












































































































































































































































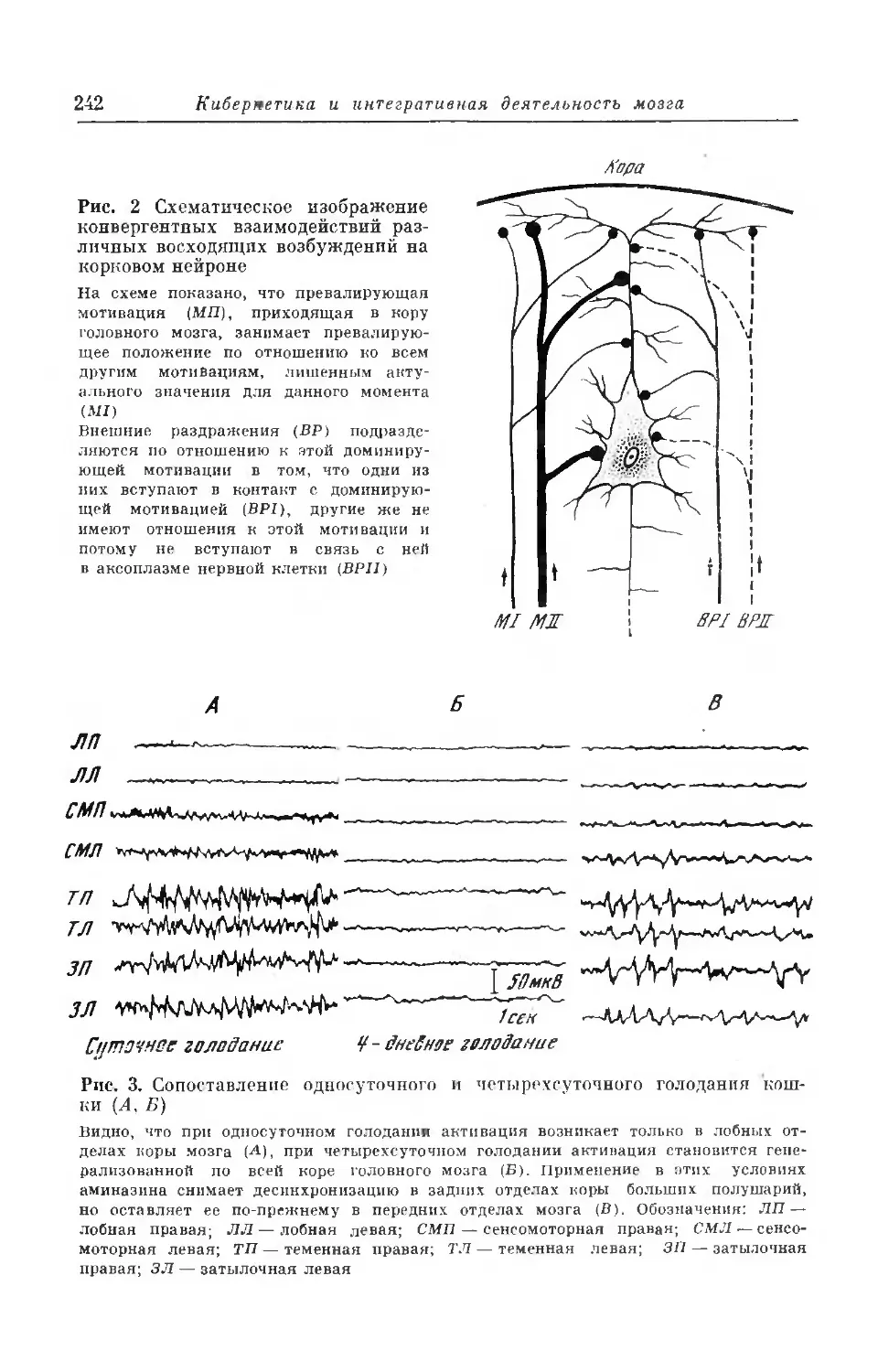


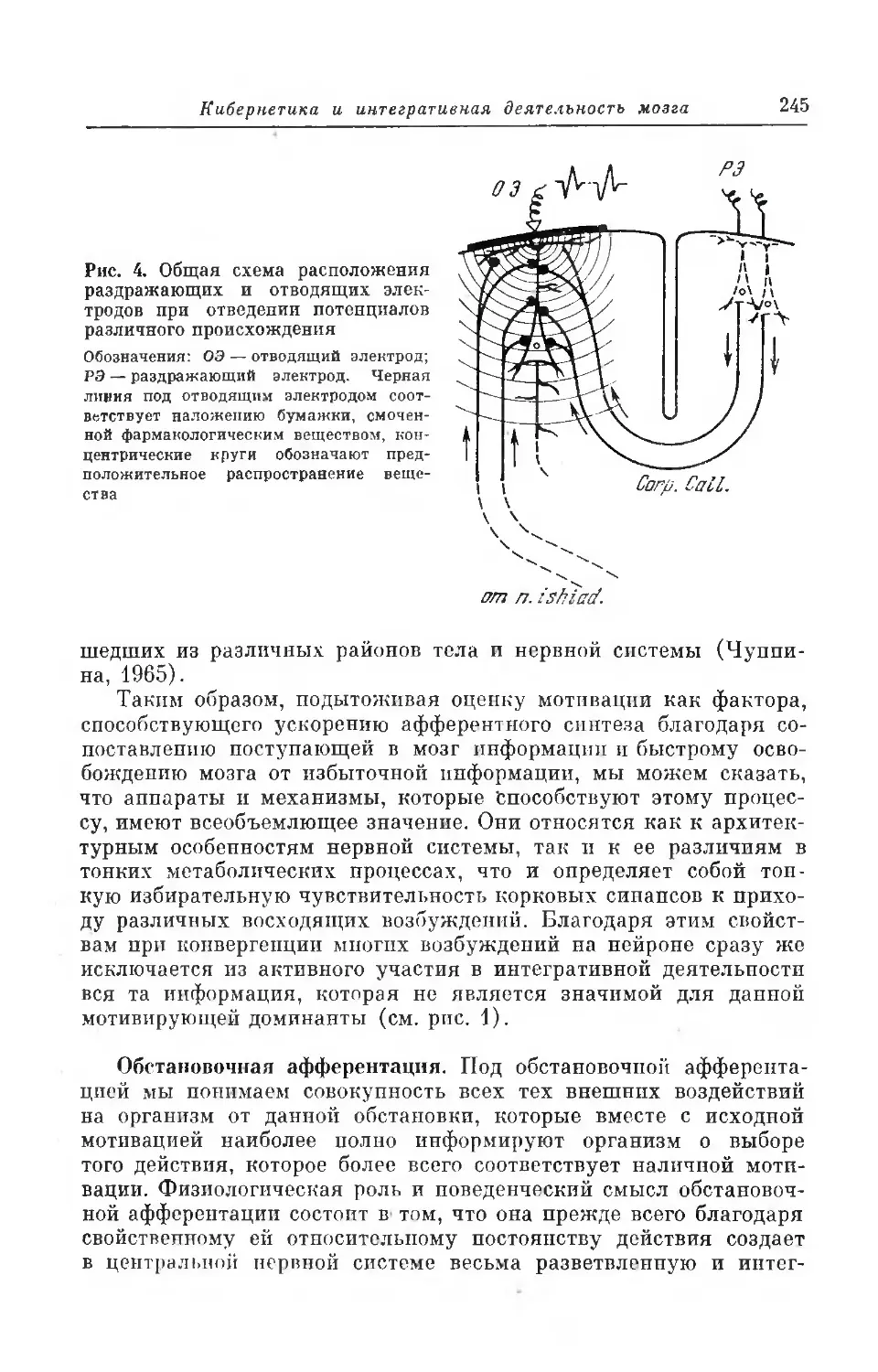







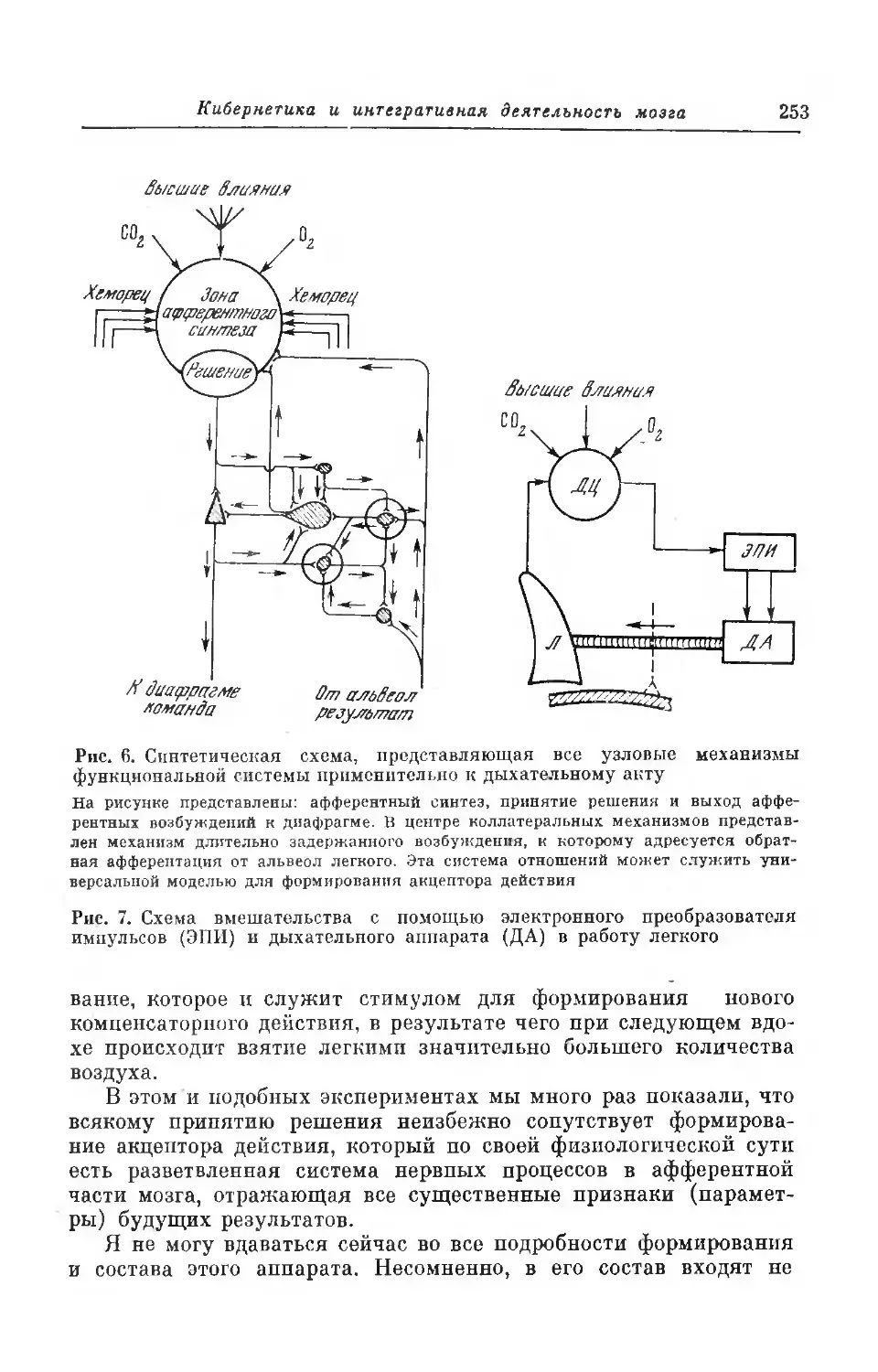




















































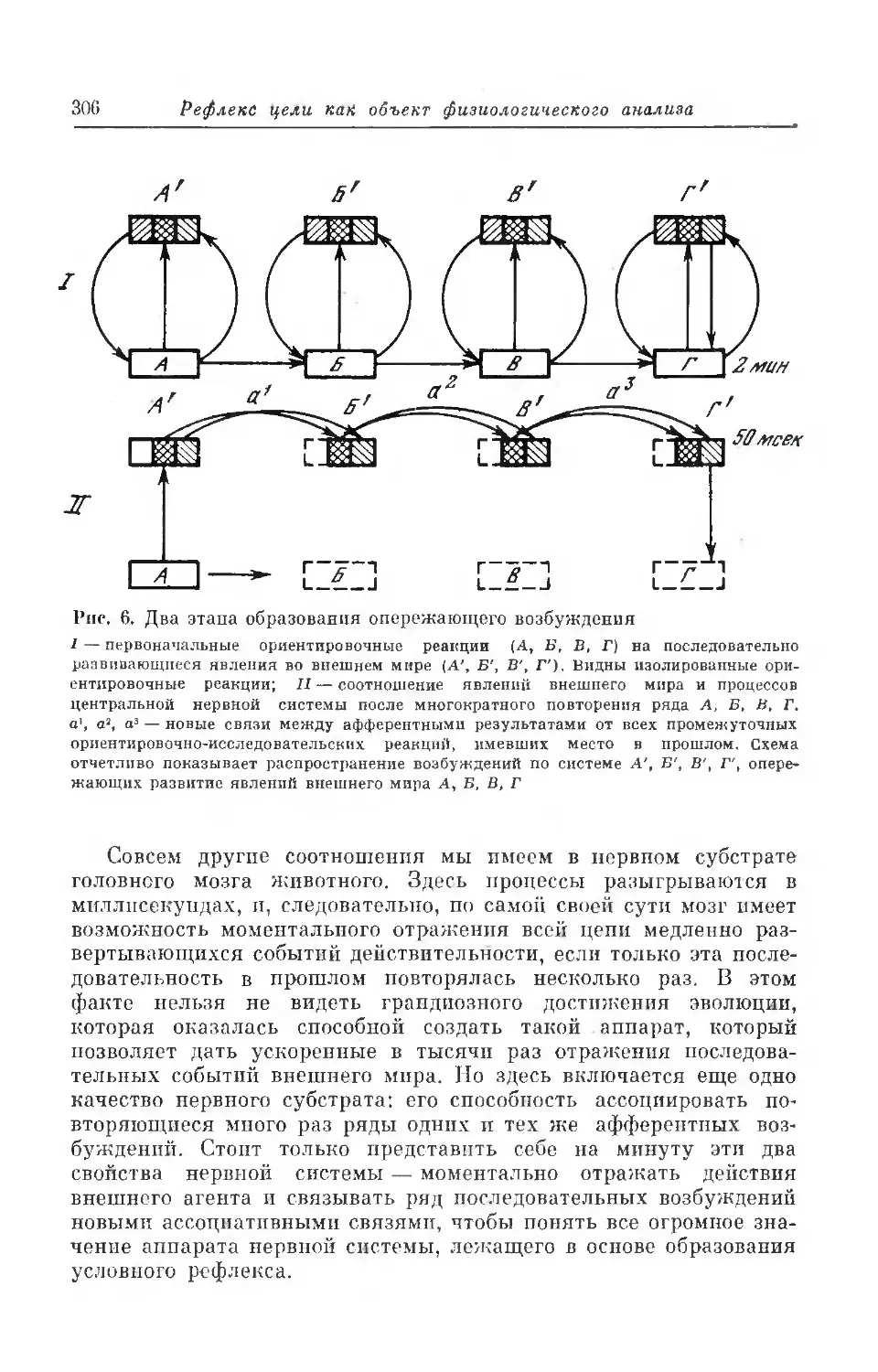













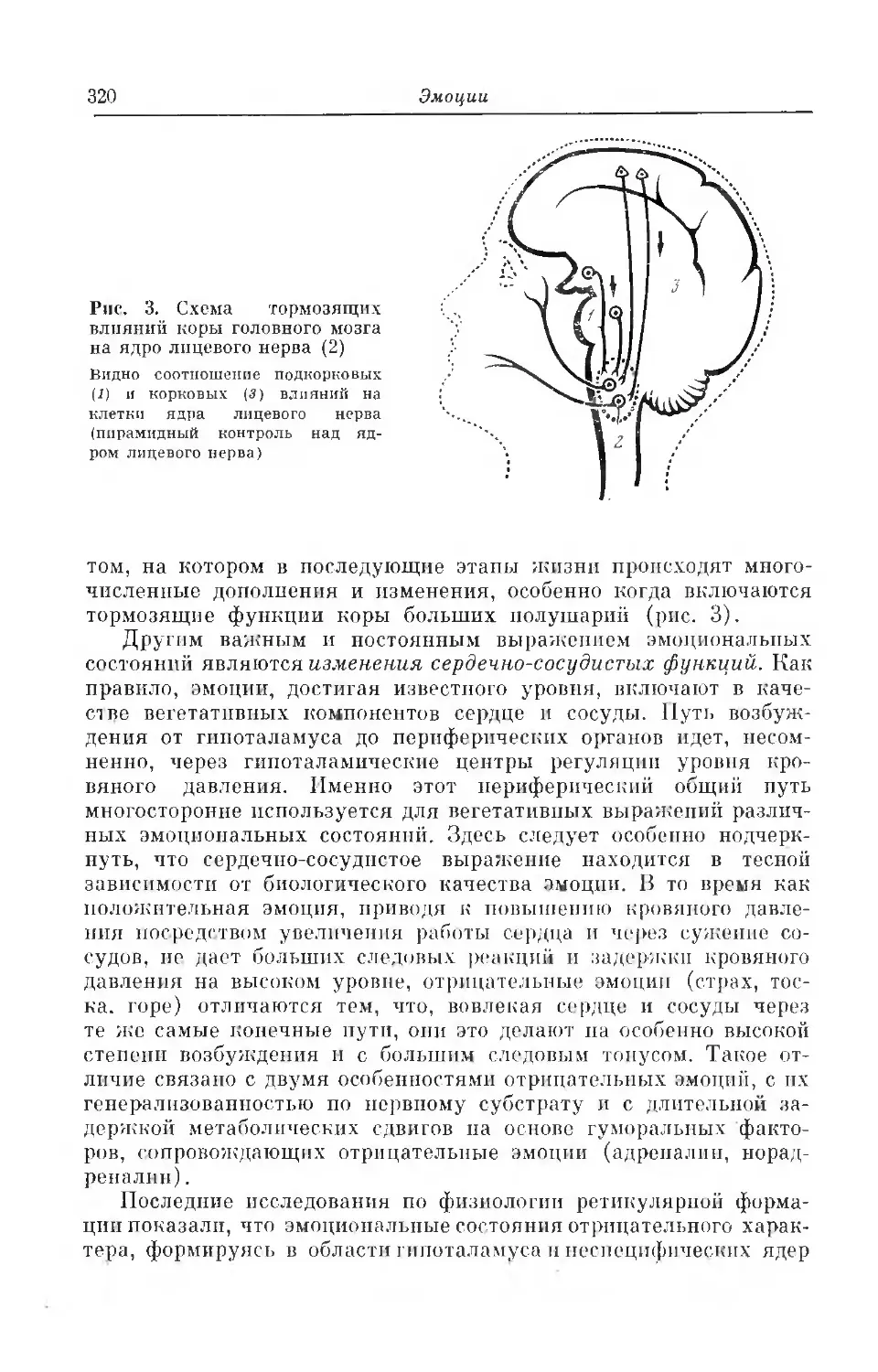

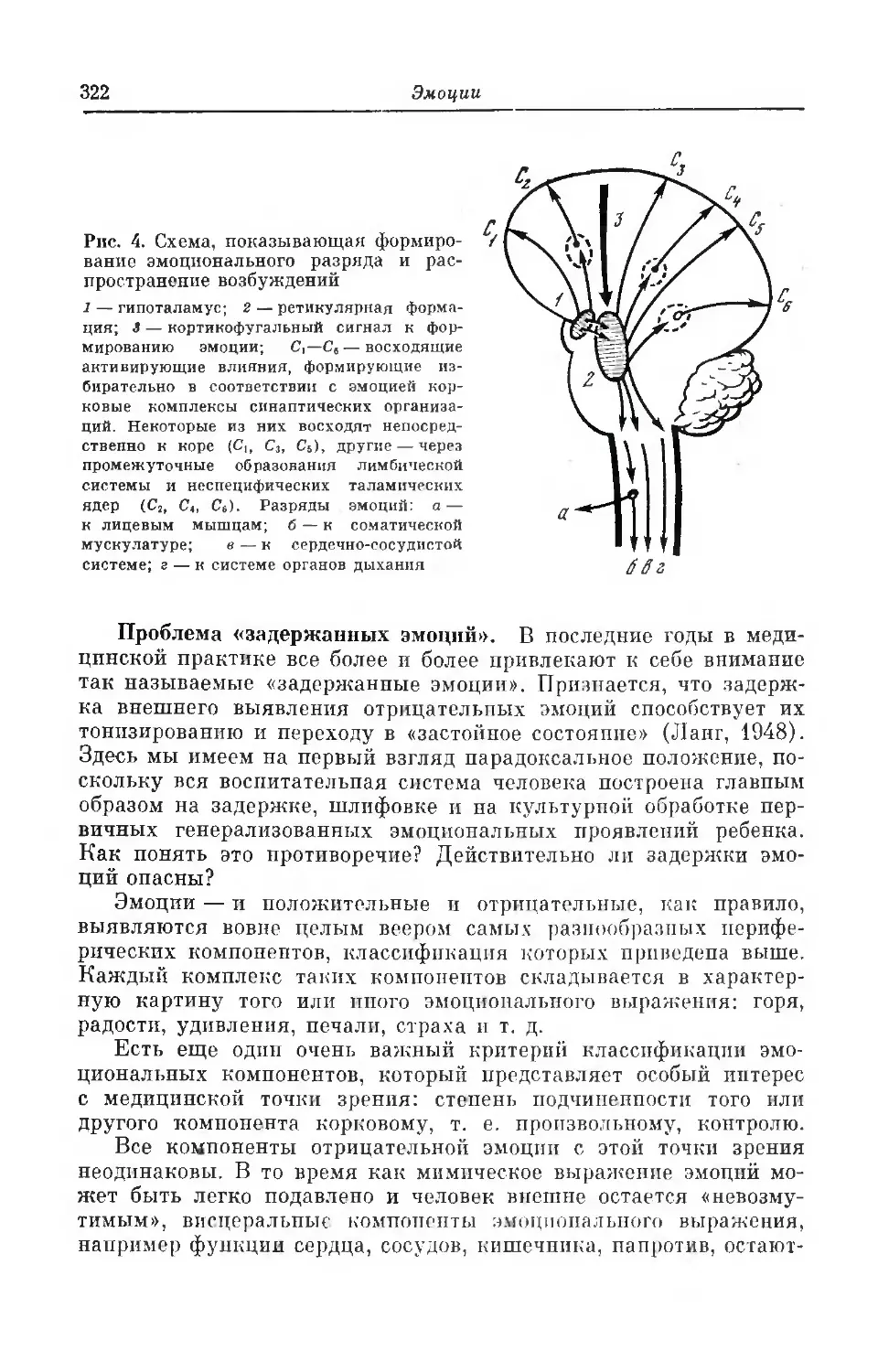

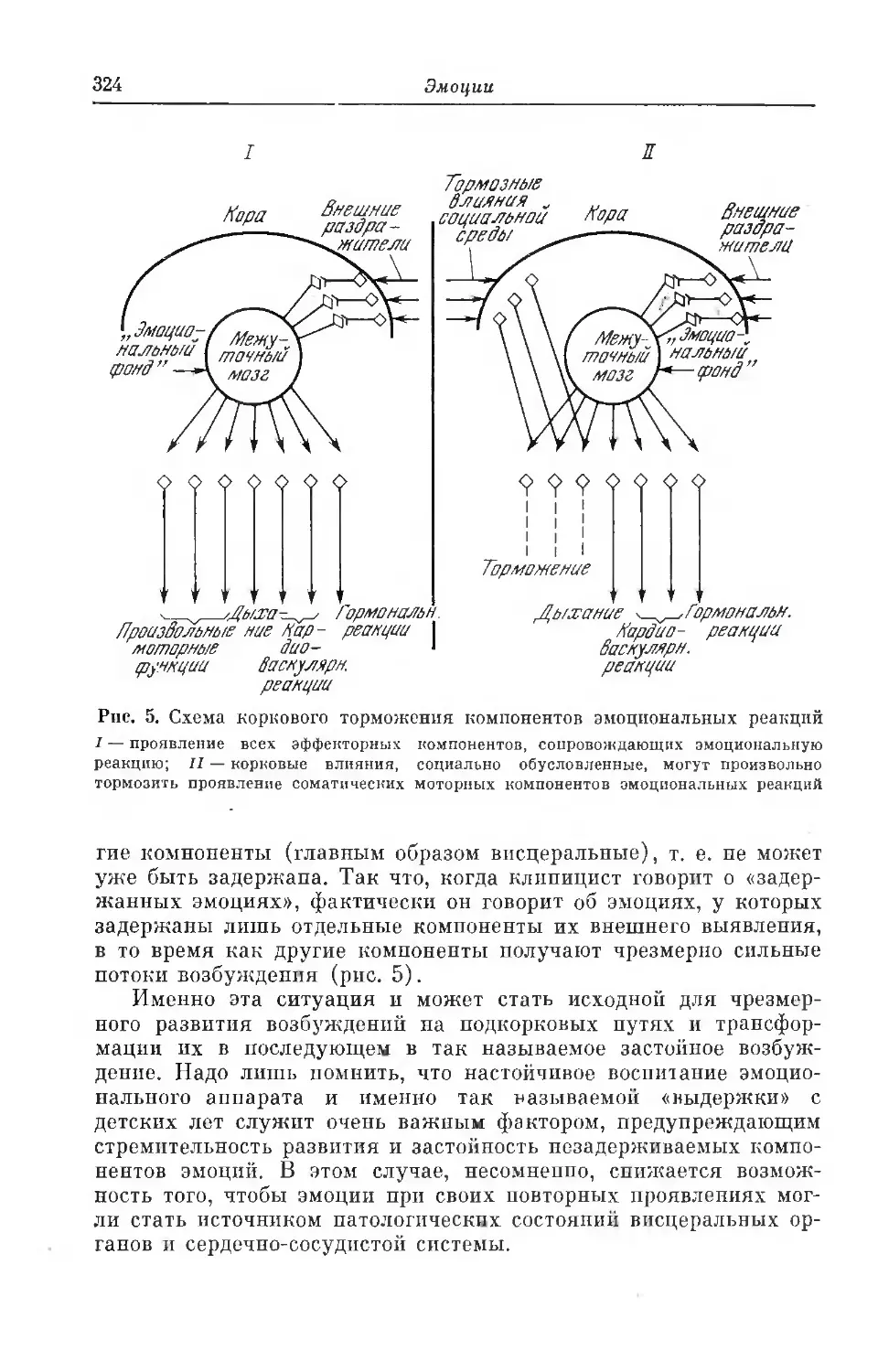


















































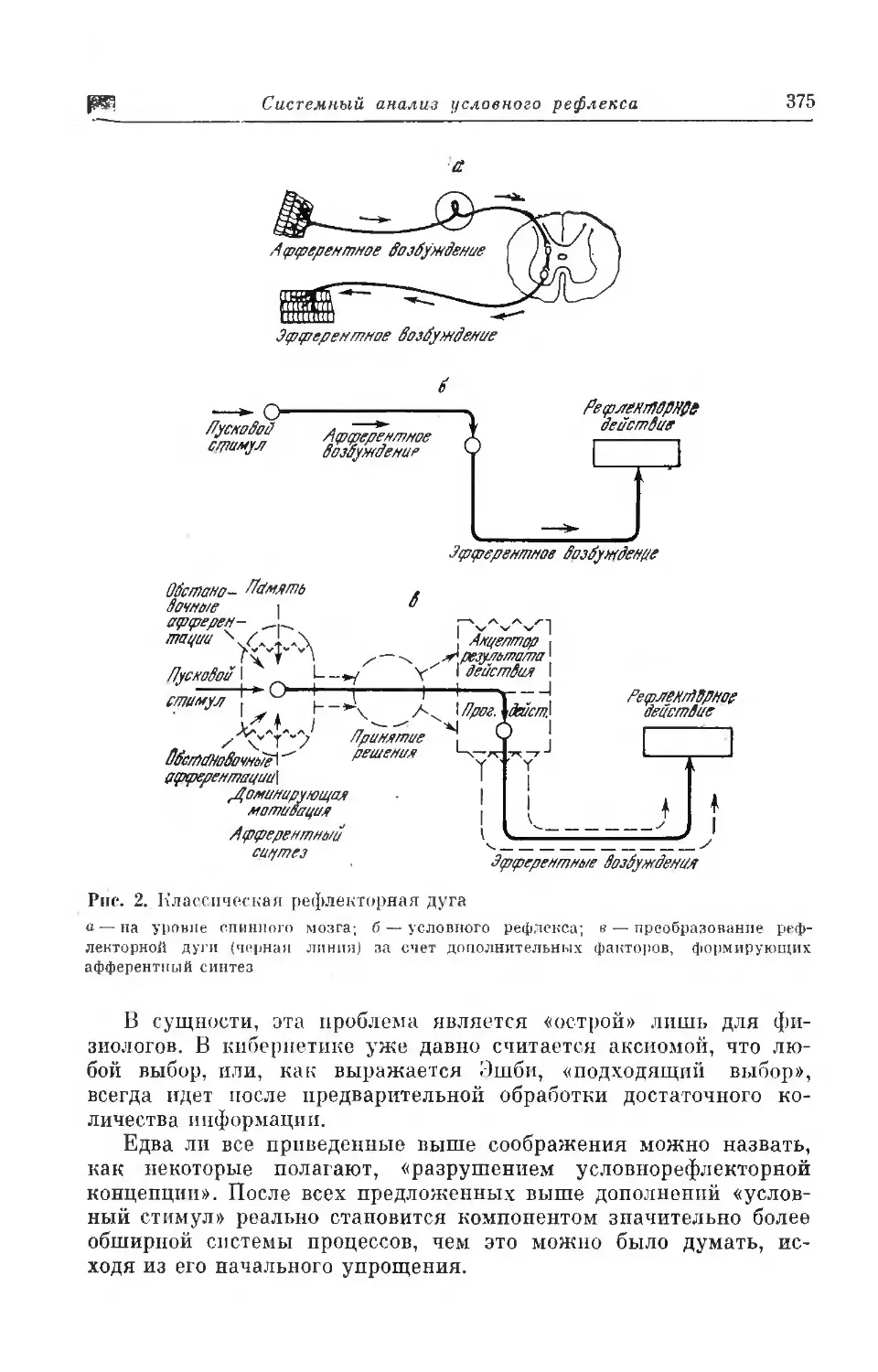




























Текст
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ ПСИХОЛОГИИ
6!
К.АНОХИН
Избранные
труды
ФИЛОСОФСКИЕ АСПЕКТЫ
ТЕОРИИ
ФУНК1 (ИОНАЛЬНОЙ
СИСТЕМЫ
И 3 Д А '1 Е Л К С Г I! О «ПЛ У К А»
МОСКВА 1978
Книга содержит избранные'работы академика П. К. Анохина,
посвященные философским и методологическим проблемам тео-
рии функциональной системы. Большой интерес представляет
разработка автором таких актуальных тем, как опережающее
отражение действительности, естественный и искусственный
интеллект, физиология и кибернетика, рефлекс цели, эмоции,
принятие решения и др.
ОТВЕТСТВЕННЫЕ РЕДАКТОРЫ:
ф. В. КОНСТАНТИНОВ, Б. Ф. ЛОМОВ, В. Б. ШВЫРКОВ
10508-232
042(02) —78 177—78
© Вступительная статья и составление.
Издательство «Паука», 1978 г.
ОТ РЕДАКТОРОВ
fl АМЫ СССР Петра Кузьмича
постановлению Президиума АН
было весьма многогранным. На
болезни, разработка проблем
многие другие. Эрудиция и
Анохина были действительно
огромного фактического мате-
он заявил о себе как о фило-
Настоящее издание избранных трудов лауреата Ленинской
премии академика АН СССР
Анохина предпринимается по
СССР.
Творчество П. К. Анохина
протяжении всей своей научной деятельности он оставался иссле-
дователем-экспериментатором, разрабатывавшим конкретные во-
просы медицины, физиологии и психологии. С его именем связа-
ны теория компенсации нарушенных функций и представления
о единстве центра и периферии в нервной деятельности, теория
системогеиеза и разработка проблем электрогенеза коры голов-
ного мозга, развитие теории высшей нервной деятельности и тео-
рия патогенеза гипертонической
нейрохимии и теории нейрона и
шпрота научных интересов П. К.
всеохватывающими: только синтез
риала, полученного в различных областях, мог позволить охва-
тить единым пониманием всю слояшость деятельности мозга. Его
заслуги в развитии конкретных областей науки нашли широкое
признание как у нас в стране, так и за рубежом.
К необходимости широких философских обобщений П. К. Ано-
хин пришел в самом начале научного пути. Уже в первой своей
философской статье «Диалектический материализм и проблемы
психического», вышедшей в 1926 г
софе. Вся его последующая обширная и разнообразная экспери-
ментальная работа была подчинена общей задаче создания единой
картины деятельности мозга и шла параллельно с глубокой тео-
ретической разработкой философских, общебпологических, кибер-
нетических и психологических проблем работы мозга.
Работы П. К. Анохина философского и методологического со-
держания, как и его конкретные исследования, посвящены раз-
витию теории функциональной системы, которая сформулирована
пм впервые в 1935 г. и постоянно совершенствовалась в течение
всех последующих лет. В настоящее издание избранных работ
включены его широко известные работы, оказавшие наибольшее
влияние на развитие философских и методологических основа-
ний современного естествознания.
При всей многогранности творчество П. К. Анохина было уди-
вительно целостным, поэтому проведенное в настоящем издании
6
От редакторов
распределение его работ по четырем разделам — философские,
биологические, кибернетические и психологические аспекты —
достаточно условно.
Первый раздел открывается статьей «Опережающее отраже-
ние действительности», в которой показывается творческое раз-
витие ленинской теории отражения; в статье «Философские
аспекты теории функциональной системы» вводятся понятия:
«континуум поведения» и «химический континуум» мозга. Обе
эти статьи сыграли значительную роль в устранении казавшего-
ся неразрешимым противоречия между принципами причинности
и целенаправленности в объяснении поведения живых организ-
мов. Наиболее полным изложением его теории функциональной
системы и философских представлений является статья «Прин-
ципиальные вопросы общей теории функциональных систем».
В статье «Философский смысл проблемы естественного и ис-
кусственного интеллекта» излагаются результаты разработки
П. К. Анохиным общих проблем соотношения среды и организма.
В первой статье второго раздела — «Системогенез как общая
закономерность эволюционного процесса» — раскрываются основ-
ные положения созданной П. К. Анохиным теории системогепеза.
Вторая статья этого раздела — «Методологический анализ узло-
вых проблем условного рефлекса» — обобщает результаты твор-
ческого развития П. К. Анохиным идей своего учителя И. П. Пав-
лова, ярко выявляет преемственность между учением о высшей
нервной деятельности и теорией функциональной системы.
В следующий раздел включены три статьи. Первая из них —
«Физиология и кибернетика» — интересна тем, что в ней прово-
дится сравнительный анализ понятия «обратная связь» в кибер-
нетике и в теории функциональной системы. В статьях «Кибер-
нетика и интегративная деятельность мозга» и «Методологическое
значение кибернетических закономерностей» отражаются этапы
разработки и модификации теории функциональной системы и
утверждения ее в качестве основного принципа функционирова-
ния биологических систем.
В последнем разделе помещены статьи, имеющие непосредст-
венное отношение к анализу механизмов поведенческого акта и
деятельности мозга как органа психики. Некоторые работы да-
ются с небольшими редакционными сокращениями.
В публикуемых в сборнике работах II. К. Анохина подняты и
оригинально решены те проблемы, которые интенсивно разраба-
тываются в философии естествознания в настоящее время.
Академик Ф. В. КОНСТАНТИНОВ
Член-корр. АН СССР Б. Ф. ЛОМОВ
Канд. мед. наук В. Б. ШВЫРКОВ
ОПЕРЕЖАЮЩЕЕ ОТРАЖЕНИЕ
ДЕЙСТВИТЕЛЬНОСТИ *
В настоящее время развитие ряда конкретно-научных дисцип-
лин выдвигает задачу исследования специфических форм про-
странственно-временной структуры мира на различных уровнях
организации материи, в частности на уровне живой материи.
Пространственно-временная структура мира является тем
фундаментом, на котором первичная жизнь приобретала свои
основные свойства, а живые существа — приспособительные каче-
ства в процессе своей эволюции вплоть до высшего ее этапа — че-
ловека.
Мы. несомненно, встречаемся здесь с новыми свойствами вре-
мени, которые никогда не оценивались в аспекте их биологиче-
ского значения. С зарождением жизни на Земле материя обога-
тилась принципиально новым фактором — активным отношением
живой материи к всевозможным превращениям пространственно-
временной структуры неорганического мира, и, следовательно,
время для животного приобретало свое специфическое значение
К сожалению, время как фактор эволюции живых существ
на земном шаре и как важнейший параметр развития их нерв-
ной деятельности никогда не подвергалось серьезному анализу.
Этот важнейший для физиологии высшей нервной деятельности
вопрос фактически надо исследовать заново. Возникают пробле
мы: в каких формах время как универсальный фактор мире
могло оказать влияние на развитие живой материи, как организ-
мы приспособились к временным факторам движения материи и
какие именно параметры времени оказались наиболее решающи-
ми для живых существ, обеспечили им выживание и прогрес-
сивную эволюцию?
Попытаемся выяснить, что является для нас наиболее важ-
ным во временных соотношениях живой и неживой природы?
Первым и существенным моментом является то, что основные
формы движения материи в пространственно-времепных рамках
существовали в неорганической природе до появления первых
живых существ на земном шаре. Это очень важный момент. Жи-
Опубликовано в журнале «Вопросы философии» (19G2, № 7).
8
Опережающее отражение действительности
LH
вая материя, «вписавшись» в уже готовую пространственно-вре-
менную систему мира, не могла не отразить ее свойства, ее архи-
тектуру, если только эти свойства имели отношение к основному
свойству самой живой материи — выживаемости.
С того момента, когда на земле возникла жизнь, отношение
к отдельным параметрам пространственно-временной структуры
со стороны этой качественно новой формы материи стало суще-
ственно иным. Гора, например, как форма неорганической мате-
рии не относится избирательно к климатическим и метеорологи-
ческим факторам, которые на нее действуют. Для горы не суще-
ствует той проблемы, которая возникла с появлением живой
материи и которую можно формулировать двумя словами: «при-
способиться и выжить». Для живых существ с момента их фор-
мирования в процессе эволюции весь внешний неорганический
мир со всеми его многообразными воздействиями стал «взвеши-
ваться» только на этих весах прогрессивной эволюции, только в
сопоставлении с конечным эффектом этих воздействий.
Появилось активное отношение к внешним неорганическим
факторам и неизбежно последовало за этим разделение всех
факторов на две большие категории — вредные для жизни и спо-
собствующие ее сохранению.
Начиная с этого момента все разнообразные формы движения
материи в рамках временной структуры мира стали распреде-
ляться первичными живыми существами по этой реципрокной
схеме, а живая материя начала отражать и закреплять в
структуре эти универсальные закономерности.
Из всего сказанного выше следует, что мы прежде
Н
своей
F
всего
должны установить, с какими конкретными временными парамет-
рами движения материи, уже существовавшими в неорганическом
мире нашей планеты, столкнулась первичная и примитивная
жизнь. Иначе говоря, мы должны дать точную оценку времен-
ной структуре мира, т. е. тех разновидностей пространственно-
временных движений материи, которые могут стать факторам!
формирующими приспособительные реакции живой материи.
Сейчас на этот вопрос мы не имеем ответа ни в философской,
ни в биологической литературе, поскольку гносеологическое
абстрагирование фактора времени оттеснило на задний план раз-
работку временной структуры неорганического мира именно в
том аспекте, который помог бы выявить значение этой структуры
для прогрессивной эволюции живых организмов.
Это ставит нас перед необходимостью вычленить те особен-
ные черты временной структуры материального мира, которые
явились своего рода «категорическим императивом» для разви-
тия жизни на Земле.
При любой попытке конкретизировать пространственно-вре-
менную структуру материального мира мы неизбежно встречаем-
Опережающее отражение действительности
9
ся с последовательным перемещением тел в пространстве, после-
довательными воздействиями одного тела на другое, последова-
тельным развитием
аз движения и преобразования материи.
н
Можно сказать, что с точки зрения диалектического материа-
лизма наиболее существенной чертой пространственно-времен-
ной структуры мира, определяющей временное отношение пер-
вичных организмов к внешнему неорганическому миру, является
последовательность воздействий внешнего мира на эти организмы
независимо от интервала этих
воздействий
от качества
их
энергии. Этим самым мы вычленяем временной параметр отно-
шений организма к неорганическому миру как в какой-то степени
самостоятельный фактор в смысле значения его для приспособи-
тельных превращений.
Мы поставили перед собой конкретные вопросы: с какими
формами и вариантами этого основного временного фактора —
последовательности воздействий — встречается организм? Каким
образом дробится всеобъемлющее движение материи в рамках
абсолютного временного параметра — последовательности явле-
ний? Иначе говоря, с какой уже готовой временной структурой
неорганического мира, вынуждающей на приспособление к ней,
встретились первичные организмы на пашей планете.
Нам кажется, что мы имеем полную возможность вычленить
некоторые временные параметры из основной формы — последо-
вательности и проанализировать то значение, которое они могли
иметь для приспособительных отношений первичных организмов.
Первый вариант — существование ряда таких последова-
тельных явлений, каждое из которых никогда затем не повто-
ряется на протяжении всей жизни данного организма. Символи-
чески этот ряд мог бы быть изображен так: а, б, в, ...
Само собой разумеется, конечно, что факторы неорганическо-
го мира а, б, в и т. д. должны быть связаны между собой при-
чинно-следственными отношениями и развиваться в последова-
тельном порядке. Несомненно также, что все они или некоторые
из них могут оказывать последовательное воздействие на орга-
низм. Однако но условию ни одно из этих воздействий никогда
не должно иметь повторного действия на организм. Конкретно
это может быть солнечное затмение, гроза или ураган необычай-
ной силы, прохождение кометы вблизи Земли и т. д. Конечно,
неповторяющиеся воздействия могут исходить и от непосредст-
венно окружающих его факторов. Некоторые из этих неповторяю-
щихся воздействий внешнего мира могли иметь какое-то суще-
ственное значение для организма — или разрушительное, или
благотворное. Первичный организм мог либо противостоять этому
действию с помощью уже имевшихся у пего возможностей, либо
разрушиться. Однако эти воздействия для пего являлись всегда
внезапными, новыми, никогда им не испытывались.
10
Опережающее отражение действительности
Возможна и другая временная структура в последователь-
ности внешних воздействий на организм: некоторые факторы
внешнего мира могут возвращаться и возобновлять свое действие
через определенные промежутки времени, а следовательно, могут
быть названы повторяющимися рядами последовательных воз-
действий. Символически эта вторая возможность может быть изо-
бражена так: (абвг), (абвг), (абвг) ... (абвг).
Приведенная схема показывает, что мы можем иметь в не-
органическом мире последовательность таких событий, которые
ритмически повторяются в том же составе и в том же порядке
следования компонентов. Пусть это будут, например, день, вечер,
ночь, утро или, например, лето, осень, зима, весна, или, напри-
мер, туча, молния, гром. Эти ряды последовательно развиваю-
щихся и ритмически или апериодически повторяющихся явлений
неорганического мира имеют абсолютную направленность и свя-
заны с астрономическими, метеорологическими и физическими
закономерностями, хотя длительность ритмов различна, и, зна-
чит, повторяемость этих циклов может иметь различное значе-
ние для организмов с различной длительностью жизни.
До сих пор мы брали в качестве примеров такие ряды после-
довательно развивающихся событий, которые обладают свойством
абсолютной устойчивости. Но мыслимы также и такие последо-
вательные явления, которые, обладая тем же параметром повто-
ряемости или возвращаемости
вместе с тем имеют свойство
относительной устойчивости. Так, например, дерево, растущее на
берегу реки и имеющее определенный возраст, может быть мес-
том обитания для различных видов животных только на протя-
жении определенного срока, после которого оно будет заменено
другими такими же или отличающимися от него деревьями.
Однако на протяжении своей жизни это дерево будет представ-
лять собой для его обитателей серию последовательных, ритми-
чески повторяющихся явлений в зависимости от различных вре-
мен года, особенно для северных широт.
Таким образом, как в первом случае, так и во втором мы
имеем один главнейший временной признак последовательно раз-
вивающихся событий — повторяемость, абсолютно устойчивую и
относительно устойчивую.
Конечно, само свойство повторяемости явлений неорганиче-
ского, а позже и органического мира как частный параметр
абсолютной пространственно-временной последовательности мо-
жет иметь бесконечные вариации в длительности самих ритмов,
в их устойчивости, в составе и продолжительности существова-
ния отдельных компонентов, в их пространственной локализации
и т. д.
Однако при всех этих вариантах именно параметр повто-
ряемости будет наиболее характерным и специфическим.
Опережающее отражение действительности
11
До сих пор мы разбирали последовательность явлений неорга-
нического и отчасти органического мира только с одной целью —
выделить различные временные параметры неорганического мира,
которые неизбежно должны были воздействовать на органиче-
скую материю уже при самом ее возникновении. Эти времен-
ные параметры (неповторяемость, повторяемость, длительность,
устойчивость, изменчивость и т. д.) были нами изложены безотно-
сительно к другому их важному качеству, которое они стали при-
обретать специально при взаимодействии с живой природой.
Таким новым качеством для этих временных параметров стали
существенность или несущественность их для поддержания и
закрепления жизненного процесса.
Несомненно, что уже на самых первых этапах организации
живого вещества (может быть, еще на стадии первичных коацер-
ватов) изменения внешних материальных условий «классифици-
ровались» этими зачатками живого вещества как «вредные» и
«полезные» именно по критерию их «существенности» для стаби-
лизации многомолекулярных систем. Этот вид отношений первич-
ных форм живой материи к внешним воздействиям особенно
полно характеризует Л. И. Опарин в своей книге «Возникнове-
ние жизни па земле» (1957), к которой мы и отсылаем читателя.
Несмотря на относительное постоянство неорганических усло-
вий, в которых зародилась жизнь (возможно, это были глубины
первородных океанов), даже в тех условиях осуществлялась по-
следовательная смена внешних воздействий на кусочки первич-
ного живого существа (смена температур, реагирующих веществ
химической среды, течений, приливов и отливов и т. д.), кото-
рая способствовала своего рода первичному естественному «от-
бору» наиболее устойчивых образований типа «открытых много-
молекулярпых систем».
Эта последовательная смена внешних воздействий приобрела
особенно большое значение тогда, когда у первичных живых
существ появилась способность размножаться и рассепваться по
водным пространствам с различным составом экологических фак-
торов.
Еще большее количество вариаций повторяемости возникло
с того момента, когда первичные организмы приобрели способ-
ность к активному передвижению. Тогда всякое повторное дви-
жение около неподвижного неорганического объекта с широкой
изиологической точки зрения приводило к повторным более или
менее однотипным воздействиям. Исключительное значение этот
тип повторного воздействия внешнего мира имеет в жизни
человека с развитием его широчайших возможностей к переме-
щению.
В данный момент нам важно одно — что и в этот период вре-
менная структура окружающего организм неорганического мира
i2
Опережающее отражение действительности
передвижении
осталась той же и включала в себя по крайней мере четыре
основные формы.
1. Действие относительно постоянных факторов.
2. Последовательные ряды внешних воздействий, повторяю-
щихся ритмически или апериодически.
3. Ритмические или апериодические воздействия относительно
постоянных внешних факторов при активном
живых существ.
4. Действие последовательных рядов никогда
щихся факторов.
На этот последний вид внешних воздействий,
и на все другие виды, правило существенности для сохранения
жизни также распространяется.
Хотя эти последние воздействия и являлись только «вредны-
«полезными» и «нейтральными» неповторяющимися эпизо-
не повторяю-
л
как, впрочем
ми»
дами для живой материи, сами по себе они, естественно, всегда
являлись звеньями непрерывных п последовательно развиваю-
щихся явлений в общем потоке эволюции пространственно-вре-
менной структуры материального мира. Они встречались с жиз-
ненным циклом организмов, так сказать, по касательной.
Из этого можно сделать вывод, что эти никогда не повторяв-
шиеся воздействия не могли оказать какое-либо решающее влия-
ние на эволюцию высших форм приспособления живой материи
к окружающим условиям и, следовательно, не могли стать фак-
торами организации протоплазматической структуры живых
организмов.
Вообразим себе на минуту, что временная структура неорга-
нического мира была бы представлена только этой последней
формой движения материи, т. е. рядами последовательных и ни-
когда не повторяющихся явлений.
Могла ли при этом условии развиться на Земле жизнь в ее
стабилизированных, имеющих всегда приспособительное значе-
ние структурах?
Нет, не могла. Живой организм не мог бы иметь устойчивой
и прочной структуры, ибо последняя может появиться только
как результат отражения ритмически и апериодически повторяю-
щихся воздействий неорганической природы. И даже само поня-
тие «приспособление» потеряло бы всякий смысл в таком мире
всегда новых, никогда прежде не испытывавшихся организмом
воздействий. Совершенно очевидно, что из общего потока собы-
тий пространственно-временной структуры мира только ритмиче-
ски и апериодически повторяющиеся явления могли стать вре-
менной основой для развития приспособительных реакций пер-
вичных организмов.
Итак, мы пришли к решающему выводу относительно эволю-
ции организмов в зависимости от постоянных свойств времен-
Опережающее отражение действительности
13
ной структуры неорганического мира. Этот вывод можно форму-
лировать следующим образом: основой развития жизни и ее
отношений к внешнему неорганическому миру являлись повто-
рявшиеся воздействия этого внешнего мира на организм. Именно
эти воздействия как результат изначальных свойств пространст-
венно-временной структуры неорганического мира обусловили
собой всю анатомическую организацию и приспособительные
функции первичных живых существ. В этом отношении органи-
зация живых существ представляет собой в подлинном смысле
слова отражение пространственно-временных параметров их кон-
кретной среды обитания.
Проделанный выше анализ приводит нас к заключению, что
именно эта форма временных соотношений — повторяющиеся воз-
действия — представляет собой универсальную форму связи уже
сложившихся и индивидуализировавшихся живых существ с
окружающей их средой.
Такое заключение немедленно ставит перед нами вопросы:
с помощью каких же конкретных механизмов эти жизненно важ-
ные, повторяющиеся воздействия делают организм приспособлен-
^«ым к ним? В каких интимных процессах живой протоплазмы
могли отражаться эти повторные воздействия внешнего неоргани-
ческого мира па уже оформившиеся живые существа?
Для ответа на этот вопрос мы можем использовать общепри-
нятое представление, что простейшие живые существа представ-
ляли собой «открытые системы», которые были связаны с окру-
жающей средой через ряд химических превращений, начинав-
шихся на границе живой миогомолекулярной протоплазмы,
продолжавшихся в форме целой цепи отдельных реакций и закан-
чивавшихся либо вредным, либо полезным для жизни итогом.
Таким образом, каждое из внешних по отношению к организ-
му последовательных явлений благодаря особенным свойствам
живой протоплазмы отражается в вей в форме более или менее
длинных цепей химических превращении. Эта цепь реакций
может иметь обменный характер, поддерживающий жизнь орга-
низма, его самовоспроизведение, но может возникнуть и такая
цепь превращений, которая окажется вредной для этого обмена
веществ и, следовательно, не поддерживающей жизненный про-
цесс. Одпако, несмотря на эти два биологически противополож-
ных исхода, и то и другое воздействие вызывает ряд последова-
тельных и вполне закономерных химических превращений. Здесь-
то и выступает на первый план тот основной параметр
пространственно-временной структуры неорганического мира,
который всегда играет доминирующую роль в развитии живой
материи,— повторяемость явлений и воздействий.
Сейчас уже нетрудно установить, к каким реальным послед-
ствиям могли привести те воздействия на первичный организм,
14
Опережающее отражение действительности
которые имели существенное значение для его выживания и мно-
гократно повторялись или были постоянными на протяжении
миллионов лет. По общему признанию биохимиков любое внеш-
нее воздействие на первичные организмы неизбежно служило
отправным пунктом для образования цепей химических процес-
сов и взаимодействий, кончавшихся или разрушением организма,
или, наоборот, укреплением его метаболической стабильности.
Вместе с тем повторявшиеся много раз последовательные ряды
воздействий неизбежно приводили к облегченному и ускоренно-
му катализаторному типу развития этих цепных процессов
(Опарин, 1957). Так создавались в конце концов доминирующие
линии цепных химических реакций.
Мне трудно себе представить, чтобы вне повторяющихся воз-
действии внешнего неорганического мира на организм существо-
вали какие-то иные пути, которые смогли бы создать преимуще-
ственные цепи обменных и других химических реакций. Именно
повторяемость последовательно развивающихся внешних воздей-
ствий, существенных в том или другом отношении для жизни
организма, смогла создать непрерывную и последовательную цепь
химических реакций в протоплазме организма.
На этой основе в эволюции жизни па Земле произошло одно
весьма знаменательное событие, которое в дальнейшем на мно-
гие миллионы лет определило ведущий признак приспособитель-
ных реакций организмов. Я имею в виду вытекающую из основ-
ных свойств первых живых существ (как «открытых образований»
с многомолекулярным составом протоплазмы) способность реаги-
ровать па изменения в окружающем мире более или менее об-
ширными химическими перестройками, если только эти измене-
ния находят физико-химический контакт с этими образованиями.
И потому естественно, что если во внешнем мире последова-
тельно развивается несколько специфических событий (напри-
мер, сезонные ритмы, смены температур, течения в океанах),
то организм должен отразить каждое из них в специфических
химических перестройках своей протоплазмы, если только эти
события достигают известного порога действия.
Специфические химические перестройки протекают своеобраз-
но в зависимости от физических свойств неорганических фак-
торов.
Интересно, что Л. II. Опарин, характеризуя эти соотношения
первичной живой материи с окружающей неорганической приро-
дой, также считает, что последовательность и быстрота реакций
протоплазмы являются решающими факторами материальной
организации первых живых существ (Опарин, 1957).
Знакомство с теориями происхождения жизни на Земле пока-
зывает, что и с широкой биологической точки зрения, так же как
и с точки зрения философского анализа роли пространственно-
Опережающее отражение действительности
15
временной структуры мира, движение материи ио последователь-
ным, ритмически повторяющимся
•азам является универсальным
законом, определяющим основную организацию живых сущест»
на нашей планете.
Появление первичных белковых тел, приобретших в дальней-
шем ферментативную функцию, радикально изменило весь про-
цесс совершенствования жизни. Создалась возможность развития
отдельных преимущественных, т. с. с избирательным каталити-
ческим ускорением, цепей реакций, и, конечно, в первую очередь
тех реакций, которые, являясь существенными для сохранения
жизни, повторялись множество раз под влиянием внешних воз-
действий. Эти воздействия могли быть весьма различными, на-
пример периодические смены в поступления веществ, необходи-
мых для первичных организмов как «открытых систем». Однако
остается несомненным, что именно специфический катализ
создал возможности преимущественных условий для развития тех,
а не других цепей реакций.
Это ускорение реакций!, как показывает анализ некоторых
широко распространенных ферментативных комплексов, может
достигать величины в сотни миллионов и даже миллиардов раз.
Во всяком случае па первых стадиях развития жизни этот «прин-
цип максимальной скорости» дал возможность па фоне в какой-то
степени гомогенных коацерватных образований сформировать
пути преимущественных цепей реакций, развивающихся с огром-
ной скоростью.
II здесь, в этом узловом пункте развития живых организмов,
было сделано одно замечательное приобретение, которое в даль-
нейшем оказало решающее влияние на весь исторический путь
развития животного мира на земном шаре.
Дело в том, что между последовательными воздействиями
внешнего мира и реакциями на эти воздействия со стороны
живого вещества существовала принципиальная разница. Внеш-
ние воздействия на организм могли совершаться в самых разно-
образных временных интервалах и исходить из самых различных
источников (их объединял лишь принцип последовательности).
Вместе с тем все эти воздействия конвергировали к одной и той
же протоплазме живого существа, вызывая различные по каче-
ству химические реакции, развертывающиеся в пределах одного
и того же протоплазматического образования.
Таким образом, создались все условия для формирования
одного из решающих процессов живого вещества, который опре-
делил пути дальнейшего развития животного мира и
юрму его
приспособительных реакций по отношению к внешнему миру.
В самом деле, если в небольшом комочке живого вещества,
представляющего собой многомолекулярную открытую систему,
стали возможными многообразные химические реакции да еще
16
Опережающее отражение действительности
с весьма большими скоростями, то некоторые последовательно
повторявшиеся ряды внешних воздействий, пусть даже разделен-
ных большими интервалами, получили возможность отражать себя
в быстрых химических превращениях этого вещества в соответ-
ствии с физическим или химическим качеством этих воздействий.
Постепенно складывались такие соотношения, при которых
все более и более выявлялось различие временных параметров
для событий, протекавших во внешнем неорганическом мире,
и для их отражения в химических перестройках живого су:
ства. В результате этих совершенно реальных взаимодействий
уже иа этой стадии сложилась одпа универсальная закономер-
ность в приспособлении организма к внешним условиям, которая
в дальнейшем бурно развивалась на протяжении всей эволюции
животного мира: в высшей степени быстрое (в ценных химиче-
ских реакциях) отражение медленно развертывающихся событий
внешнего мира.
Для более четкой формулировки этого положения я попытаюсь
проиллюстрировать эту закономерность в схематическом виде.
Допустим, что во внешнем мире развивается последователь-
ный ряд явлений —Л, Б, В, Г, Д. Эти явления действуют на орга-
низм па значительных временных промежутках в течение нолу-
суток. Пусть каждое из этих явлений вызывает в протоплазме
живого существа последовательный ряд химических превраще-
ний, который мы обозначим соответственно символами а, б, в, а, д
(рис. 1, 2).
Допустим, что ряд внешних воздействий па организм (Л, Б,
В, Г, Д) систематически повторяется на протяжении многих лет и
имеет существенное положительное значение для его обменных
процессов, устанавливая более совершенные цепи химических
реакций, т. е. помогая стабилизации жизненного процесса. В ре-
зультате длительного и многократного воспроизведения в прото-
плазме живого существа этого определенного ряда химических
реакций между отдельными звеньями устанавливается органиче-
ская связь, делающая весь ряд превращений а — б—в — г — д не-
прерывной и быстро развертывающейся цепью химических реак-
ций (см. рис. 1, 2).
Несомненно, что решающее значение в образовании этих
цепей реакций имело формирование специфических катализато-
ров, определяющих преимущественное течение цепных химиче-
ских реакций именно в данном направлении, а не в другом.
Все эти условия, создавшиеся уже на самых ранних этанах
эволюции живой материи, привели к тому, что протоплазма по-
лучила возможность отражать в микроиптервалах времени своих
химических реакции те последовательные события внешнего ми-
ра, которые по самой своей природе могут развертываться, наобо-
рот, в макроинтервалах времени.
Опережающее отражение действительности
17
Рис. 1. Соответствие протоплазматических реакций внешним событиям
А, Б, В, Г, Д — последовательно развивающиеся внешние события на различных
интервалах; а, б, в, г, 0 — протоплазматические реакции, возникающие от индиви-
дуальных энергетических особенностей каждого отдельного внешнего воздействия
Рис. 2. Опережающее отражение в состоянии протоплазмы медленно раз-
вертывающихся событий внешнего мира
Видно, что после многочисленных повторений последовательного ряда явлений А,
Б, В, Г, Д в протоплазме образовалась непрерывная цепь химических превращений,
возникающих только в ответ на первое событие (А) во внешнем мире. Процесс
отражения в протоплазме опережает ход последовательных событий во внешнем
мире. Реакция протоплазмы (б) уже произошла, в то время как событие (Д) на-
ступит еще только в будущем
Так постепенно формировалась способность первичных орга-
низмов отражать внешний неорганический мир не пассивно, а ак-
тивно, с опережением в своей протоплазме последовательно и по-
вторно развертывающихся явлений внешнего мира.
В самом деле, в протоплазме живого организма установилась
единая цепь химических реакций, ранее вызывавшаяся последо-
вательным действием внешних факторов А, Б, В, Г, Д. И если эти
факторы разделены значительными интервалами времени, то те-
перь уже действие только одного первого
актора (Л) способно
начать и привести в активное состояние всю последовательную
цепь химических реакций.
Исторически в живой протоплазме неизбежно должны были
создаваться эти условия, при которых быстрота химических ре-
акций протоплазмы обеспечивала опережение организмом развер-
тывания последовательных, много раз повторявшихся внешних
воздействий.
Таким образом, с пашей точки зрения, эволюция уже очень
рано сформировала этот универсальный и единственно возмож-
18
Опережающее отражение действительности
ный путь приспособления организма к внешнему миру. С помо-
щью процесса опережения уже самые начальные живые организ-
мы, отгородившиеся от внешнего мира простейшими мембранами,
могли с огромной быстротой своих химических превращений
строить цепи химических реакций по первому звену много раз
повторявшегося последовательного ряда внешних воздействий.
Основой для построений таких цепей химических реакций явля-
лись многомолекулярпость и в высшей степени сложная по строе-
нию протоплазма первичных живых существ.
Живая организация получила от такой формы реагирования
на повторные ряды последовательных воздействий огромные
преимущества. По сути дела без этого опережающего разверты-
вания цепей химических реакций трудно представить себе суще-
ствование даже такого простого процесса, как, папример, погло-
щение и переваривание амёбой какого-либо кусочка пищи или
захватывание и переваривание бактерий бактериофагом.
Вся история животного мира служит наглядным примером
усовершенствования этой универсальной и самой древпей законо-
мерности, которую можно было бы назвать опережающим отра-
жением действительности, т. е. ускоренным в миллионы раз раз-
витием цепей химических реакций, которые в прошлом отражали
последовательные преобразования этой действительности.
Возвращаясь к исходной предпосылке этой проблемы, мы мог-
ли бы сказать, что опережающее отражение действительности
есть основная форма приспособления живой материи к простран-
ственно-временной структуре неорганического мира, в котором по-
следовательность и повторяемость являются основными времен-
ными параметрами.
С этой точки зрения фактор Д, являясь каким-либо полезным
для жизни фактором, делает весь ряд воздействий, много раз
предшествовавший ему в истории развития, сигнальным по от-
ношению к нему как к конечному звену этого ряда, а сами цепи
последовательных химических реакций, образовавшиеся на опи-
санной выше основе, предстанут перед нами как временные связи.
Следовательно, самый факт появления «сигпалыюсти» и «вре-
менных связей» может быть признан одной из древнейших зако-
номерностей развития живой материи. Именно в этом смысле на-
до понимать выражение И. П. Павлова «временные связи есть
универсальное явление природы». Сопутствуя самому происхож-
дению и совершенствованию живой материи и являясь естест-
венным следствием пространственно-временной структуры мира,
эта закономерность закреплялась все глубже и глубже естествен-
ным отбором в стабилизированных структурах протоплазмы.
Следовательно, принцип опережающего отражения мира в прото-
плазматических процессах организма может считаться основой
для создания и закрепления всех тех структур организма, кото-
Опережающее отражение действительности
19
рые целесообразно приспосабливают его к внешнему миру на ос-
нове принципа сигналыюсти и временных связей. Эта способность
организма, открывающая ему широчайшие возможности приспо-
собления и прогресса, стала быстро специализироваться в особом
субстрате, основной функцией которого стало быстрое химическое
сцепление всех одновременно и последовательно развивающихся
химических реакций. Таким субстратом и стала нервная систе-
ма.
С этой широкой точки зрения первичное нервное вещество и
вообще центральную нервную систему можно рассматривать как
субстрат высокой специализации, который развивался как аппа-
рат максимального и быстрейшего опережения последовательных
и повторных явлений внешнего мира.
В одной из наших прежних работ мы показали, что реальный
факт приближения служителя с кормом к собаке, направляюще-
гося с первого этажа на третий, дает определенную последова-
тельность стимулов (Анохин, 1962). Такое Продвижение служи-
теля с кормом требует значительного времени, ибо оно соверша-
ется в рамках физико-механических возможностей пространства.
Совсем другие соотношения мы имеем в нервном субстрате го-
ловного мозга животного. Процессы в рецепторах и в нервном
веществе развертываются в течение миллисекунд, и, следова-
тельпо, по самой своей сути мозг имеет возможность уже при
действии первого агента всего ряда событий (стук дверью ниж-
него этажа) моментально воспроизвести всю цепь химических
реакций, зафиксированных в прошлом медленно протекавшими
событиями действительности, если только одна и та же последо-
вательность этих событий в прошлом повторялась много раз.
В этом факте нельзя не видеть грандиозного достижения эво-
люции, которая оказалась способной на основе первичных про-
топлазматических цепей химических реакций создать такой ап-
парат, который позволяет дать в тысячи раз ускоренное отраже-
ние действительности, во много раз опережающее последователь-
ный ход реальных явлений внешнего мира. Именно различие в
скоростях течения явлений в самой натуральной действительно-
сти и в нервном веществе мозга животного создает предпосылки
для распространения такого опережающего возбуждения.
Если сопоставить разобранные нами выше столь различные
уровни организации живой материи, то мы увидим, что на всем
длинном пути эволюции живой материи принцип опережающего
отражения внешнего мира является неотъемлемой стороной жиз-
ни, ее приспособления к окружающим условиям. У мозга как
специализированного в этом направлении органа нет границ для
такого опережающего отражения действительности. Он обладает
возможностью отражать в микроинтервалах времени цепь собы-
тий, которые могут длиться целые годы.
20
Опережающее отражение действительности
Должен заметить, что когда я сопоставляю ту высокоспе-
циализированную форму опережающего отражения внешнего ми-
ра, представленную условным рефлексом у высших животных,
с опережающим отражением у примитивных живых существ,
то я отнюдь не снимаю качественной разницы между тем и дру-
гим явлением. Необходимо лишь помнить, что всякое сопоставле-
ние многих феноменов и их классификация неизбежно требует
ведущего критерия для такого сопоставления.
Поэтому, естественно, мы должны поставить перед собой
вопрос: какой признак условного рефлекса является наиболее для
него характерным, определяющим? Из всех возможных его ка-
честв качество «упреждения», пли «сигнализации», о предстоя-
щих событиях внешнего мира («предупредительная деятельность»,
по И. П. Павлову) является наиболее решающим его качеством.
Ни изменчивость, ни прочность, пи нриобретешюсть и т. д. не
могут сравниться по значению с этим его биологическим каче-
ством. Именно потому, что животные имеют возможность подго-
товиться по сигналу к еще только предстоящим звеньям после-
довательно развивающихся событий, он стал центральным пунк-
том прогрессивной эволюции. И открытие его — величайшая за-
слуга нашего учителя И. П. Павлова.
Но если этот основной признак (параметр) разбираемых про-
цессов является древнейшим, то, следовательно, по этому приз-
наку мы вполне можем сопоставлять приспособительные воз-
можности низших и высших животных. Естественно, что струк-
турные и количественные усложнения самих аппаратов, обеспе-
чивающих более обширную сигнализацию, привели к дальнейшим
качественным изменениям в поведении животпых, однако сигпаль-
ность как универсальный параметр приспособления к внешнему
миру не утратила от этого своего принципиального значения.
Следовательно, условный рефлекс высших
ниваемый по параметру сигнальности, есть только частный слу-
чай высокоспециализированпых форм опережающего отражения
действительности, т. е. приспособления к будущим, но еще не на-
ступившим событиям (Анохин, 1949).
В результате произведенного нами анализа исторических за-
кономерностей формирования биологических основ условнореф-
лекторной деятельности становится очевидной искусственность
целого ряда вопросов, относящихся к условному рефлексу: ког-
да в эволюции появился условный рефлекс? Есть ли условный
рефлекс у простейших и у растений? Может ли врожденная дея-
тельность быть сигнальной? и т. д.
С изложенной выше точки зрения универсальным принципом
приспособления всех форм живого к условиям окружающего ми-
ра является опережающее отражение последовательно и повтор-
но развивающихся событий внешнего мира, «предупредительное»
животных, оце-
Опережающее отражение действительности
21
приспособление к предстоящим изменениям внешних условий,
или в широком смысле — формирование подготовительных из-
менений для будущих событий.
Как мы видели, этот принцип начинает действовать уже с
первых этапов формирования живой материи. Поэтому вопрос мо-
жет быть только о форме и о конкретных аппаратах, в каких
этот принцип опережающего отражения внешнего мира представ-
лен на данном уровне развития. У одноклеточных он представлен
в форме цепей химических преобразований протоплазмы, опере-
жающих развитие последовательного ряда внешних событий.
У высших животных он проявляется в форме участия специали-
зированных нервных аппаратов, дающих огромный выигрыш в
охвате внешнего мира и в быстроте опережения. Одпако во всех
случаях эта форма опережающего отражения имеет одну и ту
же решающую характерную черту — сигнальность. Для живот-
ных, имеющих нервную систему, это и будет условный рефлекс.
В свете сказанного теряет смысл и вопрос о том, можно ли
выработать временную связь у простейших и растений?
Поскольку те и другие представляют собой живые образова-
ния со сложным молекулярным и многоклеточным строением и
поскольку они подвержены постоянным рядам последовательных
и повторяющихся воздействий (особенно последних), постольку
можно с определенностью утверждать, что в тех и других имеют
место натуральные сигнальные изменения или что эти изменения
можно выработать. При этом только должны быть соблюдены
указанные выше требования: длительность повторения и доста-
точно удачно выбранный существенный фактор для жизни дан-
ного растения или простейшего.
Принципиально же следует признать, что ни одно растение
не смогло бы существовать и немедленно погибло бы в результа-
те естественного отбора, если бы оно реагировало только на налич-
ные факторы среды, т. е. только на то
момент, и не реагировало бы по принципу опережающего отра-
жения.
Так, например, сезонные изменения температуры обычно
таковы, что могут иметь место самые различные перепады тем-
ператур, следовательно, внезапный мороз мог бы полностью на-
рушить протоплазматические свойства растительной клетки.
Совершенно отчетливо решается также и кардинальный для
физиолога высшей первной деятельности вопрос о врожденных
реакциях сигнального типа. Мы уже говорили, что сигнальность
является наиболее характерной чертой для условного рефлекса.
Именно эта черта — опережающее отражение действительно-
сти — приобрела в условном рефлексе наиболее выраженный и
наиболее специализированный вид. Однако, по общепринятой
классификации, врожденные, т. е. безусловные, рефлексы не мо-
, что действует в данный
22
Опережающее отражение действительности
гут иметь этого самого характерного свойства условного рефлек-
са — сигнального значения.
Фактический материал нашей лаборатории по изучению эко-
логической обусловленности первых поведенческих актов ново-
рожденных свидетельствует о том, что этот вопрос не может
быть решен так просто. Исследования нашего сотрудника
Я. А. Милягипа показали, что сразу же после вылупления из
яйца птенец грача безотказно реагирует поднятием головы и рас-
крытием клюва, т. е. пищевой реакцией, на такие раздражители,
которые сами по себе не имеют никакого пищевого значения (дви-
жение воздуха, звук «кар-р-р», сотрясение гнезда). Анализ нату-
ральной экологической ситуации, в которой живут грачи в первые
дни после рождения, показал пам, что все три приведенных выше
агента имеют экологическое значение и служат сигналами пред-
стоящего вкладывания пищи отцом-грачом в раскрытый клюв
птенца (Милягин, 1967).
Таким образом, на основе исторической повторяемости последо-
вательных изменений нервного субстрата, предшествующих момен-
ту кормления, закреплялись наследственностью те структурные
связи, которые обеспечивают теперь опережающее отражение
внешних событий, т. е. с принципиальной стороны здесь происхо-
дит то же самое, что и у первичных протоплазматических обра-
зований. И потому, хотя само раздражение кожи, например, не
является пищевым раздражением, возбуждение от него благодаря
многовековым предшествованиям кормлению с огромной скоро-
стью (30 миллисекунд) и по готовым (врожденным) синаптиче-
ским связям доходит до пищевого центра — птенец поднимает го-
лову и раскрывает клюв. Здесь природа дает исключительно на-
глядный пример опережающего отражения внешних последова-
тельных событий и его приспособительного значения в эволюции.
Интересно, что тщательные электрофизиологические иссле-
дования слуховых способностей уха грача показали поразитель-
ную приспособленность эмбрионального развития к его постна-
тальпой экологии. Оказалось, что в слуховом рецепторе к момен-
ту вылупления из яйца созрели лишь те клетки, которые способ-
ны воспринять звук «кар-р-р».
Можно привести поразительные примеры опережающих отра-
жений внешних событий из различных экологий, причем в каж-
дом отдельном случае это имеет огромный приспособительный
смысл и точно соответствует экологическим факторам данного
вида птицы, т. е. характерным для нее именно последовательно-
стям явлений, предшествующих кормлению.
Так, например, мухоловка-пеструшка выводит своих птенцов
в дупле, куда лучи света поступают лишь через то маленькое
отверстие, через которое мать и отец должны непременно про-
лезть, прежде чем они смогут кормить птенцов. Естественно, что
Опережающее отражение действительности
когда мать или отец просовывают голову в это маленькое отвер-
стие, то исходная небольшая освещенность дупла исчезает и на
какой-то момент воцаряется полная темнота. И как раз именно
эта временная темнота и служит сильнейшим стимулом для птен-
цов дуплянки: они немедленно всей компанией вытягивают шеи,
раскрывают клювы и, таким образом, оказываются готовыми к
приему будущей пищи. После этого мать вкладывает каждому из
них соответствующую порцию (исследования нашего сотрудника
С. И. Хаютпиа, проведенные в натуральных условиях).
Во всех случаях с различной экологией (я мог бы привести
значительно большее количество вариантов) проявляется один
и тот же закон.
Таким образом, опережение событий внешнего мира является
универсальным явлением жизни, которое определило собой все
формы приспособительного поведения животного: врожденное,
условнорефлекторное. С точки зрения этой закономерности любой
член последовательного ряда воздействий внешнего мира может
стать сигналом той конечной жизненно важной реакции, которой
замыкается этот ряд последовательных воздействий.
Для избежания недоразумений в понимании нашей точки зре-
ния следует более подробно остановиться на характеристике сход-
ства и различия двух уровней ассоциативной деятельности: на
ступени примитивной жизни и в форме условного рефлекса. Здесь,
так же как и раньше, важно точно формулировать тот критерий
или параметр этих двух категорий явлений, по которому они
сравниваются.
Основным параметром для сопоставления мы берем решающий
биологический критерий: формы и механизмы отражения в обоих
феноменах последовательно развивающихся внешних событий, по-
скольку сама последовательность является универсальным зако-
ном неорганического и органического мира. Как известно, раз-
витие органов чувств у животпых связано с необходимостью бо-
лее совершенного приспособления к внешнему миру. Органы
чувств нереводят различные но качеству внешние энергии в еди-
ный нейрохимический процесс, смысл которого состоит в том,
что даже в ответ на одиночный раздражитель создается опреде-
ленная система нейрохимических реакций на различных уровнях
центральной нервной системы. Уже один такой раздражитель
обусловливает систему специализированных химических реакций,
состоящую по крайней мере из миллионов точек. Эта система
имеет определенную архитектуру и дает обширную информацию
целому организму не только о том, «как» и «какое» раздражение
было нанесено, но также и о том, «где» и «когда» было нанесено
это раздражение.
Таким образом, уже на самом первом этапе контакта различ-
ных организмов с внешними факторами сказывается огромная
24
Опережающее отражение действительности
разница в объеме той информации о внешнем мире, которую они
получают даже от одиночного раздражения. Поэтому по парамет-
ру сложности распределения микроочагов химических реакций
мозг, несомненно, не идет ни в какое сравнение с химическими
последствиями раздражения, например, уже оформившегося, по
еще примитивного живого существа. В этом последнем случае
химическое раздражение элементарно, оно разыгрывается в об-
щей протоплазме, и лишь только специфика химических валент-
ностей и катализа удерживает эту химическую реакцию в преде-
лах более или менее ограниченных путей.
Если же на минуту представить себе, сколь сложны эти ар-
хитектуры микрохимических очагов в сложнейших специальных
условиях при образовании огромных количеств связей, включаю-
щих в себя отложенный в памяти опыт прошлого, то при сопо-
ставлении опережающего отражения действительности у прими-
тивного живого существа и у человека только по одному парамет-
ру сложности и утонченности станет очевидным, что они не-
сравнимы.
Исторически накапливавшиеся количественные усовершен-
ствования реакции привели к совершенно очевидному новому
качеству приспособительных функций мозга. Это качество мозга
выражается в его способности строить грандиозные планы па
будущее.
Однако, если эти явления (т. е. опережающее отражение дей-
ствительности у примитивных существ и условный рефлекс у выс-
ших животных) мы будем сопоставлять строго по параметру от-
ражений последовательно развивающихся явлений внешнего
мира, мы придем к выводу, что механизм формирования опере-
жающего отражения действительности и там и здесь в точности
соответствует той схеме, которая нами была дана выше.
Химическое объединение раздельно, последовательно и по-
вторно возникающих химических реакций и запуск в последую-
щем всей этой цепи реакций с первого звена — вот что является
принципиально общим параметром для обоих столь различных
по уровню феноменов приспособлений. Вместе с тем это и есть
историческая основа для развития всех видов сигнальных при-
способлений, т. е. подготовки организма к будущим событиям.
Возможен еще и другой, более широкий философский аспект
этой проблемы. До II. П. Павлова физиология животных как
наука знала две категории времени — прошлое и настоящее. Обе
эти категории были представлены соответствующими процессами,
хорошо укладывающимися в паше понимание реакций животных.
Все, что касалось третьей категории времени — будущего, отно-
силось к целесообразному поведению человека и было предметом
почти исключительно идеалистической психологии. Приспособле-
ние к будущим событиям — «предвосхищение будущего» было
Опережающее отражение действительности
25
м
LIE
достоянием человека и являлось признаком наиболее высокоцеле-
сообразпых процессов мозга.
Формулируя условный рефлекс как «предупредительную»
функцию, т. е. функцию приспособления животного к грядущим
событиям, И. П. Павлов тем самым ввел в физиологию третью
категорию времени — будущее. Отныне будущее в жизни орга-
низмов стало предметом такого же объективного научного иссле-
дования, как и две другие категории времени. Таким образом,
открытие условного рефлекса было событием огромной важности
для физиологии. В этом лежит одна из причин столь огромного
успеха изучения поведения животных под этим углом зрения. На-
копление материалов на этом пути все больше расширяло рамки
условного рефлекса, и в настоящее время мы можем твердо ска-
зать, что И. П. Павлов вскрыл кардинальную и всеобщую черту
исторического развития и усовершенствования животного мира.
Условный рефлекс есть снециализированная форма того само-
го процесса, который в цепях элементарных химических реакций
первичной протоплазмы утверждал жизнь на первых этапах ее
развития в форме опережающего отражения пространственно-вре-
менного движения материи в неорганической природе и образо-
вания стабильных структур организма на этой основе. Понадоби-
лись миллионы лет, чтобы эта примитивная форма цепных хими-
ческих реакций фиксировалась н развивалась в самой совершен-
ной форме живой материи — в нервном веществе.
Однако, несмотря на весь грандиозный путь этой эволюции и
на значительные преобразования самих форм опережения, ее ос-
новной закон — опережающее отражение последовательно повто-
ряющихся рядов внешних явлений — остался в силе. Следует от-
метить, что на основе этой формы отражения сформировался и
специализировался и сам мозг как орган психической деятельно-
сти, т. е. орган всеобщего отражения мира в мыслительной дея-
тельности человека. Опережающее отражение действительности,
раскрытое нами здесь па ряде примеров, является лишь одной
из форм отражательной способности живой материи.
Теперь мы должны показать, какое место занимает данная за-
кономерность в универсальной теории отражения, сформулиро-
ванной В. И. Лепиным. Общеизвестно, что теория В. И. Ленина
дает материалистическое объяснение всех процессов организма и
особенно его психической деятельности в зависимости от первич-
но существующих материальных воздействий внешнего мира на
организм, па его мозг. В гносеологическом смысле теория отра-
жения В. И. Лепина является основой для понимания матери-
альной обусловленности психики человека
философская черта этой теории. Она прежде всего утверждает
объективное существование мира и столь же объективное и законо-
мерное отражение этого мира в психической деятельности чело-
это главнейшая
26
Опережающее отражение действительности
века. Отражение это адекватное, и оно является определяющим
фактором для всей мыслительной деятельности человека.
Теория отражения В. И. Ленина, являясь общей философской
основой, открывает широчайшие возможности для исследования
различных форм отражательной деятельности, конкретных физио-
логических механизмов этого отражения и развития в эволюции
различных уровней отражательной деятельности. Она включает в
себя все виды отражения объективной действительности в живой
материи вообще и в психической деятельности в частности. Сюда
входит прежде всего энергетическое отражение, которое благода-
ря последним работам по теории информации стало наилучшим
доказательством точности и адекватности отражения объектив-
ного мира в нашем сознании. Теория отражения В. II. Лепина
включает в себя также и историческое отражение внешнего ми-
ра, т. е. адекватное отражение всей совокупности экологических
►акторов в организации формы и в динамике приспособительной
деятельности животных.
Из сказанного следует, что лепппская теорпя может стать ис-
ходным пунктом для глубоких исследований всех уровней отра-
жательной деятельности живых организмов до высших форм пси-
хической деятельности человека включительно.
Настоящая работа является лишь скромной попыткой рас-
крыть исторические корни отражательной деятельности организ-
мов в отношении временной структуры мира. Целью этой работы
было доказать, что опережающее отражение действительности
как одна из форм отражения организмом объективного мира яв-
ляется историческим фундаментом для появления всех высших
форм предупреждения будущих событий, и прежде всего для по-
явления условного рефлекса.
Л И ТЕГА Т У Г А
Анохин И. К. Узловые вопросы в изучении высшей нервной деятельности.—
В кн.: Проблемы высшей нервной деятельности. М., 1949, с. 9.
Анохин П. К. Рефлекс цели как объект физиологического анализа.— Журн.
высш, нервн. деят., 1962, т. 12, № 1, с. 7.
Милягин Я. А., Лебедева А. 1\, Бурсина М. //. Роль экологических факто-
ров в развитии врожденных форм интегративной деятельности моз-
га.— В кн.: Интегративная деятельность мозга. М., 1967, с. 58.
Опарин Л. //. Возникновение жизни на Земле. М., 1957.
ФИЛОСОФСКИЕ АСПЕКТЫ
ТЕОРИИ ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ*
Общая оценка
опережающего отражения действительности
После того как нами была опубликована статья, в которой под-
робно аргументировалась наша концепция об «опережающем от-
ражении действительности» (см. «Вопросы философии», 1962,
№ 7), в литературе появился ряд монографий, статей и замеча-
ний, разбирающих эту концепцию под различными углами зре-
ния. В основном авторы склонны признать «опережающее отра-
жение действительности» одним из путей развития фундамен-
тальных проблем, поднятых ленинской теорией отражения
(Т. Павлов, Б. С. Украинцев, М. Ф. Веденов, Г, А. Югай и др.).
В самом деле, представление о том, что отражение действи-
тельности может быть активным и, так сказать, «забегать вперед»
на основании использования прошлого опыта, находится в русле
идей ленинской теории отражения и показывает, насколько эф-
фективным может быть применение теории к объяснению явле-
ний отражения внешнего мира.
Опережающее отражение является одной из возможных форм
отражения, в процессе которого, используя опыт прошлого, ор-
ганизм активно приспосабливается к предстоящим событиям.
Условный рефлекс является наивысшей формой этой изначаль-
ной закономерности всего живого.
Эта закономерность формирования живого и приспособления
его к внешнему миру лишний раз убеждает нас в несостоятельно-
сти критиков ленинской теории отражения, утверждающих, что
само понятие «отражение» говорит лишь о пассивном (?!) отно-
шении организма к окружающему миру. Применительно к эволю-
ции организмов мы можем сказать, что отражение действительно-
сти всегда было активным процессом деятельности самого
организма и на более высоких уровнях эволюции вылилось в не-
посредственное воздействие на внешний мир с целью его преоб-
разования (употребление орудий и приспособление к внешним
условиям)...
Опубликовано в книге ((Философские проблемы биологии. Труды Второго
Всесоюзного совещания по философским вопросам современного естест-
вознания, посвященного 100-летию со дня рождения В. И. Ленина. Мо-
сква, декабрь 1970 г.» (М., «Наука», 1973).
Философские аспекты теории функциональной системы
Особенно наглядно этот процесс опережения событий как при-
способительный был продемонстрирован в последние годы на при-
мере куколок некоторых насекомых, которые по условиям разви-
тия вынуждены бывают зимовать на открытом воздухе (напри-
мер, паразитическая оса Вагсоп). Этот факт вызывал немало
удивления в среде ученых и долгое время оставался интригую-
щей загадкой. Действительно, как может куколка осы, содержа-
щая достаточное количество воды в своей протоплазме, устоять
против зимних морозов? Настойчивые исследования в этом на-
правлении ряда ученых привели к поразительным открытиям, ко-
торые полностью объяснили эту загадку. Оказалось, что уже пер-
вые осенние похолодания стимулируют в протоплазме клеток, со-
ставляющих тело куколки, особый процесс: быстрое образование
глицерина. Таким образом, секрет был раскрыт, поскольку изве-
стно, что глицерин, представляет собой вещество, значительно
снижающее криоскопическую температуру клеточных масс.
Конкретный эксперимент показал, что уже ранней осенью
(или в условиях искусственного содержания куколок при темпе-
ратуре 5° ниже пуля) куколки приобретают способность благода-
ря накоплению глицерина перенести мороз в 40—70°. Стоит по-
местить куколок в нормальную температуру летнего дня — и гли-
церин из протоплазмы клеток немедленно исчезает (через 3 дня).
Особенно поразительно и демонстративно это появление и устра-
нение глицерина у пенсильванских муравьев-древоточцев. С ними
можно проделывать подобные опыты несколько раз. И каждый
раз в зависимости от смены температуры глицерин то появляет-
ся (до 100%), то пропадает.
Эта способность куколок в ответ на первые похолодания на-
капливать глицерин, который будет необходим еще только в де-
кабре, является наглядным подтверждением опережающего отра-
жения действительности, выработанного на протяжении миллио-
нов лет.
Из дальнейшего изложения и особенно из разбора специфи-
ческих механизмов мозга можно будет видеть, каковы механизмы
реализации этого опережающего отражения действительности.
Проблема «вписанности» живого
в абсолютные законы неорганического мира
Перед биологами и физиологами возникает новая и интерес-
ная проблема, которую можно было бы охарактеризовать как
проолему «вписанности» живого в фундаментальные законы неор-
ганического мира. Можно выделить весьма большое количество
процессов и явлений неорганического мира, к которым живые ор-
ганизмы приспособились в ходе эволюции но принципу volens no-
lens.
Философские аспекты теории функциональной системы
29
В самом деле, возьмем для примера силу тяжести. Она имела
место задолго до появления жизни, а в масштабе Галактики и
всей Вселенной, конечно, представляла собой изначальное свойст-
во материи.
И действительно, наличие силы тяжести послужило причиной
того, что у разнообразных животных были выработаны вынуж-
денные приспособления самого удивительного характера. Так, на-
пример, у пресмыкающихся сила тяжести определяет своеобраз-
ный характер передвижения, у птиц — многообразные способы
полета, а у рыб — развитие органов и систем регуляции, позво-
ляющих им по-разному использовать «тяжесть» тела в зависимо-
сти от удельного веса тела и воды, и т. д.
Мы можем дополнительно сказать, что развитие всего живо-
го, его самых существенных черт структурно-функциональной
организации, образование самых прочных межнейрональных свя-
зей определяются действием законов всемирного тяготения. По
сути дела начало этого процесса восходит к периоду зарождения
жизни, или, точнее, к периоду формирования предбиологических
систем.
Точно так же
все детали организации
разнообразнейших
структур органов чувств точнейшим образом «пригнаны» к энер-
гетическим свойствам внешнего мира. В более широком плане
мы имеем подлинно активное отражение законов неорганического
мира, с которыми взаимодействовали все живые организмы на-
шей планеты. Мы можем сказать, что активное отражение изна-
чальных свойств внешнего неорганического мира в основных
структурных формах животных является абсолютным законом
жизни.
На фоне этой фундаментальной закономерности движения ма-
терии пространство и время являлись особенно фундаментальны-
ми постоянными факторами, которые уже с момента зарождения
жизни воздействовали на все живое. Только благодаря приспо-
соблению к пространственно-временным взаимодействиям жизнь
могла сохраниться на нашей планете. Постараемся представить
себе, к каким фундаментальным приспособлениям животных по-
вели пространственно-временные факторы.
Пространственно-временной континуум
как фактор биологической адаптации
В основе пашей оценки приспособительной деятельности жи-
вотных лежит необходимость признания искусственного выделе-
ния отдельных важных моментов в их жизни. Так, например,
говоря о действии стимула на организм, мы считаем это искус-
ственное допущение основополагающим в понимании деятельно-
сти организма. Именно оно позволило сформулировать представ-
30
Философские аспекты теории функциональной системы
ление о дискретных актах животного и о дискретном действии
внешних факторов на организм.
Успехи последних лет в области физики и других наук все
более и более позволяют нам сделать вывод о том, что развитие
событий в мире идет па основе непрерывно-прерывного движения
материи. Это представляет собой абсолютный закон, в одинаковой
степени неизбежный как для неорганического, так и для живого
мира.
Однако «биологический экран» здесь вносит существенное до-
полнение, создавая прерывность в этом движении материи в
соответствии с биологической значимостью различных компонен-
тов континуума.
В самом деле, совсем не все компоненты континуума необходи-
мы, например, животному с точки зрения выживания. Некоторые
из них могут иметь решающее значение для жизни животного,
тогда как другие не играют для него никакой роли. Появление,
папример, крупного хищника является жизненно важным собы-
тием, в то время как движение листьев на дереве или движение
воды в реке может остаться незамеченным. Именно эти факторы
биологической значимости отдельных событий создали для живот-
ного своеобразную прерывность пространственно-временных отно-
шений. Однако об этом далее.
Как известно, имеется несколько типов пространственно-вре-
менного континуума. В первом, наиболее простом случае, в так
называемом одномерном континууме, любое положение точки,
движущейся в одном направлении от исходного пункта, может
быть определено координатами пространства и времени, которое
понадобилось для продвижения точки к данному пункту. Суть
этого типа континуума состоит в том, что любому возможно ма-
лому показателю пространственного продвижения точки может
соответствовать столь же малый интервал времени. Мы можем
эти отрезки пространства и времени сделать максимально малы-
ми и, таким образом, на этом микроуровне получить фактически
пространственно-временной континуум, составляющий как бы
индикатор материальных процессов.
Но мы можем оторвать эту точку от одномерного конти-
нуума и поместить ее па какой-либо плоскости, придав ей еще
одну степень свободы (движение в различных направлениях) в
пределах ограниченной плоскости. Тогда положение точки на
каждый данный момент может быть определено уже по двум
осям координат. Следовательно, куда бы и как бы эта точка ни
передвигалась, ее положение всегда может быть «засечено» и вы-
ражено двумя величинами, фиксирующими ее положение по
двум координатам.
Для пас в этом втором типе пространственно-временного кон-
тинуума важно то, что передвижение точки и здесь может быть
Философские аспекты теории функциональной системы
31
представлено в микрорасстояниях и мнкроинтервалах времени,
что позволяет выразить ее различные положения в координатах
двумерного континуума.
Еще дальше усложняя данный пример, можно поместить нашу
точку таким образом, чтобы она продвигалась в трех измерениях.
Тогда, следовательно, ее положение в каждый данный момент
может быть оценимо по отношению к трем перпендикулярным
плоскостям. Однако наряду с этим такая точка может передви-
гаться и менять свои положения и время продвижения также в
зависимости от интервалов пространства и времени. Поскольку
эти интервалы могут быть взяты в мыслимых микроразмерах
пространства и времен:
мы опять-таки,
как и в первом случае,
7
будем иметь континуум, однако более сложного характера, чем
это мы разбирали до сих пор. Это будет уже трехмерный конти-
нуум.
Любое тело, живое или неживое, находится в непрерывно ме-
няющемся трехмерном пространственно-временном континууме.
Это составляет существенную сторону бытия всего живого на
земном шаре. Но именно поэтому мы полагаем,
что отношение
живого к внешнему миру нужно понимать как непрерывную пере-
работку информации в протоплазме или — па высших уровнях
эволюции живого — в его нервной системе, как обработку конти-
нуума воздействий, не имеющего скачкообразного разрыва в про-
странстве и во времени.
Как нейрофизиолог, я могу настаивать па этом положении, по-
тому что практически все временные параметры, в которых функ-
ционирует нервная система, оказываются значительно более дли-
тельными п значительно более компактными, чем минималь-
ные интервалы в течение событий пространственно-временного
континуума внешнего мира. Вместе с тем явления внешнего ми-
ра, развивающиеся в пространственно-временном континууме, мо-
гут находиться в большом удалении друг от друга, и тем не ме-
нее они непрерывно фокусируются в одном и том же мозге,
в одних и тех же нервных клетках.
Так, например, реактивный самолет, летящий по небосклону
перед нашими глазами, может, продвигаясь в пространственном
континууме, за несколько минут преодолеть десятки километров,
и тем не менее все его положения от начальной и до конечной
точки видения, как в фокусе, отражены в микроинтервалах про-
странства и времени в форме непрерывного течения нейрональ-
ных процессов мозга.
Именно этот чудесный механизм живого образования, который
окусирует в микроскопическом пространстве грандиозные интер-
валы пространственно-временного континуума внешних явлений,
стал центральным пунктом всей эволюции жизни на Земле. Толь-
ко благодаря такому биологическому экрану, фокусирующему
32
Философские аспекты теории функциональной системы
огромные масштабы событий внешнего мира в молекулярных ре-
акциях мозговой ткани, стало возможно «объять мир» во всем
его разнообразии и во всей его грандиозности небольшим кусоч-
ком мозгового вещества. Неспециалисты часто забывают об этом
грандиозном достижении эволюции, благодаря которому мы, лю-
ди, на высшем ее этапе приобрели способность в короткие времен-
ные интервалы охватить явления и события, протекавшие в мас-
штабах целого земного шара. Этому способствовали молекуляр-
ные процессы в нервных клетках, принимающие па себя и отра-
жающие грандиозные масштабы Вселенной с помощью непрерыв-
ных процессов, проходящих в самых разнообразных органах
чувств. Ниже мы увидим, какие специфические особенности ор-
ганизма и мозга помогают приспособиться к этому безграничному
пространственно-временному континууму мира.
Понятие о химическом континууме
в мозговых процессах
Достижения
илософии,
lie
изики и математики последних лет
н
показали, что традиционный подход к изучению нервной деятель-
ности и поведения животных уже не может удовлетворить ней-
рофизиологов. Стало очевидным, что искусственное дробление
нервных процессов и в особенности поведепческих актов живот-
ного не может удовлетворить исследователя, который хочет по-
нять смысл этих явлений в аспекте совершенно достоверной ар-
хитектуры мира, складывающейся п развивающейся в рамках
закона пространственно-временного континуума.
В самом деле, если я начинаю изучать животное и исчислять
его способности к поведенческим актам только с момента начала
опыта, то это является совершенно явным изъятием животного
из сложного континуума его жизни и во многом подчиняет ис-
следование процессов, протекающих в его мозгу, концепциям и
замыслам исследователя. Эта искусственная дискретизация явле-
ний внешнего мира особенно значительно проявляется в области
нейрофизиологии в изоляции стимула, который, по представле-
нию физиологов, занимает исключительное положение в форми-
ровании поведепческих актов животного.
В последние годы на основании новейших успехов нейрофизио-
логии становится все более и более ясным, что стимул является
лишь толчком к раскрытию и выявлению того, что создавалось
в мозгу под влиянием многих факторов как нечто целостное, ин-
тегрированное. Стимул же является лишь толчком, раскрываю-
щим эту подготовленную интеграцию. В свое время эту скрытую
и ни в чем себя не проявляющую внутреннюю мозговую интегра-
цию мы назвали «предпусковой интеграцией» (Анохин, 1949).
Но что такое «предпусковая интеграция»? Это — состояние,
выраженное в многочисленных связях нервных элементов, кото
Философские аспекты теории функциональной системы
33
рое складывалось в непрерывном континууме в течение, напри-
мер, дня, когда проводился эксперимент. В самом деле, уже од-
ного прихода служителя в виварий за экспериментальным живот-
ным в определенное время дня достаточно, чтобы в мозгу сфор-
мировался комплекс нервных процессов, отражающий все детали
будущего эксперимента и будущего подкрепления животного
жизненно важными стимулами (корм, боль и т. д.). Практически
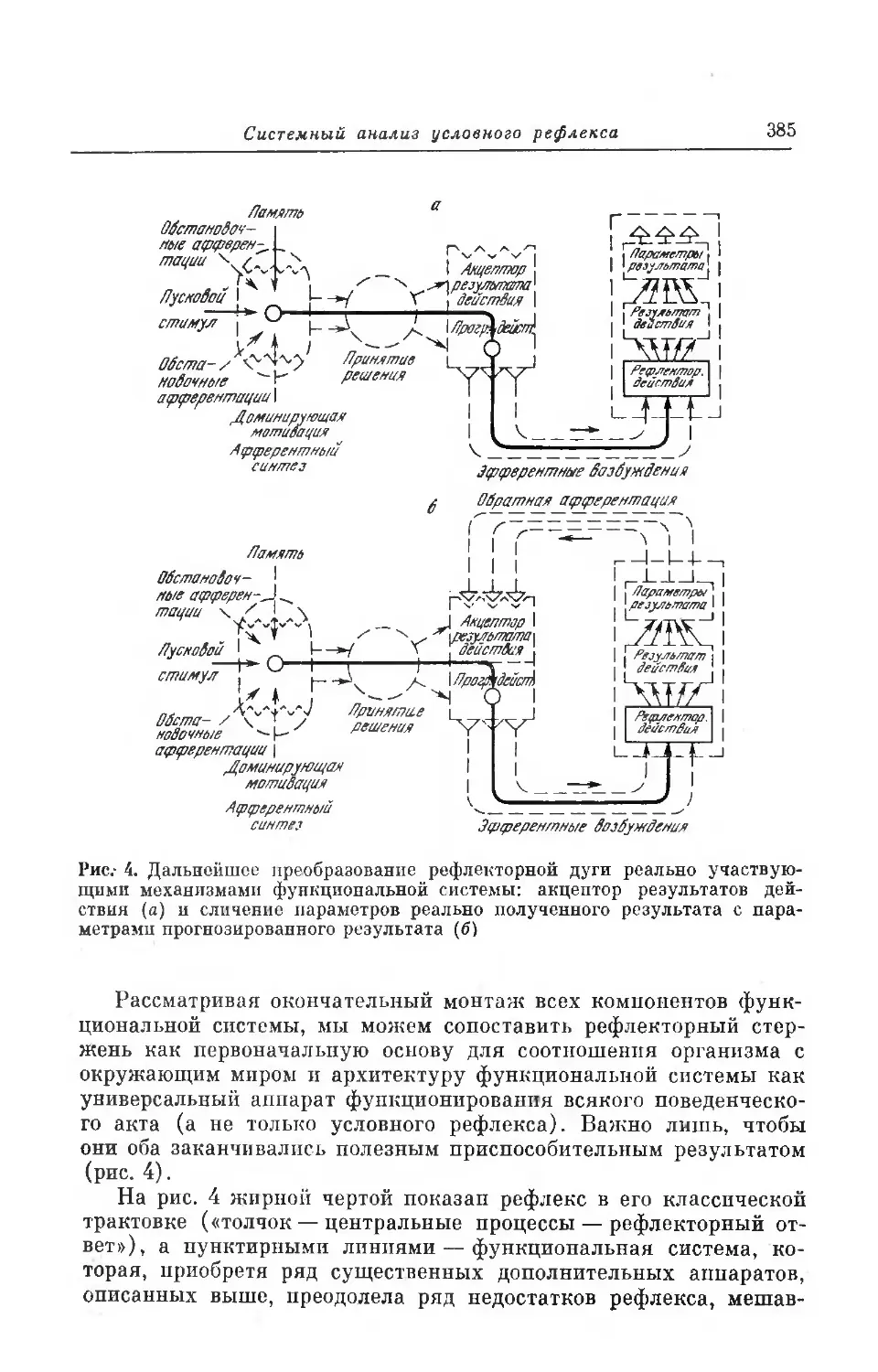
между всеми деталями этого непрерывного сцепления явлений мы
не найдем какого-либо разрыва, который мог бы нарушать кон-
тинуум. Совершенно очевидно, все явления внешнего мира только
искусственно могут быть подвергнуты какой-то дискретизации,
на самом деле они развиваются в подлинном пространственно-
временном континууме.
Таким образом, жизнь животного в целом мы могли бы оха-
рактеризовать как «дискретное» проявление жизненно важных
узловых событий, возвышающихся над реальным континуумом
незаметных для него явлений жизни. Мы так привыкли, что в
нашу жизнь вторгаются отдельные события, которые существенно
затрагивают наше состояние, вызывают эмоциональную реакцию,
что существование истинного пространственно-временного конти-
нуума представляется чем-то нереальным, неощутимым. Тенден-
ция развития биологического объекта такова, что организм неми-
нуемо «засекает» все, составляющее существенные стороны его
жизни, его достижений и его неудач. И тем не менее все эти
островки значимых для нас событий соединены между собой
реальным континуумом явлений и процессов, которые, хотя и не
имеют решающего жизненного значения, по все же являются сое-
динительными звеньями в континууме мира. Этим самым дости-
гается универсальный охват всех компонентов континуума, одни
из которых жизненно важны, а другие служат сигналом их по-
явления.
Современная нейрофизиология па основе нейрохимических мо-
лекулярных процессов доказывает, что жизненная значимость от-
дельных событий представлена в мозгу даже в специфических
химических процессах мозга, которые как бы засекают «шаги»
этих жизненно важных событий. Так, например, мы имеем раз-
личную химию страдания, тоски, страха, радости и других су-
щественных эмоциональных переживаний и событий в жизни жи-
вотных и человека. Существенность всех этих явлений в жизни
человека и эпизодический прорыв их в сознание приводит к тому,
что истинный пространственно-временной континуум нашего по-
ведения субъективно не воспринимается нами как континуум.
Если взять бесконечное количество примеров нашего поведе-
ния, то при тщательном анализе можно убедиться, что раздраже-
ние нашей нервной системы происходит в совершенно непрерыв-
ном континууме.
2 П. К. Анохин
Философские аспекты теории функциональной системы
Попробуем теперь с помощью воображения перенестись внутри
Мозга, в бесконечное множество его процессов, отражающих дей-
ствительность. Современная техника исследования мозга сделала
возможном не только изучение грубых, суммарных электрических:
явлений в мозгу, имеющих место во время его деятельности..
Электроника и ее тончайшие приборы дают возможность проник-
нуть в деятельность любой нервной клетки из всех четырнадцати
миллиардов клеток мозга. Такой подход значительно расширил
наши представления о топких процессах, развивающихся в нерв-
ной клетке в момент ее деятельности под тем или иным воздей-
ствием внешнего мира. Каждая нервная клетка имеет в среднем:
около 5 тыс. контактов с другими нервными клетками и с орга-
нами чувств. Кроме того, каждая нервная клетка может испыты-
вать по крайней мере 6 разнообразных общих состояний. Очень»
трудно представить себе то количество возможных специфиче-
ских состояний, которые может испытать мозг при наличии тако-
го огромного количества переменных.
Мы можем поставить вопрос: как же вся эта необъятная ком:-
бинация нервных процессов, происходящих в мозгу, отражает не-
прерывное течение внешних событий в различных пространствен-
ных и временных комбинациях? «Мудрость мозга» определяется
не только тем, сколько количественных показателей он способен
обработать, но и тем, как мозг отражает все свойства простран-
ственно-временного континуума внешнего мира в его динамиче-
ских процессах и в качественной особенности его структурных
взаимоотношений.
Представим себе на минуту, что непрерывный ряд внешних
событий последовательно действует на первную систему челове-
ка, и это действие осуществляется с интервалами, которые могут
быть выражены в миллисекундах. В таком допущении пет ниче-
го невероятного. Паша жизнь заполнена такого рода действиями.
Можно взять любой пример из нашего повседневного поведения и
показать, что оно складывается из непрерывных, одпо в другое
переходящих действий и из таких же непрерывных афферентных
возбуждений нервной системы.
Фактически мы ни в одном нашем акте не можем найти какую-
либо дискретность, достаточную для того, чтобы мы могли кон-
статировать наличие интервала в виде полного «молчания» нерв-
ной системы между какими-либо достаточно отдаленными воздей-
ствиями пространственно-временного континуума внешнего мира.
Паши представления о дискретных действиях различных сти-
мулов на организм и о дискретных процессах мозга в значитель-
ной степени павеяпы формой искусственного эксперимента —
внезапным применением нужного нам стимула. Однако, как я
уже упоминал, даже и в этом случае процессы, происходящие в
мозгу, например, экспериментального животного, текут в той же
Философские аспекты теории функциональной системы
35
непрерывности, как и описанные выше, в естественном пове-
дении.
Очевидно, мы должны будем принять в качестве основного за-
кона работы мозга следующее: абсолютный и универсальный за-
кон неорганического мира — развитие явлений в пространственно-
временном континууме — в процессе эволюции живого привел к
тому, что мозг животных как специальный орган отражения и
приспособления приобрел свойство непрерывного течения его
процессов в полном соответствии с компонентами этого контину-
ума в пространстве и во времени.
Все формы нашего поведения, все вариации его и, как увидим
ниже, активный выбор, оценка и классификация живыми орга-
низмами отдельных этапов и компонентов этого континуума впи-
саны в этот великий закон природы.
Можно сказать, что все относительное, меняющееся в зависи-
мости от совокупности многих условий жизни, всегда вписано в
этот абсолютный закон.
Следовательно, мы должны искать лишь условия и механизмы
того, как появляется и как ведет себя относительное и меняющее-
ся в пределах этого абсолютного закона. Именно здесь лежит, по
нашему мнению, «точка отсчета» для изучения работы мозга во
всем ее многообразии.
Разбирая достоинства теории относительности А. Эйнштейна,
Макс Планк следующим образом выражает ее отношение к аб-
солютным законам мира: «В основе так называемой теории от-
носительности заложено нечто абсолютное; таковым является оп-
ределение меры пространствепно-времеппбго континуума, и как
раз особенно привлекательная задача состоит в том, чтобы разы-
скать то абсолютное, которое придает относительному его под-
линный смысл. Мы можем исходить всегда только из относитель-
ного. Все наши измерения имеют относительный характер... Речь
идет о том, чтобы во всех этих данных обнаружить то абсолют-
ное, общезначимое, инвариантное, что в них заложено» (кур-
сив.— П. А.).
В этих словах создателя квантовой теории дана программа для
исследования любого^ даже весьма незначительного, явления на
фойе и в рамках абсолютных законов мира. В особенности это
положение применимо и даже настоятельно необходимо для изу-
чения эволюции жизни на Земле, которую Планк, конечно, мень-
ше всего имел в виду.
В эволюции живого все относительное и вариантное «пришито»
весьма «откровенными» нитями к абсолютным законам мира. Осо-
бенно отчетливо это иллюстрируется эволюцией мозга.
В дальнейшем изложении мы увидим, насколько тонко все
структуры и механизмы мозга оказались приспособленными к
максимальному охвату того многомерного континуума мира, ко-
2*
36
Философские аспекты теории функциональной системы
торый способствовал развитию многих специальных приспособле-
ний в работе мозга, обеспечивающих организму выживание.
Однако сейчас я верпует» к тому, что составляет саму природу
этих приспособлений,— к деятельности одиночной нервной
клетки, к процессам, происходящим в ее протоплазме. Само при-
способление всегда имеет интегральный, системный характер, од-
нако элементарным процессом в этих системах является разряд-
ная деятельность нейрона.
Выше мы установили, что практически при любом движении
животного или при выполнении им какого-либо поведенческого
акта пространственно-временной континуум внешнего мира дей-
ствует в мпкроинтервалах времени на его органы чувств и, сле-
довательно, на нейрональные элементы мозга. Как же сами
нервные элементы реагируют на эти воздействия компонентов н
ас
$аз пространственно-временного континуума?
Для того чтобы ответить на этот вопрос, мы должны
обратиться к физиологическим свойствам нейронов мозга. Одной
из самых замечательных особенностей нейрональных элементов
мозга является разнообразие их индивидуального поведения, раз-
нообразие реакций на приходящие возбуждения.
Так. например, одни из них переходят на режим повышенной
активности в момент действия приходящего возбуждения. Дру-
гие, наоборот, находясь в постоянной фоновой активности, пере-
ходят во время действия раздражителя в состояние торможения.
Одни нейроны прекращают свою ответную повышенную разряд-
ную деятельность с прекращением раздражения, другие, наобо-
рот, еще очень долго в виде «следовой деятельности» продолжа-
ют разряжаться. У различных нейронов эта так называемая
следовая активность может продолжаться различное время. В то
время как у одних нейронов она длится всего лишь несколько
миллисекунд, у других она может продолжаться секунды и даже
минуты. Ясно, что все это разнообразие индивидуального поведе-
ния нейронов мозга в ответ на раздражения является выражени-
ем их места, их функционального значения в обширных интег-
ративных образованиях, складывающихся в процессе формирова-
ния сменяющихся функциональных систем организма.
Однако сейчас мы не будем рассматривать эти проблемы с точ-
ки зрения нейрофизиологии. С этим можно подробно ознакомить-
ся по ряду солидных монографий (Берптов, 1969).
Сейчас же хотелось бы обратить внимание читателей именно
па те нейроны, а их в головном мозгу большинство, которые
имеют следовую активность, простирающуюся на целые секунды
после прекращения раздражения. Как должны были бы вести
себя эти клетки в случае непрерывной смены внешних раздраже-
ний в микроинтервалах, т. е. по крайней мере в интервалах, оп-
ределяемых миллисекундами?
Философские аспекты теории функциональной системы
3?
Прямые эксперименты с употреблением парных
раздражени
пли целого ряда раздражений показывают, что происходит неиз-
бежное наложение возбуждений, их суммация и значительное
пролонгирование активного состояния нервной клетки. Эта «следо-
вая деятельность» может приобретать различный характер, глав-
ным же образом изменяется конфигурация разрядов.
Однако остается бесспорным, что имеет место совершенно оче-
видное наложение последующих разрядов клетки, возникших в
ответ па каждое предыдущее раздражение или на последействие
от предыдущей деятельности. Такой процесс, несомненно, про-
исходит ежеминутно и в естественных условиях: при смене, на-
пример, многообразных впечатлений некоторые зрительные клет-
ки с выраженным следовым эффектом находятся в непрерывном
перекрытии разрядов, вызванных различными и последователь-
ными зрительными раздражениями. Стоит лишь каждому из нас
припомнить переход из одной комнаты в другую или выход па
улицу.
Факт перекрытия активностей определенных нервных клеток
мозга не подлежит сомнению. Отдельные компоненты внешнего,
пространственно-временного континуума могут действовать на
слуховой или зрительный аппарат на еще более коротких интер-
валах, чем те, которые указывались выше. Следовательно, пере-
крытие деятельности нервных клеток остается постоянным и су-
щественным фактором в работе мозга. Причем важно отметить,
что это перекрытие компонентов континуума может быть одно-
родным, т. е. подряд могут следовать два различных, например,
зрительных раздражения. Но оно может заключаться и в том,
что на одну и ту же нервную клетку падают в небольшом интер-
вале два различных возбудителя,— например, звуковой, а потом
зрительный.
Это значит, что определенные типы клеток нервной си-
стемы по самой своей сути «ведут» в мозгу непрерывную «мело-
дию» пространственно-временного континуума внешнего мира.
Очевидно, именно им принадлежит прерогатива создания абсо-
лютной основы для жизненно важных эпизодов нервной дея-
тельности.
Именно они, по-видимому, поддерживают непрерывную впи-
санность всего относительного и вариантного в абсолютный закон
пространственно-временного континуума мира.
Здесь мы подошли к последнему ряду аргументов, направлен-
ных в защиту положения о наличии нейрональной деятельности,
обеспечивающей постоянный контакт мозга, а следовательно, и
организма со всеми процессами, происходящими на разных эта-
пах развертывания пространственно-временного континуума.
Но рано или поздно у нас должен возникнуть вопрос: что же
представляют собой по своей глубокой сути, т. е. по природе, тс
Философские аспекты теории функциональной системы
клеточные разряды, которые нами улавливаются сейчас с помо-
щью электронной техники как электрические взрывные про-
цессы?
Сейчас не остается сомнений в том, что разряд нервной клет-
ки — это ее «голос», «крик», и в него вложена вся история данно-
го живого существа, обладателя данного мозга. Он отражает
многочисленные влияния, пришедшие к данной нервной клетке.
Однако по своей глубокой природе этот разряд есть производный
эффект весьма специализированных химических процессов, вы-
званных к жизни пришедшим к нейрону возбуждением.
Итак, все импульсы, приходящие к нервной клетке, трансфор-
мируются в ее протоплазме в химический процесс, который вто-
рично ведет к электрическому разряду.
Любые влияния или возбуждения, действующие на нервную
клетку, неизбежно проходят стадию химической трансформации,
которая и определяет кодирование интегративного результата в
нервной деятельности клетки в каждый данный момент в форме
того или иного рисунка нервных импульсов. Это положение при-
водит нас к окончательному заключению, которое и составляет
основную цель этого раздела работы.
Сам факт наличия следовых разрядов у некоторых видов нерв-
ных клеток неизбежно приводит к тому, что химические процес-
сы нейрона, вызванные предыдущими влияниями, перекрываются
новыми химическими процессами, вызванными последующим ком-
понентом пространственно-временного континуума.
Иначе говоря, за перекрытием нервных импульсов или клеточ-
ных разрядов мы всегда должны видеть перекрытие и химиче-
ских процессор нейрона в целом. Однако континуум событий
внешнего мира не имеет перерыва и не имеет конца. Следователь-
но, подавляющее большинство клеток мозга непрерывно пребы-
вает в химическом континууме тех реакций, которые вызваны
постоянно п повторно раздражающими факторами.
Следовательно, в протоплазме нервных клеток мозга имеет
место реальный химический континуум, отражающий непрерыв-
ность событий внешнего мира, т. е. его пространственно-времен-
ной континуум.
Это и есть то абсолютное, та основа, на которой строится
все грандиозное разнообразие структурных и функциональных
проявлений мозга.
Поведение как континуухм результатов
Из предыдущего видно, что мозговые процессы связаны меж-
ду собой и представляют непрерывную цепь химических процес-
сов, флуктуирующих непрерывно в зависимости от действия тех
или иных раздражений. Мы установили, что именно след от
Философские аспекты теории функциональной системы
39
этих флуктуаций возбудимости, выраженный в разрядной дея-
тельности нейрона, является тем связующим звеном, которое
предыдущее в жизни мозга цементирует с последующим. В этом,
па наш взгляд, и состоит глубокая нейрофизиологическая приро-
да химического континуума мозга.
Однако такое описание процессов мозга должно вызвать есте-
ственный вопрос читателя: как же этот, казалось бы, монотонный
процесс химических последовательностей может сконструировать
все разнообразие поведенческих актов животного и человека, так
тонко и динамически приспосабливающих организм к изменениям
внешнего мира? Как может сам поведенческий акт, включающий
на разных этапах разнообразные анатомические структуры, осу-
ществить этот химический континуум?
При глубоком и внимательном анализе существа дела мы
можем вступить в область исключительно интересных закономер-
ностей, где эволюция структуры нервной системы замечательным
образом приспособлена к абсолютному закону пространственно-
временного континуума мира, создавшего континуум химиче-
ских процессов мозга. Этот анализ убеждает пас также в том,
что мозг в процессе эволюции выработал специальные структуры
и их соотношения, которые специально предназначены для вос-
произведения прострапствеппо-времепибго континуума внешнего
мира и для включения в него жизненно важных моментов.
В самом деле, развитие структур мозга, обладающих способ-
ностью к генерализованной активации, т. е. к распространению
активирующих влияний на весь мозг, обеспечило одну из самых
важных функций поведенческих актов — их сигнальный харак-
тер. В настоящее время в высшей степени важно разобрать конк-
ретный механизм этого действия.
Допустим, что во внешнем мире в короткие интервалы вре-
мени развивается действие ряда раздражителей на мозг. Как мы
знаем, огромное количество клеток мозга непрерывно соединяет
химические последствия этих раздражителей.
Допустим также, что все эти раздражители не имеют ника-
кого существенного значения для организма, т. е. они не грозят
ему несчастьем, не несут ему радости.
Теперь сделаем другое допущение, что в этом ряду простран-
ственно-временного континуума внешних раздражений один из
раздражителей оказался чрезвычайно опасным для жизни или
целостности организма, т. е. вызвал сильнейшее болевое ощуще-
ние, Тем самым конкретный раздражитель возбудил весь комп-
лекс врожденных биологически значимых процессов в области
ствола мозга и гипоталамуса. Эти возбуждения восходят до
самой коры, п в данном случае они возбудят положительно все
клетки коркового уровня. Во всяком случае трудно найти такую
клетку коры больших полушарий, которая бы нс отреагировала
40
Философские аспекты теории функциональной системы
учащением или торможением своих разрядов па возбуждение
ретикулярной формации и гипоталамуса.
Постараемся представить себе, что же происходит во всех
следах, которые остались от предшествовавших, последовательно
действовавших па организм раздражителей перед тем, как он
получил болевое раздражение. Некоторые из корковых клеток,
несомненно, сохранили следы только что подействовавших на
них возбуждений, т. е. находились в состоянии затухающих раз-
рядов. Совершенно естественно поэтому, что чем ближе было
раздражение к моменту сильного болевого возбуждения, тем от-
четливей был след от этого возбуждения, совпавший с моментом
болевого состояния. Для теории условных рефлексов это был бы
«коротко отставленный сигнал».
Благодаря этому совпадению следов от предшествовавших ин-
дифферентных раздражений с генерализованным возбуждением
от ретикулярной формации и гипоталамуса происходит интерес-
ная картина: где бы ни был предшествовавший раздражитель, он
неминуемо будет «настигнут» генерализованным болевым возбуж-
дением и произойдет в такой же степени неминуемая встреча
двух химических процессов в протоплазме одной и той же нерв-
ной клетки. Это и есть момент химического сцепления всех пред-
шествовавших «химий» с «химией» болевого возбуждения.
К чему поведет такая встреча и, главное, каков химический
результат этой встречи? В каждом случае, когда повторяется
последовательность тех же самых внешних воздействий, процесс
пойдет по нервным связям, химически уже фиксировавшимся до
момента формирования реакций на болевое раздражение, хотя
вся последующая цепь возбуждений, предшествующих болевому
раздражению, не была подкреплена соответствующими им конк-
ретными воздействиями. Иначе говоря, мы имеем подлинное опе-
режение компонентов пространственно-временного континуума
внешнего мира специфическим химическим, мозговым процессом.
В поведенческом смысле это и будет сигнальная, пли предупре-
дительная, реакция, по И. П. Павлову.
Наличие химического континуума в мозгу животных и чело-
века при постепенном приближении к узловому пункту прост-
ранственно-времепнбго континуума внешнего мира, имеющему
существенное биологическое значение, является неоспоримым и
реальным фактом. Нам сейчас важно лишь установить, какое
влияние этот континуум имеет на формирование всех поведенче-
ских актов.
Мне хотелось бы отметить во избежание непонимания один
важный этап развития, связанный с приобретением живыми су-
ществами способности к свободному передвижению. Не изменяя
принципиального содержания взаимозависимости химического
континуума от пространственно-временного континуума внешнего
Философские аспекты теории функциональной системы
41
мира, свободное движение создает тем не менее условия для
огромного скачка в развитии приспособительного поведения у
животных.
В самом деле, теперь уже само животное, двигаясь самостоя-
тельно в различных направлениях, создает континуум внешних
раздражений своих анализаторов, как бы подставляя себя под
их действие. Континуум уже непосредственно начинает влиять
на построение целевого или целенаправленного поведения, сво-
дящегося к активному поиску таких компонентов внешнего кон-
тинуума, которые определяют удовлетворение каких-либо потреб-
ностей организма. Организм как «открытая система» активно
ищет для своих «входов» точно запрограммированные ее обме-
ном веществ недостающие компоненты.
