
































































































































































































































Автор: Елинов Н.П.
Теги: биология клетки и субклеточных частиц цитология микробиология химия биология
ISBN: 5-06-000089-3
Год: 1980




Текст
ЭВОЛЮЦИЯ
МИКРО-
ОРГАНИЗМОВ
Нефотосинтетические
аэробные
эукариоты
* Animalia
► Protozoa
► Mycetalia
Дробные
пркариоты
Анаэробные
протоэунариоты
ЛУИ ПАСТЕР
(1822-1895)
Фотосинтетичские
анаэробна
пронаркон
Зеленые и пурпурные
► фотобактерии
—►Цианобактерии
Фотосинтетические
аэробные
эукариоты
Фотосинтетические
анаэробные
эукариоты
Аэробные
’ бактерии
11 Plantes
! Algae
Неклеточные
формы
1
Предковые
анаэробные
пронариоты
wAh
а
АрхеЗ
4 млрд, лет назад
- Фаги ——
—►Вирусы
около
3 млрд, лет назад
.около
^1(5 млрд, лет назад
ЛИСТОК СРОКА ВОЗВРАТА
КНИГА ДОЛЖНА*ВЫТЬ
ВОЗВРАЩЕНА НЕ ПОЗЖЕ
УКАЗАННОГО ЗДЕСЬ СРОКА
/?2
01'46- f А/
Гб sty
Н.П. БЛИНОВ
Химическая
микро/
биология
Допущено Государственным комитетом СССР
по народному образованию в качестве
учебника для студентов
высших учебных заведений, обучающихся
по специальности «Биотехнология»
МОСКВА «ВЫСШАЯ ШКОЛА» 1989
ББК 28.4
Е51
УДК 576.8
Рецензенты:
кафедра микробиологии Московского государственного университета
им. М. В. Ломоносова (зав. кафедрой проф. Н. С. Егоров) и проф.
Н. Б. Градова (ВНИИсинтезбелок)
I ЕГЯШИЯА i
им. Н. И. ЛоБачзвсного
|*зшквго гес. ужерсятеп
Блинов Н. П.
Е51 Химическая микробиология: Учеб, для студентов химико-
технол., технол., фармац. и др. ин-тов, аспирантов и практ.
работников. — М.: Высш, шк., 1989. — 448 -е.; ил.
ISBN 5-06-000089-3
Учебник включает материалы о химическом составе, строении, топологии и
функции микро- и макромолекул в микробных клетках. Изложены химические
основы процессов роста, развития, размножения, дифференцировки, обмена ве-
ществ и патогенности микроорганизмов, а также противомикробного иммуни-
тета.
1905000000(4309000000)—407
001(01)—89
137—89
ББК 28.4
57.04
ISBN 5-06-000089-3
© Издательство «Высшая школа», 1989
ПРЕДИСЛОВИЕ
В планах экономического и социального развития СССР до
2000 г. намечены грандиозные задачи, требующие неотложного
решения. Немалая роль в этих планах отводится использованию
микробиологических процессов, объектами которых являются
природные (естественные) и целенаправленно измененные клетки
(мутантные формы, гибридные клетки, а также клетки, несущие
искусственно заданную генетическую информацию). Указывает-
ся на необходимость дальнейшего многопланового развития
фундаментальных и прикладных исследований в области моле-
кулярной биологии, иммунологии, генетики и физиологии микро-
организмов, генной и клеточной инженерии. Для этого необходи-
ма подготовка соответствующих кадров биотехнологов, владею-
щих знаниями в области химико-биологических, химико-техно-
логических, биотехнологических и других смежных с ними дис-
циплин.
Существующая форма преподавания микробиологии не отве-
чает современному уровню развития науки и техники. В связи
с этим назрела необходимость подготовки специального учебни-
ка «Химическая микробиология».
При написании учебника автор использовал опыт препода-
вания курса «Молекулярно-генетические основы микробиологии»
в Ленинградском химико-фармацевтическом институте (ЛХФИ).
Для обеспечения преемственности, главным образом курсов «Ор-
ганическая химия» и «Микробиология», учебник «Химическая
микробиология» разделен на три части и восемь глав. В первой
части (главы 1—3) изложены химические основы структурно-
функциональной организации микроорганизмов. Вторая часть
(главы 4 и 5) посвящена химическим основам жизнедеятель-
ности микробов, третья часть (главы 6—8) — прикладному зна-
чению химической микробиологии.
В подготовке материалов по иммунохимии существен вклад
старшего научного сотрудника ЛХФИ М. А. Кашкиной, любезно
согласившейся участвовать в написании восьмой главы.
У читателя могут возникнуть критические замечания, реко-
мендации и пожелания. Они будут приняты автором с призна-
тельностью.
Автор благодарен рецензентам проф. Н. С. Егорову (МГУ
им. М. В. Ломоносова) и проф. Н. Б. Градовой (ВНИИсинтез-
белок Минмедбиопрома), чьи замечания способствовали улучше-
нию книги.
Автор
ВВЕДЕНИЕ
Химическая микробиология — наука о химии и биохимии микро-
организмов, находящихся в конкретных условиях среды обита-
ния. Химическая микробиология является составной частью мик-
робиологии — науки о микроорганизмах. Она базируется на зна-
нии структуры и функций различных молекул в индивидуальном
и интегрированном виде, т. е. в живой клетке, обитающей в
различных условиях существования (включая человека, живот-
ных и растения).
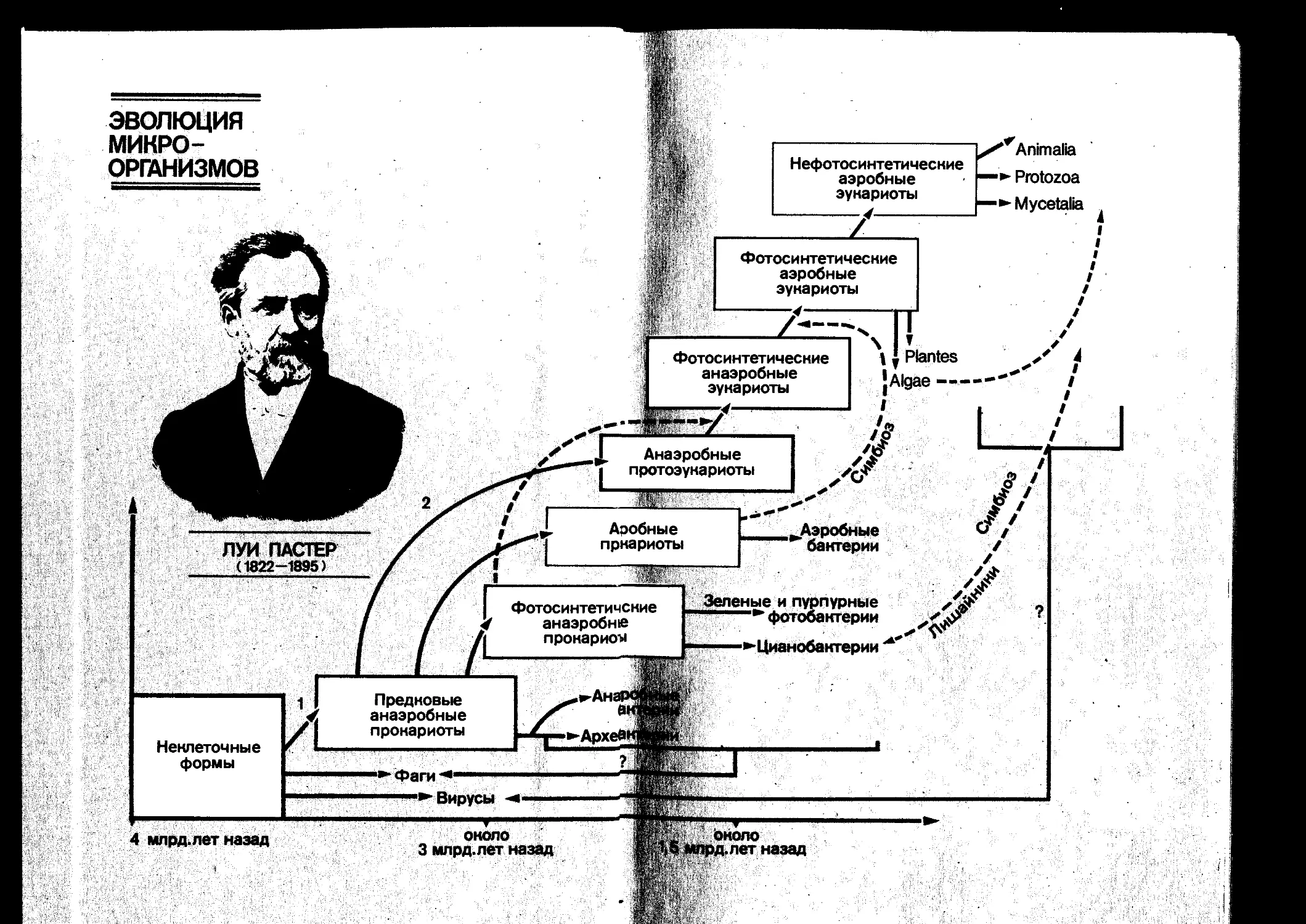
Великий французский ученый Луи Пастер (1822—1895) —
основоположник научной микробиологии — внес химический под-
ход к раскрытию отдельных процессов, совершающихся при
участии микроорганизмов. Разделив с помощью плесневого гриба
право- и левовращающие оптические изомеры винной кислоты,
он заложил основы стереохимии. Пастер открыл микробы, жи-
вущие в бескислородных условиях, внеся тем самым неоценимый
вклад в понимание процессов биоэнергетики и биохимической
эволюции живых организмов. Им было доказано, что некоторые
(инфекционные) заболевания передаются с помощью микроорга-
низмов.
Около 40 лет своей научной деятельности Пастер посвятил
изучению процессов брожения, в результате которых под дей-
ствием микроорганизмов образуются спирты или органические
кислоты при распаде растительных материалов. Он показал,
что каждый химический тип брожения, сопровождающийся
образованием главного конечного органического вещества, свя-
зан с развитием определенного микроба.
Важной вехой в понимании биохимических реакций, осуще-
ствляемых микробными клетками, было открытие ферментатив-
ной активности дрожжевого сока (М.. Манасеина, 1871), а затем
подтверждение его Э. Бюхнером (1897). Это открытие считается
началом становления биохимии. Теперь самые сложные физиоло-
гические процессы в клетке могут быть объяснены с физико-
химических позиций.
Честь выяснения роли микробов в геохимии принадлежит
С. Н. Виноградскому (1856—1953), который установил участие
микробов в круговороте углерода, азота и серы на Земле.
В конце XIX и начале XX в. были подвергнуты глубокому
изучению возбудители инфекционных болезней, иммунитет микро-
организмов к указанным возбудителям, химическая активность
различных микробов, получила развитие химиотерапия ин-
фекционных заболеваний. Сформировались самостоятельные
4
научные дисциплины как части общей микробиологии: промыш-
ленная микробиология, медицинская микробиология, инфекцион-
ные болезни, сельскохозяйственная микробиология, микробиоло-
гия пищевых продуктов и др. Микробиология воды, воздуха,
почвы и пищевых продуктов составила основу санитарной микро-
биологии. В то время понятие «химическая микробиология» еще
не приобрело самостоятельного значения, так как лишь к 40-м
годам текущего столетия ученые окончательно утвердились во
мнении, что все живые системы в основе своей сходны на биохи-
мическом уровне. Кроме того, определенное отставание в разви-
тии генетики микроорганизмов в немалой степени сдерживало
наше продвижение к более глубокому познанию отдельных сто-
рон химии и биохимии функциональной активности этих существ,
в том числе — последствий наследуемых изменений (мутаций).
Материализация гена шм.исе, мжиьим •.:! ..«.w ......
исследований в области юнетикн живых ср. Jfrs лил
всего микроор! аннзмив, сблир-и = ,► <
ей генома и способности.) быстро' • рззмво/м нм.' б г. . . у.;у--.
когда конечным пр'КТ'>М би П'.АН ,Л- \ ! С: . у У'.. гУ.
ется биомасса к.ж'-> ку, - ‘ ч ижу-оу у у- дарчу! .ч/л .) ун-
циальных условиях обе^всчивжн наю чуйте о- г, 'юлмпю хо/ш
чествах за короткий лоршн: времени; (ч-ти же • г.-нгчшм .н'-г-лу*'
том является нервинный и :н птощс-ный мсчабоии-: о •: и ; жа-
ние его можно измени1 уятчиво щей', гвся иг. фишелши
ческие и рсгуляюрные •>,,-< я у ; н сиик.оа мегшй . i и • ншяы.ч
микроорганизмом Р. об чн . ..в-шях ад/бода"’- i нсбос/ч.- яошё
ние фиЗИОЛО! ИИ МИлр;,О!>1-Ш;-'-iMO’’ И О'В'.ХиМИ'КЧ К--)? 'ИЧНОСТИ
реакций, проигходяших В СО-.КрС! НЫ'.- .".ТВИН' М ЛЬ! PiH-iPO! ;иия
продуцентов.
К середине 40-х годов произошло сближение микробиологии,
генетики, химии и биохимии, что породило молекулярную биоло-
гию и затем углубило дифференциацию дисциплин, обогащен-
ных накоплением фактического материала в последующие годы.
К этим дисциплинам относятся фармацевтическая, химическая,
космическая микробиология и биотехнология. Фармацевтическая
микробиология связана с изучением микрофлоры лекарственных
средств, механизма ее взаимодействия с компонентами лекар-
ственных форм; источников и причин попадания микробов в
лекарства в заводских и аптечных условиях, а также с выра-
боткой мер, предупреждающих недопустимую обсемененность
ими лекарственных средств.
Космическая микробиология впервые возникла в нашей стра-
не в годы подготовки и первого запуска космического спут-
ника (1957). Это наука об изучении влияния космических усло-
вий на свойства микроорганизмов, находящихся в различных
средах существования (в том числе — в организме человека), о
предотвращении заноса микробов в космос и о борьбе с микро-
организмами, занесенными в космические аппараты, снаряжение,
оборудование и др.
5
В современном понимании биотехнология -- это наука об ис-
пользовании биологических процессов в технике и промышлен-
ном производстве. В виде суммы эмпирических технологических
процессов биотехнология существует с библейских времен, од-
нако ее научные основы были заложены Луи Пастером. Второе
рождение биотехнологии связано с внедрением глубинных мето-
дов крупномасштабного культивирования микроорганизмов (ко-
нец 30-х — начало 40-х годов XX в.), в том числе — с промыш-
ленным производством пенициллина и других антибиотиков.
Третье рождение ее связано с возникновением генной инжене-
рии (1972) и внедрением методов генной инженерии в производ-
ство.
Из изложенного выше видно, что микробиология в своем
развитии постоянно опиралась на химическую и биохимическую
основы, и теперь, когда мы можем вторгаться в статические
и кинетические процессы и структуры, стало необходимым изу-
чать молекулярные и генетические механизмы роста, развития,
размножения и метаболизма микроорганизмов с химических и
физико-химических позиций. В этой связи и подготовлен настоя-
щий учебник «Химическая микробиология». Данная дисциплина
переживает свой расцвет и ее успехи внесут несомненный вклад
в общее благосостояние человечества.
Часть I
Химические основы
структурно-
функциональной
организации
микроорганизмов
Глава 1
ПРЕДМЕТ, ОБЪЕКТЫ. П.ЕЛИ И ЗАДАЧИ
X ИМ И Ч ЕС К О И М И К Р О Б И OJIО к' И И
Химическая микробиология — самостоятельная научная дис-
циплина, сформировавшаяся к началу 50-х годов и представляю-
щая собой ветвь общей микробиологии. Химическая микробиоло-
гия является наукой о химии и биохимии микроорганизмов, изу-
чающей химический состав, строение, топологию, функцию сво-
бодных и конъюгированных молекул, а также обмен веществ
прокариот и эукариот (от лат. pro — вместо, греч. xapicov —
ядро, еи — хорошо).
К прокаршл НЩ-Ч ’И"-’ П:Ш. 1Ч11НЖ' Ж. (ЙН.
не имеют оформлении! о ядре И ня ныи жжди.1 . •• жж ,
НИХ ОТ ЦИ1 (ИI.i<i Жи Прон ,.!№: Р фор Ж >- >' »|.те
ВОЙ спирали ДЖЖжСИ|Ш'йци члгж.Ч-ЖоЙ Ж'-ЖД"
К Э^карио! ПЧ('( I.HM У!ЖрЖ5(>! ЯНИ’-W-.- ЧП,,ЖЖ! I ertso; ' • >||ро
ЛИ, лишайники И Про'; Ж '!'!,( ( )1<ч ,ор. р Ч НД('О , !
деленное о> цигом- <лчмы ядеждой ж миражж. а ж пле.; а рг
делена на гнепшин ;;|ро.нчн!>ые номпир; менш д-гс.•>.и , ,л;ржь-
ч ен н ы е мембранами-.
Концепция о прокариотах и эукариотах, сложившаяся к сере-
дине XX в., оказалась весьма плодотворной. Она помогла решить
ряд проблем в области дифференциации определенных клеточ-
ных структур, генетики и систематики микробов, а также внесла
соответствующий вклад в понимание эволюции жизни на Земле.
Используя эту концепцию, удалось более точно соотнести накоп-
ленный фактический материал, например, о топологии молеку-
лярных и надмолекулярных структур, о химических механизмах
регуляции роста, развития и размножения микроорганизмов
и т. д.
7
Химическая микробиология во многом базируется на данных
пограничных наук, с которыми имеет тесную связь. К их числу
относятся неорганическая, органическая, физическая, коллоид-
ная и биологическая химия, генетика, микробиология, иммуно-
логия, биотехнология и др. Более того, химическая микробиоло-
гия, особенно ее объекты, нередко используются в смежных
дисциплинах для выявления и подтверждения некоторых обще-
биологических закономерностей (например, о строении и функ-
циях мембран, генетического аппарата, обмене веществ и др.).
1.1. ОБЪЕКТЫ ХИМИЧЕСКОЙ МИКРОБИОЛОГИИ
Объектами химической микробиологии являются микробы, а так-
же химические компоненты микробных клеток и вирусов. Микро-
бы отличаются своими малыми размерами; они измеряются в
микрометрах (мкм) (например, бактерии) и в нанометрах (нм)
(например, вирусы). Так, размеры некоторых энтеробактерий
соответствуют (0,5...2,0)Х(3,0...15,0) мкм; поперечник нитей
актиномицетов равен 0,5...1,5 мкм; грибы — микромицеты замет-
но больше бактерий (например, клетки дрожжевых организмов
в — 10 раз крупнее стафилококков). Отдельные грибы — макро-
мицеты образуют плодовые тела, измеряемые сантиметрами
(белые грибы, подберезовики и др.), хотя поперечник нитей гриб-
ницы этих же форм находится в пределах 3...7 мкм. Размеры
отдельных водорослей составляют от 1,0 до 17-103мкм. Но не
все водоросли могут быть отнесены к микроорганизмам, на-
пример ламинария, длина которой может достигать 50 м. Ве-
личина простейших колеблсзся в среднем от 3,5-103 до
60,0-103 мкм. Некоторые данные о величине различных микро-
бов приведены в табл. 1.
Таблица I. Сравнительные размеры микробов
Микробы Диаметр, нм
Некоторые водоросли Algae 1000...17-Ю6
Простейшие Protozoa 3500... 60000
Мукор Mucor species (поперечник гиф) - 10000
Пеницилл Penicillium species (конидии) -4000
Палочка чудесной крови Serratia marcescens 750
Риккетсии Провачека Rickettsia prowazekii (попе-
речник) 300-600
Актиномицет Streptomyces species (поперечник ни-
тей) 300
Вирус оспы Poxvirus 210X260
Вирус бешенства Rhabdovirus 125
Вирус гриппа Orthomyxovirus (Influenza virus) 85
Аденовирус Adenovirus 75
Бактериофаг Т3 Bacteriophag Т3 45
Вирус полиомиелита Poliovirus 27
Вирус японского энцефалита Togavirus 18
Вироиды Viroidi 15
8
Исключения из правил, касающиеся размеров, имеются в
каждой группе микроорганизмов. Например, известны нитчатые
и стебельковые бактерии, заметно превышающие по длине боль-
шинство других бактерий; размеры вириона полиэдроза тутового
Шелкопряда Bombix mori составляют 30,0...50,ОХ200,0...320,0 нм.
’* Микробный мир, включающий вироиды и вирусы, бактерии,
грибы, водоросли, лишайники и протозойные организмы, систе-
матика которых представлена в табл. 2, характеризуется своими
особенностями.
Таблица 2. Систематическое положение микробов
Надцарства Царства Отделы
Vira Protovira Euvira Ribovira
Deoxyvira
Procaryotae Bacteria Gracilicutes
Firmicutes
T enericutes
Mendosicutes
Eucaryotae Mycetalia (My cot a) Myxomycota
Eumycota Lichens*
Plantae Algae
Animalla Protozoa
* Лишайннкн — продукт симбиоза некоторых видов грибов и водорослей (или циа-
нобактерий) .
Вирусы — наименьшие организованные формы жизни в виде
частиц (не клеток!), не обладающие собственным обменом ве-
ществ и стоящие на грани между живой и неживой природой.
Это безусловные или облигатные паразиты, содержащие какой-
либо один тип нуклеиновой кислоты — дезоксирибонуклеиновую
(ДНК) или рибонуклеиновую (РНК). К вирусам относятся ви-
роиды и бактериофаги.
Вироиды — инфекционные агенты, вызывающие заболевания
людей, животных и растений. Они представляют собой нуклеино-
вые кислоты, состоящие из коротких однонитевых циркулярных
молекул рибонуклеиновой кислоты, лишенных капсида,
с молекулярной массой (М) 1ОО...12О.кДа*. Вторичная структура
их может быть двухнитевой с короткими однонитевыми петлями.
Возбудители болезни скрейпи у овец, раннего старческого сла-
* 1 дальтон (Да) — 1/12 массы чистого изотопа |2С, 1 килодальтон
(кДа) = 1000 Да.
О
10 нм
MC\k\t«™KTeP"C™Ke ВИРУСОВ: ВИрИ0Н (а); спиРальный тип укладки
Да (о), кубический тип укладки капсида (в);
тип укладки капсида (в); суперспирализация вирусной
ДНК, некоторых фагов (а); форма некоторых фагов (<?):
1 — фаги Тки Т5 ; 2 — стафнлофаг 6; 3 — фаг Т, 4 — фаг Т2
10
Продолжение рис. 1
боумия у людей и другие напоминают вироиды. Это молекулы
ДНК е молекулярной массой порядка 70...100 кДа.
Вирусы, вызывающие заболевания или обитающие в орга-
низме человека и животных, относятся преимущественно к РНК-
типу. К этому типу принадлежит и подавляющее большинство
растительных вирусов.
Молекулярные массы вирусных РНК. находятся в пределах
1 • 106,..15-10* Да, вирусных ДНК — 1,5-106...160-106 Да. Цирку-
лярные ДНК нередко суперспирализованы (рис. 1, г).
Инфекционные частицы (единицы) вирусов называют вирио-
нами. Каждый вирион имеет капсомеры, представляющие собой
морфологические единицы, состоящие из одной или нескольких
белковых молекул. Сумма капсомеров, уложенных в определен-
ном порядке, составляет капсид, который вместе с нуклеиновой
кислотой образует нуклеокапсид (рис. 1,а).
По характеру укладки капсидов судят о типах симметрии:
спиральном (у вирусов гриппа, парагриппа, бешенства и др.);
кубическом (у вирусов герпеса, аденовирусов, ветряной оспы, по-
лиомиелита, некоторых фагов и др.), смешанном (у ряда фагов).
Кубический тип симметрии может быть представлен тетра-
эдром, октаэдром, икосаэдром (правильным двадцатигранником)
с осями симметрии соответственно 2:3, 4:3:2 и 5:3:2 и минималь-
ным числом структурных единиц — 12, 24, 66. Икосаэдрический
кубический тип симметрии наиболее распространен у вирусов.
Некоторые типы укладки капсидов показаны на рис. 1,6, в.
Вирусы выращивают на куриных эмбрионах или на культу-
рах клеток различных животных, растений (растительные ви-
русы) .
Бактериофаги — вирусы бактерий содержат ДНК, однако
имеются фаги, содержащие РНК. Форма некоторых фагов при-
ведена на рис. 1,6. Вирусы цианобактерий называют цианофага-
н
ми, актиномицетов — актинофагами. Все бактериофаги класси-
фицируют по бактериям-хозяевам и типу содержащейся нуклеи-
новой кислоты. Структурно в них выделяют головку, содержа-
щую ДНК, и хвостик. Хвостик имеет воротничок, чехол, стер-
жень, базальную пластинку с шестью шипами и шестью фибрил-
лами. При адсорбции фагов на чувствительных клетках проис-
ходит инъекция нуклеиновой кислоты, после чего клетка может
направленно синтезировать фаговую нуклеиновую кислоту, вновь
формируя 200...300 фаговых частиц к концу своего деления.
•',! •• ’ liafi й ~~ 4- lh ;• и ч.'ржиг одну молекулу
! УКДС Иг: • >'< • J,'; UP кД.|), Ч'О " г'1 ;»>!,:> НУТ 50 %
T из,---.- cvv'ii ЧЛ 1ШГ е 4"') МНИМ ма.Ч'Ы фги ОМОН ЧЗеТИПЫ
H’-MV ..(.jm н>-!ЛП.)ПЬН' Др) ИГ Ю'ИПОНГН гы '!'пги так-
на У МНОЖУ -
чаи вреза
. В перво,у
вы-
!<) при
Бактерии, грибы, водоросли,
низмы морфологически (от греч. цорур — вид, форма) отлича-
ются друг от друга, причем протозойные организмы являются
лишайники
и протозойные Орга-
микроскопическими животными существами.
По организации ядерного аппарата грибы, водоросли и про-
тозоа стоят выше бактерий и, следовательно, эволюционно они
12
появились позднее. Накопленные материалы о каждой группе
микробов настолько обширны, что это давно послужило основой
\для выделения самостоятельных научных дисциплин: бактериоло-
гии, микологии, альгологии, лихенологии и протистологии.
I Бактерии, согласно определителю Д. X. Берги (1896), под-
разделены на четыре отдела: грациликуты (от лат. gracilis —
стоойный, cutis — кожа), фирмикуты (от лат. firmus — проч-
ный), тенерикуты (от лат. tener — нежный) и мендосикуты (от
лат. mendosus — ложный). К двум первым отделам отнесено
большинство бактерий, в том числе фотобактерии и скотобакте-
рии. Так, фотоавтотрофные цианобактерии лишены полного
цикла трикарбоновых кислот и дыхание у них осуществляется
через окислительный пентозофосфатный цикл. Цианобактерии
могут расти на свету в минеральной среде, используя в качестве
источника азота молекулярный азот, а в качестве источника
углерода — диоксид углерода.
Азотфиксирующие бактерии являются обычно нитчатыми
формами, образующими так называемые гетероцисты.
Пурпурные и зеленые бактерии являются аноксигенными фо-
травтотрофными и фотогетеротрофными анаэробами (фотосин-
тез у них не сопровождается выделением кислорода), их под-
разделяют на серные и несерные. Серные накапливают элемен-
тарную серу в результате окисления сероводорода, несерные —
не накапливают.
Скотобактерии (от греч. охютсоо — мрак, темнота), включаю-
щие примерно 20 основных групп, можно условно подразделить
на бактерии без клеточной стенки (микоплазмы), бактерии с
однослойной клеточной стенкой (грамположительные) и бакте-
рии с двухслойной клеточной стенкой (грамотрицательные).
В 1978 г. было предложено выделить из прокариот царство археобактерий
Archaeobacteria, представляющих своеобразные формы жизни, куда включают
три группы микроорганизмов: метанобразующие, облигатные галофильные и
термоацидофильные бактерии. Они обитают в экстремальных природных усло-
виях. От обычных бактерий отличаются по внутриклеточным, специфичным для
них, транспортным и рибосомальным 16S, 18S рРНК, а также тем, что не содер-
жат муреинового каркаса (см. гл. 3) в клеточной стенке. Тем не менее многие
геиосистематики не считают правомерным выделять археобактерии из прокариот
В виде отдельного царства, а в определителе Д. X. Берги (1986) они составляют
единственный класс в отделе мендосикутов.
Бактерии (прокариоты) весьма разнообразны по форме и чис-
ленности. На рис. 3 изображены представители всех основных
групп бактерий, включая археобактерии. Они имеют сфериче-
скую, овальную, палочковидную, нитевидную и другие формы.
Сферическими являются микрококки (от греч. pixpwo — малый,
хоххыо — кокк), располагающиеся в виде изолированных клеток;
стафилококки — гроздевидные кокки (от греч. атадчАл] — вино-
градная гроздь); диплококки, имеющие форму сдвоенных клеток
(от греч. бглХохоо — двойной), например гонококки, менинго-
кокки и пневмококки; стрептококки, располагающиеся короткими
или длинными цепочками (от греч. отрцлтош — витой, круче-
ный); тетракокки, располагающиеся по четыре клетки (от греч.
i-rqTOa — четыре); сарцины, или пакеты (от лат. sarcina), при-
сущие, например, виду Sporosarcina urea, разлагающему моче-
вину.
\ Палочковидные бактерии (от греч. paxvqpicov — палочка) мо-
гугг располагаться изолированно, попарно (диплобактерии), в
циючках (стрептобактерии), как сферические или овальные
фсюмы.
нитевидным бактериям относятся, например, актиномицеты.
Изогнутыми и извитыми бактериями являются вибрионы (на-
пример, холерные), спириллы (возбудитель болезни содоку),
лептоспиры (возбудитель желтушного лептоспироза), трепонемы
(возбудитель сифилиса).
Ряд палочковидных бактерий способен образовывать внутри-
клеточные споры (одна спора на клетку). Такие клетки чаще
называют бациллами Bacillus (Вас.) или Clostridium (от лат.
Clostridium — веретенообразная бактерия). Клостридиальные
формы, как правило, имеют утолщение в середине либо в конце
клетки в форме головки, или булавы (от греч. —
булава), что видно из рис. 3, 13a, б. Некоторые актиномицеты так-
же образуют споры. Они формируются у них на воздушных ни-
тях —- спороносцах, которые могут быть прямыми, изогнутыми и
спиралевидными. У отдельных представителей актиномицетов
могут образовываться подвижные споры в так называемых
актиноспорангиях. Кокки, как правило, не образуют спор и не-
подвижны. Исключение представляет названная выше споросар-
цииа, которая подвижна и образует споры.
Многим бактериям присуща подвижность благодаря наличию
у них жгутиков — органов локомоции. В качестве примера мож-
но назвать кишечную палочку Escherichia coli, сенную палочку
Вас. subtilis, синегнойную палочку Pseudomonas aeruginosa,
холерный вибрион Vibrio cholerae, спирохету (трепонему) сифи-
лиса Treponema pallida и т. д.
Другой вид подвижности — скольжение — проявляется лишь
при контакте клеток с твердыми субстратами. Такой подвиж-
ностью обладают, например, многие цианобактерии, некоторые
водоросли.
В 1884 г. X. Грам предложил метод окраски бактерий, соглас-
но которому все бактерии подразделяют на грамположительные
Рис. 3. Морфология бактерий:
1 археобактерии (а — метанобактерии, вегетативные и б — покоящиеся формы, в —
галофилы, живущие в средах с высокими концентрациями некоторых солей, г — термо-
ацидофильные бактерии); 2 — цианобактерии (а — б); 3 — микоплазмы; 4 — микрококки;
“-“Диплококки; 6 — тетракокки; 7— сарцины; 8 — стрептококки; 9— стафилококки;
10 бактерии; 11 — диплобактерии; 12 — стрептобактерии; 13 — бациллы (а — клостри-
дии, б— плектридии); 14 — диплобациллы; 15 — стрептобациллы; 16 — вибрионы; 17 —
спириллы; 18 — спирохеты (а — трепонемы, б — боррелии, в — лептоспиры, г—кристи-
спиры, д — спирохеты); 19 — актиномицеты (а — микобактерии, б — нокардии, в —
стрептомицеты); 20— риккетсии; 21 — миксобактерин; 22 — стебельковые бактерии
(a Nevskia, б — Gallionella, в — Caulobacter, г — Rhodomicrobium, д — Hyphomicro-
и грамотрицательные. К грамположительным относят большин-
ство кокков (кроме гонококков и менингококков), споровых
бактерий — бацилл (кроме Desulfotomaculum) к неспоровые
молочнокислые бактерии Lactobacterium species. К грамотрица-
тельным относят не образующие спор бактерии (кроме молочно-
кислых), гонококки, менингококки, микоплазмы, L-формы бакте-
рий, риккетсии. При оценке того или иного вида бактерий учи-
тывают морфологические признаки клеток: величину; характер
концов (закругленные у Вас. subtilis, обрубленные у Вас.
anthracis и т. д.); изогнутость; наличие или отсутствие спор и
капсул и пр. В специальных опытах выявляют- тип дыхания,
или биологического окисления (в присутствии кислорода живут
аэробы, в его отсутствие — анаэробы), способность вызывать
заболевание у экспериментальных животных, т. е. являются ли
они болезнетворными (патогенными) или неболезнетворными
(сапрофитами), при необходимости оценивают их рост на средах
с повышенными концентрациями солей (выявление галофилов).
Грибы (от лат. fungus — гриб, губка; от греч. цихт]О —
гриб) — обширная группа эукариотических микроорганизмов,
насчитывающая свыше 100 000 видов. Грибы были известны
научному миру раньше бактерий и изучались в курсе ботаники.
Их подразделяют на микромицеты (например, плесневые нит-
чатые или мицелиальные грибы, дрожжи) и макромицеты (на-
пример, шляпочные -и пластинчатые съедобные и несъедобные
грибы) (рис. 4).
О,е.,ь<.ыб ।,,>и.<;к ?л>лbufи<‘। грибов образование
|л;и щгч-й. л-ц j шЬ. знылн: г.лзл.!ънги.< ы.дщ щ-ptнлетающи.м.я
в пл;' ру-о; ц • * \ <у-Ды < hjj !ЗЛ,ы.-д (г-т ; иеч. ii4AA<i>0"
м-щ-.дой л.ыч они -j шиш ча щ; = а• щьшп щн-дах и<кам>-
л. л-.н.но [>лл>яЪ|д ль- , фор-.н,.- и >ации, особенно --
1 ill [Щ->А ’> (’' 1ОВ-Ш-СНМ и >, lepUjK. I»'; оЧ.ЧЫе формы Грибов
• н.!! 1 р и мер, ’луюц.ы. лрР.т.д<: . однако гщльши.щтъо видов много
1..ееТ1,<В1>,1 ( ciepcvi в; PObiSiUiC Вл, a v.li'P! И л.: i.l. фр ЩрИУМЫ И 1 Д.).
CpViiBi.iie • очные Ы- ЫЫ1Г eol’Pei ЫЧе.ы с ыыш«. i дгда как
щ юг-.ж,тс ночные а ,-.avi.ic ! щ 5ч<, имени-, нары ородки. или
вечны I or ъ.н зерД:’л ш <ыы,'ро;оыд , г. ю.ы дюз;.
!фибы. нздтд нс '.'р-ще--- ро-.-ыыы называкл совер-
t'./ee,'/ы,«д !-I ни a it'' > р. м дворовые rpi-e'.j, ..'г,.., ,.’ья); не имеющие
иа-оаиге не. -AH-piuemibiми ;и'Пример, альтернация).
Все грибы подразделяют на два подотдела — грибы-слизеви-
ки Myxomycetes и настоящие грибы Eumycetes. Миксомицеты
занимают промежуточное положение между Eungi и Protozoa,
так как их ассимилятивная стадия развития в виде плазмодия
является бесклеточной, многоядерной, амебоидной массой, окру-
женной слизистым чехлом, варьирующим от микроскопических
размеров до обширной системы из ветвящихся и анастомози-
рующих сплетений, через которые отмечается ритмичный и обра-
тимый поток протоплазмы. При благоприятных условиях плаз-
модий трансформируется в одно или большее число плодовых
16
Рис. 4. Морфология некоторых представителей грибов:
а — дрожжи-сахаромицеты; б— сапролегння (/ — зооспорангий, 2 зооспоры); в
мукор (/'— спорангий, 2'— спорангиоспоры, 3—колонка, 4 — спорангиеносец); г
аспергилл и пеницилл (/ — головка, 2 — конидненосец, 3 — стеригмы, 4 — конндиоспо-
ры, 5—опорная клетка); д — базнднальный гриб (/ — плодовое тело, 2 базидня с
базидиоспорами, 3 — мицелнй); е — несовершенный гриб (альтернарня) конидненосец
с коиидиоспорами
тел — спорофор, обеспечивающих размножение организма с по-
мощью спор. Миксомицеты — преимущественно сапрофиты, не
вызывают заболеваний у человека и животных.
Эумицеты (микро- и макромицеты) обладают чрезвычайно
разнообразными морфологическими формами, особенно это
касается зародышевых клеток — спор и конидий.
У низших грибов споры располагаются в так называемых
спорангиях, это эндоспоры. У большинства высших грибов споры
располагаются открыто — экзоспоры, или конидии (рис. 4). Все
споры образуются на спороносцах (спорофорах), или конидие-
носцах (конидиофорах).
Известны грибы, способные образовывать нитчатые и дрожжевые формы,
например болезнетворный гриб Histoplasma capsulatum и сапрофитный Аигео-
basidium (PuUularid) pullulans. Некоторые мукоры образуют дрожжевые клетки
при увеличении в среде выращивания парциального давления диоксида угле-
рода. Это так называемый диморфизм, являющийся отражением зависимости
формы клеток от условий выращивания организма.
17
Мицелий грибов подвержен видоизменениям, играющим важную роль в оп-
ределении видов, выделяемых, например, из природных субстратов. К таким
видоизменениям мицелия относят: хламидоспоры (от греч. хЛацтхт — накид-
ка)— концевые (терминальные) и срединные (интеркалярные); геммы (от лат.
gemma — почка); гаустории, или присоски (от лат. haustor — пьющий); тяжи
и шнуры, ризоиды (от греч. ptga — корень); пряжки (рис. 5). Например, хла-
мидоспоры у Candida albicans только концевые, у Coccidioides immitis имеются
и интеркалярные; геммы присущи A. pullulans; гаустории — ржавчинным гри-
бам; тяжи, шнуры, пряжки — базидиальным грибам; ризоиды — низшим грибам
фикомицетам.
С учетом морфологических особенностей и размножения грибов их подразде-
ляют на семь классов: Chytridiomycetes, Hyphochytridiomycetes, Oomycetes,
Zygomycetes, Ascomycetes, Basidiomycetes, Deuteromycetes.
Большинство бактерий и грибов культивируют на естественных и искусствен-
ных питательных средах. В то же время, например, лепрозные бактерии не
удается вырастить в лабораторных условиях, а в эксперименте заражаются
лепрой лишь броненосцы.
Водоросли Algae (от лат. alga — морская трава) — преиму-
щественно автотрофные фотосинтезирующие талломные организ-
мы, содержащие разной формы специальные органеллы — хро-
матофоры (хлоропласты) и живущие, как правило, в воде и
почве. Хроматофоры содержат хлорофилл и другие пигменты, а
также особые белковые тельца — пиреноиды (от греч. mpiyv —
косточка), вокруг которых откладываются крахмал и другие гли-
каны. У многих водорослей зеленая окраска от хлорофилла
перекрывается пигментами иного цвета.
Рис. 5. Видоизменения мицелия:
хламидоспоры (/ — терминальная, 2— интеркалярная, 3— геммы); гаустория, нлн
присоска (4); тяж (5); шнур (6); пряжки (7); ризоиды (8); столон (9)
18
формы клеток водорослей весьма разнообразны: сферические,
Грушевидные, яйцевидные, спиралевидные, цилиндрические, вере-
^ообразные и др. Известны водоросли — строгие фотосинте-
•йиКИ (например, зеленые хламидомонады и хлорококки), равно
к*к и водоросли, приспособленные к фотоавтотрофному и гетеро-
трЪфному питанию (например, пиррофитовые).
’^Наиболее широко принято деление водорослей на 9 типов:
пИрЬофитовые, золотистые, диатомовые, желто-зеленые, бурые,
кржйые, эвгленовые, зеленые и харовые. Несмотря на большое
разнообразие форм водорослей, их можно свести к следующим
осрбйным структурам: монадная, амебоидная (ризоподиальная),
пальмеллоидная, коккоидная, нитчатая, пластинчатая (тканевая)
и сифоновая.
Монадиая структура характеризуется подвижностью клеток с помощью
двух, реже одного или нескольких жгутиков. Она наблюдается в течение всей
вегетативной жизни более просто устроенных водорослей (например, эвглено-
вых), тогда как у более высокоустроенных водорослей (например, золотистых)
она типична для зооспор при бесполом или для гамет при половом размно-
жении.
./ Для амебоидных структур характерно передвижение с помощью псевдопо-
дий (ложноножек); такие клетки имеются, например, у отдельных представи-
телей красных водорослей, находящихся на определенных стадиях размножения.
Пальмеллоидная структура — скопление нескольких или многих независимых
друг от друга клеток, погруженных в общую слизь (например, у зеленых
водорослей). Коккоидная структура характеризуется неподвижными клетками
различной формы, одиночными или соединенными в различные колонии (напри-
мер, у, золотистых водорослей). Нитчатая структура — это соединение одного
или нескольких рядов неподвижных клеток в простые или разветвленные нити
(например, у желто-зеленых водорослей). Пластинчатая структура представля-
ется в виде узких или широких плоских шнуров, формирующихся в результате
продольного и поперечного деления клеток (например, у зеленых водорослей).
Сифоновая структура, присущая, например, отдельным представителям зеленых
водорослей, проявляется в виде одной многоядерной большой клетки.
Лишайники Lichenes — симбионты, обладающие специфиче-
скими морфологическими признаками и строением. Они включа-
ют трибы (фикомицеты, аскомицеты или базидиомицеты) и циа-
нобактерии (носток, глеокапса, хлорококк) или зеленые водорос-
ли (цистококк, хроококк, хлорелла, реже — кладофора, паль-
меЛла). Например, гриб окружает цианобактерии или клетки во-
дорослей и даже проникает в них, этим обеспечивается своеоб-
разие физиологии таллома лишайника. Приспособляемость ли-
шайников к внешним условиям исключительна, поэтому их
можно встретить повсеместно на различных почвах, скалах,
деревьев, в воде, на металлических конструкциях, на бетоне,
стекле и т. д.
Известно около 300 родов лишайников. Размножаются они
либо вегетативно — фрагментами слоевища, изидиями и сореди-
ями, либо половым и бесполым путем за счет спор. Изидии —
бородавчатые палочковидные или коралловидные выросты на по-
верхности таллома, покрытые корой и содержащие клетки орга-
низмов-симбионтов. Соредии — микроскопические клубочки из
грибных гиф с окруженной ими одной или несколькими клетками
19
водоросли или цианобактерии. Соредии, как правило, многочис-
ленны и образуют порошковидную беловато-желтоватую массу.
По внешнему виду различают накипные (корковые), листо-
ватые и кустистые лишайники.
Протозойные организмы (Protozoa')—нефотосинтезирующие,
одноклеточные эукариоты. Предполагается, что Protozoa про-
изошли от различных водорослей, в большинстве своем потеряв-
ших хлоропласты, а с ними — способность использовать энергию
солнечных лучей. Наиболее примитивные из них — жгутиковые
формы, из которых затем произошли амебоидные и реснитчатые
типы; промежуточные типы в определенные фазы развития имеют
жгутики, в другие фазы — псевдоподии.
Все Protozoa подразделяют на плазмодромные (классы —
мастигофоровые Mastigophora, саркодовые S ar coda, споровики
Sporozoa) и реснитчатые (Ciliophora).
Среди протозойных микроорганизмов известны патогенные
для человека виды: вызывающие лямблиоз Giardia lambda, три-
хомоноз Trichomonas vaginalis, трипаносомоз Trypanosoma
gambiense, лейшманиоз Leishmania donovani, относящиеся к
жгутиковым; к саркодовым относится возбудитель амебной ди-
зентерии Entamoeba histolytica', к споровикам — возбудители
малярии Plasmodium sp., к реснитчатым — возбудитель балан-
тидиоза Balantidium coli (рис. 6).
В природных условиях микроорганизмы находятся в составе
биогеоценозов (от греч. ршс — жизнь, уесо— земля, xwivwg —
община). По определению В. Н. Сукачева биогеоценоз — это
участок земной поверхности, на котором в тесном взаимодей-
Рис. 6. Некоторые протозойные организмы:
/ — лямблия; 2— трихомонада; 3— трипаносома; 4 — лейшмання; 5 — дизентерийная
амеба; 6 — малярийные мерозоиты; 7 — кишечная балантидия
20
efsiiH развиваются однородная по составу и производительности
растительность; однородный комплекс животных и микроорга-
низмов; однородная по физико-химическим свойствам почва;
ррплерживается однородная газовая и климатическая ситуация
цШгтанавливается однородный по масштабу и ритму материаль-
н^эйергетический обмен между всеми составляющими биогео-
цеЙоза. Частью биогеоценозов являются микробоценозы, главная
bofj» которых заключается в минерализации органических ве-
щдав.
'Основной резервуар сапрофитных микробов — почва, где они
мбгут выживать какое-то время, находясь в состоянии анабиоза
или метабиоза. Анабиоз — приостановка жизнедеятельности ор-
ганизма при неблагоприятных условиях существования с после-
дующим восстановлением нормальной жизнедеятельности при
наступлении благоприятных условий. Метабиоз — рост, развитие
и размножение организма (см. гл. 4) при определенном уровне
обменных процессов, или метаболизма, в соответствующих усло-
виях среды обитания. Под метаболизмом понимают все химиче-
ские реакции, имеющие место в живых клетках. Метаболизм
складывается из катаболизма — разложения химических ве-
ществ в целях получения энергии, и анаболизма — синтеза ве-
ществ клетки. Реакции промежуточного обмена называют амфи-
болизмом.
При анабиозе микроорганизмы выживают, но не размножа-
ются. При мггабио-.е « природных условиях микробы постоянно
взаимодействую' сп'ЖД} -)бой и другими организмами. Эти
взаимоотношения иоцр;,! •*( ляют i»a симбиотические. или симбиоз
(коменсализм, мутузлч >, наразизн гм); антагонистические, или
антибиоз (одно и тп =р- гетсроантагонизм; изоантаго-
ннзм; виолентоангаынн- с или насилье гневный антагонизм).
В природных и ж c i витых условиях микроорганизмы
подвержены из.м<'не”-гзv ц<. форме, типам размножения, об-
менным процесса г- ' табильпыс формы называют поли-
морфными (от л - лзого, i реч щорфг • форма, вид), а
само явление - /. ,-л. • -сссмои [реже плеоморфными (от греч.
nletov — больше) и. .нх/’с'р'пизмом ( К таким микробам отно-
сят холерный вибри !' > учбы дермдюфиты и др, особенно, если
они длительно храня; в табор.)сорных условиях.
Методы химической микробиологии специфичны, но их ис-
пользуют как в микробиологии, так и в химических науках.
Это различные виды микроскопии, культивирование на питатель-
ных средах в лабораторных и производственных условиях;
разрушение микробных клеток в дезинтеграторах; центрифуги-
рование (аналитическое и дифференциальное); выделение компо-
нентов клеток с помощью смешивающихся с водой органиче-
ских растворителей, диализа, гель-фильтрации, электрофореза
И др. Для анализа элементного состава клеток и вирусных
частиц применяют методы неорганической химии. Методические
приемы, используемые в биохимии животных и растений, в рав-
ной мере применимы к микробным объектам (анализ фермен-
тов и др.)- Методы органической и физической химии необходи-
мы при анализе состава и строения микробных белков, углево-
дов, липидов, различных конъюгатов и т. д.
Целями и задачами химической микробиологии является изу-
чение химического состава, строения и физиологической актив-
ности микроорганизмов.
Химическая микробиология имеет важное теоретическое и
прикладное значение. Исследование химического состава и строе-
ния микроорганизмов, их первичных и вторичных метаболитов
помогает проникнуть в организацию и функциональную актив-
ность живой материи и получать необходимые для практики
вещества (пищевые, лечебные, вспомогательные и т. д.). Изу-
чение организации и работы генетического аппарата вирусов и
микробных клеток вносит неоценимый вклад в понимание зако-
номерностей наследования признаков и свойств организма в ряду
поколений, в использование быстрорастущих и развивающихся
видов, в генно-инженерных работах, в становление и развитие
новых производств.
бурное развитие химии иммуншета, или иммунохимии, преж-
де всею ююано с изучением патогенных микроорганизмов.
Системагичсское и глубокое изучение ответных реакций макро
организма при инфекционных процессах было положено в основу
пс только иммунитета нря заразных заболеваниях, но и в фун-
дамент современной иммунологии и иммунохимии.
Перспективы дальнейшего развития химической микробиоло-
гии многопрофильны и заманчивы. Она находится на стыке
биологических и химических дисциплин, где больше возможно-
стей для прогнозируемых открытий и свершений.
1.2. СТРОЕНИЕ КЛЕТОК ПРОКАРИОТ И ЭУКАРИОТ
Основные химические вещества, входящие в состав живой ма-
терии, тождественны для одноклеточных и многоклеточных су-
ществ, тогда как специфические структуры у тех и других суще-
ственно различны. Например, такие полимеры, как протеины,
нуклеиновые кислоты, полисахариды, содержатся в клетках лю-
бых живых существ, но они специфичны для каждого вида. Так,
гликолактиловые кислоты имеются лишь в клеточных стенках
определенных бактерий; белки клеток нитчатых грибов отличны
от белков клеток животных организмов, несмотря на то, что
включают однотипные аминокислоты. ДНК, определяющая спе-
цифичность наследственных характеристик вида, также различна
у разных организмов, хотя состоит из одних и тех же кирпичи-
ков-блоков — дезоксирибозы, фосфорной кислоты, двух пурино-
вых (аденин, гуанин) и двух пиримидиновых (тимин, цитозин)
оснований.
Таким образом, на фоне величайшего разнообразия форм
живых организмов в природе не менее удивительным представ-
22
длегся химическое (точнее биохимическое) однообразие струк-
•рувиых блоков у всех этих форм. Очевидно, однообразие строи-
тельного материала является отражением эволюционных процес-
в ходе которых происходил отбор наиболее универсальных
с4®«кгур для организации, выживания и приспособления наибо-
совершенных форм жизни, возникшей на Земле около
4 йирд- лет назад.
ЛТредковые прокариоты были исходными формами для разви-
тиЙвсех других организмов, в том числе фототрофных, способ-
^Дгиспользовать энергию солнечного света (рис. 7). И если у
всех организованных частиц и клеток наследственными веще-
ствами выступают нуклеиновые кислоты, то можно предполагать,
что эти структуры сформировались в числе первых полимеров
в ходе многовековой эволюции жизни. Так, чтобы закодировать
всю информацию о 4 тыс. белков в бактерии и о 100 тыс. белков
у человека, необходимо соответственно Н0~11 и 6-10-12 г ДНК
на клетку (диплоидную у человека). Следовательно, в бакте-
риальной клетке ДНК меньше примерно в 600 раз. Число нуклео-
тидных пар в этих ДНК также различно. Например, в ДНК
кНШечной палочки определено примерно 4 млн. нуклеотидных
иар< в ДНК из клеток, млекопитающих — около 5,5 млрд. пар.
В атмосфере 21%
кислорода ——.
В атмосфере
более 0,2%
кислорода
В атмосфере
иет кислорода
В атмосфере
0.2% кислорода
В атмосфере
менее 0,2%
кислорода
В атмосфере
нет Кислорода
Конец кембрия
(500 млн лет назад)
Цианобактерии
Рис. 7. Спираль эволюции жизни на Земле (схема)
Настоящее время
Эукариотические клетки
(1,4 млрд, лет назад)
' - (2 млрд, лет назад)
--------------------------(2,8 млрд, лет назад)
Проводоросли Анаэробные аутотрофы
- (более 3,5 млрд, лет назад)
Возникновение жизни
(анаэробы)—(около 4 млрд, лет назад)
Рис. 8. Строение микробной клетки в обобщенном
виде:
1 — гранулы поли-р-окснмасляной кислоты; 2— жировые
включения; 3 — капельки серы; 4 и 5 — тилакоиды; 6-
газовые пузырьки; 7 — хроматофоры; 8 — нуклеоид; 9 —
рибосомы; 10 — цитоплазма; 11 — базальное ядро (блефа-
ропласт); 12— жгутики; 13— капсула; 14 — клеточная
стенка; 15 — клеточная мембрана; 16 — мезосома; 17 — га-
зовые вакуоли, 18 — пластинчатые структуры; 19 — углевод-
ные гранулы; 20 — метахроматин
Чтобы записать буквенную последовательность нуклеотидов одной хромосомы
кишечной бактерии, надо заполнить 2000 страниц размером 16X25 см, а для
такой же записи последовательности нуклеотидов в ДНК одной клетки человека
потребуется уже 1 млн. страниц.
Таким образом, для живых организмов характерно не только
эволюционирование во времени, но и формирование исключи-
тельно компактных структур в одиночных клетках любых пред-
ставителей микро- или макромира.
На рис. 8 схематично представлено строение клетки. Основ-
ные структурные компоненты таких клеток: клеточная стенка
(ее нет у микоплазм и протозоа), клеточная (плазматическая)
24
^выбрана, цитоплазма, ядро, митохондрии (или их аналоги),
рибосомы. Принципиальные различия между клетками прокариот
д’эукариот показаны в табл. 3.
S. || ц а 3. Сходство и различие в структурных компонентах
Ж прокариот и эукариот
Ж Структурный компонент Прокариоты Эукариоты
“ •*‘‘^ЙЙ>рмленное ядро +
'./^даточная стенка ±* ±**
Кдёгочная мембрана + +
Эндоплазматический ретикулум — +
Аппарат Гольджи и его производные — +
Мятохоидрии — +
Мезосомы 1 »»» —
* Некоторые прокариоты лишены клеточной стенки, например микоплазмы.
** Клеточная стенка отсутствует у простейших.
Тилакоиды у цианобактерий.
^УКлеточная стенка. Микроорганизмы обитают в различных
уровнях, поэтому они должны быть устойчивыми к изменениям
температуры, pH, давления, состава сред. В значительной сте-
пени такая устойчивость клеток обеспечивается клеточной стен-
кой, имеющей достаточно жесткую (ригидную) структуру. Бла-
годаря ей клетка выдерживает высокое внутреннее осмотиче-
ское давление (5...20 МПа). За счет этой жесткости клетки со-
храняют форму. В то же время существуют микробы, которые
совсем утратили клеточную стенку, — это микоплазмы [ранее
Язывали Pleuro-pneumonia-like organisms (PPLO)]. Они
еризуются большой плеоморфностью, репродуктивностью
на питательных средах (наименьший размер их репродуктивных
единиц 125...250 нм), устойчивостью к пенициллинам, неспособ-
ностью переходить в какие-либо бактериальные родительские
формы. Микоплазмы по форме могут быть кольцевидными, нит-
чатыми, палочковидными, спиральными, гранулярными.
? Отсутствие клеточной стенки может быть временным явле-
нием, возникающим под влиянием каких-то внешних факторов
(Ф&рменты, антибиотики и др.). При этом из грамотрицательных
wtepHft образуются протопласты, полностью лишенные клеточ-
ной стенки. Если стенки остаются хотя бы частично, то говорят
О сферопластах. L-Формы бактерий (по названию института
ИЙ.- Листера в Шотландии) являются дефектными по клеточной
стенке; они размножаются и образуют колонии на уплотненных
питательных средах, некоторые из них стабилизируются, а другие
ревертируют (от лат. reversio — возврат, возвращение) в исход-
ное родительские формы, особенно в присутствии 15...30 %
желатина или 2,5 % агар-агара. Такую реверсию блокируют
ингибиторы белкового синтеза, например антибиотики из группы
тетрациклинов.
25
Таким образом, микоплазмы нельзя отождествлять с прото-
пластами, сферопластами и L-формами бактерий, так как между
ними нет генетического родства. Дефектные по клеточной стенке
бактерии в отличие от микоплазм способны продолжать синтез
некоторых клеточно-стеночных веществ, присущих родительским
клеткам (например, капсульный полисахарид и М-протеин у
L-форм стрептококков).
. Установлено, чн; ригидный ьаркас г.щточной стенки мноп
слоен у (рамноложительных б.чкгерий и <> ; югюеь у fpaMojpc
дательных бактерий Толщина щенок у гамиоложиюльн).'
бактерий достигает 15...80 нм, у ; рамотрицагеллны< н\
У грибов (более выражений, чем у бактерий! к т.' точна я степь
является природным КОМПОЗИТОМ, COClOHIHliM И :<v\ фаз. ОДП
из которых выполняет роль микрофибрилляргой сирины, В1-
рая -- роль аморфного наполни для матрикса I <>.:чюпа кле^н
ной стенки у грибов достигает 1 мьм. Исполыпя фщ.мевтативны
гидролиз, можно получить протопласты из клеток . рибов. Григ
ные клеточные стенки более ригидны, чем у бактерий и пред’та:
лены в значительной части углеводными комщин'нп1чи.
В клеточной стенке грамположительных бактерий сравнитель-
но больше углеводсодержащих полимеров, чем в клеточной стен-
ке грамотрицательных бактерий, однако архитектоника ее у
грамотрицательных видов от этого не становится проще, а скорее,
наоборот, — сложнее (рис. 9, а).
Поверхность микроорганизмов, представленная клеточной
стенкой, не бывает гладкой. Это хорошо видно на снимках,
выполненных с помощью сканирующего электронного микроскопа
Рис. 9. Приближенная архитектоника клеточной оболочки грамотрицательных
бактерий (в):
1—липополисахарид, 2 — двухслойная фосфолипидная мембрана, 3 — внешний слой,
4 — оболочка, 5 — клеточная мембрана, 6 — периплазматическое пространство, 7 —
пептидогликан (ригидный слой), 8 — липопротеин. Поверхность клеток Candida lipoly-
tica в сканирующем электронном микроскопе X 12000 (б)
26
9,6). Особым разнообразием поверхности отличаются
з&р^иышевые клетки-споры.
Клеточная, или цитоплазматическая, мембрана (плазмалем-
|кикрасполагается сразу же за клеточной стенкой (см. рис. 8).
той и другой имеется так называемое периплазматиче-
ЯиРпространство, в котором могут находиться тейхоевые кис-
'лИ У грамположительных бактерий. Можно предполагать, что
ДИгяу клеточной мембраной и клеточной стенкой у микроорга-
димт отсутствует какая-либо прочная связь. Доказательством
ЛЬт относительная легкость получения протопластов у грам-
йииЬятельных бактерий и сферопластов у грамположительных
<ЯИврий. окруженных клеточной или цитоплазматической мем-
Поперечник (толщина) клеточной мембраны достигает
7,5...8 нм. Инвагинации (внутренние складки) ее у неко-
торых видов бактерий образуют специализированные структу-
ры — мезосомы. Мезосомы бывают двух типов: перегородочная
И боковая, из которых первая участвует в образовании перего-
родки, формирующейся в процессе деления клетки, и к ней при-
ЫЙЙляется нуклеоид, а вторая локально связана с клеточной
мдораной где-то сбоку и не принимает участия в образовании
Ймегородки. Функционально мезосомы отдаленно напоминают
((фтрхондрии у эукариот. У большинства бактерий складчатость
шгораны (кроме мезосом) выражена слабо. Исключение пред-
ставляют, например, фотобактерии и азотофиксаторы, у которых
осуществляется более активный транспорт электронов в процессе
биологического окисления (дыхания).
. Мембранными образованиями у прокариот являются тилакои-
d&,J)UKобилисомы, аэросомы, хлоросомы (хлоробиум-везикулы)
Л$/£боксисомы. Происхождение мембран у трех последних орга-
не^ окончательно не выяснено. Тилакоиды представляют собой
систему элементарных мембран, к внешней стороне которых при-
креплены фикобилисомы (см. рис. 8).
Дэросомы, или газовые вакуоли, имеются у фототрофных и
хемотрофных водных бактерий (цианобактерий, пурпурных серных
бактерий, зеленых бактерий, у некоторых археобактерий и др.).
Оед состоят из газовых пузырьков, представляющих собой полые
цМЙИйдрики с конусовидными концами, размерами 75Х(200...
*0Ov) нм, окруженные мембраной толщиной 2 нм. Пузырьки, как
Правило, собираются в регулярные ряды субъединиц, и возобнов-
ЩНОтся они заново, а не путем заполнения газом ранее запустев-
ших пузырьков. Через мембрану пузырьков проникают все обыч-
ные газы, поэтому поддержание их в заполненном состоянии все-
цело зависит от растворенных в среде газов. Газовые вакуоли обес-
печивают и регулируют плавучесть водных микроорганизмов.
Хлоросомы (хлоробиум-везикулы) в виде сигарообразных
пузырьков [50Х (100.. .150) нм] обнаруживают у зеленых фотосин-
"^ирующих бактерий. Располагаются они непосредственно под
клеточной мембраной. Хлоросомы окружены однослойной мембра-
ной толщиной 3...5 нм. В них сосредоточена лишь часть фотосинте-
27
тического аппарата, а именно — пигменты, выполняющие функ-
цию антенны.
Карбоксисомы, или полиэдральные тела (50...500 нм), обнару-
живают у фотосинтезирующих, у некоторых хемолитотрофных
(например, нитрифицирующих) бактерий. Они окружены одно-
слойной мембраной толщиной 3,5 нм. В них содержится рибулозо-
дифосфаткарбоксилаза, или карбоксидисмутаза, выполняющая
ключевую роль в процессе фиксации СОг.
У эукариотических микроорганизмов клеточная мембрана те:
но связана с эндоплазматическим ретикулумом (от лат. retie;,
lum — сеточка). Мембрана и ретикулум трехслойны. На наружно,
слое, обращенном в периплазматическое пространство, имею; .
одиночные субъединицы диаметром до 11 нм; на внутреннем, обрл
щенном к цитоплазме, эти субъединицы сгруппированы по 2..
и более. Предполагают, что они являются комплексами ферменте;.
Эндоплазматический ретикулум бывает гладким и шероховаты,'
В последнем случае к нему тесно примыкают риоосомы - бело;
синтезирующие образования.
За счет ретикулума клетка подразделяется на множество свое-
образных отсеков (компартментов). Эндоплазматический ретику-
лум примыкает к ядру и за счет его элементов образуется ядерная
мембрана. Он же формирует систему канальцев, пузырьков и цис-
терн, наблюдаемых в клетках под микроскопом. Количество и ка-
чество их стоит в прямой зависимости от функциональной актив-
ности клеток. Цистерны могут формировать вакуоли, в том числе
центральную, где откладываются запасные питательные вещества.
Мембрану вакуолей иногда называют тонопластом.
Мембрана хлоропластов у водорослей двухслойна. Наружный
слой ее имеет сходство с наружной мембраной митохондрий. Внут-
ренняя мембрана укладывается в форме параллельных слоев
нерегулярными группами вдоль длинной оси хлоропласта (ламел-
лы).
Хотя эндоплазматический ретикулум у эукариотических микро-
организмов развит слабее, чем в клетках растений и животных,
тем не менее он достаточно выражен у них и с ним связан аппарат
Гольджи, участвующий в образовании первичных лизосом, фаго-
сом и сегресом. Аппарат Гольджи включает параллельно упако-
ванные дисковидные пластинки-диктиосомы с валикообразными
расширениями и пузырьками по периферии.
Лизосомы бывают первичные и вторичные. Однослойные пер-
вичные лизосомы с ферментами-гидролазами формируются из
элементов аппарата Гольджи в нормальных условиях роста и
развития клеток эукариот до времени их проникновения в цент-
ральную вакуоль. Как только произошло это проникновение и
запасные вещества подверглись гидролизу, центральная вакуоль,
содержащая первичные лизосомы, становится вторичной лизо-
сомой.
Если клетка оказывается в экстремальных условиях (от лат.
extremum — крайность), т. е., когда ее физиологические функции
28
.• I мкм
Рис. 11. Простеки
0. Пероксисомы у Hansettula
jrpha
перенапрягаются, в ней образуются фагосомы и сегресомы,
ивающие кислую фосфатазу. Известны еще гидрогеносомы
ксисомы (рис. 10), окруженные однослойной мембраной
ржащие специфические ферменты.
У грибов имеются хитосомы и ломасомы. Хитосомы — это
Ждизикулы (40...70 нм в диаметре), содержащие хитинсинта-
дитосомах происходит синтез микрофибрилл хитина. Лома-
ср»йН —это производные клеточной мембраны. Формируясь на
стороне внутреннего слоя мембраны в виде
ДДЙий с пузырьками и гранулами, они затем
ЯЙну периплазматического пространства. В
««мнении ломасомы мало изучены.
4ж®яеточная мембрана и клеточная стенка у
куполообразных
выпячиваются в
функциональном
микроорганизмов
ВДЙосте составляют оболочку, переходящую у некоторых грамотри-
ЦДХедьных бактерий в простеки (от греч. лрыо — пре, пред, при,
про;.0тр<г] — вместилище, капсула, оболочка), например у Prosthe-
icrobium pneumaticum (рис. 11). У ряда микробов с оболочкой
ДНО образование капсул, жгутиков, пилей и фимбрий (от лат.
| — волосок, fimbria — ворсинка, волокно).
капсулы формируются такими видами прокариот, как лейко-
Дки (Leuconostoc mesenteroides), пневмококки (Streptococcus
fnoniae), различные бациллы (например, Вас. licheniformis),
«Которые свободноживущие в почве азотофиксирующие бактерии
V^ptobacter chroococcum) и др., а также клетками многих эука-
®ДЛ. pullulans, Cryptococcus neoformans, Lipomyces lipofer,
otorula glutinis и др.). В природных условиях некоторые пред-
«ОВКгели нормальной микрофлоры человека вырабатывают так
^“^•Ваемый гликокаликс, состоящий из длинных полисахаридных
фИррилл и обеспечивающий клеткам тесное прилипание к поверх-
»встн органа или ткани (например, Streptococcus mutans к зубной
29
Жгутики являются органами локомоции (движения) у отдель-
ных представителей прокариот и эукариот. Они характерны для
вибрионов, многих энтеробактерий, спирилл, спирохет, трипано-
сом, грибных зооспор и др. Жгутики начинаются крючком и бле-
фаропластом, или базальным телом, связанным с клеточной мем-
браной (рис. 12, а). У грибов блефаропласт часто связывается
с ядром при помощи нити (ризопласта). Блефаропласт у микробов
отождествляют с центриолей в клетках высших организмов. Как
видно из рисунка, базальное тело включает набор колец и стер-
жень. У грамположительных бактерий имеется два кольца (М и
S), у грамотрицательных — четыре (М, S, Р, L).
Число жгутиков различно у разных видов микроорганизмов;
один полярный (монотрих) у Vibrio metschnikovii, пучок жгутиков
на одном полюсе клетки (лофотрих) у Ps. aeruginosa, пучок жгути-
ков на обоих полюсах клетки (амфитрих) у Spirillum species,
по всей поверхности клетки (перитрих) у Proteus vilgaris. Диаметр
жгутиков— 12...30 нм, иногда 60 нм (например, у Bdellovibrio
bacteriovorus); длина — от 5 до 50 мкм.
У ряда низших грибов жгутики характерны для зооспор и под-
вижных гамет (иногда их называют ресничками); они имеются
также у отдельных видов протозойных организмов. Жгутики пред-
ставляют собой однокомпонентную систему с завитками и волнооб-
разными изгибами. Длина волны строго постоянна для каждого
вида, например 2,5 мкм для жгутиков Salmonella typhimurium.
У таких извитых форм, как спирохеты, подвижность обеспечивает-
ся за счет осевой нити, состоящей из двух рядов жгутиков, окру-
женных клеточной стенкой.
Если у бактерий жгутики однообразны по форме, то у грибов
выделяют два типа их: бичевидный (плетевидный), или колеблю-
щийся, и сверкающий, или блестковидный. Клетки со жгутиками
подразделяют на четыре типа: с одним задним жгутиком (колеб-
лющийся тип), с одним передним жгутиком (сверкающий тип),
с двумя жгутиками обоих типов, с двумя жгутиками колеблющего-
ся типа (рис. 12, б).
У живых клеток жгутики представляют собой полужесткие
спиралевидные роторы, приводящие клетки во вращательное дви-
жение благодаря подвижному прикреплению S-кольца к внутрен-
ней поверхности клеточной стенки и фиксации М-кольца в клеточ-
ной мембране. Вращение S-кольца относительно фиксированного
М-кольца обеспечивает жгутику вращательное движение. Причи-
на движения S-кольца окончательно не выяснена. Направленное
движение клеток (таксис) связано с функцией жгутиков, хотя,
например, в случае хемотаксиса пищевые рецепторы располагают-
ся на клеточной мембране.
Пили, или фимбрии (рис. 12, в), короче и тоньше жгутиков.
Они обнаруживаются у многих грамотрицательных бактерий и
других микроорганизмов. Их диаметр может достигать 8,5...
12,0 нм; длина — до 10,0 нм (у некоторых до 2 мкм). Пили могут
быть у жгутиковых и безжгутиковых организмов. Различают прос-
зо
Рис. 12. Жгутики:
а— часть бактериального жгутика — крючок (кольца: М — в мембране, S —
стабилизирующее надмембранное, Р — в пептидогликановом слое, L — в ли-
пополисахаридиом слое); б — жгутики грибных зооспор (1 — задний колеблю-
-. - щегося типа, 2 — передний сверкающего типа, 3 — жгутнки обоих типов,
4 — два жгутика одного типа); в — sex-пили и фимбрии у Salmonella
Species
Ми половые волоски, или sex-пили, являющиеся полыми труб-
ками. Число последних обычно не более двух на одну клетку. Пили
Участвуют в адгезии (прилипании) и в половой конъюгации кле-
У0К?»(см. гл. 4).
Цитоплазма (гиалоплазма) более гомогенна у прокариот,
ч®И. у эукариот. Например, в растущем мицелии верхушки гиф
ЧЗДйостью заполняются цитоплазмой, тогда как на определенной
Дистанции от них формируются вакуоли, особенно в период ста-
₽«Вйя клеток. Вакуоли заполняются клеточным соком, в них откла-
дываются резервные вещества — фонды пуринов и пиримидинов,
аминокислот, полифосфатов, некоторых витаминов. Вакуоли обес-
31
печивают тургор клетки, необходимый для ее роста и поддержа-
ния внешней формы, а также частично выполняют роль лизосом
в клетках высших организмов. В цитоплазме происходит сборка
рибосом, синтезированных в ядре.
Цитоплазма — гетерогенная система, в которой различают
дисперсионную среду и дисперсную фазу. Дисперсионной средой
является вода с растворенными в ней низкомолекулярными ве-
ществами, а дисперсной фазой — высокомолекулярные соедине-
ния: белки, липиды, углеводы и их конъюганты. Водные (гидрат-
ные) оболочки вокруг белковых и углеводных молекул стабилизи-
руют эти молекулы и цитоплазму в целом. Соотношение свободной
и связанной воды неодинаково у разных микроорганизмов и ме-
няется у одного и того же штамма с возрастом, в зависимости от
условий выращивания. Если связанная вода удерживается моле-
кулами белков и углеводов преимущественно за счет водородных
связей, то свободная вода — капиллярными силами в тончайших
канальцах эндоплазматического ретикулума и в трубчатой системе
разного рода мембран. Ацетон, метанол, этанол легко отнимают
свободную воду и воду гидратных оболочек, приводя к коагуляции
макромолекулярные структуры.
В цитоплазме одни вещества находятся в виде истинно ;
раствора (неорганические соли, витамины, аминокислоты, мот
зы), а другие — в коллоидном состоянии (белки). Внутриклеп
ное осмотическое давление, например, бактериальной кле: '
эквивалентно 10...20 %-ным растворам сахарозы (такие ра-
воры сахарозы используются для стабилизации сферопластсг ;
протопластов in vitro).
Многие молекулы в цитоплазме заряжены, из них белковые
несут положительный и отрицательный заряды одновременно.
Следовательно, гидратные оболочки, коллоидные свойства и элек-
трические заряды обеспечивают высокую стабильность цитоплаз-
мы в целом. В интактных (от лат. intactus — нетронутый) клетках
вязкость цитоплазмы близка вязкости воды, но при делении клеток
она резко возрастает — возможен переход золя в гель — тиксот-
ропия (отлат. tixis — прикосновение, trepo — изменяться), напри-
мер при формировании нитей ахроматинового веретена в размно-
жающихся клетках эукариот.
Свободные белки цитоплазмы имеют обычно коэффициент се-
диментации между 3S и 6S (1S = 1 сведберг-единица =
= КГ13 см/с/ед поля). С белками связана основная ферментатив-
ная активность.
Такие вещества, как Н2О, СО2, N2, SO«~, NCXf, РО3-, НРО< ,
Н2РОг, NHt и др., поступают из внешней среды и являются акти-
ваторами обменных процессов в клетках, а также предшествен-
никами молекул ацетата СН3СОО~, карбамоилфосфата H2N—
—СО—О—РОГ, а-кетокислот (например, R—СО—СОО-), мала-
та ~ООС—СНг—СНОН—СОО~, малоната ~ООС—СНг—СОО .
рибозы НОН2С—(СНОН)з—СНО, фосфопирувата НзС—СО—
—СО—ОРОГ- Эти молекулы оказываются промежуточными в об-
паэбйании строительных блоков, к числу которых относятся ами-
нокйёлоты, глицерин, жирные кислоты, простые сахара (монозы
я олигозы), мононуклеотиды.
Д Вещества-предшественники имеют молекулярную массу от 18
Ж 150 Да, промежуточные молекулы — от 50 до 250 Да, строитель-
нйе блоки — от 100 до 350 Да. Из строительных блоков в цитоплаз-
ме синтезируются главнейшие макромолекулы, в основном обес-
дечивакиЦие структурную организацию микробной клетки. Это
белки, липиды, нуклеиновые кислоты, полисахариды, молекуляр-
ная масса которых достигает 1О3...1О9 Да (или 1...106 кДа). Из
макромолекул формируются надмолекулярные ферментные, рибо-
сбмальные и другие комплексы с М = 103...106 кДа, обесиечива-
Ющие многие функции клетки.
.Некоторые надмолекулярные комплексы объединяются в виде
Понятие органеллы или органоиды достаточно условное,
ТЙК’йак надмолекулярные комплексы входят в состав не только
достоянных, но и временных структурных компонентов клетки. До
яёДайнего времени к ним относили лишь реплицирующиеся внутри-
клеточные мембранные образования эукариот (ядро, митохондрии,
хлфропласты), полагая, что их нет у прокариот, клетка которых
сайа является как бы единичной органеллой. В настоящее время,
ШЗфдя из положения о том, что клеточная стенка вместе с просте-
камгй и капсулой может быть потеряна клетками микробов в опре-
делённых условиях и что клеточная мембрана выступает основой
ЖЙ* форм ирования ряда мембранных структур, к органеллам сле-
относить постоянные внутриклеточные образования. В клет-
'эукариот такими образованиями являются клеточная мембра-
на, верный аппарат, аппарат Гольджи и его производные — ли-
и хитосомы, пероксисомы, митохондрии, гидрогеносомы,
ГЙдаэсомы; в клетках прокариот — клеточная мембрана, ядерный
яйшрат, мезосомы, тилакоиды и фикобилисомы, хлоросомы,
гДЗбвые пузырьки-аэросомы, карбоксисомы; жгутики и волоски
(У Прокариот и эукариот).
В самом деле, ядро эукариотической клетки имеет свою обо-
лочку, а ядро прокариотической не имеет ее, однако от этого не
изменяется роль ядра как главенствующей органеллы. В равной
Степени аналогия возможна между тилакоидами у цианобактерий
и хлоропластами в растительных клетках, где осуществляется
процесс фотосинтеза.
Свободные липиды, гранулы ?гр.--з. Kf :дму .
Гликоген), полифосфатсо.-щржзн. ИГ : ТШМЫ (н. к,:',! у
хроматин), у некоторых бщ-тущ-ш чрщтал’-ы.
могут накапливаться и цятопл.ч « «кле -и-
чё'ний. Они выявляются, как правяит при выоа.щивш-и'- V'-
брРйнизмов в средах .тщ» t «•г.-чь'* ц..(
йвЩбства, а исчезают ы-'- г,хч.-.
ц Жировые включения у ряда бактерий представлены в виде
гранул поли-0-гидроксимасляной кислоты, а также другими липи-
дами. Поли-Р-гидроксимасляная кислота синтезируется из угле-
2-1617 33
родсодержащих соединений при дефиците источников азота в сре-
де. Гранулы ее покрыты мембраной (см. рис. 8), но их не относят
к числу органелл.
Включения располагаются либо в цитоплазме (у прокариот),
либо в вакуолях (у эукариот). Поскольку ядро у прокариот не
имеет своей оболочки (нуклеолеммы, или кариолеммы) и ядерной
цитоплазмы (кариоплазмы), цитоплазму клетки можно отожде-
ствлять с протоплазмой.
Ядерный аппарат у микроорганизмов. В ядре клеток микробов
(как и в клетках других организмов) содержится вся основная
наследственная информация в форме ДНК. Эта информация реа-
лизуется в синтезе специфических белковых молекул. Данный
процесс разобщен во времени и связан с различными структурами,
которые можно отнести к ядерному аппарату: у прокариот — нук-
леоид, рапидосомы (у некоторых видов), перегородочные мезосо-
мы, рибосомы (их рассматривают также в качестве самостоятель-
ных органелл, а не в составе ядерного аппарата); у эукариот —
ядро, ядрышко, нуклеолемма, поросомы, центриоли, митотический
аппарат, рибосомы.
Ядро. В клетках прокариот в цитоплазме ядро (нуклеоид)
представлено в виде области с высоким содержанием ДНК (см.
рис. 8) в виде замкнутой нити. Размеры этой нити, например,
у кишечной палочки—1,4-106ХЗ нм, масса 1-Ю-14 г и М =
— 6-106 кДа. Такую ДНК длиной ~ 1,4 нм в разомкнутом состоя-
нии (в ~ 500 раз длиннее бактериальной клетки, вмещающей эту
ДНК) рассматривают единственной хромосомой. Она прикреп-
ляется в одной точке к перегородочной мезосоме, играющей ключе-
вую роль в репликации двух сестринских хромосом.
Саморепродуцирующимися автономными (внехромосомными
генетическими элементами в клетках многих микроорганизмов
являются плазмиды. К плазмидам относят F-факторы (от англ,
fertile — плодородный); R-факторы (от англ, resistance — сопро
тивление); Col-факторы (от англ, colicinogenicity — колициноген
ность); профаги; плазмиды деградации некоторых органически?
соединений (камфоры, салициловой кислоты, октана и т. д.); крип
тические, или скрытые, плазмиды. Все известные плазмиды
двухцепочечные ДНК (М ~ Г-104...7- 104 кДа), содержащие д;,
100 генов. Исключение представляет киллер-плазмида дрожжей
являющаяся двухнитевой РНК. Плазмиды не являются необходи
мыми структурами для клеток-хозяев и могут быть ими потерян:-
или, напротив, приобретены извне.
Плазмиды могут существовать в клетке и в интегрированном
состоянии с бактериальной хромосомой, сохраняя при этом способ-
ность переходить к автономии. Будучи интегрированными, они не
являются частью линейной структуры хромосомы. Автономность
и интегрированность плазмид исключают друг друга. Особенности
отдельных классов плазмид выявляются по характеру их поведе-
ния в автономном состоянии и фенотипическому выражению этого
состояния.
34
IМирной структурой для ядра у некоторых бактерий (Proteus,
'^bacterium, Pseudomonas, Saprospira) являются рапидосомы
j^T. rapidus — быстрый) или микротрубочки (микротубули).
Цосомы могут быть в виде полого и неполого цилиндров раз-
М 33X225 нм.
Викробные клетки эукариот содержат оформленное ядро, от-
Виое от цитоплазмы двухслойной мембраной (нуклеолеммой,
Ьомембраной), связанной с эндоплазматическим ретикулу-
Ядро содержит ДНК и основные белки — гистоны. Хромо-
щй материал в виде компактных гранул, включающий по од-
ыолекуле ДНК на хромосому, погружен в ядерную. основу.
В Шйрфазный период хромосомы мало различимы, хотя они не
мышр свою индивидуальность. Митотический аппарат в это
ним*- не обнаруживается. Молекулярная масса ДНК одной
хммйсомы (из 17) у Saccharomyces cerevisiae составляет
вИГ кДа.
эукариотических клетках форма ядра чаще округлая, его
в среднем 1...6 мкм в диаметре. В ядре имеется ядрыш-
место образования рибосом и рибосомальных РНК. Оно
Явонт из плотных гранул (около 15 нм в диаметре) и тонких
толщиной 4...8 нм, погруженных в виде сеточки в аморф-
1|Ьодцество.
* Оболочка ядра включает внешнюю и внутреннюю мембраны,
в^Ци^йе поросомы, или ядерные поры, диаметром 10... 12 нм. Они
эщэдкняются аморфно-фибриллярной диафрагмой-пробочкой,
«меняющей над плоскостью поры в обе стороны на
® нм. На 1 мкм2 ядерной оболочки дрожжей приходится
iQt>>15 пор.
й^йИйсло ядер в клетках весьма различно. Так, кокковые фор-
Mli-бактерий содержат, как правило, по одному нуклеоиду, па-
ЭДЙЖОВИдиые формы — два и кратное двум число ядер как след-
ИМбе десинхронизации скорости роста и скорости деления клет-
|Цн11?гетативные клетки высших грибов могут содержать одно
ЯЙИмыпе ядер. Низшие грибы полинуклеарны (от лат. poly —
В nucleus — ядро). Такие грибы называют еще ценоцит-
ериод митотического деления, когда разрушаются яд-
и ядерная мембрана, в эукариотических клетках образует-
'отический аппарат — биполярная структура в форме
а. Эта структура состоит из микротрубочек 20...30 нм в
давМетре, включающих глобулярные белковые субъединицы
жг 50...60 кДа). Микротрубочки имеют упорядоченную струк-
Wh обладают сократительными свойствами. Они выходят из
ИФЧфиолей, расположенных на полюсах клетки. Центриоли так-
ЯНЙЙ^еют цилиндрическое строение и состоят из девяти пар
^•ЯфОтрубочек. Очевидно, распределение хроматид по полюсам
*яегок индуцируется в центриолях и осуществляется с помощью
«ЙИ^ена. При формировании дочерних ядер веретено распа-
Щгся.
35
30S
а бе
Рис. 13. Рибосома (а) и ее субчастицы — большая (б),
малая (в)
Рибосомы. В клетках прокариот и эукариот содержится боль-
шое количество нуклеопротеиновых частиц, или рибосом (у бак-
терий — в среднем 104, у грибов и других эукариот — 105 — 106).
Размер бактериальных рибосом находится в пределах 20Х30Х
ХЗО нм; рибосомы в клетках эукариот крупнее — 20X40X40 нм,
их молекулярная масса 2,7-1(г — 4,4-103 кДа (рис. 13). Бакте-
риальные рибосомы (равно как и рибосомы митохондрий и хло-
ропластов у эукариот) в большинстве своем относятся по кон-
станте седиментации к типу 70S (для малых субъединиц — 30S,
для больших — 50S); грибные — к типу 80S (для малых субъе-
диниц — 40S, для больших — 60S).
На рибосомах осуществляется синтез специфического белка.
В этом случае они агрегируются в полисомы с более высокими
константами седиментации. У эукариот рибосомы образуются в
ядре клетки.
Митохондрии. Основные реакции, сопровождающиеся высо-
коэнергетическим выходом, протекают у эукариотических организ-
мов в органеллах, называемых митохондриями (от греч. щтсоо —
нить; xtovSpwo — зернышко, крупинка). Предполагают, что около
2,5...3 млрд, лет назад некая более крупная анаэробно живущая
клетка поглотила более мелкую клетку — бактерию, каким-то
образом видоизменившуюся в направлении возможности исполь-
зования кислорода в дыхании. Возникло эндосимбиотическое
взаимоотношение, при котором, например, фотосинтетические
предки эукариот стали получать больше энергии, чем при ана-
эробных процессах, осуществляющихся у них до симбиоза. Такие
клетки стали прародителями всех современных клеток эукариот,
в которых симбионтные в прошлом аэробные бактерии трансфор-
мировались в митохондрии, но сохранили свои гены.
Митохондрии (рис. 14) имеют удлиненно-овальную форму 2...
5X0.5 мкм. На поперечном разрезе видны внутренняя и внеш-
няя мембраны. Первая из них сильно складчатая, и складки ее
пронизывают жидкое содержимое (матрикс); тогда как вторая —
гладкая. Она сходна с наружной мембраной хлоропластов у
водорослей.
В митохондриях происходит собственный синтез белка, однако
большая часть их структурных и некоторых других белков за-
36
рована в ядре эукарио-
жой клетки, т. е. функ-
’митохондрий в значи-
зй мере зависит от ге-
клетки.
:ы
1. Что такое «Химиче-
{ая микробиология» и какое
есто она занимает в ряду
икробиологических и хими-
гских дисциплин?
2. Какие объекты (кле-
зчной и неклеточной орга-
йЗацйи) подлежат изучению
курсе химической микро-
иологии?
3. В чем принципиальное
кристы
межмембранное
пространство
матрикс
Рис. 14. Митохондриальная гранула в
разрезе
наружная
мембрана
внутренная мембрана
... з. В чем принципиальное различие между прокариотами и эукарио-
тами? К какой группе из них относятся археобактерии, фото- и скотобакте-
" рии, микро- и макромицеты, трипаносомы, водоросли и лишайники?
-уу 5. Что представляют собой вироиды, фаги, вирусы? Почему они отно-
сятся к микробам?
6. Какое теоретическое и практическое значение имеет химическая
микробиология?
и 7. Что такое органоиды (органеллы) в клетках микроорганизмов и
. какие структурные компоненты клеток к ним относятся?
н 8. Какое строение имеет клеточная- стенка у прокариотических микро-
организмов?
9. Какое строение имеет клеточная стенка у эукариотических микро-
• организмов?
. 5 10. Какова архитектоника мембран
микроорганизмов?
11. Что такое компартментализация
’ микроорганизмов она имеется?
V 12. Что представляют собой ядро и
иизмов?
13.
14.
15.
и мембранных образований у
клеток, ее значение и у каких
ядерный аппарат у микроорга-
в
Каковы химический состав цитоплазмы и ее функции?
Что такое плазмиды? Какие виды плазмид известны?
Имеется ли ядро у вирусов?
а
2
ИЯ МИКРООРГАН
•АЛЫЕ И БОЛЬШИЕ МОЛЕКУЛЫ КЛЕТОК
•шкробы, как и все прочие организмы, возникли в процессе эво-
из элементов, широко представленных на Земле. Меньшее
ИйЖ’болылее количество этих элементов обычно обнаруживается
•'ЖИКробных клетках. К основным элементам (органогенам) от-
веятся азот, водород, кислород и углерод, на них приходится от
атомных долей, %.
АгГруппу микроэлементов составляют калий, кальций, магний,
Натрий, сера, фосфор, хлор; на них приходится от 0,02 до 0,1
Дойных долей, %. Ультрамикроэлементы — бор, ванадий, желе-
37
зо, кобальт, кремний, марганец, медь, молибден, цинк — состав-
ляют менее 0,001 атомных долей, %.
Основные элементы обладают рядом примечательных свойсть
!) они самые легкие среди элементов, способных образовывав
прочные ковалентные связи благодаря спариванию электроне:
(прочность ковалентной связи находится в обратно пропорции
нальной зависимости от атомных весов элементов); 2) легк ,
взаимодействуют друг с другом; 3) азот, кислород и углерод м<:
гут образовывать ординарные и двойные связи, благодаря kots
рым резко приумножается число образуемых ими соединений
4) углерод способен образовывать тройные связи с другими атс
мами углерода и азотом; 5) углерод может образовывать угле
род-углеродные связи, благодаря чему возможно формирован»*
бесчисленного количества разнообразных органических молекул.
6) соединения углерода обладают различной трехмерной структх
рой благодаря образованию тетраэдрической конфигурации вс
круг каждого атома углерода за счет спаренных электронов.
Существует еще понятие универсальные элементы. К ним от-
носят азот, водород, железо, калий, кислород, натрий, серу, угле-
род и фосфор. Они называются универсальными потому, что
имеют структурно-функциональное значение для всех видов жи-
вых клеток. Другие элементы (микро- и ультрамикроэлементы)
рассматривают как минорные: Си, Со, Na, Cl, Мп, Mo, Zn, Se,
Ni, W. В процессе эволюции жизни на Земле ионные состав и ба-
ланс в живых системах отличались (отличаются и теперь) своим
постоянством. За миллиарды лет со времени возникновения жиз-
ни на нашей планете во всех клетках сохраняется более или ме-
нее постоянный ионный баланс, характерный для морской воды
(при меньших концентрациях). Это, очевидно, и есть доказатель-
ство того факта, что жизнь впервые возникла в воде (признак,
сохранившийся даже у человека).
Все перечисленные выше элементы образуют молекулы, кото-
рые можно подразделить на две условные группы: малые и боль-
шие молекулы. К первым относят молекулы-предшественники,
поступающие в клетку извне: Н2О, СО2, N2, Mg2+, Са2+, NOT,
SO2-, РО2-, С1_, К+ и другие ионы; промежуточные молекулы
органических кислот, рибозы; молекулы таких строительных бло-
ков, как аминокислоты, мононуклеотиды, монозы и низшие оли-
гозы (простые сахара), глицерин, жирные кислоты. К большим
молекулам или макромолекулам относят нуклеиновые кислоты,
белки, полисахариды и липиды. Все они являются молекулами
полимеров.
На примере Е. coll установлено, что малые молекулы состав-
ляют 73 % от общей массы клетки; из них на воду приходится
70 %, на неорганические ионы — 1, на прочие малые молекулы —
2 %. Макромолекулы составляют 27 % от общей массы клетки;
из них на белки падает — 15 %, на РНК — 6, на углеводы — 3,
на жиры — 2, на ДНК — 1 % массы клетки. Такое соотношение
молекул обеих групп характерно для большинства типов клеток.
38
молекул каждого вида, приходящееся на одну клетку,
оно следующее: неорганических ионов — 12, молекул про-
бочных соединений и строительных блоков — 500, белков —
РНК — 1000, углеводов — 50, жиров — 40, ДНК — 1 [т. е.
'малых молекул (без воды) —512, макромолекул — 4091].
этих цифр можно сделать следующие выводы: вода как
•сионная среда количественно преобладает в клетках микро-
омов (как и других организмов); на макромолекулы при-
fl большая доля массы клетки, чем на малые молекулы, не
воды; белки как продукт функции генов преобладают
угими макромолекулами; молекулы ДНК, хранящие и пе-
дие генетическую информацию, составляют наименьший
ft среди макромолекул.
Малые молекулы
ММ, Молекула воды относится к малым молекулам, ее значение
^Жизнедеятельности микроорганизмов огромно. Количество воды
|aзныx клетках колеблется от 60 до 90 %, даже в спорах бакте-
I она содержится до 18...20 %. Молекулы воды легко взаимо-
'Ствуют, образуя межмолекулярные водородные связи. Для
дой воды (льда) известно не менее 10 модификаций: I, 1с,
ПП и стеклообразный лед, в которых молекулы воды соеди
пены водородными связями О—Н...0 и четырьмя соседними мо-
40даами так, что атомы кислорода располагаются в углах и
центре тетраэдра. Следовательно, все формы воды представляют
собой огромные макромолекулы. В этом кроется диалектическое
единство противоречия понятий микро- и макромолекула воды.
ЭЙёргия водородной связи равна 88,89-103 Дж/моль, что значи-
tWtbHO меньше энергии ковалентной связи между кислородом и
ородом (377,1 • 103Дж/моль), но больше индукционных ди-
ьяых и дисперсионных сил (или ван-дер-ваальсовых сил).
5>То важно для структуризации определенных молекул и их функ-
Ц1К>нирования в живых системах микробных клеток. Например,
ЦЦИ1Де гидратных оболочек вода экранирует противоположно
«ряженные молекулы и способствует их стабилизации в диспер-
«Юнной среде. Это в равной мере относится к гидратационной
|Яве, располагающейся вокруг ионов. Вода имеет высокие пока-
Цубли диэлектрической проницаемости, поверхностного натяже
>и теплоемкости, благодаря которым она выступает раство-
ЙЙЙВДем для многих веществ, электростатическим и тепловым
дИВЙ>ОМ. Как диэлектрик вода растворяет неорганические соли,
фЙРЮЦИирующие на ионы; она растворяет алифатические спирты
К» многие ароматические и гетероциклические произ-
окое поверхностное натяжение воды отражает существен-
^^^Йологический эффект живой материи — способность проти-
резким колебаниям внутренней среды при попадании
внесении) в нее каких-либо других веществ. Если такие
39
вещества заметно и резко изменяют поверхностное натяжение
воды, то они, как правило, губительно действуют на микроорга-
низмы (катионные и анионные поверхностно-активные вещества)
или вызывают какие-либо эффекты, не проявляющиеся в физио-
логически нормальных для клетки условиях (неионогенные по-
верхностно-активные вещества). Высокая теплоемкость воды
предохраняет клеточные структуры от термической инактивации
в допустимых, пределах. Известно, что микроорганизмы приспо-
собились жить в определенных температурных интервалах и, сле-
довательно, в показатели теплоемкости их дисперсионной (вод-
ной) среды природа внесла соответствующие коррективы (мута-
ции) за счет дисперсной фазы. Большинство микроорганизмов
относят к группе мезофилов. Оптимум температуры для них
20...45 °C. При температуре выше 45 °C лучше растут термофилы,
а ниже 20 °C — психрофилы.
Приведенное в табл. 4 деление микробов в отношении темпе-
ратур достаточно условное, так как указанные интервалы часто
перекрываются.
Таблица 4. Разделение микроорганизмов по отношению к температуре
Группа
Представители
Интервалы температур (°C)
Психрофилы
Мезофилы
Термофилы
Вас. globisporus
Xanthomonas
phormiicola
Candida species
E. coli
Lactobacterium
lactis
Neisseria gonor-
rhoeae
Вас. subtilis
Вас. coagulans
Вас. stearother-
mophilus
* До 25 °C; ** до 55 °C; *»• до 65 °C; *«• от 35 до 75 °C.
Как следует из табл.. 4, можно выделить факультативные
психрофилы, для которых верхний температурный предел превы-
шает 20 °C; облигатные психрофилы, не растущие выше 20 °C;
стенотермофилы, не растущие ниже 37 °C; эвритермофилы, расту-
щие ниже 37 °C. Снижение количества воды в некоторых клеточ-
ных образованиях (споры) повышает их термоустойчивость.
Термофилы в сравнении с мезофилами содержат в клетках
меньше денатурирующихся белков и больше насыщенных жир-
ных кислот, чем ненасыщенных.
40
образом, теплоемкость клеток разных микробов явля-
Зийготносительно стабильным свойством и в значительной мере
дх^поотся содержанием в них воды.
^Е^соответствии с законом действия масс константа диссоциа-
ЯК^иды равна 1-10^ 14 при 25 °C: Кд= Hg» , в чистой
ДрН+1 = [ОН“] = 1 • 10~7, pH = — 1g [Н+] = 1g-^= 7,0.
ЗКичения pH цитоплазмы интактных клеток микроорганизмов
ЗИГбыть выше 7 (у бактерий) и ниже 7 (у грибов), однако
ДКиения от нейтральной зоны не столь значительны. Цито-
клеток различных микробов обладает выраженными бу-
ыми свойствами, поэтому возможные колебания pH во
ней среде не очень заметно отражаются на показателях pH
нищфазмы. Это связано с тем, что вода усиливает диссоциацию
электролитов или полиэлектролитов, каковыми являются
макромолекулы живых клеток. Большинство бактерий
пфйдночитает среды с pH около 7,3...7,5, большинство грибов —
ftvu.6,5, хотя исключения имеются в разных группах микроорга-
fhj$MOB. Например, для роста холерного вибриона наиболее опти-
Шдоым является pH 8,00...10,0, для отдельных видов Candida
>,0...5,6.
(^Диоксид углерода. Из числа малых молекул (кроме воды)
$$№,.£02 значительна. В молекуле СОг все три атома приобре-
тают октет электронов
20: + .с.—► :д::С::6:
^‘Цианобактерии, зеленые и красные водоросли используют
В качестве единственного источника углерода. Вода в реак-
является донором электронов, а СОг — их акцептором
4Й1Г ' 6Н2О + 6СО2--------------. (СН2О) 6 + 6О2
цианобактерии
{следствии происходит синтез глюкозы, вступающей в соот-
ггвующие реакции обмена веществ.
(1л я пурпурных и зеленых бактерий в отличие от цианобакте-
"донором электронов выступают сероводород и газообразный
^>од; или для пурпурных — органические соединения (но не
Й). В результате, например, из H2S образуется элементарная
i. накапливающаяся в форме капелек внутри клеток (у Ch.ro-
iutn species) при последующем выталкивании их (экзоцитоз)
„В клетках не откладывающаяся (у Chlorobium species):
• 2H2S 4- CO2---------------------CH2O + H2O + 2S
IV пурпурные серные и зеленые
, ’ серные бактерии
затем окисляется до сульфата. Суммарная реакция будет
Следующей:
H2S 4-2СО2-р2Н2О— (CH2O)2 + H2SO4
Схематичная реакция с газообразным водородом — донором
электронов — может быть представлена в следующем виде:
2Н2+со2--------------------► СН2О4-Н2О
пурпурные несерные и
зеленые несерные бактерии
У пурпурных несерных бактерий донором водорода являются
какие-либо органические вещества, например изопропанол:
2СН3СН (ОН) СНз + СО2----------► СН2О + 2НзС—СО—СНз + Н2О
пурпурные несерные
изопропанол бактерии ацетон
Все указанные реакции фиксации СОг осуществляются авто-
трофными (от лат. autos — сам, trophe — питание) организмами;
однако известны также реакции гетеротрофной (от лат. heteros —
другой) фиксации, например в анаплеротической (возмещаю-
щей) реакции Вуда и Беркмана у некоторых пропионовых бак-
терий:
Пируват + СО2 4- АТФ Оксалоацетат -|- АДФ 4- Фн
Приведенные примеры доказывают огромную роль СОг в жиз-
недеятельности различных групп микроорганизмов, а способность
использовать СОг в качестве простейшего источника углерода
сохранилась с древних времен до наших дней у фототрофов
(в том числе — анаэробных).
Азот. Для синтеза жизненно важных молекул (нуклеиновых
кислот, белков) микроорганизмы нуждаются в азоте. В качестве
малой молекулы N2 в свободном состоянии находится в воздухе
(~78 %) и эволюционно в числе первых его потребителей стали
цианобактерии. В молекулярном азоте каждый атом приобре-
тает октет электронов, причем шесть из них являются общими
для двух атомов :N: -N: , за счет такого октета молекула стаби-
лизируется в целом. Оказалось, что связанного азота в виде
аммонийных и нитратных солей в биосфере недостаточно для
покрытия дефицита его в образовании биомассы. На первых эта-
пах эволюции не было и не могло быть того круговорота азота
в природе, который происходит в настоящее время, так как тогда
отсутствовали растения и животные организмы. Лишь позднее
цианобактерий возникли свободноживущие микроорганизмы и
микробы-симбионты, способные усваивать молекулярный азот.
К свободноживущим аэробным азотофиксаторам относятся азо-
тобактеры, актиномицеты, некоторые вибрионы, микобактерии,
спириллы, спирохеты. Из анаэробов азотофиксаторы известны
среди клостридий, цианобактерий, фотосинтезирующих бактерий.
Азотофиксаторы—симбионты, ризобактерии и актиномицеты —
выявлены в клубеньках бобовых растений, болотного мирта,
лоха, облепихи, ольхи.
Представителями симбионтов являются лишайники, в которых
цианобактерии и зеленые водоросли фиксируют молекулярный
42
IВ трансформируют его в более сложные азотистые соеди-
1етки.
ганические соли — нитраты и нитриты — могут потреб-
иногими грибами и бактериями в качестве источников
[ри этом нитратный азот восстанавливается при участии
а нитратредуктазы, а затем при участии нитритредук-
грит восстанавливается до NH3.
ация свободного азота из воздуха и восстановление
j (нитритов) до аммиака осуществляется с участием
ов, содержащих в своем составе молибден и железо или
юлибден.
другие малые молекулы-предшественники (неорганиче-
;оли магния, кальция и калия, фосфаты, сульфаты и др.)
Одимы для синтеза молекул — строительных блоков типа
(уклеотидов, серосодержащих аминокислот, а также для
уздврения разнообразных каталитических процессов. Магний,
цвиример имеет отношение более чем к 20 ферментативным
«исциям, он же необходим для функционирования рибосом; ка-
важен для синтеза белка; кобальт входит в состав витамина
который может содержаться в ферментах; кальций — важ-
ЙМ компонент эндоспор у бактерий; медь и железо имеют пря-
цюе отношение к цитохромам, некоторым пигментным структурам
фйгёнанинам, гемопротеинам и др.). Замечено, что металлы с
Мюмным весом более 55 не активируют деятельность биокатали-
ЗМГГОров. Катионы ртути и серебра токсичны для большинства
ММфобов. Так называемое олигодинамическое действие некоторых
фйаллов (например, серебра) связано с токсическим действием
ЖЛЫх катионов, тогда как тяжелые металлы необратимо блоки-
руют белковые молекулы, содержащие сульфгидрильные груп-
WSH
По отношению к неорганическим солям в среде микроорга-
низмы подразделяют на крайние галофилы, умеренные галофилы,
1рские организмы и негалофилы, которые выдерживают соот-
гственно следующие концентрации NaCl: 5...36 % (насыщен-
Й раствор); 0...20,5 %; 0,1...5 % и 0...4 %. Например, к первым
росится Halobacterium salinarium, ко вторым — Paracoccus
lodenitrif leans, к третьим — Pseudomonas marina, к четвер-
М, — Е. coll.
Небольшие молекулы и ионы поддерживают оптимальный
Уровень обмена веществ в клетках, они вносят определенный и
‘Существенный вклад в осморегуляцию микроорганизмов. Если
®1Утриклеточное осмотическое давление меньше внешнего, то
•ада
выходит из клетки, объем цитоплазмы уменьшается и кле-
•УНИая мембрана повреждается. У грамположительных бактерий
У^^очная стенка при этом отстает от клеточной мембраны. Та-
процесс называют плазмолизом. Вещества (электролиты и
^Ййектролиты) в водных растворах проявляют различную осмо-
Лйцтость. Осмолярным называют одномолекулярный раствор
•Жального неэлектролита; осмотическое давление его равно
43
2986,4 Па при О °C, а температура замерзания воды в нем сни-
жена на 1,86 °C.
Как правило, внутриклеточная осмолярность всегда превы-
шает осмолярность среды, что зависит от накопления в клетках
ионов калия. Морские бактерии нуждаются в высоких концентра-
циях ионов натрия, который обеспечивает нормальную функцию
транспортных механизмов, поддерживает стабильность и актив-
ность ферментов. Постоянство внутриклеточной концентрации
даже в условиях низких концентраций ионов калия во внешней
среде обеспечивается за счет действия специфических активных
транспортных систем, локализующихся в мембранах.
В практике выращивания микроорганизмов в лабораторных
и производственных условиях в водопроводную воду перед сте-
рилизацией вносят основные элементы и микроэлементы, будучи
уверенным, что ультрамикроэлементы всегда находятся в воде
или в других неорганических солях, используемых в этих слу-
чаях. Лишь в специальных опытах все элементы вносят в дис-
тиллированную воду в виде химически чистых ингредиентов. Из
рассмотренных малых молекул-предшественников (НгО, СО2, N2,
неорганические соли) образуются промежуточные соединения и
строительные блоки, также относящиеся к разряду малых мо-
лекул.
Органические кислоты как промежуточные соединения явля-
ются исходным материалом для биосинтеза ряда строительных
блоков (аминокислот, глицерина, жирных кислот, мононуклеоти-
дов, моноз и низших олигоз, или простых сахаров). Так, уксусная
и малоновая кислоты используются в клетках для синтеза глице-
рина и жирных кислот; пальмитиновая кислота становится родо-
начальником всех других жирных кислот (олеиновой, стеарино-
вой, лауриновой и др.) или их альдегидов; глицерин входит в
состав липидов.
-Ou;
10!
имеет
Атом серы в некоторых аминокислотах первично
генное происхождение из малых серосодержащих молекул неор-
ганического ряда. В последующем он может переноситься с по-
экзо-
44
-льмцл ферментативных реакций с одних молекул аминокислот
5адл»ругие. Например, цепь цистеина происходит из серина, а
из метионина.
Ж^ировиноградная кислота является основным продуктом в об-
ЯЬшх процессах, связанных с превращением глюкозы как ис-
|Кдоа углерода для многих гетеротрофных микроорганизмов,
идет прежде всего о фосфоенолпирувате с макроэргической
Ku (~):
Н2С=С—соон
О~РО3
qHMtftp в него включается из АТФ, а в АТФ — экзогенно из не-
«ф^анического фосфата. Первичный синтез пирувата из простей-
ШМ малых молекул у ряда организмов происходит путем обра-
зования его из многих аминокислот у гетеротрофных видов:
L-Треонин
л
о
Ж ;
lii
Глицин
L- Серин -•-
I
н3с—со—СООН
пируват
СН3—СН(ОН)—СН—СООН
| nh2
сн2—соон
NHj
!
НО—СН,—CH—СООН
NH2 |
HS—CH(NHj)—СООН
L- цистеин
S—СН2—СН—СООН
I
S—СН2— СН—соон
nh2
£-цистин
Н3С—СН (NH,)—СООН
аланин
<10. л. о
малым молекулам промежуточных соединении относится
«Кронная кислота НООС—СНг—СН(ОН)—СООН, или малат.
ЖЗсфоенолпируват и малат участвуют в синтезе углеводов через
ИХ называемые центральные и обходные пути. Из органических
Л«слот большое значение имеет мевалоновая кислота — пред-
шественник изопреноидных структур у многих микроорганизмов,
относится к разряду веществ, называемых факторами роста:
Ж
НОН2С—СНг—С (ОН)— СНг—СООН
сн3
Ий промежуточных малых молекул важное место занимают кар-
ОДМоилфосфат и рибоза как исходные вещества для синтеза
Мононуклеотидов. Так, из карбамоилфосфата и аспарагиновой
кислоты осуществляется биосинтез оротовой кислоты — пред-
шественника уридина:
nh2
H2N—CO—OPO3 + НООС—CH2—CH—COOH--------
карбамонлфосфат аспарагиновая кислота
оротовая
кислота
Синтез карбамоилфосфата (например, у энтеробактерий, дрож-
жей и др.) осуществляется из аммиака, диоксида углерода и
фосфата, т. е. из малых молекул неорганической природы.
।____________I
рибоза
I___________________I
мононуклеотид
уридин-5'-фосфат
Рибоза образуется из глюкозы в гексозомонофосфатном шун-
те у гетеротрофных организмов. Имеются микроорганизмы, син-
тезирующие необходимые углеводы (в том числе рибозу) из СО2,
например водородные бактерии. D-Рибоза является структурной
единицей не только нуклеиновых кислот, но и ряда важнейших
коферментов: АТФ, НАД, НАДФ, флавопротеинов. Отсюда ста-
новится понятной ее роль в ключевых процессах роста и разви-
тия микроорганизмов.
К малым молекулам относятся также аминокислоты, простые
сахара (низшие олигозы и монозы), мононуклеотиды, витамины,
молекулярная масса которых чаще не превышает 350 Да. Они
являются строительными блоками, лежащими в основе макро-
молекул.
Аминокислоты. Важнейшие макромолекулы клеток любъл
организмов (в том числе микроорганизмов) — белки, они со
стоят из мономерных единиц — аминокислот, определяющих ос-
новные свойства полимеров. Большое значение при этом имеет
|ин аминокислот, порядок их соединения и пространственно
взаимодействие между аминокислотами.
Известно около 300 аминокислот, из них живая природа ото-
46
ta ДЛЯ формирования белковых макромолекул лишь 20 (при-
£-а-аминокислоты) (табл. 5). Этого оказалось вполне
«точно для того, чтобы образовать множество различных
В у всех видов организмов. Например, в каких-то белках
ёя по 100 аминокислот, т. е. каждая из 20 (или часть из
повторяется не один раз. Тогда возможно 2О100 вариантов
вых молекул, в меньшей или большей степени отличающих-
уг от друга. Белок, содержащий последовательность ами-
лот в порядке 1, 2, 3, 4, 5, 6, 7, 8, 9, ..., 20, отличается от
в в которых последовательность аминокислот будет, на-
p. 1, 3, 4, 2, 5, 7, 6, 8, 9, ..., 20 и 3, 2, 1, 4, 5, 6, 7, 9, 8, ...,
, 20 и т. д. Приводим общую формулу для аминокислот,
1дикал R может быть полярным или неполярным:
н
R—C“-NH2
I
С ООН
, В белках найдены также и редкие производные указанных 20
аминокислот, но свыше 200 других известных аминокислот,
являющихся, как правило, производными а-аминокислот, никогда
не обнаруживаются в природных белках, они находятся в свобод-
ВИЛи связанном состоянии в других структурах. Кроме того,
Т быть 0-, у-, б- и е-аминокислоты и аминокислоты с Р-кон-
фЯГурацией. Так, в клеточных стенках бактерий найдены Р-аланин
у Р «
И у*Р-глутаминовая кислота НООС—СН—СН2—СН2—СООН,
NH2
о-в-мезодиаминопимелиновая кислота
НООС—СН—(СН2)з—СН—СООН и др.
i' . nh2 nh2
i
Все аминокислоты (кроме глицина) являются оптически актив-
ин, поскольку содержат хотя бы один хиральный. (асимметриче-
1Й) атом углерода. Гидроксилизин, 4-гидроксипролин, изолей-
<ДИЩ и треонин имеют по два хиральных атома углерода, поэтому
КВЖДЫЙ из них может существовать в 4 изомерных формах (звез-
Дочкой помечены хиральные атомы углерода) :
СООН СООН • СООН СООН
H2N—с*—н и—с*—nh2 h2n—с*—н н—с*—nh2
H—С*—ОН НО—с*—н но—с*—н н—с*—он
11 II
СНз СНз СНз СНз
Ь-треоиин D-треонин L-аллотреоннн D-аллотреонин
47
о©
Таблица 5. Природные аминокислоты в белках микроорганизмов
Группа R Название Символы Строение R Полярность
Алифатическая цепь Глицин* Аланин Валент Лейцин Изолейцин Гли Gly Ала Ala Вал Vai Лей Leu Иле Не н- СНз— (СНз)2=СН— (СНз)2=СН—СН2— (C2H5)—СН— Полярный Неполярный » »
сн3
С гидроксилом Серин Треонин Сер Ser Тре Thr НО—СНг— СНз—СН(ОН) — Полярный »
С атомом серы Цистеин** Метионин Цис Cys Мет Met HS—CH2— НзС—S—(СН2)2— » Неполярный
С карбоксильной или имидогруппой Аспарагиновая кислота Аспарагин Глутаминовая кислота Глутамин Асп Asp Аси Asn Глу Glu Глн Gin НООС—СНз— H2N—ОС—СН2— НООС—(СН2)2— H2N—ОС— (СН2) 2— Полярная » » »
С основной группой Аргинин Apr Arg H2N—С—NH—(СНг) з— »
Лизин*** Гистидин Лиз Lys Гис His II NH H2N—(СН2)з— |==|—СНз— » »
HN N
ы; —>—_. .Д. Ч .XT.lffv-r
Группа R Название Символы Строение R Полярность
С бензольным и другими циклами Фенилаланин Тирозин Триптофан Фен Phe Тир Туг Три Тгр СбНз— СН?— НО—С6Н4—СН2— Шсн- NH Неполярный Полярный Неполярный
Иминокислота Пролин’!** Про Pro ^Р^соон Н Неполярный
* Глицин не содержит хирального атома углерода, поэтому он не относится ни к D-, ни к t-ряду аминокислот; радикал R представлен
водоводом, который эффективно не заслоняет полярные группы у а-углеродного атома. с с
Д ** два остатка цистеина входят в состав аминокислоты цистина, где они соединяются^днсульфиднои связью S S .
«‘Известны гидроксилнзин H2N—СН—(СН2)2—СН—СООН и 4-гидроксипролнн Т^ХСООН- Они образуются соответственно
ОН NH2 Й
из лизнна и пролина, которые гидратируются после включения в полипептидные молекулы.
СООН
H2N—с*—н
Н—С*—СНз
сн2
СНз
£-нзолейцин
СООН
н—с*—nh2
НзС—с*—н
СН2
I
СНз
Р-изоленцнн
СООН СООН
h2n—с*—н н—с*—nh2
I I
НзС—С*—Н Н—С*—СНз
I I
СН2 СН2
СНз СНз
£-аллонзолейцин О-аллонзолейцин
Однако аллотреонин и аллоизолейцин не обнаружены в природ-
ных белках.
Большинство аминокислот растворяется в полярных раствори-
телях (воде, спирте) и не растворяется в неполярных растворите-
лях (бензоле, гексане, эфире). Гистидин, тирозин, триптофан и фе-
нилаланин поглощают ультрафиолетовые лучи (>240 нм), при-
чем больше всего триптофан.
Наличие карбоксильной и аминной групп обеспечивает амино
кислотам способность взаимодействовать с соответствующими
ингредиентами в реакциях ацилирования, солеобразования и эсте
рификации. Главнейшая реакция аминокислот в живой клетке
образование пептидной связи. В отдельных случаях некоторые
аминокислоты служат исходными продуктами для синтеза органи-
ческих аминов, пуринов и других веществ. Например, из аргинина
образуются полиамины спермин и спермидин — факторы роста
для иерсиний и нейссерий.
Глицерин НО—СНг—СН(ОН)—СН2ОН относится к простей-
шим сахароспиртам и является составной частью липидов. Глице-
роальдегид — родоначальник L- и D-рядов моноз. Наличие одного
хирального атома углерода в его молекуле создает возможность
существования D- и А-форм глицероальдегида:
Н—С=О
Н—С—ОН
СН2ОН
D-глицероальдегнд
н—с=о
но—с—н
<^Н2ОН
/.-глицероальдегид
При восстановлении глицероальдегид превращается в оптиче-
ски неактивный глицерин. Глицероальдегид выполняет ключе-
вую роль в превращении углеводов в клетках различных орга-
низмов.
Жирные кислоты являются компонентами липидов в клеточных
мембранах и в виде свободных липидов (включений) могут нахо-
диться в клетках различных микроорганизмов.
Углеродные атомы в формулах жирных кислот нумеруют арабскими цифрами,
начиная с карбоксильного (1-й); второй углеродный атом обозначают как а, тре-
тий — р, а концевой углерод в метильной группе — о>. Двойные связи обозначают
знаком А (например, А7 — это двойная связь, расположенная между 7-м и 8-м
углеродными атомами). Обычно указывают общее число углеродных атомов, число
и позицию двойных связей.
50
Например, пальмитоолеиновая кислота
. U 13 12 И Ю 9 8 7 6 5
СНг-СНг-СНг—СНг—СНг—СН^Н—СНг-СНг-СНг—СНг-
г 3 4 5 6 ;
4 3 2 1
СНг—СНг—СНз—СООН
iV
быть условно обозначена как 16: 1,9, т, е. она содержит 16 углеродных
одну двойную связь между С9 и СЮ. В другом варианте эту же кислоту
ачают таким образом: ® = 7, С 16:1, т. е. двойная связь располагается
С7 и С8, начиная счет их с метильного конца (при наличии 16 углеродов
‘.х-лК насыщенным жирным кислотам относят пальмитиновую
С|»Нл СООН, лауриновую СцНгзСООН, миристиновую С13Н27СООН,
стеариновую С17Н35СООН, арахиновую С19Н39СООН, лигноцери-
новую С23Н47СООН; к ненасыщенным —
пальмитоолеиновую СНз(СН2)5СН=СН(СН2)?СООН,
олеиновую СНз (СН2)?СН=СН (СНгЬСООН,
линолевую СН3(СН2)4СН=СНСН2СН=СН(СН2)7СООН,
линоленовую
СНзСН2СН=СНСН2СН=СНСН2СН=СН(СН2)7—СООН,
81хидоновую СНз (СН2) 4СН==СНСН2СН=СНСН2СН=
СН2СН=СН(СН2)зСООН. К необычным жирным кислотам мо-
ДОо отнести туберкулостеариновую СНз(СН2)7СН—(СНг^СООН,
I
•Ж" СНз
ЩШстобактериновую СНз(СН2)бНС—СН(СН2)эСООН и др.
\/
' сн2
Наличие двойных связей в молекулах ряда жирных кислот
ддределяет возможность их цис- и транс-конфигураций. В транс-
чформе жирные кислоты более стабильны. Липиды с большим со-
держанием ненасыщенных жирных кислот будут менее вязкими
..Я, наоборот, липиды с большим содержанием насыщенных кислот
Кудут более вязкими (сравните масла и жиры).
Простые сахара, или монозы, и низшие олигозы. Первичными
.Амлыми биомолекулами являются такие монозы, как триозы,
Тетрозы, пентозы, гексозы, гептозы, октозы, декозы. К простей-
шим триозам относят глицериновый альдегид; к тетрозам —
Эритрозу и треозу с общей формулой НО—НгС—(СНОН)г—
*^СОН, к пентозам — рибозу, арабинозу, ксилозу, ликсозу с об-
Шей формулой НО—Н2С—(СНОН)з—СОН; к гексозам — алло-
'•Шеальтрозу, глюкозу, маннозу, гулозу, идозу, галактозу, талозу
в общей формулой НО—НгС—(СНОНН—СОН и т. д.
•’ У некоторых моноз первичная спиртовая группа замещается
водород. Многие бактерии и некоторые грибы в составе поли-
амидов содержат фукозу, или 6-дезоксигалактозу. Примеры
моноз:
51
:0
•О
। \н | \н
с< СС н—с—он н—с—он
рн I \н I I
Н— с— он 1 Н— С —ОН 1 но—с—н но—с—н
Н— С — он н—с—ОН 1 н—с—он 1 но—с—н 1
1 СН2ОН 1 н—С—ОН н— с — он 1 н—с—он 1
1 СН2ОН 1 СН2ОН СН2ОН
D-эритроза D-рибоза D-глюкоза D-галактоза
с< сн2он
рн I
н—с—он с=о
НО—С—Н НО—с—н
но—с—н н—с—он
I I
н—с—он н—с—он
СНз СН2ОН
фукоза D-фруктоза
Все моносахариды (полиоксиальдегиды и полиоксикетоны)
подразделяют на альдозы (например, глюкоза) и кетозы (напри-
мер, фруктоза). Это кристаллические бесцветные вещества, хо-
рошо растворимые в воде и обладающие сладким вкусом. Боль-
шинство моносахаридов содержит хиральные атомы углерода.
Как видно из табл. 6, число N оптически активных изомеров для
альдоз и кетоз определяется правилом 2"; у кетоз того же ряда,
как и у альдоз, число хиральных атомов на один меньше.
