвления, боли подложечкой, тифоподобное состояние и т. д. Иногда ли-
чинки сырной мухи выходят в живом состоянии с рвотой.
ГЛАВА III
НАСТОЯЩИЕ МУХИ (СЕМЕЙСТВО MUSCIDAE)
Некоторые представители семейства Muscidae играют первостепенную
роль как механические переносчики различных микроорганизмов.
£1. НАРУЖНОЕ СТРОЕНИЕ
К этому семейству относятся мухи средней и малой величины (рис. 225),
реже — крупные. Все тело мух густо покрыто щетинками и волосками
разной величины.
а) Голова и ее придатки
Голова (рис. 226) выпуклая спереди, плоская сзади. По бокам головы
находятся большие сложные глаза. На средней линии головы, за слож-
ными глазами, расположено три простых глазка. На голове находится
большое число щетинок. Для мух этого семейства характерны щетинки,
расположенные на лбу между глазами, так называемая «лобная аллейка».
358
У самцов лоб узкий и глаза сближены или даже соприкасаются; у самок
лоб широкий и глаза широко расставлены.
На передней поверхности головы в углублении находятся усики.
Они состоят из трех члеников, из которых последний длиннее двух пер-
вых; на нем расположены чувствительные ямки — органы обоняния.
На спинной стороне 3-го членика сидит трехчлениковая щетинка, у неко-
торых видов голая, у других — усаженная волосками.
Хоботок* От нижней поверхности головы отходит хоботок; его строе-
ние различно в зависимости от способа питания мухи. У большинства ви-
Рис. 225. Musca dom^stica — ком-
натная муха, самка (общий вид со
спинной стороны).
Рис. 226. Голова Musca domestica (вид
спереди).
1— глаза; 2— глазки 3— «лобная аллейка»; 4—
усики (левый усик не нарисован); 5 — щетинка на
спинной стороне 3-го членика усикь: 6— хоботок.
дов семейства Muscidae он приспособлен только для подлизывания свобод-
ных жидкостей, у кровососущих форм служит для нанесения раны.
Хоботок лижущего типа (рис. 227) — мягкий, широкий; когда муха
не питается, он складывается под углом и втягивается в углубление на
нижней поверхности головы.
Хоботок состоит из двух частей: основной и нижней. В основной ча-
сти — rostrum расположена глотка; всасывание пищи происходит при
растягивании глотки мышцами, расположенными поперек rostrum. К этой
части хоботка спереди прикреплены щупики. Нижняя часть хоботка —
haustellum, включает в себя ротовую полость, которая заключена между
надглоточником (labrum-epipharynx) и подглоточником (hypopharynx).
На конце хоботка находится ротовое отверстие, прикрытое с боков
подушковидными образованиями—ротовыми лопастями; обычно они спу-
скаются с боков ниже ротового отверстия, и мухи при питании приклады-
вают к пище эти лопасти. На своей поверхности приротовые лопасти несут
359
органы осязания и вкуса. На нижней и внутренней (обращенных друг
к другу) сторонах находятся узкие канальцы, которые носят название
псевдотрахей. Псевдотрахеи лежат параллельно друг другу и ведут к ро-
товому отверстию. По длине псевдотрахей расположены мельчайшие от-
верстия, через которые пища поступает в эти канальцы, а по ним перехо-
дит в рот. Ширина отверстия на псевдотрахеях равна 0,03—0,04 мм. Та-
ким образом, в них может поступать лишь жидкая пища. Однако мухи
могут питаться и твердыми веществами благодаря тому, что у них имеется
внекишечное пищеварение. В хоботке открывается проток слюнных же-
лез, из которого слюна выпускается наружу на твердые питательные ве-
Рис. 227. Хоботок ком-
натной мухи (А— вид
сбоку) и приротовые ло-
пасти (В — вид снизу).
1—основная часть хоботка (rostrum); 2—щупики; 3—нижняя
часть хоботка (haustellum); 4 — ротовые лопасти; 5 — надглоточной;
— подрлоточник; 7 — проток слюнных желез; 8 — псевдотрахеи;
9 — ротовое отверстие; 10 — приротовые зубы. 1
щества и растворяет их; растворенная таким образом пища может всасы-
ваться псевдотрахеями. Иногда питание твердой пищей происходит и иным
путем. По бокам ротового отверстия, с внутренней стороны приротовых
лопастей, находятся хитиновые отростки — приротовые зубы. В неко-
торых случаях муха при питании отгибает лопасти кверху, и тогда приро-
товые зубы соприкасаются с пищей, скоблят ее, и отдельные части посту-
пают, минуя псевдотрахеи, непосредственно в рот. Ротовое отверстие ши-
ре отверстий псевдотрахей, оно равно 0,45 мм.
К основанию хоботка прикреплены одночлениковые щупики, на ко-
торых расположены органы чувств. Хоботок лижущего типа имеется
у Musca, Muscina, Fannia.
Хоботок колющего типа—длинный, твердый и тонкий (рис. 228).
В спокойном состоянии не втягивается и сильно выступает вперед; при пи-
тании опускается вниз. Приротовые лопасти очень малы. Приротовые зу-
бы большие и сильно хитинизованы. При соприкосновении хоботка с ко-
жей ротовые лопасти раздвигаются, приротовые зубы приближаются
к коже и расцарапывают ее, образуя ранку; кровь из ранки поступает
в хоботок.
На хоботке у основания расположены одночлениковые щупики.
Колющий хоботок имеется у Stomoxys и некоторых других мух.
360
б) Грудь и ее придатки
Грудь я ее придатки. Переднегрудь очень мала (рис. 229). Сзади нее
ио бокам расположены передние грудные дыхальца; к переднегруди при-
крепляется первая пара ног.
Рис. 228. Хоботок жигалки Stomoxys calcitrans (вид сбоку)
1—основание хоботка; 2—щупики; 3— подглоточник; 4 — над-
глоточник; 5 — приротовые лопасти; 6 — проток слюнных желез.
Большая часть грудного отдела занята среднегрудыо. На спинной
стороне она разделена швом на переднюю и заднюю половины; от послед-
ней глубоким швом отделяется небольшой отдел, имеющий треугольную
форму; он нависает над брюшком и носит название щитка. К среднегруди
прикреплена вторая пара ног и крылья.
Заднегрудь — очень узкий отдел груди. К нему прикрепляется
третья пара пог и жужжальца — остатки второй пары крыльев.
Рис. 229. Схема грудного отдела Musca domestica (вид сбоку
и со спинной стороны).
1 —переднегрудь; 2— среднегрудь; 3—заднегрудь; 4— переднее
и 5 — заднее грудные дыхальца; 6 — поперечный шов среднеспинки;
7 — щиток; 8—место прикрепления крыльев; 9— жужжальце;
10 — основания ног.
Ноги. Ноги мухи (рис. 230) состоят из следующих отделов: тазик,
прилегающий к груди, вертлугв виде небольшого кольца, два длин-
ных отдела—б едро и голень; затем идет пятичлениковая лапка;
на конце последнего членика лапки (рис. 231) сидит пара коготков и под
ними — две подушечки — пульвиллы, которые служат присосками и по-
могают мухе при ползании по отвесным гладким поверхностям, например,
по стеклу. На пульвиллах расположены органы осязания и вкуса. Когда
361
муха ползает по различным веществам, то вкус этих последних воспри-
нимается прежде всего пульвиллами; если вещества оказываются съедоб-
ными, муха выпускает хоботок; в противном случае хоботок остается
втянутым.
Крылья. Крылья мух (рис. 232) перепончатые с продольными и по-
перечными жилками. По переднему краю крыла идет костальная
( С) жилка, следующая продольная жилка — субкостальная (SC )—
впадает в костальную. Далее кзади расположен радиальный
ствол жилок; у Muscidae он состоит из трех ветвей. Первая радиаль-
ная ( R±) впадает в костальную позади субкостальной. За ней идут две
Рис. 230. Нога Musca domestica
(вид сбоку).
1 — тазик; 2 — вертлуг; 3 — бедро;
4 — голень; 5 — лапка.
Рис. 231. Пятый
членик лапки Mus-
ca domestica (вид с
брюшной стороны).
1— коготки; 2— по-
душечки.
прямые жилки, каждая из которых слилась из двух, — радиальная
2 4-3 (Т?2+з) и радиальная 4 4- 5 (Я44-5). За радиальными жилками
следует медиальная (М ). У некоторых мух она идет прямо к вер-
шине крыла (Fannia), у других образует дуговидный изгиб по направле-
нию к переднему краю крыла (Muscina, Stomoxys), у третьих она изогнута
под углом (Musca). Далее кзади идет кубитальная (Си) жилка;
последняя продольная жилка анальная (Ап), у большинства видов
не доходит до края крыла.
Из поперечных жилок следует упомянуть рад по -меди аль-
н у ю (гт), соединяющую последнюю радиальную с медиальной, и з а д-
нюю поперечную, соединяющую медиальную с кубитальной.
Участки крыла, ограниченные жилками, образуют ячейки; главные
из них носят следующие названия. Между радиальными жилками лежат
радиальные ячейки (г), У основания крыла между R и М и гт находится
передняя основная (6а); между М, Си и задней поперечной —
задняя основ п а я (Ьр). Ближе к переднему краю крыла к послед-
ней примыкает диско и да льна я (d) ячейка, лежащая между М и
36 2
Си. Между Си и Ап у основания крыла лежит анальная (ап) ячейка.
По заднему краю крыла расположены заднекрайние ячейки.
Жужжальца. Жужжальца представляют собой булавовидные образо-
вания и являются остатками второй пары крыльев. Жужжальца служат
органами управления полетом и равновесия; перед полетом они приходят
в движение, и только после этого муха взлетает.
Рис. 232. Крылья мух семейства Muscidae.
Л — Musca; Б—Muscina; В — Fannia: 1—костальная жилка;
? —’субкостальная; 3—радиальные; 4—медиальная; 5—куби-
тальная; 6 — анальная; 7 — радиомедиальна;!.
в) Брюшко и его придатки
Брюшко состоит из 5 члеников; сверху каждый членик покрыт спин-
ной пластинкой — тергитом, снизу — брюшной пластинкой — с т ер-
нит о м. На боковых краях тергитов расположены брюшные дыхальца;
на переднем тергите их две пары, так как он слился из 1-го и 2-го тер-
гитов. Отдельные хитиновые пластинки брюшка соединены между собой
тонкими перепонками, благодаря чему брюшко напившейся мухи или
у самки со зрелыми яичниками может сильно растягиваться.
На конце брюшка расположен наружный половой аппарат: гипо-
пигий у самцов и яйцеклад у самки.
Гипопигий самцов и яйцеклад самки. Гипопигий состоит из парных
крючков, при помощи которых самец придерживает самку. Между ними
расположен penis.
Яйцеклад (рис. 233) трубчатый, состоит из нескольких последних
сильно суженных члеников брюшка (у Musca domestica из четырех); на по-
следнем членике находятся два отверстия — половое и анальное. На конце
яйцеклада находятся парные придатки — церки. Отдельные сегменты
яйцеклада могут вкладываться друг в друга, как труба телескопа. В та
ком сложенном виде яйцеклад обычно втянут в брюшко. При кладке яиц
он выходит из брюшка, и у комнатной мухи длина его равна длине
остального брюшка.
363
2. ВНУТРЕННЕЕ СТРОЕНИЕ
а) Пищеварительный аппарат
Пищеварительный аппарат (рис. 234) состоит из передней, средней и
задней кишки. Первые отделы передней кишки — ротовая полость и глот-
ка, как уже говорилось выше, находятся в полости хоботка.
Передняя кишка. В хоботок впадает проток слюнных желез. Они
представляют собой пару тонких и очень длинных трубочек, лежащих
в брюшке и груди. В передней части груди
в тонкий проток; далее оба протока со-
единяются в непарный, впадающий в
хоботок.
Из глотки пища поступает в пище-
вод, проходящий через голову в груд-
ной отдел. Здесь пищевод впадает
в преджелудок — proventriculus. Пе-
каждая железа переходит
Рис. 234. Пищеварительный аппэра
комнатной мухи.
J—хобогон; 2—пищевод; 3 — слюнной
проток; 4—слюнные железы; 5—пред-
желудок; 6 — проток зоба; 7 — зоб:
8 — средняя кишка; о — задняя кишка;
10—прямая кишка; 11—клапан задней
кишки; 12 — ректальные железы.
Рис. 233. Брюшко комнатной мухи
с вытянутым яйцекладом.
ред впадением пищевода в преджелудок от него отходит тонкий проток,
на конце которого расположен зоб. Последний представляет собой двух-
лопастной мешок. У Musca зоб, наполненный пищей, велик и занимает зна-
чительную часть полости брюшка. Он служит резервуаром, из которого
пища постепенно переходит через преджелудок в среднюю кишку. У кро-
вососущих видов (Stomoxys) зоб невелик и редко используется. Преджелу-
док несет две функции. Во-первых, он служит клапаном, не допускающим
обратного возвращения пищи из среднего отдела кишечника. Во-вторых,
он вырабатывает перитрофическую перепонку — тонкую оболочку, кото-
рая обволакивает пищу, находящуюся в средней и задней кишке.
Средняя кишка. За преджелудком начинается средняя кишка. Первый
ее отдел расположен в груди и представляет собой тонкую прямую труб-
ку. В брюшке она значительно расширяется и образует желудок. В зад-
ней части брюшка желудок делает двойную петлю, после чего переходит
в третью часть средней кишки, более узкую и прямую. Эта последняя со-
единяется с задней кишкой в точке впадения мальпигиевых сосудов.
Задняя кишка. Задняя кишка состоит из тонкой передней части и зад-
ней расширенной прямой кишки. По середине тонкой части находится
клапан, не допускающий возвращения переваренных остатков пи-
щи в переднюю часть кишечника. В прямой кишке находится четыре со-
скообразные выроста — ректальные железы, вероятно, играющие роль
при всасывании воды из переваренных остатков пищи.
Мальпигиевых сосудов с каждой стороны имеется по два; оба сосуда
каждой стороны впадают в кишечник общим протоком.
б) Дыхателъный^аппарат\
Трахейная система мух построена в основных чертах так же, как и
у комаров (стр. 214). Однако у мух в некоторых частях тела трахейные
стволы сильно расширяются и образуют воздушные мешки (рис. 235),
Рис. 235. Воздушные мешки комнатной мухи (вид сбоку).
1 — головные воздушные мешки; 2 — грудные мешки; 3 — брюшной
мешок; 4 — брюшные дыхальца; 5 — трахеи, отходящие от брюш-
ных дыхалец.
от которых отходят мелкие ветви трахей. В грудном отделе Musca лежит
пара воздушных мешков удлиненной формы, которые берут начало от перед-
них грудных дыхалец. От этих же дыхалец отходит пара трахей, идущих
в брюшко и здесь образующих большие воздушные мешки, занимающие
часть полости брюшка.
в) Органы кровообращения
Трубчатое сердце, разделенное на камеры, лежит на спинной стороне
брюшка; спереди, в грудном отделе, оно переходит в аорту, из которой
кровь поступает в полость тела. Между камерами сердца расположены
клапаны, не допускающие обратного тока жидкости. С боков каждой
камеры имеются отверстия (остии), через которые кровь поступает из
полости тела в сердце. Эти отверстия также снабжены клапанами.
г) Нервный аппарат
Центральный нервный аппарат мух отличается очень сильной кон-
центрацией. Кроме хорошо развитых подглоточного и надглоточного
365
узлов (рис. 236), на брюшной стороне грудного отдела лежит грудной
узел, образовавшийся из слияния трех пар грудных и всех брюшных
узлов. От него отходят нервы к органам груди и брюшка.
д) Половой аппарат}
Яичники (рис. 237) состоят из яйцевых трубочек; их число в каждом
яичнике колеблется от 17 до 90 (в зависимости от вида и размеров самки).
На конце каждой яйцевой трубочки нахо-
дится концевая камера, ниже которой рас-
положены в виде цепочки фолликулы в чи-
сле не более пяти (комнатная муха имеет
не более трех фолликулов), которые нахо-
дятся на различных фазах развития: более
молодые у концевой камеры, более зрелые
на противоположном конце трубочки. Каж-
Рис. 236. Нервный аппарат
комнатной мухи (вид со
спинной стороны).
1 — надглоточный узел (голов-
ной мозг); 2—зрительные до-
ли; 3 — отверстие, через кото-
рое проходит пищевод; 4 —
грудной узел; 5 — брюшной
нерв; 6 — грудные нервы.
Рис. 237. Женские органы размно-
жения осенней жигалки.
*— яичник; 2 — отдельные яйцевые тру-
j с развивающимися фолликулами;
— парный яйцевод; 4— непарный яйце-
вод; 5—влагалище; 6—придаточная
железа; 7 — семяприемники.
дый фолликул состоит из 16 клеток, из которых одна является яйцевой,
остальные 15 — питательными. Снаружи фолликул одет однослойным фол-
ликулярным эпителием. Все яйцевые трубочки каждого яичника впадают
в парные яйцеводы, последние впадают в непарный яйцевод, конечная часть
которого носит название влагалища. Перед переходом яйцевода во влага-
лище в него впадает три семяприемника — два с левой стороны и один •— с
правой (осенняя жигалка имеет два семяприемника). Каждый семяприем-
ник состоит из небольшой хитинизованной капсулы и протока. Рядом с
семяприемниками в яйцевод впадает пара трубчатых придаточных желез.
Рядом с ними имеется пара прозрачных придаточных пузырьков. В се-
мяприемниках хранится сперма, полученная при копуляции. Когда зре-
лое яйцо, выходя из яйцевой трубочки наружу, проходит через непарный
яйцевод, из семяприемников выделяется часть спермы и оплодотворяет
яйцо. Придаточные железы вырабатывают клейкое вещество, которое вы-
366
пускается на яйца во время их откладки и служит для приклеивания их
к субстрату и друг к другу.
Органы размножения самца. Семенники (рис. 238) имеют грушевид-
ную форму и окрашены в красновато-коричневый цвет. От них отходят
парные тонкие семяпроводы, впадающие
в непарный семяизвергательный про-
ток; последний открывается в совоку-
пительный орган.
3. ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
СЕМЕЙСТВА MUSCIDAE, ИМЕЮЩИХ
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
(В ПРЕДЕЛАХ СССР)
1 (10). Медиальная (четвертая продоль-
ная) жилка изогнута по направлению к пе-
реднему краю крыла.
2 (3). Медиальная жилка изогнута под
углом Musca L.
3 (2). Медиальпая жилка изогнута ду-
гообразно.
4 (5). Хоботок мягкий; когда муха не пи-
тается. втянут в углубление головы, вершина
щитка красновато-желтая..Muscina R. D.
5 (4). Хоботок сильно хитинизовап, не
втягивается, в спокойном состоянии направ-
лен вперед. Щупики хоботка короткие, значи-
тельно короче самого хоботка -........
Рис. 238. Мужские органы размно-
жения комнатной мухи.
1 — семенники; 2 — семяпроводы; з —
семяизвергательный канал.
................... Stomoxys Geofl г.
6 (1). Медиальная жилка прямая. Средние голени с внутренней стороны в корот-
ких и густых волосках........................................Fannia В. D.
4. РОД MUSCA L.
а) Комнатная муха (Musca domestica L.)
1. Подвиды комнатной мухи
Род Musca содержит много видов, но наибольшее эпидемиологическое
значение имеют два подвида комнатной мухи — Musca domestica domes-
tica L. (рис. 225) и Musca domestica vicina Macq.
Первый подвид встречается в умеренном климате и на севере, второй—
в жарких странах. В СССР он известен в Закавказье, в среднеазиатских
республиках и в Уссурийском крае.
Оба подвида комнатной мухи очень тесно связаны с человеком: во взрос-
лом состоянии они обитают в жилищах человека; их развитие протекает
во всевозможных отбросах поблизости от человеческого жилья; в дикой
природе эти мухи обнаружены не были, — они являются ярко выражен-
ными синантропами (сожителями человека). Биология обоих подвидов
очень сходна, поэтому в дальнейшем они рассматриваются совместно.
2. Стадии развития
Яйца. Зрелая самка откладывает яйца на те вещества, в которых бу-
дет происходить развитие личинок. Яйца (рис. 239) белого цвета, продолго-
вато-овальные, около 1 мм длины. Вдоль спинной стороны по бокам идут
два валика, соединяющиеся спереди. При вылуплении личинки из яйца
пластинка, находящаяся между валиками, приподнимается, и в образовав-
шееся отверстие личинка выползает из хориона — оболочки яйца.
367
Развитие яйца длится несколько часов. Сроки развития изменяются
в зависимости от температуры (табл. 11).
Личинка. Только что вылупившаяся личинка I возраста имеет длину
2 мм. Она имеет вид белого, почти прозрачного червячка. Тело личинки
(рис. 240) состоит из 13 члеников. Передний ее конец заострен, задний
расширен и заканчивается тупо. На переднем конце помещается ротовое
Рис. 239. Яйца комнатной мухи.
А — сильно увеличенные; Б — отложенные на навоз.
Рис. 240. Взрослая личинка комнатной
мухи.
А — вид сбоку; I — XIII ’— сегменты тела;
1—’головной конец; 2 —передние дыхальца;
м — брюшные валики.
отверстие, ведущее в глотку. Над глоткой и по бокам ее расположен так
называемый головоглоточный хитиновый скелет; передняя часть этого
скелета представлена крючками, способными выдвигаться наружу; они
служат для размягчения пищи; кроме того, личинка пользуется ими при
передвижении, упираясь ими в субстрат. На брюшной стороне каждого
из восьми задних члеников тела имеется валик, покрытый щетинками,
служащий личинке для ползания. На заднем конце тела располо-
жена пара задних дыхалец. У ли-
чинки I возраста они имеют вид
сердцевидных щелей (рис. 241, А).
Развитие личинки I возраста
занимает от 20 до 36 часов (в зави-
симости от температуры). После
этого личинка линяет и переходит
во II возраст. Личинки II возра-
ста отличаются от личинок I воз-
раста следующими признаками:
каждое заднее дыхальце состоит
из двух продолговатых щелей.
По бокам 3-го членика тела имеются передние дыхальца, которых еще не
было у личинок I возраста. Передние дыхальца представляют собой вы-
росты, снабженные на конце пятью-семью пальцеобразными отростками.
При 25—30° развитие личинок II возраста занимает около 24 часов. При
низких температурах оно может растянуться на несколько дней. После
второй линьки личинка переходит в III возраст. В этом возрасте личинки
особенно усиленно растут и к концу его достигают длины 12—13 мм; с воз-
растом, вследствие сильного развития жирового тела, личинки постепенно
приобретают желтоватый оттенок. Личинки III возраста отличаются от
предыдущего по строению задних дыхалец. Каждое дыхальце (рис. 241, Г)
состоит из трех сильно извитых щелей, расположенных на дыхальцевой
пластинке; пластинка окружена сильно хитинизованным кольцом. По
мере роста личинки пластинка хитинизуется все сильнее, темнеет
и у взрослых личинок щели становятся почти незаметными.
Рис. 241. Задние дыхальца личинки комнатной мухи.
А—задний конец личинки I возраста (оригинал); Б—задний конец личинки
П возраста; В—задний конец личинки III возраста; Г — заднее дыхальце ли-
чинки III возраста; 1—задние дыхальца; 2 — сильно извитые щели.
Рис. 242. Пупа-
рий комнатной
мухи.i
Передний конец
цупария отделен
круговой щелью
и разделен про-
дольно после вы-
хола^мухи.
Рис. 243. Среднее число
личинок комнатной му-
хи в 250 см3 субстрата.
1 — свиной навоз; 2 — кон-
ский навоз; 3 — кухонные
отбросы; 4 — коровий на-
воз.
Окукление. Когда развитие личинки мухи закончено, то она переходит
в следующую фазу развития — предкуколку (ргаерира). На этой фазе
идет подготовка к окукливанию, в первую очередь выбор места, где будет
24 Учебник медицинской энтомологии
369
извиваться куколка. Личинка при переходе в предкуколку перестает пи-
таться, кишечник ее освобождается от экскрементов. Вначале предкукол-
ка быстро ползает, но, выбрав место для окукливания, остается здесь непо-
движной. Превращение предкуколки в куколку внешне выражается в том,
что она сокращается в длину и из червеобразной превращается в яйце-
видную, затем она линяет, не сбрасывая шкурку. Эта старая шкурка об-
разует собой покров куколки (пупарий) (рис. 242). Вначале он такого же
желтоватого цвета, как и личинка, затем постепенно темнеет ц через не-
сколько часов становится темнокоричневым.
3. Экология личинки
Местообитание личинок. Личинки комнатной мухи обитают в толще
скоплений разных гниющих веществ растительного или животного проис-
хождения. Они встречаются в кухонных отбросах в мусорных ящиках или
на полях минерализации, в фекалиях уборных или в скоплениях навоза
домашних животных в хлевах и на дворе.
Число личинок в этих веществах может быть огромно. Личинками
М. domestica особенно сильно бывает заселен свиной навоз. На рис. 243
приведена диаграмма среднего числа личинок этого вида в 250 см3 раз-
ных сортов навоза и кухонных отбросов. Для М. domestica vicina в табл. 10
приводится макс и м а л ь н о е число личинок в 1 кг вещества.
Таблица 10
Максимальное число личинок М. domestica vicina в 1 кг вещества
(из Зимина и Тетеровской)
Конский Коровий Свиной Фекалии Кухонные отбросы
навоз помет навоз помет навоз помет с земли из убор- ных
12 900 6 500 1280 48 1 | 7 800 i 8 400 16 200 4 7 700
Все те вещества, в которых личинки обитают, служат им пищей.
Личинки не могут заглатывать твердую пищу, но у них имеется внеки-
шечное пищеварение — в их экскрементах содержатся ферменты, растворяю-
щие твердую пищу. Благодаря этому в тех участках, где находится
основная масса личинок, субстрат более разжижен, чем там, где личинок
нет; это бывает хорошо видно, например, на таких веществах, как карто-
фельные очистки или мясные отходы.
Продолжительность развития. Сроки развития личинок зависят от
внешних условий. Особенно большое влияние имеет температура. В табл. 11
приведено время развития личинок в зависимости от температуры.
Таблица 11
Сроки развития комнатной мухи в зависимости от температуры
Температура
16° 18° 1 | 20° 25° 30° 36° 40° 43°
Яйцо Личинка Куколка 36 ча- сов 18 суток 19 » 27—30 часов 12 суток 13 » 25 часов 3 суток 10 » 18 часов 7 суток 8 » 10 часов 5 суток 5 » 7—8 ча- сов 3 суток 8—9 ча- сов 4 суток 6 » Неиз- вестно 5 суток Все по- гибли
370
Наиболее быстро, в 3 дня, личинки развиваются при 36°. Как пони-
жение, так и повышение температуры задерживают развитие. Особенно
сильно оно затягивается при низких температурах: так, при 16° оно длится
2х/2 недели.
Температурные условия личиночных местообитаний. Личинки ком-
натной мухи теплолюбивы; в естественных условиях личинки М. domes-
tica часто встречаются при температурах 40—46°, а М. domestica vicina—
при более высоких температурах до 55°. Такая температура создается
в скоплениях органических веществ благодаря происходящим в них
процессам гниения. В разных участках скопления температура бывает
различна. Например, в большой куче конского навоза самая высокая тем-
пература (70—80°) бывает в центре; в глубину температура постепенно
понижается, так как внизу бывает обычно более старый навоз, уже пере-
гнивший и остывший; кверху от центра температура постепенно падает
благодаря охлаждающему влиянию внешнего воздуха. В жаркие летние
дни температура 40—46° встречается на глубине 2—3 см, и здесь обнару-
живается самое большое скопление личинок. В более холодную погоду верх-
ние слои навоза охлаждаются и личинки переползают на глубину
Ю—12 см, где температура оказывается выше, чем вблизи поверхности.
В табл. 12 показано расположение личинок в конском навозе в за-
висимости от температуры воздуха.
Таблица 12
Распределение личинок в конском навозе
(на свалке) при разной температуре воз-
духа
Температура
воздуха
Глубина на-
хождения
личинок в см
Температура на-
воза, при кото-
рой встречались
личинки
12°
17°
20°
21,5°
22°
23,5°
24°
26°
26,5°
32,5°
12—13
8—9
5—6
1—2
3—4
2—3
2—3
2—3
1—2
2—3
46°
46°
45°
45°
44°
45°
42°
46°
45°
44°
Благодаря способности к миграции (передвижениям) в теплые участ-
ки, личинки имеют возможность почти всегда развиваться при высокой тем-
пературе, а потому их развитие протекает быстро. В естественных усло-
виях в конском навозе развитие личинки занимает около 3 дней. Таковы же
сроки развития и в скоплениях других веществ. Этот факт имеет большое
практическое значение. При установлении сроков вывоза нечистот или об-
работки их химическими веществами необходимо учитывать, что личинки
развиваются не под влиянием воздушной температуры, а при более высо-
кой температуре скоплений гниющих веществ, которая значительно со-
кращает сроки развития.
Личинки комнатных мух не только теплолюбивы, но и теплоустойчи-
вы, особенно в III возрасте. Личинки М. domestica, попадая из свойствен-
ных им температур 40—45° в температуры 50—60°, погибают не сразу.
24*
371
Они могут выползти из этих горячих участков и перебраться в более хо-
лодные. Личинки М. domestica vicina способны переживать еще более
высокую температуру — выше 63,5°.
Ь? Эти данные надо иметь в виду при проведении так называемых «биотермических»
методов борьбы с мухами, имеющих целью уничтожение личинок с помощью высокой
температуры. Для этого навоз и отбросы складывают в особые биотермические камеры
или ямы, компостируют или же свежий навоз закапывают в глубокие слои разогретого.
Все эти способы достигают цели только в том случае, если в субстрате достаточно бы-
стро достигается температура 60° для М. domestica и выше 65° для iM. domestica vicin«.
Действие на личинку влажности, газов, света. Распределение личи-
нок в субстрате зависит не только от температур, но также и от влажности.
Рис. 244. Взрослые личинки комнатной мухи из навоза.
В подсохшем субстрате личинки мух не встречаются; также не встречаются
они и в очень влажных, полужидких веществах.
Несмотря на то, что личинки комнатной мухи развиваются в толще
гниющих веществ, они очень чувствительны к содержанию газов в суб-
страте. При отсутствии притока кислорода личинки быстро погибают.
Этим объясняется, что в таких веществах, как свиной навоз, личинки дер-
жатся, как и в конском навозе, вблизи поверхности, хотя свиной навоз
никогда не разогревается так, как конский, и личинки могли бы спустить-
ся и на большую глубину. В мусорных ящиках личинки сосредоточивают-
ся главным образом в тех участках, где между стенкой ящика и неровной
поверхностью отдельных частей мусора создаются большие воздушные
пространства.
Личинки мух не переносят света и не выползают на поверхность суб-
страта. Если снять верхний слой, под которым находятся личинки, то
они моментально скрываются в глубину.
4. Экология предкуколки
Продолжительность фазы предкуколки. Продолжительность фазы пред-
куколки у комнатной мухи незначительна. При благоприятных условиях
она длится не долее суток. Но низкая температура (ниже 12°), излишняя
372
влажность или, наоборот, слишком большая сухость могут растянуть ее*
на несколько дней. Однако в некоторых случаях неблагоприятные условия
действуют в обратном направлении, вызывая преждевременное окукли-
вание личинок, еще не перешедших в фазу предкуколки. Например, это
имеет место при воздействии на личинок слабых, несмертельных, доз
отравляющих веществ; это же явление наблюдается в естественных, усло-
виях при наступлении дождливой погоды. В этих случаях предкуколоч-
ный период совсем выпадает и личинки переходят в куколку, не достиг-
нув окончательного роста.
Миграция предкуколки. С наступлением предкуколочного периода
у личинок изменяется отношение к температуре и влажности; они стре
Рис. 245. Куколки комнатной мухи на бумаге из мусорного ящика.
мятся покинуть те места, где питались, и переползают в более холодные*
и сухие места; иногда они мигрируют при этом на значительные расстояния.
В естественных условиях окукливание происходит в земле, иногда
на значительной глубине (до 60 см). Перед окукливанием личинки способ-
ны не только к вертикальными миграциям, но и к горизонтальным — от му-
сорных ящиков или навозных куч они уползают на расстояние более
чем в 50 см.
Нередко окукливание происходит не в земле, а в субстрате (рис. 245),
например, у основания навозных куч, под тонким слоем подсохшего наво-
за. В мусорных ящиках куколок можно обнаружить в участках, где нет
гниющего мусора, например, в комках бумаги или тряпках, иногда на дне
ящика или в щелях между дном и стенками.
Предкуколка М. domestica не обязательно ползет перед окукливанием
вниз: если условия более подходящи в верхних слоях, го окукливание
происходит в них. Например, если мусор или навоз засыпать землей или
торфом, то куколки будут встречаться в этой покрышке.
£73
Во всех местах, где происходит окукливание, температура и влаж-
ность ниже, чем в тех участках субстрата, где происходило питание личи-
нок. Чаще всего окукливание имеет место при^ температурах не выше 25°.
5. Экология куколки
Сроки развития куколки зависят от внешних факторов, в первую оче-
редь от температуры. Эти данные приведены в табл. 11. Наиболее быстрое
развитие куколки, 3—4 дня, имеет место при 36°. При 16° оно затяги-
Рис. 246. Вы-
ход имаго ком-
натной мухи пз
пупарпя.
I— лобный пу-
зырь в выпячен-
ном состоянии.
вается почти до 3 недель.
Уход личинок в предкуколоч-
ный период из теплых .и влажных
мест связан с тем, что куколки пло-
хо переносят и высокую темпера-
туру, и высокую влажность. При
42°, т. е. при той температуре, при
которой происходит нормальное раз-
витие личинок, погибает 50% куко-
лок, а при 43° погибают все ку-
колки .
В естественных условиях довольно ча-
сто можно наблюдать гибель куколок от
высокой температуры. В компостах, напри-
мер, окукливание происходит в земляном
или торфяном слое на поверхности кучи;
когда затем сверху накладывается слой
гниющих нечистот, то температура в том
слое, где находятся куколки, поднимается
и они гибнут.
Рис. 247. Новорож-
денная муха.
Крылья находятся опте
в сложенном состоянии.
Плохо действует на куколок и
высокая влажность: так, при влаж-
ности песка 0,2%; 5,5%; 10%; 16%;
процент гибели куколок?; 14; 40; 100.
В’естественных условиях погибшие от высокой влажности куколки встречаются
иногда после сильных дождей около уборных или свинарен, где почва долго не про-
сыхает.
Если высокая влажность действует непродолжительное время*(24—48
часов), то часть куколок заканчивает свое развитие и дает нормальных
мух.
6. Вылупление имаго
Выход из пупария. Перед выходом мухи из куколки на пупарии по-
является круговая щель, отделяющая передний конец в виде крышечки,
кроме того, сама крышечка разделяется второй щелью на две половинки —
спинную и брюшинную. Через это отверстие муха и выходит наружу
(рис. 246).
Новорожденные мухи (рис. 247) отличаются по внешнему виду or
более зрелых мух. Хитин у них еще совсем мягкий и светлый, брюшко
гораздо короче и уже нормального, крылья находятся в сложенном
состоянии.
Вышедшая из пупария муха быстро и суетливо бегает в течение 10—12
минут, иногда это продолжается даже около часа. После этого срока
муха усаживается неподвижно и начинает приобретать нормальный вид:
у нее расправляется брюшко и крылья, а затем начинает постепенно твер-
374
деть и приобретать темную окраску хитин. Только после этого муха ста-
новится способной к полету и переходит к нормальному образу жизни.
Весь период от момента выхода из куколки и до начала нормальной по-
движной жизни занимает при температуре 20° около 2 часов. Все это вре-
мя муха проводит в непосредственной близости от мест выплода: на крыш-
ке и стенках мусорного ящика или на стенках строений около скопления
навоза. Этот факт можно использовать для борьбы с мухами, применяя
в местах выплода контактные яды.
Выход на поверхность. Как уже говорилось выше, куколки комнатной
мухи в большинстве случаев лежат под слоем земли, навоза или отбросов;
поэтому новорожденная муха для выхода на поверхность должна пройти
этот слой субстрата. При этом она пользуется так называемым «лобным
пузырем». Выпячиваясь, пузырь раздвигает частицы почвы, затем он втя-
гивается, и перед мухой открывается небольшое свободное пространство,
на которое она передвигается. Далее все повторяется сначала: выпячивание
пузыря, раздвигание почвы, втягивание пузыря и продвижение самой
мухи вперед. Таким способом мухи могут преодолевать большие простран-
ства. Например, все мухи, вышедшие из куколок, которые были засыпаны
слоем песка в 50 см, пробрались через этот слой на поверхность. Даже
через слой песка в 1 м проходило больше половины новорожденных мух.
Земля труднее проходима для мух, и все же около 80% мух проходит через
слой земли толщиной в 30 см.
Это показывает, что метод борьбы с мухами при помощи засыпки мусора рыхлым
слоем земли в 5—10 см предохраняет только от свежей откладки яиц на отбросы,
но не может служить препятствием для выхода па поверхность тех мух, которые раз-
вились из находившихся в мусоре куколок.
Уплотненная или утрамбованная земля непроходима для мух, особенно если трам-
бовке подвергается влажная почва: чем почва влажнее, тем опа сильнее уплотняется
и тем труднее мухам выйти па поверхность.
Молодые мухи способны проходить через слой мусора толщиной
более 30 см.
7. Экология взрослых мух
Соотношение полов и копуляция. При выведении мух из куколок,
полученных из нормально питавшихся личинок, число самцов и самок
приблизительно одинаково. Но если личинки питались недостаточно, то
самок получается меньше, чем самцов. Из кладки, отложенной одной сам-
кой, самцы вылетают раньше самок.
Самки комнатной мухи копулируют один раз. Жизнеспособность спер-
матозоидов сохраняется у них в семяприемнике и они в течение всей жи-
зни откладывают оплодотворенные яйца. Копуляция может происходить
в первый же день по выходе мух из куколки, по иногда наступает значи-
тельно позднее, через 5—7 дней. Соединенная пара может сидеть на месте,
ползать или летать.
Продолжительность жизни. Продолжительность жизни мух в есте-
ственных условиях летом неизвестна. В лаборатории, при температуре
20—25°, мухи живут около месяца. При низких температурах мухи жи-
вут дольше, чем при высоких. В неактивном состоянии, вызванном низ-
кой температурой, мухи могут прожить гораздо дольше, чем при самых
благоприятных условиях в активном состоянии. Например, при темпе-
ратурах от 0 до 6—7° мухи переживают всю зиму, т. е. в умеренном кли-
мате живут 6—7 месяцев.
Влияние температуры на взрослых мух. Влияние температуры преж-
де всего отражается на степени активности мух. При температурах ниже
7—8° М. domestica совсем неподвижны, при 9° начинают слегка ползать,
375
при 10,5° еще не летают по собственному почину, но, будучи вспугнутыt
взлетают; добровольный полет начинается с 12°. С 15° мухи начинают нор-
мально питаться, с 17—18°— откладывать яйца. При дальнейшем повыше-
нии температуры до 30° мухи становятся все активнее— чаще перелетают
с места на место, чаще питаются и т. д.
Этим объясняется, что уловы мух на липкие листы или в мухоловки в жаркие
дни бывают обильнее (рис. 248), чем в холодные дни, даже в том случае, если число мух
в помещении остается неизменным.
Мухи охотно скапливаются в местах, где температура бывает 42—43°, сидят
на горячей поверхности печки, на горячей крышке кастрюли и т. д.
Отношение мух к свету. Комнатные мухи светолюбивы. На открытом
воздухе они встречаются в хорошо освещенных местах. Из очень темных
помещений они стремятся вылететь
на свет.
Н а этом основаны следующие меры
борьбы с мухами. В помещении, где мно-
го мух, затемняют все окна, за исключе-
нием одной форточки, в которую встав-
ляется мухоловка, сделанная из метал-
лической сетки: все мухи из темного
помещения устремляются к свету, про-
ходящему через сетку, и таким образом
вылавливаются в мухоловку. На све-
толюбивости мух основано и применение
в помещениях пищевой промышленности
окраски оконных стекол в синий цвет,
что создает в помещении полумрак. Мухи
привлекаются запахом, доносящимся из
этих помещений, но темнота их отталки-
вает, и в тех цехах, где окна окрашены
синей краской, мух не бывает.
Рис. 248. Влияние температуры на улов
комнатной мухи.
По вертикали — среднее число мух на одну му-
холовку за сутки; по горизонтали — темпера-
тура
Вместе с тем комнатная муха, залетая с улицы в комнату, способна
в ней свободно передвигаться — она не устремляется на свет, исходящий
от окна, как это бывает со многими насекомыми (бабочки, осы, шмели)г
которые, попав случайно в помещение, сразу устремляются к окну и бьют-
ся о него, если оно закрыто. Зрелые самки М. domestica в поисках мест
для откладки яиц летят с освещенной солнцем улицы в темные или полу-
темные помещения для животных, забираются в мусорные ящики и т. д.
При искусственном освещении мухи ведут себя так же, как и при естест-
венном, и не бьются о лампу, как это делают бабочки и другие насеко-
мые. Цвета мухи, повидимому, различать неспособны.
Питание взрослых мух. Молодые, только что вышедшие из куколки
мухи не сразу начинают питаться. В течение первого часа после расправ-
ления крыльев мухи никакого внимания на корм не обращают. Большая
часть мух начинает принимать пищу через 2 часа после перехода к по-
движному образу жизни, поэтому первое питание происходит обычно не
около мест выплода, а в помещении, куда мухи обычно устремляются„
получив способность летать.
Голодная муха, придя в соприкосновение с кормом, выпускает хобо-
ток и принимается сосать. Очень редко муха насасывается не отрываясь.
Обычно, немного пососав, она меняет положение и начинает сосать в дру-
гом участке; часто она совсем отходит или отлетает от корма с тем, чтобы
через несколько секунд вновь к нему вернуться. Заглатывание жидкого
корма происходит очень быстро: муха насасывается менее чем в одну ми-
нуту (в среднем через 43 секунды).
Первые глогки пищи попадают в зоэ, а затем пища начинает попадать и в же-
лудок, так что после запотнепия зоэа заполняется и желудок. С началом пищева-
рения пища постепенно начинает переходить и заполнять заднюю часть средней
376
кишки и одновременно с этим из зоба переходит в желудок. При 28° через 2 часа’после
принятия пищи зоэ оказывается уже совсем’пустым, это же отпосигся и к желудку,
но зато вся остальная часть кишки бывает заполнена к этому времени переварива-
ющейся или переваренной пищей.
У мухи наблюдается явление отрыжки. Наевшаяся хмуха отрыги-
вает пищу, которая выступает в виде капли прозрачной жидкости на кон-
чике хоботка, а затем втягивается обратно. Муха может отрыгивать и втя-
гивать каплю несколько раз подряд, но часто выпускает ее на поверх-
ность, на которой сидит. Эти капли подсыхают и образуют пятна, которые
вместе с пятнами испражнений составляют всем хорошо известные «муши-
ные пятна» на стеклах, посуде, сахаре и т. д.
Первые испражнения появляются через 2 часа после еды. В течение
суток муха может оставить до 50 пятен испражнений. Так как обычно
и испражнения, и капли отрыжки содержат живых микробов, то понятно,
что даже одна муха может сыграть значительную роль в распространении
инфекции.
Нормально муха приступает к еде вторично до того, как первая пор-
ция пищи переварится. Часто она начинает вновь принимать пищу в то
время, когда старая пища имеется не только в кишке, но еще и в зобе.
Эта черта биологии мух имеет огромное эпидемиологическое значение: чем
чаще мухи соприкасаются с разного рода пищей, тем легче они могут распространять
инфекцию, так как они могут питаться зараженными веществами — отбросы, фекалии
больных людей, остатки пиши после больных и т. д., и веществами, которые они могут
заражать, — пищей человека.
Взрослые комнатные мухи употребляют в пищу самые разнообраз-
ные вещества как растительного, так и животного происхождения. Преж-
де всего питанием для них служит пища человека, которую они находят
в жилище, в магазинах, предприятиях общественного питания и т. д.,
а также отбросы в мусорных ящиках и помойках. Комнатная муха исполь-
зует в качестве питания также экскременты животных и человека. В жар-
кие дни М. domestica может быть очень надоедлива, стремясь сесть на
человека, на котором она охотно подлизывает пот, выделения слизистых
оболочек и ран. Но этот характер питания имеет для Хмух данного вида
второстепенное значение. Первое место в их питании занимает пища чело-
века, фекалии и кухонные отбросы.