В этом случае мы сталкиваемся с новой закономерностью,
которая стала, так сказать, сквозной для всех этапов развития
жизни на Земле. Продвижение вперед живого существа создало
условия для развития поведенческого континуума животпых, по-
скольку в последовательном действии внешних агентов на орга-
низм в процессе его активного продвижения мы опять-таки не
имеем перерыва в химически обусловленной разрядной деятель-
ности нейронов.
Однако здесь появилось нечто новое. Это — полезный резуль-
тат деятельности животного, который непременно возникает в
процессе его активного отношения к пространственно-времен-
ному континууму внешнего мира. Можно на любом поведенче-
ском акте показать, что, будучи вписан в пространственно-вре-
менной континуум, о котором мы говорили выше, он вместе с
тем состоит из цепи полезных результатов, переходящих один в
Другой.
С точки зрения более общей оценки поведения мы видим, что
речь здесь идет о все той же закономерности — поведение, чело-
века в пространственно-временном континууме предстает перед
нами как континуум больших и малых результатов с непремен-
ной оценкой каждого из них с помощью обратной афферен-
тации.
Итак, мы определили в приведенной
ормуле закон конст-
руирования поведения живых существ, по которому оно пред-
ставляет непрерывную цепь результатов с последующей оценкой
их достаточности.
Здесь-то и вмешивается роль «биологического экрана», клас-
сифицирующего значимость результата и вводящего тот элемент
генерализующего подкрепления, который перекрывает все преды-
дущие этапы раздражений, оставивших след в центральной нерв-
ной системе.
Именно биологически значимые или эмоцпогенные раздраже-
ния, скрепляющие следовые реакции от прежних раздражений,
Философские аспекты теории функциональной системы
составляют необходимое условие, подготавливающее путь для
будущих опережающих реакций под действием какого-то отдален-
ного звена пространственно-временного континуума.
Итак, непрерывность явлений внешнего мира 'отражается на
«биологическом экране» как «прерывность» жизненно важных
событий, отставленных друг от друга во времени, но связанных
континуумом малозначащих результатов целого поведенческого
акта животных и человека.
Заключение
В настоящей работе я сделал попытку на основе теории отра-
жения показать, что основной, или, по выражению Планка, «аб-
солютный закон» неорганического мира — пространственно-вре-
менной континуум определял собой все формы поведения живого
от примитивных его форм до человека включительно.
Появление жизни на Земле позволило в микромире молеку-
лярных реакций живой материи отразить все макропроцессы,
протекающие с огромными интервалами времени. Создались ус-
ловия для «забегания в будущее», а следовательно, появилось
в высшей степени приспособительное поведение.
«Биологический экран» не отражает пассивно континуум внеш-
него мира. Внося в него «потребности жизни», обеспечивая со-
хранение ее, он связал получение любых результатов с этими
основными требованиями. Так организовался континуум резуль-
татов, в котором «мелкие» результаты, лежащие на пути полу-
чения «больших» и «грандиозных результатов», сливаются в еди-
ный поведенческий континуум результатов и сопровождают жизнь
от ее зарождения до ее уничтожения.
Я хотел бы заранее оговорить, что все высказанные здесь
общие положения о структуре взаимоотношения неорганическо-
го п органического мира не являются лини» чисто теоретически-
ми построениями, а имеют существенное практическое значение.
Едва ли надо говорить о том, что такого рода обобщения,
значительно меняющие сам исследовательский подход, должны
быть доброжелательно обсуждены всеми теми, кому дороги проб-
лемы, поднятые в этой статье: философами, физиологами, биоло-
гами и др. Такое обсуждение, я надеюсь, поможет нам посмот-
реть с высоты философских обобщений на работу мозга, объект,
наших каждодневных исследовательских усилий.
Выступление П. К. Анохина па Совещании *
Ученые многих направлений науки в наши дни с большим
вниманием изучают возможности системного подхода, который
позволил бы осмыслить то, что нельзя понять при аналитическом,
* Опубликовано в журнале «Вопросы философии» (1971, № 3).
Философские аспекты теории функциональной системы
43
или, как выразился В. А. Энгельгардт, «редуцированном накоп-
лении материала». Этот подход является в какой-то мере жестом
отчаяния. Совершенно очевидно, что мы сейчас стоим перед
опасностью утонуть в обилии материала, накопившегося по «ча-
стным» проблемам, не связанным в систему, и это грозит поте-
рей общего направления.
Один из крупных нейрофизиологов, получивший недавно Но-
белевскую премию, Дж. Экллс, выразил эту мысль так: надо стоять
в центре сражения за проблему мозга, чтобы увидеть, что по
крайней мере 95% публикуемых исследований не годится для
построения большой теории. И это относится не только к ней-
рофизиологии, но и к другим областям — биологии и т. д.
Таким образом, поиски ключа, который соединил бы уровень
целостности и аналитический уровень получения деталей,— это
и есть та кардинальная проблема, которая продолжает стоять не
только в мировоззренческом плане, но и носит чисто практиче-
ский характер.
Естественно, необходимы обобщения и частных аналитических
материалов, которые сами по себе могут быть успешно исполь-
зованы и в практической деятельности и при больших теорети-
ческих обобщениях. В настоящее время этот поиск системы ве-
дется в высшей степени интенсивно. Достаточно сказать, что
целый ряд обществ, журналов, симпозиумов занят изучением си-
стем — словом, система стала тем маяком, который непосредст-
венно освещает путь к эксперименту и всем другим видам науч-
ного исследования. Хотя тенденция к тому, чтобы связать це-
лостный уровень с уровнем частичным, с уровнем результатов
аналитических исследований, значительно усилилась, к этому
явлению надо подойти критически; следует сказать, что все это
научное движение пока еще не имеет строгой логики, которая
необходима для того, чтобы соединить оба уровня исследований.
Отсутствие строгой логики в постановке вопроса о том, что
ищется и для чего ищется, завело в тупик целый ряд исследо-
вателей и мыслителей, в том числе и самого Л. Берталанфи.
Поэтому прежде всего необходимо решить, для чего мы ищем
формулировку системы, которая могла бы быть приемлема для
многих. Эта основная цель поиска в высшей степени важна для
того, чтобы конечный итог был действительно продуктивным.
В самом деле, задача заключается в том, чтобы заполнить
ту пропасть, которая разделяет еще во многих науках уровень
целостного и уровень частного, аналитически полученного
результата. Заполнить эту пропасть нельзя словами, ее нельзя
заполнить формулировками. Нужно найти действенные принципы
работы в этом направлении. Решение этих двух задач поможет
ликвидировать пропасть, выработать конкретные принципы ра-
боты.
44
Философские аспекты теории функциональной системы
1
этом. Закрыть эту про-
значит сформулировать принцип работы, который, с од-
, находится в области целостности, т. е. носит
а с другой — в аналитической
и т. д.). Но эти исследования находятся в тесном единстве
Тем не менее эти вопросы еще не решены. Значит, логика
развития самой проблемы требует прежде всего их решения.
Если мы их не решим, то будем вынуждены остаться на уровне
словесных формулировок и не достигнем того конкретного ре-
зультата для науки, который может дать формулировка понятия
системы в истинном смысле.
Задача, которую мы ставили в своей практической и теоре-
тической работе, состояла именно
пасть
ной стороны, находится в области целостности,
черты интегративного целого
области. При данном условии этот принцип выступает как бы
приводным ремнем между целым и его частями.
Концепция функциональной системы представляет для нас
именно такой принцип. Функциональная система позволяет осу-
ществлять исследование в любом заданном участке целого с
помощью любых методов (и микроэлектронного, и электрофоре-
за,
благодаря функциональной системе, которая показывает, где и
как ведутся данные исследования.
Такая возможность возникла в результате решения двух во-
просов, которые не были решены в системном подходе, но без
которых двигаться вперед нельзя.
Первый вопрос, который мы поставили уже давно и который
на данном этапе получил удовлетворительное решение, заключа-
ется в следующем. Все известные формулировки систем по сути
дела имеют своим лейтмотивом принцип взаимодействия (это —
взаимодействие множества компонентов, взаимодействие и отно-
шение между компонентами и т. д.).
Если просто рассчитать количество клеток, количество синап-
сов, т. е. возбуждений, которые получает каждая отдельная
клетка, то количество взаимодействий, возможных в мозгу, долж-
но быть
единица
9,5 млн.
природа
ды, этих
хаос
в единую систему. Вот почему слово «упорядочение» начинает
все больше и больше проникать в формулировку систем. Однако,
если мы введем этот термин, но не раскроем фаз, по которым
множество упорядочивается, мы будем недалеки от понятия эн-
телехии, т. е. от утверждения того, что кто-то упорядочивает,
какая-то сила вмешивается в систему и упорядочивает. Поэтому
необходимо проанализировать конкретную систему и посмотреть,
что же упорядочивает это множество степеней свободы.
выражено огромным по величине числом, это число —
с нулями, где нули выписываются на ленте длиной в
км. Это даже не астрономическая цифра! И ясно, что
развивалась в сторону увеличения этих степеней свобо-
возможных взаимодействий. Но возник бы невероятный
если бы все эти взаимодействия были организованы сразу
ЭЙ
Философские аспекты теории функциональной системы
45
Многие работы, проведенные в нашей лаборатории, показали,
что таким упорядочивающим фактором является результат дея-
тельности системы. Только он может через обратную связь (как
называют кибернетики афферентацию) воздействовать на систему,
перебирая все степени свободы и оставляя только те, которые
содействуют получению данного результата.
Результат господствует над системой, и над всем формирова-
нием системы доминирует влияние результата. Результат имеет
императивное влияние на систему: если он недостаточен, то
немедленно эта информация о недостаточности результата пере-
страивает всю систему, перебирает все степени свободы, и в
конце концов каждый элемент вступает в работу теми своими
степенями свободы, которые способствуют получению результата.
Введение результата в интегрирующий фактор позволило рас-
крыть пути упорядочения в системах.
И вторая важная проблема, которая не решена системным
подходом в западных странах, особенно в США, состоит в сле-
дующем: до тех пор, пока скачки будут делаться от целого прямо
к аналитической детали, трудно ожидать положительного резуль-
тата в употреблении системных идей как инструмента повсе-
дневного исследования. Функциональная система заполняет и
этот пробел, так как в ней разработана совершенно определен-
ная операциональная рабочая архитектоника со специфичным ме-
ханизмом и со специфичными свойствами, которая и допускает
постановку вопроса в аналитическом плане. В функциональной си-
стеме есть стадия афферентного синтеза, и на этой стадии решает-
ся вопрос, какой результат должен быть получен.
Здесь введение физиологических детерминистических понятий
сняло видимость телеологичности целого. Целое — это есть нечто,
запрограммированное в конкретных афферентных параметрах бу-
дущего результата. Мы считаем, что, раз весь процесс подходов
к целому виден на нейрофизиологическом материале (что хорошо
показал И. Т. Фролов), проблема эта целесообразна и никакой
телеологичности вкладывать в нее не надо и невозможно.
В этом пункте мы столкнулись с необходимостью решения
философской проблемы, вернее, с необходимостью сделать фило-
софское обобщение, которое разъяснило бы весь процесс, проис-
ходящий в функциональной системе до получения результатов.
Акцептор действия — это аппарат, который примет решение,
получив какой-то результат, закодирует все свойства этого ре-
зультата до того, как он будет реализован. И это обстоятельство
заставило нас задуматься.
Мы не могли не поставить вопроса: как же так? Основным
законом рефлекторной теории является поступательный ход воз-
буждения. Закон поступательности есть основной принцип реф-
лекторной теории.
46
Философские аспекты теории функциональной системы
момент принятия решения идет процесс,
намного опере-
жающий события. Он формируется не поступательно. Он уже
имеет закодированный в ведущих параметрах результат, который
может быть получен через минуты, часы, а может быть, и
годы.
Как формируется опережение событии? Я думаю, что многие
знакомы с законом опережающего отражения действительности.
Опережение событий есть прежде всего активное поддержание
поставленной цели до момента ее реализации.
В последнее время в пашей лаборатории нолучен факт ней-
изиологического
порядка, па основе которого удалось «под-
смотреть» и выявить ту «хитрость», которую создала природа
эффектом длительного поддержания цели.
Какие силы сохраняют цель до момента ее реализации? Ока-
зывается, рефлекторный аппарат. Наши действия дают ответвле-
ния возбуждения, которые заряжают части мозга, и эти импуль-
сы идут обратно в мозг, в кору больших полушарий, и там они
все время дают энергию для формирования цели.
Таким образом, раскрыта физиологическая природа поддержа-
ния цели в активном виде на протяжении большого периода
времени.
Как это опережение происходит? Мы неизбежно должны были
прийти к формулировке пространства и времени в философском
плане. И тут мы столкнулись с важным обстоятельством, которое
пришлось продумать.
Как физическая, так и философская формулировка простран-
ства и времени учитывает то, что главным параметром здесь яв-
ляется последовательность. В статье В. II. Лебедева и В. С. Сте-
пина (Лебедев и Степин, 1970) говорится, что последовательность
есть главный параметр времени. Мы поставили этот вопрос не-
сколько иначе, и именно этот аспект мне бы хотелось здесь
обсудить.
Физическая теория времени исходит из действительного аб-
солютного времени, т. е. того, что имеется вне органического
мира и что существовало и до формирования жизни на Земле.
Но появилась жизнь, и мы знаем, что она вписана многочислен-
ными механизмами во все законы мира, которые существовали
до происхождения жизни.
Поэтому был бы законным вопрос, который мы поставили:
а как же организм вписался в фундаментальный закон мира,
в пространство? Оказалось, что возбуждающий акцептор действия
в пределах функциональной системы — это один из признаков,
который существует па протяжении миллионов лет эволюции
живых существ.
Может ли организм вписываться во время, которое имеет
только один параметр — последовательность? Нет. Никакой жиз-
Философские аспекты теории функциональной системы
47
ни на нашей планете пе было бы и, наверное, ие было бы и
этого симпозиума, на котором я выступаю.
Что такое последовательность? Это — следование одного за
другим событий внешнего мира, которые могут не повторяться,
и возможность устойчивости жизни вне органического мира в
этом случае полностью была бы исключена. Однако другие па-
раметры времени (повторяемость и ритмичность событий внеш-
него мира) оказались тем компонентом времени, который спо-
собствовал развитию жизни. Почему это так? В самом деле,
если пространство, в которое мы вписаны, которое ежесекундно
действует на нас, составляет континуум, а мозг есть орган при-
способления, то ни одна минута этого континуума не может
пройти мимо мозга. Иначе приспособление к событиям внешнего
мира было бы невозможно. Организм не мог бы выжить, и га-
рантия естественного отбора была бы исключена.
Мы рассмотрели мозг именно в плане того, как он мог бы
отразить пространственный континуум.
Определенные категории нервных клеток продолжают свои
реакции на внешние раздражения и после прекращения действия
непосредственного раздражителя, оставляя определенный «след».
Этот «след» выражается в разрядах нервных клеток, которые
основаны на химических процессах самой клетки, па ионных,
метаболических процессах. Но континуум внешнего мира таков,
что события следуют в нем гораздо быстрее и непрерывнее, чем
может работать отдельная клетка. 11 поэтому естественно, что
каждое новое событие, как показали все проводившиеся экспе-
рименты, рассмотренные под этим углом зрения, постоянно на-
слаивается одно на другое.
Мы не можем представить себе сейчас, уже после того, что
мы знаем о мозге, пи одного внешнего впечатления, которое пе
оставило бы своего «следа» в химических процессах.
Здесь перед нами открывается новая область исследований,
на первый взгляд необъятных. Как может мир с этим непрерыв-
ным течением событий, вызывающим непрерывное течение хими-
ческих процессов в мозгу, восприниматься разумно?
Эту проблему мы решили в теоретическом плане; весь вопрос
теперь состоит в том, по какому пути идти в дальнейших иссле-
дованиях.
В неорганических предметах физическая последовательность
внешних событий представляет собой только последовательность
в пространстве и во времени. Для человека же не все элементы
этого пространственного и временного континуума одинаково
важны. Вот в чем ключ к объяснению того, почему организм
преодолел этот кажущийся хаос в реакциях на все внешние со-
бытия. Специфичность системы работы мозга состоит в том, что
он пе только отражает пространственно-временной континуум,
48
Философские аспекты теории функциональной системы
но благодаря особой способности живого вещества, кроме всего
прочего, накапливает опыт прошлого. Это свойство человеческого
мозга выражается в способности через континуум строить опе-
режение событий. И эта способность живого была очень широко
использована на всем протяжении эволюции.
Проблемы, которые я здесь эскизно набросал, являются для
нас ведущими, и в какой-то степени они революционизируют
наш исследовательский процесс. Сейчас не только многие его
формы могут и должны быть пересмотрены, но п весь процесс
исследования. Это не только наша задача; возникает огромная
возможность комплексной работы, и прежде всего с
философа-
ми, но и не только с философами.
Это объединение обнадеживает. Мы можем дать ряд обобще-
ний, идей, без которых идти вперед "нельзя. Правда, иногда обоб-
щения, идеи, концепции разрушаются и строятся новые, но это
ученый с самых первых шагов своей исследовательской работы
должен считать непреложным фактом. Гипотезы стареют, а если
они сохраняются, то это вызывает сомнение в их правомочности.
И я хочу закончить словами Клода Бернара: «Какое счастье
разрушить теорию, в особенности свою собственную».
ЛИТЕРАТУРА
Беритов И. С, Структура и функции коры большого мозга. М., 1969.
Лебедев В. П., Степин В. С. Гносеологический аспект понятия времени.—
Вопросы философии. 1970, № 10.
Планк М. Единство физической картины мира. М., 1960.
ПРИНЦИПИАЛЬНЫЕ ВОПРОСЫ
ОБЩЕЙ ТЕОРИИ
ФУНКЦИОНАЛЬНЫХ СИСТЕМ *
Общие предпосылки
Трудно найти в истории цивилизации такой момент, о кото-
ром можно было бы сказать, что именно тогда возникла идея о
целостности, о единстве мира. Вероятно, уже при первой попытке
понять мир мыслящий человек столкнулся с поразительной гар-
монией между целым, «универсумом», и отдельными деталями,
частями. Однако по самой сути человеческого ума он всегда
имеет дело с непосредственным, с его конкретным окружением,
с явлениями изолированной «ниши», и это коренным образом
повлияло на весь ход его познавательной деятельности.
Непосредственное, частичное имело прямое практическое
значение для усовершенствования приспособительной деятельно-
сти человека, и потому абстрактное «целое» пришло в его по-
знавательную деятельность значительно позднее.
От диффузных и недифференцированных форм целое посте-
пенно приобретало значение чего-то организованного, с постоян-
ным гармоническим взаимодействием своих частей, подчиняю-
щимся своим специфическим законам, не свойственным частям,
деталям целого. Так постепенно подготавливалось то научное
движение, которое в настоящее время получило широкое назва-
ние «системный подход».
Если внимательно проанализировать успехи современной био-
логии и вдуматься в попытки обобщить и осмыслить значение
этих успехов, то, пожалуй, системный подход представляет со-
бой наиболее выраженное и широко принятое направление во
всех этих поисках.
Такой успех системного подхода вполне понятен. Исключи-
тельно бурный рост числа новых публикаций в области конкрет-
ных естественных наук приводил к столь же бурному росту чув-
ства беспомощности исследователя неред половодьем аналитиче-
ских фактов. По этому поводу красочно выразился крупнейший
физик нашего времени Оппенгеймер, говоря, что в настоящее
время «исследователь ощущает свое невежество тем больше, чем
больше он знает...» (цит. по Юнгу, 1960).
♦ Опубликовано в книге «Принципы системной организации функций»
(М., «Наука», 1973).
50 Принципиальные вопросы общей теории функциональных систеМ
, . , -—-В-*-*
н
Стало очевидным, что только какой-то более высокий прин-
цип поможет разобраться в логических связях между отдельны-
ми фактами и что только этот принцип позволит более успешно
и па более высоком уровне проектировать новые исследования.
Сторонники системного подхода все более и более настойчи-
во подчеркивают, что именно система является тем изоморфным
принципом, который проникает через все границы, исторически
сложившиеся между различными науками, несмотря на то, что
эти науки изучают как будто качественно различные классы
явлений: организмы, общество, машины.
Однако из сказанного видно, что поиски «системы» как более
высокого и общего для многих явлений принципа функциониро-
вания могут дать значительно больше, чем только один анали-
тические методы при изучении частных процессов. Благодаря
такой оценке роли системного подхода возникли самые различ-
ные направления поиска. С одной стороны, делаются попытки
объяснить организацию больших биологических систем, с тем
чтобы связать поведение организма с молекулярным уровнем
процессов, включенных в это поведение. С другой стороны, идет
настойчивый поиск законов формирования больших систем для
социально-экономических явлений (В. Г. Афанасьев, Б. С. Ук-
раинцев и др.), машинных устройств и т. д. В зависимости от
общей концепционной обстановки к этим классам явлений при-
бавляется . также и трактовка обширных машинных устройств,
которые способны к самоорганизации. Все это направляет мысль
на новые поиски, на формулировку п раскрытие новых законо-
мерностей, и именно это обстоятельство сейчас является наибо-
лее впечатляющим в области нового и прогрессивного движения
в науке, получившего название системного подхода.
Развитие этого нового научного движения в последние десять
лет ознаменовалось весьма радикальными выступлениями, в ко-
торых роль системы в развитии наук и общества поднимается
настолько высоко, что некоторые энтузиасты этого направления
стали говорить о наступлении в науке «эры системы» (Эллис).
Вместе с тем наметилась тенденция к выделению системного
подхода в особую науку—«системологпю» (Г. Паск, В. М. Глуш-
ков и др.), а коллектив авторов из NASA1
выделить специальную науку о «биологических системах» («Bio-
logikal Systems Science»,
авторов этого издания опубликована специальная монография
«Toward a general science of viable systems», в которой делается
попытка сформулировать некоторые основные положения этой
науки (Iberall, 1972).
предлагает вообще
1971). В последнее время одним из
1 National Aeronaftik and Space Administration (Национальное управление
по исследованию космического пространства, США).
Принципиальные вопросы общей теории функциональных систем 51
Таким образом, внимание к системному подходу очепь велико,
хотя Калмап, Фалб и Арбпб в своей книге «Очерки по математи-
ческой теории систем» (1971) назвали системный подход «об-
ластью увлекательной, но неупорядоченной». Как мы увидим
ниже, в какой-то степени с ними можно согласиться.
Трудно сказать, когда возникла необходимость введения це-
лостного подхода при объяснении функций организма. Эта по-
требность фактически ощущалась всеми исследователями, но ре-
шалась ими различным образом. Одни ученые полностью отрица-
ли что-либо специфическое в целостной организации и делали
попытку объяснить ее, исходя только из свойств элементов, вхо-
дящих в целостные образования. Так возник механистический
подход к объяснению целого.
Другие допускали, что множество элементов организма при-
водится в организованное целое при помощи некоторой неорга-
нической силы, которая, находясь все время в «падорганическом
состоянии», обладает специфическим качеством «одухотворения»
и формирования организованного целого (например, «энтелехия
Дриша», «мнема Блейлера», «руководящая сила Клода Бернара»
и др.). Такую позицию заняли виталисты всех направлений, хотя
некоторые из них давали объективные оценки целостности.
Преимущество изучения целостного организма с особой силой
выражено уже в очепь раппих работах И. П. Павлова. Так, на-
пример, еще в конце прошлого столетия он выдвинул идею, что
наиболее нормальные фупкцтш организма можно изучать не у
ограниченного в подвижности животного, т. е. в условиях виви-
секции, а у целостного, пенаркотизировапного животного. Так
появились его знаменитые методы исследования: изучение кро-
вяного давления, фистульные методы при изучении пищеваре-
ния и, наконец, изучение высшей нервной деятельности.
Однако физиология XIX и XX вв. характеризуется именно
тем, что в ней сосуществовали противоречивые тенденции. Наря-
ду с совершенно четко ощущаемой необходимостью изучения
целостного, динамически функционирующего организма физио-
лог-экспериментатор находился все еще под опьяняющим влия-
нием анализа рассечения организма и часто ограничивался ис-
следованием лишь его тонких механизмов, не относя резуль-
татов к более высоким принципам организации.
В последние годы теидепцпя к изучению специфических за-
кономерностей именно целостного организма стала уже совер-
шенно непреодолимой в связи с нарастающим разочарованием
исследователей в продуктивности одного аналитического подхода
к исследованию и к объяснению полученных результатов. Ста-
новится все более очевидным, что необходимо сформулировать
такой рабочий принцип, который мог бы перебросить «концеп-
туальный мост» между теми фактами, которые получаются при
52 Принципиальные вопросы общей теории функциональных систем
изучении явлений у целостного животного, и теми, которые по-
лучаются при тонком аналитическом эксперименте.
В сущности, эта проблема актуальна не только для исследо-
вателей-биологов, но и для физиков, экономистов и других спе-
циалистов. Достаточно проследить крайне интересную дискуссию
между Нильсом Бором и Альбертом Эйнштейном, чтобы понять,
какой животрепещущей проблемой является выработка этих
новых принципов целостного подхода к изучаемому объекту.
В этой дискуссии столкнулись два подхода к вопросу, как обе-
спечить уровень специфически целого в научном исследовании,
например, организма и в то же время не потерять огромных пре-
имуществ уровня тончайшего анализа.
Как известно, Нильс Бор выразил свою точку зрения в кон-
цепции, известной как «принцип дополнительности». Согласно
этой концепции
целое и целостный подход должны дать иссле-
дователю возможность найти дополнительные характеристики
изучаемого объекта, более или менее отражающие специ
ические
черты именно целого. Он считает, что условия наблюдения, т. е.
ракурс, в котором мы наблюдаем изучаемый объект, могут из-
меняться в процессе исследования и это дает дополнительные
опорные пункты для всестороннего познания объекта. По своей
сути этот подход, расширяя возможности изучения целостного
объекта, ставит исследователя в положение наблюдателя, опре-
деляющего, как складываются наблюдаемые явления.
Альберт Эйнштейн, наоборот, стремился найти такой подход,
который заменил бы феноменологический подход Бора динамиче-
ским подходом, позволяющим проникнуть в природу внутреннего
взаимодействия в каком-либо сложном целостном
еномене.
В то время как при помощи феноменологического подхода к
целостному процессу можно определить, как устроены изучаемые
объекты, подход Эйнштейна заключается в стремлении эмпириче-
ски найденную закономерность представить как логическую не-
обходимость.
Нам сейчас важно подчеркнуть, что дискуссия между двумя
величайшими учеными нашего времени принимает как нечто
определенное и обязательное поиски переходного принципа це-
лостного подхода к явлениям. Речь идет лишь о том, в какой
LH
£орме этот принцип мог бы быть наиболее эффективным для
конкретной научно-исследовательской работы, поскольку целост-
ный подход вообще, оставаясь мечтой исследователя, не давал в
то же время никаких конструктивных решений для
ормулиров-
ки задач повседневного исследования. «Целое» п аналитическое
экспериментирование по-прежнему уживались в двух параллель-
ных плоскостях, не обогащая друг друга.
Появление системного подхода дало ученым некоторую на-
дежду на то, что, наконец, «целое» из диффузной и неконструк-
Принципиальные вопросы общей теории функциональных систем 53
тивной формы примет четкие очертания операционального иссле-
довательского принципа. Однако, прежде чем анализировать при-
чины, по которым этот принцип не был найден,
краткую оценку развития системного подхода в
мы хотим дать
различных
энологических школах.
Термин «система» имеет весьма древнее происхождение,
и едва ли есть какое-либо научное направление, которое его не
употребляло. Достаточно вспомнить «систему кровообращения»,
«систему пищеварения» и т. д., которые до сих пор некоторыми
исследователями принимаются за выражение системного подхо-
да. Большей частью термин «система» употребляется там, где
речь идет о чем-то собранном вместе, упорядоченном, организо-
ванном, но, как правило, не упоминается критерий, по которому
компоненты собраны, упорядочены, организованы.
Эти общие недостатки естественны. Мы не должны забывать,
что последовательное приложение системного принципа к явле-
ниям различного класса (организму, машинам, обществу) не яв-
ляется простой сменой терминологии, перестановкой лишь поряд-
ка исследовательских приемов. Системный подход к исследованию
является прямым следствием перемены теоретического подхода
к пониманию изучаемых объектов, т. е. в какой-то степени след-
ствием изменения самой формы мышления экспериментатора.
Естественно, что такой процесс не может быть одномоментным.
Как будет показано ниже, наиболее характерной чертой си-
стемного подхода является то, что в исследовательской работе
не может быть аналитического изучения какого-то частичного
объекта без точной идентификации этого частного в большой си-
стеме. Таким образом, со стратегической и практической точек
зрения исследователь должен иметь прежде всего конкретную
концепцию системы, которая должна удовлетворять основным
требованиям самого понятия системы, и лишь после этого форму-
лировать тот пункт системы, который подлежит конкретному ис-
следованию.
В области физиологических исследований И. П. Павлов, пожа-
луй, первым употребил выражение «система» для некоторых спе-
циальных случаев своей экспериментальной работы. Речь идет
п
прежде всего о
ормировании динамического стереотипа.
Как
известно, эта система создается тем, что изо дня в день повторя-
ется стереотипный порядок одних и тех же условных раздражи-
телей. В результате длительной тренировки этот порядок раздра-
жителей, обнаруживаемый по специфическому для данного раз-
дражителя количеству слюны, проявляет себя даже в том случае,
когда применяется один и тот же раздражитель.
Прямые электроэнцефалографические исследования мозга в
момент создания такого динамического стереотипа, проведенные
в на
:ей лаборатояии А. Д. Семененко, показали весьма инте-
II
54 Принципиальные вопросы общей теории функциональных систем
ресные свойства мозга как целого. Так, например, оказалось,
что к каждому предстоящему раздражителго в случае укреплен-
ного динамического стереотипа мозг автоматически, т. е. только
на основе прежней тренировки и независимо от реального внеш-
него раздражителя, готовит состояние, качественно отражающее
именно тот раздражитель, который применялся на этом месте
много раз в прежних тренировках. Конечно, создаваемые таким
образом стереотипные состояния мозга, отражающие комплекс
условных раздражителей данного опытного дня, далеки по своему
физиологическому смыслу от того системного подхода, который
бурно развивается в последнее время. И все же эти эксперимен-
ты показали, что мозг может на основе приобретенного опыта
создавать некоторые целостные состояния, объединяющие раз-
дражители целого экспериментального дня и раскрывающиеся
независимо от реальной экспериментальной обстановки.
Мы пе будем приводить здесь все те случаи, в которых поня-
тие «система» привлекалось для обозначения чего-либо упорядо-
ченного, ибо хорошо известно, как широко этот термин употреб-
ляется для обозначения подобного рода феноменов. Пас интере-
суют здесь по преимуществу системы, обладающие способностью
экстренной самоорганизации, динамически и адекватно приспо-
сабливающие организм к изменению внешней обстановки. Иначе
говоря, нас интересуют натуральные функциональные системы
организма.
Системный подход в форме теоретической концепции под наз-
ванием «общая теория систем» возник как реакция па исключи-
тельно бурный рост аналитических подходов в науке, все более
и более удаляющих творческую мысль от того, что длительное
время называлось проблемой целостного организма.
Общая теория систем как новая форма мышления, несмотря
на свои глубинные дефекты, сразу же привлекла внимание за-
падных ученых и получила особенно широкий отклик в среде
теоретиков. Этому содействовала также организация регулярного
ежегодного издания «Общая теория систем», которое стало свое-
образным дискуссионным клубом и для сторонников и для про-
тивников системного подхода как универсальной концепции, объ-
единяющей интересы самых разнообразных наук.
К этому времени и в среде биологов-экспериментаторов наз-
рела необходимость целостного подхода к объяснению материала,
и
накопленного в результате аналитического подхода к предмету.
В отчетливой и красочной форме эту потребность выразил Уод-
дингтон в предисловии к книге Гудвина «Временная организация
клетки» (1966), Подчеркивая необходимость разработки теоре-
тической биологии, направленной на понимание принципов орга-
низации биологических систем, он пишет: «Мы можем восхищать-
ся теориями, говорящими нам о назначении структуры в про-
Принципиальные вопросы общей теории функциональных систем 55
стекших живых объектах, таких, как например, вирусы, которые
почти целиком состоят из нуклеиново-кислотного стержня, за-
ключенного в белковую оболочку, но мы пе можем удовлетворить-
ся ими. Пам необходимо развить, исходя из этого, надструктур-
ную теорию, которая позволила бы нам понять организацию
высших, наиболее сложных форм жизни. Однако разработка та-
кого базиса, необходимого биологии для того, чтобы проделать
путь от вируса до мыши, является, вероятно, еще более гранди-
озной задачей, чем та, которую решала физика па пути от
атомного ядра к молекуле, полупроводнику и звезде» [Гудвин,
1966, с. 14-15].
Нельзя не согласиться с Уоддингтоном в том, что этот путь
действительно труден, однако он не был бы таким, если бы с
самого начала изучения биологических систем было решено, что
целое, система, при своем становлении приобретает собственные
и специфические принципы организации, не переводимые па
принципы и свойства тех компонентов и процессов, из которых
формируются целостные системы (например, возбуждение и тор-
можение) .
Значительное влияние па развитие системного подхода оказа-
ло интервью «отца кибернетики» Норберта Винера. Отвечая на
вопрос корреспондента о том, какой будет паука в 1984 г., он
сказал: «Главные проблемы биологии также связаны с системами
и их организацией во времени и пространстве. II здесь самоорга-
низация должна играть огромную роль. Поэтому мои предполо-
жения в области наук о жизни касаются не только их постепен-
ной ассимиляции физикой, но и обратного процесса — постепен-
ной ассимиляции физики ими» (Винер, 1964).
Сразу же после этого интервью общая теория систем и си-
стемный подход возбудили особый интерес среди исследователей
разных специальностей. Прежде всего было организовано не-
сколько центров ио изучению системы: при Кливлендском и Окс-
фордском университетах («Исследование систем») и др.
Во Франции недавно был создан специальный Институт выс-
шего синтеза, организаторы которого, в сущности, также сосре-
доточили свое внимание на изучении принципов организации
целостных образований в природе и обществе. Его центральным
направлением является формулировка «идей науки» для раскры-
тия высшего синтеза в явлениях природы и общества. В 1971 г.
этот Институт провел III Международный конгресс, посвящен-
ный искусственному и естественному интеллекту (Institut des
Hautes Synthes. Nice, France).
В последние годы один за другим стали организовываться
симпозиумы, конференции и другие встречи, посвященные из-
учению системы. Особенно большую активность в этом направ-
лении (с 1950 г.) проявили Берталанфи и Научное общество по
Принципиальные вопросы общей теории функциональных систем
общей теории систем, а также и Кливлендский центр ио изуче-
нию систем. Последний уже провел три международных симпо-
зиума по «системному подходу в биологии» и осуществил ряд
изданий, посвященных этой проблеме (например, «Теория си-
стем и биология»). Естественно, должен возникнуть вопрос: как
далеко продвинулись биологи и физиологи после этих многосто-
ронних попыток приблизиться к пониманию системы и исполь-
зованию ее как методологического инструмента в формулировке
новых задач исследования и понимании уже накопленных мате-
риалов? Можем ли мы сейчас с большим успехом сформулиро-
вать само понятие системы, чем раньше? Несмотря на то что об-
щие задачи системного движения в науке сформулированы были
достаточно правильно, результаты исследований, особенно кон-
кретизация понятия системы и формулировка ее конкретных и
специфических только для нее биологических свойств, остаются
очень неутешительными. Не будет преувеличением сказать, что
дело остановилось на подборе определений, формулировок, ко-
торые охарактеризовали бы систему и выделили бы ее из кате-
гории несистем.
Из всех определений системы, которые даются участниками
дискуссии по общей теории систем (см. ниже), видно, что чисто
теоретический подход к проблеме практически не сдвинул ее с
места. В качестве аргумента можно привести достаточно красно-
речивую характеристику, данную одним из прогрессивных биоло-
гов Гудвином в книге «Временная организация клетки» (1966).
Он писал: «Центральное место в биологической науке занимает
концепция организации, хотя само понятие организации и не
имеет четкого определения». И это верно. Можно взять десяток
определений системы как у самого Берталапфп, так и у его по-
следователей и увидеть, что ни одно из них не дает возможности
активно использовать понятие «система» как инструмент для
более усовершенствованной исследовательской работы.
Действительно, если подвести итоги поисков системного под-
хода и приложить их к пониманию всего накопленного материа-
ла в биологических и
изиологических
исследованиях, то сразу
же обнаруживается их неспособность хоть в какой-либо мере
помочь конкретному исследованию. Мы не сможем каждый
конкретный результат, полученный при аналитическом экспери-
менте, поставить в определенное место системы, чтобы он при-
обрел свое реальное значение органического компонента системы,
содействующего своими степенями свободы получению резуль-
тата системы. Вспомним, что одной из главных целей поисков
системы является именно ее способность объяснить и поставить
на определенное место даже тот материал, который был задуман
и получен исследователем без всякого системного подхода.
В чем причина этого очевидного неуспеха в таких оживленных
Принципиальные вопросы общей теории функциональных систем 57
поисках системного подхода при достаточно правильно сформули-
рованной исходной цели? Почему при правильно сформулиро-
ванных недостатках теоретических обобщений в биологии и фи-
зиологии и при широком внимании к этим недостаткам мы тем
не менее не имеем конструктивных итогов, которые своей полез-
ностью заслужили бы широкую популярность у эксперимента-
торов — биологов и физиологов? Нам кажется, что одной из глав-
ных причин такого печального положения с поисками конкретных
качеств системы является излишнее теоретизирование всей проб-
лемы в целом. Действительно, необходимость начать системный
подход возникла у специалистов главным образом в результате
смутного ощущения неправильного развития биологии и физио-
логии, т. е. на основе чисто теоретических соображений. Достоин-
ства системного подхода мыслились теоретически, но они еще не
имели соответствующего, уже найденного в конкретном исследо-
вании эквивалента.
Сложилось курьезное положение: с одной стороны, не было
поддержки конкретных биологических и физиологических наук
в виде открытия конкретных, специфических только для системы
механизмов, а с другой — непомерно разрослась часть теорети-
определений, часто украшенных обширны-
то
ческих поисков и
мп математическими выкладками. Именно этим можно в како:
ч
степени объяснить парадоксальное явление: у всех теоретиков
системы и у философов поразительно схожи сами определения
понятия системы, хотя ни у тех, пи у других это понятие не
имеет действенного значения как инструмент, облегчающий кон-
кретную исследовательскую работу.
Особенности отдельных тенденций
в разработке теории систем
Интересно выяснить, какие же цели ставят перед собой раз-
личные группы исследователей, какими методами опп осущест-
вляют настойчивый поиск системного подхода?
При первой же попытке понять эти цели по имеющимся ли-
тературным источникам мы встречаемся по крайней мере с че-
тырьмя оформившимися тенденциями.
Первая состоит в том, чтобы сделать системный подход до-
статочно попятным с точки зрения формулировки и направить
все внимание на философский смысл и объем системного подхо-
да, примененного, в частности, к социальным явлениям. В этих
поисках система выступает как научная и философская катего-
рия, ведущая к усовершенствованию познавательного процесса.
Такое направление, в основном разрабатываемое философами и
историками, развивается в настоящее время по пути исключи-
тельно теоретического поиска и теоретической оценки, при кото-
58 Принципиальные вопросы общей теории функциональных систем
рых трудно пока говорить о каком-либо контакте с практи-
ческими запросами исследовательской работы.
Представителями второй тенденции являются сторонники ма-
тематической формализации системы, или математической теории
систем (Месарович, Раппопорт, Кухтин и др.).
Последователи системного подхода, поддерживающие третье
направление, считают, что теория систем должна вырасти из
изучения натуральных систем п затем стать конкретным инстру-
ментом исследования. Такие системы должны помогать исследо-
вателю ставить новые, более прогрессивные задачи научных ис-
следований, быть способными объяснять накопленный ранее
материал и преодолеть только аналитический подход к исследо-
вательской работе. Эта категория ученых обычно ожидает, что
теория систем проложит «концептуальный мост» через пропасть,
пока еще разделяющую синтетический и аналитический подходы
к объектам исследования.
Можно считать, что к последнему направлению примыкают
сторонники системного подхода (четвертая тенденция), которые
под этим углом зрения анализируют социально-экономические
системы.
В последние годы становится особеппо популярной проблема
«больших систем», объединяющая все виды организации промыш-
ленных, производственных и обслуживающих систем (В. А. Тра-
пезников, В. Г. Афанасьев, Д. М. Гвишиани, Хапике, Б росс
п др.).
Как видно из этой краткой характеристики, исследования
м
различных ученых значительно отличаются по своим подходам
и целям, что по-разному влияет па репутацию самой полезности
разработки теории систем.
Естественно, что исследователь, работающий в конкретной
области науки (биологии, физике, физиологии, медицине), заин-
тересован прежде всего в том, чтобы теория систем вошла в его
интеллектуальный статус как вполне попятное научное движение,
значительно обеспечивающее прогресс его конкретной научной
работы. Обычно эта категория исследователей мало обращает вни-
мания на общефилософское и методологическое обсуждение тео-
рии систем, поскольку такое обсуждение, как правило, не уста-
навливает «концептуального моста» между философией системы
и ее применением к какому-либо изучаемому объекту.
Такое отношение к философскому разбору системного подхода
отнюдь нельзя назвать узким прагматизмом, как это может пока-
заться на первый взгляд. Скорее, эту позицию можно определить
как склонность изучать не «методологию вообще», а «методоло-
гию моего дела».
Если представленные выше особенности разработки различных
аспектов теории систем затрагивают интересы различных сто-
Принципиальные вопросы общей теории функциональных систем 59
1 1 Ч » i j I ^MI 1| ,
рондиков системного подхода, то фактор, который мы собираемся
обсудить ниже, является общим и даже обязательным для лю-
бого исследователя теории систем с любыми требованиями к ее
практическому эффекту.
Решающая роль системообразующего фактора
Таким обязательным положением для всех видов и направле-
ний системного подхода является поиск и формулировка системо-
образующего фактора. Эта ключевая проблема определяет как
само понятие системы, так и всю стратегию его применения в
исследовательской работе. Иначе говоря, принесет ли пользу
конкретным наукам системный подход или не принесет, будет
зависеть от того, насколько успешно мы выделим системообра-
зующий фактор и насколько полно будет описапо его операцио-
нальное значение для формирования системы. Только при этом
условии мы можем применить принципы системообразования для
всех тех классов явлений, в которых происходит упорядочение.
Между тем вокруг этой проблемы сложилась крайне странная
ситуация. Почти все сторонники системного подхода и общей
теории систем подчеркивают как центральное свойство системы
«взаимодействие множества компонентов» (Берталанфи, Раппо-
порт и др.). Близким является «упорядоченное взаимодействие»
или «организованное взаимодействие». По сути дела именно на
этих определениях понятия системы и покоится все обсуждение
системного подхода. Хотя весь успех понимания системной дея-
тельности, особенно у организмов, зависит от того, определим ли
мы, какой именно фактор упорядочивает «беспорядочное мно-
жество» и делает это последнее функционирующей системой,
вопрос о системообразующем факторе просто никогда не был
поставлен в отчетливой форме системологами.* Он не ставится
ни главным идеологом «общей теории систем» Берталанфи, ни
группой его последователей (Акоф, Раппопорт, Месаровпч, Уотер-
ман и др.). Пожалуй, так же дело обстоит н у советских теоре-
тиков системы, объединенных Институтом истории естествозна-
ния и техники и Институтом философии АН СССР (В. И. Са-
довский, И. В. Блауберг, Э. Г. Юдин, АГ И. Уемов, К. М. Хай-
лов и др.).
В результате этого коренного недостатка — отсутствия си-
стемообразующего фактора все имеющиеся сейчас определения
системы случайны, не отражают ее истинных свойств и поэтому,
естественно, неконструктивны, т. е. не помогают ставить новых,,
более широких вопросов для исследования (рис. 1).
Ознакомившись подробно со всеми публикациями Общества
общей теории систем (Society of General Systems Theory), можно
с уверенностью утверждать, что теоретическая неопределенность,
ЙО Принципиальные вопросы общей теории функциональных систем
Рис. 1. Схематическое изображение «концептуального моста» между си-
стемным уровнем и тонкими аналитическими процессами
а — уровень целостной системной деятельности; б — уровень тонких аналитических
процессов; в — путь обычных корреляционных отношений; г — включение системо-
образующего фактора, который объясняет процесс упорядочивания между множе-
ством компонентов системы; д — операциональная архитектоника системы и ее уз-
ловые механизмы.
Схема демонстрирует непрерывность исследовательского процесса, обеспечивающего
непосредственный переход от системного уровня к тонким физиологическим дета-
лям системы до молекулярного уровня включительно
отсутствие связи с конкретными научными дисциплинами п не-
конструктивность основных положений непосредственно для
исследовательской работы являются следствием игнорирования
основной проблемы системологип — раскрытия системообразую-
щего фактора. Без определения этого фактора пи одна концепция
теории систем не может быть плодотворной. Трудно допустить
без него существование какой-либо теории систем, и прежде
всего общей теории систем. Отсюда возникают и терминологиче-
ские вопросы. Так, например, можно утверждать, что термин
общая, примененный к теории систем Берталаифи, пе имеет
достаточного логического обоснования. Именно это чрезвычайно
ограничивает ее конструктивное использование в научно-иссле-
довательском процессе.
Постараемся произвести строгий логический анализ этой проб-
лемы.
В каком случае мы могли бы говорить именно об общей тео-
рии систем? Только в том случае, если бы были даны убедитель-
ные доказательства того, что она может быть отнесена к самым
разнообразным классам явлений, т. е. выявляет какие-то общие
черты в разнообразных классах явлений, например в неорга-
нической природе, организме, машинах, обществе. Так клеточная
Принципиальные вопросы общей теории функциональных систем 61
теория является, несомненно, общей теорией для всего живого на
земном шаре, поскольку клеточное образование является общим
и изоморфным фактором для всех организмов независимо от
уровня их развития и положения па биологической лестнице. Зна-
чит, растения и животные именно по этому критерию оказывают-
ся изоморфными образованиями.
Исходя из всего сказанного, мы можем построить следующий
ряд логических положений.
1. Теория может получить право стать общей только в том
случае, если она вскрывает и объединяет собой такие закономер-
ности процессов или механизмов, которые являются изоморфны-
ми для различных классов явлений.
2. Изоморфизм явлений различных классов может быть выяв-
лен только в том случае, если мы найдем достаточно убедитель-
ный критерий изоморфности. Чем более значимым является этот
критерий для разбираемых явлений, тем более выраженным яв-
ляется их изоморфизм.
3. Для принятия «общей теории систем», пригодной для раз-
личных классов явлений, наиболее важным критерием изоморф-
ности, естественно, является изоморфность системообразующего
фактора.
Достаточно внимательно проанализировать эти три положе-
ния, чтобы увидеть, в чем состоят конструктивные трудности
«общей теории систем», выдвинутой Берталанфи.
Эта теория не вскрыла того фактора, который из множества
компонентов с беспорядочным взаимодействием организует «упо-
рядоченное множество» — систему.
Это обстоятельство, т. е. отсутствие системообразующего фак-
тора, не дает возможности установить изоморфность между яв-
лениями различного класса, а следовательно, и не может сделать
теорию общей. Именно этот недостаток бросается в глаза при
изучении аргументов сторонников общей теории систем. И это
же обстоятельство неизменно препятствует общей теории систем
стать инструментом конкретного научного исследования.
Математическая теория систем
Оценивая все стороны системного движения, которые являют-
ся тормозящим фактором в быстром и широком использовании
теории систем в повседневном научном исследовании, мы не мо-
жем оставить в стороне так называемую математическую теорию
систем.
Едва ли приходится сомневаться в том, что моделирование в
настоящее время стало всеобщим и неоспоримым помощником в
теории и практике различных областей деятельности. Большую
пользу приносит и математическое моделирование.
62 Принципиальные вопросы общей теории функциональных систем
Однако вряд ли то же самое можно сказать и нро «мате-
матическую теорию систем», и именно потому, что ее общая стра-
тегия вступает в противоречие с конкретной пользой от ее при-
менения там, где речь идет о приложении математической тео-
рии систем к биологическим системам.
Прежде всего — и это, пожалуй, самое главное — сторонники
математической теории совершенно радикально решают вопрос о
соотношении этой теории с биологией. Так, например, Месарович,
апологет математической теории систем, пишет: «Мы будем
рассматривать системный подход как использование теории си-
стем для изучения и объяснения биологических явлений. Таким
образом, эта статья будет посвящена теории систем пли, более
конкретно, рассмотрению вопроса о том, может ли эта теория
служить одновременно и принципиальной основой, и практиче-
ским методом для научного объяснения биологических явлений»
(1970, с. 137).
И еще более демонстративно эта парадоксальная, с точки зре-
ния биолога, последовательность разработки биологических си-
стем выражена в тезисах Месаровича, регламентирующих при-
менение математической теории систем к биологическим явле-
ниям: «После того как построена система (математическая мо-
дель) и определено конструктивное задание, задача теории систем
сводится к изучению свойств данной системы (методами мате-
матической дедукции или путем машинного моделирования).
Таким образом, методология системного подхода в биологии сла-
гается из следующих этапов:
а)
ормалпзацпя
(абстрагирование) — построение системы S
и определение для нее конструктивного задания;
б) дедукция — исследование свойств системы S с использова-
нием дедуктивных методов;
в) интерпретация — изучение смысла найденных (дедуктив-
ными методами) свойств в контексте рассматриваемого биологи-
ческого явления» (Месарович, 1970, с. 141).
По сути дела той же точки зрения придерживаются Налман,
Фалб, Арбнб — авторы известной монографии «Очерки по мате-
матической теории систем».
Среди советских исследований теории систем особенно выде-
ляются работы проф. А. И. Кухтенко, который, применяя мате-
матическую теорию систем к «большим системам» промышлен-
ного типа, принципиально так же начинает процесс изучения
производственных явлений после предварительной формулиров-
ки математических моделей систем.
Итак, мы видим, что во всех видах применения математиче-
ской теории систем декларируется один и тот же принцип ее
использования. Сначала на чисто теоретическом основании форму-
лируется математическая теория систем, и только после этого
Принципиальные вопросы общей теории функциональных систем 63
ее «задания» начинают применяться к объяснению тех пли иных
биологических явлений.
Для биолога и физиолога такая последовательность приме-
нения математической
теории и
ормалпзашш
кажется весь-
ма странной. Как можно чисто математическую модель, разрабо-
танную уже заранее в обход всех современных знаний об осо-
бенностях именно биологической организации, применить к
объяснению и формулировке биологических закономерностей?
Так, например, в результате многолетней практики такого
подхода пи одна из тысяч математических моделей нейрона
абсолютно не отразила истинные особенности нейрона и ни на
один шаг не продвинулись вперед наши знания о действительных
законах его функционирования. Более того, можно с уверенно-
стью утверждать, что исследование нейрона с применением
электронной микроскопии, микропонофореза, ультрацентрифуги-
рования, культуры нейронов in vitro и нейрохимических иссле-
дований всегда на несколько десятков лет опережает довольно
простенькие математические модели нейрона.
Не считаясь с действительными биологическими и физиоло-
гическими свойствами систем, математическая теория систем
фактически переводит вопрос в плоскость настолько запутанного
теоретизирования, что практически до сих пор опа мало помогла
разработке системного подхода в области биологических явлений.
В этом вопросе, по нашему мнению, предпочтительнее рас-
сматривать в обратном порядке взаимодействие этих двух обла-
стей знаний. Нет сомнения, что реальные «системные закономер-
ности» могут быть почерпнуты и разработаны только на основе
конкретного материала биологии и физиологии последних дней.
Именно этот материал и должен стать реальной основой форма-
лизации. Благодаря строгому и быстрому математическому ос-
мысливанию этих закономерностей более реальной станет и пер-
спектива развития наших знаний о биологических системах.
Крайняя трудность заимствования результатов исследователь-
ской работы теоретиков математических систем состоит именно
в этом. Придавая первичное и решающее значение именно ма-
тематической обработке биологических явлений, сторонники это-
го подхода крайне затрудняют использование их разработок био-
логами и физиологами — профессионалами.
Практически же сторонники математической теории систем
пе могут, на наш взгляд, формализовать истинную биологическую
систему хотя бы уже по одному тому, что они, как это видно по
формулировкам Месаровича, не исходят из наиболее существен-
ных критериев оценки именно биологических систем.
В самом деле, можно ли математически определить биологи-
ческую систему, если мы пе можем наделить эту модель системы
самыми важнейшими свойствами живой системы: формпрова-
64 Принципиальные вопросы общей теории функциональных систем
нием потребности получить тот, а не другой результат и опре-
деленной целью, которую обычно ставит перед собой биологи-
ческая система уже в самом начале формирования поведенче-
ского акта. Ни одна из известных нам вариаций математической
теории систем не решает этого кардинального вопроса, а это
значит, что мы не можем признать полезными и любые матема-
тические выкладки, если они сформулированы без учета таких
важных системообразующих факторов. Образно говоря, с точки
зрения биолога и физиолога, математическая модель дает нам
в какой-то степени приемлемую форму кузова автомобиля, одна-
ко она неспособна наградить его мотором и топливом...
Формируясь в пределах самой биологической системы на ос-
нове ее потребностей, внешних факторов и памяти, цель всегда
опережает реализацию ее организмом, т. е. получение полезного
результата.
Интересно, что Месарович, наиболее солидный теоретик био-
логических систем, в конце концов приходит к признанию, что
математическая теория именно биологических систем не может
быть построена без привлечения целенаправленного поведения:
«Существует важный класс ситуаций, в которых эффективное
конструктивное задание системы удается получить только при
помощи описания, основанного на понятии целенаправленности
(т. е. телеологического описания); при таком описании основ-
ной характер системы как некоторого (математического) отноше-
ния остается неизменным.
Под описанием, основаниььм на понятии целенаправленности,
здесь подразумевается вся совокупность системных описаний,
представленных с помощью понятий, которые выражают цели в
поведении системы (такие, как адаптация, эволюция, управле-
ние, гомеостаз и т. п.)».
Однако при попытке представить себе эту «целенаправлен-
ность» в поведении систем математически Месарович приходит к
таким упрощениям (стимул-ответам), что вообще теряется само
явление целенаправленности. Так, одно из самых специфических
свойств целенаправленности — принятие решения и предсказа-
ние результата — оказывается полностью не представленным в
его рассуждениях.
Как из приведенного выше высказывания Месаровича, так и
других его работ яспо, что он не видит своеобразных черт био-
логической системы, которые только искусственно, а потому и не
конструктивно могут быть превращены в математическую модель.
В самом деле, своеобразие биологической системы состоит в том,
что потребность в каком-либо полезном результате и цель полу-
чения этого результата зреют внутри системы, в глубине ее ме-
таболических и гормональных процессов, и только после этого
по нервным «приводным ремням» эта потребностг реализуется
Принципиальные вопросы общей теории функциональных систем 65
в поведенческих актах, допускающих в какой-то степени матема-
тическую формализацию. Этот путь возбуждений от реализации
метаболической потребности в адекватных мотивационных струк-
нейрофгтзиологпческой точностью (К.
В. Судаков,
:еппо попятно поэто-
турах мозга до выявления первых попыток удовлетворения этой
потребности изучен в настоящее время в нашей лаборатории с
большой
Б. В. Журавлев, Л. В. Котов и др.). Совер^
му (это отмечает п Месарович), что математическая модель си-
стемы накладывает ограничения пменно на эти свойства биоло-
и
гической системы, т. е. па то, что и составляет самую суть меха-
низмов биологических систем.
Таким образом, вопрос о применимости и эффективности ма-
тематической теории систем должен быть подвергнут специаль-
ной дискуссии.
Вопрос о том, какую помощь развитию системного подхода
приносит математическая теория систем, становится особенно
острым, если мы применим к пей наиболее важный для биолога
и физиолога критерий: прокладывает ли математическая теория
систем тот «концептуальный мост», который должен соединить
два края пропасти — синтетический уровень подхода исследова-
теля к биологическим объектам и аналитический уровень изуче-
ния этих объектов. Этот чисто аналитический уровепь исследо-
вания в биологии и физиологии становится все более и более
опаспым и угрожает утонпть пас в половодья разрозненных и
часто не объединенных ничем фактов.
Самый смысл «концептуального моста», как мы увидим ниже,
состоит в том, что система должна объединить непрерывной де-
терминистической логикой оба края этой пропасти, что дало бы
возможность исследователю всегда видеть тот район целой систе-
мы, в котором ведутся его тончайшие аналитические исследования.
Само собой разумеется, что этот «мост» должен быть пост-
роен на материале фактов, терминов п понятий конкретных наук.
Приходится глубоко сомневаться в том, что математическая мо-
дель системы обеспечит построение такого «моста» и значитель-
но расширит творческие возможности исследователя. Тем не ме-
нее, поскольку этот вопрос является дискуссионным, оставим
то или иное решение его специалистам этой области системного
подхода.
В связи со сказанным выше совершенно особо выглядит точ-
ка зрения современной общей теории систем на ведущие зако-
номерности системы, а именно па взаимодействие множества
неупорядоченных компонентов п перевода их в ранг упорядочен-
ной системы. Изучая систему как совокупность математически
сформулированных заданий, математик — теоретик системы обыч-
но не идет дальше своих излюбленных формулировок о «взаимо-
действии множества компонентов» при формировании системы.
3 П. К. Анохин
66 Принципиальные вопросы общей теории функциональных систем
Понятие системы
Как показано в одной из последних работ (Анохин, 1970),
взаимодействие как таковое не может сформировать систему, по-
скольку анализ истинных закономерностей функционирования с
точки зрения функциональной системы раскрывает скорее меха-
низм «содействия» компонентов, чем пх «взаимодействие». Это
серьезный вопрос, связанный с выработкой самого понятия си-
стемы, и он требует специального и подробного разбора.
Возникает вопрос: может ли взаимодействие компонентов,
взятое само по себе, создать что-то системное, т. е. что-то упо-
рядоченное? Неопределенность и неконструктивность всех приве-
денных выше формулировок делает понятным тот удивительный
факт, что, несмотря на многолетнюю пропаганду, системный под-
ход, в особенности общая теория систем Берталанфи, не стал
достаточно популярным среди исследователей конкретных орга-
низмов п не привел к значительному преобразованию самой ис-
следовательской практики.
В самом деле, что специфически системного может извлечь
исследователь-физиолог, например, из выражения «система — это
комплекс взаимодействующих компонентов», если взаимодействие
частей организма даже для начинающего исследователя являет-
ся аксиоматическим фактором жизни?
Мне кажется, что именно здесь тот узел, не развязав кото-
рого исследователь никогда не приблизится к истинным механиз-
мам системы и потому, естественно, не сможет использовать ее
в своей конкретной работе.
Попытаемся провести более глубокий анализ этих ходячих оп-
ределений, чтобы вскрыть их недостаточность как для форму-
лировки понятия системы, так и для утверждения ее как фактора
научного прогресса. Прежде всего поставим перед собой вопрос:
может ли вообще «взаимодействие компонентов» быть основой
какого-то системного процесса? Мы даем совершенно определен-
ный ответ: нет, не может. И этот ответ легко аргументировать
на любом примере взаимодействия.
Для характеристики понятия множеств и для подсчета чис-
ла возможных степеней свободы для взаимодействия в этом мно-
жестве приведем пример расчета, сделанного Эшби. Он берет
пло:
[адку с 400 лампочками
(20X20) и делает расчет Возмож-
к
ного количества комбинаций взаимодействий, которые можно
составить из этих лампочек. Оказывается, что этих взаимодейст-
вий такое огромпое количество (1010120), что они превосходят
общее количество атомов, содержащихся в видимой нами Вселен-
ной (10 73).
Но ведь эта квадратная площадка с 400 лампочками в коли-
чественном отношении совершеппо ничтожна по сравнению с го-
Принципиальные вопросы общей теории функциональных систем 67
ловным мозгом. Для того чтобы применить такое же вычисление
по отношению к мозгу, мы должны взять в качестве исходного
количества «лампочек», т. е. первных клеток, по крайней мере
количество в 14 млрд.
Кроме того, известно, что соединения между этими «лампочка-
ми» идут через синаптические контакты, так что каждая распо-
лагает не двумя возможными состояниями, как в примере Эшби,
а ъ среднем по крайней мере 5000 возможных состояний в зави-
симости от приходящих к синапсам импульсаций. Причем надо
номпить, что каждый из этих контактов может придать состоянию
нейрона особое качество.
Однако для полной характеристики множества взаимодействий
на примере мозга даже и этого количественного расчета недоста-
точно. Мы непременно должны учесть также и те общие состоя-
ния каждого отдельного нейрона, которые определяют характер
его участия во взаимодействиях нейронального множества.
Так, например, Буллок указывает, что имеется по крайней
мере пять возможных изменений в градации состояний нейрона,
а следовательно, и синаптического образования: возбуждение или
торможение, облегчение или депрессия, положительное или отри-
цательное последействие (или оба вместе), спонтанное расслабле-
ние или тонизация нейрона, градуированные ответы спайкового
или неспайкового характера.
Важно, что каждое из этих взаимодействующих множеств
(нейрон, синапс, градуированное состояние нейрона и др.) может
создать условие, при котором деятельность элемента в таком об-
ширном «множестве» может радикально измениться, а это значит,
что конечный результат деятельности мозга может быть иным.
Трудно решить даже с помощью воображения задачу подсчета
того количества комбинаций взаимодействия в целом мозге, ко-
торое может быть выведено нз указанных выше цифр.
Однако, предложив решить эту задачу опытным математикам,
мы получили совершенно фантастическую цифру. Оказалось, что
число степеней свободы нервных клеток мозга с учетом всех тех
переменных, которые были разобраны выше, может быть выраже-
но единицей с таким количеством нулей, что они могут уместить-
ся только па ленте длиной... в 9 500 000 км. Стоит только предста-
вить себе это «множество», чтобы понять, что человек практиче-
ски никогда не сможет использовать всех грандиозных резервов
своей мозговой деятельности.
Представим себе, какой хаос сложился бы в нервной системе,
если бы все это множество стало взаимодействовать и взаимовли-
ять друг па друга! Ясно, что этот хаос не допустил бы никакого
организованного поведения целого организма. II тем не менее
взаимодействие вообще все-таки непременно входит во все фор-
мулировки понятия системы как решающий критерий.
3*
68 Принципиальные вопросы общей теории функциональных систем
Нам кажется, что такое положение наблюдается потому, что
этот важнейший вопрос никогда пе был серьезно проанализиро-
ван до конца, по крайней мере для биологических систем, и имен-
но поэтому мы не имеем исчерпывающей и научно обоснованной
формулировки системы.
Применительно к поведенческому акту мы должны прежде
всего исходить из абсолютно достоверного нейрофизиологического
факта, что каждый отдельный нейрон потенциально имеет огром-
ное число степеней свободы как объект взаимодействия с другими
нейронами.
Следовательно, говоря о взаимодействии вообще, мы тем са-
мым неизбежно допускаем одновременное и неорганизованное
использование всех этих степеней свободы нейрона.
И это действительно так, поскольку все формулировки поня-
тия системы, делая акцепт на «взаимодействии», не содержат в
себе и не имеют даже в виду какие-либо факторы, ограничиваю-
щие многочисленные возможные степени свободы взаимодействия
данного компонента с другими. Практически это касается не толь-
ко нервной системы, где этот процесс особенно отчетлив и много-
образен. Это же наблюдается и в мышечной системе, где малей-
шее отклонение в механических соотношениях между сокращаю-
щимися мышцами ведет к хаосу и потере целенаправленности
движения, и во многих химических констелляциях.
Итак, мы пришли к очень важному выводу; взаимодействие,
взятое в его общем виде, не может сформировать системы из
«множества компонентов». Следовательно, и все формулировки
понятия системы, основанные только на «взаимодействии» и на
«упорядочении» компонентов, оказываются сами но себе несостоя-
тельными.
существующих подходах к выработке общей
Совершенно ясно, что именно в этом пункте мы имеем корен-
ной недостаток в
теории систем. Становится очевидным, что в проблему понятия
системы необходимо ввести некоторые дополнительные аспекты,
которые придали бы этому понятию конкретные механизмы орга-
низованного целого, детерминистически достоверного и логически
попятного. Точнее говоря, мы должны вскрыть те детерминирую-
щие факторы, которые освобождают компоненты системы от избы-
точных степеней свободы.
Внесение в формулировку системы выражения «упорядоченное
множество» не исправляет исходного дефекта и, пожалуй, даже,
наоборот, вносит в проблему некоторый привкус телеологического.
В самом деле, кто «упорядочивает» распределение множества ком-
понентов в системе? По какому критерию производится это
«упорядочение»? Не может же какое-либо множество стать «упо-
рядоченным» без критерия (1) этой «упорядоченности». Должен
быть конкретный фактор, который «упорядочивает» систему.
Принципиальные вопросы общей теории функциональных систем 69
Ясно, что, не имея четкого ответа на эти вопросы, мы факти-
чески продолжаем стоять на месте. Примером такой нечеткости
может служить выражение Эшби, который определил существо
«самоорганизующихся систем» как «изменение от неорганизован-
ной системы к организованной». Все сказанное выше заставляет
нас признать, что необходимо более глубоко проанализировать
некоторые до сих пор еще пе вскрытые детерминанты, направ-
ляющие взаимодействия компонентов в системе.
Пока системологи не определят точно фактор, который ради-
кально ограничивает степени свободы участвующих в данном мно-
жестве компонентов, все разговоры о системе и ее преимуществах
перед несистемным подходом будут столь же неплодотворны, как
до сих пор была неплодотворной в конкретной исследовательской
работе и сама общая теория систем.
Ниже и будет сделана попытка вывести системообразующий
фактор из свойств живого организма п обсудить изоморфность
этого фактора для различных классов явлений (организма, ма-
шины, общества).
Конкретный результат деятельности системы
как системообразующий фактор
Достаточно понаблюдать после каких-либо нарушений за вос-
становлением какой-нибудь простой и очевидной функции с весь-
ма четким результатом, например за удержанием тела человека в
вертикальном положении, чтобы ответить с определенностью на
поставленные выше вопросы.
Таким императивным фактором, использующим все возмож-
ности системы, является полезный результат системы, в данном
случае вертикальная поза, и формируемая им обратная афферен-
тация. Именно достаточность или недостаточность результата оп-
ределяет поведение системы: в случае его достаточности организм
переходит на формирование другой функциональной системы с
другим полезным результатом, представляющим собой следующий
этап в универсальном континууме результатов (Анохин, 1970).
В случае недостаточности полученного результата происходит
стимулирование активирующих механизмов, возникает активный
подбор новых компонентов, создается перемена степеней свободы
действующих синаптических организаций и, наконец, после не-
скольких «проб и ошибок» находится совершенно достаточный
приспособительный результат.
Мы не вскрываем всех детальных механизмов, при помощи
которых система подбирает необходимый на данный момент ре-
зультат. Это будет сделано в одном из последующих разделов
статьи. Сейчас же нам необходимо лишь дать исчерпывающую
формулировку системы, в которой результат является ведущим
компонентом.
70 Принципиальные вопросы общей теории функциональных систем
Включение в анализ результата как решающего звена системы
значительно изменяет общепринятые взгляды на систему вообще
и дает новое освещение ряду вопросов, подлежащих глубокому
анализу.
Прежде всего оказывается возможным как всю деятельность
системы, так и ее всевозможные изменения представить в терми-
нах результата, что еще более подчеркивает его решающую роль
в поведении системы. Эта деятельность может быть полностью
выражена в вопросах, отражающих различные этапы формиро-
вания системы.
1. Какой результат должен быть получен?
2. Когда именно должен быть получен результат?
3. Какими механизмами должен быть получен результат?
4. Как система убеждается в достаточности полученного ре-
зультата?
По сути дела эти четыре вопроса решаются основными узло-
выми механизмами системы. Вместе с тем в них выражено все
то, ради чего формируется система.
Возьмем для примера последний вопрос. Центральная нервная
система должна непременно иметь информацию о полученном
результате. Мы знаем, что в механических системах эта инфор-
мация называется «обратная связь». В нашей лаборатории она
получила название «обратная афферентация», или «санкциони-
рующая афферептация», поскольку она может санкционировать
последнее распределение в системе эфферентных возбуждений,
обеспечивших достижение полезного результата.
Все это будет четко показано в дальнейшем изложении на
основе конкретных физиологических механизмов. Сейчас же вам
необходимо отметить одно решающее обстоятельство: результат
обладает императивными возможностями реорганизовать распре-
деление возбуждений в системе в соответствующем направлении.
Итак, мы видим, что формирование системы подчинено полу-
чению определенного полезного результата, а недостаточный ре-
зультат может целиком реорганизовать систему и сформировать
новую, с более совершенным взаимодействием компонентов, даю-
щим достаточный результат.
Что может быть более убедительным для доказательства
справедливости положения о том, что результат является в самом
деле центральным фактором системы? И вместе с тем очевидно,
что пе может быть понятия системы без ее полезного результата.
Правда, нам приходилось не раз слышать замечания, что си-
стема с результатом — это «специальный случай» системы. Но
тогда очень важно было бы узнать, что является у системы без
результата тем фактором, который обеспечивает переход, выра-
жаясь языком Эшби, «от неорганизованного к организованному»,
г. е* от хаоса взаимодействия к системе.
Принципиальные вопросы общей теории функциональных систем 71
* —
Такое положение радикально меняет и наше отношение к по-
нятию «взаимодействие», которое, как мы видели, будучи основ-
ным критерием для определения понятия системы у многих со-
временных исследователей, определило общий неуспех всего на-
правления.
Показанная выше роль результата во всех превращениях си-
стемы делает невозможной какую-либо формулировку системы,
не основанную па роли результата в ее деятельности, ибо, как
мы видели, только он может «изменить неорганизованное множе-
ство в организованное».
Важным последствием включения результата в систему как
решающего операционального фактора системы является то, что
сразу же делаются попятными механизмы освобождения компо-
нентов системы от избыточных степеней свободы. А ясность в
этом вопросе — это серьезный шаг в разрешении противоречий,
возникших в связи с недостаточностью понятия «взаимодействие».
Следует отметить, что, говоря о «степопях свободы» централь-
ной нервной системы как важнейшего звена функциональной
системы, мы имеем в виду степени свободы, определяемые коли-
чеством участвующих пе только нейронов, но и синапсов из
всех синапсов, имеющихся на каждом из 14 млрд, нейронов.
В самом деле, допустим, что какая-либо система имеет в
своем составе а, Ъ, с, d, е — компоненты. Возникает вопрос,
какие факторы устанавливают вполне определенные системные
(!) взаимоотношения, например, между компонентами Ъ и е. Что
может вообще установить между всеми компонентами системы
такие взаимоотношения, которые устранили бы хаос всеобщего
взаимодействия, т. е. одновременной реализации всех степеней
свободы каждого компонента?
Для пас ответ па этот вопрос является вполпе определенным:
упорядоченность во взаимодействии множества компонентов си-
стемы устанавливается па основе степени их содействия в полу-
чении целой системой строго конкретного полезного результата.
Степени же свободы каждого компонента системы — нейрона, пе
помогающие получению полезного результата, устраняются пз
активной деятельности.
Таким образом, к системе с полезным результатом ее деятель-
ности более пригоден не термин «взаимодействие», а термин
«взапмосодействпе». Опа должна представлять собой подлинную
кооперацию компонентов множества, усилия которых направле-
ны па получение конечного полезного результата. А это значит,
что всякий компонент может войти в систему только в том слу-
чае, если он вносит свою долю содействия в получение запрограм-
мированного результата.
Именно по этому принципу, как мы увидим ппже, п проис-
ходит вовлечение всякого нового компонента в трудных условиях
72 Принципиальные вопросы общей теории функциональных систем
ь — * II , I —
функционирования для получения полезного результата. Компо-
нент при своем вхождении в систему должен немедленно исклю-
чить все те степени своей свободы, которые мешают или не по-
могают получению результата данной системы. Наоборот, он
максимально использует именно те степени свободы, которые в
той или иной мере содействуют получению конечного полезного
результата данной системы.
Главное качество биологической самоорганизующей системы и
состоит в том, что опа непрерывно и активно производит перебор
степеней свободы множества компонентов, часто даже в микро-
интервалах времени, чтобы включить те из них, которые прибли-
жают организм к получению полезного результата.
Возвращаясь к спору Бора и Эйнштейна о наиболее эффек-
тивном подходе к изучению целостных организаций, мы бы ска-
зали, что термин «взаимодействие» звучит более феноменологи-
чески, нейтрально, в то время как термин «взаимосодействие»
звучит более каузально, активно, поскольку в создание этого взаи-
модействия включаются многие новые специализированные меха-
низмы.
Несколько удивляет то обстоятельство, что, несмотря на весь-
ма широко развивающийся в настоящее время системный подход
к биологическим и техническим объектам, результат системной
деятельности как решающий самооргапизующий фактор системы
не был взят в качестве основного операционального фактора,
а соответственно с этим не были даны формулировки и сделаны
те преобразования в наших поисках, которые вынуждают ста-
вить многие вопросы по-новому.
Суммируя все сказанное выше, мы можем теперь легко дать
ту формулировку понятия системы, которая, с пашей точки зре-
ния, наиболее полно отражает ее суть.
Системой можно назвать только комплекс таких избирательно
вовлеченных компонентов, у которых взаимодействие и взаимоот-
ношения принимают характер взаимосодействия компонентов для
получения фокусированного полезного результата.
Конкретным механизмом взаимодействия компонентов являет-
ся освобождение их от избыточных степеней свободы, не нужных
для получения ‘ данного конкретного результата, и, наоборот,
сохранение всех тех степеней свободы, которые способствуют
получению результата.
В свою очередь, результат через характерные для него пара-
метры и благодаря обратной афферентации имеет возможность
реорганизовать систему, создавая такую форму взаимодействия
между ее компонентами, которая является наиболее благоприят-
ной для получения именно запрограммированного результата.
Таким образом, результат является неотъемлемым и решаю-
щим компонентом системы, инструментом, создающим упорядо-
Iрезультат JLрезультат ЛГрезультат
Рис. 2. Схематическое изображение упорядочивающего действия результата системы на реорганизацию
ее степеней свободы
Сплошные линии — взаимодействия между нервными эле-
ментами только на основе общего числа их степеней сво-
боды (4) и ограничение избыточных степеней свободы
системы благодаря обратной афферентации, формирую-
щейся на основе различных параметров результата (2—4).
Пунктирные линии — исключенные степени свободы. В ка
честве одиночной степени свободы для центральной нерв
ной системы предполагается одиночное синаптическое об
разование
74 Принципиальные вопросы общей теории функциональных систем
ценное взаимодействие между всеми другими ее компонентами.
Теперь мы сможем вполне определенно ответить на постав-
ленный выше вопрос: какой фактор упорядочивает множество
компонентов системы? Таким решающим и единственным факто-
ром является результат, который, будучи недостаточным, активно
влияет на отбор именно тех степеней свободы у компонентов
системы, которые при их интегрировании определяют в дальней-
шем получение полноценного результата (рис. 2).
Именно потому, что в нашей концепции результат оказывает
центральное организующее влияние на все этапы формирования
функциональной системы, а сам полезный результат является,
несомненно, функциональным феноменом, мы и назвали всю ар-
хитектуру функциональной системой. Ниже будет дана более
подробная аргументация этого понятия. Сейчас же мы считаем
нужным разобрать одно принципиальное следствие нашей кон-
цепции для общепринятой кибернетической терминологии.
Имеется в виду прежде всего весьма распространенное выра-
жение «управляющая система», которое ни семантически, ни ло-
гически не может быть принято теорией функциональной систе-
мы. В самом деле, что означает это выражение? Ничего, кроме
традиционного игнорирования результата системы при обсужде-
нии кибернетических закономерностей.
Действительно, выражение «управляющая система» по своей
сути предполагает, что управляемый объект не является компо-
нентом управляющей системы, т. е., попросту говоря, он нахо-
дится за пределами (!) самой управляющей системы.
Из самого выражения «управляющая система» следует, что
она является полноценной, несмотря на то что управляемый объект
находится вне ее. С точки зрения всех решающих влияний
результата на систему, которые были изложены выше, такое
положение является неприемлемым.
В самом деле, стоит лишь поставить вопрос, что произойдет
с управляющей системой,
если результат, т. е. управляемый
объект, окажется не совпадающим с запрограммированным ре-
зультатом, т. е. если попросту результат окажется неполно-
ценным.
На основании предложенных нами выше представлений о си-
стеме результат должен сам немедленно сигнализировать о своей
недостаточности и стимулировать управляющую систему на ре-
организацию, которая временами заходит так далеко, что может
привести к полной перемене установившихся взаимоотношений
между компонентами системы.
Можно нойти еще дальше и поставить чисто риторический
вопрос: откуда управляющая система «узнает», каким именно
объектом ей надо управлять, если она уже система? Ведь, чтобы
управлять чем-то, надо иметь весьма адекватные связи и соот-
Принципиальные «опросы общей теории функциональных систем 75
ношения с управляемым. И не обстоит ли дело так, что уже на-
чальные процессы формирования самой управляющей системы
находятся полностью под управлением будущего, необходимого в
данный момент организму результата?
Именно так позволяет смотреть на этот вопрос теория функ-
циональной системы, которая вгупочает приспособительный ре-
зультат функционирования системы как органическую составную
часть системы. Только в этом случае можно уйти от бесплодных
терминов, которые, уводя мысль в сторону, несомненно, наносят
ущерб пониманию самой системы. Именно к этому ведет чрез-
вычайно широко распространенная тенденция употреблять такие
выражения, как «управляющая система», «управляемый объект»,
«управляемая система», «биоуправление» и т. д.
Как мы видели выше, при более глубоком анализе все эти
понятия превращаются в научную фикцию, поскольку они не
соответствуют истинным соотношениям в действительной регуля-
ции, т. е. просто-напросто не имеют реального содержания.
Однако эта тенденция оперировать понятиями, не имеющими
конкретной основы, отразилась и на построении моделей систем.
Они все, как правило, имеют input u output, т. е. вход и выход.
Большей частью выход замыкается на вход дополнительной пет-
лей обратной связи.
Для примера можно указать на недостаточность схем Винера,
Гродииса, Мплсума, Эшби, Мак Кея, Паска и многих других,
хотя они и внесли весьма полезный, а часто и решающий вклад
в теорию регуляции.
Но что значит «выход» с точки зрения биологического объ-
екта, папрпмер развитого организма? Выходом может быть и ре-
альное возбуждение, выходящее всего лишь на эфферентные
пути к рабочим органам; выходом может быть также и работа
этих органов, т. е. то, что соответствует физиологическому терми-
ну «рефлекторное действие». Но этот выход никогда фактиче-
ски не связывается с результатом функционирования системы,
конкретные и измеримые параметры которого, как правило, оп-
ределяют все дальнейшее поведение системы.
Традиция избегать результат действия как самостоятельную,
физиологическую категорию не случайна. Она отражает тради-
ции рефлекторной теории, которая заканчивает «рефлекторную
дугу» только действием, не вводя в поле зрения и не интерпре-
тируя результат этого действия. Между тем сейчас становится
все более и более очевидным, что именно результат функциони-
рования системы является движущим фактором прогресса всего
живого на пашей планете.
Еще одно замечание. Часто приходится встречаться с поня-
тием «состояние системы», которое при неправильном егф при-
менении может повести мысль в ненужном направлении^ Так,
76 Принципиальные вопросы общей теории функциональных систем
например, Хапике говорит: «Каждый раз, когда возникают воз-
мущения равновесия, система стремится пайти устойчивое состоя-
ние» [1969, с. 331]. Было бы совершенно непрогрессивным для
живой природы, если бы система «стремилась» пайти лишь устой-
чивое состояние.
Если и употреблять термин «стремиться», то наиболее пра-
вильно такое выражение: система «стремится» получить запро-
граммированный результат и ради этого результата может пойти
на самые большие возмущения во взаимодействиях своих компо-
нентов. Следовательно, центральным пунктом, ради которого про-
исходят всякого рода «изменения состояний системы», является
опять-таки результат. Именно он в случае затрудненного его полу-
чения может привести всю систему в крайне беспокойное и не-
устойчивое состояние. Итак, вся трактовка состояния системы
радикально меняется, как только мы пойдем естественным путем,
приняв в качестве центрального фактора получение системой по-
лезного результата.
Но так как организм живет в среде непрерывного получения
результата, в подлинном континууме результатов, то после до-
стижения определенного фазного результата начинается «беспо-
койство» по поводу последующего результата.
Надо обратить внимание па одну особенность функциональ-
ной системы, по укладывающуюся в обычные физиологические
представленпя. Речь идет о том, что содержание результата,
т. е., выражаясь физиологическим языком, параметры результа-
та, формируется системой в виде онределенпой модели раньше,
чем появится сам результат. Именно этот чудесный и реальный
подарок всему жпвому на Земле, имеющий характер предсказа-
ния, отпугнул от себя даже гениальных экспериментаторов.
Вряд ли кто-либо из теоретиков павловского учения о высшей
нервной деятельности обратил внимание па один эпизод в генеа-
логии творческих приемов И. П. Павлова. Как известно, в 1916 г.
смелый гений ученого замахнулся па самое тонкое и самое со-
кровенное в работе головпого мозга человека — на цель поведе-
ния. И. П. Павлов пазвал свое известное выступление по этому
вопросу «Рефлекс цели». Казалось бы, с этого момента должна
была развиваться бурная исследовательская деятельность павлов-
ской лаборатории по этому физиологически, психологически и
идеологически важному вопросу. Однако нам хорошо известно,
что И.
дальнейшей жизни к этому вопросу не возвращался. Почему?
Нам кажется, что причина ухода И. П. Павлова от важнейшей
проблемы — деятельности мозга заключается в том, что сам факт
возникновения цели для получения того или иного результата
вступает в принципиальное противоречие с основными чертами
рефлекторной теории. И. П. Павлов, несомненно, думал и об
. Павлов никогда больше па протяжении всей своей
рефлекторной теории. II. П. Павлов,
Принципиальные вопросы общей теории функциональных систем 77
н
Такая грандиозная роль ре-
является
этом, видел и то, что, поставив проблему цели, он вынужден был
бы значительно перестроить то грандиозное здание, которое с
такой гениальной смелостью и настойчивостью строил всю свою
жизнь.
Как известно, представление о рефлекторном процессе постро-
ено на нерушимом принципе поступательного хода возбуждения
от пункта к пункту по всей рефлекторной дуге.
армирования цели И. П. Павлов, наоборот, встретплся с
совершенно неожиданным принципом работы нервной системы.
Здесь модель конечного результата данного акта создается уже
на начальных этапах распространения возбуждения, т. е. раньше,
чем закончится весь процесс формирования поведенческого акта,
и раньше, чем будет получен сам результат.
Действительно, совершенно ясно, что цель к получению дан-
ного результата возникает раньше, чем может быть получеп сам
результат. Причем интервал между этими двумя моментами мо-
жет равняться и минуте, и годам
зультата во всех поведенческих актах животных и человека,
естественно, не может быть игнорирована, если мы хотим сформу-
лировать системный подход и построить модель системы.
Сейчас, пожалуй, единственной областью, где результат, «по-
лезность результата» и проблема оценки этого результата стано-
вятся почти центральным фактором исследования
область промышленно-экономических систем. Такой успех и зна-
чение результата пмеппо в этой области спстемпого подхода по-
нятны, ибо здесь полезность деятельности настолько очевидна,
что игнорировать ее было бы просто неразумно. В самом деле,
если мы имеем «большую систему» в виде производственно свя-
занных заводских агрегатов, например в нефтеперерабатывающей
промышленности, то игнорирование полезности результата на
уровне каждой субсистемы этой большой системы привело бы к
расточительности и полной нерентабельности всего предприятия.
Именно состояние ценности и полезности результата в каждой
субсистеме этого предприятия и сочетание их с окончательным
результатом могут дать решающее суждение о том. насколько
полезен конечный результат и в какой степепи выгодно все об-
ширное предприятие.
Одни из теоретиков полезности экономист Bross (1953) так
определяет значение пользы: «Суд последней инстанции — это не
блестящий словесный аргумент, не солидно звучащий абстракт-
ный принцип и даже пе ясная логика пли математика,— это ре-
зультат в реальном мире». Данное высказывание весьма показа-
тельно для мышления теоретиков управления предприятиями. По
сути дела и большие системы организма также сотканы из малых
субсистем. Наконец, результат деятельности целого организма
также является «судом последней инстанции».
78 Принципиальные вопросы общей теории функциональных систем
Мы уже видели, что именно отсутствие результата во всех
формулировках системы и делает их неприемлемыми с операцио-
нальной точки зрения. Этот дефект полностью устранен в разви-
ваемой нами теории функциональной системы.
1. В функциональной системе результат представляет собой
ее органический фактор, оказывающий решающее влияние как на
ход ее формирования, так и на все ее последующие реорганизации.
2. Наличие вполне определенного результата как решающего
компонепта функциональной системы делает недостаточным по-
нятие «взаимодействия» в оценке отношений компонентов систе-
мы между собой. Именно результат отбирает все адекватные для
данного момента степени свободы компонентов системы и фоку-
сирует их воздействие.
3. Если деятельность системы заканчивается полезным в ка-
ком-то отношении результатом, то «взаимодействие» компонентов
данной системы всегда будет протекать по типу их взапмосодей-
ствпя, направленного на получение результата.
4. Взаимосодействие компонентов системы достигается тем,
что каждый из них под влиянием афферентного синтеза или об-
ратной афферентации освобождается от избыточных степеней
свободы и объединяется с другими компонентами только на основе
тех степеней свободы, которые вместе содействуют получению
надежного конечного результата (рпс. 3).
5. Включение результата в функциональную систему исклю-
чает необходимость применять как несовершенные формулировки
самой системы, так и многие другие («управляющая система»,
«управляющий объект», «биоуправление» п т. д.).
Состав функциональной системы
и иерархия систвхМ
После того как мы определили само понятие системы, другим
важным общим вопросом характеристики системы является вы-
яснение ее состава и роли отдельных компонентов функциониро-
вания системы. Определение состава необходимо еще и потому,
что именно в этом пункте происходит довольно часто интерферен-
ция нового представления о системе со всем тем, что в прежнее
время свободно определялось термином «система» без каких-либо
строгих ограничений формулировок и понятий.
Критериями употребления термина «система» являлось все,
что представляло собой нечто упорядоченное по сравнению с дру-
гими явлениями, относящимися к иным классам (например, си-
стема кровообращения, система пищеварения, мышечная систе-
ма и др.). Ясно, что здесь термин «система» употребляется в
смысле принадлежности данного феномена к определенному типу
анатомических преобразований, объединенных типом функциони-
Принципиальные вопросы общей теории функциональных систем 79
Рис. 3. Схематическое изображение соотношений возбужденных и невоз-
бужденных синапсов на нейроне. В зависимости от этих соотношений одна
и та же клетка может выдавать разную активность
Черные линии и точки — возбужденные в данный момент волокна и синапсы;
пунктир и белые кружки — невозбужденные
роваппя. Еще и до сих пор имеются исследователи, которые ви-
дят в употреблении термина «система» только этот смысл.
Говоря о системе в этом последнем смысле, мы выделяем из
целого организма какую-то часть, объединенную типом анатоми-
ческого строения или типом функционирования, и по сути дела
исключаем возможность понимания этих выделенных структур в
истинном системном плане. Совсршеппо очевидно, что система
кровообращения никогда не выступает как нечто отдельное, ибо
это было бы нонсенсом в физиологии. В полноценном организма
80 Принципиальные вопросы общей теории функциональных систем
* .... 11 1 '1 1 1 ' 1
кровообращение всегда ведет к получению какого-то приспособи-
тельного результата (уровня кровяного давления, скорости кро-
вотока и т. д.). Однако ни один из этих результатов нельзя по-
лучить только за счет системы кровообращения. Сюда непремен-
но включаются нервная и эндокринная системы и др. И все эти
компоненты объединены но принципу взаимосодействия. Поэтому
при новом системном подходе вопрос идет об акцепте пе па ка-
ком-либо анатомическом признаке участвующего компонента, а на
принципах организации многих компонентов (из многих анатоми-
ческих систем) с непременным получением результата деятельно-
сти этой разветвленной гетерогенной системы.
Очень часто приходится встречаться с каким-то нарочитым
подчеркиванием именно структурпо-анатомической принадлеж-
ности компонентов системы (например, «структурно-системная
организация», «структурные уровни» и т. д.). Это, однако, ведет
к неправильной ориентации читателя. В самом деле, что значит
«структурно-системный»? Разве может быть какая-то система ор-
ганизма, например, дающая полезный приспособительный резуль-
тат, бесструктурной, т. е. функционировать без структуры?
С самого начала надо подчеркнуть, что функциональные си-
стемы организма складываются из динамически мобилизуемых
структур в масштабе целого организма и на их деятельности и
окончательном результате не отражается исключительное влия-
ние какой-нибудь участвующей структуры анатомического типа.
Более того, компоненты той или иной анатомической принадлеж-
ности мобилизуются и вовлекаются в функциональную систему
только в меру их содействия получению запрограммированного
результата.
Как мы уже видели из всего изложенного, эти компоненты,
входя в систему, теряют свои избыточные степени свободы; оста-
ются лишь те из них, которые содействуют получению именно
данного полезного результата, поскольку поведение в целом пред-
ставляет собой истинный континуум результатов. Едва ли поэто-
му будет разумным то терминологическое усложнение, которое
вводится термином «структурно-системные отношения».
Кроме того, нельзя пе отметить и того, что введение понятия
структуры в формулировку системы привносит привкус чего-то
тем одним из
самых характерных свойств функциональной системы является
жестко структурно детерминированного. Между
именно динамическая изменчивость входящих в нее структурных
компонентов, изменчивость, продолжающаяся до тех пор, пока пе
будет получен соответствующий полезный результат. Ясно, что на
первый план в формировании истинно функциональных систем
выступают законы результата и динамической мобилизуемое™
структур, обеспечивающие быстрое формирование функциональ-
ной системы и получение данного результата.
Принципиальные вопросы общей теории функциональных систем 81
Нам хотелось бы отметить одно важное обстоятельство, кото-
рое проходит мимо внимания исследователя. Это свойство внезап-
ной мобилизуемое™ структурных элементов организма в соответ-
ствии с непрерывными функциональными требованиями, которые
функция предъявляет к структуре. Под свойством мобилизуемое™
мы понимаем возможность моментального построения любых
дробных комбинаций, обеспечивающих функциональной системе
получение полезного приспособительного результата.
Практически, если бы не было этой потенциальной способно-
сти структур к внезапной мобилизуемое™, причем в любой аран-
жировке, моментальная организация функциональных систем
была бы просто невозможна и, следовательно, приспособление
было бы несовершенным.
Так, например, сидя за письменным столом, мы можем взять
с него предмет правой рукой, но если нам в этот момент необхо-
дим другой предмет, лежащий позади нас, то мы с такой же лег-
костью, разворачивая руку в сторону, можем взять и этот пред-
мет. Ясно, что только способность иннервациопных аппаратов
мышц к внезапной перестройке и к выборочной организации от-
дельных дробных компонентов и обеспечивает формирование
функциональных систем, полезных в данный момент.
Таким образом, существование результата системы как опре-
деляющего фактора для формирования функциональной системы
и ее фазовых реорганизаций и наличие специфического строения
структурных аппаратов, дающего возможность немедленной моби-
лизации объединения их в функциональную систему, говорят о
том, что истинные системы организма всегда функциональны по
своей сути. Это значит, что функциональный принцип выбороч-
ной мобилизации структур является доминирующим. Потому
вполне естественно назвать такую систему «функциональной», что
мы в свое время и сделали (Анохин, 1935).
В связи с вопросом структурного состава функциональной си-
стемы возникает также и вопрос об иерархии систем, который
становится в последние годы все более и более актуальным. Мы
никогда не имеем по-настоящему изолированные
н
ункцпональ-
ные системы организма, можно только с дидактической целью
выбрать определенную систему, обеспечивающую какой-то резуль-
тат на данном уровне иерархии систем (рис. 4).
Так, например, соотношение актина и актомиозина, конечно,
составляет по своей операциональной архитектонике вполне очер-
ченную функциональную систему, заканчивающуюся положитель-
ным результатом, который можно было бы сформулировать как
сокращение мышечной фибриллы. Но такая функциональная си-
стема представляется лишь промежуточной между еще более
тонкими молекулярными соотношениями протоплазмы мышечной
клетки и между движением (например, движением охотника по
82 Принципиальные вопросы общей теории функциональных систем
Рис. 4. Схема иерархических объеди-
нений функциональных систем раз-
личной сложности, но построенных
на одном пейцмекерном физиологи-
ческом образовании, например на
возбуждениях пищевого центра. Схе-
ма дает возможность сопоставить
реальную организацию функцио-
нальных систем 1, 2, :::6, имеющих
общий пейцмекер (Р)
Пунктирные линии показывают, что
функциональные системы такого типа
не могут быть поняты из концепции
уровня, так как любой уровень органи-
зации неизбежно связан с энергетиче-
ским пунктом, питающим системы раз-
личных уровней
Z
лесу в поисках дичи), поскольку это движение осуществляется в
конце концов также с помощью актина и актомиозина. Но как
обширен диапазон, в который включено множество функциональ-
ных систем, составляющих эту грандиозную иерархию систем!
Естественно поэтому, что, говоря о составе функциональной
системы, мы должны иметь в виду, что каждая функциональная
система, взятая для исследования, неизбежно находится где-то
между тончайшими молекулярными системами и наиболее высо-
ким уровнем системной организации в виде, например, целого
поведенческого акта.
Нетрудно понять, что, раскрыв реальные физиологические
механизмы объединения функциональных систем различных уров-
ней, мы приблизились бы к решению проблемы органического
объединения анализа и синтеза в самом исследовательском про-
цессе.
В самом деле, сокращенпе мышечного волокна представилось
бы нам в двух аспектах: с одной стороны, как процесс вообще
сократительных структур, а с другой — сокращение, составляю-
щее какую-то очень дробную субсистему, например, в спортив-
ном прыжке. Ясно, что мы имеем огромное различие в составе
этих систем и, может быть, более грандиозную задачу опреде-
ления места компонента в большой системе.
В последние годы вопрос об «уровнях» организации больших
систем особенно подчеркивается рядом советских (Кремянский,
Введенов и др.) и зарубежных авторов (Джерард, Браун, Нови-
Принципиальные вопросы общей теории функциональных систем 83
ков и др.). Наиболее полный обзор этого направления в поисках
интегративных закономерностей дан в книге и статьях В. И. Кре-
минского. Это направление выступает под различными названия-
ми: «интегративные уровни», «структурные уровни», «иерархия
систем» п др.
Однако опять-таки в силу того же излишнего теоретизиро-
вания ни один из авторов не дает ничего конструктивного для
конкретного исследовательского процесса и пе отвечает па основ-
ные вопросы иерархии.
В сущности, па это указывает и употребление термина «уровни»,
который находится в абсолютном противоречии с понятием
«система». Главное же то, что ни в одной концепции уровни не
обладают какой-либо функциональной архитектоникой. Следова-
тельно, как способ соединения уровней
так и механизмы, удер-
живающие единство всей архитектуры целого, естественно, не
могут быть найдены.
Все сказанное выше с совершенной очевидностью убеждает
пас в том, что перед исследователем стоят по крайней мере два
кардинальных вопроса, пе решив которые он не может надеяться
на понимание тонких механизмов сложных функциональных си-
стем организма.
Несколько лет назад эти вопросы были сформулированы пами
в следующем виде.
1. Какими конкретными механизмами соединяются между со-
бой субсистемы при образовании суперсистемы? Учитывая нали-
чие в функциональной системе определенного количества специ-
фических для пее узловых механизмов, вопрос можно поставить
более конкретно: какими именно узловыми механизмами своей
архитектуры соединяются субсистемы, чтобы образовать суперси-
стему?
2. Различается ли чем-либо принципиально архитектура функ-
ционирования как весьма элементарных, так и сложных субси-
стем? Иначе говоря, функционируют ли системы всех уровней по ,
одной и той же архитектуре, которая характерна для функцио-
нальной системы вообще, или эти архитектуры чем-то отличают-
ся друг от друга?
Не объединяются ли по этому же принципу и «большие си-
стемы» промышленных предприятий? Мы предоставляем право
судить об этом соответствующим специалистам. Но если бы это
было так, перед нами встал вопрос о замечательной гармонии в
организации всех тех систем, где результат является решающим
фактором системообразования.
Мы допускаем, что при анализе такого существенного вопроса,
как объединение субсистем в суперсистему, могут возникнуть
другие вопросы, однако несомненно одно, что без решения двух
поставленных выше вопросов нельзя надеяться на решение всей
84 Принципиальные вопросы общей теории функциональных систем
проблемы иерархии систем в целом. Они принципиальны по своей
5
сутз
Отвечая па первый вопрос, пужпо исходить из того вывода,
к которому мы пришли при формулировке самого понятия «си-
стема». Центральным моментом для системы является резуль-
тат, так как любой комплекс и любое множество становятся си-
стемой только благодаря результату. Вместе с тем система пе
может быть стабильной, если сам результат своими существен-
ными параметрами не влияет на систему обратной афферента-
цией. А если это так, то любая система, какой бы значительной
она ни была в иерархическом ряду, должна подчиняться этим
правилам.
Все эти соображения приводят пас к окончательному и фун-
даментальному выводу о составе иерархии: все функциональные
системы независимо от уровня своей организации и от количест-
ва составляющих их компонентов имеют принципиально одну и
ту же функциональную архитектуру, в которой результат явля-
ется доминирующим фактором, стабилизирующим организацию
систем.
Решение второго из поставленных выше вопросов облегчает-
ся принятием положения, что архитектуры систем принципиаль-
но тождественны.
Если допустить, что какие-то субсистемы соединяются меж-
ду собой, вступают в контакт с помощью каких-то промежуточ-
ных механизмов, ведущих к получению полезного результата,
то сразу же будет видно, что такое допущение пе может быть
сделано. Тогда какие-то субсистемы пе смогут развить своего
основного функционального значения, т. е. получения результа-
та, и, таким образом, сама система не может быть названа си-
стемой. Поэтому наиболее вероятно, что именно полезный резуль-
тат системы, какой бы малой она ни была, представляет тот
реальный вклад, который она может сделать при образовании су-
персистемы, пли «большой системы».
Отсюда следует, что при образовании иерархии систем всякий
более низкий уровень систем должен как-то организовать кон-
такт результатов, что и может составить следующий, более вы-
сокий уровень систем и т. д. Очевидно, организм формирует
своп системы именно таким образом и только при этом возможно
организовать системы с обширным количеством компонентов. Ес-
тественно, что в этом случае «иерархия систем» превращается в
иерархию результатов каждой из субсистем предыдущего уровня.
Другой важный вопрос, возникающий при образовании иерар-
хии систем, состоит в следующем: как действует эта субордини-
рованная иерархия, когда ей надо выступать как целое?
Хороший пример представляет соотношение уровня кровяно-
го давления и какого-либо эмоционального состояния, возник-
Принципиальные вопросы общей теории функциональных систем 85
tero под влиянием внешних воздействий. Мы знаем, что при
возникновении эмоции, например страха, происходит быстрый
подъем кровяного давления, что имеет несомненное приспособи-
тельное значение. Но в то же время мы знаем, что постоянный
уровень кровяного давления представляет собой результат само-
стоятельной разветвленной функциональной системы, независи-
мой от эмоционального разряда (Анохин, 1960). Как эмоциональ-
ный разряд, возникший по внешнему поводу, находит доступ к
функциональной системе кровообращения? На какие компоненты
этой более низко организованной системы действует эмоциональ-
ный разряд?
Кора головного мозга и вызванная ею эмоция нуждаются в
поддержании высокого уровня кровоснабжения и метаболиче-
ских процессов в условиях стрессового состояния целого орга-
низма. Поэтому ясно, что эмоциональный разряд должен оказать
свое действие на «результат» функциональной системы — на уро-
вень кровяного давления. Но этот уровень — физическая вели-
чина.
Таким образом, эмоциональный разряд должен действовать
прежде всего на эфферентные механизмы, определяющие уровень
кровяного давления. Следовательно, в нисходящем направлении
эмоциональный разряд должен подействовать на эфферентное зве-
но системы, определяющее уровень давления, т. е. па сосудосу-
живающий центр.
Обращает па себя впимание один весьма интересный факт:
в этом случае уровень кровяного давления не зависит от того
афферентного синтеза, который производится ежесекундно сосу-
досуживающим центром па основе барорецепторной афферент-
ной сигнализации. В случае эмоционального разряда возбуждение
суперсистемы прямо занимает эфферентные пути субсистемы и
устанавливает нужный уровень кровяного давления, минуя аффе-
рентный синтез «хозяппа» субспстемы. На долю же этой послед-
ней остается лишь сопротивляться при помощи барорецепторов
чрезмерному давлению, оказываемому на ее сосуды...
Подводя итог сказанному, следует заметить, что главной чер-
той каждой функциональной системы является ее динамичность.
Структурные образования, составляющие функциональные систе-
мы, обладают исключительной мобилизуемостью. Именно это свой-
ство систем и дает им возможность быть пластичными, внезап-
но менять свою архитектуру в поисках запрограммированного по-
лезного результата.
86 Принципиальные вопросы общей теории функциональных систем
Внутренняя операциональная архитектоника
функциональной системы
Не будет преувеличением сказать, что трудность развития си-
стемного подхода вообще и «общей теории систем» Берталанфи
в частности состоит именно в том, что обсуждение ведется на
уровне глобальных свойств системы, так сказать, обсуждения
системы «черного ящика». Подавляющее большинство исследова-
телей не делают попытки проникнуть во внутреннюю архитек-
тонику системы и дать сравнительную оценку специфических
свойств ее внутренних механизмов. При таком подходе обсуж-
даемая система всегда выглядит как нечто гомогенное, в кото-
ром клетки одинаковы, все компоненты равноценны и все меха-
низмы равнозначны.
В действительности же дело обстоит совсем наоборот. Функ-
циональная система всегда гетерогенна. Опа всегда состоит из
определенного количества узловых механизмов, каждый из кото-
рых занимает свое собственное место и является специфическим
для всего процесса формирования функциональной системы. Ста-
новится очевидным, что, не вскрыв этих своеобразных механиз-
мов, составляющих внутреннюю операциональную архитектонику
системы, мы не приблизимся к самой решающей цели системно-
го подхода вообще — обеспечению органического единства в ис-
следовательском процессе системного уровня функционирования
с индивидуальной характеристикой каждого дробного элемента
или механизма, принимающего участие в этом функционирова-
нии (рис. 5).
Смысл системного подхода состоит именно в том, что элемент
или компонент функционирования не должен пониматься как
самостоятельное и независимое образование, он должен понимать-
ся как элемент, чьи оставшиеся степени свободы подчинены об-
щему плану функционирования системы, поправляемому получе-
нием полезного результата. Компонент должен быть органическим
звеном в весьма обширной кооперации с другими компонентами
системы. Естественно, что это правило относится к любому ком-
поненту любой субсистемы, как бы элементарна она пи была.
Одним из существенных п даже, пожалуй, решающих отли-
чий теории функциональной системы от всех предлагаемых к
обсуждению системных моделей является наличие в пей четко
отработанной внутренней операциональной архитектоники. Такая
внутренняя архитектоника, выраженная в физиологических поня-
тиях, является непосредственным инструментом для практиче-
38
ского применения
ункцпопальпой системы в исследовательской
работе, если даже опа касается молекулярного уровня исследуе-
мого объекта.
Практически система может стать методологическим принци-
Принципиальные вопросы общей теории
ункциональных
систем
Обстановоч-
ные а Фре-
рен тации
Обстано-
вочные
Пуснабой
стимул^
шж
та^аи Доминирующая
мотивация ~
Афферентный
синтез
Обратная афрерентацая
Орфе рентные бвзбужбения
Параметры
результата
шп
Результат
деистбая
ШЕ
,Деистбие
Рис. 5. Общая архитектура функциональной системы, как логической моде-
ли поведенческого акта, представляющая собой основу «концептуального
моста» между уровнями системных и аналитических процессов
пом исследования и перебросить «концептуальный мост» от син-
тетических обобщений к аналитическим деталям только в том
случае, если опа будет иметь четко очерченную,
изиологически
достоверную и логически оправданную внутреннюю архитекто-
нику.
Внутренняя архитектоника функциональной системы выража-
ет собой дальнейшее развитие идеи взаимосодействия компонен-
тов системы, она раскрывает ее топкие механизмы, при помощи
которых компоненты системы освобождаются от избыточных сте-
пеней свободы, чтобы установить взаимосвязь с другими компо-
нентами па основе императивного влияния результата на всю си-
стему.
В одной из своих работ Эшби очень разумно говорит о том,
что сама множественность компонентов системы и их потенци-
ально безграничное взаимодействие должны быть упрощены в
соответствии с требованиями анализа, поскольку, как мы видели
на примере с площадкой с 400 лампочками, совершенно невоз-
можно эффективно анализировать хаотические «взаимодействия»
этого множества. Именно это обстоятельство заставило его гово-
рить об упрощении и об «улучшенной логике механизма». Он
выразился даже еще более радикально, говоря, что «теория си-
88 Принципиальные вопросы общей теории функциональных систем
стем должна строиться на методах упрощения и что она пред-
ставляет собой науку упрощения» (Эшби, 1962).
Однако опять-таки он не указывает самого важного: па ос-
нове какого критерия должна быть построена «логика механиз-
ма» и должно быть произведено «упрощение множества»? Теория
функциональной системы решает этот вопрос четко и обоснован-
но. Центральным критерием упрощения множества является ре-
зультат системы, который, как мы видели, предъявляет решаю-
щие требования к определенным степеням свободы компонентов
системы. С точки зрения теории функциональной системы, «улуч-
шенная логика механизма» есть не что иное, как внутренняя
операциональная архитектоника системы, отвечающая на все тре-
бования тончайшего физиологического анализа механизмов си-
стемы до молекулярного уровня включительно.
Ниже мы приводим узловые специфические механизмы, пред-
ставляющие собой внутреннюю архитектонику системы и вместе
с тем обеспечивающие объединение в одной исходной концеп-
ции и высшего синтеза, и тончайшего анализа.
I til ।
ерентный синтез
для функциональ-
результат должен
основе ее законо-
н
Как мы уже установили в начале раздела,
ной системы характерно то, что вопрос, какой
быть получен, решается внутри системы и на
мерных механизмов. Это обстоятельство радикально отличает био-
систему от самых сложных машинных устройств автоматической
регуляции. Практически для всех машин цель поставлена за пре-
делами машины и для нее допускается лишь некоторая способ-
ность самоорганизации в процессе получения запрограммирован-
ного не ею результата. Биосистема даже очень простой иерархии
сама, на основе своих внутренних процессов, принимает решение
о том, какой результат нужен в данный момент ее приспособи-
тельной деятельности. Вопрос этот решается именно в стадии
афферентного синтеза.
Поскольку этот процесс много раз описывался нами в различ-
ных публикациях, мы дадим здесь лишь краткую формулировку
его особенностей. Прежде всего вопрос о его составе. Какие имен-
но афференты синтезируются в этой стадии?
Мы предложили четыре решающих компонента афферентного
синтеза, которые должны быть подвергнуты одновременной обра-
ботке с одновременным взаимодействием на уровне отдельных
нейронов: доминирующая на данный момент мотивация, обстано-
вочная афферентация, также соответствующая данному моменту,
пусковая афферентация и, наконец, память (рис. 6).
Основным условием афферентного синтеза является одновре-
менная встреча всех четырех участников этой стадии функцио-
Принципиальные вопросы общей теории функциональных систем 89
&СМ76
Рис. 6. Различные типы конвергенции возбуждений на одном и том же неЗ
Л
роне
1 — мультисенсорная конвергенция — встреча возбуждений от различных рецеп-
торных поверхностей; II — мультибиологическая конвергенция — возможность кон-
вергенции от различных биологических состояний, например голода, боли, ориенти-
ровочно-исследовательской реакции; III — сенсорно-биологическая конвергенция,
благодаря которой различные сенсорные возбуждения имеют возможность вступать
в контакт с биологическими состояниями организма, широко генерализованными по
огромному количеству корковых и подкорковых нейронов; IV — аксонально-сен-
сорно-биологическая конвергенция, благодаря которой на одном и том же нейроне
или на комплексе нейронов создается возможность встречи не только сенсорных,
биологических, антрацентральных возбуждений, но и тех возбуждений, которые
выходят на конечный нейрон в виде эфферентных. Эти возбуждения отходят по
коллатеральным ответвлениям от основного аксона. Такая конвергенция возбужде-
ний с полифункциональным содержанием представляет собой подлинный интегра-
тивный блок
нальной системы. Микроэлектродный и микрохимический ана-
лизы и другие формы аналитического исследования нейрона в
момент встречи на нем всех упомянутых выше типов возбужде-
ний показали, что этот процесс поддерживается и облегчается
рядом динамических процессов нервной системы. К ним относят-
ся прежде всего восходящая активация, вызванная ориентиро-
вочно-исследовательской реакцией, как правило, сопутствующая
¥ ¥
ЭНН
А А
ерентному синтезу и предшествующая принятию решения,
90 Принципиальные вопросы общей теории функциональных систем
процесс корково-подкорковой реверберации и процесс центробеж-
ного повышения возбудимости вовлеченных в афферентный син-
тез рецепторов.
Своеобразие состоит в том, что этот синтетический процесс,
если его отнести к масштабам нейрона, совершается на основе
центральной закономерности интегративной деятельности мозга,
именно на основе конвергенции возбуждений на одном п том же
нейроне.
Нам хотелось бы обратить внимание на следующее: одновре-
менность обработки всех четырех типов возбуждений в стадии
афферентного синтеза основана на том, что каждый из них при-
обретает свои особенные физиологические свойства именно в про-
цессе симультанной обработки. Именно здесь происходит освобож-
дение нейрона от избыточных степеней свободы благодаря при-
ходу к нему именно тех, а пе других возбуждений.
В связи с этим уместно отметить, что существующая в ней-
рофизиологии тенденция изучать и рассматривать такие компо-
ненты афферентного синтеза, как мотивация, память, стимул
и т. п., в качестве отдельных, самодовлеющих, проблем неверна
п малоэффективна.
Возьмем, например, проблему памяти. Взятая в отдельности,
сама по себе, как самодовлеющая проблема, опа может повести
мысль исследователя совершенно в другую сторону. Действитель-
но, в каком аспекте сейчас развивается проблема памяти? По-
давляющее большинство исследователей проблемы все внимание
сосредоточивают на моменте фиксации пережитого опыта живой
системы. Это, несомненно, важная сторона памяти, но совсем по-
другому выглядит весь вопрос о пей, как только мы будем ее
рассматривать как один из компонентов, органически включен-
ных в проблему принятия решения. Здесь сразу же центр собы-
тий перемещается с фиксации опыта иа динамическое извлече-
ние этого опыта из молекулярных агрегатов большой системы.
Этот процесс извлечения из памяти является еще более пора-
зительным, если вспомнить, с какой легкостью он ежесекундно
и безошибочно помогает принять нужное решение в конце аффе-
рентного синтеза. Обратим внимание, с какой легкостью мы из-
влекаем из памяти самые тончайшие нюансы нашей мысли, раз-
говора и всего того, что было накоплено за всю нашу жизнь.
Совершенно очевидно, что эта чудесная способность памяти быть
готовой ежесекундно отдать то, что было накоплено за много лет
и что требуется в данной стадии афферентного синтеза, не мо-
жет быть изучена иначе, как на основе полного контакта п взаи-
модействия всех четырех компонентов афферентного синтеза.
Значительное облегчение при решении проблемы афферентно-
го синтеза пам принесла разработка концепции об интегратив-
ной деятельности нейрона. Она дала возможность сформулировать
Принципиальные вопросы общей теории функциональных систем 91
положение, на основе которого строится динамическое участие
памяти в афферентном синтезе: извлечение прошлого опыта из
памяти происходит но топ же нейрохимической трассе, по ко-
торой он был зафиксирован в момент приобретения опыта (Ано-
хин, 1974).
Таким образом, афферентный синтез, приводящий организм к
решению вопроса, какой именно результат должен быть получен
в данный момент, обеспечивает постановку цели, достижению ко-
торой и будет посвящена вся дальнейшая логика системы.
Нетрудно видеть, что афферентный синтез, являющийся абсо-
лютно необходимым этапом формирования функциональной си-
стемы, содержит все необходимое для постановки цели, которая
так долго пугала исследователя-материалиста и так долго нахо-
дилась в безраздельном владении идеализма.
Все детали афферентного синтеза в нашей лаборатории изу-
чаются на протяжении многих лет с помощью экспериментально-
физиологических методик. Так, например, неразделимое единство
пусковой и обстановочной афферентации было показано на основе
опытов с экстирпацией лобных отделов коры мозга собаки и по-
следующего изучения реакции выбора на различные условные
раздражители (Шумилина, 1949).
В последнее время были подробно изучены различные виды
восходящих активирующих влияний, которые своеобразно помо-
гают афферентному синтезу. Здесь прежде всего надо указать на
чрезвычайное увеличение дискриминационной способности корко-
вого нейрона к частоте импульсаций под влиянием «мотивацион-
ных» раздражений гипоталамуса (Хаютип, 1971). Другой важный
механизм, выявляющийся при раздражении лимбической систе-
мы,— это перевод нейронов из категории мопомодальпых в муль-
тиконвергептпую категорию (Макаров, 1970). Легко видеть, что
уже только эти два механизма могут значительно повысить ин-
формационную емкость нейрона и, следовательно, вместе с дру-
гими описанными выше механизмами сделать особенно эффек-
тивной стадию афферентного синтеза. К разряду механизмов,
облегчающих афферентный синтез, надо отнести также детектор-
ные свойства периферических рецепторов и побочное вытормажи-
ваиие «шумовых процессов» при прохождении афферентной ин-
формации по центральной нервной системе.
Особенно подробно (до микроэлектродной и до микрохимиче-
ской методик включительно) была изучена роль доминирующей
мотивации в установлении синтетических взаимодействий в этой
стадии на поверхности одиночного нейрона. Само собой разуме-
ется, что возможность такого тонкого анализа отдельных про-
цессов, составляющих систему, появилась только тогда, когда
была выработана «улучшенная логика механизма», т. е. внутрен-
няя операциональная архитектоника системы.
92 Принципиальные вопросы общей теории функциональных систем
В управленческом деле, как и в биологических системах, так-
же следует поставить вопрос: какой результат должен быть по-
лучен системой? Иначе говоря, совокупность всех предваритель-
ных расчетов и соображений должна составить (например, в тор-
говом деле) основу афферентного синтеза, приводящего к решению
и выбору действия с наилучшим результатом. Так, например,
Хапике, касаясь проблемы управления, пишет: «Цель заключает-
ся в том, чтобы обеспечить принятие решений па основе систе-
матического анализа» [1969, с. 41].
Насколько отчетливо именно в проблеме управления пред-
приятиями встают вопросы о необходимости афферентного син-
теза, совершенно схожие с биологическими проблемами, видно из
того удивительного совпадения, которое получилось в аргумента-
ции Ханике и нашей собственной по поводу роли афферентного
синтеза.
Для выявления наличия широкого афферентного синтеза у че-
ловека перед принятием решения рассмотрим пример, когда пе-
шеход переходит улицу. Перед тем как принять решение о пере-
ходе, человек должен тщательно оценить довольно большое коли-
чество компонентов этого афферентного синтеза (число машин,
скорость движения, ширину улицы, своп силы и др.). Аргумен-
тируя наличие этой стадии в системной деятельности человека,
Ханике также приводит именно этот пример, перечисляя все па-
раметры, которые должны быть учтены для принятия решения о
переходе.
Принятие решения
1£
Принятие решения — один из самых интересных моментов в
развертывании системных процессов. Тот факт, что само понятие
принятия решения долгое время изгонялось так называемой чи-
стой наукой из области научных исследований,— лучшее доказа-
тельство того, что оно происходит на основе весьма комплексных
процессов мозга.
В настоящее время, как видно из предыдущего изложения,
теория функциональной системы сделала «принятие решения»
полноценным участником объективного процесса формирования
системы, критического процесса, благодаря которому афферент-
ная оценка всех условий завершается доступным исследованию
механизмом. Афферентный синтез, подчиняясь доминирующей в
данный момент мотивации и под коррекцией памяти, ведет та-
кой подбор возможных степеней свободы, при котором возбуж-
дения избирательно направляются к мышцам, совершающим нуж-
ное действие.
Возникает весьма существенный для всей нашей концепции
вопрос, когда и как происходит принятие решения, связанного
с получением именно этого, а не другого результата?
Принципиальные вопросы общей теории функциональных систем 93
Последние данные сотрудников нашей лаборатории (А. И. Шу-
милина, В. Б. Швырков) заставляют думать, что оценка воз-
можных результатов при конкретной доминирующей мотивации
происходит уже в стадии афферентного синтеза. Однако эти ре-
зультаты не получаются реально, а следовательно, их оценка
происходит при помощи какого-то пока не изученного нами ме-
ханизма. То же, что происходит в процессе «принятие решения»,
является уже результатом выбора па основе длительной оценки
различных, внутренне (!) формирующихся результатов.
Иначе говоря, любое принятие решения, после того как за-
кончится афферентный синтез, является выбором наиболее под-
ходящих степеней свободы в тех компонентах, которые должны
составить рабочую часть системы. В свою очередь, эти оставшие-
ся степени свободы дают возможность экономно осуществить
именно то действие^ которое должно привести к запрограммиро-
ванному результату.
Как происходит это освобождение от избыточных степеней сво-
боды? Почему момент принятия решения часто имеет характер
внезапной интуиции?
В настоящее время мы еще не можем ответить на эти вопро-
сы, но выработанный подход к ним дает полную гарантию того,
что принятие решения в биологических системах с большой и
малой иерархией является вполне анализируемым и доступным
для объективной пауки феноменом.
Опыты А. 11. Шумилиной, которая удаляла лобные отделы у
собаки, находящейся в условиях активного выбора одной из сто-
рон станка, убедительно показали, что в этот момент, т. е. в мо-
мент принятия решения, вся обрабатываемая информация инте-
грируется именно
команда к подбору наиболее оптимальных аппаратов поведения.
Интересно отметить, что, являясь наиболее требовательным к
объему афферентной информации, процесс принятия решения
страдает в первую очередь после различных вмешательств в цент-
ральную нервную систему. Весьма доказательной иллюстрацией
являются результаты опытов Gambaryan (1971). Разрушая неко-
торые подкорковые аппараты (бледное тело и др.), он нашел, что
прежде всего длительно страдает активный выбор стороны стан-
ка, на которой должно быть сделано пищевое подкрепление. На-
оборот, казалось бы, более сложный «произвольный акт» — нажа-
тие педали для подачи корма — остается неразрушенным или
мало пострадавшим.
Попытаемся понять сам момент принятия вполне определен-
ного решения с точки зрения общефизиологических закономер-
ностей. Его механизмы станут ясными, если мы представим себе
на минуту, какое количество возможных действий может сделать
человек, употребляя хотя бы только один мышечный аппарат.
в области лобных отделов, откуда и идет
94 Принципиальные вопросы общей теории функциональных систем
Мы уже знаем, что вся эфферентная часть организма должна
обладать свойством экстренной мобилизуемости. Малейшего из-
менения в распределении мышечных усилий достаточно для того,
чтобы было произведено вполне целенаправленное движение ко-
нечности или целого организма.
Принципиально таких возможностей движения у организма
существует необозримое множество,— вероятно, не меньше, чем
количества комбинаций взаимодействия на площадке с 400 лам-
почками. Вместе с тем мы знаем, что в каждом отдельном слу-
чае (в соответствии с принятым решением) возбуждения выхо-
дят на совершенно определенные мышечные группы. Иначе гово-
ря, происходит очень быстрое отбрасывание всех тех степеней
свободы в наших движениях, которые не соответствовали бы при-
нятому решению. Таким образом, сама реализация принятого
решения в физиологическом плане должна объяснять два важных
вопроса.
1. Какие механизмы производят отбор нужных в данный мо-
мент степеней свободы и вытормаживают все те степени свободы
на моторных нейронах и мышечных аппаратах, которые не име-
ют отношения к получению данного результата?
2. Где может быть преимущественная локализация этого в
высшей степени конденсированного процесса, который одновре-
менно обрабатывает на основе доминирующей мотивации всю при-
шедшую в мозг афферентную информацию, производит непре-
рывное сопоставление этих результатов с прошлым опытом и,
наконец, переводит результаты этой обработки на эфферентные
пути, точно соответствующие распределению возбуждений для со-
вершения нужного акта, обеспечивающего получение нужных ре-
зультатов?
Поставив эти два вопроса, мы переходим в область, абсолют-
но новую для нейрофизиологии и потому, естественно, совсем
неразработанную. Насколько интересны те новые вопросы иссле-
дования, которые при этом возникают, показывает, например,
роль доминирующей мотивации в отборе необходимой в данный
момент афферентной информации на уровне коры, на ее нейронах.