Разнообразие моносахаридов возрастает за счет различных
типов изомерии (табл. 7). Изомерия моноз существенно прояс-
нилась с введением понятия о циклических структурах их и не-
плоскостных конформациях типов С, В, Т и Е (от англ, chair —
Таблица 6. Число оптических изомеров для некоторых моносахаридов
Моносахариды
Число хиральных
атомов углерода
п
Число оптических
изомеров 2"
Альдотриозы
Альдотетрозы и кетопентозы
Альдопентозы и кетогексозы
Альдогексозы и кетогептозы
Таблица 7. Основные типы изомерии
Изомеры
Структурные (скелетные)
Геометрические (£>-, L-,
цис-, транс-, эпимеры)
1
2
3
4
2
4
8
16
Признаки, по которым различаются
молекулы изомеров
Расположение атомов в углеродном скелете
Расположение заместителей вокруг атомов угле-
рода в пространстве
52
Продолжение табл. 7
Изомеры
сложения (а- и р ано-
Признаки, по которым различаются
молекулы изомеров
Положение полуацетального гидроксила
гл0итические (энантио-
формационные (кон-
еры)
Стереоизомерные молекулы, относящиеся друг к
другу как предмет и его зеркальное отражение
Степень поворота вокруг простой связи
о, boat — лодка, twist — скрученный, envelope — конверт):
он
носн2
СН2ОН
н
он
/J-D-фруктофураноза
Н
Я-D-глюкопираноза
В-форма
a-D-глюкозы
Т-фуранозные циклы
Е- фуранозный цикл
Моносахариды (гчюкогамин. галактозамин, мурамовая, или
Миколацтиловая, мтic-та) нередко обнаруживаются у микро-
организмов в виде ам.иноса ха роз и их производных. Водород
Аминогруппы в пминмсахаридах нередко оказывается замещен-
ным, например на а не" и. i. За счет полуацетального гидроксила
Моносахариды обра.ук.- :'л>ч озз структуры как с аналогич-
*ными молекулами тих л,(гомо- и гегерот.чиканы), так
Я с другими молем не-лл ,например, нуклеозиды. некоторые
Антибиотики). По правилам номенклатуры моносахаридов можно
ЯФИЮЬзовать символы вмел го формул (табл. 8). Указанная сим-
$№&Ика в равной мере применима и к L-ряду моносахаридов.
»<. К простым сахарам относятся и низшие олигосахариды (оли-
Таблица 8. Названия и символы некоторых моносахаридов D-ряда
Название Символы
русские латинские
D-Абеквоза D-Абе D-Abe
D-Арабиноза D-Apa О-Ага
D-Рибоза D-Риб D-Rib
D-Ксилоза D-Кси D-Xyl
D-Ликсоза D-Лик D-Lyx
D-Аллоза О-Алл D-All
D-Альтроза D-Альт D-Alt
О-Глюкоза D-Глю D-Glc
D-Манноза D-Ман D-Man
D-Г улоза ОТул D-Gul
D-Идоза D-Идо D-Ido
D-Галактоза D-Гал D-Gal
D-Т алоза О-Тал D-Tal
D-Фукоза D-Фук D-Fuc
D-Г алактопираноза D-Г алр D-Galp
D-Идофураноза D-Haof D-Idof
D-Маннопирануроновая кислота D-МанрУК D-ManpUA
2-Амино-2-дезокси-0-глюкоза (глюко- D-ГлюГЧ D-GlcN
замии)
2-Ацетамидо-2-дезокси-О-галактоза О-Гал^ц D-GalNAc
(ацетилгалактозамин)
2,3,4-три-О-метил-р-О-арабииоза 2,3,4-три-О-метил- 2,3,4-tri-O-methyl-
p-D-Apa p-D-Ara
гозы), включающие от 2 до 5 моноз. Например, низший гомооли-
госахарид-биоза — мальтоза, состоит из 2 остатков глюкозы:
К гетероолигосахаридам относится сахароза, состоящая из глю-
козы и фруктозы:
Биозы, или дисахариды, такие, как лактоза и мелибиоза, со-
стоящие из глюкозы и галактозы, относятся к гетеробиозам;
трегалоза, ламинарибиоза, целлобиоза — к гомобиозам, включа-
ющим только остатки глюкозы:
54
CH2OH
-t* Трисахариды (триаозы), тетраозы, пентаозы, а также высшие
ОйИгозы (с 6—10 остатками моноз) у микроорганизмов не обна-
Вжены, тем не менее монозы и олигозы играют огромную роль
к энергосубстраты и структурные блоки полисахаридов и поли-
*&Йхаридсодержащих молекул в микробных клетках. Классифика-
*Ля углеводов и углеводсодержащих полимеров, или гликоконъю-
гатов, приведена в табл. 9. Полисахариды и гликоконъюгаты отно-
сятся к макромолекулам, включающим моносахаридные строи-
тельные блоки.
ица 9. Классификация углеводов и гликоконъюгатов
Группа Подгруппа Название Число моно- мерных единиц
Моно- «СИфиды и произ- водные 1. Альдозы и кетозы Монозы (триозы, тетрозы, пен- тозы, гексозы, гептозы, октозы, нонозы, декозы) 1
Продолжение табл. 9
Г руппа Подгруппа Название Число моно- мерных единиц
2. Ангидриды сахаров Ангидромонозы (1,6-ангидро-Д- D-глюкопираноза; 1,6-ангидро-0- D-глюкофураноза; 3,6-ангидро-0- D-глюкопираноза и т. д.) 1
3. Уроновые кислоты 4. Гликозиды с О-, N-, S- или С- типами гликозид- ных связей (аль- дозиды, кетози- ды): Галактуроновая, глюкуроновая, идуроновая кислота и т. д. 1
а) голозиды* (см. гр. II и III. Содержат О-гликозидные связи) 1*
б) гетерозиды Галактозиды, гликозиды, рамн- озиды, рибозиды, фукозиды и др. Ангидропиранозиды (метил-3,6- ангндро-а-О-глюкопиранозид и др.), ангидрофуранозиды (метил- 3,6-ангидро-а-О-глюкофураиозид и т. д.) 1 и >**
5. Ангидрогли- козиды 1
6. Аминосахара Монозамины, ацетилмонозами- ны (галактозамин, глюкозамин, N-ацетилглюкозамин и др.) 1
7. Гликолакти- ловые (мурамо- вые) кислоты Глюколактиловая (глюкомура- мовая) кислота, галактолактило- вая (галактомурамовая) кислота и т. д. 1
II. Олиго- сахариды 8. Дезоксиса- хара Гомо- и гетеро- олигосахариды (голозиды, со- держащие О-гли- козидные связи) Моио-, ди-, тридезоксиозы (6- дезокси-О-галактоза, или О-фу- коза; 3,6-дидезокси-О-ксилогексо- за, или абеквоза; 3,6-дидезокси- /.-ксилогексоза, или колитоза; 3,6-дидезокси-О-рибогексоза, или паратоза; 3,6-дидезокси-О-араби- ногексоза, или тивелоза; 6-дезок- си-£)-манноза, или О-рамноза, и др.) 1
а) низшие Олигозы (дисахариды, или био- зы; трисахариды, или триаозы; тетрасахариды, или тетраозы; пентасахариды, или пентаозы) 2—5
б) высшие Олигозы (гексаозы, гептаозы, октаозы, ионаозы, декаозы) Полиозы-гомогликаны (глюкан, 6—10
111. Поли- а) Гомополиса- >10
сахариды, или глика- ны хариды (голози- ды, содержащие О-гликозидные связи) маннан и др.)
56
Продолжение табл. 9
•О?."* ;
ед»*
Ч'-*.
«Йм.
(IgfeX
tP
•MP-
Глико-
каты
Подгруппа Название Число моно- мерных единиц
б) Гетерополи- сахариды (голо- зиды, содержа- щие О-гликозид- ные связи) Полиозы-гетерогликаны (ксан- гаи, ксилоглюкурономаннан и др.) >10
I. Гликопро- теины Аллергены (трихофитины, кан- дидины), ферменты (амилаза, карбоксипептидаза, рибонуклеаза и др.), иммуноглобулины (IgM, IgG, IgA, IgE, IgD) и т. д. Разное
2. Пептидогли- каны, или протео- гликаны Муреин, полипептидилбиозами- нилуронил (гепарин, хондроитин- сульфаты и др.) »
3. Гликозами- ногликаны Гиалуроновая кислота, хитии, хитозан, N-ацетилхитобиозилман- нан и т. д. »
4. Гликолипи- ды Бактопренол (ундекапренол), долихол (Glc-NAc-пирофосфо- рилдолихил), липотейхоевые (гли- цериитейхоевые) кислоты, корд- фактор туберкулезных микобак- терий и др. »
5. Липоглика- ны Липополисахариды, или эндо- токсины, грамотрицательных бак- терий О-Антигены грамотрицательных бактерий, полученные по методу Буавена и Месробяну (это липо- полисахарид + протеин) »
6. Гликолипо- протеины »
7. Тейхоевые*** кислоты (!?) Рнбиттейхоевые кислоты »
г&^оносахаридные остатки в олиго- и полисахаридах соединяются О-глнкозидными
1Ями и число моноз в них всегда больше единицы; они выделены в самостоятельные
н III.
* * В некоторых гетерозидах содержится, например, по 2 остатка моноз(ы) (в анти-
«Мтике стрептомицине, в коферменте НАДФ и др.).
* ** Липотейхоевые кислоты относятся к гликолипидам; рнбиттейхоевые кислоты
time связываются с пептидогликаном.
Мононуклеотиды. Важной группой малых внутриклеточных
•Кмекул, участвующих в формировании структур, хранящих и
Явредающих наследственную информацию, являются мононуклео-
*ЯДЫ общей формулой
о3р—о—сн2
ISMsfc 4'
ОН ОН(Н)
R — один из остатков пуриновых (аденин, гуанин) или пири-
“Диновых (тимин, урацил, цитозин) оснований:
57
пурин
пиримидин аденин
(6-аминопурин)
гуанин
(2-амино-6-оксипур ин)
О
тимин
(2,4-диокси-5-метил-
пиримидин)
урацил цитозин
(2,4-Диоксипиримидин)
(2-окси-4-аминопиридин)
Дезоксирибонуклеиновая кислота (ДНК) включает аденин,
гуанин, тимин, цитозин, дезоксирибозу и фосфорную кислоту.
Следовательно, она состоит из четырех мононуклеотидов: адени-
ловой кислоты (дезоксиаденозин-5:фосфат), гуаниловой кис-
лоты (дезоксигуанозин-5'-фосфат), тимидиловой кислоты (дезо-
кситимидин-5'-фосфат) и цитидиловой кислоты (дезоксицити-
дин-5'-фосфат), или соответственно из аденозинмонофосфата
(АМ.Ф), гуанозинмонофосфата (ГМФ), тимидинмонофосфата
(ТМФ) и цитидинмонофосфата (ЦМФ). Все они являются силь-
ными кислотами.
Рибонуклеиновая кислота (РНК) включает такие мононук-
леотиды, как адениловую, гуаниловую, уридиловую (вместо ти-
мидиловой в ДНК) и цитидиловую кислоты, а также рибозу
(вместо 2-дезоксирибозы в ДНК) и фосфорную кислоту.
Дезоксирибоза (рибоза) соединяется с пуриновыми и пири-
мидиновыми основаниями посредством N-гликозидных связей
соответственно в позициях СГ->С9 и СГ->СЗ. Природные
нуклеотиды чаще находятся в анти-, чем в си«-конформационном
состоянии. Эти состояния показаны на примере гуанозина:
сип -конформация
гликозидной связи
в гуанозине
анти -конформация
гликозидной связи
в гуанозине
58
1-Конформация гликозидной связи приобретает важное
е в наведении водородных связей между пуриновыми и
циновыми основаниями в двойной спирали ДНК (см.
.12). N-Гликозидные связи характерны для всех моно-
идов, независимо от того, в свободном или связанном
ми они находятся. Однако имеются исключения. Напри-
некоторых типах РНК (тРНК) остаток рибозы может
ггься с С5 урацила. В этом случае вместо N-гликозидной
уется С-гликозидная связь. Этот нуклеотид назван псев-
в которой ф— псевдоуридин:
о
.иловой кислотой (ф-МФ),
но—р—о—сн2
Ан
NH
1*г X)
Н
о
ж
С5' дезоксирибозы
статки
ибозы
псевдоуридиловая кислота
фосфорной кислоты в позиции
в нуклеотидах служат местом присоединения других
идов по СЗ' позиции пентозы. Следовательно, моно-
иды, явдяясь структурными блоками нуклеиновых кислот,
ются в виде нитей или цепей по СЗ' и С5' позициям пен-
) освобождается около 3768 кДж
угорые нуклеотиды в соответствующих реакциях высво-
•т большое количество энергии. К таким нуклеотидам от-
АТ Ф — универсальный источник энергии в клетках раз-
микроорганизмов, который не входит в состав нуклеино-
!лот. При гидролизе 1 моль АТФ до аденозиндифосфата
; энергии.
о
о-4
О" О- О н
nh2
О—Р~О—Р—о-
он
он
нуклеозид-аденозин
АМФ
АДФ
АТФ
Ж
О о
59
Следовательно, мононуклеотиды могут находиться как в по
мерном (в нуклеиновых кислотах), так и в мономерном (свосч
ном) .состоянии. Свободными нуклеотидами (кроме АМФ, А.,
и АТФ) являются также аденозинцикло-3', 5'-фосфаг или ник,
веский АМФ (цАМФ); циклические гуанозин- и цитозинмонофг
фаты (цГМФ и цЦМФ соответственно); ааенозин-3'-фосфа?
фосфосульфат; S-аденозилметионин и др.
цАМФ принимает участие в регуляции отдельных реакций
обмена углеводов. Установлено, что, например, клетки Е. coli
содержат специальный белок, названный белком — активатором
катаболизма (БАК) [ от англ—catabolism activatoring protein
(САР)], который связывается с цАМФ и в таком виде стимули-
рует транскрипцию (считывание) Lac-мРНК (матричная РНК,
кодирующая синтез фермента, катализирующего реакцию гидро-
лиза лактозы):
При недостатке простых сахаров (катаболитов) концентрация
цАМФ в клетках повышается, при достаточном или избыточном
количестве таких сахаров концентрация цАМФ снижается. Роль
цАМФ особо выражена при катаболитной репрессии. Кроме
цАМФ в клетках образуются также цГМФ и цЦМФ, различа-
ющиеся по типу основания, связывающемуся с С1' рибозы.
В некоторых случаях цГМФ выступает антагонистом цАМФ.
Аденозин-3'-фосфат-5'-фосфосульфат относится к числу ак-
тивных сульфатов, принимающих участие в синтезе, например,
сульфатированных полимеров:
аденозиН'3'-фосфат-5'-фосфосульфат
Активным метионином является S-аденозилметионин, высту-
пающий в качестве донора метильных групп в реакциях трансме-
тилирования и как источник пропиламина в синтезе полиаминов:
60
ДФ и ГТФ так же, как АДФ и АТФ, имеют макроэргические
н и участвуют в реакциях, требующих затрат энергии, а так-
s синтезе белков и углеводов:
Кроме того, в цикле трикарбоновых кислот на стадии окисле-
1*Я в-кетоглутаровой кислоты до сукцинил-КоА сопряженно
Происходит окислительное фосфорилирование с передачей фосфа-
Т» на ГДФ. При этом образуется ГТФ.
^‘Предшественником всех пуриновых нуклеотидов, синтезиру-
МКихся заново, является гипоксантиновый рибонуклеотид —
МЦозиновая кислота. В некоторых реакциях фосфорилирования
Принимают участие инозиндифосфат (ИДФ) и инозинтрифосфат
дИТФ), отличающиеся от инозинмонофосфата (ИМФ) возраста-
числа фосфатных групп соответственно до двух и трех.
ИМФ
61
Уридин-5'-дифосфат (УДФ) и уридин-5'-трифосфат (УТФ),
цитидин-5'-дифосфат (ЦДФ) и цитидин-5'-трифосфат (ЦТФ)
относятся к пиримидиновым нуклеотидам:
УДФ
УТФ
Важным исходным продуктом в биосинтезе углеводов у многих
микроорганизмов является УДФ-глюкоза:
62
Н2ОН
о
о
- >~О—Р—о—сн2
О- 0“
н
он
о
н
о
УДФ-глюкоза
выше свободные мононуклеотиды относят-
* перечисленные
1акроэргическим соединениям. Это объясняется резонансной
явностью их неорганического фосфата:
о
О~
о-
О~ « СТ—Р— О- — О~—Р=О
О-
он
он
Г : .
влением свойств ангидридов фосфорной кислоты:
ff-. О О Основание
« и ।
ЗГ О —Р—О—Р —О—Рибоза
I И I
feu. О-у 0~
атомные (ангидридные) связи
IF1'
ДЕрбностью к ионизации и внутримолекулярному электростати-
ММЙУ отталкиванию.
^Возникновение ангидридных связей сопряжено с уменьшением
ИР** резонансных групп в молекуле, а ее термодинамическая
|ИУичивость определяется способностью принимать большее или
уьшее число резонансных форм. Поэтому макроэргические
иШ^УРЫ менее стабильны и при их гидролизе выделяется
иое количество энергии.
(рнонуклеотиды могут быть структурной частью ряда вита-
|.и коферментов. Так, витамин Bi2 (цианкобаламин), пред-
ающий собой циклическую корриновую систему, содержит
нуклеотид (5,6-диметилбензимидазол-рибонуклеотид).
; выступает в роли кофермента, то в такое соединение
to CN-группы включается нуклеозид Б'-дезоксиаденозин:
63
он н
CN~
1\_*
5'-дезокси-
аденозин-R
Витамин В2 (рибофлавин) также относится к нуклеотидам.
В его структуре рибоза заменена пятиатомным спиртом рибитом,
или рибитолом. Таким образом, В2 представляет собой флавин-
мононуклеотид (ФМН)
н3с
Н3С
6,7-диметилизоаллаксазин
СН2—(СНОН)3—СН2—О—РО3Н2
ФМН
64
t витаминам группы В относится и никотиновая кислота
СООН
который входит в
состав никотинамидмононуклеотида
о—РО3Н2
аденилов® кислота
кофермент А
рмент А переносит ацильные группы, присоединяющиеся
-Шожноэфирной связью к тиоловой группе. Итак, ФМН, НМН
Й&оА — мононуклеотидные коферменты.
. Известны также и динуклеотидные коферменты: флавинаде-
нуклеотид (ФАД), никотинамидадениндинуклеотид (НАД),
тинамидадениндинуклеотидфосфат (НАДФ), последний от-
я от НАД лишь тем, что в рибозе АМФ находится фос-
ая группа по месту С/2.
ибофлавнн (витамин В2)
Адении
9
етилизоаллаксазин-рибит—Р—О—Р—О—5'—Рибоза
j°H
аденозин
ФМН
АМФ
ФАД
'I
о
о
65
Никотинамид Аденин
1 9
Г О О
II II
Рибоза—5'—О—Р—О—Р—О—5'—Рибоза
।_________А।
НМН АМФ
Динуклеотидные коферменты играют важную роль в цепи пере-
носа электронов при окислительном фосфорилировании.
Витамины. К ним относят витамины группы В (водораствори-
мые) и витамины A, D, Е, К (жирорастворимые).
Витамин В\ (тиаминхлорид)
сн2он
1АНЛХН1
Cl-
витамин В^тиаминхлорид)
достаточно устойчив в кислой среде, но быстро инактивируется
при нагревании в нейтральных и щелочных растворах. В виде
кофермента тиаминпирофосфата (кокарбоксилазы) витамин В|
о~ о-
о
о
H3C'NjxN'!!5YxNH2 сн2—о—р—о—р—о~
^Х^^СН2—Ni-------“—СН3
тиаминпирофосфат
участвует в окислительном декарбоксилировании а-кетокислот
и в транскетолазной реакции. Витамин Bi могут синтезировать
многие микроорганизмы, но некоторые из них не способны осу-
ществлять такой процесс и нуждаются в добавлении витамина
в питательные среды (Leuconostoc species, Candida species и др).
Витамин В2 (рибофлавин) относится к светочувствительным
соединениям. При облучении его ультрафиолетом в нейтральном
или кислом растворе отщепляется остаток рибита и образуется
6, 7-деметилаллоксазин (люмихром), а в щелочной среде — 6,7,9
триметилизоаллоксазин (люмифлавин):
66
it люмихром
мьшинство бактерий и грибов синтезирует витамин Bj.
Кд некоторые мутантные дрожжи, отдельные бактерии (на-
ер, Lactobacterium species) нуждаются в добавлении его
», Рибофлавин служит предшественником коферментов ФМН
Д. Ферменты, содержащие ФМН и ФАД, участвуют в реак-
' окисления — восстановления и, в частности, в переносе
'ронов при окислительном фосфорилировании.
итамин Вь (пантотеновая кислота, пантоил-0-аланин) обра-
я большинством микроорганизмов:
СНз
I
НОСНг—С—СНОН—СО—NH—СНг—СООН
I
СНз
пантотеновая кислота
ХНая связь в структуре витамина В5 устойчива в кислой и
Мной средах. Пантотеновая кислота входит в состав кофер-
а А и ацилпереносящего‘белка (АПБ), производные которых
гвуют более чем в 70 ферментативных реакциях. АПБ, на-
юр у Е. coli, содержит 4'-фосфопантетеин, присоединенный
рйновому остатку фосфодиэфирной связью и выступающий
^связывающим звеном:
пептид
В—CHsr-O— Р—О—СН2—С—СН ОН—С О—N Н— (СН2) г—СО—N Н
-II I
3 О СНз HS—(СН2)2
ептид
4х-фосфопантетеин
участвует в биосинтезе жирных кислот.
67
Холин* (гидроксид ₽-оксиэтилтриметиламмония) — сильное
основание, полностью диссоциирующееся в воде на ионы холина
и гидроксила:
+
(H3C)a=N—СН2—СН2ОН + ОН~
Холин может выступать субстратом для некоторых флавиновых
ферментов в митохондриях у отдельных эукариотических орга-
низмов:
Холин-----> Убихинон----> ФАД
(акцептор
электронов)
Холин входит в состав соответствующих фосфатидов и обнару-
живается в мембранах многих микроорганизмов.
Витамин В6 (пиридоксин) является предшественником пири-
доксальфосфата — кофермента всех трансаминаз, который вы-
ступает переносчиком аминогрупп:
пиридоксин пиридоксальфосфат
(пиридоксол)
Витамин Be образуется большинством микроорганизмов. Лишь
отдельные мутанты являются ауксотрофами по данному витами-
ну, т. е. нуждаются в добавлении его в питательные среды извне.
Витамин Н** (биотин) — производное имидазола; в виде про-
стетической группы входит в состав ряда ферментов, катализи-
рующих реакции включения СО2 в органические соединения (аце-
тил-СоА-карбоксилазы, метилмалонил-транскарбоксилазы, пиру-
ваткарбоксилазы и др.):
—(СН2)4—соон
биотин
* Ранее холин называли витамином В4, а витамином В6 — НАД.
** Ранее называли витамином В?.
68
вызование комплекса биотин=СО2 происходит по следующей
;еме:
Mg'
АТФ + HCO3
Ж'
о
о
Hbr нН
биотин-фермент
АДФ
о=с
углеродно-
фосфорный
ангидрид
карбоксибиотин-фермент
биотин образуется многими микроорганизмами, включая некото-
рых представителей нормальной микрофлоры кишечника чело-
века. Вместе с тем известны виды, ауксотрофные по данному
витамину, например Candida sp. и др.
Витамин В» (инозитол) — производное циклогексана. Он мо-
жет находиться в виде семи оптически неактивных и двух опти-
чески активных форм. Биологически активна лишь одна оптиче-
ская форма — лшо-инозитол, который в виде составной части
липидов обнаруживается у кислотоустойчивых бактерий:
л/ио-инозитол
Инозитол входит в состав фосфолипидов, локализующихся
В мембранах микроорганизмов, и в форме инозитолгексафосфата
(фитиновой кислоты) связывает двухвалентные катионы. Маг-
ниево-кальциевая соль фитиновой кислоты названа фитином.
Инозитол участвует в липидном и углеводном обменах.
Витамин Вс (фолиевая кислота, фолацин) — олигопептид с мо-
лекулярной массой более 350 Да. Синтезируется одними микро-
организмами и необходим извне для других, например для Strep-
tococcus faecalis. Фолиевая кислота участвует в переносе одно-
69
углеродных фрагментов при метаболизме аминокислот, а также
в образовании пуринов и тимина:
птероевая кислота
ПАЕК*
Глу
2-амино-4-окси-6- Go 1 ' I
фолиевая кислота из бактерий
(птероилглутаминовая кислота)
Глу
СН—соон
(СН2)2
соон
Фермент-дигидрофолат—редуктаза бактерий (А4Г~ 15-103...
18-103) восстанавливает фолиевую кислоту до тетрагидрофолие-
вой кислоты в позициях С5, С7 и С8.
При гидролизе Ns, ю-метилентетрагидрофолата образуется
Nio-формилтетрагидрофолат, выступающий донором формильной
группы во многих реакциях биосинтеза у микроорганизмов:
Чн2
1 10
CH2~^>N +НОН
{-------сн2
10
Ч'-СН2
I ю
НС—СН—N—
С=О
i i
N-формилтетрагидрофолат
Витамин С (аскорбиновая кислота) проявляет окислительно-
восстановительные свойства. В щелочной среде дегидроаскорби-
новая кислота гидролизуется с образованием А-дикетогулоновой
кислоты НООС—СО—СО—СНОН—СНОН—СН2ОН. Уксусно-
о=с
но—с
но—с
Н—С'
+ 2Н+ +2е-
но—с—н
СН2ОН
но—с—н
I
СН2ОН
L-аскорбиновая Г-дегидроаскорбиновая
кислота
кислота
* ПАБК — парааминобензойная кислота.
70
МИиииые бактерии трансформируют многоатомный спирт сорбитол
здв&прбозу. являющуюся исходным продуктом при синтезе L-ac-
аЬфбиновой кислоты. Однако непосредственное участие витами-
ЯБ G в окислительно-восстановительных реакциях у микроорга-
яКвмов остается проблематичным.
Витамин В3 (никотиновая кислота, витамин РР, ниацин)
Щ^ляется производным пиридина и синтезируется подавляющим
4£мьщинством микроорганизмов. Это термостабильное соедине-
ние с температурой плавления 232 °C, поэтому оно устойчиво
-СООН
Я
никотиновая
кислота
Ви разных видах стерилизации. В виде нуклеотидов НАД+ и
ЩФ+ выступает коферментом во многих окислительно-восста-
4 Довительных реакциях, катализируемых оксидоредуктазами. Это
Sfo многом зависит от способности пиридинового кольца нахо-
диться во множественных резонансных формах, обеспечивающих
дему относительную стабильность.
ж Парааминобензойная кислота (ПАБК) — предшественник
‘.фолиевой кислоты. Например, фекальные стрептококки, нуждаю-
щиеся в фолиевой кислоте, легко синтезируют ее в случае вне-
^«ения в " *""
?•-
питательную среду ПАБК и аденина:
соон
nh2
ПАБК
Для многих микроорганизмов ПАБК выступает фактором
проста и относится к разряду водорастворимых витаминов груп-
пы В. ПАБК — антагонист сульфаниламидов (антиметаболит) и
•Поэтому снимает их губительное действие в силу структурного
Сходства молекул.
Витамин В\2 (кобаламин) обычно выделяют из культураль-
Мюй жидкости в виде цианкобаламина, способного восстанав-
j ЛИваться в добаламин — сильный нуклеофил, выступающий пере-
* Носчиком алкильных групп. Витамин В12 как кофермент необхо-
•; Дйм при образовании СН4 различными анаэробными бактериями;
’ он также участвует в реакциях трансметилирования. Кобаламин
^Синтезируется исключительно бактериями, для человека он ва-
Цйен как противоанемический фактор (в печени он находится в
$ форме метилкобаламина, аденозилкобаламина и гидроксокоба-
$,О!амина). Наиболее устойчивая форма — цианкобаламин, в такой
2£-форме витамин выпускается промышленностью. Алкилкобалами-
ны относят к разряду природных металлорганических соединений.
Витамин Bi3 (оротовая кислота) — производное пиридина
(см. разд. 2.1.1). Синтезируется в клетках различных дрожжей.
Оротовая кислота стимулирует рост некоторых бактерий и дру-
гих микроорганизмов. В виде оротата калия находит применение
в медицине.
Витамин В\5 (пангамовая кислота, £)-глюконодиметилаце-
тат) — эфир D-глюконовой кислоты и полностью алкилирован-
ной аминоуксусной кислоты. Обнаружен в клетках многих эука-
риот и, в частности, пивных дрожжей:
HOOC—(СНОН)4—СН2—ОСО—СНг—№=(СН3)2
пангамовая кислота
Синтетическим путем возможно получение 4-, 8- и 12-метиль-
ных производных пангамовой кислоты соответственно:
R—ОСО— СН2—N= [СН (СНз) 21 г R—ОСО—С Н—N= [СН (СНз) 2] 2
N=[CH(CH3)2]2
N=[CH(CH3)2]2
R—ОСО—С—N= [СН (СНз) 2] 2
N=[CH(CH3)2]2
Пангамовая кислота содержит лабильные метильные группы и
может принимать участие в реакциях метилирования и трансме-
тилирования. Кальциевая соль пангамовой кислоты рекомендо-
вана для использования в практике здравоохранения (пангамат
кальция устойчив в кислой среде и неустойчив в щелочной):
[ (СНз) 2=N—СНг—ОСО—СН2—(СНОН) 4—COO] f • Са • Н2О
пангамат кальция
Витамин N (липоевая кислота) образуется многими про-
кариотическими и эукариотическими микроорганизмами. Как ко-
фермент липоевая кислота ковалентно связывается амидной
связью с е=МНг-группой лизина в составе апофермента:
СН2 СООН
_ <1 СН2 сн2
липоевая кислота
Участвует в окислительном декарбоксилировании а-кетокислот.
Применяется в медицине.
Витамины А и D жирорастворимые, построены из блоков изо-
преноидного типа (равно как и витамины Е и К). Они образу-
ются в клетках некоторых микроорганизмов в форме провитами-
нов, например у пигментных коринебактерий и дрожжей. В мемб-
ранах клеток облигатных галофилов Halobacterium sp. изопре-
ноиды защищают клетки от фотохимического повреждения. Вита-
мины Е и К не типичны для микроорганизмов.
72
ЖкЖ; Большие молекулы, или макромолекулы
яИВГ У-' —
шэдшие молекулы, или макромолекулы, — это полимерные
^ЯиУКтУРы с большой молекулярной массой (в среднем 103...
1|?Да). К ним относятся нуклеиновые кислоты, белки, полисаха-
Ь1ды и гликоконъюгаты, липиды и липоконъюгаты.
•^Нуклеиновые кислоты. Известно два типа нуклеиновых кис-
Двг: ДНК и РНК.
Дезоксирибонуклеиновая кислота состоит из мононуклеотидов
фениловой, гуаниловой, тимидиловой и цитидиловой кислот),
соединенных в полимерную нить через 3', 5'-фосфодиэфирные
дгостики:
•УХе R и Ri — пуриновые или пиримидиновые основания, соеди-
ненные N-гликозидными связями с дезоксирибозой.
Информационная сущность ДНК заключается в последова-
тельности оснований. Например, последовательность их в отрез-
ке цепи ДНК: аденин (А) — цитозин (Ц) — аденин (А) — ти-
Нин (Т) —гуанин (Г) —тимин (Т), т. е. А — Ц—А — Т — Г — Т,
не идентична последовательности Г — Ц--А..А —Т — Т, хотя ка-
чество и количество оснований одинаково в том и другом отрез-
ках.
^ Полимерная цепь ДНК обладает полярностью. Один конец ее
^Одержит 5'-гидроксил или фосфат, другой — З'-фосфатный или
•Ядроксильный остаток:
Й
73
Полярность цепи ДНК важна для формирования второй анти-
параллельной цепи ДНК при образовании двойной спирали.
Поскольку генетическая информация заключена в порядке
расположения оснований в полимерной цепи ДНК, постольку
должны существовать механизмы воспроизведения или реплика-
ции этой специфической информации с исключительно высокой
точностью. При анализе состава молекул ДНК различных орга-
низмов было установлено, что количество адениновых нуклеоти-
дов равно количеству тимидиновых нуклеотидов (т. е. А=Т),
такое же отношение было установлено для гуаниновых и цити-
диновых нуклеотидов (т. е. Г=Ц) (табл. 10).
Таблица 10. Состав молекул ДНК у некоторых микроорганизмов
Источник ДНК Молярное отношение А + T г + Ц*
А т г Ц
Saccharomyces cerevisiae 10 10,5 6 5,5 1,79
Proteus vulgaris 10 9,8 6,6 6,9 1,47
Aspergillus niger 10 10 10 10 1
Escherichia coll 10 9,6 10,5 10,4 0,93
Pseudomonas aeruginosa 10 9,6 19,6 20,2 0,49
Фаг фХ174 10 13,1 10,6 8,2 1,23
* Цитознн нли 5-метилцитозин (5СНзЦ).
Отношения А-фТ к Г + Ц могут быть положены в Основу под-
разделения ДНК на 3 группы: в одной из них (А-|-Т) >(Г + Ц),
во второй — (А + Т) == (Г + П) ив третьей — (A-f-T) < (Г + Ц).
74
^отдельных молекулах ДНК различия в молярных отношениях
ценина к тимину и гуанина к цитозину весьма заметны (фаг
Ml74)-
JJp Выявленное количественное равенство А=Т и Г=Ц было
.^вложено Дж. Уотсоном и Ф. Криком в основу предложенной
ййи в 1953 г. модели двойной спирали ДНК (рис. 15,1). Две
в 1953 г. модели двойной спирали ДНК (рис. 15,1). Две
2,0 нм
Рис. 15. Двойная спираль
ДНК (I), водородные свя-
зи между пуриновыми и
пиримидиновыми основа-
ниями в ДНК (П), уча-
сток двойной спирали с
антипараллельиой на-
правленностью углеводно-
фосфатных цепей (Ш)
75
комплементарные нити спирали закручиваются вправо (форма В)
и удерживаются рядом благодаря наведению водородных связей
между пуриновыми основаниями в одной нити с пиримидиновыми
основаниями — в другой. Вследствие ограничений, накладывае-
мых фосфодиэфирной связью, в частности образование пред-
почтительной анти-конформации гликозидной связи (см. разд. 2.1.1)
и присутствие оснований в кетоформе (а не в енольной фор-
ме), формирование пар возможно лишь из А и Т, а также Г
и Ц.
Связи между парами оснований весьма специфичны. Учитывая
тот факт, что между А и Т наводятся две водородные связи, а
между Г и Ц — три, связь Г=Ц примерно на 50 % прочнее связи
А=Т и, следовательно, плотность ДНК, содержащей большее
количество пар Г=Ц, будет выше (рис. 15,11). Содержание
Г + Ц у микроорганизмов имеет таксономическое и филогенети-
ческое значение.
В двойной спирали ДНК цепи идут антипараллельно, т. е.
одна нить следует направлению 5'—►З', а другая — направлению
3'—>-5'. Как видно из рис. 15, Ш, пуриновые и пиримидиновые
основания находятся между углеводно-фосфатными цепями, т. е.
внутри спирали, защищающей их от возможных дополнительных
влияний извне. Кроме того, размеры пар А=Т и Г^Ц близки и
поэтому последовательность оснований в цепи не налагает каких-
либо ограничений на структуру ДНК в целом. Вот почему архи-
тектура ДНК у разных организмов тождественна и различа-
ется лишь по специфической последовательности пар основа-
ний.
Наследственная информация хранится в обеих цепях ДНК, *
не только в одной, так называемой смысловой. гледовательш
комплементарная ей антипараллельная цепь лишь условно явля
ется антисмысловой. Однако считывание информации нроисхо
дит только с одной цепи из двух (несимметричная транскрип
ция). Если бы обе цепи ДНК использовались в качестве матрш
одновременно, то на одном транскриптоне лпш. шровались
две комплементарные мРНК, заключающие информацию дл1-
двух разных белков.
ДНК, схематично изображенная на рис. 15, принадлежит
клеткам эукариот. У бактерий, большинства фагов и ДНК-содер-
жащих вирусов животных ДНК не имеет 3'- или 5'-концов и у
них она кольцевидная, хотя и не лишенная полярности. В ДНК,
выделенной из различных источников, основания внутри спирали
располагаются на расстоянии 0,34 нм друг от друга, на шаг спи-
рали (3,4 нм) приходится 10 пар оснований; диаметр спирали
равен 2,0 нм. Фосфатные группы располагаются на наружной
стороне спирали на расстоянии 0,9 нм от ее оси.
Расстояние между гликозидными связями, соединяющими
пары оснований с углеводно-фосфатным кором (от англ, core —
стержень, скелет); одинаковы для каждой пары и равны
1,085 нм:
76
$ Данная характеристика приведена для спирали ДНК в фор-
W В. Эта форма может трансформироваться в форму А при сни-
^цёнии влажности препарата ДНК ниже 75 %. В форме А пары
•’Оснований располагаются неперпендикулярно оси спиралей, а с
Наклоном под углом около 20°. В итоге уменьшается шаг спирали
'Др 2,8 нм, на виток приходится 11 пар оснований и сама цепь
укорачивается примерно на 25 %.
г Известна С-форма ДНК с 9 парами оснований в одном витке
И шагом спирали, равном 3,3 нм. Описаны еще Z- и SBS-формы
ДНК. Первая из них представляет собой левую спираль с 12 па-
рами оснований, приходящимися на один виток. Z-форму, или
форму зигзага, приобретает углеводно-фосфатный остов в участ-
ках, содержащих чередующуюся последовательность Г и Ц.
SBS-Форма ДНК (от англ, side by side — бок о бок) характери-
. зуется отсутствием взаимозакрученности цепей в двойную спи-
раль, что, очевидно, приобретает важное значение для разделе-
,ния цепей при биосинтезе ДНК.
V , Жесткая двуспиральная структура ДНК поддерживается в
1 Нормальных условиях не только за счет водородных связей, но
; Н за счет гидрофобных взаимодействий между расположенными
«стопкой» основаниями. Тем не менее двухнитевая ДНК может
Подвергаться плавлению при повышенных температурах или при
уменьшении концентрации солей в растворах — при этом нити
расплетаются и нарушается парность оснований, хотя сами осно-
’ Вания остаются связанными с углеводно-фосфатными цепями.
Это так называемая денатурация ДНК, которая может быть пол-
яой и неполной. Денатурация возможна также и от действия
сольватирующих агентов, например формамида или мочевины.
Сольватация (от лат. solvo — растворяю) означает взаимодейст-
вие молекул растворителя с молекулами растворенного вещества.
Сольватация в водных растворах называется гидратацией. При
' Денатурации не происходит разрыва ковалентных связей. Денату-
рированные молекулы ДНК дают гиперхромный эффект, т. е.
‘ Наблюдается увеличение поглощения света в ультрафиолетовой
• области (в норме максимум поглощения порядка 260 нм); умень-
шение положительного вращения плоскости поляризованного
»Света; уменьшение вязкости.
Если, например, нагретые водные растворы ДНК подвергать
медленному охлаждению при поддержании в течение какого-то
времени температуры несколько ниже температуры плавления
(отжиг), то может происходить ренатурация ДНК, когда компле-
ментарные нити ее воссоединяются (хотя и не всегда точно по
исходному плану). При быстром охлаждении нагретых водных
растворов ДНК ранее комплементарные цепи ее не воссоединя-
ются, и она остается в денатурированном состоянии. Степень
сплавляемости ДНК разных видов характеризует и степень их
гомологии, что широко используют в геносистематике микроор-
ганизмов. При этом проводят гибридизацию молекул ДНК, при-
надлежащих разным видам.
Размеры молекул ДНК у разных организмов варьируют в
широких пределах (табл. 11).
Таблица 11. Размеры молекул ДНК различных организмов
Организм Хромосомы ДНК
число в гаплоид- ной клет- ке форма длина, см среднее число пар основании на хромо- сому (X 103)
Saccharomyces 17 Незамкнутая 0,33 1000
cerevisiae Escherichia coli 1 Кольцевая 0,13 4000
Фаг фХ174 1 Кольцевая, одноце- 0,00018 5,4
Вирус SV4o 1 почечная; двойная реп- ликативная Кольцевая 0,00017 5,1
обезьяны Человек 23 Незамкнутая 4,1 125 000
В ДНК эукариотических организмов содержатся обращенные
повторы, или палиндромы (от греч. — перевертыш),
т. е. последовательности, повторяющиеся в обратном порядке
А—Т
Г Г
5' - ЦТТЦГАТ Г ГААГ - 3' Ц • Г
................... * Т • А
3' - Г А А ГЦТ АЦЦТТЦ - 5' Т • А
< ' Г ! . Г _ 1'
В состав линейной ДНК палиндромы не влияют на ее вторич
ную структуру, однако в суперспирализованном состоянии (кон-
формации) длинные палиндромы (10...20 и более пар оснований)
образуют крестообразные структуры, которые, как предполагают,
служат сигналами для узнавания определенных участков ДНК
ферментами (рестриктазами, метилазами) и регуляторными бел-
ками, регулирующими действия генов.
78
. двухспиральной ДНК
тся большая и малая
+Н1
гистоновый
октамер
-Н1
Рис. 16. Нуклеосомы (с гистоном, +Н1)
и нуклеосомный кор (без гистона, —Н1)
„вки по длине углеводно-
фатного кора. У эукарио-
еских организмов в этих
авках специфические бел-
взаимодействуют с моле-
рами ДНК. В прокариоти-
—р.ких клетках ДНК может
связана с незначитель-
ном количеством белковых
Молекул, а с учетом того факта, что на каждый межнуклеотидный
фосфат приходится один анион с рКа —1, очевидно, что при нейт-
«кльном pH нуклеиновые кислоты находятся в виде натриевых,
Магниевых и других солей. У некоторых вирусов и фагов ДНК
«Нейтрализована органическими аминами (кадаверином, путресци-
«ёЙм, спермидином, спермином).
Ядерная ДНК у более высокоорганизованных эукариот неко-
Кентно связана с основными белками (гистонами), которых
ержнтся около 50 %. У высших эукариот (животные и чело-
) в ДНК найдены еще негистоновые белки (кислые и ббль-
Вго размера, чем гистоны) и небольшое количество РНК. Все
1 вместе составляет хроматин, наблюдаемый в электронном
«Микроскопе в виде сферических частиц — нуклеосом (12,5 нм
^диаметре), схематично представленных на рис. 16. В связи с
(Ййтерпретацией рисунка приведена номенклатура гистонов
Ъабл. 12).
Таблица 12. Номенклатура гистонов
J Гистоны 1 ; -,« Соотноше- НИЯ Лиз Apr Обозначения по
первоначальной, схеме новой схеме
Обогащенные лизином 19,0 fl (12С) 1в (V) Hl H5
Слабо обогащенные лизином 1,1 2,0 f2a2 f2B IIbI IIb2 H2A H2B
Умеренно обогащенные аргинином И содержат цистеин 0,7 f3 III H3
Обогащенные аргинином и глици- ном 0,7 12a 1 IV H4
Гистоны не найдены в хромосомах разных бактерий, однако
; из их клеток выделены фракции, обогащенные лизином и арги
ннном. Детали структуры ДНК эукариотических мицелиальных
•грибов, дрожжей и других организмов остаются противоречи
79
s'
3'
Рис. 17. Шпилька РНК, формирующаяся из одной
цепи при внутримолекулярном спаривании основа-
ний
выми. Несомненно лишь одно, что различные химические модг
фикации в хроматине имеют отношение к образованию структу;
высшего порядка и к изменению взаимоотношений между гистс
нами и ДНК во время репликации и транскрипции.
Рибонуклеиновая кислота — полимер, состоящий из пурино-
вых и пиримидиновых нуклеотидов, связанных 3',5'-фосфоди-
эфирными мостиками (как в ДНК). В отличие от ДНК рибонук-
леиновая кислота содержит урацил вместо тимина и рибозу
вместо дезоксирибозы. Нативная РНК преимущественно однони-
тевая. Однако свойство комплементарности оснований с противо-
положной полярностью присуще и РНК при складывании ее цепи
в виде шпильки (рис. 17). В отдельных случаях существуют при-
родные двухнитевые РНК, например киллер-плазмида (от англ,
killer — убийца) у дрожжей, двухнитевая РНК у человека —
индуктор противовирусного белка интерферона. В однонитевых
РНК существует количественное равенство между аденином и
урацилом, между гуанином и цитозином.
РНК всех прокариотических и эукариотических организмов
можно разделить на три основных класса: матричная, или инфор-
мационная (мРНК); рибосомальная (рРНК); транспортная
(тРНК) РНК каждого класса отличается размером, функцией и
общей стабильностью. мРНК наиболее гетерогенны по размеру
и стабильности, так как они выступают посредниками между
ДНК и рибосомами (рис. 18). Вследствие того, что информация
считывается с одной нити ДНК (смысловой), то мРНК сходна по
строению с антисмысловой цепью ДНК (только тимин в мРНК
заменяется на урацил).
ДНК
ТАГГЦТЦЦЦТТТЦГАГАТААГТАТЦЦТАТГ
АТЦЦГАГГГАААГЦТЦТАТТЦАТАГГАТАЦ
---Антисмысловая
цепь
---Смысловая цепь
мРНК (п) УАГГЦУЦЦЦУУУЦГАГАУААГУАУЦЦУАУГ (и) ----Транскрипт
Рис. 18. Транскрипт мРНК, комплементарный смысловой цепи ДНК:
и — инициация, п — пунктуация
80
i клетках прокариот мРНК образуется сразу в процессе транс-
дции, тогда как в клетках эукариотических организмов мРНК
рплазмы являются продуктами предварительной обработки
•рогенных ядерных РНК (гяРНК) или про-мРНК, которые
[чно больше по размерам, чем мРНК цитоплазмы. Молекуляр-
t масса гяРНК достигает 107 Да, тогда как молекулярная мас-
мРНК обычно менее 106Да. Поэтому гяРНК должна пройти
специальную обработку, или процессинг (от англ, process — про-
^исс). В ходе обработки первичные транскрипты (РНК, обра-
зующиеся при транскрипции) превращаются в мРНК и стано-
Еся в цитоплазме матрицей для синтеза полипептидных молекул
ка. У прокариот мРНК менее стабильна, чем у эукариот.
рРНК, имеющая форму V или Y, составляет каркас рибосом,
^клетках некоторых бактерий на нее приходится 80 % общей РНК.
(f. бактерий рРНК обычно существует в виде трех компонентов,
дцеющих соответственно следующие коэффициенты седимента-
ции: 5S, 16—18S и 23—28S. У эукариот она представлена че-
-*вд>ьмя видами молекул, на которые приходится менее 100 специ-
фических молекул белка на рибосому. Причем в более крупной
Субъединице рибосомы (60S) содержатся 5S и 5,8S (прежде была
обозначена как 7S) и 28S рРНК, а также около 40 специфиче-
ских полипептидов. В меньшей субъединице рибосомы (40S) со-
держится одна 18S рРНК и примерно 30 полипептидных цепей
' (табл. 13). 5S рРНК, прокариот гомологичны 5,8S рРНК эука-
риот, но в составе митохондрий клеток эукариот 5S рРНК не
обнаружена (она содержится в хлоропластах).
Таблица 13. Типы РНК в рибосомах клеток прокариот и эукариот
Клетки Субъединицы рибосом микроорганизмов РНК
константа седи- ментации S М, Да константа седи- ментации S М, Да
Прокариот 30 1,0-106 16 0,56-106
50 1,8-106 23 1,1-106
5 4,1-104
Эукариот 40 1,3-106 18 750-103
60 2,7 • 10б 5 35-Ю3
5,8 45-Ю3
28 1,5 • 106
Все рибосомальные молекулы РНК у эукариот, за исключе-
нием 5S рРНК, образуются в ядрышке из единого предшествен-
ника— 45S РНК. Очевидно, рРНК типа 5S имеет своего соб-
ственного, независимо транскрибирующегося предшественника.
В ядрышке упаковываются высокометилированные молекулы
рРНК со специфическими рибосомальными белками основного
характера. У Е. coli белки для малой субъединицы рибосом обо-
значают как SI, S2, ..., S21 (от англ, small — маленький), для
81
большой субъединицы — L1, L2, ..., L34 (от англ, large — боль-
шой). Рибосомы микроорганизмов в цитоплазме остаются доста-
точно стабильными и готовыми к многократным трансляциям.
Каждая молекула транспортной РНК (тРНК) состоит при-
мерно из 75...90 нуклеотидов и, следовательно, молекулярная мас-
са ее порядка 23 000...30 000 Да; тРНК составляют 10...20 % и
являются как бы итогом ядерного процессинга молекул-пред-
шественников. В каждой клетке прокариот и эукариот имеется
по крайней мере до 60 разных видов тРНК, каждый вид пере-
носит в рибосому только одну какую-либо аминокислоту.
Порядок нуклеотидов в каждой тРНК различен, хотя класс
тРНК в целом имеет много общего. Молекула тРНК представ-
ляет собой одну цепь, закрученную «на себя». Первичная струк-
тура, зависимая от последовательности соединения нуклеоти-
дов, обеспечивает всем молекулам тРНК возможность формиро-
вать складки и соответственно внутрицепочечную комплемен-
тарность. Это в свою очередь генерирует вторичную структуру
в виде клеверного листа, показанную на примере аланиновой
тРНК на рис. 19, а, на котором демонстрируется ее первичная
структура, во многом сходная с тРНК, изолированными из дру-
гих источников.
Все молекулы тРНК имеют общие структурные части: 1) ан
гикодоновую петлю, содержащую 7 неспаренных нуклеотидов
из которых 3 терминальных признают кодон на мРНК (анти
кодоновая петля тРНК и кодоновая петля мРНК антипараллель
ны но своей комплементарностиV. 2) тимин-псевдоуридин-ци-
гидиновую петлю, участвующую в. связывании аминоацил-тРНК
с поверхностью рибосомы на месте синтеза белка; 3) дигидро
уридиновую пеглю. играющую важную роль для правильное»
признания «своим» ферментом - аминоацил-тРНК-синтетазой.
4) АЦЦ-последовательность на З'-конце углеводно-фосфатногс
кора, где через З'-гидроксильную группу аденозильного остатка
присоединяется (через карбоксил) соответствующая аминокисло
га (рис. 19. б).
тРНК, изолированные из клеток разных микроорганизмов,
характеризуются консервативностью и, как следствие, большой
структурно-функциональной общностью.
Белки (протеины) — это высокомолекулярные полипептиды
и их производные. Белки, состоящие только из аминокислот,
называют простыми. Молекулярная масса простых белков нахо-
дится в пределах 8 000... 10 000 Да, молекулы с меньшей моле-
кулярной массой оценивают как малые, а с большей — как боль-
шие полипептиды. Если в состав белков входят небелковые
(простетические) группы, то их называют сложными. К слож-
ным белкам относятся гемопротеины, гликопротеины, нуклео-
протеины, металлопротеины, липопротеины, которые содержат
соответственно, гемоглобин, углеводы, нуклеиновые кислоты,
металлы и липиды.
Белки различают по растворимости, форме, функции и трех-
82
[ОЙ структуре. Напри-
f. эуглобулины нера-
римы в воде, тогда как
умины и псевдоглобу-
ны растворимы в ней.
лки могут быть глобу-
сными и линейными; по
&3ункции — каталитически
Дивными, структурными
транспортными. Оцен-
трехмерной структуры
Щелков полезна для выво-
Сда — имеет ли данный бе-
Ъугок надмолекулярную
^структуру. Для этого про-
;'водят кристаллографиче-
ские исследования с по-
' мощью рентгеновских лу-
дЧей. В формировании трех-
дверной структуры белков
Важную роль играет отсут-
-^'фгвие свободного враще-
''•Ййя вокруг С—N-связи в
"вёптидных цепях.
W
к
Рис. 19. Дрожжевая аланиновая тРНК (а), связывание анти-
кодона тРНК с кодоном мРНК в рибосоме (б)
» В основе первичной структуры протеинов находится пептид-
I пая связь, образующаяся между карбоксильным остатком одной
;' Аминокислоты и аминогруппой другой:
________ пептидная связь
*hN— СН—COiOH + HjN—СН—СООН Н2О + H2N СН—СО—NH—СНСООН
>. ' I ---------------il I I
HR, R Ri
i ' ' дипентид
83
Пептидная связь является планарной и обладает некоторым
характером двойной связи, поэтому нет свободного вращения
молекулы или части молекулы вокруг нее. Это обстоятельство
приобретает важное значение, для формирования структуры бо-
лее высокого уровня:
Некоторые размеры, характеризующие полностью растянутую
молекулу полипептидной цепи, дают возможность заключить,
что: 1) атомы, образующие пептидную связь О=С—N—Н,
копланарны, т. е. они находятся в одной плоскости; 2) амидная
группа имеет трамс-конфигурацию, т. е. два асимметрических
атома углерода располагаются в противоположных углах от нее
и свободное вращение может иметь место вокруг связей, соеди-
няющих а-углерод с а-азотом и а-карбонилом (показано на схе-
ме круговыми стрелками); 3) растянутая полипептидная цепь
представляет собой полужесткую структуру, в которой 2/3 атомов
находятся в фиксированном плоскостном взаимоотношении друг
с другом.
Различают олигопептиды, включающие ди-, три-, тетра-,
пента-, гекса-, гепта-, окта-, нана- и декапептиды, и полипепти-
ды, включающие свыше 10 аминокислот:
трипептид
Названия всех пептидов прочитывают в направлении от
N-конца к С-концу, т. е. слева направо (указанный трипептид
представляет собой глицил-цистеил-валин). Таким образом, ли-
нейная последовательность аминокислот в олиго- или полипеп-
тидах определяет первичную структуру пептидных молекул. Если
полипептид включает большое число аминокислотных остатков,
84
$' прибегают к сокращенному написанию линеинои последо-
1ательности (см. табл. 5): Гли-Цис-Вал.
Пептидная (амидная) связь не несет заряда при любых
аизиологических значениях pH, хотя сами пептиды заряжены
на N- и С-терминальных группах и на функциональных группах
полярных аминокислотных остатков в позиции а-С:
н
Н3С—С—СН3
н в । 9 н
H2NX I /С. С“ Н /N4 I СООН
i-O Н н L__
Гсн2| гснл
Il I
। I I ( Полярные I I ।
[_SHj группы при [OH_J
а-С в трнпеп-
тнде
Значение рК для С-терминальной группы полипептида выше,
•чем значение рА для карбоксила соответствующей аминокисло-
•ты, т. е. пептидный карбоксил относится к разряду слабых кис-
лот. Для пептидов, так же как и для аминокислот, существует
$ изоэлектрическая точка (р7), определяемая исходя из значения
,р/С
”8 Микроорганизмы имеют кроме белковых молекул достаточно
большое разнообразие пептидов, содержащих от 3 до 100 ами-
<нокислотных остатков. Например, циклопептидный антибиотик
^грамицидин С включает 5 пар аминокислот, причем некоторые
.'.'ИЗ них (например, фенилаланин) относится к P-ряду (аминокис-
л-лота орнитин не встречается в природных белках):
; Вал Орн Лей О-Фен Про
t 4
Про D-Фен Лей <- Орн Вал
грамицидин С
' Близким, но не тождественным грамицидину С является анти-
- биотик тироцидин А. К циклопептидным антибиотикам относятся
также полимиксин, бацитрацин, включающие Р-аминокислоты
; Или другие аминокислоты, не встречающиеся в естественных
_ белках, например L- и Р-диаминомасляную кислоту (ДАМ).
'¥-- Тетра- и пентапептиды входят в состав пептидогликана кле-
Точной стенки грамположительных и грамотрицательных бакте-
Г рий, где он обеспечивает ригидность (жесткость) форм клеток
( * целом. Приводим строение тетрапептида, в котором R—оста-
С* Ток гликолактиловой кислоты, связанной с N-ацетилглюкозами-
ДАП — диаминопимелиновая кислота:
Ж 85
£-Ала D-Глу /.-ДАН D-Ала
R—NH—CH—CO-1-1NH—CH—COi-'NH—CH—CO-'^NH—CH—COOH1
СНз (СН2)2 ((^Н2)з СНз
I ।
СООН h2n—сн—соон
Названные пептиды синтезируются вне рибосом при действии
специфических ферментов, определяющих порядок чередования
аминокислот (нематричный синтез).
Молекулы белков стабилизируются двумя типами сильных
связей — пептидной и дисульфидной, а также тремя типами
слабых связей — водородными, гидрофобными и электростатиче-
скими. Все слабые связи при денатурации белков разрушаются.
Сильная дисульфидная связь соединяет две части одной поли-
пептидной цепи. Она образуется за счет остатков цистеина:
ОС
НС—Н2С—S—S—СНг—С
Эта связь может быть разрушена восстанавливающими (на-
пример, р-меркаптоэтанол HS—СН2—СН2ОН восстанавливает
SH-группы в молекулах цистеина) или окисляющими (напри-
мер, надмуравьиная кислота НОС—О—О—СОН окисляет связи
—S—S—) агентами. Первичная структура полипептидных цепей
при этом не нарушается:
+ 2HS—СН2—СН2ОН
ОС^
_^2н\:—ch2sh +(сн2)г—он
NH i
>° i
(<!:н2)2—он
86
ОС'
СО о
о
;со
ос
.—s—сн
,NH
ОС.
о
2Н,С—CH2SO3H + 2НСН
NH
^СО
‘А’Р
В полипептидах наводится множество слабых водородных
4 .£Вязей благодаря расположению атомов водорода между азо-
£-тЪм и карбонильным кислородом различных пептидных связей
.• в одной и той же или разных полипептидных цепях:
$
R
-N—
i.
Водородная
связь------
О
Н
н
11
н
Ri
1
В поддержании структуры нативной белковой молекулы за-
метную роль играют гидрофобные взаимодействия неполярных
боковых цепей нейтральных аминокислот. Если же в боковых
заряженные группы, то при
разноименных зарядов возни-
например, между вторым кар-
и'второй МНф-группой лизина:
Цепочках аминокислот имеются
противоположном расположении
кают электростатические связи,
боксилом глутаминовой кислоты
— [—ОС—СН-(СН2)2—СОО ~] |Н>N - (СН2)з-сн—NH—] —
Г
—NH
Глу
—СО
Лиз
’f'' Водородные связи, возникающие между всеми СО- и
.г ин-группами в молекулах белков, стабилизируют формирую-
Йгуюся правую а-спираль. На один виток а-спирали приходится
3,6 аминокислотных остатков; шаг спирали составляет 0,54 нм,
4,'Аиаметр вместе с боковыми цепями— 1,05 нм и 3 аминокислот-
87
ных остатка в одной петле, скрепленной водородными связями:
формула одной петли
а-спирали белковой
молекулы
Таким образом, а-спираль представляет собой вторичную струк-
туру белковой молекулы. Эта структура может быть представ-
лена также складчатыми слоями белковых молекул 0-структу-
рой, или «складчатым листком». В таких белках сближенные
полипептидные цепи прилегают друг к другу и идут параллель-
но (во флаводоксине) или антипараллельно (в супероксиддис-
мутазе Е. coll). Обе цепи стабилизируются многими водородны-
ми связями в виде сети:
₽сн^ н,ср рсн^
с=о • • н—n jc=d
Н—N/ \С=О---Н—N-/
антипараллельный
«складчатый листок»
параллельный «складчатый листок»
Водородные связи могут возникать между боковыми цепя-
ми (R) аспарагиновой кислоты и серина, гистидина и тирози-
на (1). Ионные связи устанавливаются между противополож-
но заряженными боковыми цепями, например аспарагина, ги-
стидина и лизина, с одной стороны, и глутаминовой и аспараги-
новой кислот — с другой (2). В неполярных гидрофобных взаи-
модействиях участвуют R-цепи валина, лейцина, изолейцина,
фенилаланина и триптофана (3):
88
а- и 0-Спирали в белках могут взаимодействовать между
’.'^Обой и друг с другом, образуя сверхвторичные структуры бел-
з^овой молекулы, или ансамбли, которые энергетически наиболее
|о|ыгодны. Короткие участки таких ансамблей обнаружены, на-
пример, в микробном глобулярном белке — бактериородопсине.
Вторичная структура полипептидной цепи включает еще и
(от англ, domain — область), представляющие собой
®«убобласти молекулы, соединенные шарнирными участками.
^Структурные домены обнаружены, например, в глутатионредук-
^.'уазе дрожжей.
Известны также третичные и четвертичные структуры белков.
Третичная структура определяется ковалентными дисульфидны-
ми связями и нековалентными взаимодействиями между участ-
г'гхами полипептидной цепи (дополнительные водородные и ион-
з'Щяе связи), а также взаимодействиями гидрофобных R-rpynn.
1 'В глобулярных белках до 30 % аминокислотных остатков имеют
Гидрофобные R-группы и до 50 % содержат полярные или ионные
R-группы. Дисульфидные связи могут возникать при сближении
Полипептидных цепей, содержащих цистеин, однако они форми-
руются не во всех белках. Например, в белках кишечной палоч-
практически отсутствуют дисульфидные связи. Следовательно,
вклад их в третичную структуру отдельных белков отсутствует
Или ничтожен. Но в тех белковых молекулах, где имеются
дисульфидные связи (например, четыре —S—S—связи в лизо-
циме), разрыв их сопровождается утратой биологических свойств
молекул.
Известно, что многие белки включают больше, чем одну по-
•’ липептидную цепь, причем и сами цепи могут быть различными.
В этом случае, исходя из данных о функциональной активности
какого белка, обладающего специфической биологической ак-
тивностью, его расценивают как одну молекулу. Две и более
. Полипептидных цепей (субъединиц) в составе такого белка
•*' ассоциируют на основе полярных и гидрофобных взаимодействий
•£. 6 последующим формированием четвертичной структуры моле-
кулы (табл. 14).
(г Например, четвертичная структура 0-галактозидазы кишеч-
ной палочки объединяет 4 тождественные субъединицы — поли-
89
Таблица 14. Четвертичная структура белков некоторых микроорганизмов
Белок Молекулярная масса, Да Число субъединиц
Алкогольдегидрогеназа Saccharomyces cerevisiae 150 000 4
Аспартат-транс-карбамоилаза Е. coli 50 500 6+6
P-Галактозидаза Е. coll 540 000 4
Енолаза Saccharomyces cerevisiae 67 000 2
Рибонуклеаза Т1 Aspergillus niger 11 100 1
пептидные цепи, состоящие каждая из 1173 аминокислотных
остатков. Триптофансинтаза того же микроба также включает
4 субъединицы, подразделяющиеся на две пары разных поли-
пептидных цепей, одна из которых содержит 268 аминокислотных
остатков, вторая — 450. Из данных внутримолекулярных и меж-
молекулярных взаимодействий следует, что вторичная, третичная
и четвертичная структуры белков всецело определяются их пер-
вичной структурой.
Полисахариды и гликоконъюгаты. К большим молекулам
относят углеводные полимеры, включающие свыше 10 остатков
моносахаридов, и гликоконъюгаты, в которых углеводные цепи
ковалентно связаны с белками или липидами.
Полисахариды (гликаны, полиозы) (см. табл. 9) — самые
распространенные в природе биополимеры и микроорганизмы
занимают важное место как источники различных углеводов.
Гликаны — продукты конденсации многих молекул моносаха-
ридов. Например, эмпирическая формула для полимерной гек-
созы (гексозан) — (СбНюО5)п, для полимерной пентозы (пен-
тозан) — (CsHgO^n, т. е. при конденсации п молекул монозы
происходит отнятие п молекул воды по схеме:
(С6Н12Об)„ + CeHiaOe —(CeHioOsjn+i + лНгО
Гомогликанами являются, например: глюкан (аубазидан),
образуемый диморфным грибом Aureobasidium pullulans\ ман-
нан (родэксман), продуцируемый некоторыми видами дрожже-
вых организмов рода Rhodotorula; глюкан (декстран), образуе-
мый бактерией Leuconostoc mesenteroides. К гетерополисахари-
дам относятся, например, капсульные полисахариды дрожже-
вых организмов из рода Cryptococcus. Некоторые из гетеро-
полисахаридов включают остатки маннозы, галактозы, ксилозы
и глюкуроновой кислоты (полиурониды). Сложный гетерополи-
сахарид ксантан образуется бактерией Xanthomonas campestris.
Звено ксантана состоит из целлобиозного кора и триаозной
боковой цепочки, включающей 4,6-пируваткеталь-маннозу, глю-
куроновую кислоту и ацетил-6-маннозу.
Все названные выше полисахариды содержат гликозидные
90
зи между моносахаридными остатками и поэтому являются
озидами. Если гликозид образуется, за счет С1 глюкозы, то
называют глюкозидом, а все С1-производные других угле-
,ов — гликозидами. Гликозидные связи могут быть между
,, и С2, С1 и СЗ, С1 и С4, С1 и С6 углеродными атомами.
Гликозидные связи более устойчивы к кислотному гидролизу,
м a-связи, особенно связь |3-С1->-СЗ. Фуранозиды, дезокси-
хариды и ангидропроизводные углеводов в составе полимеров
устойчивы к кислотам. В полиуронидах гликозидные связи
обще устойчивее к кислотному гидролизу, чем в других поли-
жаридах.
grafa. Полисахариды могут включать аминосахаридные остатки,
1!&йк. например, в хитозане у некоторых нитчатых грибов; если
Едаиногруппа его замещена ацетильным радикалом, то полисаха-
рид называют хитином:
могут быть нейтральны-
от свойств функциональ
или меньшем количестве
Полисахариды по своим свойствам
I, кислыми и основными, что зависит
х групп, содержащихся в большем
иблекуле полимера. Из числа названных полисахаридов нейт-
г льными являются аубазидан, родэксман, декстран; кислыми —
^йрнелан и ксантан; основным — хитозан. Низкомолекулярные
жЫи относительно низкомолекулярные полисахариды растворимы
Коде, но нерастворимы в большинстве органических раствори-
ей. Гидратные оболочки вокруг молекул полисахаридов раз-
;';.ф-ушаются при добавлении таких водоотнимающих средств, как
этанол и ацетон. Полисахариды, как правило, оптически активны.
Ж*;.1 Химические и физические свойства полисахаридов опреде-
лдяются строением, конфигурацией и конформацией составляю-
JglUHx их моносахаридов. Под термином строение понимают при-
-%реду и последовательность связей между атомами в молекуле;
йод термином конфигурация — определенную группировку
дтомов заместителей вокруг атомов углерода (сравнить галакто-
зу и маннозу); под термином конформация — различное распо-
ложение атомов молекулы в пространстве вследствие вращения
вокруг одной или нескольких простых связей (С- и В-формы
^.^иранозного цикла).
9k Зная строение, конфигурацию и конформацию, можно гово-
^МТь о структуре полисахарида. В настоящее время доказано
^существование полисахаридов на различных уровнях организа-
-жЧИМ молекул — первичном, вторичном, третичном и четвертич-
ном (сравните с белками). Первичная структура их определяет-
ся природой, последовательностью и характером связи моноз в
полисахариде; вторичная — относительной ориентацией полиса-
харидных цепей в пределах возможностей для ковалентно соеди-
ненных моноз; третичная — энергетически благоприятными
взаимодействиями между цепями полисахарида; четвертичная —
взаимодействиями полисахаридных «глобул» между собой или с
другими полимерами с компактной третичной структурой. При-
менительно к номенклатуре полисахаридов выработан следую-
щий единый подход. Названия гомополисахаридов слагаются из
названий входящих в их состав редуцирующих моносахаридов,
в которых суффикс «оза» заменяется на суффикс «ан» (арабан,
глюкан, маннан и т. д.). Если полисахарид представлен раз-
ветвленной молекулой гетерогликана, в основной цепи которого
находятся, например, остатки глюкозы, а в боковых цепочках —
остатки маннозы, то его называют манноглюканом; в случае
обратного распределения моноз — глюкоманнаном.
Иногда полисахариды называют по продуценту, сохраняя
суффикс «ан», например ксантан (продуцент — Xanthomonas
campestris), аубазидан и пуллулан (в обоих случаях ‘ проду-
цент — Aureobasidium pullulans). Лишь некоторые тривиальные
названия сохранились за отдельными полисахаридами — крах-
мал, гликоген, инулин, гранулеза.
Полисахариды, образуемые микроорганизмами, различаются
как по составу, так и по молекулярной массе. Например, фос-
фоманнан, образуемый Hansenula holstii, имеет молекулярную
массу, равную 157 000 Да, а некоторые декстраны— 1-108 Да.
Гликоконъюгаты — ковалентно связанные молекулы угле-
водов с белками, липидами и другими веществами. Различают
следующие гликоконъюгаты: гликопротеины, пептидогликаны
(протеогликаны), гликолипиды, липополисахариды (липоглика-
ны), гликолипопротеины, тейхоевые кислоты.
Глипопротеины широко распространены у представителей
микробного мира. Им присущи структурные, транспортные,
каталитические функции, а также функции узнавания и при-
соединения. Они входят в структуру иммуноглобулинов. В гли-
копротеинах наиболее часто выявляются галактоза, манноза,
фукоза, N-ацетилгалактозамин, N-ацетилглюкозамин, реже —
арабиноза, ксилоза, сиаловая кислота. Приводим формулу сиа-
ловой (N-ацетилнейраминовой) кислоты:
сиаловая кислота
92
•(Фукоза и сиаловая кислота всегда находятся в дистальном
йожении к полипептидной цепи, тогда как ацетилглюкозамин
^галактоза — в проксимальном положении и часто образуют
едзь с аминокислотами в молекулах белков, в частности с
тарагином, серином, треонином, гидроксилизином и гидрокси-
юлином. N-Гликозидная связь образуется исключительно меж-
г N-ацетилглюкозамином и аспарагином. Гликозилирование
риновых и треониновых остатков объясняется тем, что обра-
тощийся гликоконъюгат достаточно легко проникает, напри-
*р, через мембрану эндоплазматического ретикулума:
W
§
полипептидные цепи
остаток
ацетилгалактозамина
ч Гликопротеин, содержащий N-гликозидную связь, имеет
Iстержневую часть, или кор, который, в свою очередь, включает
Маннопиранозил-fJ-l4-М-ацетилглюкозаминил-|3-1 4-N-ane-
-->.уилглюкозаминил-0-аспарагин. Последовательность N-ацетил-
£$ЯЮКОзаминил-|3-14-ЬГацетилглюкозаминил называют ди-N-
5‘|Ц1|етилхитобиозной, поскольку она соответствует последователь-
ности N-ацетил-глюкозаминил в хитине.
Ж*-. К гликопротеинам, например, относится карбоксипептидаза
„г^фожжей с молекулярной массой 100 000 Да. Приводим схе-
IМатичное строение дрожжевой карбоксипептидазы:
V-
П
О
л
и
п
Е
п
т
и
д
%
Асн—(Тлю N Ац)2—(Ман)13
Асн—(Тлю N Ац) 2—(Мал)] 3
Асн—(Тлю N Ац)2—(Ман)13
Асн—(Тлю N Ац)2—(Ман)13
Асн—(ГлюК Ац)2—(Ман)13
^Гликопротеинами являются также отдельные ферменты микро-
-^Организмов (амилаза, рибонуклеаза, плазмокоагулаза), аллерге-
из патогенных грибов и др.
Пептидогликаны (протеогликаны). В них углеводная часть
$1 Состоит из повторяющихся дисахаридных звеньев, в которых
Жреегда присутствуют D-галактозамин или D-глюкозамин, уроно-
Т'вая (глюкуроновая, или L-идуроновая) кислота. Муреиновый
93
каркас клеточной стенки бактерий является типичным пептидо-
гликаном. В нем тетра- (три-, пента-) пептид связан с углевод
ной частью молекулы через посредство гликолактиловой (мура-
мовой) кислоты. Гликановые цепочки у разных микробов вклю-
чают от 10 до 65 дисахаридных единиц:
В результате гидролиза пептидогликана из него можно полу-
чить изолированные протеиновую и углеводную (гликозамино-
гликан) части.
Гликолипиды — полимеры, в которых углеводные части ко-
валентно связаны с липидными частями. Ключевыми глико-
зиллипидами мембран прокариотических и эукариотических мик-
роорганизмов являются соответственно бактопренол и долихол,
относящиеся к полиизопреноидным структурам (см. липоконъю-
Бактопренол (ундекапренол) содержит две транс- и 9 цис-
связей. Бактопренол и долихол участвуют в сборке гликолипи-
дов. Изопреноидами являются каротиноиды. Отдельные из них
представляют собой гликолипиды, например миксоксантофилл.
Иные гликолипиды содержатся в хроматофорах цианобактерий,
например моно- и дигалактозилглицериды, сульфохиновозилди-
94
,ерид с общей формулой R—Н2С—СН—СН2—OCORi, где
ocor2
и R2 — жирные кислоты, a R — углеводный компонент.
К гликолипидам относятся сфинголипиды (см. липиды и ли-
конъюгаты), корд-фактор туберкулезных микобактерий, опре-
1яющий их вирулентность (см. гл. 7), а также гликозилдиа-
1глицерины, обнаруженные в микроорганизмах. Сфинголипиды
•ут относиться и к гликолипидам и к липоконъюгатам, причем
>еброзиды как сфинголипиды или липоконъюгаты, содержа-
еся в нервных клетках высших организмов, здесь не рассмат-
1аются. Однако в тех же нервных клетках, например мозга,
(ержатся гликолипиды-гамглиозиды, присущие также и бакте-
^м, например нейссериям. Данные полимеры находятся у них
Пилях и определяют специфическое прилипание бактериальных
(вток к другим поверхностям:
•fc-t.
Л’’
:н2он
>н
Гал
ООН
H3COCNH
агон
н,он
но.
'Н.н-
остаток
N-ацетил-
нейраминовой
кислоты
со
Гал
неполярная часть
(хвост)
н2он
(СНОН)2
Н NHCOCH
Гал N Ад
.СН—снон
вдАн
(СН2)16 (СН2)12
Н3С сн3 .
:н,он
СН
НС
полярная часть (голова)
ганглиозид
он
«i I
,?,1НО)Н122СбТ-НС—НС—ОС—!
It С24Н49 Н
H
но
1122С61—НС—НС—ОС—I
С24Н49
ОН
О
н ОН
корд-фактор Mycobacterium tuberculosis
95
моногалактозилдиацилглицерин
Липополисахариды (липогликаны) обнаружены у грамотри-
цательных бактерий. Липополисахариды весьма разнообразны
по составу моноз в концевом блоке, тогда как стержневая
часть (кор) более постоянна, что показано ниже для Salmonella
newington:
।-------------------------------------------------------------------------------------------------------------------------------------и___________________________________________________________________________________________________________________________________и________________________________________________________________।
липид A
кор
терминальный
блок триаозы,
повторяю щий ся
до 25 раз
липополисахарид
где Р — пирофосфат; Си — миристиновая кислота; ЖК —
другие жирные кислоты; КДО-2-кето-З-дезоксиоктулонат; Геп —
Ь-глицеро-В-манногептоза; Глю — глюкоза; ГлкМАц— N-аце-
тилглюкозамин; Гал — галактоза; 3tNH2 — этаноламин; Рам - -
рамноза; Ман — манноза; R — один из остатков КДО может
иметь боковую цепь — [KDO-P-StNHj] .
КДО содержится не во всех типах липополисахаридов. Так,
его нет у многих цианобактерий, отдельных ксантомонасов и
других скотобактерий:
96
КДО
Липополисахариды обладают свойствами эндотоксинов. Ток-
1еские свойства их обусловлены липидом А, а полисахарид
ределяет специфичность О-антигенов клеток.
.Гликолипопротеины содержатся в нативном виде в клеточ-
X стенках и мембранах грамотрицательных бактерий. При
пользовании методов кислотного гидролиза (например, с по-
щью трихлоруксусной кислоты, фенола) удается выделить
иколипопротеиновый комплекс, ранее называемый полноцен-
IM антигеном. Данный комплекс можно разделить на составные
сти (углевод, липид, белок, липид-белок, углевод-липид).
Тейхоевые кислоты — полимерные структуры, занимающие
граничное положение между гликоконъюгатами и фосфоли-
дами. Так, к гликоконъюгатам относятся глицеринтейхоевые
„рибиттейхоевые кислоты, в которых углеводные компоненты
язаны с другими частями молекул ковалентными связями,
дример в глицеринтейхоевых кислотах, из клеток Staphylo-
ccus lactis и Streptomyces kanamyceticus, в рибиттейхоевой
слоте из клеток Bacillus subtillis:
звено глицеринтейхоевой кислоты
из клеток Staphylococcus lactis
звено глицеринтейхоевой кислоты
из клеток Streptomyces kanamyceticus
1 fi 1 7
D-Ала
О Глю—О—СН
Н2С
сн2
НО—СН
НО—СН
О
звено рибиттейхоевой кислоты
из клеток Bacillus subtilis
Тейхоевые кислоты найдены только у грамположительных бакте-
рий, у которых они либо связаны с клеточной мембраной, с поли-
сахаридами клеточных стенок, или локализуются в периплаз-
матическом пространстве.
Липиды и липоконъюгаты. Липиды представляют собой ге-
терогенную группу молекул, в большей или меньшей степени
родственных жирным кислотам. Те и другие нерастворимы в
воде, но растворимы в неполярных растворителях (бензин,
хлороформ, эфир). К липидам относят жиры, масла, воска и
родственные им вещества.
Липиды подразделяют на простые и связанные. Первые
из них называют липидами, вторые — липоконъюгатами, в кото
рых все составные части молекулы соединены ковалентными
связями. Липиды (простые липиды) включают жиры, масла и
воска. Жиры и масла представляют собой эфиры жирных кислот
и глицерина. Кроме глицерина обнаружены такие спирты, как
цетиловый, холестерин и др. Масла можно характеризовать как
жидкие жиры. В жирах обычно содержатся жирные кислоты с
четным числом атомов углерода, поскольку синтез их базируется
на двухуглеродных структурах. Воска — эфиры жирных кислот
и высокомолекулярных одноатомных спиртов (пчелиный воск,
ланолин). У микроорганизмов они не обнаружены.
Жирные кислоты могут быть насыщенные, ненасыщенные к
полиненасыщенные (полиеновые). Полиеновые кислоты не пай
дены в липидах у бактерий.
Некоторые грибы образуют простагландины (PG), которые
были впервые выделены из предстательной железы. Они синте-
зируются из 20 углеродных полиненасыщенных жирных кислот,
например дигомо-у-линоленовой, арахидоновой и тимнодоновой.
и образуют серию PG1 —PG3 соответственно:
98
>н
он
PGF,.
ОН
юон
он
СООН
20
PGF3a
PGF2a
биоло-
Простагландины обладают высокой и разносторонней
еской активностью, представляющей интерес для медицины.
Среди жиров и масел известны нейтральные жиры (ацил-
,церины), фосфоглицериды (глицерофосфаты) и фосфорибити-
. (рибитофосфаты). Нейтральные жиры (ацилглицерины) под-
,зделяют на три-, ди- и моноацилглицерины, когда гидроксилы
ицерина этерифицированы жирными кислотами соответственно
юностью, два и один:
О—СО—R,
R—О—СО—СНз—СН—СН2—О—СО—r2
триацилглицерин
О—СО—Ri
1,2-диацилглицерии
R—О—СО—СН2—СН (ОН) — СН2ОН
1-моиоацилглицернн
Триацилглицерины могут быть простыми и смешанными,
первом случае R—R2 представляют собой один и тот же оста-
к жирной кислоты (например, триолеин), во втором — разные
одному, двум и трем остаткам кислоты, например 2-стеароил-
пальмитоилглицерин (2-стеародипальмитин):
Нз 1С1 з—О—С О—С Н 2— С Н—СН 2—о—С О—С1 зН з 1
отличие от жиров масла содержат больше ненасыщенных жир-
IX кислот. Например, тристеарин — твердое вещество, а три-
еин — жидкое. В случае выращивания микробов при понижен-
ие температурах в них повышается содержание липидов с не-
сыщенными жирными кислотами. При щелочном гидролизе
мыление) ацилглицеринов образуются соответствующие со-
— мыла жирных кислот и глицерин:
+ 3NaOH
R—ОСО—СНз—СН—СН?—ОСО—R2 --------►
| Омыление
ОСО—R,
НОН2С—СН—СН2ОН + 3R (Rb R2)— COONa
ОН
грментативное расщепление нейтральных жиров липазами
оисходит по типу кислотного гидролиза:
99
+ЗН2О
R—ОСО—СНг—СН—СНг—ОСО—R2 —------НОН2С— СН—СН2ОН +
। Лнпаза ।
ОСО—Ri ОН
+ 3R(Rb R2)—СООН
Фосфоглицериды и фосфорибитиды содержатся в клеточных
мембранах. В их структуре одна из первичных спиртовых
групп глицерина или рибита этерифицирована фосфорной кис-
лотой, поэтому они называются еще фосфолипидами. Фосфоли-
пиды подразделяют на фосфатидную кислоту и фосфатидил-
глицерины, фосфатидилхолин (лецитин), фосфатидилэтаноламин
(кефалин), фосфатидилинозит, фосфатидилсерин, лизофосфоли-
пиды, плазмалогены, тейхоевые и липотейхоевые кислоты:
СН2—ОСО—R
I
Ri—ОСО—СН О
I II
СНг—О—Р—ОН
ОН
фосфатидная кислота
СН2—ОСО—R СН2ОН
Ri—ОСО—СН О Н^ОН
I II I
СНг—О—Р—О—СН2
ОН
фосфатидил глицерин
Если к фосфатидилглицерину присоединяется еще одна молекула
фосфатидной кислоты, то образуется кардиолипин, обнаруживае-
мый в клеточных мембранах бактерий и в мембранах митохонд-
рий эукариотических клеток. Фосфатидилглицерин не обнаружи-
вается (или обнаруживается в следовых количествах) в клеточ-
ной мембране эукариот, но является распространенным липидом
у прокариот. Фосфоглицерины относятся к полярным соедине-
ниям, так как остаток фосфорной кислоты полярен:
Rj—ОСО—СН—СН2—OCOR
I °
СН2—О—Р—он
о
I
носн
I
сн2
ОСО—r2 о
I I
R3—ОСО—Н2С—НС—сн2—О—Р=О
он
кардиолипин
СН2—ОСО—R
R,—ОСО—СН О
СН2—О— >—(СН2)2—N=(CH3)3
о-
фосфатидилхолин
(лецитин)
CH2OCOR
Ri—ОСО—СН О
1 и
СН2—О—Р—О—(CH2)2—NH 3
О-
фосфатидилэтаноламин
(кефалин)
100
t
CH2—ОСО—R
СН,—ОСО—R
Г*-. I
но—сн
R]
I—ОСО—сн
О—Р—О—(СН2)2—№(СН3)3
О
сн2—о—р—о
он
н
он
он
н
О”
лизофосфолипид
(лизолеицитин)
фосфатидилинозит
СН2—О—СН==СН—R
СН,—ОСО—R
•ОСО—СН
Rj—ОСО—СН
NH3
СН2—О—Р—О—(СН 2 )2—NH з
снг-о-
-О-
о_
плазмалоГен
он
фосфатидилсерин
о—сн2
Н3с—СН—ОСО—СН Р
NH-
н2с/
звено глицеринтейхоевой
кислоты из клеток
Lactobacterium casei
IH
Н—С—OR
н—с—он
н—с—он
Н2С-
0--СН2
,р-
IH
п
звено рибиттейхоевой кислоты
из клеток Streptococcus faecalis
(R=H, D-Ала или остаток
янтарной кислоты)
он
H ОН
липоглицеринтейхоевая кислота из клеток Streptococcus pyogenes
(R-остаток жирной кислоты)
о
О
О
о
3 всех перечисленных выше фосфолипидов только тейхоевые
липотейхоевые кислоты обнаруживаются в клетках грампо-
ЭЖительных бактерий; другие фосфолипиды широко распростра-
&ны в мембранах других прокариот и эукариот.
Липоконъюгаты представляют собой вещества, в составе ко-
фых имеются -ковалентно связанные жирные кислоты, спирты
ие всегда и не глицерин) и какие-либо другие компоненты,
йпример белки, углеводы. В этой связи они подразделяются
а сфинголипиды, микостерины, терпены, гликолипиды, липо-
I
101
протеины и липогликаны (гликолипиды и липогликаны см. ра-
нее).
Сфинголипиды не содержат глицерина, но включают спирт
дигидросфингозин и его ненасыщенный аналог сфингозин. Пред-
ставителем сфинголипидов является сфингомиелин, обнаружи-
ваемый в мембранах эукариот:
остаток сфингозина
Н Н ОН
Н3С—(Н2С)|2—= С—СН—СН—NH—СО—R
транс । I____I
СНг остаток
жирной
кислоты
остаток
I +
L о— (сн2)2—n=(CH3)3
остаток холина
сфингомиелин
К сфинголипидам относят цереброзиды, содержащие монозу,
спирт сфингозин и жирные кислоты, хотя по наличию углевода
они также могут быть включены в группу гликолипидов (см.
гликоконъюгаты). Цереброзиды не содержатся в клетках микро-
организмов, однако некоторые их производные церемиды (це-
реброзиды, лишенные углеводного компонента) обнаружены в
пилях некоторых штаммов Е. coli (например, серовар К 88).
Микостерины не содержат глицерина. Они часто ассоциируют
с другими липидами. Найдены у нитчатых грибов и дрожжей.
Микостерины являются производными пергидроциклопентанфе-
нантренового ядра, лежащего в основе стероидов, а именно —
спирта стерина (характеризуется наличием гидроксила у СЗ и
разветвленной алифатической цепи у С17 ядра):
пергидроциклопентанфенан-
треновое ядро
В клеточных мембранах эукариотических клеток содержится
эргостерин. Поэтому, например, дрожжи используются для полу-
чения эргостерина как провитамина D:
102
эргостерин кальциферол (витамин D2)
[удельные стероиды могут быть факторами роста для мико
Казм.
г Терпены включают изопреноидные структуры:
Г Н2С=С—СН=СН2
I- I
fc СНз
Пч изопреи
Й;, (2-метил-1,3-бутадиен)
Терпены с двумя изопреновыми группами называют монотер
(Внами; с тремя — сесквитерпенами; с четырьмя, шестью, во
0<ыо и десятью — соответственно ди-, три-, тетра- и пентатер
игами.
? У всех аэробных микроорганизмов важную роль в цепи пе
Ьйоса электронов играет убихинон, или кофермент Q. Полиизо
феиоидные структуры обнаружены в клетках прокариот и эука
Йот. Они участвуют в синтезе бактериальных клеточных сте
0к, гликопротеинов и протеогликанов в клетках эукариот. У бак
£рий полиизопреноидом является бактопренол, в клетках эука
йот — долихол:
ОН
он
убихинон (п = 6... 10)
f К изопреноидам относятся также каротиноиды, синтезирую
(неся многими пигментными микроорганизмами (Corynebacte
him, Pseudomonas, Rhodotorula, Sporobolomyces и др.) и имею
отношение к фотосинтетическим процессам клеток:
* Л — постоянная Планка, v — длина волны.
У некоторых цианобактерий обнаруживаются зеаксантин, мик-
соксантофилл (см. гликолипиды) и эхинон:
зеаксантин
эхинон
Липопротеины являются составной частью клеточной стенки
грамотрицательных бактерий (см. рис. 9, а). Их белковый ком-
понент, включающий 57 аминокислотных остатков в форме по
вторяющихся блоков, связан с остатками диаминопимелиновой
кислоты в пептидогликане — муреине. Следовательно, липопротеи-
ны у грамотрицательных бактерий, как правило, входят в состав
гликолипопротеинов, из которых могут быть выделены в виде
индивидуальных компонентов.
Макромолекулярные вещества клеток микроорганизмов (нук-
леиновые кислоты, белки, полисахариды и гликоконъюгаты,
липиды и липоконъюгаты) — основа надмолекулярных комплек-
сов, дающих в свою очередь начало органеллам и другим
структурам, обеспечивающим цитоархитектонику и функциональ-
ную активность прокариот и эукариот.
2.2. ЛОКАЛИЗАЦИЯ НИЗКОМОЛЕКУЛЯРНЫХ И ПОЛИМЕРНЫХ
ВЕЩЕСТВ В КЛЕТКАХ И ОРГАНИЗОВАННЫХ ЧАСТИЦАХ
Структурно-функциональная иерархия клеточных компонентов и
органелл обеспечивается малыми и большими молекулами хими-
ческих веществ. Одни из этих веществ занимают ведущее место
в формировании жестких или лабильных структур (пептидо-
гликаны клеточных стенок, липопротеины клеточных и митохонд-
104
ЙЛьных мембран); другие — в обеспечении обменных процес-
в, в которых из низкомолекулярных веществ возникает боль-
ннство первичных и вторичных метаболитов, накапливается и
сходуется энергия макроэргических связей АФТ и других мо-
‘ ул (ферменты, коферменты, кофакторы) ; третьи — в сохране-
I и передаче в последующих генерациях наследственной ин-
>мации (нуклеиновые кислоты) и т. д.
Компартментализация, присущая клеткам эукариот, вносит
дественные коррективы в распределение и локализацию мно-
веществ по «отсекам», тем самым оптимизируется синхро-
ш>ация процессов катаболизма и анаболизма. Тем не менее
ивая клетка любого микроорганизма является открытой систе-
й, поэтому в процессе ее роста, развития и размножения одни
щества постоянно должны поступать извне, а другие являются
юсительно устойчивыми структурно-функциональными компо-
итами метаболизирующих клеток.
Низкомолекулярные вещества (молекулы-предшественники,
эмежуточные вещества и молекулы строительных блоков) с
пекулярной массой не более 350 Да локализуются в клетках
>авнозначно. Вода с растворенными в ней веществами, вклю-
< некоторые макромолекулы белков, занимает основной кле-
1НЫЙ объем. Она как дисперсионная среда составляет зна-
ггельную часть цитоплазмы, вакуолей, лизосом, микросом, ядер
кариот. Следовательно, любая молекула клетки находится в
дной среде или связана с водой (например, наличие гидратных
олочек вокруг молекул). Вода, кислород, иногда натрий пас-
вно диффундируют в клетки до установления равновесных
нцентраций их внутри и снаружи. Для большинства других
зкомолекулярных веществ существуют специальные транспорт-
le системы в мембранах, обеспечивающие перенос этих ве-
еств из среды в клетку. Таким способом переносятся моно-
гхариды у Е. coli и Staphylococcus aureus. Следовательно,
шрерывность метаболических процессов базируется на непре-
явном поступлении низкомолекулярных веществ в клетку, рас-
здуемых в конструктивном и энергетическом обменах. Конструк-
1вный обмен (транспорт питательных веществ и синтез био-
элимеров) сопровождается расходованием энергии, накапли-
гемой и выделяемой при энергетическом обмене (брожение,
Ихание). Вот почему в цитоплазме прокариот и матриксе эука-
sot постоянно находятся низкомолекулярные вещества, необ-
"димые клетке. Отдельные малые молекулы, например пурины
пиримидины, аминокислоты, полифосфаты, некоторые вита-
ны (Вг), сера в виде клеточных фондов, или пулов (от англ.
о1 — фонд), накапливаются в вакуолях. Поступившие в клетку
тионы и анионы могут становиться составными частями мак-
молекул, включаясь преимущественно в белки — ферменты
и ассоциируются с ними, выступая в качестве кофакторов,
к, железо содержится в цитохромах, ферредоксинах; магний —
бактериохлорофилле; цинк — в алкогольдегидрогеназе, щелоч-
ной фосфатазе, альдолазе, РНК- и ДНК-полимеразах; марга-
нец — в супероксиддисмутазе; молибден — в нитрогеназе и
нитратредуктазе; медь — в цитохромоксидазе и оксигеназах; ни-
кель — в уреазе и т. д. Локализация катионов в отличие от
содержащих их солей может быть совершенно иной. Например,
они могут фиксироваться мембранами или включаться у эукарио-
тических клеток в органеллы типа митохондрий, лизосом, аппа-
рата Гольджи и др.
Многие гетеротрофные организмы используют малые моле-
кулы органических веществ в качестве источников углерода, на-
пример моносахариды — глюкозу и фруктозу, дисахарид —
лактозу и т. д. В подобных случаях названные вещества транс-
портируются в клетку без какой-либо предварительной деграда-
ции молекул. Так, лактоза переносится извне в Е. coll с по-
мощью лактозотрансферазы, а внутри клетки биоза расщепля-
ется с помощью 0-галактозидазы до глюкозы и галактозы. Ма-
лые молекулы используемых углеводов находятся в растворен-
ном состоянии в цитоплазме прокариот или достигают, напри
мер, митохондрий у эукариот, а затем подвергаются превра
щениям в энергетическом обмене. Поэтому потребляемые клет-
ками моносахариды рассматриваются транзиторными вещества-
ми, срок пребывания которых в неизмененном виде весьма огра-
ничен.
Организованные частицы — вирусы бактерий, растений и жи
вотных — не имеют собственного обмена веществ, поэтому пр>.
репродукции они используют строительный материал клеток
У них нет транзиторных веществ, но имеются макромолекулы
организующиеся в белки капсомеров (капсидов) и нуклеиновьг
кислоты (РНК или ДНК).
Белки многих вирусов представлены ферментами инвазив-
ности (от лат. invasio — вторжение); у некоторых РНК-вирусов,
вызывающих опухолевые процессы, имеется фермент обратная
транскриптаза, катализирующая реакцию транскрипции ДНК
по РНК-матрице вирусов. В вирусных частицах могут содержать-
ся липиды, входящие в структуру вирусов, такие вирусы чувст-
вительны к обработке эфиром и другими органическими раство-
рителями. В вирусных оболочках найдены гликопротеины.
Фаги содержат лишь два типа макромолекул: белки и нуклеи-
новые кислоты (обычно ДНК). Белки формируют головку, стер-
жень хвостика, сократительный чехол хвостика, фибриллы. Фаг
PSM 2 (или фаг Pseudomonas marine) имеет, как исключение,
двухнитевую ДНК и сама частица вириона окружена мембраной
липопротеина. Кроме того, фаг содержит ферменты эндонуклеазу
и ДНК-зависимую РНК-полимеразу.
Макромолекулы могут быть постоянными структурами на про-
тяжении всей жизни клеток или временными. Из них первые мо-
гут иметь строгую локализацию или топологию (от греч. тсолсоо—
место). Так, хлорофилл у цианобактерий является неотъемлемой
частью фотосинтетических пластинок, или тилакоидов, у водорос-
106
s— частью хлоропластов; белки-ферменты, катализирующие
кции цикла Кребса, находятся в митохондриях эукариотиче-
х микроорганизмов; ДНК как составная часть ядра форми-
тся в виде хромосом и плазмид у эукариотических и прока-
тических микроорганизмов или в виде самостоятельной струк-
_>ы входит в состав вирионов.
<Строго приписана» топологически, например хитинсинтаза
хитосомах грибов. У других полимеров такая «строгость» не
наруживается, но это не исключает постоянства их локализа-
। в определенных структурах клетки (ДНК, ферменты-гидро-
ы и т. д.).
В вакуолях микроорганизмов временно могут накапливаться
каны, липиды, белковые кристаллы, ферменты-гидролазы,
ютиноиды; в цитоплазме — гликоген, гранулеза, поли-0-гид-
кимасляная кислота, тРНК, ферменты — гидролазы и оксидо-
уктазы, каротиноиды и т. д.
Необходимо иметь в виду, что, несмотря на структурно-меха-
ческую и химико-физиологическую дифференциацию микробных
'ток, отсутствует выраженная разобщенность их обменных
щессов в пространстве и времени. Напротив, все интегриро-
о в одной системе-клетке, где процессы осуществляются
имосвязанно.
Вопросы
1. Что представляют собой основные элементы, микроэлементы и
ультрамикроэлементы в микробных клетках и вирионах?
2. Что означают понятия «универсальные» и «минорные» элементы
в клетках и организованных частицах (вирусах)?
3. Назовите молекулы химических веществ, относящихся к разряду
малых в клетках и вирусах.
4. Что такое большие молекулы или макромолекулы в микробных
клетках и вирусах?
5. Какие молекулы составляют наименьший процент среди макромо-
лекул у микроорганизмов и почему?
6. Какие физико-химические свойства воды определяют ее исключи-
тельную роль в микробных клетках и вирусах?
7. Какие микроорганизмы используют СОг в качестве единственного
источника углерода и какие субстраты являются донорами электронов
в ходе трансформации углерода из диоксида в органическую молекулу?
8. В чем сущность реакции Вуда и Беркмана у пропионовых бакте-
рий?
9. Какие органические кислоты являются предшественниками изопре-
ноидов, уридина, пальмитиновой кислоты?
10. Почему пируват относят к разряду ключевого продукта в процес-
сах превращения глюкозы у гетеротрофных микроорганизмов?
11. Какие аминокислоты входят в состав микробных белков? Какие
из них не являются оптически активными?
12. Какими свойствами обладает пептидная связь в белках?
13. Родоначальником каких химических структур является глицерино-
вый альдегид? В какие вещества у микроорганизмов он входит в состав?
14. Какие углеводы относят к монозам и низшим олигозам?
15. Назовите основные типы изомерии, присущие моносахаридам.
16. На какие группы подразделяют углеводы и углеводсодержащие
полимеры?
107
17. Какие пуриновые и пиримидиновые основания входят в состав
нуклеиновых кислот у микроорганизмов?
18. Какие типы гликозидных связей имеются в нуклеозидах, и что озна
чают син- и антиконформании гликозидной связи в них?
19. Каковы химическая структура цАМФ и его роль в углеводное
обмене у микроорганизмов?
20. Какие мононуклеотиды относятся к макроэргическим соединениям
и почему?
21. Какие витамины входят в состав нуклеотидов (каких)?
22. Какие витамины являются составными частями коферментов (ка
ких)?
Глава 3
химия И ФУНКЦИИ ОСНОВНЫХ СТРУКТУР
И КОМПОНЕНТОВ МИКРООРГАНИЗМОВ
Клетки микроорганизмов могут находиться в живом (биотиче
ском) или мертвом (абиотическом) состоянии. В последнем слу-
чае они попадают в общий круговорот веществ в природе, рас-
падаясь вначале до таких конечных продуктов, как NH3, N2,
H2S, СО2, Н2О, фосфатов и других солей. Поэтому в случае рас
смотрения связей химического состава, строения и функций клеток
имеют в виду живые микроорганизмы. Эти последние, в свою
очередь, могут быть в состоянии анабиоза или метабиоза. Ана-
биоз — состояние клеток, при котором резко снижена или при
остановлена жизнедеятельность организма в неблагоприятных
условиях его существования. В благоприятных условиях вое
станавливается нормальная жизнедеятельность. Метабиоз — -
рост и развитие организма при определенном уровне метаболиче
ской активности в соответствующих условиях среды обитания
В зависимости от целей и задач при исследовании химии и
функциональной активности используют живые или убитые (в
том числе — дезинтегрированные) микробные клетки или искус-
ственно смоделированные структуры, помогающие проникнуть
в сущность их деятельности.
3.1. КЛЕТОЧНЫЕ СТЕНКИ И КАПСУЛЫ
Клеточная стенка возникла на каком-то определенном этапе
эволюции микроорганизмов. У археобактерий отмечено существен-
ное различие химического состава ее от клеточной стенки других
прокариот. Условно принимая эволюцию микроорганизмов как
универсализацию организации и дифференциации клеточных
структур в ряду: фаги и вирусы------► бактерии----► дрожжи -►
-►мицелиальные грибы------► водоросли (растения)----►простей-
шие, отметим, что фаги, вирусы и простейшие не имеют клеточной
стенки (фаги и вирусы не способны синтезировать ее, а простей-
шие утратили эту способность). Археобактерии (метанобразу-
108
le бактерии, облигатные галофильные бактерии и термоаци-
йльные бактерии) лишены муреинового каркаса — пептидо-
дана, типичного для бактерий. Лишь отдельные виды из них
ёржат ацетилсахара и /.-аминокислоты вместо ДАП; у них
О-аминокислот.
При переходе от бактерий к мицелиальным грибам отмечается
ощение молекулы пептидогликана. Если в клеточной стенке
мположительных бактерий пептидогликан многослоен, то у
мотрицательных бактерий он однослоен. С пептидогликаном
зана окраска бактерий по Граму. Дрожжевые организмы и
чатые грибы не имеют муреинового каркаса. Вместо него у
появляется своеобразный маркер — хитин. Причем у ди-
|фных грибов хитина больше в клетках мицелиальных вариан-
. Это свидетельствует в пользу целесообразности более жест-
универсальной структуры, способной противостоять неблаго-
[ятным внешним воздействиям. Тем более, что многие грибы
ешли из водной среды к наземному существованию. Хитин
только остался в растительных организмах, но и сохранился
ке у некоторых животных (членистоногие, рыбы).
У нитчатых грибов появляется в клеточной стенке клетчатка,
;орая по массе становится ведущей структурой у растений,
клеточных стенках грибов, представляющих собой фибрилляр-
аморфные матрицы из углеводных полимеров, обнаружены
хгеины, гликопротеины, липиды (реже) и липопротеины. Так,
Saccharomyces cerevisiae имеется фермент «большая инвер-
а» и даже такие органеллы, как хитосомы, транспортирующие
крофибриллы хитина в клеточную стенку. Фосфорилированные
шгалактозамины найдены в клеточных стенках Aspergillus
„ег, Botrytis cinerea, Neurospora crassa, N. sitophila\ гликопро-
ины обнаружены в клеточных стенках, например у патогенных
ибов, индуцирующих выраженную аллергию у больных людей,
глактано-маннано-пептид из дерматофитов выступает аллерге-
»м при немедленном и замедленном типах аллергии. Причем
леводная часть пептида имеет отношение к первому типу, а бе-
к— ко второму типу аллергии (см. гл. 8). Липиды и липо-
гьюгаты в клеточных стенках грибов выявляются в весьма
1ых количествах, а у некоторых видов отсутствуют (у ряда
>жжевых организмов).
Клеточная стенка у мицелиальных грибов представлена труб-
ой структурой, внутри которой находится цитоплазматиче-
е содержимое, одетое клеточной мембраной. Функция клеточ-
стенки преимущественно защитная и опосредованно связа-
с обменными процессами. Клеточная стенка бактерий — мно-
омпонентная система, в которой взаимодействуют гидрофоб-
i и гидрофильные, заряженные и незаряженные молекулы.
Пептидогликан построен по единому принципу у грамположи-
ьных и грамотрицательных бактерий. В -основе его находится
рамовая, или гликолактиловая (глюкомурамовая), кислота
lyp NAn)
109
СН2ОН
н\/н \z°H
Н(У\р. у\н
Н NHCOCH3
о
Н3С—<1н—СООН
глюкомурамовая кислота
звено тетрапептида (пептидогликана) с биозой
(например, у E.coli)
которая поочередно связывается с N-ацетилглюкозамином и в
виде биозы соединяется в цепочку из 10...65 повторяющихся
блоков, а по месту карбоксила молочной кислоты присоединяется
тетра-(три-, пента-) пептид из L- и Р-аминокислот.
В изображенном звене пептидогликана пятый (концевой)
остаток P-Ала обнаруживается в остатке гликоконъюгата не
у всех бактерий. А-ДАП является связывающим местом с кон-
цевым Р-аланином соседней субъединицы тетрапентида
L-ДАП
----------!
—ГлюЫАц—MypNAu—L-Ала—D-Глу—NH—СН—СО—D-Ала
(СН2)з D-Ала
СН—NH—СО—СН—СНз'
I I
СООН NH
ГлюИАц—MypNAu—L-Ала—Р-Глу
или формирует интерпептидный мостик. У некоторых бактерий
L-ДАП заменена на L-лизин (у Aerococcus viridans) или L-
орнитин (у Spirochaeta stenostrepa) и т. д.
но
Таким образом, в пептидогликанах различают кор, тетра-
Ш-, пента-) пептидные цепи, поперечные сшивки между леп-
ными цепями. В изображенной выше схеме строения звена
[тидогликана Е. coli поперечная сшивка имеется между L-
Л одного тетрапептида с £)-Ала другого тетрапептида.
В пептидогликане Staph, aureus поперечная сшивка пред-
авляет собой мостик из пентапептида (пентаглицина):
фрагмент пептидогликана Staphylococcus aureus
химиче-
Кор пептидогликана имеет в основном однотипное
Кое строение у всех бактерий, тогда как тетра- (три-, пента-)
ептидные боковые цепочки и интерпептидные мостики неодина-
овые у разных видов. Например, .иезо-ДАП в тетрапептиде ти-
ична для пептидогликанов из всех грамотрицательных бакте-
й, грамположительных лактобактерий, коринебактерий, споро-
разующих бацилл, актиномицетов, миксобактерий. У бифидо-
1Ктерий и некоторых спирохет вместо л<езо-ДАП находится L-op-
111
нитин; у стрептококков, стафилококков и лейконостоков —
А-лизин и т. д.:
1. /.-Ала (Тли, /.-Сер)
I
2.(3-гидроксиГлу) D-Глу - NH2 (Тли, Гли NH2, D-Ала NH2)
t
3. мезо-ДАП[(£-Лиз, £-Орн, LL-ДАП, мезо-гидрокси-ДАП,
Л-ДАМ, 1-гидрокси-Лиз);
у
иногда N- »» ацетил-Л-Д АМ, £-гомосерин
£-Ала, £-Глу]
4. D-Ала
।
।
!
5. D-Ала
В приведенной схематичной формуле в скобках указаны ами-
нокислоты (или некоторые пептиды) — возможные заместители
соответствующих аминокислот в тетра- (три-, пента-) пептиде.
Например, глицин и Л-серин могут быть вместо L-аланина (по-
зиция 1); глицин, глициламид и D-аланиламид— вместо D-глу-
таминовой или З-гидрокси-Р-глутаминовой кислоты (позиция 2).
Заметная вариабельность заместителей вместо мезо-ДАП наблю-
дается в позиции 3. Указанные в круглых скобках заместители
имеют вторую аминогруппу. Поэтому они являются якорными,
способными перекрестно связывать пептидные субъединицы в
пептидогликанах. Якорную функцию в некоторых случаях выпол-
няет трифункциональная (по числу NH2- и НООС-групп) D-глу-
таминовая кислота. T-Амино- и у-карбоксигруппы ее участвуют
в построении пептидных связей, тогда как а-карбоксигруппа
может формировать поперечную связь в случаях, когда ДАП или
ее заместители не способны выполнять подобную роль:
Г ₽ а
НООС—СН2—СНг—С Н—СООН
I
nh2-
D-Глу
Тогда, например, в тетрапептиде с N-ацетил-ДАМ в цепи (вмес-
112
1АП) образование интерпептидных связей берет на себя
утаминовая кислота:
—MypNAu—
4
(1)4. -Ала
4
со
$
уСН2
(2)О-Глу
₽СН2
аСН—СО-—«-интерпептидная сшивка
NH
(3) N-Ацетил-ДАМ
СО
СН—СН2—СН2—NH—СОСНз
NH
| ч- место действия О-Ала-карбоксипептидазы
(4)О-Ала
якорная функция О-глутамнновой кислоты
онцевой £>-аланин может отщепляться за счет действия £>-Ала-
зрбоксипептидазы, в других случаях к концевому D-Ала при-
чиняется еще один остаток D-Ала. В первом варианте обра-
ется трипептид, во втором — пентапептид.
Все пептидогликаны бактерий подразделяются на две боль-
це группы — А и Б. В группе А пептидогликановые субъедини-
£ связываются между собой за счет е-аминогруппы диамино-
РЧ0™
(позиция 3) и карбоксигруппы 2)-Ала (позиция 4):
—MypNAu
L-Ала(1)
I
О-Глу(2)
I
мезо- ДАП (е — NH2)(3)
(ОС)О-Ала(4)
—MypNAu.—
I
(1)£-Ала
4
(2)0-Глу
4
(З)лето-ДАП(е — NH2>
4
(4)О-Ала
4
звено пептидогликана типа А
4>' В группе Б пептидогликана субъединицы соединяются за счет
Шрбоксигрупп /)-глутаминовой кислоты (позиция 2) и D-ала-
113
нина (позиция 4). Поскольку взаимодействия между двумя кар-
боксилами не происходит, в интерпептидном мостике должна всег-
да присутствовать диаминокислота:
— МурИАц —
— МурМАц —
| NH
W 1?И СН- СН3
I I
(2)0- Глу (T-COOH)(NH-CH2-CO)2-HN-(CH2)2-CHNHCO
1 1-----------1 I
, ’ Гли соон
(3) Z— Лиз —
I * —Гли, О—Глу, Z—Лиз опущены
(4) О-Ала (1) (2) (3)
О - Ала (4)
Пептидогликаны типа Б встречаются реже, чем пептидогликаны
типа А. Они обнаружены, например, у фитопатогенных корине-
бактерий. Существуют многие варианты интерпептидных связей в
обоих типах гликоконъюгатов. Если интерпептидный мостик
включает остатки какой-либо одной аминокислоты, то его относят
к разряду регулярных (например, у Staph, aureus); если разные
аминокислоты, то — к разряду нерегулярных. Интерпептид-
ная прямая связь исключает наличие аминокислотного мос-
тика.
Пептидогликан у грамположительных бактерий составляе:
30—90 % сухой массы клеточных стенок, у грамотрицательны
бактерий — в среднем около 10%. Хотя этот гликоконъюгат or
носится к ригидным структурам, он динамичен в своих конфор
мационных состояниях. Пептидогликан в клеточной стенке баг
терий выполняет роль каркаса, достаточно легко синтезирующе
гося и разрывающегося (с последующей репарацией) в ход-
деления клеток; он прямо или косвенно участвует в обменных
процессах клетки, обеспечивая поступление в клетку и выход и •
нее различных веществ (здесь важны плотность интерпептидны
мостиков у грамположительных и грамотрицательных бактерий
а также слоистость молекул пептидогликана). У грамположг.
тельных бактерий пептидогликановый слой пронизывают лило
тейхоевые кислоты, идущие от клеточной мембраны. В свою оче
редь с пептидогликаном могут связываться рибиттейхоевые
114
оты, образующие фосфодиэфирные связи с С6 гликолактиловой
ютой:
' Тейхоевые кислоты связывают ионы магния, необходимые
(етке; вносят определенный вклад в формирование общего отри-
йтельного заряда микробной клетки, образуют часть рецептор-
ах мест для бактериофагов. В качестве углеводного компонента
некоторых грамотрицательных бактерий найдены сиаловые ки-
тоты — производные нейраминовой а-кето-6-аминопентаокси-
йрбоновой, или нонулозаминовой) кислоты. Нейраминовую
кислоту можно рассматривать продуктом альдольной конденса-
ции маннозамина и пировиноградной кислоты.
Полимер N-ацетил-О-ацетилнейраминовой кислоты опреде-
ляет антигенную специфичность менингококков группы С. Липо-
Чфотеины входят в состав клеточной стенки грамотрицательных
бактерий, относящихся к родам Escherichia, Klebsiella, Proteus,
.Pseudomonas, Salmonella, Serratia и др., у которых они построе-
ны по единому плану. Так, липопротеин Е. coli включает 58 ами-
нокислот, среди которых нет гистидина, глицина, пролина, трип-
тофана и фениланина. Молекулярная масса полипептида порядка
?000 Да, т. е. это маленькая протеиновая блочная структура, на
Терминальном конце которой находится глицерилцистеин, этери-
1ированный остатками жирных кислот (Ri — R3). Из 58 ами-
ислот 9 приходится на аланин; по 7< — на аспарагиновую
лоту и аспарагин; 6 — на серин; 5 — на лизин; по 4 — на
мн, лейцин и аргинин; 3 — на глутамин; по 2 — на глутамино-
> кислоту, треонин и метионин, по одной — на изолейцин, ти-
ин и цистеин:
115
ocor2
СН2—S—СН2—СН—СН2—OCOR,
I 5
Н N—НС—С О—С ер—Сер—Аси—Ал а—Л из—Ил е—Асп—Г л у>
COR3
чЛей 10
Асн—Лей—Тре—Глн—В ал—Асп—Сер—Сер-
Ал
20 25
Лиз—Вал—Аси—Глу—Лей—Сер—Аси—Асп
Ала
35 30
Глн—В ал —Асп—Сер—Apr—Мет—Ала—Аси
Вал
40 45
Ал а—Л из—Асп —Асп—Ал а—Ал а—Apr—Ал а-
>Асн
50
Тре—Ала—Мет—Аси —Асп—Лей—Apr—Глн-
55Лиз
58
Тир—Apr—Лиз—СООН
eNH
D-Ал а—ДАП—Глу—Ал а
...ГлюЫАц—MypNAu—ГлюЫАц—MypNAu—EnioNAu—MypNAu—ГлюЫАц-
строение липопротеина клеточной стенки Е. coli
Часть липопротеина связана с пептидогликаном (порядка
2,4-105 молекул на клетку), но большая его доля находится в
свободном состоянии (4,8-105 молекул на клетку). Функциональ-
но липопротеины обеспечивают интегративность клеточной стенки
как ригидной структуры в сохранении формы клеток и в транс-
порте различных веществ, а также выступают своеобразными
посредниками между пептидогликановым и фосфолипидным слоя-
ми (рис. 20, а, б).
Фосфолипиды топологически располагаются снаружи от ли-
попротеинового слоя (см. рис. 9, а), т. е. между липопротеином
и липополисахаридом. На 1 мкм2 условной поверхности прихо-
дится 106 молекул фосфолипидов. Фосфолипиды представлены
в клеточной стенке в виде двухслойных мембран, нековалентно
связанных с липидными компонентами липопротеинов. Внешний
слой фосфолипидной мембраны граничит с липополисахаридом
(гидрофобное взаимодействие), а между слоями располагаются
молекулы белков. Фосфолипиды обладают заметной способ-
116
пептидогликан
L
g.f.:
Полисахарид
Липид А
Выводящие поры
Липопрси еиь
фИКСНЫИ (Н'ЛОК
1ПИД
11
из белковых гранул в клеточ-
20. Структурные слои в оболочке (а), каналы
гстенке (б) грамотрицательных бактерий:
6—липополисахарид; ВМ — внешняя мембрана;
периплазматическое пространство; ЦМ — цитоплазматическая мембрана
ПГ — пептидогликановый слой;
Г9ШМ
фгыо к боковому смещению. Основными компонентами фосфо-
пидного слоя (равно как и клеточной мембраны) являются
ффатидилглицерины и фосфатидилэтаноламин, в присутствии
Дорых проявляется активность некоторых ферментов. К числу
1ких ферментов относится галактозилтрансфераза Salmonella
'phimurium, катализирующая реакцию переноса галактозы в
)р липополисахарида.
Количество фосфолипидов в клеточной стенке бактерий обыч-
о незначительно, лишь у коринебактерий и кислотостойких
икобактерий их достаточно много — 30 и 60 % соответственно,
составляющих их жирных кислотах имеется не более одной
Войной связи. В бактериальных фосфолипидах выявляются
?Ирные кислоты с 0-окси- и циклопропановыми группами:
н н
•Н3С—(СН2)„—£н—ЙНг—СООН СНз—(СН2)5—С —С —(СН2)9—СООН
I \/
он сн2
Таким образом, фосфолипидный слой полярен, его гидрофиль-
на части обращены кнаружи, у грамотрицательных бактерий
выступает своего рода подложкой для липополисахаридного
оя. Белковые глобулы, распределенные в фосфолипидном слое,
Дают своеобразные каналы для поступающих в клетку гид-
)ильных веществ (рис. 20, б). Эта фосфолипидная мембрана
яется внешней по отношению к внутренней, т. е. клеточной
117
мембране, в которую заключена цитоплазма. Внешняя мембрана
вносит определенный вклад в устойчивость клеток к неблаго-
приятным воздействиям.
Белки у грамположительных кокков располагаются между
пептидогликаном и капсульным веществом. На 1 мкм2 клеточной
поверхности приходится около 105 белковых молекул. Протеин
А стафилококков имеет выраженное родство к Fc-фрагменту
иммуноглобулинов. У стрептококков различают М-, Т- и R-npo-
теины клеточной стенки. С М-протеином связана вирулентность
стрептококков группы А. Роль Т- и R-протеинов, ассоциирован-
ных с М-протеином, еще не определена. У грамотрицательных
бактерий это белковые глобулы — матричный протеин (см. фос-
фолипиды), формирующий каналы для гидрофильных веществ.
Выделяют мажорные и минорные протеины. Первые по кодиру-
ющим структурным генам подразделяют на отрА, отрС и ompF,
их молекулярная масса 30 000...42 000 Да. Мажорный отрА-про-
теин выступает акцептором при конъюгации с помощью F-пилей;
ompF- и отрС-протеины (порины) образуют комплексы с пепти-
догликанами, подтверждая тем самым интегральность структур в
клеточных стенках грамотрицательных бактерий, они же прини-
мают участие в формировании гидрофильных пор в клеточной
стенке. Между белками ompF и отрС имеется иммунологическое
родство.
У бактерий присутствует от 10 до 20 минорных протеинов.
Так, у Е. coll известны минорные белки bfe6, cir, cit, D, tonAb и
др. (в клеточных стенках найден также лектин, специфичный к
маннозе). Минорные белки связывают различные вещества или
каким-то образом включаются в репликативный процесс ДНК
при делении клеток; некоторые из них рецептируют колицины,
четные и нечетные Т-фаги; а белок tonAb — дополнительно анти-
биотик альбомицин.
Среди белков в клеточных стенках грамотрицательных бакте-
рий выявлено много ферментов: аспарагиназа, аденозин-5'-фос-
фатаза, АДФ-глюкозопирофосфатаза, глюкозо-6-фосфатаза, де-
зоксирибонуклеаза, кислая фосфатаза, лактат-дегидрогеназа,
5'-нуклеозидаза, рибонуклеаза, сукцинат-дегидрогеназа, уридин-
5'-фосфатаза, фосфоглюкоизомераза, фосфодиэстераза, фосфо-
трансфераза, щелочная фосфатаза, эндонуклеаза. Таким обра-
зом, условно выделяемый протеиновый слой в клеточных стенках
бактерий исключительно разнообразен по входящим в его струк-
туру белковым молекулам.
Липополисахарид (липогликан)—гликоконъюгат, содержа-
щийся в клеточной стенке грамотрицательных бактерий. Тополо-
гически он находится кнаружи от фосфолипидного слоя (см.
рис. 9, а и 20, а). Липополисахарид состоит из трех частей: липи-
да А, кора и терминального триаозного блока. Через фосфоди-
эфирные связи липид А соединен с пептидогликаном. Десять
моноз кора в основном однотипны (например, у бактерий из се-
мейства Enterobacteriaceae и Pseudomonadaceae), триаозные
118
3rNHj
I
РР АраИАц
I I
КДО Р₽
I I
|оки видоспецифичны и являются детерминантными группами
►антигенов кишечных бактерий.
» Липополисахарид Salmonella tyrhimurium схематично пред-
ъявляют в следующем виде:
' гг*
Ман—Абе—ОСОСН3
I
: Рам
I
Гал
| Ман —Абв-ОСОСН3
О ।
Рам ГлюМАц Гап Гап
I I I I
Гал---- Глк>*Гал*Глю--*Геп-*Г0п-*ЧКДО]2—*ГлюМН-*-ГлюМН—РР —ЭгЫН2
L U U-__________I I ----------.—r—» । -----,------
внешняя часть внутренняя часть [ЖК]в
< । ________________________ - « < , ,, —I
опигосахаридный кор лнлид А
U---------------------------------------------------------'
липополисахарид
ел (£-глицеро-О-манногептоза) и КДО постоянно обнаружи-
аются в липополисахаридах. У энтеробактерий находятся
,6-дидезоксисахар а: абеквоза (3,6-дидезокси-Ь-ксилогексоза,
ии 3,6-дидезоксигалактоза); колитоза (3,6-дидезокси-А-ксило-
йксоза, или 3,6-дидезоксигалактоза); паратоза (3,6-дидезокси-
|грибогексоза, или 3,6-дидезоксиглюкоза); тивелоза (3,6-диде-
ркси-О-арабиногексоза, или 3,6-дидезоксиманноза):
S" н
ОН2С I он
Липополисахариды грамотрицательных бактерий являются
Идотоксинами, причем токсические свойства определяются липи-
Дм А. Повторяющиеся концевые блоки в О-цепи могут включать
>Иозу, триаозу, тетраозу, пентаозу или гексаозу. Повторяющийся
Йок у Serratia marcescens: —£->-ЗГлЮ1—&->2PaMi-^> ; у К/е-
Siella aerogenes: —> 3 Гал1 —3 Гал( —3 ГлЮ1 j > . В ос-
<№е структуры О-антигенов А-, В-, D- и Е-серологических групп
119
(от лат. serum — сыворотка) рода Salmonella находится триао-
за:
где Y обозначает какую-либо монозу, а знак — необязатель-
ность присоединения моноз. В разных серологических группах
Salmonella найдены 3,6-дидезоксигексозы: паратоза — у S. para-
typhi (серогруппа А), абеквоза у S. typhimurium (серогруппа В)
и S. newport (серогруппа С), тивелоза — у S. typhl и S. enteriti-
dis (серогруппа D).
