























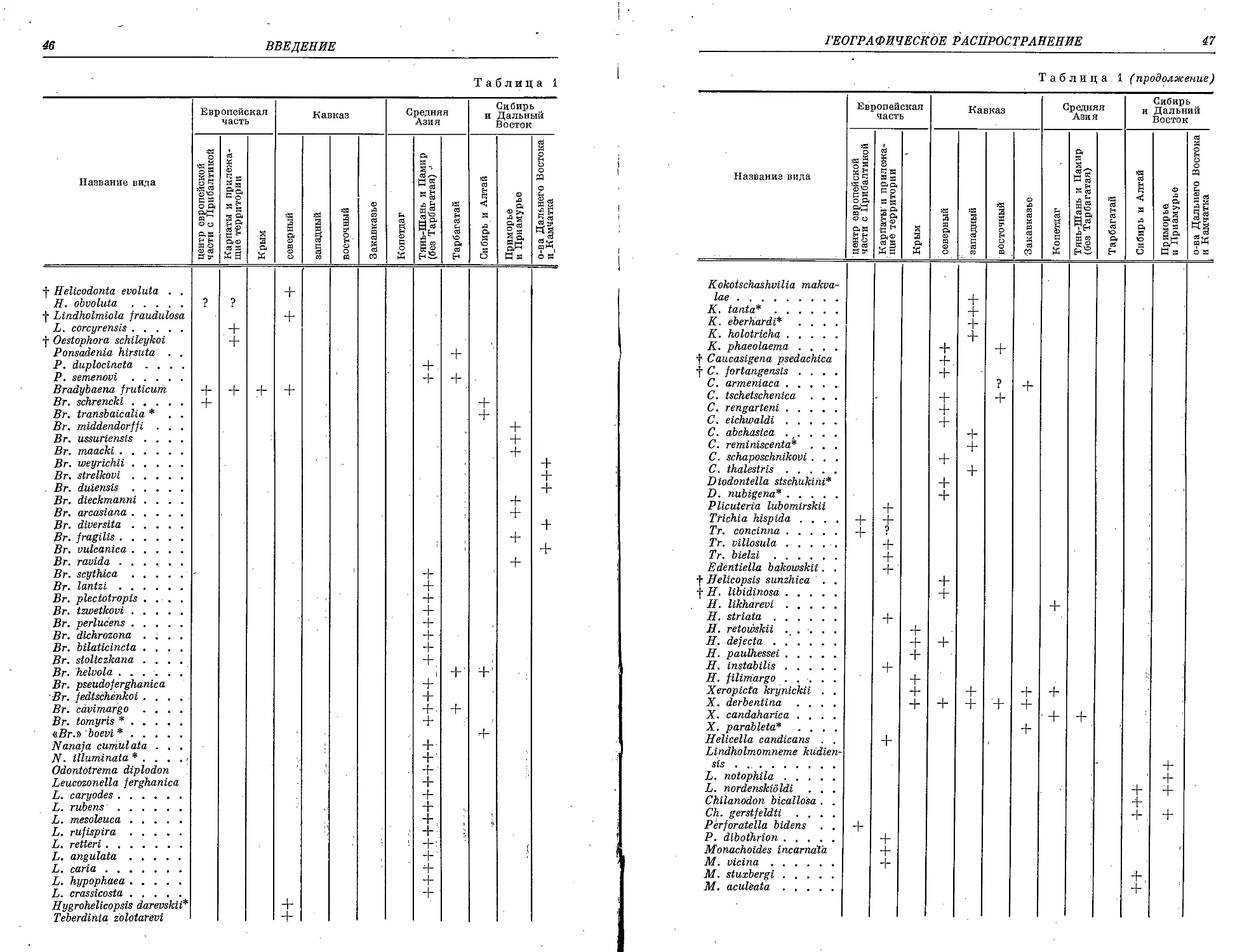

































































































































































Текст
ФАуНА
СССР
МОЛЛЮСКИ
ш
выпуск
6
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ, № 117
ФАУНА СССР
МОЛЛЮСКИ
Том III, вып. 6
А. А. ШИЛЕЙКО
НАЗЕМНЫЕ МОЛЛЮСКИ НАДСЕМЕЙСТВА
HELICOIDEA
Под редакцией
| А. А. СТРЕЛКОВА
ЛЕНИНГРАД
«НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1978
УДК 594.38
Наземные моллюски надсемейства Helicoidea. Шилейко А. А.
(В серии: Фауна СССР. Моллюски. Т. Ш, вып. 6). Л., «Наука»,
1978. 384 с.
Систематический обзор всех видов Helicoidea, обитающих на территории
СССР, как современных (166 видов), так и ископаемых (15 видов). Общий
очерк организации представителей надсемейства. На базе морфо-функ-
ционального анализа сделана попытка реконструировать ход историче-
ского развития группы в целом и проведена таксономическая ревизия;
в результате система таксона сильно перестроена. Таблицы для определе-
ния семейств, подсемейств, родов, подродов и видов. По каждому виду,
за незначительным исключением, приведены фотографии раковин и изо-
бражения внешнего вида и внутреннего отроения полового аппарата.
Лит. — 386 назв., ил. — 471 рис. и 21 табл, вкл., табл. — 2.
Главный редактор серии «Фауна СССР»
директор Зоологического института АН СССР
О. А. Скарлато
Редакционная коллегия:
А. А. Стрелков | (отв. ред. серии), И. М. Лихарев (зам. отв. редактора),
И. М. Громов, В. Ф. Зайцев, О. Л. Крыжановский, Л. /1. Кцтикова,
Г. С. Медведев, М. Е. Тер-Минасян
21008-538
Ш055(02)-78527'7/
<С Изчательство «Наука», 1978
ПРЕДИСЛОВИЕ
«Was ist Helix?»
H. Iliering, 1892
Среди основных крупных группировок наземных легочных моллюсков надсемей-
ство Helicoidea — одно из самых заметных и самых известных. Такая популярность
связана с широким распространением группы, ее большим объемом (в мировой фауне
насчитывается более 2500 видов), а также с тем, что представители Helicoidea, отлича-
ясь зачастую большими размерами, образуют порой значительные скопления и бро-
саются в глаза даще неискушенному наблюдателю. Этими же обстоятальствами опре-
деляется и большая роль названных моллюсков в общей экономике природы. Кроме
того, отдельные виды с давних пор — излюбленные объекты изучения для биологов
самых различных специальностей, благодаря чему по ним существует обширная
литература. Особенно это касается крупных видов, таких как Helix pomatia, Cryptom-
phalus aspersa, Eobania vermiculata, видов рода Cepaea и др.
Многие виды надсемейства — серьезные вредители сельскохозяйственных культур
и важные переносчики различных паразитических нематод и трематод.
Вместе с тем существует резкое противоречие между широкой известностью и зна-
чительностью этой группы для человека и практически полным' отсутствием попыток
разработать, систему группы. Точнее, такие попытки предпринимались (Iliering, 1892,
1909; Lindholm, 1927а), но они не пошли дальше построения формальной классифика-
ции по ограниченному набору разрозненных признаков, установления границ отдель-
ных родов и составления их диагнозов.
Несовершенство наших представлений о систематической структуре надсемейства
в большой степени связано с крайне бедной признаками раковиной, характерной для
его членов. Трудности усугубляются широкой внутривидовой изменчивостью боль-
шого числа видов и наличием многочисленных случаев конвергентного сходства вйдов
из разных таксонов. В результате составить дифференциальный родовой диагноз
сплошь да рядом невозможно. Даже в тех редких случаях, когда раковина отдельных
видов отмечена ярким своеобразием, не исключено конвергентное сходство: европей-
ский Isognomostoma и североамериканские Tiiodopsis; северо-западноафриканские
Tingitana и филиппинские Pyrochilus.
Нет нужды лишний раз повторяться, пропагандируя преимущества анатомиче-
ского метода в изучении системы брюхоногих моллюсков. перед психологическим.
Однако стоит подчеркнуть, что так называемый анатомический метод в. понимании
многих, даже современных малакологов заключается во внешнем и часто довольно
поверхностном осмотре полового аппарата. Вместе с тем раскрытие путей эволюции
множества рецентных таксонов должно быть основано не только на детальном исследо-
вании внешнего строения гениталий, но и на выяснении их внутренних структур.
Такое исследование позволяет нам, не ограничиваясь регистрацией наличия или от-
сутствия определенных морфологических элементов, выявить характер взаимоотноше-
ний между таксонами, точнее, определить таксономический вес отдельных признаков
или состояний признака, и, что особенно важно, выявить «векторизованные» черты,
позволяющие определить тенденции в морфолого-историчесйом развитии группы.
Углубленное изучение организации животных, и в первую очередь полового аппарата,
дает возможность установить, на какие признаки можно опереться, разрабатывая
систему конкретного таксона.
В связи со сказанным возникает настоятельная необходимость максимально
точной графической регистрации изучаемых структур: важность тонких деталей строе-
ния вызывает стремление к передаче их на рисунке с применением приемов светотени.
К сожалению, подавляющее большинство рисунков анатомии является, по существу,
схемами, к тому же часто пе совсем точными. Необходимо оговориться, что изучение
внутреннего строения гениталий, часто хорошо иллюстрированное, осуществлялось
4
ПРЕДИСЛОВИЕ
более 70 (Wiegmann, 1900) и 40 лет назад (Hesse, 1931, 1934, и др.; Pilsbry, 1939, 1940,
и др.) либо путем поперечных сечений соответствующих отделов, либо путем прямого
их вскрытия. Однако на таксономических построениях полученные факты никак не от-
ражались или отражались неадекватно; к тому же данные такого рода носили фраг-
ментарный характер, а их функциональный аспект оставался нерасшифрованным.
Что касается видов, обитающих на территории нашей страны, то по большинству
из них сведения о внутреннем строении гениталий отсутствуют. Исключение состав-
ляют лишь Bradybaenidae Средней Азии, детально исследованные П. В. Матёкиным
(1972).
При написании настоящей работы мы ставили себе двоякую цель: ревизию фауны
Helicoidea, обитающих в СССР, и разработку принципиальных основ системы и фило-
гении надсемейства в целом. Последнее оказалось возможным благодаря хорошей изу-
ченности моллюсков таких регионов, как Европа, Средиземноморье и Северная Аме-
рика; в то же время изученность азиатских Camaenidae и Bradybaenidae недостаточна;
даже по тем районам, из которых эти моллюски анатомически изучены, мы не распола-
гаем сравнительным материалом (Филиппинские о-ва, Центральная Азия). Неясно,
что собой представляют так называемые африканские Bradybaenidae (Н aioli mnohelix
S. lat., Haplohelix, Urguesella),
В систематической части приводится описание 181 рецентного и ископаемого вида
(в известной сводке Лихарева и Раммельмейер, 1952 — 128 рецептных видов); из них
два не обнаружены на территории СССР, но не исключено их присутствие у наших
западных границ (скорее всего, на территории Белоруссии). Ископаемые (15 видов)
приведены только те, по которым существуют хорошие четкие описания: в старых
работах (Синцов, 1876; Богачев, 1936) описан еще ряд видов «Helix», но описания на-
столько несовершенны, что трудно себе представить, с чем имел дело автор.
В определительную таблицу включены только рецентные виды.
По всем видам, где это оказалось возможным, указаны лектотипы или обозначены
голотипы. В этом отношении неоценимую помощь нам оказали материалы И.М. Ли-
харева, который указал лектотипы видов, описанных Вестерлундом и хранящихся
в музеях Стокгольма, Гетеборга и Хельсинки.
Изложение ведется по определенной схеме; все виды описаны на основе
личного знакомства с материалом. Фактическая основа монографии составлена нашими
сборами, осуществленными в средней полосе европейской части СССР, на Карпатах,
в Крыму, на Кавказе, в Средней Азии и на Дальнем Востоке, а также обширные кол-
лекции Зоологического института АН СССР. Кроме того, частично использованы кол-
лекции следующих учреждений: Биолого-почвенного ин-та ДВНЦ АН СССР (Влади-
восток), Геологического ин-та А.Ц'СССР (Москва), Государственного природоведче-
ского ин-та (Львов), Зоологического ий-та Польской.АН (Варшава), Зоологического
музея МГУ (Москва), Ин-та зоологии "§Н. Армянской ССР (Ереван), Ин-та зоологии
АН Грузинской ССР (Тбилиси), Калифариййской АН (Сан-Франциско), Музея естест-
венной истории (Лейден), Палеонтоло^ческого ин-та АН СССР (Москва). Мы выра-
жаем глубокую благодарность администрации названных учреждений.
При написании этой работы мы пользовались любезным содействием многих кол-
лег, но чувство особой признательности мы выражаем сотрудникам Зоологического
ин-та АН СССР И. М. Лихареву и Я. И. Старобогатову за постоянную и всестороннюю
помощь.
Ценные советы и коллекционные материалы Мы получали от Н. Н. Акрамбвского
(Ереван,) Р. Я. Братчика (Владивосток), Р. Я. Бутылина (Ош), Е. Гиттенбергера
(Лейден), В. В. Горохова (Москва), В. Гудеца ^Прага), А. С. Довгялло (Ленинград),
Э. Клаусса (Кведлинбург), Г. И. Лежавы (Тбилиси), Ю. А. Мешкова (Москва),
С. М. Паттерсон (Энн Эрбор, Мичиган), А. Риделя (Варшава), Д. А. Сахарова
(Москва), А. Смита (Сан-Франциско) и К. К. Увалиевой (Алма-Ата); считаем прият-
ным долгом поблагодарить перечисленных лиц.
Мы благодарны также директору Биолого-почвенного ин-та ДВНЦ АН СССР
Н. Н. Воронцову за возможность в течение длительного срока продуктивно работать,
всецело отдавая время написанию настоящей монографии.
Наконец, мы выражаем самую глубокую признательность Л. В. Шилейко
и Ю. П. Губарю за постоянное внимание и помощь в нашей работе.
Фотографии раковин и монтаж таблиц, а также подавляющее большинство рисун-
ков выполнены автором; в немногих случаях, когда иллюстрации заимствованы из ли-
тературных источников, это оговорено.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
I. Семейство HELICODONTIDAE Hesse
I. Подсемейство Helicodontinae Hesse
1. Род Helicodonta Fer.
Стр.
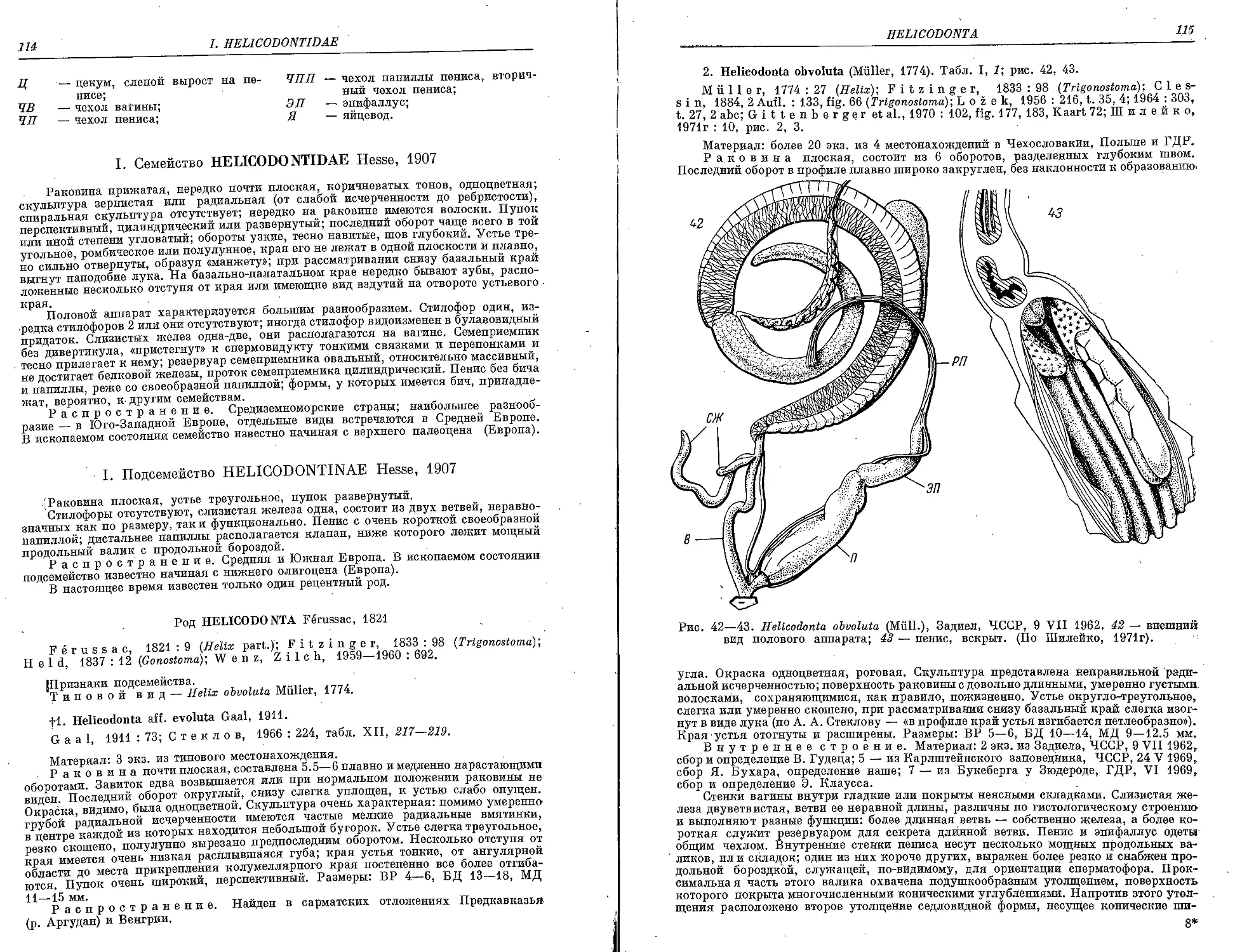
fl. ff. evoluta Gaal ................................................... 114
2. H. obvoluta (Miill.) ........................................... • 115
II. Подсемейство L i n d h о 1 in i о 1 i n a e Schileyko, subfam. nov.
1. Род Lindholmiola Hesse
fl, L. fraudulosa (Steklov) ............................................ 116
2. L. corcyrensis (Fer.) ............................................... 116
2. Род Oestophora Hesse
tO. schileykoi (Prysjazhnjuk).......................................... 118
II. Семейство BRADYBAENIDAE Pilsbry
1. Род Ponsadenia Schileyko, gen. nov.
1. Подрод Tarbagat a ja Schileyko, subgen. nov.
1. P. (T.) hirsute, (Matiokin)............'............................. 119
2. Подрод M e s a s i a t a Schileyko, subgen. nov.
2. P. (M.) duplocincta (Mart.).......................................... 120
3. Подрод Ponsadenia s. str.
3. P. (P.) semenovi (Mart.)............................................ 121
2. Род Bradybaena Beck
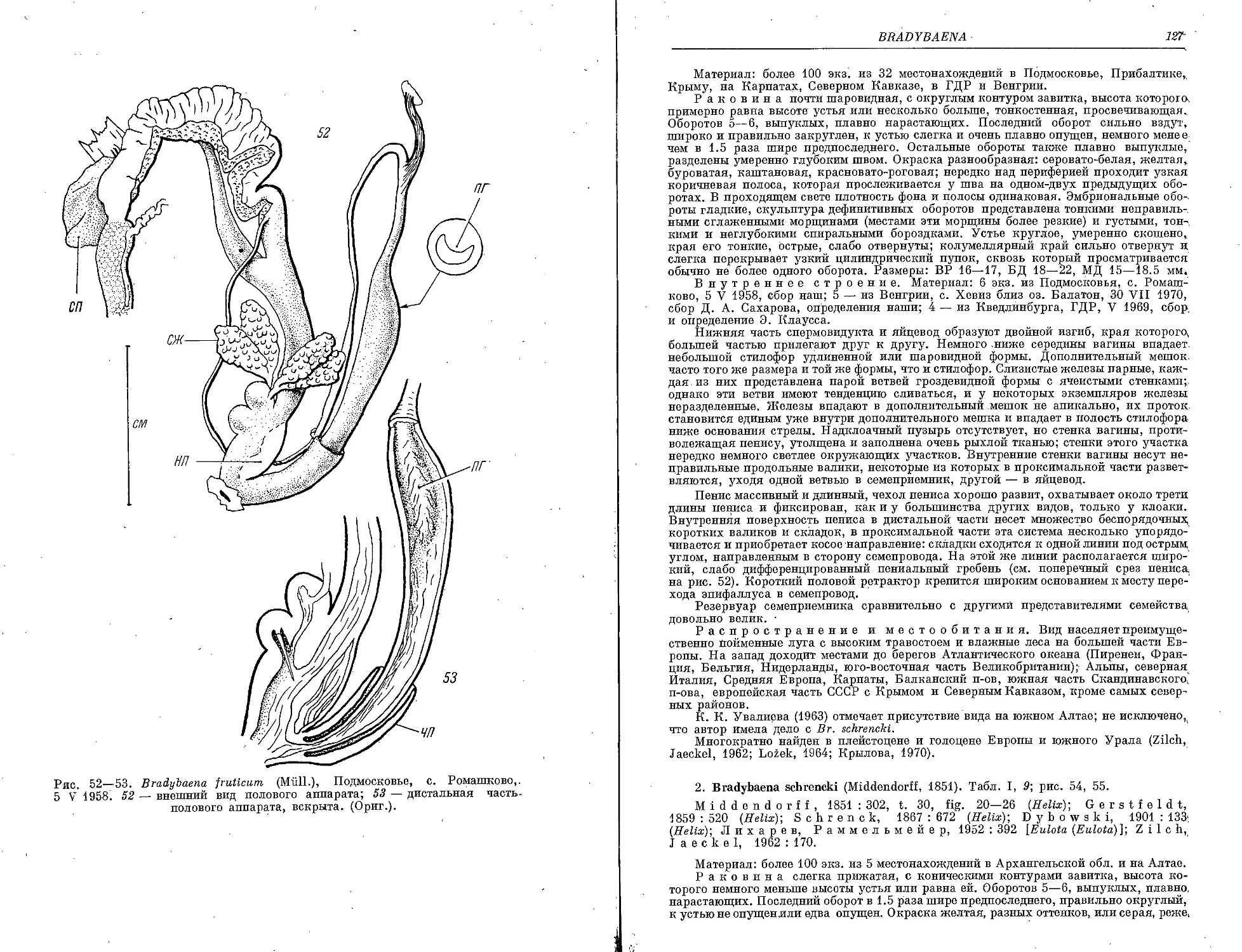
1. Br. fruticum (Miill.)............................................. 125
2. Br. schrencki (Midd.)...... . ................................ 127
3. Br. transbaicalia Schileyko, sp. nov.............................. 128
4. Br. middendorfji (Gerstf.)...................'...................... 130
5. Br. ussuriensis (West.)........................................... 131
6. Br. maacki (Gerstf.) ............................................... 132
7. Br. weyrichii weyrichii (Schrenck) ................................ 133
7a. Br. weyrichii bocageana Crosse...................................... 135
8. Br. strelkovi Lich. et Ramm.......................................... 135
9. Br. duiensis (West.) ............................................. 137
10. Br. dieckmanni (Mouss.)............................................. 137
11. Br. arcasiana (Crosse et Deb.) . ................................. .139
12. Br. diversita Schileyko et Bratschik, sp. nov...................... 140
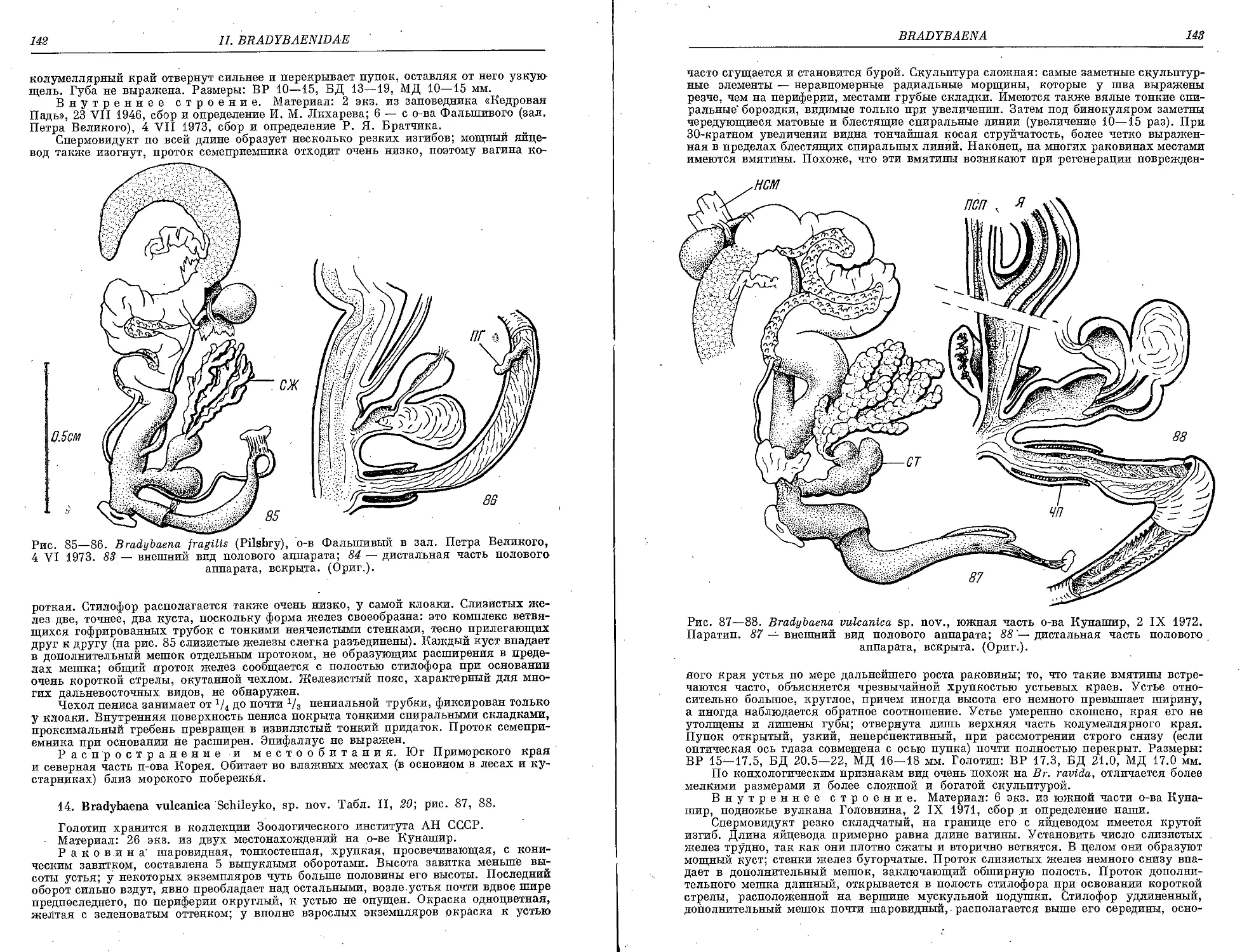
13. Br. fragilis Pilsbry................................................ 1H
14. Br. vulcanica Schileyko, sp. nov. ................................ .142
б СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
15. Br. ravida (Benson)............................................................. 144
16. Br. scythica (West.)..................................... . . ................ 145
17. Br. lanizi (Lindh.) ............................................................. 147
18. Br. plectotropis (Mart.).............'........................................... 148
19. Br. tzwetkovi Uvalieva et Soboleva .............................................. 151
20. Br. perlucens (Rosen) .......................................................... 151
21. Br. dichrozona (Mart.)........................................................... 153
22. Br. bilaticincta (Mart.)....................................................... 154
23. Br. stoliczkana (Nevill)......................................................... 155
24. Br. helvola (L. Pfr.) . ......................................................... 157
25. Br. pseudoferghanica Schileyko, sp. nov.......................................... 158
26. Br. fedtschenkoi (Mart.)............... . ................................... 160
27. Br. cavimargo cavimargo (Mart.).................................................. 161
27a. Br. cavimargo tarbagataica Matiokin . ............................................ 162
28. Br. tomyris (Lindh.).......................................................... 152
29. «Br.» boevi Uvalieva ............................................................ 153
III. Семейство HYGROMIIDAE Tryon
I. Подсемейство Trichiinae Zilch et Jaeckel
1. Род Nanaja Schileyko, gen. nov.
1. N. cumulata Schileyko, sp. nov.................................................. 164
2. N. illuminata Schileyko, sp. nov................................................ 165
2. Род Odontotrema Lindh.
O. diplodon Lindh........................................................... 167
3. Род Leucozdnella Lindh.
1. Подрод Leucozonella s. str.
1. L. (B.) ferghanica (Lindh.)............................................ 169
2. L. (B.) caryodes (West.)................................................ 170
3. L. (L.) rubens (Mart.).................................................. 171
4. L. (B.) mesoleuca (Mart.).............................................. 172
5. L. (L.) rufispira (Mart.).............................................. 173
6. L. (L.) retterl (Rosen) ............................................... 175
7. L. (L.) angulata (West.)................................................ 176
8. L. (L.) caria Schileyko............................................... 177
2. Подрод N a r i n a Schileyko, subgen. nov.
9. L. (TV.) hypophaea (Lindh.)..................................................... 178
10. L. (N.) crassicosta Schileyko, sp. nov........................................... 180
4. Род Hygrohelicopsis Schileyko
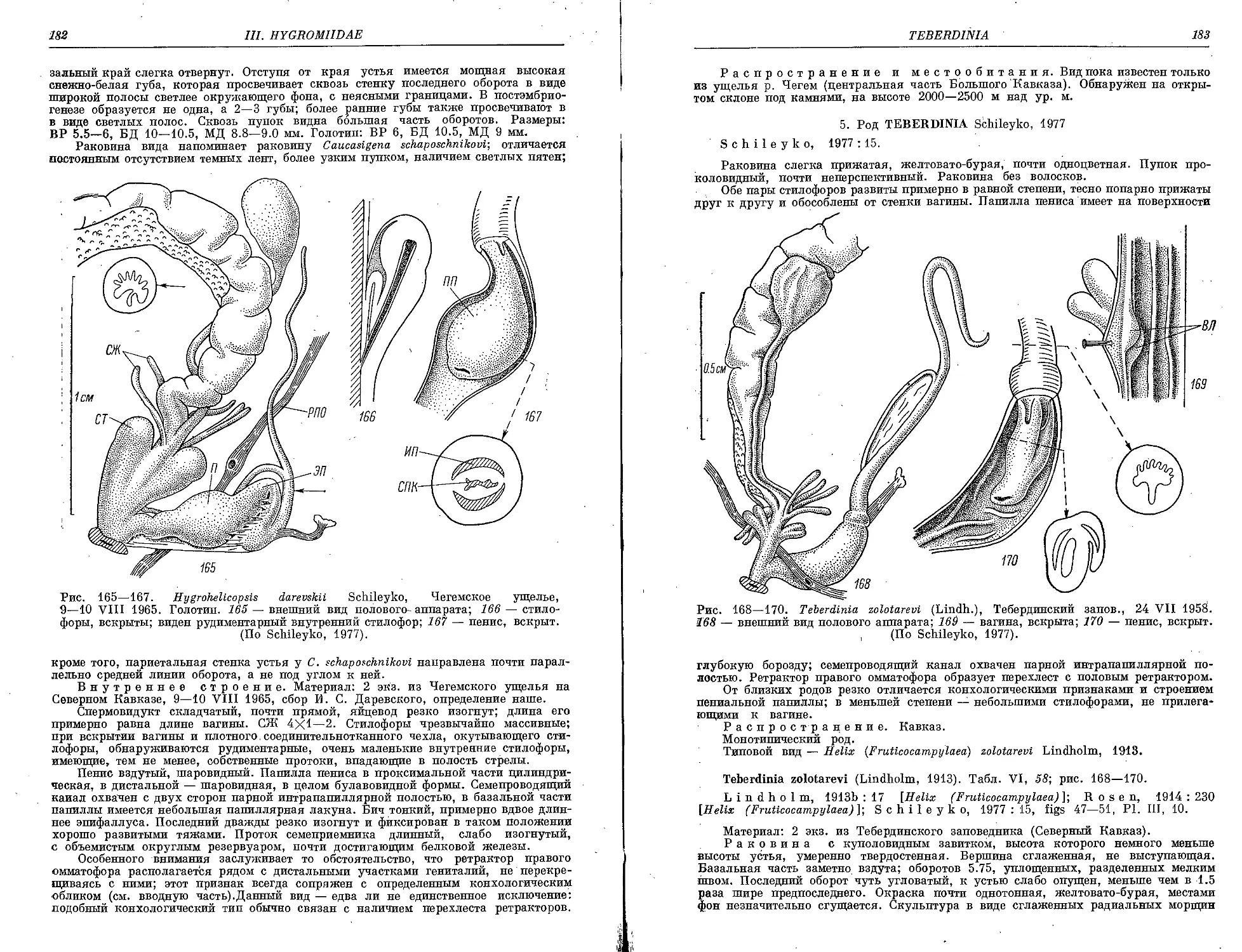
H. darevskii Schileyko 181
5...............................Род Teberdinia Schileyko
T. zolotarevi (Lindh.) ......... 183
6. Род Kokotschashvilia Hudec et Lezhawa
1. K. makvalae (Hudec et Lezhawa)............................................... 184
2. K. tanta Schileyko ......................................................... 186
3. K. eberhardi Schileyko......................... . . :...................... 187
4. K. holotricha (Bttg.) ....................................................... 158
5. K. phaeolaema (Bttg.)..................................................... 190
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ' 7
7. Род Caucasigena Lindh.
1. Подрод Caucasigena s. str.
fl. С. (C.) psedachica (Steklov)................................................ 192
f2. С. (C.) fortangensts (Steklov)......................' . . .'................ 192
3. С. (C.) armeniaca (L. Pfr.)................................................. 192
.4. С. (C.) tschetschenlca (Ret.).............................................. 194
5. С. (C.) rengarteni (Lindh.) ................................................. 195
6. С. (C.) eichwaldi (L. Pfr.)................'................................ 196
7. С. (C.) abchasica (Lindh.).................................................- 198
8. С. (C.) reminiscenta Schileyko, sp. nov...................................... 199
2. Подрод Anoplitella Lindh.
9. C. (A.) schaposchnikovi (Rosen).............................................. 200
3. Подрод Dioscuria Lindh.
10. C. (D.) thalestris (Lindh.)............................................... 201
8. Род Diodontella Lindh.
1. D. stschukini Lindh......................................................... 203
2. D. nubtgena Lindh.......................................................... 203
9. Род Plicuteria Schileyko
Pl. lubomirskii (Slos.) 204
10. Род Trichia Hartmann
1. Tr. hispida (L.)........................................................... 205
2. Tr. concinna (Jeffr.) ............................................... . . . 207
3. Tr. villosula (Rssm.) •...................................................... 208
4. Tr. bielzi (A. Schm.) ....................................................... 209
11. Род Edentiella Polinski
E. bakowskii (Poliftski) 210
12. Род Helicopsis Fitz,
fl. H. sunzhica (Steklov) 212
f2. H. libidinosa (Steklov) .................................................. 212
3. H. likharevi Schileyko, sp. nov............................................. 212
4. H. striata (Miill.)........................................................ 213
5. H. retowskii (Cl.) 215
6. H. dejecta (Cr. et Jan)..................................................... 216
7. H. paulhessei (Lindh.) .................................................... 216
8. H. instabilis (Rssm.)....................................................... 217
9. H. filimargo (Kryn.)........................................................ 217
13. Род Xeropicta Mont.
1. X. krynickii (Kryn.) ......'................................................ 219
2. X. derbentina (Kryn.)....................................................... 220
3. X. candaharica (L. Pfr.).................................................... 222
4. X. parableta (Bttg.)......................................................... 223
14. Род Helicella Fer.
H. candlcans (L Pfr.) ......................................................... 224
8
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
II. Подсемейство Hygromiinae Tryon
1. Род Lindholmomneme Haas
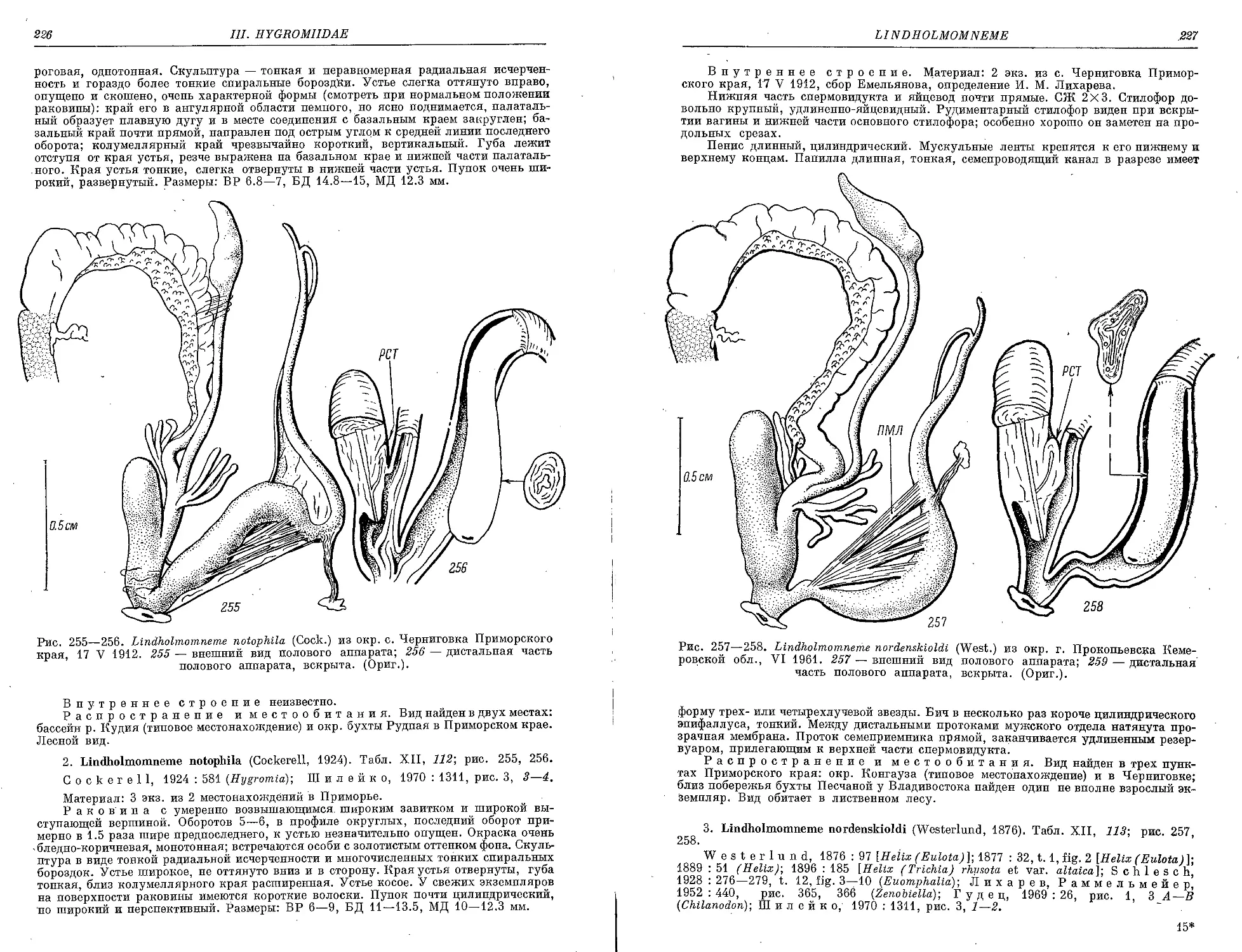
1. L. kudiensis (Cock.)........................................................ 225
2. L. notophila (Cock.) . ................................................... 226
3. L. nordenskioldi (West.)................................................... 227
2. Род Chilanodon West.
1. Ch. bicallosa (L. Pfr.)..................................................... 228
2. Ch. gerstfeldti (Dyb.)........................................................ 229
3. Род Perforatella Schluter
1. P. bidens (Chemn.) ........................................................... 231
2. P. dibothrion (Kim.) ......................................................... 232
4. Род Monachoides Gude et Woodward
1. M. incarnata (Mull.) ........................................................ 234
2. M. vicina (Rssm.)........................................................... 235
3. M. stuxbergi (West.) ........................................................ 237
4. M. aculeata (Uvalieva) .................................................... 238
5. Род Pseudotrichia Licharev
P. rubiginosa (A. Schm.) . ....................................................... 238
6. Род Urticicola Lindh.
