




































































































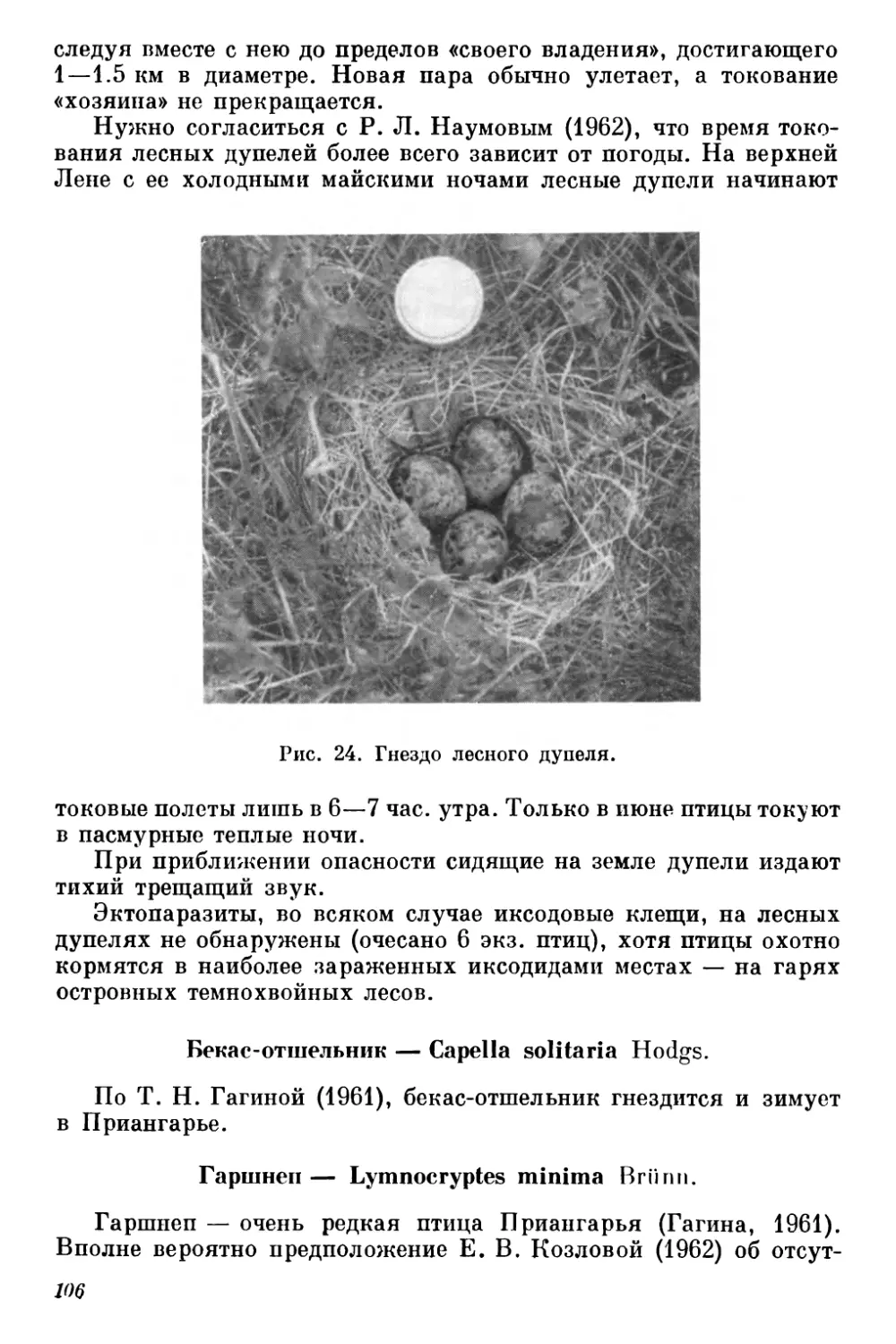



















































































































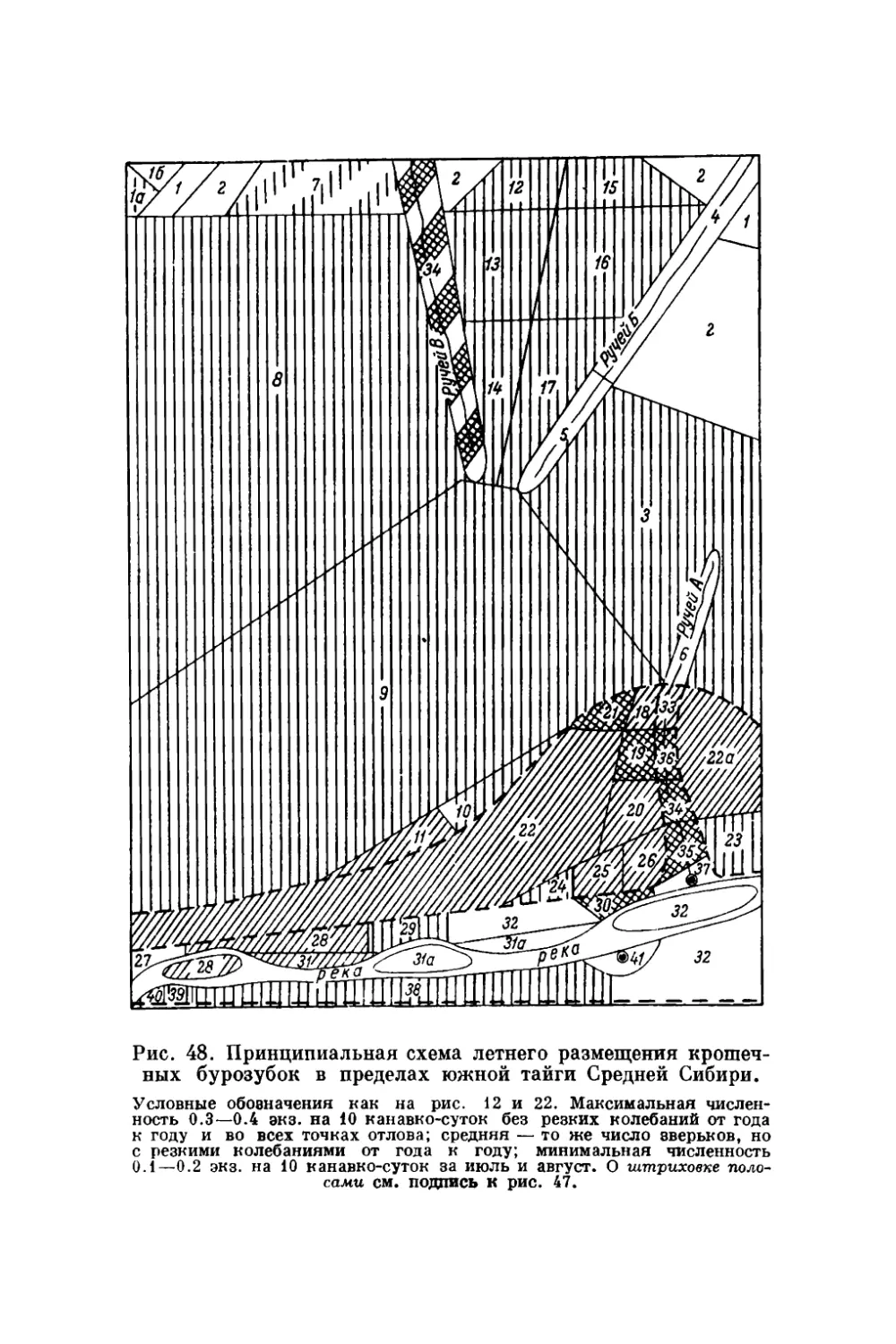








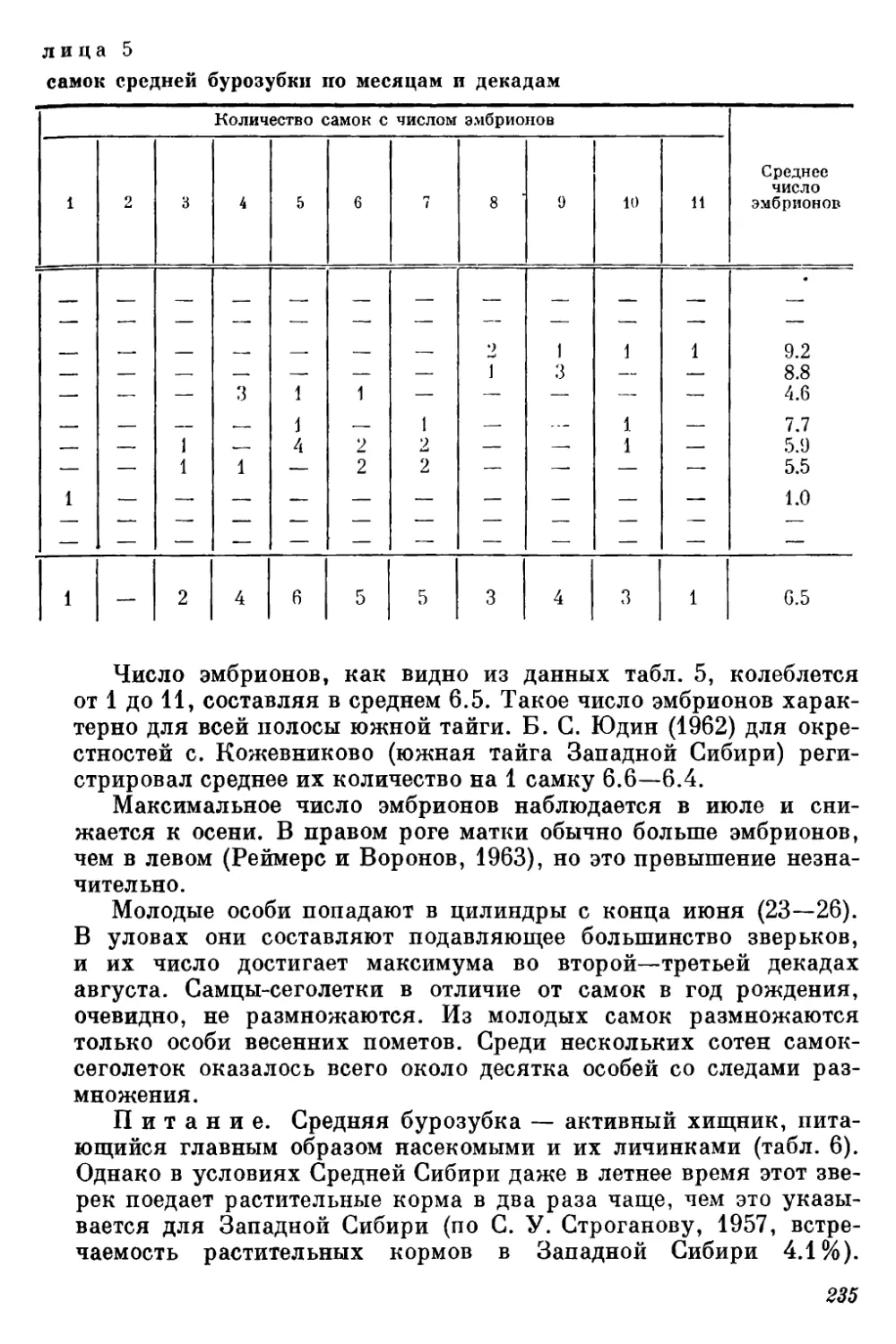
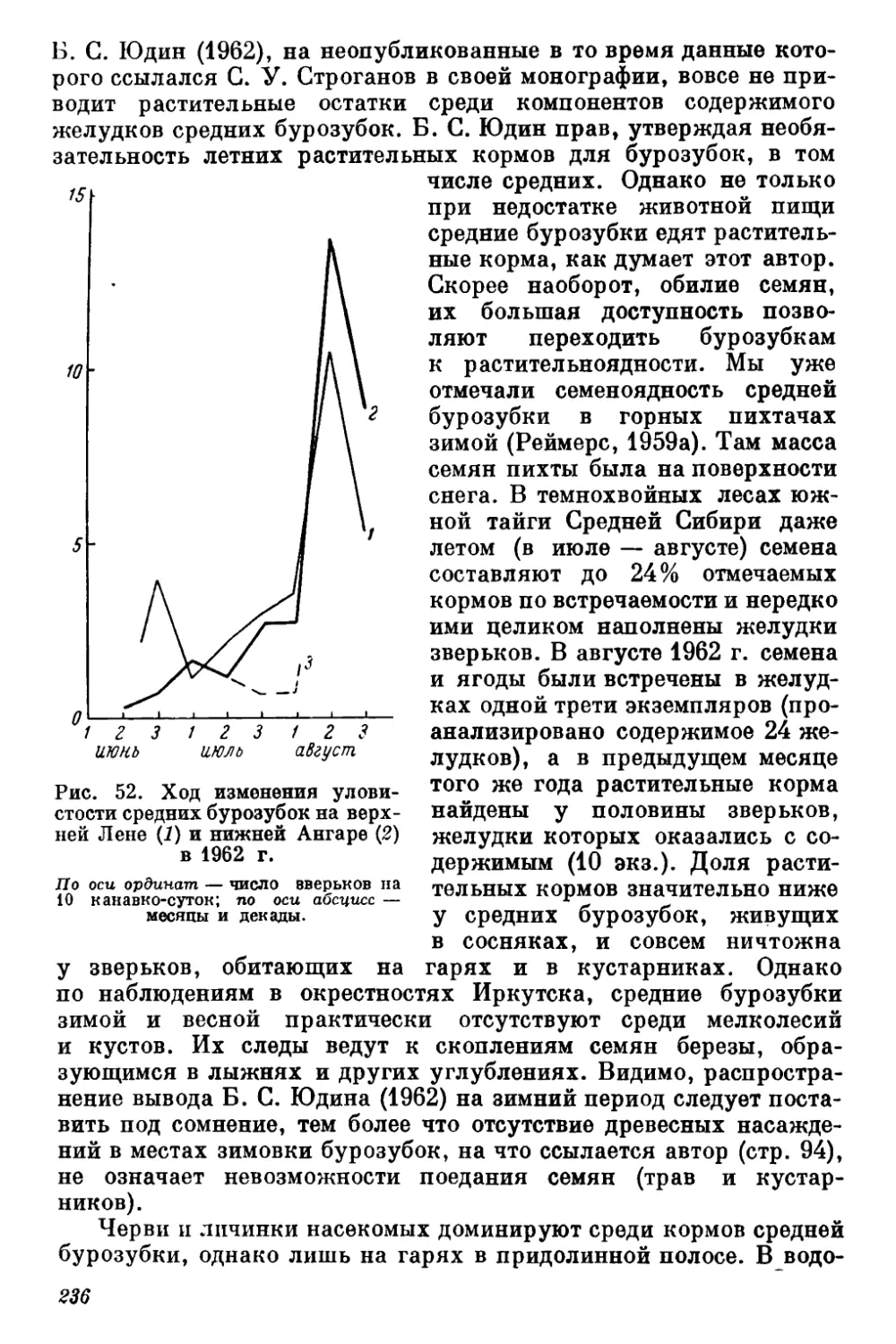









































































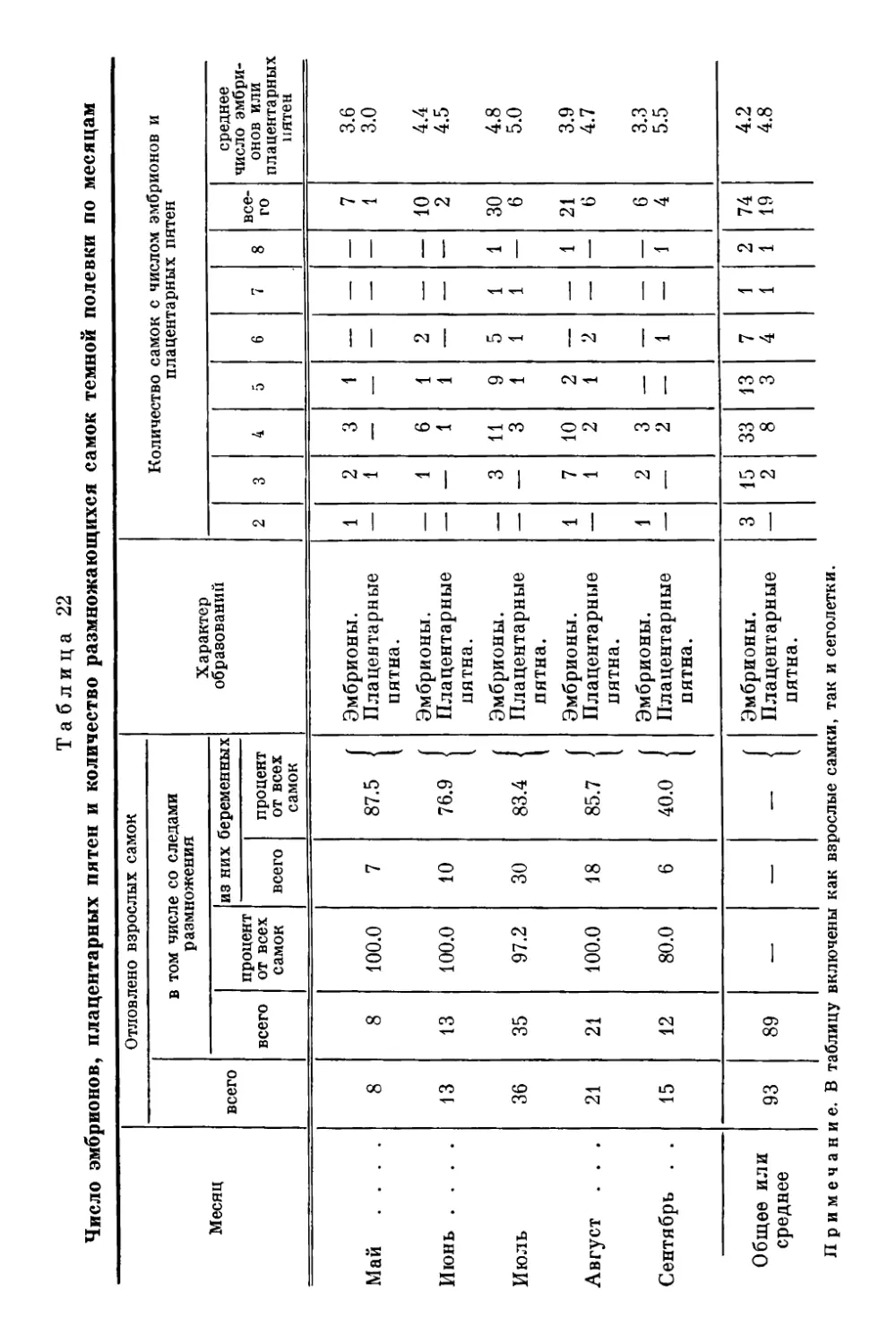
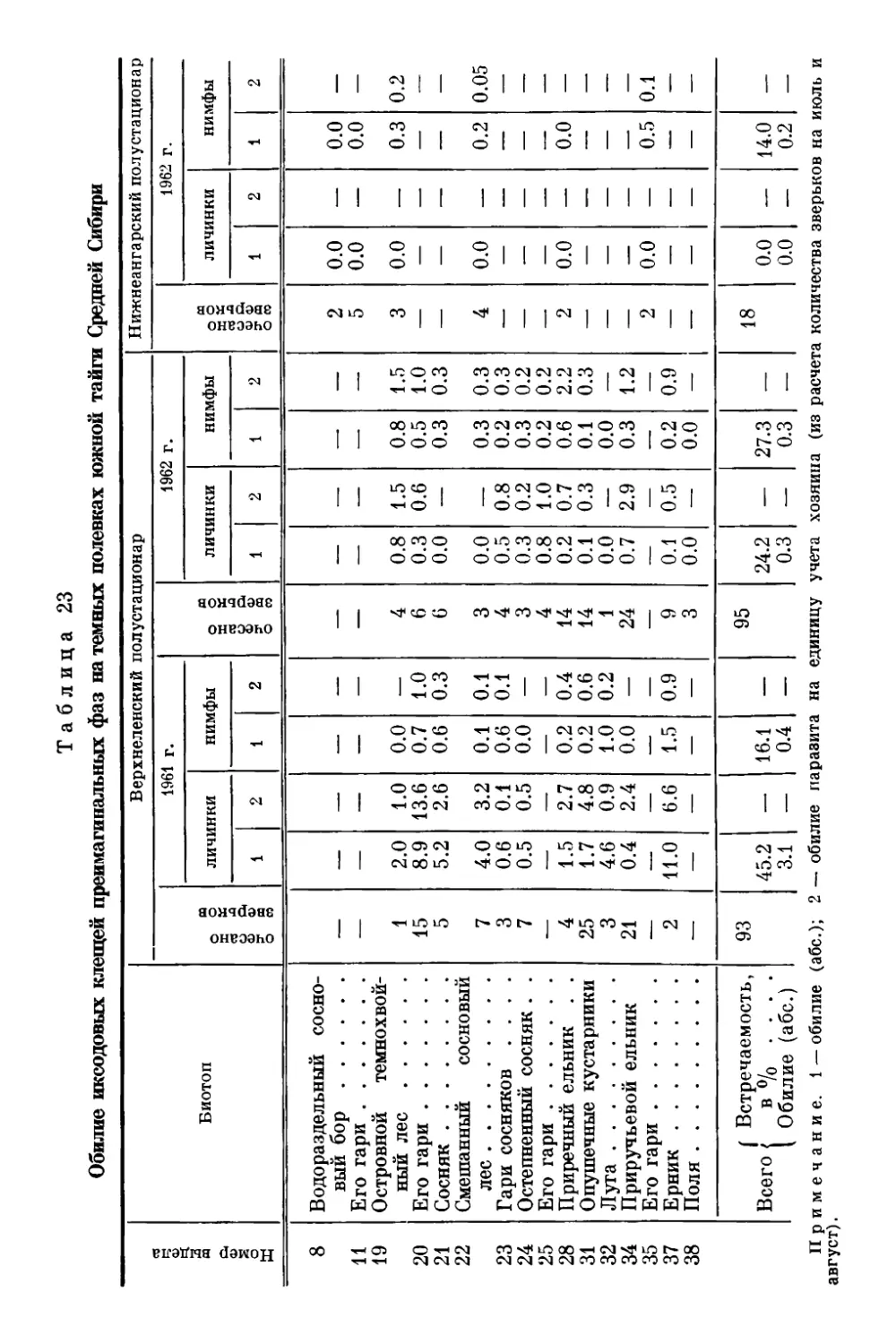



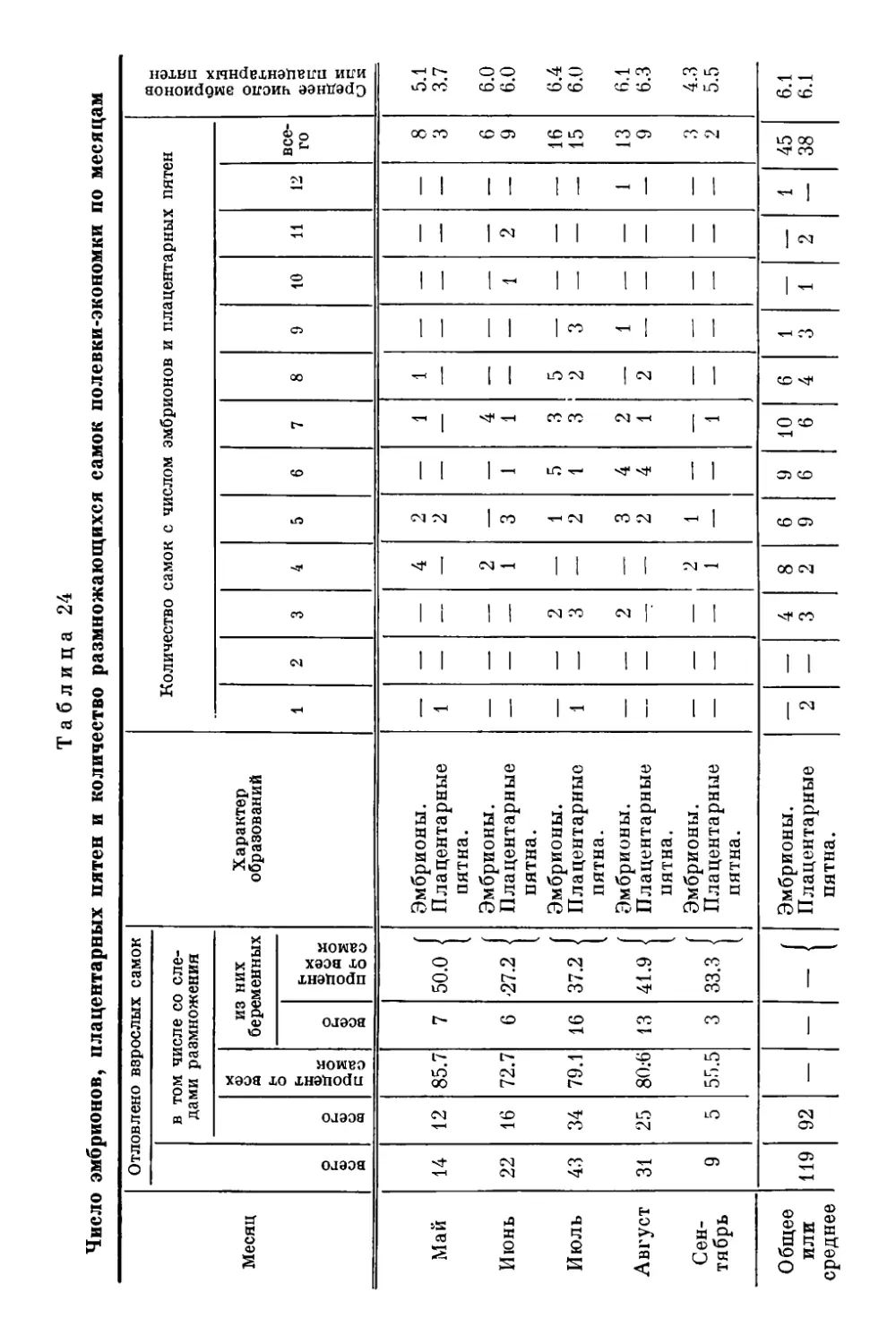



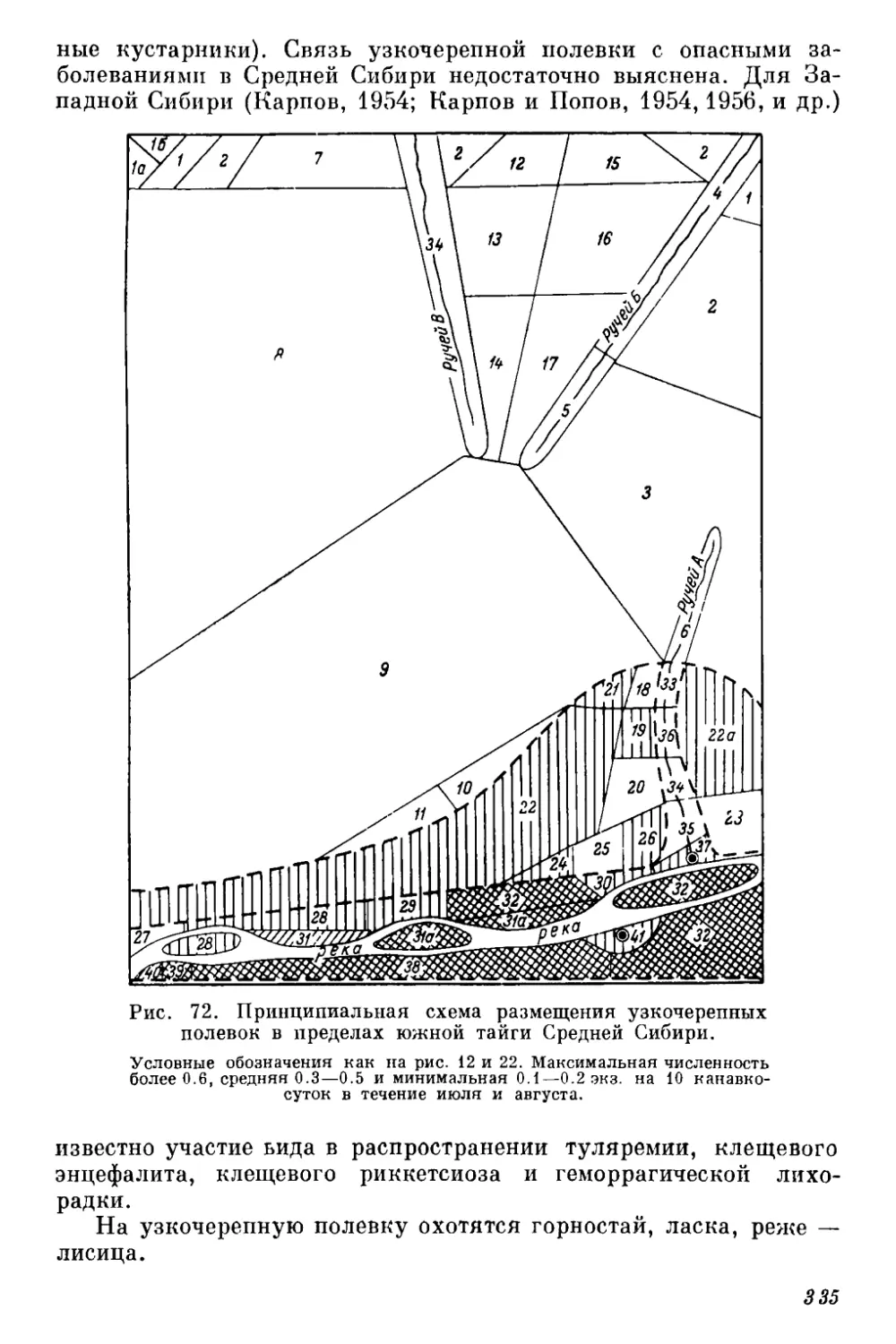

































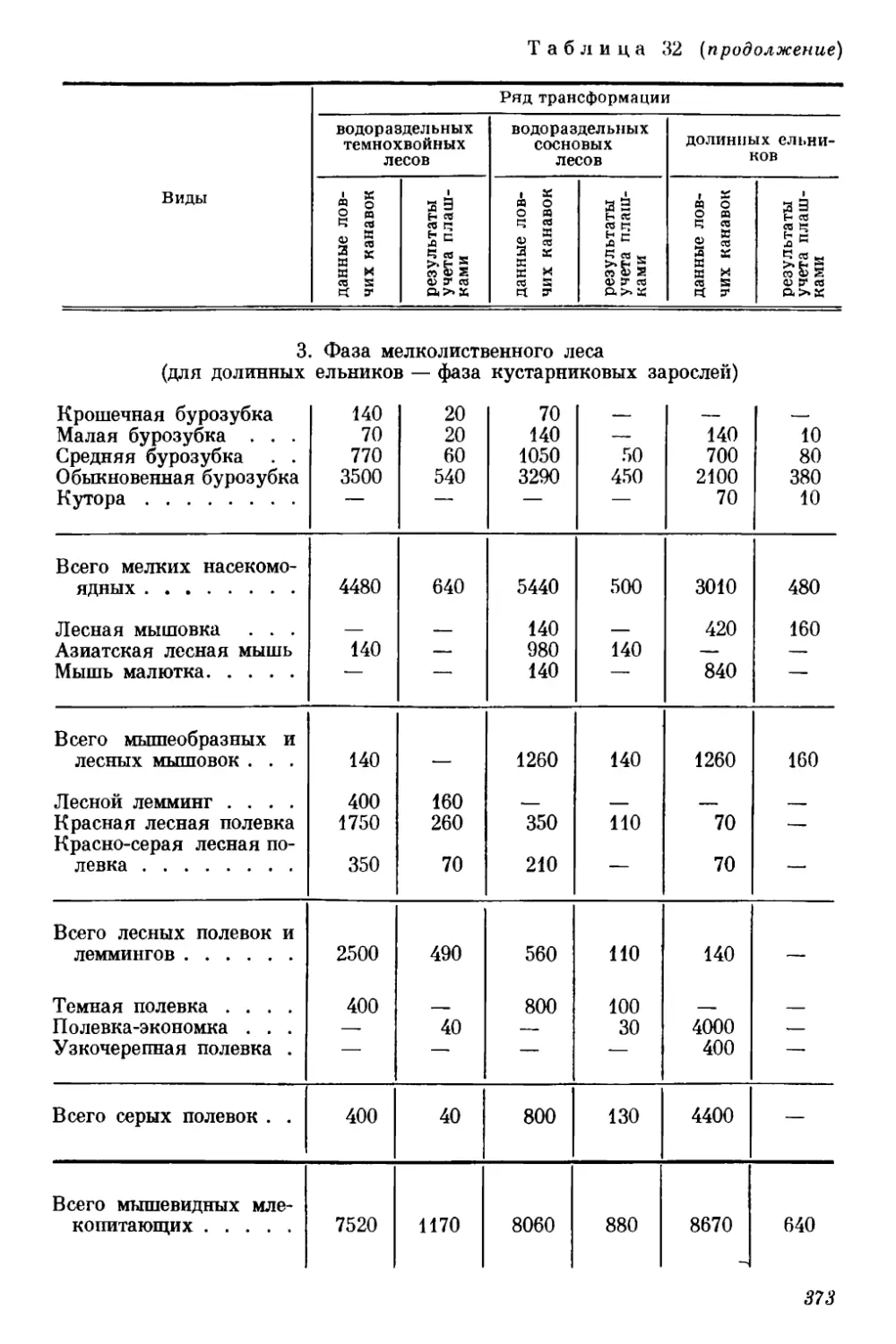
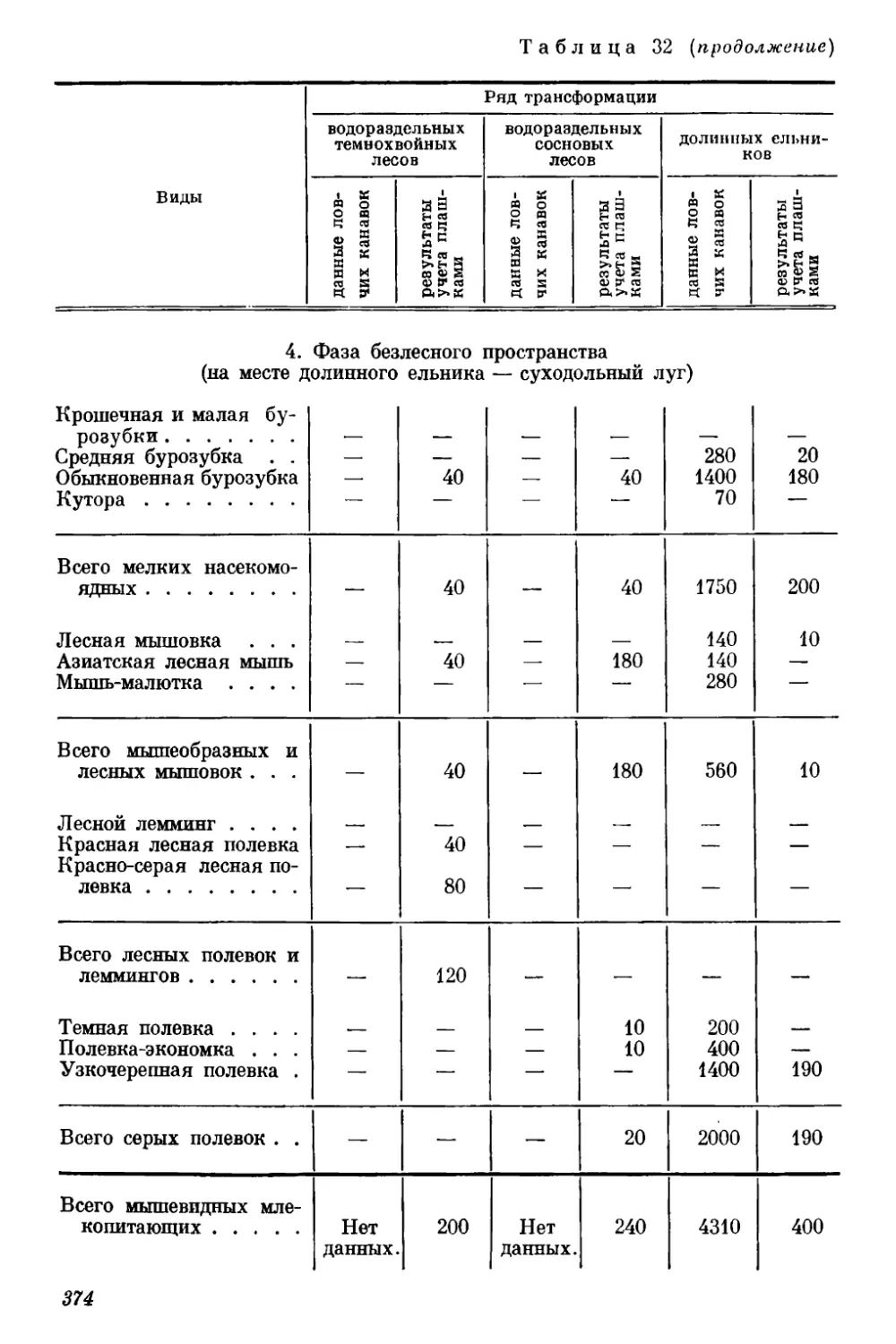

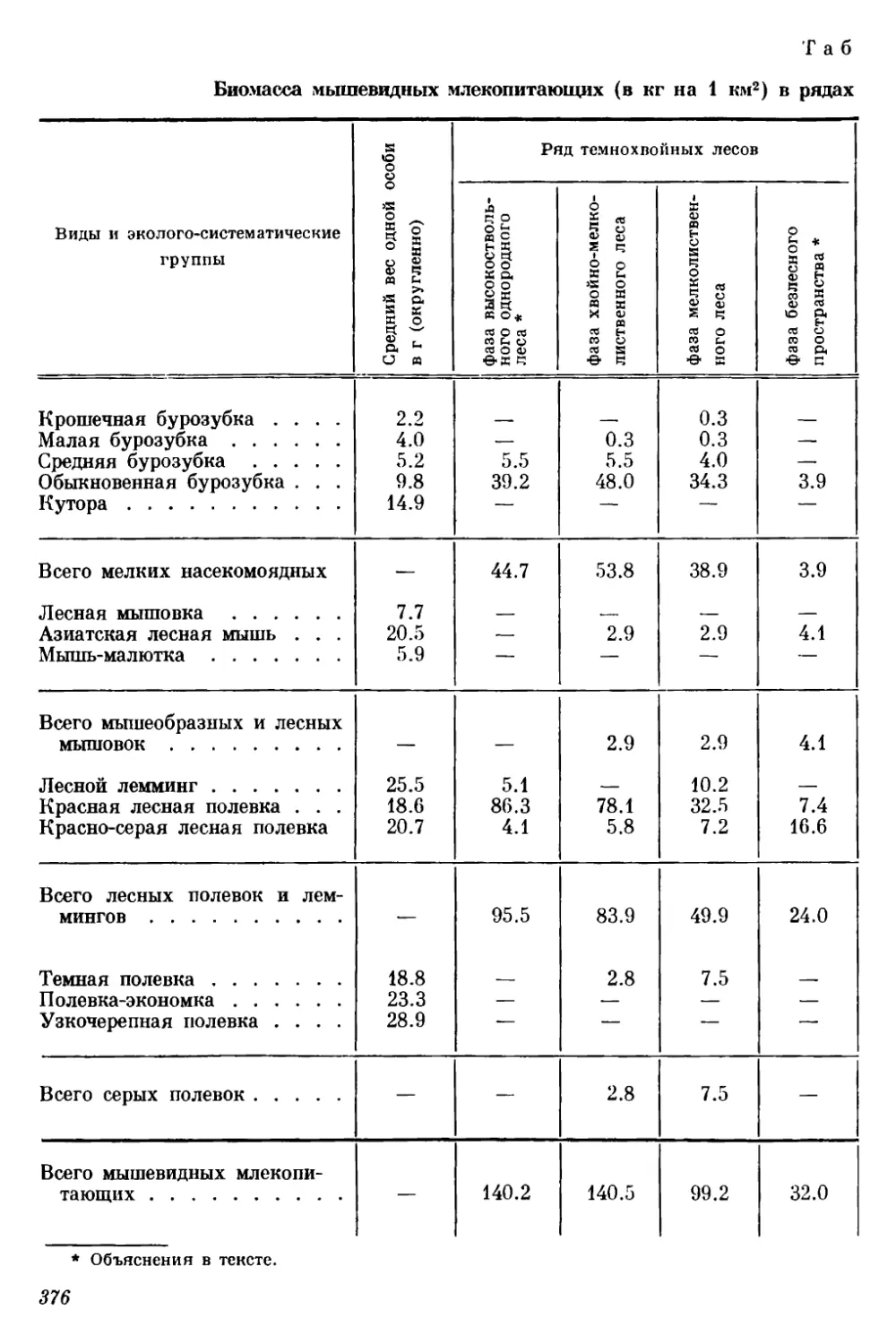
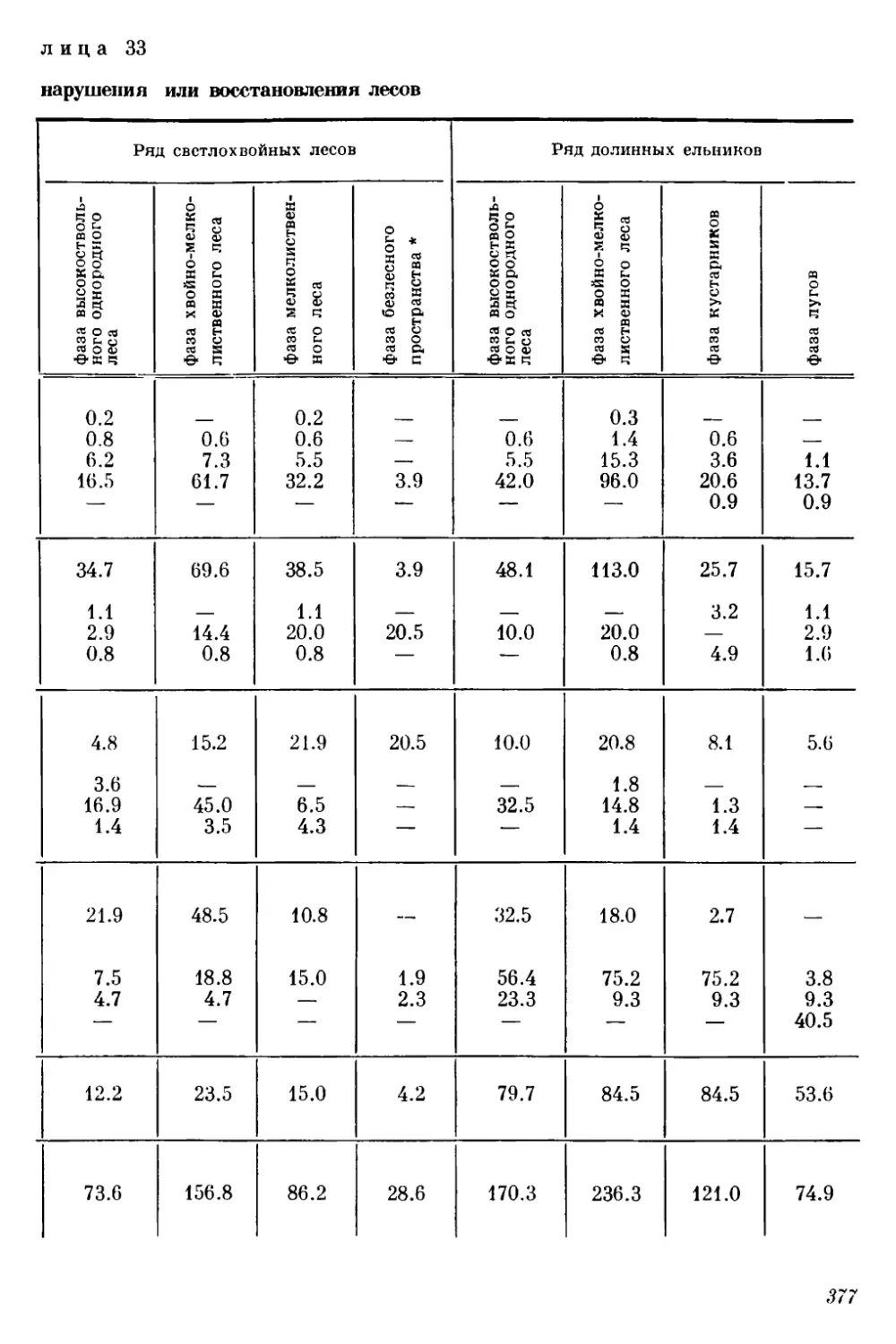












































Текст
ПТИЦЫ
и МЛЕКОПИТАЮЩИЕ
ЮЖНОЙ ТАИТИ
СРЕДНЕЙ СИБИРИ
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
САХАЛИНСКИЙ КОМПЛЕКСНЫЙ
НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ
Н. Ф. Ре Vim ерс
птины
МЛЕКОПИТАЮЩИЕ
ЮЖНОЙ ТАЙГИ
СРЕДНЕЙ СИБИРИ
о о
в
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА — ЛЕНИНГРАД
19 6 6
Самоотверженному помощнику — моей жене
Анне Васильевне Алиной
ВВЕДЕНИЕ
История современного освоения человеком территории нашей
страны сложилась таким образом, что обширные таежные простран-
ства Сибири оставались долгое время вне интересов земледелия
и промышленности. Однако развитие индустрии, особенно
в период развернутого строительства коммунизма, потребовало
мобилизации колоссальных природных ресурсов сибирской
тайги.
В процессе приобщения таежных богатств к интенсивному
использованию потребовалось изучение многих вопросов, свя-
занных со слабым знанием тайги, ее животного населения.
Первоочередной оказалась проблема борьбы за жизнь человека,
которому угрожали трансмиссивные заболевания, передаваемые
паразитическими членистоногими от диких животных.
Люди пошли по пути коллективной и индивидуальной защиты
от паразитов, по пути предохранительных прививок от болезней.
Однако пассивная оборона никогда не была лучшим средством
ведения войны, а вакцинация против всей огромной массы болез-
ней просто невозможна.
Самый правильный метод борьбы с силами природы, когда они
угрожают человеку, — метод глубокого познания естественных
процессов, расшифровки их кодов, отыскание слабых звеньев
в цепи явлений, нарушение этих цепей (но без вредных побочных
последствий!), и, таким образом, нейтрализация губительного
влияния природных факторов.
Среди частных вопросов естествознания, тесно связанных
с проблемой сохранения здоровья людей и их хозяйством, немалое
место занимает изучение животного мира. Позвоночные животные
служат резервуаром многих опасных инфекций, вредят сельскому
и лесному хозяйству и вместе с тем являются объектом промысловой
и спортивной охоты. Знание деталей жизни позвоночных живот-
ных помогает борьбе с трансмиссивными инфекциями, вредите-
1*
3
лями полей и огородов, позволяет наметить пути к расширению
и сознательной регулировке охотничьего промысла.1
Южная тайга Средней Сибири — край первоочередного освое-
ния. Это заставляет обратить внимание прежде всего на этот регион.
Определение четких границ территории, принятой для изуче-
ния, всегда бывает затруднено, подобно проведению любой разгра-
ничительной линии в природе. Исследователи нередко выходят
из положения, принимая за пределы изучаемой площади админи-
стративные границы различного ранга. Такой подход привычен,
оправдан интересами региональной экономики, удобен автору
работы, обычно не вызывает возражений, однако едва ли научно
состоятелен. Административные границы искусственны. Это поло-
жение не требует доказательств. Значительно правильнее ограни-
чивать район исследований какими-то естественными рубежами.
Однако определение этих рубежей не простое дело. Физико-
географами, биогеографами и, уже, ботанико- и зоогеографами
предложено огромное количество схем районирования. Эти схемы
не совпадают друг с другом ни в границах, ни даже в рангах
выделяемых регионов. Нет нужды останавливаться на разборе
существующих шкал районирования. Отметим только, что при
выборе площади для исследования обязательно необходим учет
того, насколько степень однородности совпадает с дробностью
исследования, т. е. насколько масштаб исследования совпадает
сего глубиной. Определяя границы нашего региона, мы не смогли
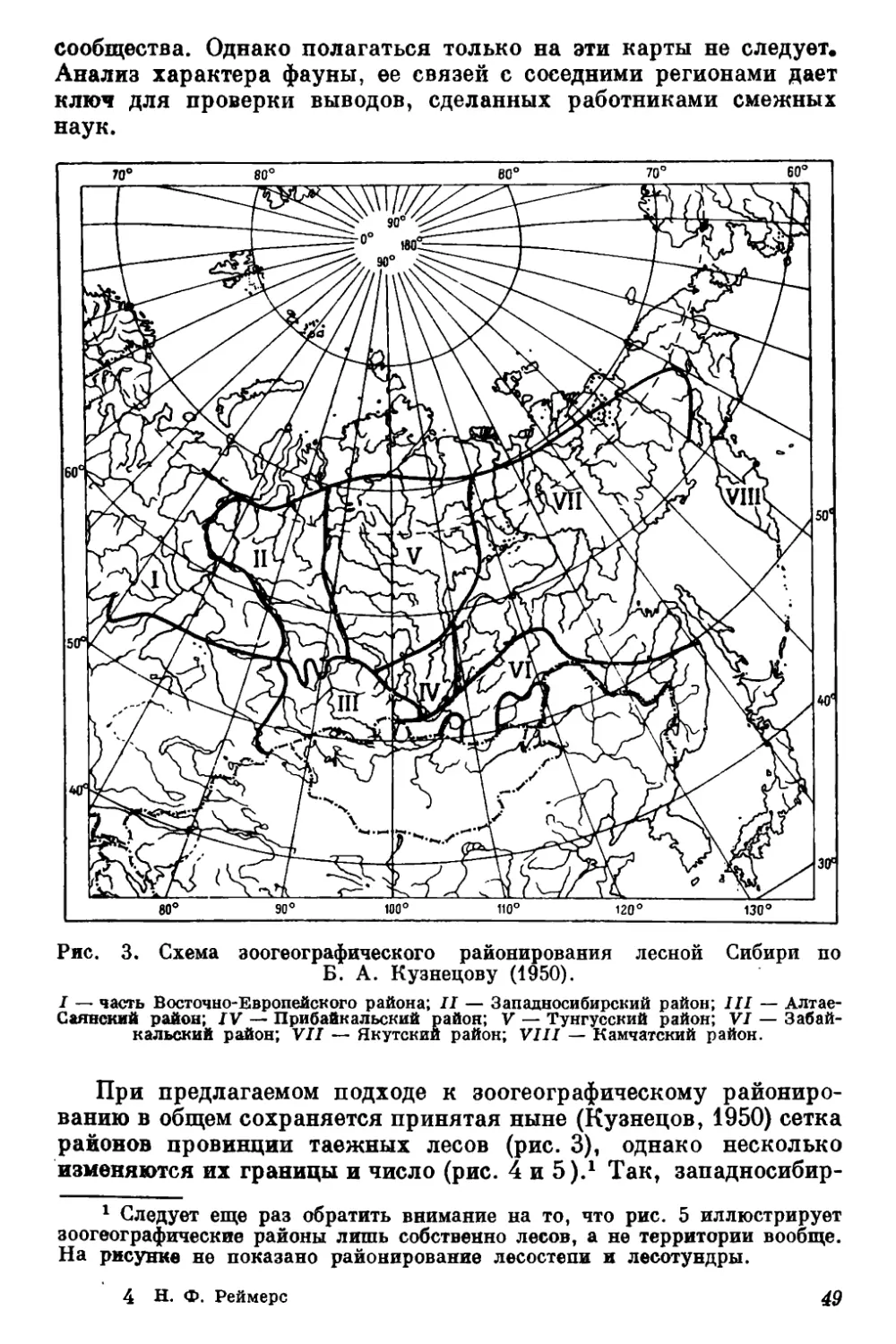
удовлетвориться имеющимися зоогеографическими схемами.
В первых главах работы поэтому кратко изложены типолого-
зоогеографические воззрения автора, а затем в новых границах
очерчен Прибайкальский зоогеографический район (южной тайги
Средней Сибири). Схематично он занимает предгорья Саян,
южную половину Средне-Сибирского плоскогорья, поднимается
языком вдоль р. Лены, оставляет вне своих границ Прибайкаль-
ские хребты, но включает в себя их низкогорные части, в том
числе Онотскую возвышенность? 1 2
1 Следует отметить, что роль насекомоядных видов и вообще косвен-
ная польза позвоночных животных выяснена в условиях таежной Сибири
очень слабо. Первые опыты изучения этого вопроса показывают, что
птицы и млекопитающие как регуляторы численности насекомых-вреди-
телей имеют второстепенное или даже третьестепенное значение. Даже
в условиях массового размножения такого вредителя, как сибирский
шелкопряд, птицы уничтожают очень незначительную часть его популя-
ции (Реймерс, 1958в, 1959д; Рожков и Реймерс, 1958; Реймерс и Рожков,
1960). Очевидно, роль этих животных должна быть изучена на широком
биоценотическом материале с тщательной расшифровкой трофических
цепей. Это не входило сейчас в нашу задачу.
2 Таким образом, очерченные нами границы южнотаежной части
Средней Сибири, очевидно, не отличаются от принятых в новейшей
(вышедшей после окончания работы над книгой) монографии «Средняя
Сибирь» (1964). К сожалению, в этой фундаментальной сводке освещена
лишь западная часть рассматриваемой территории, поэтому в полном объеме
сравнение произвести нельзя.
4
Строго говоря, наша задача — освещение фаунистических
закономерностей лишь собственно южной тайги Средней Сибири,
т. е. ее лесов. На лесные формы мы прежде всего и обращали
внимание. Ряд обстоятельств: а) отсутствие новой фаунистической
сводки для юга Средней Сибири («новейшей» до сих пор остается
монография В. К. Тачановского, изданная в 1893 г. на француз-
ском языке), б) проведенные и ведущиеся в настоящее время интен-
сивные исследования в соседних районах (Якутия, Приенисейская
полоса), оставляющие очерченный нами участок почти без вни-
мания, в) тот факт, что строго лесные участки охватывают коль-
цом лесостепь Средней Сибири и, наконец, г) весьма условный
характер границ между тайгой, «подтайгой» и вторичной лесо-
степью (особенно среднесибирского типа) — заставляют нас вклю-
чить в обзор некоторые формы, мало характерные для лесных
биоценозов. Многие из этих видов мы не изучали специально,
наблюдения за ними случайны, описаны они конспективно, часто
по литературным материалам.
Все теоретические построения в работе основаны на исследова-
нии лесных биоценозов. Особое внимание уделялось непромысло-
вым видам, так как они хуже изучены.
У нас не было стремления создать полную региональную
сводку или монографию, завершающую цикл работ соответству-
ющего профиля. Однако мы надеемся, что по результатам нашего
исследования можно будет оценить как то, что уже сделано, так
и то, что еще предстоит сделать. Исходя из таких воззрений, мы
старались не просто перечислить известные факты, но пытались пх
осмыслить, имея в виду практическую значимость животных для
человека. Однако было бы неверно приводить лишь общие рас-
суждения. Теория развивается. Факты остаются фактами вечно,
давая пищу для новых обобщений.
Всякая научная работа обесценивается, если не определено
ее главное направление. Для нашего исследования — это изуче-
ние динамики природы в различных ее проявлениях с тем, чтобы
представить себе изменения, происходящие в природе под влиянием
естественных и антропогенных факторов в настоящее время,
и попытаться приподнять завесу будущего. Без понимания сук-
цессионно-исторических процессов в природе, на наш взгляд,
невозможно управлять ее силами, предвидеть те полезные и вред-
ные изменения, которые происходят вслед за освоением новых тер-
риторий.
В работе нет обширных разделов о физико-географических
условиях района исследований. Это не означает недооценки роли
климата и орографии в жизни животных. Сложившаяся практика
помещения в изданиях значительных вкраплений из других
отраслей, часто без достаточной связи с основным повествованием,
нам кажется нецелесообразной. Мы привлекали материалы смеж-
ных дисциплин лишь в тех случаях, когда требовалось что-либо
5
расшифровать, т. е. когда уже улавливалась логическая связь
между явлениями.
Специально оговорим отсутствие исторического очерка. Исто-
рия исследований — особая ветвь науки. Знание ее помогает
понять пути изучения и полноту изученности района. Однако мы
сознательно опустили традиционный раздел о трудах предыду-
щих поколений, тем более что для большей части региона южной
тайги Средней Сибири недавно составлена полная библиография,
выдержанная в хронологическом плане (Библиография Иркут-
ской области: Биология, Иркутск, 1956; Физическая геогра-
фия, Л., 1957; Лесное хозяйство и лесные промыслы, Иркутск,
1962).
Несколько слов о структуре книги. Логически кажется
более правильным изложение частных вопросов и материалов
перед главами, освещающими общие проблемы. Однако познание
частностей настолько тесно связано с формированием представле-
ний о целом, что чисто техническое разделение того и другого
вызывает затруднения. Еще больше осложнений возникает, когда
трактуются вопросы, выходящие за рамки конкретного материала
региона: частное и типичное настолько переплетаются, что стано-
вятся неотделимыми.
Исходя из сказанного, первый раздел, включающий три главы
(1—3), основан на более широких материалах (в значительной ча-
сти литературных), отчасти предвосхищает конкретные факты из
второго раздела и рассматривается нами как теоретико-методологи-
ческое введение. Второй раздел (главы 4—6) касается непосредствен-
но животного мира южной тайги Средней Сибири. Такая схема из-
ложения не бесспорна, и можно предвидеть возражения, особенно
в отношении обоснованности выводов первого раздела. Однако
перенесение материалов из повидовых очерков в общие разделы,
во-первых, не позволило бы изложить многие факты, непосред-
ственно не касающиеся теоретических вопросов (что при слабой
изученности территории нежелательно), во-вторых, лишило бы
книгу характера фаунистического справочника. И вместе с тем
такой перенос все равно не восполнил бы недостатка оригиналь-
ных материалов из других районов Сибири. Заполнение же этого
пробела потребовало бы детального изучения всей территории,
о чем мы лишь ставим вопрос. Одновременно нужно обратить
внимание на право авторов решать некоторые общие вопросы без
детального обоснования каждой частности в тексте. Иначе дока-
зательство очень широких задач, идущее «от печки», потребует
колоссальных нагромождений фактов.
Предлагаемая работа — результат экологических и зоогеогра-
фических исследований, проведенных Ангаро-Ленским комплекс-
ным физико-географическим отрядом Среднесибирской южно-
таежной экспедиции Института географии Сибири и Дальнего
Востока СО АН СССР, которым в течение пяти лет руководил
автор. Вошли в нее и более ранние наблюдения, сделанные нами
в составе коллективов существовавших тогда Восточно-Сибир-
ского и Западно-Сибирского филиалов АН СССР. Специальные
и попутные исследования в южной тайге заняли в общей сложности
12 лет (1950—1963) с двухлетним перерывом.
Общая длительность непосредственных полевых работ автора
составила несколько лет (экспедиционный период в году коле-
бался от 8 до 2 месяцев и чаще всего составляй пол года). Были
заложены 34 ловчие канавки для ловли мышевидных зверь-
ков, проработавшие 65 канавко-сезонов, в общей сложности
4500 суток (22 500 цилиндро-суток). Число ловушко-суток пре-
высило 40 000. Проведен длинный ряд количественных учетов
птиц на постоянных, специально размеченных (пронумерован-
ных) через 25 м, пятикилометровых маршрутах. В ходе этих
учетов каждый раз составлялись кроки маршрута, на которых по-
мечались (с ориентацией на номер ближайшей цифровой отметки)
встречи поющих самцов, отдельных особей и стаек птиц. Так как
различные виды птиц особенно заметны в разное время весны и лета
(в пределах гнездового периода), учеты неоднократно повторя-
лись. Двойная повторность учета считалась достаточной для
составления одной схемы кроков. На более длинных маршрутах —
конных и пеших — также производилось элементарное картиро-
вание. Сопоставление картосхем помогало выяснению характера
размещения птиц. В ряде биотопов мы применяли учет численно-
сти птиц на площадках. Выделенный для этой цели прямоугольник
приблизительно (измерение производилось шагами) 200 на 500 м
рассекался маршрутами через 50 м. Вне гнездового периода учеты
проводились для выяснения изменений в размещении видов птиц
и концентрации пролетных стай. Широко применялась визуаль-
ная оценка числа птиц.
Одновременно с учетом птиц выяснялось обилие млекопита-
ющих. Для этого на кроках мы помечали встречи зверей или их
следов (на снегу).
Зимние наблюдения, помимо кратковременных выездов, были
организованы в окрестностях Иркутска в сезоны 1950—1951
и 1959—1960 гг. В пределах южной тайги было добыто более
2000 экз. птиц и свыше 10 000 млекопитающих.3 Кроме того,
привлечены небольшие музейные материалы.
Нами применялись главным образом полустационарные мето-
ды, и в исследованиях мы опирались на два основных полуста-
ционара: Верхнеленский (Якимовский) в Жигаловском районе
Иркутской области, на р. Тутуре — правом притоке верхней
Лены и Нижнеангарский (Яркинский) в Богучанском районе
Красноярского края, на р. Чадобец — правом притоке нижней
3 Вместе с материалами, переданными нам Г. А. Вороновым и собран-
ными паразитологами, более 11 500 зверей и 2200 птиц.
7
Ангары. Оказались полезными некоторые данные, полученные
при работе (с зимовкой) на полустационаре в Хамар-Дабане
(1955—1956 гг.), а также при маршрутах в смежных с южной тай-
гой Средней Сибири регионах — Хамар-Дабане (1951—1956 гг.),
Саянах (1952—1954 гг.), Алтае (1957 г.), Туве (1958 г.), южной
тайге Западной Сибири (1957—1958 гг.) и Горной Шории
(1957 г.).4
В пределах собственно южной тайги Средней Сибири были
также совершены пешие и автомобильные маршруты: дважды
с лошадьми пересечен Око-Ангарский водораздел (1959 г.), три-
жды осмотрено Ангаро-Ленское междуречье в районе Малышовка—
Жигалово (1960—1961 гг.), сделаны маршруты по Верхоленью
(Качуг—Жигалово, Качуг—Шевыкан, Жигалово—Чикан) и бо-
лее поверхностно произведено ознакомление по маршрутам Ир-
кутск—Тулун—Братск—Дубинине, Иркутск—Канск—Чунояр—
Богучаны—Яркино, Иркутск—Качуг, Иркутск—Черемхово—
Тальники, Иркутск—Тулун—Аршан (на р. Ие) и по другим,
более коротким. Кратковременные полустационары (не менее
10 дней) были организованы в пределах лиственничной лесо-
степи Средней Сибири (б. Баяндаевский район Иркутской обла-
сти, улус Байша, 1959 г.), пади «Глубокая» (между Иркутском
и Слюдянкой, 1952 г.) и Онотской возвышенности (1954 г.). Окрест-
ности Иркутска (в широком понимании — в радиусе до 80 км)
были ареной наблюдений в 1951—1953 и 1959—1960 гг. Для
общего ознакомления с южной тайгой совершены авиационные
экскурсии на самолетах местной авиации (АН-2 и ЯК-12) по марш-
рутам: Иркутск—Жигалово—Киренск—Непа—Ербогачен, Ир-
кутск—Братск и Красноярск—Канск—Богучаны—Яркино. Места
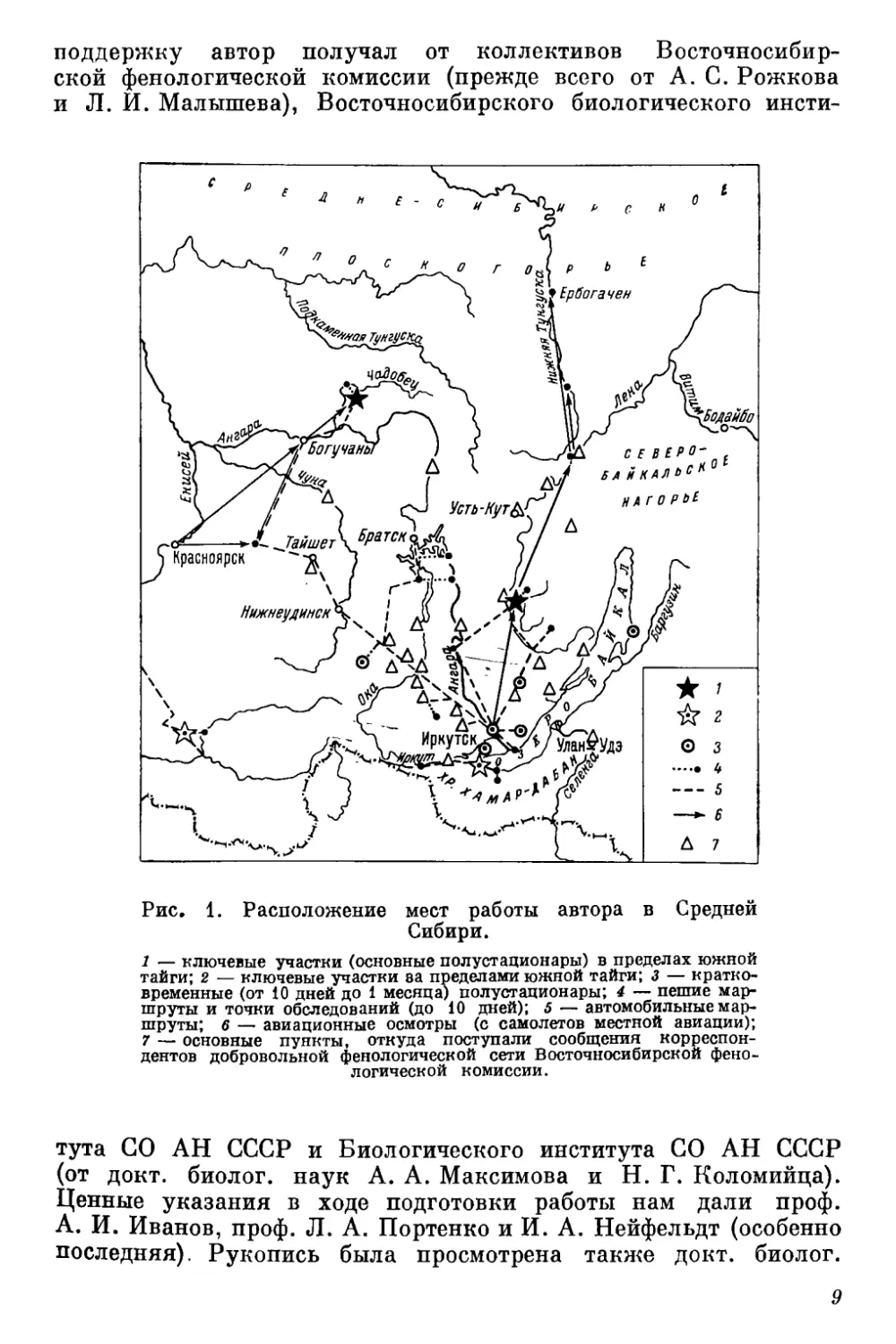
работы автора показаны на рис. 1.
Некоторые дополнительные материалы были получены
с помощью корреспондентов добровольной фенологической сети
Восточносибирской фенологической комиссии, организованной
и возглавлявшейся автором в 1959—1963 гг. Этих энтузиастов —
любителей сибирской природы автор всегда вспоминает с осо-
бой теплотой.
В ходе полевых работ автор неоднократно пользовался това-
рищеской помощью ландшафтоведа В. А. Ряшина, ботаников
В. Я. Балаганова и А. В. Белова, паразитолога Б. В. Вершин-
ского, врача И. Т. Меркачева и студентов иркутских и столичных
вузов. Значительную часть технической работы выполнил зоолог
Г. А. Воронов, подготовивший кандидатскую диссертацию по
части материалов, собранных на верхней Лене. Неоценимую
4 Интересные сопоставления удалось сделать в ходе работы на Сахалине
(1963 г.). Особенно ценной оказалась проверка и подтверждение многих
теоретических положений автора в новых для него условиях Дальнего Вос-
тока.
8
поддержку автор получал от коллективов Восточносибир-
ской фенологической комиссии (прежде всего от А. С. Рожкова
и Л. И. Малышева), Восточносибирского биологического инсти-
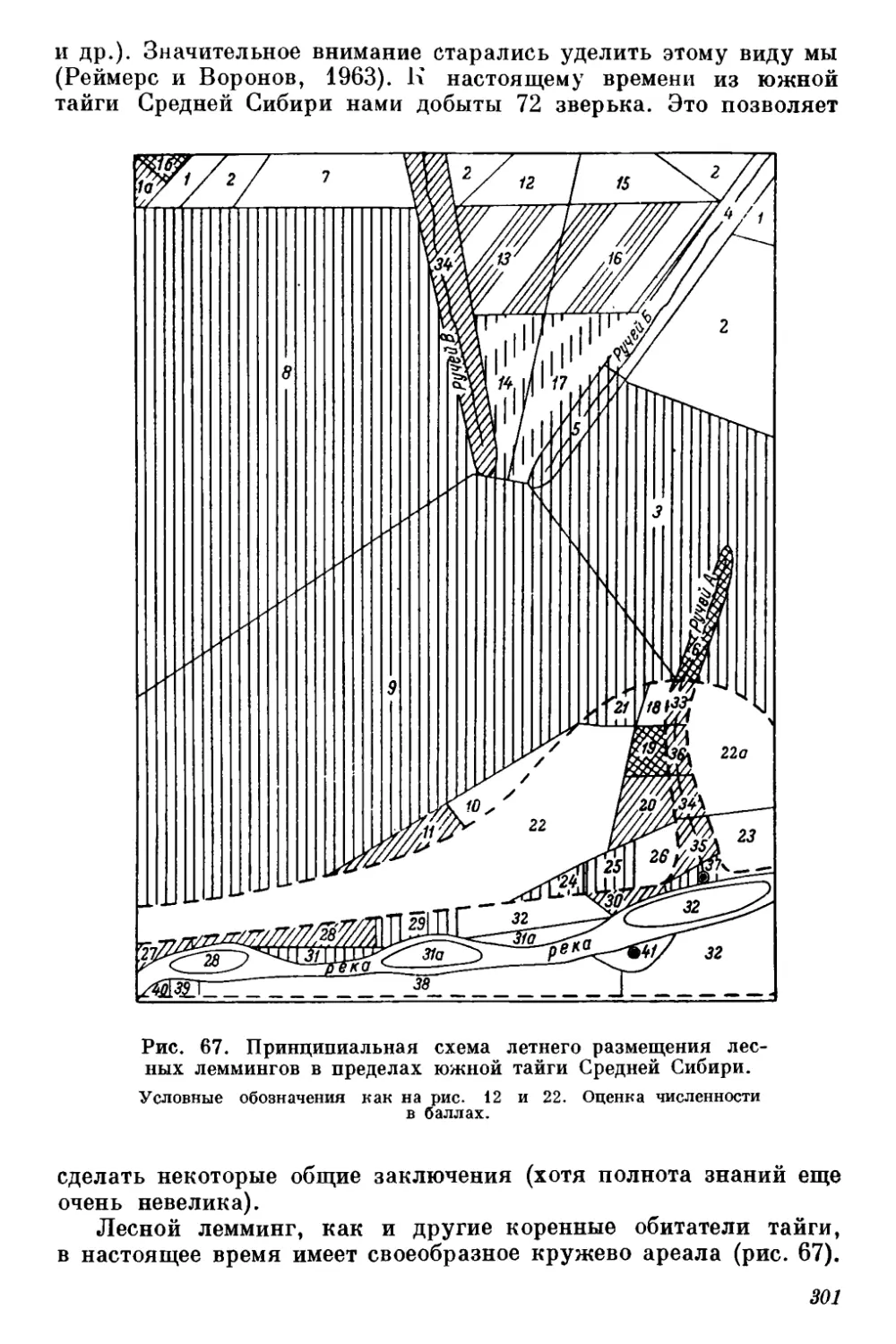
Рис, 1. Расположение мест работы автора в Средней
Сибири.
1 — ключевые участки (основные полустационары) в пределах южной
тайги; 2 — ключевые участки ва пределами южной тайги; 3 — кратко-
временные (от 10 дней до 1 месяца) полустационары; 4 — пешие мар-
шруты и точки обследований (до 10 дней); 5 — автомобильные мар-
шруты; 6 — авиационные осмотры (с самолетов местной авиации);
7 — основные пункты, откуда поступали сообщения корреспон-
дентов добровольной фенологической сети Восточносибирской фено-
логической комиссии.
тута СО АН СССР и Биологического института СО АН СССР
(от докт. биолог, наук А. А. Максимова и Н. Г. Коломийца).
Ценные указания в ходе подготовки работы нам дали проф.
А. И. Иванов, проф. Л. А. Портенко и И. А. Нейфельдт (особенно
последняя). Рукопись была просмотрена также докт. биолог.
9
наук А. А. Максимовым и доц. К. И. Шурыгиной. Замечания
перечисленных лиц позволили исправить ряд погрешностей.
Искренне благодарю всех лиц и коллективы, которые помогли
мне в работе.
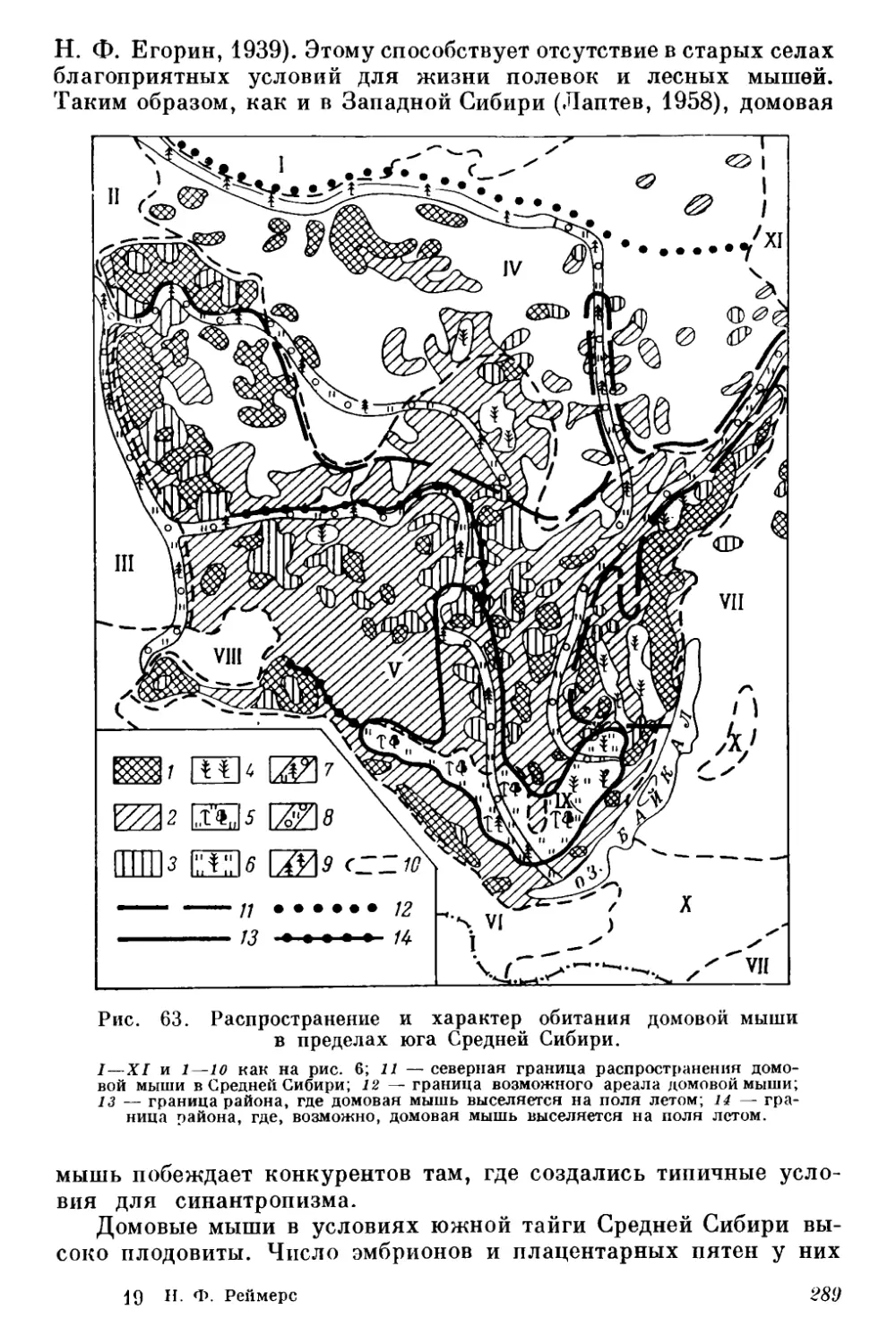
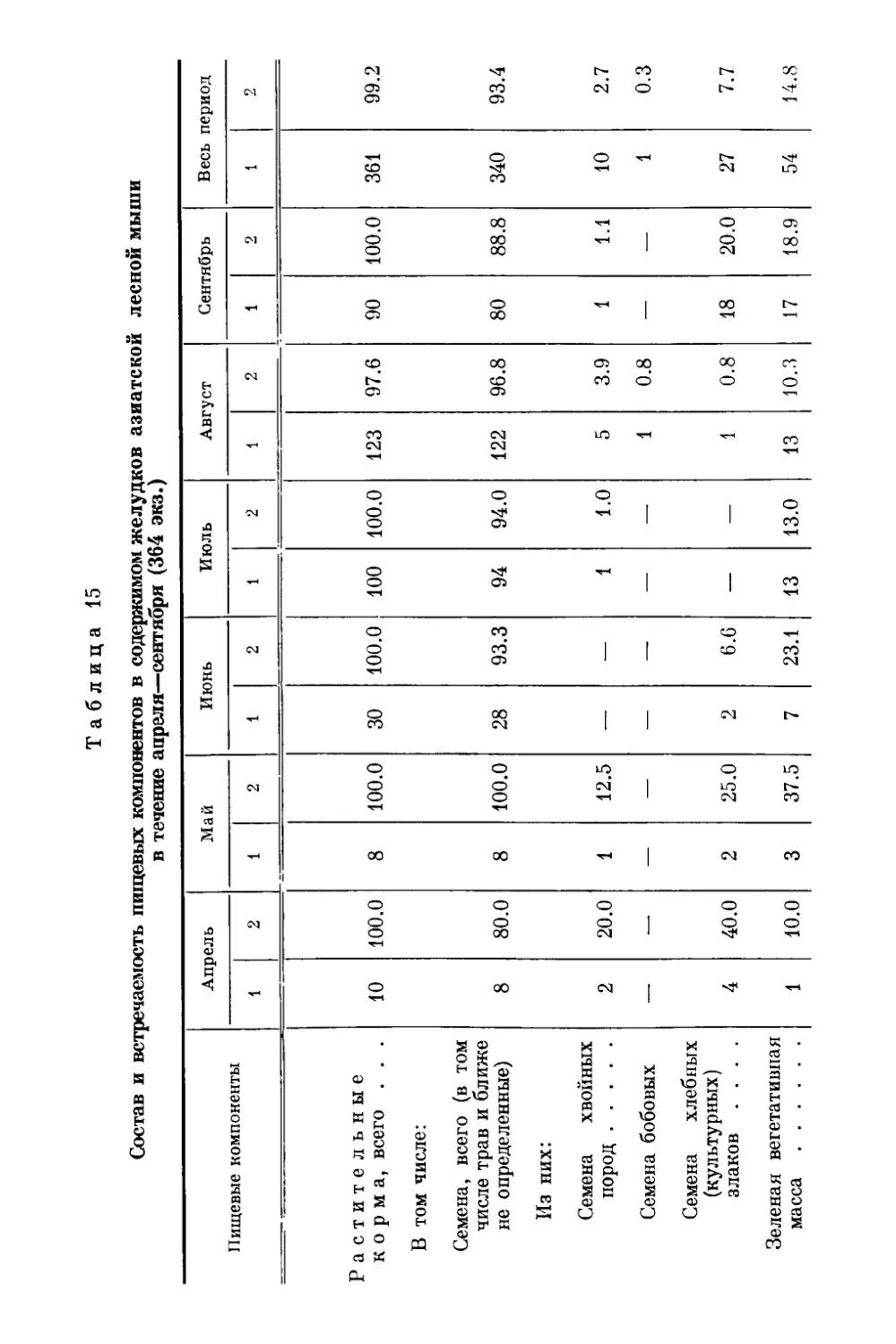
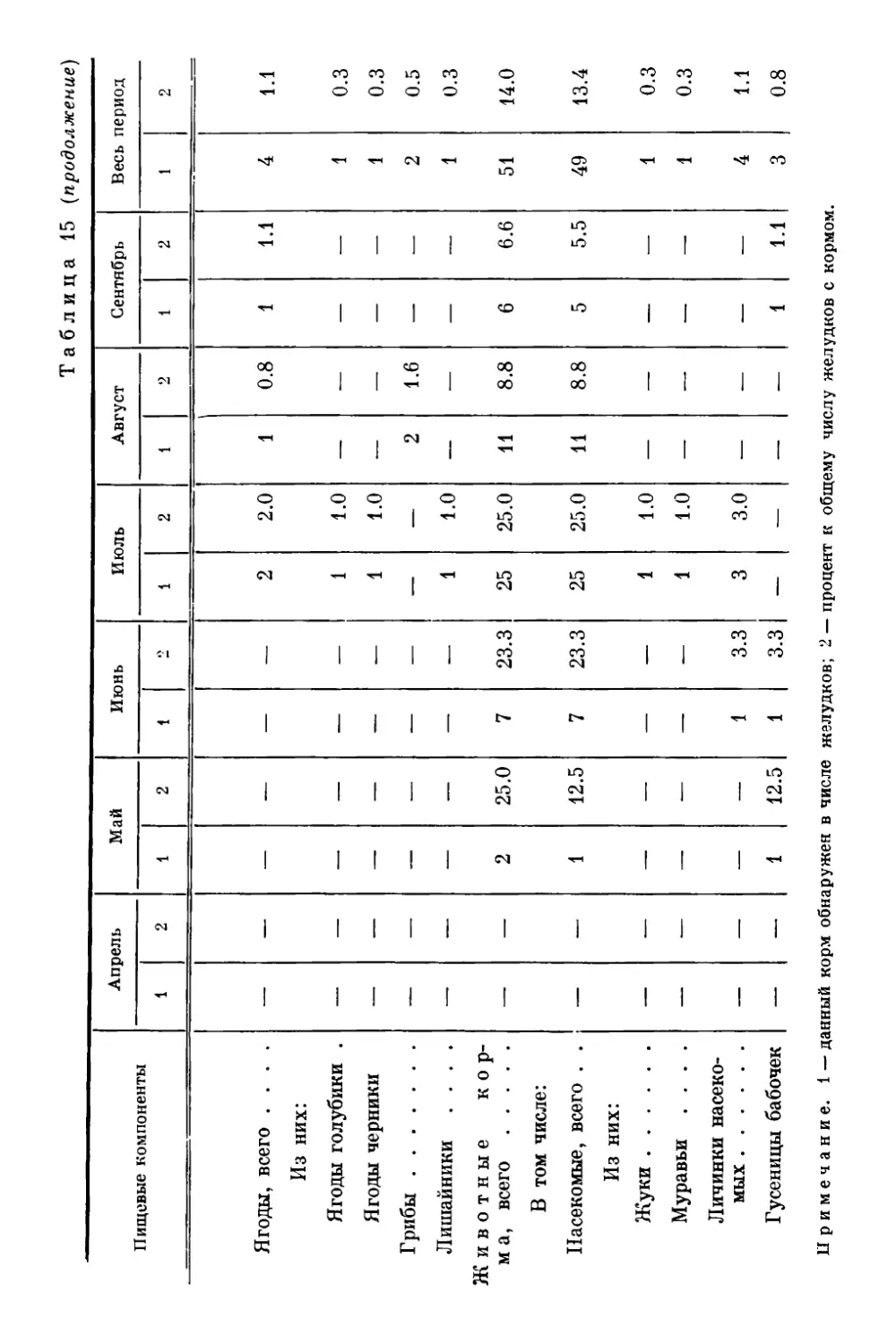
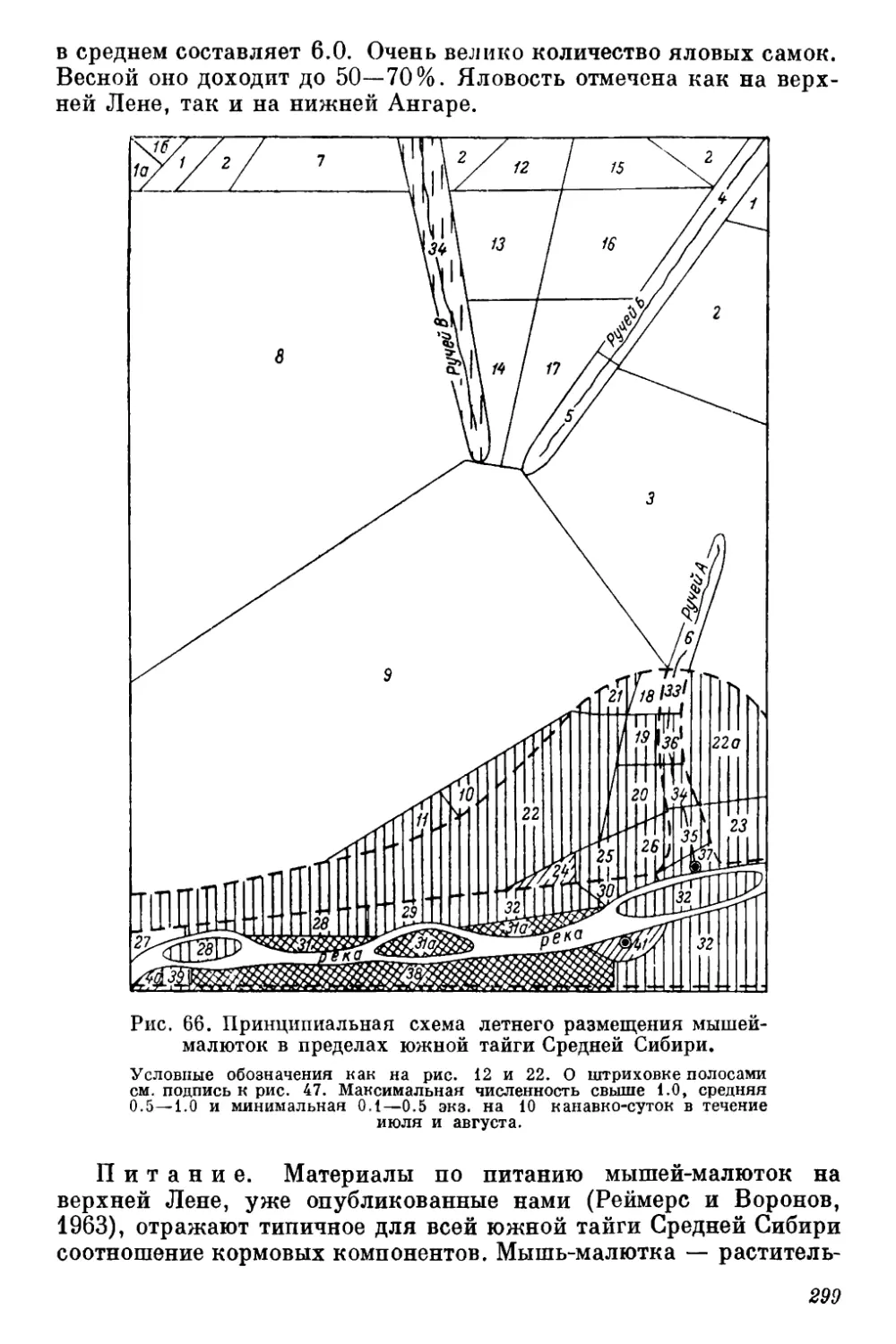
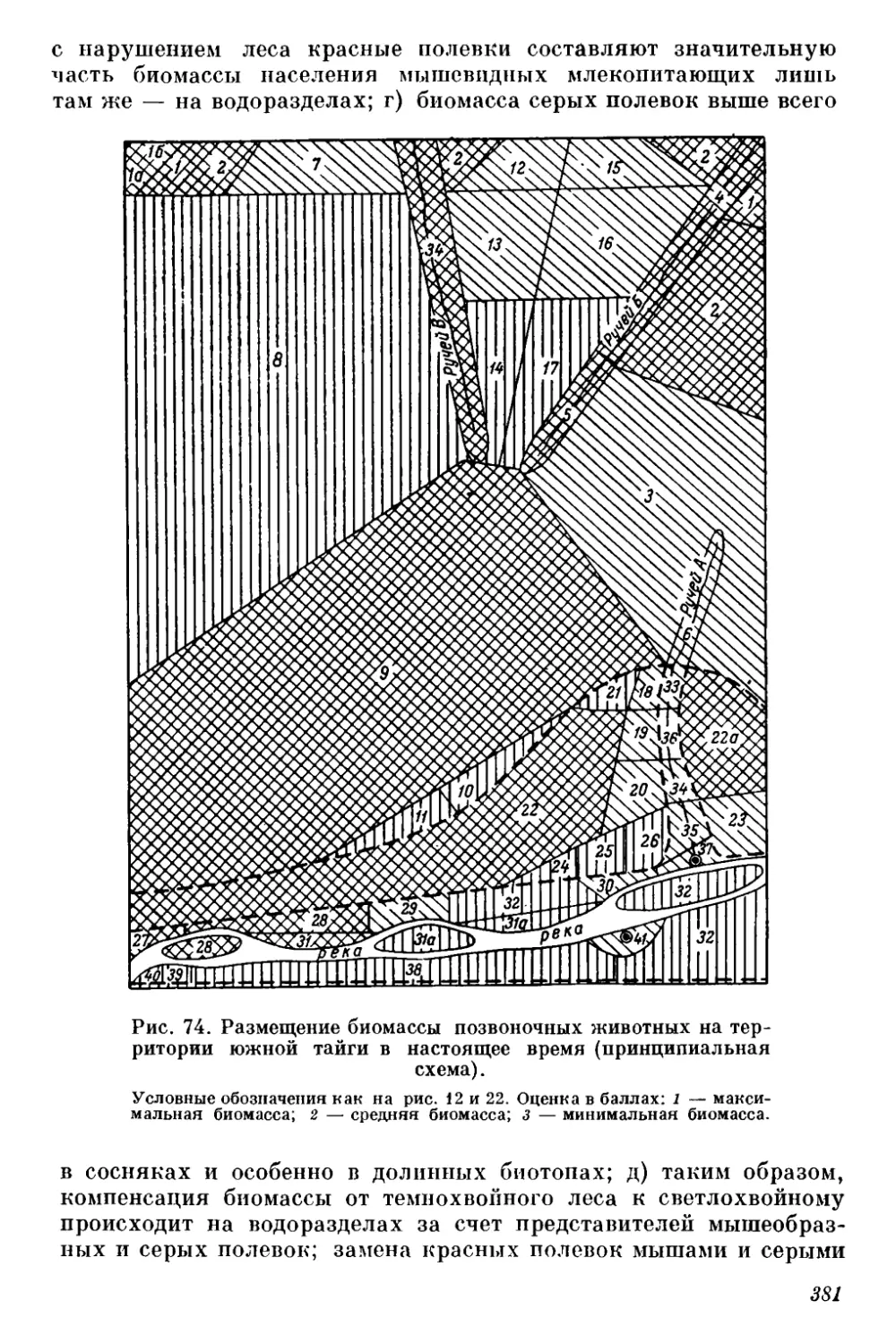
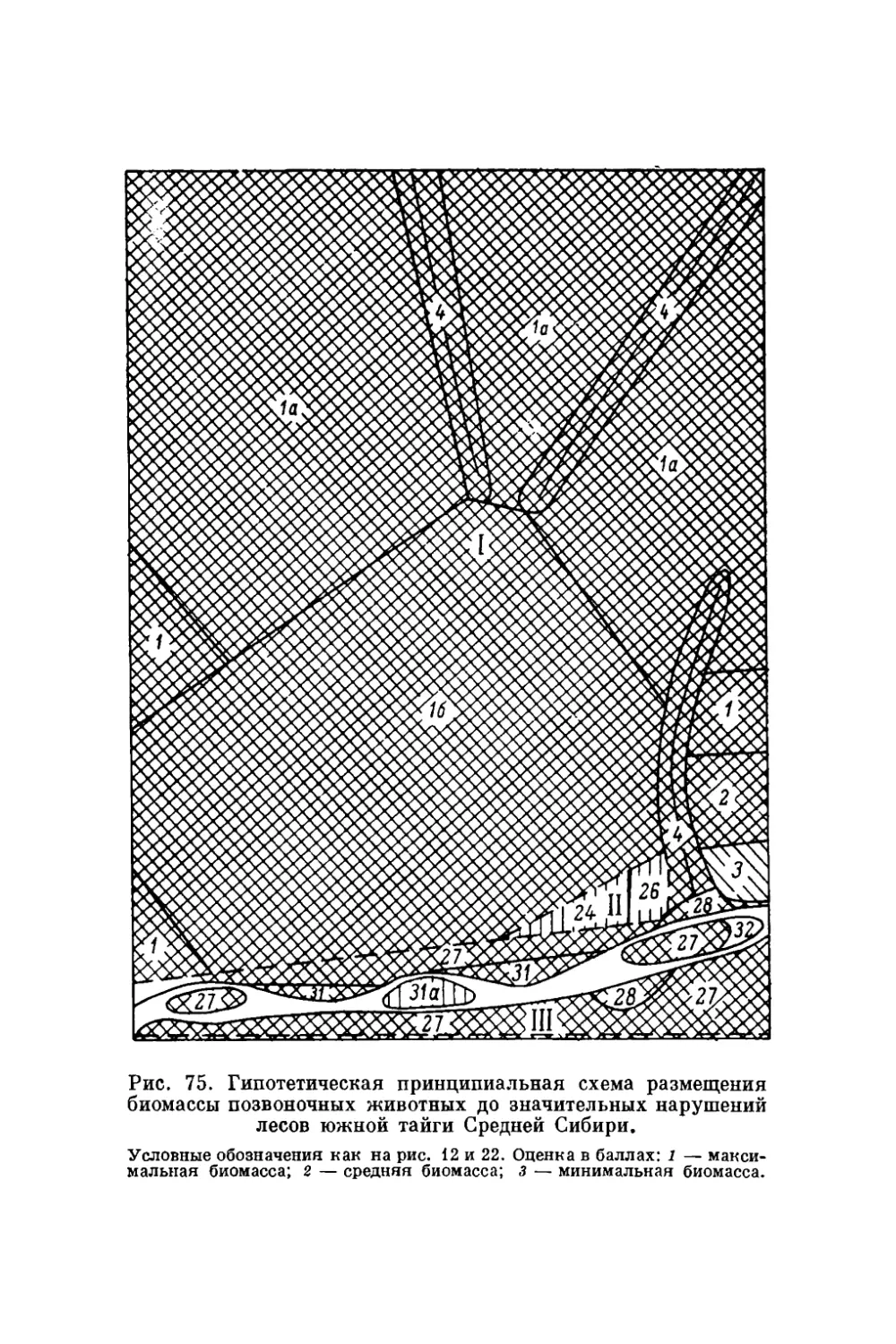
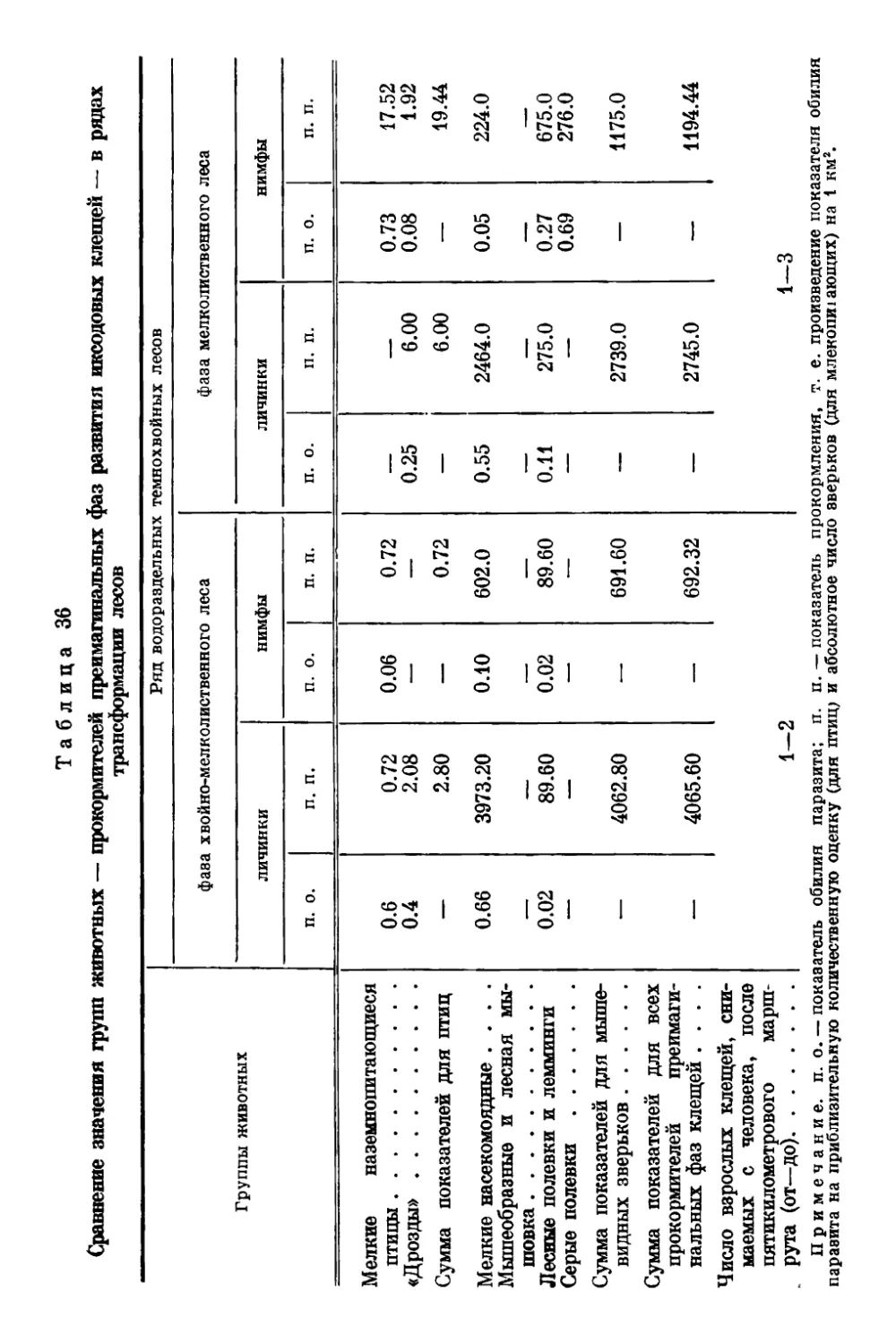
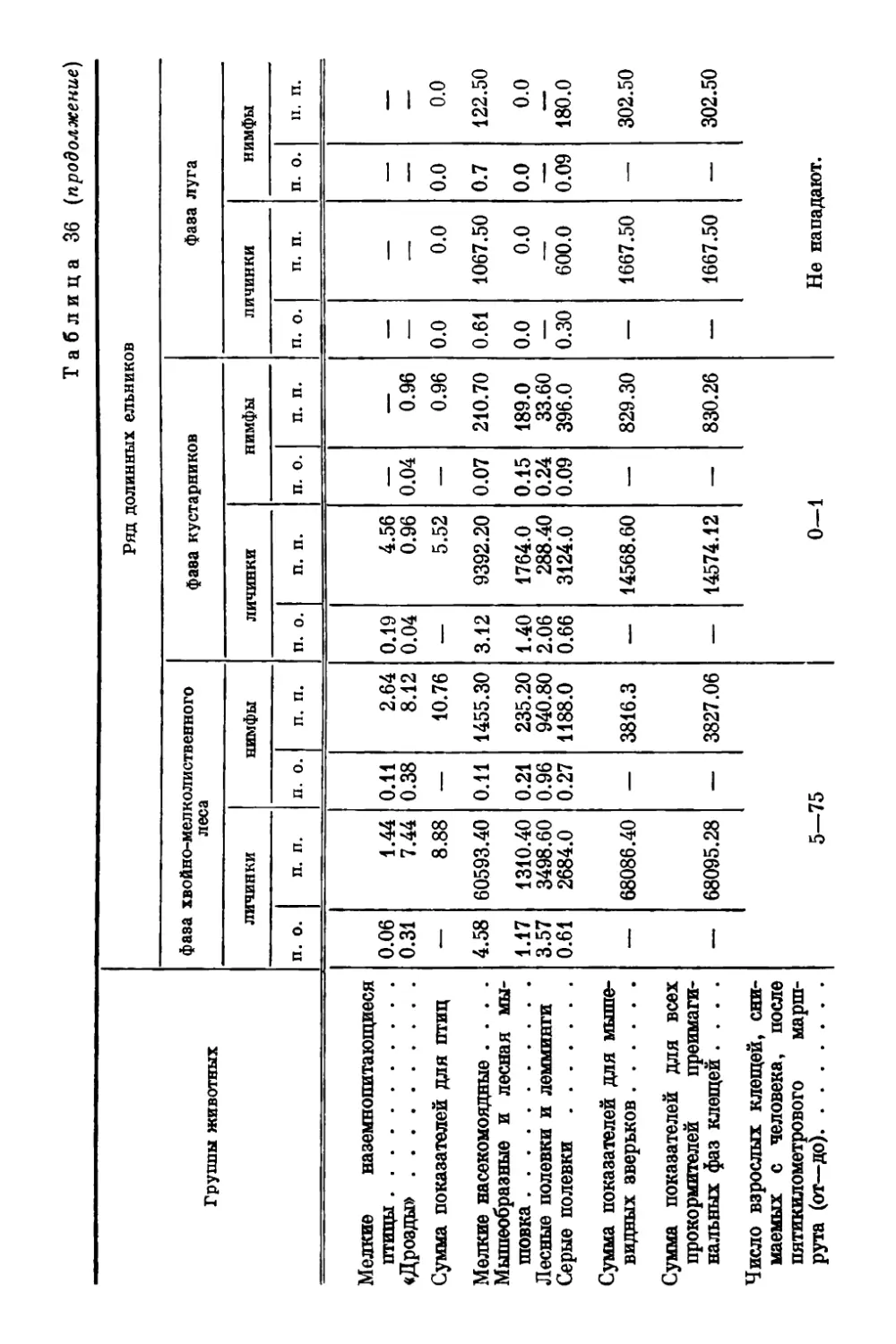
gg Оригиналы всех рисунков в книге принадлежат автору.
РАЗДЕЛ I
ЭКОЛОГО-ЗООГЕОГРАФИЧЕСКАЯ ТИПОЛОГИЯ
И МЕЛКОМАСШТАБНОЕ ЗООГЕОГРАФИЧЕСКОЕ
РАЙОНИРОВАНИЕ ТАЕЖНОЙ СИБИРИ1
Глава 1
ЭКОЛОГО-ЗООГЕОГРАФИЧЕСКАЯ ТИПОЛОГИЯ
В практической работе мы столкнулись с трудностями двоякого
рода: во-первых, нужно было определить границы той территории,
на которую можно экстраполировать получаемые данные, во-вто-
рых, приходилось искать общие типологические принципы, кото-
рые бы позволили рассматривать южную тайгу как некоторое
метамерное образование. Так перед нами встали вопросы зоо-
географической типологии и районирования. Взаимосвязь их
очевидна.
Вопрос о природных элементарных единицах сейчас разраба-
тывается большим числом авторов. Предложено немалое количе-
ство систем и методических подходов (экосистемы американских
авторов, биогеоценозы лесоводов, фации географов и т. д.). Все
эти системы довольно близки между собой и не будут служить
предметом нашего разбора. Отметим лишь, что зоология позво-
ночных, имея дело с особым («крупным» и подвижным) материалом,
вынуждена создавать свою сетку элементарных единиц. Законо-
мерность и правомочность такого подхода отмечает Н. В. Тимо-
феев-Ресовский (1961). Необходимость его признают многие уче-
ные, разрабатывающие природно-типологическую систему в при-
менении к миру позвоночных животных.
1 В данном случае под названием Сибирь понимается вся зауральская
часть СССР (без Казахстана), включая Дальний Восток. Такая расшиф-
ровка имеет довольно широкое хождение: Сибирское отделение АН СССР
включает Дальневосточный филиал, а также расположенные на Сахалине,
Камчатке и в Магадане институты, многотомная книга «Звери Сибири»
С. У. Строганова охватывает чуть ли не всю азиатскую часть СССР, и т. д.
11
Первые такие схемы были предложены С. В. Кириковым
(1952, 1956), Т. Б. Вернандер с соавторами (1959), Л. П. Никифо-
ровым (1959) и рядом других авторов. Наиболее полно разработал
зоогеографическую типологию Л. П. Никифоров. Характеризуя
Тоболо-Ишимское междуречье, этот исследователь делит его на
четыре природных района (южная, средняя и северная лесо-
степь, притобольские боры), затем внутри природных районов
производит деление на категории угодий (лесные и кустарнико-
вые, травянистые, водные и болотные, населенные пункты). Кате-
гории угодий разделены на типы угодий: леса и кустарники на
1) сосновые леса, 2) суходольные березняки, 3) низинные колки,
4) ивовые кустарники; травянистые угодья подразделяются на
1) луга, примыкающие к колкам, 2) солончаковые луга, 3) солон-
цовые степи и т. д. Типологическая система Л. П. Никифорова
хорошо продумана, но страдает тремя недостатками: 1) она не
универсальна, 2) в ее подразделения входят разномерные образо-
вания и 3) построена она на эклектическом ландшафтно-геоботани-
ческом принципе, без применения зоогеографических критериев.
По сути дела, это разработка ландшафтно-геоботанической типо-
логии для целей экстраполяции зоогеографических данных —
как раз то, к чему и стремился автор, создавая свой интересный
зоогеографический профиль.
Г. А. Зиновьев (1959), разбирая методические приемы соста-
вления карты энтомокомплексов лесных биоценозов, подчеркивает
насущную необходимость сопоставления материалов геоботаниче-
ской основы с данными непосредственной съемки энтомокомплек-
сов. На это замечание следует обратить особое внимание. Дей-
ствительно, как мы уже отмечали (Реймерс, 1965), при картографи-
ческой методике выявления природных закономерностей многие
авторы следуют используемой основе, невольно забывая, что они
работают с зоологическим материалом.
При применении картографических методик наиболее трудно-
понимаемым оказывается вопрос о соотношении биотопа, биоце-
ноза и, более узко, — типологических единиц животного населения
позвоночных. Не углубляясь в вопросы биоценологии, отметим, что
наиболее сложно соотношение зоокомплексов с геоботаническими
или иными типологическими единицами. Действительно, очень
трудно вычленить из биоценоза как целого животное население.
Еще труднее привыкнуть к мысли, что это выделенное есть лишь
органическая часть целого. Картографически (и в своей сущности)
единые биотоп и биоценоз совпадают в изображаемых выдел ах.
Также совпадает выдел местообитания вида с распространением
организма. Следовательно, смысл биогеографической карты всегда
кодирован: по ее контурам нельзя определить, что она изобра-
жает — расшифровка таится в легенде. Если рассматривать тер-
риторию с точки зрения условий существования видов и ком-
плексов, карта будет изображать распределение биотопов. Ставя
12
во главу угла животное население, получаем карту комплексов
животного населения, заполняющего взятые биотопы. Это поло-
жение очень легко воспринимается при взгляде на природу в «мел-
ком масштабе». Н. П. Наумов (1955) совершенно прав, полагая,
что «в понятие „степь“, „пустыня“, „тундра“ и „луг“ необходимо
включать не только растительность, но и животное население не
потому, что они живут совместно, но потому, что они обоюдно
необходимы» (стр. 163). Однако в «крупном масштабе» при рас-
смотрении элементарных выделов у зоологов возникает стремление
к расшифровке кода геоботанического термина путем прибавления
видовых названий доминирующих форм (Иоганзен, 1953; Кузя-
кин, 1962). Возникают непривычные и комические сочетания слов,
необходимость которых сомнительна. Упоминание одного-двух
доминантов из мира позвоночных животных (почему не беспозво-
ночных?) не раскрывает кода называемого выдела и не помогает
пониманию карты.
Перечисление только руководящих форм, на наш взгляд, также
ничего не дает. Во-первых, название характерных видов ничего
не прибавляет для лиц, не знающих фауны, во-вторых, три-четыре
перечисленных вида даже в слабой степени не расшифровывают
схемы кода. Между тем создается ложное впечатление, что наз-
ванные виды — самые основные, важнейшие в данной фаунисти-
ческой группировке. Однако большей частью это не так. Число
массовых видов обычно больше, чем то, которое можно ука-
зать в легенде карты, а зооэдификаторы (Кучерук, 1959) чаще
всего еще неизвестны. Относительно малочисленные или мало-
специфичные виды могут играть в биоценозе настолько замет-
ную роль, что ими ни в коем случае нельзя пренебрегать. Наконец,
не следует забывать, что раритеты указывают на исторические тен-
денции развития биоценоза в прошлом или будущем, что имеет
большое значение для прогнозирования и расшифровки динамики
биоценозов и ландшафтов. В природе нет ничего маловажного,
несущественного, а именно на обратное намекает перечисление
нескольких характерных форм.
Кажется более правильным в этом отношении подход А. И. Ку-
ренцова (ряд работ, в том числе 1959а, 19596, 1963а), рассматрива-
ющего фаунулы различных лесов как отдельные образования,
несовместимые зоогеографически. Вместе с тем этот автор говорит
о фаунуле в целом как о некоторой исторически сложившейся
системе, не отрывая ее части друг от друга и от населяемых ею
биотопов.
Таким образом, мы отдаем предпочтение собирательным назва-
ниям, взятым из арсенала смежных наук. Практически будем
называть биотопы, подразумевая животное население, как говорят
о фауне степи, тундры и т. д.
Возвращаясь к сущности вопроса, следует сформулировать
прежде всего основные требования к типологическим единицам
13
в зоогеографии позвоночных животных. Очевидно, следует исхо-
дить из специфического материала науки, а именно: из фауны
и животного населения. В основу типологии должна быть положена
единица, имеющая достаточно четкие не только количе-
ственные, но и качественные показатели. Такая
единица должна быть природной в своей основе, т. е. быть
адекватной среде и исторически обусловленной, так сказать, корен-
ной для природы. Очевидно также, что наибольшее преимущество
имеет основная таксономическая единица, единая не только в зоо-
географическом, но и в биогеографическом смысле.
Типологии необходимо быть единой снизу доверху и при-
годной для использования в любых картографических масшта-
бах. Типологические единицы зоокомплексов должны быть внут-
ренне относительно однородны, характеризоваться опреде-
ленным (но не обязательно строго стабильным) набором
индивидуальных участков особей (а следова-
тельно, и составом фауны), отражать одновременно струк-
турные особенности кружева ареалов2 видов, входящих
в комплекс животного населения.
Таким образом, мы считаем, что фаунистический комплекс
(фауна или фаунула) и животное население есть различные пока-
затели единых градаций животного мира, первый — качества,
второй — количества.3 Изучение того и другого в отдельности
возможно и целесообразно для специальных целей, но в типоло-
гии показатели качества и количества должны рассматриваться
в единстве.
В связи с тем что животные крайне трудны для наблюдения,
а их территориальные комплексы не поддаются непосредственному
изучению методами аэрофотосъемки и т. п., типологические еди-
ницы зоогеографии должны опираться на какие-то таксоны смеж-
ной науки, позволяющие делать достаточно широкую экстрапо-
ляцию данных.
Как мы уже показали (Реймерс, 1965), для лесных участков
ведущими факторами следует считать растительность, местополо-
жение и хозяйственный режим.
Следует подчеркнуть, что практически для большинства лес-
ных позвоночных животных именно лес, растительность — среда
жизни. Почти все подлинно таежные животные непосредственно
никак не связаны с почвой: не в почве, а в лесной подстилке и под
корнями деревьев проходит жизнь большинства лесных землероев
2 Термин «кружево ареала» с легкой руки А. Н. Промптова (1934)
прочно вошел в отечественную литературу: Ю. А. Исаков (1952а, 1957),
А. Г. Воронов (1958), А. Н. Формозов (1959) и др.
8 Несколько специальных наших работ, находящихся в печати, посвя-
щены доказательству этого положения. О соотношении фаунистического
комплекса и животного населения говорится также в кандидатской диссер-
тации Г. А. Воронова.
14
из мира позвоночных.4 * Лес создает необходимый биоклимат,
доставляет зверям и птицам пищу и дает укрытия. И даже там, где
почвенные или литологические факторы выступают на первый
план, растительность остается лучшим индикатором особенностей
местности, тем более что без растений, без растительных кормов
зооценоз вообще не может существовать.
Выше мы определили требования к основной зоотипологиче-
ской единице. Этим требованиям отвечает комплекс животного
населения таксономического уровня, равного формации геобота-
ников, а в приложении к позвоночным животным называемой нами
(Реймерс, 1963г, 1965) зооформацией.
Зооформация — сообщество животного
населения определенного фаунистическо-
го состава размножающихся, кормящих-
ся и зимующих видов, сходных количеств
особей отдельных видов, заселяющее
одну растительную формаци ю.6 Действитель-
но, фауна и животное население соснового леса во все времена
года весьма четко отличаются от зоокомплекса темнохвойной
тайги (Реймерс, 1963г, 1965).
На уровне формации контуры геоботанической и зоогеогра-
фической карт максимально приближаются друг к другу. Ряд
сходных зооформаций мы предлагаем объединять и называть ана-
логично объединениям растительных формаций (Реймерс, 1963г):
группы зооформаций (животное население темнохвойных лесов,
животное население светлохвойных лесов), классы зооформаций
(животное население хвойных лесов, животное население широко-
лиственных лесов), наконец, классы зооформаций образуют тип
фаунистического комплекса, или тип зооформаций — животное
население лесного типа растительности.
Основная типологическая единица не есть в то же время наи-
меньшая. А эта наименьшая имеет неодинаковый территориаль-
ный объем в применении к различным эколого-систематическим
группам животных. Так, мышевидные зверьки и вообще млеко-
питающие дают значительно более однородную картину размеще-
ния (Снигиревская, 1959), чем многочисленная и мобильная группа
птиц; качественные отличия еще резче и дробнее выступают
в группе открыто живущих насекомых и т. д.
Количественные изменения в расселении позвоночных живот-
ных можно отметить даже при переходе от одного типа леса (ассо-
циации) к другому (Никсо-Никочио, 1956), т. е. минимальные
4 Виды, связанные с почвой (крот) или делающие в ней норы (барсук,
серые полевки и т. д.), либо выходцы из лесостепи и лесотундры (т. е. их
родина — открытые пространства), либо связаны с широколиственными
лесами.
6 Границы растительной формации рассматриваются нами как условные,
тем более, что биоценозы — саморегулирующиеся, но не замкнутые системы.
15
сдвиги в среде, отличающие один биогеоценоз от другого, приводят
к улавливаемым отличиям в животном населении. Однако если
условия меняются коренным образом для растений (другой тип
леса) и для многих беспозвоночных животных, то в среде более
эврибионтных птиц и млекопитающих они не всегда приводят
к качественным сдвигам (фаунистический состав может оставаться
постоянным, а население меняться в пределах, не выходящих
за рамки типичного для данной фаунистической разности). Нао-
борот, внутри одного типа леса (даже в узком понимании школы
В. Н. Сукачева, не говоря уже о широком школы П. С. Погреб-
няка) могут быть различные фаунистические образования, вызван-
ные к жизни незначительными для лесной типологии изменениями
в составе леса, например минимальной примесью другой лесной
породы, возрастом насаждения. Для практических целей (напри-
мер, борьба с вредителями и паразитами) важно знать наименьшие
единицы среды, которые бы четко отличались по животному насе-
лению (фаунистическому составу) всех позвоночных животных
или одной эколого-систематической группы. Существенно такое
знание и для изучения размещения отдельных видов животных,
выяснения кружева их ареалов, популяционной экологии и т. д.
Очевидно, что наименьшей типологической единице должно
быть присуще единство качественных и количественных отличий.
Специальные полевые исследования показывают, что элемен-
тарный фаунистический комплекс поз-
воночных животных отличается от сосед-
него наличием или отсутствием хотя бы
одного постоянно присутствующего раз-
множающегося вида (редко) или группы
(чаще) постоянно присутствующих видов.
Такой фаунистический комплекс населяет лес приблизительно
одной возрастной фазы, одной формации, состоящей из одной или
нескольких ассоциаций с относительно одинаковыми кормовыми
и защитными условиями.
Отличие хотя бы на один постоянно присутству-
ющий размножающийся вид обычно сопровождается заметным
различием в количестве особей других видов, поэтому его можно
характеризовать как качественное изменение — перерыв посте-
пенности.6 Эти дифференциальные виды — основные элементы
математического анализа, которые отвечают схеме «да — нет»,
«плюс — минус».
6 Элементарный фаунистический комплекс позвоночных животных —
искусственно отчлененный компонент биоценоза и так же, как он, представ-
ляет собой низшую единицу, имеющую устойчивый состав видов и их сред-
нюю численность (Hesse, 1924; Friederichs, 1927, и др.). Мы лишь повторяем
известное правило в применении к пространственному размещению группы
позвоночных животных и придаем ему методическое значение как критерию
при выделении территориальных разностей.
16
Внутри одной зооформации, непрерывно развивающейся, но
существующей сравнительно долгое время, элементарные фауни-
стические комплексы значительно более эфемерны и возникают
под влиянием природных и антропогенных причин, действующих
как раздельно, так и в совокупности. Чаще всего мы имеем дело
не с природными комплексами, а с образованиями, слагающимися
под влиянием той суммы факторов, которую В. Я. Балаганов
(1962) назвал природно-антропогенным режи-
мом. Однако в Сибири и сейчас еще есть клочки относительно
нетронутой природы, развивающейся, условно говоря,7
под влиянием природных, вековых динамических процессов.
С другой стороны, природно-антропогенный режим поддержи-
вает некоторые биоценозы в состоянии относительного равновесия
долгое время. Вот эти коренные, узловые, по терминологии
П. Д. Ярошенко (1949, цит. по 1961), растительные сообщества
населены и сравнительно постоянным комплексом животных
(как качественно, так и количественно). Внутри одной зооформа-
ции возможно наличие нескольких таких узловых сообществ —
экологических вариантов зооформации.
Животному населению не безразличен возраст насаждений
и та сукцессионная фаза, в которой находится растительность.
По этому вопросу накапливается все большая литература (Adams,
1908; Строгий, 1911; Поспелов, 1957; Turcek, 1957; Данилов,
1958, и мн. др.). Возражение, обычно выдвигаемое против зоо-
географического исследования сукцессий, заключается в том, что
динамическая карта отражает краткий миг в жизни природы.
Однако исследования последних лет показывают, что изменения
ареалов отдельных видов животных и даже групп видов проис-
ходят с быстротой, не уступающей частоте смен сукцессионных
фаз. Изменение лика природы вообще столь стремительно, что
любая карта живой природы, кроме исторических, устаревает
чрезвычайно быстро. Мнение, что следует изучать не динамику
реально существующих разностей в природе, а лишь то, что дол-
жно быть на месте этих разностей в идеале, очевидно, ошибочно.
Природу нужно изучать во всех аспектах — как в прошлом, так
в настоящем и будущем.
Антропогенные нарушения узловых сообществ — экологиче-
ских вариантов зооформаций — приводят к образованию а н-
тропогенного сукцессионно-возрастного
ряда, или ряда трансформации, состоящего из нескольких
элементарных фаунистических комплексов, например населяющих
гарь темнохвойного леса, хвойно-мелколиственный лес как про-
межуточную фазу сукцессии и, наконец, темнохвойный лес.
7 На самом деле и эти участки испытывают сильное влияние деятель-
ности человека, опосредованное через общее им изменение климата, гидро-
логического режима, состава воздуха и т. д.
2 н. Ф. Реймерс
17
Природные смены, идущие в сотни раз медленнее, приводят
к образованию другого сукцессионного ряда элементарных ком-
плексов, например сомкнутого темнохвойного леса, редкостойного
темнохвойного леса, заболоченного редкостойного темнохвойного
леса и т. д. Этот ряд принципиально отличается от антропоген-
ного своей «обратимостью» (а не односторонней направленностью
к сравнительно устойчивому экологическому варианту зооформа-
ции).
Следовательно, элементарный фаунистический комплекс —
производное образование, существующее сравнительно короткое
время. Чаще всего он является этапом в процессе антропогенной
трансформации или природной сукцессии биоценоза, слагающе-
гося в определенных условиях природно-антропогенного режима
(в этом биоценозе позвоночные животные составляют «экологи-
ческий вариант зооформации»).
Таким образом, вся предлагаемая нами таксономическая
система для животного населения позвоночных лесной Сибири
отображается схемой рис. 2.
Элементарный фаунистический комплекс отличается от другого
прежде всего строго привязанными (стенобионтными)8 видами-
индикаторами, достигающими наибольшей своей численности
в этом комплексе (по сравнению с другими биотопами) и нередко
практически отсутствующими в других комплексах.9
Между понятиями «вид-индикатор», «верный вид» школы Браун-
Бланке и «детерминант», по Л. Г. Раменскому (1938), можно поста-
вить знак равенства, т. е. вид-индикатор — верная форма, детер-
минирующая зоокомплексы. Очевидно, среди животных возможны
все степени верности, различаемые Браун-Бланке (1928 и 1951,
цит. по Ярошенко, 1961): виды верные, привязанные, свободные,
бродячие и случайные. Для различения зоокомплексов важны,
8 Необходимо подчеркнуть, что один и тот же вид может быть в одном
зоогеографическом районе привязан к одному биотопу, в другом — к дру-
гому, а в третьем вести себя как форма широкой экологической валентности.
Например, альпийская пищуха (Qchotona alpina Pall.) в южной тайге Средней
Сибири привязана к долинным темнохвойным лесам водораздельного типа,
в горах Южной Сибири вид хотя и тяготеет к каменным россыпям, но жи-
вет в самых разнообразных условиях темнохвойной тайги; на Сахалине же
пищуха связана с высокогорьями, поросшими кедровым стлаником, и прак-
тически не встречается в темнохвойных лесах южной тайги острова. Таких
примеров можно привести много, некоторые из них указаны в гл. 5,
9 Отличие лишь в количественных характеристиках дает внутренние
подразделения элементарного фаунистического комплекса (в идеале — части
биоценоза), называемые К. В. Арнольди и Л. В. Арнольди (1963) динамиче-
ским вариантом биоценоза. Этот термин явно неудачен, поскольку слово
«динамика» может быть понято различно: вековая динамика, антропогенная
динамика и т. д. Подразделения биоценоза, имеющие различные количествен-
ные показатели, лучше называть до предложения специального термина его
экологическими вариантами, так как сгустки населения могут быть вызваны
самыми различными факторами, в том числе антропогенными, микроклима-
тическими и т. д.
18
помимо форм высшей степени верности, прочие характерные виды—
привязанные и свободные. В связи с тем что среди животных
редко встречаются формы абсолютно верные, группа привязан-
ных видов практически может быть объединена с верными под
названием «виды-индикаторы». Свободные из характерных форм
мы предлагаем, в отличие от видов-индикаторов, называть просто
верными видами.
Антропогенный.
сукцессионно-
возрастной ряд
(трансформации)
элементарных
фаунистических
комплексов
Рис. 2. Таксономическая система эколого-зоогеографиче-
ской типологии.
Постоянно присутствующие в биотопе виды низкой верности
подобны сопровождающим видам Браун-Бланке, но термины «со-
провождающий», а тем более «бродячий» мало удачны в применении
к зоологии. Нельзя согласиться и с расшифровкой этих терминов:
«виды, не предпочитающие какой-либо определенный тип сооб-
ществ». Таких видов среди животных нет, все они предпочитают
одни и избегают другие биоценозы. Мы предлагаем для третьей
степени верности термин «постоянный вид». Формы,
заходящие в биотоп из соседних, временно включающиеся в эко-
логические связи биоценозов, можно называть случайными.
Таким образом, можно различать четыре основных градации
-верности видов в зоокомплексе: 1) виды-индикаторы, 2) верные
виды, 3) постоянные виды, 4) случайные виды.
Понятие «вид-индикатор» определено выше. Это стенобионтная
в данной местности форма. Так, в южной тайге Средней Сибири
только по приручьевым темнохвойным лесам водораздельного
2*
19
типа живут пятнистые коньки и альпийские пищухи, лишь на над-
пойменных лугах — степные коньки, по приречным ивнякам —
ширококлювые мухоловки; на южном Сахалине только в темно-
хвойной тайге обитают длиннохвостые мышовки и красные полевки.
Верные виды в отличие от видов-индикаторов населяют
несколько природных разностей, но в ряде их достигают наи-
высшей для себя численности. Например, красная полевка —
верный вид всего ряда трансформации водораздельных темнохвой-
ных лесов юга Средней Сибири, а средняя бурозубка занимает
такое же положение в темнохвойных лесах южного Сахалина.
Постоянные виды населяют данный биотоп с малой для себя
численностью или живут в широком круге биотопов. Так, обыкно-
венная бурозубка в южной тайге Средней Сибири или когтистая
бурозубка на Сахалине входят во все биоценозы. Почти повсеме-
стен в южной тайге Средней Сибири пухляк. Красная полевка
населяет, помимо водораздельных темнохвойных лесов, приреч-
ные и приручьевые ельники, светлохвойные леса различных раз-
ностей, но в них не достигает той численности, которая характерна
для вида на водоразделах.
Нередко констатируется случайное обитание животных в не-
свойственных им стациях. Очень яркий пример такого рода —
гнездование дрофы в забайкальском лесу (Измайлов и Стариков,
1961) или приводимый в настоящей работе случай нахождения
альпийской пищухи в подвале жилого дома. Более обычно посе-
ление серых полевок в темнохвойных лесах водоразделов юга
Средней Сибири, где, как правило, представители этого рода
не живут.