При отыскании пищи мухи очень охотно летят на запахи бродящих
веществ. Поэтому они могут быть особенно многочисленны на хлебозаво-
дах и в пекарнях, куда их привлекает запах теста. На этоАм основании
в мухоловки в качестве приманки рекомендуется наливать хлебный квас
или класть закваску.
Для нормальной жизнедеятельности мухи необходимо, чтобы в состав ее
пищи входили углеводы и белки. Первые служат для поддержания общего обмена ве-
ществ, вторые используются для развития яичников.
Мухи охотно пьют теплую пищу, подогретую до 38—48° и даже до 55°. Холодная,
ниже 10°, пища их не привлекает.
Развитие яичников и размножение. Развитие яичников у мух (рис. 249)
протекает сходно с тем, что имеет место и у комаров. Деление процесса
развития яичников на 7 фаз можно применить и к мухам (стр. 222).
При температуре около 20° созревание первого фолликула занимает
около 5 дней. Таким образом, при нормальном питании на 6-й день после
вылупления самка может отложить свою первую кладку. Во время разви-
тия первого фолликула в яйцевой трубочке образуется сначала второй,
а затем и третий фолликул. До откладки яиц первого ряда второй фолли-
кул достигает фазы ПЬ, а третий — фазы I. После откладки первой порции
377
г
Рис. 249. Развитие яйцевых фолликулов у комнатной мухи.
А — фаза No; Б — фаза I; В—фаза На; Г — фаза ПЬ; Д— фаза III; Е — фаза IV;
Ж—фаза V. 1—яйцевая клетка; 2—питательные клетки; 3—фоллику-
лярный эпителий; 4—концевая камера; 5—концевая нить; 6—начало образо-
вания „следующего фолликула; 7 — зерна желтка; 8 — скопления желтка;
9 — остатки распавшихся питательных клеток.
Рис. 250. Расположение трахеол на фолликулах новорожденной (Л) и
клавшей (Б) самок комнатной мухи. Фаза развития фолликула— 111
(оригинал Кавериной).
I — трахеолы свернуты в клубочки; 2 — трахеолы растянуты по фолликулу.
Рис. 251. Желтые тела в яйцеводах клавших самок комнатной мухи (оригинал
Казериной).
А — у однократно клавшей самки; Б — у многократно клавшей самки. 1 — желтые тела.
зрелых яиц второй фолликул достигает V фазы через 2 дня; поэтому вто-
рая и последующие кладки могут происходить через каждые 2—3 дня.
Определение возраста. Молодые, не клавшие, самки мух отличаются
от клавших следующими признаками. К фолликулам подходят мелкие
трахеи (рис. 250, Л), ветвящиеся на трахеолы. У новорожденных самок
эти трахеолы свернуты в клубочек, у клавших самок они веерообразно
растянуты по фолликулу (рис. 250, Б). Кроме того, у клавших самок в яй-
цеводе заметны «желтые тела», представляющие собой остатки питатель-
ных клеток, застрявших в яйцеводе
после откладки зрелых яиц. У самки,
отложившей один раз, желтые тела
мелкие (рис. 251, Л) и бледно окра-
шены. У самки, многократно клав
шей (рис. 251, Б), они крупные у
окрашены в яркожелтый цвет.
Плодовитость. Число одновре-
менно созревающих яиц у самкг-
комнатной мухи в среднем равно
109. Однако у отдельных особей оно
сильно колеблется, что зависит от
личиночного питания (рис. 252).
Если личинка питалась хорошо, то
из нее впоследствии выходит круп-
ная муха с большим числом яйце-
вых трубочек (до 186). Наоборот,
при плохом питании личинки по-
является мелкая муха с небольшим
числом яйцевых трубочек (38 штук).
Число кладок, откладываемых
одной мухой, зависит от продолжи-
Рис. 252. Зависимость между средним
весом куколки и средним числом яиц,
отложенных мухами, выведенными из
этих куколок.
По вертикали—среднее число яип; по гори-
зонтали — средний вес куколки в милли-
граммах.
тельности ее жизни: чем дольше она проживет, тем больше отложит
кладок. Одна муха, прожившая в лаборатории 33 дня, отложила
11 кладок, большинство же погибает раньше — после 4—6-й кладки.
Откладка яиц. Яйца откладываются самками М. domestica непосред-
ственно на те субстраты, которые будут служить для жизни и питания
личинок. При откладке яиц самки забираются в разные щели и трещины,,
имеющиеся в субстрате, или засовывают в них вытянутый яйцеклад. Чаще
всего яйца откладываются на глубину 1—2 см, благодаря чему они ока-
зываются защищенными от различных неблагоприятных внешних воздей-
ствий: подсыхания, смачивания дождем, действия прямых солнечных лу-
чей. Отложенные яйца склеены между собой в виде одной кучки выде-
лениями придаточных желе^.
Выбор субстрата. Очень многие вещества растительного и животного
происхождения, находящиеся на разных стадиях гниения, могут служить
источником выплода комнатных мух. В сельских условиях на первом
месте стоит навоз разных домашних животных; в городах основными ме-
стами размножения служат кухонные отбросы, содержащие гниющие
овощи и разлагающиеся остатки рыбы и мяса.
Помимо этих ботее или менее постоянных субстратов, самки Musca могут откла-
дывать яйца и в случайные вещества, например, в мокроту, смешанную с песком в пле-
вательницах, на бумагу или тряпки, испачканные испражнениями, на содержимое
детских матрацов, прочитанное мочой. Яйца часто откладываются в силос, разбросан-
ный окото силосных башен, в гниющую шелуху семян хлопка, в отруби в кормушках
домашних животных. В Средней Азии немалое значение как место выплода комнатной
мухи имеют кизяки во время их приготовления.
380
Нередко самки комнатной мухи откладывают яйца на скопления муки
и остатков пищи в щелях пекарен и кухонь. Число мух, выплачиваю-
щихся в этих условиях, довольно велико. Например, из 250 см3 мусора,
взятого из щелей пола пекарни, было выведено более 250 мух.
Кроме того, самки М. domestica, привлекаемые запахом гноя, могут
откладывать яйца на незащищенные раны человека и животного.
Местом откладки яиц могут служить и скопления фекалий в уборных,
но это происходит не всегда. Уборные не заселяются комнатной мухой,
если их содержимое очень жидко, что имеет место, когда выгреб вырыт
в глинистой почве или когда он цементирован. В условиях такой жидкой
среды личинки комнатной мухи суще-
ствовать не могут. Они не встреча-
ются также и в тех случаях, когда фе-
калии заселены хищными личинками
других видов мух (Hydrotaea, Ophy-
га), уничтожающих личинок М. do-
mestica. Возможно, что и в том, и в
другом случае самки откладывают
яйца в фекалии, но личинки поги-
бают, и поэтому выплода мух не про-
исходит.
При выборе места для откладки
яиц большую роль играет' выделяе-
мый субстратом запах. Яйцекладу-
щие самки особенно охотно летят
на запах аммиака; все । вещества,
в которых обычно размножается ком-
натная муха, выделяют этот запах.
На этом свойстве мух основан один
метод борьбы. Для того чтобы отвлечь мух
от обычных субстратов, где уничтожить
яйца было бы трудно, рядом с этими суб-
стратами расставляют чашечки с отрубя-
ми, смоченными 10—20% раствором угле-
кислого аммония. Самки откладывают
яйца в эти чашечки, содержимое которых
легко может быть уничтожено.
телячий навоз
ЕЗ норовой - • -
3 телячий помет
О КОHCh'U и HC6JJ
овечий - -
Щ свиной - • -
Рис. 253. Степень заселенности разных
сортов навоза комнатной мухой в раз-
ных условиях (оригинал).
А— в среднем по нескольким пунктам; Б —
в колхозе под Москвой: В—в колхозе
Кабарде; Г— в колхозе Архангельской
области.
Но, кроме запаха, М. domestica при выборе места для откладки яиц
руководствуются также температурой и влажностью поверхности суб-
страта. Для комнатной мухи очень большое значение имеет характер
скопления вещества. Одно и то же вещество в небольших скоплениях
совсем не привлекает самок, в то время как в больших массах оно очень
охотно используется ими для откладки. Причиной этого является то, что
небольшие скопления быстро подсыхают и процессы гниения в них не раз-
виваются. Наоборот, в больших скоплениях, благодаря интенсивным про-
цессам гниения, создается высокая температура и высокая влажность,
которые и привлекают самок. Этим объясняется, например, тот факт, что
в умеренном климате самки М. domestica, охотно откладывая яйца в скоп-
ления навоза разных домашних животных, почти никогда не отклады-
вают их на помет тех же животных. В более теплых местностях, где похмет
нагревается сильнее, М. domestica vicina чаще использует его для кладки.
Нои большие скопления субстратов играют в выплоде мух не всегда
одинаковую роль, что хорошо видно при сравнении разных сортов навоза.
Мухи могут разхмножаться в конском, свинОхМ, телячьем, овечьем и даже
381
коровьем навозе, но который из них играет основную роль,_______за-
висит от его наличия в данном населенном пункте и от способа его хране-
ния (рис. 253).
Для самки комнатной мухи не имеет большого значения, находится ли
субстрат в помещении или на открытом воздухе. Если навоз убирается
из конюшни рано утром, до начала дневной активности мух, то откладка
будет происходить на кучи, сложенные во дворе. Наоборот, если навоз
остается лежать в помещениях все утро и даже весь день, то он тут и бу-
дет заражаться. Место откладки в кухонные отбросы зависит от степени
их разложения. На совершенно свежие отбросы самки комнатной мухи
яиц не откладывают: их привлекают только те отбросы, в которых нача-
лось гниение. Это может произойти в мусорном ящике или уже на полях
минерализации. Бывают случаи заражения отбросов яйцами мух еще
в квартире, в помойном ведре, если последнее выносится нерегулярно или
даже и при регулярной очистке не промывается и от него исходит гни-
лостный запах, привлекающий мух.
Приуроченность к жилью человека. Musca domestica — обитатель
человеческого жилья. Большую часть времени мухи этого вида проводят
в помещении человека. На открытом воздухе они встречаются только
вблизи разного рода отбросов. В теплые солнечные дни мухи этого вида
многочисленны около мусорных ящиков, грязно содержащихся дворовых
уборных, кормушек для животных и всюду, где они могут найти питание.
Среди незагрязненной растительности они почти не встречаются. На-
пример, среди 10 000 мух, выловленных в течение лета в саду, было всего
лишь 10 экземпляров комнатной мухи.
Круговорот мух в жилье. Но и вблизи отбросов на открытом воздухе
мухи остаются лишь временно: как только эти места оказываются в те-
ни, мухи оттуда исчезают. В помещении мухи проводят больше времени,,
но и здесь продолжительность их пребывания зависит от погоды. В жар-
кие и ясные дни они все время перелетают с улицы в комнату и обратно.
В холодную и пасмурную погоду, залетев в помещение, мухи остаются
в нем. Это бывает хорошо заметно осенью, когда мухи совершенно исче-
зают с улицы и из холодных построек и скапливаются в значительном
числе в теплых помещениях.
Повышенная активность мух в жаркую погоду имеет существенное
эпидемиологическое значение: благодаря постоянным перелетам с улицы
в дом и из одного дома в другой, облегчается распространение мухами
инфекций.
Места кормежки. Среди разных типов помещений наиболее привле-
кательными для них являются те, откуда исходят запахи пищи. Поэтому
мухи особенно многочисленны в пекарнях, общественных кухнях и сто-
ловых, а также в квартирах, особенно в часы приготовления пищи.
Большие скопления мух наблюдаются также в свинарнях, если в них
происходит варка корма. В конюшнях и других помещениях для животных,
а также в уборных они встречаются в меньшем числе.
Таким образом, мухи многочисленнее в тех помещениях, где они
находят питание. Места выплода привлекают меньшее число мух — сюда
залетают только зрелые самки, но и они здесь не задерживаются, а, вы-
брав место и отложив яйца, быстро вылетают.
Число мух в тех помещениях, куда они залетают в поисках питания,
зависит от двух факторов: близости места выплода и привлекательности
для мух самого помещения.
Рассеяние мух в деревне. Зависимость между числом мух, залетевших
в помещение, и его близостью к источнику выплода особенно хорошо вид-
382
на в условиях колхозной деревни (рис. 254). Здесь обычно имеется один
ующпый очаг выплода, сосредоточенный на колхозном дворе в виде скоп-
лений навоза. Обычно все жилые помещения деревни по своему санитар-
ному состоянию, а следовательно, и по своей привлекательности для мух,
очень сходны; поэтому мухи, летящие из места выплода, задерживаются
в домах, ближайших к колхозному двору; чем дальше отстоит дом от
конюшен и свинарника, тем меньше в нем мух (рис. 254). Обычно
мухи бывают многочисленны в домах, расположенных на рассто-
янии не далее 150—200 м от источника
выплода. Исключение составляют поме-
щения, которые оказываются почему-либо
особенно привлекательными для мух, на
пример, те дома, в которых происходит
варка корма для животных. В этих домах
число мух бывает очень велико даже в том
случае, если они находятся на рассто-
янии более 400 м от источника выплода.
Рассеяние мух в городе. В городах
мы встречаемся с иными санитарными
условиями, а поэтому и с иным распреде-
лением мух по помещениям. В условиях
города мы имеем дело с мелкими очагами
выплода в виде мусорных ящиков при
каждом домовладении. Кроме того, сани-
тарное состояние жилых помещений в
городе очень различно. Близость очага
выплода сказывается только в тех квар-
тирах, окна которых выходят непосред-
ственно на мусорный ящик, отстоящий от
них не далее 10—20 м. В таких кварти-
рах мухи обычно многочисленны. Но их
число зависит гораздо больше от санитар-
Рис. 254. Кривая средних уловов
мух в домах небольшого колхоза
в зависимости от расстояния от
колхозного двора (оригинал).
По вертикали — среднее число мух в
мухоловке за сутки; по горизонтали—
расстояние дома, где производился
улов, от конюшни ’в метрах.
ного состояния самого помещения: в грязных квартирах, где на столе
остается неубранная пища и крошки, их значительно больше, чем в чи-
стых, независимо от расстояния до мусорного ящика (табл. 13).
Таблица 13
Средний улов мух за сутки в квартирах разного санитарно-
го состоянии
Помещение Санитарное состояние . Расстояние от мусорно- го ящика в м Среднее число мух
ДворА
Квартира № 1 . . . Грязная 100 332
» № 2. . . Чистая 60 27
Д в о р Б
Квартира № 1 . . . Грязная 80 169
» № 2. . . Чистая 40 37
Кроме того, в городах имеются предприятия общественного питания,
которые особенно сильно привлекают мух, и в них мух может быть больше,
чем в жилых квартирах, даже в тех случаях, когда очаг выплода распо-
383
ложен ближе к жилой квартире, чем к помещению общественного питания.
Таким образом, мухи минуют менее привлекательные точки и устрем-
ляются к более дальним, но более привлекательным (табл. 14).
Табл и’ц а 14
Средний улов мух за сутки в’местах обществен-
ного питания и в жилых квартирах
Помещение Расстояние от очага вы- плода в м Среднее число мух
Квартира,№ 1 . . . 25 ! 145
» № 2 ... ' 150 30
Общественная кухня i 355 1 275
Перелеты мух. Musca domestica способна к значительным перелетам.
Отдельные экземпляры специально окрашенных и выпущенных мух на-
ходили на расстоянии 13 и даже 21 км от места выпуска. Такое про-
странство они покрывают постепенно, перелетая от одной привлекатель-
ной точки к другой и из одного населенного пункта в другой. Но такие
дальние перелеты совершаются лишь отдельными особями.
В распространении мух немалую роль играет пассивная перевозка
их разными видами транспорта. На машинах, отвозящих мусор, а также
на таре с различными съестными припасами можно постоянно видеть мух.
Такая транспортировка мух происходит не только в пределах населен-
ного пункта, мухи могут перевозиться на большие расстояния в поездах ,
на пароходах и в самолетах.
8. Годовой цикл вида
Зимовка. У М. domestica отсутствует диапауза, и при благоприятных
внешних условиях она может быть активна круглый год: в течение всего
года у нее может продолжаться нормальное размножение и развитие.
Фактически это имеет место у М. domestica vicina в тропиках и неко-
торых субтропических странах. В странах с холодной зимой мухи про-
водят ее в неактивном состоянии. Чем длительнее период низких темпе-
ратур, тем дольше продолжается зимовка мух. На юге СССР (Ереван,
Тбилиси) М. domestica vicina бывает неактивна 4—5 месяцев. Под
Москвой зимовка М. domestica продолжается около 7 месяцев.
Но и в холодном климате мухи могут оказаться в условиях, допускаю-
щих их размножение зимой. Это бывает, когда с осени они попадают в ота-
пливаемое помещение, в котором постоянно имеется подходящий суб-
страт для размножения. Такими помещениями бывают иногда пекарни или
кухни, где под полом, как об этом говорилось выше, происходит развитие
личинок мух. В таких помещениях мухи активны всю зиму, и их числен-
ность в течение этого периода не только не падает, а даже возрастает.
Поэтому весной эти помещения становятся источником рассеяния мух по
всему населенному пункту.
Кроме описанного способа переживания зимнего периода, у М. do-
mestica имеются и другие. Если наступление осеннего похолодания за-
стает мух в помещениях с низкой температурой, то под ее воздействием
они впадают в неактивное состояние и остаются там до наступления тепла.
Обычно местами зимовок мух служат подвалы, чердаки, утепленные хле-
84
ва и конюшни. Часто мухи зимуют между оконными рамами. На зимовках
самки встречаются чаще, чем самцы. Яичники могут быть у них на любой
фазе развития. Если самок с зимовки перенести в теплую комнату, то
они становятся активными, начинают питаться и откладывать яйца. Од-
нако среди зимующих имаго наблюдается большая гибель, доходящая
иногда до 85—88%.
Кроме того, М. domestica может зимовать и на неполовозрелых ста-
диях развития. Зимующие личинки и куколки этого вида остаются в земле
под мусорными ящиками, около навозохранилищ и в толще мусора и на-
воза. В земле наибольшее число особей встречается на глубине до 20 см. Зи-
мующие куколки встречаются чаще, чем личинки. Чтобы иметь представле-
ние о числе мух, зимующих на предимагинальных стадиях, можно привести
следующие цифры. Из 150 проб (величиной 0,5 м3) земли вокруг мусор-
ных ящиков было получено 300 личинок и 600 куколок. Такова же чи-
сленность зимующих личинок и куколок М. domestica и в толще мусора.
Очень важно знать, какой способ зимовки имеет преимущественное место в каж-
дом населенном пункте В одном и том же климате, в зависимости от условий челове-
ческого хозяйства, способ переживания зимы мухами может быть различен- Например,
если в населенном пункте имеются антисанитарно содержимые предприятия обще-
ственного питания, то значительный процент популяции будет зимовать в активном со-
стоянии. Это встречается в городах или крупных поселках. В деревнях, где предприя-
тий общественного питания нет, этот способ зимовки обычно не встречается.
От способа переживания зимнего периода зависит, во-первых, время
появления мух весной: они появляются раньше в том случае, если
переживали зиму в активном состоянии или зимовали в виде има-
го; при зимовке личинок и куколок весной требуется еще время на окон-
чание их развития. Во-вторых, способ зимовки отражается на численно-
сти мух весной: мухи, перегнивающие зиму в активном состоянии, в теп-
лых помещениях, меньше подвергаются неблагоприятным воздействиям,
чем мухи, зимующие в неактивном состоянии. Поэтому в тех населенных
пунктах, где распространена зимовка мух в активном состоянии, весной
численность их будет выше, чем там, где такого способа переживания
зимы нет.
Зпапие способов зимовки в каждом случае необходимо для разработки мер борьбы
с мухами. При зимовке мух в теплых помещениях необходимо с осени принимать меры
для недопущения размножения их в течение зимы — уничтожение щелей в полах, при-
менение приманок с углекислым аммонием и т. д.
Для борьбы с зимовками на стадиях развития рекомендуется пе оставлять осен-
ние скопления навоза в поселках, а вывозить их на поля до выплода мух весной. Земля
после вывоза этих скоплений должна перекапываться и обрабатываться отравляющими
веществами.
Сезонный ход численности. Зимой размножение мух, за некоторым
исключением, прекращается, по смертность в этот период продолжается;
потому весной, до появления нового поколения, численность мух находится
на самом низком уровне. Затем, с повышением температуры до степени,
допускающей размножение и развитие, численность мух начинает посте-
пенно возрастать, и этот рост продолжается до наступления неблаго-
приятных условий осенью, когда под влиянием пониженной температуры
начинается задержка в откладке и развитии яиц; с дальнейшим похолода-
нием оба эти процесса приостанавливаются до весны.
Характер сезонного хода численности может изхменяться в зависимо-
сти от разных факторов.
Климат влияет на продолжительность сезона размножения: на юге
последний длиннее, чем на севере. Например, в Тбилиси и Ереване пер-
вые кладки встречаются в конце апреля, а последние — в конце ноября.
25 Учебник медицинской энтомологии оос
В Москве первые кладки удается находить лишь в середине мая, а по-
следние — не позднее октября.
В местностях с жарким и сухим летом, как, например, в Ереване и
Ашхабаде, сезонная кривая имеет две вершины: в начале (июнь) и в конце
(сентябрь) сезона с провалом посредине (июль, август); последний указы-
вает на сильное снижение численности мух в этот период. Это падение
объясняется сокращением мест выплода вследствие быстрого подсыхания
отбросов и навоза. Примеры сезонных кривых численности мух для раз-
Рис. 255. Кривые сезонпого хода
численности комнатной мухи (ори-
гинал).
1—в колхозе под Москвой; 2—в Тби-
лиси; ‘ 3 — в ’Ашхабаде; 4 — в Томске.
По вертикали—месячные уловы (в процен-
тах к максимальному месячному улову);
по горизонтали — месяцы.
ных климатов приведены на рис. 255.
Сезонные кривые отражают и' из-
менения погоды. Во-первых, погода
изменяет активность мух: в теплые дни
они более подвижны и имеют больше
шансов попасть на липкий лист или в
мухоловку, при помощи которых ве-
дется обычно учет численности мух.
В холодную погоду уловы всегда пони-
жены не потому, что мух в эти дни
меньше, а потому, что они под влия-
нием холода сидят неподвижно на по-
толке и стенах и в уловы не попадают.
Кроме того, погода может отра-
жаться и на самой численности мух;
например, продолжительное похолода-
ние среди лета задерживает процессы
их размножения и развития. Поэтому
вслед за холодной погодой число мух
падает, так как нового вылупления в
этот период не было. Выше уже гово-
рилось, что характер сезонного хода
численности зависит от способа зимовки
мух. В городах, где имеются помещения
с зимним выплодом мух, последние появляются весной рано, например,
в Томске (рис. 255) и в большем числе, чем в небольших поселках. Такой
пример разной численности мух в начале лета приведен в табл. 15.
Таблица 15
Средний улов мух на липкий лист за сутки
в городе и поселке Московской области 5 июни
Помещение I Город Поселок
Столовые 200 8
Жилые квартиры 60 0
Состояние очагов выплода также отражается на сезонном ходе чи-
сленности мух. Примером этого могут служить местности с двувершинной
сезонной кривой, где падение численности мух летом объясняется сокра-
щением очагов выплода. Хозяйственная деятельность человека тоже может
оказывать влияние на очаги выплода комнатной мухи. Такой факт, как
перевод лошадей летом из конюшни на подножный корм, ведет к сокра-
щению скоплений навоза в поселках, а следовательно, и к сокращению
численности мух (рис. 256).
386
Изучение сезонного хода численности мух представляет двойной ин-
терес. С одной стороны, на основании этих данных планируются сроки
мероприятий по борьбе с мухами, с другой — противоэпидемические ме-
роприятия.
Роль мух в распространении инфекций тем больше, чем выше их
численность и их активность.
б) Эпидемиологическое значение муэс
М. domestica занимает одно из первых мест как распространитель ин-
фекционных заболеваний.
Это объясняется следующими причинами: 1. Комнатная муха очень
тесно связана с человеком и всей
окружающей его обстановкой.
2. Она питается всякого рода отбро-
сами, с одной стороны, и пищей че-
ловека — с другой. 3. Благодаря
частым приемам пищи, мухи все
время приходят в соприкосновение
то с зараженными, то с незаражен-
ными веществами и поэтому рас-
пространяют инфекции. 4. Микро-
бы, простейшие и яйца глистов
не подвергаются воздействию пи-
щеварительных соков мухи и про-
ходят через ее кишечник в живом
состоянии. 5. Благодаря частым
испражнениям и отрыжке, мухи
всюду рассеивают живые микро-
организмы. 6. Все тело мухи гу-
сто усеяно щетинками и волоска
ми, к которым легко пристают
частички грязи и отбросов, кото-
Рис. 256. Влияние уничтожения очага вы-
плода на сезонный ход численности комнат-
ной мухи в одном из ко 1хоюв Кабарды,
1939 г. (оригинал).
рые она оставляет затем на про-
дуктах питания или посуде.
7. Постоянные перелеты с улицы в
помещение и из одного помещения
1—сезонная кривая для дома, около которого
очаг существовал все лето; 2— сезонная кривая
для дома, около которого очаг перестал суще-
ствовать с июля. По вертикали—среднее число
мух на мухоловку за сутки; по горизонтали—
даты.
в другое тоже способствуют рас-
пространению инфекций. 8. В населенных пунктах легко созда-
ются условия массового размножения комнатной мухи (скопления от-
бросов, навоза и т. д.), благодаря чему она часто достигает большой
численности.
Количество микроорганизмов на поверхности тела и в кишечнике
мухи огромно— оно достигает 6 млн. в первом случае и 28 млн. во втором.
Число бактерий, переносимых мухами, зависит от санитарного состоя-
ния окружающей обстановки. Так, в чистых районах города число микро-
организмов на каждой мухе может быть в 500 раз меньше, чем в неблаго-
устроенных районах. На наружных покровах бактерии гибнут под влия-
нием солнечных лучей, в кишечнике они сохраняются дольше. Однако и
в кишечнике мухи могут развиваться факторы, неблагоприятно дей-
ствующие на бактерии. Так, например, в некоторых местностях к кон-
цу лета в кишечнике мух накапливается дизентерийный бактериофаг
(Змеев), уничтожающий дизентерийных микробов, заглоченных мухой.
25*
387
Ниже мы приводим основные виды бактерий, вредных для человека,
которых находили на мухах, пойманных в естественных условиях —
в помещениях и на открытом воздухе (свалки, рынки и т. д.): бациллы
брюшного тифа, бациллы паратифа А, бациллы паратифа В, дизентерийная
палочка,холерный вибрион,туберкулезнад палочка, дифтерийная палочка,
споры сибирской язвы и многие другие. Всего на мухах было обнаружено
44 вида микробов. Кроме того, мухи способны переносить некоторых про-
стейших возбудителей болезней. Неоднократно находили дизентерийную
амебу, лямблий, кокцидий и др. В табл. 16 приводятся сроки существо-
вания бактерий на поверхности тела, в пищеварительном тракте и в ис-
пражнениях мух.
Таблица 16
Срок существования бактерии
Бактерии Лапки Крылья Голова Зоб 1 Кишеч- ник Faeces
Бациллы брюшного тифа __ 1 6 дней 2 дня
Бациллы паратифа В 7 дней — 7 дней <8 дней 7 » ; —
Туберкулезная палочка — — — 3 дня 16 » 13 дней
Холерный вибрион 30 часов 5 часов 5 часов 2 » 2 дня 30 часов
Споры сибирской язвы 20 дней 20 дней 20 дней 13 дюй 20 дней 1 13 дней
Хотя мухи способны переносить различные инфекции, но их основ-
ное эпидемиологическое значение заключается в распространении летних
кишечных заболеваний. Среди факторов распространения кишечных эпи-
- Мухи_________Дизентерия______Ьрмщной тиф
Рис. 257. Численность мух и заболеваемость дизентерией
и брюшным тифом в 1938 г. в Москве (по Ванской).
По вертикали—число мух, число заболевании дизентерией и число за-
болеваний брюшным тифом (все кривые в процентах к максимальному
числу); по горизонтали — месяцы.
демий мухи стоят на одном из первых мест. Именно благодаря участию
мух кишечные эпидемии носят сезонный характер: падают на конец лета—
начало осени. Кривые этих заболеваний вполне соответствуют кривым се-
зонного хода численности мух. При этом кривые хода дизентерии совпа-
дают с кривыми увеличения численности мух (рис. 257): максимум числа
заболеваний совпадает с периодом максимальной численности мух. Это
388
ооъясняотся тем, что дизентерия имеет очень короткий инкубационный пе-
риод, т. е. признаки болезни проявляются через 1—2 дня после заражения.
Болезни с длительным инкубационным периодом, например, брюшной
тиф, который проявляется лишь через 15—20 дней после заражения, имеют
кривую, отстающую от «мушиной» кривой, т. е. максимум заболеваний от-
мечается приблизительно через месяц после максимума численности мух.
Это означает, что наибольшее число
большей численности мух, но, бла-
годаря длительной инкубации, за-
болевания проявились позднее
(рис. 257).
Комнатная муха принимает
участие и в рассеянии гельминтов:
когда муха питается на испражне-"
ниях, зараженных яйцами, послед-
ние пристают к поверхности ее
тела. Если после этого муха садит-
ся на пищу, она заносит на нее
и яйца глистов.
Диаметр ротового отверстия ком-
натной мухи меньше диаметра боль-
заражений происходило в период наи-
Рпс. 258. Musca sorbens—базарная муха,
самка.
Общий вид со спинной стороны.
шинства яиц глистов, поэтому опа их не
заглатывает и в испражнениях комнат-
ной мухи яйца большинства человече-
ских гельминтов обнаружены не были.
М. domestica может быть возбудителем случайных и факультативных
миазов. Первые наблюдаются, когда человек заглатывает вместе с пи-
щей личинок мух, вторые, —когда самки откладывают яйца в гноящиеся
раны, в которых и происходит развитие личинок.
в) Базарная муха
(Musca sorbens W. D.)
Musca sorbens (рис. 258) несколько мельче, чемM. domestica. На спин-
ке груди она имеет только две темные полоски. Это южный вид, который
в СССР встречается в Средней Азии и в Азербайджане.
Развитие происходит главным образом в фекалиях человека на
земле и реже в свином помете; в уборных, как правило, личинки не
встречаются. Обилие личинок в фекалиях бывает очень велико и дохо-
дит до 42 000 особей в 1 кг субстрата.
При температуре 20—25° развитие яйца занимает б1/^—7 часов,
развитие личинки—около 4 дней и куколки—около 8 дней. Взрослые
мухи особенно многочисленны на базарах, в ларьках и других местах
открытой продажи пищи. В помещения они залетают редко и на ночь
никогда в них не остаются. На базарах они в 42 раза многочисленнее,
чем в жилых квартирах.
В Таджикистане первые мухи этого вида появляются в мае — июне,
количество их достигает максимума в августе; встречаются они до де-
кабря.
М. sorbens очень охотно нападает на человека, особенно на откры-
том воздухе. Опа питается потом, выделениями слизистых оболочек носа
и рта, кровью и гноем ран. Но особенно настойчиво она нападает на глаза
и является переносчиком возбудителей конъюнктивитов. В Сталийабаде
ежегодные эпидемии этого заболевания совпадают с сезоном лёта базар-
389
ной мухи. Не исключена возможность, что М. sorbens, будучи, с одной
стороны, тесно связана с фекалиями, с другой — с местами продажи пищи,
может принимать участие и в распространении летних кишечных заболе-
ваний.
б. РОД FANNIA R. D.
МАЛАЯ КОМНАТНАЯ МУХА (FANNIA CANICULARIS L.)
Взрослые мухи (рис. 259) значительно мельче, чем М. domestica.
У самцов брюшко удлиненное, серое, с черной продольной полоской на
спинной стороне. На передних члениках брюшка с боков просвечивающие
желтоватые пятна. Голова белая, с черным лбом, ноги черные. У самки
все тело черноватое, брюшко у основания
желтоватое, голова серая.
Самка откладывает около 80 белых
яичек; по бокам они имеют крыловидные
отростки; яйца откладываются разбро-
санно по поверхности субстрата.
Рис. 260. Взрослая ли-
чинка малой комнатной
мухи.
1—передние дыхальца;
2—задние дыхальца; 3 —
отростки.
Рис. 259. Fannia canicu-
laris—малая комнатная
муха, самец.
Общий ви i со спинной
стороны.
Развитие F. canicularis обычно происходит в фекалиях в уборных,
но личинки встречаются и в полужидком свином навозе. Личинки (рис. 260)
коричневатого цвета, плоские; на спине и по бокам они имеют довольно
длинные отростки. Окукливание происходит в земле или в подсохшем
субстрате. Все развитие от яйца до взрослой мухи при 21—26° занимает
около 10 дней.
Во взрослом виде эта муха встречается в хлевах и наружных убор-
ных, а также и в человеческих жилищах. В квартирах этот вид занимает
по численности второе место после М. domestica. Чаще всего их можно
видеть летающими под потолком или около спускающихся с потолка ламп.
Основным питанием взрослых мух слу?кат, с одной стороны, экс-
кременты, а с другой — продукты питания человека; отсюда следует, что
они могут играть значительную роль в распространении инфекций.
F. canicularis менее теплолюбивы, чем М. domestica, и на открытом воз-
духе встречаются больше в тени, чем на солнце.
390
Зимовка происходит на стадии куколки. Этот вид обладает диапаузой:
у особей, окуклившихся осенью, наблюдается задержка в развитии ку-
колки на несколько месяцев даже в условиях высокой температуры.
Сезонный ход численности характеризуется тем, что Fannia много-
численнее весной и осенью, чем летом; в наиболее жаркий период числен-
ность их понижается, а ранней осенью опять возрастает.
6. РОД MUSCINA R. D.
ДОМОВАЯ МУХА (MUSCINA STABULANS FLLN.)
Взрослая муха (рис. 261) крупнее, чем М. domestica. Отличительной
особенностью является красновато-желтый цвет на кончике щитка. Ноги
и щупики желтые. Хорион — оболочка яйца — несет продольные реб-
рышки. Отличительной особенностью личинок является форма щелей
задних дыхалец (рис. 262):
они не извитые, как у
М. domestica, а лишь слегка
изогнуты под углом.
Рис. 261. Muscina stabulans — домовая муха,
самка. Общий вид.
Рис. 262. Задние дыхальца
личинки домовой мухи
III возраста.
Питание личинок М. stabulans отличается исключительным разно-
образием. Они могут паразитировать в живых растениях и в живых насе-
комых (кузнечиках), а также и на теплокровных, в том числе иногда вы-
зывают миазы и у человека. Личинки Muscina встречаются и на трупах
позвоночных. Наряду со всем этим, они часто развиваются в кухонных
отбросах, навозе и особенно фекалиях. Чаще, чем в уборных, этот вид
встречается в отдельных порциях фекалий в хлевах или на дворе. Ли-
чинки I и II возраста питаются экскрементами, а в III возрасте часто
прибегают к хищничеству, поедая личинок других видов, например,
М. domestica.
Яйца и личинки М. stabulans очень влаголюбивы и встречаются в вы-
гребах с жидким содержимым. Наиболее обычным местообитанием взрос-
лых мух данного вида являются неканализованные уборные. Эти мухи
бывают многочисленны и в хлевах, особенно в тех, где имеются челове-
ческие faeces. Реже М. stabulans встречается в жилых домах и здесь по
численности во много раз уступает М. domestica.
В табл. 17 показан средний улов за сутки домовой и комнатной мухи
в помещении Кабардинского колхоза.
Нередко домовая муха встречается и на рынках. Основным пита-
нием ей служат фекалии, но опа охотно садится и на пищу человека.
Нередко у особей, пойманных в домах, кишечник содержит экскременты.
Отсюда ясно, что, несмотря на свою немногочисленность в домах, эта муха
может иметь существенное эпидемиологическое значение, так как, с одной
391
Таблица 17
Средний улов за сутки М. stabulans и М. dome-
stica в разных помещениях
,, Musca dome-
Характер помещения | stica
- ।
Muscina
stabulans
Дома............. S3 I
Хлева.............. 1 I 10
Уборные............. 3,5 , 17
стороны, садится на продукты питания, а с другой — очень тесно свя-
зана с faeces. Число одновременно созревающих яиц у М. stabulans равно
122—123. Яйца разбрасываются по
поверхности субстрата, а иногда
даже над субстратом — на стенке
мусорного ящика.
Рис. 263. Кривая сезонного хода числен-
ности домовой мухи.
домовая муха; 2 —комнатная муха. По вер-
тикали — среднее число мух в мухоловке за сут-
ки, подекадно (в процентах к среднему суточ-
ному улову мух в той декаде, на которую па-
дает максимальный вылов); по горизонтали —
декады и месяцы.
Эта способность Muscina откла-
дывать яйца не на субстрат имеет прак-
тическое значение, так как даже при-
крытые продукты могут оказаться зара-
женными, если запах от них проникает
наружу; в таком случае самка кладет
яйца около крышки, а личинки поль-
зуются мельчайшими щелями, чтобы
переползти на продукты.
Повидимому, этот вид может
зимовать как на стадиях личинки
или куколки, так и на стадии
имаго.
Весной М. stabulans по-
является раньше, чемМ. domestica.
Сезонный максимум (рис. 263) на-
блюдается приблизительно на ме-
сяц раньше, чем максимум ком-
натной мухи, и в тот период, когда
числоМ. domestica достигает сво-
его максимума, численность попу-
ляции М. stabulans находится уже
на очень низком уровне. Эту разницу в сезонном ходе численности
комнатной и домовой мухи можно видеть на рис. 263.
7. РОД STOMOXYS GEOFFR.
ОСЕННЯЯ ЖИГАЛКА (ST. CALCITRANS L.)
По своим размерам взрослая муха-жигалка почти не отличается от
комнатной мухи. В спокойном состоянии она держит крылья более ши-
роко расставленными, чем комнатная муха. Главным отличительным при-
знаком осенней жигалки является ее твердый, длинный хоботок с корот-
кими и тонкими щупиками (рис. 228 и 264).
Яйца Stomoxys несколько мельче яиц комнатной мухи. Отличительной
особенностью личинок является форма и расположение задних дыхалец.
Расстояние между дыхальцевыми пластинками значительно больше, чем
у Musca, они мелкие и почти треугольной формы. Изгиб щелей несколько
напоминает латинскую букву S.
392
Развитие жигалки происходит в скоплении гниющих растений и
в навозе разных домашних животных, смешанном с соломенной или травя-
ной подстилкой. В чистом навозе или помете домашних животных личин-
ки этого вида встречаются очень редко. Иногда личинки обнаруживаются
и в мусорных ящиках, если в них присутствует гниющая трава или со-
лома. Личинки S. calcitrans даже в основных очагах выплода не бывают
так обильны, как личинки М. domestica; максимальное число личинок
жигалки в 250 см3 навоза, смешанного с соломой, — 200 штук.
Рис. 264. Stomoxys calcitrans — осенняя жи-
галка, самка. Общий вид.
Рис. 265. Личинка осенней
жигалки.
Задний конец; дыхальца широко
расставлены; справа — увеличен-
ное заднее дыхальце.
Развитие S. calcitrans идет медленнее, чем развитие М. domestica.
В табл. 18 приводятся сроки развития от яйца до имаго в зависимости от
температуры.
Таблица 18
Продолжительность развития S. calcitrans от яйца до имаго при разных температурах
Температура 1 1« = 19э 22° | | 25° | 29э 33^ 35°
Число дней. . 42 30 2L 16 12 10 .2
Личинки жигалки не так теплолюбивы, как личинки комнатной му-
хи, и в скоплениях гниющих веществ выбирают температуру от 20° до 30°
и не встречаются при температуре выше 36°. Окукливание Stomoxys проис-
ходит в тех местах, где и питание личинок; в землю, как правило, они не
уползают.
Все стадии развития Stomoxys очень влаголюбивы. Личинки этого
вида встречаются иногда в навозе такой жидкой консистенции, в какой ли-
чинки Musca существовать не могут. Имаго Stomoxys кровососущи. Кровью
питаются и самцы, и самки. Основной Добычей им служит рогатый
скот и лошади; иногда они нападают и на человека. Кроме крови, они
могут пить и другие жидкости, например, навозную жижу, но прибегают
к этому питанию редко.
Кровь, получаемая путем прокола кожи животного, поступает в кишечник, а
жидкость, выпиваемая без прокола, — в зоб. За один раз муха выпивает количество
крови, превышающее первоначальный вес ее тела в 1х/2—2 раза.
393
Переваривание крови при 26° длится около 30 часов.
В отличие от комаров и слепней, которым одной порции крови доста-
точно для созревания одной порции яиц, самкам Stomoxys для
созревания первой порции яиц необходимо пить кровь не ме-
нее трех раз. Развитие яичников протекает так же, как и у Musca,
но у жигалок в каждой яйцевой трубочке развивается одновременно по
пяти фолликулов. Среднее число яиц в кладке — 104. Кладок в течение
Рис. 266. Кривая сезонного хода численности
Stomoxys в окрестностях Москвы (оригинал
Кузиной).
до 100 м от хлевов, служащих местом
жизни самки несколько, и число
их зависит от продолжитель-
ности ее жизни.
Для откладки яиц самка
осенней жигалки выбирает гни-
ющие растения; даже при раз-
множении в навозе она откла-
дывает яйца не на сами экскре-
менты, а на кусочки растений,
примешанных к навозу. Яйца
откладываются небольшими
кучками или более или менее
разбросанно.
В условиях умеренного
климата S. calcitrans встречает-
ся главным образом в челове-
ческих поселках и большую
часть времени проводит в ко-
ровниках, реже в конюшнях и
свинарниках. Здесь их можно
видеть на животных или на сте-
нах и потолке. На пастбища
вслед за скотом Stomoxys ле-
том, как правило, не летит;
это имеет место иногда поздней
осенью. В жилище человека
жигалка — случайный посети-
тель, хотя в тех домах, кото-
рые находятся на расстоянии
выплода этих мух, они довольно
многочисленны.
Массовые нападения на животных наблюдаются при температуре
выше 15°. Жигалка светолюбива и питается на животных только днем.
Жигалка может зимовать на стадии личинки, куколки и имаго и при
благоприятных внешних условиях может размножаться круглый год.
Сезонный ход численности жигалки (рис. 266) своеобразен. Название
«осенняя жигалка>> эта муха получила благодаря тому, что в заметных
количествах появляется очень поздно, только в июле, и своего макси-
мума достигает в сентябре.
Жигалка изредка является механическим переносчиком ряда ин-
фекций. Перенос этот осуществляется в том случае, если муха, не насо-
савшись полностью на больном животном или человеке, нападает на здоро-
вых. В это время болезнетворные микроорганизмы на хоботке мухи еще
живы и, попав в кровь, могут вызвать заболевание.
Жигалка может заражать человека сибирской язвой, очень редко —
кожным лейшманиозом, возвратным тифом, полиомиелитом и, пови-
димому, туляремией.
394
8. РОД GLOSSINA WIED.
МУХА ЦЕ-ЦЕ
Род Glossina (рис. 267) имеет много видов. Три из них играют основную роль как
переносчики возбудителя африканской сонной болезни (возбудитель—Trypanosoma
gambiense). Все они—обитатели Центральной Африки, и поэтому только здесь встре-
чается и это заболевание. Муха це-це передает также возбудителя (трипаносому) на-
ганы— тяжелого заболевания домашних животных, которое в огромном большинстве
•случаев заканчивается смертью.
Так Кс1к виды Glossina — единственные переносчики сонной болезни, борьба с
ними является насущной необходимостью. Это заболевание имеет обычно массовый ха-
рактер и ведет иногда к вымиранию целых туземных селений.
ГЛАВА IV
СЕМЕЙСТВО LARVIVORIDAE
Отличительным признаком этого семейства по сравнению с семей-
ством Muscidae является наличие щетинок на бочках груди перед зад-
ними дыхальцами (над тазиками задних ног). Медиальная жилка на
крыле изогнута под углом, как у Musca. Рассмотрим представителей
двух подсемейств: Calliphorinae и Sarcophaginae.
1. ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПОДСЕМЕЙСТВА CALLIPHORINAE,
ИМЕЮЩИХ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
1 (2). Средняя пара рядов щетинок на спинке груди перед швом отсутствует. Все
тело металлически синее, с зеленоватым оттенком....................