Гетерополисахаридная часть липополисахарида формирует
нечто подобное микрокапсул толщиной ~ 0,2 мкм. Бактерии в
этом случае вырастают на уплотненных питательных средах в ви
де гладких или S-форм колоний (от англ, smooth — гладкий).
Если же клетки не содержат ферментов, катализирующих синтез
О-специфических цепей липополисахаридов, то они вырастают
в виде шероховатых, или R-форм, колоний (от англ, rough -
грубый, шероховатый). Если в липополисахариде отсутствует
только видоспецифический олигосахаридный блок, то такой мик
рооргаиизм относится к RII мутанту, если же при этом отсутст
вует и N-ацетилглюкозамин, то это будет Rl-мутант. В целом
30...40 % наружной поверхности бактериальной клетки покры-
вается липополисахаридом. На 1 мкм2 наружной мембраны при-
ходится около 105 молекул липополисахарида. Во всей массе
клеточного липополисахарида имеется 3,4-10® остатков жирны:<
кислот на одну клетку, что сравнимо с фосфолипидными остат
ками (2,9-10® на клетку). При отсутствии липополисахарида
в клеточной стенке наблюдается замещение его фосфолипидами.
Все это обеспечивает клеткам более высокую приспособляемость
к изменяющимся внешним условиям. Липополисахаридные мо-
лекулы обладают способностью бокового смещения, хотя и мень
шей, чем это присуще фосфолипидам. Допускается транслокация
их в клеточную стенку из клеточной мембраны (рис. 21).
Липополисахариды грамотрицательных бактерий кроме ток-
сичности обладают пирогенной активностью. На практике нашли
применение пирогеналы из определенных видов энтеробактерий
и псевдомонасов. Пирогеналы являются многоканальными эф-
фекторами в макроорганизме, где они вызывают повышение тем-
120
Рис. 21. Возможная транслокация липополисахарида из внутрен-
ней мембраны в наружную
•ратуры тела, вазомоторные (от лат. vasomotor — сосудодвига-
!ль) расстройства, нарушение обмена веществ, геморрагии (от
1т. haemorrhagia — кровотечение) во внутренних органах, изме-
;ние устойчивости к возбудителям инфекционных заболеваний
т. д.
Капсулы микроорганизмов различны по форме и химическому
ставу. В соответствии с организацией их можно подразделить
। три группы: 1) микрокапсулы — переходные структуры от
(еточной стенки к капсуле, они типичны для грамотрицательных
щтерий и состоят из липогликанов (липополисахаридов) и бел-
>в, не выявляются светооптическими методами;
2) макрокапсулы — переходные структуры от микрокапсул
обильно ослизненным, хорошо выявляются при негативной ок-
1ске по методу Р. Бурри или Р. Бурри — X. А. Гинса;
3) слизи структурно напоминают макрокапсулы, но более
1ффузные, легко накапливаются в питательной среде в форме
щивидуального продукта.
Капсулы состоят из водной фазы и полимеров, преимущест-
!нно углеводной природы. Потенциально капсульный материал
дбл. 15) образуется различными микроорганизмами — бакте-
Гблица 15. Химический состав капсул некоторых микроорганизмов
Микроорганизм Природа капсулы Субъединицы капсульного полимера
Acetobacter xylinum Целлюлоза* Глюкоза
Aureobasidium pullu- lans Глюкан Глюкоза
Azotobacter vinelandii Полиуронид Глюкуроновая н ман- нуроновая кислоты
Вас. anthracis Полипептид D-Глутаминовая кисло- та
121
Продолжение табл. 15
Микроорганизм Природа капсулы Субъединицы капсульного полимера
Вас. licheniformis Полипептид D-Глутаминовая кисло- та
Cryptococcus neofor- mans Г етерополисахарид Манноза, ксилоза, глю- куроновая кислота
Отдельные виды из се- мейства Enterobacteria- сеае Многие типы сложных полисахаридов, колано- вая кислота $ Галактоза, глюкоза глюкуроновая кислота, пи- ровиноградная кислота фукоза и др.
Klebsiella pneumoniae Л-63 Сложный полисахарид Галактоза, галактуро- новая кислота, фукоза
Leuconostoc mesenteroi- des Глюкан (декстран) Глюкоза
Pseudomonas aerugi- nosa Полиуронид или дру- гие полисахариды Глюкуроновая, манну- роновая кислота и др.
Streptococcus haemolyti- cus Гиалуроновая кислота N-Ацетилглюкозамин, глюкуроновая кислота
Streptococcus pneumo- niae Многие типы сложных полимеров, например Тип I Тип II 3-Дезоксигалактоза, галактуроновая кислота Глюкоза, глюкуроновая кислота
Streptococcus saUva- rius Фруктан (леван) Фруктоза
* Целлюлоза у A. xylinum выступает больше связующим веществом для клеток,
нежели капсульным.
риями (клебсиеллами, пневмококками, стрептококками и др.),
дрожжевыми организмами, макромицетами.
Химическое строение полисахаридов, как правило, блочное,
независимо от характера организации капсул. Универсальная
роль капсул — защитная. У патогенных микроорганизмов капсу-
лы выявляются чаще in vivo или в особых условиях культивиро
вания in vitro. Например, пневмококки и сибиреязвенный микроб
формируют выраженные капсулы при попадании в макроорга
низм.
У сапрофитных видов капсулы помогают клеткам выживать
при неблагоприятных условиях существования (при пониженных
температурах, в засушливых географических регионах и т. д.).
В связи с главной функцией соподчиненная роль их сводится к
удержанию влаги (образование гидратных оболочек), к раство-
рению некоторых солей и поступлению их в клетки, к узнаванию
определенных структур, например, у растений, с которыми они
находятся в симбиотических взаимоотношениях, и др.
122
КЛЕТОЧНЫЕ МЕМБРАНЫ И МЕМБРАННЫЕ
ГАЗОВАНИЯ
•окариотические клетки не содержат компартментов, характер-
ах для эукариотических микроорганизмов, у них имеется одна
£тема элементарных мембран. Исключение составляют циано-
^ктерии, у которых фотосинтезирующий аппарат сосредоточен
t уплощенных слоистых мембранных мешках, или тилакоидах,
доминающих тилакоиды хлоропластов зеленых растений. Но
Ян в хлоропластах тилакоиды заключены в специальные граны,
,,у цианобактерий их нет. По химическому составу мембраны
сток всех микроорганизмов состоят преимущественно из белка
липидов. На долю белков в мембранах эукариот приходится от
<до 80 % ее массы, на долю липидов — около 30 %. Благодаря
вей специфичной организации природные мембраны обладают
особностью к самоорганизации, вследствие чего ни в каких
(Пробных клетках не обнаруживаются мембраны со свободными
аями — они всегда смыкаются и формируют замкнутое прост-
Иство.
•' Выявляемые у некоторых бактерий мезосомы и сходные с ни-
I образования являются своеобразным продолжением клеточ-
й мембраны (исключение — тилакоиды у цианобактерий).
йможно, и такие органеллы, как хлоросомы (у фотосинтези-
ющих зеленых бактерий), аэросомы (у водных прокариот),
[рбоксисомы (у многих фототрофов и хемоавтотрофов), обра-
ются из клеточной мембраны, утратившей трехслойность. Все
и органеллы покрыты однослойной мембраной толщиной 2...
нм, тогда как клеточные мембраны у разных микробов имеют
лщину от 6 до 10 нм. Принципиальная жидкостно-мозаичная
вдель клеточной мембраны у прокариот и эукариот однотипна,
ггя у прокариот она проявляет большую избирательность к
ществам внешней среды (рис. 22). Для оценки различий кле-
чных мембран прокариот- и эукариот используют данные по их
одическому составу и функции (табл. 16).
Клеточная мембрана у различных микроорганизмов выпол-
ет разнообразные функции, связанные с метаболизмом. Через
е поступают различные
щества в клетку и выхо-
т из клетки, поэтому мем-
ана проявляет выражен-
ie осморегулирующие и
анспортные функции. Транс-
рт веществ через мембра-
может осуществляться
тем простой диффузии,
легченной диффузии и ак-
вного транспорта, а у неко-
рых эукариот — с помощью
Доцитоза. В нормальных
Рис. 22. Жидкостно-мозаичная модель
клеточной мембраны
Таблица 16. Химический состав и функции клеточных мембран
у микроорганизмов
Общие и дифференциаль- ные признаки клеточной мембраны У прокариот У эукариот
Соотношение между лн- Генетически детерми- Генетически детермини-
пидными компонентами нировано ровано
Гликопротеины Имеются Имеются
Транспортные системы » »
Различные ферменты » »
Кофермент Q »
Жирные кислоты
а) насыщенные и мо-
ноненасыщенные » Отсутствуют
б) полнненасыщен- Отсутствуют* Имеются
ные
в) простагландины Имеются у некоторых
грибов
Ацилглицерины Имеются Имеются
Фосфолипиды » »
а) кардиолипин Отсутствует »
б) фосфатиднлглице-
рин Имеется Отсутствует или имеет-
ся в следовых количе-
ствах
в) липотейхоевые Имеются (у грамполо- Отсутствуют
КИСЛОТЫ жительных бактерий)
Гликолипиды Имеются Имеются
а) моногалактознл- Имеются (у зеленых Отсутствуют
диглицериды бактерий и цианобакте-
Рий)
б) дигалактозилди- Имеются (у цианобак-
глицериды терий)
в) сульфохиновознл- Имеются (у цнанобак-
диглицернды терий)
г) ундекапренол Имеется
(бактопренол)
д) долихол Отсутствует Имеется
Липоконъюгаты Имеются Имеются
а) сфингомиелин Отсутствует Имеется
б) стерины Отсутствуют** Имеются
Связь с ядерной ДНК Имеется Отсутствует
Функция барьера про- Осуществляется Осуществляется
ннцаемостн
Экзоцнтоз и эндоцитоз Не осуществляется
Участие в энергетиче- Участвует Не участвует
ском обмене
* Кроме цианобактерий, способных синтезировать полинеиасыщенные жирные
кислоты.
** Микоплазмы могут включать экзогенные стерины в состав мембраны.
124
овиях мембрана непроницаема для большинства полярных
1екул. Однако некоторые газы (N2, СО2) и водные растворы
>гих веществ свободно диффундируют через мембрану. Веду-
fl роль здесь отводится концентрационному или электрохими-
скому градиентам*. При этом не затрачивается энергия клеткой,
юрость диффузии ингредиентов через мембрану не лимитируется
1сецело зависит от пропорциональной растворимости их в гидро-
>бном коре бислойной мембраны. Электролиты медленно прохо-
т через мембрану, и чем больше заряд вещества, тем транспорт
о меньше. При пассивной диффузии молекула растворенного
щества не изменяет свою первоначальную молекулярную форму.
В природных мембранах имеются каналоподобные структуры,
и поры, генерирующие ион-проводящие пути. Отдельные мик-
бные пептиды, или ионофоры, формируют также ионные кана-
1, которые действуют в прямом и обратном направлениях
ис. 23, а). Эти ионофоры содержат гидрофильные центры,
торые связывают специфические ионы, находясь сами в гидро-
>бном окружении. К числу таких ионофоров относится: цикли-
ский декапептид— антаманид из базидиомицета, переносящий
ны натрия, грамицидин С, ускоряющий поступление через
.мбрану таких ионов, как литий, калий, натрий, рубидий и це-
й; валиномицин, обладающий таким же эффектом в отношении
лия, рубидия и цезия (но не Н+, Na+ и Li4"); нонактин —
отношении калия и др.
Белковые поры, через которые осуществляется транслокация
ецифических молекул, изменяют свою форму (рис. 23, б). Пе-
1ферические и интегральные мембранные белки могут действо-
ть взаимосвязанно, чтобы осуществилась облегченная диффу-
Я. При этом периферические белки могут проявить специфич-
сть благодаря стереоспецифическому связыванию лиганда, тог-
как интегральный протеин формирует трансмембранную пору.
Составной частью системы активного транспорта является
грмент АТФаза, проявляющая активность в присутствии фос-
алипидов. Энергия для первичного активного транспорта пос-
еляется одним из двух способов: 1) когда транспорт сопряжен
одновременным прохождением через мембрану второго вещест-
, движущемуся по градиенту концентрации и в том же направ-
нии, что и первая молекула, — симпорт, или в обратном на-
бавлении — антипорт, &G этого процесса большая и отрица-
льная величина (см. разд. 5.1); 2) когда энергия поставляется
i счет сопряженного гидролиза АТФ или какого-либо другого
гсокоэнергетического соединения, этот способ называют насо-
За счет функционирования систем первичного активного
•анспбрта на мембране создается градиент ионов (прежде все-
* Градиент концентраций — это разность концентраций; диффузия — сум-
фное перемещение вещества, зависимое от градиента концентраций; электро-
Мическнй потенциал — сумма зарядов диффундируемых веществ.
125
го — протонный АрН+), который индуцирует вторичный актив-
ный транспорт — перенос с помощью белков-переносчиков ами-
нокислот, аминов, сахаров и других веществ. Из белковых сис-
тем активного переноса сахаров в мембранах микроорганизмов
известен, например, М-белок, переносящий 0-галактоз иды у
Е. coli и являющийся компонентом трансферазной системы.
У других бактерий имеются белки, специфичные для каждого
сахара, транспортируемого в клетку, для системы переноса суль-
фатов и многих других веществ, а также для системы переноса
ряда веществ из клетки.
Функции систем активного переноса следующие: а) поддер-
жание оптимальной и постоянной концентрации неорганических
ионов, важных для активирования внутриклеточных энзиматиче-
ских реакций и других процессов; б) поддержание стабильных
концентраций метаболитов независимо от колебаний веществ во
внешней среде; в) извлечение из окружающих клетки питатель-
ных сред необходимых веществ даже при низкой их концентра-
ции; г) регуляция метаболизма.
126
Иембранные системы переноса у микроорганизмов генетиче-
детерминированы и синтез этих мембран генетически сцеплен
нтезом ферментов. Мембраны действуют как барьеры с селек-
дой проницаемостью, которая обеспечивается порами, насоса-
И рецепторами для ферментов, субстратов и сигналов (напри-
I, гормональных). Мембраны являются также местом превра-
1ия энергии при фотосинтезе, окислительном фосфорилиро-
ии.
Имеющиеся в жидкостно-мозаичной мембране (см. рис. 22)
и представляют собой нековалентно собранные термодинами-
ки стабильные, но метаболически активные структуры. Белко-
! молекулы как бы заякорены в мембране и несут специфичес-
функции. Одни гликопротеины в мембране не сопряжены с
гими компонентами и могут диффундировать латерально в
скости мембраны; другие, напротив, интегрированы с матрик-
; мембраны прочно.
Кроме регуляции избирательного транспорта веществ мембра-
и мембранные образования участвуют в процессах фотосин-
а и биологического окисления. У цианобактерий через посред-
о тилакоидов мембрана участвует в оксигенном фотосинтети-
ком процессе, сопровождающемся выделением кислорода.
1ущая роль здесь принадлежит хлорофиллам, весьма эффек-
тно поглощающим видимый свет (проявляется в виде спектров
лощения). Известны хлорофилл а и бактериохлорофиллы а, Ь,
1, е (табл. 17).
„блица 17. Характеристика радикалов в бактериохлорофиллах
? V Радикалы в циклах R
I II III IV при С7 V
при СЗ при С4
—СО-СНз —СНз —С2Н5 —СНз Фи- тнл или гера- нил-ге- раннл —ОСОСНз —Н
=СН—СНз Фи- тил »
£ —СНОН—СНз —с2н5 —С2Н5 Фи- тил, сте- арил, фарне- зил —н —СНз
» —Н
—сон —СНз
У пур-
У цианобактерий содержится хлорофилл а, тогда как
ных — бактериохлорофиллы а и Ь, у зеленых — а (минор-
i), с, d и е (основные):
127
В отличие от хлорофилла а цианобактерий, водорослей и
высших растений винильная группа в пиррольном кольце I бак-
териохлорофилла а замещена на ацетильную и связь между СЗ
и С4 насыщена.
Функцию светопоглощения несут и другие пигменты циано-
бактерий — фикобилинпротеины, сосредоточенные в фикобилисо-
мах — органеллах, располагающихся на внешней поверхности
тилакоидов. Фикобилисомы имеют упроленную форму, размеры
их порядка 30X50 нм. Подсчитано, что одна фикобилисома
приходится на 2000 молекул бактериохлорофилла. Пигменты в
этих органоидах (фикоэритробилин, фикоцианобилин, аллофи-
коцианобилин) ковалентно связаны с белком и представляют
собой фикобилинпротеины, сходные по химическому строению
и организации в клетке:
Фикоцианобилин (синий) отличается от фикоэритробилина
наличием СН3—СН2-группы вместо СНг=СН-группы в IV пир-
128
|»ном кольце; он же является хромофором и в аллофико-
нобилине. Фикобилинпротеинами называют фикоэритрин, фи-
ианин и аллофикоцианин (у цианобактерий преобладают два
ледних). Энергия в фикобилисоме переносится в такой по-
!овательности: Фикоэритрин----> Фикоцианин----->- Алло-
социанин----->• Бактериохлорофилл.
Другие фототрофные прокариоты (пурпурные и зеленые бак-
ии) отличаются от цианобактерий тем, что они не имекуг
;акоидов и фикобилисом. У пурпурных серных (Chromatium,
ocystis, Thiospirillum и др.) и несерных (Rhodomicrobium,
odopseudomonas, Rhodospirillum) бактерий фотосинтезирую-
J аппарат* локализован в клеточной мембране; у зеленых
терий серных (Chlorobium) и несерных (Chloroflexus) пиг-
[ты, улавливающие свет, находятся в хлоросомах, а пигменты
кционных центров сосредоточены в клеточной мембране,
жционные центры у пурпурных и зеленых бактерий представ-
ит собой нуклеопротеины, небелковая часть которых пред-
влена бактериохлорофиллами (см. табл. 17), ионами двух-
(ентного железа, убихиноном (кофермент Q) и каротинои-
ли.
У Chromatium и
>ероиден
каротиноид —
Rhodospirillum имеется
н3со
сфероиден
зеленой несерной бактерии Chloroflexus — 0- и
леных серных — хлоробактин
у-каротины, у
н,с
н,с
сн3 сн, сн
сн3
хлоробактин
Функция каротиноидов в реакционных центрах — защита
тивированного бактериохлорофилла от фотоокисления. Они
личаются от других мембранных каротиноидов своим отчужде-
ем от функции светопоглощения, как это присуще другим
:мбранным каротиноидам. Некоторые представители археобак-
рий (Halobacterium species) содержат в мембране красные
ротиноиды. Эти галофильные бактерии относят к группе
•тосинтезирующих; они синтезируют АТФ с помощью механиз-
1 фотофосфорилирования, не связанного с участием хлорофилл-
* Фотосинтезирующий аппарат состоит из системы улавливания световой
!ргин (содержит пигменты), реакционного центра фотосинтеза (содержит
1сленный хлорофилл) и цепи переноса электронов (обеспечивает образование
1617
129
содержащих реакционных центров. Однако каротиноиды нужны
им и для предохранения клеток от губительного фотодинами-
ческого действия солнечного света.
Хромофором в мембранном хромопротеине — бактериоро-
допсине галобактерий выступает ретиналь (каротиноид С20).
Бактериородопсин является протонным насосом, работающим
под действием света с образованием АТФ. Он сходен с родопси-
ном в сетчатке глаза позвоночных (зрительный пигмент):
НзС/^СНз
ретиналь
Таким образом, каротиноиды в клеточной мембране и мем-
бранных образованиях могут принимать или не принимать не-
посредственное участие в фотосинтезе. Однако во всех случаях
они защищают клетки от фотодинамической гибели.
Фотосенсибилизатор Р -f- hv -» Триплетное состояние фотосенсибилнзатора Р*
Р* 4- Ог -► Р + Оз (синглетное состояние кислорода)
Синглетный кислород — сильный окислитель. Каротиноиды
гасят синглетное состояние кислорода ("Ог), возникающее под
влиянием пигментов — фотосенсибилизаторов (например, хлоро-
филлов, цитохромов).
Клеточная мембрана эукариот, имеющая принципиально
сходные химический состав и организацию с клеточной мембра
ной прокариот, приобрела черты более высокой специализа
ции. В частности, ее компартментализация за счет эндоплазма
тического ретикулума создает возможность локализации реак
ций биосинтеза и распада соответствующих веществ в разны-
отсеках, а также оптимизировать энергоснабжение таких реакций
Клеточная мембрана проявляет свойства барьера, дифферент:
рующего поступление в клетку малых и больших молекул. Син
тезируемые рибосомами белковые молекулы поступают в каналь
цы эндоплазматического ретикулума и транспортируются в др\
гие участки клетки.
Через эндоплазматический ретикулум выделяются продукты
обмена клетки, следовательно, транспортировка веществ в эука-
риотических клетках имеет преимущественно направленный ха-
рактер. Элементы ретикулума располагаются вокруг ядра, фор
мируют ядерную мембрану, которой нет у прокариот. С эндо
плазматическим ретикулумом связаны ферменты, катализирую
щие реакции синтеза липидов, гидроксилирование и другие ре-
цепторы для внешних возбудителей (химических веществ при
хемотаксисе, слабых электрических зарядов, света при фототак
сисе и т. д.).
Структурно-функциональные компоненты мембраны эукариот
во
Зержены заметным изменениям и сдвигам благодаря ее
коподобной мозаичной организации, которая зависит от ко-
;ства входящих в мембрану ненасыщенных жирных кислот.
..ротив, стерины и насыщенные жирные кислоты увеличивают
кость мембраны и ограничивают боковое смещение молекул
:ков в плоскости мембраны. Этому же способствуют и микро-
бочки, содержащие белок тубулин (Af ~ 60 000 Да), укла-
квающийся спирально. Микротрубочки постоянно разрушаются
.самособираются. В сборке и дезинтеграции их регуляторную
(ль выполняют внутриклеточные ионы кальция. При митозе
разующееся митотическое веретено в основе своей имеет
даротрубочки. Они же входят в состав рапидосом, ресничек
Исгутиков.
Клеточная мембрана таких эукариотических микроорганиз-
9, как Protozoa, участвует в процессах фагоцитоза (напри-
3, бактерий и других относительно небольших плотных объек-
>), пиноцитоза и экзоцитоза. Фагоцитоз — захват частиц,
раничение их мембраной в виде мешочка, последующая транс-
рмация мешочка в вакуоль (пищеварительную); пиноцитоз —
Ьдный с фагоцитозом процесс захвата капелек жидкости,
агоцитоз и пиноцитоз чаще объединяют в одно название —
•доцитоз. При этом в обоих случаях мембрана клеток форми-
гет эндоцитотический пузырек (рис. 24). С эндоцитоза в при-
даных условиях начинается эндосимбиоз. Обратный эндоцитозу
•оцесс называют
юстейших могут
ества.
Комплекс, или
снимающий участие в построении клеточной мембраны, а так-
е в формировании лизосом, вакуолей и хитосом, имеет непо-
зедственное отношение к секреции
ликов и полисахаридов. В нем они
одвергаются биохимической обра-
отке, приобретая при этом необхо-
нмую завершенность в структуре,
ак, с помощью специфичных глико-
<лтрансфераз углеводные участки-
;реносятся на полипептидные цепи
образованием гликопротеинов и
д. Следовательно, аппарат Гольд-
и содержит различные ферменты,
i же выполняет функцию транспор-
i секретируемых
|астки клетки и
кзоцитоз).
Л изосомы, —
экзоцитозом. С помощью экзоцитоза клетки
выводить образовавшиеся внутри них ве-
аппарат Гольджи, эукариотических клеток,
веществ в другие
к ее поверхности
органеллы, окру-
енные однослойной мембраной и
|еющие 0,5Х (2...3) мкм в диамет- Рис 24 Эндоцитотический
содержат различные гидролазы: рек
131
арилсульфатазы, гликозидазы, липазы, нуклеазы, протеазы, фос-
фолипазы и фосфатазы. Кислая фосфатаза — один из маркерных
ферментов — накапливается также в фагосомах и сегресомах.
Лизосомальные ферменты активируются во вторичной лизосоме.
Хитосомы грибов содержат фермент хитинсинтазу (хитинсин-
тетазу), катализирующий синтез микрофибрилл хитина. Хито-
сомы осуществляют функции переноса микрофибрилл к клеточ-
ной стенке, где происходит сборка макрофибрилл хитина. В них
нет АТФ (в отличие от клеточной мембраны). Хитосомы обра-
тимо ассоциируются. Субъединицы их имеют молекулярную мас-
су около 500 000 Да.
Пероксисомы — одномембранные образования эукариотиче-
ских клеток, где имеет место биохимический процесс, в котором
основным промежуточным продуктом является пероксид водоро-
да (см. рис. 13, г). Пероксисомы, очевидно, более древние по
происхождению, чем митохондрии, успешно приспособились к
появлению молекулярного кислорода в атмосфере нашей плане-
ты, имея примитивную дыхательную цепь:
SH2
°2----
z оксидазы
s
—* Н2О2
hmSH2 нм S
--------------► 2Н2О
' А 1 ► 2Н2О + О2
Субстрат SH2 окисляется с помощью одной из оксидаз, передаю-
щей электроны водорода к кислороду. Образующаяся Н2О2 вос-
станавливается каталазой до воды. Необходимые для этого
электроны используются из низкомолекулярных субстратов (на-
пример, из нм ДНг, в случае отсутствия таковых — из пероксида
водорода). Пероксисомы у некоторых инфузорий (Tetrahymena
pyriformis) называют глиоксисомами. В них содержится лишь
два фермента глиоксилатного пути окисления веществ (изоцит-
рат-лиаза и малат-синтаза), обеспечивающих глиоксилатный
шунт. Изоцитрат поступает в пероксисомы из митохондрий, где
он превращается в малат, возвращающийся в митохондрии:
СООН
но—(i—н соон
Из митохондрий ->-Н—с:—СООНСНОСН2 —
I I I
СН2 СООН Н—С—ОН
СООН СООН
изолимонная глиокси- яблочная
кислота ловая кислота
кислота
превращение в пероксисомах
Поступление в митохондрии
132
Пероксисомы играют важную роль в обмене липидов — в них
(сходит так называемое р-окисление жирных кислот. Следо-
ушно, при наличии реакций (5-окисления и глиоксилатного
па повышается эффективность биосинтеза углеводов. В перо-
гомах разных микроорганизмов содержится неодинаковое
ю ферментов (всего их описано более 40). В пероксисомах
ферментов цикла Кребса и цитохромов.
У трихомонад (Trichomonas vaginalis*) имеются гидрогено-
ы — одномембранные органеллы, в которых происходит окис-
[ие пирувата до ацетата и диоксида углерода, сопряженное
интезом АТФ. В аэробных условиях роста трихомонад элек-
•ны при окислении субстрата передаются на кислород, а при
пробных условиях переходят к протонам Н+ и образуется
1ород Нг. Следовательно, в гидрогеносомах имеется соответ-
ующая гидрогеназа, а также ферредоксин — железосодержа-
й белок (молекулярная масса 5 000...10 000 Да) с низким
[слительно-восстановительным потенциалом (—417 мВ), спо-
рный переносить электроны. В частности, в гидрогеносомах
принимает электроны, возникающие в реакции, катализируе-
i ферментом пируватдегидрогеназой:
Н2 + НАД+ гидрогеназа НАД Н н+
Гликосомы — также однослойные органеллы, в которых про-
ает гликолиз. Они открыты у ряда протозойных микроорганиз-
1 (возбудителей сонной болезни, болезни чагаса, лейшманио-
). Очевидно, клетки, содержащие гликосомы, таким путем
еличивают свой запас прочности. При аэробных условиях гли-
сомы как бы помогают митохондриям в выработке дополни-
йьной энергии. Но гликосомы действуют и в анаэробных усло-
ях или в условиях избирательного блокирования митохондрий
помощью салицилгидроксамовой кислоты:
_ /ОН
салицилгидроксамовая
кислота
Митохондрии — органеллы, в которых происходит окисли-
льное фосфорилирование. Благодаря этому процессу клетка
еспечивается основной массой энергии, необходимой для ее
ста, развития и размножения. Количество митохондрий, при-
•дящихся на одну клетку, широко варьирует (от единиц до
ен) и зависит от интенсивности обменных процессов. Посту-
ощие в матрикс митохондрий вещества подвергаются окисле-
о в реакциях цикла Кребса, а освободившиеся при этом
ктроны передаются в цепь переноса электронов, расположен-
о на внутренней мембране митохондрий (рис. 25). Указанная
* У 77. vaginalis есть пероксисомы, но нет митохондрий.
133
митохондрии
кристы
мембрана
Рис. 25. Криста митохондрии в разрезе (схема)
субмитохондриальные
частицы
передача электронов сопряжена с фосфорилированием АДФ и
превращением его в макроэргическую молекулу АТФ, т. е. низко-
калорийное «топливо» в виде окисляемых веществ трансформи-
руется в высококалорийное топливо — АТФ. Иными словами, ми-
тохондрии — трансформаторы энергии и выступают как бы элек-
тростанциями клетки.
Схема реакций сопряженного окисления и фосфорилирования
выглядит следующим образом:
1) АН2 + В + X А + ВН2 ~ X
2) ВН2 ~ X + Y « X <х> Y + ВН2
3) Х~ Y + <P„^Y^<P„ + X
4) Y <х> Ф„ + АДФ Y + АТФ
где А и В — переносчики электронов; X и Y — условные посред-
ники в процессе сопряжения; Ф„ — фосфат неорганический.
Митохондрии имеют собственные ядро и рибосомы, в них
может осуществляться синтез белка. В митохондриях сосредото-
чены ферменты, катализирующие реакции цикла Кребса, в кото-
ром на одну молекулу окисляемой глюкозы образуется 24 моле-
кулы АТФ, многие оксидоредуктазы, некоторые лигазы, трансфе-
разы и др.
Наружная мембрана митохондрий относительно легко прони-
цаема для разных молекул с массой до 10 000 Да. Внутренняя
мембрана состоит из гидрофобного липидного бислоя, содержа-
щего преимущественно фосфоглицериды. Жирные кислоты в ли-
пидах мембран составляют около 60 %, из них лишь 20 % прихо-
дится на наружную мембрану. На внутреннюю мембрану прихо-
дится почти весь клеточный кардиолипин (~ 20 % всех липидов).
Если в ней сосредоточены ферменты цепи переноса электронов,
то в межмембранном пространстве находятся аденилаткиназа,
нуклеозиддифосфокиназа, сульфитоксидаза; в матриксе — пиру-
ватдегидрогеназа, цитратсинтаза, аконитаза, изоцитратдегидро-
геназа, фумараза, а-кетоглутаратдегидрогеназа, малатдегидроге-
наза, система окисления жирных кислот, фосфоенолпируваткар-
боксилаза, глутаматдегидрогеназа, аспартат-глутамат-амино-
трансфераза, орнитин-карбомоилтрансфераза, НАД.
На клеточную мембрану, митохондрии и другие мембранные
134
зования падает огромная функциональная нагрузка в пе-
, жизнедеятельности микроорганизмов при нередко изменя-
хся внешних условиях. Несмотря на кажущуюся лабиль-
ь клеток прокариот и эукариот, их мембранные образования
бильно функционируют в достаточно широком диапазоне ко-
>аний различных факторов (температура, концентрация пита-
ьных веществ, pH и др.).
ЖГУТИКИ и пили
угики (у некоторых видов Protozoa, например у инфузо-
I Tetrahymena pyriformis, их называют ресничками) своей
ювной массой выступают за пределы клетки, однако они явля-
:я органеллами, поскольку структура, инициирующая движе-
! жгутика, находится внутри клетки и тесно взаимодействует
щеточной мембраной. Ресничка у простейших (например, у
pyriformis) начинается кинетосомой (аналогичной базальной
1стинке у бактерий), каждая из которых соединена фибриллой
шетодесмой) с соседней. Все вместе формируют так называе-
е кинетии. При утрате клеткой ресничек кинетии сохраняются.
Жгутики состоят из спиральных нитей белка-флагеллина, к
юрому могут присоединяться углеводы и липиды. Белок пред-
елен субъединицами, имеющими молекулярную массу в сред-
। 51 000 Да. Число субъединиц в белке может быть разным,
це 8...9 у бактерий, 11 у грибов и простейших (у простейших
и распределяются по принципу «9 + 2», т. е. 9 субъединиц
ставляют стенку цилиндра, а две субъединицы находятся в
нтре).
Жгутики у бактерий (2 % от веса сухой клетки) различаются
। размерам, что зависит от величины шага спирали и от коли-
ства субъединиц, которых может быть меньше, чем 8—9. Виде-
ние их на всем своем протяжении возможно лишь после по-
льзования щадящих методов разрушения клеток (осмотическое
створение, обработка поверхностно-активными веществами),
других случаях жгутики обрываются на уровне наружной по-
рхности клетки.
Углеводная фракция прочно связана с флагеллином и в целом
от комплекс мало растворим в воде и солевых растворах, ус-
йчив к действию различных денатурирующих агентов (щелочи,
фменты). Особенно устойчивы жгутики, снабженные липидным
хлом, стабилизирующим гликопротеиновый комплекс. Флагел-
н включает 14 аминокислот: аланин, аргинин, аспарагиновую
слоту, валин, глицин, глутаминовую кислоту, изолейцин, лей-
,н, лизин, метионин, серин, тирозин, треонин, фенилаланин.
1стидин и пролин обнаруживают редко; цистин и триптофан —
когда; метионин и фенилаланин — в небольших количествах;
парагиновая и глутаминовая кислоты, напротив, — в больших
личествах. У некоторых энтеробактерий в жгутиках содержит-
редко встречающаяся в природе аминокислота e-N-метилли-
135
зин. Флагеллин склонен к агрегации и, как следствие, к рекон-
струкции in vitro. Однако для этого необходима затравка в виде
коротеньких фрагментов жгутика. Субъединицы белка синтезиру-
ются внутри клеток и передвигаются через полый цилиндр к вер-
хушке, где происходит наращивание органеллы.
Полисахариды и липиды остаются мало изученными и меха-
низм их встройки в жгутики неизвестен.
Жгутикам (как и клеточной мембране) присуща АТФазная
активность, с которой отчасти связывают их подвижность. Оче-
видно, в механохимическом движении жгутиковых форм аэроб-
ных бактерий существенную роль играет генерация трансмем-
бранного электрохимического потенциала.
Пили, или фимбрии, грамотрицательных бактерий, ряда
дрожжей и грибов остаются мало изученными. Они начинаются
от мембраны клетки. Различают простые и половые пили. Пер-
вые, например у Е. coli, представляют собой относительно жест-
кие скрученные структуры с диаметром осевого пространства
2...2,5 нм; у других бактерий они являются агрегатами или ли-
нейными образованиями. Классификация пилей еще преждевре-
менна, хотя имеются некоторые попытки к их систематизирова-
нию (табл. 18).
Таблица 18. Классификация и характеристика простых бактериальных
пилей (выборочно)
Вид или род бактерий Типы no Пили
Дью- гиду Брин- тону постоян- ство у вида, % число на клет- ку длина, им ширина, мкм
Corynebacterium 4 III 1...100 10...100 0,2...0,3 2...6
Enterobacter liquefaciens 1 I 10 — 1,3 7,9+ 0,3
Escherichia coli 1 I 3...100 50...400 0,2...2 7
Klebsiella 3 II 80 400... 700 0,2... 1,5 4,8
Neisseria gonorrhoeae — — 90 20...200 0,5...4 7
Neisseria meningitidis — — 5...80 50 0,5...4,3 7
Proteus mirabilis 4 Ill 10 50...500 0,2...6 3,4...4
Pseudomonas aeruginosa — — 4...100 1...25 1...5 6 + 2,8
Pseudomonas echinoides — IV — 1...50 1...10 5
Salmonella 1 I 5...90 10...300 0,1...1,5 7
Serratia 1 I 50...90 100...400 0,2...1,5 6,5...7
Shigella 1 I — 100...250 0,3...2 7
Vibrio cholerae — — 10 9 1 7;2±0,3
Vibrio El-Tor — — 50 50 3,3 7,9+ 0,2
Среди половых, или sex-пилей, различают типы F и I. Первые
из них имеют диаметр 7,5...13,5 нм и длину до 20 нм, вторые —
(6...12) Х2000 нм. Морфологическая структура их сходна с тако-
вой простых пилей, однако они длиннее и заметно выделяются
на фоне других фимбрий (см. рис. 12, в).
Обычно гаплоидные клетки любого скрещивающегося типа
образуют редкие короткие пили, часть из которых (как и среди
136
овых пилей) образует терминальные луковицы, например у
Ькжей Saccharomyces и ржавчинного гриба Ustilago. Пили —
^имущественно белковые структуры, состоящие в основном из
рка пилина, к которому могут присоединяться углеводный и
видный компоненты (табл. 19). В пилине содержатся неполяр-
fe аминокислоты: аланин, валин, глицин, изолейцин, лейцин,
1ГИОНИН, пролин, триптофан, фенилаланин. Повторяемость
Йинокислот в полипептидной цепи пилина различна.
'блица 19. Физико-химическая характеристика некоторых простых пилей
J Виды бактерий Пили
молеку- лярная масса, ДаИО3 изоэлек- триче- ская точка (Р/) остатки аминокис- лот, мае. доля, % ЧИСЛО фосфат- ных групп углеводы, мае. доля, %
Corynebacterium renale 19...19,4 4,35 44,7 — —
Escherichia coli Т1 16,6 4,5...5,1 54,0 — —
Moraxella nonliquefaciens 17 — 50,2 — —
Neisseria gonorrhoeae 19±2,5 5,3 43,8 ±45,8 1,0...2,0 1,3...2,0
Pseudomonas aeruginosa 17,8±0,3 3,9 53,7 — —
пилине Е. coli наибольшее количество аминокислотных остат-
№ приходится на аланин, аспарагиновую кислоту, треонин, гли-
IM; в пилине С. renale — на глутаминовую и аспарагиновую
(слоты, аланин.
Организация пилей происходит в связи с цитоплазматической
ьмбраной. Исходный фонд веществ в клетках для образования
гутиков и пилей один. Из этих веществ образуются отдельные
бъединицы их, а затем формируются органеллы. Информация
i этом закодирована в хромосомах и плазмидах.
' Функция простых пилей у бактерий — обеспечивать их адге-
гю (прилипание) к поверхностям других тел вместе с вещества-
И-адгезинами, локализующимися на поверхности клеток, на-
)имер липопротеиновый антиген К-88 у Е. coli. У дрожжей и
>угих грибов пили гаплоидных клеток скрещивающихся типов
’рают определенную роль в конъюгации, а также в процессах
покуляции (от лат. flocculus — клочок) дрожжевых организ-
Эв. В пилях высокоспецифичные структуры, определяющие
(очную адгезию клеток, относятся к разряду углеводов, глико-
(отеинов и гликолипидов. У энтеробактерий — это D-манноза и
юботетразилцерамид (Е. coli К-88); у нейссерий — гликопро-
мны и гликолипиды (в частности, ганглиозиды); у вибрионов —
‘фукоза и манноза.
Бактерии с пилями обладают гемагглютинирующей актив-
ные. В ряде случаев пытаются вывести зависимость виру-
(тности от наличия пилей. Например, около 80 % штаммов
менингококков, выделенных от больных людей, лишь 5 % лабора-
торных штаммов несут пили. Однако у сальмонелл даже непато-
генные штаммы могут иметь пили.
Во время процесса конъюгации клеток бактерий половые
пили как трубчатые волоски переносят соответствующие плаз-
миды, называемые фактором плодовитости, или F-фактором {Fer-
tility factor)-, фактором устойчивости, или R-фактором (Resis-
tance factor), и колициновым фактором, или Col-фактором (Cod-
eine factor). Причем первые два из них переносятся с участием
F-пилей, а третий — с участием I-пилей. Через эти же пили про-
исходит заражение клеток соответствующими фагами. F- и 1-во-
лоски (по одному или вместе) обнаруживаются числом 1...2 на
клетку, иногда — больше, но свыше 20 — никогда. Они обладают
специфическими антигенными детерминантами (от лат. determi-
nans — определяющий). Доказано, что для клеточного контакта
sex-пилей существуют высоко специфичные рецепторы у грам-
отрицательных бактерий.
3.4. ЯДЕРНЫЙ АППАРАТ
В широком понимании ядерный аппарат включает все структур!.,!
клетки, которые взаимосвязанно функционируют в процесс!
реализации генетической информации. Главенствующую роль пр:-,
этом играет ядро или его аналог у бактерий и условно у ви
русов.
Постоянным компонентом ядра (или нуклеоида) является
ДНК, входящая в состав хромосом (от лат. chroma — окрашен-
ный, цветной, греч. о<оца — тело). Каждая хромосома содержит
только одну молекулу ДНК, в которой в линейном порядке сгруп-
пированы самовоспроизводящиеся дискретные (от лат. discre-
tus — раздельный) единицы, или гены (цистроны), обладающие
элементарной биохимической функцией. Определенный ген в хро-
мосоме может занимать один локус (от лат. locus — место) со
своим аллеломорфом. Аллеломорфный ген — другая форма
одного и того же гена (от греч. aAXqXtov — взаимно). Аллели
одного локуса различаются тем, что один из них доминантен, а
другой рецессивен. Обычно аллель дикого (нормального) типа
доминантен по отношению к мутантному (мутация — скачкооб-
разное наследуемое изменение гена).
Генетические символы обозначают буквами латинского алфа-
вита, причем заглавной обозначают доминантный ген, строч-
ной — рецессивный ген. Два гена, обозначенные одинаковой бук-
вой, являются аллеломорфными. Разные гены, оказывающие
тождественные влияния на внешние проявления признака (или
признаков), могут обозначаться одинаковой буквой, но с разны-
ми индексами (например, W, и W2 или w~/w+).
Ген состоит не из одной тысячи пар нуклеотидов. Причем
один нуклеотид или сотни и тысячи их могут мутировать, явля-
ясь единицей мутации, или мутоном. Единица рекомбинации (ре-
139
Про О |
bio
Не
ind-
руг Е
mtl
cvs *
(а); группа сцепления
оперона на хромосоме
►
[?0.с
карта хромосомы
генетических элементов лактозного
аЛ*с
Лс>>
руг C,D,F
аго А
pure Ip
cySB
г'У —+с
Tlrec ГВ
6
Рис. 26. Генетическая
генов и
£. colt (б):
Р — ген-регулятор; Про — инициатор, или промотор; О — оператор; С —
структурные гены; Т — терминатор
и) — отдельный нуклеотид. Различают: структурные гены, от-
•ственные за синтез белковых молекул; гены-регуляторы, опре-
[яющие синтез цитоплазматических продуктов-репрессоров, от
'явности которых зависит функционирование структурных ге-
операторы — участки, локализованные на хромосомах по
едству со структурными генами, эти генетические элементы
(яются местом приложения действия репрессоров; оперон, или
ппу сцепления структурных генов, находящуюся под контро-
J оператора (рис. 26, а). Другие генетические элементы —
ициаторы, или промоторы, располагаются перед операторами
являются начальными точками переписывания (транскрипции)
формации; терминаторы блокируют транскрипцию и преры-
139
вают синтез белка. Смысл указанной группировки генов в хро-
мосомах заключается в обеспечении митотического перемещения
генетического материала в процессе деления клеток.
У бактерий и вирусов не доказано существование хромосом
и, следовательно, не доказан митотический механизм распреде-
ления генетического материала. Вот почему термины «бакте-
риальная хромосома», «хромосомы фагов» весьма условны. Они
имеют кольцевидные формы и состоят из одной молекулы ДНК
(рис. 26,6). Поэтому каждая такая ДНК принимается за одну
хромосому. Установлено, например, что длина разомкнутой ДНК
Е. coll равна 1,3 мм, т. е. в 500 раз длиннее самой клетки кишеч-
ной палочки. На каждые 3,4 нм приходится десять пар оснований
в двухцепочечной ДНК. Следовательно, хромосома Е. coll содер-
жит 3 823 530 пар оснований, т. е. ~ 3,824 • 106 или 3824 кв (от англ.
। -I . -. 1,3 мм (1 300 000 нм) • 10 ооо л
kilobases — килоосновании): — -1——---------= 3824 кв.
Кроме лактозного оперона Е. coli (рис. 26, а) изучены гисти-
диновый оперон у Salmonella typhimurium (его 15 генов коорди-
нируют биосинтез 10 ферментов), у некоторых грамотрицатель-
ных бактерий — опероны биосинтеза аргинина, валина, галакто-
зы, изолейцина, лейцина и др. Однако lac-оперон Е. coll иссле-
дован с большей полнотой. Доказано, например, что 1ас-репрес-
сор является белком-протеином, включающим 250 аминокислот-
ных остатков. На одну бактериальную клетку приходится всего
5—10 молекул этого белка, его молекула состоит из 4 идентич-
ных субъединиц с 44 = 30 000 Да. Оператор включает 27 пар
оснований ДНК с двухскладчатой поворотной симметрией (отме-
чено сплошными и прерывистыми линиями вокруг пунктирной
оси на схеме) в регионе из 21 пары нуклеотидов по длине ДНК:
I
5-ААТТГТГАГЦ^ГАТ ААЦААТТ
3ТТ ААЦАЦ ТЦГЦЦТАТ ТГТТАА
Минимальный эффективный размер оператора для связывания
1ас-репрессора составляет 17 пар оснований (на схеме буквы
набраны жирным шрифтом). Каждый раз только 2 субъединицы
репрессора присоединяются к оператору, и в пределах 17 пар
оснований по крайней мере одно основание из каждой пары под-
ключается к узнаванию 1ас-репрессора и связыванию с ним. Ами-
нокислотные остатки (позиции 1...52) в субъединицах репрессора
узнают ДНК неспецифически, в то время как аминокислотные
остатки в позициях 53...58 специфически связываются с регионом
из 17 пар оснований (6...7 нм длиной). Аминокислотные остатки
в позициях 74...75 предназначаются для присоединения индукто-
ра к молекуле репрессора.
На рис. 26, б представлена генетическая карта хромосомы
140
yli Hfr. Кольцевая ДНК образует порядка 50 петель, скреп-
ых общим «остовом» из РНК, а каждая петля свернута в
аль, содержащую около 200 витков. Поэтому вся ДНК нахо-
дя в суперспирализованном состоянии. С нею связывается
чно небольшое количество белка и катионы (чаще двухва-
гные).
Эчевидно, протеиновые рапидосомы, или микротубули, выяв-
ше у некоторых бактерий (Proteus, Pseudomonas, Sapro-
а и др.), представляют собой якорную ось, вокруг которой
ерспиральный нуклеоид вращается во время репликации,
фтав ДНК будет медленно изменяться, если, например, прояс-
нит замена отдельных оснований в результате мутаций. Эф-
КТ таких мутаций носит кумулятивный, или накопительный,
рактер. Подсчитано, что для существенного изменения ДНК
(Слетке бактерии должно смениться 2-107 их генераций, и если
’ >мя одной генерации равно 1 ч, то для такого изменения по-
буется 2000 лет. Вот почему организмы довольно стойко со-
1няют свои признаки, особенно в понятиях человека, средняя
•должительность жизни которого менее ста лет. Неконтроли-
мые мутации происходили и происходят постоянно, без них
знь на Земле могла бы остановиться (например, на уровне
ггерий, амеб) и не было бы истории эволюции видов вплоть
человека, а точнее — жизни не было бы вообще.
< В клетках эукариот содержится определенное число хромосом
аще всего между 10 и 50). Хромосомы всегда парны (гомоло-
чны) и каждая пара морфологически отличается от других пар.
ядрах клеток многих грибов имеется по 8 хромосом (для срав-
мия — в соматических клетках человека по 46 хромосом). В це-
дящихся клетках эукариот хромосомы, как правило, не видны,
<к как ДНК деспирализована. У некоторых жгутиковых прое-
вших, например динофлагеллят, хромосомы остаются видимы-
I в интерфазном ядре. В делящихся эукариотических клетках
омосомы выявляются в виде определенных структур (ДНК
бализована).
Опираясь на генетические закономерности, общие для эука-
отических клеток, можно предполагать, что микробам-эукарио-
м присущи подвижные, или «прыгающие», гены. Они оказы-
ют влияние на другие гены, находясь от них на значительной
’станции. Здесь, очевидно, велика роль суперспирализации ДНК
хромосомах. Благодаря мутациям, названным «транспозицион-
1ми взрывами», возможно массовое и в известной мере направ-
иное перемещение генетических элементов. Известны также
»астки ДНК-усилители, активирующие работу генов. Многие из
х способны активировать ген, находясь на дистанции в не-
лько тысяч нуклеотидов от места считывания гена или даже
;ализуясь на хромосоме после гена. Прыгающие гены нередко
дны с участками ДНК-усилителями по последовательности
ований, однако механизм действия усилителей остается пока
ясным.
141
Половые клетки эукариот располагают одинарным, или гапло-
идным, набором из п хромосом. При оплодотворении женской
половой клетки, или гаметы (—), мужской ( + ) образуется зиго-
та с двойным, или диплоидным, набором хромосом (2п). Таким
образом, у микроорганизмов, размножающихся половым путем,
наблюдается чередование фаз — гаплофазы и диплофазы. Если
при оплодотворении сливаются три и более гаплоидных ядер, то
говорят о полиплоидии. Как уже было сказано, ДНК в клетках
эукариот связана с основными белками — гистонами, т. е. она
является нуклеопротеином, называемым хроматином.
По современным воззрениям в хроматине содержатся моно-
мер Н1, димер (Н2А и Н2В) и тетрамер из двух молекул гистона
НЗ и двух молекул гистона Н4. Один тетрамер и два димера
взаимодействуют с 200 парами оснований ДНК на участке дли-
ной II нм, образуя нуклеосомы или НЮ-тела (см. рис. 16), моле-
кулярная масса каждой нуклеосомы 200 000 Да. Гистон Н1 рас-
полагается в разделяющих бусинки секторах, ДНК — на поверх-
ности белковых частиц с радиусом вращения 3 нм. Гистоны
имеют важное значение в упаковке молекул ДНК в хромосоме и
выступают регуляторами генетической активности. Эти белки не-
редко содержат модифицированные аминокислоты: моно-, ди-,
три-е-М-метиллизины; (o-N-метиларгинин; 3-метилгистидин; a-N-
ацетилсерин; e-N-ацетиллизин; О-фосфосерин, О-фосфотреонин
и др. Причинность модификаций аминокислот в гистонах оста-
ется невыясненной, однако несомненно, что она имеет отношение
к взаимодействию белков с ДНК и модулированию активности
генов.
Несмотря на определенное постоянство хромосом, они подвер-
жены структурным изменениям, в частности, можно наблюдать
фрагментацию, делеции, инверсии, дупликации и транслокации
хромосом. Фрагментация — разрыв хромосом в одном или не-
скольких местах, например, при воздействии рентгеновских лу-
чей, некоторых химических веществ и пр. Эти же факторы могут
вызвать делецию — выпадение (утрату) части хромосомы. Если
участок внутри хромосомы поворачивается на 180°, то имеет мес-
то инверсия. Перестройка хромосом, противоположная делеции,
когда какой-то участок хромосомы удваивается, названа дупли-
кацией. Обмен местами участков разных хромосом называют
транслокацией.
Ранее было отмечено, что генетическая информация, зало-
женная в ДНК (РНК), реализуется в процессе синтеза специфи-
ческих белковых молекул. Специфичность белков определяется
строгой последовательностью аминокислот, что в свою очередь
зависит от последовательности нуклеотидов в ДНК- Поэтому при
оценке качества белка более важно определить не качественный
состав аминокислот, а их последовательность в молекуле, так же
как и последовательность нуклеотидов в ДНК (РНК). Физиоло-
гический смысл генетической информации, содержащейся в нук-
леиновых кислотах, заключается в переводе ее на «язык белка».
142
>то связано с проблемой кодирования, с генетическим ко-
Генетический код — это система эквивалентности в последо-
ельности нуклеотидов и последовательности аминокислот,
орая позволяет установить соответствие между полипептид-
: цепью и геном (цистроном). Набор из трех нуклеотидов —
плет, или кодон, соответствует определенной аминокислоте,
актически оказалось, что конкретной аминокислоте может со-
«тствовать более чем один кодон, в этом случае код назы-
,от вырожденным. Вырожденность кода проявляется в созда-
и запаса генетической информации, страхующей клетку от слу-
Йных изменений в структуре ДНК. Так, известно, что в состав
НГК входит 4 нуклеотида — адениловая, тимидиловая, гуанило-
Я и цитидиловая кислоты, т. е. АТГЦ. Если бы одно основание
дировало включение в белок лишь одной аминокислоты, то
шекула протеина состояла бы только из 4 аминокислот. Кодон
трех оснований (43) полностью удовлетворяет всем требова-
ям. Кодирование в этом случае с избытком перекрывает (64)
обходимость включения в полипептид 20 природных аминокис-
1Г. Это объясняет факт существования нескольких разных ко-
нов, соответствующих одной аминокислоте.
В то же время код является неперекрывающимся, т. е. при
итывании генетической матрицы любое нуклеиновое основание
ляется членом лишь одного кодона:
—А—Г—Ц—Г—Ц—Т-
кодон
кодон
ли бы код был перекрывающимся
,-А-Г-Ц-
,-г-ц^т-,
при считывании генетической матрицы каждое нуклеиновое
нование принадлежало бы двум и более кодонам. Расшифров-
[ генетического кода стала возможной после открытия мРНК и
(яснения ее роли (табл. 20). Неоспоримо доказано, что код в
«овном универсален, т.е. клетки различных видов организмов
Организованные частицы — вирусы имеют один и тот же «кодо-
й словарь», хотя в него внесены некоторые дополнения. На-
имер, в митохондриях дрожжей и некоторых нитчатых грибов
'pergillus, Neurospord) триптофан кодируется не только трип-
‘ом УГГ, но еще и УГА; у дрожжей триплет АУА кодирует
гионйн, а четыре триплета с двумя первыми нуклеотидами ЦУ
даруют лейцин. Благодаря универсальности кода получила
ретическое и практическое развитие генная инженерия.
из
Таблица 20. Генетический код:
Первая Вторая буква кодона Третья
буква буква
кодона У ц А Г кодона
У УУУ] Фен УЦУ1 УАУ] Тир УГУ] Цис У
УУЦ УЦЦ УАЦ УГЦ ц
УУА Лей УЦА Сер УАА' 1 А УГА ( + ) А
УУГ уцг] УАГ] (+) УГГ] Три Г
ц ЦУУ ЦЦУ ЦАУ ЦГУ" У
ЦУЦ Лей ццц Про ЦАЦ 1 ИС ЦГЦ Apr ц
ЦУА ЦЦА ЦАА Глн ЦГА А
ЦУГ. ццг ЦАГ цгг. Г
А ' АУУ1 АЦ.У1 ААУ АГУ У
АУЦ Иле АЦЦ ААЦ Асн АГЦ Сер ц
АУА. АЦА 1 ре ААА Лиз АГА' А
АУГ+|Мет АЦГ J ААГ АГГ Apr Г
Г ГУУ ГЦУ] ГАУ] ГГУ] У
ГУЦ Вал ГЦЦ ГАЦ Асп ГГЦ ц
ГУА ГЦА rtjld ГАА" Глу ГГА 1 ли А
ГУГ+ гцг] ГАГ] ггг] Г
* В матричной РНК имеется У вместо Т в ДНК; + кодон — инициатор; ( + ) кодон ---
терминатор.
Из табл. 20 видно, что одна и та же аминокислота кодируется
во многом сходными триплетами. Например, все 4 кодона, на-
чинающиеся с дублета ЦЦ, кодируют пролин, а с дублета АЦ —
треонин. Дублетное правило правомочно для всех аминокислот,
кроме аргинина, лейцина и серина. Во всех других случаях
аминокислота определяется двумя первыми основаниями кодона.
Следовательно, третий нуклеотид в кодоне менее важен, чем два
других, для включения специфической аминокислоты в белковую
молекулу — в этом главное объяснение вырожденности кода.
Код не бывает двусмысловым, данный специфический кодон ко-
дирует одну-единственную аминокислоту. Кодоны УАА, УАГ и
У ГА, ранее считавшиеся бессмысленными, как оказалось, явля-
ются кодонами С-пунктуации, или терминации, т. е. они приоста-
навливают процесс сборки полипептидной цепи. Генетический
код сложился около 4 млрд, лет назад, он выдержал натиск при-
родных воздействий. Происшедшие в нем возможные перестройки
нелегко распознать вследствие существенных изменений, имев-
ших место в процессе эволюции живых организмов. Функцио-
нальная активность ядерного аппарата и уровень метаболиче-
ской активности клетки взаимосвязаны. С учетом скорости раз-
множения микроорганизмов (особенно прокариотических) можно
утверждать, что в благоприятной среде обитания синтез бел-
ков у них протекает в исключительно короткий интервал вре-
мени.
144
Реплика
Реплика
Родительская цепь
Рис. 27. Репликация ДНК
.« В соответствии с центральным постулатом моле-
Влярной генетики, сформулированным Ф. Криком, генетическая
(формация переносится по схеме ДНК—РНК—белок. Допус-
вется перенос информации от РНК к ДНК (например, у ретро-
русов), но никогда — от белка к РНК. Все виды передачи ге-
мической информации основаны на матричном механизме. Мат-
1ей служит одна из цепей ДНК — прямая транскрипция (или
К у вирусов—обратная транскрипция). Перенос генетиче-
й информации от ДНК к ДНК (или от РНК к РНК у виру-
) называется репликацией или самоудвоением.
Репликация ДНК осуществляется полуконсервативным спо-
'Ом, т. е. двухспиральная молекула деспирализуется и каждая
'ь индуцирует синтез комплементарной себе нити при участии
>мента ДНК-полимеразы (рис. 27).
В процессе репликации ДНК выделяют три этапа: инициа-
о; элонгацию, или рост цепи; терминацию.
' Репликативный биосинтез ДНК начинается лишь после того,
ж заново образуются короткие праймерные цепочки РНК (при-
фно по 10 нуклеотидов в длину) под действием ферментов
*имаз (специальные РНК-полимеразы):
145
К З'-ОН-группе праймерной РНК присоединяется дезоксирибо-
нуклеозидмонофосфат, в такой форме она готова к нуклеофиль-
ной атаке на следующий присоединяющийся дезоксирибонуклео-
зидтрифосфат (на его а-фосфатный остаток) с последующим
отщеплением пирофосфата. Отбор собственного дезоксирибо-
нуклеотида, у которого должна быть атакована З'-ОН-группа,
зависит от собственного спаривания с матричной нитью ДНК
(согласно правилу комплементарное™). Например, в соответству-
ющей позиции на матричной ДНК стоит остаток гуаниловой кис-
лоты, тогда к нему присоединится цитидилтрифосфат и его а-фос-
фатный остаток будет атакован З'-ОН-группой дезоксирибонук-
леозидмонофосфорила, только что присоединившегося к полиме-
146
Таким ступенчатым процессом матрица (исходная нить ДНК)
ует комплементарность дезоксирибонуклеозидтрифосфата и
сживает его на месте водородными связями, в то время как
Н-группа растущей нити атакует и включает новый нуклео-
в полимер:
матричная ДНК
У-
5'
5'р.
Т Ц А г
А Г У Ц
''ОН ''ОН ''ОН '
РНК-праймер
д г т т
Т Ц А А
г г
Ц
Ц
г
'он Нон* I * 3 * 5'
растущая нить Р
комплементарной ДНК |
Р
-------------------------входящий
иуклеозид-
трифосфат
Эти фрагменты ДНК, присоединенные к РНК-инициатору,
ли названы фрагментами Оказаки (по фамилии исследовате-
; впервые описавшего их). Указанные фрагменты ДНК затем
иваются с помощью ДНК-лигазы.
В процессе репликации точно воспроизводится исходная струк-
а ДНК. Но если произошла ошибка (мутация) в процессе
ирования, то она будет с предельной точностью копироваться
I последующих репликациях изменившейся ДНК. Показано,
' участки ДНК, содержащие скопления нуклеотидов, обладают
ышенной склонностью к мутациям.
В настоящее время установлено, что последовательность нук-
тидов в области начала репликации кольцевых хромосом бак-
ий и фагов узнается не ДНК-полимеразой, а РНК-полиме-
ой. РНК-транскрипт области начала репликации служит за-
1вкой, которая затем удлиняется с помощью ДНК-полимеразы.
а последняя катализирует синтез дезоксиполинуклеотидной
пи, следуя нитям-матрицам расплетенной спирали ДНК-
Репликация ДНК у прокариот и эукариот происходит одно-
еменно на обеих нитях. Однако у микробов нет фермента, спо-
5ного катализировать реакцию полимеризации в направлении
—5', поэтому обе заново реплицирующиеся нити ДНК не могут
сти в одном и том же направлении одновременно. Один и тот
5 фермент (репликаза) обеспечивает репликацию одной нити
идирующая нить) в направлении 5' —► З', и он же обеспечивает
ерывистую репликацию второй нити (запаздывающая нить) с
зоротом ее назад. Разделение нитей обеспечивается специфи-
:кими белковыми молекулами, стабилизирующими однонитевые
>уктуры в виде репликационной вилки (Y), не подавляя при
iM матричной функций нитей.
147
По расчетным данным, репликация ДНК у прокариот (вме-
сте с раскручиванием молекулы по 10 нуклеотидных пар одно-
моментно) должна происходить со скоростью 400 000 оборотов
в секунду, что явно превышает реальную скорость репликации.
Вот почему обоснованным было предположение о существовании
фиксаторов, включенных в молекулы ДНК всех организмов.
Функцию фиксатора обеспечивает специфический фермент —
ДНК-топоизомераза. Некоторые топоизомеразы катализируют
реакции объединения молекул ДНК в зацепленные кольца - -
катенаны. Топоизомераза I способна разрывать одну из цепей
суперспирализованной ДНК, после чего цепи раскручиваются,
уменьшается число супервитков, затем этот же фермент устра-
няет разрыв в нити ДНК. Топоизомераза II, или гираза, катали-
зирует суперспирализацию ДНК.
Определенные участки ядерной ДНК прокариот и эукариот
(транспозоны) могут реплицироваться автономно и копии их
способны встраиваться в геном в разных случайных точках. Пре-
обладающая часть транспозонов бактерий кодирует фермент
транспозазу, катализирующую встраивание транспозона в ДНК.
Геномы бактерий и фагов реплицируются как единое целое,
т. е. как организованные единицы репликации, или репликоны.
Репликон имеет начало ori (от англ, origin — начало) — ориен-
тированное направление и в некоторых случаях концевой (тер-
минальный) участок ter. Репликация бактериальной ДНК проте-
кает почти непрерывно в течение периода генерации клеток, и
скорость синтеза ДНК удваивается примерно к середине цикла
генерации. Поэтому завершение репликации кольцевой родитель-
ской молекулы ДНК и начало репликации двух дочерних моле-
кул произойдут задолго до разделения родительской клетки.
Окончание процесса репликации изучено недостаточно, особенно
у эукариотических микроорганизмов. В ДНК кишечной палочки
имеется специальный участок — терминатор. У многих бактерио-
фагов терминаторы отсутствуют и, как предполагают, реплика-
ция, например, у бактериофагов Е. coll (Р2, Р22, Т4, X) осуще-
ствляется по типу «катящегося кольца». Двигателем в распреде
лении реплицированных молекул ДНК по дочерним клеткам
выступает клеточная мембрана, к которой прикреплена ДНК нук-
леоида (рис. 28).
Транскрипция—процесс переписывания закодированной в
ДНК информации и перенос ее к месту синтеза белка, т. е. на
рибосомы, находящиеся в цитоплазме. Матричная, или информа-
ционная, рибонуклеиновая кислота (мРНК) способна образовы-
вать цепь, комплементарную одной нити ДНК. Две такие цепи
дают гибрид РНК-ДНК, устойчивый к ферменту РНКазе. мРНК
определяет (закодированную в ДНК) последовательность амино-
кислот в будущих молекулах белка. Цепь ДНК, комплементар-
ную мРНК, называют кодирующей, вторую цепь — замыкающей.
Все стадии ДНК-зависимого синтеза РНК составляют цикл
транскрипции: инициацию, элонгацию, терминацию. Эти стадии
148
Рост
Рис. 28. Прикрепление ДНК к клеточной мембране в период
деления клетки
Гаточно подробно изучены у бактерий. На стадии инициации
исходит взаимодействие РНК-полимеразы с ДНК-матрицей.
К-полимеразы имеются у вирусов (фагов), бактерий, эука-
гических микро- и макроорганизмов. Мелкие РНК-полиме-
ы фагов содержатся в клетках прокариот, где они на матри-
ДНК фагов синтезируют мРНК, кодирующие фаговые белки.
К-полимераза бактерий — протеин, состоящий более чем из
Э аминокислот, организующихся в 5 субъединиц в четвертич-
структуре: две а-субъединицы, р-, и о-субъединицы. а-
ъединицы состоят примерно из 365, р-субъединица — из 1365,
149
Рис. 29. Синтез мРНК при участии
РНК-полимеразы
Р'-субъединица — из 1450 и
о-субъединица — из 780 ами-
нокислотных остатков. С ДНК
связывается р'-субъединица,
обеспечивающая присоедине-
ние всей РНК-полимеразы к
матрице.
Синтез мРНК на ДНК ядра
(или нуклеоида), осуществляе-
мый при участии фермента
РНК-полимеразы, показан на
рис. 29. Сборка молекул мРНК
происходит последовательно в
направлении 5'->3' (элонга-
ция). Скорость элонгации в
клетках составляет 45...50 ну-
клеотидов при 37 °C в 1 с.
Каждая молекула мРНК 10...20 раз используется в белковом
синтезе, затем она расщепляется при участии фермента РНК-азы.
Например, мРНК Вас. sultilis функционирует как матрица в пре-
делах 2 мин. Остановку синтеза мРНК вызывает терминатор, или
стоп-сигнал, представляющий собой определенную последова-
тельность нуклеотидов в ДНК- У Е. coli терминаторы эффектив-
ны только в присутствии р-фактора (белок с Af = 50 кДа, не
связанный с РНК-полимеразой). Другой белок, участвующий в
терминации, назван х-частицей. Как только участок мРНК отделя-
ется от ДНК-матрицы, то сразу же к нему присоединяются рибо-
сомы, способствующие отделению мРНК от матричной ДНК и
начинающие синтез белка, — образуется единый транскрипци-
онно-трансляционный комплекс.
В отличие от прокариот, у которых транскрипция и трансля-
ция сопряжены, у эукариот эти процессы разобщены. Разобщи-
телем аппаратов транскрипции и трансляции выступает ядро
клетки. У эукариот не вся синтезированная в ядре ДНК-подобная
РНК (Д-РНК) переходит в цитоплазму, часть ее разрушается
в ядре.
Синтезированные в ядре молекулы Д-РНК связываются с
белками информатинами (М = 800 000 Да), образуя нуклеопро-
теины, или информоферы, которые подвергаются действию нук-
леаз. В результате отщепляются избыточные нуклеотиды и обра-
зуется мРНК, которая транспортируется в цитоплазму и связы-
вается там с белками. Так формируются комплексы — инфор-
мосомы, они связываются с рибосомами, образуя полисомы.
Полисомы обычно включают 4... 12 рибосом, связанных с мРНК
через 30S (40S) субъединицы.
Известно, что не все молекулы РНК, находящиеся в цито-
плазме клетки, являются продуктами непосредственного синтеза
на матрице ДНК. Они вначале проходят процессинг, или сплай-
синг (от англ, processing — продвижение вперед, splicing — спле-
150
е нитеи), т. е. в них вырезаются некоторые нуклеотиды с по-
ующим сшиванием разрывов, а затем уже РНК оказываются
топлазме.
Следовательно, в ядре эукариотической клетки ядерная РНК
НК) составляет 4...10 % от суммарной РНК клетки.
3 ядре синтезируются короткоживущие РНК-затравки, необ-
1мые для синтеза ДНК, высокомолекулярные предшествен-
г (пре-РНК) рибосомных 28S, 18S РНК и 5S РНК; тРНК;
[К- Высокомолекулярные предшественники рРНК и тетеро-
не низкомолекулярные РНК (нмРНК) локализуются в яд-
ке. От 2 до 10% всей яРНК составляет особая гетерогенная
К-подобная по нуклеотидному составу ядерная РНК (гяРНК)
онстантой седиментации в среднем 25...30S. гяРНК является
дшественником цитоплазматической мРНК. Молекулярная
ха гяРНК может достигать 107 Да. Наибольшая часть транс-
бированной гяРНК деградирует в ядре.
Оказалось, что кодирующие области ДНК, транскрипты кото-
[ появляются в цитоплазме как одиночные молекулы мРНК,
рываются большими некодирующими ДНК-последовательно-
ми (100...2000 нуклеотидов) —интронами. Кодирующие уча-
и ДНК называют экзонами или доменами. Интроны обычно
много раз больше экзонов, они разделяют функциональные
оны, облегчая быструю и необходимую генетическую пере-
ойку, направленную на синтез специфических белковых моле-
. Поэтому можно считать, что сплайсинг завершается в ядре
олько после этого сформированная мРНК поступает в рибо-
!ы, перемещающиеся затем в цитоплазму.
Важным было открытие у ретровирусов фермента, катализи-
эщего реакции синтеза однонитевых, а затем двухнитевых
1екул ДНК на однонитевой матричной РНК. Это РНК-зависи-
я ДНК-полимераза, или обратная транскриптаза (ревертаза).
ачале синтезируется гибридная молекула ДНК-РНК на РНК-
тричном геноме. Специфический фермент РНаза Н разрушает
ъ РНК, а остающаяся нить ДНК служит матрицей для био-
теза двухнитевой молекулы ДНК, содержащей информацию,
'воначально имевшуюся в РНК-геноме вируса. Этапы биосин-
а здесь следующие:
ГЧ11Г. обратная транскриптаза т.тгтг „ РНаза И
РНК —----------------► ДНК-РНК-гибрид--------
-----днк - лн—и-а-а— ДНК-ДНК
(одно- (двухните-
нигезая) вая(
Трансляция — перевод закодированной информации, заклю-
ной в мРНК, в последовательность аминокислотных остатков
[ипептидных цепей. Организующими центрами процесса тран-
ции являются рибосомы. Как и другие матричные процессы,
нсляция включает инициацию, элонгацию и терминацию. На
вой стадии аминокислоты активируются с помощью фермен-
151
тов аминоацил-тРНК-синтетаз (АРСазы)
энергии АТФ:
при использовании
R—СН—СО—АМФ +
NH2
R—СН—СООН + АТФ
nh2
Активирующий фермент
+ ФФ„ (пирофосфат)
Далее активированная аминокислота присоединяется к 3'—ОН-
группе аденозина в триплете ЦЦА (—ЦЦА—3х—ОН) на конце-
вом участке тРНК (см. разд. 2.1.2) с образованием комплекса
аминоацил-тРНК, который и является промежуточным продуктом
в ходе синтеза специфичной белковой молекулы («узнавание»,
или рекогниция):
+ АМФ + 29,2 кДж/моль
Для сборки полипептидной молекулы из аминокислот (элон-
гация) кроме аминоацил-тРНК, мРНК, рибосомных 30S- и
SOS-субъединиц необходимы еще белки, называемые факторами
элонгации (от англ, elongation factor — фактор удлинения):
1) у прокариот — EF-Tu, EF-Ts и EF-G; 2) у эукариот — EF-1
и EF-2. Эти факторы не входят в структуру рибосом, а присоеди-
няются к ним на определенных этапах. Роль факторов элонгации
в процессе сборки полипептидной молекулы у прокариот пока-
зана на схеме:
EF-Tu + EF-Ts + ГТФ + Аминоацил-тРНК—I-*- EF-Tu-аминоацил-тРНК
EF-Ts
I
ГТФ
EF-G + ГДФ + Фн ------
и 1
[Рибосома-мРНК-пептидил-тРНК] ------1
।--------------------- EF-Tu + ГДФ + Ф„
[Рибосома-мРНК-пептидил-тРНК-аминоацил-тРНК]
удлиненная на одну амино-
кислоту полипептидная
цепь перемещается
в пептильный центр
новообразованная полипептидная
цепь удлиняется на одну
аминокислоту
------------------------------------— тРНК
[Рибосома-мРНК-удлиненная пептидил-тРНК]
EF-G + ГТФ
152
!десь I—аминоацил-тРНК активируется, чтобы присоеди-
ся к аминоацильному центру рибосомы; 2 — аминоацил-тРНК
юединяется к аминоацильному центру рибосомы. Наличие
' (но не его гидролиз) необходимо для связывания EF-Tu
босомой, тогда как гидролиз, очевидно, необходим для осво-
дения фактора элонгации из рибосомы после завершения
гесса удлинения. Это дает основание предполагать, что ГТФ
рупает здесь в качестве аллостерического эффектора.
J бактериальной клетке около 80 % суммарной РНК состав-
• рРНК и лишь около 5 % — мРНК. Одна молекула мРНК
ходит через несколько рибосом (полисом). Поступающие к
осомам аминокислоты располагаются в молекуле белка в со-
етствии с расположением кодонов, детекторами которых явля-
.я антикодоны в составе тРНК-
В клетках прокариот имеется тРНК, несущая на своем
-ОН-конце N-формилметионин (у эук-ариот— Мет-тРНК):
Н(НзС)— S— СН2—СН2—СН—соо
HN- C=0
H
и аминокислоты вместе с тремя факторами инициации (IF-1,
2, IF-3) необходимы для начала сборки полипептидной цепи.
1<есте с мРНК, 305-субчастицей рибосомы и гуанозилтрифосфа-
л (ГТФ) они образуют комплекс инициации, соединяющийся
iOS-субчастицей рибосомы и формирующий в итоге функцио-
йьную 705-рибосому. При этом гидролизуется одна молекула
Ф с образованием ГДФ и неорганического фосфата и осво-
«дением IF-факторов. Такая последовательность реакций
Одна с образованием и диссоциацией комплекса EF-Tu при
астии ГТФ (рис. 30, а).
Завершение синтеза и отделение полипептидной цепи условно
Дразделяют на три стадии: 1) ориентация молекулы амино-
Ил-тРНК. вдоль мРНК и их взаимодействие с участком А рибо-
йы (рис. 30,6); 2) образование пептидной связи под дейст-
пептидилтрансферазы; 3) перемещение пептида с помощью
1нслоказы и перемещение рибосомы.
При каталитическом действии фермента пептидилтрансферазы
разуется первая пептидная связь. При участии фермента
Знслоказы и ГТФ образовавшийся дипептид перемещается из
Детка А в участок П рибосомы, вытесняя освободившуюся от
Шнокислоты тРНК. Рибосома сдвигается на следующий трип-
t и против участка А становится очередной кодон. Цикл по-
Оряется, и цепь полипептида продолжает расти до момента
квления стоп-кодона, или кодона пунктуации. Биосинтез завер-
ется, и цепь белковой молекулы отделяется, приобретая соот-
Ствующие вторичную и третичную структуры. Рибосомы осво-
кдаются от мРНК и диссоциируют на субъединицы.
©елки вирусных капсидов в клетках синтезируются по анало-
153
EF-Tu <-----ГТФ
EF-Ts
EF-G ----->.ГДФ+ФН
Факторы
освоЬождения
—Завершение < — Синтез "
полипелтидной полипептидной
цепи цепи
Белковый
продукт
Рис. 30. Рибосомные частицы в циклическом процессе синтеза белка (а)
и место связывания fMet-TPHK(6):
П — полипептидный и Л — аминокислотный участки в большой субъединице
(50S) рибосомы
гичным путям. У прокариот, а также в митохондриях и хлоро-
пластах эукариот донором формила для образования формил-
метионина выступает 10-формил-тетрагидрофолиевая кислота.
Для метионина кодоном в мРНК выступает АУГ, а для фор-
милметионина — АУГ и ГУГ. Если триплет АУГ находится в са-
мом начале или почти в начале цепи мРНК, то его опознает фор-
мил-тРНК и полипептид начинается с формилметионина. Удален-
ный от начала цепи кодон АУГ опознается обычной Мет-тРНК,
которая включает метионин в соответствующее место полипептид-
ной цепи. Триплет ГУГ в начале цепи также определяет начало
синтеза с формилметионина, но, будучи в середине цепи, он коди-
рует включение валина. Общая схема синтеза N-формилметио-
нил-полипептида показана на рис. 31.
У эукариотических клеток Мет-тРНК также является инициа-
торной, но в отличие от Мет-тРНК прокариот она не формили-
руется, а взаимодействует с факторами инициации eIF-5, eIF-2,
eIF-З с 405-субчастицей рибосомы и мРНК- Последующие реак-
ции протекают однотипно у эукариот и прокариот.
Не все белки остаются по истечении времени в том виде, в
каком они были синтезированы рибосомальным аппаратом клет-
ки. В некоторых случаях отдельные белки претерпевают так на-
зываемые посттрансляционные (вторичные) модификации, т. е.
изменения в ковалентных связях после синтеза белков на рибосо-
мах. Так, например, в белке вируса табачной мозаики в цито-
хромах с-типа и других концевые а-аминогруппы (рК 8) вторично
модифицируются присоединением ацетила. Вероятное биологи-
ческое значение процесса маскировки а-аминогрупп заключа-
154
в защите белков
Рис. 31. Схема синтеза N-формилметионнл-поли
пептида
гаки аминопепти-
авно как и в защи-
рбоксигрупп амм-
онием (р/< около
т карбоксипепти-
возможным пост-
сляционным изме-
ни белков относят-
юдификации боко-
цепей в ферментах,
шмер, аденилиро-
е тирозина в глу-
нсинтетазе транс-
щрует зависимость
[ентативной актив-
fl от Mg2+ на зави-
сть от Мп2+; фосфорилирование серина или треонина перево-
неактивную форму фосфорилазы в в активную форму фосфо-
ры а и т. д. Любые модификации белков расширяют спектр
войств.
'егуляция генной экспрессии. В ходе приспособления микро-
низмов к изменяющимся условиям существования при на-
ении и сохранении энергии возникает ответ на внешние
алы. В общем плане ответ может проявиться в положитель-
или отрицательном смысле. Когда выражение (экспрессия)
'ической информации количественно возрастает вследствие
утствия специфической регуляторной молекулы, регуляцию
уют положительной; если же экспрессия генетической ин-
1ации при таких же обстоятельствах уменьшается — отри-
льной. Регуляторную молекулу в первом случае называют
жительным регулятором, во втором — отрицательным. Но
которых случаях имеет место как бы удвоение отрицатель-
эффекта с положительным конечным результатом. Напри-
вещество (эффектор) тормозит функцию отрицательного
аятора — эффект, сходный с положительной регуляцией.
•егуляторные сигналы могут вызывать у микроорганизмов
типа ответов: присутствие; транзиентный, или преходящий;
ределенно долгий (рис. 32). Первый тип ответа характерн-
ей возрастанием темпа генной экспрессии, которая зависит
продолжающегося наличия (присутствия) индуцирующего
ала. При устранении сигнала темп генной экспрессии возвра-
тен к исходному уровню. Если сигнал возобновляется, то и
ессия вновь увеличивается (кривая /). Транзиентный тип
а проявляется в возрастании скорости генной экспрессии,
•ая быстро уменьшается даже при продолжающемся дей-
сигнала (кривая 2). В случае исключения действия сиг-
и полного восстановления клетки (возврат к исходному
155
Рис. 32. Типы ответов и характер генной
экспрессии на специфические регуляторные
сигналы:
VGE—характер генной эспрессии; Т—время;
1 — тип присутствия; 2 — транзиентный тип; 3 -
неопределенно долгий тип
состоянию — до действия
сигнала) можно получить
второй транзиентный от-
вет на регуляторный сиг-
нал. В таком случае спе-
цифическая белковая мо-
лекула (продукт гена) яв-
ляется преходящей и пос-
ле ее устранения нет экс-
прессии, хотя сигнал еще
продолжает действовать.
Неопределенно долгий тип
ответа характеризуется
тем, что возросший темп
генной экспрессии на регу-
ляторный сигнал продол-
жается неопределенно
долго даже в случае пре-
кращения действия сигнала (кривая 3). Здесь сигнал действует
как пусковой (триггерный) механизм. Такой ответ характерен для
процессов дифференциации. Раз начавшись в клетке, генная экс-
прессия не заканчивается даже в дочерних клетках, поэтому она
необратима и наследуется (например, при метилировании осно-
ваний в ДНК).
Наименьшей единицей генной экспрессии является цистроп.
Это генетическая единица кодирования субъединицы белковой
молекулы. Если ранее последовательно провозглашали посту
латы: «один ген — один фермент» и «один ген — одна белковая
молекула», то в настоящее время более резонной представляется
формулировка: «один ген — одна субъединица белковой мол*
кулы».
Весь комплекс регуляторных механизмов экспрессии генов
можно подразделить на репрессию — подавление синтеза бел-
ка — и индукцию — стимуляцию синтеза белка. Основные меха-
низмы регуляции генной экспрессии у микроорганизмов (прока-
риот и эукариот) во многом аналогичны, однако у эукариотиче-
ских видов они сложнее. Для тех и других существуют зависи-
мые механизмы регуляции скорости синтеза белка на разных ста-
диях от воздействия различных факторов (внутренних и внеш-
них). У прокариот к регуляторным генам относятся те, продукты
которых (белковые молекулы) вовлекаются в регуляцию других
генов, включая гены-репрессоры. В регуляции экспрессии генов
участвует определенная (регуляторная) последовательность ДНК
(например, промоторы, операторы).
Белок, связывающийся с регуляторной последовательностью
ДНК (оператором) и в таком состоянии блокирующий транс-
крипцию гена, называют репрессором. Вещества, переводящие
репрессор из неактивного состояния в активное, называются ко-
репрессорами, а вещества, инактивирующие репрессор, — ин-
156
срами. Из этих определений следует, что репрессор имеет
тки связывания корепрессора и индуктора. Репрессорный
•к кодируется регуляторным геном (рис. 33) . Этот аллостери-
ий белок в активной форме ингибирует синтез определенных
центов (репрессия). Отдельные низкомолекулярные вещества
(укторы) взаимодействуют с репрессором, блокируют или де-
ивируют его и после этого он не может тормозить синтез
ментов. Следовательно, это и есть индукция или дерепрессия
геза белков (ферментов). Дерепрессия может быть координи-
Ьнной и некоординированной.
Небольшие молекулы (пурины, аминокислоты) выступают в
естве корепрессоров. Они могут блокировать синтез всех фер-
тов, вовлеченных в биосинтез данных малых молекул. Напри-
, добавление гистидина при выращивании Salmonella typhi-
turn репрессирует синтез всех ферментов биосинтеза этой
нокислоты. Подобная репрессия конечным продуктом, назы-
вая координированной, не является общей для всех путей
синтеза. Рассмотренные примеры репрессии — дерепрессии
эсятся к разряду регуляторных механизмов по типу обратной
ш (от англ, feedback — repression).
Ген, экспрессия которого возрастает на индуктор — специфи-
<ий регуляторный сигнал, называют индуцибельным. Если же
дая экспрессия достаточно высока в отсутствие любого спе-
жческого регуляторного сигнала, то такие гены называют
ститутивными. Некоторые, ранее индуцибельные, генные про-
ты (белки) становятся конститутивно экспрессивными в ре-
ьтате мутаций (конститутивные мутации).
Первичная структура белков кодируется генетическим кодом.
1езультате мутаций код изменяется и, как следствие, модифи-
(уется первичная структура белка (фермента). Мутации могут
вести к частичной и полной утрате ферментом каталитической
ивности и реже к возрастанию ее. Таким образом, мутация —
ова регуляции генной экспрессии.
Другой механизм регуляции метаболизма — индукция. При
,авлении в среду некоторых субстратов (индукторов) происхо-
существенное возрастание синтеза ферментов — это адап-
ные, или индуцибельные, ферменты, каковыми являются р-га-
дозидаза Е. coll, пенициллиназа Bacillus cereus.
Хотя многие индукторы выступают субстратами для соответ-
ующих индуцируемых ими ферментов у прокариот, есть индук-
ы, структурно сходные с субстратом, но не являющиеся тако-
«и. Их называют беспричинными индукторами. Напротив, не-
орые вещества могут быть субстратами, но не индукторами.
>едко один какой-то компонент индуцирует различные фер-
ты катаболитного пути (например, лактоза индуцирует (3-га-
дозилтрансферазу и 0-галактозидазу).
Фермент может быть конститутивным для одного штамма,
1уцибельным для другого и отсутствовать у третьего. Клетки,
собные к индукции такого фермента, всегда содержат некий
157
измеряемый основной уровень его, даже когда они растут без
добавления индуктора. С помощью индукции количество фер.
мента может возрастать в 2...1000 раз.
Качество и величина ответа на индуктор определяются гено-
мом клетки. Когда все структурные гены, кодирующие синтез
группы катаболитных ферментов, составляют оперон и все эти
ферменты данного оперона индуцируются одним индуктором, то
индукцию называют координированной, т. е. оперон представляет
собой систему координированной регуляции.
Строение лактозного оперона Е. coli было рассмотрено ранее
в этом разделе. Здесь приведен схематичный механизм его регу-
ляции на основе репрессии и дерепрессии (рис. 33). В отсутствие
индуктора P-ген определяет конститутивный синтез белковой
молекулы репрессора, который присоединяется к оператору в
ДНК и тем самым предупреждает связывание фермента РНК-по-
лимеразы на этом же участке ДНК. Таким образом, исключается
транскрипция структурных генов. Если же индуктор присутству-
ет, то он инактивирует репрессорные белковые молекулы — про-
дукты гена регулятора, и оператор остается незанятым. В при-
сутствии циклического аденозинмонофосфата (цАМФ) и связы-
вающего его белка — активатора катаболизма (КАП, или САР)
РНК-полимераза может транскрибировать структурные гены и
катализировать реакцию синтеза полицистронной молекулы
мРНК, используемой затем для трансляции при синтезе молекул
Рис. 33. Схематичный механизм регуляции лактозного оперона на основе репреС'
снн — дерепрессни
158
истозидазы, пермеазы и ацетилазы, участвующих в катабо-
лактозы.
бстраты, используемые клеткой под влиянием конститутив-
(ерментов, вызывают катаболитную репрессйю индуцибель-
[етаболических путей. Это могут быть глюкоза, фруктоза,
зо-6-фосфат, /.-глутамат и другие вещества.
микробных клетках цАМФ и КАП могут образовывать
екс, соединяющийся с промотором в /ас-опероне. Только
сутствии данного комплекса становится возможной ини-
я транскрипции этого оперона. Следовательно, комплекс
>—КАП действует как положительный регулятор, так как
жсутствие необходимо для генной экспрессии.
эукариотических микроорганизмов не обнаружены регуля-
,е белки типа репрессоров у бактерий; опероны у них обыч-
ноцистронные с обширными регуляторными зонами, а струк-
ie гены разбросаны по геному (у прокариот сосредоточены
ом опероне). У эукариот больше развита групповая регуля-
активности структурных генов, чем у прокариот и вирусов,
е регуляции генной экспрессии существует регуляция фер-
ггивной активности, а все биохимические процессы в живых
ах становятся возможными лишь при согласованном дей-
ферментов и ферментных систем. К тому же продуктами
й экспрессии в значительной мере являются молекулы бел-
ферментов (см. гл. 5).
>менчивость микроорганизмов. Под изменчивостью пони-
разнообразие организмов, ныне существующих и вновь
кающих. При работе с микроорганизмами в качестве объек-
шользуется не одна особь, а популяции.
опуляция — многочисленные клетки (особи) одного вида,
ающие в ряду поколений в определенных условиях, где нет
гных преград для случайного свободного скрещивания этих
Ж. Микроорганизмы в популяции характеризуются общно-
генетической программы и возможностью свободного обме-
(нетической информацией. Для популяции характерны такие
йатели, как плотность — число особей на единицу объема
Площади; прирост — число новых особей, возникающих в еди-
' времени; гибель — число особей, гибнущих в течение опре-
шого промежутка времени. Изменение численности популя-
мяражается кривыми размножения или кривыми выживания.
тура микроба называется чистой, если родительские и до-
ае особи практически неразличимы в ней и между ними нель-
тановить родственные связи. Чистые культуры (как популя-
мало или совсем непригодны для генетических исследова-
Генетические исследования выполняются на клоновых куль-
Клон — культура, состоящая из наследственно однород-
иеток, возникающих в результате бесполого размножения.
:и грибов могут быть многоядерными (например, у однокле-
ix низших грибов), поэтому получить их клоновые культуры
а трудно.
159
Ненаправленные (спонтанные) мутации у разных микроорга-
низмов происходят с частотой 1 • 10 ~4...1-10~ ° на одну особь в
единицу времени или на одну генерацию. Это значит, что в дан-
ный период изменяется одна клетка из 10 000 или 10 млрд, кле-
ток, и время существования клоновой культуры ограничено.
Однако по каким-либо специфическим признакам (например, об-
разование антибиотика) наследственная однородность культуры
может поддерживаться при рассевах в течение длительного вре-
мени. Такую культуру называют штаммом. Он может сохранять
специфический признак при размножении половым и бесполым
путем.
Из множества признаков, используемых для учета изменчиво-
сти микробов, — морфология, структурно-химические особенности
клеток, культуральные свойства, антигенность (см. гл. 8) — наи-
большую ценность для генетики представляют стабильные изме-
нения, наследуемые в потомстве клеток. Различают наследуемые
мутации, ненаследуемые модификации и наследуемые длитель-
ные фенотипические модификации.
Мутация — генотипическое и необратимое изменение нормы
реакции организма. Норма реакции организма — проявление
фенотипа в разных условиях существования вида, а фенотип —
определенная сумма признаков организма в конкретных внешних
условиях. Факторы, обусловливающие мутации, называют мута-
генными или мутагенами. Известны химические мутагены — про-
изводные этилена, уретаны, алкилсульфонаты, иприты, этилени-
мины и др.; физические — излучения (ионизирующие, ультрафио-
летовые), ультразвук, повышенная температура; биологиче-
ские—вирусы бактерий (фаги).
Клетки, наследственно изменившиеся под влиянием мутаге-
нов, называют мутантными, а культуры, получаемые из них, —
мутантами. Мутации могут быть индуцированные (контролируе-
мые) и спонтанные (неконтролируемые). Последние имеют место
в естественных условиях без вмешательства экспериментатора.
Мутации проявляются, например, в изменении устойчивости к
антибиотикам, в потребности в каких-либо факторах роста (аук-
сотрофы), в дефектности по синтезу отдельных ферментов и т. д.
Адекватность генотипического изменения клеток условиям среды
обитания оценивается в ходе естественного отбора при адапта-
ции, так как мутанты могут быть жизнеспособными и нежизне-
способными. Если мутация произошла в эукариотической клетке
с диплоидным набором хромосом в ядре, то такая мутация может
нивелироваться доминантным геном и ее выявление будет затруд-
нено или невозможно без расхождения ядер по разным клеткам-
Ненаследуемые модификации — это смена фенотипов при из-
менении условий существования клона. Например, потеря пиг-
ментации у Serratia marcescens при повышении температуры вы-
ращивания и возврат к пигментообразованию при снижении тем-
пературы. Конкретный фенотип воспроизводится в совершенно
определенных условиях существования вида и зависит всецело
160
аследственной конституции, или генотипа клетки. Генотип
диняет ядерный аппарат — геном и цитоплазматические на-
ственные структуры — плазмон. К ненаследуемым модифика-
[ относят, например, индуцибельный синтез ферментов. Под
нием специфического субстрата-индуктора образуется спе-
1ческий фермент. При удалении из среды индуктора клетки
ращаются в исходное репрессированное по ферменту состоя-
Индуцируемый синтез ферментов иногда называют адаптив-
синтезом, а образующие ферменты — адаптивными. Адапта-
' (в широком понимании) — приспособление наследственно
йетентной клетки к изменившимся условиям существования.
Длительность модификационных изменений обусловливается
зчением стабильных метаболических циклов. В процессе раз-
кения родительские клетки передают дочерним поровну
ктуры генотипа и другие клеточные компоненты, которые
определенных условиях существования микроорганизма ав-
ггически поддерживают наведенный цикл обменных реакций
:чезают из клетки лишь спустя несколько генераций. На
,ых порах они передаются потомству как бы по наследству —
длительные наследственные фенотипические модификации.
присущи только клеточным структурам, но не вирусам.
Рассмотренные виды изменчивости определяют и соответст-
вий ее анализ. Так, генетический анализ у микроорганизмов
ючает мутационный и гибридологический анализы. Первый из
предусматривает точное количественное изучение процесса
никновения мутаций и разнообразия мутантов. С его помощью
ется устанавливать существование наследственного различия
пределить фенотипические особенности мутанта. Гибридоло-
еский анализ предусматривает количественное изучение на-
девания родительских свойств в ряду поколений. Здесь по-
но также использовать гибридизацию протопластов клеток
ариотических микроорганизмов. С помощью такого анализа
>ется выявлять генетическое детерминирование признака и
анавливать локализацию генов в геноме и локализацию мута-
1нного изменения внутри гена.
Микробы, как и высшие организмы, способны собирать и пе-
аспределять уже имеющуюся наследственную информацию
<ду родственными, но генотипически неидентичными клет-
и. Процесс комбинирования в одной клетке мутировавших
эв из двух разных клеток называют генетической рекомбина-
й. Она наблюдается при трансформации, трансфекции, конъю-
ии и трансдукции. При этом не происходит истинного слия-
клеток и лишь часть генетического материала переходит
летку-реципиент. Образуется неполный, или частичный, дип-
д — мерозигота, или частичная зигота (от греч. цершо —
ть, oiycoTwo — спаренный). Геном реципиентной клетки назы-
»т еще эндогенотой, а привносимый в нее фрагмент ДНК —
огенотой. Экзогенота может в клетке гидролизоваться (рест-
ция) с помощью фермента рестриктазы или эндонуклеазы;
1617
161
сиариваться с эндогенотой, образуя рекомбинантную хромосому;
сохраняться в индивидуальном состоянии и реплицироваться^
формируя в последующем клон частично диплоидных клеток; со-
храняться без репликации лишь в одной клетке потомства в виде
мерозиготы (абортивная трансдукция).
У эукариот имеет место истинное слияние половых клеток
(гамет) с последующим образованием диплоида (зиготы). Ре-
комбинация у эукариот имеет место при кроссинговере хромо-
сом (от англ, crossing — скрещивание; over — на, поверх), а у
прокариот — при интеграции двух разных репликонов (репли-
кон — единица репликации), например в случае с фагом А.,
интегрирующимся с бактериальной хромосомой.
Трансформация — процесс, при котором осуществляется пере-
нос информации химически чистой ДНК от бактериальной клет-
ки-донора в бактериальную клетку-реципиент, где в результате
рекомбинации замещается специфическая последовательность
генома. ДНК при этом выступает трансформирующим агентом
без каких-либо посредников. Она чувствительна к ферменту
ДНК-азе. Процесс трансформации известен для грамположи-
тельных и грамотрицательных бактерий, он подразделяется на
пять фаз:
1) обратимая фаза контакта ДНК с клеткой-реципиентом,
ДНК здесь чувствительна к ДНКазе;
2) необратимая фаза проникновения ДНК в клетку-реци-
пиент, ДНК донора здесь не чувствительна к ДНКазе и не изме-
няется при обработке левомицетином;
3) фаза спаривания ДНК донора с хромосомой реципиента,
ДНК не чувствительна к ДНКазе, последующие события инги-
бируются левомицетином;
4) фаза интеграции ДНК донора с ДНК реципиента в ре-
зультате рекомбинации, ДНК не чувствительна к ДНКазе, вве-
денный маркер реплицируется при репликации ДНК хромосомы
реципиента;
5) фаза репликации интегрированной в хромосоме реципиента
новой информации, ДНК не чувствительна к ДНКазе, развива-
ется трансформированный клон.
ДНК с молекулярной массой 1-107 Да обладает высокой
трансформирующей активностью, особенно двухцепочечная, хотя
в клетку проникает лишь одна нить. Трансформация по времени
протекает в секунды (у пневмококков) или в минуты (у гемо-
фильных палочек’и других бактерий).
Трансфекция — процесс, при котором информация перено-
сится в компетентную бактериальную клетку при поглощении
ею ДНК фага. Различают пять этапов трансфекции: 1) адсорб-
ция клеткой ДНК фага, 2) поглощение клеткой адсорбирован-
ной ДНК фага, 3) фрагментация поглощенной фаговой ДНК
с помощью ферментов эндонуклеаз, 4) первичная рекомбинация,
сопровождающаяся воссоединением фрагментов фаговой ДНК,
5) образование зрелых фаговых частиц.
162
роцесс поглощения ДНК фага компетентной клеткой при
гфекции сходен с процессом трансформации бактериальной
, однако они существенно различны, поскольку фрагмен-
1 ДНК происходит лишь при трансфекции. Конечная фаза
иинает процесс, происходящий при заражении клеток фа-
ми частицами, т. е. естественным путем. Таким образом,
трансфекции компетентная клетка поглощает фаговую ДНК,
варительно освобожденную из фаговой частицы. Трансфек-
проходит с большей эффективностью посредством ДНК
тагов, выделенных из лизогенных клеток. Она осуществи-
на сферопластах компетентных бактерий.
конъюгация — процесс частичного или полного переноса
гического материала из мужской особи в женскую при уста-
енном клеточном контакте между разнополыми клетками
ерий. Конъюгация проходит четыре фазы: 1) конъюгиро-
е клеток противоположного типа спаривания (+ и —),
Перенос генетического материала через конъюгационный
•ик из мужской клетки в женскую, 3) включение всего или
И переданного генетического материала в хромосому клетки-
пиента, 4) сегрегация (от лат. segregare — разделять)
зовавшейся рекомбинантной хромосомы.
1ол у бактерий определяется переносимым (трансмиссибель-
) генетическим элементом, называемым F-фактором (от
; fertility — плодовитость). Женская клетка (F-), получив-
в процессе конъюгации F-фактор, становится мужской (F+).
shoc хромосомных маркеров при конъюгации бактерий —
айное событие. Кроме того, оно является вторичным ре-
татом переноса F-фактора, зависимым от интеграции хро-
мы с фактором в донорной клетке.
'-фактор относится к внехромосомным наследственным
рминантам — плазмидам, переносимым чаще через F-пили
через Р-пили (у псевдомонасов). В популяции Р+-клеток
грация F-фактора с хромосомой в результате кроссинговера
сходит примерно в одной из 100 000 клеток в каждой гене-
:и. Клетки, в которых произошла интеграция, и образован-
ими клоны называют клонами высокой частоты рекомби-
1Н Hfr (от англ. High frequency of recombination). Процесс
грации обратим в популяции клеток Hfr. Исключение
жтора в результате второго кроссинговера происходит при-
ю с такой же частотой, как и интеграция в популяции
Клеток. На примере Е. coli К12 направленность рекомбина-
5 применительно к половым типам клеток выглядит следую-
й образом:
шчество генетических рекомбинантов в направлении 1 на
|Цнной выше схеме не превышает 0,001% от числа бактерий-
163
партнеров, тогда как в направлении 2 их образуется в 100...1000
раз больше).
При реверсии Hfr-клеток -в F+-клетки иногда появляются
такие варианты, у которых F-фактор включает часть бакте-
риальной хромосомы. Подобный фактор называют промежу-
точным (F'). При его передаче от F+- к Р“-клеткам образуются
мужские клетки с признаками F+- и Hfr-бактерий. Перенос
генетического материала бактерий при конъюгации с помощью
F'-факторов называют сексдукцией или F'-дукцией.
Трансдукция — процесс переноса ДНК из бактериальной
клетки-донора в фагочувствительную клетку-реципиент с по-
мощью фага. Фаг выступает посредником между донорной и ре-
ципиентной клетками. Среди различных фаговых частиц лишь
немногие могут быть трансдуцирующими (от 1 • 10~5 до
1- КГ8). Различают два вида трансдукции: неспецифическую
(общую) и специфическую (ограниченную). При общей транс-
дукции любой хромосомный маркер включается в фаговую
частицу примерно с равной вероятностью. При специфической
в состав фага включаются только локусы, непосредственно при-
легающие к местам-сайтам (от англ, site — место) интеграции.
Так, фаг X трансдуцирует один признак — способность фер-
ментировать галактозу. В ДНК Е. coli имеется лишь один сайт,
в котором он может рекомбинировать с ДНК фага X. В ре-
комбинации участвуют «липкие концы» ДНК, поскольку после-
довательность оснований на двух одноцепочечных концах ли-
нейной ДНК фага А комплементарна. При нагревании раствора
ДНК фага X при 60 °C и последующем медленном охлаждении
липкие концы соединяются друг с другом за счет комплемен-
тарного взаимодействия оснований.
Нельзя смешивать трансдукцию с фаговой, или лизогенной,
конверсией (нехромосомной лизогенией), когда свойства бакте-
риальных клеток определяются только фаговыми частицами.
Такие свойства присущи лишь лизогенным или зараженным
фагом штаммам и никогда не появляются в клетках штаммов,
свободных от фагов. Например, клетки Corynebacterium diphthe-
riae образуют токсин только при заражении определенным фа-
гом. Без подобного фага клетки не приобретают этого свойства
даже в результате мутации. То же можно сказать о токсигенных
стафилококках. Фаговая конверсия может быть причиной виру-
лентности вследствие образования соответствующих ферментов
патогенности (см. гл. 7) — гиалуронидазы, коагулазы, гемоли-
зинов, лецитиназы, фибринолизина.
3.5. ВНЕХРОМОСОМНАЯ ДНК
К внехромосомным дезоксирибонуклеиновым кислотам относят-
ся митохондриальная ДНК и ДНК хлоропластов в клетках
аэробных эукариот, двухмикронная ДНК (плазмида) у некото-
рых дрожжевых организмов и плазмиды бактерий.
164
Аитохондриальная ДНК (мт ДНК) относительно полно
ена у ряда дрожжей, мицелиальных грибов, трипаносом
юропластов водорослей. Она сходна с хромосомой бактерий
>едставляет собой двухспиральную кольцевидную ДНК, не
эанную с белками (у некоторых Protozoa митохондриальная
К разомкнута). Однако геном митохондрий существенно
ьше бактериального и приближается по размерам к вирус -
у. Он включает более 78 000 нуклеотидов с большими интро-
и внутри генов. Функционально мтДНК служит матрицей
рРНК рибосом митохондрий и тРНК, а также мРНК субъ-
иц комплексов дыхательных ферментов внутренней мембра-
Однако большинство рибосомных белков митохондрий и
юпластов импортируется из цитоплазмы клеток. Все другие
и внутренней мембраны, матрикса, внешней мембраны зако-
ваны в ядерной ДНК клетки. Следовательно, функция
гического аппарата митохондрий в значительной мере за-
т и обеспечивается ядерным аппаратом клетки; оба они
ггают согласованно. Установлено, что, например, гены мито-
дрий дрожжей похожи на гены митохондрий человека (!).
различаются они по своей организации. На рис. 34, а пред-
длена карта мтДНК дрожжей, на которой имеются длинные
кодирующие участки — интроны. При генетической экспрессии
;ой ген транскрибируется целиком. Образованная гяРНК
(вергается сплайсингу (или процессингу), т. е. интроны выре-
зтся, а экзоны сшиваются с помощью лигаз в зрелую мРНК.
В хромосомах ядра интроны не кодируют синтез никаких
шов, а в митохондриях они кодируют синтез отдельных белков,
орые сами могут участвовать в -йоследующем вырезании
тронов. Некоторые молекулы ферментов (белков) кодируются
повременно экзонами и интронами. Один из таких ферментов
тохондрий назван матуразой (от англ, mature — созревать).
зможно, что для каждого интрона существует своя матураза
Ис. 34, б). Интроны митохондриальной ДНК похожи на вак-
хальные транспозоны; предполагается, что матуразы произо-
щ от транспозаз. В ДНК митохондрий дрожжей открыто мно-
ство точек инициации синтеза РНК, что свидетельствует о
}можности в них контроля процесса транскрипции.
j У некоторых эукариот в ядерной ДНК обнаружены сегменты
ДНК- В этой связи очевиден факт переноса генов из орга-
ри в ядра клеток. Открытие интронов в генах археобактерий
гранило одно из последних возражений против эндосимбио-
ческой гипотезы о происхождении митохондрий.
Двухмикронная ДНК дрожжей — кольцевая двухнитевая
йзмида, содержащая 6 300 пар нуклеотидов. Найдена у ряда
дов рода Saccharomyces. В диплоидной клетке может содер-
ггься 60... 100 копий этой плазмиды. Функционально двух-
Кронная плазмида расценивается цитоплазматическим элемен-
я, хотя в действительности она локализуется в нуклеоплазме,
етки не имеют барьеров, предотвращающих их размножение,
165
В1 /1 В2 12 ВЗ /3
ДНК ямввжвммямамммшямм1
РНК
кольцевая РНК
В5 /5
дальнейший |
прессинг |
Bl В2 ВЗ В4 В5 В6
фермент,
закодированный
в ядре
Рис. 34. Карта митохондриальной ДНК у дрожжей (а), образование матуразы
через РНК-матуразу, необходимой для вырезания интрона 12 (б)-.
В1...В6 — экзоны, 11...15 — интроны
если двухмикронные плазмиды попали внутрь. Эти плазмиды
образуют гибриды с ДНК хромосом и могут рекомбинировать
с ними. Двухмикронная ДНК дрожжей (но не мтДНК) упа-
166
деается в нуклеосомы, со-
[ащие нормальный состав
ных гистонов и активно
скрибируется преимущест-
во РНК-полимеразой II.
рхмикронная плазмида (рис.
по крайней мере часть вре-
де находится в нуклеоплаз-
она ковалентно не связы-
ся с хромосомной ДНК.
жце плазмиды имеются об-
де А, В и С, кодирующие
•ез трех белковых молекул
молекулярной массой 35 000,
Э00 и 48 000 Да соответствен-
1 Допускается наличие в
дмиде двух меньших откры-
X областей, кодирующих белок с молекулярной массой ~ 18 000
* 12 000 Да. Все открытые области транскрибируются в форме
|бильной полиаденилатной РНК-
Плазмида способна автономно реплицироваться в клетке,
отличие от бактериальных плазмид двухмикронная дрожже-
плазмида синхронизирована с клеточным циклом в период
‘й репликации. Несущие их клетки фенотипически не отли-
тся от клеток, не содержащих двухмикронные плазмиды,
ледние составляют лишь около 2 % от генома клетки. Функ-
1альная роль двухмикронной дрожжевой плазмиды интен-
ю изучается. С ней связывают устойчивость клеток к анти-
гику олигомицину и возникновение секторной летальности в
ониях на агаризованных средах.
Плазмиды бактерий — автономно реплицирующиеся молеку-
ные структуры. Плазмиды, находящиеся в интегрированном
актериальной хромосомой состоянии, иногда называют эпи-
ами, это частный случай плазмид. Между эписомными эде-
мами, находящимися в интегрированном состоянии, и сосед-
де областями хромосомы бактерий могут происходить реком-
деции. К плазмидам относятся умеренные фаги, факторы
ггериоциногенности, F- и R-факторы. Все плазмиды пред-
вляют собой кольцевидные двухцепочечные молекулы ДНК
ключение — киллер-плазмида у дрожжей с молекулярной
:сой от 2-104 до 3,2-108 Да). Мелкие плазмиды содержат
етическую информацию примерно для двух белков, крупные —
I 200 белков.
Лизогенные бактерии (см. разд. 1.1), несущие умеренные
Ш (профаги), устойчивы к заражению теми же или близко-
1ственными фагами. Лизогенизация может проявиться к 1...5
:ам фагов. Фагоустойчивость культур имеет большое практи-
ков значение в производстве, например, кисломолочных
Щуктов, антибиотиков и других веществ. Профаги привносят
167
свою собственную информацию в клетки-реципиенты, которые
могут приобрести способность вырабатывать токсины, изменить
устойчивость к химиотерапевтическим агентам и т. д. Это так
называемая лизогенная или фаговая конверсия (см. транс-
дукция).
Факторы бактериоциногенности впервые были найдены у
кишечной палочки Е. coli и названы колицинами. Генетические
детерминанты, контролирующие синтез специфических бактерио-
цинов, сходны с умеренными фагами. В отличие от последних
бактериоцины убивают чувствительные бактерии, не размно-
жаясь в них. Некоторые бактериоцины адсорбируются на спе-
цифических рецепторах, которые могут быть утеряны при мута-
ции клеток. Бактериоцины найдены у многих грамположительных
и грамотрицательных бактерий. Их называют по происхождению,
например, альвецины из Hafnia alvei, каротоворицины из Erwinia
carotovora, морганоцины из Proteus morgani, колицины из Е. coli
и некоторых других видов (А, В, С, D, Е1 — Е4, G, Н, I, К, V,
SI, S4, L, М, О, Р, Q и др.).
Бактериоцины подразделяют на два класса. К первому клас-
су относят те из них, которые напоминают бактериофаги или
их фрагменты; ко второму — бактериоцины, которые являются
либо простыми белками, либо белками, связанными с компо-
нентами клеточной стенки. В последнее время некоторые бакте-
риоцины, например колицины, классифицируют по их активно-
сти в отношении набора штаммов (производных из одного
чувствительного штамма), каждый из которых устойчив лишь к
одному какому-либо классу колицинов.
Бактериоцины придают штамму-носителю (и популяции плаз-
мид, обитающих в этом штамме) селективное преимущество
перед родственными штаммами, которых он убивает. Колицины
могут убивать родственные клетки из-за разобщения энергоза-
висимых процессов, протекающих в клеточной мембране; вслед-
ствие деградации ДНК и подавления деления клеток; за счет
расщепления 16Б-рибосомной РНК и прекращения синтеза
белка.
F-фактор, или половой фактор (см. конъюгацию), контроли-
рует процесс конъюгации и перенос генетического хромосомного
материала из бактериальной клетки-донора (F+) в клетку-ре-
ципиент (F-). На одну хромосому приходится один F-фактор.
R-фактор, или фактор множественной устойчивости к хи-
миотерапевтическим веществам (от англ. Resistance — устойчи-
вость), представляет собой трансмиссибельную плазмиду. Он
никогда не возникает в результате мутации, а передается через
F-пили (как и F-фактор) от одних клеток к другим или посред-
ством трансдукции, будучи интегрированным с хромосомными
ДНК (например, у стафилококков). На одну хромосому в клетке
приходится обычно одна или две копии крупной плазмиды, но
могут формироваться неконъюгирующие миниплазмиды (от 5 до
30 копий на клетку).
168
R-плазмида имеет кольцевидную структуру, в которой нет
I содержится сегмент RTF (от англ. Resistance transfer fac-
— фактор, передающий устойчивость) и r-детерминанты ус-
чивости. Молекулярная масса RTF-сегментов равна ~ 6- 107Да,
-детерминант ~ 1 • 107 Да. Между RTF и детерминантами ус-
чивости нет ковалентной связи. R-факторы регулируют образо-
1ие sex-пилей. Клетки с R-фактором образуют половые пили
Йгановятся клетками-донорами данной плазмиды (R+). R-плаз-
щы могут теряться клетками спонтанно при хранении рези-
|Нтных культур или при обработке химическими веществами,
стягивающими молекулу ДНК (бромистый этидий и акрифла-
дЙ) или ингибирующими синтез РНК (антибиотик рифампицин):
бромистый этидий
акрифлавин
Клетки, несущие R-плазмиды, могут быть избирательно убиты
кими веществами, как этилендиамин-тетрацетат, натрия доде-
илсульфат. R-плазмиды приобрели большое значение в инфек-
юнной патологии (см. гл. 7), поскольку клетки патогенных
жробов, содержащие R-факторы, устойчивы ко многим извест-
1м лечебным химиопрепаратам. Такая устойчивость, обуслов-
нная плазмидами, чаще всего является результатом модифи-
ции химиотерапевтических средств (сульфаниламиды, антибио-
ки) ферментами, которые вырабатываются патогенными мик-
сами, содержащими R-факторы.
Известны и другие двухспиральные ДНК-содержащие плаз-
1ды. Например, плазмиды биодеградации в штаммах Pseudo-
onas putida (табл. 21). Такие штаммы бактерий играют опре-
йенную роль в разложении названных веществ в природных
1И иных условиях. В клетках дрожжей Saccharomyces cerevi-
169
Таблица 21. Некоторые плазмиды деградации
Деградируемый субстрат Число фермен- тативных стадий Плазмида Штамм Размеры плаз- миды, МДа
Камфора 15...20 САМ PgGl 165
Ксилол 1 1 XYL Рху ?
Нафталин >11 NAH Pg G7 49
Салицилат 8 SAL RI 51
siae обнаружена двухнитевая РНК-плазмида, называемая кил-
лер-плазмидой, индуцирующая синтез белка-токсина, убиваю-
щего чувствительные клетки того же рода. Клетки-продуценты
токсина устойчивы к такому белку. Механизм действия токсина
окончательно не выяснен, но конечным результатом выступает
повреждение клеточной мембраны, например, киллер-плазмида
имеет существенное значение в повреждении дрожжей, исполь-
зующихся в виноделии.
Вопросы
I В чем заключается ипформационная сущность ДНК?
2 Канон механитм формирования нар из нуклеотидов в пукле ио
кислотах? Какие формы ДНК известны?
3 Чю такое дещиурация, ренжурация и гомология ДНК?
! 'Ы, ню»' палиндромы в ДНК и какова их роль?
> Какие 'иетоны входя! г. состав хроматина клеток эукариоте
микроорганизмов?
h. Что такое хромосомы и какое число их может быть у прокарио,:
гких и эукариотических микроорганизмов?
7 . Какие |ины РНК имеются в клетках микроорганизмов? Каков
ф\ чкиии?
Я Что огначаст процетсинг РНК в клетках эукариотических микро •
ни тмивJ
ч I'.iKiie pas.THHiin имеются между рибосомами у прокариотиче, >
г, парно: и чес них микроор! а имамов?
10 . Чю означает первичная, вторичная, третичная и четвертичная
тхры молекул полисахаридов (н сравнении с белками)?
II Какие основные структурные блоки имеются в нснгидсилт .< :
к.пне Hi них важны для дифференциации нентидо!ликанов?
12 Какое строение имеют липополисахариды i рамотрицательных
при и ?
1' г Какой химичеткин состав капсул у сине! войной палочки и на • т
сибирской язвы?
14 Каковы химический состав и архитектоника клеточной (цитоила
тической) мембраны?
io. Назоните орышеллы. являющиеся ирон [водными клеточной мет
пы Какова их функция в клетке?
16 Дайтч определения понятиям кодон, ген. цистрон, геном.
17. Что такое репликация ДНК и каков ei механизм?
13. В чем заключает я механизм 1|>анскрннции?
I’.l Чю i.'ikoc трансляция мРНК и каков ее механизм?
26 Китие составные части входят в комплекс инициации?
1:1 Что отн,ищет ностгранс.'тяциониая (вторичная) модификация бел
примеры ее?
Каковы о, сонные .механизмы регуляции генной эксирестии у ' 1
Юрию '
’3. Что пре 1СТ.Ш.1ЯЮТ собой плазмиды и какова их функция в к,в
прокарио! и эукариот?
21. Что понимают под терминами: клон, штамм, чистая культур.’
170
но
0
мические
новы
1знедеятельности
нроорганизмов
ава 4
[МИЧЕСКИЕ ОСНОВЫ ПРОЦЕССОВ РОрТА
РАЗВИТИЯ МИКРООРГАНИЗМОВ
оцессы роста, развития, размножения, морфогенеза и диффе-
щировки микроорганизмов взаимосвязаны (рис. 36). Напри-
>, трудно уловить какие-либо качественные изменения в клет-
i бактерий после размножения, так как переход их в зрелое
в
. 36. Примеры роста, развития, размножения и дифференцировки микроор-
1змов:
рост нитчатого гриба (/ — ростковая трубочка проросшей споры, 2 — образование
!Й мицелия); Б — размножение коккобактерии; В — развитие и дифференцировка
>икового гриба из класса Basidiomycetes во время жизненного цикла (а — образова-
днкариона, б—начало развития, в—образование базидиокарпа, г—образование
1ДИЙ и базидиоспор, д — гаплоидные базидиоспоры, е — развитие монокарнонов, ж —
•Могамия, з — миграция ядра)
171
состояние осуществляется в пределах минут. У грибов (микро- и
макромицетов) такие изменения проследить легче, поскольку
жизненные циклы их сложнее, чем у бактерий, и разграничены
во времени. Тем не менее в каждое из этих понятий вклады-
вается четко определенный смысл. Рост — это координирован-
ное увеличение размеров и веса клетки. Под термином разви-
тие понимают совершенствование во времени структур и функ-
ций клетки, унаследованных потомками от родительских форм.
При увеличении во времени числа клеток популяции определен-
ного вида или штамма в питательной среде говорят о раз-
множении микроорганизма. Под морфогенезом понимают
процесс образования соответствующих форм микроорганизмов в
конкретных условиях их существования. Дифференциров-
ка — это дифференциальное выражение генов, содержащихся в
геноме клетки.
4.1. РОСТ И РАЗВИТИЕ МИКРООРГАНИЗМОВ
Со времени образования новой генерации клеток все особи про-
ходят определенный период роста. Если время генерации какой-
то бактерии в оптимальных условиях занимает 30 мин, время ге-
нерации дрожжевого организма — 40 мин, то до наступления
очередного периода размножения проходит приблизительно та-
кой же временной интервал, в течение которого клетки растут
в размерах и по массе. Размеры и масса микробных клеток
обусловлены достаточным поступлением в клетку ингредиентов,
необходимых для конструктивного и энергетического обменов.
Особенно велика роль источников азота, углерода, микроэлемен-
тов для некоторых микроорганизмов — ряда витаминов, амино-
кислот и других веществ.
Размножение и рост микроорганизмов существенно зависят
от условий культивирования их (состав и консистенция пита-
тельных сред, поверхностное и глубинное культивирование на
жидких средах, с подачей или без подачи кислорода воздуха
или иных газов, и т. д.). Например, при недостатке источника
углерода (лимитирующий фактор) некоторые дрожжи в первые
минуты размножаются почти с такой же скоростью, как и в ус-
ловиях отсутствия лимитирующего фактора. Однако при дефи-
ците источника углерода клетки становятся сильно вакуоли-
зированными и меньше в размерах.
Прокариотические организмы на жидких средах могут обра-
зовывать осадок, поверхностную пленку (это нередко типично
для подвижных клеток) , кольцо по стенкам сосуда на поверхно-
сти среды (иногда ползущее вверх), равномерную суспензию
(мутность). Подобная картина присуща и многим дрожжевым
организмам. Грибы-аэробы образуют различной толщины плен-
ки на поверхности жидких сред в стационарных условиях.
Среды могут быть при этом разной степени мутности или про-
зрачными, содержать или не содержать осадки. При выращива-
172
3456789 10 W
питательный агар
I
ife
Рис. 37. Профиль колоний:
^—полусферический, 2— конусовидный (сопковидный), 3 — седловидный, 4 —
уплощенно-выпуклый, 5 — каплевидный, 6 — росовндный, 7 — выпукловолнистый,
$— сосцевидно-выпуклый, 9 — сферический, 10 — выпукло-сосцевидно-вдавлен-
<НЫЙ, 11— кратеровидный, 12—прямоугольный, 13 — срезанный обращенно-кону-
совидный, 14 — срезанный конусовидный, 15 — блюдцевидный, 16 — стелющийся
?(а); /, /' — неправильные, 2, 2'— врастающие в среду, 3, <3'— ворсинчатые;
—игольчатые; 5 — зубчатые; 6 — шиповидные (б); форма краев колоний: ров-
ная (/), неровная (2—6), 2 — волнистая, 3 — фестончатая, 4 — лопастная, 5 —
уэваиая, 6 — реснитчатая (а)
I грибов в погруженных условиях (в ферментаторах) рост
развитие их может быть в виде гомогенной суспензии, зер-
тости, хлопьев или комочков, напоминающих вату.