U. umbrosa (C. Pfr.) 240
7. Род Fruticocampylaea Kobelt
1. Fr. narzanensis (Kr.yn.) ..................................................... 241
2. Fr. kobiensis (Bttg.)........................................................ 243
8. Род Shileykoia Hudec
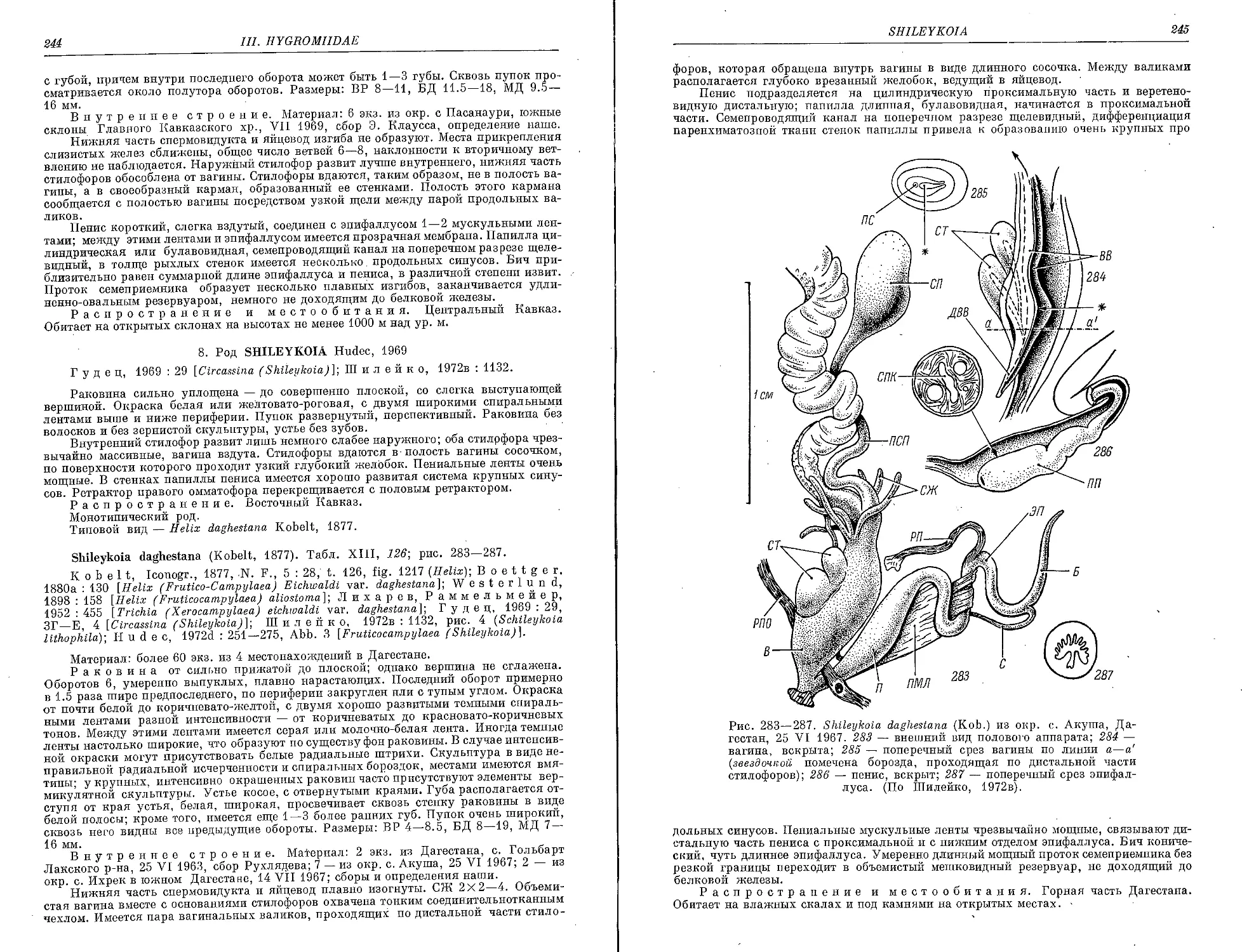
Sh. daghestana (Kobelt) 244
9. Род Circassina Hesse
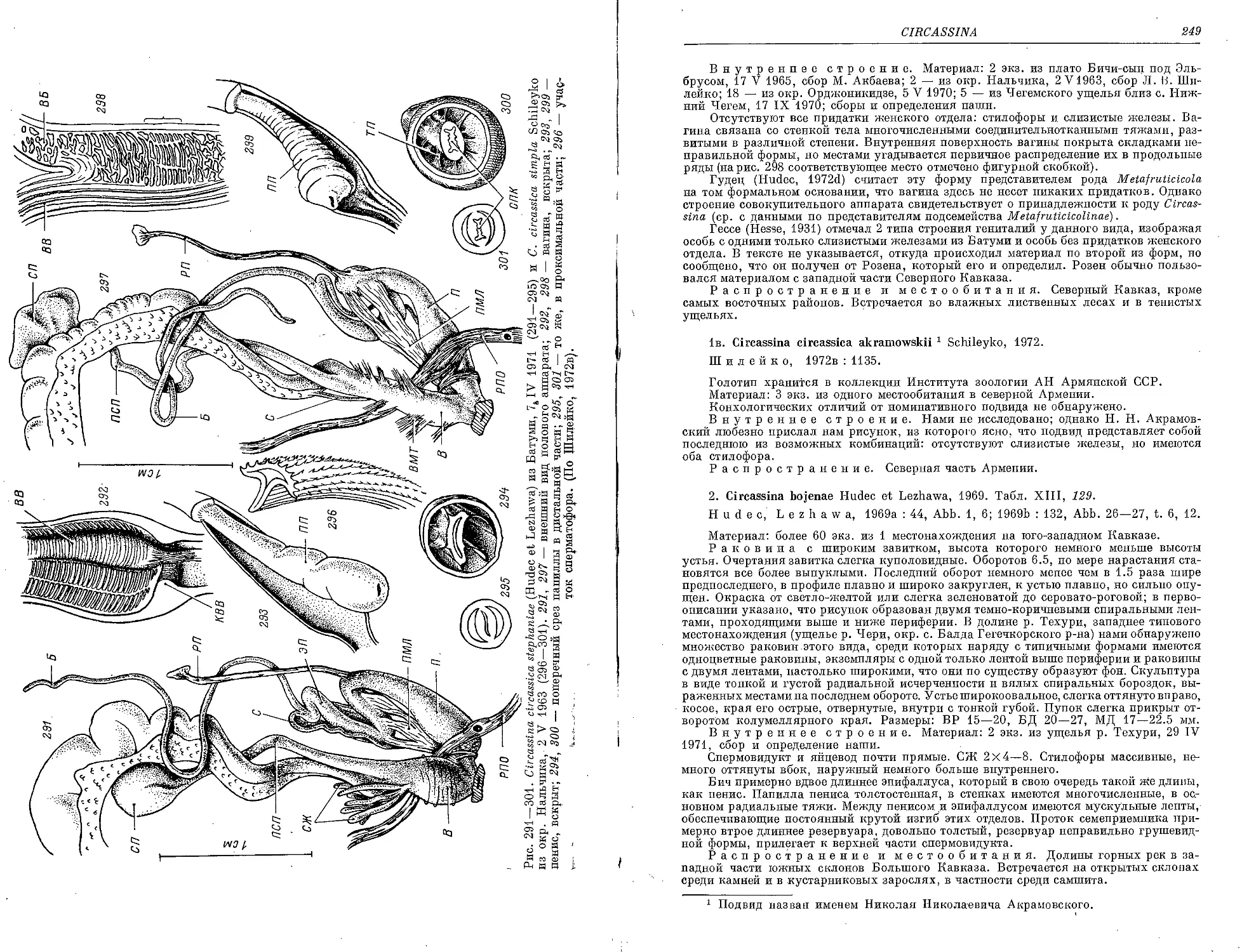
1. C. circassica circassica (Mouss.) .......................................... 246
la. C. circassica stephaniae (Hudec et Lezhawa)................................ . . 247
lb. C. circassica simpla Schileyko...........................................'. 247
1c. C. circassica akramowskii Schileyko .......................................... 249
2. C. bojenae Hudec et Lezhawa . .............................................. 249
3. C. frutis (L. Pfr.)......................................................... 250
10. Род Cernuella Schliiter
1. C. virgata (Da Costa) 250
2. C. tiflisiana (Lindh.) ...................................................... 252
11. Род Xerosecta Mont.
1. X. schelkovnikovi (Bogatschev) ........................................... 254
2. X. crenimargo (L. Pfr.) ...................................................... 254
3. X. arcadlana (Schileyko)...................................................... 255
III. Подсемейство Arc h a i c i n a e Schileyko, subfam. nov.
1. Род Archaica Schileyko
1. Подрод Euarchaica Schileyko
1. A. (E.) heptapotamica (Lindh.)................- . . .. ....................... 257
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 9'
2. Подрод А г с h a i с a s. str.
2. A. (A.) apollinis (Mart.)........................................... 259
3. Подрод U g a m a Schileyko, subgen. nov.
2. A. (U.) labianix Schileyko, sp. nov................................. 261
IV. Подсемейство Euomphaliinae Schileyko, subfam. nov.
1. Род Karabaghia Lindh.
K. bituberosa (Lindh.) .................................................. 262
2. Род Hesseola Lindh.
1. H. solidior (Mouss.)................................................. 263
2. H. transcaspia (Bttg.) ............................................. 265
3. Род Euomphalia West.
1. E. aristata (Kryn.) ...........'..................................... 266
2. E. appeliana (Mouss.)................................................ 268
3. E. strlgella (Drap.) ............................................... 269
4. Род Platytheba Pilsbry
1. Подрод J as о n ell a Lindh.
1. P. (/.) mingrelica (Hesse)............................................ 271
2. Подрод Platytheba s. str.
2. P. (P.) prometheus (Bttg.) ........................................... 273
5. Род Oscarboettgeria Lindh.
0. euages (Bttg.)..................................................... 273
6. Род Stenomphalia Lindh.
1. Подрод Stenomphalia s. str.
1. S. (S.) selecta (Klika) . ;.......................................... 275
2. Подрод В atumica Schileyko, subgen. nov.
2. S. (B.) maiae (Hudec et Lezhawa)..................................... 278
3. Подрод Diplobursa Schileyko
3. S. (D.) pisiformis (L. Pfr.)........................,................ 278
4. Подрод Harmozica Lindh.
4. S. (H.) ravergieri (Fer.)............................................ 280
7. Род Monaeha Fitz.
1. Подрод P a r a. t h e b a Hesse
1. M. (P.) fruticola (Kryn.) ....................................... . 282
2. M. (P.) caucasicola (Lindh.) ....................................... 283
3. M. (P.) talyschana (Mart.).......................................... 285
10
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
2. Подрод Monacha s. str.
-j-4. M. (M.) externa Steklov ........................................... 286
5. M. (M.) carthusiana (Miill.)......................................... 286
............. 3. Подрод Metatheba Hesse
6. M. {Met.) samsunensis (L. Pfr.)........................................ 288
4. Подрод В о e m i c a Schileyko, subgen. nov.
•j-7. M. (B.) praeorientalls Steklov...................................... 289
8. M.(B.) subcarthusiana subcarthusiana Lindh. . . . .................. 289
8a. M. (B.) subcarthuslana scrobiculosa Lindh............................. 291
V. Подсемейство Paedhoplitinae Schileyko, subfam. nov.
1. Род Angiomphalia Schileyko, gen. nov.
1. Подрод Angiomphalia s. str.
1. A. (A.) copiosa Schileyko, sp. nov. .................................. 292
2. A. (4.) caelesttmontana (Tzvetkov) ................................... 293
3. A. (4.) exasperata Schileyko et Uvalieva, sp. nov..................... 295
4. 4. (A.) regeliana (Mart.) ............................................ 296
5. 4. (4.) almalensis Schileyko et Uvalieva, sp. nov..................... 297
6. 4. (4.) seducttlis (West.) ........................................... 298
2. Подрод L e n t i g a Schileyko, subgen. nov.
7. 4. (L.) lentina (Mart.)............................................... 300
8. 4. (L.) buamica (Tzvetkov et Tzvetkova)............................... 301
2. Род Paedhoplita Lindh.
1. P. laminata Lindh............................................... . . 302
2. P. lindholmi Tzvetkov et Tzvetkova.................................... 303
3. P. kirgisensis Lich. et Ramm. . . . , ............................... 304
VI. Подсемейство Metafruticicolinae Schileyko
1. Род Caucasocressa Hesse
1. C. pratensis (L. Pfr.)................................................. 305
2. C. joannis (Mort.)................................................... 306
IV. Семейство HELICIDAE Rafinesque
I. Подсемейство Ariantinae Morch
1. Род Faustina Kobelt
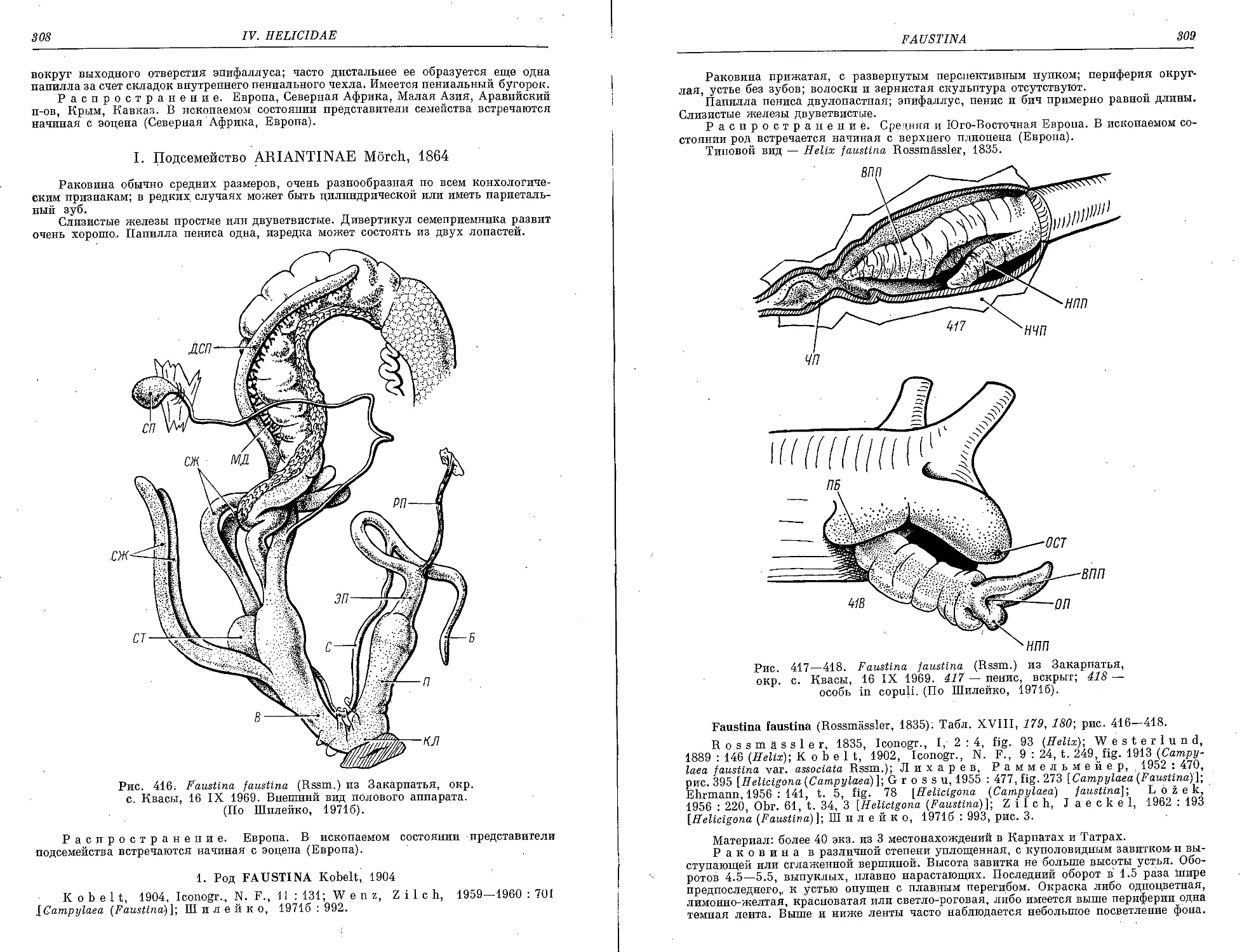
F. faustina (Rssm.)....................................................... 309
2. Род Arianta Turton
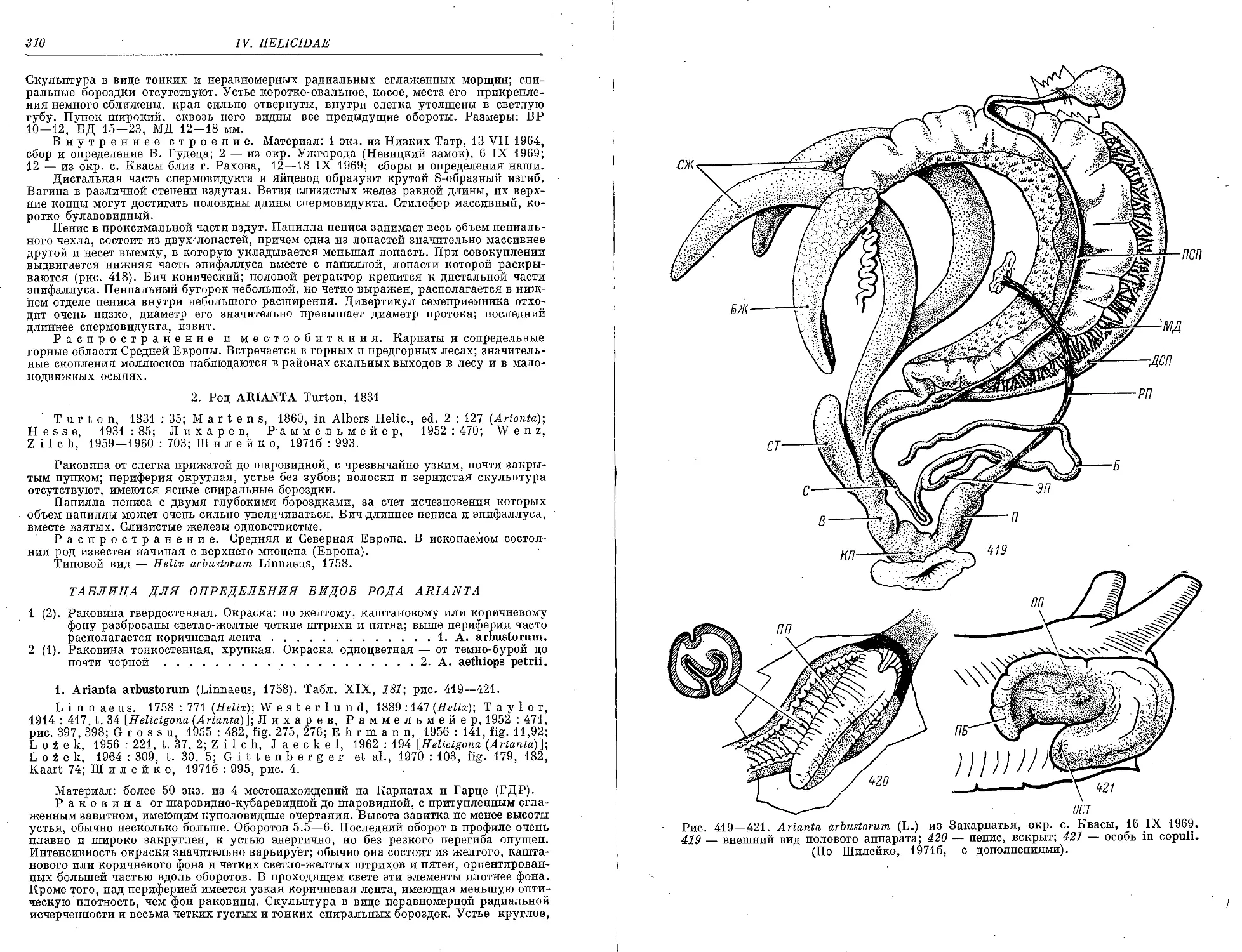
1. 4. arbustorum (L.).................................................... 310
2. 4. aethlops petrii Kim................................................ 313
3. Род Helicigona Fer.
H. lapicida (L.)..................................................... . 313
4. Род Isognomostoma Fitz.
I. personatum (Lam.).................................................... 315
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 11
5. Род Drobacia Brusina
О. banatica (Rssm.) ?............................................ 317
6. Род Causa Schileyko
C. holosericum (Stud.) 317
7. Род Klikia Pilsbry
j-K. giengensls zamechovense Prysjazhnjuk et Schileyko......................... 319
II. Подсемейство H e 1 i c i n a e Rafinesque
1. Род Cepaea Held
1. Подрод Cepaea s. str.
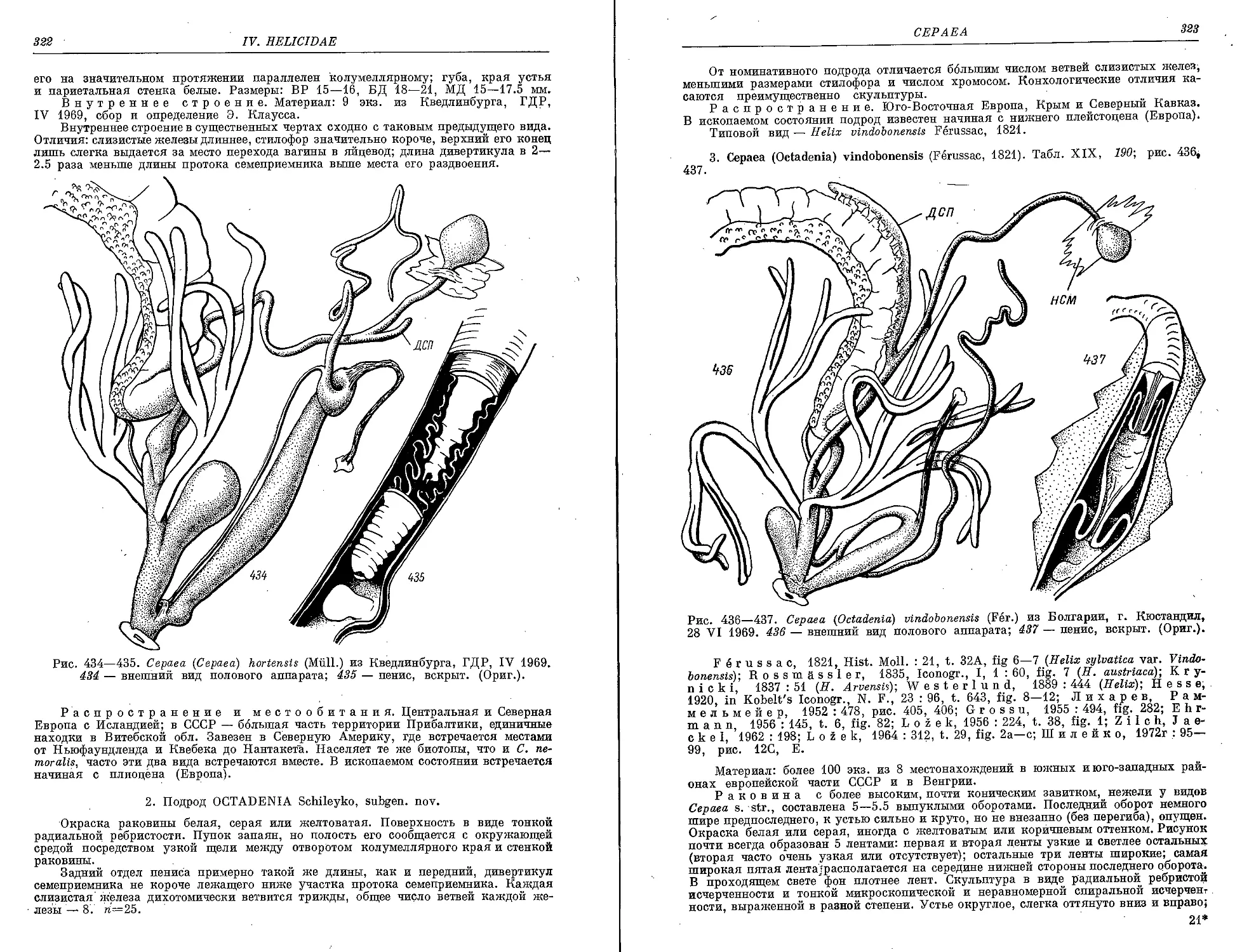
1. С. (C.) nemoralts (L.)..................................................... 320
2. С. (C.) hortensis (Miill.) . . ........................................... 321
2. Подрод Octadenia Schileyko, subgen. nov.
3. C. (0.) vlndobonensis (Fer.)............................................... 323
2. Род Cryptomphalus Charp.
Cr. aspersa (Miill.)........................................................... 324
3. Род Eobania Hesse
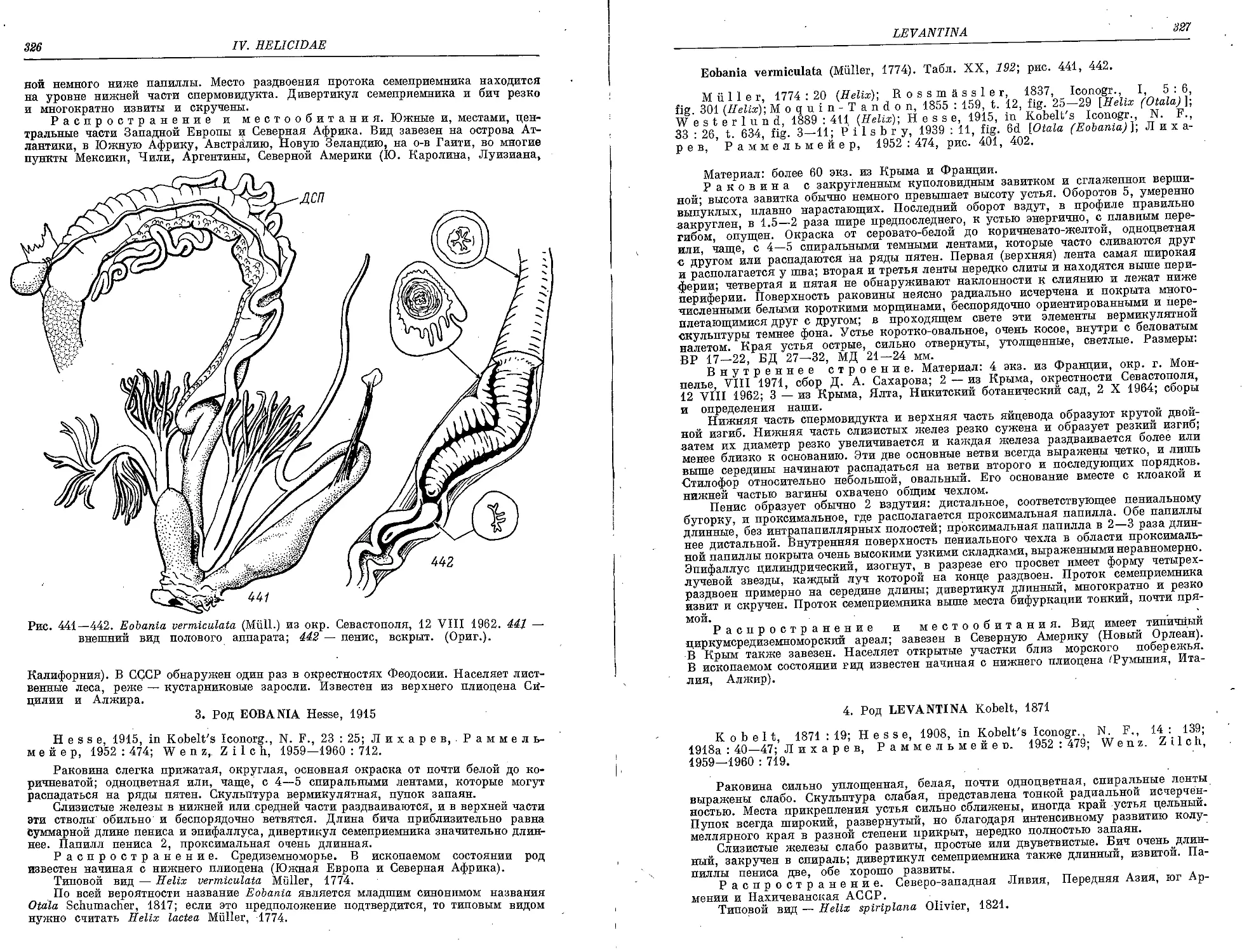
E. vermiculata (Miill.) 327
4. Род Levantina Kobelt
1. L. djulfensis (Dubois de Mont.)............................................ 328
2. L. escheriana (Bourg.)..................................................... 328
5. Род Caucasotachea C. Bttg.
fl. C. andrussovi Steklov ..................................................... 331
f2. C. jortangensis Steklov ................................................... 331
f3. C. kubanica Steklov ....................................................... 332
4. C. lenkoranea (Mouss.).................................................... 332
5. C. atrolabiata (Kryn.) ............................................... . 332
6. C. calligera (Dubois de Mont.)............................................ 335
6. Род Steklovia Schileyko, gen. nov.
f5. maslovae (Steklov) 337
7. Род Helix L.
fl. H. maeotica Steklov '........................................ 338
2. H. nordmanni Mouss......................................................... 338
3. H. christophl Bttg....................................................... 340
4. H. albescens Rssm....................................'..................... 340
5. H. lucorum L............................................................... 342
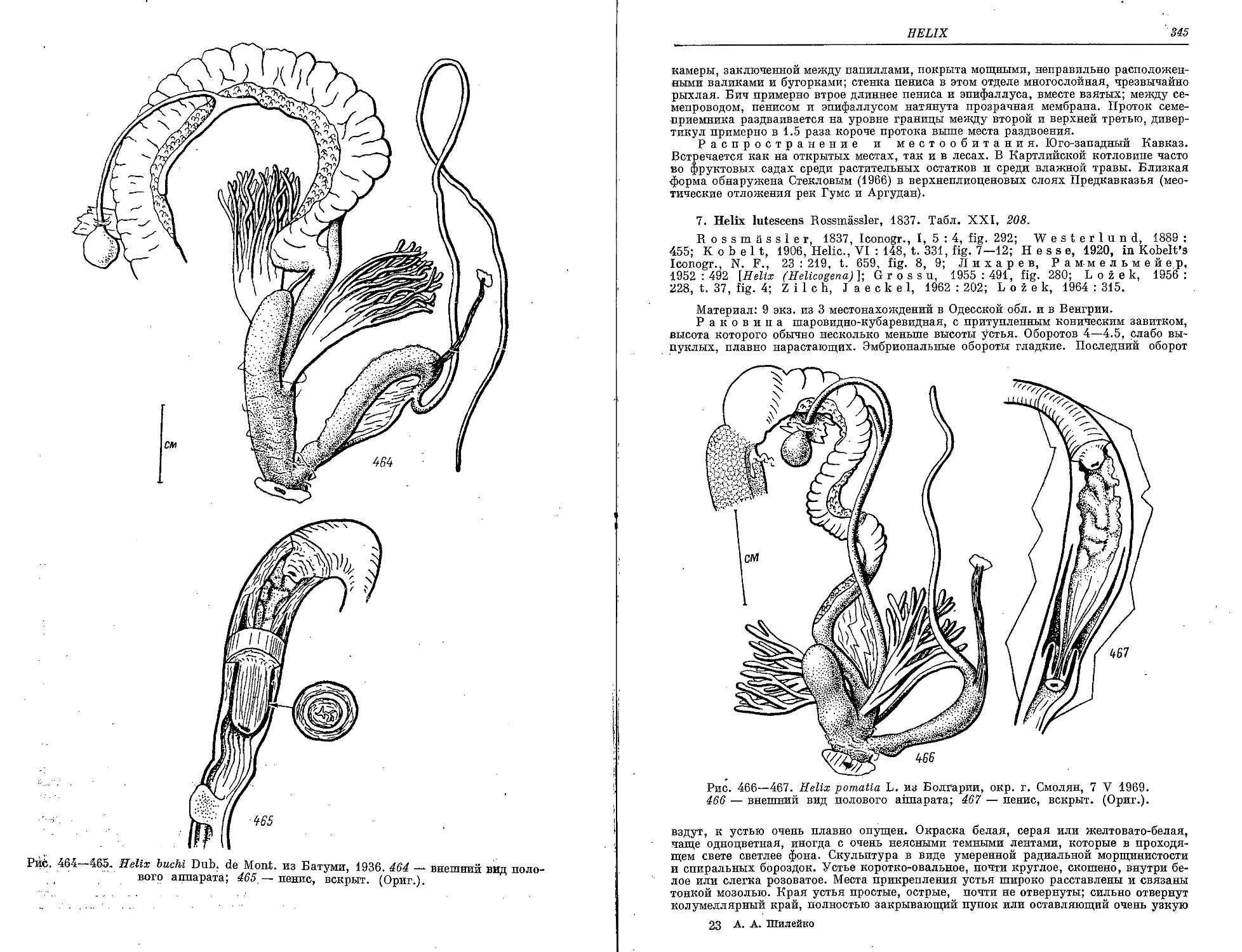
6. H. buchl Dubois de Mont.................................................. 343
7. H. lutescens Rssm......................................................... 345
8. H. pomatia L. . . . .............,........................................ 346
ПРИЛОЖЕНИЕ
Надсемейство POLYGYROIDEA
Семейство POLYGYRIDAE Pilsbry
Подсемейство Triodopsinae Pilsbry
Род Triodopsis Rafinesque
Tr. subpersonatum (Midd.)...................................................... 34S
ВВЕДЕНИЕ
ОБЩАЯ ХАРАКТЕРИСТИКА МОЛЛЮСКОВ
НАДСЕМЕЙСТВА HELICOIDEA
Животные с наружной раковиной, никогда не имеющей склонности к редукции..
Оборотов чаще всего 4—7. Раковина правозакрученная (декстральная, дексиотроп-
ная), значительно реже левозакрученная (синистральная, леотропная). Форма рако-
вины — от плоской до шаровидной, очень редко цилиндрическая (Cylindrus) или в виде
высокого конуса (Cochlicella, Buliminopsis), тогда число оборотов может достигать 12.
Последний оборот в профиле закругленный, угловатый или с килем; к устью чаще
всего в разной степени опущен. Окраска
разнообразная, некоторые тропические-
виды окрашены ярко за счет спираль-
ных лент. Скульптура может быть почти
любого типа, на поверхности раковины
часто присутствуют волоски. Устье чаще
всего с губой, неармированное; реже-
имеются зубы в виде утолщений ба-
зального и палатального краев, «на-
плыва» на колумеллярной стенке, а
также складок и бугорков на париеталь-
ной стенке. Пупок от полностью запа-
янного до очень широкого, разверну-
того, перспективного.
Тело явственно подразделяется на
голову, ногу и внутренностный мешок.
Голова несет две пары щупалец и три
околоротовые лопасти. Подошва цель-
ная, не разделена продольными бороз-
' дами; однако на фиксированных эк-
земплярах, предварительно расправлен-
ных, часто видно три продольных поля:
гладкое или покрытое продольными мор-
щинами центральное и два боковых,
смятых в поперечные бугры и складки
(рис. 1). Перипедальная борозда одна,
проходит по краю подошвы. Хвостовая
ямка отсутствует.
Рис. 1. Подошва фиксированного экземп- Кожа от очень тонкой и почти глад-
ляра Leucozonella rubers. (Ориг.). кой до грубой, сильно мускулизованной,
покрытой короткими грубыми морщи-
нами.
Край мантии (мантийный валик) несет дыхательное отверстие (пневмостом), аналь-
ное отверстие и отверстие мочеточника; два последних сильно сближены. На мантийном
валике всегда имеются 3 лопасти: анальная (верхняя) и две боковые — правая и левая.
Почка характерная для подотряда Sigmurethra, с вторичным мочеточником. Форма
легкого зависит от формы последнего оборота, степень его васкуляризации стоит
в прямой зависимости от размеров животного.
Челюсть всегда хорошо развита, представляет собой изогнутую пластинку, клино-
видную в поперечном сечении, без срединного выступа (исключение: у Sphincterochila.
челюсть со срединным выступом). Челюсть несет поперечные ребра, иногда чрезвы-
чайно резкие, выступающие за края челюстной пластинки.
Нервный аппарат характеризуется высокой степенью концентрации, состоит
из 7 ганглиев, плотно слитых в окологлоточное кольцо.
ОБЩАЯ ХАРАКТЕРИСТИКА HELICOIDEA
13
Для полового аппарата характерно наличие. 4, 2 или 1 стилофора («сумки любовных
стрел») и функционально связанных с ним слизистых желез; иногда наблюдается пол-
ная редукция всех придатков женского отдела. Пепис в виде простой трубки или с па-
пиллой и бичом. Семеприемник первично с дивертикулом или без него. Половой рет-
рактор крепится к дну мантийпой полости, в редких случаях отсутствует.
Осевая мускулатура представлена колумеллярным мускулом, который распада-
ется на пару глоточных ретракторов, пару ретракторов омматофоров (глазных щу-
палец) и множество педальных мускульных пучков, группирующихся более или менее
отчетливо в два продольных ряда. Половой ретрактор паллиального происхождения,
но в редких случаях отходит от колумеллярного мускула.
При втягивании животного в раковину устье замыкается не ногой, а краем мантии.
Ареал надсемейства охватывает всю Палеарктику,(кроме самых северных районов),
Северную li частично Восточную Африку, Малайский архипелаг с о-вом Новая Гвинея,
северное побережье Австралии, Северную и Центральную Америку, северную и северо-
западную части Южной Америки; учитывая, что многие виды завезены в ряд пунктов
Западной Африки, Южной Америки, Австралии, Новой Зеландии и Океании, можно
сейчас говорить о всесветном распространении надсемейства.
В ископаемом состоянии представители Helicoidea встречаются начиная с верх-
него мела.
В мировой фауне насчитывается более .2500 видов Helicoidea, в фауне СССР заре-
гистрировано 166 рецентных видов.
Надсемейство Helicoidea входит в состав отряда Geophlla1 подкласса Pulmonata
класса Gastropoda.
МОРФОЛОГИЯ И АНАТОМИЯ ПРЕДСТАВИТЕЛЕЙ
HELICOIDEA С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
ВНЕШНИЙ ВИД И РАКОВИНА
Наружная морфология тела моллюсков данной группы описывалась в литературе
но раз и хорошо известна (Moquin-Tandon, 1855; Lehmann, 1877; Иванов, 1946; Ki-
lias, 1960, и др.). Более или менее ясно отграниченная затылочной складкой голова
несет 3 околоротовые лопасти (нижнюю и пару боковых) и две пары щупалец; на кон-
цах каждого из верхней пары располагаются глаза, заметные в виде темных точек.
Под ротовым отверстием заметна поперечная щель, на дне которой открывается педаль-
ная железа. Позади и несколько ниже основания правого (у синистральных форм —
левого) омматофора заметно округлое или щелевидное отверстие половой клоаки.
Система бугорков, складок и борозд на поверхности тела ориентирована вдоль спины
продольно, на боках — поперечно и немного косо, вокруг полового отверстия либо
пропадает, либо располагается концентрически. Наиболее постоянны две борозды,
идущие вдоль средней линии тела от головы к мантийному валику. Задний конец ноги
заострен пли закруглен или в разной степени раздвоен.
Окраска тела — от светло-серой или почти белой до угольно-черной и почти крас-
ной, причем изменчивость окраски в пределах вида — явление обычное. В этом случае
носветление начинается с мантийного валика; в последнюю очередь теряют пигмента-
цию покровы головы. Подошва обычно гораздо светлее верхней стороны тела. У многих
видов мантия покрыта узором, состоящим из черных пятен, точек и размытых линий.
Хотя нередки случаи, когда животные с толстостенной раковиной имеют темную ок-
раску тела, имеется общая тенденция к усилению пигментации мантии у видов с тон-
костенной просвечивающей раковиной.
Мантийный валик светлый, реже — пигментированный, несет 3 лопасти: аналь-
ную и 2 боковые; между анальной и правой лопастями располагается дыхательное от-
верстие (пневмостом), анальное и отверстие мочеточника..
Что касается формы раковины, то, если взять Helicoidea в полном объеме, гораздо
легче отдифференцировать немногие семейства, раковина которых не может быть спу-
тана с раковиной представителя настоящего надсемейства, чем описать признаки,
по которым можно всегда отличить раковину представителей этого таксона; по су-
ществу, таких признаков нет..
Анатомический метод в разработке системы Helicoidea играет большую роль, не-
жели в большинстве других крупных таксонов Pulmonata именно в силу чрезвычайной
расплывчатости и бедности конхологических признаков. Если раковина некоторых так-
сонов ранга семейства обладает характерным общим габитусом, замыкательным ап-
паратом плп хорошо развитой устьевой арматурой, к которой подчас добавляется
система продольных складок внутри последнего оборота, то раковины представителей
1 О разделении группы Stylcmmatophora см.: Миничев, Славошевская. 19.'I.
14
ВВЕДЕНИЕ
обсуждаемой группы большей частью различаются скульптурой, опушенностью,
формой, наличием или отсутствием губы и ее расположением, степенью перспективности
пупка и некоторыми другими признаками, из которых большинство носит качествен-
ный характер и не выражается измеряемыми величинами. Благодаря этому различе-
ние многих видов доступно лишь наметанному глазу; дело осложняется еще и тем,
Рис. 2. Основные типы раковин представителей Helicoidea. А — плоская; Б — линзо-
видная; В—.Г — прижатая; Д—3 — кубаревидная; И — широко кубаревидная;
И — шаровидная; Л~М — шаровидно-кубаревидная. (Ориг.).
что многие признаки обнаруживают значительную изменчивость в пределах одного
вида и даже в пределах одной популяции.
Наиболее широко распространен типичный «хеликоидный» облик раковины:
диаметр 10—30 мм, форма более или мепее прижатая, округлая, с постоянной кривизной
последнего оборота и небольшим открытым пупком; обычно имеется губа или утолща-
ется сам край устья’— перистом. Отклонения от этого типа касаются величины (5—
50 мм в диаметре); формы — от совершенно плоской до почти башневидной; развиваю-
щейся угловатости последнего оборота вплоть до появления резкого, грубо зазубрен-
ного киля; присутствия устьевой арматуры в виде бугорков, пластинок и складок.
На рис. 2 изображены основные типы раковин, между которыми имеются много-
численные переходы. Если раковина плоская, все обороты лежат в одной или почти
в одной плоскости (рис. 2, Л). Чаще бывает так, что завиток более или менее выдается'
ВНЕШНИЙ ВИД И РАКОВИНА
15
над последним оборотом. В этом случае периферия может быть угловатой или нести
киль, что придает раковине характерную форму двояковыпуклой линзы (рис. 2, В);
если же периферия с небольшим углом или плавно закруглена, различаются два типа:
прижатая раковина, если высота завитка меньше высоты устья (рис. 2, В—Г), и кубаре-
видная, если высота завитка равна или превышает высоту устья (рис. 2, Д—3). Раз-
новидность второго типа — ширококубаревидная раковина — последний оборот вздут,
с выпуклой базальной частью (рис. 2, И). Далее можно выделить шаровидную форму,
название которой говорит само за себя (рис. 2, К), и шаровидно-кубаревидную с резко
вздутым последним оборотом и большим устьем, несколько оттянутым вниз и в сто-
рону (рис. 2, Л—М). 'Более дробная классификация в применении к данной группе
не оправдана, так как самая общая характеристика раковины уже достигается употреб-
лением приведенных терминов.
Число оборотов колеблется от 3.5 до 12; в фауне СССР отсутствуют виды, раковина
которых состояла бы более чем из 7 оборотов. Эмбриональные обороты в числе 1.25—
2.0 чаще всего полупрозрачные, гладкие или с неясной пунктировкой, коричневые,
розоватые или черные. Эмбриональные обороты формируют вершину раковины, ко-
торая может быть закругленной (рис. 3, А) или выступающей (рис. 3, В). В первом
Рис. 3. Типы вершин раковины и положение угла по периферии
относительно средней линии последнего оборота. А — не вы-
ступающая вершина, угол на периферии совпадает со средней
линией; В — выступающая вершина, угол на периферии сдвинут
вверх относительно средней линии. (Ориг.).,
случае завиток имеет конические или куполовидные очертания, во втором — немного
вогнутые. Дефинитивные обороты, формирующиеся в постэмбриогенезе, в профиле
чаще округлые, но молодь большей частью имеет угловатый последний оборот.
Периферией раковины мы именуем часть последнего оборота, наиболее удаленную
от столбика, т. е. по существу от оси навивания; если периферия не округлая, обычно
отмечается, совпадает ли киль или угол со средней линией последнего оборота или
периферия сдвинута вверх (рис. 3). В очень редких случаях выше угловатой периферии
наблюдается еще один угол.
У разных видов обороты имеют разную скорость нарастания. Когда они нарастают
стремительно, т. е. диаметр оборотов от вершины к устью увеличивается быстро, пос-
ледний оборот вздут, а число оборотов невелико — 3.5—4.5; если же диаметр оборотов
увеличивается медленно, то и число оборотов больше — до 7.
Исходный вариант окраски раковины — вероятно, роговой фон со светлой полосой
по периферии. Этот тип окраски у большинства видов в той или иной степени сохраня-
ется в пределах большинства таксонов нашей группы. Одним из широко распростра-
ненных путей изменения окраски является общее посветление фона и вторичное появ-
ление узора, состоящего обыкновенно из спиральных темных лент. Рисунок из не-
скольких широких спиральных полос (лент) независимо появляется в разных группах
Helicoidea, причем связан преимущественно с крупными размерами раковины (Helix,
Humboldtiana, Ponsadenia duplocincta, Bradybaena lantzi). У ксерофильных видов
из семейства Hygromiidae наблюдается возникновение тонких спиральных темно-
коричневых или черных лент в числе до 12, причем степень их выражен-
ности и количество очень непостоянно в пределах вида. У ряда видов из разных так-
сонов наблюдается независимое возникновение двух спиральных лент — выше и ниже
периферии — на светлом фоне. Эти ленты обычно в разной степени выражены в пре-
делах даже одной популяции; в первую очередь всегда исчезает нижняя полоса.
Радиальное расположение элементов окраски — явление более редкое; радиаль-
ные цветовые полосы никогда не бывают у представителей Helicoidea четкими, напро-
тив, границы их всегда размыты, сами они очень нерезкие, непостоянны и разной
ширины: такие полосы наблюдаются только на светлых раковинах и имеют светло-
коричневую, бурую, редко — почти черную окраску.
Скульптура поверхности раковины очень разнообразна. Наиболее обычный скульп-
турный элемент — радиальная исчерченность или морщинистость, связанная с не-
равномерным ростом раковины. В случае усиления радиальных морщин возникает
ребристость, причем в отдельных случаях имеет место формирование очень грубых
16
ВВЕДЕНИЕ
резких ребер, которые, доходя до шва, придают ему зазубренный облик. Иногда на по-
верхности раковины имеются короткие радиальные морщины, несущие у некоторых
видов кожистые шипики или чешуйки. Если наличие чешуек — явление сравнительно
редкое, то присутствие волосков на поверхности раковины наблюдается часто, причем
такие волоски порой можно заметить у отдельных экземпляров, относящихся к видам,
для которых наличие волосков не характерно. Волоски бывают достаточно длинными
и, располагаясь часто, создают определенный «шерстистый» облик раковины. Нередко
бывает, что молодь имеет волоски, утрачивающиеся с возрастом. В том случае, если
волоски опадают, остаются более или менее ясно выраженные рубцы. Волоски появ-
ляются в эмбриогенезе как составная часть верхнего рогового слоя раковины — перио-
стракума. Каждый волосок формируется в специализированной сумке, закладываю-
щейся в борозде железистого эпителия мантийного валика (см. ниже). Осевая часть
волоска вначале гомогенна и очень эластична, затем она обволакивается конхиолино-
вым слоем, а позже — вторым таким же слоем. Наблюдаемое обычно изгибание во-
Рис. 4. Характер опущенности последнего оборота к устью. А — последний оборот
прямой; В — последний оборот плавно и постепенно спускается к устью (оползает);
В — последний оборот плавно опущен близ устья; Г — последний оборот оЦущен энер-
гично, с перегибом. (Ориг).