...............................Phormia R. D. — весенняя падальная муха.
2 (3). Средние щетинки на спинке груди перед швом имеются по меньшей мере
в числе одной пары.
3.....(4). Тело металлически зеленое, щиток с каждой стороны с четырьмя щетинками
...... Lucilia R. D. — зеленая падальная муха.
4 (3). Тело синее, с беловатым налетом, щетинка усиков покрыта длинными во-
лосками .......................Calliphora R. D. — синяя падальная муха.
395
2. РОД PHORMIA R. 1).
Phormia terrae-novae И. D.
Основные места развития этого вида — трупы рыбы и рыбные отбросы. <)т?
муха нередко приносит большой вред на промыслах, откладывая яйца па рыбу, при
готовленную для обработки. Развитие этого вида может также происходить и на тру-
пах млекопитающих, а также в мясных отбросах и в гниющих скоплениях других орга-
нических веществ. Phormia в изобилии встречается в городах, где ее местами выплода
являются мусорные ящпки. В сельских условиях численность этого вида значительно
меньше, так как навоз им почти не используется. Исключение составляет свиной навоз
в тех случаях, когда к нему примешано много кухонных отбросов, служащих кормом
для свиней. Окукливание часто происходит не в земле, а на подсохших остатках
субстрата, которым питались личинки. Например, на мясных или рыбных костях,
иногда в комках бумаги или тряпок в мусорных ящпках можно найти массу плоти с
прилипших' к этим предметам куколок Phormia.
Рис. 268. Calliphora
erythrocephala. Общий
вид.
Рис. 269. Задние дыхальца личинки
синей падальной мухи.
1 — прямые щели.
Отличительными признаками личинок Phormia является грязнобелый цвет тела,
мясистые отростки на заднем конце и прямые щели задних дыхалец. Во взрослом состо-
янии Phormia многочисленны около мусорных ящиков па рынках, особенно в мясном
и рыбном ряду. Весной они облепляют стволы деревьев, если из них сочится сок. Число
яиц, откладываемых за один раз, около 300. В течение жизни самка откладывает не-
сколько кладок. Phormia обладает имагинальной диапаузой; мухи зимуют под корой
деревьев, в щелях деревянных построек и т. д. Весной они появляются очень рано:
в конце марта или начале апреля их можно видеть на пригретых солнцем стенах
строений. Phormia имеет значение как механический переносчик инфекционных забо
леваний и возбудитель факультативных миазов.
3. СИНИЕ ПАДАЛЬНЫЕ МУХИ (РОД CALLIPHORA В. D.)
a) Calliphora erythrocephala Мегд.
Места выплода — трупы млекопитающих и мясные отбросы в мусор-
ных ящиках; иногда яйца откладываются на изготовленную пищу. Окук-
ливание происходит в земле на различной глубине — в плотной почве'
до 6 см, в рыхлой — на глубине более 20 см. Местообитание имаго — от-
крытые места продажи пищи; они встречаются на мясных, рыбных, молоч-
ных продуктах и на фруктах. В большом числе эти мухи залетают в по-
мещения боен, в колбасные цехи и т. п. Самки откладывают за один
раз около 200 яиц. В течение жизни кладок бывает 4—5.
У С. erythrocephala имеет место диапауза на предкуколочной фазе.
Зимовка происходит в земле. Этот вид холодолюбив: весной и осенью имаго
многочисленнее, чем летом. Сезонный максимум в Москве падает па
октябрь. Эпидемиологическое значение — механический перенос микро-
организмов и яиц гельминтов. Размеры ротового отверстия этой мухи
допускают заглатывание яиц глистов. Последние проходят через ее ки-
шечник неповрежденными. Поэтому в фекальных пятнах Calliphora мож-
396
но обнаружить живые япца аскарид, остриц, широкого лентеца и др.
С. erythrocephala является возбудителем факультативных миазов, откла-
дывая яйца в незащищенные раны. Неоднократно отмечались случаи и
кишечных миазов, возникающих в тех случаях, когда яйца или личинки
Calliphora заглатывались с пищей.
б) Calliphora uralensis Vill.
Отличается от предыдущего вида более желтой окраской передней
части головы и очень слабым белым налетохм на брюшке.
Основными местами выплода служат скопления фекалий в уборных.
В остальном аналогичен предыдущему виду. Отсюда вытекает, что эпиде-
миологическое значение С. uralensis больше, чем С. erythrocephala.
4. ЗЕЛЕНЫЕ ПАДАЛЬНЫЕ МУХИ (РОД LUCILIA R. D.)
LUCILIA SERICATA MEIG.
Рис. 270. Задние дыхальца
зеленой падальной мухи.
К роду Lucilia принадлежит несколько синантропных видов, близ-
ких между собой по своей биологии и экологии. В качестве примера
дается описание вида L. sericata.
Места выплода — трупы животных, кухонные
отбросы, иногда фекалии и свиной навоз. Личинки
этого вида (рис. 270) часто встречаются в ранах че-
ловека и животных. Окукливание происходит на
земле.
L. sericata встречаются в местах открытой про-
дажи пищи, па бойнях, па экскрементах человека
и животных на открытом воздухе. Для откладки
яиц предпочитают свежие, еще не разлагающиеся
трупы. Число яиц, откладываемое самкой, около 200.
Кладок бывает несколько (4—5). Зимовка происходит на стадии личинки. Осенью впав-
шие в диапаузу личинки уходят в землю на большую глубину, а весной, выйдя из со-
стояния диапаузы, передвигаются ближе к поверхности, где и происходит окукливатше.
Виды Lucilia имеют большое значение как механические переносчики микробов
и яиц гельминтов.
5. ПОДСЕМЕЙСТВО SARCOPHAGINAE. ВОЛЬФАРТОВА МУХА
(WOHLFAHRTIA MAGNIFICA SCHIN)
Из многочисленных представителей подсемейства Sarcophaginae
(рис. 32) наибольшую роль в патологии человека играет вольфартова муха
(рис. 271)—Wohlfahrtia magnifica—возбудитель облигаторных миазов. Вид
этот обычен в скотоводческих районах. В СССР распространен в степной
полосе, но встречается и за ее пределами. Самка откладывает не яйца,
а рождает личинок, по 120—160 штук за один раз. Новорожденные ли-
чинки 1 мм длины.
Имаго светлосерого цвета, на груди имеет три темные продольные
полоски. Брюшко яйцевидное, с черным блестящим рисунком — посре-
дине черная полоса с неровными краями, по бокам — круглые пятна.
Все тело личинки (рис. 272, Л) покрыто рядами мелких темных шипи-
ков, направленных назад. Приротовые крючки сильно развиты. Передние
дыхальца с 5—6 отростками. Задние дыхальца (рис. 272, Б) расположены
в углублении, которое образовано выступающими краями заднего чле-
ника. Задние дыхальца имеют три прямые щели, окруженные незамкну-
тым хитиновым кольцом.
Самки вольфартовой мухи не нуждаются в белковом питании для
созревания яиц и развития личинок. Основным их питанием является нек-
тар, поэтому они обычно встречаются на цветах. Когда личинки готовы
397
к откладке, самка начинает отыскивать животных, на которых их и
откладывает. Вольфартова муха заражает главным образом копытных,
однако нередко откладывает яйца в глаза
Рис. 271. Wohlfahrtia magnifica — вольфартова
муха, самка.
Общий ВИД.
нос, уши и на раны спя-
щего человека. Проникая
вглубь тканей через царапи-
ны или ранки на коже, ли-
чинки выедают все мягкие
ткани вплоть до костей, мо-
гут попадать в носовую и
гайморову полость, десны
и т. д. Когда личинки прокла-
дывают новые ходы в тка-
нях, то ощущается сильней-
шая боль. Части тела, пора-
женные личинками Wohb
fahrtia, воспаляются, появ-
ляются нагноения, гангре-
нозные процессы. Описано
несколько случаев, когда
миазы (рис. 273), вызванные
личинками вольфартовой му-
хи, заканчивались смертью.
Развитие личинок W. magnifica занимает 3—4 дня. За этот период они
могут произвести большие разрушения, поэтому обнаруженные в тканях
личинки должны быть немедленно удалены. Для этого употребляют
Рис. 272. Личинка вольфартовой мухи.
А — общий вид сбоку; Б — заднее дыхальце.
хлороформную воду (1 : 100), которой промывают рану; личинок уда-
ляют пинцетом.
Перед окукливанием личинки покидают рану; окукливание происхо-
дит в земле.
Основная масса миазов наблюдается с июля по сентябрь.
ГЛАВА V
ЖЕЛУДОЧНЫЕ ОБОДЫ (СЕМЕЙСТВО GASTROPHILIDAE)
Из видов этого семейства рассмотрим один—Gastrophilus intestina-
lis Deg—большой желудочный овод лошади (рис. 274).
Мухи крупной величины; все их тело покрыто желтоватыми и сероватыми волос-
ками, крылья желтоватые со сьетлобурыми пятнами. Potoi ые органы не развиты,
так как имаго не питается.
398
Рис. 273. Миаз, вызванный личинками
вольфартовой мухи.
Самка G astrop hilus intestinal is откладывает яйца, приклеивая их к волосам ло-
шади. Развившаяся личинка выходит из яйца при прикосновении к нему влажного
предмета и сейчас же внедряется в эпидермис кожи, проделывая в ней извилистые
ходы и вызывая при этом сильный зуд. Лошадь расчесывает ходы и проглатывает
находящихся в них личинок. Дальнейшее развитие личинок протекает в желудке
лошади. Взрослые личинки выходят
наружу с испражнениями лошади и
окукливаются в земле.
Случаи миазов, вызванных
личинками Gastrophilus, описаны и
у людей, главным образом у лиц,
имевших дело с лошадьми. Личинки
I возраста внедряются в эпидермис
кожи человека, не проникая в глуб-
жележащие ткани. Описан случай,
когда миграция личинки под кожей
мальчика 21 /2 лет продолжалась в
течение двух месяцев; ход’начинался
от лобка и через весь живот проходил
на грудь, где он был очень изви-
лист; длина хода равнялась прибли-
зительно 190 см. Он имел вид тонкой
* буроватой полоски, похожей на след
подживающей царапины (рис. 275).
Личинка I возраста около 1 мм
длины. Спереди тело личинки не-
сколько шире, чем сзади. На голов-
ном конце много шипиков. Сегменты
соскообразных шипов. На заднем конце тела два коротких выроста с дыхальцами
на концах. Ротовые крючки большие (рис. 276).
с 3-го по 11-й несут по переднему краю ряды
Рис. 274. Gastrophilus intestinalis — желу-
дочный овод лошади, самка
Общий вид.
Рис. 275. Миаз, вызванный
личинками желудочного овода.
Извлечение личинки производится хирургически (вскрывается конец хода и ли-
чинка вынимается) или же путем наложения повязки с 5% креолиновой мазью.
Г Л А В А VI
КОЖНЫЕ ОВОДЫ (СЕМЕЙСТВО HYPODERMIDAE)
Из видов этого семейства рассмотрим Hypoderma bovis L., кожного
овода рогатого скота.
Крупные мухи черного цвета; спереди средпеспинка в желтовато-серых, сзади
в черных вотосках с черными блестящими продольными полосками; брюшко у осно-
вания в беловатых, посредине в черных, на конце в красноватых волосках. Ротовые
органы не развиты. Самки откладывают яйца па шерсть задних ног животного. Выхо-
дящие из яиц личинки проникают под кожу, а затем по тканям постепенно мигрируют
399
почти через все тело животного и заканчивают свое развитие под кожей на спинной
стороне. Окукление происходит в земле. t
Описаны случаи нахождения личинок Hypoderma у человека. Невидимому,
самка откладывает яйца на части тела, покрытые волосами. Вышедшая личинка про-
никает под кожу, мигрирует и затем, уже будучи во II возрасте, обнаруживается где*
Рпс. 276. Личинка
I возраста желу-
дочного овода-
1— передний конец;
2—заднее дыхальце.
Рис. 277. Hypoder-
ma bovis — кож-
ный овод. Взрос-
лая личинка (вид
сбоку).
1 — передний конец;
2—спинная сторона.
Рис. 278. Личинка I возраста Oest-
rus ovis — овечьего овода.
А—общий вид с брюшной- стороны:
Б— задний конец. 1— ротовые крючки:
2—когтеобразпые шипы.
либо под кожей, например, па спине, на* руке, на лице. В том месте, где находится
личинка, образуется почти безболезненная опухоль.
Брюшная сторона тела личинки (рис. 277) плоская, спинная выпуклая. Все
тело покрыто мельчайшими шипиками. Ротовые крючки небольшие.
Личинки удаляются хирургическим путем.
ГЛАВА VII
ПОЛОСТНЫЕ ОВОДЫ (СЕМЕЙСТВО OESTKIDAE)
Рассмотрим двух представителей этого семейства: Oestrus ovis L.—
овечий овод и Rhinoestrus purpureus Br.—лошадиный, или русский, овод.
Самки рождают личинок I возраста. Подлетев к животному, самка выпускает
в его ноздри моточнобелую жидкость, содержащую личинок. Развитие'личинок проте-
кает.в носовых и глоточной полостях и в лобных пазухах.
Иногда самка полостного овода выпускает личинок в глаз человека. Глазные
миазы, вызванные личинками полостных оводов, могут быть наружными, когда
личинка прободает веко, слезный мешок или слизистую оболочку, или внутренними,
когда личинка проникает внутрь глазного яблока.
У личинки Oestrus ovis I возраста (рис. 278) па каждом сегменте тела располо-
жено 3—4 ряда шипиков. По бокам тела имеются пучки длинных волосков. На заднем
конце тела 20—22 темпожелтых когтеобразных шипа. Ротовые крючки большие.
Для удаления личинки глаз анестезируют раствором кокаина и промывают бор-
ной кислотой. Если промыванием личинку удалить не удается, ее вынимают пинце-
том.
400
1ЛАВА VIII
КРОВОСОСКИ (СЕМЕЙСТВО IIIPPOBOSCIDAE)
Из представителен этого семейства рассмотрим Hippobosca equina L.,
лошадиную кровососку (рис. 279).
Тело плоское, покрыто очень толстым хитином; антенны скрыты во впадинах го-
ловы; коготки на лапках сильно развиты, с одним-двумя дополнительными зубцами.
Рис. 279. Hippobosca equina — лошадиная
кровососка.; Общий вид.
Рпс. 280. Пу парий
кровососки.
Взрослые мухи питаются кровью и ведут образ жизни постоянных паразитов,
проводя почти все время на теле лошади или других млекопитающих. Иногда нападаю!
и на человека. Самка рождает по одной личинке, находящейся в предкуколочной ста
дии, совершенно неподвижной. Самка, в теле которой находится готовая к рождению
личинка, покидает хозяина и отыскивает место для откладки, обычно в расщелинах
стен или почвы. Сейчас же после откладки предкуколка превращается в куколку
(ряс. 280), а самка возвращается на хозяина.
Другая обычная форма, Melophagus ovinus L., овечий рунец, совершенно лишена
крыльев. Живет на овце в качестве постоянного паразита. Пупарии находятся в шер
сти хозяина.
2б Учебник медицинской энтомологии
РАЗДЕЛ Д
БЛОХИ (отряд Aphanlptera)
1. ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Блохи — весьма важные в эпидемиологическом отношении насеко-
мые, являющиеся переносчиками такого опасного заболевания, как чу-
ма. Они являются также основными переносчиками одного из риккетсио-
зов — крысиного сыпного тифа, которым могут заболевать и люди. Ве-
роятно, блохи участвуют и в распространении туляремии среди грызунов
(Тифлов). Блохи паразитируют на млекопитающих и птицах, как прави-
ло на тех из них, которые живут в гнездах или норах. Особенно большое
значение имеют те виды блох, которые живут в жилищах людей и напа-
дают на них (блоха человека Pulex irritans, блоха собаки Ctenocephalides
canis, блоха кошки Ctenocephalides felis, блохи крыс Xenopsylla cheo-
pis и Ceratophyllus fasciatus и др.).
2. НАРУЖНОЕ СТРОЕНИЕ
Строение блох приспособлено к пребыванию и свободному передви-
жению в шерсти или перьях хозяина: сжатое с боков тело (рис. 281),
гладкость и твердость хитинового покрова, направленные назад щетинки
и шипы, нередко собранные в гребенки или ктенидии, отсутствие
крыльев, хорошо развитые коготки на лапках.
Голова. Хорошо развитая передняя часть головы, прикрывающая
ротовые органы, служит насекомому как бы тараном, при помощи кото-
рого оно продвигается сквозь гущу волос. Антенны, состоящие из двух
основных члеников и обычно девятичлениковой булавы, спрятаны в осо-
бые ямки, что предохраняет их от поломки и облегчает ползание в шер-
сти. Сложные глаза отсутствуют, простые глазки развиты в различной
степени у разных видов, но могут также совсем отсутствовать.
Ротовой аппарат. Ротовой аппарат состоит из следующих важнейших
частей. Имеется один непарный колющий орган, стилет, с внутренней сто-
роны выемчатый, происходящий из верхней губы (надглоточника). Под-
глоточник (hypopharynx) сильно редуцирован, имеет вид треугольной
пластинки, расположенной у основания верхних челюстей. На его сво-
бодней конце открывается проток слюнных желез. Верхние челюсти
(mandibulae) — хорошо развитые, узкие, длинные пластинки с зазубрен-
ными краями у вершины и желобками на внутренних поверхностях. Ман-
дибулы примыкают с боков к стилету надглоточника и образуют канал,
по которому кровь устремляется в глотку. Зазубренные же края служат для
402
разрезания кожи при уколе. Таким образом, по своим функциям верхние
челюсти играют весьма важную роль в ротовОхМ аппарате блохи. Нижние
челюсти (maxillae) развиты слабее и не принимают участия в проколе;
к ним причленяются щупики (palpi maxillares), состоящие из 4 члеников.
Весь ротовой аппарат снизу прикрыт нижней губой (labium), которая не-
сет на конце пару длинных нижнегубных щупиков, или пальп, выемча-
тых с внутренней стороны. У различных видов блох пальпы имеют раз-
Рис. 281. Человеческая блоха (Pulex irritans) (вид сбоку).
I — глаза; 2 — антенны; 3 — щетинки; 4 — 1—3-й грухтные сегменты;
5 — грудные стигмы; 6— тазик; 7 — вертлуг; 8 — бедро; 9 — голень;
10 — лапка; 11— 1—8-й сегменты брюшка; 12—пигидий;
13 — брюшные стигмы.
ное число члеников, образующих так называемый хоботок. В состоянии
покоя в выемки щупиков вкладывается верхняя губа и верхние челюсти.
Грудь. Грудной отдел состоит из трех члеников, подвижных отно-
сительно друг друга. В ногах сильно развиты тазики (coxae), в которых
помещается мощная прыгательная мускулатура. Величина прыжка че-
ловеческой блохи около 32 см в длину и 9 см в высоту. Лапка пятичлени-
ковая. Последний членик несет пару крепких коготков.
Брюшко. Брюшко состоит из 7 члеников обычной формы и нескольких
измененных члеников, форма которых представляет важнейшие признаки
для распознавания видов блох. Позади 8-го членика на спинной стороне
расположено чувствительное поле, покрытое тонкими чувствительными
волосками, так называемый пигидий, который служит, повидимому, для
восприятия движения воздуха. 9-й и 10-й членики сильно видоизменены.
У самцов они превращены в мощные клешни, по которым самцов и отличают
от самок; у последних конец брюшка овальной формы и несет небольшие
церки. Грудь имеет две пары дыхалец, брюшко — восемь пар. Обычные
размеры блох от 1 до 5 мм, ио крупные «беременные» самки алакурта
(доркадий) (см. ниже) достигают 16 мм в длину, мелкие Echidnophaga —
0,7э мм. Самки крупнее самцов.
26*
403
3. ВНУТРЕННЕЕ СТРОЕНИЕ
Пищеварительный аппарат. Пищеварительный аппарат блох состоит
из следующих отделов. От мускулистой глотки отходит тонкий пищевод
(рис. 282), впадающий в преджелудок. Последний густо выстлан направ-
ленными назад хитиновыми щетинками, не мешающими прохождению
пищи, но являющимися в тоже время как бы клапаном, задерживающим
обратный ток содержимого желудка. Средняя кишка, или желудок, начи-
Рис. 282. Внутренняя анатомия самки блохи.
1 — пищевод; 2 — преджелудок; 3 — желудок; 4 — мальпигиевы со-
суды; 5 — задняя кишка; 6 — прямая кишка; 7 — слюнные железы;
8—слюнной насос; 9— яичники; 10—парный яйцевод; И—не-
парный яйцевод (матка); 12 — семяприемник; 13 — придаточные
железы.
нается от преджелудка; она мешковидной формы, стенки ее выстланы ци-
линдрическим эпителием. Позади средней кишки впадают четыре мальпи-
гиевых сосуда, за которыми следует довольно длинная задняя кишка.
Задняя кишка переходит в прямую, несущую ректальные железы. По
обеим сторонам кишечника лежит по две слюнные железы, состоящие
из небольшого числа гигантских клеток. Протоки обеих желез каждой
стороны вскоре соединяются между собой, а в общий непарный слюнный
проток они объединяются в области шеи. В голове проток расширяется
в так называемый слюнный насос.
Половой аппарат. Половой аппарат самки состоит из двух яичников,
заключающих в себе по нескольку яйцевых трубочек с концевыми каме-
рами. Яйцевые трубочки впадают в парные яйцеводы, которые
объединяются в области задней кишки в непарный яйцевод, или матку.
Имеется один или два семяприемника; в них отличают резервуар и при-
даток, форма которых имеет значение для систематики блох. Тонкий про-
ток соединяет семяприемник с совокупительной сумкой (bursa copulatrix),
которая открывается в матку в месте ее перехода во влагалище. Мужской
половой аппарат состоит из семенников, семяпроводов с придаточными
железами, непарного выводного канала, совокупительного органа с хи-
тиновой арматурой (вооружением) и половой клешни.
4. РАЗВИТИЕ
Блохи относятся к насекомым с полным превращением (рис. 283).
Яйца, как правило, белые, длиной от 0,4 до 0,65 мм. Личинки
блох стройные, червеобразные. Тело состоит из головы и 13 члеников
404
из которых три первых — грудные. Длина тела около 5 мм. Личинка
блох безногая и лишена глаз, антенны короткие, ротовой аппарат жева-
тельный. На каждом членике тела имеется несколько щетинок. На заднем
конце тела расположен «анальный гребешок», состоящий из одного-двух
рядов щетинок. Кишечник личинок длиннее, чем у взрослых. Он состоит
из двух отделов: переднего, выстланного плоским эпителием, и заднего,
Рис. 283. Развитие блох.
1— яйцо; 2— личинка; 3 — куколка; 4— кокон, очищенный от по-
сторонних частей; 5 — кокон (нормальный вид).
выстланного высоким эпителием. В месте перехода одного отдела в дру-
гой имеются слепые выросты. Личинки блох проходят три возраста. Ку-
колки лежат в коконе.
5. ЭКОЛОГИЯ
По характеру своего паразитизма блохи представляют различ-
ные переходы между гнездовыми, или норовыми, паразитами, с одной
стороны, и настоящими эктопаразитами — с другой. Они никогда не уда-
ляются далеко от хозяина или его гнезда. В тех случаях, когда по своему
образу жизни хозяин сравнительно мало связан с гнездом, блохи вынуж-
дены проводить большую часть времени на хозяине. Таким образом, по
образу жизни блохи делятся на две основные группы: блохи гнезда и бло-
хи шерсти (Пофф). Каждый вид блох имеет своего специфического хозяина,
но при отсутствии последнего легко переходит на других животных. Так,
например, Pulex irritans, будучи первоначально паразитом какого-то
другого млекопитающего, приспособился к человеку и в то же время
с успехом нападает на свиней, а также на многих хищников. Способность
блох питаться на различных животных очень важна в эпидемиологическом
отношении. Как правило, блохи легко переходят на тех животных, кото-
рые тесно соприкасаются с их хозяином. Так, например, блоха тотчас же
покидает мертвое животное и переходит на хищника, убившего ее хо-
зяина. Следует отметить, однако, что блохи большинства видов вполне
нормально чувствуют себя лишь в том случае, если питаются на своих
специфических хозяевах. Так, например, при кормлении на необычных
хозяевах наблюдается сокращение продолжительности жизни блох,
прекращение откладки яиц и т. д.
Блоха питается только кровью. Желудок крысиной блохи вмещает
до 0,5 мм3 крови, но во время сосания блоха выпивает иногда много боль-
ше, так как во время еды у многих видов из анального отверстия неодно-
кратно выделяются капельки свежей крови, быстро высыхающей среди
405
шерсти хозяина. Человеческая блоха (Pulex irritans) за первые полчаса
после принятия пищи испражняется до 10—20 раз. Поскольку блохи
питаются только стерильней пищей в виде крови, их кишечник не содержит
дсстаточного количества активных бактерицидных веществ (т. е. веществ,
убивающих попавших в него бактерий), и в тех случаях, когда те пли иные
микробы все же попадают в кишечник блохи, некоторые виды бактерий
могут там поселиться. Чумные палочки, попав в кишечник блохи, начи-
нают в нем размножаться и своей массой закупоривают кишечник; когда
такая блоха с закупоренным преджелудком пытается пить кровь, чумные
палочки отрыгиваются в ранку, что возможно лишь при нарушении функ-
ции клапанного аппарата в преджелудке, о которОхМ упоминалось раньше.
Экскременты блох, состоящие из темных комочков, остаются на платье
человека, в шерсти животных или прямо на подстилке гнезда. Блохи
могут пить кровь ежедневно, но при содержании в неволе их достаточно
кормить каждые 2—3 дня. «Блохи шерсти», как правило, питаются чаще,
чем «блохи гнезда». Сосание длится от одной минуты до многих часов.
Копулируют блохи многократно. Спустя несколько дней после пре-
бывания на хозяине самка начинает откладывать яйца в гнезде послед-
него. Блохи человеческого жилья откладывают япца в щели полов и дру-
гие укромные уголки. В течение жизни человеческая блоха (Pulex ir-
ritans) откладывает до 450 яиц. Личинки вылупляются из яиц прп помощи
яйцевого зуба. Они питаются всевозможными органическими частицами.
Личинки некоторых видов не могут обходиться без питания кровью, ис-
точником которого для них являются экскременты взрослых блох. Разви-
тие личинок блох протекает обычно в норах при достаточно высоко!! влаж-
ности, не ниже 80%, и сравнительно небольших колебаниях температуры.
Поэтому пределы температуры, благоприятной для развития личинок,
довольно узки: для Pulex irritans 18—27°, минимум 13°, для Xenopsylla
chaeopis 21—27°, минимум 15°, для Ceratophyllus fasciatus 13—16°.
Высокие температуры при недостаточной влажности действуют на
блох губительно. Температура выше 40° вызывает их быструю гибель.
Взрослая личинка делает себе кокон, в котором покоится некоторое
время. После этого она линяет в третий раз и превращается в куколку,
остающуюся в этом же коконе. Интересно, что вылупившаяся из куколки
молодая блоха также задерживается в коконе, дожидаясь сотрясения, что
обычно совпадает с появлением в норе хозяина, который сразу обеспечи-
вает ей пищу. Вылупление из кокона может быть связано также с рез-
кими изменениями температуры и влажности, но чаще всего бывает при-
урочено к определенному сезону. Сроки пребывания в коконе различны
и колеблются от 7 до 373 дней.
Продолжительность жизни блох зависит, с одной стороны, от климати-
ческих факторов (повышение температуры и понижение влажности сокра-
щают их жизнь) и, с другой стороны, от питания: нормально питающиеся
блохи живут дольше, чем те, которым приходится голодать, но и без пищи,
при благоприятных условиях, некоторые блохи могут жить очень долго —
до многих месяцев. Конечно, разные виды блох в различной мере выносят
голод; более стойки в этом отношении те из них, которые более связаны
с гнездом, чем с хозяином. Самцы переносят голод хуже самок.
Сезонный ход численности блох различен для разных видов и для
разных местностей. Так, например, южная по своему происхождению
Xenopsylla cheepis становится у нас наиболее многочисленной к осени,
максимум их падает на сентябрь. Pulex irritans также многочисленны
осенью, но максимум их более растянут, занимая август и сентябрь.
Минимум их приходится на январь.
406
6. РАЗЛИЧНЫЕ ФОРМЫ ПАРАЗИТИРОВАНИЯ СРЕДИ БЛОХ
На примере блох можно проследить все переходы от периодически питающихся
норовых паразитов до постотнных эктопаразитов (Vermipsylla, Tunga), у которых вы-
работался целый ряд характерных приспособлений, как, например, увеличение пло-
довитости, способность личинок жить в открытой природе, приспособления для под-
стерегания хозяина и т. д.
К периодически нападающим паразитам относятся те блохи, которые сильнее -
связаны с гнездом хозяина, проводящего в нем значительную часть времени. В том
случае, если животпое бездомно, как, например, дикие копытные, блохи приспосаб-
ливаются к более долгому пребыванию на хозяине. Так, папример, у паразитов ко-
пытных Vermipsylla alacurt и V. dorcadia, живущих в горах Тянь-Шаня, молодые, не-
больших размеров самки (до 2—3 мм длины) присасываются осенью к хозяину, пара-
зитируют почти всю зичу и по мере созревания яиц увеличиваются до 8—16 мм
в длину и 3—7 мм в ширину (Пофф).
Другой пример перехода к постоянным паразитам представляет Echidnophaga
gallinacea, живущая в теплых странах, но встречающаяся и у нас в Средней Азии. Эти
блохи нападают на различных животных,в том числе и на домашних животных,и птиц, и
иногда на человека. Они плотно присасываются к хозяину, но в отличие от нижеупоми-
наемой Tunga в кожу не вбуравливаются. Если у большинства блох готова вытянута
и хоботок направлен назад, то у этого вида голова несколько уплощена и хоботок на-
правлен вперед. Прикрепление к хозяину совершается при помощи мандибул. Гребешки
и щетинки у этих блох сильно редуцированы, а прыгательные ноги значительно ослаб-
лены. Плодовитость Echidnophaga раза вЗ—4 превышает плодовитость обычных блох,
причем яйца откладываются преимущественно ночью (40 яиц откладывается ночью
и 3—4 яйца днем),
Tunga - (Sarcopsylla) penetrans, живущая в тропиках, не только присасывается
к коже, но и целиком входит в нее, вбуравливаясь при помощи верхних челюстей;
наружу остается торчать лишь копчик брюшка со стигмами. При этом брюшко
разрастается до сравнительно огромных размеров и в нем созревает громадное
количество яиц. Яйца после созревания выталкиваются наружу или же выпадает
вся блоха. Личинки живут в земле. Молодые блохи присасываются главным обра.зохМ
на ногах, часто между пальцами и, растут до размера горошины. Пребывание блохи
в коже вызывает зуд и воспалительную реакцию и нередки случаи нагноений и зара-
жения крови, вследствие загрязнения расчесанной кожи. Самцы в кожу не вбуравли-
ваются и к хозяину не прикрепляются.
РАЗДЕЛ Е
ВШИ (подотряд Siphunculata отряда Anoplura)
1. ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Все вши являются постоянными эктопаразитами млекопитающих,
причем каждый данный вид вши придерживается определенного вида хо-
зяина. Паразитами человека являются вши из семейства Pediculidae:
платяная вошь — Pediculus humanus corporis, головная — Р. humanus
capitis и лобковая — Phthirus pubis.
2. НАРУЖНОЕ СТРОЕНИЕ
Вши — постоянные паразиты, которые всю жизнь проводят на чело-
веке или на его одежде. Строение вшей (рис. 284) хорошо приспособлено
к паразитическому образу жизни. Тело их сильно сплющено в дорсо-
вентральном направлении. На теле имеются щетинки. Их число и места
прикрепления служат систематическими признаками. Окраска вшей
4- зависит от выпитой крови, которая просвечивает сквозь тонкий хитин,
а также от цвета хитина. Последний зависит от цвета фона, на котором
жила вошь. Известно, что у людей с темной кожей вши темнее. Самцы
немного мельче и стройнее самок.
Голова суживается кпереди. Сложные глаза отсутствуют, имеется два
простых глазка. Короткие антенны состоят из 5 члеников. Колющий ро-
товой аппарат состоит из пары максилл, гипофаринкса со слюнным про-
током и нижней губы; в состоянии покоя он глубоко втянут в голову, по-
мещается в особом футляре. Голова соединяется с грудью довольно тол-
стой шеей.
Грудной отдел не расчленен на отдельные кольца, членики его лишь
иногда намечаются. Ноги развиты хорошо, состоят из пяти обычных отг
делов. На дистальном конце голени, с внутренней стороны, имеется загну-
тый выступ. Лапка одночленистая, заканчивается коготком, который,
соединяясь с выступом голени, образует нечто вроде клешни, при помощи
которой вошь может крепко держаться за волосы. На груди Ихмеется толь-
ко одна пара стигм.
Яйцевидное брюшко состоит из 10 члеников. 1-й и 2-й членики силь-
но редуцированы. Членики брюшка с 3-го по 8-й несут стигмы. Женское
половое отверстие лежит у заднего края 8-го членика, а мужское — у 9-го.
Самку вши легко отличить по раздвоенному на конце брюшку. У самца
задний край брюшка ровный. Сквозь три последних сегмента просвечивают
две сходящиеся под острым углом полоски—мышцы дилятаторы, имеющие
значение при копуляции.
408
Рис. 284. Вши.
А — платяная вошь (Pediculus humanus corporis); Б — головная вошь
(Р. hum anus capitis самец и С'мка, сильно увеличены); В^-площица
(Phthirus pubis).
3. ВНУТРЕННЕЕ СТРОЕНИЕ
Пищеварительный аппарат. Пищеварительный аппарат вши начи-
нается ротовой полостью (рис. 285), которая ведет в глотку, снабженную
насосом, с сильной кольцевой мускулатурой. Глотка переходит в пище-
вод, который в свою очередь впадает в обширный желудок. В месте впаде-
ния пищевода желудок образует два слепых мешка, направленных вперед.
Желудок занимает пространство до 6-го членика брюшка, где постепенно
переходит в петлеобразно изогнутую тонкую кишку. На ее границе с зад-
ней кишкой впадают четыре мальпигиевых сосуда. Задняя кишка направ-
ляется вперед, образует петлю и поворачивает назад, переходя затем в пря-
мую кишку с шестью ректальными железами. Вши имеют две пары слюн-
ных желез. Выводные протоки желез постепенно объединяются в общий
проток, открывающийся в гипофаринкс.
Половой аппарат. В мужском половом аппарате имеется две пары
семенников: небольшие протоки, отходящие от каждого семенника, объе-
409
дйняясь, образуют с каждой стороны по тонкому семенному протоку, впа-
дающему в семенной пузырек, на конце которого расположены придаточ-
ные железы. Семенные пузырьки на дистальном конце соединяются в не-
парный семяизвергательный проток (ductus ejaculatorius). Яичники состоят
каждый из пяти яйцевых трубочек, впадающих в парные яйцеводы, в каж-
дом из которых имеется вздутие, так называемая яйцевая ампула. Пар-
Рис. 285. Внутренняя анатомия вшей (Pedicuius).
А — самка; Б — половые органы самца. 1 — глоточный насос;
2 — пищевод; 3 — желудок; 4 — мальпигиевы сосуды; 5 — задняя
кишка; 6 — слюнные железы; 7 — яичник; 8 — парный яйцевод;
9—яйцевая ампула; to—матка; 11—семенники; 12—семенные
протоки; 13 — семенные пузырьки; 14 — выводной проток.
ные яйцеводы впадают в непарный яйцевод, неясно разделенный на матку
и влагалище. С каждой стороны непарного яйцевода имеется по трехлопа-
стной придаточной железе; секрет этих желез служит для приклеива-
ния яиц.
4. РАЗВИТИЕ
Вши относятся к насекомым с неполным превращением. Яй-
ца, так называемые гниды, удлиненно-овальной формы (рис. 286),
бело-желтого цвета, покрытые продырявленной крышечкой, на которой
помещается возвышенная площадка, образованная камерами хориона.
Высота этих площадок различна у разных видов вшей и является систе-
матическим признаком. Япца приклеиваются самкой к волосам или вэр-
синка.м ткапи. Клейкое вещество после затвердевания не уступает в проч-
ности хитину, что весьма затрудняет удаление гнид. Вылупившиеся мо-
лодые вши сразу по вылуплении начинают сосать кровь. После трех линек
вошь становится взрослой.
410
5. ЭКОЛОГИЯ
В отличие от периодически нападающих на хозяина паразитов,
стремящихся получить за один прием возможно большую порцию крови,
еши питаются часто и небольшими порциями (в неволе их надо кормить
2__3 раза в день). Количество выпиваемой крови зависит в большой мере
от того, насколько голодна вошь. Если она питается один-два раза в день,
то за один прием может выпить от 0,7 до 1,1 мг крови — до появления
красной капли у анального отверстия. Самцы пьют несколько меньше.
Переваривание крови длится
около 5 часов. В отличие от
блох, вши не могут долго голо-
дать. Они легче выносят го-
лод при низких температурах:
при оптимальной температуре
28—30°—не более 2 дней, при
10—20°—7 дней, при 0—10°—10
дней. При трех кормлениях в
день и температуре около 35°
молодая вошь линяет в первый
раз через 3 дня, во второй раз
через 5 дней и в третий раз че-
рез 8 дней. Копуляция повтор-
ная. В первый раз она может
происходить через 10 часов пос-
ле третьей линьки. Яйца от-
кладываются через 24—48 ча-
сов после последней линьки.
За всю свою жизнь самка
может отложить до 300 яиц,
Рис. 286. Гниды различных видов вшей.
А — гнида площицы; Б — гнила головной вши;
В—гнида платяной вши. 1— яйцо; 2— приклеи-
вающее вещество; 3—крышечка.
откладывая в день от 5 до 14 штук. Оптимальная температура для от-
кладки 28—30°; при 25° кладка немного подавлена, а ниже 15° вообще не
имеет места. Сроки вылупления из яиц также зависят от темпе-
ратуры.
От температуры зависит также и дальнейшее развитие. Приведем
сроки личиночного развития при разных температурах.
46° 37° 34° 30° 27° 22-24°
12—13 дней 10 дней 11—12 дней 15—16 дней 20—21 день 22—23 дня
Таким образом, полный цикл развития от яйца до яйца занимает
по меньшей мере 15 дней. Продолжительность жизни вшей от 40 до 50 дней.
В связи с жизнью на теле при определенной температуре вши довольно
чувствительны к ее изменениям. Оптимальной температурой для них
является 28—30° х, и всякое повышение температуры выше 30° заставляет
вшей уходить в более прохладное место. Вши, как и многие кровососу-
щие насекомые, не одинаково охотно нападают на разных людей, руковод-
ствуясь, очевидно, обонянием. Обоняние их действует на сравнительно не-
большом расстоянии, в несколько сантиметров.
Таким образом, при тесном контакте вши могут переползать с одного
человека на другого, запах которого оказался для них более привлека-
тельным. Такое поведение вшей очень важно с точки зрения распростра-
нения тифа.
То есть та температура, которая бывает под бельем у здорового человека.
411
При экспериментальных работах, помимо человека, вшей можно на-
кормить на обезьяне, свинье, но весьма трудно накормить их на обычных ла-
бораторных животных, например, на морских свинках, кроликах. Реак-
ции людей на укус вши различны. Для одних укус остается почти не-
заметным, у других он вызывает сильный зуд, который может повторяться
в течение 10 дней. Расчесывание кожи может привести к экземе, к появле-
нию корост и т. д. Заражение вшами может происходить при непосред-
ственном контакте людей, через белье, а при сильной завшивленности вши
могут переползать по полу.
6. СИСТЕМАТИКА ВШЕЙ ЧЕЛОВЕКА
Платяная и головная вошь. Головная вошь отличается от платяной
меньшим размером, брюшко ее меньше и уже; границы члеников высту-
пают яснее. Щетинок на теле головной вши больше и сами они крупнее,
чем у платяной вши. Имеются отличия в строении голеней. По окраске
эти вши темнее платяных, но, так же как у последних, окраска их в боль-
шой мере зависит от цвета кожи человека. Головная вошь селится в воло-
сах головы и лишь очень редко попадается на теле. Отличия Р. h. corporis
и Р. h. capitis невелики, что дает возможность не выделять их в два само-
стоятельных вида, тем более что удалось найти промежуточную расу между
обоими. Образ жизни Р. h. capitis достаточно сходен с таковым Р. h.
corporis. Самка менее плодовита: она откладывает всего 140 яиц — не
более 4 яиц в день.
Гниды имеют несколько иной вид (рис. 286, Б) и приклеиваются
к волосам. Средняя продолжительность развития до 17 дней. Головная
вошь встречается также и во влажном тропическом климате, в котором
условия жизни для платяной вши весьма неблагоприятны. Вред, при-
носимый головной вошью, заключается главным образом в ранении
кожи при расчесывании с последующими заражениями (экзема, струпья,
язвы, колтун).
Лобковая вошь. Лобковая вошь, или площица (Phthyrus pubis),
меньше размерами, чем вши рода Pediculus. Тело короткое, грудь и брюш-
ко неясно отграничены. Живет эта вошь на участках тела человека, слабо
покрытых волосами, главным образом в волосах лобка, но также может се-
литься на волосах подмышек и на частях лица, покрытых растительностью
(борода, брови, ресницы), иногда забирается в волосы на голове, но там
ей трудно пробираться из-за широкого размаха ног. Так же, как и другие
вши, самки площиц откладывают яйца, приклеивая их к волосам. Пер-
вая откладка имеет место через 24—48 часов после последней линьки. Пло-
довитость лобковой вши очень невелика: за всю свою жизнь самка отклады-
вает до 26 яиц. Через 5—8 дней из гнид вылупляются личинки. Личи-
ночный период занимает 15—17 дней, продолжительность жизни имаго
до 26 дней.
Лобковая вошь гораздо менее подвижна, чем вши рода Pediculus.
Молодые личинки по нескольку дней остаются на месте, погрузив
хоботок в кожу человека и сося кровь с частыми промежутками. Взрослая
площица также длительное время принимает кровь. В связи с таким спо-
собом питания, площица вне тела человека чувствует себя очень плохо и
уже через 10—12 часов погибает. Переход их с одного человека на другого
возможен при тесном контакте (главным образохм половом).
Не являясь переносчиком какого-либо заболевания, лобковая вошь
причиняет большее беспокойство человеку, вызывая сильнейший зуд, а
при сильных расчесах могут возникать различные заболевания кожи.
412
ЭПИДЕМИОЛОГИЧЕСКАЯ РОЛЬ ВШЕЙ РОДА PEDICUWS
Вши рода Pediculus, главным образом платяные, играют огромную
роль как переносчики сыпного и возвратного тифа.
Сыпном тиф. Возбудитель сыпного тифа, риккетсия (Ricketsia рго-
wazeki) присутствует в крови больного. Вошь заражается, напившись
зараженной крови.
Риккетсии поселяются в клетках эпителия желудка и тонкой кишки
вши, здесь же размножаются, сильно повреждая эпителий, и в громад-
ном числе выделяются испражнениями вши. Вошь становится заразной
через 4 дня после заражения и остается заразной до конца своей жизни.
Человек заражается, втирая в расчесы испражнения зараженной вши
или раздавливая ее самое. В испражнениях вшей риккетсии остаются
живыми и заразными очень долго, при благоприятных условиях — ме-
сяцами.
Через яйца, от самки к потомству, риккетсии у вшей не передаются.
Первоначально сыпной тиф являлся, быть может, болезнью диких
морских свинок Южной Америки, среди которых он существует и сей-
час и передается свойственными этим зверькам власоедами и блохами.
Возвратный тиф. Возбудитель возвратного тифа — спирохета
(Spirochaeta recurrentis). Во время приступов они массами встречаются
в крови больного. Попав с кровью в желудок вши, спирохеты там быстро
исчезают, но через 8 дней появляются в полости тела насекомого и зара-
жают его внутренние органы. Человек заражается, раздавливая вошь,
занося спирохет из ее гемолимфы в расчесы или на слизистые оболочки
своего тела (рот, веки).
г*
РАЗДЕЛ Ж
КЛОПЫ (подотряд Hemiptera отряда Rhynchota)
1. ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
В отряде клопов представители двух семейств нападают на человека:
Triatomidae nCimicidae. Триатомиды — крупные, ярко окрашенные кры-
латые формы (рис. 26, В), свойственные тропикам; в Южной Америке они
являются переносчиками болезни Чагаса — тяжелого заболевания чело-
Рис. 287. Клопы.