Микрофизиологическое исследование нейронов вентро-меди-
ального гипоталамуса показало, что уровень их возбудимости за-
висит от начального возбуждения латерального ядра гипоталаму-
са «голодной кровщо».
Дозируя это возбуждение в натуральных условиях сроками
голодания (1—4 дня), мы показали, что количество доминирую-
щего возбуждения меняется на соответствующих синапсах корко-
вых нейронов,
и в связи с этим значительно меняется интегри-
рующее влияние доминирующей мотивации на соотношение дру-
гих синаптических возбуждений данных кортикальных нейронов.
Есть основание предполагать, что этот механизм сосредоточен
Принципиальные вопросы общей теории функциональных систем 95
в наиболее интегрирующих и компактных структурах головного
мозга. Одпако вопрос физиологической интерпретации принятия
решения, возникший благодаря системному подходу к предмету,
должен быть фактически разработан заново.
Формирование акцептора результатов действия
Аппарат акцептора результатов действия — один из самых
интересных в деятельности мозга. Практически он так же универ-
сален и «вездесущ», как обратная афферентация и афферентный
синтез. Формирование этого механизма нарушает устоявшееся
представление о поступательном ходе возбуждений по централь-
ной нервной системе согласно рефлекторному процессу. Акцеп-
тор результатов действия па основе многостороннего механиз-
ма афферентного синтеза не является выражением последова-
тельного развития всей цепи явлений поведенческого акта. Он
«предвосхищает» афферентные свойства того результата, который
должен быть получен в соответствии с принятым решением, и,
следовательно, опережает ход событий в отношениях между орга-
низмом и внешним миром.
Акцептор результатов действия является весьма сложным ап-
паратом. По сути дела оп должен сформировать какие-то тон
кие нервные механизмы, которые позволяют не только прогнози-
ровать признаки необходимого в данный момент результата, но и
сличать их с параметрами реального результата, информация о
которых приходит к акцептору результатов действия благодаря
обратной афферептацпи. Именно этот аппарат дает единственную
возможность организму исправить ошибку поведеппя или довести
несовершенные поведенческие акты до совершенных. Здесь еле-
дует также подчеркнуть, что различного рода «поиски» и компен-
сации также могут повести к полезному результату через такого
рода оценку обратной афферептацпи. Циркулярное развитие этих
возбуждений при «узнавании» н «поиске» может быть столь бы-
стрым, что каждый блок этой функции, состоящей из компонен-
тов: результат, обратная афферептацпи, сличение п оценка ре-
альных результатов в акцепторе результатов действия, коррекция
новый результат и т. д
кунды. Особенно быстро этот процесс протекает в условиях «ска-
нирования» и «слежения».
Факт организации этого аппарата непосредственно после при-
нятия решения был доказан в нашей и других лабораториях как
в обычном эксперименте с условными рефлексами, так и при по-
мо:
может развиться буквально в доли се-
и
щ тонких электрофизиологических приемов.
Возможно, сам момент встречи опережающего комплекса воз-
буждений с информацией о реально полученных результатах
происходит па основе каких-то весьма тонких признаков воз-
96 Принципиальные вопросы общей теории функциональных систем
Suga (1964). Он изучал получение обратного, отражен-
объекта, сигнала при ультразвуковой локации у летучих
Оказалось, что при посылке ультразвукового поискового
в пространство в мозге летучей мыши формируется комп-
возбужденных клеток с различными латентными перио-
песколько сенсорных компо-
сложным. Наши последние
результатов действия как
показали, что эти средства
1Ш1
буждения вообще, допускающих количественное и композицион-
, пое сличения.
Одна из интересных форм сличения в акцепторе результатов
действия, хотя ее автор и не был знаком с общей архитектурой
функциональной системы, была недавно представлена японским
ученым
ного от
мышей,
сигнала
леке из
дами, периодом торможения и частотной конфигурацией. Имен-
но эти комплексы в зависимости от фазы, в которой происходит
встреча их с отраженным от объекта ультразвуковым сигналом,
решают вопрос о расстоянии, на котором паходится преследуе-
мый объект.
Конечно, этот аппарат оценки результата действия сравни-
тельно простой, но оп подсказывает нам, какими возможностями
и параметрами нервное возбуждение решает проблему сличения.
В более сложных формах результата, например при очинке ка-
рандаша, этот аппарат, включающий
нептов, является, песомпенпо, более
исследования по составу акцептора
комплекса разнородных возбуждений
сличения могут быть очень различны.
Что может входить в состав этого аппарата? Совершенно оче-
видно, что существенные признаки будущего результата динами-
чески формируются благодаря многосторонним процессам аффе-
рентного синтеза с извлечением из памяти прошлого жизненно-
го опыта и его результата.
Некоторые последние данные заставляют, однако, думать, что
в стадии афферентного синтеза складывается несколько возмож-
ных результатов, по они пе выходят па эфферентные пути и по-
этому пе реализуются. Решение же совершается после того, как
произведен выбор наиболее адекватного результата по отноше-
нию к данной доминирующей мотивации.
Этот комплекс возбуждений — в подлинном смысле слова аф-
ферентная модель будущего результата, и именно эта модель, яв-
ляясь эталоном оценки обратных афферентация, должна направ-
лять активность человека и животных вплоть до получения за-
программированного результата.
В последнее время были получены дополнительные сведения о
том, что в этот нервный комплекс, обладающий высокой степе-
нью мультиконвергентного взаимодействия, приходит еще одно
возбуждение совершенно иной, не афферентной, а уже эффе-
рентной природы. Речь идет о коллатеральных ответвлениях пи-
рамидного тракта, отводящих ко многим межуточным нейронам
II1II
Принципиальные вопросы общей теории функциональных систем 97
«копии» тех эфферентных посылок, которые выходят на пирамид-
ный тракт. Интересно, что эти эфферентные возбуждения кон-
вергируют на те же межуточные нейроны сенсомоторной обла-
сти, куда поступают и все те афферентные возбуждения, которые
могут составить параметры реального результата.
Таким образом, момент принятия решения и начала выхода
рабочих эфферентных возбуждений из мозга сопровождается
формированием обширного комплекса возбуждений, состоящего
из афферентных признаков будущего результата и из коллате-
ральной «копии» эфферентных возбуждений, вышедших на пери-
ферию по пирамидному тракту к рабочим аппаратам. В зависи-
мости от интервала между постановкой цели и ее реализацией
к этому же комплексу возбуждений через определенное время
приходят возбуждения и от реальных параметров полученного
результата. Самый процесс оценки полученного реального резуль-
тата осуществляется сличением прогнозированных параметров и
параметров реально полученного результата. Именно здесь, в этом
пупкте, осуществляется таинство оценки полученного результата,
которое, как мы видели па примере с летучей мышью, произ-
водится специальным динамически формирующимся аппаратом.
Оценка же и ее результат определяют дальнейшее поведение
организма. Если результат соответствует прогнозированному, то
организм переходит к следующему этапу поведенческого конти-
нуума.
Если же результат не соответствует прогнозу, то в аппарате
сличения возникает рассогласование, активирующее ориентиро-
вочно-исследовательскую реакцию, которая, поднимая ассоциатив-
ные возможности мозга па высокий уровень, тем самым помогает
активному подбору дополнительной информации. Все эти процес-
сы поведения акцептора действия достаточно хорошо изучены в
нашей лаборатории.
Так, например, было показано, что в момент выхода пз коры
головного мозга нпрамидное возбуждение отдает «копию» эффе-
рентного возбуждения не только в комплексы оценки результа-
та, т. е. в кору мозга, но и в ретикулярную формацию. Рети-
кулярная формация в обратном кортикопетальпом направлении
имеет возможность оказать дополнительное энергетическое воз-
действие на те циркулярные возбуждения («круги ожидания»),
которые должны удержать свою активность до момента прихода
информации о получении полезного результата. Как мы знаем,
в некоторых случаях комплекс акцептора результатов действия
должен очень долго быть в напряженном состоянии, прежде чем
будет получен реальный запрограммированный результат.
4 П. К. Анохин
98 Принципиальные вопросы общей теории функциональных систем
Заключение
Большая часть раздела посвящена тем разграничительным ли-
ниям, которые отделяют общий системный подход от разработан-
ной в пашей лаборатории теории функциональной системы, имею-
щей и иное начало, и иной путь отработки принципиальных
позиций и понятий.
В одной из своих работ Берталанфи пишет, что идея «орга-
низменного» или «целостного» подхода родилась у него очень
рано, кажется, в 1937 г., однако интеллектуальный климат того
времени не был подходящим для восприятия «организменной»
идеи, которую он предложил. Действительно, интеллектуальный
климат того времени был воспитан па подражании законам фи-
зики и механики и потому считалось научным только то, что
могло быть переведено на принципы и понятия этих наук.
И только в 1950 г. Берталанфи решился сделать официальные
шаги в защиту своей идеи, ибо в это время, т.
ные годы, резко изменился сам интеллектуальный климат. Весь-
ма быстро и настоятельно проявился интерес к абстрактным фор-
мулам, моделям, синтетическим понятиям и вообще к иптердис-
е. в нослевоен
циплияарным взаимоотношениям между учеными, и только в этот
период стало возможным говорить об общей теории систем.
Теория функциональной системы претерпела также немало
изменений с момента ее первой формулировки, однако условия
1932—1933 гг. и была
ее возникновения и самые первые шаги в ее развитии принци-
пиально отличались от путей развития общей теории систем. Как
. концепция она зародилась примерно в
сформулирована в достаточно развитой форме уже в 1935 г. в
сборнике «Проблемы центра и периферии в физиологии нервной
деятельности».
Однако в первых же
стема как интегративное образование организма включала все те
компоненты, которые характерны для нее и сейчас, в том числе и
обратную, или санкционирующую, афферентацию. В отличие от
общей теории систем, которая до сих пор пребывает в стадии
исканий конкретных и полезных путей, теория функциональной
системы в первые же годы после ее формулировки явилась для
нашей лаборатории толчком к бурному развитию именно кон-
кретной исследовательской работы, которая нашла в функцио-
нальной системе совершенно очевидный конструктивный стимул
к формулировке новых задач исследования.
Эта особенность теории функциональной системы как конкрет-
ной конструктивной теории резко отличает ее от судьбы общей
формулировках функциональная си-
теории систем, которая даже к настоящему времени практиче-
ски еще нс имеет связи с конкретной исследовательской рабо-
той.
Принципиальные вопросы общей теории функциональных систем 99
Достаточно указать, что уже в 1937 г. была опубликована
кардинальная для наших исследований работа «Функциональная
система как основа интеграции нервных процессов в эмбриогене-
зе». Фактически это было зарождением той эволюционной кон-
цепции, которая в 1945 г. была сформулирована как теория си-
стемогенеза.
Очень часто задают вопрос: к каким принципиальным обоб-
щениям привели исследования на основе теории функциональной
системы? Системогенез может служить самым демонстративным
примером конструктивной продуктивности теории функциональ-
ной системы как методологического инструмента, помогающего
поставить новые вопросы исследования.
Системогенез существенно изменил имевшие представления о
путях эмбрионального развития. Это стало возможным только по-
тому, что теория функциональной системы оказалась конструк-
тивным принципом, подсказывающим в каждой области новый
подход, новые трактовки и новые формы экспериментирования.
Мы не можем здесь перечислять все те направления исследо-
ваний в различных областях биологии и физиологии, которые
получили совершенно новое освещение при использовании теории
функциональной системы как рабочего принципа в повседневном
исследовании.
Казалось бы, общая теория систем в соответствии с ее весь-
ма радикальными и даже эволюционными стремлениями в борь-
бе с механистическим аналитизмом должна была бы вызвать не-
медленное преобразование самой логики научного исследования
и, несомненно, приобрести всеобщую симпатию реально мысля-
щих ученых. Однако этого не произошло. Почему?
В этом разделе мы пытались выявить причины этого пара-
доксального явления. Нам кажется ясным, что имеются две при-
чины того ничтожного результата, который был получен при об-
суждении «общей теории систем».
Первая состоит в том, что авторы ее пошли по неправильно-
му пути как в поисках понятия системы, так и в общей тактике
ее разработки.
Как можно было видеть, при всякой попытке сформулировать
само понятие системы, т. е. дать принципиальное кредо этому
новому интересному направлению, положительно все исследовате-
ли попадают в заколдованный круг традиционных понятий. Здесь
часто употребляются «целостность», «организменность», «взаимо-
действия», «организованная сложность», «упорядоченное множе-
ство» и другие подобные термины, которые становятся даже
центральными критериями понятия системы. Ясно, что все эти
термины по своей сути являются лишь вариациями понятия це-
лостности. Именно поэтому, не выходя за пределы понятийного
доля целостности, все эти определения не дают какого-либо су-
4*
100 Принципиальные вопросы общей теории функциональных систем
щественного скачка как в понимании системы, так и в конкрет-
ной разработке ее.
Между тем главный смысл системного подхода состоит имен-
но в том, что любая деталь наблюдения или экспериментирова-
ния должна быть неизбежно вписана в какой-то из узловых ме-
ханизмов внутренней архитектоники системы. Практически ни-
какая новая научная тема не могла бы быть сформулирована
вне конкретной внутренней архитектоники системы, где эта тема
только и может получить широкий познавательный смысл. Точно
так же и трактовка полученных результатов даст наибольший
эффект, если она будет построена на основе системных меха-
низмов.
Вторая причина состоит в том, что интеллектуальный кли-
мат для принятия системного воззрения действительно изменил-
ся. Однако большинство ученых с трудом отказываются от усто-
аналитическом аспекте. Такой переход — не простая
явшихся традиции рассматривать все накопление научных дости-
жений в
перемена названий и выражении, как сначала думали некото-
рые исследователи; он требует радикального изменения самих
принципов подхода к элементарным процессам и общей тактике
исследования.
Действительно, для исследователя,
использующего теорию
•Е
ункциональной системы как методологический инструмент экс-
периментирования, не может быть «возбуждения вообще», «аф-
ферентации вообще», «мотивации вообще» и даже «памяти вооб-
ще». Конструктивная роль этих привычных понятий выявляется
благодаря их положению в качественно своеобразном механизме
внутренней архитектоники функциональной системы?
Возьмем для примера зрительную афферептацию. Она обычно
определяется как сенсорная модальность, и с точки зрения этой
аналитической характеристики специфической чертой этой аффе-
рептации будет именно ее оптический характер. Но так ли об-
стоит дело, если оценивать зрительную афферептацию но ее ме-
сту в архитектонике функциональной системы?
Зрительная афферентацпя может быть «пусковой афферента-
цией», как это, например, наблюдается в случае условного зри-
тельного стимула. Но та же зрительная афферептация в других
условиях может составлять и обстановочную афферентацию, оп-
ределяющую совсем другой механизм системы — предпусковую
интеграцию нервных процессов.
И даже более того, зрительная афферептация может иметь
еще третью и совершенно иную функциональную значимость:
она может участвовать в оценке полученйого системой резуль-
тата. Таким образом, сформулировав внутреннюю операциональ-
ную архитектонику системы, мы тем самым изменили подход к
привычным для нас физиологическим понятиям и процессам.
Принципиальные вопросы общей теории функциональных систем 101
Еще более заметной становится роль функциональной систе
мы, когда надо, так сказать, «анатомировать» какое-то сложное
явление в работе мозга животных или человека. Так, например,
оценка изменения психической деятельности человека после опе-
рации на мозге производится обычно при помощи стандартных
тестов, а в последние годы — при помощи оценки различных
электрофизиологических показателей в работе мозга (ЭЭГ, вы-
званного потенциала, активности нейронов).
Функциональная система вносит определенный порядок и ло-
гическую последовательность в эту оценку. Можно задать вопрос:
изменился ли афферентный синтез у данного больного при вы-
работке определенного решения? Если он изменился, то больной
не может принять адекватного решения. Тогда сейчас же возни-
кает следующий вопрос: какой из компонентов и какой из нейро-
динамических процессов, определяющих успех афферентного син-
теза, является нарушенным? По мы знаем, что может оказаться
ненарушенным ни один из компонентов афферентного синтеза и
больной принимает вполне адекватное решение и выполняет со-
ответствующее движение. Однако у этих больных может быть на-
рушен очень важный завершающий механизм системы: оценка
результатов действия в акцепторе результатов действия. В этом
случае мы видим, что, правильно выполняя инструкцию, больной
пе может остановиться на правильно выполненном результате и
продолжает его «выполнять» повторно. Физиологически это зна-
чит, что больной не может оборвать действие при правильно полу-
ченном результате только потому, что аппарат сличения и, сле-
довательно, торможения дальнейшего повторения оказывается не-
полноценным (систематические исследования А. Р. Лурия).
Можно назвать многие другие области научного знания, кото-
рые получили с применением теории функциональной системы
иное и более конструктивное решение, чем при обычном подхо-
де. Так, например, в корне изменилась расшифровка компенса-
торного процесса при восстановлении нарушенных функции, бо-
лее глубоко понят генез гипертензивных состояний вообще п в
частности гипертонической болезни, изменилась расшифровка по-
нятий заболевания и выздоровления и т.
формы нарушений и нормальные функции организма значительно
более легко расшифровываются с точки зрения теории функцио-
нальной системы, чем при обычном подходе. II здесь, конечно,
на первом плане стоит расшифровка самого поведения животного
и человека. Каждый поведенческий акт, приносящий какой-то
результат, большой или малый, неизбежно формируется по прин-
ципу функциональной системы.
И, наконец, решительно изменилось понимание онтогенетиче-
ского развития функций и механизмов отбора на протяжении фи-
логенеза животных.
д. Практически все
102 Принципиальные вопросы общей теории функциональных систем
Приведенные выше примеры являются далеко не единствен-
ными, и заметный практический успех во всех этих случаях де-
лает особенно необходимой глубокую и всестороннюю разработку
теоретических основ функциональной системы.
Обратимся к вопросу о развитии системного подхода в Совет-
ском Союзе. Нам кажется, что попытки популяризировать си-
стемный подход как новую форму трактовки и, вообще, подхода
к научным фактам весьма полезны. Мы должны вооружить нашу
научно-исследовательскую молодежь этим методом и дать ей воз-
можность работать в ускоренном темпе, с более рациональным
исходом. Одпако надо отметить, что публикации последнего года,
относящиеся к этой проблеме, весьма мало сдвигают проблему
системного подхода именно в сторону конструктивных методов
исследования. Виной этому, как мы уже отмечали, излишнее тео-
ретизирование конкретных биологических, машинных или обще-
ственных систем.
Общая теория систем может быть интересна исследователям
в области конкретных наук только в одном плане, именно в обо-
гащении приемов и подходов к конкретному научному исследо-
ванию. В познавательном же отношении максимальный интерес и
максимальное внимание должны заслужить те концепции, гипо-
тезы и теории, которые показали совершенно очевидный конст-
руктивный эффект в конкретном научном исследовании
Можно только вкратце отметить важнейшие обобщения, ко-
торые нами были разработаны с помощью общей теории функ-
циональных систем и не могли быть получены иначе. Прежде
всего это — концепция системогенеза, которая сейчас помогла по-
пять основной закон эмбрионального созревания функций, обе-
спечивающих выживание новорожденного (Анохин, 1937, 1948,
1964, 1968).
Концепция системогенеза по своей сути вступает в противо-
речие с общепринятым понятием органогенеза и дает реальное
объяснение многому, что не находило до этого достаточно чет-
кой расшифровки. Многочисленные работы наших сотрудников,
начиная с 1932 г., охватывают эмбриогенез животных самых раз-
нообразных видов, включая и изучение живых плодов (рыб, ам-
фибий, птиц, млекопитающих, человека) (Я. А. Милягин,
К. В. Шулейкина, Е. Л. Голубева и др.).
Особенно интересно подчеркнуть, что приложение новой кон-
цепции — системогенеза к онтогенетическому развитию животных
разных видов дало возможность по-новому подойти к некото-
рым нерешенным проблемам теории естественного отбора и
«биогенетического закона» Мюллера — Геккеля (Ата-Мурадова,
1970).
Эта широкая перспектива приложения системогенеза к рас-
крытию некоторых сторон
ундаментальных проблем биологии
Принципиальные вопросы общей теории функциональных систем 103
сделала эту концепцию одним из отправных пунктов для построе-
ния теоретической биологии (Анохин, 1970).
Однако все сказанное выше о системогепезе касается целиком
той стадп созревания плода, когда отдельные фрагменты функ-
циональной системы (мышцы, нервные центры, кости и др.) за-
кладываются раздельно и развиваются, так сказать, навстречу
друг другу до момента их функциональной консолидации. По-
следняя обычно происходит за несколько дней или недель до
рождения, и к моменту рождения система готова функциониро-
вать и обеспечить новорожденному выживание.
Рано или поздно должен возникнуть вопрос о том, какие фак-
торы в раннем эмбриогенезе направляют возникновение и разви-
тие так гармонически сочетающихся впоследствии компонентов
системы. Этот вопрос подвел нас вплотную к закономерностям
общей генетики, ибо только она может помочь решить карди-
нальные вопросы развития: как закодированы функциональные
системы в геномных организациях и какой фактор в дальней-
шем дирижирует таким синхронным объединением всех компо-
нентов функциональной системы (гетерохрония в закладках ком-
понентов и синхронность в консолидации к моменту рождения)?
Для решения этих вопросов в настоящее время мы вступили
в рабочий контакт с
надеемся, что новый
Институтом общей генетики АН СССР и
вопрос о генетике конкретных
и
рупкпио-
нальпых систем организма будет в какой-то степени ос. вешен.
Другим значительным обобщением, вытекающим из физиоло-
гической архитектоники функциональной системы, является Фор-
мулировка интегративной деятельности нейрона, построенная на
совершенно иных основаниях, чем это принято в современной ней-
рофизиологии.
Эта новая концепция возникла как следствие топкого анализа
механизмов афферентного сиптеза, являющегося аванпостным уз-
ловым механизмом в .развертывании функциональной системы.
В самом деле, суть афферентного синтеза состоит в том, что
процессы возбуждения различного функционального значения и
пространственного рецепторного происхождения должны быть не-
минуемо обработаны совместно и часто одновременно.
Возникает критический вопрос: где может быть организована
эта «встреча» возбуждений, составляющих афферентный синтез
функциональной системы? Ответ может быть только один: такая
встреча может произойти па одном и том же нейроне, сколько
бы ни было переключений и обогащений этих возбуждений на
пути к коре головного мозга. Отсюда возникла целая серия ра-
бот, которая привела вначале к формулировке идеи о гетерохи-
мпческом характере субсппаптических образований п новой кон-
цепции об интегративной деятельности нейрона (Апохин, 19743
Сейчас можно лишь перечислить те новые направления, кото-
104 Принципиальные вопросы общей теории функциональных систем
рые возникли у нас в лаборатории и которые, как правило, по-
вели мысль по новому пути.
Совсем другой аспект возник в объяснении природы вызван-
ного потенциала, и, что особенно важно, он перестал быть только
электрическим феноменом с изменяемыми «параметрами»: ампли-
тудой, частотой, длительностью латентного периода и др. Благо-
даря системному подходу вызванный потенциал стал инструмен-
том нейрофизиологического анализа субкортикальных взаимо-
действий, а вместе с тем дал возможность сделать оценку
восходящих возбуждений этих подкорковых образований
(Ф. А. Ата-Мурадова).
Можно назвать области физиологии, которые получили благо-
даря общей теории функциональных систем повое направление и
в объяснении и в разработке. Сюда относятся компенсация на-
рушенных функций, гипертоническая болезнь, эмоциональные
стрессы и т. д.
Если к этому прибавить использование теории функциональ-
ных систем педагогами, медиками, музыкантами и многими дру-
гими специалистами, то можно достаточно уверенно утверждать,
что в общей теории функциональных систем были пайдепы уни-
версальные черты функционирования, изоморфные для огромного
количества объектов, относящихся к различным классам явлений.
Это обстоятельство полностью удовлетворяет тем требованиям,
которые мы предъявили в самом начале этой статьи к теории си-
стем вообще и общей теории функциональных систем в частности.
ЛИТЕРАТУРА
Акоф Р. Л. Системы, организации и междисциплинарные исследования.—
В кп.: Исследования по общей теории систем. М., 1969. с. 143—144.
Анохин П. К. Проблема центра и периферии в современной физиологии
нервной деятельности.— В кн.: Проблема центра и периферии в нерв-
ной деятельности. Горький, 1935, с. 9—70.
Анохин II. К. Системогенез как общая закономерность эволюционного про-
цесса.—Бюл. экспер. бпол., 1948, т. 26, вып. 2, № 8, с. 81—99.
Анохин П. К. О физиологическом субстрате сигнальных реакций.— Жури,
высш. нерв, деят., 1957, т. 7, № 1, с. 39—48.
Анохин П. К. Методологическое значение кибернетических закономерно-
стей.— В кн.: Материалистическая диалектика и методы естественных
наук. М, 1968, с. 547-587.
Анохин И. К. Функциональная система
биологического и физиологического исследования.
организация Y ¥
Анохин Л. К. Теория функциональной системы.—Успехи физиол. наук,
1970, т. 1, № 1, с. 19—54.
Н
как методологический принцип
,— В кн.: Системная
изпологических функций. М., 1968, с. 5—7.
Анохин П. К. Системный анализ интегративной деятельности нейрона.—
Успехи физиол. наук, 1974, т. 5, № 2, с. 5—92.
Анохин П. К., Стреж Е. Изучение динамики высшей нервной деятельно-
Принципиальные вопросы общей теории функциональных систем 105
сти. 6. Характеристика рецептивной функции коры больших полушарий
в момент действия безусловного раздражителя.— Физиол. журн. СССР,
1934, т. 17, № 5, с. 1225-1237.
Ата-Мурадова Ф. А, Анализ вызванных ответов коры мозга с помощью
локального теплового блока ее поверхностных слоев.— Физиол. журн.
СССР, 1966, т. 52, № 6, с. 696—708.
Ата-Мурадова Ф. А. Гетерохронное созревание отрицательных компонен-
тов вызванного потенциала зрительной коры в онтогенезе кролика.—
В кн.: Электрофизиологические исследования центральной нервной
системы позвоночных. Л., 1970, с. 85—92.
Афанасьев В. Г. Управление обществом как социологическая проблема —
В кн.: Научное управление обществом, вып. 2. М., 1968, с. 183—224.
В кн.: Научное управление обществом, вып. 2. М
Берталанфи Л. Общая теория систем — критический обзор.— В кн.: Иссле-
дования по общей теории систем. М., 1969, с. 23—24.
Вейсс II. Динамика клетки.— В кн.: Современные проблемы биофизики,
т. 1. М., 1961, с. 21—34.
Гвишиани Д. М., Мику лине кий С, Р., Ярошевский М. Г. Социальные и
психологические аспекты изучения деятельности ученого.— Вопр. фи-
лософии, 1971, № 3, с. 83—92.
Глушков В. М. Информация по кибернетике. Киев, 1970.
Гудвин Б. Временная организация клетки. М., 1966, с. 251.
Дубинин II, II. Общая генетика. М., 1970, 487 с.
Журавлев Б. В. Особенности реакций нейронов гипоталамуса у голодных
кроликов на пищевое подкрепление,— Труды Ин-та норм, и пат. физиол.
АМН СССР, 1969, т. 12, с. 194-196.
Зачиняева II. А, Выработка динамического стереотипа на двигательных
условных реакциях.— В кн.: Проблемы высшей нервной деятельности.
М., 1949, с. 521-539.
Калман Р., Фалб П., Арбиб М. Очерки по математической теории систем.
М., 1971, 400 с.
Кузнецов Б. Г. Этюды об Эйнштейне. М., 1970.
Кухтин В. С. Системно-структурный подход и специфика философского
знания.— Вопр. философии, 1968, № 41, с. 47—58.
Ланге О. Целое и развитие в свете кибернетики.— В кн.: Исследования по
общей теории систем. М., 1969, с. 181—251.
Лобные доли и регуляция психических процессов. Нейропсихологические
исследования. М., 1966, 740 с.
Макаров В. А. Роль миндалевидного комплекса в механизме конвергенции
возбуждений различной сенсорной модальности на нейронах коры боль-
ших полушарий.— Докл. АН СССР, 1970, т. 194, № 6, с. 1454—1457.
Месарович М. Д, Теория систем и биология: точка зрения теоретика.—
В кн.: Системные исследования. М., 1970, с. 137—163.
Милсум Дж. Анализ биологических систем управления. М., 1968.
Павлов II, П. Рефлекс цели,— Поли. собр. трудов, т. 3. М.— Л., 1949, с. 242—
Павлов II, П. Динамическая стереотипия высшего отдела головного мозга.—
Поли. собр. трудов, т. 3. М.— Л., 1949, с. 496—499.
Паск Г. Естественная теория цепей.—В кн.: Саморегулирующая система.
М., 1964, с. 318-357.
Полянцев В, А. Методика самоуправляемого искусственного дыхания.—
Пат. физиол. и экспор. терапия, 1961, т. 5, № 4, с. 77—79.
Правдивцев В. А, Эфферентио афферентная конвергенция на нейронах
зрительной коры.— В кн.: Интегративная деятельность мозга. М. 1967,
с. 67-69.
Раппопорт А. Математические аспекты абстрактного анализа систем. М.,
1969, с. 83—105.
106 Принципиальные вопросы общей теории функциональных систем
Садовский В. II., Юдин Э. Г. Задачи, методы и приложения оощей теории
систем.— В кн.: Исследования по общей теории систем. М., 1969, с. 3—
Синичкин В. В. Особенности распространения возбуждения, вызванного
антидромным раздражением пирамидного тракта на нейроны зритель-
ной области коры головного мозга.— В кн.: Интегративная деятельность
мозга. М., 1967, с. 72—74.
Системные исследования. Ежегодник. М., 1970. 208 с. (Институт истории
естествознания и техники АП СССР).
Судаков К. В. Нейрофизиологические механизмы пищевого возбуждения.
Докт. ди с. М., 1965, 488 с.
Судаков /V. В. Пейсмекерным механизм пищевой мотивации.—Рефераты
докл. II Всесоюз. съезда физиологов, т. 1. Л., 1970, с. 187—192.
Судаков К. В., Котов А. В. Взаимодействие мотивационного и подкрепляю-
щего возбуждений на нейронах коры мозга как основа пищевого услов-
ного рефлекса.— Материалы симпозиумов 22-го совещания по пробле-
мам физиол. высшей нервной деятельности. Л., 1969, с. 20—21.
Судаков К. В., Фадеев Ю. А. Особенности восходя
головного мозга в состоянии физиологического голода и при болевом
раздражении.— Физиол. журн. СССР, 1963, т. 49, № 11, с. 1310—1917.
Трапезников В. А. Человек в системе управления.— Тезисы 5-го Всесоюз.
совещания по проблемам управления. М., 1970.
Уоддингтон К. Предисловие.— В кн.: Б, Гудвин, Временная организация
клетки. М., 1966.
Урсул А. Д. Природа информации. М., 1968.
Ухтомский А. А. Собр. соч., т. 2. Л., 1951.
Хаютин С. II. Активность одиночных нейронов зрительной области коры
в условиях выраженной пищевой
ункциональная и нейрохимическая
с. 175—179.
Ханике Ф. Новые идеи в области управления. М., 1969.
Холл А. Д., Фейджин Р. Е. Определение понятия системы.— В кн.: Иссле-
дования по общей теории систем. М., 1969, с. 252—282.
Шумилина А. И. Функциональное значение лобных областей коры голов-
ного мозга в условнорефлекторной деятельности собаки,
блемы высшей нервной деятельности. М., 1949, с. 561—627.
Шумилина А. И. Экспериментальный анализ методом вызванных потенци-
алов корково-подкорковой реверберации возбуждений.— В кн.: Интег-
ративная деятельность нервной системы в норме и патологии. М., 1968,
с. 276-287.
Эйнштейн А. Физика и реальность. М., 1965, 359 с.
Функциональная система как основа интеграци нервных процессов в эм-
бриогенезе.— Сборник докл. 6-го Всесоюз. съезда физиологов, биохими-
ков и фармакологов. Тбилиси, 1937, с. 148—156.
Эшби У. Р. Конструкция мозга. М., 1964, 411 с.
Эшби У. Р. Общая теория систем как новая научная дисциплина.— В кн.:
Исследования по общей теории систем. М., 1969, с. 125—142.
Юнг Р. Ярче тысячи солнц. М., 1960.
Bross J. D. Design for decision. New York, 1953.
Bullock T. H. Parameters of integrative action of the nervous system at the
neuronal level.— In: The submicroscopic organization and function of ner-
vous cells, 1958, p. 323.
Macting D. C. La experimentacion instrumental a la luz de conception de
Anojin.— Bol. Inst, neurofisiol. psychoL, 1966, v. 2, p. 14.
Suga N. Recovery cycles and responses to frequency modulated tone pulses
in auditory neurones of echo-locating bats.— J. Physiol., 1964, v. 175,
p. 50-80.
ЗЕ
it
геи активности коры
31
мотивации.— В кн.: Структурная,
организация эмоций. Л
1971
В кн.: Про-
ФИЛОСОФСКИЙ смысл
ПРОБЛЕМЫ ЕСТЕСТВЕННОГО
И ИСКУССТВЕННОГО ИНТЕЛЛЕКТА*
Вступление
Трудно назвать более интересную научную проблему, чем про-
блема познаппя мозга, его фундаментальных механизмов и его
молекулярной природы. Прямым следствием развития этой сферы
знания должно быть разумное управление мозгом в будущем,
а также использование законов его деятельности для конструи-
рования различных механизмов, составляющих основу техниче-
ского прогресса в пашу эпоху.
Когда один из корреспондентов спросил «отца кибернетики»
Норберта Випера, допускает ли он возможность того, что высо-
коорганизованные «интеллектуальные машины» смогут в буду-
щем поработить человека, Винер пе без иронии ответил: «Если
это и произойдет, то только по вине человека...»
И вопрос и ответ подчеркивают крайнюю злободневность про-
блемы естествсппого и искусственного интеллекта. Дело в том,
что некоторые кибернетики полагают, будто можно все смодели-
ровать и даже создать машины, которые по своим интеллекту-
альным качествам превзойдут человека. Такие надежды широко
распространены среди физиков, математиков, электроников. Од-
нако здесь допускается серьезная логическая ошибка, суть кото-
рой будет разъяснена в данной статье.
Несмотря на важность проблемы интеллекта, переход от изу-
чения фундаментальных проблем мозговой деятельности на уров-
не интеллекта к использованию результатов исследований в тех-
нической кибернетике пока еще наталкивается на серьезные пре-
пятствия: отсутствует достаточно полная модель искусственного
интеллекта, соответствующая современным представлениям о дея-
тельности мозга в естественных условиях.
Проблема естественного п искусственного интеллекта ставит
перед нами также и целый ряд вопросов философского характе-
ра. Действительно, тезис материалистической философии — «ма-
терия первична, сознание вторично» — устанавливает органиче-
скую историческую связь между этими феноменами, поскольку
мы знаем, что неорганический мир существовал задолго до по-
явления жизни па нашей планете, и, следоватольпо, интеллект
должен был неизбежно отразить законы неорганического мира и
Опубликовано в журнале «Вопросы философии» (1973, № 6).
108
Философский смысл проблемы интеллекта
«вписаться» в них. По если это так, то все свойства интеллекта
должны были развиться па базе предшествующих органических
форм и, естественно, должны быть приспособлены для опериро-
вания объектами внешнего мира.
Иначе говоря, естественный интеллект (в примитивной фор-
ме — интеллект животных и в высшей форме — интеллект чело-
века) неизбежно должен действовать па основе объективно по-
знаваемых процессов и механизмов. Рассматривая этот вопрос в
философском аспекте, мы можем сказать, что изучение искусст-
венного интеллекта является одним из важнейших этанов позна-
ния материальной природы психических явлений и, следователь-
но, способствует дальнейшему развитию диалектического мате-
риализма.
Ясно, что надеяться па создание искусственного интеллекта
можно только после создания достаточно солидного «концепту-
ального моста», который даст возможность максимально исполь-
зовать наши фактические знания о принципах работы мозга.
Важнейшие черты интеллекта
и их характеристика
В последние годы в связи с попытками конструирования ис-
кусственного интеллекта исследователи столкнулись с необходи-
мостью определения самого интеллекта и выявления его харак-
терных черт. Без этого невозможен контакт между нейрофизио-
логами, психологами и специалистами по технической реализации
основных черт интеллекта в моделях и рабочих конструкциях.
Успех Маккаллока в создании искусственной нейронной сети
объясняется как раз тем, что оп наиболее четко выделил неко-
торые характерные логические черты мозговой деятельности и
использовал их для конструкции распознающего и «мыслящего»
устройства (Маккаллок, Питтс, 1956*1, McCulloch, 1968; МсСау,
1959; Минский, 1961). Благодаря этим исследованиям проблема
искусственного интеллекта стала широко разрабатываться имен-
но нейрокибернетиками, а пе нейрофизиологами. Последние про-
должали оставаться па позициях классической нейрофизиологии
с господствующей в пей «рефлекторной» манерой мышления, пе
дающей возможности понять решающие свойства, характерные
именно для интеллектуальной деятельности. Естественным след-
ствием этого была неопределенность в понимании нейрофизиоло-
гических свойств интеллекта и отсутствие научно обоснованных
формулировок. Это обстоятельство значительно затруднило кон-
такт между психологами, нейрофизиологами и кибернетиками.
Конструкторы искусственного интеллекта все чаще и чаще
подходили вплотную к изучению как раз тех свойств мозга, ко-
торые физиологи в своих исследованиях даже нс затрагивали.
Философский смысл проблемы интеллекта
109
Пожалуй, Марон был первым из кибернетиков, кто пришел к
выводу, что пе может быть и речи о понимании интеллекта во-
обще и о конструировании «интеллектуальной машины», если
эта система не будет обладать способностью к предсказанию. Со-
поставляя человеческий мозг с наиболее совершенными машина-
ми кибернетического характера, он особенно отчетливо сформу-
лировал их различие: способность к предсказанию у мозга и
отсутствие этой способности у машин.
Условный рефлекс, по И. П. Павлову, несомненно, основыва-
ется на предсказании, поскольку условная реакция имеет «пре-
дупредительный» характер. Как показал наш анализ, в составе
условного рефлекса имеется аппарат, который создается в про-
цессе формирования рефлекса для оценки предстоящей ситуации,
т. е. акцептор результата реакции (Павлов, 1947).
Однако нейрофизиологи совсем не затрагивают в своих рабо-
тах этой проблемы, поскольку господствующий принцип в пони-
мании нервной деятельности — принцип рефлекторной дуги пол-
ностью исключает саму возможность предсказывать будущее.
Суть дела в следующем: нервное возбуждение, вызванное
раздражением какого-либо рецептора, согласно рефлекторной тео-
рии распространяется по «рефлекторной дуге» на основе линейно-
поступательного принципа, т. е. от пункта к пункту. Между тем
по своей сути предсказание предполагает «забегание вперед», опе-
режение хода возбуждений: в самом начале появляются процес-
сы и физиологические аппараты, которые должны проявить себя
лишь в заключительной стадии рефлекторного действия (Анохип,
1935, 1949).
На этот вид нервной деятельности указывал О. Аттли. Опи-
сывая конструкцию управляющих механизмов, он отмечал: «Вы-
числитель может непрерывно подсчитывать для каждого управ-
ляющего движения, которое опробовалось раньше, вероятность
того, что оно приведет к цели» (Аттли, 1964, с. 430). Говоря о
цели, к которой ведут все движения, Аттли тем самым подчер-
кивает ее направляющее влияние па те действия, которые при-
ближают будущие события.
Наиболее полно роль целп и предсказания рассмотрена в кни-
ге Л. Фогеля, А. Оуэнса и М. Уолша (1969).
Определяя понятие «искусственный интеллект», авторы стре-
мятся найти те характерные признаки, которые могли бы быть
общими и для естественного и для искусственного интеллекта.
Они совершенно правильно, па паш взгляд, сосредоточивают вни-
мание не па топкости, четкости и быстроте выполнения отдель-
ных операций, а па логике механизмов, составляющих интел-
лект. Среди этих механизмов на первый план они ставят механиз-
мы «принятия решения» и «предсказания», т. е. формирования
цели. Авторы считают, что будет «...более содержательным оп-
110
Философский смысл проблемы интеллекта
ределять интеллект в терминах поведения некоего стремящегося
к цели существа и измерять степень его интеллекта по адекват-
ности принимаемых им решений» (Фогель и др., 1969, с. 19).
Таким образом, мы видим, что определение интеллекта охваты-
вает наиболее сложные формы поведенческой деятельности: цель,
«принятие решения», предсказание. По сути дела это верно. Од-
нако недостаток такого определения состоит в том, что сущест-
венные и характерные для интеллекта факторы просто перечис-
ляются, а пе даются в той логической связи и последовательно-
сти, которая соединила бы их прочной питью системного
детерминизма.
И действительно, в приведенных выше определениях, как,
впрочем, и во многих других, «цель» выступает как нечто напе-
ред данное. А дальше начинает рассматриваться цепь поведен-
ческих актов, направленных на достижение этой цели. Но как
возникла сама цель? Какие факторы и какие материальные про-
цессы предшествовали ее появлению и создали из пее матери-
альный аппарат, направляющий специфическое стремление орга-
низма? Упомянутые выше авторы, а также многие другие (Меса-
рович, 1970; Уотермап, 1970; Sadovsky, 1971) эту фазу «предцелп»
совершенно пе рассматривают.
То же самое можно сказать и о принятии решения. Какие
факторы толкают организм па принятие именно этого, а пе дру-
гого решения? Ясно, что в процессе принятия решения происхо-
дит непрерывный подбор наиболее адекватного для данной ситуа-
ции решения. Но как это происходит? На основе каких конкрет-
ных нейрофизиологических механизмов выбирается одиа-едипст-
венная поведенческая степень свободы из миллионов возможных
степеней?
Обычно все эти вопросы исследуются отдельно, впе их логи-
ческой связи в масштабе целого поведенческого акта, а потому
иногда и правильно выделенные факторы естественного интел-
лекта, как, например, «предсказание» (Фогель и др.), цель и
принятие решения (Аттли), остаются изолированными фрагмен-
тами интеллекта, пе связанными между собой логикой функцио-
нирования.
Оценивая нынешний этап в исследовании основных специфи-
ческих характеристик естественного и искусственного интеллек-
та, мы можем сказать, что главным недостатком этих исследова-
ний является отсутствие универсальной модели, которая логиче-
ски связала бы все этапы формирования интеллектуальных актов.
Естественно, что эта модель должна достаточно полно отражать
и нейрофизиологические механизмы каждого фрагмента интел-
лектуальных процессов.
Анализируя значение и содержание всех попыток моделиро-
вания интеллектуальных процессов па базе перцептронов, Ф. Ро-
Философский смысл проблемы интеллекта
111
зенблатт очень ярко отразил стремления тех, кто занят исследо-
ванием процессов «принятия решения». «В ближайшем
будущем,— писал он,— потребуется, по-видимому, разработать це
лую программу психологических экспериментов с животными или
людьми для пополнения наших сведений о характеристиках мо-
делей. Когда это произойдет, модели фактически станут исполь-
зоваться как спреде называющие^ устройства, способные
вырабатывать определенные данные (вначале, возможно, доволь-
но грубые), которые у людей до сих пор не наблюдались. Ко-
нечным использованием модели мозга с точки зрения психоло-
гической ценности как раз и являются эксперименты такого
рода, в которых модель правильно предсказывает явления,
не открытые в биологических системах» (Rosenblatt, 1962).
и
Как видим, Розенблатт возлагает весьма большие надежды па
модели мозга типа «предсказывающих» устройств. Однако, чтобы
эти модели правильно предсказывали и правильно ориентировали
в будущих явлениях, они должны заимствовать у мозга те его
свойства и механизмы в форме акцептора результатов действия,
которые дадут им возможность формировать цель поведения, пред-
сказывать сам результат поведения и ежесекундно контролиро-
вать и сличать полученный результат с поставленной заранее
целью. Именно этими-то свойствами не обладает ни одна из су-
ществующих моделей мозга, т. е. искусственного интеллекта.
Ясно, что такая удовлетворяющая исследователя модель может
быть построена только при непременном использовании данных
нейрофизиологии для постоянной коррекции работы этой модели.
Несмотря на то, что проблемам принятия решения, формули-
ровки цели и предсказания в последнее десятилетие уделяется
очень много внимания, все попытки создания искусственного
интеллекта находятся лишь в самой начальной стадии.
Как известно, при изучении биологических систем в силу
устоявшихся традиций принято считать, что любой поведенческий
акт заканчивается действием. При этом полезный результат
действия фактически никогда не включается в процесс как само-
стоятельная физиологическая категория. А между тем именно в
этом пункте лежит причина трагического взаимонепониманпя
между представителями весьма близких областей пауки — нейро-
физиологии и психологии. Для последней, как известно, цель
и принятие решения стали необходимыми факторами в изучении
интеллектуальных процессов.
Кибернетика внесла в психологию целый ряд смелых идей,
которые заставили ее принять такие синтетические понятия, как
цель, польза, предсказание и т. д.
Можно было бы назвать большое число исследований послед-
них лет, в которых детальнейшим образом разрабатывается проб-
лема «принятия решения» в самых разнообразных ситуациях.
112
Философский смысл проблемы интеллекта
Наиболее полные обзоры результатов изучения этой темы были
сделаны на специальных симпозиумах *.
Одним из значительных событий в исследовании характерных
свойств интеллекта явилась организация во Франции Институ-
та «высшего синтеза», занимающегося изучением «принятия ре-
шения» и пути построения «искусственного интеллекта» (Insti-
tut des Hautes Syntheses Nice, France). На одной из последних
сессий института (1971 г.) специально обсуждался вопрос о со-
отношении естественного и искусственного интеллекта.
XX Международный психологический конгресс (Токио, 1972)
провел особый симпозиум по проблематике «принятия решений»
(Dynamic* Aspect of «Decision Making») 1 2. В докладах У. Эдварса,
Г. Экеля, М. Тода, М. Месика были еще раз поставлены вопросы
об основных признаках принятия и выполнения решений. Экель
акцентировал внимание также па физиологических коррелятах
принятия решения и соответствующих ему положительных и от-
рицательных эмоциональных состояниях.
Все сказанное выше можно резюмировать следующим образом.
а) Большое внимание к проблемам искусственного и естест-
венного интеллекта привело специалистов различных областей
науки к актуальным задачам по изучению характерных черт
естественного интеллекта и применению результатов этой работы
к построению искусственного интеллекта. Сама возможность удач-
ного решения этого вопроса сулит широкие перспективы прогрес-
сивного развития многих областей экономики и промышленности.
Использование результатов таких исследований в электрони-
ке, медицине, педагогике и в других областях может привести к ре-
волюционным сдвигам в этих пауках. Поэтому весьма важно
правильное понимание сути такого прогресса.
б) Несмотря на значительность проблемы, ее нынешнее со-
стояние нельзя считать удовлетворительным. Нет четкого опреде-
ления самого понятия интеллекта, его состава и решающих меха-
низмов его отдельных операций. Наиболее синтетические и ха-
рактерные для интеллекта узловые механизмы, такие, как «при-
нятие решения», «цель», «предсказание», не только пе изучены
в их глубоком нейрофизиологическом содержании, но даже не
вскрыты их операциональные взаимодействия в момент осуществ-
ления интеллектуальных актов.
Пожалуй, одним из самых существенных пробелов в изучении
искусственного и естественного интеллекта является то, что не
выявлено логическое единство в самой архитектуре интеллекту-
альных актов, не установлена детерминистическая связь между
указанными выше узловыми механизмами интеллекта. Каждый
1 См., например: Decision Processes. Ed. by R. M. Thrall. N. Y., 1954.
2 См.: XX Congress International de Psychologie. Guide Resume. Tokyo, 1972.
Философский смысл проблемы интеллекта
113
из них берется для исследования как нечто отдельное, независи-
мое от других свойств и механизмов интеллекта,
В последующих разделах статьи мы попытаемся применить
для исследования этой проблемы системный подход в виде теории
функциональных систем, разрабатываемой в нашей лаборатории
на протяжении последних сорока лет.
Функциональная система
как логическая модель искусственного интеллекта
Как мы уже видели, одной из существенных тенденций со-
временной нейрофизиологии является изоляция отдельных меха-
низмов мозга для удобства экспериментирования над ними в це-
лях изучения их свойств. Этот аналитический прием, общий для
многих биологических наук, уже дал значительные результаты.
Однако прием этот полезен и хорош только па определенной ста-
дии научно-исследовательского процесса: при сборе первичных
материалов и в период подготовки к широким обобщениям.
Все функции организма и особенно функции его нервной си-
стемы по своей природе являются логически целостными, и по-
тому понимание их биологического смысла зависит от того «выс-
шего синтеза», в котором выявится реальная роль каждого ме-
ханизма в образовании целого. Теория функциональной системы
как раз и имеет своей целью выявить органическое единство ме-
ханизмов, которые обычно исследуются в отдельности.
Много лет назад, изучая процесс компенсации нарушенных
функций, мы увидели, что все сложные факторы деятельности,
такие, как память, эмоция и цель, выступают в органическом
единстве и только это единство способно восстановить нарушен-
ную функцию (Анохин, 1935). Такое функциональное единство
в силу era системного характера было названо нами функцио-
нальной системой. Она представляет собой законченную единицу
деятельности любого живого организма и состоит из целого ряда
узловых механизмов, обеспечивающих логическое и физиологиче-
ское формирование поведенческого акта.
Так как характеристика теории функциональной системы неод-
нократно приводилась нами в различных публикациях, здесь бу-
дет дана лишь краткая характеристика ее узловых механизмов
с точки зрения их важности для построения «искусственного ин-
теллекта».
Функциональная система устраняет дефект имеющихся схем
интеллекта. Как мы уже отмечали, механизм «принятия решения»
большинством авторов рассматривается как нечто первичное и
исходное для всех других процессов интеллектуального акта.
Такой подход не может удовлетворить объективно мыслящего
исследователя, поскольку принятию решения должен предшество-
114
Философский смысл проблемы интеллекта
вать весьма сложный процесс обработки многообразной информа-
ции.
Эта стадия интеллектуального акта была названа нами «а
и
ферентным синтезом» в связи с тем, что в процессе этого синте-
за происходит одновременная обработка самой разнообразной
информации, поступающей в центральную нервную систему из
внешнего и внутреннего мира. На этой стадии «предрешения»
синтезируется целый ряд возбуждений. Как п весь поведенче-
ский акт в целом, стадия «предрешения» формируется на основе
доминирующей в данный момент эмоции или мотивации. Послед-
няя, говоря психологическим языком, представлена желанием или
потребностью. Такое доминирующее возбуждение, как показыва-
ют эксперименты на простых формах потребности (голод, жажда,
половая потребность и т. д.), обладает способностью извлекать из
многочисленных синаптических образований мозга все то, что было
связано в прошлом с удовлетворением или разрешением именно
этой, доминирующей в данный момент, потребности (исследова-
ния наших сотрудников Судакова, Котова, Журавлева и др.).
В процессе распространения возбуждения по нейронам мозга
неизбежно привносятся и другие возбуждения — от совокупности
факторов внешней обстановки.
Таким образом, и это было показано в эксперименте, на каж-
дом нейроне коры головного мозга одновременно обрабатываются
возбуждения трех различных источников: внутреннее возбужде-
ние, связанное с формированием той или иной доминирующей
мотивации, внешние возбуждения, представленные содействием
данной обстановки, и возбуждения памяти, извлеченные как мо-
тивацией, так и данной обстановочной афферентацисй. Только
одновременная обработка этих возбуждений и сопоставление всех
комбинаций возбуждений с прошлым опытом дают возможность
организму принимать то или иное решение для получения полез-
ного результата.
Экспериментальные исследования свидетельствуют о том, что
все упомянутые возбуждения, а иногда дополнительно и специ-
альный пусковой фактор (например, условный сигнал) должны
одновременно встретиться на одном и том же нейроне, или, пра-
вильнее говоря, на каждом из миллионов нейронов.
В стадии «предрешения», т. е. афферентного синтеза, во всех
случаях формирования поведенческого акта решается главней-
ший вопрос: какой полезный результат должен быть получен в
данной ситуации и при данной комбинации составных возбужде-
ний этой стадии?
Мы видим, что только строго научное исследование стадии
«предрешения» может привести к совершенно четкому детермини-
стическому объяснению и самого процесса принятия решения.
Действительно, микроэлектродное исследование отдельных ней-
Философский смысл проблемы интеллекта
115
роиов коры головного мозга показало, что этот процесс обработки
всей исходной информации совершается при помощи многих ди-
намических механизмов, биологический смысл которых состоит в
том, чтобы выработать наиболее адекватное решение для данной
ситуации и обеспечить наиболее точное его исполнение.
Так, например, активизирующие аппараты подкорковой обла-
сти (гиноталамус, ретикулярная формация) обеспечивают образо-
вание ассоциаций и извлечение информации из памяти. Эти же
активирующие возбуждения значительно повышают различные
способности нервных элементов коры мозга, и в частности способ-
ность к конвергенции па них разнородных возбуждений. К этому
надо добавить еще усиление реверберации возбуждений между
корой и подкорковыми областями, благодаря чему осуществляется
поиск наиболее продуктивного синтеза для предстоящего принятия
решения (опыты Шумилиной).
Таким образом, нам необходимо представить себе все то, что
могут дать нам нейрофизиологические эксперименты для понима-
ния механизмов принятия решения как одного из главнейших
факторов формирования интеллекта.
Пе йрофизиологпческие предпосылки
принятия решения
Для понимания этого ответственного синтетического процесса
в интеллектуальной деятельности мы должны представить себе
отдельный нейрон и миллионы нейронов как образования, обла-
дающие бесчисленным количеством степеней свободы, которые
обусловлены способностью нейрона производить самые разнооб-
разные конфигурации первных разрядов.
Простой математический расчет показывает, что количество
степеней свободы в масштабе целого мозга с трудом может быть
записано цифрой длиной в 9,5 млн. км!.. Именно это количество
степеней свободы головного мозга и представляет собой ту бес-
конечную клавиатуру, на которой разыгрываются сотни мил-
лионов различных мелодий — поведенческих и интеллектуальных
актов.
Итак, мозг и организм в каждый данный момент обладают
необъятным количеством степеней свободы, одновременный запуск
которых привел бы к чудовищному хаосу в поведении организма.
Организованное поведение человека и животпых предполагает
неизбежное ограничение этого огромного разнообразия. Следова-
тельно, принятие решения по своей сути представляет собой
выбор одной степени свободы, наиболее адекватно удовлетворяю-
щей требованиям данной ситуации. Суть проблемы заключается
в том, каким образом мозг осуществляет выбор из миллиардов
116
Философский смысл проблемы интеллекта
возможных одной-единственной степени свободы, дающей полез-
ный эффект именно в даппой ситуации.
Здесь необходимо коснуться вопроса, который обычно усколь-
зает от исследователя, но который неизбежно возникает, если
придерживаться системной точки зрения, т. е. рассматривать весь
процесс построения поведенческого акта в аспекте
н
>ункциональ-
ной системы.
При внимательном изучении общей схемы последовательного
действия узловых механизмов функциональной системы можно
увидеть, что принятие решения ориентировано на тот результат,
который соответствует доминирующей в данный момент мотива-
ции. Наблюдения последних лет свидетельствуют, однако, о том,
что в стадии афферентного синтеза из памяти извлекаются не
только общие афферентные черты той или иной внешней ситуа-
ции, но и признаки тех результатов, которые когда-то получались
при подобных мотивационных и эмоциональных состояниях.
Иначе говоря, наш мозг обладает поразительной способностью
к системной генерализации возбуждения, способностью охваты-
вать не только частные признаки каких-либо событий, но и сте-
пень успешности и полезности тех результатов, которые были
получены в аналогичных ситуациях в прошлом. Результаты прош-
лого могут последовательно извлекаться из памяти и сопостав-
ляться с потребностью данной ситуации до тех пор, пока домини-
рующая ныне мотивация не станет вполне соответствовать одному
из результатов прошлого. Пожалуй, это одна из самых замеча-
тельных способностей нашего мозга, которую можно было бы
назвать перебором мнимых результатов прошлого и сопоставлени-
ем их с потребностью данного момента.
Возникнув в эмоциональных структурах мозга (гипоталамус,
лимбическая система и ретикулярная формация), мотивационное
возбуждение иррадиирует даже в те структуры мозга, которые
хранят в памяти результаты различных удовлетворений именно
данной мотивации в прошлом.
Например, состояние аппетита зависит от того, что латеральное
ядро гипоталамуса непрерывно раздражается «голодной» кровью.
Это возбуждение, поднимаясь в кору мозга в восходящем
направлении, мобилизует здесь элементы прошлого опыта, отно-
сящегося именно к данной мотивации. Мы начинаем перебирать
возможность ее удовлетворения в соответствии с данной ситуа-
цией. Попросту говоря, ищем
где бы мы могли закусить. При
этом часто говорим, что в такой-то ресторан мы не пойдем, «по-
скольку там плохо кормят».
Что значит такое решение с нейрофизиологической точки
зрения? Оно означает, что, перебирая многие возможности
удовлетворения пищевой мотивации, мы не только извлекаем
из памяти сведения о посещении этого ресторана в прошлом,
Философский смысл проблемы интеллекта
117
но также и результат посещения
т. е. вспоминаем саму еду и
вкусовые ощущения от пищи, полученные когда-то именно в
этом ресторане.
Под влиянием доминирующей мотивации в процесс воспоми-
нания включается практически вся функциональная система со
всеми ее механизмами, в том числе и механизм оценки получен-
ного результата.
Поразительная вещь! Интеллект оперирует гармоническим
сочетанием главнейших факторов нейрофизиологической основы,
необходимых для принятия решения,—всей настоящей ситуацией
(голод, обстановка) и всем многообразием опыта прошлого, также
связанного с удовлетворением пищевой мотивации. Казалось бы,
эта система взаимодействия так далека от реальной мозговой
структуры. Однако мы видим, что каждый элемент нашей интел-
лектуальной деятельности имеет вполне определенную нейрофи-
зиологическую основу. Составляющие ее механизмы функциональ-
ной системы детально изучаются в нашей лаборатории (Анохин,
Судаков, 1971).
Возвращаясь к принятию решения, которое согласно нашей
схеме, является результатом предшествующего афферентного син-
теза, мы должны на основании всего предыдущего признать, что
на стадии афферентного синтеза осуществляется универсальный
перебор извлеченных из памяти всех прежних результатов дейст-
вия и всех прежних оценок этих результатов в соответствии с
наличной доминирующей мотивацией. Именно для этой ответст-
венной стадии, очевидно, и нужен процесс реверберации, мобили-
зующий все сокровища кладовых нашей памяти. Следовательно,
«принятие решения» является той процедурой, результат кото-
рой был признан после перебора «мнимых» результатов наибо-
лее адекватным для данной обстановки.
С нейрофизиологической точки зрения этот процесс выбора
единственной степени свободы состоит, очевидно, в непрерывном
сканировании различных результатов, а эталоном для этого ска-
нирования служит наличная в данный момент доминирующая
мотивация. Экспериментами нашей лаборатории показано, что в
ряде специфических случаев кортико-гипоталамическая ревербера-
ция может быть весьма отчетливой (Каграманов, 1965).
Акцептор результатов действия
В этом разделе мы подходим к анализу такого нейрофизиоло-
гического аппарата, в котором скрестились главнейшие пути
исторических поисков отгадки тайн человеческой психики («цель»,
«предсказание», «ошибка», «память», «ожидание» и многое дру-
гое). Как оказалось, все эти факторы имеют единый нейрофизио-
118
Философский смысл проблемы интеллекта
логический стержень, совершенно четко формирующийся в момент
(или несколько позднее) принятия решения. Прежде всего я имею
в виду нейрофизиологический аппарат предсказания, названный
нами «акцептором результатов действия». Что это за аппарат?
Какова его природа и каковы функции? Благодаря перебору всех
признаков прошлых результатов и сличению их с данной доми-
нирующей мотивацией этот аппарат сосредоточивает в себе все
афферентные признаки того конечного результата, по поводу ко-
торого было принято решение.
Разберем пример. Если принято решение взять со стола ста-
кан, то в появляющемся акцепторе результатов будут сконден-
сированы все относящиеся к действию признаки стакана: его
внешний вид, вес, тактильные особенности, воспринимаемые кож-
ными рецепторами, температура и т. д. Смысл этого аппарата,
опережающего и предсказывающего свойства будущего результа-
та, состоит в том, что в конце действия, т. е. после взятия ста-
кана, должна быть получена вся информация о параметрах этого
действия. Именно в этот момент в центральной нервной системе
и происходит сличение результата, который прогнозировался в
акцепторе результатов действия (взять стакан), с параметрами
реально полученного результата (Анохин, 1949).
В момент сличения двух комплексов возбуждений наша нерв-
ная система осуществляет контроль результатов произведенного
действия. Если сличение показало, что прогнозированные пара-
метры (предсказание) в акцепторе будущего результата полностью
совпадают с параметрами реально полученного результата, то
данное действие заканчивается, а его результаты получают «санк-
цию» и используются для формирования следующего этапа пове-
дения. Если же выявляется несовпадение параметров реально по-
лученного результата действия с запрогнозированнымп, то это
рассогласование стимулирует построение и подбор новой програм-
мы действия, более точно обеспечивающей получение запрогнози-
рованных результатов.
Поскольку все наше поведение представляет собой подлинный
континуум результатов, больших и маленьких (Анохин, 1971),
то практически такого рода сличения происходят в нервной си-
стеме непрерывно. Так, например, даже результаты таких незна-
чительных действий, как открывание двери на лестницу, спуск
по лестнице, посадка в автобус и т. д., оцениваются и формируют
цепи получения последующих результатов. Однако и они могут
быть дискретизированы на еще более мелкие результаты: напри-
мер, постановка ноги на первую ступеньку автобуса, на вторую
и т. д. Поразительно, что от каждого такого «маленького» резуль-
тата наша нервная система должна непременно получить инфор-
мацию, которая обрабатывается в соответствующем акцепторе
результатов действия. Малейшее несоответствие результата прог-
Философский смысл проблемы интеллекта
119
позу (мы оступились) — и мозг немедленно подбирает новое дви-
жение.
Из этой краткой характеристики функций аппарата, прогнози-
рующего результаты, становится понятным и его название
«акцептор результатов действия». Латинское слово acceptare со-
держит в себе два смысла — «принимаю» и «одобряю», которые
представлены в функциях акцептора результатов действия (Ано-
хин, 1949; Анохин, 1955).
В нашей лаборатории были проведены многочисленные экспе-
рименты па клеточном и нейрохимическом уровнях для исследо-
вания того, как создается этот аппарат предсказания в каждом
отдельном случае и каковы его функции в масштабе целой функ-
циональной системы.
Теперь мы можем оценить значение этого аппарата для ин-
теллектуальной функции человека и животных и определить его
роль в изучении искусственного интеллекта. Прежде всего он
является аппаратом предсказания, поскольку в нем прогнозируют-
ся свойства будущего, еще пе полученного результата. Так как
во всех наших действиях получение того пли иного результата
связано с заранее поставленной целью, то совершенно очевидно,
что аппарат акцептора результатов действия практически явля-
ется и аппаратом цели. Из этого положения вытекает, что цель
в нашем понимании и в наших экспериментах не является чем-
то изначальным, а подготавливается сложной работой нервной
системы в стадии афферентного синтеза. Именно это обстоятель-
ство позволяет выразить цель как психологическое понятие па
языке нейрофизиологических механизмов и объективных при-
чинных связей между процессами, происходящими в головном
мозге.
Необходимо подчеркнуть, что прп рассмотрении вопросов о
предсказании и цели перед нами особенно четко вырисовывается
философская сторона проблемы интеллекта и ее решения па ос-
нове концепции функциональной системы.
Действительно, совсем недавно одно лишь произнесение таких
слов, как «предсказание», «цель», «целесообразное поведение»,
грозило физиологу обвинением в идеализме, в отрыве от материа-
листических принципов. Такая ситуация в физиологии исторически
вполне объяснима, поскольку еще ие был подготовлен соответ-
ствующий «научный климат» для материалистического разрешения
проблемы цели и предсказания. Такой климат был создан в фи-
зиологии мозга в основном рефлекторной теорией.
И потому, естественно, мы мирились с поразительным пара-
доксом: каждый мыслящий человек прекрасно осознавал, что он
ставит цель сделать «что-то» значительно раньше, чем реализует
это «что-то». Но вместе с тем физиология мозга ие имела средств
для объяснения тех механизмов, с помощью которых мозг ставит
120
Философский смысл проблемы интеллекта
перед человеком «цель» и с помощью которых он может пред-
сказывать реализацию этой «цели».
С выявлением объективных нейрофизиологических законо-
мерностей, обеспечивающих высшие функции интеллекта, отно-
шение к этой проблеме радикально изменилось. В настоящее
время, как можно было видеть, она успешно разрабатывается на
основе принципов и категорий диалектического материализма,
что приближает пас к реальному моделированию интеллекта.
Это становится возможным благодаря тому, что концепция
функциональной системы позволяет охватить все те основные
механизмы, которые помогают попять естественный интеллект.
Как показывают электрофизиологические исследования форми-
рования подобного аппарата у человека, мы можем искусственно
вводить и выводить из акцептора результатов действия любые
новые компоненты (так называемое обогащение акцептора ре-
зультатов действия), что значительно расширяет пашу власть,
нашу способность воздействовать на интеллектуальную деятель-
ность, в частности на процессы обучения.
Эволюция основных свойств интеллекта
Возникает, однако, один вопрос: являются ли все описанные
выше принципиальные механизмы интеллекта специфическими
только для высших уровней развития животных или даже только
для человеческого мозга? Это вопрос краеугольный, поскольку он
очень тесно связан с другими вопросами, например, с такими:
есть ли интеллект у животных, когда и у каких животных он
появляется в процессе эволюции?
Интересную попытку сделали Фогель и другие, попробовав
построить эволюционную модель «искусственного интеллекта»,
которая должна совершенствовать свои основные свойства путем
разрастания примитивных механизмов в последующих «поколе-
ниях» (Фогель и др., 1969). Следует отметить, что в эволюцион-
ном моделировании накопление опыта осу:
^ествляется несколько
и
иным способом, чем в интеллекте животных в процессе эволюции.
Отвечая на поставленные выше вопросы, мы должны прежде
всего высказать основное положение, сложившееся у нас в ре-
зультате многих лет работы над описанными выше свойствами
интеллекта: ни одно из тех свойств мозговой деятельности, ко-
торые мы выше рассматривали как характерные признаки интел-
лекта, не появлялось внезапно, на каком-то «рубпконе», до ко-
торого этого свойства не было и после которого оно появилось.
Все эти свойства возникли уже па заре зарождения жизни,
и все они являлись уже тогда частью динамической физиологи-
ческой архитектуры. Более того, они явились conditio sine qua
non самого развития живых существ.
Философский смысл проблемы интеллекту
121
Эю может показаться странным, поскольку мы всегда описы-
ваем интеллект как выдающееся свойство живого, присущее по
крайней мере самым высшим и совершенным представителям жи-
вотного царства.
Однако паше недоумение немедленно рассеется, если мы
представим себе, как происходило формирование интеллекта.
Возьмем для примера предсказание будущих событий или резуль-
татов какой-либо деятельности, выполняемой некоторой четко
очерченной функциональной системой.
При каких условиях внешнего и внутреннего мира'животного
возможно предсказание? Главным условием предсказания являет-
ся то, что цепь событий, по поводу которой осуществляется пред-
сказание, неоднократно повторялась в прошлом в определенных
местах пространства в определенные моменты времени. Наш ин-
теллект может предсказать, что после дня последует вечер, а пос-
ле вечера ночь, только потому, что этот нерушимый ход внешних
событий повторялся миллионы лет, и тогда, когда были живые
существа, включая человека, и даже тогда, когда не было намека
на их появление.
Здесь потребуется несколько отойти от конкретных нейрофи-
зиологических процессов и механизмов и обратиться к обобщени-
ям более широкого характера.
Пространственно-временной континуум движения материи, как
справедливо отмечал Планк, является абсолютным законом мира.
Но этот закон действовал задолго до появления жизни на Земле.
Иначе говоря, жизпь, т. е. живые существа, должна была volens
nolens «вписаться» в рамки, задаваемые этим фундаментальным
законом, и только при этом условии им было обеспечено выжива-
ние. Этот факт и привел к тому, что именно «вписанность», или
отражение живыми существами пространственно-временного кон-
тинуума, стала совершенно неизбежной предпосылкой предска-
зания.
Рассмотрев этот вопрос применительно к целому ряду биоло-
гических явлений, мы в свое время сформулировали принцип
опережающего отражения мозгом действительного ряда событий
во внешнем мире. Именно это свойство является изначальным
свойством протоплазматических процессов даже у низших живот-
ных, для которых смена, например, сезонных явлений (лето —
осень — зима — весна — лето) была неизменным условием жизни
на протяжении миллионов лет.
Можно привести десятки примеров поразительной точности и
целесообразности приспособления к этим абсолютным законам
неорганического мира.
В сущности, открытие И. П. Павловым условного рефлекса
было открытием опережающих отражений внешнего мира в вы-
сокоспециализированном субстрате — в нервной системе. В самом
122
Философский смысл проблемы интеллекта
деле, когда в ответ на звонок у собаки выделяется слюна, то это
происходит совсехм не потому, что слюна должна «переваривать
звонок», а потому, что в будущем появится пища, которую надо
переваривать. Следовательно, в силу повторения последователь-
ности определенных воздействий внешнего мира мы создали ли-
нию облегченных реакций, в которых достаточно первого толчка,
чтобы химическая реакция протоплазмы, подобно реакции бикфор-
дова шнура, распространилась по нервной системе в будущее,
опережая последовательное развитие внешних событий.
Из разнообразных примеров и приведенных рассуждений ста-
новится ясным, что «предсказание» как феномен изолированного
интеллектуального акта имеет глубокие исторические корни. На
высшем этапе эволюции органом этого опережающего процесса
стала нервная система. Именно она в сотни раз обострила и уско-
рила опережающие процессы, и именно благодаря этому мы мо-
жем совершить почти фантастическое путешествие в будущее в
ответ на какой-либо толчок или сигнал из внешнего мира.
Эволюция, начавшаяся с примитивного протоплазматического
«предсказания», усовершенствовала этот процесс в материальных
явлениях мозга до такой степени, что мозг стал органом, который
в каждый данный момент своей деятельности сочетает в себе
прошлое, настоящее и будущее.
Все это не является фантазией нейрофизиолога. Микроэлект-
родный метод дает нам возможность установить, что некоторые
нейроны, испытывая настоящее раздражение, включают накоп-
ленный в прошлом опыт и одновременно с этим формируют про-
цессы, содержащие в себе качества того результата действия,
который будет получен только в будущем. В нашей лаборатории
эти нервные клетки мозга мы назвали «нейронами трех времен».
Резюмируя обсуждение проблемы «предсказания», мы должны
подчеркнуть, что «предсказание», выявляемое па высших этапах
интеллектуальной деятельности, есть продукт наиболее совершен-
ного развития того прототыпного процесса, который проявляет-
ся уже в опережающем протоплазматическом отражении действи-
тельности.
Однако возвратимся к естественному развитию событий при
формировании поведенческих актов па высшем уровне. Процесс
афферентного синтеза, как мы уже говорили, закапчивается
принятием решения, являющегося итогом перебора возможных
результатов, органически связанных в прошлом с данной моти-
вацией. Таким путем осуществляется одно из самых замечатель-
ных явлений в активности мозга: формирование на уровне нерв-
ной системы модели всех признаков и свойств будущего полезного
результата, в связи с которым и ради которого развивались про-
цессы афферентного синтеза. Это и есть цель.
Философский смысл проблемы интеллекта
123
Заключение
Приведенные в этой статье соображения убеждают нас в том,
что для познания основных свойств естественного и искусствен-
ного интеллекта необходимо идейное перевооружение всей совре-
менной нейрофизиологии, выработка новых методических и мето-
дологических подходов. И действительно, поиски наиболее харак-
терных черт искусственного интеллекта показали, что нейрофизио-
логия, строящаяся только на традиционной, по преимуществу
аналитической, основе, не может надеяться на успех в решении
данной проблемы.
В свое время в связи с возникновением кибернетики крупней-
ший французский физиолог А. Фессар писал, что наука вступила
в эпоху обратных связей (Fessard, 1953—1954). Это было совер-
шенно верно. По к этому надо добавить вступление нейрофизио-
логии в эпоху более широкого и глубокого синтеза нейрофизио-
логических и поведенческих исследовательских задач, нового син-
теза данных, добытых в отдельных биологических науках. Преж-
де всего здесь следует сказать об усовершенствовании системного
подхода, который содержал в себе все возможности для изучения
самых высших форм деятельности мозга: «принятия решения»,
«цели»,, «предсказания», «интеллекта» и др.
Предлагаемый в данной статье подход, опирающийся па тео-
рию функциональной системы, как нам кажется, приближает нас
к решению все еще загадочных проблем интеллекта. Во всяком
случае, нам стали ясны некоторые общие аспекты проблемы,
открылся доступ к конкретному научному п экспериментальному
исследованию тех аспектов, которые еще совсем недавно были
прерогативой психологии, а часто оказывались основой для идеа-
листических интерпретаций.
И здесь при изучении исторических предшественников интел-
лекта были сделаны серьезные обобщения, позволившие понять
непрерывную эволюцию интеллекта. Главнейшие свойства интел-
лекта — афферентный синтез, постановка цели, принятие реше-
ния и оценка полученного результата, предсказание и обратная
афферентация, санкционирующая получение полезного результа-
та. Эти синтетические процессы имели большую предысторию:
они развивались из тех примитивных форм, которые сложились
уже на заре жизни на нашей планете.
Эта общность архитектуры поведенческого акта и есть тот
исторический фактор, благодаря которому жизнь и мозг разви-
лись до высшего этапа — человеческого интеллекта.
Исследования последних лет еще более убеждают нас в ус-
пешном применении теории функциональной системы к решению
вопросов интеллекта. Ее конкретные синтетические узловые ме-
ханизмы, выросшие на системной основе, дают возможность пере-
124
Философский смысл проблемы интеллекта
кинуть «концептуальный мост» между нейрофизиологией, психоло-
гией и теми проблемами, которые возникают на путях познания
тайн интеллекта.
Итак, диалектический материализм еще раз получает доказа-
тельство того, что интеллект, сознательная деятельность и актив-
ное преобразование самого приспособления к внешним факторам
являются истинным продуктом исторического развития от материи
к сознанию на основе фундаментальных законов неорганическо-
го и живого мира на нашей планете.
ЛИТЕРАТУРА
Анохин //. К. Проблема центра и периферии в современной физиологии
нервной деятельности. В кн.: Проблема центра и периферии в физио-
логии нервной деятельности. Горький, 1935, с. 9.
Анохин 11. К. Узловые вопросы в изучении высшей нервной деятельности.
В кн.: Проблемы высшей нервной деятельности. М., 1949, с. 9.
Анохин П. К. Особенности афферентного аппарата условного рефлекса и
их значение для психологии.— Вопр. психол., 1955, № 6, с. 16.
Анохин П. К. Принципиальные вопросы общей теории функциональных си-
стем. М., 1971.
Анохин П. К., Судаков К. В. Нейрофизиологические механизмы голода и
насыщения.— Успехи физиол. наук, 1971 т. 1, № 11, с. 3.
Аттли О. М. Механизмы процессов мышления (заключительная речь). В кн.:
Самоорганизующиеся системы. М., 1964, с. 430.
Каграманов К. М. О холинергических и адренергических механизмах дея-
тельности головного мозга. Автореф. канд. дис., М., 1965.
Маккаллок У., Питтс У. Логическое исчисление идей, относящихся к нерв-
ной активности. В кн.: Автоматы. М., 1956.
Месарович М. Теория систем и биология: точка зрения теоретика. В кн.:
Системные исследования. М., 1970, с. 137.
Минский М. На пути к искусственному интеллекту., 1961, 49.
Павлов И. П. Лекции о работе больших полушарий головного мозга.— Поли,
собр. трудов, т. 4. М.— Л., 1947.
Уотерман Т. X. Теория систем и биология: точка зрения биолога. В кн.:
Системные исследования. М., 1970, с. 164.
Фогель Л., Оуэнс А., Уолш М. Искусственный интеллект и эволюционное мо-
делирование. М., 1969.
Fessard A. Points de contact entre neurophysiologie et cybernetiqne.— In:
Structure et evolution des techniques, Paris, 1953—1954, N 35—36.
McCay D. M. Operational aspect of intellect.— In: Mechanization of thought
processes, 1959, vol. 1.
McCulloch W. S. Logic and closed loops for a computer junket to mars.—
In: Neural Networks. Symposium. Ed. by E. R. Caianiello. N. Y., 1968.
Rosenblatt F. Principles of neurodynamics. Washington, 1962.
Sadovsky V. N. The history and perspectives of the systems approach deve-
lopment and general systems theory.— In: International congress of the
history of sciences (18—24 August, 1971).
СИСТЕМОГЕНЕЗ
КАК ОБЩАЯ ЗАКОНОМЕРНОСТЬ
ЭВОЛЮЦИОННОГО ПРОЦЕССА*
Если провести оценку направлений современной физиологии,
то можно увидеть, что у ряда представителей физиологической
мысли выявляется отчетливая тенденция понять механизм фи-
зиологической организации наиболее комплексных процессов
организма, иначе говоря, имеется стремление осмыслить физио-
логические закономерности интеграции. Становится все более и
более ясным, что каждый новый частный факт, получаемый в
изолированном эксперименте, делается все менее и менее пригод-
ным для использования его в объяснении функций целого орга-
низма. Из этого и вытекает необходимость преодолеть традицию
простого суммирования физиологических результатов, ежегодно
получаемых в огромном количестве на путях аналитических ис-
следований. На последний путь исследователи становились не
однажды, но оп мало помог выработке именно руководящих прин-
ципов для изучения регуляторных приспособлений целого орга-
низма. Можно привести пример Шеррингтона (Sherrington), Эд-
риана (Adrian), Гесса (Hess) и др.
Известно, что еще Шеррингтон в конкретной форме поставил
вопрос о необходимости изучения интегративных процессов нерв-
ной системы. Шеррингтон не сделал главного: оп пе вскрыл тех
принципиально новых свойств, которые приобретает целостная
организация, как только она становится таковой.
Книга Шеррингтона также говорит более о средствах интегра-
ции, чем о ее специфических принципах, свойственных только
интеграции и пе свойственных тем частям, на основе которых она
строится.
Этот дефект в общей концепции Шеррингтона неизбежно дол-
жен был дать себя знать. Отсутствие руководящих принципов в
понимании физиологической интеграции привело Шеррингтона в
последнее время к высказыванию чисто идеалистических представ-
лений о нервной интеграции.
Только знание общих принципов физиологической организа-
ции приспособительной деятельности организма приводит к проч-
♦ Опубликовано в «Бюллетене экспериментальной биологии и медицины*
(1948, № 8, т. 26).
126 Системогенез как общая закономерность эволюционного процесса
ному материалистическому мировоззрению о целостном организ-
ме. Примером такого общего принципа является закон образова-
ния условного рефлекса или вообще закон временных связей,
развитый нашим великим соотечественником И. II. Павловым.
Этот закон лег прочным камнем в фундамент материалистического
понимания сложных форм поведения животных и человека до
психической деятельности включительно.
Чем мы можем объяснить такой успех и такое универсальное
значение принципов условных связей? Главным образом тем, что
II. II. Павлов, не отрываясь от конкретных физиологических
закономерностей нервной системы, указал принцип, который
имеет свои собственные характерные черты, отличающие его от
других тем, что он охватывает более высокую форму деятельно-
сти организма. Этим же свойством отличаются и другие направ-
ления русской физиологии (Введенский, Ухтомский, Орбели,
Быков и др.).
Наряду с этим русская паука в лице школы А. И. Северцова
располагает огромным опытом в изучении
ских закономерностей
рило-эмориогенетиче-
который значительно обогащает наши
зё
представления о материальных основах целостности организма.
Однако наряду с хорошо оформленной концепцией о корреля-
тивных связях организма, характеризуемых с морфологической
точки зрения, до последнего времени пе было сделано сколько-ни-
будь существенной попытки представить физиологический аспект
этой целостности. Структура и функция оказывались еще вне ор-
ганической связи и не составили того диалектического единства,
которое помогло бы нам на основе общих законов развития по-
нять прогрессивную эволюцию организмов.
Характеризуя соотношение морфологических и физиологиче-
ских понятий при изучении эволюционных закономерностей,
А. Н. Северцов писал: «Для данного вида безразлично, выполня-
ется ли данная функция у потомков тем же органом, что и у
предков, или же она выполняется каким-либо другим органом,
лишь бы биологически, т. е. с точки зрения выживания в борьбе
за существование, опа выполнялась лучше, чем у предков. Этим
собственно и отличается наша точка зрения от точки зрения спе-
циалистов-физиологов: они исследуют функции органов как тако-
вые, мы же рассматриваем их как средство, с помощью которого
виды сохраняются и выживают в борьбе за существование. Изме-
няющиеся органы являются лишь орудиями, благодаря которым
у потомков данпой формы образуются биологически важные ак-
тивные или пассивные приспособления» (Северцов, 1939).
Легко видеть, что в этих формулировках понятию функции
придан биологический масштаб, т. е. указано как раз именно на
то, что в природе каждодневно и происходит. Обстановка жизни
ставит перед организмом одну-едпнствеыпую задачу — выжить
Системогенез как общая закономерность эволюционного процесса 127
любыми средствами. Отсюда, действительно, безразлично, какая
комбинация органов выполняет данную
п
ункцию. Важно лишь
одно, чтобы эта комбинация органов и взаимодействие частей
этих органов приводили к благоприятным результатам в смысле
выживания вида. Нетрудно видеть, что при таком подходе поня-
тие «функция» приобретает очепь общий биологический смысл и,
несомненно, в какой-то мере надстраивается над понятием конк-
ретного органа пли отдельных тканевых образований.
Пятнадцать лет назад я сделал попытку представить себе,
каким образом частные физиологические процессы при выполне-
нии каких-либо приспособительных функций организма образуют
единое целое, обладающее своеобразием связей, отношений и вза-
имных влияний именно в тот момент, когда все эти части орга-
низма мобилизованы па выполнение функции. Этот комплекс
объединений мной был назван «функциональной системой». Идея
функциональной системы как единицы физиологической интегра-
ции возникла в нашей лаборатории на основе изучения цент-
рально-периферических закономерностей в деятельности организ-
ма. Производя различного рода гетерогенные анастомозы нервных
стволов, мы делали физиологический анализ тех механизмов, с по-
мощью которых извращенные вначале соотношения центра и пе-
риферии создавали повое единство, возвращая организму его при-
способительные способности. Изучая эти механизмы постепенных
«перестроек» нервных центров, мы обнаружили, что ни «центр»,
ни «периферия» не определяют этого успеха перестройки, если их
взять только в чисто анатомическом понимании. Выяснилось, что
любая перестройка нервных центров после наложения гетероген-
ных анастомозов первов находится в зависимости от того, в какой
большой системе отношений находится сам перестраивае-
мый центр. Во всех наших опытах эта система отношений оказа-
лась четко очерченной единицей, имеющей определенный приспо-
собительный эффект. Так мы должны были неизбежно подняться
до понятия физиологической интеграции, а в качестве единицы
ее приняли функциональную систему (Анохин, 1935).
В процессе изучения физиологических особенностей организа-
ции функциональной системы мы пришли к убеждению о необхо-
димости охарактеризовать ранний онтогенез функциональной си-
стемы.
Всякая функциональная система, входящая в фонд приспосо-
бительных способпостей организма, должна была обязательно
возникнуть как определенное следствие непрерывно развиваю-
щихся структур эмбриона. Как складывается функциональная
система в том периоде, который получил не совсем правильное
название дофункциональпого периода и по традиции стал приви-
легией механики развития? Постановка этого вопроса неизбежно
должна была привести нас на путь объединения общей физиоло-
128 Системогенез как общая закономерность эволюционного процесса
гии нервной системы с той огромной областью исследования, ко-
торая была намечена в основном работами русских и советских
эволюционистов-морфологов. Ниже мы даем краткое содержание
результатов наших исследований, которые были проведены на
этом пути. Вместе с тем я считаю нужным дать и формулировку
тех общих закономерностей, которые нами были вскрыты в этих
исследованиях и которые направляют развитие любой
нальной системы у любого представителя животного
н
< л ож Л
»ункцио-
мира.
Функциональная система
как единица физиологической интеграции
Прежде чем перейти непосредственно к описанию эмбриогене-
тических закономерностей, я считаю нужным дать общую форму-
лировку тех физиологических свойств, которые характеризуют
функциональную систему как самостоятельную организацию, об-
ладающую специфическими, только ей присущими свойствами.
Иначе говоря, мы должны рассмотреть функциональную систему
как правомерное понятие физиологии, как понятие, обладающее
своими специфическими чертами в качестве метода исследования
и своими физиологическими свойствами в качестве объекта ис-
следования,
1. Как единица интеграции функциональная система представ-
ляет строго очерченную группу процессов и структур, объединен-
ных для выполнения какой-либо определенной качественно свое-
образной функции организма или акта его неведения. К таким
функциональным системам, заканчивающимся определенным эф-
фектом приспособления, можно отнести, например, локомоторные
акты любого назначения: прыжок, хождение, плавание, перевора-
чивание со спины, кашель, дыхание и т. д.
Все эти акты, состоящие из отдельных компонентов, пред-
ставляют собой хорошо интегрированную систему процессов, пе-
реходящую в такое распределение возбуждений па перифериче-
ских органах, которое приводит к осуществлению полезных для
организма функций.
Этим самым определяется как своеобразное целостное качест-
во функциональной системы, так и объем участвующих в пей
структур и процессов.
2. Исследования моих сотрудников показали, что имеются все
физиологические основания для классификации функциональных
систем организма по их более или мепее тесному отношению к
структурам организма. В самом деле, ряд функциональных си-
стем, о которых главным образом и будет идти речь ниже, скла-
дывается на основе морфогенетических закономерностей уже в
ранних стадиях развития эмбриона. Эти системы имеют вполне
определенные структурные основы и вступают в действие автома-
Системогенез как общая закономерность эволюционного процесса 129
тически, как только раздражается рецептивное поле системы.
Такие функциональные системы
составляют фонд врожден-
ных реакций организма и чрезвычайно разнообразны у различных
животных. Они точно отражают экологические особенности данно-
го вида животного и потому можно вполне обоснованно утверж-
дать, что своеобразных эмбриогенезов функциональных систем
имеется столько же, сколько различных видов животных.
Наряду с этими функциональными системами, структурная
основа которых морфогепетически подготовлена в условиях эмб-
рионального развития, в жизни высших животных в каждый от-
дельный момент их активной деятельности могут складываться
самые разнообразные, так сказать, эпизодические функциональ-
ные системы, которые приспосабливают организм к какой-то экст-
ренно сложившейся ситуации. Так, например, животное, достаю-
щее корм в условиях какого-то специально затрудненного поло-
жения, мобилизует в этот момент все ресурсы своей нервной и
моторной активности, которые имеют конечным эффектом доста-
JIIH
ванне пищи.
Благодаря этой способности организма в каждый данный мо-
мент мобилизуются все те аппараты, которые приводят к дости-
жению определенного приспособительного эффекта. Своеобразие и
четкая очерчешюсть функциональных систем в таких случаях ха-
рактеризуется еще и тем
ный момент функциональная система является единственной. Она
что каждая развертывающаяся в дан-
целиком занимает интегративные аппараты организма и исключа-
ет возможность сосуществования с другими функциональными
системами. Так, например, в момент прыжка лягушки, который
развивается как вполне очерченная функциональная система,
исключается возможность вступления в деятельность других
функциональных систем организма.
Если нормальной лягушке положить па область бедра смочен-
ный серной кислотой кусочек бумажки, то опа отвечает на это
несколькими формами движения, которые могут сосуществовать
только в том случае, если они являются развитием или вариан-
том одной и той же функциональной системы. Если же движения
относятся к различным функциональным системам, то они неиз-
бежно взаимно исключают друг друга. Вот почему такая лягуш-
ка в момент осуществления самого прыжка не реагирует на при-
ложение кислотной бумажки. Однако стоит закончиться прыжку
(в момент посадки), как все моторные аппараты лягушки пере-
страиваются и устанавливаются па сбрасывание этого кусочка бу-
мажки. Исключительность, или взаимоисключение функциональ-
ных систем,— одно из самых характерных свойств, подчеркиваю-
щих значение функциональной системы как целостной
физиологической организации, экстренно складывающейся в при-
способительном поведении животного.
5 П. К. Анохин
130 Системогенез как общая закономерность эволюционного процесса
;ью в себе», если оно не выявляется в своеобразных
н
ин
3. Состав функциональной системы не ограничивается лишь
нервными структурами, которые, естественно, выполняют наибо-
лее тонкую интегрирующую роль в ее организации, придавая ей
биологическое качество.
Надо помнить, что качество центральной интеграции остается
лишь «ве'
комбинациях рабочих усилий периферического аппарата. Эта реа-
лизация происходит прежде всего по закономерностям централь-
но-периферических соотношений в нервной деятельности, благо-
даря которым рабочая периферия может адекватно отразить
особенности данной центральной интеграции. Только совместно
все эти структуры составляют очерченную функциональную си-
стему со своими собственными закономерностями.
Так, например, плавание лягушки как биологическая форма
приспособления к внешней среде является результатом детальных
и избирательных связей в нервных центрах, которые и определяют
ее облик как функциональной системы. Однако эта центральная
интеграция оказывается бесполезной, если уменьшить до извест-
ного предела общую афферептацию системы, исходящую от кож-
ной поверхности и мьг
В этом эксперименте никак не затрагивается центральная ин-
теграция функциональной системы сама по себе. Здесь лишь под-
черкивается ее зависимость от наличия известной суммы
периферических афферентаций. Детальнейшие исследования этой
зависимости (М. Ф. Чепелюгипа) показали, что каждая функ-
циональная система имеет свои специфические количествен-
ные требования к этой афферептацпи. Так, например, максималь-
ную потребность в афферентирующих импульсах имеет та
функциональная система, которая складывается в момент перево-
рачивания лягушки на спину и заключается в мобилизации мо-
торных усилий для достижения нормального положения. Кроме
лабиринтной афферентации, которая в данном случае создает тол-
чок к появлению содействующих друг другу усилий, афферентные
импульсы от кожи и мышц вносят свой существенный вклад в об-
щую сумму координированных попыток перевертывания.
Нарастающая при перерезке задних корешков деафферента-
ция лягушки говорит о том, что именно эта функциональная си-
стема разрушается прежде всего и лягушка оказывается беспо-
мощной в достижении нормального положения. Однако, если ля-
гушку посадить в нормальное положение, она совершенно
свободно может совершать прыжок, ходить при реципрокном уча-
стии всех четырех конечностей и нормально плавать в бассейне.
Наиболее резистентной в отношении деафферентации, т. е. наиме-
нее требовательной к объему афферентных импульсаций, являет-
ся функциональная система плавания. В тех случаях, когда про-
изведена максимальная деаффереитацпя, охватывающая большую
Системогенез как общая закономерность эволюционного процесса 131
sc
часть поверхности кожи и мышечных групп и никакие моторные
акты невозможны, лягушка еще достаточно легко может совер-
шать плавательные движения и пользуется ими как в водном бас-
сейне, так и на столе.
Совершенно очевидно, что* из общего комплекса функциональ-
ных систем этого животного, обеспечивающих ему локомоторные
приспособления, только одна из них может проявить свою цент-
ральную интеграцию в условиях минимальных афферентных им-
пульсаций с периферии. Этот факт различных специфических
требований разных функциональных систем к периферическим
импульсациям еще больше подчеркивает своеобразие каждой
функциональной системы как некоторой самостоятельной едини-
цы интеграции.
Исследования этой закономерности показывают, что потреб-
ность к афферентным импульсациям увеличивается соответствен-
но тому, как функциональная система, удаляясь от стандартного
типа приспособительной реакции, все больше приближается к си-
стемам, возникающим экстренно в условиях данной ситуации.
Такой структурный состав функциональной системы, ее тес-
нейшая зависимость от объема и качества афферентных импуль-
саций делают ее в собственном смысле целостным проявлением
организма. Этим самым выдвинутый нами принцип функциональ-
ной системы коренным образом отличается от всех тех попыток
представить интегративную деятельность, которые ставят акцент
на той или
попытка
нервной
вопросы
что все
к нашей
инои степени сложности центральных процессов
(«интеграция» Шеррингтона, «констелляция центров» Ухтомско-
го, «рабочее объединение» всякого рода и т. д.). Всякая
представить себе совокупную работу тканей, органов и
системы неизбежно в той или иной степени затрагивает
интеграции процессов организма. Естественно поэтому,
эти попытки в той или иной степени имеют отношение и
концепции. Однако все они не удовлетворяют основным требова-
ниям физиологического исследования целостных процессов орга-
низма: они пе указывают подлинной единицы интеграции функ-
ций организма, не отмечают специфических свойств, присущих
только этой физиологической единице и не присущих ее частям,
и, наконец, самое главное, эти представления о «рабочих объеди-
нениях» оторваны от биологического смысла таких объедине-
ний. Они не исходят в оценке интеграции от конечного приспо-
собительного эффекта в интересах целого организма, в то время
как именно только последний определяет всевозможные
регулятивных изменений в пределах системы.
4. Состав функциональной системы не определяется топогра-
фической близостью структур или их принадлежностью к какой-
классифпкации (вегетативной
примеры
либо существующей анатомической
или соматической).
5*
132 Системогенез как общая закономерность эволюционного процесса
31
31
н
В нее могут быть избирательно вовлечены как близко, так и
отдаленно расположенные структуры организма. Она может во-
влекать дробные разделы любых цельных в анатомическом отно-
шении систем и даже частные детали отдельных целых органов.
Единственным фактором, который определяет эту избиратель-
ность различных комбинаций структур и процессов, является био-
физиологическая архитектура функции, а в отдельных случаях
даже и ее механика. Так, например, биомеханические свойства
прыжка заключаются именно в том, что вся тяжесть тела благо-
даря отталкивающим действиям задних конечностей перемещает-
ся в переднем направлении. Эта физическая конструкция прыж-
ка с неизбежностью приводит к распределению процессов в систе-
ме тела, обусловливает их удельпое значение в различные фазы
прыжка и т. д., что в эволюции в конце концов ведет к своеоб-
разной анатомической организации таких животных (например,
тушканчик). Наоборот, животные, вынужденные по особенностям
их экологии передвигаться в средах с большим физическим со-
противлением, вырабатывают в эволюции такие функциональные
системы, в которых действующие силы и компоненты располага-
ются в обратном направлении. В качестве примера обладателей
этой второй категории функциональных систем можно привести
крота, у которого наряду с мощным развитием когтей, лап и
мышц передних конечностей совсем незначительно развиты зад-
ние конечности.
Следовательно, говоря о структурном составе
систем, мы не можем отрешиться от экологической роли той или
иной функциональной системы, а вместе с тем от того, что в каж-
дом отдельном случае именно экология диктует избирательное объ-
единение тех или иных структур организма в жизненно важной
функциональной системе. Если единственным фактором, опреде-
ляющим эту избирательность, является архитектура функции, то
единственным критерием полноценности этих объединений явля-
ется конечный приспособительный эффект для организма в целом,
который наступает при развертывании процессов данной
опальной системы. Каждая функциональная система обязательно
приводит к приспособительному эффекту. Этим самым понятие
функциональной системы непосредственно смыкается с темп био-
логическими понятиями и факторами развития, которые опреде-
ляют прогрессивную эволюцию организмов. Вместе с тем концеп-
ция функциональной системы отличается от традиционных фи-
зиологических концепций, которые, характеризуя изолированный
физиологический процесс, мало интересуются тем, какое место он
займет в системе отношений, определяющих приспособительный
эффект организма в целом. Почти совсем не уделяется внимания
вопросу, что делается с элементарным процессом, когда он входит
в совокупность отношений, дающих организму приспособление,
функциональных
t э <_Л
31
31
»ункци-
л я
Системогенез как общая закономерность зеолюционного процесса 133
т. е. меняет ли он свои свойства при вхождении в эту систему
отношений?
5. С точки зрения физиологической
ункциональная система
обладает одним замечательным свойством. На основе известных
физиологии закономерностей (иррадиация, «рекрутирование»,
торможение и т. д.) функциональная система приобретает внут-
реннюю регулятивность, т. е. она может выбирать, перебирать и
комбинировать количество участвующих в ней структур. Она мо-
жет динамически изменять характер протекающих в ней процес-
сов, расширяя его в сторону других, не относящихся к данной
»♦
к этим видам афферентных
н
системе структур организма до тех пор, пока организм как целое
не получит приспособительного эффекта.
Подробный анализ всех видов афферентных имиульсаций орга-
низма (пусковая, регулирующая, ведущая, санкционирующая)
привел нас к выводу, что одним из самых выразительных фи-
зиологических свойств функциональной системы как целостной
организации служит ее отношение
воздействий. Функциональная система может уменьшить объем
афферентных воздействий («сужение афферентацип»), однако
это «сужение афферентации» касается только функциональной
системы и служит ее интересам. Только функциональная система
в своей физиологической архитектуре определяет, какая именно
необходима афферентация и насколько она может быть устране-
на из общей регуляции единства функциональной системы при
переходе на стандартное течение.
С тех пор как мы встали на путь изучения физиологической
основы функциональных систем организма, мы много раз видели
это поразительное ее свойство подчинять своим интересам участ-
вующие в ней отдельные частные процессы.
Так, например, с общефизиологической точки зрения наличие
проприоцептора какой-либо мы
[,ы делает само по себе разуме-
1111
ющимся участие этого проприоцептора в деятельности организ-
ма. Да и на самом деле это предположение имеет все основания.
Если животное производит какое-либо движение конечностью, то
проприоцептор в силу одних своих структурных особенностей
должен неизбежно продуцировать центростремительные импульсы
и вряд ли можно представить себе, что нрп растяжении мышцы
или при ее сокращении эти импульсы отсутствовали. Но эта
чисто периферическая закономерность возникновения проприо-
цептивных импульсаций еще не определяет их назначения в смы-
сле активного участия в той или иной функциональной системе.
Функциональная система, например, прыжка почти нацело ис-
ключает эту проприоцептивную афферентацию и пе включает ее
в систему аппаратов своей центральной интеграции. Здесь полу-
чается явное противоречие между общефизиологическими пред-
ставлениями о роли проприоцептора и его конкретным использо-
134 Системогенез как общая закономерность эволюционного процесса
ванием в интегративных единицах приспособительной деятельно-
сти организма.
Такое избирательное использование и перераспределение важ-
ности афферентных импульсаций в пределах данной функцио-
нальной системы определяется отношением организма к внешне-
му миру, которое диктуется экстренно сложившейся ситуацией.
Я считаю нужным подчеркнуть здесь, что наш опыт изучения
интегративной деятельности организма на основе общефизиологи-
ческих закономерностей дал определенный положительный ре-
зультат. Оп показал возможность использования методических
приемов общей физиологии в решении задач целостной физиоло-
гии.
Функциональная система как основа развития
нервной деятельности в эмбриогенезе
После ряда исследований в этой области (Голубева, Милягин,
Цинде и др.) мы пришли к окончательному заключению, что
только своеобразие дефинитивных приспособительных функций
новорожденного данного вида животных может служить критери-
ем оценки эмбриогенетического созревания его функций. Как по-
рядок, так и темпы развития отдельных структур находятся в
прямой зависимости от того, что именно новорожденный данного
вида животных должен использовать в момент рождения, чтобы
добиться, как выражается А. Н. Северцов, своей «нормы выжи-
вания».
Возьмем пример эмбриогенеза сумчатой крысы (Opossum Vir-
ginia). Весь эмбриогенез у этого животного продолжается 12,5 су-
ток. С точки зрения обычных физиологических понятий не толь-
ко периферические нервы и мышцы, но и задние конечности
этого животного в момент рождения оказываются совершенно не-
созревшими. Однако все то, что необходимо для продвижения
новорожденного к сумке матери, оказывается в полной функцио-
нальной готовности. Его продвижение к сумке матери связано с
тонкими движениями передних конечностей, цеплянием за шерсть
и волнообразными движениями туловища. С помощью этих дви-
жении он добирается до сумки матери, немедленно присасывается
там к соску и продолжает своё «эмбриогенетическое развитие» на
протяжении последующих 20 с лишним дней.
Можно привести много положений как из работ самого Север-
цова, так и его учеников, в которых отчетливо выявлена эта за-
кономерность эмбрионального развития, приводящая к своевре-
менному обеспечению организма необходимыми приспособитель-
ными реакциями. Естественный отбор ежесекундно работает над
тем, чтобы все особи, которые по тем или иным условиям раз-
вития не обеспечили себе этого будущего, немедленно устраня-
лись из жизни.
Системогенез как общая закономерность эволюционного процесса 135
Допустим, что у эмбриона белой крысы почему-то произошла
задержка в развитии тончайших синаптических связей между ра-
но закладывающимися в эмбриогенезе ядрами лицевого и трой-
ничного нерва. Пусть это будет какая-то мутация или вредное
действие внешних условий. В результате этой задержки развития
сосательный акт неизбежно окажется дезинтегрированным, а со-
сательная
ункция в первые минуты после рождения окажется
неосуществимой. И так как природа пе знает искусственного
вскармливания, то данная генерация белых крыс с замедленны-
ЗЕ
ми темпами развития некоторых нервных элементов окажется
элиминированной. Мы видим, что в природе идет непрерывная
борьба за своевременное созревание сложных функциональных
систем, и потому, естественно, что у эмбриона эти системы
имеют смысл только для будущей жизни.
Необходимо поэтому более точно установить место функцио-
нальной системы в организации эмбриональных процессов и ее
особенности по сравнению с другими принципами эмбриологиче-
ского исследования. Прежде всего надо формулировать ее отно-
шение к понятию «функция».
Когда биолог говорит о функции животного, особенно ново-
рожденного, он понимает под этим словом окончательный при-
способительный эффект в жизнедеятельности животного, позволя-
ющий последнему выжить в данной комбинации внешних усло-
вий. Для биолога функция является формой приспособления жи-
вотного к внешним условиям, дающей ему возможность успешно
бороться за су
it
чествование. С этой точки зрения можно говорить
о функции дыхания, о функции пищеварения, о функции движе-
ния п т. п. Биологу в широкой степени безразлично, каким со-
четанием структур и какой архитектурой физиологических про-
цессов обеспечивается данная функция, лишь бы только она
успешно приспосабливала животное к внешним условиям и позво-
ляла ему идти по пути прогрессивной эволюции. В этом заклю-
чается качественная сторона деятельности биолога, определяющая
уровень его операций с физиологическими понятиями.
Для физиолога, вступившего па путь изучения эволюционных
проблем, только с этого момента и начинается как постановка
вопроса, так и само исследование.
Его прежде всего интересует, с помощью каких структурных
аппаратов и с помощью каких сочетаний детальных физиологи-
ческих процессов обеспечивается успешное выполнение данной
функции.
Здесь необходимо понятие, которое охватило бы эти структу-
ры и процессы в целом, подчеркнуло бы их интегративный ха-
рактер. Таким понятием для нас является понятие функциональ-
ной системы.
Став па позицию изучения системной деятельности, физиолог
136 Системогенез как общая закономерность эволюционного процесса
неизбежно должен заинтересоваться закономерностями биологи-
ческого порядка. Он уже не может ограничиться исследованием
какого-то частпого процесса; он должен обязательно учесть биоло-
гический эффект функции, ее роль в прогрессивной эволюции
данного вида. II на этом пути для пас возникает прежде всего
вопрос: какими путями и с помощью каких механизмов к моменту
рождения животного складываются полноценные функциональ-
ные системы?
Наши исследования но эмбриогенезу убедили нас, что разви-
тие функциональных систем животного не может оцениваться
только с точки зрения непосредственных взаимодействий в преде-
лах данной стадии развития. Оно может быть понято в каждом
отдельном случае только с точки зрения окончательного функцио-
нального эффекта, к которому ведут морфогенетические процес-’
сы. Точнее говоря, в оценку и анализ морфогенетических про-
цессов надо вложить физиологический смысл, который может
быть совершенно полно определен только функциональными си-
стемами новорожденного и взрослого животного.
Все приведенные выше соображения дали мне основание
предложить новый подход к исследованию эмбрионального разви-
тия, в основу которого была положена изложенная выше концеп-
ция функциональной системы. Поскольку для изучения эволюци-
онных закономерностей этот подход является повым, а совокуп-
ность процессов, подлежащих исследованию
, иная, нежели это
было принято в классической эволюционной морфологии, я пред-
ложил ввести понятие системогенеза (Анохин, 1943). В это поня-
тие я вкладываю совокупность эмбриональных процессов, которые
через посредство морфогенетических закономерностей созревания
отдельных структурных элементов ведут к избирательным систем-
ным связям. Это избирательное созревание структур протекает не-
зависимо и от топографической близости субстратов. Конечным
этапом такого последовательно развивающегося объединения все-
гда является та или иная функциональная система, обеспечива-
ющая выживание данного новорожденного. Понятие системогене-
за принципиально отличается от понятия морфогенеза и органо-
генеза, которые являются в настоящее время в значительной
степени руководящими. Такое объединение не относится к какой-
либо одной ткапи зародыша, оно выборочно включает в себя эле-
менты всех ткапей организма, принимающих участие в осуществ-
JE
лении данной функции. Кроме того, созревание функциональной
системы происходит в некоторой степени независимо от созрева-
ния того или иного органа как целого. Последняя особенность
представляет собой также существенное отличие теории системо-
генеза от коррелятивных теорий, имеющих в виду только органы
или отдельные тканевые образования, не объединенные физиоло-
гическим единством функции.
Системогенез как общая закономерность эволюционного процесса 137
Подробный анализ литературы с этой точки зрения и ряд спе-
циальных экспериментов, поставленных моими сотрудниками,
убедили меня, что экспериментальная разработка системогенеза
является крайне необходимой и своевременной.
Достоинство концепции оценивается по плодотворности тех
исследований, которые на ней основываются. К настоящему вре-
мени понятие системогенеза помогло нам вскрыть некоторые но-
вые закономерности, которые направляют морфогенетические
процессы в «дофункциональной» стадии развития зародыша и
приводят к появлению полноценной функции новорожденного.
Вскрытые нами закономерности являются закономерностями
функциональной системы и не могут быть отнесены к созрева-
нию какого-либо отдельного специфического тканевого образова-
ния, как, например, нерва, мышц или железы.
Принцип внутрисистемной гетерохронии
Учение о гетерохрониях вообще возникло в биологии давно.
Уже Геккель, пытаясь обосновать установленный им биогенети-
ческий закон, встретился с рядом затруднений, которые мешали
признать полностью его положение о рекапитуляции признаков
предков в эмбриональном развитии современных животных.
В числе этих препятствий и была гетерохрония в развитии от-
дельных органов.
Гетерохрония в буквальном смысле слова — это сдвигание
во времени закладки какого-либо органа или изменение темпов
развития у эмбриона данной особи по сравнению с закладкой
этого органа у предков. Из самого понятия следует, что сдвига-
ния во времени возможны или в сторону более ранней закладки
органа, или, наоборот, в сторону запаздывания закладки. Эмбрио-
генез вы с:
и
:их животных дает много примеров подобных «сдвига-
ний во времени». Так, например, мозг млекопитающих, хотя и
развивается на более поздних стадиях эволюции, закладывается,
однако, очень рано. В данном случае мы имеем дело с «онтогене-
тическим ускорением» развития (акселерация). Первоначально
все исследования гетерохронии проходили исключительно под
знаком ее отношения к биогенетическому закону.
Мейнерт был первым эволюционистом, который рассматривал
вопрос о гетерохрониях с точки зрения их приспособительного
значения в процессе эволюции целого организма. Сделав попытку
объяснить причину возникновения гетерохроний, он при:
и
:ел к вы-
воду о решающей роли упражнений в индивидуальной жизни
предков данного животного.
Более полную характеристику роли гетерохроний для жизни
животных дает в своей классической монографии A. II. Северцов.
Указывая на то, что гетерохронии эмбрионального развития, по-
138 Системогенез как общая закономерность эволюционного процесса
явившиеся в филогенезе, должны быть полезными и для эмбрио-
на и для взрослого, он пишет, что «...нам приходится поставить
вопрос о функциональном значении гетерохроний», «...завершение
морфогенеза того или другого органа зависит от того, в какой
именно период жизни животного, в зависимости от общих условий
его существования, требуется функционирование данного органа»
(Северцов, 1939).
В приведенных формулировках Северцова отчетливо выраже
на роль гетерохроний, заключающаяся в закреплении наилучше-
го приспособления животного к окружающей обстановке, т. е.
в прогрессивной эволюции его организации. Особенно большое
значение в этом смысле имеет ускорение эмбрионального разви-
тия органов. Можно указать на интересные исследования
Б. С. Матвеева и В. С. Емельянова, которые, изучая развитие
позвоночника некоторых костистых рыб (Cyprinidae и др.), пока-
зали, что костные тела, позвонки и ребра у них развиваются
прямо из соединительной ткани, минуя необходимую для других
видов хрящевую стадию.
Это ускорение способствует, в свою очередь, более раннему
превращению мальков в рыбок, т. е. в общем приводит к прогрес-
сивному развитию данного вида.
последние годы вопрос о гетерохрониях нашел отражение в
ряде работ школы А. II. Северцова, в особенности в систематиче-
ских исследованиях Б. С. Емельянова.
Исследуя гетерохронные закладки и различные темпы течения
морфогенетических процессов, В. С. Емельянов пришел к выводу,
что гетерохрония морфологических образований меньше всего за-
висит от перехода органов в рудиментарное состояние, как это
думал Мейнерт, и связано совсем с другими факторами.
На основании своих исследований он считает, что при всякого
рода изменениях внешних обстоятельств, требующих более ранне-
го вступления в функцию какого-либо морфологического образо-
вания, «подгонка» морфологического субстрата сначала происхо-
дит путем ускорения темпов развития без сдвигания на ранние
сроки самой закладки этого образования. Только в том случае,
когда одного ускорения темпов развития оказывается недостаточ-
но, чтобы удовлетворить требованиям естественного отбора в
данной экологической обстановке, вступает новая закономер-
ность — сдвигание закладки морфологического образования на бо-
лее ранний срок.
Анализируя современное состояние учения о гетерохрониях,
мы можем отметить, что оно еще далеко пе использовано даже в
исследованиях морфологов-эволюционистов, не говоря уже о том,
что физиологи совершенно не касались этой закономерность эм-
брионального развития. А между тем гетерохропное развитие раз-
личных структур эмбриона представляет собой именно ту около-
Системогенез как общая закономерность эволюционного процесса 139
►
гическую закономерность, которая вызвала огромное разнообразие
точек зрения и подходов у современных представителей эмбрио-
физиологических школ. Я постараюсь это доказать несколько
ниже.
В чем заключаются недостатки современного состояния проб-
лемы гетерохроний?
Рассуждения различных авторов — Оппель (Oppel), Мар-
шалл (Marschall), Кейбель (Keibel), Мейнерт, Северцов, Емелья-
нов — касаются в основном гетерохронии в развитии органов и
«морфологических образований», которым придается самодовлею-
щее значение. Однако, если сопоставить весь ход рассуждений,
приведенный нами в предыдущих разделах, с этим традиционным
подходом, то станет совершенно ясно, что ни орган сам по себе,
ни какие-либо морфологические образования, какими бы важны-
ми они ни были, не могут определить собой ни приспособления, ни
выживания животного. Это можно доказать разбором хотя бы од
ного из примеров, который приводит А. Н. Северцов.
Анализируя явление ускоренного развития плавников у Holo-
stei и Tellostei, которое происходит у них даже без промежуточ-
ных стадий, он пишет: «Для нас важно то, что в результате этой
акселерации у Holostei и у Tellostei брюшные плавники начина-
ют функционировать раньше, чем у предков, и функционируют
лучше
берущего в качестве показателя гетерохронии какой-либо отдель-
ный орган животного или даже только отдельный его признак, та-
кое заключение вполне достаточно. Однако для физиолога с этого
момента только и начинается постановка вопроса о механизмах и
чем у них». С точки зрения мо{
олога-эволюциониста
•in
путях этого ускорения.
В самом деле, что может дать полезного ускоренное созрева-
ние плавника целому животному? Ясно, что ускоренное созрева-
ние (акселерация) должно обязательно охватить также и мышцы,
с помощью которых этот плавник может двигаться, нервы и ней-
ромускулярные синапсы, с помощью которых возбуждение пере-
дается на мышцы. И, наконец, самое главное — должны ускорен-
но созревать и нервные центры, которые должны войти в слож-
ную интеграцию всего плавательного акта, т. е. установить с ним
адекватные и во всех отношениях совершенные синаптические
связи. Только при этих условиях рано созревшие плавники при-
обретают какое-то приспособительное значение для целого живот-
ного и определяют его успех в борьбе за существование. Любой
дефект, например запаздывание в развитии синапсов, благодаря
этой системе отношений, делает бесполезным для животного вся-
кое ускорение в развитии отдельного «органа»: приспособитель-
ный эффект этого органа будет равен нулю.
Из всего сказанного можно сделать вывод, что проблема ге-
терохроний в том смысле, как она была поднята Мейнертом и
140 Системогенез как общая закономерность зволюционного процесса
разработана школой Северцова, является, в сущности, не гетеро-
хронией органов, а гетерохронией функциональных систем, в ко-
торых те или иные анатомические части лишь наиболее выделя-
ются и потому становятся заметными для исследователя.
Развивая эту мысль дальше, мы можем сказать, что эмбрио-
нальное созревание животного есть прежде всего подготовка тех
ункциональных систем
которые необходимы новорожденному
непосредственно после рождения, и только затем уже функцио-
нальных систем, необходимых на различных этапах постнатально-
го развития.
Таким образом, «гетерохрония в развитии органов» есть толь-
ко наиболее бросающийся в глаза частный признак гетерохронно-
го развития тех функциональных систем, которые приспосаблива-
ют животное к внешним условиям на всем протяжении его жиз-
ни — от рождения до старости.
Для иллюстрации случая внутрисистемной гетерохронии могу
указать па особенности развития функции четвероногого хожде-
ния у хвостатых амфибий, представителем которых является ак-
солотль, и у рептилий. В то время как первый крайне замедлен-
но приобретает в своем онтогенезе конечности, у рептилий конеч-
ности становятся органом хождения сразу же после вылупления
из яйца.
Если мы посмотрим па эти факты в аспекте закладки и раз-
вития только конечностей, то здесь не возникает никакой мор-
фофизиологической проблемы. Но процесс усовершенствования
локомоторной функции есть процесс системного характера, и оп
не ограничивается одними лишь конечностями. С точки зрения
биологической, акселерация в закладке и развитии только конеч-
ностей не имела бы никакого смысла, если бы вместе с этим пе
происходили акселерация в закладке и развитии нервных центров
спинного мозга, их синаптическое объединение и полпая переме-
на в последовательности и темпах развития центров ствола мозга.
Только при этих условиях к моменту вылупления зародыша реп-
тилий функциональная система, обеспечивающая хождение, ока-
зывается физиологически совершенно интегрированной. Но тогда
закладка отдельных нервных соотношений моторной функции на
уровне спинного мозга должна у рептилий происходить значи-
тельно раньше по сравнению с аксолотлем.
Это и есть внутрисистемная гетерохрония в ее филогенетиче-
ском значении. Она ведет к перестановке времени закладки и
темпов развития отдельных компонентов в такой функциональной
системе, которая имеет одно и то же биологическое значение и
у высших
и у низших животных. Она позволяет любой функцио-
нальной системе переставлять время закладки и темпы развития
отдельных своих компонентов.
Главные факторы, направляющие
ункциопальную
систему
Системогенез как общая закономерность эволюционного процесса 141
н
на путь гетерохронпого развития отдельных ее компонентов, за-
ключаются прежде всего в длительных изменениях внешних усло-
вий. Последние через интенсификацию определенных свойств
функциональной системы, подхватываемых естественным отбо-
ром, способствуют возникновению гетерохроний.
Если рассматривать в пределах разобранного нами случая
возникновения локомоторного акта, то можно указать на интерес-
нейшие примеры внутрисистемной гетерохронии, имеющие место
у одного и того же класса животных. Если мы возьмем, напри-
мер, грачонка и цыпленка, то мы увидим, что каждый из них пос-
ле вылупления из яйца попадает в совершенно различные усло-
вия внешней среды.
В то время как грачонок долгое время обречен оставаться в
сидячем положении, цыпленок уже в первые часы способен к са-
мостоятельному передвижению. Такое различие в постнатальном
поведении связано с биологическими особенностями существова-
ния. Грачонок получает корм, сидя на месте, а цыпленок с пер-
вого часа начинает сам клевать крошки и зерна. Если на это
явление посмотреть с точки зрения эмбрионального созревания
центров и клеточных элементов спинного мозга, то мы можем
предполагать, что закладка люмбальных сегментов спинного моз-
га и темпы развития клеточных элемептов, обеспечивающих ра-
боту конечностей, у обоих эмбрионов должны протекать по-раз-
ному.
Прямые морфофизиологические исследования обоих зароды-
шей, проведенные в моей лаборатории, показали, что это имеет
место па самом деле (Милягин, Пушкарева).
Подводя итог всему изложенному, мы можем уточнить общую
формулировку понятия внутрисистемной гетерохронии.
Внутрисистемная гетерохрония — это процесс сдвигания во
времени закладки и изменения темпов развития отдельных фраг-
ментов функциональной системы по сравнению с закладкой этих
же фрагментов и для этой же функциональной системы у пред-
ков, Внутрисистемная гетерохрония является одним из наиболее
действенных средств организма, с помощью которых он отражает
всякое длительное изменение в окружающей его обстановке; при
условии, если это изменение протекает на протяжении значитель-
ных геологических эпох. С точки зрения классификации форм
приспособления животных в эволюции,
внутрисистемная гетерохрония является прямым следствием от-
бора.
Таким образом, оценивая время закладки отдельных фраг-
ментов данной функциональной системы в сравнении с закладкой
у предков в историческом плане, мы обращаемся к механизму
возникновения данной функции в филогенезе. Проводя же срав-
нительную оценку закладки и развития отдельных фрагментов в
н
мы можем сказать, что
142 Системогенез как общая закономерность эволюционного процесса
пределах функциональной системы данного животного, мы приме-
няем принцип внутрисистемной гетерохронии для изучения меха-
низмов возникновения функций в онтогенезе особи. Первый под-
ход представляет собой лишь общий фон для наших дальнейших
исследований, в то время как второй — применение внутрисистем-
ной гетерохронии — является предметом всего последующего
изложения материала данной статьи.
В направлении изложенных выше положений мои сотрудники
провели ряд исследований, посвященных созреванию функцио-
нальных систем у различных животных. Нами были использова-
ны эмбрионы аксолотля, грача, цыпленка, морской свинки и чело-
века. В каждом отдельном случае проводились тщательные мор-
фологические исследования на самой ранней стадии закладки
фрагментов соответствующих функциональных систем. В боль-
шинстве случаев это было связано и с внешними приспособи-
тельными качествами созревающей функциональной системы (Го-
лубева, Милягин, Вайнштейн, Шулейкина, Пушкарева, Резничен-
ко и др.).
Особенно тщательно в порядке морфофизиологической корре-
ляции были изучены в Московском институте мозга мной сов-
местно с II. И. Цинде эмбрионы морской свинки и Я. А. Миля-
гипым — эмбрионы грачей и цыплят.
Во всех случаях брались те жизненно важные функциональ-
ные системы, которые обеспечивают новорожденному выживание.
Это были функциональные системы питания (грач, человек),
функциональные системы специфических форм передвижения
(прыжок, летание, хождение, плавание) и специальная форма
раннего постнатального приспособления обезьян, представленная
в рудиментарном виде у человека (удерживание тела в подве-
шенном состоянии с помощью хватания).
Во всех случаях неизменно констатировались различия в сро-
ках развития отдельных фрагментов упомянутых выше функцио-
нальных систем. Так, например, нейромускулярные синапсы, фор-
мирующиеся на нервных окончаниях нижней ветви лицевого нер-
ва, заполняют к четвертому месяцу эмбриогенеза человека все
мышцы, участвующие в сосании (orbicularis oris). Наоборот, ней-
ромускулярные синапсы для мышц лобной мускулатуры не толь-
ко не созревают к этому моменту, но едва лишь начинают фор-
мироваться.
Эта межсистемная гетерохронность идет совершенно парал-
лельно с внутрисистемной гетерохронией. Ядра лицевого нерва и
тройничного нерва в продолговатом мозгу начинают группиро-
ваться уже в нейробластической стадии, когда нет еще и речи о
соотношении нерва и мышцы на периферии, да и сами мышцы
еще находятся на стадии ранних миобластов (Голубева, Вайн-
штейн) .