Развитие клеток прокариотических и эукариотических микро-
анизмов на стандартных уплотненных средах отличается
бильностью и специфическим проявлением. Клетки форми-
5т колонии, характеристика которых может быть сведена к
дующим показателям: I — форма профиля (выпуклые, плос-
, стелющиеся, неправильные, врастающие в среду, ворсинча-
, игольчатые, зубчатые, шиповидные); II — рисунок (вид
>ху) [гладкие, шероховатые, узорчатые (кружевные, мозго-
вые, паутинные, смешанные)]; III — окраска (окрашенные,
сращенные); IV — форма края колонии [ровный, неровный
1нистый, фестончатый, лопастный, рваный), опушенный];
- консистенция (сметанообразные, крошковатые, комковатые,
зчатые, сухие, слизистые); VI — прозрачность (прозрач-
, полупрозрачные, непрозрачные); VII — размер (очень ма-
ше; маленькие, или карликовые; крупные, или большие;
173
очень крупные, или очень большие); VIII — характер блеска
(глянцевые, матовые).
Своеобразие колоний можно дополнительно описать, исполь-
зуя какие-то другие характеристики. При этом следует помнить,
что генетически неоднородная популяция микроорганизмов одно-
го и того же вида может формировать колонии нескольких
морфологических типов одновременно. Так, среди выпуклых
колоний можно выделить 11 вариантов, тогда как плоские
(12... 15) и стелющиеся (16) колонии менее разнообразны
(рис. 37, а); профили и форма других колоний показаны на
рис. 37, б, в. Колонии неправильной формы присущи старым
бациллярным и некоторым грибным культурам.
Окраска колоний микроорганизмов может включать весь
спектр цветов или комбинацию цветовых гамм. Например, коло-
нии Serratia marcescens приобретают кроваво-красный цвет,
некоторые виды сарцин — лимонно-желтый, грибы — от белого
(дрожжи) до черного (аспергиллы). Наиболее распространенны-
ми пигментами являются каротины, ксантофиллы и меланины.
Ксантофиллы являются кислородсодержащими аналогами каро-
тинов:
виолаксантин (ксантофилл)
p-Каротин содержится, например, в дрожжевых организмах
из группы Rhodotorula, а виолаксантин — в мембране тилакои-
дов цианобактерий в виде эпоксида, где он синтезируется под
действием фермента зеаксантин-эпоксидазы.
Меланины — нерастворимые, полимерной природы пигменты
(черные, коричневые и красные), синтезирующиеся из феноль-
ных соединений. Их подразделяют на эумеланины (черные
тирозиновые полимеры у животных), феомеланины (желтые и
красные тирозиновые полимеры волос животных и перьев птиц),
алломеланины (черные безазотистые, чаще пирокатехиновые пиг-
менты растений и микроорганизмов). Меланины обычно связаны
с белками (меланопротеины) или с гликопротеинами (мелано-
гликопротеины). Части молекул меланинов включают преимуще-
ственно остатки 5,6-индолхинона (а) и пирокатехина (б):
174
S—СН,—CH(NH,)—соон
соон
а
б
Меланины наряду с ферментами — супероксиддисмутазой,
талазой и пероксидазами — обладают функцией протекторов
кислородных радикалов (Ог~) и синглетного кислорода
'Ог), являющихся сильными окислителями. Клетки, образую-
<е меланины, быстро погибли бы без соответствующих про-
кторов.
Некоторые пигменты хорошо растворимы в воде, что обу-
овливает окраску уплотненных питательных сред вокруг вы-
вших колоний. Не требуется дополнительных пояснений к при-
Денной выше характеристике колоний по консистенции, про-
ачности и блеску. В стандартных условиях выращивания
кроорганизмов они достаточно стабильны, равно как и разме-
1 колоний, измеряемые по диаметру. Так, диаметр очень ма-
тьких не превышает 1 мм; маленьких, или карликовых, нахо-
гся в пределах от 1 до 3 мм; крупных, или больших, — от 3
10 мм; очень крупных, или очень больших, — более 10 мм.
Таким образом, рост и развитие микроорганизмов зависят
многих факторов. Причем различия в потребностях могут
ть существенными не только между родами и видами, но и
‘аммами одного и того же вида, что необходимо учитывать
й оценке роста соответствующих культур на естественных и
Кусственных питательных средах.
‘ Важное значение приобретает также концентрация пита-
«льных веществ, вносимых в искусственные среды для выращи-
!йия микроорганизмов. При составлении таких сред рекомен-
тот следующие ориентировочные концентрации (г/л): доноров
акцепторов водорода ~ 2,0; источника азота ~ 1,0; источника
йерода ~ 1,0...2,0; микроэлементов (сера, фосфор, магний)
0,05 каждого; ультрамикроэлементов ~ 0,0001...0,001 каждого;
ютовых факторов: витаминов ~ 0,0001...0,001 каждого; амино-
|слот, пиримидинов; пуринов и др. ~ 0,05 каждого.
175
Наличие в среде одновременно различных источников угле-
рода, азота и других веществ может по-разному сказываться'
на росте и развитии микроорганизма. Тем не менее клетки мик-
робов в большинстве своем достаточно лабильны и легко при-
спосабливаются к окружающим условиям. Их популяции в
конкретных участках биосферы (экологических нишах) находят-
ся в тесной взаимосвязи. Эта взаимосвязь называется синтро-
фией (от греч. 0iv —вместе, трохрт] — питание). Как правило,
микроорганизмы могут использовать не один какой-либо тип
источников углерода, азота и других веществ, а разные. На-
пример, кишечная палочка может расти и развиваться в средах,
содержащих в качестве источников углерода глюкозу, глицерин,
уксусную кислоту или этанол; в качестве источников азота —
разные аминокислоты, аммиак, пиримидины, пурины и т. д. Если
в среде будет содержаться смесь таких веществ, то клетки
микроорганизма будут использовать прежде всего более доступ-
ные, например аминокислоты, а не аммиак. Это связано с тем,
что аминокислоты могут непосредственно включаться в обмен
веществ и в клетках выключается процесс синтеза ферментов
(репрессия), катализирующих реакции образования аминокис-
лот из аммиака.
Репрессия возможна и применительно к потреблению источ-
ников углерода. Например, в присутствии арабинозы, галак-
тозы, лактозы или мальтозы глюкоза тормозит активность фер-
ментов, необходимых для сбраживания упомянутых углеводов
клетками Е. coli. Причем непосредственно глюкоза не ингиби-
рует ферменты, их ингибирует продукт ее расщепления, или
катаболит. Подобный эффект глюкозы называют катаболитной
репрессией.
У всех организмов цАМФ образуется из АТФ под действием
фермента аденилатциклазы с выделением неорганического пи-
рофосфата. Если, например, рост клеток Е. coli лимитирован
источником углерода и энергии, то в них отмечается высокий
уровень содержания цАМФ, а при наличии в среде глюкозы —
низкий.
В клетках Е. coli комплекс цАМФ-БАК контролирует транс-
крипцию (считывание) лактозного оперона (1ас-оперона) в хро-
мосоме клетки. Такой контроль может быть положительным,
когда комплекс с цАМФ-БАК обеспечивает возможность РНК-
полимеразе связываться с матрицей перед началом транскрип-
ции, или отрицательным, когда этот комплекс не обеспечивает
такой возможности. На рис. 38 схематически указана последо-
вательность участков ДНК в lac-опероне кишечной палочки, в
том числе участок взаимодействия с комплексом БАК-цАМФ.
В ряде биотехнологических процессов необходимо накапли-
вать биомассу клеток в качестве конечного продукта (произ-
водство вакцин, кормовых и пищевых дрожжей и пр.), доби-
ваясь хороших показателей роста и развития соответствующих
культур грибов или бактерий.
176
lacP
1
। lacO
lacZ
J
6
a
, 38. Схема части хромосомы E. coli:
:тки взаимодействия с БАК-цАМФ (а) и РНК-полнмеразой (б); Lad — ген-регу-
ф; LacP — промотор; LacO — ген-оператор; LacZ — ген 0-галактозидазы
начальный период после засева среды, например, грибковыми спорами
1й из них образует ростковую трубочку, т. е. в определенный отрезок вре-
клетка лишь растет, но не размножается. Однако позднее, при созре-
, она будет размножаться. Иначе говоря, размножение является следствием
. И если рост микроорганизма происходит в течение значительного вре-
то масса клеток его в различные ростовые периоды будет неодинаковой.
микробов можно определять либо по массе клеток в весовых единицах
У= Ктах- Ко,
— урожай; К— клетки (в момент засева среды — Ко, в момент съема —
либо по их линейному росту во времени, например, применительно к нит-
грибам;
£ — Li — Lq,
— прирост грибной нити в единицах длины; Lo — длина нити в начале
• а времени роста, Lt — длина нити через какое-то время t.
\ожно определять экономический коэффициент (ЭК) по отношению урожая
у
к (У) к потребленному субстрату (С„): ЭК=-~~, или молярный экономи-
С п
й коэффициент (МЭК) по отношению урожая в граммах (Уг) к числу молей
у
бленного субстрата (Л4С„): МЭК = .
/ИСП
ели известен путь превращения (расщепления) какого-либо субстрата с
;ом АТФ, то можно рассчитать энергетический коэффициент (ЭнК), выражен-
В граммах на 1 моль АТФ; ЭнК= • Когда говорят о росте
фиальной массы, то имеют в виду размножающиеся клетки и определяют
исло N = Nt — No, где N — урожай клеток, No — количество засеянных
ок, Nt — количество клеток через какое-то время t.
Клеточную массу определяют также по занимаемому объему влажных кле-
после центрифугирования, по сухой массе, по количеству азота на определен-
биомассу, по мутности (для бактерий и дрожжей), регистрируемой с по-
ЬЮ спектрофотометра или фотоэлектрокалориметра.
Со временем клеточная популяция отмирает, если ее частично или пол-
ью не переносят в свежую питательную среду. В каждый данный момент в
содержатся разнокачественные клетки — более старые и более молодые. Mo-
te отличаются наибольшими потенциями к последующему выживанию, они
одны, менее вакуолизированы.
[ля микроорганизмов, как и для более высокоорганизован-
существ, характерна общебиологическая закономерность —
итие в течение роста, т. е. совершенствование структур и
5м. У бактерий развитие плохо поддается учету из-за боль-
скорости их роста и размножения. Тем не менее ясно, что в
развития бактериальных клеток в них воспроизводятся и
ршенствуются нуклеоиды, рибосомы, клеточные стенки,
177
3) 61
Рис. 39. Развитие Aspergillus species (а) и почкование Saccharomyces cerevisiae
(б) в направлении а-*-з:
/ — конидия; 2 — прорастание конидии; 3,4 — формирование конидиеносца; 5 — образо-
вание головки со стеригмами; 6 — продукция конидий
мембраны, капсулы. У грибов удается проследить некоторые эта-
пы процесса развития, например, от одноклеточной споры (ко-
нидии) до сформировавшегося многоклеточного организма
(рис. 39, а).
У дрожжей почки, как правило, меньше родительской клет-
ки, после отделения они продолжают расти и развиваться опре-
деленное время (рис. 39,6). У нитчатых грибов, вырастающих
в виде колоний на поверхности питательных сред, перифериче-
ская зона состоит из более молодых нитей мицелия, центральная
зона — из более старых, рост которых заметно ограничен. У ди-
морфных грибов, например, из рода Candida существенно изме-
няется химический состав клеточной стенки при переходе дрож-
жевой формы в мицелиальную. В частности, у них в 1,5...2 раза
увеличивается при этом количество хитина.
Фазовое развитие протозойных организмов также четко вы-
ражено и может быть прослежено во времени, например, у маля-
рийного плазмодия.
Таким образом, в растущей популяции чистой культур.. ик-
роорганизмов клетки могут находиться и рашой степени ю-
сти, и любые аналитические приемы с ними будут прик. ми
лишь для получения приближенных результатов. Если ж< об-
ходимо оценить свойства одиночной клетки (например ез
ДНК при делении; чувствительность к химиотерапевта -им
агентам и т. д.). то прибегают к методу синхронизации ста
культуры. Это особенно важно примениюльно к бакгер. У
которых невозможно изучать многие процессы на изолирог -их
клетках.
Такой метод впервые предложил Р. Д. Хочкисс (1954). При
этом культуру микроорганизма вначале выдерживают при пони-
женной температуре, а затем — при оптимальной. Так, например,
выдерживая клетки Streptococcus pneumoniae вначале при 25 °C
в течение 15 мин, а затем при 37 °C, удается достичь синхро-
низированного (одновременного) удвоения числа клеток и во вре-
мени тем меньшем, чем длительнее период генерации культуры.
Синхронизация ауксотрофных мутантов доступна при их вы-
держивании на средах, лишенных продукта-маркера, с последуй'
178
.переносом на полные среды. Используют для синхрониза-
какже метод фильтрации культуры через мембранные фильт-
ямеющие поры строго определенного диаметра. Этим пу-
Йолучают клетки одинакового размера, деление которых
входит затем синхронно. Синхронизированные культуры со-
ат клетки равной степени развитости, но синхронизация
'ся не более 2...4 генераций, после чего культуры переходят
щхронному (неодновременному) росту, развитию и размно-
ю.
УМНОЖЕНИЕ ПРОКАРИОТ И ЭУКАРИОТ
уже было сказано ранее, микроорганизмы обладают спо-
стью передавать генетический материал от одной клет-
г) к другой (—), т. е. для них характерен половой процесс,
ге с тем известно, что, например, гаплоидные клетки дрож-
одиночные бактериальные клетки, различные грибы (на-
»р, дейтеромицеты) размножаются бесполым путем. Очевид-
многие микроорганизмы наиболее приспособлены к вегета-
®>му размножению, чем более высокоорганизованные особи,
Имер из царства животных. При размножении одиночных
ж прокариот и эукариот образуются соответствующие
1яции чистых культур микроорганизмов в конкретных усло-
среды обитания. Изменение численности популяции выра-
ся кривыми размножения или кривыми роста. Кривые раз-
:ения и роста одноклеточных и многоклеточных микроорга-
эв разнопрофильны. Так, при засеве ими питательных сред
ледующим периодическим определением числа клеток в 1 мл
.о наблюдать закономерность, представленную на рис. 40.
.енительно к одноклеточным (кривая /) внесенные в пита-
ую среду клетки адаптируются к условиям существования
:менно сохраняются в том же числе.
длительность первой начальной стационарной фазы po-
ll) зависит от организма, его возраста и потенциальных
ожностей, от качества среды и других факторов. Вторая
' ускоренного размножения (2) наступает, когда возрастает
>та клеточного деления и нарастает число клеток. В третьей
">ифмической, или экспоненциальной, фазе размноже-
(3) наблюдается постоянная продолжительность существо-
ч возникающих друг за другом поколений клеток. Если на
рдинат отложить число клеток, а на оси абсцисс — время,
висимость получится в виде прямой линии (рис. 41). Четвер-
аза — уменьшение скорости размножения (см. рис. 40, 4) —
агеризуется увеличением продолжительности существования
ющих поколений клеток. Скорость размножения падает
истощения компонентов питательной среды, появления
'Ирующих факторов роста, накопления продуктов обмена и
х факторов. В пятую заключительную стационарную
размножения (см. рис. 40, 5, 6) общее число и масса живых
к достигают своего максимума. После этого наступает шес-
179
In N
Рис. 40. Кривые возрастания биомассы
одноклеточных (/) и мицелиальных (//)
микроорганизмов:
log N — логарифм числа клеток; Md
масса сухого мицелия; t — время
Рис. 41. Экспоненциальный
рост в полулогарифмическом
выражении
Рис. 42. Двухфазный рост (диа-
уксия) клеток Е. coli в питатель-
ной среде, содержащей глюкозу
и сорбит:
р— плотность; t — время
тая фаза — отмирание, когда в основном за счет автолити-
ческих процессов биомасса клеток уменьшается. В случае ориен-
тировочной оценки кривой I (см. рис. 40) выделяют только че-
тыре фазы: лаг-фазу (соответствует начальной стационарной
фазе и фазе ускоренного размножения), логарифмическую,
стационарную (соответствует заключительной стационарной фа-
зе) и фазу отмирания. Для бактерий фазы размножения часто
(но менее точно) называют фазами роста. При графическом
изображении процесса размножения клеток прокариот в ряде
случаев могут выявляться две лаг-фазы, предшествующие ло-
гарифмической фазе. Это связано с регуляторными процессами
в клетках, селективно и последовательно потребляющих внача-
ле более доступный субстрат, а затем — менее доступный. Так,
клетки Е. colt вначале потребляют глюкозу из смеси ее с сорби-
толом, и лишь после исчерпания моносахарида используют мно-
гоатомный спирт. Это явление называют диауксией (от греч.
6i(o) — дважды, ai)£av(o — увеличиваю, расту), двухфазным или
двуцикличным ростом (рис. 42).
У нитчатых грибов отсутствует
логарифмическая фаза, хотя она не
исключается для растущих верхушек
гиф (см. рис. 40, кривая II). В по-
верхностных культурах выявляется
прямолинейная зависимость между
возрастом культуры и кубическим
корнем из показателя массы мице-
лия. У таких организмов растущие
мицелиальные верхушки конкуриру-
ют за питательные вещества более
выраженно, чем погруженные куль-
туры одноклеточных организмов.
180
д., т. е. раз-
прогрессии.
прямо про-
1И размножении в благоприятных условиях из каждой жизнеспособной
сальной клетки (и в приближении — из клеток немицелиальных дрожже-
)ганизмов) возникают две новые клетки, далее — четыре и т.
ние происходит экспоненциально, или в геометрической
ный конкретный момент скорость размножения культуры
жальна числу клеток:
i ^=ЧГ-
зеле интегрирования уравнение (4.1) имеет следующий вид:
(4.1)
(4.3)
(4.2)
, — число клеток в нулевой точке отсчета времени; — число их в любое,
позднее время /; р — индекс скорости размножения, или удельная ско-
уазмножения.
йшая уравнение (4.2) для ц, получаем:
In (JV/JV0)
f ----
р отражает скорость, при которой натуральный логарифм числа клеток
стает со временем и может быть определен графически.
,аждая генерация бактерий (и приближенно одноклеточных немицелиальных
жей) в экспоненциальную фазу роста удваивается в сравнении с предыду-
Следовательно,
N = ЛД,2«.
имер, g = 8, тогда ^ = 256(1 Х28 = 256). Подставляя вместо его значе-
В уравнения (4.2), получаем:
'J Nf = N02s.
(4.4)
(4.5)
швнения (4.5) следует:
Г
g м
t 1п2 ’
(4.6)
(4.7)
значения
цавнивается g/t (генерации/час) с константой размножения р.
Используя уравнение (4.4), можно рассчитать время генерации:
1 logiV— log Д'г,
8 1Бр
(апример, при экспоненциальном росте посевной материал от
врастает до 109 клеток, тогда
log(109)-log(103) 9-3 оп
8 log2 0,3
Для 20 генераций потребовалось, например, 10 часов, то скорость размно-
была 20 : 10 = 2 генерации в час.
гстота смены генераций зависит от многих факторов (pH,
:ратуры, концентрации питательных веществ, аэрации, пере-
вания). В отдельных случаях кинетика размножения бак-
оказывается линейной (как и для нитчатых грибов с вер-
ным ростом). Это, например, можно наблюдать в опытах
‘антами, дефектными по синтезу какого-либо необходимого
в определенных условиях. Тогда скорость прироста клеток
Постоянна: -4^-= С, а при интегрировании N = Ct, т. е. чис-
» dr
1еток N (а не логарифм N) является линейной функцией
гни t.
181
Формы размножения микроорганизмов весьма разнообразны
и зависят от особенностей вида. Большинство прокариот размно-
жается путем изоморфного деления клеток, или цитокинеза
(см. рис. 3). В таком случае круглые формы бактерий, или кокки
могут образовывать скопления разобщенных клеток — микрококки,
соединенные попарно — диплококки, по четыре — тетракокки, в
пакеты — сарцины, в грозди — стафилококки, в цепочки —
стрептококки. Палочковидные формы при делении могут дать
в последующих генерациях изолированные бактерии, диплобакте-
рии, стрептобактерии. Спирохеты размножаются поперечным де-
лением.
Присущие родительским клеткам формы и типы деления сох-
раняются в последующих генерациях по наследству. Следователь-
но, конкретный тип цитокинеза прокариотной клетки обусловлен
ее геномом. Замечено, что, например, холерный вибрион легко
изменяет форму запятой на палочковидную при размножении на
питательных средах в лабораторных условиях. Очевидно, форма
данного микроба является фенотипическим признаком в конкрет-
ных условиях его обитания. События, развивающиеся в этом слу-
чае, обусловлены биохимическими реакциями, связанными с про-
цессами синтеза клеточной стенки.
У некоторых бактерий можно наблюдать гетероморфные клет-
ки, соединенные в одной цепочке, и, очевидно, у них происходит
гетероморфное деление. В качестве примера можно назвать
аэробные грамотрицательные бактерии Streptobacterium (Strep-
tobacillus) moniliformis, обнаруживаемые в глоточной области
зева у крыс. Они полиморфны, располагаются нерегулярными
цепочками, состоящими из перемежающихся веретенообразной
формы клеток и относительно больших округлых тел. Стимулом
изоморфного или гетероморфного размножения является зре-
лость выросших клеток.
Большинство бактерий размножается путем бинарного ='•
ния. При этом клетки удлиняются, формируются поперс
клеточная мембрана и новая клеточная стенка, растущие нс- ’Ь.
В этом процессе участвуют перегородочные мезосомы. Ну к. ч
удваивается перед делением и поровну распределяется в дь< -о-
разующиеся клетки. Таким образом, удвоение клеток при ра во-
жении в оптимальной питательной среде сопровождается
нием других учитываемых параметров это так назына ,е
сбалансированное размножение, типичное для бактерий в '<'•
ненциальной фазе. При таком размножении скорость при;' ;а
любого учитываемого компонента популяции одна и та же ’ ”'0
не наблюдается при несбалансироетнн'>м размножении в ст • °’
нарную фазу.
Деление клетки Е. coli при 37 °C совершается с определенным
постоянством, в среднем через 20 мин, когда завершается ранее
начавшаяся репликация (удвоение) хромосомы. Такая законо-
мерность в основном не зависит от скорости роста клетки. В свою
очередь полная репликация хромосомы занимает примерно
182
2
3
А
н
s
в
43. Размножение Е. coli:
г/—5); Б — (/—5); 1 — интерфазная клетка (нулевое время отсчета); 2 — удвоение
фсомы и места ее прикрепления к мембране; 3 — начало репликации новообразован-
;ромосомы; 4 — начало формирования межклеточной стенки; 5 — полная реплика-
Еромосомы и завершение формирования межклеточной стенки; 6 — образование
ювых клеток из одной родительской
1ин при той же температуре (рис. 43). Клетка не делится,
репликация хромосомы не закончилась.
'ост, развитие и деление (А) могут превышать по времени
репликации ДНК (Б), тогда А — Б=В, гли В — время,
1ение которого не наблюдается синтеза ДНК. Как видно из
43, А равно 45 мин, Б — 40 мин и, следовательно, В —
н; продолжительность собственно деления клетки охватывает
рвал в 20 мин, т. е. от 25 до 45 мин (рис. 43).
•епликация останавливается в точке терминации; заново син-
рованные молекулы ДНК расходятся или сегрегируют (от
^segregere — отделять, удалять) в образующиеся новые клет-
(Процесс такого деления клеток при размножении характерен
многих бактерий. Однако известны прокариотические микро-
|Иизмы, размножающиеся другими путями. Так, хламидии
К5удители трахомы, пситтакоза) — облигатные внутриклеточ-
! паразиты, близкие к грамотрицательным бактериям, прохо-
своеобразный цикл размножения и развития. Благодаря
цитозу хламидии, или элементарные тельца (0,3 мкм в диа-
«), оказываются внутри клеток организма-хозяина, где в
олях трансформируются в вегетирующие формы — ини-
гьные, или ретикулярные, тельца (0,5...1,5 мкм), потенциаль-
Пособные к делению. После ряда делений они превращаются
омежуточные формы, дающие начало очередному поколению
1Идий. Описанный цикл роста, размножения и развития длит-
среднем 40...48 ч.
Эблигатный паразитизм хламидий накладывает определенную
йфику на строение их клеток, потерявших способность синте-
Вать АТФ. Хламидии — это энергетические паразиты, поль-
1иеся энергией организма-хозяина.
Микоплазмы, по размерам близкие к крупным вирусам
меньшие репродуктивные единицы их имеют диаметр около
.250 нм), отличаются исключительным разнообразием мор-
»гических форм (полиморфизм). Однако наиболее частыми
«ами их являются яйцевидные (овоидные) или сферические,
оторых вначале возникает несколько нитевидных выростов
ой до 10 мкм. В этих нитях формируются затем сферические
зования, которые располагаются цепочкой и полностью вы-
ождаются после фрагментации истонченных нитей. Описан-
Цикл роста, размножения и развития характерен, например,
183
для Mycoplasma mycoides. У других (Mycoplasma pulmonis)
в нитях формируются сегменты, из которых концевой отделяется
в виде бактериальной почки (сравните размножение микоплазм
с размножением актиномицетов). Наблюдаются и другие процес-
сы размножения микоплазм — поперечным делением при синхро-
низации репликации ДНК нуклеоида с делением клетки; почко-
ванием крупных шаровидных тел, предварительно возникших из
клеток микоплазм.
Для актиномицетов характерны мицелиальные формы клеток,
что является отличительным признаком лишь данной группы
грамположительных прокариот (см. рис. 3). Актиномицеты под-
разделяют на проактиномицеты и эуактиномицеты. У первых ми-
целиальные формы ограничены и временны; размножаются они
фрагментацией мицелия на короткие, палочковидные клетки.
Проактиномицеты близки к бинарно делящимся полиморфным
коринебактериям. К проактиномицетам относят виды из родов
Actynomyces, Bifidobacterium, Geodermatophilus, Mycobacterium,
Nocardia.
Эуактиномицеты растут и развиваются исключительно в виде
мицелия, а размножаются с помощью одноклеточных спор, обра-
зующихся при сегментации или фрагментации спороносцев
(рис. 44), или заключенных в специальные образования — спо-
Рис. 44. Streptomyces
globisporus (фрагмен-
тация спороносцев,
X 22 000)
рангии (см. размножение низших грибов).
Лишь немногие представители из них, напри-
мер Thermoactinomyces sacchari, образуют
исключительно термоустойчивые эндоспоры
(выдерживают нагревание при 100 °C более
1 ч). Следовательно, актиномицеты как про-
кариоты по характеру размножения зани-
мают промежуточное положение между бак-
териями и настоящими эукариотическими
грибами.
Размножение грибов весьма разнообраз-
но по форме, что объясняется более выра-
женной дифференциацией их структур, чем,
например, у большинства бактерий. Так,
клетки дрожжей из рода Schizosaccharomy-
ces размножаются простым делением, а гри-
бы из рода Aspergillus имеют специальные
органы размножения. У некоторых низших
грибов вегетативные клетки представлены
в виде протоплазматических образований,
превращающихся на определенных стадиях
в споры — холокарпные организмы (от лат.
holo — врожденный, carpus — кисть). Низ-
шие и высшие грибы с дифференциро-
ванными вегетативными органами и орга-
нами размножения относят к эукарпным
организмам (от греч. еи— правильный)-
184
. этой связи различают вегетативное и репродукционное
ножение грибов. В первом случае грибы не имеют специаль-
органов и размножаются участками грибницы, почкованием,
зованием яйцевидных клеток (оидий), или хламидоспор,
тым делением. Оидии возникают вследствие распада гиф на
явные овальные клетки, из которых затем возникает мице-
и формируются грибницы. Репродукционное размножение
ов происходит с помощью спор, т. е. организм формирует
иальные органы размножения с многочисленными спо-
I. Репродукция осуществляется бесполым или половым
м.
1есполое репродукционное размножение сопровождается об-
ванием эндогенных или экзогенных спор. Эндоспоры типич-
например, для некоторых низших грибов (Mucor species,
rolegnia species и др.). И если какие-либо представители
I грибов приспособились обитать в водной среде (Saproleg-
, то они могут образовывать подвижные эндоспоры — зоос-
я, лишенные оболочки и являющиеся голыми комочками про-
1азмы с 1...2 жгутиками. Через какое-то время они теряют
тики, окружаются оболочкой и прорастают в мицелий. Вмес-
ища с эндоспорами соответственно называют спорангиями и
порангиями, а эндоспоры — спорангиоспорами и зооспорами
рис. 4).
Г наземных низших грибов спорангии образуются на споран-
ippax, или спорангиеносцах, которые имеют терминальные
тирения различной формы — колонки. Наружные споры, или
споры, образуются у представителей низших и высших гри-
их чаще называют конидиями, а несущие конидии гифы —
'.диофорами или конидиеносцами. Споры и конидии грибов
ма разнообразны по форме, цвету и размерам. Они могут
• гладкими, шероховатыми или бугристыми, прямыми, и изог-
1ми, одно-, двух- и многоклеточными, беспигментными и
шенными.
1а каждом конидиеносце может быть по одной или несколько
дий. Иногда конидии образуются одна за другой — в виде
чки. В тех случаях, когда верхняя конидия является самой
ой, а нижняя (базальная) — самой молодой, способ их фор-
>вания называют базипетальным (от лат. basis — основание,
-е — искать). Если верхняя конидия оказывается самой мо-
й, а базальная — самой старой, такой способ называют
петальным (от греч. ахрсоо — верхушка, крайний) или ба-
нальным (от лат. fugas — изгоняющий).
шорангиеносцы и конидиеносцы могут быть простыми и раз-
ленными. Различают следующие типы ветвлений: монопо-
ьный, симподиальный, дихотомический, мутовчатый. Кони-
осцы возникают на грибнице одиночно или группами. Харак-
группирования определяет те образования, которые принято
йвать коремиями, ложами, пикнидами (рис. 45). Коремия —
пучок тесно прижатых друг к другу конидиеносцев, несущих
185
\s
Рис. 45. Группы конидиеносцев:
I — коремия; 2 — ложе; 3 — пикнида
на своих вершинах конидии (у Penicillium species, Chaetomium
species, Stysanus species и др.).
Ложе — образование из плотно переплетенных гиф, развива-
ющееся внутри или на поверхности субстрата. На внутренней
поверхности его сосредоточиваются конидиеносцы с конидиями
(например, у Coryneum species).
Пикнида — округлой формы вместилище из плотно перепле-
тенных гиф. Внутри пикниды располагаются конидиеносцы с ко-
нидиями, которые при созревании выходят наружу через вывод-
ное отверстие (например, у Cytospora species).
У некоторых грибов концы конидиеносцев расширены в виде
булавы, шара или других форм (например, у аспергиллов). Такой
терминальный пузырек называют головкой (отсюда происходит
название «головчатая плесень»). Все аспергиллы обычно имеют
опорную клетку, из которой выходит конидиеносец; из терминаль-
ного пузырька формируются бутылочковидные стеригмы, верхуш-
ки которых являются ростковыми точками, где в последователь-
ном порядке образуются конидии. Их иногда называют катену-
лятными (от лат. catena — цепь), поскольку они остаются
соединенными в цепочки.
У пенициллов нет опорной вегетативной клетки, из которой
возникает конидиеносец, и нет терминального пузырька (голов-
ки), а путем ветвления формируется кисточка (отсюда происхо-
дит название «кистевик» или «леечная плесень»). Особенности
строения аспергиллов и пенициллов показаны на. рис. 4.
Отдельные представители грибов образуют более чем один
тип конидий на одной и той же грибнице (Fusarium species,
Microsporum species и др.). Одни конидии одноклеточные неболь-
ших размеров — микроконидии, другие многоклеточные боль-
шие — макроконидии. Конидии могут возникать непосредственно
на гифах без каких-либо поддерживающих структур — это сидя-
чие (сесильные, или латеральные) конидии. Они могут распола-
186
ся и на боковых веточках гиф (не на конидиеносцах) в виде
Ерадных гроздей (у Fusarium species) или одиночно (алей-
Й дерматофитов— возбудителей кожных заболеваний). Если
|дии образуются на конце коротких конидиофор (стебель-
li то их называют стебельчатыми или педицеллятными.
При репродукционном половом процессе, или митозе, у гри-
к существенные изменения претерпевает ядерный аппарат,
зечается слияние цитоплазмы и ядер (плазмогамия и карио-
вя). В общей схеме митоза выделяются две главные стадии:
Пирование митотического аппарата и перемещение хромосом
уносам с помощью этого аппарата. Митотический аппарат —
ароматические и ахроматические структуры клетки, подвер-
мой митотическому делению (от лат. chroma — цвет, краска,
отрицание «не»). Различают 4 фазы, связанные с измене-
и ядра: профазу, метафазу, анафазу и телофазу. Длитель-
рь митоза в зависимости от вида организма от 30 мин до 3 ч.
Каждую репродуктивную клетку грибов, несущую определен-
число хромосом, называют гаметой. Она соответствует яйце-
гке или сперматозоиду высших организмов. Гаметы морфоло-
Вски трудо дифференцировать по половому признаку, поэтому
Обозначают как (+) мужская и (---) — женская клетки
типы спаривания, т. е. они гетероталличны. Число хромосом
»Ибов весьма различно, например: у сумчатых - 4, у ржав-
ных — 8 (реже 6, 10). Характерное для вида число хромосом,
|ржащихся в гамете, называют гаплоидным (от греч.
|»<оа — одинарный) и его обозначают как п или х (или соот-
|гвенно 1 п и 1х). При слиянии однополых гамет (изогамия)
|зуется гомозиготный диплоид, при слиянии гетерополых га-
|(гетерогамия) — гетерозиготный диплоид. Диплоиды обозна-
(T как 2п или 2х. Большинство пивных и хлебопекарных дрож-
^являются гетерозиготными динлоидами, Такие же дрожжи-
^оиды, образующие. 4 половые споры (аскоспоры), обычно
|ируют из природных субстратов.
гомозиготные диплоиды более стабильны, чем гетерозигот-
(„так как они обычно не образуют аскоспор и размножаются
тативно до тех пор, пока не произойдет слияния гаплоидных
хж, возникших на каком-то этапе их жизненного цикла.
ГДа гаплоидные гаметы становятся стабильными и тогда раз-
Нощиеся из них организмы являются аспорогенными (многие
^матики такие формы относят в группу несовершенных гри-
|так, митоз характерен для диплоидных клеток, т. е. клеток
|йным набором хромосом. Поскольку в процессе митоза про-
Щит точное копирование наследственного материала, переда-
тося от родительского организма потомству, постольку для
1зования гамет с гаплоидным набором хромосом необходим
юй процесс, в ходе которого восстанавливается исходное
to хромосом п. Этот процесс называется мейозом или редук-
шым делением. Сущность мейоза состоит в том, что ядро
187
праевает два последовательных деления, сопровождающихся
лидним делением хромосом.
щесс мейоза подразделяют на две фазы: первое деление
ме1 и второе деление мейоза. Итогом первого деления являет-
ся 1зование гаплоидного набора хромосом в ядре, где каж-
дыен этого одинарного набора представлен парой сестрин-
скфомосом; итогом второго — формирование и созревание
га» гаплоидным набором хромосом, когда разделяется общая
цемера сестринских хромосом и эти сестринские хромосомы
оттаются к противоположным полюсам с последующим обра-
зовм мужских или женских половых клеток. В первом деле-
нийоза выделяют: профазу, включающую лептонему, зигоне-
му,инему и диплонему (от лат. leptos — узкий, zygote — сое-
дини в пару, pachis — толстый, diplos — двойной, пета —
нити, как при митозе, метафазу, анафазу и телофазу. После
завения первого деления мейоза наступает короткая интер-
фазосле которой происходит второе деление мейоза, во мно-
гоядное с митотическим делением диплоидных клеток. При
пер делении мейоза могут наблюдаться внутрихромосомные
кохации, выявляемые по образованию так называемых хиазм
(щрме греческой буквы (хи) %). Обмен генами, связанный
с пением хиазм, называют кроссинговером.
овая репродукция у разных грибов определенно имеет
свообенности, но в большинстве случаев грибам свойствен-
на .ксуальность, т. е. они являются двудомными видами, спо-
соба в зависимости от условий образовывать гаметы, функ-
цисующие то как мужские, то как женские образования.
Мий вида может относиться одновременно к плюс- и минус-
варгам, способным давать зиготы. Установлено, что путем
сек|и половых гормонов (гамонов) у ряда грибов осуществляет-
ся роль и направленность сексуальных процессов. Так, грибы
из । Allomyces образуют сиренин, индуцирующий тактильное
двиие (от лат. tactilis — осязательный) и относящийся к раз-
рядародышевых аттрактантов (от лат. attractio — притяже-
ниеготение) — эротактинам. Антеридиол из Achlya species
и поровые кислоты из Mucor species и Blakeslea species,
индоующие формирование и дифференциацию сексуальных
стрр, относят к группе эрогенов:
сиренин
188
'соон
эиспоровая кислота А (С18Н2бО4)
он
о
СООН
триспоровая кислота В (С18Н24О4)
н
ООН
триспоровая кислота С (С]8Н2бО4)
юны, контролирующие хемотаксис, в частности сиренин, от-
г к эротропинам. \
азвитие плодовых тел у базидиальных грибов находится
:е под контролем специальных гормонов. Имеются данные о
что некоторые метаболиты, образуемые, например, актино-
ггами, могут влиять на проявление половой репродукции у
атых грибов. С этим, возможно, связаны определенные
моотношения микроорганизмов в природных условиях,
значительное число грибов размножается лишь бесполым ре-
укционным путем с помощью конидий или вегетативно,
юму их выделили в искусственную группу Deuteromycetes,
Fungi imperfedl — несовершенные грибы. У ряда грибов с
[альным половым процессом размножения, а также у многих
еромицетов имеется так называемый парасексуальный цикл,
а рекомбинация осуществляется в митозе и не связана с
дотворением и мейозом (рис. 46), но связана с возникнове-
Рис. 46. Парасексуальный цикл развития у Penicillium
chrysogenum
189
нием анастамозов между клетками, через которые происходит
обмен ядрами. Образующийся мицелий с гаплоидными ядрами
разных генотипов (гетерокарион) распадается на два исходных
мутантных генома, дающих одноядерные конидии. В гетерокарио-
тическом мицелии возможна также диплоидизация ядер с после-
дующим развитием диплоидных гетерозиготных гиф. При пара-
сексуальном цикле постепенно уменьшается число хромо-
сом в ряду митотических делений вплоть до их гаплоидного
числа.
Из приведенных данных очевидно, что в процессе размноже-
ния грибы образуют самые различные споры, возникающие в
результате бесполого и полового процессов размножения. Так
называемые бесполые споры образуются в результате митоза,
они могут быть гомокариотическими или гетерокариотическими.
Половые споры возникают в результате мейоза, и на их основе
создаются новые генетические рекомбинации. С другой стороны,
споры иногда обозначают как мемноспоры (от лат. memnonius —
коричнево-черный) и ксеноспоры (от греч. grjvcDcr — чужой).
Мемноспоры не перемещаются с места своего возникновения и
выполняют функции сохранения видов в неблагоприятных усло-
виях (при высушивании, голодании). К мемноспорам относятся
хламидоспоры, зигоспоры. Ксеноспоры отделяются от родитель-
ского мицелия и активным или пассивным путями распространя-
ются во вне на большие и малые расстояния, способствуя
экспансии вида во внешней среде (конидии, многие аско-
споры).
Одноклеточные, нефотосинтезирующие, подвижные протисты
(Protozoa) размножаются, как правило, делением: продольным
(жгутиковые — Mastigophora), бинарным (амебоидные — Rhizo-
poda), множественным (споровики — Sporozoa), поперечным
(инфузории — Ciliata). Некоторые протисты размножаются и по-
ловым путем, например, инфузория Tetrahymena pyriformis имеет
клетки двух различных типов спаривания, способных конъюги-
ровать, претерпевать мейоз и митоз с последующим образова-
нием двух дочерних клеток, содержащих по одному макро- и
одному микронуклеусу.
Промежуточной группой между простейшими и настоя' чи
грибами являются миксомицеты, или грибы-слизевики. ГЬ ль
тивной структурой у них выступает многоядерный недифу- н-
цированный плазмодий, из которого в определенных у< • ях
развивается плодовое тело. В этот период одноядерные хи
плазмодия покрываются клеточной стенкой и трансформиг .-я
в одноядерные споры, находящиеся на плодовом теле. Отт !>,ь
от плодового тела, споры прорастают, формируя одножгутш
амебоидные клетки-гаметы. Эти последние сливаются, оО.
двухжгутиковые зиготы, трансформирующиеся после утери
тиков в новый плазмодий. Диплоидные ядра растущего пл °’
дия претерпевают мейоз перед самым образованием ее в
плодовом теле.
190
ЭРФОГЕпЕЗ И ДИФФЕРЕНЦИРОВКА У ПРОКАРИОТ
;ариот
сроорганизмов (как и у других организмов) имеют место
.еленные жизненные циклы. Причем у высокоорганизован-
видов эти циклы включают чередование мейоза и митоза,
гаплофазы и диплофазы. Иначе говоря, жизненный цикл —
!сс морфологической дифференциации клеток от любой ста-
одительского организма до такой же стадии дочернего орга-
1. Жизненные циклы неравнозначны, схематично они изоб-
ды на рис. 47.
•е организмы имеют форму, которую они проявляют в про-
бстве в данный конкретный период времени. Это и есть так
даемый морфогенез, определяемый генотипом. Под контро-
Генотипа находятся фенотипические признаки вида. Фено-
- это все параметры и характеристики организма, которые
одаются в различных условиях существования вида и кото-
«огут быть измерены или учтены с использованием различ-
иетодов, включая оценку химического состава всех клеточ-
«жпонентов. Генотип постоянен в течение жизненного цик-
фенотип варьирует. До тех пор, пока окружающие условия
отся соответствующими для роста и развития микроорганиз-
шд всегда проявляет присущие ему характерную форму и
цию. Например, грибы из родов Aspergillus и Rhizopus
зуют бесполые споры на питательном агаре с картофельным
(актом, но морфогенез их совершенно и постоянно различен,
ичие морфологических структур, присущих одному и тому
рганизму, свидетельствует об их дифференцировке. Очевид-
то споры у бактерий или сумки у дрожжей отличаются по
е от нормальных вегетативных клеток. Эта дифференциация
!нная, присущая клеткам, находя-
я в различных стадиях их жизнен-
(иклов. Другую дифференцировку
р определить как стабильное раз-
организма с измененной струк-
: и функцией (клетки эукариот
шлофазы до гаплофазы). Прехо-
е нестабильные изменения явля-
результатом адаптации или при-
бления организма к изменившим-
ловиям существования. Следова-
Ь, главный критерий дифферен-
»ки — необратимость процесса.
Ьрфогенез и дифференцировка
ны с контролируемыми изменения-
мического состава и функций кле-
! прежде всего с изменениями,
ицимися ферментов, находящихся
рямым генетическим контролем
му центральной проблемой диф-
5
Рнс. 47. Схема жизненных
циклов у микроорганизмов:
1 — бесполый; 2 — гаплоидный;
3 — гаплоидно-дикариотический;
4 — гаплоидно-диплоидный; 5 —
диплоидный; М — мейоз; жир-
ные линии — диплоидная фаза,
спаренные тонкие линии — ди-
кариотическая фаза
191
ференцировки является изучение генетических и биохимических
механизмов, имеющих непосредственное отношение к регуляции
дифференциальной генной экспрессии и биосинтезу информацион-
ных макромолекул (ДНК, РНК, белки). С дифференцировкой свя-
зана продукция вторичных метаболитов (см. гл. 5), многие из ко-
торых имеют важное народнохозяйственное значение (см. гл. 6).
Дифференцировку условно подразделяют на морфологиче-
скую и биохимическую, из них морфологическую уловить труднее
(например, у многих неспорообразующих бактерий). Прокарио-
тические микроорганизмы обладают слабо выраженной морфо-
логической дифференцировкой. Так, у Е. coli при размножении
амитотическим делением вновь образующиеся клетки морфологи-
чески идентичны родительским клеткам. Если в те же клетки вне-
сена R-плазмида, то они биохимически (по устойчивости к ряду
химиотерапевтических веществ) отличаются от клеток Е. coli,
не содержащих данной плазмиды. Морфогенез у спорообра-
зующих бактерий-бацилл более выражен в период спорообра-
зования, когда вегетативная клетка приобретает иную форму.
Биохимическое выражение указанной дифференцировки заклю-
чается в таких характеристиках, как низкое содержание пар ГЦ,
особенно у клостридий (22...28 %); отсутствием л/езо-диаминопи-
мелиновой кислоты (л/езо-ДАП) в муреине большинства видов
из родов Bacillus и Clostridium; наличием специфического пеп-
тидогликана в кортексе спор у большинства спорообразующих
бактерий:
аланиновая субъединица тетрапептадная субъединица
I------------------II----------------------1
фрагмент пептидогликана в кортексе спор бацилл
Многие аэробные и анаэробные бациллы в связи со споруля-
цией синтезируют особые пептиды с М ~ 1400 Да, обладающие
противомикробным действием (антибиотики циклопептиды: ба-
цитрацины, грамицидины, микобациллин, полимиксины, тироци-
дины), и накапливают до 10...15 % от сухого веса дипиколината
кальция; культура Вас. thuringiensis образует по соседству со
спорой бипирамидальный белковый кристалл, токсичный для
гусениц различных чешуекрылых насекомых и т. д.
Дифференцировка спорообразующих бактерий на примере
Вас. cereus показана на рис. 48, на котором видны все этапы
формирования, высвобождения и прорастания спор по схеме:
вегетативная клетка--> спора-* вегетативная клетка.
192
48. Морфогенез и дифференцировка Bacillus cereus:
(ёгетативиая клетка с двумя нуклеоидами; 2—конденсация нуклеоидов в палочко-
гю структуру; 3 — начало образования поперечной перегородки; 4 — завершение
юваиия поперечной перегородки и отграничение споры от вегетативной клетки; 5 —
(Ование новой оболочки вокруг новой проспоры; 6—завершение образования обо-
I вокруг проспоры; 7 — созревание споры; 8 — высвобождение зрелой споры; 9 —
астаиие споры; 10 — образование вегетативной клетки
Биосинтез дипиколиновой кислоты связан с биосинтезом трео-
ia, изолейцина, метионина, лизина или лезо-ДАП:
о—с
с=о
о
о
дипиколинат
кальция
НООС— СН2—СН—СООН
I—СН—СООН
он
NH2
гомосерин
NH2
аспарагиновая кислота
н nh2
полуальдегид Asp
О—С—СН2—СН—СООН
СН3—СО—СООН
пируват
НООС
ООН
дигидродипиколиновая
кислота
Н |С—СН—СН—СООН
ОН NH;
треонин
—СН2—СН—СН—СО0Й
СН3 NH2
изолейцин
HS—СН2—CH(NH2)—СООН
метионин
ноос—нс:—(сн2)3—сн—соон
nh2 nh2
.«я»-dap
Н2С—(СН2)3— СН—СООН
NH-
NH2
лизин
(только
в спорах)
дипиколиновая
кислота
Для вида Вас. subtilis построена генетическая карта сцепле-
I, на которой выделены гены, определяющие споруляцию. Эти
1617
193
гены располагаются группами, захватывающими примерно пять
сегментов хромосомы.
У эукариотических микроорганизмов морфогенез и диффш • ;t.
цировка более выражены. Важными вегетативными структур,,
например у грибов, являются нити мицелия или их эквива.-ц нц
у одноклеточных форм (дрожжи и хитридиевые грибы). Фп ,,,а
большинства грибных клеток обеспечивается ригидной клетс, !<>й
стенкой и изучение ее структуры может дать ключ к более по. ио.
му пониманию морфогенеза соответствующего вида и даже ж L (.
сказать направленность проявлений его в конкретных уело; ;,чх
существования данного организма. Лишь амебоидные или ж- , <ь
ковые стадии при развитии некоторых миксомицетов и ф;жо-
мицетов существуют без ригидной клеточной стенки в определен,
ные периоды жизненных циклов. Клеточные стеНки грибов
строятся из переплетенных микрофибрилл с погруженным в них
(или «зацементированным») аморфным матриксным веществом.
Скелетная часть клеточных стенок у нитчатых грибов состоит из
хитина (хитозана) или целлюлозы, у дрожжей она формирует,-я
из друитх глюканов. Вещества матрикса представлены главным
образом полисахаридами тина галактана, глюкана, маннан.- и
гетерополисахаридов, конъюгированными с белками. На подж а-
хариды приходится 80...90 % массы сухих грибных клето-шых
стенок, структурными единицами в них выступают гексозы и ;• ш-
ногексозы. 1екеуроновые кислоты, пентозы и метилпени-.ы.
Выявлена определенная корелляция химического состава кле;"Ч-
ных стенок грибов и их систематического положения (табл. ')
Состав и реконструкцию клеточных стенок удобно изучать
на клетках грибов, относительно легко трансформируемых в
протопласты. Регенерация протопластов в исходные нитчатые
формы создает исключительные возможности для изучения син-
теза компонентов клеточных стенок и факторов контроля диффе-
ренциации клеточных форм. В любой нити гриба на всем ее про-
тяжении достаточно легко отдифференцировать три зоны: апи-
кальную (или верхушечную), субапикальную и вакуолизирован-
ную. В апикальной зоне скапливается большое количество вези-
кул почти при полном вытеснении рибосом и каких-либо других
структурных элементов в цитоплазме клетки. В отдельных случаях
удается наблюдать слияние везикулярных мембран с цитоплаз-
матической мембраной. В субапикальной зоне концентрируются
ядра, рибосомы, митохондрии, эндоплазматический ретикулум,
микротельца, микротубули и т. д.
Субапикальная зона переходит в зону вакуализации. Послед-
няя тем выраженнее, чем дальше расположена от верхушки.
Параллельно с возрастанием вакуолизации увеличивается содер-
жание липидов в этой же зоне. Апикальные везикулы участвуют
в синтезе клеточной стенки. Они образуются в субапикальной
зоне и перемещаются к верхушке, где секретируют свое содержи-
мое или сливаются с мембраной. В них также могут быть экзо-
ферменты, которые выходят на уровне гифальной верхушки. Ве-
194
,ица 22. Таксономическое положение некоторых грибов в связи
щеским составом их клеточных стенок
Неводные компоненты ; клеточных стенок Род грибов Таксономическая группа
1ГИН — р-глюкан Allomyces, Blastocladiella Chytridiomycetes
Aspergillus, Histoplasma, Neurospora Euascomycetes
Fames, Polyporus, Schi- zophyllum Homobasdiomycetes
ялюлоза — хитин Rhizidiomyces Hyphochytridiomy- cetes
^люлоза — р-гликан Phytophthora, Pythium, Saprolegnia Oomycetes
ггнн — хитозан Mucor, Phycomyces, Zy- gorhynchus Zygomycetes
|ниан-р-глюкан Е Candida, Saccharamyces Ascomycetes
Ьииан — хитин Rhodotorula, Sporobolo- myces Heterobasidiomyce- tes
димеры, содержащие ^тозамин — галактозу Amoebidium Trichomyces
^люлоза — гликоген Dictyostelium, Polysphon- dylium Acrasiales
ли, возникающие из эндоплазматического ретикулума, сли-
лся и образуют цистерну внутренней (проксимальной) части
!иосомы; содержимое цистерны и мембраны последовательно
[Сформируется, перемещаясь к наружной (дистальной) части
мосомы в связи с продолжающимся образованием новых
•ерн. Здесь цистерны превращаются в пузырьки, а затем —
Креторные везикулы, которые мигрируют к верхушке гифы,
эторые везикулы увеличиваются в размере, другие сливаются
ду собой и таким образом формируются большие секретор-
везикулы. Отдельные везикулы непосредственно достигают
Очной мембраны и сливаются с ней. Слившиеся с мембраной
кулы освобождают свое содержимое в периплазматическую
гсть апикальной части гифы, где осуществляется синтез
Очной стенки и сбалансированный лизис скелетных микро-
)илл.
г всех высших грибов в апикальных частях мицелиальных
й имеется так называемое верхушечное тельце сферической
ы, которое исчезает при остановке роста гифы. Положение
la на верхушке служит направляющим вектором простран-
ного роста гифы. Например, эксцентрическая позиция его
*вает на то, что верхушка изогнется в ту же сторону.
195
Хотя гифы всех грибов растут за счет апикальных частей,
утолщение клеточных стенок может наблюдаться и около верху-
шек, а иногда — ив дистальных частях, например при образо-
вании хламидоспор. Тем не менее если ограниченный рост ми-
целия в длину потенциально возможен, то увеличение ширины
его весьма лимитировано. При поверхностном культивировании
грибов в культуре образуются: субстратный (поверхностные и
погруженные в среду гифы) и воздушный мицелий, имеющий
свое морфологическое и физиологическое выражение.
Визуально наблюдаемое скопление гиф представляется в виде
паутинного налета, войлока, пленок, шнуров. Если множествен-
ные гифы не срастаются своими стенками и остаются свободны-
ми, то грибницу называют плектенхимной тканью. Напротив,
если гифы срастаются и теряют свой первоначальный вид, то
образованную ткань называют псевдопаренхимой или ложной
паренхимой. Она находится в покровах плодовых тел и выпол-
няет защитную функцию.
Мицелий ряда грибов претерпевает своеобразные видоизме-
нения. Морфогенез возникающих форм и функция их специфич-
ны (см. рис. 6).
Фенотипическая двойственность клеточных форм у ряда гри-
бов проявляется также в виде ранее упоминавшегося плеснеш.-
дрожжевого диморфизма. Подобный диморфизм иногда называ-
ют примитивным морфогенезом. Доказано, что существуют три
главные группы диморфных грибов: температурозависимые (на-
пример, Blastomyces dermatitidis), зависимые от температуры
и питательных веществ (Histoplasma capsulatum), зависимые
лишь от питательных веществ (Candida albicans).
Многообразны проявления морфогенеза и дифференцировки
вегетативных форм у дрожжевых организмов. Для сравнения
можно указать на дрожжи из родов Candida, Endomycopsis, Sac-
charomyces, Schizosaccharomyces, Cryptococcus, Sporidiobolus,
Trigonopsis и др. У филаментирующих дрожжей преобладание
мицелиальных (псевдомицелиальных) форм или дрожжевых во
многом зависит от окружающих условий (питательная среда,
аэрация). При ограничении азотного питания в аэробной культу-
ре клетки S. cerevisiae приобретают удлиненную форму; при
ограничении доступа кислорода и глюкозы С. albicans образует
больше псевдомицелия и хламидоспор; удлинение клеток дрож-
жей Hansenula schnegii стимулируется триптофаном. Очевидно,
многие грибы синтезируют из триптофана индолуксусную кисло-
ту, которая индуцирует удлинение клеток, например у S. cere-
visiae:
триптофан
индолуксусная кислота
196
Змесь метионина и холина стимулирует образование треу-
>ной формы клеток Т rigonopsis variabilis.
вегетативный рост большинства эукариотических микроорга-
мов сопровождается митотическим делением ядер с последу-
ем образованием новых клеток. Этот митотический цикл под-
Деляют на четыре фазы: Gi — фаза роста в период до синтеза
К (от англ, gap — пробел); S — фаза синтеза ДНК (от англ,
thesis — синтез); G2 — вторая фаза роста в период после син-
а ДНК; М — фаза митоза (от англ, mytosis — митоз). У Schi-
accharomyces pombe на Gi-, S- и Ga-фазы приходится 150 мин.
Цикл деления ДНК и цикл роста — независимо контролируе-
; процессы в клетках; этим обеспечивается потенциальная
можность функционирования механизма контроля за разме-
1И клеток. В клеточном цикле происходит последовательное
азование ферментов и, следовательно, состав их непрерывно
1Яется, что отражается на качестве и уровне контроля мета-
ических процессов, протекающих в клетке. Каждому классу
,бов присущ свой жизненный цикл. Например, слизистые гри-
Dictyostelium mucoroides и Polysphondylium violaceum, вы-
яющие разные аттрактанты (от лат. attractio—притяжение,
•отение), имеют сходную морфологию и жизненные циклы,
итая в виде отдельных клеток, любой из них поглощает все
{терии в сфере своего обитания, после чего начинается фаза
)щественного» развития. Клетки гриба собираются в одной
<ке и сливаются (сотнями и тысячами) в единую слизистую
огоклеточную массу. Затем наступает процесс дифференциров-
складывающийся из образования выпячивания на фронталь--
1 части слизистой массы — это начало закладки и формиро-
1ия стебля, растущего верхушечной частью; клетки всползают
стеблю вверх, накапливаются там, теряя подвижность. Так
>азуется плодовое тело, состоящее из разветвленного или не-
шетвленного стебля (остатки целлюлозных оболочек от отмер-
..х клеток) с шаровидной головкой — спорангием со спорами
нкапсулированные амебы). Как только споры рассеиваются
юдой или при механическом соприкосновении с другими объек-
ми, из них после разрыва оболочек высвобождаются амебы, и
ui начинается в той же последовательности. Положительный
ютаксис клеток у этих двух видов грибов индуцируется разны-
веществами — акразинами*. У Dictyostelium акразином вы-
дает цАМФ. В фазу агрегации миксамебы секретируют цАМФ
(мерно в 100 раз больше, чем в предшествующую фазу изоли-
анного существования. У Р. violaceum акразин — аттрактант
дставляет собой дипептид глорин**, состоящий из производ-
: глутаминовой кислоты и орнитина:
* Acrasia — имя колдуньи из сказки Спенсера «Королева фей», которая
лекала к себе людей и превращала их в животных.
** Название глорин происходит от первоначальных двух букв: Глу и Орн с
влением окончания «ин».
197
остаток этилового эфира
СН3—СН2—О
'Ч’С=О g лактам
Н3С—(СН2)2—CO-j-NH—СН—(СН2)2—CQ— NH—СН—(СН2)2—СН2
остаток пропионовой остаток глютаминовой остаток орнитина
кислоты (пропионил) кислоты
Глорин
Глорин оказался еще более активным аттрактантом, чем цАМФ,
для Dyctyostelium.
Хитридиальные грибы образуют подвижные клетки (зооспо-
ры) с одним задним бичевидным жгутиком. Они дают начало
гаметоталлусу. Причем у некоторых видов, например у Allomyces
macrogynus, образующиеся женские гаметы секретируют зароды-
шевый аттрактант — сиренин.
На примере Mucor mucedo (рис. 49) показаны морфогенез
и дифференцировка клеток в зависимости от стадии жизненного
цикла. В жизненном цикле М. mucedo дважды отмечается обра-
зование спорангиев. Однако в гаплофазе спорангии содержат
однотипные спорангиоспоры, тогда как в диплофазе спорангии
одновременно содержат ( + ) и (—) спорангиоспоры. Гаплоидные
споры при прорастании образуют мицелий, в клеточной стенке
которого синтезируется хитин (у некоторых видов — хитозан).
В клеточных стенках гаплоидных спорангиоспор откладываются
глюканы и пигмент меланин. При взаимодействии ( + ) и (—)
нитей мицелия выделяются триспоровые кислоты, возрастает син-
тез каротинов, а под влиянием зиготропных гормонов происходит
слияние зигофор. На стадии формирования гаметангия наступает
лизис стенки и слияние содержимого ранее отграниченных клеток
гаметангия (плазмогамия). Каротины, образующиеся в ходе по-
ловой репродукции, полимеризуются до спорополленина (оксиге-
нированный полимер p-каротина), входящего вместе с мелани-
ном в состав высокоустойчивой стенки зигоспор.
Из рассмотренных жизненных циклов у низших грибов видно,
что в проявлении морфогенеза и дифференцировки клеток су-
щественную роль играют половые гормоны (табл. 23). Очевидно,
что в природных условиях гормональная активность грибов имеет
важное значение во взаимоотношениях культур не только одного,
но и разных родов и видов, особенно с учетом тех экологических
ниш, в которых ассоциативные взаимоотношения микроорганиз-
мов установились более прочно. Жизненные циклы сумчатых и
базидиальных грибов также характеризуются различной диффе-
ренциацией клеток на разных стадиях их развития. Эти циклы
более простые у ряда дрожжевых организмов и более сложные —
у нитчатых форм. В период спорообразования в клетке снижает-
ся уровень биосинтеза белка и РНК и возрастает содержание
ДНК, число ядер после первой метафазы и число зрелых сумок.
198
49. Жизненный цикл Mucor mucedo'.
гаплоидные спорангиоспоры; 2 — вегетативный мицелий; 3 — спорангии, обра-
щиеся на вегетативном однодомном мицелии; 4 — образование зигофор в резуль-
> взаимодействия ( + ) и (—) нитей мицелия; 5 — слияние зигофор; 6 — формиро-
не парагаметангия; 7 — образование отграниченного гаметангия с суспензорами;
молодая зигоспора; 9— зрелая зигоспора; 10 — прорастание зигоспоры
екоторых дрожжевых организмов гаплоидные клетки могут
5илизироваться и размножаться впоследствии бесполым пу-
„ (Candida, Zygosaccharomyces и др.).
^Многоступенчатую дифференцировку проходит в своем жизнен-
цикле сумчатый гетероталличный гриб — хлебная плесень
trospora crassa, образующий плодовые тела — перитеции с
ками, в которых содержится по 8 аскоспор. Формирование
199
итециев связано с биогенезом меланинов и возрастанием
вня тирозиназы. При 35 °C на минимальных средах задер-
гаются процессы синтеза меланина и образования перитециев;
Алогичный эффект проявляют высокие концентрации азота,
акже ингибиторы тирозиназы.
На примере гомобазидиальных агариковых грибов показано,
образование плодовых тел у Agaricus bisporus стимулируют
юторые грамотрицательные бактерии из рода Pseudomonas,
фимер Ps. putida, и родственные ей виды. Экстракты из этих
стерий, являющиеся стимуляторами спорообразования, отно-
>ся к группе железосодержащих хелатирующих агентов.
(.Таким образом, процессы морфогенеза и дифференцировки
йкариотических и эукариотических микроорганизмов прослеже-
। у многих видов и штаммов, однако биохимическая сущность
остается далеко не изученной. Несомненно лишь одно, что для
фференциального выражения генов необходимо не только их
исутствие в геноме клетки, но и создание соответствующих
повий, в которых они проявят свою активность.
Вопросы
h
1. Какова химическая структура микробных пигментов виолоксаитина
' и фрагментов молекул меланина? Какой цвет они придают колониям
| микроорганизмов на питательных средах?
2. Что такое синтрофия и в чем она проявляется у микроорганизмов?
? 3. Что обозначает термин «энергетический коэффициент» и как его оп-
ределяют?
fc 4. Что такое популяция микроорганизмов и каковы ее показатели?
Г 5. Какими методами добиваются синхронизации роста культур микро-
it . организмов и в каких случаях необходима синхронизация?
р 6. На какие фазы подразделяют размножение (рост) одноклеточных
с микроорганизмов и как характеризуются эти фазы?
if 7. Каким образом определяют удельную скорость размножения одно-
з клеточных микроорганизмов?
I' 8. Каким образом определяют рост и развитие мицелиальных грибов?
I 9. Какие принципиальные различия между прокариотами и эукариота-
ми по характеру размножения?
10. Какую химическую структуру имеют половые гормоны низших гри-
F бов — антеридиол, сиренин и триспоровые кислоты? На какие группы их
* подразделяют?
11. Что такое мемноспоры и ксеноспоры и каковы их функции?
/ 12. Что такое жизненный цикл микроорганизма (на примере различ-
J них классов грибов)?
й 13. В чем сущность морфогенеза и дифференцировки микроорганиз-
S мов?
14. Какую химическую структуру имеет пептидогликан в кортексе
' спор у бацилл?
15. Каковы химическая структура дипиколипата кальция, его рать в
дифференцировке бацилл и путь биосинтеза в клетках?
16. Что такое диморфизм грибов и какие группы диморфных грибов
известны?
17. Какие известны фазы роста эукариотических микроорганизмов в
1. митотическом цикле? Средняя продолжительность их.
с 18. Каковы химическая структура и функция акразинов у Dictiostelium
I* muc'oroiiies и Polijsphoudylum violaceum?
I
t 201
200
Глава 5
Ш.мгн HI ОМ'С'.ТВ V МИКРООРГАНИЗМОВ
Все реакции жизнеобеспечения, происходящие в микробной
клетке и катализируемые ферментами, составляют обмен ве-
ществ, или метаболизм. Промежуточные или конечные продукты,
образующиеся в соответствующей последовательности фермента-
тивных реакций, в результате которой разрушается или синте-
зируется ковалентно связанный скелет конкретной биомолекулы,
называют метаболитами.
Существуют понятия о реакциях первичного и вторичного об-
мена или о первичном и вторичном метаболизме. Реакции пер-
вичного обмена сходны для всех живых организмов — это образо-
вание и расщепление нуклеиновых кислот и оснований, белков
и их предшественников, а также большинства углеводов, липи-
дов, некоторых карбоновых кислот. К реакциям вторичного обме-
на относят те из них, в результате которых образуются несущест-
венные для микроорганизма вещества в период так называемой
идиофазы; образование этих веществ (антибиотики, гибберелли-
ны, триспоровые кислоты, алкалоиды и др.) свойственно лишь
некоторым видам.
Реакции первичного и вторичного обмена трудно дифферен-
цируемы, даваемые им определения неточны; расчленение мета-
болизма на первичный и вторичный обмен условно, но в учебных
целях имеет смысл сохранить понятия о первичных и вторичных
метаболитах. Тогда, принимая во внимание последовательность
развивающихся событий в живой клетке, первичными метаболи-
тами являются белки, синтезирующиеся по схеме: ДНК -► РНК ~>
-+ Белок. Включение ферментов — первичных метаболитов —
в последующие реакции обмена сопровождается образованием
вторичных метаболитов. Это можно представить в виде схемы:
Простые питательные вещества, поступающие извне — преметаболиты
(углекислота, аммоний, фосфаты, сульфаты, ионы металлов и др.*)
Эндогенные промежуточные продукты (интермедиаты) — прометаболиты
(простые сахара, аминокислоты, нуклеиновые основания и др.)
ДНК= РНК
I
Эндогенные первичные метаболиты
а) ферментные белки---------------------------->
б) неферментные белки
Вторичные метаболиты, эндо-
генной и(или) экзогенной лока-
лизации (алкалоиды, антибиоти-
ки, липиды, полисахариды, гор-
моны и др.)
* Например, для гетеротрофов.
202
шной схеме образование нуклеиновых кислот как бы изоли-
1но от других процессов. В этом видна условность понятий
!ичные и вторичные метаболиты, так как синтез ДНК и РНК
(лизируется ферментами, т. е. первичными метаболитами. Вот
му при рассмотрении образования обоих типов метаболитов
сообразно выделять в особую группу так называемые инфор-
ионные молекулы (нуклеиновые кислоты и специальные бел-
особенности которых определяются непосредственно генети-
ки материалом клетки. В этом случае приведенную выше
iy можно изобразить следующим образом:
Преметаболиты
Интермедиаты, или про-
метаболиты-*--------
.нформационные молекулы
рвичные метаболиты
а) ферментные белки
б) неферментные белки
Вторичные метаболиты
Становится очевидным, что синтез информационных молекул
же катализируется первичными метаболитами, или фермен-
[и. Более того, распад (деградация, разложение) вторичных
аболитов до исходных продуктов возможен под действием
>ментов одного и того же микроорганизма. Наконец, образо-
ие прометаболитов, т. е. интермедиатов, или промежуточных
дуктов, осуществляется при действии первичных метаболитов-
•ментов. Поэтому можно заключить, что биосинтез вторичных
аболитов не кодируется ДНК хромосом или плазмид.
Процесс выращивания того или иного продуцента подразде-
эт на трофофазу (от греч. трохр») - питание, пища) и илио-
>у (от греч. i6io>o - свой, специфичный). В период трофофазы
рость роста организма высокая, а продукция вторичных
'аболитов низкая. Напротив, в период ндиофазы скорость
:та микроба низкая, а продукция вторичных метаболитов вы-
:ая.
Наиболее важной регуляторной особенностью вторичного ме-
тодизма клеток является его компартментализация (табл. 21).
Вторичные метаболиты классифицируются по группам на ос-
днии механизма их синтеза в клетках. Они являются произ-
,ными ацетил-КоА и жирных кислот, мевалоновой кислоты,
Инокислот, углеводов, шикимовой или ароматических аминокис-
203
лот, интермедиатами цикла трикарбоновых кислот, продуктами
различных других метаболических путей.
НОж /СН;
г'
сн2он соон
мевалоновая кислота
Таблица 24. Вторичные соединения и мембраносвязанные
первичные метаболиты
Вторичный метаболит Первичный метаболит (фермент)
название локализация в клетке
Целлюлоза р-С1 -> С4-Глюкансин- тетаза (глюкансинтаза) Плазмалемма, эндоплазмати- ческий ретикулум и его произ- водные, периплазматическое пространство
Хитин Хитинсинтетаза Хитосомы, плазмалемма
Полисахариды матрикса клеточ- ной стенки 0-С1 -> СЗ-Глюкансинте- таза и другие синтетазы углеводов Плазмалемма и ее произ- водные
Каротиноиды Ферменты биосинтеза ка- ротиноидов Хлоропласты, эндоплазмати- ческий ретикулум и другие структуры
Как было сказано, в период трофофазы микроорганизмы
активно растут (имея в виду их рост, развитие и размножение).
В этот период питательные вещества среды активно транспор-
тируются в клетку и осуществляется конструктивный и энергети-
ческий обмен; синтетические процессы в клетках преобладают,
в них возрастает количество первичных метаболитов.
Обмен веществ и энергии микроорганизмов приводит к воз-
никновению разности концентраций веществ внутри и вне клеток,
что может быть лишь при условии отделения содержимого клеток
от окружающей среды мембранами с различной степенью про-
ницаемости для разных веществ. Растворенные вещества посту-
пают внутрь клетки через клеточную стенку (у отдельных видов
микроорганизмов — еще и через капсулу) и клеточную мембрану.
Следовательно, химические молекулы из среды проникают либо
через три барьера — капсула, клеточная стенка, клеточная мем-
брана (например, у Leuconostoc mesenteraides, Aureobasidiutn
pullulans); либо через два — клеточная стенка и клеточная мем-
брана (например, у Streptomyces griseus, Saccharomyces cerevi-
siae); либо через один — клеточная мембрана (например, у My-
coplasma species).
Капсулы микроорганизмов, преимущественно углеводной при-
роды, имеют рыхлую структуру, проницаемую для многих ве-
204
гв. Однако некоторые микроорганизмы образуют капсулы,
эчающие, например, уроновые кислоты (Cryptococcus spe-
. В этом случае активность их в отношении, например, ка-
юв металлов существенно возрастает и, очевидно, увеличи-
ся вклад полиуронидов в регулирование поступления в клет-
лектролитов.
клеточная стенка различных микроорганизмов также не яв-
-ся инертной структурой, в ней имеются поры, диаметр кото-
; у бактерий колеблется от 9 до 74 нм, у грибов — от 4,0 до
?нм. Поэтому молекулярные массы веществ, проникающих че-
‘ клеточные стенки, могут быть достаточно большими (10...
И 4,5...4,7 кДа соответственно).
Даким образом, клеточные стенки дрожжевых и нитчатых
фоорганизмов менее проницаемы для крупных молекул, чем
точные стенки бактерий. Вместе с тем наличие у бактерий
!колактиловых и тейхоевых кислот заметно ограничивает дос-
электролитов в клетку. Следовательно, одни размеры пор
леточных стенках микроорганизмов не могут рассматриваться
ущим показателем в оценке проникновения веществ внутрь
яки — важна интегральная структура в целом.
Благодаря метаболизму микроорганизмы получают энергию
нечного света или химическую энергию из субстратов пита-
ьных сред; превращают экзогенные вещества в строительные
>ки, из которых синтезируют полимерные структуры (белки,
еводы, нуклеиновые кислоты, липиды или их конъюгаты) и
мие компоненты клеток, а также осуществляют другие спе-
зические процессы, например синтез и разрушение молекул,
бходимых для каких-либо определенных функций (синтез
оторых каротиноидов и последующий частичный распад их
5разованием гамонов у низших грибов). Метаболизм подраз-
,яют на энергетический и конструктивный:
• АТФ
окение
хание
>
1 к
АДФ
Синтез нуклеиновых кислот,
белков, углеводов, липидов, их
конъюгатов
Активный транспорт питатель-
ных веществ
Биолюминесценция
Тепло
(ргетический обмен
акции, сопровождаю-
8ся выделением энер-
Конструктивный обмен
(реакции, сопровождающиеся
расходованием энергии)
Все микроорганизмы, использующие энергию солнечного све-
называют фотосинтезирующими или фототрофными. Микро-
'анизмы, использующие энергию при окислении химических
205
веществ, называют хемосинтезирующими или хемотрофными.
Если при этом учесть и потребляемый источник углерода, т0
микроорганизмы можно подразделить на группы (табл. 25).
Таблица 25. Деление микроорганизмов по способам питания
в зависимости от источников углерода и энергии
Группа Используемый источник Подгруппа
углерода энергии
Автотро- фы Неорганиче- ский (СО?) Фотохимическая ре- акция (фототрофы) Неорганические веще- щества (хемотрофы) Фотосинтезирующие (фотолитотрофы) Хемосинтезирующие (хемолитотрофы)
Гетеро- трофы Органиче- ский Фотохимическая ре- акция (фототрофы) Неорганические ве- щества (хемотрофы) Органические веще- ства (хемотрофы) Фотосинтезирующие (фотоорганотрофы) Хемосинтезирующие (хемолитоорганотрофы) Хемосинтезирующие (хемоорганотрофы)
Пути использования потребляемой энергии у фототрофов и
хемотрофов сходные. Они включают реакции промежуточного об-
мена, или амфиболизма, и реакции синтеза веществ клетки, или
анаболизма. Высвобождение энергии, связанное с расщеплением
химических веществ, называют катаболизмом.
Химические реакции, сопровождающиеся выделением энергии,
называют экзергоническими, а реакции, протекающие с поглоще-
нием энергии, — эндергоническими. В биологии стандартным вы-
ражением энергии является внесистемная единица килокалория
(ккал). Одна килокалория — это количество энергии (или тепло-
ты), необходимое для повышения температуры одного килограм-
ма воды на Г; 1 ккал = 4,1868 • 103 джоулям (СИ) или 1 Дж—
= 2,388-10~4 ккал.
5.1. ФЕРМЕНТЫ МИКРООРГАНИЗМОВ—БИОКАТАЛИЗАТОРЫ
ОБМЕННЫХ ПРОЦЕССОВ
Многие реакции в живых организмах сопровождаются выделе-
нием энергии и в то же время самопроизвольно они не протека-
ют. Для того чтобы вещества взаимодействовали, их необходимо
привести в состояние критического энергетического уровня, при
котором могло бы происходить эффективное столкновение моле-
кул, приводящее к химической реакции. Энергию, необходимую
для этой цели, называют энергией активации (Еа). Следователь-
но, самопроизвольные экзергонические реакции возможны, если
206
льшое число активированных молекул окажется инициато-
реакции, в результате которой освобождается достаточное
чество энергии для активирования соседних молекул.
(ля микроорганизмов (как и всех других живых систем)
е общим методом активации реагирующих молекул является
«огический катализ. Метаболическая активность живых кле-
проявляется при относительно низких температурах, и поэто-
они используют экзергонические реакции (Дб<0). Кроме
>, при отсутствии в среде источника энергии активации реаги-
щих молекул в клетках используется биокатализ. В качестве
логических катализаторов выступают ферменты (энзимы).
[ ускоряют химические реакции, но не расходуются при этом
• входят в состав конечных продуктов:
Е + S ES EX EY , ► EZ Е - Р Р + Е
Е — фермент; S — субстрат; Р — конечный продукт; ES —
»мент-субстратный комплекс; X, Y, Z — переходные формы
страта перед образованием конечного продукта Р, который
обОждает фермент, вступающий затем в очередную специфи-
кую реакцию.
Часть потенциальной энергии реагирующих веществ, которая
жет быть использована для осуществления полезной работы,
)ывается свободной энергией G, выражаемой в Дж/моль:
G = Н - TS,
! Я — энтальпия; Т — абсолютная температура; S — энтропия.
Энтальпия — количество общей энергии системы (теплосодер-
ние), Дж; энтропия — мера разупорядоченности системы, или
га вероятности данного состояния системы, обладающей энер-
!Й (S = felnT), Дж/град. Чем больше степень упорядоченности
ггемы, тем меньше ее вероятность и энтропия.
Изменение свободной энергии ДО служит критерием само-
эизвольного протекания химических реакций — спонтанные
акции возможны лишь при ДО < 0 (экзергонические реакции)
невозможны при Д0'>0 (эндергонические реакции):
, \G = \H-T\S или ДЯ = ДО + 7’Д5.
(и ДЯ со знаком (—) — реакция экзергоническая, со зна-
4 ( + ) —эндергоническая. При равновесных реакциях ДО =
>©. Изменение стандартной свободной энергии в биологических
Сектах зависит от pH, при pH = О ДО обозначают как ДО0,
| pH 7,0-до0:
В энергетическом обмене центральную роль играет АТФ, па-
янный Ф. Липманом (1941) высокоэнергетическим фосфатом,
Держащим высокоэнергетическую фосфатную связь ~ Р: Аде-
Ьин —Р~Р~Р.
207
Наряду с АТФ и другими высокоэнергетическими фосфа-
тами имеются низкоэнергетические фосфаты, что зависит от зна-
чения AG°' их гидролиза (кДж/моль):
ДО”',
кДж моль
Фосфоенолпируват* —61,9
Карбамоилфосфат —51,4
1,3-Дифосфоглицерат —49,3
Креатинфосфат —43,1
Ацетилфосфат —42,3
АТФ -> АДФ + Ф„ -36,8
Аргининфосфат —32,2
Глюкозо-1-фосфат —20,9
Фруктозо-6-фосфат —15,9
Глюкозо-6-фосфат —13,8
Глицерол-1, или 3-фосфат —9,2
Система АТФ — АДФ потому и занимает промежуточное по-
ложение, что она выполняет посредническую роль в переносе
фосфатных групп от высокоэнергетических фосфатов (располо-
жены выше АТФ) к акцепторным молекулам (расположены ниже
АТФ).
На рис. 50 показано энергетическое соотношение реакции в
присутствии и отсутствие фермента, где энергетический выигрыш
ферментативной реакции весьма значителен.
Ферхменгы в клетках находятся в очень малых концентраци....
по все они обладаюг достаточно высокими числами оборачиз
мости, которые указывают на то, что молекула фермента моз.. г
катализировать следующие одна за другой реакции тысяч м<>
кул субстрата в минуту (молекулярная активность).
В процессе жизнедеятельности микроорганизмов постоянно
происходит трансформация энергии, например: химической в теп-
ловую у гетеротрофов, световой в химическую у аутотрофов, хи-
мической в световую у светящихся бактерий. Указанная транс-
формация осуществляется при обязательном участии ферментов.
Ферменты — глобулярные белки, молекулярная масса кото-
рых колеблется от 15 до нескольких тысяч килодальтон. Они
являются простыми и сложными белками. Так, например, уреаза,
пепсин, трипсин — простые, а карбоксипептидаза, амилаза, рибо-
нуклеаза— сложные белки (точнее — гликопротеины), которые
могут быть двух- или многокомпонентными. Следовательно, фер-
менты (холоферменты) наряду с белком (апоферментом, феро-
ном), содержат еще термостабильную небелковую часть (агон).
Если эта часть лабильно связана с ферментным белком и легко
диссоциирует из холофермента и, следовательно, легко может
быть отделена, например, при диализе через полупроницаемые
мембраны, то ее называют коферментом. Если же связь между
белковой и небелковой частями фермента прочная, то кофермент
называют простетической группой. В качестве простетических
* При 37°С, pH 7,0, избытке ионов Mg2+ и 1 м концентрации исходных
веществ и продуктов.
208
8
Ход реакции
Рис. 50. Энергетическое соотношение
реакций в отсутствие и в присутствии
фермента:
S — субстрат; Р — продукт реакции; AG—
изменение свободной энергии; ДЕ + Е —
энергия активации ферментативной реак-
ции; ЛЕ — Е — энергия активации нефер-
ментативной реакции
коферментных групп могут
> относительно несложные
нические молекулы, напри-
витамины или комплексно
энные металлы: железо —
талазе, пероксидазе, цито-
шой системе, медь — в со-
е полифенолоксидазы и
рбиноксидазы, молибден
ггратредуктазе, цинк — в,
.ьной ангидразе, кальций —
блочной протеиназе. Кроме
зрментов известны еще ко-
коры* — вещества или ионы
щлов, активирующие дея-
ность ферментов. В норме
не входят в состав фер-
ов.
’еакции изомеризации, оки-
ия—восстановления, пере-
i и реакции, сопровождаю-
ся образованием ковалентных связей, катализируются слож-
и ферментами, тогда как реакции гидролиза — простыми или
жомпонентными ферментами.
<ак правило, одинаковые химические связи в живой и нежи-
природе обладают равной прочностью. Так, энергия расщеп-
1Я связи Н—Н (длина 0,074 нм) составляет 422,9 кДж/моль,
101 ккал; связи С=О (длина 0,122 нм) —607,1 кДж/моль,
145 ккал; связи С-—N (длина 0,147 нм) —205,2 кДж/моль,
49 ккал. Для разрыва самой слабой из них необходимо в
раз больше той энергии, количество которой необходимо
>рме для разрушения связи в биологических системах. Подоб-
количество энергии не вырабатывается в живых системах,
гому одним из главных путей снижения энергии активации
яется наведенная асимметрия реагирующих молекул с по-
1ью ферментов. При этом возрастает энергия электронов в
страте, они переходят на более высокий уровень, а связи ста-
ится менее прочными.
Дестабилизация субстрата обеспечивается за счет преиму-
ггвенной связи его в нескольких точках с активным центром,
( каталитическим сайтом фермента, представляющего собой
сток фермента, построенный за счет определенных амино-
лот с возможным участием простетической группы или кофер-
1та. В активных центрах ферментов чаще находятся гистидин,
ин, тирозин и другие, боковые цепочки которых обычно иони-
ованы и обладают электронодонорными и электроноакцептор-
Некоторые авторы отождествляют кофакторы с коферментами.
209
Рис. 51. Модель каталитического сайта:
схема трехточечного присоединения субстрата (а)
к плоскости активного сайта фермента (б)
(a)
ферментный сайт
ними свойствами. Элек-
тронодонорные группы
могут быть связаны с
ионами магния, мар.
ганца и цинка.
На рис. 51, а изобра-
жена модель каталити-
ческого сайта, отра-
жающая возможность
индуцированной асим-
метрии фермента. Если
связывание фермента с
субстратом изобразить
в пространстве, то ката
литический сайт в плоскости, как правило, соединяется тремя
точками (рис. 51, б).
В противоположность небелковым катализаторам, ускоря-
ющим многие химические реакции, каждый фермент катализиру-
ет чаще только одну реакцию. В этом проявляется специфичность
ферментов и определяется их особая роль в метаболизме клеток
живых организмов, поскольку именно специфичность лежит в ос-
нове тончайшей регуляции скорости метаболических процессов.
Ферменты обладают абсолютной оптической специфичностью, по
крайней мере для части молекулы субстрата. Например, мальта-
за катализирует гидролиз а-, но не р-гликозидной связи; фермен-
ты прямого окислительного пути катализируют конверсию D-,
а не А-фосфосахаров.
Ферменты, катализирующие реакции гидролиза, действуют на
особые химические группы субстратов. Следовательно, множест-
во субстратов с подобными связями не предопределяет такого же
множества ферментов (природа экономит набор ферментов).
В отдельных случаях некоторые гидролазы проявляют более
высокую групповую специфичность. Например, карбоксипептида-
зы и аминопептидазы отщепляют аминокислоты ступенчато с
карбоксильного или аминного конца соответственно.
Все ферменты подразделяют на 6 главных классов, имеющих
строго определенные номера: 1 — оксидоредуктазы; 2 — трансфе-
разы, 3 — гидролазы, 4 — лиазы, 5 — изомеразы и 6 — лигазы.
В основу такой классификации ферментов (КФ) положен тип
катализируемых ферментами реакций.
Оксидоредуктазы катализируют окислительно-восстановитель-
ные реакции между двумя субстратами:
S red + S ОХ -------- S ох + S red
К ним относятся НАД, НАДФ, каталаза и др.
Трансферазы катализируют реакции межмолекулярного пере
носа различных химических групп (не водорода) и одноуглероД
210
остатков, альдегидной и кетонной групп, ацила, гликозила,
|>орсодержащей группы и др.:
S—G + S' = S'G + S
G — переносимая группа.
Гидролазы катализируют реакции гидролитического расщеп-
1Я внутримолекулярных связей (эфирных, пептидных, глико-
<ых, С—С, в ангидридах кислот, R—N):
СН2—СН—СН2 +3H29 3RCOOH + сн2—СН—СН2ОН
I I । Липаза •
OCOROCOROCOR
он он
-СН—(СО—NH)„—СН—R, +п11гО „ nR—СН—СООН + nR,—СН—NH2
। Протеиназа
СООН
‘nh2
nh2
соон
C0Hn O5-(O-CeHl0O4)„-O-C6H„O5 Гл + оГидОаза
Чиазы (от франц, liaison — соединение) катализируют реак-
присоединения групп по двойным связям и обратные реакции
лва таких групп:
X Y
с-
НОСН2 (СНОН) 3—С—СН2—О—Р
Альдолаза НОСН2—С—СН2—О—Р +
О
фруктозо-1 -фосфат
+ носн2—снонс^
н
глицероальдегид
О
дигидрооксиацетон-
фосфат
О
JHOOC—CH2—СНОН—СООН -Раз>а НООС—СН=СН—СООН + Н2О
яблочная кислота фумаровая кислота
Гласно КФ альдолазу соответственно называют кетозо-1-фос-
f-альдегид-лиазой, а фумаразу — L-малат-гидролиазой.
Изомеразы катализируют реакции изомеризации, т. е. внут-
1юю конверсию оптических, геометрических изомеров или
1еров положения (позиционных). К ним относятся рацемазы
имеразы, цис-транс-изомеразы и др.:
6 5 4 3 2 1
Р—О—С—С—С—С—С—с=о
Глюкоз о-6-фосфатизомераза
6 5 4 3 2 1
Р—О—С—С—С—С—С—С—
II
о
211
R' R'
HQ—С—н Рацемаза H-C-OH
I I
R" R"
L-форма D-форма
Лигазы (синтетазы) катализируют реакции соединения (си-
лат. ligare — связывать) двух молекул, сопряженные с расщеп-
лением пирофосфатной связи в молекуле АТФ или аналогичного
нуклеозидтрифосфата. При этом образуются связи С—О, С—N,
С—С, С—S, например:
НООС—(СН2)2—СН—СООН + АТФ + NHt г\1.!отаминсинтетаз>а
I
nh2
L-глутаминовая кислота
—^О=С—(СН2)2—СН—СООН + АДФ + Фн
I I
nh2 nh2
глутамин
Согласно КФ глютаминсинтетазу называют Д-глютамат-аммо-
ний-лигазой (АДФ).
По КФ каждый фермент имеет шифр, включающий 4 цифры,
из которых первая указывает на класс, вторая — на подкласс,
третья — на подподкласс и четвертая — на порядковый номер
фермента в данном подподклассе. Так, шифр 3.5.1.5 принадлежит
карбамидамидогидралазе (уреазе), которую относят к третьему
главному классу — гидролазам. Она действует на С—N-связь,
отличающуюся от пептидной (шифр 3.5); встречающуюся в ли-
нейных амидах (шифр 3.5.1); порядковый номер уреазы 5.
Скорости реакций, катализируемых ферментами, различны и
зависят от количества и активности ферментов, концентрации
субстрата, pH, температуры, присутствия в среде активаторов
и ингибиторов. Количество фермента не всегда удается опреде-
лить в биологических системах, хотя его присутствие доказано
с помощью специфической реакции. Поэтому было введено поня-
тие активность фермента, приложимое к очищенному и неочи-
щенному ферментам. Активность измеряют в международных,
единицах (ME) : 1 ME соответствует количеству фермента, прев-
ращающему 1 микромоль (мк/моль; 10-6 моль) субстрата в
1 мин в стандартных условиях.
Единицы активности определяют также с учетом превращения
1 нано- (10~9) и 1 пико- (10-12) моль субстрата в 1 мин. Тогда
обозначения международных единиц микро-, нано- и пико- буду1"
соответственно мкЕ, нЕ и пЕ. Удельную активность выделенного
фермента выражают в единицах на 1 мг белка. В 1973 г. была
введена единица ферментативной активности катал (кат), соот-
ветствующая количеству катализатора, способного превращать
1 моль субстрата в продукт за 1 с. Катал связан с ME следуюши-
212
^равенствами: 1 кат = 1 моль субстрата-с- ==60 мольХ
:ин 1 = 60-106 мкмоль-мин”1 == 6 -107 ME, 1 ME = 1 мкмольХ
[ин-1 = 1/60 мкмоль-с1 — 1/60 мккат = 16,67 нкат =
16670 пкат.
Если ферменты состоят из двух и более полипептидных цепей
отомеров, мономеров, или субъединиц), то их именуют олигоме-
tu. Олигомерные ферменты с несходными протомерами могут
чествовать в различных формах, называемых изоферментами
изозимами.
Например, известные изоферменты лактатдегидрогеназы раз-
аются по четвертичной структуре. Активная молекула лактат-
Идрогеназы (Af = 130 000 Да) состоит из четырех протомеров
£ типов — Н и М, молекулярная масса которых около
Ю0 Да. Только тетрамерные молекулы обладают каталитиче-
й активностью. Независимо от порядка расположения прото-
ов они комбинируются следующим образом: НННН (Е),
НМ (I2), ННММ (1з), HMMM (Е), ММММ (15). Синтез
(томеров Н и М контролируется различными генетическими
усами. Концентрация ферментов и изоферментов активно
рируется в клетке главным образом в результате репрессии
теза ферментов.
Синтез, сопряженный с использованием энергии, предотвра-
щен эндогенно образующимися веществами-репрессорами.
: репрессоры инактивируются субстратами ферментов или
цными с ними компонентами. Индуцибельные ферменты назы-
>т адаптивными, тогда как ферменты, образуемые клеткой
отсутствие субстратов-индукторов, именуют конститутивными.
зестны также ферменты, активность которых на каталитиче-
м сайте модулируется аллостерическими эффекторами (от
». aZZtoff — другой, (Нортроп — пространственный), распо-
сенными на аллостерическом сайте. Последний физически
ичен от каталитического сайта (рис. 52). Эффекторы могут
ъ положительными и отрицательными. Аллостерические фер-
ты также являются олигомерами, состоящими из двух и бо-
протомеров. Они могут быть в неактивном и активном состоя-
х. В процессе аллостерической активации четвертичная струк-
а фермента остается неизменной.
По числу взаимодействующих молекул ферментативные реак-
(как и другие химические реакции) подразделяют на реак-
первого, второго или третьего порядков в соответствии с
( как при данных условиях скорость реакции зависит от кон-
трации реагирующих веществ. При малых концентрациях суб-
ата ферментативная реакция протекает в соответствии с ки-
йкой реакций первого порядка, т. е. со скоростью, прямо про-
циональной концентрации реагирующего вещества в каждый
ный момент времени: А -► Р.
Такие реакции описывают уравнением
213
QO
on
1
Рис. 52. Аллостерическая активация (схема):
1 — положительный аллостерический эффектор; 2— молекула аллостерического фер-
мента в неактивном состоянии (а — г — протомеры); 3—аллостерический сайт;
4 — каталитический сайт; 5 — активированный фермент; 6 — субстрат; 7 — активное
состояние аллостерического фермента в отношении субстрата; 8 — продукты транс-
формации субстрата
где [А] — молярная концентрация вещества A; t — время; k —
константа скорости (размерность с_| или мин-1). Если такая
реакция протекает в воде, то химические изменения не должны
зависеть от наличия воды.
Скорость реакции второго порядка пропорциональна произ-
ведению концентраций двух реагирующих веществ или второй
степени концентрации одного из них. Схема биомолекулярных
реакций имеет вид: А + В Р или А + А -► Р, их описывают
следующими уравнениями:
-^T- = ^ar- = fe[A] 1В1 или = [А] =*[А]2.
Размерность константы скорости реакции второго порядка —
моль”’с-1. Скорости распада А и В должны быть равны, по-
скольку одна молекула А реагирует с одной молекулой В.
В реакции третьего порядка скорость пропорциональна про-
изведению трех концентраций. Она имеет следующий вид: А +
+ В + С Р. Реакции третьего и более высокого порядков
очень редки, так как вероятность столкновения трех, четырех и
т. д. молекул ничтожно мала.
Выделяется еще реакция нулевого порядка, когда в единицу
времени превращается постоянное количество вещества вне зави-
симости от его начальной концентрации (—d[S]/df = k), т. е.
при реакциях нулевого порядка фермент насыщен своим суб'
стратом и при снижении концентрации фермента не происходит
изменения скорости реакции.
Каждая ферментативная реакция характеризуется константой
214
аэлиса Кт- Она обозначает концентрацию субстрата, при
рой ферментативная реакция протекает со скоростью, равной
>вине максимальной, ее размерность моль/л. Кт может быть
делена экспериментально (графически — как функция ско-
и). Уравнение Михаэлиса — Ментен описывает поведение
•их ферментов в зависимости от изменений концентрации
тратов ([S] в молярных концентрациях):
Vma,[S]
ym+ [S] •
1ля удобства графического изображения экспериментальных
1ых уравнение Михаэлиса — Ментен алгебраически преобра-
г в другие формы (например, уравнения Лайнуивера — Бэр-
Эди — Хофсти).
Активность ферментов зависит от температуры, pH и других
горов. Подобно всем химическим реакциям, ферментативные
дии ускоряются с повышением температуры. Так, возраста-
температуры на 10° усиливает активность биокатализаторов
и в 2 раза. Но белки являются термочувствительными струк-
,ми и при высоких температурах они денатурируют. При
’С каталитическое действие большинства ферментов необра-
) прекращается. Для каждого фермента имеется оптимум pH,
пение которого на 2 единицы в кислую или щелочную сторо-
риводит почти к полной потере активности. Многие вещества,
являющиеся субстратами ферментов, могут активировать
цваторы), другие, напротив, тормозить их (ингибиторы).
5олее часто свойства активаторов проявляют такие кофакто-
как ионы кобальта, магния, марганца, цинка. Аллостериче-
ферменты активируются преимущественно органическими
фелн inn ибиторов фермен тов известны конкурентные и не-
ypenoi.!te. Структура конкурентных ингибиторов подобна
ктурс субстрата и поэтому они конкурируют с субстратом за
ыванш с активным центром фермента. Конкурентные инги-
ры выг-.сняются избытком субстрата. Неконкурентные инги-
ры ре;и ируют с другими важными частями молекул фермен-
те с активными центрами). Такой частью может быть
фгидрильная группа SH. Подобные ингибиторы не вытес-
"ся избытками субстрата.
Микроорганизмы отличаются, как правило, высокой приспо-
яемостью к изменяющимся условиям среды обитания и, сле-
тельно, их согласованные процессы анаболизма и катаболиз-
йходятся под постоянным контролем регуляторных механиз-
(рис. 53). В ходе так называемой амфиболической стадии
завершается деградация макромолекул до ССЬ, и в эту же
вю поставляются низкомолекулярные субстраты — пред-
венники для анаболических процессов.
; микроорганизмов регуляция обменных процессов осущест-
ГСя с помощью ферментов (см. разд. 5.1). Выделяют три
215
Ill
Рис. 53. Три стадии анаболизма и катаболизма, регулируемых ферментами:
пунктирные стрелки — анаболические пути; непрерывные стрелки — катаболические пути
уровня регуляции ферментативных реакций с учетом потребности
клеток в энергии: изменение абсолютного количества конкретного
биокатализатора; изменение фонда реагирующих веществ, иск-
лючая фермент; изменение каталитической активности фермента.
Абсолютное количество конкретного фермента определяется ско-
ростью его синтеза (Ks) и скоростью распада (Kd) по схеме:
К, Л
Фермент Аминокислоты
As
Комбинированный процесс синтеза и распада фермента назы-
вают оборотом фермента (не путать с числами оборачиваемости
216
ента!). Количество фермента может увеличиться за счет
щтания Ks либо уменьшения Kd или того и другого вместе,
отив, уменьшение количества фермента может происходить
ет снижения Ks или возрастания Kd либо за счет того и другого
ге. Следовательно, в этом проявляется относительно независи-
регуляция синтеза и распада ферментов.
•а каталитическую активность и регуляцию активности фер-
ов существенное влияние оказывают: 1) ксмпартментализа-
ферментативных процессов у эукариотических организмов;
бъединение ферментов в макромолекулярные комплексы,
хинирующие их деятельность и направление интермедиатов
ответствующему метаболическому пути; 3) ингибирование по
обратной связи — Feedback inhibition (не смешивать с
back — репрессией), когда тормозится активность фермента
в начале биосинтетического пути конечным продуктом (Д):
a в-!дг-^д
ри биосинтезе Д из А в реакциях участвуют ферменты Ei — Е4,
'да концентрация Д становится значимой для клетки, то Д
Зирует превращение А в Б. Конечный продукт при этом свя-
ётся с аллостерическим сайтом на Е| и ингибирует фермент,
уная в роли отрицательного аллостерического эффектора,
[бирование по типу обратной связи регулирует синтез Д.
(бирование по типу обратной связи наблюдается и в тех слу-
, когда метаболические пути оказываются разветвленными:
А, Б, В — предшественники четырех конечных продуктов
р4; Г — предшественник Р2 и Р3; Д — предшественник Р4.
о существу, последовательности реакций в разветвленном
5олическом пути имеют линейный характер, и они ингибиру-
по типу обратной связи конечными продуктами:
В——♦'р1, Г —♦Д’2’ >3, В^^д—^>4
ге с тем при разветвлении метаболических путей фермента-
ie реакции могут регулироваться за счет возникновения
:ественных петель по типу обратной связи:
217
Таким образом, клетки микроорганизмов располагают меха-
низмами грубого и тонкого контроля регуляции синтеза и актив-
ности ферментов, катализирующих соответствующие реакции об-
мена первичных и вторичных метаболитов. Например, ингибиро-
вание синтеза фермента конечным продуктом относится к меха-
низму грубого контроля регуляции. Ранее образовавшиеся моле-
кулы фермента продолжают выполнять свою роль до тех пор, пока
они не будут разбавлены при дальнейшем клеточном росте, хотя
ненужный синтез белка прекращается немедленно. С другой
стороны, ингибирование ферментативной активности тем же конеч-
ным продуктом относится к разряду тонких механизмов регуля-
ции, так как, например, поток источника углерода на конкретном
пути постоянно и точно регулируется.
Feedback-ингибиция у микробов — один из- самых действенных
путей регуляции «прохождения» источников углерода через био-
синтетические пути. Конечный продукт аллостерически тормозит
активность лишь первого фермента, но все последующие реакции
в конкретном метаболическом пути прекращаются, т. е., хотя
ферменты в последующих реакциях и не подвержены действию
конечного продукта, активность их не проявляется.
В ряде случаев клеткам микробов выгоднее использовать
конечный или промежуточный продукт в качестве активатора, а не
ингибитора какого-либо особого фермента. Это аллостерическая
активация. Например, на пути превращения глюкозы в глюко-
зо-6-фосфат и фосфоенолпировиноградную кислоту у Е. coli из-
быток их служит сигналом переключения части глюкозы на путь
синтеза гликогена, что сопровождается аллостерической актива-
цией фермента, катализирующего реакцию превращения глюко-
зо-1-фосфата в АДФ-глюкозу (рис. 54).
Нередко в мире микроорганизмов наблюдается кооператив-
ность как способ тонкой регуляции метаболизма. Здесь речь идет
о кооперативном взаимодействии фермента и субстрата. Поли-
мерные энзимы, обладающие более чем одним местом связывания
с субстратом, соединяются с ним одним каталитическим центром,
чем создается повышенное родство других мест для дополнитель-
ных молекул субстрата. Суммарный эффект такого воздействия
должен выражаться в геометрическом возрастании каталитиче-
ской активности по отношению к арифметическому повышению
концентрации субстрата.
В тонкой регуляции метаболизма важную роль играют фер-
менты, выступающие в качестве аллостерических белков, когда
каждый энзим имеет не только каталитический центр, соединяю-
щийся с субстратом, но также один или более других мест, к кото-
рым присоединяются небольшие регуляторные молекулы — эффек-
торы. Присоединяясь к аллостерическому белку в своем месте,
эффектор вызывает конформационные изменения в энзиме — роД'
ство каталитического центра фермента к субстрату уменьшается
(наступает аллостерическая ингибиция) или, напротив, возрастает
(наступает аллостерическая активация).
218
Г лю
I
Г лю-6-Ф-----*- г лю-1-Ф----*- АДФ-Г лю------Гликоген
I * *
I . I 1 .
Фру-6-Ф 'J
I ~ — — АДФ — —j ।
* Фру-1,6-ди-Ф | |
З-Фосфоглицери- I
новая кислота I
------ФЕП-------------------и
«U--------------------------!
Пируват
I
54. Аллостерическая ингибиция (1) и аллостерическая активация (2)
регуляторные механизмы использования глюкозы клетками Е. coli
I учетом времени размножения, роста, развития и отмирания
юбных клеток энзиматические реакции в них протекают в
елах минут, секунд или их долей. Следовательно, клетка —
гма динамичная, представляющаяся в каждый момент с но-
стороны. Вот почему любой шаг (реакция) метаболизма
ит свой вклад в процессы регуляции. Это наипростейший ме-
зм регуляции, известный у всех организмов.
Открытие гормоноподобных структур у некоторых грибов слу-
основанием для предположения о существовании у данных
«организмов гормональной регуляции отдельных метаболиче-
процессов.
1ЕТАБОЛИЗМ БЕЛКОВ
1ервичные метаболиты в клетках микроорганизмов являются
ами, большинство из которых обладает каталитическим дей-
iM (ферменты). Поэтому обмен белков непосредственно свя-
' функцией генетического аппарата клеток. Механизм синтеза
ифических белков рассмотрен в гл. 2. В данном разделе из-
отся основные сведения о метаболизме аминокислот, являю-
‘я структурными элементами всевозможных белков.
Метаболизм аминокислот
«кислоты находятся в центре процессов, обеспечивающих
су белками, амфиболическими интермедиатами и другими спе-
изированными продуктами, поэтому важно знать, каким обра-
зом синтезируются аминокислоты у прокариотических и эукарио-
тических микроорганизмов.
Особое место занимает синтез аминокислот у микроорганиз-
мов, способных фиксировать свободный азот из воздуха. Прежде
чем такой окисленный элемент станет составной частью амино-
кислот, он должен быть восстановлен. Микроорганизмы — фикса-
торы молекулярного азота — подразделяют на свободноживущие
в природе и находящиеся в симбиозе с растениями или другими
микробами, например, некоторые грибы с отдельными водоросля-
ми или цианобактериями — лишайники. Число азотофиксаторов
невелико, но от них в буквальном смысле слова зависит жизнь на
нашей планете. С помощью микробов-азотфиксаторов ежегодно
фиксируется около 17,5-107 т молекулярного азота из воздуха, из
которых 10...15 % приходится на цианобактерии. Схема фиксации
азота представляется в следующем виде:
Основными компонентами системы фиксации азота являются
сильные восстановители: АТФ, ферредоксин или флаводоксин и
нитрогеназа (ферментный комплекс). У азотобактеров, клубень-
ковых бактерий Rhizobium species и других микроорганизмов АТФ
образуется в ходе обмена углеводов в реакциях с участием НАДФ.
У аэробных свободноживущих и симбионтных бактерий восста-
новители также образуются в ходе углеводного обмена в реак-
циях с НАДФ, у фото- и цианобактерий сильные восстановители
образуются фотохимически.
Бактериальные ферредоксины, содержащие один (Fe2S2) или
два (Fe4S4) так называемых железосероцентра (молекулярная
масса их от 6,3 до 9,6 кДа), имеют во многом гомологичные амино-
кислотные последовательности. Каждый атом железа в них скоор-
динирован с четырьмя атомами серы четырех цистеиновых остат-
ков. Окислительно-восстановительный потенциал ферредоксинов
порядка —400 мВ.
У ряда фотобактерий, анаэробных бактерий и цианобактерий
имеются флаводоксины (М — 15...21 кДа), содержащие по одному
ФМН (флавинмононуклеотиду). Окислительно-восстановитель-
ный потенциал для пар ФМН/ФМН-Н и ФМН-Н/ФМН-Н’
—100...200 мВ и 400 мВ соответственно, т. е. это самый низкий
потенциал, известный для какого-либо флавопротеина и поэтому
флаводоксины могут заменять ферредоксины в фотохимических
реакциях.
220
Дитрогеназа — двухкомпонентный ферментный комплекс, каж-
компонент которого (большой или малый) неактивен в отдель-
*и. Молекулярные массы компонентов различны, что зависит от
э микроба — источника нитрогеназы (М большого компонен-
- 150...270 кДа, малого — 50...70 кДа). В большом содержатся
езо, молибден и лабильно связанный сульфид, в малом —
езо и лабильный сульфид. Следовательно, большой компо-
— железо-молибден-протеин (молибдоферредоксин), ма-
— желез о-протеин (азоферредоксин). На большой компонент
годится два малых.
восстановление Na до NH3 с участием нитрогеназы происходит
ледующей суммарной реакции:
N2 + 6Fe2+ + 6H+ + 12АТФ -* 2NH3 + 6Fe3+ + 12АДФ + 12Ф
:тановительные эквиваленты (Н+) от НАДФ через флаво-
'еины поступают к ферредоксину (флаводоксину), который
танавливает азоферредоксин в нитрогеназе, а последний вос-
[авливает молибдоферредоксин. Азоферредоксин связывает
'Ф на один атом Fe; 4 АТФ гидролизуются в присутствии обоих
тонентов фермента при условии переноса пары электронов
ферредоксина к молибдоферредоксину. Ингибитором нитроге-
1 является кислород, очевидно поэтому, например, у циано-
герий стенка гетероцисты надежно отделяет фермент от вы-
1емого ими кислорода в процессе фотосинтеза.
Образование аммиака из молекулярного азота с участием нит-
иазы можно представить следующим образом:
n2
h2n — nh2
2Н| | 2Н2О
H3N-«-2NH2OH
Лногие микроорганизмы способны восстанавливать нитраты до
иака. В этом случае нитрат вначале восстанавливается в
>ит с помощью фермента нитратредуктазы. Донором электро-
у бактерий выступает НАД-Н, а у грибов — НАДФ-Н.
ратредуктаза (М = 800 кДа) — тетрамер, состоящий из двух
₽ров, одна субъединица димера имеет молекулярную массу
кДа, другая — 50 кДа. Это также молибденсодержащий фер-
г. Реакция восстановления нитратов в присутствии нитрат-
чстазы протекает в две стадии:
НАДФ-Н + Н+ + NO3~ NOf +НАДФ+ + Н2О
NOr + 8Н + 6е~ -> NHt + 2Н2О
221
Непосредственным восстановителем нитрита выступает сиро-
гем — железотетрагидропорфирин:
соон соон
сиротам
Аммиак, образующийся различными путями в природе (в том
числе у микроорганизмов), включается в состав органических
соединений и прежде всего в состав аминокислот благодаря трем
главным реакциям:
Г лутаматдегидрогеназа
1) NHt + НООС—С— (СН2) 2—СООН + НАДФ Н НООС—СН— (СН2) 2СООН +
A nh2
а-кетоглутаровая глутаминовая
кислота + НАДф + Н2О кислота
2а) НООС—СН—(СН2)2—COOH+NH3+AT®r----------► НООС—СН—(CH2)2CONH2 +
I Глутамат- .. i
NH2 синтетаза .
глутаминовая глутамин
кислота ддф _|_ фи
26) НООС—С— (СН2)2СООН + НООС—СН—(CH2)2CONH2 + НАДФ-Н + нт-
i bffl2
а-кетоглутаровая
кислота
------------► 2НООС—СН—(СН2)г—СООН 4-НАДФ+
Глутаматсинтаза ।
NHo О
За) NH:i + СО2 + 2АТФ Карбамоилфосфатсинтетаза +
ОН
-р 2АДФ I Фн карбамоилфосфат
36) НООС—СН— (СН2)2СОМН2 + СО2 + 2АТФ Карбамоилфосфатсинтетаза
nh2
- НООС—СН— (СН2) 2СООН + H2N—СО—ОРОзНг + 2АДФ + Ф„
NH2
222
1утаминовая кислота и глутамин фактически являются ис-
1ком всех атомов азота, входящих в состав любых амино-
>т или других органических соединений. Карбамоилфосфат
(дуется либо на синтез аргинина, либо на синтез пиримиди-
ногие микроорганизмы синтезируют все природные амино-
1ты, но есть виды, нуждающиеся в большинстве амино-
>т (например, молочнокислые бактерии). Концентрация каж-
1минокислоты в фонде клетки (или пуле) определяет скорости
нтеза и расходования в синтетических процессах.
1. Биосинтез аминокислот
Мн образуется из пировиноградной кислоты путем трансами-
вания, или переаминирования, т. е. переноса аминогруппы
есто кислорода кетогруппы в кетокислотах. Донором амино-
П является глутаминовая либо аспарагиновая кислота:
1) СНз—СО—СООН + НООС—СН—СНг—СООН Дансаминаза
пировиноградная
кислота
nh2
аспарагиновая
кислота
СНз
соон
СН—nh2
+ со
СООН
аланин
сн2
соон
ща в елево-
уксусная кислота
2) СНз—СО—СООН + НООС—(СН2) 2—СН—СООН 2РансамииаД
nh2
Глутаминовая кислота
СНз
соон
СН—nh2
+ сн2
соон
сн2
аланин
со
соон
а-кетоглутаровая
кислота
иридоксальфосфат — важная часть каталитического сайта
!аминаз, как и многих других ферментов, субстратами кото-
223
рых выступают аминокислоты. Во всех пиридоксаль-зависимых
реакциях биосинтеза аминокислот первым интермедиатом стано-
вится основание Шиффа, связанное с ферментом:
R—СО—СООН
а-кетокислота
(субстрат)
Аланин в заметных количествах входит в состав пептидогли-
канов бактериальных клеточных стенок (частично как О-изомер),
где содержание его от общей массы пептидогликана, например у
Streptococcus faecalis, составляет 39...40 %; у Staphylococcus
aureus — 67 %. 0-Аланин является составной частью витамина —
пантотеновой кислоты.
Аргинин синтезируется из орнитина, который, в свою очередь,
образуется из глутаминовой кислоты:
I. НООС—(СН2)г—СН—СООН + АТФ АцетилКоА.
| Ацетилаза
NH2
глутаминовая кислота
----------► НООС—(СН2)2—СН—СООН —Тф.
NHCOCH3
N-ацетилглутаминовая кислота
----- Н2О3РО—СО—(СН2)2—СН—СООН .
I НАД-Н-1-Н+
NHCOCH3
а-N-ацетил глутамил фосфат
О
-----н—с— (сн2)2— снсоон 2рансам_нназ!
| Аминокислота
NHCOCH3
N-ацетилглутамил-у-полуальдегид
224
Н2С—(СН2)2—СН—СООН АцетилоРнитиназа
NH2 NHCOCH3
N-ацетилорнитин
h2n—сн2—(сн2)2—сн—соон
NH.
ориитин
J. Н2С—(СН2)2—СН—СООН + H2N—СО—О-РО3Н2
nh2
nh2
карбамоил фосфат
орнитин
NH2
Фн
АТФ
СО
Аспарагиновая
кислота
СООН
НС—N=C—NH— (СН2)з—СН—СООН-
сн2
nh2
nh2
NH
СООН
(СН2)з
аргининсукцинат
СН—nh2
соон
цитруллин
соон
HN=C—NH—(СН2)3
СН
nh2
chnh2
4-
НС
соон
аргинин
СООН
фумаровая кислота
не входит в состав i
формамидина при синтезе стрептомицина культурой
ИТИН
ф°М
ptomyces griseus. Бактерии превращают аргинин в полиами-
- спермин и спермидин, использующиеся как факторы роста
ггорыми штаммами Neisseria и Yersinia-.
природных белков. Аргинин является
17
225
HN==C—NH—(СН2)з—CH—СООН
h2n
nh2
4 аргинии 4
HN—(CH2)r—NH HN—(CH2)4—NH2
(<^h2)2 (<!:h2)2 (ch2)2
h2n^h2 ch2nh2 h2nch2
спермин
спермидин
Аспарагин у микроорганизмов синтезируется из аспарагиновой
кислоты и аммиака за счет энергии АТФ при участии фермента
аспарагинсинтетазы и в присутствии ионов Mg .
О
Mg2+ II
НООС—СНг-СН—СООН + АТФ + NH3 *H2N—С—СНг—СН—СООН 4-
I . I
nh2 nh2
аспарагиновая аспарагин
кислота
+ АМФ 4-ФФ„
Бактерии не используют вместо МНз какие-либо другие доноры
аминогрупп (например, глутамин).
Аспарагиновая кислота синтезируется всеми формами микро-
организмов путем трансаминирования щавелевоуксусной кислоты:
НООС—(СН2)2-СН—СООН + НООС—СО—СНу—соон ->-
I
nh2 щавелевоуксусная
глутаминовая кислота
кислота
НООС—СН—СНг—СООН + НООС—(СН2)2—С—соон
I II
NH2 о
аспарагиновая а-кетоглутаровая кислота
кислота
Аспарагиновая кислота входит в состав белков бактерий, грибов,
водорослей, протозоа, а также служит исходным сырьем для био-
синтеза аспарагина.
Валин синтезируется из двух молекул пировиноградной кис-
лоты при участии четырех ферментов: ацетолактатсинтазы (1),
диоксикислота-редуктоизомеразы (2), диоксикислота-дегидрата-
зы (3), аминотрансферазы аминокислот с разветвленной
цепью (4). В биосинтезе участвуют также тиаминпирофосфат
СНз СНз СНз
2(io 1ПЯ со ”АД-Н > н3с—(i—сн—соон —Я
| I 2' Mg2+ н+ I I 3
СООН НзС—с—соон он он
I
он
пировииоград- ацетолактат
ная кислота
а,р-диоксиизовалериа-
новая кислота
226
СНз СНз
----- НзС—СН—С—СООН пеРеаминир°ваниеНзС_JH_СН—СООН
II 4 I
о nh2
e-кетоизовалериановая кислота валин
мстидин образуется многоступенчато. При глубоком изуче-
механизма биосинтеза гистидина у Salmonella species и Е. coli
о установлено, что гистидиновый оперон в хромосоме бакте-
содержит 9 генов, кодирующих синтез соответствующих фер-
гов. Первая реакция биосинтеза, катализируемая аллостериче-
4 ферментом АТФ-фосфорибозилтрансферазой, специфически
4озится конечным продуктом, т. е. гистидином:
И'-(5'-фосфорибозил) АМФ К'-(5'-фосфорибозил, Формимино-
-5-амииоимидазол-4 -карбокса мид-
рибонуклеозид
—Рибозофосфат
Глн
5
N—Рибозофосфат
СН2О—Р
имида зол-
глицерофосфат
Ы'~(5'-рибулозил)-формимино-
-5-аминоимидазол-4-карбоксамид-
рибонуклеотид
имидазол-
. ацетолфосфаз
L - тис I и л и ноя - L гистидинол
фосфат
СН
I
hcnh2
соон
L-i исгидин
227
В реакции участвуют: 1 — АТФ-фосфорибозилтрансфераза
2 — пирофосфогидролаза, 3 — фосфорибозил-АМФ-циклогидр0’
лаза, 4 — фосфорибозилформимино-5-аминоимидазолкарбокс-
амидрибонуклеотид-изомераза, 5 — глутамин-амидотрансфераза
6 — имидазолглицерофосфат-дегидратаза, 7 — А-гистидинолфос-
фат-глутамат-аминотрансфераза, 8 — гистидинолфосфатфосфата-
за, 9 — гистидинолдегидрогеназа. В девятистадийной реакции
имеется несколько точек контроля биосинтеза гистидина.
Глицин образуется из серина под каталитическим действием
пиридоксальфосфата и переноса его 0-углеродного атома на
тетрагидрофолиевую кислоту
(UI Н
СО—NH—СН—(СН2)—СООН
н соон
I
н
тетрагидрофолиевая кислота (ТГФК)
в которой N5 и N10 принимают участие в переносе одноуглеродных
групп:
\ 10
НОСНг—СН—СООН + НС—CHg—NH—
I I
nh2 сн
серин ТГФК
—N5 --------СН2
\ \
СН2—СООН + НС—СН2—N —
nh2 <^н2
глицин N5,N10-метилентетра гидрофолат
Глицин необходим не только для биосинтеза белков, он сам яв-
ляется источником и поставщиком активных одноуглеродных
соединений (СН3ОН, НСООН, НСНО — в зависимости от уровня
окисления).
Глутамин и глутаминовая кислота синтезируются всеми фор-
мами микроорганизмов. Они составляют фундамент процессов
биосинтеза аминокислот, поскольку, приводимые ниже реакции
являются основным путем образования а-аминогрупп непосред-
ственно из аммиака. Синтез глутамина катализируется глутамин-
синтетазой при обеспечении реакции энергией АТФ:
НООС— (СН2) 2—СН—СООН H2N—СО—(СН2)2—СН—СООН +
I NH.+ |
nh2 nh2
L-глутаминовая +АДФ + Ф„ L-глутамин
кислота
Синтез глутаминовой кислоты катализируется L-глутаматде-
гидрогеназой:
228
ЭОС—(СН2)2—СО—СООННАДФ'Н’Н> НООС—(CH2)2—CH—соон +
NH^ I
жетоглутаровая кислота
L-глутамино-
вая кислота
+ НАДФ + Н2О
Дрожжи и нитчатые грибы содержат две глутаматдегидроге-
ы, специфичные для НАД+ или НАДФ+. НАД-связанные
идрогеназы катализируют реакции окисления-восстановления
путях окислительного метаболизма, особенно при гликолизе,
Икле трикарбоновых кислот и в дыхательной цепи митохондрий;
ДФ-связанные дегидрогеназы постоянны для восстановитель-
; синтезов (например, в синтезе жирных кислот и стероидов
митохондрий), как коферменты дегидрогеназ в гексозомоно-
:фатном шунте и т. д.
Бактерии содержат лишь НАД+-зависимую дегидрогеназу и
тому могут синтезировать глутаминовую кислоту с помощью
тих реакций, например из глюкозы и аммония. В этом случае
г превращении глюкозы в цикле трикарбоновых кислот в га-
гве интермедиата образуется а-кетоглутаровая кислота, из
орой синтезируется глутаминовая кислота по указанной выше
кции с участием НАДФ, т. е. восстановительным аминиро-
ием. Образовавшаяся глутаминовая кислота широко исполь-
гся затем в реакциях переаминирования или трансаминирова-
Глутамин служит донором аминогрупп при биосинтезе гистиди-
глюкозамин-6-фосфата, пуринов, триптофана, а также карба-
,лфосфата из предшественников производных пиримидина.
Изолейцин синтезируется по механизму, сходному с синтезом
ина. Общими для обоих механизмов являются четыре фер-
та, находящиеся в клетке в форме мультиферментного комплек-
Это, кроме треонин-дезаминазы (1), ацетилолактатсинтета-
(2), диоксикислота-редуктоизомераза (3), диоксикислота-де-
ратаза (4) и аминотрансфераза аминокислот с разветвленной
ью (5):
г—сн—сн—соон .снз—сн2—с—соон Триа»»н”ир°фосфа;
|| 1 II СНзСНО, 2
он nh2 о
треоиин а-кетомасляная кислота
ОН СН3
:н3—СНг—А—СООН НАД,Н- Н+' М-С СНз—СНг—С—СН—COOH -^/Q
io 3 Ан Ан
СНз
а-ацето-а-окси- о-рдиокси-р-метил-
масляиая кислота валериановая кислота
229
СНз СНз
СН3—СНг—СН—СО—СООН ПеРеамиииР0Ван^ снз—СНг—^н—сн—соон
5 I
а-кето-₽-метилвале- NHz
риановая кислота
изолейцин
Источником четырех (из шести) углеродных атомов в изолей-
цине является треонин. На первой стадии катализатором реак-
ции выступает треониндезаминаза, коферментом которой служит
пиридоксальфосфат. На уровне этого же фермента осуществляет-
ся эффективная специфическая регуляция реакции изолейцином
(ингибирование и репрессия).