А — самка постельного клопа (Cimex lectularius); Б— задний конец тела самца постель-
ного клопа С. lectularius; В — задний конец тела самки. 1 — влагалище; 2 — выемка,
ведущая в совокупительную сумку; 3 — совокупительный орган.
века, возбудитель которого, Schizotrypanum cruzi, относится к трипано-
сомам (класс Flagellata). Второе семейство кровососущих клопов, Cimi-
cidae, включает постельного клопа, Cimex lectularias, а также ряд дру-
гих видов, являющихся гнездовыми паразитами птиц и летучих мышей.
Постельный клоп подозревался в передаче различных инфекций,
но до сих пор не удалось установить переноса им какого-либо заболева-
ния. Тем не менее клопы настолько досаждают человеку своими укусами,
что иногда заставляют его покидать дом.
414
2. НАРУЖНОЕ СТРОЕНИЕ ПОСТЕЛЬНОГО КЛОПА (РОД С1МЕХ)
Рис. 28S. Голова и ротовые части постель-
ного клопа.
1—глаза; 2— антенны; 3—верхняя губа;
4 — верхние челюсти; 5 — нижние челюсти;
6 — нижняя губа.
Тело клопа (рис. 287) сплющено в дорсо-вентральном направлении,
что позволяет ему пролезать в узкие щели. Голова широко причленяется
к груди и несет два сложных глаза. Усики четырехчлениковые.
Ротовой аппарат. Ротовой аппарат (рис. 288) состоит из полукруглой
короткой верхней губы и мандибул, т. е. верхних челюстей — тонких ще-
тинок, зазубренных на конце; на
их внутренней поверхности имеет-
ся два желобка — верхний, бо-
лее широкий, и нижний — узкий.
Тесно примыкая одна к другой,
мандибулы образуют два канала—
один, широкий, для принятия
крови и другой, узкий, для введе-
ния слюны в ранку. Нижние че-
люсти, в виде колющих щетинок,
расположены по бокам верхних
челюстей; они не участвуют в об-
разовании каналов. Мандибулы и
максиллы вкладываются, как в
ножны, в нижнюю губу, состоя-
щую из 4 члеников. В состоянии
покоя хоботок подвернут на
брюшную сторону. Щупиков, ни
челюстных, ни губных, у клопов
не бывает.
Грудь. Переднегрудь сильно
развита и несет по бокам два вы-
роста, прикрывающих глаза. Менее развитая среднегрудь несет ру-
дименты передней пары крыльев. Заднегрудь развита также довольно
слабо. Ноги умеренной длины. Голени несут на концах щеточки из креп-
ких щетинок. Лапки четырехчлениковые с двумя коготками на конце
последнего членика.
Брюшко. Брюшко состоит из 8 члеников. У самца конец брюшка асим-
метричен вследствие наличия расположенного с левой стороны сильно изо-
гнутого совокупительного органа. Самку можно узнать по выемке заднего
края 4-го членика брюшка, которая ведет в совокупительную сумку (bur-
sa copulatrix). При копуляции сюда вводится сперма; сперматозоиды че-
рез протоплазматические стенки мешка проникают в полость тела, откуда
пробираются в семяприемники и, наконец, добираются до яичщтков, но
не через полость яйцеводов, а через эпителий. Неприятный запах клопов
обязан выделению секрета пахучих желез, которые у взрослых насеко-
мых открываются позади тазиков третьей пары ног, а у личинок—на спин-
ке 3—5-го члеников брюшка. У клопов имеется две пары грудных и семь
пар брюшных дыхалец.
3. ВНУТРЕННЕЕ СТРОЕНИЕ ПОСТЕЛЬНОГО КЛОПА
Органы пищеварения начинаются сосательной трубкой в хоботке,
переходящей в глотку; глотка снабжена мышцами и работает, как насос;
за глоткой следует пищевод в виде тонкой, нежной трубки; он впадает
в среднюю кишку, так называемый желудок, удлиненной формы. Корот-
кий последний отдел кишечника — задняя кишка — незаметно переходит
415
в прямую кишку, или rectum. В начале задней кишки открываются маль-
пигиевы сосуды. Маленькие круглые слюнные железы расположены по
обеим сторонам передней части желудка. От них отходят протоки, соеди-
няющиеся в общий проток, немного не доходя слюнного насоса.
Половые органы самца состоят из двух семенников, построенных каж-
дый из семи почковидных долек. Выводные протоки после веретеновид-
ного расширения соединяются в непарный семяизвергательный проток (due
tus ejaculatorius), соединяющийся с совокупительным органом. Пара раз-
ветвленных придаточных желез открывается в пузыреобразные расшире-
ния, которые в свою очередь открываются с каждой стороны в дистальных
частях парных выводных протоков.
У самки каждый яичник состоит из семи яйцевых трубочек, впадаю-
щих в парные яйцеводы, объединяющиеся в непарный яйцевод, по обеим
сторонам которого имеется по семяприемнику.
4. РАЗВИТИЕ ПОСТЕЛЬНОГО КЛОПА
Яйца овальные, с заостренными, косо срезанными концами, покрыты
крышечкой, которая сбрасывается вылупляющейся личинкой. Яйцо сна-
чала белое, позже оно желтеет. Самка откладывает яйца ежедневно в ко-
личестве от 1 до 12 штук в день. В лаборатории от одной самки удалось
получить 541 яйцо; обычно это число значительно меньше. Яйца отклады-
ваются в те же места, которые служат убежищем для клопов: различные
щели, складки матрацев и т. д. Они приклеиваются по всей длине к суб-
страту, но клеящее вещество не столь прочно, как у гнид вшей, и легко
растворяется в воде. Вылупление из яиц при комнатной температуре
от 14 до 18° имеет место через 21—22 дня, при 22—26° — через 8—9 дней,
при 35—37°— через 5—6 дней. Температура ниже 14° тормозит развитие.
Клопы имеют неполное превращение. На протяжении своего развития
они линяют 5 раз. Личинка отличается закругленным брюшком, вслед-
ствие отсутствия наружных половых органов, неясной сегментацией и
двухчлениковыми лапками. Антенны у личинок короче и толще, чем у
взрослых. С каждой линькой антенны по строению приближаются к та-
ковым взрослых клопов. При особо благоприятных условиях питания и
температуры (30э) личинка линяет в первый раз через 6 дней, затем че
рез 5, третий раз через 5, четвертый раз через 5 и, наконец, в последний
раз она линяет через 6 дней, так что в общей сложности развитие ее за-
нимает 28 дней. При умеренном питании и комнатной температуре разви-
тие может продолжаться 6—8 недель. Личинки и нимфы для перехода в еле
дующую стадию непременно должны каждый раз выпить полную порцию
крови. Размеры различных стадий личинок, а также и взрослых зависят
в большей мере от степени наполнения кишечника, но в среднем они сле-
дующие: I стадия — 1,3 мм, II— 2 мм, III — 3 мм, IV — 3,7 мм и V ста-
дия — 5 мм.
5. ЭКОЛОГИЯ ПОСТЕЛЬНОГО КЛОПА
Клоп — ночное животное. На всех стадиях развития питается только
кровью. Как правило, клоп нападает на добычу в темноте, но, если он очень
голоден, то нападает и при свете. Продолжительность сосания для взрос-
лых клопов — 15 минут, для личинок — 1 минута и даже меньше. Взрос-
лая самка может выпить до 7 мг крови, личинка I стадии — х/3 мг. Клопы
нападают главным образом на человека, но они нападают также и на
других животных. Они кусают преимущественно участки тела, не при-
крытые одеждой или одеялом. Реакция на укус клопа у различных лю
416
дей различная. Сам укус почти нечувствителен. Ощущение укуса появ-
ляется после того, как насекомое убежит. Обычно клоп колет несколько
раз, прежде чем найдет подходящее место для сосания.
Продолжительность голодания зависит от температуры. Так,
при низких температурах клоп может голодать год, при умеренных тем-
пературах — несколько месяцев, при высоких температурах — значи-
тельно меньшие сроки. В тропиках он голодает от 19 до 34 дней. Если кло-
пам нечего есть в данном помещении, они покидают его, часто совершая
в поисках добычи большие переходы. Взрослый клоп переползает в 1 ми-
нуту 1,25 м, молодая личинка — до 25 см. Взрослые клопы очень долго-
вечны, — они могут жить до 14 месяцев.
Ввиду того что быстрота размножения и развития клопов при высо-
ких температурах значительно больше, максимальное размножение их
происходит либо в летнее время, либо в очень хорошо отапливаемых по-
мещениях.
27
Учебник медицинской энтомологии
РАЗДЕЛ 3
ТАРАКАНЫ (подотряд Blattodea отряда Orthoptera)1
Эпидемиологическое значение. Тараканы (семейство Blattidae) —
широко распространенные домашние насекомые. Чаще всего в домах
встречается черный таракан (Blatta orientalis) (рис. 289) и рыжий тара-
Рис. 289. Черный таракан. *
А—самец; Б—самка черного таракана Blitta orientalis сверху;
1— голова; 2— глаза; 3— антенны; 4 — переднеспинка; 5— сред-
неспинка; 6 — надкрылья; 7 — заднеспинка; 8— брюшко: 9 — бед-
ро; 10— голень; 11— лапка; 12— церки; 13— грифзльки.
кан, или прусак (Blattella germanica). Тараканы могут служить механи-
ческими переносчиками различных инфекций, так как встречаются на
фекалиях, мокроте и т. п., после чего переползают на продукты питания.
Кроме того, они могут нападать на спящего человека и обгрызать кожу.
Строение тела. Голова. Тело таракана сплющено, что позволяет ему
забираться в узкие щели. Голова подвижно сочленена с грудью; по бокам
головы расположены сложные фасеточные глаза; имеется также пара
простых глазков. К передней поверхности головы прикреплены ните-
видные антенны, состоящие у взрослых тараканов из 75—90 члеников.
Ротовые органы таракана (рис. 290) грызуще-жующего типа. Они состоят из сле-
дующих частей: 1. Подвижная верхняя губа. 2. Пара верхних челюстей, представлен-
ных двумя массивными пластинками, снабженных по внутреннему краю крепкими зуб-
цами. 3. Нижние челюсти, основная часть которых состоит из двух члеников короткого,
примыкающего к головной капсуле (cardo), и длинного стволика (stipes); к стволику
примыкают две подвижные лопасти: наружная (galea) и внутренняя (lacinia), которая
1 В настоящее время тараканы обычно рассматриваются как особый отряд
Blattodea.
418
несет на конце два зубца и участвует в разжевывании пищи; к стволику причленяются
пятичлениковые щупики. 4. Нижняя губа, состоящая из следующих отделов: подпод-
бородок (submentum), подбородок (mentum), который несет наружные и внутренние ло-
пасти (paraglossa и glossa) и пару нижнегубных щупиков. Лопасти служат для слизы-
вания пищи. 5. Подглоточник (hypopharynx) распою кен у основания нижней губы.
Он принимает участие в слизывании жидкой пищи и главным образом в протал-
кивании пищи в глотку.
Грудь и брюшко. Ноги резко выраженного бегательного типа, лапка
состоит из 5 члеников. Последний членик лапки несет пару коготков и
присоску. Крылья у обоих полов прусаков развиты одинаково. У сам-
у-
Рис. 290. Ротовые органы черного таракана.
А — верхняя губа; Б— верхняя челюсть; В—нижняя челюсть;
Г—нижняя губа. 1—основание; 2—стволик нижней челюсти;
.7 —- наружная лопасть и 4 — внутренняя лопасть нижней челюсти;
5 — нижнечелюстной щупик; б — подподбородок; 7 — подбородок;
8 — наружная лопасть и 9— внутренняя лопасть нижней губы;
10— нижнегубной щупик.
цов черных тараканов крылья развиты лучше; передняя пара, обычно
кожистая и укороченная, носит название надкрылий, или элитр. Вторая
пара тонкая, перепончатая, с прямыми жилками.
Брюшко состоит из 10 члеников. Каждый членик построен из спинной
и брюшной пластинки — тергита и стернита. Снаружи у самцов видно
9 члеников, а у самок — только 7, так как два последних погру-
жены внутрь и образуют род кармана, в котором образуется яйцевой па-
кет. 9-й стернит самца несет грифельки (styli). На конце брюшка у обоих
полов имеются хорошо развитые церки (cerci), служащие органами осяза-
ния.
Пищеварительный аппарат. Пищеварительный аппарат (рис. 18)
начинается ротовым отверстием, ведущим в глотку, которая в области
шеи переходит в узкий пищевод, постепенно расширяющийся в широкий
мешковидный зоб. После заметного сужения зоб переходит в мускулистый
жевательный желудок, впадающий затем в среднюю кишку. От передней
27* 419
части средней кишки отходит восемь слепых выростов. Граница между
средней и задней кишкой определяется местом впадения мальпигиевых со-
судов, состоящих из многочисленных тонких трубочек. У взрослого тара-
кана число их достигает 80—100. Задняя кишка переходит в прямую,
снабженную ректальными железами.
Половой аппарат. Женский половой аппарат состоит из двух яични-
ков, по восемь яйцевых трубочек в каждом. Концевые нити яйцевых тру-
бочек соединяются в одну общую концевую нить. Яйцевые трубочки от-
крываются в парные яйцеводы, соединяющиеся в непарный яйцевод или
матку, которая открывается в наружные половые органы, впяченные
внутрь в виде кармана. На спинной стороне этого кармана открывается
отверстие семяприемника. В матку впадают придаточные железы, секрет
которых идет на образование яйцевой капсулы.
Мужские половые органы состоят из семенников, построенных из
многочисленных мелких фолликулов, расположенных на концах пар-
ных семяпроводов. Семяпроводы дают расширение — семенные пузырьки,
после чего объединяются в непарный семяизвергательный канал, идущий
к наружным совокупительным органам. В семенные пузырьки впадают
многочисленные придаточные железы.
Развитие. Оплодотворенная самка прусака откладывает яйца (от 28
до 56) в капсулу, которая по мере накопления яиц начинает высовываться
наружу. Всего за свою жизнь самка откладывает до четырех таких кап-
сул. Она носит капсулу с собой в течение 15—40 дней, в зависимости от
температуры, но кладет ее только тогда, когда личинки готовы к вылуп-
лению.
Тараканы относятся к насекомьш с неполным превращением. Малень-
кие личинки лишены крыльев, число члеников антенн у них меньше, чем
у взрослых. С последовательными линьками число члеников антенн уве
личивается и отрастают крылья. Всего личинки таракана линяют 6 раз.
Продолжительность жизни личинки, начиная с момента вылупления
из кокона и кончая последней линькой, колеблется в зависимости от тем-
пературы. При 20° превращение длится около б месяцев, при 30° этот срок
сокращается до 21/2 месяцев.
Самка черного таракана носит капсулу до 5 дней, после чего кладет
ее в теплое место. Вылупление личинок происходит в различные сроки:
от нескольких дней до года. Метаморфоз черного таракана длится до 5 лет,
но при благоприятных условиях этот срок значительно сокращается.
Биология. Тараканы — ночные существа: днем они сидят, укрыв
шись в щелях, а с наступлением темноты переходят к активному образу
жизни. Условиями их существования являются тепло, влажность и изо-
билие пищи; вот почвхму они селятся в кухнях, пекарнях, столовых и т. д.
Пищей им служит хлеб, овощи, сахар и другие продукты питания. Голо-
дать тараканы могут довольно долго. Так, личинка голодает от 9 до 22 дней,
а самка 30—40 дней.
РАЗДЕЛ И
КЛЕЩИ (отряд Acarina класса Arachnoidea)
ГЛАВА I
ПОНЯТИЕ О КЛЕЩАХ
1. ОБЩИЕ ЗАМЕЧАНИЯ
Характеристика отряда клещей
Клещи (Acarina) представляют один из отрядов класса паукообраз
ных (Arachnoidea) (см. стр. 31). Наиболее характерным признаком кле
щей, отличающим их от всех остальных паукообразных, является нал®
чие у них шестиногой личинки (рис. 291). В то время как все остальны
паукообразные выходят из яйца с полным числом ног (четыре пары'
клещи покидают яйцо, имея только три пары ног; четвертая пара в эт
время отсутствует. Столь резкое отличие I стадии послезародышевог
развития клещей от всех последующих стадий развития заставляет рас
сматривать ее как личинку, а самое развитие клещей— как метаморфоз
После первой же линьки личинка получает четвертую пару ног и пре
вращается в нимфу.
Второй отличительный признак клещей — отсутствие видимой чле
нистости (рис. 292). Наружный скелет их не распадается на отдельны
тергиты и стерниты, как у большинства насекомых и у многих паукооб
разных (скорпионы, бихорки и пр.). Вместо этого хитин, покрывающи
их тело, либо сплошь тонкий, либо в его состав входят крупные щитке
иногда частью соответствующие нескольким тергитам или стернитал
Однако этот признак свойствен не только клещам, но и большинству naj
ков (отряд Araneina); в то же время существуют клещи (правда, их немног
и они встречаются редко) с более или менее выраженной членистостьк
Третий отличительный признак — отсутствие у многих клещей тог
разделения тела на отделы, которое мы видим у большинства паукообраз
ных. Этот признак также свойствен не всем клещам (рис. 298), но он хз
рактерен для целого ряда широко распространенных и важных форм этог
отряда.
Практически отличать клеща от всех других членистоногих удобв
именно по слитому в одну сплошную массу телу при наличии четыре
пар ног (исключением являются личинки, имеющие слитное тело и тр
пары ног).
Отряд клещей весьма богат видами; виды отличаются большим разн<
образием строения, что связано и с образом жизни клещей, и с эколоп
ческой обстановкой, в которой они живут. Так, клещи, живущие в воде
конечно, сильно отличаются от наземных, паразитические формы устрое-
ны иначе, чем свободноживущие, и т. д.
Клещи — по большей части мелкие, иногда микроскопические жи-
вотные. Однако крупнейшие из них (некоторые паразитические формы)
могут в сытом, раздутом состоянии достигать нескольких (3—4) сантиметров
в длину.
Рис. 291. Ixodes riclnus.
А—личинка сверху; Б—личинка снизу; В—нимфа сверху:
Г—нимфа снизу. 1—спинной щиток; 2—перитрема со стшмой:
3—половая бороздка; 4— анальное отверстие; 5—анальная бороздка.
2. СТРОЕНИЕ КЛЕЩЕЙ
Тело клещей обычно шаровидное или продолговато-овальное, реже
иной формы. Хитиновая кутикула иногда местами уплотнена и образует
твердые щитки разной формы. Ноги на вершинах часто вооружены ко-
готками или подушечками-присосками, помогающими клещу ползать по
поверхности субстрата и цепляться за проходящее мимо животное в поис-
ках хозяина для паразитирования.
422
Ротовой аппарат. Ротовые органы устроены сложно и разнообразно,
применительно к способу питания клеща. Существуют колющие, режущие,
грызущие, сосущие и хватательные ротовые органы. Сосущие ротовые
органы обозначаются как хоботок.
Ротовые органы всех клещей образованы двумя первыми парами их
конечностей: хелицерами и педипальпами. Хелицеры у многих клещей
имеют клешневидное строение, подобно хелицерам многих других пауко-
образных. Клешневидные хелицеры
могут быть приспособлены для хвата-
ния, грызения, разрезания, прокола.
У некоторых паразитических клещей
хелицеры теряют клешневидную
форму и _ превращаются в более со-
вершенные органы прокола и проре-
зания. Коксы педипальп обычно сли-
ваются между собой и образуют
нижнюю стенку ротовой полости; са-
ми педипальпы играют роль щупалец
(пальпы). За счет кокс педипальп и
стенок ротовой полости у многих кле-
щей образуются различные парные
и непарные пластинки, также участ-
вующие в принятии пищи. Ротовой
аппарат наиболее для нас важных
иксодовых клещей (рис. 293) состоит
из пары режущих хелицер и из
гипостома (hypo — под, stoma—рот,
гипостом — подротовой придаток).
Последний представляет длинный и
массивный вырост нижнего края ро-
тового отверстия, покрытый обратно
направленными зубчиками. При со-
сании крови на животном клещ вво-
дит гипостом в ранку кожи и с его
помощью удерживается на хозяине
до тех пор, пока не насосется. Пе-
редний конец тела, на котором рас-
положены ротовые органы, у иксо-
довых клещей подвижно сочленен
с остальным телом и носит название
основания хоботка.
Дыхательный аппарат. Органы
дыхания у клещей могут вообще от-
сутствовать. В таком случае дыхание
осуществляется через покровы тела
(кожное дыхание). У наиболее высоко
Рис. 292. Ixodes ricinus, самка.
А — слабо насосавшаяся (вид сверху); Б—то
же (вид снизу); В—сильно насосавшаяся
(вид сверху). 1—хоботок; 2—основание
хоботка; 3 — спинной щиток; 4 — половое
отверстие; 5—перитрема; 6—половая бо-
роздка; 7 — анальное отверстие; 8 — аналь-
ная бороздка.
организованных клещей дыхательные органы представлены системой
дыхательных трубочек — трахей, открывающихся наружу дыхальцами—
стигмами. Дыхальца иногда располагаются на специальных пластинках—
перитремах — и могут быть снабжены сложно устроенным замыкающим
аппаратом. В разных группах клещей дыхальца открываются на разных
частях тела. На различиях в устройстве дыхательного аппарата и
в местоположении дыхалец построена классификация подотрядов
клещей.
Пищеварительный аппарат. Пищеварительные органы у крупных кро-
вососущих клещей надсемейства Ixodoidea (рис. 294; рис.7, Б) состоят из
короткого пищевода, объемистой средней кишки, способной вместить много
пищи, тонкой кишки и задней кишки. К последней примыкает особый рек-
* 6
Рис. 293. Ixodes ricinus. Хоботок самки (Л — вид сверху;
Б — вид снизу).
1—пальпа; 2— гипостом; 3— хелицеры; 4—футляр хелицер;
5 — основание хоботка; 6 — поровые поля.
Рис. 294. Ixodes ricinus. Внутренние органы' самки.
1 — пальпы; 2 — основание хоботка; 3—поровые поля; 4 — орган Женэ, 5 —слюн-
ная железа; 6— отростки средней кишки; 7— средняя кишка; 8—ректаль-
ный пузырь; 9 — мальпигиевы сосуды; 10 — яичник; 11 — яйцевод; 12 — трахеи;
13— центральный отдел нервного аппарата; 14— пищевод.
424
тальный пузырь, в котором скопляются продукты выделения (гуанин).
Анальное (выделительное) отверстие замыкается специальным клапаном.
Имеются п слюнные железы. Они вырабатывают секрет, впрыскиваю’
щийся в ранку хозяина при питании на нем. Секрет препятствует сверты-
ванию крови хозяина и обезболивает укус. Последнее имеет для клеща
важное значение, так как защищает его от уничтожения хозяином. Вме-
сте с тем слюна клещей ядовита и, попадая в ранку, может вызвать
различные болезненные явления у хозяина клеща.
Выделительный аппарат. Выделительный аппарат состоит из одной
пары мальпигиевых сосудов, впадающих в ректальный пузырь. Кроме
того, у быстро напивающихся кровососущих клещей (семейство Arga-
sidae) имеется еще одна пара сегментальных выделительных органов —
коксальные железы. Они служат для выделения из организма избытка
непитательной жидкой составной части всосанной клещом крови. Клещу
важно избавиться от обременительного избытка всосанной жидкости,
так как он часто насасывается до состояния, в котором затруднительно
передвижение. Выделив через коксальные железы избыток жидкости, клещ
вновь приобретает подвижность и становится способным уползти в защи-
щенное место.
Нервный аппарат и органы чувств. Центральный отдел нервного ап-
парата в высшей степени концентрирован: вся брюшная цепочка слилась
в один узел, лежащий на брюшной стороне тела. Из органов чувств могут
быть развиты глаза; их бывает одна пара или больше. Лишенные глаз
клещи (например, многие аргазиды) все же хорошо ощущают свет, пови-
димому, всей поверхностью тела, и способны ориентировать по отношению
к нему свои движения (например, уползать от яркого света в затемненное
место). Иногда бывает развит особый орган обоняния (орган Галлера),
располагающийся на вершине первой пары ног. Клещ, передвигаясь в
поисках хозяина для присасывания, размахивает по воздуху этой парой
ног, улавливая запахи. Кроме того, у всех клещей имеется множество
осязательных волосков и других кожных органов чувств.
Половой аппарат. Клещи раздельнополы. Внешние различия между
самцами и самками иногда резко выражены (половой диморфизм), иногда же
сводятся лишь к мелким признакам (меньший рост самца, форма полового
отверстия и др.). Половые органы у крупных кровососущих форм состоят
у самцов (рис. 7, Б) из одного-двух семенников, семяпроводов и набора при-
даточных желез. Из секрета придаточных железу многих клещей (например,
у Ixodoidea) образуется особая сумка сперматофор, в которой заключены
мужские половые продукты. Сперматофор при оплодотворении прикреп-
ляется к половому отверстию самки, и содержимое его переходит во внут-
ренние половые пути последней. Женские половые органы состоят из не-
парного яичника, пары яйцеводов, более или менее развитой матки, вла-
галища, добавочных желез и иногда семяприемника. У Ixodoidea имеется
еще так называемый орган Женэ, играющий важную роль при откладке
яиц. Орган этот представляет собой железистый мешочек, расположенный
в передней части тела и нормально ввернутый внутрь тела. Во время от-
кладки яиц (рис. 295) он через особую щель выворачивается наружу, охва-
тывает яйцо и покрывает его смазкой, препятствующей высыханию яйца.
3. БИОЛОГИЯ, ЭКОЛОГИЯ И РАСПРОСТРАНЕНИЕ КЛЕЩЕЙ
Жизненный цикл. Клещи имеют сложное превращение. Из яйца, от-
ложенного самкой, вылупляется шестиногая личинка; личинка после
линьки превращается в нимфу, имеющую уже 4 пары ног, но еще не обла-
425
дающую половым отверстием. Нимфальных стадий бывает одна или не-
сколько, т. е. нимфа I возраста линяет или на нимфу II возраста, или же
сразу на взрослого клеща. Точно так же и нимфа II возраста переходит
или в нимфу III возраста, или же в имаго и т. д. Нимфальных стадий может
быть до семи. У большинства видов клещей число нимфальных стадий
бывает строго постоянным; у других клещей число нимфальных стадий
непостоянно, и в потомстве одной
Рис. 295. Ixodes ricinus. Момент от-
кладки яйца: выпяченный орган Же-
нэ охватывает выделенное из поло-
вого отверстия яйцо.
1—орган Женэ; 2—ноги; 3—яйцо;
4 — пальпа; 5 — гипостом; 6 — половое
отверстие.
и той же самки половозрелые особи полу-
чаются после разного числа нимфаль-
ных стадий. У некоторых свободно-
живущих клещей имеется особая поко-
ящаяся стадия (гипопус), способная
выдерживать неблагоприятные внеш-
ние условия. Эта стадия появляется
в случае ухудшения условий существо-
вания, и в ней клещи пережидают до
момента наступления более благопри-
ятных условий жизни. Эта же стадия
служит для расселения клещей: она
переносится ветром или различными
животными, к поверхности тела кото-
рых прикрепляется. Сравнительно не-
многие клещи живородящи.
Распространение. Клещи на земном
шаре чрезвычайно многочисленны; не-
которые свободноживущие формы при
подходящих условиях встречаются
миллионами в пространстве нескольких кубических дюймов. Парази-
тические формы подчас также размножаются в большом числе.
Распространены клещи во всех частях света — и на севере, и на юге.
Имеются они и под тропиками, и на островах Ледовитого океана. Но в
основном клещи все же теплолюбивые формы, и преобладающее большин-
ство их живет в теплых странах.
Образ жизни. По образу жизни клещи делятся на свободноживущих
и паразитических. По роду пищи они бывают растительноядные, сапро-
фаги (т. е. питающиеся различными мертвыми остатками), хищники я
паразиты.
Паразитические клещи могут быть в различной степени связаны с хо-
зяевами, за счет которых существуют. Наименее прочны связи с хозяевами
у клещей — периодически питающихся кровососов: они соприкасаются
с хозяином лишь на время питания (например, большинство аргасовых
клещей). Иногда этот контакт длится всего несколько минут. Периоди-
чески питающиеся кровососы, напившись крови, покидают хозяина и
остальное время, т. е. большую часть жизни, проводят во внешней среде.
Совершенно иначе связаны с хозяином клещи — накожные паразиты
(клещи, вызывающие накожниковую чесотку). Эти клещи всю свою жизнь
не покидают хозяина, живут на поверхности его тела, питаются корочками
и выделениями кожи и здесь же откладывают яйца. Еще далее зашел па-
разитизм у клещей — внутрикожных паразитов (например, чесоточные
клещи). Клещи эти живут в глубоких слоях эпителия кожи млекопитаю-
щих и человека, проделывают в них ходы, питаются тканевой жидкостью
и тут же размножаются. Наибольшую связь с хозяином мы нахо-
дим у клещей-паразитов полостей и внутренних органов хозяина.
Таковы клещи, обитающие в воздушных мешках птиц, в печени, брю-
шине и т. д.
426
Клещи, которые тесно связаны с хозяином, обычно встречаются
всюду или почти всюду, в тех же странах, где живут их хозяева, например,
некоторые чесоточные клещи, расселившиеся вместе с человеком и его
домашними животными почти по всей земле. Клещи, значительную часть
жизни проводящие в свободном состоянии во внешней среде, сильно под-
вергаются воздействию внешних условий; поэтому они могут жить лишь
там, где эти условия им подходят. Иными словами, такие клещи связаны
в своем распространении с определенными ландшафтными зонами.
4. ЗНАЧЕНИЕ КЛЕЩЕЙ ДЛЯ ЧЕЛОВЕКА
Сельскохозяйственное значение. Клещи имеют очень большое прак-
тическое значение. Некоторые растительноядные свободноживущие фор-
мы являются вредителями культурных растений, например, паутинный
клещик хлопка. Другие являются важными амбарными вредителями, по-
вреждающими зерно, муку, сыры, сухие фрукты и т. п. Размножаясь в скла-
дах в несметном количестве, они портят продукты и вредят человеку, со-
прикасающехмуся с зараженными материалами. Другие клещи вредят
сельскохозяйственным животном, высасывая у них много крови (некоторые
клещи высасывают крови в 200 раз больше, чем весят сами до питания),
отравляя их своей слюной и, самое главное, иногда передавая им крово-
паразитов — возбудителей заболеваний (пироплазмозы, тейлериозы, спи-
рохетозы и др.).
Медицинское значение. Медицинское значение клещей велико и раз-
нообразно. За последние десятилетия каждый год открываются новые
связи клещей с различными заболеваниями человека. Самые разнообраз-
ные клещи способны вызывать у человека при соприкосновении с ним
раздражение кожи — дерматиты; чесоточные клещи вызывают различные
виды чесотки; интоксикация от укусов клещей может повести к явлениям
паралича, иногда смертельного; наконец, клещи переносят человеку воз-
будителей заболеваний.
Заболевания, передаваемые клещами, относятся к разным катего-
риям. Во-первых, среди них имеются вирусные, т. е. такие, возбудителями
которых являются фильтрующиеся вирусы. Таковы таежный клещевой
энцефалит, шотландский энцефалит, крымская геморрагическая лихо-
радка. Во-вторых, риккетсиозы, т. е. заболевания, вызываемые риккет-
сиями. К этой категории относится большой список клещевых сыпнотифоз-
ных лихорадок, распространенных во всех частях света (лихорадка Ска-
листых гор в Северной Америке, средиземноморская, или марсельская,
лихорадка на юге Европы, цуцугамуши в Японии, ряд заболеваний в на-
шем Союзе и др.). В-третьих, бактериальные болезни (туляремия, чума,
бруцеллез) и, наконец, в-четвертых, группа спирохетозов — клещевые
возвратные тифы, возбудителями которых являются спирохеты.
5. КЛАССИФИКАЦИЯ КЛЕЩЕЙ
Для практических целей удобно пользоваться простой классифика-
цией, делящей отряд клещей на пять подотрядов, преимущественно по
устройству дыхательного аппарата.
Подотряд I. Brachypoda. Клещи первого подотряда вовсе
не имеют трахей; тело их микроскопически хмалой величины, червеобраз-
ной формы, с короткими, недоразвитыми ножками. К этому подотряду
относятся клещи-возбудители железничной чесотки (сехмейство Demo-
dicidae).
427
Рис. 296. Dem ode х folliculorum—
железница.
А — общий вил; Б — железяицы в
волосяной сумке собаки (сильно уве-
личено).
Подотряд II. Astigmata. Также бестрахейные клещи, но
имеющие тело округлое или продолговатое и относительно хорошо раз-
витые ноги. Это паразитические чесоточные клещи (надсемейство Sar-
coptoidea) и некоторые свободноживущие клещи, среди которых стоят
и амбарные вредители (надсемейство Tyroglyphoidea).
Подотряд III. Heteros tigm a t а. У представителей третьего
подотряда самцы без трахей, самки же имеют трахеи, связанные с хели-
церами (надсемейство Tarsonemoidea). Среди этих клещей имеется хищный
клещик, иногда нападающий на человека
и сосущий его кровь (Pediculoides ventri-
cosus).
j Подотряд IV. Prostigma-
ta. У клещей этого подотряда трахеи
всегда развиты и открываются в хелице-
рах или у их основания. Сюда относятся
преимущественно свободноживущие кле-
щи, наземные и водные. Среди них одно
надсемейство краснотелок (Trombidoidea)
имеет большое медицинское значение.
Подотряд V. Mesostigma-
t а. Сюда относятся клещи с трахеями,
открывающимися на боках тела. Клещи
эти образуют два надсемейства (Parasitoi-
dea и Ixodoidea), представляющие наи-
больший интерес как вредители здоровья
человека.
ГЛАВА II
ВАЖНЕЙШИЕ В МЕДИЦИНСКОМ
ОТНОШЕНИИ КЛЕЩИ ФАУНЫ СССР
1. ЧЕСОТОЧНЫЕ КЛЕЩИ
Различают чесотку железпичную,
зудневую и накожниковую. Последняя
встречается только у животных, почему
мы ее касаться не будем.
а) Железница (JDemodex folliculorum Simon)
Железничную чесотку вызывают мельчайшие (0,3 X 0,04 мм)
клещики железницы, или угрицы (Demodex folliculorum) (рис. 296),
относящиеся к надсемейству Demodicoidea (подотряд I Brachypoda).
Клещики эти живут в сальных железах и волосяных сумках кожи
лица, ушей, живота и т. д. В одной железе “может быть 10—15 штук
клещей. Все развитие клещи проходят в железе. В состав жизнен-
ного цикла входит яйцо, личинка, две нимфальные стадии и взрослые
клещи.
Находясь в железе, клещи эти могут не вызывать никаких болез-
ненных явлений и оставаться незамеченными. Но иногда их присут-
ствие сопровождается закупоркой прохода железы, воспалением, обра-
зованием угрей или сыпи. Заражение происходит путем соприкосно
428
вения с зараженным человеком. Вне хозяина клещики эти быстро
погибают. Железница животных, видимо, не способна переходить на
человека.
б) Чесоточиый зуденъ (Acarus siro L.)
Зудневая чесотка есть результат паразитирования в коже клещиков
семейства Acaridae (подотряд II Astigmata), или чесоточных зудней. У че-
ловека чесотку вызывает чесоточный зудень Acarus siro (рис. 297). Кле-
Рис. 297. Acarus siro var. equi, чесоточный зудень.
А — самец, вид с брюш к>й стороны; Б — самка, вид со спинной
стороны (сильно увеличено).
щик этот малой величины, со складчатьш телом, покрытым направлен-
ными назад щетинками, препятствующими обратному движению клеща
в ходах, проделанных в коже. Клещ, попадая на тело человека, вне-
дряется в толщу эпидермиса в местах с более нежной кожей: между паль-
цами, подмышками и т. д. В коже клещ проделывает ходы и постепенно дви-
жется вперед, питаясь тканевой жидкостью и оставляя позади себя
экскременты и, если это самка, — яйца. Самка дает около 20—50 яиц за
всю жизнь. Из яиц выходят личинки, которые начинают проделывать
новые ходы. В дальнейшем они слиниваютна нимф (две нимфальные стадии
у самки и одна у самца) и далее на имаго. Взрослый клещ способен жить
40—50 дней, а цикл развития может завершиться в 2—3 недели. Несмотря
на столь быстрое размножение, чесоточные клещи обычно не бывают у
одного и того же человека особенно многочисленными.
Движущийся в коже клещ раздражает нервные окончания и вызывает
нестерпимый зуд. Зуд этот особенно усиливается ночами, когда человек
согревается в постели. Расчесы зудящих мест вызывают собственно кар-
тину чесотки. Картина эта держится еще долго и после того, как^клещей
429
Рис. 298. Tyroglyphus farinae, мучной
клещик.
А—самец, вид снизу; Б — самка, вид снизу;
В— самка, вид сверху (сильно увеличено)'.
уже нет в коже человека. В расчесы могут попадать различные бактерии
и вызывать осложнение болезни.
Диагноз чесотки особенно легко установить, если удается найти
в коже самих клещей. Ходы их бывают заметны в виде тонких темных
полосок; на конце хода иногда бывает пузырек, в которохм находится клещ.
Клеща можно извлечь из пузырька тонкой иголкой. При расчесывании
пузырьки сдираются, клещи из них могут быть выдавлены и перенесены
на другое место тела или на другого человека. Заражение происходит или
при непосредственном контакте с
больным, или через общие с ним
предметы обихода (постель, поло-
тенце и т. д.).
Чесотка — мучительная бо-
лезнь. Местами, при низком куль-
турном уровне населения, опа
очень распространена. Лечение
удается втиранием специальных
мазей, например, вилькинсонов-
ской, содержащих зеленое мыло,
препарата К и др. В качестве
профилактики рекомендуется чи-
стота, внедрение санитарных на-
выков и общее повышение куль-
турного уровня населения.
Зудневая чесотка некоторых
животных, называемая особыми
видами чесоточных клещей, мо-
жет иногда переходить на чело-
века (например, с собак, лошадей,
верблюдов). Но обычна для чело-
века только описанная форма че-
сотки.
2. АМБАРНЫЕ КЛЕЩИ (НАДСЕМЕЙ-
СТВО TYROGLYPHOIDEA)
Мелкие бестрахейные клещи,
во взрослом состоянии не веду-
щие подлинно паразитического
образа жизни. Тело разделено на
два отдела насечкой, проходящей
между второй и третьей парой ног.
Имеется большое число видов. Живут в зерне, муке, сырах, сухих
фруктах, сене, луковицах, высохших трупах и т. д.
Мучной клещик. Представитель (рис. 298) — мучной клещик (Tyrogly-
phus farinae de Geer), важный амбарный вредитель. Мучной клещ, сырный
клещ и др. в массе размножаются в запасах продуктов и портят их,
выедая, заполняя экскрементами и трупами погибших. Могут с продуктами
перевозиться на большие расстояния. Во множестве носятся с пылью в воз-
духе в складах. При соприкосновении с кожей человека могут вызывать
раздражение ее (дерматиты); попадая при дыхании в его дыхательные
пути, способны вызывать их воспаление и общую болезненную реакцию
со стороны организма (нервные явления и т. п.). В результате перевари-
вания и всасывания проглоченных с продуктами клещей и их экскрементов
430
могут возникать расстройства пищеварения, катарр желудка и кишок
и общие явления отравления. Часть проглоченных клещей может целиком
проходить через кишечник человека и выделяться с испражнениями;
в исключительных случаях (при поносе) клещи могут оказаться в испраж-
нениях даже живыми. Однако это не служит показателем паразитирования
их в кишечнике человека, так как они могут только проходить через него
транзитпо, но не паразитировать в нем (лжепаразиты, псевдопаразиты).
Существует еще целый ряд свободноживущих клещей, относящихся к разным
подотрядам и семействам, способных при соприкосновении с человеком вызывать раз-
дражения кожи, а иногда и сосать его кровь. Последствием могут быть зуд, жжение,
сыпь, иногда даже лихорадочные явления.
3. КРАСНОТЕЛКИ (СЕМЕЙСТВО TBOMBIDIIDAE)
Небольшие клещи, относящиеся к подотряду IV Prostigmata. Снабже-
ны трахеями, открывающимися у ротовых органов. Хелицеры их воору-
жены коготками. Тело обычно выглядит пушистым от покрывающих его
5
Рис. 299. Trombicula akamushi, краснотелка.
А — личинка, вид сверху; Ь’— личинка, вид снизу (сильно увеличено).
густых и длинных щетинок. Личинки (рис. 299), а иногда и взрослые,
яркокрасного цвета, откуда их название — краснотелки. Взрослые ведут
свободный образ жизни, личинки же паразитируют на членистоногих, на
диких и домашних позвоночных и на человеке.
Тромбидиоз в Европе. Особенно много личинок появляется осенью.
Они сидят обычно кучками на растительности, кочках, почве, ожидая до-
бычи. Нападают на посещающих такие места животных и человека. При-
сасываясь, вызывают сильное раздражение и воспаление кожи, иногда
сопровождающиеся лихорадкой. Болезнь носит название тромбидиоза,
осенней, или крыжовниковой, болезни (совпадает по времени со сбором уро-
жая). Поражаются ею люди, связанные с садовыми и полевыми работами,
посещающие леса и т. д.
В Японии имеются виды краспотелок (например, Trombicula akamushi) (рис. 299),
передающие через укус речную лихорадку, или цуцугамуши. Болезнь эта относится
к числу сыпнотифозных лихорадок (риккетсиозов), местами дает до 30—70% леталь-
ности.
Болезньэта — зооноз, т. е. нормально, в природе, ею болеют животные (полевки,
крысы и др.). Личинки Trombicula, кормясь на больных животных, воспринимают
от них возбудителя; когда вылупившиеся из этих личинок самки созревают, они пере-
дают риккетсий своему потомству. Личинки, происходящие от зараженной матери, за-
ражены от рождения и при питании передают болезнь новому животному. Человек
431
заболевает, если посещает места обитания клещей и оказывается покусанным заражен-
ными личинками.
Кроме Японии, цуцугамуши широко распространена и в других странах юго-
востока Азии, на островах Индо-Малайского архипелага, а также на севере Австралии.
Рис. 300. Dermanyssus gallinae,
птичниковый клещ, самка, вид сверху
(сильно увеличено).
4. КЛЕЩИ НАДСЕМЕЙСТВА
PARASITOIDEA
Относятся к подотряду V Mesostig-
mata. Имеют трахеи, заканчиваю-
щиеся дыхальцами на боках тела, и хе-
лицеры в форме щипчиков или острых
колющих щетинок. Ротовой аппарат
сложного устройства. Богатое видами
надсемейство. Многие представители —
паразиты беспозвоночных и позвоноч-
ных животных. Часть из них имеет то
или иное хозяйственное, медицинское
или ветеринарное значение.
Относящийся к этому надсемейству
птичниковый клещ (Dermanyssus gallinae de
Geer) (рис. 300) достигает размеров около
0,75 х0,4мм. Живет в стенах и насестах курят-
ников. Ночами нападает на кур и сосет их
кровь, после чего вновь прячется в щели
помещения. Цикл развития проходит в этих
щелях. Вредит птицеводству. При случае
кусает и человека, вызывая зуди папулки на
коже. Борьба ведется заделкой щелей в сте-
нах, окуриванием курятников серой, опрыс-
киванием, например, керосином с нефтью, и
т. д. В Америке является переносчиком энце-
фалита Сан-Луи — болезни кур, опасной и для
человека, которому она передается комарами.
б
Рис. 301. Bdellonyssus bacoti, крысиный клещ.
А — самка, вид_сверху; Б — самка, вид снизу (сильно увеличено).
432
Крысиный клещ. Крысиный клещ (Bdellonyssus bacoti Hirst, рис. 301)
связан в своем жизненном цикле с крысами. Живет в крысиных норах,
а также в трещинах стен, фундаментов домов и т. д. Иногда размножается
в жилье в больших количествах. Охотно нападает на человека, вызывая
сильное раздражение кожи. Может передавать укусом от крысы крысе
или человеку крысиный сыпной тиф и чуму.
Другие клещи надсемейства Parasitoidea. В составе надсемейства имеются и дру-
гие клещи — паразиты грызунов, видимо имеющие эпидемиологическое значение. Так
некоторые из них (род Laelaps), постоянные эктопаразиты грызунов, подозреваются
в распространении средп водяных крыс туляремии. Однако человека клещи роде
Tsaelaps не кусают.