И
Системогенез как общая закономерность эволюционного процесса 143
а |» >» в» !
по сравнительном
ЭЕ
При морфологической оценке темпов миелинизации лучевого
нерва и отдельных ветвей срединного нерва у зародыша челове-
ка можно обнаружить, что уже в стволе самого срединного нер-
ва, обеспечивающего функцию мышц-флексоров, происходит за-
метное ускорение развития миэлина в волокнах, иннервирующих
сгибатели пальцев, по сравнению с волокнами, иннервирующими
межкостную мышцу (Шулейкина).
В развитии спинного мозга морской свинки очень рано мож-
но констатировать, что отдельные скопления нейробластов на
люмбальных и плечевых сегментах начинают ускоренно диффе-
ренцироваться и образовывать аксоны передних корешков задол-
го до того, как эти отдельные очаги гетерохронного развития
будут объединены в функциональную систему. Интересно, что в
одном из случаев нам удалось показать значение мейнертовско'го
положения о рудиментарности органов для замедленного их раз-
вития (ретардация). В работе Пушкаревой
оценке темпов развития плечевых сегментов у зародышей птиц,
имеющих различную экологию (грач и курица), было пайдепо,
что клеточные столбы передних рогов спинного мозга у грача
имеют непрерывный ход от верхних к нижним сегментам, в то
время как на этих же сегментах у зародышей курицы происхо-
дит группировка моторных нейронов, или, правильнее сказать,
нейробластов, только на уровне выхода передних корешков. Меж-
сегментарное же пространство спинного мозга оказывается ли-
шенным этих клеток. Я не могу разбирать здесь все эти иссле-
дования в отдельности, ибо к настоящему времени нами получен
огромный материал, который подчеркивает совершенно несомнен-
ную гетерохронность в закладке и развитии отдельных фрагмен-
тов функциональной системы. Следует отметить, что из того опре-
деления состава функциональной системы, которое нами было
дано в начале этой статьи, следует, что оценку функциональной
системы в целом надо производить на всех уровнях ее органи-
зации — от центральной нервной системы до периферического ра-
бочего аппарата включительно. Это требование не всегда можно в
достаточной степени выполнить, хотя постепенно уточняющаяся
техника наших исследований дает нам возможность надеяться,
что это затруднение будет преодолено. Все полученные нами в
процессе исследования факты не оставляют сомнения в том, что
функциональная система как целостное структурное образование
созревает путем внутрисистемной гетерохронии, являющейся пря-
мым следствием длительного естественного отбора в эволюции
данного вида животного.
Вместе с тем в наших фактах отчетливо выступает и еще од-
но важное обстоятельство. Гетерохронность (ускорение) в разви-
тии отдельных фрагментов функциональной системы находится в
зависимости от степени сложности и тонкости организации этого
144 Системогенез как общая закономерность эволюционного процесса
фрагмента и его значения в выполнении функции в целом. Гак,
например, синаптические образования центральных частей систе-
мы, как правило, требуют длительного созревания. Если учесть,
что самый характер функциональной системы, ее механика и ка-
чество интеграции определяются тонкостью этих центральных со-
отношений, то станет понятно то «забегание вперед», которое
мы имеем во всех случаях созревания нервных центров для тех
пли ипых функциональных систем. Нельзя, очевидно, отрицать
в данном случае и роли того
актора, который особенно тща-
Я
тельно был изучен В. С. Емельяновым, именно фактора морфо-
генетических воздействий в пределах данного формообразователь-
ного аппарата. Однако для жизненно важных функциональных
систем, с которыми новорожденные появляются на свет, эти фак-
торы имеют скорее вторичное, чем первичное значение.
Применяя тот способ иллюстрации, который был особенно ши-
роко использован А. Н. Северцовым, мы можем процесс внутри-
системной гетерохронии изобразить в следующем виде.
Допустим, что А, Б, В, Г и Д представляют собой различ-
ные компоненты какой-нибудь функциональной системы, жизнен-
но необходимой для новорожденного данного вида животпых.
Пусть это будут, например, мышцы, синапсы, центры, нервы, ре-
цепторы. Как было вы]
и
е показано, формула АБВГД к моменту
рождения должна быть совершенно готовой. Но отдельные компо-
ненты системы имеют различную сложность. Следовательно, со-
зревание всей системы в целом может быть достигнуто только в
том случае, если соответственные закладки а, б, в, г, д произой-
дут гетерохронно в отношении друг к другу и будут иметь раз-
личные темпы морфогенеза.
CL CL CL? CLCL *
б, б, 6) 6, 6 .
в, в, в, а, в .
Z Zj Z •
д, д3 д, д, d .
рождение
Из приведенной схемы видно, что независимо от времени за-
кладок и темпов созревания все компоненты функциональной
системы должны оказаться готовыми к моменту рождения.
Внутрисистемная гетерохрония представляет собой только
один из частных случаев в общей закономерности системогенеза.
Она распространяет свое влияние на тот период, когда будущие
функции новорожденного едва только начинают приобретать свой
нервный и рабочий субстрат. Гетерохронность этого рода прояв-
ляется именно в тот момент, когда более древний тип развития
по градиенту постепенно сменяется более поздним типом разви-
тия, именно системным типом развития, в соответствии с особен-
ностями внешних условий жизни каждого вида животпых. Однако
Системогенез как общая закономерность эволюционного процесса 145
системогенез как закономерность, охватывающая все этапы раз-
вития животного, не ограничивается только стадией внутриси-
стемной гетерохронии. Каждый из следующих этапов представ-
ляет собой специфическое явление в онтогенезе и поэтому тре-
бует специальной характеристики.
Внутрисистемная гетерохрония как закономерность, имеющая
значение для самых ранних стадий развития зародыша, являет-
ся для пас особенно важной потому, что опа вводит нас во все
последующие стадии и делает понятным их переход из одной в
другую. Именно поэтому опа памп и была подробно разобрана.
Чтобы составить представление о дальнейшем развитии функцио-
нальной системы, я вкратце остановлюсь еще па нескольких
принципах системогенеза, которые имеют вполне специфическое
морфофизиологическое значение для понимания эмбриогенеза
функции. Прежде всего я остановлюсь на том моменте, когда
гетерохронию возникшие и развивающиеся отдельные фрагменты
функциональной системы вступают между собой в тонкие синап-
тические соотношения и образуют тот комплекс структур, кото-
рый мы называем функциональной системой.
Принцип консолидации функциональной системы
Момент консолидации функциональной системы составляет
критический пункт ее развития. Как показали наши исследова-
ния иа эмбрионах морской свинки, для прыжка он заключается
в том, что стволовая часть головного мозга, установив тончайшие
синаптические связи, в которых отражена архитектура будущей
функции, постепенно завладевает нижележащими сегментами
спинного мозга. Нисходящие пути постепенно устанавливают
синаптические связи выборочно для отдельных моторных нейро-
нов спинномозговых сегментов. Этим самым создаются условия,
при которых сложившиеся до того на сегменте отношения ради-
кально. изменяются. Компоненты, которые имели определенную
последовательность в развитии, могут вопреки своему различному
возрасту поменяться местами в функциональной системе. Так,
например, при развитии спинальных сегментов морской свинки
соблюдается до некоторой степени правило осевого градиента,
т. е. плечевые сегменты закладываются и дифференцируются не-
сколько ранее, чем люмбальные сегменты, хотя и те и другие
гетерохронии ускоряются в своем развитии по сравнению с дру-
гими структурными образованиями спинного мозга. Однако, как
только нисходящие пути спинного мозга, берущие свое начало от
стволовых центров (ядро Даркшевича и др.), доходят до плече-
вых и люмбальных сегментов, значение последних сразу резко
изменяется. Люмбальные сегменты приобретают особую силу и
ведущую роль в функциональной системе прыжка. Такая резкая
146 Системогенез как общая закономерность эволюционного процесса
переоценка отдельных фрагментов функциональной системы в мо-
мент консолидации ее оказывается прямым следствием архитек-
турных особенностей данной функциональной системы.
Этот пример, обнаруживающий решающую роль стволовых
интеграций, конечно, пе единственный. Уже из исследований на-
шей лаборатории мы можем привести их несколько.
Так, например, начальная стадия (8 недель) развития челове-
ческого эмбриона характеризуется тем, что все нейроны, выра-
стающие через плечевое сплетение к верхней конечности, доходят
до мышц в точном соответствии с законом проксимо-дистального
развития, т. е. раньше иннервируется мышца плеча, затем мыш-
ца предплечья и, наконец, в последнюю очередь мышца дисталь-
ного расположения. Однако, как только наступает более поздняя
стадия развития (12—14 недель), когда нейроны стволовой ча-
сти мозга устанавливают свой контроль над моторными нейрона-
ми плечевых сегментов, происходит радикальная перемена этих
проксимо-дистальных отношений. Мышца предплечья и ее иннер-
вация приобретают сразу ведущее значение, благодаря чему про-
исходит спастическое удерживание предмета сжатыми пальцами
при еще недостаточной функциональной зрелости плечевой и ту-
ловищной мускулатуры.
Приведенных примеров, заимствованных мной из работ нашей
лаборатории, достаточно для того, чтобы оценить значение такого
критического положения, когда все фрагменты
А
ункциональной
системы объединяются к моменту рождения. Мы считаем поэто-
му, что
дующие
самая закономерность консолидации и немедленно сле-
за этим перемены в значимости отдельных фрагментов
системы должны стать предметом специального исследования.
Принцип минимального обеспечения
функциональной системы
Эта закономерность, подмеченная нами на примере развития
ряда функциональных систем, нредставляет огромное достижение
эволюции и, вероятно, одну из самых совершенных форм дости-
жения успеха в борьбе за «норму выживания». Она заключается
в том, что функциональная система, пройдя период консолида-
ции, становится в какой-то степени полноценной задолго до того,
как все ее звенья получат окончательное оформление и дефини-
тивное состояние. Функциональная система благодаря этому при-
обретает приснособленную роль в жизни новорожденного раньше,
чем она окончательно созреет. Нельзя не видеть в этой законо-
мерности важнейшего дополнительного фактора созревающих жи-
вотных в борьбе за существование. В самых разнообразных ус-
ловиях жизни различных животных могут быть созданы такие
условия, при которых спасение от врагов или добывание пищи
Системогенез как общая закономерность эволюционного процесса 147
становится необходимым задолго до того, как все детали какой-
либо функциональной системы получат окончательное оформ-
ление.
Применяя морфологический и физиологический методы иссле-
дования, нам удалось показать, что эта тенденция функциональ-
ной системы «выйти в жизнь», прежде чем созреют все ее ком-
поненты, проявляется в самых разнообразных формах. Так, на-
пример, малёк аксолотля, выходя из икринки, попадает в
условия свободной жизни, где ему со всех сторон может грозить
опасность. Сильным средством, спасающим его от врагов, служит
плавание, ы мы видим, что этот малёк способен производить до-
вольно быстрый плавательный бросок вперед, состоящий из ряда
волнообразных движений туловища. Казалось бы само собой ра-
зумеющимся, что все нейромускулярные синапсы туловищной му-
скулатуры к этому моменту окончательно созрели и только при
этом условии могут довольно быстро проводить возбуждение при
плавании.
Однако специальные исследования моей сотрудницы
Т. Т. Алексеевой показали, что если па этой стадии мальков ак-
солотля поместить в раствор кураре, то они продолжают свободно
плавать; это говорит о том, что кураре оказалось недеятельным.
Можно было бы думать, что кураре просто не проникает к ней-
ромускулярпым синапсам малька аксолотля, однако на самом
деле это пе так. Если взять малька аксолотля более поздней
стадии, поместить его также в раствор кураре, достаточный для
того, чтобы полностью обездвижить взрослую особь, то получает-
ся крайне интересное явление: передняя часть тела и жабры
оказываются парализованными, в то время как хвостовая часть
туловища не подвергается действию кураре, В результате такой
комбинации условий отравленный маленький аксолотль продви-
гается вперед только одними волнообразными движениями хво-
стовой части туловища.
Этот
акт
убеждает нас
в том, что
и
сложная химическая система протоплазмы синапса созревает,
проходя ряд промежуточных стадий, а организм в целом исполь-
зует все минимальные фазные способности развивающейся про-
топлазмы проводить возбуждение для консолидации первичной
функциональной системы. Можно указать еще на один пример
подобного рода. В опытах Е. Л. Голубевой на зародышах мор-
ской свинки было показано, что прыжок эмбриона осуществляет-
ся уже на 25-й день беременности. Он заключается в коорди-
нированном соотношении передних и задних конечностей, причем
последние делают сильно отталкивающие движения.
Для нас ясно, что это соответствует как раз моменту кон-
солидации
ункциональпой системы. Однако, если взять попереч-
ные срезы спинного мозга над люмбальными сегментами и обра-
ботать их по Вольтере — Кульчицкому, то можно увидеть, что
148 Системогенез как общая закономерность эволюционного процесса
ни одно из нисходящих волокон спинного мозга еще не миэли-
низировано.
Каким же образом возбуждение от вышележащих отделов голов-
ного мозга распространяется до люмбальных сегментов и вызы-
вает в конце концов довольно мощные движения задней конеч-
ности?
Если стать на точку зрения тех авторов, которые считают,
что миэлип абсолютно необходим для проведения возбуждения, то
перед нами возникает непреодолимое затруднение. Вместе с тем,
если предположить, что миэлин совсем не нужен для координа-
ции процессов в довольно сложной функциональной системе, мы
должны прийти к другому противоречивому заключению. Тща-
тельный микроскопический анализ всех уровней нервной системы
зародыша этого возраста убеждает нас в том, что ни то, ни дру-
гое заключение неправильно. Стволовая область головного мозга,
где лежат основные аппараты, интегрирующие прыжок в целом,
оказывается полностью миелинизированной, и, таким образом,
процесс распределения возбуждения в самом ответственном фраг-
менте функциональной системы является совершенным. Послед-
ний случай особенно убеждает нас в том, что вопрос о соотно-
шении миэлинизации и функции нервного волокна не может быть
решен в масштабе нервного волокна, а только в масштабе функ-
циональной системы в целом. Именно функциональная система
в целом определяет, какие структуры и ее фрагменты должны
быть к моменту функции обязательно созревшими, а какие могут
вступить в эту функцию даже и не совсем дифференцированными.
Из этого видно, что принцип минимального обеспечения является
подлинным принципом функциональной системы как целостной
организации и использован эволюцией как одно из средств в
борьбе за выживание новорожденного.
Заключение
В заключение считаю необходимым отметить ту роль, которую
должна приобрести концепция системогенеза в частных эмбрио-
физиологических исследованиях, и в особенности в подходе к эво-
люционным проблемам, изучаемым в плане эволюционной морфо-
логии.
Весь изложенный выше материал убеждает пас в том, что он-
тогенетическое созревание отдельных функциональных систем
осуществляется независимо от созревания целого органа и нахо-
дится в прямой связи с выборочным оформлением необходимых
в первую очередь структур. Только лишь в силу единства фор-
мообразовательного аппарата мы можем допустить влияние рано
созревающей части органа на поздно созревающие его части.
В качестве примера можно указать на раннее созревание двух
Системогенез как общая закономерность эволюционного процесса 149
мышц конечностей аксолотля (mm. anconeus communis coracob-
rachialis), которые опережают все остальные мышцы передней
конечности и служат для прижатия конечностей в момент пла-
вания личинки аксолотля. В этой стадии конечность как целый
орган еще не дифференцирована, однако в выборочном порядке
развитие отдельных ее структур гетерохронно ускоряется и, опе-
режая остальные структуры, уже включается в жизненно важную
функциональную систему эмбриона — плавание.
Можно привести много примеров, которые убеждают нас в
том, что в процессе онтогенеза происходит постепенная функ-
циональная фрагментация органа. Рассматривая это явление в
масштабе только органа, мы должны отметить, что оно целиком
совпадает с тем, что эволюционисты-морфологи определяют как
«полифункциональность» органа (Северцов, Матвеев, Емелья-
нов). Однако, с пашей точки зрения, это явление должно рас-
сматриваться в масштабе организма, т. е. в нлапе созревания его
жизненно важных функциональных систем. А это значит, что мы
должны рассматривать созревающий в эмбриогенезе орган как
арену, на которой разыгрывается постепенный и гетерохронный
захват отдельных его структур в интересах отдельных функцио-
нальных систем, в целом находящихся далеко за пределами этого
органа.
Это явление, названное нами функциональной фрагментацией
органа в онтогенезе, в значительной степени изменяет отноше-
ние эволюциониста-физиолога к органу как к объекту эволюцион-
ных исследований. Принятие концепции системогенеза расширяет
возможности оценки эволюции изолированного органа и, что осо
бенно важно, делает понятным его трансформацию в зависимо-
сти от внешних факторов. При оценке этой трансформации осо-
бенно важно помнить, что внешние факторы, как бы они ни
изменялись (внезапно или медленно), всегда оказывают влияние
на функциональную систему в целом, а не на изолированный
орган.
Следовательно, во всех случаях трансформации органа ис-
следователь неизбежно должен пройти этап оценки специфиче-
ских особенностей соответствующей функциональной системы, ее
состава и ее реакций на соответствующие изменения внешней
среды.
Эволюционирует и изменяется под воздействием внешних
факторов всегда только функциональная система, а морфологи-
ческое изменение внешних органов есть только отдаленное и до
некоторой степени отраженное следствие этого изменения. Следо-
вательно, встав на позицию системогенеза и рассматривая от-
дельные органы только как фрагменты обширных функциональ-
ных систем, исследователь приобретает новое средство более со-
вершенного анализа морфологической трансформации органов.
150 Системогенез как общая закономерность эволюционного процесса
Значение концепции системогенеза для чисто эмбриофизиоло-
гических исследований очевидно само по себе. С точки зрения
широкого понимания эмбрионального развития нет, например,
нерва вообще, так же как нет и мышцы вообще как каких-то
определенных субстратов любого эмбриона. Точно так же нет и
какого-то вполне определенного, нервного центра. Есть только
структуры, которые с той или иной степенью гетерохронности
приближаются к организации жизненно важных функциональных
систем, с которыми эмбрион должен появиться на свет.
Становится ясным, что такое выражение, например, как «хро-
наксия нерва зародыша», является, в сущности, физиологической
фикцией. В самом деле, из всего приведенного материала сле-
дует, что нерв, принадлежа какому-то определенному животному,
имеет специфическое отношение к функциональным системам
именно данного животного, а последние, созревая с известной
гетерохронией, определяют неравномерное созревание отдельных
волокон в пределах одного и того же ствола. Следовательно, фи-
зиолог, раздражая какой-то нервный ствол эмбриона, вмешивает-
ся в систему волокон, находящихся в динамическом процессе
различного и постепенного созревания. Благодаря этому всякое
указание на «возбудимость» нервов в данном случае теряет ка-
кое-либо специальное познавательное значение. Это есть возбуди-
мость каких-то волокон, обеспечивающих какие-то функциональ-
ные системы, находящиеся в какой-то стадии гетерохронного
развития.
Конечно, тип развития, например, синаптической передачи в
нейромускулярном синапсе будет более или менее одинаков для
всех моторных нейронов, имеющих отношение к поперечнополо-
сатым мышцам, и, следовательно, только в этом смысле можно
поставить вопрос для исследования. Это будет вопрос о стадиях
созревания моторного синапса вообще или приобретения возбу-
димости каким-то нервным волокном вообще.
Все это говорит о том, что если не исходить из общеэволю-
ционной точки зрения в оценке созревания функции, то любое
исследование специальных тканевых образований дает лишь весь-
ма ограниченный материал для суждения о стадиях созревания
данной ткани.
Учитывая особенности эмбриогенеза, т. е. движения от эквипо-
тенциальных образований ко все более и более дифференциро-
ванным, можно заранее сказать, что каждый исследователь част-
ного процесса тканевого созревания неизбежно будет иметь дело
со все более и более возрастающей способностью дифференциро-
ванно и максимально быстро отвечать на нанесенное раздра-
жение.
Изучение этих свойств созревающих в онтогенезе структур яв-
ляется совершенно необходимым, поскольку они составляют осно-
Системогенез как общая закономерность эволюционного процесса 151
“ “ *' 1,1 ' 1 ._х' “
ву для более высокой интеграции функций. Однако само по себе
изучение эмбриогенетических превращений частных возбудимо-
стей и реакций в свете широкого эволюционного аспекта имеет
смысл только тогда, когда будет точно установлено, в пределах
какой функциональной системы и в какой стадии ее развития
включается изучаемая морфологическая структура или реакция
зародыша.
ч В этом состоит смысл системогенеза как ведущей закономер-
ности эмбрионального развития. Именно он определяет смысл,
тип развития и быстроту созревания какой-либо изолированной
частной деятельности организма.
МЕТОДОЛОГИЧЕСКИЙ АНАЛИЗ
УЗЛОВЫХ ПРОБЛЕМ
УСЛОВНОГО РЕФЛЕКСА *
Вступительные замечания
Хорошо известно, что учение об условных рефлексах в послед-
ние десять лет широко распространилось по всему миру, про-
никло даже в те нейрофизиологические лаборатории, которые
традиционно занимались только тонким анализом самого нервного
процесса (Fessard, Bremer, Morell, Magoun и др.). Это привело к
тому, что в настоящее время учение об условных рефлексах и
общая нейрофизиология начинают объединять всю работу по изу-
чению мозга.
Такое широкое обращение к методу условных рефлексов не
случайно. Оно детерминировано историческими закономерностями
развития самой общей физиологии нервной системы, которые мы
постараемся показать в дальнейшем. Девятнадцатый век не ста-
вил перед нейрофизиологом-аналитиком вопрос: для чего нужны
те топкие процессы, которые он изучает? Считалось само собой
разумеющимся, хотя бы в общем натуралистическом плане, что
аналитически изучаемые нервные процессы п есть те самые про-
цессы, которые непосредственно участвуют в работе центральной
нервной системы, создавая приспособительные реакции целого ор-
ганизма.
Развитие электронной техники направило усилия нейрофизио-
логов на изучение еще более тонких процессов нервной системы,
протекающих уже в одной нервной клетке и даже на молеку-
лярном уровне. Однако это прогрессирующее утончение аналити-
ческой стороны исследования таило в себе принципиальные про-
тиворечия, которые п пе замедлили выявиться. Оказалось, что,
чем более глубокие и топкие процессы исследует нейрофизио-
лог, тем дальше он уходит от того, что является конечной целью
всякого нейрофизиологического исследования, а именно понима-
ние работы целого мозга, целостного поведения, психической дея-
тельности, а затем и управление работой мозга в условиях нормы
и патологии.
Естественно, что мысль прогрессивных нейрофизиологов обра-
тилась в сторону комплексных актов поведения животных, закоп-
* Опубликовано в книге «Философские вопросы физиологии высшей нерв-
ной деятельности и психологии» (М., 1963).
Методологический анализ узловых проблем условного рефлекса 153
ченных нейрофизиологических единиц интегративной деятельно-
сти организма. Вот здесь-то и выявилось, что единственным
физиологическим принципом деятельности целого мозга, объяс-
няющим целостное поведение животного
принцип условного рефлекса.
Стало общепризнанным положение, что
и человека, служит
тончайшие процессы
нервной системы приобретают свой высший смысл и занимают
свое истинное место в архитектуре целостных поведенческих ак-
тов только в том случае, если само изучение их непрерывно со-
относится с образованием и проявлением условного рефлекса.
Так подготавливался и возник современный этап изучения
высшей нервной деятельности, когда тонкое нейрофизиологиче-
ское исследование и изучение условного рефлекса органически
объединились в одной лаборатории.
Условный рефлекс стал рассматриваться как самый необходи-
мый фактор деятельности мозга. Именно этим объясняется то,
что традиционные международные симпозиумы по изучению моз-
га получили название «Механизмы мозга и обучение».
Совершенно очевидно, что мы, ученики и последователи
И. П. Павлова, не можем пе заметить и недооценить этот прин-
ципиально новый этап в изучении глубинных нейрофизиологиче-
ских законов формирования условного рефлекса как единицы це-
лостного поведения в связи с изучением его архитектурных осо-
бенностей.
Кроме того, параллельно с ростом популярности условного
рефлекса в собственно нейрофизиологических лабораториях нара-
стало внимание к нему и среди клиницистов и, что особенно
важно, в среде фрейдистов. Эти последние или полностью игно-
рируют значение условного рефлекса как фактора высшей нерв-
ной деятельности человека, или делают попытку привлечь его к
психоаналитическим приемам лечения, которые по своей сути не-
совместимы с материалистическим подходом к изучению челове-
ческой психики.
Отсюда появляется необходимость установления тесного кон-
такта физиологов высшей нервной деятельности с психологией
как наукой, охватывающей наиболее общие и наиболее тонкие
проявления человеческого поведения. Формулируя пограничные
проблемы и устанавливая взаимный контакт, мы сможем найти
точки соприкосновения для совместной разработки наиболее
принципиальных и решающих проблем психической деятельности
человека. Это, в свою очередь, позволит нам заполнить тот раз-
рыв, который создавался на протяжении ряда лет между физиоло-
гией высшей нервной деятельности и психологией.
Все отмеченные выше факты приводят к выводу, что Совеща-
ние по философским вопросам учения о высшей нервной дея-
тельности вполне своевременно. Оно должно дать философский
154 Методологический анализ узловых проблем условного рефлекса
анализ нового уровня и нового этапа в развитии учения об ус-
ловных рефлексах в связи с богатейшими достижениями в обла-
сти общей физиологии нервной системы и тем самым послужить
толчком для дальнейших, более эффективных исследований.
Трудно представить философский анализ какого-либо научно-
го направления, тем более такого разветвленного, как учение
И. П. Павлова о высшей нервной деятельности, без конструк-
тивных предложений конкретного его развития и формулировки
ближайших исследовательских задач.
Методология диалектического материализма тем и сильна, что
позволяет перейти на более высокий уровень обобщений и напра-
вить научное исследование по более эффективным путям, веду-
щим к разрешению проблем.
Очень часто приходится встречать в общем правильную фор-
мулировку, что философское осмысливание результатов исследо-
вания и анализ существующего уровня науки под углом зрения
методологии диалектического материализма предупредят ошибоч-
ное отклонение ученого в его собственной работе и позволят ему
правильно разобраться во всех неприемлемых для нас (с идеоло-
гической точки зрения) научных теориях.
Все это так и абсолютно верно. Но все же, с конструктив-
ной точки зрения, это только половина дела. Деятельность не
ошибающегося, но и пе развивающего науку исследователя едва
ли может служить показателем того, какая огромная движущая
сила скрыта в диалектико-материалистическом методе.
В этом смысле настоящее совещание должно дать большой эф-
фект, поскольку в изучении высшей первной деятельности назре-
ло много противоречий и новых принципиальных вопросов, по ко-
торым совершенно необходима взаимная консультация специали-
стов по физиологии высшей первной деятельности. Без
разрешения этих вопросов, как я думаю, мы не сможем устано-
вить правильного пути для более эффективной разработки всем
нам близкой проблемы условного рефлекса.
Мы не должны забывать одного важного факта, что в школе
И. П. Павлова в годы ее бурного роста по необходимости было
высказано много рабочих гипотез и предположений. Это составля-
ет естественный признак роста школы. Однако логика научного
прогресса такова, что в процессе исследовательской работы все
большее и большее количество «вероятных» положений получает
свою проверку, в результате которой они или переходят в раз-
ряд достоверных и прочно доказанных истин, или, наоборот, не
выдерживают экспериментальной проверки и заменяются более
приемлемыми рабочими гипотезами.
Чрезвычайно тяжелым обстоятельством для развития науч-
ной школы выступает такое* положение, когда все многочислен-
ные гипотезы
1
и достоверные, и вероятные, н даже сомнитель-
Методологический анализ узловых проблем условного рефлекса 155
ные, вдруг сразу приобретают значение нерушимых догм, абсо-
лютно достоверных истин. История науки показывает, что с этого
момента обычно задерживается прогресс научных исследований,
прекращаются поиски новых путей, начинается рост вширь, воз-
никает бесконечное дублирование и варьирование незначитель-
ных экспериментов без заметных признаков обобщений и движе-
ния вперед.
Хотя в школе И. П. Павлова и имеются все условия для про-
грессивного развития его идей в большом количестве лаборато-
рий его учеников, тем не менее мы должпы сказать, что целый
ряд важнейших вопросов, составляющих фундамент общего на-
правления, не подвергался серьезному анализу и не получил раз-
вития в связи с бурным ростом смежных дисциплин, и в особен-
ности в связи с развитием общей физиологии нервной системы.
Так, например, за последние 15 лет в связи с развитием тон-
ких электронных методов исследования нервной системы, допу-
скающих вмешательство в любые области мозга и любые клетки
нервных центров, совершенно изменили свое лицо прежние ней-
рофизиологические концепции об интегративных процессах мозга
и природе основных его процессов — возбуждения и торможения.
В настоящее время мы — свидетели того, как рухнули прежние
представления Шеррингтона об интегративной деятельности моз-
га и на совершенно новых материалах закладывается фундамент
современных представлений о деятельности целого мозга.
Хорошо известно, что Павлов, сделав гениальное открытие
условного рефлекса, был поставлен в начале нашего века перед
тяжелой необходимостью использовать современную ему, весьма
бедную, нейрофизиологию для выработки представлений о кон-
кретных нервных механизмах как самого условного рефлекса, так
и многих форм его проявления (иррадиация процессов возбуж-
дения и торможения, отрицательная индукция, корковое торможе-
ние и т. д.). И действительно, только гений мог на основе аб-
солютно несовершенной нейрофизиологии того времени построить
такие рабочие концепции, которые послужили толчком к продук-
тивным научным исследованиям па протяжении десятков лет. Но
это все же были только гипотезы о механизмах корковой дея-
тельности, которые с разных пунктов «входа» и «выхода» засека-
ли таинственные механизмы «черного ящика».
В настоящее время полностью изменила свое лицо прежняя
нейрофизиология.
Разработаны электронные
методы прямого
внедрения в «черный ящик», открыты совершенно новые законы
в деятельности мозга и вырастает совершенно новая концепция
о механизмах интегрирования многочисленных процессов мозга.
Наряду с этим все то, что было вложено И. П. Павловым в трак-
товку механизмов высшей нервной деятельности 50 лет назад, по
сути дела остается в силе и сейчас и во многих случаях про-
156 Методологический анализ узловых проблем условного рефлекса
должает еще служить основой для понимания условного рефлекса
и его физиологических превращений.
Возникает естественный вопрос: имеют ли какое-либо отноше-
ние к процессам высшей нервной деятельности все современные
достижения по изучению физиологии нервной системы? Можно
ли установить логический контакт между этими достижениями в
области нейрофизиологии и руководящими концепциями павлов-
ского учения?
Я считаю, что такой контакт должен быть установлен немед-
ленно; чем дальше, тем труднее будет создать органическое
единство между нейрофизиологией и физиологией высшей нерв-
ной деятельности. Мы должны помнить, что овладение современ-
ным уровнем нейрофизиологии потребует чрезвычайно напряжен-
ного труда. Поэтому естественно, что уже сейчас должны быть
намечены те принципиальные и узловые вопросы учения о выс-
шей нервной деятельности, которые требуют первоочередного ана-
лиза их на основе последних успехов нейрофизиологии.
Все, что говорилось выше, касается контакта высшей нерв-
ной деятельности с «низшими» закономерностями в работе цент-
ральной нервной системы. Они должны создать естественный
фундамент для высшей нервной деятельности, не снимая, однако,
специфических черт, определяющих ее отношение к высшим фор
мам поведения животных и человека. И даже больше того, фи-
зиолог высшей нервной деятельности должен иметь свою собст-
венную специфическую тактику в использовании современного
нейрофизиологического материала. Оп должен помнить, что ус-
ловный рефлекс есть высшая интегративная закономерность в
деятельности мозга, подчиняющая себе и приспосабливающая к
себе соответствующим образом все отдельные процессы нервной
системы, которые обычно служат объектом изучения тонкими
аналитическими приемам и.
Необходимо, однако, оговорить связь физиологии высшей
нервной деятельности с психологией, с изучением психических
явлений и высших форм сознательной деятельности. К сожале-
нию, приходится отметить, что искусственно поддерживаемый на
протяжении ряда лет разрыв между физиологией и психологией
оказал огромное отрицательное влияние на прогресс пашей со-
ветской паукп в изучении психической деятельности па основе
общепризнанных законов высшей нервной деятельности. Я хочу
сказать, что при тесном контакте высшей нервной деятельности
с психологией мы пошли бы вперед к изучению «психического»
гораздо более быстрыми шагами, чем идем сейчас.
Достаточно указать, что уже Павлов паметил целый ряд про-
блем пограничного значения, к которым мы могли бы «прокла-
дывать туннели», идя навстречу друг другу. Можно указать, на-
пример, на «рефлекс цели», вторую сигнальную систему и др.
Методологический анализ узловых проблем условного рефлекса 157
В самом деле, разве цель, формирующая в деталях предстоя-
щее действие, цель, без которой вообще не совершается пи одно
действие человека, разве опа не достойна того, чтобы объединить
физиолога и психолога в их исследовательской работе? В этом
смысле данное совещание должно помочь и физиологам и психо-
логам сформулировать пограничные проблемы, которые, являясь
для физиологов в какой-то степени психологическими, а для пси-
хологов — физиологическими, закрыли бы пропасть, незаконно
существующую между нашими науками уже много лет.
В настоящем докладе будет сделана попытка па основании
анализа существующих данных сформулировать некоторые новые
положения относительно некоторых затронутых выше методологи-
ческих проблем павловского учения. При этом надо иметь в виду,
что многие частные, хотя бы и в высшей степени актуальные,
вопросы не могут быть здесь, естественно, разобраны, как, напри-
мер, отдельные рабочие гипотезы о механизмах образования ус-
ловного рефлекса, представления о соответствии новых данных
нейрофизиологии главным теориям механизма движения нервных
процессов и т. д. Я обращу внимание по преимуществу на основ-
ные методологические проблемы, от правильного разрешения ко-
торых зависит само понимание частных вопросов.
Прежде всего я остановлюсь на исторических корнях самого
рефлекса и попытаюсь показать, что он представляет собой вы-
ражение универсальных законов, поддерживающих пространст-
венно-временную структуру материального мира.
Этот анализ происхождения условного рефлекса неизбежно
должен закончиться преобразованием и расширением общеприня-
той рефлекторной теории, так называемой классической, ведущей
свое начало от Декарта. Эта теория давно уже стала, по выра-
жению И. II. Павлова, «первобытной», и в настоящее время она
в своем прежнем архитектурном выражении («рефлекторная
дуга») представляет собой несомненную помеху для дальнейшей
разработки нейрофизиологического фундамента целостного пове-
дения животного.
Едва ли надо говорить, что рефлекторная теория, сформули-
рованная Декартом, сыграла революционную роль в развитии
А
экологии нервной системы. В течение 300 лет она успешно на-
правляла мысль в тысячах исследований. Но вместе с тем на
современном уровне пауки она значительно ограничивает воз-
можности объяснения именно целостного поведения животных и
человека.
Мы уже давно проводим работу по расширению классической
рефлекторной теории. Теория функциональной системы, сформу-
лированная нами еще в 1935 г., в настоящее время является
пока наилучшей формой физиологического преодоления недостат-
ков рефлекторной теории. Однако только через 11 лет после
158 Методологический анализ узловых проблем условного рефлекса
опубликования принципа функциональной системы все основные
физиологические черты этого принципа были развиты в кибер-
нетическом направлении и в настоящее время разрабатываются
в самых различных областях физиологии, психологии, лингвисти-
ки и т. д.
Естественным развитием этих двух разделов доклада служит
формулировка тех физиологических основ и процессов, которые
предшествовали в истории живого мира появлению психических
функций. Именно в первичных проблесках психического и их
развитии до высших форм мыслительной деятельности человека
надо искать фактическое объединение условного рефлекса и
психического, которое составляет для нас, физиологов высшей
нервной деятельности, одну из важнейших философских проблем.
Условный рефлекс как отражение
пространственно-временной структуры мира
Диалектический материализм
время как формы существования
чит, что только через изменение
и законов их взаимодействия мы
рассматривает пространство и
и движения материи. Это зна-
материальных физических тел
можем познать пространство и
время.
На разных этапах развития науки представления о простран-
стве и времени базировались на различной материальной основе
в полной зависимости от успехов физики. Так, например, несколь-
ко десятков лет тому назад, когда основные представления физи-
ки покоились на взаимодействии твердых тел, были созданы кон-
цепции о пространстве и времени в полной зависимости от их
свойств, со всеми вытекающими отсюда ограничениями и недо-
статками (прерывность, противоречивость законов).
С успехами физики в области изучения электромагнитной ос-
новы всех видов материальных тел на сцену выступила электро-
магнитная «матрица», связывающая все формы движения и со-
стояния материального мира. В частности, вполне обоснованная
теория относительности могла возникнуть только с развитием в
науке представлений об универсальности этих электромагнитных
процессов, заполняющих собой мировое пространство, и благо-
даря этому удалось сформулировать новые способы связей между
всеми материальными явлениями Вселенной (А. Эйнштейн).
В аспекте интересов настоящего доклада важно установить,
что пространство и время, взятые в отдельности, не являются
тем абсолютным законом Вселенной, каким служит комплекс
«пространство — время». Однако для оценки составляющих
торов очень важно знать, что оба они почти никогда не играют
одинаковую роль в тот пли иной момент. Мы всегда сталкиваем-
ся с тем, что какой-то отдельный параметр этой абсолютной
Методологический анализ узловых проблем условного рефлркса 15
в масштабе нашей планеты они, есте-
основы материального мира приобретает на данный момент ве-
дущее значение.
В этом смысле надо различать две совершенно различные
плоскости оценки пространства и времени — общефилософскую,
гносеологическую и соотносительную, т. е. пространство и время
как факторы приспособления живой природы.
Оценивая гносеологическое значение пространства и времени,
А. Д. Александров пишет: «Основываясь на законах движения
материи, теория пространства и времени отвлекается от конкрет-
ных свойств предметов и процессов, с тем чтобы выделить имен-
но общие законы пространственных и временных отношений и
свойств. Она необходимо абстрагирует пространственные и вре-
менные формы от их конкретного материального содержания, так
как иначе она и не была бы теорией пространства и времени»
(1959, с. 94). (Курсив мой.— П. А.).
Эта формулировка, совершенно приемлемая в общем фило-
софском плане, оказывается недостаточной для наших целей.
В самом деле, поскольку пространство и время как абсолютные
факторы материального мира имели место во все времена преоб-
разования Вселенной, то
ствеппо, были изначальной структурой материального мира и
имели свои абсолютные свойства задолго до появления и разви-
тия живой материи на Земле. Уже первые формы живого ве-
щества и примитивные живые существа были «вписаны» в
основные законы пространственно-временных соотношений, и эти
последние стали абсолютными факторами приспособления живой
материи к внешнему миру, определившими ее выживание.
Этот кардинальный факт эволюции корепным образом изме-
няет наше отношение к теории пространственно-временных соот-
ношений. Мы не можем остаться только на уровне гносеологи-
ческого абстрагирования его параметров, а должны оценить все
разнообразие временной структуры мира в аспекте прогрессив-
ной эволюции живой материи. Мы, несомненно, встречаемся здесь
с новыми факторами материального мира, в котором пассивное
отношение к параметру времени в процессе бесконечного движе-
ния материи обогатилось новым качественным фактором —
активным отношением живой материи к всевозможным превра-
щениям пространственно-временной структуры неорганического
мира.
К сожалению, мы не нашли в современной философской ли-
тературе анализа этого узлового пункта эволюции. Время как
фактор эволюции живых существ на земном шаре и важнейший
параметр развития их нервной деятельности никогда не под-
вергалось серьезному анализу, и потому этот важнейший для
физиологии высшей нервной деятельности вопрос фактически
приходится разрабатывать заново.
160 Методологический анализ узловых проблем условного рефлекса
Что для нас наиболее важно во временных соотношениях
неживой и живой природы?
Первым и существенным моментом является то, что основные
формы движения материи в рамках пространственно-временных
факторов существовали и в неорганической природе задолго до
возникновения первых живых существ на земном шаре. Это, ко-
нечно, важный фактор. Он сам по себе делает очевидным, что
живая материя, «вписавшись» в уже готовую пространственно-
временную систему мира, не могла пе отразить ее свойства, ее
архитектуру, если только эти свойства имели отношение к основ-
ному свойству самой живой материи — выживаемости.
С того момента, когда па Земле возникла жизнь, отношение к
отдельным параметрам пространственно-временной структуры со
стороны этой качественно повой формы материи кардинально из-
менилось. Гора, например, как форма неорганической материи,
не относится избирательно к климатическим и метеорологическим
факторам, которые на нее действуют. Для горы не существует
той проблемы, которая возникла заново с появлением живой ма-
терии и которую можно формулировать двумя словами: «приспо-
собиться и выжить». Для живых существ с момента их форми-
рования в процессе эволюции весь внешний неорганический мир
со всеми его многообразными воздействиями стал «взвешиваться»
только на этих весах прогрессивной эволюции.
Появилось активное отношение к внешним неорганическим
факторам и неизбежно последовавшее за этим их разделение па
две большие категории — вредных для жизни и способствующих
ее сохранению.
Начиная с этого момента, все разнообразные формы движения
материи в рамках временной структуры мира стали распреде-
ляться первичными живыми существами по этой реципрокной
схеме, а живая материя начала отражать и закреплять в своей
структуре эти универсальные закономерности.
С какими временными параметрами движения материи в неор-
ганическом мире нашей планеты столкнулась первичная и при-
митивная жизнь? А еще точнее, в рамках каких временных па-
раметров формировалась сама живая материя?
К сожалению, на этот вопрос мы пе имеем ответа в современ-
ной философской и биологической литературе, поскольку гносео-
логическое абстрагирование фактора времени оттеснило па зад-
ний план разработку временной структуры неорганического мира
именно в том аспекте, который помог бы нам выявить значение
этой структуры для прогрессивной эволюции живых организмов.
Это ставит нас перед необходимостью вычленить те особенные
черты временной структуры материального мира, которые яви-
лись своего рода «категорическим императивом» для развития
жизни на Земле.
Методологический анализ узловых проблем условного рефлекса 161
Как разрешается сейчас проблема пространства и времени в
философском аспекте?
Обсуждая проблему пространства и времени с точки зрения
диалектической методологии в связи с анализом особенностей
элеатской философской школы, Гегель пишет: «Сущность време-
ни и пространства есть движение потому, что оно всеобще; по-
нять его, значит высказать его сущность в форме попятия». Раз-
бирая эти положения Гегеля, В. II. Лепин отмечает па полях сво-
их заметок в «Философских тетрадях»: «верно!»
При любой попытке конкретизировать пространственно-вре-
менную структуру материального мира, мы неизбежно встречаем-
ся с последовательным перемещением тел в пространстве, после-
довательными воздействиями одного тела на другое, последова-
тельным развитием фаз движения и преобразования материи.
Резюмируя эти рассуждения, мы можем сказать, что, с точки
зрения диалектического материализма, наиболее существенной
чертой пространственно-временной структуры мира, определяю-
щей временное отношение первичных организмов к внешнему
неорганическому миру, служит последовательность его воздейст-
вий независимо от качества энергии этих воздействий. Тем са-
мым мы вычленяем временной параметр отношения организма к
неорганическому миру как в какой-то степени самостоятельный
фактор для понимания его приспособительных превращений.
Мы поставили перед собой конкретный вопрос: с какими фор-
мами и вариантами этого основного временного фактора — после-
довательности воздействий — встречается организм? Каким обра-
зом дробится всеобъемлющее движение материи в рамках абсо-
лютного временного параметра — последовательности явлений?
Иначе говоря, с какой уже готовой временной структурой неор-
ганического мпра, вынуждающей к приспособлению, встретились
первичные организмы па пашей планете?
Мы постараемся вычленить некоторые временные параметры
из основной формы — последовательности и проанализировать то
значение, которое они могли иметь для приспособительных отно-
шений первичных организмов.
Первая возможность этого — существование ряда таких по-
следовательных явлений, каждое из которых никогда пе повто-
ряется на протяжении жизни данного организма. Символически
этот ряд мог бы быть изображен так: а, б, в...
Несомненно, факторы неорганического мира а, б и т. д. свя-
заны между собой причинно-следственными отношениями и раз-
виваются в последовательном порядке; несомненно также, что они
могут оказывать при этом последовательные воздействия па ор-
ганизм. Однако ни одно из этих воздействий никогда не должно
иметь повторного действия па организм. Конкретно это могут
быть солнечное затмение, необычайной силы гроза или ураган,
6 П. К. Анохин
162 Методологический анализ узловых проблем условного рефлекса
прохождение кометы вблизи Земли и т. д. Но могут быть, ко-
нечно, неповторяющиеся воздействия и более близких к организ-
му окружающих его факторов.
Конечно, некоторые из этих пеповторяющихся воздействий
внешнего мира могут иметь какое-то существенное значение для
организма — разрушительное или благотворное — и первичный
организм мог или противостоять этому внезапному действию с
помощью уже имеющихся у пего возможностей, или разрушить-
ся. Однако эти воздействия для него были бы всегда внезапными.
Возможна, однако, другая временная структура в последова-
тельности внешних воздействий на организм: некоторые отдель-
ные факторы внешнего мира могут повторять свое действие через
определенные промежутки времени и, следовательно, могут быть
названы «повторяющимися рядами последовательных воздейст-
вий». Символически эта вторая возможность может быть изобра-
жена так: (абвг), (абвг), (абвг) ...(^бвг).
Мы можем иметь в неорганическом мире последовательность
событий, которые ритмически повторяются в том же составе и в
том же порядке следования компонентов. Пусть это будут, напри-
мер; день, вечер, ночь, утро или лето, осень, зима, весна.
Оба эти ряда последовательно развивающихся явлений неор-
ганического мира имеют абсолютную направленность и связаны
с астрономическими закономерностями, хотя длительность ритмов
в обоих случаях различна, и, следовательно, повторяемость этих
циклов может иметь разное значение для организмов с различной
длительностью жизни.
До сих пор мы брали в качестве примеров такие ряды после-
довательно развивающихся событий, которые обладают свойством
абсолютной устойчивости. Но мыслимы также и такие последова-
тельные явления, которые, обладая тем же параметром повторяе-
мости или возвращаемости, вместе с тем имеют свойство относи-
тельной устойчивости. Так, например, дерево, растущее па берегу
реки и имеющее определенный возраст, может быть местом оби-
тания для различных видов животных только па протяжении оп-
ределенного срока, после которого оно будет сменено другими
такими же или отличающимися деревьями. Однако на протяже-
нии срока своей жизни это дерево будет представлять собой се-
рию последовательных, ритмически повторяющихся явлений,
имеющих место в различные времена года, особенно для северных
широт.
Таким образом, как в первом случае, так и во втором мы име-
ем один главнейший временной признак последовательно разви-
вающихся событий — повторяемость, абсолютно устойчивую и от-
носительно устойчивую в масштабе жизни организма.
Конечно, само свойство повторяемости явлений неорганическо-
го, а позже и органического мира как частный параметр абсолют-
Методологический анализ узловых проблем условного рефлекса 163
ной пространственно-временной последовательности может иметь
бесконечные вариации в длительности самих ритмов, их устой-
чивости, составе и продолжительности отдельных компонентов,
их пространственной локализации и т. д., однако при всех этих
вариантах параметр повторяемости будет наиболее характерным
и специфическим.
До сих пор мы разбирали последовательность явлений неорга-
нического и отчасти органического мира только с одной целью:
выделить различные временные параметры неорганического мира,
которые неизбежно должны были воздействовать на органическую
материю уже при самом ее возникновении. Однако эти временные
параметры (неповторяемость, повторяемость, длительность, устой-
чивость, изменчивость и т. д.) нами были изложены безотноси-
тельно к другому их важному качеству, которое они приобретают
специально при взаимодействии с живой природой; это — свойст-
ва существенности и несущественности данных параметров для
поддержания жизненного процесса
Несомненно, уже на самых первых этапах организации живо-
го вещества, еще на стадии первичных коацерватов, изменения
внешних материальных условий «классифицировались» зачатками
живого вещества как «вредные» и «полезные» именно по крите-
рию «существенности». Эту форму отношения первичных форм
живой материи к внешним воздействиям особенно полно характе-
ризует А. И. Опарин (Опарин, 1957).
Несмотря на относительное постоянство неорганических усло-
вий, в которых зародилась жизнь (глубины первородных океа-
нов), даже и в той ситуации имелась последовательная смена
внешних воздействий на первичные живые тела, которая могла
производить своего рода первичный естественный «отбор» наи-
более устойчивых образований типа «открытых многомолекуляр-
ных систем» (смена температур, реагирующих веществ, течений,
приливы и отливы и т. д.).
Однако это последовательная смена внешних воздействий при-
обрела особенно большое значение тогда, когда первичные живые
существа получили способность размножаться и рассеиваться по
различным водным пространствам с разным составом экологи-
ческих факторов.
На этом этапе последовательность как универсальное свойст-
во движения неорганической материи, составляющее существен-
ное условие для выживания, приобрела исключительное значе-
ние. Сезонные, суточные ритмы, эпизодические, неповторяющиеся
воздействия в самом океане, как п огромное количество ритмиче-
ски повторяющихся воздействий, связанных с астрономическими
факторами,— все это составило основной фонд факторов «окру-
жающей среды», которые приобрели избирательное жизненное
значение для различных организмов.
6*
164 Методологический анализ узловых проблем условного рефлекса
Еще большее количество вариаций повторяемости возникло с
того момента, когда первичные организмы приобрели способность
к активному передвижению. Тогда всякое повторное движение
около неподвижного неорганического объекта приводило к пов-
торным более или менее однотипным воздействиям. Исключитель-
ное значение этот тип повторного воздействия внешнего мира
имеет в жизни человека с развитием его широчайших возмож-
ностей к перемещению.
В данный момент нам важно одно, что и в этом периоде вре-
менная структура окружающего организм неорганического мира
осталась той же и включила в себя три основные формы:
1) действие относительно постоянных факторов,
2) действие последовательных рядов повторяющихся факто-
ров (ритмически пли апериодически);
3) действие последовательных рядов никогда не повторяю-
щихся факторов.
Все эти факторы могли быть ближайшими и отдаленными воз-
действиями, также, конечно, передающимися через непосредствен-
ную водную среду.
Мы уже отмечали, что последовательный ряд никогда пе пов-
торяющихся воздействий практически имеет одно существенное
значение для жизни организмов: отдельные звенья этой цепи
внешних явлений могут быть несовместимы с жизнью в силу
своего экстремального значения. Они уже самим своим единст-
венным и внезапным воздействием могут нарушить устойчивость
организовавшейся живой материи п привести ее к распаду. Ина-
че говоря, на этот вид внешних воздействий, как, впрочем, и на
все другие, распространяет свое влияние правило существенно-
сти для сохранения жизни.
Хотя эти последние воздействия и оказывались только «вред-
ными» пли «полезными» неповторяющимися эпизодами для жи-
вой материи, сами по себе они всегда были звеньями непрерыв-
ных и последовательно развивающихся явлений в общем потоке
эволюции пространственно-временной структуры материального
мира. Опп встречались с жизненным циклом организмов, так
сказать, по касательной.
Из этого яспо следует, что эти никогда не повторяющиеся воз-
действия не могли оказать какое-либо решающее влияние па
эволюцию высших форм приспособления живой материи к окру-
жающим условиям и, следовательно, пе могли стать факторами
организации самой протоплазматической структуры живых орга-
низмов.
Представим себе на минуту, что временная структура неорга-
нического мира состояла бы только из первой формы движения
материи — из ряда последовательных и никогда пе повторяющих-
ся явлений. Смогла бы при этом условии развиваться на Земле
Методологический анализ узловых проблем условного рефлекса 165
жизнь в ее стабилизованных, имеющих всегда приспособительное
значение структурах? Нет, не могла. Живой организм не мог бы
иметь устойчивой п прочной структуры, ибо последняя может
появиться только как результат отражения ритмически и апе-
риодически повторяющихся воздействий неорганической приро-
ды. Само понятие «приспособление» потеряло бы всякий смысл в
этом мире всегда новых, никогда пе испытывавшихся организмом
воздействий.
Совершенно очевидно, что из общего потока событий прост-
ранственно-временной структуры мира только ритмически и апе-
риодически повторяющиеся явления смогли стать временной ос-
новой для развития приспособительных реакций первичных ор-
ганизмов.
Итак, мы пришли к решающему выводу относительно эволю-
ции организмов в зависимости от постоянных свойств временной
структуры неорганического мира.
Этот вывод можно формулировать следующим образом: осно-
вой развития жизни и ее отношения к внешнему неорганическо-
му миру явились повторяющиеся его воздействия на организм.
Именно эти воздействия, как результат изначальных свойств
пространственно-временной структуры неорганического мира,
обусловили собой всю анатомическую организацию и приспособи-
тельные функции первичных живых существ. В этом отношении
организация живых существ представляет собой в подлинном
смысле слова отражение пространственно-временных параметров
их конкретной среды обитания.
Естественно, что относительное постоянство пространственно-
временных соотношений организма с внешним миром может быть
рассматриваемо в качестве вариаций повторяющегося воздейст-
вия на организм. Все дело в том, какие конкретные временные
соотношения имеются между продолжительностью жизненного
цикла организма и длительностью внешнего воздействия.
Постоянным (относительно) воздействием условно можно счи-
тать такое, которое перекрывает по своей длительности миллионы
жизненных циклов, как, например, наличие кислорода в атмосфе-
ре; относительно менее постоянным можно считать такое, кото-
рое перекрывает лишь тысячи жизненных циклов, как, например,
природные факторы данной местности; и, паконец, еще менее по-
стоянным,— например, влияние какого-либо мелиоративного или
водного преобразования, которое может по своей длительности
перекрыть всего лишь сотни жизненных циклов.
Кроме того, ритмические и апериодические воздействия внеш-
него мира уже на первичные организмы могли быть двух
видов — тех самых, которые обязательны и для высших живот-
ных. Одни из них — это ритмические воздействия внешних усло-
вий на организм, находящийся в неподвижном состоянии. Другие,
166 Методологический анализ узловых проблем условного рефлекса
или апериодические, зависят, однако, от
также ритмические
повторяющихся движений самого организма. Одпако в обоих слу-
чаях мы имеем принципиально тождественный конечный про-
цесс — повторяющееся воздействие каких-либо внешних факторов
на организм.
Итак, проделанный выше анализ приводит нас к выводу, что
именно эта форма временных соотношений — повторяющиеся воз-
действия — представляет собой универсальную форму связи уже
сложившихся и индивидуализировавшихся живых существ с ок-
ружающей их средой.
Такой вывод немедленно ставит перед нами вопрос: с помощью
каких же конкретных механизмов эти жизненно важные повто-
ряющиеся воздействия делают организм приспособленным к
ним?
В каких внутренних процессах живой протоплазмы могли бы
отражаться эти повторные воздействия внешнего неорганического
мира на уже оформившиеся живые существа?
Для ответа мы можем использовать общепринятое представ-
ление, что простейшие живые существа — это «открытые систе-
мы», которые связаны с окружающей средой через ряд химиче-
ских превращений, начинающихся на границе живой многомоле-
кулярной протоплазмы и продолжающихся в форме целой цепи
отдельных реакций, которые закапчиваются вредным или полез-
ным для жизни итогом. Таким образом, каждое из внешних по от-
ношению к организму последовательных явлений благодаря осо-
бенным свойствам живой протоплазмы отражается в протоплазме
самого живого существа в форме более или менее длинных цепей
химических превращений.
Эта цепь реакций может иметь обменный характер, поддер-
живающий жизнь организма, его самовоспроизведение, но может
оказаться вредной, не поддерживающей жизненный процесс. Од-
пако, несмотря па эти два биологически противоположных исхо-
да, и те и другие воздействия вызывают ряд последовательных п
вполне закономерных химических превращений. Здесь-то и вы-
ступает на первый план тот основной параметр пространствен-
но-временной структуры неорганического мира, который всегда
играет доминирующую роль в развитии живой материи, а именно
повторяемость явлений и воздействий.
Сейчас нетрудно установить, к каким реальным последствиям
могли привести те воздействия па организм, которые имели су-
щественное значение для его выживания и повторялись или были
постоянными на протяжении миллионов лет.
По общему признанию биохимиков, любое внешнее воздейст-
вие на первичные организмы неизбежно служило отправным
пунктом для образования целых цепей химических процессов п
взаимодействий, кончавшихся или разрушением организма, или,
Методологический анализ узловых проблем условного рефлекса 167
наоборот, укреплением его метаболической стабильности. Но вме-
сте с тем повторяющиеся много раз одни и те же последователь-
ные ряды воздействий неизбежно приводили к облегченному и
ускоренному катализаторному типу развития этих цепных про-
цессов. Создавались доминирующие линии цепных химических
реакций.
Таким образом, оценка всех условий развития первичных ор-
ганизмов под воздействием постоянных п много раз повторяющих-
ся воздействий приводит пас к выводу, что именно эти последо-
вательные ряды воздействии составляли основную причину
первичной структурной организации живых существ.
Вне повторяющихся воздействий внешнего неорганического
мира на организм нет других путей, которые смогли бы создать
доминирующие цепи обменных и других химических реакций.
Именно повторяемость последовательно развивающихся внеш-
них воздействий, существенных в том или другом отношении для
жизни организма, смогла создать непрерывную и последователь-
ную цепь химических реакций в протоплазме организма.
Па этой основе в эволюции жизни па Земле происходит одно
весьма значительное явление, которое в дальнейшем па многие
миллионы лет определило ведущий признак приспособительных
реакций организму, его способность реагировать на изменения в
окружающем мире более или менее обширными химическими пе-
рестройками.
И потому естественно, что если во внешнем мире последова-
тельно развивается несколько специфических событий (напри-
мер, сезонные ритмы, смены температур, течения в океанах), то
организм должен отразить каждое из них в специфических хими-
ческих перестройках своей протоплазмы, если только они дости-
гают известного порога действия.
Эти специфические химические перестройки могут иметь свое-
образие в зависимости от физических свойств каждого неоргани-
ческого фактора из длинного последовательного ряда этих пос-
ледних.
Интересно, что А. И. Опарин, характеризуя эти соотношения
первичной жизни материи с окружающей неорганической приро-
дой, также считает, что последовательность и быстрота реакций
протоплазмы служат решающими факторами материальной орга-
низации первых живых существ.
Знакомство с теориями происхождения жизни на Земле пока-
зывает, что и с широкой биологической точки зрения, так же как
и с точки зрения философского анализа роли пространственно-
временной структуры мира, движение материи по последователь-
ным, ритмически повторяющимся фазам представляет собой уни-
версальный закон, определявший основную организацию живых
существ на нашей планете.
168 Методологический анализ узловых проблем условного рефлекса
Появление первичных белковых тел, приобретших в дальней-
шем ферментативную функцию, радикально изменило весь про-
цесс совершенствования жизни. Создалась возможность развития
отдельных цепей реакций с избирательным каталитическим ук-
реплением, и, конечно, в первую очередь тех реакций, которые,
будучи существенными для сохранения жизни, повторялись мно-
жество раз под влиянием внешних воздействий. Эти воздейст-
вия могли быть весьма различными, как, например, периодиче-
ские смены в поступлении веществ, необходимых для «открытых
систем» первичных организмов. Одпако остается несомненным,
что сам специфический катализ создал возможности благоприят-
ных условий для развития тех, а не других цепей реакций.
Это ускорение реакций, как показывает анализ некоторых ши-
роко распространенных ферментативных комплексов, может до-
стигать сотен миллионов и даже миллиардов раз. Легко предста-
вить, какое огромное преимущество перед другими приобрели те
цепи химических реакций, которые в силу частой повторяемости
сформировали в себе какой-либо каталитический комплекс.
Мы видим, что уже здесь, на этой стадии развития жизни,
возникла та «борьба за быстроту», которой Дори придавал такое
большое значение во взаимоотношении организма л внешней
среды.
Во всяком случае па первых стадиях развития жизни этот
«принцип максимальной скорости» дал возможность на фопе в
какой-то степени гомогенных коацерватных образований сформи-
ровать пути доминирующих цепей реакций, развивающихся с ог-
ромной скоростью.
И здесь, в этом узловом пункте развития живых организмов,
было сделано одно замечательное приобретение, которое в даль-
нейшем оказало решающее влияние па весь исторический путь
развития животного мира на земном шаре.
Дело в том, что между последовательными воздействиями
внешнего мира и реакциями па них со стороны живого вещества
существовала одна принципиальная разница. Внешние воздейст-
вия на организм могли совершаться в самых разнообразных вре-
менных интервалах и происходить из самых различных источни-
ков (их объединял лишь принцип последовательности). Вместе с
тем все эти воздействия конвергировали к одной и той же прото-
плазме живого существа, вызывали различные по качеству хими-
ческие реакции, разыгрывающиеся в пределах одного и того же
протоплазматического образования.
Здесь создались все условия для формирования одного из ре-
шающих процессов живого вещества, который определил пути
дальнейшего развития животного мира и форму его приспособи-
тельных реакций по отношению к внешнему миру. В самом деле,
если в небольшом комочке живого вещества, представляющего
Методологический анализ узловых проблем условного рефлекса 169
* — ~ ш — » Mi* , || ill Hiii । 1
собой многомолекулярную открытую систему, стали возможными
многообразные химические реакции, да еще с весьма большими
скоростями, то некоторые последовательно повторяющиеся ряды
внешних воздействий, пусть даже разделенных большими интер-
валами, получили возможность отражать себя в быстрых хими-
ческих превращениях этого вещества в соответствии с физиче-
ским или химическим качеством этих воздействий.
Постепенно складывались такие соотношения, при которых
все более и более выявлялось различие временных параметров для
событий, протекающих во внешнем неорганическом мире, и их
отражения в химических перестройках живого существа.
Так, например, внешние факторы могли воздействовать на
организм в непрерывной последовательности на протяжении су-
ток (степень инсоляции, приливы и отливы и т. д.), и, следова-
тельно, различные химические реакции в живых организмах, со-
ответствовавшие каждому из этих воздействий, также развива-
лись на протяжении суток. По именно то, что эти отдельные хи-
мические реакции последовательно разыгрывались в небольшом,
но весьма комплексном многомолекулярном образовании, при-
вело к теснейшим взаимодействиям этих реакций и образованию
непрерывных и взаимозависимых цепей реактивных изменений.
Следовательно, одни и те же свойства первичных живых об-
разований — мпогомолекулярность, каталитическое ускорение
реакции и открытые возможности для отражения внешних воз-
действий обеспечили прогрессивное развитие живых существ и
отражение ими в химических перестройках своей протоплазмы
последовательно повторяющихся воздействий внешнего мира.
В результате этих совершенно реальных взаимодействий уже
на данной стадии сложилась одна универсальная закономерность
в приспособлении организма к внешним условиям, которая в
дальнейшем бурно развивалась на протяжении всей эволюции жи-
вотного мира.
Проиллюстрируем схемой эту закономерность. Допустим, что
во внешнем мире развивается последовательный ряд явлений
А, Б, В, Г. Пусть эти явления действуют на организм через зна-
чительные временные промежутки на протяжении полусуток и
каждое из этих явлений вызывает в протоплазме живого сущест-
ва последовательный ряд химических превращений, который мы
обозначим соответственно символами а, б, в, г. Допустим, что ряд
внешних воздействий иа организм Л, Б, В, Г систематически пов-
торяется па протяжении многих лет и имеет существенное поло-
жительное значение для его обменных процессов, устанавливая
более совершенные цепи химических реакций. Тогда в результа-
те длительного и многократного воспроизведения в протоплазме
живого существа этого определенного ряда химических реакций
между отдельными звеньями пх установится органическая связь,
170 Методологический анализ узловых проблем условного рефлекса
пассивно,
с опережением в своей протоплазме последовательно
•I
делающая весь ряд превращений а-+б-+в^г непрерывной и быст-
ро развивающейся цепью химических реакций.
Несомненно, что решающее значение в образовании этих це-
пей реакций имело формирование специфических катализаторов,
определяющих преимущественное течение цепных и химических
реакций именно в данном направлении, а не в другом.
Все эти условия, создавшиеся уже на самых ранних этапах
эволюции живой материи, привели к тому, что протоплазма по-
лучила возможность отражать в микроинтервалах времени своих
химических реакций те последовательные события внешнего
мира, которые по своей природе могут развертываться, наоборот,
в макроинтервалах времени.
Так постепенно формировалась способность первичных орга-
низмов отражать внешний неорганический мир
а активно
и повторно развертывающихся явлении внешнего мира.
В самом деле, поскольку в протоплазме живого оргапизма
установилась единая цепь химических реакций, ранее вызывав-
шаяся последовательным действием внешних факторов А, Б, В,
Г, и если эти факторы разделены значительными интервалами
времени, то теперь уже действие только одного первого фактора
А способно начать и привести в активное состояние всю после-
довательную цепь химических реакций.
Исторически в живой протоплазме неизбежно должны были
создаваться эти условия, при которых быстрота ее химических
реакций обеспечивала опережение организмом развертывания
много раз повторявшихся внешних воздействий.
Таким образом, с нашей точки зрения, эволюция очень рано
«напала» на этот универсальный и единственно возможный путь
приспособления организма к внешнему миру. С помощью про-
цесса опережения уже самые начальные живые организмы, от-
городившиеся от внешнего мира простейшими мембранами, мог-
ли с огромной быстротой своих химических превращений пост-
роить цепи химических реакций но первому звену много раз
повторявшегося последовательного ряда внешних
Основой для построений таких цепей химических
лись мпогомолекулярность и
строению протоплазма первичных живых существ.
Живая организация получила от такой формы
на повторные ряды последовательных воздействии огромные
преимущества. По сути дела без этого опережающего действи-
тельность развертывания цепей химических реакций трудно
представить себе существование даже таких простых процессов,
как, например, поглощение и переваривание амёбой какого-либо
кусочка пищи или захватывание и переваривание бактерий бак-
териофагом.
воздействии,
реакций яви-
сложпая по
высшей степени
реагирования
Методологический анализ узловых проблем условного рефлекса 171
Вся история развития животного мира есть наглядный пример
усовершенствования этой универсальной и самой древней законо-
мерности, которую можно было бы назвать «опережающим отра-
жепиелМ действительности», т. е. ускоренным в миллионы раз раз-
витием ценой химических реакций, которые в прошлом отражали
последовательные преобразования этой действительности.
Возвращаясь к исходной предпосылке этой проблемы, мы
могли бы сказать, что опережающее отражение действительности
есть основная форма приспособления живой материи к простран-
ственно-временной структуре неорганического мира, в котором
последовательность и повторяемость служат основными времен-
ными параметрами.
Попробуем рассмотреть описанные выше закономерности под
другим углом зрения, близким к задачам изучения приспособи-
тельного поведения в физиологии высшей нервной деятельности.
Если посмотреть на схему, изображающую последовательное раз-
витие событий А, Б, В, Г, то можно видеть, что это только огра-
ниченный ряд последовательных воздействий па организм, ис-
кусственно вырванных из непрерывного развития и движения
внешнего мира. Конечно, и перед А были нескончаемые ряды по-
следовательных изменений в неорганической прпроде, точно так
же как и после Г непременно должны следовать бесконечные
преобразования внешнего мира.
Таким образом, выделение
оправданным только тем, что
особое существенное значение
ряда А, Б, В, Г могло бы быть
эти
акторы имеют какое-либо
для сохранения и поддержания
жизни.
Пусть с этой точки зрения
к
актор Г — какой-либо полезный
для жизни фактор, который поддерживает метаболический про-
цесс и тем самым стабилизирует живой организм. Предполагает-
ся при этом, что естественный отбор уже проделал ту работу,
благодаря которой выжили организмы с наиболее быстрыми и
наиболее специализированными цепями химических реакций и
хорошо дифференцированной структурой протоплазмы. Тогда
весь ряд воздействий, много раз предшествовавший жизненно
полезному фактору Г, неизбежно становится сигнальным по от-
ношению к конечному звену этого ряда, т. с. к Г, а сами цепи
последовательных химических реакций, образовавшихся на опи-
санной выше основе, предстанут перед нами как временные
связи.
Следовательно, самый факт появления «спгналыюсти» и «вре-
менных связей» может быть признан одной из древнейших зако-
номерностей развития живой материи. Именно в этом смысле
надо понимать выражение И. П. Павлова: «временная нервная
связь есть универсальнейшее физиологическое явление в жи-
вотном мире и в нас самих».
172 Методологический анализ узловых проблем условного рефлекса
Сопутствуя самому происхождению и совершенствованию
живой материи и будучи естественным следствием простран-
ственно-временной структуры мира, эта закономерность выявля-
лась естественным отбором и закреплялась все глубже и глубже
в стабилизированных структурах протоплазмы. Следовательно,
принцип опережающего отражения мира в протоплазматических
процессах организма может считаться основой для создания и
закрепления всех тех структур организма, которые целесообразно
приспосабливают его к внешнему миру на основе принципа сиг-
нальности и временных связей.
Едва ли поэтому следует удивляться, что вся эта функция ор-
ганизма, предоставляющая ему широчайшие возможности при-
способления и прогресса, стала быстро специализироваться в
особом субстрате, основной функцией которого стало быстрое хи-
мическое сцепление всех одновременно и последовательно разви-
вающихся химических реакций. Таким субстратом и стала нерв-
ная система.
С этой широкой точки зрения первичное нервное вещество и
вообще центральную нервную систему можно рассматривать как
субстрат высокой снецпализации, который развивался как аппа-
рат максимального и быстрейшего опережения последовательных
и повторных явлений внешнего мира.
У мозга как специализированного в этом направлении орга-
на нет границ для такого опережающего отражения действитель-
ности. Оп обладает возможностью отражать в мпкроинтервале
времени цепь событий, тянущуюся целые годы.
Должен заметить, что когда я сопоставляю ту высокоспециа-
лизировапную форму опережающего отражения внешнего мира,
какая представлена условным рефлексом у высших животных,
с опережающим отражением у примитивных живых существ,
я отнюдь не исключаю качественной разницы между тем и дру-
гим явлением. Но всякое сопоставление многих феноменов неиз-
бежно требует ведущего критерия для их оценки. Поэтому,
естественно, мы должны поставить перед собой вопрос: какой при-
знак условного рефлекса наиболее для него характерен?
Из всех его качеств «предупредительность», пли «сигнализа-
ция» о предстоящих событиях внешнего мира,— самое характер-
ное. Ни изменчивость, ни прочность, ни приобретенность и т. д.
не могут сравниться по значенпю с этим его биологическим каче-
ством.
Именно потому, что животные имеют возможность подго-
товиться по сигналу к еще только предстоящим звеньям после-
довательно развивающихся событий, условный рефлекс стал
центральным пунктом прогрессивной эволюции, и именно в этом
своем качестве он был открыт И. П. Павловым. Но если этот ос-
новной признак (параметр) — древнейший, то, следовательно,
по
Методологический анализ узловых проблем условного рефлекса 173
нему мы вполне можем сопоставлять приспособительные возмож-
ности низших и высших животных.
Естественно, что структурные и количественные усложнения
самих аппаратов, обеспечивающих более обширные сигнализации,
привели к дальнейшим качественным изменениям в поведении
животных, однако сигнальность как универсальный параметр
приспособления к внешнему миру пе теряла от этого своего прин-
ципиального значения.
Следовательно, условный рефлекс высших животных, оцени-
ваемый по параметру сигнальпости, есть только частный случай
высокоспециализированных форм онережающего отражения дей-
ствительности — приспособления к будущим, но еще не насту-
пившим событиям («предупредительная деятельность», по
И. П. Павлову) (Апохпп, 1949).
В свете разобранных выше исторических закономерностей
формирования биологических основ условнорефлекторной деятель-
ности становится очевидной вся искусственность ряда вопросов,
относящихся к условному рефлексу: когда в эволюции появился
условный рефлекс? Есть ли «условный рефлекс» у простейших
растений? Может ли врожденная деятельность быть сигнальной?
и т. д.
С изложенной выше точки зрения универсальным принципом
приспособления животного к условиям окружающего мира слу-
жат опережающее отражение последовательно и повторно разви-
вающихся событий внешнего мира, «предупредительное» приспо-
собление к предстоящим изменениям внешних условий, или в ши-
роком смысле — формирование подготовительных изменений для
будущих событий.
Как мы видели, этот принцип имеет силу уже с первых эта-
пов формирования живой материи. Поэтому вопрос может быть
только о форме, в какой этот принцип опережающего отражения
внешнего мира представлен на данном уровне развития. У одно-
клеточных оп выражен в форме цепей химических преобразова-
ний, опережающих развитие последовательного ряда внешних со-
бытий. У высших животных он проявляется в форме участия
специализированных нервных аппаратов, дающих огромный выиг-
рыш в быстроте опережения. Однако во всех случаях эта форма
опережающего отражения имеет одну и ту же решающую харак-
терную черту — сигнальность. Для животных, имеющих нервную
систему, это и будет условный рефлекс.
В свете сказанного теряет смысл и второй вопрос: можно ли
выработать временную связь у простейших и растений?
Поскольку и те и другие представляют собой живые образо-
вания со сложным молекулярным и многоклеточным строением и
поскольку они подвержены постоянным рядам последовательных
и повторяющихся воздействий (особенно последних), постольку
174 Методологический анализ узловых проблем условного рефлекса
существенный фактор для жизни данного
в работах мичуринской школы
известно
примеры того, как «толчок», представленный
что действует в данный мо-
можно с определенностью утверждать, что и те и другие уже име-
ют натуральные сигнальные изменения или эти изменения мож-
но выработать заново. При этом только надо соблюсти изложен-
ные выше требования: длительность повторения и достаточно
удачно выбранный
растения.
Насколько мне
имеются наглядные
осенними факторами, дает в некоторых зернах и растениях на-
чало таким процессам, которые являются, по существу, уже за-
щитой против зимних условий. Мне кажется, что вопрос выработ-
ки временных связей у растений — прежде всего вопрос подбора
адекватных условий выработки опережающего отражения.
Принципиально же следует признать, что ни одно растение не
смогло бы существовать и немедленно было бы отброшено естест-
венным отбором, если бы оно реагировало только на наличный
фактор среды, т. е. только на то
мент, а не реагировало бы по принципу опережающего отраже-
ния. Так, например, сезонные изменения температуры обычно
таковы, что могут быть различные перепады, следовательно,
внезапный мороз мог бы полностью нарушить протоплазматиче-
ские свойства растительной клетки.
Совершенно отчетливо решается также и третий, кардиналь-
ный для физиолога высшей нервной деятельности, вопрос о врож-
денных реакциях сигнального типа. Мы уже говорили, что сиг-
нальность — наиболее характерная черта для условного рефлек-
са. Именно эта черта — опережающее отражение действительно-
сти — приобрела в условном рефлексе наиболее выраженный и
специализированный вид. Однако, по общепринятой классифи-
кации, врожденные, т. е. безусловные, рефлексы пе могут иметь
самого характерного свойства условного рефлекса — сигнального
значения.
Фактический материал нашей лаборатории по изучению эко-
логической обусловленности первых поведенческих актов ново-
рожденных показал, однако, что этот вопрос не может быть ре-
шен так просто. Исследования нашего сотрудника Я. А. Миляги-
на показали, что, вылупившись из яйца, птенец грача сразу же
безотказно реагирует поднятием головы и раскрытием клюва,
т. е. пищевой реакцией, на такие раздражители, которые са-
ми по себе не имеют никакого пищевого значения (движение
воздуха, звук «кар-р-р», сотрясение гнезда). Анализ натуральной
экологической ситуации, в которой живут грачи в первые дни
после рождения, показал, что все три приведенных выше аген-
та имеют экологическое значение и служат сигналами предстоя-
щего вкладывания пищи отцом-грачом в раскрытый клюв птен-
ца (Милягин, 1957).
Методологический анализ узловых проблем условного рефлекса 175
f
Таким образом, совершенно очевидно, что мы имеем в этом
примере парадоксальный случай — «врожденный условный реф
леке», который обладает всеми чертами сигнальной деятельности.
Если рассмотреть этот феномен под углом зрения общебиоло-
гпческой закономерности «опережающего отражения действитель-
ности», то случай с птенцом окажется особенно важным для
понимания эволюции этой закономерности от низших
живых
существ к высшим, имеющим хорошо сформированную нервную
систему.
Мы отмечали, что длительная повторяемость последователь-
ных рядов внешних воздействий вызывает в протоплазме живого
существа специфическую для каждого воздействия химическую
реакцию. Эти реакции, если они повторяются на протяжении
многих веков, фиксируются в конце концов естественным отбо-
ром в стабильных протоплазматических структурах данного
организма, закрепляются огромным ускорением с помощью спе-
цифических катализаторов п отбираются как наиболее быстрая
и совершенная форма приспособления животного к внешним воз-
действиям. С этой широкой биологической точки зрения все
организованные протоплазматические структуры могут быть
признаны результатом фиксации постоянных последовательных
внешних воздействий на живой организм, если только они имели
существенное значение для сохранения жизни.
Так, например, в жизни многих животных, особенно птиц,
воздух и его движение — почти абсолютные факторы внешнего
мира, сопровождавшие эволюцию всей жизни на Земле. Неуди-
вительно поэтому, что в процессе развития приспособительных
реакций грача движение воздуха как неизбежное следствие дви-
жения крыльев матери включено было в процесс исторического
формирования такого специализированного лабильного субстрата,
как нервная система.
II поскольку не отмечалось еще случая, чтобы в истории
нашей планеты с момента зарождения жизни не было атмосфер-
ного воздуха, постольку рецепторы, воспринимающие его движе-
ние, и соответствующие реакции нервной системы вошли в длин-
ный ряд тех последовательных изменений нервного субстрата,
которые предшествуют моменту кормления. Образовались закреп-
ленные наследственностью структурные связи, обеспечивающие
опережающее отражение внешних событий принципиально так
же, как это нами было показано выше и для первичных прото-
плазматических образований.
В самом деле, движение воздуха — не «пищевой продукт», оно
не является безусловным раздражителем в том смысле, как мы
его принимаем в обычной трактовке павловской школы, но это —
истинный сигнал кормления. Все структурные предпосылки для
восприятия и реализации этого раздражения вполне созрели уже
176 Методологический анализ узловых проблем условного рефлекса
только па одном морфогенетическом основании в периоде эмбрио-
генеза.
Здесь возникает естественный вопрос: почему это возбужде-
ние кожных рецепторов птенца движущимся воздухом идет имен-
но к центральному ядру пищевой функциональной системы и
формирует уже описанную выше реакцию подготовки птенца к
приему пищи? У нас пет сомнения в том, что мы имеем здесь
наглядный пример фиксации в лабильном нервном субстрате той
цепи последовательных химических реакции, которые отражают
собой реальную последовательность воздействия внешнего мира
на нервную систему птенца в характерной для него экологии,
заканчивающейся кормлением.
Эта последовательность представлена следующими факторами:
1) вылет матери из гнезда и обнажение птенцов; 2) повторный
вылет; 3) обмахивание крыльями птенцов, лежащих в гнезде;
звук «кар-р-р»; 4) прилет отца и резкое сотрясение гнезда, когда
он садится на край; 5) вкладывание пищи в раскрытый клюв;
6) возбуждение вкусовых рецепторов и рецепторов пищевари-
тельного тракта; 7) выделение соков и переваривание пищи.
Я не отметил еще многих других факторов, которые реально
предшествуют этому ряду явлений. Дело в том, что отец приле-
тает с кормом и садится на отдаленное дерево так, чтобы ока-
заться в пределах зрительного поля матери, сидящей на птенцах.
Мать, завидя отца-грача, поднимается с гнезда и, размахивая
крыльями, дает начало описанному выше ряду последовательных
явлений.
Совершенно очевидно, что раздражение кожи птенца движу-
щимся воздухом — лишь один из промежуточных этапов этого
длинного ряда последовательных агентов, но такой, который воз-
действует на нервную систему птенца наряду с другими факто-
рами (звук «кар-р-р», сотрясение гнезда). II хотя само раздра-
жение кожи не является пищевым, возбуждение от него благо-
даря многовековым предшествованиям кормлению с огромной
скоростью (30 миллисекунд) и по готовым синаптическим свя-
зям доходит до пищевого центра. Поэтому птенец поднимает
голову и раскрывает клюв.
Здесь природа дала исключительно яркий пример опережаю-
щего отражения последовательных событий внешнего мира и его
приспособительного значения в эволюции. Такое опережающее
действительность распространение возбуждения было зафиксиро-
вано наследственностью в нервных структурах и приобрело важ-
нейшее приспособительное жизненное значение для птенцов
грача. Стоит только на минуту представить себе, что к моменту
вылупливания птенца у него недостаточно созрела нервная связь
между раздражением воздухом и пищевым центром (хотя бы
один-два синапса оказались неготовыми), как сейчас же станет
Методологический анализ узловых проблем условного рефлекса 177
ясным, что такой птенец должен быть элиминирован естествен-
ным отбором.
Интересно, что тщательные электрофизиологические исследо-
вания слуховых способностей уха грача показали поразительную
приспособленность эмбрионального развития к его постнатальной
экологии. Оказалось, что в слуховом рецепторе к моменту вылуп-
лпвания из яйца созрели лишь те клетки, которые способны вос-
принять звук «кар-р-р».
Опережающее отражение внешнего мира, будучи универсаль-
ным явлением жизни, определило собой все формы приспособи-
тельного поведения животного: «врожденное», «сигнальное»,
«условнорефлекторпое». С точки зрения этой закономерности
любой член последовательного ряда воздействий внешнего мира
может стать сигналом той конечной, жизненно важной реакции,
которой замыкается этот ряд последовательных воздействий.
Примеры, подобные разобранным, можно привести и из общей
физиологии мозга как субстрата быстрейших ассоциаций после-
довательно протекающих химических реакций. Так, например,
хорошо известно (Джаспер), что если длительное время давать
последовательно звук и свет, то в коре мозга животного эта по-
вторяемая последовательность фиксирует отношения типа опере-
жающего отражения действительности, т. е. наносимых раздра-
жений. После целого ряда применений пары «звук — свет» стоит
дать только один звук, как в мозгу, именно в том его отделе,
который воспринимает раздражение света, развиваются явления
точно такие, какие возникают, когда дан свет, хотя последний
будет включен только через несколько секунд.
Здесь совершенно очевидно опережающее отражение внешних
явлений благодаря тому, что возбуждение из слуховой области
коры больших полушарий распространилось в зрительную рань-
ше, чем туда пришел свет. В этом элементарном физиологическом
факте с исключительной отчетливостью проявляется истинная
функция центральной нервной системы: ассоциировать такие
молекулярные цепи, которые по описанной выше закономерности
способствуют предвосхищению наступающих событий. Нервный
же субстрат с точки зрения этой закономерности выступает как
субстрат, специализировавшийся па быстрое «химическое сцепле-
ние» тех молекулярных процессов, которые последовательно воз-
никают от действия последовательных агентов внешнего мира.
Одна из моих сотрудниц изучала с помощью электроэнцефа-
лографической записи, как протекает электрическая реакция
мозга па стандартную тройку раздражителей «звук — сирена —
свет». После десятков раз применения этого стереотипного ряда
раздражений можно видеть, что характерная для света реакция —
подавление альфа-ритма в зрительной области — наступает даже
в том случае, если вместо света применить звонок, который
178 Методологический анализ узловых проблем условного рефлекса
обычно не вызывает такого подавления. Это электроэнцефалогра-
фическое выражение того, что 11авлов назвал «динамическим
стереотипом». Однако в данном эксперименте мы наблюдаем не-
посредственно процессы самого мозгового субстрата. Исследова-
ния показали, что в мозге после многократного повторения одного
и того же последовательного комплекса раздражений устанавли-
ваются такие прочные цепи нейронных контактов, что даже на-
личный раздражитель (звонок) не может преодолеть их инерцию,
и потому в данном пункте стереотипа развивается реакция на
отсутствующий раздражитель — свет.
Можно показать также,
что эта способность мозга — фикси-
ровать в новых молекулярных связях последовательно повторяю-
щиеся воздействия внешнего мира — идет еще дальше.
Однако для избежания недоразумений в понимании нашей
точки зрения следует более подробно остановиться па характе-
ристике сходства и различия этих двух уровней ассоциативной
деятельности.
Логика учит нас, что при сопоставлении и сравнительной
оценке сложных феноменов (особенно биологических и физиоло-
гических) прежде всего важно точно формулировать тот крите-
рий сложных явлений, по которому они сравниваются. Опере-
жающее отражение действительности и условный рефлекс, не-
сомненно, представляют собой явления со многими параметрами,
и поэтому уже один правильный выбор критерия для их сопо-
ставления служит залогом будущего успеха.
Основным параметром для сопоставления мы берем решаке
щий биологический критерий: формы и механизмы отражения
в обоих феноменах последовательно развивающихся внешних
событий, поскольку сама последовательность является универ-
сальным законом неорганического мира.
Как известно, развитие органов чувств у животных связано
с необходимостью более совершенного приспособления к внеш-
нему миру. Органы чувств переводят различные по качеству
внешние энергии в единый нейрохимический процесс, смысл ко-
торого состоит в том, что в ответ даже на одиночный раздражи-
тель создается определенная система нейрохимических реакций
на различных уровнях центральной нервной системы. Уже один
такой раздражитель создает систему специализированных хими-
ческих реакций, состоящую по крайней мере из миллионов точек.
Она имеет определенную архитектуру и дает обширную инфор-
мацию целому организму не только о том, как и какое раздраже-
ние было нанесено, но также и о том, где и когда оно было
нанесено.
и
Таким образом, уже на самом первом этапе контакта различ-
ных организмов с внешними факторами сказывается огромная
разница в объеме той информации о внешнем мире, которую они
Методологический анализ узловых проблем условного рефлекса 179
получают даже от одиночного раздражения. Поэтому по парамет-
ру сложности распределения микроочагов химических реакций
мозг, несомненно, пе идет ни в какое сравнение с химическими
последствиями раздражения, например, уже оформившегося, но
еще примитивного живого существа. В этом последнем случае
химическое последствие раздражения элементарно разыгрывает-
ся в общей протоплазме и лишь только специфика химических
валентностей и катализа удорживаег эту химическую реакцию
в более или менее ограниченном виде.
Исторически накапливавшиеся количественные усовершенст-
вования реакции привели к совершенно очевидному новому
качеству приспособительных функций мозга. Однако, если эти
явления, т. е. опережающее отражение действительности у при-
митивных существ и условный рефлекс у высших животных, мы
будем сопоставлять по параметру отражений последовательно
развивающихся явлений внешнего мира, мы придем к выводу,
что механизм формирования опережающего отражения действи-
тельности в обоих случаях идет в точности по той схеме, кото-
рая была нами дана выше.
Химическое объединение раздельно, последовательно и повтор-
но возникающих химических реакций и «запуск» в последующем
всей этой цепи реакций с первого звена — вот что является прин-
ципиально общим параметром для обоих, столь различных по
уровню феноменов приспособления. Вместе с тем это
историческая основа для развития всех видов сигнальных приспо-
соблений, т. е. подготовка организма к будущим событиям.
Разобранные выше закономерности и иллюстрации к ним
делают еще более убедительным то, что условный рефлекс в его
основпых биологических и нейрофизиологических механизмах
представляет собой частный и весьма специализированный слу-
чай той универсальной закономерности, которая в примитивной
форме стабилизации цепных химических реакций появилась уже
на стадии примитивных живых существ.
До сих пор мы рассматривали условный рефлекс как высоко-
специализированную форму деятельности нервной системы, даю-
щую чрезвычайно ускоренное опережение событий, последова-
тельно и повторно развивающихся во внешнем мире. Это и
составляет высочайшее приспособление высших животных к
окружающим условиям.
Но возможен еще и другой, более широкий, философский
аспект этой проблемы. До Павлова в физиологии животных были
известны две категории времени — прошлое и настоящее. Обе
они были представлены соответствующими процессами, хорошо
укладывающимися в наше понимание реакций животных. Все,
что касалось третьей категории времени — будущего, относилось
к целесообразному поведению человека и было предметом почти
есть
180 Методологический анализ узловых проблем условного рефлекса
н
£орме опережающего отражения пространствепно-вре-
движения материи в неорганической
н
исключительно идеалистической психологии. Приспособление к
будущим событиям, «предвосхищение будущего» рассматривались
только как достояние человека и считались признаком наиболее
высокоцелесообразных процессов мозга.
Формулируя положение об условном рефлексе как «предупре-
дительной» функции приспособления животного к грядущим
событиям, И. П. Павлов тем самым ввел в физиологию третью
категорию времени — будущее. Отныне «будущее» в жизни орга-
низмов стало предметом такого же объективного паучпого иссле-
дования, как две другие категории времени. Таким образом,
открытие условного рефлекса было событием огромной важности
для физиологии, и в этом лежит одна из причин столь огромного
успеха изучения поведения животных под этим углом зрения.
Накопление материалов па этом пути все больше расширяло
рамки условного рефлекса, и в настоящее время мы можем твер-
до сказать, что Павлов вскрыл кардинальную и всеобщую черту
исторического развития животного мира.
Условный рефлекс есть специализированная форма того про-
цесса, который в цепях элементарных химических реакций пер-
вичной протоплазмы утверждал жизнь на первых этапах ее раз-
вития е
меннбго движения материи в неорганической природе и
образования стабильных структур организма на этой основе.
Понадобились миллиарды лет, чтобы эта примитивная форма
цепных химических реакций фиксировалась и развивалась в
самой совершенной форме живой материи — в нервном веществе.
Однако, несмотря на весь грандиозный путь этой эволюции
и значительные преобразования самих форм опережения, ее
основной закон — опережающее отражение последовательно по-
вторяющихся рядов впешних явлений — остался в силе. Он при-
обрел лишь специальный аппарат — мозг, вещество которого
высоко специализировано именно на «химическое сцепление»
последовательных и повторных воздействий внешнего мира. Это и
обусловило столь широкое и совершенное овладение будущим,
какое мы видим в поведении высших животных и человека. Так,
изучение энергетической стороны отражения, т. е. трансформа-
ции организмом внешних энергий (свет, тепло, химические агенты
и т. д.) в адекватную информацию мозга о материальных свой-
ствах внешнего мира, несомненно, является одной из очередных
задач нейрофизиологии. В настоящее время этот первичный узел
отражения может быть изучен на очень высоком уровне, если
только для этой цели будет применен весь арсенал современных
достижений (биофизика, теория информации, теория кодирова-
н
Но есть и другая форма отражения — отражение в свойствах
организма временной структуры мира, которая и была предме-
Методологический анализ узловых проблем условного рефлекса 181
сформулированной В. И. Лениным. Общеизвестно, что
•t
том обсуждения настоящей главы нашей работы. Мы видели,
что лейтмотивом этой формы отражения служит опережающее от-
ражение, что дает мозгу на высшем этапе развития организмов
совершенно адекватную информацию о последовательных и повто-
ряющихся рядах явлений внешнего мира. Здесь следует отметить,
что на основе этой формы отражения был сформирован и специа-
лизировался и сам мозг как орган психической деятельности, т. е.
орган всеобщего отражения мира в мыслительной деятельности
человека.
Все сказанное выше убеждает нас, что опережающее отраже-
ние действительности, раскрытое нами в этой главе на ряде при-
меров,— лишь одна из форм отражательной способности живой
материи.
После изложения биологической и философской аргументации
о наличии па всем протяжении развития жизни опережающего
отражения действительности мы должны определить, какое место
занимает данная закономерность в универсальной теории отра-
жения,
эта теория дает материалистическое объяснение всех процессов
организма, особенно его психической деятельности, в зависимости
от первично существующих материальных воздействий внешнего
мира на организм.
В гносеологическом смысле ленинская теория отражения —
основа для понимания материальной обусловленности психики
человека, что составляет главнейшую философскую черту этой
теории. Она прежде всего утверждает объективное существование
мира и столь же объективное и закономерное отражение его в
психической деятельности человека. Это отражение адекватное,
и потому оно служит исходным достоверным фактом для позна-
ния мира, мыслительной деятельности человека.
Теория отражения В. И. Лепина, давая общую философскую
основу, открывает широчайшие возможности для исследования
различных форм отражательной деятельности, конкретных физио-
логических ее механизмов. Так, например, уже сейчас можно
указать по крайней мере несколько форм отражательной деятель-
ности, связанных именно с временной структурой движения
материи в неорганическом мире, которые должны стать предме-
том тщательного исследования.
Марксистско-ленинская теория отражения может стать исход-
ным пунктом глубокого исследования всех уровней отражатель-
ной деятельности, от низших живых организмов до высших форм
психической деятельности человека включительно.
Настоящая глава представляет собой лишь скромную попытку
раскрыть исторические корни одной из форм отражательной дея-
тельности — условного рефлекса и временной связи.
н
182 Методологический анализ узловых проблем условного рефлекса
Опыт преобразования
рефлекторной теории Декарта
анализ рефлекторной
«классической»,— это
ней, но и поставить
принципиальную ли-
которая называется
Едва ли надо говорить, что рефлекторная теория для физио-
лога высшей нервной деятельности имеет принципиальное мето-
дологическое значение. Дать философский
теории Р. Декарта
значит не только определить отношение к
новые проблемы исследования, установить
нию в повседневной работе самого исследователя-физиолога.
На протяжении многих лет мы получали экспериментальные
результаты, которые не совпадали с общими положениями реф-
лекторной теории Декарта и потому заставляли нас создавать
более подходящие и более эффективные представления о первной
деятельности животных и человека. Уже И. П. Павлов много лет
назад называл классическую рефлекторную теорию «первобыт-
ной».
Однако в физиологии нервной системы не было ни одной
серьезной и направленной попытки устранить ее недостатки и
построить более совершенную концепцию о деятельности цент-
ральной нервной системы целостного организма.
Следует отметить при этом, что отдельные критические вы-
ступления против теории Декарта со стороны психологов не идут
здесь в расчет. Эта критика рефлекторной теории не имела своей
целью развитие и усовершенствование ее основ как теории мате-
риалистического детерминизма, что особенно ценил в рефлектор-
ной теории Павлов. Наоборот, ее критика со стороны психологов
вела или к отрицанию этого детерминизма и замене его представ-
лениями об «эквипотенциальности» мозгового вещества, или к
«растворению» его в субъективной, т. е. идеалистической, психо-
логии (Лешли и др.). В этом смысле полный «ответ физиолога
психологам» был дан самим Павловым.
Прежде чем приступить к анализу рефлекторной теории в том
ее виде, как она была дана Декартом и сохранилась до наших
дней, мне хотелось бы оговорить один важный пункт, касающий-
ся рефлекторной теории Павлова. В последние годы в многочис-
ленных публикациях, посвященных анализу павловского учения
об условном рефлексе, фигурирует такая «рефлекторная теория
Павлова», которая, на мой взгляд, по общей трактовке
ски не соответствует тому, чем обогатил эту теорию сам Павлов
и что составляет фундамент нового направления в современной
физиологии нервной системы.
В качестве исчерпывающей характеристики рефлекторной
теории Павлова некоторые авторы перечисляют те три методо-
логические принципа, которые он привел в своем знаменитом
ответе психологу Лешли. Эти принципы широко известны: прин-
рактиче-
Методологический анализ узловых проблем условного рефлекса 183
цип детерминизма, принцип единства динамики и конструкции,
принцип единства анализа и синтеза. Очень часто характеристика
«рефлекторной теории Павлова» ограничивается разбором имен-
но этих трех принципов.
Но разве они отражают действительный смысл научных до-
стижений И. П. Павлова, его революционное влияние на физио-
логическую мысль и именно на рефлекторную теорию Декарта?
Мне кажется, что такое ограничение в характеристике роли
Павлова в развитии рефлекторной теории чрезвычайно обедняет
научный смысл павловского учения об условных рефлексах и
сводит все его достоинство к формулировке общепринятых поло-
жений, обязательных для любого диалектика-материалиста, неза-
висимо от того, что он рассматривает — рефлекторную деятель-
ность или процесс клеточного деления. В самом деле, мы не
можем считать, что эти три принципа составляют то оригиналь-
ное, в чем заключается суть учения об условных рефлексах.
Фактически любой материалист-исследователь, в особенности
стоящий на позициях диалектического материализма, принимает
и в такой же форме выражает свое отношение к рефлекторным
нервным процессам организма. Разве Энгельс иначе смотрел на
развитие нервных процессов? Разве другой какой-либо исследо-
ватель-материалист, работавший задолго до Павлова, не прини-
и
что именно указанные три прин-
мал полностью этих трех принципов как руководящих для пони-
мания материальных процессов вообще всей живой природы?
Конечно, мы должны всегда подчеркивать тот огромной важ-
ности факт, что Павлов, сложившийся как исследователь в эпоху
революционных исканий русской передовой интеллигенции, сде-
лался подлинным материалистом и даже диалектиком-материа-
листом. И все же утверждать,
цппа отличают павловскую рефлекторную теорию от классиче-
ской рефлекторной теории Декарта, значило бы весьма обеднить
научный смысл и эпохальное значение всех открытий ученого
в этой области. Поэтому я хочу прежде всего отметить, что нель-
зя подменять материалистическим подходом. Вклад Павлова в
классическую рефлекторную теорию — отправной пункт для пре-
образования рефлекторной теории Декарта.
Что же внес нового И. П. Павлов в рефлекторную теорию,
определившую тип физиологического функционирования?
вестно, «рефлексом» был назван отражательный акт, в
нервный процесс проходит двойной путь от рецептора
ральной нервной системе и от центральной нервной системы к
екторным органам. Именно этот путь в силу его внешнего
сходства с дугой и был назван «рефлекторной дугой», и это
название закрепилось за ним на протяжении 300 лет в истории
физиологии, и именно эта схема составляет основу рефлекторной
теории Декарта.
Как из-
котором
к цент-
। ш ।
184 Методологический анализ узловых проблем условного рефлекса
Следовательно, так называемая классическая рефлекторная
теория — прежде всего теория о вполне определенной архитектуре
физиологического функционирования, предусматривающая впол-
не определенный путь распространения нервного возбуждения,
заканчивающийся формированием ответного акта животного.
Именно эта архитектурная часть и составляет логическую суть
рефлекторной теории Декарта.
Когда Декарт впервые сформулировал свои представления о
таком архитектурном
•I
изиологнческом
принципе, он оказал
исключительное революционное влияние на ход исследования раз-
личных поведенческих актов животных и человека. И это понят-
но. Вместо схоластически понимаемых причин поведения, вместо
спонтанных сил и всякого рода «одухотворенных» начал Декарт
совершенно отчетливо поставил вопрос о начальном материаль-
ном стимуле всякого поведения. Именно благодаря этой материа-
лизации и детерминированию поведения Декарт подвергался
большой опасности разделить судьбу Джордано Бруно и должен
был долгое время скрываться от преследовавших его церковни-
ков. Именно эта сторона учения Декарта о рефлексе особенно
привлекала Павлова, и он много раз говорил о том, что введение
понятия материального стимула послужило источником для раз-
вития физиологического детерминизма в области изучения слож-
ных поведенческих актов. Достаточно указать на то, что на про-
тяжении 300 лет с лишним и до настоящего времени эта идея
организует мысль в оценке основного закона существования
животных — рефлекторного отражения внешних воздействий.
Я не собираюсь хоть в какой-нибудь степени отрицать все
грандиозное значение рефлекторной теории Дскатра для разви-
тия физиологии! Об этом достаточно полно я говорил в своей
книге «От Декарта до Павлова», где подробно охарактеризовал
все исторические этапы развития этой идеи. Вместе с тем, как
показали открытия самого И. А. Павлова, а также богатейшие
научные материалы последних лет, теория пе может остаться
в настоящее время в ее «первобытном» виде.
Что же внес Павлов в эту трехчленную дугу рефлекса? В чем
изменилась основная физиологическая архитектура рефлекторного
акта в результате последних достижений нейрофизиологии?
Павлов внес в «первобытную» декартовскую схему два суще-
ственных дополнения: он раскрыл в рефлексе его сигнальное
значение и преодолел ограниченность декартовской теории тем,
что ввел подкрепляющий фактор как естественное продолжение
рефлекторного акта в сторону его результатов.
Это огромной важности дополнение и создало законную славу
И. П. Павлову, оно именно и составляет те отличительные черты
павловской рефлекторной теории, которые только и могли создать
основу для развития оригинальной научной школы. Такое пони-
Методологический анализ узловых проблем условного рефлекса 185
мание павловской рефлекторной теории целиком сохраняет ориги-
нальное научное значение всех его грандиозных открытий в обла-
сти физиологии мозга.
Значение этого факта выступит более отчетливо после того,
как мы произведем философскую диалектико-материалистическую
оценку декартовской рефлекторной теории.
Мне всегда казалось странным, что некоторые авторы, желая
продемонстрировать свою преданность декартовской рефлектор-
ной теории, объявляют ее чуть ли не универсальным принципом
диалектического материализма. Здесь допускается явное преуве-
личение ее философского значения и, что особенно важно, этим
самым закрепляются те недостатки рефлекторной теории, которые
уже на примере самого Декарта не замедлили дать свой отрица-
тельный философский результат.
Попытаемся разобраться более глубоко в философской сути
рефлекторной теории Декарта и значении для науки о мозге ее
конкретного рабочего принципа, получившего название «рефлек-
торной дуги».
Понятие «прерывность» отнюдь не означает остановку разви-
тия материи, абсолютную изоляцию какого-либо фрагмента из
пространственно-временного последовательного движения мате-
рии, из всеобщей взаимозависимости явлений материального
мира.
Говоря о развитии материи в связи с разбором диалектиче-
ского метода Гегеля, В. П. Ленин пишет: «С ,,принципом разви-
тия44 в XX веке (да и в конце XIX века) „согласны все44.—
Да, по это поверхностное, непродуманное, случайное, филистер-
ское „согласие44 есть того рода согласие, которым душат и опо-
шляют истину.— Если все развивается, значит, все переходит из
одного в другое, ибо развитие заведомо пе есть простой, всеобщий
и вечный рост, увеличение (respective уменьшение) etc.— Раз
так, то во-1-х, надо точнее понять эволюцию как возникновение
и уничтожение всего, взаимопереходы.— А во-2-х, если все
развивается, то относится ли сие к самым общим понятиям и ка-
тегориям мышления? Если нет, значит, мышление пе связано с
бытием. Если да, значит, есть диалектика понятий и диалектика
познания, имеющая объективное значение» (Ленин, с. 229).
Конечно, в целях исследования какого-либо феномена приро-
ды мы часто изолируем его для удобства изучения и тогда он ка-
жется как бы «изъятым» из всеобщего движения материи. Одна-
ко это только технический прием исследования, допустимый в
исследовательской работе, и оп, конечно, не равноценен призна-
нию реального существования даппого явления в изолированном
виде.
Следует специально подчеркнуть, что, изолируя явление из
непрерывного движения материи в интересах его изучения, мы
186 Методологический анализ узловых проблем условного рефлекса
допустили бы весьма серьезное отступление от принципов диа-
лектического материализма, если бы уже в самом процессе иссле-
дования не расширяли время от времени свой кругозор и не охва-
тывали причинно-следственных отношений разбираемого феноме-
на с предшествующими и с последующими этапами движения
материи.
Эту особенность познавательного процесса очень хорошо выра-
зил В. И. Ленин в своей реплике па гегелевскую характеристику
мышления: «Мы не можем представить, выразить, смерить, изоб-
разить движения, не прервав непрерывного, не упростив, угрубпв,
не разделив, не омертвив живого» (Ленин, с. 233).
Особенно действенно это «угрублепие» истинного непрерывно-
го движения материи тогда, когда в процессе исследования раз-
рывается логическая завершенность какого-либо этана в прост-
ранственно-временных преобразованиях живой природы.
В данном случае речь идет именно о тех особенностях взаимо-
действия живой материи с окружающим миром, которые мы оха-
рактеризовали избирательным отношением живых организмов к
«существенным» и «несущественным» факторам. Причем крите-
рием существенности в данном случае служит поддержание жиз-
ни, способствование ее прогрессивному развитию.
Описанная выше особенность отношения живых организмов к
окружающим условиям создает и особую форму непрерывности
движения материи — она должна иметь фазовую завершенность,
как только данный фактор окружающего мира реализовал свое
значение в обменно-жизненных циклах организма. Такая завер-
шенность отдельных этапов в непрерывном движении материи по
вносит, естественно, разрыва в эту непрерывность, но создает ка-
чественную очерченность отдельных этапов движения живой ма-
терии, которые имеют оформленную логическую архитектуру на
основе удовлетворения определенной потребности.
В жизни уже оформившихся организмов, особенно высших,
такими «узлами» в непрерывном процессе приспособления несом-
ненно являются отдельные поведенческие акты. Как сами эти по-
веденческие акты, так и их отдельные фрагменты могут быть, ко-
нечно, временно изолированы с целью их изучения. Однако часть
какой-то архитектуры поведенческого акта не может остаться на-
всегда вырванной из непрерывного потока преобразований живой
материи.
Можно проиллюстрировать эту мысль сложным жизненным
примером. Человек сидит за письменным столом и работает. Но
вот у пего появилось чувство жажды, которое оказалось доста7
точно сильным, чтобы прервать его работу. Он встает из-за стола,
идет на кухню, ищет там воду, наливает в стакан, подносит ко
рту, выпивает воду и чувствует удовлетворение жажды. Конечно,
с точки зрения общей непрерывности движения материи, имелась
Методологический анализ узловых проблем условного рефлекса 187
огромная цепь внешних явлении и ощущении, которые предшест-
вовали появлению у человека ощущения жажды. Точно так же и
после появления у него чувства утоления жажды последуют це-
лые цепи поведенческих актов.
Принципиально в жизни человека нельзя найти «перерыва не-
прерывности». Однако если мы изолируем этап, начинающийся
появлением жажды и кончающийся чувством ее утоления, мы по-
лучим из общего и непрерывного потока поведенческих актов че-
ловека вполне определенный этап, имеющий логически завершен-
ную архитектуру.
Представим себе теперь, что, начав с возникновения жажды,
мы взяли только ту цепь поведенческих актов, которая оканчива-
ется на моменте взятия в руку стакана. С точки зрения схемы
рефлекса мы имеем уже рефлекторное действие — взятие стакана.
Но с точки зрения завершенности определенного этапа поведения
мы разорвали непрерывное в таком пункте, который делает осо-
бенно трудным понимание целостного акта поведения.
Итак, «прерывание непрерывного», разрыв отдельных завер-
шенных этапов в движении живой материи как временный прием
исследования может быть допустим. Однако с точки зрения диа-
лектического материализма нельзя допустить, когда этот «пере-
рыв непрерывного» ил;
исследований логически узаконивается как ведущий принцип, как
теория и кладется в основу научного исследования природы.
Исторически именно так и произошло с рефлекторной теори-
ей Декарта, и это составляет ее «ахиллесову пяту».
Ниже я постараюсь более подробно обосновать фактическую
сторону этого положения.
Весьма прогрессивные для своего времени представления Де-
карта о рефлекторном акте поставили стимул к действию и «дуго-
образное» движение процессов по нервной системе на уровень ре-
шающих материальных факторов в развитии поведения животных
и человека. Это — то бессмертное, что всегда будет связано с
именем Декарта *.
Однако Декарт вместе с тем совершил просмотр, который
сыграл огромную роль как в деятельности его самого, так и в ра-
боте целого ряда поколений физиологов. Анализируя дугообразное
движение процессов, он довел их только до ответного действия и
на этом остановился. Как известно, в этой же форме идея рефлек-
са дошла и до пас. В действительности же на этом пи один ответ-
ный акт животных и человека никогда не кончается. Он естест-
венным образом всегда переходит в оценку нервной системой жп-
: укорочение пространственно-временных
♦ Правда, недавно один исследователь доказал, что принцип рефлекса был
сформулирован еще за 100 лет до Декарта испанским врачом Перейра.
Но для нас это сейчас не имеет значения.
188 Методологический анализ узловых проблем условного рефлекса
вотного тех результатов, которые получены были при совершении
данного действия.
Вот здесь-то и произошел тот критический «перерыв непре-
рывности», который на много лет приостановил материалистиче-
ское исследование целостного поведения. Целостное поведение
связывает все моменты деятельности в единую завершенную цик-
лическую систему, и никогда самый момент ответного действия не
может иметь места без немедленной посылки информации о ре-
зультатах этого действия, т. е. о его достаточности или недоста-
точности для животного в приспособительном смысле. Никакое
координированное поведение высших животных не было бы воз-
можным, если бы от каждого этапа сложных поведенческих актов
пе поступало информации о том, что они выполнены в соответст-
вии с исходным замыслом.
Так, например, вам понадобился карандаш и вы протягиваете
руку, чтобы его взять. Налицо все элементы декартовской «дуги
рефлекса». Был стимул — вид карандаша. Это зрительное раздра-
жение перебросилось на эффекторные пути, что вызвало соответ-
ствующее сокращение мышц — рука приблизилась к карандашу.
С точки зрения классической рефлекторной теории рефлектор-
ный акт закончился действием, т. е. «рефлекторная дуга» сформи-
рована. Однако центральная нервная система должна получить
информацию о том, что ваша рука прикоснулась к карандашу и
взяла именно его. В самом деле, вы могли, отвернувшись па
секунду, взять в руку по ошибке и другой предмет. Обычно вы
сейчас же исправляете ошибку. Однако классическая рефлектор-
ная теория не дает нам оснований для понимания этого момента.
Опа с самого начала оставила в стороне вопрос, что же происхо-
дит в конце самого действия, поскольку она искусственно выр-
вала понятие «дуги» из циклически непрерывного, последователь-
но развивающегося ряда физиологических явлений.
Между тем окончательным моментом, завершающим действие,
служит прикосновение к карандашу и получение от него суммы
своеобразных раздражений, в комплексе определяющих все пара-
метры этого предмета (вес, температура, форма и т. д.). Сюда же
надо отнести и зрительное раздражение в виде приближения ру-
ки к карандашу. Мы видим, что в этом пункте целостного поведе-
ния происходит стык между тем, что было, и тем, что будет, по-
скольку в конце действия к центральной нервной системе неиз-
бежно направляется целый поток разнообразных афферентных
возбуждений от различных рецепторных областей. Именно прихо-
дом в центральную нервную систему этого сложного интеграла
обратных афферентных импульсаций от результатов данного
действия и сопоставлением его с основным намерением к со-
вершению данного действия и завершается рефлекторная ре-
акция.
Методологический анализ узловых проблем условного рефлекса 189
Перед нами уже давно возник вопрос: является ли эта аффе-
рентная сигнализация о результатах действия нужной для даль-
нейшего поведения животного и в какой степени? Опыты показа-
ли, что получение такой афферентация, которую мы назвали «об-
ратной», или «санкционирующей афферентацней», абсолютно
необходимо для того, чтобы окончить данный поведенческий акт
или, наоборот, сформировать другой, с более успешным результа-
том.
Доведя рефлекс только до действия, Декарт остановился в не-
решительности перед проблемой всегда целесообразных рефлек-
торных актов животного, но, оборвав развитие целого процесса и
потеряв таким образом материальный аппарат целесообразности,
он вынужден был пойти для ее объяснения на целый ряд допуще-
ний вроде «высшего разума» и других нематериальных факторов.
Таким образом, начав с совершенно материалистического понятия
стимула, Декарт не мог объяснить всегда целесообразных ответ-
ных действий животного, и это сделало его дуалистом.
Впервые с этой проблемой мы встретились в 1932 г. при изу-
чении компенсаторных приспособлений животного к тому или
иному нарушению его функций.
Производя разнообразные операции с перекрестом нервных
стволов, мы столкнулись с фактом, что отнюдь не всегда нарушен-
ная функция может восстановиться немедленно. Оказалось, что
прежде чем восстановить полностью какую-либо функцию, живот-
ное должно несколько раз произвести разнообразные движения,
пока какое-либо из них не окажется приемлемым; оно и закрепля-
ется. Мы, конечно, пе могли пе задуматься над физиологическим
механизмом, который руководит подбором этих движений и, про-
ведя ряд контрольных опытов, увидели, что от каждого этапа вос-
становления нарушенной функции в центральную нервную систе-
му идут потоки обратных афферентных возбуждений, сигнализи-
рующих о результатах совершенного действия. И только после
того, как эта обратная афферентация окажется достаточной и
адекватной для поставленной цели движения, данный этап пове-
дения закапчивается и животное переходит к следующе-
му этану.
Таким образом, мы показали, что «рефлекторная дуга» не
кончается действием, а результаты данного действия возбуждают
определенный интеграл афферентных импульсаций. Эти обратные
афферентные воздействия идут в центральную нервную систему
и информируют ее о достаточности или недостаточности совер-
шенного акта.
Мне понятно, почему на протяжении 300 лет этот, казалось
бы, простой факт — информация о результатах действия — не был
специально обработан и не повлиял на представление физиологов
того времени об общей архитектуре рефлекса. Дело в том, что фи-
190 Методологический анализ узловых проблем условного рефлекса
зиологи XVIII и XIX вв., воодушевленные возможностью дробить
центральную нервную систему на отдельные детали и рефлектор-
ные дуги, почти все свое
внимание направили на острые экспери-
менты, вивисекцию. Но имеино в этих-то условиях, под наркозом
и на операционном столе, животное никогда не нуждается в об-
ратной афферентации, т. е. в оценке совершенного действия. Имен-
но поэтому вивесекционная методика отвлекла внимание иссле-
дователей от разработки продолжения классического рефлекса в
сторону обратной афферентации о результатах действия.
Рефлекторная теория Декарта разорвала естественную группу
последовательно развивающихся явлений и сделала изолирован-
ный фрагмент этого целого циклического комплекса в форме «ду-
ги рефлекса» принципиальным теоретическим постулатом. Она
изолировала действие от его результатов и, разорвав единый ло-
гически оформленный и завершенный комплекс действий, на це-
лое столетие скрыла от взора исследователей материальный ме-
ханизм целесообразного поведения животного. Этим самым вся
проблема целесообразности поведения, особенно врожденного, бы-
ла отдана на долгие годы идеализму и витализму.
Возникает естественный вопрос: как сам II. II. Павлов, введя
хронический эксперимент для изучения поведения животного,
смотрел на «первобытную» дугу рефлекса? Здесь мы встречаемся
с одним досадным обстоятельством, которое, несмотря на огром-
нейшие оригинальные достижения школы Павлова, неоправданно
укрепило в физиологии высшей нервной деятельности позиции
классической, т. е. декартовской, теории рефлекса.
Ввиду особой важности этого момента в жизни нашей школы
я позволю себе более подробно остановиться на нем. При обсуж-
дении недостаточности рефлекторной теории Декарта мне не раз
приходилось слышать замечания такого рода: «так никто теперь
и не думает», «Павлов уже давно отказался от нее» и т. д. Сло-
вом, создается впечатление, что уже нет «дуги рефлекса» и уже
нет всех тех трактовок нервной деятельности, которые на ней
построены. На самом деле, как это, впрочем, ясно каждому рабо-
тающему в области физиологических наук, дело обстоит совер-
шенно иначе. «Рефлекторная дуга» и по настоящее время служит
основой всей педагогической работы в области физиологии в ме-
дицинских и биологических вузах, так же как и основой общих
лредставлений о нервной деятельности в физиологических лабора-
ториях. Я не хочу сказать, что это неправильно. Нет, это необхо-
димый этан изучения реактивных особенностей нервной системы
в условиях острого эксперимента. Но понятия «рефлекторная
дуга» абсолютно недостаточно, когда мы подходим к изучению
физиологического фундамента поведенческих актов.
Именно в связи с этим нам важно привести точку зрения на
этот счет самого И. II. Павлова.
Методологический анализ узловых проблем условного рефлекса 191
Приступая к своим систематическим лекциям по физиологии
высшей нервной деятельности, которые мне посчастливилось слы-
шать, И. П. Павлов говорил следующее о своем отношении к
рефлекторной теории Декарта: «Основным исходным понятием у
нас является декартовское понятие, понятие рефлекса. Конечно,
оно вполне научно, так как явление, им обозначаемое, строго де-
терминируется. Это значит, что в тот или другой рецепторный
нервный прибор ударяет тот или другой агент внешнего мира или
внутрепнего мира организма. Этот удар трансформируется в нерв-
ный процесс, в явление нервного возбуждения. Возбуждение по
нервным волокнам, как проводам, бежит в центральную нервную
систему и оттуда благодаря установленным связям но другим
проводам приносится к рабочему органу, трансформируясь в свою
очередь в специфический процесс клеток этого органа. Таким
образом, тот или другой агент закономерно связывается с той или
другой деятельностью организма, как причина со следствием»
(Павлов, т. 4, с. 22).
Приведенная цитата ясно показывает, что в основу своих
представлений о сущности приспособления животного к внешнему
миру И. П. Павлов положил идею рефлекса именно в его класси-
ческом выражении, поскольку в его формулировке имеются три
звена и сам рефлекс заканчивается действием.
Но тогда возникает прежний вопрос. Как мы уже указывали,
введя подкрепляющий фактор как обязательное условие выработ-
ки условпого рефлекса, Павлов, несомненно, преодолел ограничен-
ность декартовской схемы. Именно это создало то новое и ориги-
нальное, что было положено в основу всего учения о высшей
нервной деятельности. Однако это несомненное расширение де-
картовской схемы рефлекса Павлов пе сформулировал как преодо-
ление ее важнейшего недостатка, а рассматривал как «прибавку»,
которая лежит в пределах той же «дуги рефлекса».
Отвечая Лешли, он писал: «Учение об условных рефлексах
бесспорно утвердило в физиологии факт временной связи всевоз-
можных (а пе определенных только) как внешних, так и внутрен-
них раздражений с определенными единицами деятельности орга-
низма, т. е. рядом с проведением нервных процессов в высшей
центральной станции точно констатировало также замыкание и
размыкание их. Через эту прибавку, конечно, никакого сущест-
венного изменения в понятии рефлекса не произошло» (т. 3, кн.
2, с. 170) (курсив мой.— П. Л.).
Итак, мы можем теперь заключить, что. открыв закономерно-
сти образования условных рефлексов у животного в естественных
условиях, которые явно преодолевают ограниченности декартов-
ской схемы («дуга рефлекса»), Павлов тем по менее стремился
включить это открытие в рамки существующей классической реф-
лекторной теории.
192 Методологический анализ узловых проблем условного рефлекса
Именно это и определило парадоксальное положение классиче-
ской рефлекторной теории в высшей нервной деятельности. Из
всего смысла открытия условного рефлекса следовало, что эта тео-
рия уже не могла существовать в прежнем виде, тем пе менее
официально опа считалась основой всего учения о высшей нерв-
ной деятельности.
Прямым следствием такого сочетания противоречивых тенден-
ций было то, что условный и безусловный рефлексы еще до сих
пор рассматриваются как две отдельные разомкнутые рефлектор-
ные дуги, а выработка условного рефлекса — как ассоциация та
ких же разомкнутых рефлексов.
В результате такого положения непрерывный, циклически раз
вивающийся поведенческий акт, начинающийся с условного сти
мула и заканчивающийся в качестве обратной афферентация по
лучепием пищи, фактически стал рассматриваться как комплекс
нескольких отдельных рефлекторных дуг.
Мы видели уже, что в данном случае речь идет об обратной
афферентация, приходом которой в центральную нервную систе
му пепременно завершается каждый поведенческий акт, и имен
но только после этого завершения животное может перейти к сле-
дующему этапу своего поведения.
В этом случае безусловный раздражитель в виде действия пи-
щевого вещества (подкрепление) становится источником двух
воздействий. С одной стороны, это возбуждение направляется к ап-
паратам сопоставления, завершающим условный акт, с другой —
к подкорковым пищевым центрам, начинающим весь необходи-
мый процесс пищеварения. Этот факт дает нам основание счи-
тать, что и в данном случае единицу интегративной деятельности
целого организма составляет также циклическое образование,
в которое рефлекс вписан лишь как наиболее важный стержневой
его
н
рагмент.
При сопоставлении данных, полученных при изучении поведе-
ния собак, птиц и других животных, мы все время видели, что
обратная афферентация о результатах действия — абсолютно не-
пременное составное звено любого приспособительного акта. Од-
нако уже па первых шагах наших исследований неизбежно воз-
ник вопрос, что же является «адресатом», получающим обратную
афферентацию с периферии в конце рефлекторного действия? Ка-
кой аппарат в центральной нервной системе получает ее, где и
как происходит оценка этой обратной афферептацпи и сопостав-
ление ее с тем, что предназначалось делать, прежде чем началось
формирование самого действия?
Вот здесь-то, в этом узловом пункте, и замкнулась вся цепь
фактов, начиная от опережающего отражения последовательности
внешних явлений у примитивных живых существ и кончая целе-
направленным, так называемым произвольным действием высших
ЗЕ
Методологический анализ узловых проблем условного рефлекса
193
животных и человека, В ряде экспериментов вновь подтверди-
лась идея о распространении опережающего отражения действи-
тельности. Было показано, что в самом начале действия различ-
ных условных раздражений возбуждение от них стремительно
распространяется по всем тем афферентным следам, которые
остались от прежних повторений данного раздражения и тех аф-
ерентных (всегда множественных) возбуждений, которые явля-
лись следствием полученных результатов. Все мельчайшие аффе-
рентные признаки прошлых результатов от данного действия не-
н
медленно воссоздаются в ответ на условный раздражитель,
и раньше, чем сформировалось и закончилось само рефлекторное
действие, уже наперед формируется афферентный аппарат оцен-
ки возможных результатов еще только предстоящего действия»
Этот своеобразный афферентный аппарат мы назвали «акцепто-
ром действия», что означает аппарат, который принимает обрат-
ную афферентацшо и сопоставляет ее с тем, что составляло цель
данного действия, и в зависимости от результата этого сопоставь
ления может начаться формирование нового, более точного, ответ-
ного действия.