лосков в сторону устья объясняется более интенсивным выделением секрета желези-
стых клеток в задней части мантийной борозды. Возникновение очередного ряда во-
лосков предшествует суточному росту раковины и образованию очередного слоя на-
растания. После образования всего комплекса волосков железистый эпителий, участ-
вующий в его образовании, очень быстро дегенерирует. При повреждении раковины
волоски регенерируют в последнюю очередь, постепенно восстанавливается располо-
жение рядов волосков параллельно линиям нарастания (Polinski, 1928—1929; Kai-
ser, 1966).
Функция волосков может быть различной в зависимости от их характера. Надо
обратить внимание, что раковины таких видов, как Рseudotrlchia rubiginosa, Trichia,
villosula и др., нередко обитающих на влажной липкой глинистой почве, не загрязня-
ются глинистыми частицами. Густота и изогнутость волосков позволяют предполагать,
что они играют роль своеобразных пружинок, образующих- в совокупности слой, от-
талкивающий капли воды и частицы почвы от поверхности раковины. Когда волоски
короткие, редкие и прямые, они могут выполнять обратную функцию, т.е. способствуют
удержанию на поверхности раковины комочков почвы в целях маскировки (молодь
некоторых видов Monacha, Euomphalta aristata).
Спиральная исчерченность раковины — явление гораздо более обычное, чем принято
считать: у большинства видов можно обнаружить выраженные в той или иной степени
спиральные бороздки, но лишь у немногих видов они настолько резки, что Видны нево-
оруженным глазом. То же, в несколько меньшей степени, касается и так называемой
скульптуры «удары молотка» — вмятин, которые наблюдаются на последнем обороте.
Зернистая скульптура может возникать двумя путями. Первый путь начинается
с четко выраженных спиральных и радиальных бороздок, образующих более или менее
ясно выраженную сетку; ячейки этой сетки и представляют собой гранулы. Этот путь
сравнительно редок (отдельные виды Kokotschashvilla, Caucasocressa). Более распростра-
нен второй путь, когда зернистость связана с наличием чешуек (Stenomphalia maiae,
карпатские виды Monachoides), располагающихся на возвышениях. По достижении
полного числа оборотов чешуйки чаще всего опадают, а зернистость остается.
ВНЕШНИЙ ВИД И РАКОВИНА
17
Спиральная ребристость у Helicoidea наблюдается крайне редко; все случаи такого
рода касаются неарктических групп (некоторые виды Oreohelix).
Наконец, отметим еще один тип скульптуры, так называемую вермикулятную
скульптуру, представляющую собой сочетание коротких морщин, обычно светлее фона,
ориентированных без видимой системы, переплетающихся и сливающихся друг с дру-
гом (Bradybaena scythica, Arlanta arbustorum, Eobania vermiculata, Cryptomphalus aspersa,
некоторые популяции Shileykoia daghestana).
Рис. 5. Форма базального края устья при рассмотрении снизу.
А — базальный край изогнут в виде лука; В — базальный
край плавно выгнут; В — базальный края прямой. (Орйг.).
Рис. 6. Варианты строения края устья (схематиче-
ские сагиттальные разрезы через базальную стенку,
вид справа). А — край устья умеренно отогнут, губа
уплощенная и широкая; Б — край устья не отог-
нут, губа почти не выражена; В — край устья утол-
щен, образует перистом; Г — утолщенный край
устья отогнут; Д — край устья сильно отогнут,
губа мощная и резкая; Е — край устья круто ото-
гнут по линии губы, образуя манжету. (Ориг.).
Края устья могут быть широко расставленными или в различной степени сближен-
ными; в исключительно редких случаях край устья цельный, т. е. не прерван париеталь-
ной стенкой (например, Helici gona lapicida); вместе с тем нередки случаи сильного сбли-
жения краев устья, тогда говорят о наклонности к образованию цельного устья.
Представители Helicoidea, как и подавляющее большинство других Geophila
растут не всю жизнь: по достижении половой зрелости рост раковины полностью оста!
навливается, и формируются
дефинитивные признаки. Од-
ним из таких признаков явля-
ется опущенность последнего
оборота к устью. В характере
опущенности наблюдаются 3
основных варианта: 1) послед-
ний оборот начиная с середины
или несколько позже медленно
и плавно отходит вниз от сред-
ней линии, оползает (рис. 4,
Б}-, 2) последний оборот опус-
кается плавно, без ясного пе-
региба, но непосредственно
близ устья (рис. 4, 5); 3) пос-
ледний оборот близ устья резко
перегибается; этот вариант
связан, как правило, с сильной
скошенностью устья (рис. 4, Г).
Надо заметить, что последний
оборот почти всегда в той или
иной степени опущен, но
иногда этот признак выражен
очень слабо.
Если рассматривать рако-
вину снизу, со стороны пупка,
то можно заметить, что базальный край устья иногда выгнут в виде лука и края устья,
таким образом, не лежат в одной плоскости (рис. 5, A; Helicodontidae); гораздо чаще
базальный край устья плавно и равномерно выгнут (рис. 5, Б) или прямой (рис. 5,
В); прямой край связан с тем, что базальная часть последнего оборота уплощена.
Устье половозрелых особей имеет обыкновенно в разной степени отогнутые края
и снабжено концентрическим утолщением — губой, расположенной несколько отступя
от края устья. Интенсивнее всего отворот края устья и губа развиты в колумеллярной
области; у некоторых видов Trichia s. str., Paedhoplita laminata и др. губа резко обры-
вается на стыке базального и палатального краев. Сочетание уплощенной широкой
губы й умеренно отогнутых краев устья в пределах надсемейства может рассматриваться
как исходный вариант по отношению к другим (рис. 6, А). У многих видов, в первую
очередь ксёрофильных Hygromiidae, край устья прямой, а губа не развита или выражена
2 А. А. Шилейко
18
ВВЕДЕНИЕ
очень слабо (рис. 6, В). Второй путь модификации дефинитивного устья — исчезнове-
ние участка устьевого края, лежащего кпереди от губы, утолщенным в этом случае
оказывается непосредственно край устья, именуемый в этом случае перистомом (неко-
торые Bradybaena, Helix) (рис. 6, В). Дальнейший шаг на этом пути сводится
к тому, что отгибается наружу перистом (ряд западноафриканских Heliclnae) (рис. 6, Г).
Наконец, третий путь — резкое усиление губы (некоторые Caucasigena, Archaica и др.)
(рис. 6, Д) и крутое отгибание края устья, приводящее к возникновению манжеты (тер-
мин А. А. Стеклова). Последний тип встречается среди Helicoidea редко (Isognomostoma)
(рис. 6, Е). Показательно, что среди множества особей Isognomostoma personatum
изредка встречаются экземпляры, у которых края устья отогнуты не столь круто и,
таким образом, отвечают картине, изображенной на рис. 6, Д.
Нередки случаи независимого возникновения зубов на колумеллярном, базаль-
ном и палатальном краях. Эти зубы представляют собой бугорки, возникающие в ре-
Рис. 7. Внешний вид пупка: А — запаянный; В — проколовидный, цилиндри-
ческий; В — перспективный, умеренно широкий; Г — развернутый. (Ориг.).
зультате местного впячивания участка губы; снаружи таким зубам соответствуют ямки-
Значительно более редко наблюдается возникновение зубов на париетальной стенке
устья. Из видов фауны СССР париетальные зубы имеются только у 3 видов рода
Paedhoplita и Isognomostoma personatum. Есть все основания предполагать, что у пос-
леднего поперечная париетальная пластинка сформировалась за счет разрастания па-
риетальной мозоли, связывающей края устья, видов вымершего рода Klikia', в част-
ности, К. osculum не имеет никаких серьезных отличий от I, personatum, но отличается
чрезвычайно мощной париетальной мозолью. Что касается париетальных зубов
Paedhoplita, то эта группа конхологически весьма специализирована, и вопрос о про-
исхождении париетальных зубов здесь пока неясен. Роль подобных образований, ве-
роятно, неоднозначна. Тут, по-видимому, и сужение просвета устья для уменьшения
интенсивности испарения, и содействие выделению эпифрагмы, и защита от хищных
насекомых. Солем (Solem, 1972) специально обращает внимание на роль дугообразных
париетальных складок, направленных концами наружу, в связи с проблемой защиты
от хищников. Наконец, надо отметить, что на колумеллярном крае у Helicoidea зубов
не бывает, но. нередко (особенно у Heliclnae) возникает наплыв — утолщение губы.
Пупок (рис. 7) — пространство между внутренними стенками оборотов — большей
частью открытый, т. е. сквозь него просматривается хотя бы часть предпоследнего обо-
рота. Если сквозь пупок видны внутренние стенки всех или почти всех оборотов, го-
ворят о перспективном пупке. Здесь могут быть два варианта: или пупок глубокий,
в виде колодца, или мелкий, в виде чаши. Во втором случае П. В. Матёкин предлагает
пользоваться термином «развернутый пупок» («. . .через пупочное отверстие видна
значительная часть внутренних стенок последнего и предыдущих оборотов» — Матё-
кин, 1966, стр. 100). Следовательно, пупок может быть перспективным, но не развер-
нутым. Полностью запаянный пупок — по-видимому, прерогатива Helicidae. В этом
случае колумеллярный край устья сильно расширен и плотно прирастает к нижней
стенке последнего оборота вокруг пупка; иногда между отогнутым колумеллярным краем
остается узкая щель, но называть ее пупком нельзя: это не пупок, но лишь отверстие,
посредством которого полость столбика имеет сообщение с наружной средой. Правда,
употребляемый часто термин «щелевидный пупок» на практике удобен и поэтому он
оставлен в тексте.
МАНТИЯ, НОЖА И ЭКТОДЕРМАЛЬНЫЕ ЖЕЛЕЗЫ
Мантия состоит из наружного и внутреннего эпителиальных слоев и разделяющего
их слоя субэпителиальной соединительной ткани. Клетки наружного эпителия содер-
жат пигментные гранулы (Abolins-Krogis, 1963), обеспечивающие рисунок на верхней
поверхности мантии. Форма и длина легкого зависит от формы последнего оборота,
но в общем не превышает 3/4 его длины.
Мантийный валик мощный, богат одноклеточными железами, несет три лопасти:
анальную, которая располагается наверху слева от анального отверстия и пневмостома,
и две боковые, причем левая развита лучше правой. Боковые лопасти гребневидны и
МАНТИЯ, КОЖА И ЖЕЛЕЗЫ
19
либо направлены вперед, либо прилегают к поверхности мантийного валика. Роль
лопастей хорошо видна, если, закрепив на пластилине раковину, рассиатрпвать об-
ласть устья под бинокуляром с сильным источником света. Моллюск, раздражаемый
сильным светом и тепловым потоком, начинает втягиваться в раковину. При этом ло-
пасти мантийного валика расправляются и очень плотно прилегают к ноге, отирая с ее
поверхности выделяемую в этот момент слизь. После того как задний конец ноги пол-
ностью втянут, виден мантийный валик с полностью сомкнутыми лопастями и слоем
слизи, снятой ими с поверхности тела (рис. 8). Застывая, этот слой слизи образует
эпифрагму. Блок (Block, 1971) различает два типа эпифрагмы: в виде толстой пористой
известковой пластинки или в виде тонкой полупрозрачной мембраны, состоящей из
высохшей слизи и инкрустированной гранулами углекислого кальция. Ла поверхности
Рис. 8. Стадии втягивания ноги у Kokotschashvilia phaeolaema. Объяснение
в тексте. (Ориг.)'.
эпифрагмы видны линии смыкания лопастей мантийного валика и беловатое пятно,
расположенное напротив пневмостома, которое мы предлагаем называть «отдушиной»
(Блок именует этот участок «ventilator»). Отдушина пориста, и сквозь нее осуществля-
ется газообмен. Поры отдушины можно видеть после обработки поверхности эпифрагмы
раствором нитрата серебра, который проникает довольно медленно, но делает рельеф-
ными края пор. Убедиться в функции отдушины несложно: стоит опустить моллюска
с эпифрагмой в теплую воду, и вскоре через отдушину начинается выделение пузырь-
ков воздуха.
Выявлены 3 закономерности в образовании эпифрагмы: 1) животные с пигменти-
рованным мантийным краем всегда образуют прозрачную эпифрагму, причем этот
процесс протекает с очень малой скоростью (до 14 суток); 2) если край мантии светлый,,
эпифрагма формируется быстрее (в пределах 6 суток) и относится к первому тину
(см. выше); 3) эпифрагма, образованная быстро (за 1—2 часа), всегда прозрачна, т. е.
состоит только из слизи, независимо от степени пигментации мантийного валика.
При длительной засухе может образоваться несколько эпифрагм; они могут на-
пластовываться одна на другую, но чаще разделены промежутком, причем в экспери-
ментах Блока Cryptomphalus aspersa при групповом содержании выделяли до 6 эпи-
фрагм, а при одиночном — до 16.
Функции эпифрагмы разнообразны: защита от врагов и патогенных микроорга-
низмов; защита тела моллюска от механических повреждений; прикрепление к суб-
страту; ограничение испарения; экскреция.
По поводу последней из перечисленных функций надо добавить, что в эпифрагме
отсутствует столь обычный экскреторный материал, как мочевая кислотаи ее дериваты,
но присутствуют фосфаты. Вместе с тем экскреция не может быть первостепенной функ-
цией эпифрагмы хотя бы потому, что последняя часто поедается моллюском после ос-
вобождения из раковины.
2*
so
ВВЕДЕНИЕ
.?• Блок отмечает еще одну важную подробность: все последовательно образующиеся
эпифрагмы связаны между Собой, а последняя — с телом моллюска посредством
тонкого тяжа — аристы (рис. 9). Последнюю аристу, т. е. ту, которая свя-
зывает: самую внутреннюю, последнюю по времени образования эпифрагму с мантий-
ным.валиком, обнаружить очень трудно, так как она еще не затвердела и рвется при
быстром втягивании животного, когда повреждается раковина. Химически ариста от-
личается от эпифрагмы тем, что обнаруживает кислую реакцию, а эпифрагма — основ-
ную., В связи с существованием аристы представляется вероятным, что эпифрагма
действует подобно психрометру, изменяя кривизну поверхности при повышении влаж-
ности: натяжение аристы усиливается, и животное получает соответствующий сигнал.
Вост раковины осуществляется деятельностью мантийного валика (рис. 10).
Параллельно его краю проходит мантийная борозда, кнаружи от которой эпителий об-
разует краевую складку; эпителий этой складки переходит на переднюю часть мантий-
ного края. С внутренней стороны к мантийной борозде прилегает слой высоких секре-
Рис. 9. Несколько последовательных эпифрагм у Сераеа hortensis (раковина частично
удалена) — I; эпифрагма — II. А — ариста; КМ — край мантии; О — отдушина;
звездочка — линия смыкания лопастей мантийного валика. (По Block, 1971, с из-
менениями).
торных клеток — железистая подушка. Наружный роговой слой раковины — пери-
остракум — выделяется клетками мантийной борозды, известковый слой — желе-
зистой подушкой, а самый тонкий внутренний слой — гипостракум — всей верхней
поверхностью мантии и внутренностного мешка (Иванов, 1940).
^ Пространство между верхней поверхностью ноги и нависающим передним краем
Мантии именуется супранухальной полостью, которую некоторые авторы гомологизи-
руЮт-; полностью или частично, с жаберной полостью Prosohranchia (Regondaud, 1961;
Harry,''4964).
Ма’н1ййнаЯ полость сообщается с окружающей средой посредством пневмостома,
расположенного в правом (при рассматривании раковины в нормальном положении
от наблюдателя — в левом) верхнем углу мантийного валика. Частота дыхания, о чем
можно судить по частоте открывания пневмостома, находится в прямой зависимости
от' температуры воздуха. По всей видимости, важную роль в механизме газообмена
выполняет тонкий мышечный слой, расположенный под столбчатым эпителием дна
мантийной полости (Pohunkova, 1967).
Одной из дополнительных функций легочной полости является также накопление
й соХфанение запаса воды: многие Helicoidea, сокращаясь при резком раздражении,
выбрасывают через дыхательное отверстие жидкость, содержащуюся в легком. Таким
образом; паллиальная вода играет роль депо, откуда происходит пополнение ее запа-
сов в организме по мере испарения (Blinn, 1964). Возможно также, что эта вода выпол-
ийЬт'роль своеобразной физической жабры, механизм действия которой может быть
бейовйй на разности парциальных давлений кислорода внутри легкого и в наружной
среде.
••••Jtjjjga представителей Helicoidea имеет различную толщину и окраску. Наиболее
пЙб'ХЙйё'Щокровы, выделяющие относительно небольшое количество слизи, характерны
'Ййй'Хгр^йныХ Helicinae с грубой раковиной. Очень тонкая кожа, особенно богатая сли-
'зйстымй ’одноклеточными железами, наблюдается у некоторых дальневосточных
Bfddybaena (Br. ravida, Br. vulcanica); при раздражении этих животных обильно выде-
МАНТИЯ, КОЖА И ЖЕЛЕЗЫ
21
ляется пузырящаяся слизь жидкой консистенции. Вообще надо отметить, что содер-
жание понятия «слизь» в применении к наземным моллюскам в последнее время зна7
чительно расширилось, и этим термином обозначается большой набор разных по хи-
мической природе выделений желез и органов, объединяемых только их консистенцией.
Так, у отдельных видов насчитывается до 8 сортов слизи (Campion, 1961). Одноклеточ-
ные кожные железы залегают под эпителиальными клетками. Опорожнение этих же-
лез происходит в результате сокращения мускульных фибрилл либо наружу/’ либо
внутрь, в гемоцель. Секрет этих желез состоит из мукополисахаридов, протеинов,»
СаСО3 в виде очень мелких гранул, жировых капель и пигмента (флавон). При раздра-
Рис. 10. Сагиттальный разрез через край мантии и раковину Trichia
hispida. ГС — гипостракум; Ж11 — железистая подушка; ЙС — из-
вестковый слой раковины; КВ — клетки мантийного валика; К МБ —
клетки мантийной борозды; КС — краевая складка; ИМ — поверх-
ность мантийного валика; ПМБ — просвет мантийной борозды;
ПС — периостракум. (По Муанье-де-Вильпуа, из: Иванов, 1940, с из-
менениями) .
жении моллюска, например кристалликом соли, выделяется слизь, содержащая боль-
шое количество воды, способствующей смыванию разражающего агента.
Среди желез эпидермального происхождения надо особо выделить педальную же-
лезу, залегающую в толще тканей ноги и открывающуюся под ротовым отверстием ши-
рокой щелью. Морфология педальной железы обнаруживает очень большое сходство
у видов из самых разных групп — Achatina (Ghose, 1963), Onchidiella (Binot, 1965),
Zonitoides (Шилейко, 1972a) и, как мы имели возможность наблюдать, у многих
Helicoidea. Это сходство позволяет широко экстраполировать данные по формированию
железы в эмбриогенезе. Дефинитивная непарная железа образуется парным впячива-
нием эктодермы. Дно железы несет два продольных валика, выраженных в разной сте-
пени и покрытых ресничками. Свод протока железы состоит из плоского эпителия,
часто расслаивающегося. Слизь, выделяемая этой железой, имеет кислую реакцию,
тогда как одноклеточные железы покровов продуцируют нейтральную слизь.
Что касается общей архитектоники ноги, то на поперечных срезах выделяются
3 слоя, резче выраженные в переднем и среднем отделах. Самый внутренний слой—
железистые клетки, расположенные в рыхлой соединительной ткани. Это большого
размера клетки, изливающие секрет в просвет краевой железы. Средний слой — зона
интенсивного развития собственной мускулатуры ноги, волокна которой идут в разно-
образных направлениях. Наружный слой занят одноклеточными железами, выраба-
тывающими слизь, покрывающую поверхность тела. Границы между этими зонами
нерезкие.
22
ВВЕДЕНИЕ
РЕНОПЕРИКАРДИАЛЬНЫЙ АППАРАТ1 И ЛЕГКОЕ
Ш"
Сердце и почка располагаются на крыше мантийной полости, занимая заднюю
ее часть (рис. 11). Сердце, характерное для представителей Euthyне ига, состоит из двух
камер и окружено перикардом. Желудочек отличается значительной толщиной сте-
нок. Сердце срастается с перикардом в местах входа и выхода; от предсердия начина-
ется легочная вена. Перикард представляет собой замкнутую полость с единственным
крошечным реноперикардиальным отверстием; стенки его образованы многочислен-
тп нп
Рис. 11. Участок крыши легкого Fruticocampylaea
narzanensis. ВМ — вторичный мочеточник; Ж — желу-
дочек сердца; К — кишка; ЛВ — легочная вена; МС —
мускульный слой; НП — нижняя доля печени; ОП —
отверстие между почкой и первичным мочеточником;
П — почка; ПК — перикард; ПМ — первичный мо-
четочник; ПС — предсердие; РП — реноперикардиаль-
ное отверстие; PC — ректальный венозный синус;
ТП — тонкостенный отдел почки. (Ориг.).
ными переплетающимися мускульными волокнами. Эти волокна разветвлены, синти-
циальны и не обнаруживают поперечной полосатости. На границе предсердия и желу-
дочка имеется клапан, предотвращающий обратный ток крови из желудочка в пред-
сердие; в местах входа и выхода в камеры сердца имеются мощные сфинктеры. Сокра-
щение сердца начинается в области аорты и кончается в предсердии. Газообмен в лег-
ком и деятельность сердца обнаруживают четкую взаимосвязь (Ghose, 1964). У всех
Helicoidea васкуляризована только дорсальная часть легкого, причем Дроздовский
(Drozdowski, 1970) отмечает прямую зависимость степени развития венозной системы
1 Выражения «выделительная система», «половая система» и т. и. имеют широкое
хождение в современной литературе. Мы присоединяемся, однако, к мнению В. Н. Бек-
лемишева (1964), что целесообразнее в данном случае пользоваться термином «аппарат»,
поскольку «система органов» — понятие тектологическое, т. е. обозначает совокуп-
ность органов одинаковой функции и строения (так, системой является осевая муску-
латура); аппарат же суть совокупность органов чаще всего различного строения, мо-
гущих играть различную роль, но объединенных функционально и морфологически
в единый комплекс, выполняющий единую более общую функцию.
СЕРДЦЕ, ПОЧКА И ЛЕГКОЕ
23
легкого от величины животного: в связи с тем, что площадь участвующей в газо-
обмене поверхности легкого у крупных моллюсков относительно меньше, чем
у мелких, возрастает удельная роль легкого в дыхании животного большого размера.
Выводы Дроздовского могут быть истолкованы, однако, несколько иначе (Шилейко,
1972г): вопрос о кожном дыхании наземных легочных вообще пока неясен, тем более
у крупных моллюсков с грубой кожей; более интенсивная васкуляризация легкого
у последних может стоять в простой зависимости от величины тела, содержащего боль-
шое количество жидкости. У видов небольшого размера диффузное дыхание, естест-
венно,'сильно облегчено. Кроме того, минимальный диаметр капилляра имеет предел,
Рис. 12. Поперечный разрез мантийной полости Bradybaena dichorozona.
ВМ — вторичный мочеточник; Д — диафрагма; Ж — желудочек сердца;
К — кишка; МС — мускульный слой; 11В — почечные выросты; ПК —
перикард; ПМ‘—первичный мочеточник; ПП — полость почки; PC—
ректальный венозный синус. (Ориг.).
и количество сосудов, свойственное легкому Helix, может просто не уместиться в лег-
ком животных такого размера, как например Karabaghia или Chilanodon. Добавим,
что рисунок, образуемый сосудами на поверхности легкого, подвержен значительной
индивидуальной изменчивости.
К перикардию справа прилегает почка, выделяющаяся на бесцветной прозрачной
поверхности легкого светло-желтой окраской. Почка представляет собой тонкостен-
ный мешок, стенки которого изнутри покрыты многочисленными высокими складками
и лепестками, вдающимися в полость почки (рис. 12). В стенках почки имеются тонкие
кровеносные сосуды. Проксимальный отдел почки, прилегающий к печени, выделяется
особенно тонкими стенками и отграничен от остальной части. Очень узким отверстием
на переднем крае почка сообщается с мочеточником, который, образуя очень резкий
изгиб, проходит вдоль тела почки спереди назад, тесно прилегая к нему. Этот отдел
именуется первичным мочеточником. Круто изогнувшись, первичный мочеточник пе-
реходит во вторичный, который проходит по правой стороне мантии, и, сопровождае-
мый прямой кишкой и ректальным венозным синусом, открывается рядом с анальным от-
верстием. Подобное строение почки, т. е. наличие вторичного мочеточника, свойственно
представителям подотряда Sigmurethra (Pilsbry, 1900; Taylor, Sohl, 1962).
Почечные сосуды особенно развиты в заднем тонкостенном отделе почки, куда по-
ступает венозная кровь из большого синуса тела, а также из сосудов, расположенных
на ее нижней и верхней поверхностях. Артериальное кровоснабжение осуществляется
за счет выносящих легочных сосудов правой части мантийной полости, впадающих
в почку справа.
24
ВВЕДЕНИЕ
ПИЩЕВАРИТЕЛЬНЫЙ ТРАКТ
В глубине ротового отверстия, окруженного околоротовыми лопастями, распола-
гается роговая челюсть, покрытая поперечными ребрышками и лишенная срединного
выступа; только у Sphincterochila челюсть без ребрышек и со срединным выступом.
Челюстные ребрышки особенно сильно развиваются у видов, отличающихся большими
размерами тела, а также у видов, обитающих в аридных условиях (Шилейко,
1968а).
Далее располагается ротовая полость, занятая радулярным аппаратом. Послед-
ний состоит из радулы и системы мышц и хрящей, обеспечивающих ее работу. Радула
слагается из прозрачной седловидной мембраны и расположенных на ней зубов; по
средней линии вдоль проходит ряд срединных зубов, имеющих более или менее сим-
метричное строение. Справа и слева располагается по несколько рядов боковых зубов,
боковое положение занимают ряды краевых зубов. В задней части глотки находится
радулярное влагалище, где осуществляется формирование новых зубов взамен стер-
шихся. Образование зуба начинается с закладки базальной пластинки, красящейся
по методу Риттера и Олесона (Пирс, 1962) на нейтральные и кислые мукополисахариды
в фиолетовые тона, что указывает на ее базофильный характер; пластинка образуется
одной продуцирующей клеткой, расположенной в эпителиальном слое стенки раду-
лярного влагалища. Полость влагалища заполнена соединительнотканными клетками,
образующими тело зуба, т. е. по существу одонтобластами. В формировании тела зуба
принимают участие 2 типа клеток: «ядро», т. е. центральная, внутренняя часть зуба,
образуется одной соединительнотканной клеткой, накапливающей нейтральный муко-
протеид. Эта клетка красится в интенсивный темно-красный цвет. Накопившая муко-
протеид клетка подходит к базальной пластинке и плотно с ней контактирует; при этом
клетка приобретает коническую форму и превращается в «ядро» зуба, це теряя при этом
гистохимических характеристик. Примечательно, что зубы на этой стадии окраши-
ваются также эозином. Следующая стадия заключается в окутывании «ядра» плотным
чехлом, выделяемым несколькими небольшими удлиненными соединительнотканными
клетками. На препаратах часто видны зубы, имеющие темно-красную сердцевину,
покрытую толстой прозрачной оболочкой. За исключением первых 6—8 рядов, зубы
не окрашиваются (Шилейко, 19726). Это связано со сменой химического состава зу-
бов: вначале зубы состоят из белков и хитина (возможно, из гликопротеина, подобно
хитину насекомых); позже наблюдается появление аминогрупп и материала, дающего
положительную реакцию на серебро. Наконец, в теле зуба откладываются соли же-
леза и кремния, придающие ему механическую прочность (Runham, 1962, 1963).
Издавна. принято рассматривать радулу как двусторонне-симметричный орган
с осью симметрии, проведенной по основным зубцам срединного ряда зубов. При этом
принималось, что зубы с одинаковыми порядковыми номерами в пределах одного ряда
являются зеркальными отражениями друг друга, и такое положение не меняется от
одного горизонтального (поперечного) ряда зубов к другому, за исключением несколь-
ких рядов самых молодых и самых старых (передних). Кроме того, постоянными видо-
выми признаками считаются число зубов в каждом горизонтальном ряду и число самих
рядов. В связи с этим во многих систематических работах приводятся в качестве одного'
из видовых признаков радулярные формулы, составляемые обычно так: примерно в се-
редине радулы выбирается половина одного горизонтального ряда, подсчитывается
число боковых и краевых зубов, учитывается степень их расчленения, и полученный
результат удваивается — получается формула для одного ряда, которая и приводится
как отражающая особенности радулярных признаков вида в целом. Специально пред-
принятое нами исследование показало несостоятельность принятого способа описания
радулы (Шилейко, 19726). Изучение радулярных особенностей 16 видов позволило
прийти к следующим выводам.
1. Радула представителей надсемейства асимметрична. Асимметрия проявляется
в непостоянстве границ между боковыми и краевыми зубами в правой и левой сторонах
одного горизонтального ряда; в числе зубов в правой и левой половинах одного ряда;
в разном характере расщепления зубов в правой и левой половинах одного ряда; иногда
асимметрия проявляется и в строении срединного зуба. Если имеются какие-то верти-
кальные закономерности (обнаружены у единственного вида — Monacha carthusiana), то
они касаются только одной половины радулы, либо справа и слева они касаются раз-
ного числа зубов, имеющих неодинаковые порядковые номера.
2. Радула, как правило, имеет для каждой половины каждого горизонтального .
ряда свою формулу, подчас совершенно непохожую на формулу предыдущего или сле-
дующего ряда.
3. Радулярные признаки, имея мутагенную природу, носят ярко выраженный
индивидуальный характер, и, как правило, видовая специфичность в этих признаках
не проявляется.
4. В тех редких случаях, когда радула обладает специфическими видовыми при-
знаками, такие признаки должны описываться особо; составление традиционных раду-
ПИЩЕВАРИТЕЛЬНЫЙ ТРАКТ
25
лярных формул нецелесообразно, так как такие формулы не отражают и не могут отра-
жать видовой специфики органа.
Размер базальной мембраны, отдельных зубов и плотность их расположения нахо-
дятся в прямой зависимости от размеров животного (Drozdowski et al., 1966; Isaran-
kura, Runham, 1968).
Вообще же радулы Helicoidea могут быть двух типов. Наиболее широко распростра-
нен первый тип — имеются основной и дополнительный зубцы; те и другие конической
формы. Второй тип обнаружен у некоторых Camaenidae (Papuina): все зубы примерно
одинаковы, имеют форму долота (Solem, 1972а). Интересно, что наличие таких типов
отмечено и у группы, далекой в систематическом отношении от Helicoidea — у Clau-
siliidae (Лихарев, 1962).
Радулярная мембрана располагается на одонтофоре, представляющем собощси-
стему хрящей, основу которой составляют два удлиненных хряща, ориентированных
дорсовентрально.
Рис. 13. Пищеварительный тракт Brydybaena plectotropis. ВИ — верхняя
доля печени; Г — глотка; Ж — желудок; ЖН — железа Налепа; 3 — зоб;
НП — нижняя доля печени; ПК — прямая кишка; ПРП — отверстия про-
токов печени; РГ — ретрактор глотки; СЛ — стенка легкого; СЛЖ — слюн-
ная железа. (Ориг.).
От заднего конца глотки, в которую, переходит полость рта, начинается короткий,
часто S-образно изогнутый пищевод, переходящий в объемистый тонкостенный зоб,
внутренние стенки которого покрыты тонкими редкими высокими продольными склад-
ками. На поверхности зоба лежит пара сильно расчлененных светлых слюнных же-
лез, от каждой из которых кпереди отходят тонкие протоки, впадающие в глотку;
места впадения протоков окружены так называемыми железами Налепа.
Экскреторные отделы слюнных желез содержат два основных типа клеток: про-
дуцирующие слизь — мукоциты и продуцирующие зимогенные гранулы — зимоциты.
Цитоплазма мукоцитов заполнена вакуолями, содержащими слизь, идущую на1увла-
жнение пищевого комка. Зимоциты — большие клетки диаметром более 50 мкм, их
цитоплазма включает вакуоли, митохондрии и эндоплазматическую сеть. Все экс-
креторные клетки слюнных желез имеют крупные ядра, богатые ДНК. Выводные про-
токи желез выстланы мерцательным эпителием, среди клеток которого также имеются
экскреторные элементы. Дистальные отделы выводных протоков выстланы ; только
покровным эпителием, железистых клеток здесь нет (Quattrini, 1967).
Задняя часть зоба сужается и переходит во внутренностный мешок, где впадает
в округлый желудок. С желудком сообщаются печеночные протоки, по которым пища
поступает в полость печени, точнее, гепатопанкреаса. Здесь растворенные вещества
всасываются печенью, а белковые частицы фагоцитируются, подвергаясь внутрикле-
точному пищеварению. Гликоген и фосфорнокислая известь откладываются в печени,
а жир, кроме того, в соединительной ткани по соседству с печенью. Непереваренные
частицы выводятся обратно в желудок, а затем — в тонкую кишку. Основной функцией
последней является вывод экскрементов наружу (Иванов, 1946). Длина тонкой кишки
зависит от длины легкого, поскольку сопровождает его по всей длине: у форм с неболь-
шим числом стремительно нарастающих оборотов легкое короче, чем у форм с много-
оборотной раковиной. Соответственно меняется длина тонкой кишки (Schmidt, 1936),
26
ВВЕДЕНИЕ
Печень состоит из 2 лопастей — правой и левой. Правая (верхняя) лопасть зани-
мает верхние обороты, в ткани ее погружена гонада. Левая лопасть располагается в об-
ласти желудка и тонкой кишки и подразделяется на две доли (рис. 13).
Отходящая от желудка тонкая кишка образует 2 петли в контакте с поверхностью
печени и частично даже погружаясь в нее, затем проходит по правому краю мантийной
полости, сопровождаемая тонкой мышечной лентой, и заканчивается анальным отвер-
стием. Внутри кишки продольно проходит высокая тонкая складка — тифлозоль,
хорошо выраженная у животных большого размера, но неясная у мелких видов.
МУСКУЛАТУРА
Осевая мускулатура очень однотипна и представлена колумеллярным мускулом,
крепящимся к столбику на уровне 2—4 оборотов (рис. 14). От этого мускула вперед от-
ходят две мощные ленты — ретракторы глотки. Более или менее независимо от них
отходят ретракторы омматофоров, нередко пигментированные в дистальной части,
причем у одних видов ретрактор правого омматофора проходит между пенисом и ваги-
ной, а у других видов — рядом с ними. В первом случае соответствующий участок
ретрактора часто имеет непосредственную связь с основанием пениса посредством мус-
кульных волокон и соединительнотканных перепонок. Ниже глотки и по ее бокам распо-
IW РГ ПДР
Рис. 14. Схема опорно-двигательного аппарата Helicoidea.
БПГ — боковой протрактор глотки; Д — диафрагма; ДГ —
депрессоры глотки; НДР — педальные ретракторы; ПЖ — пе-
дальная железа; ПР — половой ретрактор; РГ — ретрактор
глотки; РНЩ — ретрактор нижнего щупальца; РО — ретрактор
омматофора; СТ — столбик. (По Trappmann, 1916, с изменениями).
лагаются ретракторы губных щупалец. Если ретракторы глотки имеют антагонистов —
протракторов глотки, то ретракторы щупалец антагонистов не имеют, и выворачивание
щупалец осуществляется давлением полостной жидкости. От колумеллярного мускула
вниз отходят многочисленные короткие мощные педальные ретракторы. Антагонистом
осевой мускулатуры в целом является кожная мускулатура, особенно кольцевая; ее
сокращение вызывает вытягивание головы и туловища. Определенную роль играет
также перераспределение крови в туловищных синусах.
Помимо колумеллярной мускулатуры имеются два независимых мускульных ком-
плекса. Первый — мышечный аппарат глотки, состоящий из 4 групп мышц: глоточный
сфинктер, группа протракторов (передних, боковых и вентральных), группа леваторов
и группа депрессоров (опускателей). Второй комплекс представлен единственным мус-
кулом — половым ретрактором, связывающим место перехода эпифаллуса или семе-
провода в пенис с нижней стенкой мантийной полости, именуемой часто диафрагмой.
Нога и многие внутренние органы также имеют собственную мускулатуру. Осо-
бенно обильны мускульные волокна в составе стенок стилофора, где они образуют по-
многу кольцевых и продольных слоев.
НЕРВНЫЙ АППАРАТ
21
НЕРВНЫЙ АППАРАТ И НЕЙРОСЕКРЕЦИЯ
Для нервной системы легочных вообще, и рассматриваемого таксона в особенности
характерна высокая степень концентрации ганглиев, группирующихся в окологлоточ-
ное нервное кольцо. Однако в пределах надсемейства наряду с группами, в ЦНС ко-
торых различимы все основные ганглии, имеются таксоны, у представителей которых
субэзофагеапьный комплекс настолько компактен, что выглядит как одно целое, и на-
личие отдельных ганглиев обнаруживается лишь на срезах. Кроме того, в моменты
интенсивной нейросекреции эпиневральная оболочка сильно разбухает, придавая под-
пищеводному ганглиозному комплексу вид единой сплошной массы, где невозможно
увидеть отдельные ганглии (рис. 15, IT).
Над местом перехода глотки в пищевод располагается пара церебральных ганглиев,
соединенных короткой комиссурой, выраженной в различной степени. От церебральных
ганглиев назад и вниз отходят длинные церебро-плевральные и церебро-педальные кон-
нективы, а вперед — церебро-буккальные. Буккальные ганглии маленькие, лежат
на верхнебоковых поверхностях глотки. От латеральных выростов церебральных ган-
глиев отходят обонятельные нервы, к которым почти примыкают очень тонкие зритель-
ные; они сопровождают ретракторы омматофоров. От заднебоковых поверхностей ган-
глиев отходят наружные, средние и внутренние губные нервы, иннервирующие соот-
ветственно мускулатуру глотки, губные щупальца и стенки головного отдела вместе
с передней частью глотки. От церебральных ганглиев между церебро-плевральными и
церебро-педальными коннективами проходит очень тонкий нерв статоцистов. Наконец,
от правого церебрального ганглия начинается непарный нерв пениса (рис. 15, Z).
Подглоточная, точнее, подпищеводная нервная масса составлена 7 ганглиями:
парой педальных, парой плевральных, паллиальным, париетальным и абдоминальным.
Самые большие — педальные ганглии, они занимают всю вентральную часть подгло-
точного отдела нервного кольца и иннервируют ногу, стенки тела и дистальные части
полового аппарата. Плевральные ганглии занимают верхнее латеральное положение
и отличаются небольшими размерами. Паллиальный ганглий иннервирует посредством
левого мантийного нерва левую чаЪть мантии и располагается ближе к левой стороне.
Париетальный ганглий располагается справа почти симметрично по отношению к пал-
лиальному, иннервирует правую сторону мантии. Абдоминальный ганглий возник в ре-
зультате слияния висцерального и субинтестинапьного (правого париетального) ган-
глия переднежаберных моллюсков. От этого ганглия отходит интестинальный нерв;
сопровождая головную аорту, он входит во внутренностный мешок, где разделяется на
3 ветви. Одна из них проходит по левой стороне тепа, иннервируя часть кишки и пе-
чени, другая иннервирует реноперикардиальный аппарат, а третья — белковую
железу и гермафродитный проток. От этого же ганглия отходит анальный нерв.
Буккальные ганглии связаны комиссурой и иннервируют глотку, пищевод и
слюнные железы; кроме того, от церебр'о-буккальных коннективов отходят нервы
к стенкам глотки.
В связи со способностью церебро-педальных и церебро-плевральных коннек-
тивов заметно растягиваться положение окологлоточного нервного кольца относи-
тельно глотки может быть различным: при сильном сокращении глоточных ретрак-
торов вся глотка целиком проходит сквозь нервное кольцо, и последнее оказывается
в непосредственном соседстве с передней стенкой тела. Другими словами, глотка
имеет значительную подвижность относительно нервного кольца (а не наоборот).
Наряду с центральным нервным аппаратом, углубившимся под кожу, имеется
и периферическое нервное сплетение (рис. 16) — диффузная сеть, способная к само-
стоятельным рефлексам. Сложное сплетение имеется и во внутренних органах;
с центральным отделом нервного аппарата оно стоит в связи через буккальные и ви-
сцеральные (плевральные) ганглии (Беклемишев, 1964).
Из всех видов чувствительности у Helicoidea лучше всего развита тактильная чув-
ствительность. Особенно ярко она проявляется при раздражении придатков головы:
околоротовых лопастей и щупалец. Обоняние локализовано на щупальцах (Судзуки,
1968). Моллюски воспринимают запахи растений, на которых они обычно питаются,
проявляя направленную реакцию к источнику запаха. В экспериментах Судзуки
(Suzuki, 1967) на Ezohelix jlextbilis эта реакция исчезала после ампутации одного
глазного щупальца, но сохранялась после удаления обоих губных щупалец.
Следовательно, дистантная хеморецепция осуществляется глазными щупальцами,
а губные щупальца действуют лишь при непосредственном контакте моллюска
с источником раздражения.
На вершине верхних щупалец располагаются глаза, почему и сами щупальца
получили название омматофоров. Обонятельный нервный узел располагается по
соседству с глазом в концевом вздутии щупальца. Глаз представляет собой отшну-
ровавшееся впячивание покровов — глазной пузырь. Полость глаза занята сфери-
ческой линзой — хрусталиком; между хрусталиком и стенкой глазного пузыря име-
ется слой стекловидного тела. Вероятно, возможности зрения наших моллюсков
значительно выше, чем обычно считают; во всяком случае, Otala lactea при передви-
I
Рис. 15. Окологлоточное нервное кольцо. I — Bradybaena
fruticum (экземпляр фиксирован в августе); II — Frutico-
campylaea narzanensis (экземпляр фиксирован в мае). АГ —
абдоминальный ганглий; АН— анальный нерв; Б Г — буккаль-
ный ганглий; ГН — глоточный нерв; ИН — интестинальный
нерв; НС — нерв статоциста; ОН — обонятельный нерв;
НАГ — паллиальный ганглий; ИГ — педальный ганглий;
ПЛГ — плевральный ганглий; ПМН — правый мантийный
нерв; ПН — педальные нервы; ПРГ — париетальный ганг-
лий; С — статоцист; ЦГ — церебральный ганглий; ЦПК —
це ребро-педальная комиссура; ЦПЛК — церебро-плевраль-
ная комиссура. (Ориг.).
ПОЛОВОЙ АППАРАТ
29
Рис. 16. Периферическое
диффузное нервное спле-
тение в подошве ноги
Helix pomatia. (По Нап-
strom, из: Беклемишев,
1964).
жении обладает способностью зрительной ориентировки и может оценивать размеры
объектов (Hermann, 1968). Внутренняя роговица соприкасается с внешней, представ-
ляя собой переднюю стенку собственно глаза. Боковая и задняя стенки роговицы
представляют собой сетчатку, состоящую из пигментных и
пых клеток (Schwalbach et al., 1963; Eakin, Brandenburger,
Статоцисты — органы равновесия — располагаются
на поверхности педальных ганглиев; особенно хорошо
они заметны на тотальных препаратах ЦНС, просветлен-
ных и заключенных в канадский бальзам.
Нейросекреторный материал накапливается и в ней-
ронах ганглиев (Kuhlmann, 1963; Fernandez, 1966; Са-
darin, Beladeen, 1968; Sanchis, Zambrano, 1969). По-
мимо нервных элементов нейросекрет выделяется так
называемой нейрогемальной областью, которая распро-
странена от окологлоточного нервного кольца до пери-
ферических . нервов (губных, артериальных, комиссу-
ральных).
Нейросекреторные клетки с ацидофильным продук-
том секреции в цитоплазме обнаружены у моллюсков
из разных классов (Gabe, 1965). Однако истинные ней-
рогемальные органы, теснейшим образом связанные
-с нейросекреторной деятельностью, обнаружены только
у легочных моллюсков, в частности у Helicoidea. Несом-
ненно, имеется связь нейросекреции с годичным циклом,
с регуляцией водного баланса и с функционированием
полового аппарата. В. этой связи надо отметить, что
важную роль в механизме гормонального контроля иг-
рают глазные щупальца. Особи с удаленными щупаль-
цами продуцируют яйца с более мощной оболочкой и бо-
лее крупные ооциты (Stephens G. J., Stephens G. С.,
1966).
собственно зритель-
1967; Rogers, 1971).