Нередки случаи, когда вид размножается в одном биотопе,
а питается в другом и, таким образом, перманентно присутствует
в двух отличных биоценозах, в одном — в качестве постоянного
вида, в другом — постоянного гостя. Такие формы можно назы-
вать полистациальными в отличие от других, у которых место-
обитание включает лишь одну стацию (мы присоединяемся в пони-
мании этого термина к Н. П. Наумову, 1955). По отношению
к биоценозу, где полистациальные виды постоянно гостят, они
не могут считаться случайными, мы их предлагаем называть
гостевыми. Степень их верности кормовой стации может
быть высока и низка, поэтому гостевые виды теоретически могут
входить во все градации верности, кроме высшей — индикацион-
ной. Следовательно, необходимо различать при изучении верно-
сти форм аборигенные и гостевые виды.
Среди первых трех групп верности можно отличать две серии
форм. Одна из них — изначальные члены комплекса от природы,
другая вошла в него после воздействия человека на биоценоз.
Например, редины в лесу могут заселять случайные виды из сосед-
него комплекса, но может создаваться такое положение, что при
сохранении общей структуры комплекса в него проникают чуж-
20
дне ему формы. Так, у орехопромысловых балаганов в кедровни-
ках поселяются большие горлицы. Зимой в глухой тайге вдоль
троп обитают зайцы-беляки и т. п. Для коренных компонентов
существует термин «автохтонный» вид, для пришлых —
«аллохтонный». Этими терминами пользуемся и мы. В дан-
ном случае речь идет не о проникновении вида с другой удаленной
территории, а о расширении кружева ареала формы. В девствен-
ной природе расширение местообитания вида за счет других био-
геоценозов — явление редкое. Ему должны предшествовать веко-
вой сукцессионный процесс, изменение экологических потребно-
стей вида или образование «эрзац-биотопа» (например, сочетание
кедровых стлаников и березовых лесов на Камчатке создает
«эрзац-биотоп» темнохвойного леса). С нарушением природных
комплексов для многих видов возникают предпосылки для эври-
топности, поскольку появляется много «эрзац-биотопов». Так, сме-
шанный сосново-мелколиственный лес для многих видов темно-
хвойной тайги оказывается «эрзац-стацией» размножения. В на-
рушенной природе, таким образом, происходит смешивание
животных, а это затрудняет различение элементарных фауни-
стических комплексов.
Следующий важный критерий индивидуальности зоокомпле-
кса — иерархия доминирования. Этот вопрос
хорошо разработан в применении к зоокомплексам (Вернандер,
Викторов и др., 1959; Кузякин, 1962). Нужно лишь отметить, что
до широкого применения методик абсолютного учета животных
шкалы доминирования были очень и очень условны из-за
относительности показателей, а десятичные градации не всегда
можно использовать в практической деятельности (см. гл. 6).
Виды-индикаторы, верные и постоянные могут входить в раз-
личные градации доминирования, в том числе оказаться рарите-
тами. В связи с этим следует возразить тем авторам, которые пред-
лагают при изучении природы отбрасывать малочисленные виды
как несущественные (Кузякин, 1962).
Среди всех групп животных, населяющих биоценоз и составля-
ющих его зоокомплекс, имеются виды-эдификаторы,
определяющие физиономию комплекса, создающие его. Среди
позвоночных животных только в редких случаях можно сразу
констатировать эти формы, тем более что их влияние на зооком-
плекс обычно опосредовано. Например, копытные животные
степей — ее эдификаторы, кедровка — эдификатор кедровой тайги.
К сожалению, истинного значения большинства видов в биоценозе
мы не знаем.
Зоокомплексы изменяются в течение суток, сезонов и ряда лет.
Это приводит к еще почти совершенно не изученным в тайге пере-
стройкам в их структуре. Однако едва ли можно ожидать, что
в ходе сезонных или других изменений может происходить смеще-
ние границ участков, занимаемых комплексами. По сумме пока-
21
зателей: фауне, верности видов и шкале доминирования — зоо-
комплексы безусловно отличаются друг от друга в любой момент
(в то же время качественно и количественно отличаясь от тех,
которые существовали на их месте в течение другого сезона или
времени суток).
Единство многих показателей элементарного фаунистического
комплекса заставляет предполагать, что внутри него каждый вид
имеет определенные по размеру индивидуальные участки особей
или определенные «секторы» таких участков. Таким образом,
несмотря на слабую изученность этого вопроса, можно гипотети-
чески считать элементарные фаунистические комплексы едини-
цами типизации индивидуальных участков обитания.
Итак, основные принципы, из которых мы исходим, заклю-
чаются в следующем.
1. Фаунистический комплекс и животное население есть раз-
личные показатели градаций животного мира: первый — каче-
ства, второй — количества. Территориальные их границы совпа-
дают, хотя ареалы отдельных видов могут выходить за пределы
области распространения обоих комплексов.
2. Типология должна быть единой снизу доверху и позволять
использование ее в любых картографических масштабах и любом
месте. Вместе с тем мы допускаем возможность разработки ее при-
менительно к различным ландшафтным зонам, поскольку в каж-
дой из них могут быть свои ведущие факторы среды.
3. Основная единица типологии должна иметь четкие каче-
ственные и количественные критерии (характеризоваться опреде-
ленным фаунистическим комплексом и животным населением).
Такая единица должна быть природной в своей основе,
т. е. адекватной среде и исторически обусловленной — коренной
для природы. Она должна быть единой не только в зоогеографи-
ческом, но и в биогеографическом смысле.
4. Допускается возможность выработки типологических кри-
териев для различных эколого-систематических групп животных,
условно вычленяемых из биоценоза. В нашем частном случае
типология разрабатывается для всего комплекса наземных поз-
воночных животных — зверей и птиц.
5. Наименьшая (элементарная) таксономическая единица
должна характеризоваться минимальным перерывом качества —
отличием фаунистического комплекса хотя бы на один постоянно
присутствующий вид.
6. Таксоны нашей системы типологических единиц до зоо-
формации включительно — природные образования. Экологиче-
ские варианты зооформаций — узловые сообщества, существова-
ние которых поддерживается природно-антропогенным режимом.
Природные и антропогенные ряды сукцессий приводят к об-
разованию определенного экологического варианта зооформа-
ции.
22
7. Все таксоны отличаются друг от друга видовым составом,
рядом верности видов и всей иерархией доминирования. Практи-
чески в мелком масштабе можно пользоваться сравнением видо-
вого состава — разницей в фаунистических комплексах: уже на
уровне элементарной типологической единицы разница хотя бы
в один постоянно присутствующий размножающийся вид обычно
сопровождается заметным отличием в количестве особей других
видов. Изменение комплекса на один вид констатируется очень
редко. Это связано с тем, что, во-первых, при исследовании про-
пускаются многие малочисленные формы и потому при тщатель-
ных поисках можно найти более широкий перерыв постепенности,
во-вторых, и это основное, сам перерыв постепенности происходит
скачком, характеризующимся присутствием или отсутствием
в новом комплексе сразу нескольких видов.
8. При выделении характерных биотопов на местности в при-
менении к группе позвоночных животных следует различать лишь
те образования, которые отличаются населением (фаунистическим
комплексом) данной группы животных, т. е. искать перерывы
постепенности и в среде, и в животном населении. В этом случае
получаемые выделы имеют универсальное значение показателей
фауны, животного населения и среды.
Глава 2
ТЕРРИТОРИАЛЬНЫЕ ВАРИАНТЫ ЗООФОРМАЦИЙ
И ТИПОЛОГО-ЗООГЕОГРАФИЧЕСКАЯ КАРТА ЛЕСНОЙ СИБИРИ
Зооформация, как любое природное образование, изменяется
во времени и пространстве. Изменение во времени связано как
с сезонными ее аспектами, так и с общим колебанием условий
от года к году. В состав зооформации и ее подразделений могут
входить случайные и аллохтонные виды. Количественный состав
животного населения, как известно, постоянно изменяется. Тем
не менее основное ядро зооформации — виды-индикаторы, верные
и постоянные виды, а также их масса, сохраняют некоторую сред-
нюю величину, отличающуюся от средней другой зооформации.1
При многолетних стационарных исследованиях можно определить
эти средние величины и математически показать возможные коле-
бания биомассы животного населения внутри той или другой
зооформации, элементарного фаунистического комплекса и т. д.
Такие подсчеты могут быть сделаны как в относительном выраже-
нии (при применении методик определения относительного обилия
животных), так и в абсолютных числах.
1 Это положение для условий Средней Сибири доказывается нами в ряде
отдельных работ (Реймерс, 1965, и в печати), поэтому здесь мы не приво-
дим детальной аргументации.
23
Другая сторона вопроса — изменение зооформации в простран-
стве. При изучении любых закономерностей в природе важно
знать географические пределы возможных экстраполяций, т. е.
границы района исследований. Мы уже отмечали, что это одно из
самых больных мест современной фаунистики, да и не только ее.
Если изменение видового состава влечет за собой перемену
количественных характеристик сообщества животных в пределах
одного небольшого участка, а появление нового вида может слу-
жить сигналом изменения численности многих других форм, то
естественно ожидать проявления таких же закономерностей в про-
странстве. Нами было уже показано (Реймерс, 1965) наличие изме-
нений зооформации в пространстве, а крупные ее отдельности были
названы территориальными вариантами.
Территориальный вариант зооформа-
ции—однотипное сообщество позвоноч-
ных животных, характеризующееся в пре-
делах своего ареала относительно посто-
янным фаунистическим составом и живот-
ным населением. Критерием для деления зооформации
на территориальные варианты мы считаем сравнительно одновре-
менное изменение числа видов или замену одних видов другими
(т. е. совпадение границ ареалов нескольких видов), сопровождае-
мое переменой количественных показателей, сменой типа кружева
ареалов и других характеристик животного мира. Минимальным
одновременным изменением числа видов при выделении террито-
риальных вариантов зооформации следует считать 3—5. Эти пока-
затели числа повторностей имеют статистическую достоверность.
Поскольку характер кружева ареала большинства видов мало
известен, а результаты количественных учетов животных имеют
относительное значение и чаще всего не поддаются статистической
оценке (недостаточное число повторностей), качественный пока-
затель — состав фауны — все еще остается самым надежным кри-
терием в зоогеографии.
Таковы принципы разделения зооформации на территориаль-
ные варианты. Причины возникновения территориальных вари-
антов многообразны. Это общие климатические различия —
зональные, азональные и связанные с орографией. Это взаимное
влияние смежных зооформаций в их историческом развитии.
Наконец, это степень антропогенных воздействий на природный
комплекс.
Границы территориальных вариантов различных зооформа-
ций, расположенных бок о бок, могут как совпадать, так и не сов-
падать между собой. Так, Енисей служит достаточно четкой реги-
ональной границей внутри группы зооформаций темнохвойных
лесов, но зооформация сосновых лесов практически не имеет тут
перерыва постепенности. Он наблюдается лишь по линии Бай-
кала. Еще яснее это видно на примере лесотундры: лесные зоо-
24
формации имеют четкие региональные границы по западной окра-
ине севера Средне-Сибирской возвышенности, а тундровые — столь
же выраженные разграничения лишь восточнее Лены. Может
быть, отсюда и проистекают разногласия в районировании полосы
лесотундры.
Причина несовпадения границ территориальных вариантов
зооформаций лежит, по нашему мнению, в совокупном действии
геоисторических и антропоисторических факторов. В одних местах
зооформация развивается автохтонно и давно уже сложилась,
в других — аллохтонно, и ее комплекс, так сказать, еще не сфор-
мировался. В третьем месте уже появились предпосылки для
проникновения элементов данной зооформации — налицо расти-
тельная формация, но наблюдается влияние соседних экологи-
чески близких зооформаций, и в результате складываются
несколько иные фаунистические комплексы. В последнем случае
резко возрастает значение географических преград.
Исходя из изложенных теоретических предпосылок, мы попы-
тались произвести деление зооформаций лесной Сибири на терри-
ториальные варианты и составить типолого-зоогеографическую
карту этой территории. Для первого опыта был избран десяти-
миллионный масштаб. Большая дробность показалась нам излиш-
ней в силу слабой изученности вопроса и практического отсут-
ствия хороших основ для экстраполяции данных.
В масштабе 1 : 10 000 000 могут быть отражены не все зоо-
формации или даже их группы. Особенно это касается речных
долин, в пределах которых приходится весьма схематично и
условно выделять сочетания нескольких зооформаций (луговых,
кустарниковых и лесных) и высокогорий с их сложной форма-
ционной структурой.
Проведенное нами деление следует считать безусловно про-
визорным, тем более что мы не могли охватить своими полевыми
исследованиями и трети колоссальной территории Сибири, а в ряде
случаев вынуждены пользоваться весьма туманными литератур-
ными указаниями на приуроченность и численность тех или иных
видов.
Пример деления зооформации на территориальные варианты2
При рассмотрении фауны зооформаций и их сочетаний мы
будем вынуждены обращаться к анализу экологических потреб-
ностей видов (характер их размещения) и к сопоставлению границ
2 В связи с ограниченным объемом данной книги ниже приводится лишь
пример деления зооформации на территориальные варианты. В качестве
образца избрана группа зооформаций темнохвойных лесов, широко распро-
страненная на территории Сибири, а не отдельная зооформация. Поскольку
принципы вычленения территориальных вариантов в том и другом случае
идентичны, приводимый пример характерен и для отдельных зооформаций.
25
ареалов видов, мысленно воспроизводя кружево этих ареалов.
Следует сразу же оговориться, что наши современные знания
и того и другого вопроса далеки от совершенства. Почти неизве-
стны также древность, величина и местоположение первичных
ареалов животных, т. е. история видов. Ареалы животных, как
и растений (Толмачев, 1958, 1962), могут быть изначально лока-
лизованными или обширными, относительно молодыми, развива-
ющимися, и старыми, стабилизировавшимися. Также очевидно,
что теоретическая возможность существования вида, благоприят-
ная биотопическая обстановка еще не означает, что вид будет
обязательно тут присутствовать. Ему могут противодействовать
экологически близкие виды, наличие болезней с неминуемо леталь-
ным исходом и т. п. Эти обстоятельства нужно четко осознавать
при рассмотрении границ и кружева ареала животных. Следует
учитывать и гибкость экологии видов, их «полиландшафтность»,
по терминологии В. Ф. Палия (1958). Однако при всей эвриби-
онтности животных количественный анализ (даже относительный)
распределения вида всегда устанавливает его излюбленные био-
топы, места максимума и минимума плотности его населения, но
при этом четких количественных характеристик для больших
площадей, к сожалению, сейчас дать нельзя из-за отсутствия
материалов. Немаловажно и то обстоятельство, что почти неиз-
вестна потребность видов в отношении климатических условий.
С другой стороны, исключительно бедны наши представления
о микроклимате лесов.
Следует учитывать еще одну трудность в отнесении видов
к той или другой зооформации: антропогенные изменения в при-
роде. Если бы мы имели дело с девственными биоценозами, то рас-
пределение животных приобрело бы более четкие формы. Но в сов-
ременной природе человек создал такую мозаику рядов элемен-
тарных фаунистических комплексов, что резко сместил все
естественные границы, «перемешал» представителей различных
зооформаций. Действительно, экологический облик гари темнохвой-
ного леса, поросшей березняком с подростом темнохвойных пород,
очень близок к облику гари соснового бора, также поросшей берез-
няком, но с густым подростом сосны. По тенденциям развития
фауны и количественным характеристикам животного населения
эти участки не одно и то же и принадлежат к разным зооформациям,
а по общему составу фауны позвоночных животных в указанной
сукцессионной фазе они близки (см. гл. 6).
Списки видов, населяющих зооформацию, при современном
уровне знаний еще не могут претендовать на полноту и высокую
точность. Вместе с тем следует ожидать, что степень их несовер-
В дальнейшем все конкретные материалы, на которых основано деление»
вынужденно оставлены между строк, а текст ограничен самыми необходимыми
краткими пояснениями и выводами. Иллюстрируется эта глава цветной кар-
той.
26
шенства приблизительно одинакова для всех районов и поэтому
они в целом сравнимы.
Для примера разберем деление на территориальные варианты
группы зооформаций темнохвойных лесов. Список видов, населя-
ющих эти леса в пределах Сибири, ориентировочно включает 76
(видимо, больше) форм (табл. 1). Анализ их распространения пока-
зывает наличие минимум пяти территориальных вариантов
в совокупности зооформаций высокоствольных темнохвойных лесов
и отличной от них зооформации кедровых стлаников:
а) животное население западносибирских северных равнинных
темнохвойных лесов;
б) животное население западносибирских южных равнинных
темнохвойных лесов;
в) животное население среднесибирских среднегорных темно-
хвойных лесов;
г) животное население южносибирских горных темнохвойных
лесов;
д) животное население дальневосточных горных темнохвойных
лесов.
Зооформация кедровых стлаников в пределах Сибири, оче-
видно, едина: животное население зарослей кедрового стланика.
Каждый территориальный вариант группы зооформаций темно-
хвойных лесов населяют 45—60 видов, из которых от 18 до 23 форм
не повторяются в соседнем варианте.
Наличие или отсутствие вида в темнохвойном (как и всяком
другом) лесу определяется множеством факторов, в том числе
соседством других биотопов, вернее их сочетанием. Например,
полевка-экономка в типе не встречается в высокогорных темно-
хвойных лесах и не входит в их зооформацию, однако в-горах
южной Сибири сочетание крошечных луговин антропогенного
происхождения со сплошной кедрово-пихтовой тайгой и наличие
болотистых участков позволяют экономке селиться в тайге. Еще
один пример. Желтобровая овсянка в типе не живет в высоко-
ствольных темнохвойных лесах юга Средней Сибири. Однако на
гарях этих лесов, уже поросших молодым лесом (жердняком),
или в разреженных островных участках темнохвойных лесов
с обильным подседом, расположенным куртинами, это обычная
птица.
Формы, подобные полевке-экономке, в целом для темнохвой-
ных лесов случайны и, строго говоря, относятся к другим зоо-
формациям. В очень крупном масштабе они попали бы в соответ-
ствующие выделы. Однако в мелком масштабе их приходится
включать в общий список животного населения доминирующей
зооформации. Иное положение занимает желтобровая овсянка
и подобные ей виды. Заселяя лес в определенной фазе сукцессии
и входя в элементарный фаунистический комплекс сукцессионно-
возрастного ряда зооформаций темнохвойных лесов, такие формы
27
Таблица 1
Распределение птиц и млекопитающих в группе зооформаций
темнохвойной тайги (водоразделы)
Виды Западносибир- ская равнинная темнохвойная тайга (юг) 1 Западносибир- ская равнинная темнохвойная тайга (север) Среднесибирская среднегорная темнохвойная тайга Южносибирская горная темно- хвойная тайга Дальневосточная горная темно- хвойная тайга Заросли кедро- вых стлаников
Глухарь + + + +
Каменный глухарь . . . — + —
Рябчик + + + + + —
Дикуша + —
Большая горлица . . . + — + + + —
Ястреб-тетеревятник . . + + + + + —
Ястреб-перепелятник . . + + + + + —
Малый перепелятник . . — — + + + —
Филин + + + + + —
Мохноногий сыч .... + + +? + + —
Воробьиный сыч .... + + + + —
Бородатая неясыть . . +? + +? — —
Длиннохвостая неясыть + + +? + + —
Глухая кукушка . . . + + _? + + +
Желна + + + + + —
Трехпалый дятел . . . + + + — + —
Большой пестрый дятел + + + — —
Ворон +? +? + + +? +
Кукша + + + + +
Кедровка + + + + + +
Снегирь + + + + + —
Сибирская чечевица . . — + +
Обыкновенная чечевица — — — + —
Щур — + + + — +
Клёст-еловик + + + + +
Белокрылый клёст . . . + + —
Черногорлая овсянка . — — +
Желтобровая овсянка — — + — —
Лесной конек + + + — — —-
Пятнистый конек . . . + — +? —
Пищуха + + -? + —
Поползень + — + + + —-
Московка + + + + + —
Пухляк + + + + + —
Сероголовая гаичка . . + + — + —
Восточноазиатский сви-
ристель — — — —• + —
Мухоловка-касатка . . — — + —• + —
Малая мухоловка . . . + — + + + —
Мухоловка-мугимаки . . — + +? + —
Желтоголовый королек + — + + —
Пеночка-королек .... — + + + —
Пестрый дрозд .... + + + + —
28
Таблица 1 (продолжение}
Виды Западносибир- ская равнинная темнохвойная тайга (юг) t Западносибир- ская равнинная темнохвойная тайга (север) Среднесибирская среднегорная темнохвойная тайга Южносибирская горная темно- хвойная тайга Дальневосточ- ная горная тем- нохвойная тайга Заросли кедро- вых стлаников
Певчий дрозд + + + + — —
Темнозобый дрозд . . . + + + + —. —
Бледный дрозд .... — — + — + —
Синехвостка — + + —• + —
Синий соловей — — + + + —
Соловей-свистун .... — — +? — + —
Сибирская завирушка . — + — + +
Кроты обыкновенный и сибирский + + — +? —. —
Крошечная бурозубка + + +? + + +
Малая бурозубка . . . + + + + + +?
Средняя бурозубка . . + + + + + +
Обыкновенная и когти- стая бурозубки . . + + + + + +
Барсук + — — — — —
Соболь + + + + +
Солонгой —- — — + + —
Горностай +? — — — — +
Ласка — — ? + + +
Росомаха + + +’ + + +
Рысь + + + + + —
Бурый медведь .... + + + + + +
Благородный олень . . —' — — + + —
Северный олень .... +? + + + + +
Кабарга —. — + + + —
Заяц-беляк + + + + + +
Пищуха алтайская . . . — — + + + +
Летяга + + + + + —
Белка + + + + + —
Бурундук + + + + + +
Азиатская лесная мышь — — + + + +
Лесной лемминг .... + + + + +
Рыжая полевка .... + — — — — —
Красная лесная полевка + + + + + +
Красно-серая лесная полевка + + 4" + + +
Полевка-экономка . . . —? — —? + _? —
птиц 31 28 41 32 36 6
Всего млекопитающих 20 17 19 24 22 17
видов
(общее число . . 51 45 60 56 58 23
Негативное и пози- тивное отличие от фау- ны тайги Средней Си- бири 21 21 23 18
29
относятся к собственно зооформациям темнохвойных лесов, соста-
вляют их «видовой фонд». Следовательно, в списки попали виды,
очень различные по экологическим потребностям.3 Это вызывает
затруднения в четком определении зооиндикаторов отдельных
территориальных вариантов зооформаций или их групп, во всяком
случае при рассмотрении списков форм не ниже видового ранга.
Анализ подвидовых списков повысил бы степень различия, но
привел бы к еще большей неточности нашего разбора, поскольку
многие подвиды еще не настолько хорошо охарактеризованы
и всеми признаны как виды. Безусловно — это затруднение
временное, связанное с недостатком знаний о распределении
животных.
Следует обратить внимание (табл. 1) на приблизительно оди-
наковое количество видов во всех перечисленных участках темно-
хвойных лесов (кроме северных и стлаников). Это указывает на
близость экологических возможностей различных участков темно-
хвойных лесов и значительную древность фаунистических обра-
зований в них («полноту» фаунистических комплексов). Однако
близость количества видов не означает идентичности фаун. Так,
горные темнохвойные леса Дальнего Востока и Южной Сибири
имеют почти равное число видов, но отличаются друг от друга
21 формой видового ранга (приблизительно 35—40% от состава
фауны позвоночных животных, отмеченных в списке).
Указанные различия связаны как с экологическими особенно-
стями участков, так и с историческими факторами. Серый снегирь,
как показал еще Б. Дыбовский (Dybowski, 1874), населяет в Средней
Сибири и на Дальнем Востоке (Воробьев, 1954) главным образом
темнохвойные леса водоразделов, экологических аналогов которых
в Западной Сибири нет. Фауна дальневосточных темнохвойных
лесов в историческом развитии пополнилась за счет хвойно-широ-
колиственных формаций. Для темнохвойных лесов Средней
и Западной Сибири такое пополнение было невозможно, поскольку
они не граничат (во всяком случае в последние столетия) с хвойно-
широколиственными формациями. Однако фауна темнохвойных
лесов Средней Сибири сохранила в своем составе ряд форм релик-
тового характера (например, мухоловка-мугимаки), оставшихся
от зооформаций хвойно-широколиственных лесов, предшество-
вавших темнохвойным. В истории Западной Сибири не было
такого стыка. Сочетание исторических и экологических условий
привело здесь к некоторой обедненности животного населения
темнохвойных лесов.
Животное население зооформаций кедровых стлаников близко
к лесным фаунам группы зооформаций темнохвойных лесов, однако
3 В табл. 1 включены главным образом виды, обитающие в самих темно-
хвойных лесах или заселяющие молодняки темнохвойных пород. Большая
часть форм гарей опущена.
30
оно беднее видами и имеет некоторые специфичные черты. Попытка
дать список птиц и млекопитающих, живущих в кедровых стла-
никах, сделана в последней колонке табл. 1. Приведенный список
нужно дополнить, очевидно, несколькими видами птиц, не живу-
щих в темнохвойных лесах, но обитающих в кедровых стланиках, —
это пеночка-таловка, пеночка-зарничка (?) и темный дрозд
Науманна. Многие виды приходится включать в список под вопро-
сом, так как вообще фауна кедровых стлаников изучена крайне
плохо. Очевидно, стланики населены приблизительно 8—10
видами птиц и 15—17 млекопитающих, т. е. всего 23—27 видами
позвоночных животных.
Зооформации лесной Сибири4
I. Животное население лесотундры
Исследователи фауны лесотундры колеблются, относить ли
эту полосу к лесной зоне или тундровой. Некоторые авторы счи-
тают лесотундру особой зоной (Портенко, 1937). Другие (Успен-
ский, 1960) категорично относят лесотундру к лесной полосе.
Эколого-фаунистический анализ животного мира лесотундры
(Кузнецов, 1950; Сдобников, 1958; Успенский, 1960) показывает,
что виды, населяющие эту полосу, относятся минимум к пяти
экологическим группам: 1) тундровые, 2) виды, тяготеющие к реч-
ным уремам, 3) лесные, 4) лугово-болотные, 5) озерно-речные.
Распространение форм каждой из групп специфично. Тундровые
виды главным образом распространены севернее лесотундры —
в тундре. Виды, тяготеющие к речным уремам, встречаются как
в зоне тундр, так и южнее — в зоне тайги. Эта группа имеет много
общего с лесной. Строго лесные формы немногочисленны в лесо-
тундре. Основную массу птиц ее фауны составляют болотные,
речные и озерные виды, которые распространены также далеко
на юг от лесотундры. Своих видов, живущих только в полосе
лесотундры, выделить не удается.
Речные, лугово-болотные и озерные виды мало специфичны
и большей частью не могут служить индикаторами ландшафтной
полосы. Очевидно, лесотундру должны характеризовать прежде
всего тундровые и лесные формы (ведь сама полоса лесо-
тундр а!). При таком подходе кажется совершенно логичным
вывод Т. Н. Дунаевой и В. В. Кучерука (1941) о том, что переход-
ная полоса лесотундр тяготеет к двум соседним «типичным» зонам.
4 Как уже отмечено выше (стр. 25), в принятом нами масштабе иссле-
дования отражены не все зооформации, и, наоборот, в ряде случаев прихо-
дится говорить о сочетании нескольких зооформаций. Однако всюду при
рассмотрении лесной полосы основой для выделения территориальных ва-
риантов служит зооформация или группа очень близких зооформаций. К их
сочетаниям мы обращаемся лишь при анализе фауны речных долин, а также
нетаежных ландшафтных зон или вертикальных поясов.
31
В лесной своей части она примыкает к лесам, в тундровой —
к тундрам.
Рассмотрение ареалов тундровых и лесных видов лесотундры
выявляет различия в распространении этих экологических групп.
Почти все лесные формы лесотундры широко распространены
в пределах последней. Однако западносибирские и восточно-
сибирские участки лесотундры отличаются 2—3 видами лесных
млекопитающих и 4 видами птиц: на востоке полосы отсутствуют
крот и крошечная бурозубка, но появляется (если справедливо
выделение этого вида) колымская бурозубка; в восточносибирской
лесотундре, очевидно, нет обыкновенного снегиря, белокрылого
клеста и горихвостки-лысушки, зато появляется дрозд Науманна.
Граница всех этих изменений в видовом составе лежит на водо-
разделе между Енисеем и Хатангой у западных пределов возвы-
шенной части севера Средне-Сибирского плоскогорья. Таким
образом, лесная лесотундра имеет четкую зоогеографическую
границу по Енисею, что было уже давно констатировано (Тугари-
нов, 1927; Колюшев, 1936, и др.).
Однако Б. А. Кузнецов (1950), рассматривавший лесотундру
с несколько иной точки зрения, совершенно справедливо отри-
цает эту границу как единственную. По его мнению, лесотундра
четко делится на две разности — северосибирскую (от европей-
ской части Союза до реки Индигирки, вернее водораздела между
Леной и Индигиркой) и колымо-анадырскую (на восток от реки
Индигирки). Тундровый комплекс птиц и млекопитающих лесо-
тундры действительно имеет перерыв постепенности в указанном
Б. А. Кузнецовым регионе. Таким образом, сочетания лесных
и тундровых зооформаций возможны в трех вариантах: 1) западно-
сибирские лесные и северосибирские тундровые элементы; 2) во-
сточносибирские лесные и северосибирские тундровые элементы;
3) восточносибирские лесные и колымо-анадырские тундровые
элементы. В соответствии с естественно выделяющимися сочета-
ниями мы и разбиваем полосу лесотундр.
(1) Животное население западносибирских лесотундр из запад-
носибирских лесных и сибирских тундровых элементов.
(2) Животное население среднесибирских лесотундр из восточ-
носибирских лесных и сибирских тундровых элементов.
(3) Животное население восточносибирских лесотундр из
восточносибирских лесных и колымо-анадырских тундровых эле-
ментов.
Более дробное деление лесотундры в масштабе 1 : 10 000 000
невозможно по техническим причинам.
Факт несовпадения границ зоогеографических разностей тунд-
ровой и лесной лесотундры заставляет предложить для картогра-
фического изображения полосы лесотундры расцветку из пере-
межающихся полос, где один цвет отражает характер тундровых
участков, другой — лесных.
32
II. Животное население лесов
А. Животное население темнохвойных
лесов. К полосе лесотундр естественно примыкает обширная
область распространения северосибирских редкостойных листвен-
ничников, протянувшихся, так же как и лесотундра, вдоль всего
севера Сибири. Однако по традиции принято начинать разбор
биот таежных участков с темнохвойных лесов. Деление этой
группы зооформаций на территориальные варианты разобрано выше
в качестве примера, поэтому здесь мы ограничимся лишь упоми-
нанием, что в легенде карты эти выделы отмечены цифрами (4) — (9).
Б. Животное население лиственничных
лесов. Рассмотрение ареалов форм, предпочитающих листвен-
ничники в качестве мест обитания, позволяет различить внутри
зооформации лиственничных лесов пять территориальных вари-
антов. Северные отличаются бедным видовым составом (общее
число видов около 35) и приблизительно совпадают с редкостой-
ными лиственничными лесами Западной и Восточной Сибири.
Фауна этих лесов имеет много общего с лесотундрой (лесными эле-
ментами), но отличается большим разнообразием. Меридионально
зоокомплекс полосы редкостойных лиственничников распадается
минимум на два территориальных варианта: редкостойные листвен-
ничники Западной Сибири имеют животное население, отлича-
ющееся от фаунистического комплекса таких же лиственничников
Средней и Восточной Сибири. Граница между этими территориаль-
ными вариантами проходит приблизительно по горам Путорана,
различие фаун здесь примерно 10 видов (около 30% от всего
фаунистического состава позвоночных животных).
Полоса сомкнутых лиственничных лесов в пределах Сибири
распадается на три фаунистических территориальных варианта,
из которых в пределах каждого почти вдвое больше видов, чем
в редкостойных лесах (60—65 вместо 35). Среднесибирские, восточ-
носибирские и южносибирские горные лиственничники имеют
отличие между собой в 12—21 вид при общем почти стабильном
их числе.
Таким образом, зооформация лиственничных лесов в пределах
Сибири делится на 5 территориальных вариантов.
(10) Животное население северных редкостойных лиственнич-
ников Западной Сибири.
(И) Животное население северных редкостойных лиственнич-
ников Средней и Восточной Сибири.
(12) Животное население среднесибирских среднегорных лист-
венничников.
(13) Животное население южносибирских горных лиственнич-
ников.
(14) Животное население восточносибирских горных листвен-
ничников.
3 II. Ф. Реймерс S3
Как уже отмечено, лиственничные редколесья фаунистически
меньше отличаются от лесных участков лесотундры, чем от южнее
расположенных сплошных лиственничных лесов. Это обстоятель-
ство подчеркнуто идентичностью условного обозначения лесных
участков лесотундры и лиственничных редколесий.
К зооформации лиственничных лесов следует относить лесные
участки лиственничных лесостепей и якутских районов господ-
ства аласов. Однако в масштабе 1 : 10 000 000 лесные участки
невозможно графически отделить от степных и луговых, поэтому
лесостепь и подобные ей образования приходится рассматривать
как сочетание лесных и степных участков вместе, а в легенде
карты выделять специальные пункты.
В. Животное население сосновых лесов.
Общая конфигурация ареала сосновых лесов в Сибири напоми-
нает распространение темнохвойных лесов. Однако фауна западно-
сибирских южных сосновых лесов оказывается (в отличие от
фауны темнохвойных лесов) почти идентичной таковой средне-
сибирских сосняков. При общем количестве 55—59 видов позво-
ночных животных, населяющих эти леса, разница между ними не
более чем в 5—6 малоспецифичных форм (лесные мыши, рыжая
лесная полевка, клинтух, возможно осоед и сероголовая гаичка).
При этом не наблюдается никакого четкого перерыва постепен-
ности, образованного границами ареалов нескольких видов. Одни
формы распространены в сосняках с запада до линии Оби (клин-
тух, европейский осоед, обыкновенная лесная мышь), другие —
восточные — встречаются в таежных сосняках до Енисея (азиат-
ская лесная мышь), третьи с запада перешагнули Енисей (рыжая
лесная полевка). Более четкая зоогеографическая граница внутри
зооформации сосновых лесов проходит по линии Байкала: восточ-
носибирские сосновые леса отличаются по фауне от среднесибир-
ских на 7 видов, а от суммы видов фауны сосновых лесов Западной
и Средней Сибири на 10 форм.
Животное население северных сосняков Западной Сибири отли-
чается от животного мира сосновых лесов юга страны. Сумма
негативных и позитивных отличий в фауне составляет 17 видов.
Таким образом, зооформация сосновых лесов в пределах Сибири
может быть разделена на 3 территориальных варианта.
(15) Животное население северных равнинных сосновых лесов
Западной Сибири.
(16) Животное население южных равнинных и среднегорных
сосновых лесов Западной и Средней Сибири.
(17) Животное население восточносибирских горных сосно-
вых лесов.
Г. Животное население мелколиствен-
ных лесов. Мелколиственные леса из березы и осины зани-
мают довольно обширные пространства на юге Западной Сибири,
переходя в березовые колки южнее и перемежаясь с темнохвой-
34
ными и сосновыми лесами севернее. В Средней Сибири осиновые
леса с примесью березы и березняки весьма характерны для При-
ангарья. Далее на восток, за Байкалом, мелколиственные леса
занимают небольшие площади, которые часто невозможно отра-
зить на карте масштаба 1 : 10 000 000.
Трудность рассмотрения фауны мелколиственных лесов заклю-
чена в том, что в ряде мест березняки и осинники не представ-
ляют собой самостоятельных лесных массивов. Они лишь пере-
ходный этап быстро идущей лесной сукцессии (трансформации)
сосняков или темнохвойных лесов и поэтому не достигают в чистом
виде даже возраста жердняка. Эти зарастающие гари нередко
экологически ближе к кустарникам, чем к лесам, и физиономи-
чески их фауна смыкается с животным миром не лесных ассоциа-
ций, а кустарниковых зарослей того своеобразного комплекса,
который создан человеком и который можно назвать лесопольем
и лесолужьем. Отделить высокоствольные березняки и осинники
от березняков и осинников гарей — дело не простое, особенно
при изучении литературы. Фауна смешанных лесов средней фазы
лесной сукцессии должна рассматриваться внутри фаунистиче-
ского сукцессионно-возрастного ряда лесов коренного комплекса,
а не как производная мелколиственных лесов, доказательством
чему служит переходный характер фауны смешанных лесов
с тенденцией развития комплекса в сторону коренных образований.
Группа зооформаций мелколиственных лесов в пределах Запад-
ной Сибири имеет единый фаунистический состав. Перерыв посте-
пенности между фаунами этих лесов Западной и Средней Сибири
достигает 16 видов. Мелколиственные леса Восточной Сибири
отличаются от среднесибирских приблизительно тем же числом
форм видового ранга.
Особняком стоит в группе зооформаций мелколиственных
лесов фауна камчатских лесов из каменной березы. Близость кед-
ровых стлаников и березняков создает условия эрзаца темнохвой-
ных лесов. Очевидно, этим объясняется тот факт, что на Камчатке
многие формы, живущие обычно в хвойных лесах, поселяются
в березняках (Куренцов, 1963). Изолированность мелколиствен-
ных лесов Камчатки сказывается в бедности видового состава
фауны камчатских березняков (около 30 видов позвоночных
животных).
Можно подметить, что если число форм в фауне темнохвойных
и лиственничных лесов уменьшается с востока на запад, то фауни-
стические комплексы сосняков и мелколиственных лесов обога-
щаются в том же направлении (в мелколиственных лесах Запад-
ной Сибири 61, Средней — 50, Восточной — 46 видов). Очевид-
но, этого следовало ожидать. Мелколиственные леса Средней и
Восточной Сибири генетически моложе западносибирских, зани-
мают меньшие площади и смыкаются не с родственными
лиственными лесами Восточной Европы, а с азиатскими степями
3*
35
и хвойными лесами Забайкалья. Процесс освоения животными
мелколиственных лесов Сибири, видимо, не совсем закончен (в срав-
нении с другими, более сложившимися зооформациями). Особенно
заметно продвижение западных форм на восток в Средней Сибири.
Намеченные перерывы постепенности позволяют, таким обра-
зом, выделять среди группы зооформаций мелколиственных лесов
следующие территориальные варианты.
(18) Животное население западносибирских равнинных мелко-
лиственных лесов.
(19) Животное население среднесибирских среднегорных мелко-
лиственных лесов.
(20) Животное население восточносибирских среднегорных
мелколиственных лесов.
(21) Животное население камчатских мелколиственных лесов.
Д. Животное население хвойно-широко-
лиственных и широколиственных лесов.
Мы специально не исследуем фауны хвойно-широколиственных
и широколиственных лесов, оставляя их в рамках «Зоогеографи-
ческой карты СССР» (Осмоловская, 1960). В легенде нашей карты
животное население этих лесов отмечено, соответственно, под
номерами (22) и (23).
III, Животное население сибирских высокогорий
На современных зоогеографических картах фауну сибирских
высокогорий либо выделяют в виде единого образования (Осмо-
ловская, 1960), либо объединяют ее с лесными фаунами (Осмолов-
ская, 1962). Оба эти подхода достаточно объективно обоснованы —
гольцы Сибири поросли во многих местах кустарниками, заселен-
ными лесными и кустарниковыми видами. Мы в своем анализе
старались рассматривать лишь гольцовые формы, опуская лесные
виды, связанные с нижележащим лесом, а также виды, тяготе-
ющие к водоемам.1
Серия высокогорных зооформаций имеет четкие перерывы
постепенности в распространении видов между Средней и Восточ-
ной Сибирью на севере (около 10 видов), а также между системой
восточносибирских хребтов и алтае-саянских высокогорий. Пос-
ледние также не едины: алтае-тувинские хребты значительно богаче
видами, чем высокогорья Саян. Отличие фаун этих регионов дости-
гает 20 форм.
Фауна гор Камчатки несколько отличается от высокогорий
северо-востока Сибири, но это происходит главным обра-
зом за счет малоспецифичных для высокогорий форм (полярная
овсянка, рогатый жаворонок, серый сорокопут, пеночка-зар-
ничка и т. д.), поэтому гольцов Камчатки мы не выделяем особо.
По тем же причинам остаются не отделенными от восточносибир-
ских высокогорий гольцы Сахалина.
36
Таким образом, легенда зоогеографической карты в части,
посвященной высокогорьям Сибири, состоит из четырех пунктов.
(24) Животное население алтае-тувинских высокогорий.
(25) Животное население саянских высокогорий.
(26) Животное население высокогорий севера Средней Сибири.
(27) Животное население восточносибирских высокогорий.
IV. Животное население лесостепи и сходных участков
Лесостепь, так же как лесотундра, — переходная полоса, где
перемежаются и тесно контактируют лесные и нелесные зоофор-
мации. Для лесостепи характерно сочетание лугостепных участков
с лесными колками. Она издавна освоена человеком, во многих
местах распахана и с помощью огня и топора искусственно про-
двинута в те места, где по физико-географическим условиям сама
по себе существовать не может. Эти антропогенные лесостепи
в некоторых местах существуют теперь как длительно-производ-
ные сочетания и физиономически неотличимы от естественных;
в других — они постоянно поддерживаются хозяйственной дея-
тельностью человека. Такие участки следует отличать от лесо-
степей. В мелком масштабе это почти невозможно, поэтому мы
объединяем лесостепь, лесополье и лесолужье в одну группу.
На геоботанических картах (Лавренко и Сочава, 1956; Сочава,
1960, и др.) лесостепи рассматриваются вместе со степями. Фауна
лесостепей, взятая в целом, безусловно не идентична степной,
поскольку в нее входят лесные представители. Нам кажется, что
правильнее считать лесостепь не подзоной степей, как это делают
в последнее время фитогеографы (Лавренко, 1950, 1954; Юнатов,
1950), а во всяком случае особой переходной полосой (зоной),
как это принимали Н. А. Северцев (1877) и в наше время Б. А. Куз-
нецов (1950). Как будет показано ниже, лесостепь в отличие от
лесотундры, вопреки мнению, утвердившемуся в литературе
(Кириков, 1956, и др.),5 имеет свой специфический зоокомплекс
и потому не может считаться образованием, полностью аналогич-
ным лесотундре.
Б. А. Кузнецов (1950) совершенно законно обращает вни-
мание на то, что степи в полосе лесостепи сочетаются с лесами раз-
личного состава: дубравами и сосновыми борами в Восточной
Европе, березовыми колками в Западной Сибири, сосновыми
и березовыми колками в Средней и Восточной Сибири. М. А. Мен-
збир (1882) в соответствии с этими особенностями лесных участков
лесостепей разделил «полосу островных лесов» на три провинции:
5 Когда рукопись уже находилась в издательстве, «Бюллетень МОИП»
опубликовал материалы совещания, посвященного становлению и развитию
лесостепи как отдельной ландшафтной зоны (Совещание по вопросам лесо-
степи; Никифоров, 1964; Образцов, 1964; Тибет, 1964). С. В. Кириков (1963)
также фактически отошел от своей первоначальной позиции.
37
1) Среднеевропейскую, 2) Южно-западносибирскую и 3) Даур-
скую. Совершенно очевидно, что и лугостепи, входящие в^состав
лесостепи, также неоднородны. В соответствии со стоящей перед
нами задачей, мы не рассматриваем формы, тяготеющие к соб-
ственно степям, а следовательно, оставляем вне нашего разбора
степные зооформации лесостепей. Можно лишь отметить, что
в связи с генетическим различием фаунистических комплексов
степи и леса следует ожидать несовпадения границ распростране-
ния степных и лесных элементов животного населения полосы
лесостепи.
Степные участки, входящие в состав лесостепей, имеют много
общего с южнее расположенными степями. Мы делим их, как
общепринято, на Европейско-Казахстанские и Монгольские. К пер-
вым относятся степные участки Западной Сибири, ко вторым —
степи и степные участки лесостепей Алтая, Тувы и Забайкалья.
В островных лесах лесостепи представлены те же фаунистиче-
ские комплексы, что и в лесах севернее расположенной таежной
полосы. Число лесных видов в лесостепи нередко резко снижено
по сравнению с соответствующими зооформациями таежной
полосы. Однако очень трудно констатировать, когда это снижение
вызвано историческими или общими физико-географическими
причинами, а когда объясняется лишь экологическими отличиями
маленьких островов от больших лесных массивов. Фауна березо-
вых колков в основе все же фауна таежного мелколиственного леса,
хотя между ними и имеются существенные различия. То же можно
сказать об островных сосновых лесах и лесных участках листвен-
ничной лесостепи. В связи с этим можно было бы рассматривать
животный мир островных лесов лишь как отличающуюся часть
соответствующих зооформаций.
Все сказанное вполне справедливо для внутренних частей
сравнительно крупных участков леса среди степи. Однако в лесо-
степи очень велико число видов, тяготеющих к опушкам лесов.
Это значительно обогащает фауну лесостепи и отличает ее лесные
зоокомплексы от таежных.
Мелкие лесные острова — это, условно говоря, сплошная
опушка — пограничнай полоса между лесом и степью. Опушка
леса фаунистически уже не лес, она населена другими зоокомпле-
ксами. Но это и не степь. Степь населена своими зоокомплексами.
Таким образом, лесостепь — образование тройственное: в ней
есть элементы таежных зооформаций, собственно степные (или
луговые) участки и, наконец, то особое, не совпадающее ни с лес-
ным, ни со степным, животное население, которое фаунистически
отличает лесостепь от собственно степи и тайги. Очевидно, форми-
рование фаун указанных трех зоокомплексов лесостепи шло парал-
лельно, но за счет различных элементов и под влиянием различных
факторов. Если представить себе лесостепи совершенно безжиз-
ненным пространством, находящимся между идеальными лесом
38
и степью, в которое вдруг привнесены современные растительные
ассоциации, то строго лесные и степные формы должны немед-
ленно занять подходящие стации, став аллохтонами лесостепи.
Опушечный комплекс будет формироваться на месте автохтонно.
В природе процесс заселения происходит не так, как в идеале.
Приведенным рассуждением мы лишь подчеркиваем отличие спе-
цифической фаунистической группировки лесостепи.
Очевидно, тройственный, а в других местах (там, где нет соб-
ственно степного комплекса) двойственный характер фаун лесо-
степи должен отображаться зоогеографической картой. Введение
трех разноцветных полос равной ширины едва ли не приведет
к путанице, поэтому мы применяем одноцветное изображение
лесостепи с внемасштабными. знаками, отображающими наличие
в ней соответствующих лесных и степных зоокомплексов. Там,
где собственно степной комплекс животных отсутствует, пред-
лагается система из двух полос, одна из которых соответствует
лесному, другая — собственно лесостепному зоокомплексу.
Лесостепи Западной Сибири, характеризующиеся березовыми
колками, весьма схожи между собой по характеру лесной фауны,
которая обогащается вблизи обширных лесов и обедняется по мере
удаления от них.
Кузнецко-Минусинский участок березовых колков фаунистиче-
ски отличается от лежащих к западу. Б. А. Кузнецов (1950),
очень дробно разделивший лесостепи на основе рассмотрения
всех экологических групп лесостепных млекопитающих, выде-
ляет Кузнецкий и Минусинский лесостепные зоогеографические
районы. При нашем подходе к вопросу можно лишь констатиро-
вать некоторое отличие фауны березовых островов Кузнецкого
бассейна и Минусинской котловины от западносибирских колков
за счет внедрения таежных форм (белка, бурундук, летяга, крас-
ная и красно-серая полевки). Однако следует учесть, что полоса
лесостепей Кузнецко-Минусинского участка тянется узкой лентой
и зажата между распаханными степями и тайгой. Мы не имеем
никаких доказательств того, что на стыке лесных и лесостепных
участков в любом географическом пункте лесные формы не про-
никают в глубь островных лесов столь же далеко, как в полосу
лесостепей Кузнецкого бассейна и Минусинской котловины.
Особняком стоят ленточные боры — остепненные леса древнего
происхождения. Они давно изолированы от таежных сосняков,
и их животный мир занимает особое место в зооформации сосновых
лесов.
Лесостепи и лесопольные участки, тянущиеся в Средней
Сибири вдоль железной дороги (Красноярско-Канские и Иркут-
ско-Балаганские), в большей своей части носят отпечаток недав-
него расширения и длительной изоляции от других степных и лесо-
степных массивов. На крайнем западе у Ачинска они сходны с запад-
носибирскими березовыми лесостепями, но восточнее приобретают
39
характер соснового лесополья с фауной, характерной для сосно-
вых лесов района.
Б. А. Кузнецов (1950) не включил лесостепи Средней Сибири
ни в один из зоогеографических округов или районов. Автор аргу-
ментировал это тем обстоятельством, что фауна Ачинской, Красно-
ярской, Канской и Иркутской лесостепей состоит почти полностью
из таежных и эвритопных форм. Это не совсем так. Правильнее
было бы сказать, что среднесибирские лесостепи лишены строго
степных видов. Лесостепные формы так же характерны для лесо-
степья и лесополья в Средней Сибири, как и в других ме-
стах.
При возможных отличиях в частностях горные лесостепи Алтая
и Тувы очень сходны по характеру фауны лесных участков. Это
более или менее обширные колки из лиственницы, иногда парко-
вого характера.
Своеобразный характер имеет лиственничное лесополье верх-
ней Лены. Фауна лесных участков этой местности имеет много
общего с фауной сосновых островов среднесибирской лесостепи,
но, с другой стороны, тут можно встретить и ряд видов, характер-
ных для зооформации лиственничных лесов. Отличается листвен-
ничное лесополье верхней Лены и от лесостепей Алтая и Тувы.
Лиственничные леса лесополья верхней Лены обычно достигают
значительных площадей и могут быть объединены вместе с соот-
ветствующими таежными.
Лесные острова лесостепей Забайкалья фаунистически отли-
чаются от прибайкальских, как и животное население лиственнич-
ных, сосновых и мелколиственных лесов этих районов.
В Центральной Якутии лежит огромная страна лугов, болот
и лиственничных лесов. Лесостепи и лесолужье Якутии заняты
лесными участками с фауной, очевидно несколько отличающейся
от сплошных лиственничных лесов.
Итак, зоогеографическое деление островных лесов лесостепи
можно представить следующим образом:
а) фауна березовых колков западносибирской равнинной лесо-
степи и лесополья;
б) фауна ленточных боров западносибирских и казахстанских
равнинных лесостепи и лесополья;
в) фауна сосново-мелколиственных лесных островов средне-
сибирской лесостепи и лесополья;
г) фауна лиственничных колков южносибирской горной лесо-
степи и лесополья;
д) фауна островных сосновых лесов забайкальских горных
лесостепей;
е) фауна островных мелколиственных лесов забайкальских
горных лесостепей;
ж) фауна лесов центральноякутского лугово-лиственничного
района.
40
Выделение группы собственно лесостепных форм весьма за-
труднено тем обстоятельством, что эти виды очень глубоко проникли
в полосу лесов в связи с нарушением природных биоценозов.
Особенно заметно это проникновение вдоль речных долин, где
издавна скотоводы расчищали луга, а позже земледельцы стали
распахивать склоны. Долинные леса среди степей вместе с зональ-
ной лесостепью были местом формирования лесостепного зоо-
комплекса, поэтому у долин и лесостепи много общих видов.
Млекопитающие, распространенные значительно равномернее,
чем птицы, не дают строго специфических опушечных форм, однако
выявляют существенные отличия в числе особей. Например, лес-
ная мышовка, мышь-малютка и темная полевка явно предпочи-
тают лесополье и лесостепи. Эти виды не живут в Сибири ни в степи,
ни в девственной тайге.
Монолитный лесоопушечный комплекс Западной Сибири отли-
чается от среднесибирского приблизительно 15 формами. Южно-
сибирские горные лесостепи по опушечной фауне более сходны
с западносибирскими, но все же отличие составляет 10 видов. Забай-
кальские горные лесостепи по опушечному комплексу близки
к среднесибирским, но отличаются рядом форм, число которых
достигает 7—8. Лесоопушечное население Забайкалья еще четче
дифференцировано от южносибирского — минимум 13 отличных
видов. Особое место занимает Центральная Якутия. Опушки
лесов среди луговых степей и аласов этого района населены очень
небольшим числом характерных видов (в два раза меньшим, чем
Забайкалье или Средняя Сибирь). Отличие здешних опушечных
комплексов от других сходных участков чисто негативного харак-
тера, но выражено весьма резко.
Таким образом, мы предполагаем наличие семи разностей
лесного животного населения лесостепей, минимум двух разно-
стей (очевидно, их больше!) степной фауны и пяти разностей
собственно лесостепных комплексов. Часть из указанных разно-
стей совпадает по территории, другие — нет. Среднесибирские
и якутские лесостепи практически не имеют строго степных ком-
понентов, однако отличаются комплексами опушечных форм.
Забайкальские и западносибирские лесостепные участки, включая
в себя сравнительно однородные опушечные зоокомплексы, имеют
внутреннее различие лесных зооформаций. Все эти особенности
должны быть отражены картой населения позвоночных живот-
ных. Следовательно, в окончательном виде часть легенды карты,
посвященная лесостепям Сибири, должна иметь такой вид.
(28) Животное население западносибирских равнинных лесо-
степей и лесополья в сочетании с европейско-казахстанскими
степными зооформациями и зооформацией березовых лесов (кол-
ков).
(29) Животное население западносибирских равнинных лесо-
степей и лесополья в сочетании с европейско-казахстанскими
41
степными зооформациями и зооформацией сосновых (ленточных)
боров.
(30) Животное население среднесибирских среднегорных лесо-
степей и лесополья в сочетании с зооформацией островных южно-
сибирских сосновых (сосново-мелколиственных) лесов.
(31) Животное население среднесибирских среднегорных лесо-
степей и лесополья в сочетании с зооформацией островных южно-
сибирских горных лиственничных лесов.
(32) Животное население южносибирских горных лесостепей
и лесолужья в сочетании с монгольскими степными зооформациями
и зооформацией южносибирских горных лиственничных лесов.
(33) Животное население забайкальских горных лесостепей
и лесолужья в сочетании с монгольскими степными зооформаци-
ями и зооформацией восточносибирских островных горных сосно-
вых лесов.
(34) Животное население забайкальских горных лесостепей
и лесолужья в сочетании с монгольскими степными зооформаци-
ями и зооформацией восточносибирских среднегорных островных
мелколиственных лесов.
(35) Животное население центральноякутских лесостепей, лесо-
полья и лесолужья в сочетании с зооформацией восточносибир-
ских горных лиственничных лесов.
V. Животное население болот
По Н. Я. Кацу (1948), болота лесной зоны могут быть раз-
делены на три зональные полосы: 1) практически лишенные
древесной растительности (во всяком случае в центральной
выпуклой части) болота северного типа, 2) сосново-сфагновые
или лиственнично-сфагновые болота средней полосы, 3) травяные
и гипновые болота юга лесной зоны и лесостепи. Перечисленные
типы болот встречаются чаще всего совместно друг с другом
и между ними имеются переходы. Ботаническая типология не
учитывает (или почти не учитывает) наличия открытой воды.
Для позвоночных животных это важный фактор. Схематически
экологические разновидности болот сводятся к следующему:
а) с открытой водой
б) без открытой воды
Болота:
1) облесенные
2) кустарниковые
3) травяные
4) сфагновые и моховые
Провести достаточно резкую грань между облесенным болотом
и заболоченным лесом едва ли возможно. Болота с открытой водой
и без нее в мелком масштабе не поддаются раздельному изобра-
жению. Таким образом, остаются три возможных разновидности
болот: 1) сфагновые открытые, 2) травяные, 3) лесо-кустарнико-
вые. Последние доминируют и очень тесно сочетаются как со сфаг-
42
новыми, так и с травяными. Однако можно принять, что для севера
Западной Сибири типичны открытые сфагновые болота. Для юга
этой страны характерны кустарниковое, лесо-сфагновые и травя-
ные болота. В Средней и Восточной Сибири доминируют болота
травяно-кустарниковые с вкраплениями сфагновых участков.
Для Камчатки указываются выпуклые сфагновые болота того
же типа, что и на севере Западной Сибири. Наконец, Дальний
Восток обладает весьма полным набором болот, из которых все
же доминируют лесо-кустарниковые разновидности. Лишь на
крайнем юге Дальнего Востока широко распространены травяные
(и тростниковые) болота, которые, очевидно, составляют особую
группу. Фауна этих болот больше тяготеет к маньчжуро-китай-
скому комплексу видов.
Рассмотрение ареалов болотных видов птиц лесной Сибири
(о «болотных» млекопитающих можно говорить лишь условно)
показывает, что особые фаунистические комплексы имеют север-
ные болота Западной Сибири, открытые сфагновые болота Кам-
чатки и лесо-кустарниковые болота юга Дальнего Востока (тра-
вяные болота этой страны мы не рассматриваем). Все перечислен-
ные участки отличаются друг от друга и от соседних 5—12 видами
животных. Наиболее богата болотная фауна юга Западной Сибири.
В ее состав входит до 20 видов. Фаунистический комплекс болот
Средней и Восточной Сибири несколько беднее, но его отличие от
западносибирского невелико. Видимо, все лесо-кустарниковые
болота Сибири фаунистически сравнительно однородны. В связи
с этим восточносибирский фаунистический комплекс болот мы
выделяем сугубо условно.
Таким образом, в пределах лесной полосы Сибири в мелком
масштабе можно выделить по крайней мере три достаточно хорошо
очерченных болотных региона.
(36) Животное население открытых сфагновых болот севера
Западной Сибири.
(37) Животное население лесо-кустарниковых болот юга Запад-
ной Сибири.
(38) Животное население лесо-кустарниковых болот Средней
и Восточной Сибири.
(39) Животное население открытых сфагновых болот Камчатки.
VI. Животное население рек и речных долин
Речные долины — своеобразные «интразональные» образова-
ния. По ним на север в тундру проникают леса, а в зону лесов
внедряются лесостепные и лесопольные формы, и эти же самые
долины позволяют видам-таежникам глубоко продвигаться в глубь
зоны степей.
В пределах речных долин можно выделить две резко отличные
группы биотопов: одна — широкой полосой тянущиеся вдоль рек
43
луга, кустарники, приречные ельники и тополевые уремы; дру-
гая — значительно более узкая — ленточка речного бичевника
и крутых берегов водоемов. Эти группы не равноценны по объему
(первая значительно шире, чем вторая) и состоят из ряда зоофор-
маций, подчас совершенно не родственных друг другу ни морфо-
логически, ни исторически. Пойменные луга, заливные и неза-
ливные, различного генезиса, кустарники первичного и антропо-
генного происхождения, приречные ельники и тополевые уремы —
все это приходится объединять из-за мелкого масштаба и плохой
изученности территории.
Анализ территориального распределения видов позвоночных
животных, наиболее охотно поселяющихся в речных долинах
полосы лесотундры, лесной и лесостепной зон, показывает нали-
чие минимум пяти различных сочетаний территориальных вари-
антов зооформаций. Можно констатировать определенное разли-
чие между фаунами речных долин севера и юга (речные долины
северных рек заселяет почти вдвое меньше форм, чем долины
южных рек). Фаунистические комплексы речных долин севера
лесной полосы Западной Сибири достаточно четко отличаются
от среднесибирских. Между севером Средней и Восточной Сибири
таких различий не наблюдается, очевидно, это единый регион.
Речные долины лесной полосы юга Средней и Западной Сибири
отличаются длинным рядом видов (около 25). Несколько меньше
разница между фаунами речных долин Средней и Восточной
Сибири. Фауны речных долин этих регионов отличает короткий
ряд видов (если не считать самой южной полосы Восточной Сибири,
где много маньчжурских выходцев): коростель, турухтан, средний
кроншнеп, лесной дупель, луговой и степной луни, пеночка-вес-
ничка, садовая камышевка, славка-завирушка, варакушка, крот
сибирский, лесная мышовка и узкочерепная полевка. Следова-
тельно, в пределах лесной Сибири можно выделить следующие
характерные разности животного мира речных долин.
(40) Животное население пойменных и надпойменных лугов
в сочетании с кустарниками, приречными ельниками и тополе-
выми уремами речных долин севера лесной и лесотундровой полос
Западной Сибири.
(41) Животное население речных долин юга лесной полосы
и лесостепи Западной Сибири.
(42) Животное население речных долин севера лесной полосы
и лесотундры Средней и Восточной Сибири.
(43) Щивотное население речных долин юга лесной полосы
и лесостепи Средней Сибири.
(44) Животное население речных долин юга лесной полосы
и лесостепи Восточной Сибири.
(45) Животное население речных долин советской части Маньч-
цсуро-Китайской подобласти (включается условно без специального
разбора).
44
Речным долинам для удобства изображения на карте придана
гипертрофированная ширина. Она как бы компенсирует невозмож-
ность показа долин средних и мелких рек и речек, населенных
теми же комплексами животных.
Особое место в речных долинах занимают околоводные участки,
обрывы берегов, полосы галечника, мелкие островки и участки
водной растительности. Старичные озера как бы составляют мостик
от рек к озерам, да и сами озера нередко служат истоком рек. Пти-
цы и млекопитающие околоводных полос тесно связаны с фауной
долин, и четкую грань между этими фаунистическими комплексами
уловить подчас трудно. Однако если виды долин (пойменных и над-
пойменных лугов в сочетании с кустарниками, приречными ель-
никами и тополевыми уремами) могут в подавляющем большин-
стве жить в долине независимо от того, имеется ли в ней водоток
или нет, то многие околоводные или полуводные формы рек и озер
существовать без открытой воды не могут. На этом основано про-
водимое нами разделение.
Различия околоводной фауны лесной полосы особенно отчетливы
между Западной и Восточной Сибирью (в широком понимании пос-
леднего термина, включая Среднюю Сибирь) и между севером и югом
(приблизительно по 65° с. ш.) этих территорий. Ряд форм, живущих
на юге Сибири, отсутствует в ее средней части (малая чайка,
серощекая поганка и зимородок). Околоводная фауна Средней
Сибири отличается присутствием желтых трясогузок. Несмотря
на эти отличия, мы полагаем, что разделять район, занятый око-
ловодной фауной Заенисейской Сибири, на два участка едва ли
целесообразно, поскольку степень единства фауны выше, чем
имеющиеся различия.
Во многих отношениях особняком стоит околоводная фауна
Камчатки. Ее не удается уверенно объединить ни с севером, ни
с югом Восточной Сибири. В фауну ее околоводных пространств
входят виды то севера, то юга без видимой закономерности.
Обедненность разбираемой фауны Камчатки не столь высока,
чтобы ее считать резко отличной от других таких же фаунистиче-
ских комплексов Сибири. Мы пока не выделяем ее как особое
фаунистическое образование.
Таким образом, часть легенды, касающейся фауны рек и мел-
ких озер, имеет следующий вид.
(48)® Животное население рек и мелких внутренних водоемов
севера лесной полосы и лесотундры Западной Сибири.
(49) Животное население рек и мелких внутренних водоемов
юга лесной полосы и лесостепи Западной Сибири.
(50) Животное население рек и мелких внутренних водоемов
севера лесной полосы и лесотундры Средней и Восточной Сибири.
• (46) Монгольские степи.
(47) Европейско-Казахстанские степи.
45
(51) Животное население рек и мелких внутренних водоемов
юга лесной полосы и лесостепи Средней и Восточной Сибири.
(52) Животное население рек и мелких внутренних водоемов
советской части Маньчжуро-Китайской подобласти (включается
условно без специального разбора).
VII. Животное население побережий крупных озер и морей
В принятом нами масштабе могут быть отражены лишь немно-
гие озера типа Чаны и Байкала. Лесная полоса Сибири выходит
на побережье морей Тихого океана. Фауна позвоночных животных
озер и морей определенным образом связана с экологическими
условиями самих водоемов и их берегов. Эти условия весьма спе-
цифичны. Очевидно, зоогеографическое деление побережий круп-
ных озер и морей — особого типа фаунистических комплексов —
нецелесообразно ставить в связь с соответствующими подразде-
лениями лесной полосы, а следует проводить, исходя из разбора
особенностей водоемов более широкой территории. Подобный
разбор требует специального исследования и не входит в наши
задачи. Мы оставляем этот тип зооформаций не разделенным, отме-
чая лишь существование зоокомплексов побережий крупных озер
и морей.
В легенде карты животное население побережий не получило
отражения
VIII. .Животное население сибирских населенных пунктов
Позвоночные животные издавна заселили жилища человека.
Вместе с товарами некоторые виды распространились почти по
всему свету. Но тем не менее фаунистические комплексы населен-
ных пунктов не однородны в различных местах. И эта разница
касается не только синантропных птиц, в большей степени, чем
млекопитающие, зависящих от климатических факторов. Прежде
всего четко отличается фауна сельских и городских поселений.
Современные города с их высокими каменными домами, асфаль-
том улиц и дворов, централизованным вывозом мусора, с газо-
нами экологически резко отличаются от деревень, где дома пере-
межаются с огородами, в хозяйственных пристройках живет скот,
амбары и подвалы значительно доступнее, чем хранилища про-
дуктов в городах. Однако столь значительная разница существует
лишь между центральной частью городов и населенными пунктами
сельского типа. За редким исключением сибирские города окру-
жены поясом предместий или деревень, а в ряде случаев благо-
устроенные кварталы перемежаются с еще не реконструирован-
46
ними районами. Почти все населенные пункты Сибири располо-
жены в долинах рек, поэтому их фауна испытывает влияние долин-
ных зоокомплексов.
Следует отметить, что зоокомплексы населенных пунктов не
образуют сукцессионных рядов и вообще при всей своей близо-
сти к природным образованиям резко отличаются от них по струк-
туре и внутренним взаимосвязям. Очевидно, фауна населенных
пунктов — это особый тип фаунистических комплексов. В пре-
делах таежной Сибири этот тип представлен одной зооформа-
цией населенных пунктов лесной полосы. Географическая разница
заселенности жилищ человека и его хозяйственных построек
синантропными видами не находит себе объяснения в различных
типах сооружений. Она зависит от исторических и климатических
причин, которые из-за слабой изученности биологии видов диф-
ференцируются с большим трудом. Можно предполагать, что раз-
ница между севером и югом прежде всего вызвана факторами
климата, в то время как меридиональные различия возникают от
исторических причин. Но это лишь предположения.
В населенных пунктах Сибири живет весьма ограниченное
число форм позвоночных животных. В связи с этим мы считаем
достивэрным различие в 2—3 вида. Севернее 62—65° с. ш. насе-
ленные пункты вдвое беднее позвоночными животными, чем на
юге. Северные села Западной, Средней и Восточной Сибири отли-
чаются друг от друга на 2 вида: домовый воробей и деревенская
ласточка отсутствуют в Средней Сибири, те же виды и полевой
воробей не живут в Восточной Сибири, зато здесь в жилищах
человека обитает азиатская лесная мышь. Южная полоса имеет
перерывы постепенности от Западной к Средней Сибири в 7 видов
(различные голуби, нет белопоясого стрижа, в населенных
пунктах — азиатская лесная мышь, но нет обыкновенной, к синан-
тропному существованию приспособилась красная полевка, но
практически нет обыкновенной), а от Средней к Восточной
Сибири — в 2 вида (черный стриж и скворец).
Таким образом, зооформация населенных пунктов в пределах
лесной полосы Сибири делится на 6 территориальных вариантов.
(53) Животное население населенных пунктов севера лесной
полосы и лесотундры Западной Сибири.
(54) Животное население населенных пунктов юга лесной
полосы и лесостепи Западной Сибири.
(55) Животное население населенных пунктов севера лесной
полосы и лесотундры Средней Сибири.
(56) Животное население населенных пунктов юга лесной
полосы и лесостепи Средней Сибири.
(57) Животное население населенных пунктов севера лесной
полосы и лесотундры Восточной Сибири.
(58) Животное население населенных пунктов юга лесной
полосы и лесостепи Восточной Сибири.
47
Глава 3
МЕЛКОМАСШТАБНОЕ ТИПОЛОГО-ЗООГЕОГРАФИЧЕСКОЕ
РАЙОНИРОВАНИЕ СИБИРСКОЙ ЧАСТИ ПРОВИНЦИИ ТАЕЖНЫХ
ЛЕСОВ И ВОПРОСЫ МЕТОДИКИ ИССЛЕДОВАНИЯ КРУПНЫХ
РЕГИОНОВ
Рассматривая территориальные варианты зооформаций, их
групп или сочетаний, можно подметить приуроченность границ
этих образований к определенным переходным полосам. Это явле-
ние дает возможность проводить зоогеографическое райониро-
вание.
Разделение зоогеографических округов на районы обычно
производится чисто эмпирически при констатации отличий в фауне
соседних территорий. От этого эмпиризма, видимо, пока нет воз-
можности избавиться даже при большом желании. На нем практи-
чески основано и наше деление зооформаций на территориальные
варианты. Однако мы делим более однородные образования, чем
обычно это делают зоогеографы. Эмпирическое разделение одной
зооформации более обосновано, чем деление их сочетаний, так же
как правильнее в зоогеографическом районировании исходить из
ареалов видов, нежели из ареалов крупных систематических
групп. Тем более ошибочно ориентироваться на разницу в фауне,
происходящую от наличия или отсутствия видов совершенно
различной приуроченности, например, отличать равнинные и гор-
ные районы на основе того, что на равнине нет альпийских видов.
Очевидно, при всяком разделении территории следует условиться,
что подразумевать под той или другой единицей районирования,
какой ей придавать объем. В этом отношении еще нет единого
мнения, хотя систем районирования великое множество. Мы пред-
лагаем называть зоогеографическим районом
значительную по площади часть зоогео-
графического округа (или зоны), занятую
в основном одним, отличным от соседних,
территориальным подразделением (терри-
ториальным вариантом) зооформации
(группы зооформаций), одной, отличной
от соседних, зооформацией или не-
сколькими, отличными от соседних, тер-
риториальными вариантами зооформаций
с приблизительно совпадающими грани-
цами распространения.
При выделении района следует обращать внимание прежде
всего на коренные зооформации, развивавшиеся автохтонно и пред-
ставляющие собой «климакс» сукцессионного ряда. В этом отно-
шении имеют большое значение геоботанические карты восстано-
вленной растительности, показывающие коренные растительные
48
сообщества. Однако полагаться только на эти карты не следует.
Анализ характера фауны, ее связей с соседними регионами дает
ключ для проверки выводов, сделанных работниками смежных
наук.
Рис. 3. Схема зоогеографического районирования лесной Сибири по
Б. А. Кузнецову (1950).
I — часть Восточно-Европейского района; II — Западносибирский район; III — Алтае-
Саянский район; IV — Прибайкальский район; V — Тунгусский район; VI — Забай-
кальский район; VII — Якутский район; VIII — Камчатский район.
При предлагаемом подходе к зоогеографическому райониро-
ванию в общем сохраняется принятая ныне (Кузнецов, 1950) сетка
районов провинции таежных лесов (рис. 3), однако несколько
изменяются их границы и число (рис. 4 и 5).1 Так, западносибир-
1 Следует еще раз обратить внимание на то, что рис. 5 иллюстрирует
зоогеографические районы лишь собственно лесов, а не территории вообще.
На рисунке не показано районирование лесостепи и лесотундры.
4 Н. Ф. Реймерс
49
ская тайга не остается монолитной, а делится на три района:
западносибирское лиственничное редколесье, северная тайга Запад-
ной Сибири и южная тайга Западной Сибири. Западные границы
1 — лесотундра: 1а — Западносибирский лесотундровый подрайон, 16 — Северосибир-
ский (Среднесибирский) лесотундровый подрайон, 1в — Колымо-Анадырский (Восточно-
сибирский) лесотундровый подрайон; 2—6 — лесостепь: 2 — Западносибирский лесостеп-
ной район, з — Среднесибирский лесостепной район, 4 — Южносибирский лесостепной
район, 5 — Забайкальский лесостепной район, 6 — Центральноякутский лесолуговой
и лесостепной район.
указанных трех районов мы не определяли. Разграничительная
линия вдоль р. Оби, проведенная И. П. Лаптевым (1958), очевидно,
разделяет таежную Западную Сибирь на подрайоны. Именно такое
таксономическое значение придает этой границе упомянутый
автор. В Заенисейской Сибири при сохранении Тунгусского,
50
Прибайкальского, Якутского, Забайкальского и Алтае-Саяя-
ского районов выделяются районы лиственничного редколесья
и Приохотский (дальневосточная темнохвойная горная тайга).2
Рис. 5. Схема зоогеографического районирования лесной Сибири по основ-
ным территориальным вариантам зооформаций, их группам и сочетаниям.
I — район северного редколесья Западной Сибири; II — район северной тайги Западной
Сибири; III — район южной тайги Западной Сибири; IV — Алтае-Саянский район;
V — Прибайкальский район; VI — Тунгусский район; VII — Забайкальскйй район;
VIII — Якутский район; IX — Приохотский район; X — район северного редколесья
Средней и Восточной Сибири; XI — Камчатский район.
2 Районирование Дальнего Востока, проводимое А. И. Куренцовым
(1947, 1959а, 1963) и Ю. В. Авериным (1957), относится к иной степени
дробности — среднему или даже крупному масштабу. Деление Якутии,
построенное на другом принципе (Аверин, и др., 1962; Воробьев, 1963), так же
как и районирование Прибайкальской Сибири Т. Н. Гагиной (1958, 1961),
очевидно, впредь до детального анализа не совместимо с проводимым нами.
Это замечание относится и к другим системам зоогеографических подразде-
лении, основанных на ином, чем наш, подходе к фауне.
4*
51
Все указанные районы совпадают с фоновыми территориаль-
ными вариантами зооформаций или с самими зооформациями
и четко Видны на карте населения позвоночных животных (см. цвет-
ную карту). Переходные полосы между районами образуются
взаимопроникающими пятнами соседних зооформаций.
Следовательно, сибирская часть провинции таежных лесов
делится на 11 зоогеографических районов, 3 из которых входят
в восточное крыло Западнотаежного округа и 8 — в Восточно-
таежный округ.
Лесотундра в пределах Сибири совершенно естественно раз-
бивается на три региона: Западносибирский, Северосибирский
(или Среднесибирский) и Колымо-Анадырский (или Восточно-
сибирский). Думается, что придание этим территориальным
единицам значения округов (как делает это Кузнецов, 1950)
неправомочно, поскольку они несоизмеримы с округами провин-
ции тайги. Очевидно, не имея специфичных зооформаций, а лишь
различные сочетания тундровых и лесных комплексов, лесотундра
не может быть разделена вполне аналогично лесной полосе. Вы-
являющиеся разности оказываются типологически не равными
лесным районам. По аналогии с лесостепью (см. ниже) мы пред-
лагаем сочетаниям зооформационных комплексов лесотундры
Сибири придавать лишь ранг подрайонов.
Районирование лесостепи, в нашем понимании, также приоб-
ретает новый вид. Если признать существование в этой полосе
территориальных единиц ранга округов, то это будут Западно-
сибирский (включая горные лесостепи и лесолужье Алтая и Тувы)
и Восточносибирский округа. Фауны этих территорий отличаются
рядом видов, имеющих прямо противоположные тенденции рас-
пространения: восточные формы продвигаются на запад, запад-
ные — на восток. Ранг лесостепного района следует придать всем
пяти зооформационным подразделениям собственно фауны лесо-
степи. Это будут Западносибирский, Среднесибирский, Южно-
сибирский, Забайкальский и Центральноякутский районы. Типо-
логическим сочетаниям, имеющимся в лесостепи, можно придать
значение подрайона. Западносибирский лесостепной район, таким
образом, делится на Урало-Енисейский и Предалтайский под-
районы; Среднесибирский район распадается на Ангаро-Саянский
и Ангар о-Ленский подрайоны; Забайкальский район состоит из
Селенгинского и Даурского подрайонов.
Более мелким территориальным единицам следует придать
значение лишь зоогеографических участков. Этому таксономиче-
скому рангу соответствуют многие лесостепные районы, выде-
ленные Б. А. Кузнецовым (1950). Мы предлагаем следующую
систему деления на участки (там, где район или подрайон делится
на них). Урало-Енисейский подрайон может быть подразделен
на Урало-Иртышский, Барабинский, Кузнецкий, Минусинский
и Красноярско-Канский участки. Южносибирский район состоит
52
из Алтайского и Тувинского участков. Все указанные подразде-
ления хорошо видны на типолого-зоогеографической карте
(см. цветную карту) и показаны на рис. 4.
Наличие в лесостепи специфического зоокомплекса приводит
к мысли, что придание лесотундре и лесостепи одного таксономи-
ческого ранга, как делает это Б. А. Кузнецов, сомнительно. Либо
за лесостепью следует признать ранг подобласти, либо отнести
к более низкому рангу переходную зону лесотундры. Окончательно
решить этот вопрос можно будет лишь после генеральной ревизии
районирования Палеарктической области, что не входит в нашу
задачу.
Деление на регионы сибирских высокогорий, болот и речных
долин (так же как и лесотундры) не соответствует по точности
районированию лесов и лесостепей, поскольку из-за мелкого
масштаба карты были рассмотрены не зооформации, а лишь их
сочетания. Условно можно принять, что выделенные разности
адекватны типолого-зоогеографическим районам.
Южную тайгу Средней Сибири (равно Прибайкальский зоо-
географический район) хорошо очерчивают соответствующие тер-
риториальные варианты групп зооформаций темнохвойных и мел-
колиственных лесов, а также среднесибирский раздел сочетания
зооформаций речных долин и южносреднесибирский территори-
альный вариант зооформации населенных пунктов. В указанный
район входят восточное крыло территориального варианта зоо-
формации южносибирских сосновых лесов и западное крыло
восточносибирского раздела сочетания зооформаций околоводных
пространств. Следовательно, из общего разнообразия фаунистиче-
ских комплексов сибирской части провинции тайги в полосу южной
тайги Средней Сибири входит 6 зооформационных и комплексных
подразделений (из 56). Все они (кроме сосняков) принадлежат к
Восточнотаежному зоогеографическому округу. По расположению
среди других зоогеографических районов южная тайга Средней
Сибири занимает центральное место, не имея непосредственных
границ ни с тундрой, ни со степью. Лишь на юге она разорвана
участками среднесибирских лесостепей и лесополья. Обширных
болот в пределах района нет. Имевшийся когда-то их массив на
р. Оке ныне залит водами Братского моря. Безлесные выпуклые
болота северо-западной части южной тайги следует относить
к образованиям западносибирского типа.
На основе единства территориального варианта зооформации
нужно ожидать проявления единых закономерностей на всей
площади зоогеографического района (а внутри некоторых зоо-
формаций закономерности должны быть общи и вне района).
Следовательно, мы вправе ожидать, что по всему району видовой
состав животного населения зооформации будет единым. Близки
будут и количественные показатели по видам, а также в целом
по биомассе позвоночных животных. Нами было уже показано.
53
что это так (Реймерс, 1965), во всяком случае в пределах При-
байкальского района. Действительно, на нижней Ангаре мы
нашли все ожидаемые виды в составе тех зооформаций, в кото-
рые они должны были входить, при этом оказалось прибли-
зительно верным и созданное заранее представление о возможной
численности особей. Более четко указанная закономерность про-
явилась в наименее нарушенных человеком коренных биотопах.
Таким образом, был найден принцип для экстраполяции дан-
ных и определения границ таких экстраполяций. Одновременно
открылись большие возможности, во-первых, для использования
метода корреляций (чаще всего опосредованных, так что внешне
это простые совпадения), и, во-вторых, для практической
разработки программы и методики широкого изучения живой
природы Сибири.
Действительно, если для определенной территории известен
элементарный фаунистический комплекс, можно на основе кор-
релятивных зависимостей определить, каково здесь количество
особей видов, в том числе полезных и вредных, при этом не только
позвоночных животных, но и некоторых их паразитов. Знание
коэффициентов коррелятивных зависимостей позволит довольно
точно наметить возможные пределы распространения многих
важных для человека видов, не прибегая к первоначальным
полевым исследованиям. Все силы можно сосредоточить на уточ-
нении уже намеченных границ. С другой стороны, единство ком-
плекса и составляющих его видов позволяет при применении
предложенной типологии составлять карты кружева ареалов
отдельных видов в крупном, среднем и мелком масштабах при
сравнительно простом переходе от масштаба к масштабу на основе
геоботанических карт (Воронов, 1963). Все перечисленное чрез-
вычайно важно для эпидемиологии трансмиссивных заболеваний
и прогноза численности полезных и вредных животных в районах
нового освоения.
Вместе с тем деление огромной территории Сибири на внут-
ренне однородные части очень важно методически. Совершенно
очевидно, что изучить животный мир Сибири сплошь невозможно.
Это неосуществимо в силу огромных пространств страны. Следо-
вательно, нужно наметить пути для достаточно полного иссле-
дования с помощью сравнительно простых и малотрудоемких
методик. Наиболее подходящ в этом случае прием изучения ста-
ционарных ключевых участков (Нейфельдт, 1959, и др.).
Поскольку методические принципы исследований на ключевых
участках разработаны еще недостаточно, мы остановимся на этом
вопросе более подробно.
Беглый просмотр картографических и литературных мате-
риалов и рекогносцировочные экспедиционные маршруты обычно
выявляют два элемента в характеристике изучаемой территории:
1) степень ее зональной и меридиональной однородности в мелком
54
масштабе и 2) характерные природные разности внутри каждой
отличной пространственной единицы (в сравнительно крупном
масштабе). Очевидно, число ключевых участков должно соответ*
ствовать общему масштабу исследований, их намечаемой дроб-
ности. Для общей характеристики зоогеографического района
достаточно одного ключевого участка, для среднемасштабного
изучения района такое исследование необходимо в каждом из
подрайонов, наконец, для сравнительно дробного познания био-
логических закономерностей нужно иметь стационар в каждом
зоогеографическом участке. Однако подробное исследование дол-
жно опираться на менее дробное. Как же иначе установить деле-
ние на участки?
Слишком мелкомасштабное исследование целесообразно лишь
при изучении очень больших территорий, поскольку оно обычно
не дает ничего, кроме теоретических построений. Мы сочли наи-
более рациональным для территории южной тайги Средней Сибири
изучение животного мира в среднем масштабе, опирающееся на
ключевые участки, расположенные в каждом зоогеографическом
подрайоне. Однако определение смысловых рамок зоогеографи-
ческого подрайона не имеет общепринятого толкования, и эта
категория выделяется большей частью эмпирически. Мы пред-
лагаем учитывать при выделении подрайонов в основном два
фактора: первый — тенденция в естественном развитии биоце-
нозов, второй — влияние соседних территориальных вариантов
зооформаций и соседних зооформаций. Особенно важно прини-
мать во внимание позитивное влияние соседних фаунистических
комплексов. Всякая региональная единица, если она не ограни-
чена резкими географическими преградами, должна рассматри-
ваться как единое целое с соседними, поэтому в ней можно всегда
обнаружить элементы из соседних регионов. Если «соседи» рас-
положены слева и справа, то их влияние на центральный регион
будет идти также слева и справа до определенных рубежей, опре-
деляющихся комплексом природных и исторических условий.
Тенденция в развитии природных образований, скорость вос-
становления коренных растительных формаций также не без-
различны для фаунистических комплексов. Например, быстрое
восстановление темнохвойной тайги в присаянских районах Сред-
ней Сибири препятствует распространению сюда растительных
и животных формаций мелколиственных и сосновых лесов, а сле-
довательно, дает большую почву для влияния зооформаций гор-
ных темнохвойных лесов юга Сибири, чем для зооформаций равнин
юга Западной Сибири, где характерны смены лесных пород.
Позитивное влияние на лесные зооформации южной тайги
Средней Сибири возможно с четырех сторон — с запада, юга,
севера и востока. Однако влияние северных зоокомплексов листвен-
ничных лесов почти не распространяется на коренные экологи-
ческие варианты зооформаций южной тайги, и этим влиянием
55
при делении на подрайоны можно пренебречь. Западные выходцы
характерны прежде всего для сосновых и мелколиственных лесов,
наиболее распространенных в бассейне Ангары. В узкой полосе
присаянских лесов южного типа много выходцев с гор Сибири.
В бассейне Лены сильно ощущается влияние забайкальских
элементов (Реймерс и Воронов, 1963).
Таким образом, в пределах Прибайкальского района намети-
лись три зоогеографических подрайона: Присаянский, Ангар-
ский и Лено-Байкальский (рис. 6).
Следует особо подчеркнуть, что разница между подрайонами
не абсолютна, т. е. ощущается не во всех зооформациях. Напри-
мер, группа зооформаций темнохвойных лесов практически едина
по составу для всего района южной тайги Средней Сибири. Под-
районы отличаются, во-первых, тенденцией к господству опре-
деленной зооформации (в силу направленности процесса трансфор-
мации природных разностей) и, во-вторых, некоторыми отличиями
внутри отдельных зооформаций, чаще всего еще не сло-
жившихся или сложившихся недавно в результате антропоген-
ных воздействий. Примером второго отличия могут служить
особенности животного населения лугов речных долин, кустар-
никовых зарослей в долинах и населенных пунктах (см.
гл. 5). Именно они дифференцируют Ангарский и Лено-Байкаль-
ский подрайоны (рис. 6). Таким образом, подрайоны в отличие
от районов имеют перерывы постепенности не во всех зооформа-
циях и даже лишь в наименее физиономически существенных
для территории.
Еще слабее отличие зоогеографических участков. В южной тайге
Средней Сибири оно вызвано также влиянием соседних регионов
и обусловлено теми же факторами, что и разделение района на
подрайоны. Однако в данном случае отличия проявляются не
в фаунистическом составе коренных экологических вариантов
зооформации, а лишь в процессе трансформационных смен (внутри
элементарных фаунистических комплексов). Так, практически
лишь на гарях и вырубках живет рыжая полевка в Западнопри-
саянском участке Присаянского зоогеографического подрайона
(Штильмарк, 1963в). Также лишь на холодных заболоченных
гарях — березовых молодняках — живут северные виды внутри
Нижнеангарского участка Ангарского подрайона (см. гл. 6).
Более дробное (чем до уровня участков) зоогеографическое деле-
ние едва ли целесообразно, поскольку уже не имеет объективных
критериев даже внутри рядов элементарных фаунистических
комплексов.
Элементы районирования до подрайона включительно легко
очерчиваются на карте, поскольку различия между ними обна-
руживаются либо как территориальные варианты водораздель-
йых зооформаций, либо внутри зоокомплексов речных долин
(таким образом отличаются подрайоны речных систем Ангары
56
Рис. 6. Подрайоны и участки Прибайкальского зоогеографического
района.
I — район северного лиственничного редколесья Средней и Восточной Си-
бири; II — район северной тайги Западной Сибири; III — район южной тайги
Западной Сибири; IV— Тунгусский район; V — Прибайкальский район (южной
тайги Средней Сибири); подрайоны: Va: — Присаянский, V6 — Ангарский,
Ve — Лено-Байкальский; участки подрайонов: Vai — Западпо-Присаянский,
Va2 — Восточно-Присаянский, Vot — Нижне-Ангарский, V62 — Верхне-Ангар-
ский, Vet — Верхне-Ленский, Ve2 — Байкальский; VI — Алтае-Саянский
район; VII — Забайкальский район; VIII — Красноярско-Канский участок
Западносибирского лесостепного района (Урало-Енисейского подрайона);
IX — Среднесибирский лесостепной район; X — Забайкальский лесостепной
район; XI — Якутский район. 1 — темнохвойные леса; 2 — сосновые леса;
3 — мелколиственные леса; 4 — участки лиственничных лесов с животным на-
селением восточносибирских горных лиственничников; 5 — сосново-мелколист-
венная лесостепь Средней Сибири; 6 — лиственничная лесостепь Средней
Сибири; 7 — речные долины юга лесной полосы и лесостепи Средней Сибири;
8 — речные долины юга лесной полосы и лесостепи Восточной Сибири; 9 — реч-
ные долины севера лесной полосы и лесотундры Средней и Восточной Сибири;
Ю — границы зоогеографических районов; 11 — границы зоогеографических
участков; 12 — границы зоогеографических подрайонов.
и Лены). Иногда они отмечены географическими преградами
(Присаянский подрайон отделен от прочих полосой лесостепи).
Границы участков определить значительно труднее, тем более
что именно внутри них чаще всего происходит пульсация границ
ареалов отдельных видов животных (гл. 5).
Присаянский подрайон (вместе с более высокогорным Алтае-
Саянским районом) был объектом изучения в 1951—1956 гг.
В остальных двух подрайонах южной тайги Средней Сибири
Рис. 7. Долина р. Тутуры у с. Якимовка.
были заложены ключевые участки и проведены маршруты в 1959—
1963 гг. Таким образом, мы основываемся на данных трех ключе-
вых участков, расположенных в каждом из подрайонов: в Лено-
Байкальском — Якимовскпй (рис. 7), в Ангарском — Яркинский
(рис. 8) и в Присаянском — Хамар-Дабанский (рис. 9). Более
дробные закономерности, выраженные внутри участков, остались
вне поля нашего внимания. В связи с этим проведенные нами
на рис. 6 границы участков следует рассматривать как прови-
зорные.
Размер ключевого участка — второе важное звено в методике
работ. В каждом зоогеографическом подрайоне можно просле-
дить значительное число элементарных фаунистических ком-
плексов. Ключевой участок должен, с одной стороны, охватывать
сравнительно полную серию элементарных фаунистических ком-
плексов, с другой — отражать их площадное соотношение в при-
роде, т. е. быть как бы макетом типичных условий в зоогеогра-
фическом регионе. Очевидно, размер «ключа» определяется глав-
ным образом разнообразием природных условий во всех трех
58
Рис. 8. Долина р. Чадобца у дер. Яркино.
Рис. 9. Темнохвойная тайга (Присаянский подрайон).
измерениях и антропогенным влиянием на природу. Как показал
опыт наших работ в Западной и Средней Сибири, а также на Саха-
лине, лучшим вариантом ключевого участка будет полоса,
тянущаяся от водораздела до водораздела или от морского побе-
режья до водораздела.3 В упомянутых нами местах протяжен-
ность такой полосы колеблется от 30 до 60—70 км. Практически
такую длинную полосу стационарно исследовать невозможно,
поэтому целесообразно брать лишь один водораздел, т. е. полосу
от водораздела до середины долины или до шлейфовой части склона
противоположного водораздела. Длина такой полосы не превы-
шает 30—35 км. Ширина полосы определяется исключительно
пестротой условий. Эмпирически установлено, что обычно все
разнообразие условий бывает отражено в южной тайге Средней
Сибири при ширине полосы, равной 14—15 км. Таковы рациональ-
ные размеры большого ключевого участка, предназначенного для
исследования основных биотических закономерностей, отражае-
мых картосхемами масштаба 1 : 100 000.
Совершенно естественно, что ключевой участок площадью
в 450 км2 не может быть подробно исследован, хотя вполне реально
изучение на нем всех основных типов характерных выделов (эле-
ментарных фаунистических комплексов). Технически при нали-
чии 4—5 работников (в том числе лишь 2 квалифицированных)
приходится выделять на территории большого ключа 2—3 малых
ключа, доступных для ежедневного посещения. Конфигурация
малого ключа зависит от особенностей места работ, но, как пра-
вило, в условиях пересеченной местности целесообразно распо-
лагать малый ключ поперек большого с таким расчетом, чтобы
один малый ключ — водораздельный — охватывал все разнооб-
разие условий на водоразделе, другой — долинно-придолинный —
мог бы характеризовать основные выделы долины и придолинной
полосы (объем указанных полос см. в гл. 4). Желательнее, конечно,
иметь большее число малых ключевых участков, однако это
не всегда возможно при малом числе наблюдателей. Каждый
малый ключ для достижения полноты исследований, как ясно
из вышеизложенного, должен иметь в длину около 15 км. Ширина
его практически линейна, так как ежедневное «прочесывание»
более значительной площади невозможно, да и не нужно. При
двух наблюдателях, работающих на малом ключе, каждый из
них проходит ежедневно до 15—20 км, что достаточно утоми-
тельно. Мы практиковали раздельный подход к изучению млеко-
питающих и птиц. Конный наблюдатель объезжал все линии
3 Следует предостеречь против выбора ключевого участка в долине боль-
шой реки. Такой «ключ» будет характеризовать прежде всего зооформации
речных долин и практически не захватит полосу водоразделов, занимающую
в любом месте большую площадь, чем долины. Кроме того, речные долины —
древние торные пути, вдоль которых природа в значительно большей степени
(и раньше) трансформирована человеком.
G0
плашек и канавки на малом ключе, кроме тех, около которых
проходил короткий маршрут орнитолога. Обычно ежедневный
маршрут конного наблюдателя не превышал 40 км, пешего —
10—15 км.
Описанная система «большой ключ—малые ключи» дает возмож-
ность достаточно подробно исследовать биологические законо-
мерности в природе и получить сравнительный материал для
характеристики значительных площадей, занимаемых зоогеогра-
фическими районами, подрайонами и участками. Знание общих
закономерностей позволяет установить довольно точные границы
подрайонов (учитывая наличие широких переходных полос между
ними) и наметить провизорные грани между зоогеографическими
участками. Дальнейшие уточнения (зная, где и в чем искать
перерывы постепенности) должны производиться путем марш-
рутных исследований. Таким образом, методически целесообразно
изучать значительные регионы в три этапа: 1) маршрутное обсле-
дование, 2) многолетние стационарные исследования на базовом
(районном) ключевом участке и, по возможности, такие же по
длительности, но можно и в течение более короткого срока —
одного сезона, — на подрайонных и участковых полустацио-
нарах, и, наконец, 3) маршрутное уточнение границ географи-
ческих разностей (ареалов видов и зоокомплексов).
РАЗДЕЛ II
ПТИЦЫ И МЛЕКОПИТАЮЩИЕ
ЮЖНОЙ ТАЙГИ СРЕДНЕЙ СИБИРИ
И АНТРОПОГЕННАЯ ДИНАМИКА ИХ НАСЕЛЕНИЯ
Глава 4
КРАТКАЯ ИСТОРИКО-БИОГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
ЮЖНОЙ ТАЙГИ СРЕДНЕЙ СИБИРИ
Южная тайга Средней Сибири — один из самых лесистых краев
мира. Здесь свыше 95% площади занято различными лесными
угодьями. По приблизительным подсчетам лесоводов (Л. В. По-
пов, устное сообщение), в пределах южной тайги сосна занимает
36.0, лиственница 22.6, береза 20.4, ель 7.8, кедр 7.2, осина 3.2
и пихта 2.8% лесопокрытой площади. Хвойные породы, таким
образом, составляют по площади 76.4% лесов, лиственные —
23.6. Перечисленные соотношения будут несколько иными, если
принять положение о нехарактерности лиственничных лесов для
южной тайги Средней Сибири. Помимо этого следует учесть то
обстоятельство, что лесоводы нередко относят к лиственничникам
темнохвойные леса с лиственницей в первом ярусе и смешанные
лиственнично-сосновые субори, имеющие экологический облик
сосняков. Сами сосновые леса почти всегда имеют примесь листвен-
ницы. В некоторых случаях эта примесь бывает значительной,
но по облику фауны лес все-таки остается сосновым.
Чисто лиственничные леса южнотаежного типа встречаются
лишь на небольшом участке крайнего юго-востока Прибайкалья и
имеют весьма специфический характер лесопольных образований.
Весьма неточно охарактеризованы лесоводами площади пихтовых и
еловых лесов, поскольку эти породы часто растут вместе в раз-
личном соотношении. Учтя все эти оговорки, можно принять,
что различные леса, типичные для южной тайги Средней Сибири,
занимают площади в следующем процентном соотношении: сос-
62
новые 46.5%, березовые 26.5, елово-пихтовые 13.5, кедровые 9.0
и осиновые 4.5%, т. е. группа зооформаций темнохвойных лесов
распространена на 22.5%, а зооформация сосновых лесов на
46.5% площади южной тайги.
Группа зооформаций мелколиственных лесов должна была бы
занимать 31.0% площади, но на самом деле она распространена
далеко не так широко. Значительная часть березовых и осиновых
лесов — этапы восстановления темнохвойной тайги или сосняков.
Фауна и животное население таких участков должны быть отне-
сены не к характерным образованиям мелколиственных лесов,
а к антропогенно-сукцессионным рядам хвойных зооформаций.
Практически же отсутствие материалов делает невозможным
такое разделение.
Характер лесов южной тайги и состав их фауны в значительной
мере определяются климатическими особенностями региона. Ана-
лиз климатических карт новейших атласов (Атлас СССР, 1962;
Атлас Иркутской области, 1962) показывает, что ряд метеоро-
логических показателей меняется именно у северной границы
южной тайги (величина солнечной радиации, температурные
показатели, изолиния 220-дневного снежного покрова и т. д.).
Северный предел южной тайги совпадает с границей распрост^а-
нения^’ таких чутких климатических индикаторов, как пихта.
Тут сходит на нет распространение в Средней Сибири крупных
массивов кедровников и сосняков. Вместе с тем по той же линии
происходит стык ареалов сибирской и даурской лиственниц.
Все этр<— качественные показатели климатического единства
южной тайги Средней Сибири. Однако не только климатические
факторы регулируют распространение животных и растений.
С появлением человека и особенно с развитием современного
хозяйства природные воздействия стали проходить через призму
антропогенных влияний. Именно сочетание природных и антро-
погенных факторов создает картину современной южной тайги
Средней Сибири.
Эколого-физиологические потребности большинства животных
Сибири практически неизвестны. В связи с этим подробная клима-
тическая характеристика района южной тайги едва ли может дать
что-либо для понимания фаунистических закономерностей. Можно
лишь заметить, что животные населяют леса различных типов
не только потому, что те обладают определенными кормовыми
и защитными свойствами, но и оттого, что эти леса имеют совер-
шенно четко очерченные микроклиматические характеристики.
Говоря «темнохвойный лес», мы подразумеваем, таким образом,
не только растительность, но одновременно и сумму природных
влияний, с одной стороны, позволяющих существовать этому лесу,
а с другой — создаваемых самой темнохвойной тайгой. Следо-
вательно, разбирая смены растительности, мы неминуемо подра-
зумеваем и перемены в мезо- и микроклимате, определенные
63
изменения в неживом покрове и т. п. Количественные характе-
ристики всех этих перемен, быть может, дали бы критерии
для каких-нибудь зоогеографических построений, но всегда была
бы угроза впасть в ошибку: post hoc, ergo propter hoc. Вот поэтому
мы и предпочитаем качественные показатели, среди которых
растительность — верный индикатор.
Подавляющее большинство исследователей растительности
южной тайги Средней Сибири (Дробов, 1909; Боровиков, 1910
Рис. 10. Водораздельные сосновые леса (Ангарский подрайон).
1911, 1912, 1913; Криштофович, 1910, 1911, 1913; Ганешин, 1910,
1912; Кучеровская, 1912; Васильев, 1933; Поварницын, 1934,
1937, 1944, и др.) пришли к выводу о широком распространении
здесь темнохвойной тайги в прошлом. Сейчас площадь, некогда
занятую темнохвойными лесами, завоевывают сосна (рис. 10)
и местами лиственница. Однако возможность существо-
вания темнохвойных лесов во многих местах сохранилась: темно-
хвойные породы вновь стремятся вытеснить лесообразователей-
пришельцев. В зависимости от микроклиматических и геологиче-
ских условий местности этот процесс идет с различной скоростью.
При всей пестроте и сложности взаимоотношений лесных пород
удается проследить несколько основных рядов смен растительно-
сти. Кратко они уже были нами описаны (Реймерс и Малышев,
1963).
По характеру сукцессионных процессов южную тайгу Средней
Сибири можно схематически разделить на три региона: с полной
64
сменой лесных пород, с частичной их сменой и с отсутствием смен.
Первый из районов в указанной выше работе назван Приангар-
ским боровым, второй — Катангско-Ленским, третий — Средне-
горным районом темнохвойных лесов.
В Приангарском боровом районе прослежены следующие ант-
ропогенные изменения растительности.
1. На легких, особенно супесчаных и карбонатных, слабо-
и среднеувлажненных почвах темнохвойную тайгу непосредственно
сменяют сосновые леса с небольшой примесью лиственницы.
Новообразовавшиеся сосняки обычно имеют свои ряды транс-
формации и не сменяются вновь темнохвойной тайгой.
В ряде мест крутые склоны, ранее поросшие сосной, в резуль-
тате повторных нарушений оказываются лишенными леса. Обра-
зуются остепненные участки — степоиды по терминологии
В. Б. Сочавы и В. В. Липатовой (1960). Эти вторичные остепнен-
ные участки оказываются весьма близкими по растительности
к первичным образованиям такого же рода, возникшим на эро-
зионных поверхностях, первоначально лишенных леса.
2. На аллювиальных суглинках сгоревшие темнохвойные леса
бывают замещены лиственнично-сосновыми ассоциациями. При
отсутствии повторных нарушений светлохвойные леса этих место-
положений постепенно обогащаются темнохвойными элементами
и превращаются с течением времени в темнохвойные леса со зна-
чительной примесью лиственницы, а на более ранних фазах транс-
формации — и сосны.
3. На тяжелых, обычно переувлажненных почвах темнохвой-
ные леса сменяются мелколиственными — осиновыми и березо-
выми (рис. 11). Мелколиственные леса, особенно осиновые на сред-
ней Ангаре, достигают исключительного развития — диаметр
осин бывает до 50 см.
4. Нарушение лиственнично-сосновых лесов, несмотря на боль-
шую огнестойкость лиственницы, в Средней Сибири приводит
в конце концов к господству сосны. Везде, кроме тяжелых почв,
смены пород не происходит. На аллювиальных суглинках и дру-
гих тяжелых почвах светлохвойные массивы сменяются мелко-
лиственными, прежде всего березняками. Сосновые боры, как пра-
вило, замещаются смешанными сосново-мелколиственными молод-
няками, из которых затем береза и осина выпадают.
5. Выгорание приречных и приручьевых ельников происходит
лишь при больших лесных пожарах. Чаще эти массивы бывают
вырублены людьми при расчистке земель под луга. При повальных
пожарах ельники сменяются березняками или ерниками (из
березки низкой, кустарниковой лапчатки и таволги иволистной).
Березняки в ряде случаев оказываются весьма устойчивыми
и существуют длительное время, постепенно сменяясь вновь ель-
никами. Заброшенные луга (не подвергающиеся выпасу и сено-
косу) чаще всего зарастают сначала кустарниками (особенно в пере-
5 Н. Ф. Реймерс
65
увлажненных западинах), а затем постепенно и ельником. На воз-
вышенных участках поселяются параллельно лиственница и сосна.
Первая сохраняется в составе ельников длительное время, вторая
сравнительно скоро выпадает.
6. Ивовые заросли, тянущиеся полосами вдоль рек и стариц,
возобновляются хороню. Лишь в местах исключительно активного
истребления (около деревень) они постепенно исчезают. В дев-
ственной природе происходит постоянное наступление приречных
Рис. И. Гарь темнохвойной тайги, зарастающая мелколиственным
лесом (над ним возвышаются полузасохшие лиственницы, уцелевшие
от пожара).
ельников на ивняки. Здесь сказывается природный сукцессион-
ный процесс. Первичные луга, образовавшиеся на речных наносах,
с углублением ложа реки постепенно сменяются зарослями ивня-
ков, а эти последние, попадая под полог ельников, оказываются
оттесненными к реке. Уничтожение ельников приводит не только
к образованию березняков, но и к расширению кустарниковых
полос. Человек, раскорчевывая ельники и кустарники, создает
антропогенные луга, которые в настоящее время занимают значи-
тельные площади.
Таковы антропогенные смены, происходящие в Приангарском
боровом районе. Помимо них, в крайне редких случаях удается
наблюдать и естественные вековые сукцессионные ряды. Темнохвой-
ная тайга с возрастом самоизреживается, заболачивается, затем
вновь появляется возможность для лесовозобновления, процессы
старения и омоложения ассоциации сбалансировываются и воз-
никает разновозрастный лес, типичный для слабо нарушенных
участков.
66
В предгорьях Саян (Среднегорный район темнохвойных лесов)
на гарях сразу вырастают темнохвойные молодняки, почти без
изменений породного состава переходящие в спелые леса (Реймерс,
19586). В Катангско-Ленском районе частичной смены лесных
пород можно проследить все только что описанные процессы транс-
формации. Специфичной для него следует считать смену темно-
хвойных лесов темнохвойно-мелколиственными молодняками (сна-
чала молодняком березы, под пологом которого быстро вырастает
подрост темнохвойных пород), подобно смене сосновых боров
такими же смешанными ассоциациями (см. выше, пункт 4).
Таким образом, водораздельные темнохвойные леса вовлечены
в один из рядов трансформации растительности. Для водораздель-
ных пространств южной тайги Средней Сибири наиболее харак-
терны четыре основных типа таких смен. Первый тип: 1) темно-
хвойный молодняк, 2) средневозрастной темнохвойный лес, 3) зре-
лый темнохвойный лес; второй тип: 1) гарь, поросшая мелко-
лиственным молодняком, 2) смешанный мелколиственно-темно-
хвойный лес, 3) зрелый темнохвойный лес; третий тип: 1) гарь,
поросшая мелколиственным молодняком (березовым или осино-
вым), 2) мелколиственный лес (березовый или осиновый), 3) такой
же лес с подлеском темнохвойных пород, 4) смешанный мелко-
лиственно-темнохвойный лес, 5) зрелый темнохвойный лес; нако-
нец, четвертый тип: 1) гарь, поросшая мелколиственным лесом,
а также вырубка или залежь, занятая лиственнично-сосновым
молодняком, 2) смешанный мелколиственно-светлохвойный лес,
3) светлохвойный лес, 4) такой же лес с подростом темнохвойных
пород, 5) темнохвойный лес со значительным участием светло-
хвойных или мелколиственных пород, 6) зрелый темнохвойный
лес. При последнем типе смен не всегда бывают выражены все
этапы, в частности вырубки, поросшие изначально светлохвой-
ным лесом, переходят сразу в фазу светлохвойного леса, а эта
последняя нередко выпадает при превращении гари, поросшей
мелколиственным лесом, в светлохвойное насаждение с подлеском
из темнохвойных пород. Приручьевые участки темнохвой-
"ных лесов испытывают смены первого типа. Следовательно,
водоразделы имеют набор биотопов, показанный на рис. 12
(7-/7).
Выше уже отмечено, что все выделенные биотопы имеют спе-
цифическую фауну и животное население позвоночных.
Несколько слов нужно сказать о самой схеме на рис. 12. На
ней мы пытались отобразить не только принципиальное размеще-
ние биотопов и приблизительное их соотношение по площади,
но и совместить на одном бланке все разнообразие природных
разностей южной тайги Средней Сибири. В любом месте южной
тайги на основе лесных планов, крупномасштабных геоботаниче-
ских карт, а лучше всего динамических карт растительности можно
«развернуть» предлагаемую схему в соответствующую картосхему
5*
67
Рис. 12. Принципиальная схема размещения биотопов в полосе южной тайги
Средней Сибири (пунктир разделяет перечисляемые полосы).
I — полоса слабой и умеренной нарушенности (сравнительно редкой пов-
торяемости нарушений) лесов на водоразделах и в расчленяющих их вер-
ховьях речек и ручьев.
1 — биотопы фаунистического комплекса темнохвойного (лиственнично-кедрово-пихто-
вого или лиственнично-елово-пихтового) леса: 1а — биотоп фаунистического комплекса
разреженного перестойного темнохвойного леса с признаками поверхностного забола-
чивания; 2 — биотоп фаунистического комплекса темнохвойного леса с участием мелко-
лиственных пород (березы, осины); з — биотоп фаунистического комплекса гарей на месте
темнохвойного леса, поросших мелколиственным лесом-молодняком (березовым, осино-
вым); 4 — биотоп фаунистического комплекса приручьевых участков темнохвойного
(лиственнично-кедрово-пихтово-елового) леса; 5 — биотоп фаунистического комплекса
приручьевых участков темнохвойного леса с участием мелколиственных пород (березы);
6 — биотоп фаунистического комплекса гарей на месте приручьевых участков темнохвой-
ного леса, поросших мелколиственным лесом-молодняком (березовым); 7 — биотоп фа-
унистического комплекса сосновых лесов (лиственнично-сосновых) с подлеском темно-
хвойных пород; 8 — биотоп фаунистического комплекса сосновых (лиственнично-сосновых)
лесов; 9 — биотоп фаунистического комплекса сосновых (лиственнично-сосновых) лесов
с участием мелколиственных пород (березы, осины); 10 — биотоп фаунистического ком-
плекса гарей и вырубок, поросших сосновым молодняком; 11 — биотоп фаунистического
комплекса гарей на месте сосновых лесов, поросших мелколиственным лесом-молодня-
ком (березовым, осиновым); 12 — биотоп фаунистического комплекса березовых лесов
с подлеском темнохвойных пород; 13 — биотоп фаунистического комплекса березовых
лесов; 14 — биотоп фаунистического комплекса березовых молодняков; 15 — биотоп
фаунистического комплекса осиновых лесов с подлеском темнохвойных пород; 16 — био-
топ фаунистического комплекса осиновых лесов; 17 — биотоп фаунистического комплекса
осиновых молодняков.
II — полоса умеренной и сильной нарушенности (перманентно повторяю-
щихся нарушений) лесов на коренных берегах долин крупных и средних рек
с прилегающими низкими водоразделами.
18 — биотоп фаунистического комплекса островных участков темнохвойных (листвен-
нично-кедрово-пихтово-еловых) лесов среди светлохвойных массивов и мелколиствен-
ных лесов (в том числе гарей); 19 — биотоп фаунистического комплекса островных уча-
стков темнохвойных лесов с участием мелколиственных пород (березы, осины); 20 — био-
топ фаунистического комплекса гарей островных участков темнохвойных лесов, поросших
мелколиственным лесом-молодняком (березовым, осиновым); 21 — биотоп фаунисти-
ческого комплекса сосновых (лиственнично-сосновых) лесов; 22 — биотоп фаунисти-
ческого комплекса сосновых лесов с участием мелколиственных пород (березы, осины)
как фазы процесса трансформации (антропогенной сукцессии) соснового леса; 22а—тот же
самый биотоп, возникший в результате разреживания соснового леса рубкой и смены
в прогалах сосны березой; 23 — биотоп фаунистического комплекса гарей и вырубок
на месте сосновых лесов, поросших мелколиственным лесом-молодняком (березовым,
осиновым); 24 — биотоп фаунистического комплекса остепненных сосновых (лиственнично-
сосновых) лесов; 25 — биотоп фаунистического комплекса гарей на месте остепненных
сосновых лесов, поросших лиственнично-березово-сосновым молодняком; 26 — биотоп
фаунистического комплекса «степоидов» (безлесных гарей на месте остепненных сосновых
лесов и остепненных луговых склонов).
III — полоса сильной нарушенности (перманентных нарушений) и полного
преобразования природных комплексов в долинах крупных и средних рек
с низовьями впадающих в них речек и ручьев.
27 — биотоп фаунистического комплекса приречных ельников; 28 — биотоп фаунисти-
ческого комплекса приречных ельников с участием мелколиственных пород (березы,
тополя); 29 — биотоп фаунистического комплекса гарей на месте приречных ельников,
зарастающих мелколиственным молодняком (березой); 30 — биотоп фаунистического
комплекса березовых лесов речных долин; 31 — биотоп фаунистического комплекса иво-
вых и черемуховых зарослей в поймах рек и ручьев, примыкающих к приречным ельни-
кам; 31а — то же самое, но по границам лугов; 32 — биотоп фаунистического комплекса
суходольных и заливных пойменных лугов; 33 — биотоп фаунистического комплекса при-
ручьевых ельников; 34 — биотоп фаунистического комплекса приручьевых ельников с
участием мелколиственных пород (березы); 35 — биотоп фаунистического комплекса гарей
на месте приречных ельников, поросших мелколиственным лесом-молодняком (березовым);
36 — биотоп фаунистического комплекса березовых лесов долин ручьев; 37—биотоп фау-
нистического комплекса ерниковых зарослей.
Антропогенные участки.
38 — биотоп фаунистического комплекса возделываемых полей; 39 — биотоп фаунисти-
ческого комплекса открытых залежей; 40 — биотоп фаунистического комплекса залежей,
поросших сосновым молодняком; 41 — биотоп фаунистического комплекса населенных
пунктов (огородов и зданий).
68
в зависимости от поставленной задачи: будь то схема распростра-
нения вида (гл. 5) или комплекса животного населения (гл. 6).
Вблизи от населенных пунктов частота антропогенных нару-
шений возрастает. Это меняет не только облик леса, но и состав
его фауны. В часто нарушаемых сосняках на тяжелых почвах
увеличивается примесь мелколиственных пород, а процесс транс-
формации идет по схеме: 1) мелколиственный молодняк (рис. 13),
2) смешанный мелколиственно-светлохвойный лес (рис. 14), 3) свет-
лохвойный лес (рис. 15). Наоборот, на легких песчаных почвах
и на выходах известняков сосна получает преимущество перед
мелколиственными породами. Вместе с тем в таких местах проис-
ходит внедрение под полог сосняков степевидных ассоциаций —
возникают остепненные сосновые леса, дающие свой антропогенно-
сукцессионный ряд: 1) степоид (рис. 16), 2) светлохвойно-мелко-
лиственный молодняк, нередко с подлеском ксерофильных кустар-
ников, 3) остепненный сосновый бор.
Биотопы придолинной полосы дополняет еще один антропо-
генно-сукцессионный ряд. Его составляют фазы восстановления
островных темнохвойных лесов. Небольшие сохранившиеся уча-
стки главным образом приручьевых темнохвойных лесов в усло-
виях повышенной влажности почвы (и воздуха) стойко сопроти-
вляются наступающему на них со всех сторон светлохвойному
лесу. Однако эти участки уже ни флористически, ни фауни-
стически не идентичны темнохвойным массивам. Подобно кол-
кам в степи, они стоят в море светлохвойных лесов, и как
фауна колков отличается от лесной, так и животный мир остров-
ных темнохвойных лесов отличен от фауны сплошной тайги.
Итак, придолинная полоса (см. рис. 12), имеющая свой при-
родно-антропогенный режим, четко отличается от водораздельной.
В нее входят 9 специфичных разностей (18—26).
Приручьевые ельники (рис. 17) пронизывают придолинную
полосу, словно жилки лист. Но они, как уже отмечено, име-
ют особую историю трансформации. Близки к ним приречные
ельники (рис. 18), но судьба этих образований сильнее связана
с приречными кустами (рис. 19), лугами и самой деятельностью
реки. Они в большей степени носят характер околоводных биотопов.
Ивняки оказываются различными по количественным характе-
ристикам животного населения в зависимости от того, примыкают
ли они к лугам (рис. 20) или к приречному ельнику (31а и 31 на
рис. 12). Однако это различие не столь существенно, чтобы было
основание считать эти участки различными биотопами.
Группу приречных участков дополняют места, коренным обра-
зом преобразованные человеком. Это сельскохозяйственные угодья
и населенные пункты. Брошенные поля в южной тайге постепенно
поглощаются лесом. Прежде чем зарасти типичным лесом-молод-
няком с лесной подстилкой (чаще всего сосновым, изредка — на
средней Ангаре — березовым), заброшенное поле переживает две
70
Рис. 13. Молодой березняк на месте соснового бора.
Рис. 14. Нарушенный сосновый бор с примесью мелколиственных пород.
Рис. 15. Сосновый бор.
сукцессионные фазы, животное население которых представлено
особыми элементарными фаунистическими комплексами, —просто
залежи и залежи, поросшей сосновым молодняком.
Обрывы речных берегов, населенные пункты, речные старицы
и водные глади рек замыкают список биотопов долинной полосы.
Таким образом, в ее пределах можно наметить основные выделы,
отмеченные на рис. 12 цифрами 27—41. В дополнение нужно
упомянуть еще два выдела, не выражающиеся в принятом нами
масштабе: а) биотоп фаунистического комплекса речного бичев-
ника и обрывов речных берегов, б) биотоп фаунистического ком-
плекса речных стариц.
74
Описание приведенных антропогенно-сукцессионных смен
помогает в общих чертах представить облик территорий южной
тайги Средней Сибири в далеком прошлом (рис. 21). Водораздель-
ные темнохвойные леса в то время смыкались с при ручьевыми
Рис. 17. Приручьевой ельник.
и приречными ельниками. При этом приручьевых ельников в сов-
ременном их виде не было. Они сближались по экологическим
показателям с приручьевыми участками темнохвойных лесов.
Редкие естественные нарушения (ветровалы, стихийные лесные
пожары) приводили к кратковременному существованию мелко-
лиственных лесов, а чаще — к образованию смешанных молод-
няков.
75
Исследователи времен Переселенческого управления (Дробов,
1909; Боровиков, 1910, 1911, 1912, 1913; Криштофович, 1910,
1911, 1913, и др.) единодушно признавали, что сосновые боры по
Ангаре и ее притокам существовали издавна и до интенсивного
Рис. 18. Приречный ельник.
нарушения приангарских лесов пожарами. М. Г. Попов (1953)
предполагал наличие вдоль сибирских рек степных участков
приблизительно там, где, по другим авторам, должны были расти
первичные сосновые боры. Сейчас трудно решить вопрос о том,
каков был характер речных долин в отдаленные времена, однако
именно здесь, на песчаных дюнах речных долин Приангарья,
должны были в первую очередь закрепиться сосновые леса. Отсюда
они могли проникнуть на степные склоны выходов карбонатных
76
пород, а с приходом современного человека широко распростра-
ниться на водоразделы.
Приречные ельники сжимали речные долины. Лишь вдоль самой
кромки воды и на островах лепились кустарники и крохотные
участки первичных лугов. Изредка в узких долинах приречные
Рис. 19. Кустарниковые заросли у реки в половодье.
ельники прерывались поперечными полосками галечников времен-
ных водотоков.
Так выглядела тайга юга Средней Сибири до появления ее
основного врага — частых лесных пожаров. Следует подчеркнуть,
что горимость первобытных лесов из-за высокой влажности
воздуха в темнохвойной тайге была минимальной. 1
1 За годы работы в темнохвойных лесах мы наблюдали десятки случаев
попадания молний в сухие или суховершинные деревья. Нередко стволы при
этом расщеплялись от вершины до комля, а трещины достигали глубины
десятков сантиметров. Абсолютно сухие деревья при прохождении молний
лишь изредка начинали тлеть, однако во всех нами прослеженных случаях
77
Освоение прибайкальской тайги человеком началось в конце
палеолита — около 30 тыс. лет тому назад. В те отдаленные вре-
мена на месте современной южной тайги простирались субаркти-
ческие тундры (Окладников, 1956). Стоянки человека обнаружены
главным образом в долинах Енисея, Ангары и Лены. В неолите
(5—5.5 тыс. лет тому назад) людей уже окружала тайга. В могиль-
никах исаковской культуры найдены кости только лесных и лесо-
Рпс. 20. Выпасаемый луг и сохранившийся участок приречного
ельника.
степных форм: лося, марала, бобра и косули. Очевидно, в это
время уже сложились почти современные условия жизни. Во вся-
ком случае к бронзовому веку (около 4.5 тыс. лет тому назад)
имелась определенная разница между степными племенами и пле-
менами тайги. Однако еще раньше, в неолите, на Ангаре и Лене
существовали лесостепные или сходные с ними лесолуговые (воз-
можно, подобные центральноякутским аласам) участки. Понома-
ревские могильники с костями косуль четко указывают на это
обстоятельство. Да и лоси — основной объект охоты и искусства
древних сибиряков — едва ли могли быть столь многочисленны
в условиях слишком глухой тайги.
В бронзовом веке можно предполагать резкое увеличение леси-
стости и одновременное уменьшение числа крупных копытных
возгорания не происходило. Интересно, что специальное поджигание стоя-
щих деревьев при высокой влажности воздуха никогда не приводило к рас-
пространению огня по лесу.
78
зверей — люди перешли главным образом к рыболовству. Однако
охотничий промысел продолжал существовать, и в первом тыся-
челетии до нашей эры обрел равноправное положение с рыболов-
ством. Видимо, люди тех времен все-таки обитали не в условиях
Рис. 21. Гипотетическая схема размещения биотопов юж-
ной тайги Средней Сибири до резкого нарушения лесов
человеком.
Условные обозначения как на рис. 12.
сплошной тайги, как предполагает А. П. Окладников, на работы
которого мы опираемся (Окладников, 1948, 1949, 1956). Среди
глухих лесов едва ли могли развиваться связи между племенами,
жившими в разных речных системах, а тем более разделенными
многими водоразделами. Примитивное сухопутное сообщение
более вероятно в степных и лесостепных местностях. Между тем
у народов Прибайкалья в первом тысячелетии до нашей эры
79
обнаружены котлы скифского типа и вещи, указывающие на связь
с древним Китаем. Очевидно, так называемые лесные племена
Прибайкалья поселялись в условиях, близких к лесостепным.
Курыкане — племена первого тысячелетия нашей эры, жившие
вокруг Байкала, еще больше связаны со степными участками,
что нашло отражение в искусстве. Художники племени изобра-
жали не только коней, скачущих во весь опор, но и верблюдов.
Курыкане были скотоводами, а для скотоводства нужны паст-
бища. Трудно себе представить, чтобы племя, разводившее домаш-
них верблюдов, могло жить среди глухих лесов. Локализация
укреплений — городищ курыкан вокруг Байкала в местах, где
и ныне имеются степные участки (Поплавская, 1913, 1916; Корот-
кий, 1916; Николаев, 1949; Попов, 1953; Малышев, 1956), заста-
вляет предположить наличие в то время довольно обширных
степей. Это, конечно, не значит, что степи того времени должны
были господствовать в Прибайкалье. Люди тогда были не слишком
многочисленны и могли довольствоваться сравнительно неболь-
шими площадями.
Наш экскурс в археологию показывает, что развитое выше
представление о юге Средней Сибири как огромной лесной стране
с господством темнохвойных лесов относится к исторически корот-
кому сроку и требует дополнений. Очевидно, во времена курыкан
темнохвойная тайга была широко распространена на между-
речьях, степные же и лесостепные участки (Канские, Око-Ангар-
ские и Приленско-Балаганские) окаймлялись лесами из листвен-
ницы, сосны, но главным образом березы. Эти леса не занимали
больших площадей и были изолированы от родственных сосновых
и мелколиственно-широколиственных массивов востока и запада.
На западе их разобщали покрытые темнохвойной тайгой отроги
Саян, Кузнецкого Алатау и Салаира, смыкавшиеся с урманными
кедровниками Западно-Сибирской низменности. На востоке сто-
яли Прибайкальские хребты и пролегала водная гладь Байкала.
Южносибирские горы не были особым препятствием для культур-
ного обмена курыкан с монголами (такой обмен мог происходить
по Иркуту и Тункинской долине), но эти же самые поросшие тай-
гой горы замыкали Предбайкальские лесостепи с юга.
В последние сотни лет, видимо, наблюдалось резкое наступа-
ние лесов на степи, что следует из наблюдений за взаимоотноше-
нием леса и степных участков (Николаев, 1949; Попов, 1953; Малы-
шев, 1956; Мартынов, 1958; Пешкова, 1960, 1962). Однако при-
родный процесс на этот раз встретил серьезное сопротивление
человека. Расчищая леса под пастбища и поля, буряты, пришед-
шие на смену курыканам, создали на месте лесостепей и окраин
лесов участки лесополья и лесолужья.2 Товарищем бурят был
2 Очевидно, Б. В. Надеждин (1959) встретил именно такие участки,
поэтому его выводы и оказались прямо противоположными заключениям
других авторов.
80
огонь. И поныне буряты для улучшения лесных лугов выжигают
ветошь, а с нею часто и леса на колоссальных пространствах.
Именно благодаря такой системе земледелия и образовались
лиственничные лесостепи Средней Сибири. Это яркий пример
особого ландшафта, созданного человеком. В связи с уменьшением
значения Верхоленья как земледельческого района многие ранее
использовавшиеся поля и луга заросли лесом. Благодаря малому
количеству осадков, повышенной горимости нарушенных лесов
(Реймерс и Малышев, 1963) и все еще продолжающимся лесным
пожарам там существуют вторичные лиственничники, которые
геоботаники относят к южносибирскому типу.
На крайнем юге Средней Сибири русские встретили уже нару-
шенную тайгу. Однако нарушения не распространялись слишком
широко, особенно на водоразделы, тем более что в то время уже
была в полном разгаре фаза наступания леса на нелесные уча-
стки, а темнохвойной тайги — на сосновые боры и лиственнич-
ники. Когда в XVII в. сложился путь землепроходцев по Енисею,
Ангаре, Илиму и Лене, а также менее использовавшийся — по
Нижней Тунгуске—Лене (Покшишевский, 1951), вся северо-
западная часть южной тайги Средней Сибири, не подвергавшаяся
в историческом процессе остепнению, еще была почти не тронута
пожарами. Редкие охотничьи племена, вся жизнь которых про-
ходила в тайге и зависела от ее целостности, не могли произво-
дить значительных нарушений. Другое положение сложилось
с приходом русских. Для них лес был врагом, а тайга начала
гореть настолько интенсивно, что был издан ряд правительствен-
ных циркуляров, направленных против этого зла (Скалой,
19516).
В результате пожаров процесс наступания темнохвойных
лесов был не только остановлен, но сменился на обратный: темно-
хвойные леса начали исчезать. Местами былые их площади отме-
чены мелколиственными ассоциациями, а иногда их полностью
сменили сосняки.
Исчезновению темнохвойных лесов способствовало и то обсто-
ятельство, что в последние полвека наметилась тенденция поте-
пления климата Средней Сибири (Галазий, 1954). Оно сопровож-
далось повышением температуры воздуха и почвы, уменьшением
влажности воздуха, сокращением количества осадков, т. е. как
раз теми факторами, которые благоприятны для развития пожаров
(Мелехов, 1936, 1938; Нестеров, 1945; Анцышкин, 1952; Сныткин,
1963, и др.).
Нарисованная картина истории формирования современных
биоценозов южной тайги Средней Сибири иллюстрируется и фау-
нистическими данными. В предыдущих главах отмечено, что различ-
ные зоокомплексы сложились в пределах темнохвойных и мелко-
лиственных лесов, в то время как животный мир сосняков Средней
Сибири специфических черт почти не имеет.
6 II. Ф. Реймерс
81
История речных долин, где проходило «младенчество» чело-
вечества, ближайших от долин водоразделов, откуда черпал
человек топливо и где прежде всего он охотился, и дальних водо-
разделов, куда человек проникал лишь в поисках хороших охот-
ничьих угодий в более позднее время, складывалась различно.
Вот эти различия в истории развития вместе с климатическими
факторами и тем влиянием, которое оказывает на образование
и перемещение биотопов эрозионная деятельность крупных водо-
токов, и сложили неоднократно упоминавшиеся в этом разделе три
полосы: 1) полосу слабой и умеренной нарушенности (сравнительно
редко повторяющихся нарушений) лесов на водоразделах и в рас-
членяющих их верховьях речек и ручьев, 2) полосу умеренной и
сильной нарушенности (постоянно повторяющихся нарушений) ле-
сов на коренных берегах долин крупных и средних рек с прилега-
ющими низкими водоразделами и 3) полосу сильной нарушенности
(перманентных нарушений) и полного преобразования природных
комплексов в долинах крупных и средних рек с низовьями впада-
ющих в них речек и ручьев (рис. 12).