Этот афферентный аппарат должен отражать все мельчайшие
признаки (параметры) того результата, который раньше полу-
чался при данном действии.
Л что такое «результат» в афферентном выражении? Можно
на простом примере показать обширность и динамичность этого
нервного образования.
Давая в обычных экспериментальных условиях пищевой раз-
дражитель, например хлеб или мясо, мы обозначаем его поняти-
ем «безусловный раздражитель». Стоит, однако, посмотреть на
этот безусловный раздражитель с точки зрения результатов топ-
ких электрофизиологических исследований, как перед нами от-
кроется замечательный мир микропроцессов, определяющих в
конечном счете всю изменчивость поведения животных и чело-
века.
В самом деле, действие безусловного раздражителя складыва-
ется из раздражающего действия целого ряда специфических па-
раметров данного раздражителя: тактильный, температурный, хи-
мический и т. д. Каждое из этих свойств безусловного раздражи-
теля имеет свой рецептор, свой порог раздражения, скорость
и путь распространения и, наконец, различную локализацию в ко-
ре больших полушарий. Таким образом, безусловный раздражи-
тель для мозга — это целый уз<ор афферентных возбуждений, рас-
положенных в различных пунктах головного мозга, органически
ассоциированных данным безусловным раздражителем. Эта кор-
ковая система возбуждений является в подлинном смысле «отра-
жением» качеств данного безусловного раздражителя, отличаю-
и
[их его от другого.
7 П. К. Анохин
194 Методологический анализ узловых проблем условного рефлекса
Но мы взяли только одно действие безусловного раздражите-
ля, именно прямое действие на поверхность языка. Если же к
этому прибавить действие его «условно натуральных» качеств и
многих других, сопровождающих прямое действие, то мы должны
будем представить себе весьма разветвленную локализованную
в различных пунктах коры и подкорки систему возбуждений, ко-
торая и будет в целом представлять собой афферентный резуль-
тат действия, т. е. обратную афферептацию в случае реального:
подкрепления.
Анализ человеческого поведения, так же как и свободного по-
ведения животпых, убедил нас в том, что акцептор действия есть,
универсальный аппарат сопоставления заданного с получившим-
ся и формируется он целиком па основании опережающего отра-
жения действительности благодаря быстрому распространению-
возбуждений по сложнейшим и длинным нейрональным систе-
мам, образованным в прошлом опыте, относящимся к данной си-
туации и данным пусковым раздражителям.
Надо иметь в виду, что эта обширная система возбуждений,
представленная разными фрагментами различных анализаторов,
не могла бы сделаться единым функциональным целым, если бы
все ее многочисленные компоненты пе объединялись динамически
единым энергетическим источником.
Долгое время мы искали ту нервную организацию, которая
могла бы «сцементировать» эти многочисленные звенья, однако
только последние достижения в области общей нейрофизиологии.
именно открытие активирующего
восходящего действия па кору
мозга со стороны ретикулярной формации, заполнило тот пробел,
который мы ощущали при построении нашей концепции (Ано-
хин, 1955, с. 16).
Как показали наши исследования, вопреки концепции о «пе-
специфической активации» подкорковые аппараты производят из-
бирательную энергетическую активацию корковых клеток, опре-
деляя тем самым их объединение в большие интегративные обра-
зования, всегда имеющие функциональный смысл, например ак-
цептор действия (Апохин, 1960, 1961).
Таким образом, «первобытная» форма рефлекса в самом деле
ныне не может удовлетворить пас, и мы должны ее расширить,
введя в нее специальные механизмы, которые формируются в
нервном субстрате благодаря последовательным и повторяющимся
воздействиям на нервную систему животного и опережающим эти
воздействия на данном этапе фиксации приобретенного опыта.
В целом мы ввели в окончательную физиологическую архи-
тектуру условного рефлекса несколько новых компонентов, кото-
рые пополнили понимание условного рефлекса как универсальной
и
единицы поведения животных и человека. Мы ввели понятия а<
ферентного синтеза, обратной афферентации и акцептора деист-
Методологический анализ узловых проблем условного рефлекса 195
вия, которые все, вместе взятые, представляют собой органиче-
ские составные части не только условного рефлекса, но и любого
целостного поведенческого акта, в особенности целенаправленно-
го поведения.
Выше мы разобрали роль обратной афферентации и акцепто-
ра действия как аппаратов оценки уже совершенного действия.
Остается лишь отметить физиологическую роль и состав той ста-
дии, которую мы назвали стадией афферентного синтеза.
Но нашим обычным представлениям, навеянным понятием
«рефлекторной дуги», в ответ на раздражающий условный стимул
процесс нервного возбуждения приходит сначала в «корковое
представительство» данного анализатора и данного условного
рефлекса, затем переходит в представительство данного безуслов-
ного рефлекса и, наконец, весь процесс завершается на эффектор-
ных путях условнорефлекторным ответом.
Как показали исследования, на самом деле нет такого линей-
ного и поступательного распространения возбуждения в форме
дуги, ибо в каждом узловом пункте возбуждение обогащается
циклическими взаимодействиями с периферией, например в про-
цессе осуществления ориентировочно-исследовательских реакций
животного. При каждом поисково-исследовательском движении
животное получает огромное количество афферентных информа-
ций из внешнего мира. Эти обратные афферентации от результа-
тов ориентировочно-исследовательских реакций сопоставляются с
ностоянно действующими многообразными раздражениями от
данной обстановки,
обогащаются возбуждениями из прошлого
оныта, и на основании всего этого многоооразного и разносторон-
него в отношении состава афферентных воздействий комплекса
возбуждений происходит то, что мы назвали стадией афферентно-
го синтеза. Как мы не раз показывали, например, в опытах с уда-
лением лобных отделов коры, эта стадия совершенно необходима
в общей физиологической архитектуре поведения животных и че-
ловека.
В самом деле, только в результате этого афферентного син-
теза и могут формироваться замысел действия или цель предсто-
ящего действия, в существовании которых ни один думающий че-
ловек не сомневается. Как бы мы ни назвали этот важный этап,
после которого начинают формироваться дальнейшие этапы по-
веденческого акта, важно то, что этот вполне очерченный но фи-
зиологическому составу этап реально имеет место и только после
его окончания начинают формироваться соответствующие фраг-
менты поведения — акцептор действия, само эффекторное дейст-
вие и обратная афферентации о результатах действия.
Интересно отметить, что кибернетика, моделируя отдельные
свойства мозга, упустила из виду этот важный этап в развитии
каждого поведенческого акта. Именно он, афферентный синтез,—
196 Методологический анализ узловых проблем условного рефлекса
самый ответственный узловой пункт поведения, поскольку в этой
W
азе из миллионов возможностей действия выбирается определен-
ное действие, точно соответствующее стимулу, обстановке, прош-
лому опыту и состоянию организма в данный момент.
Если исходить из сравнительной оценки временных парамет-
ров всех фаз условной реакции, особенно поведенческого много-
этапного акта человека, то стадия афферентного синтеза оказыва-
ется максимально быстрой. В то время как поведенческий акт
(особенно целая серия их) развивается обычно на протяжении зна-
чительного времени, сам момент решения «что делать?», т. е. аф-
ферентный синтез, складывается почти моментально.
Например, переход через площадь большого города требует
вполне ощутимого времени, в то время как решение пройти эту
площадь с учетом всей сложной ситуации — потоков транспор-
та, светофоров ит. д.- происходит почти моментально.
Мы считаем, что введение понятия афферентного синтеза как
важного промежуточного этана в формировании поведенческого
акта необходимо не только потому, что оно ставит новые вопро-
сы для дальнейших физиологических исследований. Важно и то,
что это понятие необходимо также и для тесного контакта с пси-
хологией, которая в понятии афферентного синтеза получает фи-
зиологически оформленный фундамент для объяснения целена-
правленных поведенческих актов и многих понятий чисто психо-
логического характера («заключение», «интуиция» и т. д.).
Если стадию афферентного синтеза понимать в широком смыс-
ле, именно как обработку афферентных сигналов перед формиро-
ванием действия, то в разной степени сложности она оказывается
обязательной для любой деятельности, включая и деятельность на
спинальном уровне. Даже простой рефлекторный акт на спиналь-
ном уровне совершается в результате синтеза многих возбужде-
ний, конвергирующих к моторному нейрону. Дальнейшее редуци-
рование этих многочисленных возбуждений может быть представ-
лено, скажем, в рефлекторном акте, совершающемся в пределах
одного или двух сегментов. Но даже и в этом случае афферент-
ный синтез представлен суммой разных по качеству возбуждений,
приходящих с периферии по различным системам афферентных
волокон, что п определяет некоторый, весьма редуцированный,
конечно, афферентный синтез.
В этом случае понятие «синтез» служит синонимом необходи-
мого количества афферентных импульсов для того, чтобы произо-
шел разряд на конечных нейронах.
Наоборот, примером максимального объема этой стадии аффе-
рентного синтеза может служить какой-либо ответственный посту-
пок человека, которому предшествовала долгая и упорная работа
по предварительной оценке многочисленных афферентных пред-
посылок в виде поводов, причин, сопоставлений их с прошлым
Методологический анализ узловых проблем условного рефлекса 197
опытом, предвосхищений того, что может быть в будущем и т. д.
Но независимо от широты этой стадии афферентного синтеза де-
ло кончается формированием вполне определенных эффекторных
возбуждений определенного назначения.
В нашей лаборатории с помощью специальной кибернетиче-
ской модели дыхательной системы па основе электронных схем,
оценивающих состояние дыхательного центра, была показана воз-
можность конструирования на этой основе замкнутой саморегули-
рующейся системы. В этой единой системе, наполовину живой,
наполовину механической, дыхательный нервный центр своими
собственными возбуждениями может управлять работой искусст-
венного дыхательного аппарата, обеспечивающего поступление
воздуха в легкие (В. А. Полянцев).
Прежде всего необходимо отметить, что формирование ком-
плекса эффекторных возбуждений в моторной части дыхательно-
го центра есть результат синтеза многих приходящих к нему раз-
нородных афферентаций. Дыхательный центр получает афферен-
тацию от легкого, воздухоносных путей, например трахеи. От
уровня парциального давления СО2 и О2 крови оп получает так-
же гуморальное возбуждение.
Принципиально здесь также необходим какой-то афферентный
синтез, ибо только от интегрального значения всех этих аффе-
рентаций возникает очередное возбуждение вдоха сначала в клет-
ках дыхательного центра, а затем и в мышечных элементах ды-
хательного аппарата. Объем этого эффекторного возбуждения
всегда находится в прямой зависимости от потребности орАаниз-
ма в окислительных процессах на данный момент. Эти закономер-
ности были очень хорошо показаны как в эксперименте, так и в
хирургической клинике на примере пульмонэктомии. Результаты
экспериментов заставляют думать, что в тот момент, когда закан-
чивается афферентный синтез и начинает формироваться возбуж-
дение моторной части дыхательного центра, оказывается сформи-
рованным также и добавочный афферентный аппарат, играющий
роль акцептора действия.
Что же будет принимать с периферии в виде обратной аф-
ферентации этот элементарный акцептор действия? Такими об-
ратными афферентациями для него выступают афферентные им-
пульсы прежде всего от альвеол легкого, которые служат абсолют-
ным показателем количества принятого воздуха и, следовательно,
доставляют дыхательному центру самую исчерпывающую инфор-
мацию о достаточности конечного приспособительного результата,
т. е. объема вдоха.
Как показал в нашей лаборатории К. Д. Груздев, афферента-
ция от альвеол легкого в норме оказывается ведущей для интегра-
ции дыхательного акта в целом. Наоборот, афферентные импуль-
сации, например, от трахеи, слизистой поса и т. д., являясь только
198 Методологический анализ узловых проблем условного рефлекса
региональными афферентациями, не могут определить всю инте-
грацию целостного дыхательного акта.
Учитывая, таким образом, решающую роль легочной афферен-
тации, мы построили эксперимент, носящий в нашей лаборато-
рии название «внезапной подмены подкрепления (обратной аффе-
рентации)». Смысл его состоит в том, что искусственно и вне-
запно создается «рассогласование» между акцептором действия п
поступающей к нему обратной афферентацней. Такой прием,
употребленный нами впервые в обстановке работы по секреторно-
двигательной методике (подмене безусловного раздражителя),
позволил нам впервые увидеть, что данное условное возбуждение
(во время действия условного раздражителя) по своему качеству
соответствует качеству предстоящего безусловного подкрепления.
Условный рефлекс оказывается «закрытым» только в том случае,
если обратная афферентация от подкрепляющего фактора точно
соответствует акцептору действия.
В полумеханической модели дыхательной системы, разрабо-
танной В. А. Полянцевым в нашей лаборатории, постановке экс-
перимента предшествовали следующие соображения.
Как мы видели, дыхательный центр посылает к дыхательным
мышцам такое количество возбуждений, которое точно соответст-
вует происшедшему в нем интегрированию всех поступивших к не-
му афферентных возбуждений, т. е. он точно отражает потреб-
ность организма в определенном объеме воздуха (К. Д. Груздев).
А это, в свою очередь, значит, что альвеолы расширятся в пол-
ном соответствии с объемом сокращения дыхательных мышц и,
следовательно, дыхательный центр должен получить от легкого,
т. е. от результатов данного вдоха, такую обратную афферента-
цию, которая в количественном отношении точно коррелируется с
эфферентными возбуждениями, посланными самим же дыхатель-
ным центром на периферию, к дыхательным мышцам. Этим за-
мыкается круг саморегулирующейся дыхательной системы.
Но возникает естественный вопрос: какой аппарат или клеточ-
ный комплекс принимает на себя эту обратную афферептацию об
объеме принятого легким воздуха? И вообще, должен ли быть
этот промежуточный, воспринимающий обратную афферептацию
аппарат, оценивающий ее достаточность?
Есть несколько физиологических соображений и фактов, ко-
торые заставляют думать о существовании такого промежуточно-
го аппарата тина весьма элементарного акцептора действия.
Прежде всего этим аппаратом не может быть самый конечный
нейрон, поскольку он сам — исполнитель сложившегося на дан-
ный момент афферентного синтеза. Кроме того, соотношения
между обратной афферентацней от легкого и посылкой дыхатель-
ного «зална» на периферию таковы, что они как раз противоречат
тому, что можно было бы ожидать от такого предположения.
Методологический анализ узловых проблем условного рефлекса 199
В самом деле, пульмонэктомия в эксперименте и клинике по-
казала, что удаление легкого, т. е. полное устранение обратной
аффереитацип па данной стороне, приводит к резкому увеличе-
нию возбудимости и количества разрядов именно на этой полови-
не дыхательного центра. Эти явно парадоксальные отношения
(уменьшение стимуляции приводит к повышению количества раз-
рядов) говорят о том, что имеется какой-то промежуточный ап-
парат, который после получения обратной афферентацип от лег-
кого производит ее обработку, адекватную и в соответствии с
достаточностью приспособительного эффекта на периферии. Если
это так, то этот аппарат выполняет функцию, подобную функции
акцептора действия в сложных поведепческих актах.
Полуэлектроппая модель дыхательной функциональной систе-
мы дает возможность проверить это соображение постановкой
упомянутых выше опытов с впезапной подменой обратной аффе-
рентации («рассогласование» в кибернетике).
С помощью электромеханических приспособлений можно было
внезапно уменьшить количество обратных афферентных импуль-
сацпй. Этим самым дыхательный центр, пославший на перифе-
рию необходимое для данной газообмешюй ситуации количество
нервных пмпульсаций, оказывается «обманутым»: он получает от
легкого такую обратную афферентацию, которая информирует его
так, как будто бы оп не дослал необходимое количество импуль-
сов.
Опыт показывает, что па это «рассогласование» между послан-
ной от центра и обратно полученной информациями от легкого
дыхательный центр немедленно реагирует значительным увеличе-
нием посылки своих пмпульсаций к дыхательным мышцам; в ре-
зультате наступает компенсаторное увеличение вдыхаемого воз-
духа.
Такой тип эксперимента воочию убеждает в том, что одновре-
менно с окончанием афферентного синтеза и посылкой на перифе-
рию определенного количества эффекторных возбуждений дыха-
тельный центр формирует и акцептор результатов будущего дей-
ствия, который служит своеобразным эталоном для сопоставления
того, что послано на периферию, с тем, что там произошло.
Следует, однако, подчеркнуть, что это всегда динамическое об-
разование, ибо с каждой повой газообменной ситуацией этот ап-
парат будет толкать па компенсацию и при другом количестве
обратных пмпульсаций.
Итак, мы видим, что формирование аппаратов типа акцептора
действия — закономерность центральной нервной системы. Осо-
бенно выпукло она представлена, конечно, в сложном целеустрем-
ленном поведении. Там акцептор действия является основным ме-
ханизмом, входящпхм и в оценку результатов целеустремленного
поведения. Однако проблема цели в ее физиологических механпз-
200 Методологический анализ узловых проблем условного рефлекса
так и обеспечивающих все этапы
•I
мах нами была разобрана в другом месте (Анохин, 1962), поэто-
му сейчас мы оставим ее в стороне.
Сопоставляя все особенности замкнутых функциональных об-
разований как вегетативных
ормирования поведенческого акта, мы сформулировали в 1935 г.
понятие ’ функциональной системы как единицы интегративной
деятельности организма.
Как можно было видеть из разобранных выше примеров,
функциональная система представляет собой динамическое обра-
зование, в котором возбуждение циркулирует по многочисленным
циклам, обладающим своей специфической ролью и своим конеч-
ным приспособительным эффектом в осуществлении поведенче-
ского акта. Эта организация есть в подлинном смысле слова
функциональная система, имеющая непременно центрально-пери-
ферическую архитектуру и, конечно, всегда развертывающаяся на
конкретном материальном субстрате. К этой системе не приложи-
мо понятие рефлекса в форме его трехчлеппой архитектуры, хотя,
конечно, рефлекс и составляет какую-то органическую часть этой
функциональной системы. Функциональная система условного
рефлекса, например, была пополнена тремя важными факторами,
о которых речь шла выше (афферентный синтез, акцептор дейст-
вия, обратная афферентация).
Понятие функциональной системы и обратной афферентация
нами было предложено за 11 лет до развития кибернетики. Дан-
ная нами общая характеристика функциональной системы как
физиологического замкнутого образования полностью предвосхи-
тила кибернетическое понимание циклических систем организма
с «обратной связью».
Я не имею возможности разбирать здесь все общие физиоло-
гические свойства функциональной системы как целостного об-
разования, на основе которого развиваются все многообразные
деятельности организма до самых высших включительно. Для
доказательства абсолютной необходимости этого принципа при ис-
следовании интегративной деятельности целого организма приве-
ду несколько примеров.
Так, например, сумма всех аппаратов и механизмов, поддер-
живающих постоянное осмотическое давление крови, представля-
ет собой совершенно отчетливую функциональную систему с ко-
нечным приспособительным эффектом (константа осмотического
давления). Эта функциональная система действует на основе тех
же принципиальных закономерностей, как и более высоко орга-
низованные функциональные системы человека, например речь,
приспособительное поведение человека и даже его психическая
деятельность.
Почему функциональная система является столь общим прин-
ципом функционирования для столь разнообразных функций ор-
Методологический анализ узловых проблем условного рефлекса 201
ганизма? На этот вопрос ответить легко. Центральным пунктом
функциональной системы любого уровня организации служит
конечный приспособительный э
>ект. Именно он
представляет
нЦ|
А А
собой организующий фактор, поскольку всякие отклонения от его
константного или заданного уровня сейчас же с помощью обрат-
ной афферентации вызывают к жизни целый ряд новых приспо-
собительных реакций, пока пе установятся адекватные отноше-
ния между результатами акта и исходным намерением, отражен-
ным в акцепторе действия.
Можно привести пример из области более сложных форм по-
ведения. Так, например, если при произношении какой-либо фра-
зы отдельное слово, произнесенное человеком, оказалось не соот-
ветствующим уже ранее сформировавшейся мысли и фразе
уже сформировавшемуся акцептору действия, то последний бла-
годаря звуковой обратной афферентации и «рассогласованию» с
заданным афферентным аппаратом способствует исправлению
ошибки и ведет к формированию нового слова, более адекватного
исходному намерению.
Описанный выше тип «подгонки» конечного приспособитель-
ного эффекта к исходному намерению осуществляется принци-
пиально одной и той же системой отношений, хотя, конечно,
эта система в различных случаях будет иметь огромную разницу
но составу участвующих компонентов и исторической предпосыл-
ке для вхождения каждого из них.
Вторым важным признаком функциональной системы служит
наличие описанных выше аппаратов афферентного синтеза: ак-
цептора действия и обратной афферептацпи. Эти аппараты не-
прерывно поддерживают соответствие между намерением и ре-
зультатами действия, создавая меняющееся, но координированное
взаимоотношение между многочисленными компонентами каждой
функциональной системы.
Как мы уже видели выше, здесь неизбежно должен был воз-
никнуть вопрос: каким образом и на основе какого энергетиче-
ского резерва все многочисленные компоненты функциональной
системы составляют единое и всегда напряженное целое? Нату-
ральные условия деятельности целого организма убеждают нас,
что в зависимости от афферентных потребностей данного момен-
та и афферентного синтеза этих потребностей функциональные
системы организма могут меняться, мобилизуя почти мгновенно
различные составы аппаратов и механизмов для осуществления
конечного приспособительного действия. При такой моментальной
смене функциональных систем наиболее загадочным механизмом
опять-таки был энергетический базис, который мог бы поперемен-
но поддерживать доминирующее состояние этих вновь и вновь
формирующихся функциональных систем.
202 Методологический анализ узловых проблем условного рефлекса
орме восходящей активации (пшотала-
•I
В последние годы физиология получила возможность объясне-
ния этой энергетической основы функциональных систем орга-
низма. Такой основой, несомненно, является та постоянная или
перемежающаяся активация, которая восходит к коре больших
полушарий и поддерживает единство всех компонентов системы,
сколько бы их ни было в данный момент. И больше того, есть ос-
нования утверждать, что всякое новое «обрастание» ядра функци-
ональной системы новыми компонентами и расширение ее за счет
новых условпорефлекторных приобретений происходят все время
на основе той энергетической основы, которую доставляют под-
корковые образования в
мус и ретикулярная формация).
Работы нашей лаборатории, выполненные за последние 10 лет,
дают нам все больше и больше оснований думать, что функцио-
нальные системы организма формируются на основе двух различ-
ных по природе факторов — архитектурного и энергетическо-
го. Оба эти принципа работы центральной нервной системы орга-
нически объединяются при формировании самых различных
функциональных систем с разными конечными приспособитель-
ными эффектами. В эволюционном смысле оказалась более про-
дуктивной именно такая интеграция нервной деятельности, в ко-
торой ассоциативные и архитектурные факторы приобрели
собственные возможности изменяться и варьировать. Кроме того
наличие этого разделения способствует самому главному условию
изменчивой и пластичной работы целого организма — быстрой
смене действующих в данный момент и, следовательно, доминиру-
ющих функциональных систем организма.
Из огромного многообразия врожденных и созданных жиз-
ненным опытом функциональных систем организма, составляю-
щих его первейшую ценность, энергетический фактор помогает
«выходу па сцену» именно данной
В этих сложных взаимоотношениях инициативная роль в том,
когда и какую функциональную систему «выпускать на сцену»,
несомненно остается за корой головного мозга, где осуществляет-
ся афферентный синтез.
Таким образом, теория функциональной системы раскрывает
перед нами широкие конструктивные возможности в исследова-
нии именно интегративной деятельности организма.
Она позволяет нам, например, преодолеть недостатки теории
интеграции Шеррингтона, который, дав много интересных и цен-
ных фактов и соображений, не смог тем не менее вскрыть прин-
ципов интегрирования функций в масштабах целого организма.
Более подробная критика недостаточности представлений
Шеррингтона о принципах интеграции нами была дана в специ-
альной работе об интеграции физиологических функций.
В настоящее время целый ряд исследователей, работающих в
а не другой деятельности.
Методологический анализ узловых проблем условного рефлекса 203
смежных областях (психология, педагогика, физиология различ-
ных функций организма), широко использует принцип функцио-
нальной системы для объяснения и анализа различных форм
деятельности организма до психических функций
включительно.
Можно указать на значительный успех в анализе компенсатор-
ных приспособлений человека после различного рода нарушений
целости его центральной нервной системы (А. Р. Лурия). В ана-
лизе целенаправленных пли произвольных действий принцип
функциональной системы, особенно представление об акцепторе
действия как фрагменте функциональной системы, также дает по-
ложительные результаты (Е. И. Бойко и др.).
Оценивая полученные нами результаты на протяжении по-
следних лет, можно с полным убеждением сказать, что анализ
сложного приспособительного поведения животных и человека на
основании принципа функциональной системы более продуктивен,
чем анализ
осуществленный в пределах представления о «реф-
лекторпой дуге».
Это положение, конечно, ни в какой мере не устраняет «реф-
лекторной дуги» как стержневого процесса, формирующего ответ-
ное действие. Но именно потому, что сам процесс приспособле-
ния никогда не заканчивается действием, необходимы были
дальнейшие расширения и преобразования этого понятия.
С нашей точки зрения, это и есть то конструктивное, что дает
нам диалектико-материалистический метод для преобразования
рефлекторной теории Декарта.
Заключение
Разобранные выше проблемы условного рефлекса, относящие-
ся к историческому возникновению самого принципа образования
временных связей, так же как и к недостаточности классиче-
ской теории рефлекса,— несомненно, важнейшие методологиче-
ские проблемы учения о высшей нервной деятельности. Они ле-
жат в основе понимания самой природы тех явлений, которые в
о
совокупности составляют учение о высшей нервной деятельности.
В этом смысле опи могут стать отправным пунктом как для
деловой дискуссии, так и для постановки экспериментов под но-
вым углом зрения. Именно по этому пути пошла паша лаборато-
рия, и мы имеем к настоящему времени значительное количест-
во результатов, позволивших нам в целом изменять наши пред-
ставления о ряде вопросов, по которым, казалось бы, существуют
устоявшиеся взгляды.
Сюда относятся прежде всего те дополнения, которые были
нами введены в общую физиологическую архитектуру условного
рефлекса, и изменения в общепринятой концепции об активиру-
ющем действии на кору мозга ретикулярной формации.
204 Методологический анализ узловых проблем условного рефлекса
К ряду новых взглядов мы относим также и концепцию о
системогенезе как общей закономерности развития функций в эм-
бриогенезе животных, о зависимости этого развития от экологиче-
ских факторов данного вида животпых.
К этой же категории новых представлений, возникших в на-
шей лаборатории, мы относим и представления о компенсации
функций как циклическом центрально-периферическом процессе.
Сюда же надо отнести и представление о физиологических осно-
вах патогенеза гипертонической болезни.
С первого взгляда может показаться, что речь идет о совер-
шенно различных феноменах, пе объединенных какой-либо одной
общей идеей. В самом деле, что, казалось бы, может быть обще-
го между опережающим отражением действительности, наиболее
древней закономерностью развития живых существ на земном ша-
ре, и, например, активирующим действием ретикулярной форма-
ции на кору головного мозга? Или: что может быть общего меж-
ду тем же опережающим отражением действительности и пробле-
мой компенсации нарушенных функций?
И тем не менее все эти области функционирования живой ма-
терии имеют один ключевой принцип, который меняет лишь свое
и
выражение в зависимости от качества функции или историческо-
го этапа развития живых существ. Этот принцип — функциональ-
ная система, свойства которой как интегративной организации
изучались памп па протяжении последних 30 лет в самых раз-
нообразных направлениях.
В самом деле, сигнализация как принцип могла вырасти и
закрепиться в эволюции живого мира только потому, что было
о чем сигнализировать, а это последнее всегда представляет со-
бой какой-то жизненно важный фактор организма. В свою оче-
редь, эта жизненная необходимость всегда реализуется в каком-то
конечном приспособительном эффекте, а информация об этом эф-
екте теми пли иными путями поступает к основным регулирую-
щим механизмам.
Эта обратная афферентации о существенности для жизни дан-
ного приспособительного эффекта может поступать по нервным
коммуникациям, например, высших животных. Но опа может по-
ступать также и по сложной цепи молекулярных процессов, на-
пример, простейших форм живых существ (амёба), где опа стаби-
лизует тот или иной обменный процесс.
Принципиально же в обоих случаях типичная архитектура
функциональной системы как «замкнутого контура» автоматиче-
ской регуляции остается той же. Именно эта .всеобщая основа
приспособления всех' живых существ к внешним условиям дала
начало сложным вариациям в функциях самых разнообразных
живых существ.
Поэтому надо считать естественным, что эта единая законо-
Методологический анализ узловых проблем условного рефлекса 205
мерность, на основе которой строятся все приспособительные
функции всех живых существ, может быть ключом к раскрытию
многих частных узловых процессов жизни.
Тридцать лет назад мы сформулировали теорию функциональ-
ной системы как циклической единицы саморегуляции с обратной
афферентацпей и показали ее огромное конструктивное значе-
ние для исследований в различных областях физиологии. Особое
значение это имело тогда для нас в раскрытии механизмов ком-
пенсаторных приспособлений организма после нарушения его
функций. Оказалось, что никакое восстановление нарушенных
функций невозможно без постоянной обратной и санкционирую-
щей афферептацпи о результатах данной попытки восстановле-
ния утраченной функции.
Еще большее значение принцип
ункциональной
системы
имеет для раскрытия приспособительного значения условнореф-
лекторной деятельности. Здесь открываются беспредельные воз-
можности углубленного исследования и понимания узловых пунк-
тов этой высшей формы саморегуляции.
Однако многие годы принцип функциональной системы был
руководящим лишь в нашей лаборатории. Только некоторые уче-
ные как у пас, так и за рубежом увидели в нем широкие воз-
можности для понимания функционирования целого организма
(А. Сперанский. А. Лурия).
Развитие кибернетики с ее основным принципом «регуляпип
с обратной связью» коренным образом изменило положение. Ос-
новной принцип автоматической регуляции с обратной связью
как основа кибернетики поразительно совпал с нашими представ-
лениями о «замкнутой функциональной системе».
В настоящее время у пас нет колебаний в признании того,
что все формы оценки конечного приспособительного эффекта
идут через обратную афферентацию, а любая форма саморегуля-
ции развивается через разветвленный аппарат функциональной
системы.
Таким образом, с точки зрения обитой оценки развития мыс-
ли в области изучения высшей нервной деятельности, мы можем
указать прежде всего на философскую или гносеологическую
оценку учения о высшей нервной деятельности. Здесь должно
быть разобрано значение учения И. П. Павлова для материали-
стического понимания оспов психической деятельности и проис-
хождения самой психики в процессе развития животных.
Другой аспект оценки — методологический анализ узловых
проблем в развитии самого учения о высшей нервной деятель-
ности. Этот анализ, конечно, должен быть органически связан
с основами диалектического материализма, поскольку основная
гносеологическая формула — материя первична, сознание вторич-
но — остается в силе для всех этапов исследования,
206 Методологический анализ узловых проблем условного рефлекса
Мы постарались показать огромное философское значение ге-
неза временных связей и недостаточности классической рефлек-
торной теории для современного уровня наших знаний. Но, ко-
нечно, этими проблемами отнюдь не ограничивается все то ак-
туальное, что должно стать именно сегодня предметом самого
большого внимания и конструктивного развития. Из тех общих
принципов функционирования, которые были разобраны выше,
сейчас же вытекает целая серия актуальных вопросов, которые
могут стать уже предметом лабораторного исследования. Сюда
можно отнести проблему генерализации, афферентного синтеза,
физиологической основы образования временных связей и т. д.
Сейчас же важно установить, что все частные вопросы зай-
мут свое место и приобретут свой смысл, значительно более эф-
фективную форму исследования в том случае, когда будут об-
суждены фундаментальные методологические вопросы учения о
высшей нервной деятельности.
ЛИТЕРАТУРА
относитесь-
1ШК
Ленин В. И. Поли. собр. соч., т. 29.
Александров А, Д. Философское содержание и значение теории
ности.— В кн.: Философские проблемы современного естествознания.
М., 1959.
Анохин П. К. Узловые вопросы в изучении высшей нервной деятельно-
сти.— В кн.: Проблемы высшей нервной деятельности. М., 1949.
Анохин П. К. Особенности афферентного аппарата условного рефлекса и
их значение для психологии.— Вопр. психол., 1955, № 6, с. 16.
Анохин П. Е. Новые данные к характеристике специфичности восходящих
активаций.—Журн. высш, нервн. деят., 1962, т. 12, вып. 1, с. 7.
Милягин Я, А. Определяющее действие экологических факторов на эмбрио-
генез безусловных реакций. Автореф. докт. дне. М., 1957.
Опарин А. И. Возникновение жизни на земле. М., 1957.
Павлов И. П. Поли. собр. соч., т. 3, кн. 2.
Павлов И. П. Поли. собр. соч., т. 4.
ФИЗИОЛОГИЯ И КИБЕРНЕТИКА *
Бурный рост нового и своеобразного направления научного*
мышления, объединенного понятием «кибернетика», вполне за-
конно привлекает к себе пристальное внимание деятелей самых
разнообразных научных дисциплин. Успех этого направления по-
нятен... Достаточно указать на полную автоматизацию ряда про-
мышленных предприятий, расчета дальнодействующих аппаратов
п вообще всех процессов,, которые требуют коммуникаций,
т. е. отдаленных связей и управлений.
Возникла особая ветвь кибернетики — теория моделирования
биологических процессов в механических схемах и устройствах.
Кульминационного пункта это направление достигает в попытках:
моделировать мозг человека в его наиболее тонких формах дея?
тельпости — вторая сигнальная система и т. п.
Неудивительно, что именно эта сторона кибернетического на-
правления — моделирование и производство всевозможных «робо-
тов» (Винер, 1954) — приобрела особенно широкую известность.
И все же более глубокая и всеобщая суть кибернетики находит-
ся несколько в другой плоскости. Создание механических моде-
лей является лишь частным следствием этой сути.
Основная трудность состоит сейчас в формулировании того
наиболее специфического, что несет с собой кибернетика...
Для обсуждения этих вопросов устраиваются международные
конгрессы: почти в каждой стране представители различных об-
ластей знания собираются на специальные симпозиумы и т. д.
Причем наряду с совершенно законным признанием огромных
достижений, которые были получены в технике па основе при-
менения кибернетических принципов, высказываются также и
весьма разумные предостережения по поводу чрезвычайно расши-
ряющихся претензий этой повой отрасли знания. Так, например,
Луи де Бройль, очень высоко оценивая значение нового направ-
:ет: «Чувство интереса
ления научной мысли, все же пи:
и
а иног-
1
да и восхищения принимает у меня оттенок недоверия...» Даль-
ше он поясняет, на какой основе складывается этот «оттенок
недоверия»: «Мне кажется, что устремления кибернетики, столь
* Опубликовано в журнале «Вопросы философии» (1957, № 4).
208
Физиология и кибернетика
полезные в качестве стимула к исследованиям, выходят в данное
время за пределы той области, которую она может охватить с
полным правом. Некоторые же из ее соображений явно выходят
за рамки тех результатов, над которыми она может обеспечить
действенный контроль» (Луи де Бройль, 1953—1954, с. 1).
Эти сомнения становятся особенно настойчивыми и повсемест-
и
ными, когда от математического анализа «сервомеханизмов»
и нескольких удачных примеров моделирования физиологических
феноменов кибернетика делает попытки перейти к раскрытию са-
мых высших функций человеческого мозга, и в особенности об-
щественного поведения человека. Здесь оказался предел притяза-
ний кибернетики, и именно поэтому возникла широкая дискус-
сия о смысле и методах этого научного направления.
Следует повторить, однако, что никто при этом не сомневает-
ся в огромной важности кибернетики как области знания, уста-
навливающей новые перспективы исследования и обеспечиваю-
щей чрезвычайно полезные преобразования в области техники..
В этом смысле надо признать весьма ценной инициативу акад..
С. Л. Соболева, проф. А. А. Ляпунова, А. И. Китова и других,,
которые на страницах журнала «Вопросы философии» смело по-
ставили вопрос о недопустимости игнорирования кибернетики..
Опи дали возможность сделать кибернетику предметом открытой
дискуссии и этим самым прекратили невежественные попытки
объявить ее «вредным вымыслом». Именно после этого выступле-
ния в советской прессе появился ряд статей, разбирающих раз-
личные стороны кибернетики.
Оценивая все имеющиеся в настоящее время наиболее значи-
тельные высказывания зарубежных ученых, явившиеся откликом
на выступления Норберта Винера, а также ряд выступлений на-
ших советских ученых, можно сделать несколько существенных,
на наш взгляд, замечаний, которые и послужат исходным пунк-
том для дальнейших наших собственных соображений по этой
проблеме.
Постановка вопроса и ее недостатки
Можно отметить прежде всего, что почти все работы, касаю-
щиеся проблемы кибернетики, берут для обсуждения какие-либо
частные стороны этой теории, но пе касаются ее принципиально-
го содержания, ее руководящих положений. Причиной такого по-
ложения дела является прежде всего недостаточная четкость по-
становки этого вопроса и у самого П. Винера. Он пе развил
мысль о том, на каком общем основании покоится возможность
переноса, сопоставления и аналогизирования механизмов автома-
тической регуляции в машинных системах с работой таковых в
животном организме. Вернее, он недостаточно ее развил. Он пе
Физиология и кибернетика
209
указал также и топ границы, у которой должны остановиться
сопоставление и моделирование живых систем и машин.
В результате именно этой нечеткости внимание подавляюще-
го числа исследователей было направлено главпым образом в сто-
рону моделирования различных функций целого организма в ма-
шинных схемах и действующих моделях. Между тем моделиро-
вание — это далеко не все то, что содержит в себе кибернетика и
что от нее надо ожидать для взаимного обогащения различных
наук.
Что такое моделирование? Как полагают некоторые исследова-
тели (см., например, Бэлэнеску), это — «воспроизведение» (!)
в машинных устройствах какой-либо деятельности организма. Од-
нако в данном случае понятие «воспроизведение» употреблено не
в соответствии с точным смыслом, и потому оно скорее запуты-
вает вопрос, чем разъясняет его. Так как такие операции с не-
раскрытыми понятиями начинают уже в настоящее время мешать
дальнейшему прогрессу во взаимопонимании различных специа-
листов, то я считаю необходимым уделить им несколько подчерк-
нутое внимание.
Прежде всего зададим себе вопрос: что значит «воспроизве-
сти» какое-либо явление? Понятие «воспроизведение», употреб-
ленное без специальных ограничительных формулировок, всегда
предполагает идентичность явлений, процессов и предметов по
их существу. Можно, например, воспроизвести другой автомо-
биль, подобный (!) первому, можно воспроизвести какой-либо ин-
дивидуальный организм, который содержит в себе все органиче-
ские признаки своего вида, но нельзя с помощью радиолампы
или полупроводника германия «воспроизвести» межнейрональный
сипапс (Куффиньяль, 1953—1954). А между тем во всех случаях,
когда речь идет об аналогиях машинных и живых систем, поня-
тие «воспроизводить» фигурирует как нечто само собой разумею-
щееся.
Так что же все-таки «воспроизводится» в машинных моделях,
и почему они так неотразимо гипнотизируют наше внимание?
Этот вопрос может быть решен только на путях тщательного
анализа всех сторон того многообразного процесса, который ка-
жется воспроизведенным. Возьмем в качестве примера процесс
проведения тока через полупроводник германий, которы
так ши-
роко использован Эшби в его «гомеостатах». Если расположить
по отношению к германию соответствующим образом подводящие
и отводящие ток провода, то можно создать такие соотношения,
которые во многом походят на некоторые формы синаптического
проведения. Так, например, два одновременно входящих тока мо-
гут «тормозить» друг друга в полупроводнике, и благодаря этому
из кристалла германия не выходит ток па отводящие провода.
Создаются условия, которые могут быть признаны (как это и
210
Физиология и кибернетика
делают зарубежные ученые) моделью синаптического торможе-
ния, т. е, в конце концов пессималыюго торможения.
Но «воспроизвели» ли мы здесь в самом деле торможение?
И какую его сторону мы воспроизвели? Если произвести строжай-
ший логический анализ этого «воспроизведения», то окажется,
что мы имеем перед собой феноменологическую сторону «отсутст-
вия чего-то», и поскольку при торможении также в конце кон-
цов отсутствует какая-то заторможенная деятельность, то от фе-
номена отсутствия делается интерполяция и в существо процесса
1
которому и приписывается термин «торможение».
Можно было бы мне возразить, что ведь каждое моделирова-
ние предполагает получение какого-либо сходного, т. е. аналогич-
ного, феномена и что пи одна модель не претендует на тожде-
ство природы механизмов. Это действительно так. Но отсутствие
четких формулировок, определяющих, что именно моделируется и
что именно «воспроизводится» в каждом отдельном случае, ме-
шает до конца разобраться в этом запутанном вопросе, а часто
ведет к вульгаризации и самой идеи моделирования. Опасность
такой неопределенности состоит прежде всего в том, что часто за
сходными внешними феноменами лежит пропасть между принци-
пиальными качествами сравниваемых механизмов. В самом деле,
если выражаться языком математической кибернетики, то за фе-
номеном синапса лежат необъятно широкие вероятности измене-
ний, поскольку его протоплазматический и ионный состав пред-
ставляет собой структуру с бесконечно большой пластичностью.
Наоборот, возможности полупроводника чрезвычайно ограничены,
и это отнюдь пе только количественные различия.
Становится ясно, что «воспроизведение» феноменологической
стороны как одной из многочисленных сторон такого сложного
явления природы, как синаптическое проведение, отнюдь пе дает
нам оснований питать иллюзии о полном отождествлении этих
узловых пунктов в системе моделируемых процессов. Как увидим
ниже, это обстоятельство оказывается решающим в оценке кибер-
нетики вообще как перспективного направления научной
мысли.
Вместе с тем следует отметить, что весьма интересно постав-
ленный И. Бэлэнеску вопрос, несомненно, продвигает вперед всю
проблему о сходстве и различии механических и физиологиче-
ских систем саморегуляции. Однако II. Бэлэнеску довольно часто
высказывает категорическое положение о принципиальном разли-
чии физических и физиологических явлений, не указывая, где
кончается сходство между ними и начинается различие. Так, на-
пример, сопоставляя работу механических систем и работу мозгаг
оп пишет: «Процессы нервной системы, головного мозга являются
биологическими процессами, в то время как процессы машины,,
как бы сложна и совершенна она ни была, являются физически-
Физиология и кибернетика
211
ми процессами (механическими, электрическими, электронными
и т. д.)» (Бэлэпеску, 1957).
Заслуживает внимания интересная попытка И. Бэлэнеску вы-
работать классификацию тех общих законов, которые в одинако-
вой степени применимы как к механическим, так и к живым
системам. Конечно, эти законы слишком общи и не дают пока
видимой связи между заколами живой и неживой природы, тем
не менее они направляют мысль исследователя в определенную
сторону при поиске этой связи и потому могут быть припяты
за положительные факторы.
Мы переживаем самый начальный период развития киберне-
тики; я полагаю, что строгая и четкая формулировка понятий,
определяющих процессы и механизмы, подлежащие в каждом от-
дельном случае моделированию, может только помочь нам в опре-
делении грапиц и возможностей этого научного направления. Это
особенно касается области, которая мне ближе всего,— физиоло-
гии центральной нервной системы. Здесь мы имеем самые не-
ожиданные заключения, способные вселить смущение в умы лю-
дей. интересующихся кибернетикой.
Так, например, оказывается, что безусловный рефлекс можно
моделировать, условный также можно, а рефлексы второй сиг-
нальной системы нельзя (ТО. П. Фролов). Почему? Никаких убе-
дительных оснований к этому не дано. Здесь мы имеем опять тот
же результат отсутствия четких формулировок того, в чем именно
кибернетике^ находит общие основания для разнородных процес-
сов и как далеко это общее может простираться. Бросается в
глаза также и другой эпизод. И. И. Гальперин в своей интерес-
ной статье с особенной настойчивостью отстаивает возможпость
называть машины «рефлекторными». Как увидим ниже, в самом
этом требовании есть глубокая противоречивость, поскольку оп
наряду с этой тенденцией также отрицает возможпость модели-
рования второй сигнальной системы. Явный парадокс! Если вто-
рая сигнальная система («сигналы сигналов») действует так же
на основе принципа рефлекса, как и два более низких уровня
организации (первая сигнальная система, безусловный рефлекс),
то почему же вторая сигнальная система не может быть моде-
лирована «рефлекторными машинами»?
212
Физиология и кибернетика
О некоторых частных принципах кибернетики
Как было отмечено выше, в большинстве литературных пуб-
ликации и в нашей советской научной прессе, и в зарубежной в
центре внимания стоят лишь фрагменты кибернетики. Мне при-
ходилось не раз слышать доклады «по кибернетике», которые
начинались с понятия информации и заканчивались им. Но ведь
информация, как и сама теория информации, представляет со-
бой только часть кибернетического принципа, только средство,
с помощью которого создается целостная функциональная архи-
тектура, приобретающая после этого черты, обязательные для
любого другого аналогичного образования. Таким образом, весьма
важный фрагмент теоретической кибернетики приобретает как бы
самостоятельное значение и начинает обсуждаться и анализиро-
ваться без определения его места в общей системе кибернети-
ческих представлений.
Такая изоляция одного из звеньев целостной архитектуры
особенно остро сказывается на разработке соотношений киберне-
тики и физиологии нервной системы. Здесь изоляция «информа-
ции» сводится к отдельному рассмотрению процессов нервной
сигнализации. При такой редукции общей архитектуры целой
функциональной системы организма кибернетика для физиологии
теряет фактически всякий конструктивный смысл. Конечно, тео-
рия информации, чрезвычайно широко разработанная математи-
кой, рассматриваемая даже в отдельности, способна дать направ-
ляющие идеи в изучении, например, анализаторных функций
центральной нервной системы (Гершуни). Однако этот частный
процесс целой функциональной системы, сколь бы он глубоко ни
разрабатывался отдельно, в отрыве от целостной архитектуры
функции, никогда не умножит того специфического, что принесла
кибернетика в физиологию целого организма.
В последнее время, пожалуй, еще более широко разбирается
принцип «обратной связи», как обязательное включение в систе-
му автоматически регулирующихся механизмов. Здесь также име-
ются попытки поиска аналогичных соотношений и в живом ор-
ганизме. Однако и в данном случае «обратная связь» обсуждает-
ся как что-то самодовлеющее, без детального анализа ее роли
в системе целой реакции. Не ставится четко вопрос: что общего
между организмом и машиной в отношении обратной связи и
на какой основе генетически возникли эти аналогичные соотно-
шения? Положение в этом вопросе несколько усугубляется еще и
тем, что физиология целые столетия существовала без осозна-
ния обратной связи, и потому даже вполне конкретные многолет-
ние призывы пашей лаборатории к обязательному учету обратных
афферентаций (в случае восстановления нарушенных функций)
лишь с большим трудом входят в поле зрения исследователей,
Физиология и кибернетика
213
Следует отметить при этом, что абсолютная необходимость об-
ратных афферептаций при восстановлении нарушенных функций
целого организма нами была сформулирована в совершенно от-
четливой форме задолго до того, как вообще возникло само ки-
бернетическое направление (Анохин, 1935).
Только недостаточно внимательным отношением физиологов к
достижениям своей собственной науки можно объяснить то лож-
ное мнение, согласно которому идея об «обратных связях» при-
шла в физиологию якобы от кибернетики. На самом деле, повто-
ряю, она давно уже была тщательно разработана нами на основе
конкретных экспериментальных моделей. Суть дела состояла в
следующем.
После ряда перерезок нервных стволов у собаки эти стволы
перешивались друг к другу перекрестно, т, е. создавались усло-
вия так называемого гетерогенного анастомоза нервов. Впослед-
ствии эти модели получили название «химер», поскольку пере-
крестно сшитые нервные стволы проявляли целый ряд парадок-
сальных функциональных черт. Так, например, при прикосновении
рукой к коже передней ноги можно было вызвать кашель и даже
рвоту, передняя конечность проделывала вместе с диафрагмой
«дыхательные движения» и т. д.
В большой серии этих экспериментов было выяснено, что вос-
становление функций нервной системы, т. е. «переучивание цент-
ров», происходит при непрерывном обратном афферентировании с
периферии каждого этапа восстановления функций. Можно ска-
зать, что в каждый отдельный момент животное при восстанов-
лении функций только тогда переходит к следующему этапу вос-
становления, когда оно получит с периферии обратную афферен-
тацию, информирующую центральную нервную систему о
достигнутом эффекте. Без этой обратной афферентации не может
быть и не происходит никакого восстановления нарушенных
функций. Анализируя этот процесс восстановления (компенсация
функций), мы пришли к выводу, что особое значение для орга-
низма имеет та обратная афферентации, которая сигнализирует
в центральную нервную систему окончательное достижение по-
лезного эффекта. Эта обратная афферентации полностью стаби-
лизирует («санкционирует») то распределение возбуждений в ап-
паратах коры и подкорки, которое смогло дать на периферии
окончательный полезный эффект. В силу такой роли конечной
обратной афферентации она нами была названа «санкционирую-
щей афферентацней» (Анохин, 1935).
В последнее время наблюдается не совсем разумная, на мой
взгляд, тенденция — подменить физиологические термины и поня-
тия терминами, заимствованными из арсенала кибернетики. Ши-
роко вводятся понятия «информация», «кодирование», «програм-
мирование». Но особенно в этом отношении повезло понятию «об
214
Физиология и кибернетика
ратная связь». Я должен категорически высказаться против та-
кого формального переноса, ибо введенное нами понятие «обрат-
ная афферентация» п понятие «обратная связь» не только не
совпадают по своему содержанию, но, наоборот, последнее зпачи-
тельно обедняет первое!
В самом деле, говоря о регулирующем значении обратной аф-
ферентацип, физиолог немедленно же ставит перед собой вопрос
о составе этой обратной а
во времени. Иначе говоря, понятие обратной а
И1П
рерентации как в пространстве, так и
ерентации «зо-
о охи
вет» к дальнейшим поискам и ставит перед исследователем
конкретные исследовательские задачи. Наоборот, понятие «обрат-
ная связь» констатирует лишь самый
и
акт наличия связи и не
нуждается в дальнейшем уточнении. В этом смысле мне казалось
бы нерациональным стремиться к обязательному внедрению имен-
но кибернетической терминологии в специальные биологические
вопросы.
Приведенный нами материал о роли обратных афферентаций
в саморегулирующихся приспособлениях организма убеждает нас
прежде всего в том, что «обратная связь» так же, как и «инфор-
мация» не могут быть вырваны из системы циклических связей,
приводящих в конечном итоге к полезному эффекту. Наоборот,
их смысл становится совершенно ясным именно только в этой
системе циклических связей.
Сделанные в последнее время попытки приложить теорию ин-
формации к разрешению проблемы стимуляции различных форм
поведения животных показали, что работа по применению идей
кибернетики к физиологическим проблемам находится только в
начале пути.
Так, например, статистическая сложность раздражителя бе-
рется как нечто вполне определенное в смысле количества вход-
ной информации. Количество последней исчисляется по соответ-
ствующей формуле, в которой довольпо полно предусмотрены все
возможности градации частоты, силы и времени действия данно-
го комплекса раздражений. Таким образом, все, что необходимо
для исчисления и точного расчета информации, получаемой ка-
кой-либо машиной с автоматической регуляцией, здесь имеется.
По крайней мере именно на этом пути теория информации
впервые получила свое развитие п принесла богатейшие полез-
ные материалы.
Однако положение резко меняется, когда теория информации
начинает применяться к оценке и определению целостного пове-
дения животного, как это, например, сделали в последние годы
Гарнер и Хэйк (Garner and Hake, 1951).
Проблема приложения теории информации к исследованию
физиологии нервной системы,
и особенно физиологии высшей
нервной деятельности, чрезвычайно сложна и актуальна. Физио-
Физиология и кибернетика
215
информации,
логия, несомненно, должна взять от теории информации все, что
обеспечит ей более быстрый прогресс. Но вот вопрос: что именно
она должна взять?
Сделанные до сих пор попытки представляют собой только
первые шаги и пе вскрывают значительных трудностей на этом
пути. Например, так называемая статистическая сложность раз-
дражителя учитывает лишь наличное количество
нредставленпое действительными внешними и внутренними аген-
тами. Однако для организма эта статистическая сложность раз-
дражителя, хорошо рассчитываемая математически, не составляет
самой существенной информации. Для него максимальную важ-
ность имеет сигнальное значение этой информации, т. е. в ко-
нечном счете биологическая значимость того, что последует толь-
ко в будущем. Вот эту-то наиболее значительную долю инфор-
мации теория информации пока и не может рассчитать, поскольку
здесь решающим моментом в реакциях организма является пере-
житая им история.
Не удивительно поэтому, что американские авторы Гарнер и
Хэйк допустили весьма существенные ошибки в приложении тео-
рии информации к объяснению целостного поведения, взяв за
критерий получаемой информации ошибочность или правильность
Конечной реакции. Для физиолога ясно, что этот критерий никак
пе может быть использован для оценки информации. Тем более
трудно согласиться с предположением, что если возникла ошибоч-
ная реакция, то мы имеем дело с «потерей информации» (?).
Как раз, наоборот, чем более «тренируется» какая-либо ре-
, следовательно, чем большей становится вероятность ее
акция и
правильности, тем все более и более редуцируется первоначаль-
ный состав афферентных влияний. Эта закономерность нами была
названа еще двадцать лет назад «сужением афферептации». Вме-
сте с тем при всяком внезапном увеличении внешних афферент-
ных воздействий, как правило, увеличивается и вероятность оши-
бочной реакции. Таким образом, приложение теории информации
к целостному поведению животного убеждает нас, что имеющих-
ся материалов в этой области далеко не достаточно. Здесь перед
физиологами и математиками стоят крайне увлекательные про-
блемы.
Заключая разбор тех трудностей, которые встречаются в на-
стоящее время на нути развития кибернетики и именно ее свя-
зей с физиологией центральной нервной системы, мы можем ска-
зать, что весь этот вопрос находится в самой начальной стадии
развития. Не удивительно поэтому, что внимание исследователей
направлено в сторону наиболее заметных и выделяющихся фраг-
ментов общей системы циклических взаимоотношений, представ-
ляю:
тики. В противоположность этому общие принципы кибернетики
цтх собой самую характерную и решающую черту кпберне-
216
Физиология и кибернетика
что четкая формулировка универсального
вскрываются п используются с большими трудностями, а следо-
вательно, и с меньшей охотой. Однако мы уже пе раз указывали,
характера этих общих
принципов кибернетики должна быть обязательным условием
для постановки частных задач в будущих исследованиях. Именно
поэтому я считаю необходимым сделать попытку установить
генетическое родство между такими, казалось бы, разнородными
областями научной мысли, как физиология мозга и кибернетика.
Чем они объединяются? На какой основе между ними возникает
творческий контакт, который сулит все большие и большие прак-
тические результаты?
Вот вопросы, которые должны быть освещены нами в после-
дующих разделах этой статьи.
Общие принципы кибернетики
и их происхождение
е вопрос о той основе, которую дает кибернетика.
и
По общему признанию, заслуга кибернетики состоит в том,
что она установила тончайшие логические нити, связывающие та-
кие области знания, которые много лет развивались независимо
друг от друга. Именно здесь надо искать ответ па поставлен-
ный нами вы;
И здесь мы должпы обратиться прежде всего к Норберту Ви-
перу, декларировавшему общие задачи кибернетики. В предисло-
вии к своей последней книге он дает довольно отчетливое ука-
зание на то, откуда он получил первый толчок к поискам общего
у различных на первый взгляд научных направлений. Оп излага-
ет взгляды Гиббса, который поставил вопрос о возможности поис-
ков общего у различных миров явлений: насколько ответ, полу-
ченный для одного мира явлений, может быть допустим для объ-
яснения явлений других миров? Вероятность такого переноса
закономерностей одного мира па закономерности другого мира,
по Гиббсу, тем меньше, чем более пндетерминистичеп или чем
больше вероятность случайностей в каком-либо из миров.
И, наоборот, возможность переноса и установления общих зако-
номерностей между двумя мирами тем больше, чем меньше в них
вероятность случайных изменений, чем менее индетерминистичен
данный мир явлений. Так постановка вопроса об общих законах
в различных мирах явлений неизбежно должна была войти в ор-
ганический контакт с учением об энтропии.
В самом деле, с нарастанием энтропии и с уменьшением слу-
чайностей,
вптся все более и более вероятным выявление той общей основы,
которую они имеют. Математика дает нам возможность вскрыть
вероятности общей основы у различных классов явлений, и это
составляет новую черту в консолидации и в зависимом развитии
определявших индивидуальность двух миров, стано-
Физиология и кибернетика
217
различных наук нашего времени. В этом смысле кибернетика де-
лает довольно смелые попытки объединить, казалось бы, несрав-
нимые классы явлений по их общим свойствам. Так, например,
Винер, разбирая необходимость обратных коммуникаций при
управлении шлюзами Панамского канала, заключает: «Следова-
тельно, административные руководители, будь то правительства,
университеты или общественные корпорации, должны принимать
участие в двустороннем потоке коммуникаций, а не только в од-
ном, идущем сверху вниз. Иначе, высшие чиновники могут столк-
нуться с тем, что они основывали свою политику на совершен-
ном непонимании тех фактов, которыми располагают их подчи-
ненные» (Винер, 1954).
Таким образом, мы видим, что понятие «обратная связь», осо-
бенно широко распространенное применительно к механическим
системам автоматического регулирования, с одинаковым успехом
применяется и к явлениям общественного порядка. Мы хорошо
знаем, как эта «обратная связь» необходима в правильной орга-
низации обществеппо-трудовых
соотношении
и каждый может
для этого наити многие примеры в своей повседневной жизни.
Достаточно указать, что всем известный принцип проверки ре-
зультатов, получивший ходовое обозначение «контроля исполне-
ния», представляет собой типичный пример обратных коммуника-
ций.
Таким образом, первое теоретическое положение кибернетики
о наличии общих закономерностей у различных рядов явлений и
в различных науках эмпирически может быть реализовано на са-
мых разнообразных примерах.
Однако это всего лишь только иллюстрация закона. Но такие
иллюстрации далеко ие достаточны для того, чтобы ответить на
коренной вопрос о природе такой общпости закона обратных ком-
муникаций. В самом деле: почему так сложился мир физических,
живых и общественных систем, что какие-то формы функциони-
рования для них оказываются в одинаковой степени обязатель-
ными? Па этот кардинальный вопрос Випер не дает ответа. Он
даже не делает попытки и поставить его. Для нас же, с точки
зрения философской, пмеппо этот вопрос является центральным,
ибо, ответив на пего, мы легко сможем объяснить и использо-
вать все дальнейшие разветвления логических построений кибер-
нетики. Так как ответ па этот кардинальный вопрос касается
прежде всего механических и биологических аналогий, особенно
аналогии в работе машин и центральной нервной системы, то мы
считаем необходимым попытаться дать такой ответ.
Прежде всего мы должны поставить вопрос: чем вызвало по-
явление в процессе эволюции обратных афферептаций, которые
положительно у всех живых существ дают информацию о степе-
ни успешности и полезности произведенного действия? Простой
218
Физиология и кибернетика
разбор любого рефлекторного акта убеждает нас, что обратная
афферентацпя неизбежно должна была появиться и причиной
этого является обязательная биологическая полезность любой ре-
акции животного, любого приспособительного акта. Живая систе-
ма существует как целое до тех пор, пока ее жизненно важные
константы строго поддерживаются на определенном уровне. Но
вот здесь-то и должно было возникнуть препятствие для сохра-
нения жизни. Дело в том, что любая реакция животного пли
любой приспособительный эффект, с точки зрения теории вероят-
ностей, есть одна из многих возможных вариаций одной и той
же реакции.
Эта рассеянность степеней свободы реакции, зависящая от
особенностей многочисленных связей в центральной нервной си-
стеме, была бы одним из серьезных препятствий для развития
совершенного приспособления животного к внешнему миру. Мож-
но представить себе, что реакция животного А в зависимости от
условий может иметь варианты в виде целой серин близких реак-
ций — «1, а2, а2....ап. Какая из них закрепится в структурах ор-
ганизма как наиболее полезная? Как организм оценит эту степень
полезности данной реакции? Естественно, что без этой оценки по-
лезности все приспособительное поведение животного преврати-
лось бы в сплошной хаос.
И вот в процессе эволюции живых существ, вероятно, уже на
очень ранней стадии развития, возникло универсальное приспо-
собление в виде обратной сигнализации о полезности совершен-
ного действия. Этим самым был достигнут тот уровень развития,
при котором степени свободы каждой реакции (рассеянность ре-
акции) значительно уменьшились, а организм получил возмож-
ность на основе этих обратных сигналов осуществлять непре-
рывную «подгонку» своего поведения вплоть до получения мак-
симального полезного эффекта. Именно в этой форме при изучении
компенсации фупкций обратная афферептация и была впервые
открыта в нашей лаборатории. Следовательно, обратная инфор-
мация в мире живых существ является абсолютно необходимым
условием выживания и потому получила свое широкое развитие
задолго до того, как на Земле появился человек. Это положение
очень важное. Оно нам понадобится в дальнейших рассужде-
ниях .
Обращает па себя внимание один существенный момент, что в
центре этих эволюционных превращений стоят полезность эффек-
та, ради обнаружения и оценки которого появилась на сцепу об-
ратная афферентацпя. Оно и понятно: там, где вопрос стоит о
полезности эффекта, необходима и оценка степени этой полезно-
и
сти. Но удивительно поэтому, что эта дочсловеческая об:
(ая
за-
кономерность в последующем направляла и эволюцию человека.
В самом деле, первобытный человек, употребив впервые при-
Физиология и кибернетика
219
митивное орудие, свое отношение к этому последнему строил:
также на основе оценки копенного полезного эффекта. Весь вы-
бор орудии и их преобразование должны были покоиться на осно-
ве этого универсального закопа, ибо, разумеется, не оцепив по-
лученного эффекта (обратная афферептация), первобытный чело-
век не знал бы, в какую сторону изменять своп примитивные
орудия.
Положение дела не изменилось и тогда, когда человек упо-
требил более или менее сложные орудия, например, соху для об-
работки земли. Здесь, как н во всех предыдущих случаях, по-
лезный эффект пемипуемо должен был оцениваться непрерывно,
на оспове зрительных аффереитаций, получаемых от вспаханной
земли. В зависимости от характера этого конечного эффекта че-
ловек делал те или иные движения и усиливал или ослаблял
давление на поручни сохи.
Здесь важно отметить одну замечательную особенность: не-
смотря па то что человек начал употреблять орудия, принципи-
альное значение обратной афферентации в циклической системе
трудового акта нисколько пе изменилось. В цикл включилось
лишь новое звено — орудие. И хотя некоторые трудовые процес-
сы уже давно были подвергнуты автоматической регуляции (Пол-
зунов, 1763; Уатт, 1784), еще весьма долгое время человек был
центральным звеном даже самых сложных машин. Он сам оцени-
вал конечный полезный эффект машины и в зависимости от этой
оценки направлял энергетические воздействия на регулируемый
объект. Как мы знаем, еще и в настоящее время по такому спо-
собу работает множество машин. В чем состоит здесь роль чело-
века? Оп получает от полезного эффекта, производимого маши-
ной, обратную афферептацию и в зависимости от ее значения
производит какую-то настраивающую работу, приводящую в ко-
нечном итоге к сдвигу полезного эффекта в желаемую сторону.
По своей сути это — циклический процесс, и в пего человек
вставлен как «чувствительный элемепт» и как некоторое «преобра-
зующее устройство», если выражаться языком техники автома-
тического регулирования.
Современный этап в развитии техники автоматического регу-
лирования дал возможность полностью исключить человека из
циклических связей при получении какого-нибудь производствен-
ного полезного эффекта. Это значит, что сигнализацию о степе-
ни полезности эффекта машины теперь получает уже специаль-
ное механическое или электронное устройство, которое сконструи-
ровано таким образом, что служит специфическим «рецептором»
именно для данного полезного эффекта. Здесь полезный эффект
«оценивается», и в зависимости от этой оценки через целую се-
рию приборов в машину направляется регулирующее энергетиче-
ское влияние.
220
Физиология и кибернетика
Л & тематически и ре гуля тар
-*- ! л-t-A *м> w
Рис. 1. Схематическое изображение пяти стадий
( Яыаед системы - атялеяеяие
регулируя май величины)
Физиология и кибернетика
оценки полезного эффекта деятельности системы
развития циклической системы с обратной сигнализацией
/ — циклическая функциональная система животных на
ранних стадиях развития жизни на Земле (дочеловеческая
эра); II— стадия употребления примитивного орудия;
III — стадия употребления усовершенствованных орудий
труда; IV—стадия машинного производства, в котором
человек является «аппаратом» оценки полезного эффекта
машины и нового регулирующего воздействия на регули-
руемый объект; V — стадия полного устранения человека
в механизмах с автоматической регуляцией. Видна прин-
ципиальная схема цикличности, которая является спутни-
цей всего процесса в дочеловеческой и человеческой ста-
дии развития приспособительного поведения. Части цикли-
ческой системы, относящиеся к участию животного и
человеческого организма, обведены пунктирОхМ
222
Физиология и кибернетика
Если приведенные выше пять этапов развития оценки полез-
ного эффекта изобразить в общепринятых символах техники ав-
томатического регулирования, то мы получим весьма поучитель-
ный ряд схем (рис. 1).
Первая часть рис. 1 (I) дает изображение тех натуральных
отношений в нервной системе, которые соответствуют натураль-
ному приспособительному поведению живых существ. Мы видели,
что такая система отношений сформировалась в истории живых
существ как единственно возможная. Она — следствие той вари-
антности поведения, которая составляет один из кардинальных
законов мира. Эта система непременно должна быть всюду, где
речь идет о полезности приспособительного эффекта, и потому
опа неминуемо должна была сопутствовать и первым попыткам
первобытного человека ввести и в эту циклическую систему ору-
дие как средство повышения полезности натурального приспосо-
бительного эффекта (11).
В сущности, на всех последующих этапах речь шла только
о более эффективном заполнении этой циклической системы раз-
личными, сначала механическими, а затем и электрическими уз-
лами, которые все более и более освобождали человека от излиш-
них усилий (III и IV). И, наконец, на последнем этапе техниче-
ского усовершенствования регуляторная роль человека в этой
циклической системе была полностью устранена. Человечество
вступило в век автоматической регуляции полезной деятельно-
сти машин (V). На этом пути человечество произвело несколь-
ко технических революций: сначала появились паровые машины,
затем на сцену выступило электричество и сделало неузнаваемым
не только промышленность, но и быт человека. Теперь мы вступи-
ли в век радио, электроники и атомной энергии. Перед челове-
чеством открываются необъятные возможности усовершенствова-
ния его быта, коммуникаций, промышленности.
Однако, если мы сравним I и V этапы нашей схемы, то заме-
тим один поразительный факт: принципиальная схема цикличе-
ских зависимостей, определяющая получение любого полезного
эффекта, осталась извечной и сохранила именно ту форму, в ка-
кой эволюция нащупала ее на самых первых шагах развития
жизни на Земле. По-прежнему полезный эффект имеет несколько
вероятностей и потому должен быть регулируем центральной
нервном спет сноп пли электронными устройствами на основе сиг-
налов обратной связи.
Здесь мы имеем дело с одним из самых кардинальных зако-
нов жизни, который определяет собой все формы полезных при-
способлений человека до сложных ма:
и
:ин автоматической регуля-
ции включительно. Этот закон мог бы быть сформулирован сле-
дующим образом: всякая функциональная система, механическая
или живая, созданная или развившаяся для получения полезного
Физиология и кибернетика
223
эффекта, непременно имеет циклический характер и не может су-
ществовать, если не получает обратной сигнализации о степени,
полезности произведенного эффекта.
Это и есть та общая основа, которая должна была бы быть
сформулирована в четком виде в самом начале, чтобы мысль ис-
следователей различных специальностей была строго детермини-
рована в обсуждении проблем кибернетики. Последняя в самом
деле «нащупала» узловой пункт в развитии жизни и техники.
Но именно потому, что ни сам Випер, ни его последователи не
сделали этот закон очевидным и не поставили его в качестве
исходного пункта для всякого рода интерполяций, мы оказались
в настоящее время перед грандиозным «смешением языков»...
В самом деле, если познакомиться с современной, довольно
уже обширной литературой по кибернетике, то бросается в гла-
за огромное разнообразие частных интересов, вкусов и точек зре-
ния отдельных авторов. Некоторые из них все свое внимание уде-
ляют теории коммуникаций, другие — энтропии, а третьи — тео-
рии информации. По чем объединены эти искания? Из
приведенных нами схем ясно, что представленная на них цикличе-
ская система может существовать как целое только при том усло-
вии, если между ее отдельными узлами будет непрерывно цирку-
лировать адекватпая для каждого отдельного момента сигнализа-
ция. Окончательный полезный эффект такой циклической системы
будет всегда находиться в зависимости от того, какая это сиг-
нализация (или информация) в каждом отдельном звене, какой
ее состав и каковы «полосы ограничения».
Именно на этой основе, т. е. па оспове математической об-
паботкн вероятностей входной и выходной информации, возникла
теория информации. В своем теоретическом арсепале она уже
имеет много изученных закономерностей и в некоторых случаях
проявляет явную тенденцию к автономности и замкнутости. Од-
нако нельзя ни па минуту забывать, что результаты этой теории
могут быть использованы только в пределах той циклической си-
стемы, которая обеспечивает полезный эффект, и здесь не имеет
значения, какая это система — автомат, общественные коммуни-
кации или что-либо другое. Строго говоря, теория информации
есть теория о поддержании сигнализаций и связей в пределах
какой-либо циклической системы — живой или механической —
с действующей установкой на получение максимально полезного
эффекта.
Из приведенной формулировки видно, что разработка отдель-
ных частных вопросов теории информации в порядке нарастаю-
щей специализации дает, несомпенно, полезный результат только
потому, что эта работа ведется в пределах все той же целостной
ппклической архитектуры, которая памп была разобрана выше.
Следовательно, на всех этапах работы инженера, физиолога и ма-
224
Физиология и кибернетика
тематика их объединяет эта универсальная общая закономер-
ность.
Здесь может возникнуть один недоуменный вопрос: почему,
несмотря на столь глубокую древность закономерности цикличе-
ского характера при получении всякого полезного эффекта, ее
универсальные черты были вскрыты только сейчас? Иначе говоря,
почему кибернетика появилась на арене научных исканий с та-
ким большим запозданием? Ответ на этот вопрос может быть в
некоторых отношениях весьма поучительным для различных спе-
циалистов, и потому ему следует уделить особое внимание.
Достаточно взглянуть на пашу схему и сопоставить ее I и
V этапы, чтобы получить убедительный ответ на поставленный
вопрос. Механике и физике эту общую закономерность помешал
вскрыть человек, который, находясь в центре «замкнутого кон-
тура», ручной настройкой приборов и оценкой полезного эффекта
машины маскировал циклическую зависимость между отдельными
звеньями этого коптура. И только тогда, когда путем внедрения
электронных релейных установок стало возможным полностью
устранить человека из управления машинами, во свю ширь встал
вопрос об «обратных связях» и их кардинальном значении в тео-
рии автоматического регулирования.
Но почему же тогда физиология, имея дело па протяжении
столетий с функциональными системами циклического характера,
не вскрыла их истинного смысла и не «подсказала» технике идею
моделирования этих циклических зависимостей в различных ма-
шинных устройствах? Теперь уже пе подлежит сомнению, что
физиологии в этом деле помешала рефлекторная теория Декарта.
Ее чрезвычайный авторитет и та огромная польза, которую она
приносила физиологическим исследованиям, на целые столетия
скрыли от глаз физиолога ее недостаточность в объяснении пол-
ноценного натурального поведения животпых.
Рефлекс Декарта оканчивается только ответным действием,
т. е. третьим и последним звеном классической «дуги рефлекса».
По своей сути он не мог ничего дать для оценки результатов
совершившегося действия. Короче говоря, физиология на протя-
жении целых столетни не замечала наличия обратной афферен-
тации, которая служит абсолютно обязательным звеном всякого
приспособительного поведения животных,
причин этого факта убеждает нас в том,
вивисекция, которая по понятным причинам на целые столетия
заняла доминирующее место в анализе рефлекторной деятельно-
сти центральной нервной системы. В самом деле, никакая обрат-
ная афферептация о рефлекторном эффекте не может способст-
вовать наплучшему приспособлению у собаки, привязанной к опе-
рационному столу. Опа просто здесь пе пужпа, а поэтому и пе
смогла быть выявлена в многочисленных работах физиологов.
Ближайший анализ
что виной этого была
Физиология и кибернетика
225
HIH
Мне не раз приходилось указывать на необходимость расши-
рения наших обычных представлений о «дуге рефлекса», что было
в свое время подтверждено вескими экспериментальными данны-
ми. И надо сожалеть, что совершенная необходимость обратных
афферентаций в организме стала очевидной только сейчас, с та-
ким значительным запозданием и к тому же лишь в результате
изучения такой далекой области, как машины с автоматической
регуляцией. Ясно, однако, что «обратная связь» пришла в маши-
ны только потому, что ее достоинство и полезность были прове-
рены природой на протяжении сотен миллионов лет...
О пределах аналогии
между живыми и механическими системами
Установив общую основу для явлений разного порядка — жи-
вых, механических, общественных,— мы должны поставить перед
собой вопрос: как далеко это тождество общей архитектуры соог
ношений может быть распространено на свойства и сущность от-
дельных узлов и отдельных звеньев всей циклической системы
соотношений? В этом пункте как сам Винер, так и его последо-
ватели во всех странах имеют, как мне кажется, довольно опре-
деленную солидарность, хотя здесь есть много такого, с чем нель-
зя безоговорочно согласиться. Поскольку от четкого решения
этого вопроса зависит и то, насколько успешно мы можем разре-
шить проблему моделирования, я позволю себе несколько подроб-
нее представить здесь точку зрения самого Винера на этот
предмет.
Он рассуждает следующим образом: если механическая систе-
ма и живая система
ункционируют «одинаковым образом», т. е.
в обоих случаях имеются обратные связи и автоматическая регу-
ляция, и если для понимания механической системы имеется ма-
тематическая теория, то, следовательно, эта математическая тео-
рия в одинаковой степени может быть применена и к физиоло-
гическим системам. Так, например, теория информации может
быть целиком применена к исследованию получаемых организ-
мом всевозможных сигнализации из внешнего и внутреннего ми-
ра, теория коммуникаций с особенностями ее расчетов («полоса
пропускания» и т. д.) также может быть применена к изучению
проведения возбуждений по самым разнообразным проводникам
центральной и периферической нервной системы.
Особую смелость Винер проявляет в допущениях, касающих-
ся функции собственно центральной нервной системы. Если дока-
зано, что функция нервной системы в отношении ряда регу-
IE
ляций проявляется в форме сервомеханизмов, а последние уже
имеют разработанную математическую теорию, то нет никаких
препятствий, с точки зрения Винера, к тому, чтобы применить
8 П. К. Анохин
226
Физиология и кибернетика
эту математическую теорию и к объяснению деятельности нерв-
ной системы.
Таким образом, мы видим, что на основе действительного и
несомненного тождества принципиальных циклических систем, по
которым развертывается саморегуляция машин и саморегуляция
функций организма, делается определенная попытка отождест-
вить и те конкретные звенья передач «информации», которые
поддерживают единство этих систем.
Однако как далеко может быть распространен па конкретные
механизмы тезис тождества общей архитектуры циклической си-
стемы организма и системы механической? От того, как мы отве-
тим на этот вопрос, зависит и наше представление о пользе ки-
бернетики как повой формы мышления для разработки проблем
физиологии центральной нервной системы. Именно па этом пути
происходит вульгаризация самого принципа аналогий.
Можно на конкретном примере показать, к каким результатам
приводит в отдельных случаях перепое закономерностей механи-
ческих схем на физиологические явления. Допустим, что человек
полез в ящик письменного стола, чтобы взять конверт, который
там обычно находится. Однако, раскрыв ящик, он не увидел
конверта. Проделав несколько ищущих движений среди бумаг
ящика и не найдя конверта, он закрывает ящик и начинает ис-
кать конверт на столе п в шкафах, пока его пе найдет. Но может
случиться так, что конверта вообще ие окажется, и тогда чело-
век может взять его у соседа, послать кого-либо па почту или
сам пойти на почту, но в конце концов конверт оказывается при-
обретенным и письмо посланным.
Если сопоставлять, как нами дано в универсальной схеме
(см. рис. 1), все эти поведенческие акты с машиной автоматиче-
ской регуляции, то в известных границах мы найдем то же ар-
хитектурное сходство. В самом деле, человек, поставив пред собой
цель найти конверт, открывает ящик — это рефлекторное дейст-
вие. Но это действие не закончилось тем полезным эффектом,
достижение которого было поставлено как цель. Выражаясь
языком физиологии, все органы чувств человека не получили той
обратной афферентации, которая должна соответствовать нахож-
дению конверта. В результате этого в центральную нервную си-
стему поступает афферептация, пе соответствующая поставленной
цели.
В технике автоматического регулирования такое соотношение
получило название «рассогласования». Это значит, что в регули-
рующее устройство вместо заданной величины С70, которая долж-
на быть получена при устойчивой работе системы в порядке об-
ратной информации от регулируемого объекта, поступает инфор-
мация Ui. Тогда Uo будет выражать величину рассогла-
сования. До сих пор аналогия между машиной и человеком по
Физиология и кибернетика
227
общей схеме саморегуляции может быть достаточно полной» Од-
нако реакция организма и машины на рассогласование и поис-
ки решения поставленной задачи значительно разнятся и прин-
ципиально не могут быть аналогизированы»
В самом деле, рассогласование в машине автоматической регу-
ляции с заданными величинами и с известным диапазоном коле-
баний обратной информации может быть выражено математиче-
скими уравнениями и формулами. Точно так же и реакция
регулирующего устройства на рассогласование может быть в точ-
ности рассчитана как по интенсивности, так и по составу. В ре-
зультате мы имеем установку полезного эффекта па нужном
уровне, а рассогласование при этом сведется к нулю.
Если мы посмотрим, каковы реакции человека на рассогласо-
вание между целью поведения и его конечными результатами, то
мы можем с определенностью утверждать, что никакой матема-
тической формулой не охватить все те возможные условия, кото-
рые толкают человека на разнообразные поведенческие акты.
Кроме того, в машинах с автоматической регуляцией есть точ-
ная пропорциональность между величиной рассогласования и
реакцией регулируюхцего устройства на эти рассогласования. На-
оборот, в приведенном нами примере несовпадение между целью
действия и обратной афферентацней от результатов действия пе
имеет никакой пропорциональности и находится в прямой зави-
симости от побочных условий, не входящих в данную систему
циклической регуляции (пет конверта у соседа!).
Из разбора этого конкретного примера мы видим, что положе-
ние Випера о возможности перенесения математической теории
сервомеханизмов на частные механизмы человеческого поведения
неверно. Во всяком случае, это положение совершенно пока не
обосновано разбором конкретных примеров в сравнительном пла-
не. Между тем уверенность в возможности применения матема-
тической теории сервомеханизмов к человеческому поведению ве-
дет и к уверенности в возможности моделирования всех без остат-
ка поведенческих актов человека.
С этой точки зрения, надо признать совершенно правильным и
сомнение Куффипьяля, который указывает на то, что «математи-
ческий анализ показывает, что это неосуществимо... Одни лишь
сервомеханизмы сами по себе не могут создать все человеческое
мышление, и, таким образом, приходится отказаться от первой
важной части доктрины Винера» (Куффиньяль, 1953—1954, с. 3).
Разобранный пример показывает, что, хотя архитектурные
черты циклических систем саморегуляций и являются общими
для явлений различных классов, тем не менее свойства звеньев,
узлов и механизмов таковы, что математические теории механи-
ческих систем не всегда могут быть с успехом использованы для
анализа и понимания частных механизмов жнвои системы мозга
8*
228
Физиология и кибернетика
Во всяком случае на этом пути предстоит основательная и впол-
не конкретная аналитическая работа по установлению степени
приложимости математических теорий и расчетов к отдельным
конкретным формам функций и механизмов. Только
при этом ус-
ловии возможно движение вперед в этом весьма интересном,
но уже изрядно запутанном вопросе.
Условный рефлекс и кибернетика
Не случаен тот факт, что и сам Винер, и многие
его последо-
ватели, да и критически настроенные авторы при анализе пер-
спектив развития кибернетики непременно закапчивают изложе-
ние характеристикой отношения ее к условным рефлексам и к
поведению человека. Мы видели из приведенной схемы, что эта
органическая связь вытекает из самой эволюции циклических
систем с обратной связью.
Обсуждая значение обратной связи для поведения животных,
Винер называет ее «способностью живого существа устанавли-
вать свое будущее поведение па основании прошлого опыта».
Он пишет: «Обратная связь может быть такой же простой, как
при обычном рефлексе, но она может быть и обратной связью
высшего порядка, при которой прошлый опыт используется для
регулирования не только специальных движений,
но и целой
линии поведения. Такая обратная связь, определяющая поведе-
ние, может проявляться, да часто и проявляется, в виде того,
что нам известно в одном аспекте как условный рефлекс, а в
другом — как обучение» (Винер, 1954, с. 33). Луи де Бройль, об-
суждая проблемы кибернетики, пишет: «Теория счетных машин,
теория передачи сигналов... должна дать нам много сведений о
нормальной и патологической деятельности нервной системы и,
в частности, о механизме рефлексов». «Блестяще изученные
Павловым условные рефлексы также входят в эту общую схему»
(Луи де Бройль, 1953—1954, с. 48, 52).
Можно привести и еще ряд подобных высказываний, но дело
не в этом. Нам важно сейчас установить, что именно условный
рефлекс кибернетики находят удобным для своих далеко иду-
щих аналогий. Из приведенных высказываний следует, что при-
обретение опыта и использование приобретенного опыта для бу-
дущего поведения являются теми общими способностями живот-
ных и человека, подражание которым в машинных моделях за-
крыло бы принципиальную пропасть между «роботом» и челове-
ком, до сих пор существующую и осознаваемую всеми киберне-
тиками. Но именно здесь-то и надо вспомнить то сомнительное
положение кибернетики, по которому общность функциональных
проявлений должна соответствовать общности их частных меха-
низмов. При ближайшем знакомстве с этим вопросом можно ви-
Физиология и кибернетика
229
Рис. 2. Принципиальная схема условнорефлекторного ответа в новом осве-
щении
At А\ А2 — анализаторы, на которые падают раздражения от обстановки (обет.)
и от пускового условного раздражения (У. Р); Афф. с — афферентный синтез,
включающий в себя и возбуждения, поступающие через ретикулярную формацию
ствола мозга; ФР — формирование рефлекторной реакции; Обр. афф.— обратная
афферентация о результатах реакции; АД — акцептор действия
деть, что сторонники кибернетики при обращении к условным
рефлексам имеют в виду то кардинальное и принципиально но-
вое, что дало открытие условного рефлекса в начале девятисотых
годов. Однако для более детального обсуждения вопроса этого
уже недостаточно. Необходимо установить, какую внутреннюю ар-
хитектуру и какие механизмы имеет условный рефлекс, если его
рассматривать в аспекте всех последних достижении как школы
И. П. Павлова, так и вообще нейрофизиологии.
Только вскрыв глубокие физиологические механизмы условно-
го рефлекса, мы сможем установить, что именно и в каком виде
может быть заимствовано кибернетикой для моделирования и по-
строения различных аналогий. Вместе с тем глубокий физиологи-
ческий анализ механизмов условного рефлекса может показать
нам, какими конкретными средствами нервная система позволяет
животному и человеку использовать прошлый опыт для построе-
ния будущего поведения. А разобрав эти специфические особен-
ности механизмов условного рефлекса, мы, естественно, сможем
поставить вопрос и о том, имеются ли аналогичные приспособ-
ления в машинных устройствах с автоматической регуляцией.
Все эти вопросы можно разрешить только на конкретной схе-
ме механизмов условного рефлекса, которые мы можем в настоя-
щее время считать прочно установленными (рис. 2).
230
Физиология и кибернетика
Приведенная схема является синтетической. Она включает в
себя все достижения современной нейрофизиологии, помогающие
понять архитектуру условного рефлекса, и, кроме того, в ней
отражены и те данные из физиологии высшей нервной деятель-
ности, которые получены были нами за последние годы.
Первый пункт, требующий тщательного обсуждения, состоит
в особенностях соотношения двух видов афферентных раздраже-
ний, на которых строится условный рефлекторный ответ. Их
можно назвать соответственно: а) «пусковой афферентацней»,
представленной непосредственным действием самого условного
раздражителя (У. 1\), и б) «обстановочной афферентацней»,
представленной совокупностью раздражений от обстановки экспе-
римента и от всех тех предшествовавших опыту раздражений,
которые создают определенное доминантное состояние животного.
Что касается пускового раздражения, то его значение не под-
лежит сомнению, и именно оно более всего изучалось в много-
численных экспериментах школы И. II. Павлова. Значение
же обстановочных раздражителей, подготавливающих специфиче-
скую предпусковую интеграцию нервных процессов в форме
скрытой доминанты, стало ясным только в последние годы. Ре-
шающая и направляющая роль этой предпусковой интеграции
нервных процессов с особенной отчетливостью была выявлена
в двух формах эксперимента: в динамическом стереотипе (Купа-
лов, Асратян, Скипип) и после удаления лобных отделов коры
больших полушарий (Шумилина). В первом случае нарочито со-
зданная стереотипная обстановка предрешает (!), в какой форме
проявится условный рефлекс, независимо от того, какой пусковой
раздражитель будет применен. В этой форме эксперимента была
получена отчетливая диссоциация обстановочной и пусковой аф-
ферентацпй, органически объединенных при экспериментирова-
нии с условным рефлексом.
Недавние эксперименты с прямым электроэнцефалографиче-
ским анализом феномена динамической стереотипии показали,
что это предпусковое доминирование достигается тем, что к мо-
менту предстоящего действия условного раздражителя в соответ-
ствующих областях коры и подкорки происходит опережающее
повышение возбудимости (Анохин, Симоненко).
Во второй форме эксперимента, проделанного в нашей лабора-
тории, было показано, что органическое объединение обстановоч-
ных и пусковых афферентацпй и их последовательное действие
осуществляется при прямом влиянии лобных отделов коры мозга.
Удаление лобных отделов нарушает этот афферентный синтез,
и обстановочные раздражители становятся пусковыми раздражи-
телями, полностью дезинтегрируя условные двигательные реакции
животного.
Уже сейчас можно отметить, что для кибернетики во всех ее
Физиология и кибернетика
231
попытках моделировать условный рефлекс пли какие-либо его
фрагменты это органическое объединение обстановочных и пус-
ковых афферентаций, несомненно, составит «камень преткнове-
ния». В самом деле, как мы видели, совокупность обстановочных
афферентаций создает предпосылку к развертыванию той или
другой условной реакции. По состав обстановочных афферентаций
исключительно сложен и, что особенно важно, разнообразен и ди-
намичен до бесконечности. Сможет ли машина когда-либо пост-
роить свое поведение в зависимости от синтеза обстановочных и
пусковых воздействий па нее? Во всяком случае, это первая, но
еще не последняя трудность на пути подражания условному реф-
лексу. Обычно же для анализа и моделирования берется изоли-
рованное действие только какого-либо пускового раздражителя.
Из приведенной схемы условного рефлекса видно, что все разд-
ражители внешнего и внутреннего мира неизбежно должны прой-
ти стадию афферентного синтеза. Физиологический смысл вы-
деления этой стадии состоит в том, что никакое рефлекторное
действие пе может быть сформировано в эффекторных аппаратах
до тех пор, пока не закопчен синтез всех афферентных воздейст-
вий, падающих па организм в данный момент. Именно резуль-
таты афферентного синтеза определяют, какое действие должно
быть сформировано при наличии данной совокупности афферент-
ных раздражений. Важно помнить, что без этой стадии ие может
быть сформировал пи один поведенческий акт животного и чело-
века. Однако этим дело не ограничивается. Многолетние работы
нашей лаборатории показали, что точно в тот же момент, когда
заканчивается синтез всех афферентных раздражений и когда
начинается формирование в эффекторпой части соответствующего
рефлекторного действия, складывается особый аппарат, назван-
ный нами в свое время акцептором действия (см. рис. 2, А. Д.).
Физиологические свойства этого аппарата памп были подробно
описаны в специальных публикациях, и потому я сейчас пе бу-
ду па этом подробно останавливаться (см. Анохин, 1949, 1952,
1955).
Для настоящей же статьи нам важно лишь отметить два важ-
нейших свойства этого аппарата, о которых, к сожалению, пе име-
ют представления авторы, делающие попытки моделировать услов-
ный рефлекс. Прежде всего этот аппарат формируется раньше, чем
будет сформирован сам условный рефлекс. Следовательно, он на
какое-то время опережает появление ответного действия. Это об-
стоятельство определяет и его дальнейшую физиологическую
роль: принимая па себя (см. рис. 2, А. Д.) в форме обратной
афферентация (Обр. афф.) все афферептпые раздражения, кото-
рые возникают от результатов действия, аппарат акцептора дей-
ствия производит сопоставление результатов афферентного син-
теза с результатами произведенного действия. Таким образом, со-
232
Физиология и кибернетика
впадение этих возбуждений заканчивает весь циклический про-
цесс, в то время как их «рассогласование» вызывает целый ряд
новых реакций, которые должны дать в конце концов рефлектор-
ный ответ, соответствующий характеру акцептора действия. Во
всех случаях так называемого произвольного поведения, как и
при появлении условного рефлекса в ответ на условный раздра-
житель, акцептор действия является постоянным направляю-
щим фактором, дающим возможность определить соответствие
произведенного действия исходному замыслу.
Как мы уже видели из предыдущего изложения, во всех пред-
ставлениях кибернетики вопрос «обратных связей» является
центральным, поскольку благодаря им осуществляется последую-
щая подгонка соответствующих воздействий на «регулируемый
объект». Однако во всех случаях моделирования условного реф-
лекса по внешним чертам сходства авторы этих моделей никогда
не думали о том, что в головном мозгу раньше, чем появится
действие, всегда формируется аппарат оценки этого действия.
Мне представляется, что это — второе принципиальное препятст-
вие для моделирования условного рефлекса, так как без акцепто-
ра действия немыслима никакая приспособительная деятельность
животных и особенно человека. Фессар в своей недавней статье
по кибернетике указывает, что нечто подобное существует в ма-
шинах автоматического регулирования, но он пе знает аналога
этому в деятельности нервной системы. Он пишет: «Употребля-
ются также цепи «предвидения», когда рабочий механизм
предупреждается заранее о будущем приходе приказа. Очень ве-
роятно, что такие же приспособления существуют и при работе
нервной системы. Нейрофизиологам надо попытаться решить эту
трудную задачу — доказать существование таких приспособле-
ний в различных частных случаях» (Фессар, 1953—1954, с. 30)
(курс, наш.— П. Л.).
Мы уже показали выше, что аппарат «предвидения» давно на-
ми найден и, следовательно, надо только посмотреть, в какой
мере он может быть воспроизведен в соответствующих механиче-
ских моделях. Я лично сомневаюсь, чтобы он смог быть смонти-
рован в механических моделях, поскольку он всегда складывается
ex tempore и всегда на основании синтеза многообразных внеш-
них и внутренних раздражений организма. Если будущее пока-
жет, что мы можем приблизиться к этому, то уж во всяком слу-
чае с обязательным учетом факта наличия такого аппарата в
нервной системе.
Подводя итог всему приведенному материалу, мы можем вы-
сказать следующие резюмирующие положения.
1. Кибернетика — это направление мысли, стремящееся найти
общие черты у явлений различного порядка и на основе этой
общности раскрыть механизмы функционирования, в частности
Физиология и кибернетика
233
нервной системы. В этом смысле данное направление мысли за-
служивает всяческой разработки специалистами различных про-
филей.
2. Основные исходные положения кибернетики еще и сейчас
не являются четко сформулированными, а исторические основа-
ния для наличия общих черт у разнородных явлений остаются
еще не вскрытыми. Между тем только на этом пути можно отчет-
ливо представить себе, до какого предела общие черты у явлений
различного класса могут быть использованы для моделирования.
Имеющаяся литература по этому вопросу (как зарубежная, так
и наша) пе дает здесь хоть сколько-нибудь утешительного ана-
лиза.
содержит в себе все черты «обратных связей»
3. Теория обратной афферентации в физиологии, развиваемая
нами с 1935 г
кибернетики и потому может служить для расширения ее воз-
можностей при построении аналогичных механических моделей.
4. Общая архитектура циклических систем с обратной а
। hi ।
рентацией или с обратной связью применима для понимания всех
тех приспособительных актов человека, которые заканчиваются
полезным эффектом. В этом смысле вторая сигнальная система
как специфическая принадлежность человека целиком подчиня-
ется этому правилу.
5. Никакие модели условно-рефлекторных ответов не могут
претендовать даже на отдаленное сходство с действительным ус-
ловным рефлексом, если в них не будут учтены два кардиналь-
ных процесса: а) процесс афферентного синтеза и б) образование
акцептора действия как аппарата «оценки» результатов действия.
ЛИТЕРАТУРА
Бэлэнеску И. Н. Кибернетика и некоторые вопросы физиологии и психоло-
гии.— Вопросы философии, 1957, № 3.
De Broglie L. Vue generale et philosophique sur la cybernetique.— In: Structu-
re et evolution des techniques, Paris, 1953—1954, N 35—36.
De Broglie L, Sens philosophique et portee pratique de la cybernetique.— In:
Structure et evolution des techniques, Paris, 1953—1954, N 35—36.
Couffignal L. Methodes et limites de la cybernetique.— In: Structure et evo-
lution des techniques, Paris, 1953—1954, N 35—36.
Fessard A, Points de contact entre neurophysiologie et cybernetique.— In:
Structure et evolution des techniques, Paris, 1953—1954, N 35—36.
Wiener N, The human use of human beings. Boston, 1954.
КИБЕРНЕТИКА
И ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ
МОЗГА *
Соотношение кибернетики и нейрофизиологии выражается
прежде всего в том, что кибернетика стремится моделировать
основные законы деятельности мозга.
Будучи сама производной от циклических замкнутых, т. е.
автоматических саморегуляций в технических системах, киберне-
тика наложила определенный отпечаток и па подходы нейрофи-
зиологов к изучению процессов целого мозга. Нейрофизиология
широко использует в настоящее время формализацию процессов,
преобразование информации в сложных системах, математиче-
скую обработку материалов исследования и т. д. Все эти заим-
ствования из кибернетики стали в работе нейрофизиолога по-
стоянными и необходимыми приемами для расшифровки регуля-
торных приспособлений целого организма.
В свою очередь, кибернетика постепенно совершенствует своп
модели мозговой деятельности и этим самым позволяет исполь-
зовать их в конструировании различного рода технических си-
стем с повышающимся уровнем «интеллигентности», т. е. со свой-
ствами, приближающимися к работе самого мозга.
Это взаимодействие совершенно определенно показало, что ус-
пех воспроизведения мозговой деятельности в информационных
моделях находится в прямой зависимости от того, в какой сте-
пени полно отражены в каждом отдельном случае моделирова-
ния истинные законы деятельности мозга, логически формализо-
ванные нейрофизиологами. Иначе говоря, успех изучения про-
цессов мозга и особенно реализация их в поведенческих актах
зависит от того, в какой степени в этом встречном движении от-
ражены истинные архитектурные черты работы целого мозга, син-
тезированные на основе последних достижений нейрофизиоло-
гии.
В настоящее время стало очевидным, что успех такого кон-
такта зависит от того, сможем ли мы создать столь адекватную
логическую модель работы целого мозга, которая не только об-
легчила бы возможность производить анализ любого механизма
* Опубликовано в журнале «Вопросы психологии» (1966, № 3). Доклад на
XVIII Международном психологическом конгрессе в Москве.
Кибернетика и интегративная деятельность мозга
235
любого поведенческого акта как интегративного образования, но и
помогла бы формулировать и новые исследовательские работы.
Такой подход к интегративной деятельности целого мозга по-
казал, что она пе может быть изучаема без оценки конечного
приспособительного результата, в связи с которым происходит
как формирование, так и перестройка любой функциональной
системы.
В этом смысле результаты исследований школы крупнейшего
английского физиолога Шеррингтона дают возможность судить
лишь о частных механизмах интегративной деятельности, по-
скольку они не отнесены к работе целого мозга и остаются не-
зависимыми от конечного приспособительного эффекта целого ор-
ганизма (Лнохин, 1948).
Становится все более очевидным, что рабочие контакты ней-
рофизиологии и кибернетики поставили совершенно недвусмыс-
ленные требования построить такую нейрофизиологическую мо-
дель интегративной деятельности целого мозга, которая отвечала
бы двум основным требованиям: а) она должна быть достаточно
убедительно обоснована фактическими результатами современной
нейрофизиологии и вместе с тем б) быть той большой киберне-
тической архитектурой, в которую вписываются все тонкие ана-
литические результаты, полученные при изучении любых част-
ных механизмов мозга. Именно поэтому, прежде чем перейти
к копкретпым нейрофизиологическим механизмам формирования
поведенческого акта, нам нужно раскрыть состав самой архитек-
туры поведенческого акта. Я уже имел однажды счастливый слу-
чай — на Международном симпозиуме по обработке информации
в центральной нервной системе (Лейдеп, 1962)—предложить та-
кую физиологическую архитектуру, которая, как нам кажется, от-
вечает поставленным выше требованиям и которая построена на
основе нашей более равней концепции функциональной системы,
как саморегуляторпой организации с обратной афферептацпей
(рис. 1).
Однако к настоящему времени моими сотрудниками получе-
ны многие новые результаты, которые позволяют еще глубже
понять те универсальные механизмы, которые поддерживают в
единстве все многочисленные компоненты функциональной систе-
мы на всем протяжении формирования конечного полезного эф-
фекта. Интегративные механизмы в работе целого мозга, пред-
лагаемые нами для объяснения структуры поведепческих актов,
вступают в тесный контакт с принципами самоорганизации и мо-
делирования работы мозга, разрабатывающимися в настоящее вре-
мя в нейрокиберпетпке.
Описываемая ниже физиологическая архитектура интегра-
тивной деятельности целого мозга в настоящее время уже пере-
растает свое значение только как модели, объясняющей меха-
236
Кибернетика и интегративная деятельность мозга
ОР
рррерентные оазиутнаения ПА
Обратная аррерентация
О А
1 О А
Яро грамма
оеистбая
р/ебстбае
Viiiii
МША
Анцеатар
Оеистбия
/Щ/ШМе/77/76/
бебстбия
__//11111___
Результаты
Яеаетбая
О А
Оррерентные бпабуна/епня
Аррерентнь/а
синтез
РА
Обратная аррерентар/я
пара метре/
РеастРан
__Z 1.1 1 1 XV
Резуябп?ап?б/
Оеисп7Рия
р/еистбае
ОА
ОА
Акцептор
беастбия
Ярагпамма
оеастбия
РР
Аррерентне/и
синтез п.
НА
Кибернетика и интегративная деятельность мозга
237
низмы деятельности, и становится методом для формулировки
новых исследовательских задач на уровне тончайших аналити-
ческих нейрофизиологических приемов. Эта модель упорядочива-
ет многочисленные поиски в нейрофизиологических условиях и
точно определяет то частное место, которое каждое из этих ис-
следований занимает в большой архитектуре поведенческого акта.
Ниже мы последовательно разберем узловые механизмы этой
физиологической архитектуры, пригодной для любого поведен-
ческого акта.
I. Стадия афферентного синтеза
Нейрофизиологический смысл этой стадии в формировании по-
веденческого акта состоит в том, что она позволяет тщательно
обработать, сопоставить и синтезировать всю ту информацию,
которая необходима организму для совершения наиболее адекват-
ного для данных условий приспособительного акта.
Афферентный синтез является наиболее ответственным мо-
ментом интеграции, он является в полном смысле этого слова уз-
ловым механизмом функциональной системы. Он помогает реше-
нию трех основных параметров всякого поведенческого акта: что
делать, как делать и когда делать?
Эти три параметра вписывают всякий данный поведенческий
акт в гармоническую последовательность многих поведенческих
актов и устанавливают то адаптивное отношение животного к
внешнему миру, без которого невозможно было бы его выжива-
ние. Афферентный синтез позволяет животному с определенной
точностью удовлетворять его непрерывно сменяющиеся потребно-
сти и приводить их в соответствие с бесконечно разнообразны-
ми воздействиями внешнего мира.
Рис. 1. Принципиальная схема модели поведенческого акта, представлен-
ная в трех этапах развития
А — для понимания динамики нелинейных процессов функциональной системы пока-
зана стадия, когда закончился афферентный синтез (АС), произошло формирование
цели и вслед за этим акцептора действия. Эфферентные возбуждения в этой стадии
лишь вышли на афферентные периферические пути. Однако в этой стадии не полу-
чен полезный приспособительный результат.
Б — дальнейшее развертывание процессов в системе, в связи с чем был получен
конечный результат, соответствующий сходной цели действия. Видно, что резуль-
таты действия через свои параметры формируют обратную афферентацию, которая
направляется в центральную нервную систему и вступает в контакт с акцептором
действия.
В — эта часть рисунка иллюстрирует момент несовпадения обратной афферентапии
с акцептором действия. На схеме видно, что это несовпадение немедленно активи-
рует ориентировочно-исследовательскую реакцию, которая по своему значению
является фактором, облегчающим образование новой программы действия.
ОР — ориентировочная реакция; О А — обстановочная афферентация; ПА — пусковая
афферентация; РФ — коллатерали ретикулярной формации; М — мотивация; П —
память
238
Кибернетика и интегративная деятельность мозга
Сама необходимость учета невероятно большого количества
переменных параметров, определяющих поведенческий акт, исто-
рически сделала совершенно необходимым формирование стадии
афферентного синтеза. Непременное наличие многочисленных аф-
ферентных воздействий па организм из внешнего мира сделало
афферентный синтез универсальным фактором всякой жизни, по-
скольку последняя даже в самой примитивной форме должна
устанавливать адекватные соотношения между своими потребно-
стями и совокупностью последовательно развивающихся внешних
воздействий.
Другим фактором, относящимся к характеристике роли аффе-
рентного синтеза, является то, что он непременно и всегда дол-
жен предшествовать принятию решения (desision making).
Едва ли сейчас надо говорить о том, что момент «принятия
решения» является физиологическим «рубикопом» в формирова-
нии поведенческого акта.
Как уже говорилось, афферентный синтез призван решить
наиболее общие вопросы поведения: что делать, как делать и ког-
да делать? Если вдуматься в эти три вопроса, связанные с при-
нятием решения, то станет очевидным, что каждый из этих во-
просов представляет собой фактор выбора, т. е. исключения из-
быточных степеней свободы, являющихся неизбежным следствием
анатомической и физиологической композиции организма.
В самом деле, попытаемся проанализировать, какой смысл
вложен в решение вопроса: что делать? В силу особенностей
расположения и иннервации отдельных, часто весьма дробпых,
мышечных образований организм животных так сконструирован,
что практически в каждый данный момент потенциально он мо-
жет совершить миллионы возможных движений и, следовательно,
миллионы моторных актов. Иллюстративно говоря, человек мо-
жет указать пальцем на какой-то предмет, он может встать со
стула, он может повернуть голову к окну и т. д. Ясно, что та-
ких индивидуализированных моторных актов он может произве-
сти миллионы. Однако в данной ситуации, например, человек
протягивает руку именпо к карандашу и берет его.
Возникает естественный вопрос: па каком физиологическом
основании к периферическим органам, т. е. к мышечным аппа-
ратам, вышли возбуждения именно для этого моторного акта,
а все остальные миллионы степеней свободы были исключены?
Причем тот же карандаш в ряду последовательных поведен-
ческих актов данного человека должен быть взят именно в тот
момент, когда он является следствием завершенного афферентно-
го синтеза, переходящего в «принятие решения» — «взять каран-
даш». Если бы карандаш был взят значительно позднее того мо-
мента, когда должен был быть совершен акт письма, он не привел
бы к положительному приспособительному эффекту. Следователь-
Кибернетика и интегративная деятельность мозга
239
но, вопрос «когда?» также должен решаться адекватно в резуль-
тате афферентного синтеза, т. о. в результате сопоставления и
соподчинения всех действующих в данный момент афферентных
сигналов из внешнего мира поставленной цели к действию. Со-
вершенно очевидно, что это ограничение потенциальных степеней
свободы и выбор вполне определенного по значению и по времени
поведенческого акта является результатом учета основных по-
требностей организма па данный момент, возможности осущест-
вления конкретной потребности в данных условиях и т. д.
Из всех этих соображений следует, что с чпсто нейрофизио-
логической точки зрения проблема формирования афферентного
синтеза исключительно сложна и потому мы должны упорядо-
чить обсуждение конкретного материала этой решающей стадии
по следующим направлениям.
Л. Состав афферентного синтеза и его тонкие
гические и нейрохимические механизмы.
Б. Общая нейродипамика его осуществления.
Все, что говорилось выше, показало, что из
неирофизиоло-
всех узловых
механизмов поведенческого акта напоолее ответственным и наи-
более широким по охвату специфических свойств целого мозга
является афферентный синтез. Именно отсюда происходит и его
значимость для поведения вообще, и его особая чувствительность
ко всякого рода воздействиям па организм. Мне думается, что
именно эти характерные свойства афферентного синтеза застави-
ли II. II. Павлова уже очепь давно подчеркнуть, что афферент-
ная сторона в деятельности центральной нервной системы явля-
ется «так сказать, творческой», в то время как за эфферентной
функцией он признавал только «техническую» роль.
Это определение весьма красочно и в то же время наиболее
адекватно отражает роль стадии афферентного синтеза. Как мы
уже говорили, она неизбежно заканчивается «принятием реше-
ния», но это и есть, в сущности, «творческий акт», поскольку
на основании тысяч разнородных информаций принимается ре-
шение о выборе одного акта из миллионов возможных!
Бросается в глаза многообразие тех афферентных информа-
ций, которые непременно должны быть обработаны, собраны и со-
поставлены с прошлым опытом, прежде чем произойдет то кри-
тическое событие, которое получило выразительное название
«принятие решения» (Бншон, 1963; Буллок, 1959).
Мы можем высказать, как нам кажется, вполне достоверное
положение, что пи одно «принятие решения», насколько бы при-
способительный акт нп был элементарным, пе может произойти
без предварительного афферентного синтеза. Вот почему опреде •
ление его состава и анализ тончайших нейрофизиологических
механизмов является самой актуальной задачей нейрофизиологии
высшей нервной деятельности, учения о поведении, кибернетики.
240 Кибернетика и интегративная деятельность мозга
Все это должно составить материальную основу для совмест-
ного разрешения пограничных проблем нейрофизиологии, высшей
нервной деятельности и психологии (И. II. Павлов, И. С. Бери-
ташвили, А. Н. Леонтьев, А. Р. Лурия и др.).
А. Состав стадии афферентного синтеза
Каков же состав афферентного синтеза по качеству и коли-
честву всех информаций, обработка которых должна повести к
«принятию решения»?
Физиологический анализ методом условных рефлексов пока-
зывает, что в его состав входят следующие качественно различ-
ные компоненты: доминирующая мотивация; обстановочная аффе-
рентация; пусковая афферентация; память.
Только всесторонний объективный анализ всех составных ком-
понентов стадии афферентного синтеза и их интегральных взаи-
модействий поможет нам понять, каким образом разнородные
формы информации, имеющие различные по сложности струк-
туры и качественно разное значение для организма, могут иног-
да в высшей степени быстро интегрироваться, сопоставляться
друг с другом, оценивать сложившуюся в данный момент ситуа-
цию и принимать именно то решение, которое является наиболее
адекватным для всей ситуации в целом. Ниже и будет более под-
робно разобрана физиологическая роль перечисленных составных
компонентов афферентного синтеза.
Доминирующая мотивация. Состояние доминирующей мотива-
ции может быть создано или на основе исходных гормональных
и метаболических превращений организма, или па основе преж-
ней последовательности в поведенческих актах. Примером первого
типа мотиваций может служить чувство голода, примером второго
типа мотиваций — стремление поступить в высшее учебное заве-
дение, коллекционировать какие-либо предметы и другие «основ-
ные влечения», по Павлову. II та и другая мотивация, являясь
доминантной, обладает способностью обрабатывать поступающую
в мозг информацию в соответствии со своим энергетическим по-
тенциалом. Конечно, второй вид мотивации для своего формиро-
вания требует более длительной обработки огромного количества
информации, часто поступавшей на протяжении большого перио-
да времени, однако принципиальные архитектурные черты этой
стадии для того или другого случая одинаковы. Какие физиоло-
гические механизмы начинают и реализуют в коре больших по-
лушарий доминирующую мотивацию? Специальные электрофи-
зиологические эксперименты, проведенные в нашей лаборатории
К. В. Судаковым и другими сотрудниками па голодающих жи
вотных, показали, что мотивация, т. е. господствующее в данный
момент состояние, весьма интересно действует на кору головного
Кибернетика и интегративная деятельность мозга
241
мозга. Восходящие возбуждения биологически различных мотива-
ций избирательно мобилизуют в коре головного мозга именно те
синаптические организации, которые в процессе прошлого жиз-
ненного опыта животного или человека были связаны с удовлет-
ворением такого же состояния. Благодаря такому избирательно-
му распространению возбуждений по коре больших полушарий
создаются энергетические условия для ’занятия превалирующего
в функциональном отношении положения теми корковыми ас-
социациями, которые в прошлом способствовали возникновению
и удовлетворению данной потребности.
В сущности этот факт усиленного возбуждения через синап-
сы корковых клеток, принадлежащих определенной биологиче-
ской деятельности, и составляет основу для исключения той из-
быточной информации, которая в каждый данный момент в ог-
ромном количестве поступает в Ц.Н.С., т. е. главным образом в
кору головного мозга.
Информация начинает подбираться и как бы «взвешиваться»
на весах этого господствующего или доминирующего мотиваци-
онного состояния, своеобразно представленного почти на каждом
корковом нейроне.
Легко видеть, что уже одно наличие доминирующей мотива-
ции значительно облегчает организму «принятие решения». Сра-
зу же на уровне первых синаптических организаций подкорки
происходит «оттеснение» с помощью тормозного процесса всей
той информации, которая функционально не связана ни с прош-
лым опытом, ни с наличной мотивацией (рис. 2).
Перед нами встал естественный вопрос: какими средствами
каждая мотивация, имеющая определенную биологическую мо-
дальность, может осуществить свое столь избирательное влияние
на синаптические организации коркового уровня. Применив ме-
тод аппликации и внутривенной инъекции различных нейрофар-
макологических и нейрохимических средств, мы раскрыли весь-
ма интересные интимные механизмы той избирательности, с по-
мощью которой различные восходящие влияния оккупируют
корковый уровень нервной организации. Оказалось, что различ-
ные биологические модальности восходящих возбуждений опери-
руют синапсами различной нейрохимической природы, что и со-
ставляет, вероятно, инструмент первичного избирательного рас-
пространения возбуждений по коре больших полушарий. Мы были
удивлены разнообразием и спецификой тех механизмов, с помо-
щью которых осуществляется избирательное воздействие на кору
больших полушарий со стороны подкорковых восходящих возбуж-
дений.
Так, например, кошка, голодавшая всего один-два дня, имеет
заметную десипхронизацию корковой электрической активности,
по преимуществу передних отделов коры головного мозга.
Кибернетика и интегративная деятельность мозга
Рис. 2 Схематическое изображение
конвергентных взаимодействий раз-
личных восходящих возбуждений на
корковом нейроне
На схеме показано, что превалирующая
мотивация (МП), приходящая в кору
головного мозга, занимает превалирую-
щее положение по отношению ко всем
другим мотивациям, лишенным акту-
ального значения для данного момента
(MI)
Внешние раздражения (ВР) подразде-
ляются по отношению к этой доминиру-
ющей мотивации в том, что одни из
них вступают в контакт с доминирую-
щей мотивацией (ВР/), другие же не
имеют отношения к этой мотивации и
потому не вступают в связь с ней
в аксоплазме нервной клетки (BPJ7)
А'ора
Суточное голодание ¥ - днедное голодание
Рис. 3. Сопоставление односуточного и четырехсуточного голодания кош-
ки (Л, Б)
Видно, что при односуточном голодании активация возникает только в лобных от-
делах коры мозга (А), при четырехсуточном голодании активация становится гене-
рализованной по всей коре головного мозга (Б). Применение в этих условиях
аминазина снимает десинхронизацию в задних отделах коры больших полушарий,
но оставляет ее по-прежнему в передних отделах мозга (В). Обозначения: ЛП—
лобная правая; ЛЛ — лобная левая; СМП — сенсомоторная правая; СМЛ — сенсо-
моторная левая; ТП — теменная правая; ТЛ — теменная левая; ЗП — затылочная
правая; ЗЛ — затылочная левая
Кибернетика и интегративная деятельность мозга
243
В это же время остальные области коры находятся в состоя-
нии обычной, характерной для сна медленной фоновой активно-
сти (рис. 3, Л). Контрольными экспериментами (коагуляция, по-
ляризация и т. д.) было установлено, что источником этой вос-
ходящей активации является гипоталамус (К. В. Судаков).
Однако после пяти дней голодания картина радикально меняется:
теперь уже вся кора больших полушарий оказывается в состоя-
нии десинхропизации (рис. 3, Б),
Казалось бы естественно думать, что эта генерализованная
десипхронизация есть результат широкой иррадиации по коре
мозга возбуждений той же пищевой биологической модальности
и распространяется из того же подкоркового источника, т. е.
из гипоталамуса. Однако оказалось, что это не так, хотя сама
десинхропизация по своему внешнему выражению и остается оди-
наковой.
Прямые исследования показали различную нейрохимическую
природу подкорковых и корковых синаптических организа-
ций, включенных в формирование этих двух активаций. Зная
на основе прежних наших исследований (А. И. Шумилина, В. Гав-
личек), что аминазин блокирует на подкорковом уровне болевую
активацию, мы решили применить его в той стадии голодания
кошки, когда локальная активация передних отделов мозга пере-
растает в генерализованную активацию всей коры мозга. Резуль-
тат оказался весьма демонстративным. Хлорпромазин полностью
снял активацию пятидневного голодания в задних отделах коры
мозга, но оставил нетронутой локальную активацию в передних
отделах мозга, т. е. первичную активацию, возникающую в начале
голодания (рис. 3, В).
Кроме того, прямое наложение амизила (М-холинолитика) па
область первичной активации показало, что эта активация осу-
ществляется с помощью холинергических механизмов (Турепко,
1965).
После этих экспериментов стало ясно, что начальные стадии
голодания формируются преимущественно мобилизацией холинер-
гических механизмов коры и подкорки, в то время как при мно-
годневном голодании активирующая деятельность, т. е. формиро-
ванне мотивированного поведения, начинает осуществляться по
преимуществу адренергическими механизмами.
Интересно, что микроэлектродные исследования с нанесением
различных фармакологических средств на поверхность коры боль-
ших полушарий показали, что даже одна и та же клетка коры
больших полушарий может получать восходящие влияния в виде
конвергенции от различных биологических мотиваций: от пище-
вой и от оборонительной (Фадеев, 1965; Котов, 1965). Какая за-
мечательная способность мозга оперировать динамически, вклю-
чая все то, что
в той или ипои степени адекватно наличной
244
Кибернетика и интегративная деятельность мозга
мотивации, и исключая всю ту информацию, которая ей неадек-
ватна!
Возникает вопрос: с помощью каких конкретных механизмов
реализуется эта замечательная способность корковой клетки при-
нимать и классифицировать огромное количество конвергиру-
ющих возбуждений, различных по происхождению и по нейрохи-
мическим характеристикам? Как мы видели, узловым механиз-
мом этой избирательной мобилизации корковых элементов
в соответствии с господствующей мотивацией служит широкая
конвергенция возбуждений на нейроне.
Чтобы ближе подойти к этому важному механизму, мы разра-
ботали специальный прием исследования. Замысел этих исследо-
ваний был таков: если с одного и того же пункта коры мозга
снимать электрически вызванные ответы, то мы, несомненно, бу-
дем иметь дело с каким-то количеством клеток, принимающих
на себя различные возбуждения. Эти различия могут состоять
как в сенсорной модальности (Юнг, Фессар и др.), так и в био-
логической модальности (Анохин; Фадеев, 1965). Из конструкции
экспериментов следует, что мы получили возможность сопоста-
вить между собой как вызванные потенциалы, так и клеточные
разряды,1 возникающие в одном и том же пункте в ответ на
раздражения различного качества и различного происхождения.
Кроме того, замысел состоял в том,
что именно в этих условиях
мы должны были апплпцировать на поверхности коры в пункте
отведения различные фармакологические средства.
Сопоставляя теперь влияние этих средств на различные ком-
поненты вызванного потенциала и на разряды нервных клеток,
мы могли, следовательно, сделать вывод о химической специфике
синаптических образований, и в особенности субсинаптических
мембран.
Например, с одного и того же пункта коры мы отводили од-
новременно вызванные потенциалы нескольких типов: транскол-
лазального происхождения, от седалищного нерва и от других
областей мозга и организма (рис. 4). Обычно потенциалы были
характерны для каждого отдельного источника. Наложение раз-
личных фармакологических средств на пункт отведения различ-
ных вызванных потенциалов и разрядов клеток дало возможность
выяснить, что независимо от одного и того же электрического
знака компонентов вызванного потенциала они менялись по-раз-
ному при различных аппликациях. Внимательное изучение этих
изменений показало, что они связаны только лишь с химически-
ми особенностями анплицированного вещества, а не с электри-
ческой характеристикой компонентов вызванного потенциала.
Выявилось огромное различие в химической чувствительности
по внешнему виду одинаковых отрицательных компонентов вы-
званного потенциала, но возникших на основе возбуждений, при-
Кибернетика и интегративная деятельность мозга
245
Рис. 4. Общая схема расположения
раздражающих и отводящих элек-
тродов при отведении потенциалов
различного происхождения
Обозначения: ОЭ — отводящий электрод;
РЭ — раздражающий электрод. Черная
линия под отводящим электродом соот-
ветствует наложению бумажки, смочен-
ной фармакологическим веществом, кон-
центрические круги обозначают пред-
положительное распространение веще-
ства
/7/77 /7. isb tad.
шедших из различных районов тела и нервной системы (Чуппи-
на, 1965).
Таким образом, подытоживая оценку мотивации как фактора,
способствующего ускорению афферентного синтеза благодаря со-
поставлению поступающей в мозг информации и быстрому осво-
бождению мозга от избыточной информации, мы можем сказать,
что аппараты и механизмы, которые Способствуют этому процес-
су, имеют всеобъемлющее значение. Они относятся как к архитек-
турным особенностям нервной системы, так и к ее различиям в
тонких метаболических процессах, что и определяет собой тон-
кую избирательную чувствительность корковых синапсов к прихо-
ду различных восходящих возбуждений. Благодаря этим свойст-
вам при конвергенции многих возбуждений на нейроне сразу же
исключается из активного участия в интегративной деятельности
вся та информация, которая не является значимой для данной
мотивирующей доминанты (см. рис. 1).
Обстановочная афферентация. Под обстановочной афферента-
•ян
цией мы понимаем совокупность всех тех внешних воздействий
на организм от данной обстановки, которые вместе с исходной
мотивацией наиболее полно информируют организм о выборе
того действия, которое более всего соответствует наличной моти-
вации. Физиологическая роль и поведенческий смысл обстановоч-
ной афферентации состоит в том, что она прежде всего благодаря
свойственному ей относительному постоянству действия создает
в центральной нервной системе весьма разветвленную и интег-
246
Кибернетика и интегративная деятельность мозга
рированную систему возбуждений, своего рода нервную модель
обстановки. Оставаясь длительное время подпороговой, эта систе-
ма от применения «пускового стимула» (см. ниже) может перейти
из скрытого состояния в явное, г. е, сформировать поведенче-
ский акт, интегрирующий интересы мотивации и возможности об-
становки.
Значение обстановки как особого типа афферентной информа-
ции особенно отчетливо проявляется в классических эксперимен-
тах с условными рефлексами. Здесь животное, находясь много раз
в обычной для него обстановке эксперимента, фактически ока-
зывается полностью подготовленным к секреторной реакции, т. е.