Лейцин синтезируется из а-кетоизовалериановой кислоты, яв-
ляющейся также непосредственным предшественником валина,
а метилированная форма ее — предшественником изолейцина.
Следовательно, валин, изолейцин и лейцин синтезируются на пу-
тях, имеющих общие перекрестки. В реакциях биосинтеза лейцина
катализаторами выступают пять ферментов: а-изопропилмалат-
синтаза (1), а-изопропилмалатизомераза (2), изопропилмалат-
дегидрогеназа (3), декарбоксилаза (4) и аминотрансфераза
аминокислот с разветвленной цепью (5):
СНз
СНз—СН—СО—СООН + io
(Ьз S-KoA
он
—5^Д-СН3—СН—С—СН2СООН —
1 I I
НзС соон
а-кетоизовалериа-
новая кислота
Ацетил-КоА
а-изопропиляблочная
кислота
СНз—СН—СН-СН—СООН -НАД^ СНз—сн—сн—со—соон
III 3 I I
СНз СООН ОН СНз соон
р-изопропиляблочная а-кето-р-изопропил-
кислота яблочная кислота
СНз—СН—СНз—СО—СООН ЕеРеамииироваии! СНз—СН—СНг—СН—СООН
I 5 I I
СНз СНз nh2
(1'Кетоизокапроиовая кислота лейцин
Продукт первой реакции выступает индуктором синтеза фер-
ментов последующих реакций, но биосинтез самого индуктора
регулируется по механизму обратной связи (Feedback inhibition).
Лизин синтезируется двумя путями: диаминопимелатным
(у бактерий, некоторых низших грибов) и аминоадипинантныя
(у высших и ряда низших грибов). В первом случае доказано
участие следующих ферментов: дигидропиколинатсинтазы (!)•
Д'-пиперидеин-2,6-дикарбоксилатдегидрогеназы (2), N-сукцинил-
диаминопимелатглутаматаминотрансферазы (3), N-сукцинилди-
230
цнопимелатдесукцинилазы (4), эпимеразы (5) и диаминопи-
(атдекарбоксилазы (6).
СООН со СНО СНз СН k + со -не сн 1 1 1 1 chnh2 СООН СН2 1 пировиноград- MVi СООН ная кислота । ^-аспартил- СООН полуальдегид *— — -Н2О
1
2,3-дигидродипико-
лииовая кислота
Д'-пиперидеии-2,6-
дикарбоновая кислота
СООН соон
I I
СО г ./ л СО
I Сукцииил-КоА ।
1 НООС- (СН2)2СО—S-KoA I .
—* (СН2)з ------------------------ (СН2)з
<кмн2 hcnh—со—сн2
iooH соон сн2
а-амино-е-кетопиме- СПОН
линован кислота tiJUn
N-сукцинил-а-амино-е-
кетопимелиновая кислота
соон соон СООН
h2nch h2nch hcnh2 н2с—nh2
(k)3 т <к)з V 4 I 5 (СН2)з-^ее (СН2)з 1 6 1
HCNHCO—СН2 hcnh2 hcnh2 hcnh2
। соон сн2 соон СООН соон
1 соон /,,Л-а,е-ДАП мезо-а,е-ДАП Г-лизин
N -сукцииил-ТД-а.е-
диаминопимелиновая
кислота
Установлено, что у Bacillus megaterium после второй реакции
епи биосинтеза лейцина ацилирующим агентом является аце-
ьная группа вместо сукцинильной, например у Е. coll. Конеч-
23!
ный продукт биосинтеза—лейцин ингибирует первую реакцию,
т. е. и здесь имеет место регуляция по типу обратной связи.
В аминоадипинатном пути биосинтеза лизина исходные продук-
ты конденсации — ацетил-КоА и а-кетоглутаровая кислота. Часть
последующих реакций напоминает реакция в цикле трикарбоно-
вых кислот:
СНз +НООС— (СН2)2—СО—СООН J±?£L
CO-КоА а-кетоглутаровая
кислота
а цетил-КоА
ОН
— НООС—(СН2) 2—С—сн2—соон —н-'° >
I 2
соон
гомолимонная кислота
« НООС—(СН2)2—С=СН—соон
I 3
соон
цис-гомоаконитовая кислота
он
«НООС— (СН2)2—сн —сн—соон НАД+-Mg-
I 4
соон
гомоизолимониая кислота
— НООС—(СН2)2—СН—СО~СООН ~ НООС—(СН2)з—СО—СООН
СООН а-кетоадипиновая кислота
щавелевоглутаровая кислота
« НООС— (СН2)3—СН—СООН АТФ’2+АД”
I Mg2+, 6
nh2
а-аминоадипииовая кислота
О
и глутамат
— НС—(СН,)з—СН—СООнД^ДФ7+ НД
nh2
6-полуальдегид
а-аминоадипиновой кислоты
СООН
« НООС— (СН2)2—СН—NH(CH2)4—СН—СООН -АД+8 Н2°
nh2
сахарофин
[e-N (L-глутамил-2) -L-лизин]
232
^NH2CH2—(СН2)з—СН—СООН + НООС— (СН2) 2—СО—соон
I
NH2 а-кетоглутаровая кислота
лизин
цифрами обозначены гомоцитратсинтаза (1); ццс-гомоакони-
а (2); гомоаконитатгидратаза (3); гомоизоцитратдегидроге-
а (4); а-аминоадипат-глутамат-аминотрансфераза (5); а-ами-
дипилполуальдегиддегидрогеназа (6); аминоадипилполуальде-
-глутамат-редуктаза (7); сахарофиндегидрогеназа (8).
а-Аминоадипиновая кислота образуется в результате переами-
ования. Сахарофин — промежуточный продукт у дрожжей
a Sacharomyces при биосинтезе лизина. При этом сахарофин
вращается в лизин и а-кетоглутаровую кислоту после реак-
i восстановления и гидролиза. ДАП и лизин широко распростра-
ы в составе пептидогликанов бактерий, а дигидродипиколи-
в виде кальциевой соли выполняет исключительную роль в
рообразовании у бацилл.
Метионин синтезируется различными микроорганизмами. Он
<ен как донор метильных групп. Углеродный скелет метионина
>мируется из гомосерина (б), который в свою очередь обра-
тен из аспарагиновой кислоты (а):
НООС—СН2—СН—СООН Аспартилкиназа
nh2
аспарагиновая кислота q
Н2РОз—ОСО—СН2—СН—СООН с—СН2—СН—СООН НАДФ'Л
| НАДФ' I |
nh2 н nh2
Р-аспартилфосфат p-аспартилполуальдегид
НОСН2—СН2—СН—СООН
nh2
гом осерин
НОСН2—СН2—СН—СООН Сукцинид-КОА
nh2
гомосерин
С н2—О—-с о
<^Н2 (^Н2
I I
chnh2 сн2
ioOH соон
SH—СН2—CHNH2—соон
—сукцинил. 2 *
-сукцинилгомосерин
233
СН,—S—СН2
СН, CHNH, —снзСОСООН, —NH3
I I 3
chnh2 соон
I
соон
SH
(CH2)2
(^HNH2
соон
Дезоксиаденозил-
кобаламин
ЬР-Метил-ТТФК, 4
цистатионин
гомоцистеин
СНз—S— (СН2) 2—сн—соон
nh2
метионин
Ферменты, катализирующие реакции биосинтеза метионина:
гомосеринсукцинилаза (1), цистатионин-у-синтаза (2), циста-
тионин-р-лиаза (3), гомосерин-трансацетилаза и метионин-
синтетаза (4). Простетической группой последнего фермента яв-
ляется восстановленное производное кобаламина, а донором мети-
ла выступает №-метилтетрагидрофолиевая кислота (№-метил-
ТГФК). Метионин ингибирует первый фермент в цепи биосинтеза
по типу Feedback inhibition.
Пролин синтезируется из глутаминовой кислоты с расходова-
нием энергии АТФ. Пути биосинтеза его сопряжены с путями
биосинтеза орнитина — исходного продукта для образования ар-
гинина.
Глутаминовая кислота + АТФ + Ацетил-КоА ---->- N-Ацетилглутаминовая
Ацетилаза
АТФ кт л жж НАДФ-Н-1-Н .
кислота »-a-N-Ацетилглутамилфосфат—=-----М-Ацетилглутамил--у-полу-
альдегид
Далее пути синтеза орнитина и пролина расходятся (см. ар-
гинин) .
NH—СОСНз nh2
°Ч. 1 °ч. । но
>С—СН,—СН2— СН—СООН _ ?С—СН,—СН,— СН—СООН
н/ Н/ + н,о
N-a цетил глутамил-у-полу-
альдегид
/^соон
НАД-Н + Н +
НАД+ ' 1
Д'-пиррол ин-5-карбоновая
кислота
глутамин-у-полу-
альдегид
Возможен путь биосинтеза пролина и без ацетилирования
у-глутамилфосфата, однако он у микроорганизмов не доминирую-
щий. У спорообразующих клостридий имеет место прямое превра-
щение орнитина в пролин с выделением аммиака. Эта реакция
катализируется орнитинциклазой:
H2N—СН2—СН,—СН,—СН—COOH^Wh2N—СНг—СН,—СНг-СО—СООН+
I
NH, 2-кето-5-аминопеитановая
орнитин кислота
234
+ NH3
НАД-Н + H +
НООС/^Ж НООС/4' hr
I
t_I
Д'-пиррол ин-2-карбоновая п
кислота пролин
Серин синтезируется из 3-фосфоглицериновой кислоты, обра-
лцейся при метаболизме углеводов (см. гликолиз):
н А л +
{2О3Р—О—CHg—СН—СООН . . „ н2о3р—о—сн2—со—соон
। Н' + НАДН
QPI 3-фосфопировиноградная кислота
-фосфпглицериновая кислота
Псреаминирование с Глу , Н 2О3Р—О— С Н—С ООН О—С Н 2— С Н—С ООН
I I
nh2 nh2
3-фосфосерин серин
:ред образованием серина 3-фосфосерин гидролизуется серин-
сфатазой.
Серин участвует в биосинтезе пурина и пиримидина; р-углерод
> выступает источником метильных групп для тимина и холина;
ингибирует (отрицательный эффектор) фосфатазу и 3-фосфо-
ицератдегидрогеназу.
Тирозин, фенилаланин и триптофан имеют общие пути био-
нтеза, начиная с эритрозо-фосфата и фосфоенолпировиноград-
й кислоты (ФЕП):
НАД*
2 3
5-дегидрохинная
?Н
НООС—СО—CHj—СН—СН--СН—СН5—О—Р —
он Ан
3-дезокси-2-кето- D -арабинотегттулозоновая кислота-
7-фосфат
ОН
5-дегидрошикимовая
кислота
ОН
шикимовая
кислота
6н
5-фосфошикимовая
кислота
соон
ФЕП
3-енои л-5-фосфопирувил -
шикимовая кислота
235
Р—о—сн2
п -оксифенилпиро-
виноградная кислота
тирозин
Путь к триптофану начинается с 5'-фосфорибозилантранило-
вой кислоты:
индол-3-глицерофосфат
неон
I
СН2—О—Р
триптофан
глицероальдегид-
3-фосфат
Более многоступенчатый путь биосинтеза характерен для трип-
тофана, но все три аминокислоты берут свое начало от хоризмовой
кислоты (реакции 8 и 8'). Затем точкой дихотомического ветвле-
ния реакций синтеза становится префеновая кислота: в направле-
нии 9" и 10" происходит образование тирозина, а в направлении
9' и 10' — фенилаланина. Триптофан синтезируется в направлении
реакций 8...12.
236
Цо седьмой реакции включительно катализаторами выступа-
одни и те же ферменты для всех трех названных аминокис-
i 3-дезокси-2-кето-В-арабиногептулозонат-7-фосфатсинтаза (1),
адрохинатсинтаза (2), 5-дегидрохинат-дегидратаза (3), шики-
дегидрогеназа (4), шикиматкиназа (5), 3-енолпирувилшики-
-5-фосфатсинтаза (6), хоризматсинтаза (7). Реакции синтеза
озина далее катализируют: хоризматмутаза (8'), префенатде-
рогеназа (9"), тирозинаминотрансфераза (10"). Ферменты
синтеза фенилаланина: префенатдегидратаза (9') и фенилала-
аминотрансфераза (10'). Синтез триптофана катализируют:
ранилатсинтаза (8), 1-антранилат-пирофосфорилпирофосфат-
нсфераза (9), М-5'-фосфорибозилантранилат-изомераза (10),
;ол-3-глицерофосфатсинтаза (11) и пиридоксальфосфат-зави-
!ая трипТофансинтаза (12).
Тирозин у некоторых микроорганизмов образуется путем гид-
силирования фенилаланина под действием фенилаланин-гид-
силазной системы (фенилаланин-4-монооксигеназа). Реакция
текает в две стадии:
nh2
СНг—СН—СООН
н
он
/.-тирозин
h2n
О
+ Н2О
СН—СН—СНз
Он Он
дигидробиоптерин
N
I) Дигидробиоптерин -|- НАДФ-Н -f- Н+ Тетрагидробиоптерин + НАДФ+
фвая стадия катализируется фенилаланин-гидроксилазой, вто-
Я — дигидроптеридин-редуктазой. Тетрагидробиоптерин высту-
ет донором электронов, в этом он сходен с фолиевой кислотой.
Конечные продукты биосинтеза — тирозин, фенилаланин и
Иптофан — могут ингибировать синтез по типу обратной связи:
237
а) тирозин тормозит фермент 1 в цепи реакций, репрессируй!
синтез тирозин-чувствительного фермента; б) фенилаланин тормо.
зит ферменты 1, 8', 9'; в) триптофан тормозит ферменты 8 и у
Хоризмовая и префеновая кислоты ингибируют фермент 1.
Треонин, являющийся исходным продуктом для биосинтеза
изолейцина, сам образуется из О-фосфогомосерина при участия
пиридоксальфосфата:
NH2
О-фосфогомосерин
о
сн
сн2—сн—соон +
фермент (Е)-пи-
ридоксальфосфат
р—о—сн2—сн2—с—соон
кетимин
N
н
О-фосфогомосерин
в форме кетимина
сн2=сн—с—соон
N
Е
н
|Н3С—сн=с—соон|
Альдимин N
сн
Н+ (из среды)
ОН~ (из среды;
НзС
Е
238
ОН
НзС—(^Н—СН—соон
Н—С—СООН
N
СНг— О— Р
Н
СН
Н+ (из среды)
Н
N
Е
ОН
Н—СН—СООН + Е-пиридоксальфосфат
j nh2
треоиин
Модный гомосерин вначале фосфорилируется АТФ с участи-
>мента гомосеринкиназы, после чего треонинсинтаза доводит
jc до образования треонина. Другой способ биосинтеза трео-
асуществляется при участии также пиридоксаль-зависимой
•трансоксиметилферазы:
СНзСОН + H2N—СНг—СООН — НзС—сн—сн—соон
’ I I
ацетальдегид глиции 1 •'
ОН 1NH2
треоиии
гиком ацетальдегида является пировиноградная кислота,
стеин у ряда микроорганизмов синтезируется из Л-серина
водорода под каталитическим действием цистеинсинтазы,
мой от пиридоксальфосфата:
НО—СНг—СН—СООН + H2S HS—СНг—СН—СООН + Н2О
nh2
/.-серии
NH2
/.-Цистеин
о равновесие этой реакции сдвинуто влево. У других видов
организмов серин вначале ацетилируется за счет ацетил-
НО—СНг—С Н—СООН + НзС—СО-КоА^-К^
nh2
‘в НО—СНг—СН—COOhJ=^HSCH2—СН—СООН + СН3СООН
I 1
, NHCOCH3 nh2
Йализатор первой реакции — серинтрансацетилаза, вто-
* О-ацетилсерин-сульфгидролаза; оба действуют в активном
Йексе при соотношении 1 :2 соответственно.
239
Сероводород в природе в основной своей массе — конечный
продукт «сульфатного дыхания» специальных микроорганизмов,
когда происходит ассимиляционное восстановление сульфата.
Сульфат также может образовываться в результате окисления
восстановленных соединений серы, например тиобактериями,
некоторыми пурпурными несерными бактериями. Любым путем
образовавшийся сульфат используется затем в качестве источни-
ка энергии микроорганизмами — продуцентами его. В результате
ассимиляционного восстановления сульфата образуется сульфид,
который расходуется на синтез цистеина:
SO?- + 8[Н+] -> H2S + 2Н2О + 2ОН-
Цистеин — важнейшая аминокислота в природных белках,
поскольку она является местом образования дисульфидных мос-
тиков —S—S—, благодаря чему вносит свой вклад в формирова-
ние и поддержание вторичной структуры данных полимеров.
Таким образом, в биосинтез 20 природных аминокислот вовле-
каются неорганические вещества (источники азота и серы)
(рис. 55) и органические (преимущественно как источники угле-
рода) (рис. 56).
Пять аминокислот являются родоначальниками всех других
аминокислот на первых или промежуточных этапах их биосинте-
за; ведущая роль среди них принадлежит глутаминовой кислоте.
На схеме указаны исходные вещества для синтеза всех природ-
ных аминокислот из углеводов, а также вклад в этот процесс
цикла трикарбоновых кислот, являющегося поставщиком а-кето-
глутарата и оксалоацетата. Кроме того, указаны аминокислоты,
синтез которых сопряжен с расходованием энергии АТФ.
Необходимо подчеркнуть, что все аминокислоты, входящие
в состав природных белков, как правило, относятся к А-ряду.
Однако у микроорганизмов, например, в клеточных стенках
(в составе пептидогликанов) присутствуют D-аминокислоты.
Превращение D-форм в L-формы осуществляется в присутст-
вии пиридоксальфосфата. Следовательно, пиридоксальфосфат
NOJ
n2
а -кетоглутарат, НАД (О)- Н+Н > L-Гпу
нитратредукция ассимиляционная фиксация из воздуха Пируват, НАД-Н + Н+ 3, ! -А1а
NH4f Фумарат э^Г-Асп
L-Глу+АТФ / -Глн
SO32 Восстановление _
gO42— ассимиляционное
Цистеинсинтаза
----------------------L -Цис
или АцКоА, серин-
трансацетилаза, ацетил-
серинсульфгидролаза
Рис. 55. Пути биосинтеза аминокислот из неорганических источников
240
Г ексозы
СН j—СО —СООН
Пируват
S -Фосфо-рибозо-
1-дифосфат
С
(СНОН)2
АТФ
СН2— О—Р
Эритрозо-4-фосфат
Трп
Асн
ГомоСерин » Мет
ж ТиР
Фен
Вал Леи Ала
Лиз .
Дап]
АТФ
М-Тли
Сер JAHC
СО—СООН
Н2С—СООН
Оксалоацетат
Тре Иле
АТФ
Н2С—СООН
сн2
Про
со—СООН
а-кетоглутарат
Орнитин
Аргинин
Цитруллин*
Рис. 56. Пути биосинтеза аминокислот из гексоз при участии интермедиатов
углеводного обмена
зет весьма заметную роль в трех типах реакций аминокислот:
заминирования, рацемизации и декарбоксилирования (см.
(д. 5.2.1.3).
.1.2. Биосинтез некоторых олигопептидов,
риновых и пиримидиновых нуклеотидов, металлопротеинов
овь синтезированные аминокислоты включаются в белки под
ггролем генетического аппарата клетки (см. гл. 2). Однако,
к уже было показано, при биосинтезе самих аминокислот,
также других соединений аминокислоты могут быть донорами
Ноуглеродных групп —СН2ОН, —СНО (серин), аммиака (глу-
иин), метильных групп (метионин), аминогрупп при переами-
ровании (глутаминовая кислота); служить исходным материа-
и для биосинтеза амидов, олигопептидов и нуклеотидов.
Образование амидов из аминокислот рассмотрено ранее на
241
примере аспарагина и глутамина. Таким же путем синтезируются
и другие амиды. Реакции протекают однотипно: на первой стадии
образуется активированный энзимом комплекс ацилфосфата, из
которого на второй стадии после реакции с аммиаком образуется
амид:
1) RCOOH + АТФ-> E-RCOOPO3H2 + АДФ
2) E-RCOOPO3H2 + NH3 ->- RCONH2 + Н3РО4 + Е
Сложнее протекают синтезы олигопептидов и нуклеотидов.
Биосинтез олигопептидов. Глутатион — трипептид, представ-
ляющий собой у-глутамилцистеинилглицин. Он синтезируется в
соответствии со следующими уравнениями реакций:
НООС—сн—(СН2)2—СООН + HS—СН2—СН—СООН + АТФ + Mg2\ АДФ. К-
I I _ф“’1
nh2 nh2— sh
L-глутаминовая кислота L-цистеин
-> НООС—СН—(СН2)2—СО—NH—CH—СООН + Гли\+АТФ'+
| । — АДФ, — Ф„72
nh2 СН2 —SH
глутамил цистеин
-> НООС—СН—(СН2)2—СО— NH—СН—СО—NH—СН2—соон
I I
NH2 СНг—SH
глутатион
Первая реакция соединения глутаминовой кислоты с цистеи-
ном катализируется у-глутамилцистеин-синтазой (1), а вторая —
глутатион-синтазой (2) (оба фермента неспецифичны). Первый
фермент может катализировать реакцию синтеза у-глутамила-
ланина или у-глутамил-а-аминомасляной кислоты, второй —
реакции синтеза офтальмовых кислот, обнаруженных вначале
в хрусталике глаза (офтальмовые кислоты — аналоги глутатио-
на, в котором цистеин замещен какой-либо другой аминокисло-
той). Офтальмовые кислоты сильно ингибируют глиоксилазу,
катализирующую превращение метилглиоксаля в лактат, напри-
мер у Е. coli:
ОНС—СО—СН3 -> СНзСНОНСООН
метилглиоксаль лактат
Глутатион содержится в клетках бактерий и грибов, где он
выполняет роль восстановителя при синтезе дезоксирибонуклео-
протеинов (окисленный глутатион восстанавливается с помощью
глутатионредуктазы, сопряженной с НАДФ), а также участвует
в транспорте аминокислот. При взаимодействии с аминокислотой
происходит перенос глутамильного остатка глутатиона на транс-
портируемую аминокислоту:
Глутатион 4- R—СН—СООН ->-у-Глутамил—СО—СН—R
I I
nh2 nh2
+ H2N—СН—СО—NH—сн?—СООН
I
СН,—SH
цистеинилглицин
242
Указанная реакция катализируется у-глутамилтрансферазой.
глутамиламинокислотного дипептида транспортируемая ами-
дслота поступает в клетку, а глутаминовая кислота вновь
пользуется для синтеза исходного трипептида. В указанной
кции донорами глутамильной группы могут быть также и дру-
у-глутамилпептиды, а акцепторами — все аминокислоты
юме пролина).
Депсипептиды. К ним относятся циклопептидные структуры,
•являющие выраженные антибиотические свойства: грамици-
[ С, полимиксины, бацитрацины, тироцидин, микобациллин,
жулины и др., синтезируемые различными видами бацилл,
шь разные штаммы Вас. brevis образуют более 10 полипептид-
с молекул: бревиген, бревин, бреволин, бресеин, грамицидины
, В, С, CD, D), гратизин, грацейлин, комисан, тироцидины
, В, С), эдеин, эсеин. Все эти олигопептиды содержат до 34 ос-
нов аминокислот и синтез их отличается от матричного синте-
высокомолекулярных белков. У бактерий циклопептиды могут
юлнять роль ионофоров. Рассмотрим наиболее изученные цик-
’ептиды, синтез которых базируется на эдеине, включающем
ць 4 аминокислоты.
Грамицидин С включает 5 аминокислот, каждая из которых
(теряется в структуре два раза (одна из них относится к D-
iy). Следовательно, грамицидин С отличается регулярностью,
о нельзя сказать о полимиксинах и других упомянутых цикло-
тидах:
А-Орн -* А-Лей
t
А-Вал
t
А-Тир
i
A-Глн — А-Асн —
тироцидин А
А-ДАМ -> изо-Лей -*• £)-Лей
4
А-ДАМ
I
£>-Фен А-Ала А-Асп -> А-Про -*• А-Асп — А-Глу
1 t 1
L -Про А-Асп А-Тир
t 1
L -Фен А-Глу А-Асп
1
£>-Фен A-Лей — А-Асп — А-Сер- А-Тир
микобациллин
А-ДАМ — А-Тре— A-ДАМ —А-Тре —А-ДАМ — L-ДАМ—6-МОК
циркулин А
полимиксине В и других олигопептидах ДАМ означает а,у-
зминомасляную кислоту: H2N—СНг—СНг—CH(NH2)—СООН.
а находится в D- и A-формах, как и некоторые из 10 амино-
слот; 6-МОК — это 6 метилоктановая кислота:
НООС—СНг—СНг—СНг—СНг—СНг—СН —СН3
СНз
243
В бацитрацине А имеется тиазолидиновое кольцо с аминоме-
тилбутильным остатком, которое присоединено к олигопептиду
включающему 10 D- и /.-аминокислот:
£-Лей —»- D-Фен —/.-Про—«-/.-Вал—»- £-Орн
I,,-J
£-Орн -»—/-Вал—»-£-Про-«—D-Фен —L-)1ей
грамицидин С
NH2
t
/-Лей —— /-ДАМ —>- /-ДАМ—NH,
1 I
D-Фен £-Тре
f 1
£-ДАМ-«—£-ДАМ-«—Z-ДАМ—NH2
I I
NH2 £-Тре
1
H2N-«— D-ДАМ—6МОК
полимиксин В
D-Фен —«-/-Гис—«-/-Асп—«-D-Acn
f I
изо-Лей-»-D-Орн -•--/-Лиз
н3с
н3с—н2с—нс—сн—nh2
I—L-CO
—/ - изо -Лей -•— D-Глу -«— / -Лей
бацитрацин А
Непосредственные участники синтеза олигопептидов — эдеин-
синтаза и эдеин — локализованы соответственно в клеточной
мембране и в периплазматическом пространстве. Синтетаза гра-
мицидина С, бацитрацин-синтетаза, тироцидин-синтетаза и
эдеин-синтетаза представляют собой мультиферментные комплек-
сы и состоят из двух или трех комплементарных ферментных
фракций. Эти фракции могут быть далее разделены на катали-
тически еще активные простые белки с молекулярной массой око-
ло 70 кДа, каждый из которых активирует одну из аминокислот,
участвующих в биосинтезе олигопептида.
В синтетазах обнаруживаются и меньшие субъединицы (/•! =
=з 17 кДа), являющиеся пантетеин-содержащими белками-шжи-
телями, сходными с ацилпереносящими белками (АПБ). Молеку-
ла протеина имеет длину около 1,5 нм и его SH-группа может
связываться с активными центрами ферментных белков снгге-
тазного комплекса. Эти белки-носители при помощи челночного
механизма осуществляют промежуточное последовательное тр-.нс-
пептидирование активированных аминокислот. Приводим схема-
тичное изображение мультиферментного комплекса панте! eeri-
с. одержи щего белка-носителя:
244
и
белок
NH
НС О
.о-
> сн2
он
NH
^С=°
белок 1
СН NH
сн3 дн3Д
фосфопантотеновая кислота
4-фосфопантетеин
Hj
СН2
сн2
н
1,5 нм
Исходя из этого показано, что, например, тироцидин-синтетаза
цставляет собой ферментный комплекс, образованный 12 раз-
ными субъединицами, катализирующим активацию и присоеди-
ие 10 аминокислот. Указанные ферментные комплексы катали-
уют также реакции рацемизации некоторых аминокислот-пред-
твенников. Для синтеза циклопептидов необходима энергия
р. Аминокислоты при этом активируются путем образования
Ф-производных, которые трансформируются затем в тиоэфиры,
данные с ферментным комплексом. Пептид высвобождается из
ментного комплекса при циклизации. В грамицидине С, напри-
, местом циклизации являются указанные пунктиром связи.
Гомопептиды, состоящие лишь из одной аминокислоты, также
тезируются в качестве вторичных метаболитов при действии
гветствующих лигаз. К таким гомоолиго- и гомополипептидам
осятся полиглутамил-пептиды:
НООС— [ — (СН2)2—СН—СО—NH—СН— (СН2)2— ] —соон
nh2 соон
Полиглутамил-пептиды обнаруживаются в виде капсул у Bacil-
anthracis, Вас. lichenifortnis, Вас. subtilis и др. Молекулярная
са их находится в пределах от единиц до 50 кДа. Входящая
i состав глутаминовая кислота почти исключительно относится
-ряду.
Матричный синтез полипептидов рассмотрен в разделе 4.2.4.
Биосинтез пуриновых нуклеотидов. Каждая микробная клетка
гезирует пуриновые нуклеотиды, являющиеся составными час-
и нуклеиновых кислот. Но для нормального роста и размноже-
многих из них необходимо добавление к среде определенных
Инов в готовом виде. Синтез пуриновых нуклеотидов возможен
1уриновых оснований или из аминокислот, углекислоты и ам-
ка. Этот синтез у всех микробов протекает во многом одинако-
। путем. Так, адениловая кислота (АМФ) образуется из инозин-
юсфата (инозиновая кислота) при участии аспарагиновой кис-
ы и ГТФ под действием аденилосукциназы:
245
инозин-5'-фосфат
гуанозинтрифосфат
+ НООС—СНу-СН—СООН—
nh2
А сп
(ИМФ) (ГТФ)
Синтез гуаниловой кислоты или гуанозин-5'-фосфата (ГМФ)
также происходит из инозиновой кислоты под каталитическим
действием инозинатдегидрогеназы:
ИМФ
ксантозин-5'-моно-
фосфат (КсМФ)
Указанная реакция ингибируется ГМФ. Последующая реакция,
катализируемая гуанилатсинтетазой, завершается образованием
ГМФ.
КсМФ + АТФ + H2N—СО—СНг—СНг—СН—СООН Mg2-\
nh2
глутамин
О
ГМФ
+ НООС—СНг—СНг—СН—СООН + АМФ + ФФ
nh2
Рибозо-5'-фосфат
глутаминовая
кислота
246
^сли катаболизм нуклеиновых кислот и нуклеотидов в куль-
х микроорганизмов сопровождается образованием свобод-
пуринов, то развивающиеся и размножающиеся клетки
т использовать их для синтеза пуриновых нуклеотидов (за-
юй путь). Тогда свободные пурины взаимодействуют с 5-фос-
ибозил-1-пирофосфатом, образовавшимся из рибозо-5-фосфа-
ФРФФ) и АТФ, с последующим синтезом нуклеозид-5'-моно-
фатов:
1. Адении + ФРФФ»Аденозин-5'-фосфат 4-ФФИ
2. Гуанин + ФРФФ Гуанозии-5'-фосфат -|- ФФ„
.Первая реакция катализируется аденинфосфорибозилтранс-
►азой, вторая — гуанинфосфорибозилтрансферазой. Выделяю-
йся неорганический пирофосфат (ФФ„) легко гидролизуется
hvo неорганической пирофосфатазой, поэтому реакции приоб-
ают необратимый характер.
Под влиянием пуриннуклеозидфосфорилазы возможно превра-
аие свободных пуринов в нуклеозиды, а под влиянием киназ —
леозидов в нуклеотиды (это также запасной путь).
Аденин + Рибозо-1-фосфат Аденозин 4- Ф„
Гуании + Рибозо-1-фосфат Гуанозин 4- Ф„
Аденозин 4- АТФ — АМФ 4- АДФ
Гуанозин 4- АТФ —» ГМФ 4- АДФ
АМФ 4- АТФ АДФ 4- АДФ
ГМФ 4- АТФ ГДФ 4- АДФ
АДФ 4- АТФ =* АТФ 4- АДФ
ГДФ 4- АТФ ГТФ 4- АДФ
С помощью веществ, меченных изотопами, удалось расшифро-
ъ общую схему биосинтеза пуринового ядра из нуклеотидных
'дшеств енников:
Происхождение атомов в ядре следующее: азот в позиции
- из аспарагиновой кислоты, в позициях 3 и 9 — из амидного
та глутамина, в позиции 7 — из глицина; углерод в позициях
। 8—из муравьиновой кислоты (формиата), в позициях 4 и
- из глицина, в позиции 6 — из углекислоты.
(-Биосинтез пуринов из аминокислот, СО2 и NH3 однотипен у
«ариотических и эукариотических микроорганизмов. Суть его
тоит в формировании пуринового ядра у первого углеродного
ма рибозо-5-фосфата в последовательности из десяти реак-
247
ций, промежуточными продуктами которых являются: а-5-фосфо-
рибозил-1-пирофосфат (ФРФФ), 5-фосфорибозил-1-амин, гли-
цинамидрибонуклеотид, a-N-формилглицинамидрибонуклеотид
a-N-формиглицинамидинрибонуклеотид, 5-аминоимидазол-рибо-
нуклеотид, рибонуклеотид-5-аминоимидазол-4-карбоновой кисло-
ты, рибонуклеотид-5-аминоимидазол-4-М-сукцинкарбоксамида,
рибонуклеотид-5-аминоимидазол-4-карбоксамида, инозин-5'-фос-
фат (ИМФ). Как уже было сказано, ИМФ выступает затем ис-
ходным нуклеотидом для образования других пуриновых нуклео-
тидов.
Биосинтез пиримидиновых нуклеотидов. Пиримидиновые нук-
леотиды, как и пуриновые, могут синтезироваться клетками мик-
роорганизмов, использующими запасные пути, или из аминокис-
лот, углекислоты и аммиака. Так, урацил и 5-фторурацил могут
трансформироваться в УМФ и S-F-УМФ непосредственной реак-
цией с ФРФФ под действием урацил-фосфорибозил-трансферазы:
урацил
(или 5-фтор-урацил)
рибозо-5'-фосфат
уридин-5'-фосфат (УМФ) или
5-фтор-уридин-5'-фосфат (5 - F -УМФ)
Урацил может также взаимодействовать с рибозо-1-фосфа-
том в присутствии уридиндифосфорилазы с образованием уриди-
на. Биосинтез УМФ из уридина возможен в присутствии уридин-
цитидинкиназы:
О
II
|)+АТФ
Рибоза
Mg2+^
УМФ + АДФ
уридин
Подобная реакция имеет место с цитидином, фторцитидином и
фторуридином. Образование ди- и трифосфатов из УМФ проте-
кает аналогично с ди- и трифосфатами пуриновых мононуклео-
тидов.
Структурным предшественником пиримидинов (урацила, ти-
мина, цитозина и 5-метилцитозина) является оротовая кислота,
обнаруживаемая в клетках различных микроорганизмов. Она
синтезируется из карбамоилфосфата и аспарагиновой кислоты:
248
соон
оротовая кислота
Образование карбамоиласпартата — ключевая реакция в био-
тезе пиримидиновых нуклеотидов; она ингибируется ЦТФ.
(имные превращения пиримидиновых дезоксирибонуклеотидов
ут быть представлены в следующем виде:
д-ТТФ* д-УТФ д-ЦТФ
t t t
д-ТДФ д-УДФ— УДФ д-ЦДФ*— ЦДФ
|f ।
Тимин -* Тимидии д-ТМФ *-- д-УМФ--д-ЦМФ
Рибонуклеозидтрифосфаты являются предшественниками всех
ов РНК, а дезоксинуклеозидтрифосфаты — предшественника-
ДНК.
Биосинтез белков-металлопротеинов. К металлопротеинам
осят гемосодержащие белки, в которых гем представляет со-
железопротопорфирин, или ферропротопорфирин. Такие бел-
выделили в группу гемопротеинов. Негемовое железо содер-
г бактериальные дегидрогеназы, упомянутые ранее ферре-
сины и другие структуры. Гемопротеины — цитохромы, ката-
а, пероксидаза, сирогем (см. разд. 5.2.1)—обнаружены у
к аэробных микроорганизмов. Если геминовая часть у гемо-
теинов идентична, то различия их обусловлены последова-
ьностью аминокислот и конформацией белковой части.
Из всех цитохромов цитохромы b имеют самый низкий окис-
:ельно-восстановительный потенциал. Цитохромы а и аз со-
вляют цитохромоксидазу — медьсодержащий белок, соединен-
включающей
с особой формой гема, названной геМом А,
лфарнезильный остаток:
этилфариез иль-
ный остаток
к пиррольному кольцу порфина
ковый комплекс в цитохромоксидазе (М «200 кДа) содержит
гбъединиц с различной молекулярной массой. Восстановленный
охром аз легко восстанавливает молекулярный кислород Ог.
новая часть гемопротеинов синтезируется матричным спосо-
। и затем присоединяется к гему.
• Буквой «д» обозначено «дезокш».
249
5.2.1.3. Протеолиз и расщепление аминокислот
Микроорганизмы синтезируют аминокислоты или используют
некоторые из них в готовом виде из среды обитания не только
для конструктивного обмена. Во многих случаях аминокислоты
подвергаются катаболизму и потребляются как источники угле-
рода и энергии. Различные микроорганизмы как бы лавинообраз-
но осуществляют процесс круговорота азота в природе. В приво-
димой схеме стрелками снизу показаны этапы подключения
микробов, расщепляющих белки. Это расщепление (неспецифи-
ческий протеолиз) катализируется ферментами из класса гид-
ролаз:
Белки -у Полипептиды у Олигопептиды у Аминокислоты —...
С помощью эндопептидаз белки распадаются на крупные бло-
ки полипептидов разной молекулярной массы. Этот процесс осу-
ществляют гнилостные микроорганизмы, например, из родов
Bacillus, Proteus, Pseudomonas и др., а также многие грибы.
Нередко еще используют устаревшее название пептоны, которые
представляют собой обработанные пепсином белки, содержащие
полипептиды, олигопептиды и до 30 % свободных аминокислот.
Аминокислоты, образованные в процессе протеолиза, подверга-
ются дальнейшим превращениям или используются как таковые
микроорганизмами. Некоторые полипептиды и олигопептиды так-
же могут поглощаться микробными клетками с последующим
гидролизом внутриклеточными пептидазами до аминокислот.
Кроме неспецифического протеолиза известен еще и ограни-
ченный, или специфический, протеолиз. Так, специфическим рас-,
щеплением полипептидов является каскад реакций связывания
комплемента (см. гл. 8), морфогенетические процессы, прису-
щие бактериофагам. Специфическое посттрансляционное расщеп-
ление белков исключительно важно при активации и инакти-
вации внутриклеточных ферментов (табл. 26).
Таблица 26. Ограниченный (специфический) протеолиз белков
и его назначение
Биологический процесс Пример,^ ограниченного протеолиза Назначение протеолиза
Действие токсина Действие компле- мента Дифтерийный ток- син—* ферментативно ак- тивный фрагмент + про- никающий фрагмент Фактор комплемен- та -> один или два ак- тивных белка комплемен- та Протеаза пораженной клетки выделяет летальный фрагмент токсина, обладающий фермент- ной активностью Защитная пусковая (триггер- ная) система примерно из 18 белков сыворотки крови; серией специфических протеолитических реакций ограничивается дейст- вие комплемента во времени и пространстве. Растворимые бел- ки присоединяются к мембране
250
Продолжение табл. 26
шологическии
процесс
Пример ограниченного
протеолиза
Назначение протеолиза
□рфогенез ви-
ых частиц
разоваиие спе-
ческих белков
трусов живот-
Две независимые суб-
структуры вируса Ti->
образование из двух суб-
структур
Предшественник . бел-
ков вируса полиомиели-
та -> белки вируса по-
лиомиелита
Расщепление полипептидной
цепи открывает новые центры
связывания; необратимость про-
теолиза способствует упорядоче-
нию вирусных частиц
Адаптация вирусов к специ-
фике синтеза белка в клетках
животного организма
Все белковые структуры подвержены инактивации и расщеп-
ию. Принято считать, что первая реакция расщепления бел-
(одной или нескольких пептидных связей) является специ-
еской, после чего наступает неспецифический протеолиз,
ршающийся полным распадом протеина.
юльшинство белков гидролизуется с той или иной глубиной
•ими протеолитическими ферментами, или пептидазами. При-
। аминокислот, образующих пептидные связи, а также поло-
се гидролизуемых связей (например, рядом с заряженными
пами или вдали от них) являются основой для классифика-
пептидаз. Различают две группы пептидаз: 1) экзопепти-
I, для действия которых необходимо наличие либо свободной
,евой аминогруппы (аминопептидазы), либо свободной кон-
й карбоксильной группы (карбоксипептидазы), 2) эндопеп-
зы, катализирующие реакции гидролиза внутримолекуляр-
пептидных связей в белках. Экзопептидазами являются:
;инаминопептидаза, карбоксипептидазы А, В, С и многие дру-
а эндопептидазами — субтилизин, эндопептидаза гриба
trnaria species, клостридиопептидаза В и т. д.
Аминокислоты, образующиеся в результате протеолиза белков,
ут подвергаться дальнейшему расщеплению. При этом в реак-
[ катаболизма включаются азот аминогрупп и углеродный
[ет. Катаболизм подавляющего большинства аминокислот на-
.'ается с отщепления а-аминогруппы, либо с трансаминирова-
либо с окисления. В реакциях ферментативного трансамини-
ания участвуют аминокислоты: Ала, Apr, Асн, Асп, Вал, Иле,
з, Тир, Три, Фен, Цис; а также следующие кетокислоты: пиро-
)градная, а-кетоглутаровая или щавелевоуксусная. При этом
шногруппа переносится к а-углеродному атому в одной из
1анных выше кетокислот. В итоге аминогруппа меняется ме-
I с кетогруппой. Наиболее известны аланин’грансаминаза и
•аматтрансаминаза, содержащие пиридоксальфосфат в каче-
кофермента.
i результате дезаминирования аминокислот все аминогруппы
[раются в единый фонд в виде глутаминовой и аспарагиновой
ют, а также аланина. При этом конечным акцептором амино-
251
групп оказывается а-кетоглутаровая кислота, которая превращу,
ется в глутаминовую кислоту, завершающую катаболизм амино-
кислот. В итоге аминогруппы передаются соответствующим ак-
цепторам, представляющим собой конечные продукты азотистого
обмена.
Дезаминирование, при котором аммиак отщепляется от ами-
нокислоты, может быть прямым, гидролитическим и окислитель-
ным. В результате прямого дезаминирования образуются нена-
сыщенные соединения:
R—СН2—СН—СООН — NH3 + R—СН=СН—СООН
nh2
Например, прямое дезаминирование гистидина происходит
под влиянием гистидин-аммиак-лиазы Klebsiella aerogenes с об-
разованием урокановой кислоты:
р==—СН=СН—СООН
N^^/NH
В случае гидролитического дезаминирования образуются
аммиак и оксикислоты:
R—СН—СООН- +HaO . R—СН—СООН + NH3
I I
nh2 он
Однако такие реакции не характерны для микроорганизмов,
у них дезаминирование чаще происходит за счет амидной группы.
Например, пиогенные стрептококки осуществляют следующую
реакцию гидролитического дезаминирования (дезамидирова-
ния):
НООС—СН—(СН2)2—CONHj -+Н2-— НООС—СН—(СН2)2—соон + NH3
I I
NH-> NH2
глутамин глутаминовая кислота
У некоторых микроорганизмов гидролитическое дезаминиро-
вание протекает одновременно с декарбоксилированием той же
аминокислоты (Clostridium propionicutn):
ЗСНз—СН—СООН —2СНз—СНг—СООН + СНзСООН + СО2 + 3NH3
I н пропионовая уксусная
Nr*2 кислота кислота
аланин
К разряду гидролитического дезаминирования относится так-
же распад мочевины под каталитическим действием фермента
уреазы, образуемого рядом бактерий (Bacillus pasteurii, Sporo-
sarcina ureae, Proteus vulragis, Pseudomonas aeruginosa и др ):
H2N—C—NH2 + H2O — 2NH3 + CO2
252
Наиболее распространен окислительный вид дезаминирова-
I, проходящий с образованием кетокислот:
R—СН—СООН — R—С—СООН +Нг° > R—С—СООН + NH3
I II II
NH2 NH О
Биокатализаторы этих реакций — оксидазы аминокислот и
идрогеназы. Роль акцепторов водорода выполняют НАД,
Д (или ФМН). Подобному дезаминированию подвергается,
фимер, глутаминовая кислота под действием НАД-зависимой
таматдегидрогеназы:
НАД+
ООС—СН2—СНг—СН—СООН + НОН. _=±НООС (СН2) 2—С—СООН +
I II
nh2 о
а-кетоглутаровая
кислота
+ NH3 + HA4-H + H+
хтановленный НАД используется как восстановитель в соот-
ствующих реакциях биосинтеза. Глутаматдегидрогеназа —
?чевой аллостерический фермент в реакциях дезаминирования
ереноса аминогруппы.
ФАД- и ФМН-зависимые дегидрогеназы, катализирующие
1кции окислительного дезаминирования, называются оксида-
щ D- или /.-аминокислот:
ФАД+
ZJ-R—СН—СООН -l Trv— О-R—С—СООН + NH3 + ФАД-Н2
I + Н2О II
nh2 о
ФМН+
Z.-R—СН—СООН —Дц,* Z.-R—С—СООН + NH3 + ФМН-Н2
| +Н2О ||
nh2 о
ФАД- и ФМН-зависимые дегидрогеназы являются флавопро-
нами.
Аминокислоты могут подвергаться также декарбоксилирова-
с образованием СОг и биогенных аминов. Такие реакции
,е протекают в кислых средах под влиянием декарбоксилаз;
СН2— (СН2)з—СН—СООН СНг—(СН2)з—СН2 + СО2
nh2 Ah2 nh2 nh2
лизин кадаверин
триптофан
СН2—СН2
I +СО2
nh2
триптамин
253
Декарбоксилазы аминокислот имеются у сапрофитных и пато-
генных микроорганизмов. Причем у отдельных болезнетворных
видов декарбоксилазы могут быть отнесены к факторам агрессии
(см. гл. 7). Коферментом декарбоксилаз (за исключением гисти-
диндекарбоксилазы) служит пиридоксальфосфат.
Любые превращения аминокислот завершаются тем, что их
углеродный скелет оказывается в составе пировиноградной кис-
лоты либо ацетиЛ'КоА, включающихся затем в метаболизм угле-
водов и липидов. У фотосинтезирующих бактерий деградация
аминокислот играет малозаметную роль, поскольку они в соот-
ветствии с потребностями легко синтезируют необходимые ами-
нокислоты.
Отдельные аминокислоты могут использоваться, например,
псевдомонасами в качестве единственного источника углерода,
из которого синтезируются все необходимые клетке вещества.
В этом случае метаболизм аминокислот почти подобен метабо-
лизму, происходящему в печени животных, и если он сопровож-
дается образованием пировиноградной, а-кетоглутаровой и ща-
велевоуксусной кислот, то такие аминокислоты называют глико-
генными (Ала, Apr, Асн, Асп, Вал, Гис, Гли, Глу, Глн, Мет,
оксмПро, Про, Сер, Тре, Три, Цис). Если аминокислоты превра-
щаются в ацетил-КоА или ацетоацетат, то такие а-минокислоты
называют кетогенными (Лей). Известны аминокислоты, относя-
щиеся и к гликогенным, и к кетогенным одновременно (Иле, Лиз,
Тир, Фен).
Метаболизм каждой аминокислоты достаточно сложен и его
направленность зависит от ряда факторов (наличия у микроор-
ганизмов соответствующих ферментов, качества питательной
среды и др.). Например, окисление 20 природных аминокислот
разными микробами происходит по 20 различным путям с поис-
тине огромным числом биокатализаторов, участвующих в этих
реакциях, и промежуточных продуктов, которые могут быть пред-
шественниками различных клеточных компонентов. Пути ката-
болизма аминокислот переплетаются с другими метаболическими
путями, например с циклом трикарбоновых кислот, когда амино-
кислоты выступают поставщиками соответствующих углеродных
скелетов, из которых образуются определенные компоненты цик-
ла (рис. 57).
Анаэробные микроорганизмы {Clostridium botulinum, Cl-
perfringens, Cl. tetani) способны осуществлять катаболизм ами-
нокислот в парах (реакция Стикленда), когда одна из них слу-
жит донором электронов, а другая — акцептором их:
СНз—СН—СООН 4-2СН—СООН + 2Н2О - ЗСНз—СООН +3NH3 +СО2
ткн2 >jh2
аланин глицин
В результате указанного взаимного окисления и восстанов-
ления аминокислот происходит генерация АТФ и, следовательно,
аминокислоты при этом являются источниками энергии.
254
БЕЛКИ
пептидазы (протеолиз)
57. Связь катаболических путей аминокислот с циклом трикарбоновых
от
.4. Катаболизм пуриновых и пиримидиновых
еотидов
менты, катализирующие реакции гидролиза нуклеиновых
ют, полинуклеотидов и олигонуклеотидов, называют нуклеа-
и (нуклеодеполимеразами). Ферменты, специфичные лишь в
лпении ДНК или РНК соответственно, относят к ДНКазам
к РНКазам. Нуклеазы, атакующие полинуклеотидные цепи
•нцов, называют экзонуклеазами. Если же нуклеазы атакуют
ые а и б связи в схематично изображенной полинуклеотидной
J и не нуждаются в наличии свободной 3'- или 5'-гидроксиль-
группы на концах цепи, то их называют эндонуклеазами.
°/ v
. Р 3_________________
5' 6 ~а7
Р ,,
5 а/
Р 3'
б/ 3__________________
5' аТ
Р
6/
фрагмент полимерной цепи
основание
основание
основание
255
Экзо- и эндонуклеазы расщепляют фосфодиэфирные связи в
ДНК и РНК- Если гидролизуется связь а, то при этом освобож-
даются нуклеотидные единицы в виде нуклеозид-5'-фосфатов.
Если же гидролизуется связь б, то освобождаются нуклеозид^
З'-фосфаты. ДНКазы и РНКазы проявляют специфичность к
углеводным компонентам субстратов (дезоксирибозе и рибозе со-
ответственно), тогда как другие нуклеазы не обладают такой
специфичностью.
Деполимеризующие нуклеазы широко распространены среди
представителей микробного мира. Некоторые из них обнаружи-
ваются во внешней среде при росте микроорганизмов на подхо-
дящих субстратах (например, гуанил-РНКазы актиномицетов),
другие локализуются внутриклеточно.
Освобождающиеся в результате деполимеризации нуклеотиды
гидролизуются нуклеотидазами, или фосфатазами, с освобожде-
нием неорганического фосфата и нуклеозидов. На следующем
этапе нуклеозиды расщепляются нуклеозидазами до пентозы
(рибозы или дезоксирибозы) и оснований, которые могут исполь-
зоваться микроорганизмами для процессов биосинтеза; в других
случаях основания подвергаются дальнейшим превращениям.
Так, анаэробные бациллы из рода Clostridium могут расщеплять
их до аммиака, глицина, муравьиной кислоты и СО2.
Поскольку микроорганизмы обитают в природных условиях в
виде ассоциаций, образующиеся промежуточные продукты транс-
формации нуклеиновых кислот (нуклеотиды, нуклеозиды, осно-
вания, органические кислоты и аминокислоты) могут использо-
ваться ассоциантами или другими представителями биогеоцено-
зов, например растениями.
5.3. МЕТАБОЛИЗМ УГЛЕВОДОВ
В общебиологическом смысле углеводы выполняют защит <чо
роль. Это можно сказать, например, о полисахаридных капстжх
соответствующих микроорганизмов, о пептидогликанах и хитине
клеточных стенок бактерий и грибов соответственно, о поли-
цией-, дезокси-) рибозе в нуклеиновых кислотах и т. д. Даже,
будучи в виде запасного энергетического материала (глико ен,
гранулеза, крахмал), полисахариды первыми направляют1': в
«энергетическую топку» клетки при голодании микроорганп o.ia
или попадании его в экстремальные условия существовав ж-1 ”
отношении обеспечения источниками энергии. На фоне защит мй
роли углеводы выполняют многие другие соподчиненные ф'лж
ции: узнавания, транспорта, энергетические и др.
Распад отдельных, существенных для микроорганизмов угле-
водов завершается, как правило, образованием трех главных
моносахаридов — глюкозы, фруктозы и галактозы, которые слу-
жат исходным материалом для осуществления различных реак-
ций обмена веществ.
256
. Анаболизм углеводов
мотря на параллелизм путей биосинтеза и распада макро-
екул полисахаридов, эти два процесса во многом различны.
^синтез (анаболизм) начинается с образования прометабо-
ов, соединяющихся затем в полимерные углеводы — вторич-
• метаболиты. Следовательно, прометаболиты являются строи-
мыми блоками для синтеза полисахаридов. К ним относятся
жде всего фосфосахара (равно как аминокислоты для синтеза
ков, пуриновые и пиримидиновые основания для синтеза
Деиновых кислот и т. д.). Строительные блоки образуются
путях промежуточного или центрального обмена — амфибо-
ма (от греч. apupi— оба), связывающего сопряженно проте-
>щие процессы анаболизма (рис. 58) и катаболизма (распада
{еств).
Полисахариды микроорганизмов синтезируются из таких про-
куточных продуктов (прометаболитов, или интермедиатов),
3-фосфоглицериновая кислота, 3-фосфоглицериновый альде-
, глюкозе-1-фосфат. Из 3-фосфоглицериновой кислоты обра-
ггся все углеродсодержащие соединения клетки. В свою оче-
,ъ эта кислота образуется либо при катаболизме полисахари-
!, либо из углекислоты, формиата, ацетата и липидов.
Ключевым интермедиатом в процессе биосинтеза различных
еводов является также глюкозо-1-фосфат. Он образуется из
[фоенолпировиноградной кислоты (фосфоенолпирувата, ФЕП),
. из неуглеводного источника. Вот почему образование глюко-
(или полисахарида) из неуглеводных предшественников назы-
>т глюконеогенезом. Таким путем могут синтезироваться сво-
шисахариды
!«...
>угие углеводы
иокозо-6-фосфат
1/С=°
С—СН2—О—Р
(юсфоглицери-
|ВЬ1Й альдегид
Присоединение
водорода из
субстратов к НАД,
НАДФ, и ФАД
I ДЮ О
с;
Из АДФ +
цепь
переноса
электро-
нов
|>осфоглицерино-
Пя кислота
♦
’ * -,С00~
"=С^СН
Ируват
Фотосинтез'
О2 Н2О
tc. 58. Схема биосинтеза полисахаридов
617
257
бодная глюкоза, другие моносахариды, дисахариды, полисахарц.
ды клеточных стенок, запасные углеводы — крахмал, гликоген:
Пируват —► Оксалоацетат — Малат — Оксалоацетат — ФЕП —►
— 2-Фосфоглицериновая кислота —► Глицероальдегид-З-фосфат
—- Фруктозодифосфат — Фруктозо-6-фосфат —- Глюкозо-6-фосфат —►
р- Глюкозо-1-фосфат —< УДФ-глюкоза — полисахарид
I—.Глюкоза
Глюконеогенез характерен для гетеротрофных микроорганиз-
мов. У автотрофных фотосинтезирующих микробов глюкоза об-
разуется при восстановлении углекислоты по следующей реак-
ции:
6СО2 + 18АТФ + 12НАДФ • Н + 12Н+ — C6Hi2O6 + 18Ф„ + 18АДФ + 12НАДФ+
глюкоза
Эта суммарная реакция включает 15 стадий, из которых 8 на-
правлены на образование глюкозы и 7 — на регенерацию рибу-
лезо-1,5-дифосфата (цикл Кальвина).
Глюконеогенез у микроорганизмов возможен также из про-
межуточных продуктов цикла Кребса, ацетил-КоА, аминокислот
(например, аланина, аргинина, валина, глутаминовой кислоты,
изолейцина, лизина, метионина, пролина, серина, треонина
и др.).
Благодаря реакциям в цикле ди- у трикарбоновых кислот
(см. разд. 5.3.2), а также в глиоксилатном цикле, ацетил-КоА
и аминокислоты могут вносить свой вклад в биосинтез глюкозы
t
Реакции, катализируемые изоцитратазой
и малатсинтазой, не участвующих в цикле Кребса
Рис. 59. Схема глиоксилатного цикла
258
УДФ-ГалА
УДФ-ИдА
УДФ-Глю А
У^Ф-Глю _
УДФ-Кси
УДФ-L-Apa
УДФ-Гал
ГДФ-Глю
i-Ф
Глю-1-Ф
Ф
Фосфоинозит
Ф-6-Ф
♦
Ман-6-Ф
f
Ман-1—Ф
♦
ГДФ-Ман
АДФ-Г лк>
УДФ-Глю
УДф-Мур N Ас
УДФ-Глю N Ас
N Ас
Глю N Ас-6-Ф
ЦМФ Nana
ГДФ-Д-Фук
ГДФ-МанА—*- ГДФ-ГулА
•ис. 60. Основные пути превращений глюкозо-6-фосфата до нуклеозид-
(ифосфатсахаридов у микроорганизмов:
£лю — глюкоза; Гал — галактоза; ГлюА — глюкуроновая кислота; ГалА — галак-
уроновая кислота; Ара — арабиноза; ГлюЫАс — N-ацетилглюкозамин; Гa.iN Ас —
(•ацетилгалактозамин; Глю\Ас-6-Ф — N-ацетил-глюкозаминил-б-фосфат; Ман —
Мнноза; МанА— маннуроновая кислота; ГулА — гулуроновая кислота; Рам —
«миоза; Кен — ксилоза; УДФ — уридиндифосфат; Ф-6-Ф — фруктозо-6-фосфат;
tax — сахароза; Ф — фосфат неорганический; ГДФ — гуанидиндифосфат; ТДф-
ЙИиДиндифосфат; ЦМФ — цитидинмонофосфат: Nana — нейраминовая кисло-
3; Фук — фукоза; ИдА — идуроновая кислота; MypNAc — N-ацетилмурамовая
«слота
I глиоксилатном цикле на 2 молекулы потребляемого ацетил-
ft выделяется одна молекула янтарной кислоты и два атома
юрода. Янтарная кислота используется, в частности, в био-
Йгезе глюкозы, а водород включается в дыхательную цепь
ислительное фосфорилирование). Янтарная кислота транс-
иируется в щавелевоуксусную, служащую предшественником
а эта последняя — в глюкозо-1-фосфат:
Юс— (СН2)г— СООН — 2НООС—СНг—СО—СООН — 2СН2=С—СООН —
сукцинат
оксалоацетат
фосфоенолпнруват
:н—(снон)4—сн2он
О— Р
глюкозо-1 -фосфат
акне аминокислоты, как глутаминовая и аспарагиновая,
т непосредственно превращаться в а-кетоглутарат и оксало-
259
Глю —Глю-6-Ф ——» Глю-1-Ф
УТФ
УДФТлю-пирофосфорилаза
АДФ-Глю-пирофосфорилаза
АДФ-Г лн>
Липополисахарид
УДФ-Гал
Экзополисах ар ид
УДФ-Глю
УДФ-Гал-4-эпимераза.
УДФ-Глю А
УДФТлю-дегидрогеназа
Гликоген
Экзополисахарид
Экзополисахар ид
Липополисахарид
Рис. 61. Схема анаболических превращений глюкозы у микроорганизмов
ацетат соответственно. Другие аминокислоты трансформируются
в глюкозу через ацетат-КоА.
Глюкоза при необходимости может трансформироваться в
другие моносахариды, содержащие 6 углеродных атомов, напри-
мер в галактозу, маннозу, фруктозу, глюкуроновую кислоту. При
этом обязательно участие соответствующих ферментов. Так, эпи-
меризация глюкозы в галактозу протекает в 3 стадии с участием
УДФ, НАД, НАД-Н и гидролазы.
Нуклеозиддифосфатсахара могут служить источником нук-
леозиддифосфатпроизводных дезоксисахаров типа Л-рамнозы и
L-фукозы, входящих, например, в состав липополисахаридов
клеточных стенок ряда грамотрицательных бактерий (Salmonella
species). На рис. 60 приведены основные пути превращений моно-
сахаридов с участием соответствующих нуклеотидов.
В обобщенном виде анаболические превращения глюкозы до
конечных полисахаридов показаны на рис. 61.
УДФ-глюкопирофосфорилаза для многих микроорганизмов
является ключевым ферментом, катализирующим реакции сио-
синтеза экзополисахаридов и полисахаридов клеточной стенки.
Пути биосинтеза клеточных и внеклеточных полисахаридов мик-
роорганизмов неравнозначны, так как, например, внеклеточные,
внутриклеточные резервные и клеточно-стеночные углеводы от-
личаются друг от друга по строению. На основании всестсу лп
него изучения реакций, ведущих к образованию того или и.;; то
полимера у бактерий, были определены преимущественные п\ •• и
очередность синтеза полисахаридов (рис. 62).
Изопреноидные структуры с числом С-атомов от 50 до 60
(бактопренолы) у прокариот и с числом С-атомов от 80 до 100
(долихолы) у эукариот сосредоточены на уровне мембранного
аппарата клеток. Причем гидрофобная часть изопреноидов за-
крепляется в мембране, а гидрофильная — выступает в цито-
260
Ьзму. Способность до-
кола переносить, на-
ИА-киназа
вмер, маннозильный
таток при синтезе
щнана у дрожжей
ображена на рис. 63.
В цикле долихолфос-
гга переносится радикал
(ГлкМАц)2Ман9Глю3]
! аспарагин в белко-
ft молекуле с образо-
иием гликопротеина.
(икопротеин может
одить в состав каких-
бо органелл либо
кретироваться во вне
апример, фермент ин-
ртаза у дрожжей).
Муреиновый каркас
жтерий, являющийся
ИА
ИФ-фосфотаза
Гликопротеин
Липополисахарид или
тейхоевая кислота
Экзополисах арид
Рис. 62. Преимущественные пути и очередность
синтеза полисахаридов у бактерий:
ИПФ — С55-нзопреннлпнрофосфат; ИФ — С55-НЗ0-
преннлфосфат; ИА — Css-нзопреноид-алкоголь
лтидогликаном, или протеогликаном, синтезируется с участием
йтопренола. Очередность реакций при этом следующая:
1
1. УДФ-Глю\Ац-ФЕП, УДФ-ГлюЫАцЕП + Ф„
2. УДФ-ГлюЫАцЕП НАДФ-Н + Н+ MypNAu + НАДФ+
3. УДФ-MypNAu АТФ’ £~АлД УДФ-MypNAu (или УДФ-Мур\Ац-£-Ала)
О
I
ОС—СН—СНз
nh2
4. УДФ-MypNAц-^.-Aлa АТФ’ ° Е~Д yдф.MypNAц-Z.-Aлa-£)-Глy
'ис. 63. Участие долихолфосфата в синтезе дрожжевого маннана:
(ол-Ф-долнхол — монофосфат; Сер — серии; пп — полипептид
261
5. УДФ-МурЫАц-А-Ала-В-Глу АТФ' - ~Ли?УДФ-МурМАц-£-Ала-О-Глу-/.-Лиз
6. УДФ-Мур^ц-£-Ала-В-Глу-£-Лиз АТФ’ ДАла-О-Ала
— УДФ-МурМАц-£-Ала-О-Глу-£-Лиз-О-Ала-В-Ала
(или УДФ-МурЫАц-пеитапептид)
Дипептид £>-Ала-О-Ала (реакция 6) синтезируется бактериаль-
ной клеткой отдельно.
Единица синтезированного муреинового каркаса УДФ-
МурЫАц-пентапептид включается в так называемый муреино-
вый цикл, протекающий с участием бактопренол-фосфата
(Бол-Ф). Биосинтез пептидогликана клеточной стенки Staphylo-
coccus aureus показан на рис. 64.
Тейхоевая кислота грамположительных бактерий синтезирует-
ся сопряженно с синтезом пептидогликана-муреина. Оказалось,
что пути синтеза стеночных и мембранных тейхоевых кислот не-
тождественны. На путях синтеза пептидогликана имеются звенья,
подверженные ингибиторному (тормозящему) действию пяти
антибиотиков: 1) циклосерин ингибирует реакции присоединения
£>-аланина перед образованием УДФ-МурЫАц-пентапептида;
2) бацитрацин ингибирует реакции на уровне включения бакто-
Пептидогликам
пентапептидамид
пентаглицин (связующий мостик)
•Рис. 64. Биосинтез пептидогликана клеточной стеики Staphylococcus aureus
262
|лфосфата; 3) ванкомицин и ристотецин ингибируют реак-
после отделения бактопренол-фосфата на предпоследней
и формирования пептидогликана; 4) пенициллин ингиби-
последнюю стадию образования пептидогликана (перед от-
ием Д-аланина).
энорами сахарных компонентов для стеночных тейхоевых
т являются соответствующие УДФ-сахара, а источниками
фосфатных единиц —ЦДФ-рибит и ЦДФ-глицерин. Фер-
I, катализирующие синтез стеночных тейхоевых кислот и
^ящиеся в плазмалемме, нуждаются для своей деятельности
гаточно высоких концентрациях ионов магния (10...50 мМ).
LO путях биосинтеза мембранных тейхоевых кислот известно
Достаточно. Для некоторых из них донором глицерофосфатной
Дни цы выступает фосфатидилглицерин, переносящийся без ка-
|-либо предшественников. Подтверждается также факт «заин-
|есованности» полипреноид-фосфатов в синтезе тейхоевых кис-
| обоих типов. О-Антигены клеточных стенок бактерий из се-
|ства Enterobacteriaceae синтезируются также с участием изо-
^нолфосфата, или бактопренолфосфата, являющегося перенос-
ном фосфогалактозильного остатка, к которому присоединяют-
другие моносахаридные звенья в последующей цепи реакций.
Таким образом, с участием изопреноидфосфатов (бактопре
1Ы, долихолы) синтезируются многие гликопротеины, пептидо-
|жаны (протеогликаны), гликолипиды и маннаны клеточных
;нок дрожжей, но не гликаны и не хитин.
.Главный компонент клеточной стенки дрожжей рода Saccha-
*nyces — (J-CI — СЗ-глюкан, хотя в ее составе имеются также
Ьаннан и маннопротеин. Синтез хитина и глюкана катализи-
бтся хитинсинтазой и глюкансинтазой соответственно. Хитин-
Ьтаза сосредоточена преимущественно в хитосомах, глюкан-
втаза — в плазмалемме. Синтез хитина осуществляется по
“ме:
.ГлюЫАц^ + УДФ-ГлкМАц-----------------------►(ГлкМАц)п-н + УДФ
Хнтинсинтаза хитин
В процессе сборки молекул глюкана, например целлюлозы,
Юкозильные единицы переносятся глюкансинтазой от ГДФ-
Юкозы или УДФ-глюкозы:
«ю)„ + ГДФ-Глю (или УДФ-Глю)----------(Глю)„+1 + ГДФ (или УДФ)
Глюкансинтаза целлю-
лоза
По аналогичному пути идет синтез гликогена и крахмала.
Внеклеточные полисахариды синтезируются преимущественно
уровне мембран и затем выделяются в окружающую среду,
Вример в культуральную жидкость. Однако есть полисахари-
которые синтезируются внеклеточно, например декстран, об-
эуемый видами Leuconostoc.
В процессе биосинтеза полисахаридов клетками микроорга-
263
низмов полимер нередко оказывается полидисперсным, что обус-
ловлено опосредованным контролем углевода по схеме: ДНК ^
—► РНК —► Белок (фермент)—► Полисахарид. Субстрат, из кото-
рого синтезируется полисахарид, может не проникать в клетку,
однако чаще проникает в нее. Известны три механизма этого
процесса: облегченная диффузия, активный транспорт, группо-
вая транслокация.
Ряд полисахаридов содержит ацильные и кетальные группы.
Время их присоединения к полимеру точно неизвестно. Пред-
полагают, что ацилирование происходит в тот период, когда
полимер находится в связанном с липидом состоянии. Освобож-
дение полисахаридов от изопреноид-липидов, очевидно, обуслов-
лено действием ферментов.
5.3.2. Катаболизм углеводов
Реакции катаболизма углеводов относятся преимущественно к
окислительному типу. Они сопровождаютя выделением энергии,
частично накапливаемой в форме АТФ (коэффициент полезного
действия в отношении накопления энергии, например в цикле
Кребса, достигает 39%).
Процесс катаболизма углеводов условно можно подразде-
лить на три стадии:
1) распад полисахаридов на составляющие их мономерные
единицы. Например, глюкан (СбНюО5)л распадается на п моле-
кул глюкозы по схеме:
(С.„Н|„О5)„ + пН2,О—юканаза, гаСбН1гОб
' глюкоза
2) превращение моноз в глицеральдегид-3-фосфат с после-
дующим расщеплением его до ацетила:
С—н
С6Н12О6--...Н—С—ОН _ ...СН3СО-КоА
I
СН2—О—Р
ацетильный радикал чаще всего входит в состав ацетил-КоА;
3) окисление ацетила до СО2 и Н2О.
Катаболизм и анаболизм углеводов у эукариот разобщены
(компартментализованы) пространственно, энзиматически и ре-
гуляторно, но существуют метаболические перекрестки — амфи-
болические пути. Кроме того, связь катаболических и анаболи-
ческих реакций существует потому, что продукты первых из них,
как правило, служат субстратом для вторых.
Катаболизм углеводов включает:
а) анаэробные процессы — гликолиз (гликолитический рас-
пад, фруктозо-дифосфатный, или ФДФ-путь, превращение угле-
264
,ов по Эмбдену—Мейергофу—Парнасу), гексозомонофосфат-
i шунт (пентозо-фосфатный, или ПФ-путь, окисление глюкозы
Варбургу—Дикенсу—Хорекеру), кетодезоксифосфоглюконат -
й, или КДФГ-путь (превращение глюкозы в трехуглеродные
зечные продукты по Энтнеру—Дудорову, или ЭД-путь);
б) аэробный процесс — цикл ди- и трикарбоновых кислот
икл лимонной кислоты, или цикл Кребса).
Гликолиз (от греч. yXixro — сладкий; от лат. lysis — раство-
ше, распад). Это один из видов брожения, посредством кото-
'о анаэробные микроорганизмы получают химическую энергию
углеводов в форме АТФ. Наблюдаемые при этом реакции от-
:ятся к окислительно-восстановительному типу. Донором элек-
жов является углевод, акцептором электронов — какая-ни-
хь органическая молекула, образующаяся в процессе этого же
зжения. При гликолизе средняя степень окисления исходного
конечного продуктов оказываете^ одной и той же. Так, при
иртовом брожении
С6Н|2О6 — 2СО2 + 2СгН5ОН
шень окисления углерода в глюкозе равна нулю, в углекислоте
-4) и в этаноле (—2), тогда баланс окисления и восстановле-
я будет следующим: 6X0 = 2X4 +2 [2Х (—2) ] = 8 + (—8).
Синтез АТФ при гликолизе осуществляется на уровне суб-
эата. Ключевым продуктом здесь выступает пировиноградная
слота (пируват). Из одной молекулы глюкозы образуются
молекулы АТФ и 2 молекулы НАД-Н2.
Гликолитический распад глюкозы осуществляют гомофермен-
тивные молочнокислые бактерии в такой последовательности
акций:
D-фруктозо-1,6-дифосфат
265
4)
СН2—О—Р
___________ . Н—С—ОН
K+Zn2+Ca2+,Fe2+ I 0
Альдолаза
СН2—О—Р
сн2—О—р
'=0
СН2ОН
5)
6)
2
СН2—О—Р
сн,он
.0
|ХН
Н—С—ОН
СН2—О—Р
С С
Р
Т риозофосфатизомераза
Г лицероальдегид -3-
фосфат-дегидрогеназа
+2НАД++ 2ФН
'Н
Ц-глицероаль-
дегид-3- фосфат
диоксиацетон -
фосфат
с:
D-глицероальдетид-З-
- фосфат
2 Н—С—ОН +
2НАД-Н + 2Н+
СН2—О—Р
13-дифосфогицерат
.0
7)
2
н—С—ОН
Фосфоглицераткиназа
+ 2АЛФ
2 Н—С—ОН + 2 АТФ
8)
2
9)
2
10)
сн2—о—Р
СН2—О—Р
3 - фосфоглицерат
z
Ск
сС
н—С—ОН
СН2—О—Р
с;
Н—С—О—Р
СН2ОН
Фосфоглицератмутаза
+ Mg
Енолаза
+ Mg2+ или Мп2+
2 Н—С—О—Р
СН2ОН
2-фосфоглицерат
cf
2 С—О—Р + 2Н2О
сн2
фосфоенолпируват
2
С—О—Р
Пируваткиназа
+ 2АДФ, К+, Rb2+ или Cs2+,
Mg2+ или Мп2+
2 С=О + 2 АТФ
сн3
пируват
СН2
266
! И) 2 С=О
Лакгатдегидрогеназа
+ 2НАД-Н +Н +
2 Н—С—ОН + 2НАД +
сн3
реакций гликолиза
сн3
лактат
можно подразделить на
Одиннадцать
^стадии. Первая из них включает реакции 1...5, в результате
торых глюкоза превращается в глицероальдегид-3-фосфат и
5людается расход энергии 2 АТФ; вторая стадия включает
акции 6...11, когда в результате окислительно-восстановитель-
х процессов отмечается образование 4 АТФ и лактата. Следо-
гельно, в процессе гликолиза из одной молекулы глюкозы об-
язуются 2 молекулы лактата, а 2 молекулы АДФ за счет неор-
“нического фосфата превращаются в 2 молекулы АТФ. Выиг-
;ш энергии при этом составляет немногим более 120 кДж, или
tee 30 ккал (1 кал = 4,1868 Дж).
Совпадает с гликолизом спиртовое брожение, осуществляе-
>, например, дрожжевыми организмами. Разница лишь в
I, что 11-я реакция при гликолизе заменяется двумя другими
1 спиртовом брожении:
:О
На) 2С=<
СНз
Пируватдекарбоксилаза 2СН3 | 2СО2
ацетальдегид
;О
С< СН2ОН
I Nd . I
mi оАи Алкогольдегидрогеиаза
!2) 2СН3 +2наД.Н + Н+ ' 2СНз
этанол
Суммарная реакция спиртового брожения следующая:
СвН|2Об -|- 2АДФ -j- 2Ф„ —► 2СО2 -|- 2С2НзОН -|- 2АТФ -|- 2Н2О
[игрыш энергии составляет 166 кДж/моль-1.
Полисахариды, например крахмал, гранулеза, гликоген, мо-
.' использоваться многими микроорганизмами, причем началь-
;е этапы гидролиза данных полимеров протекают по схеме:
(СбНюОз)„ а-1 л Глюканфосфорилаза Глю-1-Ф + (С6Н,0О5)л_,
4~Фн
Глюканфосфорилаза атакует полисахарид с нередуцирующе-
0 конца, отщепляя один за другим глюкозные остатки, присоеди-
«ющие пирофосфат (фосфоролиз) к С1. Следовательно, каждый
аг реакции сопровождается укорочением полисахарида на еди-
267
ницу монозы. В тойках ветвления вступает в реакцию а-1,6-глю-
каназа, после чего глюканфосфорилаза вновь атакует полимер.
Образовавшийся глюкозо-1-фосфат под действием фермента
фосфоглюкомутазы трансформируется в глюкозо-6-фосфат, а
этот последний включается в цепь реакций гликолиза:
Глю-1-Ф «-М-Фосфоглюкомутаза^ Глю.6.ф
+Mg2+
Глю-6-ф -ФоЛ’оглю^ОИЗОМ1?£аза-» фру-6-ф и т. д.
Если гликолиз протекает в среде с крахмалом, то всего обра-
зуется 3 молекулы АТФ (вместо двух при сбраживании глюко-
зы); энергетический баланс реакций составляет 149,34 кДж
(AG = —35,67 ккал).
Гликолитический процесс лежит в основе других брожений.
При этом пируват как ключевой промежуточный продукт транс-
формируется в какой-либо вторичный метаболит. Возможность
образования последнего зависит от ферментативного оснащения
соответствующего организма (табл. 27).
Таблица 27. Примеры брожений, основанных на гликолизе
Вид брожения Микроорганизмы, вызывающие брожение Конечный продукт, илн вторичный метаболит
Молочное гомо- ферментативное Streptococcus sp., Lactoba- cterium sp. Молочная кислота
Спиртовое Некоторые грибы (преиму- щественно дрожжи Saccharo- myces sp.) Этанол
Пропионовокис- лое Clostridium propionicum, Propionibacterium sp., Coryne- bacterium diphtheriae, некото- рые виды из родов Micromo- nospora, Neisseria, Veillonella Пропионовая кислота, уксусная кислота, янтарная кислота
Молочнокислое гетерофермента- тивное Бактерии из родов: Esche- richia, Proteus, Salmonella, Schigella Молочная кислота, ук- сусная кислота, муравьиная кислота, этанол, янтарная кислота
Маслянокислое Butyribacterium sp., Sarcina maxima, некоторые виды из родов Clostridium, Neisseria Бутанол, масляная кисло- та, ацетон, изопропанол, уксусная кислота, этанол
Бутилен-гликоле- вое Некоторые виды из родов: Aeromonas, Bacillus (polymy- ха), Enterobacter Ацетон, бутилен-гликоль, молочная кислота, муравьи- ная кислота
Молочнокислое гетероферментативное и маслянокислое бро-
жения являются смешанными по характеру образующихся ко-
нечных продуктов и поэтому данные им названия являются до-
вольно условными.
Названные брожения используют на практике в различных
268
нах мира. В СССР особенно широко распространены спир-
ю и молочнокислое гомоферментативное брожения. Некото-
вторичные метаболиты являются диагностическими тестами
выявлении и определении отдельных видов патогенных и
овно патогенных микроорганизмов (например, относящихся
семейству Enterobacteriaceae).
Гексозомонофосфатный шунт. Данный путь распада углево-
в, включающий две фазы, заметно отличается от гликолиза
и, что реакции окисления имеют место уже на первых ступе-
[X процесса, а углекислота оказывается характерным продук-
»м, который вообще не образуется при гликолизе. Первая фаза
-пути состоит из следующих реакций:
6-фосфоглюконолактон
1 6-Фосфоглюконатдегндрогеназа I
4) НО—С—Н +НДДФ+ Mgr+. Мп2+ нлн Са2+ С=О
(НСОН)г (НСОН)2
6СН2—О—Р 6СН2—О—Р
3-кето-6-фосфоглюконо-
СООН вая кислота
неон сн2—он
5) с=о С=о
(НСОН)2 (НСОН)2
I I
СН2—О—Р СН2—О—Р
рибулезо-5-фосфат
269
Рибулезо-5-фосфат является ключевым продуктом в ПФ-пути
от которого последующие реакции превращения пентозы обра-
тимо могут протекать в различных направлениях (ветви а и б
второй фазы гексозомонофосфатного шунта). Ветвь а включает
реакции 6а 66, 7а и 76; ветвь б — реакции 8а, 86 и 9:
6а 66, 7а и 76; ветвь б — реакции 8а, 86 и 9:
СН2ОН
6а)
с=о
н—С—ОН
Рибозо-5-фосфат-кето-изомераза
С—он
(Н—С—ОН) 2
(Н—С—ОН) 2
СН2—О—Р
СН2—О—Р
промежуточная енольная
форма рибулезо-5-фосфата
СН2—он
СНг—ОН
66)
с=о
Н—с—он
с=о
Рибулезо-5-фосфат-нзомераза
7а)
н—с—он
;Нг—О—Р
н—с—он
с—он
н—с—он
н—с—он
н—С—он
ксилулезо-5-фосфат
рибозо-5-фосфат
СН2—О—Р
рибозо-5-фосфата может присоединиться еще один оста-
ток пирофосфата за счет АТФ в присутствии солей магния, тогда
образуется рибозо-1,5-дифосфат.
сн2—о—р
К С1
76)
Фосфотрансфераза
рибозо-1-фосфат
Ветвь а ПФ-пути
является важной
для синтеза
и нуклеиновых кислот в клетках микроорганизмов:
+ СН,—он
нуклеотидов
+ СН2ОН
+ с=о
.О
СН2—О—Р
8а) НО—С—Н
н—с—он
Транскетолаза
+ Mg
НО—С—н
н~с—он
(НСОН)з
’СН2—О—Р
СН5-0—р
СН2—О—Р
седогептулезо-7-
- фосфат
3-фосфоглииери-
новый альдегид
270
86)
СН2ОН
’С=О
НО—С— н
(НСОН)з
;О
Н-*С—ОН
Трансальдолаза
*СНг—О— Р
"СНг—ОН
"С=О
Н—С=О
но—с—н
н—С—ОН
н-^с—он
H-tC—он
н—с—он
*СН2—О—Р
фруктозо-6-фосфат
СН2—О—Р
эритрозо-4-фосфат
Н—С=О
СНг—ОН
9) Н—С—ОН
Н—С—ОН
•С=О
но—<!:—н
Транскетолаза
+Mg2+-"
СН2—О—Р
н—С—ОН
ксилулезо-5-фосфат
(из редакции 66)
СНг—ОН
НО—С—Н
Н—С—ОН
H—с=о
н—он
н—с—он
СНг—О—Р
фруктозо-6-фосфат
3-фосфоглицериновый альдегид
Кроме участия в реакции 9 эритрозо-4-фосфат может, напри-
ер, использоваться бактериями для синтеза ароматических ами-
окислот.
Семь реакций второй фазы ПФ-пути (ветви а и б) представ-
яют собой лишь процессы превращения пентозофосфатов в
:ксозофосфаты и обратно, тогда как первые пять реакций за-
‘ршают процесс окисления глюкозы образованием рибулезо-
фосфата.
271
Гексозомонофосфатный шунт присущ большинству микроор-
ганизмов. Он в двух точках может смыкаться с гликолизом и,
следовательно, оба этих пути взаимно дополняют друг друга при
необходимости образования таких ключевых веществ, как фрук-
тозо-6-фосфат и 3-фосфоглицериновый альдегид, которые могут
переключить ПФ-путь на гликолитический. Тем не менее гексозо-
монофосфатный шунт имеет выраженный циклический характер
(рис. 65).
Следует заметить, что кроме окислительного ПФ-пути имеют-
ся неокислительные ПФ-пути перестройки гексозофосфатов в
пентозофосфаты, эритрозо-4-фосфат и седогептулезо-7-фосфат:
2,5С6 ЗС5; 2Сб ЗС4; 3,5Сб -► ЗС7. С помощью ПФ-пути рас-
щепляется до 20...30 % гексоз, например клетками дрожжевых
организмов рода Candida. Здесь не происходит запасания энер-
гии в форме АТФ, однако накапливается восстановленный
НАДФ в форме [НАДФ-Н-|-Н+], используемый клетками в
анаболизме.
Кетодезоксифосфоглюконатный (КДФГ) путь. Ключевым
продуктом пути Энтнера—Дудорова является 2-кето-З-дезокси-
6-фосфоглюконовая кислота:
а. - D - глюкозо - 6 - фосфат
272
2)
СН2—О—Р
о
НАДФ^ Mg 2 ‘ или Са2 +
Г люкозо-6-фосфат-
дегидрогеназа
СН2—О—Р
о
о
6-фосфоглюконолактон
СН2—О—Р
о
3)
соон
Глюконолактонгидролаза
+Н2О, Mg2+, Мп2 + или Са2+
СООН
(СНОН)4
СН2—О—Р
6-фосфоглюконовая кислота
соон
н—с—он
с=о
Ъ 4) НО—С—Н
6-Фосфоглюконатдегидр атаза
фНАДФ + , Mb,iF, Мп2+ или Са1'
сн2
н—с—он
н—с—он
н—С—он
н—с—он
СН2—О—Р
СН2—О—Р
2-кето-3-дезоксн-6-фосфо-
глюконовая кислота
СООН
с=о
5)
2-кето-3-дезоксн-6-фосфо-
глюконат-альдолаз а
соон
I Н
+ н—с—он
н—с—он
н—с—он
СНз
пируват
СН2—О—Р
3-фосфоглице-
риновый альдегид
СН2—О—Р
Образовавшийся 3-фосфоглицериновый альдегид окисляется
> пирувата согласно реакциям 6... 10 пути Эмбдена — Мейерго-
Парнаса. В результате окисления глюкозы в КДФГ-пути
фазуется 1 моль АТФ. Впервые этот путь был открыт у
seudomonas saccharophila.
' В сравнительном плане ФДФ- и ПФ-пути имеют универсаль-
>е значение, так как их можно обнаружить, как правило,
^большинства микроорганизмов, хотя завершенность процессов
^разных видов выражена неодинаково. Например, Rhodotorula
facilis не содержит фермента фосфофруктокиназы (3-я реакция
|иколиза) и поэтому не может довести до конца гликолитиче-
|ий процесс — вся глюкоза расщепляется в ПФ-путях (20 % —
«окислительном и 80 % — в неокислительном). Продуцент дек-
Ьана Leuconostoc mesenteroides не содержит фермента альдо-
В 273
лазы и, следовательно, не может расщеплять ФДФ на две моле-
кулы триозофосфата. Поэтому превращение молекулы глюкозы
осуществляется по пути реакций в ПФ-шунте, смыкающимся
с гликолизом на уровне пирувата у гетероферментативных
молочнокислых бактерий.
КДФГ-путь присущ немногим микробам и главным образом
тем из них, которые способны включать в катаболические реак-
ции глюконовую кислоту. Это псевдомонады, водородные бакте-
рии и некоторые другие. Учитывая тот факт, что в природе суще-
ствует больше аэробных микроорганизмов и факультативных
анаэробов в сравнении с облигатными (безусловными) анаэро-
бами, можно не сомневаться в том, что типичный гликолитиче-
ский процесс оказывается единственным способом получения
энергии у анаэробов.
Факультативные анаэробы могут осуществлять брожение
(гликолиз) и дыхание (аэробное окисление, при котором акцеп-
тором электронов выступает кислород) одновременно. Очевидно,
что строгие аэробы используют только дыхание для получения
энергии, запасаемой в форме АТФ. Аэробное окисление эволю-
ционно более позднее, чем гликолиз (или ПФ- и ЭД-пути), и
энергетически более выгодное:
Брожение: С6Н|2Ос - ► 2С2Н5ОН + 2СО2 + 166 кДж/моль1
, глюкоза
Дыхание: СеН^Ое —> 6СО2 -J-6H2O + 2872 кДж/моль-1
глюкоза
На примере окисления глюко<ы витно. чю броженш •
чем в 17 раз энергетически менее в.+ i одно. чем дыхание
Цикл ди- и грикарбоновых KUC’ior {цикл Кребна) заве.'
окислительный катаболизм л.юбок) шеогоисiочинка в ичц •
условиях. Как уже были смвало в начале pa ; юла 5 3.2. в
стадия превращения моноз соировожт;ич<’я образование.•
тильного радикала, входящего в а че: ил КоА. На
шающей третьей стадии аиехиз <>> ю он'-пя щ. ю юющ-
воды. Точнее, вида образуется из аюмоз во нч'ода ( умм
уравнение цикла Кребса следующее:
СНзСООН +2Н2О 2СО2 +8Н
Водород или электроны водорода включаются в цепь пере-
носа электронов (дыхательная цепь), переносящую его к молеку-
лярному кислороду:
Углевод или другой
органический субстрат
НАД -------Флавопротеин----------»-
I I
(-0,32) (-0,20)
Цитохромы
I---------1
Хинон -b —► с —а ---------------О 2
I 1111
(-0,07) (+0,01) (+0,22) (+0,34) (+0,81)
274
НАД
С—СООН
СН2—СООН
оксалоацетат
НО—СН—СООН
СН —СООН
£-малат
в Н2О
Н—С—СООН
•С—С —Н
фумарат
СН2СООН
сукцинат
ФАДН2
ФАД
---@ Малонат
ГТФ
ГДФ+ФН
loA-SH
СН2
С <л S-KoA
сукцинил-КоА
Углевод
S—КоА
, (С2) KoA-SH
Н2О
НО—С —СООН
цитрат
Флуороацетат
Н2О
С—СООН
СН—СООН
uuc-аконитат
Fe
Н2О
СН —СООН
н— —СООН
изоцитрат
НАД
I 4
НАДН+Н^***|
СН—СООН
о=с—СООН
оксалосукцинат
Арсенит
СО2
а-кетоглутарат
НАДН+Н
НАД
сн2
о—с—соон
О
СО2
е. 66. Схема реакций в цикле Кребса с указанием мест ингибирования флуоро-
ггатом, арсенитом и маловатом:
- цитратсннтетаза; 2,3 — аконитаза; 4,5 — изоцитратдегидрогеназа; 6 — а-кетоглута-
/дегидрогеназный комплекс; 7 — сукцннаттиокиназа; 8 — сукцинатдегндрогеназа; 9 -
мараза; 10 — малатдегидрогеназа
275
В круглых скобках указаны значения окислительно-восста-
новительного потенциала в вольтах при стандартных условиях
(pH 7, температура 25 °C, концентрация 1 М). Следовательно,
в направлении от углевода к кислороду происходит заметное
уменьшение свободной энергии и сопряженно синтезируется АТФ
из АФД и неорганического фосфата (окислительное фосфорили-
рование) .
Изменение свободной энергии сопровождается переходом
пары электронов от НАДН кОги образованием 3 молекул АТФ.
Переносчики электронов связаны с мембраной и ориентированы
таким образом, что перенос электронов сопровождается переда-
чей протонов от внутреннего листка мембраны к внешнему. Как
только мембрана становится непроницаемой для протонов, т. е.
энергизованной, то говорят о том, что она достигла протонного
градиента.
Энергия протонного градиента может использоваться клеткой
для осуществления различных процессов: активного транспорта
других ионов и небольших органических молекул, вращения
жгутиков и 1енерации АТФ.
Окислительные реакции цикла Кребса сосредоточены в ми-
тохондриях эукариотических клеток, хотя и в цитоплазме могут
быть обнаружены некоторые ферменты, относящиеся к циклу, но
они включаются преимущественно в реакции других метаболи-
ческих путей (аконитаза, малатдегидрогеназа, фумараза). Клет-
ки прокариот осуществляют реакции цикла Кребса в цитоплазме
и в кооперации с плазмалеммой. В настоящее время все сходятся
на той точке зрения, что цикл ди- и трикарбоновых кислот одно-
типен для всех аэробов, находящихся на разных ступенях эволю-
ционной лестницы. На рис. 66 приведены реакции цикла в общем
виде.
Ацетил, образующийся при разложении какого-либо углевода
(возможно также из белков, липидов), активируется с помощью
КоА и в такой форме подвергается дальнейшим превращениям.
Во время каждого оборота двухуглеродный ацетил соединяется
с четырехуглеродной щавелевоуксусной кислотой, образуя шести-
углеродную лимонную кислоту. Эта последняя отдает две моле-
кулы СО2 и через янтарную кислоту окисляется вновь до щаве-
левоуксусной, готовой начать очередной цикл взаимодействия с
ацетилом.
В результате окисления одной молекулы ацетила, катализи-
руемого дегидрогеназами, образуется 3 молекулы НАДФ-Н и
одна молекула ФАД-Н2. Восстановительные коферменты переда-
ются в дыхательную цепь, где НАД-Н генерирует 3 высокоэнер-
гетические молекулы АТФ благодаря этерификации АДФ с обра-
зованием АТФ в ходе окислительного фосфорилирования;
ФАД-Нг в этих условиях генерирует образование двух молекул
АТФ, так как на первом этапе передачи электронов в дыхатель-
ной цепи он не принимает участия, а передает электроны непо-
средственно к убихинону Q:
276
Сукцинат
I
АТФ Флавопротеин 2
\ 1 Цитохромы
X ' I-----1
Субстрат —► НАД Флавопротеин] —Хинон ► b—с—а »- О2
(малат, X. X.
ИЗОТ" АТФ АТФ
Высокоэнергетический нуклеозидтрифосфат генерируется так-
на уровне субстрата самого цикла Кребса, например, во вре-
конверсии сукцинил-КоА в янтарную кислоту; образовавший-
ГТФ может взаимодействовать с АДФ: ГТФ + АДФ
ГДФ + АТФ. Таким образом, 12 новых высокоэнергетических
чекул АТФ образуется за каждый оборот цикла ди- и трикар-
ювых кислот (табл. 28). Если при этом учесть образование
Д-Н на стадии пируват —>- ацетил-КоА:
Пируват + НАД+ -|-КоА — Ацетил-КоА -|-НАД-Н -|-Н+ -f-COs
всего за один оборот реакций цикла Кребса образуется 15 мо-
:ул АТФ.
Главным источником внемитохондриальных редуцирующих
бивалентов является цитоплазматический 2НАД-Н, образую-
йся при гликолизе.
(ависимости от механизма поступления этого НАД-Н в дыха-
ьную цепь митохондрий образуется еще четыре или шесть
Чекул АТФ (без учета двух молекул АТФ, накапливающихся
анаэробную фазу). Следовательно, при аэробном окислении
иоль глюкозы (принимая в расчет анаэробную фазу) клетка
1учает до 38 моль АТФ с учетом образования двух молекул
)увата из одной молекулы глюкозы (6...8 моль на стадии гли-
шза, 6 моль на стадии окисления пирувата до ацетил-КоА
14 моль в цикле Кребса).
блица 28. Этапы образования молекул АТФ в цикле Кребса
№ $акции цикле Ферменты Способ образования высоко- энергетического фосфата Число образую- щихся молекул АТФ
4 Изоцитратдегидрогеназа Окисление НАД - Н в ды- хательной цепи 3
6 а-Кетоглутаратдегидро- геназа То же 3
7 Сукцинаттиокиназа Окисление на уровне суб- страта 1
t 8 Сукцинатдегидрогеназа Окисление ФАД - Н2 в ды- хательной цепи 2
* 10 Малатдегидрогеназа Окисление НАД - Н в ды- хательной цепи 3 Итого ... 12
277
Таким образом, при анаэробном и аэробном распаде у:
дов энергетически более выгодным является последний К.аг
уже сказано, эволюционный процесс ра 1вшич микрооргаг
(применительно к типам еякаппг! i ше.) в направлении
эробы -> факультативные анаэробы >- аэробы. Поэтому на-
смотренные нуги катаболшма vi.H'Bo/iob более или менее
связаны друг с другом
Необходимо учитывать еще одно обстоятельство. Промежу-
точные структуры в рассмотренных путях превращения углево-
дов могут быть исходными веществами для анаболических про-
цессов. Поэтому необходимо пополнение расходуемых молекул за
счет возмещающих или анаплеротических реакций (от греч.
ava — поровну; лХцргосио — наполнение, восстановление). Одной
из важнейших, например, является реакция Вуда и Беркмана
у псевдомонад:
„ СООН
Пируваткар- ।
СНзСОСООН +СО2+АТФ -°к-с"лаз^ СО + АДФ + Ф„ + 2,09 кДж
пируват 1..
С.Г12
соон
оксалоацетат
Образующийся оксалоацетат выступает акцептором ацетил-КоА
в цикле Кребса.
Весьма распространена и другая реакция, катализируемая
фосфоенол-пируват-карбоксилазой (ФЕП-карбоксилазой):
с^ соон
С—О—Р + СО2+Н2О ФЕП-карбоксилаза ^.=Q
II I
СН2 СН2
ФЕП х соон
оксалоацетат
Ранее рассмотренный глиоксилатный путь также относится
к разряду анаплеротических, поскольку он может быть возме-
щающим в глюконеогенезе.
У некоторых микробов пируват образуется в так называемом
метилцитратном цикле, например у Saccharomyces lipolytica
(рис. 67).
В процессе обмена веществ многие микроорганизмы образуют
вторичные метаболиты смешанного типа, т. е. когда в структуру
конечного продукта входят химически различные субъединицы,
в том числе углеводной природы. К таким метаболитам относят-
ся многие антибиотики (амфотерицин В, гентамицин, леворин,
нистатин, олеандомицин, стрептомицин, эритромицин и т. д.)1
атипичные нуклеозиды; коферменты и др. Все они являются гли-
козидами. Агликоны в них имеют разнообразную структуру и оп-
278
Пропионил-КоА
Рис. 67. Схема реакций в метилцитратном цикле:
1— метилцитрат-синтаза; 2, 3 — метилцитрат-дегидратаза; 4 — метилизоцит-
рат-лиаза; 5 — сукцинатгидрогеназа; 6 — фумараза; 7—малатдегидрогеназа
аеляют специфичность действия на соответствующие молекулы
клетках. Тем не менее при отщеплении углеводных компонен-
в активность веществ, как правило, снижается или полностью
везает (например, в случае с антибиотиками). Поэтому важ-
сть углеводных остатков исключительно высока; более того,
И могут выступать определяющим звеном в сохранении актив-
сти молекул.
. МЕТАБОЛИЗМ ЛИПИДОВ
шиды, относящиеся к гидрофобным структурам, выполняют
жнейшие функции в клетках различных организмов, включай
1кробы. Они входят в состав клеточных мембран и обеспечива-
регулируемый транспорт различных веществ в прямом и об-
тном направлениях. В ряде случаев липиды накапливаются в
де сильно преломляющих свет гранул (наблюдение в световом
кроскопе) как запасные питательные вещества. Это происхо-
т главным образом в процессе культивирования микроорганиз-
в на обогащенных углеводами питательных средах, что сви-
гельствует о трансформации части углеводов в липиды. Обра-
вание липидов (липогенез) происходит внутриклеточно, хотя
которые микроорганизмы продуцируют их внеклеточно в сре-
обитания (Rhodotorula sp. и др.).
279
5.4.1. Биосинтез жирных кислот
Природные жирные кислоты включают, как правило, четное чг-с-
.10 углеродных атомов, что подтверждает путь биосинтеза и ; из
двухуглеродных предшественников. Синтез осуществляется ;и)д
каталитическим действием мультиферментного комплекса -- син-
тетазы жирных кислот (его молекулярная масса, напри jy
у дрожжей 2,3- К)3 кДа). Этот комплекс, состоящий из 7 ферхи н-
тов, весьма устойчив у различных грибов. Если у дрожжей ..к-
тивность проявляет лишь сам ферментный комплекс (но не его
субъединицы), то у кишечной палочки активньт все 7 субтк.ш-
ниц синтетазы.
Биосинтез жирных кислот у различных микробов (как и у
всех других организмов) в основном одинаков. Инициатором,
или затравкой, при этом выступает одна молекула ацетил-КоА,
конденсирующаяся с 7 молекулами малонил-КоА, в результате
чего образуется пальмитиновая кислота (С15Н31—СООН):
Ацетил-КоА Д- 7 Малонил-КоА Д- 14 НАДФ-Н Д- 14 Н+ -►
СНз—(СН2)|4—СООН + 7СО2 + 8КоА Д- 14НАДФ+ Д- 6Н2О
пальмитиновая кислота
Ацетил в ацетил-КоА становится донором 15-го и 16-го ато-
мов углерода для СНз—СН2-концевой группы (карбоксильный
остаток для 15-го атома и метильный — для 16-го атома) в паль-
митиновой кислоте. Ацетил-КоА присоединяется к SH-rpynne
одной из субъединиц ферментного комплекса с отщеплением КоА.
Одновременно к другой субъединице комплекса — ацилперенося-
щему белку (АПБ), к SH-группе 4-фосфопантетеина, присоеди-
няется малонил-КоА также с отщеплением КоА. Следовательно,
АПБ — якорь, обеспечивающий через SH-группы ацетил-транса-
цилазы (1) и малонил-трансацилазы (2) присоединение ациль-
ных промежуточных продуктов:
1) Ацетил-S-KoA Д- HS—АПБ Ацетил-Б-АПБ Д-\КоА-БН
2) Малонил-S-KoA Д- HS—АПБ Малонил-Б-АПБ Д- KoA-SH
Из 16 атомов углерода пальмитиновой кислоты 14 ведут свое
происхождение от малонил-КоА, который образуется из гидро-
карбоната и ацетил-КоА с помощью ацетил-КоА-карбоксилазы
(Е) в присутствии Мп2+; ацетил-КоА-карбоксилаза содержит
ковалентно связанный биотин:
СНзСО—S-KoA Д- НСО3- + АТФ Б Мп? НООС—СН2—СО—S-KoA Д-
Ацетил-КоА Малонил-КоА
+ АДФ Д- Ф„
У некоторых микробов малонил-КоА образуется не карбокси-
лированием ацетил-КоА, а активацией малоната специфической
лигазой в присутствии АТФ и КоА и некоторыми другими
путями. Малонил-КоА выполняет исключительную роль в образо-
вании пальмитиновой кислоты, поэтому регуляция активности
280
тил-КоА-карбоксилазы является эффективным методом конт-
я биосинтеза жирных кислот.
Суммарная последовательность реакций биосинтеза насыщен-
[ жирных кислот из ацетил-Б-АПБ и малонил-Б-АПБ может
'Ь представлена в следующем виде:
3) Ацетил-8-АПБ + Малонил-8-АПБ Ацетоацетил-8-АПБ 4- СОг
t 4) Ацетоацетил-8-АПБ 4- НАДФ-Н 4* Н+ 3= D(—)-|5-Оксибутирил-8-АПБ +
+ НАДФ+
5) D(—)-р-Оксибутирил-8-АПБ Кротонил-8-АПБ 4- Н2О
6) Кротонил-8-АПБ + НАДФ-Н 4- HAj Бутирил-8-АПБ 4- НАДФ+
6
J 7) Бутирил-Б-АПБ + Малонил-Б-АПБ р-Кетокапроил-Б-АПБ -|- СОг
и т. д.,
> 3 — 0-кетоацил-АПБ-синтаза; 4 — р-кетоацил-АПБ-редукта-
j 5 — р-оксиацил-АПБ-гидратаза; 6 — еноил-АПБ-редуктаза;
— р-кетоацил-АПБ-синтаза.
Реакции 4...7 продолжаются в виде повторных циклов с удли-
няем цепи на два углеродных атома в каждом цикле. В ука-
нной выше последовательности седьмая реакция повторяет
етью, и четырехуглеродный бутирил присоединяет двухугле-
дный остаток малонила, трансформируясь в р-кетокапронил,
‘орый восстанавливается с помощью НАДФ-Н до оксипроиз-
щого (повторение реакции 4), затем дегидратируется до не-
дельного соединения (повторение реакции 5), которое вновь
:станавливается до остатка предельной шестичленной кислоты
овторение реакции 6). С последующим присоединением мало-
л-КоА цикл повторяется снова и Так до тех пор, пока не прои-
йдет удлинения углеродной цепи до 16 атомов углерода, т. е. до
разевания пальмитиновой кислоты СНз—(CHs)i4—СООН. На
следней стадии биосинтеза с участием пальмитоил-АПБ-деа-
лазы происходит освобождение пальмитата.
i В указанной последовательности реакций источником ацетил-
»А могут быть избыточная глюкоза в среде, полисахариды и
инокислоты, сохраняющиеся в фондах клетки и не использу-
циеся ею для других целей. Из приведенных реакций также
дно, что биосинтез жирных кислот происходит в присутствии
1ДФ-Н, который в свою очередь образуется преимущественно
И НАДФ-Н-зависимом окислении глюкозо-6-фосфата (см.
Зд. 5.3.2), когда 7 молекул его окисляются до рибулезо-5-фос-
гга и СО2, и образуется достаточное количество НАДФ-Н для
©синтеза одной молекулы пальмитиновой кислоты.
Удивителен тот факт, что в рассмотренном процессе не обра-
тен ни стеариновая СНз—(СНг)1б—СООН, ни более высоко-
Лекулярные жирные кислоты. Процесс остановки биосинтеза
пальмитате остается нераскрытым. Удлинение цепи пальмити-
вой кислоты возможно за счет ацетил-КоА или малонил-КоА,
281
но без участия синтетазы жирных кислот. У микроорганизмов
(например, у Е. coll) имеются ферментативные механизмы обра-
зования ненасыщенных жирных кислот. Известны метилирован-
ные жирные кислоты, например, туберкулостеариновая в клетках
туберкулезных микобактерий:
СНз— (СН2) 7—с (СН3)=СН (СН2) 7—соон
образующаяся из олеиновой кислоты через промежуточную ме-
тиленстеариновую кислоту СНз(СН2)7—С—(СНг)в—СООН.
II
сн2
К жирным кислотам, содержащим 20 углеродных атомов и
циклопентановое кольцо, относятся простагландины (PG) — про-
изводные простаноевой кислоты, равно как и С20-три-, тетра-
и пентаеновой жирных кислот:
простаноевая кислота
Их подразделяют на две основные группы — Е и F, каждая из
которых включает по три простагландина — Ei, Е2, Е3 и Fia, F2a,
F3a соответственно. Эти шесть простагландинов являются пер-
вичными. PG из группы Е содержат кетогруппу при С9 и гид-
роксильную группу при СП, тогда как PG из группы F содержат
гидроксильные группы в тех же положениях. Нижние индексы
PG 1, 2 и 3 в каждой группе обозначают число двойных связей
в боковых цепях Rt и R2.
В результате химического или ферментативного превращения
PGE получаются вторичные простагландины:
о
он
PGE
он
PGB
PGC PGD2
Предполагают, что биосинтез простагландинов у грибов и
более высоко организованных существ во многом сходный и на-
чинается с арахидоновой кислоты под влиянием циклооксигеназы
жирных кислот, входящей в состав простагландинсинтазного
комплекса. Ацетилсалициловая кислота (аспирин) и другие про-
тивовоспалительные лекарственные средства блокируют циклоок-
сигеназу жирных кислот, что необходимо учитывать при назначе-
нии таких лекарств больным, получающим PG, или у которых
контролируют уровень PG.
282
. Биосинтез нейтральных жиров (триацилглицеринов)
•сфатидов (фосфолипидов)
ie биосинтеза жирные кислоты сразу же расходуются на
азование триацилглицеринов и фосфатидов. Триацилглицери-
обычно накапливаются в клетках микробов в качестве запас-
L питательных веществ. Их синтез (как и некоторых фосфа-
) осуществляется через фосфатидную кислоту из КоА-произ-
|Ных жирных кислот (из АПБ-производных жирных кислот
coli).
2R—СО—КоА + НОСН2—СНОН—СН2—ОРО3Н2
L-глицерол-З-фосфат
->• 2КоА + R—СОО—СН2—СН—СН2—ОРО3Н2
I
ROOC
L-фосфатидная кислота
Глицерол-З-фосфат образуется из глицерина, который фосфо-
ируется в присутствии специфичной киназы и АТФ, или же
восстановлении диоксиацетонфосфата.
/.-Фосфатидная кислота + Н2О -+ Диацилглицерин -р Ф„
R—СО—S—КоА -р Диацилглицерин -> Триацилглицерин -р КоА—SH
Центральная роль фосфатидной кислоты видна из рис. 68.
£-Фосфатидная кислота в реакции с ЦТФ превращается в
ндиндифосфатдиацилглицерин, выступающий предшественни-
I всех фосфоглицеридов:
-Фосфатидная кислота -|- ЦТФ Цитидиндифосфатдиацил-
глицерин (ЦДФ-диацилглицерин) + ФФ„
да в соответствии с обозначениями на рис. 68:
I) ЦДФ-диацилглицерин -р Серин Фосфатидилсерин -р ЦМФ
2) ЦДФ-диацилглицерин -р Инозит Фосфатидилинозит -р ЦМФ
3) ЦДФ-диацилглицерии -р Глицерофосфат З-Фосфатидилглицерол-1'-
|>ат 4- ЦМФ
Фосфатидилсерин, содержащийся, например, в мембране
coli и в митохондриях эукариот, под действием фосфатидил-
ин-декарбоксилазы декарбоксилируется с образованием фос-
гидилэтаноламина: о
ROOC—CH
NH^
’ OCOR' О
З-Фосфатидил-глицерол-Г-фосфат является предшественни-
1 мембранных фосфатидилглицерина и кардиолипина у бак-
.нй, в мембране Salmonella typhimurium содержится порядка
% фосфатидилглицерина:
ROCO— CH2—CH—CH2—О— P—О—СНг— CH—CH2OH
OCOR' OH
фосфатидилглицерин
OH
283
R—COO—CH2—CH
ROOC
I
CH
I
CH2 — OPO3H2
£-фосфатидная кислота
CH2—О —COR
СНО—С—COR
I
СН2ОН
Серин
ЦТФ
ЦДФ-диацилглицерид
Инозит^
Путь синтеза
диацилглицерина
Фосфа ти дил-
серин
Фосфатидил-
инозит
Глицеро-
фосфат
Фосфатидил-
глицерол-
1-фосфат
-*н 1
фосфатидил-
этаноламин
Фосфатидил
глицерин
J 3(СНГ)
Фосфатидил-
холин
—глицерин^,
Кардиолипин
| СО2
Пути биосинтеза фосфоглицеридов
Рис. 68. Участие фосфатидной кислоты в синтезе различных веществ
Кардиолипин входит также в состав мембраны митохондрий
эукариот (~ 5%), он образуется в результате взаимодействия
двух молекул фосфатидилглицерина с выделением одной молеку-
лы глицерина:
О О
II II
R—ОСО—СН2—СН—СН2—О— Р—О—СН2—СН(ОН)— СН2—О— Р—О СН2—
I I I
OCOR' ОН ОН
— СН—CH2OCOR"
I
OCOR"'
кардиолипин
Предшественником фосфатидилэтаноламина является этанол-
амин, фосфорилирующийся в присутствии специальной киназы
Е и Mg2+:
НО—СН2—СН2—NH2 + АТФ Р—О—СН2—СН,—nh2 + АДФ
фосфоэтанолам ин
На фосфатидилэтаноламин приходится до 75 % сухого веса
мембран у грамотрицательных бактерий. Фосфоэтаноламин взаи-
модействует с ЦТФ и образуется цитидиндифосфоэтаноламин,
трансформирующийся затем в фосфатид:
284
+h3n—сн2—сн2—о—р—о—р—
о-
О
О”
ROCO—С—Н
Н2С—COOR
СН5-ОН
Mg'
цитидиндифосфоэтаноламин
1,2-диацил глицерин
О
Н2С—COOR
косо—c—fl о + ЦМФ
Н2С—О—Р—О—CHj—сн— nh)
фосфоати дил этаноламин
В случае последовательного переноса метильной группы
грансметилирование) от трех молекул S-аденозилметионина
фосфатидилэтаноламину образуется фосфатидилхолин-.
Н—<
соо~
I
сн2
Н2С—с—COOR
н2
н,е—s— CH2/O.
S-аденозилметионин
Н2С— COOR
RCOO—СН О
R’OCO—СН
О"
фосфатидилэтаноламин
СОО“
+ Метил трансфераза
НС—NH2 nh
СН2
сн2
Н2с—О—Р—ОСН2—СН2—N(CH3)3
HS-----СН2
I
3
о-
фосфатидилхолин
S-аденозилгомоцистеин
S-Аденозилгомоцистеин далее может расщепляться на адено-
н и гомоцистеин.
4.3. Анаболизм липоконъюгатов
1к уже было сказано (см. гл. 2), к липоконъюгатам относятся
(инголипиды, микостерины, терпены, гликолипиды и липогли-
ны (липополисахариды). Строительными блоками сфинголипи-
285
дов являются длинноцепочечные сфингозин и дигидросфингозин,
биосинтез которых происходит с участием пиридоксальфосфата:
П 2 +
Пальмитоил-КоА -|- Серин—Дигидросфингозин ->
Флавопротеин
— Восстановленный
Сфингозин
В ходе биосинтеза сфинголипидов сфингозин ацилируется в
0-положении КоА-эфиром жирной кислоты с образованием N-
ацилсфингозина (церамида), к которому затем в a-позиции при-
соединяется ЦДФ-холин:
Сфингозин + КоА-производное жирной кислоты —-^д-
-*• Церамид ~~Сфингомиелин
Церамид, соединяясь с галактозой или глюкозой, образует
цереброзид, который может сульфатироваться в позиции С6 мо-
нозы. При синтезе ганглиозидов церамид соединяется в a-поло-
жении с несколькими моносахаридными единицами — глюкозой,
галактозой, N-ацетилглюкозамином, N-ацетилгалактозамином,
фукозой и N-ацетилнейраминовой кислотой. Ганглиозиды могут
затем связываться с протеинами, образуя сложные комплексы —
гликолипопротеины, протеолипиды, фосфатидолипиды.
Микостерин, или стерин грибов (эргостерин), под действием
светового излучения трансформируется в витамин Da (кальци-
ферол), который стимулирует синтез белка в тонком кишечнике
человека, участвующего в переносе ионов кальция.
Витамины группы D образуются из ненасыщенных стеринов,
а эти последние — из ацетил-КоА. Три молекулы ацетил-КоА
конденсируются в мевалоновую кислоту, которая затем превра-
щается в сквален, относящийся к тритерпенам:
НОЧ /СНз
>с<
HOH2CZ СН2СООН
мевалоновая кислота
Из сквалена образуются четыре конденсированных кольца хо-
лестерина путем циклизации:
286
сквален
К липидам с изопреноидными структурами относятся также
1ихол, обнаруживаемый в мембранах у эукариот; бактопре-
I — в мембранах у прокариот; хиноны — в составе цепей пере-
:а электронов у бактерий и грибов. Биосинтез изопреноидов
[лючая стероиды) начинается с конденсации двух молекул
ггил-КоА, в результате чего образуется ацетоацетил-КоА, ко-
пай конденсируется с третьей молекулой ацетил-КоА с образо-
шем: p-окси-Р-метилглутарил-КоА; из этого последнего обра-
тен важное промежуточное соединение — мевалоновая кис-
а, являющаяся источником каротина, убихинона, фитола и др.
Мевалоновая кислота после двух последовательных реакций
:форилирования и декарбоксилирования трансформируется в
пентенилпирофосфат — ключевой продукт для синтеза поли-
преноидов:
н2с.
>С—СН2—СНг—О—Р—Р
H3CZ
нзопентил пирофосфат
Конденсацией нескольких молекул изопентилпирофосфата об-
уются смешанные цыс-транс-полимеры терпеноидной структу-
, например долихолы и бактопренолы. К изопреноидным
уктурам относятся коферменты Q (убихиноны). Они содержат
;овые цепи из 5... 10 изопреноидных остатков или 25...50 атомов
>ерода. Коферменты Q принимают участие в переносе электро-
I в окислительно-восстановительных реакциях, в частности при
(слительном фосфорилировании у аэробных микроорганизмов.
Хрофобными частями боковые цепи прикрепляются к липид-
»у компоненту мембраны, а хиноидное ядро остается свобод-
и для окисления — восстановления.
Убихиноны синтезируются из р-оксибензойной кислоты и соот-
ствующего пренилпирофосфата:
2-декапренилпирофосфат
287
Донором метоксильных и метильных групп выступает метио-
нин. р-Оксибензойная кислота у бактерий образуется из хориз-
мовой кислоты, которая может трансформироваться в салицило-
вую кислоту у Mycobacterium tuberculosis при дефиците железа
соон
соон
хорнзмовая кислота
При конденсации изопентилпирофосфата по типу «голова к
голове», через ряд промежуточных реакций, образуется фитоин:
(R — повторение цепи справа после прерывистой линии). Бес-
цветный фитоин подвергается ступенчатому окислению с транс-
отщеплением двух водородных атомов и последующим образова-
нием ликопина, из которого происходит семейство каротинов:
Замыкание и циклизация боковых 6-углеродных звеньев моле-
кул дает Р-каротин. К каротиноидным соединениям относятся
триспоровые кислоты (С1з-терпеноиды), являющиеся у ряда низ-
ших грибов половыми гормонами (см. гл. 4). Ковалентное свя-
зывание липидов с углеводами или белками сопровождается об-
разованием более сложных молекул гликолипидов, липогликанов,
липопротеинов, протеолипидов и гликолипопротеинов.
288
4. Катаболизм липидов и липоконъюгатов
:пад липидов и липоконъюгатов у микроорганизмов происхо-
биокаталитически. Иначе говоря, микробные клетки обла-
>т не только ферментами биосинтеза липидов, но и фермента-
их разложения. Распад нейтральных жиров катализируется
азами, относящимися к гидролазам. Активность их у различ-
; микроорганизмов выражена по-разному, однако имеются,
ример, штаммы нитчатых грибов, проявляющие высокую
азную активность. Отщепление жирнокислотных остатков от
церина происходит ступенчато. Освобождающийся глицерин
<ет включаться в процесс гликолиза или расходоваться на
гие соответствующие нужды клетки. Жирные кислоты рас-
аются за счет р-окисления до ацетил-КоА, который вклю-
гся в цикл ди- и трикарбоновых кислот или участвует в дру-
процессах биосинтеза.
р-Окисление жирных кислот — наиболее распространенный
трех известных путей расщепления. При р-окислении разры-
тся связь между а- и p-углеродными атомами в молекуле
зной кислоты:
0 а
R—СН2—СН2—СО ~ КоА -+ ... R—СО ~ КоА + СН3—СО ~ КоА
ацнл-КоА ацил-КоА ацетил-КоА
Сп — 2
Если насыщенная кислота содержит четное число углеродных
мов, то в конечном итоге она полностью расщепляется до
тил-КоА, если нечетное, то процесс заканчивается образова-
м соответствующего числа молекул ацетил-КоА и одной моле-
ы пропионил-КоА.
Энергия, высвобождаемая при расщеплении жирных кислот,
новится доступной для микроорганизма вследствие последу-
их превращений ацетил-КоА, НАД-Н и ФАД •№ Расщепле-
ненасыщенных жирных кислот также осуществляется путем
<исления, но предварительно такие кислоты превращаются
ментативным путем в соединения, которые атакуются фермен-
я р-окисления.
i-Окисление свободных жирных кислот (рис. 69) осуществ-
'ся реже, чем р-окисление, и не всеми микроорганизмами
^имущественно эукариотами). Оно протекает при участии
виновых ферментов и молекулярного кислорода с образова-
[ жирной кислоты, укороченной на один углеродный атом;
я кислота может вновь подвергнуться а-окислению. а-Окис-
е — важный процесс в обмене жирных кислот и при образо-
и, например, производных ацетилена у базидиальных грибов.
ie производные могут быть представлены неразветвленными
затическими соединениями или соединениями, содержащими
лические и гетероциклические кольца. Так, для ряда грибов
чен диатрен-. НООС—СН=СН—(CsC):i—СН2ОН.
)-Окисление жирных кислот возможно, когда происходит
оксидирование концевой метильной группы при участии ок-
1617
289
(Cn-1)
(Сл.,)
0.-D- оксижирная кислота
(Cn)
Рис. 69. а-Окисление свободных жирных кислот:
1 — ФАД-Щ-, 2 — НАД-зависимая дегидрогеназы
сигеназы со смешанной функцией. Такие оксигеназы атакуют
непосредственно электроны, участвующие в образовании связи
С—Н, и приводят к образованию «-оксижирной кислоты. После-
дующее окисление дает дикарбоновую кислоту, которая может
быть укорочена с любого конца молекулы путем р-окисле-
ния.
Образование а-, «-дикарбоновых кислот у некоторых микро-
организмов происходит путем расщепления у двойной связи,
например, олеиновой кислоты (позиции С9, СЮ). В этом случае
появляется азелаиновая кислота НООС-(СН2)7—СООН (с 9 уг-
леродными атомами), подвергающаяся затем р-окислению с
образованием предшественника биотина — пимелиновой кис-
лоты.
Образующийся из жирных кислот с нечетным числом углерод-
ных атомов пропионил-КоА может участвовать в различных про-
цессах биосинтеза, например С27-фтиеновой и С32-микоцерозовой
кислот у микобактерий Н3С—(СН2)|7—СН(СН3)—СНг—
—СН(СН3)—СН=С (СН3)—СООН, НзС— (СН2) 19—СН(СНз)~
— [СН2—СН(СН3)]з—СООН соответственно. У других микро-
организмов, например актиномицетов, пропионил-КоА участвует
в биосинтезе ряда антибиотиков из группы макролидов (эритро-
мицина, метимицина, магнамицина).
Простагландины также подвергаются р- и «-окислению. Вна-
чале происходит окисление при С15 с участием 15-оксипроста-
гландин-дегидрогеназы. Затем следует восстановление С13-
двойной связи под каталитическим действием А|3-редуктазы-
После этого имеют место реакции р- и «-окисления. Названные
превращения схематично показаны на примере молекулы
PGE2:
290
о
р-окисление
С13,14-редукция
Фосфолипиды также подвергаются ферментативному гидроли-
Ферменты, гидролизующие одну или более связей в фосфа-
,ах, называются фосфолипазами. Известно 5 фосфолипаз:
Аг, В, С и D. Каждая из них соответственно гидролизует,
|ример в фосфатидилхолине, связи А, В, АВ, С и D:
сн2------сн--------
—COR А—§—OCOR'
о
3 С 11 D +
СН2—О—Р—О—СН2—СН2—N^(CH3)3
Следовательно, фосфолипаза Ai катализирует реакцию гид-
[иза одной ацильной группы по месту связи А; фосфолипаза
— по месту связи В; фосфолипаза В — по месту связей А и В.