Смена и нарушение растительности приводили к изменению
в фауне и животном населении района. Фауна первичных темно-
хвойных лесов обогатилась за счет разнообразия видового состава
животных сукцессионных фаз, но вместе с тем резко трансформи-
ровалась из-за смены природных рядов на антропогенные. Светло-
хвойные леса с «прорывом» изоляции с запада стали обогащаться
европейскими выходцами. Общее потепление климата, как плане-
тарное, так и местное (за счет сведения лесов), дало возможность
южным формам проникнуть дальше на север. Наоборот, развитие
местами переувлажненных мелколиственных и луговых ассо-
циаций северного облика дало приют северным формам. Существо-
вание степных, лесостепных и лесопольных участков сделало
возможным распространение в южной части тайги лесостепных
(опушечных) форм. Все это создало современную физиономию
животного мира южной тайги Средней Сибири.
Подвергавшийся в прошлом значительному остепнению при-
ленский регион получил черты Лено-Байкальского зоогеографи-
ческого подрайона. Исконно таежный Ангарский подрайон в силу
внедрения западных форм распался на два зоогеографических
участка.
Тот же фактор расчленил Присаянский подрайон на два
зоогеографических участка, из которых в восточном в наименее
трансформированном виде сохранились первичные фаунистиче-
ские группировки.
Таким образом протекала история формирования биотических
закономерностей в рассматриваемом регионе, так сумма истори-
ческих и природно-антропогенных факторов складывала совре-
менные региональные и типологические взаимоотношения внутри
южной тайги Средней Сибири.
82
Перейдем теперь к изложению материалов по распространению
и размещению отдельных видов животных и биологическим замет-
кам.
Глава 5
ПОВИДОВОЙ ОБЗОР ПТИЦ И МЛЕКОПИТАЮЩИХ
ЮЖНОЙ ТАЙГИ СРЕДНЕЙ СИБИРИ
Птицы
Прежде чем приступить к изложению собранных материалов
по распространению, размещению и биологии отдельных видов,
следует напомнить, что в соответствии с нашей основной задачей —
изучением динамики животного населения — мы обращали наи-
большее внимание на характер кружева видовых ареалов и его
изменение в связи с нарушениями лесов. В своей работе мы опи-
рались на крупномасштабные карты, которые здесь заменены при-
веденной в предыдущей главе принципиальной схемой биотопов.
Напомним, что эта схема практически применима в любой точке
южной тайги Средней Сибири: если абсолютные значения баллов
численности животных могут меняться в зависимости от места
и времени, среднее их соотношение в пределах зоогеографического
района почти одинаково (Реймерс, 1965).
В ряде случаев количественная оценка птичьего населения
сделана не на основе точных учетов (об этих учетах см. «Введение»),
а приблизительно. Такой подход, на наш взгляд, оправдан тем, что
1) население птиц резко колеблется по сезонам, 2) точность разо-
вых учетных данных очень низка, особенно для скрытноживущих
видов, 3) лучше дать приблизительную оценку численности, чем
не иметь никакой.
Некоторые общепринятые методы количественного учета
птиц оказались в условиях южной тайги Средней Сибири непри-
менимыми в силу особенностей природных условий. Так, в период
глухариных токов снег еще очень глубок, ночью покрыт настом,
днем наступает «распар» (мокрое кашеобразное состояние, очень
характерное для всей полосы южной тайги нашего района и гор
юга Сибири). Наст трещит под ногами, и пробиться через мокрый
снег — дело нелегкое. Удаленность глухариных токов от дорог
усугубляет трудность учета этих куриных. Вслед за таянием снегов
и одновременно с ним вздуваются реки. Уровень средних рек
поднимается до 9—10 м. Даже маленькие речки делаются на 2—3 м
глубже. Южная тайга превращается в своеобразный лабиринт.
Маленький ручеек летом, весной — грозный поток, сбивающий
с ног. Все это делает водоразделы доступными лишь с середины
июня. К этому времени некоторые виды уже прекращают петь
и их учет по голосам технически невозможен.
6*
83
Сложность условий проведения орнитологических наблюдений
заставляет нас с большой осторожностью делать выводы об отсут-
ствии видов в составе фауны полосы южной тайги. В ряде слу-
чаев мы лишь ссылаемся на литературный источник. Включение
в список не зарегистрированных нами видов, на наш взгляд,
оправдано тем, что, оставляя вопрос об их обитании открытым,
мы указываем на возможность обнаружения животного.
Равномерное исследование всех систематических групп и видов
птиц практически невозможно. Основное внимание уделено
лесным формам, наименьшее — связанным с водоемами и откры-
тыми пространствами. Такой подход оправдан целями исследо-
вания.
Следует с огорчением отметить, что в силу ряда обстоятельств
мы не смогли обработать часть собранных материалов, чем и
объясняется крайняя фрагментарность приводимых данных о пи-
тании птиц.
Русские и латинские названия птиц даны по монографии
«Птицы Советского Союза» (1951) под редакцией Г. П. Дементьева
и Н. А. Гладкова.
Отряд КУРИНЫЕ — GALUFORMES
Белая куропатка — Lagopus lagopus L.
По А. В. Михееву (1952), в пределах южной тайги Средней
Сибири проходят границы ареалов трех подвидов белой куро-
патки: L. I. koreni Thayer et Bangs, L. I, brevirostris Hesse и
L. I. serebrowsky Doman. Однако, как отмечает К. А. Юдин (1952),
южная граница вида в тайге левобережья Енисея проходит около
62° с. ш., т. е. у северного предела южной тайги. Далее к востоку
куропатка доходит до 58° с. ш. (Гагина, 1958а). В пределах соб-
ственно южной тайги Средней Сибири нами этот вид на гнездовье
не отмечен. Зимой 1950/51 г. белые куропатки были обычны под
Иркутском. Позже мы их тут не регистрировали. Это обстоятель-
ство связано с резкими колебаниями численности вида. Т. Н. Га-
гина отмечает переселение куропаток к югу в 1953—1954 гг.
Как летом, так и зимой белая куропатка приурочена к гольцам
(она весьма обычна в западном Хамар-Дабане среди высокого-
рий, поросших водяникой), лесостепям, лесополью, болотам.
Кружево ее ареала зимой почти совпадает с вязью водотоков.
Летом его нити имеют соединения между гольцовыми высоко-
горьями на юге и водораздельными болотами на севере. Сведение
лесов благоприятно для зимовки птиц и их проникновения
в южную тайгу на гнездовья. Однако до сих пор белая куропатка
не стала аборигеном южнотаежных пространств.
84
Тундреная куропатка — Lagopus mutus Mont.
В пределах южной тайги Средней Сибири этот вид не встре-
чается. Отмечен лишь в окружающих ее высокогорьях (Гагина,
1961).
Тетерев — Lyrurus tetrix L.
Как мы уже отмечали (Реймерс, 1963г), тетерев в Средней
Сибири — типичная форма лесополья. На водоразделах тетеревов
нет даже среди обширных гарей, зато в лесостепи это весьма рас-
пространенный вид. В лиственничной лесостепи Ангаро-Ленского
водораздела вблизи населенных пунктов на 1 км можно насчи-
тать по опушкам до десятка токующих самцов. В глубине тайги
мы не встречали птиц далее 5 км от открытых пространств. Тетерев
относится к видам придолинного комплекса болыпих рек, проник-
шим в южную тайгу с развитием земледелия. На Ангаро-Под-
каменнотунгусском междуречье тетерев ранней весной настолько
привязан к полям, что буквально становится синантропом.
По малым таежным рекам, там, где нет полей, тетерева не встре-
чаются (Око-Ангарский водораздел).
Первые тетеревиные тока отмечены в окрестностях Жигалово
на Лене 19 апреля 1960 г., следующей весной —24 апреля. В 1962 г.
на р. Чадобец тетерева начали токовать также 24 апреля. На основе
наблюдений добровольной сети Восточносибирской фенологи-
ческой комиссии в 1961—1963 гг. можно составить весьма при-
близительное представление о сроках начала токования тетеревов
на территории южной тайги Средней Сибири. В районах сосново-
мелколиственной лесостепи вдоль железной дороги тетерева
начинают токовать, в зависимости от скорости хода весны, между
24 марта и серединой апреля. В лиственничной лесостепи Ангаро-
Ленского междуречья и районах лесополья верхней Лены начало
токования отмечено ранней весной 25 марта, но обычно происхо-
дит между 10 и 25 апреля. Широтных отличий в начале токования
тетеревов отметить не удалось из-за недостатка материалов.
В начале августа молодые тетерева достигают полукилограм-
мового веса.
Ранней весной и осенью тетерева обычны на полях, где питаются
зерном. В первой половине лета птицы поедают много насекомых,
во второй — питаются в основном ягодами. Питанию тетеревов
посвящена статья А. С. Фетисова (1934).
В малоснежные зимы тетерева нередко погибают, очевидно от
холода. 14 мая 1959 г. в окрестностях улуса Байша (лиственнич-
ная лесостепь, б. Баяндаевский район Иркутской области) были
найдены остатки 3 тетеревов, погибших зимой. Высохшие трупы
птиц не имели следов насильственной смерти. Предыдущая зима
1958/59 г. была очень малоснежной, а морозы в районе Байши
достигали 50°.
85
На двух обработанных паразитологами молодых птицах с верх-
ней Лены иксодовых клещей обнаружено не было.
Глухарь — Tetrao urogallus L.
Южную тайгу Средней Сибири населяет обыкновенный глу-
харь, который в части ареала, совпадающей с местами распростра-
нения каменного глухаря, дает с последним помеси (Кирпичев,
1958).
Места распространения глухарей в южной тайге охарактеризо-
вать весьма сложно. Птица связана с высокоствольными лесами,
нередко паркового характера, однако четкой приуроченности
к определенным лесным породам нет. Недаром М. А. Мензбир
(1934) писал: «К характерным же лесным животным надо причис-
лить из птиц глухаря (Tetrao urogallus), распространившегося
по Европе и Азии, но с запада ли на восток или с востока на запад—
неизвестно» (стр. 191—192). По глазомерной оценке больше всего
глухарей в водораздельных разреженных темнохвойных лесах.
На втором месте следует поставить сосняки, на третьем — листвен-
ничники. Поскольку лиственничники не характерны для южной
тайги, следует признать, что глухари должны быть довольно
многочисленны в ее пределах. Однако благодаря преследованию
человеком и общему омоложению лесов Средней Сибири (в связи
с гибелью их от пожаров и сибирского шелкопряда) резко сокра-
тилось число глухарей.
Второму фактору мы придаем значительно большее значение,
чем первому. Нередко вблизи населенных пунктов глухари бывают
весьма обыкновенны, а дальше на водоразделах, изобилующих
гарями, птиц почти нет. Приведем еще один пример. В августе
1959 г. мы дважды пересекли Око-Ангарский водораздел. В месте
южного пересечения, где много гарей, мы встретили всего одну
пару (самец и самка) птиц почти на 100 км. Северный маршрут,
проходивший по сосновым и осиновым высокоствольным лесам,
дал на том же расстоянии 7 встреч, т. е. численность птиц оказа-
лась выше более чем в три раза. Среди сплошных гарей глухари
нередко встречаются в совершенно неожиданных местах. Так,
на том же Око-Ангарском междуречье мы вспугнули упомянутую
пару из зарослей приречного ивняка.
Взрослый самец, добытый на верхней Лене 5 июля 1961 г.
в осиново-березовом лесу, весил 4 кг. Размер тестикул 10.2 X
X 6.6 мм. В зобу и желудке найден мох Polytrichum sp., несколько
семян, цветок растения из сем. сложноцветных и камешки. Моло-
дой самец 30 июля весил 1 кг, имел тестикулы 5.5 X 2.3 мм.
В его зобу и желудке найден хвощ (50% содержимого), ягоды
голубики, черники и красной смородины приблизительно в одина-
ковом соотношении.
В ранние весны глухари начинают токовать в конце марта—
начале апреля^(1961 г.). Обыкновенно же начало токов прихо-
86
дится в южной тайге Средней Сибири на середину апреля. В лесо-
степной полосе и вообще на юге птицы начинают токовать на
4—5 дней раньше, чем на верхней Лене и нижней Ангаре. При-
близительно такое же опережение в начале токов наблюдается
у птиц, живущих у юго-западных пределов южной тайги, по срав-
нению с юго-востоком.
Каменный глухарь — Tetrao urogalloides Midd.
Этот вид распространен лишь за пределами южной тайги Сред-
ней Сибири. Очевидно, С. В. Кириков (19526) ошибочно включил
истоки Ангары в пределы ареала вида. Б. К. Штегман (Steg-
mann, 1936) правильно отмечал, что каменного глухаря нет у исто-
ков Ангары, на южном Байкале и северных скатах Хамар-Дабана.
Рябчик — Tetrastes bona si a L.
Ареал рябчика полностью включает южную тайгу Средней
Сибири. Однако рябчик — отнюдь не повсеместно распростра-
ненная птица, как нередко указывается в литературе (Taczanow-
ski, 1872—1874, 1891—1893; Stegmann, 1936, и др.). Сейчас уже
имеется достаточное количество материалов для того, чтобы очер-
тить кружево ареала рябчика в Средней Сибири.
В южной горной Сибири рябчик гнездится по долинам рек
и склонам гор в смешанных лесах всегда с присутствием березы
или ольхи (Козлова, 1930; Реймерс, 1959а). Позже, летом, ряб-
чики поднимаются на хребты. Однако численность птиц в темно-
хвойных лесах ниже, чем в березняках с примесью темнохвойных
пород. По данным А. Г. Банникова (1957), полученным в Кентее,
численность рябчиков в кедровой тайге 12—36 птиц на 10 км;
в березняках по долинам рек на то же расстояние приходится
23—41 особь. В случае хорошего урожая ягод рябчики осенью
концентрируются в ягодниках. Зимой птицы населяют главным
образом менее заснеженные «упечные» склоны и старые леса, где
под выворотами корней они находят необходимые им камешки.
К северу — в собственно южной тайге — рябчики также четко
приурочены к долинам рек и ручьев. Но тут они связаны не с берез-
няками, а с ельниками (Реймерс, 1963г) и вообще приручьевыми
темнохвойными лесами. Долинных березняков рябчики в южной
тайге избегают. Летом птицы, так же как и в горах юга Сибири,
весьма обычны в хребтовых темнохвойных лесах, но их практи-
чески нет в остальных водораздельных массивах. Принципиальная
схема весенне-летнего размещения рябчиков показана на рис. 22.
Осенью рябчики выходят на ягодники гарей, встречаются в сос-
ново-березовых лесах, однако наивысшая численность птиц наблю-
дается по приручьевым ельникам. Этот же биотоп населяют ряб-
чики зимой. Нередко они в холодное время года кормятся на опуш-
ках лесов, выходящих к рекам, гарям и полянам, подчас прибли-
87
жаясь вплотную к населенным пунктам и залетая в кустарники
на огородах.
В полосе лесостепей, как горных (Тува), так и плакорных
(Ангаро-Ленский водораздел, сосново-мелколиственные лесостепи
вдоль железной дороги), рябчики малочисленны и гнездятся в лес-
ных колках не менее чем в нескольких десятках метров от опушек.
Этот вид входит в лесную фауну лесостепи.
У северных пределов южной тайги с появлением значительных
массивов лиственницы и в местах господства сосны (западные части
южной тайги Средней Сибири) рябчики выходят на водоразделы
лишь по опушкам гарей и темнохвойным участкам там, где они
имеются. Здесь вновь усиливается связь вида с березой (По-
ляков, 1873), почками и сережками которой рябчики питаются.
В ходе сукцессии водораздельных лесов южной тайги рябчики
достигают наивысшей численности в фазе разреженного пере-
стойного леса. В глухих разновозрастных древостоях рябчиков
мало. Таким образом, в исторический период происходило изме-
нение числа рябчиков дважды. С первоначальным омоложением
лесов и их нарушением число рябчиков снизилось на водораз-
делах, но повысилось в долинах (нарушенные ельники населены
рябчиками плотнее, чем глухие). Второй этап снижения числен-
ности птиц начался с развитием скотоводства и земледелия, когда
исчезли огромные массивы приречных ельников, а водоразделы
начали покрываться гарями. Вместе с тем второй этап омоложения
темнохвойных лесов привел к некоторому увеличению числен-
ности рябчиков на водоразделах. Снижение произошло лишь тогда,
когда темнохвойные леса стали здесь сменяться сосняками.
В мае рябчики начинают размножаться. В середине июня
рябчата еще не летают (в 1961 г. на верхней Лене последний неле-
тающий выводок встречен 22 июня), но в конце месяца (29—30-го)
даже у северных пределов южной тайги встречаются летающие
выводки (р. Чадобец, 29 июня 1962 г.). Однако в начале июля
птенцы по весу едва превышают 100 г. В двадцатых числах июля
молодые рябчики (самцы с р. Чадобец) весили 152.5, 192.0 г
(1962 г., придолинная полоса) и 125.0, 126.0, 177.3 г (водораздел).
Молодые самки в это же время несколько крупнее — 262.0, 223.6 г.
Такие же цифры получены и на верхней Лене. С задержкой весны
размножение у рябчиков происходит в более поздние сроки и моло-
дые поднимаются на крыло только в середине июля. Так, в 1961 г.
в двадцатых числах июля встречались особи менее 100 г (84.8)
весом, а в августе вес молодых не достиг 300 г даже у самок: 8 авгу-
ста 226.8, 18 августа 261.0 и 290.0 г. Обычно же рябчики в это
время весят более 300 г (как самцы, так и самки) и к концу авгу-
ста мало отличаются от взрослых особей.
В выводке, встреченном 23 июня 1961 г., было 6 птенцов.
Это обычная численность семьи, хотя число откладываемых яиц
часто намного больше.
88
' И2 НИШ5 О4
Рис. 22. Принципиальная схема весенне-летнего размещения
рябчиков в южной тайге Средней Сибири.
1 — максимальная численность; 2 — средняя численность; з — мини-
мальная численность; 4 — данный вид практически отсутствует. Осталь-
ные обозначения как на рис. 12. Оценка численности в баллах.
Нам удалось добыть свыше 50 рябчиков. Объекты питания их
весьма разнообразны: ягоды брусники, черники, голубики, малины,
почки и сережки березы и ольхи (зимой), гусеницы бабочек (глав-
ным образом зеленые голые) — иногда 100% содержимого зоба;
реже — жуки и другие насекомые и еще реже — семена травя-
нистых растений (например, анемоны). Только один раз были
обнаружены в зобу рябчика проросшие орехи кедра.
Зараженность рябчиков южной тайги иксодовыми клещами
невысока. На верхней Лене из 14 птиц на 4 были обнаружены
нимфы клещей (3 раза по 1 и один раз 2 экз.). Рябчики с нижней
Ангары дали еще меньший процент заражения — из 13 птиц лишь
на 1 обнаружено 3 нимфы иксодид. Следует заметить, что в южных
районах рябчики заражены и на водоразделах, где общая пора-
женность животных очень низка. Перекочевывая из долинных
лесов в водораздельные, рябчики заносят туда клещей.
Как уже упомянуто, с уничтожением девственных лесов чис-
ленность рябчиков резко снижается. В южной тайге меняется
и кружево ареала. Птицы оказываются буквально привязанными
к узким полоскам приручьевых ельников. Это в свою очередь
приводит к быстрому истреблению вида в результате неумеренного
отстрела даже вдали от населенных пунктов. Каждая геологическая
или лесоустроительная партия, а иногда и группы туристов в окре-
стностях своих лагерей хищнически истребляют рябчиков всех
возрастов. Охотники-любители нередко уничтожают рябчиков в ра-
диусе 15 км от села. Если роль рябчика как объекта охотничьего
промысла в Средней Сибири не слишком велика, то в жизни про-
мысловиков эта птица имеет безусловное значение как одна из
лучших приманок для ловли соболя. Следует организовать более
действенную охрану вида.
Обыкновенный перепел — Coturnix coturnix L.
В пределах южной тайги Средней Сибири встречается лишь
обыкновенный перепел С. с. coturnix L. Это типичный вид лесо-
лужья, обитающий на сухих гарях придолинных сосняков (нередко
остепненных). На водоразделах птица нигде не встречается.
Перепел — очень редкая птица на верхней Лене, изредка
встречается на средней Ангаре и не найдена нами на нижней Ангаре
(р. Чадобец). Однако Е. Е. Сыроечковский (1960) отмечает при-
сутствие птицы в долине Подкаменной Тунгуски и даже на Ниж-
ней Тунгуске вблизи Туры. В общем, хотя перепел и не связан
непосредственно с южной тайгой, границы его ареала близко
совпадают с северными пределами южнотаежных лесов.
Любопытно то, что хотя на р. Чадобец мы перепелов не добыли,
бой их слышали неоднократно. Крик несся из заболоченных низин
с кустарниками. Оказывается, бою перепелов очень искусно под-
ражают варакушки.
90
На верхней Лене бой перепела мы слышали в 1961 г. последний
раз 3 августа. По данным Т. Н. Гагиной (19626), перепела изредка
встречаются в полосе среднесибирской лесостепи зимой.
Серая куропатка — Perdix perdix L.
Серую куропатку для «Приангарского комплекса» указывает
Т. Н. Гагина (1961). Мы никаких материалов по этому виду не
имеем.
Бородатая куропатка — Perdix daurica Pall.
Н. Н. Карташев (1952) и Т. Н. Гагина (19586, 1961) указывают
бородатую куропатку для юга Иркутской области и долины
Ангары. Как и предыдущий вид, бородатая куропатка не связана
с тайгой и принадлежит к лесостепному и лесолуговому ком-
плексам. Новых данных о бородатой куропатке у нас нет.
Отряд ГОЛУБИ — COLUMBIFORMES
Сизый голубь — Columba livia Gm.
Т. Н. Гагина (1961) упоминает эту птицу как изредка гнездя-
щуюся в бассейне р. Ангары.
Скалистый голубь — Columba rupestris Pall.
Полуодомашненные скалистые голуби обычны на Лене и Ангаре
[в том числе значительно севернее и северо-западнее, чем это пока-
зано Р. Н. Мекленбурцевым (1951) на приложенной к очерку вида
картосхеме]. В полосе, прилегающей к предгорьям Саян (но не
в горах), скалистый голубь живет и вне поселений человека.
Однако с переходом к лесостепи, расположенной к северу, это уже
синантропный вид.
Заселение южной тайги человеком привело к широкому рас-
пространению скалистых голубей. Очевидно, вид будет продол-
жать расселяться за пределы нынешнего ареала.
Полуодомашненные скалистые голуби выводят птенцов не-
сколько раз за лето без четкой периодичности. На верхней Лене,
по наблюдениям Г. А. Воронова (личное сообщение), 19 мая
1962 г. птенцы были уже крупными (103 г). В предыдущем году
вылет птенцов первой кладки отмечен 9 июня. Интересно, что
в это время уже была полная вторая кладка, тем не менее роди-
тели продолжали кормить птенцов. Третья кладка у этой пары
птиц появилась в августе, 15 августа вывелись птенцы, 4 сентября
они вылетели из гнезда, следовательно, период выкармливания
в гнезде длился 20 дней. У другой пары 30 августа еще были
в гнезде яйца.
91
Большая горлица — Streptopelia orientalis Lath.
Большая горлица весьма характерна для южнотаежных лесов
Средней Сибири. По Лене она распространена далеко к северу
и гнездится под 63°30' с. ш. (Иванов, 1929). Нередка большая
горлица и на Ангаро-Подкаменнотунгусском междуречье, так что
Р. Н. Мекленбурцев (1951) провел границу ареала этого вида
на Ангаре неверно.
Кружево ареала большой горлицы очертить довольно сложно—
птица гнездится в разнообразных местах. Однако в гнездовой
период ее чаще всего можно встретить в приречных и приручьевых
ельниках, в прилегающих к ним сосняках и по опушкам лесов.
Как и в Саянах, горлица живет в кедрачах вблизи полян, обра-
зовавшихся вокруг охотничьих зимовий и особенно орехопромыс-
ловых балаганов. Лишь в обширных глухих водораздельных
темнохвойных лесах с малой долей участия кедра в древостое
горлинки отсутствуют. Видимо, к таким местам, а не вообще к тем-
нохвойной тайге, относится указание А. II. Шведова (1962) об
отсутствии горлинки в темнохвойных лесах среднего Приангарья.
Человек создает благоприятные условия для проникновения
горлиц в глубь южной тайги. Вне лесополья птица встречается
намного реже. В связи с этим следует предположить, что числен-
ность горлиц с нарушением лесов нарастала. Этот процесс должен
был сильнее всего выразиться при смене темнохвойных лесов
светлохвойными. Очевидно, до значительных нарушений тайги
горлицы обитали главным образом вдоль окраин степных участков
Ангаро-Ленского междуречья и верхней Ангары. С развитием
скотоводства по долинам рек, а затем и земледелия, горлицы стали
проникать к северу. Можно предполагать, что, как и у многих
других видов подобного происхождения, северная граница ареала
горлиц стабилизируется в Средней Сибири вблизи 62° с. ш.
Осенью горлица покидает приречные и приручьевые ельники,
сосредоточиваясь по окраинам полей и в местах орехового кедро-
вого промысла. Птицы делаются очень обычными по краям полей,
нередко держатся тут стаями в 5—7 экз.
Первые горлицы весной прилетают рано. На верхней Лене
в 1960 г. они появились 19 апреля (Реймерс, 1963г), в 1961 г.
птицы отмечены с 24 апреля. Прилетая рано, они, возможно,
даже откочевывают на юг во время резких похолоданий. Массо-
вый прилет горлиц начинается позже: на верхней Лене они появи-
лись в массе в 1960 г. 18 мая, в 1961 г. 9 мая; на побережье Бай-
кала в 1961 г. птицы стали очень заметными 22 мая; на р. Чадобец
в 1962 г. горлицы начали попадаться как обычные пернатые
с 25 мая. По наблюдениям Л. И. Малышева (1963), под Иркутском
горлицы появляются между 30 апреля и 12 мая (данные пяти лет).
Также постепенно покидают горлицы полосу южной тайги.
Основная масса птиц улетает в конце августа. Однако по окраинам
92
полей их можно встретить еще в сентябре. Так, на верхней Лене
у р. Чекан 14 сентября на опушке леса у скошенного пшеничного
поля была встречена стайка горлинок из 4 экз. В горах южной
Сибири горлинок мы отмечали еще позже — 27 сентября 1954 г.
(Реймерс, 1959а). Очевидно, птицы окончательно исчезают из
пределов района к октябрю. Это подтверждается данными Л. И. Ма-
лышева (1963), который встречал горлиц до конца сентября
и запоздалую особь отметил 15 октября 1954 г.
Численность горлинок в общем невысока: на маршруте 10 км
в смешанном сосновом лесу встречено 4 воркующих экземпляра
(24 мая, 5—9 час. утра). Осенью на опушках тех же лесов, выхо-
дящих к полям, можно встретить до 20—25 птиц на 5 км.
27 мая 1961 г. на верхней Лене было найдено гнездо горлинок
с 1 яйцом. Гнездо помещалось на кусте ивы среди опушки приреч-
ного ельника, выходящей к сосновому бору, на склоне. Гнездо
было столь рыхло сложено, что яйцо видно с любой точки, в том
числе снизу и сбоку наискось. 11 июня горлинка продолжала
насиживание (дату начала насиживания и появления второго
яйца мы не установили). Размер яиц 36.5 X 26.0 и 35.4 X 25.6.
24 июня птенцы были уже покрыты перьями, лишь на брюхе еще
оставался пух.
Начиная с 1—10 июля встречаются молодые, самостоятельно
живущие птицы (в 1961 г. — верхняя Лена, в 1962 г. — нижняя
Ангара). Вес их достигает к этому времени 150—200 г (взрослые
особи весят 230—320 г). В период размножения у горлиц сильно
развиты зобные железы. У молодой самки, добытой 4 июля, вместе
с другими кормами в зобе и желудке найдено «молочко».
Ассортимент кормов горлицы весьма ограничен (просмотрено
34 экз.): это семена трав и особенно культурных злаков, кедровые
орехи, ягоды (чаще всего черники), сухопутные моллюски (весьма
характерный корм птицы) и дождевые черви. В зобах найдено до
40 зерен овса, 45 орехов кедра, 36 улиток, 3 дождевых червя.
Весной и осенью в питании преобладают культурные злаки, в кед-
рачах — орехи, среди которых много проросших. Кормятся
горлицы и на полях подсолнуха.
Горлицы оказались свободными от иксодид. Пухоеды и гама-
зовые клещи — обычные их паразиты.
Отряд ПАСТУШКИ — RALLIFORMES
Лысуха — Fulica atra L.
Редкая в пределах южной тайги птица нами не встречена.
Пастушок — Rallus aquaticus L.
Вид указан Т. Н. Гагиной (19586, 1961) для долины Ангары.
93
Коростель — Crex crex L.
По E. П. Спангенбергу (19516), ареал коростеля у восточных
пределов распространения вида охватывает верхнюю Лену и Ниж-
нюю Тунгуску до ее среднего течения. С 1959 по 1961 г. включи-
тельно нам не приходилось встречать этих птиц в пределах южной
тайги Средней Сибири. В прежние годы (1950—1958) птица была
редка, и все точки ее регистрации относились к районам лесо-
степья. И вдруг в 1962 г. коростель стал самой обычной птицей
верхней Лены и Ангары. На всех мокрых лугах были слышны
голоса птиц. Была ли это пульсация границ ареала или значитель-
ное колебание числа птиц внутри него, точно установить не уда-
лось, так как мы не имеем новых сведений с Нижней Тунгуски
и средней Лены.
Погоныш — Porzana porzana L.
В связи со сказанным о предыдущем виде интересно указание
А. К. Федосенко (1958) о находке погоныша на Подкаменной
Тунгуске у Байкита. Ранее птица была отмечена лишь в окрестно-
стях Красноярска (Тугаринов и Бутурлин, 1911).
Погоныш-крошка — Porzana pusilia Pall.
Для района южной тайги указан Е. П. Спангенбергом (19516)
и Т. Н. Гагиной (1961).
Отряд ЖУРАВЛИ — GRUIFORMES
Серый журавль — Grus grus L.
Журавли связаны отнюдь не с тайгой, но в Средней Сибири
северная граница распространения серого журавля совпадает
с самыми северными форпостами тайги южного типа. В пре-
делах собственно южной тайги журавли — обитатели заболочен-
ных лугов и долинных болот, севернее они встречаются на водо-
разделах, где имеются болота, которых в южной тайге мало,
поэтому журавль тут не такая уж обычная птица. В связи с этим
данные о пролете журавлей, полученные корреспондентами-
фенологами и нами, довольно фрагментарны и сбивчивы. Наблю-
датели указывают на начало пролета с конца марта (31 марта
1960 г., Черемховский р-н, с. Голуметь, А. А. Давыдов), с начала
и середины апреля (большинство других указаний). Массовый
пролет журавлей происходит в ранние весны в двадцатых числах
апреля. Поздними веснами птицы летят большими стаями лишь
в мае. На Лене пролет проходит в более поздние сроки, чем в запад-
нее расположенных местностях, обычно не раньше середины мая.
94
Разница между сроками пролета журавлей на Ангаре и Лене очень
значительна (первые птицы: 1961 г. — Ангара 5—30 апреля,
Лена 11—26 мая; 1962 г. — Ангара 20—28 апреля, Лена 18—
22 мая). С другой стороны, журавли прилетают в Якутск раньше
(в среднем около 1—2 мая, Скалой, 1956), чем на верхнюю Лену,
Причина такого явления нам не ясна.
В Средней Сибири человек преследует серых журавлей лишь
вблизи населенных пунктов. Промысловики их не добывают.
Нужно думать, что общая численность этих птиц и характер их
распределения в пределах южной тайги Средней Сибири мало изме-
нились в историческое время.
Отряд КУЛИКИ — CHARADRIIFORMES
Очень немногие кулики связаны с лесом. В связи с этим они
не достигают большого разнообразия в пределах южной тайги
Средней Сибири. Здесь кулики —• характерные обитатели речного
бечевника, берегов озер, заливных лугов и окраин болот. Эти
биотопы занимают немного места среди обширной тайги. Однако
в западной половине и особенно на северо-западе Прибайкаль-
ского района — в низовьях Ангары и по ее притокам — проле-
гают торные пролетные пути многих видов куликов. На верхней
Лене сезонные миграции куликов почти незаметны, зато на р. Чадо-
бец в низовьях Ангары огромные стаи птиц буквально заполняют
воздух над заливными лугами, речными старицами и даже полями.
Золотистая ржанка — Charadrius apicarius L.
С 19—20 мая на р. Чадобец начался пролет ржанок. 22 мая
из большой стаи золотистых ржанок, присевшей на залитый водой
болотистый луг у околицы деревни, была добыта самка. Это уже
не первый случай добычи золотистой ржанки далеко на востоке:
Е. В. Козлова (1961) указывает на наличие в коллекции ЗИН
АН СССР экземпляра от 25 мая из Байкита (Подкаменная Тун-
гуска). Вес добытой нами птицы 205.0 г. Эктопаразиты не обна-
ружены.
Бурокрылая ржанка — Charadrius dominicus Mull.
Т. Н. Гагина (1961) отмечает этот вид как пролетный для
Ангары. Нами он не добыт.
Хрустан — Charadrius morinellus L.
Хрустан также отмечен Т. Н. Гагиной (1961) для бассейна
Ангары как пролетный вид.
95
Малый зуек — Charadrius dubius Scop.
Обычный вид галечных отмелей на берегах рек. Первая добыча
на р. Чадобец 13 мая 1962 г., на верхней Лене (р. Тутура) 17 мая
1961 г. Очевидно, эти даты близки к истинному началу весеннего
пролета птиц. На Лене летом малый зуек сравнительно редкая
птица: 29 мая 1961 г. Г. А. Воронов на 6 км русла р. Тутуры
встретил 3 экз. На Ангаре он обычен (Гагина, 1961).
Чибис — Vanellus vanellus L.
А. Я. Тугаринов и С. А. Бутурлин (1911) указывали на воз-
можность пребывания чибиса на Енисее под 64° с. ш. М. И. Тка-
ченко (1937) отметил эту птицу как редкую для верховий Нижней
Тунгуски. Однако Н. А. Гладков (1951а) и Е. В. Козлова (1961)
проводят северную границу распространения чибиса на Енисее
по 58° с. ш. Действительно, севернее этой линии чибис очень редок.
На верхней Лене мы его не встретили (хотя птица обычна на юге
Ангаро-Ленского междуречья). На р. Чадобец чибис был отмечен
весной 1962 г. 13 мая. В районе дер. Яркино птицы держались
все лето. Одна из них 29 августа была добыта. Можно предполо-
жить, что чибис гнездится на р. Чадобец.
Камнешарка — Arenaria interpres L.
Вид указан как пролетный для бассейна Ангары (Гагина,
1961).
Краснозобик — Calidris testacea Pall.
Для Ангары и Лены краснозобик упоминается как пролетный
вид (Гагина, 1961).
Чернозобик — Calidris alpina L.
Пролетный вид Ангары и Лены (Гагина, 1961).
Кулик-воробей — Calidris minuta Leisl.
Т. Н. Гагина (1961) указывает этот вид как пролетный для
Ангары.
Песочиик-красношейка — Calidris ruficollis Pall.
Как и предыдущий, этот вид Т. Н. Гагина (1961) относит к числу
пролетных видов Ангары.
96
Длиннопалый песочник — Calidris subminuta Midd.
Ареал длиннопалого песочника охватывает большую часть
района южной тайги Средней Сибири (Гладков, 1951а), однако
границы его неясны. Т. Н. Гагина (1961) указывает этот вид для
Ангары лишь как пролетный. Мы длиннопалых песочников не
встречали.
Белохвостый песочник — Calidris temminckii Leisl.
С конца мая происходит интенсивный пролет белохвостых
песочников вдоль р. Чадобец. Самая ранняя добыча — 28 мая
1962 г. Не исключена вероятность, что начало пролета вида было
зарегистрировано нами с запозданием на 2—3 дня. По какой-то
случайности все 7 птиц, добытых по 2 июня, оказались самцами.
Размер семенников достигал 8.0 X 6.0 мм. Т. Н. Гагина (1961)
отмечает, что белохвостый песочник был встречен на Ангаре
и летом. К сожалению, летние материалы у нас отсутствуют.
На Лене мы этот вид не нашли. Судя по коллекционным экзем-
плярам географического факультета МГУ, белохвостый песочник
летом обитает на северо-восточном побережье Байкала: Н. Скря-
бин добывал их там в июле и августе.
Острохвостый песочник — Calidris acuminata Horsf.
Указан как очень редкая пролетная птица Ангары (Гагина,
1961).
Турухтан — Philomachus pugnax L.
В 1962 г. турухтаны появились на р. Чадобец в середине мая.
21 мая начался их массовый пролет. С этого дня в течение недели
птицы летели огромными плотными стаями, присаживаясь на мел-
ких болотцах, поросших зеленью. Размер семенников экземпляра,
добытого 27 мая, был равен 20.0 X 11.0 мм. Птицы держатся
в окрестностях дер. Яркино ежегодно (по рассказам местных
жителей) и в течение всего лета. Весьма вероятно, что турухтаны
гнездятся на р. Чадобец. В литературе так много указаний на изо-
лированные места гнездования этой птицы, что невольно напра-
шивается мысль о более южном расположении границы ареала,
чем указано в монографии «Птицы Советского Союза».
Песчанка — Croce thia alba Pall.
Вид указан как пролетный для Ангары (Гагина, 1961).
Грязовик — Limicola falcinellus Pont.
Очень редкая пролетная птица Ангары (Гагина, 1961).
7 Н. Ф. Реймерс Р7
Щеголь — Tringa erythropus Pall.
Пролетная птица Ангары и Лены (Гагина, 1961).
Большой улит — Tringa nebularia Gunn.
По словам А. П. Шведова (1962), большой улит встречался
ему несколько раз на таежных речках. Нами этот вид был найден
на оз. Мильчиково (рис. 23), расположенном среди Око-Ангар-
ского междуречья, 6 августа 1959 г. Птицы держались парой,
однако поздняя встреча не дает оснований для предположения
о гнездовании. На старице р. Чадобец также в августе (22-го) была
добыта пара больших улитов — самец и самка. Гонады самки
развиты слабо. Семенники были размером 3.5 X 2.0 мм.
Южная граница ареала большого улита доходит до южной
части Прибайкалья, не захватывая ее. Обычно виды с таким рас-
положением ареала распространены до северных пределов лесной
зоны. Исходя из этого, следует предположить, что большой улит
будет встречен севернее ныне известной границы распространения
не только на Енисее (Сыроечковский, 1960), но и в других местах.
Черныш — Tringa ochropus L.
Чернышей нам удалось добыть лишь на верхней Лене и средней
Ангаре, где на лугах и в примыкающих к ним приречных зарослях
птица весьма обычна. Встречаются черныши и на луговых бе-
регах водораздельных озер (оз. Мильчиково, Око-Ангарское
междуречье).
С ходом антропогенных нарушений придолинных лесов
численность чернышей возрастает.
Самая ранняя добыча черныша относится к 15 мая 1961 г.,
а первая регистрация в том же году И мая (верхняя Лена).
На пролете птицы держатся берегов реки — линии бечевника.
Позже они здесь исчезают (в 1961 г. на р. Тутуре с 23 мая) и
гнездятся на заливных лугах, прорезанных мелководными реч-
ными рукавами и открытыми старицами. 28 июля в стайках птиц
молодых уже не удавалось отличить от взрослых куликов.
В желудке 1 экз. найдены остатки насекомых и несколько
улиток (23 июля 1960 г.).
Фифи — Tringa glareola L.
В противоположность чернышу фифи не найден на верхней
Лене, но очень обычен на нижней Ангаре. Стации, занятые видами,
идентичны. Первая добыча фифи на р. Чадобец в 1962 г. относится
к 22 мая, а первая встреча птицы — к 12 мая. Самый крупный
фолликул у самки, добытой 28 мая, имел диаметр 5 мм. Птицы,
отстрелянные в июне и августе, имели мелкие фолликулы.
98
Рис. 23. Озеро Мильчиково на Око-Ангарском междуречье.
Пепельный улит — Tonga incana Gmel.
T. H. Гагина (1961) указывает эту птицу как изредка гнездя-
щийся вид Ангарского бассейна. Достоверных сведений о пепель-
ном улите у нас нет. Возможно, к этому виду принадлежат три
самки, добытые 4 июня работавшими с нами студентами. Экзем-
пляров мы не видели (их не сохранили), поэтому наше предполо-
жение строится лишь на определении студентами размеров и веса
птиц.
Перевозчик — Tringa hypoleucos L.
Перевозчик — весьма обычная птица берегов рек Средней
Сибири. На верхней Лене (р. Тутура) самая ранняя добыча
птицы — 17 мая 1961 г., на р. Чадобец — 27 мая 1962 г. Послед-
няя дата близка к истинному сроку прилета перевозчиков, которые
в массе появились на р. Чадобец дня на 3 раньше их первой
добычи. До этого срока птицы были очень редки. День появления
первых птиц мы упустили. На Лене в 1961 г. первые перевоз-
чики появились 7 мая. Ко времени добычи первой птицы они уже
были в массе.
27 мая отстреляна самка с фолликулом, достигающим 10 мм
в диаметре. Фолликулы самки, добытой 17 мая на верхней Лене,
были всего 4 мм.
Эктопаразитов на перевозчиках не обнаружено (очесано 7 экз.).
19 июля 1961 г. мы проплыли на байдарке по р. Тутуре от
с. Чекан до с. Якимовки, учитывая птиц. На расстоянии в 30—
35 км (27 км по проселочной дороге, идущей значительно прямее
течения реки) было учтено 93 экз. перевозчиков. Это число следует
увеличить раза в полтора, поскольку не везде мы были в состоянии
отметить птиц на обоих берегах реки и совершенно упускали ту
часть особей, которая находилась в протоках. Таким образом,
на 1 км берегов р. Тутуры приходится 4 кулика-перевозчика.
Мородунка — Terekia cinerea Guld.
Единственная мородунка была добыта нами 1 июня 1962 г.
на р. Чадобец.
Круглоносый плавунчик — Phalaropus lobatus L.
Птица изредка бывает на пролете в бассейне Ангары (Гагина,
1961).
Большой веретенник — Limosa limosa L.
Большой веретенник не найден нами в пределах южной тайги
Средней Сибири. Т. Н. Гагина (1961) отмечает залет этой птицы
на Ангару.
100
Большой кроншнеп — Numenius arquata L.
По T. Н. Гагиной (1961), большой кроншнеп — птица, обычно
гнездящаяся на верхней Лене и Ангаре, Однако ни на р. Чадобец,
ни на р. Тутуре мы этот вид не нашли. Не встретился большой
кроншнеп нам и при более поверхностных маршрутных исследо-
ваниях на средней Ангаре и в других местах. Под Иркутском мы
наблюдали больших кроншнепов, но неизвестно, гнездятся ли
они там.
Средний кроншнеп — Numenius phaeopus L.
Ареал среднего кроншнепа весьма своеобразен. Н. А. Гладков
(1951а), составлявший картосхему распространения этой птицы
с применением принятого в монографии «Птицы Советского Союза»
метода широкой экстраполяции данных, изображает область
распространения вида с широким разрывом в области Средней
Сибири от Лены до Оби. Е. В. Козлова (1962), очень осторожно
пользовавшаяся экстраполяцией данных, в Сибири отмечает
лишь отдельные пятна распространения птицы, причем новейшие
исследования (Сыроечковский и Рогачева, 1958, и др.) показали
наличие вида на Енисее. При применении методики составления
картосхем, принятой в издании «Птицы Советского Союза», нахо-
док средних кроншнепов на Енисее достаточно, чтобы соединить
восточный и западный отрезки ареала птицы в единый, обнима-
ющий во всяком случае полосу северной тайги Сибири. Е. Е. Сыро-
ечковский и Э. В. Рогачева (1958) предполагают, что в настоящее
время происходит смыкание двух разобщенных областей обитания
вида. Вероятность такого предположения не исключена.
В южной части Средней Сибири средний кроншнеп, видимо,
не гнездится или гнездится редко, хотя, по указанию Т. Н. Гагиной
(1961), постоянно встречается летом. Зато на пролете это обыч-
нейшая птица Ангары, особенно нижней.
Первая особь была добыта на р. Чадобец 25 мая 1962 г. В сле-
дующие дни средние кроншнепы стали очень обычны, можно было
бы сказать, даже многочисленны, если бы птицы не держались
обособленно, одиночными экземплярами. Больше всего их было
на полях, засеянных пшеницей, реже они встречались на лугах
вдоль реки и у уреза воды. Самец, добытый 25 мая, имел семен-
ники 8.0 X 9.0 мм. Фолликулы самки (27 мая) были мелки. На
р. Лене среднего кроншнепа мы не отметили, но севернее у с. Ви-
тим (б. Киренского уезда) 21 мая 1901 г. был добыт экземпляр
этой птицы (Иоганзен, 1916/1917).
Трудно предположить, чтобы такая масса птиц с Ангары летела
в крошечные островки обитания на Енисее или Нижней Тунгу-
ске. Более вероятно, что птицы в подходящих местах гнездятся
всюду по лесной и лесотундровой Сибири.
101
Кроншнеп-малютка — Numenius borealis Forst.
Вид отмечен как пролетный для верхней Лены (Гагина, 1961).
Вальдшнеп — Scolopax rusticola L.
Вальдшнеп — один из лесных куликов. В свете новых мате-
риалов (Сыроечковский и Рогачева, 1958; Федосенко, 1958;
Сыроечковский, 1960) можно предполагать, что вальдшнепы насе-
ляют всю полосу тайги, а не только ее южную половину. Однако
характер' распространения и особенно размещения (кружево
ареала) вида резко меняется от юга к северу. В предгорьях Саян
и в южной тайге Средней Сибири вальдшнепы обычны как среди
темнохвойной, так и светлохвойной тайги. Тем не менее необхо-
димость для них густого леса весьма относительна. Слишком
густых многоярусных и темных лесов птица избегает. Зато она
обычна у опушек и в лесах с большой примесью березы. На Лено-
Ангарском междуречье вальдшнепы очень многочисленны в сосно-
во-лиственных лесах с большой долей участия березы, но не всюду,
а лишь в водораздельной полосе. Вообще в южной тайге Средней
Сибири вальдшнеп — птица водоразделов, лишь изредка встре-
чающаяся вблизи крупных рек. Летом после периода токовых
полетов вальдшнепов днем найти в лесу нелегко. Зато ночью птицы
часто вылетают из-под колес автомобилей, идущих по глухим
лесным дорогам.
Совсем иначе размещены вальдшнепы в полосе лесостепи и лесо-
полья на юге, а также на северном пределе распространения вида.
В лесостепи птицы живут на окраинах лесных участков вблизи
слегка заболоченных лугов, нередко поросших мелким березня-
ком. На севере вальдшнепы также птицы лесополья, весьма ред-
кие на водоразделах и живущие в светлых лесах на старых гарях
и среди редколесий (но отнюдь не в водораздельных глухих лесах).
Видимо, именно резкое нарушение тайги людьми позволило
вальдшнепам жить так далеко к северу. Вообще же птица тяготеет
к разреженным лесам южного типа.
Под Иркутском вальдшнепы весной появляются между 2 и 9
мая (Малышев, 1963). В начале—середине мая отмечают начало
токовых полетов птиц все наблюдатели фенологической сети (Рей-
мерс, 19636, 1963в). Материалы, полученные с помощью коррес-
пондентов, показывают, что вальдшнепы на южную Ангару,
и вообще в полосу сосново-мелколиственной лесостепи, прилетают
на 10—15 дней раньше, чем на Лену. На р. Чадобец в 1962 г.
вальдшнепы отмечены лишь с 26 мая, т. е. несколько позже, чем
на Лене, куда птицы обычно прилетают к 24 мая. Таким образом,
одна изолиния одновременного прилета птиц проходит вдоль
лесостепи (9—10 мая), вторая с интервалом в 10—15 дней — по
верхней Лене и нижней Ангаре.
102
Осенью вальдшнепы встречаются еще в августе и даже (под
Иркутском) в сентябре. Наиболее поздняя находка — 28 сен-
тября 1955 г. (Малышев, 1963).
Бекас — Capella gallinago L.
Обыкновенный бекас редко встречается в полосе южной тайги
Средней Сибири. Поймы рек с мокрыми лугами и травяными боло-
тами — обычное место обитания вида. Сравнительно часто встре-
чается птица лишь среди кочкарников в широких полузаболочен-
ных долинах медленно текущих ручьев. Эти долины когда-то были
расчищены от леса под пастбища и сенокосы. Обыкновенный бекас
в отличие от азиатского избегает ерников и заболоченного мелко-
лесья.
До освоения южной тайги человеком бекасы были, очевидно,
значительно более редкими птицами, чем сейчас. Ныне бекасы
занимают биотопы главным образом антропогенного происхож-
дения. В прежнее время птицы могли находить благоприятные для
жизни места на первичных заливных лугах, где они безусловно
должны были встречать мощную конкуренцию других видов этой
приуроченности.
Наиболее ранняя добыча обыкновенного бекаса на р. Чадобец—
2 июня. Однако токовые полеты птицы начали намного раньше —
16 мая, а первую птицу мы встретили 12 мая. Л. И. Малышев
(1960) встречал бекасов под Иркутском с середины мая (он пола-
гает, что все птицы принадлежали к виду С. gallinago). Всюду
в Средней Сибири токование бекасов начинается около середины
мая, в среднем на верхней Ангаре дней на 10 раньше, чем на Лене
и в Нижнем Приангарье. Так, в 1962 г. на Ангаре токование бека-
сов началось около 6—8 мая. В 1963 г. на южной покати Лено-
Ангарского междуречья бекасы начали токовые полеты 10—12
мая, а на Лене — лишь 22-го числа.
У самца обыкновенного бекаса 2 июня 1962 г. размер тести-
кул 16.2 X 10.4 мм.
Азиатский бекас — Capella stenura Bp.
Этот бекас встречается в полосе южной тайги Средней Сибири
еще реже, чем обыкновенный. Однако Е. В. Козлова (1962) про-
явила излишнюю осторожность, исключив Прибайкальский район
из зоны обитания азиатского бекаса. Распространение птицы на
верхней Лене показывает, что ареал вида сплошной, а не разор-
ванный, как это показано на картосхеме, приводимой Е. В. Коз-
ловой (стр. 248).
Поймы рек с мокрыми лугами и травяными болотами — место
обитания азиатских бекасов. Птицы часто встречаются в кустар-
никах (ерниках) и заболоченном мелколесье. Однако азиатский
ЮЗ
бекас селится по опушкам и не заходит глубоко в леса, чем заметно
отличается от лесного дупеля. На кормежке азиатские бекасы
чаще всего встречаются на слегка заболоченных лугах вблизи
приречных кустарников или в самих кустах, где птицы роются
в иле, скопившемся между стволами во время половодья.
Как и обыкновенный бекас, азиатский ныне тесно связан
с антропогенными биотопами и, очевидно, стал более многочислен-
ной птицей (или во всяком случае шире распространился в полосе
тайги). Ранее птица, видимо, обитала лишь на первичных лугах
у рек и по краям заболоченных пространств.
Наиболее ранняя добыча азиатского бекаса на р. Чадобец
25 мая, токовые полеты начались в середине месяца. Бекас (се-
голеток), весивший 117.8 г (вес взрослой птицы 123—125 г),
был найден 13 сентября 1961 г. на краю поля. Семенники ази-
атского бекаса, добытого 25 мая, 14.0 X 6.0 мм.
Дупель — Capella media Lath.
Восточная граница дупеля проходит по долине Енисея. Однако
указание К. А. Юдина, приводимое Е. В. Козловой (1962), на
нахождение дупелиных токов в Канской лесостепи заставляет
предполагать возможность распространения птицы на этом уча-
стке лесостепи или во всяком случае вероятность пульсации гра-
ницы ареала дупеля в этом районе.
Лесной дупель — Capella megala Swinh.
В 1962 г. опубликовано два исследования этого вида: превос-
ходная монография Е. В. Козловой и очень обстоятельная статья
Р. Л. Наумова. Повторять монографически сведенные данные нет
необходимости. Отметим лишь высокую точность наблюдений
Р. Л. Наумова и сделаем несколько общих замечаний.
Лесные дупели весьма обычны на Ангаро-Подкаменнотунгус-
ском междуречье и безусловно тут гнездятся. В связи с этим сле-
дует думать, что северная граница ареала птицы в Средней Сибири
очерчена Н. А. Гладковым (1951а) вернее, чем Е. В. Козловой
(1962) — лесной дупель не ограничен в распространении долиной
Ангары.
Ареал лесного дупеля, разорванный на два участка, очень
любопытен. Дупель — вид хорошо прогреваемых солнцем мелко-
лиственных и мелколиственно-светлохвойных древостоев,— при
этом едва ли можно сказать лесов, скорее гарей и редкостойников—
своеобразных «лесных степей». 1 Оба участка ареала лесного
1 Мы уже акцентировали внимание (Реймерс и Малышев, 1963) на том,
что редкостойные леса, по данным микроклиматологов (Сапожникова, 1950),
имеют условия теплообмена примерно такие же, как на степных и полевых
участках.
104
дупеля — среднесибирский и дальневосточный — лежат в преде-
лах тайги южного типа. Более того, в Средней Сибири вид
заселяет всего охотнее гари на месте темнохвойной тайги (гари
островных участков темнохвойного леса, осушенные и разрежен-
ные приречные ельники и их производные). В связи с этим наи-
более вероятно предположение о климатогенном характере раз-
рыва ареала. Границы же его должны близко совпадать с областью
распространения темнохвойной тайги, прерываемой достаточно
широкими речными долинами с четко выраженной антропогенной
придолинной полосой. На гарях водораздельных темнохвойных
лесов эти птицы также гнездятся (во всяком случае на верхней
Лене), однако лишь вблизи ручьев и в очень малом количестве.
Нарушение лесов человеком безусловно привело к расшире-
нию ареала вида и к резкому увеличению численности особей.
Звук токового полета лесных дупелей резко отличается от
бекасиного: он грубее (более низких тонов) и громче. Место тока
и характер токового полета очень точно описаны Р. Л. Наумовым
(1962). Действительно, бекасы — болотные птицы, лесные дупели—
сухолюбивые. Однако гнездовья лесных дупелей приурочены
к местам с крайне высокой влажностью воздуха, почти все время
достигающей 85—95%, так что сухолюбив лесного дупеля весьма
относительно.
Найденные нами два гнезда лесного дупеля помещались
довольно открыто, оба были под кустами (Р. Л. Наумов не нашел
гнезд с таким расположением). Диаметр лотка первого из них,
найденного 5 июня 1960 г., 134, глубина 50 мм, размер яиц 43.3 X
X 30.3, 41.1 X 30.4, 41.8 X 30.3 и 39.5 X 30.2, в среднем 41.4 X
X 30.3. Яйца темного варианта. Гнездо помещалось на сухом
склоне под кустиком жимолости.
Второе гнездо (рис. 24) было найдено 18 июня 1961 г. Оно
помещалось на кочке под кустом у небольшого, пересыхающего
летом ручейка. Лоток его имел диаметр 10 и глубину 4.5 см.
Выстилка — сухая осока. Размер яиц: 41.6 X 28.6, 41.0 X 30.5,
42.3 X 30.5 и 39.9 X 29.7, в среднем 41.2 X 29.8. Окраска яиц
также темного варианта.
20 июня 1961 г. был пойман пуховой птенец лесного дупеля.
На верхней Лене в 1961 г. лесные дупели начали токовые
полеты 17 мая; в 1962 г. на р. Чадобец первое токование птиц
отмечено 21 мая; в этом же году на верхней Лене первый экземпляр
добыт 19 мая. В окрестностях Красноярска (Юдин, 1952; Наумов,
1962) токовые полеты начинаются раньше. Токование чаще всего
происходит парами, реже — большими группами или в одиночку.
Пролетные особи, попадающие уже на занятую другими токую-
щими птицами гарь, или токующие соседи, вторгшиеся в чуж-
дые пределы, встречают сопротивление «хозяев». При этом «хозяин»,
продолжая токовать, издает особые трескучие звуки, видимо,
означающие угрозу, и летает над вновь прибывшей парой, иногда
105
следуя вместе с нею до пределов «своего владения», достигающего
1—1.5 км в диаметре. Новая пара обычно улетает, а токование
«хозяина» не прекращается.
Нужно согласиться с Р. Л. Наумовым (1962), что время токо-
вания лесных дупелей более всего зависит от погоды. На верхней
Лене с ее холодными майскими ночами лесные дупели начинают
Рис. 24. Гнездо лесного дупеля.
токовые полеты лишь в 6—7 час. утра. Только в июне птицы токуют
в пасмурные теплые ночи.
При приближении опасности сидящие на земле дупели издают
тихий трещащий звук.
Эктопаразиты, во всяком случае иксодовые клещи, на лесных
дупелях не обнаружены (очесано 6 экз. птиц), хотя птицы охотно
кормятся в наиболее зараженных иксодидами местах — на гарях
островных темнохвойных лесов.
Бекас-отшельник — Capella solitaria Hodgs.
По Т. Н. Гагиной (1961), бекас-отшельник гнездится и зимует
в Приангарье.
Гаршнеп— Lymnocryptes minima Briinn.
Гаршнеп — очень редкая птица Приангарья (Гагина, 1961).
Вполне вероятно предположение Е. В. Козловой (1962) об отсут-
10$
ствии гаршнепа в пределах южной части Средней Сибири, где нет
обширных сфагновых болот и почти не встречаются топкие тра-
вяные болота.
Отряд ЧАЙКИ — LARIFORMES
Даже на сравнительно больших реках типа верхнего течения
Лены и притоков Ангары представители отряда чаек — редкость.
Появления чаек на верхней Лене отмечаются местными жителями
как важное событие, о котором они сообщают в Восточносибир-
скую фенологическую комиссию.
Средний поморник — Stercorarius pomarinus Temm.
Зарегистрирован залет птицы этого вида на Апгару (Гагина,
1961).
Серебристая чайка — Larus argentatus Pont.
20—21 мая 1961 г. вдоль р. Лены наблюдался пролет крупных
чаек. Птицы летели группами от 3 до 7 экз. па очень большой
высоте. За точность определения ручаться не приходится, но
возможно, что это были пролетные серебристые чайки.
Сизая чайка — Larus canus L.
Вид отмечен на Ангаре. Но уже на ее притоках сизая чайка
практически не встречается.
Обыкновенная чайка — Larus ridibundus L.
Т. Н. Гагина (1961) полагает, что этот вид редок на Ангаре
и обычен на Лене. О районе наших исследований этого сказать
нельзя — даже на Лене этот вид мы не встретили.
Речная крачка — Sterna hirundo L.
Считается, что речная крачка гнездится как в бассейне Ангары,
так и Лены (Гагина, 1961). В пределах южной тайги Средней
Сибири нам не пришлось видеть крачек. Очевидно, на водораз-
дельных озерах с заболоченными берегами, расположенных среди
тайги (а только такие озера мы могли наблюдать), крачки не гнез-
дятся.
Отряд ГАГАРЫ — GAVIIFORMES
По Г. П. Дементьеву (1951а), в пределах южной тайги Средней
Сибири могут быть встречены краснозобая и чернозобая гагары
(Gavia stellata Pont, и G. arctica L.). Фактически же эти виды
107
на юге обитают лишь на высокогорных озерах и правильнее отме-
чать разрыв ареала гагар между северной тайгой и высокогорьями
Южной Сибири (за исключением района оз. Байкал).
Отряд ПОГАНКИ — COLYMBIFORMES
Чомга — Colymbus cristatus L.
В главнейших сводках последнего времени (Козлова, 1947;
Дементьев, 19516) очертания ареала чомги на приводимых карто-
схемах почти совпадают. Северная граница его минует Прибай-
калье. Т. Н. Гагина (19586) приводит чомгу как залетный вид
верхней Лены. В следующей своей обзорной работе (1961) она
уже считает большую поганку видом, гнездование которого в бас-
сейне верхней Лены лишь требует дополнительных доказательств.
Мы этого вида на верхней Лене не встречали.
Рогатая поганка — Colymbus auritus L.
По данным Т. Н. Гагиной (1961), гнездится в бассейне верх-
ней Лены и встречается на пролете по Ангаре.
Ушастая поганка — Colymbus caspicus Hab.
Малая поганка — Colymbus ruficollis Pall.
Для обоих видов отмечены залеты в бассейн р. Ангары (Га-
гина, 1961).
Отряд ГУСЕОБРАЗНЫЕ — ANSERIFORMES
Как и большинство водных или околоводных форм, гусеобраз-
ные почти не связаны с тайгой, особенно водораздельной. Кружево
ареалов большинства видов полностью совпадает с мелкомасштаб-
ным изображением гидрологической сети: большинство гусей и
уток живет в полосе южной тайги на сравнительно крупных
реках и озерах.
Лебедь-кликун — Cygnus cygnus L.
Массовый пролет лебедей-кликунов в 1961 г. на верхней Лене
отмечен 15 мая, предыдущей весной — 21 мая. Это единственные
регистрации нами вида. И. С. Поляков (1873) предполагал, что
кликуны не гнездятся на Лене, однако Т. Н. Гагина (1961) ука-
зывает, что это не так. Нельзя исключать возможности гнездования
лебедей, но для большинства мест южной тайги это все-таки про-
летный вид.
108
Тундровый лебедь — Cygnus bewickii Yarrell.
T. H. Гагина (1961) указывает на пролет тундрового лебедя
в бассейне Ангары.
Серый гусь — Anser anser L.
Очерчивая ареал серого гуся, Е. С. Птушенко (1952) включил
в район обитания вида верхние течения Ангары и Лены, однако
в монографии нет указаний на материал или источник, на которые
опирался автор. На Лене и Ангаре в их верховьях большинство
пригодных для гнездования серого гуся участков густо населено
и сравнительно хорошо обследовано. Тем не менее нам неизвестны
случаи доказанного гнездования там серого гуся.
В предгорьях Саян (р. Ия) 22 июня 1954 г. нами встречены
пуховички серого гуся, там он живет по низовьям горных речек.
Белолобый гусь — Anser aibifrons Scop.
На Ангаре изредка наблюдается пролет этого вида (Гагина,
1961). В старых работах указывается на сравнительно частую
встречу белолобого гуся под Иркутском (Taczanowski, 1872—
1874).
Пискулька — Anser erythropus L.
Г. А. Воронов, работавший вместе с нами на верхней Лене,
16 мая 1961 г. зарегистрировал стаю гусей, которых он определил
как пискулек. Т. Н. Гагина (1961) считает, что для Лены, как и
для Ангары, пискульки — очень редкий пролетный вид.
Гуменник — Anser fabalis Lath.
Гуменники весной — наиболее рано летящие гуси. На это
указывают сообщения, сделанные корреспондентами добровольной
фенологической сети (Скалой, 1956). Имеющиеся у нас данные
показывают, что весной гуменники прилетают на территорию
верхнего Приангарья в зависимости от течения весны между
15 и 25 апреля. На верхней Лене гуси появляются позже — между
концом апреля (26-го) и серединой мая. Более поздние указания
вызывают сомнения, поскольку могут относиться к другим видам
гусей. Маловероятны и очень ранние указания корреспондентов,
например 28 марта 1962 г., для окрестностей с. Икей (Б. С. Спи-
цын).
Массовый весенний пролет гусей начинается через 6—10 дней
вслед за появлением первых стай. Общая продолжительность
пролета сильно колеблется, но, так же как и в Якутии (Скалой,
1956), длится около двух недель.
109
Осенний пролет гусей начинается в последних числах сентября
(не раньше начала последней декады) и первых числах октября.
Конец осеннего отлета и его продолжительность не установлены.
Все данные о весеннем пролете приведены нами на основе
сообщений, сделанных многими корреспондентами в течение
1959—1963 гг. Время осеннего отлета указано на основе сообщений
в 1960 и 1962 гг. Часть конкретных дат опубликована (Реймерс,
19636).
Достоверных сведений о гнездовании гуменника в пределах
южной тайги Средней Сибири у нас нет. Т. Н. Гагина (1961)
указывает, что гуменники гнездятся в бассейнах Ангары и Лены.
Для нижней Ангары это вполне вероятно, поскольку там мы встре-
чали пары гуменников в начале июня. Гнездование птиц на верх-
ней Лене требует подтверждения, но отнюдь не исключено, особен-
но в местах внедрения участков северного типа в южнотаежную
полосу (по р. Киренге).
Краем озобая казарка — Rufibranta ruficollis Pall.
Отмечен залет на Ангару (Гагина, 1961).
Пеганка — Tadorna tadorna L.
Т. Н. Гагина (1961) констатирует залет в Приангарье.
Огарь — Tadorna ferruginea Pall.
Залетает в бассейны Ангары и Лены (Гагина, 1961).
Шилохвость — Anas acuta L.
На р. Чадобец шилохвости появились в 1962 г. около 5 мая.
7 мая птицы были уже в заметном количестве. 17 мая отмечена
максимальная интенсивность пролета птиц. К 20 мая пролет стал
менее заметным и в последующие дни почти полностью прекратился.
В пределах южной тайги шилохвости если и гнездятся, то
в небольшом количестве.
Чирок-свистунок — Anas сгесса L.
Чирок-свистунок — одна из самых многочисленных уток в пре-
делах южной тайги Средней Сибири. Он гнездится по ручьям,
речкам и озерам, иногда проводя птенцов через приречные лесные
участки. Л. Тачановский (Taczanowski, 1872—1874) указывал,
что прилетает чирок-свистунок во второй половине апреля и
остается до конца сентября. На р. Чадобец мы зарегистрировали
начало пролета 30 апреля — почти одновременно с появлением
ПО
крякв. Со 2 мая чирки начали лететь массами. Очень интенсивный
пролет отмечен 17—20 мая. Значительная часть особей загнезди-
лась, поэтому конец пролета четко установлен не был. Резкое
ослабление его отмечено после 25 мая.
Вдоль р. Чадобец идет миграция колоссального числа свистун-
ков. Их стаи достигают многих десятков особей. В отличие от
нижней Ангары в Среднем Приангарье свистунки — редкая птица.
Немногочисленны они и на верхней Лене. Эта разница очень су-
щественна и объясняется прежде всего тем, что долины притоков
нижней Ангары очень широки, изобилуют старицами и заболочен-
ными участками.
Чирок-клоктун — Anas formosa Georgi
Этот чирок гнездится лишь на северо-востоке полосы южной
тайги (Исаков, 1952а) и безусловно не характерен для нее. Первые
пролетные особи на р. Чадобец нами были пропущены. В ночь
на 20 мая наблюдался исключительно интенсивный пролет клок-
тун ов. На верхней Лене пролет этих птиц отмечен не был (за 3
года стационарных работ).
Касатка — Anas falcata Georgi
Прилетают касатки поздно. Начало их весенней миграции
на р. Чадобец зарегистрировано только 25 мая. Здесь — это одна
из наиболее поздно прилетающих уток.
Кряква — Anas platyrhynchos L.
Кряквы очень обычны на реках, протекающих через южную
тайгу Средней Сибири. Они прилетают раньше других уток, по-
этому именно о них сообщают корреспонденты-фенологи в разде-
лах «Пролет первых уток крякв» и «Массовый пролет уток крякв».
В 1960 г. кряквы появились в пределах южной тайги между 8
и 24 апреля (Реймерс, 19636). Весной следующего года прилет
первых особей отмечен между 1 и 24 апреля (два сообщения от-
носятся даже к 20 и 30 марта, Реймерс, 1963в). В 1962 г. в связи
с поздней весной кряквы появились между 11 апреля и 15 мая
(на Ангаре 11—25 апреля, на Лене и в нижнем течении Ангары
30 апреля—15 мая). Между 18 апреля и 3 мая прилетели кряквы
в 1963 г. Максимумы интенсивности пролета наблюдаются дней
через 5—10 после его начала.
Значительно меньше данных имеется по датировке начала осен-
него пролета уток. Самое раннее указание на начало пролета
относится к 15, большинство — к 20—25 сентября.
Кряква с шестью уже подросшими утятами встречена 18 июля
1961 г. на р. Тутуре.
111
Желтоносая кряква — Anas poecilorhyncha Forst.
Отмечен залет на Ангару (Гагина, 1961).
Свиязь — Anas penelope L.
На р. Чадобец весенний пролет свиязей начался в 1962 г.
23 мая. 29 мая 1961 г. свиязей учитывал на р. Тутуре Г. А. Воро-
нов. На маршруте в 6 км он встретил всего 1 пару этих уток.
Чирок-трескунок — Anas querquedula L.
Эта утка открытых ландшафтов распространена лишь несколько
шире, чем другие виды, проникшие в полосу южной тайги из степи
и лесостепи. В пределах таежного Приангарья трескунок немного-
числен. На верхней Лене птица обычна. В 1962 г. трескунки на
р. Чадобец появились в середине мая, 17 мая зарегистрирован их
массовый пролет.
Широконоска — Anas clypeata L.
На р. Чадобец в 1962 г. широконоски появились в середине
мая. Массовый пролет их отмечен с 20 мая. К июню пролет резко
ослабел и практически прекратился. На верхней Лене этот вид
нами не найден.
Красноголовый нырок — Aythya ferina L.
Гнездится на Ангаре и Лене (Исаков, 1952а; Гагина, 1961).
Хохлатая чернеть — Aythya fuligula L.
Гнездится повсюду в пределах южной тайги Средней Сибири.
Зимует на Ангаре. Под Киренском взрослый самец был добыт
18 мая 1901 г. (Иоганзен, 1916/1917).
Морская чернеть — Aythya marila L.
Зимует на Ангаре (Гагина, 1961).
Сибирская гага — Somateria stelleri Pall.
Одиночная пролетная самка добыта нами 21 мая 1962 г. на
р. Чадобец. Птица присела на небольшое озерко среди ерников
речной долины.
112
Горбоносый турпан — Melanitta deglandi Bp.
Гнездится в пределах южной тайги Средней Сибири (Исаков,
1952а).
Каменушка — Histrionicus histrionicus L.
Зимует на Ангаре и изредка гнездится на верхней Лене (Га-
гина, 1961).
Морянка — Clangula hyemalis L.
Зимующий на Ангаре и пролетный на верхней Лене вид (Га-
гина, 1961).
Гоголь обыкновенный — Bucephala clangula L.
Гнездится на Лене и зимует на Ангаре (Гагина, 1961).
Луток — Mergus albellus L.
Длинноносый крохаль — Mergus serrator L.
Большой крохаль — Mergus merganser L.
Все три вида крохалей гнездятся в пределах южной тайги
Средней Сибири (Исаков, 1952а; Гагина, 1961), но в местах наших
исследований оказались крайне редкими птицами. Обычны они
на Ангаре в ее верховьях и в дельте Селенги (Бакутин, 1957).
Отряд ФЛАМИНГО — PHOENICOPTERIFORMES
Обыкновенный фламинго — Phoenicopterus ruber L.
Рассказы о залетах фламинго в окрестности Иркутска и на
южный Байкал мы слышали неоднократно. Отмечает фламинго
как залетный вид и Т. Н. Гагина (1961).
Отряд ГОЛЕНАСТЫЕ ПТИЦЫ — CICONHFORMES
Эта группа, как и другие, связанные с водоемами и болотами,
лишь в крайне слабой степени зависит от характера окружающей
сухопутной растительности.
Колпица — Platalea leucorodia L.
На Ангаре случаи залета колпиц — довольно обычное явление.
Один экземпляр птицы был доставлен в Восточносибирский филиал
АН СССР в 1952 г. Имеются и более поздние сообщения о залетах
колпиц на Ангару и южный Байкал.
8 Н. Ф. Реймерс
113
Черный аист — Ciconia nigra L.
В южной тайге Средней Сибири черный аист — редкая птица.
Он отмечен нами лишь на верхней Лене.
Серая цапля — Ardea cinerea L.
Мы не встречали серых цапель в пределах описываемого района.
Т. Н. Гагина (1961) также считает эту птицу очень редкой и встре-
чающейся лишь на верхней Лене. Е. П. Спангенберг (1951а),
отмечая распространение серой цапли на верхней Лене, показы-
вает на картосхеме, приложенной к очерку вида, кроме того,
изолированный участок обитания вида в Центральной Якутии.
Здесь, по данным К. А. Воробьева (1963), серая цапля стала
обычной лишь в последние десятилетия. Однако в Центральной
Якутии участок ареала цапли не изолирован. Как указывает
К. А. Воробьев, птица встречается на Лене, Вилюе, Амге, Алдане,
Мае и Олекме. Вообще лено-якутский язык ареалов очень характе-
рен для многих видов, распространенных подобным образом.
Выпь — Botaurus stellaris L.
Отмечена нами лишь на р. Чадобец. Первый раз весной 1962 г.
ее «уханье» было услышано 28 мая.
Отряд ХИЩНЫЕ ПТИЦЫ — FALCO N1FORMES
Сокол-сапсан — Falco peregrinus Tunst.
Сапсана неоднократно наблюдали на р. Тутуре как автор,
так и работавший с ним Г. А. Воронов. Встречи птиц отмечены
в 1961 г. 31 марта, 18 и 19 июля. На р. Чадобец сокола удалось
видеть лишь во время весенних перемещений птиц.
Гнезда сапсана найдены В. Д. Сониным (1962) в лесостепных
местностях. В июле птенцы были полуоперившимися. В. Д. Со-
нин среди объектов питания сокола обнаружил пухового птенца
утки. Поскольку наблюдения за сапсанами вообще редки, со-
общения В. Д. Сонина (1962) и Л. И. Малышева (1960) заставляют
предполагать, что нападения соколов на плавающую дичь—обыч-
ное явление.
Кречет — Falco gyrfalco L.
По Т. Н. Гагиной (1961), кречеты зимуют и даже, возможно,
изредка гнездятся в пределах бассейна Ангары. Мы этот вид
не встречали зимой ни на Лене, ни в окрестностях Иркутска.
Летние (июньские) находки известны нам лишь по музейным
материалам из района Восточного Саяна. В. Н. Скалой (1934)
114
полагал, что кречет для Ангары — лишь залетная форма. В. В. Ти-
мофеев (1958) считает факт гнездования кречета не установленным
для всей Иркутской области.
Чеглок — Falco subbuteo L.
Этот лесостепной и лесопольный вид сравнительно малочислен
даже в подходящих биотопах. На 35 км по р. Тутуре мы отметили
две пары этих птиц. Вне лесополья чеглоки нам не встретились.
С развитием земледелия число птиц возрастает. До нарушения
тайги человеком чеглоки либо отсутствовали в пределах лесной
полосы, либо спорадически встречались по речным долинам.
В водораздельных лесах, даже изобилующих гарями, видеть
птиц нам не пришлось.
В полосе южной тайги чеглоки появляются в первой половине
мая или в самом конце апреля. В 1961 г. на р. Тутуре чеглок
отмечен 11 мая. Весной следующего года на р. Чадобец птицы
появились 19 мая (возможно, несколько раньше). Из Киренска
известен экземпляр от 14 мая 1901 г. (Иоганзен, 1916/1917). Пре-
делы района чеглоки покидают не раньше сентября. Наиболее
поздние даты добычи под Иркутском — 1 и 2 сентября 1952 г.,
на р. Чадобец — 21 и 24 августа 1962 г. В указанные дни под
Иркутском нами были добыты молодые самец и самка, на р. Чадо-
бец — взрослые особи.
30 мая 1961 г. Г. А. Воронов нашел яйца чеглока в гнезде
сороки на ивовом кусте в 2 м от земли (куст 3 м высотой). Выстилка
гнезда состояла из шерсти мелких грызунов. 5 яиц имели размеры
39.5 X 31.2, 39.5 X 32.2, 39.8 X 32.0, 40.0 X 31.7, 40.5 X 31.5,
в среднем 39.8 X 31.9. Первый птенец вылупился 9 июня.
На р. Чадобец чеглоки нападали на пролетные стаи рогатых
жаворонков.
Дербник — Falco columbarius L.
В южной тайге Средней Сибири дербник — лесопольный вид,
присутствующий только в освоенных человеком речных долинах.
На р. Чадобец дербники в 1962 г. появились 23 апреля и, очевид-
но, сразу загнездились (пара птиц постоянно держалась в одном
районе). Гнездо другой пары было найдено на опушке сосняка
у поля, но, к сожалению, не осмотрено. Птица слетела с него,
когда наблюдатель приблизился на несколько шагов.
На Лене дербник нами встречен не был.
Обыкновенная пустельга — Falco tinnunculus L.
Пустельга в пределах южной тайги Средней Сибири встречается
повсеместно, в том числе и на р. Чадобец, т. е. несколько севернее
отмеченной Г. П. Дементьевым (1951в) границы. Повсюду весной
8*
115
птицы появляются в последней декаде апреля и сразу же при-
ступают к гнездованию. На р. Чадобец пустельга добыта 28 ап-
реля 1962 г. 22 апреля под Иркутском Л. И. Малышев (1963)
отметил начало гнездования обыковенной пустельги, а в середине
мая им найдены полные кладки (13 мая 1957 г. — гнездо с 6
свежими и слабо насиженными яйцами и второе гнездо с 5 свежими
яйцами). Летающие молодые этим же автором встречены 19 июля
1956 г. (там же).
Объекты питания пустельги — водяная полевка, узкочерепная
полевка, лесная мышовка (Сонин, 1962) — хорошо подчеркивают
нетаежный характер этой птицы. Обыкновенная пустельга —
лесостепной вид, никак не связанный с таежными лесами.
Степная пустельга — Falco naumanni Fleisch.
По сведениям Т. Н. Гагиной (1961), отмечен залет этой птицы
на Ангару.
Кобчик — Falco vespertinus L.
Птица встречается всюду, где расчищены большие массивы
лугов и полей: на Ангаре и Лене. Добыт кобчик и на р. Чадобец,
где птицы отмечены с начала июня (прилететь они могли раньше).
13 августа была отстреляна полувзрослая самка, так что на
р. Чадобец кобчик, очевидно, гнездится. Достоверных сведений о
гнездовании кобчика на верхней Лене у нас нет. 21 мая И. С. По-
ляков (1873) подстрелил кобчика у Жигалово. Также в мае (25,
1906 г.) братья Сапожниковы (Иоганзен, 1916/1917) добыли
птицу этого вида у с. Залог б. Верхоленского уезда.
Ястреб-тетеревятник —Accipiter gentilis L.
Тетеревятники гнездятся повсеместно: и в долинах рек, и на
водоразделах. Глухой тайги, не перемежаемой гарями и полями
или каменными россыпями и гольцами, птицы избегают. Любимое
место их гнездования — 100—150 м в глубине опушки леса,
выходящей к большой поляне или любому достаточно значитель-
ному прогалу в тайге. Однако, несмотря на значительное увели-
чение числа подобных мест и общую сильную нарушенность тайги
юга Средней Сибири, тетеревятники не становятся там многочис-
ленным видом. Лишь вдоль рек — притоков нижней Ангары,
изобилующих водяной крысой (что вообще мало характерно для
южной тайги Средней Сибири), тетеревятников относительно
много — до 3 охотящихся птиц на 5 км береговой полосы (р. Чадо-
бец, май). Обычно же тетеревятники не встречаются чаще одной
пары на 10—15 км, а нередко и намного реже — пара на 30 км
и больше. Порода, составляющая лес, для птиц безразлична,
лишь бы были крупные хвойные деревья для устройства гнезда.
116
Весной ястребы медленно перекочевывают вслед за перелетными
стаями птиц. Под Иркутском они появляются обычно в начале
апреля, на верхней Лене и нижней Ангаре — лишь к середине
мая (первые птицы добыты в окрестностях дер. Яркино на р. Ча-
добец 21 мая, на Лене отмечены приблизительно в эти же сроки).
Нападают ястреба как на птиц, так и на грызунов, особенно
водяных крыс, бурундуков и северных пищух. 26 августа 1961 г.
у с. Качуг мы наблюдали тетеревятника, несшего живую гадюку.
Змея извивалась в лапах хищника. Наши непосредственные на-
блюдения подтверждают предположение В. Д. Сонина (1962) о
том, что тетеревятники разоряют гнезда птиц — врановых и
дроздов.
Малый перепелятник — Accipiter virgatus Temm.
Все указания на места находок малого перепелятника в Сред-
ней Сибири, обобщенные Г. П. Дементьевым (1951 в), относились
к горным системам Саян, Алтая и Кузнецкого Алатау. Эти со-
общения пополнились новыми данными Н. Ф. Реймерст (1959а)
и Т. А. Кима (1962). Т. Н. Гагина (19586) на основе просмотра
литературных и музейных материалов включила описываемый
вид в число редких гнездящихся птиц верхней и средней Ангары,
но не упоминает о нем в графе «Долина р. Лены». В следующей
работе этот автор (1961) относит малого перепелятника к редким
гнездящимся птицам Верхнего и Среднего Приангарья и очень
редким пернатым р. Лены.
Малый перепелятник добыт нами дважды: на верхней Лене
9 августа 1961 г. в лиственнично-осиновом лесу (самка, 145 г,
крыло 185 мм, самый крупный фолликул 1.0 мм) и в бассейне
р. Чадобец 3 июня 1962 г. в островном темнохвойном лесу с при-
месью осины и березы (самец, 100 г, крыло 164 мм, тестикулы
7.0 X 4.0 мм). В последнем случае птица была добыта из пары,
явно находившейся вблизи гнезда.
Действительно, малый перепелятник весьма обычен в долинах
горных рек Южной Сибири. Лишь в послегнездовое время птицы
появляются на хребтах (Реймерс, 1959а). Почти все виды речных
долин гор Южной Сибири распространены в пределах зоогеогра-
фического района южной тайги Средней Сибири. Для этого района
стацией малого перепелятника следует считать различные комби-
нации хвойных лесов с мелколиственными, особенно сочетание
темнохвойных пород (прежде всего пихты) с осиной. Пихта и
замещающая ее осина служат показателем определенных климати-
ческих условий и одновременно образователями микроклимата
и других экологических условий, благоприятных для жизни
малого перепелятника и многих других видов южнотаежного
характера.
Следует подчеркнуть, что в ходе антропогенных нарушений
тайги и последующих сукцессионных процессов пихтово-осиновые
117
леса закрепились лишь на низких водоразделах и отсутствуют
вблизи рек. Исследователи, концентрирующие свое внимание
лишь на изучении приречных пространств, неминуемо пропускают
ряд видов, характерных для водоразделов. Этим можно объяснить
незначительное число данных о находке многих видов, в том числе
и малого перепелятника.
Перепелятник — Accipiter nisus L.
Наиболее многочисленный из ястребов вид. Перепелятник, как
и тетеревятник, тяготеет к речным долинам, но в отличие от
своего крупного собрата никогда не встречается на водоразделах.
Наиболее благоприятны для перепелятника районы лесополья,
но не малых лесов и больших полей, а, наоборот, небольших рас-
чищенных участков в светлохвойных массивах или перемежаю-
щихся приречными ельниками лугов. Гнезда помещаются всегда
в густых зарослях сравнительно молодого леса (чаще ельника или
намного «реже — сосняка).
Так же как и тетеревятники, весной перепелятники медленно
перекочевывают на север, а осенью — на юг. На р. Чадобец
в 1962 г. первый перепелятник зарегистрирован 13 мая. Предыду-
щей весной на Лене перепелятник добыт 26 апреля. Осенний про-
лет хищников на Лене был особенно заметен в этом году 1—2 сен-
тября. Массовые перемещения перепелятников совпадают по
времени с интенсивным лётом лесных и пятнистых коньков, на
которых успешно охотится описываемый вид.
Мы уже сообщали (1963г) о находке Г. А. Вороновым
23 июля 1960 г. птенцов-перепелятников в гнезде, помещавшемся
в приречном ельнике (вес птенцов 99, 140 и 145 г).
Помимо мелких птиц, перепелятники охотятся и на четвероно-
гих, в том числе ящериц. 26 апреля 1961 г. в желудке птицы най-
ден хвост живородящей ящерицы.
Перепелятники сильно заражены гельминтами и кровососками.
Иксодовых клещей на них не было обнаружено (осмотрено 4
взрослых птицы и 3 птенца).
Полевой лунь — Circus cyaneus L.
Полевой лунь — наиболее многочисленная птица этого рода
в пределах южной тайги. Однако с лесом лунь практически не
связан. Это типичный обитатель долинных лугов, перемежающихся
ивняками и черемушниками. Именно среди лугов и кустарников,
а не долинного лесолужья (лугов среди приречных ельников),
луни наиболее многочисленны.
В период до нарушения долинных лесов человеком полевые
луни должны были либо отсутствовать в полосе тайги, либо быть
там большой редкостью.
118
Весной полевые луни появляются в середине—последней
декаде апреля (верхняя Лена: в 1959 и 1960 гг. 25 мая, в 1961 г.
18 мая; р. Чадобец 6 мая 1962 г. — возможно, дата первого по-
явления пропущена). Начало осеннего пролета не отмечено, но
к концу сентября—октябрю полевых луней уже встречать не
приходится.
В пищевом тракте самца, добытого на р. Тутуре Г. А. Вороно-
вым 19 мая 1962 г., было найдено 4 экз. узкочерепных полевок,
1 экз. малой бурозубки и 8 живородящих ящериц. Судя по на-
блюдениям за лунями в природе, мышевидные млекопитающие и
рептилии — их основной корм. На юге в полосе лесостепи полевые
луни охотно нападают на молодых длиннохвостых сусликов. От-
мечены случаи преследования степных коньков и других мелких
птичек речных долин. Приблизительно к таким же выводам раньше
нас пришел В. Н. Скалой (1934).
Степной лунь — Circus macrourus Gm.
Известны случаи залета степных луней на Ангару (Гагина,
1961).
Болотный лунь — Circus aeruginosus I.
На Ангаре болотный лунь обычен (Скалой, 1934). На Лене
мы этого вида не нашли. Т. Н. Гагина (1961) считает, что для
Лены факт гнездования болотного луня нуждается в проверке.
Черный коршун — Milvus korschun Gm.
Черный коршун — еще один типичный представитель лесо-
полья речных долин. В отличие от луней для черного коршуна
необходимо наличие леса. Однако не характер леса и близость
реки, а достаточно обширные безлесные пространства определяют
возможность существования вида. В связи с тем, что поля и другие
открытые пространства размещаются вблизи населенных пунктов,
коршун — частичный синантроп. Его гнезда нередко бывают не-
далеко от селений и со своеобразной выстилкой из старых носков,
варежек, шерсти домашних животных и т. д. (рис. 25).
На 35 км вдоль р. Тутуры мы учли 5 экз. коршунов. Истинная
численность этих птиц на этом расстоянии — 8—10 экз., т. е.
около 1 экз. на 5 км. На р. Чадобец коршуны намного малочислен-
нее. Если на каждую деревню на р. Тутуре приходится приблизи-
тельно 2 пары коршунов, то на р. Чадобец только одна пара.
Среднее расстояние между населенными пунктами на этих анало-
гичных по величине речках разнится также приблизительно в 2
раза (больше на р. Чадобец), так что коршунов на притоке Лены
раза в 4 больше, чем на р. Чадобец. На верхней и средней Ангаре
коршуны не менее многочисленны, чем на Лене.
119
По свидетельству Е. Е. Сыроечковского (1960), залетные кор-
шуны по Енисею встречаются до лесотундры. Очевидно, в ареале
коршуна можно выделить несколько полос равной численности:
максимальной, северная граница которой идет вблизи 56° с. ш.,
средней — до северных пределов полосы южной тайги (62° с. ш.),
низкой — до последних форпостов биоценозов южного типа и
случайной — севернее (в пределах северосибирских редколесий).
Рис. 25. Гнездо черного коршуна.
В пределы южной тайги Средней Сибири коршуны прилетают
в середине апреля (верхняя Лена, 1960 г. — 16 апреля, 1961 г. —
13 апреля). Однако на р. Чадобец первая встреча коршуна за-
регистрирована лишь 11 мая. Возможно, мы констатировали птицу
с запозданием.
Орлан-белохвост — Haliaeetus albicilla L.
Редкая птица южной тайги. Один молодой экземпляр был до-
быт нами на р. Чадобец 17 мая 1962 г.
Орлан-долгохвост—Haliaeetus leucoryphus Pall.
В пределах южной тайги птица неизвестна. Она добыта лишь
в окрестностях Иркутска 9 февраля 1895 г. (Скалой, 1934).
120
Бородач — БураёСив barbatus L.
В литературе имеется указание на добычу бородача у северного
предела южной тайги на Лено-Нижнетунгусском междуречье
(Бутурлин, 1928).
Беркут — Aquila сЬгуваё^в L.
В районах Верхнего Приангарья, по В. Н. Скалону (1934),
беркут обычен. Т. Н. Гагина (1961) считает этого хищника гнездя-
щейся птицей верхней Лены.
Орел-могильник — Aquila heliaca Sav.
Для Балаганской степи В. Н. Скалой (1934) считает орла-мо-
гильника обычным видом. Это утверждение повторяет Т. Н. Га-
гина (1961). Как далеко орел-могильник проникает в полосу лесо-
степи и лесополья — нам не ясно.
Степной орел — Aquila тарах ’1ешш.
Этот вид упоминает В. Н. Скалой (1934), считая его редкой
птицей Балаганской степи. Экземпляр, имеющийся в Иркутском
музее, очевидно, залетная птица, что констатировано Т. Н. Гаги-
ной (1961).
Большой подорлик — Aquila clanga Pall.
Наличие большого подорлика как гнездящейся птицы в глу-
бине южной тайги Средней Сибири вызывает сомнение. Скорей
всего все данные о гнездовании вида относятся к степным участкам
(Скалой, 1934; Гагина, 1961) и прилегающим к степи узким поло-
сам лесостепи и лесополья. Одна молодая самка была добыта
нами 7 августа 1959 г. на Око-Ангарском междуречье вблизи
р. Оки.
Орел-карлик — Aquila pennata Gm.
Сведения о добыче этого вида в пределах не горной части Сред-
ней Сибири очень туманны. 1 экз., «по слухам» (Скалой, 1934),
был подстрелен близ Иркутска Б. Дыбовским. Е. Хартерт (Наг-
tert, 1903—1922) полагает, что орел-карлик изредка гнездится
на Лене. Г. П. Дементьев (1951в) указывает на находку орла-кар-
лика у Жигалово, но на картосхеме в пределы ареала вида бас-
сейн р. Лены не включает.
Сарыч — Buteo buteo L.
Сарыч пополняет комплекс лесопольных видов южной тайги
Средней Сибири. Птица встречается здесь нечасто и лишь в окуль-
туренных районах. На р. Чадобец сарыча мы не наблюдали. Он
предпочитает речные долины лишь как лесопольный биотоп.
121
В районе Братска после расчистки водораздельных лесов под
пашни сарычи, как и коршуны, стали обычны на водоразделах.
Такое переселение было постепенным процессом, и его можно
связать с перемещением основных кормовых объектов сарычей —
серых полевок. Вообще долинные виды хищных птиц связаны
обычно с сэрыми полевками (узкочерепными и экономками) и
водяной крысой. Сарычи с охотой поедают этих последних зверь-
ков и нередко концентрируются вдоль рек и болот, населенных
ими. Красные полевки занимают в питании сарычей значительно
более скромное место (Сонин, 1962).
Указанный автор отмечает пролет канюков под Иркутском
в конце апреля 1958 г. В 1959 г. на Ангаро-Ленском водоразделе
пролет сарычей констатирован нами со 2 мая. Весной следующего
года под Жигалово канюки прилетели 25 апреля. В раннюю
весну 1961 г. там же канюки появились около середины апреля.
Мохноногий канюк — Buteo lagopus Brunn.
Мохноногий канюк зимует в Верхнем Приангарье (Скалой,
1934). Т. Н. Гагина (1961) не подтверждает сказанное В. Н. Ска-
лоном. На р. Чадобец зимняки появились во второй половине
мая (отмечены впервые 19 мая). Не исключено, что птицы гнездятся
на северных притоках Ангары (во всяком случае на р. Чадобец они
были многочисленны). На верхней Лене мы не видели этих птиц
даже на пролете.
Мохноногий курганник — Buteo hemilasius Temm. et Schleg.
Отмечен залет этого вида на Ангару (Гагина, 1961).
Степной сарыч — Buteo rufinus Cretzsch.
Т. Н. Гагина (1961) регистрирует залет степного сарыча
в бассейн Ангары.
Хохлатый осоед — Pernis ptilorhynchus Temm.
Хохлатый осоед — южная форма лиственных лесов. Данных
об этом виде так мало, что трудно делать выводы даже об ареале
птицы. Однако следует предположить, что хохлатый осоед шире
распространен на западе, чем указывает Г. П. Дементьев (1951в).
Если не отбрасывать сообщения М. И. Ткаченко (1924) о находке
вида на Нижней Тунгуске, то, очевидно, птица обитает значитель-
но севернее указанной в монографии «Птицы Советского Союза»
границы. Как и все южнолесные птицы («водораздельные»), хохла-
тый осоед может быть встречен до водоразделов Подкаменной
Тунгуски. Во всяком случае тут расположена зона теоретического
предела возможного гнездования вида.
122
26 августа 1959 г. мы добыли осоеда в осиннике на Око-
Ангарском междуречье (между реками Кежмой и Тореем). Ни-
каких данных о гнездовании вида здесь у нас нет, но вероятность
этого нельзя исключать. Дело в том, что обширные осиновые леса
водоразделов Приангарья еще ждут своего исследователя. Образо-
вавшиеся на месте темнохвойных массивов могучие осинники
могут быть гнездовым биотопом осоедов. Наличие этого вида
под Красноярском и на Алтае заставляет предполагать отсутствие
в Приенисейской Сибири каких бы то ни было непреодолимых
рубежей для распространения хохлатого осоеда. Скорее всего
кажущаяся изолированность красноярской и алтайской находок
объясняется неполнотой данных, а ареалы хохлатого и европей-
ского осоедов смыкаются.
Добытый нами экземпляр (самец?) весил 1250 г, длина его
тела 622, крыла 412, размах крыльев 1330 мм. Во рту и в желудке
обнаружена масса личинок ос (несколько десятков экземпляров).
Скопа — Pandion haliaetus L.
Весьма обычная на саянских речках, скопа в южной тайге
Средней Сибири редка. На р. Чадобец мы ее не видели. Она живет
на верхней Лене, верхней и средней Ангаре. Составить предста-
вление о численности скопы можно из того, что на 40 км течения
р. Тутуры и ее притока р. Чекан жила 1 пара птиц. Малое число
птиц можно связать с истощением рыбных запасов рек южной
тайги.
Отряд СОВЫ — STRIGIFORMES
Полярная сова — Nyctea scandiaca L.
Полярная сова, по В. Н. Скалону (1934), обычна в Верхнем
Приангарье. Однако нам не удалось ее увидеть под Иркутском,
несмотря на десятки зимних экскурсий. Не встречали мы сову
и в других местах. Местные жители не знают полярной совы и не
отвечают на вопросы о времени ее появления. Очевидно, полярные
совы не регулярно прилетают на зимовку даже в лесостепные рай-
оны юга Средней Сибири и безусловно отсутствуют в таежных
ее частях там, где слабо выражено лесополье и нет обширных бо-
лотистых открытых пространств, характерных для более северной
полосы.
Филин — Bubo bubo L.
Наблюдения над этим видом у нас отсутствуют. По литератур-
ным данным, филин населяет всю полосу южной тайги Средней
Сибири.
123
Обыкновенная сплюшка — Otus scops L.
Лесостепной вид, отсутствующий в собственно южной тайге
Средней Сибири. Однако на берегах Байкала сплюшка — не
редкость (Малышев, 1958, 1960).
Ушастая сова — Asio otus L.
В. Д. Сонин (1962) совершенно прав, относя ушастую сову
к обитателям речных долин и разреженных лесов придолинной
полосы. На водоразделах эта сова не живет даже среди горель-
ников. В соответствии с этим в питании ушастой совы доминируют
серые полевки: экономка и узкочерепная (Скалой, 1934; Сонин,
1962). В желудках 2 экз., добытых нами на рр. Чадобец и Тутуре,
были также остатки серых полевок. На первой из упомянутых
сов обнаружен один гамазовый клещ.
Болотная сова — Asio flammeus Pontopp.
Очень многочисленный вид сов на нижней Ангаре и ее прито-
ках. На Лене мы этой птицы не нашли. Болотная сова в южной
тайге еще более, чем ушастая, связана с зарослями ивняков и
других кустарников по долинам рек. Вечером можно наблюдать,
как пары хищников беззвучно кружатся над залитыми половодьем
ерниками и другими кустарниковыми зарослями. В водораздель-
ных лесах болотные совы отсутствуют.
На р. Чадобец совы этого вида появились в массе около 20
апреля (21 апреля 1962 г. добыт первый экземпляр). Наибольший
фолликул у самки, добытой 21 мая. 3 мм.
Мохноногий сыч — Aegolius funereus L.
Отнюдь не многочисленная птица. Нами добыт лишь 1 экз.
(самец) на р. Тутуре 1 апреля 1961 г. Птица присела на крышу
сарая в населенном пункте.
Принадлежность этого вида к хвойным лесам южной тайги
вызывает сомнение. Мы не знаем экземпляров, добытых тут летом.
Лишь весной мохноногий сыч отмечен в районе Красноярска (Кру-
товская, 1958).
Воробьиный сыч — Glaucidium passerinum L.
Эта маленькая сова относится к птицам водораздельных лесов
и, очевидно, гнездится в темнохвойных лесах, особенно перестой-
ных. Обладая агрессивным нравом, она сразу обнаруживает себя.
20 июня 1961 г. ее отметили на водоразделе между Леной и Ту-
турой. 23 июля 1962 г. сова была добыта в бассейне р. Чадобец
124
работавшими с нами студентами (ее вес 79.0 г, крыло 103, размах
крыльев 355, хвост 45 мм, пол определен не был, очевидно самка).
Добыча всего одного экземпляра и единичная регистрация по-
казывают крайнюю редкость вида в южной тайге.
Ястребиная сова — Surnia ulula L.
Мы не встретили этого вида в пределах южной тайги Средней
Сибири. Упоминание о летних экземплярах из южной полосы
имеется лишь у В. Н. Скал она (1934), который добыл ястребиную
сову у с. Янды на Ангаре летом 1929 г. Все остальные из извест-
ных нам регистраций вида относятся к ранней весне (марту)
или осени. Наиболее ранняя добыча — 30 сентября 1900 г. у с.
Залог б. Верхоленского уезда (Иоганзен, 1916/1917).
Бородатая неясыть — Strix nebulosa Forst.
Столь обычная в горных темнохвойных лесах Сибири (Рей-
мерс, 1959а), бородатая неясыть в южной тайге редка. За все время
мы добыли (на р. Тутуре) лишь 1 экз. 24 августа 1961 г. на поле
у леса. Это был взрослый самец.
Длиннохвостая неясыть — Strix uralensis Pall.
В горах Сибири и в полосе лесостепи обычна, в южной тайге
исследованного района редка. На верхней Лене не встречена,
ранее была там добыта братьями Сапожниковыми (Иоганзен,
1916/1917). Отстреляна на р. Чадобец 30 августа 1962 г. работав-
шими с нами студентами. Характер распределения вида в южной
тайге неясен. По свидетельству В. Д. Сонина (1962), для охоты
длиннохвостая неясыть вылетает на открытые места. Многочислен-
ные встречи в желудках длиннохвостой неясыти узкочерепных
полевок подтверждают сказанное этим автором.
Отряд КУКУШКИ — CUCULIFORMES
Обыкновенная кукушка — Cuculus canorus L.
Обыкновенная кукушка живет буквально везде — от приреч-
ных зарослей до водораздельных глухих лесов. Тем не менее —
это прежде всего долинная форма. Тут, в долинах рек и на их
склонах, гнездится огромная масса воробьиных птиц, в гнезда
которых кукушка подбрасывает свои яйца. На водоразделах ку-
кушки обычны по берегам истоков ручьев и на окраинах гарей,
но малочисленны в сплошной тайге. Практически обыкновенные
кукушки не живут в разновозрастных глухих темнохвойных лесах
125
без кустарников. В таких лесах почти нет открытогнездящихся
птиц, к дуплогнездникам кукушки яиц не подбрасывают.
Результаты учетов кукушек очень различны в зависимости
от времени их проведения, поэтому сравнение затруднено. При-
ведем лишь некоторые факты. В водораздельной темнохвойной
тайге с примесью березы в последней декаде июня 1961 г. число
птиц оказалось меньше одной на 5 км учетного маршрута. В светло-
хвойных лесах такого же типа на то же расстояние и в то же время
было 1—2 кукушки (полоса учета кукушек около 50—70 м
в каждую сторону, т. е. 100—150 м). Среди опушек сосновых лесов,
приречных и приручьевых ельников, а также в примыкающих
к ним кустарниках в начале июня 1961 г. численность обыкновен-
ных кукушек достигала 5 птиц на 1 км маршрута, а местами даже
50 экз. на 5 км. В этом году было феноменальное скопление ку-
кушек на верхней Лене. Весной предыдущего лета обилие птиц
было по крайней мере в 2—3 раза ниже.
На р. Чадобец в 1962 г. наибольшее количество кукушек отме-
чено также в приречной полосе, но их число не превышало 2—3
на 1 км. Водоразделы тут населены приблизительно с такой же
плотностью, как и на Лене.
Если по датам первого кукования обыкновенной кукушки
имеются довольно большие материалы, то о днях истинного при-
лета птиц сообщений значительно меньше. Наши наблюдения по-
казывают, что кукушки появляются нередко на 3—5 дней раньше
регистрации первого крика. Однако такое явление отмечено лишь
в очень ранние, но холодные весны. Весной с поздним течением
кукушки кукуют уже на пролете, что вообще более характерно
для вида.
Вдоль линии железной дороги в пределах Средней Сибири
обыкновенные кукушки начинают куковать в среднем около 20
мая (Корчагин, 1936; Буторина и Крутовская, 1958; Крутовская
и Буторина, 1958; Реймерс, 19606; Малышев, 1963). Любопытно,
что И.С. Поляков (1873) считал, что кукушка под Иркутск прилетает
6—9 мая. Средняя дата начала кукования обыкновенной кукушки
в Якутии, т. е. за пределами южной тайги, также отмечена 19
и 21 мая (Скалой, 1956). Такое совпадение заставляет предположить
одновременность начала кукования на всей территории Средней
Сибири. Это не совсем так. Обработка сообщений за 4 года из 16—
18 точек и сравнение средних дат с такими же показателями из
соседних районов (Улан-Удэ, Чита — Корчагин, 1936; Котов,
1957, и др.) показывают, что обыкновенные кукушки на западе
Средней Сибири начинают куковать несколько раньше, чем на
востоке (в пределах лесной, а не лесостепной полосы). Так, в тайге
бассейна р. Чуны обыкновенные кукушки начинают куковать на
неделю раньше, чем в бассейне верхней Лены [на западе часто
повторяются прилеты в первой декаде мая, на востоке — в третьей
декаде этого месяца; средние за 4 года (1960—1963) для Чуны —
126
метеопункт Баер, по сообщению наблюдателя А. И. Павловой, —
12 мая, на верхней Лене у Жигалово — 20 мая]. Однако южнее
р. Чуны птицы начинают куковать позже. Маловероятно, чтобы
их не было на юге во время начала кукования на севере. Скорей
всего в лесостепной полосе птицы чаще молчат в первые дни при-
лета.
В пределах южной тайги обыкновенные кукушки держатся
до конца августа. 22 августа 1961 г. встречены последние птицы
под Качугом. В это время под Жигалово — на 100 км севернее —
кукушек уже практически не было.
Куковать обыкновенные кукушки прекращают в конце июля.
Однако в упомянутый день 22 августа 1961 г. под Качугом ку-
кушка куковала. Это самая поздняя дата кукования, которую
нам и Л. И. Малышеву (1963) удалось зарегистрировать за 12 лет
(1951—1963).
Разгар брачных игр обыкновенных кукушек падает на конец
мая—начало июня. В 1961 г. на верхней Лене особо массовое
спаривание наблюдалось 1 июня.
Из 17 обыкновенных кукушек, обработанных паразитологами,
ни одна не была заражена иксодовыми клещами. Вообще заражен-
ность обыкновенных кукушек эктопаразитами ничтожна.
Глухая кукушка — Cuculus saturatus Bly th.
Принято считать, что глухая кукушка населяет главным обра-
зом высокоствольные участки леса независимо от породы составля-
ющих его деревьев (Судиловская, 1951). Как мы уже отмечали
(Реймерс, 1959а,) это не совсем так. Б. К. Штегман (Stegmann,
1928) наблюдал глухую кукушку даже в степи. Местами в пределах
южной тайги птица обычна в березовых молодняках вблизи на-
селенных пунктов. Однако численность вида всюду низкая. Наи-
высшего значения она достигает по южной и юго-восточной пери-
ферии южной тайги. Тут в невысоких горах, поросших темно-
хвойными лесами, глухих кукушек приблизительно столько же,
сколько и обыкновенных — около 1 птицы на 5 км учетного
маршрута. Под Красноярском численность глухих кукушек
уступает кукушке обыкновенной (Наумов и Бурковская, 1959)
В Приангарье и на верхней Лене глухие кукушки встречаются
во много раз реже обыкновенных.
Экологическая пластичность весьма характерна для кукушек.
В горах юга Сибири глухая кукушка — вид, обитающий в высоко-
ствольных лесах паркового типа, нередко в пределах подгольцо-
вого пояса. Она избегает склонов, но многочисленна в речных
долинах. В равнинных краях и на севере южной тайги Средней
Сибири долины становятся ее основным местом обитания. Для
всех этих районов общей чертой является некоторая прерывистость
127
леса: редкостойность, лесопольность или сочетание леса и кустар-
ников.
Очень трудно определить, как повлияло на популяцию глухих
кукушек нарушение лесов. Омоложение редкостойных темно-
хвойных лесов привело к невозможности существования вида;
в то же время образование прогалин, гарей и участков лесополья
благоприятно сказалось на численности птиц. Можно предполагать,
что в Сибири глухие кукушки распространены в тех же пределах
и с такой же численностью, как и до резкого нарушения лесов
человеком. Обилие птиц должно было измениться при переходе
от сплошных редкостойких лесов к более омоложенным, т. е.
на первом этапе нарушения лесов южной тайги.
Весной глухие кукушки начинают куковать позже обыкновен-
ных в среднем на 3—7 дней, однако данные о сроках их прилета
очень немногочисленны. В 1959 г. на Ангаро-Ленском междуречье
у улуса Байша глухие кукушки прилетели 22 мая (обыкновенные—
18 мая). Весной следующего года наблюдатели-фенологи отмечали
очень большой разрыв в прилете обоих видов кукушек (до 20 дней)
и появление глухих между 24 и 30 мая. В 1961 г. глухие кукушки
начали куковать в южной тайге 20—22 мая (на верхней Лене на
6 дней позже обыкновенных). Кукование глухой кукушки в 1962
и 1963 гг. не отмечено ни одним из корреспондентов. На южной
Ангаре в 1962 г. глухие кукушки появились 29 мая — на 12 дней
позже обыкновенных. У Красноярска средняя дата начала кукова-
ния глухой кукушки — 29 мая (Крутовская, 1958), т. е. на 9—
10 дней позже, чем обыкновенной. Имеющиеся данные позволяют
с некоторой степенью достоверности предположить, что чем запад-
нее пункт, тем позже прилетают глухие кукушки.
Куковать глухие кукушки перестают около середины июля.
Е. А. Крутовская (1958) среднюю дату прекращения кукования
определила как 11 мая, а наиболее поздний крик птицы слышала
24 июля 1942 г. В 1962 г. у Верхоленска мы слышали глухую ку-
кушку 27 июня. Летом 1962 г. это была последняя регистрация.
На два года раньше на Ангаро-Ленском водоразделе у дороги
Жигалово—Балаганск кукушки кричали 15 июля.
В августе глухие кукушки начинают продвигаться на юг.
Дата исчезновения последних птиц осталась невыясненной.
В горах юга Сибири (Реймерс, 1959а) глухие кукушки держатся
до конца августа.
Отряд КОЗОДОИ — CAPRIMULGIFORMES
Обыкновенный козодой — Caprimulgus europaeus L.
Как и все южнобореальные формы Северной Азии, обыкновенный
козодой широко распространен в осветленных сосновых и сосново-
лиственничных лесах южной тайги Средней Сибири и особенно
128
на их гарях, поросших мелколесьем березы. На средней Ангаре
(Шведов, 1962) и верхней Лене (наши материалы) птицы добыты.
На нижней Ангаре этого сделать не удалось, но крик их зарегистри-
рован. Можно предполагать, что козодой будет констатирован
приблизительно до 62° с. ш. — обычного предела распростране-
ния таких форм в Сибири. На верхней Лене козодои присутствуют
(и, очевидно, гнездятся) ежегодно (отмечены с 1959 по 1963 г.).
Странно, что их раньше там не отметили другие авторы.
Самец, добытый из пары 29 мая 1961 г. на р. Тутуре, имел
размер тестикул 16.2 X 8.8 мм. Эктопаразитов на нем не оказа-
лось.
Отряд СИЗОВОРОНКИ — CORACIIFORMES
Обыкновенный зимородок — Alcedo atthis L.
О присутствии обыкновенного зимородка в пределах южной
тайги мы судим лишь по литературным данным (Гагина, 1961;
Шведов, 1962).
Отряд УДОДЫ — UPUPIFORMES
Удод — Upupa epops L.
Удоды практически никак не связаны с лесами и проникают
в тайгу по лесопольным участкам. В пределах среднесибирского
лесостепья они обычны. По лесопольным участкам удод доходит
до верхней Лены (Качуг—Верхоленск) и, возможно, средней
Ангары.
Отряд ДЛИННОКРЫЛЫЕ — MICROPODIFORMES
Черный стриж — Apus apus L.
Черный стриж — форма, уплотняющая кружево ареала в пре-
делах южной тайги Средней Сибири вместе с освоением таежных
пространств человеком. Первоначально птицы были распростра-
нены, видимо, в горах и вдоль лесостепи.
У нас нет полной уверенности, что черные стрижи, как показал
Е. С. Птушенко (1951), отсутствуют в междуречье Ангары и Лены.
Тут мы наблюдали стаи птиц над водоразделами. Черные стрижи
наряду с белопоясными либо залетают на большое расстояние от
возможных мест гнездования (до 30 км), либо гнездятся на водо-
разделах. При отсутствии выходов скал птицы могут гнездиться
в колоссальных лиственницах, возвышающихся над тайгой на
многие метры.
Черные стрижи появляются в последней декаде мая—начале
июня. К сожалению, сообщения добровольных корреспондентов-
9 Н. Ф. Реймерс
129
фенологов не всегда можно использовать, так как «стрижами» и
особенно «стрижками» местные жители называют городских ласто-
чек.
В с. Якимовке на р. Тутуре 4 июня 1961 г. в сильный снегопад
у черных стрижей наблюдались случаи шока от холода. Одна
птица была подобрана нами и после обогрева благополучно уле-
тела.
Белопоясный стриж — Apus pa cificus kLath.
Восточная форма, также перешедшая к синантропному суще-
ствованию, но менее связанная с нарушением лесов, чем черный
стриж. Белопоясные стрижи чаще, чем черные, встречаются над
водоразделами и поднимаются в горы южной Сибири до самых
больших высот. Гнездятся белопоясные стрижи в каменных зда-
ниях, но при большой численности не гнушаются двухэтажных
деревянных строений. В одноэтажных деревянных домах стрижи
никогда не живут.
Сроки весеннего прилета у обоих видов стрижей очень близки —
главным образом начало последней декады мая. В Приангарье
белопоясный стриж появляется на несколько дней раньше, чем
на верхней Лене.
Иглохвостый стриж — Hirundapus caudacutus Lath.
По литературным данным (Гагина, 1961), гнездится на Ангаре
и Лене. Нам этот вид наблюдать не пришлось.
Отряд ДЯТЛЫ — PICIFORMES
Желна — Dryocopus martius L.
В пределах южной тайги Средней Сибири желна вездесуща
там, где есть высокоствольные леса с примесью сосны или листвен-
ницы. Только в этих лесных породах нам удалось видеть гнездо-
вые дупла черного дятла. Питаться же дятлы нередко предпочи-
тают в приречных и приручьевых ельниках, особенно размеща-
ющихся на речных террасах. Причина этого — большое число
муравейников, достигающих здесь огромных размеров. Вторым
источником муравьев служат трухлявые пни, обильно населенные
муравьями-компанотусами (Companotus sp.) и личинками усачей.
Последних желна легче всего достает из рыхлой древесины пихты.
Небольшая примесь светлохвойных пород всегда имеется среди
темнохвойной тайги. В связи с этим мест для устройства гнезд
у желны было всегда достаточно. Можно считать поэтому, что
повышение доли светлохвойных пород в лесах южной тайги не
привело к значительному увеличению плотности населения желны.
130
Омоложение же лесов безусловно снизило возможности для устрой-
ства гнезд, поэтому нужно думать, что с освоением тайги числен-
ность дятлов должна была снизиться.
Плотность населения желны в высокоствольных лесах всюду
«стандартна» — 1 пара на 5—7 км маршрута по хребту или неши-
рокому водоразделу, между водоразделами низшего порядка —
1 пара на 1.5 — 5—7 км, в среднем на 3—4 км. Таким образом, сред-
няя численность желны около 1 пары на 20 км2. Почти такую же
численность наблюдала А. П. Шкатулова (1963) в Читинской
области — 8 птиц на 100 км2 и Э. В. Рогачева (1962) на нижнем
Елогуе (Красноярский край) — 1 птица на 5 км маршрута
(в сосновых и сосново-лиственничных лесах).
Брачные игры желны мы наблюдали в первых числах мая
(в полосе лиственничного лесостепья). Птицы строили гнездо на
опушке лиственничника. На верхней Лене самец, добытый 11 мая
1961 г., имел крупные семенники: 12.0x7.0 мм. К концу мая
семенники у желны невелики: 5—6 мм длиной (измерены у 3 экз.).
На 5 обработанных паразитологами птицах эктопаразитов не
найдено.
Седой дятел — Picus canus Gm.
За седым дятлом установилась слава редкой птицы или во
всяком случае редко попадающейся на глаза (Гладков, 19516).
Второе определение более подходит для условий южной тайги
Средней Сибири. Седые дятлы в гнездовой период настолько скрыт-
ны, что их трудно заметить, поэтому седой дятел был впервые от-
мечен у дер. Осипово вблизи 62° с. ш. лишь совсем недавно
Е. Е. Сыроечковским (1960). Теоретически именно до этих преде-
лов и должны быть распространены седые дятлы в Средней Си-
бири.
Максимальной численности вид достигает в сосновых борах
и лиственнично-березовых лесах придолинной полосы. Тут в на-
чале—середине мая можно учесть 2 пары птиц на 5 км маршрута
(1 пару на 15—25 га). В водораздельных сосняках седых дятлов
меньше — 1 пара на 5 км (25—30 га). В темнохвойных лесах
водоразделов седые дятлы практически отсутствуют.
Следует заметить, что для учета седых дятлов пригоден лишь
период с начала и до середины мая. В июне, даже точно зная место
предыдущих встреч и расположения гнездового дерева, седых
дятлов наблюдатель не отмечает.
В послегнездовой период (с конца августа—начала сентября)
седые дятлы поселяются в темнохвойных участках леса, где бывают
очень заметны. Их в это время никак нельзя назвать молчали-
выми. С громким отрывистым криком потревоженные дятлы
перелетают с места на место. На фоне снега они кажутся очень
темными, почти черными.
9*
131
Нарушение лесов, очевидно, привело к некоторому увеличению
числа седых дятлов и безусловно сделало вязь кружева их ареала
более частой, а «нити» более «толстыми».
Седые дятлы встречаются парами уже в апреле. Их барабанный
бой значительно громче и глуше, чем у большого пестрого дятла.
Птицы токуют на сухих вершинах лиственниц и сосен. Однако
гнезда седые дятлы устраивают чаще всего в осинах. Разгар брач-
ных игр приходится на первую половину мая. В это время птицы
перелетают парами с дерева на дерево, «нападают» друг на друга.
Барабанящий дятел сидит совершенно неподвижно. Вдруг он
издает короткую, но очень громкую раскатистую «трель» и вновь
замирает.
Появление молодых отмечено 10 июня (1961 г., р. Тутура).
В это время самцы уже имеют спавшиеся семенники длиной не бо-
лее 2.5 мм. Полувзрослые птицы добыты 17 июля (1961 г.).
На 4 обработанных паразитологами седых дятлах паразитов
не оказалось.
Трехпалый дятел — Picoides tridactylus L.
Трехпалый дятел (Р. t. tridactylus L.) — прекрасней пример
того, как изменяется кружево ареала вида с характером местности.
В высокогорьях юга Сибири трехпалый дятел в гнездовое время —
птица нижнего и среднего пояса гор. Тут он достигает наивысшей
численности на гарях небольшого размера среди лиственничных
на западе (Тува) и темнохвойных на востоке (Саяны, Хамар-Да-
бан) лесов. Среди смешанных лесов Восточного Саяна и Хамар-
Дабана дятлы многочисленнее, чем в чистых насаждениях. Нако-
нец, на высотах свыше 1200 м, где в Хамар-Дабане, кроме кедра
и пихты, нет высоких деревьев, трехпалые дятлы не гнездятся
(Реймерс, 1963а). На Алтае, где живет другой подвид — тянь-
шаньский трехпалый дятел (Р. t. tianchanicus But.), птицы гнез-
дятся на значительно больших высотах, иногда, как мы наблю-
дали в районе Курайского перевала, у верхней границы леса.
Осенью в горных странах юга Сибири трехпалые дятлы по-
являются высоко в горах, но к зиме исчезают (Реймерс, 1963а).
В полосе южной тайги трехпалые дятлы достигают наивыс-
шей численности в водораздельной темнохвойной тайге и на ее
гарях. Лишь весной и осенью птицы делаются обычными в долин-
но-придолинной полосе, откуда вновь исчезают на зиму. Именно
темнохвойные, а не лиственные леса (как указывает Н. А. Глад-
ков, 19516), наиболее благоприятны для существования трехпалых
дятлов в южной тайге. Особенно заметно это на севере: небольшие
участки островных темнохвойных лесов буквально насыщены
трехпалыми дятлами.
Судя по сводкам, сделанным Н. А. Гладковым (19516),
К. А. Воробьевым (1963), и другим материалам, севернее южной
132
тайги трехпалые дятлы охотно живут в лиственничных лесах,
а еще далее на север делаются птицей урем (Михель, 1935; Пор-
тенко, 1939). Приречной птицей становятся дятлы и у южного пре-
дела распространения вида в Западной Сибири (например, на
крайнем юге Новосибирской области, где нам пришлось работать).
Таким образом, на севере и в горах трехпалый дятел — вид
более теплых (по сравнению с соседними) лесов, в пределах южной
тайги — более холодных, но вместе с тем и более кормных биото-
пов. Очевидно, такое распределение птиц связано с наличием
излюбленного корма трехпалых дятлов — личинок усачей, кед-
ровых орехов и семян пихты.
Для условий южной тайги Средней Сибири неверен вывод
Н. А. Гладкова (19516) о том, что нарушение лесов благоприятно
сказывается на популяции трехпалых дятлов. Справедливо это
замечание в пределах известных нам районов для гор южной Си-
бири и отчасти равнинной южной тайги Западной Сибири. Сведение
темнохвойных лесов в Средней Сибири безусловно снизило жиз-
ненные возможности вида.
Численность трехпалых дятлов в пределах южной тайги не-
высока и резко колеблется от сезона к сезону и от года к году.
Осенью 1960 г. на верхней Лене отмечено наивысшее число трехпа-
лых дятлов в долине р. Тутуры. Их было приблизительно в два раза
меньше, чем больших пестрых дятлов (Реймерс, 1963г). Там же и
в то же время, нов 1961 г., трехпалых дятлов было так мало, что 25
октября мы отметили присутствие вида как необычную редкость.
Разгар брачных игр у трехпалых дятлов приходится на конец
мая—начало июня. В это время у самцов наиболее крупные се-
менники: 8.Ох5.0 мм — 29 мая, 10.0x5.0 мм — 1 июня и
3.5X1.Змм в середине июня (р. Чадобец, 1962 г.).
Гнездовые дупла трехпалые дятлы делают на высоте от 2 (ред-
ко) до 5 м над землей, чаще всего в сухой пихте. Усиленно крича-
щие молодые отмечены на верхней Лене 29 июня, а нар. Чадобец
27 июня. В бассейне р. Чадобец слетки встречались в последней де-
каде июля.
На 9 осмотренных паразитологами птицах иксодовых клещей
не оказалось. С 2 экз. сняты гамазовые клещи.
Большой пестрый дятел — Dendrocopos major L.
Большой пестрый дятел в условиях южной тайги Средней
Сибири вполне может быть назван «сосновым дятлом», хотя он
не чуждается и уремного высокостволья (но не приречных ельни-
ков!), осинников и березняков с небольшой примесью хвойных
пород. В темнохвойных лесах он гнездится лишь при наличии
осины или комбинации пихты и березы. Не избегают дятлы и
лиственничного лесополья. И. С. Поляков (1873), отмечая приуро-
ченность больших пестрых дятлов к сосновым борам, считал при-
133
чиной этого явления обилие в сосняках муравьев. Действительно,
в сухих борах и парковых лиственничниках муравьи находят
благоприятные условия для жизни. В болотистых листвягах и
темнохвойной тайге муравьиных куч, как правило, нет. Однако
в водораздельных сосновых борах муравьев немного, а дятлы
обычны. Нет муравьиных куч и в осинниках водоразделов. Здесь
дятлы питаются короедами, древесными муравьями-компаноту-
сами и массой голых гусениц. Но особенно большое значение
сосна имеет для большого пестрого дятла зимой. Ее семена могут
заменить лишь орехи сибирского кедра (тоже сосны!).
Максимальной численности большие пестрые дятлы в гнездовой
период достигают в придолинных сосновых борах. Тут встречается
до 4—5 пар на 5 км учетного маршрута. В водораздельных сосня-
ках на это же расстояние встречается всего 1 пара. Столько же
птиц (1—2 пары на 5 км) в нарушенных темнохвойных лесах с бе-
резой и осинниках. Нет больших пестрых дятлов на обширных
гарях (на небольших гарях — не более 1 км в поперечнике — бы-
вает до 1 пары на 10 га), в глухих темнохвойных лесах и приречных
ельниках. Общий характер размещения вида в южной тайге ле-
том показан на рис. 26. Материалы по зимнему размещению боль-
ших пестрых дятлов уже опубликованы нами (Реймерс, 1963а).
Наибольшее число дятлов в это время держится в разреженных
сосновых лесах. Значительная часть особей в сосняках живет
оседло. Из лиственных лесов и с опушек гарей темнохвойных
лесов (где дятлы также нередко гнездятся, отсутствуя внутри об-
ширных гарей вдали от леса) дятлы откочевывают в сосняки,
в глубь темнохвойных массивов и отчасти к югу — в кедровники
и боры крайнего юга Сибири (Малышев, 1963).
Тесная связь больших пестрых дятлов с сосняками, большая
часть которых в пределах Средней Сибири антропогенного происхо-
ждения, осинниками и другими производными лесными ассоциа-
циями позволяет считать, что численность больших пестрых
дятлов резко возросла за период нарушений лесов человеком.
Выборочная рубка сосняков вторично увеличила плотность на-
селения дятлов. Однако широкое распространение пожаров при-
вело к неравномерности в распределении вида, образованию
скоплений птиц в одних местах и вторичному отсутствию в других.
На это обстоятельство следует обратить внимание в связи с про-
блемой зимней циркуляции вируса энцефалита (Краминский и
ДР., 1962).
Первая «барабанная дробь» большого пестрого дятла обычно
слышна в южной тайге около 10—15 марта. Однако в теплые дни
конца февраля дятлы также приходят в возбуждение и барабанят.
К середине апреля у больших пестрых дятлов образуются пары —
вслед за обратной кочевкой части птиц с юга на север, что про-
исходит в конце марта—начале апреля (Малышев, 1963). Разгар
брачных игр приходится на первые дни мая. В середине—конце
134
мая дятлы выдалбливают дупла. К концу июня в гнездах уже
громко кричат птенцы, а в конце июля обычны самостоятельно
живущие молодые. Возможно, бывают случаи рекордно раннего
Рис. 26. Принципиальная схема весенне-летнего размещения
больших пестрых дятлов в пределах южной тайги Средней
Сибири.
Условные обозначения как на рис. 12 и 22.
вылета птенцов из гнезда. Так, 5 июня 1961 г. был добыт молодой
пестрый дятел в темнохвойном лесу Лено-Тутурского между-
речья. К сожалению, этого экземпляра, добытого паразитоло-
гами, мы не видели, поэтому нет уверенности в правильности
определения.
Г&>
Гнездовое дупло большие пестрые дятлы делают в различных
породах, часто в осине и березе. Однажды нам встретилось не-
обычное дупло, выход из которого был расположен всего в 2.5 м
от земли (в высоком восьмиметровом пне березы диаметром 30 см).
Уровень заклещенности больших пестрых дятлов очень ни-
зок — на 12 птицах паразитологи не нашли исксодид, однако
факт вирусоносительства подтвержден многими авторами
(Р. Л. Наумов и др., 1963, и др.).
Белоспинный дятел — Dendrocopos leucotos Bech.
Т. Н. Гагина (1961) считает белоспинного дятла обычной
гнездящейся птицей бассейна Ангары и верхней Лены. Однако
мы этот вид отметили лишь на юге — в Хамар-Дабане (16 августа
1955 г.). Во всех известных нам работах авторы либо вообще
на упоминают белоспинного дятла, либо упоминают о нем как
о пролетной птице, либо говорят об этом виде весьма и весьма
неопределенно. Создается впечатление, что белоспинный дятел
распространен в Прибайкалье лишь в пределах Саян и в верховьях
Ангары.
Малый пестрый дятел — Dendrocopos minor L.
Малый пестрый дятел — птица речных долин. В горах юга
Сибири эти дятлы живут среди лиственных и смешанных лесов
по берегам рек. В южной тайге Средней Сибири они типичные
обитатели приречных ивняков. Там, где приречные ельники сво-
дят ивняки на нет, малые пестрые дятлы не гнездятся. Нам не ясна
причина этого явления, но на Лене малые пестрые дятлы встре-
чаются намного реже, чем в пределах Саян и на Ангаре, особенно
нижней.
Поскольку малые пестрые дятлы охотно поселяются в приреч-
ных кустарниках и не живут в тех же кустах среди приречных
ельников, следует думать, что человек создал этим птицам благо-
приятные условия для распространения на север и увеличения
общей численности популяций.
Брачные игры у малых пестрых дятлов начинаются в одно
время с большими пестрыми — в конце апреля—начале мая.
В это время семенники у дятлов достигают размера 10.0x4.0 мм.
В сентябре (с конца августа) молодые дятлы широко кочуют по
речным долинам, нередко встречаясь в бурьянах огородов.
На взрослой самке, добытой! сентября 1961 г. нар. Тутуре
в ивовых зарослях, найдена личинка иксодового клеща.
Вертишейка — Jynx torquilla L.
Вертишейка в южной тайге Средней Сибири — типичный оби-
татель придолинной полосы. Гнездовые дупла этой птицы бы-
вают в лиственницах и соснах, почти обязательно сухих и стоящих
136
среди открытого пространства (чаще всего на небольшой по раз-
меру гари) или редколесья. Нарушение тайги весьма благоприятно
для вида.