имеет все центральные процессы для осуществления ее. Однако
по нашему опыту мы знаем, что условной секреторной реакции у
животного в промежутках между раздражениями ие бывает до
тех нор, нока не будет введен условный стимул. Таким образом,
в момент, когда животное спокойно сидит в станке в промежутке
между применениями отдельных условных раздражителей, в его
нервной системе имеется комплекс разнородных организованных
ранее возбуждений, который мы в свое время назвали «предпу-
сковой интеграцией» (Анохин, 1949). Применение же условного
стимула выводит эту интеграцию из подпорогового состояния и
формирует активный поведенческий акт. Какие же факторы уста-
навливают такую гармонию между процессами, определяющими,
«что делать?», и процессами «как делать?»
Специальные опыты А. II. Шумилиной, проведенные в нашей
лаборатории, показали, что, действительно, в моменты покоя жи-
вотного между применениями условных раздражителей происхо-
дит сложное уравновешивание обстановочных и пусковых воз-
буждений, создающих строгое распределение во времени отдель-
ных фрагментов поведенческого акта (стояние в станке, побежки
к кормушке п условносекреторная реакция в ответ па пусковой
стимул).
Как известно, удаление лобных отделов коры мозга приводило
животное к такой форме реакций, которая свидетельствовала о
нарушении различия между обстановочными и пусковыми стиму-
лами: животное уже теперь не сидело спокойно в станке, а по-
переменно бегало то к одной, то к другой кормушке, явно пере-
путав правильный распорядок между этими двумя формами аф-
ферентных сигнализаций.
В прежнее время заметно преувеличивалась роль условного
раздражителя в условном рефлексе. Считалось само собой разу-
меющимся, что условный раздражитель служит абсолютным и
единственным поводом к возникновению условной секреции. Те-
перь, с раскрытием закономерности динамического стереотипа
(II. Павлов), становится все более и более ясным, что видимая
условная реакция является результатом обширного афферентного
Кибернетика и интегративная деятельность мозга
синтеза, в который включаются в различной последовательности
как внутренние, так и внешние факторы. Из сопоставления и
разбора предыдущих компонентов вытекает и роль третьего ком-
понента афферентного синтеза — пусковой афферентации.
Пусковая афферентация. Характерной чертой пускового сти-
мула является то, что он точно приурочивает к определенному мо-
менту реализацию того интегрированного нервного образования,
которое уже сложилось в мозгу до пускового стимула на основе
мотивационных и обстановочных афферентации.
Если доминирующая мотивация определяет собой, что именно
должен делать организм, а обстановочная афферентация опреде-
ляет, как он должен это делать в данной ситуации, то пусковой
стимул определяет третий важный параметр поведенческого акта:
когда надо делать то, что подсказано и мотивацией и обстанов-
кой. Какими конкретными физиологическими механизмами пу-
сковой стимул переводит «скрытую» интеграцию поведенческого
акта в видимую, реализованную форму? Где локализовано это
сближение возбуждений от пускового стимула с подпороговой мо-
делью возбуждений, созданных мотивацией и обстановкой,— все
это вопросы, которые должны неизбежно стать предметом иссле-
довательской работы, поскольку сам процесс афферентного синте-
за стал решающей стадией в формировании поведенческого акта.
Казалось бы, что афферентный синтез мог бы быть полностью
обеспечен разобранными выше компонентами и вся структура
поведенческого акта могла бы быть введена в действие. Однако
практически пи один поведенческий акт не может быть сформи-
рован, если из памяти организма, т. е. из его прошлого опыта,
не будут извлечены все те молекулярные следы, условнорефлек-
торные связи и ассоциации, которые накапливались на протяже-
нии всей предыдущей жизни организма в аналогичных или по-
хожих ситуациях.
Именно потому, что из памяти во время афферентного синте-
за непрерывно извлекается все то, что соответствует мотивации
и обстановке, мы и посвятим ей специальный раздел.
Память как компонент афферентного синтеза. Едва ли было
бы разумным развивать в связи с проблемой афферентного син-
теза все те представления о природе памяти, которые были раз-
виты в последние годы (Hyden, Me Conel, Gaito). Сейчас нам
важны механизмы памяти только в топ мере, в какой они обога-
щают возможности афферентного синтеза и принятие решения к
совершению одного из миллионов возможных поведепческих ак-
тов. В последние годы, основываясь на данных нейрохимическо-
го исследования, мы построили рабочую
гипотезу о так называ-
емом конвергентном замыкании условного рефлекса. Ядром этой
248
Кибернетика и интегративная деятельность мозга
Рис. 5. Схематическое изображение
двух синапсов на мембране одной и
той же клетки с различными субси-
наптическими мембранами (СМ)
Окончания приходящих аксонов обра-
зуют концевые колбы, заполненные вези-
кулами и окруженные преспнаптической
мембраной (ПМ).
Точками показана область аксоплазмы
данного нейрона, а различными стрел-
ками начинающиеся от субсинаптиче-
ской мембраны цепи специфических
ферментативных процессов (ФП1, ФПП)
гипотезы является процесс субсинаптпческой мембраны, на осно-
ве которого из молекул нервной клетки извлекаются элементы
фиксированного в прошлом жизненного опыта.
Как известно, вся проблема памяти состоит из момента запо-
минания пережитого данной физиологической системой и момен-
та извлечения этого фиксированного жизненного опыта, т. е вос-
поминания.
Если в настоящее время мы в какой-то степени и можем со-
ставить себе представление, хотя бы и гипотетическое, о процес-
се
иксации пережитого опыта
то по сути дела мы ничего пе
можем сказать о том, как формируется воспоминание, т. е. с по-
MO1
I
(ыо каких механизмов происходит извлечение фиксированно-
го опыта именно тогда, когда он способствует принятию реше-
ния, наиболее адекватного данной ситуации.
Исходя из представлений о молекулярной фиксации пережи-
того опыта, мы можем представить себе, что цепи химических
реакций, возникающие на субсинаптической мембране, продолжа-
ются и в аксоплазму. Здесь они должны обладать высокой фер-
ментативной специфичностью. Нам кажется, что именно по этой
ферментативной цепочке, начинающейся от субсипаптической
мембраны, впечатления прошлого могут «вытягиваться» в точном
соответствии с потребностями афферентного синтеза в данный мо-
мент. Только химическая специфика ферментативных процессов
может объяснить нам скорость, точность и своевременность вос-
произведения прошлого опыта из памяти в процессе формирова-
ния поведенческого акта (рис. 5).
Мы считаем вполне разумным допустить, что на каком-то эта-
пе этих процессов между субсипаптической мембраной и генера-
тором аксонных разрядов клетки в механизм воспроизведения
могут включаться и нуклеиновые кислоты (Hyden).
Надо представить себе все динамическое многообразие про-
цессов, описанных выше, чтобы понять, насколько грандиозным
является объем процессов, вовлеченных в этот синтез.
Кибернетика и интегративная деятельность мозга
249
Б. Общая динамика осуществления афферентного синтеза
Сложность афферентного синтеза состоит не только в том, что
он содержит огромное количество процессов и компонентов, но
еще и в том, как эти компоненты взаимодействуют между собой.
Все они в полном смысле этого слова производят непрерывную
«примерку» одного процесса к другим до тех пор, пока не
сформируется решение и пе сложится афферентная модель ожи-
даемых результатов. Иначе говоря, на первый план выступает
проблема нейродинамики взаимодействий между многочисленны-
ми процессами афферентного синтеза и, что особенно важно, ак-
тивная «подгонка» этих процессов к моменту принятия решения.
Всю эту работу мозга выполняют два важнейших фактора, спо-
собствующих облегчению взаимодействий между даже весьма уда-
ленными нервными элементами мозга. Эти факторы следующие:
ориентировочно-исследовательская реакция и центробежное акти-
вирующее и тормозящее влияние как на промежуточные этапы,
так и на чувствительность периферических рецепторных аппара-
тов (Мещерский, 1966).
Первый из этих факторов состоит в том, что восходящее ак-
тивирующее влияние на кору больших полушарий от гипотала-
муса и ретикулярной формации фактически не устраняется на
протяжении всего процесса афферентного синтеза. Следовательно,
корковые нейроны приобретают состояние, значительно облегча-
ющее «нащупывание» благоприятных связей между различными,
адекватными для данной мотивации синаптическими организа-
циями. Фактически та избирательная мобилизация корковых си-
наптических связей, о которой мы говорили в разделе мотива-
ции, оказывается в условиях ориентировочно-исследовательской
реакции значительно более обширной и, следовательно, вероят-
ность принятия правильного решения становится максимальной.
Афферентный синтез является взаимодействием рассмотрен-
ных выше его компонентов, но взаимодействием, происходящим
на высоком уровне энергетической активности коры мозга, под-
держиваемой гипоталамусом и ретикулярной формацией в форме
непрерывной реверберации.
Как показали работы Эрнандес Пеона и Левинстона, про-
цесс активирования периферических рецепторов также благо-
приятствует осуществлению оценки внешней информации. По ра-
ботам Гранита, Линдсли и других, центробежное влияние на ре-
цептор приводит к тому, что порог их чувствительности
снижается, а восходящая активация способствует тому, что коэф-
фициент дискриминации раздражений, приходящих в кору моз-
га, также значительно увеличивается.
Эта общая характеристика нейродинампческих ситуаций,
складывающихся во время афферентного синтеза, совершенно пол-
250
Кибернетика и интегративная деятельность мозга
но определяет роль ориентировочно-исследовательской реакции и
центробежной активации рецепторов в процессе афферентного
синтеза. Одиако для объяснения, несомненно, остается еще огром-
ное количество вопросов.
Так, например, восходящая активация, сопутствующая ори-
ентировочно-исследовательской реакции, несомненно, способству-
ет извлечению элементов прошлого опыта из кладовых памяти.
Но как она это делает? Какие топкие молекулярные процессы
разыгрываются на подступах к этим кладовым?
Мы имеем, правда, основание думать, что субсинаптические
мембраны, через которые возбуждение ориентировочной реакции
поступает в нервную клетку, имеют свои особенности, химиче-
скую специфику. Может быть, эта специфика и состоит в том,
что процесс, начинающийся на субсинаптической мембране, при-
водит к ускорению всякого рода ферментативных реакций в ак-
соплазме нервной клетки? Может быть, эта новая химическая
ситуация на мембране нервной клетки и в аксоплазме клетки и
представляет собой ту благоприятную ситуацию, которая спо-
собствует наиболее быстрому и наиболее правильному с вероят-
ностной точки зрения контакту между различными типами ин-
формации (см. рис. 5).
Все это пока еще далеко идущие гипотезы. Однако гипотезы,
уже не столь невероятные, как это могло бы показаться, напри-
мер, десять лет тому назад, т. е. до периода весьма быстрого
расширения наших знаний по нейродинамике и нейрохимии си-
наптических процессов.
II. Формирование действия
и аппарата предсказания его результатов
Афферентный синтез неизбежно приводит к принятию реше-
ния, т. е. к выбору вполне определенной деятельности из мил-
лионов возможных. Ясно, что этот процесс выбора знаменует со-
бой начало избирательного распространения возбуждений но эф-
ферентной части нервной системы. Как осуществляется это
принятие решения и выбор адекватной деятельности — это со-
ставляет один из самых загадочных вопросов для нейрофизио-
логии поведения.
Несомненно одно, что в процессе осуществления афферент-
ного синтеза непременно должен наступить момент, когда ио ка-
ким-то признакам мозг определяет, что афферентный синтез дол-
жен быть закончен и должно быть принято соответствующее ре*
иьение. Можно думать, что определение этого момента происхо-
дит благодаря широчайшей конвергенции возбуждений на корко-
вых нейронах и всестороннему сопоставлению на их поверхности
афферентных возбуждений различного происхождения.
Кибернетика и интегративная деятельность мозга
251
Можно согласиться с Бишопом, что в момент принятия ре-
шения все процессы мозга должны быть как-то особенно скон-
денсированы во времени и пространстве (Бишоп, 1963). Точно
так же утверждает и Т. Буллок, который в одной из своих ра-
бот уделил специальное внимание проблеме принятия решения
(Буллок, 1959).
Как и где происходит этот наиболее интегрированный про-
цесс формирования поведенческого акта? Ответа на эти вопросы
мы пока не имеем, да и сами эти вопросы поставлены только
в последние годы, причем в значительной степени под влиянием
кибернетики. Однако уже сам факт, что вопрос этот поставлен в
четкой форме, представляет собой огромное достижение в изуче-
нии интегративной деятельности головного мозга.
Опыты Л. II. Шумилиной с удалением лобных отделов моз-
га в какой-то степени подсказывают мысль, что наиболее инте-
грированным и экономным процессом при осуществлении выбора
является процесс, формируемый лобными отделами коры головно-
го мозга, в которых в какой-то форме должны быть представле-
ны результаты афферентного синтеза.
Значительно более определенно мы можем сказать относитель-
но формирования весьма важной стадии, которая завершается по-
строением модели
афферентных элементов,
целом составля-
ющих наиболее важные параметры будущих результатов действия.
Как показывает анализ поведенческих реакций и любой физио-
логической регуляции, этот аппарат, названный -нами акцепто-
ром действия, т. е. воспринимающим информацию о результатах
действия, является важнейшим образованием, которое упорядочи-
вает и коррегирует недостаточность поведенческих актов живот-
ных и человека.
Прямые наблюдения за обычной условной пищевой реакцией
в случае внезапной подмены хлеба мясом дали возможность
вскрыть, что условный раздражитель в совокупности с остальны-
ми компонентами афферентного синтеза действительно формирует
афферентную модель, которая по своему составу весьма точно
отражает характерные признаки будущего подкрепления. Именно
поэтому внезапная подмена хлеба, для которого уже заготовлена
афферентная модель, другим пищевым веществом — мясом созда-
ет своеобразное «рассогласование» между заранее сформирован-
ными признаками результата (подкрепление хлебом) и реальной
совокупностью афферентных признаков от подкрепления мясом
(Анохин и Стреж, 1933).
Подобные эксперименты с экстренной подменой или устране-
нием подкрепляющих факторов в других формах эксперимента,
например в условиях самораздражения, по Олдсу, неизменно по-
казывают один и тот же результат. В этом последнем случае
новая совокупность афферентных признаков от нового результата
252
Кибернетика и интегративная деятельность мозга
(выключение раздражающего тока) оказывается неадекватной для
уже ранее сформировавшейся модели аппарата предсказания,
подсказанной афферентным синтезом.
Для понимания функций аппарата акцептора действия важно
помнить, что механизм его формирования является универсаль-
ным. ;Он имеет место в нервной системе всюду там, где сло-
жившаяся функциональная система обеспечивает выход на пе-
такои
риферию командных процессов и происходит получение какого-
либо приспособительного эффекта. В этом смысле даже
автоматизированный акт, как дыхательный, формируется с несом-
ненным участием акцептора действия, хотя физиологическая ар-
хитектура здесь, конечно, своеобразная.
Так, например, было установлено, что прежде чем на перифе-
рии осуществится команда дыхательного центра о взятии четы-
рехсот кубиков воздуха, в самой структуре дыхательного центра
заведомо до реального взятия воздуха (1,5—2,0") формируется
афферентный аппарат, задачей которого является последующий
прием информации от легкого о том, взято ли действительно че-
тыреста кубиков или команда выполнена неточно (рис. 6).
Эти представления были подтверждены в пашей лаборатории
с помощью специального электронного устройства (Полянцев,
1963). Импульсации диафрагмального нерва преобразовывались
в электронном устройстве и направлялись к прибору искусствен-
ного дыхания, а этот прибор вводил в легкое воздуха ровно
столько, сколько должны были дать импульсации, направляющие-
ся от дыхательного центра к диафрагме.
Такие приспособления способствовали созданию модели само-
управляемого дыхания, благодаря которому легкое с помощью
прибора искусственного дыхания всегда получало воздуха столь-
ко, сколько этого требовалось по условиям афферентного синтеза,
т. е. при учете всех важнейших факторов дыхания (рпс. 7) —
наличия СО2 и О2, периферической хеморецепториой афферента-
ции и т. д.
Изменяя в электронном устройстве передачу информаций на
дыхательный аппарат, мы могли в любой момент ввести в легкое
столько воздуха, сколько это было необходимо по соображениям
эксперимента, а пе по тому, сколько это нужно было организму.
Так, например, вместо трехсот кубиков, которые были закодиро-
ваны в пмпульсациях дыхательного центра, в эксперименте было
введено в легкое всего лишь 100 кубиков. Альвеолярные рецеп-
торы легкого в этих условиях могли сигнализировать в дыха-
тельный центр информацию, эквивалентную взятию легкими ста
кубиков воздуха. Итак, в результате приведенных выше экспе-
риментов в дыхательном центре создаются условия явного не-
совпадения между тем, что было подсказано командой, и резуль-
татом, который получен в действительности. Налицо рассогласо-
Кибернетика и интегративная деятельность мозга
253
Дь/сшие влияния
ломанаа результат
Рис. 6. Синтетическая схема, представляющая все узловые
Дысшае влияния
механизмы
31
ункциональной системы применительно к дыхательному акту
На рисунке представлены: афферентный синтез, принятие решения и выход аффе-
рентных возбуждений к диафрагме. В центре коллатеральных механизмов представ-
лен механизм длительно задержанного возбуждения, к которому адресуется обрат-
ная афферентация от альвеол легкого. Эта система отношений может служить уни-
версальной моделью для формирования акцептора действия
Рис. 7. Схема вмешательства с помощью электронного преобразователя
импульсов (ЭПИ) и дыхательного аппарата (ДА) в работу легкого
ванне, которое и служит стимулом для формирования нового
компенсаторного действия, в результате чего при следующем вдо-
хе происходит взятие легкими значительно большего количества
воздуха.
В этом и подобных экспериментах мы много раз показали, что
всякому принятию решения неизбежно сопутствует формирова-
ние акцептора действия, который по своей
изиологической сути
есть разветвленная система нервных процессов в афферентной
части мозга, отражающая все существенные признаки (парамет-
ры) будущих результатов.
Я не могу вдаваться сейчас во все подробности формирования
и состава этого аппарата. Несомненно, в его состав входят не
254
Кибернетика и интегративная деятельность мозга
только афферентные признаки конечных результатов, которые
отражают цель данного поведенческого акта, но и все промежу-
точные этапы в совершении поведенческого акта.
И здесь также каждое несоответствие какого-либо промежу-
точного результата и его обратной афферентации с уже сфор-
мировавшимся акцептором действия ведет к исправлению этой
неточности или ошибки. Фактически имепно этот механизм де-
лает наши действия и все поведение в целом гармоничным и
целесообразным.
В последние годы, воспользовавшись особенной простотой и
моделируемостью дыхательной функции, мы попытались с по-
мощью микроэлектродпого метода охарактеризовать те процессы
дыхательного центра, которые способствуют образованию аппара-
та предсказания предстоящих результатов действия. Из сопостав-
ления деятельности отдельных нейронов дыхательного центра
можно вывести пока еще лишь предварительное соображение о
том, каким образом формируется акцептор действия.
Несомненно то, что в момент, когда формируется «принятие
решения» и оно начинает реализоваться, на эфферентных кол-
латеральных путях аксона происходит ответвление эфферентных
возбуждений, составляющих в целом своеобразную «копию ко-
манды» к действию.
Этот комплекс коллатеральных эфферентных возбуждений в
контакте с некоторыми процессами афферентного синтеза и со-
ставляет первоначальное ядро акцептора действия.
Это ядро должно находиться в непрерывном возбуждении до
тех пор, пока с периферии поступят сигналы о получении ре-
зультатов.
Конкретные микрофизиологические исследования показывают,
что в дыхательном центре имеются весьма разнообразные типы
клеток, и некоторые из них по своей деятельности вполне со-
ответствуют этой логике процесса.
Мне кажется, что конкретным нервным субстратом, с по-
мощью которого осуществляется передача в акцептор действия
«копии команды», являются многочисленные аксонные колла-
терали, особенно у корковых нейронов.
Хотя образование акцептора действия и представляет собой
узловой механизм поведенческого акта, однако он является не
единственным механизмом, формирующим закрытый циклический
процесс в конце поведенческого акта. Одним из самых принци-
пиальных вопросов построения универсальной модели поведен-
ческого акта является получение полезных результатов, которые
по своим параметрам должны удовлетворять как а<
ШОР
е рентный
синтез, так и принятое решение.
Кибернетика и интегративная деятельность мозга
255
III. Результаты действия
как самостоятельная физиологическая категория
В истории изучения мозга как интегративного образования
явно преобладала фиксация внимания на самом действии, а не
на его результатах.
Так, например, мы говорим: «чесательный рефлекс»
тельный рефлекс»,
этих выражениях? В них,
, «хватз-
«мигательный рефлекс» и т. д. Что отражено в
как правило, отражено только реф-
лекторпое действие и даже нет намека на результаты. Термин
«чесательный рефлекс» означает только, что произошло чесание,
но он совершенно не отражает, какие результаты были получе-
ны вслед за этим чесанием. Точно так же «хватательный реф-
лекс» отражает только то, что произошло хватание, но какие
результаты получил организм от этого, остается неотраженным.
Можно перебрать всю терминологию, созданную рефлекторной
теорией, и убедиться, что результаты действия были полностью
исключены из привычного мышления физиолога.
Еще и до сих пор есть много исследователей, которые огра-
ничиваются представлением о рефлекторной дуге. Однако в этом
случае, естественно, исключается и механизм афферентного син-
теза, и аппарат предсказания результатов (акцептор действия),
поскольку в рефлекторной дуге все течение нервных процессов
имеет линейный характер...
Концепция о «рефлекторной дуге» исторически выросла на
действии и именно поэтому оставалась «открытой», поскольку
«закрыть» ее могло лишь только представление о результатах
действия и представление об информации мозга о этих резуль-
татах, т. е. об обратной афферентации.
На протяжении всего исследовательского процесса «резуль-
таты действия» подразумеваются как нечто само собой разуме-
ющееся, но это «нечто» не нашло себе места в органической не-
прерывности поведенческого акта. Естественно, что отсутствие в
поле зрения результатов действия повело за собой исключение из
внимания исследователя и механизма оценки результатов дейст-
вия, т. е. обратную афферентацшо.
Используя приведенные выше примеры рефлексов, мы можем
сказать, что результатом «хватательного рефлекса» будет не
само хватание, как действие, а та совокупность афферентных
раздражений, которая соответствует признакам «схваченного»
предмета (результат действия). Движение этих возбуждений в
сторону мозга и создает конечный этап — сопоставление этих
афферентных возбуждений в акцепторе действия. Для карандаша,
например, это будет совокупность характеризующих его призна-
ков (граненность, вес, температура и т. д.).
Мне кажется, что именно в этом пункте надо искать причину
256
Кибернетика и интегративная деятельность мозга
крайней устойчивости представления о «рефлекторной дуге», ко-
торое на протяжении многих лет фиксировало внимание физио-
логов на совершившемся действии и непроницаемой ширмой
отделяло их от оценки полученных результатов, являющихся
органическим следствием действия.
IV. Обратная афферентация
Понятие обратной афферентаций «закрывает» пробел между
действием и его центральным эффектом и делает поведенческий
акт стройной архитектурой, цельным «блоком», заканчивающим-
ся оценкой результатов в акцепторе действия.
Здесь будет уместно оговорить одно неправильное выраже-
ние, которое часто применяется при анализе поведенческих ак-
тов. Некоторые авторы предпочитают вместо предложенного
нами термина «обратная афферентация» применять термин «кор-
ригирующая афферентация». Это последнее выражение непра-
вильно по своей физиологической сути. Афферентная сигнали-
зация от результатов действия, от их параметров, ничего не
может корригировать, поскольку она всегда является пассивным
следствием наличия любых, даже неправильных, результатов
действия. Обратная афферентация возникает даже в том случае,
когда совершена поведенческая ошибка, поскольку она всегда
может возникнуть как результат столкновения организма с ка-
кими-то факторами среды. ГТ только лишь акцептор действия,
включая в себя все афферентные параметры цели, подсказанной
афферентным синтезом, может иметь корригирующее влияние.
Это корригирующее влияние па полученный результат акцеп-
тор действия оказывает не прямо, не непосредственно, а возбуж-
дает все активирующие аппараты центральной нервной системы,
которые создают более тонкий афферентный синтез и более точ-
ную программу действия в виде сопоставления имеющихся уже
параметров результата с параметрами, закодированными в обрат-
ной афферентация (ориентировочно-исследовательская реакция).
HIM
Известно, что обратная афферентация является почти всегда
многопараметрпой, отражая многообразие черт самого результа-
та. Так, например, результат действия может состоять в поступ-
лении обратной афферентаций через зрительный канал, но она
может поступить также и через кожные рецепторы, через зву-
ковые рецепторы и т. д. А это значит, что каждый из параметров
результата может быть локализован в своей собственной проек-
ционной зоне и, следовательно, комплекс параметров, характе-
ризующих данный специфический результат, должен быть по-
строен по принципу весьма обширной системы связей, охваты-
вающей всю кору больших полушарий.
Возникает естественный вопрос: каким образо^м такая обшир-
Кибернетика и интегративная деятельность мозга
257
ная система может вступить в связь с акцептором действия
со-
держащим также афферентные параметры? Из поставленного во-
проса следует: если обратная афферептация проецируется в раз-
личные зоны коры мозга, часто весьма удаленные друг от друга,
то и акцептор действия должен был бы иметь такую же обшир-
ную, т. е. дисперсную структурно-
изиологическую архитектуру.
Такой вывод логичен, но едва ли мозг в действительности ра-
ботает по такому принципу. Я склонен думать, что вначале,
т. е. в первой стадии поступления в кору мозга, распыленные
афферентные возбуждения в дальнейшем на каких-то элементах
коры и подкорки собираются в более компактном виде, может
быть даже на отдельных нейронах, п уж во всяком случае на
каком-то весьма компактном ядерном образовании. Тогда
логическое противоречие между дисперсией афферентных приз-
наков результата по коре мозга, принятием решения и акцеп-
тором действия было бы устранено.
V. Рассогласование между целью
и полученным результатом
Пожалуй, одним из замечательных свойств функциональной
системы поведенческого акта является момент сопоставления того
афферентного прогноза результатов, который сконденсирован в
акцепторе действия, с темп нервными импульсами, которые при-
ходят к акцептору действия как сигналы о полученных резуль-
татах. Этот момент, несомненно, является решающим для форми-
рования актов дальнейшего поведения в непрерывном и после-
довательном ряду поведенческих актов животного и человека.
В этом сопоставлении акцептор действия выглядит как нечто
постоянное и направляющее (модель мотивации), в то время как
обратная афферентация результатов может все время меняться
в зависимости от степени успешности этих результатов. Эти
изменчивые соотношения особенно широко представлены в по-
ведении, формирующемся по типу «проб и ошибок».
Механизм сопоставления интегрированных признаков будущих
результатов с сигналами о реально полученных результатах
включает еще одно важное дополнение, а именно эмоциональный
фактор удовлетворенности или неудовлетворенности. Ясно, что
этот фактор служит дополнительным стимулом к поиску новых,
более адекватных цели программ афферентного действия. Эта
связь процессов рассогласования с эмоциональным характером
всей реакции оказывается особенно выраженной в тех случаях,
когда рассогласование связано с полным отсутствием обратной
афферентации биологически положительного характера, как, на-
пример, от акта еды. В этих случаях имеет место рассогласова-
ние между акцептором действия, включающим в себя все пара-
9 П. К. Анохин
258
Кибернетика и интегративная деятельность мозга
метры пищевого вещества
и отсутствием реальных раздражите-
лей от пищевого вещества: вкусового, обонятельного, тактиль-
ного п т. д.
Типичным примером такого рассогласования является угаше-
ние условной пищевой реакции в экспериментах с условными
рефлексами. Уже первое неподкрепление едой акцептора дейст-
вия, созданного условным раздражителем, в некоторых случаях
вызывает резкую агрессивную реакцию и весьма энергичные
попытки достать корм из кормушки (Анохин, 1925).
Можно думать, что в процессе эволюции фактор рассогласо-
вания между поставленной целью и полученным результатом был
связан с возникновением отрицательной эмоции там, где рас-
согласование создавалось на основе неполучения жизненно важ-
ных факторов. Наоборот, если отсутствует обратная афферента-
цпя, сигнализирующая о биологически отрицательном действии,
то рассогласование ведет к положительным эмоциям.
Можно думать, что такая связь механизмов оценки конечно-
го результата в поведенческих актах с эмоциональным аппара-
том является весьма прогрессивным и целесообразным приобре-
тением в широком биологическом плане.
Для оценки этого момента рассогласования и совпадения в
акцепторе действия мы должны еще раз указать на огромную
роль ориентировочно-исследовательской реакции. Во всех случаях
несовпадения акцептора действия с конкретно полученными ре-
зультатами автоматически включается механизм ориентировочно-
исследовательской реакции. Последнее ведет к немедленному рас-
ширению и мобилизации афферентных информаций за счет более
тщательного анализа окружающей среды.
Этот процесс длительный. Он связан с непрерывной цирку-
ляцией
•ин
ерентных
системах центрально-периферических
циклов возбуждений. В результате создается новая программа
действия, которая может сформировать па периферии именно те
конечные результаты, которые соответствуют сложившемуся уже
ранее акцептору действия.
Как мы видели, нейродинампка формирования афферентного
синтеза в том и состоит, что на протяжении этой стадии ориен-
тировочно-исследовательская реакция создает два основных усло-
вия, обогащающих афферентный синтез все большим и боль-
шим количеством афферентной информации.
С одной стороны, это достигается чрезвычайно мощным энер-
гетическим воздействием ретикулярной формации и гипоталамуса
на кору больших полушарий. Благодаря этим воздействиям па
корковых нейронах создаются все условия облегченного взаимо-
действия между клеточными элементами коры мозга, располо-
женными в различных, часто весьма удаленных друг от друга
пунктах коры.
Кибернетика и интегративная деятельность мозга
259
С другой стороны, та же самая ориентировочно-исследова-
тельская реакция способствует центробежным влияниям и на
периферические рецепторы. Это последнее действие приводит к
тому, что порог чувствительности включенного в данный момент
более успешной
в ориентировочно-исследовательскую реакцию анализатора зна-
чительно снижается, что еще больше соответствует успеху аф-
ферентного синтеза и формированию новой.
программы действия.
Из приведенных выше нейродипамических
условии, которые
немедленно создаются после рассогласования между реальным
результатом и акцептором афферентной модели результата, ак-
цептор действия служит весьма важным стимулом, создающим
новый поиск с включением новых афферентных аппаратов и
созданием повой программы действия.
Заключение
Приведенный выше материал дает законченную архитектуру
поведенческого акта, все узловые механизмы которого имеют
нейрофизиологическое обоснование и много раз были показаны
в специально поставленных поведенческих экспериментах (Ма-
каренко, Асмаяц и др.).
Эта физиологическая архитектура представляет собой весьма
разнородную картину по качеству интегративных процессов,
причем само взаимодействие между этими процессами является
специфической чертой именно интегративного целого, а не его
частей. Так, например, возникновение акцептора действия не-
мыслимо без предварительного афферентного синтеза и без тех
процессов, которые связаны с оценкой сигналов с периферии о
полученных результатах.
_ Совершенно очевидно, что каждый из этих узловых механиз-
мов содержит в себе бесконечное переплетение и взаимодейст-
вие возбуждений и торможений и в каждом из этих механизмов
много раз встречаются одни и те же и различные возбуждения.
Но именно в этом и состоит смысл универсальной модели пове-
денческого акта. Хотя ее архитектура по составу процессов и
механизмов весьма сложна, она представляет собой логически
законченное образование, чем и определяется место каждого
процесса и механизма в этой большой архитектуре.
С точки зрения кибернетики, такая модель работы мозга
является особенно удобной, поскольку, будучи построена на
тончайших нейрофизиологических закономерностях, вместе с тем
открывает широкие перспективы для моделирования и матема-
тической обработки ее частных механизмов. Но именно соотноше-
ние этих частных механизмов с целой архитектурой и представ-
ляет собой то, что может быть использовано при моделировании
9*
260
Кибернетика и интегративная деятельность мозга
и конструировании электронных и технических конструкций.
Именно этим и объясняется тот факт, что многие нейрофизио-
логи и кибернетики обратили специальное внимание на достоин
ства изложенной выше физиологической архитектуры целого по-
веденческого акта (Мэгун, 1963; Клаус, 1962; Паск, 1963; Мин-
ским, 1964; Фессар, 1961; и др.).
В данной работе мне интересно было продемонстрировать для
психологов физиологическую архитектуру поведенческого акта
как целого потому, что именно поведенческий акт служит свя-
зующим звеном между нейрофизиологией, высшей нервной дея-
тельностью и психологией.
Когда-то Альфред Фессар, принимая наши принципиальные
положения о физиологической архитектуре поведенческого акта,
заметил, что наступил момент, когда в тонком нейрофизиологи-
ческом эксперименте мы должны показать те механизмы и про-
цессы, с помощью которых удерживается как единое интегратив-
ное целое эта поведенческая архитектура (Фессар, 1959). Сейчас
мы уже можем сказать, что значительная доля тайн физиоло-
гической
архитектуры поведенческого
акта нами в какой-то
степени раскрыта, хотя еще многое остается исследовать, для
того чтобы сделать ясной эту удивительно целесообразную ор-
ганизацию, создававшуюся миллионы лет.
Во всяком случае, мы сейчас близки, например, к пониманию
механизмов афферентного синтеза до молекулярного уровня вклю-
чительно. Мы подошли вплотную к оценке составных физиоло-
гических компонентов акцептора действия и поняли огромную
роль результата как самостоятельного компонента всей архитек-
туры.
И лишь только два узловых механизма, с логической точки
зрения локализованные довольно точно, до сих пор еще не под-
даются тонкому нейрофизиологическому анализу и детерминисти-
ческой расшифровке. Это — «принятие решения» и «рассогласо-
вание» между реально полученными результатами и афферент-
ной моделью данных результатов, отраженные в акцепторе дей-
ствия.
Однако, основываясь на добытых нами весьма многочислен-
ных нейрофизиологических фактах, я не имею оснований думать,
что эти сложные вопросы являются неразрешимыми. В качестве
примера, оправдывающего такой оптимизм, можно указать на то,
что роль «копии» команды при совершении какого-либо аффе-
рентного приказа к действию становится все более и более ясной.
Как показывают микроэлектродные исследования, ответвляясь по
коллатералям тысяч различных аксонов, эти эфферентные воз-
буждения долгое время остаются активными в замкнутых «ло-
вушках возбуждения», открытых Лоренте де Но. Эти возбужде-
ния,
очевидно, являются
циклическими
в целом комплексе
Кибернетика и интегративная деятельность мозга
261
остаются активными до того момента, когда придет обратная
афферентация о полученных результатах.
Для меня сейчас является несомненным, что решающий акт
Hill
оценки полученных результатов происходит где-то здесь, на сты-
ке еще свежих, энергетически заряженных следов от эфферент-
ного приказа к действию с приходящими позднее сигналами о
полученном результате.
Мы прилагаем сейчас все усилия на самых различных уров-
нях исследования, от молекулярных процессов до высшей нерв-
ной деятельности и психологии включительно, чтобы объединить
в одной концепции все уровни наших знаний о мозге.
Задача, несомненно, трудная, но мы хорошо знаем, что ни-
когда еще в истории науки не было такого случая, чтобы страх
перед трудностями исследования способствовал научному про-
грессу.
МЕТОДОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
КИБЕРНЕТИЧЕСКИХ ЗАКОНОМЕРНОСТЕЙ *
Общие предпосылки
Общеизвестно, что кибернетический подход получил в настоя-
щее время наиболее широкое развитие в технических пауках и в
областях, где применяются разного рода электронные машины п
приспособления. По кибернетическая революция — это не только
технические преобразования; это прежде всего революция в
мышлении, в самом подходе к явлениям природы и в методе
анализа. Фиксация внимания только па технических аспектах
кибернетики рано или поздно станет недостаточной для научного
прогресса в целом.
Вместе с тем недостаточны и попытки философского
нпя самого существа тех кибернетических положений
пришли в науку вместе с пей. Так, до сих пор не уделено серь-
осмысле-
которые
езного внимания кардинальному вопросу: какие законы лежат в
основе того парадоксального факта, что явления различных клас-
сов, относящиеся к технике, живой природе и обществу, разви-
ваются и действуют па основе одних и тех же общих принципов
функционирования? Этот вопрос по своему характеру является
философским. И мне кажется, что корень той неопределенности,
с которой мы встречаемся при попытке определить, что такое
кибернетика, лежит именно в этом.
В самом деле, возьмем наиболее полное и наиболее устояв-
шееся определение кибернетики как науки об управлении в жи-
вых и механических системах. Если остаться па уровне частных
закономерностей жизни в видимых ее пределах, то такое опре-
деление как будто удовлетворяет нас. Однако как только мы
захотим определить в более общем аспекте само управление, то
сейчас же возникнут новые вопросы: кому необходимо это уп-
равление, как вообще могла возникнуть сама необходимость уп-
равления как организационного принципа жизни? Действительно,
с точки зрения материалистической диалектики управление не
может не иметь истории развития и, следовательно, не может не
иметь вполне закономерных причин своего возникновения.
Этот вопрос приобретает еще большую актуальность, если мы
остановимся на другом определении кибернетики, а именно, что
* Опубликовано в книге «Материалистическая диалектика и методы естест-
венных наук» (Мм 1968).
Методологическое значение кибернетических закономерностей 263
_________________•__-_ — — -- ________ - т—-
кибернетика есть наука о преобразовании информации в живых
и технических системах. Это определение также немедленно
вызывает вопросы: что такое информация и чем вызвана необ-
ходимость ее преобразования? Иначе говоря: какие более общие
законы предопределяли появление информационных зависимо-
стей, как и где родилась или возникла первичная информация,
почему она должна преобразовываться и какие законы направ-
ляют это преобразование? Философский аспект этих вопросов
гораздо более широк, чем тот, в котором принято рассматривать
их в обычных кибернетических формулировках.
То же самое относится и к центральному пункту кибернети-
ческого мышления — к наличию у всех классов явлений обрат-
ной связи.
Никто не сомневается в том, что обратная связь является
«душой кибернетики» и что только она почти нацело определила
все успехи технической кибернетики. Не случайно поэтому не-
которые исследователи считают, что мы живем в «эру обратных
связей». К этой центральной закономерности кибернетического
мышления также приложимы аналогичные вопросы, которые
были поставлены выше: почему обратная связь стала такой уни-
версальной закономерностью, пригодной и для машин, и для
организмов, и для общества; когда и при каких условиях эта
закономерность могла возникнуть в истории земного шара?
Отсутствие ответа на эти вопросы показывает, что, несмот-
ря на развитие кибернетического направления в технике, биоло-
гии и общественных пауках, все же более широкий взгляд на
особенности кибернетики, именно философский взгляд, остается
до сих пор мало разработанным.
Очевидно, что этот аспект кибернетики можно будет считать
в той или иной степени ясным только в том случае, если будет
точно определена всеобщая роль кибернетических закономерно-
стей в эволюции пашей планеты как историческая необходимость.
Ниже мы попытаемся использовать добытые нами экспери-
ментальные данные для раскрытия некоторых исторических
причин формирования основных кибернетических закономер-
ностей.
Система и результат
Одной из самых замечательных черт современного развития
наук является ноиск «ключа», который позволил бы наиболее
быстро понять огромное разнообразие
актнческих
результатов
научного исследования, добытых в различных науках. Таким
универсальным «ключом» является понятие системы.
Предсказывая в одном из своих предсмертных интервью, ка-
кой будет физика и биология в 1984 г., Порберт Винер выразил
эту мысль в весьма отчетливой форме. Он рассказывал о своей
264 Методологическое значение кибернетических закономерностей
совместной с доктором Дела Риччи работе над развитием идеи,
согласно которой в квантовой физике и в квантовой механике
должна играть большую роль организация системы. Главное
направление развития биологии также пройдет, по его мнению,
через организацию системы в пространстве и во времени. Здесь
самоорганизация должна играть основную роль. Следовательно,
полагал Винер, не только биологические науки будут сближаться
с физикой, но и физика будет ассимилировать некоторые биоло-
имепно в кибернетических
спасение от половодья раз-
степени обоснованной. Спе-
гические идеи.
Роль системы как обобщающего и организующего фактора в
развитии всех наук особенно выпукло представлена в формулиров-
ках Эллиса и Людвига, которые опубликовали недавно книгу
«Философия системы», установив, что «эпоха джетов», «эпоха
кибернетики» и «эпоха космоса» обязаны своим происхождением
появлению систем с универсальными свойствами саморегуляции;
они предлагают назвать нашу эпоху «эпохой системы» (Ellis,
Ludwig, 1962)...
Вполне естественно поэтому, что
обобщениях многие ученые увидели
розненных научных фактов.
Такая надежда была в какой-то
цпалисты различных областей знаний нашли в кибернетике об-
щий язык, и возникли широкие научные контакты, обеспечиваю-
щие взаимообогащение. В практику вошли симпозиумы и конг-
рессы, в которых наряду с математиками и физиками принимали
участие биологи, физиологи, филологи, лингвисты.
Эту роль кибернетики особенно полно и точно охарактери-
зовал в своей монографии «Кибернетика и философия» Г. Клаус:
«Кибернетика имеет большое зпачение в первую очередь потому,
что эта наука осуществляет общую тенденцию к интегральному
объединению многих наук... Опа выдвигает много проблем, об-
щих для самых различных специальных паук. Опа конструирует
понятийное поле, в котором объединяются многие элементы, при-
сущие разным специальным паукам; при помощи общей систе-
мы своих категорий и понятий она стремится подвести фундамент
под тенденцию к интеграции расчлененных конкретных наук»
(Клаус, 1963).
Возвращаясь к вопросам, которые были поставлены нами во
вступительной части этой статьи, мы могли бы сказать, что имен-
но эта «тенденция» кибернетики к обобщению и формированию
ею «поцятийного поля» и составляет одну из научных проблем
философии кибернетики. Мы должны и здесь опять-таки поста-
вить перед собой прежний вопрос: в чем состоит причина того
факта, что именно кибернетика создала такое «понятийное поле»,
которое смогло выявить нечто общее в огромном разнообразии
наук и явлений жизни?
Методологическое значение кибернетических закономерностей 265
Именно потому, что кибернетика вызвала к жизни много
частных теорий и особенно практических применений киберне-
тических правил, создалась диспропорция в развитии ее общих
принципов и частных механизмов. Стало совершенно очевидным,
что необходим тот «общий знаменатель», который помог бы вы-
разить с помощью единой основы разнообразие линий исследо-
ваний и такое же разнообразие фактов п понятий.
Можно присоединиться к Винеру, Клаусу, Эллису, и Людвигу
и к другим исследователям, которые считают, что «система»
является наиболее удобным понятием для организации научного
исследования и построения обобщающих гипотез. Однако сейчас
же встают вопросы: как определить самое понятие системы и как
выразить в четких формулировках наиболее характерные приз-
наки, отличающие ее от «несистемы»?
Несмотря на широко принятое употребление термина «систе-
ма», основные характеристики ее остаются несформулирован-
ными. Отсюда и проистекает досадное взаимонепонимание между
учеными различных специальностей, а иногда даже в пределах
одной и той же специальности.
Успех кибернетического направления определяется главным
образом тем, что кибернетика ввела в жизнь понятие системы
с весьма четкими параметрами, поддающимися математической
обработке. В сущности, это и должно служить нам отправным
пунктом для обсуждения. Другое дело в биологии. Здесь понятие
системы всегда употребляется в том случае, когда хотят отме-
тить что-то сложное, превосходящее по масштабам обычные про-
цессы, механизмы и элементы. И даже Эшби с его строгим ма-
тематическим мышлением, желая охарактеризовать системы
разного масштаба, употребляет такие неопределенные выраже-
ния, как «малая система» п «очень большая система», ставя при
этом акцент только лишь на простом увеличении количества ком-
понентов и на объеме взаимодействий между системами и суб-
системами (Эшби, 1964).
Успех обсуждения любой проблемы обычно находится в пря-
мой зависимости от того, насколько точно будут определены все
параметры объекта дискуссии и общее направление самой дис-
куссии. Именно поэтому мне хотелось бы дать характеристику
тех свойств кибернетической самоуправляемой системы, которые
относятся именно и только к системе, а не к ее компонентам.
Эта формулировка покоится на многолетней разработке в на-
шей лаборатории
упкциональных систем организма и,
следо-
вателъно, имеет экспериментальное обоснование.
1. Ни одна организация, сколь бы обширной она ни была по
количеству составляющих ее элементов, не может быть названа
«самоуправляемой», «саморегулируемой» системой, если ее функ-
ционирование, т. е. взаимодействие частей этой организации, не
266 Методологическое значение кибернетических закономерностей
заканчивается каким-либо полезным для системы результатом и
если отсутствует обратная информация в управляющий центр о
степени полезности этого результата. Только при таком условии
все части системы вступают в консолидацию, взаимную коорди-
нацию п субординацию. Благодаря этому вступление в действие
каждого компонента системы происходит в точно определенный
момент, иначе говоря, взаимодействие между ними имеет орга-
низованный и направленный характер.
У всякого другого объединения частей, хотя бы и взаимо-
действующих, по не имеющих конечного полезного результата,
нет основного условия для системной работы. Система самоуп-
равления — это не простое взаимодействие, это интегрирование
активности всех компонентов в одном-единственпом направле-
нии — на получении необходимого в данный момент и специ-
фического для системы приспособительного результата. У системы
нет другого пути для упорядочения работы ее компонентов, как
фокусирование степени их участия в конечном результате.
Посмотрите на котенка, который проделывает ритмические че-
сательные движения, устраняя какой-то раздражающий агент в
области уха. Это не только тривиальный «чесательный рефлекс».
Это в подлинном смысле слова консолидация всех частей системы
на результате. Действительно, в данном случае не только лапа
тянется к голове, т. е. к пункту раздражения, но и голова тянет-
ся к лапе. Шейная мускулатура на стороне чесания избиратель-
но напряжена, в результате вся голова наклонена в сторону лапы.
Туловище также изогнуто таким образом, что облегчаются сво-
бодные манипуляции лапой. II даже три не занятых прямо че-
санием конечности расположены таким образом, чтобы с точки
зрения позы тела и центра тяжести обеспечить успех чесания.
Как можно видеть, весь организм «устремлен» к фокусу ре-
зультата и, следовательно, ни одна мышца тела пе остается без-
участной в получении полезного результата. Мы имеем в под-
линном смысле слова систему отношений, полностью подчинен-
ную получению полезного организму в данный момент резуль-
тата.
Прямые эксперименты нашей сотрудницы М. Чепелюгиной с
перерезкой задних (и особенно передних корешков для задней
ноги) в весьма отчетливой форме показали этот факт «взаимосо-
действия» частей системы при достижении результата (Анохин,
1949).
2. Из предыдущего тезиса видно, что результат самоуправля-
емой системы приобретает центральное значение для ее органи-
зации во времени и пространстве. Именно полезный результат
составляет тот операциональный фактор, который способствует
тому, что система в случае недостаточности данного результата
может полностью реорганизовать расположение своих частей в
Методологическое значение кибернетических закономерностей 267
пространстве и во времени, что и обеспечивает в конце концов
необходимый в данной ситуации приспособительный результат
организма. В качестве примера можно указать на ампутацию ко-
нечности. Этот пример показывает, что такая система представ-
ляет собой логически оформленный комплекс процессов и меха-
низмов. Причем последнее распределение мышечных усилий
немедленно закрепляется обратной санкционирующей афферен-
тацией от полученных положительных результатов (Анохин,
1935).
Акцепт на результате саморегуляции как наиболее характер-
ном и ведущем параметре системы сделан нами не случайно. Не-
обходимость такого акцента вытекает уже хотя бы из анализа со-
временных физиологических концепций. Если посмотреть наибо-
лее важные концепции физиологии, то они сформулированы при
полном устранении именно понятия результата...
Наши наблюдения над живыми системами привели нас к пол-
ному убеждению в том, что любое физиологическое исследование
во много раз продуктивнее, если оно будет придерживаться выше-
приведенного представления о системе. Поскольку каждая такая
система несет определенную функцию с четко очерченными ре-
зультатами, мы и назвали ее в свое время «функциональной си-
стемой» (Анохин, 1935). Как было установлено, такая функцио-
н
L _ — Л
нальная система непременно содержит ряд специфических узло-
вых механизмов, принадлежащих ей только как интегральному
образованию.
Так как все дальнейшее изложение нашего представления о
философском смысле и методологическом значении основных ки-
бернетических закономерностей будет основано на теории функ-
циональной системы, я считаю совершенно необходимым дать
здесь хотя бы краткую характеристику ее основных узловых ме-
ханизмов, как они представляются нам по последним экспери-
ментальным данным.
Стадия афферентного синтеза
Эта стадия является начальной в развитии любого приспосо-
бительного акта животного. Важность этой стадии состоит в том,
что только после завершения ее наступает следующий этап в
формировании поведенческого акта. Афферентный синтез, пожа-
луй, наиболее обширный и сложный механизм функциональной
системы. Есть основание думать, что вся эволюция мозга и осо-
бенно все те усложнения, которые он приобрел па последних
этапах своего развития (кора головного мозга, п особенно ее лоб-
ные отделы), связаны были с нарастающим усложнением имен-
но афферентного синтеза. Это станет понятным, если предста-
вить себе, что именно в этой стадии любой организм решает три
268 Методологическое значение кибернетических закономерностей
важнейших вопроса своего поведения: что делать, как делать,
когда делать?
В процессе эволюции животных с прогрессирующим усложне-
нием условий их существования именно эти три вопроса требова-
ли для своего разрешения все более и более сложных мозговых
структур. В этом можно видеть смысл той высокой оценки «аф-
ферентного отдела» центральной нервной системы, которую дал
ему в свое время И. П. Павлов.
Многочисленные исследования моих сотрудников (А. И. Шу-
милина, Ф. А. Ата-Мурадова, К. В. Судаков) привели нас к вы-
воду, что афферентный синтез осуществляется на основе четырех
важнейших компонентов, имеющих весьма разнородные возбуж-
дения. Взаимодействию же этих возбуждений (и последующему
«принятию решения») помогают три нейродинамических фактора:
ориентировочно-исследовательская реакция, конвергенция воз-
буждений на нейроне и корково-подкорковая реверберация воз-
буждений. Эти механизмы способствуют объединению всех раз
нородных возбуждений, сопоставлению их и вынесению «реше
ния» об осуществлении определенного поведенческого акта
наиболее подходящего для данной ситуации.
В состав же самого афферентного синтеза у высших живот-
ных непременно входят следующие четыре формы афферентации
корковых нейронов: доминирующая мотивация, обстановочная
афферентация, пусковая афферентация, аппараты памяти. Кон-
вергенция и сопоставление всех этих возбуждений па отдель-
ных нейронах коры и последующая интеграция результатов этого
взаимодействия в масштабах целого мозга приводят в конце кон-
цов к формированию цели самого действия и к «принятию» наи-
более эффективного решения, т. е. к выбору наиболее эффектив-
ной в данных условиях программы действия.
Принятие решения
«Принятие решения» является реальным подарком кибернети-
ки физиологам, поскольку этот узловой момент в совершении лю-
бого поведенческого акта оставался скрытым из-за преимущест-
венной линейной точки зрения на формирование поведения, при-
нятой рефлекторной теорией. В самом деле, если представить себе,
что внешний стимул служит толчком к действию, то по сути
дела на протяжении всей рефлекторной дуги пет места принятию
решения как механизму, с помощью которого формируется цель
и программа предстоящего действия.
Понятие цели, как оно было представлено в статье И. П. Пав-
лова «Рефлекс цели», предполагает неизбежное опережение со-
бытий, т. е. формирование модели последнего этапа действия
раньше, чем окончились начальные его этапы. Едва ли кто-ни-
Методологическое значение кибернетических закономерностей 269
будь будет утверждать, например, что оп поставил перед собой
цель купить шляпу только в тот момент, когда он оказался в ма-
газине головных уборов. Ясно, что принятие решения и форми-
рование цели действия неизбежно опережают и даже подсказы-
вают ход будущих событий, и поэтому весь процесс не является
последовательно линейным.
Однако, несмотря на четкую постановку вопроса И. П. Пав-
ловым, проблема цели в исследовательском плане не стала объ-
ективно-научной проблемой, поскольку этому не соответствовал
общий уровень нейрофизиологических знаний того времени. По-
видимому, этим и надо объяснить тот парадоксальный факт, что
сам И. П. Павлов никогда больше не возвращался к своей сме-
лой попытке рассматривать цель как рефлекс.
Итак, принятие решения и формирование цели, предусмат-
ривающие достижение определенного результата, составляют не-
пременный результат стадии афферентного синтеза, вместе с тем
эти процессы предрешают ход событий.
Другую характерную черту принятия решения составляет его
исключающая, ограничительная роль. Благодаря принятию реше-
ния о совершении вполне определенного акта организм освобож-
дается от огромного количества потенциальных степеней свобо-
ды, которые оп мог бы реализовать в каждый данный момент
(Ухтомский, Васильев, Виноградов, 1927). Принципиально в
данный момент организм может совершить миллион различных
действий, что находится в прямой зависимости от композиции
многообразных связей в центральной нервной системе и от пла-
стичных объединений мышечных групп тела. Однако в момент
принятия решения из этих бесчисленных степеней свободы дела-
ется выбор лишь одного-единственного действия, которое наибо-
лее полно приспосабливает организм к данным условиям и в
данный момент.
Теперь уже не подлежит сомнению, что эту пригонку выбора
действия к совокупности данных условий нервная система про-
изводит именно в стадии афферентного синтеза после всесторон-
ней обработки имеющейся на данный момент внутренней и внеш-
ней информации.
Из сказанного ясно, что принятие решения представляет собой
узловой пункт всей функциональной системы, поскольку оконча-
тельный успех или неуспех приспособления будет находиться в
прямой зависимости от того, насколько точно сделан выбор необ-
ходимого поведенческого акта.
В пределах обычного поведения момент принятия решения яв-
ляется в той или иной степени замаскированным и часто совер-
шается в высшей степепи быстро. Это обстоятельство и служит
причиной того, что в физиологических исследованиях ему не уде-
ляли абсолютно никакого внимания, полагая по традиции, что
270 Методологическое значение кибернетических закономерностей
наличие одного начального стимула вполне достаточно для того,
чтобы был сформирован соответствующий поведенческий акт.
Многочисленные эксперименты показывают, что момент при-
нятия решения сопровождается немедленным возникновением
двух взаимосвязанных комплексов возбуждений: а) адекватной
программы действия, т. е. интеграла эфферентных возбуждений;
б) специфического аппарата предсказания результатов еще пе
законченного действия. Следует подчеркнуть, что этот последний
комплекс афферентных признаков результата появляется задолго
до того, как совершится само действие и будут получены реаль-
ные результаты. Оба эти комплекса возбуждений связаны тесней-
шим образом. Как показывают, например, тонкие микрофизиоло-
гические исследования, посылка эфферентных возбуждений бла-
годаря аксонным коллатералям принимает самое близкое участие
в формировании нейрофизиологических механизмов афферентной
модели будущих результатов действия (В. А. Правдивцев,
В. В. Синичкин, А. В. Котов, Б. В. Журавлев и др.).
Нейрофизиологическая природа
акцептора действия
Кажется удивительным, что проблема будущего, т. е. такие
решающие моменты пашего поведения, как постановка цели дан-
ного действия, предсказание результатов, которые должны по-
лучиться при осуществлении этой цели, пе была предметом спе-
циального объективного научного исследования. Впервые эта
проблема попала в поле зрения физиолога после того, как вы-
явилось, что внезапная подмена стандартного безусловного раз-
дражителя другими безусловными раздражителями (папример,
хлеба мясом) ведет на первый взгляд к совершенно непонятным
поведенческим проявлениям: к отказу от пищи, возникновению
исследовательской реакции и т. д. С точки зрения описательной
эти проявления нельзя было попять иначе, как «рассогласова-
ние» между тем, что «ожидалось», и тем, что «появилось» на
самом деле (Анохин, Стреж, 1933). Ясно было, что наличная, но
необычная еда, т. е. мясо, не соответствовала по своим характер-
ным признакам какому-то комплексу возбуждения, который был
уже «заготовлен» в ответ на условный раздражитель до появле-
ния самой еды. При первых попытках объяснить физиологиче-
скую сторону этого феномена мы исходили только из этого од-
ного несогласия, почему и назвали это возбуждение «заготовлен-
ным».
Дальнейшие исследования подобных фактов, названных нами
«методом сюрпризов», на человеке, на животных, а также и на
примере вегетативных функциональных систем (дыхание) .убе-
дило нас в том, что речь идет о новом феномене в деятельности
Методологическое значение кибернетических закономерностей 271
мозга. Было показано, что любые функциональные системы орга-
низма, начиная от вегетативных и кончая сложными поведенче-
скими актами, если только они заканчиваются полезным эффек-
том, непременно осуществляются через стадии афферентного син-
теза, принятия решения и предсказания результата будущего
действия в форме центрального моделирования афферентных при-
знаков этого результата. В начале действия, до его завершения,
в мозге формируется афферентная модель будущих результатов.
С точки зрения психических эквивалентов поведения этот
аппарат, вероятно, близок по своему содержанию к изучаемо-
му 11. С. Беритаипшли поведению, направляемому «образом»
(Бериташвили, 1953). Несомненный успех, который он получил
при этом подходе в расшифровке поведения, лишний раз убеж-
дает нас в том, что быстрые процессы мозга всегда опережают
медленно организующиеся отдельные этапы поведения.
В последние годы в нашей лаборатории были предприняты
шаги к дальнейшей конкретизации состава и топких нейрофизи-
ологических механизмов ацептора действия, т. е. к дальнейшей
материализации предсказания будущего. Были получены весьма
интересные результаты, раскрывающие эволюционную и нейро-
физиологическую суть функции предсказания будущего.
Мы вполне можем рассматривать механизм конвергенции
разнородных возбуждений на одном и том же нейроне как на-
чальную стадию сложной интегративной деятельности целого
мозга. Конечная же интеграция в масштабе целого мозга, несом-
ненно, рождается на основе многочисленных связей между этими
нейронами, в которых уже были обработаны па основе механизма
конвергенции разнородные возбуждения.
До последнего времени нейрофизиология отмечала только та-
кие типы конвергенции возбуждений на одном и том же нейро-
не, в которых приходящие к клетке возбуждения имели исклю-
чительно афферентный характер — зрительный, слуховой, так-
тильный и т. д. (так называемая мультисенсорная конвергенция).
Оказалось, однако, что этим видом конвергенции далеко пе ис-
черпывается афферентный синтез, начинающийся уже в масшта-
бе одной нервной клетки. В нашей лаборатории было показано,
например, что существует особая категория корковых нейронов,
вероятнее всего промежуточных нейронов, которые принимают
на себя не только разнообразные афферентные возбуждения, по,
что особенно важно, и эфферентные возбуждения, приходящие
к этим же межуточным нейронам по аксонным коллатералям от
пирамидпых клеток коры мозга.
Все эти исследования были проведены микроэлектродным ме-
тодом нри различной комбинации кортнко-петальных, кортико-
фугальных и антидромных возбуждений (Фельдшеров, 1966;
Правдивцев, 1967). Конвергенция на одной и той же нервной
272 Методологическое значение кибернетических закономерностей
клетке «копии» команды, т. е. эфферентного возбуждения, вы-
шедшего на аксоны нервной клетки, с афферентными возбужде-
ниями, приходящими от периферических рецепторных аппаратов,
представляет собой все необходимые условия для оценки полу-
ченных результатов. Этот же механизм после ряда его тренировок
сразу при выходе командных возбуждений и формирует функцию
предсказания сенсорных параметров будущих результатов.
С раскрытием описанного выше нейрофизиологического меха-
низма, удивительным образом созданного эволюцией для предска-
зания свойств будущих результатов, радикально изменилось и
наше отношение к «целесообразным актам»
к «предвидению» и
вообще к любому прогнозированию хода будущих событий.
Здесь очень важно оценить степень вероятности самого пред-
сказания. Некоторые авторы, принявшие в общем нашу концеп-
цию акцептора действия, вводят, однако, различные понятия,
как, папрпмер, «вероятностное прогнозирование» (Файгенберг),
«потребное будущее» (Бернштейн) и др.
Для успеха дела здесь уместно будет отметить, что в каждом
я
поведенческом акте мы всегда имеем два фактора: предсказанный
результат в параметрах акцептора действия и реальный результат,
полученный уже при завершении действия. Из этих двух факто-
ров наибольшей вероятностью обладает акцептор действия, по-
скольку его параметры подсказаны потребностью самого организ-
ма и всем процессом афферентного синтеза.
Наоборот, полученный результат в силу значительного разброса
эфферентных возбуждений и реальных возможностей организма
имеет значительно меньшую вероятность, которая оценивается и
повышается до уровня прогноза акцептора действия при помощи
яти
обратной афферентации путем поиска и сопоставлений — вооб-
ще формирования приспособительного поведения. Следовательно,
выражение «вероятностное (?) прогнозирование» просто не соот-
ветствует физиологическому смыслу событий п неправильно ори-
ентирует читателей, интересующихся нашими работами.
Вероятность прогноза, т. е. параметров акцептора действия,
всегда максимальна и равна единице, поскольку он отражает по-
требность организма па данный момент. Наоборот, результат всег-
да менее вероятен по отношению к акцептору действия, и, следо-
вательно, выражение «вероятностное прогнозирование» едва ли
оказывается полезным при изучении живых систем.
Нельзя не отметить также, что наконец-то стало попятным
значение аксонных коллатералей у значительного большинства
корковых, подкорковых н спинальных нейронов. Наличие их в
огромном количестве до последнего времени было абсолютной
загадкой. Зачем, казалось бы, рассылать по всему мозгу тысячи
«копий команды», посылаемой в данный момент к перифериче-
ским рабочим органам?
Методологическое значение кибернетических закономерностей 273
Не удивительно, что многие исследователи последних лет ос-
танавливались в нерешительности перед этим богатейшим нерв-
ным аппаратом, ограничиваясь при оценке его замечанием о том,
что «функция этих нервных образований до сих пор остается
таинственной» (Amassian, Patton, 1960).
Мы подвергли подробному обследованию функцию этих аксон-
ных коллатералей с помощью микроэлектродной техники и вы-
явили, что она состоит в длительном поддержании циклических
возбуждений, своеобразных «кругов ожидания». Поскольку таких
коллатералей, в общем, тысячи, а каждая из них еще ветвится на
несколько более топких окончаний, то возникает представление
о поразительной широте охвата действующими в данный момент
эфферентными возбуждениями различных районов коры и под-
корки головного мозга. Так, например, пирамидный тракт отдает
еще тысячи коллатералей па уровне ретикулярной формации. Эти
коллатерали через аппарат ретикулярной формации и через вос-
ходящие возбуждения еще раз способствуют поддержке активно-
сти в уже созданных «кругах ожидания» (В. В. Синичкин). Изу-
чение длительности возбуждения в этих «кругах ожидания»
показывает, что опа вполне достаточна для того, чтобы с перифе-
рии уже поступила информация о результатах.
Таким образом, получил функциональное объяснение массо-
вый характер, казалось бы, ненужных аксонных коллатералей у
корковых клеток.
После систематических исследований в этой области мы
можем представить себе ту картину циклических процессов
между центром и периферией, которая в целом обеспечивает
как получение полезного результата в итоге предпринятых дейст-
вий, так и оценку этого результата в кругах возбуждения, сфор-
мировавшихся до получения результата, но уже содержащих
афферентный код этого будущего результата.
Как только принимается решение о каком-либо действии и
как только возбуждение выходит на эфферентные пути, то уже
в этот самый момент коллатеральные возбуждения, распространя-
ясь но самым различным отделам мозга, создают в масштабе це-
лой коры многочисленные системы циклических возбуждений,
отражающих посланную па периферию команду и воспроизводя-
щих опыт прошлых результатов в форме акцептора действия.
Следовательно, в момент начала действия в соответствии с
, кора головного
принятым решением уже весь мозг и, конечно
мозга подготавливают модель будущих результатов — акцептор
действия. Эта модель осуществит в дальнейшем прием обратной
афферентаций от полученных результатов, произведет сопостав-
ление этой информации о реальных результатах с предсказанной
(или прогнозированной) моделью этих результатов. В сущности
это и есть конкретный субстрат предсказания, возбуждение кото-
1
274 Методологическое значение кибернетических закономерностей
рого все время опережает ход событий в функциях целого
мозга.
Благодаря этим исследованиям вопрос о приспособлении ор-
ганизма к ходу будущих событий получил в нашей лаборатории
чисто нейрофизиологическое решение.
Механизм обратной афферентации
Под обратной связью в кибернетике имеется в виду более или
менее точная информация о конечных результатах, полученных
в какой-либо саморегулирующей системе. И только в этом смыс-
ле обратная связь делается неотъемлемым фактором организации
успешного результата. Но по своей сути обратная афферентация
в биологических системах меняется в зависимости от широких
флюктуаций самого результата, и, следовательно, непременно
должен быть аппарат, который ее принимает и оценивает, т. е.
санкционирует тот приспособительный результат, о котором она
информирует.
Именно поэтому такая афферентация в нашей лаборатории
первоначально и была названа «санкционирующей», хотя впо-
следствии оказалось, что правом «санкции» обладает лишь акцеп-
тор действия (Анохин, 1935). Сама же обратная афферентация
есть пассивное следствие любого результата, успешного и неус-
пешного. Момент сопоставления всех параметров обратной аффе-
рентации, сигнализирующих о свойствах результата действия с
афферентной моделью этих результатов, закодированных в акцеп-
торе действия, завершает логику любого отдельного поведенческо-
го акта.
Если сопоставление показывает, что параметры реально полу-
ченных результатов, закодированные в обратной афферентации,
соответствуют параметрам предсказанного афферентным синтезом
результата, то данный акт завершается и организм переходит к
формированию очередного поведенческого акта. Если же оказы-
вается, что параметры результатов, закодированных в обратной
афферентации, не совпадают с тем, что было сформировано в
принятии решения и в акцепторе действия, то наступает момент
рассогласования, служащий стимулом для новой цепи реакций.
В этом последнем случае прежде всего немедленно возникает
ориентировочно-исследовательская реакция с весьма быстрой и
сильной активацией корковой деятельности (Анохин, 1958). Это
способствует активному подбору новых компонентов для аффе-
рентного синтеза и, что особенно важно, облегчает иптракорти-
кальные взаимодействия восходящих возбуждений (Lindsley,
1960).
В результате такого расширения афферентной информации
формируется в конце концов и более совершенная программа дей-
Методологическое значение кибернетических закономерностей
275
ствия и, следовательно, более успешные результаты, соответст-
вующие принятому решению и акцептору действия.
Нарисованная выше картина общей архитектуры поведенче-
ского акта выражает универсальную закономерность в построе-
нии живых систем.
Однако, если бы мы захотели сопоставить эту архитектуру
функциональной системы с любым типом саморегулирующихся
механических систем, то мы увидели бы то поразительное сход-
ство принципов функционирования, которое с такой настойчиво-
стью подчеркивал Норберт Винер. Правда, он писал об этом в
своей хорошо известной книге «Кибернетика» (1958), имея в виду
всего лишь обратную афферептацию, пли обратную связь —
«фид—бэк». Адрес этой обратной связи для биологических систем
не был еще и
звестен. Им стал для нас акцептор действия.
В чем же причина такого поразительного сходства в архитек-
турных чертах функционирования столь различного класса явле-
ний? Почему и в организме и в машинах контрольный аппарат
формируется раньше, чем появятся результаты, которые будут
подлежать контролю?
Наша концепция, по которой результат действия является
центральным пунктом любой системы отношений, позволяет от-
ветить па эти вопросы вполне четко. Наличие аналогичных черт
обеих систем диктуется тем, что и в той и в другой системе
конечным продуктом деятельности является четко очерченный
полезный результат. Именно последний представляет собой тот
уравнивающий
актор, который и в том и в другом случае не-
избежно требует одинаковой функциональной архитектуры.
И если бы получаемые результаты всегда математически точ-
но соответствовали заданным результатам, то по сути дела пол-
ностью исключалась бы необходимость контроля их и оставался
бы лишь стимул этих результатов к формированию последую-
щего звена непрерывной поведенческой мелодии. Однако в дей-
ствительности результаты всегда имеют множественный разброс
около какого-то предсказанного акцептором действия эталона,
необходимого именно в данный момент. Вот именно для устра-
нения избыточных степеней свободы в получении результатов и
служит во всех случаях контроль или управление па основе за-
готовленной модели результатов.
Где причина этого удивительного подобия функциональных
архитектур у систем различного класса? Какие исторические
факторы обусловили организующую роль результата действия?
Эти вопросы поднимают само понятие результата на ступень ис-
торического анализа и подчеркивают его универсальное и, сле-
довательно, философское значение в картине того мира, в котором
мы живем. Единственную возможность ответа на поставленные
выше вопросы я вижу в анализе того критического периода в
276 Методологическое значение кибернетических закономерностей
жизни нашей планеты, когда после длительного периода всевоз-
можных «нредбпологпческих» комбинаций материи возникла пер-
вичная жизнь.
Необходимость полезного результата как движущего и стаби-
лизирующего систему фактора возникла именно где-то здесь, на
стыке предбиологпческих и примитивных биологических комби-
наций материи. Следовательно, искать ответ на поставленные
выше вопросы об универсальной роли результата в поддержании
стабильности систем надо именно здесь. Широкую общность за-
конов и, следовательно, «философский корень» кибернетики надо
искать в том периоде развития нашей планеты, когда сложилась
первичная необходимость полезного результата.
Однако в связи с этим нам придется несколько по-новому по-
смотреть на диалектико-материалистическую формулировку жиз-
ни. Сохраняя свое принципиальное содержание, эта формула, как
нам кажется, на новом этапе развития наук должна включить в
себя все то, что создано в последние годы биологическим и фи-
зиологическим экспериментом.
Опыт расширения
диалектико-материалистической формулы жизни
Уже сам факт, что центральным пунктом прогресса жизни яв-
ляется полезный результат действия, заключенный в единую
л.
ункциональную систему, говорит о том, что для полной харак-
теристики жизненного процесса сейчас уже недостаточно только
чисто субстратного его основания. Материальный субстрат не мо-
жет быть основой жизни без того, чтобы он не составил какую-
либо систему отношений с более или менее стабильным конечным
результатом, в каком-то отношении полезным самой системе. Это
вытекает из свойств саморегуляторной динамической организа-
ции, которая была нами рассмотрена выше.
Как известно, диалектико-материалистическая формула жиз-
ни, вытекающая из общих установок «Диалектики природы» Эн-
гельса, состоит в том, что на первый план ставится специфика
субстрата, т. е. белковые тела. Несомненным остается и сейчас,
что формулировка Энгельса: «Жизпь есть способ существования
белковых тел» (с. 616),— является по своей глубокой материали-
стической сущности верной. Именно белковые тела составили
тот субстрат, без которого невозможен был прогресс жизни и
невозможно было совершенствование ее форм.
Однако, согласно последним достижениям молекулярной био-
логии, белковые тела представляют собой настолько высокую
степень организации материи, что исторический подход к разви-
тию жизни на Земле заставляет пас поставить вопросы: как про-
изошли сами белковые тела; являются ли они пассивным про-
Методологическое значение кибернетических закономерностей ЪТ1
дуктом случайных предбиологических химических комбинаций,
в миллионных количествах возникавших на основе самых разно-
образных физических и химических условий пашей планеты, или
сам белок как высокополимеризованный продукт есть продукт
каких-то активных процессов, толкнувших предбиологические
системы на путь неудержимого прогресса?
Специальная литература о происхождении жизни на Земле,
и в особенности о происхождении предбиологических систем, под-
чинена почти исключительно субстратной концепции жизни и
направляет внимание в сторону всех тех возможных физических
условий, когда-то существовавших на нашей планете, которые
способствовали формированию именно таких субстратов.
Достаточно посмотреть последнюю литературу, относящуюся
к происхождению жизни, чтобы увидеть, что все попытки разре-
шить проблему происхождения жизни на Земле связаны с поис-
ками тех возможных условий физического химического и метео-
рологического характера, которые могли бы сформировать нечто
близкое по составу какому-либо из белковых компонентов. Если
отвести в сторону вопрос о межпланетных причинах происхож-
дения жизни па Земле, то в этом смысле особенно важно отме-
тить теорию А. И. Опарина (1963), Дж. Холдейна (1966),
Дж. Бернала (1966) и др.
Характерной чертой «субстратных теорий» происхождения
жизни являются поиски тех соединений, которые, однажды сло-
жившись, могли бы в процессе дальнейших превращений приве-
сти к формированию белковой жизни. Интересно отметить, что
эти теории жизни, почти как правило, считают наиболее важным
моментом в появлении примитивной жизни ее способность к вос-
произведению, т. е. по сути дела наиболее поздний и производ-
ный фактор жизни.
Вполне естественно поэтому, что в связи с новыми открытия-
ми в области биологических наук (генетика) внимание ученых
стало сосредоточиваться на роли в этом процессе дезокси- и рибо-
нуклеиновых образований.
Однако концепция поиска случайных субстратных комбина-
ции и нланетарпых катаклизмов неизбежно должна была приве-
сти к представлению, что само возникновение жизни находится в
зависимости от случайного наличия соответствующего материа-
ла — белка или его компонентов.
Как уже говорилось выше,
это положение в своей общей фор-
ме правильно, ибо на всех этапах развития жизни именно белко-
вые полимеры оказались способными обеспечить выживание. Од-
нако последние достижения молекулярной биологии, и особенно
ее достижения па клеточном уровне, дают возможность высказать
предположение, что субстрат жизни как таковой не является
единственной и решающей причиной ее появления. Сейчас уже
278 Методологическое значение кибернетических закономерностей
невозможно обойтись без динамических характеристик первичной
организации, «предбиологической» или даже «предоргапической».
Становится все более ясным, что белковые тела, являясь вы-
сокоспециализированным и более поздним субстратом жизни, не
могут быть исключительным фактором, который подготовил жиз-
ненную организацию в ее предбиологической фазе, а может быть
даже п в предбелковой стадии.
Наоборот, последние достижения науки дают все основания
думать, что появлению белка как полимерного образования и
даже появлению одного-едииствеиного нуклеотида неизбежно
должны были предшествовать такие динамические принципы
организации материи, которые впоследствии па более высокоорга-
низованном уровне послужили своего рода «колыбелью» для по-
явления и развития самих определяющих белковых образований.
Что же это за принципы организации? И почему они оказа-
лись столь решающими в дальнейшем отборе все более и более
совершенных форм структурного развития? В разделе «Система и
результат» я уже подчеркнул то важное положение, что никакая,
даже самая обширная комбинация процессов и элементов не
способна составить саморегулирующуюся систему, если эта ком-
бинация не приводит к появлению каких-то результатов, обрат-
но влияющих на распределение взаимодействую:
щх сил в систе-
и
ме и тем самым полезных самой системе.
Следовательно, до тех пор, пока при взаимодействии процес-
сов результат деятельности этой системы не станет стабилизи-
рующим и саморегулирующим фактором, система пе может быть
устойчивой. Становится ясным, что всякий субстрат может быть
оцениваем только па фоне уже сложившейся устойчивой дина-
мической системы, т. е. только с точки зрения того, в какой сте-
пени, входя в систему, оп способствует пли совершенствует по-
лучение соответствующего результата. А это означает, что сами
поиски субстрата жизни должны быть расширены поисками той
формы устойчивого результата, который, став своеобразным фо-
кусом, стал обрастать все более и более новыми компонентами,
совершенствующими или уничтожающими уже созданные ранее
устойчивые системы.
Едва ли будет преувеличением, если я скажу, что стабилиза-
ция на основе принципов саморегулирования является самой пер-
вичной и самой решающей чертой жизненного процесса, именно
она обеспечила поступательное развитие структур в предбио-
логическом периоде. Можно говорить с уверенностью, что ника-
кие близкие к жизни субстраты, в том числе и белок, сами по
себе не могут составить жизни, если они не вовлечены в какую-
то более обширную систему, функционирующую по принципу
саморегуляторной стабилизации.
Приведенные выше рассуждения ставят перед нами в песколь-
Методологическое значение кибернетических закономерностей 279
ко новой форме те кардинальные вопросы, которые связаны с
изучением происхождения жизни на Земле. Мы неизбежно долж-
ны поставить вопрос: что произошло ранее, белковые тела и био-
генные амины, которые впоследствии составили саморегулятор-
ную систему — жизнь, или, наоборот, вначале создалась какая-то
(или какие-то) примитивная химическая стабилизация с каким-
то, может быть даже еще небиологическим, результатом? Но этой
стабильной устойчивости было вполне достаточно для того, чтобы
такая длительно «переживающая система» стала пунктом даль-
нейшего субстратного усовершенствования па основе пусть весь-
ма примитивного, по все же «естественного отбора».
В самом деле, уже факт наличия какой-то стабильности само-
регуляторпого тина и появления результата, хотя бы только в
форме самой устойчивости системы, становится неизбежным фак-
тором резистепции такой примитивной системы против всякого
рода внешних возмущающих воздействий. Следовательно, с по-
явлением устойчивых систем саморегуляции критерием допусти-
мости или недопустимости нового ингредиента системы становит-
ся его значение для ее стабилизации: если от присоединения чего-
то нового система становится еще более стабильной, то этого
достаточно, чтобы компонент нашел в ней свое место, а система
стала еще более устойчивой. И наоборот, если внешний физико-
химический фактор вредит стабилизации системы, то последняя
или преодолевает его действия, или сама перестает существовать
как стабильная система.
На все эти вопросы можно дать только один ответ: первичной
могла быть только какая-то стабильная система процессов, для
которой вначале единственно полезным результатом, очевидно,
была сама ее устойчивость.
Мы хорошо знаем нз теории ультрастабильных систем, что
каждая такая система неизбежно приобретает некоторую рези-
стентность по отношению к внешним воздействиям хотя бы уже
по одному тому, что она «стремится» сохранить стабильность.
Разбирая подробно способность стабильных и полистабильпых
систем к адаптации, Эшби, например, пишет, что повторяющиеся
воздействия на такую систему способствуют «отбору форм, обла-
дающих особой способностью противостоять ее изменяющему
действию». И дальше, говоря о «системах с миллионными пере-
менными», которые на протяжении миллионов лет па поверхно-
сти Земли претерпевают самые разнообразные комбинации взаи-
модействий, оп пишет: «Разве только чудо могло предохранить ее
(Землю.— П. Л.) от тех состояний, в которых переменные груп-
пируются в комплексы, способные к интенсивной самозащите»
(Эшби, 1964).
Я хочу еще пояснить развитую выше точку зрения. Есть все
основания думать, что эти первично стабилизировавшиеся систе-
280 М етод о логическое значение кибернетических закономерностей
мы, которые вначале могли быть весьма простыми в химическом
отношении, в последующем могли бы стать сами своеобразными
«воспитателями» сложных полимерных продуктов, которые явля-
ются уже неотъемлемым признаком более высокого уровня пред-
биологических и биологических организаций.
Изложенная нами точка зрения пе исключает грандиозной
роли белковых тел в развитии жизни, опа лишь предполагает
появление белковых тел как решающего этапа жизни в системе
уже сформированных стабильных систем, может быть даже и не-
органического или элементарного органического характера. Под-
держивая устойчивость за счет саморегуляторпых соотношений с
окружающей средой, эти системы в последующем становятся
усовершенствованной основой биологических систем в подлинном
смысле этого слова.
С этой точки зрения наиболее близкой к истине гипотезой
происхождения жизни является, как нам кажется, гипотеза
А. И. Опарина о коацерватных капельных объединениях, где
многие компоненты могли бы вступать между собой в самые раз-
личные взаимодействия. Этой гипотезе пе хватает лишь того, что-
бы разнообразные химические ингредиенты, входящие в состав
этих капель, составили какую-то саморегуляторную устойчивую
систему за счет динамических взаимодействий с внешней средой.
Ясно одно, что как бы пи была мала эта первичная саморе-
гулирующаяся система, она должна иметь стабилизирующий ее
конечный результат. Этот результат и должен служить поддержа-
нию ее стабильного состояния. При этих условиях все другие
окружающие систему субстраты и факторы среды могут или до-
ставлять ей исходный материал, или потреблять ее конечный
продукт.
Таким образом, в целом это должна быть открытая система,
которая теоретически может поддерживать устойчивость постоль-
ку, поскольку окружающая ее среда будет удовлетворять описан-
ным выше требованиям. Иначе говоря, эта гипотетическая
первичная устойчивая система в самой своей стабилизации приоб-
ретает «полезный результат» и неизбежно должна обладать спо-
собностью настойчиво «защищать» этот результат от посторонних
«возмущений».
Важно подчеркнуть, что именно в этот момент и должна была
родиться «целесообразность» в ее объективном, научном и мате-
риалистическом смысле, или, как говорил И. П. Павлов, «в дар-
винском смысле».
Поскольку у этих первичных систем устойчивость была пер-
вым полезным результатом их деятельности, мы можем сказать,
что всякая встреча этих систем со средой, приводившая к ук-
реплению этой устойчивости, была полезна для системы в целом.
Наоборот, все то, что нарушало ее устойчивость, становилось для
Методологическое значение кибернетических закономерностей 281
нее отрицательным, неполезным. Иначе говоря, устойчивость
первичных предбиологических систем стала решающим крите-
рием самых разнообразных соотношений этих систем с окружаю-
щей их средой.
При такой точке зрения неизбежно должен возникнуть вопрос,
какие же химические организации в первородном океане могли
бы вступить в такого рода стабилизированные взаимодействия и
имеем ли мы что-либо подобное в известных нам в настоящее
время химических взаимодействиях. Молекулярная биология дает
нам достаточное количество примеров для того, чтобы допущение
существования таких первичных систем стало вполне правдопо-
добным. И здесь прежде всего следует обратить внимание на те
узловые пункты химических систем разной степени сложности,
которые носят признаки самоорганизации.
Для примера можно взять хотя бы феномен так называемого
«ретроингибирования», или «торможения конечным продуктом».
Этот фепомен имеет место у таких дробных частей обширных
цеппых химических реакций, что эти части могут быть названы
узлами стабилизации. Для подобных «открытых систем» вход
формируется на основе специфических реакций с окружающей
средой, а выход, состоящий в формировании какого-то результа-
та — продукта, связан с уходом или потреблением этого продук-
та также во внешнюю среду.
Современная литература знает много примеров, когда на очень
узком отрезке какого-то обширного метаболического процесса
формируется своеобразный стабилизирующий комплекс, регули-
рующий па основе обратного влияния весь ход метаболизма в
данной области. Литература по саморегуляторным приспособле-
ниям протоплазмы па молекулярном уровне обширна.
Можно указать, например, работы Jacob и Monod (1963),
Dickson и Webb (1958), Novick и Szilard (1954) и многих других,
которые показали, что в ряде случаев определенная концентрация
конечного продукта в цепи метаболических реакций оказывает
тормозящее действие на начальную стадию всей цени реакции.
Совершенно очевидно, что здесь мы имеем элементарную, но
типичную саморегулирующуюся систему, которая может поддер-
живать стабильный уровень, например, триптофана пли изолей-
цпна в данной среде и обладает определенными чертами устой-
чивости. Если же все, что окружает узел этой химической реак-
ции, принять за среду по отношению к этой, как ее назвал Жакоб,
«аллостерической системе», то мы будем иметь нечто подобное
той предполагаемой первичной устойчивой системе, о которой мы
говорили выше. Здесь уже не только устойчивость как таковая
является результатом системы, но и сам конкретный продукт себя
поддерживает в данной среде на определенном уровне его кон-
центрации.
282 Методологическое значение кибернетических закономерностей
Пожалуй, наиболее демонстративно этот процесс развивается
в случае биосинтеза цитидин-5'-трифосфата. Здесь исходным про-
дуктом является карбамилфосфат, который с помощью целой се-
рии ферментов через стадии L-дигидрооротовой кислоты, оротовой
кислоты, оротидин-б'-фосфата, уридин-б'-фосфата приходит к об-
разованию цитидин-5'-трпфосфата. Мы имеем типичную саморе-
гулирующуюся систему с весьма четким исходным материалом и
конечным продуктом.
Само собой разумеется, что вся динамика этой системы будет
зависеть от того, в какой степени внешняя среда по отношению
к этой системе может непрерывно поставлять карбамилфосфат и
аспарагиновую кислоту и в какой степени та же среда сможет
потреблять конечный продукт этой саморегулирующейся системы,
т. е. цитпдин-б'-трифосфат.
Важно отметить одпо обстоятельство: такой универсальный
механизм создания и усовершенствования стабильных систем, как
ретроингибирование, может эволюционировать, совершенствовать-
ся и, следовательно, вступать в более комплексные отношения
с окружающей систему средой. Конечно, следует помнить, что в
данном случае мы имеем систему с органическими компонентами,
т. е. систему, уже в какой-то степени получившую свое оформле-
ние в пределах обширной живой системы. Но едва ли можно со-
мневаться в том, что в первородном океане, который содержал
миллионы различных компонентов и еще больше различных ком-
бинаций взаимодействий, пе создавались системы подобного ти-
па на примитивном органическом уровне, например па основе
фотосинтеза и с неорганическими катализаторами.
В разобранной выше устойчивой системе первый фермент —
аспартаттранскарбамилаза в процессе эволюции может стать спе-
цифическим продуктом саморегулирующейся, устойчивой систе-
мы, приспособленным к ее химическим свойствам, т. е. к вполне
определенному конечному продукту.
Этот последней факт лишний раз подтверждает вышеприве-
денные положения, что начальным этапом в прогрессе предбполо-
гических систем должна была быть первичная устойчивая систе-
ма с чертами саморегуляции. II только в процессе «защиты» этой
устойчивости и в процессе многообразных взаимодействий системы
с внешними факторами среды происходит непрерывное «обраста-
ние» ее все новыми и новыми компонентами, вступающими в
прогрессивное взаимодействие с уже имевшимися ранее компо-
нентами этой системы.
Следовательно, и первичные белковые тела, и более специа-
лизированные белки в виде ферментов явились следствием
постепенно совершенствовавшихся стабильных предбиологичес-
ких систем, послуживших основой для развития жизни на
Земле.
283
Методологическое значение кибернетических закономерностей
~ * *' * а ° а ° - а * 1 а ° °
В заключение этого раздела статьи отметим три важных об-
стоятельства.
1. Само появление устойчивых систем с чертами саморегуля-
ции стало возможным только потому, что возник первый резуль-
тат этой саморегуляции в виде самой устойчивости, способной к
сопротивлению внешним воздействиям. Следовательно, регулятор-
ная роль результата системы была первым движущим фактором
развития систем, который сопровождал все этапы предбиологи-
ческого, биологического и социального развития материи.
2. Сейчас мы не имеем возможности предположить, что бел-
ковые тела и, в частности, рибонуклеиновые кислоты возникли
как нечто отдельное и самостоятельное, на основе только чисто
субстратных перекомбипаций на нашей планете, как это, напри-
мер, полагает Ehrensvard (1962). Сама химическая структура
белковых образований, и особенно ферментов, настолько точно и
очевидно приспособлена к детальным
ункциям
саморегулятор-
ного характера у весьма различных систем, что нет никакой воз-
можности допустить, что опи могут возникнуть как бы спонтанно,
независимо от этих систем, т. е. вне принципов, удерживающих
систему в стабильном состоянии.
3. Эти новые положения, сформулированные нами, как можно
было видеть, не отрицают той огромной роли белковых тел в жиз-
ненном процессе, которая была сформулирована Ф. Энгельсом,
и, следовательно, диалектико-материалистическое понимание
основ жизни остается в силе. Однако приведенные выше рассуж-
дения, основанные на успехах кибернетического, физиологическо-
го и биохимических исследований живых систем, дают возмож-
ность представить себе развитие жизни на Земле и, следователь-
но, белковых полимеров в связи с совершенствованием результата
как центрального фактора саморегулирующихся систем.
Результат как критерий
для оценки кибернетических закономерностей
В предыдущем изложении не раз отмечалось, что существую-
щая до сих пор физиологическая терминология практически ис-
ключила результат действия из основных понятий рефлекторной
деятельности. Это обстоятельство, как мы видели, значительно
повлияло на физиологические подходы к объяснению целостных
поведенческих актов.
По сути дела то же самое мы имеем и в области кибернети-
ческих представлений. Целый ряд терминов и понятий киберне-
тики хотя и имеет широкое хождение среди исследователей, но
именно в силу того, что из них также исключено понятие резуль-
тата как основного фактора саморегуляции, эти понятия имеют
284 Методологическое значение кибернетических закономерностей
весьма ограниченное влияние па самый исследовательский про-
цесс. Ниже я постараюсь разобрать некоторые из таких кибер-
нетических понятий, чтобы показать, в какой степени меняется
их трактовка и подход к ним при внесении в кибернетические
модели понятия о результате.
«Управляющая система». В числе общих представлений о ки-
бернетике широкое хождение получил термин «управляющая
система». Мне хотелось бы здесь отметить тот факт, что это по-
нятие ни лингвистически, ни логически не соответствует тому
объему представлений, которые составляют основу кибернетиче-
ского мышления.
В самом деле, термин «управляющая» означает лишь то, что
какое-то устройство воздействует с помощью определенных сиг-
налов на управляемый объект, находящийся за пределами дан-
ного устройства. Весь смысл же кибернетических взаимоотноше-
ний состоит именно в том, что «управляемый», т. е. результат
действия системы, должен непременно сообщать о степени его до-
статочности или недостаточности туда, где используется резуль-
тат системы и где формируется программа действия.
Таким образом, выражение «управляющая система» не может
быть принято хотя бы уже в силу того, что смысл выражения
«управляющая система» не включает в себя конечный полезный
результат саморегулирующейся системы.
Предложенное нами еще в 1935 г. понятие «функциональная
и
система»,
как можно было видеть вьи
:е
наиболее полно отра-
жает основной смысл кибернетических закономерностей. Функ-
циональная система включает в себя центральную нервную си-
стому,
в которой происходят афферентный синтез, принятие
решения и складывание программы действия. Опа включает
также управляющее воздействие на периферический аппарат, т. е.
формирование самого действия, она включает формирование ре-
зультата этого действия и весьма специфическую сигнализацию
о нем, наконец, принятие и сопоставление этой информации в ак-
цепторе действия.
Все звенья этой системы связаны взаимовлиянием, которое
не происходит хаотически, или неориентированно, а развивается
таким образом, что все компоненты системы содействуют друг
другу, т. е. их усилия фокусируются на путях получения полез-
ного результата для данной системы или для организма в целом.
Следовательно, в функциональной системе результат является
универсальным организующим
фактором. Именно это и опреде-
и
ляет, в какой степени, в каком месте и в какое время включит-
ся тот или иной компонент системы.
«Иерархия систем» — иерархия результатов. Принятие поло-
жения, что результат является центральным пунктом системы,
неизбежно ведет нас к иному пониманию организации целого ор-
Методологическое значение кибернетических закономерностей 285
ганпзма, в котором происходит объединение систем и субсистем
в определенной иерархии...
Организм не мог бы существовать как гармоническое объеди-
нение систем с огромным количеством результатов, если бы объ-
единялись только системы в их целостном содержании. Биологи-
ческий смысл формирования каждой более высокой системы на
основе субсистем состоит в том, что объединяются именно ре-
зультаты субсистем, а уже это объединение результатов состав-
ляет новый уровень саморегулирующихся систем с новым конеч-
ным полезным результатом...
Из этих положений с очевидностью следует, что, когда мы го-
ворим о том, что более высоко организованная система состоит
из субсистем, то это надо понимать отнюдь не в прямом смысле
этого выражения. Правильнее было бы говорить, что всякая функ-
циональная система более высокого уровня организации состав-
ляется на основе результатов субсистем, т. е. результатов функ-
циональных систем более низкой организации. Мне кажется, что
этот подход к соотношению «систем» и «субспстем» значительно
расширяет наши представления об интегральной деятельности
целого организма, заканчивающейся полезным приспособительным
результатом.
Кибернетика и «причинные сети». Недавно на страницах на-
шей печати возникла дискуссия о том, какое содержание следует
вложить в понятие «кибернетики» как науки. Как мы уже видели,
в первом параграфе этой статьи были сделаны самые разнообраз-
ные акценты па детальных сторонах саморегуляции, обеспечи-
вающих динамическое целостное образование.
С точки зрения А. А. Маркова (1964), акцент при определе-
нии кибернетики делается на детерминистическом характере вза-
имодействия частей системы. Что такое «причинная сеть», если
к ней применить критерий результата как центрального пункта
саморегулирующейся системы? Сам А. А. Марков предлагает
рассматривать кибернетику как науку о «причинных сетях».
При ближайшем рассмотрении концепции А. А. Маркова в
аспекте кибернетических закономерностей становится совершен-
но очевидным, что причинная сеть ие является организованным
целым в кибернетическом смысле: она не имеет цели, прогноза
результатов своего функционирования и, что самое главное, не
имеет полезного результата функционирования, а следовательно,
никак не связана в своих реорганизациях с информацией о по-
лученном результате.
Если принять во внимание все приведенное нами в предыдущих
разделах статьи, то формулировка «причинной сети» А. А. Мар-
кова является наилучшим аргументом в пользу необходимости
принять результат деятельности системы в качестве основного
параметра саморегуляции. В противном случае все изменения в
г
286 Методологическое значение кибернетических закономерностей
«причинной сети» пе будут достаточно детерминированными и це-
ленаправленными. Мы должны помнить, что никаких «причин»
в этой сети не может быть вне формирования конечного полез-
ного результата, и, следовательно, только он может определить,
какая причина и какие следствия должны быть получены в каж-
дом данном участке системы.
«Надежность» как кибернетическое понятие. Хороню известно,
что надежность, пожалуй, самое устоявшееся понятие в кибер-
нетике. Мы можем часто встретить выражение, что задачей ки-
бернетики является «построение надежных систем из ненадеж-
ных элементов». Исходя из органического единства «система —
результат», мы должны сказать, что это выражение представляет
собой кибернетический нонсенс.
В самом деле, может ли быть система «падежной», если «не-
надежны» ее элементы? Говоря о ненадежности элементов, мы в
первую очередь, конечно, предполагаем или их «выход из строя»,
или их внезапную изменчивость в процессе работы системы. Но
если отдельные элементы системы оказываются изменчивыми пли
нарушенными, то может ли при этом система быть «надежной»,
т. е. не изменить и не нарушить взаимодействия всех своих ос-
тальных компонентов?
Поясню этот вопрос конкретным поведенческим примером.
В лабиринтных опытах у крысы был выработан в ответ на оп-
ределенный сигнал навык побежки к кормушке. После того как
крыса выработала этот навык и безошибочно различала все хо-
ды лабиринтов, ведущие к кормушке, у нее были ампутированы
все четыре конечности. Естественно, что теперь крыса не могла
передвигаться обычным способом. Но она нашла другой способ
двигаться к кормушкам: опа продвигалась бочкообразно, т. е.
подобно тому, как катится бочка. Благодаря этому способу она
докатывалась до кормушки и добивалась этого же результата,
т. е. того же самого пищевого подкрепления.
Постараемся понять этот эксперимент с точки зрения приве-
денных выше рассуждений. Что здесь произошло? Ясно, что из
обширной локомоторной системы животного были вырваны ее
весьма значительные компоненты — конечности. Следовательно,
компоненты большой системы, поведенческого акта, несомненно,
стали «ненадежными». Значит ли это, что вся локомоторная си-
стема осталась «надежной», т. е. неизменпвшейся и ненарушен-
ной?
Физиология нервной системы па это отвечает вполне опреде-
ленно: все элементы локомоторной системы в приведенном слу-
чае подверглись значительному изменению, произошло радикаль-
ное перемещение удельного веса отдельных оставшихся мышеч-
ных компонентов и резкое изменение распределения возбуждений
по системам в целом. Таким образом, совершенно очевидно, что
Методологическое значение кибернетических закономерностей
287
у гармонически сочетанной функциональной системы, обладавшей
всеми чертами саморегуляции, каждое нарушение или изменение
ее элементов немедленно ведет к перестройке и перераспределе-
нию роли всех других компонентов системы.
Но что же тогда в данном примере является все-таки дейст-
вительно «надежным»? Надежным является результат деятель-
ности системы, т. е. получение пищи. Именно оп остался одним
и тем же, несмотря па радикальные изменения как отдельных
элементов, так и всей системы в целом. И в этом состоит уни-
версальный закон приспособления живых систем ко всякого рода
экстренным нарушениям. Итак, приведенное выше ходячее вы-
ражение о «ненадежности» элементов и «надежности» систем яв-
ляется парадоксальным с точки зрения понятия о системе.
Во всех приведенных выше определениях надежности резуль-
тат системы не фигурирует как фактор, определяющий все воз-
можные модуляции и перестройки взаимоотношений элементов в
пределах целой системы. Модифицируя общеупотребительную
формулу надежности, мы могли бы сказать, что термин «надеж-
ность» во всех саморегулирующихся системах может быть при-
меним только к результату деятельности системы. Именно на-
дежность результата представляет собой тот primum movens, ко-
торый своими отклонениями определяет любую «надежность»,
любое изменение и любую перестройку как в элементах, так и в
целой системе.
Иначе говоря, как бы пи была изменена функциональная си-
стема и как бы ни было перераспределено участие ее компонентов,
элементы и вся система всегда будут падежными, если в этих
случаях оказался падежным результат ее деятельности. Приме-
нительно к живым системам это положение можно было сформу-
лировать следующим образом: результат действия только потому
и может стать надежным, что и элементы системы, и сама опа
в целом могут быть пластичными, т. е. «ненадежными».
Понятие «информационного эквивалента» результата. Мы уже
говорили о том, что определения кибернетики, основанные па по-
нятии преобразования информации, хотя и берут существенную
сторону саморегуляции, являются недостаточными, поскольку они
не отражают специфического характера архитектурных особенно-
стей саморегуляции. В самом деле, информация может циркули-
ровать и преобразовываться и в таких взаимодействиях отдель-
ных компонентов, которые могут быть и несаморегулируемымп.
Однако информация, циркулирующая в саморегулирующейся
системе, организует ее взаимодействия, которые оканчиваются
полезным приспособительным результатом. Именно этот решаю-
щий факт заставляет нас посмотреть и на преобразования ин-
формации с несколько другой точки зрения. В этом смысле мне
хотелось бы обратить особое внимание на высказывание Эшби
288 Методологическое значение кибернетических закономерностей
* Mil I II I
относительно того, что представляет собой циркуляция информа-
ции в системе. Давая в одной из своих последних работ матема-
тическое выражение переходных состояний системы и кодиро-
вания в ней информации, оп пишет: «Суть идеи машины, кото-
рая уже давно понималась интуитивно, будучи выражена строго,
сводится к тому, что «в машине» мы видим кодированный вариант
простой последовательности событий» (Эшби, 1964).
Едва ли можно согласиться с тем, что в «машине» (а в дан-
ном случае под машиной им подразумевается биологическая
система) кодирование подчиняется «простой последовательности
событий». Конечно, в любой системе, поскольку опа подчинена
временной структуре нашего мира, события развертываются и ко-
дируются последовательно. Однако в любом изолированном «блоке
системы», функция которого заканчивается полезным результа-
том, эта последовательность и кодирование пе могут быть безот-
носительны к этому результату.
В самом деле, фактически на любом этапе циркулирования ин-
формации в функциональной системе сама информация неизбеж-
но кодирует какую-то долю предстоящего результата, будучи под-
чинена этому конечному результату. Любой элемент системы в
какой-то степени отражает долю своего локального участия в по-
лучении конечного результата, и, следовательно, его информаци-
онные взаимоотношения неизбежно должны отражать какой-то
эквивалент его участия в получении результата.
Постараюсь пояснить все вышесказанное конкретным
Y
изио-
и
m ж
логическим примером. Дыхательная система, постоянным про-
межуточным результатом которой является поддержание устой-
чивой концентрации СО2—О2 в крови, служит хорошим приме-
ром того, что информация, па каких бы этапах системы она нп
циркулировала, не может быть безотносительна к конечному по-
лезному результату.
Допустим, что в дыхательном центре сложилась такая ситуа-
ция, что после обработки информации от хеморецепторов сосудис-
той системы, от хеморецепторов ткани, от хеморецепторов сердца
и хеморецепторов самого дыхательного центра, т. е. после аф-
ферентного синтеза, дыхательный центр даст «команду» па пери-
ферию забрать 600 см3 воздуха. Эта закодированная команда вы-
ражает потребность организма в таком именно объеме кислорода
в данный момент.
Постараемся понять, как эта информация,
или
«команда», бу-
дет распространяться по дыхательной функциональной системе
через все информационные каналы.
Из моторной части дыхательного центра возбуждение выхо-
дит по спинному мозгу и дыхательным нервным стволам на все
дыхательные мышцы: диафрагму, межреберные мы
и
;цы, брюшные
мышцы, головные мышцы и т. д. Однако эта информация дыха-
Методологическое значение кибернетических закономерностей 289
тельного центра распределяется по дыхательным мышцам отнюдь
не беспорядочно. Каждая мышца получает нервные импульсации
в таком временном распределении, что в конечном счете все мышцы
грудной клетки сокращаются ровно настолько, что в легкие вхо-
дит именно 600 см3 воздуха. Следовательно, возбуждение, распро-
странявшееся по нервам, закодировало в количестве пмпульсаций
и интервальных соотношениях именно 600 см3 воздуха. Наряду с
этим дыхательные мышцы грудной клетки в своих сокращениях
проявили тот же самый код, т. е. сократились в такой степени,
чтобы забрать именно 600 смЛ
Пожалуй, самым интересным элементом дыхательпой систе-
мы является движение информации в обратном направлении, т. е.
когда уже взято 600 см3 воздуха. Статистически выражаясь, п-е
количество альвеол растянуто в такой степени и в такой града-
ции, что онп забирают именно 600 см3 воздуха. Но тогда совокуп-
ность весьма разнообразных импульсов от растяжения альвеоляр-
ных рецепторов обоих легких будет такова, что в целом эти об-
ратные афферентации по всем волокнам блуждающего нерва дол-
жны быть эквивалентны информации именно о 600 см3 воздуха,
взятого легкими. Таким образом, круг замыкается.
Дыхательный центр после обработки всей входной информа-
ции посылает потоки эфферентных импульсов, в которых как по
частоте, так и по интервалу закодирован будущий результат —
взятие 600 см3 воздуха. Дыхательные мышцы воспринимают эту
информацию и трансформируют ее в сократительный процесс,
который также в совокупности мышечных волокон отражает ис-
ходный код дыхательного центра — расширить грудную клетку
для взятия 600 см3 воздуха. Растянутые воздухом альвеолы, не-
смотря на все многообразие степеней растяжения, в статистиче-
ской совокупности посылают от своих рецепторов в дыхательный
центр информацию о взятии именно 600 см3 воздуха, а не 500
или 400. Эта информация проходит в дыхательный центр и всту-
пает в сопоставительный контакт с уже известным нам аппара-
том — акцептором действия. Такое соотношение было показано
в целой серии специальных экспериментов. Следовательно, сам
факт получения приспособительного результата системой неиз-
бежно определяет объем и кодирование информации на всех пу-
тях ее распространения: опа всюду должна быть эквивалентна
результату действия — и на путях его получения, и па путях
сигнализации о получении в центральную нервную систему.
Таким образом, нз теории функциональной системы следует,
что циркуляция информации по компонентам системы пе может
быть «информацией вообще»; это всегда информация, которая в
специфическом коде данного конкретного элемента системы со-
держит эквивалент пли будущего, или уже полученного резуль-
тата.
Ю П. К. Анохин
290 Методологическое значение кибернетических закономерностей
Все эти соображения, разработанные нами для всех основ-
ных функциональных систем организма, дают возможность сфор-
мулпровать следующие положения:
а) наличие приспособительного результата во всякой саморе-
гулирующейся и самоорганизующейся системе радикально ориен-
тирует все потоки
б) любой элемент системы проводит или преобразует инфор-
мацию в эквивалепте какой-то доли этого результата;
в) каждый элемент системы, информация которого не отражает
информации в системе на этот результат;
параметров результата, делается помехой для системы и немед-
ленно преодолевается пластическими перестройками всей системы
в целом.
Мы считаем необходимым для более продуктивного исследо-
вания функциональных систем организма ввести понятие «инфор-
мационный эквивалент результата». Как мы видели, этот экви-
валент имеет место во всех звеньях системы, и, следовательно,
приняв, что информация, циркулирующая в системе, всегда в
какой-то степени отражает эквивалент результата, мы облегчаем
себе изучение ее природы и ее функционального смысла па всех
этапах циркуляции...
Заключение
Настоящий очерк имел целью провести анализ основных ки-
бернетических закономерностей под углом зрения более общих
законов мира, которые определили па различных этапах жизни
нашей планеты формирование этих закономерностей. Мне кажет-
ся, что современный материал, привлеченный нами в этом очер-
ке, убеждает нас в необходимости выработки новых взглядов на
некоторые как бы устоявшиеся общепринятые истины.
Задача дать философско-методологический анализ кибернети-
ческих закономерностей и с этих позиций по-повому рассмотреть
некоторые стороны самой кибернетики, несомненно, далека от
выполнения.
Как можно было видеть, я попытался вывести па свет тот
главный фактор, который, несмотря на его определяющий харак-
тер, остается как-то вне поля зрения исследователей. Этот глав-
ный фактор — результат деятельности системы.
Сопровождая жизнь на всем протяжении ее эволюции, ре-
зультат подчинил себе все свойства и конструкции функциональ-
ной системы, которые в свою очередь обеспечили ему в бе более
и более топкое приспособление к усложнявшимся условиям су-
ществования от биологического к социальному уровню. В этом
именно и состоит главный смысл наличия общих законов функ-
ционирования систем различного класса: машин, организмов и
общественных образований. Все эти классы явлений возникли,
Методологическое значение кибернетических закономерностей 291
развивались и существуют в наше время на основе получения
конечного полезного результата. Но именно этот последний ока-
зывает свое императивное влияние на всю систему и на переор-
ганизацию ее компонентов. Он и лежит в основе общих законо-
мерностей различных классов систем.
ЛИТЕРАТУРА
Маркс К., Энгельс Ф. Соч., т. 20.
Анохин П. К. Проблемы центра и периферии в современной физиологии
нервной деятельности.— В кн.: Проблема центра и периферии в первной
деятельности. Горький, 1935.
Анохин 77. Я. Внутреннее торможение как проблема физиологии. М., 1958.
Анохин П. Я. Рефлекс и функциональная система как факторы физиологи-
ческой интеграции.— Бюл. Моск, о-ва испытателей природы, 1949, т. 54,
№ 5.
Анохин П. К. Кибернетика и интегративная деятельность мозга.— Вопр.
психол., 1966, № 3.
Анохин П. К. Биология и нейрофизиология условного рефлекса. М., 1968.
Анохин П. К., Стреж Е. Ф. Изучение динамики высшей нервной деятельно-
сти. Сообщение 3. Нарушение активного выбора в результате замены
безусловного стимула.— Физиол. журн. СССР, 1933, т. 16, К® 2.
Бернал Дж. Молекулярные матрицы живых систем.— В кн.: Происхожде-
ние предбиологических систем. М., 1966.
Бунге М. Причинность М., 1962.
Винер И. Кибернетика и общество. М., 1958.
Клаус Г. Кибернетика и философия. М., 1963.
Марков А. А. Что такое кибернетика.— В кн.: Кибернетика,
жизнь. М.. 1964.
мышление
Фельшеров Ю. Особенности реакций дыхательных нейронов на антидром-
ное раздражение афферентных дыхательных путей.— Матер. 5-й Все-
союз. конф, по электрофизиологии. Тбилиси, 1966.
Холдейн Дж. Информация, необходимая для воспроизведения первичного
организма.— В кн.: Происхождение предбиологических систем. М., 1966.
Эшби У. Р. Конструкция мозга. Происхождение адаптивного поведения. М.,
1964.
Amassian V. 7., Patton С. The pyramidal tract: its excitation and function.—
In: Handbook of physiology, section I. Neurophysiology. Washington,
1960, v. 2.
Lindsley D. B. Attention, consciousness, sleep and wakefulness.— In: Hand-
book of physiology, Sec. 1. Neurophysiology, v. 3. Washington, 1960.
Novick A., Szilard L. Experiments with the honieostat on the rates of amoni
acid synthesis in bacteria.— In: Dynamics of growth processes. New Yer-
sey, 1954.
Structure et evolution des techniques 1953/1954, N 35 —36.
Webb E.f Dickson M. The enzymes. London, 1958.
10*
РЕФЛЕКС ЦЕЛИ КАК ОБЪЕКТ
ФИЗИОЛОГИЧЕСКОГО АНАЛИЗА *
Постановка вопроса
Сорок пять лет тому назад II. II. Павлов опубликовал одно
из замечательных произведений, которое, к сожалению, в даль-
нейшем не пашло достаточного отражения как в эксперименталь-
ных работах школы Павлова, так и в огромном числе публика-
ций его учеников и последователей. Я имею в виду его сообще-
ние на III съезде по экспериментальной педагогике в Петрограде
в 1916 г. па тему: «Рефлекс цели».
В этой речи он с удивительной полнотой дал характеристику
одному из самых узловых и решающих механизмов человеческого
поведения, именно формированию цели, направляющей и орга-
низующей поведение. II. П. Павлов писал: «Рефлекс цели имеет
огромное жизненное значение, оп есть основная форма жизненной
энергии каждого из пас ... Вся жизнь, все ее улучшения, вся
ее культура делается рефлексом цели, делается только людьми,
стремящимися к той пли другой поставленной ими себе в жизни
цели... Наоборот, жизнь перестает привязывать к себе, как только
исчезает цель... До какой степени у нас отсутствуют практиче-
ские сведения относительно такого важнейшего
Y актора
жизни,
И
как рефлекс цели. Л эти сведения так нужны во всех областях
жизни, начиная с капитальнейшей области — воспитания» (Пав-
лов, с. 19).
Трудно более красочно охарактеризовать ту огромную роль,
которую играют в жизни человека нервные комплексы цели,
способные стать путеводной звездой иногда даже на протяжении
всей жизни.
В качестве модельного случая И. П. Павлов взял такую устой-
чивую цель, как формирование пищедобывательпых реакций на
основе периодически сменяющихся чувств голода и насыщения.
Естественно, что это не единственные случаи целенаправленных
реакций человека. Па самом деле вся паша жизнь состоит из
непрерывной цепи формирующихся целен и их достижений. Та-
кие цели могут быть большими, малыми, а иногда и просто пу-
стяковыми, однако, только поставив перед собой такую цель,
человек может в дальнейшем формировать гармоническое, а пе
* Опубликовано в «Журнале высшей нервной деятельности» (19G2, вып. 1).
Рефлекс цели как объект физиологического анализа
293
хаотическое поведение. Сомнительно, чтобы вообще можно было
представить себе какой-либо поведенческий акт человека или да-
же высших животных, который мог бы реализоваться без пред-
варительного формирования самой цели или намерения к совер-
шению данного действия.
Вы задумали что-то купить в магазине. Это есть цель, сфор-
мированная во всех деталях па основе раздражений окружающей
нас обстановки и систем возбуждения, сложившихся на основе
прошлого опыта. II эта цель полностью определяет ваше будущее
поведение, К магазину, может быть, ведет очень длинный путь.
Может быть, по пути надо будет использовать различные виды
транспорта пли, может быть, надо долго выбирать нужную покуп-
ку. В этом примере между моментом формирования цели и конеч-
ным звеном ее осуществления включены целые цепи побочных
новых и промежуточных «целей» и новых действий, которые, од-
нако, лежат в плоскости основной и первоначальной цели —
покупки определенной вещи.
Каким образом в нервном субстрате распределяются эти про-
цессы? Откуда они черпают энергию иногда для очень длитель-
ной устойчивости, а часто и императивности в ее выполнении?
Мы не можем думать, что этот процесс формирования цели свер-
шается где-то за пределами материального субстрата мозга. Он
есть, несомненно, продукт наиболее высоких и наиболее слож-
ных комбинаций нервных потоков, развертывающихся по пре-
имуществу па путях корково-подкорковых соотношений,
Ясно, что предшествование момента формирования цели пос-
ледующему осуществлению этой цели имеет особенно выражен-
ное проявление в высшей нервной деятельности человека, в его
социальном поведении, в его грандиозных планах на будущее.
Однако принципиально мы пе можем отказать высшим живот-
ным в каких-то более простых процессах формирования цели к
действию, которые архитектурно построены таким же образом,
т. е. они предваряют само осуществление данного действия.
В качестве примера обычно приводятся человекообразные
обезьяны, которые могут проделывать весьма длинные цепи от-
дельных поведенческих актов для достижения раз возникшей
цели (например, овладение фруктами, подвешенными па высокой
перекладине).
Всем известны знаменитые опыты с человекообразными обезь-
янами, которые постепенно нагромождают целую пирамиду из
ящиков для того, чтобы достать фрукты. Здесь конечная цель
очевидна, по формирование- такой цели относилось только к выс-
шим животным, обладающим весьма развитой корой головного
мозга.
Однако это оказалось ошибочным. Недавно было показано, что
такая птица, как канарейка, может производить подобные же
Й94
Рефлекс цели как объект физиологического анализа
целенаправленные акты (Пастор, 1955). Автор так подвязывал
канерейкам крылья, что они не могли больше летать, а должны
были передвигаться только при помощи ног. Этим самым он де-
лал возможным предъявление им целого ряда поведенческих за-
дач. Он выработал у канарейки весьма разнообразные активные
навыки по добыванию пищи. Так, папример, канарейки подтяги-
вали в свою клетку корм за нитку, конец которой находился у них
в клетке. Они решали самые разнообразные дифференцировки
па цвет, на положение п т. д.
Но, пожалуй, наиболее интересным было решение канарейками
задач на сложное доставание пищи. Они могли производить под-
тягивание (при помощи веревки и крыльев) кубиков к месту
нахождения пищи и накладывать их один на один, точь-в-точь,
как это делали с ящиками шимпанзе в опытах Келлера, Павлова
и Денисова.
Трудно отрицать в этом конструктивном поведении птиц ту
же степень целеустремленности, как и у шимпанзе, хотя, конеч-
но, имеются значительные различия в технике, в нервных меха-
низмах и в объеме использования внешних условий для реали-
зации этой цели. Вместе с тем мы знаем, что строение головного
мозга у птицы и у человекообразной обезьяны принципиально
отличается. И потому наличие у этих живых существ в значи-
тельной степени сходных по общей архитектонике поведенче-
ских актов склоняет пас к выводу, что возникновение цели к дей-
ствию является весьма древней архитектурной особенностью пове-
дения. Очевидно, основные архитектурные особенности поведения
пе так уж прямо и абсолютно связаны с теми чертами в строении
головного мозга, которые мы по традиции считаем «решающими»
и «высшими». Достаточно вспомнить хотя бы совершенно полно-
ценные условные рефлексы у рыб (Фролов, 1923).
Выдвигая для научного исследования понятие «рефлекса це-
ли» при совершении сложных и многоэтапных действий,
И. П. Павлов выдвинул на первый план, с нашей точки зрения,
два важных момента: а)
м
формирование особых нервных механиз-
мов, предшествующих самому действию; б) возможность их объ-
ективно-научной физиологической трактовки.
Все сказанное выше делает несомненным, что физиологиче-
ская постановка вопроса для изучения механизмов
армирова-
ния цели в целостном поведении играет первостепенную роль в
расшифровке высшей нервной деятельности вообще и человека в
особенности. Очень важна эта расшифровка сейчас, когда пробле-
ма высшей нервной деятельности человека становится актуаль-
ной в связи с поисками новых путей ее изучения и возможности
управлять этой деятельностью. Несмотря па несомненную важ-
ность «рефлекса цели» для изучения поведения, ему не уделяет-
ся внимания в объективном научном исследовании.
Рефлекс цели как объект физиологического анализа
295
Чем можно объяснить такое парадоксальное отношение к это-
му вопросу, органически связывающему интересы физиолога
высшей нервной деятельности п психолога? Мне кажется, что
имеется несколько причин, которые исторически способствовали
такому забвению гениального взмаха павловской мысли.
Прежде всего я хотел бы указать на пе совсем справедли-
вое отношение к понятию цели, как в какой-то степени одиозно-
му только потому, что это понятие было много раз использовано
идеалистическими концепциями о поведении человека. Именно
мы — физиологи высшей нервной деятельности должны показать
всю силу физиологического, материалистического анализа этого
узлового момента поведения и тем самым осуществить мечту
II. П. Павлова о «слитии физиологического с психологическим».
Если наличие цели к действию, которая опережает само осу-
ществление действия, является для нас совершенно достоверным
фактом в природе нашего мозга, если этот факт повседневно про-
веряется в нашем собственном поведении и если, как бы пи был
сложен этот механизм, он непременно разыгрывается на матери-
альном субстрате мозга, то тогда пе должно быть никакого
места для боязпп объективного научного подхода к этому факту.
Конечный же успех дела должен будет зависеть только от того,
насколько остроумно и насколько убедительно мы сможем разра-
ботать нашу физиологическую аргументацию.
Другим большом препятствием к изучению
«рефлекса
является его исключительная физиологическая сложность
цели»
и, что
особенно важно, некоторая принципиальная новизна в самой
постановке вопроса. Большинство наших представлений о нейро-
физиологических механизмах высшей нервной деятельности осно-
вано па допущении линейного движения первногд процесса по
центральной нервной системе, т. е. поступательного хода его по
«рефлекторной дуге» до тех пор, пока оно не осуществит конеч-
ное звено поведения — работу периферических аппаратов. К та-
кому допущению нас неизбежно ведет рефлекторная теория в ее
старом варианте. Наоборот, постановка вопроса о рефлексе цели
содержит в себе совершенно противоположное допущение, имен-
но, что координированное и длительно развертывающееся дейст-
вие должпо в каких-то своих компонентах быть сформировано
уже в момент появления цели действия, т. е. раньше, чем начнет
осуществляться первый этап действия.
Совершенно очевидно, что рефлекс цели является сложным
нелинейным комплексом возбуждений и торможений, в котором
прошлое, настоящее и будущее организма составляют единое
интегративное образование. В этом и состоит исключительная ней-
рофизиологическая сложность проблемы формирования цели
поведения, которая еще ждет удовлетворительного физиологиче-
ского разрешения. Однако уже сама постановка вопроса о реф-
296
Рефлекс цели как объект физиологического анализа
лексе цели сейчас, иа новом этапе развития пауки о мозге, весь-
ма полезна. Она ставит перед исследователями вопрос о наличии
в мозговой деятельности своеобразного критического этапа, без
которого пе может пи сложиться, ни закончиться успешно пове-
денческий акт...
Здесь следует упомянуть, что в литературе последних лет
появилось много работ, относящихся к исследованию так назы-
ваемых «мотиваций». Под этим термином подразумеваются неко-
торые общие состояния организма, формирующиеся под влиянием
внутренних, по преимуществу гуморальных факторов.
Это направление психофизиологии, за вычетом всех тех нена-
учных спекуляций, к которым особенно склонны фрейдисты, со-
держит интересный фактический материал к объективному ней-
рофизиологическому объяснению так называемого спонтанного
поведения. Особенно интересны в этом направлении систематиче-
ские работы Миллера (1957).
Обычный подход к анализу поведения животных и человека
связан с выработкой условных рефлексов, созданием их различ-
ных комбинаций, воздействием па них специально подбираемых
экспериментатором каких-либо внешних агентов и т. д. Короче
говоря, при устоявшейся в настоящее время традиции центр тя-
жести исследования переместился в зону изучения механизмов
приобретения повых условных связей, в процесс накопления ин-
дивидуального опыта, в условия изменения этого опыта под воз-
действием внешних факторов. Однако другие, пе менее важные,
стороны поведения животных и человека: механизмы реализации
накопленного опыта в свободном поведении, пластическое комби-
нирование «фрагментов памяти» и построение па этой основе но-
вых форм поведения, механизмы подбора наиболее адекватных для
данной ситуации поведенческих актов и т. д.— все это в значи-
тельной степени осталось вне поля зрения исследователей! физи-
ологии условных рефлексов. Можно указать лишь па отдельные
примеры систематического изучения свободного поведения (Кери-
тов, 1923, Куналов, 1955). (йода же мы отпоенм и наш прием
изучения выбора двух противоположных кормушек.
Надо считать, что «рефлекс цели» как проблема объективной
науки лежит именно в области реализации накопленного опыта
в условиях свободного поведения. В этом пункте объединяются
три кардинальных фактора поведения, без знания которых мы пе
можем надеяться на широкое и удовлетворяющее пас незнание
работы мозга: 1) формирование «основных влечений» животпых
и человека, врожденных и приобретенных; 2) формирование це-
ли предстоящего поведения па основе синтеза всех афферентных
воздействий в данное время и в данной обстановке; 3) реализа-
ция всего прошлого опыта под влиянием поставленной цели и
формирование адекватного приспособительного поведения в па-
Рефлекс цели как объект физиологического анализа
297
стоящем. Мы еще далеки от знания детальных механизмов каж-
дого из этих факторов (этапов) формирования приспособитель-
ного поведения.