патогенных и сапрофитных клостридий и бацилл содержится
оставе а-токсина фосфолипаза С, расцепляющая гомологич-
5 связь в фосфатидилхолине. Известны также лизофосфоли-
а и фосфолипаза D. Из них первая расщепляет связь А, вто-
1 — связь D. Фосфолипаза D находится в растительных тка-
К числу гликолипидов (липоконъюгатов) относятся гликозил-
ацилглицериды. некоторых бактерий. Например, в хроматофо-
; фотобактерий содержится 6-сульфо-6-дезоксиглюкоза, или
ювоза, связанная О-гликозидной связью с диацилглицерином.
ульфохиновозилдиацилглицерин гидролизуется с помощью
козидаз и липаз. Первые катализируют расщепление О-гли-
идной связи, вторые — расщепление диацилглицерина. Обра-
>щиеся свободные жирные кислоты подвергаются окислению,
291
а глицерин включается в процесс гликолиза или в другие мета-
болические реакции. Аналогичным процессам распада могут под-
вергаться и другие гликолипиды и липогликаны. Катаболизм
прочих липоконъюгатов изучен неполностью.
5.5. МЕТАБОЛИЗМ НЕКОТОРЫХ ДРУГИХ
ВТОРИЧНЫХ СОЕДИНЕНИЙ
Из огромного числа различных конечных продуктов, образуемых
микроорганизмами, особый интерес представляют поликетиды,
среди которых найдены антибиотики, органические кислоты и
другие вещества. В поликетидах метиленовая и карбонильная
группы регулярно чередуются. Ранее их рассматривали полиме-
рами кетонов, откуда и произошло их название. Поликетиды об-
разуются микроорганизмами под действием мультиферментного
комплекса из молекулы-затравки, например из ацетил-КоА или
малонил-КоА, но в отличие от жирных кислот в ходе их. биосин-
теза нет стадии восстановления или оно осуществляется крайне
редко.
Противогрибковый антибиотик гризеофульвин образуется
нитчатым грибом penicillium griseofulvum. Он синтезируется из
одной молекулы ацетил-КоА и 6 молекул малонил-КоА:
Разнообразные моноциклические поликетиды образуются гри-
бами и лишайниками. Один из простейших поликетидов — 6-
метилсалициловая кислота (6-МСК), биосинтез которой катали-
зируется мультиферментным комплексом 6-МСК-синтетазы:
сн3
СООН
6-метилсалициловая
кислота
К поликетидным соединениям относятся также усниновая кис-
лота, обладающая противомикробным действием, которая обра-
зуется некоторыми лишайниками (например, Ramalina reticu-
lata) :
292
СОСНз
усниновая кислота
Почвенные грибы и некоторые другие микроорганизмы прев-
,ают фенолкарбоновые кислоты в другие формы фенолов с по-
р>ю реакций окисления, гидроксилирования и декарбоксили-
1ния:
соон
шикимовая
кислота
фумагатин (фумагацин, 3,5-диоксибензойная резорцин
гельволевая кислота) кислота
В галловую кислоту непосредственно может переходить ши-
иная кислота. Данная реакция присуща Phycotnyces blakes-
inus. Этот вид образует также и протокатеховую кислоту:
соон
он
протокатеховая кислота
Ряд приведенных веществ (пирогаллол, резорцин, фумига-
) обладает выраженным антимикробным действием. Триокси-
бензолы (пирогаллол и др.) проявляют высокие аутоокислитель-
ные свойства, в связи с чем могут превращаться в ортохиноны.
Наряду с этим триоксибензолы подвергаются нуклеофильным
замещениям и присоединяют другие фенолы, аминокислоты или
амины, которые могут и далее снова окисляться или замещаться.
В конечном итоге возникают окрашенные в коричневый цвет
сложные высокомолекулярные соединения — гуминовые кислоты,
являющиеся составной частью органического вещества многих
видов почв.
К поликетидам относятся антибиотики — тетрациклины, обра-
зуемые некоторыми видами актиномицетов из рода Streptomyces:
Они различаются по радикалам Ri, R2 и R3, которые соответ-
ственно представлены у тетрациклина Н, СНз, Н; у 7-бромтет-
рациклина бромом, СНз, Н; у 6-деметилтетрациклина водорода-
ми; у 5-окситетрациклина Н, СНз, ОН; у 7-хлор-6-деметилтетра-
циклина хлором, Н, Н; у 7-хлортетрациклина хлором, СНз, Н.
Биосинтез тетрациклинов происходит путем конденсации мо-
лекул малонил-КоА (но не ацетил-КоА). Донором метильной
группы для некоторых тетрациклинов выступает метионин, тогда
как аминогруппу поставляет глутамин. Фермент оксигеназа со
смешанной функцией катализирует реакцию включения гидро-
ксильной группы, берущей начало от молекулярного кислорода.
Таким образом, ступени биосинтеза тетрациклинов представля-
ются в следующем виде:
—*~4-Десдиметиламино-4-кетоангидротетрациклии —Ангидротетрациклин или 7-Хлорангидро-
тетрациклин —5а,11а-Дегидротетрациклин или 5а,11а-Дегидро-7-хлортетрациклин----------►
—► Тетрациклин или 7-Хлортетрациклин
294
Тетрациклины обладают выраженной противобактериальной
гивностью и ряд из них входит в арсенал химиотерапевтиче-
их средств.
К другим поликетидным соединениям, образуемым грибами
spergillus, Penicillium), относятся антраценовые соединения —
грахиноны и антроны. За счет действия фенолоксидазы воз-
жна их димеризация, которая приводит к образованию сле-
щих соединений:
эндокроцин-производное эмодин-производное
антрона антрона
ОН о он
о
производное
антрахинона
Из производных антрахинона синтезируются пигменты сум-
ого гриба — спорыньи (Claviceps purpurea), называемые
охромами. Участником их биосинтеза является эмодин. Био-
тез антраценовых соединений осуществляется конденсацией
юй молекулы ацетил-КоА и семи молекул малонил-КоА.
Из производных индола заслуживают внимания вторичные
аболиты — галлюциногены, образуемые рядом базидиальных
бов из родов Conocybe, Panaeolus, Psilocybe, Stropharia.
> псилоцин и псилоцибин, беоцистин и норбеоцистин. Некото-
‘ сумчатые, низшие и несовершенные грибы (Claviceps, Rhizo-
и Penicillium) образуют индольные алкалоиды — лизергино-
> кислоту и ее производные, также обладающие галлюциоген-
м действием: о
II
НО —Р—О-
н
н
псилоцибин
псилоцин
295
беоцистин
норбеоцнстин
лизергиновая кислота
Если в формуле лизергиновой кислоты R=NH2, то получают
амид D-лизергиновой кислоты (эргин); если R==NHCHOHCH3,
то N-( 1-гидроксиэтил) амид-О-лизергиновой кислоты. Известны
изоформы D-лизергиновой кислоты, т. е. амид и М-(1-гидрокси-
этил)амид. Если вместо остатка COR содержится —СНгОН, то
это будет лизергол. Сходным по структуре является клавиновый
(от лат. слова Claviceps — спорынья) алкалоид эргометрин, об-
ладающий кровоостанавливающим действием при маточных кро-
эргометрин
296
5-ОН-ТА
/ серотонин
N-Ме-серотонин Буфотенин-И-оксид
ТА
N-Me-TA
Amanata sp.
Panaeolus sp
S-OH-ТФ-----> S-OH-ИУК
Psilocybe sp.
Conocybe sp
Panaeolus sp.
Panaeolina sp.
N.N-ДМТА
5 MeO-N.N-ДМТА
ТА
1М,1М-ДМТА > 4ОН М,1М-ДМТА
Х х. псилоцин
4-Ф-ТА
норбеоцистин
4 -Ф-N-Me-TA
беоцистин
Рис. 70. Пути образования биогенных аминов у некоторых базидиомицетов:
ТФ — триптофан; ТА — триптамин; ДМ — днметил; МеО — метокси; ОН — гид-
рокси; Мт — метил; Ф — фосфорил; ИУК — нндолилуксусная кислота
течениях. У всех выше перечисленных грибов биосинтетиче-
ие пути, ведущие к образованию так называемых биогенных
[инов, во многом сходны. Это можно продемонстрировать на
дольных производных (рис. 70).
Триптофан (рис. 70) выступает родоначальником ряда других
ологически активных соединений, из которых, например, гормон
ротонин образуется и в организме человека, где он регулирует
нус почечных сосудов и кровоток через почки. Из триптофана
кже синтезируются клавиновые алкалоиды, алкалоиды типа
1зергиновой кислоты и некоторые другие соединения; например,
дельные виды Pseudomonas образуют антибиотики пирролнит-
IH, изо-пирролнитрин:
'Х+
•СН2
I
сн
' ''"соон
СООН
н
промежуточное соединение
Cl
н
триптофан
пирролнитрин
изо-пирролнитрин
297
У многих микроорганизмов имеется геминовый фермент
триптофанпирролаза — диоксигеназа, присоединяющая к субстра-
ту оба атома молекулярного кислорода. Она катализирует реак-
цию окислительного расщепления индольной циклической струк-
туры триптофана между С2 и СЗ с образованием формилкину-
ренина:
форм илкинуренин
Из формилкинуренина затем образуется кинуренин при уча-
nh2
СО—сн2—сн —соон
nh2
кннуреннн
стии фермента формамидазы. Кинуренин трансформируется в
кинуреновую кислоту, которая через ряд промежуточных про-
дуктов, включая 4,6-диоксипиколиновую кислоту, превращается
в алифатические а-кетоглутаровую и аспарагиновую кислоты.
Эти последние могут подвергаться дальнейшим превращениям
или используются в процессе биосинтеза белков.
он
соон
кинуреновая кислота
Вопросы
I. Что такое метаболизм микробов и на какие типы сто нодразделяют J
2. Каким обратом осуществляется перенос веществ в клетки микроорга: и
мов?
3 На какие группы и подгруппы подразделяют микроорганизмы в заве ”
мости от источника углерода и энергии?
4. Какое принципиальное отличие биологического катализа от химиче.т
катализа?
5. Почему система АДФ - АТФ занимает промежуточное положенщ •
\(Г ' между высокоэиергетическими и низкоэнергетическими фосфатами i
звагь примеры тех и других)?
298
16. Какими показателями характеризуется активность ферментов?
»7. Что такое холоферменты, коферменты, кофакторы и иростетические груи-
Дпримеры их)?
*8. Что означает термин «абсолии пая оптическая специфичность фермента»
чем это выражается?
J9. Какая известна общепринятая классификация ферментов и что положено
р основу?
10. Что такое конкурентные и не.юшкхрентные ингибиторы ферментов (при-
!Ы)?
11. Какие трн уровня регуляции ферментативных реакций известны с учетом
ребностей микробных клеток в энергии?
12. Что понимают под механшмами ! py6oi о и тонкого контроля регуляции
|цессон синтеза и активности фермии юв?
13. Какое различие имеется между Fcedback-ингнбицией и Feedback-pe-
ссией?
14. Какие основные компоненты составляют систему' фиксации азота из воз-
а?
15. Укажите гри главные реакции, благодаря которым аммиак включается
встав аминокислот.
16. Какие строение и функции пиридоксальфосфата в клетках микроорга-
мовэ
17. Назовите примеры биосинтеза аминокислот, которые доказывают роль
t и Глн как универсальных источников азота.
18. Какие ферменты относятся к гемопротеинам (их функция)?
19. Каковы биосинтез и функция тетрагидрофолата?
20 Почему шикимовая и .хоризмовая кислоты являются интермедиатами
Иосинтезе тирозина, триптофана и фенилаланина?
21. Каков источник серы в молекуле цистеина и роль цистеина в моле-
ах белков у микроорганизмов?
22. В чем сущность нематричного синтеза олигопептидов (примеры)?
23- Каковы пути биосинтеза пуринов?
24. Каковы пути биосинтеза пиримидинов?
25. В чем выражается ограниченный протеолиз и каково его назначение?
26- Назовите известные основные цуги катаболитма аминокислот.
27 Каковы основные характсрис!ики нуклеаз (классификация, функция)?
28 Какие пути биосинтеза ратличных полисахаридов известны?
29 Какое значение имеет глиоксилагный цикл в обмене веществ у микро-
анизмов?
30 Что такое бактоиренолы и долихолы и какое участие они принимают
иосинтезе углеводных полимеров?
31 Какие пути биосинтеза пептидогликана у золотистого стафилококка нз-
тны?
32 В чем заключается сущность первой и второй стадий гликолиза у гомо-
Ументативных молочнокислых бактерий?
33 В основе каких анаэробных процессов у микроорганизмов находится
КОЛИТ.-1
34 Какие фазы известны в (ексо гомонофосфатном шунте (химизм реакций)
X назначение?
35 В чем заключается особенность КДФГ-пути и какой ключевой интер-
нат образуется под влиянием б фосфог.пижоиаг-дегидраi;i3bi'J
36 Какие ферменты и субстраты являются участниками HTK'J
37 Сколько молекул АТФ обратуется за один оборот реакций в НТК?
38 Сколько ферментов входит в состав му.тьтиферменгного комплекса -
Ететазы жирных кислое? Каковы основные пути биосинтеза пальмитиновой
лоты?
39 Какие известны пути расщепления жирных кислот (механизм)?
40 Какие биосинтетические eivtu ведут к образованию индольных биогенных
шов у некоторых грибо>ч?
Часть
3
Прикладное
значение
химической
микробиологии
Глава 6
ХИМИЧЕСКАЯ МИКРОБИОЛОГИЯ
ОСНОВА БИОТЕХНОЛОГИИ
6.1. БИОТЕХНОЛОГИЯ, ЕЕ ЦЕЛИ И ЗАДАЧИ
Биотехнология переживает в настоящее время бурное развитие,
что связано с качественно новым подходом к решению биотех-
нологических проблем. Согласно определению Европейской феде-
рации биотехнологов (1984), биотехнология — это интегриро-
ванное использование биохимии, микробиологии и инженерных
наук с целью достижения технологического (промышленного)
применения способностей микроорганизмов, культур клеток тка-
ней и их частей. Как и любая другая наука, она базируется на
своих объектах и методах. Объекты биотехнологии — преиму-
щественно микробы (грибы, бактерии, вирусы, простейшие) или
клетки других организмов (растения, животные); реже — биоло-
гически активные вещества специального назначения (например,
иммобилизованные ферменты, катализирующие синтез или распад
полимеров).
Наиболее типичными методами биотехнологии являются круп-
номасштабное глубинное культивирование биообъектов в перио-
дическом или непрерывном режиме, выращивание клеток расти-
тельных и животных тканей в особых условиях. Биотехнологи-
ческие методы культивирования биообъектов выполняются на
специальном оборудовании. Так, для культивирования аэробных
микроорганизмов чаще всего используют ферментаторы с необ-
ходимыми контрольно-измерительными приборами и автомати-
кой, применительно к анаэробным видам в ферментаторы не
зоо
йают воздух. Клеточные культуры растении выращивают ча-
| в стационарных условиях на твердой подложке в стеклян-
(х или полиэтиленовых емкостях, для животных клеток ис-
щьзуют стеклянные емкости. В подавляющем большинстве
учаев биотехнологические процессы осуществляют в асептиче-
jjx условиях, исключающих попадание в среду выращивания
ггогенных, а также сапрофитных микроорганизмов — конкурен-
в за питательные субстраты, антагонистов, потребителей или
крушителей конечного продукта, продуцентов токсических ве-
еств.
Следовательно, биотехнология — наука об использовании
.©логических процессов в технике и промышленном произ-
дстве. Это выражение прикладная наука, использующая ре-
льтаты фундаментальных исследований в области смежных
ологических, химических и технических дисциплин (микробио-
гии, генетики и генной инженерии, ботаники и физиологии,
[мунологии и иммунохимии, органической и биологической
мии, процессов и аппаратов и др.).
Предлагаемые иные названия для данной науки достаточно близки назва-
ю «биотехнология», но не в полной мере отражают ее содержание. Это, в ча-
иости, касается «биохимической технологии», «генной и клеточной ннжене-
и, «промышленной (технической) микробиологии». Так, например, выращива-
е микробных клеток для их использования в качестве белкового продукта
орее относится к промышленной микробиологии, чем к биохимической тех-
иогии. Генная и клеточная инженерия — это методы, с помощью которых
конструируют наследственные свойства организмов. Реконструированные виды
пользуют затем в производственных условиях, т. е. в биотехнологии. Про-
пиленная микробиология — ветвь биотехнологии, причем наиважнейшая в на-
>ящее время.
Назовем основные цели и задачи, стоящие перед биотехноло-
ми:
1) получение клеток или их составных частей (главным об-
(зом, ферментов) для направленного изменения сложных моле-
л (например, изомеразы, рестриктазы и т. д.);
2) поддержание и активация путей обмена клеток, ведущих
накоплению целевых продуктов при заметном подавлении дру-
х реакций обмена у культивируемого организма;
3) совершенствование и оптимизация аппаратурного оформ-
ния биотехнологических процессов с целью достижения макси-
(льных выходов конечных продуктов при культивировании ес-
ственных
венностью
видов организмов
методами генной и
и видов с измененной
клеточной инженерии.
наслед-
Применительно к содержанию настоящего учебника в даль-
йшем будут рассмотрены биотехнологические процессы, осно-
вные преимущественно на использовании микроорганизмов
ромышленная микробиология). Следует подчеркнуть, что по-
зные микробы сегодня занимают важное место в жизнеобе-
ечении человеческого общества, а со временем их роль воз-
стет еще в большей степени.
301
6.2. БИОТЕХНОЛОГИЧЕСКИЕ ПРОЦЕССЫ, ОСНОВАННЫЕ
НА ИСПОЛЬЗОВАНИИ ХИМИЧЕСКОЙ АКТИВНОСТИ
МИКРООРГАНИЗМОВ
Человечество с незапамятных времен использовало в своих нуждах химиче-
скую активность микроорганизмов, хотя микробная природа многих процес-
сов долго оставалась нераспознанной. Археологические раскопки в Двуречье
(Тигр и Евфрат) помогли найти сохранившиеся остатки пекарен и пивоварен,
которые были построены соответственно за 6000 и 2000 лет до н. э. С давних пор
люди используют также молочнокислые продукты (простоквашу, кефир, кумыс,
лебен, мацун, чортан, танапур, сыр), виноградные вина и т. д. Лишь после
открытий великого французского ученого Луи Пастера — основоположника
микробиологии — удалось распознать роль многих микробов в природе. С этого
времени заложенные основы технической микробиологии получили мощные им-
пульсы в виде замечательных открытий в области физиологии и генетики
микроорганизмов, процессов и аппаратов биотехнологии и других наук. К числу
таких открытий следует отнести: получение «нуклеина» (ДНК) (Ф. Мишер,
1869); установление каталитической функции ферментов (В. Оствальд, 1893);
раскрытие процессов брожения (Ц. Нейберг, 1912); разработка кинетики фер-
ментативных реакций (Л. Михаэлис и М. Л. Ментен, 1913); доказательство
мутагенного действия рентгеновских лучей (Г. А. Надсон и Г. С. Филлипов,
1925); разработка методов глубинного культивирования микроорганизмов
(А. Клюйвер и Л. X. Ц. Перкин, 1933); открытие цикла трикарбоновых кислот
(Г. Кребс, 1937). Произошло бурное развитие промышленности антибиотиков
(1940...1960 гг.), микробиологической промышленности и, прежде всего, круп-
номасштабного производства кормовых дрожжей, аминокислот, ферментов (в том
числе иммобилизованных), фруктозных сиропов и т. д. (1960...1970 гг.); в по-
следующие годы зародилась клеточная и генная инженерия, становление кото-
рой на промышленные рельсы происходит в настоящее время. На фоне исто-
щения ископаемого топлива — нефти, каменного угля и природного газа, все
большую роль для получения пищевых (кормовых) продуктов и источников
энергии приобретают воспроизводимые виды сырья — растения и микроорга-
низмы. Из последних микробы выгодно отличаются от растений быстротой
роста, метаболической лабильностью и относительно легкой приспособляемостью
к изменяющимся окружающим (внешним) условиям, а также многообразием
биохимических реакций, завершающихся образованием различных продуктов,
в том числе существенно важных для человека.
Большинство микробиологических производств до недавнего
времени основывалось исключительно на применении клеток со-
ответствующих микроорганизмов; многие такие производства су-
ществуют и в настоящее время. Это условно называемый кле-
точный уровень биотехнологических процессов. В качестве при-
мера можно назвать разные виды брожений: маслянокислое,
молочнокислое, спиртовое и др., а также окислительные про-
цессы (получение уксусной, лимонной и других кислот), произ-
водство большинства антибиотиков, дрожжей и т. д.
Немногим более десяти лет стали широко использовать ме-
тоды получения гибридных клеток на основе метода слияния
микробных протопластов (парасексуальная гибридизация). Это
уровень клеточной инженерии в биотехнологических процессах.
Таким путем удается повысить потенциальную активность исход-
ных клеток в гибридном потомстве.
Осуществлена гибридизация В-лимфоцитов и клеток миеломы
млекопитающих (рис. 71, а). Полученная гибридома способна
продуцировать так называемые моноклональные антитела (см.
302
Селезенка
Лимфоциты
Б
Антигенная
детерминанта
Смесь антител
в иммунной
сыворотке
Слияние
Клонирование
Гибридные миеломные
клетки (гибридомы)
Клетки
миеломы
•лимфоциты
Клоны
71. Схема
а
Моноклональные
антитела
получения
юклональных антител
помощью гибридомной
:ники (а); инкапсули-
занные гибридомы, об-
зующие моноклональ-
е антитела (б)
3
. 8). В Шотландии построен завод по производству таких ан-
тел, где биотехнологический процесс рассчитан на использова-
е метода выращивания гибридомных клеток в инкапсулиро-
нном виде (рис. 71,6). При достижении клетками определен-
й плотности в капсулах антитела выделяются и очищаются
помощью простейших процедур.
Еще большие возможности открывает генная инженерия
Юлекулярный уровень биотехнологических процессов), хотя
кителем измененных наследственных молекул ДНК выступают
303
клетки. Генная инженерия берет начало с 1972 г., когда П. Берг
со своими коллегами (в США) создали первую рекомбинантную
ДНК- Со времени выделения Дж. Бекуитом с сотр. химически
чистого лактозного гена Е. coll (1969) сформировавшаяся
впоследствии генная инженерия продвинулась далеко вперед.
Удалось включить в наследственный аппарат микробных клеток
гены инсулина, соматотропного гормона человека и др. Культи-
вируемые в искусственных средах клетки высших растений и
животных являются по существу микрообъектами, и к ним при-
ложимы микробиологические подходы.
При оценке любого микробиологического производства ,;а
первый план выступает многостадийность процесса, которая,
как правило, включает: подготовку питательной среды и инок-..
та, или посевного материала (от лат. inoculatio - прививани >,
засев основных аппаратов и ферментацию, сепарирование кле: . к
от культуральной жидкости, выделение целевого продукта. Э'П!
пять стадий типичны для клеточного и клеточно-инженерн,к о
уровней биотехнологических процессов, если в последнем случае
на второй стадии используют в качестве продуцента гибридш.ш
организм и не выделяют этапы его создания в отдельные гы
дии.
В тех случаях, когда конечный целевой продукт находится
в культуральной жидкости, схема выделения зависит от его
физико-химических свойств (сравнить лимонную кислоту и бен-
зилпенициллин). Если же целевым продуктом являются клетки,
то технологический процесс их дальнейшей обработки относи-
тельно несложен, хотя это зависит от предназначения клеток.
В качестве примера можно указать на мицелий гриба, исполь-
зуемый для выделения из него какого-либо антибиотика, и дрож-
жи, применяемые как белковые добавки в рационы домашних
животных. В последнем случае технологическая схема проще,
нежели в первом случае.
Как уже было сказано, биотехнологические процессы осу-
ществляются преимущественно в асептических условиях, хотя
иногда используют открытые ферментаторы для выращивания,
например, в крупнотоннажных производствах кормовых дрожжей
на жидких Н-парафинах, на гидролизатах древесины и расти-
тельных отходах сельского хозяйства. Однако дальнейшее совер-
шенствование аппаратурного оформления этих процессов позво-
лит в будущем осуществлять соответствующие производства
в асептических условиях. Глубинное культивирование клеток
оказывается предпочтительным перед поверхностным. Тем не
менее выращивание клеток многих растительных тканей удается
лишь в стационарных условиях поверхностным способом.
Стабильность химической активности микроорганизмов зави-
сит от генома клеток и условий их культивирования (состав
питательных сред и методы их подачи, pH, температура, аэра-
ция, перемешивание, окислительно-восстановительный потенциал
и др.). При стандартных условиях выращивания и системати-
1
304
ком отборе нужных вариантов из популяции микроб-проду-
кт образует с достаточным постоянством соответствующий
вечный продукт.
.1. Брожения и окислительные процессы
ифатические спирты, органические кислоты и многие другие
рдукты относятся к разряду вторичных метаболитов (см.
5). Для прокариот характерно четыре типа катаболизма
окозы (гликолиз, пентозный цикл, кето-дезокси-фосфоглюко-
|-ный цикл и цикл трикарбоновых кислот); три типа фотосин-
18 (с участием хлорофилла у цианобактерий, бактериохлоро-
Лла у зеленых серных бактерий и бактериородопсина у гало-
Ктерий); два типа дыхания или биологического окисления
наэробное и аэробное) и два способа синтеза белков (мат-
»1ный, например ферментов, и нематричный, например тетра-
ента-)-пептидов в пептидогликане муреине). Эукариотические
рроорганизмы обладают большей специализацией, для них ти-
чны: один гликолитический путь разложения глюкозы, один
я' фотосинтеза с участием хлорофилла, аэробное дыхание,
тричный синтез пептидов. Однако среди эукариот имеются
ды, способные жить при ограниченном притоке кислорода,
птример, для роста и развития Candida krusei необходима
раженная аэрация, тогда как для С. albicans этого не требует-
' особенно в период образования хламидоспор. Следовательно,
1ницы изложенной биохимической типизации прокариот и
сариот не являются резкими, но выявляются они достаточно
'ко и несложными методами.
; Катаболические превращения углеводов в анаэробных усло-
ях (брожения) завершаются образованием различных конеч-
1х продуктов, что зависит от вида микроорганизма и глубины
ансформации углевода (см. разд. 5.3.2). Г липолитический про-
се осуществляют гомоферментативные молочнокислые бак-
рии:
СбН,2О6 ... 2СНз—СО—СООН Лактатдегидрогеназа, 2НАД -Н+H^
глюкоза пируват
-> 2СНз—СНОН—СООН + 2НАД+
Ь лактат
’ При спиртовом брожении пируват вначале трансформируется
^ацетальдегид, который восстанавливается до этанола:
СНз-СО-СООН Пируватдекарбоксилаза, Mg^ СНз_сон _
-со2
Алкогольдегидрогеназа _
----НАД Н^Н+ > СНз-СН2-ОН
Пропионовокислое брожение лежит в основе созревания сы-
)в. Вкусовые качества и рисунки сыра (пустоты, или глазки)
[Висят от конечных продуктов брожения, образующихся в про-
305
цессе жизнедеятельности пропионовокислых бактерий. Химизм
реакций здесь сложнее, чем при молочнокислом и спиртовом
брожениях, хотя центральное место также принадлежит пиру-
вату:
С6Н|2О6 - ... СНз—СО— СООН ——... СНз—соон
глюкоза пируват | ацетат
СН3—СНг—СООН
пропионат
В данной схеме образования пирувата опущено 9 реакций,
при образовании ацетата опущена одна реакция и, наконец, при
образовании пропионата опущено 8 реакций. Однако из этих
схематичных реакций видно, что наряду с пропионатом обра-
зуются уксусная кислота и углекислота. Суммарно реакции
пропионовокислого брожения могут быть представлены в следую-
щем виде:
ЗС6Н12О6 4СНз—СН2—СООН + 2СН3—СООН Д- 2СО2 -f- 2Н2О Д- «кДж
При маслянокислом брожении кроме масляной кислоты обра-
зуются различные другие продукты:
1. C6Hi2O6 -> ... 2СН3—СО—СООН ——2СН3—СОН
СНз—СОН + НСН2—СОН СНз—СНОН—СН2—СОН -> СНз—(СН2)2—соон
ацетальдоль бутират
2. СНз—СО—СООН СНз—СОН СН3—СНг—ОН
2 ацетальдегид этанол
3. СНз—СО—СООН ... СНз—СО—СНз СНз—сн—он
ацетон |
СНз
изопропанол
4. СНз—СО—СООН ... СНз—(СН2)г—СО — КоА ... СН3—(СН2)2—ОН
бутанол
Возможно также образование каприловой и капроновой кис-
лот:
СНз—(СН2)6—СООН СНз—(СН2)4—соон
каприловая кислота капроновая кислота
Капроновая кислота может накапливаться в качестве основ-
ного продукта при капронном брожении, вызываемом Clostri-
dium kluyveri.
Смешанные конечные продукты образуются при условно
называемых ацетоноэтиловом и ацетонобутиловом брожениях:
2С6Н12О6-^ СНз—СО—СНз + 2С2Н5ОН + 5СО2 + 4Н2 + п кДж
ацетон этанол
12СбН|20б С4Н9ОН 4СНз—СО—СНз С2Н5ОН Д- С3Н7СООН Д
бутанол ацетон этанол бутират
+ 13Н2 4- 28СО2 -Н 2Н2О + п кДж
Возбудители этих брожений — Clostridium butyricum, Cl. асе-
tobutylicum и др. Ацетоноэтиловое и ацетонобутиловое броже-
306
по продуктам относятся к разновидностям маслянокислого
кения.
^родильными процессами являются анаэробное разложение
,чатки и брожение пектиновых веществ. В анаэробных ус-
йях целлюлозу гидролизуют многие миксобактерии, мезофиль-
и термофильные клостридии. Продукты сбраживания цел-
озы — молочная, муравьиная и уксусная кислоты, водо-
, углекислота.
Р рубце желудка жвачных животных содержатся анаэроб-
! целлюлозоразрушающие бактерии (109...10 0 клеток в 1 мл
щовой жидкости), осуществляющие распад полимерных угле-
ов до ацетата (50...70 %), пропионата (17...21 %), бутирата
...20%), молекулярного водорода и метана (всех газов до
л в сутки), валерианата и формиата (небольшие количест-
. Пектиновые вещества находятся в листьях растений, мя-
и плодов, ягод, корнеплодов, в кожице цитрусовых и пр.
ктин представляет собой метиловый эфир полигалактуроновой
лоты вместе с арабаном и галактаном. Нерастворимый пек-
, называемый протопектином, находится в виде межклеточ-
о вещества и в молодых первичных стенках растительных
ток. В соках растений присутствуют растворимые пектино-
: вещества. Наряду с £)-галактуроновой кислотой, L-арабино-
и D-галактозой в пектинах находят D-ксилозу, L-рамнозу,
>укозу и др. С помощью ферментов (пектазы, пектиназы и
ггопектиназы) пектиновые вещества гидролизуются микроор-
измами. Протопектиназа катализирует трансформацию нера-
оримого протопектина в растворимый пектин; пектинэстераза,
I пектаза, катализирует реакцию гидролиза сложноэфирных
пп с образованием полигалактуроновой кислоты; полигалак-
оназа, или пектиназа, катализирует реакции гидролиза всех
козидных
связей:
место действия
пектинэстеразы
coo-
J----0.
COOCHj
COOCH,
coo
d
о
л место действия
полигалактуроназы
От ферментативной активности микроорганизмов зависит глу-
на катаболических процессов при использовании пектина. На-
имер, Clostridium pectinovorum вызывает маслянокислое бро-
!ние пектиновых веществ, a Cl. felsineum — преимущественно
гуснокислое брожение.
Аэробные микроорганизмы превращают углеводы, как пра-
по, в кислые продукты, эти ферментативные процессы называют
делением, а не брожением. Наиболее простым и давно изу-
307
ценным окислительным процессом является получение уксусной
кислоты из этанола:
СНз—СНз—ОН + О2 СНз—СООН + Н2О + 740 кДж
При скисании вина и пива также образуется уксусная кислота
из спирта. Уксуснокислые бактерии (Acetobacter species) могут
окислять не только этанол, но и другие субстраты, включая
моносахариды, пропанол, глицерин, маннит. В этом случае, на-
пример, сорбит трансформируется в сорбозу, глицерин — в ди-
оксиацетон, маннит — во фруктозу, арабиноза — в арабановую
кислоту, глюкоза — в глюконовую кислоту и т. д. Превращение
сорбита в сорбозу лежит в основе промышленного синтеза
L-аскорбиновой кислоты из £)-глюкозы:
А-аскорбиновая кислота
Окисление углеводов, сопровождающееся образованием ли-
монной кислоты и ранее неточно называемое лимоннокислым
брожением, вызывается многими грибами из родов Aspergillus,
Penicillium, Rhizopus и др. Наилучшим источником углерода для
образования цитрата мицелиальными грибами является сахаро-
за при pH 2—4. Промышленные способы получения лимонной
кислоты базируются на использовании нитчатого гриба Asper-
gillus niger. Мукоровые грибы — активные продуценты янтар-
ной и фумаровой кислот.
6.2.2. Аминокислоты, белки, клеточная биомасса
Получение некоторых аминокислот в промышленных условиях
стало важной отраслью народного хозяйства в ряде стран. Речь
идет прежде всего о незаменимых аминокислотах валине, изо-
лейцине, лейцине, лизине, метионине, треонине, триптофане и
фенилаланине. Отдельные аминокислоты используют в пищевой
308
(омышленности в качестве питательных (лизин), ароматических
дутаминовая кислота, глицин) и вкусовых веществ (глицин),
негодное производство, например, глутаминовой кислоты в ми-
> достигло 300 000 т, лизина — 200 000 т.
г Значительное развитие получила ферментная промышлен-
ють. В качестве первичных метаболитов ферменты синтези-
'ются под контролем генетического аппарата клеток. Каталити-
ские функции многих из них приобретают важное значение
i практике. Из 6 классов ферментов гидролазы занимают пер-
•е место по объему производства, а из гидролаз — пептидазы.
В экспериментальных исследованиях и для генной инженерии
тылую роль играют рестрикционные эндонуклеазы, или рест-
1ктазы. Их выделяют из бактерий, в которых они выполняют
нцитную роль, разрушая чужеродную ДНК. Эти ферменты
нают специфические симметричные последовательности нуклео-
[дов в молекулах ДНК и соответственно катализируют рас-
епление фосфодиэфирной связи на специфических симметри-
ских сайтах в пределах специфических последовательностей,
иедовательно, рестриктазы приобретают важное значение в
хнологии рекомбинантных ДНК- Например, необходимо встро-
ъ в циркулярную плазмиду специфический сегмент ДНК.
>гда очередность путей изменения ДНК будет следующей
•ис. 72). Специфичность действия рестриктаз приведена в
йг- Hr Т- Т- LT 3‘
I I
липкий конец
5- Г
липкий конец
III ।-----------1
3ta Ц“ Т- Т- А— А
фрагмент чужеродной ДНК
Рис. 72. Схема получения рекомбинантной плазмиды:
I — плазмида; II — линейная ДНК; III — рекомбинантная плазмидная ДНК
309
табл. 29, из которой видно, что ферменты различаются не только
по специфичности к субстратам, но и по числу расщепляющихся
сайтов.
Таблица 29. Субстратная специфичность некоторых рестрикционных
эндонуклеаз
Название Расщепление в последователь- ности Число расщепляю- щихся сайтов в ДНК Продуцент фермента
фага Л адено- виру- са 2 виру- са SV40
Alu I АГ*ЦТ >50 >50 32 Arihrobacter luteus
Bam HI Г-ГАТЦЦ 5 3 1 Вас. amyloliquefaciens Н
Bglll А'ГАТЦТ 5 10 0 Вас. globiggi
Bsu I гг*цц >50 >50 18 Вас. subtilis х5
Bal I цггЩцг г’ааттц 15 17 0 Brevibacterium albidum
Eco RI 5 5 1 Е. coli RY13
EcoRII •ццтгг >35 >35 16 Е. coli R245
» . *ЦЦАГГ >35 >35 16 Е. coli R245
Hap II Ц'ЦГГ >50 >50 1 Haemophilus aphrophilus
Hha I гцгД >50 >50 2 Haemophilus haemolyticus
Hind III А‘АГЦТТ 6 11 6 Haemophilus influenzae Rd
Hpa I гттЧац 11 6 5 Haemophilus parainfluen- zae
Mbol ТАТЦ 50 50 6 Moraxella bovis
Sma I цццДгг 3 12 0 Serratia marcescens Sbb
Из более чем 3000 выделенных ферментов только около 20
внедрены в производство. Следовательно, освоение биотехноло-
гии подавляющего большинства ферментов — дело будущего.
Уже сегодня амилазы применяют в хлебопекарной и текстиль-
ной промышленности, пептидазы — в мясомолочной и коже-
венной промышленности, ряд полиферментных препаратов ис-
пользуют в сельском хозяйстве (глюкаваморин, пектаваморин
и др.); здравоохранении (биодиастаза и др.). Большие количе-
ства протеолитических ферментов используются в моющих сред-
ствах.
Пластеины — новые формы продуктов питания — также по-
лучают с помощью ферментов. Для этого какой-либо исходный
белок вначале гидролизуют специфичной пептидазой, а затем
осуществляют сшивку пептидных связей соответствующей лига-
зой, но в иной аминокислотной последовательности или даже с
включением новой, ранее не содержавшейся в исходном белке
аминокислоты.
Практически важным оказался метод иммобилизации фер-
ментов, благодаря чему удается существенно продлить работу
биокатализаторов в стандартных условиях. При иммобилизации
ферментов используют методы адсорбции, ковалентного связыва-
ло
й
j (с использованием или без использования промежуточного
йа), ковалентной межмолекулярной сшивки, заключения в
уктуру геля и в полые волокна, проточных реакторов с полу-
|иицаемой мембраной. Иммобилизованные ферменты не только
йываются фиксированными, но и нерастворимыми. В качест-
: носителей ферментов используют микрокристаллическую,
(окнистую и пластинчатую целлюлозу, диэтиламиноэтилцел-
йозу (ДЭАЭ-целлюлозу), карбоксиметилцеллюлозу, амино-
Йцеллюлозу, р-аминобензилцеллюлозу, л-аминобензил-окси-
гиловый эфир целлюлозы, крахмальный гель, диальдегидный
|хмал, агар (агарозу или сефарозу), декстран (сефадекс),
)АЭ-сефадекс, оксид никеля, дакрон, стекло и пр.
?В последнее десятилетие реализовано производство глюкозо-
уктозного, или инвертного, сиропа, получаемого из сахарозы
•омощью фермента инвертазы, или глюкозоизомеразы. Инвер-
а катализирует реакцию гидролиза дисахарида сахарозы на
тавляющие — глюкозу и фруктозу. Иммобилизованная ин-
таза, полученная, например, из нитчатого гриба, сохраняет
ойчивость длительное время (период полуинактивации —
сут при 60°C). Следует заметить, что фруктоза слаще глюко-
и немного слаще исходной сахарозы. Вот почему производ-
о глюкозо-фруктозных сиропов особенно выгодно для стран,
котирующих сахар. В промышленно развитых государствах
шзводят миллионы тонн такого сиропа, в которых соотноше-
! глюкозы и фруктозы равно 1:1. В этой связи заманчивым
[зывается использование микроорганизма, одновременно несу-
го гены а-амилазы, глюкоамилазы и глюкозоизомеразы. Тогда
[учение глюкозо-фруктозных сиропов, например, из крахмала
'ло бы быть одностадийным процессом. Промышленное зна-
:ие приобрели иммобилизованные 0-галактозидаза (лактаза),
иноацилазы, фумараза, пенициллинамидаза, некоторые гид-
ссилазы, дегидрогеназы и другие ферменты. В первом случае
помощью 0-галактозидазы получают диетический продукт,
збходимый по жизненным показаниям многим лицам, не спо-
>ным усваивать лактозу, или молочный сахар.
Аминоацилазы катализируют реакции избирательного отщеп-
1ия ацилов только из L-аминокислот, оставляя незатрону-
ии £)-аминокислоты в их исходной рацемической смеси (на-
1мер, лишь £)-фенилаланин необходим для синтеза антибио-
зов ампициллина и цефалоспорина). Фумараза обеспечивает
ансформацию фумаровой кислоты в яблочную, а эту послед-
ую используют в качестве заменителя лимонной кислоты в пи-
авой и медицинской промышленности. Пенициллинамидаза
ггализирует реакцию образования 6-амино-пенициллановой кис-
1Ты (6-АПК) из бензилпенициллина. 11-0-Гидроксилазу и Де-
гидрогеназу используют в синтезе гормона преднизолона. На
Иове литических ферментов стрептококков созданы некоторые
(чебные препараты, например фибринолитики, катализирующие
Дролиз внутрисосудистых тромбов. Таким препаратом яв-
зн
ляется стрептодеказа — продукт соединения стрептазы с моди-
фицированной полисахаридной водорастворимой матрицей сши-
того декстрана. В США широко используется тромболитик
стрептокиназа. Более перспективным для растворения внутри-
сосудистых тромбов оказывается активатор тканевого плазмино-
гена, получаемый с помощью метода генной инженерии.
В последние годы важное значение приобрело использование
иммобилизованных микробных клеток как моделей полифермент-
ных систем. Иммобилизованными являются клетки, фиксирован-
ные на твердых носителях или включенные в матрицу. Из мето-
дов иммобилизации известны: физические (адсорбционные) с
использованием в качестве носителей активированного угля,
оксида алюминия, силикагеля, полиуретанов, производных чет-
вертичных пиридиниевых оснований, ДЭАЭ-целлюлозы и др.;
механические, когда клетки включают в гель; получаемый поли-
меризацией ненасыщенных мономеров (например, сополимериза-
цией акриламида и метилен-бис-акриламида и др.); поликон-
денсацией (например, в процессе синтеза пенополиуретана и
других полиуретанов); включением клеток в гель из полимеров,
способных к пространственной структуризации (например, водо-
растворимые и вододиспергируемые смолы с ненасыщенными
группами), или в гели, получаемые на основе неструктуриро-
ванных природных и синтетических полимеров (например, колла-
гена, казеина, полифенилаланина, политирозина, каррагинана,
желатина, альгинатов, агарозы, поливинилового спирта и др.);
химические, например присоединение (связывание) клеток к по-
верхности зерен сефадекса с помощью глутарового альдеги-
да, и др.
Использование иммобилизованных клеток микроорганизмов
весьма перспективно для создания различных технологических
процессов. Иммобилизация ферментов и клеток как полифер-
ментных систем стала основой развивающейся технологической
дисциплины — инженерной энзимологии. Иммобилизация фер-
ментов наряду с иммобилизацией нативных микробных клеток
заметно расширяет пути использования микроорганизмов и вы-
рабатываемых ими биокатализаторов в различных технологиче-
ских процессах. В разных странах выпускаются в продажу
ряд иммобилизованных ферментов: алкогольдегидрогеназа, а-
амилаза, бромелин, папаин, глюкозооксидаза, лейцинаминопеп-
тидаза, пероксидаза, РНКаза, субтилопептидаза, трипсин, уреа-
за, цитохром С и др.
Из других первичных метаболитов (белков) давно известны
микробные экзотоксины. Их производство осуществляется на
предприятиях, выпускающих вакцино-сывороточные препараты
(см. гл. 8).
Из числа бактериальных экзотоксинов можно назвать ста-
филококковые, стрептококковые, сибиреязвенный, дифтерийный,
ботулинический, столбнячный, дизентерийный, коклюшный, ган-
гренозные, токсины тюрингских бацилл и др. Бактериальные
312
зтоксины в большинстве своем используются на практике
]>орме обезвреженных препаратов (токсоидов) в качестве
игенов для профилактики соответствующих заболеваний.
Bacillus thuringiensis — грамположительная спорообразую-
я палочка, впервые изолированная С. Исивата (1902) из
ибшей личинки шелковичного червя, была названа Вас. sotto.
щнее Е. Берлинер (1915) описал ее под названием Вас. thu-
liensis. В клетке данного микроорганизма образуется харак-
ное ромбовидное параспоральное тело, представляющее собой
ковый кристалл, состоящий из 18 аминокислот (с преобла-
шем аргинина, аспарагиновой кислоты, глутамина, лейцина).
, локализации — это эндотоксин, или 6-токсин. Вместе с тем
с. thuringiensis продуцирует и а-у-экзотоксины. Известны
(личные серовары Вас. thuringiensis, например: berliner, fini-
us, alesti, kenyae, sotto, dendrolimus, galleriae, subtoxicus,
prison и др.
Промышленность различных стран производит соответствую-
е энтомопатогенные препараты для народного хозяйства,
меняющиеся главным образом для борьбы с насекомыми из
гада чешуекрылых (Lepidoptera). Токсины различных грибов
производят в промышленных масштабах, но они важны с
хицинской точки зрения и являются факторами агрессии у ряда
кро- и макромицетов (см. гл. 7).
Клетки микроорганизмов широко используют как источники
иродных белковых продуктов. В 1966 г. было введено понятие
чок одноклеточных [по англ. — single cell protein (SCP)]. По
цеству, SCP представляет собой инактивированные цельные
зтки бактерий, дрожжей и нитчатых грибов. Этот белок ре-
иендован для использования в рационах сельскохозяйствен-
х животных и птиц.
В Англии, например, в 1980 г. разрешено использовать в питании людей
юк Fusarium graminearum. Следует иметь в виду, что в белках микробных
еток часто недостает тех или иных незаменимых аминокислот. Поэтому
будущем важно получение более перспективных штаммов микроорганизмов
-одами генной инженерии, что позволит скорректировать качественные и ко-
(ественные характеристики продуктивности соответствующих видов. Сырьем
। получения микробного белка являются отходы сахарного и крахмало-па-
ного производств, деревообрабатывающей и целлюлозной, нефтеперерабаты-
)щей, молочной и газовой промышленности. Крупнотоннажное производство
(ровного белка для животноводства основано сегодня на использовании
имущественно дрожжевых организмов из рода Candida, утилизирующих са-
>а гидролизатов, углеводсодержащие отходы, Н-алканы и этанол, а также
илотрофных бактерий, утилизирующих метанол и метан из природного газа,
цианобактерий давно используют Spirulina maxima как древний традицион-
й продукт питания у жителей Северного Чада. Эти цианобактерии содержат
нее 60 % белка. Перспективны и микроводоросли, например, из родов Scendes-
us, Chlorella.
Роль дрожжей в питании человека давно известна и как таковые они служат
Лавкой к некоторым продуктам в отдельных странах Юго-Восточной Азии
Др. Также известен их лечебный эффект при ряде патологических состояний.
Бурное развитие биотехнологии некоторых белковых продук-
>в основано на методах клеточной и генной инженерии (моно-
313
клональные антитела, интерфероны, гормональные препараты,
ферменты).
Получение моноклональных антител базируется на методах
клеточной инженерии. Моноклональные антитела идентичны
одной антигенной детерминанте и образуют потомство (клон)
одной клетки В-лимфоцита. Г. Кёлер и К- Милстайн (1975)
впервые получили так называемые гибридомы — слившиеся
клетки мышиной миеломы с В-лимфоцитами из селезенки мышей.
Слияния клеток достигают, например, с помощью полиэтиленгли-
коля. Миеломы — злокачественные опухоли иммунной системы,
клетки которых быстро растут и размножаются, образуя до-
статочно большие количества аномальных иммуноглобулинов,
или миеломных белков.
Клетки гибридов сочетают в себе способность миеломных
клеток непрерывно размножаться (бессмертие в идентичном
потомстве) и способность В-лимфоцитов вырабатывать специфи-
ческие антитела. Имея в распоряжении клон гибридных клеток,
образующих антитела к одной антигенной детерминанте, их мож-
но использовать для получения моноклональных антител в ис-
кусственных условиях (в культуре) или в организме животных,
которым вводят такие гибридомы (см. рис. 71, а). Клональную
природу гибридомной клетки необходимо подтвердить анализом
продуцируемых ею антител с помощью физико-химических ме-
тодов, по антигенности и специфичности. Нередко такие клетки
вводят в брюшную полость гистосовместимых мышей, предвари-
тельно стимулированных в направлении активизации роста
гибридомных клеток, которые впоследствии размножают в виде
асцитных опухолей. Возникающая при этом асцитическая жид-
кость содержит большое количество моноклональных антител.
Гибридомы удается получать на основе гибридизации клеток
миеломы человека и В-лимфоцитов человека, выделенных из
периферической крови, миндалин или селезенки. Подобные гиб-
ридомы также можно культивировать в производственных усло-
виях и масштабах.
Согласно требованиям Комитета экспертов ВОЗ (Всемирная организация
здравоохранения) по стандартизации биологических препаратов необходимо
четко указывать: источник получения гибридомного клона; его стабильность;
историю пассирования; условия хранения; проводимые тесты для доказательства
специфичности клона; доказательства моноклональных антител с помошью
физико-химических методов (изоэлектрофокусирование, электрофорез и пептид-
ное картирование фрагментов, полученных обработкой ферментами и бромциа-
ном); характеристику изотипа тяжелой и легкой цепей и аллотипов монокло-
нальных антител.
Моноклональные антитела производятся теперь в производ-
ственных условиях. Они находят широкое применение в фунда-
ментальных и прикладных исследованиях (при определении
групп крови, пассивной иммунизации соответствующих больных
людей, для выделения и очистки отдельных белковых продуктов,
например интерферона, и т. д.).
В настоящее время исследуют различные аспекты клеточной
314
<енерии, в том числе — возможность получения гибридов
еду микробами-продуцентами белка и галофильными бакте-
ми, живущими за счет солнечной энергии, и др.
Получение интерферона, гормональных и других белков так-
базируется на методах генной инженерии. В настоящее время
чедователи могут целенаправленно изменять генетическую
юрмацию в клетках прокариот и эукариот, заставляя их та-
: образом производить ранее не свойственные им белки. Такие
етки являются рекомбинантами. В этом плане особенно пер-
ективны бактерии и дрожжи, легко и быстро размножающиеся
лабораторных и производственных условиях. Чтобы ввести и
стельно поддерживать новую генетическую информацию в
етке, необходимы так называемые направляющие молекулы
НК (векторы). Другие молекулы чужеродной ДНК подвер-
дотся в клетке деградации за счет действия собственных
НКаз. Векторные молекулы ДНК способны автономно сущест-
>вать и реплицироваться в клетке в форме кольцевых мини-
НК. Репликатор, включающий реплицирующий аппарат клетки,
’ветствен за репликацию вектора. Этот последний, в свою оче-
!дь, обеспечивает клеткам-носителям маркерный признак, по
второму возможен отбор их, например, по какой-либо специ-
Ической ферментативной активности. Наиболее широко исполь-
гёмыми векторами являются многокопийные плазмиды, содер-
жите от 3 до 40 тыс. оснований (3...40 кв).
Применительно к интерферону удалось клонировать в клетки
i coli гены лейкоцитарного (а), фибробластного (0) и Т-лим-
эцитарного (у) интерферонов. Известно около 20 генов, коди-
ющих синтез 20 разновидностей а-интерферона, один ген —
!я у-интерферона и несколько генов — для 0-интерферонов.
ятерфероны 0- и у-типов — гликопротеины, тогда как а-интер-
:рон — «чистый» протеин. Все они содержат 144...166 остатков
[инокислот. Интерфероны проявляют противовирусное и проти-
сни исключительно важны для
опухолевое действие, поэтому
рапии соответствующих за-
деваний.
В СССР под руководством
;ад. Ю. А. Овчинникова полу-
пи штаммы Е. coli, несущие
комбинантную плазмиду и
нтезирующие интерферон че-
>века (рис. 73). В настоящее
ремя гены интерферона кло-
ированы в дрожжевые клетки,
жже перспективные для полу-
дня этого ценного белка в
юизводственных условиях.
Методами генной инжене-
1И удалось создать гибридные
ны, кодирующие синтез пеп-
ген интерфе
Рис. 73. Рекомбинантная плазмида,
обеспечивающая синтез интерферона
(И) человека клетками Е. coli
315
Рис. 74. Клетки Е. coli (а), образующие человеческий инсулин (б)
тидных и белковых гормонов, клонировать их в микробные клетки и
получать на практике эти гормональные регуляторы.
На примере инсулина можно показать успех генной инжене-
рии для биотехнологии данного гормона, образуемого кишечной
палочкой (рис. 74, а, б).
Из ферментных белков методом генной инженерии полу-
чают активатор тканевого плазминогена (АТП). Он образуется
внутренней оболочкой кровеносных и лимфатических сосудов,
а также сердца (эндотелий). Молекулярная масса его около
50 кДа. АТП активирует плазминоген, трансформирующийся
в плазмин, а этот последний растворяет внутрисосудистые тром-
бы. Для получения АТП применили метод встройки генов, коди-
рующих его продукцию, в клетки животных организмов. АТП
более селективен в отношении внутрисосудистых тромбов, чем
стрептокиназа или стрептодеказа. Он обладает меньшим побоч-
ным действием и быстро (в пределах нескольких минут) инакти-
вируется в печени.
В перспективе обнадеживающими представляются исследова-
ния: по переносу генов азотофиксации из бактерий в раститель-
ные организмы с целью увеличения в них содержания азота, что
имеет большое значение для обеспечения населения высокока-
чественными продуктами питания; по созданию вакцин против
многих вирусных, протозойных и других инфекционных заболева-
ний; по биосинтезу кейлонов, нейропептидов и других пептидов,
являющихся регуляторами физиологических процессов у различ-
ных организмов.
6.2.3. Углеводы
Применительно к углеводам известны биотехнологические про-
цессы получения циклодекстринов и разнообразных микробных
полисахаридов. Еще Ф. Шардингером (1903) было установлено,
что амилаза Вас. macerans катализирует образование цикло-
декстринов Из крахмала (циклодекстрины Шардингера). Извест-
316
a, p- и у-циклодекстрины, включающие соответственно шесть,
ь и восемь остатков ангидроглюкозных единиц. Следователь-
они представляют собой глюкогексаозы, глюкогептаозы и
•кооктаозы. Некоторые из них приобрели важное значение в
1евой и медицинской промышленности. Так, циклодекстрины
собны инкорпорировать (от лат. in — в, согроге — тело) дру-
молекулы, т. е. при этом наблюдается своеобразное моле-
ярное инкапсулирование. В циклодекстринах первичные
роксильные группы располагаются на внешней поверхности
:роцикла, а вторичные гидроксильные группы — внутри его,
создается аполярное пространство за счет своеобразного
пределения атомов водорода.
0-Циклодекстрины нашли наиболее широкое применение. Это
словлено прежде всего их размерами (наружный диаметр
J нм, внутренний — 0,78 нм). 0-Циклодекстрины используют
включения многих летучих и ароматизирующих вкусовых
редиентов, лекарственных веществ; рекомендуют для стабили-
ии неустойчивых структур, для трансформации жидких соеди-
ий в кристаллические или таблетированные формы, для уве-
ения растворимости мало растворимых в воде веществ и т. д.
Р -циклодекстрин
В промышленности используют различные микроорганизмы
я получения многих типов полисахаридов, из которых из-
стны клеточные, внеклеточные и капсульные бактериальные,
'также грибные полисахариды. К бактериальным относятся:
Еьгинаты, декстраны, ксантан, курдлан, полисахариды клеб-
317
сиелл, менингококков и пневмококков; к грибным — аубазидац
кориолан, лентинан, пахиман, пахимаран, пуллулан, родэксман
склероглюкан, шизофиллан и др. Названные выше полисахариды
производятся промышленностью многих стран.
Альгинаты, образуемые некоторыми водорослями, применяют
в пищевой промышленности с 1935 г. В настоящее время на
подходе к внедрению биотехнология бактериальных альгинатов,
образуемых Azotobacter vinelandu и Pseudomonas aeruginosa.
Эти альгинаты сходны по строению с альгиновой кислотой из
водорослей, обладающей блочной структурой и включающей
а-С1 —С4 линейный сополимер ^-О-маннуроновой кислоты и ее
5-эпимер a-L-гулуроновой кислоты:
[поли-МанУК 1
поли-ГулУК Х0,6ОСОСН3
полн- (ГулУК-МанУК) 1
блок альгиновой кислоты
Декстран, продуцируемый некоторыми видами лейконостоков,
например Leuconostoc mesenteroides, нашел широкое применение
в медицинской и пищевой промышленности. Это один из первых
плазмозаменителей крови, производящийся ныне в больших ко-
личествах. На его основе получены ферриглюкин, реоглюкин и
другие препараты, использующиеся при различных патологиче-
ских состояниях людей. В молекуле декстрана 95 % связей при-
ходится на а-С1 -> Сб-связи и 5 % на а-С1 ->СЗ-связи:
— [—- 6Глю1 —> 6Глю1 —► 6Глю1——
3
t
1
Глю-R
где R — Н или остаток глюкозы; С1 -> СЗ-связи оказываются
местом формирования боковых цепочек, состоящих примерно на
85 % из одного или двух остатков глюкозы; 15% боковых цепо-
чек могут неравномерно распределяться по длине скелета макро-
молекулы. Такой декстран относительно хорошо растворим в во-
де, тогда как другие декстраны, содержащие в боковых цепочках
С1 С2-, С1 -> СЗ- и С1 -> С4-связи в разных количественных
соотношениях, плохо или совсем нерастворимы в воде.
Декстран синтезируется из сахарозы под действием индуци-
бельного фермента — декстрансахаразы. Синтез осуществляется
с редуцирующего конца в присутствии двухвалентных ионов:
(С|2Н22О,,)„ Д^тр^еахараза (СбН10О6)„ + пС6Н12О6
декстран фруктоза
Фруктоза как отход производства может быть очищена и исполь-
зована для пищевых целей. В настоящее время известно произ-
водство декстрана с помощью ферментного метода. На первой
стадии выращивают лейконосток, образующий активную дек-
странсахаразу, затем клетки отделяют, а на второй стадии ис-
318
Йьзуют фермент в культуральной жидкости для биосинтеза
[страна. Следует заметить, что декстраны продуцируют и дру-
I бактерии: L. dextranicum, Betacoccus arabinosaceus, Betaba-
Ifium vermiforme, Streptococcus mutans, Str. viridans, Str.
jcosus.
tC 1960 г. Швеция производит так называемые сефадексы—
растворимые сшитые декстраны, нашедшие широкое примене-
е в технике разделения различных полимерных веществ. Суль-
фированные декстраны применяют при сердечно-сосудистых за-
деваниях, например в качестве антикоагулянтов крови (гепа-
доподобное действие). С 1963 г. известно крупномасштабное
Ьизводство ксантона — бактериального экзополисахарида, об-
^уемого фитопатогенными бактериями Xanthomonas campest-
X. phaseoli, X. juglandis. Эти виды вызывают пожелтение
стьев фасоли, увядание и опадание листьев моркови, пятнис-
сть хлопчатника. За счет выраженной продукции полисахарида
онсходит закупорка сосудов в растениях-хозяевах, которые
итоге погибают. Ксантан — сложный, высоко разветвленный
дисахарид. В небольших концентрациях ксантан придает вод-
IM растворам высокую вязкость, мало зависящую от нагрева-
я в широком диапазоне температур, от колебаний pH (в пре-
лах 6...9). Полисахарид имеет блочную структуру, включаю-
ую целлобиозо-подобный кор, к которому присоединяется боко-
я цепочка, состоящая из остатков маннозы, а также пирови-
градной, глюкуроновой и уксусной кислот. Причем содержа-
е пирувата в молекуле может колебаться в пределах 2,5...
1%, т. е. не каждый остаток маннозы соединен с пируватом,
тесте с тем водород карбоксила может замещаться каким-либо
тионом. Поэтому, несмотря на блочную структуру ксантана,
ми блоки не всегда имеют полную аналогию в строении. В при-
димой формуле ксантана пунктирные линии означают, что не
ждый блок в молекуле полимера содержит пируваткетальную
уппу. Ксантан производится в мире тысячами тонн для нефте-
бывающей, пищевой, медицинской промышленности, для сель-
ого и лесного хозяйства.
— [ -_^4Глю1А 4 Глю 1-И-]
3
t «
1
ацетил-6-Ман
2
t ₽
1
Me-Глю УК
4
t 3
,----------------, ]
4,р-пируват-кеталь-Ман
ксантан
319
Большой интерес для практики представляет курдлан (от
англ, curdle — коагулировать, уплотнять), образуемый Alcali-
gens faecalis var. myxogenes, сходные полисахариды образует
и Agrobacterium radiobacter:
— ( -i 4Глю1 Л ЗГлю1 Л ЗГлю1 Л 6Глю1 А 4Гал1 Л
->-ЗГлю1 Л ЗГлю1 Л ЗГал1 Л-ЗГлю1 Л-4Гал1 —)„^
курдлан
Курдлан благодаря реологическим (от лат. rheo — течь, logos —
учение) свойствам оказался полезным в хлебопечении, изготов-
лении кондитерских, колбасных и других изделий, в приготовле-
нии ряда лекарственных форм медицинских препаратов и т. д.
Для его получения оказалась перспективной двухстадийная не-
прерывная ферментация.
Капсульные полисахариды в составе гликоконъюгатов, изоли-
руемых из микроорганизмов рода Klebsiella, используют в ка-
честве иммуноадъювантов, например, в рибосомальной вакцине
рибомунил (Франция) против бронхопневмонии у людей.
Как правило, капсульные полисахариды Klebsiella содержат
пируваткетальные группы. Это можно показать на примере полиса-
харида, характерного для серовара Г.
— -----&4ГлюУК-1 -Д 4Ь-Фук1 -Д ЗГлю1— Д-
3\ /2
НООС/'С\сНз п
повторяющийся блок капсульного полисахарида Klebsiella К-1
Капсульные полисахариды пневмококков входят в состав так
называемой полисахаридной вакцины, выпускающейся в произ-
водственных условиях в ряде промышленно развитых стран. Вак-
цина поливалентна и включает углеводные полимеры 18 наиболее
распространенных сероваров пневмококков (Streptococcus pneu-
moniae) из 85 известных. На примере наиболее патогенного
серовара III показана особенность химического строения поли-
сахарида, состоящего из блоков целлобиуроновой кислоты, сое-
диненных fl-Cl -> СЗ-связью:
пневмококковый капсульный полисахарид серовара 111
320
Группоспецифические капсульные полисахариды восьми серо-
ов менингококков (А, В, С, X, Y, Z, 29Е и 135) являются
ерминантами вирулентности. Наиболее важное значение в
нической практике приобрели полисахаридные вакцины серо-
ов А, В и С как профилактические лечебные препараты при
тветствующих менингитах. Полисахарид А состоит из N-аце-
-О-ацетилманнозаминилфосфатных остатков; полисахариды В
С — из ацетил-О-ацетилнейраминовой кислоты, которая час-
1но ацетилирована в С-антигене.
Из числа гликоконъюгатов в производственных условиях по-
чают диагностические и лечебные липополисахариды из неко-
рых бактерий семейств Enterobacteriaceae и Pseudomonada-
ie. Разрабатываются промышленные биотехнологические про-
гсы получения ряда грибных экзополисахаридов. Так, разные
аммы диморфного гриба Aureobasidium pullulans продуциру-
1 аубазидан и пуллулан. Первый из них имеет а- и 0-конфигу-
ции гликозидных связей, тогда как второй — лишь а-конфигу-
цию связей:
(Глю1 А- 4Глю)л
1
!₽
^ЗГлю! АзГлю! А ЗГлю1—
блок аубазидана
Пуллулан содержит а-гликозидные связи преимущественно
»жду триаозными блоками, хотя в структуре полисахарида
«еются и тетраозные блоки, являющиеся местом взаимодейст-
[я с ферментом эндоамилазой:
I
«I
Глю1 А 4Глю1 А 4Глю1 Место Действия
, амилазы
1“ I
6 I
Enrol А 4Глю1 А 4Глю1 А 4Глк>1,
блок пуллулаиа ।__ п
4
Аубазидан — вспомогательное вещество для создания различ-
йх лекарственных форм медицинских препаратов, пуллулан на-
ел применение в пищевой промышленности как упаковочное
>едство, легко разрушаемое микробами во внешней среде.
Кориолан, лентинан, пахиман и шизофиллан вырабатываются
полупроизводственных условиях (Япония), их используют для
:чения некоторых онкологических больных. Все эти полисахари-
J получают из биомассы базидиальных грибов: Coriolus verst-
dor, Lentinus edodes, Porta cocos, Schizophyllum commune
ответственно. Они представляют собой (3-глюканы, содержащие
321
1617
в основной цепи С1->-СЗ-связи. Пахимаран — производное па-
химана, лишенного боковых цепей.
Родэксман — внеклеточный полисахарид, образуемый Rhodo-
torula rubra. Это маннан с чередующимися fJ-Cl -> СЗ- и 0-С1 ->
С4-СВЯЗЯМИ. Родэксман и его полусинтетические производные
представляют интерес в качестве самостоятельных лечебных
средств при некоторых заболеваниях сердечно-сосудистой сис-
темы.
Склероглюкан продуцируется несовершенным грибом Sclero-
tium glucanicum. Это в основном линейный нейтральный глюкан,
во многом сходный с аубазиданом:
— Г-1зГлю1 А ЗГлю1 Л- ЗГлю1——
t ₽
1
Глю
Склероглюкан рекомендован к использованию в здравоохра-
нении. Кроме перечисленных полисахаридов изучены многие дру-
гие грибные углеводы, часть из которых близка к внедрению в
производство. Некоторые полисахариды, детерминирующие анти-
генную специфичность условно патогенных дрожжей рода Candi-
da, нарабатываются в небольших количествах, например в СССР,
для диагностических целей (постановка аллергических проб и се-
рологических реакций).
6.2.4. Антибиотики
Промышленное получение антибиотиков занимает особое место в биотехнологии.
Развитие производства антибиотических веществ позволило не только открыть
многие новые природные химические структуры, но и создать мощные, современ-
ные по технической вооруженности производства. Сегодня антибиотическая про-
мышленность — одна из наиболее развитых отраслей биотехнологии. Ею выра-
батывается около 100 антибиотиков (всего известно более 6000), относящихся
главным образом к пенициллинам, цефалоспоринам, аминоглйкозндам, тетрацик-
линам и макролидам. К началу 80-х годов мировая промышленность выпускала
в год порядка 25 000 т антибиотиков: пенициллинов - 17 000 т, цефалоспори-
нов — 1200 т, тетрациклинов — 5000 т, аминогликозидов и др. — 1800 т. В СССР
производится около 50 антибиотических веществ, из которых готовят свыше
200 лекарственных форм.
Пенициллины составляют большое семейство антибиотических
веществ, родоначальником которых был пенициллин, образуемый
отдельными видами Penicillium и впервые полученный в кристал-
лическом виде X. Флори и Е. Чейном (1940):
боковая амидная
г-це-пь | /связь XSX сн3
R—COj—NH—СН—СН cf
j?-лактамное- t , | з* | ж тиазолидиновое
кольцо ; о=с---------N-СН—СООН кольцо
I_________________________I
6-аминопенициллановая кислота
(6-АПК)
I_____________________________________________I
пенициллин
322
В пенициллинах R может быть представлен различными
руктурами. Наиболее известным и широко используемым пре-
ратом был бензилпенициллин, или пенициллин G, в котором
представляет собой бензильный остаток (СвН5—СНг—); в пе-
циллине V радикалом является феноксиметил (СбНб—О—
СНг—) и т. д.
Позднее было установлено, что различные микроорганизмы
особны вырабатывать гидролазы, катализирующие реакции
дролиза 0-лактамного кольца и амидной связи. Первый фер-
рит был назван пенициллин-р-лактамазой (пенициллиназой);
орой — пенициллинамидазой (пенициллинацилазой). В резуль-
те действия пенициллин-^-лактамазы образуется неактивная
нициллоиновая кислота. Под действием пенициллинамидазы
разуется антибиотически мало активная 6-АПК, которая ока-
лась исключительно удобным объектом для введения радика-
в вместо одного водорода в аминной группе.
Пенициллиназу и пенициллинамидазу образуют многие мик-
организмы. Так, спорообразующие виды Вас. cereus, Вас. те-
terium, Вас. mesentericus, Вас. subtilis, неспорообразующая
coli и другие продуцируют активную пенициллиназу. Зараже-
е такими микробами культуральной среды, на которой выра-
1вался продуцент пенициллина, может быть причиной заметной
и полной инактивации конечного целевого продукта.
Пенициллинамидаза образуется многочисленными видами
'кроорганизмов из числа бактерий (палочек, кокков, актиноми-
тов) и грибов (дрожжей, нитчатых грибов), в том числе —
миМи продуцентами пенициллина. В то же время имеются мик-
бы, способные продуцировать оба фермента одновременно —
coll, Streptomyces lavendulae, некоторые виды Nocardia.
настоящее время в разных странах (в том числе в СССР) соз-
ны биотехнологические процессы с использованием иммобили-
ванной пенициллинамидазы для выделения 6-АПК из бензил-
нициллина.
Полученная 6-АПК затем используется для химического син-
за других пенициллинов, например, созданы кислотоустойчи-
ie препараты: a-феноксиэтилпенициллин (фенетициллин), име-
Щий радикал СеНб—О—СН(СН3)—; а-феноксипропилпеницил-
н (пропициллин) с радикалом С6Н5—О—СН—СН2—СН3; а-
локсибензилпенициллин (фенбенициллин) с радикалом
Н5—О—СН—СбНб. 6-АПК является также основой пеницил-
назоустойчивого 2,6-диметоксифенилпенициллина< (метицилли-
) с радикалом СбНз(ОСН3)2—, кислотоустойчивого и широ-
спектрального a-D-(—)-аминобензилпенициллииа (ампицилли-
) с радикалом CeHs—СН—NH2, кислото- и ленициллиназо-
тойчивых 5-метил-3-фенил-4-изоксиазолилпенициллина (окса-
ллина), 2-этокси-1-нафтилпенициллина (нафциллина), 3-0-
373
хлорфенил-5-метил-4-изооксазолила (клоксациллина) и др.
радикал
N С—СН,
оксациллина
радикал нафциллина
радикал клоксаииллина
Цефалоспорины — семейство антибиотиков, близкое по хими-
ческому строению к пенициллинам:
/?-лактамное
кольцо
I /SX
НООС—СН—(СН2)3— со—NH—СН4-СН сн2
1 Г I I дигидротиазиновое
' 1 ~~ | кольцо
с—сн,—осо—сн3
cZ
а - О-аминоадипиновая
кислота
nh2
о=с---N
соон
7-аминоцефалоспорановая кислота
В радикале цефалоспорина N, который относится к пеницил-
линам, содержится О-4-амино-4-карбокси-п-бутил:
/s\ /СН3
НООС—CH—(СН2)3—СО—NH—СН—СН с<
I I I ПСНз .
nh2 О=С N СН—соон
цефалоспорин N (пенициллин N)
Цефалоспорины ингибируют грамположительные и грамот-
рицательные бактерии, хотя по активности в отношении чувстви-
тельных микробов они уступают пенициллинам. Продуцируют
цефалоспорины нитчатые грибы из рода Cephalosporium.
По аналогии с 6-АПК путем химического синтеза удалось
трансформировать 7-АЦК (7-аминоцефалоспорановую кислоту)
в иные производные, некоторые из них — цефалоглицин, цефа-
лексин, цефалотин и цефалоридин (цепорин) — нашли практи-
ческое применение в химиотерапии инфекционных заболеваний.
Цефалоглицин активен в отношении грамотрицательных бакте-
рий, цефалексин — кислотоустойчив и подавляет пенициллиназо-
образующие стафилококки; цефалотин и цепорин (равно как и
цефалоглицин) активны против стафилококков, продуцирующих
пенициллиназу; они неустойчивы в кислой среде.
324
с6н5—сн—co—nh—сн—с
СН2
nh2
о=='
соон
цефалоглицин
—СНг—СО—NH—СН—Cf-f ^СНг
цефалексин
0='
ОСОСНз
:С—СНз
СООН
—СН2—<
н
цефалотин
0=С--N.
s\
хсн2
,С—СНгОСОСН
соон
—СН2—СО—NH—СН—С'
цефалоридин
0=С--N.
S\
ХСН2
СН
СООН
Родственными пенициллинам и цефалоспоринам являются
тибиотики — монобактамы — моноциклические р-лактамы, у
торых структурным ядром выступает р-лактамное кольцо. Мо-
бактамы активны против возбудителей госпитальных инфек-
;й:
R,— СО— NH
О'
/R2
N
soy И *
монобактамы
Аминогликозидные антибиотики включают ряд противомик-
обных
веществ,
в химической структуре которых имеются глико-
дные связи: стрептомицины, неомицины, канамицины, гента-
325
мицины, гигромицин, сизомицин, амикацин и др. Продуцируют
их различные виды актиномицетов. Аминогликозиды — антибио-
тики широкого спектра действия в отношении чувствительных
грамположительных и грамотрицательных бактерий. Все они тор-
мозят белковый синтез благодаря взаимодействию с 30S суб-
частицами бактериальных рибосом и нарушению их функций.
Названные антибиотики более активны в щелочной среде. Устой-
чивые к аминогликозидам бактерии инактивируют антибиотики
с помощью ферментов, образующихся при наличии в клетках
R-плазмид.
Стрептомицины — стрептомицин, маннозидострептомицин,
гидроксистрептомицин образуются актиномицетами Streptomyces
griseus, Str. bikinensis, Str. olivaceus, Str. mashuensis, Str.
humidus. Стрептомицин — это основание, состоящее из стрепти-
дина, стрептозы и №метил-£-глюкозамина. При гидролизе его
растворами кислот может быть выделена биоза — стрептобиоза-
мин (Н-метил-а-Ь-глюкозаминидоф-Ь-стрептоза). Стрептидин
представляет собой 1,3-дигуанидино-2,4,5,6-тетраоксициклогек-
сан. Ни стрептидин, ни стрептобиозамин не обладают антибак-
териальным действием. Стрептомицин легко образует соли (хлор-
гидрат, сульфат, рейнекат и др.), которые в кристаллическом
виде флуоресцируют в ультрафиолетовом свете. На практике,
как правило, используют стрептомицин-сульфат (C21H39O12N7) X
X3H2SO4:
стрептобиозамин
326
-Маннозидострептомицин — это а-£>-маннозидо-М-метил-а-£-
(>козаминидо-|3-£-стрептозидострептидин.
Гидроксистрептомицин во многом аналогичен стрептомицину.
Шь в стрептозе вместо метильной группы находится оксиме-
’ьная. Дигидрострептомицин может быть получен путем био-
теза культурой Str. humidus или при каталитическом гидри-
,ании стрептомицина. В нем альдегидная группа стрептозы
[енена на оксиметильную.
* Как было указано, стрептомицины нарушают синтез белка на
овне 30S рибосом. Если эти последние диссоциировать в гра-
енте плотности хлористого цезия на кор-частицы (16S РНК +
15 белков) и на отделяющиеся белки (6 белков), то при реас-
циации 30S рибосом доказано, что чувствительность к анти-
отикам определяется кор-частицами. Каждая рибосома может
взывать 1...2 молекулы антибиотика в присутствии ионов маг-
fl и уридин- или цитозин-содержащих полинуклеотидов. При
ом отмечается не только ошибочное считывание кода с мРНК,
। и необратимое подавление синтеза белка.
1 Сагамицин (микрономицин сульфат) —новый аминоглико-
дный антибиотик. Химическое название его О-2-амино-2,3,4,6-
^радезокси-б- (метиламино) -а-О-эритро-гексапиранозил- (1--*
4) -О-З-дезокси-4-С-метил-З- (метиламино) -(З-А-арабинопира-
^ил-(1-> 6)-2-дезокси-Л-стрептамин гемипентасульфат.
юти к широкого спектра действия. Отделение одной или
леводных частей сопровождается потерей активности. По
гзму действия сходен со стрептомицином:
Анти-
обеих
меха-
Неомицин образуется Streptomyces fradiae и некоторыми дру-
ми актиномицетами. Известны неомицины А (неамин), В и С
I и С изомеры неомицина А). К этой группе антибиотиков
носятся паромомицины I и II. Синонимы неомицина — коли-
щин, мицерин, фрамицин. Неомицины представляют собой оп-
чески активные основания, образующие соли с кислотами:
327
Мономицин В: R = Н, Ri = CH2NH2, R2 — СН2ОН, R3 — NHAc
(возможно, MOHo-N-ацильная группа находится в третьем поло-
жении того же кольца); неомицин А: часть изображенной моле-
кулы справа от пунктирной линии; неомицин В: R = Н, Ri =
= R2 = CH2NH2, R3 = NH2; неомицин С: R = R2 = CH2NH2,
Ri = Н, R3 = NH2; паромомицин I и мономицин A: R = Н, Ri =
= CH2NH2, R2 = СН2ОН, R3 = NH2; паромомицин II: R =
= CH2NH2, Ri = Н, R2 = СНгОН. Отделение любой углеводной
части молекулы лишает антибиотик противомикробной актив-
ности.
Канамицин — противобактериальный антибиотик широкого
спектра действия из группы аминогликозидов. Будучи основа-
нием, он легко образует соли с неорганическими и органическими
кислотами; оптически активен ([а]о для основания = 146°).
Продуцент канамицина — Str. kanamyceticus-. В культуральной
жидкости после роста актиномицета образуются сходные по
строению канамицины А, В и С. Канамицин А состоит из 3-амино-
3-дезокси-£)-глюкозы, 6-амино-£)-глюкозы и дезоксистрептамина:
328
рамицин С отличается от канамицина А тем, что вместо 3-Z)-
^козамина в нем содержится обычно 2-О-глюкозамин. По ме-
лизму действия канамицины относятся к антибиотикам, вызы-
ющим ошибочное считывание кода на уровне 30S субчастиц
босом в чувствительных бактериальных клетках.
Гентамицин — образуется Micromonospora purpurea. Извест-
несколько близких по составу и строению гентамицинов:
Bl, С1, С1а и С2. На практике используют препарат, содержа-
1Й 40 % фракции С1, 20 % — фракции С 1а, 40 % — фракции
I и лишь в виде незначительной примеси могут быть фрак-
й А и В.
Гентамицин — основание, образующее соли с кислотами,
медицине применяют его в виде сульфата. В щелочной среде
тибактериальная активность препарата выше, чем в кислой,
составе антибиотика имеются три цикла, два из которых в ген-
мицине А принадлежат a-D-глюкозамину и З-метиламино-З-
зокси-a-D-ксилозе (а); в гентамицине С1—3,4,6-тридезо-
и-6-метил-6-метиламино-а-£)-глюкозамину и З-дезокси-З-мети-
мино-4-метил-а-£-арабинозе (б); в гентамицине С2 — 3,4,6-
идезокси-6-метил-6-амино-а-О-глюкозамину и той же а-А-ара-
нозе, которая имеется в гентамицине С1 (б). Наконец, в гента-
щине С1а содержатся 3,4,6-тридезокси-6-амино-а-£)-глюкоза-
н и та же a-L-арабиноза
(б):
б
гентамицины: А(а);С1,С2 и С1а(б)
(в Cl R и R]-CH3, в С2 R-CH3, R,-H, вС1а R и Rj-H)
По механизму действия гентамицин сходен с канамицином.
отивомикробная активность его исчезает, если отделить от мо-
кулы
один или оба остатка углевода.
329
Касугамицин — аминогликозидный антибиотик, продуцируе-
мый Streptomyces kasugaensis и подавляющий синтез белка, но
не вызывающий ошибок трансляции. Он связывает фенилала-
нил-тРНК с комплексом рибосомы — полиурацил, а также фос-
фометионил-тРНК с 30S рибосомами. Касугамицин более акти-
вен в отношении грамположительных бактерий, чем грамотри-.
нательных; не действует на грибы. В составе касугамицина нахо-
дится 2,3,6-тридезокси-4-иминоглицил-а-О-глюкозамин. Отделе-
ние последнего при гидролизе сопровождается потерей антибио-
тической активности препарата:
Гигромицин (гомомицин) — также аминогликозидный анти-
биотик, являющийся производным дигидроксицинамановой кис-
лоты, инозамина и кетодезоксигексозы. Дезоксигексоза представ-
ляет собой 5-кето-6-дезокси-а-£)-арабиногексозу. Антибиотик об-
ладает широким спектром .действия и подавляет рост грамполо-
жительных, грамотрицательных и кислотоустойчивых бактерий,
а также эффективен против гельминтов.
6
гигромицин
Из других аминогликозидных антибиотиков известны амика-
цин, блуэнсомицин, бутирозин А, ливидомицины, рибостамицин,
соннамицин А, спорарицин' А, тобрамицин, фортимицин А. Так
же, как и предыдущие аминогликозидные препараты, все они
содержат углеводные компоненты, при отделении которых анти-
биотическая активность теряется.
ззо
Тетрациклины составляют группу антибиотиков, образуемых
1ом актиномицетов (см. гл. 5). Эти вещества (прежде всего
'рациклин и окситетрациклин) нашли широкое применение
медицинской практике благодаря своей активности против
пиположительных и грамотрицательных бактерий, риккетсий
сламидий. Кроме тетрациклина и окситетрациклина известны
другие тетрациклины: 6-деметил-тетрациклин, 7-бромтетрацик-
н и т. д. Получены также многие полусинтетические тетрацик-
ны: миноциклин, рондомицин, доксициклин, представляющие
(дктический интерес. Радикалы у этих антибиотиков показаны
габл. 30 исходя из структурной формулы, изображенной в виде
сти колец А, В, С, D:
?4 Rj R2 N(CH3)2
Id с в а
Таблица 30. Радикалы (R) у некоторых
тетрациклинов
R Доксициклин Метациклии (рондомицин) Миноциклин
Ri он он н
r2 СН3 1 сн2 н
R3 н н
R< н н N(CH3)2
Антибиотики-макролиды — противомикробные вещества, со-
ржащие в своей структуре макроциклическое лактонное
льцо, связанное с остатками углеводов. В качестве таких
татков обычно выступают аминосахара. В класс макролидных
руктур входят и полиеновые антибиотики, имеющие несколько
пряженных двойных связей, и в зависимости от этого деля-
иеся на диены, триены, тетраены, пентаены, гексаены и геп-
:ены.
Амфотерицин В продуцируется Streptomyces nodosus. Анти-
1отик относится к неароматической группе полиенов. Это ам-
отерный гептаен, содержащий углевод микозамин; одновремен-
э он является аминогликозидом, поскольку в молекуле имеется
-С—О—С-гликозидная связь между остатком 3-амино-3,6-ди-
езокси-£)-маннозы (микозамина) и остальной частью антибио-
за, представляющую собой макролактонное кольцо. В кольце
Меется гидроксилированный участок и система незамещенных
войных связей с транс-конфигурацией:
331
тельных клеток, и они утрачивают свои барьерные функции.
Антибиотик оптически активен ([а] о35 в 0,1 М НС1 в метано-
ле— 33,6°); губительно действует на ряд возбудителей грибко-
вых заболеваний и в том числе глубоких микозов (Coccidioid.es
immitis, Cryptococcus neofortnans, Blastomyces dermatitidis,
Candida species, Histoplasma capsulatum). Ацилирование амфо-
терицина В по аминогруппе микозамина переводит этот амфо-
лит в Ъещество кислого характера, легко образующего водо-
растворимые соли, но существенно теряющего при этом свою
биологическую активность. К гептаенам также относятся: кан-
дицидин и кандидин, леворин, микогептин, нистатин, перими-
цин, трихомицин; к тетраенам — римоцидин.
Неполиеновую группу макролидных антибиотиков составляют
ланкамицин, магнамицин (карбомицин), метимицин, олеандо-
мицин, спирамицины, тилозин, халкомицин, эритромицин. Их
пространственные структуры имеют некоторое подобие, поэтому
в качестве примера остановимся на олеандомицине и эритро-
мицине.
Олеандомицин (амимицин) продуцируется Streptomyces anti-
bioticus. В состав молекулы антибиотика входят два углевода —
p-D-дезозамин и a-L-олеандроза, соединенные —С—О—С-гли-
козидными связями с макроциклическим лактонным кольцом-
агликоном:
332
Олеандомицин — основание с [а] о в метаноле —65°. На
1ктике чаще используют в виде соли с ортофосфатом. Если
делить от антибиотика один или оба углеводных остатка, то
почти полностью теряет свою противомикробную активность,
иносахар всегда располагается ближе к макролактонному
1ьцу, чем нейтральный углевод:
дезозамин
схема пространственной модели
агликона олеандомицина
Олеандомицин активен против грамположительных бактерий,
кобактерий, риккетсий и хламидий. Механизм его действия
условлен подавлением синтеза белка благодаря связыванию
:тибиотика с 50S субчастицами рибосом.
Эритромицин продуцируется актиномицетом Streptomyces
ythreus. По химическому строению эритромицин — макролид,
геющий макролактонное кольцо и два углеводных остатка —
D-дезозамин и а-А-кладинозу, соединенных —С—О—С-глико-
дной связью с агликоном-эритронолидом (обведен жирными
щиями). На практике антибиотик используют в виде основания.
|1дролитическое отщепление углеводных остатков приводит к
езкой потере активности эритромицина. Максимум поглощения
^ультрафиолете 278 нм; [а] д’ в метаноле —73,5°.
; Как и в других макролидах, аминосахар в эритромицине
асполагается ближе к агликону, чем кладиноза, и простран-
твенно его молекула сходна с олеандомицином. Эритромицин
ктивен против стрептококков и стафилококков, гемофильных
алочек, бруцелл, легионелл, спирохет, дизентерийных амеб,
'икоплазм и L-форм грамотрицательных бактерий. По механиз-
iy действия он сходен с другими антибиотиками-макролидами:
ззз
Из других антибиотиков применяются грамицидин С, поли-
миксин и другие полипептидные вещества, образуемые некоторы-
ми спорообразующими бактериями; новобиоцин, рифамицины,
продуцируемые актиномицетами; гризеофульвин, образуемый от-
дельными видами пенициллов, и др.
Новобиоцин образуется Streptomyces spheroides и некоторы-
ми другими видами актиномицетов. Он представляет собой
кислородсодержащий гетероцикл, проявляющий активность глав-
ным образом против грамположительных бактерий:
Рифамицины (А, В, С, D, Е, S, SV) образуются Streptomyces
(Act.~) mediterranei (см. разд. 3.5). В этом комплексе антибиоти-
ческих веществ наиболее стабильны рифамицины В, S и SV.
Из них практическое значение приобрел последний, который в
результате химической модификации удалось превратить в ри-
фампицин (римактан), обладающий многими лучшими каче-
ствами в сравнении с рифамицинами. Он активен против грам-
положительных бактерий и туберкулезной палочки (в том чис-
ле—устойчивых против других антибиотиков), несколько сла-
бее действует в отношении грамотрицательных бактерий.
Гризеофульвин (см. разд. 5.5) образуют Penicillium urticae,
Р. nigricans и др. Он активен в отношении нитчатых патогенных
и сапрофитных грибов. Его широко применяют на практике для
лечения различных микозов, а также в сельском хозяйстве для
борьбы с некоторыми фитопатогенами.
Линкомицин образуется Streptomyces linkolnensis. Это метил-
6,8-дидезокси- [1-метил-4- (/г-пропил)-2-пирролидин-карбоксами-
до] -1-тио-£)-эритро-/)-галакто-октопиранозид:
н3с—н2с—н2с
линкомицин
334
Если гидроксил в позиции С7 заменить на хлор, то получа-
гя полусинтетический антибиотик клиндамицин, лучше линко-
щина всасывающийся в кровь из кишечника. Линкомицин и
индамицин проявляют активность преимущественно против
амположительных кокков. Механизм их противобактериального
Йствия в основном связан с нарушением синтеза белка.
Гужеротин продуцируется Streptomyces gougerotii, химиче-
ая формула— 1-(цитозинил)-4-саркозил-Д-сериламино-1,4-ди-
£Зокси-0-Д-галактопиран-уронамид:
штибиотик широкого спектра действия, но активность его не-
елика. Это ингибитор синтеза белка на рибосомах. При этом
арушается перенос аминокислот с аминоацил-тРНК на пепти-
рл-тРНК, т. е. гужеротин является ингибитором трансферной
Ьакции.
Известны антибиотики из группы ауреоловой кислоты. К ним
тносятся вещества гликозидной природы, образуемые актино-
ицетами. Молекулярная масса их колеблется в пределах 0,8...
,3 кДа, спектры Amax поглощения имеют 3 максимума — 225...
30, 275...280 и 410...430 нм. Действуют губительно на грамполо-
сительные микроорганизмы и быстро пролиферирующие нор-
1альные клетки макроорганизма. Поэтому они обладают проти-
Ьопухолевой активностью.
В группу входят ауреоловая кислота (митрамицин), оливо-
1ицин, хромомицетин и хромоцикломицин. Все они, кроме хро-
юцикломицина, имеют агликоны, состоящие из трех шестичлен-
ых колец. Углеводная часть включает различные сахара. Ме-
анизм действия их на чувствительные клетки аналогичен и
водится к подавлению синтеза ДНК и РНК (сильнее подав-
яется образование РНК). Необходимое условие для связывания
казанных антибиотиков с ДНК — спирализация последней и на-
ичие ионов магния.
Ауреоловая кислота (митрамицин) — родоначальница ве-
хеств этой группы, продуцируется Streptomyces species. Она
остроена по гликозидному типу. Агликоном является хромо-
ицинон, сахаристая часть состоит из p-микарозил-(С1—>-С4) -
335
а-олиозил-(С1->СЗ)-0-оливозила и
возила, т. е. из триаозы и биозы,
С6 агликона:
р-оливозил- (С 1->СЗ) -р-оли-
присоединяющихся к С2 и
Следовательно, ауреоловая кислота представляет собой
2- [Р-микарозил- (С1->-С4) -а-олиозил- (С 1->СЗ)-р-оливозил] -6- [р -
оливозил-(С 1->-СЗ)-р-оливозил] -хромомицинон. Триаоза, при-
соединенная к С2 позиции агликона, содержит р-Р-микарозу,
а-Р-олиозу и р-Р-оливозу. Биоза в Сб позиции агликона содер-
жит две р-Р-оливозы:
Р - D -микароза
(ЗС-метил-2,6-ди-
дезокси- О-рибогек-
соза)
Вариамицин образуется Streptomyces olivovariabilis. В состав
молекулы вариамицина входят агликон-хромомицинон и 3 типа
дезоксисахаров — олиоза, оливоза и вариаоза:
336
вариамицин
Между биозой и С6 агликона, а также между триаозой и С2
дикона образуется —С—О—С-гликозидная связь. Вариамицин
тивен против грамположительных и грамотрицательных бакте-
й, проявляет цитотоксическое действие на культуру опухоле-
IX клеток HeLa и эффективен в отношении ряда эксперимен-
льных опухолей.
Оливомицин продуцируется Streptomyces olivoreticuli. Этот
тибиотик — гликозид, в котором сахаристая часть представ-
ка триаозой и биозой; агликоном выступает оливин:
337
В состав оливомицина входят три монозы: оливомикоза,
оливомоза и оливоза (или соответственно ЗС-метил-2,6-диде-
зокси-А-арабиногексоза, 4-0-метил-2,6-дидезокси-£)-ликсогексо-
за, 2,6-дидезокси-£)-арабиногексоза):
сн3
оливомикоза
(ЗС-метил-2,6-
•дидезокси-Z,-
-арабиногексоза)
дидезокси- D -
-ликсогексоза)
оливоза
(2,5-дидезокси-
-D -арабиногек-
соза)
Причем оливомицин имеет остатки изопропионила в позиции С4
оливомикозы и ацетила — в позиции СЗ оливозы.
Хромомицин продуцируется Streptomyces griseus. Антибиотик
образует комплекс близких гликозидов, названных вариантами
А1...АЗ. Наиболее изучены хромомицины А2 и АЗ. Хромомицин
А2 полностью тождествен оливомицину, тогда как хромоми-
цин АЗ несколько отличен от него:
хромомицин АЗ
Сахаристая часть в виде биоз занимает соответственно по-
зиции С2 и С6 в агликоне. Биоза в позиции С2 представляет
собой хромозы В и С, соединенные С 1-<-СЗ-связью. Биоза в
позиции С6 включает хромозы А и D, соединенные С1-<-С4-
связью. Хромозы напоминают сахарные остатки в оливоми-
цине:
338
Нз
н3'
он
ОН
хромоза А
СН,
ОН
ОН
Streptomyces
продуцируется
species
ауреоло-
позициях
хромоза С
L Хромоцикломицин
К-7017. Антибиотик напоминает другие производные
Л кислоты. Он содержит две гликозидные связи в
J и С12а агликона. В первом случае сахаристой частью явля-
ся моноза (микароза), во втором — триаоза, состоящая из
<карозы, олиозы и оливозы:
хромоцикломицин
собой 8- fa-
представляет
ледовательно, хромоцикломицин
Икарозил] -12а-[a-микарозил- (С1->-С4)-р-олиозил-(С1->СЗ)-р -
Пивозил] -хромоциклин.
В агликоновой части антибиотика имеется четвертое сопря-
женное кольцо, отсутствующее у всех описанных выше произ-
дных ауреоловой кислоты. Сахаристая часть состоит из двух
татков микарозы и остатков олиозы и оливозы.
Антибиотики-антрациклины обладают противомикробной (не-
торые — противоопухолевой) активностью. По своей химиче-
339
ской структуре они — гликозиды, хромофорная группа которых
представлена антрациклинонами — веществами красного цвета,
нерастворимыми в воде. Антрациклиноны соединены —С—О—С-
гликозидной связью' с амино- или дезоксисахарами. К группе
антрациклинов относятся дауномицин, рубомицин, родомицины
и другие антибиотики, образуемые некоторыми актиномицетами
(табл. 31). Из таблицы видно, что рубомицин тождествен
дауномицину. Антрациклины нарушают обмен РНК в клетках.
В агликоне-хромофоре антрациклинов указаны заместители на
Таблица 31. Антибиотики-аитрациклины и их компоненты
Антибиотик Агликон Углеводный компонент
Дауномицин Дауномицинон Даунозамин (2,3,6-тридезокси-3- аминоликсоза)
Карминомицин Карминомицинон Даунозамин
Пирромицин в-Пирромицинон 1 моль родозамина
Родомицин А 0-Родомицинон 2 моль родозамина
Родомицин В » Родозамин
у-Родомицин-1 у-Родомицинон >
у-Родомицин 2 » 2 моль родозамина
у-Родомицин 3 » 2 моль родозамина и 1 моль 2-де- зокси-А-фукозы
у-Родомицин 4 » 1 моль 2-дезокси-£-фукозы и 1 моль родинозы
Рубомицин Тетраценхинон Даунозамин
Цинерубин А в-Пирромицинон 1 моль родозамина, 1 моль 2-дезок- си-£-фукозы и 1 моль дидезоксиаль- догексозы
Цинерубин В »
Алкарубицин Алкацинон Диметилдаунозамин илн дауноза- мин 2-дезокси-£-фукоза и 4-кето-роди- ноза
Таблица 32. Характер заместителей в хромофоре антрациклинов
Хромофор Заместители на месте
Ri r2 Rs r4 Rs Rs Ry
Дауномиценон Н ОСНз ОН он Н ОН СОСНз
Карминоми- цинон Н ОН ОН он Н он СОСНз
е-Пирромици- нон ОН ОН Н он С ОСН3 он с2н5
0-Родомицинон н он он он ОН он С2Н5
у-Родомицинон или ОН н или Н ОН он он он н с2н5
Тетраценхинон или ОН Н илн Н ОСНз он он н он СОСНз
340
£тах Ri—R? (табл. 32).
аиции Re агликона:
' R,
Гликозидная связь формируется в
о
Ra
Rs
R7
ОН
О
R<
Re
R2
хромофор-агликои антрациклинов
Приводим углеводные компоненты антрациклинов:
н3с
н,он
нб N(CH3)2
родозамин
н3с
н,он
он
2-дезокси-1 -фукоза
н3с
н,он
он
родиноза
н3с
н,он
NH,
ОН
даунозамин (2,3,6-тридезокси-
-3-амино-ликсоза)
Н3С
н,он
НО । СНз
СН3
диметил-даунозамин
(2,3,6- тридезокси -3-
диметиламино-ликсоза)
н3с
н,он
4-кето-родиноза
'• К антибиотикам-циклопентитам относится ристомицин (ристо-
гтин), который продуцируется Proactinomyces fructiferi var.
stomycini и др. Известны ристомицины А и В, аналогичные
*стоцетинам А и В. Их химическая структура окончательно не
:тановлена. Однако доказано, что, например, ристомицин А
леет несколько остатков углеводов, соединенных гликозидными
Вязями с циклопептидным агликоном.
В отличие от многих других антибиотиков, содержащих угле-
воды, отщепление части или всех сахаров от агликона ристоми-
Ина существенно не сказывается на его противомикробной
|стивности, направленной против грамположительных бактерий.
Механизм его действия связывают с нарушением синтеза пепти-
ргликана клеточной стенки бактерий:
341
p -D~ Г люкоза -•— a - L-Рамноза
I
a-D-Манноза
^-D-Арабиноза
^-p-D -Манноза
-*—Ристозамин
приближенная структура ристомицина А
Ванкомицин, образуемый Streptomyces orientalis, близок к
ристомицину по строению и свойствам. Антибиотик левомицетин
под названием хлорамфеникол производится в некоторых стра-
нах путем биосинтеза в виде D-трео-формы. Продуцент левоми-
цетина — Streptomyces {Act.} venezuelae.
Н NHCOCHCb
O2N-</ у- с — С —СН2ОН
Ч=/ I I
он н
А)-трео-хлорамфенокол
В СССР левомицетин получают химическим синтезом при разде-
лении рацемата, называемого синтомицином. Левомицетин —
антибиотик широкого спектра действия. Он подавляет грампо-
ложительные и грамотрицательные бактерии, риккетсии и хла-
мидии вследствие блокирования синтеза белка за счет связыва-
ния с пепхидил-трансферазным центром рибосом и создания
конкурентных помех для взаимодействия этого центра с соответ-
ствующими акцепторами.
6.2.5. Нуклеозиды, нуклеотиды, витамины и др.
Кратко рассматриваемые в этом разделе вещества (кроме неко-
торых витаминов) образуются всеми микроорганизмами, однако
биотехнологические процессы разработаны и внедрены в про-
342
шленность лишь для немногих из них. Тем не менее, используя
1етико-селекционные приемы, удается получать такие штаммы
одуцентов, которые могут быть весьма перспективными.
Кордицепин, или аденин-нуклеозид, содержащий 3-дезокси-
нтозу (кордицепозу), продуцируется Cordiceps militaris,
ipergillus nidulans. Следовательно, кордицепин — это 3-дезок-
аденозин. Он проявляет антибиотические свойства и тормозит
звитие различных грамположительных и грамотрицательных
жтерий в случае, если нуклеозид оказывается фосфорилиро-
;нным (т. е. нуклеотидом). В таком состоянии он препятствует
цтезу РНК.
Нуклеоцидин образуется Streptomyces calvus. Это эфир суль-
>миновой кислоты и пуринового нуклеозида. Обладает широ-
IM спектром действия. Механизм противомикробной активности
язан с нарушением
штидной связи:
деятельности рибосом при образовании
NH,
осн2
•OSO2NH2
кордицепин
Псикофуранин продуцируется Streptomyces higroscopicus
ir. decoyicus. Псикофуранин — нуклеозид, представляющий со-
эй 6-амино-9-£)-псикофуранозилпурин, обладающий антибиоти-
*ской активностью против грамположительных и грамотрица-
гльных бактерий. Он ингибирует ксантозин-5-фосфатаминазу.
уанин-содержащие структуры ревертируют подавленные псико-
уранином бактерии:
псикофуранин
ангустмицин А
343
Str. higroscopicus var. angustmyceticus образует псикофура-
нин и нуклеозид — ангустмицин А, в котором псикофураноза
имеет одну двойную связь С5—С6. Следовательно, ангустми-
цин А представляет собой 9-Р-(5,6-псикофуранозоенил)-6-амино-
пурин. Он менее активен против бактерий, чем псикофуранин.
Оба препарата сходны по механизму действия.
Пуромицин. Продуцент — Streptomyces alboniger. Пуромицин
представляет собой структурный аналог аминоацил аденозина —
это 6-диметиламино-9- (З'-дезокси-З'-и-метокси-А-фенил-аланил-
амидо-0-£)-рибофуранозил) -пурин. Гликозидная связь в нем рас-
щепляется при действии 1 М. НС1 (100 °C) за 10 мин, амидная
связь в аминокислоте при этих условиях остается стабильной.
Максимум поглощения для пуромицина в ультрафиолете Zmax
267,5 нм (в 0,1 М НС1) и 275 нм (в 0,1 М NaOH); [а] Ь5 -1Г.
Ингибирует многие микроорганизмы, включая трипаносомы, од-
нако терапевтическое значение его невелико.
Пуромицин приобрел важное значение в расшифровке син-
теза полипептидной цепи на рибосомах. Он подобен З'-концу
аминоацетил-тРНК, связывающемуся с определенным участком
рибосомы, и действует как аналог аминоацил-тРНК, участвуя
в синтезе пептидной связи. Пуромицин слабо связывается с рибо-
сомами типа 70S/80S и с субчастицами рибосом типа 50S/60S.
Образующаяся молекула пептидилпуромицина легко отделяется
от рибосомы. Следовательно, принимая на себя растущую по-
липептидную цепь, предназначенную для переброски на следую-
щую аминоацил-тРНК, пуромицин препятствует дальнейшему
удлинению полипептидной молекулы.
Туберцидин продуцируется Streptomyces tubercidicus. Туберци-
дин относится к числу пиррол-пиримидиновых рибозидов-нуклео-
344
дов с выраженным цитотоксическим действием. Сходными с ним
азались актиномицетные нуклеозиды — тойокамицин и сан-
вамицин, подавляющие Mycobacterium tuberculosis
туберцидин тойокамицин саигивамицин
токсичные для многих эукариотических клеток. Как аналоги
.енозина они включаются в РНК и тем самым нарушают
нтез белка.
В медицинской практике широко используют инозин (рибок-
н), его продуценты—Вас. subtilis, Вас. putnilis, Brevibacte-
urn ammoniagenes и др. Инозин назначают при заболеваниях
‘рдца (инфаркте, дистрофиях и пр.), так как он является при-
рдным метаболитом, использующимся клетками для биосинтеза
ТФ, нуклеотидов и нуклеиновых кислот
Микроорганизмы оказываются ценными продуцентами алка-
•идов (Claviceps species), а также ряда витаминов. В СССР
других странах освоена биотехнология рибофлавина (В2),
(анкобаламина (Bi2), эргостерина (D3), каротина (А). Эти
[тамины образуются соответственно Candida guilliermondii и
remothecium ashbyi, Propionibacterium species и Streptomyces
»., Saccharomyces cerevisiae, Rhodotorula sp. и Blakeslea trispo-
i. Витамины используются как предшественники соответствую-
их ферментов.
Гиббереллины — производные дитерпенов, выступающие тор-
онами растений. В основе их химической структуры находится
;тракарбоциклическое ядро гиббана:
345
4
5
Известна серия гиббереллинов, обозначаемых заглавной бук-
вой А (АьА2и т. д.). На практике чаще используют гибберел-
ловую кислоту, продуцируемую сумчатым грибом Gibberella
fujikuroi (его конидиальная стадия относится к Fusarium moni-
liforme). Гибберелловую кислоту используют в сельском хозяй-
стве как стимулятор роста растений.
Вопросы
1. Что такое биотехнология? Каковы этапы ее развития?
2. Какова сущность условных уровней биотехнологических пронес; щ
3 Какой ключевой продукт лежит в основе биохимических пронес.
ведущих к чолхченпю этанола, масляной и молочной кислот, бутанола, -
го: i.iJ
-1 Что представляют собой иммобилизованные ферменты и клеи’, i
какие методы используют для их иммобилизации?
5 Каков механизм действия реетриктаз на молекулы ДНК? Ира» и
четкое значение ресгрикгат.
6, Какой основной недостаток присущ микробному белку в виде на :
пых клеток, используемых в рационах сельскохозяйственных живо,и - -
". Что такое гибрндомы, как их получают и используют на пракн
8. Что такое векюрны.е молекулы ДНК и для каких целей их -
ио.тьтсюг на практике?
9 Какие вещества получают в настоящее время, используя м, ,
генной инженерии?
10. Что представляют собой циклодекетрипы и какое практи'ч, о
щачеиие они имеют?
11. Назовите известные крупнотоннажные производства микроб1:./ >
полисахаридов.
12 Какие стрхктхры имеют 6-ЛИК и 6-ЛЦК? Их практическое значащ.
13. Какие аминогликози.тные антибиотики приобрели наибольшее ••л
че.чие в медицине?' (Их (Троение и механизм действия.)
И Что такое гиббереллины и где они применяются?
15. Какие витамины получают путем биологического синтеза и г;п
применяют на практике?
16. Каковы химическое строение и свойства антибиотиков - полю и
17. Каковы химическое строение и свойства антибиотиков тетращч :
hob?j '
18. Что такое моноклональные антитела и методы их получения?
19. Дайте определение иммунобиотсхио.тогии. Каковы ее объекты,
годы и задачи?
20 Используются ли нуклеозиды в качестве лекарственных вещее:
Их строение и механи:м действия.
346
Рава '
|МИЧЕСКИЕ ОСНОВЫ ПАТОГЕННОСТИ
Микроорганизмов
h ПАТОГЕННОСТЬ И ВИРУЛЕНТНОСТЬ
ЙКРООРГАНИЗМОВ
Йкроорганизмы подразделяют на две большие группы — сапро-
цтные (неболезнетворные) и патогенные (болезнетворные) (от
IT. saprotes — гниль, phyton — растение; saprophyti — произ-
(Гстающие за счет мертвого субстрата; pathogenes — болезне-
(Орный, патогенный). К сапрофитным относятся большинство
Вкробов, обитающих в различных почвах и водоемах, на пок-
ойных тканях и слизистых оболочках открытых полостей у хо-
(днокровных и теплокровных животных; микробы, находящиеся
I растениях (эпифиты), и т. д. За счет химической деятельности
(болезнетворных микробов (прежде всего в почве) происходит
Внерализация органических веществ, поступающих в огромных
Оличествах во внешнюю среду. Глобальное значение такого про-
веса минерализации очевидно, без него жизнь давно бы прекра-
Иась на нашей планете. Существуют микроорганизмы (сапро-
Йты), которые трансформируют сложные органические соедине-
1я в более простые, как бы подготавливая субстраты, пригодные
1Я питания других микробов. Таким образом, все виды превра-
ений органических веществ главным образом до СО2, Н2О и
H3(N2) осуществляются с подключением в химические процес-
I различных групп микроорганизмов. Сапрофитными микроба-
I являются Вас. megaterium, Вас. subtilis, Е. coli (за исклю-
мием определенных сероваров), Lactobacterium species, Saccha-
myces cerevisiae, Aspergillus niger, Penicillium notatum и др.
В процессе эволюции происходило приспособление микроор-
низмов к макроорганизмам — растениям, животным и челове-
!. Возникли условно патогенные и патогенные виды микроорга-
Вмов. Патогенность означает потенциальную способность мик-
!юрганизма вызывать заболевание или приводить к возникнове-
|ю прогрессирующих повреждений. Болезни, вызываемые пато-
чными микроорганизмами, относят к разряду инфекционной
(тологии (от лат. infectiosus — заразный; от греч. латсоо —
клезнь, Хсоусоа — учение). Сформировались специальные науч-
ie дисциплины: инфекционные болезни человека и животных,
также фитопатология, или инфекционные болезни растений,
зависимости от природы возбудителей заболевания различают
(русные, бактериальные, грибковые и протозойные инфекцион-
ie болезни. В природе и теперь можно найти непатогенные
Юйники у патогенных микроорганизмов. Например, известны
Черный и холероподобные вибрионы, туберкулезные и сапро-
бные микобактерии, дифтерийная и ложнодифтерийная палоч-
Г, патогенный и непатогенные криптококки и т. д.
347
Условно патогенные микроорганизмы являются как бы пере-
ходными формами между патогенными и непатогенными; они
способны вести сапрофитный и паразитический образ жизни.
К ним относятся некоторые бактерии {Pseudomonas aeruginosa,
Proteus vulgaris), дрожжеподобные грибы рода Candida (С. al-
bicans) и др.
В последние годы в инфекционной патологии существенно
возросла роль условно патогенных микроорганизмов, прояв-
ляющих выраженную устойчивость к большинству известных
химиотерапевтических агентов.
К так называемым безусловным, или облигатным, паразитам,
не способным развиваться вне живой ткани, относятся возбуди-
тели вирусных заболеваний (герпес), риккетсиозов (сыпной тиф,
лихорадка Ку) и некоторых венерических заболеваний (сифи-
лис). Есть виды, серовары и штаммы более патогенные, чем дру-
гие болезнетворные виды, серовары и штаммы. Для примера
можно сравнить особо опасный возбудитель чумы Yersinia pestis
и менее патогенный возбудитель мелкоочаговой бронхопневмонии
Staphylococcus aureus или возбудители крупозной пневмонии
Streptococcus pneumoniae III и VIII сероваров (III серовар более
патогенный). Поэтому было введено понятие вирулентность,
обозначающее меру патогенности, т. е. более вирулентные и ме-
нее вирулентные микробы. Вирулентность (как и патогенность)
определяют в опытах на чувствительных экспериментальных жи-
вотных, чаще белых мышах и крысах, морских свинках, кро-
ликах.
С учетом вирулентности и степени опасности работы с ними
все болезнетворные виды подразделены на 5 групп: I группа —
возбудитель чумы; II группа — возбудители холеры, натуральной
оспы, бруцеллеза, туляремии, сибирской язвы, сапа, мелиоидоза,
эпидемического сыпного тифа, эпидемических энцефалитов, ге-
моррагических лихорадок, желтой лихорадки, лихорадки Ку, ли-
хорадки цуцугамуши, лихорадки скалистых гор, орнитозов, гис-
топлазмоза, кокцидиоидоза, токсин ботулиновый; III группа —
возбудители бактериальных (брюшной тиф, дизентерия, дифте-
рия, туберкулез, актиномикоз, сифилис и др.), риккетсиозных
(болезнь Брилля, клещевые тифы и др.), вирусных (бешенство,
полиомиелит, грипп, корь и др.), протозойных (малярия, лейш-
маниоз, трихомоноз, балантидиоз и др.), грибковых (кандидоз,
криптококкоз, дерматомикозы и др.) инфекционных заболеваний,
выделенных в самостоятельные нозологические формы (от лат-
nosologia — учение об отдельных болезнях); IV группа — возбу-
дители токсикоинфекций и острых бактериальных отравлений
(сальмонеллы, стафилококки, вибрионы, клостридии), септице-
мий и пневмоний (стафилококки и стрептококки, ринегнойная
палочка и др.); V группа — облигатная непатогенная микрофло-
ра, населяющая слизистые и кожные покровы человека, а также
микроорганизмы — показатели санитарного состояния внешней
среды (энтерококки, эшерихии, перфрингенс и др.). Министер-
348
юм здравоохранения СССР утверждены специальные режимы
5оты с микроорганизмами, относящимися к каждой из пяти
щп.
ХИМИЧЕСКИЕ ВЕЩЕСТВА, ОПРЕДЕЛЯЮЩИЕ
ОЙСТВА ПАТОГЕННОСТИ МИКРООРГАНИЗМОВ
(тогенным микробам присуща инвазивность (от лат. invasio —
зржение), т. е. способность проникать в ткани макроорганиз-
, размножаться там и распространяться. Отдельные возбуди-
1И инфекционных заболеваний могут проникать через непов-
кденные слизистые оболочки и кожу (гемолитические стрепто-
кки, стафилококки и др.). Если же патогенные микроорганизмы
реносятся какими-либо переносчиками (например: комарами,
дами, клещами), то возбудители болезни попадают непосред-
венно в кровь или лимфу (риккетсии, плазмодии малярии),
го трансмиссивные инфекции. Известно также, что отдельные
(тогенные микробы способны прилипать к эпителиальным клет-
м слизистых оболочек, например, за счет протеинов (энтеро-
догенные кишечные палочки), гликоконъюгатов (стрептококки
уппы А, некоторые дрожжеподобные организмы рода Сап-
В тесной связи с инвазивностью находится инфекциозность,
е. способность патогенного микроба (микроба-паразита) взаи-
(действовать с макроорганизмом, в который он проник через
дхательные пути, пищеварительный тракт, кожу и слизистые
юлочки (особенно поврежденные). Такой микроб, проявивший
:вазивность и инфекциозность, вызывает, как правило, инфек-
юнное заболевание. Например, у токсинобразующих микробов
ифтерийная палочка, возбудители столбняка, газовой гангрены
др.) инфекциозность в значительной степени определяется
зотоксинами, взаимодействующими in vivo с определенными
руктурами и тканями. Инвазивность и токсигенность (способ-
сть продуцировать токсические вещества) могут находиться
д дифференцированным генетическим контролем.
Патогенные свойства соответствующих микроорганизмов,
лк>чая их способность к инвазии и инфекциозности, определя-
ли так называемыми факторами агрессии и факторами пото-
чности, представляющими собой продукты обмена (метаболиз-
1) развивающихся и размножающихся клеток либо их струк-
рными компонентами.
Факторы агрессии — вещества, с помощью которых патоген-
1й микроб проникает в соответствующие ткани. К ним можно
нести преимущественно гидролитические ферменты (например,
молизины, гиалуронидазу, ДНКазы и РНКазы, пептидазы),
.гезивные вещества и пр. Факторы патогенности (гликоконъю-
ты, токсины и др.) приобретают важное значение в развитии
Секционного заболевания. Однако резкую грань между факта-
ми агрессии и факторами патогенности проводить нельзя, так
349
как факторы агрессии всегда участвуют в соответствующих пато-
логических реакциях в течение развития заболевания. В ряде слу-
чаев агрессивность микроба проявляется уже в ходе возникшего
инфекционного заболевания (например, действия пептидаз при
газовой гангрене, возбудители которой образуют сильные токси-
ны) . Поэтому все факторы агрессии и патогенности можно под-
разделить на 7 групп: белки, полисахариды, гликоконъюгаты,
липопротеины, фосфатиды, фосфопротеины и другие субстанции.
К первой группе факторов (белкам) относят ферма-
ты, неферментные полипептиды и токсины (токсические белюд.
Среди ферментов прежде всего следует назвать гидролазы, ю;-
ляющиеся преимущественно факторами агрессии. Так, лецитина-
за, или лецитовителлаза [КФ.3.1.4.3(4) ], в виде внеклеточного
фермента образуется некоторыми клостридиями, холерным виб-
рионом, коклюшной палочкой, листериями. Лецитиназа гидро.:: н-
з у от ф а с ф а т и ди л х ол и н
н>С-- осок
[TOGO—СН О
II! „
Ц..С—О—!>—(>•-(СН2) n^-hch,) л’1"™"3:’:!
। ’ Н,0
о
фоефсти,'i.h.ixo он!
НоС—OCOR О
I I!
- КОСО—СН + но— Р—О— (СИ,о—N=s(CH,))
I I
СН—он о
Стафилококки, стрептококки, дифтерийные палочки, некото-
рые клостридии, вульгарный протей продуцируют ДНКазу, или
срептодорназу [КФ-3.1.4.5(6) ], катализирующую реакцию гидро-
лиза ДНК. Микрококковая эндонуклеаза из золотистого стафи-
лококка катализирует реакции распада ДНК и РНК на олиго-
нуклеотиды и мононуклеотиды с З'-фосфатного конца. Многие ви-
ды патогенных бактерий (в том числе микоплазмы) образуют
нейраминидазу, или сиалидазу (КФ.3.2.1.18), гидролизующую
соответствующие гликозидные связи в олигосахаридах, коломи-
новой кислоте, гликоконъюгатах. Гиалуронидаза [КФ.3.2.1.35(36)[
как фактор инвазии известна сравнительно давно. Ее образуют
бруцеллы, стафилококки, стрептококки, дифтерийные бактерии,
холерные вибрионы, протей, синегнойная палочка и др. Гиалу-
ронидаза расщепляет гиалуроновую кислоту, хондроитинсульфа-
ты А и С и, в частности, эндоацетилгексозаминные связи в их
структурах. Стафилококки, холерные вибрионы, некоторые пато-
генные грибы (например, Candida species) образуют эндо- и
экзоклеточную коагулазу (КФ.3.4.11.1), сходную с лейцинамино-
пептидазой (а-аминоацил-пептидгидролазой). Коагулаза катали-
350
>ует гидролиз пептидных связей. К подобным гидролазам от-
:ят также фибринолизин, или стрептокиназу (3.4.21.7), — вне-
‘точный фермент, образуемый стрептококками, стафилококка-
холерными вибрионами, иерсиниями, синегнойной палочкой
ДР-
К внеклеточным ферментам, проявляющим свойства факторов
рессии за счет гидролиза белков, относят клостридиальную
ютеиназу В (КФ З.4.22.8); стрептококковую протеиназу
<Ф.3.4.22.10); коллагеназу (КФ.3.4.24.3) отдельных клостридий
дерматофитов; нейтральную протеиназу (КФ.3.4.24.4) сине-
рйной палочки и других бактерий; иные протеиназы [КФ.3.4.99.5
Ц2,15,20,22) ], образуемые некоторыми клостридиями, стафило-
кками, дерматофитами. Ряд болезнетворных микробов обра-
;ёт протеазы, катализирующие реакции гидролиза иммуногло-
булинов. Например, виды Neisseria и Streptococcus sanguinus мо-
»т расщеплять местно накапливающиеся секреторные антитела
•па IgA (см. гл. 8), предотвращая тем самым прилипание, опсо-
(зацию и фагоцитоз бактерий. Из числа гидролаз заслуживает
кже внимание уреаза — внеклеточный фермент, образуемый
уцеллами, некоторыми криптококками, протеем. Уреаза ката-
зирует реакцию распада мочевины до диоксида углерода и ам-
:ака.
Среди лиаз можно отметить внутри- и внеклеточные карбок-
лиазы (4.1.1), продуцируемые различными энтеробактериями
псевдомонадами. Эти ферменты декарбоксилируют кислоты,
лючая аминокислоты, и выступают преимущественно фактора-
агрессии:
R—CH(NH2)—СООН Карбоксилиазы Со2 + R—CH2—NH2
Лиазы оксокислот (КФ.4.1.3), образуемые внутри и вне кле-
: некоторыми энтеробактериями, гидролизуют 3-гидроксикис-
ы:
R—СН(ОН)— СН2—СООН _Лназы ^кислот R Ацетил-КоА +
+ НООС—СОН + Н2О
Из других ферментов можно назвать эндо- и экзоклеточные
молизины стафилркокков, стрептококков, некоторых клостри-
(й и грибов, они катализируют реакции распада эритроцитов.
Гемолитические стрептококки (наряду со стрептококками из
упп А, С и G по Лэнсфильд) образуют стрептолизины О и S
молекулярной массой около 10 кДа. Лейкоцидины стафило-
ikkob, стрептококков, пневмококков, псевдомонад лизируют тка-
вые клетки и лейкоциты. Так, мишенью для стафилококкового
йкоцидина выступает трифосфоинозитид, локализующийся
мембране лейкоцитов. Лейкоцидин состоит из двух белковых
мпонентов F и S, каждый из которых в отдельности не прояв-
ит токсических свойств.
351
он
трифосфоннозид
Названными ферментами, принимающими то или иное участие
в инвазивности и инфекциозности патогенных микробов, а также
в развитии патологических процессов, не исчерпывается весь их
арсенал. Так, например, карбогидразы (КФ-3.2.1) патогенных
микроорганизмов могут расщеплять углеводы, входящие в струк-
туру гликопротеинов крови, гликолипидов нервных клеток, имму-
ноглобулинов.
Из неферментных полипептидов факторами патогенности мо-
гут быть капсульный £)-глутамил-полипептид возбудителя сибир-
ской язвы, цетил-полипептид (протеин А стафилококков), а также
М-протеин стрептококков и пневмококков. Первый из них состав-
ляет капсульный материал Вас. anthracis, второй содержится
в клеточной стенке Staph, aureus, третий находится в клеточной
стенке и накапливается в среде обитания Streptococcus species.
Все эти полипептиды проявляют антифагоцитарное действие,
т. е. ингибируют или полностью подавляют фагоцитарную актив-
ность сегментоядерных лейкоцитов.