Наибольшей численности вертишейки достигают в парковых
лиственничниках лесостепи. На верхней Лене и на Ангаре обилие
птиц зависит от наличия благоприятных мест для гнездования.
Вдоль придолинной полосы вертишейки встречаются не чаще
одной пары на 3—5 км маршрута, главным образом на гарях
у границы между сосновыми лесами и приречными или приру-
ч ьевыми ельник ами.
Под Иркутском, по данным Л. И. Малышева (1963), вертишей-
ки появляются между 9 и 18 мая. В б. Баяндаевском районе в 1959 г.
птицы прилетели 21 мая. На р. Чадобец в 1962 г. первые пролет-
ные особи отмечены 17 мая. На пролете вертишейки держатся
разреженных приречных и приручьевых ельников, в лесостепи —
опушек лесов с редкими деревьями.
24 мая в бассейне р. Чадобец вертишейки держались парами.
Размер семенников добытой птицы 7.0x4.Омм. 10 июня на верхней
Лене гнезда были уже заняты, но что было в них — мы выяснить
не сумели.
2 экз., добытых на р. Чадобец, были свободны от эктопара-
зитов.
Отряд ВОРОБЬИНЫЕ —PASSERIFORMES
Сем. ВОРОНОВЫЕ — CORVIDAE
Ворон — Corvus corax L.
Ворон в пределах южной тайги гнездится повсеместно, предпо-
читая участки, где открытые места перемежаются высокостволь-
ными лесами. В связи с этим вороны обычны в лесостепи, лесополье
и вблизи гольцов или горных редколесий. Благоприятны для
гнездования опушки лесов, выходящие к большим, плохо зараста-
ющим гарям. Осенью вороны охотно держатся в кедровниках,
где питаются орехами кедра. Летом птицы также посещают кед-
ровники, собирая орехи в отходах кедрового промысла. Пару
воронов в таком месте мы ежедневно наблюдали в течение июня
1961 г. на Лено-Тутурском водоразделе. Сильные птицы во время
своего пребывания изгоняют с куч отходов обычных тут горлинок.
Численность воронов, как и всех крупных птиц, невысока.
По подсчетам, сделанным нами в лесостепном районе близ улуса
Байша весной 1959 г., пара птиц (и соответственно их гнездо)
приходится приблизительно на 140—150 км2. Гнезда воронов
нам не приходилось встречать ближе 10 км друг от друга. Иногда
они делают гнезда в оживленных местах. Например, пара свила
гнездо в пади Топка вблизи Иркутска (1953 г.). Гнезда мы видели
только на лиственницах и соснах в 12—15 м от земли.
137
Весенние эволюции птиц в воздухе можно наблюдать уже
в конце января. В начале мая в окрестностях Байши у воронов
были еще яйца (1959 г.). В то же время (1953 г.) под Иркутском
в гнезде было 4 пуховых птенца. К сожалению, в начале июня
гнездо кто-то разрушил. В последующие годы вороны в Топке
не гнездились.
Численность воронов, очевидно, не изменилась в результате
освоения местности человеком. Правда, вороны оттеснены к водо-
разделам, но наличие большого количества гарей позволяет
птицам находить удобные места для гнездовий. В историческом
прошлом вороны гнездились в естественно разреженной перестой-
ной водораздельной тайге. Резкое омоложение лесов на больших
пространствах приводит к сокращению числа воронов. Особенно
пагубными бывают для птиц огромные лесные пожары, приводя-
щие к омоложению леса на колоссальных пространствах.
Ворона — Corvus согопе L.
Восточная черная ворона (С. с. orientalis Ev.), обитающая
в южной тайге Средней Сибири, — птица культурного ландшафта.
Вдали от населенных пунктов этот вид отсутствует и поднимается
на водоразделы только вслед за человеком. Также благодаря
человеку ворона получила возможность зимовать в пределах
Средней Сибири (Вержуцкий, 1963).
Чем выше степень освоенности территории, тем больше число
гнездящихся ворон и тем плотнее кружево ареала вида. Однако
при полном заселении территории (что в Средней Сибири бывает
редко) вороны все же предпочитают для гнездовий днища речных
долин и речные террасы.
В начале сентября вороны откочевывают к югу и возвра-
щаются назад в середине—конце марта. Постройка гнезд на-
чинается обычно 13—17 апреля, однако 26 марта 1955 г. под Ир-
кутском была найдена полная кладка из 5 яиц (Малышев, 1963).
В отличие от окрестностей Ленинграда (Мальчевский, 1959),
где вороны устраивают гнезда чаще всего на соснах, в Средней
Сибири подавляющее большинство гнезд вороны строят на елях,
реже — на соснах и лишь иногда — на березах. На соснах и
особенно на березах вороны вьют гнезда лишь тогда, когда побли-
зости нет елей. Размеры двух гнезд, найденных близ с. Якимовки
в апреле 1961 г.: высота 25 и 26, ширина гнезда 43 и 50, ширина
лотка 22 и 25, глубина лотка 7.5 см у обоих гнезд. Выстилка
этих двух гнезд состояла из шерсти кабарги. В других гнездах
мы находили шерсть домашнего скота, нежные травинки осок,
серую вату и т. д?
В конце апреля у большинства ворон бывают полные кладки
из 4—5 яиц. Окраска их очень различна — в одних случаях
светлая голубовато-зеленая, в других — темно-зеленая. Размеры
яиц упомянутых двух кладок: 1) 43.4 X 31.1, 41.7 х 31.4, 42,3 X
X 31.2 и 44.1 X 31.9 мм; 2) 45.8 X 31.0, 46.0 X 29.9, 45.4 х
X 31.0, 47.9 X 30.0 мм.
В первом гнезде 17 мая было 2 птенца и 2 яйца. На следующий
день вылупился еще 1 птенец и 19 мая — последний. 18 мая вес
птенцов из первого гнезда был 21.7, 29.2 и 37.2 г, яйцо весило
18.5 г (за день до вылупления птенца). Начало вылупления птен-
цов во втором гнезде мы пропустили. Окончилось оно 17 мая.
Птенцы из этого гнезда весили 34.0, 50.7, 53.0 и 51.5 г. Таким
образом, птенец при вылуплении имеет вес около 20 г. В первые
дни разница между птенцами, вылуплявшимися ежедневно по
одному, была велика, позже они сравнялись по весу и развитию.
Птенцы первого из упоминавшихся гнезд вылетели 14 мая, т. е.
через 25 дней после вылупления последнего птенца и 28—29 дней
со времени начала вылупления птенцов в гнезде. Выкармлива-
ние в среднем длилось 26—27 дней.
Несмотря на вылет птенцов в середине мая, летающие молодые
начинают попадаться на глаза лишь в июне. Л. И. Малышев (1963)
под Иркутском видел их 8 июня 1955 г. Мы отметили массовое
появление слетков 16 июня 1960 г. на р. Тутуре. На р. Чадобец
из одного гнезда птенцы вороны слетели уже 12 мая, а молодых
не было заметно до конца первой декады июня.
В 28 погадках, собранных нами и Г. А. Вороновым 13 и 23 мая
1961 г., были найдены (в скобках указано число погадок, в кото-
рых обнаружены данные включения): обломки крупных костей
(9), кости мелких грызунов (10), кость зайца-беляка (1), кости
ящериц (3), рыбьи позвонки (1), растительные остатки, точнее
не определенные (12), семена растений, ближе не определенные
(4), скорлупа кедровых орехов (5), семена пшеницы, составляющие
основное содержимое погадок (22), и камешки (6). Корма вороны
в основном антропогенного происхождения. Почти все компо-
ненты могли быть собраны на окружающих деревню свалках.
В желудках 3 экз., добытых в августе 1960 г., были семена расте-
ний, ближе не определенные, зерна овса, ягоды черемухи, косточ-
ки бурозубки и насекомые, в том числе навозники и личинка
стрекозы.
На 2 экз. птиц, обработанных паразитологами, иксодовых кле-
щей не оказалось. Очень многочисленны были вши и кровососки.
Грач — Corvus frugilegus L.
Грач — редкая птица лесостепи Средней Сибири (Гагина,
1961). В пределах лесополья нам видеть его не пришлось.
Галка — Corvus monedula L.
Даурская галка — обитатель лесостепи и лесополья юга
Средней Сибири. На верхней Лене она отмечена до дер. Понома-
реве (Реймерс, 1963). 26 апреля 1962 г. обыкновенные галки
199
были встречены в окрестностях дер. Яркино на р. Чадобец. Ви-
димо, это были залетные экземпляры. По рассказам местных жи-
телей, которые хорошо знают галок, птицы бывают на р. Чадобец
не каждый год, но «часто гнездятся в долине р. Чадобец у полевых
станов».
Под Иркутском прилет даурских галок происходит 17—26
марта (Малышев, 1963). По сообщениям корреспондентов-фено-
логов, в 1962 г. галки появились 13—26 марта, весной следующего
года — около 15 марта. Небольшой материал не позволяет вы-
вести никаких закономерностей.
Галка может проникать далеко на север. М. И. Ткаченко (1924)
указывал ее для дер. Верхнее Карелино на Нижней Тунгуске.
Очевидно, как и все другие южные виды, птица в Средней Си-
бири может быть встречена на гнездовье до 62° с. ш.
Сорока — Pica pica L.
Сорока — удивительнейший вид. Загадка причин отсутствия
ее на значительной части Сибири, на Сахалине и на востоке Се-
верной Америки пока не поддается иному объяснению, чем пред-
положение о существовании в прошлом двух изолированных оча-
гов распространения: в Европе и в пределах гипотетической Бе-
рингии. Ныне восточная часть сплошного когда-то ареала распа-
лась на три участка — Юго-Восточной Азии, Северо-Восточной
Азии и Северной Америки. Вероятность такого объяснения весьма
невелика, как и вообще всех историко-фаунистических построений.
К сожалению, мы не знаем генетической стойкости видов, а без
этого опасно углубляться в вопросы причинности явлений.
Объяснение характера распространения вида, данное в свое
время И. С. Поляковым (1873) и заключающееся в том, что сороке
необходимы сухие места, изобилующие лугами, очень заманчиво,
но не всеобъемлюще. Нам приходилось видеть гнезда сорок на
кустах среди речного разлива и посредине обширных заболоченных
пространств. В это же время птицы не живут в ряде районов с ин-
тенсивным скотоводством. Скорее всего современное распростра-
нение сороки обусловлено сочетанием исторических и экологи-
ческих причин, которые пока еще не раскрыты.
В Средней Сибири сорока — лесостепная форма, проникающая
в тайгу вслед за образованием лесополья вдоль рек. Кружево
ее ареала в южной тайге совпадает с очертанием днищ долин и
частично нижних речных террас. Лишь там, где поля выходят
на водоразделы, сороки живут по хребтам. Северная граница рас-
пространения сорок в Средней Сибири совпадает с последними
форпостами южной тайги. Е. Е. Сыроечковский и Э. В. Рога-
чева (1958) сообщают о регулярном гнездовании сороки у пос.
Комса (на Енисее), но замечают, что тут сорока уже откочевы-
вает на зиму к югу. Поскольку за границей регулярных гнездовий
обычно следует полоса периодического гнездования, достигающая
14Q
у некоторых видов птиц ширины 300—400 км с юга на север (вдоль
географической широты протяженность таких полос обычно
значительно меньше), нет ничего удивительного, что вид может
быть встречен на гнездовье намного севернее обычных пределов
распространения. В связи с этим отнюдь не парадоксально, что
А. Я. Тугаринов и С. А. Бутурлин (1911) наблюдали сороку
между 64° и 65° с. ш. Однако еще севернее X. Сибом (Seebohm,
1880) скорей всего видел залетных птиц.
Можно предполагать также, что сорока гнездится по речным
долинам бассейна Витима, хотя бы изредка уже в настоящее время,
и в будущем должна заселить их. Сможет ли сорока «освоить»
Якутию, нам не ясно.
Численность сорок очень высока у населенных пунктов с оби-
лием пашен и падает вдали от деревень. При расположении сел
в 5—-10 км друг от друга и ширине освоенной долины реки в 1—
1.5 км 1 пара сорок гнездится от другой на расстоянии 2 км.
Прих более широкой долине (2—3 км) частота встреч удваивается.
Из этого можно заключить, что пара птиц на Ангаре и Лене за-
нимает участок в среднем около 2 кмI 2. В Забайкалье плотность на-
селения обыкновенных сорок вчетверо меньше (Шкатулова, 1963).
В подавляющем большинстве мест южной тайги Средней Си-
бири сорока оседла. Первые яйца птицы откладывают под Иркут-
ском в конце апреля или первые дни мая. Полная кладка состоит из
6—8, очень редко (Юдин, 1952) 9 яиц. Полные кладки зарегистри-
рованы на верхней Лене в середине, а на р. Чадобец уже в начале
мая (3—4). Насиживает сорока полную кладку не менее 18 дней
(например, с 15 мая по 3 июня 1961 г.). Птенцы во многих гнездах
вылупляются почти одновременно (в пределах недели), но бывают
случаи позднего вылупления. Особенно часты такие опоздания на
севере южной тайги и в холодные снежные весны. Вылетают соро-
чата из гнезд через 27—28 дней (под наблюдением было 2 гнезда).
Массовое появление слетков повсюду происходит в конце мая-
первых числах июня. Однако в это время (27 мая, р. Чадобец)
можно еще найти и птенцов с маховыми в чехлах (длиной 3—4 см).
Гнезда сороки чаще всего устраивают в ивовых кустах на
высоте около 4 м от земли. Одно гнездо найдено на сосне с диамет-
ром ствола (на уровне груди) около 30 см на той же высоте. Лишь
однажды мы нашли гнездо сороки на высоте 6 м. Оно было построе-
но над старым гнездом вороны, которое служило основанием
шарообразной постройки сороки. Глубина лотка этого гнезда 9,
ширина 11 см. Размеры гнезда на сосне: шар диаметром 60,
ширина собственно гнезда 28, высота его 18, ширина лотка 20,
глубина 10.5 см. Размер яиц двух кладок (в мм):
I (р. Чадобец)
35.9X25.8 35.3X26.0
35.6X25.6 34.7X25.9
36.1X25.9 35.4X26.2
II (р. Тутура)
32.5X24.0 31.0X25.6
31.6X24.2 32.7X24.0
31.2X25.7 32.9X23.6
141
Одно яйцо из гнезда, где уже вывелось 7 птенцов (29 мая), 36.0 х
X 20.4 мм (р. Тутура). Обычная окраска яиц: сероватые или
коричневато-серые с коричневыми крапинками различной интен-
сивности.
Значительную часть кормов сороки находят на окраинах на-
селенных пунктов и скотных дворов. Однако птицы ловят
полевок-экономок и крупных насекомых.
Голубая сорока — Cyanopica cyana Pall.
Голубая сорока изредка залетает на Ангару, но никогда не
достигает ее среднего течения.
Сойка — Garrulus glandarius L.
Рыжеголовая сойка (G. g. brandtii Ev.) гнездится по лиственным
лесам южной тайги, достигая на севере 61—62° с. ш. Немного-
Рис. 27. Рыжеголовая сойка.
численная в южной тайге птица, здесь, у северного предела рас-
пространения, гнездится спорадически и не ежегодно. В 1962 г.
сойки, очевидно, не гнездились в районе дер. Яркино, находящей-
ся даже несколько южнее 60° с. ш. Возможно, это связано с от-
сутствием на р. Чадобец высокоствольных березняков и осинни-
142
ков. Прежде всего этим обстоятельством объясняется редкость
соек на Лене. В окрестностях пос. Жигалово сойки обычны во
время весенних (конец апреля) и осенних (конец августа—сен-
тябрь) кочевок, когда птицы весной парами, а осенью стайками
кормятся в сосняках и на проселочных дорогах. Летом число
соек резко падает (на р. Тутуре не более 1 пары на 5—7 км при-
речной полосы), а осторожное поведение птиц почти совершенно
скрадывает их от глаз наблюдателя. Мы зарегистрировали соек
на р. Тутуре лишь 24—25 мая 1961 г. К этим датам и относится
упомянутая оценка численности птиц.
Осенью сойка весьма обычна в темнохвойных водораздельных
лесах (рис. 27) и вблизи населенных пунктов. В окрестностях
с. Якимовки птицы отмечены как многочисленные в конце октя-
бря 1961 г. Встречены они были и 20—22 декабря этого года.
С августа появились они на р. Чадобец (первая птица добыта
16 августа 1962 г.). Но особенно заметны сойки на средней Ан-
гаре. Здесь, в районе изобилия осинников на Око-Ангарском между-
речье, 8—24 августа мы насчитывали у таежных речек 1 выводок
на 1—2 км маршрута. В то же время водораздельные леса были
пусты.
Сойки питаются крупными насекомыми.
2 экз. птиц, обработанные паразитологами, оказались свобод-
ными от паразитов.
Кукша — Cractes infaustus L.
Южную тайгу Средней Сибири населяет байкальская кукша
(С. inf. siblricus Bodd.). Исходя из наших представлений о един-
стве южной тайги как фаунистического образования, мы предпола-
гаем, что западные притоки средней Лены должны быть населены
якутским подвидом, а к северу от Байкала живет южнотаежный
подвид. А. К. Рустамов (1954) в первом из упомянутых мест от-
мечает байкальскую кукшу, а во втором (на картосхеме) оста-
вляет свободное поле. Забайкалье должен населять другой под-
вид.
В Средней Сибири, начиная с ее южных хребтов, кукша всюду
приурочена к темнохвойной тайге и особенно к ее приручьевым
участкам. Обычны кукши в приручьевых ельниках. Численность
гнездящихся пар по ручьям достигает одной на 2—3 км. В темно-
хвойных лесах водоразделов кукши также приурочены к ручьям,
хотя бы к их сухим долинкам или седловинам у их истоков. Густые
сосновые молодняки иногда заменяют обычную гнездовую ста-
цию. Но особенно охотно кукши с выводками молодых держатся на
гарях, поросших сосновым мелколесьем и молодняком лиственных
пород. Именно в этих местах птицы находят обильную пищу.
Приречных ельников вблизи лугов кукши в противоположность
другим врановым избегают.
143
Как и все политопные виды, кукши в меньшей степени, чем
другие формы, страдают от нарушений в тайге. Численность кукш
пока сохраняется приблизительно на одном уровне. Однако
кружево ареала постепенно меняется. Кукши оттеснены из ни-
зовий долин речек и ручьев к водоразделам, а на водоразделах
уничтожение темнохвойной тайги и образование огромных го-
рельников привело к флуктуациям и разрежениям в населении
вида. Такое «квантовое» размещение, «узлы» на вязи кружева
ареала, характерно для многих видов животных. Островные темно-
хвойные леса населены кукшами с исключительной плотностью.
Так, на участке елово-пихтового леса размером 200 на 200 м
вблизи дер. Яркино на р. Чадобец жили 2 пары птиц, в то время
как в ее окрестностях среди сосновых и березово-осиново-сосно-
вых лесов можно было не встретить ни одной кукши на 5 км учетного
маршрута.
Кукши весной разбиваются на пары уже в марте. В это время
семенники едва достигают размера 3x2 мм, а фолликулы микро-
скопичны. Птицы подолгу сидят на верхушках небольших деревь-
ев, изредка тихо «переговариваясь». Спаривание кукш можно на-
блюдать в апреле. Позже кукши исчезают с глаз наблюдателя.
Найти птиц в это время тем более трудно, что происходит таяние
снегов, делающее тайгу непроходимой. Для учетных целей можно
использовать речные наледи, а лучше производить подсчет пар
в мае, когда птицы уже кормят птенцов.
Гнездо кукши нам посчастливилось найти лишь однажды —
7 мая 1961 г. на р. Тутуре (рч. Черемшанка). Это была задержав-
шаяся или повторная кладка. Гнездовое дерево — сухая елочка
всего 5—6 см толщиной (на уровне груди) — стояло в редкОхМ
заболоченном приручьевом ельнике (с примесью лиственницы)
вблизи границы ельника с сосновым бором на склоне. Несмотря
на такую незначительную толщину деревца, гнездо помещалось
в 3.5 м от земли и было мало заметно. Птицу, сидящую в гнезде,
с земли не видно: хвост ее напоминает в это время сучок.
Размеры гнезда: ширина 20, высота 12, ширина лотка 9, глу-
бина лотка 6 см. Материал гнезда состоит из сухих веточек, вхо-
дящих в состав крепления стенок гнезда, а не только подстила-
ющих его. Утеплено гнездо древесными гнилушками, сухим вей-
ником, кусочками листовых лишайников и главным образом бо-
родачом. Внутренняя выстилка гнезда состоит из нежных птичь-
их перышек, средн которых удалось определить перья самой
кукши и рябчика. Структура гнезд кукш чрезвычайно напоминает
построение гнезд кедровок (Реймерс, 1959г): в его стенках лишь
меныпую роль играют древесные гнилушки, да выстилки из перьев
в гнездах кедровок мы не находили.
Полная кладка у кукши состоит из 3 яиц. Такое количество
их было в найденном автором гнезде, столько же бывает молодых
в выводке (как большая редкость — 4). Яйца, к сожалению, мы
144
измерить не успели. Они голубоватые, слабо заостренные, с не-
сколько большей пятнистостью у тупого конца (рис. 28), очень
похожи на яйца кедровки, но несколько мельче.
Птица (рис. 29) сидит в гнезде очень крепко (это отмечает и
К. А. Воробьев, 1963). Ее можно трогать руками. При попытке
вытащить ее из гнезда она цепляется за его выстилку и не улетает.
Пугает птицу появление опасности сверху.
Рис. 28. Гнездо кукши с^яйцами.
Б Насиживает одна и та же птица (мы ее пометили штемпельной
краской), очевидно самка. Другая птица ее кормит, однако вне
гнезда. Эта другая подлетает с кормом на ближние деревья и
издает звуки, очень напоминающие кедровичий полусвист-полу-
повизгивание у гнезда. В ответ на этот «разговор» (издаваемые
звуки безусловно имеют какое-то «смысловое» значение) насижи-
вающая птица слетает с гнезда и активно преследует принесшую
корм. При этом она громко кричит, крылья ее трепещут, она мелко
дрожит и ерошит темную шапочку в хохолок. Крик птицы, прося-
щей корм, напоминает крик испуга, но отличается от него трудно
уловимыми деталями. После кормления насиживающая птица
подлетает к деревьям, стоящим рядом с гнездовым, затем переле-
тает на гнездовое, садится у гнезда, осматривается и лишь затем
садится в гнездо.
Бывает, что вторая птица подлетает без корма. В этом случае
она издает «разговорные» звуки, на которые получает ответ.
Ю П. Ф. Реймерс
145
Насиживающая птица не покидает в этом случае гнезда, а прилетев-
шая через^короткое время улетает.
О частоте кормления можно судить по следующему наблюде-
нию. 9 мая птица прилетела с кормом в 11 ч. 20 м., затем в
12 ч. 10 м. без корма и в 12 ч. 30 м. с кормом (между кормлени-
ями прошел 1 ч. 10 м.). 10 мая кормление произошло в 14 ч. 30 м.
и не повторялось до 16 час., когда мы ушли.
Рис. 29. Кукша на гнезде.
К сожалению, 15 мая гнездо было разорено: из него исчезли
яйца. Птица, помеченная нами, продолжала сидеть на пустом
гнезде два дня, после чего покинула его. Гнездо передано на
кафедру биогеографии МГУ.
14 мая 1961 г. уже можно было наблюдать слетков кукш,
усиленно преследовавших родителей с требованием корма. Вы-
водки кочуют чаще всего по мелколесью и гарям. Кукши тут
питаются на земле. Вообще кукши собирают корм летом главным
образом на земле. Осенью и зимой они охотно ищут насекомых в ниж-
них частях крон и лишь ранней весной охотятся в верхних частях
крон и на крайних концах веток. Видимо, весной эти части деревь-
ев лучше прогреваются и населены большим числом беспозвоноч-
ных. Питаются кукши жуками, клопами, гусеницами бабочек,
пауками, а иногда и грызунами.
Собирание кукшей корма на земле приводит к высокой заражен-
ности птицы иксодовыми клещами. Среди врановых кукши —
146
самые зараженные клещами птицы. Правда, на р. Чадобец, где
число клещей очень мало, на 13 осмотренных кукшах клещей не
было обнаружено, как и на 7 птицах с Тутуро-Ленского водораз-
дела (темнохвойных лесов). Зато из 9 кукш, добытых в придолин-
ной полосе р. Тутуры, 4 оказались зараженными нимфами и 1 —
личинками. Максимальное число паразитов было 2—3. Обильно
заражены кукши и кровососками.
Кедровка — Nucifraga caryocatactes L.
Кедровка (TV. с. macrorhynchos Вг.) — обычнейшая птица
темнохвойных лесов южной тайги. В условиях исчезновения
кедровников численность птиц резко сокращается, они предпри-
Рис. 30. Копанка кедровки в снегу.
нимают местные перекочевки (Реймерс, 1954), приспосабливаются
к гнездованию в приручьевых ельниках и других мало характер-
ных для их существования местах. Можно с достаточной степенью
точности считать, что кружево ареала кедровок в южной тайге
совпадает с распространением темнохвойной тайги. При этом мак-
симальная численность наблюдается в лесах с участием кедра.
Слабый урожай кедра приводит к уменьшению численности
кедровок, в том числе к почти полному исчезновению их из приру-
чьевых ельников. На Ангаро-Подкаменнотунгусском междуречье,
10*
147
где кедра очень мало и нет кедрового стланика, кедровки практи-
чески не гнездятся.
В пределах южной тайги нам удалось найти лишь одно гнездо
кедровки. 7 мая 1961 г., когда оно было обнаружено, птенцы
уже вылетели. Помещалось гнездо на ели (16 см диаметром) с
очень густой кроной (в «ведьминой метле») на высоте 8 м и в 3 м
от вершины дерева. Гнездо с выстилкой из сухих злаков оказалось
идентичным описанным нами ранее (Реймерс, 1959г). Любопытно,
что гнездовые деревья кедровок и кукш были расположены в 20 м
друг от друга.
22 мая мы добыли кедровку, продолжавшую еще кормить
птенцов очищенными от скорлупы ядрами орехов. Несколько
ядер было найдено в подъязычном мешке самки.
Кедровки уверенно находят свои кладовые. С веток дерева
птица безошибочно садится к месту захоронения орехов и несколь-
кими сильными ударами клюва достает орехи даже из мерзлой
земли. Зимой птица также безошибочно роет в снегу копанку,
сильно напоминающую беличью (рис. 30), слегка под углом к по-
верхности снега. Копанки кедровка делает лишь в неглубоком
снегу — не более 30 см, потому в многоснежных районах их на-
блюдать не удается.
Орехи кедра, составляющие основу в питании кедровок,
птицы начинают доставать из шишек целиком с середины августа
(14-го, 1959 г.), до этого разбивают недозревшие шишки (и орехи)
на деревьях.
Несмотря на то что кедровки весной достают «клады» орехов,
присаживаясь на землю, на всех 16 обработанных паразитологами
(верхняя Лена) птицах иксодовые клещи не найдены.
Сем. СКВОРЦОВЫЕ — STURNIDAE
Скворец — Sturnus vulgaris L.
Е. П. Спангенберг (1954) оставил открытым вопрос о северо-
восточной границе распространения скворца в Средней Сибири.
Е. Е. Сыроечковский (1960) нашел птицу в поселках Комса и
Верхне-Имбатское (на 400 и 220 км севернее, чем указывали Ту-
гаринов и Бутурлин, 1911).
Мы констатировали скворца как обычную гнездящуюся птицу
в дер. Яркино на р. Чадобец. По рассказам, есть скворец и в
пос. Кежма. Очевидно, птица населяет весь бассейн нижней Ан-
гары и, как все лесостепные и лесопольные виды, может гнездиться
до 62° с. ш.
В окрестностях дер. Яркино скворцы появились И апреля
1962 г. К 20 апреля они стали очень обычны и заселили все скво-
речники в деревне. Еще 6 мая большие стаи (до 20—25 экз.) сквор-
148
цов продолжали лететь вдоль долины р. Чадобец с общим напра-
влением на северо-восток.
В лесостепных районах Средней Сибири скворцы появляются
в первой декаде—середине апреля (в 1960 г. около 15 апреля,
в 1961 г. на грани марта и апреля, весной 1962 г. около 1—2 ап-
реля, в 1963 г. в очень растянутые сроки — между 30 марта и
27 апреля, но главным образом 8—12 апреля).
На двух птицах из пролетной стаи паразитологи не обнаружили
эктопаразитов.
Сем. ИВОЛГОВЫЕ — ORIOLIDAE
Обыкновенная иволга — Oriolus oriolus L.
Иволга — редкая птица крайнего западного крыла южной
тайги Средней Сибири. Если права Т. Н. Гагина (1962а), что
ареал иволги в Средней Сибири сокращается и граница его отсту-
пает на запад, то иволга станет почти единственным исключением
такого рода в общем процессе распространения западных форм
на восток. Исключение это будет тем более парадоксальным,
что благоприятные для иволги биотопы в Средней Сибири не умень-
шаются, а увеличиваются по площади.
Сем. ВЬЮРКОВЫЕ — FRINGILLIDAE
Дубонос — Coccothraustes coccothraustes L.
В Средней Сибири дубонос — лесостепная форма, отлетающая
на зиму в юго-восточном направлении и частично зимующая (Га-
гина, 1961) на Ангаре и Лене. Нам зимой дубоносов встречать
не приходилось. Огромные массы птиц мы наблюдали на осеннем
пролете в западном Хамар-Дабане (Реймерс, 1959а), а Н. Г. Скря-
бин и К. И. Филонов (1962) — в Баргузинском заповеднике.
Не нашли мы дубоносов и в окрестностях Жигалова. Однако нали-
чие птиц на верхней Лене констатирует Т. Н. Гагина (1961).
На р. Чадобец 31 мая 1962 г. был добыт из стаи дубонос самец.
Возможно, это была залетная стая, но не исключена вероятность
периодического гнездования дубоносов значительно севернее гра-
ницы, указанной в сводке «Птицы Советского Союза» (Горчаков-
ская, 1954). Дубоносы появились на р. Чадобец около 20 мая
и жили на опушке березово-соснового леса в кустарниках у реки.
Щегол — Carduelis carduelis L.
Щеглы залетают на Ангару и, возможно, местами даже гнездят-
ся (Гагина, 1961). Тут же они зимуют, достигая Иркутска. В ок-
рестностях этого города мы встречали щеглов зимой 1951/52 г.
149
В зиму 1959/60 г. птиц под Иркутском мы не нашли. Видимо,
численность зимующих особей бывает резко различной от года
к году.
Чиж — Carduelis spinus L.
Чижи изредка залетают при осенних кочевках в верховья
Ангары. Мы наблюдали этих птиц осенью 1959 г. под Иркутском.
Т. Н. Гагина (1961) полагает, что чижи даже гнездятся на Ангаре.
С востока или с запада прибывают чижи в южную тайгу Средней
Сибири — мы судить не беремся.
Чечетка — Carduelis flammea L.
Южная тайга Средней Сибири — район периодического гнездо-
вания чечеток. Вообще же эти птицы — обитатели северной тайги
и лесотундры. Судя по немногочисленным сообщениям корреспон-
дентов-фенологов, чечетки исчезают в поздние весны в начале
мая, а в ранние — в конце апреля. Л. И. Малышев (1963) отме-
чает наличие «урожайных» и «неурожайных» лет — с большим
и малым числом пролетающих или зимующих птиц. «Урожайными»,
по его мнению, были 1953/54 и 1954/55 гг. Много было птиц
в 1959/60 г., а следующий сезон ознаменовался очень малым числом
зимующих и пролетных чечеток. Если весной 1960 г. чечетки на
верхней Лене летели сотенными стаями (Реймерс, 1963г), то
в 1961 г. они встречались единицами. На нижней Ангаре, так же
как и на верхней Лене, интенсивный пролет чечеток был отмечен
с середины апреля и закончился к 7 мая. В общей сложности сроки
весенних массовых кочевок птиц весьма близки по всей южной
тайге Средней Сибири.
Осенью чечетки под Иркутском и во всей полосе вдоль желез-
ной дороги появляются в конце сентября или начале октября.
На верхней Лене птицы отмечены раньше — 4 сентября 1961 г.
Все приведенные данные относятся главным образом к обыкно-
венной чечетке (С. /. flammea L.). Л. И. Малышев пишет (1963),
что в стайках обыкновенных чечеток нередки их сородичи —
пепельные чечетки (С. /. exlltpes Cones). При всей субъективности
понимания термина «нередок» мы не можем согласиться с Л. И. Ма-
лышевым. Пепельных чечеток совсем можно не увидеть в десятках
стай обыкновенных. Они достаточно редкие птицы зимней лесо-
степи Средней Сибири. В пределах южной тайги этот подвид встре-
чается зимой далеко не всюду.
Нарушение тайги человеком и образование на ее месте больших
массивов плодоносящих березняков — весьма благоприятные фак-
торы для успешной зимовки чечеток в южной тайге. Возможно,
это обстоятельство играет определенную роль и во время спора-
дического гнездования птиц в пределах южной тайги Средней
Сибири.
150
Длиннохвостый снегирь — Uragus sibiricus Pall.
Длиннохвостый снегирь — яркий пример того, насколько еще
плохо известен зоологам север южной тайги Средней Сибири. На
р. Чадобец — это обычнейший вид приречных ивняков. Встре-
чается птица и на верхней Лене. Однако в окрестностях Жигалова
длиннохвостые снегири бывают не ежегодно и лишь во время
весенних кочевок (добыты 17 апреля 1961 г.). В связи с этим нельзя
отрицать спорадического гнездования тут длиннохвостых снегирей,
но рискованно считать этот вид укоренившимся.
На р.Чадобец длиннохвостые снегири появились 2 мая (1962 г.).
До этого дня птиц мы не регистрировали, несмотря на экскурсии
в течение нескольких недель. Очевидно, на зиму длиннохвостые
снегири отсюда откочевывают к югу. Миграционная волна снегирей
была особенно заметна до двадцатых чисел мая. Позже птицы
встречались лишь парами, а не стайками. В это время и немного
раньше у длиннохвостых снегирей отмечен максимальный размер
семенников 5.0 X 7.0 мм (15 мая).
5 птиц с р. Чадобец, осмотренные паразитологами, оказались
свободными от эктопаразитов.
Снегирь — Pyrrhula pyrrhula L.
В пределах южной тайги Средней Сибири встречаются две
формы снегирей: восточноевропейский (Р. р. pyrrhula L.) и
серый (Р. р. cineracea Cab.). Их ареалы обнимают весь этот
зоогеографический район, но кружево ареала снегирей раз-
лично. На этот факт обращал внимание еще Б. И. Дыбовский
(Dybowski, 1874). Многие исследователи рассматривают формы
снегирей как самостоятельные виды.
Восточноевропейский снегирь летом при-
держивается мелколесья и кустарниковых зарослей, в связи
с этим сведение лесов для него весьма благоприятно. Численность
особей формы, очевидно, увеличивается, а кружево ареала делается
более плотным.
Весенняя прокочевка восточноевропейских снегирей наблю-
дается в апреле—мае и не носит характера массового пролета.
Осенью волна кочующих красных снегирей констатирована нами
(Реймерс, 1963а) в октябре—начале ноября.
В желудках обыкновенных снегирей, добытых 27 апреля
1960 г. на верхней Лене, обнаружены орехи кедра и семена пше-
ницы. Зимой птицы кормятся по окраинам полей и в кустарниках
долин ручьев.
На 3 снегирях паразитологи не нашли иксодид.
Серый снегирь гнездится почти исключительно в темно-
хвойной тайге и в замещающих ее осинниках (как редкость — на
севере Прибайкальского зоогеографического района в сосново-бе-
151
резовых лесах с примесью темнохвойных пород). Широкое распро-
странение формы в горных темнохвойных лесах Средней Сибири
и Дальнего Востока (Воробьев, 1954, 1963) и отсутствие серого
снегиря в равнинной тайге Западной Сибири заставляют считать
эту птицу специфичной для темнохвойной тайги горных местно-
стей.
Нарушение лесов весьма пагубно для серых снегирей — они
лишаются мест гнездовий и сокращаются в числе. В водораздель-
ных темнохвойных лесах на верхней Лене обилие серых снегирей
достигает 1 пары на 1 км учетного маршрута. В осинниках средней
Ангары, возникших на месте существовавшей когда-то здесь
пихтово-кедровой тайги, численность птиц не превышает 1 пары
на 10 км учетного хода. В березняках, сосновых лесах и листвен-
нично-сосновых суборях без примеси темнохвойных пород, также
занявших место темнохвойных ассоциаций, птицы отсутствуют
вовсе.
Серые снегири прокочевывают на север несколько раньше во-
сточноевропейских — в марте-апреле (Реймерс, 1963а, 1963в)
и интенсивность этого процесса у них выше. Можно говорить даже
о пролете птиц. В мае снегири приступают к размножению. В июне
они кормят птенцов. Так, 24 июня 1961 г. была добыта самка,
имевшая в подъязычном мешке корм — какие-то мягкие семена.
27 июня мы обнаружили в мешке двух пауков. Последний раз
паук обнаружен у птицы, добытой 1 августа. В то же время был
наполнен подъязычный мешок и у снегирей с нижней Ангары.
Из 18 обработанных серых снегирей на 1 найдена нимфа иксо-
дового клеща (Лено-Тутурский водораздел). На серых снегирях
найдены также гамазовые клещи.
Сибирская чечевица — Erythrina rosea Pall.
В распространении сибирской чечевицы много неясностей. Судя
по сводке «Птицы Советского Союза», ареал птицы охватывает
значительную часть Средней и Восточной Сибири и включает
южную тайгу. Однако за 10 лет в пределах южной тайги мы ни
разу не встречали вид летом. Интенсивный пролет сибирских
чечевиц также наблюдается не ежегодно. Весной 1960 г. птицы
на верхней Лене летели с 24 апреля по 8 мая, а наиболее интен-
сивный пролет отмечен 27 апреля—1 мая (Реймерс, 1963г).
В следующем году там же не было встречено ни одной особи вида.
Сибирские чечевицы не всегда заметны и под Иркутском, особенно
на осеннем пролете (5 ноября 1955 г. их отметил тут Л. И. Малы-
шев, 1963).
Есть все основания предполагать, что сибирские чечевицы
гнездятся севернее южной тайги и по окружающим ее горным систе-
мам. Однако в Хамар-Дабане (Реймерс, 1959а) и в юго-западном
Забайкалье (Козлова, 1930) птица на гнездовье не найдена.
152
Сибирские чечевицы характерны для Якутии (Воробьев, 1963),
где гнездятся в зарослях кедрового стланика. Нельзя категорично
отрицать вероятность спорадического гнездования птиц и в упомя-
нутых выше районах юга Сибири, тем более что несколько север-
нее, на Баргузинском хребте, сибирские чечевицы — обычные
гнездящиеся птицы (Скрябин и Филонов, 1962).
У всех 3 исследованных нами птиц в желудках обнаружены
остатки кедровых орехов.
Обыкновенная чечевица — Erythrina erythrina Pall.
Обыкновенная чечевица в южной тайге Средней Сибири —
птица опушек сосняков, гарей этих лесов, поросших молодым
березняком или осинником, и приречных кустарниковых зарослей.
В отличие от гор юга Сибири, где обыкновенная чечевица характер-
на для полян в водораздельных лесах и даже для бадановых кед-
ровников (Реймерс, 1959а), в пределах южной тайги вид населяет
только долинную и придолинную полосы, не выходя к водоразде-
лам даже по долинам мелких речек.
Наивысшей численности вид достигает на гарях сосновых
лесов, поросших молодой березой. Здесь на участке около 1 га
могут гнездиться две пары птиц. Одинаковые по местополо-
жению, но более влажные гари островных темнохвойных ле-
сов, также поросшие березой, обыкновенная чечевица не засе-
ляет. ,
Человек безусловно способствовал распространению обыкно-
венных чечевиц в южной тайге Средней Сибири. В мало нарушен-
ных его деятельностью местностях чечевица гнездится лишь
вдоль уреза воды рек в черемушниках. В Нетронутой природе
такие места — редкость, поэтому первоначальное кружево аре-
ала вида в южной тайге было подобно отдельным обрывкам четок.
Четковидность «нитей» ареала сохранилась и ныне, но в целом
кружево ареала уплотнилось и стало «грубее» — птицы заселили
более широкую полосу вдоль водотоков.
На верхней Лене передовые стайки чечевиц отмечены в 1961 г.
19 мая, однако первые поющие самцы появились 27 мая. Пролет
в этом году достигал максимума между 28 мая и 13 июня. Улетели
обыкновенные чечевицы в 1960 г. 11 августа, осенью следующего
года птицы исчезли после 18 августа. В этот день 1961 г. молодая
птица еще пыталась петь. На нижней Ангаре в 1962 г. обыкновен-
ные чечевицы прилетели 1 июня. 28 августа работавшие с нами
студенты добыли птицу, определенную ими как «обыкновенная
чечевица». Экземпляра мы не видели. Если при определении не
было ошибки (т. е. птица не принадлежала к сибирскому виду),
эта дата — наиболее поздняя осенняя находка обыкновенной
чечевицы на р. Чадобец в 1962 г.
153
Максимальный размер тестикул у чечевиц — в начале июня:
1 июня 7.0x6.0 мм, 6 июня 8.0x7.0 мм, а 15 и 22 июня уже
5.0x3.8 и 4.2x3.7 мм.
Все 9 обыкновенных чечевиц, обработанных паразитологами,
были свободны от иксодид.
Щур — Pinicola enucleator L.
Щур — эндемик темнохвойных лесов. В Средней Сибири для
него обязательно сочетание кедра и пихты, восточнее — присут-
ствие кедрового стланика. Эти леса либо изначально горные,
Рис. 31. Щур в кедровнике на куче отходов от орехового промысла.
либо (в пределах южной тайги) оттеснены человеком на хребты.
В Прибайкальском зоогеографическом районе щуры гнездятся
только в водораздельной темнохвойной тайге (рис. 31). На нижней
Ангаре по р. Чадобец, где нет кедра, щуров мы не нашли. Запад-
нее эта птица населяет лишь тайгу северного типа (севернее 60—
62° с. ш.). Результат ли это нарушений темнохвойной тайги (южнее
щуры держатся летом лишь в обширных лесных массивах),
или следствие экологических потребностей вида — судить трудно.
В пределах южной тайги Средней Сибири, где ныне высоко-
ствольные темнохвойные леса становятся редкостью, щур — исче-
зающий вид. Сейчас общая численность птиц во многие десятки
раз ниже, чем до освоения южнотаежной полосы человеком. Она
154
стала снижаться с тех пор, как у первобытного населения страны
скотоводство сменило охоту и начался процесс выжигания лесов
под луга (в том числе лесные).
Щур — периодически кочующая птица, зимой появляющаяся
в сосновых лесах. Осенние кочевки наиболее заметны в сентябре-
октябре, весенние перемещения начинаются с конца февраля
(Реймерс, 1959а, 1963а; Малышев, 1963).
В начале июля у щуров крупные семенники (9 июля 1961 г.
8.3x6.2 мм), фолликулы у самок достигают 2 мм. Молодые
зарегистрированы на верхней Лене 1 августа 1961 г.
Объекты питания щуров — кедровые орехи и семена пихты.
4 щура, обработанные паразитологами, оказались свободными
от эктопаразитов.
Клёст-еловик — Loxia curvirostra L.
Летом и зимой еловик — птица приречных и приручьевых
ельников, сосняков (особенно придолинной полосы) и намного
реже елово-пихтовых лесов водоразделов. Кедровой тайги клесты
избегают, хотя в их желудках мы находили остатки кедровых
орехов.
Сведение водораздельной темнохвойной тайги, очевидно, не
отражается пагубно на численности клестов, нарушение при-
речных и приручьевых ельников разрывает кружево ареала вида,
делает его четковидным.
Сроки размножения у клестов южной тайги неопределенны.
В 1960 г. на верхней Лене максимальные размеры семенников
зарегистрированы в мае—июне (5.9x4.8, 6.0 X4.0 мм). На
р. Чадобец 2 июля 1962 г. добыта самка с фолликулами, дости-
гающими 6.0 мм, а самцы в это время имели семенники крупнее
5 мм (5.0x4.7, 6.6x6.0, 5.0x4.5 мм).
7 экз. клестов с р. Чадобец, обработанных паразитологами,
были свободны от эктопаразитов.
Белокрылый клёст — Loxia leucoptera Gm.
Белокрылый клёст — северная форма, становящаяся обыкно-
венной лишь у северных пределов южной тайги. На верхней Лене
нами был добыт лишь 1 экз. 25 апреля 1960 г. В следущие годы
мы этот вид не отмечали. На р. Чадобец белокрылый клёст весь-
ма обычен в смешанных лесах обязательно с присутствием листвен-
ницы и в водораздельных темнохвойных лесах также с листвен-
ницей, но без кедра.
Нужно думать, что белокрылый клёст теснее всего связан с
лиственницей, хотя в его питании существенную роль играют се-
мена ели. Современное знание ареала вида явно недостаточно.
Мы полагаем, что белокрылый клёст должен гнездиться значитель-
155
но севернее, чем это обычно считается (Птицы Советского Союза,
т. 5).
Самка с наседным пятном добыта 28 июля 1962 г. в сосново-
лиственнично-березовом лесу водораздельного типа (бассейн
р. Чадобец).
На 13 белокрылых клестах, подвергшихся паразитологической
обработке, иксодовые и гамазовые клещи найдены не были.
Зяблик — Fringilla coelebs L.
В последние годы литература наполнена сообщениями о про-
движении зябликов на север и восток Сибири (Юдин, 1952; Ким,
19576; Сыроечковский и Рогачева, 1958; Наумов и Бурковская,
1959; Сыроечковский, 1960, и др.). На наш взгляд, этот процесс
весьма естественен.
Ныне сосновые боры Средней Сибири, разреженные и «оте-
пленные» общим нарушением таежных лесов, почти не имеют
видов птиц среднего яруса. Вообще сосновые леса и их производ-
ные в Средней Сибири носят следы недавнего образования: био-
ценозы этих лесов «не укомплектованы» — имеют свободные
экологические ниши, и в них мало специфических форм. В связи
со сказанным нас ничуть не удивила встреча зябликов на р.Ча-
добец (далеко за пределами рамки картосхемы, приведенной
в сводке «Птицы Советского Союза»!). Нет никакого сомнения,
что вид достигнет Байкала.
Зяблики появились в окрестностях дер. Яркино очень рано:
впервые они отмечены 19 апреля 1962 г. К 25—26 апреля птицы
стали встречаться значительно чаще. С 9 мая зяблики сделались
малозаметными. Самцы в мае усердно пели в елово-лиственничных
редколесьях по слегка заболоченным долинкам ручьев при впаде-
нии их в реку. Самки отмечены в таких же местах и на опушках сос-
няков, выходящих к речным зарослям. Нередко на таких опушках
присутствует ель. Значительно реже зяблики встречались на
опушках сосняков у полей. Во всех случаях птицы держатся не
на самой опушке леса, а метрах в ста от нее в глубине леса. За-
регистрированы зяблики в разреженном рубками сосновом бору.
Встречаются они и в долине реки среди сильно изреженных кур-
тин приречных ельников. С переходом к лесам водораздельного
типа, границу которых на р. Чадобец легко определить в связи
с тем, что березово-сосновые леса сменяются лиственнично-сосно-
во-осиновыми, зяблики исчезают.
15 мая 1962 г. размер семенников зяблика был 7.0X5.0 мм.
12 мая была добыта самка с крупными фолликулами. Этих данных
мало для уверенного утверждения, но есть много оснований пред-
полагать, что зяблики гнездятся на р. Чадобец.
На одном из 4 обработанных паразитологами зябликов обна-
ружен клещ, систематическая принадлежность которого осталась
неясной.
156
Юрок — Fringilla montifringilla L.
T. H. Гагина (1961) считает юрка гнездящейся и пролетной
птицей Ангары и полагает, что гнездование вида на верхней
Лене требует подтверждения. Наши материалы и наблюдения
Л. И. Малышева (1963) свидетельствуют, что юрки гнездятся в юго-
восточной части Саян (в лиственничниках) и отсутствуют в гнездо-
вое время на Ангаро-Ленском междуречье, на верхней Лене и
средней Ангаре. Ими населен лишь западный край южной тайги
Средней Сибири и ее север. Следовательно, для верхней Лены
это, как правило, пролетный вид (спорадическое гнездование не
исключено).
В связи с общим нарушением таежных лесов и образованием
обширных сосняков на востоке южнотаежной полосы Средней
Сибири вид может заселить всю описываемую полосу сплошь.
Разреживая сосняки и приречные ельники, человек способствует
росту численности юрков, сводя эти леса — уменьшает населе-
ние вида,
На верхней Лене массовый пролет юрков в 1961 г. отмечен
10 мая. 11 мая птицы держались уже стаями в 100—150 особей.
Юрки летят вдоль лесных опушек, редколесья и гарей. Они нередки
в густом сосновом жердняке, где останавливаются на отдых.
Пролетные стаи кормятся на кустах, деревьях и земле. 14 мая
интенсивность пролета ослабела и через несколько дней птицы
исчезли. Первые юрки на нижней Ангаре появились 4 мая (нам-
ного позже зябликов), 6 мая интенсивность пролета значительно
возросла и достигла максимума 10—12 мая; 17-го пролет резко
ослабел и 22 мая полностью прекратился. Осенью пролет на верх-
ней Лене отмечен в конце сентября 1961 г.
С середины мая юрки занимают гнездовые участки. Максималь-
ная численность птиц (до 25 пар на 5 км маршрута) отмечена
в разреженных сосняках. Почти такая же численность птиц зареги-
стрирована в разреженных сухих приречных ельниках с березой.
Раза в полтора меньше птиц в водораздельных сосновых лесах
и совсем немного их в темнохвойной тайге (в последних двух
местах количественные учеты не производились).
В самом конце мая юрки беспокоились так, как обычно делают
эти птицы вблизи гнезд. Размер семенников достиг максимального
значения (у 2 экз. 9.0 X6.0 и 10.2x7.4 мм) в последней декаде
мая—начале июня. Молодые отмечены с 7 июля.
На 22 птицах, обработанных паразитологами, иксодовых и
гамазовых клещей найти не удалось.
Сибирским вьюрок — Leucosticte arctoa Pall.
Т. Н. Гагина (1961) отмечает для Ангары этот вид как залет-
ный и зимующий.
157
Сем. ТКАЧИКОВЫЕ — PLOCEIDAE
Домовый воробей — Passer domesticus L.
Домовый воробей населяет всю южнотаежную полосу Средней
Сибири, но встречается далеко не во всех населенных пунктах.
Воробьи проникли на верхнюю Лену в 1710 г. (Миддендорф,
1869), а к 1781 г. заселили ее вплоть до Витима. Поскольку на
Лене сейчас гнездятся оба вида воробьев, то не исключено, что
они проникали туда бок о бок. На средней Ангаре почти всюду
присутствуют как домовые, так и полевые воробьи. Однако на
северных притоках нижней Ангары домовых ворьбьев нет
во многих населенных пунктах. Мы не отметили вида даже в боль-
шой дер. Яркино.
Весеннее возбуждение у домовых воробьев отмечено с середины
февраля—начала марта. В это время птицы таскают материал
(перышки, бумажки), но, очевидно, не строят еще гнезд. Истинное
гнездостроение происходит лишь в конце марта—начале апреля.
К концу месяца имеются уже кладки. На верхней Лене в течение
лета 1961 г. воробьи вывели птенцов из 3—4 кладок. В июньской
кладке было 6 яиц размером 15.4x21.2, 15.5x21.2, 15.5х
Х20.0, 14.5x19.8, 16.0x21.6 и 14.3x20.5 мм. Первый пте-
нец вывелся из этой кладки 6 июля, 8 июля оставалось 1 яйцо,
9 июля вылупились уже все птенцы. 21 июля птенцы вылетели
из гнезда (общее время кормления — 12 дней со дня вылупления
последнего птенца). В то же время (17 июля) в трех других гнездах,
найденных Г. А. Вороновым в дер. Чертовской (на р. Чекан),
оказалось в одном гнезде 2 птенца, в другом — 3 птенца и в треть-
ем — 1 птенец и 1 яйцо (болтун).
22 августа у пары, выведшей птенцов в июле, уже снова были
птенцы, но на этот раз лишь 4. В других гнездах в это время от-
мечены яйца. Птенцы из гнезда, за которым велось наблюдение,
вылетели лишь 6 сентября, т. е. не покидали гнезда более
15 дней (они были найдены в возрасте около 1 дня).
На нескольких осмотренных воробьях иксодовых клещей най-
дено не было.
Полевой воробей — Passer montanus L.
Полевой воробей гнездится повсеместно, однако интересных
данных по этому виду у нас нет.
Высшая степень синантропности обоих видов воробьев пред-
полагает их продвижение в тайгу вместе с человеком.
Сем. ОВСЯНКОВЫЕ —EMBERIZIDAE
Обыкновенная овсянка — Emberiza citrinella L.
Е. П. Спангенберг и А. М. Судиловская (1954) не включили
верхнюю Лену в ареал обыкновенной овсянки. Т. Н. Гагина
(19586, 1961) отмечает этот вид как редкий и воздерживается от
158
констатации факта гнездования. Обыкновенную овсянку нам уда-
лось наблюдать в 1960 г. (Реймерс, 1963г) лишь в период с 16 ап-
реля по 7 мая. Весной 1961 г. птицы появились 21 апреля. С на-
чала мая интенсивность пения их ослабла и вновь усилилась
к 28 мая. Птицы держались здесь все лето и встречались с кормом
в клюве, следовательно, не лишено вероятия, что они гнездились.
На р. Чадобец 19 апреля 1962 г. уже шел массовый пролет
обыкновенных овсянок. Его интенсивность заметно ослабела
ко 2 маю. В это время птицы заняли гнездовые участки и пели
на одних и тех же местах.
Излюбленная стация обыкновенной овсянки — берега речек
и ручьев при впадении в реки. Изредка птицы остаются зимовать
в сосново-мелколиственной лесостепи, где держатся по полям
и опушкам лесов (наши наблюдения; Гагина, 19626).
Максимальный размер тестикул у овсянки с верхней Лены
22 апреля: 10.0x8.0 мм. На нижней Ангаре крупные семенники
измерены лишь у июньских экземпляров: 2 июня 11.0x7.0 и
9.2хб.9 мм.
На 5 птицах, обработанных паразитологами, иксодовых клещей
не найдено.
Белошапочная овсянка — Emberiza leucocephalos Gm.
В отличие от обыкновенной овсянки белошапочная не только
гнездится вблизи водотоков, но и занимает гари лесов придолинной
полосы. На гари водораздельных лесов белошапочная овсянка
не проникает. В связи с тем что придолинную полосу ныне зани-
мают главным образом сосновые боры, птица сделалась характер-
ным видом их горельников и вырубок. Гнездится она также среди
приречных кустарников, на опушках сосняков и приречных ель-
ников, на гарях ельников, поросших березовым мелколесьем,
и даже в кустах среди огородов. Однако ее нет в ерниках. Распре-
деление вида в южной тайге показано на рис. 32.
В глубь мало нарушенной тайги птицы на гнездовье не прони-
кают далее сотни метров и собирают корм вне леса. Что ограничи-
вает распространение овсянок в сторону водоразделов, неизвестно.
Для вида необязательна близость полей. Трудно предположить,
чтобы продвижение вида останавливали метеорологические фак-
торы или влажность почвы. Белошапочные овсянки гнездятся
и далеко на севере, а гнезда их приходилось находить даже на
болотных кочках. Однако факт налицо: вне участков водоразделов,
коренным образом преобразованных человеком (лесополья), бело-
шапочные овсянки — лучший индикатор границы придолинной
полосы, показатель многократности нарушения лесов и нередко
относительной необратимости антропогенного сукцессионного про-
цесса (например, замены темнохвойных лесов светлохвойными).
Распашка водоразделов и резкое изменение характера лесов
159
приводят сначала к тому, что белошапочные овсянки поселяются
на опушках у полей, а затем широко заселяют образовавшееся
лесополье. При этом можно почти наверняка говорить о том,
Рис. 32. Принципиальная схема весенне-летнего размещения
белошапочных овсянок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22.
что на этих гарях не произойдет восстановления темнохвойной
тайги, а вырастет светлохвойный лес придолинного типа (главным
образом с примесью березы и обилием видов животных придолин-
ного комплекса).
160
Человек дал возможность белошапочной овсянке — типично
лесостепному виду — проникнуть далеко в глубь таежной полосы.
Изначальный ареал птицы в Средней Сибири, видимо, обнимал
Лено-Якутские лесостепи.
Численность белошапочных овсянок местами достигает значи-
тельных величин. На опушках сухих сосняков у полей и лугов
пары гнездятся на расстоянии 50—70 м друг от друга. В глубине
разреженного пожаром или рубкой леса расстояние между парами
также не превышает по прямой 50—70 м, т. е. минимальная
площадь, занимаемая одной парой, 0.25—0.5 га. Обычно же в сто-
рону глухого леса белошапочные овсянки совсем не летают,
и их индивидуальный участок похож на веер с рукояткой у гнезда
и опахалом на поле, гари или участке луга. Указанная высокая
численность птиц отмечается на очень незначительной площади
лесных опушек, выходящих к открытым местам. Обычно же птицы
не населяют даже гари сосняков плотнее, чем 1 пара на 4—6 га
(200 X 200 или 250 X 250). Влажные гари придолинной полосы
населены с плотностью не более 1 пары на 9—10 га, остальные
обитаемые участки — еще в два-три раза реже.
Л. И. Малышев (1963) наблюдал под Иркутском пролет обык-
новенных и белошапочных овсянок в одно и то же время (1 апреля
1955 и 1956 гг., 31 марта 1957 г.). Как мы уже отмечали (Реймерс,
1963г), в 1960 г. на верхней Лене белошапочные овсянки начали
пролет раньше обыкновенных (в конце марта). Весной 1961 г.
первые белошапочные овсянки в окрестностях Жигалова появи-
лись 27 марта (вслед за появлением первых проталин на открытых
склонах южной экспозиции). 29 марта птицы летели небольшими
стайками по 2—7 экз. 31 марта была услышана первая песня
белошапочной овсянки. Одновременно появились первые пары.
Большие стаи птиц в несколько десятков особей отмечены лишь
с 13 апреля. Обыкновенных овсянок в это время еще не было,
следовательно, в течение двух лет на верхней Лене белошапочные
овсянки прилетали раньше обыкновенных.
Интенсивность пролета белошапочных овсянок на верхней
Лене весной 1961 г. заметно снизилась 21 апреля. К этому времени
местные особи заняли гнездовые участки и с начала мая пение их
ослабело.
На нижней Ангаре в середине апреля 1962 г. уже шел интен-
сивный пролет белошапочных овсянок. Со 2 мая он ослабел и
часть птиц заняла гнездовые участки. Тем не менее белошапочные
овсянки летели стаями еще до 6 мая, когда обыкновенных уже
на пролете заметно не было.
Таким образом, первые белошапочные овсянки появляются
в пределах южной тайги весной раньше обыкновенных и их пролет
продолжается дольше. Следует заметить, что часть птиц в теплые
зимы остается в пределах ангарского лесополья до весны (наши
наблюдения; Гагина, 1962а). Осенний отлет белошапочных овся-
Ц Н. Ф. Реймерс
161
нок на верхней Лене происходит с середины августа до конца
сентября. Пик его интенсивности в 1961 г. отмечен 18 сентября.
К концу мая самцы белошапочных овсянок вновь начинают
интенсивно петь, но самки из других пар в это время уже насижи-
вают яйца (рис. 33). В конце первой декады июня в гнездах по-
являются птенцы (13 июня 1960 г. — трехдневные, 13 июня
1961 г. — двухдневные). Массовое появление слетков в 1960 г.
Рис. 33. Гнездо белошапочной овсянки. Лесостепь.
отмечено 4—6 июля (Реймерс, 1963 г). Часть птиц гнездится вто-
рично, другая (очевидно, молодые из выводков прошлого года) —
запаздывает и выводит птенцов лишь к середине июля. Так, в
1961 г., когда на верхней Лене было отмечено позднее гнездо-
вание белошапочных овсянок, 13 июля найдено гнездо с не выле-
тевшим еще единственным птенцом. В южной тайге Средней Сибири
птенцов бывает 4 (1960 и 1961 гг., верхняя Лена — 5 гнезд), юж-
нее в лесостепи — 5 и нередко 6 (под Иркутском от 3 до 6). 5 и 6
яиц мы находили у белошапочных овсянок в Туве (Реймерс,
1960в).
Из 29 очесанных паразитологами белошапочных овсянок лишь
на одной была найдена личинка иксодового клеща. Видимо,
роль овсянок (Наумов, 1963) в цикле развития южнотаежных
иксодид Средней Сибири невелика. На очень немногих птицах
найдены и гамазовые клещи (1 экз. на 20). Белошапочные овсянки
заражены вшами и пухоедами.
162
Рыжая овсянка — Emberiza rutila Pall.
T. H. Гагина (1958а, 1961) полагает, что рыжие овсянки
гнездятся в бассейнах верховий Лены и Ангары. Мы не получили
никаких доказательств в пользу этого утверждения. Даже в рай-
оне Жигалова птицы встречены лишь дважды: осенью 1960 и 1961гг.
(10 августа и 10 сентября), явно на пролете (Реймерс, 1963г).
Не отметили мы птиц и для средней Ангары. Также лишь осенью
констатированы птицы в Хамар-Дабане (Реймерс, 1959а).
На нижней Ангаре в бассейне р. Чадобец работавшими с нами
студентами добыты 3 овсянки, отнесенные коллекторами к описы-
ваемому виду. Поскольку среди птиц был взрослый самец, воз-
можно, определение было сделано правильно, но ручаться мы не
можем, ибо экземпляры не сохранились. 20 июля добыта молодая
птица, значит, если определение было сделано верно, рыжая ов-
сянка гнездится на нижней Ангаре.
Следует отметить, что в распространении и размещении рыжей
овсянки остается больше неясностей, чем выясненных вопросов.
Дубровник — Emberiza aureola Pall.
Дубровник — лесопольный и лесолуговой вид, сравнительно
недавно завоевавший обширные пространства таежной полосы.
Если верить Г. Радде (Radde, 1862—1863), то в Саянах дубровник
поднимается до высот свыше 1500 м. Боюсь, что это недоразумение,
связанное с тем, что птицы в послегнездовой период широко ко-
чуют и действительно достигают высот 1200—1300 м, а местами,
где местность освоена человеком, и выше (пределы нам не ясны).
Однако гнездится птица на более низких уровнях.
В полосе южной тайги Средней Сибири дубровник — долинная
птица, гуще всего населяющая кустарники вдоль рек (но не при-
речные ивняки) и вдоль лесных опушек (особенно ельников),
выходящих к полям и лугам. Чем шире полоса освоения, тем толще
нить кружева ареала дубровника. Вид поднимается и на водо-
разделы, но лишь вслед за их распашкой. Следует полагать,
что дубровники заселили тайгу Средней Сибири уже после того,
как там появился человек.
В долинных разреженных черемушниках дубровники гнездятся
всего в 50 м друг от друга. Здесь численность птиц макси-
мальна (пара на 0.25 га). В 70—100 м гнездится пара от пары
на опушках лесов у полей (обязательно с кустарниками). На
верхней Лене в таких местах доминируют белошапочные овсянки,
а дубровников меньше, последние более характерны для средней
и нижней Ангары.
Весенний прилет дубровников на верхней Лене отмечен
в 1960 г. 1 июня по первой песне. В этот день птицы появились
сразу в массе. Весна 1960 г. была ранняя, тем не менее пролет
11* 163
птиц явно запоздал. Более поздней весной 1961 г. там же дубров-
ники в заметном количестве появились 27 мая, а передовые птицы
отмечены 24-го числа. На р. Чадобец дубровники появились
28 мая 1962 г.
Пролет дубровников проходит стремительно. Так, в 1961 г.
на верхней Лене его удалось проследить лишь с появления первых
птиц до 2—3 июня. На р. Чадобец в 1962 г. пролет длился бук-
вально 2—3 дня. Любопытно, что под Иркутском дубровники
появляются 15—21 мая (данные за 5 лет, Малышев, 1963). Прибли-
зительно в те же сроки (17 мая 1959 г.) встречены птицы в лесо-
степном районе окрестностей Байши (б. Баяндаевский район Иркут-
ской области). А на Лене у Жигалова прилет дубровников реги-
стрируется лишь после 25 мая, т. е. на 3—10 дней позже, чем под
Иркутском.
Осенью 1960 г. последние дубровники на верхней Лене отме-
чены 15 августа, в следующем году около 10-го, а на нижней Ангаре
в 1962 г. птицы держались минимум до 24 августа, когда была
добыта последняя птица работавшими с нами студентами.
Гнездо дубровника, найденное на р. Тутуре у опушки ельника
12 июля 1961 г., помещалось на земле у небольшого пенька ели.
Размер гнезда: ширина лотка 71, глубина лотка 42 мм. Размер
яиц: 15.0 X 19.8, 15.5 X 19.8, 15.3 X 20.0 и 14.8 X 20.1 мм.
Птенцы вывелись в ночь с 23 на 24 июля. 31 июля гнездо было
кем-то разорено. Описанное гнездо, видимо, имело запоздав-
шую или повторную кладку, потому что 19 июля было уже много
слетков дубровников, которые совместно с родителями кочевали
по лугам с кустарниками.
Паразитологами были обработаны 22 дубровника из наших
сборов на верхней Лене и 19 птиц с р. Чадобец. Иксодовые клещи
на них не обнаружены. На р. Чадобец с дубровников дважды
сняты гамазовые клещи. Птицы заражены также блохами, пухо-
едами и вшами.
Седоголовая овсянка — Emberiza spodocephala Pall.
Седоголовая овсянка — вид южно-восточносибирского долин-
ного комплекса, зашедший в Центральной Якутии далеко на север,
а по освобожденным от приречных ельников водотокам юга
Средней Сибири — на запад. Как мы и ожидали, седоголовая
овсянка проникает на нижнюю Ангару и ее северные притоки.
Вид, очевидно, будет найден на Подкаменной Тунгуске в освоен-
ных человеком участках долины реки (в местах ее расширения).
В долинах, где высокоствольные приречные ельники подхо-
дят почти к урезу воды, седоголовые овсянки не живут, хотя на
опушках молодых ельничков могут гнездиться. Сравнительно
редки птицы в черемушниках среди лугов, где отмечается макси-
мальная плотность населения дубровников. Зато в приречных
164
ивняках, тянущихся вдоль берегов речек, ручьев и старичных
озер седоголовая овсянка достигает наивысшей численности.
Таким образом, кружево ареала вида совпадает с той частью
гидрологической сети, берега которой освобождены от глухих
ельников, при этом максимум населения птиц приходится почти
на урез воды. Здесь же в ивняках седоголовая овсянка добывает
корм, состоящий из насекомых.
Прилетают седоголовые овсянки в пределы южной тайги поздно.
В б. Баяндаевском районе Иркутской области у улуса Байша пер-
вые седоголовые овсянки добыты весной 1959 г. 17 мая. На верхней
Лене в 1961 г. они впервые встречены 28 мая. Пролет продолжался
до 13 июня. Весной следующего года на р. Чадобец седоголовые
овсянки отмечены с 1 июня. В начале августа на Лене седоголовые
овсянки уже держатся стайками, но улетают поздно. Заметные
осенние передвижки птиц осенью 1961 г. отмечены 5—7 сентября,
интенсивный пролет продолжался 17—18 сентября, а последние
птицы исчезли в начале октября. Под Иркутском Л. И. Малышев
(1963) встретил птицу еще 5 октября (1955 г.).
Молодые на верхней Лене отмечены в 1961 г. 27—28 июля —
приблизительно в те же сроки, что и под Иркутском.
С 9 седоголовых овсянок не удалось собрать иксодовых
клещей.
Красноухая овсянка — Emberiza cioides Вг.
Возможна зимовка этого вида в пределах южной тайги Средней
Сибири (Гагина, 19626). На гнездовье мы птицу тут не встретили,
хотя под Красноярском она гнездится и зимует (Юдин, 1952).
Вообще сибирская красноухая овсянка (Е. с. cioides Вг.) предпо-
читает сухие лесостепи, прежде всего горного типа.
Садовая овсянка — Emberiza hortulana L.
Птица гнездится в Красноярско-Канской лесостепи (Юдин,
1952). В южную тайгу не проникает.
Горная овсянка — Emberiza cia L.
Встречена осенью в бассейне Ангары (Гагина, 1958а, 1961).
В теплые зимы вид остается в среднесибирской лесостепи на зи-
мовку (Гагина, 1962а).
Овсянка-ремез — Emberiza rustica Pall.
Для этой овсянки в условиях юга Средней Сибири очень бы
подошло название «болотная». В связи с этим сухое Верхоленье
ею, как правило, не населено. Среди наших материалов есть ви-
довая карточка, заполненная Г. А. Вороновым, с указанием на
165
добычу овсянки-ремеза на верхней Лене 23 июля 1962 г. Птица
(взрослый самец) попала в цилиндр для ловли грызунов. Очевидно,
ремезы, во всяком случае иногда, держатся на верхней Лене все
лето. Не исключено и спорадическое гнездование птиц. Лишь на
пролете ремез посещает бассейн верхней Ангары. Но начиная
со среднего течения реки, птица летом иногда обитает и, возможно,
гнездится. На нижней Ангаре, где уже много северных форм,
ремез — обычная птица заболоченных березняков.
Численность овсянки-ремеза даже на северных притоках
нижней Ангары в благоприятных биотопах невысока. Объективно
ее оценить очень трудно, потому что ремезы держатся в почти
нейроходимых местах. Вдоль тропы мы встречали пары прибли-
зительно через 1 км (видимость в густых березняках по болотам
очень ограничена — не далее 10—15 м, поют же птицы в период,
когда тайга становится доступной, нерегулярно).
Весной 1961 г. ремезы появились одновременно с обыкновен-
ными овсянками 21 апреля. Эта дата была днем появления ве-
сенних всходов растений на солнцепеках. Птицы летели большими
стаями, кочуя по сосновому мелколесью и среди небольших куп
елей в долине реки. Максимальной интенсивности пролет достиг
25 апреля и продолжался до 12 мая. 14 мая число летящих птиц
резко сократилось, и в следующие дни они исчезли. В 1962 г.
ремезы на нижней Ангаре появились 30 апреля. К 13 мая, когда
многие птицы держались уже парами, пролет прекратился. Оба
года мы наблюдали одновременное появление самцов и самок,
что бывает не всегда (Малышев, 1963).
На осеннем пролете в районе верхней Лены ремезы осенью
1961 г. появились около 20 сентября (23-го добыта первая птица).
В начале октября птиц уже не было видно.
Материалов о гнездовом цикле ремезов у нас нет. Максималь-
ный размер фолликул отмечен 11 мая 1962 г.—8.0 мм. Размер
семенников в это время 5.0 ХЗ.5—4.0 мм.
На 4 птицах иксодиды не найдены, но гамазовые клещи встре-
чены (последние на особях с верхней Лены). Вирусоносительство
энцефалита констатировано почти для всех овсянок (Наумов
и др., 1963), а для описываемого вида не только осенью, но и вес-
ной (Феоктистов, 1962).
Овсянка-крошка — Emberiza pusilia Pall.
Овсянка-крошка очень обычна на весеннем и осеннем пролетах
во всех участках лесостепи и лесополья Средней Сибири. Под
Иркутском, по Л. И. Малышеву (1963), весенний пролет проходит
в сжатые сроки между 11 и 29 мая, осенний — между 9 и 19 сен-
тября. Весной 1961 г. на верхней Лене птицы летели с 12 по 29 мая.
В эти же дни огромные стаи крошек летели в б. Баяндаевском
районе Иркутской области весной 1959 г.
166
Осенний пролет на верхней Лене в 1960 г. продолжался до
23 сентября. В следующем году птицы летели с 30, возможно,
25—27 августа до 16 сентября. На р. Чадобец в 1962 г. крошки
отмечены 29 августа, когда проходил осенний отлет птиц.
Все 6 пролетных птиц с верхней Лены были свободны от иксо-
ДИД.
Желтобровая овсянка — Emberiza chrysophrys Pall.
Конфигурация ареала желтобровой овсянки, очерченного
Е. П. Спангенбергом и А. М. Судиловской (1954), весьма странна.
По этим авторам, вид гнездится в средней Якутии на части бассей-
на верхней Лены и в верховьях Нижней Тунгуски, но отсутствует
на всей Ангаре и Подкаменной Тунгуске. Такого типа ареал дол-
жен был бы быть уникальным не только для птиц, нои для зверей.
Это зародило в нас сомнение в справедливости принимаемых ныне
границ распространения желтобровой овсянки.
Действительно, вид был найден нами в 1959 г. и А. П. Шведовым
в 1960 г. (Шведов, 1962) на средней Ангаре (на Око-Ангарском и
Ангаро-Илимском междуречьях). В 1962 г. мы установили, что
желтобровая овсянка весьма обычна на нижней Ангаре (р. Чадо-
бец) и Ангаро-Подкаменнотунгусском междуречье. Таким образом,
в Средней Сибири ареал вида приобретает более естественные очер-
тания, характерные для других форм с якутско-среднесибирским
эндемизмом (сибирская чечевица, лесной дупель и т. д.).
Желтобровая овсянка отнюдь не редкая, но спорадически
распространенная птица, что, впрочем, характерно для подавля-
ющего большинства видов животных. В отличие от прочих овсянок
Средней Сибири (кроме ремеза) желтобровая овсянка принадлежит
не к долинному, а к придолинно-водораздельному комплексу.
На Лене она населяет сосновые и лиственнично-сосновые леса
с подростом темнохвойных пород и такие же гари. Наличие под-
роста ели или пихты для этого вида в южной тайге абсолютно
обязательно. В связи с этим на средней Ангаре и в бассейне ее
нижнего течения желтобровые овсянки гнездятся лишь в темно-
хвойных лесах среди мелколесья, выросшего в «окнах» тайги, и
на участках успешного восстановления темнохвойной тайги.
Однако в собственно водораздельный комплекс темнохвойных
лесов желтобровые овсянки не входят, поскольку избегают об-
ширной и дремучей тайги. Для темнохвойной тайги это вид
ее островных участков среди светлохвойных массивов.
О численности желтобровых овсянок можно судить по следу-
ющему. В островке темнохвойной тайги вблизи р. Чадобец раз-
мером 100 на 200 м держалось 4 пары птиц (1 пара на 0.5 га).
На приблизительно таком же участке гари с подростом темнохвой-
ных пород на верхней Лене зарегистрировано 2 пары овсянок
(1 пара на 1 га). В светлохвойной тайге с подростом темнохвойных
167
пород встречено 1—2 пары на 5 км маршрута. Эти данные показы-
вают, что желтобровая овсянка отнюдь не редка.
На верхнюю Лену желтобровые овсянки прилетели весной
1961 г. 12 мая, п пролет их продолжался приблизительно до 20-го
числа. На р. Чадобец весной 1962 г. вид отмечен позднее —
23 мая, но вполне возможно, что первые прилетевшие особи были
пропущены. Осенний отлет не прослежен. Последние птицы на
нижней Ангаре добыты 7 августа. Очевидно, позже желтобровые
овсянки стали малочисленны (работы продолжались до конца
августа).
В верхнеленских лесах желтобровая овсянка начинает петь
20—22 мая (1961 г.). Самец поет свою несложную, но очень звуч-
ную песенку (что-то вроде «фи-и ци-ци-цп-ци-ци фи-и»), сидя на
верхушке березки, реже елки (всегда невысоко — в 3—4 м
от земли). В то же время у овсянок начинаются брачные игры,
причем птицы держатся на земле пли у земли. Максимальный
размер семенников отмечен на р. Лене 20 и 25 мая (14.0x8.0 и
10.2x8.3 мм), на нижней Ангаре на месяц позже — 9 и 25 июня
(11.5x7.5 и 10.5x7.2 мм). Первые молодые появились 5 июля.
Питается желтобровая овсянка в основном насекомыми.
Зараженность вида эктопаразитами, и в том числе иксодидами,
сравнительно высока. На 1 из 4 обработанных овсянок с верхней
Лены найдена нимфа иксодового клеща. С 1 из 27 птиц, добытых
в бассейне р. Чадобец, где уровень заклещенности очень низок,
также была снята 1 нимфа из семейства иксодид. Обе зараженные
птицы добыты во время пролета (Лена — 25 мая, Ангара —
9 июня), поэтому возможно, что нимфы были занесены овсянками
из более южных районов.
Рыжешейная овсянка — Emberiza yessoensis Swinh.
Т. Н. Гагина (1961) отмечает залет этого вида в бассейн Ан-
гары.
Полярная овсянка — Emberiza pallasi Cab.
Встречается полярная овсянка в пределах южной тайги лишь
на пролете. На верхней Лене птица добыта 22 апреля, на нижней
Ангаре особи, условно отнесенные к этому виду (сильно раз-
биты), — 28 мая. Возможно, из полярных овсянок состояла не-
большая стайка, встреченная на р. Тутуре 25 октября 1961 г.
Камышевая овсянка — Emberiza schoeniclus L.
Т. Н. Гагина (1961) отмечает для бассейна Ангары пролет
трех подвидов Камышевой овсянки. Нами встречена камышевая
овсянка лишь однажды в кустах на берегу старицы р. Чадобец
1 мая 1962 г. Была ли это отставшая пролетная птица или же
168
загнездившаяся, судить не беремся. Размер семенников 5.0 X
хЗ.О.
Лапландский подорожник — Calcarius lapponicus L.
Пролетный вид полосы южной тайги Средней Сибири. На
весеннем пролете в 1959 г. встречен 18 мая (улус Байша). Экзем-
пляр добыт из огромной стаи. Весной 1962 г. на р. Чадобец по-
дорожники появились 22 мая. С 25-го птицы летели большими ста-
ями. К 1 июня пролет резко ослабел и вскоре прекратился. Осен-
ний отлет лапландских подорожников, по литературным данным,
наблюдается в сентябре (Юдин, 1952; Малышев, 1963).
Подорожники летят главным образом над значительными от-
крытыми пространствами, поэтому обычны по долине Лены, но,
как правило, отсутствуют в более узких боковых долинах ее при-
токов.
Пуночка — Plectrophenax nivalis L.
Пуночка зимует в пределах южной тайги Средней Сибири.
В окрестностях Иркутска пролет идет 16—31 марта (Малышев,
Рис. 34. Пролетная стайка пуночек.
1963). На р. Чадобец пролет в 1962 г. начался 25 марта (рис. 34)
и продолжался до 23—25 апреля. На р. Тутуре, да и вообще на
верхней Лене, пролет пуночек мало заметен.
169
Сем. ЖАВОРОНКОВЫЕ — A LAU DI DAE
Полевой жаворонок — Alauda arvensis L.
В распространении полевого жаворонка в пределах Средней
Сибири много неясностей. Жаворонок почти не проникает в полосу
тайги и не гнездится (или очень редок) ни в долинах притоков
верхней Лены, ни нижней Ангары. Е. Е. Сыроечковский и
Э. В. Рогачева (1958) отмечают практическое отсутствие птицы
на Енисее. И вместе с тем пролет жаворонков наблюдается и на
Лене, и на нижней Ангаре. Обычны птицы на пролете в окрестно-
стях дер. Яркино на р. Чадобец.
В полосе лесостепи Средней Сибири жаворонки всюду гнездят-
ся. На Лене они распространены приблизительно до Верхоленска,
но, как правильно отмечал И. С. Поляков (1873), наиболее за-
метны до Качуга. По Ангаре жаворонок проникает, как регу-
лярно гнездящийся вид, до ее среднего течения (приблизительно
до г. Братска). Далее на север жаворонки обычны в Якутии и,
очевидно, спорадически гнездятся на Подкаменной и, возможно,
даже (под большим вопросом) Нижней Тунгуске. Наше предполо-
жение основано на факте пролета птиц, наблюдавшемся на Ан-
гаро-Подкаменнотунгусском междуречье. По сообщениям кор-
респондентов-фенологов, в верхнем течении Нижней Тунгуски
жаворонка нет.
Первая песня жаворонка под Красноярском в среднем за 27
лет отмечена 8 апреля, в окрестностях Головинской метеостанции
(Аларский аймак Усть-Ордынского Бурятского национального ок-
руга) 31 марта (средняя дата за 7 лет, Реймерс, 1960а), под Иркут-
ском, по новым данным, 27 марта — наблюдения Л. И. Малышева
(1963) за 1954, 1955, 1956 и 1960 гг. и наши за 1959, 1961, 1962 и
1963 гг. Таким образом, на востоке полосы южной тайги пролет
начинается несколько раньше (21—26 марта). Это в общем подтвер-
ждается и регистрацией первой песни жаворонка наблюдателями
фенологической сети. Данные корреспондентов-фенологов приве-
дены в табл. 2.
Под Качугом жаворонки появляются приблизительно на 1—3
дня позже, чем под Иркутском, но уже в окрестностях Жигалова
птицы прибывают лишь около 10—12 апреля. Пролет длится
недолго и прекращается приблизительно через 10 дней (в 1961 г.
21 апреля). При этом летят одиночные особи или пары поющих
птиц. В Якутии прилет регистрируется лишь в конце апреля—
начале мая (Скалой, 1956; Воробьев, 1963).
На р. Чадобец в 1962 г. жаворонки прилетели 19 апреля.
В сравнительно большом числе (парами) жаворонки держались
на полях до 23 апреля, но потом исчезли. Одиночная самка, правда,
добыта 2 июля, но это исключение.
Весной самцы, особенно старые, прилетают раньше самок.
Они держатся по полям на проталинах, но не поют. Пение обычно
170
Таблица 2
Даты первой песни жаворонков в Средней Сибири
(Иркутская обл.)
Пункт наблюдения Годы
I960 1961 1962 1963
с. Тунка 28 III 8 IV
дер. Духовщина 25 III 20 III 7 IV 7 IV
Ольхонский р-н, дер. Косая Степь . 1 IV 30 III — 10 IV
Усть-Кутский р-н, с. Марково . . . — 1 IV (?) 10 IV 11 IV
Аларский р-н, с. Иваническ .... 21 III — — —
Черемховский р-н, с. Голуметь . . . 27 III — — —
Черемховский р-н, с. Гамыль , . . — 15 III — —•
г. Зима 8 IV — 5 IV —
пос. Куйтун — — 4 IV 6 IV
Куйтунский р-н, с. Барлук .... 25 III 26 III 26 III —
Усольский р-н, с. Болыпе-Жилкино — 20 III — —
Тулунский р-н, с. Шерагул .... — 27 III 9 IV 4 IV
начинается через несколько дней после появления передовых
птиц.
Осенний пролет жаворонков на р. Чадобец отмечен в послед-
ней декаде августа. 22 августа добыта 1 птица. Под Иркутском
пролет заканчивается в конце сентября—начале октября. От-
мечена (Гагина, 1962а) задержка жаворонков на зиму.
Едва ли птицы могли успешно перезимовать под Иркутском.
Скорей всего Т. Н. Гагиной зарегистрирован факт не зимовки,
а фенологической аномалии, вызвавшей угасание инстинкта
перелета. Как отмечает упомянутый автор, птицы были сильно
истощены. Вероятно, они погибли уже в середине зимы.
На двух пролетных жаворонках, добытых в окрестностях
дер. Яркино, иксодовых и гамазовых клещей не оказалось.
Рогатый жаворонок — Eremophila alpestris L.
Пролетный вид полосы южной тайги. На верхней Лене рогатые
жаворонки в 1961 г. появились 6 мая. Весной 1962 г. на р. Чадо-
бец они отмечены с 12 мая, и их пролет продолжался до 25 мая. На
Лене стайки были невелики и редки, на Ангаре пролетные рогатые
жаворонки многочисленны как по числу стай, так и по коли-
честву птиц в стае (до нескольких сотен птиц). Ранние стайки были
по 30—40 экз. и лишь с 17 мая число птиц в них резко увели-
чилось.
Любопытно, что в лесостепи под Иркутском рогатые жаворонки
исчезают в конце марта (не позже 31-го, — Малышев, 1963). Разрыв
171
на целый месяц между исчезновением птиц в полосе лесостепи
и появлением их в более северных местах заставляет предполагать,
что в южной тайге регистрируются не те птицы, которые встре-
чаются в окрестностях транссибирской железной дороги.
Сем. ТРЯСОГУЗКОВЫЕ — MOTACILLIDAE
Белая трясогузка — Motacilla alba L.
Белая трясогузка распространена по всей южной тайге, дости-
гая большой численности в расположенных ниже широтах. Это
связано с тем, что она чаще, чем другие трясогузки, гнездится в
крупных населенных пунктах и различных строениях. Вне по-
строек человека гнездо белой трясогузки нам удалось видеть лишь
один раз — оно помещалось под нависшей дерновиной обвалив-
шегося берега реки среди переплетения корней дерева (самого
дерева на берегу не было: его давно уже спилили).
В связи с приуроченностью белых трясогузок к населенным
пунктам численность этих птиц установить трудно. Очень при-
близительно можно считать, что на верхней Лене одна гнездящаяся
пара приходится в среднем на 3 деревенских двора, но местами
в каждой постройке есть «своя» пара. На р. Чадобец белые трясо-
гузки малочисленнее — приблизительно 1 пара на 7—10 дворов.
В августе после периода гнездования белые трясогузки кочуют
вдоль крупных рек, достигая плотности (на Лене) одной стайки
(выводка?) на 1—3 км береговой полосы (одного берега).
Согласно многолетним данным, белые трясогузки прилетают
в окрестности Красноярска 11 апреля (средняя дата за 28 лет) и под
Иркутск — 7 апреля (средняя за 21 год). В последние годы под Ир-
кутском птицы появлялись также 7—9 апреля (Малышев, 1963).
В пределы Якутии белые трясогузки прибывают лишь в конце
апреля со средней датой между 18-м и 27-м (Скалой, 1956). В 1960 г.
трясогузки прилетели в южную тайгу Средней Сибири между 12 и
21 апреля. Весной 1961 г. 3—18 апреля, в 1962 г. между 2 и 21 апреля,
а в 1963 г. птицы появились 8—25 апреля. Даты прилета в очень
большой степени зависят от мезоклиматических особенностей мест-
ности (больше, чем от ее географической широты). Так, обычно
на верхнюю Лену белые трясогузки прилетают позже, чем на
верхнюю и среднюю Ангару (иногда задержка достигает, как
в 1962 г., 10 дней: Ангара — 8—16, Лена — 23—25 апреля).
Но в окрестностях с. Марково (севернее Усть-Кута) белые трясо-
гузки нередко появляются раньше, чем у Жигалова, расположен-
ного намного южнее. В 1962 г. на нижней Ангаре (дер. Яркино)
и на верхней Лене (Жигалово, Усть-Кут) белые трясогузки поя-
вились одновременно 25—26 апреля.
Осенний пролет белых трясогузок происходит на верхней
Лене с конца августа—начала сентября. Около середины месяца
172
птицы летят уже большими стаями и в двадцатых числах сентября
исчезают из пределов верховий Лены. Под Иркутском трясогузки
держатся в теплые осени до середины октября.
Сразу по прилете трясогузки занимают места гнездования и от-
гоняют «чужаков» с облюбованных крыш и чердаков. В 1960 г.
такое поведение птиц мы наблюдали на верхней Лене уже 22 ап-
реля (прилетели трясогузки 13-го). 24 мая была добыта птица с 2
яйцами в яйцеводе, одно из которых было вполне сформировав-
шимся. В 1961 г. трясогузки прилетели также 13 апреля, а 14
июня у них были птенцы 2—3 дней от роду. Найденные гнезда
помещались главным образом на полого (но не горизонтально)
поставленных балках чердачных переплетов и были сложены из
шерсти домашних животных и кабарги. Тут же ниже были гнезда
прошлых лет. В гнездах 5—6 птенцов. Они часто сильно поражены
различными эктопаразитами. Из одного гнезда с 6 птенцами
молодые вылетели через 12 дней после вылупления, но тут же 2
птенца, особенно зараженные паразитами, погибли. Видимо/
гибель птенцов — обычное явление. С родителями в летающих
выводках бывает 3—4 и редко больше птенцов. Массовое появле-
ние хорошо летающих выводков белых трясогузок отмечено 2—3
августа. Птицы кормились на лугах.
Горная трясогузка — Motacilla cinerea Tunst.
Горная трясогузка в отличие от белой связана с человеком
лишь косвенно. По берегам рек и озер она гнездится независимо
от того, нарушены ли леса человеком. Лишь по глухим таежным
речкам и ручьям условием для поселения вида служит рас-
чистка значительной поляны. Необходимо горным трясогузкам
и наличие мелкощебенистых отмелей или нагромождений из круп-
ных камней.
Численность горных трясогузок: 1 пара на 1 км береговой
полосы р. Тутуры (29 мая 1961 г.).
Прилетают горные трясогузки поздно. На верхней Лене в
1961 г. они появились впервые 15 мая. Весной следующего года
на р. Чадобец птицы появились 14 мая. Осенний пролет на верхней
Лене отмечен с 1—2 сентября. 5—7 сентября птицы летели боль-
шими стаями (1961 г.). Под Иркутском, по данным Л. И. Малы-
шева (1963), горные трясогузки задерживаются до 15 октября
(1955 г.).
Слетки горных трясогузок отмечены 25 июня.
Желтоголовая трясогузка — Motacilla citreola Pall.
По картосхеме распространения подвидов желтоголовой тря-
согузки, составленной Н. А. Гладковым (1954), значительная
часть южной тайги Средней Сибири, и в том числе все Верхоленье,
173
лишена описываемого вида. Это не так. Птица гнездится в южной
тайге повсеместно, но не достигает высокой численности. Вместе
с желтой трясогузкой она занимает мокрые луга и побережья
речных стариц. В глубине тайги она не встречается, за исключе-
нием луговин вблизи от человеческих построек (только по берегам
речек или у крупных озер; вообще же все трясогузки на водораз-
делы почти не проникают).
Весной 1961 г. в окрестностях Жигалова на Лене желтоголовая
трясогузка отмечена 15 мая, вблизи дер. Яркино на р. Чадобец
в 1962 г. — 21 мая. Под Иркутском птица появляется раньше —
в конце апреля—начале мая и улетает отсюда в конце августа
(Малышев, 1963). Приблизительно в это же время происходит
прилет и отлет под Красноярском (Юдин, 1962).
Максимальный размер семенников отмечен у особей с р. Лены
17 мая: 10.0x7.5 мм.
3 экз. желтоголовых трясогузок, обработанных паразитоло-
гами на р. Чадобец, оказались свободными от иксодовых и гама-
зовых клещей.
Желтая трясогузка — Motacilla flava L.
Желтая трясогузка — немногочисленная птица в полосе юж-
ной тайги Средней Сибири. Характерно, что под Иркутском про-
лет этого вида не наблюдается. На р. Тутуре 1 пара приходится
приблизительно на 5 км береговой полосы.
На р. Лене у Жигалова весной 1960 г. трясогузки этого вида
отмечены 19 мая, а в 1961 г. — 15 мая. На р. Чадобец вид зареги-
стрирован не был.
24—25 мая 1961 г. птицы были парами и, очевидно, загнезди-
лись.
Осенний отлет на Лене констатирован в первых числах сен-
тября.
Степной конек — Anthus richardi Vieill.
Обычнейшая птица приречных лугов. Прилетает степной конек
поздно: на Лену 1 июня (1961 г.), на северные притоки нижней
Ангары также в это время (3 июня 1962 г., р. Чадобец). Пролет
идет в очень сжатые сроки. Через 1—2 дня после прилета коньки
уже разбиваются на пары.
Фолликулы до 19 мм в диаметре отмечены 27 июня (р. Чадобец,
дер. Яркино). Хорошо летающие выводки степных коньков встре-
чены 2 августа 1961 г.
Осенний отлет птиц на р. Лене в 1961 г. наблюдался 5—7
сентября. Лишь на несколько дней позже птицу можно было уви-
деть под Иркутском.
На 14 осмотренных птицах иксодиды не обнаружены, обычны
пухоеды.
174
Лесной конек — Anthus trivialis L.
Лесной конек — вид, характерный для лесополья, сосновых
боров и нарушенной темнохвойной тайги, изобилующей полянами.
Птица избегает лишь мест с влажной почвой и поэтому отсут-
ствует в заболоченных долинах таежных речек и ручьев. В южной
тайге Средней Сибири лесной конек практически повсеместен
(кроме указанных долин и дремучих лесов).
Своим широким распространением лесные коньки безусловно
обязаны человеку, однако сосновые боры были заселены видом
издавна. Можно предполагать, что до значительного нарушения
тайги человеком птица обитала лишь в первичной лесостепи (в
том числе в Якутии) и по некоторым притокам Ангары (например,
Оке) доходила до средней Ангары. Перестойные темнохвойные
парковые леса водоразделов на определенной фазе сукцессии
(осушения) также могли быть заселены лесными коньками, но
не севернее 60° с. ш. (тут фаза осушения уже не наблюдается).
Дальнейшее продвижение вида на север происходило под воздей-
ствием антропогенных факторов. На Енисее конек достиг очень
высоких широт (Сыроечковский, 1960).
В 1960 г. коньки прилетели на верхнюю Лену в заметном
количестве 16 мая, следующей весной они появились намного
раньше — 27 апреля, но затем долго не были видны. Лишь 5—6
мая их стало много. Интенсивный пролет наблюдался до 14 мая.
На два года раньше, весной 1959 г., в б. Баяндаевском районе у
улуса Байша лесные коньки появились 9 мая. Все даты весеннего
появления коньков, отмеченные Л. И Малышевым (1963) для
окрестностей Иркутска, также относятся к лесному коньку (пят-
нистый конек, разбираемый автором совместно с лесным, никогда
не прилетает около 1 мая). На нижней Ангаре в 1962 г. лесные
коньки появились 7—8 мая (рис. 35). Это обычная дата появления
птиц под Красноярском (Крутовская, 1958). Массовый пролет
лесных коньков на р. Чадобец продолжался до середины мая.
Осенний пролет лесных коньков наблюдается на Лене с 10—
22 августа (1960 г.). Самая поздняя дата добычи лесного конька
на верхней Лене в 1961 г. 3 сентября, следующей осенью — 23
августа. На р. Чадобец последняя птица этого вида была добыта
20 августа 1962 г.
Птенцы у лесных коньков найдены в гнезде 18 июня. Массовый
вылет молодых наблюдался 25—26 июля, но первые полувзрослые
особи добыты 15 июля (1961 г.).
Лесные коньки — одни из наиболее важных прокормителей
иксодовых клещей. Они заражены преимагинальными фазами
развития паразита даже там, где общая численность имаго очень
мала. На р. Чадобец из 28 птиц долинно-придолинной полосы
зараженной оказалась одна. 24 особи с водоразделов были сво-
бодны от паразита. На р. Тутуре в 1961 г. было очесано 13 лесных
175
коньков долинно-придолинной полосы, 4 оказались зараженными
иксодидами. Среди них были один птенец и два слетка. Макси-
мальная зараженность отмечена у молодого самца 31 июля:
5 личинок и 2 нимфы (на остальных — 1—2 личинки или нимфы).
Из 14 птиц, добытых на водоразделе, заражены были 5 особей
(для этой полосы самая высокая зараженность!). Максимум
Рис. 35. Лесной конек.
обилия составили 3 нимфы (5 июля). Лесной конек так сильно
поражен иксодидами из-за наземного гнездования и обычая до-
бывать пищу на земле. Вид — вирусоноситель энцефалита (Нау-
мов и др., 1963).
Пятнистый конек — Anthus hodgsoni Riclim.
В среднесибирской лесостепи пятнистый конек, видимо, не
гнездится. Во всяком случае под Иркутском он не обнаружен.
На Лене, как мы указывали (Реймерс, 1963г), птица занимает
очень ограниченные биотопы — долины глухих таежных ручьев
и речек. В бассейне р. Чадобец пятнистый конек гнездится в ана-
логичных биотопах и заселяет островные темнохвойные леса.
Изредка вид встречается в водораздельных сосняках с примесью
лиственных пород.
Можно предполагать, что пятнистый конек был оттеснен к се-
веру в связи с общим потеплением, вызванным расчисткой при-
речных и приручьевых ельников, заменой темнохвойных лесов
176
светлохвойными, а также распашкой и превращением в лесо-
польс больших площадей южной тайги.
Прилетают весной пятнистые коньки поздно. В 1959 г. в ок-
рестностях улуса Байша птицы появились 17 мая. У Жигалова
весной 1961 г. они стали заметны 22 мая. На р. Чадобец в 1962 г.
мы пятнистых коньков отметили лишь 1 июня (очевидно, с запозда-
нием), а на верхней Лене Г. А. Воронов добыл птицу 14 мая.
Осенний отлет пятнистых коньков происходит в сентябре
и выражен очень четко. Именно пятнистые коньки (а не лесные,
как предполагает Л. И. Малышев, 1963) задерживаются под Иркут-
ском до начала октября. На Лене пролет пятнистых коньков идет
с середины августа, достигает максимума в первых числах сен-
тября и длится по 20 сентября, после чего ослабевает и к концу
месяца прекращается (1960 и 1961 гг.).
Максимальная численность пятнистых коньков на верхней
Лене — пара на 1—1.5 км маршрута вдоль таежного ручья
(Реймерс, 1963г).
В конце июня птицы выкармливают птенцов (Лена). Молодые
добыты начиная с 10 августа.
Даже на осеннем пролете в сентябре пятнистые коньки зара-
жены иксодидами. На 13 птицах с верхней Лены паразиты най-
дены дважды (по разу нимфа и личинка). 8 птиц, обработанных
паразитологами на р. Чадобец, были свободны от иксодид.
Горный конек — Anthus spinoletta L.
Отмечен как пролетная и, возможно, гнездящаяся птица бас-
сейна Ангары (Гагина, 1961). В окрестностях Иркутска он, ви-
димо, действительно гнездится. Л. И. Малышев (1963) находил
там слетков 9—14 июля. В полосе южной тайги нами вид не кон-
статирован.
Сем. ПШЦУХИ — CERTH11DAE
Обыкновенная пищуха — Certhia familiaris L.
В Средней Сибири пищуха — редкая птица. Среди лесостеп-
ных колков полосы железной дороги и лиственничной лесостепи
Ангаро-Ленского междуречья за дневную экскурсию можно встре-
тить 1—2 пары птиц. Но в пределах южной тайги за все время
работ мы видели пищуху лишь 2—3 раза на Око-Ангарском водо-
разделе, одна сомнительная встреча зарегистрирована на верхней
Лене (вне пределов лесостепи и лесополья окрестностей Качуга,
где пищухи более обычны) и 3 экз. пищух были добыты на р. Ча-
добец. Последняя находка показывает, что потенциально ареал
пищух, как и всех других птиц южной полосы, может достигать
62° с. ш. Подтверждением могут служить пищухи, добытые
Е. Е. Сыроечковским (1960) на Енисее у Комсы.
12 Н. Ф. Реймерс 177
О приуроченности вида говорить трудно, но на р. Чадобец
пищуха — типичный обитатель темнохвойного леса островного
типа. В связи с этим птица населяет в большом числе водоразделы.
Самец, добытый 23 мая 1962 г., имел размер семенников 6.0X
Хб.Омм. 7 июля была отстреляна молодая птица, что дает ос-
нование считать пищуху гнездящимся видом Ангаро-Подкаменно-
тунгусского междуречья.
Все 3 пищухи были свободны от иксодовых и гамазовых кле-
щей.
Сем. ПОПОЛЗНИ — S1TTIDAE
Обыкновенный поползень — Sitta europaea L.
Поползень живет по всей полосе южной тайги. Для него важно
лишь наличие высокоствольного леса.
В сосновых лесах, в том числе остепненных — почти без под-
леска, численность поползней достигает 8—9 птиц на 5 км (май
1961 г., р. Тутура). В водораздельной темнохвойной тайге мы
насчитывали от 6 до 12 птиц на том же расстоянии (июнь 1961 г.).
Смешанные сосново-лиственные леса населены приблизительно
вдвое реже — 3 птицы на 5 км (июнь 1961 г.). На нижней Ангаре
численность птиц значительно ниже: 1—2, редко 3—4 особи на
5 км маршрута. При этом поползень предпочитает темнохвойные
леса и участки сосняков и листвягов (рис. 36) с большим участием
лиственных пород.
Поползень — периодически кочующая птица. На севере эти
кочевки приобретают характер перелетов. Весной на верхней
Лене прокочевка малозаметна и происходит в конце марта. Осенью
перемещение птиц выражено четко и происходит с начала сентября.
На р. Чадобец мы не встречали поползней до 8 мая. 2 С отмечен-
ного срока поползни встречались регулярно. Это наводит на
мысль о том, что поползни в пределы Ангаро-Подкамениотунгус-
ского междуречья прикочевывают с юга в сравнительно поздние
сроки.
Питается поползень летом различными насекомыми, собирая
их с земли, веток и из щелей коры. Выдалбливает он лишь тех
беспозвоночных, которые живут неглубоко в коре.
Молодые отмечены с 5 июля на нижней Ангаре (1962 г.) и
с 13 июля на Лене (1961 и 1962 гг.).
Несмотря на то что поползни нередко кормятся на земле (на-
пример, расклевывают опавшие листья берез, извлекая из них
насекомых), все 14 птиц, добытых на Лене и Ангаре, были сво-
бодны от иксодид.
2 Едва ли такая заметная птица могла быть пропущена во время наших
экскурсий с 17 апреля, а наших сотрудников — с 22 марта.
178
Рис. 36. Поползень.
Сем. СИНИЦЕВЫЕ — PARIDAE
Большая синица — Parus major L.
Как и все виды южнодолинного комплекса Средней Сибири,
большая синица на севере почти не выходит за пределы полосы
южной тайги. Птица принадлежит к лесопольному комплексу
и, помимо приречных кустарников, встречается лишь на опушках
сосняков и ельников, прилегающих к реке и полям.
В остепненных сосняках верхней Лены, тянущихся вдоль
южных скатов речных долин, численность больших синиц дохо-
дит до 1 птицы на 5 км маршрута. Среди приречных кустарников
большая синица многочисленнее: на р. Лене до 2 пар на 1 км,
а на р. Чадобец даже до 3. Однако такая высокая плотность насе-
ления вида характерна лишь для участков, расположенных вблизи
от населенных пунктов. В удалении от них число птиц падает
в 2—3 раза, а на таежных речках и на неокультуренных водораз-
делах вид совсем не встречается.
Существуют исключения из указанного правила. Вблизи
сравнительно крупных водораздельных озер (типа оз. Мильчиково
на Око-Ангарском междуречье) большая синица находит благо-
приятные места для гнездования, но только там, где лес резко
трансформирован человеком. Гнездится большая синица и на
водоразделах, но всегда вблизи населенных пунктов, т. е. ведет
себя здесь как лесопольная форма.
Очевидно, вид с нарушением южной тайги стал более много-
численным и широко распространенным. Первоначальная плот-
ность населения синиц была очень низка, вид концентрировался
лишь в полосе лесостепи и по долинам рек, дисперсно проникая
на север. В Средней Сибири большие синицы изначально едва ли
были распространены севернее 56° с. ш.
Большая синица совершает в Средней Сибири регулярные
перелеты: в марте—апреле на север и в октябре на юг. Длинный
перечень дат, констатированных под Иркутском, можно найти
в работе Л. И. Малышева (1963). В пределах южной тайги время
перелетов близко к наблюдаемому под Иркутском. Отставание
и форсирование не превышает одной недели даже на севере ре-
гиона.
Крупные фолликулы (до 2 мм) найдены у птицы, отстрелян-
ной 2 июня (Лена). Самец, добытый на р. Чадобец 21 апреля,
имел еще мелкие семенники: 3.0 X 2.0 мм.
4 очесанные большие синицы оказались свободными от иксо-
довых клещей.
Белая лазоревка — Parus cyanus Pall.
Т. Н. Гагина (1961) считает белую лазоревку очень редкой,
но гнездящейся птицей бассейна верхней и средней Ангары. Нам
не пришлось встречать этой птицы.
180
Московка — Parus ater L.
Московка — экологический антипод большой синицы. Обе
птицы распространены в Средней Сибири до 62-й параллели, но
если большая синица прежде всего вид лесополья, то московка —
птица еловых лесов и водораздельной темнохвойной тайги. В не-
сколько меньшем количестве московки гнездятся в смешанных
лесах, и лишь в числе 1—2 пар на 5 км маршрута ее можно найти
в сосняках (на их опушках московки живут в большем числе и
в этом отношении сходны с большой синицей).
Смена темнохвойных лесов светлохвойными нс приводит к ис-
чезновению вида, но резко сокращает его численность.
Московки южной тайги Средней Сибири совершают регулярные
и очень четко выраженные перелеты. Под Иркутском весенние
пролеты отмечены с конца марта—начала апреля по начало—ко-
нец мая, а осенние откочевки — с начала сентября по конец ок-
тября (Малышев, 1963). На верхней Лене весенний пролет выра-
жен слабо и происходит в те же даты, что и под Иркутском. Осен-
ний отлет на верхней Лене крайне интенсивен. Птицы летят стайка
за стайкой непрерывным потоком с первых дней сентября до его
середины. На нижней Ангаре весенний пролет идет до середины
мая. В 1962 г. московки особенно интенсивно летели в первую
декаду месяца.
Вслед за прилетом московки разбиваются на пары (7 мая 1961 г.,
верхняя Лена). Размер семенников достигает максимума в июне
(р. Чадобец) —10.0 X 8.0мм. В это же время у самок найдены круп-
ные фолликулы (2 мм). Однако уже 4 июля на р. Чадобец были
слетки. На верхней Лене они констатированы 27 июля.
В состав кормов московки входят как насекомые, так и семен-
ные корма.
С 13 очесанных московок, добытых на верхней Лене и на ниж-
ней Ангаре, иксодиды не были собраны.
Черноголовая гаичка — Parus palustris L.
Констатирована Т. Н. Гагиной (1961) как редкая гнездящаяся
птица бассейна верхней и средней Ангары. Нами в южной тайге
не встречена.
Пухляк — Parus atricapillus L.
Пухляк столь широко распространен в лесной полосе, что
практически вездесущ.
В водораздельных темнохвойных лесах численность пухляков
достигает максимума на фазе смешанных насаждений: 12—15
птиц на 5 км маршрута. На гарях этих птиц нет, за исключением
опушек и мест с сохранившимися отдельными деревьями. Такой
же численности, как и на водоразделах, пухляки достигают в со-
181
сновых лесах придолинной полосы (с примесью лиственных
пород). Водораздельные же леса такого типа населены намного
реже. Общий характер размещения пухляков ясен из рис. 37.
Легко можно подметить, что некоторое нарушение лесов бла-
приятно для пухляков, а лишение местности «настоящих» лесов
и превращение ее в гари почти так же неблагоприятно для вида,
как раскорчевка леса и распашка полей. Небольшие по размеру
гари заселены пухляками с большой плотностью — до 2 птиц
на га. Но это только в том случае, если на участке есть уже моло-
дой лиственный лес и сохранились стволы или хотя бы пеньки круп-
ных деревьев.
Пары у пухляков образуются в конце марта—начале апреля.
С конца апреля—начала мая пухляки строят гнезда, при этом
нередко сами изготавливают себе дупла в гнилых березах, в по-
раженных грибками пихтах, наконец просто в торцах обломан-
ных толстых сучьев с гнилой серединой. Работают обе птицы по-
очередно.
В конце первой декады мая пухляки откладывают яйца и на-
чинают сильно беспокоиться у гнезд. Около середины июня боль-
шинство пухляков занято кормлением птенцов и их часто при-
ходится видеть с кормом в клювах. В 1961 г. на верхней Лене
первые слетки появились 25—26 июня.
С конца августа—начала сентября птицы начинают усиленно
кочевать. Видимая численность их возрастает. Прокочевка пух-
ляков на верхней Лене продолжается до конца сентября. Под
Иркутском, т. е. на 400 км южнее Жигалова, прокочевка пухля-
ков становится заметной лишь в середине сентября. Продвиже-
ние птиц на север весной совпадает по времени с образова-
нием пар.
Все 42 пухляка, обработанные паразитологами, оказались
незараженными иксодовыми клещами. Редки на пухляках и дру-
гие эктопаразиты.
Сероголовая гаичка — Parus cinctus Bodd.
Сероголовая гаичка отсутствует в полосе южной тайги всюду,
за исключением высокогорий и, возможно, Енисейского кряжа,
который к высокогорьям не может быть отнесен, но тем не менее
обладает рядом фаунистических элементов, характерных для
гор. Не нашли мы сероголовую гаичку в верховьях Лены, во вся-
ком случае до высоты 950 м над ур. м. (наличие вида в более вы-
соких поясах вполне возможно, но леса таких уровней уже от-
носятся не к южнотаежным, а к горным). В Присаянье гаичка
обычна лишь на высотах свыше 1200 м.
На нижней Апгаре сероголовые гаички отмечены с начала
августа. При этом обе добытые птицы оказались молодыми. Наши
материалы, таким образом, не позволяют ни утверждать, ни от-
182
рицать возможность гнездования гаичек на Ангаро-Подкаменно-
тунгусском водоразделе: молодые особи могли быть местными
и прикочевавшими с севера. Исходя из теоретических предпосы-
Рис. 37. Принципиальная схема весенне-летнего^ размеще-
ния пухляков в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22.
лок, полоса между 60 и 62° с. ш. должна быть областью споради-
ческого гнездования описываемого северного вида.
Обе отстрелянные птицы не были заражены иксодовыми или
гамазовыми клещами.
Длиннохвостая синица — Aegithalos caudatus L.
Для Средней Сибири длиннохвостая синица — вид южно-
таежной полосы. А в гнездовое время даже в южной тайге, где
нет березы, там нет и опсловника. В связи с этим сведение хвойных
лесов нужно считать благоприятным фактором существования
вида.
Длиннохвостая синица — долинно-придолинная птица. На во-
доразделах даже в березняках ее нет.
Численность длиннохвостых синиц в южной тайге очень низка:
около 1 пары на 5 км маршрута в сосново-березовых лесах и при-
близительно вдвое больше в почти чистых березняках на месте
лиственничников (такие участки очень редки в южной тайге).
Осенью ополовники усиленно кочуют, и большие стайки птиц
нередко перелетают на значительные расстояния. Эти перемеще-
ния иногда принимают характер откочевок осенью к югу, а вес-
ной к северу. Так, в конце сентября 1960 г. на верхней Лене
наблюдались явно направленные кочевки длиннохвостых си-
ниц. Однако в другие годы такое явление не повторилось. Не
отмечено увеличение численности птиц и осенью 1959 г. под Ир-
кутском (Реймерс, 1963а). Л. И. Малышев (1963), наоборот, счи-
тает, что в этом году откочевка синиц окончилась 4 ноября. В дан-
ном случае наши наблюдения строились на более точной мето-
дике и их достоверность выше. Вывод Л. И. Малышева о регу-
лярной сезонной перекочевке ополовников мог быть получен
в результате случайного стечения обстоятельств и требует про-
верки.
Из 6 очесанных длиннохвостых синиц 1 оказалась зараженной
гамазовыми клещами. Иксодид на птицах не обнаружено.
Сем. СОРОКОПУТОВЫЕ — LAN1IDAE
Жулан — Lanius cristatus L.
Сибирский жулан (L. с. cristatus L.), как правило, птица
лесостепи и лесополья. Лишь в мелколиственных лесах и на га-
рях, поросших мелколесьем, он выступает в роли лесного вида.
Наивысшей численности жуланы достигают на небольших влаж-
ных гарях островных темнохвойных лесов или приручьевых
ельников с отдельными сухими деревьями. Здесь число птиц на 1 га
достигает 2 пар. Сухие гари сосняков населены значительно
реже, как и гари водораздельных темнохвойных лесов. При вос-
становлении сосняков без смены пород на их гарях и вырубках
сорокопуты не живут. Мало птиц и в мелколиственных лесах
водоразделов. На втором месте по численности жуланов после
гарей островных лесов следует поставить приречные ивняки и че-
ремушники. Однако тут число птиц не превышает нескольких
пар на 5 км маршрута.
184
Рис. 38. Остатки слетка мелкой птички, наколотые сороко-
путом-жуланом .
До резкого нарушения южной тайги сорокопуты-жуланы мог-
ли находить благоприятные условия лишь по берегам рек и, оче-
видно, были очень малочисленны. В дальнейшем птицы сначала
заняли придолинную, а затем и водораздельную полосу. У се-
верной границы южной тайги жуланы концентрируются в речных
долинах, почти не’ выходя на водоразделы.
Весной они прилетают около 1 июня (верхняя Лена 1961 г.
и р. Чадобец 1962 г.). Осенью жуланы собираются в стаи уже в кон-
це июля. В 1960 г. они исчезли с верхней Лены около 15 августа,
в 1961 г. — 15—17 августа. Наиболее поздняя добыча жулана
на р. Чадобец в 1962 г. 24 августа.
Разбивка на пары у жуланов происходит сразу после при-
лета. 10 июня мы наблюдали сцены ухаживания. Одна из птиц
извивалась, вращая головой и поводя хвостом. Танцующим ша-
гом она приближалась к самке, сидящей совершенно непод-
вижно. Вся сцена происходила на вершине сухой лиственницы или
другом заметном возвышении. В это время (10—25 июня) семен-
ники птиц достигают наибольшего размера (10.2 X 8.6 мм) и, оче-
видно, происходит спаривание. Молодые в заметном ’Количестве
встречаются с 20—25 июля. Вскоре после их вылета из гнезд
и мужания сорокопуты собираются в кочующие стаи и улетают.
Питаются жуланы крупными насекомыми, главным образом
жуками, которых ловят нередко на земле. Бывают случаи напа-
дения на мелких воробьиных птиц (рис. 38).
Из 14 птиц, обработанных паразитологами на верхней Лене,
3 оказались зараженными личинками иксодид (по 1 экз.); 5 птиц
с р. Чадобец паразитов не имели. Помимо иксодовых, жуланы
заражены гамазовыми клещами.
Серый сорокопут — Lanius excubitor L.
Серый сорокопут в южной тайге Средней Сибири не гнездится.
На верхней Лене птица отмечена поздно осенью во время кочевок.
На р. Чадобец серый сорокопут был добыт 17 августа 1962 г.
В северо-западном Хамар-Дабане он отмечен нами (Реймерс, 1959а)
23 ноября 1955 г. и более не встречен ни в этой горной стране,
ни в предгорьях Восточного Саяна. Очевидно, серый сорокопут
гнездится на юге Средней Сибири лишь в высокогорьях и в не-
большом количестве зимует в ее пределах.
Сем. СВИРИСТЕЛЕВЫЕ — BOMBYCILLIDAE
Свиристель — Bombycilla garrulus L.
Как и другие северные виды, свиристели не характерны для
южной тайги и редки в ней. На верхней Лене они гнездятся не
ежегодно и лишь на глухих сырых гарях приручьевых ельни-
186
ков и приручьевых участков темнохвойных лесов. Эти гари имеют
северный облик из-за холодной заболоченной почвы. Не исклю-
чено, что по таким участкам свиристели проникают далеко на
юг по всей полосе южной тайги, а не только в верхнеленской ее
части. Во всяком случае на нижней Ангаре пары их отмечены
23 мая, когда обычно у птиц бывают гнезда с яйцами.
Холодные березняки по гарям, как мы уже отменали^выше,
заселены овсянками-ремезами и некоторыми другими видами
севера. Таким образом, нарушение тайги не во всех случаях при-
водит лишь к одностороннему проникновению южных форм на
север, но иногда, наоборот, позволяет северянам жить на юге. Это
еще раз подчеркивает сложность и неоднозначность антропоген-
ных влияний на природу.
Массовая весенняя прокочевка свиристелей на верхней Лене
наблюдалась в двадцатых числах апреля 1961 г. 1 мая встречены
птицы парами. В июне мы свиристелей не видели, но 23 июля
они появились небольшими стайками (выводками?), представляв-
шими большую редкость. Лишь в середине сентября птицы стали
обычны в связи с откочевкой на юг.
Свиристели охотно питаются ягодами жимолости татарской.
Паразитологами обработана лишь 1 птица, оказавшаяся не
зараженной иксодидами.
Сем. МУХОЛОВКОВЫЕ — MUSCICAPIDAE
Серая мухоловка — Muscicapa striata Pall.
Подобно большинству других видов, проникающих с запада
на восток, серая мухоловка населяет нижнюю Ангару (в моно-
графии «Птицы Советского Союза» северная граница почему-то
проходит от устья Ангары к ее истокам, минуя нижнее и среднее
течение реки), и, по свидетельству Е. Е. Сыроечковского (I960),
на Енисее гнездится вплоть до пос. Комсы. Т. Н. Гагина (1961)
отмечает серую мухоловку как редкий вид верхней Лены.
Встречается птица почти исключительно в сосняках (чистых
и смешанных с мелколиственными породами), как придолинных,
так и водораздельных. В последних птица многочисленнее.
На р. Чадобец серые мухоловки появились в 1962 г. 28—29 мая.
Первые молодые добыты 15 июля. Наиболее поздняя добыча серой
мухоловки отмечена 14 августа.
С 7 мухоловок этого вида не удалось снять иксодовых или га-
мазовых клещей.
Мухоловка-касатка — Muscicapa sibirica Gm.
Мухоловка-касатка встречается в пределах всей южной тайги
Средней Сибири. Однако птица не достигает высокой численности.
В водораздельных темнохвойных лесах с мелколиственными по-
187
родами мы учитывали не более 1 пары на 5 км маршрута (верхняя
Лена). Приблизительно такая же численность касаток характерна
для приречных ельников с мелколиственными породами (мухо-
ловки охотно селятся на их опушках, поросших кустами).
У северо-западного предела распространения — на Ангаро-Под-
каменнотунгусском водоразделе — мухоловки-касатки еще более
малочисленны и предпочитают уже не темнохвойные леса, а смешан-
ные светлохвойные или даже чистые сосняки. Любопытно, что та-
кое предпочтение биотопов весьма закономерно. На южном пределе
распространения в горах Кентея (Козлова, 1930) касатка также
гнездится в смешанных насаждениях и речной уреме, а не в тем-
нохвойных лесах.
В связи с исчезновением темнохвойных лесов, и особенно
приречных ельников, число мухоловок-касаток снижается.
В 1961 г. мухоловки-касатки появились на верхней Лене
2 июня. Эта дата не отличается от наблюдаемых под Иркутском
(30 мая—6 июня, Малышев, 1963). Последняя регистрация вида на
Лене осенью 1960 г. — 12 сентября.
5 июля добыта самка с готовым к откладке яйцом. Это
была запоздавшая или повторная кладка, так как подрос-
шие касатки встречались с 28 июля. Вместе с тем молодые птицы
добыты и 12 сентября.
Среди насекомых, поедаемых мухоловкой-касаткой, видное
место занимают двукрылые, в том числе слепни.
На 6 экз. мухоловок-касаток иксодовые клещи обнаружены
не были.
Ширококлювая мухоловка — Muscicapa davurica Pall.
Один из характерных видов приречных кустарников. Лишь
на средней Ангаре, где широко распространены мелколиственные
леса, эта птица, по свидетельству А. П. Шведова (1962), обычна
вне полосы долины в светлохвойных лесах с обильным подлеском,
а также на вырубках и гарях, поросших мелколиственными по-
родами и кустарниками. На верхней Лене и р. Чадобец вид лишь
слегка выходит за пределы долинной полосы, поселяясь в лесах
с обильным кустарниковым ярусом, но не проникая на водораз-
делы.
Нарушение лесов трансформировало кружево ареала этого
вида, позволив ему расселиться несколько шире, чем изначально.
Но вместе с тем исконные его биотопы — приречные кустарники —
во многих местах сведены, и их место заняли поля, подходящие
вплотную к урезу воды. Тут ширококлювые мухоловки ис-
чезли.
В окрестностях Жигалова ширококлювые мухоловки появи-
лись в 1961 г. 4 июня. 19 июля встречены первые молодые
особи. В двадцатых числах июля многочисленные стайки птиц
188
кочевали по приречным кустарникам вдоль р. Тутуры. В августе
мы птиц уже не встречали.
2 из 6 ширококлювых мухоловок с верхней Лены оказались
зараженными личинками иксодовых клещей. На каждой из птиц
было по 1 экз. паразита.
Малая мухоловка — Muscicapa parva Bech.
Малая мухоловка в пределах южной тайги—один из самых
обычных видов рода. В горах юга Сибири она занимает придолин-
ную полосу лесов с высоким участием лиственных пород. Нарушен-
ные приречные и приручьевые ельники или эти же ельники, пере-
межающиеся с кустарниками, — исконные стации вида. Нару-
шение лесов, образование смешанных ассоциаций позволило ма-
лым мухоловкам широко заселить светлохвойно-лиственные леса,
а затем проникнуть на водораздельные гари. То же самое обстоя-
тельство привело к расселению мухоловок на север, где они
нашли благоприятные для себя условия в смешанных лесах и ос-
тровных темнохвойных участках. В силу не вполне ясных об-
стоятельств (возможно, отсутствия конкурентов) малая мухолов-
ка на северо-западе южной тайги Средней Сибири более много-
численна, чем в приленской и присаянской.
Наивысшей численности малые мухоловки достигают в лесах,
богатых подлеском, у истоков ручьев, особенно на гарях приру-
чьевых ельников и на приручьевых участках темнохвойных
лесов водоразделов. Здесь число птиц иногда превышает даже
их обилие в указанной выше исконной стации вида — приреч-
ного ельника, перемежающегося кустами, вдоль стариц. Среднее
число малых мухоловок констатируется в смешанных лесах,
низкое — в сосняках. В глухих водораздельных лесах вида нет.
Как ясно из нашего изложения, антропогенные нарушения
тайги благоприятны для вида.
В 1961 г. малые мухоловки на верхней Лене появились 1 июня,
а летом следующего года на р. Чадобец — лишь в середине ме-
сяца. Тестикулы у малых мухоловок достигают наибольшего раз-
мера (7.6 X 4.6 мм) 10—15 июня. Около 10 июля появляются
слетки малых мухоловок (р. Чадобец). Осенний отлет происходит
очень незаметно. Последняя птица на р. Лене была добыта 8 сен-
тября 1961 г.
На 10 экз. с верхней Лены и 23 птицах с р. Чадобец иксодо-
вые клещи найдены не были.
Мухоловка-мугимаки — Muscicapa mugimaki Temm.
Распространение мухоловки-мугимаки проследить трудно, так
как вид имеет резко колеблющиеся по годам границы ареала,
а численность птиц подвержена столь же заметным изменениям.
189
В 1961 г. нам не удалось обнаружить их на верхней Лене, несмотря
на самые тщательные поиски. Летом предыдущего года птица
была весьма обычна. Не представляла она редкости и в 1962 г.
Это заставляет нас с осторожностью упоминать о том, что мухо-
ловку-мугимаки мы не нашли на Ангаро-Подкаменнотунгусском
междуречье у дер. Яркино на р. Чадобец. Можно предполагать,
что, как и другие южнотаежные птицы, вид может в благоприят-
ные годы заселять все пригодные для гнездования участки, вплоть
до северных форпостов южной тайги (приблизительно до 62° с. ш.).
Мухоловка-мугимаки практически живет лишь в приручьевых
ельниках и приручьевых участках темнохвойной тайги. Доста-
точно одинокой ели среди кустарниковых зарослей вдоль ручья,
чтобы на этом дереве пел самец мухоловки.
В годы высокой численности малых мухоловок мухоловки-му-
гимаки почти, а иногда полностью исчезают из приручьевых ель-
ников и встречаются лишь во вторых биотопах. Численность
птиц выше всего в приручьевых участках темнохвойных лесов —
самцы поют тут на расстоянии от 150—200 м до 1.5 км друг от
друга (Реймерс, 1963г). Нарушение тайги крайне неблагоприят-
но для мухоловки-мугимаки. Можно предположить, что вид бу-
дет в конце концов вытеснен малой мухоловкой.
В 1960 г. мухоловки-мугимаки на верхней Лене зарегистриро-
ваны с 29 мая. Приблизительно с этого же времени (30 мая —
9 июня) они встречаются под Иркутском (Малышев, 1963). Осен-
ний пролет прослеживается с большим трудом и, видимо, проис-
ходит в конце августа—начале сентября. Исключительно поздняя
задержка одиночного самца мухоловки-мугимаки зарегистриро-
вана 23 октября 1959 г. под Иркутском. Птица держалась в кустар-
никах сухой пади, окруженной сосновым лесом.
Сем. СЛАВКОВЫЕ — SYLVIIDAE
Пеночка-весничка — Phylloscopus trochilus L.
Этот северный для Азии вид практически отсутствует в южной
тайге Средней Сибири. За все годы у нас имеется лишь одна весьма
сомнительная летняя находка птицы 25 июня 1962 г. на р. Чадо-
бец. 1 сентября 1961 г. взрослая самка добыта на верхней Лене.
Нет уверенности, что это была пролетная, а не залетная птица.
Пеночка-кузнечик — Phylloscopus collybitus Vieill.
Сибирский подвид теньковки Ph. collybitus tristis Blyth.
по пению очень мало похож на известную европейскую птичку
прежде всего потому, что отсутствует тот слог, который дал попу-
лярное название виду (теньковка). Распространен кузнечик
в южной тайге Средней Сибири очень широко, и число птиц в пре-
190
делах района лишь немного уменьшается к востоку. Восточную
границу вида составляет не р. Лена, а, очевидно, Прибайкаль-
ские хребты.
Наивысшей численности кузнечики достигают на опушках
приречных ельников, выходящих к кустарниковым зарослям
и ерникам. В этих кустарниках пеночки также живут в заметном
количестве и выходят на гари, поросшие березовым мелколесьем.
Чистых сосновых боров кузнечики избегают, нет их в темнохвой-
ных водораздельных лесах, но вид присутствует в смешанных
светлохвойно-мелколиственных древесных ассоциациях с обиль-
ным подлеском.
Численность птиц в наиболее благоприятных для вида биотопах
определить довольно трудно из-за концентрации пернатых в узких
полосах леса и кустарников. На 1 км опушки приречного ельника
бывает до 3 поющих самцов. Все данные по численности вида
получены на верхней Лене. В маленьком еловом островке пло-
щадью около 1 га гнездилась 1 пара теньковок. Гнездо помеща-
лось внутри островка, но на опушке, выходящей к речной старице,
которую окружал ельник. На гари сосняка, поросшей березовым
мелколесьем, площадью в 9—10 га зарегистрирована 1 пара птиц
описываемого вида. В смешанных светлохвойно-мелколиствен-
ных лесах придолинной полосы на 5 км маршрута встречалось
до 4 поющих самцов. В почти чистых сосняках, даже с обильным
подлеском, птиц было не более 1 пары на маршруте той же про-
тяженности.
Весенний пролет на верхней Лене зарегистрирован 2 мая
1961 г., 6 мая кузнечики начали петь. На р. Чадобец птички появи-
лись 1 мая 1962 г. и 2 мая они уже пели. Осенний отлет на Лене
наблюдается в начале—середине сентября, почти в то же время,
что и в окрестностях Иркутска.
В конце мая все кузнечики уже держались парами, хотя
пролет длился до первых дней июня. У птицы, добытой
29 мая 1962 г. на р. Чадобец, в яйцеводе было готовое к от-
кладке яйцо.