Нейрофизиологические механизмы
формирования «основных влечений»
Пожалуй, наиболее выгодными для анализа этих влечений
будут два широко известных и каждодневно испытываемых вле-
чения — жажда и голод. Современные успехи в изучении механиз-
мов формирования шпцедобывательпого и вододобывателыюго
поведения сходятся в одном пункте — центральная нервная систе-
ма, а именно гипоталамическая область, имеет тончайшие чувстви-
тельные аппараты, отвечающие на обеднение организма животных
водой или питательными веществами. Явелл (1953) показал,
например, что в гипоталамусе имеются нервные клетки, которые
всей своей конструкцией служат чувствительным аппаратом, реа-
гирующим на изменение осмотического давления крови (осморе-
цептор). Как известно, поддержание постоянства этого давления
представляет собой одно из самых строгих и наиболее жестко
регулируемых констант организма.
Согласно Явеллу, некоторые нервные клетки гипоталамуса
построены таким образом, что па одной из сторон имеется нечто
подобное маленькому пузырьку, мембрана которого тесно сопри-
касается с близлежащими капиллярами. Любое изменение осмоти-
ческого давления крови может менять патяженпе этой мембраны
в ту или другую сторону, будет ли осмотическое давление крови
гипертоническим или гипотоническим по отношению к этому пу-
зырьку (рис. 1).
Получается нечто, подобное рецептору растяжения, имеюще-
муся в других органах. Колебания мембраны клетки являются
триггерным механизмом, освобождающим в форме разрядов воз-
буждения этих клеток. Затем это возбуждение по многочисленным
связям и разветвлениям аксона клетки распространяется по мно-
гим направлениям, и, падо думать, как это показано будет даль-
ше на примере пищевого возбуждения, оно распространяется до
коркового уровня включительно.
Создается весьма интересный передаточный механизм, кото-
рый построен по типу механизмов, известных в современной тех-
нике под именем «сервомеханизмов». Благодаря этим механизмам
весьма малое количество начальной энергии через мобилизацию
корковых функциональных систем может привести впоследствии
к проявлению огромного количества энергии, например к мышеч-
ным усилиям по доставанию воды из колодца.
Центральным пунктом в описанном выше механизме для нас
в данный момент является факт возникновения сложных корко
208
Рефлекс цели как объект физиологического анализа
Рис. 1. Схематически показано
формирование одного из силь-
нейших влечений человече-
ского организма — жажды
Гипертоническая кровь через
капиллярную стенку К раздра-
жает осморецептор ОР, находя-
щийся в гипоталамусе Г. КР —
кора, ПК — подкорка, РФ — рети-
кулярная формация. Клетки по-
добного типа имеют широко раз-
ветвленные связи, которые фор-
мируют все последующие этапы
вододобынательного поведения
во-подкорковых систем возбуждений, которые формируют спе-
цифические цели поведения, направленного на устранение жаж-
ды. Схематически весь этот механизм возникновения «влечения»,
формирующего цель вполне специфического поведения, можно
изобразить, как показано на рис. 1. Схема пригодна для объясне-
ния всех влечений и сформированных на их основе «рефлексов
цели». Эта схема имеет своим началом перемены какого-либо
константного гуморального фактора крови. Таким фактором может
быть изменение осмотического давления, обеднение питательны-
ми веществами, изменение содержания того пли другого гормо-
на и, наконец, колебание содержания углекислоты. Все перечи-
сленные факторы, так же как и многие другие, неупомянутые
здесь, формируют специфическое для них поведение через со-
здание цели к этому поведению по тому типу, который дан па
приведенной схеме.
Однако «удовлетворение» этого «основного влечения», возник-
шего на гуморальной основе, может иметь отнюдь пе гумораль-
ный характер. В самом деле, как известно, мы утоляем жажду
раньше, чем вода дойдет до крови, часто держа еще в руках
недопитый стакан воды. Точно так же мы испытываем чувство
насыщения уже за обеденным столом, хотя принятая нами пища
еще очень далека от кровяного русла. Очевидно, существуют два
механизма удовлетворения этих влечении: один быстрый, основан-
ный на нервных механизмах (первичное насыщение), и другой —
медленный, связанный с поступлением конкретного материально-
го субстрата в кровяное русло (вторичное насыщение).
В этом мы видим замечательное приспособление организма к
внешнему миру, благодаря которому интервал между формирова-
Рефлекс цели как объект физиологического анализа
299
нием цели к многоэтапным поведенческим актам и фактическим
устранением исходных побуждений для этих действий сокращает-
ся до минимума и делается независимым в какой-то степени от
длительного химического преобразования добытых пищевых ве-
ществ.
В особенно отчетливой форме эта закономерность была вскры-
та в экспериментах нашего сотрудника К. В. Судакова. Он пока-
зал, что даже в условиях общей анестезии гипоталамическая об-
ласть, побуждаемая «голодной кровью», непрерывно активирует
лобные отделы коры головного мозга, что и создает в них высоко-
активное состояние, выраженное в сильной десинхронизации
электрической активности (рис. 2).
Электроэнцефалографический анализ лобных и других отделов
коры и заднего гипоталамуса показал, что, несмотря на глубо-
кий наркотический сон кошки, гипоталамус непрерывно активи-
рует лобные отделы коры, между тем как другие области коры
(теменная, височная, затылочная) проявляют обычную медленную
электрическую активность, характерную для спящего животного.
Мы уже имели в прошлых работах нашей лаборатории один
замечательный пример усиления электрической активности коры
кролика под уретановым наркозом. Если на заднюю ногу кро-
лика нанести ноцицептивное раздражение в виде горячей воды,
то у кролика, находящегося в наркотическом состоянии, появля-
ется отчетливо выраженная и генерализованная по всей коре
головного мозга десинхронизацпя электрической активности. При
этом сам кролик продолжает находиться в наркотическом сонном
состоянии. Один этот факт достаточно хорошо иллюстрирует на-
шу точку зрения о наличии многих и всегда специфических ви-
дов восходящего активизирующего воздействия ретикулярной
формации па кору головного мозга (Агафонов 1956, Анохин
1959).
В опытах К. В. Судакова и Ю, Урываева был получен новый
и поразительный пример натуральной активации коры головного
мозга, а именно ее лобных отделов от подкорковых ядер, также
в условиях наркотического сна. Необходимо было проверить наши
соображения в специальном контрольном эксперименте. Если
наша догадка об активизирующем действии специфического
раздражения подкорки «голодной кровью» была правильной, то
устранение в какой-то степени этой «голодности» в крови должно
было бы изменить электрическую активность лобных отделов —
от десинхронизации к такой же медленной активности, которая
имелась в остальных зонах коры мозга. Была избрана для инъек-
ции в кровь глюкоза как вещество, наиболее значительное для
состава «голодной крови», тем более, что в литературе уже име-
лись указания на то, что глюкоза, инъекцированпая в область
гипоталамуса, значительно меняет пищевое поведение животных.
300
Рефлекс цели как объект физиологического анализа
Рис. 2. Электроэнцефалограмма, записанная от мозга кошки
1 — лобная правая; 2 — лобная левая; 3 — зрительная правая; 4 — зрительная ле-
вая; 5 — отметка времени 1 сек. Видна отчетливая десинхрониэация лобных отделов
коры мозга и обычная для сна медленная активность затылочной области
Электроэнцефалографический анализ коры мозга животного пока-
зал, что немедленно носле инъекции глюкозы в кровь десипхро-
низацпя лобных отделов в какой-то степени устраняется, хотя и
не создается полная картппа медленной активности, характерной
для сна. Однако, если к этой инъекции прибавить натуральное
орошение полости рта спящего животного молоком и введение
молока в желудок, электрическая активность лобпых отделов
очень быстро приобретает тип медленной электрической актив-
ности спящего животного с характерными ритмическими «вспыш-
ками веретен» (рис. 3). Наибольший эффект в этих эксперимен-
тах получается в том случае,
когда
раздражение
пищеваритель-
ного пути пищевым веществом идет в нормальном порядке: рот
пищевод — желудок.
Совершенно очевидно, что эти опыты раскрывают перед нами
механизм формирования и «удовлетворения» одного из самых
универсальных и самых категорических влечепий человеческого
тела — голода и аппетита. В физиологическом смысле мы должны
допустить, что потоки афферентных возбуждений, возникающих
в рецепторах пищеварительного тракта (особенно па вкусовых
рецепторах языка!), должны быть всегда достаточны по количе-
ству, чтобы затормозить гуморально возникшее возбуждение
центров гипоталамуса, а может быть, и какого-либо другого зве-
на лимбической системы.
Прежними исследованиями нашей лаборатории было показано,
что подкорковая активация коры мозга при пищевом и оборони-
тельном безусловных рефлексах весьма избирательно вовлекает в
реакцию и нервные клетки коры мозга (Шумилина 1961, Гав ли-
чек 1958). Следовательно, нейрофизиологическая основа для фор-
Рефлекс цели как объект физиологического анализа
301
Рис. 3. Отрезок электроэнцефалограммы из того же самого опыта, что и на
рис. 2, записанный после искусственного «насыщения» кошки под наркозом
мирования цели поиска пищи построена таким образом, что спе-
цифическая субкортикальная часть обширной функциональной
системы питания определяет и условия специфических взаимо-
действий па уровне коры больших полушарий. Таким образом,
пищевые поисковые поведенческие акты животного или человека,
которым предшествует формирование цели поиска, складываются
как интегративное целое в результате избирательного распростра-
нения биологически специфических подкорковых возбуждений
по системам связей коры мозга. Следует лишь только подчерк-
нуть, что в данном случае специфический «рефлекс цели» опре-
деляется сложным пейрогуморальпым началом всего процесса и
еще более сложным по архитектуре объединением нервных кле-
ток на уровне коры («основное влечение», по И. П. Павлову).
Отличительная особенность данного поискового поведения за-
ключается в том, что здесь нет налицо обычного «пищевого разд-
ражителя», наоборот, животное совершает весьма многообразные
поведенческие акты, которые сводятся к тому, чтобы самому до-
быть этот пищевой раздражитель, часто находящийся вне поля
его зрения. Мы считаем, что, такая форма поведения способствует
активному подбору афферентации, адекватпой для данного
стояния животного или человека. Не будь этого механизма,
жизнь животных превратилась бы в сплошной хаос.
В разобранном примере особенно отчетливо выявлена та
ганическая основа, на которой формируется решающий этап
со-,
вся
ор-
по-
ведения — цель действия. Вместе с тем здесь, как было показа-
но, имеется также и весьма быстрый механизм, прекращающий
формирование этих целевых поисков пищевых веществ. Уже
только одно раздражение пищей вкусовых рецепторов полости
302
Рефлекс цели как объект физиологического анализа
5
Рис. 4. Схема формирования голода и насыщения
А — схематическое изображение двух типов воздействия на клетки пищевого цен
тра гипоталамуса: голодная кровь (ГК) действует как возбуждающий фактор (+j
и пищевая афферентная импульсация от языка, пищевода и желудка (ПА), дей-
ствующая как тормозящий фактор (—) на ту же самую клетку гипоталамуса (Г).
ВК — восходящая коллатераль; Б — общая схема формирования голода («основ-
ное влечение»), пищедобывательного поведения (ПД) и насыщения в две стадии:
а) первичное насыщение через афферентные импульсации от пищепроводнык путей
и б) вторичное насыщение через сытую кровь (СК) после всасывания пищевых
веществ
рта, пищевода и слизистой оболочки желудка оказывается вполне
достаточным для того, чтобы прекратить формирование «повых
целей» поведения, повых пищедобывательных актов. Этот двойной
механизм представлен на рис. 4.
В приведенной схеме видны два механизма возбуждения ис-
ходных пищевых центров, которые так хорошо были охарактери-
зованы в работах Апдерсопа (1955) и Ананда (1951). Бросается
в глаза определенное конкурирование двух процессов, один из
которых по врожденному механизму должен быть всегда победи-
телем, а именно импульсация от вкусовых рецепторов языка и
желудка. В самом деле, только при том условии, что эти импуль-
сации в сумме достаточно сильны, чтобы затормозить возбужден-
ные голодной кровью комплексы гипоталамуса, можно попять,
Рефлекс цели как объект физиологического анализа
303
почему в момент еды исходное возбуждающее действие еще не
изменившейся «голодной крови» теряет свою силу. Оно может
быть заторможено благодаря приходящим сюда возбуждениям
от вкусовых рецепторов.
Такой тип взаимодействий в организме мы имеем и в случае
других функциональных систем. Например, подавление возбужде-
ний сосудосуживающего центра нарастающими импульсациями
от барорецепторов дуги аорты и синокаротидной области. Здесь
также должна быть зафиксирована врожденными структурными
соотношениями возможность торможения сосудосуживающего
центра барорецепторными возбуждениями. В противном случае
мы все стали бы немедленно гипертониками (Шумилина, 1961,
Анохин, 1961).
Правда, в случае градуального подавления возбужденного пи-
щевого центра импульсациями, идущими в основном от рецепто-
ров языка, может быть еще один дополнительный механизм, об-
легчающий это торможение пищевого центра в гипоталамусе.
Я имею в виду рефлекторное выбрасывание в кровь глюкозы из
печени уже в момент еды, т. е. в момент раздражения вкусовых
рецепторов языка пищей (Быков 1953; Анохин 1958).
До сих пор мы обсуждали тот вид «основных влечений», ко-
торый И. П. Павлов сопоставлял с развитием других влечений и
целей, возникающих на более высоком, социальном уровне, где
и причина и следствие строится па уровне социальных интересов...
Рефлекс цели и акцептор действия
В ряде наших публикаций, начиная с 1933 г., мы делали по-
пытки построить общую архитектуру условного рефлекса с уче-
том его неизбежного звена в виде обратной афферентации. Эти
попытки, основанные на ряде экспериментальных материалов на-
шей лаборатории, привели нас к выводу, что в любом поведен-
ческом акте, прежде чем начнет складываться эффекторная часть
этого акта, создается специальный афферентный аппарат, вклю-
чающий на основе следов прошлых возбуждений все афферентные
признаки возможного результата предстоящего поведенческого
акта (Анохин, 1955, 1958, 1961).
Этот афферентный аппарат, который должен принять обратное
афферентное возбуждение от результатов данного действия, был
нами назван «акцептором действия». В настоящее время мы
имеем все больше и больше доказательств, что этот аппарат
реально руководит оценкой результатов всех совершаемых нами
поведенческих актов и исправляет эти поведенческие акты, если
их результаты не соответствуют его афферентному составу, т. е.
исходной цели данного акта.
304
Рефлекс цели как объект физиологического анализа
Нам кажется, что нейрофизиологическая основа этого аппара-
та имеет самое ближайшее отношение к нейрофизиологическому
механизму формирования цели действия и
следовательно, к фор-
мированию целеустремленного поведения животных и человека.
Мы даже думаем, что после того, как происходит синтез всех
многочисленных афферентаций и рождается цель действия, одно-
временно формируется и аппарат акцептора действия.
Смысл формирования и того и другого механизмов состоит в
том, что нервные процессы, развивающиеся в основном в коре
больших полушарии, по самой своей природе приобрели в эво-
люции возможность опережать течение внешних событий и на
основании прошлого опыта предвосхищать те реальные афферент-
ные воздействия, которые еще только должны будут поступить в
центральную нервную систему в конце всего ряда целеустремлен-
ных действий, В 1956 г. в докладе па павловской сессии я назвал
этот тип возбуждения «опережающими возбуждениями». Однако
общая идея о значении этого опережающего возбуждения для фор-
мирования сложных приспособительных актов человека и живот-
ного, как мне кажется, не была мною изложена с достаточной
ясностью.
Я постараюсь на наиболее простом примере, реально имевшем
место в истории возникновения самого учения об условных реф-
лексах, показать смысл этого огромного приобретения в эво-
люции животного мира.
Допустим, что служитель, ежедневно кормящий собаку, при-
ходит к ней с пищей по всегда вполне определенному пути: сна-
чала он хлопает дверью первого этажа, затем он постепенно под-
нимается по лестнице в характерном для него ритме шагов, затем
он открывает дверь третьего этажа, где в стайке стоит собака и,
наконец, проходя через большую комнату, он дает собакам при-
несенный корм.
С точки зрения реакции «что такое?», каждый из этих этапов
продвижения служителя вызывал когда-то у животного в первые
разы кормления соответствующую ориентировочно-исследователь-
скую реакцию.
Следовательно, все звенья этой развивающейся цепи явлений
действительности имеют или имели своп афферентные возбужде-
ния в центральной нервпой системе.
Конечно, явления действительности всегда развиваются в не-
прерывном потоке, из которого только искусственно можно выч-
ленить какие-то наиболее заметные эпизоды. Но для иллюстрации
идеи мы можем принять, что для животного, стоящего в станке
на третьем этаже, наиболее значительными моментами являются
стук нижней дверью, шаги поднимающегося по лестнице человека,
открывание верхней двери, продвижение служителя по комнате
и, наконец, кормление.
Рефлекс цели как объект физиологического анализа
305
Рис. 5. Отдельный «блок» ориентировочно-
исследовательской реакции на определен-
ный внешний раздражитель (жирная
(стрелка)
ИЗ — проекционная зона раздражителя; ЗАО
эффекторный аппарат ориентировочной реак-
ции в центральной нервной системе; ЗВ —
эффекторное возбуждение; О А—обратная аф-
ферентация, возникающая как результат все-
стороннего обследования раздражителя jk и пот-
ным; ЦРО — центральный результат ориенти-
ровочной реакции как система разнообразных
афферентных представительств
ЗАО ПЗ ЦРО
Таким образом, с учетом ориентировочной реакции мы можем
сказать, что каждый из этих моментов может быть представлен
в виде типичного блока, имеющего некоторый приспособительный
результат. Стук дверью вызывает ориентировочно-исследователь-
скую реакцию, а результаты последней в форме обратной аффе-
рентации о результатах этой ориентировочно-исследовательской
деятельности приходят в центральную нервную систему. Схема-
тически этот блок можно представить в следующем виде (рис. 5).
Следовательно, весь прежний опыт знакомства животного с
данным рядом явлений действительности можно представить в
виде схемы (рис. 6, I). Естественно, что конечное звено этого
ряда явлений действительности — раздражение пищей вкусовых
рецепторов языка ведет к особенно мощному возбуждению
подкорковых аппаратов. Благодаря этому возникает сильнейшая
активация коры головного мозга, которая вместе с предшествую-
щей активацией ее от ориентировочно-исследовательских реак-
ций создает особенно благоприятные условия (в виде облегчения)
для последующей ассоциации этого, вначале разрозненного, ряда
последовательных возбуждений центральной нервной системы.
Таким образом, вначале мы имеем как бы противопоставлен-
ные друг другу два ряда явлений. Первый ряд — это отдельные
факты действительности, действующие как раздражители и после-
довательно развивающиеся в определенных временных парамет-
рах, свойства которых определяются физическими возможностями
внешнего мира. В самом деле, служитель, стукнув нижней дверью,
не может немедленно оказаться около собаки. Для преодоления
расстояния между нижней дверью и собакой, находящейся на
третьем этаже, неизбежно надо потратить определенное время.
Это время, конечно, в зависимости от возраста и способности
служителя, может в каких-то пределах варьировать, однако даже
самая минимальная длительность этого продвижения всегда будет
достаточно большой и измеряется или минутами, или долями ми-
нут.
306
Рефлекс цели как объект физиологического анализа
Рис. 6. Два этапа образования опережающего возбуждения
1 — первоначальные ориентировочные реакции (А, Б, В, Г) на последовательно
развивающиеся явления во внешнем мире (А', Б', В', Г'). Видны изолированные ори-
ентировочные реакции; II — соотношение явлений внешнего мира и процессов
центральной нервной системы после многократного повторения ряда А, В» В» Г.
а1, а2, а3 — новые связи между афферентными результатами от всех промежуточных
ориентировочно-исследовательских реакций, имевших место в прошлом. Схема
отчетливо показывает распространение возбуждений по системе А', Б', B't Г', опере-
жающих развитие явлений внешнего мира А, Б» В, Г
Совсем другие соотношения мы имеем в нервном субстрате
головного мозга животного. Здесь процессы разыгрываются в
миллисекундах, и, следовательно, по самой своей сути мозг имеет
возможность моментального отражения всей цепи медленно раз-
вертывающихся событий действительности, если только эта после-
довательность в прошлом повторялась несколько раз. В этом
факте нельзя не видеть грандиозного достижения эволюци:
5
которая
оказалась
способной создать такой аппарат,
который
позволяет дать ускоренные в тысячи раз отражения последова-
тельных событий внешнего мира. Но здесь включается еще одно
качество нервного субстрата: его способность ассоциировать по-
вторяющиеся много раз ряды одних и тех же афферентных воз-
буждений. Стоит только представить себе на минуту эти два
свойства нервной системы — моментально отражать действия
внешнего агента и связывать ряд последовательных возбуждений
новыми ассоциативными связями, чтобы понять все огромное зна-
чение аппарата нервной системы, лежащего в основе образования
условного рефлекса.
Рефлекс цели как объект физиологического анализа
307
Если посмотреть на приведенные схемы, то можно увидеть,
что при наличии указанных выше двух свойств нервного субст-
рата достаточно возникнуть уже первому раздражителю всей цепи
явлений, как процесс возбуждения нервной ткани, распростра-
няющийся со значительно большей скоростью, немедленно дохо-
дит до последнего звена этой цепи создававшихся в прошлом воз-
буждений. Именно различие в скоростях течения явлении в самой
натуральной действительности и в нервном веществе мозга жи-
вотного создает предпосылки для такого распространения воз-
буждения, которое опережает во много тысяч раз течение дейст-
вительных событий.
При таких соотношениях скоростей развертывания явлений в
действительности и в нервной системе мы всегда будем иметь
возбуждение в нервной системе уже в последнем звене, в то
время как в действительности наступило только первое событие
из всего длинного ряда (рис. 6, II),
Аппарат акцептора действия, опережающий своим появлением
осуществление самого действия и его конечных результатов,
и есть то последнее звено, которое когда-то закономерно создава-
лось последовательно развивающимися событиями внешней дей-
ствительпости и фиксацией в нервной системе афферентных воз-
буждений от конечного результата этого ряда действий — резуль-
тата, адекватного поставленной до этого цели действия.
Таким образом, если какой-то ряд явлений действительности
оказывается весьма длинным, то мы можем принять, что каждое
звено этого ряда имеет вначале выработки реакцию нервной
системы только па себя, причем это будет всегда реакция типа
ориентировочно-исследовательской реакции. Следовательно, до
установления ассоциативных связей между последовательными
возбуждениями центральной нервной системы
любой
внешний
агент формирует ряд процессов, каждый из которых, как мы
видели, имеет циклический характер и представляет собой нечто
вроде отдельного «строительного блока» в архитектуре большого
здания (рис. 6).
Особенность нервных механизмов, определяющих формирова-
ние цели действия, с пашей точки зрения, и состоит в том, что
многочисленные процессы, формировавшиеся в прошлом по от-
дельности, могут в долях секунды построить такую систему воз-
буждений, в которой предвосхищаются афферентные элементы
всех последующих этапов действия до конечного этапа включи-
тельно. Следовательно, с физиологической точки зрения, рефлекс
цели — прежде всего механизм формирования возбуждений, опе-
режающих течение и развитие явлений самой действительности,
и только поэтому каждый этап последующих целеустремленных
действий при помощи обратной афферентаций может контроли-
роваться и выправляться в соответствии с исходной целью.
308
Рефлекс цели как объект физиологического анализа
Из сказанного видно, что мы имеем явное сходство, а, мо-
жет быть, в некоторых случаях и почти полное перекрытие ме-
ханизмов рефлекса цели и механизмов формирования акцептора
действия. Дальнейший анализ этих двух феноменов позволит
углубить их физиологические механизмы, отдифференцировать
характерные для каждого из них черты и сопоставить их роль
л
юрмировании целенаправленного поведения животных и чело-
века. Достаточно указать, например, что оба эти феномена (реф-
лекс цели и акцептор действия) неминуемо имеют в качестве
предшествующей стадию афферентного синтеза.
Только после всестороннего синтеза всех многочисленных
внешних и внутренних афферентных возбуждений может сфор-
мироваться акцептор действия. Точно так же цель действия мо-
жет диктоваться только той же совокупностью афферентных раз-
дражений настоящего и прошлого.
Заключение
Изложенные выше факты приводят нас прежде всего к од-
ному важному выводу: проблема «рефлекса цели», поднятая
И. П. Павловым, может и должна изучаться на основе современ-
ных достижений высшей нервной деятельности и общей физио-
логии центральной нервной системы.
Однако нельзя думать, что имеющийся в пашем распоряже-
нии фактический материал и подходы к его пониманию вполне
достаточны для изучения такого сложного и решающего феноме-
на в деятельности мозга, какими являются постановка цели и
реализация ее в соответствующих действиях. Как мы видели,
некоторые подходы должны быть изменены. Ясно одно, что
игнорирование этой проблемы как предмета научного и объек-
тивного исследования в настоящее время совершенно пе обо-
сновано.
Успехи изучения высшей нервной деятельности и общей
физиологии мозга в настоящее время таковы, что они вполне
позволяют сделать попытку физиологического изучения такой
сложной архитектуры целостного поведения, какими являются
рефлекс цели, целеустремленное поведение.
Мы не можем, например, при решении проблемы физиологи-
ческого субстрата цели исключить такие важные факторы пове-
дения, как «основные влечения», которые формируются на со-
вершенно реальных материальных путях, но по своей физиоло-
гической архитектуре значительно отличаются от тех реакций,
которые возникают в ответ на внешний стимул, например, иа
условный раздражитель.
Еще несколько лет назад нас вполне удовлетворяло выраже-
ние «повышенная пищевая возбудимость», потому что она при-
Рефлекс цели как объект физиологического анализа
309
звана была объяснить нам лишь только то, почему на предъяв-
ленный в данный момент животному условный стимул выделя-
лось слюны больше, чем обычно.
Сейчас же требуется понять: по каким механизмам эта повы-
шенная пищевая возбудимость превращается во «влечение, тре-
бующее удовлетворения». По каким механизмам это влечение
формирует цель определенного поведения и реализацию этой цели
в конкретных поведенческих актах? Мы должны также понять
физиологические механизмы формирования того, что II. П. Пав-
лов назвал «основными влечениями».
Нельзя не признать огромным недостатком нашей работы то,
что этот основной двигатель, толчок к целеустремленным дейст-
виям, остается вне поля зрения наших исследовательских инте-
ресов. Такое явление можно объяснить только тем, что И. П. Пав-
лов смело поставил вопрос о рефлексе цели тогда, когда еще
физиология нервной системы в целом не была готова к его раз-
решению. Мы не знали, например, на основе каких конкретных
и
изиологпческих процессов в течение длительного времени под-
держпвается на высоком энергетическом уровне какая-либо жиз-
ненная цель человека, поглощающая, а часто и подчиняющая
себе все его другие деятельности.
В настоящее время мы знаем эту физиологическую основу.
Она обладает универсальным активирующим действием на выс-
шие уровни нервной системы и может энергетически обеспечи-
вать все более и более широкие ассоциативные связи. Как теперь
известно, субстрат этой «слепой силы», по выражению И. П. Пав-
лова, заложен в гипоталамусе и ретикулярной формации ствола
мозга (Мэгоун и Моруцци 1942, Бремер 1953 и др.). Таким обра-
зом, чисто энергетическая сторона длительного удержания в до-
минирующем состоянии раз поставленной цели действия имеет
в настоящее время достаточно убедительное объяснение. В этом
пункте проблема рефлекса цели весьма тесно соприкасается с про-
блемой доминанты, разрабатывавшейся в нашей отечественной
физиологии А. А. Ухтомским.
Л И Т Е Р А Т У Р А
••III
Агафонов В. Г. Тормозящее действие аминазина на центральный эффект
болевого раздражения.— Журн. невропат, и психиатр., 1956, № 12, с. 94.
Анохин П. К. Изучение динамики высшей нервной деятельности. I. Актив-
ный секреторно-двигательный метод изучения высшей нервной деятель-
ности.— Нижегородский мед. журн., 1932, № 7—8, с. 42.
Анохин П. К. Особенности афферентного аппарата условного рефлекса и
их значение для психологии,— Вопр. психол., 1955, № 6, с. 16.
Анохин П. К. Внутреннее торможение как проблема физиологии. М., 1958.
Анохин П. К. Новые данные о функциональной гетерогенности ретикуляр-
ной формации ствола мозга.— Журн. высш, нервн. деят., 1959, т. 9, № 4,
и
310
Рефлекс цели как объект физиологического анализа
Беритов II. С. Индивидуально приобретенные рефлексы. М., 1923.
Быков К. М. Кора и внутренние органы. М., 1953.
Гавличек В. Электроэнцефалографическая характеристика условнорефлек-
торной оборонительной доминанты.— Физиол. журн. СССР, 1958, т. 44.
Кз 4, с. 305.
Ку палов П. С. Общие результаты изучения процессов торможения в коре
больших полушарий.— Журн. высш, нсрвн. деят., 1955, т. 5, Ns 2, с. 157.
Павлов И. П. Поли. собр. трудов, 1949, т. 3.
Шумилина А. И. Сравнительная характеристика электрической активности
сетчатого образования и коры головного мозга при выработке условно-
го оборонительного рефлекса.— Физиол. журн. СССР, 1959, т. 45, № 10,
1176.
Шумилина А. II. Экспериментальный анализ электрической активности сет-
чатого образования и коры головного мозга при выработке условной
пищевой реакции.— Физиол. журн. СССР, 1961, т. 47, № 1, с. 3.
Anand В. K.f Brobeck I. В. Hypo-thalamic control of food intake in rats and
cats.— Yale J. Biol. a. Med., 1951, vol. 24, N 2.
Andersson B., McCann S. M. A further study of polydipsia evoked by hypotha-
lamic stimulation in the goat.—Acta physiol, scand. 1955, vol. 33, N 4.
Anochin P. K. New conception of physiological architecture of the condition
reflex.—^ in: Brain mechanisms and learning symposium. Blackwell scien-
tific publ., 1961, p. 189.
Bremer F. Some problems in neurophysiology, London, 1953.
Juvell P. A. The occurence of vesiculated neurones in the hypothalamus of
the dog.— Physiol. J., 1953, vol. 121, N 1.
Miller N. F. Experiments on Motivation.—Science, 1957, vol. 126, N 3286.
эмоции
Эмоции (фр. emotion от лат. emovere — возбуждать, волно-
вать) — физиологические состояния организма, имеющие ярко
выраженную субъективную окраску и охватывающие все виды
чувствований и переживаний человека — от глубоко травмирую-
щих страданий до высоких форм радости и социального жизне-
ощущения.
Общая оценка эмоций
В связи с тем, что эмоциональные состояния человека носят
подчеркнуто субъективный характер, они долгое время были пре-
рогативой психологии и вследствие этого значительно задер-
жали изучение физиологических механизмов эмоций и анализ
(с чисто медицинской точки зрения) воздействия эмоций па
организм человека. Вместе с тем некоторые физиологи из-за
субъективного характера эмоций дошли до абсурда, вообще отри-
цая существование эмоциональных состояний как специфическо-
го и реального факта живой природы. В последние годы эмоции
становятся все более и более важным объектом медико-физиоло-
гического исследования.
Так, например, с полной определенностью установлено, что
эмоции, и именно длительные отрицательные эмоции (тоска, страх,
боль и др.), играют решающую роль в развитии так называемых
неврогенных заболеваний.
Огромная роль эмоциональных состоянии в жизни человека
и необходимость всесторонне учитывать их во врачебной прак-
тике совершенно очевидны хотя бы на основании того
акта, что
врачам прежде всего приходится изучать жалобы больного, а эти
жалобы по своей глубокой физиологической сути есть не что
иное, как словесный отчет больного о его эмоциональном состоя-
нии. Наряду с этим в жалобах больного содержатся и его субъек-
тивные суждения о локализации исходного стимула для этого
эмоционального состояния.
♦ Опубликовано в Большой медицинской энциклопедии (т. 35. М., 1964).
312
Эмоции
Роль эмоций в жизни человека весьма велика. Уже класси-
ки философии и физиологии отметили их огромное значение для
всех видов чисто человеческой деятельности. «Без «человеческих
эмоций»,— писал В. И. Ленин,— никогда не бывало, нет и быть
ие может человеческого искания истины» (с. 112).
Существует ошибочное мнение, что II. II. Павлов- отрицал
эмоции и вообще субъективные состояния. Бак раз напротив, па
протяжении всей своей творческой жизни И. II. Павлов не раз
подчеркивал огромную организующую роль субъективных пере-
живаний в поведении человека. В 1923 г. в докладе на юбилее
института им. Лесгафта он сказал: «Конечно, психология, касаю-
щаяся субъективной части человека, имеет право на существо-
вание, потому что ведь наш субъективный мир есть первая реаль-
ность, с которой мы встречаемся». Того же вопроса П, II. Павлов
коснулся в 1934 г.: «Глупо было бы отрицать субъективный мир.
Само собой разумеется, он, конечно, есть. Психология, как фор-
мулировка явлений нашего субъективного мира,— совершенно
законная вещь, и нелепо было бы с этим спорить. Па этой основе
мы действуем, на этом складывается вся социальная и личная
жизнь, об этом речи быть пе может. Речь заключается в анализе
этого субъективного мира».
Специально характеризуя роль эмоций в процессах высшей
нервной деятельности, он сказал: «Эмоции — источник силы для
корковых клеток». II. П. Павлов считал, что если исключить
эмоции, то кора лишится главного источника силы. Достаточно
сопоставить это заключение с работами школы Гельхорпа, прове-
денными па протяжении последних 20 лет, чтобы убедиться, как
эти работы полностью подтвердили положения И. П. Павлова.
Таким образом, эмоциональные состояния являются реальным
физиологическим фактором в природе человека. Они составляют
неотъемлемую часть его приспособительных реакций и потому
должны быть изучаемы объективными научными методами.
Эволюция эмоций. Если исходить из того, что эмоции пред-
ставляют собой такую форму реакций, которые непременно охва-
тывают весь организм и приобретают ярко выраженный субъек-
тивный характер, то мы должны поставить вопрос: каково про-
исхождение эмоций, когда и как они появились в процессе
эволюции?
На основании дарвинского понимания эволюции приспособи-
тельных реакций организма можно утверждать, что эмоциональ-
ные состояния сыграли когда-то положительную роль, создав
условия для более широкого и более совершенного приспособле-
ния животных к окружающим условиям. Первичные ощущения
примитивных животных не могли бы удержаться в процессе эво-
люции и развиться в столь многогранные и утонченные эмоцио-
нальные состояния человека, если бы они не послужили прогрес-
Эмоции
313
су в приспособительной деятельности животных. В противном
случае они давно были бы устранены естественным отбором.
В чем же состоит это более совершенное приспособление?
Решающей чертой эмоционального состояния является его
интегралыюсть, его исключительность по отношению к другим
состояниям и другим реакциям. Эмоции охватывают весь орга-
низм, они придают состоянию человека определенный тип пере-
живания. Производя почти моментальную интеграцию (объеди-
нение в единое целое) всех функций организма, эмоции сами по
себе и в первую очередь могут быть абсолютным сигналом полез-
ного или вредного воздействия на организм, часто даже раньше,
чем определены локализация воздействия и конкретный механизм
ответной реакции организма. Именно это свойство организма —
определять благодаря эмоции качество воздействия с помощью
самого древнего и универсального критерия всего живого на
Земле — выживаемости — и придало эмоции универсальное зна-
чение в жизни организма. Вместе с тем организм оказывается
чрезвычайно выгодно приспособленным к окружающим условиям,
поскольку он, даже не определяя форму, тип, механизм и дру-
гие параметры тех или иных воздействий, может со спаситель-
ной быстротой отреагировать на них с помощью определенного
качества эмоционального состояния, сведя их, так сказать,
к общему биологическому знаменателю: полезно пли вредно для
него данное воздействие.
Насколько важна эта интеграция организма в эмоциональном
состоянии, можно видеть па примере простой болевой эмоции,
которая в той или иной мере каждым человеком испытывалась
в жизни.
Допустим, что имеется какое-то травматическое поражение
суставной сумки. Как известно, такое поражение создает крайне
тягостное болевое ощущение, болевую эмоцию. Какие средства
может употребить наш организм для построения новой моторной
координации с исключением пораженного сустава? Практика по-
казывает, что в таких случаях создаются исключительно широкие
возможности вовлечения новых мышц, новых суставов и даже
целых конечностей. Однако во всех этих случаях ограничиваю-
щим фактором служит только ощущение боли. Организм челове-
ка осуществляет в этих случаях многочисленные попытки дви-
жения, «обходя» болевое ощущение, как только оно возникает.
В данном случае болевая эмоция помогает организму воздержи-
ваться от несовместимых с жизнью движений («щажение» в
медицинской практике).
Стоит только представить себе, что этот подбор не вредящих
здоровью движений производился бы на основе каких-либо дру-
гих критериев, например степени натяжения мышцы, угла откло-
нения в суставе и т. д., как сразу же станут ясными все совер-
314
Эмоции
шенство универсального характера эмоциональных состояний и
крайняя выгодность их в приспособительном смысле. Практиче-
ски весь жизненный опыт человека, иачипая с первых дней жиз-
ни, помогает ему избегать вредных воздействий не на основе
учета объективных параметров вредящего агента (например, остро-
та повреждающего предмета, глубина погружения его в кожу
и т. д.), а на основе именно того «общего знаменателя», кото-
рый выражается эмоциональным состоянием: «больно», «непри-
ятно».
Все вышесказанное приводит к выводу, что с эволюционной
точки зрения первичные эмоциональные состояния животных как
своеобразные интегральные состояния, однажды появившись,
были немедленно включены в фонд приспособительных реакций
организма. Они давали возможность производить более быструю
оценку внешнего воздействия и более широкие возможности
выбора оптимальных условий для выживания. Именно это при-
способительное значение ощущений и эмоций позволило им в
процессе - эволюции закрепиться наследственно и достичь той
широты и изощренности, какую мы наблюдаем на стадии обще-
ственного развития человека.
Можно сказать, что всем без изъятия жизненным потреб-
ностям и отправлениям, включая и проявления интеллектуаль-
ной деятельности, сопутствует соответствующий эмоциональный
тонус, благодаря которому организм непрерывно остается в русле
оптимальных жизненных функций.
Многочисленные факты, установленные психологами и физио-
логами, демонстрируют некоторый единый план в архитектуре
живого организма, благодаря которому все разнообразные его
функции санкционируются или отвергаются на основании одного
и того же принципа — принципа их соответствия сформировавше-
муся в данное время определенному эмоциональному состоянию.
Нервный субстрат эмоций. Универсальное значение эмоцио-
нальных состояний, их руководящая роль в формировании пове-
дения уже давно привлекли к себе внимание. Первоначально
попытки определить субстрат эмоциональных состояний были
сделаны в клинике, где изучались корреляции различного рода
нарушений мозгового вещества с расстройствами поведения
больных.
Было установлено, что эмоциональная жизнь человека почти
никогда не была сколько-нибудь заметно нарушена, если разру-
шение мозгового вещества касалось новой коры головного мозга
неокортекс (некоторое исключение из этого правила составляют
лобные отделы коры). II в полную противоположность этому эмо-
циональная жизнь человека оказывается радикально нарушенной,
если в повреждение вовлечены подкорковые образования, особен-
но таламуса и гипоталамуса. Так постепенно сложилась концеп-
Эмоции 315
ция, что эмоциональная жизнь человека определяется функцией
подкорковых образований (Шарко, Монаков, Бехтерев и др.).
Экспериментальную ясность в эту проблему внесли работы
нейрофизиологов, которые, используя технику прямого вмеша-
тельства в подкорковые образования, показали, что разрушение
указанных выше подкорковых образований приводит к наруше-
нию прежде всего внешнего выражения эмоциональных состоя-
ний. Особенно много в этом направлении сделано школой
В. Р. Гесса (W. IL Hess), сотрудники которой впервые разрабо-
тали тонкую технику попадания в область гипоталамуса с целью
раздражения его и коагуляции. Было с достоверностью показано,
особенно в опытах с вживленными электродами и на свободном
поведении животных, что всякое раздражение пли разрушение
области таламуса и гипоталамуса ведет к нарушению эмоциональ-
ных реакций, изменению всего поведения животного.
В дополнение к этим экспериментам и параллельно с ними
развивалась другая линия исследований — удаление коры боль-
ших полушарии п получение «таламических» и «гипоталамиче-
ских» животных (Бард, 1950). Эти последние опыты показали,
что при наличии только таламуса и гипоталамуса и, конечно,
стволовой части мозга болевое раздражение дает отчетливое внеш-
нее выражение эмоции ярости и эмоции гнева («псевдоаффект»).
Клинические наблюдения над анэпцефалами также показали, что
эти существа способны к довольно широкой гамме эмоциональ-
ных реакций со всеми характерными признаками для этих
эмоций.
Так постепенно установилась определенная и достоверная
концепция о подкорковом генезе выражения эмоции. Как извест-
но, И. П. Павлов па основе работ по условным рефлексам также
пришел к выводу, что локализация эмоциональных выражений
имеет место прежде всего в «ближайшей подкорке», и особенно
в гипоталамусе.
В последние годы участие подкорковых отделов головного
мозга в формировании эмоции было с большой наглядностью
продемонстрировано в экспериментах, проведенных с примене-
нием электронной техники и стереотаксического метода, помогаю-
щего точно попадать в различные пункты подкорковых образова-
ний (об этих экспериментах см. ниже параграф «Теории эмо-
ций»).
Новейшими данными были подтверждены прежние опыты,
показавшие, что гипоталамус имеет непосредственное отношение
к формированию эмоции, п, кроме того, было установлено уча-
стие в этом процессе еще и амигдалярного комплекса, гиппокам-
па, поясной извилины коры, орбитальной коры и других областей
головного мозга. Таким образом, между гипоталамусом и корой
были выявлены «посредники». Была выделена специальная си-
316
Эмоции
Рис. 1. Развитие главных обо-
нятельных путей у амфибий
(вверху) и у человека (внизу)
Первичные и вторичные пути
(тонкие линии) связывают обоня-
тельный эпителий с обонятель-
ной луковицей и соответственно
с полушариями: 1 — обонятель-
ная луковица; 2 — таламус; 3 —
перегородка; 4—гиппокамп; 3 —
гипоталамус; 6 — свод; 7 — мозо-
листое тело. Пунктиром обозна-
чены проекции соответствующих
областей (по Эллиоту)
стема нервных образований, которая получила название «лимби-
ческой системы». В нее входят все те промежуточные образова-
ния, которые, являясь посредниками между гипоталамусом и корой
в формировании эмоций, представляют собой остаток весьма древ-
них нервных образований, эволюционно связанных с обонятельной
системой.
Для того чтобы показать, как складывалась циркуляция нерв-
ных возбуждений по подкорковым образованиям, Эллиот (Elliot,
1954) предложил схему (рис. 1), па которой видно, что первич-
ная обонятельная область доминирует над первичными интракор-
тикальными объединениями и над теми субкортикальными объ-
единениями, которые играют решающую роль в формировании
эмоций.
Описанные подкорковые образования, состоящие из частей
лимбической системы и развившиеся па основе первичных обоня-
тельных систем, связаны между собой теснейшими узами непре-
рывной циркуляции возбуждения. Есть основание думать, что
эта непрерывная циркуляция импульсов служит своеобразным
приводным ремнем между гуморальными раздражениями, кото-
рые содержатся в кровп и действуют па область гипоталамуса,
и высшими кортикальными функциями — Папес (J. W. Papez),
1958; Мак-Лин (Р. D. Mac Lean), 1954. В этом смысле подкор-
Эмоции
317
новая область может быть признана своеобразным энергетическим
резервуаром, который в зависимости от качества возникшей в
данный момент эмоции вовлекает в деятельность огромные обла-
сти коры больших полушарий. Весьма отчетливо эта идея Напеса
о циркуляции импульсов в лимбической системе выражена Мак-
Лином в общеизвестной схеме, очень удобной для понимания роли
лимбической системы (рис. 2). Достоинства этой схемы несомнен-
ны, однако она не показывает источники
афферентных притоков
и конкретный конечный механизм последующего вовлечения коры
Рис. 2. Круг Папеса
Затемненные поля коры представ-
ляют собой лимбическую долю
Брока или висцеральный мозг
Мак-Лина: АТ — переднее тала-
мическое ядро; М — мамилляр-
ное тело
больших полушарий, без которой, естественно, не может сфор-
мироваться осознаваемое эмоциональное ощущение.
В последние годы всеобщее внимание привлекает амигдаляр-
ное тело, которое чрезвычайно широко связано почти со всеми
решающими образованиями подкорковой области и коры больших
полушарий. В настоящее время сложилось представление, что оно
играет роль облегчающего или потенцпирующего образования,
способного снизить или повысить общий тонус эмоционального
состояния независимо от его происхождения и биологического
качества. Есть основание думать, что амигдалярное тело имеет
особенно тесную связь с областью висцеральных афферептацпи
и это в какой-то степени может совпадать и с приписываемой
ему потенцирующей ролью.
Несмотря па то что область лимбических образований бесспор-
но принимает участие в общем оформлении эмоционального со-
стояния организма, физиологическая характеристика эмоции по
современным воззрениям связана прежде всего с распростране-
нием возбуждений из области гипоталамуса на все эффекторные
органы.
Область гипоталамуса определяет первичное биологиче-
ское качество эмоционального состояния, его характерное внеш-
нее выражение.
518
Эмоции
Внешнее выражение эмоций. Наиболее известны и хорошо
изучены уже со времен первых древних наблюдателей внешние
выражения эмоции. Признаки этих выражений настолько обще-
человечпы и каждодневно наблюдаются людьми, что в специаль-
ной описании не нуждаются.
Исследования так называемых псевдоа
эпи
юктов, появляющих-
ся после удаления больших полушарий, показали, что выраже-
ние эмоций имеет стандартный характер и не зависит от высших
отделов головного мозга и без них оно реализуется на периферии
в десятках нервных и гуморальных компонентов. Достаточно
возникнуть эмоциональному возбуждению в области гипоталаму-
са, как немедленно весь организм со всеми его системами п
периферическими органами оказывается вовлеченным в эмоцио-
нальное выражение. Интересно, что любой отдельно рассматри-
ваемый компонент эмоционального выражения независим от друго-
го компонента; они выступают в некотором роде радиальными
проявлениями центрально интегрированного нервного комплекса.
Иначе говоря, каждый компонент эмоционального выражения —
вынужденный компонент системы, фрагмент интегративной моде-
ли и неизбежно обнаруживается, если даже все другие перифе-
рические компоненты будут исключены. Эта особенность выраже-
ния эмоции приобретает исключительно большое значение для
медицины, поскольку ведет к целому ряду трудно устранимых
патологических состояний па отдельных путях периферического
выражения эмоций.
Для оценки выражений эмоций прежде всего следует перечис-
лить те компоненты, которые именно для человека наиболее ха-
рактерны и постоянны. На первом месте надо поставить мимиче-
ское выражение эмоций, поскольку оно отчетливо отражает
качество эмоционального состояния, служит сигналом для окру-
жающих и, следовательно, определяет общественную роль эмо-
циональных выражений. Тонкость и вариабельность мимических
выражений эмоций таковы, что малейшего перераспределения
возбужденных мышечных волокон на лице достаточно для того,
чтобы окружающие восприняли совсем другое настроение челове-
ка. Такая изощренность лицевой мускулатуры в выражениях
эмоции, несомненно, является достижением социального существо-
вания в развитии человека. Только став приспособительным фак-
тором, именно фактором сигнального общения с окружающими,
эмоциональное выражение приобрело характер наследственно
закрепленных нервных связей.
Едва ли кто может смешать мимическое выражение горя или
радости с каким-либо другим эмоциональным выражением лица.
Все это подчеркивает, что
ских соотношений, первичная интеграция эмоций очень давно в
истории развития человека закрепилась постоянными связями с
ормируясь в области гипоталамиче-
и
Эмоции
319
ядром лицевого нерва. Последнее, находясь в области продолго-
ватого мозга, фактически служит неизбежным «конечным путем»
реализации любого мимического выражения состояний человека.
Именно здесь разыгрывается та тончайшая избирательность в
синаптических передачах, которая, мобилизуя также избирательно
мимические мышцы, формирует образ того или иного эмоциональ-
ного выражения.
При оценке мимического выражения следует иметь в виду,
что на основе врожденных' связей в области ядра лицевого нерва,
о которых говорилось выше, жизненный опыт формирует чрез-
вычайно обширную шкалу мимических нюансов, которыми чело-
век может выразить свои многообразные и тончайшие эмоцио-
нальные переживания, служащие средством общения с окружаю-
щими. Было бы неправильно думать, что эмоциональные
выражения человека целиком приобретены им в жизни. Доста-
точно вспомнить выражения первичных эмоций (плач, улыбка)
у новорожденного ребенка, чтобы попять врожденный характер
этих выражений. В связи с этим возникает весьма интересный
вопрос: до какой степени и что именно в эмоциональных выраже-
ниях человека врожденно, т. е. закреплено в готовых нервных
связях? На этот вопрос дают некоторый ответ прямые исследова-
ния эмоциональных выражений лица у плодов человека на раз-
личных стадиях их развития.
Было показано, например, что плод человека, преждевремен-
но изъятый по медицинским показаниям и помещенный в благо-
приятные условия циркуляции и питания, может выразить мими-
кой довольно разнообразные эмоциональные оттенки. Так, напри-
мер, на прикосновение щетинкой к верхней губе оп может
ответить грима сой, удивительно сходной с теми мимическими
реакциями, которыми взрослый человек выказывает отвращение,
брезгливость, чувство неприятного. При более сильном раздра-
жении плод может выразить на лице чрезвычайно наглядно
скорбь с характерным опусканием углов рта. Он даже может
выразить плач и легко всхлипывать, реагируя таким образом на
более сильное раздражение в области, иннервируемой тройнич-
ным нервом (Голубева и Шулейкина, 1956).
Сопоставляя весь приведенный материал, мы можем заклю-
чить, что мимика — это чисто человеческое выражение эмоцио-
нальных состояний — в своих основных контрастных проявлениях
(страдание, удовольствие) в значительной степени сформировала
уже на стадии эмбриогенеза. Таковы «исходные позиции», на
основе которых с момента рождения развивается и совершенст-
вуется во всем своем диапазоне человеческая мимика. В нейро-
физиологическом аспекте это означает, что первичные, подготов-
ленные эмбриогенезом связи области гипоталамуса и четверохол-
мия с ядром лицевого нерва служат тем врожденным фундамеп-
320
Эмоции
Рис. 3. Схема тормозящих
влияний коры головного мозга
на ядро лицевого нерва (2)
Видно соотношение подкорковых
(1) и корковых (5) влияний на
клетки ядра лицевого нерва
(пирамидный контроль над яд-
ром лицевого нерва)
том, на котором в последующие этапы жизни происходят много-
численные дополнения и изменения, особенно когда включаются
тормозящие функции коры больших полушарий (рис. 3).
Другим важным и постоянным выражением эмоциональных
состояний являются изменения сердечно-сосудистых функций. Как
правило, эмоции, достигая известного уровня, включают в каче-
стве вегетативных компонентов сердце и сосуды. Путь возбуж-
дения от гипоталамуса до периферических органов идет, несом-
ненно, через гипоталамические центры регуляции уровня кро-
вяного давления. Именно этот периферический общий путь
многосторонне используется для вегетативных выражений различ-
ных эмоциональных состояний. Здесь следует особенно подчерк-
нуть, что сердечно-сосудистое выражение находится в тесной
зависимости от биологического качества эмоции. В то время как
положительная эмоция, приводя к повышению кровяного давле-
ния посредством увеличения работы сердца и через сужение со-
судов, пе даст больших следовых реакций и задержки кровяного
давления на высоком уровне, отрицательные эмоции (страх, тос-
ка. горе) отличаются тем, что, вовлекая сердце и сосуды через
те же самые конечные пути, они это делают па особенно высокой
степени возбуждения и с большим следовым тонусом. Такое от-
личие связано с двумя особенностями отрицательных эмоций, с их
генерализованностыо по нервному субстрату и с длительной за-
держкой метаболических сдвигов па основе гуморальных факто-
ров, сопровождающих отрицательные эмоции (адреналин, норад-
реналин) .
Последние исследования по физиологии ретикулярной форма-
ции показали, что эмоциональные состояния отрицательного харак-
тера, формируясь в области гипоталамуса п песпецифических ядер
Эмоции
321
таламуса, вовлекают адренергический субстрат гипоталамуса и
ретикулярной формации. Этот субстрат, являясь местом форми-
рования всех реакций напряжения организма, попадающего в
трудные условия, воздействует на периферию через целый ряд
вегетативных органов. При этом один путь формирования, осо-
бенно вовлечение симпато-адреналовой системы на периферии,
идет через ретикулярную формацию в продолговатый мозг и по-
граппчиые симпатические стволы до периферических оргапов
включительно. Другой путь при возбуждении адренергического
субстрата формируется через связи гипоталамуса с гипофизом и
приводит к выделению адренокортикотропного гормона, благодаря
чему происходит еще более сильное подстегивание аппарата сим-
пато-адреналовой системы на периферии (Н. Selye, 1962).
Избыточное выделение в кровь адреналина, норадреналина и
адренокортикотропного гормона приводит к весьма устойчивым
гипертензивным состояниям и может служить исходным момен-
том для хронического подъема кровяного давления.
Нельзя не упомянуть и то обстоятельство, что адренергиче-
ский субстрат как важнейший фактор отрицательных эмоций
оказывает влияние не только на периферическое системы, но и
на подкорковый аппарат мозга, избирательно создавая очаги тор-
можения в целом ряде функциональных объединений.
В эффекторных выражениях эмоции, кроме мимической
мускулатуры и сердечно-сосудистой системы, принимают также
участие и другие вегетативные и соматические системы. Таков,
например, дыхательный компонент выражения эмоций. Этот ком-
понент выражается в учащении экскурсий грудной клетки, углуб-
лении дыхания. Чрезвычайно чувствительным и постоянным
компонентом выражения эмоций является сокращение гладкой
мускулатуры желудка и кишечника, особенно их сфинктерных
образований. Кардиальный, пилорический и сигмоидальный
сфинктеры служат довольно часто пунктами периферической раз-
рядки центрального эмоционального возбуждения. Иногда чрез-
мерная тонизация этих сфинктеров приводит к патологическим
и порой весьма затяжным состояниям (нилороспазм, кардиоспазм
и др.).
Итак, любое эмоциональное состояние всегда выявляется на
периферии в форме многочисленных компонентов, каждый из ко-
торых имеет свою физиологическую специфику и специальный
нуть для периферического выражения. Из этого следует, что эмо-
циональный разряд интегрирован центрально, как некоторая
модель с автоматическим и вынужденным выходом возбуждений
в виде отдельных компонентов (рис, 4). Эти компоненты форми-
руются как проявление целостного нервного комплекса и не зави-
сят друг от друга ни в степени выражения, ни в качественной
окраске.
11 П. К. Анохин
322
Эмоции
Рис. 4. Схема, показывающая формиро-
вание эмоционального разряда и рас-
пространение возбуждений
1 — гипоталамус; 2 — ретикулярная форма-
ция; 3 — кортикофугальный сигнал к фор-
мированию эмоции; С,—С6—восходящие
активирующие влияния, формирующие из-
бирательно в соответствии с эмоцией кор-
ковые комплексы синаптических организа-
ций. Некоторые из них восходят непосред-
ственно к коре (С,, С3, С5), другие — через
промежуточные образования лимбической
системы и неспецифических таламических
ядер (С2, С4, С6). Разряды эмоций: а —
к лицевым мышцам; б — к соматической
мускулатуре; в — к сердечно-сосудистой
системе; г — к системе органов дыхания
Мг
Проблема «задержанных эмоций». В последние годы в меди-
цинской практике все более и более привлекают к себе внимание
так называемые «задержанные эмоции». Признается, что задерж-
ка внешнего выявления отрицательных эмоций способствует их
тонизированию и переходу в «застойное состояние» (Лаиг, 1948).
Здесь мы имеем на первый взгляд парадоксальное положение, по-
скольку вся воспитательпая система человека построена главпым
образом на задержке, шлифовке и на культурной обработке пер-
вичных генерализованных эмоциональных проявлений ребенка.
Как понять это противоречие? Действительно ли задержки эмо-
ций опасны?
Эмоции — и положительные и отрицательные, как правило,
выявляются вовне целым веером самых разнообразных перифе-
рических компонентов, классификация которых приведена выше.
Каждый комплекс таких компонентов складывается в характер-
ную картину того или иного эмоционального выражения: горя,
радости, удивления, печали, страха и т. д.
Есть еще один очень важный критерий классификации эмо-
циональных компонентов, который представляет особый интерес
с медицинской точки зрения: степень подчиненности того пли
другого компонента корковому, т. е. произвольному, контролю.
Все компоненты отрицательной эмоции с этой точки зрения
неодинаковы. В то время как мимическое выражение эмоций мо-
жет быть легко подавлено и человек внешне остается «невозму-
тимым», висцеральные компоненты эмоционального выражения,
например функции сердца, сосудов, кишечника, напротив, остают-
Эмоции
323
ся в этих условиях подверженными общему эмоциональному раз-
ряду. Такое состояние центральной нервной системы, когда об-
щему эмоциональному разряду поставлен произвольный предел
на одних периферических нутях и оставлен открытым выход на
других периферических путях, может быть названо «конфликт-
ным». Здесь напряженное возбуждение коркового характера
(по преимуществу социального происхождения) преграждает путь
другому возбуждению, возникшему на уровне гипоталамуса и
ретикулярной формации. Можно указать на конкретный случай
подавления мимического выражения эмоции. Поскольку абсолют-
но неизбежным конечным путем для мимического выражения
эмоций является ядро лицевого нерва, то затормозить выход эмо-
ционального разряда на лицевые мышцы можно только здесь,
именно в ядре лицевого нерва. Фактически это и делает корко-
вый нейрон пирамидного тракта, проводящий влияние из коры
больших полушарий к ядру лицевого нерва. Этот нейрон оказы-
вается для клеток ядра лицевого нерва тормозящим в подлин-
ном смысле слова. Однако это торможение мимического выраже-
ния эмоций не исключает выхода возбуждений на сердце, сосуды,
кишечник и эндокринные железы. В результате создается положе-
ние, при котором организм получает значительно более сильное
возбуждение па висцеральных органах, чем если бы пе было за-
держки в выражении эмоций па других периферических компо-
нентах. Следовательно, крупнейшие клиницисты, утверждавшие,
что «задержанные эмоции» являются патогенными, были в какой-
то степени правы. Однако вся проблема «задержанных эмоций» с
физиологической точки зрения не была разработана, и поэтому
возникли приведенные выше парадоксальные противоречия.
ЭЙ
изиологической точки зрения выражение «задержанные
эмоции» ошибочно. Эмоцию можно назвать «задержанной» лишь
в том случае, если она не выходит потоками возбуждения ни
на один свой периферический компонент. А это означает, в сущ-
ности, предотвращение эмоций как субъективного состояния либо
путем исключения ситуаций, способных вызвать в качестве ис-
ходного стимула отрицательную эмоцию, либо путем включения
между стимулом и эмоцией интеллекта, логики, устраняющих са-
мую возможность появления вредной эмоции. Именно в этом по-
следнем случае решающее влияние оказывает воспитательная ра-
бота — культура эмоций.
Если же аффект как субъективное состояние успел сложить-
ся, то говорить о «задержанной эмоции» уже нельзя. По физио-
логическим особенностям эмоций здесь речь может идти только
о подавлении каких-то периферических компонентов эмоций,
главным образом внешнего выражения этих эмоций — оживлен-
ных движений, мимических и речевых проявлений и т. д. Однако
сама эмоция, раз возникнув, неизбежно распространяется на дру-
11*
324
Эмоции
Тормозные
ч v ' Гормонален.
Гроаздолоне/е нас Кар- реакции I
моторные дао- 1
функции даскулярн.
реакции
Дыхание ч_^_/ Гормонален.
Нарда о- реакции
васкулярн.
реакции
Рис. 5. Схема
коркового торможения
компонентов эмоциональных реакций
I — проявление всех эффекторных компонентов, сопровождающих эмоциональную
реакцию; II — корковые влияния, социально обусловленные, могут произвольно
тормозить проявление соматических моторных компонентов эмоциональных реакций
гие компоненты (главным образом висцеральные), т. е. пе может
уже быть задержана. Так что, когда клиницист говорит о «задер-
жанных эмоциях», фактически он говорит об эмоциях, у которых
задержаны лишь отдельные компоненты их внешнего выявления,
в то время как другие компоненты получают чрезмерно сильные
потоки возбуждения (рис. 5).
Именно эта ситуация и может стать исходной для чрезмер-
ного развития возбуждений на подкорковых путях и трансфор-
мации их в последующем в так называемое застойное возбуж-
дение. Надо лишь помнить, что настойчивое воспитание эмоцио-
нального аппарата и именно так называемой «выдержки» с
детских лет служит очень важным фактором, предупреждающим
стремительность развития и застойность незадерживаемых компо-
нентов эмоций. В этом случае, несомненно, снижается возмож-
ность того, чтобы эмоции при своих повторных проявлениях мог-
ли стать источником патологических состояний висцеральных ор-
ганов и сердечно-сосудистой системы.
Эмоции
325
Значение эмоций в генезе патологических состояний. Разбирая
физиологические механизмы эмоциональных состояний, мы уже
касались отчасти и вопроса перехода нормальных эмоций в пато-
логические. Однако актуальность столь важного раздела медицин-
ской патологии требует специального и подробного обсуждения
этой проблемы.
Если исходить из павловского понятия «предела работоспо-
собности» нервных клеток, то следует сказать, что ничто в дея-
тельности человеческого организма, и особенно его мозга, не под-
ходит так близко к этому пределу работоспособности нервной
системы, как эмоциональные аффекты. Именно эмоциональные
состояния в силу их указанной выше биологической роли (моби-
лизация и напряжение сил организма) придают нервным клет-
кам чрезвычайно высокий стимул деятельности. Вместе с тем это
своеобразие эмоциональных состояний мало анализировалось под
углом зрения способностей нервной клетки на определенный диа-
пазон деятельности. Лучше всего эту способность нервной клетки
назвать «пределом работоспособности» (И. П. Павлов), что соот-
ветствует биологическому оптимуму активности. Даже в самых
сильных эмоциональных состояниях здорового человека при обыч-
ных оптимальных условиях его жизни клетки подкорковых аппа-
ратов, формирующих эмоции, и корковые клетки, мобилизующие
все отношения с внешним миром на службу данной эмоции,—
все они не преодолевают верхней границы биологического опти-
мума нервных элементов. Решающим
актором в преодолении
этой верхней границы является длительное эмоциональное состоя-
ние отрицательного характера. В связи с этим можно указать
четыре главных фактора, которые образуют обязательное физио-
логическое условие перехода эмоции в длительное «застойное»
состояние на высоком уровне возбудимости, т. е. на границе па-
тологии.
1. Преодоление биологического оптимума возбудимости дан-
ных нервных клеток и поломка их триггерных механизмов. «За-
щелкивание» возбудимости на этом высоком уровне и переход ее
в хроническое состояние.
2. Все более и более широкое вовлечение нервных структур
в функцию первично и чрезмерно возбудившихся нервных эле-
ментов. Это расширение вовлеченных элементов подчиняется
принципу доминанты, сформулированному А. А. Ухтомским.
3. Стремительное нарастание тонизирующих гормонов в кро-
вяном русле, что создает дополнительное «подстегивание» пер-
вично перевозбужденных нервных элементов и переводит тече-
ние патологической эмоции в хроническую форму по типу «по-
рочного круга».
4. Формирование утонченных и весьма расширенных сигналь-
ных связей этого патологического подкоркового комплекса с фак-
326
Эмоции
торами окружающего мира через опосредующие влияния коры го-
ловного мозга*
С этого момента патологическая эмоция становится психопа-
тологическим комплексом, элементом структуры психического
расстройства.
Постараемся разобрать физиологическую сущность каждого из
этих факторов. Естественно, решающим моментом служит пере-
ход нервной клетки от биологического оптимального функциони-
рования к функции повышенного, или, как выражался И. П. Пав-
лов, «запредельного», характера. Если исходить из того факта,
что диапазон импульсовой деятельности и общего тонуса нерв-
ной клетки определяется метаболическими процессами этой клет-
ки, которые ставят естественный предел ее импульсовой деятель-
ности, то мы должны прежде всего думать о том, что возбуди-
мость клеток, перешагнувшая предельные оптимальные границы,
есть результат в какой-то степени «поломанного» метаболизма
этих клеток. В настоящее время широко исследуются некоторые
метаболиты, имеющие специальное отношение к эмоциональной
жизни, особенно группа катехоламинов. Постоянное состояние по-
вышенной возбудимости уже само по себе является нарушением
нормы, устанавливает новый и в количественном и в качествен-,
ном отношениях тип метаболизма. В клиническом смысле это
«застойное возбуждение» раскрывается в целом ряде перифериче-
ских симптомов, состав и характер которых определяются каче-
ством функции, первично вовлеченной в аффект.
В самом деле, эмоциональный разряд, как правило, построен
по такому типу, что кора головпого мозга в своей интегратив-
ной функции отражает ту подкорковую систему, которая создает
биологическое качество эмоции. Вместе с тем эта весьма ком-
пактная интегрированная эмоциональная система подкорки разре-
шается на периферии в виде функций огромного количества
разнообразных соматических и висцеральных органов. Таким об-
разом, если подкорковые структуры, формирующие эмоции,
перешли на положение стационарно возбужденных станций,
то висцеральные органы немедленно ощущают на себе «этот по-
стоянный повышенный приток возбуждения из центральной
нервной системы. Кроме того, сама чрезмерная возбудимость под-
корковых аппаратов определенного знака эмоций все более и бо-
лее расширяет поле своего влияния путем вовлечения повых
нервных элементов, избирательно пронизывая мозг в целом. Ко-
нечно, это вовлечение идет строго в функциональном плане,
по принципу обрастания доминирующей функциональной систе-
мы все большим и большим количеством добавочных поводов и
звеньев, причем сама эта эмоциональная доминанта становится
«ружьем с легко спускающимся курком». Достаточно даже незна-
чительного повода в виде внешнего раздражения, чтобы весь эмо-
Эмоции
327
циональный комплекс «взорвался» и неизбежно разрешился в по-
вышенном тонусе висцеральных органов.
Этому патологическому состоянию способствует чрезвычайное
накопление тонизирующих веществ в крови (адреналин, норад-
реналин, адренокортикотропный гормон и др.). Находясь в крови
и омывая область гипоталамуса и ретикулярной формации, эти
гормональные вещества дополнительно воздействуют на метабо-
лизм адрепергических структур и тем самым повышают застой-
ность возбуждения в подкорковых структурах. Создается пороч-
ный круг, из которого выход делается возможным только через
воздействие па обмен веществ соответствующих нервных элемен-
тов. Психофармакологические и седативные средства оказывают
нормализующее влияние на эмоциональное состояние больного
главным образом посредством нормализации обмена веществ в
клетках упомянутых нервных элементов.
Чрезвычайно важпым свойством таких застойных отрицатель-
ных эмоциональных состояний является то, что они обладают
способностью «обрастать» бесконечным количеством поводов для
возникновения, и потому практически эмоциональное состояние
становится постоянным. Это «обрастание» происходит по принци-
пу условного рефлекса, и потому могут создаться даже отдален-
ные поводы и сигналы к провокации «взрывной» отрицательной
эмоции. Прямым доказательством того, что многие висце-
ральные заболевания возникают в результате повышения цент-
рального тонуса, служит успешное лечение гипертонического
синдрома резерпином. Последний, нормализуя обмен веществ и
синаптические передачи, приводит к устранению гипертензивного
состояния через устранение отрицательных эмоций, приобретших
застойный характер.
На примере развития гипертонической болезни невроген-
ного происхождения можно хорошо видеть все стадии транс-
формации эмоциональных состояний до патологического состоя-
ния включительно (рис. 6). Как известно, сосудосуживающий
центр продолговатого мозга является общим конечным нейроном
для многочисленных состояний организма, требующих участия
этого центра. Таким образом, смена уровней его возбудимости в
значительной степени зависит от нисходящих влияний, приходя-
щих к этому центру от коры, гипоталамуса, таламуса, ретику-
лярной формации и от всей лимбической системы — главного суб-
страта, формирующего наши эмоции.
Казалось бы, при первом же аффекте кровяпое давление мог-
ло бы подняться вверх и «защелкнуться» на высоком уровне.
Однако этого не происходит, поскольку повышающемуся давле-
нию также градуально противопоставлена нарастающая импуль-
сация от дуги аорты и синокаротидной области. Потоки этих
афферентных импульсаций направляются также к сосудосужп-
328
Эмоции
Рис. 6. Схема саморегуляции кровяного давления под влиянием нисходя-
щих кортико-гипоталамических механизмов
I — норма. Афферентные депрессорные влияния (жирные линии) на вазомоторный
центр всегда сильнее, чем возбуждающие влияния высших центров (пунктирные
линии). U — гипертензивное состояние. Нисходящие возбуждающие влияния из
коры, гипоталамуса и ретикулярной формации (жирные линии), постоянно возбуж-
дающие вазомоторный центр. Депрессорные импульсы (пунктирные линии) не могут
затормозить активность этого центра
[им центрам и благодаря своей частоте приводят их в
тор-
ваю:
мозное состояние.
Таким образом, в сосудосуживающих центрах все время
исходит борьба двух сил. Одна сила — нисходящие корковые
влияния — стремится поднять возбудимость сосудосуживающего
II
про-
1
центра до предела, в то время как другая — потоки депрессор-
ных импульсаций с периферии — стремится снизить это тонизи-
рующее действие и возвратить всю систему па уровень физио-
логического оптимума. Практически эта функциональная система
работает четко и биологически построена па постоянном преобла-
дании депрессорных влияний с периферии. Тем пе менее из этого
в действительности не вытекает, что человек гарантирован от ги-
пертонии, и главным образом потому, что «застойные» отрица-
тельные эмоциональные влияния сверху создают по всей нисхо-
дящей магистрали до сосудосуживающего центра включительно
чрезвычайно высокий уровень возбудимости. Постоянство этого
эмоционального состояния и его градуальное нарастание создают
условия, при которых потоки депрессорных импульсаций с пери-
ферии уже не могут больше устранить повышенную возбуди-
мость сосудосуживающего центра. С этого момента происходит
«защелкивание» кровяного давления на высоком уровне и орга
низм в целом вступает уже в патологическое состояние, неустра-
нимое без терапевтического вмешательства. Однако исходным
Эмоции
329
пунктом для развития этого патологического сипдрома служит
первичное изменение тех нервных клеток подкорки, которые сме-
стили свой обмен веществ п распространили свою патологию на
другие клетки под действием непрерывных отрицательных аф-
фектов.
Теории эмоций
Воспроизведение переживаний человека в творениях искус-
ства было в течение многих веков истории человечества единст-
венным способом осмысления эмоций. Впоследствии, с развитием
науки, начались исследования самой природы эмоциональных со-
стояний. Поскольку эмоции являются для человека прежде всего
ярко окрашенным субъективным состоянием, все первоначальные
попытки объяснить их природу были целиком психологическими,
субъективными и в большинстве своем идеалистическими.
Ниже рассмотрены лишь те
изпологические теории, которые
имеют значение и поныне. Прежде всего надо выделить теории
эмоциональных состояний Джемса — Ланге, Кеннона — Барда,
Мак-Дауголла. Эти теории получили в настоящее время призна-
ние многих физиологов и психологов.
Согласно теории психологов Джемса и Ланге, эмоциональное
состояние является производным от состояния периферических
органов: именно периферические органы определяют, в каком
эмоциональном состоянии будет находиться в данный момент че-
ловек.
Хорошо известен афоризм Джемса: «Мы чувствуем печаль,
потому что мы плачем». Из этой формулировки следует, что
первично развивается какая-то периферическая реакция и только
через короткий промежуток времени создается соответствующее
эмоциональное состояние. Указывая на то, что мы обычно берем
сначала эмоциональное состояние, а затем уже действие, Джемс
считает такую последовательность ошибкой нашего сознания.
По его мнению, какое-либо внешнее воздействие создает прежде
всего и непосредственно рефлекторный акт. Однако от органов,
принимающих участие в данном рефлекторном ответе, немедлен-
но направляются сигналы в центральную нервную систему, и вот
именно они и создают то субъективное ощущение, которое мы
называем «эмоциональным». Джемс делит эмоции па «низшие»
и «высшие». Что касается «низших» эмоций, связанных с состоя-
нием гнева, страха, радости и т. д., то их возникновение на
основе периферических сигнализаций для Джемса не составляет
сомнения. Однако относительно некоторых «высших» эмоций,
например любознательности, различного рода эстетических по-
требностей, он допускает, что непосредственно телесных причин
здесь может и пе быть, однако когда-то, в далеком прошлом,
330
Эмоции
эти эмоции через целую цепь промежуточных этапов, несомнен-
но, развились из первичных телеспых состояний.
Теория Джемса — Ланге имеет некоторое основание для объ-
яснения целого ряда эмоциональных ощущений висцерального
генеза. Однако в целом она исходит из неправильных физиоло-
гических предпосылок. В противовес теории Джемса — Ланге
была выдвинута центральная теория эмоциональных пережива-
нии, которая получила название «таламической теории Кенно-
на — Барда». Согласно этой теории основным процессом, рождаю-
щим эмоции, является процесс, развивающийся па основе прихо-
дящих в таламус афферентных импульсов. Здесь на уровне
таламокортикальных связей и формируется эмоциональное
состояние. Кеннон и Бард различают обычный поток сенсорных
возбуждений, идущих от сигнала к коре больших полушарий,
и поток других, так называемых таламических, процессов, кото-
рые охватывают более широкие подкорковые области и совмест-
но с корой формируют соответствующую эмоциональную окраску
воспринятого раздражения из внешнего мира.
Впоследствии теория Кеннона — Барда была уточнена при
помощи прямых экспериментов с раздражением гипоталамуса и,
таким образом, субстрат формирования эмоций был из области
таламуса перемещен в область гипоталамуса (Гесс). В экспери-
ментах с электродами, вживленными в область гипоталамуса,
Гесс показал, что прямое раздражение гипоталамуса у бодрствую-
щих кошек приводит к выражению ряда эмоциональных состоя-
ний (гнев, ярость, страх и др.). В последние годы эксперименты
с телестимулированием вживленных в гипоталамус элек-
тродов были проведены на обезьянах. Вся аппаратура, восприни-
мающая по радио стимул и трансформирующая его в электриче-
ское раздражение гипоталамуса, помещалась па ошейнике обезья-
ны. Это давало возможность в любой момент нахождения
животного в различных жизненных ситуациях посылать раздра-
жение издалека (из лаборатории или с наблюдательного пун-
кта) .
В последнее время экспериментальная техника была еще более
усовершенствована, благодаря чему создалась возможность произ-
водить телеинъекции любых фармакологических веществ и гормо-
нов в область гипоталамуса обезьян. Эти опыты, подтверждаю-
щие значение гипоталамуса в генезе эмоций, были проделаны и
развиваются в настоящее время Дельгадо (J. М. Delgado) в США.
Исключительно наглядное доказательство того, что именно об-
ласть гипоталамуса является областью формирования эмоциональ-
ных состояний, было получено в экспериментах с так называе-
мым самораздражением. Дж. Олдс (J. Olds) применил методиче-
ский прием, когда электрическое раздражение гипоталамуса давал
не экспериментатор, а сама крыса с вживленными электродами,
Эмоции
331
нажимая на определенную контактную педаль. Благодаря такому
приему был установлен интересный факт, что при нахождении
электродов в определенном пункте мозга крысы с исключитель-
ной силой, преодолевая даже опасные препятствия, тянутся к
площадке, которую опи сами нажимают и тем самым включают
раздражающий ток. Если это самораздражещге не прекращать,
то крыса может давать себе самой до 2000 включений электри-
ческого тока в минуту. Это послужило прямым доказательством
того, что электроды находились в каком-то «центре удовольст-
вий». Опыты Олдса показали, что эмоции положительного и от-
рицательного характера могут быть вызваны прямым централь-
ным раздражением мозга крысы. Впоследствии оказалось, что
подобные опыты самораздражения можно поставить и на других
м
животных, например кошках, кроликах.
Некоторые дополнения к центральной теории эмоций дал про-
фессор Д. Линдсли (D. В. Lindsley), который связал эмоциональ-
ное состояние с возможностью активирующего воздействия со
стороны ретикулярной формации.
С развитием учения о лимбической системе этот комплекс,
промежуточный между древними подкорковыми образованиями и
новой корой, имеющий непосредственное отношение к эмоциям
и висцеральным органам, получил название «висцерального моз-
га», или «эмоционального мозга». По теории эмоций Папеса, Мак-
Лина, Клювера и Бьюси (Н. Kluver, Р. С. Вису), эта промежу-
точная область и составляет тот субстрат, который, получая
через ретикулярную формацию и гипоталамус уже обработанные
каким-то образом сигналы, формирует и окраску эмоционально-
го состояния и посылает своп возбуждения в кору больших по-
лушарий. Здесь эмоциональное состояние в целом приобретает
свою конечную осознанную форму.
Клинические данные, приведенные авторами всех рассмотрен-
ных выше теорий, подтверждают локализацию субстрата эмоций
в приведенных выше подкорковых образованиях. Если опухоли
и разрушения области ретикулярной формации приводят к обще-
му коматозному состоянию и потере сознания, то нарушения в
области амигдалярного тела и гиппокампа ведут к созданию вы-
раженного эмоционального состояния (тоска, половые влечения)
или, наоборот, к полной потере этих эмоций.
Несомненно одно: эмоциональные состояния могут быть вы-
званы различными стимулами кортикального происхождения, од-
нако сама эмоциональная окраска и формирование отдельных
компонентов в ее выражении связаны с процессами в тех под-
корковых образованиях, которые И. П. Павлов именовал «бли-
жайшей подкоркой». Среди них, несомненно, решаюшую роль
играет гипоталамус — область интегрирования жизненно важных
функций.
332
Эмоции
ункции организма.
и
Биологическая теория эмоций. Приведенные выше теории эмо-
циональных состояний отличаются одной чертой, которая и слу-
жит причиной их недостаточности: они не рассматривают эмоцио-
нальные состояния как закономерный факт природы, как продукт
эволюции, как приспособительный фактор в жизни животного
мира. Исходя из дарвинской точки зрения на эволюцию полезных
приспособлений, надо считать, что эмоциональные состояния,
удержавшиеся в процессе эволюции и развившиеся до своего
тончайшего проявления у человека, не могли бы ни возникнуть,
ни сохраниться, а тем более закрепиться наследственностью, если
бы они в какой-то степени были либо вредны либо бесполезны
для жизненно важных функций животного. Вопрос сводится лишь
к тому, в чем состоит биологическая и физиологическая полез-
ность эмоций и эмоциональных ощущений в осуществлении жиз-
ненно важных
Если дать общую характеристику поведения живых существ,
и человека в частности, то оно может быть грубо разделено на
две стадии, которые, непрерывно чередуясь, составляют основу
жизнедеятельности. Первую стадию можно было бы назвать «ста-
дией формирования потребностей и основных влечений», а вто-
рую — «стадией удовлетворения этих потребностей». Вниматель-
ный анализ поведения животных п человека показывает, что та-
кая классификация вполне приемлема для всех видов влечений
и для любого вида удовлетворения потребности. Есть все основа-
ния думать, что эмоциональные состояния при их первичном
появлении в животном мире включились именно в этот цикл
смены двух кардинальных состояний организма. Все виды по-
требностей приобрели побудительный характер, создающий беспо-
койство в поведении животного и формирующий различные типы
добывательного или, наоборот, отвергающего поведения. Эти по-
требности связаны с определенным эмоциональным бостоянием,
большей частью тягостного, беспокойного характера. Наоборот,
удовлетворение потребности или выполнение какой-то функции,
которая устраняет назревшую потребность, связано с чувством
удовольствия, приятного и даже иногда с чувством гедонического
характера (наслаждения).
Таким образом, если проблему эмоций рассматривать с биоло-
гической точки зрения, то падо будет признать, что эмоциональ-
ные ощущения закрепились как своеобразный инструмент, удер-
живающий жизненный процесс в его оптимальных границах и
предупреждающий разрушительный характер недостатка или из-
бытка каких-либо факторов жизни данного организма.
Обычно удовлетворение какой-либо биологической потребно-
сти человека не является только простым устранением этой по-
требности, имеющей тягостный характер. Как правило, удовлет-
ворение, устраняя потребность, сопровождается самостоятельным
Эмоции
333
подчеркнутым положительным эмоциональным переживанием.
Следовательно, с широкой биологической точки зрения удовлет-
ворение можно рассматривать как конечный подкрепляющий фак-
тор, который толкает организм на устранение исходной потреб-
ности,
С точки зрения физиологической перед нами стоит задача:
раскрыть механизм тех конкретных процессов, которые в конеч-
ном итоге приводят к возникновению и отрицательного (потреб-
ность), ги положительного (удовлетворение) эмоционального со-
стояния. И прежде всего необходимо проанализировать вопрос:
в силу каких именно конкретных механизмов удовлетворение
потребности создает положительное эмоциональное состояние.
Биологическая теория эмоций (Анохин, 1949) построена на
основе представления о целостной
•£
изиологическои архитектуре
любого приспособительного акта, каким являются эмоциональные
реакции.
Основным признаком положительного эмоционального состоя-
ния служит его закрепляющее действие, как бы санкционирую-
щее полезный приспособительный эффект. Такое закрепляющее
действие проявляется только в определенном случае, именно ког-
да эффекторный акт, связанный с удовлетворением какой-то по-
требности, свершился с абсолютным полезным эффектом. Только
в этом последнем случае формируется и становится закрепляю-
щим фактором положительная эмоция. Можно взять для примера
такой грубый эмоциональный акт, как акт чихания. Всем изве-
стен тот гедонический и протонатический характер ощущения,
которое человек получает при удачном чихательном акте. Точно
так же хорошо известно п обратное: неудавшееся чихание созда-
ет на какое-то время чувство неудовлетворенности, неприятное
ощущение чего-то незаконченного. Подобные колебания в эмо-
циональных состояниях присущи абсолютно всем жизненно важ-
ным отправлениям животного п человека. Именно для подобных
жизненно важных актов эмоция и сложилась в процессе эволю-
ции как фактор, закрепляющий правильность и полпоту совер-
шенного акта, его соответствие исходной потребности.
Как же центральная нервная система «узнает» о том, что ка-
кой-либо жизненно важный акт совершен на периферии в надле-
жащей последовательности и полноценном виде (утоление голода,
утоление жажды, опорожнение тазовых органов, кашель, чихание,
половой акт и т. д.)? Для ответа на этот вопрос необходимо
ввести два понятия:
«э<
юрентный интеграл» и «афферентный
интеграл». Каждому акту периферического удовлетворения ка-
кой-либо потребности предшествует формирование центрального
аппарата оценки результатов и параметров будущего действия —
«акцептора действия» и посылка множества эфферентных воз-
буждений, идущих к самым различным органам и частям той си-
334
Эмоции
стемы, которая должна выполнить акт удовлетворения потребно-
сти. Об успешности или неуспешное™ такого акта сигнализирует
поступающая в головной мозг а
HIM
ерентная импульсация от всех
рецепторов, которые регистрируют последовательные этапы вы-
полнения функции («обратная афферентации»).
Оценка акта в целом невозможна без такой точной информа-
ции о результатах каждого из посланных по эфферентной систе-
ме возбуждений. Такой механизм является абсолютно обязатель-
ным для каждой функции, и организм был бы немедленно раз-
рушен, если бы этого механизма не существовало.
Каждый на своем опыте знает, что всякая неудача в выпол-
нении тех или иных актов создает ощущение неудовлетворенно-
сти, трудности и беспокойства. Это и есть последствие того, что
вторая часть периферического акта, именно результативный аф-
ферентный интеграл, не была создана па периферии и потому не
произошло адекватного ее сочетания с центральной посылкой.
Суть биологической теории состоит в следующем: она утверж-
дает, что положительное эмоциональное состояние типа удовлет-
ворения какой-либо потребности возникает лишь в том случае,
если обратная информация от результатов происшедшего дейст-
вия точнейшим образом отражает все компоненты положительно-
го результата и потому точно совпадает с аппаратом акцептора
действия. Биологически этой эмоцией удовлетворения и закреп-
м
ляются правильность любого функционального проявления и пол-
ноценность его приспособительных результатов. Наоборот, несо-
впадение обратных афферентных посылок от неполноценных ре-
зультатов акта с акцептором действия ведет немедленно к
беспокойству животных и человека и к поискам той повой ком-
бинации эффекторных возбуждении, которые привели бы к фор-
мированию полноценного периферического акта и, следовательно,
к полноценной эмоции удовлетворения. В этом случае полноцен-
ное эмоциональное состояние достигается способом пробных посы-
лок различных эфферентных возбуждений.
Из
V
at
ормулировки биологической теории эмоции явствует, что
А
Джемс и Ланге в своей теории уловили правильный момент в
развитии эмоциональных состояний, во всяком случае в появле-
нии тех эмоциональных состояний, которые связаны с жизненно
важными отправлениями. Однако, фиксируя внимание на перифе-
рии, Джемс и Ланге, естественно, не могли раскрыть тот истин-
ный центральный механизм (совпадение с акцептором действия),
который служит обязательным и решающим условием для возник-
новения положительной эмоции (Анохип, 1949).
Вместе с тем Кеннон и Бард, фиксируя внимание па цент-
ральном субстрате, в какой-то степени были также правы, по-
скольку конечный момент формирования эмоционального состоя-
ния совпадает именно с областью подкорковых аппаратов. Однако
Эмоции
335
фиксация ими внимания только ла центральном механизме не
давала им возможности раскрытьистиппый механизм центрально-
периферических взаимоотношений в формировании эмоций, ощу-
щений. Эмоции висцерального происхождения не покрывают, ко-
нечно, всех тех эмоциональных состояний, которые составляют
область эмоциональной жизни человека. Многие из этих эмоций,
особенно болевая эмоция, не требуют постепенного увеличения
потребности, они возникают внезапно, и только простое устране-
ние такой эмоции создает положительное ощущение. Однако, не-
смотря па все эти изменения в деталях развития различных эмо-
ций, архитектурный план возникновения эмоции остается тем же.
Он распространяется также на потребности интеллектуального
характера. Потребности, возникшие, например, от каких-лпбо со-
циальных факторов (пусть это будет потребность в разрешении
какой-то сложной технической проблемы, потребность в коллек-
ционировании),
завершаются уже высшей положительной эмоцией
удовлетворения, как только результаты долгих и трудных ис-
каний совпадут с исходным замыслом (акцептор действия).
Есть основание полагать, что во всех эмоциях, начиная от
грубых низших эмоциональных состояний и кончая высшими со-
циальными эмоциями, используется одна и та же физиологиче-
ская архитектура. Однако эта физиологическая архитектура мо-
жет рассматриваться только как общая закономерность, прису-
щая жизненным явлениям различной степени сложности, но ни
в коей мере не делающая их тождественными по их качествен-
ному характеру, по их содержанию. Подобные общие закономер-
ности уже известны из области кибернетики. Именно эта
закономерность — закономерность формирования аппарата оцен-
ки результатов действия до совершения самого действия и получе-
ния его результатов — выступает общей для различных классов
явлений у живых организмов, в авторегулпрующихся машинах и
в общественных взаимоотношениях. Не удивительно поэтому, что
на основе именно этой закономерности и по этой универсальной
архитектуре произошло и развитие эмоциональных состояний че-
ловека — от первичных примитивных ощущений до высших форм
удовлетворения в специфически человеческой деятельности.
ЛИТЕРАТУРА
Ленин В. И. Поля. собр. соч., т. 25.
Анохин П. К. Узловые вопросы в изучении условного рефлекса.— В кн.'
Проблемы высшей нервной деятельности. М., 1949.
Вильям Д. Научные основы психологии. Пер. с англ. СПб., 1902.
Кеннон В. Физиология эмоций. Пер. с англ. Л., 1927 (библиография).
Cobb S. Emotions and clinical medicine. N.Y., 1950.
Gellhorn E.t Loofbourrow G. N, Emotions and emotional disorders. N. Y.— L.,
1963.
ПСИХИЧЕСКАЯ ФОРМА
ОТРАЖЕНИЯ ДЕЙСТВИТЕЛЬНОСТИ *
Общие предпосылка
Едва лп кто решится отрицать ту огромную роль, которую
теория отражения сыграла в прогрессе материалистического по-
знания природы, в понимании этой последней как объективно су-
ществующего движения материи, адекватно отражаемого в мозгу
и сознании человека. Огромная литература, которая посвящена
этой проблеме,— прямое доказательство философской силы этой
теории, позволившей классикам марксизма убедительно и успеш-
но полемизировать с представителями всех оттопков философско
го и научного агностицизма.
На протяжении многих лет вплоть до наших дней марксист-
ская философская наука изучает и вскрывает все новые и но-
вые стороны процесса отражения, показывая несостоятельность
таких идеалистических концепций о внешнем материальном мире,
которые ведут к агностицизму, солипсизму и к многим другим
философским течениям, гносеологически пе допускающим самую
возможность существования объективного внешнего мира и по-
знания его с помощью тончайших мыслительных операций чело-
веческого мозга.
Ясно, что реальное существование объективного внешнего
мира и возможности познания его свойств средствами человече-
ского мозга представляют собой центральное гносеологическое по-
ложение диалектического материализма. Это положение ведет нас
к ряду дальнейших выводов, которые имеют огромное философ-
ско-теоретическое значение, прежде всего — к правильному пони-
манию как биосоциальной сущности человека, так и его практи-
ческой целенаправленной деятельности.
Такое широкое внимание к проблеме отражения служит зало-
гом того, что она будет обогащаться новыми материалами и ар-
гументами, что сделает ее конкретным методологическим пособи-
ем исследователей конкретных наук, особенно в области биологи-
ческих знаний.
Нельзя не отметить один существенный недостаток в этих об-
щих усилиях: они направлены почти исключительно на гносеоло-
гическую сторону проблемы и, вместе с тем, нет сколько-нибудь
♦ Опубликовано в книге «Ленинская теория отражения и современность»
(София, 1969).
Психическая форма отражения действительности
337
значительного количества работ, ставящих акцент на том, какими
материальными средствами организм, и особенно его мозг, досто-
верно отражают в мозгу и сознании внешний объективный мир.
До какой степени и в чем именно проявляется это достовер-
ное отражение объективного мира? Как индивидуализировано это
отражение в отдельном организме и, наконец, в чем же смысл
отображаемого объективного мира в конкретном поведении каж-
дого отдельного животного? Иначе говоря, современная разработ-
ка теории отражения должна максимально полно вскрыть кон-
кретные механизмы отражательного процесса, его формы и его
роль в жизни организма. II на этом пути теория отражения долж-
на использовать все современные достижения самой первной дея-
тельности, работу целого мозга и т. д.
Я глубоко убежден в том, что для подавляющего большинства
исследователей, да и для многих философов, вопрос о гносео-
логической роли отражения объективного внешнего мира в созна-
нии в связи с блестящими достижениями нейрофизиологии, ней-
рохимии и молекулярной биологии уже перестал быть спорным.
Наоборот, ощущается настоятельная необходимость разработ-
ки теории отражения в области самих средств отражательной
деятельности. Именно здесь надо искать наиболее значительные
аргументы и иллюстрации, подтверждающие всеобщее значение
теории отражения и в философии, и в исследовании, и в прак-
тической деятельности.
Вся сила марксистско-ленинского анализа действительности
должна быть направлена на использование всего того, что дает
нам для разработки теории отражения современная наука. Под-
линная наука не может пе укреплять основной тезис диалектико-
материалистической гносеологии о реальном существовании объ-
ективного мира.
Психическое отражение и информация
Па пути глубокого научного исследования процесса отраже-
ния,
свойственного высшим животным
мы встречаемся с весьма
важной проблемой и философии и пауки о мозге. Я имею в виду
формирование в процессе эволюции организмов идеальной формы
отражения — сознания.
Сознание и субъективная жизнь человека, несомненно, явля-
ются закономерным продуктом эволюционного прогресса организ-
мов и специально эволюции мозга. В этом смысле основная фор-
мула гносеологии диалектического материализма «материя пер-
вична — сознание вторично» остается в силе. Но при этом
неизбежно возникает ряд вопросов: как осуществляется этот ре-
шающий переход от материального к идеальному? Какие естест-
венные процессы живой протоплазмы способствуют этому скачку
338
Психическая форма отражения действительности
н
на высшем пункте эволюции? Имеет ли материальная основа
психики — нервные процессы — какие-либо определенные и зако-
номерные причинно-следственные отношения с идеальным, т. е.
с отражением внешнего объективного мира в сознании человека?
Отражательный процесс развертывается таким образом, что
внешний объект через непрерывный ряд физических и физиоло-
гических процессов как бы ассимилируется организмом, т. е. отра-
жается сначала в его структурах, а потом и в сознании. Такой
порядок развития процессов отражения приводит к естественному
выводу, что по сути дела этот процесс от этапа к этапу фор-
мируется в соответствии с теорией передачи информации. Такой
подход к проблеме отражения уже был применен не раз (Т. Пав-
лов, 1965; Украинцев, 1960; Медведев, 1963; и др.). Однако я
считаю полезным еще раз возвратиться к нему с приложением
всех тех последних нейрофизиологических данных, которые осве-
щают в этом аспекте распространение возбуждений, вызванных
внешними объектами, по нервной системе.
Приступая к разбору отражательного процесса в аспекте тео-
рии информации, нам важно установить, как оценивали сами
классики теории отражения точность соответствия отражаемого
объекта и того образа объекта, который создается в конце концов
в сознании человека. Конечно, в этом отношении нам прежде
всего интересны высказывания В. II. Ленина.
К сожалению, привлечение теории информации к разрешению
или даже к иллюстрации теории отражения всегда было недо-
статочно глубоким. Между тем именно теория информации в на-
стоящее время дает пам наиболее реальпую возможность рас-
шифровки нейрофизиологических механизмов отражения и осо-
бенно осуществления перехода материальных нервных процессов
в субъективный образ, в сознание.
Роль теории информации состоит не только в том, что она
еще раз подтверждает реальное существование действительности.
Главная роль теории информации состоит в том, что опа полно-
стью исключает необходимость говорить о «копиях», «символах»,
«иероглифах» и т. д. Благодаря ей весь процесс отражения мо-
жет быть представлен в достаточно четких, научно обоснованных
понятиях и в хорошо поддающихся изучению механизмах.
Передача информации в живых и технических объектах обыч-
но происходит с исключительно большим количеством специфи-
ческих звеньев, однако она подчиняется одному важному закону:
между начальным и конечным звеном этой передачи должна быть
точная и адекватная информационная эквивалентность.
Так, например, когда мы просим ребенка изобразить звук
«а», соответствующий зрительному изображению буквы «а»,
информационный процесс охватывает огромное количество звень-
ев. Звуковые волны, продуцируемые моим аппаратом речи, пе-
Психическая форма отражения действительности
339
рекодированные барабанной перепонкой в механические колеба-
ния, попадают на кортиев орган слушателя и производят здесь
избирательное возбуждение отдельных рецепторных клеток этого
органа. Избирательность здесь весьма тонкая, она может касать-
ся частоты волнообразных колебаний воздуха, продуцируемых
мною, она может отражать интенсивность, тембр и другие пара-
метры звуковой речи.
Все эти параметры с информационной точки зрения могут
служить информационным эквивалентом воздушных колебаний,
соответствующих звуку «а». Так же развивается и следующий
этап — движение нервных импульсаций от кортиева органа по
направлению к мозгу. Объем этих импульсаций, их частота и
рисунок разрядов в нерве оказываются информационно эквива-
лентными звуку «а», хотя, конечно, в слуховом нерве нет ни
единой «звуковой волны».
Нервные импульсации возникают одновременно в тысячах во-
локон слухового нерва, но можно с достоверностью утверждать,
что ни одно слуховое волокно с точки зрения общей конфигура-
ции нервных разрядов не похоже на другое. Таким образом,
в целом этот множественный поток нервных импульсаций, бегу-
щий к мозгу по тысячам волокон слухового нерва, является
истинным информационным эквивалентом звука «а», поступивше-
го на барабанную перепонку.
Можно ли из этих потоков импульсов, которые по своей фи-
зической природе ничего общего не имеют с воздушными звуко-
выми волнами, извлечь каким-нибудь способом тот первичный
натуральный «объект», который определил весь путь формиро-
вания отражательной деятельности? Оказалось, что можно. Это
было хорошо показало в опытах Уивера и Брея. Если на слухо-
вой нерв, проводящий потоки нервных импульсаций, в которых
эквивалентно закодирован звук «а», наложить электроды и если
получающиеся при этом биотоки нервных волокон преобразовать
соответствующим образом, то на репродукторе можно услы-
шать именно звук «а». Замечательный факт! Он убеждает пас
в том, что по слуховому перву в самом деле бежит звук «а», но
лишь только в другом коде. Центральная же нервная система
делает все то, чтобы с исключительной бережливостью передать
звук «а» в специфическом информационном эквиваленте на каж-
дом звене всей длинной цепи перекодирований и достаточно точ-
но расшифровать его на конечном этапе.
Исключительно интересен в этом смысле один из эксперимен-
тов, проделанных в нашей лаборатории. Речь идет о передаче
информации в пределах дыхательной системы с сохранением ис-
ходного эксперимента на каждом этапе передачи.
Учтя все сигнализации от различных частей организма о по-
требности в кислороде, дыхательный центр дает периферическим
340
Психическая форма отражения действительности
рабочим аппаратам приказ — «взять в легкие 500 кубиков возду-
ха». Эта команда, как известно, дается по диафрагмальному нер-
ву к дыхательным мышцам.
Следовательно, команда о взятии 500 кубиков воздуха, содер-
жащаяся в потоках нервных импульсаций, теперь уже переко-
дирована в силу мышечных сокращений. Однако, несмотря па
отличие самого явления сокращения от нервных импульсаций,
в этом сокращении имеется тот же информационный эквива-
лент — «взять 500 кубиков воздуха». Расширение легкого и
растяжение альвеол — следующий этап перекодирования пер-
вичной команды центра. Однако, несмотря на различие природы
этих явлений, взятых в отдельности, они принципиально несут
тот же информационный эквивалент — «взять 500 кубиков воз-
духа».
Но вот теперь по чувствительным нервам (блуждающий нерв),
идущим от альвеол легкого, в мозг бежит обратный поток нерв-
ных импульсаций, которые в сумме несут информацию «взять
500 кубиков воздуха».
Таким образом, на всем пути передачи информации в орга-
низме, несмотря на принципиально различные средства переко-
дирования, информационный эквивалент сохраняется.
В нашей лаборатории был проделан следующий эксперимент:
было решено сопоставить и заменить информацию о взятии
500 кубиков воздуха с командой для взятия этого воздуха. Это
было осуществлено с помощью посылки командных нервных им-
пульсаций по каналу извещения о взятии воздуха в количестве
500 кубиков. Технически это достигалось тем, что командные
нервные импульсации, минуя дыхательные мышцы, легкие, аль-
веолы, преобразуются через стимулятор, переходят прямо на
блуждающий нерв, который должен был бы информировать уже
о взятии воздуха. Иначе говоря, мы хотели решить задачу: могут
ли нервные импульсации, в которых закодирована команда о
взятии 500 кубиков воздуха, подменить информацию об уже взя-
тых 500 кубиках воздуха.
Такое сопоставление показало, что центральная нервная си-
стема не принимает по каналу обратной афферентаций «коман-
ду» вместо «результата». Происходит полпая дезинтеграция ды-
хания, и животное может погибнуть.
Однако для того, чтобы понять инструмент информационного
анализа, надо прежде всего попять, какими средствами теория
информации определяет законность и научность выводов и при-
менения ее аналитического аппарата.
Кибернетика стремится ввести в обсуждение общий язык, тер-
мины и понятия, которые могут быть подвергнуты измеритель-
ной и математической проверке. Я согласен с теми авторами-фи-
лософами, которые утверждают, что нельзя в понятиях и теориях
Психическая форма отражения действительности
341
математики объяснить субъективно-психический образ объектив-
но существующего внешнего мира. Но вместе с тем нельзя п
надеяться на то, что мы сможем раскрыть материалистическую
природу психического феномена без того, чтобы на подступах к
нему, т. е. в самом процессе отражательной деятельности, мы не
изучили и не поняли материального существа передачи инфор-
мации.
Мы не должны забывать того, что, когда мы измеряем куба-
туру комнаты, то наши измерительные приборы одновременно
скользят и по объекту, и по нашему «образу комнаты».
Так, например, в теории информации существует представле-
ние о точности передачи информации о каком-либо объекте не-
зависимо от перекодирования. Я назвал бы эти этапы передачи
информации информационным эквивалентом объекта. Это значит,
что процесс информации, в каком бы звене передачи мы его ни
уловили, принципиально содержит в себе все то, что составляет
наиболее характерные черты исходного объекта, однако эти приз-
наки могут быть представлены в разных кодах. Иначе говоря,
исходный объект мог бы быть воспроизведен во всех своих чер-
тах, если бы мы нашли способ декодировать поток информации
именно в данном звене ее передачи.
Здесь возникает необходимость разобрать некоторые базис-
ные понятия теории информации, поскольку все эти понятия
полностью применимы и к процессам отражательной деятельно-
сти организмов до формирования субъективных образов включи-
тельно.
Прежде всего мы должны указать па значение понятия пара-
метра и параметризации, которые лежат в основе оценки всяко-
го объекта или явления. Параметр — это какое-либо элементар-
ное свойство или признак данного объекта, который в сумме с
другими параметрами составляет исчерпывающую характеристи-
ку данного объекта. Параметры должпы быть измеримы или по
крайней мере выражены в каких-то общепринятых единицах.
Так, например, стоящая на столе чайная чашка имеет цвет,
форму, сюжет росписи, ручку и т. д. Все эти параметры воздей-
ствуют па зрительный аппарат. Но эта же чашка имеет пара-
метры веса, температуры, форму, оцениваемую по мышечным
рецепторам, и т. д. К этим же параметрам чашки можно приба-
вить ее емкость. Все эти параметры являются индивидуальной
характеристикой этой чашки и при наличии достаточного коли-
чества способствуют вопроизведеншо в нашем мозгу субъективно-
го образа этой чашки. Конечно, не все параметры являются оди-
наково важными по отношению к основной
ункции чашки,
как,
Н
например, цвет, однако сравнительная оценка этих параметров
нервной системой лежит уже в плоскости других форм деятель-
ности.
342
Психическая форма отражения действительности
Значимость отдельных параметров чашки может в некоторой
степени меняться в зависимости от данной ситуации. Если мы
испытываем сильпую жажду, то ведущим ее параметром будет
емкость, если же мы покупаем чашку для преподнесения ее в
качестве подарка, то ее форма, антикварность и другие парамет-
ры приобретают главнейшее значение.
Художественное течение под именем «импрессионизм» хорошо
показало, как нарочитым подчеркиванием каких-лпбо весьма
ограниченных, но решающих параметров такого сложного объек-
та, как лицо человека, можно сформировать полный и, главное,
желаемый образ человека.
Итак, для оценки отдельных этапов отражательного процесса
объект должен быть параметризован, ибо в дальнейших пре-
образованиях в пределах центральной нервной системы отража-
тельный процесс будет находиться в прямой зависимости от этих
параметров.
Первой стадией, где происходит перекодирование параметров,
например, зрительного объекта, является сетчатка глаза.
В результате фотохимического процесса здесь возникают нерв-
ные импульсы, которые в самых разнообразных комбинациях и
аранжировках во времени бегут по зрительному нерву в цен-
тральную нервную систему.
В современной нейрофизиологии хорошо известно, что в этой
стадии формирования отражательного процесса полностью отсут-
ствуют те волновые колебания, которые соответствовали зритель-
ным параметрам наблюдаемого нами объекта. И тем не менее
мы имеем в этом интеграле импульсаций, развивающихся с раз-
личной скоростью, амплитудой и частотой, эквивалентный образ
наблюдаемого нами объекта, точнее говоря, эквивалент его зри^
тельных параметров.
Однако самое главное здесь заключается в том, что этот поток
импульсов несет ту же информацию об объекте, какую несли до
этого и различные волновые колебания, вызванные различными
зрительными параметрами самого объекта. Преобразованные бла-
годаря фотохимическому процессу в нервные импульсации, эти
волновые колебания были лишь перекодированы без потери точ-
ности информации об объекте.
Мы указали лишь только на первый этап в формировании
отражательного процесса. На последующих путях распростране-
ния зрительной информации об объекте по центральной нервной
системе она будет обрабатываться и перекодироваться много раз
и с различным результатом. Некоторые из нервных импульсаций
оторвутся от общего нервного потока и пойдут через филогенети-
. чески самые древние зрительные центры (нанример, через перед-
ние холмы четверохолмия и через ретикулярную формацию
мозга).
Психическая форма отражения действительности
343
Другие потоки нервных импульсаций пойдут по более моло-
дым зрительным центрам подкоркового аппарата (напри-
мер, наружные коленчатые тела). Естественно, что ни в одном
из этих центров, взятом в отдельности, мы уже не имеем того кода
зрительного объекта, который мы имели па уровне оптического
нерва — все импульсации нерва получили разнообразную в со-
ответствии с историей развития самого мозга обработку. И тем
пе менее совокупность всех нервных импульсаций во всех под-
корковых центрах, вовлеченных в этот зрительный поток, пред-
ставляет собой более высокую степень организации отражатель-
ного процесса построения субъективного зрительного образа ви-
димого объекта.
Как хорошо известно теперь в нейрофизиологии, все эти раз-
нообразные компоненты зрительного потока, в целом сохраняю-
щие информационную точность о всех реальных параметрах или
свойствах объекта, поступают в кору головного мозга совершенно
различными нервными путями. Здесь, в коре мозга, эти потоки
информации вновь объединяются и подвергаются значительной
обработке перед решающим моментом построения субъективного
образа наблюдаемого объекта. На этом высшем этапе первичная
зрительная информация еще много раз перекодируется по раз-
личным клеткам и по различным синаптическим организациям
коры мозга.
Конечно, здесь и речи быть не может о том, чтобы увидеть
первичные колебания — световые волны, которые передали сово-
купность зрительных параметров наблюдаемого объекта глазу,
сетчатке, ее фотохимическим элементам. II тем не менее все
свойства реального объекта точнейшим образом и в совершенно
измеримых нервных процессах были переданы в эту высшую ин-
станцию без потери информационной эквивалентности между на-
блюдаемым объектом и тем конечным образом, который склады-
вается на конечной стадии зрительного восприятия.
Приходится удивляться тому, с какой старательностью эво-
люционный процесс «охранял» точность передачи всех детальных
параметров наблюдаемого
мозга. Нет опасности в том,
объекта в высшие
инстанции
что первичные свойства предмета
«исказятся»,
например
па
уровне
отохимических
процессов
м
Y
А
сетчатки или в процессе распространения нервных импульсаций
по отдельным волокнам зрительного нерва. Их неприкосновен-
ность гарантирована самой структурой волокон, изолированных
от различных помех. Но совершенно по-другому распространение
зрительной информации происходит на высших инстанциях —
при обработке зрительной информации на уровне подкорковых
центров и самой коры мозга. Здесь в возбуждение втягиваются
миллиарды нервных клеток, объединенных в так называемую
нервную сеть. При этом каждый этап таит в себе опасность иска-
344
Психическая форма отражения действительности
женин первичных и, следовательно, самых верных информаций
о реальных физических параметрах объекта.
По эволюция «обошла» эту трудность самыми разнообразными
и всегда остроумными механизмами. Уже на первом этапе вступ-
ления нервных импульсов в центральную нервную систему вклю-
чаются аппараты так называемого «побочного торможения». Эти
механизмы с «хладнокровной жестокостью» отметают все побочные
«шумы», все возможные вмешательства, рождаемые «нервной
сетью» и пе относящиеся к первичному воздействию натурального
объекта. Таким образом нервная система достигает поразительной
точности информации мозга о первоначальных воздействиях
внешних объектов. Насколько важно дойти до высших центров
всему своеобразию именно первичных, натуральных, параметров
объекта, видно из последних данных нейрофизиологии. У многих
животных уже периферические рецепторные процессы переработ-
ки внешней информации об объекте насыщены биологическим
смыслом, вырабатываемым естественным отбором на протяжении
многих миллионов лет эволюции.
Так, например, было доказано, что сетчатка глаза лягушки
имеет специальные ганглиозные элементы, высокочувствительные
к весьма тонким нюансам выпуклостей, т. е. как раз к тому, что
составляет специфическую черту в «образе» насекомых, являю-
щихся добычей для лягушек. У млекопитающих животных, на-
пример, движущийся предмет служит раздражителем не только
в пунктах своего реального движения, но имеются специальные
элементы сетчатки, экстраполирующие будущие возможные пере-
движения предмета.
Приведенные примеры свидетельствуют с определенной оче-
видностью, что набор параметров покоящегося и движущегося
объекта чрезвычайно разнообразен и потому так важно донести
до окончательной обработки первоначальную, непосредственно
полученную от объекта информацию. Именно чрезвычайно важ-
ные в биологическом отношении параметры первичного воздей-
ствия из внешнего мира и должны быть точно отражены на са-
мых высших этапах обработки информации,
что, несомненно,
повышает вероятность наиболее точного отражения объекта и в
психике животных, и в сознании человека.
Можно привести многочисленные примеры того, как перифе-
рическая натуральная информация от внешнего объекта на всем
дальнейшем пути формирования отражательной деятельности ох-
раняется с максимальной бережливостью от нарушения чистоты
и точности передачи отражаемого объекта. Рассмотрим в качестве
примера летучих мышей, которые при помощи ультразвуковой
локации с поразительной точностью засекают съедобную для них
ночную бабочку.
Как известно, суть этой «погони» за жертвой состоит в том,
Психическая форма отражения действительности
345
что летучая мышь с помощью специального органа производит
специальные ультразвуковые сигналы, которые распространяются
в различные стороны. Там, где эти ультразвуковые волны не
встречают препятствий, они рассеиваются на огромные простран-
ства. Там же, где они встречают твердый предмет, например
бабочку, куст, дерево или что-нибудь другое, они отражаются и
направляются обратно, т. е. к самой летучей мыши.
Летучая мышь обладает довольно тонким органом слуха, ко-
торый получает это обратное ультразвуковое отражение и «чи-
тает» по его колебаниям ту информацию, которую оно приносит.
Стоит представить себе на минуту, какое огромное разнообразие
ответных потоков ультразвуковых волн имеет мышь при погопе
за добычей, чтобы попять тот «образ внешнего мира», на осно-
вании которого опа строит свое поведение. Она имеет в подлин-
ном смысле слова картину внешнего мира, составленную из пу-
стот, откуда мышь пе получила отражения, и из разнообразных
модуляций исходного ультразвукового сигнала в зависимости от
того предмета, от которого отразился этот сигнал.
Более того, эта способность «читать» обратную, т. е. отражен-
ную ультразвуковую посылку по частотному и амплитудному
коду столь совершенна, что мышь может вполне различать по-
роду бабочек (!), съедобную или несъедобную для нее.
Однако и жертва пе остается невооруженной. Опа также
«использует» информационные закономерности и довольно успеш-
но ими защищается. В процессе эволюции и на основе естест-
венного отбора мотыльки вырабатывают специальные рецепторы,
способные воспринимать именно ультразвуковые сигналы. Получив
сигнал, предназначенный летучей мышью для поимки жертвы,
мотылек немедленно или падает, или зигзагообразно начинает
менять уровни своего полета, спасаясь от нападения.
Некоторые же мотыльки поступают еще «хитрее». Они поль-
зуются тем, что различные породы мышей издают различные по
спектру ультразвуковые сигналы. Этого достаточно, чтобы моты-
лек с поразительной точностью различал те породы мышей, для
которых он как раз и не является съедобным. Естественно, что в
этом случае он не спешит с защитой.
Однако имеются породы мотыльков, которые решают вопрос
своей защиты опять-таки па основе естественного отбора еще
более радикально: у них вырабатывается такой густой и пуши-
стый покров, который полностью погашает все ультразвуковые
сигналы, в результате чего летучая мышь остается без отражае-
мого ультразвука, чем имитируется для нее полное отсутствие
жертвы. ।
Мы знаем сейчас, что летучая мышь посылает различные
сигналы, а для улавливания отраженных сигналов от жертвы
имеет специальные аппараты.
346
Психическая форма отражения действительности
Весь этот материал показывает огромную роль информацион-
пых взаимоотношений в животном мире и вместе с тем убеждает
нас, что отражение действительности в мозгу животных всегда
соответствует реальной объективной действительности, что под-
тверждается практикой точного приспособительного поведения.
Однако, если бы мы попытались попять, в каком виде «образ
мыши» отражается в нервной системе мотылька, какие «парамет-
ры мыши» кодируются в его нервной системе, то мы увидели бы,
что этот «образ» может быть попят только па основе учета ин-
формационных кодов. В самом деле, вероятнее всего, что мотыль-
ки никогда на получали каких-либо других стимуляций от мыши,
кроме стимулов от ее страшного оружия — ультразвука. Те же
мотыльки, которым «посчастливилось» увидеть или тактильно
ощутить мышь, несомненно, были съедены и, следовательно, уже
давно были элиминированы естественным отбором.
Итак, «образ мыши» представлен в нервной системе мотылька
почти исключительно комбинацией частоты и специфичностью
испускаемых ею ультразвуковых колебаний. Однако эта весьма
редуцированная сигнализация приводит в действие только такие
комплексы нервных возбуждений в нервной системе, которые
совершенно точно отражают объективные предметы и их глубо-
кое биологическое значение.
Итак, теория информации показывает, что любой отражаемый
в нервной системе внешний объект через ряд перекодирований
первичного сигнала на конечном этапе совершенно точно отра-
жает главнейшие, биологически важные параметры отражаемого
объекта. Эти параметры во всех своих комбинациях могут дать
и пе обязательно «зрительный образ» реального объекта, как это
мы вообще принимаем для первичных зрительных возбуждений.
Как мы видели, у мотылька этот образ отражается почти це-
ликом в сфере всевозможных модуляций ультразвуковых колеба-
ний. Однако этот «образ», будучи выражен в звуковых кодах,
всегда отражает тончайшие нюансы физических параметров ре-
ального мира. Эти звуковые модуляции с большой точностью пере-
кодированы в механические комбинации уже на стадии восприни-
мающего слухового аппарата мыши. На дальнейших этапах
отражательного процесса в нервной системе эти потоки ультра-
звуковой информации будут перекодированы еще несколько раз.
Однако конечный образ реального объекта сохраняет свой
информационный эквивалент, в точности соответствующий исход-
ному воздействию реального объекта.
Возьмем для примера крота. Для него «образ» камня, кото-
рый он встречает в своем продвижении под землей, представлен
только тактильно-обонятельными и, может быть, вкусовыми па-
раметрами. Однако окончательное отражение этой своеобразной
для крота действительности в его мозгу всегда с точностью соот-
Психическая форма отражения действительности
347
ветствует важнейшим параметрам внешнего и важного для него
объекта. Надо помнить лишь, что значимость этих параметров
находится в прямой зависимости от своеобразных экологических
факторов данного вида животных..
Убедительным примером того, что образ внешнего мира может
отразиться в мозгу и в сознании набором самых разнообразных
параметров действительного мира, является поведение слепо-глу-
хонемых людей. Замечательный внутренний мир и отражательная
деятельность этих людей представляет собой весьма яркое дока-
зательство двух положений: а) внешний мир может быть отра-
жен в мозгу человека после целого ряда перекодирований вполне
реальными параметрами, точно характеризующими этот внешний
мир, причем каждый из этих параметров может приобрести веду-
щее значение, если почему-либо устранены другие; б) наличие
точного отражения параметров внешнего мира даже при исклю-
чении важнейших из них доказывается тем, что поведение чело-
века находится в полном соответствии с теми физическими пара-
метрами внешнего мира, которые в данной ситуации оказывают-
ся жизненно важными.
Итак, заключая этот раздел нашей работы, мы можем сказать,
что современные успехи иейроморфологии, нейрофизиологии,
кибернетики и особенно теории информации не оставляют сомне-
ния в том, что окружающий нас мир является реальным миром.
Вся же эволюция живого, и особенно эволюция мозга, была на-
правлена именно на то, чтобы на всех уровнях дешифровки
первичной информации, т. е. «впечатления от самой действитель-
ности», этот реальный мнр получил бы наиболее точное отраже-
ние.
Для решения вопроса о том, в каком именно виде окончатель-
но синтезируется вся информация, обработанная на многочислен-
ных узлах нервной системы, мы должны помнить, что кора голов-
ного мозга и сознание производят окончательный синтез всей
этой обработки и формируют образ отражения, не упустив из
него более или менее важные параметры объекта внешнего мира.
Здесь следует отметить, что решающим условием охвата пара-
метров внешнего мира служит пропускная способность наших ор-
ганов чувств, которые имеют вполне определенный диапазон чув-
ствительности.
Так, например, мы не воспринимаем той части спектра, ко-
торая соответствует инфракрасному излучению. Мы не можем
видеть в темноте. Однако научные исследования различных жи-
вых объектов убеждают нас в том, что этого сделать мы не мо-
жем лишь потому, что сетчатка нашего глаза пе способна начать
отохимический процесс под влиянием именно этой волны ин-
ракрасного
излучения.
Однако мы знаем, что черный таракан
имеет такой глаз, в котором находится весьма чувствительная
348
Психическая форма отражения действительности
субстанция по отношению к инфракрасному излучению, что дает
ему высокую чувствительность к инфракрасной картине мира,
поэтому он хорошо видит в темноте.
Итак, внешний реальный мир имеет гораздо больше пара-
метров, из которых можно было бы составить (отразить) реаль-
ный образ внешнего мира, но экологические особенности каждого
животного и его индивидуальная эволюция способствовали раз-
витию тех или других датчиков, воспринимающих те или другие
параметры. Этим набором параметров и определяются в конечном
счете отраженные образы действительности у различных живот-
ных: ночной мотылек «видит» летучую мышь с помощью нерв-
ных процессов, вызванных своеобразным ультразвуковым коле-
банием, образ от общей обстановки у таракана в темноте опреде-
ляется различной температурой тел, и, следовательно, различным
инфракрасным излучением.
Что касается человека, то отображаемый им мир является
гораздо более богатым с точки зрения количества физических па-
раметров внешнего мира, и это обеспечивает создание более
полноценного и детализированного образа внешнего мира. Имен-
но этот процесс формирования последнего синтетического этапа
и составляет предмет особого внимания при гносеологическом под-
ходе к теории отражения. Однако, зная теперь, как от этапа к
этапу преобразуется (перекодируется) первичная натуральная
информация и с какой настойчивостью эволюционный процесс
удерживал па этих этапах все важные в приспособительном смыс-
ле параметры внешних объектов, мы пе можем хоть в какой-
нибудь степени сомневаться в реальности таковых параметров у
внешних объектов и, следовательно, в точности отражения их в
информационном эквиваленте на различных этапах распростра-
нения возбуждений по нервной системе.
Пусть отдаленным, но с информационной точки зрения весьма
аналогичным процессу отражения внешнего мира в организмах
является механизм телевизионной передачи.
Для тех, кто часто бывал в телевизионной студии, является
вполне обычным факт, весьма интересный для разбираемой нами
проблемы. Допустим, что мы видим и слышим в этой студии
непосредственно поющую певицу. Мы знаем, что и зрительный
образ певицы, и звук ее голоса воспринимаются соответствующи-
ми датчиками (кинескоп и микрофон) и поступают в цепь раз-
личных преобразователей. На каком-то этапе преобразования воз-
никают электромагнитные колебания, которые широко распрост-
раняются в нашей атмосфере, а может быть даже и в космосе.
Строго говоря, волнообразные колебания, полученные от вида
певицы и от ее голоса, с рассмотренной выше точки зрения
представляют собой информационный эквивалент реального обра-
за певицы.
Психическая форма отражения действительности
349
Несмотря иа несомненную точность перекодирования внешних
параметров певицы, мы не видим ее ни в атмосфере неба, ни у
себя в комнате. Нужны еще дальнейшие преобразования этих
электромагнитных колебаний, чтобы образ певицы целиком вос-
становился на экране телевизоров. В ряде случаев уже в самой
студии вы можете одновременно видеть и реальную певицу, и тут
же, недалеко от нее, телевизионное изображение этой певицы
(монитор). Однако мы знаем, что отраженный на экране теле-
визора образ мог быть получен только потому, что физические
параметры этого образа, несмотря на многоступенчатые преобра-
зования, смогли быть вновь синтезированы физическими средства-
ми в экранный образ.
Наш мозг и сознание представляют собой как бы специаль-
ный аппарат, который может сравнить реальный образ внешнего
мира (ноющая певица) и его отражение через многие этапы
перекодирования исходных параметров реального объекта.
Представим себе на минуту, что паше сознание, приобретен-
ное нами па протяжении сотен миллионов лет эволюции живого,
как раз и есть этот последний синтезирующий фактор, собираю-
щий всю ту информацию об объекте, которая по путям передачи
претерпела и дисперсию, и редукцию, и синтез. И теперь на
последней инстанции все воспринятые параметры вновь воссоз-
даются в том образе, который мы видим в своем сознании. Тог-
да сознание было бы лишь совершенным продуктом эволюции
нашего мозга, который под стать последнему каскаду телевизора
вновь восстанавливает исходный объективный мир во всей его
красочной полноте.