5. КЛЕЩИ НАДСЕМЕЙСТВА JXOBOIDEA
а) Характеристика надсемейства
В надсемейство Ixodoidea входят самые крупные виды клещей. Имеют
трахеи, открывающиеся дыхальцами на боках тела. Ротовые органы при-
способлены для разрезания кожи хозяина и укрепления на ней на время
кровососания.
Все представители надсемейства — временные наружные паразиты,
Многие имеют важнейшее медицинское значение как переносчики воз-
будителей разнообразных заболеваний.
Надсемейство состоит из двух семейств: семейство Argasidae, арга
зиды, или аргасовые клещи, и семейство Ixodidae, иксодиды, или иксо
довые клещи.
б) Аргасовые клещи (семейство Argasidae)
1) Наружное " строение аргазид
Тело аргасовых клещей (рис. 302) имеет продолговатую, реже диско-
видную форму. Спереди оно чаще приострено, сзади тупо скруглено. У ис-
тощенных клещей тело сплющено,
собрано в складки, по краям под-
час загнуто кверху. У сытых тело
раздувается, иногда становясь
почти шарообразным. Окраска у
живых—сероватых и коричнева-
тых тонов, под цвет окружающей
их естественной обстановки. По-
кров кожистый, гибкий, способ-
ный к равномерному растяжению
по всему телу. Уплотненных щит-
ков не имеется. Структура по-
крова складчатая, бугристая, зер-
нистая.
Рис. 302. Внешнее строение аргазид.
А — вид сбоку; Б — вид снизу (оригинал)
1—клюв; 2—щека; 3—основание хоботка
4 — глаз; й — шея; 6 — вульва; 7— кокса пер
вой ноги; 8 — стигма; 9 — дорсо-вентральна!
бороздка; 10 — преанальпая поперечная бороз
дка; 11—анальное отверстие; 12—аномарги
нальная бороздка; 13 — постанальная попереч
ная бороздка.
На поверхности всегда имеются
те или иные постоянные складки и бо-
роздки. Растяжение тела при напиты-
вании клеща происходит за счет раз-
глаживания этих складок и расплю-
щивание бугорков покрова. На теле ме-
стами располагаются участки с особой
структурой покрова — диски. Они яв-
ляючся точками, к которым изнутри
прикрепляются мышцы. По краю тела тянется рант (рис. 303) — шов и прилежаща?
к нему со спинной и брюшной стороны линия табличек или узких, взаимно параллель
28 Учебник медицинской энтомологии
43.
° * 2__пант* 3 — спинная пластинка
А самка (сверху); Б — личинка увеличана^личинка^лльао увеличена).
ных полосочек. Ра пт препятствует растяжению края тела, почему имеющие его клещи
и в сытом состоянии остаются по краям сплюснутыми. Устройство покрова, располо-
жение дисков и наличие тех или иных бороздок используются для целей систе-
матики аргазид.
Половое отверстие лежит па брюшной стороне па уровне чаще всего
первой-второй пары ног. По форме оно у самцов и самок различно:
у самцов оно прикрыто уплотненной пластинкой, у самок имеет вид по-
перечной щели, окаймленной растяжимой складкой покрова. Анальное
отверстие располагается на брюшной стороне позади последней пары пог.
Дыхальца открываются на маленьких пластинках.
Рис. 304. Аргасовый клещ, самец.
А — ттррелирбрюшная часть; Б— хиботок; В— нога первой пары
(оригинал). /— клюр; ?— камеростом; 3— шек;*; 4— хоботог;
6 — кокс<|; 6 — половое отверстие самца; 7 — гипостом; 8 — пальпг;
9—основание хоб. тка; >0— коготки; Л— п« д лпечка; 12 — орган
Галлера, /3— л шка; 14—дорсальные бугорки; 15— не ре дне ладна;
10 — голень; 17 — бедро; 18 — ьертлуг; 19— кокса.
В состаго их имеется обычно полулуттпая. ситогидгая, гзрешечеппая отгерстиями
пластинка и «макула» — крышечка, прикрывающая соСстгелво стигму. Воздух про-
пикте г в трахеи как через ситовидную пластинку, так и через собственно шоль стш мы.
Трахеи могут быть изолированы от окружающего воздуха особым замыкающим аппа-
ратом.
Ротовые органы (рис. 304), как правило, помещаются па брюшной
поверхности тела и не видны со спинной стороны. Лишь изредка они лежат
близко кпереди и могут даже торчать из-под переднего края тела. У личи-
нок они всегда располагаются или на самом переднем конце тела, или близ
пего. Весь набор ротовых органов лежит в особом углублении, носящем
название камеростома.
Камеростсм может быть более плп менее глубскгм. Иногда по бокам ого имеются
валпки, иногда подвижные лспас,’п- иски. Если шоки рошо развиты, они могут пол-
ностью прикрывать ротовые органы, встав; енные в какеростсм. С i среди камерост см
иногда защищен свисающим книзу от переднего выступа те; а выростом — клювом.
Основанге хоботка соединяется с телом посредством особой складки — <шеи», одетой
ножным покровом и способной вытягиваться, благодаря чему хоботок может выдви-
гаться из камеростома.
28* 435
Хелицеры вооружены набором ножичков, загнутых вершинами в сто-
роны и способных свободно двигаться вперед, назад и в стороны в особом
футляре хелицер. Клещ, присасываясь к хозяину, вначале делает про-
кол поверхности его кожи, затем поочередно вставляет в ранку то одну,
то другую хелицеру и, вытаскивая их обратно немного наискось, углуб-
ляет надрез. В надрез затем вставляется гипостом, препятствующий своими
обратно направленными зубцами вытаскиванию хоботка из ранки. Встав-
ленные в ранку и раздвинутые в стороны, наподобие якоря, хелицеры также
способствуют укреплению клеща на коже хозяина во время питания.
В самом начале процесса присасывания, еще до прокола кожи хозяина,
клещ выпускает каплю слюны. Поэтому при первом же поранении кожи
слюна попадает в ранку и, действуя анестезирующе, обезболивает укус.
Рис. 305. Alectorobius asperus.
А— личиньа (вид сверху); Б—личинка (вид снизу) (оригинал).
Гипостом аргазид имеет вид плоского цилиндрического или конического выроста,
приостренного или притупленного, иногда с выемкой на вершине. Зубцов бывает
разное число и они могут быть сильно или слабо развиты, в зависимости от того, долго
ли остается клещ на хозяине для питания; у клещей, питающихся долго, органы при-
крепления к хозяину развиты сильнее. В исключительных случаях, если та или иная
стадия клеща вообще пе питается, гипостом оказывается недоразвитым и лишенным
зубцов.
Пальпы (щупальца) состоят из четырех свободных цилиндрических
члеников. Служат для осязания, имея на вершине пучок чувствительных
щетинок.
Ноги (рис. 304) состоят из 6 члеников. Основные членики — коксы—
плотно прикреплены к телу. На конце каждой пары ног, как правило,
имеется два коготка. Присоски, за немногими исключениями, встречаются
только у личинок и то не всегда хорошо развиты.
Самцы отличаются от самок обычно меньшим ростом, формой полового
отверстия и некоторыми другими деталями строения. Нимфы в общем по-
хожи на взрослых, лишь не имеют полового отверстия; структура по-
крова у них менее отчетливо выражена, щеки развиты слабее. Чем старше
нимфальная стадия, тем более походит она на взрослых. Личинка (рис. 305)
имеет нежный покров, обычно из мелких складочек, растягивающихся
при напитывании. Изредка (в фауне СССР у рода Alveonasus) наспинной
436
стороне личинки встречается уплотненный участок покрова овальной фор-
мы — спинная пластинка. Яйцо округлой или продолговатой формы.
Различия между семействами Argasidae и Ixodidae сопоставлены в
табл. 19.
Таблица 19
Различие между семействами Argasidae^n Ixodidae
Призван* Семейство Argasidae Семейство Ixodidae
Расположение хоботка у взрослых На брюшной стороне тела На переднем конце тела
Спинной и брюшные щитки Отсутствуют, лишь у личи- нок изредка есть спин- ная пластинка Спинной щиток имеется всегда, брюшные — ино- гда
Край тела Иногда имеется рант иной структуры, чем прочая поверхность тела; насе- чек по заднему краю те- ла, обрисовывающих фе- стоны, не бывает Ранта ле бывает; обычно есть фестоны
Расположение глаз (если имеются) На боках тела По краям спинного щитка
Перитрема Маленькая, с макулой у края ситовидной пластин- ки Крупная, с макулой близ центра ситовидной пла- стинки
Присоски на лапках Отсутствуют или слабо раз- виты Хорошо развиты
Отличие самцов от самок Слабое Резкое
Количество нимфальных стадий в цикле развития Несколько Одна
Местообитание Закрытые убежища Свободные пространства, в виде исключения закры- тые убежища
Быстрота насасывания За редкими исключениями насасывается быстро (в несколько минут или часов) Насасывается медленно (за несколько дней или не- дель)
2) Роды семейства Argasidae
В составе семейства аргазид имеется несколько родов, из которых
в фауне СССР представлены четыре: Alectorobius, Alveonasus, Argas и
Carios. Два первых рода старыми авторами соединялись в один род Orni-
th odor os, два последних—в род Argas. Различия между этими четырьмя
родами приведены в табл. 20.
Наибольшее медицинское значение имеет род Alectorobius. Он пред-
ставлен на территории Советского Союза семью видами, группирующими-
ся в три подрода. Род Alveonasus имеет у нас одного представителя —
Alveonasus canestrinii (Ornithodorus lahorensis прежней терминологии).
В роды Argas и Carios входит несколько видов, часть которых еще мало
изучена.
437
Таблица 20
Отличительные признаки родов семейства Argasidae фауны СССР
Признан Alectorobius Alveonasus Arg s Corios
Форма тела Продолгова- тая, приострсн- ная кпереди Продолгова- тая, п| иостроп- ная кпереди Продэлгога- тая, притуп- ленная сп реди Дисковидная
Покров Бугорчатый, бугорки могут быть разных размеров, фор- мы и располо- жения В ямках, ок- руженных та- ликами из из- 1итых, ради- ально паправ- Л( н ]ых гре- бешков Складчатый или в малких бородавочках Складчатый или в мелких бородавочках
Диски Лежат сим- метричным, не- р а ди ал ьным рис\нсом, ча- стью по дну бо- роздок Лежат сим- метричным, не- радиальным рисунком Расположены более пли м •- Нее радиально Расположены четко радиаль- но
Край тела По структуре пэ отличается от прочего по- крова По структуре п? отличается от прочего по- крова Несет рант, состоящий из ш--а и приле- жащих к лому с дорсальной и вентральной стороны квад- ратных табли- чек или узких взаимно парал- лельных поло- сочек Несет рапт, состоящий из ш а и приле- жащих к нему с дорсальной и вентральной сторспы узких, взаимно парал- лельных поло- сочок
Бороздки дорсо-вентраль- ная и попе- речны? — пре- анальная и постанальная Имеются Отсутствуют Отсутствуют Отсутствуют, за исключением особой парной постапальной
Щеки Имеются Отсутствуют Отсутствуют Отсутствуют
Паразиты Преимущест- венно млекопи- тающих Млекопитаю- щих Преимущест- венно птиц Летучих мы- шей
Отношение к распростране- нию спирохето- зов Изученные все переносят гозбуди1елсй спирохстозов млекопитаю- щих и человека Не передают гозбудп гелей спирохетозов Передают возбудителей спирохетозов птиц Но изучены
3) Жизненный цикл аргазид
В жизненном цикле аргазид всегда имеется несколько нимфальных ста-
дий (2—6 у видов нашей фауны). После меньшего числа нимф слинивают
преимущественно самцы, после максимального их числа получаются почти
исключительно самки. Каждая стадия, как правило, питается, прежде чем
слиняет на следующую стадию. Размножение у большинства аргазид нашей
фауны имеет сезонный характер: яйцекладка происходит у них один раз
438
Рис. 306. Пещера — убежище орнитодорин
(оригинал).
Рис. 307. Норы черепах и ежа — убежище орнитодорин (оригинал).
Рис. 308. Жилье и скотпик—убежище орнитодорин
(оригинал).
в год, в конце лета — начале осени. Зимой клещи питаются реже, с вес-
ной становятся деятельнее и напитываются дружнее. Все развитие при
благоприятных условиях температуры, влажности и питания может
закончиться в 1х/2—2 месяца. Но обычно оно значительно затягивается.
4) Типы местообитаний (убежища) аргазид
Аргасовые клещи требовательны к условиям окружающей среды;
поэтому они являются обитателями закрытых убежищ, предоставляющих
им более благоприятные условия для существования. Закрытые убежища,,
обитаемые аргазидами, бывают естественные, природные, создавшиеся и
существующие независимо от человека, и искусственные, построенные
человеком или появившиеся в результате его деятельности. К первой
категории относятся пещеры (рис. 306), гроты, промоины и ниши средн
скал, образовавшиеся под действием ветра и воды, дупла, а также норы
и логовища, вырытые или устроенные разными животными (рис. 307).
Во вторую категорию входят сооружения человека, преимущественно тем-
новатые и сыроватые, глинобитные или сложенные из камня (рис. 308).
Аргазиды могут обитать как в жилье, так и в скотных помещениях и
даже в необитаемых постройках (мечети, клубы, склады и т. п.). Заселены
клещами бывают и искусственно вырытые пещеры. Европейского типа
постройки, светлые, сухие, хорошо вентилируемые, менее подходя’ для
аргазид, и последние в них встречаются реже и в меньшем числе.
Микроклимат убежищ. К 'какой бы категории ни относилось закрытое убежище,
оно всегда отличается своеобразным микроклиматом, имеет температуру летом более
низкую, зимой более высокую, чем в окружающей открытой природе. Суточные коле-
бания температуры в убежище также сглажены и тем более, чем лучше изолировало
убежище от внешней среды. Влажность в закрытых убежищах обычно несколько
повышена и также подвергается меньшим колебаниям, чем в открытом ландшафте.
Иными словами, микроклимат закрытых биотопов (убежищ) самостоятелен и отли-
чается постоянством, эвстатичпостью. Это первое, что благоприятствует обитанию
аргазид в таких местах.
Защитная роль убежищ. Кроме того, в закрытом убежище аргазиды всегда нахо-
дят затемнение от прямого солнечного света, который для них может оказаться губи-
тельным. Убежище защищает клещей и от неблагоприятных механических воздействий
(водяные потоки, ветер и т. п.) Наконец, в закрытых убежищах, где клещи всегда скры-
ваются в глубине трещин и щелей или в сыпучем субстрате дна убежища, они находят
укрытие от различных врагов, которые могли бы их уничтожить (пауки и другие хищ-
ные членистоногие, ящерицы, птицы, млекопитающие, например, мыши, ежи'.
5) Характер паразитизма аргазид
иг
Плодовитость. Почти все аргазиды проводят в закрытых убежищах
всю свою жизнь. Обнаружение их вне таких убежищ составляет редкое
исключение. В связи с постоянными благоприятными условиями существо-
вания и защищенности от врагов у аргазид наблюдается относительно
малая смертность. Поэтому у них не выработалось способности давать боль-
шое потомство. Аргасовые клещи всегда откладывают относительно мало
яиц (за всю жизнь несколько десятков, сотен или немного более тысячи)
по сравнению с иксодидами, дающими тысячи и десятки тысяч яиц.
Питание. С другой стороны, обитание в закрытохм убежище лишает кле-
щей возможности активно отыскивать хозяина для прокормления. Клещи
вынуждены питаться лишь на тех хозяевах, которые сами входят в их
убежище. Отсюда у аргазид выработался ряд приспособлений, характер-
ных для временных кровососов типа «подстерегающих пещерных кро-
вососов».
440
Прежде всего аргазиды неразборчивы в отношении источника пропи-
тания: они, как правило, способны напитываться на любом позвоночном—
от рептилий до человека (полифагия). Исключения встречаются редко
(например, некоторые аргазиды, паразитирующие на летучих мышах,
питаются исключительно на этих последних). Правда, кровь не всякого
животного одинаково полноценна для них, но за неимением лучшего кле-
щам приходится напиваться на первом попавшемся животном, хотя бы
для поддержания существования.
С другой стороны, клещу, обитателю закрытого убежища, важно,
встретившись с хозяином, накормиться на нем возможно скорее, используя
время, пока хозяин находится в убежище. Если же клещ не успеет за это
время напитаться и отпасть, хозяин вынесет его на себе наружу, где усло-
вия могут оказаться для клеща губительными. Поэтому у аргазид выработа-
лась способность напитываться в самые сжатые сроки: они кормятся от
трех минут до получаса или, реже, до немногих часов. В связи с этим для
аргазид не обязательно, чтобы в их убежище постоянно жило позвоночное—
их прокормитель. Очень часто они заселяют никем не обитаемые убежища,
используя для прокормления случайных посетителей.
Для быстрого пакармливания аргазиды обладают специальными приспособле-
ниями. Тело их способно сразу сильно растянуться, желудок велик и единовременно
вмещает относительно большие порции пищи. Непомерно раздувшийся клещ тут же
освобождается от избытка жидкости из всосанной крови через коксальные железы.
Анэстезирующие свойства слюны клеща, обезболивая укус, способствуют благополуч-
ному накармливанию на бодрствующем хозяине.
Все же при однократном быстром накармливапии аргасовые клещи не
могут взять столько питательного материала, сколько получают длительно
питающиеся клещи. Это обусловливает относительно малое число откла-
дываемых ими яиц и компенсируется повторностью питания самок. Яйце-
кладка обычно также происходит в несколько приемов. Строгой гонотро-
фической гармонии, какая существует, например, у комаров, у аргазид
не наблюдается.
Питание на случайных посетителях убежища всегда может повести
к тому, что клещу долго не представится возможности кормиться. Поэтому
у аргазид выработалась способность длительно голодать. Все стадии их
способны долго обходиться без пищи в ожидании встречи с хозяином. Чем
старше стадия, тем дольше она может голодать. Так, личинки некоторых
аргазид могут голодать более года, взрослые — более 9 лет (Павловский).
Продолжительность жизни. В связи с этим весь жизненный цикл кле-
ща может затягиваться на очень длительное время. У Alectorobius tholo-
zani papillipes, например, он, вероятно, может тянуться до 20 и 25 лет
(Павловский).
Понятно, что клещам, отличающимся столь длительным жизненным цик-
лом, нужны прочные, долговечные убежища. С такими убежищами и свя-
заны в действительности этого рода клещи. Непрочные, временные убежища
или не успевают заселиться аргазидами, или последние не успевают в них
размножиться.
Привязанность к убежищам и расселение. Раз заселенные аргазидами
убежища обыкновенно надолго остаются пораженными ими. Так, нередко
мы находим этих клещей в заброшенных, наполовину разрушенных домах
и хлевах. Зато заселение аргазидами новых построек происходит мед-
ленно, так как клещи эти вообще мало способны к расселению: активно
они не переползают, а на животных остаются недолго и имеют мало шансов
быть занесенными ими в новые места обитания. Тем не менее расселение
происходит, очевидно, именно последним путем — с хозяевами-прокор-
441
мителямп. В частности, наиболее вероятными заносителями клещей
Alectorobius tholozani papillipes в хозяйственные местообигааия являются
крысы и собаки (Кешишьян, Исаев).
Бяологич?ский цикл A! veouasus canestrinii. Отктопопио от описанпой жпзпепной
схема представляют в нашей фауне клещи Alveonasus canestrinii. Клещи эти во взрос-
лой фазе ведут такой же обр>з жизни, как пр >чио аргазиды. Ио молодые их стадии,
личинка и три нимфальпые стадии, ведут себя иначе. Личинка, присосавшись и напив-
шись, не отпадает, а тут же линяет па нимфу I возраста; нимфа зга немедленно присасы-
вается и, напившись, тоже по отпадает. Нимфа II возраста также всю жизнь остается
на хозяине и лишь сытая нимфа 111 возраста отпадает, прячется и вне хозяина линяет
на имаго. Все рьзпитие от литшки до сытой старшей нимфа приурочено к зимнему вре-
мени; линьк! на имаго происходит весной, и весь жи шепн >1й никл заканчивается в один
год (Магикашвиай). Животными-хозяевами (верб поды, овцы) этот клещ легко разно-
сится с места на место я потому широко расселился.
6) Распространение аргазид в СССР
Распространение аргазид в пределах Советского Союза ограничивает-
ся южными районами: они обитают в среднеазиатских республиках и на
Кавказе. Лишь представители рода Argas расселены шире пэ югу Евро-
пейской части СССР и на север заходят до Саратова.
Ландшафтная и вертикально-зональная приуроченность и связанность
с определенными биотопами. Несмотря па связанность с закрытыми убежи-
щами, имеющими более пли менее самостоятельный микроклимат, каждый
вид аргазид приурочен все же к определенной ландшафтной зоне (Поспе-
лова-Штром). Большая часть видов нашей фауны обитает в степных и пу-
стынно-степных предгорьях, но некоторые виды живут и на равнине, дру-
гие же заходят высоко в горы. В пределах области своего распространения
каждый вид засоляет только свои излюбленные типы убежищ (Поспелова-
Штром): одни живут в норах мелких млекопитающих, другие — в логовах
крупных зверей, третьи — в пещерах с летучими мышами, четвертые спо-
собны, наряду с природными убежищами, заселять и хозяйственные.
Очаги распространения в природной и хозяйственной обстановке.
Первоначально аргазиды были распространены в природной обстановке,
поселение их в хозяйственной обстановке — явление вторичное. Найдя
в соседстве с человеком благоприятные условия для существования, не-
которые аргазиды размножались здесь в массе и местами заселяли сплошь
все постройки поселка. Таковы, например, клещевые очаги в некоторых
поселениях Памира, Ферганы и т. д. Расселению клещей там особенно спо-
собствует крайняя скученность построек, жилых и скатных,
7) Клещевой возвратный тиф
Медицинское значение аргазид фауны СССР велико, так как многие
представители этого семейства являются перодатчикахми возбудителя кле-
щевого возвратного тифа. Возбудителем этого заболевания являются
спирохеты, которых передают клещи рода Alectorobius. Род Alveonasus
не причастен к распространению спирохет, в роде же Argas некоторые
виды переносят спирохет, не болезнетворных для человека.
Передатчики клещевою возвратного тифа. Шесть видов рода Alecto-
robius из семи известных в СССР передают спирохет клещевого возврат-
ного тифа, седьмой (Alectorobius coniceps) в этОхМ отношении еще но изучен.
Так как каждый вид аргазид нашей фауны имеет свой ареал (т. о. сваю
область обитания), связан с определеннЫхМ типом .местности (ландшафтом),
типом убежищ, животными-прокорхмителями и отличается деталями об-
442
раза жизни и склонностей, то и условия распространения клещевого воз-
вратного тифа не повсюду одинаковы.
Характеристика заболевания. Клещевой возвратный тиф характеризуется пере-
межающимися приступами лихорадки и безлихорадочных периодов. Спирохеты обычно
могут быть обнаружены в крови больного, но их бывает почти всегда немного. На на-__
шей территории заболевание является доброкачественным, смертельных исходов, как
правило, не бывает; но оно изнурительно и нередко надолго лишает трудоспособ-
ности.
В СССР имеется несколько, повидимому, самостоятельных форм клещевого воз-
вратного тифа. Каждая форма передается определенным видом клещей рода Alectorobi us.
Формы клещевого возвратного тифа различаются и по восприимчивости к ним разных
лабораторных животных (например,к одной форме более восприимчивы морские свинки,
к другой — белые мыши), и по другим признакам.
Клещевой возвратный тиф в прежнее время часто смешивали с маля-
рией. Диагноз с уверенностью ставится по нахождению в крови больного
спирохет. Распознавание заболевания облегчается обнаружением у па-
циента следов укусов клещей.
Реакция кожи человека на укус аргазид. Укус аргасового клеща сопровождается
местной реакцией кожи. Вначале появляется багровое или синеватсе пятнышко разной
величины. Через несколько часов синяк исчезает, а па его месте образуется узелок —
папулка. При этом место укуса, ранее совершенно не чувствительное, начинает зудить.
Зуд приступами длится несколько дней, иногда недоль или даже месяцев. Зуд может
быть нестерпимым, что приводит к расчесам, а нередко и к внесению в ранку различных
бактерий, в результате чего могут образоваться долго не заживающие язвы. Реак-
ция кожи человека па укус аргазид настолько характерна, что по ней можно точно оп-
ределить, какой паразит нападал на человека (Павловский).
Механизм передачи клещевого возвратного тифа. Клещевой возврат-
ный тиф по своему происхождению является зоонозом (стр. 148). В природе
им болеют различные млекопитающие: крысы, мыши, песчанки, летучие
мыши, ежи, дикобразы, лисы, шакалы и т. д. Питающиеся на них клещи
всасывают с их кровью спирохет; спирохеты в клещах размножаются и
проникают в разные их органы, в том числе часто в слюнные и коксальные
железы. Проникая в яйца, спирохеты передаются потомству клеща. За-
разившийся спирохетами клещ остается зараженным всю свою дальней-
шую жизнь. При последующем питании на свежем животном он может вне-
сти в его кровь спирохет со слюной или (у некоторых зарубежных аргазид)
с коксалыюй жидкостью, если она выпускается на хозяине и затекает
в ранку от укуса. В результате животное заболевает. Таким путем спиро-
хеты циркулируют в природе между клещами и дикими млекопитающими,
вне зависимости от человека, и клещевой спирохетоз может существовать
в местах обитания клещей в совершенно пустынной, не обитаемой челове-
ком местности (природные очаги клещевого спирохетоза, Павловский).
Заражение клещевым возвратным тифом в природной обстановке.
Все Alectorobius нашей фауны более или менее склонны нападать на чело-
века; поэтому, когда последний входит в соприкосновение с природными
местообитаниями этих клещей, он подвергается их укусам, а если среди
клещей были носители спирохет, то человек заболевает клещевым возврат-
ным тифом. Наиболее опасны те виды клещей, которые более активны и
агрессивны по отношению к человеку. Если клещи голодают, они иногда
способны нападать даже у входа в свое убежище, выходя навстречу рас-
положившемуся здесь человеку.
Заражению клещевым возвратным тифом в его природных очагах
подвергаются в первую очередь те категории населения, которые вхо-
дят в соприкосновение с. этими очагами. Пастухи, охотники, полевые
рабочие, сборщики орехов, топлива и пр., военные на полевых тактиче-
443
ских занятиях, работающие в геологоразведочных партиях и т. п. рискую)
заболеть, если, скрываясь от дождя и солнца, заходят в пещеры, гроты*
ниши среди скал, ночуют в них или же ложатся у входа в норы и убежища
разных животных. Опасности быть покусанными клещами подвергаются
также и люди, разбившие свои палатки в местах, где имеются убежища
клещей. Известны случаи заражения клещевым возвратным тифом даже
купальщиков на пляже, отдыхавших у прибрежных скал, под которыми
ютились клещи Alectorobius (северный берег Апшеронского полуострова
близ города Баку — Поспелова-Штром).
Заражение в хозяйственной обстановке. Однако заражения в природ-
ных биотопах относительно редки и встречаются обычно лишь время от вре-
мени. Иное наблюдается, если клещи Alectorobius живут вместе с челове
ком—в его жилье и надворных постройках. Здесь клещи находят постоян-
ную и легко доступную добычу в виде домашних животных (рогатый скот,
собаки и т. п.) и домовых грызунов. Поэтому клещи быстро размножаются
и достигают большой численности.
Люди, живущие в пораженных клещами усадьбах, регулярно подвер-
гаются укусам клещей, которые ведут себя наподобие клопов, скрываясь
днем и нападая ночью. В темноватых помещениях нередки случаи укусов
клещами и днем. В результате люди переболевают клещевым возвратным
тифом и вырабатывают к нему невосприимчивость (иммунитет),^которая
поддерживается постоянными повторными укусами зараженных клещей.
Местные жители обычно жалуются лишь на укусы как таковые и, стремясь
избавиться от них, спят на крышах и т. д.
Пришлое население, поселяясь в заклещевелых постройках и подвер
гаясь впервые укусам клещей, как правило, заболевает. В итоге нередко
наблюдаются массовые вспышки клещевого возвратного тифа, связанные
с прибытием в район партий свежего, неиммунного населения.
Борьба. Борьба с клещевым возвратным тифом может проводиться
путем уничтожения клещей-переносчиков (см. Курс борьбы с насекомыми
и клещами) и диких животных—прокормптелей клещей, а также путем
общественной и личной профилактики заболевания.
Культурная реконструкция поселков. Надежные результаты дает полное уничто-
жение (сжигание) всех старых, заклещевелых помещений и сооружение вместо них
культурных построек, мало пригодных для обитания клещей и мало доступных для
их проникновения. Практика показывает, что таким путем возможна полная очистка
поселка от Alectorobius и ликвидация клещевого возвратного тифа. Параллельно надо
проводить борьбу с крысами и другими животными — прокормптелями и разносите
лями клещей, особенно имея в виду, что эти же животные являются и резервуаром
спирохет.
Индивидуальная профилактика. В тех случаях, когда неизбежна ночевка или рас-
квартирование в заклещевелых поселках, следует спать не в помещениях, а на открытом
воздухе или в палатках, в стороне от подозрительных па присутствие клещей мест.
Если использования помещений нельзя избежать, рекомендуется изолировать кровати,
поместив их ножки в сосуды с керосином или обмазав их мушиным липким клеем или
дегтем. Кровать должна быть отставлена от стены и защищена густым пологом, хо-
рошо подобранным под матрац. Постельное и нательное белье полезно пропитывать
дегтярной эмульсией или мятной настойкой, отпугивающими клещей. Важно не допу-
стить укуса Alectorobius, так как минута его сосания уже может вызвать заражение
клещевым возвратным тифом (Павловский).
8) Обзор видов аргазид фауны СССР
Alectorobius tholozani papillipes Birula (=Ornithodorus papillipes Birula прежней
номенклатуры) (рис. 309). В фауне СССР важнейшим переносчиком клещевого возврат-
ного тифа является Alectorobius tholozani papillipes.Клещ этот одет бугорчатым покро-
вом; бугорки довольно мелки, звездчатой формы, мало разнятся по размерам и распо-
ложены обычно довольно равномерно, не образуя четких узоров. Диски на спинной
444
стороне лежат симметрично, поодиночке или группами, а на брюшной стороне разме-
щены по дну бороздок. Хорошо развиты продольные и поперечные бороздки вентраль-
ной стороны и бороздка, идущая на боках тела со спинной стороны на брюшную
(дорсо-вентральная бороздка). Постанальная (задняя) поперечная вентральная бо-
роздка отодвинута далеко кзади и лежит почти прямо поперек тела. Клюв и щеки хо-
рошо развиты; последние — цельные, морщинистые, слегка неровные по свободному
краю. Гипостом цилиндрический. Лапки первой пары ног с параллельными дорсаль-
ными и вентральными краями; по спинному краю они волнисты, имеют четыре-пять
бугорков, из которых лишь надвершинный развит хорошо (рис. 310, Л). Лапки чет-
в 11 тых ног гладкие, с одним хорошо развитым надвершинным бугорком.
Встречается во всех среднеазиатских республиках, начиная от юга Казахстана.
Особенно многочислен в Узбекистане и Таджикистане; в Туркмении относительно ре-
док. За рубежом имеется в Иране, Афганистане, Индии и других странах (другие под-
виды). ^Обитает преимущественно в предгорной и горной полосе; на Памире заходит до
высоты 2800 м (Павловский). Живет как в природных, так и в хозяйственных убежищах.
Рис. 309. Alectorobius tholozani papillipes.
А — самка (вид сверху); Б — самка (вид снизу)4
8 первом случае клещи чаще населяют пещеры и поры крупных животных( дикобразов);
а норках мелких зверьков (некоторых песчанок, ежей) они встречаются реже; оби-
тают преимущественно в горной полосе или в более северных районах. В хозяйственных'
убежищах местами очень многочисленны; заселяют жилье, надворные постройки, пу-
стующие здания. В цикле развития имеют 3—6 нимфальных стадий. Типичный поли-
фаг, способный сосать кровь самых разнообразных позвоночных. На человека напа-
дает очень охотно.
Передает спирохет среднеазиатского клещевого возвратного тифа. В природе
часто в большом проценте заражен спирохетами. Является наиболее частым виновником
заболевания человека. Из лабораторных животных к передаваемой им форме клещевого
возвратного тифа наиболее чувствительны морские свинки. В природе естественно зара-
жаются этой формой возвратного тифа крысы, ежи, шакалы, собаки и другие млеко-
питающие. Быть может, играет роль в распространении и хранении вируса лошадиного
энцефаломиэлита (Петрищева и Левкович).
Alectorobius asperus Warburton (=Ornithodorus verrucosus Olenev, Sassuchiu
et Feniuk) (рис. 311, В). Похож на предыдущий вид. Отличается узорчатостью покрова
вследствие расположения бугорков цепями и кругами. К заднему концу тела бугорки
заметно укрупняются. Постанальная поперечная бороздка несколько угловата.
Лапка первой ноги с параллельными дорсальным п вентральным краем и с четкими
бугорками, из которых лежащий посредине выше прочих (рис. 310, Б).
Распространен на Северном Кавказе, в Дагестане и в Восточном Закавказье (Азер-
байджан, Восточная Грузия, /Хрмения). Придерживается как предгорий и нижней гор-
ной полосы (преимущественно северный подвид), так и равнинной (преимущественно
южные подвиды). Живет почти исключительно в природных убежищах — пещерах и
норах таких крупных млекопитающих, как лисы, волки (северный подвид), или в неглу-
боких убежищах под каменными глыбами и в норках мелких животных — хомячков,
445
Рис. 310. Лапка первой пары пог.
А — Alerforobius 1ho|< z ni ] aj I —Alert ( jobius rspouF; В—AlectrrrbluB
cholodkoTskyi; Г—Alectorobius t?i t k< vsh > 1: Л —Alccturcbiue lci tends; L—Alvto-
uasus canestrinii.
Г ММ
В Г
Рис. 311. Обшип игд клешей.
А — Alectorobius chclcf'kovskji; Б — Al i to robins в r - nsis; В — Abetorobks rsprrusj
Г — Alectorobius ecniceps. uiuu (ci ГЗ3) (iudh h;> « ojh шоь части рис^ы.оь дедностъю влд
частично удалены).
птиц, черепах (южные подвиды). Несколько раз в небольшом числе найден в по-
стройках (подвал мечети, массивное старинное здание монастыря и др.). Имеет
от 3 до 5 стадий нимф. Ведет типичный образ жизни «подстерегающего кровососа»,
питающегося за счет всякого, кто войдет в его убежище. Отличается особой актив-
ностью — более других видов способен выходить из убежища и нападать на жертву,
расположившуюся у входа.
Встречается исключительно высокая природная зараженность этого вида спиро-
хетами. В связи со всем сказанным особенно часто повинен в заражении человека кле-
щевым возвратным тифом в природной обстановке (Поспелова-Штром). Передавае-
мая пм форма клещевого возвратного тифа, повидимому, своеобразна; к ней восприим-
чивы и морские свинки, и белые мыши.
Alectorobius cholodkovskyi (Pavlovsky) (=Ornithodorus cholodkovskyi Pavlovsky)
(рис. 311,А). Вид, близкий к A. tholozani. Отличается более мелкими и более резко
звездчатыми бугорками, равномерно покрывающими тело. Щеки цельные, но малень-
кие. Спинной и брюшной края лапки первой ноги не параллельны одип другому, по
спинной стороне этой лапки малоотчетливая волнистость (рис. 310.В). Лапка^четвертой
пары ног гладкая, с небольшим надвершинным бугром. Известен по отдельным на-
ходкам из разных частей Средней Азии (Узбекистан, Туркмения). Может быть связан
с пещерами, заселенными летучими мышами. Распространяет среди последних спиро-
хетоз. Человек, посетивший такую пещеру и покусанный клещами, может заболеть
спирохетозом.
Alectorobius tartakovskyi (Olenev) (=Ornithodorus tartakovskyi Olenev). Мелкий
вид, с равномерным мелкобугорчатым покровом. Бугорки более округлых очертаний,
чем у A. tholozani и у A. cholodkovskyi. Постанальная бороздка резко угловата и
расположена относительно ближе кпереди, чем у указанных видов. Щеки довольно
крупные, обычно цельные, но иногда глубоко рассеченные или приросшие впереди
к клюву. Лапка первой пары ног с тремя буграми (рис. 310,Г), лапкг1 последней пары
с одним небольшим надвершинным выступом.
Широко распространен по среднеазиатским республикам, преимущественно
по низким пустынно-степным предгорьям, реже на равнине. Обитатель исключительно
нор малого диаметра (песчанок, сусликов, черепах), вырытых в грунте или расположен-
ных под прикрытием каменных глыб. В крупных порах попадается редко, в хозяй-
ственных убежищах не встречается. Относительно менее долговечен, чем предыдущие
виды, чаще требует*питания, мепее способен голодать. Имеет 2—4 нимфальные стадии.
Не выходит активно из нор и мало агрессивен по отношению к человеку. Распростра-
няет среди песчанок спирохетоз, возбудителем которого является особая спирохета.
Человек в условиях опыта способен болеть этой формой спирохетоза (Латышев), но,
в связи с малой вероятностью укусов этого клеща, может естественно заражаться ею
лишь в исключительных условиях (раскопка нор и т. п.).
Alectorobius alactagalis (Issaakjan) (—Ornithodorus alactagalis Issaakjan)и’Alec-
torobius nereensis (Pavlovsky) (=Ornithodorus nereensis Pavlovsky) (рис. 311,Ь). Два
близких вида, имеющих крупнобугорчатый покров с резко увеличивающимися кзади
бугорками. Постанальная поперечная бороздка слегка угловатая или волнистая.
Щеки из отдельных долек. Передняя пара лапок тонкая, слабоволпистая по спинному
краю, почти без надвершинного бугра (рис. 310, Д), Лапка задней пары ног тонкая,
длинная, без всяких бугров. Между собой виды различаются мелкими признаками.
Первый вид обитает в Армении и Азербайджане, второй известен только из Ка-
ракалинского района Туркмении. Заселяют норы мелких животных в грунту п под кам-
нями. Довольно охотно нападают при случае на человека. Передают оолезнетворных
для человека спирохет. В лаборатории эта форма спирохетоза поддерживается на мышах.
Alectorobiusconiceps(Canestrini) (^Ornithodorus coniceps Canestrini) (рис. 311,Г).
Известен в СССР лишь по единичным находкам (Закавказье и берег Аральского моря).
Отличается грубым покровом в очень крупных бугорках, достигающих максимальных
размеров на заднем конце тела. Постанальная поперечная бороздка прямая, ото-
двинутая далеко кзадп. Щеки цельные, очень крупные, способные прикрыть весь
хоботок. Лапки все гладкие, без бугорков. Судя по зарубежным данным, должен быть
связан с птицами. Эпидемиологическое значение не изучено.
Alveonasus canestrinii (Birula) (—Ornithodorus canestrinii Birula = Ornithodorus
lahoresnis Neumann) (рис. 312). Крупный клещ с покровом в ямках, окруженных ра-
диально исчерченными валиками. Бороздки дорсо-вентральная и поперечные вент-
ральные отсутствуют, Клюва и щек нет. На лапках цервой пары по три крупных
бугорка (рис. 310,Е).
Широко распространен по всем среднеазиатским республикам п на юге Кавказа
вплоть до Дагестана. Живет в скотных помещениях, в исключительных случаях в пе-
щерах п порах крупных животных. Не передает спирохет, но по зарубежным данным
может предавать туляремию. Вызывает у овец явления паралича, иногда смертельного,
и в экспериментальных условиях может передавать некоторых кровопаразитов этих
448
животных (Растегаева). В Узбекистане подозревается в переносе лошадиного энцефало-
миэлйта (Петрищева и Левкович).
Представит ели родов Argas и Carios резко отличаются от всех прочих аргазид на-
шей фаупы по наличию ранта по краю тела и по радиальному расположению дисков.
Некоторые из этих клещей (Argas persicus, Argas reflexus и др.), кусая человека, могут
вызывать раздражение кожи, сыпи и как последствие расчесов — язвы па местах укусов.
А Б
Рис. 312. Alveonasus canestrinii, самка.
А — вид сверху;* Б — вид снизу (оригинал).
в) Иксодовые клещи (семейство Ixodidae)
1) Наружное строение иксодид
Тело голодных иксодовых клещей бывает продолговато-овальной фор-
мы, сужено кпереди. Кутикула кожистая, местами несет твердые, нерастя-
жимые участки — щитки (рис. 313). На спинной стороне щиток присут-
ствует всегда. У самцов он прикрывает всю спинную сторону, иногда же
и на брюшной стороне самца имеются отдельные щитки или даже вся
брюшная сторона одета щитками. В связи с этим тело самцов мало растя-
жимо: оно может увеличиваться в объеме лишь за счет растягивания мяг-
кого покрова брюшной стороны или только боков тела. Это связано с тем,
что самцы иксодид всасывают относительно мало пищи, а некоторые об-
ходятся совсем без питания. Самки, нимфы и личинки имеют лишь не-
большой щиток в передней части спинной стороны. Все остальное их тело
одето мягким, растяжимым покровом, что допускает очень большое увели-
чение объема тела и всасывание больших порций пищи. По заднему краю
тела обоих полов обычно имеются насечки, обрисовывающие несколько
фестонов.
Цвет тела голодных иксодид — коричневатых и желтоватых тонов;
у некоторых видов по основному фону щитка имеется рисунок из белого
эмалевого пигмента (род Dermacentor). На краях щитка у некоторых ро-
дов располагается пара глаз.
Половое отверстие расположено либо впереди, на уровне первой-вто-
рой пары ног, либо значительно отодвинуто кзади. Анус лежит иногда
очень далеко кзади. Около него обычно хорошо заметна бороздка (аналь-
29 Учебник медицинский энтомологии 449
Рис. 313. Внешнее строение иксодид. Hyalomma marginatum impressum, самец.
А—вид сверху. Б—вид снизу. 1—футляр хелицер; 2—пальпа; 3— гипостом; 4—о. иовани*' хоботка:
5— дорсальный щитот; *— скапула; 7— цервикальная бороздка; — глаз; 9—латеральная бороздка;
10—фестоны; 11—половое отверстие; 12—половая бороздка; 13—перитрема; 14— анальное отверстие;
15—анальная бороздка; 16—аданальпый щиток; 17—акцессорнальный щигок; 18 — субанальный щи-
ток; 19—кокса первой ноги; 20—вертлуг; 21—бедро; 22—голень; 23—передиелапка; 24— лапка;
25— орган Галлера; 26— коготки и подушечка,
ная бороздка), которая охватывает его или спереди (род Ixodes), или
сзади. У рода Boophilus эта бороздка часто вовсе незаметна. Дыхальца
открываются на больших, разнообразной формы пластинках—перитремах.
Ситовидное поле перитремы охватывает макулу со всех сторон. Форма
перитремы играет роль в систематике.
Хоботок помещается на переднем конце тела в особой вырезке. Основа-
ние хоботка может быть со спинной стороны прямоугольным, треугольным
или шестиугольным. У самок на нем сверху помещается пара пористых чув-
ствительных областей — поровых полей. Гипостом и пальпы могут иметь
разнообразную форму. Четвертый членик пальп очень мал, помещается
в углублении третьего и иногда несет на вершине пучок чувствительных
щетинок. Первый членик часто тоже мало развит. Пальпы часто имеют
выступы, зубцы, иногда резко выдаются в стороны за основание хоботка.
Ноги сильные, хорошо развитые. Их первый членик, кокса, нередко
снабжен разной формы зубцами. На первой паре ног зубцы иногда имеют
вид клешни. Зубцы на коксах помогают клещам удерживаться на хозяи-
не, так как волоски шерсти хозяина заклиниваются между зубцами коке.
Лапки па концах часто имеют зубчик и всегда вооружены парой коготков
л сильной присоской. Присоска особенно сильна на первой паре ног,
так как этой парой клещ цепляется за проходящее мимо животное.
Морфология фаз развития. Самцы резко отличаются от самок по ве-
личине дорсального щитка, иногда и по наличию вентральных щитков, и по
другим признакам. Нимфы большей частью похожи на самок, личинки
часто несколько своеобразны по деталям строения. Яйца обыкновенно
довольно мелкие.
2) Роды семейства Ixodidae
Семейство Ixodidae представлено в СССР шестью родами и многочи-
сленными видами (около 60 видов и подвидов). Важнейшие отличительные
признаки родов видны из таблицы определения родов семейства.