Лишь 2 птицы с верхней Лены были очесаны паразитологами.
На одной из них найдены 2 нимфы иксодид (27 мая 1961 г.). На
8 экз. с р. Чадобец эти паразиты найдены не были.
Пеночка-таловка — Phylloscopus borealis Blas.
Таловки редки в районе наших работ. В пределах южной тайги
Средней Сибири мы эту птицу не нашли на гнездовье, а пролет
констатирован лишь в предгорных районах Саян и Хамар-Дабана.
Т. Н. Гагина (1961) считает таловку обычной гнездящейся птицей
верхней Лены и бассейна Ангары. Пролет этих пеночек хорошо
прослеживается под Иркутском (Малышев, 1963).
191
Зеленая пеночка — Phylloscopus trochiloides Sund.
Зеленая пеночка — сравнительно обычная птица заболочен-
ных ерников среди приручьевых еловых лесов. Редкость (по пло-
щади) подобных биотопов даже при высокой степени обилия птиц
(до 3 пар на 1 га ерников на средней Ангаре, 1959 г.) делает зеле-
ную пеночку малозаметным видом, общая численность которого
низка. Видимо, это обстоятельство не позволило нам получить
данные о сроках пролета вида.
Паразитологи очесали 5 пеночек, определенных работавшими
с нами студентами как Ph. trochiloides. Поскольку экземпляры
не были сохранены для просмотра автором, мы не уверены в пра-
вильности определения. Эти птицы были свободны от иксодовых
и гамазовых клещей.
Пеночка-зарничка — Phylloscopus inornatus Bly th.
Зарничка в южной тайге Средней Сибири — птица березняков
и сосново-березовых лесов. В долинных темнохвойных лесах
этот вид встречается лишь при условии очень высокого участия
мелколиственных (не менее 5Б) пород и чаще всего по опушкам.
На водоразделы зарничка проникает по гарям, поросшим берез-
няком, но в очень небольшом числе.
На нижней Ангаре пеночки-зарнички в 1962 г. появились
24 мая. Пролет шел очень интенсивно и продолжался до первых
чисел июня. Также 24 мая прилетели зарнички на верхнюю Лену
предыдущей весной. Птицы сразу появляются в массе, довольно
большими стайками. К концу августа зарнички исчезают из пре-
делов южной тайги.
Тестикулы у самцов достигают наибольшего размера (6.0 X
X 5.5 мм) в первой декаде июня. 17 июля встречены самостоя-
тельно живущие молодые (р. Чадобец).
На 22 зарничках иксодиды обнаружены не были.
Корольковая пеночка — Phylloscopus proregulus Pall.
Корольковая пеночка — характерный вид водораздельных
темнохвойных лесов. Тут на высочайших лиственницах и кедрах
самцы поют все лето. В пределах южной тайги Средней Сибири
корольковая пеночка нигде, кроме водораздельных темнохвойных
лесов, не встречается. Численность птиц — не более 3—4 пар
на 5 км маршрута. Такая же плотность населения вида была кон-
статирована нами в кедровых лесах Хамар-Дабана (Реймерс,
1959а).
Исчезновение темнохвойных лесов, а также их омоложение
резко сокращает численность корольковых пеночек и «утон-
чает» кружево их ареала. Во многих местах вид исчез совсем.
192
Так же как в Восточном Саяне и Хамар-Дабане, корольковая
пеночка начинает петь в южной тайге Средней Сибири с 1—2 июня
даже в таком сравнительно северном месте, как Ангаро-Подка-
меннотунгусское междуречье (1962 г.).
Самцы 25 и 27 июня имели наибольшие семенники (6.0 X 4.4
и 5.2 X 3.9 мм). Молодого нам удалось добыть 9 августа 1961 г.
На 4 корольковых пеночках иксодиды найдены не были.
Бурая пеночка — Phylloscopus fuscatus Blyth.
Встречена лишь среди ерников по таежным речкам. В пределах
южной тайги Средней Сибири птица всюду довольно редка.
Молодой экземпляр добыт 21 августа 1961 г.
Голосистая пеночка-камышевка — Herbivocula schwarzi Rad do
Г. А. Воронов добыл 28 августа 1960 г. в районе Жигалова
молодую птицу, отнесенную к этому виду. Полной уверенности
в правильности определения ни у коллектора, ни у нас нет. Экзем-
пляр был в очень плохом состоянии (птица попала в заполненный
водой цилиндр для ловли грызунов).
Малая пестрогрудка — Bradypterus thoracicus Bly th.
T. H. Гагина (1961) считает этот вид изредка гнездящимся
в верховьях Ангары. В пределы южной тайги малая пестрогрудка,
так же как и сибирская пестрогрудка (В. taczanowskius Swinh.),
не проникает.
Толстоклювая камышевка — Phragmaticola a6don Pall.
В южной тайге Средней Сибири нами этот вид не найден.
Т. Н. Гагина (1961) считает толстоклювую камышевку птицей,
редко гнездящейся на Ангаре и, возможно, на Лене.
Пятнистый сверчок — Locustella lanceolata Temm.
В полосе южной тайги Средней Сибири пятнистый сверчок,
как и остальные виды этого рода, живет почти исключительно
в кустарниках по опушкам полей, особенно вблизи ручьев. Вне
культурного ландшафта вида практически нет. Изначально свер-
чок мог гнездиться в кустах на берегах пересохших стариц и по
кустарниковым опушкам между остепненными участками и лесами.
Прилетают сверчки на верхнюю Лену в конце первой декады
июня (1960 г. 8 июня, 1961 г. между 6 и 9 июня). Улетают птицы
поздно: взрослая самка была добыта между 28 и 30 сентября
1961 г. В это время на верхней Лене уже выпал снег. Предыдущей
осенью последний сверчок был добыт 24 сентября.
2 экз., обработанные паразитологами, были свободны от иксодид.
13 Н. Ф. Реймерс
193
Певчий сверчок — Locustella certhiola Pall.
Этот вид значительно более редок в пределах южной тайги,
чем предыдущий. Нам удалось добыть всего 1 экз. вне периода
гнездования (10 сентября 1961 г.) в приречном ивняке. Иксодид
на нем не оказалось.
Таежный сверчок — Locustella fasciolata Gray
1 экз. этого вида был добыт на гари соснового леса, поросшей
березовым мелколесьем, 19 августа 1961 г. (р. Тутура).
Вся группа сверчков связана с нарушенными лесами. Перво-
начально они были распространены в лесостепной полосе Средней
Сибири и вдоль рек в первичных кустарниковых зарослях.
Дроздовидная камышевка — Acrocephalus arundinaceus L.
Т. Н. Гагина (1961) констатирует залет дроздовидной камы-
шевки в бассейн верхней Ангары.
Садовая камышевка — Acrocephalus dumetorum Blyth.
2 экз., с сомнением отнесенные к этому виду Г. А. Вороновым
(личное сообщение), были добыты им 18 июня 1962 г. на р. Тутуре
в приречном ельнике. Птицы оказались зараженными гамазовыми
клещами и на одной из них были 2 вши. Т. Н. Гагина (1961)
считает садовую камышевку изредка гнездящейся птицей Ангары
и верхней Лены.
Бормотушка — Hippolais caligata Licht.
Птица известна лишь из крайних западных пределов южной
тайги Средней Сибири. Т. Н. Гагина (1961) предполагает очень
редкое гнездование вида в бассейне верхней и средней Ангары.
Садовая славка — Sylvia borin Bodd.
Этот вид доходит с запада до пределов южной тайги Средней
Сибири, но в ней, очевидно, не распространен.
Серая славка — Sylvia communis Lath.
Хотя в монографии «Птицы Советского Союза» ареал серой
славки показан с охватом присаянских районов, ни нам, ни
Т. Н. Гагиной (1961) этого вида тут наблюдать не удалось.
194
Славка-завирушка — Sylvia curruca L.
Единственная славка, широко распространенная в полосе
южной тайги Средней Сибири, населяет приречные черемушники,
кустарники по опушкам лесов у полей и лугов и участки мелко-
лесья на вырубках приручьевых и приречных ельников. Сведение
лесов благоприятно для вида и позволяет ему занять придолин-
ную полосу.
В 1961 г. завирушки прилетели на верхнюю Лену 22 мая.
Весной следующего года на р. Чадобец птицы появились 15 мая.
Интенсивный пролет в обоих местах продолжался до начала июня.
С конца мая завирушки начинают разбиваться на пары. Фолли-
кулы самки, добытой 22 мая, достигали 2 мм в диаметре. Макси-
мальный размер семенников зарегистрирован 3 июня 1962 г.
(10.8 X 6.4 мм).
На весеннем пролете славки-завирушки бывают заражены
иксодовыми клещами. Так, 22 мая 1961 г. на р. Тутуре была
добыта самка, с которой были сняты 2 нимфы иксодид. Три осталь-
ные птицы были свободны от паразитов. Также не были заражены
14 птиц с р. Чадобец, добытых в 1962 г. с 15 мая по 8 августа.
Сем. ДРОЗДОВЫЕ — TURDIDAE
Пестрый дрозд — Turdus dauma Lath.
Пестрый дрозд в полосе южной тайги Средней Сибири отно-
сится к числу типичных птиц водораздельного темнохвойно-
лиственного леса. Возможной заменой обычной стации может
быть смешанный сосново-мелколиственный лес и как исключение
сосновый бор или мелколиственное насаждение. Схема рас-
пределения этой интереснейшей птицы показана на рис. 39.
Предпочтение, которое пестрый дрозд отдает темнохвойным
лесам, заставляет предположить, что общая численность вида
резко уменьшилась в ходе нарушения лесов человеком и смены
темнохвойной тайги светлохвойной.
Сосново-мелколиственные леса населены дроздами намного
реже, чем темнохвойная тайга. Вместе с тем в смешанных темно-
хвойных лесах число пестрых дроздов выше, чем в слабо на-
рушенной тайге. Это особенно заметно в приленской части
южной тайги Средней Сибири.
В горах юга Сибири распределение плотности населения пест-
рых дроздов несколько иное. Там наивысшей численности вид
достигает в светлохвойно-лиственных лесах, на втором месте
стоят нарушенные темнохвойные леса, на последнем — сравни-
тельно нетронутые насаждения.
В ходе естественного сукцессионного процесса численность
пестрых дроздов также меняется. Редкостойные леса населены
13*
195
видом плотнее, чем сомкнутые многоярусные с разновозрастным
древостоем.
Пестрого дрозда нельзя отнести к многочисленным птицам,
но он не принадлежит и к раритетам. Количественный учет вида
Рис. 39. Принципиальная схема весенне-летнего размещения
пестрых дроздов в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22.
крайне затруднен тем обстоятельством, что к тому времени, когда
на водоразделах сходит снег, а разлившиеся реки позволяют
свободно проникать в глубь тайги, дрозды перестают свистеть
и становятся очень скрытными. Оценку численности мы сделали
196
на основании частоты добычи (а значит, и встреч), при учете,
конечно, и степени посещаемости лесов. Из 17 птиц 10 было
добыто в темнохвойном лесу со значительной примесью мелко-
лиственных пород, 4 дрозда отстреляны в сосново-мелколиствен-
ных лесах водораздельного типа, главным образом с примесью
темнохвойных пород, остальные птицы добыты в других биотопах.
Даты прилета пестрых дроздов в пределы южной тайги нами
не были установлены. Наиболее поздняя добыча вида 23 августа
1962 г. По Л. И. Малышеву (1963), под Иркутском вид наблюдается
весной с середины мая, а осенью пролет происходит в конце сен-
тября—начале октября.
Единственное гнездо пестрого дрозда было найдено работав-
шим с нами студентом В. Лямкиным 5 июля 1961 г. в темнохвой-
ном лесу. Гнездо помещалось 3 в углублении трухлявой вершины
высокого березового пня. Высота пня 3—3.5 м. Подстилка гнезда
состояла из сухих веточек кедра и его хвои, в глиняном основании
было много зеленых мхов. Размеры гнезда, к сожалению, не были
установлены. Когда наблюдатель стукнул по пню, птенцы выле-
тели из гнезда.
Молодые в массе появились на верхней Лене в 1961 г. после
10 июля, на р. Чадобец в 1962 г. после 5 июля.
Среди 12 обработанных паразитологами пестрых дроздов
с верхней Лены 2 оказались зараженными иксодидами (5 и 10
июля). С молодой самки 10 июля были сняты 3 нимфы иксодид.
3 осмотренных дрозда с р. Чадобец были свободны от иксодовых
клещей.
Сибирский дрозд — Turdus sibiricus Pall.
Никто из исследователей не наблюдал сибирского дрозда в пре-
делах южной тайги летом (Гагина, 1961). Мы не встретили вид
даже на пролете. Очевидно, сибирский дрозд не гнездится в полосе
наших исследований, хотя и обычен в горах юга Сибири.
Рябинник — Turdus pilaris L.
Рябинник — крайне стенотопный вид в южной тайге. Единствен-
ное место его гнездования — куртины приречных ельников среди
пойменных лугов и кустарников. В сплошных приречных ельни-
ках мы птицу не нашли, зато в небольшой еловой куртине была
обнаружена колония из 4—5 гнезд.
В связи с такой спецификой распределения рябинник немного-
числен, в особенности при отнесении количества птиц к общей
территории южной тайги. Сведение лесов было, очевидно, благо-
3 Описание дается со слов зоолога В. Лямкипа. Автор гнезда не видел,
поскольку место, где обнаружили гнездо, было утеряно.
197
приятно для вида, хотя уничтожение ельников приводит к сокра-
щению числа этих птиц.
Весной 1961 г. прилет рябинников на верхнюю Лену был
зарегистрирован 6 мая и длился до середины месяца. Следующей
весной на р. Чадобец рябинники достоверно были отмечены 10 мая.
Птицы держались среди залитых половодьем приречных ель-
ников, поэтому даты прилета могли быть зарегистрированы с запоз-
данием. Весенний пролет рябинников происходит весьма интен-
сивно. За 4 часа в островном приречном ельнике на р. Тутуре
побывали 2 крупных пролетных стаи птиц.
Осенний пролет птиц происходит (верхняя Лена) поздно
(в конце сентября) и в южной тайге мало заметен.
7 мая 1961 г. фолликулы у рябинников достигали 5—11 мм.
19 мая в одном из гнезд рябинников было 3 яйца, и 21 мая уже
полная кладка (6 яиц). 22 мая в 3 других гнездах было 1,3 и 6 яиц.
Расположены были гнезда следующим образом. Одно помещалось на
горизонтальной ветви ели в 1 м от ствола на высоте 3.5 м от земли.
Второе было свито вплотную к стволу небольшой елки (5—6 м
высотой) в 2.5 м от земли. Третье помещалось на приблизительно
таком же дереве с очень тонкими ветвями, но на 1 м выше от
земли. Материал гнезд: сухие стебли трав (выстилка без глины),
сухие листья, отдельные стебли мхов. Размеры одного из гнезд:
высота 125, ширина 163, диаметр лотка ПО, глубина 65 мм. Раз-
меры 3 яиц: 29.3 X 20.7, 29.3 X 20.5, 29.6 X 20.8. Со времени
откладки последнего яйца в одном из гнезд — 21 мая — до появле-
ния птенцов (4 из 6) — 2 июня — прошло 12—13 суток. Время
вылета птенцов не удалось зарегистрировать. 12 июня птенцы,
вылупившиеся 2 июня, уже оперились. Слетки в массе появились
на р. Тутуре 17—18 июня (как в 1961, так и в 1962 г.).
Кормятся рябинники почти исключительно на лугах и в кустар-
никах (местные жители на Лене считают их луговой птицей).
На верхней Лене паразитологами было обработано всего 2
рябинника. На одном из них были обнаружены пухоеды. 5 очесан-
ных птиц с р. Чадобец также были свободны от иксодид.
Деряба — Turdus viscivorus L.
Судя по литературным данным (Гагина, 1961), деряба должен
изредка гнездиться в пределах бассейна Ангары. Однако мы этот
вид не сумели обнаружить в южной тайге Средней Сибири.
Певчий дрозд — Turdus ericetorum Turt.
Южная тайга Средней Сибири — восточный форпост распро-
странения певчего дрозда. Здесь он обитает на гарях островных
темнохвойных лесов с кедрово-елово-пихтовым подростом, в сме-
шанных мелколиственно-сосновых лесах, главным образом при-
долинной полосы, и как исключение в сосновых борах с темно-
хвойным подростом на низких водоразделах и в приручьевых
ельниках, особенно молодых и разреженных. Практически певчий
дрозд почти не выходит на водоразделы, обитая лишь в долинно-
придолинной полосе. Нарушение лесов благоприятно сказалось
на численности популяции вида.
Весной певчие дрозды прилетают в южную тайгу в конце
апреля—начале мая. В 1961 г. отмечено исключительно раннее по-
явление птиц на верхней Лене: у Жигалова они зарегистрированы
25 апреля, но основная масса птиц появилась 27—29-го числа.
На р. Чадобец певчие дрозды прилетели 3 мая 1962 г. Из районов
верхней Лены эти дрозды исчезают к 20-м числам сентября.
Наиболее поздняя добыча птиц 18 сентября 1961 г.
Единственное гнездо певчего дрозда было найдено 16 июня
1960 г. Оно помещалось на елочке (в 2.2 м от земли) у дороги
в густом молодом ельнике. Диаметр гнезда 170, высота 100, диаметр
лотка 102, глубина 63 мм. 18 июня вывелось 2 птенца, 19-го числа
их было уже 5. 29 июня птенцы покинули гнездо, т. е. выкарм-
ливание их родителями в гнезде длилось 12 дней. Корм состоял
из дождевых червей (подавляющая масса) и зеленых гусениц.
Более подробно поведение родителей у гнезда нами описано ранее
(Реймерс, 1963г).
Найденное гнездо принадлежало к повторной или задержав-
шейся кладке. Слетков на р. Чадобец наблюдали в 1962 г. уже
24 июня. Однако в массе они появлялись во все годы лишь в сере-
дине—конце июля.
Певчий дрозд — один из заметных прокормителей иксодид.
Из 6 обработанных паразитологами дроздов с верхней Лены 3 ока-
зались зараженными личинками и нимфами таежных клещей.
Обилие паразитов, правда, не превышало 2 экз. на 1 птицу.
Из 4 певчих дроздов, осмотренных на р. Чадобец, 1 птица была
заражена иксодовыми клещами. Помимо клещей, с певчих
дроздов снимали кровососок и пухоедов.
Белобровик — Turdus musicus L.
Белобровик — северный вид, крайне редко встречающийся
в пределах южной тайги. Гнездится этот дрозд в пойменных лесах
и березовом мелколесье северного типа (типично высокое увлаж-
нение). Молодые наблюдались на верхней Лене 20 июля 1962 г.
Г. А. Вороновым. На р. Чадобец их появление зарегистрировано
лишь в начале августа. Пролет белобровиков на верхней Лене
отмечен 26 апреля 1961 г.
Паразитологами были осмотрены 2 белобровика с верхней
Лены и 1 с Ангары. На 1 экз. (с Лены) найдено 2 нимфы иксодид.
Обе ленские птицы были заражены кровососками.
199
Дрозд Науманна — Turdus naumanni Ternm.
В южной половине Предбайкалья рыжий дрозд Науманна нам
известен лишь на пролете. На гнездовье ни мы, ни Л. И. Малышев
(1963) этого подвида тут не находили. Правда, в 1962 г. 19 июля
Г. А. Воронов добыл на р. Тутуре 1 взрослого самца. Это
может указывать на возможность гнездования этого вида. Темный
дрозд Науманна нам не встречался и на пролете. В противовес
этому Т. Н. Гагина (1961) считает, что темный дрозд — гнездя-
щаяся птица бассейна верхней Ангары. Скорей всего случаи гнез-
дования могут относиться к северу указанного Т. Н. Гагиной уча-
стка (расшифровки автор не дает).
На нижней Ангаре, Ангаро-Подкаменнотунгусском между-
речье и в верховьях Нижней Тунгуски оба подвида (?) дрозда Нау-
манна (Г. n. naumanni Temm. и Т, п. eunomus Temm.) распростра-
нены совместно. При этом в отличие от обыкновенного и серого
снегирей их стации почти идентичны. Это березняки, возникшие
на месте приречных ельников, пойменные кустарники и как исклю-
чение смешанные светлохвойно-мелколиственные леса главным
образом придолинной полосы. Во всех случаях в местах добычи
дроздов Науманна наблюдалась хотя бы мелкая поросль
ели. Идентичность стаций и совместное обитание заставляет
предполагать видовое, а не подвидовое, как это принято в сводке
«Птицы Советского Союза», различие рыжего и темного дроз-
дов.
Оба эти вида или подвида (решающее слово для решения этого
вопроса мы оставим за систематиками) не характерны для южной
тайги и появляются как фоновые формы лишь у крайнего север-
ного ее предела. В южной тайге как типологическом образовании
дрозд Науманна отсутствует, и гнездование вида в ее полосе,
очевидно, происходит не ежегодно.
Рыжий дрозд прилетает несколько раньше темного. Под Иркут-
ском Л. И. Малышев (1963) обнаружил его между 9 и И мая,
в то время как в окрестностях Красноярска К. А. Юдин (1952)
видел темных дроздов лишь 27—28 мая. На р. Чадобец рыжий
дрозд прилетел весной 1962 г. 14 мая, а темные дрозды — лишь
2 июня.
Крупные фолликулы (3 мм) и максимальный размер семенни-
ков (12.0 X 6.4 мм) у рыжих дроздов отмечены в начале июня
(и те и другие измерены 3 июня) в то же самое время, что и у темной
формы (2 июня размер семенников темного дрозда 12.0 X 8.2 мм,
6 июня фолликулы 3 мм).
С рыжего дрозда, добытого на р. Тутуре Г. А. Вороновым,
были сняты 1 личинка таежного клеща и 18 пухоедов. 5 птиц
с р. Чадобец были свободны от иксодид. Из 8 темных дроздов,
отстрелянных на р. Чадобец и ее притоках, с 2 птиц снято по
1 нимфе иксодовых клещей.
200
Темнозобый дрозд — Turdus ruficollis Pall.
Верхнюю Лену и всю южную часть Прибайкалья населяет
краснозобый дрозд (Т. г. ruficollis Pall.). На Нижней Тунгуске
и к северу от нижней Ангары живет чернозобый дрозд (Г. г. atro-
gularis Jarocki). Между этими подвидами существуют помеси.
Поскольку в пределах Верхнеленского полустационара встре-
чается исключительно краснозобый, а на р. Чадобец — только
чернозобый дрозд и именно к этим точкам приурочена большая
часть наших наблюдений, мы будем излагать материалы, касаю-
щиеся обоих подвидов, раздельно.
Краснозобый дрозд. Гари, поросшие мелколесьем
и обязательно с куртинами или отдельными деревьями, сохранив-
шимися от хвойного леса, — любимая стация краснозобых дроз-
дов. Однако птицы не чуждаются довольно глубоких окраин темно-
хвойной тайги, будь то водораздельный или пойменный лес. Схема
распределения вида показана на рис. 40.
Численность краснозобых дроздов подвержена значительным
колебаниям, однако она всегда наивысшая на придолинных гарях.
Тут, на гари островного темнохвойного леса, перемежающейся
с темнохвойными «колками», на 1 км2 можно встретить до 10—15
пар дроздов на гнездовье. Водораздельные гари и другие биотопы
этой полосы слабее заселены краснозобыми дроздами.
Нарушение лесов крайне благоприятно для вида. Для красно-
зобых дроздов важно сочетание обширных гарей и хотя бы отдель-
ных высоких деревьев независимо от их породы. Если это одиноч-
ные кедры, то дрозды устраивают гнезда на них (подробно см.
Реймерс, 1963г), если это лиственницы, то на них поют самцы.
На гарях же краснозобые дрозды находят свой основной корм
и пищу птенцов, находящихся в гнезде, — дождевых червей.
Чем больше дождевых червей, тем больше дроздов. Глухие темно-
хвойные леса, как и сосняки на каменистой почве, совсем лишены
краснозобых дроздов. Поскольку сосняки вообще бедны дожде-
выми червями, то смена темнохвойных лесов светлохвойными
(не смешанными) резко сокращает число птиц.
Таким образом, можно нарисовать такую историю подвида
в пределах южной тайги Средней Сибири. Изначально красно-
зобые дрозды населяли первичные редкостойные леса и высоко-
горные редколесья в тех местах, где были кустарники с обилием
голых гусениц или масса червей (там, где нет вечной мерзлоты).
Нарушение темнохвойной тайги приводило к широкому рассе-
лению темнозобых дроздов по гарям. Вид заселил всю придолин-
ную и часть долинной полосы. Смена темнохвойных лесов светло-
хвойными там, где господствовали беглые палы, не приводящие
к гибели светлохвойных насаждений, привела к снижению числа
птиц. Однако всюду, где верховые и повальные пожары создавали
обширные гари сосняков, численность птиц осталась на высоком
201
уровне. В ходе описываемого процесса произошла резкая концен-
трация птиц в одних местах и исчезновение в других.
Рис. 10. Принципиальная схема весенне-летнего размеще-
ния темнозобых дроздов в пределах южной тайги Средней
Сибири.
Условные обозначения как на рис. 12 и 22.
Весной 1960 г. краснозобые дрозды на р. Тутуре были отме-
чены с 1 мая. Следующей весной птицы появились 26 апреля.
Пролет продолжался всю первую декаду мая. Сначала прилетели
одиночные птицы и лишь затем дрозды стали появляться неболь-
шими стайками.
202
В середине—конце мая краснозобые дрозды строят гнезда —
в клювах птиц нередко можно видеть строительный материал.
Гнезда птиц располагаются на высоте от 0.5 до 3 м от земли. Рас-
положение гнезд самое разнообразное: между скрещенными вет-
ками (рис. 41), на горизонтальных «лапах» кедров, на елях у ство-
лов и т. п. Размер яиц (рис. 42): 1) 27.7 X 21.5, 26.4 X 20.6,
26.5 X 21.2, 26.8 X 21.0 мм; 2) 28.0 X 20.0, 27.9 х 20.0, 27.8 X
Рис. 41. Гнездо краснозобого дрозда в развилке веток сухого куста
в 70—75 см от земли.
X 20.3, 28.2 X 20.5, 28.0 X 20.5, 27.7 X 20.3 мм. Подробно
наблюдения за гнездами нами уже описаны (1963г). Массовое
появление слетков зарегистрировано в середине июля (верхняя
Лена).
Осенний пролет краснозобых дроздов начинается рано. Уже
в середине августа птицы собираются в стаи и откочевывают к югу.
В 1960 г. на верхней Лене такие стаи отмечены с 10—12 августа,
предыдущей осенью на средней Ангаре интенсивный пролет наблю-
дался 13—14 августа, а осенью 1961 г. на р. Тутуре краснозобые
дрозды почти исчезли к концу августа. В сентябре мы этих птиц
не добывали. Однако под Иркутском Л. И. Малышев (1963) отме-
тил вид 4 октября 1954 г.
Краснозобые дрозды прокармливают большое число личинок
и нимф таежного клеща. При этом заражены не только птицы
203
придолинной, но и водораздельной полосы (правда, в значительно
более слабой степени). Из 13 птиц придолинно-долинной полосы
на р. Тутуре в 1961 г. 9 оказалось зараженными. При этом макси-
мальное число личинок на 1 экз. было 11, а нимф — 2. Общая
зараженность не была выше 11 личинок и нимф в сумме (нимфы
найдены на 3 дроздах). В том же году из 9 дроздов с водоразделов
Рис. 42. Гнездо краснозобого дрозда с яйцами (неполная кладка).
зараженными оказались 2: один 2 личинками, другой 1 нимфой.
Летом 1962 г. из 3 краснозобых дроздов придолинной полосы
зараженным был один (1 нимфа).
В силу сосредоточения дроздов на гарях, где клещей вообще
очень много, и большой подвижности птиц, краснозобые дрозды
играют значительную роль как в циркуляции инфекций,
так и в прокормлении преимагинальных фаз развития клещей.
Чернозобый дрозд. Размещение чернозобых дроздов
по биотопам совпадает с размещением предыдущего подвида.
Как и краснозобый дрозд, чернозобый связан с гарями.
На р. Чадобец в 1962 г. чернозобые дрозды прилетели 26
апреля. Интенсивный пролет продолжался до 6—8 мая. К сере-
дине мая дрозды заняли гнездовые участки. 20 и 21 мая из яйце-
водов чернозобых дроздов были вынуты готовые к откладке яйца.
204
Гнездо было найдено 29 мая в березово-осиновом лесу с подседом
из ели (рис. 43 и 44). Оно располагалось в куртине мелких елей,
окружающей большую лужу. Дрозды поместили гнездо на упав-
Рис. 43. Расположение гнезда чернозобого дрозда.
шей, но застрявшей на соседних деревьях ели (наклон 15—20°).
Диаметр дерева 15—17 см (на уровне груди). В месте расположения
гнезда ствол имел диаметр 12 см. Расположено гнездо было над
водой. Среди тех же елок было обнаружено еще два прошлогод-
них гнезда, причем одно помещалось на той же упавшей ели, но
205
ниже свежего. Выстилка гнезда — слой сухой травы (осока и вей-
ник) толщиной 12 мм. Второй изнутри слой — трава с глиной.
У верхних краев толщина слоя 10, у дна 15 мм. Изнутри этот слой
нигде не был виден, кроме верхних краев гнезда. Третий внешний
слой состоял из сухого хвоща, осоки и тонких веток ели. Гнездо
было прилеплено к стволу ели и на дне не имело слоя глины,
Рис. 44. Гнездо чернозобого дрозда с яйцами.
а лишь внутреннюю выстилку. Размеры: ширина гнезда 165,
высота 119, ширина лотка 103, глубина 92 мм. Размер 6 яиц:
29.4 X 19.3, 27.0 X 19.8, 28.0 X 20.4, 27.4 х 19.9, 27.1 X 19.5,
27.9 X 20.5 мм. Цвет голубой с коричневыми крапинками по всему
яйцу, кроме острого конца. У тупого конца небольшое скопление
пятнышек.
Слетки появились 29 июня. В это же самое время у самки
(с выводком почти выросших птенцов) в яйцеводе найдено готовое
к откладке яйцо. Таким образом, темнозобые дрозды имеют две
кладки за лето.
Основными объектами питания чернозобых дроздов, как и крас-
нозобых, следует считать дождевых червей и голых гусениц бабо-
чек.
Паразитологами было обработано 6 чернозобых дроздов
с р. Чадобец. На них иксодид не обнаружено.
206
Бледный дрозд — Turdus pallidus Gm.
Ареал бледного (оливкового — Т. р. obscurus Gm.) дрозда
обнимает всю южную тайгу Средней Сибири. В ее пределах, как
Рис. 45. Принципиальная схема весенне-летнего размещения
бледных (оливковых) дроздов в пределах южной тайги Средней
Сибири.
Условные обозначения как на рис. 12 и 22.
и в большинстве других случаев, численность вида в аналогичных
биотопах сохраняется на одном уровне. У северных пределов
южной тайги население оливковых дроздов довольно резко со-
кращается, хотя вид распространен и севернее.
20 7
Так же как и пестрый дрозд, оливковый — обитатель в основ-
ном темнохвойных лесов (главным образом водораздельных)
с примесью лиственных пород. Но в отличие от пестрого дрозда
описываемый вид охотно селится по гарям темнохвойных лесов.
Обитает оливковый дрозд и в приручьевых ельниках, смешанных
лесах и даже сосняках водоразделов. Схема размещения вида
в пределах южной тайги показана на рис. 45.
Рассматривая приведенную принципиальную схему, нетрудно
сделать вывод о влиянии нарушений леса на популяцию оливко-
вых дроздов: омоложение и разреживание темнохвойных лесов
приводит к резкому увеличению числа дроздов, смена же темно-
хвойных лесов светлохвойными снижает обилие птиц. Можно
полагать, что общая плотность населения оливковых дроздов
достаточно резко возросла со времени постоянных нарушений
тайги и теперь кривая численности птиц пошла книзу.
На верхнюю Лену оливковый дрозд прилетает поздно: весной
1961 г. птицы отмечены с 24 мая. На р. Чадобец оливковые дрозды
встречены весной 1962 г. еще позже (1 июня). Осенний отлет про-
исходит, наоборот, рано. Пролетные стайки на средней Ангаре
в 1959 г. встречены с 3—4 августа. В нашем материале нет птиц,
добытых позже конца второй декады августа.
У только что прилетевших оливковых дроздов уже крупные
фолликулы (31 мая 1961 г. 7.1 мм), а слетки встречаются с двад-
цатых чисел июля.
Из 7 обработанных паразитологами дроздов из придолинной
полосы р. Тутуры 2 оказались зараженными иксодидами. Все
14 птиц с водоразделов были свободны от этих паразитов. Из 12
очесанных оливковых дроздов с р. Чадобец 1 был заражен нимфой
иксодового клеща и 2 птицы — гамазидами. Нимфа была снята
с дрозда из придолинной полосы (сосняк).
Каменка — Oenanthe oenanthe L.
Каменка не относится к собственно южнотаежным (лесным)
птицам и гнездится по остепненным склонам на Лене, среди кам-
ней по лугам и полям на Ангаре.
В 1959 г. каменки в пределах б. Баяндаевского района Иркут-
ской области появились 20 мая. Весной 1961 г. они были зареги-
стрированы на р. Тутуре 6 мая. А в 1962 г. на р. Чадобец птицы
стали обычными И мая. Маловероятно, чтобы прилет вида в 1959 г.
был отмечен с запозданием. Район вблизи улуса Байша (Кырмен-
ский сельсовет) представляет собой любопытную фенологическую
«яму», где большинство природных явлений происходит с довольно
большим запозданием по сравнению с местами, удаленными всего
на 25—30 км.
Интенсивный осенний пролет на верхней Лене в 1961 г. наблю-
дался 13—14 сентября.
208
2 июня 1961 г. у обыкновенных каменок были уже птенцы,
громко верещавшие из-под камня, где находилось гнездо. 12 июня
удалось камень убрать. В гнезде оказалось 3 птенца, готовые
к вылету. Птенцы сидели прямо на земле без всякой подстилки,
но под камнем было очень чисто.
Ни на одной из 10 каменок иксодиды обнаружены не были.
Плешанка — Oenanthe hispanica L.
Вид нами не встречен. Т. Н. Гагина (1961) считает его гнездо-
вание в бассейне верхней и средней Ангары недоказанным.
Каменка-плясунья — Oenanthe isabellina Temm.
Т. Н. Гагина (1961) указывает, что на Ангаре этот вид гнез-
дится. Мы его не констатировали.
Черноголовый чекан — Saxicola torquata L.
В пределах юга Прибайкалья как сравнительно редкая птица
гнездится восточносибирский черноголовый чекан (S. t, stejne-
geri Parrot.). Этот же подвид, а не 5. t. тайга Pall., как указано
в монографии «Птицы Советского Союза», встречается по кустам
на полях по береговым террасам нижней Ангары и ее северного
притока р. Чадобец. На верхней Лене черноголовый чекан нами
не встречен.
Прилетели черноголовые чеканы нар. Чадобец 15 мая (1962 г.).
До 24 мая продолжался интенсивный пролет птиц. Кустарники
буквально кишели чеканами. Однако позже птицы стали очень
редкими. Скорей всего черноголовые чеканы обычны лишь для
северной полосы южной тайги.
4 пролетных чекана не были заражены иксодидами.
Синехвостка — Tarsiger cyanurus Pall.
Синехвостка способна жить в самых разнообразных лесах,
лишь бы были кустарники (но не сплошь!) и подходящая сухая
кочка для устройства гнезда. Однако нельзя считать вид безраз-
личным к условиям лесной растительности.
В горах южной Сибири синехвостки наиболее обычны вдоль
шлейфовых частей склонов или занимают переходную полосу
от днища долины к ее склону. В горы вид поднимается лишь до
тех же пределов, что и березово-темнохвойныо леса (т. е. в усло-
виях Восточного Саяна приблизительно до 1200 м). В лесостепи
Средней Сибири синехвостка гнездится внутри лесных участков
по долинам ручьев и на склонах увалов, но лишь в том случае,
если границы лесного участка позволяют расположить гнездо
14 Н. Ф. Реймерс
209
в 100 м и более от опушки. Среди южной тайги Средней Сибири
кружево ареала вида практически совпадает с гидрологической
сетью, вернее со всеми приручьевыми и приречными участками.
Нет синехвосток тут лишь в ерниках и спелых березняках. В бере-
зовом мелколесье синехвостки гнездятся. Как гнездовую стацию
птицы используют и густое сосновое мелколесье. На его опушках
можно слышать поющих самцов.
Наибольшей численности синехвостки достигают в молодых
ельниках с примесью мелколиственных пород, особенно у малых
ручьев в верхней трети их течения или там, где водоток еще отсут-
ствует на поверхности. В наиболее благоприятных местах число
поющих самцов достигает 2 на 1 км течения ручья. В водораздель-
ной темнохвойной тайге при движении по хребту (т. е. пересекая
любимые места обитания вида) можно учесть на 5 км 3—4 поющих
самца. Среди мелколиственно-светлохвойных лесов, череду-
ющихся остатками когда-то существовавших приручьевых ель-
ников, на 5 км учетной полосы приходится 1—2 поющих самца
(все данные приведены по наблюдениям в районах верхней
Лены).
На огромных пространствах светлохвойной тайги Приангарья
любая полоса ельника и остров темнохвойной тайги служат сиг-
налом, что тут будут встречены синехвостки. В связи с этим сле-
дует считать, что смена темнохвойных лесов светлохвойными резко
снизила численность птиц. Вместе с тем произошел процесс
уплотнения населения вида в одних местах и резкое разрежение
в других. В мало нарушенных лесах распределение вида было более
равномерным.
Весенний прилет синехвосток в район р. Тутуры в 1960 г. был
зарегистрирован 17 апреля, но массовое появление началось 26-го
(Реймерс, 1963г). В следующем году там же синехвостки были за-
регистрированы 25 апреля, 27-го шел уже массовый пролет. В отли-
чие от весны 1960 г., когда первые синехвостки пели «вполголоса»,
весной 1961 г. птицы сразу стали петь громко. На р. Чадобец
синехвостки в 1962 г. появились 2 мая. Весной 1959 г. в б. Баян-
даевском районе Иркутской области массовый пролет синехвосток
отмечен 6 мая.
Подготовка к осеннему отлету у синехвосток происходит
в августе. Уже в середине месяца начинается подвижка стаек
птиц. В конце месяца (26—27 августа 1959 г., 30—31 августа 1961 г.)
наблюдается массовый пролет синехвосток (средняя Ангара и верх-
няя Лена), а в сентябре регистрировались лишь отдельные экзем-
пляры, исчезнувшие после второго периода валового пролета
17—20 сентября. В 1960 г. на верхней Лене пролет синехвосток
принял массовый характер лишь 25—28 сентября, а в августе
происходила только местная подвижка птиц.
5 мая 1961 г. синехвостки держались уже парами и заняли гнез-
довые участки. Максимальный размер семенников отмечен 21 июня
210
(9.0 X 5.8 мм). Подросшие молодые стали попадаться с двадца-
тых чисел июля.
На всех подвергшихся осмотру 24 синехвостках с верхней
Лены и нижней Ангары иксодиды обнаружены не были.
Обыкновенная горихвостка — Phoenicurus phoenicurus L.
Встречаясь по всей южной тайге, обыкновенная горихвостка
приурочена всего лишь к двум местообитаниям. Она чаще всего
встречается в мелколиственно-сосновом лесу придолинной полосы
и на опушках приручьевых ельников, выходящих к светлохвой-
ному лесу или березнякам. На водоразделах вида нет. Наруше-
ние лесов, очевидно, благоприятно отражается на численности
птиц.
В смешанных лесах придолинной полосы можно встретить до
5 поющих самцов на 5 км маршрута. Птицы обычно распевают
на вершинах сухих лиственниц.
На р. Тутуре весной 1961 г. обыкновенные горихвостки появи-
лись 6 мая. Весной следующего года на р. Чадобец они отмечены
с 11 мая. Самый поздний срок добычи вида — первая декада октя-
бря (1961 г., р. Тутура). Молодая обыкновенная горихвостка
добыта 31 июля 1962 г. (р. Чадобец).
На р. Чадобец было очесано 5 обыкновенных горихвосток.
Они оказались свободными от иксодид. Одна птица с р. Тутуры
от 22 июля была заражена 2 иксодовыми клещами (личинкой
и нимфой).
Сибирская горихвостка — Phoenicurus auro reus Pall.
Сибирская горихвостка найдена лишь в юго-восточной части
полосы южной тайги. На верхней Лене птица обитает только на
опушках приручьевых и приречных ельников, выходящих на поля
и к кустарниковым зарослям долин. Тут в кустарниках птицы
неоднократно отмечены на кормежке.
В 1960 г. прилет сибирских горихвосток на р. Тутуре был
отмечен с запозданием (18 мая). Весной следующего года птицы
появились 3—5 мая (первая дата указана по рассказу местного
охотника). 9 мая сибирские горихвостки начали петь. 18 мая у птиц
начались брачные игры. 27 мая размер фолликул сибирских гори-
хвосток равнялся 4 мм. 14 августа 1960 г. мы наблюдали, как самец
кормил слетка. В 1961 г. самостоятельно летающие молодые по-
явились в заметном количестве 14 августа. Возможно, в течение
лета бывает две кладки.
На осеннем пролете в 1960 г. сибирские горихвостки отме-
чены до 24 сентября. Осенью следующего года массовый пролет
шел еще 17—18 сентября, после чего птицы исчезли.
Все 14 очесанных сибирских горихвосток оказались свобод-
ными от иксодид.
14*
211
Восточный соловей — Luscinia luscinia L.
Известны лишь залеты восточного соловья в бассейн Ангары
(Гагина, 1961).
Соловей-красношейка — Luscinia calliope Pall.
Соловей-красношейка — типичная птица приречных кустар-
ников, ерников и придолинных гарей, поросших березовым мелко-
лесьем. Поскольку ерники и гари в большинстве своем антропо-
генного происхождения, нарушение лесов благоприятно для вида.
Сначала птица была распространена лишь по кустарникам вдоль
рек и их стариц.
На р. Чадобец красношейки появились в 1962 г. 1 июня.
Под Красноярском пролет этих птиц происходит на 7—10 дней
раньше (Юдин, 1952), однако в заповеднике «Столбы» средняя
многолетняя (за 25 лет) дата прилета 3 июня (Крутовская, 1958).
Под Иркутском пролет красношеек происходит с двадцатых чисел
мая (Малышев, 1963). Молодые красношейки отмечены с конца
первой декады августа (8 августа 1962 г., р. Чадобец).
Пять красношеек с р. Чадобец не были заражены иксодидами.
Птицы с р. Тутуры (осмотрено 11 экз.) оказались зараженными
(3 случая). Однако достоверность этих материалов низкая —- все
зараженные красношейки были добыты либо плашками-давил-
ками, либо попали в ловчие цилиндры, где не был исключен пере-
ход личинок и нимф с других объектов.
Соловей-свистун — Luscinia sibilans Swinh.
Этот вид нами не встречен. Т. Н. Гагина (1961) и Л. И. Малы-
шев (1963) выражают сомнение в том, гнездится ли соловей-сви-
стун в районах южной тайги Средней Сибири.
Синий соловей — Luscinia cyane Pall.
Синий соловей — характерный обитатель приручьевых уча-
стков темнохвойного леса, особенно разреженных с высоким уча-
стием кустарников. Среди водораздельных лесов на 5 км можно
слышать 1—2 поющих самцов. Птицы очень осторожны, и добыть
их трудно.
Два самца — один из бассейна р. Тутуры, другой из бассейна
р. Чадобец — были отстреляны соответственно 25 и 26 июня 1961
и 1962 г. Размер семенников 6.4 х 4.1 и 9.8 X 5.0 мм. Вес 14.5
и 13.0 г.
Оба самца были свободны от иксодид.
212
Варакушка — Luscinia svecica L.
На верхней Ангаре варакушки не редкость и прилетают 6—7
мая (Малышев, 1963). Однако на верхней Лене вид ни нами, ни
другими исследователями не найден. В ерниках и пойменных кус-
тарниках нижней Ангары и р. Чадобец варакушка (рис. 46) —
одна из самых многочисленных птиц. До сведения приречных
ельников и образования обширных ерниковых зарослей кру-
жево ареала варакушки в пределах южной тайги было более
ажурно и численность птиц ниже, чем сейчас.
Весной 1962 г. варакушки прилетели 14 мая, и кусты напол-
нились их голосами. Птички подражали крику малых пестрых
дятлов, чирков, песням больших синиц и даже бою перепелов.
Крупные фолликулы (3 мм) отмечены 18 июня.
Все 9 очесанных птиц не были заражены иксодидами.
Сем. ЗАВИРУШКОВЫЕ — PRUNELLIDAE
Сибирская завирушка — Prunella montanella Pall.
Сибирскую завирушку на Ангаре встречали осенью, на Лене —
лишь во время пролета (Гагина, 1961). Л. И. Малышев (1963)
отмечает как весенний (7—14 апреля), так и осенний (25 сентября)
пролет этих птиц под Иркутском.
213
Альпийская завирушка—Prunella collaris Scop.
Обычна на гольцах Хамар-Дабана (добыта нами 3 июля 1954 г.),
но не встречена в пределах более низко расположенных поясов.
Гималайская завирушка — Prunella bimalayana Bly th.
Л. И. Малышев (1963) наблюдал вид на гнездовье в горах
южного Прибайкалья. В пределах собственно южной тайги гима-
лайская завирушка нами не встречена.
Сем. ОЛЯПКИ — CINCLIDAE
Обыкновенная оляпка — Cinclus cinclus L.
Обыкновенная оляпка — весьма обычная птица горных рек.
Не исключено, что она гнездится среди наиболее пересеченных
местностей южной тайги Средней Сибири. Однако мы вид в районе
наших работ не наблюдали.
Сем. ЛАСТОЧКОВЫЕ — HIRUNDINIDAE
Касатка — Hi run do rustica L.
Несмотря на то что ареал касатки очень далеко идет на север,
в Средней Сибири эта ласточка распространена лишь немного
севернее последних форпостов южной тайги. В населенных пунк-
тах, расположенных среди лесов и не окруженных обширными
полями и лугами, касатка не гнездится. Вид, таким образом,
в районе наших наблюдений безусловно связан с человеком, и его
благополучие зависит, с одной стороны, от обилия построек,
пригодных для гнездостроения, а с другой — от величины расчи-
щенной от леса площади, особенно лугов, летая над которыми
птицы кормятся.
Всюду в южной тайге мы встречали один и тот же сибирский
подвид касатки — Я. г. tytleri J ord. Поскольку указания на даты
прилета и исчезновения сибирских касаток в Средней Сибири
до сих пор немногочисленны, приведем собранные нами материалы,
основанные на сообщениях корреспондентов-фенологов в 1962
и 1963 гг. (табл. 3). Новые данные, не приводимые здесь, содер-
жатся также в ряде уже опубликованных статей (Малышев, 1963;
Реймерс, 19636, 1963в).
На р. Чадобец касатки в 1962 г. прилетели 24 мая.
Анализ всех имеющихся материалов показывает, что в запад-
ное крыло южной тайги Средней Сибири касатки прилетают раньше
чем в верхнее Приангарье, и позже всего птицы появляются на
верхней Лене. Проведение изофен пока затруднительно из-за
неполноты данных.
214
Таблица 3
Даты прилета и отлета ласточек-касаток в пределах южной тайги
Средней Сибири (Иркутская обл.) в 1962—1963 гг.
Пункт наблюдения
Усольский р-н, с. Раздолье . . .
б. Баяндаевский р-н., с. Баяндай
б. Баяндаевский р-н, с. Духовщина
Ольхонский р-н, с. Онгуреи . .
Зиминский р-н, с. Зулумай . . .
Усть-Удинский р-н, с. Усть-Уда
Куйтунский р-н, пос. Куйтун . .
Тайшетский (б. Чунский) р-н,
с. Баер.....................
Тайшетский р-н, с. Сосновка . .
Нижнеилимский р-н, с. Невон . .
Жигаловский р-н, с. Якимовка .
Усть-Кутский р-н, с. Марково . .
Прилет Отлет
1962 1963 1962
26 мая.
— 26 мая. —
20 мая. 19 мая. —
— 28 мая. —
13 мая. — 3 сентября.
— — 27 сентября.
— 30 мая. —
10 мая. — —
— 15 мая. —
— 5 июня. —
31 мая. — 21 августа.
28 мая. 9 июня. —
Осенний отлет происходит в резко различные сроки без какой-
нибудь определенной закономерности. Например, из с. Якимовки
(р. Тутура) касатки улетели 23 августа 1960 г. Следующей осенью
несколько пар касаток очень поздно вывели птенцов, и птицы
исчезли лишь 13 сентября, в то время как основная масса особей
вида покинула р. Тутуру 27—28 августа.
Касатки ежегодно занимают одни и те же гнезда, выбрасывая
старую подстилку и натаскивая новую. В 1961 г. подновление
одного из гнезд было окончено около 20 июня (прилетели птицы
31 мая). Яйца были отложены в следующие сроки: 1-е — 24 июня,
2-е — 26, 3-е — 27, 4-е — 29, 5-е — 30 июня и 6-е — 1 июля.
Размер яиц: 14.5x17.7, 14.4x17.4, 14.5x17.7, 14.4х
Х17.4, 14.3x17.4 и 14.6x17.7 мм. Насиживание началось
с первого яйца. Первый птенец появился в ночь с 12 на 13 июля,
причем за эту ночь вылупились сразу 3 птенца. 16 июля вылупи-
лись уже все птенцы, и одно яйцо (болтун) было выброшено (14—15
июля наблюдения не проводились). В двух других гнездах 16
июля еще были яйца, но в одном из них 17-го уже было 3 птенца,
т. е. вылупление нескольких птенцов сразу — явление, распро-
страненное у касаток. 19 июля в этом гнезде вывелись все птенцы
(также был 1 болтун). В третьем гнезде 17 июля было 2 яйца
и 1 птенец. Четвертое гнездо было размыто дождем, и птицы немед-
ленно приступили к строительству нового. В нем появились яйца:
1-е — 27 июля, 2-е — 28, 3-е —29, 4-е — 30 июля, 5-е — 1 ав-
густа.
215
Птенцы из первого гнезда вылетели 1 августа (находились
в нем с 13 июля). В гнезде, сделанном взамен размытого, птенцы
(3 экз.) вывелись 10 августа, т. е. кратчайший срок нормального
развития птенца был равен 12 суткам (в гнезде № 1 также 12—13
дней). Два оставшихся яйца были выброшены. 1 сентября вылетел
из гнезда первый птенец, на следующий день — еще два. Таким
образом, длительность гнездового развития птенцов была равна
20—21 дню, в то время как в гнезде № 1 птенцы оставались всего
17—18 суток. Как уже отмечено, эта пара продолжала кормить
слетков до 13 сентября, после чего птицы исчезли. В ночь с 12 на
13 сентября мороз достигал —5°, однако ласточки, очевидно, уле-
тели благополучно. Они улетели утром, когда потеплело, и не воз-
вратились вечером, хотя в следующую ночь мороз был слабее (—3°).
В предыдущем 1960 г. деревенские ласточки в одном из гнезд
отложили яйцо 28 июня и каждый последующий день отклады-
вали по 1 яйцу. 3 июля было 6 яиц. 17 июля вывелся 1-й птенец,
а 18-го числа в гнезде были уже все 6 птенцов (насиживание про-
должалось 15—17 дней). Выкармливали ласточки птенцов мелкими
гусеницами и сетчатокрылыми насекомыми. 28—29 июля выле-
тели 2 птенца, 3 августа — 3-й, 5 августа — 4-й, и 6 августа,
наконец, вылетел последний.
В заключение очерка о деревенских ласточках остановимся
на крайне любопытном случае, рассказанном А. В. Алиной, про-
водившей наблюдения за этими птицами. В одном гнезде сохра-
нился единственный птенец. По каким-то причинам родители его
бросили (возможно, они погибли). В другом гнезде было 4 значи-
тельно менее развитых птенца. Позже одинокий птенец был пере-
несен во второе гнездо, но каким образом — установить не уда-
лось. Приемные родители усердно его кормили. Через несколько
дней он вылетел. Выкармливали ли его приемные родители в даль-
нейшем — неизвестно.
Воронок, или городская ласточка — Delichon urbica L.
В южной тайге гнездится сибирский воронок (D. и. whitheleyi
Swinh.). Воронок распространен шире деревенской ласточки
и меньше связан с обширными лугами. Птица может кормиться,
летая над деревенскими улицами и вдоль берега реки. Чаще, чем
деревенскую ласточку, воронка (в ненастную погоду) можно видеть
на опушках леса или даже над ним. В такую погоду касатки летают
только низко над лугами.
Местные жители называют воронков «стрижками» и путают
их с белопоясыми стрижами. Это заставляет с осторожностью
подходить к датам, проставленным корреспондентами-феноло-
гами. В 1960 г. на Лену воронки прилетели 21 мая. Весной 1961 г.
они появились (Реймерс, 1963в) в полосе южной тайги между 17
мая и 5 июня (последняя дата сомнительна). В 1962 г. птицы заре-
гистрированы корреспондентами-фенологами 21—25 мая. На верх-
216
ней Лене воронки появились 18 мая. Весной 1963 г. городские
ласточки были замечены нашими корреспондентами между^22
и 27 мая. Всюду в Прибайкалье воронки прилетают в очень близ-
кие сроки, причем на востоке южной тайги появляются несколько
раньше, чем на западе.
Интересно, что в Якутске воронки, по многолетним наблюде-
ниям И. Ф. Охлопкова (Скалой, 1956), прилетают не позже 16 мая,
т. е. раньше, чем на юге Средней Сибири.
Осенью 1960 г. воронки покинули южную тайгу 23—26 августа.
С верхней Лены в 1961 г. эти ласточки улетели к 23 августа, и их
пролет наблюдался с 16-го по 23-е. Осенью 1962 г. наблюдатели-
фенологи перестали замечать птиц с 23—24 августа. Таким об-
разом, воронки отлетают из Средней Сибири дружно и каждый
год в приблизительно одинаковые сроки.
В 1960 г. на р. Тутуре воронки начали строить гнезда
9—11 июня — через 18—20 дней после прилета. Птенцы выле-
тели 28—30 июля, а 14—15 августа большинство воронков уже
улетело.
Весной 1961 г. городские ласточки появились в Якимовке
17 мая во второй половине дня. 26 мая у них начались брачные
игры и птицы стали поправлять гнезда. Однако лишь 14—15 июня
в гнездах появились первые яйца, откладываемые ежесуточно
по одному (например, с 15 по 18 июня — 4 яйца, т. е. полная
кладка). Размер яиц: 14.6x18.6, 14.6x18.1 и 14.3x18.6 мм
(одно было разбито).
Насиживание кладки, отложенной 15—18 июня, началось лишь
23 июня. И июля вывелся первый птенец (насиживание длилось
18 дней). Долгое время он был единственным. Лишь 22 июля
вывелся второй, но вскоре погиб. Выброшено было и оставшееся
яйцо-болтун. Единственный птенец вылетел из гнезда 1 августа,
т. е. находился в гнезде 20 дней.
В гнездах №№ 2 и 3 16 июля были полные кладки. 1 августа
во втором гнезде были обнаружены 2 подросших птенца (оперен-
ные), а в третьем, залитом дождем гнезде, — 3 погибших. Из
гнезда № 2 птенцы вылетели лишь 15 августа, а 23 августа во-
ронки исчезли из пределов верхней Лены.
17 июля в 8 гнездах воронков были найдены: 1) 2 яйца,
2) 2 яйца, 3) 2 птенца и 1 яйцо, 4) 1 птенец и 2 яйца, 5) 4 яйца,
6) 2 яйца (в том числе один болтун), 7) 3 яйца, 8) 3 яйца.
Очевидно, смертность птенцов у воронков в Средней Сибири
даже в пределах южной тайги очень высокая.
Береговая ласточка — Riparia riparia L.
Мы не добыли ни одного экземпляра береговых ласточек,
поэтому не можем судить о подвидовой принадлежности птиц
из полосы южной тайги Средней Сибири. Т. Н. Гагина (1961)
относит их к R. г. riparia L.
217
В 1961 г. береговушки прилетели на р. Тутуру 23 мая. На
р. Чадобец весной 1962 г. они были отмечены впервые 12 мая
(первые птицы) и появились в массе 19 мая. Осенний отлет птиц
не прослежен.
На р. Тутуре 11 колоний береговых ласточек было насчитано
от с. Чекан до с. Якимовки (35 км береговой полосы). Колонии
были довольно крупные, и мы приняли среднюю численность птиц
в одном гнездовье в 50 экз. Следовательно, на 35 км насчитывалось
550—600 береговушек.
Хотя человек непосредственно почти никак не воздействует
на колонии береговых ласточек, однако нарушение лесов приво-
дит к более сильной эрозии речных берегов под влиянием мощных
весенних половодий, а это создает для птиц благоприятные условия.
Расчистка лугов, оголение от леса речных стариц и озер также
благоприятно сказываются на численности вида, поскольку улуч-
шает условия питания: ласточки охотно кормятся не только над
водой, но и над сырыми лугами.
1 июня 1961 г. мы наблюдали, как береговушки усиленно
ловили выплаживавшихся комаров над большой лужей на лугу.
Вместе с касатками (и больше их) береговушки уничтожают этих
кровососущих насекомых.
Млекопитающие
При изучении животного мира трудно рассчитывать на одина-
ковое знакомство со всеми его представителями. Одни формы
оказываются более изученными, другие менее. Так же как и в раз-
деле «Птицы», повидовые очерки млекопитающих не равновелики.
Особое внимание уделено наиболее массовым непромысловым
видам, играющим значительную роль в биоценозах и жизни чело-
века. Эти животные очень плохо изучены не только в Средней
Сибири, но и в других местах. О ряде промысловых зверей мы
сделали лишь отдельные замечания. Такой подход оправдан тем,
что нашей задачей не было создание всеобъемлющей моногра-
фии.
В отличие от схем распределения птиц, при составлении кото-
рых количественные учеты играли подсобную роль, картосхемы
размещения мелких млекопитающих основаны на числовых пока-
зателях, полученных с помощью канавок и иногда учетов с помо-
щью плашек. Во всех случаях предпочтение дано методике отлова
канавками, ибо она позволяет более полно характеризовать
Фауну.
Поскольку канавки в различное время года дают резко отлич-
ные показатели численности зверьков, мы выбрали для сравнения
период июль—август. При этом за минимум принят срок отлова
в 50 дней. Выбор июля и августа как времени, для которого вычис-
лены значения уловистости за 10 канавко-суток, наиболее целе-
215
сообразен по следующим обстоятельствам. Во-первых, в августе
почти все виды мелких зверьков достигают наивысшей численно-
сти и активности, так как пик уловистости обычно совпадает со
второй—третьей декадами этого месяца. Во-вторых, именно
в июле и августе технически наиболее осуществим отлов с помощью
ловчих цилиндров: оттаивает сезонная мерзлота и оканчивается
период выпирания цилиндров грунтовыми водами.
Как и во всех случаях, оценка численности млекопитающих
произведена индивидуально (т. е. дает градации верности) для
каждого вида по скользящей четырех- или шестибалльной шкале.
Даже при низкой точности методик относительного учета приме-
нение однородных шкал для характеристики численности всех
без исключения видов (например, десятичной шкалы, которую
предложил как основную А. П. Кузякин, 1962) мало целесооб-
разно: либо вообще не улавливается разница в животном населе-
нии биоценоза, либо виды целиком оказываются в одном раз-
ряде «малочисленных», хотя могут играть в жизни природы весьма
существенную роль. Однородные шкалы получат распространение
с применением методик абсолютных учетов, когда на результатах
в меньшей мере будет сказываться активность зверьков. В каж-
дом отдельном случае мы приводим числовое выражение градаций
балльной оценки численности мышевидных млекопитающих.
Как и в разделе «Птицы», отмечены биотопы, максимально насе-
ленные данным видом, со средней численностью и т. д. Еще раз
говорится об этом, чтобы не создалось впечатление о сравнении
численности данного вида с численностью других. Такие сопо-
ставления мы делаем ограниченно в синэкологических разделах
гл. 6. Напомним, что низший балл численности вида (1) нередко
означает лишь более или менее регулярное проникновение особей
вида из других биотопов (часто одних самцов) или периодическое
заселение выдела рассматриваемой формой («пульсация» кружева
ареала).
Объем систематических рангов млекопитающих, за редкими
исключениями, мы принимаем по «Определителю млекопитающих
СССР» первого издания (1944) и по книге Б. С. Виноградова и
И. М. Громова «Грызуны фауны СССР» (1962). Исключения редки
и наиболее значительное из них — принятие зайцеобразных за
особый отряд. Другие отступления от системы, принятой в упо-
мянутых изданиях, оговорены в тексте.
Специально следует остановиться на проявленном нами «кон-
серватизме» в отношении систематики землероек. Он объясняется
тремя обстоятельствами: во-первых, мы начали работу до выхода
в свет монографии С. У. Строганова (1957), в которой произведена
полная ревизия рода Sorex, во-вторых, при массовых сборах
часть полевых определений приходится доверять студентам,
которые обычно не в состоянии за короткое время овладеть
довольно сложными определительными таблицами указанной
219
монографии. Наконец, в годы наших исследований объем видов
насекомоядных в трактовке С. У. Строганова отнюдь не был
общепризнан.4
Отряд НАСЕКОМОЯДНЫЕ — INSECTIVORA
Крот сибирский — Talpa altaica Nik.
С. У. Строганов (1957), считая сибирского крота отдельным
монотипическим родом Asioscalops, относит особей южной тайги
Средней Сибири к двум подвидам — саянскому и салаирскому.
При этом область распространения саянских особей лежит между
Западным Саяном и Байкальским хребтом, а салаирский подвид
занимает в Средней Сибири бассейн Вилюя, Подкаменной и Ниж-
ней Тунгуски. Действительно, размер кротов изменяется от юга
к северу. Для присаянских районов С. У. Строганов считает
типичной длину тела 163—232 мм при средней 180 мм. Экземпляры
с верхней Лены (7 взрослых самцов) оказались длиной 155—
180 мм (М 167). Измерение 9 половозрелых самцов с р. Чадобец
показало колебание их длины в пределах 129—158 мм при средней
141 мм. Общее измельчание особей приводит к перемене многих
морфологических показателей. Можно ли придавать всем этим
изменениям систематическое значение, нам не ясно.
Обосновывая родовую специфику сибирского крота, С. У. Стро-
ганов подчеркивал наличие у него латентной фазы беременности.
По свидетельству И. П. Копылова (1948), П. И. Худяков «Крот
в Восточной Сибири», 1937—1938, рукопись) наблюдал в одном из
присаянских районов течку кротов в конце апреля—начале мая
и через месяц — рождение молодых. Если это не ошибка, то либо
в Средней Сибири, помимо сибирского, живет европейский крот
(что маловероятно), либо латентная фаза беременности не обяза-
тельна для сибирского вида.5 6
Распространение, размещение, чис-
ленность. Фрагментарность указаний на находки крота
в Средней Сибири (Наумов, 1934; Копылов и др., 1940; Тимофеев,
1949; Реймерс и Воронов, 1963) затрудняет расшифровку харак-
тера его распространения. В. Н. Скалой в «Атласе Иркутской
4 В монографии «Млекопитающие фауны СССР» (Громов и др., 1963)
практически без ревизии приняты концепции С. У. Строганова (1957) и им
дано право гражданства. Ко времени выхода указанной монографии руко-
пись нашей книги была уже готова, а трудоемкий пересмотр всего материала
технически не осуществим. Со временем такая работа будет сделана автором
или его сотрудниками (землеройки хранятся в коллекционных фондах
Биологического института СО АН СССР).
6 В последнем случае нет уверенности в самостоятельности сибирского
крота как отдельного вида и тем более — рода.
220
области» (1962) изобразил область обитания алтайского крота на
юге Средней Сибири двумя полосами (вдоль Саян и по Ангаре—
Чоне—Вилюю) и несколькими пятнами (верхняя Лена, Илим,
север междуречья Оки и Ии). Наши маршрутные исследования
показывают, что для утверждения пятнистости в распространении
вида нет достаточных оснований, тем более что В. Н. Скалой ряд
мест, где констатировано присутствие крота, пропустил, напри-
мер, бывший Киренский округ (Погудин, 1930). Трудность наблю-
дений за кротом в полосе южной тайги Средней Сибири состоит
в том, что этот зверек в лесах не делает характерных выбросов.
Сбору опросных сведений мешает существование множества народ-
ных названий. В Средней Сибири «кротом» называют водяную
крысу, а крота во многих местах зовут «медведкой». Видимо, крот
распространен на всей территории южной тайги Средней Сибири,
но обитает в ограниченном числе биотопов.
На рис. 47 представлена принципиальная схема размещения
крота в южной тайге. Кружево его ареала долинного типа. До-
вольно много кротов на гарях и вырубках.
К осени—зиме кроты концентрируются в приречных и при-
ручьевых ельниках. Здесь в наименьшей степени промерзает
почва. При рытье ловчих канавок и вкапывании цилиндров в при-
речных ельниках выяснилось, что здесь почва промерзает до
глубины 40—50 см (1962 г., окрестности дер. Яркино на р. Чадо-
бец). Во всех других биотопах весной нам не удавалось достиг-
нуть слоя талой почвы — промерзание было глубже. Сезонная
мерзлота в Средней Сибири достигает больших мощностей и в ряде
мест смыкается с вечной, так что зимнее существование кротов
возможно лишь в особо благоприятных условиях. Обильные
наледи и весенние половодья нередко приводят к массовой гибели
зверьков. Весной трупы их нетрудно найти на поверхности,
а собаки часто вырывают мигрирующих кротов из-под снега.
Зимняя жизнь кротов все еще остается вне поля зрения иссле-
дователей.
Антропогенные нарушения южной тайги Средней Сибири при-
водят к изменению «вязи» кружева ареала кротов. С одной сто-
роны, вырубки и гари позволяют зверьку селиться на водораз-
дельных пространствах, с другой — уничтожение приречных
и приручьевых ельников лишает кротов зимних стаций. В усло-
виях южной тайги, таким образом, происходит «распыление»
популяции кротов с одновременным увеличением численности
в одних местах и сокращением в других. Осветляя приречные
ельники и тем их осушая, человек «помогает» кроту заселять
северные районы, вместе с тем полосой сосновых боров «отрезая»
кротам путь на водоразделы. В случае полного сведения приреч-
ных и приручьевых ельников крот может исчезнуть полностью.
Летнее расселение в сторону водоразделов, очевидно, возможно
в пределах около 1 км. На такое расстояние, по наблюдению
221
В. И. Павлинина (1947), способны уходить кроты (европейские)
за сезон, а следовательно, и возвращаться в зимние стации.
Рис. 47. Принципиальная схема летнего размещения крота
в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
свыше 0.4, средняя 0.2—0.4 и минимальная 0.1 и меньше экземпляров
на 10 канавко-суток. Штриховка полосами означает предполагаемую воз-
можность указанной численности (без экспериментальной проверки).
Численность кротов в южной тайге невелика и постепенно
снижается к северу. Колебания числа кротов от года к году весьма
значительны (Реймерс и Воронов, 1963).
Сезонные колебания численности кротов прослеживаются
с трудом. При отлове зверьков канавками, как это делали мы,
222
можно говорить об интенсивности расселения молодых особей
и активности взрослых. Наивысшие показатели отловов были
получены в первой—второй декадах августа. В июле—августе
отмечена наибольшая активность самцов.
Размножение. Из 115 кротов, бывших в нашем распо-
ряжении, не оказалось ни одной беременной самки. Две весенние
особи не имели следов размножения (3 апреля и 12 мая). Наиболее
крупные семенники у взрослых самцов констатированы в июле
и августе (максимумы 21.5x12.4 и 22.0x19.0 — 9 июля и 22 ав-
густа). Это время указано как период течки (Бородулина, 1951,
1953; Казанская, 1952, 1953; Строганов, 1957). Однако в бывшем
Голуметском районе Иркутской области (предгорья Саян) П. И. Ху-
дяков наблюдал течку у кротов в конце апреля—начале мая
(Копылов, 1948). Число молодых, по этому автору, 2—7, редко
8—9. Рождаются они в июне—июле. К самостоятельной жизни
молодые кроты способны с месячного возраста.
Т. Л. Бородулина (1953) имела экземпляр крота, родившегося
зимой. Под Томском и Новосибирском (Строганов, 1957) рождение
детенышей у крота происходит в мае, а в июне молодые ведут
самостоятельный образ жизни. Первых молодых на верхней Лене
нам удалось отловить 9 июля (1961 г.), а на р. Чадобец — только
в конце июля. Очевидно, роды у южнотаежных кротов происхо-
дят в июне, как правильно указывал П. И. Худяков. Остается
неясным время гона. Высокая активность самцов в июле—августе
заставляет предполагать летний период гона, однако опровергать
П. И. Худякова мы не беремся.
Питание. Как и всюду, кроты южной тайги Средней
Сибири питаются в основном дождевыми червями. Эти животные
были обнаружены во всех 54 желудках с кормом. Почвенные
личинки насекомых найдены 6 раз (11.1 %), взрослые насекомые —
3 раза (около 6% от всех желудков с кормом), в том числе один
раз — остатки жужелицы.
Паразиты. На кротах найдены гамазовые и иксодовые
клещи, однако число последних очень мало: на 32 кротах обна-
ружено всего 4 личинки, по одной на зверьке. Все находки иксо-
дид относятся к июлю (4—20). Роль кротов в распространении
природноочаговых инфекций неясна и из-за малой численности
зверьков едва ли может быть значительной. Настораживают лишь
следующие факты. Как показали исследования в Западной Сибири
(Карпов и Попов, 1956), крот может быть резервуаром лептоспи-
роза. В пределах южной тайги Средней Сибири — это обычное
заболевание свиней и крупного рогатого скота (Краминская,
1962). Колхозники имеют обычай весной выпускать свиней в при-
речные ельники, где, как мы указали, нередки трупы погибших
кротов; голодные свиньи охотно их поедают. Это один из воз-
можных путей заражения. Летом телят можно видеть в загонах
для свиней. Таким образом эпизоотия распространяется и на этих
223
животных. Тем же путем — от кротов — может происходить
заражение свиней трихинеллезом.
В свое время (Шрейнер, 1916) разгорелся спор о вреде и пользе
кротов в сельском хозяйстве. Зверька «обвиняли» в уничтожении
полезных дождевых червей. В южнотаежных районах Средней
Сибири кроты на пахотных землях почти отсутствуют, однако
пашни крайне бедны дождевыми червями. Мало крот поедает
и полезных насекомых. Местные жители часто жалуются на «крота»
как на вредителя огородов. На самом деле вредит водяная крыса
(«крот» по-местному).
Значимость крота как объекта промысловой охоты в Средней
Сибири ничтожна. Уровень его добычи не превышает одной деся-
той процента по отношению ко всей пушной продукции, загота-
вливаемой в Иркутской области. В тех местах, где крот дости-
гает промысловой плотности, значительно выгоднее охотиться
на более ценных зверей.
Крошечная бурозубка — Sorex tscherskii Ogn.
Крошечные бурозубки Средней Сибири, по С. У. Строганову
(1957), принадлежат к номинальному подвиду.
Размещение и численность. Крошечная буро-
зубка немногочисленна, но в южной тайге Средней Сибири распро-
странена повсеместно. Редкость указаний на поимку зверьков
в этом районе (Thomas, 1915; Строганов, 1957) объясняется тем,
что исследователи пользовались для отлова в основном плашками-
давилками, куда крошечные бурозубки практически не попа-
дают.
Характер размещения вида еще не ясен. В Западной Сибири
наибольшая плотность населения крошечных бурозубок отмечена
в темнохвойных лесах (Юдин, 1962), где показатель отлова дости-
гает 0.5 зверька на 10 канавко-суток (1.0 на 100 «ловушко-суток»,
по терминологии Б. С. Юдина, т. е. на 100 цилиндро-суток).
По свидетельству того же исследователя, крошечные бурозубки
концентрируются у кромок болот. Сведения о размещении зверь-
ков в южной тайге Средней Сибири отсутствуют. В Монголии
(Банников, 1954) и на Нижней Тунгуске (Наумов, 1934) вид отме-
чен в сухих лиственничниках, лиственнично-кедровых лесах
и в болотистых низинах, поросших лиственницей, ольхой и ивня-
ком.
Наши материалы позволяют заключить, что крошечная буро-
зубка предпочитает сухие места или биотопы с сухими микро-
участками, но обычно вблизи воды. На водораздельных простран-
ствах зверьков удалось отловить только на сухих гарях, порос-
ших редким березняком-жердняком, и в сосняках. В темнохвойной
тайге вид найден не был. В островных же темнохвойных лесах
и на их гарях, как и во всех лесах придолинной полосы, характе-
224
ризующейся пестрыми экологическими условиями, крошечные
бурозубки сравнительно многочисленны (рис. 48).
В сухих долинных ельниках и на их гарях, поросших березой,
вид также обычен. В кустарниках, на лугах и в дремучих забо-
лоченных ельниках крошечная бурозубка не живет.
Нарушение темнохвойных лесов приводит к некоторому уве-
личению числа крошечных бурозубок. Замена темнохвойной тайги
сосновыми борами повышает численность зверьков. Однако нару-
шение сосновых боров (в том числе весенними палами) снижает
обилие крошечных бурозубок. Практически исчезают зверьки
при преобразовании приречных и приручьевых лесов в луга
и кустарниковые заросли. К тому же ведет распашка. В период
расселения молодых крошечные бурозубки иногда забегают на
поля (пойман 1 экз.), но, как правило, тут отсутствуют.
Как мы уже отмечали (Реймерс и Воронов, 1963), численность
крошечных бурозубок стабильна от года к году. Сезонные изме-
нения активности зверьков не укладываются в какую-нибудь
определенную закономерность, но пик обилия всегда приходится
на июль—август. В пределах южной тайги Средней Сибири кро-
шечные бурозубки становятся обычнее от юга к северу.
Размножение. Данные о размножении крошечных
бурозубок весьма фрагментарны. По Б. С. Юдину (1957, 1962)
и С. У. Строганову (1957), эти зверьки размножаются в течение
теплого периода года по крайней мере дважды. Беременные самки
в Западной Сибири найдены 18 мая и 21 августа. На верхней Лене
(Реймерс и Воронов, 1963) самка с увеличенной маткой была
добыта 3 июля 1961 г. На р. Чадобец 14 июля 1962 г. была отло-
влена беременная особь с 7 эмбрионами. Молодые зверьки попа-
дали в цилиндры с 6—9 июля. Очевидно, С. У. Строганов (1957)
прав, что у крошечных бурозубок за лето бывает минимум два
помета. В условиях южной тайги Средней Сибири сроки их при-
ходятся на май—июнь и на июль—август.
Сеголетки участия в размножении, видимо, не принимают.
Питание. В 16 желудках крошечных бурозубок были
найдены почвенные личинки насекомых (6 случаев), остатки взрос-
лых насекомых, главным образом их жировые тела (11 случаев)
и семена (2 случая). 42 экз. зверьков имели пустые желудки.
Семена найдены в желудках зверьков с верхней Лены и с р. Чадо-
бец (14 и 26 июля). Очевидно, крошечная бурозубка не такой сте-
нофаг, как предполагает это Б. С. Юдин (1962).
Паразиты, значение. Паразитологи очесали 21 кро-
шечную бурозубку. На зверьках обнаружены вши, иксодо-
вые и гамазовые клещи. Личинки иксодид зарегистрированы 7 раз
и все в конце июня и июле. Обилие их невысокое — 0.7. Макси-
мальное число личинок на 1 зверька 4 экз.
► Ничтожная численность крошечных бурозубок заставляет
предполагать их малое значение в биоценозах и хозяйстве.
15 Н. Ф. Реймерс
225
Рис. 48. Принципиальная схема летнего размещения крошеч-
ных бурозубок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная числен-
ность 0.3—0.4 экз. на 10 канавко-суток без резких колебаний от года
к году и во всех точках отлова; средняя — то же число зверьков, но
с резкими колебаниями от года к году; минимальная численность
0.1—0.2 экз. на 10 канавко-суток за июль и август. О штриховке поло-
сами СМ. подпись к рис. 47.
Малая бурозубка — Sorex minutus L.
Среднесибирские особи этого вида С. У. Строганов (1957)
отнес к номинальному подвиду S. т. minutus L.
Распространение, размещение и числен-
ность. В пределах южной тайги Средней Сибири малая буро-
зубка распространена повсеместно и не проявляет тенденции
к сокращению численности на севере района.
Зверек явно предпочитает приречные и приручьевые леса,
достигая здесь наивысшей численности по опушкам и рединам,
особенно примыкающим к водоемам (рис. 49). Водораздельные про-
странства заселены малой бурозубкой несравненно реже. На верх-
ней Лене в 1962 г. на водоразделе этот вид не был отловлен, хотя
в работе было 5 канавок. В 1961 г. зверьки ловились единицами.
В ряду трансформации сосновых боров малая бурозубка дости-
гает наивысшей численности на фазе смешанного леса. Превра-
щение приречных и приручьевых ельников в смешанные насаж-
дения и затем в березняки сначала несколько повышает число
малых бурозубок, затем численность вида резко падает и дости-
гает нуля при полном преобразовании долинных лесов в луга
и поля (на полях вид отмечен лишь осенью). Приречные кустар-
ники, смыкающиеся с лесом, обладают высокой численностью
малых бурозубок; лугово-кустарниковые заросли, отрезанные
от леса значительными луговинами или полями, бедны ими.
Влияние человека на популяцию малых бурозубок противо-
речиво. С одной стороны, разрежение лесов приводит к увеличе-
нию численности вида, с другой, уничтожение лесов, замена их
гарями, лугами и полями — к сокращению числа зверьков. Оче-
видно, изначальными биотопами малых бурозубок были кромки
речных берегов, поросшие кустарниками, и опушки долинных
ельников, т. е. те места, где и сейчас вид достигает максимальной
численности. С нарушением тайги полотнище кружева ареала
малой бурозубки расширилось, а «вязь» его стала плотнее. Общая
численность вида при этом должна была увеличиться.
В настоящее время малая бурозубка — немногочисленный,
но все-таки нередко встречающийся зверек, намного, правда,
уступающий в количестве особей средней и обыкновенной бурозуб-
кам. Нам за все время удалось отловить 262 экз. описываемого вида.
Численность зверька от года к году колеблется весьма резко.
В аналогичных биотопах это колебание достигает 3—5 раз, в то
время как в течение одного сезона разница между результатами
работы канавок-дублеров6 не превышает 2 раз. Депрессия числен-
ности малых бурозубок на верхней Лене была отмечена в 1961 г.
(Реймерс и Воронов, 1963). В 1962 г. обилие зверьков возросло.
6 В ряде случаев в одном биотопе были заложены 2 канавки одновре-
менно.
15*
227
Рис. 49. Принципиальная схема летнего размещения малых
бурозубок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
0.5—1.0 и более экземпляров на 10 канавко-суток ежегодно и во всех
точках отлова, средняя 0.5—1.0 экз. не постоянно и не во всех точках,
минимальная численность не более 0.1—0.4 экз. на 10 канавко-суток
В течение июля и августа. О штриховке полосами см. подпись
к рис. 47.
Рис. 50. Ход изменения уловистости
малых бурозубок на верхней Лене (7)
и нижней Ангаре (2) в 1962 г.
По оси ординат — число зверьков на 10 ка-
навко-суток; по оси абсцисс — месяцы и
декады.
Уловистость малых бурозубок при отлове канавками (в плашки
эти зверьки практически не попадают) достигает максимума в тре-
тьей декаде августа. Второй пик обилия зарегистрирован в конце
июня—начале июля. Любопытно, что ход сезонного изменения
уловистости зверьков (рис. 50) на территории двух стационаров
(Верхнеленского и Нижнеангарского) очень близок по характеру.
Размножение. Взрослые особи редко попадают в лов-
чие канавки. В нашем распоряжении^оказалось всего 17 самцов
и 16 самок. Беременные особи
пойманы четырежды: 6 июля
1962 г. 9 эмбрионов (3.1 мм),
18 июля 1962 г. 3 эмбриона
(2.0 мм), 19 июля 1963 г. 7 эм-
брионов (8.2x7.5 мм) и 24 ав-
густа 1960 г. 9 эмбрионов
(10x8 мм). 16 августа 1960 г.
был отловлен кормящий экзем-
пляр. В течение лета размеры
семенников у самцов остава-
лись относительно стабильными
(5.0 X 4.0 мм в среднем).
Первые поимки молодых
особей зарегистрированы на
верхней Лене в 1961 г. 29 июня,
следующим летом — 28 июня.
На р. Чадобец первый молодой
экземпляр отловлен 4 июля.
Последние молодые особи в
1961 г., когда проводились поз-
днеосенние наблюдения, пой-
маны в последних числах сен-
тября.
Все указанные даты размножения малых бурозубок очень
близки к констатированным Б. С. Юдиным (1962) в Западной
Сибири, Е. М. Снегиревской (1947) в Башкирии, В. А. Поповым
(1960) в Татарии, а также С. Боровским и А. Денелем (Borowski,
Dehnel, 1952) в Польше. Очевидно, географическая изменчивость
фенологических сроков в малой степени влияет на ритмику жизни
этого вида.
Растянутость сроков размножения малых бурозубок заста-
вляет предполагать несколько пометов за лето. Наличие минимум
двух пиков обилия молодых особей наталкивает на мысль, что
малая бурозубка имеет два основных периода размножения —
в конце мая и конце июля. Поимка беременной самки 24 августа
указывает на возможность августовско-сентябрьского помета, если
это не отклонение, вызванное общим поздним ходом всех при-
родных явлений в 1960 г. Молодые самки дают приплод в июле.
229
Питание. Среди экземпляров, подвергшихся просмотру,
только 44 зверька имели в желудках корм. Результаты анализа
содержимого желудков этих особей приведены в табл. 4.
В отличие от зверьков из Западной Сибири (Юдин, 1962) малые
бурозубки южной тайги Средней Сибири больше поедают личинок
насекомых. Это, видимо, вообще характерно для вида в условиях
Таблица 4
Состав и встречаемость пищевых компонентов
в содержимом желудков малой бурозубки (44 экз.)
Пищевые компоненты Число же- лудков с дан- ным кормом Процент к общему ко- личеству же- лудков с кор- мом Время обнаруже- ния корма
Животные корма, всего 44 100.0
В том числе: Взрослые насекомые 26 59.1 Равномерно
Личинки насекомых 28 63.6 летом. То же
Из них: Почвенные личинки .... 27 61.9 » »
Гусеница бабочки (зеленая) 1 2.3 28 августа.
Дождевые черви 1 2.3 18 августа.
Не определенный корм живот- ного происхождения .... 1 2.3 —
Растительные корма (трава) 1 2.3 28 сентября.
лесов. По данным упомянутого автора, именно в лесах малые
бурозубки поедают максимум личинок насекомых.
Паразиты, значение. На малых бурозубках встре-
чаются гамазовые и иксодовые клещи, но общий уровень закле-
щевения невысок — значительно ниже, чем в Западной Сибири
и особенно на Урале (Дроздова, 1959). На верхней Лене (Реймерс
и Воронов, 1963) из 35 осмотренных зверьков оказались зара-
женными личинками иксодовых клещей 11 экз. (около 30%) и ним-
фами 3 бурозубки (около 9%). На 17 осмотренных малых буро-
зубках с нижней Ангары личинок иксодид не оказалось. Гама-
зиды также поражают этот вид бурозубок редко и в небольшом
числе (1—3 экз.).
Общая оценка роли вида в биоценозах и хозяйстве на терри-
тории южной тайги Средней Сибири сходна с данной нами для
Верхоленья: малая бурозубка — несущественный компонент ланд-
шафтов.
230
Средняя бурозубка — So rex macropygmaeus Mill.
Южная тайга Средней Сибири — практически белое пятно
в познании этого вида. С. У. Строганов (1957) не имел отсюда
экземпляров и на прилагаемой к монографии карте отметил лишь
точки находок по р. Ангаре с ссылкой на В. Н. Скалона (1933),
А. Л. Чекановского7 и О. Томаса (Thomas, 1911).
Поскольку мы пользовались систематическими критериями,
предложенными Н. А. Бобринским и А. П. Кузякиным (Бобрин-
ский и др., 1944), в описываемую группу попали, очевидно, формы,
называемые С. У. Строгановым (1957) Sorex daphaenodon Thomas
и S. arcticus Kerr. Поименованные виды из южной тайги Средней
Сибири также не были в его распоряжении.
Экземпляры средних бурозубок (или общей сходной группы)
с верхней Лены по кондило-базальной длине черепа (16.5—17.9,
М 17.4 мм) не отличимы (сличались 30 взрослых особей) от ниж-
неангарских (16.6—18.0, М 17.3 мм).
Распространение, размещение и числен-
ность. Средняя бурозубка не только повсеместно распростра-
нена в южной тайге Средней Сибири, но практически нет такого
участка, кроме водной поверхности, свежих паров и крупных
городов, где бы этот вид полностью отсутствовал. На севере, уже
за пределами южной тайги, по Н. П. Наумову (1934), это также
самый обычный зверек.
С наибольшей плотностью (рис. 51) средняя бурозубка заселяет
островки темнохвойных лесов (до 11.4 зверька на 10 канавко-
суток в июле—августе), приречные и приручьевые ельники (до 8.5
зверька на 10 канавко-суток), а также влажные гари (до 9 зверьков
на 10 канавко-суток в июле и августе). Наиболее благоприятны
для вида в южной тайге осветленные, слегка заболоченные леса.
В глухой тайге средних бурозубок меньше, но в сильно нарушен-
ных лесах также немного.
Сообщение Н. А. Бобринского и А. П. Кузякина, что средние
бурозубки предпочитают заросли у берегов рек, для Средней
Сибири не совсем точно. В кустарниках, удаленных от леса, этот
вид малочислен. Помимо прочих условий, важно то обстоятель-
ство, что половодья заливают кустарниковые заросли. В местах,
где кусты примыкают к лесу, бурозубки могут найти спасение
на деревьях или по обломкам валежника, которые несет с собой
половодье, пробраться к берегу. Такой возможности нет у зверь-
ков кустарниково-луговой поймы.
Смена темнохвойных лесов сосняками несколько снижает
численность средних бурозубок. Еще резче она падает при све-
дении ельников. Очевидно, в первобытной тайге до нарушений
ее человеком зверьков было меньше, чем в последующий период
7 Коллекционные материалы.
231
первичных нарушений лесов. Разреживание ельников и водораз-
дельных лесов привело к увеличению обилия средних бурозубок —
в редкостойных насаждениях особенно много этих зверьков.
Рис. 51. Принципиальная схема летнего размещения средних
бурозубок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
более 6.0, средняя 3.0—6.0 и минимальная 0.5—3.0 в среднем на
10 канавко-суток в течение июля и августа.
Позже, с развитием земледелия, число средних бурозубок начало
падать. Этот процесс продолжается и поныне в связи со все более
широким сведением лесов и прежде всего приречных ельни
232
ков. Вместе с тем в островках темнохвойной тайги создались
сгущения населения средних бурозубок, неестественные для не-
тронутой природы. Кружево ареала как бы становится реже и
вместе с тем «нити» его утолщаются, образуются толстые
«узлы».
Средняя бурозубка — многочисленный зверек. По уловисто-
сти она занимает в зависимости от года первое или второе (после
обыкновенной бурозубки) место. Возможно, что средняя буро-
зубка доминирует во влажные годы (1960 и 1963 — на Лене, 1962 —
на Ангаре) и уступает по численности обыкновенным бурозубкам
в сухие годы (1961 и 1962 — на Лене). Всего нам удалось отловить
1980 средних бурозубок, так что этот вид может считаться вторым
по численности среди насекомоядных.
Численность средней бурозубки, так же как и малой, испы-
тывает резкие колебания от года к году. Число зверьков в анало-
гичных биотопах в сходные сроки может увеличиваться в 5 раз.
В пределах одного биотопа (но на разных участках) обилие сред-
них бурозубок приблизительно одинаково. Приведем данные,
полученные с помощью канавок-дублеров и на разных стацио-
нарах. На водораздельной гари, поросшей березняком, две
канавки дали в 1962 г. 2.8 и 2.4 зверька (все данные взяты на 10
канавко-суток в июле и августе). Островной темнохвойный лес
на верхней Лене и нижней Ангаре имел показатель 11.0 и 11.4
зверька. Два участка гари островного темнохвойного леса дали
сходное число зверьков — 8.7 и 8.6; сосновый бор на Лене
и Ангаре — 4.8 и 3.1 зверька, смешанный сосново-мелколиствен-
ный лес — 4.6—6.1 зверька. В последнем случае регистрируется
максимальное различие данных канавок-дублеров по числу сред-
них бурозубок. Ельники дали соответственно числа 8.5—7.5 для
Лены и Ангары, а кустарники в пределах одного Верхоленского
стационара оказались совершенно идентичными в смысле числа
зверьков — 5.3 зверька на 10 канавко-суток.
Сезонная динамика численности средних бурозубок характе-
ризуется наличием двух пиков обилия (рис. 52). На рисунке
пунктиром показано продолжение падения числа зверьков на тер-
ритории Нижнеангарского полустационара в придолинной по-
лосе. Повышение уловистости в третьей декаде июля и первой
декаде августа происходило на водоразделе, а затем во вто-
рой декаде августа — всюду. Менее выраженный пик обилия в
начале июля был отмечен нами и в 1961 г. (Реймерс и Воронов,
1963).
Размножение. Как и все другие бурозубки, средние
размножаются только в теплое время года. В условиях южной
тайги Средней Сибири первые оплодотворенные самки отмечены
в третьей декаде марта. Последняя беременная особь всего с одним
эмбрионом отловлена 1 сентября 1960 г. В более позднее время
Таб
Число эмбрионов и количество размножающихся
Месяц Декада Отловлено взрослых самок
всего в том числе со следами размножения
всего процент от всех самок из них беременных
всего процент от всех самок
Нюнь .... | 2 3 6 5 1 1 16.6 20.0 — —
1 10 6 60.0 5 50.0
Июль .... | 2 9 7 77.7 4 44.4
3 16 14 87.5 5 31.3
1 17 16 94.1 3 17.6
Август .... | 2 28 26 92.8 10 35.7
3 27 25 92.6 6 22.2
1 27 22 81.4 1 3.7
Сентябрь . . | 2 20 18 90.0 — —
3 И 10 90.0 — —
Всего или сред-
нее — 178 146 — 34 —
встречались только кормящие зверьки. Таким образом, период
размножения в Средней Сибири несколько короче, чем в Баш-
кирии (Снегиревская, 1947), где беременные средние бурозубки
найдены (правда, в исключительный год) 16 октября.
В табл. 5 сведены данные о размножении средних бурозубок.
Процент размножающихся8 самок закономерно растет к осени,
однако количество беременных особей колеблется, достигая
максимумов в июле и августе. Очевидно, первый помет происхо-
дит в апреле (продолжительность беременности у бурозубок около
20 дней, Dehnel, 1952), второй — в июле и третий — в августе.
При этом в июле включаются в размножение и самки-сеголетки,
которых мы не всегда дифференцировали от перезимовавших
особей. Самки текущего года рождения имели следы размноже-
ния со второй декады июля. Эти особи затушевывают картину
периодичности размножения, тем более что они, видимо, способны
производить потомство дважды за лето.
Активность способных к размножению самцов, как и взрослых
самок, довольно четко возрастает (повышается обилие зверьков
в уловах) с периодичностью в 20 дней. Следует предположить, что
оплодотворение наступает непосредственно вслед за родами.
8 К размножающимся особям отнесены самки в течке, беременные и
кормящие.
234
лица 5
самок средней бурозубки по месяцам и декадам
Число эмбрионов, как видно из данных табл. 5, колеблется
от 1 до 11, составляя в среднем 6.5. Такое число эмбрионов харак-
терно для всей полосы южной тайги. Б. С. Юдин (1962) для окре-
стностей с. Кожевникове (южная тайга Западной Сибири) реги-
стрировал среднее их количество на 1 самку 6.6—6.4.
Максимальное число эмбрионов наблюдается в июле и сни-
жается к осени. В правом роге матки обычно больше эмбрионов,
чем в левом (Реймерс и Воронов, 1963), но это превышение незна-
чительно.
Молодые особи попадают в цилиндры с конца июня (23—26).
В уловах они составляют подавляющее большинство зверьков,
и их число достигает максимума во второй—третьей декадах
августа. Самцы-сеголетки в отличие от самок в год рождения,
очевидно, не размножаются. Из молодых самок размножаются
только особи весенних пометов. Среди нескольких сотен самок-
сеголеток оказалось всего около десятка особей со следами раз-
множения.
Питание. Средняя бурозубка — активный хищник, пита-
ющийся главным образом насекомыми и их личинками (табл. 6).
Однако в условиях Средней Сибири даже в летнее время этот зве-
рек поедает растительные корма в два раза чаще, чем это указы-
вается для Западной Сибири (по С. У. Строганову, 1957, встре-
чаемость растительных кормов в Западной Сибири 4.1%).
235
Б. С. Юдин (1962), на неопубликованные в то время данные кото-
рого ссылался С. У. Строганов в своей монографии, вовсе не при-
водит растительные остатки среди компонентов содержимого
желудков средних бурозубок. Б. С. Юдин прав, утверждая необя-
зательность летних растительных кормов для бурозубок, в том
Рис. 52. Ход изменения улови-
стости средних бурозубок на верх-
ней Лене (7) и нижней Ангаре (2)
в 1962 г.
По оси ординат — число зверьков на
10 канавко-суток; по оси абсцисс —
месяцы и декады.
числе средних. Однако не только
при недостатке животной пищи
средние бурозубки едят раститель-
ные корма, как думает этот автор.
Скорее наоборот, обилие семян,
их большая доступность позво-
ляют переходить бурозубкам
к растительноядности. Мы уже
отмечали семеноядность средней
бурозубки в горных пихтачах
зимой (Реймерс, 1959а). Там масса
семян пихты была на поверхности
снега. В темнохвойных лесах юж-
ной тайги Средней Сибири даже
летом (в июле — августе) семена
составляют до 24% отмечаемых
кормов по встречаемости и нередко
ими целиком наполнены желудки
зверьков. В августе 1962 г. семена
и ягоды были встречены в желуд-
ках одной трети экземпляров (про-
анализировано содержимое 24 же-
лудков), а в предыдущем месяце
того же года растительные корма
найдены у половины зверьков,
желудки которых оказались с со-
держимым (10 экз.). Доля расти-
тельных кормов значительно ниже
у средних бурозубок, живущих
в сосняках, и совсем ничтожна
у зверьков, обитающих на гарях и в кустарниках. Однако
по наблюдениям в окрестностях Иркутска, средние бурозубки
зимой и весной практически отсутствуют среди мелколесий
и кустов. Их следы ведут к скоплениям семян березы, обра-
зующимся в лыжнях и других углублениях. Видимо, распростра-
нение вывода Б. С. Юдина (1962) на зимний период следует поста-
вить под сомнение, тем более что отсутствие древесных насажде-
ний в местах зимовки бурозубок, на что ссылается автор (стр. 94),
не означает невозможности поедания семян (трав и кустар-
ников).
Черви и личинки насекомых доминируют среди кормов средней
бурозубки, однако лишь на гарях в придолинной полосе. В водо-
236
Таблица 6
Состав и встречаемость пищевых компонентов в содержимом желудков
средней бурозубки (542 экз.)
Пищевые компоненты Число же- лудков сдан- ным кормом Процент к общему ко- личеству же- лудков с кор- мом Время обнаружения корма
Растительные корма, всего 42 8.0
В том числе: Семена хвойных 4 0.8 30 III, 22 IV, 9 VI,
Семена, ближе не определенные 20 4.0 24 IX И юль—сентябрь.
Ягоды брусники 1 0.2 16 IX
Ягоды, ближе не определенные 8 1.5 Август—сентябрь.
Зеленая вегетативная масса 3 0.6 10 и 30 VIII, 19 IX
Остатки растений, ближе не определенных (случайные примеси) 4 0.8
Грибы 1 0.2 5 VIII
Животные корма, всего 506 93.3 —
В том числе: Насекомые, ближе не опреде- ленные 319 59.0 Весь теплый период.
Личинки насекомых, всего . . 185 34.1 » » »
Из них: Почвенные личинки насеко- мых, ближе не определен- ные 175 30.4 » » »
Гусеницы бабочек 10 1.9 Июль—сентябрь.
Дождевые черви 87 16.0 Июнь—сентябрь.
Моллюски, ближе не определен- ные 1 0.2 16 VII
Остатки позвоночных живот- ных 4 0.8 И, 14, 16 VIII
Молоко (у молодых) 3 0.6 26 VI, 15 и 18 VIII
Остатки животных, ближе не определенные 33 6.0 —
раздельных лесах эти компоненты не составляют более 20%,
а дождевые черви не встречаются вовсе.
Паразиты, значение. На средних бурозубках пара-
зитируют различные виды блох, вшей, гамазовых и иксодовых
клещей. Наибольшее практическое значение придается прокорм-
лению на бурозубках иксодид. В 1961 и 1962 гг. паразитологами
обработано на двух полустационарах 435 средних бурозубок.
Часть материалов за 1961 г. опубликована нами ранее (Реймерс
и Воронов, 1963). В табл. 7 приведены сводные данные о зара-
237
Таб
Зараженность личинками и нимфами иксодовых клещей
Номер выдела Биотоп Верхнеленский
1961 1 г.
очесано зверьков индекс встречаемости, в % обилие (абс.). индекс верности по оби- лию, в % обилие пара- зита на еди- ницу учета хозяина (из расчета количества зверьков на июль и август)
личинки нимфы
2 Смешанный водораздельный 16 0.0 0.0 0.0 0.0 0.4
темнохвойный лес. (6.2) (0.1) (25.0)
3 Его гари. 25 12.0 0.6 0.7 3.4 0.3
(4.0) (0.0) (16.6)
5 Смешанный приручьевой — — — — —
темнохвойный лес.
8 Водораздельный сосновый — — — — — —
бор.
11 Его гари. — — — — — —
19 Островной темнохвойный лес. И 81.8 12.5 16.0 56.2 —
20 Его гари. 27 77.7 10.9 13.9 62.5 —.
21 Сосняк. 7 71.4 6.1 7.9 7.9 —
22 Смешанный сосновый лес. И 82.0 3.0 4.0 6.0 0.2
(9.1) (0.1) (37.5)
23 Гари сосняков. 1 100.0 11.0 14.0 13.2 —
24 Остепненный сосняк. 10 50.0 4.1 5.2 10.2 —
25 Его гари. — — — — —
28 Приречный ельник. 16 93.7 5.3 6.7 29.1 —
31 Опушечные кустарники. 19 84.2 7.8 10.0 46.8 0.6
(5.3) (0-1) (20.9)
31а Ивняки у лугов. — — — — —
32 Луга. — — — — — —
34 Приручьевой ельник. 7 28.5 6.4 8.2 21.7 —
35 Его гари. — — —- — — —
37 Ерник. 4 100.0 10.5 13.4 18.9 —-
38 Поля. —
41 Огороды. — — — — — —
Всего / Личинки . . . 150 58.0 5.90 100.0 —-
1!сег0 1 Нимфы . . . 150 2.7 0.02 100.0 — —
Примечал и е. В скобках указаны числа, относящиеся к нимфам клещей.
238
лица 7
средних бурозубок южной тайги Средней Сибири
полустадионар Нижнеангарский полу стадион ар
1962 г. 1962 г.
о" обилие пара- о" обилие пара-
№ S зита на еди- И S зита на еди-
§ ° о ницу учета s ю о ницу учета
ф Е хозяина (из расчета Ь О о о к хозяина (из расчета
§ ф § и количества п о S количества
к сб зверьков на к Е- зверьков на
а Ф ф ь о о И а июль и август) д ft ф ф е. о ф X июль и август)
§ W аз. ф о я©- S д со я ю се Ф О Д о'' S
Д св 8 ф к ф S S индекс ЛИЮ, в X д S д <мфы гесанс о X 1 ф S ч S о а X л 11 1ЧИНК1 1МфЫ
о S о д о S ю о S ч йч д
3 33,3 0.3 1.6 0.7 — — — — — — —
5 20.0 0.2 1.1 0.7 — — — — — — —
1 0.0 0.0 0.0 — — — — — — — —
— — — — — — 14 0.0 0.0 0.0 — —
— — — — — 21 0.0 0.0 0.0
14 50.0 1.0 5.5 11.0 — 35 0.0 0.0 0.0 — .—.
28 32.1 0.8 4.4 6.9 —
16 50.0 1.9 10.5 11.4 0.6 11 0.0 0.0 0.0 — —
(6.2) (0.1) (5.8)
8 37.5 2.6 14.3 12.0 0.5 14 14.2 0.2 66.7 1.2
(12.5) (0-1) (5.8)
6 50.0 0.7 3.8 2.0 — — — —.
3 66.7 1.7 9.4 3.0 —
9 55.5 1.1 6.0 4.5 — — —
6 33.3 0.7 3.8 4.0 1.1 14 0.0 0.0 0.0 —
(W.7) (0.2) (И.9)
10 50.0 1.4 7.7 7.4 2.1 — — — .
(10.0) (0.4) (23.6)
4 0.0 0.0 0.0 — .— 4 0.0 0.0 0.0 — —
5 60.0 0.6 3.3 0.6 — -
21 47.6 3.5 19.3 22.0 0.6 —
(4.8) (0.1) (5.8)
— — — — —• — И 9.1 0.1 33.3 0.2 °:2
17 47,1 1.4 (9.1) (0.1)
7.7 7.6 0.5 — — —
(5.8) (0.1) (5.8)
2 0.0 0.0 0.0 — — — —
3 33.3 0.3 1.6 0.1 0.3 — — —
(66.6) (0.7) (41.3)
161 42.2 1.41 100.0 — — 124 2.3 0.03 100.0 __
161 4.9 0.08 100.0 — — 124 0.8 0.01 100.0 — —
239
женности клещами без деления зверьков на половозрастные группы
(что сделано в указанной работе). Судя по полученным цифрам,
роль средних бурозубок в прокормлении иксодовых клещей (в на-
шем случае, очевидно, Ixodes persulcatus) резко меняется от года
к году. В 1961 г. наивысшие показатели встречаемости и обилия
личинок были зарегистрированы в островных темнохвойных
лесах, их гарях и в группе долинных темнохвойных лесов (ельни-
ков). На следующее лето зараженность личинками резко упала
и прежде всего на гарях. Объяснить это явление нетрудно: лето
1961 г. было очень сухим, гари сильно высохли и стали неблаго-
приятным местом обитания иксодид. Наибольшее обилие пара-
зита на единицу учета хозяина сохранилось в долинных
ельниках, а в чистых и смешанных сосняках оно даже повыси-
лось.
Нижнеангарский полустационар расположен у северной гра-
ницы ареала таежного клеща. Здесь он паразитирует на сред-
ней бурозубке только в наиболее «теплых» биотопах там,
где зверек больше всего роется в подстилке, добывая дожде-
вых червей, — на гарях и в смешанных сосново-мелколиственных
лесах.
В годы высокой численности средняя бурозубка, очевидно,
прокармливает большое число личинок и сравнительно немного
нимф иксодовых клещей. Максимальное число личинок, снятых
с одного зверька, 55, нимф — 4.
Средние бурозубки, прокармливая паразитов, приносят пас-
сивный вред. Некоторый ущерб наносят они и уничтожением
семян, полезных насекомых и дождевых червей. Однако зверьки
поедают много и вредных насекомых, а высокая агрессивность
бурозубок заставляет предположить, что они играют определен-
ную санитарную роль, уничтожая больных грызунов и соплемен-
ников (случаи каннибализма очень обычны при попадании буро-
зубок в цилиндры). Общеизвестна роль бурозубок в уничтожении
трупов мелких зверьков.
Хищные птицы, особенно совы, охотно поедают средних буро-
зубок (Реймерс, 1959а). В питании ценных млекопитающих они
занимают меньшее место. Однако соболи, по данным А. С. Фети-
сова (1947), В. В. Тимофеева и В. Н. Надеева (1955), охотно пое-
дают бурозубок. Эти зверьки найдены указанными авторами в 25
и даже 30% желудков соболей.
Вместе с другими бурозубками средние разрыхляют почву
и тем способствуют лесовозобновлению (Юдин, 1962, и др.).
Роль средних бурозубок в природе и хозяйстве выяснена
далеко не полностью. До сих пор еще мало материалов об эпиде-
миологическом значении зверьков. Между тем констатация их
максимума в долинных ельниках и проникновение зверьков на
поля и огороды указывает на возможность тесных контактов
этих млекопитающих с человеком и домашним скотом.
240
Обыкновенная бурозубка — Sorex araneus L.
Под названием «обыкновенная бурозубка» в данном случае
вместе с S. araneus объединяется также S. vir Gl. Allen., экзем-
пляры которой могли попасть в наши довольно многочисленные
сборы (свыше 3500 экз.). Судя по описанию Б. С. Юдина (1962),
упомянутые две формы довольно четко отличаются местами оби-
тания: обыкновенная бурозубка предпочитает сухие участки,
плоскочерепная — влажные. Возможно, это обстоятельство не-
сколько нивелирует закономерности биотопического распределе-
ния обыкновенной бурозубки в принятом нами объеме. Биологи-
ческие особенности обыкновенной и плоскочерепной бурозубки
весьма сходны, поэтому мы считаем, что практическая ценность
наших материалов сохранится даже в том случае, если будет
полностью доказана видовая специфика этих двух форм.
Распространение, распределение и чис-
ленность. Обыкновенная бурозубка распространена в южной
тайге Средней Сибири повсеместно, к северу несколько умень-
шаясь в числе. В 1962 г. средняя численность этого вида на тер-
ритории вер хне ленского полустационара была приблизительно
в 5—6 раз выше, чем в районах Ангаро-Подкаменнотунгусского
междуречья. Сравнение различных лет рискованно, однако в 1963 г.
на верхней Лене наблюдалась численность зверьков, равная их
обилию на нижней Ангаре летом 1962 г. (к 1963 г. она могла
понизиться). Под Иркутском обыкновенная бурозубка — один
из самых многочисленных видов. Здесь ее отмечали многие авторы
(Огнев, 1928; Скалой, 1933, и др.).
О распределении этого вида по биотопам известно очень мало.
Для верхней Лены мы уже дали подробное описание (Реймерс
и Воронов, 1963), которое отражает и положение, существующее
во всей южной тайге. На рис. 53 приведена принципиальная схема
размещения обыкновенных бурозубок по биотопам с применением
шестибалльной шкалы оценки численности (этот вид нигде не
отсутствует полностью). Легко можно заметить, что обыкновенная
бурозубка при всей своей эврибионтности предпочитает места
с высокой влажностью воздуха: придолинные леса (до 23.8 зверька
на 10 канавко-суток в июле—августе), темнохвойную тайгу (до 21.5
зверька на то же число канавко-суток) и т. д., но избегает заболо-
ченных участков (не более 10.2 'зверька в тот же период).
Открытые биотопы, как и кустарники, неблагоприятны для
вида (1.0—7.6 экз. на 10 канавко-суток в июле и августе). Это
типично лесной зверек. ОднаКо и очень глухие леса населены
слабее, чем слегка разреженные. Больше всего обыкновенных
бурозубок ловилось в кустарниках у реки в местах, где кустар-
никовые заросли соприкасаются с приречными ельниками (до 30.2
зверька на 10 канавко-суток в июле и августе). Такие же ивняки
у лугов бедны зверьками (не более 11.7 экз.).
16 Н. Ф. Реймерс
241
И' VZ& ШШР
•) о;
Рис. 53. Принципиальная схема tлетнего размещения обык-
новенных бурозубок в пределах^ю/кной тайги Средней Сибири.
ДцдОры в скобках — количество зверьков.на 10 канавко-суток в июле и
августе; 1—первая максимальная ’/радиация численности (более 20);
2 — вторая максимальная градация -численности (15—20); з — первая
средняя градация численности (10—15); 4 — вторая средняя градация
численности (5—10); 5—минимальная численность (1—5). Остальные
обозначения как па рис. 12.
Сведение лесов приводит к сокращению излюбленных стаций
обыкновенной бурозубки. Уменьшается число зверьков и при
выгорании темнохвоиных лесов.
Однако разреживание сосняков
вновь увеличивает численность
обыкновенных бурозубок. Сплош-
ная вырубка или гарь соснового
леса так же бедны зверьками, как
и сам бор. В общей сложности вид
достигает наивысшей численности
на средних фазах лесной сукцес-
сии.
Общие закономерности рас-
пределения обыкновенных буро-
зубок в южной тайге Средней
Сибири весьма схожи с наблюдае-
мыми в Татарии (Попов, 1960),
конечно с учетом особенностей
этих территорий.
От 1960 до 1962 г. на верхней
Лене число обыкновенных буро-
зубок оставалось почти стабиль-
ным (в пределах биотопа не изме-
нилось больше чем в 2 раза).
В 1963 г. обилие зверьков резко
снизилось (в 5—8 раз). Таким
образом, численность вида под-
вержена значительным колебаниям
от года к году.
В течение лета можно наблю-
дать два пика активности обык-
новенных бурозубок (рис. 54).
Первый из них обычно малозаме-
тен и приходится на третью декаду
июня—первую декаду июля. Вто-
рой очень резко выражен: во вто-
рой—третьей декадах августа
обыкновенные бурозубки дости-
гают наивысшей численности, ре-
гистрируемой ловчими канавками.
Повышение обилия происходит
за счет появления большого коли-
июпь июль август, сентябрь
Рис. 54. Ход изменений улови-
стости обыкновенных бурозубок
на верхней Лене и нижней Ан-
гаре.
1 — Лена, 1960 г.; 2 — Лена, 1961 г.;
5,4 — Лена и Ангара, 1962 г. По оси
ординат — число зверьков на 10 ка-
навко-суток; по оси абсцисс — месяцы
и декады.
чества молодых особей. Число взрослых самцов в уловах по-
степенно падает к осени. Взрослые самки, наоборот, осенью
достигают наибольшего обилия (особенно в сентябре).
Размножение. В конце марта у обыкновенных буро-
зубок юга Средней Сибири наблюдаются первые признаки гона.
16*
243
Та б
Число эмбрионов и количество размножающихся
Месяц Декады Отловлено взрослых самок
всего в том числе со следами размножения
всего процент от всех самок из них беременных
всего процент от всех самок
Июнь . . . . j 2 5 2 40.0 1 20.0
3 7 4 57.1 3 42.8
1 18 6 33.3 5 27.7
Июль .... | 2 13 10 76.1 8 61.5
3 28 19 67.8 13 46.4
1 39 25 64.1 18 46.2
Август .... | 2 46 22 47.8 И 23.9
3 46 23 50.0 16 34.7
1 28 И 39.3 3 10.7
Сентябрь . . | 2 9 1 11.1 1 11.1
3 5 1 20.0 — —
Всего или сред- нее — 245 — — 79 —
В теплые весны резкое увеличение активности зверьков можно
отметить с середины месяца (под Иркутском). На верхней Лене
и нижней Ангаре половое возбуждение наблюдается несколько
позже — не раньше 25 марта и обычно уже в апреле. В среднем
апрель — начало брачной поры у обыкновенных бурозубок. По
данным различных авторов (Бобринский и др., 1944; Dehnel,
1952; Попов, 1960, и др.), продолжительность беременности у обык-
новенных бурозубок от 16 до 20 и даже 28 дней. Следовательно,
в мае должны быть уже молодые особи. По данным А. Денеля
(1952), зверьки развиваются в гнезде 21 день, по 3. А. Соколовой
(1962) — 20 дней. Действительно в первой декаде июня в уловах
появляются молодые зверьки.
Взрослые самки, кормящие молодых, нередко бывают уже
беременны вновь и в июле—августе приносят еще один помет.
Следующий (третий) помет бывает не у всех особей. Наше указа-
ние о существовании четвертого помета у взрослых самок (Рей-
мерс и Воронов, 1963) в свете новых материалов следует подверг-
нуть сомнению.
Молодые самки с признаками беременности появляются в сере-
дине июня (1961 г.) или начале июля (1962, 1963 гг.). В июле-
августе встречаются молодые кормящие самки. Часть из них дает
второй помет, отличающийся малым числом детенышей.
Общее число генераций, как правильно отметил В. А. Попов
(1960), не менее трех, но наиболее выражены две, которые обычно
244
лица 8
самок обыкновенной бурозубки по месяцам и декадам
и указываются для этого вида (Лаптев, 1958; Межжерин, 1961;
Hellwing u. Schnapp, 1962, и др.).
Количество взрослых самок, участвующих в размножении,
обычно велико (табл. 8) и сходит на нет к октябрю. Молодые самки,
как мы уже отмечали (Реймерс и Воронов, 1963), участвуют в раз-
множении не все и главным образом из ранневесенних пометов.
То же относится и к молодым самцам. В. А. Межжерин (1961)
даже полагает, что все прибылые самцы размножаются лишь
после зимовки.
Число эмбрионов у обыкновенных бурозубок южной тайги
Средней Сибири колеблется от 1 до 12, составляя в среднем около 7,
чаще всего встречаются самки с 6—8 эмбрионами. В пределах
южной тайги не наблюдается изменения числа эмбрионов от юга
к северу, во всяком случае уменьшение их числа не констатиро-
вано. Не смогли мы подметить и особых отличий в числе эмбрионов
у самок водораздельной полосы и особей долин. Несмотря на
разницу в биотопах и полукилометровый перепад высот, среднее
число эмбрионов было практически равным: водораздел (9 экз.) —
7.3, долино-придолинная полоса — 7.5 (верхняя Лена, 1962 г.).
Аналогичными оказались и крайние варианты числа эмбрионов —
обычно от 5 до 12.
Питан и е. Кормовые объекты сибирских обыкновенных
бурозубок изучены в настоящее время уже достаточно полно
(Юдин, 1956, 1962; Лаптев, 1958; Реймерс и Воронов, 1963).
245
Хорошо известно и питание зверьков в других районах (Фоли-
тарек, 1940; Лавров, 1943; Формозов, 1948; Тупикова, 1949; По-
пов, 1960; Hellwig u. Schnapp, 1962, и др.). В Средней Сибири,
по нашим данным, животные корма составляют в питании обык-
новенной бурозубки 83.4—99.0%, доля растительных кормов
в теплый период года не превышает 16.6% (Реймерс и Воронов,
1963). Среди объектов питания нами указаны семена растений
(в том числе кедра и пшеницы), ягоды брусники и земляники,
насекомые и их личинки, пауки, дождевые черви и позвоночные
животные (мелкие грызуны).
Почти у 1500 обыкновенных бурозубок из Западной Сибири
В. С. Юдин (1962) практически не нашел в желудках раститель-
ных кормов. В европейской части СССР многие из упомянутых
авторов констатировали поедание семян. Л. В. Заблоцкая (1955,
1957) полагает, что бурозубки в растаскивании семян сосны
играют большую роль, чем мышевидные грызуны. Нами было
проанализировано содержимое желудков свыше 3500 обыкновен-
ных бурозубок из южной тайги Средней Сибири. Даже в летние
месяцы (июль и август) бурозубки поедают семена и ягоды, при-
чем в августе, когда семян и ягод больше, эти корма встречаются
значительно чаще, чем в июле (доля растительных кормов в июле
около 1%, в августе — несколько более 4%). Резко возрастает
роль семян и ягод в холодный период года. Зимой по следам не-
трудно видеть, как бурозубки кормятся семенами хвойных пород
и березы.
Следует заметить, что характер питания обыкновенной буро-
зубки, как и всех мелких зверьков, в сильной степени зависит
от наличия кормов, от «кормового спектра» в биоценозе. Это хо-
рошо показывают материалы анализа содержимого желудков
обыкновенных бурозубок двух водораздельных участков на верх-
ней Лене: гарей, поросших березовым мелколесьем, и темнохвой-
ной тайги (табл. 9).
В последней колонке таблицы помещены данные анализа со-
держимого 500 желудков обыкновенных бурозубок, отловленных
на всей территории Верхнеленского полустационара в тот же
период времени (сравниваются материалы за июль и август).
Видно, что имеется довольно значительная разница в питании
бурозубок леса и его гари, и вместе с тем на водораздельных про-
странствах обыкновенная бурозубка питается резко отлично от
зверьков долин и придолинных полос (средний процент колеб-
лется именно поэтому). На водоразделе мало дождевых червей,
меньше насекомых и их личинок, зато много семенных кормов.
Все это отражается на «спектре питания» обыкновенной буро-
зубки.
Состав кормов обыкновенной бурозубки стабилен на террито-
рии южной тайги Средней Сибири. Так, на нижней Ангаре летом
насекомые найдены в 87 % желудков с кормом, черви обнаружены
246
Таблица 9
Сравнение состава и встречаемости основных пищевых компонентов
в содержимом желудков обыкновенных бурозубок,
отловленных в различных биотопах водораздельной полосы,
июль—август 1961—1962 гг.
Пищевые компоненты Темнохвийпая тайга Гари темнохвойной тайги, поросшие березовым мелко- лесьем Средний процент встречаемости корма
число желуд- ков с данным кормом процент к общему ко- личеству же- лудков с кор- мом число желуд- ков с данным кормом процент к общему ко- личеству же- лудков с кор- мом
Насекомые Личинки насекомых . . . Гусеницы бабочек .... Дождевые черви Семена Г рибы Ягоды Зелень Животные корма, ближе не определенные Всего желудков с кормом 102 18 1 14 32 2 15 143 71.3 12.5 0.7 9.8 22.3 1.4 1.4 10.5 100.0 125 12 8 23 2 1 3 20 164 76.2 7.3 5.6 14.0 1.2 0.6 1.8 12.2 100.0 96.2 47.8 2.2 63.6 4.0 0.8 1.0 0.6 16.4
в 35% желудков, семена — в 8.7 ?о желудков и т. д. Наблюдае-
мая разница (табл. 9) не выходит за пределы возможной в одной
и той же местности, но в разные годы.
Паразиты, значение. Как показали исследования
Б. С. Юдина (1962) в Западной Сибири, обыкновенные бурозубки
весьма сильно заражены гельминтами (3 вида трематод, 7 форм
цестод и 8 видов нематод) — до 40 ?6 зверьков поражено этими
паразитами. Различные гельминтозы отмечались и нами при вскры-
тии бурозубок в Средней Сибири. Виды паразитических червей
определены не были, но можно констатировать, что уровень за-
раженности гельминтами в Средней Сибири во время проведения
наших работ был ниже, чем отмеченный для Западной Сибири
Б. С. Юдиным.
Обыкновенные бурозубки заметно поражены блохами, вшами
и особенно гамазовыми клещами. Многие виды этих паразитов
свойственны и другим мелким млекопитающим. Блохи обыкно-
венных бурозубок на человека не нападают (Попов и Казанская,
1958).
Бурозубки могут служить резервуаром туляремийной и энце-
фалитной инфекции (Дунаева, 1954; Карпов, 1954; Карпов и
Попов, 1956). Степень зараженности этого вида представителями
иксодид в южной тайге показана в табл. 10. Поражаемость
247
Та б
Зараженность личинками и нимфами иксодовых клещей
Номер выдел Биотоп Верхнеленский
1961 г.
очесано зверьков индекс встречаемости, в % обилие (абс.) индекс верности по оби- лию, в % обилие пара- зита на еди- ницу учета хозяина (из расчета количества зверьков на июль и август)
личинки нимфы
2 Смешанный водораздельный 33 27.2 1.1 1.2 18.5 1.7
темнохвойный лес. (3.0) (0.1) (10.9)
3 Его гари. 50 32.0 0.7 0.8 9.8 0.3
(2.0) (0.02) (2-2)
5 Смешанный приручьевой — — — — — —
темнохвойный лес.
8 Водораздельный сосновый — — — — — —
бор.
11 Его гари. — — •— — — —
19 Островной темнохвойный лес. 36 91.6 17.9 19.3 384.8 —
20 Его гари. 58 84.5 11.2 12.4 112.0 1.0
(3.4) (0.1) (10.9)
21 Сосняк. 21 66.6 4.2 4.7 25.2 —
22 Смешанный сосновый лес. 51 86.2 8.8 9.7 136.4 1.6
(7.8) (0.1) (10.9)
23 Гари сосняков. 22 86.3 5.4 6.0 49.1 —
24 Остепненный сосняк. 33 81.8 5.3 5.9 55.6 0.6
(6.0) (0.06) (6.6)
25 Его гари. — — —. — — —
28 Приречный ельник. 68 80.8 9.4 10.4 223.7 2.4
(2-9) (0.1) (10.9)
31 Опушечные кустарники. 78 79.4 5.2 5.8 128.4 0.7
(2-5) (0.03), (3.3)
31а Ивняки у лугов. — — — — —- —
32 Луга. 26 69.2 5.0 5.6 28.5 —
34 Приручьевой ельник. 35 62.8 5.8 6.4 73.3 5.4
(2.8) (0.4) (44.3)
35 Его гари. — — — — — —
37 Ерник. И 90.9 10.6 11.8 98.6 —
38 Поля. — — — — — —
41 Огороды. — — — — — —
Rrprn f Личинки .... 552 72.2 7.1 100.0 — —
Всег0 1 Нимфы 552 2.7 0.10 100.0 — —
Примечание. В скобках указаны числа, относящиеся к нимфам клещей.
248
лица 10
обыкновенных бурозубок южной тайги Средней Сибири
полустационар Нижнеангарский полустационар
1962 г. 1962 г.
« о х ф го X о S Ф го о ft о )Н0СТИ по оби- обилие пара- зита на еди- ницу учета хозяина (из расчета количества зверьков на июль и август) д о X д ft ф 'речаемости, в % 6 шести по оби- обилие пара- зита на еди- ницу учета хозяина (из расчета количества зверьков на июль и август)
и го 2 о д ко го, ф ф о О Д X X д го о д ко го, ф % я 1эя 0 X X
го о ф П X п к . к X е S го о ф X S Ч X X 5 В X X X *9* 2
ЕС ко X X S ым X X КО х а X X
о X о X X ч X о X о х ч ч X
7 0.0 0.0 0.0 — — •— — — — —
31 16.1 0.2 1.4 2.5 1.2 — — —. — — —
17 (9.7) 11.8 (0.1) 0.2 (9.7) 1.4 2.1 2.1 -
— (11-8) (0.2) (19.6) — — 8 0.0 0.0 0.0 — —
и 0.0 0.0 0.0 __
20 45.0 1.6 11.4 18.7 0.6 8 0.0 0.0 0.0 — —
33 (5.0) 54.5 (0.05) 1.8 (5.0) 12.9 22.0 0.4 — —
25 (3.0) 32.0 (0.03) 0.5 (3.0) 3.6 3.4 0.3 3 о-.о 0.0 0.0
20 (4.0) 40.0 (0.04) 0.8 (4.0) 5.7 12.2 3.0 15 0.0 0.0 0.0 0.2
28 (15.0) 32.1 (0.2) 0.7 (19.6) 5.0 5.6 0.3 (6Л>) (0.07) (100.0)
25 (3.8) 36.0 (0.04) 0.9 (4.0) 6.4 9.7 0.4 — —
6 (4.0) 33.3 (0.04) 0.7 (4.0) 5.0 5.2 — —. .
45 46.6 0.9 6.4 14.7 0.3 8 0.0 0.0 0.0 — —
53 (2.2) 30.2 (0.02) 0.5 (2.0) 3.5 9.0 1.8
13 (9.4) 7.7 (0.1) 0.08 (9.7) 0.5 0.9 — 3 0.0 0.0 0.0 — —
3 33.3 0.3 2.1 0.3 — — — — — — —
42 52.4 1.9 13.7 19.6 1.0 — — — — — —
(7Л) (0Л) (9.7) 8 50.0 3.6 100.0 6.8
19 68.4 1.9 13.7 14.4 0.8 — — — — — —
13 (10.5) 46.1 (0.1) 1.0 (9.7) 7.1 4.3 — —. — — —
3 0.0 0.0 0.0 — — — — — —— -— —
403 37.4 0.9 100.0 — 64 6.2 0.4 100.0 — —
403 5.9 0.06 100.0 — — 64 1.6 0.01 100.0 — —
249
обыкновенных бурозубок иксодовыми клещами колеблется в за-
висимости от условий года, но всегда остается наибольшей
в островных темнохвойных лесах, смешанных сосняках, в при-
речных и приручьевых ельниках, а также на гарях ельников
и островных темнохвойных лесов.
Нами, как и Л. П. Никифоровым (1961а), констатирована
более высокая зараженность личинками иксодид самцов по срав-
нению с самками (Реймерс и Воронов, 1963).
В северных районах южной тайги Средней Сибири личинки
и нимфы иксодид обнаружены на обыкновенных бурозубках лишь
в смешанных сосново-мелколиственных лесах и на гарях при-
ручьевых ельников. Здесь же констатированы и максимумы имаго
паразита.
Общая зараженность личинками и нимфами средних и обык-
новенных бурозубок сходна. Близки показатели встречаемости
и обилия, полученные нами и Л. П. Никифоровым в Западной
Сибири (по Никифорову, встречаемость личинок клещей на обык-
новенной бурозубке 25.0%, нимф — 5.0%, обилие личинок 1.40,
нимф 0.1).
В наших материалах имеется непроверенное и сомнительное
указание на то, что со взрослой обыкновенной бурозубки был
снят взрослый иксодовый клещ.
Роль обыкновенной бурозубки как объекта питания ценных
животных аналогична значению средней бурозубки. Общая
оценка деятельности этих двух видов зверьков сходна.
Обыкновенная бурозубка, как и средняя, проникает в слу-
жебные постройки при усадьбах, но непосредственно в домах
эти зверьки зарегистрированы не были.
Кутора — Neomys fodiens Pennant.
По С. У. Строганову (1947), куторы из Средней Сибири отно-
сятся к восточному, или короткоухому подвиду N. f. orientis
Thomas.
Распространение, размещение и ч и с лен-
ное т ь. Кутора распространена в южной тайге Средней Сибири
повсеместно, вернее этот вид обитает во всей полосе, занятой
комплексом зооформаций речных долин юга Средней Сибири.
С. У. Строганов (1957) ссылается на очень небольшое число на-
ходок кутор в пределах нашего района, сделанных на Ангаре и
ее южных притоках (Middendorf, 1853; Radde, 1862; Скалой,
1933), а также в лесостепной полосе (сборы С. С. Турова под Ир-
кутском; не оговоренное, но отмеченное на картосхеме место
находки у Нижнеудинска). Однако мы нашли вид в ряде более
северных районов. Кутора оказалась весьма обычным зверьком
на верхней Лене (Реймерс и Воронов, 1963) и найдена на р. Чадо-
бец в окрестностях дер. Яркино (около 59° с. ш.).
250
На верхней Лене кутора обитает главным образом в приреч-
ных и приручьевых кустарниках, не избегая, однако, долинных
ельников. 1 экз. пойман на выпасаемом лугу в 100 м от водоема.
2 зверька, в том числе взрослый самец, были отловлены еще
дальше от постоянного водотока — в островном темнохвойном
лесу, однако вблизи этого участка имелся пересыхающий ручей.