Мы видим, что при таком подходе в материалистической кон-
цепции отражения нет места допущению чего-то нематериаль-
ного и несуществующего. II только лишь конкретные механизмы
рождения субъективного в сознании не поддаются пока аналити-
ческому описанию, однако их точная информационная связь с
исходными параметрами объективного мира не может быть под-
вергнута сомнению.
Сущность психической формы
отражения действительности
Как можно было видеть из предыдущего изложения, теория
информации, приложенная к исследованию отражательного про-
цесса, не оставляет сомнений в том, что главнейшие параметры
внешнего мира, значимые для данного животного или человека,
несмотря на многочисленные перекодирования, практически соз-
дают точное отражение действительности, хотя и в различных
кодах. Для нейрофизиолога не может быть сомнения в том, что
эта смена кода от звена к звену служит лишь специальным
350
Психическая форма отражения действительности
инструментом нервной системы, сохраняющим в точности инфор-
мационный эквивалент внешнего объекта от периферических ор-
ганов до момента появления образа в сознании. И даже более
того, само сознание представляет собой, несомненно, наиболее
совершенную форму кодирования параметров внешнего мира,
т. е. инструмент создания образа внешнего мира.
Однако сам момент превращения последних кодов внешнего
мира в нервных процессах, неизбежно переходящий в сознание,
в «субъективный образ объективного мира», остается еще мало-
известным в онтологическом аспекте, хотя его гносеологическая
сущность и ясна. Формирование специфических черт сознания,
фокусирующего весь организм иа определенных субъективных
состояниях, по-прежнему остается «крепким орешком». У нас пет
сомнения в том, что сознание является вторичным продуктом
исторического развития приспособительных способностей живот-
ных и человека. А если это так, т. е. что историческому разви-
тию сознания предшествовало прогрессивное развитие нервной
организации, то, значит, к развитию сознания во всем полном
объеме приложимы все дарвинские законы развития, включая и
естественный отбор. Именно под углом зрения исторического раз-
вития сознание обычно не обсуждается. Л между тем при оценке
отражательного процесса как феномена естественной природы
прежде всего и необходимо вскрыть биологические корни
сознания...
Мы не должны забывать также, что богатейшие исследования
в области условных рефлексов и общей нейрофизиологии делают-
ся оправданными только в том случае, если они хоть в малой
степени приближают нас к пониманию природы человеческого
поведения, психической деятельности человека. С этой точки зре-
ния вряд ли можно надеяться, что кто-либо в одиночку может
разрешить всю проблему психического со всеми его биологиче-
скими корнями и высшим развитием у человека. Мы должны
быть довольны скромными успехами каждого из нас, лишь бы
только все многочисленные вариации мысли, оттенки формули-
ровок и объяснений материальной основы психического при-
внесли хоть немножко нового в эту огромную проблему. Джеймс
был прав, когда говорил: «Мы все знаем, что такое сознание,
пока не пытаемся формулировать его...»
Диалектический материализм указывает точное гносеологи-
ческое определение сознания, считая его функцией высокоорга-
низованной материи — мозга. Наряду с этим мы не имеем к этой
формулировке соответствующей физиологической расшифровки,
основанной на работах по высшей нервной деятельности и по фи-
зиологии нервной системы.
Именно эти трудности, как мне кажется,
и создали целый
ряд противоречивых, а часто и конъюнктурных формулировок
Психическая форма отражения действительности
351
психического. В этом смысле мы должны дружными усилиями
найти те наиболее важные «параметры» психического, которые
могли бы пас приблизить к общеприемлемой формулировке пси-
хического.
Конечно, нельзя принять весьма упрощенных формулировок
психического, когда хотят просто отождествить условный рефлекс
и психическое. На этой почве вырастает представление, что ус-
ловный рефлекс — это именно и есть психический акт, в то время
как безусловный рефлекс является «беспсихическим» феноменом.
Едва ли что-либо большее может нам дать, когда психическое
отождествляется с принципом сигнальности и появляется тен-
денция отождествить психическое и сигнальное, хотя у высоко-
организованных животных они объединены в пределах одной и
той же реакции.
Мне кажется, эти попытки не учитывают одного важного
обстоятельства, что условный рефлекс и психическое — это раз-
личные параметры поведения человека, выросшие исторически
из различных потребностей организма и с удовлетворением со-
вершенно различных затруднений организма в приспособлении
к внешнему миру.
В самом деле, мы знаем, что сигнальность, т. е. деятельность
организма, предвосхищающая будущее, сформировалась в резуль-
тате непрерывного отражения абсолютной временной структуры
внешнего мира и в предвосхищении именно этого потока непре-
рывных явлений. Это — приспособление организма именно к дан-
ному своеобразному параметру неорганического мира. Мы знаем,
также, что принципиально уже примитивная протоплазма и осо-
бенно примитивные организмы прошли этот этап опережающего
отражения явлений внешнего мира на очень ранних ступенях
развития.
Едва ли кто-либо решится признать наличие психического как
сумму внутренних переживаний сложных нервных процессов уже
у первичных организмов. Между тем эти первичные организмы,
ймея элементарную раздражимость,
очень хорошо справлялись с
функцией сигнальности. Таким образом, говоря об отражении
внешнего мира в процессах живого организма и в сознании, мы
должны помнить одно основное правило для такого отражения:
неорганический мир уже в рапние эпохи своего формирования
был многопараметрным и потому, естественно, его отражение в
свойствах организма шло по пути охвата того или иного пара-
метра неорганического мира в различных процессах живого орга-
низма и в различные периоды эволюции. Во всяком случае сиг-
нальность как способность живой материи к опережающему
отражению внешнего мира развивалась под влиянием простран-
ственно-временной системы мира. И если бы живой организм эво-
люционировал только с этим свойством своей организации — сиг-
352
Психическая форма отражения действительности
нальностыо, «предупредительным поведением», то можно теоре-
тически представить, что такое поведение могло бы быть развито
и без участия психического. Конечно, если бы только организм
в процессе эволюции не встретился с другими препятствиями,
разрешаемыми на других путях его развития.
Итак, мы столкнулись с вопросом, который неизбежно возни-
кает из приведенных выше рассуждений: какой же параметр
внешнего мира и какое свойство самой живой организации исто-
рически стали толчком к проявлению первых проблесков ощуще-
ния и в какой форме впервые появилось психическое? Мы не
делаем акцепта на вопросе «когда», начиная с какого живого
существа появилось психическое, но мы ставим вопрос «почему»,
под влиянием каких движущих факторов и каких параметров
жизни появилась необходимость у организмов развивать это уди-
вительное свойство психического?
Знакомясь с многочисленными объяснениями психического,
я к удивлению своему не нашел того подхода, который мпе как
физиологу и эволюционисту всегда казался решающим. В самом
деле, если мы все согласны, что психическое подчиняется прпн-
ципу развития и должно было появиться па каком-то этапе
усложняющегося строения нервной системы, то, казалось бы,
первым и естественным вопросом должен был быть вопрос в дар-
винском смысле.
Если какое-то свойство организма появилось в начальной при-
митивной форме и если это свойство удержалось в процессе мно-
говековой эволюции и, наконец, если это свойство пе только удер-
жалось, по и развилось до тех высших этапов, какие мы имеем
в психической деятельности человека, то совершенно естественно
можно поставить вопрос: мог ли естественный отбор закрепить и
сделать фактором прогрессивной эволюции какое-либо свойство
нервного вещества, которое не определяло бы значительно боль-
ший успех выживания и приспособления и пе давало бы более
совершенное вооружение организму в борьбе за существование?
С точки зрения эволюционной теории Дарвина ответ может быть
только один: конечно, это свойство должно было бы элиминиро-
ваться естественным отбором уже в первые же периоды своего
появления, и едва ли кто-либо будет спорить с этим положе-
нием.
Но если это так, если первичные проблески ощущений и пси-
хического способствовали более успешному выживанию организ-
ма и укрепляли его в борьбе за существование, то мы непремен-
но должны выявить то свойство живого и те внешние факторы,
которые оправдывали появление и сохранение психического.
Итак, чем обогащался организм в своей отражательной деятель-
ности, т. е. в своем приспособлении к внешнему миру с появле-
нием первичных признаков психического?
Психическая форма отражения действительности
353
Какие потребности организмов способствовали в процессе эво-
люции закреплению первичных форм ощущения и развитию их
до тех высших форм психического, какие мы наблюдаем у совре-
менного человека?
Еще в 1926 г. в статье «Диалектический материализм и про-
блема психического» я поставил эти вопросы в общефилософском
плане. Однако после этого ни я сам, ни другие исследователи
психического пе ставили и не развивали этой проблемы в прямом
дарвинском смысле.
Прекрасная книга Л. Н. Леонтьева о развитии психического
ставит этот вопрос в более общепсихологическом плане и дает
много новых и интересных характеристик психической деятель-
ности. Одпако в этой книге мы также пе имеем общеэволюциоп-
ной оценки возникновения первичных форм ощущений и психи-
ческого.
Ввиду особенной важности такого именно дарвинского под-
хода к оценке возникновения психического я позволю себе сде-
лать более выпуклой и наглядной мою мысль, главным образом
в отношении определяющих факторов, под влиянием которых
могла возникнуть необходимость в развитии и усовершенствова-
нии этой высшей формы отражения и приспособления к внеш-
нему миру.
В качестве примера можно рассмотреть развитие локомотор-
ной функции у позвоночных животных. Хорошо известно, что
плавание было общей формой передвижения позвоночных до вы-
хода их па сушу. Но именно выход животных па сушу в силу
новых биомеханических особенностей потребовал от них новой
формы приспособления — хождения с помощью конечностей.
Таким образом, определяющим фактором для подобного пре-
образования формы передвижения животных явилось существо-
вание животных в таких неорганических условиях, где только
реципрокная форма движения конечностей способствовала более
успешному выживанию.
Но именно этот пример, хорошо изученный в свое время в
нашей лаборатории, может служить показателем того, как раз-
личные детерминирующие факторы внешнего мира толкают орга-
низм на преобразование его организации и отражательной дея-
тельности, определяя само направление этих преобразований.
В личиночном развитии амфибий, например аксолотля, есть
стадия, когда передние конечности имеют вид маленьких дву-
пальцевых выростов, торчащих в стороны и пока еще не достаю-
щих земли. В этой стадии развития аксолотль передвигается
только с помощью плавания. Одпако выросты еще несозревших
конечностей имеют явно регрессивное значение для аксолотля в
данной стадии его развития. Увеличивая значительно сопротив-
ление передвижений в жидкости, они должны были бы умень-
12 П. К. Анохин
354
Психическая форма отражения действительности
шить скорость движения и этим самым значительно снизить
выживаемость данного вида. Однако эмбриологическое развитие
в соответствии с принципом системогенеза, т. е. избирательного
и ускоренного развития функций, необходимых молодому орга-
низму в первую очередь, исправляет этот жизненно важный
дефект.
Уже в самом начале формирования передних конечностей
ускоренно созревают именно те мышцы, которые в момент пла-
вания (!) прижимают еще не созревшие конечности к туловищу
и тем самым обеспечивают ему вполне обтекаемую форму.
Таким образом, мы видим, что эволюция какого-либо прогрес-
сивного свойства организма расширяет его возможности приспо-
собления и «выходит на сцену» через целый ряд интересных ком-
промиссных приспособлений, которые все вместе обеспечивают
один нерушимый закон жизни: приспособиться и выжить.
Подобные примеры из эволюции приспособительных форм по-
казывают, что для появления какого-либо нового признака в мор-
фофизиологической организации животного необходимо наличие
нескольких закономерностей, которые стимулируют и направля-
ют усовершенствование этого признака. Эти закономерности сле-
дующие.
1. Необходимо наличие какого-либо вполне определенного фак-
тора внешнего мира, имеющего четко очерченные
изические
константы, например вынужденная необходимость передвижения
по суше анаботид. Этот фактор становится определителем как
возникновения, так и дальнейшего развития данного морфофизио-
логического признака.
2. У каждого такого признака неизбежно должен быть его
исторический предшественник, обеспечивавший в прошлом менее
широкие приспособительные возможности.
3. В процессе эволюции имеется специальный переходный
период от менее прогрессивной к более прогрессивной форме при-
способления, который большею частью формируется на базе все-
возможных модификаций прежних форм приспособлений (напри-
мер,
«смена
н.
ункций» Дорна, «полифункциональность органа»
А. Н. Северцова). Этот переход осуществляется в различных слу-
чаях по-разному, в зависимости от характера определяющих фак-
торов эволюции, или, образно выражаясь, в зависимости от физи-
ческих и биологических свойств самой потребности, которая тол-
кает организм на прогрессивное развитие.
Мы видим, что все эти три пункта получают вполне четкий
ответ в приведенном нами примере перехода от плавания к хож-
дению у амфибий.
В самом деле, прямым определителем этого перехода являют-
ся свойства суши как твердой опоры и широчайшие возможности
полезного приспособления. Хождение как биомеханический аппа-
Психическая форма отражения действительности
355
рат оказалось наиболее благоприятной формой для использования
всех приспособительных возможностей на суше. Но оно по ряду
признаков есть только в определенном направлении усовершен-
ствованное плавание (реципрокпость, функция передних плавни-
ков и т. д.).
Приведенные выше общебиологические примеры целиком при-
менимы к процессу возникновения первичного ощущения, прими-
тивных психических способностей, и наконец, широкой и совер-
шенной психической деятельности человека.
Желая охарактеризовать психическое, мы часто говорим, что
сначала появилось примитивное, или элементарное ощущение,
а затем оно развилось до более высоких форм психической дея-
нашим
тельности.
Но у последовательного эволюциониста, стоящего на позициях
диалектического материализма, немедленно должен возникнуть
вопрос: какая «нужда» организма толкнула его на развитие этого
первичного ощущения? И в чем это первичное ощущение более
совершенно приспособило организм к внешнему миру, поскольку
оно пе только закрепилось естественным отбором, но претерпе-
вало и огромный прогресс в процессе эволюции?
После многолетнего анализа этого вопроса я пришел к вполне
определенному ответу, который хорошо согласуется
материалистическим пониманием всей истории развития жизни
на Земле.
Так, например, развитие сигнальной деятельности у высших
животных, как мы увидим ниже, имело своим источником первич-
ное отражение пространственно-временной структуры мира.
Именно временная структура мира, существовавшая и до разви-
тия жизни, вынудила организм отразить ее в своеобразных мор-
фофизиологических конструкциях, которые стали аппаратом опе-
режающего отражения действительности. Это опережающее отра-
жение уже на ранних этапах развития жизни служило основному
требованию всего живого — приспособиться и выжить.
Таким же категорическим определителем для появления и раз-
вития первичных ощущений был критерий «вредности» или «по-
лезности» данного воздействия на организм в смысле сохранения
или разрушения жизни.
Можно привести много доказательств того, что первичные
ощущения и их дальнейшее развитие в форме субъективных ощу-
щений и, наконец, их психических коррелятов, несомненно, разви-
лись под давлением именно этой жизненной необходимости, и по-
тому они обусловили значительный качественный скачок в про-
грессивном развитии приспособительных возможностей организ-
мов. Последние приобрели в этом свойстве ощущения мощное ору-
дие почти моментального и глобального «опознания» жизненного
значения данного воздействия на организм, а это обстоятельство
12*
356
Психическая форма отражения действительности
значительно ускоряло формирование соответствующих данному
моменту приспособительных реакций.
Здесь следует отметить одпу принципиальную особенность
первичных ощущений или даже предощущений. Они приобрели
глобальность, фокуспрованность целого организма на одном каком-
то качестве состояния, которое уже пе локализовалось только по
какому-либо отдельному органу. Это и была самая высшая инте-
грация всех частей организма на одном фокусе — ощущении.
Одновременно с этой интеграцией приобреталось и качесто этого
ощущения, особенно отрицательное, подобное болевому, которое
служило сигналом к смертельному разрушению организма.
Отныне это новое свойство живой материи дало возможность
организму классифицировать все факторы внешнего мира на две
биологически важнейшие категории — отрицательные и положи-
тельные. Прогрессивно развиваясь до самых высших форм пси-
хической деятельности
животным в форме извечной антиподной пары ощущений
дания и удовольствия». В ощущении организм получил свою выс-
шую интеграцию, всегда формирующую адекватную реакцию в
эти два состояния перешли к высшим
«стра-
соответствии с жизненным значением данного воздействия.
Теперь мы можем уже ответить па поставленный нами выше
вопрос: чем же более совершенно это новое свойство материи при-
спосабливало организм к условиям его существования, поскольку
такой прогресс является необходимым условием для закрепления
его естественным отбором?
Для ответа на этот вопрос лучше всего взять такие примеры,
которые повседневно проверяются нашим опытом и особенно
медицинской практикой.
Допустим, что в кожу руки погружается игла. С помощью
каких критериев человек мог бы определить вредность этого воз-
действия для своей жизни? Он может, конечно, рассчитать в
долях миллиметра погружение иглы, ее диаметр, толщину эпидер-
миса кожи, предвидеть разрушение сосудов и, наконец, он может
подождать некоторое время, чтобы убедиться в безвредности или
вредности данного воздействия для его организма — заболел оп
или не заболел?
Все перечисленные выше критерии вредности являются совер-
шенно ненужными у обычного нормального человека, поскольку
при определенной степени погружения иглы он начинает вдруг
ощущать чувство боли, интегрирующее весь организм на форми-
рование оборонительных мероприятий.
Врачам хорошо известно, как трудно предупредить разруше-
ние той части организма человека, которая почему-либо потеряла
болевую чувствительность («пролежни»), и как много требуется
расчетов, чтобы вовремя перевернуть больного пли не положить
ему на анестезированную кожу слишком горячей грелки.
Психическая форма отражения действительности
Таким образом, мы видим, что субъективное ощущение, раз
появившись, в процессе эволюции определило самую высшую
интегрированность организма и вместе с тем организм получил
наиболее совершенную и экономичную оценку степени вредности
и полезности внешних воздействий. Несомненно, это были пер-
вые в истории живого мира отражения внешних воздействий в
примитивном субъективном состоянии. С этого момента в субъек-
тивном психическом состоянии могли быть собраны все деталь-
ные процессы организма. Все компоненты таких состояний ока-
зались в «снятом виде», и только фокус субъективного состояния,
являясь теперь представителем целого организма, стал руководя-
щим фактором.
С дарвинской точки зрения, с появлением первого субъектив-
ного ощущения была достигнута паилучшая форма приспособле-
ния к внешнему миру. Достаточно стало появления лишь опре-
деленно окрашенного субъективного состояния, чтобы организм
получил точнейшую информацию о том, как ему поступать и
строить свое поведение в данной ситуации. Этим самым доказы-
вается, что сознание как фактор эволюции целиком подчиняется
ее основному требованию — выживает и прогрессирует только то
свойство или признак организма, который обеспечивает наилуч-
шее приспособление данных живых существ к окружающей обста-
новке. Сознание же как прямое производное точного отражатель-
ного процесса в субъективном ощущении получает прямую про-
верку практикой, результатом приспособления.
Пожалуй, наиболее наглядным примером огромного приспосо-
бительного значения первичных ощущений является роль эмо-
ционального фактора в осуществлении висцеральных и вегета-
тивных функций человека. Каждому взрослому человеку хорошо
известно, что только успешный вегетативный акт сопровождает-
ся положительным субъективным эмоциональным ощущением и,
наоборот, неуснешпость или ненормальность его выполнения ведет
к тягостному эмоциональному ощущению.
Допустим, что у человека оказался неудачным акт мочеис-
пускания или даже нарушилось координированное соотношение
между отделами кишечника. Человек ие может сказать, какие
нервные клетки, какие мышцы... пе выполнили успешную функ-
цию, но тем ие менее общее эмоциональное состояние, т. е, от-
ражение этого неудачного объективного процесса в субъективном,
с поразительной точностью дает знак о неблагополучии.
И даже неудачное чихание, т. е. начавшийся, но не закон-
чившийся чихательный акт, оставляет тягостное эмоциональное
ощущение. Мы знаем, сейчас отчего это происходит: централь-
ная нервная система в случае неуспешного акта пе получает
гармонического интеграла афферентных сигналов от исполнитель-
ных органов — она пе получает «образа» успешного результата.
358
Психическая форма отражения действительности
Итак, уже на ранних этапах появления первичных субъек-
тивных ощущений они включились в важнейшие функции целого
организма: благодаря субъективному отражению организм стал
получать сигналы о благополучии там, где он никакими други-
ми средствами пе мог бы оценить жизненную важность этого не-
благополучия.
Так эволюция защитила наиболее существенные функции ор-
ганизма. Вместе с тем последний получил способность отражать
в первичном субъективном состоянии самые разнообразные, но
всегда жизненно важные внешние воздействия. Этот факт слу-
жит наилучшим доказательством материальной природы отража-
тельного процесса и теснейшей зависимости психического образа
от реального объекта действительности. Мы знаем сейчас, что
уже в момент учета организмом всего окружающего его мира со-
поставление внешних объектов с его собственными потребностя-
ми происходит при широкой мобилизации отражательной дея-
тельности и тех образов, которые опа формирует. Мы знаем так-
же, что это происходит главным образом в той стадии формирова-
ния поведенческого акта, когда происходит обширная мобилиза-
ция афферентных информаций, т. е. когда произвольный акт со-
вершается впервые (решение выносится впервые). II мы знаем
также, что сознание как высоко активный и интегрированный
акт выключается, как только данный поведенческий акт ста-
новится автоматическим п пе нуждается в непрерывной аффе-
рентной коррекции, по оно немедленно появляется и вмешивает-
ся как корригирующий фактор
неуспешным. Это еще одно доказательство того,
является оперантным фактором, реально отражающим реально
существующий мир со всем разнообразием его предметов.
Мы не можем оставить в стороне вопрос о том,
ставляет
действительности. Является ли он «копией» отражаемого объек-
та, является ли оп «образом»?..
Обсуждая эту проблему в начале нашего столетия, В. И. Ленин
знал, конечно, что уже на уровне сетчатки и оптического нерва
зрительный образ превращается в нервные импульсации и, сле-
довательно, прекрасно осознавал, что в конечном пункте отража-
тельных процессов, т. е. в сознании
верным документом действительности, тем пе менее чем-то отли-
чается от нее.
«Познание
природы. Но это не простое, пе непосредственное, не цельное
отражение, а процесс ряда абстракций, формирования, образова-
ния понятий, законов etc., каковые понятия, законы etc... и охва-
тывают условно, приблизительно универсальную закономерность
вечно движущейся и развивающейся природы».
как только акт оказывается
что сознание
что пред-
собой внутренний субъективный образ отражаемой
, он, хотя и является досто-
пишет В. И. Ленин,— есть отражение человеком
пе непосредственное
Психическая форма отражения действительности
359
Мне кажется, что уже сам факт появления в ходе эволюции
живых организмов психической деятельности и наличие у нее
способности фокусировать все состояние целого организма в од-
ном-едппствеипом сознательном акте дает нам возможность до-
пустить и возникновение в этом сознании синтеза подлинных п
важнейших параметров наблюдаемого объекта.
Пожалуй, паилучшее доказательство того, что формирование
образа в сознании требует максимального вовлечения нервных
элементов,— потеря сознания при приеме некоторых наркотиче-
ских веществ, действие которых состоит в блокаде определенных
нервных центров. Еще более наглядным является отделение со-
знания и психической деятельности от болевых ощущений в опе-
рации с применением, например, современных наркоанальгетиков.
Известно, что люди под этим наркозом абсолютно не испытыва-
ют болевых ощущений во время операции и даже, более того,
могут свободно и разумно разговаривать с хирургом и с окру-
жающими. Этот случай показывает, что отражательный процесс
обладает высокой степенью точности и реальности и имеет своп
собственные, химически своеобразные нервные элементы и синап-
тические организации.
Опережающее отражение действительности
Из предыдущего изложения следует, что отражение внешне-
го мира как универсальный процесс взаимодействия организма и
окружающей обстановки имел большую историю развития и на
протяжении этой истории совершенствовался и механизм переда-
чи информации. Особенно это относится к способам кодирова-
ния и декодирования информации по различным этапам централь-
ной нервной системы.
Однако на протяжении всего изложения мы держались в пре-
делах, так сказать, одномоментного воздействия внешнего мира
на организм и его одномоментного отражения в системах орга-
низма. Мы сознательно исключали один из важнейших факто-
ров внешнего мира — пространственно-временной континуум
мира, в котором развивалась первичная жизнь и который оказал
решающее влияние па все последующие этапы эволюции жизни
на нашей плапете.
В какой форме пространственно-временной континуум неорга-
нического мира и в особенности время отразились в конструк-
ции и деятельности живых организмов? Ведь эти последние vo-
lens nolens должны были «вписаться» в эту всеобщую законо-
мерность неорганического мира. Отвечая на этот вопрос, мы
должны, как и раньше, прежде всего сформулировать и описать
те параметры времени, которые являются его специфическими
чертами и которые приобрели жизненно важное значение для
360
Психическая форма отражения действительности
организма. Это сделать надо уже тго одному тому, чго мы па всем
протяжении этой работы подчеркивали, что параметризация внеш-
него мира, т. е. выделение в нем значимых и незначимых па-
раметров, служит абсолютно необходимым условием для научного
понимания материальной основы отражательной деятельности.
Несколько лет тому назад я уже изложил пашу концепцию
относительно «опережающего отражения действительности» в бо-
лее подробном виде. Именно к этой работе я и отсылаю читате-
лей для более детального ознакомления с этой концепцией.
Сейчас же я лишь вкратце изложил те новые соображения, ко-
торые мы развивали в этом направлении па протяжении послед-
них лет.
Наличие прострапствепно-времеппого континуума во всех яв-
лениях природы неизбежно должно было сказаться на деятель-
ности организма. В частности, человек отразил временные пара-
метры мира в своей организации и в своей деятельности с исклю-
чительной точностью. Анализ роли временных параметров мира в
развитии жизни на Земле показывает, что и зарождение жизни,
и ее прогресс находились в прямой зависимости от приспособле-
ний к этим параметрам.
Можно утверждать, что одномоментное отражение внешнего
мира в деятельности животных организмов представляет собой
и
только отдельный фрагмент из огромной
бесконечной
цепи
внешних событий, развивающихся во времени. Возникает вопрос,
какие же параметры временной структуры мира приобрели осо-
бое значение для отражательной деятельности и для выживания
организмов?
Самым естественным было бы думать, что последовательность
внешних явлений, которая представляет собой conditio sine qua
non пространственно-временного континуума, и должпа была бы
иметь решающее значение. Однако более глубокий анализ этого
вопроса показывает, что на самом деле этот параметр временной
структуры мира не смог бы обеспечить пи появления жизни, ни
ее эволюционного прогресса.
В самом деле, представим себе па минуту, что внешняя по-
следовательность явлений представляет собой бесконечный ряд
переходящих одно в другое событий, никогда неповторяющихся
и не похожих друг на друга. При такой предпосылке сохраня-
лась бы важнейшая черта пространственно-временного континуу-
ма, т. е. последовательность. По сути дела такой мир, имеющий
только простую последовательность событий, практически мог
бы существовать, если бы этот мир был только неорганическим.
В самом деле, камню и морю совсем безразлично, в какой по-
следовательности развиваются внешние события. Фигурально вы-
ражаясь, неорганический объект не имеет специальных «претен-
зий» к параметрам времени. Однако отношение к этому парамет-
Психическая форма отражения действительности
361
ру времени радикально меняется, как только на Земле, в ее
«первородном океане», появились первые зачатки жизни. Для
развития жизни кардинальным условием стала не простая
следовательиость событий, а периодически или ритмически
по-
по-
вторяющаяся последовательность отдельных фрагментов из бес-
конечного континуума явлений в общей картине мира. Только
при этих условиях стали возможными и
приспособление живого
к внешним событиям, и прогресс его материальной организации.
Естественно, и отражательная деятельность организма приобрела
от ритмически повторяющихся явлений специальный стимул для
формирования стабильных структур.
Временная структура мира фактически стала таковой, что по
своей сути, имея более короткие и более длинные последователь-
но повторяющиеся явления, опа снособствовала возникновению
самой жизни. Такие постоянные циклы, как осень, зима, весна,
лето или утро, день, вечер, ночь, были фундаментальными раз-
вивающимися циклами, и это способствовало приспособлению и
закреплению первичных живых организаций. Если проанализиро-
вать всю пашу жизнь, то по сути дела процесс повторных воз-
действий событий мира на нашу нервную систему окажется са-
мым существенным временным фактором. Мы много раз ходим по
одной и той же улице, па нас повторно действуют многие и
именно одни и те же объекты, и все это составляет основной
временной фактор отражательной деятельности нашего мозга.
Этот же параметр времени — повторяющиеся ряды явлений —
послужил толчком и к развитию исключительной способности жи-
вых организмов, которая определила весь прогресс природы и
человеческого общества. Представьте себе, что какая-то последо-
вательность явлений внешнего мира с определенными большими
интервалами времени действует на уже сформировавшуюся жи-
вую клетку. В этих случаях создаются совершенно исключитель-
ные условия для своеобразной формы отражения действительно-
сти в протоплазме этой клетки.
В самом деле, как мы хорошо знаем, внешние явления пред-
ставляют собой пространственно-временной континуум (например,
сезонные температурные смещения). Эти явления действуют на
клетку, протоплазматическое содержимое которой имеет микро-
размеры, и, следовательно, последовательные химические измене*
нпя клетки от последовательных явлений внешнего мира также
совершаются в микроинтервалах пространства и времени. Други-
ми словами, с появлением живой субстанции возник принци-
пиально новый вид отражательной деятельности — макромир че-
рез целую серию преобразований в больших интервалах времени
молекулярных пере-
стройках протоплазмы живой клетки. Отсюда следует вывод, что
отражается в микромире, т. е. в быстрых
362 Психическая форма отражения действительности
••
живая протоплазма приобретает свойство накоплять информа-
ционные посылки от внешнего мира, конденсируя их в микро-
интервальных связях.
Однако это еще не все то принципиально новое в отраже-
нии внешнего мира, что появилось с формированием живой суб-
станции.
Именно потому, что явления внешнего мира развиваются в
форме пространственно-временного континуума, живая прото-
плазма также отражала этот континуум в непрерывной цепи мо-
лекулярных перестроек. Л это значит, что пространственно-вре-
менной континуум внешнего макромира трансформируется в хи-
мический континуум молекулярных процессов микромира живых
существ. Такому отражению внешнего мира чрезвычайно способ-
ствовали следовые реакции химических процессов в протоплаз-
ме, которые возникали от непрерывного действия внешних яв-
лений. Так создавались все условия отражения внешнего конти-
нуума в химическом микроконтинууме протоплазменных
процессов.
Надо представить себе на минуту мир химических процессов
протоплазмы, в которых непрерывность внешних явлений отра-
жается в форме непрерывных цепей химических перестроек, пе-
рекрывающихся друг другом и взаимодействующих друг с другом.
Именно иа этой химической основе и родилось то «чудо», кото-
рое в дальнейшем определило весь прогресс живых организмов.
Благодаря многократному, а для некоторых внешних явле-
ний, может быть, и многомиллионному повторению того же самого
ряда молекулярных перестроек создались весьма благоприятные
условия для воспроизведения всего ряда молекулярных перестро-
ек в протоплазме под действием только начального звена в цепи
внешних событий. Благодаря химическому сцеплспию целого ряда
процессов создались условия для воспроизведения всей цени про-
топлазматических процессов от начального толчка из внешнего
мира. Протоплазма приобрела способность благодаря развитию
своих молекулярных процессов опережать во времени и простран-
стве закономерное течение последовательных событий внеш-
него мира.
Несколько лет тому назад мы назвали это свойство живых
организмов опережающим отражением действительности (Ано-
хин, 1962), а констатированная значительно ранее эта способ-
ность нервной системы была названа «опережающим возбужде-
нием» (Анохин, 1957).
Широко оценивая это свойство живой субстанции, мы должны
сказать, что организмы, приобретя способность опережать ход
внешних событий, тем самым стали с наибольшей выгодой при-
спосабливаться к будущим часто опасным явлениям внешнего
мира задолго до того, как эти явления будут иметь место.
Психическая форма отражения действительности
363
Живой мир дает, огромное количество примеров такого при-
способления к будущим явлениям, где повторения происходят
на протяжении миллиардов лет, как, например, последователь-
ность осень — зима. В результате мы видим, как весь расти-
тельный мир значительно опережает события, развивая под влия-
нием первых осенних похолоданий такую цепь химических ре-
акций, которая нужна будет для предотвращения вредных воз-
действий первых зимних морозов только в будущем. Благодаря
этой способности опережать ходом своих протоплазматических
процессов реальные внешние температуры растение спасает себя
также и от возможных внезапных заморозков...
Много миллионов лет ритмически повторяющееся в природе
замерзание привело через естественный отбор к выживанию та-
ких мутантов, которые хоть в какой-то степени могли снизить
точку замерзания протоплазмы по первому сигналу холода. Это
блестяще иллюстрирует правило опережающего отражения деист-
вительности...
Итак, существование опережающей формы отражения
объективной действительности — несомненный факт. Это — про-
дукт жизни. Развиваясь в микропнтервалах пространства и вре-
мени. опережение приобрело огромное преимущество перед собы-
тиями, медленно развивающимися во внешнем мире.
Я не буду здесь останавливаться па том, что наиболее со-
вершенной формой этого своеобразного «заглядывания» в буду-
щее является деятельность нервной системы. Именно в пей по
преимуществу стала развиваться способность организма к опере-
жению событий. Наиболее наглядной формой этого является ее
способность образовывать условный рефлекс. Мозг и стал тем ор-
ганом, который благодаря высокоразвитым рецепторам ежесекунд-
но трансформирует пространственно-временной континуум внеш-
них явлений в химический континуум, выражающийся в непре-
рывной разрядной деятельности нейрональных элементов мозга.
Способность мозга фиксировать повторяющиеся последовательно-
сти внешних событий, развившаяся на основе примитивных форм
отражения, определяет ту изумительную способность человече-
ского мозга, которая выражается в предвидении будущего в фор-
мулировке гипотез и вообще в различных прогностических оцен-
ках событий.
Именно благодаря этим мозговым,
ским механизмам, отражение внешнего мира в сознании стало
высшим пунктом отражательной функции организма. Это то отра-
жение действительности, о котором говорили классики диалекти-
ко-материалистической философии. Нейрофизиологический анализ
работы мозга показывает, что отражение внешнего мира фак-
тически и не может быть иным, пеопережающим, ибо с ин-
формационной точки зрения каждое внешнее воздействие
в основном неирохимиче-
it
на
364
Психическая форма отражения действительности
организм непременно мобилизует в нервной системе также и мо-
лекулярный опыт прошлого, связанный с данной ситуацией или
с данным раздражителем — стимулом. Это обстоятельство дает
и
возможность мозгу объединить прошлое с настоя
(ИМ
и на этой
основе предугадать в детальных параметрах наступление буду-
щих событий. Мы полагаем поэтому, что концепция опережаю-
щего отражения действительности — лишь творческое развитие
основных положепий ленинской теории отражения.
Вместе с тем сам отражательный процесс, передающий точ-
нейшим образом все параметры окружающего нас объективного
мира, является продуктом эволюционного развития животной ор-
ганизации на протяжении многих миллионов лет. И потому в
каждом акте отражения взрослого человека мы имеем подлинный
органический сгусток отражательного опыта поколений и настоя-
щей действительности. Нами были вскрыты те закономерности,
на основе которых весь опыт отражения внешнего мира организ-
мами, закрепленный в филогенезе наследственностью, вновь воз-
вращается к этому же внешнему миру через развитие зароды-
шей животных. Эту закономерность «подгонки» отражательных
способностей организма под его экологию мы в свое время назва-
ли «системогенезом».
Системогенез служит подлинным инструментом эволюции, ко-
торый является как бы посредником в передаче исторического
накопления отражательной деятельности предков их потомкам
в точном соответствии с их специфическими экологическими фак-
торами. Здесь мы встречаемся с одним из сильнейших аргумен-
тов теории отражения: с отражением объективной действитель-
ности в структуре и функции бесчисленного количества поко-
лений животных, т. е. с историческим накоплением отражатель-
ного опыта (поколения).
Системогенез как инструмент реализации
исторического накопления отражательных механизмов
Концепция системогенеза возникла в нашей лаборатории на
основе наблюдений, показавших, что у эмбриона различных жи-
вотных развитие функций происходит задолго до рождения на
основе избирательного созревания именно тех структур, которые
необходимы для осуществления этих функций. Речь идет о функ-
циях, которые должны быть обязательно готовы к моменту рож-
дения животных, ибо они обеспечивают выживание новорож-
денного (Анохин, 1937). Механизмы системогенеза обеспечивают
ускоренное созревание некоторых структур в то время, когда
еще другие структуры весьма далеки от завершения своего роста.
Так, например, круговые мышцы рта (orbicularis oris) полу-
чают иннервацию и развивают нейромускулярные синапсы весь-
Психическая форма отражения действительности
365
ма ускоренно и задолго до того, когда другие мышцы лица по-
лучают эту иннервацию. Причем такое ускоренное развитие пре-
терпевает не только круговая мышца рта, но и все другие мышцы,
особенно связи в центральной нервной системе, которые в сово-
купности обеспечивают осуществление сосательного акта. Как из-
вестно,
и
ункция сосания должна осуществляться немедленно после
рождения, в противном случае новорожденный не выживает п
устраняется естественным отбором.
Стоит только посмотреть па различных животпых, чтобы уви-
деть, как разнообразен мир адаптивного отражения внешнего
мира, и понять, каким сложным должен быть процесс подготов-
ки в эмбриогенезе специфических для каждого вида животпых
функциональных систем. Для развивающейся из икринки рыбки
необходимы определенные формы приспособления именно к вод-
ной среде.
Однако во всех случаях эмбриогенез животных, т. е. его раз-
витие до момента рождения, непременно должен обеспечить
полноценное действие тех функциональных систем, которые при-
спосабливают новорожденного к его специфическим экологиче-
ским факторам.
В нашей лаборатории па протяжении 40 лет мы изучали эмб-
рионы самых разнообразных видов животных. У пас были рыбы,
амфибии, рептилии, птицы, млекопитающие, наконец, наивысший
п совершенный организм среди млекопитающих — живые плоды
человека, И у всех изученных нами видов мы наблюдали один
и тот же закон развития функций в онтогенезе: задолго до того,
как созревают все органы и структуры зародыша, можно наблю-
дать, как ускоренно и избирательно созревают те структуры ор-
ганизма, и особенно его нервной системы, которые абсолютно не-
обходимы для жизненно важных функциональных систем именно
в момент рождения.
Для нас сейчас важно подчеркнуть, что историческое накоп-
ление отражательных процессов, обеспечивающих адекватное от-
ражение внешнего мира, у каждого новорожденного находится в
точном соответствии с теми внешними факторами, которые на
протяжении тысячелетий были решающими для жизни предков
данного животного. Этот факт замечателен в том смысле, что
история обеспечивает детальное отражение внешнего мира в моз-
гу задолго до того момента, когда внешний мир начнет действо-
вать на новорожденного. Другими словами, закрепленный в
структурах исторический опыт отражения внешнего мира предка-
ми созревает избирательно, «навстречу» специфическим факто-
рам внешнего мира, с которыми впервые встречается новорож-
денная особь. Генетическая информация развертывается в соот-
ветствии с будущими условиями...
Можно привести тысячи примеров, которые могут иллюстри-
366
Психическая форма отражения действительности
ровать одип и тот же закон: внешний мир через серию кодиро-
вания информационных процессов отбирает в теле и мозгу соот-
ветствующих животных такие структуры, которые в подлинном
смысле слова являются протоплазматическим сгустком, тончай-
шим образом отражающим свойства этого внешнего мира. Отра-
жающий аппарат мозга как бы «спешит» в своем эмбриональ-
ном развитии «навстречу» тем параметрам внешнего мира, кото-
рые на протяжении миллионов лет формировали этот отражающий
субстрат.
Поразительный пример этой закономерности дает нам сопо-
ставление развития зародыша птиц с различной экологией. Мы
сопоставляли поведение новорожденного грача, находящегося, как
известно, в гнезде йа верхушке дерева, с поведением птенца
мухоловки-пеструшки (Muscicapa Hypoleica), который выводит-
ся и живет первое время после вылупления в дупле дерева.
Сравнение дало настолько поразительный результат, что было
бы просто наивным считать, что мозг животных развивается
по каким-то своим имманентным законам, независимым от отра-
жения действительности. Птенец дуплянки также раскрывает
клюв при приеме пищи, которую приносит мать, однако, стиму-
лом для раскрытия клюва для него служит затемнение одпого-
единственного места, на которое он ориентируется, а именно от-
верстия в дупле.
Можно ли найти еще более убедительный пример, доказы-
вающий, что вся история развития животных — тончайшее отра-
жение важных внешних условий существования и фиксация этих
избранных параметров внешнего мира в специфических струк-
турах мозга. И может ли мозг человека с его выс]
ей формой —
и
сознанием, подчиняющийся в своем развитии абсолютно тем же
историческим закономерностям, каким-то образом «выпасть» из
этого закона и не служить для нас высшим отражательным
прибором, фиксирующим все детали объективно существующего
мира?
Приведенные здесь, да и многие другие результаты строго
научного исследования убеждают нас в том, что вся история
развития живой материи до ее самого высшего этапа — мысля-
щего человека подчиняется одному и тому же закону: приспо-
собительное поведение организмов, сохраняющее им жизнь и ве-
дущее их
к прогрессу, возможно только потому, что внешний
мир через разнообразнейшие параметры своего воздействия «вхо-
дит» в организм в форме тончайших информационных процес-
сов, весьма точно отражающих основные параметры этого объ-
ективного внешнего мира.
СИСТЕМНЫЙ АНАЛИЗ
УСЛОВНОГО РЕФЛЕКСА*
нейрофизио-
н
Общие замечания
На протяжении ряда последних десятилетий вопрос о доста-
точности рефлекторной теории для объяснения сложных поведен-
ческих актов принимал все более и более острую форму. Ста-
новилось очевидным, что достижения современной
логии и кибернетики вступают в противоречие с понятием реф-
лекса, ограниченным узкими рамками афферентно-эфферентных
взаимодействий.
Наиболее популярная формула поведенческого акта «сти-
мул — ответ» стала явно недостаточной, чтобы понять все раз-
нообразие тончайших приспособлений животного к окружающим
условиям при построении поведенческих актов. Совсем не укла-
дывается в схему рефлекса, например, поиск, направляемый внут-
ренне созревшими мотивациями, совершенно очевидное «приня-
тие решения», которому не было никакого эквивалента в реф-
лекторной теории, и тем более выходила далеко за пределы
рефлекса формулировка цели и целенаправленного поведения в
целом. С этой последней широко распространенной формой пове-
дения нейрофизиология, стоящая на позициях рефлекторной тео-
рии, «разделывалась» очень легко: она просто устраняла цель и
целенаправленное поведение из своего поля зрения как не отно-
сящуюся к точным наукам.
Между тем накапливающийся материал по изучению слож-
ных форм поведения и в особенности выход нейрофизиолога па
путь изучения системных закономерностей делали совершенно
необходимым выработку какого-то нового представления о пове-
денческих актах, преодолевающего жесткие рамки «рефлектор-
ной дуги».
Учение об условных рефлексах И. П. Павлова, сделавшего
исключительно важный шаг в дальнейшем развитии рефлектор-
ной теории, оказалось стоящим на грани двух «физиологиче-
ских эпох»: с одной стороны, была выявлена новая грандиозная
закономерность мозга —
ляющие успех приспособительного поведения, с другой — трак-
товка и объяснение экспериментальных результатов всецело дер-
ормировать временные связи, опреде-
н
♦ Опубликовано в «Журнале высшей нервной деятельности» (1973, вып. 2).
368
Системный анализ условного рефлекса
жались в рамках уже устоявшихся рефлекторных терминов и
понятий. Поведение животного, получающего в ответ на условный
раздражитель какой-либо подкрепляющий фактор, по-прежнему
изображалось в форме трехчленной дуги рефлекса или в виде
какой-либо суммации этих рефлексов.
Так, например, в основу теории условных рефлексов вошли
два основных рефлекса: условный и безусловный,— причем ус-
ловный рефлекс как самостоятельная физиологическая катего-
рия отграничивается от рефлекса безусловного, толчком к разви-
тию которого служит подаваемая нища (безусловный раздра-
житель). Таким образом, логически целостный поведенческий акт,
представляющий собой блок или систему отношений, включаю-
щую и обратную информацию о результате, по своей глубокой
сути был полностью подчинен первоначальной декартовской кон-
цепции о «стимуле — реакции». Это одна из причин тех глубин-
ных противоречий, которые неизбежно должны были возникнуть
при расширении идейных позиций, связанных с развитием си-
стемного подхода к анализу поведения животных.
Подчинение всех проявлений условного рефлекса принципам
рефлекторной теории прошлых веков неизбежно должно было при-
вести к противоречию с накопленными результатами экспери-
ментальной нейрофизиологии и поведенческих наук.
Попытаемся понять само существо этих противоречий и по-
строить модель поведенческого акта, которая не только включила
бы в себя условный рефлекс как форму приспособления живот-
ного к окружающему миру, но и объяснила философскую суть
развития первичных предупредительных реакций живых существ.
Постараемся понять, как И. П. Павлов подходил к объясне-
нию условного рефлекса и, главное, в какую связь оп ставил
условный рефлекс с рефлекторной теорией Декарта. Отмежевы-
вая свою позицию физиолога от бесплодных поисков субъектив-
ной психологии, он писал: «Следовательно, физиолог должен идти
своим путем. И этот путь намечен уже давно. Считая деятель-
ность животных, в противоположность человеческой, машинооб-
разной, Декарт триста лет тому назад установил понятие реф-
лекса как основного акта нервной системы. Та или другая дея-
тельность организма есть закономерный ответ на тот или другой
внешний агент, причем эта связь деятельного органа с данным
агентом, как причины со следствием, устанавливается при помо-
щи определенного нервного пути. Таким образом изучение дея-
тельности нервной системы животных было поставлено на твер-
дую естественнонаучную почву... Основным исходным понятием
у нас является декартовское понятие, понятие рефлекса. Конеч-
но, оно вполне научпо, так как явление, им обозначаемое, стро-
го детерминизируется. Это значит, что в гот или другой рецеп-
торный нервный прибор ударяет тот или другой агент внешнего
Системный анализ условного рефлекса
369
мира или внутреннего мира организма. Этот удар трансформи-
руется в нервный процесс, в явление нервного возбуждения.
Возбуждение по нервным волокнам, как проводам, бежит в цент-
ральную нервную систему и оттуда, благодаря установленным
связям, по другим проводахМ приносится к рабочему органу, транс-
формируясь, в свою очередь, в специфический процесс клеток
этого органа. Таким образом, тот или другой агент закономерно
связывается с той пли другой деятельностью организма, как при-
чина со следствием» (Павлов, т. 4, с. 22)
В этих цитатах довольно отчетливо выражен тот факт, что
концепция рефлекса ставит поведение в зависимость от внешних
факторов. В самом деле, когда причинами поведения животных
и особенно человека считались самые различные мистические фак-
торы вроде высшего разума, то введение Декартом материально-
го стимула («толчок к действию») было огромным достижением
науки, и прежде всего пауки о мозге, которая до этого находи-
лась во власти схоластического и религиозного мышления.
Именно это обстоятельство, которое всегда отмечал И. П. Пав-
лов, было одним из важных признаков материальной причин-
ности, вызывавших у него расположение к рефлекторной тео-
рии.
Однако сейчас одного этого достоинства рефлекторной кон-
цепции, как оказалось, далеко недостаточно. Современные дости-
жения нейрофизиологических паук о поведении, кибернетике и
особенно системный подход в биологических науках дали много
оснований к преобразованию рефлекторной концепции. К этому
призывают как достижения школы самого И. П. Павлова, его
учеников, так и его последователей во всем мире.
Постараемся попять, какие признаки рефлекса являются фун-
даментальными для всей концепции рефлекторной теории, в ка-
кой мере и как они могут быть видоизменены в соответствии
с более широким подходом к поведенческому акту, например,
в аспекте теории функциональной системы... Этими признаками
считаются следующие: 1) исключительность пускового стимула,
2) поступательный ход процесса возбуждения по всей рефлектор-
ной дуге, 3) завершение рефлекса «рефлекторным действием»
(«ответом»).
Исключительность стимула как фактора,
определяющего возникновение
ответной реакции
История физиологии свидетельствует, что все виды нервных
реакций начинались с действия стимула, а сама внутренняя цент-
ральная организация рефлекса расценивалась с точки зрения
свойств примененного стимула.
13 П. К. Анохин
370
Системный анализ условного рефлекса
По сути дела во всех случаях изучения условпорефлектор-
ной деятельности центральным моментом являются именно дей-
ствие условного стимула и последующая трактовка центральных
явлений. Все вариации опытов па протяжении многих лет ра-
боты школы имели дело или с различными стимулами по клас-
сификации органов чувств, пли с различными комбинациями
этих стимулов, пли с различными их интенсивностями.
Такие факторы, как обстановка и пищевая возбудимость, рас-
сматривались большей частью как дополнительные стимулы,
внешние по отношению к условному рефлексу, И поэтому они
учитывались только в особых случаях экстремального воздейст-
вия.
Эксперименты, проведенные в нашей лаборатории, с двусто-
ронним кормлением по секреторпо-двигательпой методике пока-
зали, однако, что «обстановочный раздражитель» выступает ор-
ганической частью афферентного комплекса, который формиру-
ется на основе конвергенции возбуждений па одном и том же
нейроне.
Предпосылкой к такому представлению послужили системати-
ческие многолетние эксперименты нашей лаборатории (Шумили-
на, 1966). Опыты проводились по секреторно-двигательному ме-
тоду условных рефлексов на станке с противопоставленными кор-
мушками в условиях активного выбора стороны безусловного
подкрепления. Находясь в условиях одинаковой возможности по-
лучить подкрепление как из правой, так и из левой кормушки,
собака в межсигнальных промежутках располагалась па середине
станка. При действии условного раздражителя опа направлялась
к сигнализируемой кормушке и стояла до подачи подкрепления
а после еды возвращалась на середину стайка, где находилась
до применения следующего условного раздражителя.
После удаления лобпых долей мозга у собаки резко наруши-
лось поведение. Оказавшись на станке, она маятпикообразпо
на протяжении всего опыта перебегала от одной кормушки к
другой независимо от действия условного раздражителя. Разно-
образные варианты опытов показали, что удаление лобных до-
лей привело к нарушению процессов афферентного синтеза —
распаду сложной интеграции обстановочной и пусковой аффе-
рентации. Смысл этих нарушений состоит в том, что обстано-
вочные раздражители, создающие скрытую предпусковую инте-
грацию. становятся и пусковыми стимулами. Обе афферентации
удерживаются в строгом единстве лобными отделами коры голов-
ного мозга.
Если посмотреть на начальный этап действия пускового раз-
дражителя с этой точки зрения, то становится очевидным, что
условный раздражитель имеет решающее значение для формиро-
вания реакций только при том условии, если он объединен с
Системный анализ условного рефлекса
371
обстановочной афферентацней и с пищевым возбуждением в одно
органическое целое.
Кроме этого, известны еще два фактора, без которых условный
стимул не может реализовать своего приспособительного значе-
ния: доминирующая мотивация (безусловное возбуждение) и па-
мять. Едва ли надо доказывать широко известный лаборатор-
ный факт, что применение условного стимула к сытому животно-
му не дает условной секреторной реакции. Следовательно,
условный стимул приобретает свое действие только в том слу-
чае, если он будет иметь непосредственную встречу и взаимо-
действие с генерализованным восходящим возбуждением латери-
алыюго центра гипоталамуса. Как показано экспериментами на-
шей лаборатории, эта встреча должна произойти прежде всего на
одном и том же нейроне.
То же самое мы можем сказать и о памяти. Условный сти-
мул становится деятельным в том случае, если его возбуждение,
суммируясь с предпусковой интеграцией, формирует избиратель-
ное и облегченное распространение афферентных возбуждений.
К этому следует добавить, что условный раздражитель воз-
буждает огромное количество путей, по которым распространя-
ется возбуждение к синапсам многих корковых клеток. А это
еще более усложняет вхождение стимула в обширную интегра-
цию разнородных процессов. Вероятно, надо представить себе,
что распространение условного возбуждения по уже готовой пред-
пусковой интеграции должно что-то извлечь из кладовых памяти
для формирования целой условной пищевой реакции.
Таким образом, для того чтобы сформировалась полноценная
условная реакция в центральной нервной системе, должны инте-
грироваться па субстрате памяти по крайней мере три типа раз-
личных возбуждений, и стимул вскрывает уже заготовленную
до этого предпусковую интеграцию, поэтому его исключительная
роль в
и
ормировании реакции только кажущаяся.
Возникает вопрос, где же происходит встреча и взаимодейст-
вие всех рассмотренных выше возбуждений? Микроэлектродные
исследования последних лет убедили нас в том, что такая ре-
шающая встреча и взаимодействие многих возбуждений может
произойти только на одном и том же нейроне на основе прави-
ла конвергенции разнородных возбуждений. Только при этом ус-
ловии и возможно использование информационных особенностей
каждого из перечисленных выше возбуждений, приходящих к од-
ному и тому же нейрону.
Конкретные исследования этого процесса убеждают нас, что
эта суммационная обработка разнородных возбуждений на ней-
И
роне осуществляется при помощи целого ряда новых механиз-
мов, физиологический смысл которых состоит в том, что они обе-
спечивают возможность одновременной обработки этих возбужде-
13*
372
Системный анализ условного рефлекса
We-
Обратная афферентация
Пусковой
Эфферентные возбуждения
чин.
ла д Память
Постановоч-
ные арфе-
рентации
шп
Результат
действия
VW
Действие
решения
Рис. 1. Функциональная система с ее узловыми механизмами, обеспечи-
вающими полезный результат
Обстано-
вочные
ерен-
тации Доминируннцая
мотивация
Принятие
Акцептор<
взультата
дебет Зия <
Программа
действия
Параметры
результата
ний. Можно указать па механизм корково-подкорковой ревербе-
рации возбуждений (Шумилина, I960), на повышение порога в
частотной дискриминации приходящих к нейрону возбуждений
(Хаютин, 1972), на увеличение конвергентной емкости каждого
отдельного нейрона (Макаров, 1970). Без этой неизбежной стадии
синтеза охарактеризованных выше возбуждений действие любого
условного стимула, взятого в отдельности, не может быть успеш-
ным. Поскольку в этой стадии происходит активный выбор но-
вой информации и обеспечивается симультанный синтез разно-
родных информаций, мы назвали ее «стадией афферентного син-
теза» (рис. 1).
Таким образом, афферентный синтез как начальная и решаю-
щая стадия организации любой условной реакции представляет
собой органическое объединение на отдельных нейронах следую-
щих типов возбуждений: доминирующая мотивация, обстановоч-
ная афферентация возбуждений, извлекаемых из памяти, и, на-
конец, специально в обстановке работы с условными рефлекса-
ми — наличие пускового возбуждения. Я говорю «специально»
потому, что в сугубо естественной обстановке достаточно моти-
вационного возбуждения, чтобы оно, поднимаясь до коры голов-
ного мозга и пользуясь механизмом избирательного извлечения
из нейронов прежнего опыта, сформировало организованное целе-
направленное поведение.
Системный анализ условного рефлекса
373
Итак, организация всех этих разнородных возбуждений на
стадии афферентного синтеза обеспечивает в дальнейшем один из
самых решающих механизмов поведения — механизм принятия
решения, т. е. выбор наиболее адекватной для данной ситуации
степени свободы. Именно этот последний механизм убеждает нас
в том, что в формировании условного рефлекса нет линейной
причинно-следственной зависимости, поскольку мы не можем ни
от одного компонента афферентного синтеза построить прямой
путь к принятию решения и к результату реакции. Пожалуй,
наиболее наглядным примером, отрицающим исключительное
действие стимула, является формирование динамического стерео-
типа.
Как известно, динамический стереотип формируется в резуль-
тате каждодневного повторения одного и того же порядка услов-
ных раздражителей. Впоследствии любой раздражитель, приме-
ненный па местах различных по специфике условных стимулов,
будет вызывать именно тот эффект, который характерен для этих
условных раздражителей.
Удивительно то, что этот факт не был в свое время оценен
по его глубокой нейрофизиологической сути как иллюстрация
амбивалентной роли условного стимула. В самом деле, как надо
понимать возникновение условно-секреторной реакции, характер-
ной, например, при звонке, но возникающей в ответ на примене-
ние света как условного стимула?
Постараемся понять,
суть явления динамического стереотипа. Дается условный (или
даже посторонний) раздражитель, который должен был бы вы-
звать условную реакцию в соответствии со своим прямым значе-
нием. И вдруг он дает условно-секреторную реакцию, которая в
прошлом была выработана на совершенно другой по модально-
сти раздражитель, адресовавшийся к совершенно другой проек-
ционной корковой зоне.
Явный нейрофизиологический парадокс! Он прошел без ней-
рофизиологического анализа только
исследователей доминировала
появлении условной реакции.
Действительно, если свет
нейрофизиологическая
в чем состоит
потому, что
сознании
исключительная роль стимула в
дается вместо звонка и вызывает
«звонковый» секреторный ответ, то этот феномен должен был
бы немедленно вызвать вопрос: как может свет, адресуясь к зри-
тельной зоне, вызвать условный секреторный эффект, для которо-
го началом является возбуждение слуховой области коры?
Что же имело место на самом деле? Главную роль в этом
феномене сыграла предпусковая интеграция. Она содержала в
себе все качества ранее применяемого на этом месте раздражи-
теля, включая и специфику его результата (акцептор результа-
тов действия) (Анохин, 1933). А это значит, что условный раз-
31
374
Системный анализ условного рефлекса
дражитель, который давался только один в пределах целого опы-
та, проявил лишь одну сторону своей физиологической сути —
пусковое значение, между тем как предпусковая интеграция вно-
сит в этот ответ качественные признаки, т. е. эффект звукового
раздражителя.
Итак, мы видим, что в условиях автоматизации динамиче-
ского стереотипа выявляется доминирующая роль предпусковой
интеграции. Приближение времени применения очередного разд-
ражителя сопровождается скрытым доминированием опережаю-
щего возбуждения тех структур, которые должны быть возбуж-
дены этим раздражителем.
Таким образом, условный стимул только тогда приобретает
свое приспособительное значение, когда ои применяется на фоне
уже готовой предпусковой интеграции, для которой нужен только
момент пуска, чтобы она стала законченной реакцией. Если же
исключить этот момент пуска, то скрытая интеграция может
остаться невыявленной или проявиться в довольно большом ко-
личестве внешних признаков.
Описанная выше расшифровка двух компонентов в стимуле —
пускового и определяющего качества реакции весьма убедитель-
но была показана Сименепко. Опа выработала у человека сте-
реотипную последовательность реакций на звонок, сирену, свет.
Причем одновременно она регистрировала и электроэнцефало-
грамму.
Из применявшихся раздражителей только свет вызывал за-
метную десипхронизацию медленной электрической активности.
Но как только применили «прием сюрприза» — на месте света
внезапно включили звонок — получилась совершенно парадок-
сальная реакция: звонок вызвал десипхронизацию электрической
активности в зрительной области, чего никогда не было.
Если же при такой сюрпризной пробе вообще пе применять
некоторое время очередного света, то можно заметить, что десинх-
ронизация наступит в то же самое время, когда должен быть
свет. Этот эксперимент показывает, что непрерывный континуум
мозговых процессов подготавливает к моменту пробы условного
стимула ту скрытую предпусковую интеграцию, которая была на
этом месте (Анохин, 1957).
Из сказанного следует, что начало условнорефлекторной ре-
акции представлено обширным комплексом нервных процессов,
разнородных по своему биологическому смыслу, но интегриро-
ванных уже задолго до того, как эту интеграцию раскроет ус-
ловный пусковой стимул. Кроме того, стадия афферентного син-
теза настолько многообразна по составу возбуждений — и основ-
ных, и обеспечивающих успех самого афферентного синтеза,—
что говорить об исключительной роли стимула просто невозмож-
но (рис. 2).
Системный анализ условного рефлекса
375
£
Эфферентное возбуждение
Лускодод
стимул
Афферентное
Зозоужденир
РефЛЫтМрЯфй
аеистЗие
Эфферентное дозбужденце
Обстано- Обмять *
Зочные I °
ар щерен-
W4W Чу<А 1 _
Лускобоа I L -*у Y
стимул L_J 1
!х| ]
/ \?v Y м 7 принятие
Обстмбочные^
арферентацииХ
Доминирующая
мотивация
Афферентный
синтез
| 7 ~ ~ I
Акцептор ।
Д'результата'
I ЗеистЗия I
ЭеряенФврное
ЗеистЗие
Эфферентные Зозбуждения
Рис. 2. Классическая рефлекторная дуга
а — на уровне спинного мозга; б — условного рефлекса; в — преобразование реф-
лекторной дуги (черная линия) за счет дополнительных факторов, формирующих
афферентный синтез
13 сущности, эта проблема является «острой» лишь для фи-
зиологов. В кибернетике уже давно считается аксиомой, что лю-
бой выбор, или, как выражается Эшби, «подходящий выбор»,
всегда идет после предварительной обработки достаточного ко-
личества информации.
Едва ли все приведенные выше соображения можно назвать,
как некоторые полагают, «разрушением условнорефлекторной
концепции». После всех предложенных выше дополнений «услов-
ный стимул» реально становится компонентом значительно более
обширной системы процессов, чем это можно было думать, ис-
ходя из его начального упрощения.
376
Системный анализ условного рефлекса
что условный рефлекс существенно помог мате-
Поступательный ход возбуждения
по дуге рефлекса
Эта сторона рефлекторной деятельности имеет, в сущности,
философский характер. Однако по традиции философский ана-
лиз рефлекторной теории никогда не доводился до конца. На ос-
новании того
риалистическому подходу к проблемам психики, было принято
неверное положение, что рефлекс — «категория диалектического
материализма». К сожалению, это ошибочное допущение очепь
сильно повлияло па прогресс наших знаний в этой области.
Рефлекс — это прежде всего категория научной теории и как
таковая должна подвергнуться развитию и усовершенствованию,
как и всякая другая категория.
Теперь уже после ряда работ видных философов — диалекти-
ков-материалистов (Украинцев, 1973) стало очевидным, что ме-
ханический детерминизм, сыгравший известную историческую роль
в истории материалистического мировоззрения, становится все
более и более недостаточным там, где исследователь и мысли-
тель стараются познать реальную действительность сложных био-
логических явлений.
Упрощая явления жизни, механический детерминизм создал
представление о линейном развитии в живых процессах. Каж-
дое явление есть следствие предыдущего звена — причины, а само
оно, в свою очередь, служит причиной последующего следствия.
На этой детерминистической основе образуется причинно-следст-
венная цепь с жесткими линейно-поступательными последова-
тельностями: одна причина, одно следствие.
Характерной чертой механической цепи причинно-следст-
венных отношений является тот факт, что ни одно последующее
звено цепи не может сформироваться раньше, чем предыдущее.
Иначе говоря, никакое очередное звено такой детерминистиче-
ской цепи процессов не может опередить ход событий и сформи-
ровать уже на ранних этапах то, чего еще не было, но что не-
пременно совершится в ближайшем или отдаленном будущем.
Представление о поступательном ходе событий по детерминисти-
чески определенной цепи по своей сути исключает возможность
какого-либо предсказания будущих событий.
В связи с таким представлением в исследовательских зада-
чах обычно стремятся наиболее топко вскрыть эти контакты от
звена к звену всей цепи причинно-следственных отношений.
Однако это не всегда осуществимо. Приведем пример с нерв-
ной клеткой: выходящее на аксон возбуждение не может быть
поставлено в зависимость от какого-либо одного из тех многочис-
ленных возбуждений, которые приходят в один и тот же момент
ко многим синаптическим образованиям нейрона.
Системный анализ условного рефлекса
377
и
Надо считать поразительным тот факт, что рефлекторная тео-
рия, несколько сот лет направлявшая исследовательские процес-
сы в нейрофизиологии, весьма легко сосуществовала с совер-
генно несовместимым с ней повседневным предсказанием буду-
щих событий нашим мозгом.
Целый ряд специальных исследований этого процесса показал,
что практически в любом поведенческом акте, который заканчи-
вается полезным приспособительным этапом, непременно должны
быть элементы предсказания будущего результата. На основе ка-
ких механизмов это происходит? Уже в процессе формирования
первой стадии поведенческого акта, т. е. в процессе афферент-
ного синтеза, на основе всестороннего перебора прошлого опыта
создается весьма обширный аппарат, включающий в себя а<
• рентные признаки будущего результата.
Состав этого афферентного аппарата таков, что в нем пред-
ставлены все наиболее характерные афферентные признаки буду-
HIU
• 1Ц|
щего и еще не полученного результата, подсказанного домини-
рующей мотивацией в процессе афферентного синтеза.
В свое время этот аппарат мы назвали «акцептором резуль-
татов действия». Правда, это название было дано не сразу. Сна-
чала в Горьковском медицинском институте мы назвали его «за-
готовленным возбуждением» (Анохин, 1933), затем «опере-
жающим возбуждением» и, наконец, «акцептором результатов
действия» (Лпохин, 1955) (см. рис. 2). Его главные функции
можно охарактеризовать следующим образом.
1. Прежде всего, являясь эквивалентом цели, он определяет
все этапы целенаправленного поведения, корректируя ежесекунд-
но наиболее успешное достижение необходимого полезного ре-
зультата. Действительно, искусственное «обогащение» акцептора
результатов действия промежуточными дополнительными компо-
нентами показывает, что в его состав входят не только
рентные признаки конечного результата, но также и признаки
всех подступов к конечному результату. В последнее время в
нашей лаборатории было показано, что энергию для непрерывного
управления поведенческими актами, составляющими целенаправ-
ленное поведение, акцептор результатов действия получает от ги
поталамуса и от ретикулярной формации, когда они стимулиру-
ются коллатеральными ответвлениями пирамидного тракта
(Правдивцев, 1967; Синичкин, 1967).
2. Включая в себя афферентные признаки, наиболее полно
удовлетворяющие наличную доминирующую мотивацию, акцеп-
тор результатов действия имеет возможность на основе обратной
афферентации сопоставить эти признаки с теми признаками, ко-
торые поступят через некоторое время от реального результата.
Этим сопоставлением, как правило, заканчивается каждый
дробный этап поведения, как бы ни были ничтожны его резуль-
378
Системный анализ условного рефлекса
таты, что дает нам возможность говорить о поведении как о
континууме результатов.
Анализ этого феномена был осуществлен в нашей лаборато-
рии на различных уровнях. Исторически акцептор результатов
действия был обнаружен раньше всего на уровне условнорефлек-
торного поведения при внезапной подмене безусловного подкреп-
ления. Этот прием, впервые примененный в Горьком, названный
впоследствии в нашей лаборатории «сюрпризной пробой», показы-
н
вает, что качество подкрепляюгцего
актора со
всеми его ха-
рактерными параметрами предсказано мозгом значительно раньше,
чем появится само это подкрепление. Действительно, всякое рао
согласование, т. е. внезапное появление иного подкрепляющего
фактора с иными афферентными параметрами (например, мяса
вместо обычных сухарей), немедленно вызывает бурную ориен-
тировочную реакцию и даже временный отказ от еды.
Этот афферентный аппарат, образующийся сразу же после
дачи условного раздражителя, а может быть и раньше, и пред-
сказывающий качества будущего подкрепляющего агента, был
изучен в нашей лаборатории на многих формах поведенческих
актов.
Так, например, осуществляя различную мотивационную под-
готовку животного (предварительное голодание, жажда), можно
намеренно сформировать афферентный синтез, который ведет к
вполне определенному выбору адекватного подкрепляющего аген-
та, особенно в том случае, когда животному предоставлена сво-
бода получения полезного результата.
Подмена подкрепляющего агента в кормушке, которая рань-
ше имела адекватное для акцептора подкрепление, например
воды пищей, немедленно вызывает рассогласование и многосто-
ронний поиск подкрепляющего агента (воды), адекватного для
его состояния жажды (Асмаяп, Голицын, 1971).
Эти эксперименты убеждают пас в том, что акцептор резуль-
татов действия создается уже в стадии распространения мотива-
ционного возбуждения из гипоталамических центров в кору го-
ловного мозга, а весь дальнейший поиск адекватного подкрепляю-
щего агента сводится к «перебору» результатов прошлого опыта,
вызванного к жизни доминирующей мотивацией, и к сличению
его с акцептором результатов действия, сложившимся в данной
ситуации (Асмаян, 1969; Журавлев, 1971; Судаков, 1971). Спе-
циальные электрофизиологические опыты с «опережающим воз-
буждением», т. е. практически с формированием акцептора ре-
зультатов действия при выработке классического и инструмен-
тального условных рефлексов, показали, что в случае применения
условного раздражителя процесс формирования акцептора ре-
зультатов действия происходит уже в пределах первой секунды
(Швырков, Гринченко, 1972).
Системный анализ условного рефлекса
379
Вместе с тем, разработав методику произвольного обогащения
акцептора результатов действия дополнительными компонента-
ми, мы показали, что уже с самого начала формирования услов-
ного ответа возбуждение распространяется до последнего звена,
как бы пи был сложен состав акцептора результатов действия.
Иначе говоря, возбуждение занимает и промежуточные акцепто-
ры результатов действия, находящиеся на подступах к конечно-
му результату. Для этих промежуточных результатов также не-
обходимы механизмы сличения.
Иначе говоря, если па подступах к результату имеется не-
сколько инструментальных актов животного, то каждый из них
должен непременно получать обратную афферентацию и «санк-
цию» данного фрагмента поведения прежде, чем животное достиг-
нет пищевого подкрепления из кормушки. Поясняю это конкрет-
ным экспериментом, проделанным в пашей лаборатории
В. Б. Швырковым и Ю. В. Гринченко (1972).
У животного было выработано инструментальное поведение в
следующем составе: кролик потягивал кольцо, подвешенное пе-
ред ним, и кормушка немедленно выходила в положение, гото-
вое для еды. После установления этого навыка в целях расши-
рения подступов к еде был введен дополнительный компонент —
свет. Таким образом, теперь потягивание кольца пе открывало
кормушку, а включало свет, и только потом происходила подача
кормушки. После тренировки этого ряда актов и их укрепления в
одном из предъявлений была выключена линия света, и теперь
животное, потянув кольцо, пе получило света. Кролик не пошел
к кормушке. Не получив света (рассогласование!), он продол-
жал энергично дергать за кольцо, добиваясь появления света, ко-
торое в данном варианте эксперимента должно было закончить
формирование малой субсистемы.
Едва ли можно сомневаться в том, что в данном случае по-
тягивание кольца уже было предварено в механизмах мозга соз-
данием акцептора результатов действия в виде параметров пред-
стоящего света. Отсутствие реального света при потягивании
кольца привело к нарушению феномена совпадения в акцепторе
результата действия. Таким образом, па пути приближения к
конечной цели данного поведенческого блока было введено рас-
согласование и все дополнительные реакции животного были
направлены па активный подбор дополнительных афферептаций
и на устранение рассогласования, поскольку для кролика появ-
ление света служило абсолютным условием достижения пищевого
результата.
Этот экспериментальный эпизод фактически представляет со-
бой наиболее частый случай также и в нашей повседневной жиз-
ни. Нередко в цепи явлений, развивающихся на пути к получе-
нию полезного результата, появляется ряд неожиданных помех.
380
Системный анализ условного рефлекса
Анализ таких феноменов убеждает в том, что аппараты ак-
цепторов промежуточных результатов действия, складывающиеся
по поводу даже самого незначительного промежуточного резуль-
тата, служат своеобразными «гидами» поведенческого континуума
(Анохин, 1970).
Мы сделали попытку проанализировать более тонкие меха-
низмы формирования этого аппарата и посмотреть, каким обра-
зом формируется его состав. В качестве модели мы взяли форми-
рование акцептора результатов действия в функциональной си-
стеме дыхания, существование которого было показано в экспе-
риментах В. А. Полянцева. Речь идет о заборе в данном дыха-
тельном акте того количества воздуха, которое необходимо
именно на данный момент организму.
Эксперименты показали, что любая команда дыхательного
центра о заборе легкими определенного количества воздуха пред-
варяется формированием контрольного аппарата, который должен
проверить, забрано ли именно то количество, которое подсказа-
но афферентным синтезом, т. е. обработкой всех нужд организма
в кислороде на данный момент. Именно к этому контрольному
аппарату придет обратная афферептация от альвеол обоих лег-
ких, информирующая о количестве реально взятого воздуха. Для
того чтобы понять более точные соотношения в деятельности ней-
ронов дыхательного центра, в ряде экспериментов был применен
микроэлектродный метод. Было показано, что на некоторые ней-
роны конвергируют возбуждения различного функционального
смысла: возбуждения, представляющие собой копию афферент-
ной команды и идущие по коллатералям нейрона, а также воз-
буждения, идущие с периферии по ветвям блуждающего нерва.
Мы сочетали момент включения этих нейронов с фазами ды-
хательного цикла. Оказалось, что вовлечения некоторых нейро-
нов соответствовали по времени и по характеру механизму конт-
роля полученного результата (Карпухина, 1967; Фельдшеров,
1967). Особенно подробно возможность участия коллатеральных
возбуждений пирамидного нейрона в формировании акцептора
результатов действия была изучена на примере физиологических
свойств возбуждений, распространяющихся по пирамидному ней-
рону (Правдивцев, 1967).
На основании всестороннего анализа этого феномена я могу
сказать, что в нашем поведении нет аппарата более универсаль-
ного, более быстродействующего и более точно направляющего
поведение, чем аппарат акцептора результатов действия. Поэтому
обычный вопрос, с которым мне приходится встречаться («где
локализован акцептор результатов действия?»), является физио-
логическим нонсенсом. Акцептор результатов действия как ши-
рокая система связей содержит в своем составе не только аффе-
рентные параметры конечного полезного результата данной си-
Системный анализ условного рефлекса
381
стемы, но и различно локализованные рецепторы для всех тех
обратных афферентаций, которые формируются на путях подсту-
па к полезному результату. Пример со светом и дезориентацией
поведенческого акта при устранении света достаточно убедитель-
но иллюстрирует это положение (см. выше).
Из всего сказанного о формировании акцептора результатов
действия следует, что этот аппарат представляет собой явное на-
рушение закона поступательного хода возбуждений по рефлек-
торной дуге, поскольку свойства еще не полученного результата
формируются в органический комплекс до получения самого ре-
зультата. Этот факт вынудил нас построить некоторую новую мо-
дель распространения и организации нервных процессов при фор-
мировании функциональных систем вообще и условного рефлекса
в частности (см. рис. 2).
Реализация полезного результата
Из предыдущего изложения ясно, что полезный результат си-
стемы, представленный всеми его афферентными признаками в
акцепторе результатов действия, проходит красной нитью через
всю функциональную систему, от стадии афферентного синтеза
до ее конечного момента — оценки полученного результата на
основе обратной афферентаций. Предсказанный результат реали-
зуется преимущественно через моторную сферу.
Как может акцептор результатов действия сформировать
именно те моторные возбуждения и именно на тех моторных ней-
ронах, которые в их интегративном соотношении дадут на пери-
ферии возбуждение нужного комплекса мышечных единиц?
Надо представить себе на минуту, сколь сложен этот выбор,
например, пирамидных нейронов, их синаптических образований и
интенсивностей возбуждения на каждом синапсе, чтобы понять,
когда достижение результата называется «рефлексом» по внеш-
нему описанию. И в то же время это движение во всех своих де-
талях должно соответствовать основной цели и принятому реше-
нию. Только при такой постановке вопроса можно видеть, сколь
упрощенными оказываются такие, например, выражения, как
«двигательный рефлекс», «рефлекс на мышцы» и т. д.
Действительно, допустим, что вы берете правой рукой стакан,
стоящий на письменном столе справа от вас, а затем через не-
сколько секунд протягиваете эту же руку к чайной ложке, нахо-
дящейся слева от вас. Почти безошибочно можно утверждать, что
в том и в другом движении участвуют одни и те же мышцы руки.
Но как тонко распределено участие отдельных мышечных воло-
кон этих мышц в том и другом движении. Можно без преувеличе-
ния считать, что каждое мышечное волокно этого многомиллион-
ного множества и по интенсивности, и по фазности сокращения
382
Системный анализ условного рефлекса
участвует в общем движении руки в своем индивидуальном свое-
образии. И все-таки в сумме все это необозримое разнообразие
производит движение, точно соответствующее тому акцептору ре-
зультата действия, который как «цель движения» сформировался
еще во время афферентного синтеза.
Какими средствами посылка возбуждений со всех участков
коры мозга распределяется так точно по нейронам пирамидной
зоны? Назовем этот комплекс чрезвычайно разнообразных воз-
буждений пирамидной области «командой» к совершению опре-
деленного движения. Тогда совокупность многих комплексов та-
ких же возбуждений, отходящих по впутрикорковым коллатера-
лям пирамидного тракта, совершенно определенно может быть
названа «копией команды» к совершению действия.
Этот механизм, не замеченный нейрофизиологией, несомненно,
способствует формированию акцептора результатов действия,
ибо он может поддерживать в длительном топическом возбужде-
нии всю совокупность его возбуждений до свершенпя реального
действия.
Вероятность такого энергетического питания дополнительными
реверберационными возбуждениями была показана в нашей ла-
боратории.
С помощью перерезки пирамидного тракта и электрического
раздражения этого тракта ниже места перерезки В. Правдивцев
и В. Сипичкип вскрыли один замечательный механизм (Правднв-
цев, 1967; Синичкин, 1967).
Они показали, что возбуждения, распространяющиеся но кол-
латеральным ответвлениям пирамидного тракта к ретикулярной
формации и к гипоталамусу, дают начало сильным потокам вос-
ходящих возбуждений, направляющимся при раздражении све-
том к зрительной зоне коры и к комплексу возбуждений на межу-
точных нейронах сенсомоторной коры. Как раз именно эти межу-
точные нейроны получают также «копию команды» по началь-
ным, т. е. корковым коллатералям пирамидного тракта (Нравдпв-
цев, 1967; Синичкин, 1967).
Создается совершенно определенное впечатление о всей ком-
позиции произвольных или особенно топких моторных актов,
осуществляемых, как известно, большей частью с помощью пира-
мидного тракта. Становится ясным, что «программа действия» —
это пе только просто выход эфферентного возбуждения па рабо-
чую периферию, как мы обычно понимаем. Нет, это активное
«вмешательство» потоков эфферентного возбуждения во все цент-
ральные интегративные процессы, благодаря чему создается воз-
можность как реализации, так и оценки будущего результата
действия.
На оспове этих механизмов само действие благодаря обрат-
ному влиянию через энергетически заряженные подкорковые об-
Системный анализ условного рефлекса
383
разовапия заряжает на длительное время аппараты цели дейст-
вия. И пе поэтому ли, часто несмотря на большой интервал вре-
мени, который проходит между постановкой цели получения
необходимого результата и самой реализацией этого результата,
последний оценивается всегда адекватно?
Попытаемся представить себе, что произошло бы, если бы ре-
зультат был получен с запозданием по отношению к цели, а ап-
парат оценки его, т. е. акцептор результатов действия, к этому
времени уже распался?
Вряд ли можно сомневаться, что при таких условиях невоз-
можно было бы какое-либо координированное поведение живот-
ных и человека. Весьма возможно, что некоторые формы так на-
зываемой апраксии связаны с дефектом именно этого энергети-
ческого обеспечения двигательной функции.
Вопрос о реализации запрограммированного результата имеет
самое близкое отношение к проблеме условного рефлекса, ибо он
дает возможность обсудить сравнительную характеристику
условного рефлекса и функциональной системы.
Как известно, по своей сути рефлекторная теория не предпо-
лагает и не учитывает результата действия (см. выше). Резуль-
тат отсутствует или большей частью полностью поглощается вы-
ражением «рефлекторное действие», или «рефлекторный ответ».
Как можно было видеть, это весьма существенное отличие. Со-
гласно же теории функциональной системы «рефлекторное дей-
ствие» непременно переходит в результат деятельности системы
и определяет собой всю архитектонику функциональной системы,
ибо уже первый узловой механизм системы (афферентный син-
тез) должен определить решение: какой результат должен быть
получен системой в данный момент. Именно в силу этого мы в
свое время и предложили ввести в физиологию результат дейст-
ствия в качестве самостоятельной физиологической категории
(Анохин, 1962; Анохин, 1968). По сути дела он и является тако-
вым во всех работах пашей лаборатории, что открыло перед нами
широкие перспективы по исследованию системной архитектоники
условного рефлекса.
Действительно, к чему обязывает принятие положения, что
рефлекс закапчивается пе действием, которое имеет только про-
межуточный характер, а пепримепно получением результата!
Классическая физиология, развивая представление о рефлексе,
никогда не анализировала этого ответственного пункта. В самом
деле, сказав «чесательный рефлекс», мы на этом и останавлива-
лись, поскольку классическая «дуга» этим завершалась. Но даже
в этом простом акте мы обычно не анализировали последующего
этапа — прекращения раздражающего действия на кожу,— что и
служит результатом, т. е. главным организующим фактором все-
го этого поведенческого акта.
384
Системный анализ условного рефлекса
Усладный
раздражитель
ь-
Сепреция
деззсладныи
раздражитель
(подкрепление)
Рис. 3. Рефлекторные дуги условного п безусловного рефлексов
Кроме того, если бы мы сказали, что в рефлексе имеется «ре-
зультат действия», мы немедленно должны были бы поставить
вопрос, какой аппарат призван оценить и скоррегировать ре-
зультат?
Результат есть конец деятельности функциональной системы
и с помощью обратной афферентаций, содержащей информацию
о его наиболее существенных параметрах, контролируется в ра-
нее сложившемся акцепторе результатов действия. В этом
пункте происходит логическое завершение системы как логиче-
ского блока, и никакой подменой понятия «подкрепления» на
«результат» здесь не выйти из затруднения.
Основой классической концепции об условном рефлексе яв-
ляется взаимодействие двух дуг рефлексов — условного и без-
условного, а безусловный раздражитель (подкрепление) служит
началом второй рефлекторной дуги безусловного рефлекса
(рис. 3).
Понятию «результат системы» в этом случае пет места. Имен-
но поэтому оно никогда не употреблялось, да и пе могло быть по
своей сути употреблено, поскольку результат, как было показано
выше,— неотъемлемая принадлежность системы, а информация
об этом результате возвращается к тому аппарату, который был
создан в начале формирования всей системы (акцептор резуль-
татов действия).
Функциональная система объединяет все процессы, входящие
в условный и безусловный рефлекс, в единый, органически свя-
занный блок, для которого характерна логическая поведенческая
законность.
Иначе говоря, результат выступает органическим узловым
механизмом функциональной системы как целостного образова-
ния. Системный анализ преодолевает представление о соотноше-
нии двух рефлексов — условного и безусловного и заканчивает
функциональную систему оценкой полученного результата.
Системный анализ условного рефлекса
385
Линять
Обстановоч-
ные афферен-
тации
I Акцептор
^результата'
действия I
Рефлектор,
действия
। Параметры ।
'результата^
Лу скобой ।
стимул ।
^Результат1.
1 действия I
^УМ/Z-
принятие
решения
\p£3yJW7KIfnQ I
, действия
Обета- Я
ковочные
афферентации I
Доминирующая
мотивация
Афферентный
синтез
Ламять
Обстанобоч- ।
ные афререн-Д
Лускобой ।
стимул I
побочные к-
афферентации |
Доминирующая
мотивация
Афферентный
синтез
Лринятие
решения
Эфферентные возбуждения
Обратная афферентация
\ ЛараметрыХ
'результата ।
Результат i
действия
\w/
Рнрлектор.
действия
Эфферентные возбуждения
Рис.- 4. Дальнейшее преобразование рефлекторной дуги реально участвую-
ункциональной системы: акцептор результатов дей-
щими механизмами
ствия (а) и сличение параметров реально полученного результата с пара-
метрами прогнозированного результата (б)
Рассматривая окончательный монтаж всех компонентов функ-
циональной системы, мы можем сопоставить рефлекторный стер-
жень как первоначальную основу для соотношения организма с
окружающим миром и архитектуру функциональной системы как
универсальный аппарат функционирования всякого поведенческо-
го акта (а не только условного рефлекса). Важно лишь, чтобы
они оба заканчивались полезным приспособительным результатом
(рис. 4).
На рис. 4 жирной чертой показан рефлекс в его классической
трактовке («толчок — центральные процессы — рефлекторный от-
вет»), а пунктирными линиями — функциональная система, ко-
торая, приобретя ряд существенных дополнительных аппаратов,
описанных выше, преодолела ряд недостатков рефлекса, мешав-
386
Системный анализ условного рефлекса
ших употребить его для расшифровки сложных поведенческих
актов.
В частности, такие поведенческие акты, как доставание пи-
щевой приманки с помощью накладывания ящиков один па дру-
гой (опыты И. П. Павлова с Розой и Рафаэлем), целиком и объ-
ективно расшифровываются на основе операциональной архитек-
тоники функциональной системы и не могут быть поняты только
па основе классического условного рефлекса. Именно в этом
смысле надо понимать знаменитое замечание II. И. Павлова по
поводу сложного поведенческого акта шимпанзе: «По это уже
условным рефлексом назвать нельзя ... Это уже не условный
рефлекс...» (Павловские среды, 1949).
Сделав попытку расшифровать сложные поведенческие акты
шимпанзе и оставаясь в пределах условпорефлекторных принци-
пов, И. П. Павлов увидел, что обычно принятые механизмы ус-
ловного рефлекса не могут объяснить многообразных форм ме-
няющегося поведения с испробывашгем многочисленных приемов
укладки ящиков в пирамиду.
В самом деле, при укладке даже первых ящиков обезьяна
неизбежно должна была, по описанию И. П. Павлова, оценить
самые разнообразные параметры своих «проб». Это мог быть
ящик под приманкой или в стороне от нее. Это могло быть самое
различное положение второго ящика па первом и т. д. Производя
эти многочисленные движения, шимпанзе должен был непрерывно
сличать полученный результат с конечной целью, уже сложив-
шейся в нервной системе животного еще до первой «пробы».
Выражаясь кибернетическим языком, мы можем сказать, что
на всем протяжении своих «проб» животное сканирует результат
по степени его приближения к конечному полезному — съедание
приманки. Совершенно очевидно, что на путях к конечному ре-
зультату поведения животное имело огромное количество проме-
жуточных результатов, каждый из которых был получен на осно-
ве полноценной архитектуры функциональной системы.
Трудность расшифровки этого поведения состоит и по сию
пору в том, что не был использован системный анализ поведения.
С точки зрения последнего у животного не может быть «хаоти-
ческого поиска», поскольку во всех случаях до первой пробы
произошел процесс афферентного синтеза и был сформирован
акцептор результатов действия. Налицо были все компоненты
афферентного синтеза. Именно все эти механизмы уже сложи-
лись в нервной системе Розы и Рафаэля еще задолго до первой
«пробы». Следовательно, никакого «хаоса» здесь не было, это
был в подлинном смысле целенаправленный поиск.
К сожалению, до сих пор не была сделана попытка систем-
ной расшифровки ставших уже лишними понятий «проба»,
«ошибка», хотя и не раз в своих статьях я указывал на возмож-
Системный анализ условного рефлекса
387
ность их расшифровки на объективной нейрофизиологической
основе.
Никакая «проба» в достижении какого-либо результата пе
может быть сделана без предварительного афферентного синтеза
и без предварительного формирования акцептора результатов
действия, ибо «проба», т. е. действие, должна быть оценена не-
медленно по своим результатам.
Точно так же любая «ошибка» может быть исправлена только
в том случае, если обратная афферентация от ошибочного ре-
зультата претерпела сличение в заранее запрогнозированном ак-
цепторе результатов действия.
Следовательно, системный анализ дает возможность с совер-
шенно очевидной легкостью преодолеть все те трудности, с кото-
рыми встретился в свое время II. П. Павлов при рефлекторном
объяснении сложных поведенческих актов. II. П. Павлов встре-
тился с цепью результатов, которые не могли быть объяснены с
точки зрения рефлекса, пе имеющего результата и пе имеющего
аппарата для оценки этого результата. Приходится просто удив-
ляться тому, как некоторые физиологи нрп разработке условного
рефлекса не могут заметить этого несоответствия. Вся эта чрез-
вычайно важная и поучительная проблема нашла широкий от-
клик в нашей научной прессе (Асратян, 1970; Бассин, 1971; Дуб-
ровский, 1971).
Заключение
Все рассмотренное выше представляет собой практическую
реализацию почти сорокалетпего опыта изучения системных за-
кономерностей многими сотрудниками нашей лаборатории. При
поверхностном подходе ко всем дополнительным узловым меха-
низмам, которые качественно преобразовали условный рефлекс
и в целом составляют операциональную архитектонику функцио-
нальной системы, может показаться, что они не изменяют реф-
лекторного подхода как к условному рефлексу, так и к поведен-
ческим актам вообще.
Однако легко видеть, что на самом деле это не так: описанные
выше и проиллюстрированные в рисунках дополнения радикаль-
но меняют качественную сторону исследовательской работы и
дают возможность построить более богатую перспективу изучения
как синтетических, так и топких нейрофизиологических законо-
мерностей условного рефлекса.
Как я показал выше (см. рис. 1 и 2), речь идет о принципи-
альном изменении традиционных черт рефлекса, о полном пере-
смотре начальной части реакций, где изолированный стимул пре-
вращается в многосторонний синтез основных компонентов и в
непрерывный перебор степеней свободы в поисках наиболее а дек-
388
Системный анализ условного рефлекса
ватного для данного момента «принятия решения». Это новый
мир взаимоотношений и закономерностей, включающий такие
основные нейромеханизмы, как конвергенция возбуждений на
нейроне, нейрохимическая специфика синаптических процессов у
различных нейронов, феномен реверберации кортико-подкорко-
вых возбуждений и др.
Еще более радикальным и, несомненно, качественным изме-
нением рефлекторной концепции является «принятие решения»
и формирование акцептора результатов действия как материаль-
ного эквивалента цели получения вполне определенного резуль-
тата. Этот последний аппарат формируется в нарушение одного
из самых основных показателей условного рефлекса — поступа-
тельного хода возбуждения по рефлекторной дуге.
И, наконец, получение полезного результата системы и актив-
ное включение его в операциональную архитектонику всей
и.
•унк-
циональной системы полностью порывает с классической трех-
членной дугой рефлекса. Этот решающий фактор любой систем-
ной организации является уже совершенно новым по отношению
к рефлекторному принципу деятельности и, в сущности, системо-
образующим фактором.
Однако решающий признак функциональной системы, прин-
ципиально отличающий ее от рефлекса, состоит в том, что услов-
ный рефлекс является лишь частным случаем для операциональ-
ной архитектоники функциональной системы, которая по своей
сути выступает универсальной закономерностью, распространяю-
щейся как в сторону наиболее высших функций мозга, так и в
сторону элементарных жизненных функций до молекулярных
процессов включительно,
В самом деле, как мы видели, теория
ункциональной систе-
мы раскрыла системную организацию дыхательной функции,
функции поддержания постоянства кровяного давления и т. д.,
в чем и состоит универсальный характер функциональной си-
стемы.
Этот факт, много раз реализованный как в концепции систе-
могенеза, так и в системном анализе многих вегетативных
и
унк-
ций (система кровяного давления, дыхания, осмотического давле-
ния и т. д.), выражает принципиальную черту функциональ-
ной системы, как общей теории для различных классов явлений.
В заключение мне хотелось бы ответить на одип вопрос, ко-
торый довольно часто нам задают исследователи, изучающие ме-
ханизм условного рефлекса: в чем теория функциональной систе-
мы обогащает само исследование условного рефлекса?
Все приведенные выше исследования, радикально изменившие
подход и трактовку условнорефлекторных процессов, совер-
шенно недвусмысленно отвечают на этот вопрос. Действительно,
уже сама стадия афферентного синтеза открывает необходимые
Системный анализ условного рефлекса
389
перспективы новых, совершенно оригинальных исследований для
оценки начального этапа формирования условного рефлекса.
В самом деле, вся проблема конвергенции разнородных воз-
буждений па одном и том же нейроне стала центральным меха-
низмом взаимодействия основных процессов, обеспечивающих
«принятие решения» животным. Одпако па уровне конвергенции
вопрос не заканчивается. Он неизбежно перерастает в проблему
химической интеграции, которая составляет суть работы отдель-
ного нейрона.
Как помогла нам функциональная система проникнуть в этот
уровень, мною уже не раз говорилось. Пришлось отказаться от
общераспространенного мнения, что центральным процессом ней-
рона является «электрическая суммация», т. е. сумма процессов,
возникших в силу мембранных закономерностей на поверхности
нейрона.
Сейчас же мне хочется лишь отметить те новые аспекты мик-
рофизиологических исследований, которые помогают нам попять
глубокую природу условнорефлекторных закономерностей, на-
ложенных в виде аналитических результатов на узловые меха-
низмы функциональной системы.
В качестве опорных пунктов мы взяли следующие факторы
(Швырков, Александров, 1971; Швырков, Безденежных, 1973):
а) конфигурацию вызванного потенциала на вспышку света
до выработки на него оборонительного условного рефлекса и
после выработки и уточнения этого рефлекса; б) различные типы
реагирования нейронов в момент развертывания условной реак-
ции в сопоставлении их с развитием узловых механизмов функ-
циональной системы; в) электрограмму реагирующих мышц как
показатель прихода условного возбуждения на периферию. Время
от предъявления вспышки света до появления миограммы прини-
малось по существующей традиции за «скрытый период» услов-
ной реакции; г) все моменты, изложенные в трех предыдущих па-
раграфах, были наложены на принятую в нашей лаборатории ар-
хитектуру функциональной системы.
Такое наложение было особенно интересным в том отношении,
что оно давало возможность определить время формирования
каждого узлового механизма функциональной системы... в ка-
кой-то степени неожиданно раскрыли неизвестные нам свойства
нервной системы, которые позволяют «вписать» довольно обшир-
ные системные процессы в весьма малые интервалы времени
Как мы показали, опыты с классическими условными рефлек-
сами имеют то характерное качество, что задолго до применения
условного стимула в нервной системе животного складывается
уже полностью готовая предпусковая интеграция, включающая в
себя все компоненты афферентного синтеза за исключением пу-
сковой афферентаций, т. е. возбуждений от пускового стимула.
390
Системный анализ условного рефлекса
Стоит лишь на минуту представить себе, сколько новых и ин-
тересных вопросов ставит это сопоставление. Мы можем искать
решающие механизмы выхода предпусковой интеграции из
«скрытого» состояния в активное, а также поставить вопрос: на
каких уровнях и на каких нейронах происходит это принятие ре-
шения?
Именно для этого и было произведено сопоставление компо-
нентов вызванного потенциала со стадиями развития архитекту-
ры функциональной системы. На данном последнем пути получи-
лись поразительные результаты: первые компоненты, т. е. поло-
жительно-отрицательный комплекс, сопутствуют развертыванию
всех первых фаз функциональной системы, в то время как позд-
ние компоненты вызванного потенциала соответствуют самому
действию и, вероятно, последующим фазам функциональной си-
стемы: получению результата, формированию обратной афферента-
цпи и сличению ее в акцепторе результатов действия (Швырков,
Александров, 1971; Швырков, Безденежных, 1973).
Если ко всему сказанному добавить теперь, что на всем про-
тяжении развития узловых механизмов функциональной системы
включаются то одни, то другие нейроны в различных корковых и
подкорковых образованиях, то едва ли можно сомневаться в том,
что функциональная система служит подлинной матрицей для
идентификации всех тонких процессов, разыгрывающихся при вы-
работке условных рефлексов,,.
ЛИТЕРАТУРА
Анохин П. К., Стреж Е, Ф. Изучение динамики высшей нервной деятель-
ности. Ш. Нарушение активности выбора в результате замены безуслов-
ного стимула.— Физиол. журн. СССР, 1933, т. 16, № 2, с. 280.
Анохин П. Я. Особенности афферентного аппарата условного рефлекса и
их значение для психологии.— Вопр. психол., 1955, т. 6, с. 16.
Анохин П. К. О физиологическом субстрате сигнальных реакций.— Журн.
высш, нервн. деят., 1957, т. 7, № 1, с. 39.
Анохин II. II. Рефлекс цели как объект физиологического анализа.—Журн.
высш, нервн. деят., 1962, т. 12, № 1, с. 7.
Анохин П. К. Биология и нейрофизиология условного рефлекса. М., 1968.
Анохин П. К. Химический континуум мозга как механизм отражения дейст-
вительности.— Вопр. филос., 1970, № 6, с. 107.
Асмаян II. В. Регуляция поведения собак при наличии у них голода и жаж-
ды.— В кн.: Проблемы лечебного голодания. М., 1969, т. 57, с. 47.
Асмаян Н. В., Голицын Г. А. О принятии решения в ситуации выбора.—
Вопр. психол., 1971, № 1, с. 72.
Асратян Э. А. Каузальный условный рефлекс.— Вопр. филос., 1970, № 10,
с. 112.
Басин Н. Ф. Открытое письмо Э. А. Асратяну.— Вопр. филос., 1971, № 4,
с. 159.
Дубровский Д. II. Психические явления и мозг. М., 1971.
Журавлев Б. В. Изучение нейронов гипоталамуса как центрального пункта
кортико-висцеральных взаимоотношений при пищевых реакциях,—-
Системный анализ условного рефлекса
391
Материалы 9-й Всесоюзной научной конференции по проблемам кор
тико-висцеральной физиологии. Баку, 1971, с. 91.
Карпухина Л. М. Электрическая активность нейронов дыхательного центра
в бионическом аспекте: реакция дыхательных нейронов на рассогласо-
вание команды с результатом действия.— В кн.: Некоторые проблемы
биокибернетики, применение электроники в биологии и медицине. Киев,
1967, в. 1, с. 56.
Лаптев И. И. Выработка пищевой и оборонительной условной реакции на
один и тот же раздражитель.— 3-е совещание по физиологическим проб-
лемам. М.—Л., 1938, с. 34.
Макаров В. А. Роль миндалевидного комплекса в механизме конвергенции
возбуждений различной сенсорной модальности на нейронах коры боль-
ших полушарий.— Докл. АН СССР, 1970, т. 194. № 6, с. 1454.
Павлов И. 11. Поли. собр. трудов. М.— Л., т. 4, 1957.
Павловские среды. М., 1949, с. 261.
Полянцев В. А. Изучение деятельности дыхательного центра как экспери-
ментальной модели нервного центра.— В кн.: Вопросы физиологии и
патологии нервной системы. М., 1962, с. 26.
Правдивцев В. А. Эфферентно-афферентная конвергенция на нейронах зри-
тельной коры.— В кн.: Интегративная деятельность мозга. М., 1967, с. 67.
Синичкин В. В. Особенности распространения возбуждения, вызванного ан-
тидромным раздражением пирамидного тракта на нейроны зрительной
области коры головного мозга.— В кн.: Интегративная деятельность
мозга. М., 1967, с. 72.
Судаков К. В. Биологические мотивации. М., 1971.
Украинцев Б. С. Целеполагание и целеосуществление как один из принци-
пов самодвижения функциональных систем.— В кн.: Принципы систем-
ной организации функций. М., 1973, с. 62.
Фельдшеров Ю. 11. Характеристика участия дыхательных нейронов в инте-
гративной деятельности дыхательного центра.— В кн.: Юбилейный
сборник научных работ аспирантов и ординаторов I ММИ им. Сеченова.
М., 1967, с. 35.
Хаютин С. 11. Характеристика активности одиночных нейронов зрительной
коры в условиях выраженной пищевой мотивации.— Журн. высш,
нервн. деят., 1972, т. 22, № 6, с. 1248.
Швырков В. Б., Александров Ю. 11. Связь вызванных потенциалов и разря-
дов нейронов в соматосенсорной коре кролика.— Материалы 6-й Все-
союзной конференции по электрофизиологии центральной нервной си-
стемы. Л., 1971, с. 291.
Швырков В. Б., Гринченко Ю. В. Электрофизиологическое изучение акцеп-
тора результатов действия в инструментальном поведении.— Журн.
высш, нервн. деят., 1972, т. 22, № 4, с. 792.
Швырков В. Б., Безденежных Б. И. Роль анализаторов условного и безуслов-
ного раздражений в функциональной системе условнорефлекторного
поведенческого акта.— Журн. высш, нервн. деят., 1973, т. 23, № 1, с. 15.
Шумилина А. И. Экспериментальный анализ корково-подкорковых ревер-
бераций возбуждений методом вызванных потенциалов при угашении
условной оборонительной реакции.— Труды Института нормальной и
патологической физиологии. М., 1966, т. 9, с. 5.
Шумилина А. 11. Функциональное значение лобных областей коры головно-
го мозга в условпорефлекторпой деятельности собаки.— В кн.: Лобные
доли и регуляция психических процессов. М., 1966, с. 61.
Эшби У. Р. Конструкция мозга. М., 1962.
ПРОБЛЕМА ПРИНЯТИЯ РЕШЕНИЯ
В ПСИХОЛОГИИ И ФИЗИОЛОГИИ *
Понятие «принятие решения» стало популярным в последние
годы, хотя (надо отдать справедливость) под сильным давлени-
ем нейрофизиологии. Часто это понятие переводится в область
терминологическую, в область попятий, более близких к житей-
ским, чем к научным. Обязательная необходимость ввести науч-
ное понятие «принятие решения» появилась в процессе разработ-
ки различных, больших и малых, систем, когда стало важным
определить этап, на котором заканчивается формирование и начи-
нается исполнение какого-либо акта, т. е. можно сказать, что си-
стема приняла решение.
В физиологии поведенческий акт долгое время рассматривал-
ся как прямая связь: стимул на входе определяет реакцию на
выходе. Если подходить к развитию событий с точки зрения ме-
ханистического детерминизма (т. е. звено к звену), то по сути де-
ла нет и не нужно никакого решения — ему не было места. Тем
не менее наблюдения показывали, что может быть много входов,
а выход выбирается только один — и этот выход детерминистиче-
ски, причинно ие может быть связан пи с каким отдельным вхо-
дом. Становилось яспым, что какая-то система интегрирует все
входы и принимает решение о выходе на основе учета всех вхо-
дов. Так возникло убеждение, что механистический подход не смо-
жет объяснить ту сложность, которая имеется в организме, и осо-
бенно на высшем уровне, где решение делается постоянным эта-
ном в поведении человека.
Следовательно, первый и важный момент: решение есть неиз-
бежный предмет интеграции и предмет выбора — из многих воз-
можностей выбирается одна, которая способна обеспечить орга-
низм или машипу наилучшим эффектом.
И тут появилось некоторое взапмонепонимание между специ-
алистами разных наук, поскольку в проблему решения включи-
лись специалисты всех видов (см., например, материалы, издан-
ные Институтом кибернетики АН Грузинской ССР после об-
♦ Опубликовано в журнале «Вопросы психологии» (1974, № 4). Стенограм-
ма последнего выступления акад. П. К. Анохина в Институте психоло-
гии АН СССР 23 января 1974 г.
Проблема принятия решения в психологии и физиологии
393
стоятельной, очень представительной конференции по принятию
решений).
Оказалось, что почти при всяком обсуждении понятия «при-
нятие решения» начинают с решения как такового, т. е. со струк-
туры решения как соответствующего начала для каких-то дея-
тельностей и т. д. Но решение является не началом, а результа-
том какой-то очень тонкой и обширной работы, проделанной моз-
гом. Поэтому выявилась необходимость вовлечь в сферу внимания
не только само принятие решения, но и стадию «предрешения»,
которое формирует решение и предопределяет его направ-
ленность. И тут оказалось, что принятие решения невозможно без
этой стадии предрешения, которая делает самую «культурную»,
самую «интеллигентную» работу, а именно определяет: какие об-
стоятельства должны быть учтены при принятии решения, из ка-
кого набора нужно выбирать ту деятельность организма или ма-
шины, которая окажется наиболее адекватной, и какие альтерна-
тивы рассматривать. На заключительном заседании этой
конференции пришли к выводу, что в сферу внимания должно
войти и предрешение.
Поэтому выражение «принятие решения» не должно быть гип-
нотизирующим фактором, это есть конечный акт одного весьма
разветвленного процесса и начало другого.
Возникает вопрос: какой же универсальный стержень может
быть предложен, чтобы выделить принятие решения и сделать
его объективно изучаемым процессом? Другая сторона этого воп-
роса: полный ли здесь изоморфизм по отношению ко всем видам
принятия решения в различных направлениях? Мы долго над
этим думали и работали, и оказалось, что общая теория функцио-
нальных систем дает такой стержень. Это не значит, что другого
нельзя найти, но сейчас она является единственной моделью, ко-
торая позволяет изучать самые тонкие механизмы принятия ре-
шения в различных системах и охватывает как стадию предреше-
ния, так и само решение и последствия решения.
В функциональной системе принятие решения не является изо-
лированным механизмом, изолированным актом, а служит эта-
пом в развитии целенаправленного поведения.
Это очень важный переход. В теории функциональной систе-
мы мы ставим на первое место не «отдельности», например па-
мять, как что-то изолированное. Память работает в системе на
своем месте, играет свою роль и т. д. Это же относится и к мо-
тивации, и к принятию решения, т. е. система объединяет все
эти позиции.
Подходя с этой точки зрения, мы и должны поставить воп-
рос: чему служит решение, следствием чего оно является и на
что оно направляет организм?
Первый и важный момент — нельзя осуществлять решение во-
394
Проблема принятия решения в психологии и физиологии
обще, решение, не направленное на какую-то деятельность, на ка-
кой-то положительный результат.
Можно ли принять решение, которое в будущем не обещает
никакого полезного эффекта? Очевидно, нет. Следовательно, если
говорить о принятии решения, то в нашем понимании (физиоло-
гическом и психологическом) без мотивации, которая должна
быть удовлетворена (она может быть представлена в разных ви-
дах, в простом грубом виде — типа голода, жажды, по она может
быть представлена и в виде желания поступить в институт),
нельзя обойтись. Мотивация выступает обязательным фактором,
определяющим и устанавливающим форму решения, тип реше-
ния, его общие очертания.
Мы выявили очень важный момент. Мотивация начинает про-
цесс выбора из памяти, из прошлого опыта всего того, что было в
жизни данного организма связано с удовлетворением этой мотива-
ции. Мы долго бились над тем, чтобы понять, до какой степени
мотивационное возбуждение, исходя из гипоталамуса, пли форми-
руя мотивационный комплекс с участием коры,— до какой степе-
ни это возбуждение «вытягивает» прошлый опыт: извлекается ли
только сенсорная часть или и действия, которые были совершены
по поводу данной мотивации, или и результат, который был полу-
чен при удовлетворении мотивации.
Мы пришли к выводу, что мотивация действительно извлека-
ет из памяти весь прошлый опыт с результатами включительно.
Когда вы хотите есть, вы можете эту мотивацию удовлетворить
разными способами. Если вы находитесь в степи, вы откроете
сумку и достанете охотничьи сосиски; в вагоне вы удовлетворите
голод другим путем, в городе — третьим (вы можете пойти в ре-
сторан). Пути удовлетворения могут быть различны, и мотивация
выбирает все акты из памяти, которые когда-то были связаны в
прошлом с удовлетворением этой мотивации. Какой конкретный
путь будет избран — это решает обстановка.
Таким образом, извлечение определенной мотивацией опреде-
ленного жизненного опыта (с коррекцией воздействия от внеш-
ней обстановки) и приводит к принятию именно этого, а пе иного
решения, т. е. само принятие решения опосредствовано очень
сложным процессом «примерки» жизненного опыта к данной си-
туации. Если вы в открытой степи захотели пить, то не будете
искать киоск с газированной водой, потому что обстановка толка-
ет на принятие другого решения. Поэтому, если мы возьмем при-
нятие решения как процесс, изолированный от мотивации, памя-
ти и внешних воздействий, мы не сможем вскрыть его законо-
мерностей.
Этот вопрос — место принятия решепия в целом, целена-
правленном произвольном акте с участием психических процес-
сов — является, с моей точки зрения, важнейшим в познании ре-
Проблема принятия решения е психологии и физиологии
395
шения. Если бы мы только изолировали решение, мы ничего не
могли бы сказать о принятии решения. Поэтому предрешение в
стадии афферентного синтеза (т. е. объединения всех афферен-
тов, которые предшествуют решению) оказалось в высшей степе-
ни важным в психической деятельности человека, более важным
для понимания решения, чем само принятие решения, потому что
в этой стадии расценивается, сопоставляется опыт, мотивация и
обстановка, и все это предопределяет конкретное решение.
Нейрофизиологические эксперименты показали, что все компо-
ненты афферентного синтеза встречаются на одном и том же ней-
роне, Это, конечно, пе одна клетка в мозге, а миллиарды, но
оценка производится па одном и том же нейроне, и только выс-
шая интеграция соединяет все эти решения. Таким образом, хотя
общее решение принимается всем мозгом, взаимодействие компо-
нентов афферентного синтеза, приводящее к тому или иному ре-
шению, происходит на уровне отдельного нейрона.
Вот почему все это должно привести и психологов и
Н
1ИЗИО-
логов, как и других специалистов, к решению вопроса о том, как
же нейрон может это делать, как он может сопоставить все дан-
ные прошлого опыта с внешней обстановкой в данный момент, что
и определит включение этого нейрона в новую систему для реали-
зации одного-едпнственного решения.
Взаимодействие компонентов афферентного синтеза на нейро-
не не может быть простым сложениехм разнородных возбуждений,
и проблема принятия решения в значительной степени зависит
от выяснения вопроса о том, каким образом в результате конвер-
генции к отдельному нейрону различных по природе влияний осу-
ществляется выбор наиболее благоприятной в данной обстановке
формы его активности.
Однако решение в поведенческом акте принимается, конечно,
не одним нейроном, а всем мозгом. Вот почему я хотел бы, чтобы
стадия предрешения была освещена и вовлечена в обсуждение;
без этой стадии, конечно, трудно понять, как складывается ре-
шение.
Принятие решения представляет собой поворотный пункт,
в котором происходит организация комплекса афферентных воз-
буждений, способного дать вполне определенное действие.
При любых условиях мы имеем выбор одного акта и исключе-
ние всех остальных потенциальных возможностей. Выбор этого
акта есть создание эфферентного интеграла, в котором согласова-
ны, «пригнаны» друг к другу определенные формы активности
огромного числа отдельных механизмов. Принятие решения пере-
водит один системный процесс — афферентный синтез в другой
системный процесс — программу действий. Оно является переход-
ным моментом, после которого все комбинации возбуждений при-
обретают исполнительный характер.
1
396 Проблема принятия решения в психологии и физиологии
В последние годы мы пришли к очень важному выводу по
поводу принятия решения, по поводу системных процессов. Этот
вывод заключается в том, что проблема принятия решения состо-
ит из двух частей: с одной стороны, это задача нейрона, с дру-
гой — проблема интеграции нейронов в единую систему, так как в
конце концов решение принимается всем мозгом. Это совершенно
новая проблема, над ней работает лаборатория нейрофизиологии
обучения нашего института. Это вопрос о том, каким образом ин-
формация, обрабатываемая в стадии предрешения отдельными
нейронами, соединяется в стадии решения в мощный эфферент-
ный интеграл.
Здесь необходимо ввести новое понятие, очень перспективное
для психологии, понятие степени свободы нейрона. Это понятие
открывает широчайшие перспективы для понимания того, как
нейроны соединяются между собой.
Что такое степени свободы?
На нейрон поступают тысячи возбуждений, а «выпускает)) он
через аксон одно возбуждение. Если бы в любой момент любой
вход вызывал ответ нейрона, то никакой интеграции не могло бы
быть, наступил бы полный хаос. Для включения нейрона в систе-
му необходимо устранение его излишних степеней свободы и ис-
пользование только тех входов, которые вызывают совершенно оп-
ределенную форму активности данного нейрона, способствующую
успеху системы. Система создает интегративное состояние нейро-
на за счет тысячи разномодальных влияний и делает его чувстви-
тельным то к одним, то к другим входам, использует то одни, то
другие его степени свободы и устраняет избыточные.
Принятие решения освобождает организм от чрезвычайно
большого количества степеней свободы и оставляет одну, которая
и реализуется. Организм только тогда примет решение, когда ней-
роны «подгонят» друг к другу своп степени свободы, что п создаст
их интеграцию и возможность согласованной работы. Эта подгон-
ка степеней свободы элементов лежит в основе всякой интеграции
и будет, вероятно, занимать в наших экспериментах центральное
место.
Я учитываю, что психический процесс принятия решения бо-
лее сложен и требует более углубленного анализа, но мы работа-
ем, чтобы поставить мост между психическими процессами и
между нейроном, чтобы этот переход был оправдан. Для этого
перехода и для создания общей картины необходимы и точные
психологические данные.
Я думаю, что принятие решения, которое мы сделали пред-
метом обсуждения, является наиболее важной проблемой. В при-
нятии решения очень тесно соприкасаются и нейрофизиологиче-
ские основы психической деятельности, и физиологические фено-
мены, и проекции на другие дисциплины.
Проблема принятая решения в психологии и физиологии
397
Поэтому комплексное обсуждение этого вопроса должно по-
служить центральной линией Института психологии.
Посмотрим, какие будут итоги после совещания. Я думаю, что
мы не должны смущаться, если успехи будут небольшими.
Поэтому, может быть, начнем новый путь, и в этом отношении
я желаю всяческого успеха в работе семинара...
Теперь посмотрим, чем заполняется у комара и у человека эта
функциональная система. Здесь большое различие. Есть аффе-
рентный синтез у комара, который интегрирует информацию о
температуре, влажности и т. д. и «принимает решение» — лететь
к оврагу (там прохладнее). У него таких оцениваемых качеств
4—5. Это входные ориентации, которые решают выбор.
Возьмем человека, решающего, куда ему поступить учиться.
Ему предлагают в институт пищевой промышленности или стома-
тологический. Сколько он выбирает входных информаций для
того, чтобы принять решение! Архитектура та же, по содержание
и конкретные нейрофизиологические механизмы различны, пе го-
воря уже о социальных факторах, определяющих поведение чело-
века.
Это значит, что мы и то н другое должны иметь в виду при
анализе.
Теперь должно быть понятно, что такое архитектура функ-
циональной системы и ее заполненность для различных организ-
мов. Ио архитектура всегда служит базой для начала анализа.
Далее вопрос о психологии и физиологии.
Мы изучаем человека, нам важна человеческая психология.
Если бы так говорили по поводу генетики! Вся генетика разра-
батывается на дрозофиле и на бацилле Коли. Все это оказывает-
ся применимо и для человека. Поэтому при изучении любой слож-
ной человеческой деятельности надо иметь в виду прежде всего
изоморфную функциональную систему, изоморфное функциониро-
вание и специфические черты, которые в этот узловой механизм
вложены. Недавно мне пришлось участвовать в одной дискуссии
по большим системам в нефтеобрабатывающей промышленности.
Там нет ни электродов, ни нервных клеток, но я был поражен, до
чего изоморфна сама функционирующая структура. Возьмите лю-
бое учреждение. Из какого-то министерства команда посылается,
например в Оренбург. Каждое министерство обязательно оставля-
ет копию. Это универсальный принцип организма. Организму не-
обходимо, начиная действие, оставлять у себя копию команды, что
можно показать даже на уровне нейрона. Вы видите
морфны принципы функционирования. Оставляется копия коман-
ды, чтобы проверить результаты команды. Мы называем это
«контролем исполнения».
Таким образом, говоря о человеке, мы должны различать эти
ундаментальные проблемы: проблему единства функцио-
как изо-
Две
398
Проблема принятия решения в психологии и физиологии
нальной архитектуры и проблему заполненности этой архитек-
туры...
В течение многих лет мы изучаем живые плоды человека,
взятые после какой-то операции. Естественно, мы не могли не по-
интересоваться, как складывается человек. Очень часто были не-
доноски довольно ранние, один весил 520 граммов. Он выпивал
8 кубиков молока. Нормальный новорожденный выпивает 100 ку-
биков, крупный ребенок даже 150 кубиков, а он — только 8. Но
это говорит о том, что у него функциональная система сосания в
своих узловых механизмах уже была готова. Нас занял вопрос —
в какой степени предсказание является врожденным, а в какой
оно приобретается опытом? Оказывается, для формулы принятия
молока акцептор результата действия готов. Мы сделали автома-
тическую соску и изучали детей, которые не видели еще мать.
И оказалось, что сравнительный аппарат готов у новорожденных.
Стоит немного прибавить кислоты пли соли — новорожденный не-
медленно выплевывает. Значит, так называемые инстинкты (ко-
торые у нас совершенно расшифрованы под этим углом зрения)
пользуются этой «установкой», как говорят кибернетики. Я при-
веду в пример так называемых дарвинских вьюрков на Галапа-
госских островах. Па всех островах у них выработались разные
способы доставания червяка из гнилого дерева. По очень интерес-
но: вьюрок обследует дерево, видит дырку с червяком, улетает,
находит где-то кактус, обкалывает его, берет иголку в клюв, летит
к дереву и начинает иголкой вытаскивать червяка. Что он «ду-
мал», когда летел туда, что он «думал», когда летел обратно? Без
целей, без акцепторов результатов это поведение невозможно.
В. Л. Полянцев провел интересный эксперимент и открыл, что
в дыхательном центре есть акцептор результатов действия, кото-
рый оценивает, взяли ли легкие столько воздуха, сколько требует-
ся организму. Команду дыхательный центр дает тогда, когда он
оценит потребность организма по показаниям афферентаций, по-
ступающих из аорты, вен, мышц, получит данные от всех органов
и синтегрирует их. Архитектура может быть заполнена разным
содержанием, по я ни разу не видел, чтобы были основания отка-
заться от архитектуры.
Так что универсальность этих врожденных механизмов еще
больше подчеркивает, что мы должны думать об очень широком
изоморфизме. Этот вопрос у меня очень подробно разработан в
специальной книге, которая называется «Принципы системной ор-
ганизации функций». Во вступительной статье разбирается вопрос
об изоморфизме функциональной системы...
Заключительные замечания. Общие закономерности,
о кото-
рых я говорил, дают себя знать и па высшем уровне. Возьмите
наши знания о мембране нервной клетки и яйца морского ежа.
Структура мембраны морского ежа та же, что у самой «умной»
Проблема принятия решения в психологии и физиологии
399
клетки мозга. Эти закономерности имеются у самых низших жи-
вотных. Поэтому с точки зрения подхода, поисков схемы мы
должны учитывать эти закономерности. Не случайно во всем ми-
ре, во всех научных учреждениях, разрабатывающих эксперимен-
тальную психологию, исследователи не покидают проблем челове-
ческого сознания, человеческий психики. Для них это — руково-
дящий принцип работы, но они работают па крысах. Мы должны
помнить, что есть и общие закономерности, общие принципы
функционирования и специфика.
11а вопрос о том, как сочетать биологические и высшие чело-
веческие психические функции, я отвечаю таким образом: обо-
лочка клеток или количество ДНК — РНК у бактерий такое же,
как у человека. Разумное сочетание универсальных черт функци-
онирования и того, что должно быть базой для понимания специ-
фических черт, которые приобрел человек в процессе эволюции —
это будет научной психологией...
СОДЕРЖАНИЕ
От редакторов............................ . • • • •
Опережающее отражение действительности............. 7
Философские аспекты теории функциональной системы 27
Принципиальные вопросы общей теории функциональ-
ных систем................................. 49
Философский смысл проблемы естественного и искус-
ственного интеллекта...........................107
Системогенез как общая закономерность эволюционно-
го процесса.................................... . 125
Методологический анализ узловых проблем условного
рефлекса................................... 152
Физиология и кибернетика........................ 207
Кибернетика и интегративная деятельность мозга . . . 234
Методологическое значение кибернетических законо-
мерностей ....................................262
Рефлекс цели как объект физиологического анализа . . 292
Эмоции........................................... 311
Психическая форма отражения действительности .
Системный анализ условного рефлекса
336
367
Проблема принятия решения
в психологии и физио-
логии
392
ПЕТР КУЗЬМИЧ АНОХИН
Избранные труды
ФИЛОСОФСКИЕ АСПЕКТЫ ТЕОРИИ ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ
Утверждено к печати Институтом психологии АН СССР
Редакторы: В. И. ЕвсеЪичев, Н. II, Кондакое. Художник Л. С. Эрман,
Художественный редактор С. А. Литвак. Технический редактор В. Л. Кунинл
Корректор В. A, j
гварцер
ИБ № 5061
Сдано в набор 27.01.78. Подписано к печати 21.07.78. Т-10739. Формат 60 x 90»/lt
Бумага типографская № 2. Гарнитура обыкновенная. Печать высокая.
Усл. печ. л. 25. Уч.-изд. л. 27,6. Тираж 5900 экз. Тип. зак. 159. Цена 1 р. 90 к»
Издательство «Наука*, 117485, Москва, В-48Б, Профсоюзная ул
94а
2-я типография издательства «Наука», 121099, Москва, Г-99, Шубинский пер., 10