Определение родов семейства Ixodidae фауны СССР '
1(2,3). Анальная бороздкаj огибает анус спереди; глаз нет; у сам-
цов вся брюшная сторона покрыта щитками . . . ...........................Ixodes
2(1,3). Анальная бороздка неотчетлива; глаза есть; пальпы очень короткие, ребри-
стые; у самцов на брюшной поверхности 2 пары щитков...................Boophilus
3(1,2). Анальная бороздка огибает анус сзади; глаза есть или отсутствуют;
пальпы разной формы; брюшные щитки у самцов имеются или отсутствуют
4(5). Коксы 1 нерасщепленные; глаз нет; пальпы часто резко выдаются в стороны
за основание хоботка; самцы без брюшных щитков................... Haemaphysalis
5(4). Коксы 1 более или менее глубоко расщеплены; глаза есть; пальпы не вы-
даются резко в стороны за основание хоботка; у самцов брюшные щитки имеются
или отсутствуют.
6(7). Дорсальный щиток с беловатым эмалевым рисунком; основание хоботка
прямоугольное; пальпы короткие; самцы без вентральных щитков . . . Dermacentor
7(6). Дорсальный щиток без беловатого эмалевого рисунка; основание хоботка
прямоугольное или шестиугольное; пальпы длинные или короткие, самцы с вентраль-
ными щитками.
8(9). Основание хоботка прямоугольное; пальпы длинные: 2-й и 3-й членики их
длиннее своей ширины; глаза сферические, резко заметные; самцы с тремя парами
брюшных щитков....................................................... Hyalomma
9(8). Основание хоботка шестиугольное; пальпы короткие: второй и третий их
членики не длиннее своей ширины; глаза мало заметные, довольно плоские, продол-
говатые; самцы с двумя парами брюшных щитков.....................Rhipicephalus.
29*
451
3) Жизненный цикл иксодид
Большинство иксодид подстерегает свою добычу не в убежище, а
в открытой природе, на пастбище. Поэтому встреча с хозяином для них
гораздо менее обеспечена, нежели для аргазид, подстерегающих добычу
в убежищах, более или менее регулярно посещаемых или даже постоянно
населяемых хозяевами. В связи с этим стоят очень многие особенности
жизни иксодид. Вынужденные долго поджидать добычу, они обладают
способностью очень долго выносить голод. Но очень многие личинки по-
гибают от истощения. Лишь ничтожному меньшинству удается напиться
крови и слинять в следующую стадию. Многие личинки погибают и от не-
благоприятных внешних условий, и от врагов, с которыми они сталки-
ваются в открытой природе. Те же опасности угрожают и нимфам, и взрос-
лым клещам, подстерегающим свою добычу. На каждой стадии происходит
громадная гибель. В связи с этим у иксодид выработалось приспособление:
в их развитии осталась всего одна нимфальная стадия, так что иксо-
довому клещу приходится пить кровь и подстерегать добычу самое большее
только три раза в жизни: на стадии личинки, нимфы и взрослого клеща.
Взрослая самка пьет кровь тоже один раз в жизни. И все же смертность
остается большой, и до откладки яиц доживает одна особь на тысячи. По-
этому, чтобы поддержать существование вида, необходима колоссальная
плодовитость. И действительно, самки большинства иксодид откладывают
тысячи яиц (до 17 000 самки, обитающие в нашей фауне, идо 30 000 самки
некоторых тропических форм). Столь высокая плодовитость возможна лишь
при такой же большой прожорливости; и самки иксодид нередко выпи-
вают количество крови, в сотни раз превосходящее вес их тела. Очень
прожорливы и молодые стадии: раз попав на хозяина, они берут от него
как можно больше и благодаря этому имеют возможность за счет всего-
навсего двух питаний закончить свое развитие. Принятие таких больших
порций крови требует времени, чем и объясняется длительное пребывание
иксодовых клещей на хозяине: обычно они сосут каждый раз, не отры-
ваясь, в течение нескольких дней.
Таков типичный жизненный цикл иксодовых клещей. Как мы
увидим дальше, в этом семействе имеется ряд форм, значительно укло-
няющихся по своему образу жизни и жизненному циклу от этого основ-
ного типа.
Развитие. Яйца типичных пастбищных иксодид откладываются в за-
щищенных, влажных местах: под лесной подстилкой, в трещинах почвы,
под корнями растений и т. д. Обычно они не выносят сухости и прямых
солнечных лучей.
Шестиногая личинка вылупляется в разные сроки (через неделю —
месяц), в зависимости от вида клеща и от окружающих микроклиматиче-
ских условий.. Когда хитин личинки окрепнет, она выходит па поиски хо-
зяина, всползает на возвышенные предметы—на камни, кусты и, поводя
лапками первой пары ног, снабженными обонятельным органом Галлера,
поджидает хозяина.
При приближении животного личинка цепляется за него первой
парой ног, взбирается па него, отыскивает место для присасывания и
присасывается. У разных стадий и разных видов клещей есть свои из-
любленные места присасывания (например, на голове, ушах, подгрудке,
у корня хвоста животного и т. д.). Чаще личинка нападает на более доступ-
ных ей и более многочисленных мелких животных (млекопитающих, птиц,
рептилий), но иногда она нападает и на крупных животных (например,
на крупный рогатый скот), и на человека.
452
Личинка насасывается в 3—5—7 дней, отпадает, прячется в защищен-
ное место и линяет на нимфу. Развитие нимфы может длиться от несколь-
ких дней до месяца. Нимфа, окрепну в, таким же путем, как личинка, подсте-
регает хозяина, нападает на него, сосет в среднем3—10 дней и отпадаетг
прячась для следующей линьки. Хозяевами для нимф служат обычно те же
животные, что для личинок. Линька на имаго наступает через промежуток
времени от нескольких дней до нескольких месяцев.
Самцы и самки, вылупившись и окрепнув, подстерегают или активно
преследуют обычно более крупных животных. Напав на них, самки при-
сасываются п начинают питаться, самцы же, иногда не присасываясь, чаще
пососав немного, находят самок и оплодотворяют их. Оплодотворенные
самки быстро допитываются и отпадают. Общая длительность паразити-
рования половозрелых клещей па животных около недели, но иногда зна-
чительно долее.
Трех-, двух- и однохозяинный тип развития. Описанный тип развития
носит название треххозяинного, так как при нем клещ сменяет последо-
вательно трех хозяев. В других случаях, у других видов иксодид,
личинка может, напитавшись, не отпадать, а на животном перелини-
вать на нимфу. Нимфа тут же присасывается и отпадает лишь в сытом
состоянии. Слинявшие имаго ищут себе нового, второго в цикле, про-
кормителя, почему такой тип развития называется двуххозяинным. Нако-
нец, и личинка, и нимфа могут, напившись, оставаться для линьки на
хозяине. Вышедшие на животном из нимф взрослые клещи напиваются
на том же животном, и самка отпадает лишь для откладки яиц. Здесь
все три фазы развития используют одного и того же хозяина, почему
такой тип развития называется однохозяинным.
Преимущества разных типов развития при тех или иных условиях.
Однохозяинный тип развития дает клещам некоторые преимущества, так
как на поверхности «ожи животного всегда имеются благоприятные тепло
и влажность и развитие клещей здесь идет быстрее, чем вне хозяина. Кро-
ме того, выпадает необходимость поисков защищенного места для линьки
и, главное, необходимость подстерегания нового хозяина, что всегда
влечет за собой гибель значительной части популяции клещей. При
однохозяинном типе развития на юге клещи успевают пройти за год
несколько генераций (поколений); на севере однохозяпнпые клещи
приспособились проводить на животном неблагоприятный зимний сезон
(например, Hyalomma scupense). Но для однохозяинного развития кле-
щей могут быть использованы лишь крупные, выносливые животные, на
которых личинке клещей попасть значительно труднее, чем на мелких 1
зверьков.
При известных условиях двух- и треххозяинные типы развития имеют
свои преимущества, почему они и установились у клещей. Так, Hyalom-
ma marginatum, паразитирующий в молодых стадиях на птицах, приспо-
собился использовать одну и ту же птицу для личинки и нимфы, так как
попадание на новую птицу для нимфы нелегко и мало вероятно. Полово-
зрелые клещи крупны, требуют много пищи и не могли бы прокормиться
на той же птице; они, слиняв из нимфы во внешней среде, отыскивают себе
прокормителя среди крупных млекопитающих (крупный рогатый скот,
лошади).
Треххозяинный тип развития установился, например, там, где
имеется всегда благоприятный микроклимат и доступные для присасывания
мелкие животные. Так, например, Ixodes persulcatus, типичный пастбищ-
ный подстерегающий клещ, живущий в условиях сырой, тенистой тайги,
питается в стадии личинки и нимфы последовательно на двух различных
45а
мелких животных. Животные эти недостаточно выносливы, чтобы вы-
держать длительное непрерывное питание обеих неполовозрелых стадий
клеща. Вместе с тем таких животных в тайге очень много, и нимфе
сравнительно нетрудно отыскать себе нового прокормителя. Взрослые
клещи подстерегают для питания крупных животных.
Другой треххозяинный клещ, Haemaphysalis numidiana, обита,* в пустыне,
приспособился к жизни в закрытом убежище — норе грызуна (песчанки, суслика).
В глубине поры клещ всегда имеет благоприятную прохладу и влажность, но для пита-
ния он вынужден использовать лишь хозяев норы. Прерывистое питание различных фаз
развития клеща оказывается полезным, так как дает хозяевам передышку и возмож-
ность оправиться после каждого паразитирования клещей. К тому же период размно-
•жения у этого вида клеша, благодаря подходящим условиям микроклимата в норе,
растянут па несколько месяцев и потомство разных самок паразитирует неодновре-
менно. Наконец, клещи эти мелки и высасывают относительно мало крови, не слишком
обременяя хозяина. Соответственно они дают и мало яиц, но их оказывается доста-
точно для поддержания существования вида, так как в норе, закрытом местообитании,
гибель клещей относительно мала.
Haemaphysalis numidiana представляет собой пример перехода иксодового клеща
к полностью норовому образу жизни.Другой,также треххозяинный обитатель пустыни,
Hyalomma asiaticum, имеет лишь частично норовой жизненный цикл. Его личинки
и нимфы нежны и в условиях пустыни могут существовать только в закрытом убежище.
Самка откладывает яйца в норе грызуна. Личинки и нимфы последовательно питаются
на хозяине поры. Взрослые клещи крупны и стойки: они выходят в открытую пустыню
и, не боясь жары и сухости, активно добираются до хозяев — крупных млекопитаю-
щих (верблюда, джейрана и пр.). Высасывают очень много крови. Самый трудный мо-
мент наступает, когда сытая, с трудом передвигающаяся самка должна попасть в под-
ходящее для развития потомства убежище. При этом погибает большое число клещей.
Потери эти покрываются множеством яиц (до 17 000), откладываемых самками, кото-
рым удается достичь подходящего биотопа. Множество вышедших из яиц личинок и
нимф не может, не рискуя погубить хозяина поры, проходить па нем развитие, не да-
вая ему передышки (по двуххозяинному типу). Только при условии перерыва между
паразитированием личинок и нимф хозяин успевает оправиться и выдерживает питание
обеих фаз. ..
Приведенные примеры иллюстрируют многообразие приспособлений к условиям
существования у иксодовых клещей.
Питание. По сравнению с аргазидами иксодиды сосут очень долго, но за-
то взрослая самка питается только один раз в жизни. Взрослыйиксодовый
клещ, нападающий на животное, имеет некоторые органы недоразвитыми
(например, слюнные железы, некоторые мышцы тела). Во время сосания
клещ не только растягивается, но и развивается, растет, некоторые
клетки его тела размножаются. Это обстоятельство имеет определенное эпи-
демиологическое значение, так как у зараженных вирусами или риккет-
сиями клещей, при длительном питании на хозяине, вместе с ростом слюн-
ных желез и слюна, и находящиеся в ней возбудители во все возрастаю-
щих количествах поступают в кровь хозяина. Поэтому впервые часы после
присасывания иксодовый клещ обычно еще не так опасен для хозяина,
и своевременное удаление даже заведомо зараженного иксодового клеща
нередко может предотвратить заболевание хозяина. На этом основана про-
филактика передаваемых иксодидами заболеваний, требующая периодиче-
ского, через известные промежутки времени, осмотра тела и удаления с него
клещей (Миронов, Павловский). Здесь имеется принципиальное отличие
от аргасовых клещей, которые, как говорилось, могут заразить спиро-
хетозом в первую минуту сосания на хозяине.
Если иксодовому клещу не удается найти хозяина, он способен доволь-
но долго голодать. Хотя в стойкости к отсутствию пищи эти клещи значи-
тельно уступают аргазидам, но некоторые из них все же могут голодать
до 2 лет.
454
4) Распространение иксодид СССР
Распространены иксодиды шире аргазид и встречаются на террито-
рии СССР от Крайнего Севера до южных границ. Преобладающее боль-
шинство видов населяет южные районы. Отдельные виды связаны с разны-
ми типами местности (ландшафтами): лесами, степями, пустынями, горами
и т. д. Каждый вид придерживается определенного ландшафта. Питаются
на разнообразных позвоночных, от рептилии до человека, но полифагия
выражена значительно менее, чем у аргазид, а некоторые виды довольно
крепко связаны с определенным кругом хозяев.
5) Роль иксодид в сельском хозяйстве
Иксодиды играют очень большую роль в сельском хозяйстве и как массовые
изнурительные эктопаразиты, и, главное, как переносчики возбудителей протозойных
заболеваний (пироплазмозы, тейлериозы и пр.), губительных для наших домашних
животных. Вызванное этими болезнями снижение количества скота нередко является
косвенной причиной резкого увеличения малярийной заболеваемости среди людей, по-
скольку уменьшение скота усиливает связь между малярийными комарами и чело-
веком (стр. 264).
6) Медицинское значение иксодид
Медицинское значение как переносчики возбудителей заболеваний
человека имеют отдельные представители всех родов наших иксодид
(кроме рода Boophilus).
Таежный энцефалит. Из вирусных заболеваний, передаваемых иксо-
дидами, важнейшее значение имеет клещевой, таежный, или весенне-лет-
ний, энцефалит (воспаление мозга). Заболевание это открыто и изучено
за последнее десятилетие (Зильбер, Павловский и др.); существовало оно
и ранее, но его не различали как самостоятельную болезнь. Является тя-
желым поражением нервной системы, часто приводящим к смерти пли
оставляющим последствия в виде стойких параличей отдельных групп
мышц, глухоты и т. д. Излечение без последствий наблюдается относи-
тельно редко.
Распространение заболевания и переносчики. Болезнь — зооноз.
В природе болеют некоторые млекопитающие — грызуны, насекомояд-
ные, хищные, а также и птицы. Переносчик для типичной формы клеще-
вого энцефалита практически один — Ixodes persulcatus (Миронов).
Некоторые другие иксодовые клещи (Dermacentor silvarum, Haemaphy-
salis concinna) могут передавать эту форму заболевания в эксперимен-
тальных условиях, но в природе роли, повидимому, не играют (Шубладзе,
Сердюкова, Рыжов, Скрынник, Козлова, Соловьев).
Механизм распространения заболевания. Ixodes persulcatus черпает
возбудителя болезни от болеющих в природе животных. При укусах в сле-
дующих фазах клещ передает возбудителя со слюной другим животным
или человеку. Он передает его и через яйца своему потомству, видимо
в ряде поколений (Шубладзе, Сердюкова, Чумаков). Вирус передается
не всему потомству клеща, а лишь части его, почему очаг заболевания
постепенно затух бы сам собой, если бы клещи но заражались вновь от
больных животных.
Ixodes persulcatus очень требователен к влаге (требует около 100%
относительной влажности); поэтому он живет на Дальнем Востоке в
глухой, не тронутой человеком сырой тайге. Такая тайга богата как мел-
кими, так и крупными животными. Первые служат прокормителями для
личинок и нимф клеща (треххозяинный тип развития), вторые — для
взрослой фазы.
455
Пути заболевания человека. Человек может заболеть, проникая
в природные очаги энцефалита в необжитой тайге в сезон парази-
тирования взрослых Ixodes persulcatus (преимущественно весенне-летний
период). Клещ этот охотно нападает на человека: в Уссурийской тайге
до 95,7—99,7% всех присасывающихся к человеку клещей принадлежит
к этому виду. Если присосавшийся клещ будет своеврвхменно снят, зара-
жение человека не произойдет (недоразвитые в начале питания слюнные
железы у клеща).
Но не всюду клещевой энцефалит связан с необжитыми районами.
На Урале, где переносчиком энцефалита является тот же Ixodes persul-
catus, заболевания встречаются в разреженном лесу полукультурных при-
городных участков и сельских местностей. Объясняется это тем, что кли-
мат на Урале обеспечивает клещу высокую влажность в главный период
его активности, весной, даже и в разреженном лесу. Здесь же клещ нахо-
дит достаточно прокормителей как для личинок и нимф (мелкие животные),
так и для взрослой фазы (пасущийся домашний скот). Поэтому Ixodes
persulcatus на Урале может размножаться в изобилии и вблизи от поселе-
ний человека, а вместе с клещом-переносчиком и очаги заболевания рас-
полагаются здесь же (Миронов, Чумаков).
Профилактика. Личная профилактика имеет большое значение, когда человеку
необходимо проникнуть в очаги заболевания. Сводится она к ношению закрытых
комбинезонов или к специальной заправке одежды, а также к применению отпугиваю-
щих веществ (см. Курс медицинской дезинсекции).
Шотландский, или овечий, энцефалит. На западе Европейской части
СССР, в Белоруссии, у людей встречается заболевание, очень похожее
на таежн ый энцефалит. Переносчиком этого заболевания является клещ
Ixodes ricinus. Весьма вероятно, что это заболевание тождественно с шот-
ландской вертячкой овец, или шотландским энцефалитом.
Крымская геморрагическая лихорадка. Это заболевание открыто и
описано во время Отечественной войны (Чумаков, Петрова-Пионтковская
и др.). Заболевание тяжелое; характеризуется разнообразными кровоте-
чениями (носовыми, кишечными, маточными и т. п.). Возбудитель—филь-
трующийся вирус. Переносчик — клещ Hyalomma marginatum, который
во взрослой фазе иногда присасывается к человеку. Заболевание передается
укусом; вирус черпается в природе, очевидно, от животных—прокормителей
молодых фаз (Чумаков и др.).
Сходное заболевание существует и в степной полосе Западной Сибири,
где переносчиком является Dermacentor pictus (Чумаков, Нецкий
и др.).
Клещевые риккетсиозы. С клещами Ixodidae связано также рас-
пространение большой группы риккетсиозов, или клещевых сыпных ли-
хорадок. Эти заболевания встречаются во всех частях света и местами
значительно распространены.
Течение клещевых риккетсиозов обычно тяжелое, сопровождается высокой тем-
пературой и характерной пятнистой сыпью. В отличие от эпидемического сыпного тифа,
передаваемого вшами, сыпь при этих заболеваниях бывает и на ладонях рук, и
на подошвах ног. Заболевание типично связано с летним сезоном, периодом активности
тех клещей, которые нападают на человека. Заболевают люди, посещающие местно-
сти, заселенные клещами. Некоторые риккетсиозы отличаются особенно тяжелым те-
чением и дают до 70—80% летальности, другие доброкачественны и не дают смертель-
ных исходов.
Все клещевые риккетсиозы являются зоонозами и связаны с живот-
ными, болеющими в природе (грызуны, хищные и др.). Клещи заражаются
°т этих животных. В клещах возбудитель размножается и находится в ак-
тивном состоянии, но наверняка клещ заражает хозяина лишь в тОхМ слу-
чае, если ему удастся сосать хотя бы несколько часов. Клещ, восприняв
инфекцию в одной фазе своего развития, передает ее во всех последующих
и (по крайней мере в большинстве случаев) передает возбудителя своему
потомству.
Марсельская пятнистая лихорадка. Марсельская или средиземномор-
ская пятнистая лихорадка распространена по берегам Средиземного моря.
У нас она найдена в Крыму (Алымов) и на Кавказе (Здродовский). Ве-
роятно, может встречаться и в других местах в связи с широким распро-
странением клеща-переносчика. Болезнь имеет благоприятный исход:'
смертных случаев, как правило, не бывает.
Природный резервуар болезни еще недостаточно изучен. Болеют некоторые сус-
лики; ряд других грызунов и домашних животных оказался невосприимчивым. Собаки
болеют лишь в очень молодом возрасте.
Переносчик — собачий клещ Rhipicephalus sanguineus. В молодых фазах может
кормиться на метких диких животных, по может быть связан с собаками и во всех
возрастах. Возбудителя заболевания он черпает, очевидно, или от грызунов, или от мо-
лодых болеющих собак. Передает инфекцию потомству. Приспособился все развитие
проходить в помещениях, где держат собак, и размножается там иногда в большом
числе. На человека нападает менее охотно, чем на собак, и поэтому люди болеют срав-
нительно редко. Заразиться можно и при снимании с собак и раздавливании заражен-
ных клещей, если загрязненными руками коснуться слизистых оболочек глаз, носа
и т. п. Случаи заболевания совпадают с теплым временем года. Профилактика —
борьба с к тещ эм-пере носчиком на собаках, недопущение заноса клещей собаками в по-
мещения (например, с охоты) и уничтожение клещей, обосновавшихся в помещениях.
Североазиатский клещевой сыпной тиф. Другая форма клещевого
сыпного тифа была описана первоначально в степной части Сибири (Крон-
товская и др.). Эта болезнь также доброкачественна, хотя достаточно
изнурительна. Заболевания приурочены к ранней весне, которая являет-
ся главным временем активности взрослой фазы клеща-передатчика. Пере-
носчик — степной клещ Dermacentor nuttalli. Зараженные клещи оста-
ются заразными до конца своей жизни и через яйца передают возбуди-
теля своему потомству. Заражаться и заражать могут все фазы клеща.
Резервуаром заболевания в природе служат восприимчивые гры-
зуны— полевые мыши, суслики, служащие прокормителями для молодых
фаз клеща. Взрослые клещи питаются на крупных домашних млекопи-
тающих. Человек подвергается укусам взрослых клещей и заражается в
природной обстановке. Сам он распространению заболевания не способст-
вует, д^к как уничтожает присосавшихся к нему клещей и не допускает их
оставить после себя зараженное потомство.
Клещевой сыпной тиф распространен полосой от Алтая до Приморья.
Заболевания приурочены к весеннему и летнему сезону. Вирус в природе
выделен, кроме Dermacentor nuttalli, также от Haemaphysalis concinna
и от Dermacentor silvarum (Жмаева, Савицкая п др.).
Клещевой сыпной тиф. Клещевой сыпной тиф встречается и в Кир-
гизии. Встречается в апреле. ПереносчикОхМ на человека считают клеща
Haemaphysalis punctata, для которого установлена и передача возбуди-
теля потомству (Квитницкая).
Бактериальные инфекции. Известное значение имеют иксодовые
клещи и как передатчики возбудителей бактериальных инфекций. На пер-
вом месте среди них стоит туляремия.
Туляремия. Возбудителем туляремии является Bacterium tularense. В природе ею
болеют разнообразные животные (в СССР до 20 видов). Болеют и домашние животные,
например, кошки. Восприимчивы также птицы.
457
Туляремия связана в природе с разнообразными ландшафтами: полями, лугами,
кустарниковыми опушками леса и пр. Пути заражения человека от природных очагов
многообразны и сводятся к непосредственному соприкосновению с болеющими живот-
ными или к механическому внесению заразного начала в кровь человека кровососущи-
ми двукрылыми (слепнями, комарами). Повидимому, и некоторые иксодовые клещи
(Ixodes ricinus, Dermacentor pictus), изредка кусая человека, могут передавать ему
туляремию. Но важнейшую роль играют эти клещи в поддержании туляремии в
природе (Олсуфьев).
Клещи воспринимают возбудителя туляремии от больных животных;
раз заразившись, они стойко сохраняют возбудителя в себе и при мета-
морфозе—на следующих фазах развития, а также передают через яйца по-
томству. Зараженные клещи при питании распространяют туляремию среди
животных. Причастные к передаче возбудителя туляремии иксодиды имеют
растянутый жизненный цикл. Так, у Dermacentor pictus взрослые клещи
могут оставаться неактивными до 2 лет, не питаясь и дважды зимуя.
В этих клещах Bacterium tularense может сохраняться в неблагоприятные
годы, когда грызунов мало и молодые стадии клещей гибнут от бескор-
мицы. При наступлении благоприятного периода с массовым размноже-
нием полевок сохранившийся в клещах запас возбудителя может давать
начало новым крупным вспышкам туляремии на месте старых, казалось
бы, затухших очагов. Таким образом, важнейшая роль иксодовых кле-
щей заключается именно в поддержании очагов туляремии.
Чума. Весьма вероятна роль некоторых иксодовых клещей также
в поддержании и распространении в природе чумы (Засухин).
Так, Rhipiccphalus schulzei, обычный паразит мелких млекопитающих (сусликов,
песчанок, хомяков, хорьков и пр.) Нижнего Поволжья, Закавказья и Средней Азии,
повидимому, способен распространять болезнь среди животных-прокормителей. Этот
вид может нападать и па человека.
Бруцеллез. Наконец, бруцеллы, бактерии возбудители бруцеллеза (мальтийской
лихорадки) в недавнее время были выделены из разных клещей (Dermacentor margi-
natus, Boophilus calcaratus, представителей родов Hyalomma nRhipicephalus), снятых
с больных домашних животных, коров и овец, и собранных на овечьем пастбище
(Галузо и др.). Это говорит, быть может, о причастности иксодовых клещей к распро-
странению также и этого заболевания.
Клещевые параличи. Некоторые иксодиды зарубежной фауны, приса-
сываясь, вызывают параличи животных и человека, преимущественно де-
тей (Ixodes holocyclus, Dermacentor andersoni и др.). Причина параличей
неясна. При быстром удалении клеща наступает немедленное выздоров-
ление; в противном случае паралич может окончиться смертью. Предпо-
лагается, что паралич есть реакция организма на присасывание клеща.
В фауне СССР имеются некоторые виды иксодид, подозреваемые в вызы-
вании параличей (Ixodes ricinus, Haemaphysalis sp.).
7) Важнейшие иксодовые клещи фауны СССР
Ixodes ricinus L. Некрупный клещ; самец около 2,5 х 1,5 мм, самка в голодном со-
стоянии около 4 X 3 мм, а в сытом около 11X7 мм. Щиток темнокоричневый, глянцевый,
у самок приближающийся к пятиугольной форме с выпуклыми боковыми сторонами.
На щитке мелкая точечность и топкие щетинки. Переднебоковые выступы щитка (ска-
пулы) хорошо развиты. Глаз нет. Анальная бороздка огибает анус спереди. У самца
вся брюшная поверхность одета щитками. Половое отверстие на уровне третьей-четвер-
той пары кокс; у самки оно дугообразной формы. Перитрема крупная, неправильно
округлая, немного вытянутая. Основание хоботка на вентральной стороне имеет пару
широких, в виде навеса, выступов. Поровые поля вытянутой, грушевидной формы.
На коксах первых ног у обоих полов довольно длинный, острый зубец. У самцов по зад-
нему краю этих кокс имеется слабо выраженный отдел иной структуры, чем прочая по-
верхность кокс; у фиксированных сухих клещей он бывает беловатого цвета.
458
Распространен в Европейской части СССР (на восток примерно до Волги),
Крыму и на Кавказе. Связан с ландшафтом разреженных лесов и кустарников,
луговыми пастбищами. На севере держится там, где суше, на юге, наоборот, — где
сырее.
у Треххозяинный клещ. Хозяевами для взрослых служат преимущественно круп-
ные млекопитающие, в особенности домашние животные; встречаются и на мелких
зверях (ежи, зайцы и др.), птицах, рептилиях. Личинки и нимфы паразитируют на мел-
ких позвоночных (млекопитающие, птицы, рептилии).Сезон паразитирования взрослых
на юге осенне-зимне-весенний; на севере паразитируют весной и осенью. Молодые ста-
дии питаются летом.
Передает возбудителя одной из форм клещевых энцефалитов (шотландская вер-
тячка овец); причастен к распространению туляремии. Переносит группу возбудите
лей северных пироплазмозов крупного рогатого скота.
Рис. 314. Ixodes persulcatus.
А — самка (сверху); Б— самец (сверху).
Ixodes persulcatus Р. Sell. (рис. 314). Клещ внешне очень похож на предыдущего.
У самки отмечается более округлый щиток, слабо развитые скапулы. Половое отвер-
стие самки в виде прямой или волнистой щели. На вентральной стороне основания
хоботка самки пара зубцевидных, обратно направленных выступов. Поровые поля
округлой формы. Зубец на первых коксах у обоих полов значительно короче и тупее,
чем у предыдущего вида. У самца эти коксы одинаковой структуры по всей поверхности.
Обитает в южной части таежной полосы—от Камчатки и Сахалина до Карело-
Финской ССР, на юге — до Московской, Брянской и Орловской областей, на Алтае и
в горах Семиречья. Типичный лесной обитатель.
Треххозяинный клещ. Во взрослой фазе питается па диких и домашних жи-
вотных, преимущественно крупных; охотно присасывается к человеку. Молодые фазы
паразитируют па мелких млекопитающих и птицах. Сезон питания взрослых — весна
и лето. Переносит возбудителя клещевого весенне-летнего, или таежного, энцефалита.
Haemaphysalis punctata Can. et Fanz. (рис. 315). Небольшой клещ с коричневатым
дорсальным щитком, покрытым тонкими точками. У самца этот щиток вытянутый, про-
долговатый, у самок почти сердцевидной формы. Брюшных щитов нет. Глаз нет. Поло-
вое отверстие на уровне кокс второй пары. Анальная бороздка огибает анус сзади.
Перитрема самца вытянута в виде запятой с широким отростком, направленным
к спинной стороне. У самки перитрема округлая со смотрящим в сторону спины
коротким отростком. Основание хоботка прямоугольное, у самки без дорсальных, на-
правленных кзади выступов и с нерезко очерченными поровыми полями. На вентральной
стороне основания хоботка также не имеется зубцов. Пальпы у обоих полов выдаются
своим заднебоковым краем в стороны за основание хоботка. Третий членик их несет на
вентральной стороне зубец. Кокса четвертой пары ног у самца вооружена длинным
459
зубцом в виде шпоры, вершиной направленным к средней линии тела; у самки этот
зубец лишь слегка крупнее, чем на прочих коксах. Вертлуг первой пары ног на
спинной стороне имеет обратно направленный треугольный зубец.
Распространен в южных районах Европейской части СССР, в Крыму, па Кавказе,
в среднеазиатских республиках, в лесостепной, степной, полупустынной предгорной
и нижней горной полосе; ландшафты, создавшиеся под влиянием человека.
Треххозяинный'клещ. Взрослые питаются на крупных домашних и диких млекопи-
тающих, реже на птицах; нападают и на человека. Молодые очень часто кормятся па
птицах: кроме того, встречаются на мелких млекопитающих, реже на рептилиях. Сезон
паразитирования взрослых дважды в год: весной и осенью. В южных районах изредка
встречается на животных и всю зиму. Молодые фазы паразитируют летом.
Переносчик возбудителя клещевого сыпного тифа в Киргизии» Может передавать
пироплазмоз крупного рогатого скота.
Рпс. 315. Haemaphysalis punctata.
А — самец (вид сверху); Б—то ?ке (вид снизу); В—то же, перитрема; Г— то же, лапка
четвертой (ноги; Д—самка (вид сверху); Е— то же, коксы; Ж—то же. лапка четвертой
ноги; 3— то же, перитрема.
Haemaphysalis concinna Koch. (рис. 316). Дорсальный щиток у самца широкий
овальный, у самок округлый. Цвет щитка желтовато-коричневый, точки многочислен-
ные, мелкие. Перитрема самца вытянута вдоль продольной оси тела, широкая, с корот-
ким, тупым дорсальным отростком; у самки перитрема неправильно округлая, немного
вытянутая вдоль тела, с выемкой па дорсальной стороне.Основание хоботка по заднему
краю дорсальной стороны имеет пару острых, направленных назад злбцов. У самца
зубцы эти значительно сильнее, чем у самки.На вентральной стороне зубцов ге имеется.
Поровые поля овальные, четко ограниченные. Пальпы выдаются в стороны за осно-
вание хоботка. Третий членик их песет па брюшной стороне четкий з^бец. У самцов
вершины третьих члеников пальп вытянуты в острия, загнутые кнутри, которые захо-
дят одно за другое,когда пальпы приложены к хоботку.Коксы первой пары ног у самца
и самки несут зубец более крупный, чем на прочих коксах. •
Имеет прерывистый ареал, обитая двумя пятнами: 1) южные районы РСФСР,
Крым и Закавказье (преимущественно западное), и 2) Дальний Восток до Тихого
океана, на север до устья Амура. Из промежуточной области отмечены лишь редкие,
единичные находки в Узбекистане, Казахстане, Киргизии и близ Иркутска. Обитатель
влажных лесов южных типов и сырых кустарниковых зарослей.
Треххозяинный клещ. Взрослые питаются на домашних и диких копытных, хищ-
ных, насекомоядных грызунах; присасываются и к человеку. Молодые фазы питаются
на тех же животных, по предпочтительно на более мелких, а также на птицах и репти-
лиях. Взрослые паразитируют, в противоположность предыдущему виду, в теплое
время года — смарта по август. Молодые питаются также летом и иногда встречаются
в одних сборах со взрослыми.
Передает возбудителя клещевого сыпного тифа на Дальнем Востоке.
Dermacentor pictus Herm. (рис. 317). Тело крупное. Дорсальный щиток с хорошо
развитыми белыми пятнами. Вентральных щитков нет. Глаза имеются, но они мало
460
выпуклы и слабо заметны. Анальная бороздка позади ануса. Основание хоботка пря-
моугольное, у обоих полов со спинными, обратно направленными выростами, у самца
выраженными лучше, чем у самки. На вентральной стороне выступов не имеется.
Поровые поля четкие, круглые. Пальпы короткие, несут на задне-боковом углу вто-
Рис. 317. Dermacentor pictus.
А — хоботок самца (вид "сверху); Б — то же (вид снизу); В —'щиток самца; Г — хобото к
самки (вид сверху); Д—то же (вид снизу); Е— щиток самки.
рого членика со спинной стороны острые, направленные обратно зубцы, также более
сильные у самцов. Боковыми сторонами слегка выступают за основание хоботка. На
брюшной стороне третьего членика обратно направленный зубец. Коксы первой пары
ног глубоко расщеплены наподобие клешни. На спинной стороне вертлуга первой ноги
сильный, обратно направленный зубец.
Распространен в южной половине полосы лесов, от западной границы СССР
до Красноярского края; кроме того, в горах Крыма, Кавказа, Казахстана, Киргизии,
Западного Алтая. Связан с полосой лесов, но придерживается часто сырых, разрежен-
ных и кустарииково-луговых участков.
Треххозяинный клещ. Взрослые сосут на крупных домашних и диких млеко-
питающих, реже на зайцах и ежах. Личинки и нимфы питаются па мелких млекопи-
тающих. Имаго встречаются на животных рано весной, изредка осенью; ^молодые*
кормятся исключительно летом.
Хранит в себе и передает бактерий туляремии. Переносит возбудителей пиро-
плазмозов лошадей и собак. Переносит, повидимому, и возбудителя западносибирской>
геморрагической лихорадки.
Рис. 318. Dermacentor nuttalli.
Л — самка (вид сверху); Б—[самец (вид сверху).
Рис. 319. Rhipicephalus sanguineus, самка (вид сверху)*
Dermacentor nuttalli Olen. (рис. 318). Отличается от’предыдущего более округ-
лыми и выпуклыми глазами, перитремой с узким коротким отростком, слабее разви-
тыми задне-дорсальными выступами основания хоботка, почти полным отсутствием»
зубца на спинной стороне второго членика пальп и слабо развитым зубцом на вертлуге
первой ноги.
Живет в степях Предбайкалья, Забайкалья, Минусинского и Красноярского края.
Типичный степной клещ. В природе развитие проходит треххозяинным порядком.
462
Взрослые кормятся на крупном и мелком рогатом скоте, лошадях, собаках; охотно на-
падают на человека. Молодые питаются на сусликах и полевках. Массовое паразити-
рование взрослых происходит с ранней весны до лета; единичные клещи попадаются
на скоте и зимой. Личинки и нимфы питаются последовательно, начиная с июля.
Переносчик возбудителя клешевого сыпного тифа. Причастен к передаче крове-
паразитарных болезней домашних животных.
Dermacentor si!varum Olen. Похож на предыдущие виды. Отличается от Derma-
centor pictus, во-первых, пальцами, почти не выдающимися в стороны за основание
хоботка и несущими на спинной стороне второго членика лишь ничтожный зубчик,
во-вторых, овальными поровыми полями самки и, в-третьих, слабо развитыми задне-
дорсальными выступами основания хоботка. От Dermacentor nuttalli отличается пло-
скими и узкими глазами, перитремой с широким отростком п хорошо развитым дор-
сальным зубцом на вертлуге первой пары ног.
Обитает па Алтае, по берегам Байкала, в Забайкалье, на Дальнем Востоке, в При-
морье. Придерживается кустарниковых участков тайги. Треххозяинный клещ.Взрослые
кормятся па крупных домашних п дикпх животных (копытные, хищные). Нападают
и на человека. Молодые используют для питания мелких, млекопитающих и птиц.
Сезон паразитирования взрослых — весна с максимумом во второй декаде мая.
Переносчик возбудите..я клсщсгого сыпного тифа.
Rhipicephalus sanguineus Latr. (рис. 319). Небольшой клещ с темпокоричневым
дорсальным шитком. покрытым редкими крупными и более частыми мелкими точками.
У самца две пары щитков па вентральной стороне. Глаза имеются, довольно плоские,
более светлого цвета, чем щиток. Анальная бороздка проходит позади ануса. Пери-
трема самца в форме удлиненной запятой с узким отростком; у самки перитрема оваль-
ная, снабженная маленьким узким отростком, направленным к спинной стороне и
поставленным перпендикулярно к перитреме. Основание хоботка шест ид голый е с вы-
дающимися в стороны углами. Пальпы довольно короткие, не выступают в стороны за
основание хоботка. Коксы первой пары ног глубоко клешневидно расщеп, юны.
Обитает в Крыму, Закавказье; на восток известен до Красповодска.Треххозяяп-
пый клещ, тесно связанный с собаками, способный кормиться на них во всех фазах и
жить в помещениях, где есть собаки. Личинки и нимфы иногда паразитируют и па мел-
ких диких животных.
Передает возбудителя марсельской пятнистой лихорадки. Распространяет пиро-
плазм т.з собак.
Hyalomma marginatum Koch. Тело крупное, крепкое; дорсальный щиток очень
темный, коричневый. Ноги со светлыми перевязками у сочленений. На щитке многочи-
сленные крупные, грубые точки. У сампа три пары брюшных щитков. Центральный фе-
стон на заднем краю щитка самца уже, чем прочие, и обычно не отличается от них по
цвету. По боковым краям щитка самца бороздки, доходящие впереди почти до глаз.
Щиток самки широко округлен сзади. Глаза резко заметные, сферические. Перитрема
самца в форме реторты, овальная, с длинным отростком; у самки — в виде короткой
запятой. Основание хоботка прямоугольное. Пальпы длинные, узкие, не выступают
в стороны за основание хоботка. Коксы первых ног расщеплены.
На территории СССР живет па Украине, в Крыму, в Нижнем Поволжье, в средне-
азиатских республиках,в степной зоне и полупустыне.Развитие проходит по двуххозяин-
ному типу. Взрослые питаются па крупных домашних животных (лошади, крупный
рогатый скот и др.); нападают и па человека. Личинки и пимфы кормятся па птицах
и зайцах, реже—на других мелких млекопитающих. Сезон питания имаго — все лето,
с ранней весны до глубокой осени. Молодые формы обнаруживаются на животных
с середины лета до осени.
Является переносчиком возбудителя крымской геморрагической лихорадки. Пе-
редает возбудителей некоторых кровепаразитарных болезней лошадей.
Список рекомендуемой литературы
Беклемишев В. Н., Экология малярийного комара, 1944.
Догель В. А., Зоология беспозвоночных, 1946.
Догель В. А., Курс общей паразитологии, 1948.
И о ф ф И. Г., Вопросы экологии блох в связи с их эпидемиологическим значением,
1941.
Кожевников В. П., Д о б р о т в о р с к а я Н. В. и Латышев Н. И.,
Учение о кожном лейшманиозе, 1947.
Кулагин Н. М., Зоология, 1938.
Матвеев Б. С., Курс зоологии, 1948.
Мончадский А. С., Личинки комаров, 1936.
Набоков В. А., Руководство по борьбе с малярийным комаром, 1940.
Набоков В. А., Справочник бонификатора, 1943.
Олсуфьев Н. Г., Слепни (Tabanidae). Фауна СССР. Насекомые двукрылые, т. VII,
в. 2, 1937.
Павловский Е. Н., Учебник паразитологии человека с учением о переносчиках
трансмиссивных болезней, часть I, 1945; часть II, 1948.
П а в л о в с к и й Е. Н., Наставления к собиранию и изучению блох (Aphaniptera),
1927. -
Павловский Е. Н., Методы изучения кровососущих комаров (Culicidae), 1935.
ПавловскийЕ. Н., Клещевой возвратный тиф, 1944.
Павловский Е. Н., Лихорадка паппатачи и ее переносчик, 1947.
Паразитология Дальнего Востока. Под редакцией Павловского, 1947.
Перфильев II. П., Москиты. Фауна СССР. Насекомые двукрылые, т. 1, в. 2,
1937.
Померанцев Б. И., Клещи семейства Ixodidae СССР и сопредельных стран, 1946.
Поспелов В. П., Энтомология, 1935.
Практикум медицинской паразитологии. Под редакцией Павловского, 1935.
Холодковский Н. А., Энтомология, ч. 1, 1927; ч. II, 1929.
Штакельберг А. А., Кровососущие комары Палеарктики, 1937.
Штакельберг А. А., Определитель мух, 1933.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
А
Абхазия, комары 161, 288, 300
Автогенность комаров 303, 304, 331
Аданальный щиток клещей, рис. 313
Адлер, колары 161, 301
Азербайджан, клещи 445, 448
— комары 288, 289, 295, 316
— мухи 389
Аир 248
Акцессориальный щиток клещей,
рис. 313
Алма-Ата, клещи 445
— комары 306
Алтай, клещи 457, 459, 461, 463
— комары 287
Амебы 90, рис. 51
— дизентерийные 145
Аму-Дарья, комары 132, 300, 301, 313,
317
Амур, комары 300, 309
Амфибииды 112, 248, 249
Анальная лопасть членистоногих 15,
рис. 1
Анальный сегмент личинок комаров 173
Антенны см. усики членистоногих
Антропонозы 148
Апшеронский полуостров, клещи 444
Аральское море, клещи, 448
— — комары 163, 301
---мокрецы 352
Араратская долина, комары 289, 295,
299
Аргазиды 7, 146, 147, 425, 426, 427, 433,
437, 438, 439, рис. 302, 304
— защитная роль убежищ 439
— ландшафтная приуроченность 442
Аргазиды, микроклимат убежищ 439
— очаги распространения в природе 422
—------в хозяйственной обстановке
442
— питание 431, 439
— плодовитость 439
— привязанность к убежищам 441, 442
— продолжительность жизни 441
— реакция кожи человека на укус 443
— расселение 441, 442
— связь с биотопами 442
30 Учебник медицинской энтомологии
Ареал географический 13
Армения, клещи 445, 448
— комары 316
Архангельская область, комары 285
— мухи, рис. 253
Арцела 90, рис. 51
Астрахань, комары 301
Ашхабад, москиты 346
— мухи 389
Б
Бабочки 50, рис. 29
Бабочницы 53, 335
Байрам-Али, комары 311
Бактерии 72, 74, 80, 81, 96. 144. 146,
231, 243, 387, 388
Баку, комары 303, 321, 328
Бартонеллы 144
Бедро насекомых 36
Беззубка 100, рис. 58
Белокопытник 248
Белоруссия, клещи 456
Бентос 82, 97
Биогеография 12
Биология 12
Биотоп 64
Биохронология 12
Биоценоз 64
Битиния 100, рис. 58
Бихорки 32, 153, 421, рис. 14
Бич усиков комаров 182
Благовещенск, комары 300, 311, 312
Блоха кошачья 402
— собачья 402
— человеческая 141, 142, 146, 402, 405.