Микробные токсины (от лат. toxinum — яд) являются основ-
ными факторами патогенности, определяющими патогенные свой-
ства токсигенных бактерий (табл. 33). Их можно подразделять
по локализации на экзо- и эндотоксины (не отождествлять с ли-
пополисахаридными эндотоксинами).
Таблица 33. Белковые токсины бактерий
Название Продуцент Локализация
Ботулинический Гистотоксин А Дифтерийный Нефритогенный Сибиреязвенный Стафилококковый а-ток- син Столбнячный (нейроток- син, тетаноспазмин) Энтеротоксины (холеро- ген и др.) Эксфолиатин Эритрогенин (скарлатинозный) Коклюшный термолабиль- ный Риккетсиозный Энтеротоксин клостри- диальный Clostridium botulinum Pseudomonas aeruginosa Corynebacterium diphtheriae Streptococcus haemolyticus Bacillus anthracis Staphylococcus aureus Clostridium tetani > Vibrio cholerae, E. coli, Kleb- siella spp., Shigella spp. Staphylococcus aureus Streptococcus haemolyticus Bordetella pertussis Ricketsia spp. Clostridium perfringens Внеклеточный » » » » » » » » Внутриклеточ- ный » Внутриспоровый
352
' Ботулитическии токсин избирательно поражает так называе-
те а-моторные нейроны передних рогов спинного мозга, блоки-
ет бактерицидные свойства сегментоядерных лейкоцитов; гис-
гоксин А и дифтерийный токсин блокируют синтез белка. Неф-
тогенный токсин поражает почки, а сибиреязвенный — цент-
льную нервную систему (ЦНС). Стафилококковый а-токсин
оявляет дермонекротическую, летальную и цитотоксическую
тивности; столбнячный — задерживает торможение импульсов
синапсах спинно-мозговых нейронов.
Энтеротоксины стимулируют аденилатццклазу, связанную с
:мбраной кишечника; эксфолиатин поражает кожу, а эритро-
нин обусловливает воспаление. Три эндотоксина, приведенные
табл. 33, существенно различны по действию на макроорга-
[зм. Так, термолабильный коклюшный эндотоксин вызывает
моррагии (от лат. haemorrhagia — кровотечение), дегенерацию
некроз клеток печени, почек и селезенки; риккетсиозный ток-
н поражает мелкие кровеносные сосуды и ЦНС. Клостридиаль-
ый энтеротоксин поражает кишечник.
5 Различные патогенные микроорганизмы образуют один, два
Ли более токсинов. Например, у стафилококков найдены а-, аг-,
», 6-токсины и А-, В-, Ci-, Сг-энтеротоксины. С а-токсином свя-
ывают летальное действие (от лат. letalis — смертельный),
^рмонекроз (от лат. derma — кожа, necrosis — омертвение),
емолиз (от греч. аща — кровь, Хклсг — растворение) и повреж-
ение лейкоцитов (а-, аг-, у- и 6-токсины для кролика, р-токсин
ля барана). Энтеротоксины содержат в своем составе по 18
минокислотных остатков в разных количественных соотноше-
иях. Терминальные аминокислоты в них следующие: энтероток-
<н А содержит аланин в качестве N-терминальной аминокисло-
>i и серин — в качестве С-терминальной аминокислоты; энтеро-
эксин В содержит соответственно глутаминовую кислоту и ли-
<н; энтеротоксины Ci и Сг — глутаминовую кислоту и глицин.
сибиреязвенном токсине, образуемом Bacillus anthracis, содер-
жится 3 компонента, обозначаемых EF (отечный фактор), РА
Защитный антиген) и LF (летальный фактор); каждый из них
бладает антигенными свойствами (см. гл. 8).
Дифтерийный токсин, образуемый Corynebacterium diphthe-
iae, изучен достаточно полно. Это белок с молекулярной массой
5,5 кДа. Clostridium botulinum образует один из наиболее силь-
Ых ядов (по расчетам 180 г этого токсина достаточно, чтобы
бить 1 млрд, людей). Известно 6 антигенноактивных ботулини-
еских токсинов — А, В, С, D, Е и F.
Столбнячный токсин продуцируется Clostridium tetani. Это
лассический экзотоксин-белок, выделяемый из культуральной
жидкости осаждением метанолом на холоду. Сила кристалличе-
кого токсина выражается до 100-106 минимальных смертельных
ЮЗ для белых мышей на 1 мг азота. Молекулярная масса ток-
йна около 67 кДа. Он состоит из 13 аминокислот с преоблада-
ием остатков аспарагиновой и глутаминовой кислот. Кристал-
1—1617
353
лический продукт нестабилен и самопроизвольно превращается
в высокоантигенный токсоид (см. гл. 8).
Большинство видов Salmonella и Shigella образуют только
эндотоксины, определяющие патогенность энтеробактерий. Одна-
ко Sh. disenteriae образует также и экзотоксин, часто называе-
мый нейротоксином. Это белок с молекулярной массой 82 кДа.
При его введении животные погибают в течение 3...4 дней от па-
ралича гладкой и поперечно-полосатой мускулатуры, у чело-
века неврологические осложнения отмечаются исключительно
редко.
Коклюшная палочка Bordetella pertussis образует ряд токси-
ческих и биологически активных субстанций: протективный (за-
щитный) антиген, гистамин-сенсибилизирующий фактор (HSF),
эндотоксин, термолабильный токсин, агглютининоген, гемагглю-
тинин. Все эти вещества определяют агрессивные свойства мик-
роба. Однако термолабильный токсин-белок не образуется вне-
клеточно. Он накапливается внутри клеток, из которых выде-
ляется во внешнюю среду после лизиса или искусственной их
дезинтеграции. В целях очистки и получения в гомогенном сос-
тоянии токсин адсорбируют на колонке с диметиламиноэтилцел-
люлозой с последующей элюцией раствором натрия хлорида.
Трипсин разрушает коклюшный токсин.
Гангренозные экзотоксины образуются различными видами
микроорганизмов, учитывая возможную полиэтиологическую
роль заболевания. Газовую гангрену чаще вызывают Clostridium
perfringens (Cl. welchii), Cl. novyi (Cl. oedematiens) и Cl. septi-
cunv, реже Cl. bifermentas (Cl. difficile), Cl. hystolyticum, Cl. ram-
nosum, Cl. sordellii, Cl. sporogenes и др. Названные клостридии
продуцируют разные токсины. Многие из них обладают леталь-
ным, некротизирующим и гемолитическим свойствами, которые
могут принадлежать одному и тому же токсину, иногда разным
токсинам. Наиболее полно изучен Cl. perfringens, особенно его
серовары А, В, С и D. Эти серовары образуют а-, е-, О-, х-, н- и
другие экзотоксины, из которых первый рассматривается как
наиболее важный в развитии патологического процесса у чело-
века. а-Токсин А обладает лецитиназной, гемолитической и
летальной активностями; он гидролизует лецитин на фосфорилхо-
лин и диглицерид. 0-Токсин сходен с а-токсином, но он не обла-
дает лецитиназной активностью. Некоторые штаммы Cl. .perfrin-
gens продуцируют сильный энтеротоксин (М = 90 кДа), отли-
чающийся от других клостридиальных токсинов способностью
вызывать профузный понос (диаррею). Следует подчеркнуть, что
экзотоксины патогенных бактерий проявляют ферментативные
свойства, например а-токсин Staph, aureus, а-токсин A Clostri-
dium perfringens и др. Однако не только экзотоксины, но и эндо-
токсины некоторых микроорганизмов обладают ферментативной
активностью. Например, кислый термолабильный белок — эндо-
токсин из клеток условно патогенного дрожжеподобного гриба
Candida albicans состоит из четырех субъединиц, две из которых
354
ладают карбоксипептидазной активностью и одна — фосфомо-
эстеразной.
Токсические циклопептиды (М да 2,6 кДа) образуют циано-
ктерии,. например Microcystis aeruginosa. Отдельные плесневые
шляпочные грибы образуют ядовитые циклопептиды. Penicil-
ит islandicum продуцирует хлорированный циклопептид высо-
ой токсичности — циклохлоротин (LDtb 475 мкг/кг для мышей
ри подкожном введении). Этот продукт состоит из остатков
,-аминобутиловой и р-амино-р-фенил-пропионовой кислот, серина
в соотношении 1:1:2 или 1:1:3) и карбоксипиррола. Другой
Иклопептидный токсин, образуемый Р. islandicum, — исланди-
оксин во многом сходен с циклохлоротином, но имеет другие
оследовательности и число аминокислотных остатков, Dim** или
Игпюо — 3 мг/кг для мышей при подкожном введении. Оба типа
оксинов поражают печень.
исландитоксин
* LDso — количество испытуемого вещества, приводящее к гибели 50 %
взятых в опыт животных.
** Dim — количество испытуемого вещества, приводящее к гибели 100 %
Взятых в опыт животных.
12* 355
Тентотоксин в виде циклотетрапептида образуется несовер-
шенным грибом Alternaria tenus. Токсин состоит из глицина
N-метил-дегидрофенилаланина, лейцина и N-метилаланина:
тентотоксин
Базидиальные грибы из рода Amanita образуют сильные ток-
сины, вызывающие отравление у человека. Так, бледная поган-
ка— A. phalloides продуцирует циклопептиды фаллотоксины
(табл. 34) и антитоксин антаманид:
сн2— R,
Н3С—СН-^СО—NH—СН— СО—NH—СН—СН2—С—СН2— R2
r5 NH—СО —СН—NH—СО
СНОН
I
r4
фаллотоксины'
Таблица 34. Фаллотоксины
Название Радикалы
Ri R2 Rs R, Rs
Фаллоидин а ОН Н СНз СНз ОН
Фаллоин b Н Н СНз СНз он
Фаллизин с ОН ОН СН(СН3)2 СНз он
Фаллицидин d ОН н СН(СНз)2 соон он
Фаллин В е Н н С6Н5СН2 СНз н
356
к. Антаманид — циклический декапептид, состоящий из четырех
Остатков различных L-аминокислот (аланина, валина, пролина
| фенилаланина в соотношении 1 : 1 :4 :4). В дозе 0,5 мг/кг он
полностью обезвреживает смертельную дозу для белых мышей
раллоидина (5 мг/кг).
/СбН
сн2
/СИ'"’
ZNH 9
,со
^н8
с6н
сн2
•СО—NH^,H
10
CO
7—н 7
СО
Н5С — Н2С—сн6
NH
\о
н5с6—н2с—сн5
4
СН3
NH-_CO.
•со
NH
1 СН—СН(СН3)2
со
2 Н
со
зн N
^co
-NH
антаманид
| Токсинами небелковой природы являются некоторые вторич-
|ные метаболиты условно патогенных грибов.
I Вторую группу факторов, главным образом факторов
(патогенности у болезнетворных микроорганизмов, составляют
|полисахариды, или гликаны. Сюда относятся капсульные поли-
всахариды пневмококков и стрептококков, проявляющие антифа-
|.гоцитарное действие. Полисахариды капсул и клеточных стенок
(бактероидов, гемофильных палочек, гонококков, менингококков,
(микобактерий туберкулеза, криптококков, францизелл, энтеро-
I бактерий и др. обладают противофагоцитарным действием, а не-
I которые из них индуцируют гиперчувствительность немедленного
Етипа (см. гл. 8). Полисахаридные капсулы не всегда могут быть
(факторами патогенности. Например, криптококкоз, или Европей-
ский бластомикоз Буссе — Бушке, вызывается патогенным
; Cryptococcus neoformans, образующим мощную капсулу. Другие
[виды криптококков (кроме Cr. neoformans) не способны вызы-
вать криптококкоз у человека или животных, хотя они также
' образуют сходные по химическому составу и строению полиса-
[харидные капсулы. Следовательно, химически и архитектурно
{сходные капсулы могут не быть факторами патогенности видов.
|Но Cr. neoformans, вызывая заболевание, обеспечивает себе луч-
I шие условия выживания, формируя капсулу in vivo, которая
357
здесь опосредованно выступает фактором патогенности, защищая
клетки, например, от фагоцитоза.
К третьей группе факторов (преимущественно факто-
ров патогенности) относятся гликоконъюгаты, или углеводсодер-
жащие полимеры: гликолипиды, гликолипопротеины, глико-
протеины, липополисахариды (липогликаны) и пептидогликаны
(муреины). Все они содержатся в клетках и не выделяются в куль-
туральную среду. Гликолипиды туберкулезных микобактерий и
псевдомонасов различаются по механизму действия на макро-
организмы. Первый из них (корд-фактор, представляющий собой
дитрегалозо-6,6-димиколат) задерживает миграцию лейкоцитов,
индуцирует развитие хронических гранулем, тогда как второй
гемолизирует эритроциты.
Гликолипопротеины, или О-антигены (например, иерсиний),
вызывают многие реакции в организме. Гликолипопротеины хла-
мидий поражают эпителиальные клетки дыхательных путей
и лимфоидной ткани. К разряду гликопротеинов относятся гриб-
ные аллергены, индуцирующие измененные реакции организма,
или аллергии.
Типичные эндотоксины бруцелл, гонококков, синегнойной па-
лочки, холерных вибрионов, энтеробактерий представляют
собой липополисахариды (липогликаны). Их молекулярная мас-
са порядка 100...900 кДа. Липогликаны термостабильны и сос-
тоят из трех частей: повторяющихся олигосахаридных остатков-
гаптенов (детерминантные группы), полисахаридного кора и ли-
пида А с кетодезоксиоктулонатом. При парентеральном введении
эндотоксинов отмечается повышение температуры, лейкопения
и гипогликемия, гипотензия и шок, нарушение перфузии важ-
нейших внутренних органов, активация С3-комплемента и всего
комплементного каскада реакций, внутривенный тромбоз со смер-
тельным исходом.
Пептидогликаны (муреины) могут вносить определенный
вклад в развитие очагов воспаления.
Четвертую группу факторов составляют липопротеи-
ны микобактерий туберкулеза и патогенных иерсиний. Первые
из них индуцируют чувствительность к туберкулину, вторые под
названием антигенов V и W обеспечивают выживаемость возбу-
дителей заболевания внутри клеток системы мононуклеарных
фагоцитов. Липопротеины содержатся и в других клетках пато-
генных микроорганизмов, но они изучены недостаточно.
К пятой группе факторов относятся фосфатиды, типич-
ные, например, для микобактерий туберкулеза. Они вызывают
клеточные реакции и казеозный (творожистый) распад тканей.
Эти фосфатиды — глицеринсодержащие липиды, включающие фос-
фатидилсерин, фосфатидилхолин, фосфатидилэтаноламин. Фос-
фатидилхолины называют еще лецитинами. Все фосфатиды пред-
ставляют собой диполярные ионы, способные вступать в соот-
ветствующие химические реакции in vivo. Однако фосфолипазы
микроорганизмов мало активны в отношении фосфатидов мико-
358
жтерии, поскольку в интактных клетках они находятся в связи
другими структурами, обеспечивающими таким комплексным
одествам достаточную устойчивость к действию специфических
грментов.
Шестую группу факторов составляют фосфопротеины,
которым относится относительно изученная фракция F—1,
держащаяся в клетках патогенных иерсиний и тормозящая
агоцитарный процесс.
Седьмую группу факторов составляют другие вещест-
а, не вошедшие в состав шести рассмотренных групп. Прежде
сего здесь следует назвать токсины грибов небелковой природы,
акой способностью обладают аспергиллы, пенициллы и др. Сре-
и аспергиллов наибольшую известность приобрел вид Asper-
illus f lav us — продуцент флавотоксинов, или афлатоксинов:
И, В2, В2а, Gl, G2a, Ml, М2. Афлатоксин Bl продуцируется
виде свободного гомологичного продукта (С17Н12О6), наиболее
асто он обнаруживается в зараженных Asp. flavus продуктах
кормах. По механизму действия напоминает актиномицин D,
. е. подавляет ДНК-зависимый синтез РНК вследствие образо-
ания комплекса с ДНК:
остатки)
Другие афлатоксины сходны по строению с В1, но менее ток-
сичны для человека и животных. Например, для новорожденных
тят LDioo составляет 15...20 мкг для В1, 85 мкг — для В2,
.0 мкг — для G1 и 175 мкг — для G2. Афлатоксин М2 — токси-
ческий метаболит афлатоксина В2.
359
Афлатоксины могут вызывать у чувствительных животных и
человека острый и хронический токсикозы (афлатоксикозы). При
острой интоксикации у пострадавших отмечается рвота, конвуль-
сии, или судороги, кома (от лат. сота — глубокий обморок),
отек мозговых оболочек, при этом в крови выявляется свободный
аммиак. При хроническом афлатоксикозе у чувствительных жи-
вотных (крысы, утята и др.) возникают опухоли печени, реже —
почек, иногда с метастазами в легкие. Первичный рак печени у
человека возможен в тех случаях, когда афлатоксин регулярно
поступает в организм с пищей в дозе до 500 мкг/кг веса тела.
Установлено, что Asp. parasiticus также может продуциро-
вать афлатоксины. Из других микотоксинов, образуемых разны-
ми аспергиллами, известны глиотоксин, охратоксины А, В, С,
патулин, стеригматоцистин, треморген, цитохалазины, цитринин
(табл. 35).
Таблица 35. Токсины, образуемые некоторыми аспергиллами
Название Продуцент Действие
Афлатоксины Asp. flavus Asp. parasiticus Гепатотоксичность, канцеро гениость, мутагенность, терато генность
360
Продолжение табл. 35
, Название Продуцент Действие
! Глиотоксин Asp. giganteus Asp. fumigatus Нефротоксичность
Охратоксины Asp. ochraceus Нефротоксичность, нейроток- сичность, жировая инфильтра- ция печени
Патулин* Asp. terreus Asp. niveus Asp. candidum Гепатотоксичность, нефроток- сичность, канцерогенность, отеки легких, антндиуретическое дей- ствие
Стеригматоцистин Asp. versicolor Asp. nidulans Нефротоксичность, гепатоток- сичность, канцерогенность
Треморген Asp. clavatus Asp. flavus Asp. candidum Нейротоксичность, индукция сарком в подкожных тканях
Цитохалазины Asp. clavatus Нервно-мышечные поражения, торможение фагоцитоза и пино- цитоза, энуклеация клеток
Цитринин** Asp. terreus Asp. niveus Asp. candidum Нефротоксичность, нейроток- сичность, гипотензия
* Образуется также некоторыми видами пенициллов (Р. expansum, Р. patulum и др.).
** Образуется также пенициллами (Р. citrinum, Р. lividum и др.).
Охратоксины незначительно различаются между собой. Так,
охратоксин В в отличие от типа А не содержит хлора; охраток-
син С — это этилохратоксин А.
охратоксин А
патулин
361
цитохалазин A, R-O ;
цитохалазин В, R = Н,ОН
Из других грибных токсинов следует отметить цитреовири-
дин, образуемый Renicillium citreo-viride:
При употреблении риса, пораженного данным грибом, могут воз-
никать острые отравления, сопровождающиеся в тяжелых слу-
чаях нисходящими параличами с возможным смертельным исхо-
дом за счет нарушения дыхательного центра.
Из семи пигментов, образуемыех Р. islandicum (иридоскирин,
исландицин, катенарин, лютеоскирин, руброскирин, скирин, эрит-
роскирин), лишь лютеоскирин является микотоксином. Данный
гриб, как и Р. citreo-viride, способен поражать рис. Лютеоски-
рин оказался фотохимическим реагентом, трансформирующимся
в люмилютеоскирин под влиянием прямых солнечных лучей и
ультрафиолетового облучения в течение 10 ч при 295 нм. Другие
названные выше пигменты сходны по строению с лютеоскирином,
но не идентичны с ним:
(—) лютеоскирин люмилютеоскирин
Р. islandicum кроме описанного токсичного пигмента продуци-
рует ядовитые циклопептиды (см. разд. 7.2).
362
t Известен ряд других нитчатых грибов, образующих токсиче-
ские небелковые продукты, например грибы, относящиеся к ро-
(ам Alternaria, Chaetomium, Cladosporium, Fusarium, Gibberel-
a, Helmintosporium, Sclerotinia, Sporodesmium и др. Химическое
Строение токсических продуктов, образуемых этими грибами,
фиведено ниже:
н н соон
альтернаровая кислота из Alternaria kikuchiana
СН3—(СН2)9—СН=СН—(СН2)9—С—SH
фагикладоспоровая кислота из Cl. fagi
NH—СО—СН
ацетил-у-амино-
Д“,₽-бутенолнд из
F. nivale
СН3(СН2)9—СН=СН— (СН2)13—<
о
SH
эпикладоспоровая кислота из
С/, epiphyllum
споридесмины А и В из
Sporodesmium bakerii
8-метоксипсорален из
Sclerotinia sclerotiorum
A:Rj — R2-ОН,
B:R[— OH, R2-H
363
Токсические продукты небелковой природы образуются также и
бактериями, например токсин VFDF из цианобактерий АпаЬепа
jlos-a.qu.ae-.
Факторами патогенности могут быть свободные липиды, неко-
торые органические амины, ионы металлов. Свободные липиды,
или нейтральные жиры, освобождаются из микробных клеток
в результате их автолиза и, например, в очаге воспаления они
могут блокировать фагоциты. В этом случае свободные липиды
проявляют свойства поверхностно-активных веществ, способных
образовывать мультислои на границах раздела фаз.
Органические амины — путресцин и кадаверин — образуются
столбнячной палочкой, холерным вибрионом, некоторыми гриба-
ми и др. Оба диамина относят к числу трупных ядов. Они прояв-
ляют общетоксическое действие на макроорганизм. Эти амины
возникают в инфицированных очагах под действием ферментов
декарбоксилаз на аргинин- и лизинсодержащие белки:
H2N—С—NH—(СН2)з—СН—СООН
II I
NH аргинин NH2
СО2 + CO(NH2)2 + H2N— (СН2)4—nh2
путресцин
НООС—СН—(СН2)3—СН2—NH2 ДекаЕб°ксила1!
nh2
лизин
СО2 + H2N— (СН2)5—nh2
кадаверин
Базидиальные грибы образуют различные азотистые (небел-
ковые) ядовитые продукты типа гиромитрина (протоплазмати-
ческий яд), мускарина и мускаридина (взаимодействуют с М-хо-
линорецепторами нервной системы человека) и др.:
сн,—ch=n—n—сн=о
I
сн3
гиромитрин из
Helvetia species
н3с—сн—сн—(СН2)3—№(СН3)
он он
L(+)мускарин из Amanita muscarina
мускаридин из
Amanita muscarina
364
Некоторые ионы металлов могут усиливать вирулентность
тдельных патогенных бактерий, например железо. В этом слу-
ае, очевидно, улучшается синтез железосодержащих ферментов
увеличивается выживаемость клеток возбудителя инфекцион-
ого заболевания в макроорганизме.
.3. ХИМИОТЕРАПИЯ ИНФЕКЦИОННЫХ БОЛЕЗНЕЙ
Химиотерапия — это теория и практика использования химиче-
ских средств специфического действия в комплексном лечении
заболеваний, вызванных соответствующими возбудителями. Если
такими возбудителями являются патогенные микробы, то речь
идет о химиотерапии инфекционных заболеваний. Клетки опухо-
лей могут быть приравнены к возбудителям инфекционных за-
болеваний, хотя они лишены одного из главных свойств таких
заболеваний — заразительности. В этих случаях говорят о хи-
миотерапии опухолей. Наконец, возбудителями могут быть гель-
минты — многоклеточные организмы. Тогда говорят о химиотера-
пии гельминтозов.
В данной книге представлены краткие сведения о химиотера-
пии инфекционных заболеваний. Основоположником химиотера-
пии является П. Эрлих, который сформулировал (1885) идею
химиотерапии. Он доказал антипротозойное действие бензиди-
новых красителей, впервые синтезировал и ввел в практику лече-
ния сифилиса мышьяковистые препараты — вначале сальварсан,
затем неосальварсан или новарсенол (смесь соединений I и. II):
бензидин
сальварсан
NH—-СНг—OSONa
неосальварсан
—ОН
NH—СН2—OS ON а
365
С этого времени выделяют первый период научной химиотера-
пии (ему предшествовал период эмпирической терапии). Знаме-
нательной эрой в развитии химиотерапии инфекционных заболе-
ваний было открытие красного (пронтозил), а затем белого
(пронтальбин) стрептоцидов (Г. Домагк, 1935). Начался второй
период-сульфаниламидотерапии.
H2NO2S——N=N-x^ NH2 H2NO2S—y~NH2
NH2
красный стрептоцид белый стрептоцид
Белый стрептоцид, как оказалось, является составной частью
красного; его структура лежит в основе сульфаниламидных пре-
паратов, широко используемых до настоящего времени для лече-
ния заболеваний, вызванных преимущественно стафилококками,
стрептококками, гонококками и некоторыми другими патогенны-
ми бактериями.
Третий период научной химиотерапии связан с открытием
пенициллина (А. Флеминг, 1929) и получением его в виде инди-
видуального вещества (X. Флори и Б. Чейн, 1940). Началась эра
антибиотикотерапии. В течение 15 лет были открыты стрептоми-
цины, тетрациклины, хлорамфеникол, антибиотики-макролиды и
многие другие (см. гл. 6).
С начала 60-х годов XX в. утверждается четвертый период —
период комбинированной химиотерапии, т. е. когда используют
химиотерапевтические вещества в различных сочетаниях между
собой и с другими лечебными средствами.
Введенные П. Эрлихом такие понятия, как максимально пере-
носимая доза [Dosis tolerata (DT)] и минимальная лечебная
доза [Dosis curativa (DC)], химиотерапевтический индекс 1 =
= DT/DC не потеряли своего значения. В настоящее время из-
вестно и другое определение химиотерапевтического индекса —
как отношение усредненной летальной дозы (LD50) к усреднен-
ной эффективной дозе (ED&), т. е. I = LD50/ED50.
Малотоксичный и нетоксичный химиотерапевтический препа-
рат характеризуется достаточно большим значением I (1>1).
Если I 1, то препарат токсичен или высокотоксичен. Химиоте-
рапевтический индекс — абстрактная величина, так как на прак-
тике, как правило, приходится переносить на людей результаты
экспериментов, полученные на животных. Вот почему данные
о токсичности того или иного препарата не имеют абсолютного
значения.
В практическом здравоохранении используют многие сотни
лекарственных веществ и тысячи их лекарственных форм (таб-
* Доза лекарства, излечивающая 50 % взятых в опыт экспериментальных
животных.
366
ётки, мази, суспензии, порошки, инъекционные растворы
I Т. Д.).
Практика использования индивидуальных химиотерапевтиче-
ких средств со временем стала убедительным доказательством
епригодности следующей формулы: «Если одно лекарство хоро-
ю, то два будут лучше, а три должны излечивать любое заболе-
дние». Эмпирические комбинации химиотерапевтических веществ
ыли одной из причин нарастания числа устойчивых микроорга-
низмов, особенно в клинических условиях, так как при этом не
считывалась чувствительность возбудителей инфекционных забо-
леваний к рекомендуемым лекарственным средствам. Тем не ме-
лее комбинированная химиотерапия стала велением времени.
.Необходим был научно обоснованный выбор химиотерапевтиче-
ских средств, используемых в комбинациях. Подход к такому
выбору базируется на следующих рекомендациях.
I 1. Перед началом химиотерапии инфекционного заболевания
^необходимо проверить образцы патологического материала на
'наличие возбудителя заболевания и подтвердить клинический
диагноз этиологическим диагнозом. Определить чувствительность
^возбудителя к выбранным химиотерапевтическим средствам.
5 2. При наличии предполагаемого инфекционного заболева-
ния у безнадежно больных пациентов желательно им вводить
гболее чем одно противомикробное средство в надежде на то, что
удастся в кратчайший срок подавить предполагаемого возбуди-
теля. Лекарства вводят с целью ожидаемого губительного эффек-
та на микроорганизм до тех пор, пока не будет идентифицирован
возбудитель заболевания. После этого назначают специфическое
химиотерапевтическое средство.
3. Если инфекционное заболевание вызвано не одним пато-
- генным микробом (смешанная инфекция), то возможно примене-
ние двух или более химиотерапевтических средств, каждый из
которых действует на какого-либо одного представителя микроб-
ной флоры; подобная комбинация может быть более эффектив-
ной, чем один препарат. Так иногда поступают при инфекцион-
ных заболеваниях кожи, открытых полостей тела, инфицирован-
ных ранах, когда местно используют слабо адсорбирующиеся
антибактериальные средства узкого спектра действия, например
полимиксины, некоторые аминогликозиды, бацитрацин. Анало-
гичные ситуации возможны в случаях смешанных инфекций при
поражении сердечно-сосудистой, дыхательной и мочевыводящей
систем.
4. Когда одновременное применение двух лекарств обеспечи-
вает лучший эффект, чем действие одного лекарства, в этом слу-
чае возможно повышение антимикробной активности первого
средства назначением второго против специфического возбуди-
теля. Такой синергизм химиотерапевтических веществ не всегда
предсказуем, но он характерен, например, при терапии бакте-
риальных эндокардитов, вызванных энтерококками Streptococcus
faecalis. Лечение их одним пенициллином всегда менее надежно,
367
чем лечение комбинацией препаратов пенициллин + аминогли-
козид.
5. При быстром нарастании устойчивости возбудителя к од-
ному лекарственному средству совместное использование препа-
ратов оказывается необходимым. Добавление второго препарата
в ряде случаев задерживает нарастание резистентности. Такие
ситуации имеют место при туберкулезе, и это служит основой
для рекомендации использовать при данном заболевании комби-
нации изониазида, этамбутола, стрептомицина, рифампицина
и др. Аналогичный подход целесообразен и при других хрони-
ческих инфекционных заболеваниях в затяжные периоды, когда,
например, относительно легко может нарастать устойчивость
возбудителей к одному из следующих антибиотиков: стрептоми-
цину, карбенициллину, эритромицину и др. Поэтому, например,
для борьбы с инфекцией, вызванной Pseudomonas aeruginosa,
рекомендуют комбинацию карбенициллина с гентамицином.
6. Химиотерапевтические средства в комбинациях могут
снимать частоту и интенсивность побочных реакций. Так, если
возбудитель чувствителен лишь к такой дозе одного лекарства,
от которой возникают нежелательные побочные реакции, а в комби-
нации его вполне достаточно в половинной дозе, то такая комби-
нация оправдана и обоснована (например, левомицетин с поли-
миксином против Ps. aeruginosa).
7. Комбинирование химиотерапевтических средств с иммучо-
препаратами возможно и необходимо по специальным показа-
ниям: для активации иммунной системы при хронических, вяло
текущих инфекционных заболеваниях; для более эффективного
губительного воздействия на возбудителей заболеваний. Актива-
ции иммунной системы добиваются введением вакцин пациентам
(иногда аутовакцины, приготовленной из штамма патогенного
микроорганизма, выделенного от того же больного).
В качестве средств губительного воздействия на возбудителя
заболеваний используют иммунные сыворотки или у-глобулины.
Если они антитоксические, то при их комбинировании с химио-
терапевтическими средствами преследуют цель обезвредить ток-
син, вырабатываемый патогенным микробом, и убить или задер-
жать развитие этого же возбудителя.
Динамика комбинированного использования химиотерапевти-
ческих средств может проявиться в форме синергизма, антаго-
низма или индифферентности. Синергизм — возрастание противо-
микробного эффекта при комбинации двух лекарств, когда сум-
марное действие лекарственных средств такой комбинации
превышает действие каждого из них, примененного в отдельности
и в более высоких дозах, а также превышает ожидаемые эффек-
ты в случае последовательного использования тех же лекарств.
Антагонизм — снижение антимикробной эффективности одного
лекарственного средства при добавлении другого. Индифферент-
ность — явление более частое, чем синергизм и антагонизм. При
этом химиотерапевтические средства в комбинациях по эффек-
368
явности не превышают эффектов каждого из них, используемого
отдельности, или действие выражается в арифметической сум-
е эффектов индивидуальных лекарств в назначенных дозах.
На практике иногда используют химиопрофилактику, т. е.
рименение химиотерапевтических средств для предупреждения
чедрения и развития возбудителей инфекционных заболеваний
i vivo. Она нацелена, как правило, на специальный круг лекар-
венных веществ и специфических возбудителей. Так, для спе-
ифической химиопрофилактики ревматизма и гломелуронефри-
эв, являющихся осложнениями стрептококковых инфекций, при-
1еняют препараты пенициллина и реже сульфаниламиды. Здесь
ледует иметь в виду, что у р-гемолитических стрептококков ре-
истентность к пенициллинам, как правило, не развивается. Для
даления стрептококков из глотки й зева в целях предупрежде-
ия постстрептококковых осложнений кроме пенициллина можно
спользовать эритромицин. Для химиопрофилактики менингокок-
овых заболеваний используют пенициллины, рефампицин, а так-
се некоторые препараты тетрациклинового ряда (например, ми-
юциклин); в отношении некоторых венерических заболеваний
гонорея, сифилис) рекомендуют бензилпенициллин, а для преду-
беждения бленнореи новорожденных используют 1 %-ный раст-
ор нитрата серебра. В угрожающих случаях контакта по чуме
водят сульфадиазин per os\ для химиопрофилактики сыпного
ифа используют левомицетин или тетрациклины. Химиопрофи-
лактике могут подвергаться лица, чувствительные по наследст-
венным или приобретенным характеристикам к возможному зара-
жению какими-либо возбудителями инфекционных заболеваний.
Например, люди с пороками сердца подвержены большему риску
заболеть инфекционным эндокардитом (возбудители Streptoco-
ccus viridans. Str. faecalis), чем другие индивидуумы. В подоб-
ных ситуациях эффективным средством профилактики является
бензилпенициллин.
Пневмококки и гемофильная палочка являются наиболее час-
тыми виновниками хронических бронхитов у лиц с бронхоэкта-
зиями, эмфиземой и другими легочными нарушениями. В ка-
честве химиопрофилактических средств здесь рекомендуют ампи-
циллин, тетрациклины или препараты цифалоспоринового ряда.
У некоторых лиц, страдающих тяжелыми заболеваниями типа
туберкулезной волчанки или пемфигуса (пузырчатка) и длитель-
но получающих контролируемые дозы кортикостероидных гормо-
нов, заметно снижается устойчивость к возбудителям бакте-
риальных инфекций. Таким больным можно назначать соот-
ветствующие химиотерапевтические средства с учетом наличия
резистентной микрофлоры в конкретных клинических условиях.
Иногда прибегают к предоперационной и послеоперационной
химиопрофилактике хирургических больных, подготавливаемых
для оперативных вмешательств на нижних отделах кишечника,
на сердце и на других частях тела (глаза, большие суставы).
Химиопрофилактику протозойных инфекционных заболеваний
369
осуществляют с помощью тех же препаратов, которые рекомен-
дованы для терапевтических целей. Противовирусную химиопро-
филактику используют ограниченно. Однако, например, аманта-
дин рекомендуют для предупреждения вирусного гриппа типа А.
Возможность использования иммунопрепаратов в качестве
средств профилактики инфекционных заболеваний отмечена в гл. 8.
Вопросы
I. Какие микроор!.''.hh.wi.i опкх;г, i, »г, inrannAM пара.»!гам и к
па гогенным ( примеры) ’
2. Какие группы химических веществ, образуемых патогенными м-•
организмами, отопят к разряду факторон агрессии и факторов tn.m
HOC ! И?
3. Какие виды микробов ошосяп'я к гоксигсицым и какие виды гои.,
они образуют'^
4. Какие известны <i’a,’i.ioToKcnin,i, г поя их раепгчие и какая их фх.
у микробов-продупешон3
5. Каковы химическая структура >1 механи >м действия афлатоксинов
6. Какие извести периоды в развита химио,ераиии инфекциоины •
летней, с практическим использованием каких веществ они связаны?
7. Что такое индифферентность, синергизм и антагонизм лекаре1'
комбинированном использовании их на праюике (примеры)?
8. Какие необходимы рекомендации для обоснованного выбора Д'
терапевтических средств и комбинациях?
9. Что такое па roiенноегь и вирулентность. И11вазивность и инфе >"
ноегь? Какими факторами они обусловливаются?
10. Какие ферменты могут быть факторами агрессии у .миквоощ :•
мои?
Глава 8
ХИМИЯ ПРОТИВОМИКРОБНОГО ИММУНИТЕТА
(ИММУНОХИМИЯ)
8.1. ИММУННАЯ СИСТЕМА И ИММУНИТЕТ
В обеспечении постоянства внутренней среды организма, или
гомеостаза, наряду с другими системами (нервной, эндокринной)
важная роль отводится иммунной системе, компонентами которой
являются специальные клетки, молекулы и гены. Клеточный ком-
понент включает макрофаги, лимфоциты, вспомогательные денд-
ритные клетки [иммунокомпетентные клетки (ИКК), или иммуно-
циты]; молекулярный — антитела и растворимые факторы, секре-
тируемые ИКК; генетический — гены в хромосомах. ИКК лока-
лизуются в тимусе, селезенке, лимфатических узлах, пейеровых
бляшках кишечника, костном мозгу, крови. Иммунная система —
одна из самых гетерогенных и сложных систем организма. У че-
ловека, например, она включает 2ХЮ12 лимфоцитов и 1О20 моле-
кул антител, обладающих, с одной стороны, общностью органи-
зации, а с другой — различной специфичностью.
370
В циркулирующей крови* находятся макрофагальные клетки,
[и моноциты (имеются фиксированные макрофагальные клетки
печени, соединительной ткани, лимфоузлах и других тканях
органах); лимфоциты; полиморфноядерные, или сегментоядер-
ie (зернистые) лейкоциты, называемые еще гранулоцитами или
!крофагами; эритроциты и кровяные пластинки — тромбоциты,
з них белые клетки лейкоциты — моноциты, лимфоциты, грану-
>циты; красные — эритроциты и тромбоциты.
Моноциты крови достаточно крупные по размерам (10...
! мкм в диаметре), на их долю приходится 4...8 % от общего
(личества белых клеток. Основнай функция моноцитов — обра-
>тка антигенной информации и представление ее лимфоцитам,
реди лимфоцитов различают Т- и В-клетки, которые подробно
ассмотрены в последующих разделах. Количество лимфоцитов
крови составляет примерно 30 % от общего количества лейко-
цитов. Лимфоциты по размерам подразделяют на большие
Z> Ю мкм в диаметре), средние (7...10 мкм) и малые (5...6мкм).
На долю гранулоцитов приходится-большая часть белых кле-
ок (~80%). Размеры их находятся в пределах 7... 10 мкм.
•азличают нейтрофильные (60...70 %), эозинофильные (2...4 %)
базофильные (0,5... 1 %) гранулоциты; аналоги базофилов —
каневые тучные клетки. При окраске гранулоцитов смесью ос-
овного и кислого красителя (гематоксилин + эозин) в их цито-
плазме выявляются неравнозначная по окраске, размерам и ко-
ичеству зернистость (отсюда и происходит название этих кле-
ок) и сегментированное ядро. Основные функции нейтрофи-
ов — фагоцитоз (преимущественно микроорганизмов); эозино-
филов — участие в обмене гистамина и фагоцитоз; базофилов —
акопление антикоагулянта гепарина, гистамина, кининов и дру-
их биологически активных медиаторов (см. разд. 8.6).
Эритроциты (диаметр 7...8 мкм) — переносчики кислорода
ткани и углекислоты из тканей; тромбоциты содержат фермент
ромбопластин, или тромбокиназу, участвующую в реакциях
вертывания крови.
Следовательно, среди большого разнообразия клеток и бел-
:ов лишь немногие являются компонентами иммунной системы.
Интегральное функционирование этих компонентов и иммунной
системы в целом обеспечивает иммунный статус каждого инди-
вида. Они кооперативно взаимодействуют между собой за счет
поверхностных антигенов, кодируемых генами главного комплек-
са гистосовместимости.
Условно в главном комплексе гистосовместимости можно вы-
делить три группы генов. Первая группа кодирует синтез
* Кровь, лишенную эритроцитов и тромбоцитов, называют лимфой; если
удалить из крови еще и белые клетки, то это будет плазма. Дефибринированная
плазма представляет собой сыворотку крови. Некоторые сывороточные белки (гло-
булины) представляют особый интерес и о них будет подробнее сказано в после-
дующих разделах этой главы.
371
антигенов гистосовместимости — крайне полиморфных гликопро-
теинов поверхностной мембраны всех ядерных клеток. Эти глико-
протеины одинаковы у клеток только одного животного, но отли-
чаются у организмов даже внутри вида, определяя антигенную
индивидуальность каждого организма. Структура этих антигенов
распознается цитолитическими лимфоцитами, и поэтому они
участвуют как мишени в узнавании и разрушении чужеродных
трансплантатов, злокачественных и трансформированных вируса-
ми клеток. Первичная структура и конформация некоторых из
трансплантационных антигенов уже известны или интенсивно
изучаются. У человека различают HLA — А и HLA — В антиге-
ны (от англ, human leucocyte antigen — человеческий лейкоци-
тарный антиген), их гены локализованы на 6-й хромосоме. Как
правило, они содержат тяжелую и легкую цепи. Тяжелая цепь
(Л4 = 45 кДа) состоит из небольшой фосфорилированной внут-
риклеточной С-концевой части молекулы, в которой выделяют
al-, a2- и аЗ-домены (см).
Вторая группа генов комплекса (D-локус у человека)
играет важнейшую роль в регуляции иммунного ответа. Это так
называемые 1г-гены (от англ. Immune response — иммунный от-
вет). Их продукты находятся на поверхности лимфоцитов и мак-
рофагов и реализуют эффекторные (хелперные, или супрессор-
ные, от англ, helper — помощник, suppress — подавлять) функ-
ции лимфоцитов, часто определяя, будет ли антиген распознан
как «чужое» или нет. Многие болезни человека связаны с D-
локусом комплекса, структура антигенов которого определяет си-
лу и направленность иммунного ответа против антигенов виру-
сов, бактерий и т. д.
Третья группа генов кодирует структуру некоторых
компонентов комплемента.
Иммунитет (от лат. immunitas — освобождение от чего-
либо) — врожденная или приобретенная способность макроорга-
низма к защите, специфически направленная против любых ге-
нетически чужеродных для него агентов. Такими агентами явля-
ются разнообразные клетки микро- и макроорганизмов (в том
числе измененные клетки собственного организма), а также их
структурные компоненты — белки, высокомолекулярные полиса-
хариды, гликоконъюгаты (гликопротеины, пептидогликаны, липо-
полисахариды и пр.), проявляющие свойства антигенов (см.
разд. 8.2).
В этой связи различают следующие типы иммунитета: проти-
вомикробный, антитоксический, антитрансплантационный. В пер-
вом случае он может быть направлен против вирусов, клеток
бактерий, грибов или протозойных организмов; во втором — про-
тив токсинов, вырабатываемых некоторыми бактериями (Clos-
tridium tetani, Cl. botulinum, Corynebacterium diphtheriae и др ),
растениями (клещевина Rhicinus communis), пресмыкающимися
животными, например змеями (гадюка обыкновенная Vipera
berus, гадюка степная Vipera Rennardi и др.), и т. д.; в третьем —
372
ютив чужеродных тканей при их пересадке во время оператив-
IX вмешательств (пересадка кожи, почек, костного мозга, серд-
i) или при переливании крови. Иммунитет функционально
правлен на уничтожение клеток и веществ, генетически отли-
ющихся от собственных, и на сохранение постоянства внутрен-
й среды макроорганизма в течение всей его жизни.
Основоположники иммунологии — науки об иммунитете —
И. Мечников и П. Эрлих (лауреаты Нобелевской премии
11 г.). Благодаря бурному развитию иммунологии в последние
сятилетия ее обоснованно стали подразделять на самостоятель-
ie дисциплины: общую иммунологию, иммунохимию (молеку-
ярную иммунологию), иммуногенетику, иммунопатологию, им-
[унобиологию. Современная иммунология, представляющая со-
ой науку о координированном взаимодействии генов, молекул
клеток — компонентов иммунной системы, является в то же
ремя ветвью химии, получившей название иммунохимии (тер-
[ин впервые был введен С. Аррениусом в 1907 г.). Основополож-
иком иммунохимии — науки о химии иммунологических процес-
се — был К. Ландштейнер, работы которого по изучению спе-
ифичности антигенных детерминант искусственных антигенов,
о структуре и функции гаптенов стали классическими.
Благодаря исследованиям, проводимым в течение последних
...8 десятилетий, был дан ответ на основные вопросы, выдвину-
ые при зарождении иммунохимии о структуре антигенов и анти-
ел, о сущности реакций антиген-антитело и химических процес-
се, лежащих в основе их взаимодействия.
С помощью иммунохимических методов продолжают раскры-
ать механизмы иммунологических процессов, связанных с синте-
ом иммуноглобулинов в бесклеточных системах, изменениями
нтигенных детерминант на основе полусинтеза, клеточными ре-
.епторами, медиаторами клеточного и гуморально! о иммуни -
ета.
Иммунохимия представляет собой и область иммунологии,
которой находят применение преимущественно химические и
иохимические методы исследования, в том числе высокочувст-
ительные и специфичные (иммунофлюоресцентные, радиоимму-
ологические, рентгеноиммунологические, иммуноферментные и
. д.), Иммунохимия формировалась на базе так называемой
инфекционной иммунологии (равно как иммунология в целом),
т. е. на выяснении механизмов противомикробного иммунитета.
Противомикробный иммунитет выражается в защите организма
от чужеродных инфекционных агентов независимо от того, будут
ли это собственно клетки патогенных микроорганизмов или раз-
личные составные их компоненты. Формирование противомикроб-
ного иммунитета сопровождается активной гуморальной (от
лат. humor — жидкость) и клеточной перестройкой организма,
в ходе которой синтезируются специфические антитела как сво-
бодно циркулирующие, так и связанные с мембранами лимфо-
цитов и макрофагов (см. разд. 8.3), способные уничтожать мик-
373
ровные клетки или нейтрализовать выделяемые ими токсины
(антитоксический иммунитет).
Различают видовой (естественный, наследственный) и инди-
видуальный (приобретенный) противомикробный иммунитет.
Видовой иммунитет генетически детерминирован. Он присущ ка-
кому-то одному виду живых существ и характеризуется невос-
приимчивостью этого вида к микроорганизмам, вызывающим
заболевания у других видов. Человек, например, невосприимчив
ко многим инфекционным заболеваниям животных (чуме собак,
рогатого скота и др.), тогда как животные нечувствительны
к возбудителям венерических болезней, эпидемического цереброс-
пинального менингита, дифтерии и других заболеваний человека.
Индивидуальный иммунитет приобретается в ходе развития инди-
видуума и характеризуется невосприимчивостью к заболеваниям,
вызываемым какими-либо патогенными микроорганизмами, и
строгой специфичностью. Виды противомикробного иммунитета
представлены на схеме:
Видовой
(наследственный)
Иммунитет
Индивидуальный
(приобретенный)
Естественный
Искусственный
Активный
(постинфекционный)
Пассивный Активный Пассивный
(плацентарный) (сывороточный)
Антимикробный Антитоксический Антимикробный
(поствакцинальный)
Стерильный
Антитоксйческий
(после введения
анатоксина)
Нестерильный
Иногда выделяют коллективный (приобретенный) иммунитет,
когда осуществляют прививочные мероприятия против инфек-
ционных заболеваний в целых коллективах. Тем не менее такой
иммунитет по существу является индивидуальным. Естественный
активный иммунитет возникает после перенесения того или иного
инфекционного заболевания, поэтому его называют также пост-
инфекционным. Антимикробный иммунитет в свою очередь под-
разделяют на стерильный и нестерильный (инфекционный). Им-
мунитет против микроорганизмов, сохраняющийся после их ис-
чезновения, обозначается как стерильный, или полный, иммуни-
тет. Нестерильный иммунитет обусловлен наличием в организме
возбудителей заболевания (хронически протекающие бактериаль-
ные и протозойные инфекции). В это время организм обладает
повышенной устойчивостью к тем же возбудителям, т. е. реин-
фекция здесь не наступает. Инфекционный иммунитет поддер-
374
1вается ограниченное время — в период размножения возбуди-
ли в организме и играет решающую роль при туберкулезе, сифи-
се. Естественный иммунитет может быть и пассивным, когда
передается эмбриону через плаценту с кровью матери (пла-
нтарный иммунитет) или новорожденному с грудным молоком.
Искусственный приобретенный иммунитет так же, как и ес-
ственный, бывает активным и пассивным. Активный выраба-
[вается после введения в организм вакцин в виде взвесей уби-
х или живых ослабленных возбудителей болезни — вирусов,
ктерий и анатоксинов (обезвреженных токсинов). Пассивный
ывороточный) иммунитет появляется в организме в результате
едения готовых антител (иммуноглобулинов, иммунных сыворо-
к). Он быстро возникает и быстро исчезает в отличие от актив-
но противомикробного и антитоксического иммунитета, сохра-
ющегося иногда в течение всей жизни идивидуума (например,
>и оспе, полиомиелите, кори, паротите, дифтерии, брюшном и
шном тифах и др.).
В формировании рассмотренных видов иммунитета принимает
астие вся иммунная система организма, поэтому их можно
ъединить под названием общий иммунитет. Различают также
>стный иммунитет, имеющий отношение не только к поддержа-
нию постоянства внутренней среды организма, но прежде всего
защите его поверхностей (кожи и слизистых) от возбудителей
инфекционных заболеваний. Важную роль в индукции местного
ммунитета играют секреторные антитела класса IgA (см.
азд. 8.3), доминирующие в секретах человека: молозиве, слюне,
лезной жидкости, носовой слизи, секретах бронхов, кишечника
> др. Организм, снабженный секреторными антителами, выпол-
[яющими защитные функции, представляет собой «тупик» для
нфекций, при которых поверхность слизистых оболочек служит
>е только входными воротами для возбудителя, но и местом его
окализации (при ряде вирусных респираторных и бактериаль-
ных кишечных инфекций). Секреторные антитела имеют большое
начение в коллективном иммунитете. Образование и функцио-
(ирование этих антител в данном случае мешает вирусам при
«спираторных заболеваниях выделяться из полости носа и из
рганизма в целом. За формирование противомикробного имму-
нитета ответственны неспецифические и специфические клеточные
н гуморальные механизмы.
Клеточные неспецифические факторы защиты. Кроме неспеци-
шческой защитной функции кожи и слизистых оболочек к этой
<е группе факторов относится воспаление — комплекс реактив-
ых изменений преимущественно сосудов микроциркуляторного
уела и соединительной ткани, возникающий в ответ на местное
ействие болезнетворного раздражителя (микроорганизма).
Воспалительный процесс прежде всего следует рассматривать
ак защитно-приспособительную реакцию клеток организма на
.ействие микробов, продуктов их жизнедеятельности и других
(акторов. Основным механизмом воспалительного процесса,
375
обеспечивающим борьбу организма с микробами, является фаго-
цитоз. Фагоцитоз, или «пожирание клетками» (от греч. срауоо —
поедание), представляет собой процесс поглощения корпускуляр-
ного материала (в том числе микроорганизмов) фагоцитами.
Явление фагоцитоза было открыто И. И. Мечниковым (1892),
им же были установлены роль и значение фагоцитирующих
клеток (микро- и макрофагов) в процессе защиты организма от
инфекций. Микрофаги, или полиморфноядерные нейтрофильные
гранулоциты, а также макрофаги в крови (моноциты), в соеди-
нительной ткани (гистиоциты), в печени (звездчатые клетки
Купфера), в селезенке, лимфатических узлах, костном мозге,
плевре, брюшине (свободные и фиксированные тканевые макро-
фаги) и костях (остеокласты) — все они представляют собой
фагоцитирующие клетки. Л. Ашофф (1924) объединил их под
названием ретикулоэндотелиальная система (РЭС). В 1961 г.
было предложено новое наименование — система мононуклеар-
ных фагоцитов (СМФ), включающая все фагоцитирующие клет-
ки, за исключением полиморфноядерных лейкоцитов.
Фагоциты устраняют из крови, лимфы и тканей микроорга-
низмы и другие чужеродные продукты, включая продукты рас-
пада, а также короткоживущие, отмирающие клетки собствен-
ного организма (эритроциты, лейкоциты). Фагоцитоз протекает
в несколько стадий.
1. Позитивный хемотаксис — направленное движение лейко-
цитов к очагу воспаления.
2. Аттракция, или прилипание, — адсорбция подлежащей фа-
гоцитозу частицы клеточной мембраной фагоцитов. Контакт меж-
ду фагоцитами и микроорганизмами, усиливающийся благодаря
наличию на мембране фагоцитов рецепторов для Fc-фрагмента
антител, Сзь- и Сзь-фракций комплемента, носит название имму-
ноприлипания, а фагоцитоз, осуществляемый более активно при
посредничестве антител и комплемента, — иммунного фагоци-
тоза.
3. Поглощение путем инвагинации цитоплазматической мем-
браны, в результате которой образуется вакуоль с заключенными
в нее микроорганизмами (фагосома). Фагосомы, сливаясь с лизо-
сомами, образуют фаголизосомы или вторичные лизосомы, где
микроорганизмы умерщвляются веществами, содержащимися в
лизосомах, и продуктами обмена из цитоплазмы.
4. Внутриклеточное переваривание, или внутриклеточная'сек-
вестрация (инкапсулирование), в случае неперевариваемости ма-
териала.
Фагоциты, как правило, реагируют на патогенные микроорга-
низмы, но заключительная стадия фагоцитоза может протекать по-
разному. Например, возбудители гнойных воспалений быстро
умерщвляются и перевариваются гранулоцитами и мононуклеара-
ми (завершенный фагоцитоз), а туберкулезные палочки, бруцел-
лы, гонококки, грибы родов Cryptococcus, Histoplasma могут не
только сохранять свою жизнеспособность, находясь в микро- и
376
щрофагах, но и активно размножаться в них, приводя фагоциты
гибели. Такой процесс называют незавершенным фагоцитозом.
В подавлении вирусных инфекций фагоцитоз играет соподчи-
нную роль. Он носит незавершенный характер из-за высокой ус-
йчивости вирусов к лизосомальным протеазам и другим фермен-
м фагоцитирующих, клеток. Захват и переваривание микроорга-
(змов при фагоцитозе сопровождается активными биохимиче-
ими процессами. Возрастает потребление кислорода и выделение
:роксида водорода Н2О2, усиливается анаэробный гликолиз
рез гексозомонофосфатный шунт. Активно действуют ферменты
дролазы, освобождающиеся при разрыве лизосом и поступаю-
ие в фаголизосомы. Увеличивается активность мембранной аде-
глатциклазы, изменяется активность клеточных ферментов (лак-
ггдегидрогеназы, p-глюкуронидазы, кислой фосфатазы, р-галак-
>зидазы и др.). В ходе обменных процессов в клетках накапли-
1ется токсичный супероксидный радикал Ог, переводимый с по-
эщью медь-цинк-содержащего фермента — супероксиддисмута-
1 в Н2О2.
? Фагоцитированные клетки грибов погибают в нейтрофилах бла-
Одаря воздействию ферментов системы миелопероксидазы, а за-
йм перевариваются с помощью лизосомальных ферментов. Анти-
(унгальная активность миелопероксидазы проявляется в присут-
ствии Н2Ог и ионов иода, хлора и брома. Фунгицидная активность
Иркулирующих моноцитов, так же как и гранулоцитов, опреде-
яется наличием в этих клетках системы миелопероксидазы и неко-
орых других ферментных белков.
Гуморальные неспецифические факторы защиты. Значительное
оличество патогенных микроорганизмов не попадает внутрь орга-
изма, так как они не могут преодолеть кожные и слизистые барье-
ы, где проявляют свое действие неспецифические гуморальные
(акторы местного иммунитета, обладающие бактерицидной актив-
остью. При вирусной инфекции, например, в местном воспали-
ельном очаге создаются многие неблагоприятные для вирусов
Словия, среди которых существенную роль играет усиленное обра-
бвание кислых продуктов (ацидоз) при задержке их разрушения
выделения. Образованию кислых продуктов способствуют также
копление лейкоцитов и понижение парциального давления кисло-
ода. В результате этого в очаге воспаления наблюдается анаэроб-
ый гликолиз, способствующий повышению содержания молочной
«слоты, которая снижает pH окружающей среды ниже 6,8. Бакте-
'Ицидными свойствами обладают и другие жирные кислоты, выде-
яемые потовыми и сальными железами.
В секретах слизистых оболочек дыхательного, желудочно-ки-
1ечного и мочеполового трактов содержится фермент лизоцим
КФ. 3.2.1.17), оказывающий бактерицидное и фунгицидное дей-
твия. Лизоцим вырабатывается хрящевой тканью, слезными,
люнными и молочными железами, моноцитами крови и тканевыми
(акрофагами. Защитное действие лизоцима особенно выражено
ри местном иммунитете.
377
Эпителий желудочно-кишечного тракта проявляет барьерные
функции для микроорганизмов; этому способствуют также гидро-
литические ферменты, входящие в состав слюны, желудочного и
кишечного соков. Антибактериальное действие их настолько вели-
ко, что заражение через слизистую оболочку, особенно толстого
кишечника, буквально наполненного микроорганизмами, происхо-
дит крайне редко.
К гуморальным факторам неспецифического общего иммуните-
та относят бактерицидные вещества крови. Они выделены из лей-
коцитов (лейкины), эритроцитов (эритрин), но большая часть их
обнаруживается в сыворотке крови: нормальные антитела, комп-
лемент, пропердин, трансферрин и др.
Нормальные антитела — у-глобулины сыворотки крови индиви-
дуумов, не болевших и не подвергавшихся иммунизации. В срав-
нении с иммуноглобулинами их бактерицидная активность значи-
тельно ниже.
Комплемент был обнаружен как неспецифический фактор сы-
воротки крови, дополняющий литическое действие антибакте-
риальных антител (от лат. complementum — дополнение). Компле-
мент — сложная система сывороточных белков, обладающих фер-
ментативными свойствами, способная вызывать растворение, или
лизис, микробных и других клеток. Комплемент состоит из 9 компо-
нентов, обозначаемых Cl, С2, СЗ, ..., С9. Причем отдельные
фракции, в частности С1, неоднородны и включают несколько
субъединиц (Clq, Clr, Cis и Clt). Фракции комплемента, нахо-
дящиеся в обычных условиях в неактивном состоянии, могут ак-
тивироватья специфически комплексом антиген — антитело (клас-
сический путь активации) или неспецифически — липополисаха-
ридами и полисахаридами (альтернативный путь актива-
ции).
Классический путь активации комплемента начинается со
связывания С1 по меньшей мере с двумя Fc-фрагментами анти-
тел (см. разд. 8.3). После этого совершается каскад последова-
тельных реакций, в результате которых активируются все компо-
ненты системы комплемента вплоть до С9, и эти процессы, проис-
ходящие, например, на поверхности эритроцитов, сопровождают-
ся активацией гидролитических ферментов, что приводит к раз-
рушению клеточной мембраны, а затем и самих клеток.
Для альтернативного пути активации комплемента характер-
на укороченная цепь событий, начинающаяся с активации мик-
робными липополисахаридами и полисахаридами фракции СЗ
через систему пропердина (белкового компонента нормальной
сыворотки крови, способного образовывать с липополисахарида-
ми и полисахаридами комплексы, имитирующие комплексы анти-
ген— антитело). Именно этот путь обеспечивает быстрый и эф-
фективный неспецифический способ защиты против чужеродных
агентов. Липо- и полисахариды в данном случае являются индук-
торами фракции СЗЬ, которая используется для построения и
последовательного присоединения фракций С5—С9. Конечный
378
зультат альтернативного и классического путей активации
•мплемента одинаков.
Пропердин (от лат. pro и perdere — подготавливать, разру-
ать), имеющий прямое отношение к бактерицидной активности
шоротки крови и играющий значительную роль в неспецифиче-
:ой активации комплемента при соединении с полисахаридами и
шополисахаридами, представляет собой белок, состоящий из
*тырех нековалентно связанных субъединиц. По электрофорети-
ской подвижности пропердин (М = 22 кДа) соответствует у-
юбулиновой фракции сыворотки крови. Термостабильные бак-
рицидные вещества сыворотки крови (р-лизины) могут раство-
|ть патогенные микробы, в частности аэробные спорообразую-
ие бактерии.
Противомикробную активность сыворотки крови здоровых
ивотных и человека связывают также с действием трансферри-
I, присутствующего в р-глобулиновой фракции сыворотки крови
участвующего в транспорте железа. Трансферрин — гликопро-
ин (М — 90 кДа), содержащий 5% углеводов. Одна молекула
оансферрина связывает два иона Fe3+. Благодаря связыванию
елеза трансферрин обладает бактериостатической и фунгиста-
ической активностью in vivo и in vitro. При этом возникает де-
фицит свободного железа в среде и вследствие этого уменьшение
ферментативной активности (каталазы, пероксидазы) клеток
шкроорганизмов. В сыворотке крови здоровых людей имеется
акже фактор, называемый хлопьеобразующим, он способствует
юявлению из клеток С. albicans ростковых трубок и «комкова-
нию» клеток гриба, но не губит их.
Антифунгальную активность сывороток крови людей связы-
»ают еще с действием на гриб сывороточных липопротеинов и
J-глобулинов (против Rhizopus rhisopodiformis), а также а2-
уг-глобулинов (против Cr. neof ormans). Антикриптококковую
сгивность сывороточных Я2-глобулинов объясняют взаимодей-
вием полисахаридной капсулы гриба с С-реактивным а2-глобу-
IHOM.
К числу гуморальных неспецифических защитных факторов
1ри вирусных инфекциях относятся интерферон и вируснейтра-
щзующие ингибиторы. Продукция интерферона является уни-
версальной, эффективной и немедленной ответной реакцией лю-
5ых систем чувствительных клеток позвоночных животных на
шедрение вирусов. Наибольшее количество интерферона образу-
ется лимфоцитами. Он проявляет строгую видовую клеточную
специфичность в сочетании с подавлением репродукции большин-
ства известных вирусов человека и животных в клеточных куль-
турах и в организме. При естественной вирусной инфекции син-
•ез интерферона индуцируется вирусной нуклеиновой кислотой,
I в искусственных условиях — также с помощью ряда синтетиче-
:ких полинуклеотидов и двунитевых рибонуклеиновых кислот,
остойчивых к клеточным нуклеазам.
- Защитное действие интерферона направлено на прекращение
379
процесса трансляции вирусной мРНК и синтеза вирусного белка.
Предполагают, что подавление синтеза вируса осуществляется не
самим интерфероном, а индуцируемым им другим клеточным бел-
ком. При заражении вирусами лабораторных животных и при
естественном вирусном заболевании человека происходит регу-
лярное образование интерферона в местах репродукции возбуди-
телей (при введении осповакцины — в коже, вируса гриппа —
в легких, вирусов энцефалита — в мозгу). Отсюда интерферон
резорбируется и поступает в общую циркуляцию и распределяет-
ся по всему организму. Неспецифическая защита восприимчиво-
го организма с помощью интерферона дополняется и усиливает-
ся за счет температурного фактора вирусных ингибиторов крови
и барьерных жидкостей.
Повышенная температура тела способствует продукции интер-
ферона и усилению специфических реакций иммунитета. Кроме
того, репродукция большинства вирусов подавляется при 39...
40 °C (оптимум репродукции вирусов обычно соответствует
37 °C). Термолабильные ингибиторы ((3-ингибиторы) утрачивают
свою активность при 60...62 °C, умеренно термостабильные (а-
ингибиторы) инактивируются при 75 °C, устойчивые (у-ингиби-
торы) выдерживают нагревание до 100 °C. Активность термоста-
бильных ингибиторов связана с (3-липопротеинами и в отноше-
нии вируса гриппа обусловлена высокомолекулярными (а2-мак-
роглобулины) и низкомолекулярными (трансферрин-альбумин)
белками. Механизм действия ингибиторов сходен с действием
антител. Ингибиторы, так же как и антитела, соединяются не-
посредственно с вирусом, блокируют его рецепторы, в результате
чего вирус утрачивает способность фиксироваться на поверхно-
сти чувствительной клетки и проникать в нее.
Очень часто продукты распада патогенных и непатогенных
бактерий вызывают после парентерального введения повышение
температуры тела. Такие вещества называют пирогенами, они
оказывают лечебный эффект. Обычно это эндотоксины грамотри-
цательных бактерий, термостабильные вещества, являющиеся
липополисахаридами. Экзогенный пироген вызывает лихорадку
через 60...90 мин после введения или начала действия в организ-
ме. В этот период в крови отмечается уменьшение лейкоцитов
(лейкопения). При повторном введении организм становится не-
чувствительным к эндотоксину.
В отличие от экзогенных пирогенов известен также эндоген-
ный пироген, выделенный из зернистых лейкоцитов инфицирован-
ного организма. Это термолабильное вещество, вызывающее ли-
хорадку, длящуюся в пределах нескольких минут, и на его по-
вторное введение не возникает толерантности. По химическому
составу эндогенный пироген — белок с молекулярной массой
10...20 кДа. Он инактивируется при pH > 8, стабилизируется
при pH < 4, а также сульфгидрильными реагентами, блокирую-
щими SH-группы.
В воспалительных реакциях участвуют медиаторы — химиче-
380
;ие вещества, которые могут вызывать клинические проявления
,болевания и как гуморальные факторы неспецифически дей-
вовать на иммунологические реакции. К медиаторам воспали-
льных реакций относятся простагландины, гистамин и др.
ростагландины влияют на иммунологически опосредованное
•спадение, изменяя реакцию лейкоцитов, мигрирующих под
сиянием хемотаксических стимулов. Воспалительный процесс,
ызванный микроорганизмами, сопровождается расширением
местных артерий, капилляров и выделением гистамина, стимули-
ующего активность фагоцитоза, действующего на клеточные
•ецепторы и т. д.
Таким образом, разнообразные неспецифические клеточные и
уморальные факторы иммунитета взаимодействуют друг с дру-
ом, а также со специфическими компонентами иммунной систе-
[ы, что обеспечивает достаточную резистентность организма к
•азличным инфекционным агентам. Нарушение функций четко
координированной системы защиты хотя бы по одному из пока-
ателей повышает восприимчивость человека и животных к ин-
)екционным заболеваниям.
Специфические факторы защиты. Специфический иммунный
твет у человека и животных осуществляет иммунная система,
бладающая уникальной способностью распознавать множество
азнообразных микроорганизмов, других чужеродных агентов и
юлекул, называемых антигенами (см. разд. 8.2), и вырабаты-
ать в ответ на это распознавание специфические антитела и
енсибилизированные (от лат. sensibilis — чувствительный) лим-
юциты. Когда антиген распознается иммунной системой, это зна-
•ит, что он связываете^ с комплементарными к его простран-
твенной структуре антителами или подобными антителам ре-
цепторами поверхности лимфоцитов. Это связывание приводит к
ктивации лимфоцитов, выполняющих разнообразные эффектор-
ные функции в иммунитете в зависимости от того, к какой суб-
опуляции лимфоцитов они относятся. Субпопуляции лимфоци-
ов различают также по способности связывать на своей поверх-
ности лектины или активироваться ими.
В настоящее время известны две линии лимфоидных кле-
ок — Т- и В-лимфоциты, возникающие из кроветворных стволо-
вых клеток костного мозга. Первые дифференцируются под влия-
нием тимуса (от лат. Glandus thymus — зобная, или вилочковая,
железа); вторые—под влиянием фабрициевой сумки (бурсы)
у птиц или ее окончательно неустановленного аналога у млеко-
питающих (рис. 75). Популяции В- и особенно Т-льмфоцитов
Достоят в свою очередь из субпопуляций, которые можно разли-
чать по функциям, выполняемым данной субпопуляцией в им-
мунном ответе, или по поверхностным антигенам (для каждой
^стадии дифференциации клеток характерен свой уникальный
’Набор дифференцировб’чных антигенов).
С В-лимфоциты — предшественники плазматических клеток
(плазмобластов и плазмоцитов) — продуцентов иммуноглобулинов
381
Костный Кровь Центральные Кровь Периферические Кровь
мозг лимфоидные лимфоидные органы
органы
Рис. 75. Возникновение и развитие субпопуляций Т- и В-лимфоцитов
разных классов: М, G, А, Е, D. Субпопуляции Т-лимфоцитов
различаются по своим функциям. Клетки-помощники коопери-
руют с В-лимфоцитами при образовании гуморальных антител
против тимусзависимого антигена. Клетки-супрессоры регули-
руют активность специфического гуморального иммунного отве-
та, стимулируя или подавляя его. Эффекторные клетки клеточ-
ного иммунитета подразделяются на долгоживущие клетки памя-
ти (от англ. — memory) и клетки киллеры, или убийцы (от англ,
killer), обладающие цитотоксической активностью против чуже-
родных для организма клеток-мишеней и способные, выделяя
разнообразные медиаторы иммунного ответа (лимфокины), сти-
мулировать и вовлекать в борьбу против чужеродных антигенов
различные клетки крови и СМФ.
Реакция гиперчувствительности замедленного типа (ГЗТ)
развивается также благодаря участию Т-лимфоцитов, активация
которых антигеном сопровождается освобождением из них лим-
фокинов. При контакте с антигеном В- и Т-лимфоциты, наиболее
родственные своей рецепторной структурой к пространственной
структуре антигена, превращаются в юные, бластные формы,
делятся и, таким образом, быстро обеспечивают нужду организ-
ма в реагирующих с данным антигеном В- и Т-лимфоцитах.
Для обеспечения специфического иммунного ответа щи
большинства так называемых Т-зависимых антигенов необходим'’
взаимодействие (кооперация) различных субпопуляций ИКК : I •
В-лимфоцитов и макрофагов. Центральную роль в этой коопера
ции играют поверхностные антигены ИКК, кодируемые
главного комплекса гистосовместимости.
Как же происходит взаимодействие клеток в иммунном отве-
те? Попадающий в организм антиген сам по себе или чаще в ви-
382
е иммунных комплексов с предсуществующими антителами свя-
ывается с макрофагами (см. разд. 8.6). Иммунные комплексы
вязываются за счет имеющихся на макрофагах рецепторов для
с-фрагментов антител. Взаимодействие макрофагов с иммунны-
и комплексами активируют эти клетки, что выражается в секре-
йи макрофагами факторов, активирующих Т-клетки, называе-
мых интерлейкинами-1. Интерлейкины-1 в свою очередь стимули-
уют Т-клетки-помощники, которые выделяют факторы (в част-
ости, BRMF-фактор репликации и созревания В-клеток), спо-
обствующие пролиферации и дифференциации В-лимфоцитов;
(-лимфоциты размножаются и превращаются в антителосекре-
ирующие клетки. Однако для функционирования последова-
ельности макрофаг — Т-лимфоцит-помощник—В-лимфоцит не-
бходимо выполнение ряда условий: 1) Т-лимфоцит-помощник,
частвующий в кооперации, должен обладать антигенраспознаю-
хим рецептором против иммуногенной части антигена; 2) Т-лим-
юцит-помощник должен уметь распознать антигены на В-клетке
на макрофаге (кооперация эффективна только в случае их
дентичности); 3) В-клетка при распознавании антигена свя-
ывается своим иммуноглобулиновым антигенраспознающим ре-
ептором с соответствующей антигенной детерминантой, которая,
ак правило, не совпадает с участками антигена, распознавае-
мыми Т-клеткой.
Тем не менее взаимодействие трех типов ИКК, которое при-
одит к синтезу антител в результате действия антигенов, зна-
ительно сложнее. Так, Т-клетка-помощник, активирующая В-
летки, находится под контролем нескольких субпопуляций Т-
леток-супрессоров, которые в свою очередь контролируются по
ипу обратной связи факторами, образуемыми В-клетками и Т-
летками-помощниками. Супрессорные Т-клетки осуществляют
трицательную регуляцию Т-клеток-помощников, эффекторных Т-
имфоцитов, В-клеток и антителосекретирующих плазматиче-
ких клеток. Они специфически для данного антигена угнетают
ролиферацию и дифференциацию указанных выше клеток, тор-
мозят их эффекторные функции (биосинтез антител и различных
(акторов).
Таким образом, разные субпопуляции Т-лимфоцитов, выделяя
нтиген-специфические и неспецифические растворимые факторы,
существляют сложнейшую регуляцию ИКК или самостоятельно
ыполняют эффекторные, например цитолитические функции в
ммунитете.
Клеточный и гуморальный специфический противомикробный
иммунитет. Если первой линией защиты против инфекции служит
Ьеспецифическая противомикробная резистентность, то вторая
Линия защиты связана с появлением специфического клеточного
ja гуморального иммунитета. Клеточный иммунитет обеспечивает-
ся различными клеточными факторами, гуморальный — циркули-
рующими антителами. В целом иммунный ответ зависит от химиче-
ского состава и строения микробных клеток.
383
Доля участия клеточного и гуморального факторов в иммуни-
тете зависит от вида инфекции. Клеточный иммунитет — основа
резистентности при инфекциях, возбудители которых способны к
внутриклеточному паразитированию (туберкулез, бруцеллез, лис-
териоз, сальмонеллез). Имеются доказательства участия кле-
точных факторов в проявлениях иммунитета при туляремии,
коклюше, стафилококковой и менингококковой инфекциях. Анти-
тела, введенные животным, дают защитный эффект только на
ранних сроках заражения. Последующее развитие иммунитета
(особенно к высоковирулентным штаммам) зависит от клеточных
факторов. Даже при инфекциях, вызванных токсигенными воз-
будителями, возникающая устойчивость к заражению связана не
только с появлением антитоксинов, но и с развитием клеточного
иммунитета. Противоинфекционный иммунитет при висцеральных
микозах больше связывают не с активностью антител, накапли-
вающихся в крови у больных лиц и опытных животных, а с кле-
точными механизмами защиты. Специфическая резистентность к
возбудителям в подобных случаях обеспечивается главным обра-
зом Т-лимфоцитами. При клеточном иммунитете способность к
иммунной ответной реакции может быть передана другому ор-
ганизму только при помощи реактивных лимфоцитов, получен-
ных от предварительно сенсибилизированного донора. При гумо-
ральном им1цунитете эта способность может быть передана с по-
мощью сыворотки донора или популяции лимфоцитов, получен-
ных от сенсибилизированного донора. Гуморальный иммунитет
создается при помощи иммуноглобулинов-антител.
Тем не менее понятия клеточный и гуморальный иммунит<ч
относительны. Так, В-лимфоциты, составляющие основу гумо-
рального иммунитета, способны вырабатывать некоторые медиа
торы клеточного иммунитета; в отдельных случаях образование
медиаторов в В-клетках происходит более интенсивно, чем в Г-
клетках. С другой стороны, Т-клетки (хелперы, супрессоры) при-
нимают участие в образовании циркулирующих антител. Поэто-
му деление иммунитета на клеточный и гуморальный хотя и
удобно, но довольно условно в классификации иммунных реак
ций. Клеточный иммунитет обусловлен Т-лимфоцитами и макро-
фагами. Он развивается только при условии тесного взаимодеи
ствия этих клеток. Макрофаги нужны не только для инактивации
и расщепления возбудителя, их присутствие необходимо для син-
теза в лимфоцитах растворимых факторов (медиаторов) клеточ-
ного иммунитета (лимфокинов или интерлейкина-2), они являют
ся клетками-мишенями лимфокинов и сами активно вырабаты
вают медиаторы (монокины или интерлейкин-1), влияющие на
функцию других клеток, в том числе и тимфоцитов (табл ЗСо.
В ответ на микробные антигены образуются различные анти-
тела, обеспечивающие высокий уровень гуморального иммуните-
та. Одни антитела (агглютинины, лизины, опсонины) реагируют
с поверхностными антигенами бактерий, другие (антитоксины,
преципитины) нейтрализуют экзотоксины. Гуморальный иммуни-
384
Роль медиаторов Влияют на передвижение имму- нокомпетентных клеток, обеспечи- вая их локализацию вблизи места воздействия антигенного стимула (сенсибилизирующего агента) Активируют или подавляют функциональную активность имму- нокомпетентных клеток -
Группы медиаторов, участвующие в реализации Реакций ГЗТ • Иммуннорегу- ляторных меха- низмов ।
। Клетки- продуценты L . Лимфоциты Л А Т-лимфоциты Лимфоциты Макрофаги, Т- лимфоциты Т-лимфоциты Лимфоциты, макрофаги Лейкоциты Лимфоциты, макрофаги
М, кДа 12...25, 50...80 70 14, 70, 150 1,5...2,5 0,5 1 50...80 13, 85 25...30 30...50 10 1
Факторы Клеточный иммунитет Угнетающий миграцию клеток (MIF, ФУМ) Агглютинирующий макрофаги макрофагов эозинофилов нейтрофилов Кожной реактивности Активирующий макрофаги Активирующий лимфоциты Митогенные (бластогенные) Колониестимулирующий (CSF) Переноса (transfer factor) Угнетающие рост колоний, пролифера- цию клеток, синтез ДНК, стволовые клет- ки
Хемотаксиса
385
Продолжение табл. 36
Роль медиаторов Вызывают лизис клеток-мише- ней Влияют на интенсивность анти- телообразования (оказывают хел- периое и супрессорное влияние на иммунный ответ г——— Вызывают анафилактический шок, аллергический насморк и другие реакции ГНТ
Группы медиаторов, участвующие в реализации Цитотоксиче- ской (кнллерной) активности Т-лим- фоцитов Межклеточной кооперации при распознавании антигена и при запуске иммун- ных реакций Реакций ГНТ*
Клетки- продуцеиты Лимфоциты Т-Лимфоциты Клетки костно- го мозга Незрелые клет- ки В-ряда Тучные клетки, базофилы, ней- трофилы, тром- боциты, моноци- ты, макрофаги Лейкоциты, макрофаги
М, кДа 70...90, 35...50, 10...20 30...55 20...70 о со Малая Большая
Факторы Лимфотоксические, илн лимфотоксины Гуморальный иммунитет антигеиспеци- фические антигеннеспе- цифические Стимулирующий антителопродукцию (САП) Супрессорный медиатор костного мозга Гистамин, серотонин, кинины и др. Лизосомальные ферменты, катионные белки, гепарин и др.
Хелперные и супрес- сорные
5
ф
S
о
X
ч
о
о
Ьет, как правило, сочетается с появлением клеточной устойчиво-
кти, не зависящей от циркулирующих антител. С помощью раз-
личных клеточных реакций (розеткообразования, бласттрансфор-
иации, торможения миграции макрофагов и др.), широко исполь-
Ьуемых в настоящее время в клинической практике, установлено,
ито при ряде инфекций происходит подавление клеточного имму-
нитета. Снижение уровня Т-лимфоцитов и их функциональной ак-
тивности наблюдается при стафилококковой, пневмококковой,
Вгуберкулезной, менингококковой инфекциях; при некоторых вис-
церальных микозах — криптококкозе, гистоплазмозе, южно-аме-
мшканском бластомикозе, кокцидиоидозе.
| В механизме клеточных реакций инактивация возбудителя
|без помощи циркулирующих или фиксированных на фагоцитах
|антител может происходить в результате прямого действия кле-
|ток на объект либо с участием медиаторов клеточного иммуните-
та. Лимфокины, образующиеся под влиянием микробных антиге-
рое, действуют не только на макрофаги, но и на гранулоциты,
|фибробласты, Т- и В-лимфоциты. Фиксируясь на клеточных ре-
Гцепторах, лимфокины вызывают следующие явления, способст-
вующие иммунному ответу и развитию иммунитета: 1) стимули-
|руют или подавляют отдельные функции клеток (фактор тормо-
икения миграции макрофагов, хемотаксический фактор, фактор
[фагоцитоза и др.); 2) вызывают пролиферацию клеток или спо-
гсобствуют ей (митогенный фактор, факторы стимуляции синтеза
[ДНК и др.); 3) стимулируют дифференцировку клеток (факторы
[тимуса, фактор переноса и др.).
[ Полных сведений о структуре клеточных рецепторов для лим-
[фокинов пока не существует. Известно, что они отличаются от
[рецепторов для антител, являясь гликолипидами или гликопро-
теинами, и их специфичность связана с углеводной частью: для
[рецепторов фактора ингибирования миграции макрофагов — с
[L-фукозой; для рецептора фактора аггрегации макрофагов — с
[£)-галактозой; для рецепторов хемотаксического фактора — с L-
[фукозой и Л-рамнозой; для рецепторов фактора, активирующего
[миграцию макрофагов, -- с £)-маннозой.
F Бактериальные продукты в основном стимулируют развитие
[иммунитета, но некоторые возбудители (стафилококки, коклюш-
Гные бактерии) и их фракции могут подавлять иммунный ответ
[к неродственным антигенам. Это связано не только с токсиче-
скими свойствами таких продуктов, но и с выделением из лимфо-
цитов под их влиянием неспецифических супрессорных факт-
оров, которые способны подавлять как гуморальный, так и клеточ-
кный иммунитет.
8.2. ХИМИЯ АНТИГЕНОВ
[Понятие об антигенах. Долгое время считалось, что антигены
Цот греч. avTi—против, 'Yrjvrpo — порождать) при парентераль-
ном введении в организм, т. е. минуя пищеварительный тракт,
386
387
способны только вызывать образование специфических антител,
с которыми они могут реагировать in vivo и in vitro. Позднее
было показано, что иммунный ответ организма на антигены
заключается не только в выработке антител, но и в формирова-
нии состояния повышенной чувствительности замедленного типа
или в приобретении иммунологической толерантности (от лат.
tolerantia — терпение), характеризующейся утратой способности
иммунологически отвечать на данный антиген. Кроме того, какое-
либо вещество может быть антигенным для данного организма
лишь в том случае, если оно является чужеродным ‘для макроор-
ганизма. Таким образом, антигены — вещества, несущие призна-
ки генетически чужеродной информации, вызывающие при введе-
нии в организм его иммунную перестройку и специфически взаи-
модействующие с лимфоцитами и антителами in vivo и in vitro.
Иммунная перестройка организма в ответ на введение антигенов
может проявляться как накоплением в организме специфических
иммунокомпетентных клеток и антител, так и специфическим ос-
лаблением и подавлением иммунного ответа. Другими словами,
антигены способны in vivo и in vitro взаимодействовать с лимфо-
цитами и антителами, а при попадании в организм — «запустить»
процессы иммуногенеза (иммунологическая активность антиге-
нов) или вызвать состояние иммунологической толерентности
(толерогенная активность антигенов). При формировании специ-
фической иммунологической толерантности другие виды иммун-
ного ответа на данный антиген исключены.
Если антиген попадает в организм, предрасположенный, т. е.
имеющий наследственную склонность, к избыточному иммунному
ответу (гиперчувствительность), то его называют аллергеном,
а специфическое изменение состояния организма, являющееся
предпосылкой возникновения патологического иммунологического
процесса, — аллергией. Структура аллергенов в настоящее вре-
мя малоизвестна, поэтому их стандартизацию проводят по таким
малоинформативным показателям, как общий или белковый
азот.
Термин антиген в практической иммунологии несет в себе
двоякий смысл: с одной стороны — это очищенные от приме-
гомогенные вещества определенной химической природы (белки,
сложные полисахариды, липополисахариды, а также высокополя
мерные препараты нуклеиновых кислот); с другой (в собиратель
ном смысле) — это целые клетки, ткани или сложные их компе
ненты, содержащие большое количество отдельных антигенны
веществ. Так, если говорят, что в качестве антигена были испол;
зованы те или иные клетки микроорганизмов, имеется в виду, ч ’
речь идет о целом наборе антигенных веществ данных клекл
В зависимости от иммунологической активности антигены
подразделяют на полноценные антигены, гаптены и полугаптены.
Полноценные антигены индуцируют иммунный ответ организма и
специфически реагируют с иммунокомпетентными клетками и ан-
тителами. Гаптены (от греч. уалтщу — связывать, привязывать)
388
взаимодействуют in vivo и in vitro co специфическими антитела-
ми и лимфоцитами и могут вызывать иммунный ответ организма
ришь в случаях, когда они искусственно или естественно конъю-
гированы с другими крупными молекулами, выступающими в ро-
ди носителей гаптенов. Полугаптены при введении животным
даже в комплексе с молекулой носителя не индуцируют иммун-
ный ответ, а в серологических реакциях in vitro способны только
блокировать взаимодействие антител с полноценными антигена-
Ми или гаптенами (визуально не наблюдаемая реакция).
Существует несколько понятий, характеризующих вещество
как антиген: чужеродность, антигенность (антигенная специфич-
ность) и иммуногенность. Чужеродность необходима, чтобы ве-
щество выступало антигеном в отношении данного организма.
Например, белок плазмы лошади не будет для нее антигенен,
так как этот белок для лошади генетически не чужероден. Если
же белок лошадиной плазмы ввести кролику, для которого он
чужероден, то будет наблюдаться иммунный ответ. Значит, в
последнем случае белок плазмы лошади — антиген.
Антигенность, или антигенная специфичность, — способность
антигена взаимодействовать с антигенспецифическими рецепто-
рами Т- и В-лимфоцитов и с такими продуктами их жизнедея-
тельности, как антитела и антиген-специфические регуляторные
факторы. Это взаимодействие отличается уникальной специфич-
ностью, так что лимфоциты способны различать особенности
строения четвертичной и более низких структур молекул, осу-
ществляя распознавание различных антигенных детерминант,
т. е. химических групп, определяющих специфичность антигена.
Иммуногенность — способность антигена in vivo или in vitro
вызывать иммунный ответ (продукция антител, формирование
толерантности, накопление антиген-специфических Т-эффектор-
ных клеток). Детерминанты, обусловливающие антигенную спе-
цифичность молекул антигена, могут не совпадать с участками,
определяющими их иммуногенность. Иммуногенная активность
антигена зависит от его физико-химических свойств и от способ-
ности иммунизируемого животного отвечать на данный антиген.
По способности вызывать иммунный ответ антигены можно под-
разделить на сильные и слабые. Среди веществ определенной
химической природы наиболее активными иммуногенами, или
сильными антигенами, являются белки, хотя полисахариды, син-
тетические полипептиды и другие полимеры при определенных
условиях также могут становиться иммуногенами. Нуклеиновые
кислоты в чистом виде неантигенны, но, будучи искусственно
конъюгированными с белками, вызывают образование антител
при введении животным определенными способами.
Липиды и стероиды,неантигенны. Предполагают, что жирные
кислоты, составляющие основу липидов, не обладают достаточ-
ной жесткостью структуры молекулы.
Среди клеточных и корпускулярных антигенов хорошими
иммуногенами, как правило, выступают чужеродные эритроциты,
389
лейкоциты, тромбоциты, различные микроорганизмы. Иммуноген-
ность одного и того же антигена может быть неодинаковой при
введении животным разных видов, при разных способах иммуни-
зации, при изменении иммунизирующей дозы и других условиях.
Она зависит от молекулярной массы и структуры антигена, до-
ступности его гидролитическим ферментам макроорганизма
и т. д.
Достаточно высокая молекулярная масса — условие хорошей
иммуногенности антигенов. Например, если молекулярная масса
химического соединения ниже 10000 Да, то оно, как правило,
слабо иммуногенно. Большинство высокоиммуногенных белков
имеют молекулярную массу около 100 000 Да и выше. Иммуно-
генность низкомолекулярных антигенов можно повысить путем их
искусственной агрегации, адсорбции или присоединения к разно-
образным молекулам-носителям (к сорбентам или клеткам). По-
лисахарид декстран с молекулярной массой до 100 000 Да не
вызывает образование антител у людей, эта способность появ-
ляется у данного полимера лишь по мере дальнейшего увеличе-
ния его молекулярной массы. С уменьшением размеров и
молекулярной массы антигенов теряется индивидуальность их
строения, снижается чужеродность и иммуногенная активность.
Отсюда многие низкомолекулярные соединения, состоящие из
аминокислот или сахаров, не присоединенные к белкам, лишены
чужеродности и не являются иммуногенами.
Замечено, что чем сложнее строение молекулы ан-
тигена, тем он более иммуногенен. Эту закономерность можно
проследить, сопоставляя иммуногенность синтетических поли-
пептидов. Если полипептид построен из остатков только одной
аминокислоты, он, как правило, слабо иммуногенен. Если же по-
липептид состоит из остатков двух или трех аминокислот, он
приобретает иммуногенные свойства. Наличие в синтетических
полипептидах ароматических аминокислот (например, тирозина)
также обеспечивает иммуногенность молекулы. Доказано, что
способностью вызывать образование антител в большей степени
обладают вещества, имеющие на поверхности заряженные
группировки, чем антигены, лишенные заряда.
Иммуногенность веществ повышается, если сделать их струк-
туру более жесткой. Данное положение можно проиллюстри-
ровать на примере желатина, в состав которого входит много
остатков глицина — аминокислоты, не имеющей боковых групп в
a-положении. Именно поэтому в структуре желатина отсутствуют
ригидные боковые цепи и связи, молекула желатина не обладает
прочным внешним строением и является слабым иммуногеном.
Если стабилизировать структуру желатина искусственным вве-
дением в его молекулу циклических аминокислот (тирозина,
триптофана или фенилаланина), иммуногенность этого белка уве-
личится.
Одно из условий иммуногенной активности антигенов —
«перевариваемость» молекулы антигена гидролитиче-
390
|скими ферментами клеток макроорганизма. Практика показы-
вает, что отсутствие у животных ферментов, способных расщеп-
лять те или иные полимеры, делает их неиммуногенными. Так,
йиолекулы, построенные из остатков правовращающих амино-
кислот, неиммуногенны, они выводятся из организма во много
|раз медленнее, их содержание в лимфоидных органах животных
к сотни и тысячи раз выше, чем содержание полимеров левовра-
кцающих аминокислот, для которых известны расщепляющие их
ферменты. Деградация антигенов в фагосомах макрофагов зна-
чительно повышает их иммуногенность. Извлекаемые из макро-
фагов деградированные под влиянием лизосомальных ферментов
^антигены и их комплексы с РНК оказались особенно высоко ак-
тивными иммуногенами по сравнению с нативными антигенами.
। Сила иммунного ответа зависит от вводимой дозы антиге-
[на. Она возрастает пропорционально увеличению дозы. Однако
кта закономерность сохраняется в определенном диапазоне доз.
?При введении слишком большой дозы антигена у животных утра-
чивается способность к образованию антител, возникает так на-
зываемый «иммунологический паралич», который исчезает в те
же сроки, что и состояние иммунитета.
Способ введения антигенов также влияет на проявление
иммунного ответа, сила которого тем больше, чем дольше задер-
; живается антиген в месте введения и пребывает в организме,
пролонгируя иммунное раздражение. Применение в качестве
. иммунизирующих агентов антигенов в смеси с такими депонато-
, рами и наполнителями, как гидроксид алюминия, фосфаз алюми-
ния, водно-масляные эмульсии и др., вызывают более медленно
развивающуюся, но в то же время более сильную иммунную
реакцию. Введение антигена с депонаторами позволяет увеличить
его разовую дозу, так как постепенное поступление антигена из
• места введения уменьшает вероятность формирования иммуноло
гической толерантности из-за избытка антигена.
Один и тот же организм может быть высокореагирующим на
один антиген и низкореагирующим на другой. Наоборот, иммун-
ный ответ на один и тот же антиген может быть разной высоты у
организмов разных генотипов в зависимости от его набора генов
иммунного ответа (1г-генов). А это значит, что если по каким-то
эволюционным причинам большая часть человеческой популя-
ции несет гены слабого типа в гомозиготном положении по отно-
шению к какому-либо антигену, например к вирусу гриппа, то
никакая вакцинация этим вирусом или выделенными из него
фракциями не обеспечит развитие иммунитета у большинства
лиц. Действительно, если по своему генотипу организм не в со-
стоянии развить иммунный ответ против указанного вируса или
его продуктов, то никакая вакцина, приготовленная из этого ви-
руса путем его ослабления или извлечения из него высокоочи-
щенных отдельных субстанций, не сможет стимулировать им-
мунный ответ. Принцип конкретности иммунного ответа, сформу-
лированный акад. Р. В. Петровым, породил новый путь поиска —
391
путь фенотипической коррекции, т. е. нахождения способов пре-
вращения генетически низкореагирующих особей в высокореаги-
рующие.
В области фенотипической коррекции существуют два конку-
рирующих направления. Оба связаны с созданием искусственных
антигенов нового типа. Принципиальная сущность построения та-
ких антигенов состоит в том, чтобы в одной искусственной моле-
куле совместить два функциональных компонента. Один компо-
нент — антиген, против которого надо создать иммунитет, вто-
рой — несущая часть комплексной молекулы, которая могла бы
обеспечить фенотипическую коррекцию, т. е. обойти генный конт-
роль реакции со стороны Ir-генов. Задача вполне реальная, ибо
генный контроль реализуется через Т-систему лимфоцитов, а ан-
титела — главный защитный аппарат — вырабатываются В-лим-
фоцитами. Придание искусственной антигенной молекуле способ-
ности миновать Т-лимфоцитарный контроль или введение в моле-
кулу такого компонента, который бы замещал вспомогательную
функцию Т-клеток, может обеспечить фенотипическую коррек-
цию.
Направление по созданию искусственных антигенов нового
типа наиболее интенсивно разрабатывается М. Села с сотр.
(США) и коллективом советских ученых, руководимых Р. В. Пет-
ровым и В. А. Кабановым.
Принцип Села, по его собственному выражению, — копирование природы
не in toto (от лат. in toto—в целом), а частями. В связи с этим ведется рас-
шифровка структуры естественных белковых и полисахаридных антигенов,
ресинтез их частей, ответственных за антигенную специфичность, и сборка этих
частей на единой несущей молекуле. Несущей молекулой могут быть естествен-
ные белковая и полисахаридная молекулы или построенная по их подобию
искусственная.
Принцип Р. В. Петрова и В. А. Кабанова иной, он предполагает реализа-
цию двух условий: во-первых, комплексирование антигенной детерминанты с не-
сущей частью из так называемых неприродных полиэлектролитов, т. е. в полном
смысле искусственных макромолекул; во-вторых, использование таких несущих
макромолекул, которые обеспечивают стимуляцию иммунного ответа даже у ге-
нетически низкореагирующих особей. Этот принцип возник из исследований влия-
ния простейших полианионов, поликатионов и их сополимеров на иммунный
ответ. Было показано, что поли-4-винилпиридин и его производные, несущие
радикалы разной длины, обладают способностью замещать некоторые функции
Т-лимфоцитов и стимулировать иммунный ответ у генетически низкореагирую-
щих особей. Именно с этими искусственными молекулами были соединены гап-
тенная (тринитрофенильная) или антигенная (альбуминовая) структура. В пер-
вом случае был получен искусственный антиген, при введении которого живот-
ным развивался тимусзависимый иммунный ответ. Во втором случае было полу-
чено 50...100-кратное усиление иммуногенности. Существенно отметить, что
альбумин — слабый антиген и для стимуляции иммунного ответа против него
требуется многократная иммунизация с применением стимуляторов типа стиму-
лятора Фрейнда, представляющего собой смесь водного раствора антигена
(убитых микобактерий туберкулеза) с минеральным маслом и ланолином. Соз-
данная комплексная структура стимулировала выработку антител при однократ-
ном введении и без всяких дополнительных стимуляторов. Специальные иссле-
дования показали, что чем более прочен комплекс альбумин-полимер, тем он
более иммуногенен. Ковалентное соединение молекул дает очень высокий эф-
фект.
392
Толерогенная активность антигенов. Полисахаридные и бел-
ковые антигены при введении в больших количествах могут вы-
зывать у животных состояние специфической иммунологической
толерантности. Это явление обнаружил впервые Л. Д. Фелтон
(1949), иммунизируя мышей пневмококковым полисахаридом.
Оказалось, что инъекция 0,5 мкг полисахарида иммунизирует
мышь, т. е. заставляет ее производить специфические антитела,
а инъекция значительно больших доз (0,5...5,0 мг) не только не
иммунизирует, но даже на долгие месяцы делает животное не-
способным иммунизироваться (к данному типу пневмококков).
Фелток с сотр. назвали этот феномен иммунологическим пара-
личом.
Все формы толерантности независимо от того, какими антиге-
нами (толерогенами) они вызваны, сходны и являются след-
ствием перегрузки антигеном антителообразующих клеток. Толе-
рогены — плохо перевариваемые белки. Так, белок жгутиков бак-
терий флагеллин в больших дозах вызывает у животных состоя-
ние иммунологической толерантности. Опыт показывает, что то-
лерогенность того или иного вещества связана с наличием в его
молекуле линейно расположенных и периодически повторяющих-
ся идентичных антигенных детерминант. Полноценные антигены
могут утратить иммуногенность и стать толерогенами под влия-
нием некоторых физико-химических или биологических воздей-
ствии
(ультрацентрифугирование, ферментативное расщепление,
фракционирование с помощью перитонеальных макрофагов).
В результате этих
фракции
белковых
воздействий образуются
антигенов (альбумина,
низкомолекулярные
миелина), которые
при введении животным вызывают у них состояние иммунологи-
ческой толерантности к нативным антигенам. Толерогенная ак-
тивность антигенов влечет за собой блокаду продукции антител,
подавление
клеточных реакций
иммунитета или ингибирование
этих процессов, вместе взятых.
Специфичность антигенов. Специфичность взаимодействия
антител или иммуноглобулиновых рецепторов клеток с антигеном
обусловлена их высоким сродством (комплементарностью) к оп-
ределенным участкам молекулы антигена, называемым детерми-
нантными участками или антигенными детерминантами. Число
детерминант на молекуле антигена меняется в забисимости от
размера молекулы и сложности ее строения. Чем больше детер-
минант имеет данный антиген, тем больше молекул антител он
может связать и тем выше его валентность. Валентность анти-
гена возрастает пропорционально молекулярной массе антигена.
Так, валентность сывороточного альбумина (М = 70 кДа) рав-
няется 6, дифтерийного токсина (М = 70 кДа) —
лина
230.
(М
650 кДа)
40, а гемоцианина (М =
8, тиреоглобу-
6 500 кДа) —
При иммунизации одним и тем же антигеном животных одно-
го или разных видов в качестве детерминант могут выступать
различные участки молекул антигена, так как каждый индиви-
393
дуум отличается способностью иммунокомпетентных клеток спе-
цифически распознавать лишь те или иные антигенные детерми-
нанты. В динамике иммунного ответа могут вырабатываться
антитела против все новых и новых детерминантных участков
той же молекулы антигена. Эти вначале скрытые детерминанты
выявляются у антигена в результате его деградации в организме
под влиянием клеточных ферментов. Поэтому результаты оценки
валентности антигенов с помощью иммунных сывороток, получен-
ных от животных разных видов на различных этапах иммуниза-
ции, могут значительно варьировать.
Основы учения о структуре и свойствах антигенных детерми-
нант заложил в начале XX в. К- Ландштейнер своими исследова-
ниями антигенных свойств низкомолекулярных гаптенов, присое-
диняемых с помощью диазосвязи к лизиновым, тирозиновым и
гистидиновым остаткам молекул белков. С помощью этих экспе-
риментов была установлена роль гаптена и белка-носителя в
индукции иммунного ответа. Кроме того, были охарактеризованы
особенности строения и размеры детерминантных участков анти-
генов, обеспечивающих их специфическое взаимодействие с анти-
телами или рецепторами иммунокомпетентных клеток. Ландштей-
нер и его последователи путем перекрестных иммунных реакций,
применяя большое количество гаптенов, белков-носителей и им-
мунных сывороток против них, а также с помощью задержки
серологических реакций гаптенами показали, что антигенными
детерминантами белковых молекул являются олигопептиды из
5...8 аминокислот, а полисахаридных — олигосахариды из 5...
7 остатков моносахаридов. У разных антигенов при взаимодей-
ствии с иммунными сыворотками разного видового происхожде-
ния размер антигенных детерминант может изменяться. У ди-
нитрофенолполилизина, например, антигенная детерминанта об-
разована динитрофенолом, присоединенным к трем остаткам ли-
зина. Антигенная специфичность белка вируса табачной мозаики
(одного из наиболее хорошо изученных белковых антигенов)
определяется его С-концевым трипептидом (Тре-Арг-Ала), хотя
иммунные сыворотки некоторых кроликов, морских свинок и мы-
шей реагируют только с С-концевыми полипептидами из 5, 6 и
даже 7 аминокислот. Поданным Е. А. Кэбота, антигенные'детер-
минанты имеют размеры, близкие к 3,4 X 1,2 X 0,7 нм. Таким об-
разом, антигенными детерминантами являются определенные
участки полипептидной или полисахаридной цепочки антигена,
обладающего специфичной первичной структурой молекулы.
Специфичность антигенов может быть связана с особенно-
стями вторичной, третичной и четвертичной структур молекул
белка.
Антигены естественного происхождения могут иметь детерми-
нанты, специфические для представителей определенного вида
(видовая специфичность)-, группы (групповая специфичность,
обусловливающая различия среди особей одного вида организ-
мов); типа (типоспецифичность). Типоспецифичность — понятие,
394
; чаще всего имеющее отношение к микробным видам. Например,
: пневмококки по своим полисахаридным антигенам делятся на
типы (серовары), количество которых насчитывает свыше 80.
Возбудители ботулизма по характеру образующегося токсина
подразделяют на типы: А, В, С, D и Е. Антигены, благодаря
которым разные особи одного вида различаются между собой,
называют изоантигенами. Некоторые антигены могут обладать
детерминантами, характерными только для данного индивидуума.
У антигенов животного происхождения удается также различать
специфичности, характерные для клеток разных органов (орган-
ная специфичность) и для отдельных субклеточных структур
(ядро, митохондрии, микросомы и др.) разных клеток (органоид-
ная специфичность).
В некоторых случаях идентичные антигенные детерминанты
могут быть обнаружены у антигенов клеток организмов, филоге-
нетически далеко стоящих друг от друга. В таких случаях приня-
то говорить о гетероспецифичности и о гетероантигенах. Приме-
ром такого рода антигенов может быть антиген Дж. Форсмана,
встречающийся в составе гликолипидов клеточных мембран и
гликопротеинов различных жидкостей любого организма и вызы-
вающий при введении кролику образование антител, гемолизи-
рующих бараньи эритроциты. Антиген Дж. Форсмана представ-
лен в эритроцитах овец, лошадей, собак, кошек и ряда других
животных, а также в клетках растений (шпинат) и у микроорга-
низмов разных видов (сальмонеллы, шигеллы, пневмококки,
стафилококки, дрожжи и др.). К гетероантигенам можно отне-
сти и широко применяемый в настоящее время для идентифика-
ции антител протеин А стафилококков.
Если антигенная специфичность обусловлена той или иной
гаптенной группировкой, употребляют понятие гаптеноспецифич-
ность. В настоящее время в качестве гаптена часто используют
динитрофенильную группировку, которую присоединяют к белко-
вым или липополисахаридным носителям, получая так называе-
мые конъюгированные антигены, обладающие новой иммуноло-
гической специфичностью. Гаптеноспецифичность могут приобре-
тать белки, конъюгированные с лекарственными веществами,
выступающими в роли гаптенов. Этим могут объясняться различ-
ные лекарственные аллергии, в том числе аллергические реакции
на антибиотики, которые сами по себе неантигенны.
Антигены — индукторы иммунного ответа. Полифункцио-
нальные антигены, способные вызывать гуморальный иммунный
ответ без участия в нем Т-клеток («помощников»), были названы
тимус-независимыми, или TI-антигенами (от англ, independent —
независимый).
Тимус-независимые антигены отличает многовалентность,
т. е. многократная повторяемость в молекуле одних и тех же ан-
тигенных детерминант, достаточно высокая молекулярная масса,
способность выступать в качестве поликлональных митогенов
В-лимфоцитов и способность активировать СЗ компонент компле-
395
мента. TI-антигены подразделяют на две категории: TI-1 и TI-2.
К TI-1 антигенам отнесены конъюгаты, в которых гаптены
конъюгированы с липополисахаридом микробов кишечной груп-
пы, Brucella abortus, водорастворимым экстрактом Nocardia.
К Т1-2 отнесены конъюгаты, приготовленные на основе капсуль-
ных полисахаридов пневмококка-Ш, фиколла, полиакриламид-
ных частиц, декстрана. TI-1 и TI-2 антигены способны взаимо-
действовать с различными популяциями В-лимфоцитов.
Антигены, при взаимодействии с которыми В-клетки для
дифференцировки в антитела-секретирующие клетки нуждаются
в кооперации с Т-лимфоцитами, называют тимус-зависимыми
или TD-антигенами (от англ, dependent — зависимый). Установ-
лено, что молекула TD-антигена для индукции гуморального им-
мунного ответа должна, как минимум, быть бифункциональной
и иметь: а) участок взаимодействия с антиген-специфическим
рецептором В-клетки (антиген-специфическая детерминанта гап-
тена, или гаптенная группа); б) участок взаимодействия с Т-
клеткой (детерминанта носителя). Именно носитель обеспечивает
кооперацию В- и Т-клеток в иммунном ответе В-лимфоцитов на
TD-антигены. Моновалентные антигены, как, например, гаптен-
ные группировки динитрофенола, тринитрофенола, фосфорилхо-
лина, пенициллоила и др., используемые для иммунизации без
носителя, обычно не вызывают иммунного ответа. Однако конъю-
гация их с белковым, полипептидным иммуногеном или с корпус-
кулярными носителями позволяет получать против этих гаптенов
В-клеточный ответ.
При изучении особенностей структуры антигенов, обеспечи-
вающей кооперацию Т- и В-лимфоцитов, выявлена важная роль
участков молекулы, связывающих отдельные детерминанты. Так,
конъюгированные бифункциональные антигены вызывали гумо-
ральный иммунный ответ, если две их детерминанты были ригид-
но связаны полипептидом из десяти аминокислот. Уменьшение
длины связки или замена ее более гибким звеном снижает
эффективность кооперации Т- и В-лимфоцитов и продукцию соот-
ветствующих антител. Бифункциональные антигены с гибкой
связью между идентичными детерминантами стимулировали про-
лиферацию лимфоцитов в 2... 15 раз более активно, чем антигены
с ригидной связью. По-видимому, связанные лабильно идентич-
ные детерминанты антигена могут взаимодействовать с несколь-
кими тождественными рецепторами той же лимфоцитарной клет-
ки, усиливая ее митотическую активность.
Белковые антигены разного происхождения обычно вызывают
как клеточный, так и гуморальный иммунный ответ. Химические
модификации антигенов могут изменить их способность стимули-
ровать тот или иной ответ. Например, сывороточные белки, на-
груженные остатками тринитрофенола, ацетильными или этокси-
метилен-фенил-азолоновыми группами, сохраняют способность
индуцировать у животных клеточные реакции гиперчувствитель-
ности замедленного типа, но утрачивают способность вызывать
396
; образование антител. Карбоксилирование лизоцима повышает
способность его вызывать у животных клеточный и снижать гу-
моральный иммунный ответы и т. д.
Таким образом, иммуногенность, способность индуцировать
клеточный или гуморальный иммунный ответ зависят от особен-
ностей и физико-химических свойств антигена, размеров его мо-
лекулы, характера, количества и локализации антигенных детер-
минант в молекуле антигена. Среди различных антигенных детер-
минант одни из них определяют взаимодействие антигена с ре-
цепторами специфических иммунокомпетентных клеток, стимули-
руя пролиферацию ИКК. Другие детерминанты помогают Т- и В-
клеткам удерживаться на одной и той же молекуле антигена и
кооперироваться. В результате такой кооперации Т-лимфоциты
дифференцируются в клетки-эффекторы иммунного ответа. Третьи
детерминанты антигена выступают структурами, с которыми мо-
гут взаимодействовать макрофаги; в результате кооперации с
лимфоцитами у макрофагов активируется фагоцитарная и био-
химическая активность. Искусственное изменение заряда, блока-
да детерминант антигена позволяют увеличивать либо уменьшать
активность антигена и таким образом при введении измененного
антигена избирательно стимулировать (подавлять) в организме
клеточный и гуморальный иммунный ответ.
Антигены микробного происхождения достаточно сложны для
моделирования, так как их строение во многом остается неизу-
ченным. К тому же антигенный состав и свойства микроорга-
низмов, циркулирующих в природе, подвержены заметным изме-
нениям. Так, штаммы стафилококков, выделенные в 50-х годах и
используемые для приготовления вакцин, обладали чувствитель-
ностью к большинству антибиотиков, в то время как штаммы,
выделенные в конце 70-х годов, приобрели резистентность к ним.
У таких штаммов возникли дополнительные белковые антигены,
не имеющие общих детерминант с антигенами чувствительных
штаммов.
У микроорганизмов антигены связаны с клеткой (эндоанти-
гены) или выделяются ею во внешнюю среду (экзоантигены).
Среди эндоантигенов различают антигены жгутиков (Н-антиге-
ны), ресничек, капсульные антигены (К-антигены), антигены кле-
точной стенки (О-антигены), внутриклеточные антигены. Среди
экзоантигенов — экзотоксины, гемолизины, фибринолизины, фер-
менты (гиалуронидаза, протеиназы и др.). Поверхностные эндо-
антигены (жгутиковый, капсульный и клеточной стенки) харак-
теризуются большей антигенностью, чем внутриклеточные. Имму-
ногенность биополимеров, полученных из микробных антигенов,
после выделения и очистки значительно ослабевает.
Жгутиковые Н-антигены (от нем. hauch—дыхание)—тер-
молабильные белковые антигены, в которых носителем антиген-
ности является флагеллин. При ферментативном расщеплении
он распадается на фрагменты А, В, С и D. Молекулярная масса
фрагментов — 4,5...18 кДа, и расположены они в молекуле фла-
397
геллина в последовательности BACD. Фрагмент А обладает наи-
большей иммуногенностью.
Капсульные К-антигены, разделяющиеся по чувствительности
к температуре на А- (наиболее термостабильные), В- (менее тер-
мостабильные) и L- (термолабильные) антигены, в большинстве
случаев состоят из кислых полисахаридов, в состав которых
входят уроновые кислоты, и частично из белков. Капсульные
антигены изучены у бактерий кишечно-тифозной группы, клеб-
сиелл, пневмококков. У сальмонелл среди капсульных антигенов
выделяют Vi-антиген — полимер N-ацетиламиногексуроновой кис-
лоты, называемый также антигеном вирулентности.
Антигены клеточной стенки. Это прежде всего О-антиген
(от нем. ohne hauch — без дыхания), локализующийся в клеточ-
ной стенке грамотрицательных бактерий семейства Enterobacte-
riaceae. О-антиген представляет собой комплекс липидов (в ос-
новном фосфолипидов), белков и углеводов, обозначаемый как
эндотоксин. По современным представлениям, белок обеспечи-
вает иммуногенность О-антигена, липид отвечает за острые не-
специфические токсические эффекты, а полисахарид определяет
антигенную специфичность. Полисахарид и липид относительно
прочно связаны и образуют так называемый липополисахарид
(ЛПС) (см. гл. 3).
Внутриклеточные антигены — это антигены цитоплазма-
тических мембран, цитоплазмы, рибосом, митохондрий, ядра и
др., определяющие органоидную специфичность. Антигены цито-
плазмы — обычно белки, антигены ядра — нуклеиновые кислоты.
В состав антигенов цитоплазматической мембраны входят бел-
ки, липопротеины, гликолипиды. Для стрептококков группы D
характерна локализация на цитоплазматической мембране груп-
поспецифического полисахарида, состоящего из углеводного ядра
с полисахаридными боковыми цепями, несущими сильную имму-
ногенную группировку. Внутриклеточные антигены наиболее изу-
чены у грамположительных микроорганизмов (стрептококков,
стафилококков и др.).
Бактериальные белковые токсины относят к экзотоксинам,
которые обладают антигенными свойствами. Химическая структу-
ра токсических групп окончательно не выяснена. Большая часть
токсинов — ферменты, обладающие протеолитической, липолити-
ческой или коагулирующей активностью. Для стрептококков
группы А, например, известно порядка 20 экзотоксинов (экзо-
энзимов): стрептолизины О и S, гиалуронидаза, протеаза, А-,
D-стрептодорназы, стрептокиназа, рибонуклеаза, липопротеаза,
глюкуронидаза и др. (см. гл. 7).
Среди антигенов микроорганизмов выделяют еще протектив-
ные антигены, представляющие собой группу иммунологически
активных веществ патогенных микроорганизмов, включающую
макромолекулы различной химической природы. Их объединяет
одно общее свойство — способность вызывать продукцию за-
щитных антител, предохраняющих макроорганизм от развития
398
патологического процесса. По локализации протективные анти-
гены включают эндо- и экзоантигены. Они локализуются либо
на поверхности клетки (клеточная стенка, клеточная мембрана),
например у коклюшных микробов, стрептококков групп А и В,
реккетсий, либо в цитоплазме у стафилококков.
Антигены вирусов. Антигенный состав вирусов сложен и за-
висит от структуры вирусов. Вирусы, как правило, — сильные
иммуногены. Носителями их антигенности чаще всего высту-
пают белки и комплексы белков с другими соединениями. У ви-
русов различают антигены, имеющие первостепенное значение
для формирования активного иммунитета (поверхностные анти-
гены) , и антигены, играющие роль в патогенезе инфекции, кото-
рые являются чужеродными, аллергенными и нередко токсиген-
ными веществами (внутренние антигены). Те и другие являются
вирусоспецифическими антигенами. К вирусоспецифическим по-
верхностным антигенам вируса гриппа относятся гликопротеин
гемагглютинин (Л4 ~ 150 кДа) и фермент нейраминидаза
(М ~ 250 кДа). Эти антигены у вирусов гриппа типа А претерпе-
вают выраженные изменения. Внутренний (стабильный) антиген
вируса гриппа представляет собой рибонуклеопротеин.
Кроме вирусоспецифических антигенов у вирусов формируют-
ся антигены клетки-хозяина — клеточноспецифические антигены
вириона. Они образуются во время созревания и «отшнуровыва-
ния» вируса из ядра и клеточной мембраны. Способность вирусов
включать в структуры своих наружных оболочек антигенные
компоненты клеток, в которых они репродуцируются, является
особенностью вирусов, отличающей их от бактерий и всех других
живых существ. В клетках, инфицированных вирусами, могут
возникать и новые антигены, отличающиеся как от антигенов са-
мого вируса, так и от антигенов незараженных клеток. Такие анти-
гены называют вирусиндуцированными антигенами, они харак-
теризуются специфичностью, определяемой вирусом, а не видовой
принадлежностью клеток. Примером вирусиндуцированного анти-
гена может служить антиген Томсена — Фриденрейха, открытый
впервые О. Томсеном (1927) в эритроцитах, инфицированных
нейраминидазоактивными бактериями. Этот антиген возникает
как новый и чужеродный для организма в наружных оболочках
эритроцитов, пораженных вирусами гриппа и парагриппа, обла-
дающими нейраминидазной активностью. В результате от муко-
протеиновых веществ наружных оболочек инфицированных эрит-
роцитов отщепляется нейраминовая кислота, наступает инакти-
вация определенных групповых антигенов, в структуру детерми-
нанты которых входит нейраминовая кислота.
Вирусиндуцированные антигены в зависимости от времени
появления в клетке хозяина, локализации и других свойств име-
ют разные названия: ранние, неструктурные, ядерные, поверх-
ностные, трансплантационные и т. д. Механизм их возникновения
изучен пока недостаточно. Поскольку многие из этих антигенов
возникают на самых ранних этапах вирусной инфекции, их мож-
399
но рассматривать как вещества-предшественники, необходимые
для запуска синтеза компонентов вируса. Вирусиндуцированные
антигены имеют большое значение для иммунитета, так как, бу-
дучи чужеродными для организма, они стимулируют возникнове-
ние и развитие клеточных и гуморальных защитных реакций.
8.3. ХИМИЯ АНТИТЕЛ
Антитела — это гликопротеины, образующиеся в ответ на введе-
ние антигена. Они относятся к классу иммуноглобулинов, обла-
дают общностью строения и отличаются по способности связы-
вать различные антигены. Их основная структурная единица —
мономер, состоящий из двух одинаковых тяжелых Н (М 50...
77 кДа) и двух одинаковых легких L (М = 22 кДа) полипептид-
ных цепей (рис. 76). Легкие цепи бывают двух типов: х и X. Тя-
желые цепи относятся к пяти классам: у, р, а, е, б; они опреде-
ляют классовую принадлежность антител (иммуноглобулинов
Рис. 76. Строение мономера иммуноглобулинов (1g):
1—легкая цепь (х, А); 2 — активный центр антитела; 3— тя-
желая цепь (у, ц, а, е, 6); Vl, Cl — домены легкой цепи; Vh,
Сн1, Сн2, СнЗ — домены тяжелой цепи; Fab, Fc, Fd, Fv —
фрагменты молекулы иммуноглобулинов (схема)
400
Таблица 37. Иммуноглобулины* человека
Обозначения, принятые
в настоящее время
Сокращенное
обозначение
Старое обозначение
Иммуноглобулин G
Иммуноглобулин А
Иммуноглобулин М
Иммуноглобулин D
Имунноглобулин Е
IgG
IgA
IgM
IgD
IgE
y-G-Глобулин, 78-у-глобулин
•у-А-Глобулин, 02-А-глобулин
•у-М-Глобулин, 198-у-глобулин
Реагины, IgND
* Термин введен впервые Дж. Ф. Хереманс (1959).
Ig): IgG, IgM, IgA, IgE и IgD. В табл. 37 приведена современ-
ная номенклатура иммуноглобулинов человека, рекомендованная
Всемирной организацией здравоохранения (ВОЗ).
Антитела синтезируются в значительных количествах лимфоид-
ными плазматическими клетками, а обнаруживаются в сыворотке
крови и продуктах секреции различных желез. Они главным обра-
зом определяют гуморальный иммунный ответ, но небольшая часть
молекул антител прочно прикрепляется к мембранам поверхности
таких иммунокомпетентных клеток, как лимфоциты й макрофаги,
и тогда эти иммуноглобулины играют роль антигенных рецепторов,
имеющих существенное значение в клеточном иммунитете.
Первые структурные исследования антител были затруднены
в связи е их физико-химической гетерогенностью. Поэтому боль-
шинстводанных о структуре иммуноглобулинов было получено при
исследовании так называемых миеломных белков, выделенных из
сыворотки крови преимущественно мышей, крыс и человека, по-
раженных миеломной болезнью, характеризующейся генерализо-
ванной неопластической пролиферацией плазматических клеток,
образующих патологически повышенное количество монокло-
нальных Ig или их фрагментов. Миеломные белки — удобная мо-
дель для структурного изучения антител, так как они представ-
ляют собой структурно-гомогенные антитела, вырабатываемые
опухолевыми плазматическими клетками — потомками одной зло-
качественно-перерожденной клетки.
При анализе аминокислотной последовательности полипептид-
ных цепей иммуноглобулинов оказалось, что в каждой цепи су-
ществуют участки длиной около ПО аминокислотных остатков,
обладающих высокой степенью подобия первичной и простран-
ственной структур, стабилизированных дисульфидной связью.
Такие участки цепей иммуноглобулинов были названы доменами.
Каждая из легких цепей (к и Z) состоит из двух доменов (около
220 остатков), а тяжелые у-, а-цепи — из четырех доменов, ц-,
б- и е-цепи — из пяти доменов. При сравнении первичной струк-
туры антител, относящихся к одному и тому же классу и типу,
оказалось, что они обладают практически одинаковой аминокис-
лотной последовательностью, за исключением N-концевых доме-
401
нов в тяжелой и легкой цепях. Эти домены были названы вариа-
бельными (V) в отличие от других, так называемых константных
(С). Между первым Сн1 и вторым Сн2 константными доменами
тяжелой цепи есть так называемый шарнирный участок полипеп-
тидной цепи, по своей структуре непохожий на другие структуры
иммуноглобулиновой молекулы. В у-цепях этот участок обычно
содержит около 10 аминокислотных остатков, из которых полови-
на приходится на пролин, определяющий гибкость молекулы ан-
титела, т. е. изменение угла между ее Fab-фрагментами. 6-Цепи
содержат значительно больший шарнир, а р,- и a-цепи вообще
его не имеют.