Всюду места отлова расположены не далее 1 — 1.5 км от реки.
Исключение составила поимка молодого самца куторы в при-
ручьевом ернике, сравнительно удаленном от значительного водо-
тока — более чем на 4 км.
К северу численность кутор резко падает. На р. Чадобец
зверьки найдены лишь у самой воды в приречных ивняках. Зи-
мой местные жители нашли мертвую кутору на льду. Возможно, что
она погибла по вине людей, безжалостно глушащих рыбу на реке.
При наблюдениях на Верхнеленском полустационаре в 1961 —
1963 гг. резких колебаний численности кутор не отмечено.
Размножение, питание. Молодые зверьки по-
падают в канавки с середины июля. В это же время найдены
только что родившие самки. В начале августа отловлены две
самки в начальной фазе беременности. Эти факты заставляют пред-
положить возможность 3 генераций в течение лета.
Как мы уже отмечали (Реймерс и Воронов, 1963), возможно
размножение кутор-сеголеток. Во всяком случае размер их
семенников (6.1x3.4 мм) равен величине тестикул взрослых
экземпляров.
В желудках кутор найдены насекомые, мелкая рыбешка,
водные личинки насекомых и мясо лягушки.
Паразиты, значение. На куторах обнаружены
блохи, гамазовые и иксодовые клещи (последние лишь в одном
случае — 1 личинка). По свидетельству Т. Н. Дунаевой (1954),
куторы могут заражаться туляремией, а по данным С. П. Карпова
и В. М. Попова (1954), зверьки служат объектом резервации ма-
лярийного плазмодия. Последний факт заслуживает внимания,
поскольку куторы довольно многочисленны на берегах водохра-
нилищ (в частности, Иркутского «моря»). Остается только неяс-
ным, каково отношение найденного возбудителя к тому, который
паразитирует в человеке. Малярия — весьма специфичная бо-
лезнь.
Общая роль кутор в наземных биоценозах и как истребителя
мальков рыб невелика. Зверьки ловят главным образом так назы-
ваемых сорных рыбешек.
Отряд РУКОКРЫЛЫЕ — CHIROPTERA
Летучие мыши в пределах южной тайги Средней Сибири —
редкие зверьки, чаще всего встречающиеся в долинах рек у насе-
ленных пунктов и на опушках водораздельных лесов, выходя-
251
щих к обширным гарям. Образ жизни и эпидемиологическое зна-
чение летучих мышей почти совершенно не известны. На рис. 55
мы постарались с наиболее возможной полнотой нанести все
Рис. 55. Места находок летучих мышей в нределах южной тайги
Средней Сибири.
I—XI и 1—ю как на рис. 6; 11 — прудовая ночница; 12 — водяная ночница;
13 — ночница Наттерера; 11 — ночница Иконникова; 15 — усатая ночница;
16 — длиннохвостая ночница; 17 — обыкновенный ушан; 18 — северный
кожан; 19 — двухцветный кожан; 20 — сибирский трубконос.
известные находки летучих мышей в пределах южной тайги
и в пограничных полосах смежных районов. Следует обратить
внимание на то, что в пределах Средней Сибири летучие мыши не
обнаружены севернее полосы господства южнотаежных лесов»
252
Едва ли тут можно усмотреть прямую связь с темнохвойной тай-
гой и ее производными. Скорее эти леса служат индикатором при-
родных условий, необходимых для существования рукокрылых.
Прудовая ночница — Myotis dasycneme Boie.
В пределах южной тайги Средней Сибири вид не отмечен.
Единственная находка относится к долине Енисея (Кузякин,
1950).
Водяная ночница — Myotis daubentonii Kuhl.
Водяная ночница найдена главным образом в лесостепных
районах в окрестностях Иркутска, Канска и Красноярска.
Однако встречается на верхней Ангаре (Кузякин, 1950) и в Вос-
точном Саяне (Ким, 1961).
В окрестностях Красноярска вид зимует (Емельянова и Вы-
соковский, 1962) в Караульской пещере.
По свидетельству Т. А. Кима (1961), водяная ночница питается
в основном комарами. Кормежка длится 30—40 мин. Те же са-
мые корма найдены нами в желудке экземпляра (взрослого сам-
ца), добытого в с. Якимовка 24 июля 1961 г. Помимо комаров,
в желудке оказались мелкие перепончатокрылые. Размер тестикул
самца 6.1x4.5 мм. На зверьках паразитируют гамазовые клещи
Spinturnix vespertilionis и Hirstionyssus sp.
Ночница Наттерера — Myotis nattereri Ogn.
Этот вид найден лишь южнее исследуемого района — в Тун-
кинской долине (Кузякин, 1950).
Ночница Иконникова — Myotis ikonnikovi Ogn.
В пределах южной тайги Средней Сибири эта летучая мышь
отмечена в долине Иркута и на верхней Лене. Встречается вид
в Хамар-Дабане. А. П. Кузякин (1950), ссылаясь на А. С. Фе-
тисова, считает характерным местообитанием вида горную тайгу.
Очевидно, эта мышь была отмечена нами на реках Ие и Малой
Белой в предгорьях Саян в 1952 и 1954 гг. Мышей мы не сумели
добыть и предположение основываем на порхающем и небыстром
характере полета зверька. Вылетали мыши поздно вечером, дер-
жась у опушки большой гари кедрового леса.
Усатая ночница — Myotis mystacinus Kuhl.
Этот вид широко распространен в пределах южной тайги Сред-
ней Сибири. Он найден в верховьях Подкаменной Тунгуски и на
р. Муре (Наумов, 1934), на северном Байкале (Radde, 1862), на
253
Енисее (Огнев, 1928) и Иркуте (Кузякин, 1950). Под Краснояр-
ском вид зимует в Торгашинской и Караульской пещерах (Ким,
1961; Емельянова и Высоковский, 1962).
На усатой ночнице из Караульской пещеры найдены блохи
Myodopsylla trisellis и гамазовые клещи (Емельянова и Высоков-
ский, 1962).
Длиннохвостая ночница — Myotis longicaudatus Ogn.
В пределах рассматриваемого района вид добыт однажды
под Красноярском — у дер. Додоновой 7 VII 1905 А. Я. Тугари-
новым (Кузякин, 1950). Зверек очень редок: известно всего не-
сколько экземпляров в пределах СССР.
Обыкновенный ушан — Plecotus auritus L.
Обыкновенный ушан широко распространен в пределах
южной тайги Средней Сибири. Самые северные находки его отно-
сятся к долине Подкаменной Тунгуски (Наумов, 1934). Известен
он из Киренска на Лене. Специальные поиски, предпринятые,
нами в окрестностях Жигалова на Лене, результатов не дали.
Вид зимует в пещерах окрестностей Красноярска (Емельянова
и Высоковский, 1962).
На обыкновенном ушане в условиях зимовки найдены блохи
Ischnopsyllus hexactenus kolenatii^ личинки краснотелок ТготЫ-
cula pavlovskyi, гамазовые клещи Spinturnix vespertilionis и
Hirstionyssus sp. (там же). Одна из этих форм — краснотелка
Т. pavlovskyi — паразитирует и на грызунах.
Северный кожан — Vespertilio nilssonii Keys, et Blas.
Вид широко распространен в лесостепи Средней Сибири и до-
лине Енисея. Н. П. Наумов (1934) указывает на устье р. Виви
(северный приток Нижней Тунгуски) как на самую северную
точку распространения вида. Под Красноярском северный кожан
зимует в Торгашинской и Караульской пещерах (Емельянова и
Высоковский, 1962).
Летом зверек обитает в населенных пунктах — найден
Т. А. Кимом на чердаке деревянного дома окраины г. Канска.
На зимующих зверьках были найдены 2 вида блох Ischnop-
syllus hexactenus kolenatii и Ischnopsyllus needhami (Емельянова и
Высоковский, 1962). Паразитируют на северных кожанах гамазо-
вые клещи Spinturnix vespertilionis и Hirstionyssus sp.
Двухцветный кожан — Vespertilio murinus L.
Для южной тайги Средней Сибири неизвестен, хотя найден
в Саянах (Кузякин, 1950), в том числе вблизи Красноярска.
254
Сибирский трубконос — Murina hilgendorfi Peters.
Л. П. Кузякин (1950) указывает единственное место регистра-
ции этого вида в Средней Сибири — р. Казыр-Сук, приток вер-
ховий Енисея, и полагает, что вид крайне редок. Однако
н. д. Емельянова и Н. И. Высоковский (1962) нашли этого
зверька в довольно большом количестве (5 самок и 4 самца)
в Караульской пещере под Красноярском. На зимовке в этой
пещере он оказался вторым по численности после обыкновенного
ушана. Те же авторы нашли на сибирском трубконосе два вида
блох (/schnopsyllus hexactenus kolenatii и Myodopsylla trisellis) и
гамазовых клещей.
Отряд ХИЩНЫЕ — CARNIVORA
Представители отряда хищных изучены в пределах южной
тайги Средней Сибири намного лучше зверей других групп.
В связи с этим мы либо только упоминаем формы хищников,
либо ограничиваемся фрагментарными замечаниями, сопрово-
ждая их минимальными библиографическими ссылками.
Соболь — Martes zibellina L.
Соболю Восточной Сибири посвящена колоссальная литера-
тура, неоднократно подвергавшаяся критическому разбору и
монографическому обобщению (Фаворский, 1935; Тимофеев, 1951;
Тимофеев и Надеев, 1955; Дулькейт, 1957; Сыроечковскип и Рос-
солимо, 1960; Строганов, 1962, и мн. др.).
Несмотря на известную лабильность в выборе биотопов, зверек
в южной тайге Средней Сибири должен считаться типичным оби-
тателем темпохвойных лесов. Здесь с древнейших времен конста-
тировалась наивысшая численность зверьков. Ниже мы покажем
(см. также: Реймерс и Воронов, 1963), что кормовые возможности
темнохвойной тайги для этого хищника наиболее благоприятны,
поэтому даже теоретически плотность соболиного поголовья в свет-
лохвойных лесах не может достигнуть того уровня, который ха-
рактерен для кедрово-пихтовых и елово-пихтовых массивов.
Гари и вырубки темнохвойпых лесов населены значительно
реже, чем лесные чащобы. Подчас хищник на гарях отсутствует
вообще (Соколов и Хлебников, 1962; Штильмарк и др., 1963).
Степной хорек — Putorius eversmanni Less.
Степной хорек чужд южной тайге — это зверь лесостепи и
степи. Северная граница вида довольно четко совпадает с пре-
делами широкого распространения лесостепи. В настоящее время
вид почти не выходит на водоразделы, придерживаясь не засе-
255
ленных людьми речных долин. Следует ожидать, что с дальней-
шим сведением тайги степной хорек будет широко проникать
в полосу южной тайги и достигать тут более высокой численности.
Американская норка — Putorius vison Schr.
Американская норка была завезена в районы южной тайги
Средней Сибири в 1936—1940 гг. (Лавров, 1946; Копылов, 1948).
Несмотря на первые обнадеживающие результаты (Владимиров,
1940), норка прижилась только в районах лесополья и не достигла
ожидаемой высокой плотности поголовья. Очевидно, этот вид не
сможет влиться в околоводные биоценозы южной тайги. В Сибири
реки замерзают надолго, а норки, не имея доступа к воде, гибнут
(Собанский, 1958).
Колонок — Mustela sibiricus Pall.
Колонок широко распространен в южной тайге Средней Си-
бири. К северу, уже за ее пределами, численность зверька посте-
пенно уменьшается. Вид тесно связан с долинами рек и ручьев.
Вне долин колонок всюду редок. Кружева ареалов колонка, гор-
ностая и ласки очень близко совпадают и могут служить негати-
вом схемы распространения соболя. Водораздельных темнохвой-
ных лесов колонок избегает, но селится по их приручьевым
участкам. Сведение леса не приводит к уменьшению поголовья
колонков. Гари, поросшие осиново-березовым лесом, благоприятны
для вида (Дулькейт и Козлов, 1958). В общей сложности, ги-
бель темнохвойного леса несколько «размывает» кружево ареала
вида, а численность зверька увеличивается. К такому же вы-
воду пришли Ф. Р. Штильмарк, Г. А. Соколов и их соавторы
(1963), исследуя млекопитающих лесов среднегорной полосы За-
падного Саяна.
Солонгой — Mustela altaica Pall.
Этот вид не живет в пределах южной тайги Средней Сибири,
однако может быть констатирован у ее юго-восточных окраин.
В Хамар-Дабане, по нашим наблюдениям, солонгой очень обы-
чен и спускается в низкогорья, поэтому возможно наличие
зверька в переходной полосе от горной тайги к плакорной.
Горностай — Mustela erminea L.
Горностай — обычнейший вид речных долин южной тайги
Средней Сибири. Зверьки, особенно молодые, нередко попадают
в цилиндры, установленные в канавки для отлова грызунов
(нами было отловлено 14 экз.: 12 молодых зверьков и 2 взрослых
256
самца). Зверьки на р. Чадобец весной, сразу после того как
стаял снег, вели себя очень смело, перебегая на глазах у людей.
Позже они стали скрытными.
Наиболее многочисленны горностаи летом по приречным ку-
старникам-ивнякам и ерникам. Реже зверьки встречаются в до-
линных ельниках и островных темнохвойных лесах. Изредка горно-
стаев удается констатировать в придолинных сосняках.
Среди типичной южной тайги горностаи значительно много-
численнее, чем колонки, и почти так же обычны, как ласки. Все
эти виды практически отсутствуют на водоразделах. Горностай
больше других мелких куньих привязан к кустарниковым за-
рослям. Сведение долинных лесов благоприятно для вида. Чис-
ленность горностаев, очевидно, увеличивается с нарушением
тайги, однако кружево ареала остается стабильным.
Горностаи исследованной полосы тайги питаются в основном
серыми полевками. По данным Г. Д. Дулькейта и В. В. Козлова
(1958), в заповеднике «Столбы» значительную часть рациона хищ-
ника составляют красные полевки.
Из 14 зверьков, отловленных на верхней Лене, подавляющее
большинство оказались хорошо упитанными. Единственный
взрослый самец с нижней Ангары (отстрелянный) был средней
упитанности.
Семенники взрослого самца, добытого 17 мая, имели размер
17.0x11.0 мм.
Все зверьки были поражены гамазовыми клещами. На 7 из
10 очесанных паразитологами экземпляров оказались иксодовые
клещи, главным образом нимфы. Обилие иксодид на горностаях
невелико: 1.2 личинки и 0.7 нимфы. Максимальная зараженность
1 зверька — 2 нимфы и 6 личинок.
Роль горностая, как и прочих куньих, в хозяйстве человека
следует признать положительной: зверек дает пушнину и уничто-
жает вредных грызунов, часто вблизи населенных пунктов.
Ласка — Mustela nivalis L.
Как отмечает С. У. Строганов (1962), ласки исследованного
нами района дают столь великое число вариаций, что их подви-
довую специфику он не взялся определить.
Ласки весьма обычны в Средней Сибири, тем не менее законо-
мерности их распространения отражены в литературе весьма проти-
воречиво. Для Саян наличие ласки во всех ландшафтных зонах
отмечают А. И. Янушевич и К. Т. Юрлов (1949). И. П. Копылов с
соавторами (1940) считает, что наиболее многочисленны зверьки
в верхнем течении Нижней Тунгуски, в Приангарье и Присаянье,
а на Лене и Витиме редки. Между тем в низовьях Нижней Тун-
гуски ласка редка (Наумов, 1934). К этому можно добавить, что
при многочисленности ласки к югу от Ангары (б. Киренский
17 Н. Ф. Реймерс
257
округ, Погудин, 1930) на междуречье Ангары и Подкаменной
Тунгуски мы отметили низкую численность зверьков. На верхней
Лене и средней Ангаре ласка весьма обычна и хорошо ловится
в цилиндры. Все эти указания не составляют четкой схемы рас-
пространения вида. Поэтому, опираясь на свои наблюдения, мы
склонны нарисовать следующую картину.
В горах южной Сибири ласки наиболее многочисленны вдоль
горных речек. По их долинам (практически не поднимаясь на
облесенные водоразделы!) ласки выходят к гольцам, где обычны
в зарослях кедровых стланцев на востоке и в ерниках на западе.
Зимой они широко кочуют по гольцам, а из долин выходят на
облесенные склоны гор. Летом, насколько можно судить по сле-
дам на последних летних и ранних осенних порошах, кружево
ареала вида в горах более четко и редко («нити» его приурочены
к водотокам). В полосе южной тайги Средней Сибири ласки жи-
вут по долинным кустарникам и придолинным лесам (рис. 56).
Зимой зверьки продвигаются несколько дальше к водоразделам,
проникая в их леса, особенно на гари. На грани южной тайги
Прибайкальского зоогеографического района с лесами Тунгус-
ского района численность ласок резко сокращается и вместе
с тем меняется кружево арела. Максимумы населения ласок здесь
регистрируются по опушкам леса, выходящим к болотистым
пространствам. Вдоль рек численность зверьков достигает наи-
высшего значения лишь вблизи жилья человека.
Молодые ласки попадали в цилиндры с конца июня.
Питаются ласки главным образом серыми полевками (на верх-
ней Лене, видимо, экономками). Однажды (11 августа 1961 г.)
в желудке зверька обнаружены остатки обыкновенной землеройки.
От года к году численность ласок изменяется под влиянием
нам неясных причин (видимой связи с кормовыми условиями
нет). Максимальное обилие зверьков с 1960 по 1963 г. на верхней
Лене отмечено в 1960 г.
В годы наших исследований ласки были слабо заражены экто-
паразитами. Иксодовых клещей на них не обнаружено.
Ласка безусловно весьма полезный зверек, уничтожающий
вредных грызунов вблизи жилья человека.
Росомаха — Gulo gulo L.
Росомаха очень и очень редка в южной тайге Средней Сибири.
Вне горных районов, где имеются гольцы и редколесье, и мест,
где леса перемежаются болотами северного типа, этого зверя
практически нет. Очень редко вид констатируется в лесостепье
(Погудин, 1930; Троицкий, 1930). Росомаху нельзя считать ха-
рактерным видом южной тайги. Судя по литературным данным,
этот вид вообще нужно с осторожностью относить к лесному ком-
255
плексу млекопитающих. Скорей росомаха — зверь горных и рав-
нинных лесотундр, а также аналогичных таежно-болотных мест-
ностей.
Рис. 56. Принципиальная схема летнего размещения ласок
в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Оценка численности
в баллах.
Барсук — Meles meles L.
Барсуки обычны в лесостепи и лесополье, где их отмечали
многие авторы (Поляков, 1873; Копылов и др., 1940, и др.). Се-
верный предел распространения зверя в Средней Сибири совпа-
17*
259
дает с границей южной тайги. Однако в самой этой тайге звери
очень редки. За пределами лесостепи и лесопольных участков
мы нигде не встречали следов деятельности животного. В горных
лесостепях юга Сибири барсук очень обычен. Среди лиственничной
лесостепи Ангаро-Ленского водораздела зверь немногочислен.
Темнохвойных лесов барсук совершенно избегает.
Выдра — Lutra lutra L.
Зверь широко распространен в пределах южной тайги Сред-
ней Сибири (Копылов и др., 1940). Мы этот вид не изучали, по-
этому ограничиваемся только упоминанием.
Бурый медведь — Ursus arctos L.
Медведи, по мнению многих авторов (Наумов, 1934; Юрген-
сон, 1938, и др.), тесно связаны с темнохвойными лесами. С пере-
ходом к тайге северного типа их число резко снижается (Наумов,
1934). В южной тайге Средней Сибири максимальное число мед-
ведей отмечено в районах, где сохранились темнохвойные леса:
в Присаянье, на верхней Лене, особенно между Леной и Байкалом
(Петри, 1930), в верховьях Нижней Тунгуски. Между прочим,
следы весенних медвежьих отметин (медвежьи «метки») чаще всего
можно наблюдать на пихтах.
На средней Ангаре (Око-Ангарский водораздел) медведей
немного. Особенно резко сократилось их число в 1960—1963 гг.,
когда был катастрофический неурожай кормов (семян и ягод).
Шатуны нападали на людей, много геологов и туристов стало
жертвой изголодавшихся хихцников. Зимой 1961/62 г. погибло
8 чел. (Гудритис, 1963). В низовьях Ангары и на верхней Лене
случаев бродяжничества медведей было намного меньше: на Ан-
гаре медведи вообще не слишком многочисленны (имеется неко-
торый запас кормовых ресурсов), на Лене больше сохранилось
кедрачей, давших в описываемые годы небольшой урожай ши-
шек. Вторым положительным для популяции медведей фактором
была возможность вертикальных миграций. В гористых местно-
стях юга Сибири медведи, как правило, совершают периодические
перемещения: весной кормятся в долинах, летом поднимаются
в подгольцовый пояс на черемшанники, затем спускаются в плодо-
носящие кедрачи и, наконец, направляются к местам берлог.
В наиболее выровненных местах южной тайги происходит пере-
мещение из придолинных ягодников (голубичников) в сосняки-
брусничники и плодоносящие кедровники. Совпадение неурожая
всех ягод и нынешнее отсутствие темнохвойных лесов на средней
Ангаре привело к жесточайшей бескормице.
Столь же сильная бескормица наблюдалась в северных райо-
нах за пределами южной тайги Средней Сибири. Шатуны были
260
обычны по Нижней Тунгуске (Гудритис, 1963). Здесь особенно ска-
залось отсутствие урожая ягод. По сообщению L. Kelso (in litt.),
та же причина и в те же годы вызвала появление шатунов в Се-
верной Америке. Они были зарегистрированы на Аляске, в Ка-
наде и на севере США.
Медведи южной тайги залегают в берлогу почти одновременно
с исчезновением бурундуков (Реймерс, 19636) — во второй-тре-
тьей декадах октября. Также почти одновременно они появля-
ются (в апреле). Авторы, исследовавшие биологию медведей в гор-
ных районах Восточной Сибири — южном Забайкалье, Баргу-
зинском хребте (Лавов, 1955; Устинов, 1960), указывают, что
зверь залегает в берлогу в первой половине октября — несколько
раньше, чем в южной тайге. Однако в горах южной Сибири, по
нашим наблюдениям, даты залегания медведей близки к отмечае-
мым корреспондентами-фенологами для южной тайги. Время вы-
хода медведя из берлоги в большой степени связано с ее место-
положением и климатическими особенностями года.
По всей южной тайге появление медведей происходит с не-
сколько меньшим колебанием в сроках, чем выход бурундуков.
При дружной весне медведи просыпаются почти одновременно
в течение 5—6 дней. В горах выход из берлог задерживается
приблизительно на неделю. Рождение медвежат, по данным кор-
респондентов Восточносибирской фенологической комиссии
А. И. Солдатова, А. И. Павловой и А. И. Арбатского, происходит
в пределах южной тайги Средней Сибири между 1 и 10 апреля
(чаще всего 5—10-го).
В девственных лесах медведи не приносят вреда человеку.
Однако со сведением этих лесов и заменой их малокормными био-
топами учащаются случаи полных неурожаев семян, что прину-
ждает медведей к активному хищничеству. Они делаются опас-
ными для хозяйства человека, а в отдельных случаях — и для
его жизни.
Волк — Canis lupus L.
Серый волк — обитатель открытых пространств и лесов в мало-
снежных районах — встречается на юге Средней Сибири в лесо-
степи и лесополье (Худяков, 1937). Практически его ареал совпа-
дает с районом распространения косуль. В облесенных горах
южной Сибири, вопреки утверждению Д. К. Соловьева (1920),
волки очень редки. Севернее, в полосе южной тайги (за пределами
лесостепи), зверь встречается спорадически вдоль освоенных
земледелием долин рек, почти не заходя на водоразделы.
Чем больше степень освоенности тайги, тем значительнее
потенциальные возможности расселения волков, однако успешная
охота на хищника резко снижает его численность. Последний раз
мы встретили следы волчьей стаи вблизи г. Иркутска зимой
261
1950/51 г. Во время Великой Отечественной войны, по свиде-
тельству местных жителей, волки были многочисленны во всех
земледельческих районах.
Лисица — Vulpes vulpes L.
Типичный лесопольный вид южной тайги Средней Сибири.
Новые материалы об этом виде у нас отсутствуют.
Песец — А1 орех lagopus L.
По долинам рек иногда забегают в пределы северной части
южной тайги Средней Сибири. Новые факты заходов нам не
известны.
Красный волк — Си on alpinus Pall.
С. У. Строганов (1962), ссылаясь на П. С. Палласа (Pallas,
1811), отмечает красного волка у истоков Лены. Позднейших
подтверждений обитания здесь этого хищника нет. Утверждения
о наличии красного волка на северных скатах Саян, видимо,
мало обоснованны. В свое время А. С. Фетисов сообщил нам ряд
фактов захода красных волков по долине Иркута. Тункинская
долина — один из немногих возможных каналов проникновения
красных волков в пределы южной тайги Средней Сибири. Если
сообщение П. С. Палласа о наличии волка в Верхоленье не осно-
вано на недоразумении, зверя здесь следует считать очень интерес-
ным реликтом центральноазиатского происхождения.
Енотовидная собака — Nyctereutes procyonoides Gray.
В пределах южной тайги Средней Сибири (Качугский район
Иркутской области) небольшая партия енотовидных собак
(17 экз.) с колхозной зверофермы была выпущена в 1942 г. Зверь
не прижился (Лавров, 1946).
Рысь — Lynx lynx L.
Нам не пришлось встречать этого весьма редкого в южной
тайге зверя. По литературным данным (Копылов и др., 1940;
Тимофеев, 1949, и др.) составить точного представления о раз-
мещении рыси не удается. Одни авторы утверждают, что рысь
придерживается глухих участков тайги, другие, наоборот, по-
лагают, что излюбленные места рыси — разреженные участки
тайги и гари. Возможно, что правы и те и другие. По опросным
данным, рыси обычны в местах, где тайга нарушена, но жилье
человека далеко. В таких районах сравнительно много копытных
и зайцев ~ основных объектов питания рыси.
262
Отряд ПАРНОКОПЫТНЫЕ — ARTIODACTY LA
Кабан — Sus scrofa L.
По свидетельству В. Г. Гептнера, А. А. Насимовича и
А. Г. Банникова (1961), южную тайгу Средней Сибири населяет
забайкальский кабан S. s. sibiricus Staffe. Эти авторы отмечают,
что ареал кабана узким клином вдается в полосу южной тайги
по рекам Оке и Чуне и тянется вдоль северных предгорий Восточ-
ного Саяна. В. Н. Скалой (1962) указывает более узкую область
распространения животного в пределах южной тайги Средней
Сибири: лишь самая восточная часть предгорий Восточного Саяна.
Согласно имеющимся у нас сведениям, регулярное обитание ка-
бана отмечено еще дальше от железной дороги, чем показывает
В. Н. Скалой.
Кабан живет в предгорьях Саян — лишь по долинам рек со
сравнительно неглубоким снежным покровом, поэтому на южном
берегу Байкала его нет (во всяком случае зимой).
В Средней Сибири кабан всюду связан с кедровыми лесами.
Очевидно, граница восстановленного ареала кабана в Средней
Сибири, весьма провизорно проведенная указанными тремя авто-
рами, требует уточнения. Исходя из палеоэкологических крите-
риев (непосредственных свидетельств практически нет), следует
предполагать, что кабан мог более широко заселять Верхоленье,
проникая из Прибайкалья (а не только из Забайкалья) в район
Бодайбо. На наличие кабана в Верхоленье указывал И. С. По-
ляков (1873).
По всей вероятности, кабан никогда не заселял южной тайги
Средней Сибири сплошь. Это вид речных долин предгорий с их
влажным умеренным климатом, обильным плодоношением кедров-
ников и ягодных кустарников, а также луговинами, поросшими
буйными корневищными травами.
Кабарга — Moschus moschiferus L.
Сибирская кабарга (Гептнер и др., 1961) заселяет всю южную
тайгу, однако не сплошь. Кабарга распространена отдельными
пятнами (Копылов, 1950). Пятнистость в распространении свя-
зана в южной тайге не столько со скальными выходами, сколько
с островами темнохвойной тайги. Особенно отчетливо эта законо-
мерность проявляется в пределах лиственничного лесостепья
юга Ангаро-Ленского междуречья. В условиях сглаженного рель-
ефа зверь встречается зимой почти исключительно по полоскам
приручьевых ельников (в верхнем течении ручьев), отсутствуя
или почти отсутствуя в лиственничниках и сосняках. На скаль-
ные отстой кабарга выходит, главным образом спасаясь от беспри-
зорных собак (рис. 57). Особенно много зверей гибнет от собак
в период образования наста.
263
У северного предела южной тайги — севернее 60° с. ш. —
кабарга очень редка (опросные сведения; Копылов и др., 1940;
Копылов, 1950). Еще севернее вид распространен спорадически
и численность его ничтожна. В пределах южной тайги Средней
Сибири кружево ареала кабарги в связи с исчезновением темно-
хвойных лесов изреживается, хотя границы области распро-
странения зверя сохраняют свое положение.
Пятнистый олень — Cervus nippon Temm.
Пятнистые олени жили в условиях полувольного содержания
в питомнике на Байкале (Дорогостайский, 1930; Подаревский,
1931).
Благородный олень (марал) — Cervus elaphus sibiricus Sev.
Маралы — обитатели горной тайги. Эти животные страдают
от постоянного преследования, но их наличие также зависит от
нарушений природных условий. В южной полосе горной тайги
они придерживаются наиболее возвышенных районов (Копылов,
1950). У северной границы распространения, где олени вообще
редки, они сохранились по наименее освоенным охотниками водо-
разделам (Чуно-Ангарскому, Ангаро-Ленскому, Лепо-Кирепг-
скому).
Следует отметить определенное совпадение мест обитания ма-
ралов с сохранившимися массивами темнохвойной тайги южного
типа. О распространении марала на Око-Ангарском водоразделе
говорил И. П. Копылов (1950), ссылаясь на рукопись В. В. Ти-
мофеева 1949 г. В 1959 г. в этом районе мы оленя не нашли. Этот
факт можно объяснить тем, что Око-Ангарское междуречье ока-
залось совершенно лишенным темнохвойных лесов. Вот почему
численность маралов там очень низкая или они даже полностью
отсутствуют.
Очевидно, область возможного спорадического распростране-
ния марала совпадает с границами южной тайги, т. е. маралы
в принципе могут обитать до Подкаменной Тунгуски. По Енисею
звери зарегистрированы еще севернее — у 63° с. ш. близ Боро-
дино и Искуп (Сыроечковский и Рогачева, 1961).
При сохранении границ ареала вида число маралов в южной
тайге непрерывно падает, а кружево ареала изреживается.9
9 В горной тайге юга Сибири охранные мероприятия привели к поло-
жительным результатам; в южной тайге, очевидно, основная причина сни-
жения численности — не в преследовании зверя человеком, а в биотопиче-
ских особенностях местности.
264
Рис. 57. Кабарга, загнанная собаками на отстой.
Косуля — Capreolus capreolus L.
Ареал сибирской косули (С. с. pygargus Pall.) в южной тайге
Средней Сибири может служить негативом ареала благородного
оленя. В противоположность таежному маралу лесостепная ко-
суля занимает наиболее нарушенные леса. Однако северные пре-
делы распространения обоих зверей почти совпадают. Косуля
ограничена в распространении изолинией 50-сантиметровой вы-
соты снежного покрова. Это легко вывести из сравнения кли-
матических карт «Атласа Иркутской области» (1962) с «Карто-
граммами распространения марала в Иркутской области» (Ко-
пылов, 1950). В непойменных темнохвойных лесах косуля прак-
тически не встречается. Среди приречных и приручьевых ельников
зверь находит надежное убежище. По подсчетам В. В. Тимофеева,
приводимым А. С. Фетисовым (1953), средняя плотность населе-
ния косуль в Иркутской области 0.56 особи на 1 км2. Местами
численность зверя значительно выше (до 3 особей на 1 км2).
С нарушением лесов ареал косули расширяется, а кружево
его «уплотняется» и занимает водораздельные пространства.
Вместе с тем намечается разрежение популяции косули по доли-
нам рек, где ее преследуют люди и особенно бродячие собаки
по весеннему насту.
Лось — Alces alces L.
Восточносибирский лось A. a. pfizenmayeri Zukowski (Гепт-
нер и др., 1961) населяет всю полосу южной тайги Средней Си-
бири, достигая высокой численности на севере и практически
отсутствуя в лесопольной части, примыкающей к железной до-
роге. Очень мало лосей в многоснежных предгорьях Саян. Хотя
лось поднимается во время кочевок до высот 2000—2200 м (Фе-
тисов, 1957), он обитатель низинных угодий.
Условия глухих темнохвойных лесов, глубокие снега и пре-
следование человеком — сумма факторов, отрицательно влияю-
щих на численность лося. Однако и резкое нарушение тайги,
особенно большие пожары, снижают поголовье лосей. Все пере-
численное приводит к тому, что лось в Средней Сибири оттеснен
к северным пределам южной тайги и стал весьма малочисленным
зверем в долине верховий Ангары и Лены. Раньше эти реки были
заселены большим количеством сохатых — любимым объектом
охоты древних жителей Приангарья и Верхоленья (Окладников,
1956). Таким образом, за последние сотни лет произошла пере-
дислокация лосей: с выгоранием темнохвойной тайги звери в боль-
шей степени заселили водоразделы, а преследование человеком
привело к обеднению лосями речных долин.
266
Северный олень — Rangifer tarandus L.
Сибирский лесной олень (/?. t. valentinae Flerov.) в пределах
южной тайги Средней Сибири распространен почти повсеместно,
по имеет весьма сложное кружево ареала. Оленя нет лишь в по-
лосе вдоль железной дороги и в лиственничной лесостепи Ангаро-
Ленского междуречья. Однако в зимнее время звери преодоле-
вают этот «пустой» коридор, и, очевидно, происходит обмен между
северной и южной популяциями.
В летнее время северная и южная группы оленей приурочены
к различным местам обитания. Горные олени юга Сибири летом
не распространены в южной тайге — они обитают в полосе гор-
ного редколесья и горных лесотундр. Олени северной популяции
связаны с редкостойными лиственничниками и лишайниковыми
сосняками — образованиями, также не характерными для южной
тайги. Следовательно, северный олень в южной тайге Средней
Сибири оказывается интразональным элементом и заселяет ее
почти сплошь лишь зимой. Однако в разнотравных и бруснични-
ковых сосняках южного типа, так же как среди лиственничного
лесополья и лесостепи, северные олени не встречаются и зимой.
Нужно думать, что область распространения северных оле-
ней в южной тайге Средней Сибири за последние столетия изме-
нилась несильно, хотя, как правильно отмечает В. В. Тимофеев
(1949), численность оленей упала (в связи с их преследованием
и выгоранием ягельников). Претерпело изменение и кружево ареала:
олени были оттеснены из речных долин.
Отряд ГРЫЗУНЫ — RODENTIA
Летяга — Pteromys volans L.
Южную тайгу Средней Сибири населяют, очевидно, две формы
этого зверька — Р. v. turovi Ogn. и Р. и. incanus Mill., несколько
отличающиеся друг от друга окраской (Огнев, 1940). На это об-
стоятельство указывал еще А. А. Черкасов (1867), хотя относил
эти различия к особям Забайкалья («бусенькие» и «красненькие»
летяги).
Распространение, размещение и ч и с лен-
нос т ь. Летяга заселяет юг Средней Сибири сплошь, однако
далеко не везде достигает высокой для вида численности. И. П. Ко-
пылов, А. В. Добровольский и И. А. Шергин (1940) указывают,
что на р. Лене население летяг реже, чем в бассейне Нижней
Тунгуски, в Приангарье и в Присаянье. Судя по нашим наблю-
дениям, это не совсем так. В Среднем и Нижнем Приангарье
летяги оказались значительно более редкими зверьками, чем
в районах верхней Лены. Весьма обычна летяга в предгорьях
267
Саян. Обширные сосняки западной части южной тайги Средней
Сибири практически лишены этого вида.
С. В. Кириков (1952а) совершенно справедливо причислил
южиоуральских летяг к характерным представителям фауны
березовых лесов. В более северных районах связь летяги с бе-
резой выражена намного слабее. Так, в Кировской области зве-
рек обитает в долинных елово-пихтовых лесах (Плесский, 1952).
Изменение приуроченности летяг четко прослеживается и в Сред-
ней Сибири. В горах юга Сибири летяга обитает в редкостойных
старых кедровниках, избегая пихтово-кедрового мелколесья и
глухих долин ручьев. В предгорьях Саян по всей полосе южной
тайги грызун обычен в водораздельных кедровниках, часто
с примесью березы и осины, но его население достигает наивысшей
плотности, как и в Кировской области, в приручьевых ельниках.
Намного реже встречается здесь летяга в березово-сосновых
лесах. В лесостепи Средней Сибири зверек живет почти исклю-
чительно в долинах ручьев, поросших березовым и осино-
вым лесом. На севере, как указывает Н. II. Наумов (1934),
летяга селится исключительно в долинных лесах (не листвен-
ничных).
Кружево ареала летяги в южной тайге Средней Сибири резко
трансформировалось в ходе антропогенных нарушений лесов.
Изначально в темнохвойных лесах водоразделов этот зверек
достигал наивысшей численности в редкостойных участках, не-
редко заболоченных (7, 1а и 16 нашей схемы, см. гл. 4). Смешан-
ные леса с участием березы и осины практически отсутствовали.
Они стали основным местом обитания летяг лишь с повсемест-
ным распространением пожаров. Дремучая долинная тайга также
не характерна для вида, если она естественно или искусственно не
разрежена, поэтому следует предположить, что в этих участках
летяга не достигала высокой плотности. В целом распространение
вида было более равномерным без наблюдаемых ныне флуктуа-
ций и «пустых» мест.
Оценить численность летяг весьма трудно из-за скрытности
зверьков. По рассказам охотников, в приручьевых участках темно-
хвойных лесов и приручьевых ельниках южной тайги летяга
встречается почти столь же часто, как и белка, а иногда и намного
чаще. За день охотничья собака облаивает в этих местах по
10 зверьков.
Размножение, питание, образ жизни.
О размножении летяг в Средней Сибири известно очень мало.
Самка, пойманная нами в Хамар-Дабане 23 мая 1956 г., не имела
следов беременности. А. А. Черкасов (1867) указывал на наличие
в выводке 1—2 молодых. Г. М. Хабаева (1959) встретила самку
с 3 детенышами. То же число указано для окрестностей Томска
Г. Э. Иогапзеном (1930), С. В. Кириковым (1952а) для южного
Урала и В. А. Поповым (1950) для Волжско-Камского края.
268
В течение лета бывает один помет, достоверных сведений о вто-
ром помете нет.
Вне полосы темнохвойной тайги летяга питается в основном
почками березы, осины, лиственницы и других древесных пород
(Паллас, 1786; Черкасов, 1867; Кириков, 1952а; Егоров, 1961,
и др.). Эти же корма она заготавливает впрок (Дулькейт и Коз-
лов, 1958). В темнохвойной тайге летяге служат пищей семена
хвойных деревьев: кедра, ели, пихты. Сережки берез, ягоды
и грибы отступают в питании на второй план (Копылов и др.,
1940). Содержавшиеся нами в неволе зверьки ели орехи, почки
и молодые побеги кедра, кору с однолетних побегов и почки ив,
почки и ягоды рябины, ягоды черники. Отказывались летяги от
цветочных бутонов золотистого рододендрона и цветочных буто-
нов ольхи, сливочного масла, хлеба и колбасы (наши наблюдения
над зверьком из кедровников Хамар-Дабана). На верхней Лене
(Реймерс и Воронов, 1963) летяга поедала траву, молодые побеги
кустарников, грибы, ягоды, но в неволе предпочитала кедровые
орехи, фрукты, ела различные крупы и жареное (но не сырое) мясо.
Гнезда летяг найдены в дуплах деревьев — осин (Мартынов и
др., 1960) и сосен (Хабаева, 1959), в искусственных дуплянках
(Дулькейт и Козлов, 1958) и гнездах белок (Доброхотов, 1953).
Под зимнее убежище, как уже нами отмечено (Реймерс и Воронов,
1963), летяга способна перестроить гнездо дрозда (краснозобого),
утеплив его сухим мхом.
Летом летяга активна только ночью. Зверьки лишь изредка
появляются в пасмурные дни. В холодное время года летяг можно
видеть и днем. Видимо, к прохладным дням относится замечание
А. А. Черкасова (1867), что летяга встречалась ему «совершенно
неожиданно среди белого дня» (издание 1962 г., стр. 250). Зим-
няя дневная активность зверька была известна И. С. Полякову
(1873).
Зверьки опускаются на землю и некоторое расстояние пере-
двигаются по ней: на верхней Лене мы добыли 2 экз. летяг в ме-
таллические цилиндры, врытые в дно канавок для отлова мелких
грызунов.
Значение. Специфическая роль летяги в лесах пока
еще не совсем ясна. Много общих черт у этого вида с белкой.
Летяга — конкурент белки, однако объекты питания этих видов
становятся предметом борьбы лишь в малоурожайные годы. Диа-
пазон кормов у летяги шире, чем у белки, и, очевидно, она более
конкурентноспособна. Имеются указания на вытеснение белок
летягами из гнезд (Доброхотов, 1953).
Сетования охотников на то, что летяга «выгоняет» белку, не
следует принимать на веру. Промысловиков очень раздражает,
что летяга отвлекает собак при белковании. Поэтому летяги и
считаются виновниками многих бед, в том числе оскудения бе-
личьих запасов.
269
Белка — Sciurus vulgaris L.
Этому хорошо изученному виду посвящена обширная литера-
тура, одно перечисление которой заняло бы несколько страниц.
Экология и биология белки южной тайги Средней Сибири освещены
В. Н. Троицким (1930), Н. Г. Грудининым (1939), И. П. Копыло-
вым с соавторами (1940), В. В. Тимофеевым (1947, 1949), а в более
северных районах Н. П. Наумовым (1930, 1934), А. Н. Формозовым
с соавторами (1934), О. В. Егоровым (1961) и многими другими
исследователями. Фрагментарные материалы о белках верхней
Лены приведены нами (Реймерс и Воронов, 1963). Повторять уже
опубликованные данные мы не считаем уместным. Остановимся
лишь на одном вопросе — общем изменении численности белок.
В последние годы, помимо периодических колебаний количе-
ства зверьков, отмечается постепенное падение численности вида,
которое с первого взгляда не коррелируется пи с урожаем хвой-
ных пород и других кормов белки, ни с каким-либо массовым
«мором». Одни авторы связывают это явление с хищничеством
соболя (Павлюченко, 1958; Лавов, 1959, и др.), другие эту связь
отрицают (Надеев, Тимофеев, 1955; Тимофеев, 1962), третьи
в этом вопросе придерживаются нейтральных позиций (Егоров,
1961).
Нам падение численности белок кажется весьма закономерным.
Как неоднократно было уже показано, в том числе нами (Реймерс
и Малышев, 1963), лучшие угодья белки — темнохвойные леса —
непрерывно сокращаются по площади и омолаживаются, что
крайне неблагоприятно для этого зверька. По данным Ф. Р. Штиль-
марка (1963г), на верхней Лене численность белки в водораздель-
ных кедровниках 1.5—5.6 (или 0.9—3.0) зверька на 1 км мар-
шрута; в приречных ельниках на том же расстоянии встречается
также 1.2—3.0 зверька. В лиственничниках же число белок ко-
леблется лишь от 0.1 до 1.5 экз. на 1 км маршрута. Сокращение
массивов темнохвойной тайги уже привело к тому, что наиболее
продуктивные угодья вдоль рек вообще исчезли. Нам не удалось
в полосе южной тайги Средней Сибири увидеть даже небольшой
участок нетронутой природы у реки. Это относится и к рекам,
вовсе не имеющим прибрежного населения, но судоходным хотя
бы для лодок. Темнохвойные леса вдоль рек полностью выгорели
(за исключением долинных ельников).
Берега таежных ручьев служат обычно трассами охотничьих
троп. Еще и сейчас, при общем сокращении области промысла, охот-
ники заходят в тайгу на 50—60 км. В недалеком прошлом они
осваивали почти все массивы, строя зимовья даже в 100—120 км
от ближайшего жилья. Еще дальше, нередко по звериным тропам,
также тянущимся вдоль ручьев, уходят экспедиции. Неосторож-
ное обращение с огнем привело к выгоранию лесов и по долинам
ручьев.
270
В связи с тем что темнохвойная тайга сохранилась главным
образом по гребням водоразделов, резко изменился баланс уро-
жайности семян хвойных пород. При равномерном распределении
леса в силу неоднородности мезо- и микроклиматических условий
местности не равновелик урожай семян: на склоне одной экспо-
зиции он больше, другой — меньше, на гребне семена могут во-
обще отсутствовать. Это явление создает благоприятные условия
для существования таежных семеноедов — полного неурожая
в больших массивах тайги практически не бывает. Современные
острова темнохвойного леса, расположенные главным образом
по гребням водоразделов, в большей степени подвержены повсе-
местным неурожаям.
Белка вынуждена менять биотопы. В годы урожаев семян ели
зверьки переходят в приручьевые и приречные ельники. Бывает,
как говорят охотники, «еловая» белка. Концентрация зверь-
ков в узких полосах таких ельников позволяет охотникам
резко увеличить добычу. Создается иллюзия общего высокого
урожая белки. Вместе с тем скопление зверьков в ельниках,
более частое соприкосновение их с родственными видами млеко-
питающих, болеющими теми же болезнями (кокцидиозом, туляре-
мией и др.), что и белка, неблагоприятно сказывается на популя-
ции белок. Происходит резкое сокращение числа зверьков.
Повторные неурожаи в водораздельной темнохвойной тайге
и долинных лесах заставляют белку заселять сосняки. «Сосновая»
белка, как утверждают охотники, удобный объект промысла,
но предвестник неурожая промыслового вида в будущем году.
Вредность смолистых сосновых шишек для белки общеизвестна.
В лучшем случае зверьки питаются в сосновых борах ягодами и
грибами.
Хорошие урожаи семян лиственницы в южной тайге Средней
Сибири — редчайшее явление. В эти годы белка может находить
корм повсеместно, поскольку лиственница входит в состав всех
лесов района.
Изменение кормового режима в сибирских лесах резко ме-
няет соотношение конкурирующих видов. В годы неурожаев
семян темнохвойных пород межвидовая борьба достигает наивыс-
шего напряжения. Соболь и летяга могут оказывать резкое по-
давляющее влияние на популяцию белки. Белка попадает в «ле-
тяжьи» угодья. А соболь безусловно конкурентноспособнее белки.
Серьезно изменились взаимоотношения белки и соболя в исто-
рическом процессе освоения тайги человеком. На этом вопросе
мы подробно остановимся в следующей главе, а сейчас заметим,
что соболь в силу совершенно объективных факторов должен все
сильнее преследовать белку — меньше в тайге стало других жи-
вотных кормов, доступных этому хищнику.
Следовательно, численность белки уменьшается, так как сокра-
щаются площади темнохвойной тайги и урожай семян темнохвой-
271
ных пород, во-вторых, повышается напряженность конкуренции
и становится возможным более широкое распространение заболе-
ваний. Все это кажется нам закономерным процессом. Причины
явления кроются в общей трансформации тайги, которая в ряде
мест стала значительно резче сказываться на благополучии бе-
личьего поголовья в связи с восстановлением запасов соболя.
Дальнейшее уменьшение массивов темнохвойных пород
в южной тайге Средней Сибири приведет к еще большему падению
урожаев белки. Такое падение будет продолжаться до уровня,
определяемого кормностью сосновых и сосново-мелколиственных
лесов и напряженностью конкуренции в них. Вырубка этих мас-
сивов, как показывают наши наблюдения на территории присаян-
ских леспромхозов, ведет к почти полному исчезновению вида.
Бурундук — Eutamias sibiricus Laxm.
Бурундуки Средней Сибири относятся к подвиду Е. s. sibi-
ricus Laxm. (Огнев, 1940; Зубчанинова, 1962).
Фенология. В условиях южной тайги Средней Сибири
бурундуки появляются, в зависимости от района и главным обра-
зом микроклиматических условий расположения норы, в конце
марта, начале—середине апреля. Собранные нами опросные све-
дения, полученные корреспондентами-фенологами (Реймерс, 19636,
1963в), позволяют отметить, что на верхней Лене выход бурунду-
ков происходит несколько позже, чем в Приангарье, и близок
по срокам к наблюдавшемуся в горах Хамар-Дабана (Реймерс,
1960а). В среднем в полосе лесостепи бурундуки появляются из
нор на грани марта и апреля, в Приангарье — в течение первой
декады апреля, в районах верхней Лены и на Ангаро-Подкаменно-
тунгусском междуречье — в конце второй декады апреля, а в Сая-
нах и Хамар-Дабане — лишь в середине последней десятидневки
апреля; в мае они обыкновенно уже выходят из нор, хотя в пер-
вой половине мая на хорошо знакомом нам Лазурьском хребте
М. П. Тарасов (1962) их не видел. На грани апреля и мая появ-
ляются бурундуки за пределами северной границы южной тайги
на р. Туре (Наумов, 1934). Имеющиеся в нашем распоряжении
данные (за 1960—1963 гг.) позволяют составить приблизитель-
ную схему изофен появления бурундуков на территории юга
Средней Сибири (рис. 58).
Исчезают бурундуки около середины октября, практически
одновременно во всей полосе южной тайги Средней Сибири.
Н. П. Наумов (1934) в это же время констатировал исчезновение
бурундуков в низовьях Нижней Тунгуски. Несколько раньше —
в конце сентября — перестают выходить из нор бурундуки гор
юга Средней Сибири (Реймерс, 1960а).
Как мы отмечали (Реймерс и Воронов, 1963), погода мало
влияет на время появления бурундуков. Очевидно, зверьки при-
способлены к средней фенологической ритмике местности.
272
Первые выходы бурундуков весной обычно не связаны с по-
исками пищи. Зверьки либо греются на солнышке, либо сидят
у входа в нору, почти не бегая по снегу.
Рис. 58. Изофены весеннего выхода бурундуков из нор.
I—XI и 1—10— как на рис. 6. Жирные сплошные линии — изофены.
Распространение, размещение и ч и с лен-
ное т ь. Бурундук распространен повсеместно, но далеко не
равномерно. В присаянских районах зверек очень многочислен,
значительно меньше бурундуков на верхней Лене и в Приангарье,
особенно Нижнем.
18 Н. Ф. Реймерс
273
В пределах зоны низкой численности бурундуки заселяют био-
топы с однообразной плотностью. Например, на верхней Лене при
учетах без собаки в различных лесах констатированы 2—6 встреч
зверьков на 5 км маршрута (Реймерс и Воронов, 1963).10 Прибли-
зительно такая же встречаемость бурундуков на р. Чадобец.
В Нижнем Приангарье и на Лене население бурундуков дости-
гает высокой плотности лишь на опушках леса у полей (до 12 зверь-
ков на 5 км маршрута) и среди темнохвойных лесов у орехопро-
мысловых балаганов. Некоторая концентрация бурундуков от-
мечена на пограничных участках между долинными ельниками
и сосново-лиственничными суборями (Воронов и Реймерс,
1962).
По материалам Л. А. Тибет и И. Ф. Кузьмина (1963), число
бурундуков в темнохвойных (елово-пихтовых) лесах северного
Заангарья в июле—августе достигает 100 экз. на 1 км2, в сосняках
и лиственничниках отмечена средняя плотность, а в березняках
минимальная — 25 бурундуков на 1 км2. Эти данные очень близки
к полученным нами более примитивным способом — с помощью
учетов без собаки. Л. А. Тибет и И. Ф. Кузьмин указывают, что
визуальный учет без собаки снижает результаты учета в 1.8 раза,
таким образом, вычисленное нами число бурундуков — 40—
50 на 1 км2 темнохвойного леса, оказывается при увеличении
почти вдвое весьма близким к 100, констатированным указан-
ными авторами.
В сосновых борах бурундуков очень мало, если в лесу нет
примеси лиственницы,— не больше 10—12 на 1 км2.
В Присаянье численность бурундуков заметно выше. В июле—
августе, когда выходят молодые, в кедровниках бывает до 19.3
зверька на 1 га (Штильмарк, 1963а). Даже если это число
завышено (существование на1 км2 1930 бурундуков можно поста-
вить под сомнение), количество бурундуков в кедрачах Присаянья
по крайней мере в несколько раз больше, чем в остальных участках
Средней Сибири. Для взрослых зверьков Ф. Р. Штильмарк приводит
показатели 7.1, 4.6 и 8.2 экз. на 1 га. По нашим данным (Рей-
мерс, 1958а), пересчитанным на площадь учетной полосы, весной
численность бурундуков в кедровниках достигает 275 зверьков на
1 км2 (или 2.75 на 1 га).
Вблизи орехопромысловых балаганов количество бурунду-
ков очень велико, однако число 40 зверьков на 1 га (4000 экз. на
1 км2), указываемое Ф. Р. Штильмарком, кажется нам несколько
завышенным. Обычно возле одного балагана, окруженного
кучами отходов орехового промысла (рис. 59), держится 7—8 зверь-
ков. Они собираются с площади не менее 0.3—0.5 га, так что
наивысшая численность может быть оценена самое большее в 25
10 Ф. Р. Штильмарк (1963г) с собакой учитывал до 9 зверьков на 1 км
пути в водораздельных кедровниках.
274
бурундуков на 1 га. Такой численности зверьки достигают на
небольших участках и в исключительных случаях, собираясь
сюда из окрестных лесов. Наша глазомерная оценка почти совпа-
дает с максимальным числом, полученным Ф. Р. Штильмарком на
пробных площадках (26.6 экз./га). Данные Р. В. Йорджера (Yer-
Рис. 59. Бурундук на куче отходов орехового промысла.
ger, 1953), на которые ссылается указанный автор, относятся
к другому виду и к весьма продуктивным кленово-буковым лесам,
имеющим богатый подлесок.
Не располагая достаточным количеством числовых дан-
ных, мы все же решили показать распределение бурундуков по
глазомерной оценке на схеме (рис. 60). Составлена эта схема,
подобно прочим, на основе четырехбалльной шкалы. Некоторые
числовые данные могут быть привлечены из указанной уже работы
Л. А. Тибет и И. Ф. Кузьмина (1963). Они приведены нами
выше.
Размножение. Гон у бурундуков происходит вслед
за их выходом из нор: при дружном появлении в теплые весны
сразу же, а при затяжной пасмурной весне через 7—10 дней после
первых «разминок». Ф. Р. Штильмарк (1963а) высказывает сом-
нение в правильности нашего указания (Реймерс, 1960а) на более
18*
275
ранний выход из нор самцов бурундуков. Однако ранней весной
нами осмотрены в бинокль многие десятки зверьков. Все они
были самцами. Они намного активнее самок, и их следы часто
Рис. 60. Принципиальная схема размещения бурундуков в пре-
делах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Штриховка полосами ука-
зывает места осеннего скопления зверьков (на полях). Оценка числен-
ности в баллах.
ведут к норам последних. Допустимо, конечно, что спаривание
происходит не в норе, а лишь под поверхностью снега, но не исклю-
чена возможность полового акта и в норе. Первые добываемые
самки бывают уже беременными.
276
В горах юга Сибири гон происходит в краткие сроки. Этого
нельзя сказать о южной тайге Средней Сибири. Тут период гона
растянут и заканчивается лишь в июне. Иногда июньский гон
можно наблюдать и в горах. Такое явление было нами отмечено
23—26 июня 1951 г. в Хамар-Дабане. Однако, следовала ли затем
беременность, мы не установили. В южной тайге вторая беремен-
ность — редкое явление, отмеченное лишь на верхней Лене и лишь
в одном 1961 г., когда была очень ранняя весна, а затем период
длительного похолодания, вызвавший ряд фенологических ано-
малий. Б. С. Виноградов и С. И. Оболенский (1927) полагали,
что два помета за лето для бурундуков юга Иркутской области —
правило.
Одна из самок, имевшая плацентарные пятна двух порядков
(2 августа 1961 г.), принесла в первом помете 6 и во втором 5
молодых. Видимо, это максимально возможная плодовитость
бурундуков за сезон.
Среднее число молодых в помете, как показывает анализ
количества эмбрионов и плацентарных пятен, 4.7 (табл. 11).
Довольно много самок не участвует в размножении. Причины этого
явления неясны.
Молодые зверьки начинают вести самостоятельный образ
жизни в возрасте около 1 месяца при весе 35—50 г. В это
время они очень доверчивы, их легко поймать и приручить.
Питание. В работе, посвященной насекомоядным и гры-
зунам верхней Лены (Реймерс и Воронов, 1963), мы привели ма-
териалы анализа содержимого 120 желудков бурундуков. Спектр
кормов, констатированный для верхней Лены, типичен для всей
южной тайги. Данные, полученные на нижней Ангаре и в других
районах, очень близки к собранным на Лене, поэтому в настоящей
работе мы питание бурундука подробно не рассматриваем. Можно
лишь упомянуть, что основу питания зверька составляют расти-
тельные корма (встречаемость 100%), а из них — семена (74.4%).
С неубранных полей бурундуки охотно собирают урожай как
осенью, так и весной. 2 мая 1962 г. близ дер. Яркино на р. Чадо-
бец добыт зверек, имевший в защечных мешках семена овса (22 шт.)
и «дикой гречихи» (12 шт.).
Паразиты, значение. Бурундуки поражены бло-
хами (до 12 экз. на 1 зверьке), гамазовыми и иксодовыми клещами.
Работавшие с нами паразитологи очесали в условиях полуста-
ционаров свыше 100 бурундуков (табл. 12).
Личинок на бурундуках немного, нимф — значительно больше,
и они встречаются намного чаще. На бурундуков нападают также
комары.
Бурундуки могут служить резервуаром энцефалитной (Бром
и др., 1961), туляремийной (Попов и Иголкин, 1956; Дунаева,
1963) и, возможно, токсоплазмозной инфекций. Последняя найдена
пока лишь у сусликов Средней Сибири (Тарасова и Кудинова, 1961).
277
Таблица И
Число эмбрионов и количество размножающихся самок бурундука по месяцам
Месяц Отловлено взрослых самок Характер образований Количество самок с числом эмбрионов и плацентарных пятен Сред- нее число эмб- рионов
всего в том числе со следами размножения 2 3 4 5 6 7 8 9
всего про- цент от всех самок из них беремен- ных
всего процент от всех самок
Май 13 9 69.2 8 61.5 Эмбрионы. — — 1 6 1 — — — 5.0
Эмбрионы. 1 2.0
Июнь 24 21 87.5 1 4.2 |
Плацентарные пятна. 3 2 4 3 4 2 — 1 4.7
Июль 7 6 87.5 — 0.0 » » 1 1 1 1 1 — — 4.6
Август 10 6 60.0 — 0.0 » » — 1 — 3 1 — — — 4.8
Эмбрионы. 1 — 1 6 1 —. — — 4.7
Всего или среднее . . 54 42 — 9 — |
Плацентарные пятна. 5 4 5 13 7 3 — 1 4.7
Таблица 12
Зараженность личинками и нимфами иксодовых клещей различных половозрастных групп бурундуков
Половозрастные группы Фаза развития паразита Верхнеленский полустационар Нижнеангарский полустацио- нар, 1962 г.
1961 г. 1962 г. всего ис- следовано (ЭК8.) индекс встречае- мости, в % обилие (абс.)
всего ис- следовано (экз.) индекс ВСТреЧае- МОСТИ, в % обилие (абс.) всего ис- следовано (экз.) индекс встречае- мости, В °/о обилие (абс.)
Взрослые 1 Личинки 18.8 0.8 14.3 0.1
16 5 - 7
самцы | Нимфы 37.5 0.8 — — 14.3 1.0
Взрослые 1 Личинки 11.1 1.1 — — — —
27 3 8
самки | Нимфы 37.0 0.8 33.3 0.3 12.5 0.5
Молодые 1 Личинки 14.3 1.1 28.6 1.4 — —
7 7 3
самцы | Нимфы 28.6 1.0 7J.4 3.8 66.6 1.0
Молодые 1 Личинки 8 — — 8 12.5 0.4 1 — —
самки I Нимфы 12.5 0.1 62.5 1.4 — —
Всего | Личинки 58 12.1 0.9 23 13.0 0.6 1.7 19 5.3 22.2 0.05 0.7
Нимфы 32.8 0.7 47.8
В сельском хозяйстве бурундук — второстепенный вредитель
полей (Виноградов и Оболенский, 1927; Скалой, 1931; Плятер-
Плохоцский, 1932, 1936, и др.), в лесном — наносит значитель-
ный урон почвенному запасу семян (Формозов и др., 1934; Рей-
мерс, 1956а, 19566, 1960а; Лащинский и Реймерс, 1959; Хлопов
и Реймерс, 1960, и др.). Особенно много зверьки уничтожают
орехов кедра (Телегин, 1961; Штильмарк, 1963а, и др.). В остров-
ных лесах бурундуки вместе с другими грызунами и птицами
уничтожают урожай орехов полностью, препятствуя восстано-
влению кедровников. В силу этого при многократном выгорании
лесов кедр постепенно выпадает из состава древостоя и леса из
кедрово-елово-пихтовых превращаются в елово-пихтовые. Это
следует учитывать при оставлении семенных куртин на вырубках
темнохвойных лесов.
Рациональных способов борьбы с бурундуками в лесопитом-
никах еще не разработано (Штильмарк, 19636).
Длиннохвостый суслик — Citellus undulatus Pall.
Вид в Средней Сибири представлен номинальной формой С. и.
undulatus Pall. (Виноградов и Громов, 1952).
Длиннохвостый суслик не относится к таежным животным.
Он в южной тайге обитает лишь по луговым долинам рек. В Чуно-
Ангарском крае — на западе южной тайги Средней Сибири —
суслик в пределы тайги не заходит (Троицкий, 1930). По Ангаре
зверек распространен сплошь в пределах лесостепи, а затем встре-
чается по лугам отдельными колониями до Братска. В лесостепной
части длиннохвостый суслик заселяет весь Ангар о-Ленский водо-
раздел, но уже по линии Малышевка(Балаганск)—Жигалово
встречается лишь по ангарской покати междуречья. Вблизи Ка-
чуга зверек обычен, но на север не идет дальше окрестностей этого
поселка и во всяком случае, как и другие лесостепные формы, не
распространен севернее Верхоленска. На отрезке Жигалово—
Киренск, по опросным данным, его нет, за исключением непрове-
ренных сведений из окрестностей Усть-Кута (с. Марково).
И. К. Лавров (1962) указывает, что сусликов в Усть-Кутском
районе нет.
Весной раньше всего просыпаются суслики типично степных
мест на западном берегу Байкала (Косая Степь). Здесь зверьки
появляются из нор в середине марта (все данные корреспондентов-
фенологов, частично опубликованные нами, 19636, 1963в). Однако
на самом побережье Байкала суслики просыпаются на месяц
позже. В остальной части лесостепи зверьки выходят из нор
с конца марта до середины апреля в зависимости от экспозиции
и прогреваемости склонов и течения весны. В ранние весны (1960,
1961 гг.) суслики появляются на 10—15 дней раньше, чем в позд-
ние (1962, 1963 гг.).
280
Осенью суслики исчезают в конце сентября. Момент ухода
их с поверхности трудно уловим, как и все периоды окончания
фенологических явлений.
Длиннохвостые суслики могут служить источником заражения
бруцеллезом овечьего типа (Пинигин, 1954). Они — основные
прокормители пастбищных клещей (Лавров, 1960).
Бобр — Castor fiber L.
В 1950 г. начаты работы по реакклиматизации .бобров в южной
тайге Средней Сибири. По литературным данным (Копылов, 1962),
опыт расселения бобров оказался удачным. Места выпуска бобров
и районы их натурализации показаны И. П. Копыловым и В. Н.
Скалоном на карте, помещенной в «Атласе Иркутской области»
(1962).
Прежнему распространению бобров в Северной Азии посвя-
щена монография В. Н. Скалона (1951а).
Степная мышовка — Sicista subtilis Pall.
Зверек в южную тайгу совершенно не проникает, даже по
сильно остепненным участкам. Населяет он лишь микроучастки
типичных луго-степей, избегая лесополья.
Лесная мышовка — Sicista betulina Pall.
Вид, очевидно, монотипичен (Виноградов и Громов, 1952).
Распространение, размещение, числен-
ность. Лесная мышовка при отлове грызунов плашками обычно
ускользает от внимания исследователей, поэтому данные об этом
виде крайне скудны. Сделанные до сих пор сборы в Средней Си-
бири создавали иллюзию, что зверек распространен только в лесо-
степи. Именно так описывали ареал вида Б. С. Виноградов и
С. И. Оболенский (1927). Только в лесостепной полосе отмечены
находки лесной мышовки «Определителем млекопитающих СССР»
(Бобринский и др., 1944). Между тем северная мышовка живет
в полосе лесотундры и практически по всей таежной зоне Западной
Сибири (Лаптев, 1958). Наши исследования показали, что лесная
мышовка распространена в южной тайге Средней Сибири очень
широко. Она нередка на верхней Лене (Реймерс и Воронов, 1963),
найдена в довольно большом количестве на р. Чадобец. Ее зоофор-
мационная принадлежность к долинам южных среднесибирских
рек заставляет предположить, что вид распространен по всей
Ангаре (у Иркутска высокая численность зверьков констатиро-
валась Н. И. Литвиновым, 1960а), в южнотаежной части бассейна
Лены, по Подкаменной Тунгуске и, возможно, по значительной
части Нижней Тунгуски. На последних двух реках лесная
281
мышовка должна быть редким зверьком. Подтверждением ска-
занного может служить добыча лесной мышовки в северной тайге
(Рогачева, 1959).
Рис. 61. Принципиальная схема летнего размещения лесных
мышонок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
0.7—1.1, средняя 0.4—0.6, минимальная 0.1—0.3 экз. на 10 канавко-
суток .
В пределах южной тайги лесная мышовка — представитель
долинно-придолинного комплекса (рис. 61). Она достигает наи-
высшего обилия в приречных кустарниках среди лугов, островных
приречных ельниках (только на опушках) и на гарях остепненных
сосповых боров. Много лесных мышовок и на гарях сосняков при-
долипной полосы. С созреванием урожая большое (для вида)
число мышовок концентрируется на полях. Интересно, что лесная
мышовка при высокой численности в долинных нарушенных ель-
никах редка на их гарях, поросших мелколесьем березы. Очевидно,
здесь ей холодно и слишком сыро.
Рис. 62. Лесная мышовка в неволе.
Распространение лесной мышовки тесно связано с нарушением
лесов человеком. Остепненные гари, нарушенные приречные
ельники, луга, перемежающиеся кустарниками, — все это вто-
ричные образования. Однако вид мог быть распространен в южной
тайге издавна, занимая узкую полосу первичных лугов у уреза
воды и приречные ивняки. На Лене для лесной мышовки было
благоприятно сочетание остепненных склонов с приречными ель-
никами. Безусловно, лесная мышовка будет расширять свой ареал,
а кружево его должно становиться плотнее. Особенно заметно
это явление в районах нового освоения.
Как мы уже отмечали (Реймерс и Воронов, 1963), уловистость
лесных мышовок возрастает за лето дважды: в июне (период размно-
жения) и в августе (период расселения молодых). Для целей сопо-
ставления учетных данных лучше сравнивать майско-июньские, а
283
не июльско-августовские показатели обилия. Однако в Средней
Сибири в это время земля еще не оттаивает и отлов ловчими ка-
навками возможен лишь при наличии зарытых с осени цилиндров.
Показатели обилия мышовок в аналогичных элементарных фау-
нистических комплексах южной тайги Средней Сибири очень близки
на севере и юге. Повсюду заселены одни и те же биотопы. В пре-
делах изученной нами территории численность лесных мышовок
довольно стабильна. От года к году происходят некоторые ее
колебания. На верхней Лене наибольшее обилие мышовок отме-
чено в 1962 г. Величина колебаний численности мала — не более
1.5—2 раз, судя по показателям обилия в июле—августе.
Размножение. Лесные мышовки размножаются один
или два раза за лето. При двукратном размножении рождение
детенышей происходит в первой половине июня и середине июля.
Однократное рождение молодых (при затянувшейся весне) при-
урочено к началу—середине июля. Число новорожденных от
3 до 7, в среднем 4.8 (вскрыто 3 беременных самки и 4 со свежими
плацентарными пятнами).
Таблица 13
Состав и встречаемость пищевых компонентов в содержимом желудков
лесной мышовки (75 экз.)
Пищевые компоненты Число желудков с данным кормом Процент к общему количеству желудков с кормом Время обнаруже- ния корма
Растительные корма, всего 72 96.0 Преобладают
В том числе: Семена трав . . - 52 69.3 осенью. Осенью.
Ягоды, ближе не определенные . 1 1.3 16 VIII
Зеленая вегетативная масса . . . 30 40.0 Преобладает
Среди нее: Цветы прострела 1 1.3 весной. 24 V
Цветы горицвета 1 1.3 28 V
Лишайники 1 1.3 26 VI
Животные корма, всего . . 39 52.0 Преобладают
В том числе: Насекомые, ближе не определен- ные 31 41.3 весной и летом. Преобладают
Личинки насекомых, ближе не опре- деленные 5 6.6 летом. Июнь.
Остатки животных, ближе не опре- деленные 5 6.6 —
284
Первые молодые в уловах появляются при двукратном летнем
размножении — в начале июля, при доминирующем однократном
половом цикле — лишь в августе.
Как мы уже отмечали (Реймерс и Воронов, 1963), сеголетки
участия в размножении не принимают.
Мышовки, даже взрослые, легко приручаются (рис. 62).
Питание. Лесная мышовка питается в основном семенами
трав, мелкими насекомыми и вегетативными частями растений
(табл. 13). Каких-нибудь местных особенностей в питании зверьков
различных районов мы не отметили. Сходен состав кормов и по
годам. Весной значительную долю кормов составляют зелень
и насекомые, осенью — семена.
Паразиты, значение. Лесных мышовок поражают
блохи, гамазовые и иксодовые клещи преимагинальных фаз раз-
вития. Зараженность личинками и нимфами иксодид колеблется
по годам сравнительно мало (меньше, чем зараженность других
грызунов) и не достигает высокого значения (табл. 14). Вызывает
интерес высокая зараженность мышовок нимфами. Этот зверек
служит объектом нападения очень большого числа нимф, хотя
он по величине значительно уступает многим другим грызунам.
Как видно из материалов, приведенных в табл. 14, самцы лесной
мышовки заражены иксодидами сильнее самок. Это, видимо,
связано с тем, что они более подвижны.
Таблица 14
Зараженность личинками и нимфами иксодовых клещей различных
половозрастных групп лесных мышовок
Половозраст- ные группы Фаза развития паразита Верхнеленский полуста- ционар, 1961—1962 гг. Нижнеангарский полу- стационар, 1962 г.
всего ис- следовано (экз.) о S - gSs к о о Зд Sffl обилие (абс.) всего ис- следовано (экз.) О 5 - § ад S ад обилие (абс.)
Взрослые ( Личинки 1R 40.0 1.8 11
самцы 1 Нимфы 33.3 0.4 j. 1 9.9 0.09
Взрослые ( Личинки 2 — — 0 — —
самки 1 Нимфы — — 16.6 0.16
Молодые ( Личинки Z 50.0 0.5 4 — —
самцы ( Нимфы 25.0 0.2 1 — —
Молодые ( Личинки £ 20.0 0.2 — —
самки 1 Нимфы О 20.0 0.2 — —
Всего { Личинки Нимфы 26 34.6 23.1 1.1 0.3 18 11.1 0.1
285
В целом в южной тайге Средней Сибири зверек не может
считаться важным вредителем — лесная мышовка здесь слитком
малочисленна для этого.
Тушканчик-прыгун — Allactaga saltator Ev.
По рассказам охотников и жителей деревень, расположенных
на р. Тутуре, в верховьях этой реки был добыт зверек, по опи-
санию весьма похожий на тушканчика-прыгуна. На западном
берегу Байкала найдены многие реликтовые формы степной флоры
и фауны. В связи с этим на указание охотников стоит обратить
внимание. К собственно южной тайге вид, конечно, не имеет
никакого отношения.
Крыса амбарная, серая, или пасюк — Rattus norvegicus Berk.
Восточному подвиду пасюка (/?. п. caraco Pall.) уделено вни-
мание в обширной литературе, весьма полно использованной
А. П. Кузякиным (1951). Специально крысе карако посвящена
небольшая книжка А. С. Фетисова (1945). Более поздняя литера-
тура (Бром, 1959; Литвинов, 1960а; Реймерс и Воронов, 1963,
и др.) не внесла значительных корректив в существующие знания.
Особняком можно поставить работу Т. А. Кима и В. М. Лихашер-
стова (1961), где на основе большого материала, собранного
в г. Красноярске, описывается размножение пасюка.
Не повторяя упомянутых исследователей, мы сделаем лишь
несколько замечаний о современном распространении и размеще-
нии крыс в южной тайге. В работе А. П. Кузякина (1951) приве-
дена карта распространения пасюка в СССР. Автор правильно
отмечает, что в Средней Сибири имеются значительные «пустоты»
внутри ареала вида. Практически кружево ареала серой крысы
можно было бы в мелком масштабе картографически изобразить
в виде гидрологической сети колонизированных человеком рек,
опуская те водотоки, которые с большой плотностью заселены
водяной крысой. У нас нет материалов, доказывающих наличие
конкуренции между этими видами, но обычно в Средней Сибири,
там, где много водяной крысы, мало пасюков в природе и наоборот.
В связи с этим к южной тайге Средней Сибири неприложим тезис,
выдвинутый А. П. Кузякиным, что пасюк предпочитает равнинные
реки с тихим течением. Здесь очень много водяной крысы и почти
нет серой. Между тем слабо меандрирующие реки холмистой
Средней Сибири заселены пасюком сравнительно плотно.
Крыса карако Средней Сибири хотя и живет летом
в природе — частичный синантроп. Наивысшей численности
пасюк достигает в свинарниках и вообще в населенных пунктах.
Здесь, как показали исследования Т. А. Кима и В. М. Лихашер-
286
стова (1961), крыса размножается круглогодично. Этого нельзя
сказать о природной популяции (Реймерс и Воронов, 1963).
На севере Средней Сибири серая крыса распространена шире,
чем показано на карте А. П. Кузякина (1951). Еще И. С. Поляков
(1873) говорил об этом виде как обычном на Лене, в том числе
в нынешнем Бодайбинском районе Иркутской области. На Ан-
гаро-Подкаменнотунгусском междуречье мы крыс в природных
условиях не добыли. По рассказам местных жителей, крыса в се-
лах есть, во всяком случае иногда появляется. Очевидно, участок
между Ангарой и Нижней Тунгуской — полоса пульсации гра-
ницы ареала этого животного.
Таким образом, крысы живут в природе постоянно лишь в са-
мой южной части Средней Сибири (Иркутско-Балаганская лесо-
степь). Севернее они связаны зимой с населенными пунктами
(средняя Лена, низовья Ангары, Енисей). Наконец, у северного
предела южной тайги расположена зона пульсации ареала крысы.
Вид может постоянно существовать лишь в крупных населенных
пунктах.
Серая крыса — источник многих инфекций, в том числе ши-
роко распространенного в Средней Сибири лептоспироза (Крамин-
ская, 1962). Возможна передача ею трихинеллеза и других заболе-
ваний. Роль вида в распространении эпизоотий до сих пор иссле-
дована слабо, хотя, как показывают исследования в Западной
Сибири (Траханов, 1962), численность крыс может достигать фан-
тастических размеров.
Черная крыса — Rattus rattus L.
Наличие «пустот» внутри ареала серой крысы позволило ее
менее конкурентноспособному родичу появиться после Великой
Отечественной войны в полосе южной тайги Средней Сибири
(Бром, 1961). Черная крыса заселила с большой плотностью
скотные дворы в Тайшетском районе Иркутской области. По
предположению И. П. Брома, зверька завезли с товарами из
Северо-Восточного Китая. Автор делает, на наш взгляд, пра-
вильный вывод о том, что черная крыса может появиться и в дру-
гих районах Восточной Сибири, тем более, добавляем мы, что
имеется немало мест, где серая крыса отсутствует.
Домовая мышь — Mus musculus L.
По Б. С. Виноградову и И. М. Громову (1952), ссылающихся
на работу А. И. Аргиропуло (1940), домовые мыши южной тайги
Средней Сибири относятся к номинальному подвиду М. т. muscu-
lus L.
Как и в отношении серой крысы, мы можем опираться на тща-
тельно подобранную сводку литературы, сделанную Н. В. Ту-
287
пиковой (1947). Новые материалы по Средней Сибири довольно
фрагментарны (Дулькейт и Козлов, 1958; Бром, 1959; Марты-
нов и Зонов, 1960; Фирстов, 1960; Реймерс и Воронов, 1963,и др.).
Здесь мы сделаем некоторые поправки и добавления к суще-
ствующим данным.
Н. В. Тупикова разделила ареал домовой мыши на уча-
стки, исходя из характера связи зверька с жилищами чело-
века. Экологические подразделения ареала показаны на карто-
схеме. Как и у серой крысы, кружево распространения домовой
мыши напоминает мелкие бусы, нанизанные на гидрологическую
сеть. Северная граница этих бусинок, очевидно, расположена
несколько дальше, чем полагала Н. В. Тупикова, — домовая мышь
очень обычна по северным притокам Ангары и, видимо, более ши-
роко распространена по Подкаменной и Нижней Тунгускам.
По опросным сведениям, мышь есть в Ербогачене, т. е. в среднем
течении Нижней Тунгуски. В низовьях этой реки Н. П. Наумов
(1934) мышей не нашел, но предсказал их скорое появление.
В лесостепной полосе и частично в полосе лесополья домовая
мышь в условиях Средней Сибири летом выселяется в поля. Такие
факты указаны для района Иркутска Е. П. Мартыновым и
Г. В. Зоновым (1960), для Качугского лесополья — И. К. Лавро-
вым (1962), а для окрестностей Братска — Н. И. Фирстовым
(1960). На верхней Лене (за пределами лесополья) и на нижней
Ангаре выселения мышей на поля, как правило, не наблюдается
даже при очень высокой численности грызуна, констатированной
в пределах населенных пунктов Нижнеангарского полустационара
(однако здесь зарегистрирована поимка домовой мыши в приреч-
ных кустарниках 30 июня 1962 г.). Таким образом, «зона выселе-
ния мышей в поля только на лето» В. Н. Тупиковой распростра-
няется и на Среднюю Сибирь, занимая район, очерченный на
рис. 63.
Численность домовых мышей прямо пропорциональна древ-
ности человеческого поселения и степени его изолированности
от леса. Как отмечали многие авторы (Аникин, 1902; Лаптев,
1958; Реймерс и Воронов, 1963), домовая мышь, появившаяся
в Сибири вместе с первыми русскими переселенцами (Миддендорф,
1869), не выдерживает конкуренции полевки-экономки и других
факультативно-синантропных видов. С полей и огородов ее вы-
тесняют полевка-экономка и азиатская лесная мышь. Но как
только стена леса отступает от населенного пункта, а во дворах
и приусадебных участках не остается сырых луговин и огородных
залежей, поросших бурьяном, численность полевок-экономок
и лесных мышей начинает резко снижаться. Их место занимают
домовые мыши. Достигая высокой плотности, популяции домовых
мышей оказываются весьма конкурентноспособными, и зверек вы-
тесняет других мелких синантропов, проникающих в населенные
пункты с полей (такие факты в Западной Сибири констатировал
288
Н. Ф. Егорин, 1939). Этому способствует отсутствие в старых селах
благоприятных условий для жизни полевок и лесных мышей.
Таким образом, как и в Западной Сибири (Лаптев, 1958), домовая
Рис. 63. Распространение и характер обитания домовой мыши
в пределах юга Средней Сибири.
I—XI и 1—ю как на рис. 6; 11 — северная граница распространения домо-
вой мыши в Средней Сибири; 12 — граница возможного ареала домовой мыши;
13 — граница района, где домовая мышь выселяется на поля летом; 14 - гра-
ница района, где, возможно, домовая мышь выселяется на поля летом.
мышь побеждает конкурентов там, где создались типичные усло-
вия для синантропизма.
Домовые мыши в условиях южной тайги Средней Сибири вы-
соко плодовиты. Число эмбрионов и плацентарных пятен у них
19 Н. Ф. Реймерс
289
колеблется от 7 до 11 и, видимо, даже больше (в нашем распоря-
жении было всего 3 беременных самки).
С появлением домовой мыши связано в Сибири распростране-
ние ряда инфекций, впрочем мало специфичных для вида: туляре-
мии (Комарова, 1945), энцефалита (Карпов, 1955) и псевдотубер-
кулеза (Тимофеева и Головачева, 1962). На домовых мышах
паразитологами были найдены только гамазовые клещи.
Полевая мышь — Apodemus agrarius Pall.
Этот вид распространен только в полосе лесостепи и ангар-
ского лесополья, достигая на севере Братска (Фирстов, 1960).
В бассейне Лены мы этот вид не нашли ни в окрестностях Жигалова,
ни намного южнее — почти на вершине водораздела Ангара-
Лена в б. Баяндаевском районе Иркутской области. Возможно,
этот вид обитает лишь в сосново-мелколиственной лесостепи
Средней Сибири, не заходя в лиственничную. В лиственничной
лесостепи Тувы полевой мыши нет (Янушевич, 1952; Реймерс,
1959в).
Полевая мышь ныне расширяет ареал и, очевидно, в ближайшее
время заселит все ангарское лесополье, во всяком случае до уча-
стка, где Ангара резко поворачивает на запад.
Азиатская лесная мышь — Apodemus speciosus Temm.
Азиатские лесные мыши (рис. 64) из южной тайги Средней
Сибири относятся к западному мелкому подвиду A. s. major
Radde.11
Распространение, размещение, числен-
ность. Б. А. Кузнецов в «Определителе млекопитающих» (Боб-
ринский и др., 1944) проводит северную границу распространения
азиатской лесной мыши таким образом, что она перерезает по
диагонали южную тайгу Средней Сибири. В нашем распоряжении
еще нет материалов широких исследований, проводимых на Енисее
и в низовьях Ангары Институтом географии АН СССР и Институ-
том медицинской паразитологии и тропической медицины АМН
СССР, поэтому мы не можем с уверенностью констатировать при-
сутствие мыши в северо-западном углу южной тайги. На р. Чадо-
бец азиатская лесная мышь водится в заметном количестве
(в сосново-березовом лесу 0.7 зверька на 10 канавко-суток в июле
и августе). Очевидно, азиатская лесная мышь в Средней Сибири
распространена значительно севернее, чем указано в упоминав-
11 Накопленные ныне факты заставляют предполагать наличие боль-
шего числа форм азиатских лесных мышей, чем принято в новейших моно-
графиях (Громов и др., 1963, и др.). Очевидно, существуют две внутривидо-
вые экологические разности, населяющие горы юга Сибири и севернее рас-
положенный Прибайкальский зоогеографический район.
290
шемся определителе и, без сомнения, с развитием сельского хо-
зяйства заселит всю полосу южной тайги.
В Прибайкальском зоогеографическом районе азиатская лесная
мышь достигает наивысшей плотности населения у крупных рек
и практически почти не живет на водоразделах (рис. 65). В отличие
от горных местностей, где мыши многочисленны по ручьям (Кузне-
Рис. 64. Азиатская лесная мышь.
цов, 1929; Фетисов, 1942; Банников, 1954; Реймерс, 1959а, и др.),
среди водораздельной южной тайги зверьки избегают долин мелких
водотоков. Такое явление вполне объяснимо.
В горах по ручьям растут леса с обильным высокотравьем,
а хорошая дренированность не препятствует жизни сухолюби-
вого вида. В южной тайге долины ручьев резко переувлажнены,
поэтому азиатской лесной мыши тут нет. Влажных мест мышь
избегает и в горах (Тарасов, 1962). В нарушенных приручьевых
и приречных ельниках мышь находит обилие сухих островов,
поэтому она тут многочисленна. Сведение же этих лесов, образо-
вание малокормных и сырых березняков сводит население мышей
почти на нет.
19*
291
Вывод, сделанный нами ранее (Реймерс и Воронов, 1963),
о предпочтении мышами увлажненных мест ошибочен. Мы слишком
широко экстраполировали данные, полученные в хорошо дрени-
Рис. 65. Принципиальная схема летнего размещения азиат-
ских лесных мышей в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. О штриховке полосами
см. подпись к рис. 47. Максимальная численность свыше 1.0, средняя
0.5 —1.0 и минимальная менее 0.5 экз. на 10 канавко-суток в течение
июля и августа.
рованных верховьях водотоков, имеющих обильную травянистую
растительность. В этих микроучастках складывается положение,
аналогичное горным околоводным пространствам.
292
Очень важно для азиатской лесной мыши наличие засеваемых
полей. Именно у полей и на полях население мышей достигает
наивысшей плотности. Много мышей и на залежах. Гари сосновых
лесов, поросшие мелколесьем березы, и сосново-мелколиственные
леса — излюбленные видом биотопы. В общей сложности можно
констатировать, что азиатская лесная мышь селится в южной
тайге Средней Сибири почти исключительно в биотопах антропо-
генного происхождения. Нарушение лесов весьма благоприятно
для этого вида, однако до определенного предела: сведение ель-
ников и образование на их месте лугов снижает число мышей.
Таким образом, мы приходим к выводу, что азиатская лесная
мышь в южной тайге — поздний пришелец, обязанный своим
появлением человеку. Наши выводы диаметрально противо-
положны сделанным Ф. Р. Штильмарком (1963в). Причина такого
расхождения ясна. Указанный автор работал в более высокогор-
ных местах Верхоленья, заселенных другой экологической (а
может быть, и систематической) разностью вида. Как и в горах
юга Сибири, в бассейне р. Орленги обитают мыши, связанные
с темнохвойными лесами. Для них наиболее благоприятным
биотопом следует считать хорошо плодоносящие водораздельные
кедровники.12
Численность лесных мышей в целом весьма стабильна. За
годы нашей работы нам не удалось подметить ни значительного
увеличения, ни падения количества мышей в уловах (в нашей
совместной с Г. А. Вороновым работе 1963 г. мы не учли смещения
пика сезонного обилия мышей и сделали противоположный вывод).
На севере мышей меньше, на юге, особенно на верхней Лене, —
больше, но наиболее благоприятные для вида биотопы (сосново-
мелколиственные леса) заселены почти одинаково.
В течение теплого периода года наблюдается несколько пиков
обилия азиатских лесных мышей в уловах. Наибольший совпадает
с третьей декадой августа—первой декадой сентября (Реймерс
и Воронов, 1963).
Размножение. Явления, связанные с размножением,
мы по возможности подробно описали в упомянутой работе. На
верхней Лене число эмбрионов у азиатской лесной мыши 3—8.
Восемь эмбрионов зарегистрировали мы и на нижней Ангаре.
Здесь, как и на Лене,первый помет приходится намай,но первые
молодые в уловах отмечены лишь в начале июля (на Лене они
пойманы летом 1961 г. также в середине июня). Ю. В. Богород-
ский (1962) находил у азиатских лесных мышей от 5 до 9 эмбри-
онов и плацентарных пятен (обычно 6—8).
Питание. В работе о насекомоядных и грызунах верхней
Лены (Реймерс и Воронов, 1963) мы привели анализ содержимого
12 Более подробно на этом вопросе мы останавливаемся в специальной
работе, переданной для опубликования в «Зоологический журнал».
293
268 желудков азиатских лесных мышей. Здесь мы, исходя из не-
сколько более обширных материалов, по возможности рассматри-
ваем сезонную динамику в питании вида (табл. 15). Данные при-
веденной таблицы говорят сами за себя и не требуют обширных
комментариев. Азиатская лесная мышь — почти чисто семенояд-
ный вид. Большинство желудков этих зверьков наполнено на
100% семенами трав и древесных пород, однако первые всегда
доминируют в питании мыши. В общей сложности — это вид
прежде всего травяных типов леса.
Паразиты, значение. На азиатских лесных мышах
живут обычные эктопаразиты мелких грызунов — гамазовые и
иксодовые клещи, блохи и вши. По свидетельству И. К. Лаврова
(1960), мышь не играет большой роли в прокормлении пастбищ-
ных иксодид. Н. И. Литвинов (1960а) упоминает среди паразитов
азиатской лесной мыши блох Amphipsylla kuznetzovi Wagn.,
AmphaUus runatus I. et R. Вид высокочувствителен к туляремии
(Дунаева, 1954) и, очевидно, может заражаться энцефалитом.
Близость азиатской лесной мыши к желтогорлой позволяет ожи-
дать возможного носительства ею вируса лихорадки Ку.
Зараженность азиатских лесных мышей преимагинальными
фазами развития иксодовых клещей на верхней Лене в 1961 —
1962 гг. характеризуется данными, сведенными в табл. 16. На
нижней Ангаре паразитологами было обработано всего 15 ази-
атских лесных мышей. Зараженность личинками оказалась не-
значительной (встречаемость 6.6%, обилие 0.2), поражение ним-
фами, наоборот, весьма сильное (встречаемость 26.6%, обилие
0.4). Взрослый Ixodes persulcatus встречен на старом самце
лесной мыши лишь однажды.
Любопытно, что зараженность азиатской лесной мыши ним-
фами в 1961 и 1962 гг. осталась стабильной (табл. 16), в то время
как число личинок резко изменилось (в 2 раза по встречаемости,
в 10 раз по обилию). Видимо, мышь служит надежным прокорми-
телем иксодид в сосново-мелколиственных лесах и на гарях сосно-
вых лесов.
Мыши, обитающие на полях, оказались свободными от иксодид.
Не найдено этих паразитов и на экземплярах, отловленных в ого-
родах. Видимо, перенос личинок и нимф, о котором мы упоминали
(Реймерс и Воронов, 1963), не достигает значительных масштабов.
Необходимо специально остановиться на роли азиатских
лесных мышей в жизни леса. В прежних своих работах (Реймерс,
19596; Реймерс и Воронов, 1963, и др.) мы относили этот вид к не-
сомненным вредителям лесовозобновления. Действительно, азиат-
ская лесная мышь — семеноед, уничтожающий в кедровых и
лиственничных лесах много семян древесных пород. Нередко в же-
лудках мы находили прорастающие семена (что не нашло отраже-
ния в приведенной выше таблице объектов питания из-за неуве-
ренности в точном определении этих компонентов в содержимом
294
Таблица 15
Состав и встречаемость пищевых компонентов в содержимом желудков азиатской лесной мыши
в течение апреля—сентября (364 экз.)
Пищевые компоненты Апрель Май Июнь Июль Август Сентябрь Весь период
1 2 1 2 1 2 1 2 1 2 1 2 1 2
Растительные корма, всего . . . 10 100.0 8 100.0 30 100.0 100 100.0 123 97.6 90 100.0 361 99.2
В том числе: Семена, всего (в том числе трав и ближе не определенные) 8 80.0 8 100.0 28 93.3 94 94.0 122 96.8 80 88.8 340 93.4
Из них: Семена хвойных пород 2 20.0 1 12.5 — — 1 1.0 5 3.9 1 1.1 10 2.7
Семена бобовых — — — — — — — — 1 0.8 — — 1 0.3
Семена хлебных (культурных) злаков .... 4 40.0 2 25.0 2 6.6 — — 1 0.8 18 20.0 27 7.7
Зеленая вегетативная масса 1 10.0 3 37.5 7 23.1 13 13.0 13 10.3 17 18.9 54 14.8
Таблица 15 (продолжение)
Пищевые компоненты Апрель Май
1 2 1 2
Ягоды, всего .... Из них: — — — —
Ягоды голубики . — — —
Ягоды черники — — — —
Грибы — — — —
Лишайники .... Животные кор- — — — —
м а, всего В том числе: — — 2 25.0
Насекомые, всего . . Из них: — — 1 12.5
Жуки — — — —
Муравьи .... Личинки насеко- — — — —
мых — — — —
Гусеницы бабочек — — 1 12.5
Примечание. 1 — данный корм обнаружен в числе
Июнь Июль Август Сентябрь Весь период
1 - 1 2 1 2 1 2 1 2
— — 2 2.0 1 0.8 1 1.1 4 1.1
— — 1 1.0 — — — — 1 0.3
— — 1 1.0 — — — — 1 0.3
— — — — 2 1.6 — — 2 0.5
— — 1 1.0 — — — __ 1 0.3
7 23.3 25 25.0 11 8.8 6 6.6 51 14.0
7 23.3 25 25.0 11 8.8 5 5.5 49 13.4
— — 1 1.0 — — — — 1 0.3
— — 1 1.0 — — — — 1 0.3
1 3.3 3 3.0 — — — — 4 1.1
1 3.3 — — — — 1 1.1 3 0.8
желудков; 2 — процент к общему числу желудков с кормом.
Таблица 16
Зараженность личинками и нимфами иксодовых клещей различных
половозрастных групп азиатских лесных мышей на верхней Лене
(1961—1962 гг.)
Половозрастные группы Фаза развития паразита 1961 1962
всего ис- следовано (экз.) о _ X о S Q. Е-< в И О О° S m S щ обилие (абс.) всего ис- следовано (экз.) i о 2 . g S S и о о S д Sffi обилие (абс.)
Взрослые ( Личинки 39 64.1 4.0 24 29.2 0.3
самцы | Нимфы 25.6 0.4 20.8 0.4
Взрослые ( Личинки с; 40.0 0.8 5 — —
самки 1 Нимфы и 20.0 0.2 — —
Молодые ( Личинки 45.4 2.6 32 21.8 0.4
самцы 1 Нимфы 9.1 0.1 9.3 0.1
Молодые ( Личинки 24 54.1 3.3 14 21.4 0.2
самки 1 Нимфы 4.1 0.04 7.1 0.1
Всего | Личинки Нимфы 90 55.5 15.5 3.3 0.2 75 22.7 12.0 0.3 0.2
желудков). Возможно, данные таблицы о встречаемости древесных
семян значительно занижены. Однако основу питания азиатской
лесной мыши в южной тайге составляют семена трав. Именно
в травяных типах леса она многочисленнее всего в южной тайге
Средней Сибири. В связи с этим следует предположить, что в
исследованном районе мышь не оказывает серьезного влияния на
процесс естественного лесовозобновления. Вместе с тем, обладая
прекрасно выраженным ориентировочным рефлексом, этот вид бе-
зусловно вредит посевам лесных пород, уничтожая высеянные
семена.
Азиатская лесная мышь наиболее многочисленна в местах,
где практически нет ценных пушных видов, вредит полям, прони-
кает в жилые здания. Это несомненно один из вреднейших видов
мышевидных грызунов.
Мышь-малютка — Micromys minutus Pall.
Подвидовая специфика южнотаежных малюток Средней Сибири
неясна. Видимо, это М. т, batorovi Kastsch.
Распространение, размещение, чис-
ленность. Северная граница ареала мыши-малютки в Средней
Сибири показана Б. А. Кузнецовым (Бобринский и др., 1944)
ориентировочно. Она проведена значительно южнее действитель-
ного предела распространения вида (Рогачева, 1959). Есть осно-
вания считать, что малютка встречается в южной тайге повсеместно.
297
Любопытно, что в Забайкалье этот вид считается редким (Скалой,
1935, и др.). А. С. Фетисов (1936) даже не упоминает о нем в работе
о Западном Забайкалье. Нет мыши-малютки в Муйской долине
(Хабаева, 1959). Но в Восточном Забайкалье малютка в соответ-
ствующих биотопах обычна (Линник, 1936; Некипелов, 1962).
Литературные сведения о мыши-малютке Средней Сибири
весьма ограничены. Имеются упоминания о поимке зверька на
полях (Мартынов и Зонов, 1960; Фирстов, 1960). Часть матери-
алов, полученных нами на верхней Лене, также опубликована
(Воронов и Реймерс, 1962; Реймерс и Воронов, 1963). Для смеж-
ных районов данные по экологии вида сведены М. М. Слепцовым
(1947) и Лаптевым (1958).
Все авторы единодушно утверждают, что мышь-малютка —
обитатель прежде всего полей (рис. 66). Почти столь же много-
численна мышь-малютка в приречных и опушечных кустарниках
и особенно у полей. С пашен она откочевывает в приручьевые
ельники. Из природных биотопов наиболее благоприятными для
вида нужно признать остепненные сосняки, первичные приреч-
ные ивняки и опушки ельников.
Без сомнения, мышь-малютка — дитя лесополья. Она пришла
в южную тайгу вслед за человеком и благодаря ему широко рассели-
лась. Кружево ареала мыши-малютки весьма напоминает характер
распространения лесной мышовки и отчасти азиатской лесной
мыши. Последний вид лишь более широко занял водоразделы.
Видимо, все мыши (домовая, полевая, лесная и малютка) — спут-
ники освоения тайги. Между прочим, это заметно и в горах южной
Сибири. Там азиатские лесные мыши нередко тяготеют к челове-
ческому жилью и весьма обычны в таежных зимовьях.
Численность мышей-малюток подвержена заметным колеба-
ниям. За один год (1961—1962) происходит снижение их обилия
внутри одного биотопа в 2—6 раз. 1961 г. на верхней Лене был
периодом высокой численности мыши-малютки, предыдущий и
последующий — меньшей. Заметной разницы между числом мы-
шей-малюток на верхней Лене и нижней Ангаре не отмечено.
Южную тайгу Средней Сибири вид заселяет равномерно.
В течение теплого периода года мышь-малютка наивысшего
обилия в уловах достигает в последней декаде сентября. Летом
зверек ловится плохо и неравномерно (кривая обилия имеет резко
ломаную форму, — Реймерс и Воронов, 1963).
Размножение. В отличие от районов Дальнего Востока
(Слепцов, 1947), мышь-малютка Средней Сибири дает не 4, а 3
генерации в течение теплого периода года (май—июнь, июль,
август—сентябрь). Как мы уже указывали (Реймерс и Воронов,
1963), большая часть самок размножается дважды, молодые са-
мочки весенней генерации дают лишь один помет в сентябре.
Беременные особи (8) нами отмечены в начале июня, июле, конце
августа и сентябре. Число эмбрионов колеблется от 4 до 9 и
298
в среднем составляет 6.0. Очень велико количество яловых самок.
Весной оно доходит до 50—70%. Яловость отмечена как на верх-
ней Лене, так и на нижней Ангаре.
Рис. 66. Принципиальная схема летнего размещения мышей-
малюток в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. О штриховке полосами
см. подпись к рис. 47. Максимальная численность свыше 1.0, средняя
0.5 —1.0 и минимальная 0.1—0.5 экз. на 10 канавко-суток в течение
июля и августа.
Питание. Материалы по питанию мышей-малюток на
верхней Лене, уже опубликованные нами (Реймерс и Воронов,
1963), отражают типичное для всей южной тайги Средней Сибири
соотношение кормовых компонентов. Мышь-малютка — раститель-
299
ноядный вид (99.3% встреч), в питании которого основное место
составляют семена (86.0% встреч). Животные корма найдены лишь
в 8.1% желудков. Древесными семенами мышь-малютка почти
не питается, предпочитая семена трав и зерна культурных злаков.
Паразиты, значение. Мыши-малютки сравнительно
сильно заражены гамазовыми и таежными иксодовыми клещами,
блохи и вши на них — редкость. По мнению И. К. Лаврова
(1960), вид может служить второстепенным хозяином для паст-
бищных иксодид. Занимая в основном поля, огороды и кустарни-
ковые заросли, мыши-малютки прокармливают сравнительно
немного личинок иксодовых клещей — встречаемость 35.1, обилие
1.0, однако на них обнаружены нимфы — встречаемость 12.3, оби-
лие 0.1 (Реймерс и Воронов, 1963). В районе Нижнеангарского
полустационара мыши-малютки оказались свободными от иксо-
дид (очесано 22 экз.).
Мышь-малютка приносит некоторый вред на полях, служит
передатчиком туляремии (Дунаева, 1954) и восприимчива к за-
болеванию клещевым энцефалитом (Карпов, 1955). Выселяясь
осенью из природных урочищ в скирды и стога, она может пере-
носить природноочаговые заболевания в населенные пункты.
В лесном хозяйстве мышь-малютка почти не вредит. Зверек слу-
жит объектом питания горностая, ласки и лисицы. Однако роль
вида в питании этих хищников, предпочитающих охотиться на
полевок, мала. В общем балансе южнотаежной природы Средней
Сибири мышь-малютка не играет существенной роли и может
считаться пришлым и нежелательным для человека элементом.
Даурский хомячок — Cricetulus barabensis Pall.
В пределах района наших работ даурский хомячок встречается
лишь в полосе лесостепи и прилегающей к ней узкой ленте лесо-
полья, не проникая, собственно, в южную тайгу (Лавров, 1960,
1962; Лавров и Мартынов, 1960). Нам отлавливать этого зверька
не приходилось.
Лесной лемминг — Myopus schisticolor Lili.
По Б. С. Виноградову и И. М. Громову (1952), лесные лем-
минги южной тайги Средней Сибири относятся к подвиду
М. sch. saisanicus Hint. Эти авторы, очевидно, отказались от приз-
нания выделяемого ранее подвида М. sch. middendorffi Vinogr.
Распространение, размещение и числен-
ность. Практически о лесных леммингах Средней Сибири до
сих пор почти ничего не было известно. Очень небольшие мате-
риалы приведены в нашей статье (Реймерс, 19596) и в работе
М. П. Тарасова (1962). Для других мест имеются лишь упоминания
(Скалой, 1935; Дулькейт и Козлов, 1958; Мартынов и др., 1960,
зоо
и др.). Значительное внимание старались уделить этому виду мы
(Реймерс и Воронов, 1963). К настоящему времени из южной
тайги Средней Сибири нами добыты 72 зверька. Это позволяет
Рис. 67. Принципиальная схема летнего размещения лес-
ных леммингов в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Оценка численности
в баллах.
сделать некоторые общие заключения (хотя полнота знаний еще
очень невелика).
Лесной лемминг, как и другие коренные обитатели тайги,
в настоящее время имеет своеобразное кружево ареала (рис. 67).
301
Бросается в глаза парадоксальное явление: лесной лемминг
в долинно!! и придолинной полосах тяготеет к темнохвойным
лесам, а на водоразделах в них почти не встречается. Вместе
с тем зверек живет в сосновых борах, по гарям темнохвойных
лесов как влажным (приручьевым), так и весьма сухим, с чахлой
растительностью. В связи с этим закономерно предположить сле-
дующее. Лемминг в первобытных южнотаежных лесах достигал
наивысшей численности, с одной стороны, в лесах, по ручьям,
с другой — в перестойной, заболачиваемой водораздельной тайге.
Эти биотопы обладают весьма благоприятным термическим ре-
жимом: высокая влажность воздуха смягчает перепады темпера-
тур, вместе с тем условия теплообмена в редкостойных лесах
близки к степным (Сапожникова, 1950). Кроме того, эти участки
в меньшей степени, чем другие темнохвойные водораздельные
леса, заселены красными полевками — весьма конкурентноспо-
собным видом.
Нарушение лесов пожарами привело к образованию обширных
пространств сосняков и гарей с весьма изменчивыми микроусло-
виями. Ныне лемминги находят на гарях благоприятные микро-
участки, достигая довольно высокой численности в одних местах
(до 0.8 на 10 канавко-суток в июле и августе) и отсутствуя в других.
Наиболее устойчива их популяция при сочетании островов темно-
хвойного леса и березняков (выделы 19 и 20 нашей схемы био-
топов, гл. 4).
В северных лесах лесные лемминги, очевидно, более много-
численны, чем в южных. Скорее всего это объясняется меньшей
конкуренцией экологически близких форм грызунов.
При общей низкой численности лесных леммингов говорить
о ее колебаниях невозможно. В плане вековой динамики тайги
следует думать, что лемминги уменьшились в обилии как по доли-
нам рек, так и на водоразделах.
Размножение. Нами отловлены 4 беременных самки и
столько же особей со свежими плацентарными пятнами. Число
эмбрионов от 3 до 6, в среднем 4; количество темных пятен 3—5,
в среднем 4.2. Как мы уже отмечали (Реймерс и Воронов, 1963),
за лето бывает три генерации (в мае—июне, июле—августе и сен
тябре). Сеголетки весенних пометов участвуют в размножении (это
указание требует проверки). Одна самка дает, видимо, два помета.
Питание. У 27 леммингов желудки оказались с содержи-
мым. Мох и трава обнаружены в 25 из них (92.6%), в одном най-
дены цветы азиатской купальницы. Неожиданно выяснилось,
что в июле и августе лемминги довольно охотно поедают семена
растений (каких ?); они найдены в 6 желудках (22.2%). У 1 экз.
желудок был нацело заполнен ягодами, видовую принадлеж-
ность которых установить не удалось.
Паразиты, значение. На леммингах паразитируют
вши, блохи, среди которых М. П. Тарасов (1962) указывает
302
C. penicilliger, гамазовые и иксодовые клещи. Показатели встре-
чаемости и обилия личинок и нимф иксодид на леммингах с верх-
ней Лены в 1961—1962 гг. (12 экз. очесанных зверьков) следу-
ющие: личинки — 33.3%, 7.9 экз. (столь высокий показатель
обилия связан с нахождением в одном случае 91 личинки, обычно
их 1—2), встречаемость нимф 33.3%, обилие их 0.7 экз. Таким
образом, зараженность нимфами высокая. На нижней Ангаре
при очесе 22 особей найдены по одному разу нимфа и личинка,
т. е. зараженность была ничтожной.
Крайне малочисленный вид, лесной лемминг играет скромную
роль в жизни южной тайги Средней Сибири. Массовых размно-
жений зверька никто здесь не наблюдал. Эпидемиологическое
значение лемминга, видимо, качественно сходно с известным
для лесных полевок. К сожалению, эпидемиологи нередко не уз-
нают этот специфичный вид, относя пойманные экземпляры к
красно-серым полевкам.
Рыжая лесная полевка — Clethrionomys glareolus Schr.
Рыжая лесная полевка встречается в юго-западном углу нашего
района. Ее находили в окрестностях Красноярска (Дулькейт
и Козлов, 1958) и в Западном Саяне (Огнев, 1950; Соколов и Хлеб-
ников, 1962; Штильмарк, 1963в). В последнем районе на вырубках
рыжая полевка составляет до 2.2 зверька на 100 ловушко-суток
(Штильмарк, 1963в). Видимо, этот вид связан с окраинами Красно-
ярско-Канского лесополья и нарушенными лесами Западного
Саяна. В Восточном Саяне и севернее — в южной тайге Средней
Сибири — новейшими исследованиями рыжая лесная полевка
не была отмечена. Правда, О. Л. Россолимо и Е. Е. Сыроечков-
ский (1961) считают, что по Енисею вид доходит до 61—62° с. ш.,
в том числе по правому берегу реки. Однако достоверные находки
восточнее нам не известны. В наших сборах имеются особи, по
некоторым признакам напоминающие Cl. glareolus^ однако
пристальное исследование приводит к отрицательному выводу.
Красная лесная полевка — Clethrionomys rutilus Pall.
Всю полосу южной тайги Средней Сибири заселяет номиналь-
ный подвид красной лесной полевки Cl. rutilus rutilus Pall.
(Виноградов и Громов, 1952).
Размещение, численность. В. А. Попов (1960),
начиная раздел, посвященный красной лесной полевке Волжско-
Камского края, пишет: «Распространение красной полевки...
в значительной степени совпадает с распространением елово-
пихтовых лесов» (стр. 354). Н. Н. Воронцов (1961) считает зелено-
мошные ельники стацией переживания вида. Без сомнения, темно-
хвойная тайга — наиболее благоприятный для вида биотоп
303
(рис. 68). Здесь наблюдается наивысшая численность красных
полевок в южной тайге Средней Сибири (до 13.6 зверька на
10 канавко-суток в течение июля и августа). Такого обилия
Рис. 68. Принципиальная схема летнего размещения красных
лесных полевок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 53. Дополнительно введен седь-
мой балл численности (цифры в скобках — количество зверьков на
10 канавко-суток в июле и августе). Градации численности: первая
максимальная (свыше 10 экз.); вторая максимальная (6—10); первая
средняя (3—6); вторая средняя (1—3); первая минимальная* 0.6—1.0);
вторая минимальная (0.1—0.5).
вид не достигает больше нигде (наивысший индекс уловистости
в других местах 9.3). Данные, полученные с помощью ловушек,
подтверждают сказанное: в темнохвойной тайге уловистость до-
304
стигает 4.3—4.6 зверьков на 100 ловушко-суток, в то время как
максимум, полученный в других местах (в те же сроки),—27 экз.
на то же число ловушко-суток.
В старых, еще не разреженных темнохвойных лесах красных
полевок несколько меньше. В долинах таежных ручьев числен-
ность зверьков высока, но распространены полевки неравномерно,
и в силу каких-то неясных нам обстоятельств (не эпидемии ли?)
долинные популяции красных полевок испытывают большие
колебания численности.
На гарях темнохвойных лесов обилие красных полевок при-
близительно на четверть ниже, чем в лесах, и при многократном
выгорании сокращается еще значительнее. Сосновые боры на
водоразделах бедны красными полевками, если нет подроста
темнохвойных пород или примеси осины и березы.
На водоразделах в период до значительных нарушений темно-
хвойных лесов человеком красные лесные полевки достигали наи-
высшей численности в перестойных разреженных темнохвойных
лесах. Заболачивание перестойной тайги приводило к некоторому
снижению числа зверьков, но затем с естественным осушением
популяция восстанавливалась. Общая численность красных по-
левок была очень высока. В горах Хамар-Дабана мы определили
зимнюю плотность популяции красных полевок перестойного
кедровника в 50 экз./га (Реймерс, 1958а). Применяя поправоч-
ный коэффициент 3 или 4 (Никифоров, 19616, 1963) к летним
данным, опубликованным М. П. Тарасовым (1962), получаем для
кашкарникового кедровника 60 или 80 особей на 1 га. Для за-
поведника «Столбы» Г. Д. Дулькейт и В. В. Козлов (1958) ука-
зывают возможную плотность популяции красных полевок 60—
150 особей на 1 га. В южной тайге Средней Сибири примене-
ние коэффициентов Л. П. Никифорова дает наибольшее число
25—3 0 экз./га в июле и 40—45 зверьков на 1 га как макси-
мум.
Таким образом, в южной тайге Средней Сибири наивысшая
численность полевок равна 30—45 зверьков на 1 га. Это средняя
плотность популяции полевок до нарушения лесов человеком.
Ныне вид не достигает и половины возможной численности.
Смена темнохвойных лесов гарями, как мы уже упомянули,
снизила плотность населения красных полевок на 25%. В листвен-
ных лесах зверьков вдвое меньше, чем в темнохвойной тайге.
Еще резче упало число зверьков с заменой темнохвойных лесов
сосновыми борами: в сосняках их меньше почти в 5 раз. Если
считать, как это сделано нами, что южная тайга была почти
сплошным морем темнохвойных лесов, ныне превращенным в сос-
няки и гари с островами тайги, то следует признать, что общая
численность красных полевок упала в 5 раз на половине площади
южной тайги и в 2 раза еще на 30% площади. Это значит, что
если раньше на «усредненном» квадратном километре водораз-
20 Н. Ф. Реймерс
305
дельной южной тайги могло быть 2500—3000 красных полевок,
то теперь их не больше 1000.
В долинной и придолинной полосах красная лесная полевка
в настоящее время не достигает максимальной численности нигде
(рис. 68), а в резко трансформированных биотопах (лугах, полях,
огородах) это случайный гость. Речные долины в пределах
южной тайги, видимо, никогда не были богаты этим видом.
Мы уделяем вопросам размещения красной полевки большое
внимание, поскольку этот вид — основа животной части рациона
соболя. Более подробно изменение численности грызунов в связи
с трансформацией тайги будет рассмотрено ниже.
Зимой красные лесные полевки придолинной и долинной по-
лос концентрируются в стогах сена и соломы, проникают в насе-
ленные пункты. В незначительном числе живут зверьки в домах
и летом, но лишь там, где нет домовых мышей.
Значительных изменений числа красных полевок в годы
наших работ не отмечено, хотя от 1960 к 1963 г. обилие вида по-
степенно нарастало: за эти годы численность зверьков увеличи-
лась в 2—3 раза (сравнение ведется в пределах аналогичных
биотопов). Стабильность населения красных полевок была от-
мечена А. Н. Формозовым (1948). Однако под Красноярском
Г. Д. Дулькейт и В. В. Козлов (1958) наблюдали резкие измене-
ния численности вида от года к году (почти в 3 раза).
Максимального обилия в уловах (ловчими канавками) красные
полевки достигают в разное время — между первой декадой июля
и второй декадой сентября. Причины этого явления совершенно
не ясны. В связи с этим следует упомянуть, что и в плашки крас-
ные полевки попадают при всех стандартных условиях с различ-
ной степенью частоты. Более того, после исчезновения снежного
покрова в течение конца мая—июня мы в 1962 г. не смогли ловуш-
ками поймать красных полевок там, где их заведомо было очень
много — в темнохвойном лесу. Все эти факты следует учитывать
при сравнениях численности зверьков с помощью различных мето-
дик относительной оценки обилия мышевидных млекопитаю-
щих.
Размножение. Довольно значительные материалы по
размножению красных полевок были недавно опубликованы
нами (Реймерс и Воронов, 1963), поэтому здесь мы ограничимся
краткими замечаниями.
В течение лета (с апреля по сентябрь) красные полевки успе-
вают дать четыре генерации,13 причем каждая взрослая самка
дает 2—3 помета. Молодые самки, как и самцы-сеголетки, раз-
множаются не все, а лишь родившиеся в мае. Среди многих де-
13 Под «генерацией» понимается число пометов, приносимых популяцией
в целом, индивидуально же зверьки могут давать меньшее число пометов.
306
Сятков самок-сеголеток беременных оказалось всего несколько.
Они отмечены в сжатые сроки — в начале июля и августа, поэтому
следует предположить, что молодые самки не дают более 2 гене-
раций за лето. Индивидуально они имеют лишь 1 помет (случаев
нахождения плацентарных пятен двух порядков у них не отме-
чено).
Число беременных особей достигает наивысшего значения
в июле—августе (табл. 17). В это время практически все взрослые
самки участвуют в размножении. Число эмбрионов у красных
полевок не превышает 10 и составляет в среднем 6.1. Все дру-
гие авторы также указывают среднюю величину выводка у
этого вида в пределах от 6 до 7 (Егорин, 1939 — 7; Кош-
кина, 1957 — 6.45; Попов, 1960 — 6.6—6.3; Воронцов, 1961 —
6.9 + 0.31). Наиболее часто встречаются 5—7 эмбрионов. У моло-
дых особей их меньше (3—5), однако встречена молодая самка
и с 8 зародышами.
Питание. В неоднократно упоминавшейся работе о насе-
комоядных и грызунах верхней Лены (Реймерс и Воронов, 1963)
мы привели результаты анализа содержимого почти 500 желудков
красных полевок и разобрали изменения в летнем рационе поле-
вок по месяцам. Было показано, что основу питания вида соста-
вляют вегетативные части растений (45.8—88.3%) и их семена
(28.4—70.1%). Из других компонентов заметную роль играют
лишь грибы (22.4% в августе) и лишайники (до 30% весной).
Приведение дополнительных материалов (ныне в нашем распоря-
жении свыше 2000 экз.) едва ли целесообразно. Здесь мы сосредо-
точим внимание на разнице в питании вида в зависимости от
условий жизни. Для этого рассмотрим особенности питания зверь-
ков в двух водораздельных биотопах — в темнохвойном лесу и
на его гарях, а также в двух долинно-придолинных выдел ах —
сосновом бору и долинном ельнике (табл. 18).
Из материалов таблицы видно, что красная полевка во всех
лесных биотопах имеет в питании сходное соотношение семенных
и вегетативных кормов: семена составляют почти половину встреч,
зеленые вегетативные части — чуть больше 50%. Грибы и лишай-
ники играют подсобную роль. На гарях красные полевки выну-
ждены потреблять меньшее количество семян и больше вегетатив-
ных кормов. Эта разница довольно существенна. Полевки, оче-
видно, активно ищут семена, поскольку на гарях в их желудках
обнаружены орехи кедра при плодоносящих деревьях, располо-
женных не ближе 250 м от мест поимки.
В других, не лесных,биотопах (кустарниках, лугах) и на гарях
сосновых лесов уровень потребления семян еще ниже, чем на
гарях темнохвойного леса, а доля зеленых кормов в питании крас-
ной полевки — выше (по нашим материалам, в среднем встре-
чаемость семян в содержимом желудков несколько больше
30%).
20*
307
Таб
Число эмбрионов, плацентарных пятен и количество размно
Месяц Отловлено взрослых самок Характер образований
всего в том числе со следами размножения
всего про- цент от всех самок из них беременных
всего процент от всех самок
Апрель 10 4 40.0 — — —
Май 12 9 75.0 5 41.6 | Эмбрионы. Плацентарные пятна.
Июнь 21 20 95.2 И 52.4 | Эмбрионы. Плацентарные пятна.
Июль 77 76 98.7 49 63.6 | Эмбрионы. Плацентарные пятна.
Август 34 31 91.2 22 64.7 | Эмбрионы. Плацентарные пятна.
Сентябрь .... 15 13 86.6 2 13.3 | Эмбрионы. Плацентарные пятна.
Общее или сред- нее 169 — — 89 — 1 Эмбрионы. Плацентарные пятна.
В аналогичных биотопах от года к году характер питания крас-
ной полевки меняется весьма незначительно. Не улавливается
разница и в питании особей из различных частей Прибайкаль-
ского района. Красные полевки, отловленные в темнохвойных
лесах на Лене и на р. Чадобец, имели аналогичный спектр кормо-
вых объектов (с разницей в 1—2%). Вообще стабильность пище-
вого рациона красных полевок, обитающих в лесу, весьма велика.
Красная лесная полевка, как уже отмечал ряд авторов (Фор-
мозов, 1948; Воронцов, 1956, и др.), в августе приступает к за-
готовке кормов на зиму, среди которых первое место занимают
орехи кедра и различные ягоды (главным образом брусника и
черника). В иных случаях запасы уложены в короткие отнорки,
в других — в ниши под коряжинами, в третьих — беспорядочно
сложены в ходах под мхом. Бессистемно красная полевка
запасает орехи кедра: они кучками до 5—7 штук густо разбросаны
под подстилкой (всегда в пустотах в отличие от запасов кедровки,
делаемых в твердом субстрате или в сплошной дерновине мха).
Ягоды зверек укладывает вперемежку со мхом и лишайниками.
308
лица 17
жающихся самок красной лесной полевки по месяцам
Это обеспечивает их длительное хранение. Случаев заготовки
красными полевками грибов нам отметить не удалось.
Осенью красных полевок можно видеть на верхушках кедров,
где они вместе с бурундуками грызут созревшие шишки. Однако
на сосну зверек залезть не может (бурундук на это дерево ла-
зает с трудом и лишь в той части, где кора грубо шероховата).
Не видели мы, чтобы полевки лазали на лиственницы, хотя физи-
чески они могут делать это с успехом.
Зимой красные полевки выходят из-под снега неохотно. В этом
отношении наши данные не совпадают с указанием Т. В. Кош-
киной (1957). Выбегают зверьки лишь вынужденно: у троп, дорог,
наледей. При этом случаев поиска кормов зарегистрировать не
удалось. Да в этом и нет никакой необходимости, поскольку ассор-
тимент кормов красной полевки очень широк и они всегда имеются
в толще лесной подстилки. Вообще следует подчеркнуть, что все мел-
кие таежные млекопитающие неохотно выходят на поверхность
снега, что и мешает проведению их зимних учетов по следам. Следы
мышевидных делаются заметными смарта—апреля, когда зверьки
309
в теплые дни выходят из-под снега, а затем начинается половое
возбуждение. Эти выходы отнюдь не связаны с недостатком кор-
мов, хотя красные полевки в это время охотно лазают на деревья
за бородачом и другими лишайниками.
Паразиты, значение. Красная полевка — один из
многочисленных видов, обильно зараженных различными пара-
зитами (блохами многих видов, вшами, краснотелками, гамазо-
Таблица 18
Состав и встречаемость основных пищевых компонентов
в содержимом желудков красных лесных полевок, отловленных в различных
биотопах южной тайги Средней Сибири (1961—1962 гг.)
Компоненты Водораздельная полоса Долинно-придолинпая полоса
темнохвойный лес (200 экз.) гарь темно- хвойного леса (100 экз.) сосновый бор (100 экз.) приречный и приручье- вой ельник (100 экз.)
1 2 1 2 1 2 1 2
Семена 100 50.0 40 40.0 47 47.0 51 51.0
Ягоды Зеленая вегетативная 12 6.0 3 3.0 1 1.0 2 2.0
масса 114 57.0 73 73.0 52 52.0 56 56.0
Мох 8 4.0 — — — — — —
Лишайник 16 8.0 2 2.0 13 13.0 22 22.0
Грибы 9 4.5 9 9.0 17 17.0 11 11.0
Животные корма . . 1 0.5 1 1.0 4 4.0 2 2.0
Примечание. 1 — данный корм обнаружен в числе желудков; 2 — процент
к общему количеству желудков с кормом.
выми и иксодовыми клещами). В Западной Сибири паразитофауна
красной лесной полевки тесно связана с комплексом паразитов
полевки-экономки и крота (Сазонова, 1947). В условиях южной
тайги Средней Сибири паразитарные цепи еще совершенно не
выяснены.
Роль красных полевок в прокормлении преимагинальных фаз
иксодовых клещей довольно высока (табл. 19), но отнюдь не в
тех биотопах, где зверек достигает наивысшей численности. На-
оборот, в местах наивысшего обилия красных полевок — на
водоразделах — показатели численности иксодид имеют ничтож-
ные значения. Наибольшая зараженность во всех случаях кон-
статируется в островных темнохвойных лесах (лес такого типа
на территории нижнеангарского полустационара имеет переход-
ный характер к водораздельному), на их гарях, в сосново-мелко-
310
лиственных лесах и на опушках долинных ельников. В этих
биотопах красные полевки заражены личинками и нимфами всего
сильнее.
Высокий уровень зараженности красных лесных полевок
сохранился как в год, обильный закончившими цикл развития
клещами (1961), так и в лето их малой численности (1962). Но
в последнем случае возросло число нимф. Можно было предполо-
жить, что в следующем году взрослых клещей будет много. Так
и случилось: летом 1963 г. клещи были очень многочисленны.
В связи с этим следует заключить, что урожайность клещей в зна-
чительной мере определяется, четко коррелируется или только
лишь совпадает с обилием нимф на красных лесных полевках.
В любом случае наш вывод интересен для санитарно-эпидемиоло-
гической службы.
Из приведенных в табл. 19 цифр совершенно ясно, что в 1962 г.
на Нижнеангарском полустационаре отмечалась низкая числен-
ность иксодид. Таким образом, падение обилия этого паразита
в 1962 г. констатировано для всей без исключения южной тайги
Средней Сибири. Это еще одно доказательство единства рассмат-
риваемого региона. Однако на его севере клещей вообще, и
на красных полевках в частности, значительно меньше, чем на
юге.
Среди других мышевидных грызунов красная полевка выде-
ляется высокой значимостью в кормовом рационе соболя и других
ценных куньих. Рассматриваемый вид в кедрачах юга Средней
Сибири питается в значительной степени зелеными кормами
(встречаемость зелени тут 54.0, семян — 53.5,—Реймерс, 19596),
поэтому не является серьезным конкурентом белке и соболю.
Наконец, там, где кедра нет, красная полевка служит для соболя
основным промежуточным звеном между ним и продуцирующей
растительностью. Среди других факторов обилие красных по-
левок — один из решающих в определении численности собо-
лей.
Вместе с тем вблизи населенных пунктов красная лесная
полевка — опасный прокормитель многочисленных паразитов.
Ее присутствие нежелательно для человека. Достаточно беглого
взгляда на картосхему размещения красных лесных полевок
(рис. 68), чтобы убедиться, что этот зверек концентрируется близ
жилья лишь в особых условиях — в островах темнохвойного
леса, участках приречных ельников и особенно в нарушенных
сосняках, составляющих основную массу смешанных сосново-мел-
колиственных лесов придолинной полосы. Сплошное окультури-
вание долинно-придолинной полосы может привести к полному
исчезновению красных лесных полевок из окрестностей населенных
пунктов. Это вид, с которым легче всего бороться путем уборки
лесных завалов, сведения малоценного мелколесья и прочими
общепринятыми лесокультурными приемами.
311
Таб
Зараженность личинками и нимфами иксодовых клещей красных
Номер выдела Биотоп Верхи еленский
19G1 г.
очесано зверьков индекс встречаемости, в 7о обилие (абс.) индекс верности по обилию, в % обилие пара- зита на еди- ницу учета хозяина (из расчета коли- чества зверь- ков на июль и август)
ли- чинки нимфы
2 Смешанный водораздельный 75 2.7 0.03 0.0 0.2 0.1
лес. (1.3) (0 03) (0.0)
3 Его гари. 69 2.9 0.04 0.0 0.2 1.8
(7^2) (0.3) (5.3)
5 Смешанный ириручьевой тем- — — — — —
нохвойный лес.
8 Водораздельный сосновый бор. — — — — — —
И Его гари. — — — — — —
19 Островной темнохвойный лес. 29 86.2 10.9 20.9 27.2 1.7
(51.7) (0.7) (12.3)
20 Его гари. 23 86.9 14.6 28.1 26.3 1.0
(30.4) (0.6) (10.5)
21 Сосняк. 19 52.6 4.8 9.2 10.6 1.3
(31.5) (0.6) (10.5)
22 Смешанный сосновый лес. 49 42.8 5.2 10.0 14.6 0.8
(24^5) (0.3) (5.2)
23 Гари Босняков. — — — — —
24 Остепненный сосняк. 14 50.0 4.9 9.4 8.3 0.7
(2L4) (0.4) (7.0)
25 Его гари. — — — — —
28 Приречный ельник. 14 85.7 7.0 13.5 7.0 1.8
(57.1) (1-8) (31.6)
31 Опушечные кустарники. 8 75.0 3.1 6.0 1.6 0.1
(25.0) (0^2) (ЗЛ)
31а Ивняки у лугов. — — — —
32 Луга. — .—. — — —
34 Приручьевой ельник. 12 8.3 1.3 2.5 3.6 0.8
(25.0) (0.3) (5-2)
35 Его гари. •— — — — — —
37 Ерник. 4 25.0 0.2 0.4 — —.
(25.0) (0.5) (8.9)
38 Поля. 1 0.0 .— 0.0 — —
41 Огороды. — — — — --- —
Прягп 1 Личинки 317 34.1 4.0 — — —
Всего| Нимфы 317 20.0 0.4 — — —
Примечание. В скобках указаны числа, относящиеся к нимфам клещей.
312
313
t-*. t-л. СЮ 00 CO CO to 1 1 to to 1-л. *q со to -J ££ to о о 1 1 ю ОО СЛ to очесано зверьков полуста десны {5 Я 0?