ПОЛОВОЙ АППАРАТ
Общая схема строения полового аппарата представ-
лена на рис.. 17. Он состоит из гонады, гроздья кото-
рой, впадающие в гермафродитный проток, погружены
в ткани верхнего отдела печени. Гермафродитный проток, вначале прямой и тонкий,
по мере приближения к белковой железе все более круто извивается, а диаметр его
увеличивается. Близ белковой железы проток вновь сужается и, образуя крутое
колено, более или менее глубоко погруженное в ткани белковой железы, впадает
в спермовидукт (рис. 18). В колене гермафродитного протока располагается квад-
ривий (камера оплодотворения), строение которого у виноградной улитки освещено
в статье Линда (Lind, 1972) и еще у 17 видов — в работе Л. В. Шилейко и А. А. Ши-
лейко (1975). Выделены два типа строения этого участка, принципиально, впрочем,
не различающиеся. I тип — гермафродитный проток слепо замыкается, его апикаль-
ная часть представляет собой собственно камеру оплодотворения, т. е. то место, где
происходит слияние яйцеклеток и спермиев. Ниже собственно камеры оплодотворе-
ния гермафродитный проток посредством небольшого отверстия сообщается с нисхо-
дящим протоком, апикальная часть которого также слепо замкнута и служит местом
накопления спермиев партнера, т. е. заслуживает наименования сперматеки (в дан-
ном смысле — термин Линда). II тип представляет собой вариант I типа, усложненный
за счет возникновения кармана на гермафродитном протоке, придатков собственно
камеры оплодотворения и 1—3 карманов сперматеки. Последние могут быть выра-
жены в разной степени и при прямом вскрытии, как правило, не видны
(рис. 19).
Внешние различия касаются в основном степени пигментации, изогнутости и
длины всего комплекса в целом, а также степени его обособленности от тканей бел-
ковой железы, но основной принцип построения, т. е. наличие собственно камеры
оплодотворения и сперматеки, сохраняется в пределах, по-видимому, всего надсе-
мейства.
Ниже белковой железы располагается спермовидукт, который внешне доста-
точно ясно подразделяется на женский отдел (матку) и простату, окружающую се-
менной желобок. Маткой принято именовать тот отдел, который мы здесь называем
верхним отделом вагины, однако функционально маткой является именно часть жен-
ского отдела в пределах спермовидукта: здесь формируются яйца, а у живо-
30
ВВЕДЕНИЕ
родящих видов (некоторые представители Oreohelicidae и азиатские Camaenidae) —
молодь.1
Внутри спермовидукта разделения женского и мужского протоков не наблюда-
ется, точнее, эти протоки разделены неполно (рис. 20). На рис. 20, I, II изображены
поперечные срезы через спермовидукт Lindholmiola corcyrensis, собранных в ноябре,
когда они уже ушли в зимнюю спячку. Видно, что стенки матки тонкие, секретиру-
ющие клетки не функционируют, находятся в спавшемся состоянии. Срез I сделан
в дистальной части, где простата отсутствует; семенной желобок окружен желези-
стыми клетками, находящимися в разных
фазах деятельности: растущие клетки
с центральным ядром и интенсивно крася-
щейся эозином цитоплазмой; сформировав-
шиеся крупные клетки; клетки, частично
излившие секрет — с эксцентричным яд-
ром и большой вакуолью; наконец, пол-
ностью опорожненные клетки, от которых
остается лишь прозрачная бесцветная обо-
лочка. Срез II относится к той же особи,
но сделан выше первого, в зоне интенсив-
ного развития простаты, носящей ациноз-
ный характер, т. е. составленной клеточ-
ными комплексами — фолликулами, каж-
дый из которых имеет более или менее
ясно выраженную центральную полость.
Помимо этого простата имеет еще общий
щелевидный просвет, куда попадает сек-
рет, выделяемый клетками фолликулов.
Рис. 17. Общая схема строения полового
аппарата Helicoidea. ВЖ — белковая же-
леза; ВВ — верхний отдел вагины; ГЖ —
гермафродитная железа; ГН — гермафро-
дитный проток; ДСП — дивертикул семе-
приемника; К Л— клоака (половой атриум);
М — матка; НВ — нижний отдел вагины;
П — пенис; ПР — простата; РП — ретрак-
тор пениса (половой ретрактор); С —
семепровод; СОД — спермовидукт; СП —
семеприемник; Ф — флаге ллум (бич);
9П — эпифаллус; Я — яйцевод. (Ориг.).
На рис. 20, III представлен другой вид — Helicopsis striata) данная особь зафикси-
рована в августе, во время интенсивной деятельности матки. Стенки органа в этот
момент мощные, состоят из больших, слабо прокрашивающихся клеток; резко выде-
ляются интенсивно окрашенные гемалауном Майера ядра. Железистое оформление
семенного желобка носит тот же характер, что и у ноябрьских L. corcyrensis) что ка-
сается простаты, то фолликулы здесь теряют определенность, клеточные оболочки
в пределах фолликулов в значительной степени лизируются, ядра сохраняются лишь-
в отдельных клетках.
Далее вниз спермовидукт разделяется: от него отходит тонкий семепровод, впа-
дающий в эпифаллус или, в случае отсутствия такового, непосредственно в пенис.
Придатком мужского отдела является также бич, присутствующий у Hygromiidae
и Helicidae и отсутствующий у Нelicodontidae и Bradyhaenidae. Кроме того, в редких
случаях при основании пениса развивается придаток (Xeropictd).
Отдел женской части от места отхождения семепровода до основания протока
семеприемника мы именуем яйцеводом, который далее переходит в верхний отдел
вагины — участок, заключенный между основанием семеприемника и придатками
женского отдела, если таковые присутствуют. К таким придаткам относятся стило-
форы и слизистые железы. Секреторная деятельность этих желез проявляется с весны
до осени и особенно усиливается в период спаривания и откладки яиц (Bornchen,
1967). При выходе моллюсков из зимней спячки железистые клетки удлиняются и
разбухают, изменяются величина и положение ядер, усложняется аппарат Гольджи,
возникают зоны РНК. Формирование секрета в виде капелек диаметром до 4.5 мкм
1 Термин «матка» в предлагаемом смысле уже употреблялся в литературе (на-
пример: Wurtz, 1955; Miller, 1972а, 1972b, и др.).
Рис. 19. Квадривий Fruticocampylaea narzmensis. Слева — внешний
вид, справа — внутреннее строение, схематизировано. ГП — герма-
фродитный проток; КГП — карман гермафродитного протока; КО —
камера оплодотворения; НП — нисходящий проток; О — отверстие
между камерой оплодотворения и сперматекой; СТ — сперматека.
(Ориг.).
Рис. 18. Нижняя часть белковой железы, квадривий и верхняя часть спермовидукта
(схема). В Ж — белковая железа; ГП — гермафродитный проток; К — квадривий;
М — матка; ПР — простата; СКЖ — скорлуповая железа. (Ориг.).
0.5 мл?
Рис. 20. Поперечные срезы через спермовидукт. I—II — Lindholmiola corcyrensis
(экземпляр фиксирован в ноябре): I — дистальная часть, близ основания протока
семеприемника; II — немного выше середины; III — Ilelicopsis striata (экземпляр
фиксирован в августе), немного выше середины. ПК — пустая клетка; ПМ — полость
матки; СЖ — семенной желобок; СК — секретирующая клетка; СМ — стенка матки;
ФП — фолликул простаты; ЦП — центральная полость простаты. (Ориг.).
БИОЛОГИЯ, 'питание
33
с заключенными в них мукополисахаридными и белковыми гранулами происходит
в аппарате Гольджи; в образовании секрета принимает участие связанная с аппаратом
Гольджи кислая фосфатаза и зона РНК, которая в период зимнего покоя распростра-
нена по всей цитоплазме. В просвете железы помимо секреторных капель находится
богатая белком гомогенная масса, образующаяся при отшнуровывании апикальной
части удлиненных эпителиальных клеток, другими словами, выделение • секрета
осуществляется как мерокринным, так и апокринным путем. В течение зимней спячки
синтез секрета резко ослабляется, причем выделение его происходит исключительно
апокринным путем в дистальных отделах железы. При искусственном прерывании
спячки деятельность железы восстанавливается уже через сутки.
В редких случаях слизистые железы и стилофоры могут отсутствовать. Отдел,
лежащий ниже стилофоров, носит название нижнего отдела вагины, он впадает в кло-
аку, или половой атриум. Надо заметить, что оба термина не вполне точны: клоакой
обычно называют объединенный отдел выделительного и полового аппаратов,
а в данном случае этот участок есть полость, в которую впадают два отдела полового
аппарата. Возможно, более удачен термин «половой атриум», поскольку слово «atrium»
означает «преддверие» и здесь невозможна путаница с atrium cordis — предсердием..
Сперма партнера поступает вначале в семеприемник, состоящий из резервуара
и более или менее длинного протока; у некоторых групп имеется дивертикул, или
отросток, семеприемника, могущий вторично дегенерировать. В этом случае имеется
четкая закономерность: чем длиннее бич, тем длиннее дивертикул (в частности, это
убедительно показано для некоторых Helminthoglyptidae) (Miller,' 1972а). По мере
поступления длинного сперматофора он вначале проникает в дивертикул семеприем-
ника, а «хвост» сперматофора заворачивается вверх, к резервуару. Однако у ряда
групп перед совокуплением сперматофор сворачивается в клубок внутри пениса и
компактным комком подается в половые пути партнера. Понятно, что в этом случае.
надобность в дивертикуле отпадает, и он редуцируется. В целом можно сказать, что
у видов с длинным дивертикулом непременно длинный бич, но не у всех видов с длин-
ным бичом имеется соответствующей длины дивертикул.
Формирование сперматофора у виноградной улитки в деталях выяснено. весьма-
обстоятельной работой Линда (Lind, 1972). В противоположность гипотезе Ригби
(Rigby, 1963) и Смита (Smith, 1965), которая была критически разобрана нами (Ши-
лейко, 1972г), доказано, что тело сперматофора формируется в эпифаллусе, а хвосто-
вая его часть — в биче. В соответствии с этим наблюдается корреляция структур
внутрй эпифаллуса и бича с внешними особенностями оболочки сперматофора
(см. ниже). 1
Пенис может быть в виде простой трубки, внутренние стенки которой? несут более
или менее специализированные структуры (Oreohelicidae, Bradybaenidae), или. име-
ется одна или две пениальные папиллы, строение которых может быть в свою очередь
очень сложным (Trichiinae, Euomphaliinae).
Дополнительной функцией полового аппарата является участие в элиминации
чужеродных частиц: в педальный синус Helix Браун (Brown, 1967) вводил коллоидную
суспензию двуокиси тория (торотраста). Радиографически установлено, что основной
путь удаления частиц— не эпителий мантии (как у пресноводных легочных), а по-
ловой аппарат: скопления наполненных- чужеродными частицами амебоцитов наблю-
дались в стенках гениталий, особенно в выводных протоках женского отдела, и кон-
центрация их наблюдалась в . просвете вагины и пениса.
БИОЛОГИЯ ПРЕДСТАВИТЕЛЕЙ .HELICOIDEA. / . .
. ПИТАНИЕ ' - .--V.
Подавляющее большинство членов нашей группы — растительноядные поли-
фаги. Среди них совершенно неизвестны хищники, хотя некоторые1 крупные виды
нередко поедают органические вещества животного происхождения. '?- , -'-
Предпочитаемые пищевые объекты- для большинства видов—подгнивающие-
зеленые части растений, гифы грибов- и лиственный опад.: Некоторые виды — пред*
ставители родов Helix, Xeropictd, Monacha, Arianta — потребляют и'свежую зелень,
особенно сочную. Так, в лаборатории моллюски особенно охотно едят свежие- Огурцы
и проростки злаков. Учитывая, что многие виды, населяющие открытые пространства,
часто образуют большие скопления, они порой наносят значительный вред молодым:
всходам сельскохозяйственных культур-(Vappula, 1965; Bishara et-al., 1968, И’Др.)»
Основное количество лесных видов питается преимущественно за счет -граасбвр®
и лиственного опада, причем наибольшее видовое разнообразие и наибольшая'длот*
ность наблюдаются в опаде осинников и ольшаников. Подавляющее- число видов
решительно избегает хвои, лишь в виде редкого- исключения они- встречаются в /чист®
хвойных ;лесах (Pseudotricliia rubiginosa .в высокогорБИчКарпат)а.*•«*<*.*> •.««•
3 А. Шилейко
34
ВВЕДЕНИЕ
Молодь ксерофильных видов, обитающих в открытых местах, держится в дер-
новинках трав и питается отмершими частями этих растений. Взрослые особи могут
потреблять и свежие стебли.
Скаложилы, а также виды пустынь и полупустынь питаются преимущественно
за счет лишайников (Yom-Tov, Galun, 1970).
РАЗМНОЖЕНИЕ, ЖИЗНЕННЫЙ ЦИКЛ
И ДЛИТЕЛЬНОСТЬ ЖИЗНИ
Подавляющее большинство Helicoidea — яйцекладущие животные; живорож-
дение отмечено только у немногих неарктических и юго-восточноазиатских групп.
Совокупление происходит ночью либо в пасмурные дни, после дождя. Яйца от-
кладываются кучками в различные укрытия.
Скорость полового созревания связана с продолжительностью жизни. Моллюски
нашей группы могут иметь одногодичный, двух- и трехгодичный и многолетний циклы
(Deblock, Hoestland, 1967; Chatfield, 1968). По данным Бонавита (A. Bonavita, 1965),
ксерофильны'е Hygromiidae (=Helicellinae auct.) имеют 1—2-годичный жизненный
цикл. Двулетние виды живут 25—30 месяцев, размножаются либо в конце весны,
либо осенью. Популяции этих моллюсков состоят из взрослых особей, погибающих
с июля до конца года; подрастающих, которые будут размножаться ближайшей осенью,
и молодых особей нового поколения. Следовательно, зиму переживает небольшое
количество половозрелых особей и молодь, появившаяся осенью. Для двух видов —
Xerpsecta crenimargo и Helicopsis retowskii — нами Зарегистрированы случаи совокуп-
ления между представителями этих возрастных групп. Роль самцов выполняют' по-
ловозрелые особи, передающие сперматофор; у особей с не вполне сформированными
гениталиями семеприемник оказывается наполненным (рис. 21), и спермин хранятся
там до полного созревания животного.
Завезенный в Австралию европейский вид Cernuella virgata, встречающийся и
в СССР, дает там одну генерацию в год, откладка яиц происходит осенью, и к весне
особь достигает размеров взрослых животных. Пик смертности, что вообще харак-
терно для большинства наземных моллюсков, приходится на зиму. Интересно, что
с возрастанием плотности популяции уменьшается число яиц в кладке, скорость
роста и длительность жизни, т. е. происходит своеобразное самоизреживание попу-
ляции, направленное на поддержание оптимальной плотности (Pomeroy, 1968, 1969).
В целом ту же картину дает Helicella caperata на Британских о-вах (Baker, 1968) й
Xerocrassa seetzeni в Малой Азии (Yom-Toy, 1971).
Будучи формально гермафродитными животными, Helicoidea, как большинство
других наземных легочных, вероятно, в отдельные сезоны представлены фактическими
самцами и фактическими самками, о чем можно судить по внешнему виду гермафро-
дитного протока и белковой железы (рис. 22). Во время преимущественного проте-
кания оогенеза гипертрофирована белковая железа, гермафродитный же проток
тонкий; когда в основном в гонаде идет мужской цикл, наблюдается обратное со-
отношение, Картину, подобную изображенной на рис. 22, мы наблюдали у всех без
исключения видов, по которым имели материал, собранный в разные сезоны (Шилейко,
1971b).
Длительность жизни, очевидно, в значительной степени связана с размером
животных: известно, что моллюски большого размера живут по многу лет. В связи
с этим и размножение у них начинается только на 3—4-й год. Так, по наблюдениям
Е. В. Андрейкович (1968, 1969), виноградная улитка в Литве начинает размножаться
на 4-й год жизни. Откладывание яиц у этого вида начинается через 18—60 дней после
спаривания, причем кладка, как правило, только одна и содержит в среднем 49 яиц
(показано для окрестностей Вильнюса). Длительность инкубации 19—37 суток, при-
чем наблюдается высокая смертность эмбрионов (до 100%). Вообще смертность
эмбрионов— закономерное явление: прекратившие развитие или погибшие яйца в сущ-
ности те же яйца-кормилки, столь характерные для морских гастропод. Они поеда-
ются вылупившейся молодью в течение первых двух недель постэмбриогенёза; веще-
ство съеденных яиц ускоряет рост и развитие молодых моллюсков и обеспечивает
их кальцием на первое время (Kothbauer, 1972).
Здесь следует Обратить внимание на чрезвычайно широкие сроки между сово-
куплением и временем откладки яиц. Многолетние полевые И лабораторные наблю-
дения привели нас к убеждению, что подавляющее большинство видов не имеют четко
фиксированных сроков совокупления и откладки яиц (см. также: Kothbauer, 1972).
У большинства видов размножение происходит весной—в начале лета; более точные
сроки отсутствуют — все зависит от конкретных погодных условий данного года.
Совокупление происходит в сырую, но теплую погоду; сперма способна сохранятьсй
в сёмеприемнике достаточно долго, и откладка яиц происходит, как только для этого
создадутся благоприятные условия. Для некоторых видов можно считать Доказанным,
что сперма, которой Партнеры обменялись осенью, сохраняется в семеприемнике
Рис. 21. Половой аппарат Helicopsis retowskii. A — вполне
половозрелая особь, фиксирована 20 VII 1962; с'емепри-
емник пустой; Б — ювенильная особь, фиксирована
6 VIII 1962; семеприемник наполнен; В — вскрытый се-
меприемник той же особи, виден пакет со спермйями.
(Ориг.).
3*
36
ВВЕДЕНИЕ
до весны: Lindholmiola corcyrensis, собранные нами 9 XI в окрестностях Кишинева,
уже ушли в зимнюю спячку, выделив мощную зимнюю эпифрагму. Гистологическое
исследование показало наличие в семеприемнике спермиев.
Рис. 22. Внешний вид гермафродитного протока и белковой железы Trichia hispida
в разные сезоны. Римские цщфры— месяцы. Б Ж — белковая железа; ГП — гер-
мафродитный’\проток; К — квадривий. (Ориг.).
Яйца у всех Helicoidea имеют внешнюю ивнутреннюю оболочки, между которыми
располагается галерта, представляющая собой мукополисахаридный гель со свобод-
ными ионами кальция. Внешняя оболочки/у' 'многих видов достигает значительной
толщины и механической прочности. Она состоит в основном из углекислой извести.
Внутренняя оболочка содержит жидкость белковой црироды, в которой располагается
эмбрион. В состав этой жидкости входят протеины;' гликопротеины, галактоген, не-
которые свободные аминокислоты, кальций и большое количество воды (Bayne, 1968),
ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА ПОЧВ
НА РАСПРОСТРАНЕНИЕ HELICOIDEA
Основной фактор, определяющий распространение моллюсков данной группы, —
степень кальцинированности субстрата. Helicoidea зависят от этого фактора осо-
бенно в связи с тем, что у большинства видов раковина толстостенная, плотная
и сравнительно массивная. На известняках и известковых почвах при прочих равных
условиях моллюски достигают наибольшего качественного (число видов) и количе-
ственного (число особей) развития. Наряду с облигатными кальцифилами имеются
виды, в малой степени зависящие от этого фактора. Это преимущественно мелкие
лесные виды с тонкостенной раковиной — Bradybaena dieckmanni, Br. fragilis,
Br. ravida, Pseudotrichia rubiginosa, Monachoides stuxbergi и др.
Популяции необлигатных кальцифилов, обитающие на нейтральных и кислых
почвах, составлены преимущественно особями мелких размеров с тонкой раковиной.
Вес раковины Cryptomphalus aspersa, обитающих на известняках, может превышать
вес раковины животных того же вида, встреченных на нейтральных почвах, в 8 раз.
Изредка, при особо благоприятных условиях, на кислых почвах встречаются
и моллюски с массивной раковиной: завезенный в Америку европейский вид Сераеа
ЖИЗНЕННЫЕ ФОРМЫ 37
hortensis массами заселяет гранитные острова американского побережья; правда,
там улитки добывают кальций, выскребая раковины морских гастропод, брошенные
чайками (Bleakney, 1966).
Вообще надо сказать, что почти во всякой работе, освещающей отдельные сто-
роны экологии наземных моллюсков, подчеркивается зависимость этих животных
от содержания в почве СаСО3 (Atkins, 1966; Резник, 1967; Talmadge, 1967; Agocsy,
1968; Valovirta, 1968; Pomeroy, 1969; Шилейко, 19696; Lozek, 1970; Wareborn, 1970,
и др.).
Что касается путей усвоения кальция, то, во-первых, нередко улитки грызут
известняки, и известковая крошка иногда обнаруживается в Зобе; во-вторых, они
получают это вещество с пищей; в-третьих, экспериментально доказано, что виноград-
ная улитка растворяет кальций в грунте выделениями ноги. Слизь подошвы пока-
зывает слабокислую реакцию цреимущественно за счет свободной углекислоты (Frick,
1965); кроме того, выше упоминалось, что кислую реакцию имеет и секрет педаль-
ной железы.
Во время интенсивного роста, а также при регенерации поврежденных участков
раковины потребность в углекислом кальции особенно велика. Повышение концент-
рации углекислого газа в воздухе ослабляет выделение СаСО3, а понижение концен-
трации этого газа оказывает обратный эффект. Моллюски, восстанавливающие по-
врежденный участок раковины, выделяют СО2 в большем количестве, чем в норме,
а во время кальциевого голодания снижают выделение этого газа. Резервы каль-
ция в теле моллюска представлены только в виде СаСО3. Резервы, имеющиеся в пе-
чени, ноге и мантии, при разных концентрациях углекислого газа пополняются по-
разному. При мобилизации резервов в первую очередь используется СО2 тканей,
при этом важную роль играют щелочные фосфатазы и карбоангидраза. Первые
в большом количестве содержатся в печени, эпителии подошвы и мантии, вторая —
В эпителии крыши легкого, а также в эпителии подошвы и других частей мантии.
Помимо того что СаСО3 необходим моллюскам для построения раковины, из-
вестковые почвы и чистые известняки обладают рядом особенностей, благоприятных
для жизни моллюсков. К таким особенностям относится, их большая пористость,
благодаря чему мелкие виды имеют надежные укрытия; в порах задерживается влага;
пористые почвы легче аэрируются. Далее, подобные типы почв обладают большой
способностью к аккумуляции тепла, что в значительной степени сглаживает суточ-
ные колебания температуры.
В редких случаях наблюдается своеобразная инверсия: на нейтральных и кислых
почвах буковых лесов Фогельхерда в Тюрингии виды Hygromiidae достигают круп-
ных размеров и большой плотности (Matzke, 1968); аналогичную картину нам дове-
лось наблюдать на лепоритовых скалах в среднем течении р. Чегем на северном Кав-
казе. В подобных случаях такая инверсия связана с особо благоприятными условиями
другого порядка — оптимальная температура и влажность, обилие укрытий, защи-
щенность от ветра и т. п.
Засолоненных почв все Helicoidea решительно избегают.
ПОНЯТИЕ О ЖИЗНЕННЫХ ФОРМАХ
Общеизвестно, что определенный тип внешних условий часто накладывает от-
печаток на различные по систематической принадлежности группы. Отражением,
такого рода наблюдений явилась формулировка понятия «жизненная форма», которое
часто встречается в литературе по высшим хордовым, но лишь немногие малакологи
(Несис, 1973) разрабатывают это понятие. Мы представляем себе жизненную форму
как сходную фенотипическую реакцию на базе различных генотипов, возникшую
в результате воздействия сходных условий среды. Имея в виду реальное существо-
вание определенных конхологических типов, свойственных конкретным условиям,
можно наметить основные конхологические типы, которые мы и называем «психоло-
гическими жизненными формами» (Шилейко, 1971а). Рассматривая проблему под не-
много другим углом, эти жизненные формы можно охарактеризовать как основные
направления адаптивной изменчивости конхологических признаков.
Приступая к рассмотрению этих адаптивных форм, считаем нужным подчеркнуть,
что отмечаемые здесь закономерности выражают лишь преимущественные тенденции,
и, констатируя это обстоятельство, мы не склонны абсолютизировать эти законо-
мерности.
Исходным конхологическим типом в пределах по меньшей мере западнопале-
арктических групп является раковина, свойственная большинству лесных европей-
ских Hygromiidae'. это небольшая или средних 'размеров раковина в общем роговой
окраски разной интенсивности, с умеренно возвышакицимся завитком, со светлой
размытой полосой по периферии и с волосками на поверхности.
.Наиболее примитивные азиатские виды западного происхождения, как например
Leucozonella rubens, L. mesoleuca или Angiomphalia regeliana, в значительной степени
.3 8 ВВЕДЕНИЕ
сохраняют и могут даже усиливать этот тип окраски: у отдельных экземпляров,, осо-
бенно молодых, раковина может быть интенсивно темно-красного цвета с ясной, почти
не размытой светлой периферической полосой. Leucozonella caria сохраняет в полной
мере опушенность раковины. Нередки особи кавказского вида Caucasigena rengar-
teni, у которых сохраняются отдельные редкие волоски. Обитатели открытых про-
странств Средиземноморья— виды рода Xerotricha— сохранили волоски на рако-
вине, что связано с их небольшими размерами и жизнью в основном под камнями
И среди растительной трухи в припочвенном слое. У некоторых ксерофилов, рако-
вина которых во взрослом состоянии лишена волосков, молодь имеет опушенную
раковину (Chatfield, 1968).
В горных районах на открытых участках с обильным травостоем и повышенной
влажностью почвы — в поймах, в местах выхода грунтовых вод, а также на влажных,
слаб,о затененных скалах — часто встречается характерный конхологический тип —
белая раковина с двумя четкими узкими спиральными коричневыми лентами выше
И ниже периферии, которые могут вторично отсутствовать. Таковы некоторые Cau-
casigena, Caucasocressa, Xerocampylaea zelebori, F ruticocampylaea kobiensis, Semi-
fruticicola, некоторые популяции Shileykoia daghestana, Hesseola solidior (Акрамов-
ский, 1949). '
Дериватом описанной жизненной формы лесных обитателей является широко
известный конхологический тип жителей аридных пространств — Helicellinae auct.
Это небольших или средних размеров раковина светлой окраски, нередко одноцветно
белая, плотная, сильно кальцинированная, с узким пупком (большей частью, но не
всегда, иногда пупок широкий; но полностью закрытым пупок не бывает никогда).
Рисунок образован несколькими — до 12 — спиральными коричневыми или даже
черными лентами, число, ширина и интенсивность окраски которых обычно колеблется
между конспецифичными популяциями.
Вопрос о группе Helicellinae заслуживает более подробного рассмотрения в связи
с тем, что выделение этой группы в качестве подсемейства ныне ни у кого сомнения
не вызывает.
При сравнении Helicellinae и Hygromiinae (если обе группы принимать в нынеш-
нем объеме) бросается в глаза повторение основных вариантов внешнего строения
гениталий в обеих группах (Шилейко, 1972 г). Различия между названными группами
сводятся к двум признакам: разные типы раковин и разная топография правого ом-
матофора. Конхологические типы охарактеризованы выше; надо только добавить,
что дифференциальные диагнозы все равно составить нельзя, так как большая часть
Признаков широко перекрывается.
Анализ двух типов топографии правого омматофора показал, что правильнее
говорить не о том, перекрещиваются ли гениталии с этим мускулом, но о взаимном
перехлесте последнего с половым ретрактором (подробнее см. в разделе, посвященном
анализу Euomphaliinae). Но между этими двумя типами расположения глазного ре-
трактора не существует четкой границы, поскольку имеется группа (Monacha s. str.)
без полового ретрактора, и в применении к названной группе вопрос о том, где про-
ходит ретрактор правого омматофора, теряет смысл: пенис способен перемещаться
относительно этого мускула, то оказываясь в состоянии перехлеста с ним, то нахо-
дясь в свободном состоянии.
Решить проблему таксономического статуса Helicellinae auct. можно путем срав-
нительного анализа адаптивного значения отмеченных признаков. Светлая раковина —
явно адаптивный признак, позволяющий экранировать солнечные лучи; степень
развития темных полос на раковине позволяет достигать очень тонкой регулировки
количества получаемого тепла. Другими словами, все конхологические особенности
Helicellinae стоят в прямой и непосредственной зависимости от характера свойствен-
ных им биотопов.
Адаптивный смысл утери перехлеста ретракторов не выглядит столь непосред-
ственным и явным, но роль этой утери, происходившей, видимо, неоднократно в про-
цессе формирования жизненной формы Helicellinae, исключительно велика. ’
На рис. 23 схематически изображен механизм втягивания правого щупальца
У Hygromiinae auct. (А, Б) и Helicellinae (В, Г). Видно, что при вытянутом щупальце
в „той и другой группе гениталии способны функционировать совершенно сходным
образом. Представим теперь, что происходит при автономном втягивании правого
омматофора. В случае А — Б сократившийся ретрактор вытянется по касательной
к окружности столбика (поскольку, представляет собой4 одну из ветвей, колумелляр-
ного мускула) и пережмет, таким образом, гениталии в области клоаки. В этбй ситу-
ации копуляция становится невозможной в силу чисто механических причин. Про-
должающееся сокращение глазного ретрактора параллельно с сокращением тела
моллюска неизбежно влечет за собой (точнее, перед собой) и' втягивание -участка,
где располагается половое отверстие. : '
По-иному этот механизм выглядит у Helicellinae'. втягивание щупальца и даже
переднего конца тела до известных пределов не препятствует совокуплению, поскольку
происходит независимо от гениталий и полового отверстия. ’
ЖИЗНЕННЫЕ ФОРМЫ
39
Сравнивая эти факты в несколько иной плоскости, можно констатировать, что
для успешного спаривания Hygromiinae auct. должны в течение этого процесса, ко-
торый, кстати, может длиться часами, находиться в расправленном состоянии.
(Прекрасная фотография копулирующих виноградных улиток, у которых ретрак-
торы также находятся в состоянии перехлеста, приведена Килиасом (Kilias; 1967,
photo 66—68)). В то же время Helicellinae благодаря утере перехлеста ретракторов
могут продолжать спаривание даже втянувшить настолько, что тела их почти скрыты
в раковинах (рис. 24). Понятно, что появление такого прогрессивного признака со-
Рис. 23. Расположение ретрактора правого омматофора (зачернен, белый кружок —
глаз) у Hygromiinae auct. (Л, Б) и Helicellinae auct. (В, Г) и результаты его автономного
сокращения (схема). Объяснение в тексте. (Ориг.).
вдает серьезные предпосылки для перехода в засушливую среду, и эти предпосылки
•были реализованы, по нашему убеждению, неоднократно в различных' группах
Hygromiidae (в объеме, принятом здесь).
Оценивая с этих позиций таксономический статус Helicellinae, приходится при-
знать, что эта группа является сборной, полифилетической, и как таковая таксоно-
мического статуса не имеет; все Helicellinae — суть жизненная форма обитателей
открытых аридных пространств (Шилейко, 1971а); возникла она вместе с появлением
на Земном шаре подобных ландшафтов и в связи с ними. Каждый,из признаваемых
ныне таксонов Helicellinae есть дериват соответствующего таксона Hygromiidae.
Другими словами, в составе большинства подсемейств Hygromiidae имеются более древ-
ние группы родов, связанные в той или иной степени с лесом и сохранившие пере-
хлест ретракторов, и группы родов более молодые, связанные с открытыми ланд-
шафтами в значительной мере благодаря утере перехлеста ретракторов и возникно-
вению адаптивных конхопогических черт.
Утрату перехлеста можно представить себе как многоступенчатый процесс: вначале
ретрактор правого омматофора постоянно располагается между пенисом и вагиной.
40
ВВЕДЕНИЕ
Далее этот мускул, сохраняя перехлест с половым ретрактором, приобретает боль-
шую подвижность относительно гениталий (правильнее говорить о подвижности пениса
относительно глазного ретрактора) в пределах иногда даже одного вида, как это наблю-
дается, например, у Euomphalia.strigella, Hesseola,, Caucasocressa, некоторых Monacha.
Следующий этап этого процесса заключается в редукции полового ретрактора, которая
может быть стойко закреплена в генотипе и представлять собой постоянный признак
(Monacha s. str.) или спонтанно проявляться у отдельных особей (Euomphalia aristata).
Однако в некоторых случаях (Hesseola solidior), может быть, правильнее говорить о по-
степенной редукции полового ретрактора. Наконец, заключительный этап — закреп-
ление в генотипе нового варианта взаимного расположения ретракторов.
Для создания целостной картины необходимо еще объяснить, почему любому
варианту строения гениталий более древних Hygromiidae соответствует такой же ва-
Рис. 24. Xeropicta candaharica in copuli. Моллюски
почти целиком скрыты в раковинах. (По Шилейко,
1972г).
риант у более молодых, т. е. у Helicellinae auct., но у последних имеется некоторое число
вариантов, отсутствующих у Hygromiinae auct.
Выше отмечалась относительная молодость Helicellinae- заняв новые обширные
пространства на заре становления и не испытывая вначале давления конкуренции,
они начали бурную, дивергирующую эволюцию, продолжающуюся до сего времени.
Другими словами, большая часть типов строения Helicellinae — продукт эволюционных
изменений исходных вариантов, наблюдаемых ныне у более древних Hygromiidae-,
не исключено также, что некоторые Helicellinae — потомки вымерших таксонов лесных
Hygromiidae, о внутреннем строении которых можно только догадываться.
Во всяком случае, большая часть Hygromiinae auct., по-видимому, исчерпала
свои эволюционные возможности (о чем свидетельствует большое число монотипиче-
ских таксонов — см.: Гептнер, 1965; Майр, 1968) или близка к этому, и перспективы
дальнейшего развития их зиждутся на филогенетически молодых ксерофильных пред-
ставителях.
Возвращаясь к общему рассмотрению конхологических типов, заметим, что на-
меченные выше закономерности неоднократно нарушаются из-за склонности многих
групп к неопределенной изменчивости, в результате чего возникают конхологические
типы, имеющие сходство с представителями других групп наземных легочных. В наи-
большей степени это относится к Ariantinae (ср. Arianta arbustorum, Isognomostoma
personatum, Cylindrus obtusus, Helicigona lapicida, Liburnica setosa).
Усиление радиальной скульптуры вплоть до образования резких ребер наблюда-
ется во всех таксонах Helicoidea и касается в первую очередь облигатных кальцифи-
лов. Похоже, что наклонность к образованию ребер имеют в первую очередь виды не-
большого размера. Кроме того, популяции, составленные небольшими особями, имеют
более сильно исчерченную раковину, чем конспецифичные популяции, представлен-
ные более крупными животными.
Равномерное утолщение раковин, в результате чего вес ее может достигать отно-
сительно большой величины, представляется явлением редким (Yom-Tov, 1971).
ЗАЩИТА ОТ ПОТЕРИ ВОДЫ И ЛЕТНЯЯ СПЯЧКА
Все Helicoidea первично гигрофильны. В определенном смысле гигрофилами оста-
ются даже обитатели открытых ландшафтов, поскольку основные жизненные отправ-
ления их происходят ночью либо во влажный сезон. К таким формам, тем не менее,
правильнее всего применять термин «ксерофилы», поскольку адаптации к ксеротерм-
ным условиям зашли так далеко, что для успешного завершения жизненного цикла
им необходим высокий уровень инсоляции и при постоянно влажных условиях в лабо-
ратории они погибают. .
Приспособления к аридным условиям свойственны преимущественно филогене-
тически молодым группам, так как сами степные и полупустынные пространства —
исторически молодые типы ландшафта..В силу изложенных обстоятельств исследова-
ния разнообразия и характера подобных адаптаций интересны в первую очередь с фи-
логенетико-таксономической точки зрения..
ПОТЕРЯ ВОДЫ И СПЯЧКА
41
Все разнообразие адаптаций к засушливым условиям можно произвольно разделить
на две взаимосвязанные категории: ювенильные и дефинитивные. Надо обратить вни-
мание на то, что молодь лесных видов лишена специальных морфологических адапта-
ций, и постэмбриональное развитие раковины сводится к ее росту, который заканчи-
вается формированием губы и незначительным опусканием дистальной части послед-
него оборота. Что касается обитателей открытых ландшафтов, то в пределах Helicoidea
провизорных ювенильных образований, как это наблюдается у некоторых других групп
моллюсков, нет (Шилейко, 1967а, 19676). Адаптации молоди здесь находят то же
морфологическое выражение, что и у взрослых особей: у многих видов в процессе
постэмбриогенеза формируется несколько губ, из которых только последняя дефини-
тивна (Hygrohelicopsis darevskii, Teberdinia zolotarevi, Kokotschashvilia, Diodontella,
Archaica apollinis, A. labiantx и др.) (рис. 25, В); иногда (некоторые Laeocathaica)
Рис. 25. Ювенильные и дефинитивные устьевые образования.
А — Paedhoplita lindholmi, нижняя стенка раковины частично
удалена; Б — Caucasigena schaposchnikovi, видны 4 губы в виде
светлых радиальных полос; В — молодой экземпляр Laeocathaica
polytyla; Г — то же, раковина взрослой особи. (Ориг.).
ранние губы значительно отличаются от дефинитивных (рис. 25, В—Г). Подобным же
образом у Paedhoplita комплекс зубов на париетальной стенке образуется от 2 до 6 раз
(рис. 25, А), причем по мере роста раковины более ранние зубы, которые оказываются
в глубине оборотов, постепенно исчезают. По наблюдениям, сделанным в полевых усло-
виях, можно заключить, что молодые особи живут вместе со взрослыми и испытывают
воздействие тех же условий в том же количественном выражении.
У других ксерофилов молодь резко отличается от взрослых особей более темной,
иногда черной окраской раковины;-точнее, у них раковина сильно просвечивающая,
но сильно пигментирована мантия. В первую очередь это касается ксерофильных
Hygromiidae. У видов этой группы молодь обитает в существенно иных условиях:
она встречается в дерновинках трав и не поднимается по стеблям, тогда как взрослые
и полувзрослые (но не ювенильные) особи образуют скопления именно на стеблях,
заползают на ветви кустарников на высоту до 1—1.5 м. Светлая окраска раковин взрос-
лых особей отражает солнечные лучи, тогда как пигментированная мантия при про-
зрачной раковине у ювенильных моллюсков создает условия для получения необходи-
мого количества солнечного тепла животным, обитающим в значительно более затенен-
ных условиях.
Наиболее критический момент в жизни моллюсков — период летней засухи.
На это время многие виды закапываются в почву (Ponsadenia semenovi, Leucozonella
caryodes, виды рода Xerosecta и др.); другие, напротив, образуют гроздевидные скопле-
ния на растениях (Monacha s. str., Xeropicta, Xerocrassa) и т. д. Биго (Bigot, 1967—1968)
не обнаружил разницы между растениями, на которых такие гроздья образуются,
и растениями, на которые моллюски не поднимаются. Число особей в одной грозди не
превышает большей частью 100, но в отдельных случаях может доходить до 1500.
Молодые (но не ювенильные) моллюски образуют обычно собственные скопления в пре-
42
ВВЕДЕНИЕ
делах грозди, образуемой взрослыми. Закономерностей в динамике'образования грозди
Биго не обнаружил: моллюски могут висеть без движения неделями, но .могут распол-
заться по ночам, собираясь на дневное время в ту же самую гроздь, добираясь до со-
седних скоплений, или образуют новую гроздь на другом месте.
Некоторые виды ведут себя во время летней засухи аналогичным образом, но скоп-
лений не образуют: одна особь Angiomphalia regeliana, прикрепившись к большому
камню, провела без движения с 20 июня по 14 июля, после чего 15 июля утром пробу-
дилась и уползла. Несомненно, моллюски подобных экологических групп нуждаются
в определенном минимуме солнечного тепла.
В любом случае одним из важнейших способов замедления испарения является
образование эпифрагмы. При изучении в экспериментальных условиях влияния влаж-
ности и температуры на образование эпифрагмы у некоторых ксерофильных видов
Франции для разных видов выявлено 3 типа зависимостей (D. Bonavita, 1964; А. Вопа-
vita, 1965). I тип: с октября по май при влажности воздуха 100% и температуре 5° С
при двухнедельной экспозиции эпифрагма не образуется. Имеются критические со-
стоянйя температуры и влажности, необходимые для образования эпифрагмы (5° и
80%, 10° и 70%, 25° и 20%). С марта по октябрь снижение температуры при повышен-
ной влажности не вызывает образования эпифрагмы, а при 5—20° эпифрагма образу-
ется при снижении влажности до 20%. II тип (свойственный другим видам): у зимую-
щих особей при влажной атмосфере эпифрагма не образуется в широких температурных
пределах — от 5 до 30°; у летних особей эпифрагма формируется только при 25—30°.
Снижение влажности вызывает образование эпифрагмы при любой температуре. III тип
(отмечен для Eobania vermiculata): снижение влажности всегда ведет к формированию
эпифрагмы, но более эффективно при повышении температуры. Соотношение в этом
случае следующее: 25—30° и 90%, 5—10° и 20% у зимних особей и 5—10° и 40% у лет-
них (см. также: Baker, 1968).
Для некоторых австралийских видов показано, что эпифрагма способна абсорби-
ровать водяной пар из воздуха при ненасыщенной влажности, компенсируя частично
потерю воды моллюсками (Warburg, 1965). Вообще надо отметить, что об адаптациях
физиологического порядка известно немного, хотя эта форма приспособления навер-
няка играет не последнюю роль в жизни моллюсков (Hunter, 1964). В состоянии летней
спячки (или, как часто теперь пишут, эстивации) уровень мочевой кислоты остается
таким же, как и у активного животного, а содержание мочевины относительно возра-
стает. Накопление мочевины, по-видимому, связано с уменьшением выделения жид-
ких экскретов при эстивации. Помимо других эффектов, даваемых повышенным со-
держанием мочевины в теле моллюска, накопление этого вещества может регулировать
осмотическое давление в теле моллюска и уменьшать потери воды на испарение. Эти
сведения касаются представителя надсемейства Bultmuloidea — Bulimulus dealbatus
(Horne, 1971); однако думается, что подобные материалы можно экстраполировать
достаточно широко, и описанная закономерность в той или иной степени касается пред-
ставителей Helicoidea.
Физиологическую и количественную стороны испарительных процессов исследо-
вал Мэчин (Machin, 1964а, 1964b, 1964с, 1966); но этот же автор в работе 1967 г. пока-
зал, что и морфологические адаптации играют важную роль в замедлении испарения.
Сравнивая скорости испарения у трех видов Helicidae, Мэчин пришел к выводу, что
различия в скорости потери воды объясняются, помимо особенностей физиологических
адаптаций, также и размерами раковины, толщиной эпифрагмы, относительной вели-
чиной устья и тем, насколько плотно устье сцособно «притираться» к поверхности суб-
страта: у более ксерофильных видов (Otala lactea) устье прилегает к ровной поверхности
с очень маленьким зазором в колумеллярной области. Способность к перенесению удли-
ненного бездождного периода способствует освоению аридных пространств,-
В тех случаях, когда наблюдается равномерное увеличение толщины раковины
(см. выше, раздел о жизненных формах), оно связано с обитанием в очень сухих усло-
виях. Изучая экологические особенности двух видов — Sphinderochila boissieri и
Trochoidea seetzeni, населяющих полупустынные и пустынные области западной части
Аравийского п-ва, Йом-Тов (Yom-Tov, 1971) обнаружил ряд интересных закономер-
ностей. В течение летнего периода оба вида находятся на открытых местах, где темпе-
ратура почвы днем достигает 70°. Вес одной особи Trochoidea составляет в среднем
1.6 г, из которых на вес тела приходится 60%; вес одной особи Sphinderochila, около
4.3 г, причем половина этого веса приходится на раковину. Массивные раковины обоих
видов обладают светлой окраской. В период засухи Sphinderochila обычно уходят
в почву, a Trochoidea, выделяя мощную эпифрагму, прикрепляются устьем к сухим
стеблям над почвой. Оба вида — ярко выраженные ксерофилы: при 40° животные спо-
собны прожить 6 недель, при 50° — до 3 недель; при. 55° улитки живут не более
суток. Для обоих видов показана, высокая способность к отражению солнечных
лучей.
Наконец, известно, что способцость к глубокому, втягиванию моллюска в рако-
вину непосредственно связана со степенью засушлцвости условий, в которых постоянно
обитает данный вид. ; .
ЗИМНЯЯ СПЯЧКА
43
ЗИМНЯЯ СПЯЧКА
Способы перезимовывания моллюсков достаточно однообразны и сводятся чаще
всего к закапыванию животных на разную глубину в почву, что защищает их как от
чрезмерно низких абсолютных температур, так и от температурных колеба-
ний.