406, рис. 281
Блохи 8, 37, 51, 143, 144, 146, 147, 149.
402
Блохи крыс 402
Богомол 36, 46, рис. 25
Божьи коровки 150
Бокоплав 29, 100, 250, рис. 5
Болезни трансмиссивные 148, 149
Болота 124, 324
Борисоглебск, комары, рис. 174
Бороздка аномаргинальная клещей 433
— дорсовентральная 433
— постанальная 433
46S
Бороздка преанальная 433
Бризы 260, 261
Бруцеллез 427. 458
Брюшко блох 403
— клопов 415
— комаров 185, 186
— мух 363
— насекомых 19, 35, 36, рис. 5 и 17
— таракана 419
Брюшная нервная цепочка 15, 23, 24,
рис. 8
Брюшные конечности насекомых 36, 37
Брянская область, клещи 459
В
Вакуоль воздушная 82, рис. 43
— пульсирующая 91
— сократительная, рис. 50
Валдайская возвышенность, комары 301
Валлиснерииды 180, рис. 62
Ванч, комары 306
Вахта 112, 248
Верблюд 430, 442, 455
Вертлуг насекомых 36
Вертячка 49, 97, рис. 56
Верхние челюсти блох 402
---насекомых 36
Веслоногие 29, 92
Веснянки 46, рис. 24
Ветвистоусые 29, 92, 95, рис. 42 и 55
Вид 13, 14
Вирусные заболевания 144, 332, 348,
427, 455
Вирусы 144
Витамины 231
Владивосток, комары 311
Водоемы олиготрофные 77
— эвтрофные 77
Водолюб 50, 103, рис. 59
Водомерка 48, 97, 250, 282, рис. 24
Водоросли 84, 231
— вольвоксовые 86
— десмидиевые 86
— диатомовые 88, 89
— зигнемовые 86
— протококковые 88
Водохранилища 117, 118
Водяная вата 105
Водяной орех 247
— ослик 29, 97, 100, рис. 57
Водяные клопы 48, ЮЗ, 250
Воздушные мешки мух 315, рис. 235
Вольвокс 87
Волга 129, 132, 135, рис. 73
— комары 309
Волосок антенпальный, рис. 93
Волоски головы личинки 204, 205,
рис. 130
— истинные 18, рис. 2
— клипеальные, рис. 93
— ложные 18, рис. 2
— пальмовидные 171, рис. 92, 131
— рулевые 173
— теменные, рис. 93
— тра натуральные, рис. 93
— хвостовые 173
4$6
Волыно-Подольская возвышенность, ко-
мары 301
Вольфартова муха см. муха Вольфарта
Воронеж, комары, рис. 171
Воротничок личинок комаров 163, 175,
рис. 93 и 100
Вошь 147
— головная 408, 412, рис. 284
— лобковая 408, 412, рис. 284
— платяная 408, 412, рис. 284
Вредоносность паразитов 142
Временные водоемы 127, 128
— водотоки 130
Вторичноводные организмы 60
Вши 37, 43, 47.139, 140, 141, 142, 143,
144, 146, 408
Выделительные органы насекомых 38
-----членистоногих 23, 425
Выделительный аппарат клещей, 23,
425
Вылупление комаров 176
Высшие раки 29
Г
Гамазоидные клещи 146
Гамбузия 81, 251, 317, рис. 59
Ганглии нервные членистых 15, рис. 1
Ганглий надглоточный членистых 15,
рис. 1
Гекконы 282
Гемоглобин 99
Гемолимфа 39, 209
Геотаксис 254
Гетеротопность комаров 155
Гетеротопные организмы 60
Гигантолинеиды 306
Гидроморфозы 61
Гидры 97, 250
Гилян, комары 300
Гипертермический водоем 67, 240
Гипопигий комаров 176, 186, 191, 193,
рис. 115, 118, 119—125
— москитов 336, рис. 201
— мух 363
Гипопус клещей 426
Гипостом клещей 423, 435, рис. 293
Гипотермический водоем 67, 240
Гипофаринкс насекомых 36, рис. 16
Гладыш 48, 103, 250, 251, рис. 59
Глаза членистоногих 26, 27
Гликоген 39
Глотка насекомых 37, рис. 18
Гнус 7, 143
Голова насекомых 19, 34, рис. 16, 5
Голень насекомых 36
Гонотрофическая гармония комаров 156,
270
-----москитов 346, 347, рис. 212
-----мощек 352
----- слепней 356
Гонотрофический ритм 156
— цикл комаров 225, 269
Горло насекомых 34. рис. 16
Горные реки 132, 306, 313
Гребень комаров 172, рис. 98
Гребляки 48
Гречиха водяная ИЗ, рис. 63
Гречиха земноводная 249
Грифельки таракана 419
— членистоногих 17
Грудь насекомых 19, 35, рис. 5 и 17
Грузия, клещи 445
— комары 288, 289, 304, 316
Губа верхняя насекомых 36, рис. 16
— нижняя насекомых 36, рис. 16
Гудермес, комары 161, 301
Гусеницы 51
— ядовитые 51, 151, рис. 29
д
Дагестан, клещи 445, 448
— комары 288, 296, 300, 310, 313,
315
Дальний Восток 145, 149
----AeJes, 322, 324, 325
-— — клещи 456, 460, 463
---- комары 310, 311, 312
Дафния 29, 73, 96, рис. 42 и 55
Двукрылые 51, 335, 357
Дербент .комары 161
Дерматиты 427
Десятиногие раки 29, 100
Детрит 231
Джейран 455
Джер-Курган, комары, рис. 181
Джонстонов орган комаров 182
Диапауза комаров 284, 286
Диантомы 92, 94. рис. 54
Диатомовые водоросли см. водоросли
диатомовые
Диафрагма брюшная насекомых 40,
рис. 135
----комаров 211, рис. 135
Дивичи, комары 235
Дизентерия 144,, 147, 149, 388,
рис. 257
Дикобраз 343, 443
Диксы 136, 186, 200
Диски имагипальные 210
— клещей 433
Дифлюгия 90, рис. 43, 51
Длинноусые 53
Длиннохвостые раки 29
Днепр, комары, 161, 301
Днепропетровск, комары 296, 312, 331
Днестр, комары 301
Дночерпатель 59
Дождевка 353. 355. рис. 217
Долгоносик амбарный рис. 28
Дома для малярийной местности 265,
рис. 158 и 160
Дорсальные бугорки клещей, рис. 304
Дунай 132
Дупляные водоемы 128, 129, 307. 322,
328
Дыхание комаров 216
— личипок комаров 207, 208, 244
Дыхательная трубка водяных клопов
39. рис. 20
Дыхательные трубки личинок комаров
172, рис. 102
Дыхательный аппарат клещей 423
Дыхальца 21, 38, 214, 215, 317,
рис. 136
Е
Европейская часть СССР, A6des, 322,
324, 332
-------клещи, 442, 455, 456, 459, 460
-------комары, 236, 285, 297, 301,
304, 306
Еж 439, 443, 445, 459, 461
Еж оголовка 248
Ереван, мухи 384, 385, 386
Ж
Жабры кровяные насекомых 39, рис. 20
— ракообразных 21, рис. 6
— трахейные насекомых 39, рис. 20
Ж валы см. верхние челюсти
Жгутиковые 89, 145, 147, 349, 414
Жгутиконосцы 147, 231
Железница 428. рис. 296
Железы коксальные 23, 425, рис. 7
— ли ночные членистоногих 18
— паутинные 32
— сегментальные 23. рис. 1
— слюнные насекомых 37, рис. 18
Желтые тела комаров 222, рис. 143
---мух 379, 380
Живородка 100, рис. 58
Животные ядовитые 150
Жи1а жа осенняя 360, 362, 364, 367,
392, 393, рис. 228, 237 264, 265
Жизненный цикл клещей 425, 426
Жилкование крыльев насекомых 37
---комаров 185, рис. 111, 112
— — мух 262, 395, рис. 232
Жировое тело комаров 209, 219, 284,
286, рис. 140
---насекомых 39, 40
Жужелицы 151
Жужжальца мух 363
— двукрылых 37
Жуки 49
— водяные 101, 250
— пластинчатоусые 50
— ядовитые 150, 151
3
Забайкалье, клещи 462, 463
Заднегрудь насекомых 35, рис. 5 и 17
Заднеспинка комаров 184
— мух 361, рис. 229
Зайсан, комары 309
Заморы 77
Западная Сибирь, 119, 120, 456
Западно-Сибирская низменность, комары
301
Зарастание водоемов 114, 115, 124, 250
Зарастание поемных водоемов 135
Зарастание рек 131
Зародышевое развитие насекомых 42,43,
рис. 22
Затылок насекомых 34, рис. 16
Зауралье 120
Зауры 317
Заяц 461. 463
Зейско-Буреинская низменность, комары,
161, 300
30*
467
Зеленые водоросли 85
Земноводные организмы 61
Зеравшан 129
Зеравшан, комары 317
Зигота 86, 87
Зимовка комаров 284—288, 293, 302,
306, 307, 310, 314, 317, 322, 329, 330,
331, 332
— москитов 342
— мух 384, 385
Златоглазик 353, рис. 217
Зоб насекомых 37, рис. 18
— комаров 212, 213, рис. 139
— москитов 337
— мух 364
Зона тяготения комаров 262
Зоонозы 148, 149
Зоопланктон 84, 89
Зоопрофилактика 266, 269
Зооспора, рис. 48
Зрение комаров 264
Зуб яйцевой комаров 174, 175, рис. 101
— — москитов 339
— — насекомых 42
Зубцы членистоногих 18, рис. 4
И
Ижевск, комары 290
Изотермический водоем 67, 309
Иксодиды 141, 144, 146, 433, 437, 449,
рис. 313
— и бактериальные инфекции 457, 458
— морфология фаз развития 451
— питание 454
— развитие 452, 453
— типы развития 453, 454
Иксодовые клещи см. иксодиды
Имагинальные диски личинок комаров
210
Имаго 28
Иммунизация 150
Иммунитет 348
Инфузории 90, 231
Ирганай, комары, рис. 190
Иркутск, клещи, 460
Иссык-Куль, клещи 457
К
Кабарда, комары 161, 264, 265, 267, 300
— мухи 256, рис. 253, табл. 17
Кавказ 32. 119, 125, 128, 129, 149
— клещи 442, 445, 448, 457, 458, 459,
460, 461, 463
— комары 131, 161 163, 236, 241, 249,
251, 252, 253, 267, 288, 289, 291,
292, 293, 295, 297, 300, 301, 302,
304, 305, 306, 307, 309, 312, 313,
315, 322, 324, 328, 331, 332
— москиты 349, 350
— мухи 367
Казахстан 70, 79, 119
— клещи 445, 460, 461
— комары 300
— мухи 350
Калининская область, комары 287
Кама, комары 129
468
Камера питательная 41, рис. 21
Камеростом клещей 435, рис. 304
Камчатка, клещи 459
Камыш 111, 248, 302, 306, рис. 62
Кантаридин 151
Кара-Кала, комары 299, 316, рис. 190 и
192
Каракалинский район, клещи 448
Каракурт 153, 154
Карапузики 49, рис. 27
Каратал, комары 281, 282
Карело-Финская ССР, клещи 459
-------комары 296
Карпы зубастые 251
Карстовые водоемы 120
Каспийское побережье, комары 163
Катушка 100, рис. 58
Кахетия, комары 161, 300
Кзыл-Орда, комары, рис. 178
Кивсяки 19, 29, рис. 13
Киев, комары 286, 307, 331
Киевская область, комары 287
Киргизия, клещи 460, 461
— комары 300
— москиты 350
Киргизский хребет, комары 161, 163
Кишечные заболевания человека 6, 144,
357
Кишечные простейшие 145, 357
Кишка задняя насекомых 38, рис. 18
— передняя 37, рис. 18
— средняя 38, рис. 18
Класпетты 186, рис. 115
Класс 13, 14
Клетки питающие 41, 222—224, 378—
380. рис. 21, 143, 212, 249
Клещ крысиный 433, рис. 301
— птичниковый 432,рис. 300
— сырный 430
— чесоточный 142, 426, 427, 429, 430,
рис. 297
Клещевые параличи 427, 458
Клещи 7, 8, 24, 34, 140, 141. 144, 154,
269.
— амбарные 430
— — отравление ими 151, 152
— аргасовые 146, 147, 425, 426, 433,
437, 438, рис. 302, 304
— иксодовые 423, 433
— красноте л ки 146
— медицинское значение 427
— мучные 430, 431, рис. 298
— образ жизни 426, 427
— пастбищные 141, 452
Клоп постельный 37, 140, 141, 142, 147,
414, 415, рис. 8, 287, 288
Клопы 24, 43, 48, 103, 140, 146, рис. 26,
414
Клюв клещей 435, рис. 304
Кобылка 46, рис. 25
Кожееды 49, рис. 27
Коканд, комары 289, 292, рис. 171 и 177
Кокандский район, комары 316
Коксальные железы клещей 425
---паукообразных 23, рис. 7
Коловратки 73, 81, 92, 231, рис. 41,
42, 53
Колхида, комары 161
Комар-долгоножка, рис. 31
— малярийный, см. Anopheles
— толкунчик 53, 97, 99, 118, рис. 31, 57
Комары 6, 26, 37, 50, 54, 57. 62, 65, 68,
80, 91, 127, 134, 135, 141—143, 146,
149, 154, 155, 156, 158, 186, 188,
200, 202, 208, 335
Комары как переносчики болезней 146,
319, 332—334
— строение щитка 183, рис. 108
-----шушков, рис. 107
Комиссуры нервные 15
Конечности брюшные насекомых 36
— ротовые насекомых 36
-----членистоногих 17, 18
Коннективы 15, 23, 24, рис. 1 и 8
— окологлоточные 15
Конъюгаты 73, рис. 45 (см. также сцеп-
лянки)
Конъюгация 91
Копет-Даг, комары 161
Коретры 118, см. также Chaoborinae
Корненожки 90
Корова 264, 265, 399, 458
Коромысло 100, рис. 57
Короткоусые 53
Кошка 458
Крабы 29, 100
Красноводск, клещи 463
Краснодар, комары 307
Краснотелка 146, 431, рис. 299
Красноярск, клещи 457, 461, 462
Креветки 29
Кровососка лошадиная 401, рис. 279,
280
Кровососки 53, 401, рис. 32
Круглошовные 53, 357
Крыжовниковая болезнь 431
Крылья комаров 184
— мух 362, рис. 232
— насекомых 37, рис. 24, 25, 30, 32
Крым, клещи 457, 459, 460, 461,
463
— комары, 128, 322, 324
— москиты 343, 349, 350
Крыса водяная 433
— незокия 343
— туркестанская 343
Крысы 149, 402, 431, 433, 442, 443, 444,
445
Ктенидии блох 402.
К тырь, рис. 31
Кубань 132, 312
— комары 161, 300
Кубышка 108, 111, 247, рис. 62
Кувшинка 108, 111, 247, рис. 62
Кузнечики 36, 46, рис. 25
Куйбышев, комары 296
Куколка москитов, рис. 207
— насекомых 28, 43
Куриное просо 126, 249
Кура-Араксинская низменность, комары
317
Куры, 432
Кутикула членистоногих 16, 17, 18, 19,
рис. 2
Л
Лабеллы комаров 181
— мух см. лопасти ротовые
Лабиум, см. губа нижняя
Лапка насекомых 36
Ласточки 282
Лейшмании 145
Лейшманиоз 145, 149
— висцеральный 349
— кожный 349
Легочные мешки паукообразных 21
Летучие мыши 282, 442, 443, 448
Лемниды 104, 246, 249
— нейстические 104, 247, 249
— планктические 104, 105, 246, 249
Ленинград, комары 315, 318, рис. 181,
190 и 193
Ленинабад, москиты 343
Лентец широкий 29
Ленкорань, комары 161, 300, 307
Лжескорпионы 33, рис. 5
Лилия водяная см. кувшинка
Лингула комаров 181
Линеиды 111, 248, 309, рис. 62
Линька комаров 156
— поденок 45, рис. 24
— членистоногих 18, 28
Лиса 443
Листоеды 150
Листоногие раки 29, 92, 95
Литораль 118, 119
Лихорадка Западно-Сибирская геморра-
гическая 462
— клещевая возвратнотифозная 149,
427, 442, 443, 444, 445, 448
— — сыпнотифозная 149 *
— крымская геморрагическая 427, 456,
463
— мальтийская 458
— марсельская, средиземноморская,
пятнистая, 427, 457, 463
— паппатачи 144, 348, 349
— речная 431
— Скалистых Гор 427
— цуцугамуши 427, 431, 432
Личинки клещей 421, 428, 429, 431 436,
437, 441, 442, рис. 291
— москитов 339, рис. 207
— мух 368, рис. 240, 241
— насекомых 42, 44
— слепней 354, рис. 220, 221
— членистоногих 28
Лоб насекомых 34, рис. 16
«Лобная аллейка» мух 358, рис. 246
Лобный пузырь куколок мух 375,
рис. 246
Ложноножки 90
Лопасти ротовые 359, 360
Лошадь 264, 265, 333, 398, 400, 401, 430,
453, 463
Лужи 66, 67, 127, 128, 240, 322, 324,
326
Лютик водяной ИЗ, рис. 62
Лютка, рис. 24
Люточки 100
Лягушечник или лягушатник 104, 247,
рис. 60
469
Ляжка насекомых 36
Львов, комары 171
М
Магнитогорск, комары 253, 296,
Ма занда ран, комары 300
Майка, рис. 81
Майкоп, комары 306
Макроспора, рис. 48
Мак силлы см. нижние челюсти
Макула клещей 435
Малощетинковые черви 97, 99
Мальпигиевы сосуды 23, 38, рис. 7 и 18
Малярийный паразит 140, 145, 148
Малярия 6. 7, 148, 149, 150, 319, 322
— птиц 149, 331
Маньчжурия, комары 300
Мандибулы см. верхние челюсти
Махач-Кала, комары 163, 302
Мацеста, комары 275
Межреберпые перепонки яиц комаров,
рис. 88
Мезосапробные организмы 73
Мервский оазис, комары 309, 312, 319
Метаморфоз 28, 421, см. также превра-
щение
Методика гидробиологии 58
Мешки легочные паукообразных 21
Миаз 357
Миграции 140
Микропиле яиц комаров 157, 158, 159,
226. рис. 87 и 88
Минусинский край, клещи 462
Многокорепник 104, рис. 60
Многон >жки 15, 16, 29, 282, рис. 13
— ядовитые 153
Мозг см. ганглий надглоточный
— брюшной членистоногих 24, рис. 8
Мокрецы 53, 57, 146, 335, 352, 353,
рис. 216
Мокрицы 29, рис. 12
Молдавия, комары 39
— москиты 349, 330
Моллюски 100, 354 w
Молотовская область, комары 290,
рис. 167 и 173
Моль 50
Москва, комары 290, 296, 302, 324, 325,
331, рис. 171
Москиты 7, 37, 142, 144, 145, 146, 149,
156, 335, рис. 200, 201, 204, 205,
206, 207
— анатомия внутренняя 337, 339
— гонотрофические взаимоотношения,
табл. 212
— места выплода 343
— методы обнаружения 343
— наружное строение 336
— окраска 346
— определение возраста 347, рис. 213
— разведение 342, 343
— сбор и хранение 348
— типы убежищ 347. 348
— характер паразитизма 346
Московская область, клещи 459
---комары 284. 304. рис. 172 и 175
---мухи 384, 386, 396, рис. 253
470
Мотыль 73, 99, рис. 8. 31 и 40
Мошки 7, 53, 57, 65, 146, 154, 156, 335,
350. 351, рис. 36, 215
Муравьи 36, 51, 282
Мургаб (река), комары 300. 317, 319
Мутчализм 143
Муха базарная 389, 390, рис. 258
— весенняя падальная 395, 396, 397
— вольфартова 139, 140, 141, 143,
397, 398. рис. 271, 272, 273
— домовая 360, 362, 363, 367. 391, 392,
рис. 232, 261, 262, 263, табл. 17
-- зеленая падальная 139, 395, 397.
рис. 270
— комнатная 37, 142, 147, 359, 360,
362, 367, рис. 225—236, 237—365
— малая комнатная 360, 362, 390,
391, рис. 253, 260
— серая мясная, рис. 32
— синяя падальная 140, 395, 396,
рис. 268. 269
— сырттая 358. рис. 223 и 224
— цеце 146, 147, 395, рис. 267
Мухи 24, 49. 145. 147
— биотермические методы борьбы 372
— в городе 383, 384
— в деревне 382, 383
— в домах 382, табл. 13 и 14
— влажность и куколки 374
— влияние температуры 375, 376,
рис. 248
— выход личинок па поверхность 375
— действие па личинку влажности, све-
та и газов. 372
— жопские половые оргапы 366
— кровососки 53, 54, 146, рис. 32, 279,
280
— мальпигиевы сосуды 365
— местообитание личинок 370
— миграция 371, 373
— мужские половые органы 367
— определение возраста 380, рис.250
и 251
— откладка яиц 380. 381. 382
— отношение к свету 376
— отрыжка 377
— паразитизм личинок 357
— плодовитость 380
— продолжительность жизни 375
— сезонный ход численности 385, 386
— сроки развития 370, 371
— эпидемиологическое значение 387,
388. 389
Мхи ИЗ
Мыши 439. 443. 448. 457
Мышцы членистоногих 17
Н
Нагана 395
Надглоточный ганглий членистоногих
15
Надглоточник личинок комаров 167
— мух 359, рис. 227
Наездники 36, 51, 139
Наннопланктон 84
Нарывники 50, 150, рис. 19, 81
Насекомые 15, 16, 31, 45, 354
Насекомые с неполным превращением
45
— с полным превращением 45, 48
Науплии 94, рис. 84
Нейстон 82
Нервный аппарат 15
— — членистоногих 23
Низшие раки 29
Нимфа насекомых 43
— клещей 421, 425, 426, рис. 291
Нимфеиды 108, 247
Нитчатки 231, 240, 245
Ново-Ивановка, комары 264
Новый Афон, комары 308
Ноги насекомых 36
Ногощупальца паукообразных 21
Ножны комаров 181
Номенклатура зоологическая 13, 14
О
Овод желудочный 140, 275, 276, 398,
399, рис. 274
— кожный 399, 400
—г лошадиный 400
— овечий 400, рис. 278
Оводы полостные 400, 401
Овца 401, 442, 448, 456, 458, 459
Ожирение комаров 220
Озера 117
Окологлоточное нервное кольцо членисто-
ногих 15
Окологлоточные коннективы членисто-
ногих 23, рис. 8
Олигоса пробные организмы 73
Опистосома паукообразных 31, 32
Орган Галлера клещей 425. рис. 304
— Женэ клещей 425, рис. 294 и 295
— зрения 26, 27, рис. 11 и 16
— вкуса членистоногих 25
— коловраща тельный 92
Органы обоняния членистоногих 25,
рис. 10
— осязательные насекомых 24, 25,
рис. 9
— ротовые насекомых 36, рис. 16
— слуха насекомых 25, 26, рис. 10
— хордотональные 25, рис. 10
— чувств клетей 425
---членистоногих 24
Орехово-Зуево, комары 276
Орловская область, клещи 459
Орша, комары 161, 301
Осоки 111
Осмотическая роль солей 71, 242
Осморегуляция 71, 242
Осы 36, 51, 282
Остии 40
Отделы тела членистоногих 19
Отростки кардиальные 38
Отряд 13, 14
П
Пальпы клещей 436
Пальчатник 249
Памир, клещи 442, 445
— комары 306
Пандорина 87
Папцырпые коловратки 92, рис. 53
Паразитизм 138, 143
Паразиты возрастные 141
— вредоносность 142, 143
— домовые подстерегающие 140
— кругложизненные 141
— многоядные 141
— одпоядные 141
— пастбищные подстерегающие 141
— постоянные 140
— степень связи с человеком 142, 143
Паратиф 144, 388
Паук-крестовик, рис. 15
Пауки 31, 32, 33, 153, 282,г 431, 439,
рис. 15, 85
— водные 250
Паукообразные 16, 31, 421
Педипальпы паукообразных 21
Пелагиаль 118
Первичнобескрылые 31, 45
Первичноводные организмы 60
Первичнокрылатые 31
Переднегрудь насекомых 35, рис. 17 и 5
Перелеты мух 384
Перемычка поперечная см. комиссура
— продольная см. конпективы
Переносчики механические 145—147
— специфические 145—148
Перепонка соединительная 17
Перепончатокрылые 51
— жалящие 152
Перидинеи 89
Перистом, рис. 52
Перитремы клещей 423, 437, рис. 291
— комаров 215, рис. 136
Перитрофическая оболочка (перепонка) 38
---- комаров 206
---- москитов 337, 338
---- мух 364
Перифитон 82, ЮЗ. 231
Песчанки 149, 343, 349, 443, 448, 454,
458
Пигидий блох 403
Пиреноиды, рис. 46, 50
Пироплазмозы 149. 427, 460. 462, 463
Питание двойственное комаров 155,
---- москитов 346
----мошек 350
---- слепней 355
Питание мух 376. 377
Пищеварительный аппарат блох 404,
рис. 282
----вшей 109. рис. 285
----клещей 423, 424. 425
----таракана 419, 420, рис. 18
Пищевод насекомых 37, рис. 18
Пиявки 99, 250
— ложноконская 99, рис. 57
— рыбья 99, рис. 57
Плавник личинок комаров 173
Плавт 48, 103. рис. 59
Плавунец 49. 101. 250, рис. 59
Планктон 82. 231
Пластинки жевательные 36. рис. 16
Площппа см. вошь лобковая
Поверхностное натяжение воды 62, 226,
227
Поволжье, клещи 458, 463
471
Поволжье комары 236
Подвид 13, 14
Подглоточник блох 402
— мух 359, рис. 227
Поденки 37, 45, рис. 24
Подкласс 13, 14
Подотряд 13, 14
Подрод 14
Подсемейство 13, 14
Полевица земноводная 111, 249
Полевки 431, 458, 463
Полипы пресноводные, рис. 64
Полисапробные организмы 73
Половой аппарат блох 404
— — вшей 409, 410, рис. 285
----клещей 425
---- таракана 420
Полтава, комары 307
Полужесткокрылые см. клоны
Поплавки яиц, рис. 88
Поровые поля клещей, рис. 294
Поселки в малярийной местности 265,
рис. 158 и 160
Превращение насекомых 43, 44, 45, 48,
49, 50, 51 (см. также метаморфоз)
Предбайкалье, клещи 462
Преджелудок мух 364, рис. 234
Предкуколка мух 369, 370
Приазовье, комары 161, 286, 289, 301
Приамурье, комары 309
Приволжская возвышенность, комары
301
Придатки кожи членистоногих 18
Прикаспийские степи, комары 70
Приморский край, клещи 457, 463
---- комары 309, 322, 325, 326, 327,
328, 331, 332
Просома 31, 32
Простейшие 89, 145, 147, 231
Протозоология 12
Протококковые 73
Прудовик 100, 250, рис. 58
Пруды 119
Прусак 418
Прямокрылые 37, 46, 418, рис. 25
Прямошовные 53, 335, 353, рис. 31
Псевдоподии см. ложноножки
Псевдотрахеи мух 360, рис. 227
Пузырчатка 104, 246, рис. 60
Пульвиллы 52, рис. 113
Пупарий мух 357, 370, рис. 242
Пухоеды 37, 47, рис. 26
Пчела домашняя 151
Пчелы 36, 45, 51
Пятигорск, комары, 307
Р
Равноногие 29
Развитие насекомых 42, рис. 22
>---с неполным превращением 43
— — с полным превращением 43
— клещей 425. 426
— таракана 420
— комаров 222, рис. 143
— москитов рис. 212
— яйцевых фолликулов мух 377, 380,
рис. 249, 250, 251
Размножение партеногенетическое 92
Рак речной 29, 100, рис. 6 и 12
Раки высшие 29, 100
— низшие 29
Ракообразные 15, 29, 92, рис. 12
Ракушечные или ракушковые 29, 92, 96
Рант клещей 433, 435, рис. 303
Распространение клещей 426
Расходы рек 129
Рдест гребенчатый 108, 120, 247, рис. 61
— плавающий 108, 247
Рдесты 105, 108, 240, 245, 247, рис. 61
Ресничные черви 250
Риккетсии 144, 146, 413
Риккетсиозы 144, 402, 427
— клещевые 456, 457
Рис 248
Рисовые поля 125, 126, 309, 313, 317,
331, рис. 71
Ришта 148
Рогоз 111, 248, рис. 62
Роголистник 67, 73, 105, 108, 245, 247.
рис. 61
Род 13, 14
Рост членистоногих 18
Ростов, комары 171, 206, 286, 287. 307
Ротовой аппарат блох, 402
-----клопа 415, рис. 288
-----клещей 423
-----комаров 181, 182
Ротовые конечности насекомых 36
Рунец овечий 401
Русский овод, см. овод лошадиный
Ручейники 50, 97, 100, 250
Рыбы 103, 251
Рыбные пруды 122, рис. 70
Ряска малая 104, 247, рис. 60
— трехдольчатая 67, 104, 246, 247,
рис. 60
С
Сабельник 248
Сальвиния 247
Самарканд, комары 289, 295
Самаркандский район, комары 237
Самоочищение воды 74
Сапробность водоемов 70, 71
Саратов, клещи 442
Сахалин, клещи 459
Свободные кровососы 141
Связь между систематическим положением
возбудителя и переносчика 145
Севастополь, москиты 343
Седло, личинки комаров 173, рис. 99
Сезонность, эпидемиологическое значение
320
Секреты 18
Семейство 13, 14
Семиречье, клещи 459
— москиты 343
Сенокосцы 33
Сенсибилизация 150, 348
Сердце насекомых 40
— членистоногих 23, рис. 6
Сетки планктонные 59
Сеточка водяная 88
Сетчатокрылые 50
Сибирская язва 144, 147, 356, 388, 394
472
Сибирь 70. 79, 119, 149
— клещи 456
— комары 149, 236, 242. 285, 296, 297,
301, 306, 324, 332
Симбиоз 143
Симулин 58, 65
Синантроп 326
Синезеленые водоросли 84
Ситняг 248, 249
Сифилис 145
Сифон личинки комаров 39, 172
Скафолебрис 29, 196, рис. 56
Скелет наружный членистоногих 17
Склериты 17
Сколопендры 29, 153
Скорпгоя водяной 48, 250, рис. 20
Скорпионы 32, 153, 421, рис. 84
Слепень 353, 354, рис. 8, 217 и 218
Слепни 7, 53, 57, 143, 146, 147, 156, 269,
335
— борьба с ними 356
— гонотрофическая гармония 356
— куколка 355, рис. 222
— личинка 354, 355, рис. 220 и 221
— определительная таблица родов 354
— эпидемиологическое значение 356
— яйца 354
Собака 149, 282, 349, 402, 430, 442,444,
445, 457, 463
Соленость воды 70, 241, 242
Солнечники 90
Сольвычегодск, комары 284
Сонная болезнь 395
Сороконожки 29, 153
Сосальщики 145
Сперматека комаров 221
— москитов 338, рис. 206
Сперматофор клещей 425
Спинная пластинка клещей, рис. 303
Спинные придатки пищевода 212, 213,
рис. 139
Спирохетозы 413, 427, 442, 448
Спирохеты 6, 144, 147, 413, 427, 442
Споровики 145
— кровяные 145
Способы переноса возбудителей 147
Среднегрудь насекомых 35, рис. 5 и 17
Средняя Азия 32, 125, 128, 129, 131, 149
---клещи 442, 448, 458, 460, 463
---комары 163, 236, 237, 241, 249,
251, 253, 257, 288, 289, 291, 292, 293,
295, 297, 299, 301, 303, 306, 309, 311,
312, 313, 315, 316, 319, 322, 332
---москиты 343, 349, 350
---мухи 367, 380, 389
Стадии Селла 273
Сталинабад, мухи 389
Стерниты 17, *35, рис. 3 и 17
Стигмальная пластинка личинки комаров
171, 172, 207, 208
Стигмы насекомых 21, 38
Стрекозы 36, 37, 45, 100, 250, 282,рис. 24
Стрелолист 113, 248, рис. 63
Субанальный щиток, рис. 313
Сувойка, рис. 52
Сукцессия 114, 135, 250
С ул а к (река), комары 306
Сусак 248
Суслики 343, 349, 448, 454, 457, 458, 463
Сухуми, комары 289, 291, 292, 295, 296,
*304, 329, рис. 171
Сфагновые болота 124
Сфагновый мох ИЗ, рис. 64
Сфинктер преджелудка комаров 213
Сцеплянки 86 (см. также конъюгаты)
Сыр-Дарьинская, комары 318, 319
Сыр-Дарья 129, 132
----комары 301, 317
Сяжки см. усики
Т
Таджикистан, клещи 445
— мухи 389
Тазик насекомых см. ляжка.
Таракан рыжий 418
— черпый 418, рис. 17, 18, 289 и 290
— эпидемиологическая роль
Тараканы 36, 37, 46, 448
Ташауз, комары 295, 299, рис. 171
Ташкент, комары 288, 289, 296, 311
Тбилиси, комары 328
— мухи 384, 385
Теджен, комары 300, 317
Тейлериозы 427
Тека комаров 181
Температура при развитии москитов 342
Температурный режим водоемов 68, рис.38
-----луж и прудов 66
-----мест обитания личинок и куколок
мух 371, 372, 374
Темя насекомых 34, рис. 16
Тенториум 210, рис. 105
Тергит см. щитки спинные
Терек, комары 161, 300, 307, рис. 74
Термез, комары 315, 318, 319, рис. 190 и
193
Термиты 45, 46
Тип 13, 14
Тиф брюшной 144, 147, 148, 149, 388
— возвратный 145, 147, 148, 149, 413
— возвратный клещевой 427, 442, 443
— клещевой на Иссык-Куле 457
— клещевой крысиный 8, 149, 402, 433
— сыпной 144, 148, 149, 413
-----клещевой 460, 463
-----североазиатский 457
Томск, комары 161, 301
Томская область, комары 334
Торус 182
Торфяные карьеры 125
Трансмиссивные болезни 145, 149, 150
— зоонозы 149
Трансовариальная передача вирусов 349,
455
-----риккетсий 431. 457
----- спирохет 443
Трахеи клещей 21, 34, 423, 428, рис. 7
и 294
— насекомых 39, рис. 19
— членистоногих 21, рис. 7
Трахеолы 22
Триатомиды 48, 414, рис. 26
Триба 13, 14
Трипаносомы 145. 395, 414
473
Трипсы 47, рис. 26
Тритон 103, рис. 59
Трифоль см. вахта
Трихоцисты 91
Тромбидиоз 431, 432
Тростник 111, 248, 302, 310. рис. 62
Тростниковые болота 125, 306
Трубач 73, рис. 52
Трубки дыхательные насекомых 39,
рис. 2)
— яйцевые 41, рис. 21
Туберкулез 388
Туляремия 8, 144, 148, 333, 334, 402, 427,
448, 458, 462
Туркмения 129
— клегци 445. 448
— комары 257. 302, 304, 306,
307, 310, 319
— москиты 346, 349
Туфелька, рис. 52
Тянь-Шань, комары 303, 305
У
Угрица 140, 427, 428
Ужевник 247
Узбекистан, клещи 445, 448, 449, 460
— комары 304, 315, 318
Узел нервный см. ганглий нервный
Украина, 128, 131
— клещи 463
- колары 252, 309, 322
- москиты 350
Улитки 100
Урал, клещи 456
— комары 285, 301, 324, 332
— (река) 246. 247, 248. 253. 309
Ургенч, комары 315, 318, 319, рис. 193
Уруть 67, 105, 108, 240, 247, рис. 61
Усики насекомых 36
— Членистоногих 19
Уссурийский край 57, 128
----клещи 456
----комары 309, 311, 331
---- мокрецы 352
Устюг Великий, комары 291, рис. 171
Уховертка 46, рис. 25
Ф
Фазы Кристоферса 222, рис. 143
Фаланга 32, 153, 421, рис. 14
Фенология комаров 6, 288, 295, 304,
308. 311, 315, 318
.— москитов 343, 346
Фергана, клещи 442
ферганская долина 129
ферганские ворота, комары 132
физиология 12
фильтрация личинок комаров 228—231
филярии 145, 334
филяриидозы 324, 352, 353, 356
фитопланктон 84
Фолпиды 111, 112, 248
Фолликулы яичников комаров 222,
рис. 143
----москитов 344, 345
— — мух 378
474
Фолликулы яичников насекомых 41
Фотокинез 232, 254, 256
Фототаксис 69, 70, 232, 233, 255, 272,
303, 307, 328
X
Хабаровск, комары 311, 312
Хамидиэ 264
Харэвые водоросли 113, 114, рис. 64
Харьков, комары 296, 307
Харьковская область, комары 287
Хаузы 122, 249
Хачмас, комары 295
Хвощи 111
Хей лети ды 143
Хелицеры клещей 423, рис. 293
— паукообразных 21
Хищничество 139
Хламидомонады 87
Хлорофилл 85
Хоботные см. клопы
Хоботок комаров 179, рис. 106
— мух 359. рис. 227
Ход численности блох 406
----- клопов 417
— — комаров 296—299, 304, 308, 311,
315, 319, 324
----- москитов 343,
— — мух 385. 386
Холера 144, 338
Хомячки 445
Хорезм, комары 313
Хорион 42. 158, 367, 391
X роматофоры 85
Хромулина 96
Хрущ мучной 50, рис. 28
ц
Центр притяжения комаров 261, 262, 263
Центры сопряженные 262
Церки насекомых 37. рис. 17
Циклопы 29, 147, 250, рис. 54
Цуцугамуши 427, 431, 432
Ч
Чарджоу, комары, рие. 190
Частуха ИЗ, рис. 63
Челюсти верхние насекомых 36
членистоногих 19
Челюсти нижние насекомых 36
членистоногих 19
Черви дождевые 15
— круглые 145
— низшие 92
— паразитические 145
— трубочники 73, 99, 118, рис. 57
Черепахи 343, 448
Черноморское побережье, комары 161,
242. 254, 295, 301, 303, 304, 307
----- москиты 350
Чернотелки 50
Черные речки 129
Чесотка 427, 429, 430
— пакожниковая 426
Чесоточные клещи 142, 426, 427
Чесоточный зудень 142, 426, 427, 429,
430, рис. 297
Чешуйки комаров 177, рис. 103
Численность популяции комаров 296.
299, 320
Чита, комары 291, рис. 171
Членистоногие И, 12, 15
— способы переноса возбудителей 147
Чу (рока) 129
Чума, 8, 144, 148, 149, 402, 427, 433, 458
Чумные палочки, 147, 149
Ш
Шакал, 149, 349, 443, 445
Шелкопряд походный, рис. 29
Шершень или шершневая оса 152, рис. 30
Шея клещей 435, рис. 302
Шипики кутикулярные членистоногих 18,
рис. 2
— ложные членистоногих 18
Шитхала, комары 267
Шмель, рис. 83
Шов 165, 179, рис. 16, 93, 105
Шотландская вертячка овец 459
Шпанка 151, рис. 19 и 81
щ
[Цеки клещей, рис. 302, 304
Щеки насекомых 34, рис. 16
Щетинка усиков мух 359
Щетинки груди комаров 183, 184
-----мух Зэ8. 395
Щит головной насекомых 34
Щитснь 128
Щитки (склериты) 17
Щиток комаров 183
— мух 361, рис. 229
Щупальца губные насекомых 36, рис. 16
Щупики блох 403
—'комаров 179—181
— москитов 335
— мух 360
— таракана 419
9
Эвглена 96
Эвгленовые 89
Эвдорина 87
Экология 12
Экзокутпкула членистоногих, рис. 2
Экзофильные комары 296, 303, 308, 310
Экзохорион 158. 159, рис. 88
Экскреция личинок комаров 209
Эктопаразиты 140
Элодеиды 104, 105, 247, 317, рис. 61
Элодея 105, 247
Эмподий 52
Эндокутикула членистоногих, рис. 2
Эндопаразиты 140
Эндофильные комары 296, 314, 318, 331
Эпдохорион 158, рис. 88
Энтомология И, 12
Энцефалит Сан-Луи 432
— таежный клещевой 8, 114, 149, 427,
455
— механизм его распространения 455,
456
— его перепосчики 455
— — профилактика 456
— шотландский овечий 427, 456
— японский 332, 333
Энцефаломиэлит лошадиный 333, 445, 449
Эпикутикула комаров 210
— членистоногих, рис. 2
Эпителий членистоногих 16, рис. 2
Эпифитные водоемы 328
Я
Яды членистоногих 150, 154
Язва ашхабадская см. лейшманиоз
кожный
— пендинская см. лейшманиоз кож-
ный
Яичник комаров 221
— москитов 338
— мух 366
— насекомых 41, 42
Яйца вшей 410, рис. 286
— клещей 437, 451, 452, рис. 295
— клопа 416
— комаров 157
— москитов 339, рис. 207
— мух 367, 368, рис. 239
— насекомых 42
— слепней 354
Яйцевод насекомых 42
Яйцеклад мух 363, рис. 223
— насекомых 36, рис. 25 и 83
УКАЗАТЕЛЬ ЛАТИНСКИХ
НАЗВАНИЙ И ТЕРМИНОВ
А
Aedes 127, 128, 154, 159, 166, 183, 269
321, 322—329
— гонотрофическая гармония 323
— дыхальца 215, рис. 136
— откладка яиц 157, 322
— и перифитон 103
— сезонный ход численности 324
— строение яиц 159, рис. 89
— число генераций 324
— aegypti 57, 206, 264, 322, 323, 324,
328, 329, рис. 197
— — эпидемиологическое значение 144,
329
— alectorovi 322
— annulipes 324
— caspius 128, рис. 181
---caspius 323, 333
---dorsalis 322, 323, 324, 333,
рис. 194
— communis 128, 324
— cinereus 323, 324, 333, 334
— esoensis 58, 325, 326, 332,333, рис. 196
— excrucians 323, 334
— - flavescens 324
— galloisi 322
— geniculatus 307, 322
— japonicus 58, 326, 332, 333, рис. 196
— lutescens 333
— niveus 322
— pulchritarsis 307, 322
— togoi 58, 322, 323, 324, 326, 327,
332, 333, рис. 195 и 196
— vexans 323, 333
Aeschna см. коромысло
Acaridae 429
Acarina см. клещи
Acarus siro 429, 430, рис. 297
Acorus см. аир
Actinosphaerium, рис. 43
Agalenidae 33
Agrionidae см. люточки
Agriopidae 33
Agrostis stolonicens см. полевица земно-
водная
Algae см. водоросли
Alectorobius 149, 438, 442, 443, 444
— alactagalis 448
476
Alectorobius asperus 437, 445, 448,
рис. 305, 310 и 341
— cholodkovskii 448, рис. 310 и 311
— coniceps 442, 448, рис. 311
— nereensis 448, рис. 310 и 311
— tartakovskyi 149, 448, рис. 310
— tholozani papillipes 441, 442, 444,445,
рис. 309 и 310
Alisma см. частуха
Alveonasus 436, 437, 438, 442
— canestrini 437, 442, 448, 449, рис. 310
и 312
Amoebae см. амебы
Amphipoda см. бокоплавы
Anabaena, рис. 44
Annelides см. черви кольчатые
Anophelini, щиток, рис. 108
Anopheles 14, 57, 63, 65, 68, 71, 74, 148,
154, 157, 333
— imago, ампулы 225
— — враги 282
— — гетеротопность 155
— — гипопигии 186, рис. 115
— — голова 179, рис. 105
— — гонотрофическая гармония 156,
270
— — цикл 225, 269, рис. 145
— — доступность добычи 265, 266
— — желудок 213, 214, рис. 139
— жилкование крыла 185, рис. 111
— зародышевое развитие, рис. 22
— заражаемость плазмодиями 319,
320
— — значение ветра 260, 261
— — численность комаров 320
— — corpora allata 225, 146
— — определение возраста 282
— — пассивный занос ветром и транс-
портом 280
— — первоначальное дозревание са-
мок 259
— — половой аппарат 221
— — признаки новорожденной самки
259
. — — причины смерти 282
— — продолжительность пищеварения
270
Anopheles, imago, рассеяние путем повтор-
ных перелетов 278, 279
— — распределение в дневных убе-
жищах 256
— — реакция кожи на уколы 154
— — роль обоняния 260
— — — размещения людей и живот-
ных 266—269
— — семяприемник 226
— — способы охоты 321
— — способ переноса плазмодиев 148
— — степень связи с человеком 320—
321
— — суточный ход метеорологических
факторов 257
— — типы дневных убежищ 257
— — эпидемиологическое значение се-
зонности 320
— куколка 157, 175, 176, 207, 223, 224
— личинка 25, 70, 71, 78, 81, 96, 97,
162, 163, 165, 166, 170, 171, 172, 173
247, рис. 93
— — анальный сегмент 173
— — биоценозы нейстические 96, 97
— — брюшко 170
— — влияние активной реакции во-
ды 78, 79, 80
— — влияние выпаса скота 116
— — — света 236
— — 38— температуры 63, 64, рис. 35
— — — течения и колебания уровня
воды 234, 235
— — влияние водных растений 114,
244, 255
— — — водорослей 85, 86, 88, 89, 114
— — — возраста водоема 115
— — возрастные различия 173—175
— — враги 49, 81, 94, 100, 101, 103,
105, 250, 251, 252
— — голова 93, 95
— — ГРУДЬ 170
— — дыхание 207, 288
— — жизнь во временных водоемах 128
— — жизнь в литорали 119
— — значение волнения в водоемах
65, 234, 235
—• — значение продуктов распада ор-
ганических веществ 243
— — местообитания 118—125,129,130,
133, 135, 235, 240, 241, 242
— — осмотическое давление водной
среды 242
— — определение анофельной продук-
ции водоемов 253
— — питание 84, 86, 88, 89, 91, 92,
96, 97, 103, 244, 245
— — прикрепление 227, 228, 246
— — размер и состав пищи 230, 231
— — роль водной растительности
244—250, рис. 152
— — роль О2 и СО2 78, 244
— — ротовые части 165, 166, 167, 170,
рис. 95
— — сфагновые болота 114
— — фильтрация воды 228—231,
рис. 147
Anopheles, личинка, экскреция 209
— algeriensis 125, 136, 158, 159, 205,
214, 215, 237, 248, 300, 320, 321, 322,
рис. 86, 119
— bifurcatus 63, 125, 136, 158, 159,
205, 237, 244, 248, 296, 303, 304.