Молекулы иммуноглобулинов имеют участки, на которые
предпочтительно действуют папаин, пепсин, трипсин, разрушаю-
щие молекулу антител на фрагменты. Под влиянием папаина
образуется три фрагмента, их разделяют методом ионообменной
хроматографии. Два из них идентичны, их обозначают как Fab
(от англ. Fragment antigen binding — фрагмент, связывающий
антиген). Fab состоят из Vh- и СД-доменов тяжелой цепи и Vl-
и С t-доменов легкой цепи. Третий фрагмент — Fc (от англ. Frag-
ment cristalline — фрагмент кристаллический) можно получить
в кристаллическом виде, в его состав входят по два Сн2- и Сн3-
домена тяжелой цепи, соединенные дисульфидными связями. Под
влиянием пепсина и трипсина образуется Р(аЬ)2-фрагмент, ана-
логичный двум Fab-фрагментам, соединенным в районе шарнира
дисульфидными связями между двумя тяжелыми полипептидны-
ми цепями.
Обработкой антител плазмином получают Fy-фрагмент (от
англ. Fragment variable — фрагмент вариабельный), состоящий
из вариабельных доменов легкой Vl и тяжелой Vh цепей, свя-
занных нековалентным, белок-белковым взаимодействием. Важ-
но, что активность антител (т. е. локализация активного центра
антител) ассоциирована не с Fc-фрагментом, а с Fv-, Fab и
F(ab) г-фрагментами (1 активный центр на 1 моль Fab и 2 актив-
ных центра на 1 моль F(ab)2).
Структура иммуноглобулина G. Антитела IgG преобладают
в нормальной сыворотке крови человека, их содержание состав-
ляет 70...75 % от общего количества иммуноглобулинов. Моно-
мерные молекулы IgG, выделенные из сыворотки (суммарная
фракция), имеют коэффициент седиментации 6,6S (М =
= 146 кДа). У человека известны четыре подкласса IgG: IgGl,
IgG2, IgG3 и IgG4. В суммарной фракции IgG они содержатся
в количестве 66, 23, 7 и 4 % соответственно. В Сн-областях име-
ются общие для всех подклассов антигенные детерминанты и спе-
цифические для каждого из них. Физико-химические свойства
всех классов иммуноглобулинов человека представлены в
табл. 38, из которой видно, что молекулы подкласса IgG3 по раз-
мерам несколько больше, чем молекулы других подклассов IgG.
Молекулы IgG-антител при электрофорезе, а также при изоэлек-
трофокусировании и ионообменной хроматографии имеют боль-
402
Таблица 38. Физико-химические свойства иммуноглобулинов человека
Иммуногло- булин Тяжелая цепь Кон- станта седи- мента- ции (S) Общая мо- лекулярная масса, Да Молекулярная мас- са тяжелой цепи, Да Число доме- нов в тяжелой цепи Содер- жание углево- дов, %
IgGl Yi 7 146 000 51 000 4 2—3
IgG2 72 7 146 000 51 000 4 2—3
IgG3 7з 7 170 000 60 000 4* 2—3
IgG4 74 7 146 000 51 000 4 2—3
IgM 19 970 000 65 000 5 12
IgAl 41 7 160 000 56 000 4 7—11
IgA2 а2 7 160 000 52 000 4 7—11
slgA** СЦ ИЛИ d2 11 385 000 52 000—56 000 4 7—11
IgD 8 7 184 000 69 700 5 9—14
IgE е 8 188 000 72 500 5 12
* Шарнирная область IgG3, возможно, имеет внутрнцепочечную дисульфидную связь,
однако при этом не образует домен из 110 остатков.
** Секреторный IgA.
шой диапазон подвижностей: от показателей для медленных
у-глобулинов до показателей для а2-глобулинов, что во многом
зависит от содержания белков разных подклассов в суммарной
фракции. Шарнирный участок IgG особенно чувствителен к про-
теолизу из-за своей доступности для разнообразных протеаз.
В молекулах IgG всех подклассов содержатся углеводы, кото-
рые, по-видимому, расположены на поверхности Сн2-доменов.
При изучении химического строения IgGl было показано, что
молекула углевода прикреплена к аспарагину (в положении 297)
в обеих тяжелых цепях. У миеломных белков класса IgG выяв-
лено значительное разнообразие в составе углеводного компо-
нента. Однако в настоящее время нет данных о том, какой угле-
вод характерен для того или иного подкласса антител.
Наличие двух подобных Fab-фрагментов и одного Fc-фраг-
мента в нативной молекуле IgG было подтверждено (кроме фер-
ментативного расщепления) электронно-микроскопическими ис-
следованиями. Был сделан вывод, что молекула IgG имеет Y-
образную форму, причем два из трех отходящих от центра от-
ростка дают при действии папаином Fab-фрагменты, а третий —
Fc-фрагмент. Угол между Fab-областями при взаимодействии их
с двухвалентными гаптенами имеет значения от 10 до 180°. Анти-
тела IgG найдены и изучены не только у человека, но и у ряда
млекопитающих животных; у кроликов обнаружен только один
подкласс IgG.
Структура иммуноглобулина А. IgA-антитела носят назва-
ние секреторных, они обнаружены в сыворотке крови и в серозно-
слизистых секретах человека как основные иммуноглобулины.
IgA сыворотки по своему содержанию составляют 15...20 % от об-
щего количества иммуноглобулинов. 78-мономеры IgA, представ-
403
ленные у человека более чем на 80 %, состоят из двух легких и
двух тяжелых a-цепей. Другая часть IgA (~20%) относится
к полимерным молекулам с коэффициентом седиментации 10, 13
и 15S, которые стабилизированы дисульфидными связями, сое-
диняющими отдельные мономеры.
У человека обнаружено два подкласса IgA: IgAl и IgA2, из
которых в сыворотке человека преобладает IgAl (80...90 %).
Тяжелая цепь белка IgAl построена из 472 аминокислотных ос-
татков, из них первые 119 остатков образуют V-область. С-об-
ласть a-цепи представляет собой три гомологичных домена, каж-
ный из которых идентичен по аминокислотной последовательнос-
ти С-доменам легких цепей, а также С-доменам тяжелых цепей
других иммуноглобулинов. В IgAl имеется дополнительный С-
концевой октадекапептид с остатком цистеина, находящимся в
цепи предпоследним. В полимерных молекулах этот остаток кова-
лентно связан с J-цепью (от англ, joining chain — соединяющая
цепь). Эта цепь соединяет молекулы мономеров в полимерные
молекулы.
Протеолитические ферменты (папаин, стрептококковая про-
теаза) медленно расщепляют IgA сыворотки крови с образова-
нием Fc- и Fab-подобных фрагментов. Секреторный IgA находит-
ся как в наружных (слюна, отделяемое трахеи и бронхов, моло-
зиво, молоко, выделения мочеполового тракта), так и во внутрен-
них секретах (синовиальная, амниотическая, плевральная, спин-
но-мозговая жидкость и водянистая влага глаза). Его коэффи-
циент седиментации 11S и AI = 380 кДа. Молекула IgA состоит
из двух мономерных субъединиц, одного секреторного компонен-
та (М = 70 кДа) и одной J-цепи (М = 15 кДа). В настоящее
время не совсем ясно, как все эти пептидные цепи соединены
между собой. Однако есть данные, согласно которым J-цепь син-
тезируется в тех же плазматических клетках, которые продуци-
руют IgA, и их соединение происходит незадолго до выхода этих
белков из клетки.
Секреторный компонент образуется не в плазматических клет
ках, а в эпителиальных, где и происходит его присоединение г.
молекуле IgA. Он может находиться в секретах больных, у кото-
рых отсутствуют IgA в свободной форме. При этом структурных
различий между связанными и свободными формами секретер
ного компонента не обнаружено. Секреторный компонент пред
ставляет собой одну полипепгидную пень, в которой содержится
~9% углеводов. Эта цепь (приблизительно у 80 % молекул
IgA) ковалентно связана с Fc-областью «-цепей. Существуют
между ними также сильные нековалентные взаимодействия, спо
собствующпе стабилизации молекулы в целом. На рис. 77 дан<
схематическое изображение строения молекулы секреторного
IgA, в котором секреторный компонент связывается и с IgAl.
и с IgA2.
Структура иммуноглобулина М. IgM и IgM-подобные белки
содержатся в сыворотке человека и почти всех позвоночных.
404
В ходе эволюции этот класс анти-
тел возник раньше других извест-
ных •классов иммуноглобулинов.
У человека молекулы IgM присут-
ствуют в основном в виде пента-
меров (М = 950... 1000 кДа), но мо-
гут существовать и более крупные
полимеры, а также мономеры.
У млекопитающих, птиц и пресмы-
кающихся встречаются пентамер-
ные молекулы IgM-подобных бел-
ков; у большинства земновод-
ных — гексамерные, а у большей
части костистых рыб — тетрамер-
ные молекулы. Несмотря на раз-
личную полимерность IgM-ан-
тител, их аминокислотный состав
во многом сходен.
В настоящее время изучена
полная аминокислотная последова-
тельность ц-цепей двух миеломных
IgM, выделенных от больных мак-
роглобулинемией Вальденстрёма.
IgM человека представляет собой
пентамер, состоящий из пяти четы-
Рис. 77. Схема строения молекулы
секреторного IgA человека
рехцепочечных мономеров, а вся молекула построена из 10 легких
и 10 тяжелых полипептидных цепей и одной J-цепи. Домены ц-
цепей сформированы пятью внутрицепочечными дисульфидными
мостиками. К каждой ц-цепи присоединяются пять углеводных
цепочек, представляющих собой простые или сложные олигосаха-
риды, содержащие глюкозамин. Известно, что эти олигосахариды
увеличивают растворимость молекулы и влияют на ее конфор-
мацию.
В состав каждого пентамера IgM входит одна J-цепь; точных
данных о способе ее присоединения к другим полипептидам мо-
лекулы пока нет. С помощью электронно-микроскопических ис-
следований установлено, что IgM-антитела млекопитающих име-
ют характерную звездообразную форму (иногда даже удается
увидеть отростки, соответствующие Fab-субъединицам пяти мо-
номеров IgM). Трипсин, папаин, пепсин и химотрипсин расщеп-
ляют IgM на Fc-,< Fab- и F(ab') 2-фрагменты, аналогичные тако-
вым иммуноглобулина G.
Структура иммуноглобулина D. IgD обнаружен только в сы-
воротке крови человека, впервые выделен из миеломного белка,
его содержание не превышает 1 % от общего количества иммуно-
глобулинов. Молекулярная масса его равна 184 кДа, а средняя
молекулярная масса тя'желой 6-цепи составляет 69 кДа. IgD
имеет (в отличие от других иммуноглобулинов) только одну ди-
сульфидную связь между тяжелыми цепями. Характерной осо-
405
бенностью IgD является наличие в его молекуле большого коли-
чества углеводов (9...18 %), среди которых присутствует N-аце-
тилгалактозамин, содержащийся еще только в IgAl. Папаин и
трипсин также расщепляют 6-цепь IgD с образованием Fab-
фрагмента. При расщеплении Рсь-фрагмента трипсином разру-
шается шарнирная область, а также утрачиваются N-ацетилга-
лактозамин и около 10 остатков галактозы. Малая концентрация
IgD в сыворотке затрудняет его выделение, кроме того, эти анти-
тела расщепляются плазмином.
Структура иммуноглобулина Е. IgE впервые были получены
Т. и К. Исидзака и др. (1966). Концентрация IgE в сыворотке
крови очень мала, его изучение стало возможным после обнару-
жения миеломных. IgE-белков. Константа седиментации IgE око-
ло 8S; молекулярная масса е-цепи равна 72,5 кДа (без углево-
дов — 61 кДа), а всей молекулы — 180 кДа. С помощью расщеп-
ления папаином и пепсином получены Fc- и F(ab') 2-фрагменты,
а также общий для обоих фрагментов участок, обозначаемый
как Fc"..
X. Бенник и X. Бар-Линстрём (1974) установили полную ами-
нокислотную последовательность е-цепи IgE. Отличительная осо-
бенность молекулы IgE -г- наличие двух соединяющих тяжелые
цепи дисульфидных мостиков, между которыми находится един-
ственный домен. Один из этих мостиков расположен между Се1-
и Се2-доменами, а другой — между Се2- и Се3-доменами (в С81-
домене есть добавочная внутрицепочечная дисульфидная связь).
В настоящее время IgE изучены недостаточно, не известны ка-
кие-либо их подклассы, нет сведений о строении IgE животных,
хотя у крыс миеломные IgE уже обнаружены.
В исследованиях первичной структуры иммуноглобулинов по-
казано, что вариабельные домены тяжелой цепи могут объеди-
няться только с константными доменами тяжелых цепей любого
класса, но никогда с константным доменом легких цепей CL; что
вариабельные домены легких к- и Х-цепей могут быть соединены
лишь с константными доменами своего типа. На этом основании
была выдвинута гипотеза, которая подтверждена эксперимен-
тально (1976) группой исследователей во главе с С.Тонегава.
Было установлено, что существует три группы сцепления генов,
кодирующих структуру иммуноглобулинов, которые расположены
на разных хромосомах. Одна группа генов — для вариабельных
и константных доменов тяжелых цепей, вторая и третья — для
доменов легких х- и %-цепей. Каждая из групп сцеплений генов
находится на одной хромосоме и состоит из тандемно располо-
женных участков ДНК, кодирующих синтез вариабельных и кон-
стантных доменов иммуноглобулинов.
При изучении биологических функций антител оказалось,
что они имеют две пространственно разобщенные области с раз-
ными биологическими свойствами, определяемыми структурой
соответствующих участков антител. Одна из областей определяет
специфику антитела — его связывание с соответствующим анти-
406
геном, которая зависит от аминокислотной последовательности
вариабельных доменов тяжелой и легкой цепей иммуноглобули-
нов. Это Fv-область (составная часть Fab-фрагмента), распоз-
нает чужеродные вещества. Другая область отвечает за эффек-
торные свойства антител, т. е. за те свойства, благодаря которым
чужеродные вещества — антигены элиминируют из организма
или разрушаются. Основные эффекторные функции антител пред-
ставлены в табл. 39. Среди них — активация комплемента, свя-
зывание с мембранами различных клеток, перенос иммуноглобу-
линов через плаценту, связывание с тучными клетками и т. д.
Эта область локализована в Fc-фрагменте антител и определяет-
ся их принадлежностью к соответствующему классу тяжелых це-
пей. Поэтому антитела одной и той же специфичности (с одина-
ковым активным центром), но принадлежащие к разным классам
(с разными Fc-фрагментами), связываясь с одним и тем же анти-
геном, могут вызывать противоположный биологический эффект
в организме.
За последние годы было найдено различие в строении секрети-
руемых иммуноглобулинов, которые служат антителами в орга-
низме, и иммуноглобулинов, связанных с мембранами лимфоци-
тов, выполняющих роль антигенраспознающих рецепторов
В-лимфоцитов. Оказалось, что эти иммуноглобулины в одной и
той же клетке идентичны по первичной структуре, за исключе-
нием небольшого участка на С-конце, определяющего, будет ли
молекула секретирована из клетки или интегрирована на поверх-
ностной мембране клетки. Так, секретируемые IgM-антитела на
С-конце ц-цепей имеют 20 гидрофильных аминокислот; а те же
IgM, но связанные с мембраной лимфоцита, — достаточно гидро-
Таблица 39. Основные эффекторные функции иммуноглобулинов человека
Иммуно- глобулин Средняя концент- рация в сыворот- ке, мг/мл Класси- ческий путь свя- зывания компле- мента Альтер- нативный путь свя- зывания компле- мента Перенос через плаценту Связыва- ние с мо- нонукле- арными клетками Связыва- ние с тучными клетками и базо- филами Реакция со ста- филоко- кковым белком А
IgGl 9 ++ — + + — +
IgG2 3 + — + — — +
IgG3 1 +++ — + + — -—
IgG4 0,5 — ♦ + — (?) +
IgM 1,5 ++ + — — — — —
IgAl 3,0 — + — — — —
IgA2 0,5 — + — — — —
slgA 0,05 — — — — — —
IgD 0,03 — — — — — —
IgE 0,00005 — ♦ — ♦♦ + Н—F —
* Агрегированные молекулы данного иммуноглобулина способны активировать комп-
лемент по альтернативному пути.
** IgE человека может связываться с микрофагами.
407
фобный участок из 41 аминокислоты. Аналогичные различия
найдены и для других классов секретируемых и мембранных им-
м у н огл обул и н ов.
Молекулы антител чрезвычайно многообразны. В ходе иммуч
ного ответа синтезируются как гомогенные (моноклональные)
так и гетерогенные антитела, включая все классы и подклада,
иммуноглобулинов. Чаще всего иммунный ответ на разные анти
гены осуществляется стереотипно: сначала образуются IgM-au
титела обычно с низкой аффинностью (от англ, affinity — срод-
ство, т. о. с низкой прочностью соединения активного центра им
муноглобулина и антигенной детерминанты), а затем IgG-анп:
тела, как правило, с высокой аффинностью. Иммунный оты-
на белковые антигены (столбнячный и дифтерийный анатоксины
тиреоглобулин) заключается в образовании всех четырех подклас-
сов IgG, активность которых примерно соответствует процентно
му содержанию -них белков в сыворотке. Углеводные антиген:
индуцируют образование более гомогенных антител. Напримщ
при иммунизации кроликов полисахаридом стафилококке,,
(тип VIII) обнаруживается клональное доминирование, при коте
ром синтезируемые антитела характеризуются очень неболыпо!
гетерогенностью либо практически полной гомогенностью. Таки-
высокополимерные микробные полисахариды, как декстраны >
леваны, при введении человеку вызывают синтез антител преим'-
щественно I gG‘2-подкласса.
Если антиген вводят в организм естественным путем, т. е.
через рот (полиовирус) или через нос (столбнячный анатоксин),
то возникает местная иммунная реакция, при которой в секретах
образуются преимущественно IgA-антитела. Повторная иммуни-
зация тем же способом вызывает появление также и других им-
муноглобулинов.
IgM- и IgG-антитела накапливаются, если антигены, в част-
ности бактерии или простейшие (возбудители малярии и трипа-
носомоза), попадают в кровь. Эти антитела обладают высокой
комплементсвязывающей активностью и способностью к агглюти-
нации. Присутствуя в крови, они играют особую роль в выведе-
нии микроорганизмов из кровотока. Если у новорожденных детей
обнаруживаются в большом количестве IgM-антитела, то можно
предполагать внутриматочное заражение краснухой, сифилисом,
токсоплазмозом и цитомегаловирусом.
IgE-иммуноглобулины образуются при различных видах ал-
лергии, они относятся к так называемым гомоцитотропным анти-
телам, обладающим способностью связываться с мембранами
базофилов и тучных клеток. И хотя у некоторых видов животных
гомоцитотропные антитела принадлежат и к другим классам и
подклассам иммуноглобулинов (IgGl, IgG2), наиболее важную
биологическую роль играют антитела, называемые реагинами,
принадлежащие к IgE-классу.
Основная функция IgD-антител до сих пор не установлена.
В сыворотке крови в минимальных количествах с помощью высо-
408
нечувствительных методов были найдены IgD-антитела к пени-
циллину G, дифтерийному анатоксину и инсулину. На поверхнос-
ти же В-лимфоцитов эти антитела представляют собой основную
фракцию иммуноглобулинов, что позволяет отвести им роль ре-
цепторов лимфоцитов. Кроме того, молекулы IgD, возможно,
участвуют в индукции иммунологической толерантности.
Молекулы антител, образуясь в ответ на введение антигена,
сами также обладают антигенными свойствами. Если вводить их
животным других видов, например барана проиммунизировать
антителами кролика, то сыворотка крови барана будет содержать
в основном иммуноглобулины против детерминант Fc-фрагмента
кроличьих антител, т. е. в данном случае Fc-область молекулы
наиболее иммуногенна. Если же гомогенные иммуноглобулины
вводить мышам гомологичной инбредной (от англ, inborn —
врожденный) линии (миеломные белки, полученные от мышей
линии BALB/c, инъецировать другим мышам той же линии), то
в сыворотке крови будут накапливаться главным образом анти-
тела, вступающие в реакцию с Fab-фрагментами. Такие антитела
называют антиидиот ипическими, так как считается, что они спо-
собны узнавать лишь тот миеломный белок, который индуциро-
вал их образование. Антиидиотипические антитела называют
также антителами против V-областей (вариабельных областей
Fab-фрагментов). Они нашли применение в опытах по изучению
природы рецепторов Т-лимфоцитов.
Иммуноглобулины одного и того же класса могут отличаться
по антигенным свойствам. Эти различия были названы аллотипи-
ческими, а соответствующие антигенные формы — аллотипами.
В ходе изучения ревматоидного фактора, ведущего себя подобно
антителу к глобулину человека, Р. Грубб (1956) обнаружил раз-
личные антигенные детерминанты, названные им Gm-группами
(Gm-маркеры). Позже наличие аллотипии у человека было под-
тверждено другими исследователями. Для у-цепей человека в
настоящее время известно до 15 аллотипических Gm-маркеров,
встречающихся в подклассах IgGl, IgG2 и IgG3. Для антител
IgA существуют два аллотипических варианта молекул подклас-
са IgA2. Описан один аллотипический маркер IgM человека,
однако о его локализации (в отличие от других известных алло-
типов антител) еще мало что известно. Для менее изученных ан-
тител классов IgD и IgE аллотипические антигенные детерми-
нанты пока не описаны.
Большие успехи за последнее время достигнуты в изучении
антигенсвязывающей области, или активного центра антител.
Для этого обычно использовали два подхода: сравнение первич-
ной структуры большого числа антител различной специфичности
и анализ пространственной структуры индивидуальных гомоген-
ных иммуноглобулинов. Сравнительный анализ аминокислотной
последовательности вариабельных доменов тяжелой и легкой
цепей антител разной специфичности показал, что степень вариа-
бельности неодинакова на протяжении полипептидной цепи. Это
409
позволило выделить менее консервативные по структуре участки,
называемые гипервариабельными участками, определяющими
комплементарность к антигену, располагающимися на конце под-
вижной Fab-области.
Еще до проведения рентгеноструктурных и электронно-микро-
скопических исследований Е. А. Кэбот (1960) разработал ориги-
нальный метод изучения антигенсвязывающей области антител
с помощью полимерных лигандов. Для иммунизации он исполь-
зовал полисахарид микробного происхождения — декстран. При
исследовании реакции антидекстрановых антител с гомологичным
антигеном-декстраном было показано, что моно-, ди-, три-, тетра-
меры и другие олигомеры этого полисахарида способны тормо-
зить реакцию между полимерным антигеном и соответствующим
антителом. При этом степень торможения увеличивалась с воз-
растанием размера олигомера до тех пор, пока антигенсвязыва-
ющая область целиком не заполнялась данным олигомером.
Дальнейшее увеличение длины конкурирующей молекулы не ока-
зывало влияния на степень ингибирования. Таким образом, было
установлено, что в системе декстран-антидекстран наибольшим
тормозящим действием обладали пента- и гексамеры изучаемого
антигена. Наиболее точные данные об антигенсвязывающей об-
ласти антител удалось получить с помощью рентгеноструктурно-
го анализа, на основании которого было показано, что активный
центр антитела в основном формируется из гипервариабельных
участков тяжелой и легкой цепей иммуноглобулинов и имеет
различную для разных антител форму (в виде щели, углубления
или полости) и размеры (например, 0,6X1,2X1,5 нм или 1,2Х
XI,5X2,0 нм). Важным моментом в анализе взаимодействия
антител с антигеном является то, что антитела мультиспецифич-
ны. Это значит, что помимо связывания с антигенами, облада-
ющими одинаковыми или подобными антигенными детерминан-
тами (перекрестно-реагирующими антигенами), антитела могут
взаимодействовать с пространственно непохожими антигенными
детерминантами. Явление мультиспецифичности антигенсвязыва-
ющих центров антител объясняется тем, что в одном и том же
активном центре антител могут быть участки, комплементарные
разным антигенным детерминантам. Правда, при взаимодействии
этих антигенных детерминант с антителами будет наблюдаться
конкуренция за активный центр, если размер антигенсвязыва-
ющего центра недостаточен для одновременного или индуциро-
ванного связывания с антигенными детерминантами.
С помощью физико-химических методов изучения антигенсвя-
зывающего центра антител были получены данные о его глубине
и степени гидрофобности, об участии легких и тяжелых цепей
в связывании лигандов, а также о стабилизации небольшими
лигандами взаимодействий между легкими и тяжелыми цепями.
Физико-химические методы необходимы также для выявления
конформационных изменений антител после их связывания с ан-
тигенами.
410
8.4. ХИМИЧЕСКИЕ ОСНОВЫ РЕАКЦИЙ ИММУНИТЕТА
В основе реакций иммунитета лежит специфическое соединение
антигена с соответствующим антителом в комплекс (реакция
антиген — антитело). В результате взаимодействия антигена с
антителом происходят конформационные изменения молекулы
антитела, обусловленные сближением определенных областей его
молекул, возникновением новых областей или маскировкой неко-
торых из них.
Первое количественное описание реакции антиген — антитело
предложил С. Аррениус (1904), рассматривавший ее как обрати-
мую на основе закона действующих масс. Схематически эту
реакцию выражают следующим образом:
АТ + АГ & АТ - АГ,
А Д
где АТ — свободное антитело; АГ — свободный антиген; АТ —
АГ — комплекс антиген — антитело; Аа — константа ассоциации;
Ад — константа диссоциации.
Пользуясь законом действующих масс, можно видоизменить
приведенное уравнение:
Аа [АТ] [АГ] = Ад[АТ—АГ]
и рассчитать, исходя из последнего, константу равновесия А:
Аа „ [АТ—АГ]
Ая Л [АТ] [АГ] '
Константа равновесия определяет аффинность — понятие, яв-
ляющееся важным в раскрытии сущности реакции антиген —
антитело. Аффинность есть сумма сил притяжения и отталкива-
ния между молекулами, которые возникают при взаимодействии
активного центра антитела (его антигенсвязывающей области)
и гомологичной антигенной детерминанты. Термин аффинность
наиболее пригоден для системы моновалентный антиген — анти-
гаптеновое антитело, когда происходит первичное взаимодей-
ствие антигена с антителом, т. е. взаимодействие одной антиген-
ной детерминанты с одним активным центром антитела. В этом
случае принято говорить о внутренней аффинности. Однако в
природе антигены и антитела, как правило, поливалентны. Анти-
гены имеют повторяющиеся антигенные структуры (антигенные
детерминанты) на своей поверхности, а антитела — несколько
антигенсвязывающих участков: от 2 в мономерных молекулах
IgG и IgE до 10 в пентамерных молекулах IgM. Такие антигены
и антитела вступают в более сложные взаимодействия, чем толь-
ко первичное связывание. Поэтому с биологической точки зрения
маловероятно, что' внутренняя аффинность объективно отражает
реакцию между антителами, обладающими разной валентностью
и поливалентными антигенами, энергия взаимодействия которых
характеризуется термином функциональная аффинность, или
авидность (от англ, avidity — жадность, алчность). Поскольку
411
встречающиеся в природных условиях антигены (вирусы, бакте-
рии) поливалентны, очевидно, что в иммунном ответе организма
на возбудители инфекционных заболеваний более важную роль
играет функциональная, а не внутренняя аффинность реакции
антиген — антитело. Поливалентное связывание антител в энер-
гетическом отношении выгодней, чем моновалентное, поскольку
оно более прочное и имеет явные функциональные и биологиче-
ские преимущества, выражающиеся в том, что для обеспечения
активного гуморального иммунитета требуются меньшие концент-
рации поливалентных антител, чем моновалентных.
Реакция между антителом и антигеном протекает в две ста-
дии, первая из которых — специфическая (непосредственное сое-
динение активного центра антитела с антигенной детерминан-
той) , вторая — неспецифическая, когда отличающийся плохой
растворимостью иммунный комплекс антиген — антитело выпа-
дает в осадок. Неспецифическая стадия, как правило, осуществ-
ляется в присутствии растворов электролитов и визуально прояв-
ляется по-разному в зависимости от физического состояния анти-
гена. Если антигены корпускулярные, имеет место феномен аг-
глютинации — склеивания различных химических частиц, сенси-
билизированных антигенами, и клеток, в том числе микроорга-
низмов. Образующиеся конгломераты выпадают в осадок, при
этом микробные клетки морфологически заметно не изменяются.
Теряя подвижность, они остаются живыми. Антитела, участву-
ющие в реакции агглютинации, называют агглютининами, анти-
гены — агглютиногенами, а образующийся агрегированный комп-
лекс — агглютинатом. Поскольку антигенная структура микробов
разнообразна, в их агглютинации принимают участие антитела
разной специфичности. Тождественность детерминантных участков
антигенов микробов разных видов обеспечивает групповые реак-
ции агглютинации с гетерологичными иммунными сыворотками.
Когда в реакции с антителами участвуют растворимые (моле-
кулярные) антигены — белки или их комплексы с углеводами
и липидами разного происхождения, бактериальные экстракты,
лизаты и фильтраты бульонных культур, наблюдается феномен
преципитации — осаждения антигена. Образующийся осадок но-
сит название преципитата, антитела — преципитинов, а антиге-
ны — преципитиногенов.
В основе иммунологической специфичности, т. е. избиратель-
ности реакции определенного антигена с соответствующим ему
антителом, лежит структурная комплементарность (стерическое
и химическое соответствие) антитела и антигена. Эти два компо-
нента системы должны сблизиться своими реагирующими поверх-
ностями на расстояние не более 0,1...0,2 нм. Только тогда начнут
действовать силы, вызывающие их взаимное притяжение. Весьма
существенную роль в реакциях антиген — антитело на 1-й стадии
играет взаимодействие неполярных (гидрофобных) молекул
с водной средой. Контакт этих молекул с молекулами воды осла-
бевает вследствие стремления гидрофобных групп к ассоциации.
412
В результате возникает неполярное (гидрофобное) связывание
и система антиген — антитело стабилизируется, что в конечном
итоге приводит к появлению сил притяжения. Сила неполярных
взаимодействий возрастает с температурой, так как возникнове-
ние этих взаимодействий является эндотермическим процессом,
определяемым энтропией. Межмолекулярные взаимодействия ан-
тигена и антитела обусловлены также кулоновскими (ионными)
силами, т. е. взаимным притяжением положительно (группа
NH^) и отрицательно (группа СОО~) заряженных боковых це-
пей иммуногена и стимулированного им антитела. Чем больше
соответствие полярных групп реагирующих молекул, тем прочнее
оказывается комплекс антиген — антитело.
Водородные связи, гидрофобное связывание, ионные взаимо-
действия и ван-дер-ваальсовы силы в сочетании с имеющим осо-
бое значение в образовании комплекса антиген — антитело сте-
рическим соответствием компонентов реакции противодействуют
стерическим силам отталкивания. Стерические силы отталкива-
ния, как и силы притяжения, возрастают с уменьшением рас-
стояния между взаимодействующими группами. Они обратно
пропорциональны двенадцатой степени расстояния между взаи-
модействующими группами, в то время как кулоновские взаимо-
действия обратно пропорциональны квадрату этого расстояния,
а ван-дер-ваальсовы силы обратно пропорциональны шестой
степени этого расстояния. Следовательно, стерические силы от-
талкивания более чувствительны в данном отношении. Они воз-
никают между атомами при взаимном проникновении их элек-
тронных облаков. Чем комплементарней электронные облака,
тем меньше силы отталкивания. Стерические силы отталкивания
важны для сохранения прочности комплекса антиген — антитело,
так как именно они определяют степень соответствия между ан-
тигенной детерминантой и активным центром антитела.
Реакция, происходящая между антителами и антигеном и
начинающаяся быстрым соединением детерминантной группы
антигена со специфическим активным центром антитела (1-я
стадия), осложняется далее образованием длинных цепей из
чередующихся молекул антигена и антител, а также разветвле-
нием этих цепей (2-я стадия). Во второй стадии реакции проис-
ходит образование решетки антиген — антитело.
Необходимое условие образования решетки — наличие более
трех антигенных детерминант на каждую молекулу антигена и по
два активных центра на каждую молекулу антитела. На рис. 78
изображены варианты соединения молекул антител с молекулами
антигена. Молекулы антигена являются узлами решетки, а моле-
кулы антител — связующими звеньями. В зоне избытка антител
(рис. 78, а, б) часть антигенсвязывающих участков свободна
и формирование комплекса тормозится. Область оптимальных
соотношений (зона эквивалентности) концентраций антигена и
антител, когда в надосадочной жидкости после образования
осадка не обнаруживаются ни свободные антигены, ни свобод-
413
Рис. 78. Структура агрегатов АГ—АТ и их молекулярные соотношения:
а — 5 : 1; б — 3 : 1; в — 2,5 : 1; г — 1 : 1,25; <5—1:2
ные антитела, представлена на рис. 78, в. В зоне же избытка
антигена (рис. 78, г, д) комплекс антиген — антитело растворяет-
ся, так как свободные и связанные в решетку молекулы антигена
конкурируют между собой за двухвалентные молекулы антител,
в результате чего решетка распадается на ряд растворимых
комплексов типа АГ — АТ — АГ.
Окончательное формирование системы антиген — антитело
протекает намного медленнее, чем 1-я стадия реакции. Это не-
специфический процесс, в который могут включаться различные
посторонние белки, захватывающиеся решетчатой структурой ан-
тиген — антитело и неспособные диффундировать сквозь узкие
петли этой решетки в надосадочную жидкость. Скорость обра-
зования решетки зависит от температуры, ионной силы раствора
и других условий. Оптимальными условиями реакции антиген —
антитело in vitro считаются следующие: / = ()...37 °C; ионная
сила 0,05...0,1 и pH 6,4...8,6.
Механизм реакций преципитации и агглютинации, при кото-
рых иммунный комплекс выпадает в осадок, вполне объясним
теорией решетки. В ходе преципитации решетка формируется не-
посредственно в растворе, в ходе реакции агглютинации бакте-
риальных клеток решетка антиген — антитело образуется на са-
мих клетках. О характере соединения антигена с антителом
судят, применяя различные методы. Существует, в частности,
метод изучения активного центра антител (метка по сродству),
основанный на реакции гаптена со специфическими антидетер-
минантами. В результате делают заключение о том, какие пеп-
тидные цепи участвуют в построении активного центра антител.
С помощью рентгеноструктурного анализа была раскрыта струк-
тура активного центра антител, формирующаяся из гипервариа-
бельных участков вариабельных доменов. При этом не было по-
лучено прямых данных, указывающих на изменение формы ак-
тивного центра после присоединения к нему антигена. Электрон-
ная микроскопия — один из важнейших методов не только изу-
чения общей конформации молекул иммуноглобулинов, но и
исследований комплексов антител с антигенами. Этим методом
414
было показано, что комплексы антиген — антитело имеют коль-
цеобразную, перекрывающуюся или решетчатую структуру.
Антитела IgG-класса образуют с антигеном (ферритином),
как правило, кольцеобразные комплексы. Электронные микрофо-
тографии иммунных комплексов, образованных при помощи
IgA-антител, интерпретировать сложно. Однако в некоторых слу-
чаях удается различить циклические комплексы, похожие на ве-
ревочную лестницу и пропеллерообразные структуры.
Иммунохимические реакции взаимодействия антиген — анти-
тело. Реакции между антигенами и антителами in vitro, имеющие
прежде всего диагностическое значение, всегда называли сероло-
гическими (от лат. serum — сыворотка), так как источником
антител служила сыворотка крови. В настоящее время бурного
развития методов иммунохимического анализа серологические
реакции стали как бы составной частью широкого набора реак-
ций, происходящих между антигенами и антителами. Если на
заре развития иммунологии в качестве антител для постановки
серологических реакций использовали только сыворотку крови,
теперь появились возможности с помощью разнообразных мето-
дов выделять из нее высокоочищенные иммуноглобулины. Для
индикации и количественной оценки того или иного вещества
антигенной природы предложены способы получения высокоспе-
цифических (моноспецифических) антител и высокочувствитель-
ные методы регистрации концентрации отдельных компонентов
реакции антиген — антитело. Это стало возможным в связи с
разработкой способов конъюгирования антигенов и антител
с флуоресцентными красителями и ферментами, введения в мо-
лекулы радиоактивных изотопов, фиксирования компонентов
реакции антиген — антитело на различных типах носителей. Дос-
тижения современной иммунохимии позволили проводить реакции
антиген — антитело на таком уровне, на котором их вряд ли
можно назвать серологическими, хотя источником антител про-
должает оставаться сыворотка крови. Понятие серологическая
реакция окончательно отпадает, когда речь заходит о взаимодей-
ствии антигена с моноклональными антителами, получаемыми
in vitro благодаря разработке гибридомной техники (см. гл. 6),
и когда реакция антиген — антитело протекает при аллергии.
В зависимости от характера антигена и условий взаимодей-
ствия с ним антитела в серологических реакциях обладают нейт-
рализующим, иммобилизующим, коагулирующим (осаждающим)
и лизирующим (растворяющим) действиями. Первое из них четко
проявляется в реакциях нейтрализации токсина антитоксином,
второе — в иммобилизации некоторых микроорганизмов, третье —
в реакциях агглютинации и преципитации, четвертое — в реак-
циях лизиса и связывания комплемента. Каждая из разновид-
ностей серологических реакций имеет свои варианты, а их поста-
новка — различные модификации, поэтому методические подходы
к проведению реакций между антигеном и антителом довольно
разнообразны.
Реакции нейтрализации. В ходе реакций нейтрализации спе-
цифические антитела нейтрализуют вредное действие антигена,
которое проябляется при попадании последнего в макроорганизм.
Если антигеном служит микробный экзотоксин (дифтерийный,
столбнячный, ботулинический и др.), то специфические антитела
нейтрализуют его. Когда реакция протекает в организме, нейтра-
лизуется только свободный, не связавшийся с клетками токсин.
Обезвреживание токсина в ходе физико-химической реакции
нейтрализации происходит за счет связывания его свободных
аминогрупп, что приводит к потере токсичности. Если антигеном
служит вирусный материал, нейтрализующие антитела полностью
подавляют специфическую активность вируса, выявляемую в раз-
личных клеточных культурах, куриных эмбрионах и на подопыт-
ных животных. Вирус перестает размножаться, теряет свою ин-
фекционность и цитопатогенность.
Реакции иммобилизации основаны на способности антител
сыворотки крови людей, больных сифилисом, холерой, амебиазом,
останавливать движение соответствующих подвижных микробов-
возбудителей и вызывать их гибель. Свое действие иммобилизу-
ющие антитела оказывают в присутствии комплемента, однако
механизм его действия в данной реакции не ясен.
Реакции агглютинации. Различают прямую и непрямую, или
пассивную, агглютинации. В прямой агглютинации антигеном
является сама микробная клетка или структурные компоненты ее
поверхностной оболочки. Предел чувствительности реакции аг-
глютинации микробов 0,01 мкг азота белка антител/мл. Количе-
ственные результаты реакции могут быть получены химическим
определением содержания азота белка иммуноглобулинов.
Агглютинация определяется в большей степени особенностями
клеточной оболочки и ее функциями, нежели свойствами агглю-
тининов, Fab-фрагменты которых могут блокировать антигенные
детерминанты, не приводя непосредственно к склеиванию. Агглю-
тинации способствует расположение антигенных рецепторов кле-
точной поверхности в виде скоплений. Если многовалентные аг-
глютинины взаимодействуют одновременно с несколькими анти-
генными рецепторами, константа ассоциации Аа повышается по
сравнению с для случаев соединения антитела с антигеном
единичной связью.
При пассивной агглютинации растворимые антигены (белки,
полисахариды и их комплексы микробного происхождения) сое-
диняют с нерастворимым носителем, выполняющим исключитель-
но индикаторную функцию. Носителем могут быть эритроциты
(пассивная гемагглютинация), частицы латекса, полиакрилами-
да, бентонита и др. Связывание антигена с носителем происхо-
дит в результате адсорбции или химического соединения. Мик-
робные полисахариды, например, можно адсорбировать на натив-
ных эритроцитах без какой-либо их предварительной обработки.
Различные белки (в том числе микробные ферменты) возможно
присоединить к эритроцитам, только обработав их различными
416
химическими веществами, обладающими дубильным действием,—
танином, хромахлоридом, формальдегидом или бисдиазотирован-
ным бензидином. Такая обработка предотвращает разрушение
красных кровяных клеток, которое может произойти при непос-
редственном контакте антигена с эритроцитами. Когда антиген,
связанный с носителем, соответствует антителу, наблюдается
агглютинация частиц носителя. Чувствительность непрямой реак-
ции агглютинации 0,02...0,04 мкг азота белка антител/мл.
Реакции преципитации. Основная модель реакции антиген —
антитело представлена преципитацией, в ходе которой антиген из
раствора осаждается специфическими антителами. Комплекс
антиген — антитело выпадает в осадок только при определенных
соотношениях концентраций реагирующих молекул. Область этих
соотношений, при которых в надосадочной жидкости после обра-
зования преципитата не обнаруживаются ни свободные антиге-
ны, ни свободные антитела, называется зоной эквивалентности.
Вне этой зоны, когда существует избыток антител или антигена,
феномен преципитации не развивается, так как образуется раст-
воримый комплекс антиген — антитело. Если в пробы с постоян-
ной концентрацией антител добавлять возрастающие количества
антигена и в образовавшихся, отмытых физиологическим раство-
ром преципитатах определять содержание азота микрометодом
Кьельдаля, можно получить количественное выражение реакции
преципитации. Реакция преципитации менее чувствительна, чем
реакция агглютинации. В зависимости от вариантов постановки
преципитации выявляемое количество антител колеблется от 2 до
18 мкг азота белка антител в 1 мл.
Существуют различные модификации постановки реакции
преципитации. Самый простой способ — кольцепреципитация,
которую выполняют в пробирках путем наслоения на иммунную
сыворотку различных разведений прозрачного раствора антигена.
На границе раздела двух реагирующих систем через определен-
ное время появляется опалесцирующее кольцо преципитации.
Именно таким образом при изучении стерильных фильтратов
бульонных культур возбудителей холеры, брюшного тифа, чумы
и соответствующих антисывороток в конце прошлого века (1897)
были открыты преципитины.
Реакция преципитации может быть поставлена в агаровом
геле (иммунодиффузия по Ухтерлони). Принцип этого метода
заключается в следующем. Растворимые антигены и антитела
вносят в лунки, вырезанные в геле на определенном расстоянии.
Компоненты реакции диффундируют в слое агара навстречу друг
другу, образуя в зоне конфронтации преципитат, становящийся
видимой линией преципитации.
Для более детальных иммунологических исследований приме-
няют метод иммуноэлектрофореза, описанный впервые П. Граба-
ром и К. А. Уильямсом (1953). При этом вначале проводят элек-
трофоретическое разделение антигена, представляющего зачас-
тую смесь белковых или других молекул, в забуференном агаро-
417
вом геле. После разделения в канавку, которая идет в направ-
лении миграции антигенных веществ, вносят преципитирующую
иммунную сыворотку. Антиген и антисыворотка диффундируют в
геле навстречу друг другу, и в месте их взаимодействия возни-
кают дугообразные линии преципитации. Число, положение и
форма этих линий дают представление о составе исходной смеси
антигенов.
Реакции дизиса и связывания комплемента. Лизирующее
(растворяющее), действие антител наглядно проявляется в реак-
циях. лизиса и связывания комплемента. В основе реакций лизи-
са лежит взаимодействие корпускулярных антигенов со специфи-
ческими антителами, адсорбирующими своими участками, распо-
ложенными в Fc-фрагменте тяжелых цепей, комплемент, в ре-
зультате чего происходит растворение клеток. Еще раз важно
подчеркнуть тот факт, что в связывании антигена и комплемента
участвуют различные домены молекулы антитела: антиген связы-
вается с V-доменами, т. е. с N-концевыми участками тяжелых
цепей, а комплемент, наоборот, — с участками, расположенными
в С-концевых областях (Fc) тяжелых цепей молекул иммуно-
глобулинов. Когда иммуноглобулины при содействии комплемен-
та разрушают красные клетки крови — эритроциты, из последних
выходит гемоглобин и реагирующая смесь из мутной взвеси эрит-
роцитов преобразуется в прозрачную красную жидкость, так
называемую лаковую кровь. Реакцию в данном случае называют
реакцией гемолиза. Это очень чувствительная реакция, способная
выявить 0,0002 мг азота белка антител в 1 мл.
Принцип гемолиза положен в основу метода локального гемо-
лиза в геле, предложенного Н. К. Ерне и А. А. Нордин (1963) для
выявления клеток, образующих антитела. В основе метода лежит
контакт антигена (эритроцитов барана) в гелевой среде с лим-
фоидными клетками иммунизированных животных. Антитела
(гемолизины) от синтезирующих их клеток диффундируют в гель
и связываются с эритроцитами. Последующее добавление комп-
лемента обусловливает лизис эритроцитов вокруг антителопроду-
цирующих клеток, 'образуя просветы (зоны гемолиза) на розовом
фоне. В центре каждой зоны находится клетка, образующая
антитела к эритроцитам. Сосчитав число зон, можно определить
число этих клеток. Зная общее число внесенных в гель лимфоид-
ных клеток, можно рассчитать число клеток, образующих антите-
ла, на 106 или 107 клеток исследуемого органа.
Когда иммуноглобулины также при содействии комплемента
разрушают оболочку бактериальной клетки, наблюдается лизис
бактерий — бактериолиз.
Реакция связывания комплемента (РСК) по механизму дей-
ствия является более сложной в сравнении с другими серологи-
ческими реакциями. В РСК задействованы две системы анти-
ген — антитело. Для визуальной регистрации связывания компле-
мента основной системой антиген — антитело в реакцию дополни-
тельно вводят вторую, или индикаторную, систему, состоящую из
418
взвеси эритроцитов и соответствующей антисыворотки (гемоли-
тической сыворотки). Если основной комплекс антиген — антите-
ло фиксировал на себе комплемент (в случае соответствия анти-
гена антителу), то гемолиз отсутствует (положительная реак-
ция). Если в основной системе антиген и антитело не соответ-
ствуют друг другу, то комплемент фиксируется на втором инди-
каторном комплексе, вызывая гемолиз эритроцитов (реакция
отрицательная). По чувствительности РСК примерно соответст-
вует реакции пассивной гемагглютинации. Она позволяет полу-
количественно определять комплексы антиген — антитело, пол-
ностью или частично фиксирующие комплемент, находящийся
в данной системе в ограниченном количестве. Убыль комплемента
из реакционной среды пропорциональна количеству образовав-
шихся комплексов.
Феномен иммуноклеточного прилипания. Метод, разработан-
ный Г. Дж. В. Носселом с соавторами (1961), основан на способ-
ности бактерий прилипать к поверхности малых лимфоцитов —
клеток, вырабатывающих специфические антитела после иммуни-
зации животных соответствующими микроорганизмами, и полу-
чил название иммуноклеточного прилипания или скучивания.
Феномен иммуноклеточного прилипания был использован для
дальнейшего совершенствования методов выявления клеток, об-
разующих антитела к антигенам микробного происхождения.
В частности, заслуживает внимания модификация метода с ис-
пользованием в качестве антигена эритроцитов, нагруженных
различными микробными гаптенами. Такие эритроциты приоб-
ретают способность специфически прикрепляться в опытах in vit-
ro к поверхности изолированных лимфоидных клеток животного,
иммунизированного тем микроорганизмом, из которого выделен
гаптен. При этом под микроскопом удается наблюдать так назы-
ваемые розетки, каждая из которых представляет собой лим-
фоидную антителопродуцирующую клетку, окруженную прочно
приставшими к ней эритроцитами. Просмотрев в общей слож-
ности 3000...4000 лимфоидных клеток в нескольких препаратах,
определяют процент розеткообразующих клеток, адсорбировав-
ших не менее двух эритроцитов, — процент адсорбции. Описан-
ный метод был разработан Я- С. Шварцманом и А. В. Исполато-
вой (1966) и получил название непрямой гемадсорбции по анало-
гии с реакцией непрямой гемагглютинации. Реакция непрямой
гемадсорбции по чувствительности не уступает реакции прилипа-
ния бактерий и превосходит ее по специфичности, особенно в тех
случаях, когда изучают образование антител к антигенам непод-
вижных бактерий.
Иммунофлуоресцентный и иммуноферритиновый методы обна-
ружения антигенов и антител., В основу иммунофлуоресцентного
метода положено такое взаимодействие антигена с антителом,
при котором один из компонентов реакции (чаще антитело) об-
ладает способностью к вторичной флуоресценции благодаря
предварительному ковалентному соединению с флуоресцентным
419
красителем. Образовавшиеся таким образом иммунные комплек-
сы становятся хорошо видимыми, ярко светящимися структурами
на темном фоне под флуоресцентным микроскопом. В качестве
флуоресцентных красителей используют флуоресцеинизотиоциа-
нат, родамин, В-изотиоцианат, лиссамин-родамин В200-сульфо-
хлорид и др., имеющие реакционноспособные группы (сульфохло-
рид, изитиоцианат и др.). Эти группы соединяют со свободными
аминогруппами молекул антител, которые не теряют при обработ-
ке флуорохромами специфического сродства к соответствующе-
му антигену.
Прямой и непрямой флуоресцентные методы. Прямой метод
основан на непосредственном специфическом соединении антиге-
на с мечеными антителами. Непрямой метод — на поэтапном
выявлении комплексов антиген — антитело с помощью флуорес-
центных красителей. Первый этап заключается в образовании
иммунных комплексов определенного антигена (например, бак-
териальной клетки) со специфическими антителами (у-глобули-
нами) иммунной сыворотки. Второй этап — в выявлении это-
го комплекса путем обработки его меченым анти-у-глобу-
лином.
Обнаружение иммуноглобулинов иммунофлуоресцентным ме-
тодом возможно, если антиген, использованный in vivo для об-
разования антител, — растворимый белок. В этом случае он мо-
жет быть конъюгирован с флуорохромом и в таком виде исполь-
зоваться для обнаружения гомологичных антител.
Иммуноферритиновый (иммуноэлектронно-микроскопический)
метод выявления антигенов основан на применении антител, мечен-
ных ферритином. Ферритин представляет собой железосодержа-
щий белок с молекулярной массой 750 кДа, где содержание железа
может доходить до 23 %. Ферритин хорошо выявляется в электрон-
ном микроскопе и поэтому используется как маркер при иммуно-
электронной микроскопии.
Иммуноферментный метод. В основу метода положена конъю-
гация антитела или гаптена с ферментом, которая не приводит к
утрате антителом или гаптеном специфичности, а ферментом —
каталитической активности. Однако в составе комплекса анти-
ген — антитело конъюгаты гаптен-фермент или антитело-фермент
теряют свою каталитическую активность.
Соединение молекул антитела с ферментом осуществляется
с помощью преимущественно глутаральдегида — бифункциональ-
ного реагента, взаимодействующего с е-аминогруппами белковых
молекул. В качестве фермента успешно применяют в зависимости
от модификации метода либо пероксидазу, р-галактозидазу,
щелочную и кислую фосфатазы (реже ацетилхолин, глюкоами-
лазу, глюкозооксидазу), либо лизоцим, глюкозо-6-фосфат-дегид-
рогеназу и малатдегидрогеназу. Используемый для маркирования
антител фермент не должен присутствовать в исследуемых,
содержащих антиген клетках, так как эндогенная ферментативная
активность недопустима.
420
и- .
Иммуноферментные методы первоначально были разработаны
для гистохимических исследований. Принцип иммуногистохимиче-
ского анализа с использованием ферментов заключается в сле-
дующем. Антитела, маркированные ферментом, соединяются со
специфическим антигеном, фиксированным на твердом носителе
(полистироловая пластина или другие непористые полимеры),
образуя комплекс, микроскопически не определяющийся. Выявле-
ние иммунных комплексов, содержащих фермент, проводится с
помощью цветной реакции, происходящей при добавлении фер-
мент-специфического субстрата. Окрашенный комплекс анти-
ген — антитело выявляется при использовании световой или
электронной микроскопии.
В дальнейшем методы иммуноферментного анализа стали
применять для количественного определения антигенов и антител
в биологических жидкостях. Были созданы гетерогенные (твер-
дофазные) и гомогенные методы, принципиально отличающиеся
способом разделения компонентов иммунохимической реакции
(гомогенный метод характеризуется протеканием реакции анти-
ген — антитело в гомогенном растворе, и этап физического раз-
деления реагентов и продуктов реакции не обязателен). Твердо-
фазные методы основаны на применении антител (или антиге-
нов), иммобилизованных на нерастворимых носителях, гомоген-
ные методы — на эффекте модуляции антителами активности
фермента, связанного с антигеном. В основу гомогенных методов
положен принцип конкурентного взаимодействия меченого и
немеченого гаптена с активными центрами антител. Чем больше
свободного гаптена введено в реакцию до прибавления меченого
гаптена, тем меньшее количество последнего прореагирует с ан-
тителами. Соответственно и ферментативная активность в раст-
воре изменится незначительно. Наоборот, чем меньше свободного
гаптена ввести в реакцию, тем большее количество ковалентно
соединенного с ферментом гаптена уйдет в иммунный комплекс,
и ферментативная активность в растворе существенно пони-
зится.
Определение неизвестной концентрации гаптена в образце
осуществляется по заранее полученной калибровочной кривой,
для построения которой устанавливается зависимость активности
фермента в данной системе от стандартных концентраций сво-
бодного гаптена.
В твердофазном иммуноферментном анализе наряду с конку-
рентными методами распространены подходы, основанные на
последовательном взаимодействии фиксированных на носителе
иммуноглобулинов с антигеном (или наоборот), а затем с имму-
ноферментным конъюгатом, представляющим собой меченные
ферментом антитела против иммуноглобулинов (сэндвич-метод,
от анг. sandwich — бутерброд).
Радиоиммунологический анализ (РИА). В радиоиммуноло-
гическом анализе специфичность реакции антиген — антитело
сочетаете^ с высокой чувствительностью, обеспечиваемой при-
421
менением радиоактивной метки, благодаря которой можно
определить количество азота антител до 1•10-8 мг/мл. Для
проведения анализа необходимо иметь антисыворотки и гомо-
логичные антигены, маркированные каким-либо изотопом иода
(1251, |311), равно как и гаптены, маркированные 14С или 3Н.
Чувствительность метода существенно возрастает при использо-
вании высокоаффинных антител с низкой перекрестной реактив-
ностью и антигенов с высокой удельной радиоактивностью. В осно-
ву радиоиммунологического анализа положен принцип конкурент-
ного взаимодействия определяемого немеченого антигена и извест-
ного количества меченого антигена с активными центрами ан-
тител. При этом происходит вытеснение меченого антигена или
гаптена из его комплекса с антителом вследствие последующего
добавления больших концентраций немеченого антигена или гапте-
на той же специфичности. Реакция (при высоком разведении ин-
гредиентов) подчиняется закону действующих масс: происходящее
вытеснение пропорционально введенному количеству немеченого
антигена или гаптена. Конкуренцию между определяемым и меченым
антигенами можно оценить количественно с помощью радиомет-
рии, предварительно отделив образовавшиеся иммунные комп-
лексы от несвязавшегося меченого антигена. Концентрацию опре-
деляемого антигена рассчитывают, исходя из сравнения соотно-
шений свободного и связанного меченых реагентов с соответст-
вующим стандартом.
Реакции между антигенами и антителами при аллергии. Ал-
лергия (измененная реактивность организма) является неотъ-
емлемой составляющей иммунитета и развивается в организме
в ответ на повторное попадание антигена (микробных клеток,
продуктов их распада, антибиотиков, пыльцы растений, перхоти
животных и др.). Если реактивность по сравнению с нормой
(нормергия) повышена, развивается гиперергия, если пониже-
на — гипоергия и, наконец, если полностью отсутствует — анер-
гия.
Известные иммунологические реакции при аллергии разде-
ляют на два вида: гиперчувствительность немедленного типа
(ГНТ) и гиперчувствительность замедленного типа (ГЗТ). ГИТ
можно рассматривать как форму гуморального иммунитета,
обусловленную антителами класса IgE (гомоцитотропными анти-
телами, или реагинами). К ней относятся анафилактический
шок; поллинозы (от англ, pollen — пыльца), например аллерги-
ческий насморк; атопическая бронхиальная астма и др. В основе
ГНТ лежит реакция между антигеном и антителом, сопровож-
дающаяся патологическими изменениями в тканях и тканевых
жидкостях организма.
Образующиеся иммунные комплексы антиген — атитело фик-
сируются на клетках-мишенях, повреждают их, в результате
чего из клеток выбрасываются в окружающую среду медиато-
ры — биологически активные вещества (гистамин, серотонин,
ацетилхолин, брадикинин):
422
гистамин серотонин
СНз
I
Н3С—N+—СН2—СНг—О—СО—СН3 Арг-Про-Про-Гли-Фен-"Сер-Про-Фен-Арг
I
СНз
ацетилхолин брадикинин
Медиаторы изменяют обменные процессы, вызывают сокращения
гладкой мускулатуры, повышают проницаемость сосудов и нару-
шают жизнедеятельность клеток. В некоторых случаях (напри-
мер, при анафилактическом шоке) может быть летальный исход
через 5... 10 минут после введения разрешающей дозы.
ГЗТ является формой клеточного иммунитета, она обусловле-
на сенсибилизированными Т-лимфоцитами, несущими на своей
поверхности подобные антителам рецепторы, специфичные анти-
гену (аллергену), и наблюдается при туберкулезе, кандидозах,
дерматомикозах, а также при отторжении трансплантатов и при
воздействии некоторых химических элементов, например никеля.
ГЗТ не опосредована гуморальными антителами. Ведущая роль
при повышенной чувствительности замедленного типа принадле-
жит Т-лимфоцитам, которые на первичную антигенную сенсиби-
лизацию отвечают экспрессией специфических антителоподобных
поверхностных рецепторов. Попадая в организм снова, антиген
реагирует с сенсибилизированными Т-лимфоцитами, вследствие
чего через 24...48 ч развивается аллергическое воспаление.
Это воспаление обусловлено действием медиаторов — лимфоки-
нов, освобождающихся из сенсибилизированных лимфоцитов.
Лимфокины обладают способностью активировать макрофаги,
которые, в свою очередь, выделяя медиаторы — монокины, вклю-
чаются в реакции ГЗТ. Медиаторы клеточного иммунитета
принципиально отличаются от медиаторов гуморального имму-
нитета. Биохимически более подробно изучены лимфокины, или
интерлейкин-2 (ИЛ-2). ИЛ-2 представляет собой полипептидную
молекулу с молекулярной массой 19...22 кДа, обладающую гид-
рофобными свойствами..Изоэлектрическая точка — 6,5...6,8. По-
казаны: чувствительность к трипсину и химотрипсину, додецил-
сульфату натрия и отсутствие чувствительности к jl-меркапто-
этанолу, 2...4 М мочевине и нейраминидазе. ИЛ-2 стабилен при
56 °C — 60 мин, при 70 °C — 30 мин, при pH 3...10. В первичную
структуру ИЛ-2 человека входит 133 аминокислоты с одной внут-
римолекулярной S—S-связью по цистеину в позициях 58 и 105.
Эта связь играет важную роль в приобретении молекулой актив-
ной конформации.
Монокины, или интерлейкин-1 (ИЛ-1), — белок с молекуляр-
423
ной массой 11 кДа и изоэлектрической точкой 7,0. ИЛ-1 чувстви-
телен к трипсину и химотрипсину, додецилсульфату натрия, не
чувствителен к 0-меркаптоэтанолу, нейраминидазе, перйодату
натрия, иодацетату; устойчив при pH 3...10 и к нагреванию до
56 °C в течение 60 мин, но разрушается при 100 °C через 2 мин.
Итак, аллергическое воспаление, вызванное медиаторами
клеточного иммунитета, сопровождается мононуклеарной ин-
фильтрацией лимфоцитами и другими мононуклеарными клет-
ками различных тканей, которая при гиперергии может привести
к некрозу, а следовательно, и к деструкции не только отдель-
ных тканей, но и целых систем организма.
Повышенную чувствительность человека или животного можно
снизить, проведя десенсибилизацию, т. е. повторное введение
аллергена в дозе, меньшей чем разрешающая, или в небольших
дозах дробно. При этом вырабатываются неполные (одновалент-
ные) антитела. Они связывают антигены, предотвращая взаимо-
действие последних с полными антителами, и вредные последст-
вия не наступают. Если введение аллергена индуцирует обра-
зование IgM- и IgG-иммуноглобулинов или в крови присутствуют
перекрестно реагирующие с аллергеном антитела IgM- и IgG-
классов, повышенная чувствительность значительно снижается
за счет отсутствия возможности у IgE-антител соединиться с
антигеном.
8.5. ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ ИММУНОХИМИИ
Благодаря интенсивному развитию иммунохимии иммунология за
последние 20...30 лет превратилась в одну из узловых наук, ока-
зывающих влияние как на теоретические исследования в области
биологии, генетики, биохимии, микробиологии, так и на многие
клинические дисциплины, где она приобрела важное практиче-
ское значение. В частности, благодаря вакцинации удалось
резко снизить заболеваемость людей различными инфекционны-
ми заболеваниями, а такая опасная в прошлом болезнь, как
оспа, — ликвидирована повсеместно на земном шаре. В качестве
вакцинирующих препаратов применяют не только корпускуляр-
ные (от лат. corpusculum — тельце) живые или убитые вакцины,
но и анатоксины, или токсоиды, — токсины, утратившие свою
токсичность в результате обработки повышенной температурой
или химическими реагентами, например формальдегидом.
Очищенные от балластных веществ анатоксины в качестве
вакцин (дифтерийной, столбнячной, ботулинической) обычно вы-
пускают с адъювантами — веществами, усиливающими иммун-
ный ответ (оксид или фосфат алюминия, минеральные масла
и др.).
В последние годы созданы безопасные и иммуногенные вак-
цины против круппозной пневмонии, эпидемического церебро-
спинального менингита и заболеваний, вызванных синегнойной
палочкой. Они представляют собой высокомолекулярные кап-
424
сульные полисахариды, индуцирующие образование антител с бак-
терицидной активностью, и являются эффективными в предотвра-
щении и лечении указанных заболеваний.
В 60-х годах XX в. выдвинут принципиально иной подход
к созданию вакцин нового типа на основе искусственных антиге-
нов (см. разд. 8.2). Эти антигены должны обладать структурами,
обеспечивающими повышение иммуногенности вакцины и фено-
типическую коррекцию иммунного ответа макроорганизма. Выри-
совываются перспективы создания искусственных вакцин против
гриппа. Конъюгаты антигенов вируса гриппа с синтетическим
полимером названы вирогатами. Они индуцируют выраженный
специфический иммунный ответ и повышают протективный
эффект у животных, зараженных вирусом гриппа.
В соответствии с современными представлениями антигенные
детерминанты природных молекул делятся на 2 типа: детерми-
нанты, активирующие хелперные Т-лимфоциты, и детерминанты,
вызывающие появление супрессорных Т-лимфоцитов. Последние
обнаружены у антигенов Mycobact. leprae (кстати сказать, весь-
ма эффективных мер борьбы с этим возбудителем до сих пор не
существует) и представляют собой уникальный фенольный глико-
липид. В этой связи есть основания предполагать, что при
рациональном подходе к созданию вакцин важно учитывать де-
ление антигенных детерминант на указанные типы и стараться
сделать такую вакцину, чтобы было как можно меньше супрес-
сорных детерминант или их не было вообще. Применение вакцин
без супрессорных детерминант ведет к смещению баланса между
популяциями хелперных и супрессорных Т-лифоцитов в сторону
развития выраженного иммунного ответа. Несомненно, и этот
принципиально новый подход к созданию вакцин в будущем най-
дет свое дальнейшее развитие.
Кроме вакцин для профилактики и лечения инфекционных
заболеваний применяют иммуноглобулины, которые при введении
в организм предупреждают болезнь или облегчают ее течение.
С помощью иммуноглобулинов удается эффективно проводить
борьбу со столбняком, коклюшем, краснухой, корью, оспой, бе-
шенством, свинкой, гриппом, герпесом и другими инфекционны-
ми болезнями.
В настоящее время созданы возможности применения на
практике моноклональных антител (см. гл. 6). Молекулы таких
антител взаимодействуют только с одним антигеном и представ-
ляют собой идеальный по специфичности реагент на определен-
ное соединение для диагностических или лечебных целей. Моно-
клональные антитела можно получить против обширного набо-
ра антигенов. Среди микробиологических объектов ими могут
быть вирусы, бактерии, протозоа, а также индивидуальные ан-
тигенные детерминанты из сложных антигенов. С помощью моно-
клональных антител против антигенных детерминант клеток
можно выявлять их топографию, определять отдельные детерми-
нанты, связанные с дифференцировкой клеток и их функциональ-
425
ными свойствами, изучать механизмы этих процессов, разли-
чать близкородственные группы клеток.
Иммунохимические методы с участием моноклональных анти-
тел перспективны для вирусологии. Известно, что для некоторых
вирусов (гриппа, лейкоза) характерна исключительно высокая
изменчивость структуры. Получив с помощью гибридной техники
ряд антисывороток против любых вариантов таких вирусов,
можно осуществлять их идентификацию. Кроме того, эти высоко-
специфические иммуноглобулины обладают также высоким
терапевтическим эффектом.
В борьбе с инфекционными заболеваниями моноклональные
антитела могут применяться как для диагностики (например,
стрептококков разных групп, отличающихся по составу полисаха-
ридов и вызывающих трудно идентифицируемый сепсис у ново-
рожденных) , так и для лечения. Моноклональные антитела,
продуцируемые гибридомами клеток человека (что особенно важ-
но), в ближайшем будущем могут стать самыми совершенными
лечебными иммунными препаратами — высокоэффективными и
свободными от побочного действия. Моноклональные антитела,
помимо иммунодиагностических и лечебных целей, могут быть
использованы в будущем для целей иммуномодуляции, в част-
ности, как иммуносупрессоры, активаторы пролиферации Т-лим-
фоцитов, индукторы интерферона и т. д. С помощью моноклональ-
ных, или моноспецифических, антител можно определять появле-
ние циркулирующих антигенов в сыворотке крови. Использова-
ние антител имеет большое будущее и в деле изолирования
чистых биомолекул посредством иммуносорбционных методов.
Для направленной иммунокоррекции (устранения различных
иммунодефицитов), при лечении ряда инфекционных и аллерги-
ческих заболеваний, а также в диагностических целях применяют
на практике следующие медиаторы (молекулярные курьеры
иммунитета).
1. Фактор переноса, активирующий клеточный иммунитет,
выделяют из лейкоцитов здоровых людей. Он может быть неспе-
цифическим, поскольку подбор доноров, сенсибилизированных
определенными антигенами, затруднителен. Считается, что
в переработанной массе клеток есть лимфоциты нужной специ-
фичности. В случае же лечения различных инфекционных заболе-
ваний эффективней оказывается специфический фактор переноса.
2. Биологические активные препараты тимуса, или гормоны
тимуса, — тимозин, тимопоэтин, тимарин, сывороточный тимус-
ный фактор, активная фракция тимуса 6 (АФТ-6) и др. Тимиче-
ские гормоны обеспечивают созревание Т-лимфоцитов, поэтому
их применение рекомендуется для ликвидации некоторых форм
иммунодефицитов, связанных с Т-системой иммунитета.
3. Фактор, угнетающий миграцию макрофагов, регулирует
клеточный иммунитет.
4. Костно-мозговой стимулятор антителопродуцирующих кле-
ток. Он эффективен при лечении некоторых вирусных инфекцион-
426
ных заболеваний у мышей, а также увеличивает титры антител
у различных видов животных при изготовлении антисывороток
к слабым иммуногенам.
Выделение медиаторов в чистом виде и достаточных количе-
ствах на данном этапе — сложный процесс. Можно надеяться,
что успешная гибридизация миеломных клеток не только с В-
лимфоцитами, но и с другими типами клеток (например, Т-килле-
рами, Т-супрессорами, Т-хелперами, макрофагами) позволит
справиться с этой трудной задачей и в будущем поставить
производство медиаторов на промышленную основу. Направлен-
ную иммунокоррекцию можно осуществлять также с помощью
различных лечебных средств (см. 8.6).
В теоретических и прикладных работах широко используют
иммунохимические реакции взаимодействия антиген — антитело.
Например, можно определять неизвестные антитела в сыворотке
больных с помощью известного антигена, проводя таким образом
серодиагностику инфекционных заболеваний. С другой стороны,
неизвестные антигены могут быть идентифицированы с помощью
известных антител. Реакции антиген — антитело применяют так-
же для исследования антигенной структуры микроорганизмов
и изучения механизма биосинтеза антител и других глобулинов.
С помощью реакции кольцепреципитации можно выявить
нейтрализующее действие антитоксинов. При соединении токсина
с антитоксином in vitro наблюдается помутнение, а затем выпа-
дение осадка (флокулята), и реакция в данном случае носит
название реакции флокуляции. Реакция нейтрализации вируса,
основанная на высокой специфичности, пригодна для тонкого
анализа антигенных особенностей всех вирусов, включая бакте-
риофаги. Ее используют для индентификации и титрования неиз-
вестных вирусов, а также для выявления нейтрализующих анти-
тел с помощью известных вирусов.
Явление иммобилизации трепонем у больных сифилисом
послужило основанием для разработки серологической реакции
иммобилизации трепонем, применяемой в диагностике данного
венерического заболевания.
Метод агглютинации бактерий применяют для качественной и
количественной оценок реакции. Так, известны: ориентировочная
агглютинация на предметном стекле; реакция Грубера для иден-
тификации неизвестных бактериальных антигенов с помощью
известной иммунной сыворотки; реакция Видаля для обнаруже-
ния антител в сыворотке крови больных против известного
бактериального антигена; реакция Вейля—Феликса для диагнос-
тики сыпного тифа, когда используют не возбудитель болезни
(риккетсии), a Proteus vulgaris, штамм 0X19, имеющий антиген-
ную общность с риккетсиями. Такой антиген носит название
гетерофильного.
Микробные антигены зачастую обладают свойствами адсор-
бироваться на эритроцитах, поэтому в диагностике некоторых
инфекционных заболеваний используют отличающуюся высокой
427
чувствительностью непрямую реакцию гемагглютинации, с по-
мощью которой можно проводить индикацию антител против
менингококков, стафилококков, коклюшных бактерий, микобак-
терий туберкулеза и других микроорганизмов. Реакцию агглюти-
нации бактерий применяют и для диагностики таких аутоиммун-
ных заболеваний, как ревматоидный артрит. Характерный приз-
нак этой болезни — наличие в сыворотке крови так называемого
ревматоидного фактора, представляющего собой антигаммагло-
булиновые аутоантитела. Один из методов обнаружения ревмато-
идного фактора — агглютинация с протеем по Р. Отто. Реакцию
проводят, нагружая бактерии рода Proteus субагглютинирую-
щими дозами специфической антисыворотки. Агглютинация про-
исходит в тех случаях, когда к реагентам (антигену и антителам)
добавляют сыворотку больного, содержащую ревматоидные фак-
торы. Кроме протея можно использовать и другие микроорганиз-
мы например бруцеллы. Помимо диагностических целей, реакцию
агглютинации применяют для определения типовой принадлеж-
ности микроорганизмов по известным иммунным сывороткам.
Реакцию бактериолиза используют для диагностики некото-
рых инфекционных заболеваний, например холеры; а реакцию
гемолиза (индикаторная система) — для постановки РСК, отно-
сящейся к традиционным методам микробиологической диаг-
ностики и являющейся важной, в первую очередь, при распозна-
вании сифилиса (реакция А. А. Вассермана).
С помощью метода локального гемолиза в геле изучают
разные стороны процесса образования антител отдельными клет-
ками в больших клеточных популяциях. Для определения кле-
ток, образующих антитела к микробным антигенам (целым
клеткам, экстрактам бактериальных культур, отдельным про-
теиновым, полисахаридным или другим молекулам), применяют
методы адсорбции антигена на клетках, синтезирующих антитела
(феномен иммуноклеточного прилипания, розеткообразования
или скучивания).
В диагностике инфекционных заболеваний все шире исполь-
зуют методы с применением меченых антигенов и антител. Осно-
воположником иммунофлуоресцентного метода считается
А. X. Кунс, впервые применивший его для обнаружения анти-
генов пневмококков (1942). В настоящее время иммунофлуорес-
ценция методически высоко совершенна, с ее помощью осуществ-
ляют индикацию одиночных искомых микроорганизмов прямо
в мазках среди других бактерий. Предварительный посев на
питательные среды не требуется. Не требуются также меченые
атитела против множества определяемых бактерий. Необходимо
иметь набор обычных кроличьих антител против этих бактерий и
всего одну меченную флуоресцеином антисыворотку против кро-
личьих иммуноглобулинов. Последняя вызывает свечение бакте-
рий, к которым присоединились специфические кроличьи антите-
ла. Иммунофлуоресцентным методом удается не только выявить
бактерии, но и определить их форму. Вирусный антигенный
428
материал можно обнаружить только после его соответствующей
агрегации, так как нижняя граница разрешающей способности
данного метода находится в пределах 0,2 мкм. Иммунофермен-
тный метод применяют для выявления молекул антигенов, ло-
кализованных внутри или на поверхности клеток.
Иммуноферритиновый анализ (ИФА) позволяет определить не
только специфические антитела, но и микробные антигены в раз-
личных тканях и биологических жидкостях организма. Фермен-
тные метки стали применяться с 1970 г. и к настоящему
времени приобрели большое распространение. В применении
ИФА в микробиологии можно выделить следующие основные
направления: 1) ранняя и ретроспективная диагностика инфек-
ционных заболеваний; 2) эпидемиологическая оценка уровня
заболеваемости (выявление больных, носителей при массовом
обследовании здорового населения, контроль эффективности вак-
цинации населения); 3) количественное определение специфиче-
ских реагентов (антигенов и антител) при производстве вакцин и
сывороток; 4) выявление аллергенов; 5) использование ИФА
в научных исследованиях (механизм патогенеза, генная и кле-
точная инженерия, разработка новых вакцин и сывороток,
структура антигенных детерминант и т. д.). Результаты ИФА
коррелируют с результатами, получаемыми при использовании
общепринятых методик постановки РСК, реакции непрямой ге-
магглютинации, иммунофлуоресценции и др., но в десятки и сот-
ни раз превосходят их по чувствительности, сохраняя при этом
высокую специфичность. Чувствительность ИФА близка чувстви-
тельности РИА, но значительно превосходит его по простоте
постановки и безопасности, так как не требует радиоактивных
изотопов. Перечисленные преимущества позволили применить
данный метод для диагностики многих заболеваний бактериаль-
ной, паразитарной и вирусной природы.
Одним из самых распространенных вариантов ИФА, исполь-
зуемых для диагностики бактериальных инфекций, является
ELISA (от англ. Enzyme-linked imunosorbent assay — фермент-
связанное иммуносорбентное определение) — разновидность ге-
терогенного ИФА. Применяется в макро- и в микромодифика-
циях.
Радиоиммунологический анализ в диагностических целях ис-
пользуется при определении опухолевых антигенов, лекарствен-
ных препаратов, вирусов, реагиновых антител и главным образом
различных гормонов, содержащихся в жидкостях организма в
тех малых концентрациях, которые лежат в пределах чувстви-
тельности данного метода (нано- и пикограммы).
В научно-исследовательских лабораториях радиоиммуноопре-
деление применяют для изучения антигенности слабоиммуноген-
ных молекул (микробных полисахаридов и др.), которая не вы-
является широко распространенными, но менее чувствительными
методами.
Для выявления аллергического состояния организма in vivo
429
применяют кожные диагностические пробы. В качестве аллерге-
нов при инфекционной аллергии используют микроорганизмы
в виде вакцин и различные фракции (белковые, полисахарид-
ные и др.) микробных клеток. Аллергены вводят разными
способами, но наиболее распространены внутрикожные и накож-
ные пробы. В тех случаях, когда проведение кожных проб опасно
для больного, применяют методы аллергодиагностики in vitro.
Большинство таких методов основано на визуальной регистрации
степени специфического повреждения клеточных элементов крови
или клеток-мишеней тканей, возникающего при реакции анти-
ген— антитело. Так, для определения ГНТ используют базо-
фильный тест Шелли, при котором в положительном случае изме-
няется форма базофилов и полностью исчезают клеточные гра-
нулы, прямую и непрямую реакции дегрануляции тучных клеток,
для определения ГЗТ — реакции повреждения нейтрофильных
лейкоцитов по Фрадкину, бласттрансформации лимфоцитов,
торможения миграции макрофагов.
Выявить этиологию аллергического процесса оказалось воз-
можным с помощью антигенспецифических IgE, т. е. антител
против тех или иных конкретных аллергенов (пищевых, пыльце-
вых, микробных и др.). Такие антигенспецифические IgE обра-
зуются в сыворотке крови при ГНТ в очень малых количествах
и для их определения лучше всего подходит радиоиммунологиче-
ский анализ. Идентификацию специфических антител осуществ-
ляют с помощью специально разработанного радиоаллергосор-
бентного теста (РАСТ). Определение специфических IgE при
помощи РАСТ является ценным методом современной аллерго-
диагностики, так как наряду с высокой чувствительностью он
полностью безопасен для больного и его использование возможно
в период острых проявлений аллергии и во всех других случаях,
когда кожные тесты противопоказаны.
8.6. ВЛИЯНИЕ ЛЕКАРСТВЕННЫХ СРЕДСТВ
НА ИММУННУЮ СИСТЕМУ
Различные лекарственные средства могут приводить к опреде-
ленным нарушениям в деятельности иммунной системы в целом
или отдельных ее компонентов, когда опасность возникновения
инфекционных заболеваний существенно возрастает. В других
случаях некоторые препараты-иммуносуппрессоры необходимы в
трансплантационной хирургии, и риск возникновения инфекцион-
ного заболевания ставят на второй план, памятуя о возможности
применения в этом случае химиотерапевтических средств. При
оценке структур, являющихся мишенями для действия иммуно-
суппрессоров, полезна схема иммунных процессов, приведенная
на рис. 79.
Первые шаги в обработке антигенной информации выполняют
макрофаги, или мононуклеарные фагоциты, и вспомогательные
дендритные клетки, которые затем поставляют ее лимфоцитам.
430
Процесс
Участники процесса
антиген/Streptococcus species
антител с
антигеном
итоговый
антитела в сыворотке
ГЗТ и клеточный иммуннитет
Рис. 79. Схема иммунных процессов в организме
Способность лимфоцитов взаимодействовать со специфическими
антигенами генетически детерминирована. С помощью сканирую-
щей электронной микроскопии показано, что поверхность у В- и
Т-клеток различна. Долгоживущие клоны малых лимфоцитов —
клетки, признающие антиген и преимущественно связывающиеся
с ним. Они находятся под влиянием гормонов тимуса, образую-
щихся эпителиальным компонентом зобной железы. Пролифе-
рация таких клонов антиген-признающих Т-клеток завершается
развитием клеточного иммунитета, проявляющегося в ГЗТ и от-
торжении чужеродной ткани при их пересадках (за счет прямого
цитотоксического взаимодействия и освобождения различных
лимфокинов).
Лимфоидные клетки, предшественники В-клеток, происходят
431
из костного мозга. В-клетки содержат на своей поверхности
(на мембране) моноклональные иммуноглобулины, выполняющие
роль антигенных рецепторов. Они располагаются на верхушках
500... 1000 микроворсинок. Синтетическая активность В-клеток
возрастает при дифференциации в плазматические клетки, секре-
тирующие в больших количествах антитела одного из пяти клас-
сов. В нормальных условиях большинство компонентов иммунной
системы (см. разд. 8.1) находится в репрессированном (подав-
ленном) состоянии до тех пор, пока они селективно не активи-
руются для специфического иммунного ответа.
Поскольку иммунная система — главный барьер для микроор-
ганизмов (включая онкогенные вирусы), токсинов и чужеродных
клегок, генерализованная иммуносуппрессия весьма опасна для
микроорганизма. При этом возрастает опасность возникновения
инфекционных заболеваний, лимфоретикулярных и других форм
злокачественных опухолей. Следует иметь в виду, что блокиро-
вать или ослабить развившийся иммунный ответ труднее, чем
первичный иммунный ответ. Все средства, влияющие на иммуно-
компетентность макроорганизма, можно подразделить на две
группы: иммуностимуляторы и иммуносунпрессоры.
Иммуностимуляторы применяют главным образом при имму-
нодефицитах, хронических инфекционных заболеваниях и раке.
К ним 'относятся: левамизол, вакцинный штамм туберкулезных
микобактерий, некоторые коринебактерии, грибные полисахари-
ды, бактериальные липополисахариды, мурамилпептиды, тимо-
зин, трансфер-фактор, двунитевые полирибонуклеотидные комп-
лексы. Левамизол впервые использован для лечения протозойных
инфекционных и глистных
левамизол
заболеваний. Это левовращающий изомер 2,3,4,5-тетрагидро-6-
фенилимидазотиазола, или тетрамизол. Он рекомендуется как
иммуномодулятор при иммунодефицитах, связанных с нарушени-
ем функций Т-лимфоцитов, макрофагов; при аутоиммунных за-
болеваниях (ревматоидный артрит, афтозный стоматит и др.),
при некоторых хронических инфекционных заболеваниях (вирус-
ный гепатит типа В, рецидивирующие пиодермиты и пр.). Он мо-
жет снижать количество лейкоцитов в крови, поэтому при приеме
левамизола необходимо контролировать картину крови.
Вакцинный штамм микобактерий, входящий в состав живой
вакцины БЦЖ, оказался иммуностимулятором при лечении рака.
Он активирует макрофаги, усиливая их эффекты взаимодействия
с лимфоцитами и удаления иммунных комплексов (например,
антителорастворимый опухолевый антиген) из крови. Вакцину
БЦЖ с определенным успехом использовали при меланомах,
саркомах и острой лейкемии. Аналогичное действие проявляют
липидные препараты — экстракты из вакцинного штамма БЦЖ-
432
В качестве неспецифических иммуностимуляторов на клеточ-
ном уровне выступают полисахариды из грибов (лентинан, па-
химан и пахимаран, шизофиллан, родэксман и др.), бактериаль-
ные липополисахариды, а также мурамилпептиды. Некоторые
дрожжевые глюканы (зимозан), маннаны, гликоконъюгаты (про-
дигиозан и др.) стимулируют интерфероногенез в макроорга-
низме. В небольших дозах они активируют клеточную базу анти-
телообразования, повышают процентное содержание лимфоци-
тов, усиливают фагоцитарную активность лейкоцитов и функцио-
нальную активность макрофагов.
На основании изучения многих синтетических производ-
ных мурамилдипептидов установлено, что они могут быть
подразделены на 3 группы: умеренноактивные, высокоактивные
и неактивные. В основе их находится Ы-ацетил-мурамил-£-ала-
нил-Д-изоглутамин, или сокращенно МурЫАц-Л-Ала-изо-Глн.
К умеренноактивным препаратам относятся вещества, содержа-
щие вместо D-изо-Глн следующие заместители: Д-ГлуОН, Д-Глу-
ГлиОН, Д-Глу-ГлиЫНа и др.; к высокоактивным — с замести-
телями вместо D-изо-Глн: Д-цзо-Глн (ОСН3), Д-Глу (NHCH3)
и др.; к неактивным — МурЫАц, МурЫАц-Д-АлаОН, Д-лактил-
L-Ала-Д-изо-Глн и пр. Высоко- и умеренноактивные мурамил-
дипептиды в неполном адъюванте Фрейнда (без клеток мико-
бактерий) индуцируют быстрое образование антител типа IgG
в высоких титрах.
Мур МАц-£-Ала-изо-Глн
Тимозин — белковый гормон зобной железы с молекулярной
массой порядка 10 кДа. Он передает Т-клеточную специфичность
недифференцированным лимфоидным стволовым клеткам. Тимозин
повышает число Т-клеток, несущих поверхностные маркеры, и
активирует их функцию. Препарат рекомендуют при Т-клеточных
иммунодефицитах, в том числе при некоторых инфекционных
заболеваниях.
Предполагают, что фактор переноса или трансфер-фактор —
либо небольшая молекула РНК, либо пептид (А1 « 5 кДа) —
вырабатывается нормальными лимфоидными клетками человека.
Фактор переноса видоспецифичен и иммунологически специфичен
к конкретным антигенам (в том числе микробным), хотя они мо-
гут выполнять роль адъювантов или суперантигенов. Развитие
клеточного иммунитета к антигенам обусловлено наличием этого
фактора. Под его влиянием отрицательная ГЗТ переходит в по-
433
ложительную. Такой эффект одной дозы трансфер-фактора про-
должается до 6 месяцев. Однако следует помнить о неблагопри-
ятном влиянии его при кандидозах кожи и слизистых оболочек,
кокцидиоидозе, лепре, туберкулезе, остеогенной саркоме. Дозиро-
вание фактора основано на подсчете числа лимфоцитов, из
которых он был получен. За одну дозу принято количество
фактора переноса, получаемого из 109 лимфоцитов.
Двунитевые полирибонуклеотидные комплексы (ДПК) полу-
чают полимеризацией мононуклеотиддифосфауов при каталитиче-
ском действии полинуклеотидфосфорилазы Streptomyces lisodeic-
ticus. ДПК полиаденилуридил (поли-А:У) связывается с мембра-
ной кортизон-чувствительных тимоцитов и индуцирует секрецию
хелпер-фактора из Т-лимфоцитов-хелперов. Следовательно, ми-
шенью поли-А:У являются Т-лимфоциты. Этот комплекс преду-
преждает возникновение толерантности к Т-зависимому антигену.
ДПК полиинозилцитидил (поли-И:Ц) и поли-А:У выступают
митогенами для В-лимфоцитов. Первый из них также заметно
активирует киллер-клетки. Связанный с поли-Ь-лизином и карбо-
ксиметилцеллюлозой, поли-И:Ц (поли-ИЦЛЦ) проявляет выра-
женное интерфероногенное и иммуноадъювантное действие.
Вследствие этого он полезен для борьбы с вирусными и некото-
рыми другими инфекциями.
Иммуносупрессоры применяют в трех случаях — при аутоим-
мунных нарушениях, при изоиммунных нарушениях и при транс-
плантации органов и тканей. К аутоиммунным заболеваниям
относятся: идиопатическая тромбоцитопеническая пурпура, ауто-
иммунная гемолитическая анемия, острый гломерулонефрит;
изоиммунным заболеванием является гемолитическая болезнь
новорожденных.
Преднизон оказывается препаратом выбора при указанных
иммунопатологиях. В табл. 40 приведены иммуносупрессивные
агенты, используемые с хорошим эффектом при соответствую-
щих заболеваниях.
СН2—NH^ ^СН2—СН2С1
!н2 О=Р—N
СН2—О 'Х'сн2—СН2С1
6- меркаптопурин
азатиоприн (имуран)
nh2
НС—сн
СН2—СН2С1
хлорамбуцил
циклофосфамид
434
Таблица 40. Практические пути использования иммуносупрессоров
Заболевания Рекомендуемые иммуносупрессоры
синтетические биологические
Аутоиммунные идиопатическая тромбоцитопеническая пурпу- ра Преднизон (реже меркаптопурин, или азатиоприн) —
гемолитическая анемия Преднизон, цикло- фосфамид, хлорам- буцил, меркаптопу- рии, азатиоприн —
острый гломерулонефрит Преднизон, мер- каптопурин, цикло- фосфамид —-
Изоиммунное гемолитическая анемия ново- рожденных — Человеческий Rho(D) иммунный глобулин
Связанные с необ- ходимостью пересад- ки костного мозга Циклофосфамид, преднизон Антилимфоцитар- ный глобулин
почек Азатиоприн, пред- низон Антилимфоцитар- ный глобулин; дакти- номицин или актино- мицин D; блеомицин; циклоспорин А; лим- фоцитарные кейлоны
сердца и пе- чени
Антибиотики актиномицин D (дактиномицин) и блеомицин
оказывают не только иммуносупрессивное действие, но также
применяются при некоторых формах злокачественных новооб-
разований. Дактиномицин включает хромофор актиноцин и две
пептидные петли, каждая из которых представляет собой пента-
пептид. Антибиотик ингибирует ДНК-зависимый синтез РНК.
Блеомицин — многокомпонентный продукт. Относится к глико-
пептидным антибиотикам. Он ингибирует клеточный митоз и
синтез ДНК- Компонентами (R) блеомицина могут быть:
А1;H2N— (СН2)3-SO-CH3 ДМА2:H2N- (СН2)3-S-CH3
A2:H2N—(СН2)3—S+=(CH3)2 B2:H2N—(СН2)<—HN—С—NH2 и др.
II
NH
435
Циклоспорин А — антибиотик, продуцируемый Cylindrocarpon
lucidum, обладает противогрибковым действием, но, что более
важно, он оказался выраженным иммуносупрессором. Цикло-
спорин А — циклический полипептид (М = 1202 Да):
Ала—Ала—*-МеЛей—»-МеЛей
I 1
МеЛей МеВал
I I
Вал N—СН3
МеЛей-*-МеГли-*—АБу~*-СО-*-СН—СН—СН—СН,—СН=СН—СН,
ОН (L3
аминобутират циклоспорин А
436
Кейлоны (от греч. xrpXov — замедлять, ослаблять) — вещест-
ва полипептидной или гликопептидной природы, образуемые
разными клетками макроорганизма (железами внутренней секре-
ции, кожей, лимфоцитами) и способные блокировать разные фазы
клеточного цикла — либо фазу Gi (кейлоны первого типа), либо
фазу G2 (кейлоны второго типа). Тем самым они удлиняют про-
должительность митотического цикла.
Кейлон лимфоцитов с М = 30...50 кДа — гликопептид, со-
стоящий из 12 аминокислотных остатков. Кейлон из грануло-
цитов представляет собой циклопептид с М а 600 Да. Их целе-
сообразно сочетать с некоторыми иммуносупрессорами — тио-
фосфамидом, блеомицином и актиномицином D. Rho (D) имму-
ноглобулин человека представляет собой концентрированный
(15 %) раствор IgG, содержащий высокий титр антител против
Rho(D) антигена эритроцитов. Его готовят из тщательно отоб-
ранной плазмы крови, взятой от Rho — негативных матерей, сен-
сибилизированных повторными несовместимыми (по резус-факто-
ру) беременностями, или от резус-отрицательных добровольцев,
целенаправленно иммунизированных Rho(D) антигеном.
Если ввести матери Rho(D) антитела в течение 3 суток после
рождения резус-отрицательного ребенка, то собственный ответ ан-
тителообразованием на чужеродные резус-положительные клетки
суппрессируется. При последующих беременностях не наблюдает-
ся гемолитической болезни у новорожденных. Введение Rho (D)
иммуноглобулина полезно и с профилактической точки зрения ре-
зус-отрицательным женщинам, ранее не подвергавшимся иммуниза-
ции Rho(D)-фактором. Вместе с тем следует помнить, что чело-
веческий Rho(D) иммуноглобулин никогда нельзя вводить детям!
Как видно из табл. 40, перечень иммуносупрессоров огра-
ничивается незначительным набором препаратов. Необходимы
совместные усилия различных специалистов в целях создания
новых эффективных средств профилактики и лечения нарушений
иммунной системы.
Известно также, что лекарственные средства нередко активи-
руют иммунную систему в нежелательном направлении, что ведет
к развитию побочных реакции, называемых лекарственной аллер-
гией. Такие реакции проявляются в виде поражений кожи, отека,
лихорадки, анафилактоидных реакций, эозинофилии. Одни люди
отличаются необычайно высокой чувствительностью к какому-
либо лекарственному средству, тогда как другие, напротив,
устойчивы к нему. Но если доза этого лекарства будет чрезмер-
ной, то все пациенты ответят на него какой-либо реакцией.
За исключением немногих белковых препаратов, большинство
лекарств не обладает свойствами полноценных антигенов, однако
многие из них способны быть гаптенами (см. разд. 8.2), всту-
пающими в химическое взаимодействие с некоторыми гомологич-
ными белками in vivo. Измененные таким образом белки стано-
вятся антигенноактивными (чужеродными) и молекула лекарст-
венного вещества в них детерминирует специфичность. В этом
437
плане достаточно хорошо изучен пенициллин, производное кото-
рого — пеницилленовая кислота (реактивный метаболит) ответ-
ственна за антигенную специфичность комплекса пенициллено-
вая кислота — белок:
HSX
R—C=N—С=СН ХС=(СН3)2
III I
О----С HN ----СН—СООН
II
О
пеницилленовая кислота
К аллергическим проявлениям, возникающим на лекарства,
относятся следующие реакции немедленного типа: анафилаксия,
крапивница и ангионевротический отек, сывороточная болезнь и
сходные с нею состояния, астма и риниты, васкулиты, лекарст-
венная лихорадка, тромбоцитопеническая пурпура, диссеминиро-
ванная красная волчанка, гемолитическая анемия; реакции за-
медленного типа проявляются в виде различных кожных пораже-
ний (малые и большие сыпи, атопические и экзематозные дер-
матиты, эксфолиативные дерматиты), агранулоцитоза и апласти-
ческой анемии.
Реакции аллергии немедленного типа связаны с функцио-
нальной активностью IgE-антител, или реагинов (см. разд. 8.4).
В норме у здоровых индивидуумов его содержится меньше, чем
1 мкг/мл, тогда как при аллергии количество его возрастает
блокада
вазоактивные пептиды
антигистаминными
препаратами
Рис. 80. Развитие реакций аллергии немедленного типа на введение аллер-
генов (включая лекарственные средства)
438
в 10 раз и более. Эти антитела способны фиксироваться на туч-
ных клетках и на базофилах крови (тучные клетки — аналоги
базофилов в тканях), что, как правило, реализуется в острой
аллергической реакции. При повторном введении лекарства моле-
кулы IgE-антител, фиксированные на поверхности базофильных
лейкоцитов, связываются с антигенной формой лекарства. При
этом базофилы и тучные клетки дегранулируются и освобождают
ряд высокоактивных медиаторов, определяющих проявление ал-
лергии фис. 80). Процесс дегрануляции базофилов связан с воз-
растанием внутриклеточного содержания цАМФ. Вещества, тор-
мозящие процесс разрушения цАМФ (теофиллин) или активи-
рующие синтез его (катехоламины), являются блокаторами про-
цесса высвобождения гистамина из базофилов. Сущность реак-
ций замедленного типа (ГЗТ) изложена в разд. 8.4.
Таким образом, при назначении того или иного лекарствен-
ного средства (включая химиотерапевтическое) желательно
знать иммунологический статус макроорганизма, его чувстви-
тельность к лекарству и другим чужеродным агентам. Это позво-
лит сократить до минимума развитие побочных реакций, иног-
да — очень тяжелых.
Вопросы
1. Из каких компонентов состоит иммунная система (их характерис-
тика)?
2. Что такое иммунитет и как его подразделяют на типы, виды и формы?
3. Какие реакции положены в основу иммунохимии?'
4. Какие известны стадии фагоцитоза и какие биохимические процессы
. активируются при этом в фагоцитах?
5. Какие вещества относят к разряду гуморальных неспецифических
факторов защиты организма?
6. Какими свойствами обладают макрофаги, В- и Т-лимфоциты?
7. Какие известны медиаторы иммунной системы, участвующие в реа-
лизации клеточного иммунитета?
8. Какие известны медиаторы иммунной системы, участвующие в реа-
лизации гуморального иммунитета?
9. Что такое антигены и какие известны антигены? Какими характе-
ристиками они должны обладать?
10. Что представляют собой искусственные антигены и каковы подходы
к их созданию?
11. Что такое антитела, каковы их химический состав и архитектоника?
12. Какие существуют типы иммуноглобулинов, в чем их химическое
и функциональное различия?
13. Какие известны иммунохимические реакции взаимодействия антиге-
нов с антителами (их механизм)?
14. Что означает ГИТ и ГЗТ, в чем заключается их сущность?
15. В каких направлениях на практике используют иммунохимию?
16. Какие вещества относят к иммуномодуляторам и в чем выражается
механизм их действия?
* *
*
Химическая микробиология как наука сформировалась на
стыке микробиологических и химических дисциплин и, по сущест-
ву, отражает глубину проникновения человеческой мысли в мир
439
микроскопических существ. С учетом общих закономерностей
строения и функциональной активности клеток различного проис-
хождения микроорганизмы нередко используют как модельные
системы для выяснения биохимической сущности наблюдаемых
явлений в живой природе. В качестве примеров можно назвать
состав и организацию мембран в клетках эукариотических
организмов, находящихся на разных ступенях эволюционного
развития; реакции биологического окисления или дыхания в
аэробных и анаэробных условиях и т. д. Более того, фундамен-
тальные основы деятельности наследственного материала оказа-
лись принципиально сходными у различных организмов. Вот по-
чему вирусы как организованные частицы (не клетки!) по праву
относятся к разряду микроскопических существ, стоящих на
грани между живой и неживой природой; их нуклеиновые кисло-
ты «срабатывают» в основном так же, как и нуклеиновые
кислоты в клетках других микроорганизмов. Отсюда становится
понятным, почему к объектам химической микробиологии отне-
сены вирусы, бактерии, грибы, водоросли, лишайники и протозоа,
т. е. существа микроскопических размеров.
Учитывая самостоятельность таких дисциплин, как вирусоло-
гия, бактериология, микология, альгология, лихенология и про-
тистология, содержание химической микробиологии изложено
в сравнительном плане, т. е. каждая глава и каждый раздел
включают материалы о всех представителях микроорганизмов.
Это позволяет сравнить, например, химический состав, строение
и функцию соответствующих веществ, а также органелл и вклю-
чений у представителей различных систематических групп. Изло-
жение в таком виде помогает составить целостное представление
о полноте раскрытия фактических данных о химическом составе,
строении вирусов и клеток, локализации в них микро- и макро-
молекул, функциональной активности структурных компонентов,
о химических основах процессов роста, развития и дифферен-
цировки микроорганизмов, их метаболизме и его прикладном
значении в биотехнологии, о химии факторов агрессии и патоген-
ности у болезнетворных микробов, об иммунохимии.
Многолетний педагогический опыт автора — главный побуди-
тельный мотив к написанию первого учебника «Химическая
микробиология» для студентов, и, если этот учебник послужит
делу совершенствования подготовки кадров по микробиологиче-
ским дисциплинам на современном этапе, то автор будет считать
свою задачу выполненной.
литература
Биотехнология//Под ред. А. А. Баева. — М.: Наука, 1984.
Биотехнология микробного синтеза//Под ред. М. Е. Бекера. — Рига: Зинат-
не, 1980.
Болдырев А. А. Биологические мембраны и транспорт ионов. — М.: Наука,
1986.
Бутенко Р. Г. и др. Клеточная инженерия//Под ред. Н. С. Егорова, В. Д. Са-
муилова. — М.: Высшая школа, 1987.
Генетика и наследственность. — М.: Мир, 1987.
Готтшалк Г. Метаболизм бактерий. — М.: Мир, 1982.
Дубинин Н. П., Пашин Ю. В. Мутагенез и окружающая среда.—М.:
Г|аука, 1978.
Блинов Н. П. Химия микробных полисахаридов. — М.: Высшая школа,
1984.
Захаров И. А., Мацелюх Б. П. Генетические карты микроорганизмов (спра-
вочное пособие). — Киев: Наукова думка, 1986.
Зенгбуш П. Молекулярная и клеточная биология. — М.: Мир, 1982.
Иммунология (справочник)//Под ред. Г. Бундшу и Б. Шнеевайса. — Киев:
Нау кода думка, 1981.
Красильников А. П. Микробиологический словарь-справочник. — Минск:
Беларусь, 1986.
Ленинджер А. Основы биохимии. — М.: Мир, 1985. Т. 1—3.
Лукнер М. Вторичный метаболизм у микроорганизмов, растений и живот-
ных.— М.: Мир, 1979.
Льюин Б. Гены. — М.: Мир, 1988.
Молекулярные и клеточные аспекты биотехнологии//Сб. научных тру-
дов//Под ред. С. Г. Инге-Вечтомова. —Л.: Наука, 1986.
Мусил Я-, Новакова О., Кунц К. Современная биохимия в схемах.—М.:
Мир, 1981.
Основы биохимии//Под ред. А. А. Анисимова. — М.: Высшая школа,
1986.
Петров Р. В. Иммунология. — М.: Медицина, 1987.
Перспективы биохимических исследований//Под ред. Дж. Туза и С. Пренти-
са. — М.: Мир, 1987.
Рекомбинантные молекулы: значение для науки и практики//Под ред.
Р. Бирса и Э. Бэсита. — М.: Мир, 1980.
Реннеберг Р. Эликсиры жизни. — М.: Мир, 1987.
Сассон А. Биотехнология. Свершения и надежды. —М.: Мир, 1987.
Спирин А. С. Молекулярная биология. Структура рибосом и биосинтез
белка. — М.: Наука, 1986.
Справочник по биологии//Под ред. К. М. Сытника. — Киев: Наукова думка,
1985.
Стейнер Р., Эдельберг Э., Ингрэм Дж. Мир микробов.—М.: Мир, 1979.
Т. 1—3.
Стент Г., Кэлиндар Р. Молекулярная генетика. — М.: Мир, 1981.
Стимуляция жизнедеятельности микроорганизмов и вирусов//Под ред.
М. X. Шигаевой. — Алма-Ата: Наука, 1986.
Строев Е. А. Биологическая химия. — М.: Высшая школа, 1986.
Уайт А., Хедлер Ф., Смит Э., Хнлл В., Леман И. М. Основы биохимии. —
М.: Мир, 1981. Т. 1—3.
Фрейдлни И. С. Система мононуклеарных фагоцитов. — М.: Медицина,
1984.
Экологическая роль микробных метаболитов//Под ред. Д. Г. Звягинцева. —
М.: Изд-во МГУ, 1986.
441
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Авидность 411
Агглютинация 412, 416, 427—428
Адаптация 161
Адьюванты 424
Азатиоприн 434—435
Азелаиновая кислота 290
Акразины 197
Актиномицин Д 435
Актинофаг 12
Аллерген 388, 430
Аллергия 422, 438, 437
Аллостерическая активация 218
Аминокислоты 46—50, 112
— гликогенные 254
— кетогенные 254
Амины органические 364
Амфиболизм 21, 206, 257
Амфиболические пути 264
Амфотерицин В 331
Анабиоз 21, 108
Анаболизм 21, 206, 257, 264, 285
Анаплеротическая реакция 278
Анатоксин 424
Антагонизм 368
Антаманид 356—357
Антибиоз 21
Антибиотики 322
— аминогликозидные 325
— антрациклины 339
— макролиды 331
— циклопентиты 341
Антибиотикотерапия 366
Антигены 381, 387—400
— валентность 393
— внутриклеточные 398
— гетерофильные 427
— жгутиковые (Н-анТигены) 397—398
— капсульные 398
— конъюгированные 395
— протективные 398
— толерогенная активность 393
Антиидиотипические антитела 409
Антипорт 125
Антитела 400—410
Антитоксин 356
Антраценовые соединения 295
Аппарат Гольджи 131
Аргинин 48, 224
Археобактерии 13
Аскорбиновая кислота 70
Аспарагин 48, 226
Аубазидан 321
Афлатоксин 359—360
Аффинность 408, 411
Аэросомы 27
442
Бактерии 13
— азотфиксирующие 13
— грамотрицательные 16
— грамположительные 15—16
— грациликуты 13
— зеленые 13, 129
— мендосикуты 13
— пурпурные 13, 129
— тенерикуты 13
— фирмикуты 13
Бактериородопсин 130
Бактериофаг 11
Бактопренолы 260
Бациллы 14
Бацитрацин А 243
Белки 46— 50, 79, 82—90, 118
— вторичная структура 88
— индукция синтеза 156—158
— миеломные 401
— первичная структура 83
— репрессия синтеза 156—157, 176
— третичные структуры 89
— четвертичные структуры 89
Биогеоценоз 20
Биотехнология 300, 301
Брожение ацетонобутиловое 306
— ацетоноэтиловое 306
— бутиленгликолевое 268
— капронное 306
— маслянокислое 268, 306
— молочнокислое 268, 305
— пропионовокислое 268, 305
— спиртовое 268, 305
Валин 48, 226
Ванкомицин 342
Вариамицин 336
Вибрионы 14
Вирион 11
Вирогат 425
Вироид 9
Вирулентность 348
Вируснейтрализующие ингибиторы 379
Вирусы 9
Витамины 66—72
Вода 39—41
Водоросли 18
Воспаление 375
Галофилы 16, 43
Гамета 187
Гаптены 388
Гексозомонофосфатный шунт 269
Гемадсорбция непрямая 419
Гемопротеины 249
Ген 138, 157
Генетический код 142, 143
Генная инженерия 303—304
Гентамицин 329
Гены 371—372, 382
— иммунного ответа (1г-гены) 391
— три группы сцепления 406
Гетероантигены 395
Гетерокарион 190
Гиббереллины 345
Гибридома 302, 314
Гигромицин 330
Гидрогеносомы 29, 133
Гидролазы 211
Гиромитрин 364
Гистидин 48, 227
Гифы 16, 195—196
Гликозилдиацилглицериды 291
Гликозилдиацилглицерины 95
Гликокаликс 29
Гликоконъюгаты 57, 92
Гликолиз 265, 305
Гликолипиды 94—96, 358
Гликолипопротеины 97, 358
Гликопротеины 92, 358
Гликосомы 133
Глиоксисомы 132
Глицерин 50
Глицин 48, 228
Глутамин 48, 228
Глутатион 242
Глюконеогенез 257
Грамицидин С 243
Грибы 16
— вегетативное размножение 185
— парасексуальный цикл 189—190
— репродукционное размножение 185
— ценоцитные 35
— эумицеты 17
Гризеофульвин 292, 334
Гужеротин 335
Гуминовые кислоты 294
Декстран 318
Делеция 142
Депсипептиды 243
Детерминанта , антигенная 393—395
Диатрен 289
Диауксия 180
Диплоид гомозиготный 187
— гетерозиготный 187
Диплококки 13
ДНК 22—23, 73—80, 164—167
— антисмысловая цепь 76
— внехромосомная 164—167
— двухмикронная 165—167
— интроны 151
— митохондриальная 165
— ренатурация 78
— репликация 145
— транспозоны 148
— фиксаторы 148
— фрагменты Оказаки 147
— экзоны (домены) 151
Долихолы 260
Домен 401
Дублетное правило 143
Жгутики 30, 135—136
Жиры нейтральные 99
Изоантигены 395
Изолейцин 48, 229
Изо-пирролнитрин 297
Изоферменты (изозимы) 213
Иммунитет 372—375
— медиаторы 385—386, 426—427
Иммунная система 370
Иммунный фагоцитоз 376
Иммуногенность 389
Иммуноглобулины, см. антитела
Иммунология 373
Иммуностимуляторы 432—434
Иммуиоциты 370
Иммунохимия 373
Иммуносуппрессоры 434
Иммуноферментный метод 420—421,
429
Иммуноферритиновый метод 420
Иммунофлуоресцентный метод 419
Индола производные 295
Индуктор 156—157
Инженерная энзимология 312
Инозин 345
Инозитол 69
Интерферон 315, 379
Информатины 150
Информоферы 150
Информосомы 150
Ионофоры 125
Исландитоксин 355
Канамицин 328
Капсулы 29, 121
Карбоксисомы 27—28
Кардиолипин 100
Кариоплазма 34
Касугамицин 329
Катаболизм 21, 206, 255—257, 264, 289
Катенаны 148
Кейлоны 437
Кетодезоксифосфоглюконатный путь
272
Киллер-плазмида 34, 170
Кинетии 135
Кинетодесма 135
Кинетосома 135
Кислота аспарагиновая 48, 226
— ауреоловая 335
— глутаминовая 48, 228
— оротовая 72
— пангамовая 72
443
— пантотеновая 67
— парааминобензойная 71
— усниновая 292
— фолиевая 69
— фосфатидная 100
Кислоты жирные 50, 280—282, 289—290
— органические 44—46
Клеточная инженерия 302
Клеточная мембрана, см. плазмалемма
Клон 159
Кобаламины 71
Кодон 143
Компартмент 28
Комплекс инициации 153
Комплемент 378
Конидии 185, 187
Коньюгация 163
Кордицепин 343
Корд-фактор 95
Коремия 185
Корепрессор 156
Кориолан 321
Кофактор 209
Кофермент 208
Кроссинговер 188
Ксантан 319
Ксеноспоры 190
Курдлан 320
Лейцин 48, 230
Лентинан 321
Лептоспиры 14
Лиазы 211
Лигазы 212
Лизин 48, 230
Лизосомы 27, 131 —132
Ликопии 288
Лимфоциты В и Т 381—387
Линкомицин 334
Липиды 98—104
Липоевая кислота 72
Липоконъюгаты 98—104, 285, 289
Липогенез 279
Липополисахариды 96—97, 118—121
Липопротеины 104, 115—116, 358
Лишайники 19
Ломасомы 29
Лютеоскирин 362
Макроконидии 186
Матураза 165
Мейоз 187—188
Мезофилы 40
Мемноспоры 190
Мерозигота 161
Метабиоз 21, 108
Метаболизм 21, 202, 219, 256, 279
Метионин 48, 233
Микостерин 102
МикроконидиИ 186
Микрококки 13
Микроорганизмы
— анаэробы 16
— аэробы 16
— дыхание 274
— жизненный цикл 191
— изменчивость 159—162
— инвазивность 349
— инфекциозность 349
— облигатные паразиты 348
— окисление 307—308
— патогенные 16, 347
— популяция 159
— прокариоты 7
— рост и развитие 172—179
— сапрофитные 16, 347
— сбалансированное размножение 182
— условно-патогенные 348
— факторы агрессии 349
— факторы патогенности 349
— фототрофные 205
— хемотрофные 206
— чистая культура 159
— эукариоты 7, 9
Митотический аппарат 35
Митохондрии 36, 133—135
Модификации 160, 161
Монозы 51—55
Мононуклеотиды 57
Моносахариды 55—56
Морфогенез 191 —194, 198—201
Муреин, см. пептидогликаны
Мускарин 364
Муска ридин 364
Мутагены 160
Мутанты 160
Мутация 40, 157, 160
Мутон 138
Негалофилы 43
НеомиЦин 327
Неосальварсан 365
Никотиновая кислота 71
Новарсенол 365
Новобиоцин 334
Нуклеазы (экзо- и эндо-) 255
Нуклеиновые кислоты 73—82
Нуклеозидазы 256
Нуклеолемма 34
Нуклеосомы 79, 142
Нуклеотидазы 256
Нуклеотиды
— пуриновые 245—248
— пиримидиновые 248—249
Нуклеоцидин 343
Окислительное фосфорилирование 276
Оксидоредуктазы 210
Олеандомицин 332
Оливомицин 337
Олигозы 53—54
Олигопептиды 84
444
Олигосахариды 56
Охратоксины 361
Палиндромы 78
Патогенность 347
Патулин 361
Пахиман 321
Пенициллины 324
Пептидная связь 83
Пептидогликаны 93—94, 109—114, 358
Пернплазматическое пространство 27
Пероксисомы 29, 132
Пикнида 186
Пили 30, 136—138
Пиноцитоз 131
Пиреноиды 18
Пиридоксин 68
Пирогены 380
Плазмалемма 27
Плазмиды 34, 163, 167
Плазмолиз 43
Плектенхимная ткань 196
Полиизопреноиды 287
Поликетиды 292
Полимиксин В 243
Полиморфизм 21
Полипептиды 84
Полисахариды 56—57, 90—92
— капсульные 121 —122, 320
Полисомы 36, 150
Полиурониды 90
Полугаптены 388—389
Порины 118
Поросомы 35
Преднизон 434—435
Преципитация 412, 417
Примазы 146
Пролин 49, 234
Простагландины 290
Простеки 29
Протеины, см. белки
Протеолиз 250
Протозойные организмы 20
Протопласты 25
Профаги 167
Процессинг 81, 150
Псевдопаренхима 196
Псикофуранин 343
Психрофилы 40
Пуллулан 321
Пуромицин 344
Радиоиммунологический анализ (РИА)
421, 429
Рапидосомы 35
Реагины 408
Реакции иммобилизации 416
Рекомбинация генетическая 161
Рекон 138
Реп л иконы 148, 162
Рестриктазы 161, 309
Рибосомы 28, 36
Рибофлавин 66
Ристомицин 341
Рифампицины 334
РНК 80—82
Ррдэксман 322
Сагамицин 327
Сальварсан 365
Сарцины 14
Сегресомы 29
Сексдукция 164
Серин 48, 235
Серотонин 297
Сефадексы 319
Симбиоз 21
Симпорт 125
Синергизм 368
Синтрофия 176
Сирогем 222
Сквален 286
Склероглюкан 322
Скотобактерии 13
Спириллы 14
Спорангии 185
Стафилококки 13
Стенотермофилы 40
Стеригматоцистин 361
Стрептококки 13
Стрептомицины 326
Сульфаниламидотерапия 366
Сферопласты 25
Сфинголипиды 94, 102
Таллом 16
Тейхоевые кислоты 97—98, 115
Тентотоксин 356
Термофилы 40
Терпены 103
Тетракокки 14
Тетрациклин 330
Тиксотропия 32
Тилакоиды 27, 123
Тирозин 49, 235
Токсины 352, 359—365, 398—399
Токсоид 313, 424
Трансдукция 164
Транскриптаза обратная 151
Транскрипция 148
Трансляция 151
Трансмиссивные инфекции 349
Транспозазы 148
Трансфекция 162
Трансферазы 210
Трансферрин 379
Трансформация 162
Треморген 361
Треонин 48, 238
Трепонемы 14
Триацилглицерины 283
Триптофан 49, 235
Туберцидин 344
445
Убихиноны 287
Фагосомы 29
Фагоцитоз 131, 376
Фактор колициновый 34, 138, 168
— плодовитости 34, 138, 163, 168
— устойчивости 34, 138, 168
— элонгации 152
Факторы защиты 377—381
Фаллотоксин 356
Фенилаланин 49, 235
Фенотип 160
Ферменты 157, 206—219
— адаптивные 161, 213
— активный центр 209
— аллостерические белки 218
— константа Михаэлиса 215
— международная единица активности
212
— молекулярная активность 208
— олигомеры 213
— порядок реакций 213—214
— простетическая группа 208
— удельная активность 212
Ферредоксин 133
Фикобилин 128
Фикобилисомы 27, 128
Фитоин 288
Флокуляции реакция 427
Фосфатидилхолин 285
Фосфатидилэтаноламин 284
Фосфатиды 283—285
Фосфоглицериды 99
Фосфолипиды 100, 116—118, 291
Фосфопротеины 359
Фосфорибитиды 99
Химиопрофилактика 369
Химиотерапия
— комбинированная 366
— определение 365
Хитосомы 29, 132
Хлоросомы 27
Холин 68
Холокарпные организмы 184
Хроматин 79, 142
Хроматофоры 18
Хромомицин 338
Хромосома 138, 140, 142
— гаплоидный набор 141, 187
— диплоидный набор 142, 187
Хромоцикломицин 339
Центриоли 35
Церамид 286
Цереброзиды 95
Цефалоспорины 324
Цианофаги 11
Цикл Кребса 274
Циклодекстрины 316
Циклоспорин А 436
Циклофосфамид 434—435
Циклохлоротии 355
Цистеин 48, 239
Цистрои 156
Цитоплазма 31—34
Цитохолазины 361
Цитреовиридии 362
Цитринин 361
Шизофиллаи 321
Штамм 160
Эвритермофилы 40
Экзергонические реакции 206
Экзогенота 161
Экзоцитоз 131
Эндергонические реакции 206
Эндогеиота 161
Эндоплазматический ретикулум 28
Эндоцитоз 131
Эргометрин 296
Эргохромы 295
Эритромицин 333
Эротропины 189
Эукарпные организмы 184
Ядро 34—35
Ядрышко 35
ОГЛАВЛЕНИЕ
Предисловие............................................................. 3
Введение ............................................................... 4
Часть 1
Химические основы структурно-функциональной организа-
ции микроорганизмов
Глава 1. Предмет, объекты, цели и задачи химической микробиологии . 7
1.1. Объекты химической микробиологии........................ 8
1.2. Строение клеток прокариот и эукариот................... 22
Глава 2. Химия микроорганизмов...................................... 37
2.1. Малые и большие молекулы клеток........................ 37
2.1.1. Малые молекулы....................................... 39
2.1.2. Большие молекулы, или макромолекулы.................. 73
2.2. Локализация низкомолекулярных и полимерных веществ
в клетках и организованных частицах.................. 104
Глава 3. Химия и функции основных структур и компонентов микро-
организмов ......................................................'08
3.1. Клеточные стенки и капсулы............................ 108
3.2. Клеточные мембраны и мембранные образования ... 123
3.3. Жгутики и пили........................................ 135
3.4. Ядерный аппарат....................................... 138
3.5. Виехромосомиая ДНК.................................... 164
Часть 2
Химические основы жизнедеятельности микроорганизмов
Глава 4. Химические основы процессов роста и развития микроорга-
низмов ......................................................... 171
4.1. Рост и развитие микроорганизмов....................... 172
4.2. Размножение прокариот и эукариот...................... 179
4.3. Морфогенез и дифференцировка у прокариот и эукариот . 191
Глава 5. Обмен веществ у микроорганизмов.............................. 202
5.1. Ферменты микроорганизмов — биокатализаторы обмен-
ных процессов.........................................206
5.2. Метаболизм белков......................................219
5.2.1. Метаболизм аминокислот...............................219
5.2.1.1. Биосинтез аминокислот..............................223
5.2.1.2. Биосинтез некоторых олигопептидов, пуриновых и пи-
римидиновых нуклеотидов, металлопротеинов ... 241
5.2.1.3. Протеолиз и расщепление аминокислот................250
5.2.1.4. Катаболизм пуриновых и пиримидиновых нуклеотидов . 255
5.3. Метаболизм углеводов.................................. 256
5.3.1. Анаболизм углеводов.................................'257
5.3.2. Катаболизм углеводов................................ 264
5.4. Метаболизм липидов.................................... 279
5.4.1. Биосинтез жирных кислот............................. 280
447
5.4.2. Биосинтез нейтральных жиров (триацилглицеринов) и
фосфатидов (фосфолипидов)..........................283
5.4.3. Анаболизм липоконъюгатов..........................285
5.4.4. Катаболизм липидов и липоконъюгатов.............. 289
5.5. Метаболизм некоторых других вторичных соединений . . 292
Ч а с т ь 3
Прикладное значение химической микробиологии
Глава 6. Химическая микробиология — основа биотехнологии .... 300
6.1. Биотехнология, ее цели и задачи.......................300
6.2. Биотехнологические процессы, основанные на использо-
вании химической активности микроорганизмов .... 302
6.2.1. Брожения и окислительные процессы...................305
6.2.2. Аминокислоты, белки, клеточная биомасса.............308
6.2.3. Углеводы............................................316
6.2.4. Антибиотики......................................322
6.2.5. Нуклеозиды, нуклеотиды, витамины и др...............342
Глава 7. Химические основы патогенности микроорганизмов .... 347
7.1. Патогенность и вирулентность микроорганизмов .... 347
7.2. Химические вещества, определяющие свойства патогенно-
сти микроорганизмов .................................... 349
7.3. Химиотерапия инфекционных болезней............. . 365
Глава 8. Химия противомикробного иммунитета (иммунохимия) . . . 370
8.1. Иммунная система и иммунитет......................... 370
8.2. Химия антигенов...................................... 387
8.3. Химия антител........................................ 400
8.4. Химические основы реакций иммунитета................. 411
8.5. Практическое использование иммунохимии............... 424
8.6. Влияние лекарственных средств на иммунную систему . . 430
Литература........................................................... 441
Предметный указатель................................................ 442
Учебное издание
Блинов Николай Петрович
Химическая микробиология
Зав. редакцией С. Ф. Кондрашкова. Редактор С. С. Трапезникова. Мл. редак-
торы С. М. Ерохина, Л. С. Макаркина. Технический редактор 3. В. Нуждина.
Художественный редактор Е. Д. Косырева. Художник В. М. Боровков. Коррек-
тор С. К- Завьялова
ИБ № 7397
Изд. № ХИМ-837. Сдано в набор 12.10.88. Подп. в печать 20.06.89. Формат 60 X 9O‘/i6.
Бум. кн.-журн. Гарнитура литературная. Печать офсетная. Объем 28,0 усл. печ. л.+ 0,25
усл. печ. л. форзац. 56,5 усл. кр.-отт. 31,48 уч. изд. л. 4- 0,43 уч.-изд. л. форзац . Тираж
19 000 экз. Зак. № 1617. Цена 1 р. 40 к.
Издательство «Высшая школа», 101430, Москва, ГСП-4, Неглинная ул., д. 29/14.
Ярославский полиграфкомбннат Госкомпечати СССР. 150014, Ярославль, ул. Свободы, 97.