При исследовании состояния педальной мускулатуры, печени и гонады виноград-
ных улиток, находящихся в состоянии зимней спячки, отмечен относительно низкий
уровень метаболизма в тканях гонады, тогда как уровень обмена и способность к само-
регуляции у педальной мускулатуры и тканей печени незначительно отличаются от
того, что наблюдается у активных животных (Nopp, Farahat, 1968). Это понятно, по-
скольку и ткани ноги, и ткани печени должны постоянно находиться в готовности
прервать зимнюю спячку: моллюски пробуждаются в течение считанных часов; в то
же время спаривание начинается лишь спустя несколько суток после пробуждения.
Рис. 26. Последовательные стадии закапывания в почву Helix pomatia
при уходе на зимнюю спячку. (По Lind, 1968).
Зимняя эпифрагма отличается от л(тней большей толщиной и более высоким от-
носительным содержанием солей кальция, в первую очередь СаСО3. При зимовке
в почве улитки всегда ориентируются устьем вверх (Carney, 1966). Это связано с не-
сколькими причинами: контакт с почвой приводит к отсыреванию эпифрагмы, что за-
трудняет газообмен; облегчается проникновение грибков и бактерий, разрушающих
эпифрагму; при плотном соприкосновении устья с почвой нарушается вентиляция,
тогда как в нормальном положении устьем вверх между эпифрагмой и покрывающим
слоем почвы всегда имеется воздушное пространство; наконец, при пробуждении жи-
вотное может скорее выбраться на поверхность, если устье обращено вверх. Последнее
обстоятельство немаловажно, поскольку весной при обильном таянии снегов улитка
рискует оказаться затопленной.
Виды, обитающие на скалах, на зиму забираются глубоко в трещины и выделяют
особенно мощную эпифрагму.
Глубина зарывания в почву виноградной улитки различна и зависит от разнооб-
разных внешних условий, в первую очередь от плотности почвы. Сложный комплекс
поведения улитки состоит из облигатного набора действий, наблюдающихся при. всех
условиях, и из нескольких факультативных актов, обеспечивающих адекватную реак-
цию на изменение конкретных условий (Lind, 1968; рис. 26).
' Молодь многих видов обнаруживает большую резистентность к воздействию зим-
них холодов, чем взрослые особи, в результате чего зимой наблюдается наибольшая
гибель взрослых моллюсков; популяции большинства видов Bradybaenidae в Примор-
ском крае вплоть до середины лета составлены почти исключительно неполовозрелыми
Особями, так как взрослые зимой погибают. Исключение составляет лийть наиболее
массовый вид — Bradybaena maacki, взрослые особи которого встречаются и весной,
но преобладание молоди все же наблюдается до середины июня.
44
ВВЕДЕНИЕ
РИТУАЛЬНЫЕ ФОРМЫ ПОВЕДЕНИЯ
Здесь мы рассмотрим поведение при спаривании, «ухаживание» (courtship — ан-
глийских авторов); к ритуальным формам поведения можно отнести также и закапыва-
ние в почву при осеннем понижении температуры, так как, оказывается, там тоже соблю-
дается определенный «ритуал», т. е. строго соблюдаемый порядок определенных актов,
но логичнее было изложить этот материал в предыдущем разделе, посвященном зимней
спячке.
Как выяснено Зилером (Sihler, 1954), брачный танец виноградной улитки пред-
ставляет собой цепь одиночных реакций, каждая из которых вызывается строго опре-
деленным раздражителем (ключом). Определенное раздражение одной особи вызывает
двигательную реакцию другой, и наоборот; поэтому действия обеих особей строго со-
гласованны. Зависимость каждой врожденной реакции от определенного сигнального
раздражителя позволяет предполагать существование определенного нервного меха-
низма, дающего реакцию на внешний раздражитель. Он, как и сигнальный раздражи-
тель, строго специфичен для каждой реакции — как ключ к замку. Быстрым и после-
довательным прикосновением к определенным местам тела моллюска можно искус-
ственно вызывать позы и движения, наблюдаемые в брачном танце.
Специфичность сигнальных раздражителей устраняет возможность межвидового
скрещивания. Надо с сожалением заметить, что отсутствуют наблюдения, позволившие
бы сравнить поведение при спаривании двух близких видов; а такое исследование по-
зволило бы выделить видоспецифичные элементы поведения и отделить их от элементов
более общих, свойственных всему таксону (подроду, роду).
В связи с исследованием Зилера надо заметить следующее. Для легочных моллю-
сков, как уже сообщалось, показана большая роль глазных щупалец в нейрогумораль-
ной регуляции половой деятельности: при экстирпации омматофоров наблюдаются
резкие изменения в гонаде (кроме Vaginulidae — Renzoni, 1969). В частности, участие
омматофоров в механизме гормонального контроля у Cryptomphalus aspersa показано
Стефенсами (G. J. Stephens, G. С. Stephens, 1966). Расположение каких-то механизмов
гормонального контроля не случайно именно на концах омматофоров, так как во время
брачных игр моллюски прикасаются щупальцами к различным участкам тела партнера,
воспринимая и передавая сигналы, свидетельствующие о степени готовности партнера
к спариванию. Мы имели случаи наблюдать такие картины у Fruticocampylaea narza-
nensis, Kokotschashvilia phaeolaema и Eobania vermiculata.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ HELICOIDEA
Надсемейство в целом распространено в северном полушарии (рис. 27). Южнее
экватора в Южной Америке по Андам до северной части Чили и северо-запада Арген-
тины спускаются представители Helminthoglyptidae;, на востоке Африки — несколько
небольших групп неясного систематического положения; на юго-востоке Азии ниже
экватора встречаются многочисленные виды нескольких родов Camaenidae (Papui-
ninae).
Ареалы семейств выглядят следующим образом.
I. Humboldtianldae: Мексиканское плато и прилежащие районы Техаса.
II. Helminthoglyptidae-. Тихоокеанское побережье Северной Америки вплоть до
Аляски (Monadenia)-, Флорида; Мексика, Центральная Америка и Вест-Индия; Анды
от Эквадора до запада Аргентины.
III. Oreohelicidae-. от южной части Саскачевана и Британской Колумбии в Канаде
через горы запада США до запада Чиуауа (Мексика); отдельные виды проникают к за-
паду до о-ва Санта-Каталина (Калифорния), к востоку — до Южной Дакоты.
IV. Helicidae: Эфиопия, Средиземноморские страны, Передняя Азия и Кавказ;
небольшое число видов в Западной, Средней и Северной Европе.
V. Camaenidae-. ареал состоит из американского и азиатского участков. Первый
включает север Южной Америки от Ориноко до южной части Никарагуа в Центральной
Америке, затем Кубу, Гаити, Ямайку, Виргинские и Малые Антильские о-ва; виды
рода Zachrysla завезены во Флориду и зону Панамского канала. Азиатский участок
включает Южную и Юго-Восточную Азию и большую часть о-вов Малайского архипе-
лага; наибольшее разнообразие на о-ве Новая Гвинея. Несколько видов обитает в Се-
верной Австралии — Квинсленд. К северу представители семейства доходят до Филип-
пинских о-вов и южной Японии.
VI. Bradybaenidae-. главным образом Центральная и Юго-Восточная Азия; Тянь-
Шань; Приморье и Приамурье; Сахалин, Курильские очва, Япония, Филиппины. На Сей-
шельских о-вах и на востоке Северной Африки, обитает ряд форм, систематическое
положение которых неясно, но относимых по традиции к данному семейству. Огромные
пространства Европы и Сибири, а также юг Камчатки включаются в ареал семейства
за счет видов Bradybaena fruticum и Br. schrencki.
VII. Ammonitellidae-. горные районы юго-запада Северной Америки (Монтана,
Айдахо, Орегон, Калифорния).
Рис. 27. Ареал Helicoidea. (Ориг.).
46
ВВЕДЕНИЕ
Таблица 1
Название вида Европейская часть Кавказ Средняя Азия и Зибирь Дальний Зосток
центр европейской части с Прибалтикой Карпаты и прилежа- щие территории Крым северный западный восточный Закавказье Копетдаг Тянь-Шань и Памир (без Тарбагатая) •' Тарбагатай Сибирь и Алтай Приморье и Приамурье о-ва Дальнего Востока и_Камчатка
Helicodonta evoluta . . +
Н. obvoluta j- Lindholmiola jraudulosa ? ? +
L. corcyrensis +
f Oestophora schileykoi +
Ponsadenia hirsuta . . P. duplocinota .... + . +
P. semenovi Bradybaena fruticum + + + + + +
Br. schrencki + +
Br. transbaicalia * . . Br. middendorffi . . . + +
Br. ussuriensis .... +
Br. maacki Br. weyrichii . ... . + +
Br. strelkovi . ... . +
. Br. duiensis . ... . Br. dieckmanni .... + +
Br. arcasiana Br. diversita + +
Br. fragilis Br. vulcanica i + +
Br. ravida ...... Br. scythica 1 + +
Br. lantzi ...... +
Br. plectotropis .... +
Br. tzwetkovt . ... . +
Br. perlucens . ... . +
Br. dichrozona .... +
Br. bilaticincta .... +
Br. stoliczkana .... Br. helvola ...... + . +' + '
Br. pseudoferghanica +
Br. fedtschenkoi . . . . +
Br. cavimargo .... + - +
Br. tomyris * +
«.Br.» boevi * Nanaja cuniulata . . . + ' +
N. illuminata * .... , + '
Odontotrema diplodon +
Leucozonella ferghanica +
L. caryodes +
L. rubens +
L. mesoleuca . ... . + . ||
L. rufispira . ... . +
L. retteri ....... + ‘ 1'
L. angulata . ... . + '
L. caria .......
L. hypophaea +
L. crassicosta . ... . Hygrohelicopsis darevskii* + +
Teberdinia zolotarevi —1_
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
47
Таблица 1 (продолжение)
Название вида Европейская часть Кавказ Средняя Азия Сибирь и Дальний Восток
центр европейской части с Прибалтикой Карпаты и прилежа- щие территории Крым северный западный восточный Закавказье Копетдаг Тянь-Шань и Памир (без Тарбагатая) Тарбагатай Сибирь и Алтай Приморье и Приамурье 1 о-ва Дальнего Востока и Камчатка
Kokotschashvllia makva-
lae ......... +
К. tanta* +
К. eberhardi* .... +
К. holotricha К. phaeolaema .... + + +
f Caucasigena psedachica +.
-j- C. fortangensis .... +
C. armeniaca ? +
C. tschetschenica . . . + +
C. rengarteni +
C. eichwaldt C. abchastca . ... . + +
C. reminiscenta* . . . +
C. schaposchriikovi . . . C. thalestris . ... . + +
Diodontella stschukini* +
D. nubigena* . ... . Plicuteria lubomirskii + +
Trichia hisplda .... + +
Tr. concinna . ... .
Tr. villosula ..... +
Tr. bielzi +
Edentiella bakowskii. . f Helicopsis sunzhica . . + ' +
f H. libidinosa . ... . +
H. likharevi . ... . H. striata H. retowskii . ... . + + +
H. dejecta + +
H. paulhessei +
H. instabilis H. filiniargo ..... + +
Xeropicta krynickii . . + + + +
X. derbentina .... X. candaharica .... + + + + + ' + +
X. parableta* .... Helicella candicans . . Lindholmomneme kiidien- + +
sis . . - +
L. notophila +
L. nordenskioldi . . . + ч-
Chilanodon bicalldsa . . +
Ch. gerstjeldti .... P er j or atelia bidens . . + + . +
P. dibothrion +
Monachoides incarnatd +
M. vicina ...... M. stuxbergi . ... . + -к
M. aculeata . ... . -1-
48
ВВЕДЕНИЕ
Таблица if продолжение)
Название вида Европейская часть Кавказ Средняя Азия Сибирь и Дальний Восток
центр европейской части с Прибалтикой Карпаты и приле- жащие территории Крым северный западный восточный Закавказье Копетдаг ^Тянь-Шань и Памирд (без Тарбагатая). Тарбагатай Сибирь и Алтай Приморье и Приамурье о-ва Дальнего Востока и Камчатка
Pseudotrichia rubiginosa + + + + + +
Urticicola umbrosa . . Fruticocampylaea narza- +
nensis + + +
Fr. kobiensis +
Shileykoia daghestana Circassina circassica . . + + + +
C. bojenae ...... +
C. frutis +
Cernuella virgata . . . Ce. tiflisiana . ... . + + + +
Xerosecta schelkovnikovl +
X. crenimargo .... + + +
X. arcadiana ..... Archaica heptapotamica + +
A. apollinis . ... . +
A. labianix Karabaghia bituberosa + +
Hesseola solidior . . . H. transcaspia .... Euomphalia aristata' + + + +
E. appeliana E. strigella + + + +
Platytheba mingreltca Pl. prometheus .... + +
Oscarboettgeria euages + +
Stenomphalia selecta + +
S. maiae ....... +
S. pisiiormis + +
S. ravergieri . ... . Monacha fruticola . . . + + +
M. caucasicola .... +
M. talyschana .... M. externa ...... + +
M. carthusiana .... M. samsunensis .... +. + +
M. praeorientalis . . . M. subcarthusiana . . + +
A ngiomphalia copiosa +
A.’ caelestimontana . . +
A. exasperata . ... . > +
A. regeliana +
A. almalensis . ... . +
A. seductilis . ... . +
A. lentina ...... +
A. buamica +
Paedhoplita laminata ; +
P. lindholmi ' +
P. kirgisensis* . ... > Caucasocressa pratensis +. + +
C. joannis ...... ; Faustina Faustina . . . + +
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
49
Таблица 1 (продолжение)
Название вида Европейская часть Кавказ Средняя Азия Сибирь и Дальний Восток
центр европейской части с Прибалтикой Карпаты и прилежа- щие территории Крым северный западный восточный Закавказье Копетдаг Тянь-Шань и Памир (без Тарбагатая) Тарбагатай Сибирь и Алтай Приморье и Приамурье о-ва Дальнего Востока и Камчатка
Arianta arbustorum + +
А. aethiops +
Hellcigona lapicida Isognomostoma persona- + +
turn ........ +
Drobacia bdnatica . . . +
Causa holosericum . . . ?
f Kllkla giengensis . . . +
Cepaea nemoralis . . . +
C. hortensis C. vindobonensis . . . + + 4- +
Cryptomphalus aspersa ?
Eobania vermiculata Levantina djulfensis + +
L. escheriana f Caucasotachea andrussovi + +
C. fortangensis .... - +
•• C. kubanica +
C. lencoranea C. atrolabiata .... + + +
C. calligera •f Steklovia maslovae . . + + +
f Helix maeotica .... +
H. nordmanni .... +
H. christophi ? +
H. albescens + + + +
H. lucorum + + +
H. buchi H. lutescens + +
H. pomatia Triodopsis subpersona- .+ +
turn ........ +
Примечание. Крестом помечены ископаемые виды, звездочкой — виды, известные только
по одной находке.
VIII. Helicodontidae: основная масса видов в западных и южных областях Европы,
центральных и западных областях Северной Африки и на о-вах Средиземного моря.
Один вид (Helicodonta obvoluta) проникает в Среднюю и Северо-Западную Европу.
IX. Hygromiidae: основная масса видов сосредоточена в горных районах Южной
Европы, Северной Африки и на Кавказе. Представлены также в Средней Азии, Цен-
тральной и Северной Европе, на Алтае, в Забайкалье и на материковой части Дальнего
Востока. Кроме того, по территории лесной части Сибири спорадически распростра-
нены два вида: Pseudotrichia rubiginosa и Monachoides stuxbergi.
X. Sphincterochilidae'. Средиземноморье; большая часть видов — в юго-западной
части, меньшая — в восточной. Эти участки связаны между собой почти циркумсреди-
земноморским ареалом Sphincterochila candidissima.
К этим географическим характеристикам, которые отражают исторически сло-
жившееся распространение, надо добавить, что благодаря вмешательству человека
многие виды Helicoidea расселились почти по всему Земному шару. Так, ряд европей-
ских видов завезен в Австралию, Новую Зеландию, Океанию и Америку. Юго-восточ-
4 А. А. Шилейко
50
ВВЕДЕНИЕ
аоазиатский вид Bradybaena similaris обнаружен недавно в Юго-Восточной Африке,
в окрестностях Лоренсу-Маркиш (Van Bruggen, 1967). Число подобных примеров
можно значительно увеличить.
Такова общая картина распространения основных групп надсемейства. Длятха-
рактеристики географического распространения 4 семейств со всеми видами приводим
сводную табл. 1.
Из приведенной табл. 1 видно, что для Helicoidea фауны СССР характерна четкая
привязанность подавляющего большинства видов и родов к определенным районам,
таксонов с широким распространением очень мало. Это — основная зоогеографическая
особенность данной группы: ясное географическое «лицо» большинства таксонов. Как
будет видно из дальнейшего текста, эта особенность хорошо согласуется с воззрением
на филогению группы и историю фауны.
Набор видов, обитающих на территории нашей страны, сформировался за счет
двух зоогеографических центров — западнопалеарктического и ориентального, при-
чем влияние западнопалеарктического центра имеет абсолютное преимущество в евро-
пейской части и на Кавказе; в Средней Азии его влияние несколько ослабевает, но
ощущается и на Алтае, и в Прибайкалье, и даже на Дальнем Востоке.
За счет ориентального центра сформирована почти вся фауна Helicoidea Дальнего
Востока и более половины общего числа видов в фауне Средней Азии. Только два вида
восточного происхождения проникли сравнительно недавно на запад (Bradybaena
fruticum и Br. schrencki).
Однако, не оставляя сомнений в источниках происхождения, современная фауна
Helicoidea основных 4 регионов (Карпаты; Крым и Кавказ; Средняя Азия и Алтай;
Дальний Восток) формировалась в значительной степени самостоятельно, что выража-
ется в высоком уровне эндемизма. Это обстоятельство в той или иной форме'не раз отме-
чалось в отечественной литературе (Пузанов, 1925, 1926, 1927а, 19276; Лихарев, 1950,
1953, 1957а, 19576, 1958а, 19586; Стеклов, 1966, 1968; Шилейко, 19686, 1968в; Акра-
мовский, 1970, 19716; Матёкин, 1972).
ОСНОВНЫЕ НАПРАВЛЕНИЯ МОРФОЛОГИЧЕСКОГО
РАЗВИТИЯ HELICOIDEA ВО (ВРЕМЕНИ- И В ПРОСТРАНСТВЕ
ОБЩИЕ ЗАМЕЧАНИЯ. ИСХОДНЫЕ ГРУППЫ
Перед тем как начать рассмотрение групп, представленных на территории нашей
страны и изученных поэтому нами достаточно полно, представляется целесообразным
дать общее представление о таксономической структуре, происхождении и ходе разви-
тия надсемейства в целом, а также изложить теоретические предпосылки построения
системы и результаты развития их в настоящей работе.
Ряд положений, особенно касающихся взаимоотношений между группами Нового
и Старого Света, в той или иной мере гипотетичен. Тем не менее их изложение оправ-
дано тем, что существование гипотезы влечет за собой появление исследований, имею-
щих специальную направленность. Без таких исследований невозможно дальнейшее
совершенствование наших представлений.
Основная и по существу единственная особенность членов инфраотряда Holo-
poda, объединяемых в надсемейство Helicoidea, — первичное присутствие стилофоров
и связанных с ними слизистых желез (dart apparatus — американских авторов). Сле-
довательно, при решении вопроса о происхождении группы в целом видное место при-
надлежит проблеме возникновения названного комплекса. Можно предполагать, что
подобные образования могли возникнуть в результате дифференциации неспециализи-
рованного, но морфологически выраженного участка железистой ткани, расположен-
ного в дистальной части женского отдела полового аппарата. Такой участок имеется
у некоторых представителей инфраотряда Aulacopoda, в частности):/оигШаеДимеется
в виду циркумвагинальная железа).
При такой постановке вопроса определенную оценку получают особенности строе-
ния ноги Helicoidea, отмеченные выше: при фиксации животных на подошве нередко
наблюдается возникновение 3 продольных полей (см. рис. 1), что само по себе свиде-
тельствует о каких-то различиях в строении между центральным и боковыми участ-
ками подошвы. Правда, отчасти это связано с тем, что педальные ветви колумелляр-
ного мускула крепятся близ средней линии ноги и при фиксации, сокращаясь, они втя-
гивают в первую очередь центральное поле. Но диффузная нервная сеть также зани-
мает не всю поверхность подошвы — она'приурочена именно к центральному полю
(см. рис. 16). В то же время наличие на подошве трех четких продольных полей, разде-
ленных бороздами, характерно для A ulacopoda. Таким образом, здесь можно предполо-
жительно говорить о последних следах существования продольных полей на подошве
современных Helicoidea.
НАПРАВЛЕНИЯ РАЗВИТИЯ
51
Далее, у большинства Aulacopoda челюсть имеет срединный выступ, которого ли-
шены представители рассматриваемого надсемейства; но у своеобразных Sphinctero-
chilidae челюсть со срединным выступом. Несмотря на то что«названное семейство до-
статочно изолировано от остальных Helicoidea (существует даже мнение, что эту группу
надо считать самостоятельным надсемейством — Forcart, 1972, и не исключено, что
это мнение справедливо), оно произошло от той же группы, что и прочие Helicoidea.
Наконец, у наших моллюсков внутри бича первично имеется хорошо видная на
поперечных срезах продольная складка, свернутая вдоль. В связи с этим и хвостовая
часть сперматофора имеет характерную форму. Сходное строение бича и хвоста спер-
матофора отмечено и у ряда представителей Aulacopoda (Van Mol, 1968, 1971).
Совокупность перечисленных обстоятельств достаточно определенно указывает
группу, из которой логично выводить надсемейство; однако нет никаких доказательств,
что все таксоны обсуждаемой группы имели общего предка внутри Aulacopoda. На-
против, есть основания думать, что Helicoidea в принимаемом пока объеме имеют, об-
разно говоря, не менее трех корней в недрах древних Aulacopoda соответственно основ-
ным рецентным группам: Sphinctero.chilidae, Hygromiidae, остальные семейства. Ниже
мы вернемся к этому предположению, а также обсудим положение Helicodontidae,
а пока обратимся к анализу взаимоотношений таксонов внутри той части Helicoidea,
которую мы обозначили как «остальные семейства».
Использование материалов по возможным путям трансформациитразличных эле-
ментов полового аппарата порознь (как чаще всего и делается), особенно если учесть
безграничное разнообразие и беспредельную пластичность этих элементов, превращает
филогенетическое исследование в классификацию большого числа признаков, давая
широкие возможности для спекуляций, часто противоречивых; при таком подходе за-
дачи филогенетического плана имеют обычно несколько взаимоисключающих решений.
Вместе с тем организм — по меткому выражению В. Н. Беклемишева (1964) — морфо-
процесс, и это определение относится к любой из составляющих егошастей; процессы же
подчиняются определенным закономерностям, и задача сравнительно-морфологи-
ческого и филогенетического исследования —- понять эти закономерности. Иными сло-
вами, классифицировать надо не признаки, но тенденции (Миничев, 1967, 1971; Голи-
ков, Старобогатов, 1968).
Следовательно, более строгий метод разработки филогении имеет основанием учет
коррелятивных связей — в данном случае между отдельными элементами репродук-
тивного тракта. Тогда вместо раскладывания пасьянса из большого набора разрознен-
ных признаков получаем 2 основных комплекса, элементы которых, изменяясь в про-
цессе эволюционного развития, всегда закономерно взаимосвязаны: 1) весь совоку-
пительный аппарат с семеприемником и 2) совокупность
придатков женского отдела. Существенно, что насколько закономерны
взаимные изменения элементов внутри каждого из комплексов, настолько независимы
друг от друга сами комплексы.
Рассмотрим вначале совокупительный аппарат с семеприем-
ником, поскольку эти органы в первую очередь важны для понимания взаимоот-
ношений внутри надсемейства. Полный набор этого комплекса состоит из бича, эпи-
фаллуса, пениса с заключенной внутри папиллой и семеприемника с протоком, резер-
вуаром и дивертикулом. Наличие этого набора целиком определяется существованием
сперматофора, строение которого во всех группах надсемейства (где он первично име-
ется) достаточно сложно и достаточно однотипно (рис. 28, А—Б). Носителями полного
набора перечисленных признаков являются неарктические Humboldtianidae и Helmin-
thoglyptidae и средиземноморские Helicidae.
Вторичные эволюционные упрощения всегда начинаются с редукции, а затем и
исчезновения дивертикула семеприемника. Это связано с двумя альтернативными при-
чинами. В одних случаях укорочение дивертикула коррелятивно связано с укороче-
нием бича, а в других — со способностью сперматофора сворачиваться в клубок внутри
пениса.
Свернутый в тугой клубок и подаваемый в таком виде сперматофор — уже каче-
ственно иное явление, нежели постоянно вытянутый в нить: значительная часть объема
этого клубка занята веществом оболочки, которая в силу этого уже не выполняет
в полной мере первоначальной функции, а потому подвергается вначале истончению,
а затем и полной редукции — сперматофор превращается в пакет желеобразной конси-
стенции, содержащий скопления спермиев. Очевидно, что исчезновение оболочки спер-
матофора влечет за собой редукцию и бича, и эпифаллуса, и папиллы (кроме немногих
случаев, когда папилла, видоизменяясь, принимает участие в передаче пакета — неко-
торые азиатские Camaenidae}', но тогда основание протока семеприемника приобретает
вздутые очертания, или по крайней мере проток семеприемника становится скорее
коническим, чем цилиндрическим (рис. 28, Д}. Подобная картина наблюдается у аме-
риканских Oreohelicidae, у подавляющего большинства азиатских Bradybaenidae и
некоторых как азиатских, так и американских Camaenidae. Заметим, кстати, что
Oreohelicidae сближает с Camaenidae (во всяком случае, новогвинейскими) также и
то, что это единственные 2 группы, у которых наблюдается живорождение.
4*
52
ВВЕДЕНИЕ
Дальнейшее развитие основания семеприемника приводит к формированию спе-
циализированной мешковидной приемной камеры при основании протока (рис. 28, Е),
как это имеет место у A mmonitellidae. В силу неодинаковой скорости редукции пере-
численных отделов, а также в связи со способностью пениальной папиллы морфологи-
чески перестраиваться и тем отвечать на новые функциональные требования наблю-
даются варианты, когда при основании семеприемника уже развита приемная камера,
но еще остается рудиментарный бич (Л mmonitellidae), или папилла и бич еще доста-
точно хорошо развиты, но семоприемник уже имеет вздутое основание (Papuininae—
Camaenidae).
Охарактеризованная тенденция по понятным причинам свойственна большей
частью группам, сформировавшимся и поныне обитающим в условиях влажного тропи-
Рис. 28. Элементы полового аппарата Helicoidea (схема). А — поперечный срез
эпифаллуса и соответствующий участок сперматофора; Б — поперечный срез бича
и соответствующий участок сперматофора; В—Е — форма протока семеприемника:
В — цилиндрический, без дивертикула, Г — цилиндрический, с дивертикулом,
Д — конический, без дивертикула, Е — цилиндрический, без дивертикула, с при-
емной камерой при основании; Ж — способ возникновения ложной папиллы пе-
ниса; 3 — продольный разрез бича и участок сперматофора у Plectopylis. (Ориг.).
ческого климата. Однако ряд представителей позже освоили аридные пространства,
причем большей частью адаптации к новым условиям нашли конхологическое, эколо-
гическое и, вероятно, физиологическое выражение, но изредка наблюдается вторичное
появление утраченных ранее атрибутов: сперматофора с хитинизированной оболочкой и,
следовательно, бича и папиллы, например у Plectopylis (Шилейко, 1973а). Вторичное
появление можно постулировать с уверенностью, так как и сперматофор, и бич, и па-
пилла имеют здесь совершенно иное строение. Особенности сперматофора видны из
рис. 28, 5; соответственно и бич мешковидный, с характерной внутренней структурой,
не имеющей ничего общего с первичным вариантом. Далее, если папилла первично
всегда закладывается в виде разрастания дна пениса вокруг отверстия эпифаллуса,
то у Plectopylis она возникает за счет энергичного погружения эпифаллуса в просвет
пениальной трубки. Показательно, что у отдельных особей двух видов настоящих
Bradybaena (Br. ussuriensis и Br. plectotropis), по которым изучен массовый материал,
мы обнаружили временные папиллы, образованные указанным путем (рис. 28, Ж).
Своеобразным аналогом дивертикула семеприемника у Plectopylis является наличие
при основании протока специализированной камеры (как у Ammonitellidae), где про-
исходит лизирование оболочки сперматофора, которая в силу сложной формы плохо
приспособлена к передвижению по узкому протоку семеприемника.
Менее характерные вторичные структуры, подобные описанным, известны и у не-
которых других Bradybaenidae (Habe, 1955, 1963).
Обратимся теперь к рассмотрению придатков женского отдела.
Принимая во внимание принцип олигомеризации гомологичных органов и отправную
Рис. 29. Филогенетическая схема надсемейства Helicoidea. А — A mmonitellidae; В — Bradybaenidae; С — Camaenidae; Н —
Humboldtianidae; Hel — Helicidae; Hg — Helminthoglyptidae; Hyg — Hygromiidae; О — Oreohelicidae; S — Sphincterochilidae.
Объяснения в тексте. (Ориг.).
54
ВВЕДЕНИЕ
точку этого принципа — множественную закладку новообразующихся органов (До-
гель, 1954), надо признать, что первично из неспециализированного участка рыхлой
ткани возникли 4 равноценных стилофора, каждому из которых отвечала слизистая
железа, причем чрезвычайно существенно, что ныне наблюдаются 2 принципиально раз-
личных и не связанных между собой варианта, вполне отвечающих такому представле-
нию. 1-й вариант — придатки женского отдела располагаются венцом вокруг вагины
(Humboldtianidae), 2-й вариант — стилофоры располагаются двумя горизонтальными
рядами (Hygromiidae — Trichiinae).
Рассмотрим вероятные пути развития 1-го варианта. Еще раз подчеркнем, что все
4 стилофора здесь действующие, т. е. со стрелой, слизистые железы еще слабо отдиф-
ференцированы от стенок вагины, а протоки их проходят в толще стенок последней
(Pilsbry, 1928, 1939).
Рис. 30. Схема пространственных взаимоотношений между семействами Helicoidea.
I — американский участок; II — средиземноморский участок; III — азиатский
участок. Остальные обозначения те же,.что и на рис. 29. (Ориг.).
Судя по всему, далее довольно быстро прошла олигомеризация за счет редукции
сразу трех стилофоров и двух слизистых желез (к чему многие Helicoidea вообще
проявляют склонность), чем создалась привычная и столь банальная ныне картина:
один стилофор в сопровождении пары слизистых желез (Helicidae-, в типичном случае
Helminthoglyptidae-, первично Bradybaenidae). Очевидно, для надсемейства в целом пер-
вичное положение слизистых желез — на вагине (если половой аппарат Humboldti-
anidae считать наиболее архаичным); затем известны случаи впадения желез в основа-
ние стилофора и, наконец, у большинства Bradybaenidae — непосредственно в стилофор,
часто через дополнительный мешок. Поскольку слизистые железы бесконечно варьи-
руют в форме, числе, месте впадения и характере строения их стенок (Шилейко, 1971г,
1973а), они дают наименее надежный материал для разработки вопросов филогении на
уровне семейств и подсемейств.
Заключительный этап эволюции женского отдела, реализованный независимо
у ряда групп, — полная редукция всех придатков этого отдела, причем процесс редук-
ции вначале касается слизистых желез, а затем стилофора. Во всяком случае, не из-
вестно ни одного вида с развитыми слизистыми железами, но без стилофора, тогда как
обратная картина изредка наблюдается (Nesiohelix, некоторые Cathalca).
Рассмотрим филогенетическую схему (рис. 29), представляющую собой синтез
излагаемых соображений. Здесь отражены лишь те варианты строения гениталий,
которые мы считаем первичными, исходными для каждого из включенных сюда се-
мейств. Исходя из этих вариантов, можно оценить любые изменения, наблюдающиеся
в той или иной группе.
НАПРАВЛЕНИЯ РАЗВИТИЯ
55
Основу схемы составляют три последовательно ветвящихся ствола, отвечающих
трем неарктическим семействам. Helicidae — старые, но непосредственные производ-
ные примитивных Humboldtianidae. Происхождение Bradybaenidae от Helminthog-
lyptidae не вызывает сомнений, поскольку все серьезные, казалось бы, различия между
этими семействами определяются в конечном счете единственным обстоятельством: ре-
дукцией сперматофора. Ветвление ствола, отвечающего Oreohelicidae, обосновано выше.
За единственным исключением, все рассматриваемые семейства имеют ясно выра-
женный зоогеографический облик (рис. 30) и представлены только в Новом Свете, или
только в Средиземноморье, или только в Восточной Азии (в любом случае имеются не-
сомненные связи с перечисленными центрами); только семейство Camaenidae пред-
ставлено как в Новом, так и в Старом Свете. Имеющиеся анатомические данные (Wurtz,
1955; Clench, Turner, 1962, 1964, 1966, 1968) как будто не противоречат объединению
двух пространственных группировок в одно семейство. Сравнительное изучение
Pleurodonte Isabella (о-в Барбадос), Rhynchotrochus macgillivrayi (Австралия, Квинс-
ленд) и Сатаепа rug ata (Хоцзи-чоу, Китай), проведенное нами, также показало до-
статочную близость этих форм, чтобы считать их представителями одного семейства.
Выше мы кратко коснулись особенностей Sphincterochilidae, которые заставляют
предполагать, что эта группа также является дериватом Aulacopoda, но независимо
от остальных Helicoidea. Кроме своеобразной челюсти названное семейство отличается
рядом других черт, а именно: отсутствием папиллы пениса при наличии хорошо раз-
витого бича и сравнительно небольшого дивертикула семеприемника; существованием
мешковидного придатка на пенисе, причем сам пенис отличается очень малыми разме-
рами; наличием своеобразного придатка женского отдела, снабженного единственной
железой «брадибенидного» облика и связанного с мощным придатком внутри вагины
посредством узкого протока.
По всей видимости, утрата папиллы пениса у Sphincterochilidae связана с разви-
тием мешковидного пениального придатка, представляющего собой камеру для свер-
нутого в комок сперматофора, чем в свою очередь вызвана полная или частичная ре-
дукция дивертикула семеприемника. Выше отмечалось, что исчезновение оболочки
сперматофора характерно для групп, обитающих в условиях влажного климата. Дей-
ствительно, в подобных условиях сперматофор не нужен. Однако и для таких облигат-
ных ксерофилов, как Sphincterochilidae, выгода передачи комка вместо нитевидного
сперматофора тоже очевидна, так как в этом случае резко сокращается время, потреб-
ное для копуляции (кстати, тенденция к сворачиванию сперматофора наблюдается и
у некоторых Helicinae). Допустимо также, что тенденция к сворачиванию спермато-
фора проявилась раньше, в иных климатических условиях, но и в аридной обстановке
эта тенденция себя оправдала.
Таким образом, основные отличия от остальных таксонов касаются челюсти и
придатков женского отдела, гомология которых неясна. Отличия эти достаточно серь-
езны, и Форкарт (Forcart, 1972), вероятно, прав, считая эту группу независимым над-
семейством.
Что касается 2-го варианта расположения стилофоров, т.е. Hygromiidae, то,
повторяем, попытка выводить их из 1-го варианта — Humboldtianidae (или наоборот)
выглядит очень натянутой: если у Humboldtianidae стилофоры располагаются мутов-
чато, то у наиболее древних Hygromiidae — Trichiinae они сидят двумя горизонталь-
ными рядами — по два в каждом ряду; характер связи слизистых желез со стилофорами
совершенно различный. Существенно также то, что у Hygromiidae всегда отсутствует
дивертикул семеприемника; основание резервуара притянуто тонкими соединительно-
тканными связками к спермовидукту. В то же время у всех остальных семейств основа-
ние резервуара подобным же образом связано с задней частью дна мантийной полости,
прилегая всегда к границе спермовидукта и белковой железы. В настоящее время трудно
филогенетически оценить эти особенности, однако сам факт таких различий свидетель-
ствует об определенном разрыве между Hygromiidae и прочими семействами.
Первичное отсутствие дивертикула семеприемника у Hygromiidae с функциональ-
ной точки зрения объясняется тем, что резервуар здесь отличается относительно очень
большим объемом, и нам не раз доводилось видеть свернутый сперматофор именно
в самом резервуаре, чего никогда не бывает у других групп.
Следовательно, надо принять, что Hygromiidae достаточно изолированы от осталь-
ных Helicoidea, хотя'они также происходят от Aulacopoda.
Что же мешает разделить Helicoidea, в современном понимании, на три надсемей-
ства? Основная и, пожалуй, единственная причина заключается в недостаточности
знаний по строению ряда групп — в первую очередь азиатских и африканских. Если
в дальнейшем не обнаружится серьезных возражений против предложенной здесь схемы,
то придется признать необходимость такого разделения.
В схему не включены семейство Helicodontidae и группа неясного систематического
ранга и систематической принадлежности — Geomitrinae. Как видно из приводимого
ниже анализа организации Helicodontidae, это семейство, во-первых, достаточно раз-
рознено, во-вторых, своеобразно и, в-третьих, не настолько хорошо изучено, чтобы
можно было высказывать суждения об его исторических связях. Пока можно лишь кон-
56
ВВЕДЕНИЕ
статировать, что это семейство явных связей не обнаруживает ни с одним из рецепт-
ных таксонов; возможно, что в это семейство следует включить чрезвычайно своеобраз-
ную форму — Ciliella ciliata (Шилейко, 1972г).
Организация Geomitrinae более компактна (Mandahl-Barth, 1937—1938), но си-
стематическое положение этой группы неясно.
Возвращаясь к анализу надсемейства в целом, обратим внимание на другой ас-
пект, именно на существование серьезных различий в таксономической структуре трех
основных пространственных групп: американской, азиатской и средизейноморской.
Для первой группы характерны большие роды — до нескольких десятков видов в каж-
дом — при сравнительно малом их числе. В пределах средиземноморской группы кар-
тина обратная: здесь имеется множество моно- и олиготипических родов, да и общее их
количество много больше, чем в фауне Нового Света (соответствующие подсчеты
см.: Wald6n, 1963, а также: Burch, 1969). Азиатская группа занимает промежуточное
положение. Такая разница объясняется неоднозначно (в частности, разной степенью
расчленения и разъединения горных массивов и совокупностью исторических процес-
сов, приведших к формированию материков в их современном виде), но если принять
во внимание ископаемые материалы, то отмеченный факт получает вполне определен-
ное освещение.
Приведенная и разобранная филогенетическая схема разработана на морфологи-
ческом материале. Рассмотрим теперь эту схему в пространственно-временном аспекте,
в частности, насколько она согласуется с данными исторической геологии и палеокли-
матологии (Страхов, 1948; Леонов, 1956; Марков, 1960; Синицын, 1967; Такеучи
и др., 1970).
Судя по палеонтологическим находкам, Helicoidea (не принимая пока во внимание
Sphincterochilidae и Hygromiidae) оформились уже в первой половине мелового периода;
поскольку наиболее архаичные группы ныне обитают в северном полушарии на западе
Северной Америки, и все ископаемые находки сделаны в том же районе, можно заклю-
чить, что местом возникновения надсемейства была юго-западная часть Лавразии, от-
деленная Тетисом от Гондваны. Между прочим, именно поэтому в южном полушарии
Helicoidea первично отсутствуют.
Уже в верхнем мелу обнаруживаются раковины Helminthoglyptidae и Oreohelicidae.
Отсутствие палеонтологических материалов по Humboldtianidae — не доказательство
того, что этих моллюсков в прошлом здесь не было: их раковины из-за больших разме-
ров и тонких стенок плохо сохраняются. Допустимо также, что раковины того облика,
который характерен для современных видов, появились сравнительно недавно, но это
не означает, что столь же недавно возникли и особенности внутренней организации
членов семейства.
Поскольку наличие сперматофора — первичное состояние в пределах надсемей-
ства, надо полагать, что условия, в которых начала складываться обсуждаемая группа,
были достаточно аридными. В самом деле, для нижнемеловой эпохи показано существо-
вание широкой аридной зоны на юго-западе Лавразии, влияние которой к концу мела
несколько ослабело.
Распространяясь в начале—середине палеогена в восточном направлении, непо-
средственные предки Humboldtianidae дали начало современным Helicidae, именно
Ariantinae, после чего развитие этих групп осуществлялось независимо благодаря воз-
никновению морского барьера между Европой и Северной Америкой.
В то же время, т. е. в эоцене, представители Helminthoglyptidae и Oreohelicidae
имели возможность распространяться в западном направлении, на территорию ны-
нешней Юго-Восточной Азии. Азиатскими производными первых стали Bradybaenidae,
а вторых — Camaenidae. Пилсбри (Pilsbry, 1911) верно наметил пути миграции амери-
канской малакофауны в Азию, но считает, что этот процесс происходил много раньше—
еще в мезозое. Возможно, поскольку речь идет о некоторых других группах, Пилсбри
прав, но Helicoidea появились в Азии не раньше начала третичного периода.
Hygromiidae в ископаемом состоянии известны только из Европы, начиная, веро-
ятно, с олигоцена, т. е. с того времени, когда Америка уже окончательно отделилась
от Европы; во всяком случае, широкие связи между этими регионами отсутствовали
уже с начала палеогена. То же самое относится к Sphincterochilidae и Helicodontidae.
Эти факты также свидетельствуют в пользу предположения о независимом возникнове-
нии этих групп. Более надежно это доказывается в отношении Sphincterochilidae,
так как они обладают характерной раковиной, чего нельзя сказать о двух других се-
мействах.
Прямая континентальная связь Европы с Азией открылась лишь в олигоцене —
миоцене, и этим объясняется четкое различие в фауне этих районов. Присутствие
Hygromiidae в Азии связано с двумя волнами миграций. Первая волна имела место
в течение миоцена — часть Hygromiinae мигрировали из Европы на восток, достигнув
берегов Тихого океана (Lindholmomneme), но минуя Среднюю Азию. Позже — в плио-
цене — плейстоцене — по линии хребтов Эльбурса и Копетдага в Среднюю Азию
проникли ксерофильные Hygromiidae (азиатские Trichiinae проникли сюда тем же пу-
тем, но много раньше — в начале миоцена).
НАПРАВЛЕНИЯ РАЗВИТИЯ
Одновременно со второй волной миграции с запада на восток в обратном направле-
нии продвинулись два вида азиатских Bradybaenidae.
СЕМЕЙСТВО HELICODONTIDAE
Основная масса видов семейства изучена формально, т. е. известен внешний вид
их гениталий (Hesse, 1918b; Ortiz de Zarate, 1962; Gittenberger, 1968b). Однако и эти
сведения вместе с данными по 4 видам, изученным нами (Шилейко, 1971г), позволяют
сделать ряд важных в филогенетическом отношении выводов.
Семейство в настоящее время представлено фрагментарно и состоит из весьма
разрозненных остатков некогда обширной группы. Ископаемые виды известны еще
из эоцена Европы. Разрозненность эта видна уже из обилия моно- и олиготипических
таксонов. Одни группы семейства обнаруживают более или менее ясную связь с дру-
гими семействами; другие такой связи не обнаруживают и филогенетическое значение
их неясно. Не исключено также, что часть таксонов, ныне включаемых в состав этого
семейства, правильнее было бы относить к другим семействам. В первую очередь это
относится к родам Mastigophallus и Trissexodon (рис. 31, А), резко отличающимся от
Рис. 31. Основные варианты строения полового аппарата Helicodontidae.
Объяснения в тексте. (Ориг.).
всех остальных родов семейства наличием хорошо развитого бича и двух стилофоров,
расположенных на одной стороне вагины — один над другим; внутри нижнего заклю-
чена стрела. Кроме того, имеется папилла пениса, отсутствующая у большинства дру-
гих родов семейства (Gittenberger, 1968b). Эти особенности дают повод предполагать,
что названные роды — аберрантные члены Hygromiinae.
Оставим пока в стороне роды Canariella и Soosia, половой аппарат которых лишен
наружных придатков (у Canariella, правда, имеется рудиментарный бич); тогда оста-
ются 3 группы, резко различающиеся между собой и резко отграйиченные от упомяну-
тых выше родов.