320—322, рис. 38, 86, 101, 104,
119, 136, 139, 181, 182, 183
— gambiae 320, 321
— hyrcanus 125, 126, 158, 205, 220, 235,
248, 257, 264,296, 308, 312, 320, 321,
322, 333, рис. 86, 123, 187 и 188
— lindesayi 205, 306, 320,322, рис. 122
и 131
— maculatus 303
— maculipennis 63, 64, 125, 126, 132,
158, 159, 160, 161, 228, 234, 235, 241,
248, 256, 257, 264, 282, 283, 284,
287, 289, 293, 300, 301, 321, 322,
333, рис. 35, 38, 86, 118, 133,
134, 135, 136, 137, 138, 140, 151,
153, 161, 162, 163, 166, 167, 171, 172,
173, 174, 179
— — водный баланс 256
— — вылет с зимовок 288, 289
— — гибель яиц в природе 226
— — диапауза 292, 293
— — corpora allata 225, рис. 146
— — состав дневок по стадиям пищева-
рения 273
— — степень связи с человеком 142,
320, 321
— — плодовитость 271
— — уход на зимовку 293, 295
— — часы влета 273, 274
— — часы вылета 273
— — круговорот на дневках 272, 273
— — личинка, роль загрязнения воды
75, 243
— — личинка, размеры 156, 157
— — atroparvus 71, 160, 161, 241,
242, 260, 264, 272, 283, 284, 286,
287, 288, 298, 301, рис. 176
— — labranchiae 160, 264
— — maculipennis 132, 160, 161, 193,
228, 241, 242, 264, 265, 275, 284,
287, 289, 293, 301
— — melanoon 160, 300, рис. 91
— — messeae 63, 64, 71, 75, 160, 161.
214, 233, 241, 242, 255, 256, 270,
283, 284, 286, 288, 289, 300, 321,
рис. 35 и 91
— — sacharovi 158, 162, 163, 220,
237,241,242, 255, 256, 260, 264,
284, 288, 291, 293, 295, 298, 299,
301, 302, 318, 320, рис. 86, 91,
177, 178, 180
— — subalpinus 160, 161, 241, 264,
284, 287, 288, 300, рис. 91.
— — typicus см. A. maculipennis
maculipennis
— marteri 205, 320, 322, рис. 121, 131
— minimus 303
— plumbeus 75, 128, 158, 159, 244,
307, 308, 320, 321, 322, рис, 86,
120, 184
477
Anopheles pulcherrimus 125,126.128,158.Calliphora
205, 214, 244, 257, 260, 261, 304, 309,
310, 311, 317,318, 319, 320, 321,322,
рис. 86, 123, 185, 186, 192, 193
— sun laicus 71
— superpictus 126, 128, 129, 132,
136, 157, 158, 205,215.227,228.234,
235, 241,244, 260, 278, 304, 308, 312—
317, 320, 321, 322, рис. 86, 125, 136,
189, 191, 192
Anoplura 47, 408
Antennae см. усики
Anurca 92, рис. 53
Aphaniptera см. блохи
Aphanizomenon flos aquae 73, 85, рис. 41
Apidae см. пчелы
Apis mellifica 152
Apus см. щитень
Apterygogenea см. первично-бескрылые
Aranea diadema рис. 15
Araneina см. пауки
Arachnoidea см. паукообразные
Arcella см. арцелла
Argas 437, 438, 442, 449
Argas persicus 449, рис. 303
— reflexus 449
Argasidae 145, 147, 426, 427, 433, 437,
438, 439, рис. 302, 304
Argyroneta aquatica см. пауки водные
Artomia salina 120
Arthropoda см. членистоногие
Articulata см. членистые
Asellus aquaticus см. водяной ослик
Asilidae рис. 31
Asplanchna 92, рис. 53
Astacus см. рак речной
Asterionella 84, рис. 43
Astigmata 428, 429, й ,
Attidae 33
В
Bacteria см. бактерии
Bacterium tularense 333, 458
Bdellonyssus 142, 149
— bacoti 433, рис. 301
Beggiatoa 73
Blatta orientalis 418, рис. 17, 18, 289, 290
Blattella gcrmanica 418
Blattodca см. тараканы
Bombus см. шмель
Boophilus 451, 455
— calcaratus 458
Bosmina 29, 96, рис. 55
Bra ch ion us см. панцырные коловратки
Branchipus 29, рис. 12
Brachyura см. крабы
Brach ycera см. короткоусые
Brachypoda 427. 428
Bursa copulatrix блох 404
— — клопа 415
Buthus, рис. 84
But om us см. сусак
Bythinia см. битиния
C
Calla palustris см. белокопытник
Calliphorinae 395
478
erythrocephala см. муха синяя
падальная
— uralensis см. муха зеленая падальная
Carabus см. жужелицы
Cardo таракана 418, рис. 290
Carex см. осоки
Carios 437, 438, 449
Ceratophyllum см. роголистник
Ccratophyllus fasciatus 402, 406
Ceratopogon 251
Ceratium hirundinella рис. 50
Chara рис. 64
Chaoborinae 163, 165, 166, 171, 251
Chaoborus см. коретры
Chelifer рис. 5
Chaethocampa см. шелкопряд походный
Cheyletidae см. хейлетиды
Chilopoda см. сороконожки
Chlamidomonas 87
Chlorophyceae см. водоросли зеленые
Chydorus 96, рис. 55
Chromulina 96
Chrysomclidae 150
Chr>sops см. златоглазик
Cbrysozona см. дождевка
Ciliata см. инфузории
Cimex lectularius клоп постельный 37,
140, 141, 142, 147, 414, 415, рис. 8,
287, 288
Cimicidae 48, 140, 414
Cladocera см. ветвистоусые
Closterium рис. 46
Coccinellidae см. божьи коровки
Coleoptera см. жуки
Comarum palustre см. сабельник
Conjugitae 86
Copepoda см. веслоногие
Corixidae см. гребляки
Corpora allata 225
Coseinodiscus 84, рис. 43
Cosmarium рис. 46
Coxa см. ляжка
Crustacea см. ракообразные
Ctenocephalides canis 402
— folis 402
Culex 122, 128, 132, 149, 157, 159, 254,
330, 333, 353, 357, рис. 89, 90, 107,
111, 113, 153
— bitaeniarbynchus57, 332, 333, рис. 196
— fatigans 206
— modestus 333
— pipiens 75, 220, 264, 328, 330, 335,
рис. 153, 181, 198
— — molestus 331, 333
— — pipiens 331
— tritaeniarhynchus 57, 326, 331, 332,
333, рис. 196
Gulicidae 57, 146, 155, 186, 188, 200,
202, 208, 335
Gulicinae 158, 188, 200, 202 (см. также
кома ры)
Culicini 157, 159, 179, 181, 184, 185, 221,
253, 254, рис. 89, 98, 107, 108, 111,
114, 117, 126, 129
— куколка 175, 176
— личинка 163,166, 167, 170, 171, 172.
173. 207, 208, 228, рис. 96, 98, 102
Culicoides 353, рис. 57
Cyanophyceae см. водоросли синезеленые
Cyclops см. циклоп
Cyclorhapha см. круглошовные
Cymbella рис. 49
Cynodon см. пальчатник
Cypridina 29, рис. 12
Cyprinodontiformes см. карпы зубастые
D
Daphnia~CM. дафния
— longispina 73, рис. 42
— pulex 96, рис. 55
Decapoda см. десятиногие раки
Decticus см. кузнечик
Dermestes lardarius см. кожееды
Dermestidae см. кожееды
Desmidium 84, рис. 43,
Desmidiaceae см. водоросли десмидиевые
Diaptomus см. диаптомы
Diatoma 73
Diatomaceae см. водоросли диатомовые
Difflugia см. дифлюгия
Diphyllobothrium latum см. лентец ши-
рокий
Diplopoda см. кивсяки
Diptera см. двукрылые
Dixa 61, 97. 163, 166, 170, 171, 208
рис. 56, 127
Dixinae 186, 188. 200
Demodex folliculorum 428, 429, рис. 296
Demodicoidea 427. 428
Dermacentor 449, 451
— andersoni 458
— marginatus 458
— nuttalli 457, 462, 463, рис. 315
— pictus 456, 458, 460, 461, рис. 317
— silvarum 455. 457, 463
Dermanyssus gallinae 432, рис. 300
Drassidae 33
Ductus ejaculatorius вши 410
— — клопа 416
Dytiscidae 49
Dytiscus см. плавунец
E
Echidnophaga gallinacea 403, 407
Echinochloa crus galli см. куриное просо
Elodea canadensis см. элодея
Entomostraca см. раки низшие
Ephemeroidea см. поденки
Equisetum см. хвощи
Eriocheir japonicus 100
Eudorina см. эвдорина
Euglena san guinea см. эвглена
— viridis рис. 50
Euglenaceae см. эвгленовые 29, рис. 12
Euphyllopoda см. настоящие листоногие
F
Fannia canicularis 360, 362, 367, 390,
391, рис. 232, 250 и 260
Femur см. бедро
Filariidae см. филярии
Finlaya 326
Forficula см. уховертка
Formicidae см. муравьи
Flagellata см. жгутиковые
Flagellum см. бич
Fragillaria 84, рис. 43
G
Galea таракана 418, рис. 290
Galeodes araneoides рис. 14
Gambusia affinis affinis см.
гамбузии
— — holbroocki см. гамбузии
Gamasoidea см. гамазоидные клещи
Gammarus 29, рис. 5
Gastropoda см. улитки
Gastropbilidae см. оводы желудочные
Gastrophilus intestinalis см. >вод же-
лудочный
Gerris см. водомерка
Gloeotrichia echinata рис. 44
Glossa таракана 419, рис. 290
Glossina palpalis см. муха це-це
Glyceria 249
Gyrinidae см. вертячка
Н
Haemopis см. пиявка ложноконская
Haemophysalis 451, 458, рис. 7
concinria 455, 457, 46 0, рис. 316
— numidiana 454, 455
— punctata 457, 459, 460, рис. 315
Haemosporidia см. споровики кровяные
Haustellum 359, рис. 227
Heleidae см. мокрецы
Heleocharis см. ситняг
Heliozoa см. солнечники
Hemimetabola см. насекомые с неполным
превращением
Heteroptera 24, 48, 103, 146, 414, рис. 26
Heterostigmata 428
Hippoboscidae см. мухи кровососки
Hippobosca equina 401, рис. 279, 280
Hister см. карапузики
Holometabola см. насекомые с полным
превращением
Homoptera 48
Hunterellus hookeri, рис. 30
Hyalomma 451, 458
— asiaticum 455
— marginatum 453, 456
— scupense 453
Hydra см. полипы
Hydrocharis morsus ranae см. лягушечник
Hydrodyction см. сеточка водяная
Hydrophilidae см. водолюбы
Hydrophilus см. водолюб
Hydrotaea 381
Hymenoptera см. перепончатокрылые
Hypoderma bovis см. овод кожный
Hypoderm.dae 399
Hypopharynx блох 402
— мух 359, рис. 227
— таракана 419
Hypostoma клещей 423. рис. 293
479
I
Ichneumonoidea см. наездники
Imago членистоногих 27
Insecta см. насекомые
Isopoda см. равноногие
Ixodes 451
— holocyclus 458
— persulcatus 8, 149, 453, 455, 456
459, рис. 314
— ricinus 456, 458, 459, рис. 291, 292,
293 294 295
Ixodidae 8. 141, 144, 146, 433, 437, 449,
451, 452, 453, 454, 455, 456, 457,
458. рис. 313
Ixodoidea 424. 425, 428, 433
L
Labium блох 403
Labrum epipharynx см. надглоточнпк мух
Lacinia таракана 418, рис. 290
Laelaps 433
Larva членистоногих 28
Larvivoridae 395
Lathrodectes tredecimguttatus см. кара-
курт
Lemna minor см. ряска малая
— trisulca см. ряска трехдольчатая
Leishmania см. лейшмании
— donovani 349
— tropica 349
Lepidoptera см. бабочки
Leptidae, рис. 31
Leptinus рис. 27
Leptodora 96, 118, рис. 55
Libellula 100
Limnadia 29
Limnanthemum nymphaeoides см. ужовник
Limnea см. прудовик
Linyphiidae 33
Liponeura 136, рис. 80
Lithobius 29, рис. 13
Locusta см. кобылки
Lopadorhynchus, рис. 5
Lucilia sericata 139, 395, 397, рис. 27&
Lycosidae 33
Lumbricidae см. дождевые черви
Lycosa, рис. 6
Lytta vesicatoria см. шпанка
M
Macrura см. длиннохвостые раки
Malacostraca см. раки высшие
Mallophaga см. пухоеды
Mandibulae блох 402
Mansonia 159, 166, 190, 202, 332, 333,
рис. 89, 129, 199
Mantis см. богомол
Maxillae блох 403
Megalopyge, рис. 82
Meloe proscarabaeus см. майка
Meloidae см. нарывники
Melosira varians 73, рис. 49
Mentum таракана 419, рис. 290
Menyanthes trifoliata см. вахта
Melophagus ovinus см. рунец овечий
480
Mesosternum 184
Mesothorax см. среднегрудь насекомых
Mesostigmata 428, 432
Metathorax см. заднегрудь насекомых
Micrasterias, рис. 46
Microcystis 85
Moina 29, 96, рис. 55
Mollusca см. моллюски
Muscidae 358, 359, 382
Musca domestica 37, 142, 147, 359, 360,
362—367, 370—389, рис. 225—236,
237—257, 263
— — vicina 360, 362, 367, 370, 371,
372, 381, рис. 253, 260
— sorbens см. муха базарная
Muscina stabulans (см. также домовая
муха) 360, 362, 363, 367, 390, 392,
рис. 232, 261, 262,263, табл. 17
Mylabris, рис. 81
Myriapoda см. многоножки
Myriophyllum см. уруть
Myzomyia 170
N
Naucoris см. плавт
Nauplius, рис. 54
Navicula 73, рис. 41
Nematocera см. длинноусые
Nematodes см. черви круглые
Neuroptera см. сетчатокрылые
Notholca longispina 73, 84, 92, 118,
рис. 42
Notodromas 29, 97, рис. 56
Notonecta см. гладыш
Nymphaea см. кувшинки.
Nuphar см. кубышки.
О
Odonata см. стрекозы
Ochlerotatus 324
Oedogonium 88, рис. 48
Oestrus ovis см. овод овечий
Oligochaeta см. малощетинковые черви
Ophyra 381
Opiliones см. сенокосцы
Oscillatoria 73, рис. 40
Ostracoda см. ракушковые
Ornithodorini см. Alectorobius
Ornithodoros 437
— alactagalis 448
— canestrinii 448
— cholodkovskyi 448, рис. 311
— coniceps 448
— lahorensis 437, 448
— nereensis 448, рис. 311
— papillipes 444
— tartakovskyi 448
— verrucosus 445
Orthorhapha см. прямошовные
Orthopodomyia 190, 202, рис. 89
— pulchripalpis 159, 307, рис. 129
Orthoptera см. прямокрылые
P
Palpi maxillares блох 403
Paraglossa таракана 419, рис. 290
Parasitoidea 428, 432, 433
Paragonimus см. сосальщики
Paederus fuscus 49, 151, рис. 81
Pandorina см. пандорина
Paramaecium, рис. 52
— putrinum рис. 39
Panstrongylus megistus рис. 26
Pediastrum рис. 43
Pediculidae 408, 312
Pediculoides ventricosus 428
Pediculus humanus capitis 408 412,
рис. 284
— — corporis 408, 412, рис. 284
Pericoma 136, рис. 80
Peridineae 89
Phlebotomus см. москиты
— argentipes 350
— graekovi 341
— caucasicus 341, 342
— chinensis 341, 342, 343, 350, рис. 202 и
210
— clydei 341
— kandelakii 337, 341, 350, рис. 202
— keshisbiani 341, рис. 202
— major 336, 341, 350, рис. 202
— minutus 341, рис. 211
— — var. arpaklensis 341
— papatasii 336, 337, 341, 342,
349, рис. 208 и 212, табл. 212
— perfilievi 341
— — transcaucasicus 341
— perniciosus var. tobbii 341, 350,
рис. 202
— sequens 341
— sergenti 341, 342, 349,. рис. 209
— — var. alexandri 241
— — — mongolensis 241
— smirnovi 341
— sogdianus 341, рис. 203
— squamipleuris 341
— sumbaricus 341, рис. 203
— wenyoni 341
Pholcidae 33
Phorm i a terraenovae см. весенняя падаль-
ная муха
Phragmites см. тростник
Phryganea рис. 57
Phthyrus pubis 408, 412, рис. 284
Payllopoda см. листоногие
Pinnularia рис. 49
Plop h i la casei см. муха сырная
Piophilidae 358
Piscicola см. пиявка рыбная
Plasmodium см. малярийный паразит
— falciparum 306, 316, 317, 319, 320
— malariae 306
— vlvax 306, 320
Planorbis см. катушка
Plecoptera см. веснянки
Poeciliidae 251
Polyarthra 92, рис. 53
Polygonum amphibium см. гречиха водя-
ная
Polytoma uvella 73
Potamobius см. рак речной
Potamogeton см. рдесты
— crispus 108, рис. 61
— lucens 108, 247, рис. 61
31 Учебник медицинской энтомологии
Potamogeton natans 108, рис. 61
— pectinatus 108, рис. 61
— perfoliatus 108, 247, рис. 61
Potamon fluviatilis 29, 100
Porcellio см. мокрицы
Praepupa см. предкуколка мух
Prostigmata 428, 431
Prothorax см. переднегрудь
Protococcales см. водоросли протокок- *
ковые
Protomonadina 145, 349
Protozoa см. простейшие
Proventriculus см. преджелудок мух
Psychodidae 335
Pseudoscorpiones см. лжескорпионы
Pseudopodiata см. корненожки
Pterygogenea см. первичнокрылатые
Pulex irritans 402, 405, 406, рис. 281
Pulmonata 100
Puparium см. пупарий мух
К
Ranunculus aqua Lieus см. лютик водяной
Reduviidae 282
Ricketsia см. риккетсии
Rhinoestrus purpureus см. овод лошади-
ный
Rhipicephalus 451, 458
— sanguineus 457, 463, рис. 319
— schulzei 458
Rhynchota см. хоботные
Rhyzopoda см. корненожки
Rostrum 359
Rotatoria см. коловратки
Rotifer 73, 92, рис. 40
S
Sagittaria см. стрелолист
Sarcopbaga carnaria, рис. 8
Sarcoptoidea 428
Scapl olebris 29, 96, рис. 56
Scarabaeidae см. жуки пластинчатоусые
Schizotrypanum cruzi 145, 414
Scirpus см. камыш
Scolecida, см. черви низшие
Scolopendra см. сколопендры
Scorpionoidea см. скорпионы
Scutellum см. щиток
Scutigera 29, рис. 13
Simuliidae 7, 146, 335
Simulium см. мошки
Siphunculata см. вши
Solifugae см. фаланги
Salvinia natans см. сальвиния
Sparganium см. ежеголовка
Sphagnum см. сфагновый мох
Sphacrotilus natans, рис. 39
Sphex, рпс. 23
Spirochaeta rccurrentis 147, 413
Spironernacea см. спирохеты
Spirodella polyrrhyza см. многокоренник
Sporozoa см. споровики
Spirogyra 86, рис. 45
Staphylinidae 49, 151, рис. 81
Stigeoclonium 73, рис. 40
Stipes таракана 418, рис. 290
481
Stegomyia 328 (см. также Aedes aegypti)
Sten tor coeruleus см. трубач
Stomoxys calci trans 360, 362, 364, 367,
392, 393, рис. 228, 237, 264, 265
Styli таракана 419
Stylonichia рис. 52
Submontum таракана 419, рис. 290
Surirella рис. 49
Sutura см. шов
Symphoromyia, рис. 31
Synchaeta 92, рис. 43 и 53
T
Tabanidae см. слепни
Tabanus bovinus см. слепень
Та bellaria, рис. 49
Tanypus, рис. 57
Та rso ne mo idea 428
Tarsus см. лапка насекомых
Tendipedidao см. комар толкунчик
Ton di pcs plumosus см. мотыль
Tenebrio molilor см. хрущ мучной
Tenebrionidao см. чернотелки
Thcobaldia 157, 159, 166, 184, 189, 202
рис. 89, 107, 109, ИЗ, 129
Tbomisidae 33
Thrips, рис. 26
Th\sanoptera 47
Tibia см. голень насекомых
Tinea pel ionol la см. моль
Tipula см. комар-долгоножка
Tipulidae 53, рис. 31
Тгара па tans см. водяной орех
Triatomidae 48, 414, рис. 26
Triarlhia 84
Trie hop tera см. ручейники
Triturus vulgaris см. тритон
— crislalus см. тритон
Trochanter см. вертлуг насекомых
Trombidiidac см. клещи красиотелкп
Trombidoidea 428, 431
Trombicula akamushi 431, рис. 299
Trypanosoma см. трипаносома
— gambie nsc 395
Trypanosomidae 145, 349
Tyroglyphidae см. клещи мучные
Tyroglyphoidca 428, 430
Tyroglyphus farinae 430, 431, рис. 298
Typha см. рогоз
Tubifex см. черви трубочники
U
Ulotrix, рис. 48
Uranofaenia 184, 188, 202, рис. 112, 129
Utricularia см. пузырчатка
V
Vallisncria spiralis 108, рис. 62
Vespa crabro см. шершень
Vespidae 51, рис. 30
Vcnnipsylla alacurt 407
— dorcadia 407
Vivi para см. живородка
Vorticella см. сувойка
Volvocalcs см. водороелп вольвоксовые
Volvox см. вольвокс
w
Wohlfahrtia magnifica см. муха
Вольфарта
X
Xenopsylla cheopis 402, 406
Z
Zygnema 86, рис. 45
Zygnemaceae см. водоросли зигпемовые
ОГЛАВЛЕНИЕ
Предисловие........................................................ 3
Введение. Развитие медицинской энтомологии в нашей стране .... Ь
Часть 1. ОБЩАЯ ЭНТОМОЛОГИЯ
Глава I. Предмет и значение энтомологии............................... 11
Глава II. Энтомология как часть биологии. Зоологические названия . . 12
Глава III. Основные особенности членистоногих (Arthropoda) . 15
1. Положение членистоногих в системе животных..................... 15
2. Строение кожного покрова как основная особенность членистоногих . . 16
3. Рост и линька членистоногих. ... 18
4. Расчленение тела членистоногих................................. 19
5. Органы пищеварения, дыхания, выделения и кровообращения членисто-
ногих ............................................................ 21
6. Нервный аппарат и органы чувств членистоногих.................. 23
7. Половой аппарат, размножение и развитие членистоногих.......... 27
Глава IV. Классы членистоногих........................................ 28
Глава V. Наружное строение насекомых .... 34
1. Расчленение тела насекомых ... 34
2. Придатки тела насекомых........................................ 36
Глава VI. Внутреннее строение насекомых .... 37
1. Пищеварительный аппарат...................................... 37
2. Дыхательный аппарат............................................ 38
3. Полость тела и кровеносный аппарат............................. 39
4. Половой аппарат насекомых...................................... 41
Глава VII. Развитие насекомых......................................... 42
Глава VIII. Подклассы и основные отряды насекомых..................... 45
Ч а с т ь 2. ГИДРОБИОЛОГИЯ
Глава I. Введение..................................................... 57
1. Определение гидробиологии. Ее значение. Медицинская гидробиоло-
гия ............................................................... 57
2. Вода как среда обитания организмов............................. 60
3. Свойства воды, делающие ее благоприятной средой для жизни .... 61
31*
483
Глава II. Взаимоотношения между водными организмами и факторами вод-
ной среды*............................................................ 63
1. Предварительные замечания......................................... 63
2. Взаимоотношения между водными организмами и движением воды .... 64
3. Взаимоотношения между водными организмами и температурой. . . 65
4. Отношение водных организмов к свету .............................. 69
5. Взаимоотношения между водными организмами и растворенными в воде
неорганическими веществами........................................... 70
6. Взаимоотношения между водными организмами и органическими веще-
ствами воды.......................................................... 72
7. Взаимоотношения между водными организмами и растворенными в воде
газами.............................................................. 75
8. Взаимоотношения между водными организмами и активной реакцией
воды................................................ . . 78
9. Взаимоотношения водных организмов................................. 80
Глава III.; Основные типы водных биоценозов и характерные для них
животные и растения................................................... 82
1. Основные типы пресноводных биоценозов............................. 82
2. Биоценозы толщи воды (плапктические биоценозы).................... 82
а) Условия существования, особенности и состав планктона........ 82
б) Обзор характерных представителен пресноводного планктона ... 84
1) Синезеленые водоросли (тип Cyanrphyceae).................... 84
2) Зеленые водоросли (тип Chl< rophyceae) ..................... 85
3) Диатомовые водоросли (тип Diatomaceae). . . 88
4) Жгутиковые (Flagellata)..................................... 89
5) Корненожки.................................................. 90
6) Инфузории................................................... 90
7) Ко говратки (класс Rotatoria)............................... 92
8) Ракообразные (класс Crustacea).............................. 92
3. Биоценозы поверхностной пленки воды (пейстические биоценозы) ... 96
4. Биоценозы дна (бентические биоценозы)............................. 97
а) Условия существования, особенности и жизнедеятельность бентоса 97
б) Обзор характерных представителей пресноводного бентоса . 99
1) Малощетинковые черви (класс Oligochaeta)................... -99
2) Пиявки (класс Hirudinea).................................... 99
3) Личинки комаров-точкунчиков (семейство Tendipedidae) . . 99
4) Личинки ручейников (отряд Trichoptera)..................... 1Э0
5) Личинки стрекоз (отряд Odonata) ........................... 100
6) Высшие раки (подкласс Malacostraca класса Crustacea) . . . 100
7) Мотлюски (тип Mollusca).................................... 100
в) Бептолектические и понтические животные. . . . 101
1) Жуки-плавунцы (семейство Dytiscidae)....... 101
2) Жуки-водолюбы (семейство Ilydrophilidae) .... 103
3) Водяные клопы...................... ..................... 103
г) Перифитон................................................. 103
5. Высшие водные растения................................. ... 103
а) Предварительные замечания....................................103
б) Физиономические группы водных растений и их характерные пред-
ставители .......................................................104
в) Взаимоотношения между видными растениями и другими организ-
мами ............................................................114
6. Старение водоема и связанная с этим смена его биоценозов.........114
484
Глава IV. Основные типы материковых водоемов .... 116
1. Предварительные замечания . 116
2. Водоемы................... ................... 117
а) Озера.............................. .... 117
б) Пруды.................................................... 119
в) Болота и заболоченности.................................... 124
г) Временные водоемы................... ... 127
3. Водотоки.......... ... 129
а) Реки и ручьи............................................... 129
б) Ключи, или родники....... 135
в) Искусственные водотоки........ 136
Часть 3. МЕДИЦИНСКАЯ ЭНТОМОЛОГИЯ Vх
Раздел А. Введение в медицинскую энтомологию..................... .... 139
Глава I. Членистоногие как паразиты человека ... . . 139
Глава II. Членистоногие как переносчики болезней...................... 144
Г л ав аШ. Ядовитые членистоногие..................................... 150
Раздел Б. Комары (семейство Culicidae)............................... 155
Глава I. Общие замечания.............................................. 155
Глава II. Наружное строение неполовозрелых фаз комаров................ 156
1. Цикл развития комаров........ 156
2. Яйцо......................................................... 157
а) Способы огкладки яиц....................................... 157
б) Наружная морфология яиц Culicinae........................ 158
1) Яйца Anopheles........................................... 158
2) Яйца Culicini............................................ 159
в) Отличия подвидов Anopheles maculipennis но яйцам . . . 162
3. Личинка........................................................ 162
а) Наружное строение личинки.................................. 162
б) Возрастные различия личинок Anopheles.................... . 173
4. Куколка..................................................... - 175
5. Вылупление комара ................ . . . 176
Глава III. Наружное строение взрослого комара......................... 177
1. Голова........................................................ 179
2. Строение ротовых органов .... 179
3. Работа ротовых органов .... 181
4. Органы чувств головы....... 182
5. Грудь и ее придатки . . . 182
6. Брюшко......................................................... 185
Глава IV. Систематика комаров 186
1. Общие замечания................................................ 186
2. Определение подсемейств Culicidae по взрослым комарам .... 188
3. Определение триб и родов подсемейства Culicinae, распространенных в
СССР, по взрослым комарам......................................188
4. Определение видов рода Anopheles, встречающихся в СССР, но взрослым
комарам............................................................190
30* 485
5. Определение видов рода Anopheles по гипопигиям самцов ... 191
6. Описание видов рода Anopheles (взрослые комары).............. .192
7. Определение и характеристика подсемейств семейства Culicidae по ли-
чинкам ...........................................................199
а) Таблица для определения подеемеисчв семейства Culicidae ио ли-
чинкам ........................................................199
б) Характеристика подсемейств семейства Culicidae по личинкам . 200
8. Определение триб и родов подсемейства Culicidae по личинкам . 202
9. Определение видов рода Anopheles по личинкам .........202
Глава V. Внутренняя анатомия и физиология личинок и куколок комаров 206
1. Пищеварительный аппарат........................................206
2. Дыхательный аппарат ... 207
3. Органы выделения . . 208
4. Кровеносный аппарат.......................................... 209
5. Жировое тело................................................. 209
6. Центральный нервный aunapai 210
7. Половой аппарат............................................... 210
8. Имагинальные диски .... 210
Глава VI. Внутренняя анатомия и физиология взрослых комаров . . . 210
1. Кутикула и внутренний скелет...... 211
2. Мускулатура....................................................211
3. Пищеварительный аппарат....................................... 211
4. Трахейная система и дыхательный аппарат .... 214
5. Выделительные органы.......................................... 217
6. Кровеносный аппарат........ 217
7. Жировое тело........ 219
8. Нервный аппарат........ 220
9. Половой аппарат . . . 221
Глава VII. Экология водных фаз Anopheles maculipennis . . . 226
1. Условия существования яиц......................................226
2. Приспособление личинки к жизни в поверхностной пленке воды. Роль
субстрата........................................ . . 227
3. Питание личинки Anopheles . . 228
а) Способы питания........................... 228
б) Механизм действия ротового аппарата....... 229
в) Размеры частиц, вылавливаемых личинкой Anophelo . z . . 230
г) Состав пищи................................................231
4. Движение личинок. Двигательные реакции. Движения куколок .... 231
5. Влияние ветра, волнения, течения и колебаний уровня воды. Влияние
глубины водоема............................................. ... 234
6. Влияние света....... 235
7. Влияние температуры.......................................... 236
а) Верхний и нижний температурный предел жизни . . . 236
б) Влияние температуры на скорость развития . . 238
в) Температурный режим личиночных биотопов . . 239
8. Влияние химических факторов среды на личинку и куколк> . . 241
а) Влияние растворенных солей .... 241
б) Влияние органических веществ воды . . . 243
в) Значение степени насыщения воды кислородом 244
г) Действие активной реакции воды на личинок .... 244
486
9. Роль водной растительности в жизни личинок . 244
а) Роль растительности.................................. . . . . 244
б) Роль основных физиономических типов растительности ...........246
1) Лемниды — свободно плавающие, неукореняющиеся растения . * 246
2) Укореняющиеся растения.................................. , 247
3) Сезонные перемещения центров обилия личинок................249
4) Выводы для практики...................................... 249
в) Влияние сукцессионного возраста водоемов на анофельную про-
дукцию ........................................................250
10. Враги личинок.................................................. 250
11. Личиночная смертность у Anopheles maculipennis .... 252
12. Анофельная продукция водоемов................................... 253
Г лава VIII. Экология Anopheles maculipennis—взрослого комара . . . 253
1. Поведение комара и действие на него внешних факторов..............253
а) Посадка, геотаксис, движения..................................253
б) Действие света.............................................. 254
в) Действие температуры и влажности. Дневные убежища ...... 255
2. крыление и первые дни имагинальной жизни ... ...............259
3. Поиски добычи и нападение.........................................2G0
а) Поиски добычи.................................................260
б) Пищевые предпочтения и внешние факторы, влияющие па выбор добычи 264
в) Нападение на добычу...........................................269
4. Пищеварение и созревание яиц .... 269
а) Вторая фаза гонотрофического цикла............................269
б) Биологические типы убежищ; круговорот комаров на дневках . . 272
в) Некоторые эпидемиологические выводы...........................274
5. Откладка яиц................................................. ... 275
6. Дальность полета........ . . .... 277
7. Продолжительность жизни ... ............280
8. Зимовка......................................................... 284
а) Общие замечания...................................... . . . . 284
б) Зимовка A. maculipennis messeae . . .....................284
в) Зимовка A. maculipennis atroparvus .... ..................286
г) Зимовка других подвидов.......................................287
Глава IX. Экология A. maculipennis как вида в целом ................... 288
1. Фенология........................................................288
а) Служба фенологии на малярийных станциях....................288
б) Весенняя часть годового цикла A. maculipennis.................288
в) Осенняя часть годового цикла A. maculipennis................ 292
г) Число поколений.........................•................. . 295
2. Численность популяции и ее i идовоа ход........................ 296
3. Географическое распространение A. maculipennis и его подвидов . . . 299
Глава X. Экология и распространение других видов Anopheles...............302
1. Anopheles bifurcatus (L).........................................302
2. Anopheles algeriensis Theob......................................306
3 и 4. Anopheles lindesayi Giles и A. marteri bene vet..............306
5. Anopheles plumbeus Theob..................................... • 307
6. Anopheles hyrcanus Pall........................................ 308
7. Anopheles superpictus Grassi.................................... 312
8. Anopheles pulcherrimus Theob. . .............................317
487
Глава XI. Сравпепие эпидемиологического значения отдельных видов
Anopheles СССР .........................................................319
Глава XII. Некоторые важнейшие виды немалярийных комаров СССР . . 322
а) Род Aides..........................................................322
1) Aides (Ochlerotatus) communis Deg................................324
2) Aides (0.) caspius dorsalis Meigen...............................324
3) Aides (Aides) cinereus Meig......................................324
4) Aedes (Aides) esoensis Jam..................................... 325
5) Aides (Finlaya) japonicus Theob....... 326
6) Aides (Finlaya) togoi Theob. . - . . 326
7) Aides (Stegomyia) aegypti (L.) .... ... 328
б) Род Culex .........................................................329
1) Culex pipiens L...................................................330
2) Culex tritaeniorhynchus Giles.................................. 331
в) Род Mansonia.................... ... ...............332
г) Комары — переносчики болезней ... .... 332
P а з д e л В. Другие семейства кровососущих низших двукрылых (Orthorhapha) 335
Глава I. Москиты (род Phlebotomus, семейство Psychodidae) ...... 335
1. Строение.............................................................335
2. Развитие........................................................... 339
3. Систематика............. ............................................340
4. Экология..............................................................341
5. Эпидемиологическое значение.........................................348
6. Географическое распространение.......................................350
Глава II. Мошки (семейство Simuliidae).................................... . 350
Глава III. Мокрецы (семейство Heleidae) .... . . 352
Глава IV. Слепни (семейство Tabanidae).................................... . 353
1. Строение и систематика слепней........................................353
2. Биология и экология слепней...................................... . . 354
Раздел Г. Высшие двукрылые/ или круглошовные (подотряд Cyclorhapha) 357
Г л а в а I. Предварительные замечания................................357
Г л а в а II. Семейство Piophilidae.................................. 358
Глава III. Настоящие мухи (семейство Muscidae) . . . 358
1. Наружное строение............................................. 358
а) Голова и ее придатки...................................... 358
б) Грудь и ее придатки.......................». . .361
в) Брюшко и его придатки..................................... 363
2. Внутреннее строение........................................ . . 364
а) Пищеварительный аппарат................................... 364
б) Дыхательный аппарат . ................................... . 365
в) Органы кровообращения..................................... 365
г) Нервный аппарат.......................................... . 365
д) Половой аппарат........................................... 366
3. Определительная таблица родов семейства Muscidae, имеющих медицин-
ское значение (в пределах СССР)................................367
4. Род Musca L.................................................. 367
а) Комнатная муха (Musca uomestica L. • . . . 367
488
1) Подвиды комнатной мухи...................................... 367
2) Стадии развития............................................. 367
3) Экология личинки............................................ 370
4) Экология предкуколки........................................ 372
5) Экология куколки............................................ 374
6) Вылупление имаго............................................ 374
7) Экология взрослых мух . .............................. 375
8) Годовой цикл вида........................................... 384
б) Эпидемиологическое значение мух............................... 387
в) Базарная муха (Musca sorbens W.—D.)............................389
5. Род Fannia R.—D. Малая комнатная муха (Fannia canicularis L.) . . . 390
в. Род Muscina R.—D. Домовая муха (Muscina stabulans Flln.)...........391
7. Род Stomoxys. Осенняя жигалка St. calcitrans (L.)................. 392
8. Муха цеце (род Glossina Wied.).................................... 395
Глава IV. Семейство Larvivoridae • ...................395
1. О предел ительпа я таблица родов подсемейства Calliphorinae, имеющих ме-
дицинское зпачепие ••••« ............................................ 395
2. Род Phormia R.—Р................................................ 396
3. Синие падальные мухи (род Calliphora R.—D.).......................396
a) Calliphora erythrocephala Meig.................................396
b) Calliphora uralensis Vill......................................397
4. Зеленые падальные мухп (род Lucilia R.— D.) (Lucilia sericata Meig.) 397
5. Подсемейство Sarcophaginae. Вольфартова муха (Wohlfahrtia magnfica
Sehin)................................................... . • 397
Глава V. Желудочные оводы (семейство Gastrophilidae) . . . 398
Глава VI. Кожные оводы (семейство Hypodermidae) . 399
Глава VII. Полостные оводы (семейство Oestridae)......................... 400
Глава VIII. Кровососки (семейство Hippoboscidce) . 401
Раздел Д. Блохи (отряд Aphaniptcra)...................................... 402
1. Предварительные замечания..............:........................ . 492
2. Наружное строение .. 492
3. Внутреннее строение . . .......... 404
4. Развитие.......................................................... 494
5. Экология........................................................ 495
6. Различные формы паразитирования среди блох................... 407
Разд ел Е. Вши (подотряд Siphunculata отряда Anoplura)...................498
1. Предварительные замечания....................... . . 408
2. Наружное строение................................................. 408
3. Внутреннее с роение................................................409
4. Развитие...........................................................410
5. Экология....................................................... . 411
6. Систематика вн ей человека.........................................412
7. Эпидемиологическая роль вшей рода Pediculus........................413
Раздел Ж. Клопы (подотряд Ilemiptera отряда Rhynchota)....................414
1. Предварительные замечания..........................................414
2. Наружное строение посюльного клопа (род Cimex).....................415
489
3. Внутреннее строение постельного клопа .... 415
4. Развитие постельного клопа....................................... 416
5. Экология постельного клопа .... ... 416
Раздел 3. Тараканы (подотряд Blattodea отряда Orthoptera)........... 418
Раздел И. Клещи (отряд Acarina класса Arachnoidea) . . 421
Глава I. Понятие о клещах............................................... 421
1. Общие замечания.................................................. 421
2. Строение клещей.................................................. 422
3. Биология, экология и распространение клещей . 425
4. Значение клещей для человека..................................... 427
5. Классификация клещей............................................. 427
Глава II. Важнейшие в медицинском отношении клещи фауны СССР . . . 428
1. Чесоточные клещи................................................. 428
а) Железница (Demodex folliculorum) . 428
б) Чесоточный зудень (Acarus siro L.) . . . . 429
2. Амбарные клещи (надсемейство Tyroglyphoidea) . 430
3. Краснотелки (семейство Trombidiidae.............................. 431
4. Клещи надсемейства Parasitoidea . . . 432
5. Клещи надсемейства Ixodoidea .... 433
а) Характеристика надсемейства.................................. 433
б) Аргасовые клещи (семейство Argasidae) . . . 433
1) Наружное строение аргазид .... 433
2) Роды семейства ArgasiJae .... 437
3) Жизненный цикл аргазид.................................... 438
4) Типы местообитания (убежища) аргазид . . . 440
5) Характер паразитизма аргазид.............................. 440
6) Распространение аргазид в СССР............................ 442
7) Клещевой возвратный тиф .... ... 442
8) Обзор видов аргазид фауны СССР........... 444
в) Иксодовые клещи (семейство Ixodidae) . . . 449
1) Наружное строение иксодид .... 449
2) Роды семейства Ixodidae . . . 451
3) Жизненный цикл иксодид......... 452
4) Распространение иксодид СССР . . . 455
5) Роль иксодид в сельском хозяйстве . . . 455
6) Медицинское значение иксодид.............................. 455
7) Важнейшие иксодовые клещи фауны СССР...................... 458
Список рекомендуемой литературы..................................... 464
Алфавитный указатель.................................................465
Редактор Л. Б. Левинсон
Техн. редактор А. Ф. Аксенов. Коррек-
торы Т. А. Михгра и В. Ф. Ксрмер
A106G7. М-У-42. Подписано к печати
27(Х1 1948 г. Печ. л. 30,75. Уч.-изд. л.
41,35 3nai or в 1 печ. л. 59000. Формат
бум. 70X108,16 Зан. 81 74. Цепа 10 р. 30шк.
Переплет 1 р.
Набрано в Первой Образцовой типсх
графии имени А. А. Жданова треста
<Полиграфкнига» ОГИЗа при Совете
министров СССР.
Москва, Валовая, 23.
Отпечатано в типографии
Государственного издательства
медицинской литературы,
Москва. Ногатинское шоссе, д. 1
Заказ 992.
СПИСОК ОПЕЧАТОК
Страница Строка Стоит Надо
77 19 снизу амбузия гамбузия
118 12 снизу Leptodoa Leptodora
228 21 сверху „микроводоемах* населяющих „микроводое- мы44
304 рис. 181 Вылет 1-го поколения в W 15 мая, а на рисунке ун 1оскве в 1946 г. произошел :азано на месяц позднее.
343 4 снизу в конце в начале
367 13 сверху 1(10) 1(6)
109 Объяснение к рисунку А—платяная вошь (Pedi- culus humanus corporis); Б— головная вошь(Р. hu- manus capitis А—головная вошь (Pedicu- lus humanus capitis); Б платяная вошь (Р. huma- nus corporis
430 20 сверху в качестве в целях
430 26 сверху называемая вызываемая
/чебник медицинской энтомологии