со со р р Sp 1 ©° 1 О 1 2 сл о о о о о тз Р 9° сл"3^ -л соТ^ cnjis слТ^ сл"^ р СЛ to р Р р to -q *q to p 3. oj q b bcP о 3- c 4^"сл to о 00,0 Сл"С5 р р io о о о 1 1 р'р р сл,о о 0.0 индекс встречаемости, в °/0 о я я о л СО
р р 2.0 (1.0) 0.2 1 Р Р N 1 р СТ> о fe’O) 0.2) 0.3 1.7 (1-2) 1.4 1.1 (8’0) (1.9) 0.9 (1-0) 2.6 (0.8) 1.7 рр р СЛ СО М 1 1 0.2 (0.1) 0.1 1 обилие (абс.) 6961 вок южной т
1 1 1 1 1 0.8 1 S 1 оо 1 р"р 0^00 СЛ СЛ p pp p о p p p p Cpp do to.cn ЬЬ OO^CO 42*^0 р ю р (11-6) 30.0 14.8 1 1 р р р 4^ СО 00 о о индекс верности по обилию, В °/о У
1 1 р 1 1 0.6 1 О о to 1 to P p CO СЛ 3.6 16.6 1.5 3.3 17.2 17.8 1 1 р 00 сл 1 ли- чинки в « свй дйр’Ь PJ SC я п
1 1 0.2 1 1 2.7 1 3.0 0.5 p 5.1 0.7 3.7 4.4 4.3 1 1 0.8 1 нимфы н, з « ops 2£ ta*® Id SH-g’O £ er g w a> s P “О я ф де
t\5 to ! 1 1 t-л to 1 1 1 1 g 1 1 1 to 1 СЛ 1 сл со о 4^ О 1 1 1 очесано зверьков Нижнеангарский полусп п S о S
4.0 9.7 1 1 (50.0) 1 1 1 1 7.2 (28.5) 1 "to to "to to [ CO сл о о 1 о о о 1 0.0 0.0 0.0 1 1 1 индекс встречаемости, в % 45 Я
0.2 0.2 1 1 1 -° -S 1 1 1 1 1 .°P 1 1 СЛ co 1 1 I о^ср 1 00 00 to to 1 1 1 1 1 1 1 обилие (абс.) S с»
i 1 1 1 (28.6) 1 1 1 1 To I CO CO ] k>o 1 "со -q^-. । р р р р 1 со^сл *о 1 ООО ООО 1 1 1 индекс верности по обилию, в % У
1 1 1 1 1 1 1 1 1 9*0 1 4.7 р йл. 1 1 1 1 1 1 1 ли- чинки обилие пара- зита на еди- ницу учета хозяина (из расчета коли- чества зверь- ков на июль и август) о» Я S о ж в»
1 1 1 1 1 о 1 1 1 1.0 1 2.0 р н*. 1 1 1 1 1 1 1 нимфы 15
Красно-серая лесная полевка — Clethrionomys rufocanus Sund.
Красно-серая лесная полевка в южной тайге Средней Сибири
представлена формой С7. г. irkutensls Ogn. (Виноградов и Громов,
1952).
Распространение, размещение и числен-
ность. Красно-серая лесная полевка распространена в южной
тайге Средней Сибири повсеместно. Странно, что ее не отмечает
для Нижней Тунгуски Н. П. Наумов (1934). Однако на Ангаро-
Подкаменнотунгусском междуречье этот вид оказался значительно
многочисленнее, чем в более южных районах. Хорошо известна
красно-серая полевка в Якутии (Виноградов, 1927; Карасева
и др., 1960, и др.). Не указывает красно-серую полевку для сред-
ней тайги Э. В. Рогачева (1959), впрочем, этот автор производил
свои сборы в основном в долине Енисея, поэтому мог легко вид
пропустить (еще в 1927 г. Б. С. Виноградов и С. И. Оболенский
отмечали, что встретить красно-серую полевку можно лишь
в «настоящей» тайге).
Красно-серая полевка — водораздельный вид. Лишь в годы
высокой численности (1963 г., верхняя Лена) он делается заметным
в речных долинах. Б. С. Виноградов и И. М. Громов (1952) со-
вершенно справедливо относят его к характерным обитателям
горной, а И. К. Лаптев (1959) малозаболоченной тайги. Больше
того, это вид редкостойной тайги, верхнего предела древесной
растительности в горах (Тарасов, 1962), каменных россыпей и
каменистых грив (Банников, 1954; Дулькейт и Козлов, 1958),
а также сухих водораздельных боров Средней Сибири (рис. 69).
Красно-серая полевка принадлежит к числу немногочисленных
животных, однако в отличие от подавляющего большинства форм
тайги способна занимать с наивысшей для вида плотностью
значительные пространства лесных гарей. Если для многих видов
кружево ареала в мелком масштабе может быть отображено
гидрологической сетью, то места обитания красно-серой полевки
вполне отобразимы негативом гидрологической сети. При этом
в долинах вид никогда не достигает максимальной числен-
ности.
Зверек избегает как темной тайги, так и любых переувлажнен-
ных биотопов, если они не изобилуют завалами и прогалинами.
В хорошо дренированных местах у горных ручьев полевки много-
численны. В первобытной тайге, как и ныне в горной полосе Си-
бири, красносерая полевка достигала максимального обилия в
перестойных лесах. С их уничтожением она нашла благоприят-
ные условия на гарях, в различных сосняках и на их выруб-
ках. Можно полагать, что несмотря на некоторое «иссечение»
кружева ареала вида полями, лугами и остепненными участками
в целом нарушение тайги оказалось для красно-серой полевки
сравнительно безвредным. В силу ослабления конкуренции на
314
3S
№
00 ГГ
СеР«^ лесЙ,Ц«пиалЬНая
Полевок в
более Q bge °6°3fiatICH
’ средняя 0 «а
«Уток0 в8 ? -
вредней
схема лотч/ч
предела? L° раз^ешенИ(т
снбнрИ' Ю^НОЙ тайги сраси°-
Рис. 19 п Реднеи
^Ченц^^^О^^^льнан чн
'°ЛЯ " автусгаЭКЗ- "а <0ХХЬ
гарях вид получил возможность даже более широкого распро-
странения за пределы первичного кружева ареала.
Следует обратить внимание на то, что красно-серая полевка
при общей низкой численности весьма равномерно заселяет ме-
стность. Средняя численность, отмеченная на рис. 69, чаще всего
составляет 0.4—0.8 экз. на 10 канавко-суток, т. е. весьма близка
к максимальной для вида. Наоборот, разрыв между средней и
низшей градациями обилия велик — минимальная отмечаемая
численность чаще всего 0.1—0.2 экз. на 10 канавко-суток в июле
и августе.
Как мы уже отметили в начале очерка, на нижней Ангаре
численность красно-серых полевок оказалась выше, чем на Лене
и средней Ангаре. С 1960 по 1962 г. на верхней Лене происходило
неуклонное падение обилия вида. В некоторых биотопах (долин-
ных ельниках) зверек совершенно исчез, в других (водораздель-
ные темнохвойные леса, сосняки) произошло сокращение числен-
ности вдвое. Стабильнее всего держалась численность вида
на сухих водораздельных гарях. В 1963 г. обилие красно-серых
полевок резко возросло и в некоторых биотопах превысило уровень
1960 г.
В течение лета уловистость красно-серых полевок в цилиндры
неоднократно меняется, достигая максимумов в середине—конце
июня, середине июля и на грани между августом и сентябрем
(абсолютный максимум). В это время ловится много молодых осо-
бей. Заметное увеличение уловистости констатируется в мае.
Это период полового возбуждения.
Размножение. Красно-серые полевки начинают раз-
множаться рано. Уже в середине мая встречаются рожавшие
самки, а гон обычно протекает с конца марта — несколько раньше,
чем у красных лесных полевок. Следы красно-серых полевок на
снегу напоминают бурундучьи в миниатюре, поэтому легко от-
личимы от следов других мелких грызунов.
Взрослые красно-серые полевки дают за лето не меньше 3 по-
метов (найдены плацентарные пятна трех порядков): в начале
мая, июне и конце августа. Молодые самки весенней генерации
размножаются в том же году и дают один, редко два помета (раз-
множение отмечено в июле). Все данные близки к констатирован-
ным Т. В. Кошкиной (1957). Данные о числе эмбрионов и пла-
центарных пятен сведены в табл. 20. Очевидно, большее число
плацентарных пятен по сравнению с количеством эмбрионов
нужно отнести за счет их длительного сохранения от предыдущих
родов.
Среднее число эмбрионов у красно-серых полевок южной тайги
Средней Сибири близко к найденному у них Т. В. Кошкиной
в северной тайге европейской части СССР (5.49—5.25).
Как мы уже отмечали (Реймерс и Воронов, 1963), самцы-сего-
летки весеннего помета имеют крупные семенники и, очевидно,
316
участвуют в размножении.
Сеголетки (самцы и самки)
летних и осенних генераций
приступают к размножению
в следующем году.
Питание. Все авторы
отмечают строгую раститель-
ноядность красно-серой по-
левки и небольшой удель-
ный вес семян в ее питании.
В 120 просмотренных желуд-
ках зверьков нам не удалось
найти ни одного включения
животного происхождения,
так что красно-серая по-
левка в южной тайге —
абсолютный вегетарианец
(табл. 21). Зверьки сравни-
тельно плохо лазают по ку-
стам и деревьям, поэтому
кормятся всегда на земле.
Зимой следы красно-се-
рых полевок найти очень
трудно. Зверьки ведут под-
снежный образ жизни, цели-
ком питаясь травой, мхом и
лишайниками. В небольшом
количестве полевки находят
и семена. В общем красно-се-
рые полевки хорошо обеспече-
ны в тайге кормами круглый
год (Некипелов, 1941; Рей-
мерс, 19596, и др.).
Семена хвойных пород
красно-серые полевки пое-
дают редко, чаще встречаются
среди компонентов содержи-
мого желудков семена трав.
Паразиты, значе-
ние. Красно-серые полевки
сравнительно несильно * 'за-
ражены блохами и вшами,
несколько больше иксодо-
выми и особенно гамазовыми
клещами. Материалы, соб-
ранные нашими паразитоло-
гами, слишком малы, чтобы
л
Д
Я
я
СО
о га о
2 о о S СО 00 1 Ь- О °0.
Ф £ agi а о СО । Ю L- 10 сс
и * й
л
о х
ф 2 о s 'СТ сч
•V* т-1
ffl £
- 1 1 1 1 -
и 1 1
я 1 1 -4 1
а
Я
ф 0^ ! - 1 1 1
га
R
Я
Я QO Т“Н чгЧ 1 1 oq
я
га
о 1 1 1 1 1
я 1 1 1 1 | 1
о
я
а м
*£ ф
Я ь « 1 сч
о
сЗ СО 1 CSJ СО
о X 1 1 1 1
о •хз* 1 1 1 1
S
о
га т—< т—< 1 1 Т—1 т-1
о
я 1 1 1 1 I I
о 1 1 1 1 1 1
й
1 1 1 1
5 я «3 я СО я
д J н £4 н
ст S <х Я я
Д ) а я я
а ) © ф ф
га СЬ 3 3 3
с ) Я я я
с •) _• л • л
а 3 «5 3 л я
g я Й О И я 3 о я О и
га а а ф л? Я £ а 3 a. J
га ко я ю Я КО 2
к 4 фК гоВ го га
5 со i
& ф я « « я я я 2.|« §‘Н Я а я
< 2 я К Н ю
Е с и га < SsS а я
8 а S' яи
га ю О
317
Таблица 21
Состав и встречаемость пищевых компонентов в содержимом желудков
красно-серой полевки (120 экз.)
Пищевые компоненты Число желудков с данным кормом Процент к общему ко- личеству же- лудков с кор- мом Время обнаруже- ния
Растительные корма,
всего 120 100.0 —
В том числе:
Семена, всего 35 29.1 Весь теплый пе-
риод.
Семена трав, в том числе
ближе не определенные 32 26.6 —
Семена хвойных (сосны?) 3 2.5 Апрель.
Ягоды, всего 34 28.3 Август—сентябрь.
Из них:
Ягоды, ближе не определен-
ные 21 17.5 —
Ягоды жимолости 1 0.8 Август.
Ягоды красной смородины 2 1.7 »
Ягоды черной смородины 1 0.8 »
Ягоды брусники 3 2.5 »
Ягоды голубики 4 3.3 »
Ягоды княженики .... Зеленая вегетативная масса, 2 1.7 »
всего 74 61.7 Весь теплый пе-
риод.
Мох 5 4.1 То же.
Лишайники 2 1.7 16 V, 19 VIII 4 и 9 VIII
Грибы 2 1.7
судить об изменении зараженности вида от года к году. За 1961 и
1962 гг. на верхней Лене было очесано 31 и на нижней Ангаре
40 красно-серых полевок. Показатели зараженности личинками
иксодид: Лена — встречаемость 50%, обилие 2.7; Ангара— встре-
чаемость 7.5%, обилие 0.07. Те же показатели для нимф: Лена —
встречаемость 22.6%, обилие 0.6; Ангара — встречаемость 5.0%,
обилие 0.07. В общей сложности зараженность личинками и ним-
фами на Лене высока, однако сам вид очень малочислен. Обра-
щает на себя внимание факт высокого заражения вида на водо-
раздельных гарях (заражено 50% особей — 3 из 6). Другие формы
давали более низкие показатели зараженности в этих биотопах.
Красно-серую полевку в свете современных знаний следует
отнести к полезным животным, поддерживающим кормовую базу
соболя и не конкурирующим с ценными семеноедами. Ущерб,
наносимый лесовозобновлению, невелик. Отрицательная роль
вида как прокормителя иксодид сравнительно мала, тем более
что зверек избегает наиболее посещаемые человеком угодья.
318
Водяная крыса — Arvicola terrestris L.
Водяной крысе (водяной полевке) посвящены многие сотни
работ (библиографию до 1957 г. составили А. А. Максимов и
С. С. Фолитарек, 1959), неоднократно подвергавшиеся критиче-
ской переработке (Вишняков, 1957; Фирстов, 1957; Кузякин,
1959; Максимов, 1960, и др.). И хотя из этой массы литературы
крысе южной тайги Средней Сибири посвящена лишь ничтож-
ная часть, общие экологические закономерности жизни вида
не требуют слишком подробного описания.
Водяная крыса занимает в Средней Сибири долинно-речные
местообитания при ленточном размещении поселений (термино-
логия по Максимову, 1959). Очагов долинно-ручьевого типа
в возвышенных частях Среднесибирского плато мы не обнаружили,
хотя по ручьям крыса внедряется в придолинную полосу весьма
далеко, особенно на севере. В годы высокой численности она
встречается на полях, в березово-сосновых лесах склонов долин
и даже на низких водоразделах. Мы уже отметили в очерке о се-
рой крысе определенную корреляцию между наличием в природе
серых крыс и массовостью водяной крысы: там, где в долинах
рек много поселений пасюков, мало водяной крысы.
Н. И. Фирстов (1957) отмечал выход шкурок водяной крысы
со 100 км2 охотничьих угодий. Пользуясь этим критерием, автор
констатировал наивысшую плотность населения зверька на се-
вере лиственничной лесостепи, северо-западе Иркутско-Балаган-
ской лесостепи и в Катангском районе Иркутской области, вытя-
нутом вдоль Нижней Тунгуски. В. И. Троицкий (1930) считает
водяную крысу редким зверьком Чуно-Ангарского края. Однако
далее на запад по Енисею крыса очень многочисленна и прони-
кает далеко на север (Наумов, 1934). Нужно заметить, что вы-
ход пушнины с единицы площади отнюдь не отражает истинной
численности зверька в природе. В лесостепных районах нет вы-
годных и обильных лесных промыслов. Здесь крыса стала объек-
том охоты. В Среднем Приангарье численность крыс выше, чем
на верхней Лене, однако в Качугском районе добывают 41 —
50 крыс со 100 км2, а в Братском — лишь от 1 до 10 зверьков
(по Фирстову, 1957). В связи с этим для нашего района не выдер-
живают критики картосхемы количественного распределения
вида, составленные Н. В. Тупиковой (1959).
Наивысшая плотность населения водяных крыс в пределах
южной тайги Средней Сибири отмечена на северо-западе района.
Весьма обильны этим зверьком нижние течения притоков Ангары,
впадающих в эту реку на восточно-западном ее отрезке. Много
крыс по рекам бассейна Нижней Тунгуски (юго-северном ее от-
резке). Второе место концентрации зверьков — присаянские
понижения с обширными болотистыми низинами. Меньше водяных
полевок в верховьях Лены (сравнительно небольшой участок
319
у Качуга) и в среднем течении Ангары (меридиональный отрезок
реки). Мало водяных крыс в большинстве районов верхней Лены
(Верхнеленское плато) и во всех возвышенных частях полосы
южной тайги. В большинстве мест кружево ареала вида может
быть отображено в мелком масштабе гидрологической сетью.
На нижней Ангаре и Нижней Тунгуске полоса заселенности
шире и достигает 20—25 км, но не доходит обычно до водоразде-
лов первого порядка (если они не заболочены). Заболоченность
водоразделов — показатель северной тайги. Тут, в северной
тайге, крыса, вероятно, выходит на водоразделы, но это уже не
район наших исследований.
Нарушение лесов весьма благоприятно для водяной крысы.
В приречных и приручьевых ельниках этот зверек почти не встре-
чается. Зато на гарях здесь он обычен. И особенно многочисленны
водяные крысы в зарослях кустарников вдоль рек и в ерниках.
Сведение лесов резко повысило уровень половодий, привело к об-
разованию широкой полосы кустарников, перемежающихся лу-
гами. Все это — лучшие стации водяной крысы. Можно предпо-
лагать, что в значительной части южной тайги Средней Сибири
водяной полевки до нарушения лесов не было вовсе. Зверек был
широко распространен в северной тайге (в том числе в промежу-
точной полосе смешения зооформаций южной и северной тайги,
относимой ботаниками к промежуточной подзоне средней тайги)
и по периферии первичных среднесибирских степей, заходя туда из
их западносибирских аналогов. На юге по широте Байкала вид
столкнулся с мощной конкуренцией серой крысы карако. Видимо,
это обстоятельство, а не горные хребты, как полагает Н. И. Фир-
стов (1957), послужило препятствием для проникновения вида
в Забайкалье. Севернее Байкала якутские аласы были заселены
водяной крысой испокон веков. Вид здесь ограничен в распростра-
нении климатическими факторами, а на юге, кроме того, викариа-
том серой крысы.
Не останавливаясь на биологических заметках (тем более что
наши материалы невелики — 31 экз.: мы его специально не со-
бирали), отметим, что в местах высокой численности водяной
крысы в Средней Сибири зарегистрированы очаги туляремии.
Эта болезнь отмечена в Нижнеудинском районе (Присаянье),
в Нижнеилимском и Братском районах (средняя Апгара),
а также в Катангском районе (Нижняя Тунгуска) Иркут-
ской области (Алтарева и др., 1957; Клец, 1957; Анциферов, 1961,
и др.). Широко распространена туляремия в Красноярском крае,
но главным образом уже за пределами южной тайги Средней
Сибири.
Наши паразитологи не обнаружили на водяной крысе иксо-
дид (очесано 22 экз.). Однако И. К. Лавров (1960) указывает,
что зверек — один из основных прокормителей пастбищных кле-
щей. Много на водяных крысах гамазовых клещей, блох и вшей.
320
Человек, создав расчисткой лесов благоприятные условия для
сплошного расселения водяной крысы по-долинам южнотаеж-
ных рек, в настоящее время распашкой и вырубками создает
предпосылки для заселения видом водораздельных пространств.
Благоприятно для вида и создание водохранилищ. В связи с этим
можно ожидать возникновения новых очагов туляремии и вообще
увеличения вредной деятельности водяной крысы, уже сейчас
приносящей урон овощным культурам (напоминаем, что водяную
крысу во многих местах называют «кротом»). Мероприятия по
снижению численности этого вредителя или хотя бы службу учета
его обилия следовало бы организовать до проявления вредонос-
ной деятельности в угрожающих масштабах.
Обыкновенная полевка — Microtus arvalis Pall.
Множество подвидов обыкновенной полевки, выделенных
С. И. Огневым (1950) или принимавшихся ранее (Бобринский
и др., 1944), несмотря на некоторую ревизию (Виноградов и Гро-
мов, 1952), требуют пересмотра. Идя вслед за С. И. Огневым (1950)
и считая, что монгольская полевка — лишь подвид обыкновенной,
следует предполагать, что в пределах южной тайги Средней Си-
бири живут две формы М. arvalis: во-первых, та, которую
Б. С. Виноградов и И. М. Громов (кстати, эти авторы в перечис-
лении подвидов не упоминают Прибайкалья), а потом и мы оста-
вили вообще без названия, и, во-вторых, М. a. mongolicus Radd.
Граница между этими подвидами проходит, очевидно, не по
линии Байкала. Монгольская полевка найдена на Лене (Реймерс
и Воронов, 1963) и отмечена на Илиме (Е. П. Мартынов и А. П. Шве-
дов, in litt.). Верхоленье, верхняя и средняя Ангара — район
распространения земледельческой культуры на периферии средне-
сибирских лесостепей. Вполне естественно, что эти места близки
к лесолужью и лесополью Забайкалья, где монгольская полевка
распространена широко и более многочисленна.
Енисейско-Канская лесостепь — язык западносибирских лесо-
степей, место развития особой земледельческой культуры. По
Енисею и нижней Ангаре распространились в южную тайгу особи
западного по отношению к монгольской полевке подвида.
Обыкновенная полевка в южной тайге очень редка. В наших
сборах ее всего 25 экз. (21 с Лены, 4 с р. Чадобец). Однако она
распространена значительно севернее, чем показано на картосхеме
«Определителя млекопитающих СССР» (Бобринский и др., 1944),
и доходит по крайней мере до Ангаро-Подкаменнотунгусского
междуречья (очень вероятно, что будет найдена и на самой Подка-
менной Тунгуске) и до Жигалова на Лене. Исходя из значительного
единства луго-кустарниковых зооформагий долины Лены, можно
ожидать находок обыкновенной полевки до Киренска или даже
до района Бодайбо. Таким образом, обыкновенная полевка в под-
21 Н. Ф. Реймерс
321
ходящих местах заселяет всю полосу южной тайги (наличие юж-
ных форм — выходцев из лесостепи собственно и отличает многие
зооформации южной тайги от северотаежных, где велико участие
северных фаунистических элементов). Севернее она замещена
полевкой Миддендорфа и северосибирской.
Обыкновенная полевка в южной тайге живет только в доли-
нах рек. При этом вид встречает конкуренцию узкочерепной
полевки и полевки-экономки. Может быть, поэтому численность
обыкновенных полевок мала, а места их находок часто неожи-
данны: маленькие луговинки и огороды у домов, кустарники
у реки, опушка ельника и т. п. На полях и лугах полевок практи-
чески нет, хотя зимой зверьки, очевидно, концентрируются в сто-
гах и скирдах сена и соломы.
У двух самок найдено 6 и 7 эмбрионов. Питается полевка глав-
ным образом зелеными частями растений (8 случаев из 10), семена
найдены 4 раза (40%). Изредка, видимо, полевка поедает насе-
комых (обнаружены остатки гусениц бабочек).
Личинки иксодовых клещей найдены 3 раза из 6, нимфы —
1 раз. При низкой численности вида он едва ли может быть су-
щественным прокормителем иксодид. По этой же причине хо-
зяйственное значение вида ничтожно.
Полевка Максимовича — Microtus maximoviczii Sch.
Этот восточный вид на верхней Лене имеет крайний западный
предел распространения. В. Г. Гептнер и Ю. Г. Швецов (1960)
считают полевку Максимовича синонимом М. fortis Biich., и,
таким образом, Лена оказывается крайним западным форпостом
восточной полевки. При очень большой редкости полевки Мак-
симовича вполне возможно, что она будет найдена не только на
прибайкальских, но и на забайкальских притоках Лены, давая
пример викариата с полевкой-экономкой (поиски нужно вести
прежде всего на р. Витим).
Распределение полевки Максимовича на верхней Лене весьма
сходно с кружевом ареала монгольской полевки и экономки. Эко-
логически эти три вида крайне близки. Они обитают в одних
и тех же биотопах, но полевка-экономка имеет значительно боль-
шую численность.
Среди 11 отловленных нами зверьков оказалась одна беремен-
ная самка с 6 эмбрионами. Молодых зверьков добыть не удалось.
Полевка Максимовича крайне редка, но тем не менее на ее
наличие стоит обратить внимание, поскольку вообще восточная
полевка имеет большое эпидемиологическое значение (носитель
туляремии, геморрагического нефрозо-нефрита, лептоспироза
и т. д.); сводка переносимых заболеваний сделана в упомянутой
статье В. Г. Гептнера и Ю. Г. Швецова (1960).
322
Темная полевка — Microtus agrestis L.
В южной тайге Средней Сибири выделен лишь один подвид —
М. a. mongol Thom. (Виноградов и Громов, 1952).
Распространение, размещение и ч и с лен-
нос т ь. Места регистрации темной полевки на территории
Средней Сибири немногочисленны. Однако находка вида на Ени-
сее у пос. Комса (Рогачева, 1959) и наш отлов большого числа
особей на Лене и Ангаро-Подкаменнотунгусском междуречье
показывают, что вид распространен в южной тайге Средней Си-
бири повсеместно. Наличие вида в Якутии (Бобринский и др.,
1944) и на севере Западной Сибири (Лаптев, 1958) позволяет ду-
мать, что темная полевка широко распространена по всей таежной
полосе Сибири.
Темная полевка относится к видам долинно-придолинного
комплекса (рис. 70). Максимальной относительной численности
население полевок достигает в нарушенных долинных ельниках
и на влажных гарях островных темнохвойных лесов. Много тем-
ных полевок на гарях в вершинах ручьев там, где хороший дре-
наж, а также в сосняках различных типов. На водоразделы по-
левка выходит в небольшом количестве, обитая постоянно
в нарушенных темнохвойных лесах и сосновых борах и спора-
дически населяя их гари. При заселении водоразделов полевке
благоприятствуют сухие теплые годы; во влажные и холодные
летние сезоны численность вида здесь резко сокращается. Это
сокращение происходит и в долинной полосе, но не столь заметно.
Так, от 1961 к 1962 г. в придолинной полосе численность темных
полевок возросла максимально: в островном темнохвойном лесу
и приречном ельнике — в 3 раза, в водораздельном темнохвойном
лесу осталась на прежнем уровне, а на гарях этого леса (на одном
из участков) поднялась от 0 до 1.1 зверька на 10 канавко-суток.
Вопрос об изменении общего обилия темных полевок в связи
с нарушением тайги человеком решить довольно сложно. Безус-
ловно с расчисткой лесов под луга произошла концентрация по-
левок в долинах рек по опушкам ельников. Но вместе с тем луга
почти лишены темных полевок. Сосновые леса придолинной группы
возникли как результат хозяйственной деятельности человека,
он же нарушил темнохвойные леса водоразделов и дал возможность
темным полевкам занять новые места обитания. Таким образом,
кружево ареала темных полевок заняло площадь большую, чем
в первобытной тайге, повысилась численность вида на террито-
рии южной тайги, но затем в ряде мест повторные нарушения
привели к снижению обилия вида.
Восстановленное кружево ареала темной полевки выглядит
приблизительно так. Максимумы видового населения наблюда-
лись на грани между долинными ельниками и приручьевыми
кустарниками. По приручьевым ельникам, перемежающимся
21*
323
у ручьев кустами, на хорошо дренированных участках поселя-
лись полуизолированные популяции зверьков, занимавшие после
случайных нарушений вторичные сосняки, гари и опушки ерни-
Рис. 70. Принципиальная схема летнего размещения темных
полевок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
более 2.1, средняя 1.0—2.0 и минимальная 0.2—0.9 экз. на 10 канавко-
суток в течение июля и августа.
ков. В незначительном числе зверьки проникали в темнохвой-
ные леса.
Уловистость темных полевок в течение лета меняется довольно
резко, но не всегда закономерно. Максимальное обилие в среднем
324
отмечено в июле—августе, однако резкие пики в отловах наблю-
дались в 1961 и 1962 гг. в середине июня, а в 1960 г. — в сентябре.
В плашки зверьки попадают сравнительно редко, лучше весной,
пока еще есть пятна снега, хуже — летом.
Размножение. Темные полевки приступают к размно-
жению в апреле и размножаются до сентября включительно.
В связи с тем что нет определенной локализации периодов размно-
жения, создается впечатление большого числа генераций, сле-
дующих с промежутками около 25 дней (Реймерс и Воронов, 1963).
Каждая взрослая самка дает 2—3 помета, иногда 4. Этот вы-
вод сделан на основе предположения, что новая беременность
следует вслед за родами, что подтверждается почти полным от-
сутствием самок, не участвующих в размножении. Непосредствен-
ных данных о числе пометов нет, поскольку мы пользовались лишь
макрообследованием, а плацентарные пятна у темных полевок
исчезают быстро.
Молодые самки дают за лето 2, может быть 3 помета, но раз-
множаются далеко не все (не более трети). Беременные самки-
сеголетки зарегистрированы в конце июня и середине июля.
В конце августа отмечены самки в течке, поэтому можно предпо-
лагать участие молодых особей в рождении сентябрьского по-
коления.
Размер выводка, судя по числу эмбрионов (табл. 22), невелик,
в среднем (с привлечением данных, предоставленных в наше
распоряжение Г. А. Вороновым, касающихся 28 беременных са-
мок, т. е. по 102 экз.) 4.2 с колебанием от 2 до 8. В. Н. Скалой
(1933) указывает в среднем 5.8 эмбрионов, но его материалы были
весьма незначительны. На Урале (Снегиревская, 1947; Кириков,
1952а) и в Татарии (Попов, 1960) получены средние, близкие
к констатированным нами (от 3 до 7 эмбрионов, в Татарии сред-
нее 4.5). От весны к осени число эмбрионов снижается, поскольку
молодые особи приносят несколько меньше детенышей. Число
эмбрионов и плацентарных пятен у них не превышает 4, а в сред-
нем 3.7 из 6 данных.
Молодые самцы из весенних пометов участвуют в размноже-
нии, из более поздних — лишь в следующем году.
Питание. В работе о насекомоядных и грызунах верхней
Лены (Реймерс и Воронов, 1963) мы привели анализ содержимого
200 желудков темных полевок. Новые данные, полученные на Ан-
гаре и Лене, существенно не изменили представления о кормовых
объектах этого вида. Семена в питании темных полевок играют
ничтожную роль (2—3%). Основу питания составляют зеленые
корма (свыше 80%). Полевка поедает много мха, нередко лишай-
ники, иногда ягоды.
Весной и осенью полевки поедают культурные злаки, в том
числе их семена. В это время зверьки приносят некоторый вред
на полях (см. гл. 6).
325
Таблица 22
Число эмбрионов, плацентарных пятен и количество размножающихся самок темной полевки по месяцам
Месяц Отловлено взрослых самок Характер образований Количество самок с числом эмбрионов и плацентарных пятен
всего в том числе со следами размножения
всего процент от всех самок из них беременных 2 3 4 5 6 7 8 все- го среднее число эмбри- онов или плацентарных пятен
всего процент от всех самок
Май .... 8 8 100.0 7 87.5 | Эмбрионы. Плацентарные пятна. 1 2 1 3 1 — — — 7 1 3.6 3.0
Июнь .... 13 13 100.0 10 76.9 | Эмбрионы. Плацентарные пятна. — 1 6 1 1 1 2 — — 10 2 4.4 4.5
Июль 36 35 97.2 30 83.4 | Эмбрионы. Плацентарные пятна. — 3 и 3 9 1 5 1 1 1 1 30 6 4.8 5.0
Август . . . 21 21 100.0 18 85.7 | Эмбрионы. Плацентарные пятна. 1 7 1 10 2 2 1 2 — 1 21 6 3.9 4.7
Сентябрь . . 15 12 80.0 6 40.0 { Эмбрионы. Плацентарные пятна. 1 2 3 2 — 1 — 1 6 4 3.3 5.5
Общее или среднее 93 89 — — — 1 Эмбрионы. Плацентарные пятна. 3 15 2 33 8 13 3 7 4 1 1 2 1 74 19 4.2 4.8
Примечание. В таблицу включены как взрослые самки, так и сеголетки.
Таблица 23
Обилие иксодовых клещей преимагинальных фаз на темных полевках южной тайги Средней Сибири
Номер выдела Биотоп Верхнеленский полустационар Нижнеангарский полустационар
очесано зверьков 1961 г. очесано зверьков 1962 г. очесано зверьков 1962 г.
личинки нимфы личинки нимфы личинки нимфы
1 2 1 2 1 2 1 2 1 2 1 2
8 Водораздельный сосно- вый бор 2 0.0 0.0
И Его гари — — — — — — — — — — 5 0.0 — 0.0 —
19 Островной темнохвой- ный лес 1 2.0 1.0 0.0 4 0.8 1.5 0.8 1.5 3 0.0 — 0.3 0.2
20 Его гари 15 8.9 13.6 0.7 1.0 6 0.3 0.6 0.5 1.0 — — — — —
21 Сосняк 5 5.2 2.6 0.6 0.3 6 0.0 — 0.3 0.3 — — — — —
22 Смешанный сосновый лес 7 4.0 3.2 0.1 0.1 3 0.0 0.3 0.3 4 0.0 — 0.2 0.05
23 Гари сосняков .... 3 0.6 0.1 0.6 0.1 4 0.5 0.8 0.2 0.3 — — — — —
24 Остепненный сосняк . . 7 0.5 0.5 0.0 — 3 0.3 0.2 0.3 0.2 — — — — —
25 Его гари .— — — — — 4 0.8 1.0 0.2 0.2 — — — — —
28 Приречный ельник . . 4 1.5 2.7 0.2 0.4 14 0.2 0.7 0.6 2.2 2 0.0 — 0.0 —
31 Опушечные кустарники 25 1.7 4.8 0.2 0.6 14 0.1 0.3 0.1 0.3 — — — — —
32 Луга 3 4.6 0.9 1.0 0.2 1 0.0 — 0.0 — — — — — —
34 Приручьевой ельник 21 0.4 2.4 0.0 — 24 0.7 2.9 0.3 1.2 — — — — —
35 Его гари 0.5 0.2 2 0.0 — 0.5 0.1
37 Ерник 2 11.0 6.6 1.5 0.9 9 0.1 0.9 — — — — —
38 Поля — — — — — 3 0.0 — 0.0 — — — — — —
| Встречаемость, Всего । в °/0 . . . . ( Обилие (абс.) 93 45.2 3.1 — 16.1 0.4 — 95 24.2 0.3 — 27.3 0.3 — 18 0.0 0.0 14.0 0.2
Примечание. 1 — обилие (абс.); 2 — обилие паразита на единицу учета хозяина (из расчета количества зверьков на июль и
август).
Животные корма (насекомые) встречены лишь весной. Это слу-
чайные объекты (2 встречи на более чем 500 желудков).
Паразиты, значение. Темные полевки заражены
довольно сильно обычными паразитами мышевидных грызунов:
блохами, гамазовыми клещами, вшами. Известно, что вид вос-
приимчив к туляремии (Дунаева, 1954).
В условиях южной тайги Средней Сибири зверек прокармли-
вает довольно много иксодид преимагинальных фаз, в особенно-
сти нимф (табл. 23). Показатели количества нимф на темных по-
левках высоки как в годы значительной численности клещей,
так и во время их минимума. Очевидно, темная полевка (вместе
с красной) — один из видов, поддерживающих популяцию кле-
щей в наиболее зараженных биотопах — на гарях островных темно-
хвойных лесов, на опушках ельников и т. д. В ерниках темная
полевка — один из основных прокормителей иксодид. К сожа-
лению, отсутствие точного определения клещей лишает нас воз-
можности решить, какие виды прокармливает тот или другой зве-
рек. Видимо, это в основном Ixodes persulcatus.
На полях темная полевка приносит ограниченный вред, живя
в посевах осенью и забегая на края пашен весной. Заметного
скопления зверьков па полях, однако, не замечено. Наивысший
полученный показатель относительной численности — 1.7 зверька
на 10 канавко-суток в июле—августе.
Следует отметить, что темные полевки в Средней Сибири не
такой редкий вид, как это может показаться при чтении специаль-
ной литературы. Среди наших сборов оказалось более 850 экз.
темных полевок — значительно больше, чем красно-серых и
экономок. Тем не менее паразитологическое и эпидемиологиче-
ское значение вида почти неизвестно, а материалы по биологии
до сих пор весьма фрагментарны.
Полевка-экономка — Microtus oeconomus Pall.
Для южной тайги Средней Сибири указан подвид М. oec. dau-
ricus Kastsch. (Виноградов и Громов, 1952).
Распространение, размещение и числен-
ность. Ареал полевки-экономки полностью охватывает юж-
ную тайгу Средней Сибири.
На рис. 71 показано распределение полевки-экономки в южной
тайге Средней Сибири. Вид этот с максимальной плотностью за-
селяет приречные кустарники, поля и огороды, обычен на лугах,
по ручьям, в кустарниках среди ельников, а также на залежах.
Особо высокой концентрации экономки достигают на огородных
залежах, поросших бурьяном, и на межах.
Некоторое отличие констатируется в заселенности водораз-
делов бассейнов Лены и Ангары. В Верхоленье экономка практи-
чески не проникает на водоразделы (кроме приручьевых частей
328
гарей), в Среднем и Нижнем Приангарье она обычный вид на-
рушенных приручьевых ельников, и вместе с ними заходит очень
далеко в глубь междуречий. Здесь экономка в небольшом числе
Рис. 71. Принципиальная схема летнего размещения полевок-
экономок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
более 1.0, средняя 0.5—1.0 и минимальная 0.1—0.4 экз. на 10 ка-
навко-еуток в течение июля и августа.
выходит в лиственные и сосновые леса водоразделов, но только
в травяные типы леса, а не в моховые; в этом отношении мы не
можем согласиться с А. С. Фетисовым (1958), считавшим основной
стацией экономки среди прочих и мшистые типы лесов.
329
В целом кружево ареала полевки-экономки имеет долинный
тип, как и у представителей мышеобразных, других серых поле-
вок и мелких куньих. В первобытных лесах вид был обычен в ку-
старниках, проплешинах долинных ельников, на грани первич-
ных лугов у уреза воды, а также в болотистых местах. Кружево
ареала полевки-экономки похоже на «вязь» распространения тем-
ной полевки, но в отличие от нее экономка почти не проникает
в приречные ельники (тут численность вида минимальная). Обе
эти формы тундрово-лесостепные по происхождению, но темная
полевка более «лесная», а экономка более «кустарниковая».
Безусловно между этими формами должна быть межвидовая кон-
куренция, в ходе которой полевка-экономка приобрела свойства
частичного синантропа, что мало характерно для М. agrestis.
Помимо того, экономка занимает заболоченные места, которых
избегает темная полевка. Это одна из причин, почему экономка
не меняет численности к северу или ее увеличивает, а темная
полевка делается к северу более редкой.
Численность полевок-экономок сравнительно постоянна от
года к году. Колебания ее за 1 год не превышают 2 раз (в сходных
биотопах). Обилие вида было наивысшим в 1961 и 1963 гг. Интен-
сивность отлова зверьков цилиндрами резко меняется в течение
лета, достигая максимумов в июле и на грани августа и сен-
тября. Пики обилия связаны с расселением молодых полевок.
Размножение. В конце апреля у полевок-экономок
происходит гон. К началу мая большинство самок уже оплодот-
ворено, а в конце месяца происходят роды, так что в начале июня
обычно встречаются уже родившие самки. В благоприятные
годы новая беременность наступает тотчас же, на что обращал
внимание Н. П. Наумов (1934). Сухое жаркое лето пагубно ска-
зывается на интенсивности размножения — самки беременеют
лишь после окончания периода лактации — в середине июня.
Несколько позже — в конце июня — подрастают самки-сеголетки
и в июле дают помет. Второй помет молодые самки приносят в ав-
густе, но, видимо, не каждый год. Также в зависимости от усло-
вий сезона с различной интенсивностью размножаются взрослые
самки. Благоприятным летом они дают до 4 генераций (с переры-
вом в июле), принося индивидуально 3 и даже 4 помета. Однако
строгой периодичности, как и у всех серых полевок, не наблю-
дается. В среднем молодые и старые самки полевок-экономок
дают ритм размножения с периодом около месяца. Последние
беременные самки в природе найдены в сентябре. Как мы уже
указывали (Реймерс и Воронов, 1963), в жилых постройках
зверьки способны к круглогодичному размножению.
В табл. 24 сведены материалы по интенсивности размножения
и числу эмбрионов (плацентарных пятен) полевок-экономок.
В дополнение к таблице упомянем, что в апреле из 8 отловленных
самок 3 оказались оплодотворенными (последние числа месяца).
330
Таблица 24
Число эмбрионов, плацентарных пятен и количество размножающихся самок полевки-экономки по месяцам
Месяц Отловлено взрослых самок Характер образований Количество самок с числом эмбрионов и плацентарных пятен Среднее число эмбрионов или плацентарных пятен
всего в том числе со сле- дами размножения
всего процент от всех самок из них беременных 1 2 3 4 5 6 7 8 9 10 и 12 все- го
всего процент от всех самок
Эмбрионы. 4 2 1 1 8 5.1
Май 14 12 85.7 7 50.0 | Плацентарные 1 — — — 2 — — — — — — — 3 3.7
пятна.
Эмбрионы. — 2 — — 4 — — — — — 6 6.0
Июнь 22 16 72.7 6 27.2 | Плацентарные — — — 1 3 1 1 — — 1 2 — 9 6.0
пятна.
Эмбрионы. — — 2 — 1 5 3 5 — — — —. 16 6.4
Июль 43 34 79.1 16 37.2 | Плацентарные 1 — 3 — 2 1 3 2 3 —- — — 15 6.0
пятна.
Август Эмбрионы. — — 2 — 3 4 2 — 1 — — 1 13 6.1
31 25 80:6 13 41.9 { Плацентарные — — —• — 2 4 1 2 — — — — 9 6.3
пятна.
Сен- Эмбрионы. — — — 2 1 — — — — — — — 3 4.3
тябрь 9 5 55.5 3 33.3 | Плацентарные пятна. — — — 1 — — 1 — — — — — 2 5.5
Общее Эмбрионы. — 4 8 6 9 10 6 1 — — 1 45 6.1
или 119 92 — — — 1 Плацентарные 2 — 3 2 9 6 6 4 3 1 2 — 38 6.1
среднее пятна.
В октябре была поймана лишь одна взрослая самка без следов
размножения. Из данных таблицы ясно, что в годы наших иссле-
дований интенсивность размножения была для вида невелика —
значительно ниже, чем у темных полевок. Наибольшее количество
неразмножавшихся самок поймано в северных районах, а среди
особей одного района — в сосняках.
Среднее число эмбрионов у полевки-экономки, учитывая воз-
можную статистическую ошибку, одинаково во всех районах
средней полосы и, быть может, несколько выше в Якутии (Ска-
лой, 1933 — 6.4; Попов, 1960 — 5.4; Карасева и др., 1960 —
6.8—7.9; Снегиревская, 1961 — 6.5; Тарасов, 1962 — 6.0). Одно-
родна и фенология размножения вида. Наблюдаются лишь не-
значительные сдвиги в сроках начала гона и других явлений
(различие между Волгой и Средней Сибирью около 10—12 дней).
Молодые самцы обычно в год рождения не размножаются.
Однако это правило не без исключений. Летом 1962 г. на нижней
Ангаре часть самцов-сеголеток участвовала в размножении, имея
семенные пузырьки и тестикулы одного размера со взрослыми
особями. На верхней Лене с 1960 по 1963 г. размножения молодых
самцов наблюдать не удалось. Очевидно, в благоприятных усло-
виях самцы достигают половой зрелости в возрасте около 4
месяцев (Попов, 1960), а в неблагоприятных — около года
(Kratochvil, Rosicky, 1955). Особи зимних пометов (из жилых
строений) размножаются одновременно со взрослыми зверь-
ками.
Найденные нами гнезда полевок-экономок помещались на
глубине всего 10 см и имели крайне простое устройство (Реймерс
и Воронов, 1963). Обнаруженные выводки состояли из 3—8 мо-
лодых.
Питание. Рацион полевки-экономки крайне однороден
в течение лета и почти не меняется в зависимости от географиче-
ской широты места (в пределах южной тайги Средней Сибири).
Растительные корма составляют основу пищи (100%), а зелень —
наиболее существенный компонент содержимого желудков (около
85%). Поедает экономка также ягоды, мох, лишайники, грибы.
Более подробные данные (анализ содержимого 134 желудков)
приведены в нашей книге (Реймерс и Воронов, 1963). Допол-
нительные материалы (всего около 450 экз.) существенно не
меняют выводов, сделанных в указанной работе. Очень под-
робно питание экономок Восточной Сибири описано А. С. Фети-
совым (1958).
Паразиты, значение. Полевки-экономки в Средней
Сибири весьма сильно заражены гамазОвыми клещами, блохами
(до 13 видов, Федорова, 1960), вшами и значительно менее иксо-
довыми клещами. Рассматриваемый вид обитает в основном там,
где мало иксодид. Однако на опушках сосняков и на залежах,
поросших молодняком сосны, экономки могут иметь значение
332
как прокормители преимагинальных фаз паразита. Здесь заражен-
ность зверьков достигает 80%, а среднее обилие — 7 личинок и
1 нимфа на полевку.
Обилие на полевках-экономках паразитов, из которых многие
паразитируют и на других грызунах, высокая концентрация
зверьков вблизи населенных пунктов, проникновение их в жилища
человека — все это заставляет считать полевок-экономок эпиде-
миологически опасным видом. С ним связаны очаги лептоспироза
(Карасева, 1956; Карасева и др., 1960; Краминская, 1962, и др.),
туляремии (Формозов, 1947; Дунаева, 1954, и др.) и еще ряда за-
болеваний. Экономка может передавать туляремийную инфекцию
через мочу (Клейтман и Иголкин, 1956).
Полевка-экономка — излюбленный корм горностая и ласки.
Питается ею лисица, нередко давят собаки. Норы экономки легко
разрывают свиньи. Это указывает на важность вида в цепи рас-
пространения альвеококкоза.
Экономка приносит некоторый вред на полях, где достигает
численности, близкой к максимальной. Она концентрируется
в стогах и скирдах, вблизи скотных дворов, проникает в склад-
ские помещения. Это один из вреднейших видов фауны грызунов
Средней Сибири. С ним нужно вести столь же беспощадную борьбу,
как и с серой крысой, — эти два вида, очевщцЛ, основные агенты
заражения скота лептоспирозной инфекцией и другими опасными
заболеваниями.
Узкочерепная полевка — Microtus gregalis Pall.
Подвидовая специфика узкочерепных полевок из Прибайкалья
нам не ясна.
Распространение, размещение и числен-
ность. Ко времени издания «Определителя млекопитающих
СССР» (Бобринский и др., 1944) граница распространения узко-
черепной полевки в Средней Сибири не была выяснена. Вид весьма
обычен в среднесибирской лесостепи (Скалой, 1931), всюду встре-
чается на средней Ангаре (Фирстов, 1960) и нередок на верхней
Лене (Реймерс и Воронов, 1963). На нижней Ангаре он не най-
ден (во всяком случае на Ангаро-Подкаменнотунгусском между-
речье).
Узкочерепная полевка — ныне восточная форма, связанная
с тундрой, сравнительно холодными лесостепями и полосой лесо-
полья. Можно предполагать, что вид широко распространился
в Средней Сибири на памяти современного человека, а раньше
был локализован в сравнительно узкой полосе первичной лесо-
степи в восточном Предбайкалье. В верхнем плейстоцене узко-
черепная полевка занимала более обширные площади, чем теперь,
достигая высокой численности даже на Волге (Верещагин, 1953).
Таким образом, ныне мы имеем дело с «урезанным» ареалом вида,
333
который вновь расширяется в сторону таежных пространств Си-
бири. По долинам рек узкочерепная полевка достигает, по всей
вероятности, средней Ангары до места, где река резко поворачи-
вает на запад. Вид должен быть найден на р. Чуне, но оттуда
материалы у нас отсутствуют и пределы распространения по-
левки там не ясны.
Все авторы единодушно указывают, что узкочерепная по-
левка — обитатель долин, лугов и полей (Виноградов и Оболен-
ский, 1927; Скалой, 1931, 1933, и мн. др.). Однако упоминание
Н. И. Фирстова (1960), что вид встречается повсеместно в смысле
размещения по биотопам, преувеличено. Узкочерепная полевка —
долинный вид, не выходящий за пределы придолинной полосы
(рис. 72). Можно заметить, что узкочерепная полевка избегает
тех мест, где много темной полевки, но, очевидно, выдерживает
конкуренцию полевки-экономки. На полях узкочерепная по-
левка придерживается межей и окраин, но в годы высокой чис-
ленности заселяет их сплошь.
Численность узкочерепных полевок максимальна в полосе
лесостепи. В лесополье зверьков немного. Нам удалось добыть
на верхней Лене всего 104 экз. этого вида. И. К. Лавров (1960)
указывает, что южнее — в полосе лесостепи — узкочерепная
полевка составлял^ 40.8% от всех отловленных грызунов. Этот
автор (Лавров, 1962) вообще не смог отловить узкочерепных
полевок на Лене севернее с. Большое Воробьеве.
Размножение. Поскольку явления, связанные с размно-
жением вида, довольно подробно описаны нами ранее (Реймерс
и Воронов, 1963), остановимся на них конспективно. Размно-
жаются узкочерепные полевки с апреля по конец августа—сен-
тябрь. Взрослые самки дают два следующие друг за другом помета
весной и один осенью (в августе). Самки-сеголетки также уча-
ствуют в размножении. За лето они дают два или три помета.
Среднее число эмбрионов несколько превышает 7 (при колебании
их числа от 3 до И). Для района, пограничного со Средней Си-
бирью (левый берег Енисея), Ю. Н. Климов (1931) указывает на-
ходку 15 эмбрионов и среднее их число 8. Самцы-сеголетки, оче-
видно, не участвуют в размножении.
Питание. Узкочерепная полевка — исключительно зеле-
ноядный вид. Семена в питании зверьков составляют по встречае-
мости 3%. Экземпляры, живущие на опушках лесов, охотно по-
едают мох — 7 регистраций (проанализировано содержимое
100 желудков).
Паразиты, значение. Узкочерепные полевки за-
ражены обычными паразитами грызунов. В лесостепи на них
довольно много пастбищных клещей — встречаемость 38.3, оби-
лие 4.2 (Лавров, 1960). На верхней Лене участие вида в прокорм-
лении иксодид незначительно (Реймерс и Воронов, 1963), однако
на 1 экз. найден даже взрослый Ixodes persulcatus (31 мая, приреч-
334
ные кустарники). Связь узкочерепной полевки с опасными за-
болеваниями в Средней Сибири недостаточно выяснена. Для За-
падной Сибири (Карпов, 1954; Карпов и Попов, 1954,1956, и др.)
Рис. 72. Принципиальная схема размещения узкочерепных
полевок в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Максимальная численность
более 0.6, средняя 0.3—0.5 и минимальная 0.1—0.2 экз. на 10 канавко-
суток в течение июля и августа.
известно участие ьида в распространении туляремии, клещевого
энцефалита, клещевого риккетсиоза и геморрагической лихо-
радки.
На узкочерепную полевку охотятся горностай, ласка, реже —
лисица.
335
Зверьки стравливают траву на лугах, вредят на полях и хотя
не проникают, как в Якутии (Карасева и др., 1960), в строения,
но могут инфицировать убранные сено и хлеба. Вид безусловно
вреден.
Ондатра — Ondatra zibethica L.
Этому виду посвящена обширная литература (Лавров, 1947;
Слудский, 1948; Околович и Корсаков, 1951, и др.), в том числе
касающаяся специально Восточной Сибири или более узко При-
байкалья (Дорогостайский, 1927; Слудский, 1934; Доброволь-
ский, 1937, 1939, 1952; Смиренский, 1950; Шергин, 1950, и др.).
Небольшие материалы приведены нами (Реймерс и Воронов, 1963).
Зверек был завезен в Иркутскую область в 1935 г. (Копылов,
1948) и распространился в пределах южной тайги Средней Сибири,
повсеместно заселяя долины рек и изредка выходя на водораз-
делы в крупные, но мелководные озера. В Верхоленье он менее
обычен, чем в более равнинных частях Средней Сибири.
Ондатра очень сильно заражена гамазовыми клещами (до
600 экз. на 1 зверьке). Очевидно, она может быть источником
туляремийной инфекции.
Отряд ЗАЙЦЕОБРАЗНЫЕ — LAGOMORPHA
Северная пищуха — Ochotona alpina Pall.
В пределах южной тайги Средней Сибири повсеместно распро-
странена мелкая форма этого вида Och. a. hyperborea Pall.
Распространение, распределение и ч и с-
л е н н о с т ь. Северная пищуха весьма неравномерно населяет
описываемый район. Горные местности (верхняя Лена) и выходы
траппов (средняя Ангара) наиболее благоприятны для вида.
Однако не меньшее значение имеет наличие водораздельных темно-
хвойных лесов, особенно их приручьевых участков. Зверек ми-
рится с гарями, но при смене темнохвойной тайги сосновыми бо-
рами практически исчезает из полосы южной тайги. В связи с этим
на востоке южнотаежной полосы северная пищуха — обычный
вид, на западе, где доминирует сосна, сравнительно редка.
Возможно, что решающим фактором является различная захлам-
ленность этих лесов — высокая в темнохвойной тайге и обычно
низкая в сосновых борах (Штильмарк, 1963г).
Максимальной плотности население пищух достигает в вер-
ховьях таежных ручьев. Тут расположены наиболее крупные
колонии. Более дисперсно распространены зверьки по долинам
речек и ручьев. Если там есть старые деревья или выходы камен-
ных россыпей, это благоприятствует жизни пищух. Как редкость
336
пищухи встречаются по долинам больших рек. В однородных
лесах водоразделов, особенно молодых, встречать пищух нам не
пришлось. В перестойных разреженных лесах по небольшим су-
хим падям пищухи поселяются. Молодые особи при расселении
встречаются даже в приречных ивняках и населенных пунктах
(Воронов и Реймерс, 1962; Штильмарк, 1963г).
Численность северных пищух определить трудно из-за не-
равномерности распределения их населения. В среднем она не-
высока — намного ниже, чем в Саянах и вообще в горах Южной
Сибири.
Размножение. Биология северной пищухи исследована
слабо. У И. П. Копылова (1948) находим такое указание: «В те-
чение теплого времени года она дает несколько пометов, по 3—
6 детенышей в каждом» (стр. 50). Ближе к истине был В. И. Троиц-
кий (1930), указывавший на наличие двух пометов. С. С. Туров
(1936) предполагал, что пищухи приносят молодых в июле. Но-
вые данные (Мартынов и др., 1960; Тарасов, 1962, и др.) показы-
вают, что в горных областях северные пищухи размножаются
дважды: в конце мая—начале июня и в середине—конце июля.
После периода размножения начинается заготовка кормов.
В полосе южной тайги Средней Сибири бывает 2—3 помета:
в мае, в начале июля и в конце августа. Первые молодые встре-
чены 29- июня. В начале июля обычны родившие самки, но
другая часть их в это время лишь на начальных этапах первой
за лето беременности (в нашем распоряжении 16 взрослых самок).
Число эмбрионов колеблется всего от 1 до 3 и чаще бывает
равно 2 (1 эмбрион у 1 экз., 2 — у 11 иЗ — уЗ пищух).
Самцы и самки-сеголетки участия в размножении не прини-
мают, следовательно, половой зрелости пищухи достигают в воз-
расте около года. Из старых самок редкая не участвует в размно-
жении (1 из 16).
Питание. О кормах пищух существует довольно обшир-
ная литература (Скалой и др., 1936; Шарашидзе, 1951; Ким,
1957а; Тарасов, 1962, и др.). Прав М. П. Тарасов (1962), что
зверек поедает те растения, которые обычны для местности, по-
этому приведение списков кормовых растений более характери-
зует состав растительности и обилие отдельных ее видов, чем
избирательную способность пищух (хотя В. А. Шарашидзе
упоминает, что пищухи заготавливают всего 41 вид из 600). Корма
северных пищух крайне разнообразны и различаются даже в пре-
делах близлежащих падей. Это главным образом травы, листья
кустарников (например, ольхи), кустарнички и мхи. Вегетатив-
ные части растений составляют по встречаемости в желудках
81.1% (проанализировано содержимое 53 желудков). Ягоды чер-
ники, брусники, жимолости, княженики найдены в 11.3% же-
лудков. Около 40% желудков содержат семена (в том числе кедра).
Изредка северная пищуха поедает грибы (найдены в 1 желудке).
22 Н. Ф. Реймерс
337
Характер устройства запасов у пищух различен. Если в горах
среди камней пищухи заготавливают сено в нишах под камнями
или в их расселинах, то среди пологих увалов местом заготовки
стожков служат пустоты под корнями и даже места, защищенные
от дождя густыми кронами (Троицкий, 1930). Однако нам не при-
ходилось наблюдать стожков, прикрытых сверху мхом, как об
этом упоминает названный автор. В. И. Троицкий обнаружил
стог в 8.2 кг. Видимо, это исключение; 2—3 кг корма в стоге —
не редкость.
В колонии пищух существует «строгий порядок»: в определен-
ных местах имеются уборные, в других — «складские помещения».
Это, правда, не мешает зверькам менять назначение площадок,
не убрав старого помета.
Паразиты, значение. Иксодовые клещи на пищу-
хах не найдены (очесано 27 экз. зверьков). В некоторой степени
пищухи заражены блохами (для гор юга Сибири указаны Ampha-
lius runatus и Ctenophyllus armatus, — Тарасов, 1962), вшами и
гамазовыми клещами. В литературе есть упоминание о значи-
тельном заражении зверьков личинками оводов (Скалой, 1936;
Тарасов, 1962). А. А. Спасский и К. М. Рыжков (1951) упоминают
4 формы гельминтов, паразитирующих в кишечнике пищух При-
байкалья.
Большой роли в питании ценных куньих пищуха, видимо, не
играет. В Баргузинском заповеднике участие ее в кормовом ра-
ционе соболя не превышает 1.8% (Мартынов и др., 1960). Гор-
ностай и ласка малочисленны вблизи колоний пищух и предпо-
читают более мелких грызунов.
Эпидемиологическое значение вида невелико: пищухи живут
обычно далеко от населенных пунктов, а зараженность их пара-
зитами намного меньше, чем других видов.
Общая роль вида в хозяйстве, очевидно, незначительна.
Заяц-беляк — Lepus timidus L.
Любопытно, что ни в «Определителе млекопитающих СССР»
(Бобринский и др., 1944), ни в книге «Грызуны фауны СССР»
(Виноградов и Громов, 1952) не указана подвидовая специфика
зайцев из Прибайкалья. Здесь водится мелкая форма, достигаю-
щая, однако, веса около 3.5 кг (но обычно около 3.0 кг).
Уже более 30 лет назад В. Н. Троицкий (1930) четко сформу-
лировал разницу между европейскими и сибирскими беляками.
Он писал: «Беляк в европейских условиях — типичный житель
лесов. В Сибирском крае он живет больше по опушкам в мелочах
и в полях среди колков, приближаясь в этом отношении больше
к европейскому русаку» (стр. 30). В европейской части СССР и
на крайнем западе Сибири беляк викариирует с русаком. Этого нет
в остальной Сибири. Морфологически зайцы севера и юга Сибири
338
резко отличаются: северные почти вдвое крупнее южносибирских.
Очень может быть, что в пределах СССР существует всего 3 хорошо
выраженных расы вида (если не считать островных форм) — евро-
пейская, южносибирская и северосибирская.
Рис. 73. Принципиальная схема весенне-летнего размещения
зайцев-беляков в пределах южной тайги Средней Сибири.
Условные обозначения как на рис. 12 и 22. Оценка численности
в баллах.
Распространение, размещение и числен-
ность. В южной тайге Средней Сибири, как и в Западной Си-
бири (Лаптев, 1958), беляк наиболее обычен в полосе лесостепи
и лесополья. Весной зайцы собираются в островных темнохвой-
22*
339
ных лесах и долинных ельниках, где проходит линька зверька.
Позже беляки расходятся по гарям и смешанным лесам, заходя
на водоразделы, но почти не встречаясь в глухих лесах хребтов
(как в темнохвойных, так и светлохвойных). Весенне-летнее рас-
пределение зайцев иллюстрирует рис. 73. Эти же биотопы зани-
мает заяц зимой, но в холодное время года зверьки приближаются
к населенным пунктам, охотнее выходят на поля и почти начисто
исчезают из водораздельных лесов. Все это хорошо объяснимо
сменой кормовых условий: на полях зайцы находят огрехи хлебо-
уборки, а подчас и просто брошенный урожай (на севере это часто
происходит из-за того, что хлеба не успевают вызреть).
В пределах юга Средней Сибири зайцы многочисленны в полосе
лесостепи и по Лене, с одной стороны, а с другой — на севере
за пределами южной тайги. Сама южная тайга вне полосы лесо-
полья зайцами очень бедна. Это подтверждается как нашими не-
посредственными наблюдениями, так и выходом пушной продук-
ции (Копылов и др., 1940; Копылов, 1948). Очевидно, беляк,
подобно другим видам лесостепи и лесолужья Сибири, проник
в западное крыло южной тайги вместе с нарушением лесов и све-
дением приречных ельников, а в восточной части Прибайкалья
в это время вторично расширил места обитания.
В годы нашей работы численность зайцев стояла на низком
уровне, несколько колеблясь от года к году (в 1961 г. был макси-
мум численности). Общее снижение числа зайцев наметилось с на-
чала 50-х годов. Видимо, 7-8- или 9-летний цикл, указываемый
в литературе (Копылов и др., 1940; Наумов, 1947) для юга Средней
Сибири, выражен нечетко, и, подобно северной окраине южной
тайги, могут наблюдаться более длительные (12 лет) пессимумы.
Для заповедника «Столбы» Г. Д. Дулькейт и В. В. Козлов
(1958) определили численность зайцев около 1 экз. на 1000 га
(1 на 10 км2). По летней глазомерной оценке и наблюдениям
по следам зимой в придолинных полосах исследованного нами
района на 1 км2 приходится до 3 зайцев (в особо благоприятных
местах сочетания островных темнохвойных лесов, их гарей, за-
лежей и полей); на водоразделах зверьков намного меньше.
Размножение. Весенний гон у беляков происходит
с конца марта, но разгар его приходится на первую половину
апреля (в горах южной Сибири даже на начало—середину мая).
Гон происходит до начала заметной весенней линьки. Самку в бе-
лом наряде мы добыли 5 мая. Период повторного гона нами точно
не установлен. Молодые (222.6 и 262.5 г) встречены 3 и 12 августа.
По данным литературы, пометов 2 (Копылов и др., 1940; Копылов,
1948) или 3 (Троицкий, 1930). Вообще изменчивость плодови-
тости у зайцев-беляков чрезвычайно высока (литература по
этому поводу сведена С. П. Наумовым, 1947). В. Н. Троицкий
(1930) не отмечал лично более 2 детенышей в помете, мы обнару-
жили у зайчих 7 и 6 эмбрионов (5 мая и 3 июня), а 19 июля 1961 г.
340
найдено 9 плацентарных пятен, по всей видимости, от одного по-
мета. Рождение молодых первой генерации происходит в июне,
редко в конце мая. 18 июня добыта молодая самка весом около
600 г. В ее желудке оказалось лишь материнское молоко. Однако
у особи, добытой 12 августа (самец 262.5 г), помимо молока, в же-
лудке обнаружена трава (другая молодая особь — самка,
пойманная 3 августа — имела в желудке одно молоко).
Значение. Вредная деятельность зайцев в лесу невелика.
Они зимой объедают кору осин, берез и ольхи, лишь изредка
молодых лиственниц, сосен и кедров, а летом питаются в основном
травой и мхом. Вред от вида ощутим лишь на лесных питомниках
и культурах.
Наибольшую опасность беляк представляет как прокормитель
иксодовых клещей — имагинальной и преимагинальных фаз разви-
тия (Устинов, 1962а, 19626). Согласно этому автору, встречаемость
имаго доходит до 75.0%, нимф — 50%, обилие паразита дости-
гает: имаго — 53.5, нимф — 19.7 на 1 экз. зверька. Эти данные,
очевидно, относятся ко взрослым зайцам. На молодом (от 3 ав-
густа) иксодиды нами не были обнаружены. На 2 взрослых и
1 молодом в июне—июле наши паразитологи нашли: на взрос-
лом — 34 имаго клеща (все самки), на втором взрослом — 2 па-
разитов (самки), на молодом — 22 личинки и 3 имаго (2 самки и
1 самец). Клещи, снятые с зайцев, нередко (в 30% случаев) за-
ражены вирусом клещевого энцефалита (Устинов, 1962а). Заяц
болеет туляремией (Дунаева, 1954).
Безусловно, этот ценный охотничий вид может явиться ис-
точником заражения людей и домашнего скота, поскольку до-
стигает наивысшей численности вблизи населенных пунктов.
Заяц-русак — Lepus europaeus Pall.
В 1938 и 1939 гг. в полосе лесостепи Средней Сибири были вы-
пущены зайцы-русаки. Зверек прижился. По свидетельству
И. П. Копылова (1962), население зайцев имеет довольно высокую
плотность. Однако вид не проник в северную часть лесополья.
Опыт акклиматизации русака даже при его удаче нельзя при-
знать целесообразным, поскольку в Средней Сибири беляки
живут в тех же биотопах и больше приспособлены к суровым
условиям среднесибирских лесостепей. К тому же русаки в мень-
шей степени, чем беляки, способны избегать весеннего преследо-
вания (по насту) бродячих собак — подлинного бедствия зайцев
и копытных Средней Сибири (весовая нагрузка у русака выше,
чем у беляка).
341
Глава 6
ВЛИЯНИЕ АНТРОПОГЕННЫХ НАРУШЕНИЙ ТАЙГИ
НА НАСЕЛЕНИЕ ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Одна из основных задач современного естествознания — ис-
следование динамики природы под влиянием антропогенных на-
рушений. Без этого знания невозможно предугадать ход естест-
венных процессов в будущем и расшифровать те изменения, ко-
торые произошли в прошлом и наложили отпечаток на настоящее.
Динамика природы связана со многими важными для жизни че-
ловека явлениями. Нет такой отрасли хозяйства или прикладного
знания, где были бы безразличны природные смены, особенно
планетарного масштаба.
Общее изменение животного населения влияет а) на судьбу
охотничьего хозяйства, б) на медико-географическую ситуацию,
в) на процесс саморазвития природно-территориальных комплек-
сов и г) на изменение значения животных как вредителей сель-
ского хозяйства. Вместе с тем динамика животного мира — одно
из звеньев генеральной трансформации географической среды
нашей планеты.
Выше мы рассмотрели изменения, происшедшие в размещении
и численности отдельных видов позвоночных животных средне-
сибирской южной тайги. Теперь попытаемся синтезировать ре-
зультаты анализа и рассмотреть животный мир в целом.
Общие закономерности влияния нарушений тайги
на отдельные виды животных
В этом разделе мы не будем приводить дополнительных мате-
риалов, считая его заключением к предыдущей главе. Как влияет
нарушение лесов на жизнь отдельных видов? На этот вопрос не
может быть однозначного ответа. Общие сведения о передвижках
границ ареалов наиболее многочисленных видов полосы южной
тайги, изменениях кружева их распространения и перемене чис-
ленности в текущий исторический момент сведены в табл. 25.
Эти данные отражают итог современного процесса в самом общем
виде. На ряд вопросов мы ответили с неуверенностью (в таблице
поставлен вопросительный знак), другой ряд остался открытым.
Два пояснения к таблице: 1) мы приняли две градации уве-
личения плотности кружева ареала — «утолщается», т. е. «нитей
столько же, но они толще», и «становится плотнее» — «нитей
больше»; 2) указание, что кружево ареала «трансформируется»,
означает перекомбинацию занимаемых видом биотопов.
Как видно из анализа данных, приводимых в таблице, антро-
погенные изменения в лесах южной тайги Средней Сибири при-
вели к довольно значительным сдвигам в границах ареалов жи-
вотных. Ряд форм смог распространиться с юга на север.
342
Таблица 25
Изменение границ и кружева ареалов, а также общей численности
видов зверей и птиц полосы южной тайги Средней Сибири
в процессе антропогенных нарушений лесов
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Тетерев. — Утолщается. Увеличивается.
Глухарь. — Становится ре- же. Снижается.
Рябчик. — Становится ре- же. Снижается.
Перепел. Расширяются на север. Становится плот- нее. Увеличивается.
Скалистый голубь. То же. Утолщается. »
Большая горлица. » » »
Коростель. Наблюдается рез- кая пульсация. Несколько утол- щается. ?
Серый журавль. — Стабильно. Стабильна.
Турухтан. Расширяются на юг. Становится плот- нее. Увеличивается.
Черныш. — То же. »
Фифи. — »
Перевозчик. — Стабильно. Стабильна.
Вальдшнеп. Расширяются на север. Становится плот- нее. Увеличивается.
Бекас. — Утолщается.
Азиатский бекас. — » »
Лесной дупель. Расширяются. »
Чирок-свистунок. — Становится ре- же, местами четковидно. »
Кряква. —• Становится ре- же, четковид- но. Снижается.
Чирок-трескунок. Расширяются? То же. »
Серая цапля. Расширяются на север. Стабильно. Увеличивается.
Сокол. — Делается четко- видным. Снижается.
Чеглок. — Становится плот- нее. Увеличивается.
Дербник. — То же. »
Обыкновенная пустельга. Расширяются на север. Становится плот- нее. Увеличивается.
Кобчик. То же. То же. »
Я стреб-тетеревят- ник. — Стабильно. Стабильна.
Малый перепелят- ник. Стабильны или расширяются на север. Становится ре- же. Снижается.
Перепелятник. То же ?
343
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Полевой лунь. Утолщается. Увеличивается.
Черный коршун. Расширяются на север? » »
Сарыч. То же. » »
Хохлатый осоед. Стабильны или расширяются на запад. ? ?
Скопа. ? Делается четко- видным. Снижается.
Филин. — Становится ре- же. »
Воробьиный сыч. Сокращаются к югу. То же. »
Ушастая сова. — Утолщается. ?
Болотная сова. — » ?
Обыкновенная ку- кушка. — Становится плот- нее. Увеличивается.
Глухая кукушка. — Стабильно. Стабильна.
Обыкновенный ко- зодой. Расширяются на север. Утолщается. Увеличивается.
Черный стриж. То же. » »
Белопоясый » » »
стриж. Желна. — Стабильно. Снижается.
Седой дятел. Расширяются на север. Становится плот- нее, утол- щается. Увеличивается.
Трехпалый дятел. — Становится ре- же. Снижается.
Большой пестрый дятел. — Становится плот- нее, утол- щается. Увеличивается.
Малый пестрый дятел. Расширяются на север. То же. »
Вертишейка. То же? Утолщается. »
Ворон. — Трансформиру- ется. Стабильна.
Ворона. — Утолщается Увеличивается.
Галка. Стабильны или расширяются на север. ? ?
Сорока. Расширяются на север и восток. Утолщается. Увеличивается.
Сойка. Стабильны. » »
Кукша. — Трансформиру- ется. Стабильна.
Кедровка. — Становится ре- же. Снижается.
Скворец. Расширяются на север и восток. Становится плот- нее, утол- щается. Увеличивается.
344
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Иволга. Сокращаются к западу? Делается четко- видным? Снижается?
Дубонос. Расширяются на север. Становится плот- нее. Увеличивается.
Чечетка. Расширяются на юг. То же. »
Длиннохвостый снегирь. Расширяются на север. » »
Серый снегирь. Сокращаются к югу. Становится ре- же. Снижается.
В осточноев роп ей- — Утолщается. Увеличивается.
ский снегирь.
Обыкновенная че- — Утолщается. Увеличивается.
чевица.
Щур. — Становится ре- же. Снижается.
Клёст-еловик. — Стабильно. Стабильна.
Зяблик. Расширяются па север п восток. Утолщается, ста- новится плот- нее. Увеличивается.
Юрок. — То же. »
Домовой воробей. — » »
Полевой воробей. — » »
Обыкновенная ов- сянка. Стабильны пли расширяются на восток. » »
Белошапочная ов- сянка. Расширяются на север. » »
Дубровник. То же. » »
Седоголовая ов- сянка. Стабильны или расширяются на север и запад. » »
Овсянка-ремез. Расширяются на юг. » »
Желтобровая ов- сянка. Стабильны или расширяются на запад. » »
Полевой жаворо- нок. Расширяются на север. » »
Белая трясогуз- — » »
ка.
Горная трясо- — » »
гузка.
Желтоголовая тря- —- » »
согузка. »?
Желтая трясо- — »?
1уоЛа. Степной конек. Расширяются на север. Становится плот- нее, утол- щается. Увеличивается.
345
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Лесной конек. Стабильны или Становится плот- У ве личивается.
Пятнистый конек. расширяются на восток. Сокращаются к нее, утол- щается. Становится ре- Снижается.
Обыкновенная пи- северу Стабильны или же. То же. »
щуха. Поползень. сокращаются к югу. » »
Большая синица. Расширяются на Утолщается. Увеличивается.
Московка. север. Стабильны или Трансформиру- Снижается.
Пухляк. расширяются на север. ется. То же. »
Длиннохвостая Стабильны или Становится плот- Увеличивается.
синица. расширяются на нее, утол-
Жулан. север. щается. То же. »
Свиристель. Расширяются на » »
Серая мухоловка. юг. Стабильны или ? ?
Мухоловка - касат- расширяются на север. То же. Становится ре- Снижается.
ка. Ширококлювая » же. Утолщается. Увеличивается.
мухоловка.
Малая мухоловка. » Становится плот- »
Мухоловка-муги- Сокращаются к нее, утол- щается. Становится ре- Снижается.
маки. югу. же.
Пеночка-кузне- — Утолщается. Увеличивается.
чик.
Зеленая пеночка. Стабильны или » »
Пеночка-зарнич- расширяются на север. Становится плот- »
ка. Корольковая пе- Сокращаются к нее, утол- щается. Становится ре- Снижается.
ночка. югу. же.
Бурая пеночка. ? Утолщается. ?
Пятнистый свер- Стабильны или » Увеличивается.
чок. Певчий сверчок. расширяются на север. » ?
Таежный сверчок. » ?
346
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Славка-завируш- — Утолщается. У величивается.
ка.
Пестрый дрозд. Сокращаются к югу. Становится ре- же. Снижается.
Рябинник. — Стабильно. Стабильна или снижается.
Певчий дрозд. — Утолщается. Увеличивается.
Белобровик. Расширяются на юг. » »
Дрозд Науманна. То же. »
Темнозобый Дрозд. — Становится плот- нее, утол- щается. »
Бледный дрозд. — Становится ре- же. Снижается.
Каменка. — Утолщается. Увеличивается.
Черноголовый че- — » »
кан.
Синехвостка. — Трансформиру- ется. Снижается.
Обыкновенная го- — Утолщается. Увеличивается.
рихвостка. Сокращаются?
Сибирская гори- хвостка. Делается четко- видным. Снижается.
Со л овей-к расно- — Утолщается. Увеличивается.
шейка.
Синий соловей. ? Делается четко- видным. Снижается.
Варакушка. — Утолщается. У величивается.
Касатка. — Утолщается. Увеличивается.
Воронок. — » »
Береговая ла- — Стабильно. »
сточка.
Крот. Сокращаются к югу. Делается четко- видным. Снижается.
Крошечная буро- зубка. — Трансформиру- ется. ?
Малая бурозубка. —* Становится плот- нее, утол- щается. У величи вается.
Средняя бурозуб- ка. — Становится ре- же. Снижается.
Обыкновенная бу- — То же. »
розубка.
Кутора. ? Стабильно.
Ночница Икон- никова. ? Становится ре- же. Снижается.
Соболь. — Становится плот- нее, утол- щается. У величивается.
347
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Колонок. — Становится плот- нее, утолщает- ся. Увеличивается.
Горностай. — То же. »
Ласка. — » »
Барсук. — Утолщается. ?
Бурый медведь. — Стабильно. Снижается.
Волк. — Становится ре- же. »
Лисица. — Утолщается. Увеличивается.
Рысь. — ? Снижается?
Кабан. Сокращаются к югу. Делается четко- видным. »
Кабарга. — Становится ре- же. »
Марал. Стабильны. То же. Снижается.
Косуля. Расширяются на север. Утолщается. Увеличивается.
Лось. — Становится ре- же. Снижается.
Северный олень. Стабильны. То же. »
Летяга. — Трансформиру- ется. Увеличивается.
Белка. — Становится ре- же. Снижается.
Бурундук. — Трансформиру- ется. Стабильна?
Лесная мышовка. Расширяются на север. Утолщается. Увеличивается.
Крыса амбарная. То же. » »
Домовая мышь. — » »
Полевая мышь. Расширяются на север. » »
Азиатская лесная То же. » »
мышь.
Мышь-малютка. » » »
Лесной лемминг. — Трансформиру- ется. Стабильна или увеличивается.
Рыжая лесная по- левка. Расширяются на восток. Утолщается. Увеличивается.
Красная лесная — Стабильно. Снижается.
полевка.
Красно-серая лес- ная полевка. — Становится плот- нее, утол- щается. Увеличивается.
Водяная крыса. — Утолщается. »
Обыкновенная по- левка. Расширяются на север. ? »
Полевка Макси- мовича. Стабильны или расширяются на запад. ?
348
Таблица 25 (продолжение)
Виды Характер изменения
границ ареала кружева ареала общей численности вида
Темная полевка. — Становится плот- нее, утол- щается. У величивается?
П о л е вк а-экономк а. — » »
Узкочерепная по- левка. Расширяются. Становится плот- нее, утолщает- ся. Увеличивается.
Северная пищуха. — Становится ре- же. Снижается.
Заяц-беляк. — Становится плот- нее, утол- щается. Увеличивается.
Примечание. Прочерк (—) означает либо отсутствие данных, либо вызван тем
обстоятельством, что граница видового ареала проходит вне полосы южной тайги Сред-
ней Сибири.
Это главным образом опушечные (лесостепные) виды. Их слишком
много, чтобы перечислять здесь. Факты подобного рода указаны
рядом авторов (Кумари, 1957; Сыроечковский, 1960, и др.). Дру-
гая группа животных — выходцев с севера (из северной тайги и
даже тундры) — проникла в пределы южной тайги. Таких живот-
ных меньше. В южной тайге они заселили холодные заболоченные
березняки речных долин, образовавшиеся на месте приречных и
приручьевых ельников (чечетка, овсянка-ремез, свиристель,
дрозд-белобровик, очевидно дрозд Науманна, во всяком случае
темный), и сырые луга, также освобожденные из-под долинных
ельников (ряд куликов, прежде всего турухтан). Однако наруше-
ние лесов не всегда приводит к позитивным результатам: сведение
тех же приречных ельников постепенно оттесняет на юг крота и
мухоловку-мугимаки, исчезновение темнохвойных лесов у се-
верного их предела (Реймерс и Малышев, 1963) привело к отступ-
лению на юг ряда видов, входящих в зооформацию темнохвойных
лесов. Ныне они как раритеты сохранились по островным участ-
кам елово-пихтовой тайги.
Та же трансформация лесной растительности вместе с истори-
ческим процессом распространения видов и изменением климати-
ческих условий привела к расширению ареалов животных к во-
стоку. В эту сторону продвигаются скворец, зяблик, обыкновен-
ная овсянка, сорока, лесной конек и, очевидно, рыжая лесная по-
левка. Одновременно наблюдается обратный процесс — сдвиг
ареалов восточных форм на запад. Это перемещение менее заметно
в южной тайге Средней Сибири. Возможно, в него включились
349
хохлатый осоед, седоголовая овсянка, желтобровая овсянка и по-
левка Максимовича.1
Общее нарушение лесов усилило климатические влияния на
животный мир — микроклимат темнохвойной тайги резко отли-
чается от микроклимата светлохвойного или мелколиственного
леса (Китредж, 1951; Молчанов, 1960, 1961). Светлохвойные
(и особенно листопадные) леса обладают меньшей климатической
сглаживающей способностью, чем темнохвойная тайга, но они
теплее в летний период. Это, с одной стороны, летом утепляет
тайгу, что важно для перелетных форм, а с другой — охлаждает ее
зимой, что весьма существенно для оседлых видов и вынуждает их
к откочевкам на юг. Темнохвойный лес имеет более выровненный
ход температур, равномерный ход влажности и т. д. Для ряда
видов эти факторы могут играть важную роль (к сожалению, кли-
матические потребности животных слабо изучены).
Резко трансформируются в ходе лесных смен также кормовые
и защитные условия биотопов. Все эти изменения являются при-
чиной «приведения в соответствие» границ климата и жизни.
Происходит напряжение процесса эволюции.1 2
Вместе с тем тот же самый комплекс факторов переформиро-
вывает кружево видовых ареалов. Одни виды (их — длинный ряд)
заселяют большие площади внутри ареалов, чем раньше, другие,
наоборот, оказываются распространенными уже (таких видов
также много).
Любое изменение кружева ареала, истончение его или расши-
рение, приводит к неравномерности распределения вида: в одном
месте численность особей уменьшается, в другом — увеличи-
вается. Флуктуации могут быть временными в результате разового
нарушения леса пожаром или вредителями, например при отко-
чевке зверей и птиц из очагов массового размножения сибирского
шелкопряда в окрестные леса (Реймерс и Рожков, 1960), а могут
носить и перманентный характер (скопление животных в остров-
ных участках темнохвойных лесов). В обоих случаях возрастает
напряжение внутривидовых и межвидовых отношений, что ска-
зывается на благополучии отдельных форм. Происходит в какой-
то степени убыстрение процесса естественного отбора.
Мозаичность, прерывистость распространения животных со-
здает условия для образования полуизолированных популяций,
которые служат материалом для последующей селекции в ходе
естественного отбора, поскольку безусловно не однородны гене-
тически.
1 В сравнительно недавнее историческое время многократно происходило
пересечение территории южной тайги видами якутских эндемиков и выход-
цами из зооформации сосновых и мелколиственных лесов. На этом вопросе
мы не останавливаемся.
2 Мысль об изменении хода естественного отбора иод влиянием наруше-
ния лесов была высказана еще в 1914 г. Г. Ф. Морозовым.
350
Нарушение лесов через изменение кормовых условий влияет
на спектр пищевых объектов вида. Такой факт мы привели в раз-
делах о красной полевке и обыкновенной бурозубке. Это еще
один аспект изменений внутри вида, фактор, разбивающий моно-
литную форму на отдельные популяции.
Резкое изменение защитных условий заставляет животных при-
спосабливаться к новым факторам среды. Мы отмечали (Реймерс
и Воронов, 1963), что красная лесная полевка и землеройки (да и
другие виды) по-разному проводят зиму в условиях сплошных ле-
сов и лесополья. В лесах нет стогов, где бы могли концентриро-
ваться мелкие млекопитающие, зато там — сплошной покров
мхов, приподнятой лесной подстилки. Исчезновение лесов делает
животных синантропами, в корне изменяет условия их зимовки.
Некоторые птицы с трансформацией тайги приспосабливаются
к новым условиям, осваивают новые стации, строят гнезда в не-
обычных для вида местах. Эти факты также свидетельствуют о на-
пряжении естественного отбора в ходе антропогенного нарушения
лесов.
Наконец, изменение облика лесов меняет экологические ус-
ловия жизни паразитов млекопитающих и птиц. Мы в предыдущей
главе неоднократно повторяли, что на водоразделах животные за-
ражены иксодидами в меньшей степени, чем в долинно-придолин-
ной полосе, и оттеняли разницу в пораженности млекопитающих
из различных биотопов. Поскольку пестрота нынешних лесов
в значительной степени обязана деятельности человека, именно
антропогенные влияния приводят к разбивке видового населения
позвоночных животных на слабо и сильно зараженные группи-
ровки, что безусловно также ведет к каким-то эволюционным пре-
образованиям. Таковы в общих чертах теоретические выводы,
вытекающие из рассмотрения экологических особенностей таеж-
ных форм позвоночных животных.
Трансформация лесов и животное население
Возвращаясь к только что рассмотренной табл. 25, следует
заметить, что в последнем столетии тайга Средней Сибири заметно
обогащается видами. Из упомянутых в таблице форм 97 увеличи-
ваются в числе особей и лишь 52 уменьшаются по численности
или сохраняют ее на одном уровне. Кружево ареалов 100 видов
расширяется или уплотняется и лишь у 38 форм сужается (у ос-
тальных 22 видов оно остается стабильным или переформировы-
вается). Увеличение числа видов, с одной стороны, свидетельст-
вует о большем разнообразии среды по сравнению с прошлым,
с другой — указывает на какие-то динамические процессы в при-
роде. Однако упомянутые статистические показатели не отражают
формационной приуроченности наблюдаемых перемен и не рас-
крывают общих количественных изменений в животном мире
351
тайги, хотя и дают материал для некоторых выводов. Это прежде
всего — увеличение обилия долинных форм, связанных с кустар-
никами и лесопольем, и снижение количества многих водораздель-
ных лесных видов (в том числе замена лесных полевок серыми и
представителями мышеобразных), затем увеличение участия
лесостепных форм. В общей сложности, если нет лесов — нет и
их фауны. Это положение достаточно очевидно. Но вместе с тем
все перечисленное указывает на очень интересный процесс сниже-
ния числа горных (водораздельных) видов и повышения участия
здесь равнинных (долинных и лесостепных) форм. Водораздель-
ная полоса темнохвойной тайги на юге Средней Сибири постепенно
стирается. В основе этого явления лежат природные и антропоген-
ные факторы. Однако влияние человека доминирует. Он сводит
леса и тем резко убыстряет природный процесс. Естественный ход
сукцессионных смен темнохвойной тайги светлохвойными лесами
в случае продолжения общего потепления занял бы еще многие
столетия и, возможно, на современном этапе не был бы заметен
вообще. У нас нет уверенности, что фаза наступления лесов на
сибирские степи уже окончилась бы и сменилась на обратную,
не будь лесных пожаров. Для окончательного решения вопроса
нужно знать цифровые климатические показатели, необходимые
для таких смен, и числовое выражение климатообразующей спо-
собности лесов. Подобных данных пока нет, однако исходя из
косвенных наблюдений вопрос должен быть решен отрицательно.
Всюду, где влияние человека уменьшается, леса восстанавли-
ваются, а сосняки сменяются темнохвойной тайгой (на^Лене, на
средней и нижней Ангаре, в Присаянье). Заселение лесом пустырей
происходит повсеместно. Это — непреложный факт.
Наибольший интерес представляют не общие изменения жи-
вотного мира, а анализ трансформации животного населения
внутри основных рядов зооформационных элементарных фауни-
стических комплексов. К разбору подобных закономерностей на
примере мелких млекопитающих мы уже обращались ранее (Рей-
мерс и Воронов, 1963). Здесь мы затрагиваем вопрос несколько
шире.
Если рассмотрение качественных изменений в процессе лес-
ных сукцессий не вызывает особых трудностей, то количественные
характеристики дать очень нелегко. В последнее время проведены
большие работы по количественным учетам птиц в самых удален-
ных частях нашей страны (Дубинин, 1953; Дубинин и Торопанова,
1956; Рогачева, 1962; Кузякин и Второв, 1963; Шкатулова, 1963,
и др.). Эти исследования лишь начаты. В процессе сбора данных
станет возможным анализ количественных изменений в природе
на строго математической основе. Мы же вынуждены пока восполь-
зоваться методом балльных оценок. В ходе анализа общих изме-
нений в южной тайге Средней Сибири будет обращено внимание
на группу зооформаций темнохвойных лесов, зооформацию сос-
352
новых лесов и долинное сочетание зооформаций ельников, кустар-
ников и лугов. Практически рассмотрение этих трех рядов даст
возможность сделать вывод об изменениях животного населения
всей южной тайги исследуемого района.
Прежде всего обратимся к верности видов. Данные по этому
вопросу сведены в табл. 26—28. Нетрудно заметить, что коренные
природные образования или близкие к ним биотопы имеют боль-
ший набор форм высокой верности. Стенобионтные для данной
местности виды связаны чаще всего с экологическими вариантами
зооформации в динамических фазах, близких к устойчивому со-
стоянию, поддерживаемому природно-антропогенным режимом.
Вместе с тем промежуточная сукцессионная фаза смешанных
лесов во всех случаях богаче видами (для зимы это обстоятельство
подчеркнуто нами ранее, — Реймерс, 1963а). В мелколиственных
лесах видов становится меньше, но несмотря на кажущуюся бли-
зость экологических условий (во всех случаях мелколиственный
березовый или осиновый лес, чаще всего молодняк) фауна гари
темнохвойной тайги не идентична (хотя и близка) комплексу
видов мелколиственного леса, замещающего сосновый бор. Даже
фаза безлесного пространства — гари, поросшей вейниками, —
различается по фауне, в зависимости от того, возникла ли она
на месте светлохвойной или темнохвойной тайги, а тем более
фаупистически отлична от луговой, имеющей древние корни (пер-
вичные луговины берегов рек).
На указанный факт следует обратить внимание. Гарь гари
рознь, не одинаковы по фауне и вторичные мелколиственные леса.
Эта разница может достигать двукратных размеров по числен-
ности видов и быть весьма существенной по их составу.
С изменением фаунистического состава не остается стабиль-
ным и животное население. Из предложенных шкал обилия жи-
вотных нам кажутся наиболее приемлемыми градации, основан-
ные на оценке биомассы.3 К этому выводу давно уже пришли ги-
дробиологи (Зернов, 1949; Зенкевич, 1951, и др.). И действительно,
парадоксально звучит утверждение, что лисица относится к вто-
ростепенным видам потому, что численность ее ниже, чем полевок.
Также неправомерно деление на многочисленные и малочислен-
ные виды, когда это относптся к землеройкам и медведю одновре-
менно. В табл. 29—31 мы постарались сгруппировать виды на
9 Необходимо отметить, что понятие биомассы также не идеально для
определения реальной значимости вида в природе. Интенсивность обмена на
единицу веса у землеройки-бурозубки, соболя и лося отнюдь не одинакова.
Это хорошо известно физиологам и экологам. Американские авторы при-
бегают поэтому к вычислению «потребляемой биомассы» — среднему коли-
честву пищи, необходимой виду (Salt, 1957). Такой показатель тоже усло-
вен, по сравнение его с общей биомассой (деление этих чисел) дает очень
важный коэффициент продуктивности сообщества. В настоящей работе мы
не прибегаем к вычислению продуктивности зоологических комплексов из-за
громоздкости таких подсчетов.
23 Н. Ф. Реймерс
353
Таблица 26
Верность видов (без случайных форм) в антропогенном ряду
нарушенных водораздельных темнохвойных лесов
Виды-индикаторы Верные виды Постоянные виды
1. Фаза Воробьиный сыч Серый снегирь Щур Корольковая пеночка Ночница Иконникова высокоствольного однорс Рябчик Малый перепелятник Трехпалый дятел Кедровка Мухоловка-касатка Пестрый дрозд Обыкновенная буро- зубка Соболь Кабарга Красная лесная по- левка эдного леса Глухарь Большая горлица Я стреб-тетеревятник Филин Глухая кукушка Ворон Кукша Лесной конек? Пятнистый конек? Обыкновенная пищуха Поползень Московка Пухляк Темнозобый дрозд Бледный дрозд Синехвостка Малая бурозубка Средняя бурозубка Бурый медведь Марал Летяга Белка Бурундук Лесной лемминг Северная пищуха
Птиц 4 Птиц 6 Птиц 14—16
Млекопитающих 1 Млекопитающих 4 Млекопитающих 9
Всего 5 Всего 10 Всего 23—25
Итого: 38—40 видов
2. Фаза хвойно-мелколиственного леса
Серый снегирь
Щур
Рябчик
Малый перепелятник
Воробьиный сыч
Кедровка
Мухоловка-касатка
Корольковая пеночка
Пестрый дрозд
Глухарь
Большая горлица
Вальдшнеп
Я стреб-тетеревятник
Филин
Глухая кукушка
Желна
Трехпалый дятел
Большой пестрый дятел
Ворон
354
Таблица 26 (продолжение)
В иды-индикаторы Верные виды Постоянные виды
Обыкновенная буро- зубка Соболь Кабарга Красная лесная полевка Кукша Лесной конек Обыкновенная пищуха Поползень Московка Пухляк Малая мухоловка Темнозобый дрозд Бледный дрозд Синехвостка Средняя бурозубка Бурый медведь Марал Летяга Белка Бурундук Красно-серая лесная по- левка Северная пищуха
Птиц 2 Птиц 7 Птиц 20
Млекопитающих 0 Млекопитающих 4 Млекопитающих 8
Всего 2 Всего 11 Всего 28
Итого: 41 вид
3. Фаза мелколиственного леса
Хохлатый осоед
Дрозд Науманна
Синий соловей
Лесной лемминг
Красно-серая лесная по-
левка
Лесной дупель
Глухая кукушка
Большой пестрый дятел
Сойка
Кукша
Пухляк
Жулан
Малая мухоловка
Пеночка-зарничка
Темнозобый дрозд
Бледный дрозд
Средняя бурозубка
Обыкновенная буро-
зубка
Лось
Красная лесная полевка
Темная полевка
23*
355
Таблица 26 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
Северная пищуха Заяц-беляк
Птиц 0 Птиц 3 Птиц 11
Млекопитающих 0 Млекопитающих 2 Млекопитающих 7
Всего 0 Всего 5 Всего 18
Итого: 23 вида 4. Фаза безлесного пространства
Красно-серая лесная по- левка Обыкновенная буро- зубка Азиатская лесная мышь Красная лесная полевка
Птиц 0 Птиц 0 Птиц 0
Млекопитающих 0 Млекопитающих 1 Млекопитающих 3
Всего 0 Всего 1 Всего 3
Итого: 4 вида
Таблица 27
Верность видов (без случайных форм) в антропогенном ряду
нарушенных водораздельных и придолинных сосновых
и лиственнично-сосновых боров
Виды-индикаторы Верные виды Постоянные виды
1. Фаза высокоствольного однородного леса
Желна Большой пестрый дятел Барсук Глухарь Черный коршун Сарыч Филин Обыкновенная кукушка Глухая кукушка Ворон Клёст-еловик Лесной конек Поползень Пухляк Бледный дрозд Крошечная бурозубка
356
Таблица 27 (продолжение)
В иды-индикаторы Верные виды Постоянные виды
Лисица Малая бурозубка Обыкновенная буро- зубка Колонок Бурый медведь Кабарга Косуля Лось Северный олень Бурундук Лесная мышовка Азиатская лесная мышь Мышь-малютка Лесной лемминг Красная лесная полевка Красно-серая лесная по- левка Темная полевка П о левка-экономка Заяц-беляк
Птиц 1 Птиц 1 Птиц 12
Млекопитающих 0 Млекопитающих 2 Млекопитающих 20
Всего 1 Всего 3 Всего 32
2» Фаза Желна Седой дятел Обыкновенная горихво- стка Итого: 36 видов хвойно-мелколиственного Тетерев Обыкновенный козодой Большой пестрый дятел Вертишейка Кукша Белокрылый клёст Юрок Лесной конек леса Глухарь Рябчик Большая горлица Я стреб-перепелятник Черный коршун Сарыч Филин Обыкновенная кукушка Глухая кукушка Трехпалый дятел Ворон Сойка Восточноевропейский снегирь Обыкновенная чечевица Клёст-еловик Белошапочная овсянка
357
Таблица 27 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
Барсук Волк Лисица Лось Косуля Азиатская лесная мышь Заяц-беляк Поползень Большая синица Московка Пухляк Мухоловка-касатка Ширококлювая мухо- ловка Малая мухоловка П еночка-кузнечик Пестрый дрозд Певчий дрозд Темнозобый дрозд Бледный дрозд Малая бурозубка Средняя бурозубка Обыкновенная бурозуб- ка Соболь Колонок Горностай Ласка Кабарга Летяга Белка Бурундук Мышь-малютка Красная лесная полевка Темная полевка Полевка-экономка
Птиц 3 Птиц 7 Птиц 28
Млекопитающих 0 Млекопитающих 8 Млекопитающих 16
Всего 3 Всего 15 Всего 44
3. < Перепел Итого: 62 вида 1>аза мелколиственного j Тетерев Обыкновенный козодой Обыкновенная чече- вица Белошапочная овсянка юса Рябчик Обыкновенная кукушка Большой пестрый дятел Сойка Кукша Лесной конек Поползень Большая синица
358
Таблица 27 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
Горностай Ласка Косуля Лось Летяга Азиатская лесная мышь Заяц-беляк Московка Пухляк Жулан Мухоловка-касатка Малая мухоловка Пеночка-зарничка Таежный сверчок? Темнозобый дрозд Бледный дрозд Соловей-красношейка Крошечная бурозубка Малая бурозубка Средняя бурозубка Обыкновенная бурозуб- ка Колонок Барсук Волк Лисица Бурундук Лесная мышовка Мышь-малютка Красная лесная полевка Красно-серая лесная по- левка Темная полевка П о левка-экономка
Птиц 1 Птиц 4 Птиц 17—18
Млекопитающих 0 Млекопитающих 7 Млекопитающих 15
Всего 1 Всего И Всего 32—33
Итого: 44—45 видов 4. Фаза безлесного пространства
Обыкновенная буро- зубка Азиатская лесная мышь Темная полевка П о левка-экономка
Птиц 0 Птиц 0 Птиц 0
Млекопитающих 0 Млекопитающих 0 Млекопитающих 4
Всего 0 Всего 0 Всего 4
Итого: 4 вида
359
Таблица 28 Верность видов (без случайных форм) в антропогенном ряду нарушенных долинных ельников
Виды-индикаторы Верные виды Постоянные виды
1. Фаза высокоствольного однородного леса
Скопа Мухоловка-мугимаки Сибирская горихвостка Крот Рябчик Большая горлица Я стреб-перепел ятник Кукша Кедровка Клёст-еловик Московка Синехвостка Обыкновенная буро- зубка Соболь Летяга Белка Красная лесная полевка Темная полевка Глухарь Сарыч Трехпалый дятел Ворон Поползень Пухляк Бледный дрозд Соловей-красношейка Малая бурозубка Средняя бурозубка Лось Северный олень? Бурундук Азиатская лесная мышь Водяная крыса? Полевка-экономка
Птиц 3 Птиц 8 Птиц 8
Млекопитающих 1 Млекопитающих 6 Млекопитающих 6—8
Всего 4 Всего И Всего 14—16
2. Фаза Скопа Ворона Мухоловка-мугимаки Дрозд-рябинник Сибирская горихвостка Итого: 32—34 вида хвойно-мелколиственного Рябчик Большая горлица Я стреб-перепе лятник Кукша Кедровка Клёст-еловик Московка Пухляк П еночка-кузнечик Синехвостка леса Глухарь Сарыч Филин Обыкновенная кукушка Желна Трехпалый дятел Большой пестрый дятел Восточноевропейский снегирь Обыкновенная чечевица Зяблик Юрок Поползень Мухоловка-касатка Малая мухоловка Темнозобый дрозд Бледный дрозд Соловей-красношейка
360
Таблица 28 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
Крот Обыкновенная бурозуб- ка Соболь Летяга Белка Бурундук Красная лесная полевка Темная полевка Заяц-беляк Крошечная бурозубка Малая бурозубка Средняя бурозубка Колонок Горностай Ласка Лось Косуля Лесная мышовка Азиатская лесная мышь Мышь-малютка Красно-серая лесная по- левка Водяная крыса Полевка-экономка
Птиц 5 Птиц 10 Птиц 17
Млекопитающих 1 Млекопитающих 8 Млекопитающих 14
Всего 6 Всего 18 Всего 31
Итого: 55 видов
3. Фаза мелколиственного леса
Длиннохвостая синица Лесной дупель Овсянка-ремез Большая синица Свиристель Пеночка-зарничка Темнозобый дрозд Дрозд Науманна Обыкновенная буро- зубка Колонок Горностай Ласка Косуля Рябчик Азиатский бекас Обыкновенная кукушка Сорока Сойка Кукша Чечетка? Длиннохвостый снегирь Московка Пухляк Жулан Ширококлювая мухо- ловка Таежный сверчок? Соловей-красношейка Крот (летом) Крошечная бурозубка Малая бурозубка Средняя бурозубка Кутора
361
Таблица 28 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
Мышь-малютка П о левка-экономка Лось Летяга Лесная мышовка Красная лесная полевка Красно-серая лесная по- левка Темная полевка Узкочерепная полевка
Птиц 1 Птиц 7 Птиц 12—14
Млекопитающих 0 Млекопитающих 7 Млекопитающих 12
Всего 1 Всего 14 Всего 24—26
Итого: 39—41 вид
За. Фаза кустарников
Полевой лунь Ушастая сова Болотная сова Малый пестрый дятел ‘Седоголовая овсянка Варакушка Дербник Сорока Длиннохвостый снегирь Дубровник Большая синица Ширококлювая мухо- ловка Славка-завирушка Пятнистый сверчок Соловей-красношейка Обыкновенная буро- зубка Колонок Горностай Ласка Мышь-малютка Полевка-экономка Узкочерепная полевка Обыкновенная кукушка Обыкновенная чечевица Обыкновенная овсянка Белошапочная овсянка Пухляк Жулан Пеночка-кузнечик Зеленая пеночка Певчий сверчок? Таежный сверчок Черноголовый чекан Крот (летом) Малая бурозубка Средняя бурозубка Кутора Лесная мышовка
Птиц 6 Птиц 9 Птиц 10—11
Млекопитающих 0 Млекопитающих 7 Млекопитающих 5
Всего 6 Всего 16 Всего 15—16
Итого: 37—38 видов
<362
Таблица 28 (продолжение)
Виды-индикаторы Верные виды Постоянные виды
4. Фаза безле Степной конек сного пространства (сухо Каменка Мышь-малютка Узкочерепная полевка щольные луга) Крот (летом) Средняя бурозубка Обыкновенная бурозуб- ка Лесная мышовка Азиатская лесная мышь Темная полевка Полевка-экономка
Птиц 1 Птиц 1 Птиц 0
Млекопитающих 0 Млекопитающих 2 Млекопитающих 7
Всего 1 Всего 3 Всего 7
Итого: 11 видов
основе учета их биомассы. Не для всех видов это удалось сде-
лать с достаточным обоснованием, однако в целом можно считать,
что биомасса доминантов в 2—3 раза выше, чем содоминантов,
а обычные виды дают массу еще в 2—3 раза ниже.
Рассматривая приведенные в табл. 29—31 списки, можно сде-
лать ряд выводов.
1. Смена темнохвойных лесов светлохвойными приводит к ук-
рупнению условных средних особей 4 в группах доминирования
и содоминирования, особенно среди млекопитающих (в эти группы
входят более крупные представители класса).
2. Процесс смен доминантов в ряду водораздельной темнохвой-
ной тайги и долинно-придолинных лесов различен: в ходе про-
цесса трансформации или, наоборот, нарушения темнохвойного
леса от древостоев к гари происходит уменьшение размера средней
особи в группах доминантов и содоминантов, трансформация
светлохвойных боров и долинных ельников приводит к укрупне-
нию средней особи массовых форм.
3. Как и следует ожидать, при смене высокоствольных лесов
мелколесьем становится больше форм, питающихся на земле
(дрозды, коньки).
4 Речь идет о среднем размере одной «поселенной единицы» животного
населения независимо от видовой принадлежности особей. Было бы очень
интересно сравнить размеры животных из разных биотопов и внутри систе-
матических групп. Но такую задачу в данной работе мы не ставили.
363
Таблица 29
Относительное доминирование (с учетом биомассы) видов
в антропогенном ряду нарушенных водораздельных темнохвойных
лесов (балльная оценка)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
1. Фаза высокоствольного однородного леса
Рябчик Глухарь Малый перепелятник
Кедровка Большая горлица Я стреб-тетеревятник
Пухляк Трехпалый дятел Филин
Пестрый дрозд Кукша Воробьиный сыч
Бледный дрозд Поползень Московка Темнозобый дрозд Глухая кукушка Ворон Серый снегирь Щур Лесной конек Пятнистый конек Обыкновенная пищуха Мухоловка-касатка Корольковая пеночка Синехвостка
Обыкновенная буро- Средняя бурозубка Малая бурозубка
зубка Летяга Соболь
Красная лесная полевка Бурундук Белка Северная пищуха Бурый медведь Марал Кабарга Лесной лемминг
2. Фаза хвойно-мелколиственного леса
Рябчик Большая горлица Глухарь
Кедровка Большой пестрый дятел Вальдшнеп
Пестрый дрозд Кукша Поползень Пухляк Бледный дрозд Темнозобый дрозд Малый перепелятник Я стреб-тетеревятник Филин Воробьиный сыч Глухая кукушка Желна Трехпалый дятел Ворон Серый снегирь Щур Лесной конек Обыкновенная пищуха Московка Мухоловка-касатка Малая мухоловка Корольковая пеночка Синехвостка
Обыкновенная буро- Средняя бурозубка Малая бурозубка
зубка Красная лесная полевка Летяга Соболь
364
Таблица 29 {продолжение)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
Бурундук 3. Жулан Обыкновенная буро- зубка Белка Фаза мелколиственного л Пухляк Темнозобый дрозд Бледный дрозд Средняя бурозубка Красная лесная полевка Бурый медведь Марал Кабарга Лесной лемминг Северная пищуха еса Лесной дупель Хохлатый осоед Глухая кукушка Большой пестрый дятел Сойка Кукша Малая мухоловка П еночка-зарничка Дрозд Науманна Синий соловей Лось Лесной лемминг Красно-серая лесная по- левка Темная полевка Северная пищуха Заяц-беляк
4. Фаза безлесного пространства
Красная лесная полевка Т я п п п я ЯЛ Обыкновенная бурозуб- ка Азиатская лесная мышь Красно-серая лесная по- левка
Относительное доминирование (с учетом биомассы) видов в антропогенном ряду нарушенных водораздельных и придолинных сосновых и лиственнично-сосновых боров (балльная оценка)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
1. Фаза в Большой пестрый дятел ысокоствольного однород! Глухарь Желна Клёст-еловик Пухляк Поползень ного леса Черный коршун Сарыч Филип Обыкновенная кукушка Глухая кукушка Ворон
365
Таблица 30 (продолжение)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
Обыкновенная буро- зубка Средняя бурозубка Бурый медведь Лось Северный олень Красная лесная полевка Лесной конек Бледный дрозд Крошечная бурозубка Малая бурозубка Колонок Барсук Лисица Кабарга Косуля Бурундук Лесная мышовка Азиатская лесная мышь Мышь-малютка Лесной лемминг Красно-серая лесная по- левка Темная полевка П о левка-экономка Заяц-беляк
2. Фаза хвойно-мелколиственного леса
Тетерев Глухарь Желна Седой дятел Большой пестрый дятел Большая горлица Кукша Юрок Поползень Пухляк Темнозобый дрозд Я стреб-перепел ятник Черный коршун Сарыч Филин Обыкновенная кукушка Глухая кукушка Обыкновенный козодой Трехпалый дятел Вертишейка Ворон Сойка Клёст-еловик Белокрылый клёст Восточноевропейский снегирь Обыкновенная чечевица Белошапочная овсянка Лесной конек Большая синица Московка М у хо л овка-касатка Ширококлювая мухо- ловка Малая мухоловка Пеночка-кузнечик Пестрый дрозд Певчий дрозд Бледный дрозд Обыкновенная гори- хвостка
366
Таблица 30 (продолжение}
Доминанты (балл 3) Содомин анты (балл 2) Обычные виды (балл 1)
Средняя бурозубка Обыкновенная буро- зубка Лось Косуля Азиатская лесная мышь Красная лесная полевка Темная полевка Лисица Летяга Белка Бурундук Полевка-экономка Заяц-беляк Малая бурозубка Соболь Колонок Горностай Ласка Волк Кабарга Мышь-малютка Красно-серая лесная по- левка
3. Фаза мелколиственного леса
Тетерев
Кукша
Белошапочная овсянка
Обыкновенная кукушка
Сойка
Обыкновенная чечевица
Лесной конек
Жулан
Темнозобый дрозд
Бледный дрозд
Средняя бурозубка
Обыкновенная бурозуб-
ка
Косуля
Азиатская лесная мышь
Темная полевка
Заяц-беляк
Колонок
Горностай
Ласка
Лось
Летяга
Красная лесная полевка
Красно-серая лесная по-
левка
Рябчик
Перепел
Обыкновенный козодой
Большой пестрый дятел
Поползень
Большая синица
Московка
Пухляк
Мухоловка-касатка
Малая мухоловка
Пеночка-зарничка
Таежный сверчок?
Соловей-красношейка
Крошечная бурозубка
Малая бурозубка
Барсук
Волк
Лисица
Бурундук
Лесная мышовка
Мышь-малютка
Полевка-экономка
4. Фаза безлесного пространства
Обыкновенная бурозуб-
ка
Азиатская лесная мышь
Темная полевка
П о левка-экономка
367
Т а б л и ц а 31
Относительное доминирование (с учетом биомассы) видов
в антропогенном ряду нарушенных долинных ельников
(балльная оценка)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
1. Фаза высокоствольного однородного леса
Рябчик Глухарь Я стреб-перепе л ятник
Кедровка Большая горлица Кукша Клёст-еловик Московка Пухляк Поползень Мухоловка-мугимаки Сарыч Скопа Трехпалый дятел Ворон Соловей-красношейка Синехвостка Сибирская горихвостка Бледный дрозд
Крот? Средняя бурозубка Малая бурозубка
Обыкновенная бурозуб- Летяга Соболь
ка Белка Лось
Темная полевка Бурундук Красная лесная полевка Полевка-экономка Северный олень Азиатская лесная мышь Водяная крыса
2. Фаза хвойно-мелколиствеппого леса
Рябчик
Большая горлица
Ворона
Кедровка
Московка
Дрозд-рябинник
Крот
Обыкновенная бурозуб-
ка
Средняя бурозубка
Бурундук
Темная полевка
Глухарь
Обыкновенная кукушка
Большой пестрый дятел
Кукша
Клёст-еловик
Юрок
Пухляк
Поползень
Мухоловка-мугимаки
Малая мухоловка
Бледный дрозд
Сибирская горихвостка
Колонок
Горностай
Ласка
Косуля
Лось
Летяга
Белка
Азиатская лесная мышь
Красная лесная полевка
Заяц-беляк
Ястреб-перепелятник
Сарыч
Скопа
Филин
Желна
Трехпалый дятел
Восточноевропейский
снегирь
Обыкновенная чечевица
Зяблик
Мухоловка-касатка
Пеночка-кузнечик
Темнозобый дрозд
Соловей-красношейка
Синехвостка
Крошечная бурозубка
Малая бурозубка
Кутора
Лесная мышовка
Мышь-малютка
Красно-серая леспая по-
левка
Водяная крыса
Полевка-экономка
368
Таблица 31 (продолжение)
Доминанты (балл 3) Содом ин анты (балл 2) Обычные виды (балл 1)
3. Фаза мелколиственного леса
Лесной дупель Кукша Рябчик
Сорока Длиннохвостый снегирь Азиатский бекас
Жулан Московка Обыкновен па я кукушка
Темнозобый дрозд Пухляк Ширококлювая мухо- ловка Соловей-красношейка Дрозд Науманна Сойка Чечетка Овсянка-ремез Большая синица Длиннохвостая синица Свиристель Пеночка-зарничка Таежный сверчок
Крот Средняя бурозубка Крошечная бурозубка
Обыкновенная бурозуб- Колонок Малая бурозубка
ка Горностай Кутора
Косуля Ласка Лесная мышовка
Лось Летя га Лесной лемминг
Полевка-экономка Мышь-малютка Азиатская лесная мышь Темная полевка Красная лесная полевка Красно-серая лесная по- левка Узкочерепная полевка
За. Фаза кустарников
Сорока
Длиннохвостый снегирь
Дубровник
Седоголовая овсянка
Ширококлювая мухо-
ловка
Соловей-красношейка
Варакушка
Обыкновенная буро-
зубка
П олевка-экопомка
Волевой лунь
Ушастая сова
Болотная сова
Обыкновенная кукушка
Малый пестрый дятел
Белошапочная овсянка
Большая синица
Крот
Колонок
Горностай
Ласка
Мышь-малютка
Узкочерепная полевка
Дербник
Обыкновенная чечевица
Обыкновенная овсянка
Пухляк
Жулан
Пеночка-кузнечик
Зеленая пеночка
Славка-завирушка
Пятнистый сверчок
Певчий сверчок?
Таежный сверчок
Черноголовый чекан
Малая бурозубка
Средняя бурозубка
Ку тора
Лесная мышовка
Красная лесная полевка
Красно-серая лесная по-
левка
*>4 и. Ф. Реймерс
369
Таблица 3! (продолжение)
Доминанты (балл 3) Содоминанты (балл 2) Обычные виды (балл 1)
4. Фаза безлесного пространства (суходольные луга)
Степной конек Узкочерепная полевка Каменка Обыкновенная бурозуб- ка Крот Средняя бурозубка Кутора Лесная мышовка Азиатская лесная мышь Мышь-малютка Темная полевка Полевка-экономка
4. От однородного леса к гари происходит постепенное увели-
чение в составе мышевидных млекопитающих-доминаптов доли
серых полевок и мышеобразных и, наоборот, уменьшение участия
лесных полевок.
5. Как уже было отмечено нами ранее (Реймерс и Воронов,
1963), средний размер мышевидного млекопитающего от леса
к безлесному пространству увеличивается — возрастают размеры
отдельных «корпускул» биомассы этих форм.
6. В соответствии с изменениями в экологической среде (в том
числе среди кормовых объектов) меняется состав массовых хищ-
ных животных (доминантами по биомассе эти формы быть не мо-
гут — биомасса жертвы всегда должна превышать во много раз
биомассу потребителя).
Переход к расчетам абсолютной численности всегда сопряжен
с опасностью досадных ошибок. В табл. 32 сделана попытка таких
расчетов по методике, предложенной Л. П. Никифоровым (1963)
для ловчих линий, и на основе учетов канавками. В последнем
случае мы исходили из предположения, что индивидуальные
участки мышевидных млекопитающих одинаковы во всех биотопах
и что канавка отлавливает стационарно живущих зверьков. Оба
предположения безусловно неверны (Карасева и Ильенко, 1957).
Изменяется индивидуальный участок в течение лета (Н. П. Нау-
мов, 1951; Карасева и Ильенко, 1957, и др.), от года к году (те же
работы, Окулова и Дубровский, 1962, и др.), в зависимости от за-
раженности зверьков инфекциями (Карасева, 1963) и т. д. Всем
этим пришлось пренебречь и принять на основе литературных дан-
ных (перечисленные работы; Godfrey, 1954; Никитина, 1961;
Ся У-пин, 1961; Никитина и Меркова, 1962, 1963, и др.), а также
собственных визуальных наблюдений по «летней» пороше (до
мая и с конца августа), что средний «радиус разбега» землероек
и лесных полевок равен приблизительно 50, лесных мышей —
25 и серых полевок — 15 м. С учетом пятидосятпметровой длины
370
Т а б л ii ц а
Приведение данных относительной численности
мелких млекопитающих к абсолютным цифрам (на 1 км2)
Ряд трансформации
водораздельных темнохвойных лесов водораздельных сосновых лесов долинных ельни- ков
Виды 5 о §3 1 X X о о я з н I X я о о и s а
“ га S 3 га 5 Ч га о га И * X X г; га „ к £ га
2 X га <- sli > ь s га л X * Ч га га о S
Й 5 Р.> X § ® а>»х azx
1. Фаза ВЫСОКОСТВОЛЬНОГО однородного леса
Крошечная бурозубка — — 70 — — —
Малая бурозубка . . . — .— 210 — 140 10
Средняя бурозубка . . — 400 1190 40 1050 100
Обыкновенная бурозубка — 1900 1680 170 4290 490
Кутора — — — — — —
Всего мелких насскомо-
ядных — 2300 3150 210 5480 600
Лесная мышовка . . . — — 140 — — 10
Азиатская лесная мышь — — 140 90 490 —
Мышь-малютка .... — — 140 — — —
Всего мышеобразных и
лесных мышовок — — 320 90 490 10
Лесной лемминг .... — 20 140 — —. —
Красная лесная но ловка Красно-серая лесная по- — 1160 910 70 110 1750 280
левка — 20 — — —-
Всего лесных полевок
и леммингов — 1200 1120 ПО 1750 280
Темная полевка .... — — 400 10 3000 10
Полевка-экономка . . . — — 200 .— 1000 —
Узкочерепная полевка — —- — — — —
Всего серых полевок . . — — 600 10 4000 10
Всего мышевидных мле-
копитающих Нет 3500 5190 420 10720 900
данных.
24*
371
Таблица 32 (продолжение}
Ряд трансформации
водо р аз дел ь иы х темнохвойных лесов водораздельных сосновых лесов долинных ельни- ков
Виды данные лов- чих канавок результаты учета плаш- ками данные лов- чих канавок результаты учета плаш- ками данные лов- чих канавок результаты учета плаш- ками
2. Фаза хвойно-мелколиственного леса
Крошечная бурозубка Малая бурозубка . . . Средняя бурозубка . . Обыкновенная бурозуб- ка Кутора 70 1050 4900 40 600 1560 140 1400 6300 500 4000 140 350 2940 9800 60 220 1550 5850
Всего мелких насекомо- ядных 6020 3200 7840 4500 13230 7680
Лесная мышовка . . . — — — — — —
Азиатская лесная мышь 140 — 700 200 980 400
Мышь-малютка .... — — 140 — 140 —
Всего мышеобразных и лесных мышовок . . . 140 — 840 200 1120 400
Лесной лемминг .... — — — — 70
Красная лесная полевка 4200 1600 1890 780 840 800
Красно-серая лесная полевка 280 __ 140 100 70 —
Всего лесных полевок и леммингов 4480 1600 2030 880 980 800
Темная полевка .... 200 — 1000 40 4000 160
Полевка-экономка . . . — — 200 20 400 560
Узкочерепная полевка — — — — — —
Всего серых полевок . . 200 — 1200 60 4400 720
Всего мышевидных мле- копитающих 10840 4800 11910 5640 19730 9600
372
Таблица 32 (продолжение}
Виды Ряд трансформации
во дор а здел ьных темнохвойных лесов водораздельных сосновых лесов долинных ельни- ков
данные лов- чих канавок результаты учета плаш- ками данные лов- чих канавок результаты учета плаш- ками данные лов- чих канавок результаты учета плаш- ками
3. Фаза мелколиственного леса
(для долинных ельников — фаза кустарниковых зарослей)
Крошечная бурозубка Малая бурозубка . . . Средняя бурозубка . . Обыкновенная бурозубка Кутора 140 70 770 3500 20 20 60 540 70 140 1050 3290 50 450 140 700 2100 70 10 80 380 10
Всего мелких насекомо-
ядных 4480 640 5440 500 ЗОЮ 480
Лесная мышовка . . , — — 140 — 420 160
Азиатская лесная мышь 140 — 980 140 — —
Мышь малютка — — 140 — 840 —
Всего мышеобразных и
лесных мышовок . . . 140 — 1260 140 1260 160
Лесной лемминг .... 400 160 —
Красная лесная полевка Красно-серая лесная по- 1750 260 350 НО 70 —
левка 350 70 210 — 70 —
Всего лесных полевок и
леммингов 2500 490 560 110 140 —
Темная полевка .... 400 800 100
Полевка-экономка . . . — 40 — 30 4000 —
Узкочерепная полевка . — — — — 400 —
Всего серых полевок . . 400 40 800 130 4400 —
Всего мышевидных мле-
копитающих 7520 1170 8060 880 8670 640
373
Таблица 32 (продолжение)
Ряд трансформации
водораздельных темнохвойных лесов водораздельных сосновых лесов долинных ельни- ков
Виды д § § S за я п о g 3 и ё g я тьтаты i плаш-
о Й св Ч & Е Л R Я 0> § л с 2 §
а к я и я к « § R се я я я g w g> S
5 § W tr « я
4. Фаза безлесного пространства
(на месте долинного ельника — суходольный луг)
Крошечная и малая бу-
розубки <— — — — — —
Средняя бурозубка . . — — — — 280 20
Обыкновенная бурозубка — 40 — 40 1400 180
Кутора — — — — 70 —
Всего мелких насекомо-
ядных — 40 — 40 1750 200
Лесная мышовка . . . — — 140 10
Азиатская лесная мышь — 40 — 180 140 —
Мышь-малютка .... — — — — 280 —
Всего мышеобразных и 180 560
лесных мышовок . . . — 40 — 10
Лесной лемминг .... —
Красная лесная полевка Красно-серая лесная по- — 40 — — — —
левка — 80 — — — —
Всего лесных полевок и
леммингов — 120 — — — —
Темная полевка .... 10 200
Полевка-экономка . . . — — — 10 400 —
Узкочерепная полевка . — — — — 1400 190
Всего серых полевок . . — — — 20 2000 190
Всего мышевидных мле-
копитающих Нет 200 Нет 240 4310 400
данных. данных.
374
самой канавки она облавливает (мы условно принимаем, что за
два месяца в канавку попадают все оседлые зверьки) землероек
и лесных полевок с 1.5, мышей с 0.75 и серых полевок с 0.5 га,
иными словами, абсолютное число отловленных мышевидных
должно быть помножено для пересчета данных на 1 км2 соответ-
ственно на 70, 140 и 200.
Как показывает анализ данных табл. 32, коэффициент Л. П. Ни-
кифорова (1963) при применявшейся нами нестандартной методике
отловов плашками (они работали не 1—2, а 5—10 дней) дает за-
ниженные результаты. Однако они почти всегда коррелируются
с вычисленными другим способом — учетом канавками. Это дает
возможность сравнений. Числа, полученные с помощью учетов
канавками, по порядку величин, видимо, близки к истинным.
Все расчеты в табл. 32 сделаны на основе минимум двукрат-
ной повторности учетов, обычно 3—4-кратной. Часть данных от-
носительных учетов численности приведена нами выше и в преж-
ней нашей работе (Реймерс и Воронов, 1963).
Прежде чем приступить к обсуждению всех сведенных
в табл. 29—32 материалов, обратимся к подсчетам биомассы мыше-
видных зверьков (табл. 33) и еще одной серии цифр (табл. 34),
на этот раз относительной численности мышевидных млекопитаю-
щих по эколого-систематическим группам. В табл. 33 данные трех
отмеченных звездочкой граф рассчитаны из сравнения результа-
тов учетов плашками и канавками. Пересчет делался по прин-
ципу: во сколько раз больше попадает зверьков данного вида
в канавку. В среднем можно принять, что в июле и августе земле-
ройки и серые полевки ловились в канавки на гарях приблизи-
тельно в 10 раз чаще, мыши — в 5 раз. Исходя из этих показате-
лей, данные, полученные с помощью ловушек, приравнивались
к вычисленным для канавок. Такое же соотношение для землероек
и лесных полевок в темнохвойных лесах было приблизительно
1 : 2. Все примененные нами коэффициенты не имеют абсолютного
значения и могут изменяться в зависимости от года и сезона про-
ведения учетов. Биомасса мышевидных млекопитающих вычис-
лена исходя из тех же показателей среднего веса зверьков, кото-
рые мы принимали в предыдущей нашей работе (Реймерс и Воро-
нов, 1963).
Для числовых сравнений биомассы птиц по всем выделам или
по большей их части материалов у нас недостаточно, поэтому мы
в табл. 35 сравниваем лишь темнохвойно-мелколиственные и
светлохвойно-мелколиственные леса. В качестве общего положе-
ния отметим, что обычно чем выше видовое разнообразие, тем зна-
чительнее общая биомасса птиц. К этому вопросу мы уже обраща-
лись раньше (Реймерс, 1963а; Реймерс и Малышев, 1963), поэтому
воздержимся от многочисленных литературных ссылок.
Весь материал, сгруппированный в табл. 29—35, позволяет
прийти к следующим заключениям.
375
Т а б
Биомасса мышевидных млекопитающих (в кг на 1 км2) в рядах
S ю о о Ряд темнохвойных лесов
о д 6 я
о я р X «5 д
Виды и эколого-систематические группы Н 8 § Д Сч д С о S 5 g S г? | ё Е- S О о g « 8 к
’Я а § я 3 о я к я К О о Ф S я ф св
1 3 ко* X ф Ю S R °
аэ , сС О сС СО Сч о СО Q со 2 со о
О и вк Я •е § е в с
Крошечная бурозубка .... 2.2 0.3
Малая бурозубка 4.0 — 0.3 0.3 —
Средняя бурозубка 5.2 5.5 5.5 4.0 —
Обыкновенная бурозубка . . . 9.8 39.2 48.0 34.3 3.9
Кутора 14.9 — — — —
Всего мелких насекомоядных — 44.7 53.8 38.9 3.9
Лесная мышовка 7.7 — .— — —
Азиатская лесная мышь . . . 20.5 — 2.9 2.9 4.1
Мышь-малютка 5.9 — — — —
Всего мышеобразных и лесных
мышовок — — 2.9 2.9 4.1
Лесной лемминг 25.5 5.1 — 10.2 —
Красная лесная полевка . . . 18.6 86.3 78.1 32.5 7.4
Красно-серая лесная полевка 20.7 4.1 5.8 7.2 16.6
Всего лесных полевок и лем-
мингов — 95.5 83.9 49.9 24.0
Темная полевка 18.8 2.8 7.5
Полевка-экономка 23.3 — — — —
Узкочерепная полевка .... 28.9 — — — —
Всего серых полевок — — 2.8 7.5 —
Всего мышевидных млекопи-
тающих — 140.2 140.5 99.2 32.0
* Объяснения в тексте.