Первая группа включает лишь один род — Lindholmiola (рис. 31, Б), с типовым
видом L. lens, который изучен недостаточно подробно; к этому же роду относится
L. corcyrensis, беспрецедентное своеобразие которого обнаруживается при изучении
внутреннего строения мужского отдела, поэтому два указанных вида мы оставляем
в пределах одного рода провизорно. В любом случае L. с orcyrensis настолько изолирован
от остальных представителей семейства, что мы сочли целесообразным придать роду
ранг подсемейства, причем отношение его к прочим группам неясно. Можно лишь го-
ворить о том, что здесь имеется своеобразное сочетание примитивных черт с признаками
узкой специализации.
Вторая группа характеризуется отсутствием бича и эпифаллуса, а также наличием
вильчатой слизистой железы, ветви которой неодинаковы как по внешнему виду, так
и функционально (рис. 31, В). Роль собственно слизистой железы выполняет большая
ветвь, а меньшая служит резервуаром, где сохраняется секрет большой ветви. К этой
группе относятся 4 рода: Helicodonta, Drepanostoma, A tenia, Caracollina', надо добавить,
что первично, по-видимому, имеется еще булавовидный придаток, который принято
считать резко видоизмененным стилофором. Слизистая железа и придаток имеется
58
ВВЕДЕНИЕ
у Caracollina', только придаток, без желозы — у 4tenia', остальные два рода имеют
только железу, без придатка. Учитывая склонность к редукции придатков, резонно
предположить, что монотипический род Soosla также относится к этой группе.
Все эти роды составлены одним-двумя видами, что свидетельствует о большой древ-
ности и таксономической неоднородности. Кроме того, ныне представляется возмож-
ность говорить о сокращении ареала: представители некоторых родов обнаружены
в миоцене Предкавказья (Стеклов, 1966), т. е. много восточнее современного ареала.
Здесь найдены представители родов Ilelicodonta и Lindholmiola, т. е. родов, заходящих
в настоящее время на восток дальше других членов семейства.
Третья группа представлена родом Oestophora, составленным не менее чем 15 ви-
дами, причем ареал рода почти целиком приходится на Пиренейский п-ов (Ortiz de
Zarate, 1962). Воздерживаясь от обсуждения чрезвычайно своеобразного португальского
вида «О», turriplana (см.: Шилейко, 1971г), можно дать очень четкий как анатомический,
так и конхологический диагноз этой группы, которая представляет собой наиболее
гомогенный и наиболее, возможно, молодой таксон семейства. Для всех Oestophora
характерно наличие одного массивного стилофора, 2—3 длинных гофрированных
неразветвленных слизистых желез и пениса в виде простой трубки, без папиллы
(рис. 31, Г). Существенно, что в пределах рода наблюдается редукция бича: довольно
длинный бич у членов подрода Gasullia, рудиментарный — у подрода Suboestophora,
и у Oestophora s. str. бич отсутствует. Таким образом, в организации полового аппарата
имеется сочетание примитивных и прогрессивных черт. Этот род наиболее близок
к Hygromiidae по сравнению со всеми прочими таксонами семейства; возможно, обсуж-
даемую группу надлежит вывести из Helicodontidae и присоединить к Hygromiidae.
Здесь также можно говорить о сокращении ареала, так как в понтийском ярусе При-
днепровья обнаружен ископаемый представитель рода — О. schlleykoi (Присяжнюк,
1974). .
Вероятно, Canariella близки к Oestophora- ti представляют собой уклонившуюся
за счет редукции придатков женского отдела ветвь последнего. В пользу такого пред-
положения свидетельствует наличие у Canariella короткого бича; правда, внутреннее
строение пениса видов этого рода не изучено.
Оценить филогенетически группу в настоящее время трудно, однако чрезвычайно
существенно, что здесь есть виды, имеющие общие черты с Hygromiidae, что само по
себе уже дает представление о возможных исторических связях. Скорее всего, семей-
ство в настоящем объеме — категория гетерогенная, и собственно Helicodontidae
к настоящему времени осталось очень мало — всего несколько видов.
СЕМЕЙСТВО BRADYBAENIDAE
По сравнению со всеми остальными семействами Helicoidea настоящее семейство
изучено наименее полно. Внутреннее строение большинства видов до сих пор неизве-
стно; попытки разработать систему семейства в целом отсутствуют. Трудности усугуб-
ляются еще и тем, что в этом семействе более, нежели в большинстве дру-
гих, важно знать внутреннее строение гениталий, а такими данными мы рас-
полагаем по очень малому числу видов. Старые работы (Semper, 1870; Jakobi,
1898; Wiegmann, 1900) мало спасают положение, так как в этих исследованиях сравни-
мых данных немного. По существу единственной попыткой серьезно разобраться в так-
сономической структуре группы явилось исследование П. В. Матёкина (1972), которое
касается исключительно среднеазиатских видов. В связи со сказанным мы считаем
преждевременным говорить о структуре семейства в целом.
| Что касается анализа на уровне рода и ниже, то надо подчеркнуть, что Brady-
baenidae фауны СССР в целом есть результат мощного адаптивно-радиационного про-
цесса. С практической точки зрения тут можно формально выделять в отдельные так-
соны каждый второй вид или считать их всех одним родом, даже без подродов (особый
случай — группа Ponsadenia; см. ниже). Дело в том, что почти каждый вид имеет осо-
бенности, отделяющие его от любого другого вида, причем эти особенности касаются
множества разных признаков, связанных с различными отделами полового аппарата
и раковины. Приходится поэтому констатировать, что те таксономические приемы,
которые оправдывают себя при анализе взаимоотношений внутри других семейств,
в данном случае приемлемы с ограничениями: морфологически далекие на первый взгляд
формы оказываются связанными между собой посредством промежуточных вариантов
й никаких естественных группировок не обнаруживается. Эта ситуация усугубляется
широкой внутривидовой изменчивостью, которая затрагивает не только раковину,
но и признаки внутренней организации.
Наибольший таксойомический вес в данном случае имеют особенности строения
женского отдела; приводимая схема (рис. 32) отражает основные направления эволю-
ции этого отдела.
Пытаясь понять систему таксона, необходимо из всех наблюдаемых вариантов
строения животных выделить такой, который ближе всего отвечал бы исходному или
НАПРАВЛЕНИЯ РАЗВИТИЯ
59
•близкому к исходному варианту. Исходным типом строения полового аппарата в пре-
делах по меньшей мере Bradybaeninae можно считать то, что наблюдается у не-
которых видов Trishoplitcr. в стилофор впадают 2 небольших резервуара (дополнитель-
ных мешка), каждый из которых несет по 2—3 неветвящиеся трубчатые слизистые
железы. Нетрудно видеть, что подобный тип есть непосредственное производное от
Helminthoglyptidae, что лишний раз подтверждает наши предположения о филогене-
тических связях внутри надсемейства.
Первый этап дальнейшего развития — слияние резервуаров в один дополнитель-
ный мешок; однако в подавляющем большинстве случаев слизистые железы впадают
Рис. 32. Основные направления морфологической эволюции
женского отдела полового аппарата Bradybaenidae. I—III —
род Ponsadenia; IV—X — род Bradybaena. Объяснения в тексте.
(Ориг.).
в этот мешок по-прежнему двумя пучками, причем в результате увеличения общего
числа ветвей, многократного их изгибания и тесного прилегания друг к другу железа
приобретает часто вид компактного мешка с ячеистыми стенками. Такому представле-
нию отвечает картина, наблюдаемая, в частности, у Br. fruttcum.
От этого варианта можно наметить две линии развития группы; эти морфологиче-
ские линии соответствуют двум пространственным группировкам — дальневосточной
и среднеазиатской. Для первой характерно наличие продольного гребневидного валика
внутри пениса (у Br. fruttcum такой валик намечается, но слабо обособлен) и желези-
стого «пояса», вокруг базальной части женского отдела и отсутствие надклоачного пу-
зыря. Кроме /того, дополнительный мешок всегда хорошо развит и не обнаруживает
заметной склонности к редукции. Члены среднеазиатской группировки не имеют греб-
невидного валика внутри пениса, железистый «пояс» у них отсутствует и первично име-
ется надклоачный пузырь, способный к значительной трансформации или редукции.
Вместе с тем железистый «пояс» дальневосточных и надклоачный пузырь средне-
азиатских видов — гомологичные образования, развиваются они из участка рыхлой
железистой ткани, который у Br. fruttcum имеется в одной из стенок вагйны (тле.,
если эти пространственные группировки признать родами, то Br. fruticum' можно
с равным успехом отнести к любой из них).
60
ВВЕДЕНИЕ
Непосредственными производными ст Br. fruticum являются, с одной стороны,
Br. schrencki и, с другой стороны, Br. transbaicalia; участок железистой ткани в стенке'
вагины у этих видов претерпевает изменения в двух направлениях. У Br. schrencki
кольцевое разрыхление в стенке вагины гораздо более определенно морфологически,
нежели у Br. fruticum'. паренхима распадается па ряд тяжей вплоть до образования
сплошной полости, а наружные покровы соответствующего участка уплотняются,
т. е. наблюдается формирование настоящего надклоачного пузыря, свойственного
многим азиатским видам (рис. 32, IV и VI). У Вт. transbaicalia железистый участок
в значительной степени изолируется от вагины, сохраняя паренхиматозную структуру,
т. е. представляет собой, по существу, «пояс», характерный для дальневосточных ви-
дов (рис. 32, IV и V); кстати, этот вид уже по раковине во многом напоминает дальне-
восточный вид — Br. middendorffi.
Несмотря на чрезвычайно обстоятельное для своего времени исследование Виг-
манна (Wiegmann, 1900), сравнимого материала по центральноазиатским видам очень
немного, что мешает детализировать представления о характере связей между дальне-
восточными и среднеазиатскими группами. В частности, это видно из широко бытую-
щего номенклатурного заблуждения: считается, что в Средней и Центральной Азии
имеется род Cathaica; типовой вид рода — С. fasciola. Меллендорф (Mollendorff, 1899)
указывает, что в Formenkreis С. fasciola входят также С. middendorffi, С. graeseri,
С. pekinensis, С. transita,ns, С. cardiostoma', в то же время первый из перечисленных ви-
дов — представитель того же рода, что и Br. fiscina (syn. Br. weyrichii), т. e. Karafto-
helix. Другими словами, если род Cathaica и реален (что маловероятно), то этот род
восточнокитайский и дальневосточный, но никак не среднеазиатский.
Таким образом, уже в пределах трех разобранных видов намечаются две тенден-
ции морфологического развития, которые были реализованы и пространственно. Необ-
ходимо подчеркнуть, что, как и в большинстве подобных ситуаций, речь идет не о вы-
ведении всей фауны Bradybaenidae из определенного рецентного вида, а о том, что этот
вид есть прямой и непосредственный потомок формы, которая, развиваясь, дала ны-
нешнее многообразие, и организация Br. fruticum в наибольшей мере приближается
к организации этой гипотетической формы, в частности невысоким уровнем морфоло-
гической специализации.
Таксономическая структура дальневосточного комплекса видов несложна. Это
набор форм, анатомически составляющий плотное единство, но весьма пестрый конхоло-
гически, что может свидетельствовать или об очень быстрой конхологической диверген-
ции, или о высоком совершенстве данного типа строения репродуктивного аппарата в
данных условиях. Учитывая, что в ходе геологической истории на территории Дальнего
Востока отсутствовали резкие климатические и флористические изменения (Кришто-
фович, 1955), мы склоняемся к предположению, что половой аппарат не имел стимула
к резким перестройкам, а потому эволюционный изменения коснулись преимущественно
раковины.
Кратко остановимся на особенностях внутренней организации дальневосточных
видов. В свое время были предложены подродовые названия Karaftohelix и Acusta
специально для некоторых дальневосточных видов. Диагнозы этих таксонов базиро-
вались на числе, форме и характере расположения слизистых желез. Существование
этих таксонов нашими данными не подтвердилось: количество слизистых желез у тех
видов, по которым мы имели возможность просмотреть большой материал, варьировало
от 3 до 12; форма слизистых желез и их размер зависят от функционального состояния
и степени зрелости животного. Даже в том случае, когда число ветвей приближается''
к максимуму, а каждая из ветвей внешне наиболее обособлена, все они впадают в до-
полнительный мешок в конечном счете посредством парного протока; группировка
их в две грозди в этом случае слабо заметна. В целом таксономический ранг этих приз-
наков не выше вида.
Вместе с тем по двум признакам наблюдаются определенные и не связанные друг
с другом линии специализации. Во-первых, это касается усиления дифференциации
пениального гребня, вершина развития которого наблюдается у Br. diversita и Br. fra-
gilis; у этих видов гребень, отчленившись от внутренней поверхности пениса, имеет
вид червеобразного придатка. Во-вторых, характер структуры внутренней поверхности
пениса меняется от вида к виду: это или совокупность параллельных валиков, или каж-
дый из них разбит на ряд бугорков, или система тонких валиков дает впечатление
сетчатой структуры или узора типа «елочки» и т. д. Кроме того, железистый «пояс»
развит неодинаково, и у некоторых видов не обнаружен.
Дополнительный мешок стилофора у всех дальневосточных видов всегда хорошо
выражен и заключает более или менее обширную полость (рис. 32, V), поэтому можно
думать, что его роль сводится к накоплению секрета слизистых желез.
Комплекс среднеазиатских видов много сложнее. Весьма характерная особенность
этой группировки —• широкая внутри- и межпопуляционная изменчивость как конхо-
логических признаков, так и черт внутренней организации. В качестве примеров сош-
лемся на наиболее показательные в этом отношении виды. Br. stoliczkana характери-
зуется чрезвычайно мощным мускулизованным чехлом пениса; у одних популяций он
НАПРАВЛЕНИЯ РАЗВИТИЯ
61
целиком срастается со стенками пениальной трубки и морфологически как таковой не
выражен. Однако у других популяций можно наблюдать основные этапы его обособ-
ления, в результате чего можно говорить, что у данного вида конкретный признак под-
вержен значительной изменчивости, хотя у других видов пениальный чехол сохраняет
постоянство формы и степени морфологической связи с пенисом. Далее, П. В. Матё-
кин (1972) указывает, что у Br. bilaticincta имеется надклоачный пузырь и длинный до-
полнительный мешок; в исследованном материале мы не обнаружили дополнительного
мешка, а равным образом и надклоачного пузыря. Число примеров можно было бы зна-
чительно умножить, однако и этого достаточно, если добавить, что вообще и надклоач-
ный пузырь, и особенно дополнительный мешок легко редуцируются, причем степень
их развития может быть разной у членов одной популяции.
Основное отличие наших взглядов на структуру фауны Bradybaenidae Средней
Азии от представлений П. В. Матёкина (1972) заключается в том, что, по нашему мне-
нию, в составе Bradybaenidae этого региона существуют две таксономические группи-
ровки — более молодая, компактная и обширная, связанная с дальневосточными и
центральноазиатскими видами (род Bradybaena), и более древняя и разрозненная —
род Ponsadenia.
Развитие последнего представляется следующим образом. К исходному варианту
строения женского отдела близка, вероятно, картина, наблюдаемая у Р. hirsuta (нами
вид не исследован, строение его приводится в соответствии с фактами, полученными
П. В. Матёкиным (1972)). У этого вида апикальная часть стилофора вытянута в трубку,
заканчивающуюся парным резервуаром, в который впадают два пучка слизистых же-
лез. Вытянутая часть стилофора резко согнута под острым углом при основании и при-
легает верхним концом к базальной части стилофора; в результате резервуары слизи-
стых желез соприкасаются с атриальной частью (рис. 32, I). С этих позиций легко
оценивается половой аппарат Р. duplocincta, причем возникший парный резервуар сли-
зистых желез у этого вида гомологичен резервуарам слизистых желез, а не надклоач-
ному пузырю видов рода Bradybaena (вспомним, что надклоачный пузырь возникает
из разрыхления стенок вагины). Проток слизистых желез у Р. duplocincta проходит,'
таким образом, внутри своеобразного мостика, соединяющего резервуары слизистых
желез с верхним концом стилофора (рис. 32, II).
Наконец, у Р. semenovi происходит интеграция резервуаров с тканями атриальной
области, а проток слизистых желез сдвигается на середину мостика, сообщаясь как
с полостью стилофора, так и с участком очень рыхлой паренхимы — остатком резер-
вуара (рис. 32, III).
Четкому разделению родов Ponsadenia и Bradybaena мешает, на первый взгляд,
то обстоятельство, что протоки слизистых желез у Br. helvola и Br. pseudoferghanica
пронизывают надклоачный пузырь (отсутствующий у Ponsadenia) и, что самое суще-
ственное, проходят в толще стенки стилофора снизу вверх (рис. 32, VIII). Подобная
картина создает формальную аналогию, поскольку есть соблазн считать эту часть
стенки стилофора продуктом слияния собственно стенки и мостика, т. е. если мостик
будет плотно припаян к стенке стилофора, так, что снаружи виден не будет, то проток
слизистых желез будет проходить в толще стенок стилофора, как и наблюдается у назван-
ных видов. Однако это сходство — не более, чем формальная аналогия: если этот надкло-
ачный пузырь — гомолог резервуара слизистых желез Р. duplocincta, то как могут его
пронизывать протоки слизистых желез, которые у Р. duplocincta впадают в этот резер-
вуар? Очевидно, такую гомологию надо решительно отклонить.
Выше уже обсуждалось возникновение надклоачного пузыря. Он развивается из
участка железистой ткани, и у таких видов, как Br. lantzi, Br. dichrozona и Br. plecto-
tropis, часто поднимается до самого основания слизистых желез, а иногда даже чуть
выше (рис. 32, VII). Учитывая, что дополнительный мешок подвержен редукции,
легко себе представить, что если редукция достаточно полная, а покровы иадклоач-
ного пузыря распространяются достаточно высоко по поверхности стилофора, они не-
избежно будут пронизываться протоками слизистых желез, причем эти протоки будут
проходить в толще стенок стилофора — в том месте, где раньше был дополнительный
мешок..
Следовательно, мы принимаем, что для рода Ponsadenia первично расположение
слизистых желез на вершине придатка стилофора, который, в свою очередь, впадает
в стилофор почти апикально.
Рассмотрим теперь настоящих среднеазиатских Bradybaena, исходя из того, что
исходным вариантом мы считаем половой аппарат типа Br. fruticum. Модифика-
ции, наблюдаемые у Br. lantzi, Br. dichrozona, Br. stoliczkana, Br. bilaticincta, Br.
plectotropis и Br. scythica, не требуют специальных пояснений: любой из них
можно представить как результат частных изменений полового аппарата, близ-
кого к исходному типу в сторону усиления или ослабления степени раз-
вития дополнительного мешка и (или) надклоачного пузыря (рис. 30, VI,
IX, X). Вместе с тем заслуживают, повторяем, внимания случаи особенно сильного раз-
вития надклоачного пузыря, в частности у Br. lantzi, где этот пузырь покрывает не
только клоаку и вагину, но и базальный отдел стилофора и даже иногда нижний отдел
62
ВВЕДЕНИЕ
дополнительного мешка. Нетрудно понять, что при дальнейшем усилении надклоач-
ного пузыря параллельно с редукцией дополнительного мешка пузырь будет обрастать
основания слизистых желез, сидящих уже непосредственно на стилофоре, т. е. полу-
чим картину, наблюдаемую в действительности у Br. helvola и Br. pseudoferghanica.
Таким образом, здесь, как и на Дальнем Востоке, весь комплекс видов представляет
собой результат мощного адаптивно-радиационного процесса. Но в составе фауны
Bradybaenidae Средней Азии существуют две, не связанные непосредственно между
собой таксономические группировки — более молодая и ныне процветающая, связан-
ная с дальневосточными видами (Brady Ьаепа), и более древняя, представленная в на-
стоящее время фрагментарно (Ponsadenia). Подчеркивая эту фрагментарность, мы каж-
дый из трех видов, входящих в род Ponsadenia, возводим в ранг подрода. Характерно,
что все три вида очень четко очерчены и не обнаруживают того размаха изменчивости,
который характерен для видов рода Bradybaena; по существу вся изменчивость здесь
выражается в непостоянстве окраски и размеров раковины, т. е. признаков, вообще
наиболее пластичных.
Заслуживает внимания также то обстоятельство, что род Ponsadenia в целом
тяготеет к наиболее влажным северо-восточным районам Средней Азии, климат которых
не столь резко изменился по сравнению с первоначальным климатом территории (Си-
ницын, 1962, 1967). Исключение составляет ярко выраженный ксерофил Р. semenovi,
ареал которого охватывает большую часть территории Тянь-Шаня, тяготея, однако,
тоже к северо-восточным его районам.
Что касается существования в фауне Средней Азии полиморфных видов, то мы при-
соединяемся к концепции П. В. Матёкина (1960, 1966, 1972), который считает, что здесь
имеется надвид — Br. plectotropis, представленный пятью полувидами (см. системати-
ческую часть). В то же время мы не согласны с оценкой Br. lantzi как надвида — это
обычный полиморфный вид. Полувид — это вид in statu nascendi; обычно полувиды
обнаруживают ясную зависимость конхологических признаков от конкретных условий
среды (см.: Шмальгаузен, 1969). От каких конкретных условий среды зависит направ-
ление закручивания раковины у Br. lantzi? Наличие наряду с правозакрученными си-
нистральных популяций мы расцениваем как своеобразный случай полиморфизма
генетической природы.
СЕМЕЙСТВО HYGROMIIDAE
Данная группа по сравнению с прочими группами того же ранга отличается на-
ибольшим разнообразием и многоплановостью в строении полового аппарата. В то же-
время| раковина представителей семейства большей частью маловыразительна,
причем здесь особенно много случаев конвергентного конхологического сходства.
В пределах семейства намечается несколько достаточно ясных филогенетических
линий, реализованных становлением соответствующих подсемейств (рис. 33).
Характеристика и пути развития этих подсемейств даются ниже; здесь хотелось бы
остановиться на двух взаимосвязанных и наиболее спорных вопросах, которые уже
кратко обсуждались в печати (Шилейко, 1971в, 1973в). Речь идет о возможных путях
эволюции женского отдела полового аппарата. Первично заложились 4 равноценных
стилофора и дальнейшее развитие сводилось, в частности, к полной или частичной
редукции разного их числа либо к видоизменению. Trichiinae и Hygromiinae близки
друг другу, но всегда отличаются одной постоянной особенностью: у первых 4,
у вторых 2 стилофора, расположенных на одной стороне вагины. Необходимо осо-
бенно подчеркнуть, что не известно ни одного вида, у которого пара стилофоров на,
одной стороне вагины была бы развита лучше другой, а такие картины мы неизбежно
должны были бы наблюдать, если признать постепенную редукцию стилофоров на
одной стороне вагины. Этот факт наталкивает на мысль о возможности «мгновенной»
редукции- стилйфоров путем мутаций. Подтверждение такому предположению было
найдено при массовом вскрытии Circassina circassica из разных точек ареала. Было
установлено, что вид существует в форме четырех подвидов, отличающихся наличием
или отсутствием стилофоров и слизистых желез во всех четырех возможных комби-
нациях.
Здесь уместно вспомнить, что стилофоры — не столь уж обязательный атрибут:
Котбауэр (Kothbauer, 1972) специально подчеркивает, что роль стрелы в совокуплении
и сопутствующих актах факультативна; причем здесь речь идет о Helix, т. е. пред-
ставителях группы, у которой наличие одного стилофора жестко закреплено в гено-
типе.
Явление того же порядка, что и подвиды С. circassica, было зарегистрировано
при изучении большого материала по Archaica heptapotamica из ущелья р. Пскем
на западном Тянь-Шане. Среди нескольких популяций обнаружена одна смешанная,
составленная как нормальными A. heptapotamica, так и Nanaja cumulata — новым
видом; единственное: отличие которого от A. heptapotamica заключается в том, что-
здесь имеются две пары стилофоров, а не одна. Существенно, что два названных вида
НАПРАВЛЕНИЯ РАЗВИТИЯ
63
не отличаются достоверно по раковине, равно как и по другим признакам внутрен-
ней организации. Вопрос о том, как практически оценивать эти формы, стоит так:
или они относятся к разным подсемействам, или представляют собой две морфы по-
лиморфного вида. Этот вопрос уже другого порядка, и, чтобы ответить на него, не-
обходимы исследования материала из одной кладки и эксперименты по скрещиванию.
В данном случае, однако, это не столь существенно — в- любом случае перед нами
один из возможных путей эволюции Trichiinae.
Эти рассуждения заставляют принять, что Hygromiinae возникли еще в олиго-
цене от каких-то древних Trichiinae сначала путем редукции одной пары стилофоров
на одной стороне вагины, закрепления нового состояния в генотипе соответствующих
Рис. 33. Филогенетическая схема семейства Hygromiidae. А — Archaicinae;
Е — Euomphaliinae; И — Hygromiinae; М — Metafruticicolinae; Р — Paedhopli-
tinae; Т — Trichiinae. (Ориг.).
видов, а затем развивались независимо от Trichiinae. Вместе с тем надо обратить вни-
мание, что все азиатские представители семейства — наиболее примитивные пред-
ставители своих групп. Однако случай, наблюдаемый у A. heptapotamica — N. си-
mulata, заставляет предполагать, что так называемые «азиатские Hygromiinae» воз-
никли только в плейстоцене (может быть, в конце плиоцена) на месте, независимо
от настоящих Hygromiinae, но тем же путем. Подчеркивая это обстоятельство, мы
выделяем «среднеазиатских Hygromiinae» отдельным подсемейством Archaicinae и
тем самым отрываем их от настоящих западных Hygromiinae.
Подобным же образом дело обстоит и с азиатскими «Euomphaliinae», но разница
заключается в том, что стилофоры этой группы можно представить лишь как продукт
постепенных изменений стилофоров Trichiinae. Однако и эти «постепенные» изменения,
по-видимому, могут происходить достаточно быстро. В этой связи уместно вспомнить
о существовании конхологических двойников — Leucozonella ferghanica и Angiompha-
lia copiosa; эти виды различаются только формой стилофоров, хотя эти различия и
достаточно глубоки. Вероятно, здесь, как и в разобранном примере с Archaicinae,
азиатских «Euomphaliinae» правильнее считать отдельным подсемейством т- Paed-
hoplitinae, возникшим независимо от настоящих средиземноморских Euomphaliinae
сравнительно недавно (не раньше конца плиоцена) в Средней Азии.
Свидетельством сравнительно недавнего происхождения обсуждаемых средне-
азиатских подсемейств служит то, что они еще не приобрели ярких отличий от ис-
ходных азиатских Trichiinae, если не считать признаки, связанные с придатками
женского отдела.
Подсемейство Trichiinae
Анализируя факты, изложенные в систематической части, надо заключить, что
развитие анцестральных Trichiinae шло в двух направлениях. Первое направление —
восточное: мигрирующие виды продвинулись с территории Эгеиды прямым путем
64
ВВЕДЕНИЕ
до горных районов Азии, по всей видимости, в начале миоцена. Испытывая воздей-
ствие новых условий, эти виды приобрели конхологический облик, в целом харак-
терный для многих среднеазиатских Helicoidea', в этом отношении особняком стоит
Odontotrema diplodon.
У исходных форм папилла пениса, вероятно, представляла собой простую трубку
без продольной борозды на поверхности и без интрапапиллярных полостей
(рис. 34, /). У рецентных азиатских видов прослеживаются все основные этапы фор-
мирования интрапапиллярной полости (рис. 34, II—IV): продольная борозда на по-
верхности папиллы вначале закладывается только в проксимальной части (Leuco-
zonella ferghanica), затем борозда распространяется на всю длину папиллы (О. dip-
lodon), углубляется {L. сагу odes), после него наблюдается тенденция к ее замыканию
Рис. 34. Варианты строения папиллы пениса у разных групп Trichiinae и их вза-
имосвязь. I — исходный тип, представляющий собой простую трубку; II—IV —
постепенное формирование интрапапиллярной полости у азиатских видов;
V—XIII — способы образования интрапапиллярных полостей у европейских
и кавказских видов. Просвет семепроводящего канала заштрихован. Более
подробные объяснения в тексте.
(В. rubens, L. rufispira)', заключительный этап — полное замыкание борозды и обра-
зование интрапапиллярной полости, причем семепроводящий канал остается плотно
припаянным к внутренней стенке папиллы (L. retteri, L. caria, L. angulata). Остатком
этой борозды является папиллярная лакуна, посредством которой интрапапилляр-
ная полость сообщается с полостью чехла пениса. То обстоятельство, что в ряду азиат-
ских форм мы видим постепенное развитие такого важного признака, свидетельствует
о сравнительной примитивности группы. Наиболее, по-видимому, высоко развитые
представители — два вида подрода Narina — имеют папиллу, лишенную интрапа-
пиллярной полости, но с папиллярной лакуной, что свидетельствует о вторичном
исчезновении полости у этих видов. Наиболее, напротив, примитивные виды (неза-
висимо от L. ferghanica) — два вида рода Nanaja, мешковидный пенис которых не
несет еще никаких следов продольной борозды.
Второе направление развития группы нашло выражение в образовании двух
группировок — кавказской и европейской. Формирование интрапапиллярных
структур у европейско-кавказских видов пошло по принципиально иному пути, чем
у азиатских: интрапапиллярные полости закладываются попарно в стенках папиллы,
окружая семепроводящий канал; параллельно с этим формируется папиллярная
лакуна, т. е. надо признать, что папиллярные лакуны азиатских и европейско-кав-
казских видов не гомологичны друг другу. Подобный тип строения папиллы имеется
у таких явно независимых групп, как Hygrohelicopsis, Teberdinia, Plicuteria, Trichia
и некоторых видов Kokotschashvilia.
Вероятный путы возникновения наиболее специализированного варианта —
сложной системы интрапапиллярных продольных полостей, разделенных септами
(Edentiella, Caucasigena), указывает нам комплекс Trichia striolata—Tr. danubialis
НАПРАВЛЕНИЯ РАЗВИТИЯ
65
(Schileyko, 1977) — разрыхление одной из стенок, в которой закладывается, таким
образом, уже третичная полость (т. е. ( производная ют вторичной), распадающаяся
затем на несколько более узких полостей. Вместе с тем необходимо подчеркнуть, что
совокупность признаков иного порядка заставляет признать независимое происхож-
дение европейских Edentiella и кавказских Caucasigena.
В пределах рода Kokotschashvilia наблюдается ряд вариантов строения папиллы,
которые мы склонны выводить из той же отправной точки (рис. 34, X—XII). Наи-
более близко к исходной ситуации строение папиллы у К. makvalae: семепроводя-
щий канал окружен парной интрапапиллярной полостью; в стенке папиллы вдоль
одного из тяжей проходит капилляр. У К. tanta этот капилляр остается, семепро-
Рис. 35. Филогенетические и пространственные взаимоотношения между основными
группами Trichiinae и происхождение других подсемейств Hygromiidae. I — европей-
ский участок; II — азиатский участок; III — кавказский участок. А — Anoplitella;
С — Caucasigena', D — Diodontella; Di — Dioscuria; E — Edentiella', H — Helicopsis;>
Hy — Hygrohelicopsis-, К — Kokotschashvilia', L — Le.ucozonella; N — Nanaja; Na —
Narina', О — Odontotrema', P — Plicuteria; T — T eberdinia;' Tr — Trichia; X — Xero-
picta. (Ориг.).
водящий канал сдвигается к одной из стенок папиллы и плотно спаивается с ней;
на противоположной стороне остается тяж (рис. 34, X), который исчезает у К. eber-
hardi (рис. 34, XI). В результате тип строения папиллы последнего вида формально
такой же, как у азиатских Leucozonella retteri, L. caria, L. angulata; то, что это сход-
ство чисто формальное, подтверждается двумя фактами: во-первых, возникновение
одной интрапапиллярной полости, окружающей семепроводящий канал у К. eber-
hardi, прослеживается у близких видов (см. выше); во-вторых, серия вскрытий
К. eberhardi разной степени зрелости показала, что у не вполне оформившихся осо-
бей семепроводящий канал одной стороной плотно спаян со стенкой папиллы, а с другой
имеется продольный тяж, разделяющий интрапапиллярную полость на две камеры —
так, как у взрослых К. tanta. У К. holotricha семепроводящий канал в максимальной
степени обособлен от стенки папиллы, и, наконец, у К. phaeolaema возникают в стенке
папиллы дополнительные синусы.
Подобно тому как папиллярные структуры Edentiella выводятся из Trichia stri-
olata—Tr. danubialis, папиллярные структуры Caucasigena параллельно и незави-
симо выводятся из Kokotschashvilia, так как у К. eberhardi внутренняя стенка па-
пиллы, противолежащая семепроводящему каналу, также несет ясные следы раз-
рыхления.
Имеется, наконец, еще один вариант строения папиллы: у Teberdinia имеется
пара интрапапиллярных полостей и борозда на поверхности пениса, причем одна
5 А. А. Шилейко
66
ВВЕДЕНИЕ
из полостей заходит в лопасть, отчленяемую бороздой (рис. 34, VI). В настоящее-
время затруднительно дать сравнительную оценку этого варианта, так как пока не-
известны близкие типы строения папиллы. Так, можно представить себе возникнове-
ние третьей полости за счет замыкания борозды, тогда получается вариант, соответ-
ствующий европейскому виду Trichia danubialis. Можно предполагать, что с замыка-
нием борозды и одновременной редукцией тяжа, фиксирующего семепрсводящий
канал, может реализоваться вариант, отвечающий Edentiella и Caucasigena-, этот путь,
более вероятен.
Столь большое место деталям строения папиллы пениса здесь уделено в связи
с богатством и разнообразием интрапапиллярных структур, эволюция которых есть,
частное проявление общей эволюции группы. Никакие другие признаки в данном,
случае не дают столь богатого материала для. филогенетических построений. Однако,
подробно останавливаясь на рассмотрении совокупительного аппарата, мы отнюдь-
не склонны абсолютизировать рассмотренные особенности, и предлагаемая филогене-
тическая схема (рис. 35) опирается на комплекс черт, пригодных для практической
таксономии. В диагнозах родов (см. систематическую часть) указаны признаки раз-
ного порядка; здесь целесообразно остановиться на некоторых из них в порядке об-
суждения схемы.
В добавление к сказанному в отношении азиатской группы обратим внимание-
на 4 конхологических типа, представленных в этом районе. Это, во-первых Odon-
totrema diplodon, резко выделяющийся благодаря наличию устьевых зубов; во-вторых,,
почти шаровидная раковина Leucozonella caryodes, L. rubens и др.; в-третьих,
прижатая раковинаL. ferghanica, L. rufispira и др., куда примыкает и L. crassicosta
с ребристой поверхностью раковины; наконец, в-четвертых, чисто европейский
конхологический тип — L. caria.
Явно примитивным видом надо считать О. diplodon. Помимо отмеченных особен-
ностей строения пениальной папиллы заслуживает внимания и такая черта, как вы-
сокая степень обособленности стилофоров. Первым этапом олигомеризации стило-
форов анцестральных форм было исчезновение стрел во внутренних (верхних) стилофо-
рах. Дальнейшая олигомеризация придаточных органов женского отдела выражается
в исчезновении внутренних стилофоров путем их уменьшения; в возникнове-
нии тесного контакта их с нижними стилофорами и вагиной; в погружении их в про-
свет между наружными стилофорами и вагиной и в постепенной редукции (макси-
мальная степень редукции наблюдается у Hygrohelicopsis). Наличие хорошо развитых,
четко обособленных внутренних стилофоров, имеющих примерно тот же размер,
что и наружные, вновь подтверждает относительную примитивность О. diplodon-,
вместе с тем этот вид достаточно специализирован, на что, в частности, указывает
его своеобразная раковина.
Исходным типом по отношению к остальным азиатским видам, очевидно, надо
считать Nanafa cumulata в первую очередь благодаря тому, что папилла пениса
у этого вида лишена каких бы то ни было полостей или борозд; далее надо поместить
L. ferghanica: намечается продольная борозда на поверхности папиллы, а раковина
сохраняет в целом облик Н. cumulata, отличаясь в основном скульптурой; вагина
у этого вида длинная, как и у N. cumulata. Следующим членом этого филетического
ряда является L. caryodes-. борозда на поверхности папиллы гораздо длиннее, а вагина
несколько короче; раковина приобретает шаровидную форму. Далее, L. rubens со-
храняет шаровидную раковину, но намечается некоторая редукция внутренних стило-
форов и возникает их тесная связь с поверхностью вагины. Вместе с тем этот вид при-
обретает ряд частных, но достаточно серьезных отличий: укороченный бич, более
резкое ветвление слизистых желез и околопапиллярный придаток. К этому виду
очень близок L. mesoleuca.
L. rufispira приобретает отличия другого порядка, будучи (независимо от
В. rubens и L. mesoleuca) производным от L. caryodes: длина бича и степень обособлен-
ности стилофоров не меняются. Меняется форма стилофоров, но глубина и характер
папиллярной борозды соответствуют тому, что наблюдается у L. caryodes. Раковина
вновь приобретает уплощенную форму.
Судя прежде всего по раковине, L. retteri и L. angulata, производные от L. ru-
fispira, приобретают в то же время качественно новую, важную в эволюционном от-
ношении черту — окончательное формирование интрапапиллярной полости, но со-
храняют в общих чертах прочие признаки родительской формы. К этой же группе
надо отнести и обитающего на Гиссарском хребте L. caria, раковина которого, подобно
раковинам европейских видов, имеет близкий к исходному облик, характерный для
анцестральных лесных форм. Уместно еще раз напомнить, что изучение внешнего
вида гениталий не дает возможности понять истинную сущность этого вида, и, поль-
зуясь только формальными признаками, пришлось бы вид отнести к европейским
Trichia s. str., что выглядело бы крайне фантастично зоогеографически.
Два вида подрода Harina — уклонившиеся формы, произошедшие от каких-то
высших азиатских Trichiinae, подобных L. retteri-, однако их развитие пошло по иному
пути — по линии резкого уменьшения объема пениса. Характерно, что у обоих видов
НАПРАВЛЕНИЯ РАЗВИТИЯ
67
сохраняется пениальная лакуна, но интрапапиллярная полость вторично отсутствует.
Конхологически эти виды резко отличаются друг, от друга.
Заканчивая рассмотрение азиатской группы Trichiinae, необходимо указать
еще на одну явно примитивную черту, свойственную всем видам. Первичным типом
строения эпифаллуса, по крайней мере в пределах Hygromiidae, надо считать равно-
мерную правильную внутреннюю складчатость, так что в разрезе полость эпифаллуса
имеет форму правильной многолучевой звезды. Затем, преимущественное развитие
получают две соседствующие продольные складки, тогда как прочие группируются
на противоположной стороне эпифаллуса. В результате образуются два направля-
ющих сперматофор «рельса» и ряд мелких складок над ними, в промежутках между
которыми помещаются ряды шипов на поверхности сперматофора. Такое преиму-
щественное развитие двух продольных складок наблюдается у большинства кавказ-
ских и европейских видов. Что касается азиатских форм, то дифференцировка направ-
ляющих валиков внутри эпифаллуса здесь только начинается, разница между двумя
группами складок порой едва заметна: просвет эпифаллуса большей частью сохраняет
первоначальный тип многолучевой, почти правильно!! звезды.
Европейско-кавказская группа родов не представляет собой единой линии раз-
вития, так хорошо выраженной у среднеазиатских форм, но является продуктом
реализации нескольких параллельных и в известной степени независимых путей
при сохранении в той или иной форме определенного комплекса сходных черт. Мы не
находим каких-либо анатомических признаков, позволяющих судить об общих эво-
люционных тенденциях кавказских и европейских родов. Но конхологические от-
личия между европейскими и кавказскими видами выражены ясно. Для европейских
форм в общем характерна довольно хрупкая, небольшая или средних размеров тонко-
стенная, покрытая волосками раковина коричневой окраски со слабой светлой
полосой по периферии. Исключения составляют Plicuteria lubomirskii и Xerocampy-
laea zelebori; первый и в анатомическом отношении довольно резко выклинивается
из общей массы видов, относительно второго мы скажем ниже. Почти все европейские-
представители подсемейства — лесные животные.
Кавказские виды связаны преимущественно с двумя типами биотопов: со ска-
лами (Caucasigena s. str.) либо с открытыми склонами — сухими или влажными.
Чисто лесной кавказский вид только один — Kokotschashvilia holotricha; этот вид,
что очень показательно, конхологически наиболее близок к европейской группе,
отличаясь лишь большими размерами. В соответствии с двумя типами биотопов пред-
ставлены два типа раковин: либо в той или иной степени уплощенные, часто исчер-
ченные или даже ребристые; либо высокие до шаровидных, тонко скульптурирован-
ные. Для первого типа в общем характерно наличие двух спиральных коричневых
полос выше и ниже периферии последнего оборота; вторые обычно более или менее
одноцветные.
Волоски на раковине у кавказских видов первично присутствовали и постоянно
сохраняются у единственного названного лесного вида; Caucasigena rengarteni волос-
ков, как правило, не имеет, но нередко встречаются экземпляры с отдельными, очень
короткими и редкими волосками. Таким образом, утрата волосков происходила
в связи с освоением новых типов биотопов.
Среди европейских видов единственным конхологическим исключением явля-
ется трансильванский вид Xerocampylaea zelebori, обитающий на скалах, подобно
некоторым кавказским группам. Конхологическое сходство заставляло долгое время
относить Caucasigena в качестве отдельной секции к роду (или подроду) Xerocampy-
laea. Однако известно, что скальный или вообще горный тип биотопов накладывает
общий отпечаток на внешний вид раковины. Ранее (Schileyko, 1977) нами было вы-
сказано предположение, что группа Xerocampylaea не имеет непосредственного от-
ношения к Caucasigena-, в дальнейшем мы имели возможность убедиться в справедли-
вости такого предположения, воспользовавшись материалами Зоологического ин-та
Польской АН: X. zelebori ближе всего стоит, как и следовало ожидать, к настоящим
европейским Trichia.
В заключение кратко рассмотрим три рода ксерофильных Trichiinae, представ-
ленных в фауне СССР: Helicopsis, Xeropicta и Helicella. Характерно, ч:о, за исключе-
нием последнего рода, эти группы представлены, во-первых, большим числом видов
(видов рода Helicopsis насчитывается не менее 15, Xeropicta — не менее 10) и, во-
вторых, они не образуют ясных филетических линий, составляя довольно плотное
единство. Тем не менее преемственность между этими родами удалось обнаружить,
причем связующим звеном оказался крымский вид Helicopsis filimargo. Основным
различием между двумя родами является присутствие придатка пениса у Xeropicta
и отсутствие такового у- Helicop sis. Однако в дистальной части пениса у Н. filimargo
обнаружена структура, характерная для пениального придатка Xeropicta-, по суще-
ству это не что иное, как пениальный придаток, находящийся в зачаточном состоянии.
Показательно распространение этих двух родов: Средняя и Южная Европа,
восточное Средиземноморье и Северная Африка, а также Кавказ; в Средней Азии
каждый род представлен одним видом: X. candaharica, имеющим широкое расирост-
5*
j
!
68 ВВЕДЕНИЕ
ранение и заходящим на территорию Афганистана, и Н. likharevi, присутствующим
только на Копетдаге. Присутствие обоих видов здесь обеспечено плейстоценовой
волной миграции с запада. Кроме того, в ряд пунктов Средней Азии интродуцирован
X. derbentina и в историческое время на Копетдаг — X. krynickii.
Другими словами, состав Trichiinae Средней Азии имеет двойственную природу:
основная масса видов — автохтоны, населяющие эту территорию с начала неогена;
кроме того, имеются 4 ксерофильных вида, проникших сюда в плейстоцене и позже.
Оба ксерофильных рода наиболее тесно связаны с Trichia s. str. Однако помимо
признаков, свойственных им как ксерофилам, у них формируется еще одна особен-
ность, свидетельствующая об их дальнейшем эволюционном развитии. У некоторых
видов Helicopsis при основании папиллы пениса в стенке пениальной трубки заклады-
вается кольцевое разрыхление тканей, которое у Xeropicta трансформируется в сплош-
ную полость; эта полость сливается с интрапапиллярной полостью, благодаря чему
пенис приобретает значительно большую подвижность и способен менять длину (ана-
логичное явление наблюдается у некоторых Euomphaliinae).
Остается сказать несколько слов об очень своеобразной группе Helicella s. str.
Это, по всей видимости, олиготипический род, представители которого обладают
парой стилофоров, имеющих в нижней части полости, широко сообщающиеся с по-
лостью вагины, а в верхней части располагается стрела. Напрашивается предполо-
жение, что в противоположность остальным Trichiinae здесь прошла редукция не
верхней, а нижней (наружной) пары, и отмеченные полости есть остатки нижних сти-
лофоров. В любом случае Helicella достаточно изолированы от прочих родов подсе-
мейства, и в настоящее время нет возможности уверенно говорить об исторических
связях. В этом отношении картина может проясниться, когда будет осуществлено
подробное изучение Semifruticicola serbica — единственного вида Hygromiinae auct.,
у которого имеется одна пара симметрично расположенных стилофоров.
Подсемейство Hygromiinae
Как отмечалось, это подсемейство — группа, производная от Trichiinae вначале,
за счет редукции стилофоров на одной стороне вагины, а затем развивавшаяся не-
зависимо. Судя по ряду особенностей, надо признать, что Hygromiinae отделились
очень рано, т. е. в то время, когда вообще шло бурное становление основных таксо-
нов; предположительно эти процессы имели место в середине—конце палеогена.
К особенностям подсемейства относятся: 1) необычайно широкий ареал, охваты-
вающий значительную часть территории Евразии и достигающий Дальнего Востока;
2) большое разнообразие форм и обилие олиготипических таксонов; 3) существование
весьма специализированных (в основном конхологически) групп.
В свете фактов, полученных ранее (Шилейко, 1970, 1972в) и изложенных в си-
стематической части, нетрудно видеть, что подсемейство в целом — довольно компакт-
ная группа, и в большинстве случаев между родами отсутствуют резкие границы.
Например, если взять европейских Urticicola, Monachoides и Perforatella, то отличия
их от европейских же Hygromia бросаются в глаза. Но обнаружение рудиментарного
внутреннего стилофора у алтайско-дальневосточных родов (Chilanodon, Lindholmo-
тпёте) стирает это резкое различие, и все подсемейство вместе с кавказскими пред-
ставителями приобретает довольно четкий анатомический облик.
Однако в пределах, допускаемых анатомическим диагнозом, по отдельным при-
знакам наблюдаются определенные направленные изменения. Причем для' суждений
об эволюционных тенденциях имеется, казалось бы, достаточное число признаков;
но это число резко уменьшается, если проследить изменчивость в более узких преде-
лах, не выходя за рамки рода или группы близких родов: выясняется, что по боль-
шинству признаков имеются параллельные ряды линий изменчивости (Майр, 1968).
Так, бич варьирует от длинного до рудиментарного в пределах рода Hygromia, при-
чем длина его ^уменьшается в ряду Н. (Рyrenaearia) carascalensis—Н. (Н.?) fusca—
Н. (Lozekia) transsylvanica—Н. (Ganula) lanuginosa—Н. (Н.) cinctella—Н. (Riedelia)
limbata (Taylor, 1914; Forcart, 1954; Gittenberger, 1968a; Hudec, 1970; Шилейко,
1972в). Подобный же независимый ряд прослеживается в цепи североазиатских видов
(Шилейко, 1970); укорочение бича происходит у европейских видов с одним стилофо-
ром (там же). Следовательно, рассмотренный признак выпадает из числа филогене-
тически значимых, отражая лишь частные адаптации отдельных таксонов невысо-
кого ранга. .
Изменчивость большей части прочих анатомических признаков подобна измен-
чивости разобранного. Таковы форма резервуара семеприемника, степень извитости
ц длина его протока, сравнительная мощность пениальной мускулатуры и пр.
В результате остаются две группы признаков внутреннего строения, которые
могут служить целям филогенетического толка: степень и характер редукции внут-
реннего стилофора и строение пениальной . папиллы.
Первая группа признаков не требует детальных пояснений: налицо непрерывная
цепь звеньев, причем начальное звено (Hygromia') характеризуется двумя примерно
НАПРАВЛЕНИЯ РАЗВИТИЯ
69
равными по размеру стилофорами, наружный из которых заключает стрелу, а конечное
звено (Urticicola, Monachoides, Perforatella, Pseudotrichia') имеет только один стилофор.
Что касается характера редукции, то этот процесс у европейских Hygromia и у ал-
тайско-дальневосточных родов идет разными путями: если у последних редукция
осуществляется за счет погружения внутреннего стилофора в щель между наружным
и стенкой вагины и возникновения общего соединительнотканного чехла стилофоров,
то у И. transsylvanica редукция идет путем истончепия внутреннего стилофора, ко-
торый совсем не связан с поверхностью вагины (Hudec, 1970, стр. 40, рис. 6), причем
общего чехла стилофоров не возникает.
Что касается пениальной папиллы, то на признаках, с нею связанных, стоит
остановиться еще и потому, что, во-первых, эти признаки не принято принимать
во внимание и, во-вторых, настоящая работа, по существу, первая, где содержится
попытка широкого привлечения их для разработки системы. «. . . the organ of grea-
test taxonomic utility is the penis», — эта мысль Уэбба (Webb, 1947, стр. 134) заклю-
чает в себе глубокий смысл: в строении пениса могут, с одной стороны, отражаться
общие эволюционные тенденции группы, и, с другой стороны, особенности строения
папиллы могут играть важную роль изолирующих механизмов, предотвращающих
интрогрессию между близкими видами (вероятно, в сочетании с этологическими пре-
градами), на чем основано таксономическое значение этого органа.
Что касается Эволюционных тенденций, прослеживающихся по признакам стро-
ения'папиллы, то можно констатировать следующее. Судя по тому, что наблюдается
у наиболее примитивных видов — представителей рода Hygromia, надо считать, что
вначале, как и у исходных Trichiinae, стенка папиллы была заполнена умеренно
и равномерно рыхлой паренхимой, которая затем начала дифференцироваться на
отдельные комплексы. Усиление этой диффренциации приводит к возникновению
и развитию системы лакун, причем на рецентном материале этот процесс четко про-
слеживается. Завершением дифференциации тканей стенок папиллы являются картины,
независимо наблюдаемые у Circassina circassica и Perforatella dibothriorr. семепроводя-
щий канал оказывается подвешенным и растянутым с помощью множества тяжей,
представляющих остатки паренхимы.
Функциональное толкование подобного типа строения папиллы сводится, по-
видимому, к тому, что роль рыхлой ткани в стенке папиллы соответствует роли cor-
pora cavernosa у млекопитающих; кстати, и морфологически эти два типа тканей
весьма сходны.
Интересно, далее, подсемейство в свете представлений о конхологических жиз-
ненных формах. По происхождению рассматриваемая группа — обитатели влажных
третичных лесов Средиземноморья. У всех лесных современных видов имеется рако-
вина типа bidens, название которого говорит само.за себя, или типа incarnata, т. е.
раковина типичного хеликоидного облика, роговой окраски, с размытой светлой по-
лосой по периферии и узким пупком. Для этого типа первично характерно наличие
волосков, а у тех видов, у которых волоски отсутствуют, имеется бугорчатая скульп-
тура, исторически связанная, вероятно, с наличием волосков или чешуек (по всей
видимости, чешуйки представляют собой видоизмененные волоски). Вместе с тем
все виды, покинувшие лесные биотопы и заселившие открытые склоны, скалы, осыпи
и т. д., резко отличаются по раковине от обитателей леса. Таков типичный скаложил
Shileykoia daghestana, политопный кавказский вид Fruttcocampylaea narzanensis (ко-
торый тем не менее вглубь леса никогда не заходит), обитатель открытых склонов
Fr. kobiensis, очень похожий на Caucasigena abchasica и виды родов Hesseola и Саиса-
socressa. Все эти виды характеризуются светлой в общем окраской раковины и двумя
спиральными темными лентами выше и ниже периферии.
Вместе с тем кавказские Circassina не имеют принципиальных отличий от типа
incarnata, но С. bojenae, обитающая на открытых и мало облесенных склонах либо
на опушках, приобретает те же спиральные ленты, сохраняя в целом все конхоло-
гические признаки рода.
Что же касается ксерофильных представителей — видов родов Cernuella и Хе-
rosecta, то их раковины вполне согласуются с- тем, что вообще характерно для всех
ксерофильных Hygromiidae. Анатомически Cernuella — непосредственные производные
Hygromia; женский отдел Xerosecta претерпел значительно большие изменения за
счет возникновения в нижней части вагины хода для папиллы пениса и формирования
кольцевой полости, аналогичной тому, что имеется у Xeropicta из Trichiinae и неко-
торых Euomphaliinae.
Каким же образом соотносятся зоогеографические особенности подсемейства
с ее морфологическими особенностями? Существуют 3 пространственные группировки,
характеризующиеся определенными морфологическими чертами (рис. 36): 1) алтай-
ско-дальневосточная (имеются слабые рудименты внутренних стилофоров), 2) кав-
казская (внутренние стилофоры развиты достаточно хорошо), 3) европейская, одни
члены которой вовсе лишены вйутренних стилофоров, тогда как другие имеют хорошо
развитые внутренние стилофоры, хотя и не заключающие стрел. Кроме того, имеется
70
ВВЕДЕНИЕ
d вид — Pseudotrichia rubiginosa, распространенный в большей части Палеарктики,
.и 1 вид рода Monachoides — М. stuzbergi, обитающий в бассейне Енисея; оба этих вида
имеют по одному стилофору. Условно к тому же роду отнесен вид, описанный как
Zenobiella aculeata; но пока не будет известно внутреннее строение дистальной части
женского отдела полового аппарата, обсуждение систематического положения этого
вида представляется преждевременным.
Необычность ситуации заключается в том, что в Европе одновременно существуют
.две не связанные непосредственно морфологические группировки, причем самое стран-
ное на первый взгляд то, что они объединяются через цепь алтайско-дальневосточ-
ных форм, у которых наблюдается постепенная редукция внутреннего стилофора.
Объяснить этот факт можно одним из двух допущений: либо у отдельных форм в Ев-
Рис. 36. Филогенетические и пространственные взаимоотношения между основ-
ными группами Hygromiinae. I — европейский участок; II — азиатский участок;
III — кавказский участок. С — Circassina; Се — Cernuella; Ch — Chilanodon;
F — Fruticocampylaea; H — Hygromia; L — Lindholmomneme; M — Monachoi-
des; P — Perforatella; Ps — Pseudotrichia; Sh — Shileykoia; U — Urticicola;
X—Xerosecta. (Ориг.).
pone вдруг и очень быстро прошла полная редукция внутреннего стилофора (гене-
тическая склонность к подобной редукции иллюстрируется разобранными случаями
с С. circassica и с парой среднеазиатских видов — см. выше), причем параллельно
с этими формами остались и более примитивные виды с двумя хорошо развитыми
стилофорами, либо надо признать, что группа видов с одним стилофором —• потомки
видов, вторично заселивших Европу с востока.
Анализ распространения отдельных таксонов в сочетании с особенностями орга-
низации заставляет полагать, что оба допущения могут быть справедливыми, и набор
европейских видов с одним стилофором имеет смешанную зоогеографическую природу.
К сожалению, отсутствуют соответствующие палеонтологические материалы.
Так, есть основания думать, что монотипический род Urticicola — автохтонный
европейский род с восточноальпийско-карпатским ареалом: это резко выклинива-
ющийся морфологически род, не имеющий близкородственных форм ни на востоке,
ни на западе. Таким же' образом род Monachoides, видимо, исконно европейская
группа, распространившаяся, однако, далеко на восток—вплоть до Енисея (возможно
и еще восточнее) в лице М. stuzbergi. Кроме того, алтайский «Zenobiella» aculeata
также, по всей вероятности, относится к этому роду. Что касается европейских пред-
ставителей рода, то они распространены следующим образом: М. vicina — карпат-
ский эндемик, М. incamata — среднеевропейский вид; на юго-востоке Франции оби-
тает М. ventouziana,. обнаруженный пока только в одной точке (Forcart, 1946); на Ма-
НАПРАВЛЕНИЯ РАЗВИТИЯ
71
дейре встречается своеобразный, главным образом конхологически, М. undata', и,
наконец, один вид — М. memnonis — имеется даже на западном побережьи Малой
Азии. Как видно, ареал рода разорван, вернее, носит пятнистый характер, и весьма
широк.
Иначе дело обстоит с родами Perforatella и Pseudotrichia, первый из которых имеет
типичный восточноевропейский ареал, а второй, по мнению Цильха и Эккеля (Zilch,
Jaeckel, 1962), — восточноевропейско-сибирский. Однако думается, что правильнее
считать ареал этого рода палеарктическим, так как вид присутствует местами на
Кавказе, в Средней Азии и на Дальнем Востоке. Показательно также, что в пределах
первых двух регионов этот вид встречается либо в высокогорье, либо в непосредст-
венной близости от воды среди густого травостоя или под камнями на очень влажной
почве. Таким образом, конкретные микроусловия, в которых он обитает, не столь
сильно отличаются в зависимости от географической широты и климата.
Если Perforatella dibothrion — типично карпатский вид с узким ареалом, то
Р. bidens местами доходит на восток до Волги. Одновременно с этим раковина послед-
него вида чрезвычайно похожа на раковину алтайского Chilanodon bicallosa. Конечно,
здесь можно подозревать и конхологическую конвергенцию, но такое предположение
маловероятно: для данного подсемейства подобный облик раковины — редкое исклю-
чение, и трудно допустить, чтобы 2 анатомически близких вида имели сходную ра-
ковину только в силу конвергенции. Поэтому нам представляется, что европейские
Perforatella — непосредственные потомки алтайских Chilanodon, которые в свою
очередь имели предками каких-то примитивных европейских же Hygromia.
Что касается Р. rubiginosa, то вероятнее всего этот вид имеет европейское про-
исхождение, поскольку для алтайско-дальневосточных форм характерно наличие
рудиментарного внутреннего стилофора и резко укороченного бича; этот вид наиболее
близок к Monachoides.
Показательно, что нет ни одного представителя подсемейства с настоящим аль-
пийским ареалом: в Европе фактически имеются 2 центра формообразования — Пи-
ренеи (Hygromia) и Карпаты (остальные роды). Среди представителей рода Hygromia
имеются 2 вида, не обладающих строго западноевропейским ареалом: это Н. cinctella,
который вообще широко распространен по северному Средиземноморью, и Н. trans-
sylvanica — единственный карпатский вид рода. Знаменательно, что именно этот
последний вид характеризуется наибольшей степенью редукции внутреннего стило-
фора (Hudec, 1970); характер этой редукции также своеобразен (см. выше).
Резюмируя сказанное, мы полагаем, что европейские Hygromiinae имеют сме-
шанную зоогеографическую природу: род Hygromia — автохтонная группа древних
лесов северо-западного Средиземноморья; роды Urticicola и Monachoides — непо-
средственные производные от каких-то Hygromia, проникших на Карпаты или в со-
седние области; род Perforatella — потомок североазиатского рода Chilanodon.
Кавказская группа видов целиком представляет собой эндемичный комплекс,
-сформировавшийся на месте за счет анцестральных форм типа Hygromia, проникших
из Средиземноморья через Эгейскую сушу с верхнего миоцена по верхний плиоцен.
В равной степени это относится и к ксерофильным видам рода Xerosecta', представи-
тели рода Cernuella появились на Кавказе, вероятно, несколько позже; наконец,
встречающийся в Крыму средиземноморский вид Cernuella virgata появился здесь
уже в историческое время, скорее всего, завезен.
Подсемейство Euomphaliinae
Данный таксон имеет строго средиземноморско-европейский ареал с Кавказом,
Крымом и Восточной Африкой (Verdcourt, 1969), и все особенности группы сформи-
ровались именно на этой территории; нет оснований подозревать миграций предста-
вителей подсемейства за пределы современного ареала в прошлом.
, Основным результатом изучения видов Euomphalia auct. и Monacha auct. явилось
убеждение, что эти группы — родственные комплексы, образующие вместе таксон
подсемейственного ранга и возникшие в результате перестройки вначале придатков
женского отдела, а затем совокупительного аппарата каких-то древних Trichiinae.
Это убеждение базируется на том, что, во-первых, единственное по существу первич-
ное различие между названными группами сводится к разному числу вагинальных
придатков (видоизмененных, но не рудиментарных!) стилофоров: если у Euomphalia
auct. имеется пара таких придатков, то у представителей Monacha auct. придаток
лишь один или отсутствует; во-вторых, о близости этих групп говорит их ареал.
Что касается строения вагинальных придатков, то оно у представителей двух
групп ничем существенным не отличается. Придаток всегда состоит из базальной
части — ствола, с очень узким извилистым протоком и мощными мускулизованными
стенками, и концевой части, которая обычно заканчивается более или менее ясно
выраженным вздутием. Внутри концевой части располагается то или иное количество
72
ВВЕДЕНИЕ
продольных валиков. Гистологические препараты с поперечными срезами вагиналь-
ных придатков дают поразительно сходные картины. В данном случае существенно,
что стенки придатков в верхней части, близ вздутия, мускулизованы много слабее,
чем в нижней части, и мышечные элементы, входящие в их состав, имеют преимуще-
ственно аксиальное направление, а не кольцевое, что характерно для стенок насто-
ящих стилофоров. Это позволяет постулировать субституцию функций, и, следова-
тельно, существенное видоизменение стилофоров.
Числу вагинальных придатков в данном случае нецелесообразно придавать глу-
бокий филогенетический смысл, поскольку для семейства в целом вообще характерна
тенденция к утрате отдельных элементов полового аппарата (полового ретрактора,
слизистых желез, стилофоров или вагинальных придатков). Любопытно, что все
известные случаи касаются только кавказских и среднеазиатских групп, и только
представителей Hygromiidae (если не считать очень небольшого числа южноазиатских
Bradybaenidae).
Более того, имеются виды, у которых в пределах одной популяции обнаружи-
ваются особи с едва развитыми рудиментами вагинального придатка, носящими ата-
вистический характер (это наблюдается только в пределах рода Monacha).
Если настоящий стилофор призван стимулировать партнера механическим путем,
то вагинальный придаток воздействует секретом железистого эпителия, выстилающего
его стенки. В этой связи надо обратить внимание на то, что иногда (Stenomphalia)
придатки способны сильно выворачиваться при совокуплении. У Euomphalia ва-
гинальные придатки не выворачиваются, но секрет их стекает в нижнюю часть вагины,
увлажняет ее стенки и играет, помимо прочего, определенную роль в создании ус-
ловий влажности, необходимых для спаривания. Показательно, что придатки выво-
рачиваются у тех видов, которые обитают в более ксеротермных условиях. Что ка-
сается членов рода Monacha s. lat.. то они решают проблему регуляции водного режима
при спаривании иным путем, при котором роль вагинальных придатков менее зна-
чительна (см. ниже). Думается, что именно в связи с этим представители данной
группы имеют тенденцию к утрате одного или даже обоих вагинальных придатков.
Показательна и своеобразна эволюция собственно совокупительного аппарата.
В Пределах подсемейства первичен, несомненно, тот характер оформления дисталь-
ной части мужского отдела, который наблюдается у кавказского Karabaghia bitube-
rosa и который связывает между собой Euomphalia s. lat. и Monacha s. lat.: имеется
простой чехол пениса с тонкими, но плотными и нерасслаивающимися стенками.
Папилла толстостенная, заключающая узкий семепроводящий канал, при совокуп-
лении полностью выдвигается, существенно не меняя формы, т. е. так, как у Tri-
chiinae.
Далее реализуются два возможных варианта: расслаивание стенки пениального
чехла на собственно чехол пениса и чехол пениальной папиллы (Euomphalia) или
закладка кольцевого разрыхления тканей в основании папиллы (Stenomphalia), ко-
торое затем превращается в кольцевую интрапапиллярную полость (Monacha).
Механизм действия папиллы представляется, в свете изложенного, следующим
образом (рис. 37). У Euomphalia чехол папиллы пениса позволяет этому органу вы-
двигаться на определенную часть длины, причем та часть папиллы, которая не погру-
жается в вагину партнера, оказывается, в случае максимального выдвижения, по-
крытой дополнительной защитой от иссушающего действия наружной среды в виде
чехла папиллы. На рис. 37 буквой А помечено место прикрепления чехла папиллы
к чехлу пениса, а буквой В — место крепления чехла папиллы к ее поверхности.
Верхнее изображение соответствует нормальному положению папиллы; далее вниз —
три стадии ее выдвижения, причем видно смещение точки Б относительно точки А,
которая в свою очередь смещается незначительно; в результате на нижнем изображе-
нии проксимальная часть вывернутой папиллы оказывается покрытой чехлом папиллы.
Наиболее дистальное крепление чехла папиллы наблюдается у Oscarboettgeria еи-
ages: в результате папилла способна лишь немного выдвинуться, а при увеличива-
ющемся ' давлении со стороны эпифаллуса начинает выворачиваться. Подобный
тип строения папиллы обеспечивает наименьшую длину вывернутых участков сово-
купительного аппарата и наибольшую скорость обратного втягивания. Вероятно,
это можно поставить в связь с тем, что этот вид населяет наиболее ксеротермные ус-
ловия по сравнению с любым другим видом группы, чьи вагинальные придатки не-
способны выворачиваться.
Относительно простое устройство папиллы у тех форм, придатки которых вы-
ворачиваются (Stenomphalia), связано с интенсивным развитием последних, роль
которых, в частности, — регуляция водного режима при спаривании.
Что касается видов рода Monacha, то при вскрытии большого материала обнару-
живается, что каждый экземпляр обыкновенно имеет свою, отличную от других форму
папиллы. Такая особенность объясняется тем, что очертания папиллы зависят от
функционального состояния органа в данный момент. В самом деле: интрапапилляр-
ная полость (которая представляет собой дериват кольцевого разрыхления тканей
НАПРАВЛЕНИЯ РАЗВИТИЯ
73
дистальной части эпифаллуса у Stenomphalia) заходит, с одной стороны, в стенку
папиллы и, с другой стороны, в стенку эпифаллуса. При втягивании последнего па-
пилла, естественно, будет сокращаться в длину, а при выдвижении — удлиняться.
Не случайно форма выходного отверстия папиллы вполне соответствует форме про-
света эпифаллуса. Допустимо говорить не только об отсутствии папиллы как таковой,
Рис. 37. Механизм действия папиллы пениса у Euomphalia.
А — место прикрепления чехла папиллы пениса к чехлу
пениса; Б — место прикрепления чехла папиллы пениса
к поверхности папиллы. (Ориг.).
но о наличии кольцевой полости в дистальном участке эпифаллуса, за счет изменения
формы которой и может образовываться папилла как временный орган.
В дополнение к тому, что излагалось выше по поводу филогенетической значи-
мости отсутствия или наличия перехлеста ретрактора правого омматофора с половым
ретрактором, обратим внимание, что по этому признаку Euomphalia s. lat. ныне от-
носят к Hygromiinae auct., a Monacha s. lat. — к Helicellinae auct. Но из 10 вскрытых
экземпляров E. strigella из одной популяции (с. Архангельское Московской обл.)
у 2 особей мы обнаружили нормальную картину — перехлест глазного ретрактора
с пенисом, а у 8 этот перехлест касался лишь ретракторов, тогда как гениталии были
свободны от перехлеста. Далее — одни особи Е. aristata имеют половой ретрактор,
другие лишены его; единственный изученный экземпляр Е. appeliana также оказался
74
ВВЕДЕНИЕ
без полового ретрактора. То же самое происходит в роде Monacha: у тех видов, ко-
торые имеют ретрактор пениса, перехлест ретракторов имеется, хотя глазной ретрак-
тор с гениталиями не перекрещивается. Но М. carthusiana, например, полового ре-
трактора не имеет, и здесь говорить о том, где проходит ретрактор омматофора,
не приходится. Относящийся к этому же подсемейству африканский вид Lejeania
isseli лишен перехлеста ретракторов (Verdcourt, 1969).
Эти обстоятельства также свидетельствуют о том, что характер расположения ре-
тракторов должен быть исключен из числа филогенетически значимых признаков,
хотя эволюционное значение утери перехлеста в перспективном преломлении велико.
Чрезвычайно своеобразен, как морфологически, так и географически, род Нез-
seola, в состав которого входят 2 вида; один из них обитает в Закавказье,1 другой —
на Копетдаге. Это несомненно наиболее уклонившаяся ветвь подсемейства. Вагиналь-
ные придатки здесь имеют совершенно иное строение, нежели у всех прочих пред-
Рис. 38. Филогенетические и пространственные взаимоотношения между
основными группами Euomphaliinae. I — европейский участок; II — кав-
казский участок; III — азпатский участок. В — Batumica; Е — Euomphalia;
Н — Hesseola; К — Karabaghia; М — Monacha; О — Oscarboettgeria; Р —
Platytheba; S — Stenomphalia. (Ориг.).
ставителей подсемейства: они имеют вид пары удлиненно-веретеновидных тел с сильно
мускулизованными стенками, которые пронизывают покровы вагины и почти сопри-
касаются в ее полости дистальными концами. В то же время строение собственно
совокупительного аппарата не имеет существенных отличий от того, что наблюдается
у представителей родов Euomphalia и Oscarboettgeria. Резкие конхологическйе от-
личия между этими двумя родами заставляют полагать, что наличие Н. transcaspia
на Копетдаге свидетельствует не о проникновении этого вида с более западных тер-
риторий, как это имело место, например, с Xeropicta candaharica в сравнительно не-
давнее время, но о том, что ареал рода в конце, вероятно, третичного времени занимал
значительно большую площадь; собственно И. transcaspia несомненно автохтонный
для Копетдага вид.
Филогенетическая схема подсемейства представлена на рис. 38.
В результате приходим к выводу, что наиболее заметную роль в формировании
представлений об эволюции группы играют тонкие детали строения мужского отдела.
Особенности раковины и придатков женского отдела играют подчиненную роль;
то же касается полового ретрактора. Особенно это относится к роду Monacha: в боль-
шинстве других групп таких различий, которые наблюдаются в строении женского
отдела у разных видов этой группы, было бы достаточно, чтобы выделенные еще Гессе
(Hesse, 1914) подроды считать четкими самостоятельными родами. Но в данном слу-
чае важно обратить внимание на непостоянство, зыбкость, слабую закрепленность
этих признаков в генотипе (например, см. описание М. subcarthusiana). В самом деле,
первичным вариантом строения надо считать наличие ретрактора пениса, слизистых
желез и одного вагинального придатка. Вместе с тем в пределах этого рода и близких
родов (выделенных, кстати сказать, на основе признаков раковины) можно обнару-
1 Не исключено, что в Закавказье имеется еще один вид.
НАПРАВЛЕНИЯ РАЗВИТИЯ
75
жить любую комбинацию из перечисленных признаков: все три признака налицо
(Paratheba); отсутствует только ретрактор пениса (Monacha s. str.); отсутствуют только
слизистые железы (Cyrnothcba)-, отсутствует только вагинальный придаток (Meta-
theba); отсутствуют все придатки женского отдела или находятся в явно рудиментар-
ном состоянии (Boemica); все придатки женского отдела всегда полностью отсутствуют
(Ashfordia).
Подсемейства Archaicinae и Paedhoplitinae
Выше отмечалось, что та легкость, с которой Trichiinae утрачивают стилофоры
или видоизменяют их, наводит на мысль, что эти процессы могли происходить неодно-
кратно, и, следовательно, есть основания подозревать полифилию как Hygromiinae,
так и Euomphaliinae. Такое подозрение усугубляется еще и тем, что при существу-
ющих взглядах на систему семейства всех Hygromiinae и всех Euomphaliinae следо-
вало бы выводить из их рецентных азиатских представителей, что выглядело бы весьма
странным. Прежде чем подробнее обсудить этот вопрос, отметим три обстоятельства.
1. Все среднеазиатские Hygromiidae наиболее, повторяем, примитивные пред-
ставители своих таксонов.
2. Наблюдаемые случаи резких различий в числе и форме стилофоров при со-
хранении всех прочих признаков имеют место только в Средней Азии. т. е. касаются
именно примитивных слабоспециализированных форм, и ничего подобного не отме-
чается в западной Палеарктике, за исключением редукции придатков женского от-
дела у специализированных Circassina, но там налицо географическая разобщенность
всех четырех форм.
3. Неазиатские группы Hygromiinae и Euomphaliinae дают достаточно логичные
филогенетические схемы, не имеющие видимых слабых мест, т. е. эти схемы не вступают
в противоречия ни с морфологией, ни с зоогеографией, ни с палеогеографическими
материалами.
Подсемейство Trichiinae, безусловно, монофилетично, но необходимо напомнить,
что если европейские и кавказские группы достаточно высоко специализированы,
то в Средней Азии до наших дней «законсервировались» примитивные представители,
близкие, по всей видимости, к исходным формам, мигрировавшим в Среднюю Азию
из Средиземноморья в эоцене—олигоцене; климат Средней Азии не претерпевал рез-
ких и внезапных изменений, но наблюдалось очень постепенное и плавное нарастание
аридности. Благодаря этому те формы, которые успевали адаптироваться к нараста-
ющей ксерофилизации, оставались в мало измененном состоянии, не имея стимула
к бурной эволюционной дивергенции.
В западной части Палеарктики имело место образование Hygromiinae и Euom-
phaliinae не позднее начала миоцена; в Средней Азии аналогичные процессы прошли
теми же путями много позже, вероятнее всего, не раньше конца плиоцена, возможно,
в ’ течение плейстоцена, и дали те таксоны, которые именовались среднеазиатскими
«Hygromiinae» и среднеазиатскими «Euomphaliinae». Отзвуками этих процессов мы
считаем разобранные выше случаи с Nanaja cumulata—Archaica heptapotamica и Le-
ucozonella ferghanica—Angiomphalia coplosa.
В свете изложенного названия «Hygromiinae» и «Euomphaliinae» должны отно-
ситься только к западнопалеарктическим таксонам, а также к алтайско-дальневос-
точным, непосредственно связанным с европейскими. В Средней Азии же имеются 2
самостоятельных подсемейства, аналогичных (по способам возникновения) назван-
ным, а именно Archaicinae и Paedhoplitinae.
Что касается путей эволюции групп, то они еще слабо проявились в связи с не-
давним происхождением: папилла пениса, как и у наиболее примитивных Trichiinae,
таких, например, как Nanaja cumulata и Leucozonella ferghanica, представляет собой
толстостенную трубку; судя по тому, что наблюдается у наиболее специализирован-
ного вида из Archaicinae — Archaica labianix, эволюция пениальной папиллы имеет
тенденцию развиваться не путем разрыхления тканей стенок, а путем дифференци-
ровки на отдел, содержащий семепроводящий канал, и на прилегающий к нему сое-
динительнотканно-мускульный отдел; при совокуплении, таким образом, эрекция
осуществляется не за счет всех стенок, но за счет выпрямления и уплотнения этого
соединительнотканно-мускульного отдела, на поверхности которого располагается
собственно семепроводящий канал.
У более богатой видами группы Paedhoplitinae меняется от вида к виду лишь
внешняя форма папиллы и остается постоянным ее строение: имеется узкий, четко
выраженный семепроводящий канал и более или менее мощные стенки, заполненные
паренхимой. Эрекция папиллы осуществляется за счет равномерного заполнения
пустот во всех стенках жидкостью, что в свою очередь связано с механизмами на кле-
точном уровне (Jong-Brink, 1969).
В стенках пениального чехла у видов этого подсемейства располагается выражен-
ная в разной степени, изредка пигментированная жзлеза, которая иногда может быть
парной (Angiomphalia exasperata); в этой связи надо сказать, что подобная железа
76 ВВЕДЕНИЕ
начинает формироваться у многих Trichiinae, в том числе и ксерофильных, но здесь
наличие ее улавливается только гистологически,
В заключение необходимо остановиться на некоторых конхологических особен-
ностях. Archaicinae имеют малоспециализированную раковину; наиболее своеобразна
раковина A. labianix, чрезвычайно напоминающая Paedhoplita buamica. Что же ка-
сается Paedhoplitinae, то здесь имеются 2 конхологических типа. Первый тип — мало-
выразительная раковина традиционного хеликоидного облика (Angiomphalia s. str.).
. Второй тип — раковина с угловатой периферией, мощной губой и системой зубов
на париетальной стенке устья. Необходимо напомнить, что подобные образования
не характерны для надсемейства в целом, и кроме Paedhoplita париетальная пластинка
имеется лишь у Isognomostoma personatum (Helicidae, Ariantinae) и немногих Cama-
enidae.
Имеется возможность проследить формирование париетальных зубов: уплощен-
ная раковина без зубов — A ngiomphalia copiosa, чечевицеобразная раковина, также
без зубов (но с массивной губой) — А. lentina, раковина с угловатой периферией и
с первыми признаками париетальных зубов — Р aedhoplita buamica, тот же тип ра-
ковины, но со сформированными париетальными зубами — Р. lindholmt, Р. kirgi-
sensis. Что касается Р. laminata, то этот вид, скорее всего, имеет общий корень
с Р. buamica, поскольку париетальные образования у него представлены прямой
пластинкой, а не сочетанием бугорков и изогнутой пластинки.
Подсемейство Metafruticicolinae
Группа диагносцируется в основном негативными признаками — отсутствием
каких бы то ни было придатков как мужского (кроме бича), так и женского отделов.
Формально это сближает представителей подсемейства с Euomphaliinae, так как для
некоторых из них характерна тенденция к утрате придатков. Однако, используя-
помимо традиционных также признаки внутреннего строения полового аппарата,
удалось показать, что наблюдаемая морфология гениталий в пределах подсемейства —
первичное состояние, чем и обосновывается статус группы (Шилейко, 1972г, 1972д)..
Обитающий на о-ве Крит Metafrutictcola pellita сохраняет примитивный тип строения
папиллы пениса с незамкнутым семепроводящим каналом, имеющим вид борозды, —
чрезвычайно архаичная черта, редко встречающаяся в пределах надсемейства во-
обще. Бйч у этого вида длинный, резервуар семеприемника достаточно четко очер-
ченный, валики внутри эпифаллуса слабо дифференцированы. Внутренняя поверх-
ность протока семеприемника покрыта неправильно расположенными продольными
складками, между которыми имеются многочисленные аностомозы. При совокуплении
пениальная папилла распрямляется и выдвигается.
Там же, на о-ве Крит, обитает Cretigena sublecta, у которой пайилла пениса не только
замкнута, но и в достаточной мере специализирована: имеется кольцевая интрапа-
пиллярная полость; дистальный конец папиллы обособлен и резко сужен. Бич не-
сколько короче, дифференцировка валиков внутри эпифаллуса на две группы наме-
чается явственно. Внешне семеприемник не отличается от семеприемника предыду-
щего вида, но существенно то, что продольные складки внутри протока семеприемника
разбиты на ряды бугорков, местами нечетких. Таким образом, этот вид — прогрес-
сивная ступень в сравнении с предыдущим.
Кавказские виды, относящиеся к роду Caucasocressa, — результат дальнейшего
эволюционного развития подсемейства. С. pratensis имеет приблизительно ту же аб-
солютную длину бича, но дистальная часть мужского отдела заметно укорочена.
Границы резервуара семеприемника с его протоком расплываются, так что пустой
семеприемник наружно почти не выражен; когда же он наполнен, то резервуар плавно,
без резкой границы, переходит в проток. Структура внутренних стенок протока се-
меприемника несет ясные черты специализации: продольные валики правильно гоф-
рированы и отделены друг от друга. Дифференцировка продольных валиков внутри
эпифаллуса резко усиливается: на разрезе видны два более крупных соседствующих
валика, а над ними — несколько рядов более мелких. Наиболее яркие черты морфо-
логического прогресса касаются папиллы пениса и заключаются в том, что кольцевая
интрапапиллярная полость вдается в глубь стенок эпифаллуса; в результате папилла
становится очень короткой или, точнее, теряет определенную длину: при совокупле-
нии этот орган лишь слегка выдвигается, а затем начинает выворачиваться (аналогич-
ную картину мы уже не раз видели в разных группах Hygromiidae).
Другими словами, выходное отверстие эпифаллуса — одновременно и выходное-
отверстие вообще мужского отдела. В определенном смысле здесь можно говорить
о редукции папиллы: если семепроводящий канал втянется достаточно сильно, па-
пилла исчезает. Просвет семепроводящего канала, как следует из сказанного, пред-
ставляет собой дистальный участок протока эпифаллуса и в связи с этим теряет мор-
фологическое своеобразие.
С. joannis имеет очень сходное строение и отличается в основном укороченным
бичом. Правда, надо иметь в виду, что длина этого органа может значительно меняться
НАПРАВЛЕНИЯ РАЗВИТИЯ 77
в пределах одного вида у многих представителей разных групп Helicoidea (это пока-
зано, в частности, очень ярко для Theba pisana — Backhuys, 1972), причем измене-
ние длины бича может носить характер клинальной изменчивости.
Предполагаемый филетический ряд pellita—sublecta—pratensis—joannis подтверж-
дается конхологически. Имея в виду морфологию раковины гипотетических исход-
ных Trichiinae, общую для семейства в целом, обратим внимание, что у Metafruti-
cicola pellita серая раковина, покрытая волосками, которые могут с возрастом опадать;
у очень близкого вида М. freytagi волоски, кстати, остаются пожизненно. Раковина
Cretigena sublecta светло-серая или белая, без волосков, с 2—3 спиральными корич-
невыми лентами (эти ленты есть и у двух предыдущих видов, но там они очень слабо
выражены). Наконец, оба кавказских вида имеют раковину, относящуюся к распро-
страненному конхологическому типу, — дво ясные спиральные темно-коричневые
ленты на почти белом фоне. Этот конхологический тип свойствен видам, обитающим
в условиях теплого климата на открытых, но не сухих местах (заболоченные участки,
долины горных рек, мокрые скалы, луга и склоны с сочной травой и т. п.)..
Показательна оценка этой филетической линии и с пространственной стороны.
Наиболее примитивные виды обитают на островах Греческого архипелага, и есть
все основания думать, что именно северная часть Эгейской суши была местом воз-
никновения подсемейства в течение, вероятно, эоцена—олигоцена (к сожалению,
ископаемые материалы отсутствуют). Позже, в миоцене, представители подсемейства
проникли в западное Закавказье, но в продвижении на север были остановлены Глав-
ным Кавказским хребтом: эти виды теплолюбивы и не смогли адаптироваться
к климату высокогорья.
В настоящее время подсемейство имеет компактный ареал, ограниченный восточ-
ным Средиземноморьем и западным побережьем Малой Азии и западным Закавказьем.
В частности, и с зоогеографической точки зрения выглядит неправомерным отнесе-
ние Circassina circassica simpla к Metafruticicolinae, как предлагает Гудец (Hudec,
1972d). Кроме того, детальное исследование этого подвида убеждает в принадлеж-
ности его к Hygromiinae.
Уточнение таксономической структуры подсемейства можно будет сделать после
изучения на соответствующем уровне других видов, так как пока известна лишь
внешняя морфология их гениталий (Hesse, 1931; Fuchs, Kaufel, 1934, 1936).
СЕМЕЙСТВО HELICIDAE
Широко известные Helicidae — яркая группа, имеющая компактный ареал,
четко очерченная анатомически и весьма разнообразная конхологически.
История развития группы вырисовывается, в частности, из сопоставления Ari-
antinae и Helicinae. Семейство, именно Ariantinae, известно с эоцена (вымершие роды
Galactochilus — верхний эоцен—нижний миоцен, Eurystrophe — верхний эоцен—
нижний олигоцен, Klikla — ? средний эоцен—средний плиоцен). Самые ранние на-
ходки/Те/гсгиае— не раньше среднего миоцена (С. Boettger, Wenz, 1921; Wenz, Zilch,
1959—1960), причем многие миоценовые группы существуют до сих пор (Theba, Du-
potetia, Massylaea, Pseudotachea, Hemicycla, Maurohelix, Caucasotachea, Helix). По-
казателен следующий подсчет. Если взять все известные таксоны выше вида, име-
ющие формальную валидность, и, не вдаваясь пока в рассуждения о правомочности
их выделения (большое число таксонов свидетельствует как минимум о конхологи-
ческом разнообразии), сравнить число рецентных таксонов с числом вымерших от-
дельно по подсемействам, получим такие результаты: Ariantinae — рецентных 21,
вымерших 12; Helicinae соответственно 49 и 6, т. е. у первых число рецентных так-
сонов вдвое превышает число вымерших, а у вторых — в 8 раз. Другими словами,
можно говорить о расцвете Helicinae сравнительно с Ariantinae в настоящее время.
Не менее показательно в этом плане и географическое распространение этих
двух групп, как видно из приводимой табл. 2.
Следовательно, распространение семейства почти строго средиземноморское,
причем ареал Ariantinae укладывается целиком внутри ареала Helicinae, занимая
преимущественно Южную Европу и не заходя на Пиренейский п-ов, за исключением
рода Elona. Но представители последнего встречаются только на склонах Пиренеев
и не спускаются ниже предгорий.
Наибольшее разнообразие Helicinae наблюдается в двух регионах: во-первых, как
и Ariantinae, в Южной и Юго-Восточной Европе и, во-вторых — на территории горной
системы Северо-Западной Африки: хребты Большой и Средний Атлас и Антиатлас,
причем наиболее своеобразные конхологические типы свойственны именно этим рай-
онам. Слабая изученность моллюсков Марокко — очень серьезное препятствие для
анализа организации высших Helicoidea, поскольку наиболее архаичные формы оби-
тают, по всей вероятности, именно на западе Северной Африки.
Заслуживает внимания тот факт, что в Средней и Северной Европе отсутствуют
^эндемичные Helicinae и вообще это подсемейство здесь представлено тремя родами,
78
ВВЕДЕНИЕ
Таблица 2
Район Число таксонов (родов и подродов)
Ariantinae Helicinae
всего эндемичных всего эндемичных
Тунис 4
Алжир — — 13 4
Марокко — — 16 8
Северная Африка в целом — — 21 16
Канарские о-ва — — 3(1) 1
Пиренейский п-ов .... Южная и Юго-Восточная — — 10 4
Европа 19 (3) 15 (3) 28 (5) 12 (1)
Средняя и Северная Европа 18 (.9) 12 (9) 4 (1) —
Малая Азия — — 10 4
Кавказ — — 4(1) 2(1)
Примечание. Цифры
в ископаемом состоянии.
в скобках — число таксонов, известных только
имеющими достаточно широкое распространение; в то же время Ariantinae представ-
лены здесь гораздо богаче, причем имеется 12 эндемичных групп, 9 из которых вымерли.
Показателен также удельный вес монотипических групп: у Ariantinae они со-
ставляют 35% , у Helicinae 32% всех таксонов, т. е. можно говорить о заметной дискрет-
ности и, следовательно, о достаточной древности семейства.
Наконец, необходимо особо подчеркнуть, что решительно все находки ископа-
емых представителей семейства сделаны в пределах современного ареала, т. е. все
частные миграции и перемещения происходили внутри Средиземноморья; есть все
основания думать, что заселение Малой Азии и Кавказа произошло не ранее миоцена
уже представителями Helicinae; что же касается Ariantinae, то эта группа, вероятно,
никогда восточнее Балкан и Карпат не продвигалась.
Детальное рассмотрение путей эволюции семейства в данной книге невозможно,
так как на территории СССР обитает незначительная часть представителей. Наиболее
важные в этом плане группы, в первую очередь южноевропейские и африканские,
изучено слабо. В связи с этим приводимый ниже морфологический анализ не претен-
дует на полноту и касается преимущественно видов фауны СССР.
При значительном разнообразии раковин Helicidae отличаются' большим посто-
янством черт внутреннего строения. Даже столь неустойчивый в ряде других семейств
признак, как число слизистых желез, здесь жестко закреплен — слизистых желез,
всегда две, ветвящихся в разной степени — от вообще нерасщепленных до много-
вётвистых, кустовидных. Стилофор всегда один и всегда хорошо развит. Первично
имеется длинный бич и длинный дйвертикул семеприемника, диаметр которого пре-
вышает диаметр собственно протока. Дивертикул имеет толстые стенки и связан со
спермовидуктом перепонкой, в которой располагаются сосуды.
Семейство единодушно разделяется на два подсемейства — Ariantinae и Helicinae;.
для первых характерны простые или двуветвистые слизистые железы и наличие пе-
ниального бугорка (Knipper, 1939); для вторых — многоветвистые железы и отсут-
ствие пениального бугорка.
Такое разделение семейства оправдано, но не на основании указанных призна-
ков. Во-первых, для всего семейства, в том числе и для Helicinae, первично присут-
ствие пениального, бугорка. Во-вторых, существуют Helicinae (например, Levantina)
с двумя-тремя простыми слизистыми железами. В-третьих, в пределах семейства
вообще можно отыскать ряд форм с разной степенью ветвления слизистых желез.
В самом деле, если для многих Ariantinae (например, Faustina') характерно ветвление'
каждой слизистой железы один раз (т. е. число концевых ветвей каждой железы 2),
то сТойт только себе представить еще одно дихотомическое ветвление, в результате
которого общее число ветвей каждой железы окажется равным 4, мы получим то,.
что имеется у видов подрода Сераеа s. str. (Helicinae); если будет еще одна точка вет-
вления, общее число ветвей каждой железы достигнет 8, получим вариант, наблюда-
емый у подрода Octadenia (показательно, кстати, что существование этих подродов
подтверждается и кариологически (Rainer, 1967), и биохимически (Kothbauer, Schnit-
zler, 1972)).