376
co to I r и* CD p
CD to *0 Cn CD b' CD bi 30
сл to £*
co | 00 p CO Cn 1 СЛ
bo Ьл bj do Ьп О
00 l-b.
p Сл | | P P j^p |
to о о do co cn do
to to
p° 1 p* 1 III p
CD to CO CD СЛ
1 to cn p t CO I P
p | co p P 1 to 1 P
co co b* bi °
to CO 00 to
p p | CD Сл 00 p- p p p
CO bi CO to ° do do oo
to 00
rfS | p Си P p * 1 P
О Cn CO to co **
СЛ £>•
p p ppp 1 III P
CD Co cn co do
с М р оо со 1 р р р р дл to bo io фаза высокостволь- ного однородного леса Ряд свстлохвойных лесов
ppi р do is сл 0.6 7.3 61.7 фаза хвойно-мелко- лиственного леса
о о р до о сп со 1 р р р р го сл до го фаза мелколиствен- ного леса
to 1 ь1 £ 1 £1II фаза безлесного пространства *
48.1 10.0 0.6 5.5 42.0 фаза высокостволь- ного однородного леса Ряд долинных ельников
op | р до о о 1 с© | р р р О до 4^ до фаза хвойно-мелко- лиственного леса
। to р | р р О ГО ^4 ГО 1 pppp 1 СО О СЛ оо фаза кустарников
15.7 1.1 2.9 1.6 1 фаза лугов
нарушения или восстановления лесов
Таблица 34
Результаты учетов мышевидных млекопитающих ловчими канавками
(на 10 канавко-суток в июле и августе)
Номер выдела Биотоп Число канавко- сСзонов Мелкие насеко- моядные Мышеобразные и лесные мышов- ки 1 Лесные полевки и лемминги Серые полевки Всего мыше- видных млеко- питающих
2 Водораздельный темно- хвойный лес с участием мелколиственных пород 3 17.0 0.1 12.3 0.4 29.8
3 Гарь на месте водораздель- ного темнохвойного леса, поросшая березовым мелколесьем 3 15.0 7.4 0.6 23.0
6 Гарь на месте приручьевого участка водораздельного темнохвойного леса, по- росшая березовым мел- колесьем 3 20.7 0.3 8.8 0.2 30.0
8 Сосновый бор (водораздель- ный) 1 5.4 . 3.8 0.6 9.8
19 Островной участок темно- хвойного леса с участием мелколиственных пород 3 18.8 0.6 5.2 2.8 27.4
20 Гарь островного участка темнохвойного леса, по- росшая березовым мел- колесьем 6 13.8 1.2 3.3 2.7 21.0
21 Сосновый бор (придолин- ный) 5 8.7 1.2 3.3 1.0 14.2
22 Сосново-мелколиственный лес (придолинный) . . . 3 16.3 1.0 4.8 1.0 23.1
23 Гарь придолинного сосно- вого бора 3 11.2 5.5 1.5 0.9 19.1
24 Остепненный сосновый бор (у реки) 3 9.7 2.1 4.0 1.6 17.4
25 Гарь остепненного сосно- вого бора, поросшая сос- ново-мелколиственным молодняком 2 7.7 2.6 2.1 2.2 14.6
28 Приречный ельник с уча- стием мелколиственных пород 4 21.4 1.7 4.8 4.5 32.4
31 Ивняки и черемушники до- лин: а) у лугов 2 8.4 1.6 0.5 4.1 14.6
б) у лесов 4 28.1 5.5 1.2 4.8 39.6
32 Луга 2 4.1 0.6 0.2 2.0 6.9
34 Приручьевой ельник с уча- стием мелколиственных пород 5 12.5 1.2 4.7 6.2 24.6
378
Таблица 34 (продолжение)
I Номер выдела Биотоп Число канавко- сезонов Мелкие насеко- моядные Мышеобразные и лесные мышов- ки Лесные полевки и лемминги Серые полевки Всего мыше- видных млеко- питающих
35 Гарь приручьевого ельни- ка, поросшая березовым мелколесьем 2 6.9 0.4 4.8 2.1 14.2
37 Ерники 2 12.9 1.6 2.9 5.3 22.7
38 Возделываемые поля . . . 1 9.4 10.7 0.8 6.6 27.6
41 Населенные пункты (при- усадебные огороды) . . 2 1.4 3.3 0.8 4.2 9.7
1. В ходе лесной сукцессии (трансформации) или, наоборот,
нарушений лесов 5 от сплошного леса через редколесье к безлес-
ному пространству сначала происходит увеличение числа особей
и биомассы позвоночных животных, затем снижение их до очень
низкого уровня (биомасса птиц на абсолютно безлесных гарях
равна нулю, млекопитающих — минимум в 5 раз ниже, чем в од-
нородном лесу).
2. Смена темнохвойных лесов светлохвойными резко снижает
биомассу животных, однако на фазе мелколиственно-хвойного
леса обе группы зооформаций обладают одинаковой биомассой.
Уничтожение приречных ельников резко снижает общую био-
массу животного мира речных долин.
3. Биомасса животных на фазе мелколиственного леса близка
во всех рядах трансформации.
4. При сравнении животного населения мышевидных зверьков
водораздельных темнохвойных лесов с таковым долинных ельни-
ков и светлохвойных лесов легко можно подметить ряд закономер-
ностей: а) показатель биомассы землероек всюду почти равен и зна-
чительно выше лишь на фазе березово-елового леса долин; б) пред-
ставители мышеобразных проникают в леса на фазах высокой па-
рушенности и дают наибольшую биомассу в ряду светлохвойных
лесов (особенно придолинных, на водоразделах мышей нередко
меньше); мыши с высокой плотностью населяют нарушенные до-
линные ельники; в) число красных полевок выше всего в водо-
раздельных темнохвойных лесах, их биомасса достигает абсолют-
ного максимума в однородных или смешанных насаждениях;
5 Эти процессы совершенно не идентичны, но сопровождаются аналогич-
ными изменениями в животном населении — в составе элементарных фауни-
стических комплексов.
379
Таблица 35
Биомасса птиц в темнохвойно-мелколиственных
и светлохвойно-мелколиственных лесах (май—июнь)
Зарегистрированные виды Учтено осо- бей на 5 км (лента 50 м) Количество особей на 1 км2 Средний вес одной особи в г (округ- ленно) БиОхМасса на 1 км2 в кг
Темнохвойно-мелколпственный лес
Рябчик 2 8 430 3.440
Большая горлица . . . 2 8 225 1.800
Большой пестрый дятел 2 8 95 0.760
Кедровка 6 24 165 4.020
Кукша 2 8 80 0.640
Серый снегирь 6 24 25 0.600
Лесной конек 3 12 20 0.240
Поползень 15 G0 15 0.900
Московка 3 12 10 0.120
Пухляк 12 48 15 0.720
Корольковая пеночка 3 12 5 0.060
Пестрый дрозд .... 7 28 140 3.920
Бледный дрозд .... 4 16 70 1.120
Темнозобый дрозд . . . 2 8 80 0.640
Синехвостка 6 24 15 0.360
Всего 73 — 19.340
Светлохвойно-мелколиственный лес
Тетерев 1 4 * 1100 4.400
Глухарь । ** —1 3000 —3.000
Большая горлица . . . 1 4 225 0.900
Желна 2 8 340 2.720
Седой дятел 4 16 120 1.920
Большой пестрый дятел 10 40 95 3.800
Вертишейка 1 4 35 0.140
Кукша 2 8 80 0.640
Обыкновенная чечевица 4 16 20 0.320
Поползень 9 36 15 0.540
Лесной конек 2 8 20 0.160
Большая синица .... 1 4 20 0.080
Московка 3 12 10 0.120
Пухляк 12 48 15 0.720
Темнозобый дрозд . . . 2 8 80 0.640
Обыкновенная гори-
хвостка 6 24 * 15 0.360
Всего 60 — — 20.460
* Учет произведен в придолинных лесах.
** Учетный маршрут 15 км.
380
с нарушением леса красные полевки составляют значительную
часть биомассы населения мышевидных млекопитающих лишь
там же — на водоразделах; г) биомасса серых полевок выше всего
Рис. 74. Размещение биомассы позвоночных животных на тер-
ритории южной тайги в настоящее время (принципиальная
схема).
Условные обозначения как на рис. 12 и 22. Оценка в баллах: 1 — макси-
мальная биомасса; 2 — средняя биомасса; з — минимальная биомасса.
в сосняках и особенно в долинных биотопах; д) таким образом,
компенсация биомассы от темнохвопного леса к светлохвойному
происходит на водоразделах за счет представителей мышеобраз-
ных и серых полевок; замена красных полевок мышами и серыми
381
Рис 75. Гипотетическая принципиальная схема размещения
биомассы позвоночных животных у.> зна^ри НарУШеНИИ
лесов южной таиги Средней Сиоири.
м^ьная^и^асм^^—средая^^иомасса; ^—Минимальная бйомТсса.
Рис. 76. Гипотетическая принципиальная схема будущего раз-
мещения биомассы позвоночных животных после дальней-
шего нарушения лесов вырубками и расчисткой под сельско-
хозяйственные угодья (при условии сохранения хаотичной
системы землепользования).
Условные обозначения как на рис. 12 и 22. Введены дополнительно:
8а — вырубка на месте водораздельного соснового леса; 9а — вырубка
на месте соснового леса с участием мелколиственных пород; 21а — вы-
рубка на месте придолинного соснового бора; 226 — вырубка на месте
придолинного сосново-мелколиственного леса. Оценка в баллах: 1 —
максимальная биомасса; 2 — средняя биомасса; 3 — минимальная био-
масса.
полевками характерна также для долин; е) те же группы живот-
ных заселяют возделываемые поля и населенные пункты.
К близким по смыслу заключениям пришли недавно В. М. Не-
ронов (1962) и Ф. Р. Штильмарк, Г. А. Соколов и др. (1963).6
Насколько нам известно, и более северные участки тайги (по срав-
нению с южной) также заметно нарушены пожарами. В связи
с этим кажется странным утверждение Э. В. Рогачевой и
Е. Е. Сыроечковского (1962), что воздействие человека на фауну
енисейской тайги через перестройку мест обитаний невелико.
Возвращаясь к гл. 4, напомним, что южная тайга за годы
ее резких нарушений изменила свой облик. Схемы на рис. 12 и
21 можно пополнить новым содержанием — размещением биомассы
различной плотности (рис. 74 и 75). Учитывая ход современных
нарушений лесов и их восстановления, можно представить гипо-
тетическую картину общей плотности населения позвоночных жи-
вотных в будущем — в ближайшие 50 лет (рис. 76). Совершенно
ясно, что территория южной тайги Средней Сибири сначала
(с первичным нарушением лесов) обогащалась видами и особями,
затем наступила фаза количественного обеднения животного на-
селения, которая продолжается до сих пор и завершится еще бо-
лее резким спадом биомассы, если не будут предприняты шаги
по окультуриванию и реконструкции природы.
Серия выводов, сделанных на основе разбора качественного
и количественного состава животного населения, а также заклю-
чений, выведенных из исходных аутоэкологических материалов,
имеет большой прикладной смысл, помогая разобраться в ряде
вопросов практической деятельности. На этом мы и остановимся
ниже.
Животные южной тайги Средней Сибири как кормовая база
ценных хищных зверей
Возвращаясь к данным гл. 4, напомним соотношение темно-
хвойных, светлохвойных и мелколиственных лесов: 22.5, 46.5
и 31.0%. Поскольку в доле светлохвойных лесов значительную
часть составляют однопородные сосняки, можно считать, что по-
ловина всех лесов южной тайги Средней Сибири относится к биото-
пам с низкой производительностью биомассы животных. С наруше-
нием тайги на колоссальных пространствах сменился видовой
спектр мышевидных зверьков: меньше стало лесных полевок,
больше лесных мышей и серых полевок.
0 Следует заметить, что в рядах трансформации лесов европейской части
СССР наблюдается несколько иная картина изменения численности грызунов.
Там нередко наибольшее число мышевидных констатируется на вырубках и
гарях (Керзина, 1956; Поспелов, 1957; Шилова, Троицкий и др., 1958, и др.).
Этот факт еще раз подтверждает разнокачественность процессов, идущих
в пределах различных природных регионов.
384
Происшедшее изменение весьма существенно. Нами уже про-
водился подсчет разницы относительной (Реймерс и Воронов,
1963) и абсолютной (см. выше) биомассы между светлохвойными
и темнохвойными лесами и их гарями. Обратимся к этому вопросу
еще раз. Относительная биомасса (т. е. биомасса количества
зверьков, попавших в канавку в течение июля—августа) водораз-
дельных темнохвойных лесов в среднем (М±2 т) ’378 + 51 г,
долинных ельников 427 + 86 г. Тот же показатель М для срав-
нительно богатых придолинных сосняков 208 + 57 г и для гарей
водораздельных темнохвойных лесов 283 + 54 г. Констатируе-
мые различия (первая их степень tv Плохинский, 1961) досто-
верны: td между водораздельными темнохвойными лесами и их
гарями равно приблизительно 3, а между темнохвойными ле-
сами и сосняками 4.5 (п см. табл. 34).
В свете этих данных можно заново рассмотреть проблему вза-
имоотношений соболя и белки. К. Д. Нумеров и П. Н. Павлов
(1963) делают вывод, что ныне полностью восстановлены запасы
соболя Енисейской Сибири. Это очень отрадный факт. Но не сле-
дует забывать, что кормовая база соболя сейчас совсем не та,
что в XVII в. Нет дремучих кедровников, о которых писал в на-
чале прошлого века В. Дмитриев (1818), значит нет и орехов.
При первых нарушениях лесов животная часть кормовой базы
соболя либо не пострадала, либо сократилась очень мало. Со сме-
ной же лесов гарями, а тайги — сосновыми борами произошел пе-
релом: к исчезновению основных растительных кормов прибави-
лось оскудение животных ресурсов.
В коренных местах обитания соболя биомасса мышевидных
зверьков обычно намного превышает 100 кг/км2, и наиболее бла-
гоприятны для зверя биотопы с биомассой лесных полевок в 80—
90 кг/км2. В светлохвойных лесах эти показатели несоизмеримо
ниже (см. табл. 33).
Общее увеличение количества соболя неминуемо должно
было привести к сокращению числа его конкурентов, особенно
при межвидовых отношениях типа хищник—жертва. Исходя из
этих теоретических предпосылок, следует признать, что размноже-
ние соболей должно приводить к сокращению численности белок.
Вместе с тем нужно думать, что высокая плотность соболиного
поголовья должна вызывать при существующих условиях и дру-
гие явления, связанные с перенаселением, прежде всего распро-
странение болезней. В связи с этим следует предостеречь работни-
ков охотничьего хозяйства и выработать рациональные нормы на-
селенности природных угодий соболями. Совершенно ясно, что
создалось угрожающее положение.
Простейшие подсчеты показывают, что общее снижение био-
массы мышевидных зверьков в Присаянье и на верхней Лене до-
стигает 30—35%, а в Приангарье превысило половину того ко-
личества, которое было характерно для района до нарушения ле-
1/425 Н. Ф. Реймерс
385
сов. Даже не принимая во внимание снижения семенной продук-
тивности лесов, следует думать, что в настоящее время нормально
может существовать лишь 50—70% от того поголовья соболей,
которое было до резкого нарушения лесов.
Совершенно иные соотношения сложились у группы долин-
ных хищников. Обилие их кормовых объектов постоянно растет.
Количество лисиц регулируется сейчас, очевидно, климатиче-
скими факторами и степенью преследования хищника людьми
и бродячими собаками. Благоприятна обстановка для горностая,
ласки и колонка. Эти зверьки должны увеличиваться в числе
и осваивать все новые и новые угодья. Вместе с тем наметившийся
процесс уничтожения приречных кустарников и распашка долин
снижают обилие мелких куньих, прежде всего в силу сокращения
площади их стаций.
Нужно обратить внимание еще на один существенный факт.
Несмотря на увеличение площадей угодий, внешне благоприятных
для долинных видов, животные с большим трудом проникают на
водоразделы. Очевидно, это явление связано у мелких грызунов
с климатическими факторами, а для хищников основную роль иг-
рает спектр охотничьих видов. Заселение водоразделов, лишен-
ных лесов, пушными видами оказывается чрезвычайно трудным,
даже невозможным мероприятием.
В заключение этого краткого раздела отметим, что, как по-
казывают данные расчетов (Соколов и Хлебников, 1962), смена
темнохвойных лесов вырубками и гарями приводит к резкому со-
кращению производительности охотничьих угодий (в 200 раз).
При этом в 100 раз сокращается выход пушной продукции в де-
нежном выражении.
Следует заметить, что и остальная продукция таких участков
минимальна. Они в Средней Сибири дадут новую древесину через
100—120 лет (Реймерс и Малышев, 1963). Заросли вейников и кип-
реев, характерные для безлесных мест в тайге, чрезвычайно мало-
ценны как корм для скота, да и технически выпас и сенокос на
вырубках и гарях невозможны из-за высокой захламленности
порубочными остатками или завалами сухой древесины.
Образование обширной «экономической пустыни» в пределах
южной тайги Средней Сибири и еще в большей степени на Дальнем
Востоке имеет глубокие природные корни. Безлесные гари не-
способны производить достаточно биомассы для прокормления
мелких животных, не говоря уже о ценных хищниках и копытных.
В пределах южной тайги Средней Сибири «экономическая пустыня»
занимает свыше 1.5 млн га. Другие 15 млн га представляют собой
«полупустыню» — малоценные леса с низкой продуктивностью
животной биомассы. На эти обстоятельства мы уже обращали
внимание (Реймерс и Малышев, 1963) и вновь подчеркиваем: без
рационального лесного хозяйства невозможно правильное и про-
дуктивное ведение охотничьего промысла.
386
Нарушение лесов и предпосылки природноочаговых заболеваний
Не имея достаточно широких специальных знаний в области
медицины, мы не беремся рассматривать проблему в плане изме-
нения нозогеографии, а лишь кратко рассмотрим те общие изме-
нения в животном мире, которые могут интересовать санитарные
учреждения. Некоторые возможные пути возникновения природ-
ных очагов заражения указаны нами в повидовых очерках.
Общим для всей южной тайги следует признать резкое увеличе-
ние путей циркуляции инфекций в связи а) с обогащением ви-
дового состава фауны прокормителей паразитов, б) с возникнове-
нием новых связей между видами долинного и водораздельного
комплексов и в) с «синантропизацией» диких животных. Действи-
тельно, проникновение в полосу южной тайги животных лесостепи
и северных форм, резкое смещение границ ареалов западных и
восточных видов должны были привести к возникновению новых
циклов в развитии инфекций природноочагового происхождения.
Изменение кружева ареалов видов привело к широкому расселе-
нию долинных форм, и зона контактов долинных и водораздель-
ных видов безмерно возросла по площади и протяжению границ.
При этом в биоценотическое соприкосновение вошли формы,
ранее жившие раздельно. Верхом «синтеза» природно-антропо-
генного режима стали смешанные мелколиственно-светлохвойные
леса — новые стации для видов как темнохвойной тайги, так и
форм светлохвойного комплекса. Сюда же влились представители
зооформации мелколиственных лесов, а на полянах поселились
формы лесостепи и лесополья. В результате создалась очень ла-
бильная система природно-антропогенного биоценоза, не имевшая
аналогов в девственной природе Средней Сибири.
Кратко следует остановиться еще на одном, выше нами не упо-
минавшемся явлении — изменении структуры ареала, т. е. сезон-
ной и годовой динамики населения животных внутри ареального
кружева (Исаков, 1963). Размещение экологических и географиче-
ских популяций животных может меняться даже при стабильном
кружеве ареала. Так, северные кедровки, сойки, московки и дру-
гие птицы периодически вливаются в популяции местных перна-
тых темнохвойных лесов гор юга Сибири (Реймерс, 1954, 1959а).
Другая серия видов меняет кружево ареала по сезонам (переко-
чевки дятлов, соек и многих других оседлых видов). Ряд переме-
щений животных вызван антропогенными причинами (лесные по-
жары, недостаток кормов в связи с нарушением лесов и т. д.).
В ходе передвижек млекопитающих и особенно птиц возникают
новые контакты как на месте, так и за пределами некоторого очер-
ченного участка.
Проникновение зверьков (азиатская леспая мышь, красная по-
левка, полевка-экономка, обыкновенная бурозубка и даже пи-
щуха) на приусадебные участки и в постройки человека сделало
25 Н. Ф. Реймерс
387
возможной прямую передачу природноочаговых болезней от жи-
вотных человеку. В условиях Средней Сибири этот вопрос еще не
получил широкого освещения, но под Москвой такие факты отме-
чены А. Д. Лебедевым и М. А. Савиной (1963).
Возможен и обратный путь инфекции — из жилищ человека
в природу. Например, длительное сохранение в земле, сенной
трухе и навозе возбудителя бруцеллеза (Пинигин, 1957) застав-
ляет предположить возможность заражения полусинантропных
форм млекопитающих этой болезнью. Возвращаясь в природные
биоценозы, эти зверьки принесут с собой инфекцию. Таким образом,
заболевание в данном случае антропогенного происхождения ста-
нет распространяться в природе как природноочаговое. Болезнь
из населенного пункта по природной цепи может проникнуть в со-
вершенно не освоенные человеком места и локализоваться в бла-
гоприятных условиях в очаги. Наиболее насыщенные биоценозы, —
очевидно, лучшие каналы для подобной циркуляции.
Следует обратить внимание также на указанный нами факт
концентрации животных в одних местах и разреженности их на-
селения в других. Скученность животных должна приводить
к образованию первоначальных очагов инфекции.
С другой стороны, как мы уже указали в разделе о белошапоч-
ной овсянке, обогащение фауны приводит к образованию тупиков
в развитии как паразитов, так, очевидно, и болезней. Вместе
с уплотнением животного населения в одних местах наблюдаются
значительные разрежения в других. Если для лугов как природ-
ных образований характерен специфичный комплекс паразитов,
в том числе иксодовых клещей, то свежие гари в Средней Сибири
имеют обедненную паразитофауну. Образование низкопроизводи-
тельных, с точки зрения охотничьего хозяйства, сосновых боров
с их редким животным населением в какой-то степени оздоров-
ляет местность, если в них не производится регулярного выпаса
скота.
Перечисленные общие положения можно подкрепить фактами
распространения в Средней Сибири иксодовых клещей и кле-
щевого энцефалита.
Исследованиями Г. В. Сердюковой и ее соавторов (Сердюкова
и Ходаковский, 1940; Сердюкова, 1960, и др.) было показано,
что численность взрослых иксодовых клещей в смешанных лесах
низкогорий выше, чем в более высоко лежащих поясах раститель-
ности, представленных однородными хвойными формациями. Ря-
дом авторов выяснено, что гари и вырубки населены иксодидами
подчас гуще, чем леса (Шилова и др., 1956; Мальков, 1959; Вершин-
ский и Балаганов, 1962; Реймерс и Воронов, 1963). То, что клещей
в смешанных лесах больше, чем в чистых хвойных, известно всем,
кто работал в тайге. В конце мая 1954 г. на р. Большой Быстрой
в Хамар-Дабане на десятикилометровом маршруте вдоль речной
долины, поросшей смешанным лесом (500—600 м над ур. м.),
388
за два сбора было снято с наблюдателя более 150 взрослых клещей.
За такой же маршрут в чистом кедровнике на высотах 1300—
1500 мнад ур. м., но в июне, который фенологически сходен с маем
в долинах, мы собирали по 3—4 экз. паразита. На год раньше
под Иркутском в конце апреля—начале мая в березово-сосновом
лесу мы снимали с себя до 40 клещей, проходя 3—5 км. В чистом
сосновом бору на тот же маршрут приходилось уничтожать не
более 2—5 паразитов. Такие же факты приводят Ф. Р. Штиль-
марк и Л. П. Климова с соавторами (1963).
Совершенно очевидно, что разница в зараженности иксодидами
между ниже и выше расположенными участками горной местно-
сти может объясняться неодинаковыми климатическими услови-
ями. Различное обилие клещей в пределах одного вертикального
пояса зависит от каких-то сочетаний микроклимата и биотиче-
ских факторов, в том числе от обилия прокормителей. Частичный
показ роли трансформации лесов в регулировании численности
иксодид уже сделан нами (Реймерс и Воронов, 1963). Здесь мы об-
ратим внимание на значение отдельных групп животных в прокорм-
лении преимагинальных фаз развития паразита. В табл. 36, как
и прежде, мышевидных зверьков мы делим на эколого-системати-
ческие группы, птицы объединены в две группировки — «дроздов»
(дрозды и сорокопут-жулан) и мелких паземнопитающихся (и не-
редко гнездящихся на земле) видов (степной, лесной и пятнистый
коньки, славка-завирушка). Данные о птицах приводятся в сред-
нем за 1961 и 1962 гг. по двум нашим основным полустационарам
(Верхнеленскому и Нижнеангарскому), показатели зараженно-
сти млекопитающих взяты лишь по Верхнеленскому полустацио-
нару.
Рассмотрение данных табл. 36 показывает, что а) высокие
вертикальные пояса неблагоприятны для развития иксодовых
клещей, но в пределах одного пояса более сильно нарушенные леса
продуцируют клещей больше, чем первичные коренные, б) вну-
три благоприятного вертикального пояса хвойно-мелколиствен-
ные леса наиболее заражены паразитами, в) в условиях южной
тайги Средней Сибири наибольшее число личинок и нимф, как пра-
вило, прокармливают землеройки (прежде всего обыкновенные),
г) в ряду долинных ельников большую роль в прокормлении
нимф играют лесные и серые полевки (бурундуков в этих лесах
немного), д) наиболее зараженными оказываются биотопы с ббль-
шим количеством представителей всех групп мелких зверь-
ков, е) мигрирующие зверьки выносят большое число личинок
и нимф в кустарники, на луга, а значит — в огороды и здания,
ж) в молодых мелколиственных лесах на месте гарей роль птиц
как прокормителей преимагинальных фаз развития иксодид
возрастает.
К сделанным выводам следует добавить еще одно заключение.
В предыдущей главе мы обращали внимание на тот факт, что в годы
25*
389
Таблица 36
Сравнение значения групп животных — прокормителей преимагинальных фаз развития иксодовых клещей — в рядах
трансформации лесов
Ряд водораздельных темнохвойных лесов
Группы животных фаза хвойно-мелколиственного леса фаза мелколиственного леса
личинки нимфы личинки нимфы
п. о. п. п. п. о. п. п. п. о. п. п. п. о. п. п.
Мелкие наземнопитающиеся птицы 0.6 0.72 0.06 0.72 0.73 17.52
«Дрозды» 0.4 2.08 — — 0.25 6.00 0.08 1.92
Сумма показателей для птиц — 2.80 — 0.72 — 6.00 — 19.44
Мелкие насекомоядные .... 0.66 3973.20 0.10 602.0 0.55 2464.0 0.05 224.0
Мышеобразные и лесная мы- шовка
Лесные полевки и лемминги 0.02 89.60 0.02 89.60 0.11 275.0 0.27 675.0
Серые полевки — — — — — — 0.69 276.0
Сумма показателей для мыше- видных зверьков — 4062.80 — 691.60 — 2739.0 — 1175.0
Сумма показателей для всех прокормителей преимаги- нальных фаз клещей .... — 4065.60 — 692.32 — 2745.0 — 1194.44
Число взрослых клещей, сни- маемых с человека, после пятикилометрового марш- рута (от—до) 1—2 । 1 -3
Примечание, п. о. — показатель обилия паразита; п. п. — показатель прокормления, т. е. произведение показателя обилия
паразита на приблизительную количественную оценку (для птиц; и абсолютное число зверьков (для млекопюающих) на 1 км2.
Группы животных
Мелкие наземнопитающиеся
ПТИЦЫ.......................
«Дрозды»....................
Сумма показателей для птиц
Мелкие насекомоядные . . . .
Мышеобразные и лесная мы-
шовка ......................
Лесные полевки и лемминги
Серые полевки...............
Сумма показателей для мыше-
видных зверьков.............
Сумма показателей для всех
прокормителей преимаги-
нальных фаз клещей ....
Число взрослых клещей, сни-
маемых с человека, после
пятикилометрового марш-
рута (от—до)................
фаза высокоствольного одно-
родного леса
личинки нимфы
П. О. П. П. П. О. П. п.
0.03 0.12
0.23 0.92 0.23 0.92
— 0.92 — 1.04
2.31 7276.50 0.03 94.50
1.70 833.0 0.22 107.80
3.14 3516.80 0.85 952.0
2.36 1416.0 0.45 270.0
— 13042.30 — 1424.3
— 13043.22 — 1425.34
1- 3 1
Таблица 36 (продолжение)
Ряд светлохвойных лесов
фаза хвойно-мелколиственного леса фаза мелколиственного леса
личинки нимфы личинки нимфы
п. о. п. п. п. о. п. п. П. О. п. п. п. о. П. П.
0.30 2.40
— — 0.27 2.16 1.81 43.44 0.17 4.08
— — — 4.56 — 43.44 — 4.08
5.91 46334.40 0.10 7840.0 2.57 13980.80 0.02 108.80
1.53 1285.20 0.07 58.80 3.01 3792.60 0.32 403.20
4.46 9053.80 0.46 933.80 7.40 4144.0 0.60 336.0
3.17 3804.0 0.25 300.0 0.55 440.0 0.44 352.0
— 60477.40 — 9132.6 — 22357.4 — 1200.0
— 60477.40 — 9137.16 — 22400.84 — 1204.08
5— 40 2-1 .5
фаза хвойно-мелколиственного
Группы животных леса
личинки нимфы
п. о. п. п. п. о. п. п.
Мелкие наземнопитающиеся птицы 0.06 1.44 0.11 2.64
«Дрозды» 0.31 7.44 0.38 8.12
Сумма показателей для птиц — 8.88 — 10.76
Мелкие насекомоядные .... 4.58 60593.40 0.11 1455.30
Мышеобразные и лесная мы- шовка 1.17 1310.40 0.21 235.20
Лесные полевки и лемминги 3.57 3498.60 0.96 940.80
Серые полевки 0.61 2684.0 0.27 1188.0
Сумма показателей для мыше- видных зверьков — 68086.40 — 3816.3
Сумма показателей для всех прокормителей преимаги- нальных фаз клещей .... — 68095.28 — 3827.06
Число взрослых клещей, сни- маемых с человека, после пятикилометрового марш- рута (от—до) 5—' 75
Таблица 36 (продолжение)
Ряд долинных ельников
фаза кустарников фаза луга
личинки нимфы личинки нимфы
п. 0. п. п. П. О. п. п. п. о. п. п. П. О. II. п.
0.19 4.56
0.04 0.96 0.04 0.96 — — — —
1 - । 5.52 — 0.96 0.0 0.0 0.0 0.0
3.12 9392.20 0.07 210.70 0.61 1067.50 0.7 122.50
1.40 1764.0 0.15 189.0 0.0 0.0 0.0 0.0
2.06 288.40 0.24 33.60 — — —— —
0.66 3124.0 0.09 396.0 0.30 600.0 0.09 180.0
— 14568.60 — 829.30 — 1667.50 — 302.50
— 14574.12 — 830.26 — 1667.50 — 302.50
0-1
Не нападают.
минимума численности имаго клещей 3 вида — азиатская лесная
мышь, красная полевка и темная полевка — прокармливают
столько же нимф или личинок, сколько п в годы максимальной
численности паразита. Это те зверьки, которые служат надежными
хозяевами клещей и составляют важную часть комплекса поло-
жительных условий в стациях переживания иксодид. В водораз-
дельной тайге такой вид прокормителя 1 — красная полевка,
в придолинной полосе их 3 (перечислены выше) и численность
2 из них — лесной мыши и темной полевки — резко повысилась
с антропогенными нарушениями лесов. Следовательно, емкость
жизненных условий для клещей возросла.
Таким образом, нарушение лесов привело в Средней Сибири
к увеличению числа иксодовых клещей. Однако фазы крайнего
нарушения (однородные боры, абсолютно безлесные гари) и пол-
ностью трансформированные участки (луга на месте ельников,
поля) либо не заселены паразитами, либо на них (луга) происхо-
дит смена видового состава иксодид. Вот почему интенсивное
ведение хозяйства оздоровляет местность. Очевидно, с этим же
обстоятельством связано и старое сибирское поверье, что древес-
ная и кустарниковая растительность у домов несет смерть и бо-
лезни. Культурные земли, и прежде всего широкая полоса полей
и огородов, мешают проникновению в населенные пункты лес-
ных мышевидных зверьков, препятствуют образованию антропо-
ургических очагов заболеваний в самих селах.
Само по себе наличие паразитов не означает еще возможности
заражения энцефалитом. Известны сочетания высокого числа пере-
носчиков с низкой их зараженностью. Однако сейчас уже накоп-
лены факты, показывающие, что нарушенные биотопы обладают
наивысшей зараженностью вирусом энцефалита как позвоночных
животных, так и клещей (Васенин, 1962; Кулакова, 1962, и др.).
Таким образом, нарушенные леса эпидемически более опасны,
чем коренные.
Большим числом исследователей показано широкое носитель-
ство вируса энцефалита птицами (Липин, 1962; Р. Наумов и
др., 1963, и др.). Изменение структуры ареалов пернатых вместе
с передачей вируса энцефалита зимой (Краминский и др., 1962)
должно приводить к образованию «бессимптомных» очагов цир-
куляции болезнетворного начала или даже к сплошному ареалу
вирусоносительства. Действительно, для туляремии уже из-
вестны «эпидемически инертные» природные очаги (Меринов и др.,
1959; Дунаева, 1963). В связи с изменениями в природе, произво-
димыми человеком, инертные очаги могут превращаться в активно
действующие. Сейчас не ясно, какие факторы могут пробудить
очаг, но не исключено, что среди них возможны и влияния, свя-
занные с проникновением в биоценозы новых видов позвоночных
животных. Это тем более вероятно, что занос переносчиков (не
исключено, что и зараженных) птицами уже доказан (Павловский,
393
1937; Крылов и др., 1959; Чабовский, 1959; Шилова, 1959;
Горчаковская и Чунихин, 1962; Nuorteva a. Hoogstraal, 1963,
и др.).
Влияние позвоночных животных на возобновление лесов
Этот вопрос неоднократно подвергался обсуждению в наших
работах (Реймерс, 1956а, 19566, 1958в, 1959а, 19596, 1959в; Лащин-
ский и Реймерс, 1959; Хлонов и Реймерс, 1960; Реймерс и Воронов,
1963; Реймерс и Малышев, 1963). К нему мы не раз обращались
в повидовых очерках, изложенных в гл. 5. Не останавливаясь на
нем повторно, мы лишь напомним, что, как это ни парадоксально
с первого взгляда (меньше соответствующих кормов), нарушение
лесов иногда приводит к относительному увеличению числа ак-
тивных семеноедов. Для некоторых из них типично питание се-
менами трав, но не отказываются они и от древесных семян.
Вместе с тем происходит исключительно резкая концентрация вре-
дителей в островных лесах, сохранившихся от рубки или пожара.
В общей сложности, человек, нарушая гармонию природы, создает
условия, при которых позвоночные животные начинают препят-
ствовать естественному ходу восстановления лесов. Определенную
роль играют тут пришлые виды (например, лесные мыши на гарях
сосновых лесов), «поддерживающие» необходимую им среду. В про-
цессе восстановления леса преодолеваются, таким образом, не
только чуждые абиотические, но и биотические условия. На по-
следнее обстоятельство часто не обращается должного внимания,
особенно при разработке приемов посева и посадки леса. В ча-
стности, совершенно забывается тот факт, что так называемые
искусственные ранения напочвенного покрова приводят к выра-
ботке ориентировочного рефлекса у грызунов, в результате чего
посадочный материал бывает в короткое время уничтожен ими.
Животные — вредители сельского хозяйства
На полях в полосе южной тайги Средней Сибири можно встре-
тить почти всех представителей насекомоядных и грызунов. Из
них наиболее серьезными вредителями следует признать бурун-
дука, серую крысу, азиатскую лесную мышь, водяную полевку
и полевку-экономку.
Бурундуки выходят на поля лишь по опушкам лесов, а числен-
ность зверьков никогда не превышает 10—15 экз. на 1 км границы
леса. Обычно бурундуков бывает всего 1—2 на 1 га или — при
узком поле — 2—4 экз. (они не заходят в глубь полей далее 100 м
от леса). Запасая на зиму по 2.5—3 кг зерна и съедая за осень
приблизительно столько же, один бурундук уничтожает около
5—6 кг урожая, т. е. общий вред зверьков не превышает четверти
центнера на 1 га.
Серая крыса очень важна как вредитель запасов кормов на
скотоводческих фермах. Тут наблюдается скопление крыс с по-
394
головьем, достигающим нескольких сотен экземпляров. За год
эти животные съедают многие центнеры кормов и к тому же слу-
жат распространителями инфекций.
Азиатская лесная мышь, как и бурундук, не живет на полях
постоянно, но численность зверьков здесь достигает осенью
60—65 экз. на 1 га. Одна лесная мышь может съесть более 10 г
зерна за сутки. Какую-то долю зерна злаков составляют и в зим-
них запасах зверька. Очевидно, мыши съедают не менее 18—20 кг
хлебов за осень (с 1 га). Лесные мыши глубокой осенью откоче-
вывают с полей в окрестные леса и на их гари. Если на следую-
щий год пашня остается под парами, в лесах сохраняется высо-
кая численность мышей. Зверьки при этом уходят далеко в глубь
лесных массивов. Засеянные вновь, поля становятся очагом раз-
множения мышей.
Водяная полевка наносит значительный вред огородам, поедая
клубни картофеля, корнеплоды моркови и свеклы. Огородниче-
ство развито в полосе южной тайги слабо и главным образом на
приусадебных участках. Вред водяной крысы на полях в годы
низкой численности зверька мало заметен, поскольку животное
нередко не поднимается даже на высокие речные террасы, концент-
рируясь почти исключительно на днищах долин. Массовые раз-
множения крыс возможны, видимо, лишь на крайнем западе юж-
ной тайги, граничащем с Западной Сибирью.
Численность полевок-экономок на полях достигает 38—40 экз.
на 1 га. Зверьки повреждают как зеленые растения, так и колосья.
Общий ущерб от экономок можно оценить в 15—20 кг/га.
Особенно много экономок на огородах — до 100 экз. на 1 га.
Вместе с водяной полевкой экономки вредят огородным культурам
и особенно лугам, оставляемым на приусадебных участках.
В окрестностях полей экономок обычно больше, чем вдали от
них. Это заставляет предположить, что сельскохозяйственное про-
изводство поддерживает высокую численность зверьков.
Общий урон от грызунов приблизительно 1—1.5 цна 1 га. Это
15—10-ячасть среднего урожая в Иркутской области (Шоцкий, 1962).
Некоторый ущерб приносят посевам и птицы. Плохо заделан-
ные семена собирают горлицы. Они же расхищают часть урожая
осенью. Питаются на полях врановые. Однако вред от птиц мал —
они поедают в основном опавшие или рассыпанные нерадивыми
земледельцами семена.
Таким образом, позвоночные животные уничтожают в полосе
южной тайги Средней Сибири сравнительно небольшую часть уро-
жая. С расширением площади полей урон снижается, так как мно-
гие грызуны выходят на пашни лишь из леса. В тех местах, где
стоимость потребляемой животными части урожая выше экономи-
ческих затрат на проведение борьбы с вредителями, можно реко-
мендовать разбрасывание отравленных приманок по опушкам ле-
сов и краям полей.
ЗАКЛЮЧЕНИЕ
По всей территории южной тайги Средней Сибири от Енисея
до Байкала и от Саян до Подкаменной Тунгуски заканчивается
единый процесс исчезновения темнохвойной тайги и замены ее
светлохвойными лесами. Уже близок момент, когда елово-кедрово-
пихтовые и елово-пихтовые леса станут прошлым Прибай-
калья.
Поскольку фауна»этих лесов наиболее четко обрисовывает этот
зоогеографический район, с исчезновением темнохвойной тайги
сотрутся его нынешние границы. В пределах южной тайги сло-
жатся иные взаимоотношения фаунистических комплексов. Прибай-
калье все в большей степени станет приобретать облик лесо-
степи, а вернее лиственничного и сосново-мелколиственного лесо-
полья.
С изменением облика южной тайги резко трансформируются
кружева ареалов большинства видов. Опушечные формы и живот-
ные речных долин вместе с полными или частичными синантро-
пами широко расселятся в ее пределах. Сохранят или расширят
свои позиции виды сосновых и мелколиственных лесов. Обитатели
темнохвойной тайги заметно сократятся в числе и займут несвой-
ственные им ранее стации, что в свою очередь позволит некоторым
из них расселиться шире, чем раньше, а других поставит на грань
вымирания.
Уничтожение темнохвойной тайги на юге Средней Сибири зна-
менует собой конец природных смен растительности и животного
мира и подчинение всех естественных процессов природно-антро-
погенному режиму. г Вмешательство человека в Готличие от
прошлого времени стало решающим фактором жизни при-
роды.
396
До сих пор взаимоотношения сибирской тайги и человеческого
хозяйства складываются хаотично. Уже тысячи лет люди черпают
из кладовых природы, полагая, что естественные ресурсы могут
восстановиться в первоначальном или почти изначальном виде
сами по себе. Это глубокое заблуждение руководит лесозаготови-
телем, сводящим кедровники Присаянья. Та же самая уверенность
ведет к разговорам о необходимости реакклиматизации всех
исчезнувших ценных видов и обязательного доведения их числен-
ности до первоначального уровня.
Нет, и еще раз нет! Природа — не добродушный заимодавец,
а жестокий ростовщик. И следует научиться считать, что окажется
выше — польза от взятых ресурсов или процент вызванных по-
терь.
Сибири не угрожает судьба пустыни Сахары, но превращение
юга Сибири в экономическую пустыню и полупустыню вполне
реально. В плане охотничьего хозяйства — это сокращение-за-
пасов белки, весьма возможное снижение поголовья маралов, па-
дение числа лесных куриных. Нет сомнения, что в ходе лесоразра-
боток и естественных процессов изреживания упадет и общая
численность соболей.
Если не будет налажено культурное ведение лесного хозяйства,
в ряде мест нужно ожидать усиления волн природноочаговых ин-
фекций. Во всяком случае природные предпосылки этих заболева-
ний — наличие подходящих резервуаров и увеличение обилия
переносчиков — налицо. Расширение очагов сильной нарушен-
ности природных комплексов вместе с * общим потеплением
климата предполагает распространение заболеваний в новых
местах.
Антропогенные вмешательства, нарушая природное равнове-
сие, создают условия, при которых обычно быстро идущие про-
цессы восстановления лесов замедляются абиотическими и биоти-
ческими факторами, в том числе и вредной деятельностью позво-
ночных животных. Создаются устойчивые безлесные пространства,
или хозяйственно продуктивные хвойные леса сменяются малоцен-
ными березняками.
Нами уже было показано (Реймерс и Малышев, 1963), что^пе-
речисленным далеко не исчерпывается список возможных вредных
последствий антропогенных нарушений природы южной тайги
Средней Сибири.
397
Все сказанное вовсе не означает, что следует призывать к ус-
тановлению режима абсолютной или частичной заповедности.
Речь идет отнюдь не об этом. Важно четко представлять себе ход
современных процессов в живой природе для того, чтобы созна-
тельно на нее воздействовать, уметь рассчитать возможный «уро-
жай» и нормы «посева». Человек сильнее природы, но он должен
быть и умнее ее — уметь свести к минимуму потери, а если и идти
на них, то четко сознавать их размер и последствия.
ЛИТЕРАТУРА
Аверин Ю. В. 1957. Зоогеографический очерк Камчатки. Бюлл. МОИП,
отд. биол., т. 62, вып. 5.
Аверина И. М., В. Г. Агапитов, Н. А. Доронина,
Н. Г. Ш а д р и н с к а я, Ю. А. Кручинин, Г. Л. Р у ти-
ле в с к и й, Р. К. Сиско, И. В. Семенов. 1962. Северная
Якутия (физико-географическая характеристика). Тр. Всесоюзн.
арктическ. и антарктическ. н.-иссл. инет., т. 236, Л.
Алтарева Н. Д., М. И. Анциферов, Е. П. Потапова,
Л. В. Федорова, Г. И. Васильев. 1957. К вопросу о ту-
ляремии в Иркутской области. Изв. Иркутск, гос. н.-иссл. противо-
чумн. инет. Сибири и Дальнего Востока, т. 15, Улан-Удэ.
Аникин В. П. 1902. Отчет о командировке в Нарымский край летом
1900 г. Изв. Томского унив., т. 22.
Анциферов М. И. 1961. Изучение географического распространения
природных очагов туляремии в Восточной Сибири и на Дальнем Вос-
токе. Вопр. мед. геогр. Сибири и Дальнего Востока, Иркутск.
Анцышкин С. П. 1952. Противопожарная охрана леса. М.—Л.
Аргиропуло А. И. 1940. Фауна СССР. Млекопитающие, т. 3, вып. 5,
М.—Л.
Арнольди К. В. и Л. В. Арнольд и. 1963. О биоценозе как одном
из основных понятий экологии, его структуре и объеме. Зоол. журн.,
т. 42, вып. 2.
Атлас Иркутской области. 1962. М.—Иркутск.
Атлас СССР. 1962. М.
Бакутин М. Г. 1957. Водоплавающие птицы дельты р. Селенги. Уч.
зап. Бурят-Монгольск. гос. пед. инет., вып. 12, каф. матем., физ.,
хим., ботан. и зоол., Улан-Удэ.
Балаганов В. Я. 1962. Опыт построения легенды геоботанической
карты с учетом динамики растительного покрова. В сб.: Исследования
по географии Сибири и Дальнего Востока. Матер, конф, молодых геогр.
Сибири (февраль, 1962), Иркутск.
Балаганов В. Я., Б. В. Вершинский иН. Ф. Реймерс.
1964. Динамика биокомплексов верхнеленской тайги. Сиб. геогр. со.,
вып. 3, М.—Л.
Банников А. Г. 1954. Млекопитающие Монгольской Народной Рес-
публики. М.
Банников А. Г. 1957. Заметки о зимних кормах рябчика и даурской
куропатки в Кэнтее. Уч. зап. Моск. гор. пед. инет., т. 65.
Библиография Иркутской области. 1956, 1957, 1962. Биология. Сост.
Л. Е. Зубашева. Тр. Научн. библиотеки (Иркутск, гос. унив
им. А. А. Жданова), вып. 15, Иркутск, 1956; Физическая география’
Сост. Л. Е. Зубашева. Тр. Научн. библиотеки (Иркутск, гос. унив*
399
им. А. А. Жданова), вып. 13, Л., 1957; Лесное хозяйство и лесные
промыслы. Сост. Л. Е. Зубашева, В. И. Яхненко. Вост.-Сиб. биол.
инет., Иркутск, 1962.
Бобринский Н. А., Б. А. Кузнецов, А. П. Кузякин.
1944. Определитель млекопитающих СССР. М.
Богородский Ю. В. 1962. Некоторые наблюдения по биологии ази-
атской лесной мыши. Докл. Иркутск, противочумн. инет., вып. 4,
Иркутск.
Боровиков Г. А. 1910. Восточное Заангарье Енисейской губернии.
Предварит, отчет о ботан. исслед. в Сибири и Туркестане в 1909 г.,
СПб.
Боровиков Г. А. 1911. Растительность Западного Заангарья. Пред-
варит. отчет о ботан. исслед. в Сибири и Туркестане в 1910 г., СПб.
Боровиков Г. А. 1912. Очерк растительности Восточного Заангарья.
Тр. почв.-ботан. экспед. по исслед. Азиатской России в 1909 г., ч. 2,
вып. 7, СПб.
Боровиков Г. А. 1913. Очерк растительности Западного Заангарья.
Тр. почв.-ботан. экспед. по исслед. Азиатской России в 1910 г.,
ч. 2, вып. 5, СПб.
Бородулина Т. Л. 1951. О латентном периоде в развитии эмбриона
алтайского крота. Докл. АН СССР, т. 80, № 4.
Бородулина Т. Л. 1953. К биологии алтайского крота. Тр. Инет,
морфол. животных им. А. Н. Северцова, вып. 9, М.
Бром И. П. 1959. Грызуны — носители инфекций в зоне водохранилища
Иркутской гидроэлектростанции. Изв. Иркутск, гос. н.-иссл. проти-
вочумн. инет. Сибири и Дальнего Востока, т. 21.
Бром И. П. 1961. Черная крыса в Восточной Сибири. Докл. Иркутск,
противочумн. инет., вып. 2, Чита.
Бром И. П., В. А. Б у б л и е н к о, Р. Р. Ж и в о л я п и н а,
3. К. К а р а к а ш л ы, Н. Н. К р а м и н с к а я и В. А. П е ре-
возников. 1961. К характеристике очагов клещевого энцефалита
в Тайшетском районе Иркутской области. Докл. Иркутск, противо-
чумн. инет., вып. 1, Улан-Удэ.
Буторина Т. Н. иЕ. А. Крутовская. 1958. О корреляции
некоторых феноиндикаторов с температурой. Тр. гос. заповеди.
«Столбы», вып. 2, Красноярск.
Бутурлин С. А. 1928. Определитель видов птиц СССР, их подвидыt
распространение, польза и вред для хозяйства, вып. 1. Дневные хищ-
ники и совы. М.
Васенин А. А. 1962. Вирусологические исследования в очаге клеще-
вого энцефалита на территории юго-западной части Иркутской об-
ласти. Тр. Иркутск, н.-иссл. инет, эпидемио л. и микробиол., вып. 7,
Иркутск.
Васильев Я. Я. 1933. Леса и лесовозобновление в районах Братска—
Илимска и Усть-Кута. Тр. СОПС АН СССР, сер. Сиб., вып. 2 (Ангар-
ская лесная экспедиция 1921 г., ч. 1), Л.
Вержуцкий Б. Н. 1963. О зимнем питании восточной черной вороны.
Бюлл. Вост.-Сиб. фенологич. комисс., вып. 2—3. Сезонная и вековая
динамика природы Сибири, Иркутск.
Верещагин Н. К 1953. Захоронение остатков верхнеплейстоценовых
животных и растений у села Нижние Карманки на юге Татарской
АССР. Зоол. журн., вып. 5.
ВернандерТ. Б., С. В. Викторов, А. Г. Воронов,
Т. И. Дыбская, Н. П. Осадчая иА. М. Чельцов-
Бебутов. 1959. Опыт среднемасштабного биогеографического
картирования. В сб.: География населения наземных позвоночных
животных и методы его изучения, М.
400
Вершинский Б. В. и В. Я. Балаганов. 1962. Очаги таежного
энцефалита и динамика растительного покрова. Докл. Инет, геогр.
Сибири и Дальнего Востока, № 1, Иркутск.
Виноградов Б. С. 1927. Заметки о млекопитающих Якутии. 2. Ры-
жие полевки (род Evotomys). Матер. Комисе, по изучению Якутской
АССР, вып. 18, Л.
Виноградов Б. С. и С. И. Оболенский. 1927. Материалы по
фауне грызунов южной части Енисейской, Иркутской губерний и
Забайкалья. Изв. Сиб. станц. защиты растений от вредителей, № 2 (5),
Томск.
Виноградов Б. С. иИ. М. Громов. 1952. Грызуны фауны СССР. М.
Вишняков С.В. 1957. Материалы по экологии водяной крысы централь-
ных областей РСФСР. В сб.: Фауна и экология грызунов. Матер,
по грызунам, вып. 5, М.
Владимиров К. А. 1940. Американская норка в Иркутской области.
Иркутск.
Воробьев К. А. 1954. Птицы Уссурийского края. М.
Воробьев К. А. 1963. Птицы Якутии. М.
Воронов А. Г. 1958. Некоторые проблемы современной зоогеографии.
В сб.: Пробл. зоогеогр. суши, Львов.
Воронов Г. А. 1963. Распространение красной полевки в верхнелен-
ской тайге. В сб.: Вопр. зоол. картограф., М.
Воронов Г. А. иН. Ф. Реймерс. 1962. К размещению и биологии
насекомоядных и грызунов в верхнеленской тайге. Докл. Инет, геогр.
Сибири и Дальнего Востока, № 2, Иркутск.
Воронцов Н. Н. 1956. О запасании корма красной полевкой Clethrio-
nomys rutilus Pall. Бюлл. МОИП, отд. биол., т. 61, № 4.
Воронцов Н. Н. 1961. Экологические и некоторые морфологические
особенности рыжих полевок (Clethrionomys Tiles.) европейского се-
веро-востока. В сб.: Морфология и экология позвоночных животных.
Тр. Зоол. инет. АН СССР, т. 29, М.—Л.
Гагина Т. Н. 1958а. К познанию массовых непериодических миграций
диких куриных в сибирской тайге. Изв. Иркутск, с.-х. инет., вып. 8,
фак. зоотехн. и охотовед., Иркутск.
Гагина Т. Н. 19586. Птицы Байкала и Прибайкалья (список и распро-
странение). Зап. Иркутск, обл. краеведч. музея, Иркутск.
Гагина Т. Н. 1961. Птицы Восточной Сибири (список и распростра-
нение). Тр. Баргузинского гос. заповеди., вып. 3, М.
Гагина Т. Н. 1962а. Примечания и дополнения к списку птиц Восточ-
ной Сибири. Тр. Баргузинского гос. заповеди., вып. 4, Улан-Удэ.
Гагина Т. Н. 19626. Редко зимующие птицы Прибайкалья. Тр. Бар-
гузинского гос. заповеди., вып. 4, Улан-Удэ.
Галазий Г. И. 1954. Вертикальный предел древесной растительности
в горах Восточной Сибири и его динамика. Тр. Ботан. инет,
им. В. Л. Комарова АН СССР, сер. 3 (Геоботаника), вып. 9,
М.—Л.
Ганешин С. С. 1910. Ангаро-Илимский край Иркутской губернии.
Предварит, отчет о ботан. исслед. в Сибири и Туркестане в 1909 г.,
СПб.
Ганешин С. С. 1912. Растительность Ангаро-Илимского края Иркут-
ской губернии. Тр. почв.-ботан. экспед. по исслед. Азиатской Рос-
сии в 1909 г., ч. 2, вып. 5, СПб.
Гептнер В. Г. и Ю. Г. Швецов. 1960. О видовом тождестве восточ-
ной (Mtcrotus fortis Buch.) и уйгурской (М. maximoviezii Sch.) полевок.
Изв. Иркутск, гос. н.-иссл. противочумн. инет. Сибири и Дальнего
Востока, т. 23, Иркутск.
Гептнер В. Г., А. А. Насимович, А. Г. Банников. 1961.
Млекопитающие Советского Союза, т. I. Парнокопытные и непарно-
копытные. М.
401
Г и б е т Л. А. 1964. Хищные птицы лесостепи Тоболо-Ишимского между-
речья. Бюлл. МОИП, отд. биол., т. 49, вып. 4.
Тибет Л. А. и И. Ф. Кузьмин. 1963. Картирование обилия и
распределения бурундука в Северном Заангарье. В сб.: Вопр. зоол.
картограф., М.
Гладков Н. А. 1951. Отряд кулики. В кн.: Птицы Советского Союза,
т. 3. М.
Гладков Н. А. 1951а. Отряд дятлы. В кн.: Птицы Советского Союза,
т. 1. М.
Гладков Н. А. 1954. Семейство трясогузковые. В кн.: Птицы Совет-
ского Союза, т. 5. М.
Горчаковская Н. Н. 1954. Род дубоносы. В кн.: Птицы Советского
Союза, т. 5. М.
Горчаковская Н.Н.иС. II. Чунихин. 1962. О переносе клещей
дроздами в тайге и лесостепи Салаирского кряжа (Кемеровская об-
ласть). В сб.: Орнитология, вып. 5, М.
Громов И. М., А. А. Гуреев, Г. А. Новиков, И. И. Сок о-
л о в, П. П. Стрелков, К. К. Ч а п с к и й. 1963. Млекопи-
тающие фауны СССР. М.—Л.
Грудинин И. Т. 1939. Белка и ее промысел в Восточной Сибири. Ир-
кутск.
Гудритис В. Э. 1963. О медведях-шатунах в Восточной Сибири. Зоол.
журн., т. 42, вып. 6.
Данилов Н. Н. 1958. Изменения в орнитофауне зарастающих вырубок
на Среднем Урале. Зоол. журн., т. 37, вып. 12.
Дементьев Г. П. 1951а. Отряд гагары. В кн.: Птицы Советского Союза,
т. 2. М.
Дементьев Г. П. 19516. Отряд поганки. В кн.: Птицы Советского
Союза, т. 2. М.
Дементьев Г. П. 1951в. Отряд хищные птицы. В кн.: Птицы Совет-
ского Союза, т. 1. М.
Дмитриев В. 1818. Сибирский кедр. Сиб. вести., издав. Г. Спасским,
ч. 1, СПб.
Добровольский А. В. 1937. Ондатра в Восточной Сибири. Иркутск.
Добровольский А. В. 1939. Техника добывания ондатры. Иркутск.
Добровольский А. В. 1952. Экология и принципы хозяйственного
освоения ондатры Восточной Сибири. Автореф. дисс., Минск.
Доброхотов М. А. 1953. Летяга и белка. Природа, № 1.
Дорогостайский В.Ч. 1927. Американская ондатра и перспективы
разведения ее в Сибири. Охотник и пушник Сибири, № 4, Новосибирск.
Дорогостайский В.Ч. 1930. Пятнистый олень на Байкале. Охот-
ник, № 2.
Дробов В. П. 1909. Растительные формации Лено-Киренского края
Иркутской губернии Верхнеленского уезда. Тр. почв.-ботан. экспед.
Переселенч. управл., ч. 2, вып. 1. СПб.
Дроздова Ю. В. 1959. Насекомоядные млекопитающие в Пермской
области как хозяева клещей Ixodes persulcatus. Сб. научно-практич.
работ Пермск. обл. сан.-эпид. стапц., вып. 2, Пермь.
Дубинин Н.П. 1953. Птицы лесов нижней части долины реки Урал,
ч. 1. Тр. Инет, леса, т. 18, М.
Дубинин Н. П. иТ. А. Торопанова. 1956. Птицы лесов долины
реки Урал, чч. 2 и 3. Тр. Инет, леса, т. 32, М.
Дулькейт Г. Д. 1957. Вопросы экологии и количественного учета со-
боля. М.
Дулькейт Г. Д. иВ.В. Козлов. 1958. Материалы к фауне млеко-
питающих заповедника «Столбы». Тр. гос. заповеди. «Столбы», вып. 2,
Красноярск.
Дунаева Т. Н. 1954. Экспериментальное исследование туляремии у ди-
ких животных (грызунов, хищных и насекомоядных) как основа изу
4()2
чения природных очагов этой инфекции. Зоол., жури., т. 33,
вып. 2.
Дунаева Т. Н. 1963. Значение млекопитающих в эпизоотологии туля-
ремии. Зоол. журн., т. 42, вып. 5.
Дунаева Т. Н. и В. В. Кучерук. 1941. Материалы по экологии
наземных позвоночных южного Ямала. Матер, к познанию фауны и
флоры СССР (МОИП), отд. зоол., вып. 4 (19), М.
Егорин Н. Ф. 1939. Материалы к изучению грызунов таежной полосы
Западной Сибири. Тр. Биол. н.-иссл. инет, при Томск, унив., сер. Е,
биол., т. 6, Томск.
Егоров О. В. 1961. Экология и промысел якутской белки. М.
Емельянова Н. Д. иН. Н. Высоковский. 1962. К вопросу
о зимовках летучих мышей и их эктопаразитах в окрестностях Крас-
ноярска. Докл. Иркутск, противочумн. инет., вып. 3, Хаба-
ровск.
Заблоцкая Л. В. 1955. Расхищение семян хвойных землеройками-
бурозубками. Бюлл. МОИП» отд. биол., т. 60, вып. 4.
Заблоцкая Л. В. 1957. Растаскивание семян хвойных и липы земле-
ройками-бурозубками. Тр. Приокско-Террасного гос. заповеди.,
т. 1, М.
Зенкевич Л. А. 1951. Моря СССР, их фауна и флора. М.
Зернов С. А. 1949. Общая гидробиология. М.—Л.
Зиновьев Г.А. 1959. К методике составления карты энтомокомплексов
лесных биоценозов. Матер, перв. сессии научи, совета по пробл.
«Биологические комплексы районов нового освоения, их рациональ-
ное использование и обогащение», М.—Л.
Зубчанинова Е. В. 1962. О географической изменчивости бурунду-
ков (Eutamias sibiricus Laxm.) СССР. Научн. докл. высш, школы,
Биол. науки, К® 4.
Иванов А. И. 1929. Птицы Якутского округа. Матер, комисси по изу-
чению Якутской АССР, вып. 25, Л.
Измайлов И. В. и И. А. Стариков. 1961. Зоологические иссле-
дования в южной части Витимского плоскогорья в 1960 году. Краеведч.
сб. Бурятск. фил. Геогр. общ. СССР, вып. 6, Улан-Удэ.
Иоганзен Б. Г. 1953. Рыбохозяйственные районы Западной Сибири
и их биолого-промысловая характеристика. Тр. Томск, гос. унив.,
т. 125. Исслед. водоемов Сибири, вып. 4, Томск.
Иоганзен Г. 1916 (1917). Заметки о птицах Иркутской губернии. Изв.
ВСОРГО, т. 45, Иркутск.
Иоганзен Г. Э. 1930. Из жизни Томской природы. Фенологические
заметки за 1916 г. Матер, по изуч. Сибири, вып. I, Томск.
Исаков Ю.А. 1952. Подсемейство утки. В кн.: Птицы Советского Союза,
т. 4. М.
Исаков Ю.А. 1952а. Опыт изучения распространения вида внутри
ареала. Бюлл. МОИП, отд. биол., т. 57, вып. 6.
Исаков Ю.А. 1957. Некоторые вопросы изучения фауны и географиче-
ского распространения птиц. Тр. Второй Прибалт, орнитол. конф., М.
Исаков К). А. 1963. Некоторые вопросы изучения ареала. В сб.: Вопр.
зоол. картограф., М.
Казанская В. Г. 1952. Крот Томской области и его промысловое ис-
пользование. Тр. Томск, гос. унив., т. 118.
Казанская В. Г. 1953. Биология размножения сибирского крота.
Тр. Томск, гос. унив., т. 123.
Карасева Е. В. 1956. Некоторые особенности развития эпизоотии
лептоспироза у полевок-экономок, изученные методом мечения зверь-
ков. Зоол. журн., т. 35, вып. 9.
Карасева Е. В. 1963. Изучение закономерностей течения эпизоотии
лептоспироза с помощью картирования индивидуальных участков
животных. В сб.: Вопр. зоол. картограф., М.
403
Карасева Е. В. и А. И. Ильенко. 1957. Некоторые особенности
биологии полевки-экономки, изученные методом мечения зверьков.
В сб.: Фауна и экология грызунов. Матер, по грызунам, вып. 5, М.
Карасева Е. В., Э. И. К о р е н б е р г и М. А. М е р к о в а. 1960.
Мелкие млекопитающие Центральной Якутии и их значение в природ-
ных очагах некоторых болезней человека. Зоол. журн., т. 39, вып. И.
Карпове. П. 1954. Дальнейшие наблюдения по эпидемиологии клеще-
вого энцефалита в Томском очаге и мерам борьбы с ним. Тез. докл.
научи, конф., посвящ. 350-летию г. Томска, Томск.
Карпов С. П. 1955. Краевая эпидемиология клещевого энцефалита и
меры борьбы с ним. Тр. Томск, н.-иссл. инет, вакцин и сывороток,
т. 6, Томск.
К а р и о в С. П. и В. М. Попо в. 1954. Млекопитающие Западной Си-
бири — резервуары и источники инфекций человека. Тез. докл. научн.
конф., посвящ. 350-летию г. Томска, Томск.
Карпов С. П. и В. М. Попов. 1956. Млекопитающие Западной Си-
бири как резервуары и источники инфекций человека. Тр. Томск*,
гос. унив., т. 142.
К а р т а ш о в Н. Н. 1952. Род куропатки. В кн.: Птицы Советского Союза,
т. 4. М.
К а ц Н. Я. 1948. Типы болот СССР и Западной Европы и их географиче-
ское распространение. М.
КерзинаМ. Н. 1956. Влияние вырубок и гарей на формирование лесной
фауны. Матер, к познанию флоры и фауны СССР (МОИП), отд. зоол
вып. 35 (50). Роль животных в жизни леса. М.
К и м Т. А. 1957а. К экологии северной пищухи Кизир-Казырского между-
речья. Уч. зап. Красноярск, пед. инет., т. 10.
К и м Т. А. 19576. К вопросу об ареале зяблика. Уч. зап. Красноярск, пед.
инет., т. 10.
К и м Т. А. 1961. Заметки о летучих мышах Красноярского края. Уч. зап.
Красноярск, пед. инет., т. 20, вып. 2, каф. зоол.
К и м Т. А. 1962. О распространении некоторых видов птиц в Красноярском
крае. Изв. Красноярск, отд. Геогр. общ. СССР, вып. 2. Красноярский
край (матер, по геогр. и истор.), Красноярск.
К и м Т. А. и В. М. Л и х а ш е р с т о в. 1961. О размножении пасюка
в г. Красноярске. Уч. зап. Красноярск, пед. инет., т. 20, вып. 2, каф.
зоол.
К и р и к о в С. В. 1952а. Птицы и млекопитающие в условиях ландшафтов
южной оконечности Урала. М.
К и р и к о в С. В. 19526. Род глухари. В кн.: Птицы Советского Союза,
т. 4. М.
К ирик о в С. В. 1956. Животный мир. В сб.: Лесостепь и степь Русской
равнины, М.
К ириков С. В. 1963. О «смешанной» фауне в степной и лесной зонах.
Изв. АН СССР, сер. геогр., № 1.
К ирпичев С. П. 1958. О помесях между обыкновенным глухарем и ка-
менным. В сб.: Орнитология (вып. 1). Уч. зап. МГУ, вып. 197.
К и т р е д ж Дж. 1951. Влияние леса на климат, почвы и водный режим. М.
К л е й т м а н Е. И. и Н. И. Иголкин. 1956. Течение туляремии у стад-
ной полевки и полевки-экономки и их эпидемиологическое значение.
Тр. Томск, н.-иссл. инет, вакцин и сывороток, т. 7, Томск.
К л ё ц Э. И. 1957. Вспышка туляремии в Нижнеудинском районе. Изв.
Иркутск, гос. н.-иссл. противочумн. инет. Сибири и Дальнего Востока,
т. 15, Улан-Удэ.
Климов Ю. Н. 1931. К биологии узкочерепной полевки \Microtus (Ste-
nocranius) gregalis gregalis Pall. ] и приманочный метод борьбы с ней.
Тр. по защите растений Сибири, т. 1 (8), Новосибирск.
Козлова Е. В. 1930. Птицы юго-западного Забайкалья, Северной Мон-
голии и Центральной Гоби. Л.
404
К оз л оваЕ. В. 1947. Гагарообразпые, трубконосые. Фауна СССР. Птицы,
т. 1, вып. 3. М.—Л.
Козлова Е. В. 1961. Ржаикообразпые. Подотряд кулики. Фауна СССР.
Птицы, т. 2, вып. 1, ч. 2. М.—Л.
Козлова Е. В. 1962. Ржапкообразные. Подотряд кулики. Фауна СССР.
Птицы, т. 2, вып. 1, ч. 3. М.—Л.
К о л юшевИ. И. 1936. Млекопитающие крайнего севера Западной и Сред-
ней Сибири. Тр. Биол. н.-иссл. инет, при Томск, гос. унив., т. 2,
Томск.
Комарова А. Ф. 1945. Эпидемиология туляремии в Томской области.
В сб.: Эпидемпол. и профилакт. инфекц., Томск.
К о п ы л о в И. П. 1948. Охотничье хозяйство Иркутской области. Иркутск.
К о и ы л о в И. П. 1950. Дикие копытные животные Иркутской области.
Иркутск.
Копылов И. П. 1962. Охотничье хозяйство. Атлас Иркутской области,
М.—Иркутск.
Копылов И. II., А. В. Добровольский и И. А. Ш е р г и и.
1940. Промысловые звери Иркутской области. Иркутск.
Копылов И. II. и В. Н. Скалой. 1962. Обогащение охотничьей
фауны [карта]. Атлас Иркутской области, М.—Иркутск.
К о р о т к и й М. Ф. 1916. Степные явления в Баргузпнской тайге (экспе-
диция на р. Мую). Предварит, отчет о ботан. исслед. в Сибири и Тур-
кестане в 1914 г., Пгр.
Корчагин Н. В. 1936. Фенологические наблюдения в опорных пунктах
Восточной Сибири. В сб.: Природа Восточно-Сибирского края (фе-
нология), Иркутск.
К о т о в И. С. 1957. Фенологические типы весен в г. Улан-Удэ и его окре-
стностях и календарь цветения некоторых видов растений. Краеведч.
сб. Бурят-Монгольск. фил. Геогр. общ. СССР, вып. 1, Улан-Удэ.
К о пт к и и а Т. В. 1957. Сравнительная экология рыжих полевок в север-
ной тайге. В сб.: Фауна и экология грызунов. Матер, по грызунам,
вып. 5, М.
К р а м и и с к и й В. А., Н. И. К р а м и и с к а я, И. П. Бром,
Р. Р. Ж и в о л я п и и а, В. А. 11 е р е в о з и и к о в, В. А. Б у б-
л и е и к о. 1962. Выделение вируса клещевого энцефалита от зи-
мующих птиц. Матер, юбил. научн. конф. Иркутск, н.-иссл. инет,
эпидемиол. и микробпол., Иркутск.
К р а м и н с к а я Н. II. 1962. Лептоспирозы домашних животных в Иркут-
ской области. Тр. Иркутск, и.-лсел. инет, эпидемиол. и микробиол.,
вып. 7, Иркутск.
К ришт оф о вич А. Н. 1910. Ботанико-географические исследования
в области Березовского хребта и Балаганской степи в Иркутской
губернии. Тр. почв.-ботан. экспед. по исслед. колопизац. район. Азиат-
ской России, ч. 2, вып. 3, СПб.
К р и in т о ф о в и ч А. Н. 1911. Растительность Око-Апгарского края.
Предварит, отчет о ботан. исслед. в Сибири и Туркестане в 1910 г., СПб.
К рпштофович А. II. 1913. Очерк растительности Око-Ангарского
края. Тр. почв.-ботан. экспед. по исслед. колопизац. район.
Азиатской России, ч. 1, вып. 3, СПб.
Крутовская Е. А. 1958. Птицы заповедника «Столбы». Тр. гос.
заповеди. «Столбы», вып. 2, Красноярск.
Крутовская Е. А. и Т. Н. Буторина. 1958. Сезонное развитие
природы горной тайги. Тр. гос. заповеди. «Столбы», вып. 2, Красно-
ярск.
Крылов Д. Г., Б. Д. Васильев и Ю. А. Рудяков. 1959. Зна-
чение птиц в очагах клещевого энцефалита Пермской области. Сб.
научпо-практич. работ Пермской обл. сан.-эпид. станц., вып. 2, Пермь.
Кузнецов Б. А. 1929. Грызуны Восточного Забайкалья. Изв. Ассоц.
н.-иссл. инет, при физ.-матем. фак. I МГУ, т. 2, № 1, М.
26 И. Ф. Реймерс
405
КузнецовБ.А. 1950. Очерк зоогеографического районирования СССР. М.
КузякинА. П. 1950. Летучие мыши. М.
Кузякин А. П. 1951. История расселения, современное распространение
и места обитания пасюка в СССР. В сб.: Фауна и экология грызунов.
Матер, по грызунам, вып. 4, М.
Кузякин А. П. (Ред.). 1959. Водяная крыса и борьба с ней в Западной
Сибири. Новосибирск.
КузякинА. П. 1962. Зоогеография СССР. Уч. зап. Моск. обл. пед. инет.,
т. 109, биогеограф., вып. 1, М.
КузякинА. П.иП.П. Второв. 1963. К ландшафтной орнитогеогра-
фии охотской тайги. В сб.: Орнитология, вып. 6, М.
Кулакова 3. Г. 1962. Зараженность половозрелых клещей вирусом
клещевого энцефалита в Лысьвенском районе Пермской области.
Автореф. докл. на расширенном засед. комит. по борьбе с клещевым
энцефалитом. Омск.
Кумари Э. В. 1957. Динамика орнитофауны Прибалтики за последние
столетия и вероятные направления ее дальнейших изменений. В сб.:
Вопр. экологии (по матер. 3-й эколог, конф.), т. 2. Киев.
КуренцовА. И. 1947. О зоогеографических округах Приморского края.
Комаровские чтения, вып. 1, Владивосток.
Куренцов А. И. 1959а. Животный мир Приамурья и Приморья. Ха-
баровск.
Куренцов А. И. 19596. Вопросы зоогеографии южных частей Дальнего
Востока. Зоол. журн., т. 38, вып. 2.
Куренцов А. И. 1963. Зоогеография Камчатки. В сб.: Фауна Камчат-
ской области, М.—Л.
Кучеровская С. Е. 1912. Растительность Ангаро-Ленского края
близ Илимского тракта. Предварит, отчет о ботан. исслед. в Сибири
и Туркестане в 1911 г., СПб.
Кучерук В. В. 1959. Степной фаунистический комплекс млекопитаю-
щих и его место в фауне Палеарктики. В сб.: География населения
наземных животных и методы его изучения, М.
Л а в о в М. А. 1955. Медведь Забайкалья. Зам. по фауне и флоре Сибири,
вып. 18, Томск.
Л а в о в М. А. 1959. Влияние соболя на популяцию белки в Забайкалье.
Зоол. журн., т. 38, вып. 2.
Лавренко Е. М. 1950. Основные черты ботанико-географического раз-
деления СССР и сопредельных стран. Пробл. ботан., вып. 1, М.—Л.
Лавренко Е.М. 1954. Степи Евразиатской степной области, их геогра-
фия, динамика и история. Вопр. ботан., т. I, М.—Л.
Лавренко Е. М. иВ. Б. Сочава. [Ред.]. 1956. Геоботаническая
карта СССР. М. 1 : 4 000 000. М.
Лавров И. К. 1960. К экологии Dermacentor nuttalli 01. в условиях Пред-
байкалья. Тр. Иркутск, н.-иссл. инет, эпидемиол. и гигиены, вып. 5,
Иркутск.
Лавров И. К. 1962. Материалы по фауне грызунов правобережных тер-
риторий верхнего течения реки Лены. Докл. Иркутск, противочумн.
инет., вып. 4, Иркутск.
Лавров И. К. и Е. П. Мартынов. 1960. Материалы по экологии
даурского хомячка Предбайкалья. Биол. сб. (1960), Иркутск.
Л а в р о в Н. П. 1943. К биологии обыкновенной землеройки. Зоол. журн.,
т. 22, вып. 6.
Лавров Н.П. 1946. Акклиматизация и реакклиматизация пушных зве-
рей в СССР. М.
Лавров Н.П. 1947. Ондатра. М.
Лаптев И. П. 1958. Млекопитающие таежной зоны Западной Сибири,
Томск.
Лащинский Н.Н.иН.Ф. Реймерс. 1959. Роль животных в жизни
лиственничных и кедровых лесов Алтая. Изв. Сиб. отд. АН СССР, № 2.
406
Лебедев А. Д. и М. А. Савин а. 1963. Миграция грызунов и насе-
комоядных из природы в постройки человека и эпидемиологическое
значение этого явления. Геогр. сб. (Инет, научн. информ. АН СССР), М.
Линник Т. Г. 1936. Заметки по биологии Micromys minutus Pall. Изв.
Иркутск, гос. противочумн. инет. Сибири и Дальнего Востока, т. 3,
М.—Иркутск.
Липин С. И. 1962. Птицы как прокормители лесного клеща Ixodes per-
sulcatus Р. Sch. в Присаянье. Тр. Иркутск, н.-иссл. инет, эпидемиол.
и микробиол., вып. 7, Иркутск.
Литвинов Н.И. 1960а. Грызуны зоны затопления Иркутского водохра-
нилища. Биол. сб. (1960), Иркутск.
Литвинов Н.И. 19606. Заметки о фауне наземных позвоночных острова
Ольхона на Байкале. Биол. сб. (1960), Иркутск.
М аксимов А. А. 1959. Распределение водяной крысы по зонам и ланд-
шафтам Западной Сибири. В сб.: Водяная крыса и борьба с ней в За-
падной Сибири, Новосибирск.
Максимов А. А. 1960. Природные очаги туляремии в СССР. М.—Л.
Максимов А. А. и С. С. Ф о л и т а р е к. 1959. Библиографический
указатель литературы по водяной крысе (биология, вредная деятель-
ность, эпизоотологическое и эпидемиологическое значение, борьба).
В сб.: Водяная крыса и борьба с ней в Западной Сибири, Новосибирск.
Мальков Г. В. 1959. Лесные грызуны очагов клещевого энцефалита
Пермской области и борьба с ними. Сб. научно-практич. работ Перм-
ской обл. сан.-эпид, станц., вып. 2, Пермь.
Мальчевский А. С. 1959. Гнездовая жизнь певчих птиц. Размножение
и постэмбриональное развитие лесных воробьиных птиц европейской
части СССР. Л.
Малышев Л. И. 1956. Растительность лесного пояса побережий Север-
ного Байкала. Автореф. дисс., Иркутск.
Малышев Л. И. 1958. Материалы к орнитофауне Прибайкалья. Зоол.
журн., т. 37, вып. 7.
Малышев Л. И. 1960. Материалы к орнитофауне северо-западного
побережья Байкала. Тр. Вост.-Сиб. фил. СО АН СССР, вып. 23, сер.
биол., зоол., Благовещенск.
Малышев Л. И. 1963. Сезонная биология птиц южного Предбайкалья.
Бюлл. Вост.-Сиб. фенология, комисс., вып. 2—3. Сезонная и вековая
динамика природы Сибири, Иркутск.
Мартынов В. П. 1958. О солонцеватости и осолодел ости некоторых
почв Иркутской области. Изв. Сиб. отд. АН СССР, № 2, Новосибирск.
Мартынов Е. П. иГ. В. 3 о н о в. 1960. Сведения по экологии гры-
зунов долины реки Ушаковки. Биол. сб. (1960), Иркутск.
Мартынов П. И., Н. Г. Скрябин, К. П. ФилоновиС. К. Ус-
тинов. 1960. Материалы по млекопитающим Баргузинского запо-
ведника. Тр. Баргузинского гос. заповеди., вып. 2, Улан-Удэ.
Межжерин В. А. 1961. Особенности экологии бурозубых землероек
(Soricinae} и динамика их численности в лесостепи и полесьи Украины.
Автореф. дисс., Киев.
Мекленбурцев Р. Н. 1951. Отряд голуби. В кн.: Птицы Советского
Союза, т. 2. М.
Мелехов И. С. 1936. Лесные пожары и борьба с ними. М.—Л.
Мелехов И. С. 1938. Борьба с лесными пожарами. М.—Л.
Мензбир М. А. 1882. Орнитологическая география Европейской Рос-
сии. Уч. зап. Моск, унив., отд. естеств.-истор., вып. 2—3.
Мензбир М. А. 1934. Очерк истории фауны европейской части СССР
(от начала третичной эры). М.—Л.
Меринов М. П., Е. Ф. Литвиненко, И. А. Степанова,
А. С. С л е с а р е н к о, В. М. С т у п н и ц к а я, В. В. С л е-
саренко, О.П. Хижинская, С. Г. Буяло иВ.В. Ада-
мович. 1959. К вопросу о природной очаговости туляремии в Ук-
26*
407
раинской ССР. Тез. X совещ. по паразитол. пробл. и природноочаго-
вым болезням, М.
М ид ден д орф А. 1869. Путешествие на север и восток Сибири, ч. 2.
Сибирская фауна. СПб.
Михеев А. В. 1952. Род белые куропатки. В кн.: Птицы Советского
Союза, т. 4, М.
Михель Н.М. 1935. Материалы по птицам Индигирского края. Тр.
Арктическ. инет., вып. 31, Л.
Молчанов А. А. 1960. Гидрологическая роль леса. М.
Молчанов А. А. 1961. Лес и климат. М.
Морозов Г. Ф. 1914. Роль вмешательства человека в жизнь леса. Лесо-
промышленный вестник, № 18.
Надеждин Б. В. 1959. Материалы к характеристике почвенного покрова
Усть-Ордынского Бурятского национального округа Иркутской об-
ласти. Тр. Вост.-Сиб. фил. СО АН СССР, сер. биол., вып. 17 (Почво-
ведение) , Благовещенск.
Наумов Н.П. 1930. Материалы к познанию «урожая» белки. Предва-
рительное сообщение. Тр. по лесн. опытн. делу, вып. 7, М.
Наумов Н.П. 1934. Млекопитающие Тунгусского округа. Тр. полярн.
комисс., вып. 17, Л.
Наумов Н.П. 1951. Новый метод изучения экологии мелких лесных гры-
зунов. В сб.: Фауна и экология грызунов. Матер, по грызунам,
вып. 4, М.
Наумов Н.П. 1955. Экология животных. М.
Наумов Р.Л. 1962. К биологии лесного дупеля в Средней Сибири. В сб.:
Орнитология, вып. 4, М.
Наумов Р.Л. 1963. Накармливается ли таежный клещ на обыкновен-
ной и белошапочной овсянках? Зоол. журн., т. 42, вып. 4.
Наумов Р. Л. и Т. Е. Б у р к о в с к а я. 1959. Новые сведения о пти-
цах Красноярского края. В сб.: Орнитология, вып. 2, М.
Наумов Р. Л., Е. И. Л ев кович и О. Е. Р ж а х о в а. 1963.
Об участии птиц в циркуляции вируса клещевого энцефалита
(Красноярский край). Мед. паразитол. и паразитарн. болезни,
№ 1.
Наумов С. П. 1947. Экология зайца-беляка. М.
Нейфельдт И. А. 1959. Некоторые особенности картирования распре-
деления птиц по биотопам. Матер, перв. сессии научи, совета по пробл.
«Биологические комплексы районов нового освоения, их рациональное
использование и обогащение», М.—Л.
Некипелов Н. В. 1941. Новые наблюдения по биологии красно-серой
полевки. Тр. Моск, зоотехп. инет., т. I, М.
Некипелов Н. В. 1962. Распределение млекопитающих по биотопам
юго-восточного Забайкалья. Изв. Иркутск, гос. н.-иссл. противочумн.
инет. Сибири и Дальнего Востока, т. 24, Иркутск.
Неронов В. М. 1962. Особенности изменения населения мышевидных
грызунов при освоении черневой тайги в предгорьях Алтая и Салаира.
Вести. МГУ, сер. 5, геогр., № 5.
Нестеров В. Г. 1945. Пожарная охрана леса. М.
Никитина Н. А. 1961. Результаты мечения мелких млекопитающих
в Коми АССР. Бюлл. МОИП, отд. биол., т. 66, вып. 2.
Никитина Н. А. и М. А. М е р к о в а. 1962. Использование терри-
тории некоторыми видами мышей и полевок. В сб.: Вопр. экологии,
т. 6, М.
Никитина Н. А. и М. А. М е р к о в а. 1963. Использование терри-
тории мышами и полевками по данным мечения. Бюлл. МОИП, отд.
биол., т. 68, вып. 5.
Никифоров Л. П. 1959. Опыт биосъемки населения млекопитающих
Тоболо-Ишимской лесостепи. В сб.: География населения животных
и методы его изучения, М.
408
Никифоров Л. П. 1961а. Опыт зоолого-паразитологического иссле-
дования природных очагов клещевого энцефалита в Тяжинском районе
Кемеровской области. Мед. паразитол. и паразитарн. болезни,
№ 1.
Никифоров Л. П. 19616. Опыт абсолютного учета численности мел-
ких лесных млекопитающих. В сб.: Вопр. организ. и мет. учета ре-
сурсов фауны наземных позвоночных, М.
Никифоров Л. П. 1963. Опыт абсолютного учета численности мелких
млекопитающих в лесу. В сб.: Организ. и мет. учета птиц и вредных
грызунов, М.
Никифоров Л. П. 1964. Распространение млекопитающих в лесостепи
Тоболо-Ишимского междуречья. Бюлл. МОИП, отд. биол., т. 49,
вып. 4.
Николаев И. В. 1949. Почвы Иркутской области. Иркутск.
Никсо-Никочио Н. В. 1956. Особенности фауны в связи с типами
леса. Научно-методич. информация, № 12 (Моск, лесотехнич. инет.).
Нумеров К. Д., П. Н. Павлов. 1963. Прошлое и настоящее соболя
Енисейской Сибири. Уч. зап. Красноярск, пед. инет., т. 24, вып. 5,
каф. зоол., Красноярск.
Образцов Б.В. 1964. Лесостепь как ландшафтная зона с точки зрения
зоолога. Бюлл. МОИП, отд. биол., т. 49, вып. 4.
Огнев С. И. 1928. Звери Восточной Европы и Северной Азии, т. I. Насе-
комоядные млекопитающие и летучие мыши. М.
Огнев С. И. 1940. Звери СССР и прилежащих стран, т. 4. Грызуны. М.
Огнев С. И. 1950. Звери СССР и прилежащих стран, т. 7. Гры-
зуны. М.
Окладников А. П. 1948. Древняя тюркская культура в верховьях
р. Лены. Кратк. сообщ. Инет, истор. материальной культ. АН СССР,
вып. 19, М.
Окладников А. П. 1949. Древнейшие наскальные изображения Се-
верной Азии. Советская археология, № И.
Окладников А. П. 1956. Народы Восточной Сибири. В кн.: Народы
Сибири. Этнографические очерки. М.
Околович А. К. и Г. К. Корсаков. 1951. Ондатра. М.
Окулова Н.М. иЮ.А. Дубровский. 1962. О подвижности двух
фоновых видов грызунов в очаге клещевого энцефалита Салаирского
кряжа. В сб.; Пробл. зоол. исслед. в Сибири, Горно-Алтайск.
Осмоловская В. И. [Консультант]. 1960. Зоогеографическая карта
СССР. М. 1 : 10 млн. М.
Осмоловская В. И. [Консультант]. 1962. Зоогеографическая карта.
Фаунистические подразделения. Атлас СССР. М.
Павлинин В. Н. 1947. Материалы по кольцеванию крота (Talpa euro-
раса L.) на Урале. Зоол. журн., № 6.
Павловский Е. Н. 1937. Учение о биоценозах в приложении к неко-
торым паразитологическим проблемам. Изв. АН СССР, № 4.
Павлюченко В.М. 1958. Соболь и белка в восточно-сибирской тайге.
Природа, № 8.
Палий В. Ф. 1958. О типах распространения животных и характере
зоогеографического районирования. В сб.: Пробл. зоогеогр. суши,
Львов.
Паллас П. С. 1786. Путешествие по разным провинциям Российского
государства, ч. 2, кн. 2. Пер. Ф. Томанского. СПб.
П е т р и Б. Э. 1930. Описание охотничьей территории тутурских тунгусов.
Изв. биол.-геогр. н.-иссл. инет, при Иркутск, гос. унив., т. 5, вып. 2,
Иркутск.
Пешкова Г. А. 1960. Краткий анализ флоры степей Приангарья. Научн.
чтения нам. М. Г. Попова, вып. 1, Новосибирск.
Пешкова Г. А. 1962. Взаимоотношения леса и степи в Приангарье.
Тр. Вост.-Сиб. биол. инет., вып. 1 (Ботаника), Иркутск.
409
II и и иг ин А. Ф. 1954. Бруцеллезная инфекция у длиннохвостых сусли-
ков. Изв. Иркутск, гос. н.-иссл. противочумн. инет. Сибири и Даль-
него Востока, т. 12, Иркутск.
Пинигин А. Ф. 1957. Сохраняемость бруцелл во внешней среде в усло-
виях низких температур. Тез. докл. конф. Иркутск, гос. н.-иссл.
противочумн. инет. Сибири и Дальнего Востока, вып. 2, Улан-Удэ.
Плесский П. В. 1952. Животный мир Кировской области (млекопитаю-
щие). В сб.: По родному краю, Киров.
Плохинский Н. А. 1961. Биометрия. Новосибирск.
Щлятер-П лохоцкий К. А. 1932. Бурундук и борьба с ним.
М.—Хабаровск.
Плятер-Плохоцкий К. А. 1936. Вредные и полезные млекопи-
тающие в сельском хозяйстве. Хабаровск.
Що варницын В. А. 1934. Леса и лесовозобновление в бассейне реки
Большой Белой в Восточных Саянах. Тр. СО ПС АН СССР, сер. сиб.,
вып. 7 (Ангарская лесная экспедиция 1931 г.), ч. 2, Л.
Поварницын В. А. 1937. Почвы и растительность бассейна р. Верхней
Ангары. В сб.: Бурят-Монголия (Тр. СОПС АН СССР, сер. Вост.-
Сиб., вып. 4), М.—Л.
Поварницын В. А. 1944. Кедровые леса СССР. Красноярск.
Погудин А. А. 1930. Очерк охоничьего промысла Киренского округа.
Тр. по лесн. опытн. делу, вып. 7, М.
Подаревский В. В. 1931. Летние корма пятнистого оленя в условиях
Восточно-Сибирского края. Союзпушнина, № 14—16.
Покшишевский В. В. 1951. Заселение Сибири. Иркутск.
Поляков И.С. 1873. Географическое распространение животных в юго-
восточной части Ленского бассейна. Зап. Русск. геогр. общ., Отд.
общ. геогр., 3, СПб.
Поплавская Г. И. 1913. Бассейн р. Верхней Ангары. Южные откры-
тые склоны. Предварит, отчет по исслед. почв Азиатской России, СПб.
Поплавская Г. И. 1916. На северной окраине Селенгинской Даурии
(Ботанико-географический очерк). Тр. Ботан. музея АН, вып. 16, СПб.
Попов В. А. 1960. Млекопитающие Волжско-Камского края. Насеко-
моядные, рукокрылые, грызуны. Казань.
Попов В. М. и Н. И. Иголкин. 1956. Фауна блох грызунов семей-
ства беличьих (Sciuridae), обитающих в лесной полосе Западной Си-
бири. Тр. Томск, н.-иссл. инет, вакцин и сывороток, т. 7, Томск.
Попов В. М. и В. Г. Казанская. 1958. Блохи насекомоядных лес-
ной зоны Западной Сибири. Тр. Томск, н.-иссл. инет, вакцин и сыво-
роток, Томск.
Попов М. Г. 1953. Вопрос о взаимоотношении леса (тайги) и степи в Сред-
ней Сибири. Бюлл. МОИП, отд. биол., т. 58, вып. 6.
Портенко Л. А. 1937. Фауна птиц Северного Урала. М.—Л.
Портенко Л. А. 1939. Фауна Анадырского края, чч. 1—2. Л.
Поспелов С. М. 1957. Птицы и млекопитающие ельника-черничника
различных возрастов. Зоол. журн., т. 36, вып. 4.
Промптов А. Н. 1934. Об экологических факторах изоляции у птиц.
Зоол. журн., т. 13, вып. 4.
Птицы Советского Союза. 1951—1954. Тт. 1—6. М.
Птушенко Е. С. 1951. Отряд длиннокрылые. В кн.: Птицы Советского
Союза, т. 1. М.
Птушенко Е. С. 1952. Отряд гусеобразные (Характеристика отряда
и подсемейство Гусиные). В кн.: Птицы Советского Союза, т. 4. М.
Раменский Л. Г. 1938. Введение в комплексное почвенно-геоботани-
ческое исследование земель. М.
Реймерс Н.Ф. 1954. К биологии кедровок южного Прибайкалья. Зоол.
журн., т. 33, № 6.
Реймерс Н. Ф. 1956а. Роль кедровки и мышевидных грызунов в кедро-
вых лесах южного Прибайкалья. Бюлл. МОИП, отд. биол., т. 61, вып. 2.
410
Реймерс Н. Ф. 19566. Роль млекопитающих и птиц в возобновлении
кедровых лесов Прибайкалья. Зоол. журн., т. 35, вып. 4.
Реймерс Н. Ф. 1958а. О некоторых особенностях количественного
учета птиц п мелких млекопитающих в условиях горной тайги юга
Средней Сибири. Зоол. журн., т. 37, вып. 8.
Реймерс Н. Ф. 19586. Облесение гарей и лесных массивов, уничтожен-
ных шелкопрядом, в горной кедровой тайге юга Прибайкалья, и роль
позвоночных животных в этом процессе. Бюлл. МОИП, отд. биол.,
т. 63, вып. 4.
Реймерс Н. Ф. 1958в. Уничтожение сибирского шелкопряда зверями
и птицами, населяющими кедровые леса и восстановление леса на
шелкопрядниках и гарях. Тр. по лесн. хоз., вып. 4, Новосибирск.
Реймерс Н. Ф. 1959а. Птицы кедровых лесов юга Средней Сибири и
их роль в жизни кедра. Тр. Биол. инет. Сиб. отд. АН СССР, вып. 5,
зоол., Новосибирск.
Реймерс Н.Ф. 19596. Бурозубки и мышевидные грызуны кедровой тайги
Прибайкалья и их роль в жизни леса. Зоол. журн., т. 38, вып. 8.
Реймерс Н. Ф. 1959в. К вопросу о роли позвоночных животных в во-
зобновлении лиственничных лесов лесостепной Тувы. Изв. Сиб. отд.
АН СССР, № 12.
Реймерс Н.Ф. 1959г. Гнездование тонкоклювых кедровок в Средней
Сибири. Зоол. журн., т. 38, № 6.
Реймерс Н. Ф. 1959д. Птицы и млекопитающие в очагах массового
размножения сибирского шелкопряда среди лиственничных лесов
Тувы. Изв. Сиб. отд. АН СССР, № 10.
Реймерс Н.Ф. 1960а. Бурундук и северная пищуха в кедровой тайге
Прибайкалья. Тр. Вост.-Сиб. фил. СО АН СССР, вып. 23, сер. биол.,
Благовещенск.
Реймерс Н. Ф. 19606. Фенологические наблюдения, произведенные
в Аларском аймаке Усть-Ордынского национального округа (Голо-
винская метеостанция) П. В. Поповым в 1933—1939 гг. Бюлл. Вост.-
Сиб. фенология, комисс., № 1. Вопросы региональной фенологии и
биогеографии, Иркутск.
Реймерс Н. Ф. 1960в (1961). Материалы к познанию орнитофауны лес-
ных участков лесостепной зоны Тувинской автономной области.
Биол. сб. (1960), Иркутск.
Реймерс Н.Ф. 1963а. Размещение и численность лесных птиц зимой
в ландшафтах юга Средней Сибири, Сиб. геогр. сб., вып. 2, М.
Реймерс Н. Ф. 19636. Восточно-Сибирская фенологическая комиссия
в 1960 году (Обзор поступивших материалов и деятельность комиссии).
Бюлл. Вост.-Сиб. фенология, комисс., вып. 2—3. Сезонная и вековая
динамика природы Сибири, Иркутск.
Реймерс Н. Ф. 1963в. Весна 1961 года на юге Средней Сибири. Бюлл.
Вост.-Сиб. фенология, комисс., вып. 2—3. Сезонная и вековая динамика
природы Сибири, Иркутск.
Реймерс Н.Ф. 1963г. О биологии, фенологии и размещении лесных птиц
в окрестностях пос. Жигалово (р. Тутура, приток верхней Лены).
Бюлл. Вост.-Сиб. фенология, комисс., вып. 2—3. Сезонная и вековая
динамика природы Сибири, Иркутск.
Реймерс Н. Ф. 1963д. Зоогеографическая карта лесной Сибири. М.
1 : 10 000 000. В сб.: Вопр. зоол. картограф., М.
Реймерс Н.Ф. 1965. Эколого-зоогеографическая типология и райони-
рование таежной Сибири. Чтения пам. М. Г. Попова, Иркутск.
Реймерс Н. Ф. и Г. А. Воронов. 1963. Насекомоядные и грызуны
верхней Лены. Иркутск.
Реймерс Н. Ф. и Л. И. Малыше в. 1963. Нарушенность лесов
Средней Сибири. Бюлл. Вост.-Сиб. фенология, комисс., вып. 2—3.
Сезонная и вековая динамика природы Сибири, Иркутск.
411
Реймерс Н. Ф. и А. С. Рожков. 1960. Уничтожение сибирского
шелкопряда зверями и птицами (в условиях кедровых лесов Прибай-
калья). Тр. Вост.-Сиб. фил. СО АН СССР, вып. 23, сер. биол., зоол.,
Благовещенск.
Рогачева Э. В. 1959. Очерк фауны млекопитающих и птиц Енисей-
ской тайги и лесотундры. В сб.: География населения наземных жи-
вотных и методы его изучения, М.
Рогачева Э.В. 1962. Численность и размещение птиц нижнего Е логу я
(приенисейская тайга). В сб.: Орнитология, вып. 5, М.
Рогачева Э. В., Е. Е. Сыроечковский. 1962. О влиянии дея-
тельности человека на фауну енисейской тайги. Уч. зап. Моск. обл.
пед. инет., т. 109, биогеограф., вып. 1.
Рожков А. С. и Н. Ф. Реймерс. 1958. Материалы по питанию птиц
сибирским шелкопрядом (Dendrolimus sibiricus Tschtv.) в очаге мас-
сового размножения вредителя. Зоол. журн., т. 37, вып. И.
Россолимо О. Л. и Е. Е. Сыроечковский. 1961. Новые
сведения о распространении рыжей полевки (Clethrionomys glareolus
Schreb.) в Сибири. Научн. докл. высш, школы, Биол. науки,
№ 2.
Рустамов А. К. 1954. Семейство вороновые. В кн.: Птицы Советского
Союза, т. 5. М.
Сазонова О. Н. 1947. О блохах грызунов и насекомоядных низовьев
р. Иртыша. Новости медицины; Паразитология и трансмиссивные
болезни.
Сапожникова С. А. 1950. Микроклимат и местный климат. Л.
Сдобников В. М. 1958. Сравнительно-экологический анализ фауны
тундры и тайги. Зоол. журн., т. 37, вып. 4.
Северцев Н. А. 1877. О зоологических (преимущественно орнитоло-
гических) областях внетропических частей нашего материка. Изв.
Русск. геогр. общ., т. 13, вып. 3. СПб.
Сердюкова Г. В. 1960. Цикл развития клещей семейства Ixodidae.
Тр. Всесоюзн. Энтомология, общ., т. 47.
Сердюкова Г. В. иА.Н. Ходаковский. 1940. Иксодовые клещи
в Западных Саянах. Тез. докл. Втор, совещ. по паразитол. пробл.,
М.—Л.
С
С
С
С
С
с
с
с
с
к а л о н В. Н. 1931. О некоторых млекопитающих — вредителях Верх-
него Приангарья (в оглавлении: «вредящих в Восточной Сибири»)
и мерах борьбы с ними. Тр. по защите растений Сибири, т. 1 (8), Но-
восибирск.
к а л о н В. Н. 1933. К изучению фауны грызунов Восточносибирского
края. В кн.: Вредные саранчевые. Вредители огородничества. Грызуны
Приангарья. Мокрая головня. М.—Иркутск.
к а л о н В. Н. 1934. Пернатые хищники Верхнего Приангарья и их роль
в жизни человека. Изв. Иркутск, гос. противочумн. инет. Сибири и
Дальнего Востока, т. 1. Сб. работ противочумн. организ. Восточно-
сибирск. края за 1932—1933 гг., М.—Иркутск.
калон В. Н. 1935. Новые данные по фауне млекопитающих и птиц Си-
бири и Дальнего Востока. Изв. Иркутск, гос. противочумн. инет.
Сибири и Дальнего Востока, т. 2, М.—Иркутск.
а л о н В. Н. 1936. Материалы к познанию фауны южных границ Си-
бири. Изв. Иркутск, гос. противочумн. инет. Сибири и Дальнего Вос-
к
к
к
к
к
тока, т. 3, М.—Иркутск.
а л о н В. Н. 1951а. Речные бобры Северной Азии. М.
а л о н В. Н. 19516. Русские землепроходцы XVII века в Сибири. М.
а л о н В. Н. 1956. Обзор материалов о пролете птиц в Якутии. Изв.
биол.-геогр. н.-иссл. инет, при Иркутск, гос. унив. им. А. А. Жда-
нова, т. 16, вып. 1—4, Л.
а л о н В. Н. 1962. Млекопитающие (карта). Атлас Иркутской области.
М.—Иркутск.
412
Скалой В. Н., Н. В. Некипелов и В. Н. Ефимова. 1936.
Характеристика запасов пищухи. Изв. Иркутск, гос. противочумн.
инет. Сибири и Дальнего Востока, т. 3, М.—Иркутск.
Скрябин Н. Г. и К. П. Филонов. 1962. Материалы к фауне птиц
северо-восточного побережья Байкала. Тр. Баргузинского гос. за-
поведи., вып. 4, Улан-Удэ.
Слепцов М. М. 1947. К биологии уссурийской мыши-малютки (М ic го-
ту s minutus ussuricus Barr.-Ham.). В сб.: Фауна и экология грызунов.
Матер, по грызунам, вып. 2, М.
Слудский А. А. 1934. Ондатра в Восточносибирском крае. Охотник
и рыбак Сибири, № 3, Новосибирск.
Слудский А. А. 1948. Ондатра. Алма-Ата.
См и ренский А. А. 1950. Типы ондатровых угодий Восточного При-
байкалья и их хозяйственная оценка. Тр. Всесоюзн. н.-иссл. инет,
охотн. промысла, вып. 9, М.
Снегиревская Е. М. 1947. Материалы по биологии размножения и
колебаниям численности землероек в Башкирском заповеднике. Тр.
Башкирск. заповеди., вып. 1, М.
Снегиревская Е. М. 1959. Картирование распределения грызунов.
Матер, перв. сессии научи, совета по пробл. «Биологические комплексы
районов нового освоения, их рациональное использование и обогаще-
щение», М.—Л.
Снегиревская Е. М. 1961. Наблюдения над полевкой-экономкой на
островах средней Волги. В сб.: Морфология и экология позвоночных
животных. Тр. Зоол. инет. АН СССР, т. 29, М.—Л.
Сныткин Г. В. 1963. Шкалы пожарной опасности для лесов Иркутской
области. В сб.: Лесные пожары и борьба с ними. М.
Собанский Г. 1958. Норка в Красноярском крае. Охота и охотничье
хозяйство, № 12.
Совещание по вопросам лесостепи. 1964. Бюлл. МОИП, отд. биол.,
т. 69, вып. 4.
Соколов Г. А., А. И. Хлебников. 1962. Влияние концентрирован-
ных рубок в кедровых лесах Западного Саяна на производительность
охотничьих угодий. В сб.: Организация лесного хозяйства и инвента-
ризация лесов, вып. 1. Тр. Инет, леса и древесины СО АН СССР, т. 58,
Красноярск.
Соколова 3. А. 1962. Постэмбриональное развитие обыкновенной
бурозубки (Sorex araneus L.). Научн. докл. высш, школы, Биол. науки,
№ 3.
Соловьев Д. К. 1920. Саянский промыслово-охотничий район и собо-
линый промысел в нем. Отчет Саянской экспедиции 1914—1916 гг.
Тр. экспед. по изучению соболя и исслед. соболиного промысла. Сер. 2.
Саянская экспедиция. Пгр.
Сонин В. Д. 1962. Некоторые материалы по питанию и распространению
хищных птиц Иркутской области. Изв. Вост.-Сиб. отд. Геогр. общ.
СССР, т. 60, Иркутск.
Сочава В. Б. (ред.). 1960. Карта растительности СССР.
М. 1 : 10 000 000. М.
Сочава В.Б. иВ.В. Липатова. 1960. Группировки степных расте-
ний в амурской подтайге. Тр. Моск. общ. испытателей природы,
т. 3, М.
Спаигенберг Е. П. 1951а. Отряд голенастые птицы. В кн.: Птицы
Советского Союза, т. 2. М.
Спаигенберг Е.П. 19516. Отряд пастушки. В кн.: Птицы Советского
Союза, т. 3. М.
Спаигенберг Е.П. 1954. Семейство скворцовые. В кн.: Птицы Со-
ветского Союза, т. 5. М.
Спаигенберг Е.П. и А. М. Судиловская. 1954. Род овсянки.
В кн.: Птицы Советского Союза, т. 5. М.
413
Спасский А. А., К. М. Рыжиков. 1951. Гельминты пищух При-
байкалья. Тр. Гельминтология, лабор. АН СССР, т. 5, М.
Средняя Сибирь. 1964. Сер.: Природные условия и естественные ре-
сурсы СССР. М.
Стариков Г. Ф. и А. А. Степанов. 1963. О реконструкции гео-
графических ландшафтов Приамурья. Вопр. геогр. Дальнего Востока,
сб. 6, Хабаровск.
Строганов С. У. 1957. Звери Сибири. Насекомоядные. М.
Строганов С. У. 1962. Звери Сибири. Хищные. М.
Строгий А. А. 1911. О лесах Сибири. СПб.
Судиловская А. М. 1951. Отряд кукушки. В кн.: Птицы Советского
Союза, т. 1. М.
Сыроечковский Е. Е. 1960. Изменение ареалов птиц в Средней
Сибири в результате потепления климата и воздействия человека.
В сб.: Орнитология, вып. 3, М.
Сыроечковский Е. Е. и О. Л. Россолимо. 1960. Соболь в бас-
сейне Подкаменной Тунгуски. Зоол. журн., т. 39, № И.
Сыроечковский Е. Е. и Э. В. Рогачева. 1958. Новые сведе-
ния о распространении некоторых птиц в приенисейской тайге. Про-
блемы Севера, вып. 2 (отдельный оттиск).
Сыроечковский Е. Е.иЭ.В. Рогачева. 1961. Соболь и дикие
копытные Приенисейской Сибири. Природа, № 3.
С я У - п и н. 1961. Численность и участки обитания азиатской лесной
мыши (кит.). Реф. журн. Биология, Зоол. позвоночных, № 1.
Тарасов М. П. 1962. Стадиальное размещение и относительная числен-
ность массовых видов грызунов западного Хамар-Дабана. Изв.
Иркутск, гос. н.-иссл. противочумн. инет. Сибири и Дальнего Востока,
т. 24, Иркутск.
Тарасова В. Е. и 3. С. Кудинова, 1961. Предварительные дан-
ные о токсоплазмозе в Иркутской области. Докл. Иркутск, противо-
чумн. инет., вып. 1, Улан-Удэ.
Телегин В. И. 1961. О вреде бурундука в Западной Сибири. Матер,
планово-метод. совещ. по защите раст. зоны Урала и Сибири, Ново-
сибирск.
Тимофеев В. В. 1947. Белка и беличий промысел. Иркутск.
Тимофеев В. В. 1949. Звери нашей области. Иркутск.
Тимофеев В. В. 1951. Соболь Восточной Сибири. Иркутск.
Тимофеев В. В. 1958. Наши хищные птицы. Иркутск.
Тимофеев В. В. 1962. Влияние соболя на численность белки в Восточ-
ной Сибири. Тр. Всес. н.-иссл. инет, животн. сырья и пушнины,
вып. 19, М.
Тимофеев В. В. и В. Н. Н а д е е в. 1955. Соболь. М.
Тимофеев-Ресовский Н. В. 1961. О некоторых принципах клас-
сификации биохорологических единиц. Тр. Инет. биол. Уральск,
фил. АН СССР, вып. 27, Свердловск.
Тимофеева Л. А. и В. Я. Головачева. 1962. Характеристика
псевдотуберкулезных культур, выделенных от грызунов. Докл.
Иркутск, противочумн. инет., вып. 3, Хабаровск.
Ткаченко М. И. 1924. Распространение некоторых птиц по рекам
Н. Тунгуске, Алдану и Мае. Изв. ВСОРГО, т. 47, Иркутск.
Ткаченко М. И. 1937. Птицы Нижней Тунгуски. Изв. Иркутск, гос.
музея, т. 2, Иркутск.
Толмачев А. И. 1958. Ареал вида и его развитие. В сб.: Проблема вида
в ботанике, т. 1, М.—Л.
Толмачев А. И. 1962. Основы учения об ареалах. Л.
Траханов Д. Ф. 1962. Распространение и экономическое значение
мышевидных грызунов на фермах Тюменской области. Тр. Всесоюзн.
н.-иссл. инет. вет. санитарии, т. 20, М.
414
Троицкий В. Н. 1930. Охотничий промысел Чуно-Ангарского района
б. Канского округа. Изв. Сиб. краев, научн. охотн.-промысл. станц.,
Новосибирск.
Тугаринов А. Я. 1927. Птицы Приенисейской Сибири. Список и рас-
пространение. Зап. Средне-Сиб. отд. РГО, сер. 2, т. 1, вып. 1, Красно-
ярск.
Тугаринов А. Я.и С. А. Бутурлин. 1911. Материалы по птицам
Енисейской губернии. Зап. Красноярск, подотд. Вост.-Сиб. отд.
РГО по физическ. геогр., т. 1, вып. 2—4, Красноярск.
Тупикова Н. В. 1947. Экология домовой мыши Средней полосы СССР
(по материалам, собранным в Москве и Московской области). В сб.:
Фауна и экология грызунов. Матер, по грызунам, вып. 2, М.
Тупикова Н. В. 1949. Питание и характер суточной активности земле-
роек средней полосы СССР. Зоол. журн., т. 28, вып. 6.
Тупикова И. В. 1959. Опыт картирования густоты заселенности тер-
ритории водяной крысой по данным районных заготовок пушнины.
В сб.: География населения наземных животных и методы его изуче-
ния, М.
Туров С. С. 1936. Материалы по млекопитающим северо-восточного побе-
режья Байкала и Баргузинского хребта. Сб. Тр. гос. зоомузея (при
МГУ), вып. 3, М.
Успенский С. М. 1960. Широтная зональность авифауны Арктики.
В сб.: Орнитология, вып. 3, М.
Устинов С. К. 1960. Годовой цикл жизнедеятельности бурого медведя
Северо-Восточного Прибайкалья. Изв. Иркутск, с.-х. инет., вып. 18.
Устинов С. К. 1962а. Состояние и пути дальнейшего изучения роли
крупных млекопитающих в очагах клещевого энцефалита на террито-
рии СССР. Тр. Иркутск, н.-иссл. инет, эпидемиол. и микробиол.,
вып. 7.
Устинов С. К. 19626. К познанию экологии крупных млекопитающих
в первичных очагах клещевого энцефалита. Тр. Иркутск, н.-иссл.
инет, эпидемиол. и микробиол., вып. 7.
Фаворский В. П? 1935. Соболь Восточной Сибири. Иркутск.
Федорова Л. В. 1960. Материалы по фауне кровососущих членисто-
ногих Братского района Иркутской области. Биол. сб. (1960),
Иркутск.
Федосенко А. К. 1958. Орнитологические находки в Красноярском
крае. Бюлл. МОИП, отд. биол., т. 63, вып. 4.
Феоктистов А. 3. 1962. Выделение вируса клещевого энцефалита от
теплокровных ранней весной. Докл. Иркутск, противочумн. инет.,
вып. 3, Хабаровск.
Фетисов А. С. 1934. Материалы по питанию тетерева (Lyrurus tetrix L.).
в юго-восточной части Прибайкалья Зоол. журн., т. 13, вып. 2.
Фетисов А. С. 1936. Материалы по систематике и географическому рас-
пространению млекопитающих западного Забайкалья. Изв. Иркутск,
гос. противочумн. инет. Сибири и Дальнего Востока, т. 3, М.—Ир-
кутск.
Фетисов А.С. 1942. Новые исследования по фауне грызунов Западного
Забайкалья. Изв. Биол.-геогр. н.-иссл. инет, при Иркутск, гос. унив.
им. А. А. Жданова, т. 9, вып. 3—4, Иркутск.
Фетисов А. С. 1945. Крыса-карако в Восточной Сибири. Иркутск.
Фетисов А. С. 1947. Материалы по питанию соболей Восточной Сибири.
Иркутск.
Фетисов А. С. 1953. Козуля в Восточной Сибири (ее биология и промы-
сел). Иркутск.
Фетисов А. С. 1957. Материалы по экологии восточносибирского лося
в связи с воспроизводством его запасов (4 Ices alces pftzenmayerl Zu-
kowsky). Изв. Биол.-геогр. н.-иссл. инет, при Иркутск, гос. унив.
им. А. А. Жданова, т. 17, вып. 1—4, Иркутск.
415
Фетисов А. С. 1958. Материалы по экологии полевки-экономки (Micro-
tus oeconomus Pall.) в Восточной Сибири. Бюлл. МОИП, отд. биол.,
т. 63, вып. 2.
Фирстов Н. И. 1957. О восточной границе распространения водяной
крысы. Изв. Иркутск, гос. н.-иссл. противочумн. инет. Сибири и
Дальнего Востока, т. 16.
Фирстов Н. И. 1960. Грызуны района Братской ГЭС. Изв. Иркутск,
гос. н.-иссл. противочумн. инет. Сибири и Дальнего Востока, т. 23.
Фолитарек С. С. 1940. К биологии обыкновенной землеройки (Sorex
araneus L.). Зоол. журн., т. 19, вып. 2.
Формозов А. Н. 1947. Очерк экологии мышевидных грызунов, носите-
лей туляремии. М.
Формозов А. Н. 1948. Мелкие грызуны и насекомоядные Шарьинского
района Костромской области в период 1930—1940 гг. В сб.: Фауна
и экология грызунов. Матер, по грызунам, вып. 3, М.
Формозов А. И. 1959. Предисловие (к сборнику). В сб.: География на-
селения наземных животных и методы его изучения, М.
Формозов А. Н., Н. П. Н а у м о в и И. Д. К и р и с. 1934. Экология
белки. М.—Л.
Хабаева Г. М. 1959. К фауне грызунов Муйской долины. Краеведч.
сб. Бурятск. фил. Геогр. общ. СССР, вып. 4, Улан-Удэ.
Хлонов Ю. П. и Н. Ф. Реймерс. 1960. Почвенный запас семян и
роль некоторых позвоночных животных в возобновлении липового
леса Горной Шории. Изв. Сиб. отд. АН СССР, № 4.
Худяков П. И. 1937. Борьба с волком в Восточной Сибири. Иркутск.
Чабовский В. И. 1959. Роль позвоночных животных в расселении
клещей Ixodes persulcatus. Сб. научно-практич. работ Пермск. обл.
сан.-эпид. станц., вып. 2, Пермь.
Черкасов А. А. 1867. Записки охотника Восточной Сибири. СПб. (Из-
дания: 1884, СПб.; 1958, Чита; 1962, М.).
Шарашидзе В. А. 1951. К экологии северной пищухи, или сеноставки
в Красноярском крае. Природа, № 11.
Шведов А. П. 1962. К распространению птиц среднего Приангарья
(Братский и Заярский районы Иркутской области). В сб.: Орнитоло-
гия, вып. 4, М.
Шер г ин И. А. 1950. Ондатроводство в Иркутской области. Иркутск.
Шилова С. А. 1959. Значение позвоночных животных в формировании
очагов клещевого энцефалита. Тез. X совещ. по паразитол. пробл.
и природноочаговым болезням, М.
Шилова С. А., Г. В. М а л ь к о в, В. И. Ч а б о в с к и й и Е. В. М е-
щерякова. 1956. Влияние депрессии численности мышевидных
грызунов на прокормление личинок и нимф клещей Ixodes persul-
catus Р. Sch. в очаге клещевого энцефалита. Бюлл. МОИП, отд. биол.,
т. 61, вып. 3.
Шилова С. А. и Д. Г. К р ы л о в. 1957. Изменение роли птиц в очагах
клещевого энцефалита под влиянием вырубок леса. Бюлл. МОИП,
т. 62, вып. 6.
Шилова С. А., В. Б.Троицкий, Г. Б. Мальков и В.М.Бель-
кович. 1958. Значение подвижности лесных мышевидных грызу-
нов в распределении клещей Ixodes persulcatus Р. Sch. в очагах ве-
сенне-летнего энцефалита. Зоол. журн., т. 37, вып. 6.
Шкатулова А. П. 1963. К количественной характеристике орнито-
фауны некоторых физико-географических районов Читинской области.
Уч. зап. Читинск. гос. пед. инет. (секц. геогр. и биол.), вып. 8, Чита.
Шоцкий В. П. 1962. Сельское хозяйство. Атлас Иркутской области.
М.—Иркутск.
Шрейнер Я. Ф. 1916. Крот обыкновенный (Talpa еигораеа еигораеа
L.) по новейшим исследованиям и важнейшие способы борьбы с ним.
Игр.
416
Штильмарк Ф. Р. 1963а. Об экологии бурундука (Eutamias sibiricus
Laxm.) в кедровых лесах Западного Саяна. Зоол. журн., т. 42, вып. 1.
Ш т ильмарк Ф. Р. 19636. Лесохозяйственное значение позвоночных
животных Сибири и задачи исследований в этой области. Матер, по
изучению лесов Сибири и Дальнего Востока (тр. конф.) [Инет, леса
и древесины СО АН СССР], Красноярск.
Ш т ильмарк Ф. Р. 1963в. Европейская рыжая полевка в лесах Запад-
ного Саяна и Салаирского кряжа. Уч. зап. Красноярск, пед. ипст.,
т. 24, вып. 5, каф. зоол., Красноярск.
Ш т и л ь м а р к Ф. Р. 1963г. Материалы о фауне и размещении по биото-
пам грызунов верхнеленской тайги. В сб.: Защита лесов Сибири от
насекомых-вредителей, М.
Штильмарк Ф. Р., Л. П. Климова, Н. К. Носкова,
Т. А. Ким, Л. В. Скиданова и Н. П. Елистратова.
1963. Зоолого-паразитологические материалы к изучению природных
очагов инфекции в лесах Западного Саяна (сообщение первое). Уч.
зап. Красноярск, пед. инет., т. 24, вып. 5, каф. зоол., Красноярск.
Штильмарк Ф. Р., Г. А. Соколов, А. И. Хлебников и
Н. П. Елистратова. 1963. Материалы о распространении и
численности млекопитающих в лесах среднегорной полосы Западного
Саяна. В сб.: Защита лесов Сибири от насекомых-вредителей, М.
Ю д и и Б. С. 1956. Материалы по питанию бурозубок (Sorex L.) Западной
Сибири. Тр. Томск, гос. унив., т. 142.
Ю д п н Б. С. 1957. К экологии крошечной бурозубки (Sorex minutissimus
Zimm.) в Западной Сибири. Тез. докл. на 9 научн. конф. Новосиб.
с.-х. ипст.
10 д и и Б. С. 1962. Экология бурозубок (род Sorex) Западной Сибири. Тр.
Биол. инет. СО АН СССР, вып. 8, Новосибирск.
Юдин К. А. 1952. Наблюдения над распространением и биологией птиц
Красноярского края. Тр. Зоол. инет. АН СССР, т. 9, вып. 4. Сб. работ
по позвоночным животным, М.—Л.
10 п а т о в А. А. 1950. Основные черты растительного покрова Монголь-
ской Народной Республики. Тр. Монгольск. комисс., вып. 30, М.—Л.
Ю ргенсон П. Б. 1937. Материалы по экологии и питанию бурого мед-
ведя. Тр. Центр, гос. заповеди., вып. 2.
Ю р г е н с о н П. Б. 1938. Материалы к познанию млекопитающих прпте-
лецкого участка Алтайского гос. заповедника. Тр. Алтайск. гос.
заповеди., вып. 1, М.
Я и у ш е в и ч А. И. 1952. Фауна позвоночных Тувинской области. Ново-
сибирск.
Я и у ш е в и ч А. И. и К. Т. 10 р л о в. 1949. Вертикальное распростране-
ние млекопитающих и птиц в Западном Саяне. Изв. Зап.-Сиб. фил.
СО АН СССР, сер. биол., т. 3, вып. 2, Новосибирск.
Ярошенко П. Д. 1961. Геоботаника. М.—Л.
Adams С. С. 1908. The ecological succession of birds, Auk, vol. 25.
Borowski S. i A. Dehnel. 1952. Materialy do biologii Soricidae.
Annales Universitatis Mariae Curie-Skladowska, vol. 7, 6.
Braun-Blanquet J. 1951. Pflanzensoziologie. 2-e Aul‘1. Wien.
Dehnel A. 1952. Biologia rozmozania ryjowki S. araneus L. w warunkach
laboraloryjnych. Annales Universitatis Mariae Curie-Skladowska,
vol. 6, 11.
Dybowski B. 1874. Notiz liber die ostsibirischen Pyrrhula-Arten. J. fur
Ornithologie, H. 1.
Friederichs K. 1927. Gtundsatzliches liber die Lebenseinheiten ho-
herer Ordnungund den okologischen Einheitsfaktor. Naturwissensch., 15.
Godfrey G. K. (Mrs. Crowcroft). 1954. Tracing field voles (Microtus agre-
stis) with a Geiger-Mliller counter. Ecology, vol. 35, № 1.
Hartert E. 1903—1922. Die Vogel der Palaarktischen Fauna. Berlin.
Hartert E. 1904. On the Birds collected by Mr. Robert Hall, of Melbourne,
417
on the Banks of the Lena River between Gigalowa and its mouth.
Ibis.
Hell wing H. M., B. S. Schnapp. 1962. Beitriige zur Kenntniss von
Sorex araneus tetragonurus Herm. in der Rumanischen Volksrepublik
(Vorausgehende Mitt.). Trav. Museum histoire natur. «Gr. Antipa», 3.
Hesse H. 1924. Tiergeographie auf okologischer Grundlage. Jena.
Kratochvil I., B. Rosicky. 1955. Hrabos severni (M icrotus oecono-
mus) relikt zvereny z doby ledove v 6SR. Pace Sesit 1, Rocnik 27.
M id d end orf A. J. 1853. Sibirische Reise. Saiigethiere, Vogel und
Amphibien. Bd. 2. St.-Petersb.
Nuorteva P., H. H о о g s t r a a 1. 1963. The incidence of ticks (Ixodoi-
dea, Ixodidae) on migratory birds arriving in Finland during the spring
of 1962; Reprint from Ann. Med. exp, Fenn. Vol. 41, Fasc. 4.
Pallas P. S. 1811. Zoographia Rosso-Asiatica, t. 1. Petropoli.
R a d d e G. 1862—1863. Reisen in Siiden von Ost-Sibirien in Jahren 1855—
1859. St.-Petersb.
Salt G. W. 1957. An analysis of avifaunas in the Teton Mountains and Ja-
ckson Hole, Wyoming. Condor, vol. 59, № 6.
Seebohm H. 1880. Siberia in Asia. A visit to the valley of the Yenessay
in East Siberia.
Stegmann B. 1928. Die Vogel Siid-Ost Transbaikaliens. Ежегодник Зо-
ологического музея АН СССР (отдельный оттиск).
Stegmann В. 1936. Die Vogel des nordlichen Baikal. J. fiir Ornitho-
logie, H. 1.
Taczanowski L. 1872—1874. Bericht uber die Ornithologischen Unter-
suchungen des Dr. Dybowski in Ost-Sibirien. J. fiir Omithologie,
1872 : 340—365, 433—453; 1873 : 81—118; 1874 : 315-337.
Taczanowski L. 1891—1893. Faune Ornithologique de la Siberie Ori-
entate. Mem. de 1’Acad. d. Sciences de St. Petersburg, 7 serie, vol. 39.
Thomas O. 1911. New mammals from Central and Western Asia, mostly
collected by Mr. Douglas Carruthers. Ann. Mag. Nat. Hist., ser. 8,
vol. 7.
Thomas 0. 1915. On a minute shrew from Lake Baikal. Ann. Mag. Nat.
Hist., vol. 15.
Tur 6 e k F. J. 1957. The bird succession in the conifer Plantations on mat-
grass Land in Slovakia (6SR). Ibis, 99, № 4.
Yerger R. W. 1953. Home Range, Territoriality and Populations of the-
Chipmunk in Central New York. J. Mammal., vol. 34, № 4.
ОГЛАВЛЕНИЕ
Стр.
Введение......................................................... 3
Раздел I
ЭКОЛОГО-ЗООГЕОГРАФЙЧЕСКАЯ ТИПОЛОГИЯ
И МЕЛКОМАСШТАБНОЕ ЗООГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ
ТАЕЖНОЙ СИБИРИ
Глава 1. Эколого-зоогеографическая типология..................... И
Глава 2. Территориальные варианты зооформаций и типолого-зоо-
географическая карта лесной Сибири ....................... 23
Пример деления зооформации на территориальные варианты 25
Зооформации лесной Сибири.................................. 31
I. Животное население лесотундры.................... 31
II. Животное население лесов......................... 33
III. Животное население сибирских высокогорий ... 36
IV. Животное население лесостепи и сходных участков 37
V. Животное население болот......................... 42
VI. Животное население рек и речных долин............ 43
VII. Животное население побережий крупных озер и морей 46
VIII. Животное население сибирских населенных пунктов 46
Глава 3. Мелкомасштабное типолого-зоогеографическое райониро-
вание сибирской части провинции таежных лесов и вопросы
методики исследования крупных регионов ................... 48
Раздел II
ПТИЦЫ И~МЛЕКОПИТАЮЩИЕ ЮЖНОЙ ТАЙГИ СРЕДНЕЙ СИБИРИ
И АНТРОПОГЕННАЯ ДИНАМИКА ИХ НАСЕЛЕНИЯ
Глава 4. Краткая историко-биогеографическая характеристика
южной тайги Средней Сибири ............................... 62
Глава 5. Повидовой обзор птиц и млекопитающих южной тайги
Средней Сибири............................................ 83
Птицы ..................................................... 83
Отряд Куриные — Galliformes ............................ 84
Отряд Голуби — Columbiformes ........................... 91
Отряд Пастушки — Ralliformes............................ 93
Отряд Журавли — Gruiformes ............................. 94
Отряд Кулики — Charadriiformes.......................... 95
Отряд Чайки — Lariformes............................... 107
Отряд Гагары — Gaviiformes............................. 107
419
Отряд Поганки — Colymbiformes.......................... 108
Отряд Гусеобразные — Anseriformes...................... 108
Отряд Фламинго — Phoenicopteriformes.................... ИЗ
Отряд Голенастые птицы — Ciconiiformes................ ИЗ
Отряд Хищные птицы — Falconiformes .................. 114
Отряд Совы — Strigifonnes............................ 123
Отряд Кукушки — Cuculiformes ........................ 125
Отряц Козодои — Caprimulgiformes..................... 128
Отряд Сизоворонки — Coraciiformes.................... 129
Отряд Удоды — Upupiformes............................ 129
Отряд Длиннокрылые — Micropodiformes ................ 129
Отряд Дятлы — Piciformes............................. 129
Отряд Воробьиные — Passeriformes..................... 137
Млекопитающие ........................................... 218
Отряд Насекомоядные — Insectivora ..................... 220
Отряд Рукокрылые — Chiroptera.......................... 251
Отряд Хищные — Carnivora............................... 255
Отряд Парнокопытные — Artiodactyla .................... 263
Отряд Грызуны — Rodentia .............................. 267
Отряд Зайцеобразные — Lagomorpha ...................... 336
Глава 6. Влияние антропогенных нарушений тайги на население
позвоночных животных .......................................... 342
Общие закономерности влияния нарушений тайги на отдельные
виды животных ......................................... 342
Трансформация лесов и животное население................. 351
Животные южной тайги Средней Сибири как кормовая база цен-
ных хищных зверей.................................... ?84
Нарушение лесов и предпосылки природноочаговых заболева-
ний ................................................. 287
Влияние позвоночных животных на возобновление лесов . . . 394
Животные — вредители сельского хозяйства ................ 394
Заключение .................................................... 396
Литература.................................................... 399
Николай Федорович Реймерс
ПТИЦЫ И МЛЕКОПИТАЮЩИЕ ЮЖНОЙ ТАЙГИ СРЕДНЕЙ СИБИРИ
Утверждено к печати
Сахалинским комплексным научно-исследовательским институтом Сибирского отделения
Академии наук СССР
Художник В. В. Грибакин. Технический редактор Н. А. Кругликова.
Корректоры Г. А. Аухимович, А. А. Гинзбург, О. М. Лаушкина и Г. А. Мошкина
Сдано в набор 26/IV 1965 г. Подписано к печати 23/XII 1965 г. РИСО АН СССР М 25—189В.
Формат бумаги 60х90’/1в. Бум. л. 131'8. Печ. л. 2б1/4 = 261/< усл. печ. л. 4-1 вкл. Уч.-изд.
л. 28.34-|-1 вкл. (0.4). Изд. № 2580. Тип. зак. Me 196. М-60118. Тираж 1500.
Цена 1 р. 90 к., переплет 20 к.
Ленинградское отделение издательства «Наука»
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. издательства «Наука». Ленинград, В-34, 9 линия, д. 12
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Стра- ница Строка Напечатано Должно быть
17 2 снизу общее им изменение общее изменение
101 16 сверху в Сибири в Средней Сибири
131 6—7 » между водоразделами низшего порядка — 1 пара на 1.5 —5—7 км, в среднем на 3—4 км. расстояние между водоразделами низшего порядка 1.5 —5—7 км, в среднем 3—4 км.
219 12 снизу (1962). (1952).
314 22 сверху И. К. Лаптев. И. П. Лаптев.