Поэтому ясно, что степень ветвления слизистых желез не может быть исполь-
зована в таксономических целях на уровне выше рода.
НАПРАВЛЕНИЯ РАЗВИТИЯ
79
Признание существования подсемейств Ariantinae и Helicinae диктуется тем,
что в лице этих двух групп мы имеем два типа организации совокупительного ап-
парата. Пенис у Ariantinae, при всем его разнообразии (Шилейко, 19716), построен
в общем по той же схеме, что и у Hygromiidae, т. е. папилла заключает семепроводя-
щий канал, через который при совокуплении поступает в половые пути партнера
сперматофор в виде длинной нити.
Значительная перестройка этой схемы, наблюдаемая у Helicinae, позволила
усовершенствовать механизм передачи сперматофора (рис. 39, I). Внутри чехла пениса,
на границе с клоакой, имеется, как упоминалось, пениальный бугорок. Пенис со-
Рис. 39. Схема действия пениса Сераеа. I — состояние
покоя; II — в пространство между папиллами по-
ступает сперматофор, оболочки начинают выравни-
ваться; III — комок из сперматофора сформирован,
все оболочки расправлены. ДП — дистальная папилла;
ПБ — пениальный бугорок; ПИ — проксимальная па-
пилла; С — сперматофор. (По Шилейко, 1972г, с из-
менениями) .
стоит из двух чехлов и двух пениальных папилл. Наружный чехол охватывает весь
пенис; внутренний образует своими складками дистальную (ложную) папиллу,
а затем — многочисленные широкие складки и одевает проксимальную (истинную)
папиллу (рис. 39, 11).
Механизм работы мужского отдела выглядит следующим образом. В покое между
пениальным бугорком и внутренними стенками чехла пениса имеется небольшой
щелевидный просвет. Далее располагается ложная папилла, отделяющая дистальную
часть полости внутреннего' чехла от проксимальной, стенки которой широко гофри-
рованы. На дне проксимальной части располагается истинная папилла, поверхность
которой также покрыта многочисленными кольцевыми складками. Семепроводящий
канал внутри папиллы образует высокую продольную складку (нечто вроде тифло-
золя в кишечнике), за счет выравнивания которой диаметр канала способен зна-
чительно увеличиваться.
При поступлении сперматофора через истинную папиллу в просвет проксимальной
части пениса сперматофор многократно скручивается и свивается в этом отделе клуб-
ком, целиком заполняя его (рис. 39, III). В результате проксимальная часть пениса
увеличивается в объеме за счет выравнивания (г. е. исчезновения) складок, причем
80
ВВЕДЕНИЕ
«дно» и «потолок» образовавшейся камеры возникли за счет растяжения стенок обеих
папилл, которые, следовательно, в этом случае как таковые исчезают.
Одновременно пениальный бугорок наполняется жидкостью (он заполнен очень
рыхлой паренхимой), раздувается и запирает собой выходное отверстие пениса. Именно
в этом и заключается функция пениального бугорка, а не в том, что он действует как
раздражающее тельце, на то есть стилофор со стрелой. При совокуплении дистальная
часть чехла пениса выворачивается вместе с пениальным бугорком.
Наличие этого бугорка у Ariantinae заставляет подозревать, что частичное скру-
чивание сперматофора имеет место и в этом подсемействе; но нам не доводилось видеть
здесь сперматофор иначе, как уже в семеприемнике.
Описанный механизм достоверно показан для видов рода Сераеа-, по строе-
нию папиллы, он в той или иной мере характерен и для многих других Helicinae.
Эта оговорка сделана в связи с тем, что, вскрывая многочисленные экземпляры раз-
пых видов Helicinae, мы не раз находили внутри дивертикула семеприемника передний
конец сперматофора, т. е. последний при спаривании направляется передним концом
вначале в дивертикул, а затем его «хвост» заворачивается вверх к резервуару. Пилс-
бри (Pilsbry, 1939) отмечал аналогичный факт в отношении Helminthoglypta.
Подобные наблюдения позволяют сделать два вывода. Во-первых, накопление
сперматофора в пространстве между папиллами не обязательно влечет за собой пе-
редачу его в виде компактного комка. По-видимому, при совокуплении этот комок
у ряда видов может разворачиваться и подаваться все-таки в виде нити. Во-вторых,
способность передавать сперматофор в виде комка отражается на степени развития
дивертикула семеприемника; понятно, что здесь имеется обратная зависимость.
Вместе с тем нельзя забывать, что картина порой осложняется существованием прямой
корреляции между длиной бича и длиной дивертикула, причем корреляция эта
к тому же не всегда ясно выражена из-за способности сперматофора скручи-
ваться.
Имеется единственный вид, у некоторых популяций которого дивертикул семе-
приемника может превышать длину бича, — Theba pisana-, но этот вид вообще от-
личается значительным своеобразием.
Пока известен единственный случай, когда представитель Helicinae имеет только
одну папиллу, — Cryptomphalus aspersa; однако и здесь формируется подобная «на-
копительная» камера за счет развития мощного полукольцевого валика на месте ди-
стальной папиллы.
Заключая сравнительный анализ подсемейств, надо' оговориться, что и ранее
предпринимались попытки исследовать внутреннее строение пениса (Hesse, 1931,
1934; Garcia san Nicolas, 1957), но эти исследования ограничивались констатацией
факта, что папилла присутствует; к тому же разрез проводился только до дистальной
папиллы, и вопрос о существовании проксимальной оставался открытым. Тем более
отсутствуют исследования по механизму работы совокупительного аппарата..
Мы уже имели случай отметить конхологическое разнообразие семейства. Оста-
ется добавить, что в этой группе имеется один широко распространенный по Среди-
земноморью вид — Theba pisana, который принадлежит жизненной форме <tHell-
cellinae» и очень часто образует скопления, подобно многим ксерофилам из Hygro-
miidae, часто встречаясь вместе с ними.
Именно в силу разнообразия раковин среди Helicinae выделено большое число
родов; наши материалы заставляют предполагать, что истинное число родов в под-
семействе невелико.
МЕТОДЫ ИССЛЕДОВАНИЯ
СБОР МАТЕРИАЛА И ФИКСАЦИЯ
Моллюски рассматриваемой группы отличаются в целом достаточно большими
размерами. Лишь изредка встречаются виды, у которых большой диаметр раковины
меньше 7—8 мм. Поэтому собирают их вручную; лучше всего при сборе материала
пользоваться мягким (т. е. не тугим) глазным или отолярингологическим пинцетом.
Помимо пинцета надо при себе иметь баночку со спиртом или иным фиксатором, не-
сколько пустых пробирок и спичечных коробков; кроме того, желательно иметь полот-
няный мешочек.
Собирать живых моллюсков лучше всего в сырую погоду (но не во время дождя)
или ранним утром, пока не просохла роса, когда большинство моллюсков активно и
их легче заметить, а также понаблюдать за ними в природной обстановке. Обращать
внимание необходимо на стебли растений, на поверхность скал или камней и трещины
в них. Особенно богатый набор видов бывает в осыпях, окруженных кустарником или
густым травостоем; здесь можно найти комплекс видов, свойственных различным био-
топам.
МЕТОДЫ ИССЛЕДОВАНИЯ
81
Богатый конхологический материал дают выбросы, оставляемые реками и ручьями
после разлива.
В жаркую погоду большинство улиток скапливается в различных укрытиях —
под корой поваленных деревьев и пней, под валяющимися сучьями и камнями и в ниж-
них горизонтах осыпей. Некоторые виды висят на растениях, приклеившись устьем.
Надо иметь в виду, что ряд видов (особенно среднеазиатских) на сухой период зака-
пываются в почву. Поэтому, если присутствуют раковины какого-то вида, но живых
моллюсков не видно, надо отвернуть самые большие камни и коряги, которые сборщик
в состоянии одолеть, и исследовать почву под ними на глубину нескольких сантиметров,
осторожно разрыхляя ее.
Если в данном месте какой-то вид встречается в большом количестве, часто возни-
кает вопрос — сколько и каких экземпляров надо собрать. Практика подсказывает, что
лучше всего 4—5 живых взрослых экземпляров поместить непосредственно в спирт,
40—50 таких же особей взять с собой в лабораторию и присоединить к ним 8—10 ули-
ток разного возраста — от совсем маленьких до тех, которые уже почти завершили
формирование раковины.
Материал размещается по коробкам или, если виды отличаются большими размерами
и грубой раковиной, складывается в мешочек. В плотно закрытые банки живой ма-
териал сажать нельзя — животные могут задохнуться, особенно если стоит жаркая
погода. В .коробки надо положить листья, стебли травы или нарезанную полосками
бумагу, чтобы раковины не перекатывались и не ломались. Ватой для этой цели лучше
не пользоваться: она склеивается от слизи и прилипает к раковинам.
Большинство видов могут прожить в совершенно сухом, но аэрируемом помещении
много дней и недель, а отдельные виды — даже месяцев и лет. Поэтому нет необходи-
мости торопиться с обработкой материала, но надо периодически просматривать все
коробки, проветривать их и следить, чтобы в этих коробках не появились мухи (в ос-
новном Sciomysidae), личинки которых питаются моллюсками, или личинки жуков
(Lampyrus, A blattarta), которые уже могли быть в собранных раковинах (Горохов, 1970).
Часто человек стремится создать улиткам условия, приближенные к естественным;
полагая, что так они проживут дольше, т. е. периодически смачивает их и добавляет
свежих растений в качестве корма. Этого делать не следует: если моллюски активны,
они интенсивно дышат, выделяют много экскрементов и слизи и поэтому быстро
погибают.
Этикетки не надо вкладывать в коробку, где сидят моллюски, они могут ее съесть
или повредить настолько, что прочесть текст будет невозможно. Поэтому лучше пи-
сать этикетку прямо на коробке или приклеивать ее снаружи.
В лаборатории материал сортируется: сухие раковины раскладываются по ко-
робкам, а живые моллюски помещаются в банку, наполненную доверху водой, которая
затем плотно закрывается. Через 20—30 час. моллюски гибнут в расправленном со-
стоянии; если жарко, то гибель моллюсков наступает быстрее.
Обычно материал рекомендуется из воды переносить прямо в 70—75%-й спирт
(Лихарёв, Раммельмейер, 1952; Малевич, Старобогатов, 1958; Шилейко, 19736); од-
нако этот Метод лучше немного видоизменить, а именно переложить моллюсков из воды
в слабый спирт (лучше всего отработанный, т. е. слитый из банок с материалом при вто-
ричной фиксации). В этом случае процессы разложения тканей, начавшиеся тотчас
после гибели животных, прекращаются (да и то не полностью) только в тех частях тела,
которые высунуты из раковины, и спирт во внутренностный мешок проникает мед-
ленно. В первую очередь лизируется основание колумеллярного мускула — в месте
прикрепления его к столбику. Через 4—7 дней пребывания в слабом спирте материал
перекладывается в 80%-й спирт, а еще через 1—2 недели — окончательная фиксация
75%-м спиртом. Если фиксация прошла удачно, то моллюск легко вытягивается из
раковины, и при вскрытии ее не приходится разрушать, что чаще всего крайне неже-
лательно. В то же время заметных следов разрушения мягких тканей не видно.
Формалин для фиксации моллюсков крайне нежелателен, так как он, во-первых,
портит раковину и, во-вторых, сильно затрудняет вскрытие, делая ткани неэластич-
ными. Если иного фиксатора нет, можно воспользоваться формалином в сильном раз-
ведении (4%), но при первой же возможности надо заменить спиртом, причем спирт
после формалина надо сменить не менее двух раз с интервалом в 1.5—2 недели.
. Если есть возможность, часть материала рекомендуется зафиксировать специаль-
ными фиксаторами (жидкость Буэна, Шаудина, Джильсона, Флемминга, Ценкера
и др.), но надо иметь в виду, что применение гистологических фиксаторов к материалу,
убитому выдерживанием в воде, обычно нецелесообразно, так как от воды происходит
частичная мацерация тканей, расслаивание эпителиальных слоев, разрыв клеточных
оболочек. Поэтому, если нужен расправленный материал для гистологических целей,
рекомендуется один из двух способов. Моллюск заливается водой; через некоторое
время он начинает двигаться; тогда его берут за раковину и сажают на лучинку.
Когда улитка приблизится к одному концу лучинки, ее берут за другой конец и бы-
стрым движением опускают моллюска в кипящий фиксатор — он погибает, не успев
сократиться и втянуться в раковину. Второй способ используется в том случае, если
6 А. А. Шилейко
82
ВВЕДЕНИЕ
данный фиксатор не рекомендуется нагревать или температура кипения его низка.
Суть его сводится к тому же, но для умерщвления животного используют кипящую
воду, после чего немедленно фиксируют.
При длительном хранении в спирте раковины тоже портятся, особенно страдает
периостракум и тонкие элементы скульптуры. Поэтому надо стремиться, чтобы каждый
сбор содержал как сухие раковины, так и спиртовой материал. Если один из спиртовых
экземпляров подвергается вскрытию, его раковина присоединяется к сухой коллекции.
КАМЕРАЛЬНАЯ ОБРАБОТКА
Изучение собранного материала начинается с определения видовой принадлеж-
ности экземпляров. Для ознакомления с признаками раковины ее надо правильно ори-
ентировать. При нормальном положении вершина раковины смотрит вверх, а устье на-
правлено на наблюдателя (рис. 40). В таком положении раковину измеряют, а также
обращают внимание на ее форму. Схема основных промеров дана на рис. 40, I.
Далее, оценивают разницу в ширине последнего и предпоследнего оборотов.
Для этого мысленно проводят прямую линию от края устья к вершине, держа раковину
вершиной к себе, и сравнивают отрезки, расположенные на этой линии и ограниченные
периферией и швом: самый наружный и примыкающий к нему (рис. 40, II). Понятно,
что разница будет тем сильнее, чем стремительнее нарастают обороты. -
Рис. 40. Схема основных промеров раковины и конхологические
признаки. I — нормальное положение раковины; II — положе-
ние раковины вершиной к наблюдателю. БД — большой диа-
метр раковины; БК — базальный край устья; В — вершина
раковины; ВЗ — высота завитка; ВР — высота раковины;
Г — губа; КК — колумеллярный край устья; МД — малый
диаметр раковины; П — пупок; ПК — палатальный край устья;
ПР — периферия раковины; ПРК — париетальный край устья;
1П — шов; ШП — ширина последнего оборота; ШПП — ширина
предпоследнего оборота. (Ориг.).
Кроме большого диаметра (ширины) раковины старые авторы указывали малый,
диаметр (рис. 40, II) и напрасно от этого промера в дальнейшем отказались: ширина
раковины варьирует главным образом за счет последнего оборота и особенно за счет
дистальной его части. Поэтому разброс величин малого диаметра меньше, и эта величина
бывает полезной при сравнении близких видов и установлении границ размаха из-
менчивости. В связи с этим в систематической части почти по всем видам приведены и
значения малого диаметра.
Затем внимательно изучают скульптуру и рисунок на поверхности раковины.
' В определительной таблице и описаниях линиями именуются только скульптурные
элементы; цветовые спиральные полосы называются лентами. Словом «полоса» обозна-
чаются радиальные цветовые элементы рисунка раковины.
Наконец, обращают внимание на остальные признаки: глубину шва, степень и
характер выпуклости оборотов, развития губы, скошенности устья и опущенности
последнего оборота, перспективности и ширины пупка, оценивают, насколько высту-
пает вершина раковины, и т. п.
При пользовании определительной таблицей для определения видов надо иметь
в виду следующее. Раковины большинства видов бедны четкими признаками, и, кроме
того, имеется много конвергентно схожих раковин; существование различных эко-
логических морф еще больше затрудняет определение. Поэтому необходимо внимательно
и до конца прочесть тезу и антитезу и выбрать то, что больше подходит. При составле-
нии таблицы мы исходили из того, что изучаемый материал этикетирован, поэтому
МЕТОДЫ ИССЛЕДОВАНИЯ
83
широко используются географические признаки. Применение в ключах географических
признаков порой считается неправомерным, но в данном случае с этим трудно согла-
ситься, поскольку подавляющее большинство видов имеет достаточно четкий и огра-
ниченный ареал, а случаев завоза не так много; когда речь идет о видах, изменивших
ареал при участии человека, они (т. е. виды) определяются без учета их распро-
странения.
Наконец, имеются 3 пары видов, психологически почти не различающихся. Для
этих пар в таблицу включены анатомические признаки.
Рис. 41. Вскрытие представителя Helicoidea. I—IV — основные стадии вскрытия.
БЖ — белковая железа; Г — глотка; 3 — зоб; К — кишка; КМ — колумеллярный
мускул; МЛ — мускульная лента мантийной полости; МП — мантийная полость;
НСЛ — нижняя стенка легкого; ОНК — окологлоточное нервное кольцо; П — пенис;
ПО — половое отверстие; ПР — половой ретрактор; ПЧ — печень; ПЧК — почка;
РГ — ретрактор глотки; РПО — ретрактор правого омматофора; С — сердце; СОД —
спермовидукт; СТ — стилофор. Звездочка — первый разрез. (Ориг.).
Определив вид по таблице, надо убедиться в правильности определения, изучив
его описание и фотографии. Для большей надежности изучаемый экземпляр рекомен-
дуется вскрыть.
Как видно из систематической части, таксономические признаки базируются глав-
ным образом на особенностях строения полового аппарата.
Первым этапом изучения мягких частей является извлечение их из раковины.
При удачной фиксации они легко вытягиваются. Если колумеллярный мускул крепко
удерживает тело в раковине, можно, захватив пинцетом тело моллюска выше мантий-
ного валика, покачать выступающие части, одновременно осторожно вытягивая их.
Тогда дистальная часть внутренностного мешка обычно отрывается достаточно высоко,
и в оторванной части тела остаются все органы, необходимые для определения. Если же
тело нужно целиком, но его не удается вытянуть, раковину ломают и удаляют
по частям.
Далее моллюска укладывают в парафиновую ванночку (в парафин лучше под-
мешать немного воска и черного сапожного крема, так как на темном фоне удобнее вскры-
вать) и заливают спиртом. Можно использовать отработанный йрофильтрованный
спирт. Вода для вскрытия нежелательна, поскольку в ней некоторые органы, в осо-
бенност и спермовидукт, разбухают, а сама вода быстро мутнеет.
6*
84
ВВЕДЕНИЕ
Животное ориентируют правой стороной вверх, а подошвой к себе (синистраль-
ные формы — левой стороной вверх), и отыскивают половое отверстие, видное в виде
округлой впадины или поперечной щели ниже и позади основания правого глазного
щупальца.
Вскрытие проводят под бинокуляром МБС-2 или, лучше, МБС-1 при косом кон-
центрированном свете. Вначале позади полового отверстия и несколько выше него де-
лают короткий продольный разрез (рис. 41, I, II). В этот разрез вставляют половинку
маленьких ножниц или, если животное невелико, заточенную иглу и продолжают раз-
рез к переднему концу тела, обходя половое отверстие и тщательно следя за тем, чтобы
не были повреждены органы, прилегающие изнутри к покровам. Особенно удобны для
вскрытия хирургические ножницы для роговицы или пинцет-ножницы. В другую сто-
рону разрез ведется до мантийного валика (рис. 41, III).
Затем отгибают покровы спинной стороны и прикалывают их булавками ко дну
ванночки; подошву также прикалывают. Дальнейшие операции заключаются в частич-
ном удалении мантийного валика и в постепенном продолжении разреза, сквозь ко-
торый уже можно видеть все внутренние органы. Необходимо обратить внимание на
характер расположения ретрактора правого омматофора относительно гениталий или
полового ретрактора. Освобожденные гениталии вынимают из тела вместе с участком
покровов, оставшимся вокруг полового отверстия, и участком дна мантийной полости,
к которому крепится половой ретрактор, и расправляют с помощью булавок на дне
ванночки.
Для изучения внутреннего строения полового аппарата желательно иметь не ме-
нее 2 экз. Гениталии одной особи изучаются методом прямого вскрытия: в половое от-
верстие вводят половинку ножниц и, следя за тем, чтобы она не повредила внутренние
стенки, разрезают вагину до основания семеприемника, а чехол пениса — до основа-
ния папиллы. Если имеется вторичный чехол пениса (чехол папиллы), он при этом
обычно повреждается. Если папилла отсутствует, разрез проводят до места впадения
эпифаллуса или семепровода. Затем стенки этих органов отворачивают и исследуют
их внутреннюю поверхность, а также внешний вид пенцальной папиллы.
Дальше папиллу разрезают поперек в трех местах — при основании, посредине
и близ дистального конца.
Гениталии второго экземпляра служат для того, чтобы, разрезав их дистальную
часть ножницами вдоль, исследовать те структуры, которые были пропущены при изу-
чении первого экземпляра. Таким путем выявляется,, в частности, наличие кольцевых
полостей, вторичного пениального чехла и т. п.
Это лишь самые общие указания. Вообще надо иметь в виду, что каждый вид тре-
бует некоторых модификаций этих приемов, которые вырабатываются в каждом кон-
кретном случае. Для установления размаха изменчивости признаков внутреннего
строения рекомендуется вскрывать по нескольку экземпляров каждого вида.
После того как половой аппарат удален, видна глотка с отходящим от нее кишеч-
ником, окологлоточное нервное кольцо и колумеллярный мускул с отходящей от него
системой педальных, глоточных и глазных ретракторов (рис. 41, IV).
Для изучения радулы у достаточно крупных видов ее можно удалить с помощью
пинцета и тонких игл, а у более мелких глотку целиком отделяют и кипятят в 50 % -м
растворе щелочи до полного растворения тканей, после чего остается радула, соеди-
ненная с челюстью. Ее промывают холодной водой и готовят препарат. Наилучшие ре-
зультаты дает воднобальзамный метод с последующей окантовкой, очень подробно опи-
санный в работе И. М. Лихарева (1962, стр. 99, 100).
Рэдвин (Radwin, 1969) рекомендует методику, с применением которой можно по-
лучить препараты радул из высохших моллюсков. Моллюсков кипятят в насыщенном
растворе тринатрийфосфата, где ткани размягчаются. Затем в течение 3—10 сек.
объект кипятят в концентрированном растворе NaOH, после чего переносят в часовое
стекло с 70 %-м спиртом, где под бинокуляром радула'освобождается от остатков мяг-
ких тканей.
ЛИТЕРАТУРА
Абрикосов Г. Г., Цветков Б. Н. Материалы по систематике и экологии моллюсков ок-
рестностей Ашхабада и прилежащих частей Копет-Дага. — Учен. зап. Моск,
ун-та, 1945, вып. 83, с. 203—215.
А крамовский Н. Н. Наземные моллюски лавового плато в окрестностях гор. Ере-
вана. — Науч., тр. Ереван, гос. ун-та, 1943 (1947), с. 259—273.
Акрамовский Н. Н. Наземные моллюски территории селения Гнишик в Советской Ар-
мении. — В кн.: Зоол. сб. Вып. 6. Ереван, 1949, с. 127—183.
Акрамовский Н. Н. Зоогеографический облик современной фауны моллюсков Армении
и зоогеографическое районирование республики. — Биол. журн. Армении, 1970,
т. 23, цып. 11, с. 118—127.
А крамовский Н. Н. Некоторые итоги изучения современной фауны моллюсков Арме-
нии. — В кн.: Моллюски. Пути, методы и итоги их изучения. Л., 1971а, с. 108—
111.
ЛИТЕРАТОРА
85
Акрамовский Н. Н. Краткий каталог современной фауны моллюсков Советской Ар-
мении. — Биол. журн. Армении, 19716, т. 24, вып. 6, с. 3—12.
Акрамовский Н. Н., Азарян Дж. М. К вопросу о видовой самостоятельности Euom-
phalia arpatschaiana (Mousson, 1873) (Gastropoda—Stylommatophora, Helicidae). —
Изв. АН АрмССР. Биол. науки, 1964, т. 17, вып. 4, с. 69—75.
. Андрейкович Е. В. О размножении Helix pomatia (L.) в Литовской ССР. — В кн.:
Моллюски и их роль в экосистемах. Л., 1968, с. 59—61.
Андрейкович Е. В. Биология размножения виноградной улитки в Литве. — Тр. АН
ЛитССР, 1969, В, № 1 (48), с. 111—117.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. 1964. Т. I. Про-
морфология, с. 1—432. Т. II. Органология, с. 1—446.
Богачев В. В. Пресноводные и наземные моллюски из верхнетретичных отложений бас-
сейна р. Куры. — Тр. Азерб. фил. АН СССР. Геол, сер., 1936, вып. 13, с. 5—
' 96.
Величковский В. К фауне моллюсков окрестностей Никольска-Уссурийского. — Изв.
Южно-Уссур. отд-ния Рус. геогр. о-ва, 1924, т. 8, с, 153—154.
Гаузе Г. Ф., Смарагдова Н. П. Потеря в весе и смертность у правозавитых и левозави-
тых особей улитки Pruticicola lantzi. — Зоол. журн., 1939, т. 18, вып. 2, с. 154—
161.
Гептнер В. Г. Структура систематических групп и биологический прогресс. — Зоол.
жури., 1965, т. 44, вып. 9, с. 1291—1308.
Голиков А. Н., Старобогатов Я. И. К построению системы переднежаберных брюхо-
ногих моллюсков. — В кн.: Моллюски и их роль в экосистемах. Л., 1968,
с. .5—7.
Горохов В. В. Химические и биологические методы борьбы с моллюсками — проме-
жуточными хозяевами гельминтов. — В кн.: Гельминтозы с.-х. животных.
Итоги науки. М., 1970, с. 132—170.
Гудец В. О некоторых малоизвестных видах моллюсков (Pulmonata, Helicidae)
из СССР. — Биол. науки, 1969, № 9, с. 26—34.
Догель В. А. Олигомеризация гомологичных органов как один из главных путей эво-
люции животных. Л., 1954, с. 1—368.
Иванов А. В. Класс брюхоногих моллюсков (Gastropoda). — В кн.: Руководство по зо-
ологии. Т. II. М., 1940, с. 323—455.
Иванов А. В. Mollusca. — В кн.: Большой практикум по зоологии беспозвоночных.
Ч. II. М., 1946, с. 57—218.
Политика 3. И. К изучению зонального распределения наземных моллюсков северных
склонов Центрального Кавказа и Восточного Предкавказья. — В кн.: Здраво-
охранение и медицина в Сев. Осетии. Вып. VII, ч. 1. М.—Л., 1958, с. 157—181.
Палитина 3. И. Наземные моллюски Кабардино-Балкарской АССР и их роль в рас-
пространении зоонозных и антропозоонозных заболеваний. — В кн.: Антропо-
зоонозные болезни, профилактика и их искоренение. М., 1967, с. 65—73.
Прастин Н. И., Дворядкин В. А., Кухаренко Н. С. К фауне и экологии наземных мол-
люсков Амурской области. — Тр. Науч.-исслед. проблемной лаб. паразитол.
Благовещ. с.-х. ин-та, 1969, вып. 1, с. 92—96.
Приштофович А. Н. Развитие ботанико-географических областей северного полуша-
рия с начала третичного периода. — Вопр. геологии Азии, 1955, вып. 2, с. 824—
844.
Круликовский Л. К познанию фауны моллюсков России. — В кн.: Прилож. к 60 т.
Зап. Акад, наук, 1889, № 7, с. 1—35.
Крылова Л. И. Стратиграфия и малакофауна антропогеновых отложений Среднего
Урала и Южного Зауралья. Автореф. дис. Л., 1970, с. 1—19.
Леонов Г. П. Историческая геология. М., 1956, с. 1—364.
Линдгольм В. А. Заметки о палеарктических наземных и пресноводных моллюсках. —
Ежегод. Зоол. муз., 1922а, т. 23, с. 304—320.
Линдгольм В. А. Описание нескольких кавказских видов наземных моллюсков (Ga-
stropoda, Pulmonata). — Ежегод. Зоол. муз., 19226, т. 23, с. 355.—361.
Линдгольм В. А. О некоторых представителях семейства Helicidae (Mollusca, Pul-
monata) Кавказского края. — Ежегод. Зоол. муз., 1928, т. 28, с. 539—549.
Линдгольм В. А. Моллюски. — В кн..” Тр. Памир, экспедиции 1928 г. VIII. 1931,
с. 29—64.
Линдгольм В. А. Новый вид рода Helicella (Helicidae, Gastropoda) из Крыма и некоторые
наблюдения над экологией моллюсков. — Тр. Зоол. ин-та АН СССР, 1936,
т. 3, с. 439—442.
Лихарев И. М. Новый китайский элемент в фауне Приморского края. — ДАН СССР,
1949, т. 64, № 3, с. 417—420.
Лихарев И. М. Наземные моллюски [степной зоны]. — В кн.: Животный мир СССР.
Т. 3. М., 1950, с. 466—470.
Лихарев И. М. Особенности распространения моллюсков Приморского края. —
Тр. Зоол. ин-та АН СССР, 1953, т. 13, с. 277—288.
86
ВВЕДЕНИЕ
Лихарев И. М. К систематическому положению некоторых среднеазиатских наземных
моллюсков. — Тр. Зоол. ин-та АН СССР, 1955, т. 21, с. 179—185.
Лихарев И. М. Материалы к фауне наземных моллюсков островов Итуруп и Шикотан
(Южные Курильские острова). — Тр. Ленингр. о-ва естествоиспыт., 1957а,
т. 73, вып. 4, с. 77—80.
Лихарев И. М. Географическое распространение наземных моллюсков Кавказа и не-
которые пути происхождения этой фауны. — В кн.: Матер, к совещ. по вопр.
зоогеографии суши. Львов, 19576, с. 73—75.
Лихарев И. М. Наземные моллюски [Крыма]. — В кн.: Животный мир СССР. Т. 5.
М., 1958а, с. 133—136.
Лихарев И. М. Наземные моллюски [Кавказа]. — В кн.: Животный мир СССР.
Т. 5. М., 19586, с. 464—476.
Лихарев И. М. .Моллюски клаузилииды. — В кн.: Фауна СССР. Моллюски. Т. 3,
вып. 4. Нов. сер., № 83. М.-Л., 1962, а. 1-317.
Лихарев И. М., Раммелъмейер Е. С. Наземные моллюски фауны СССР. — В кн.:
Опред. по фауне СССР. Вып. 43. М.—Л., 1952, с. 1—511.
Лихарев И. М., Старобогатов Я. И. Материалы по фауне моллюсков Афганистана. —
Тр. Зоол. ин-та АН СССР, 1967, т. 42, с. 159—197.
Майр Э. Зоологический вид и эволюция. М., 1968, с. 1—597.
Малевич И. И., Старобогатов Я. И. Наземные моллюски Подмосковья как объект са-
мостоятельных работ студентов на летней практике и в зоологическом кружке. —
Учен. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, 1958, т. 84, вып. 7,
с. 269—316.
Марков К. К. Палеогеография. М., 1960, с. 1—267.
Мартенс Э. Слизняки (Mollusca). — В кн.: А. П. Ф е д ч е нк о. Путешествие в Туркестан.
Вып. 1, т. 2, ч. 1. 1874, с. 1—64.
Матёкин П. В. Систематическое положение некоторых видов наземных моллюсков
фауны Тянь-Шаня. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол., 1954,
т. 59, вып. 2, с. 90.
Матёкин П. В. Материалы по фауне наземных моллюсков Средней Азии. Дис. М.
1960, с. 1—516.
Матёкин П. В. Определитель раковинных моллюсков Средней Азии — промежуточных
хозяев гельминтов; — В кн.: Гельминты животных Киргизии и сопред. терри-
торий. Фрунзе, 1966, с. 97—137.
Матёкин П. В. Наземные моллюски семейства Bradybaenidae Средней Азии. — Сб.
тр. Зоол. муз. Моск. гос. ун-та, 1972, т. 12, с. 112—144.
Миничев Ю. С. Исследования по морфологии низших Opisthobranchia. — В кн.: Мор-
фология беспозвон. животных. М., 1967, с. 109—182..(Тр. Зоол. ин-та АН СССР,
т. 44).
Миничев Ю. С. Положение Soleolifera в системе Gastropoda. — В кн.: Моллюски. Пути,
методы и итоги их изучения. Л., 1971, с. 8—10.
Миничев Ю. С., Славошевская Л. В. Особенности эволюции реноперикардиального
комплекса наземных Pulmonata. — Зоол. журн., 1971, т. 50, вып. 3, с. 350—•
360.
Мухитдинов А. Материалы к фауне моллюсков северного Таджикистана. — Изв.
АН ТаджССР. Отд-ние биол. наук, 1972, № 3 (48), с. 39—49.
Несис К. Н. Экологическая классификация (жизненные формы) головоногих моллю-
сков. — В кн.: Промысловые моллюски. Итоги науки. Зоология беспозвоночных,
т. 2. М., 1973, с. 8—59.
Николаев В. А. Систематическое положение и изменчивость наземных моллюсков рода
Trichia (Helicidae) Среднерусской возвышенности. — В кн.: Моллюски и их
роль в экосистемах. Л., 1968, с. 32—33.
Пирс Э. Гистохимия. М., 1962, с. 1—783.
Полевийа А. А. К фауне наземных моллюсков Закарпатья. — Докл. и сообщ. Ужгор.
гос. ун-та. Сер. биол., 1959, т. 3, с. 65—68.
Присяжнюк В. А. Про наземш молюски з понтичних в!дклад1в Одесько! облает!. —
ДАН УССР, сер. Б, 1974, вып. 4, с. 317—321.
Пузанов И. И. Материалы к познанию наземных моллюсков Крыма. Ч. I. Моллюски
горного Крыма. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол., 1925, т. 33,
с. 48—104.
Пузанов И. И. Материалы к познанию наземных моллюсков Крыма. Ч. II. Моллюски
степного Крыма. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол., 1926,
т. 35, с. 84—101.
Пузанов И. И. Крымские наземные моллюски в связи с вопросом о происхождении,
фауны Крыма. — Тр. II съезда зоологов, анатомов и гистологов СССР, 4—
10 мая 1925 г. М., 1927а, с. 52—54.
Пузанов И. И. Материалы к познанию наземных моллюсков Крыма. Ч. III. Состав, рас-
пределение и генезис крымской малакофауны. — Бюл. Моск, о-ва испыт. при-
роды. Отд-ние биол., 19276, т. 36, с. 221—282.
ЛИТЕРАТУРА
87
Резник 3. В. Изучение фауны наземных моллюсков Ставропольского края. — Сб.
статей по зоологии Ставроп. пед. ин-та, 1967, с. 97—101.
Розен О. В. Материалы к познанию фауны слизняков Закаспийской области и Хоро-
сана. — В кн.: Фауна Закасп. обл. Т. II. 1894, с. 39—57.
Розен О. В. Материалы к познанию фауны слизняков Кавказа и Средней Азии. — В кн.:
Диевн. Зоол. отд-ния О-ва любит, естествозп., антропол. и этногр., 1901, т. III,
вып. 2, с. 7—13.
Рошковский В. Tachea argonautarum, nomen novum (=atrolabiata auct., non Krynicki).—•
Изв. Кавк, муз., 1919, т. 12, с. 134—136.
Синицын В. М. Палеогеография Азии. М.—Л., 1962, с. 1—267.
Синицын В. М. Введение в палеоклиматологию. Л., 1967, с. 1—231.
Синцов И. Ф. Предварительное сообщение о новых и малоисследованных формах
раковин из третичных образований Новороссии. — Зап. Новорос. о-ва естество-
испыт., 1876, т. 4, вып. 1.
Скворцов А. Л. К изучению фауны моллюсков Воронежской области в связи с их ролью
в распространении глистных заболеваний домашних животных. — Тр. Воро-
неж. зоовет. ин-та, 1938, т. 4, с. 178—188.
Скворцов Л. Л. К фауне моллюсков Южного Казахстана. — Тр. Каз. науч.-исслед.
вет. ин-та, 1940, т. 3, с. 361—378.
Стеклов А. А. Наземные моллюски неогена Предкавказья и их стратиграфическое
значение. — Тр. Геол, ин-та АН СССР, 1966, вып. 163, с'. 1—262.
Стеклов А. А. Этапы развития неогеновых наземных моллюсков Предкавказья. —
Malacologia, 1968, т. 6 (3), с. 243—251.
Страхов Н. М. Основы исторической геологии. Ч. 2. М.—Л., 1948, с. 1—396.
Судзуки Н. Ориентировка по запаху наземного моллюска Ezohelix flexibilis. — Zool.
Mag., 1968, v. 77, № 6, p. 178—184. (На япон. яз.).
Такеучи X., Уеда С., Канамори X. Движутся ли материки? М., 1970, с. 1—248.
Увалиева К. К. Фауна наземных моллюсков Южного Алтая. — Тр. Ин-та зоол.
АН КазССР, 1963, т. 19, с. 152—160.
Увалиева К. К. Новый вид наземных моллюсков Алтая (Gastropoda, Helicidae). — Тр.
Ин-та зоол. АН КазССР, 1964, т. 22, с. 199—202.
Увалиева В. В., Соболева Т. Ц. Наземный моллюск Bradybaena tzwetkovi sp. nov. —
эндемик Заилийского Алатау. — Зоол. журн., 1973, т. 52, вып. И, с. 1717—
1718.
Цветков Б. Н. Правые и левые формы моллюска Fruticlcola lantzi Lndh. и их географи-
ческое распространение. — Бюл. Моск, о-ва испыт. природы. Отд-ние биол.,
1938, т. 47, вып. 5—6, с. 414—419.
Цветков Б. Н. К познанию фауны моллюсков пастбищ Алма-атинской области. — Тр.
Каз. науч.-исслед. вет. ин-та, 1940, т. 3, с. 379—419.
Цветков Б. Н. Материалы к фауне моллюсков Средней Азии. I. Представители подрода
Pseudiberus (р. Cathaica). — Сб. тр. Зоол. муз. Моск. гос. ун-та, 1941а, т. 6,
С. 277—286.
Цветков Б. Н. Изменчивость Fruticlcola lantzi (Mollusca, Pulmonata). — Сб. тр. Зоол.
муз. Моск. гос. ун-та, 19416, т. 6, с. 287—302.
Цветков Б. Н., Цветкова Е. А. Род Paedhoplita и его представители. — Изв. Каз. фил.
АН СССР. Сер. зоол., 1943, т. 2, с. 117—121.
Цветков Б. Н., Цветкова Е. А. Фауна и распространение наземных моллюсков —
промежуточных хозяев гельминтов на юго-востоке Казахстана. — Тр. Каз.
науч.-исслед. вет. ин-та, 1950, т. 5, с. 335—366.
Шилейко А. А. О факторах изменчивости некоторых наземных моллюсков..— Вести.
Моск. гос. ун-та. Сер. VI, 1967а, вып. 2, с. 14—21.
Шилейко И. И. О биологии размножения и ювенильных адаптациях слизня Parmacella
ibera Eichw. — Зоол. журн., 19676, т. 46, вып. 6, с. 946—948.
Шилейко А. А. Замечания о системе рода Helicella Fer. (Pulmonata, Helicidae) с описа-
нием нового вида из Талыша. — Вести. Моск. гос. ун-та. Сер. VI, 1967в, вып. 4,
с. 27—34.
Шилейко А. А. Моллюски семейства Helicidae (Pulmonata, Stylommatophora) Талыша. —
Зоол. журн., 1968а, т. 47, вып. 3, с. 337—347.
Шилейко А. А. Наземная малакофауна лесного Талыша как пример фауны островного
типа. — В кн.: Моллюски и их роль в экосистемах. Л., 19686, с. 27—28.
Шилейко А. А. Зоогеографическая структура и история формирования фауны назем-
ных моллюсков Талыша. — Malacologia, 1968в, т. 7 (1), с. 117—134.
Шилейко А'. А. Эколого-фаунистический обзор наземных раковинных моллюсков гор-
ного Дагестана. — Вести. Моск. гос. ун-та. Сер. VI, 1969а, вып. 1, с. 26—31.
Шилейко А. А. Очерк экологии наземных моллюсков Талыша. — Бюл. Моск, о-ва
испыт. природы. Отд-ние биол., 19696, вып. 3, с. 80—88.
Шилейко. А. А. Объем, система и филогения группы Perforatella—Zenobiella—Chilano-
don (Pulmonata, Helicidae). — Зоол. журн., 1970, т. 49, вып. 9, с. 1306—1321.
Шилейко А. А. Конхологический и анатомический методы исследования Hellcacea
88
ВВЕДЕНИЕ
и области их применения. — В кн.: Моллюски. Пути, методы и итоги их изу-
чения. Л., 1971а, с. 104—106.
Шилейко А. А'. Сравнительноанатомическое исследование некоторых Ariantinae (Pul-
monata, Helicidae). — Зоол. журн., 19716, т. 50, вып. 7, с. 990—1001.
Шилейко А. А. Малакология: обманчивость сходства и природа различий. — При-
рода, 1971в, № 7, с. 75—80.
Шилейко А. А. Таксономический статус Helicodontinae (Pulmonata, Helicidae). — Науч,
докл. высш, школы. Биол. науки, 1971г, № 12, с. 7—16.
Шилейко А. А. Организация Zonitoides nitidus (Moll.) в связи с вопросом о таксономи-
ческом ранге Gastrodontidae (Gastropoda, Stylommatophora). — Сб. тр. Зоол. муз.
Моск. гос. ун-та, 1972а, т. 12, с. 145—156.
Шилейко А. А. Морфология радулы Helicidae и таксономическое значение радулярных
признаков. — Сб. тр. Зоол. муз. Моск. гос. ун-та, 19726, т. 12, с. 157—168.
Шилейко А. А. Таксономическая и географическая структура трибы Hygromiini
Schileyko (Pulmonata, Helicidae). — Зоол. журн., 1972в, т. 51, вып. 8, с. 1129—
1141.
Шилейко А. А. Некоторые аспекты изучения современных континентальных брюхо-
ногих моллюсков.— В кн.: Зоология беспозвоночных. Итоги науки. Т. 1.1972г,
М., с. 1—188.
Шилейко А. А. Подсемейство Metafruticicolinae Schileyko, 1972 как филетическая ли-
ния (Pulmonata, Hygromiidae). — Науч. докл. высш, школы. Биол. науки,
19729, № 12, с. 12—19.
Шилейко А. А. Сравнительная характеристика палеарктических наземных моллюсков
Helicoidea. — Зоол. журн., 1973а, т; 52, вып. 4, с. 492—506.
Шилейко А. А. Моллюски. — В кн.: Гельминтол. оценка пастбищ. М., 19736, с. 112—
168.
Шилейко Л. В., Шилейко А. А. К морфологии проксимального отдела полового аппа-
рата Stylommatophora (Gastropoda, Pulmonata). — Науч. докл. высш, школы.
Биол. науки, 1975, № 1, с. 7—13.
Шмалъгаузен И. И. Проблемы дарвинизма. Изд. 2-е. Л., 1969, с. 1—493.
Яблоков А. В., ВалецкийА. В. Об изменении ареала левозакрученных форм Eulota
lantzi в Заилийском Алатау за последние десятилетия.- — Зоол. журн., 1971,
т. 50, вып. 3, с. 436—438.
Abolini-Krogis A. The histochemistry of the mantle of Helix pomatia L. in relation to the
repair of the damaged shell. — Arkiv zool., 1963, v. 15, p. 461—476.
Adams A. On the species of Helicidae found in Japan. — Ann. and Mag. Natur. History,
1868, v. 4, N 1, p. 459—472.
Ag6csy P. Data to quantitative conditions in the mollusk faunas of two different substra-
tes in Central Hungary. — Acta zool. Acad, scient. hung., 1968, v. 14, N 1—2,
p. 1—6. .
Albers J. C. Die Heliceen nach natiirlicher Verwandtschaft systematise!! geordnet. 2-te
Ausgabe, herausgegeben von E. Martens. Leipzig, 1860, S. 1—359.
Atkins Ch. G. Factors affecting the structure and distribution of terrestrial Pulmonata. —
Proc. Iowa Acad. Sci., 1966, v. 73. Des Moines, 1968, p. 408—416.
Backhuys W. Notes on Theba pisana ustulata (Lowe, 1852), the land-snail of Salvages
Islands. — Basteria, 1972, v. 36, N 2—5, p. 117—130.
Baker R. The ecology of the wrinkled snail, Helicella caperata Mont, on the Braunton
Burrows Sand Dune System. — Proc. Malacol. Soc. London, 1968, v. 38, N 1,
p. 41-54.
Bayne Ch. Histochemical studies on the egg capsules of eight gastropod molluscs. —
Proc. Malacol. Soc. London, 1968, v. 38, N 3, p. 199.—212.
Beck H. Index molluscorum praesentis aevi musei principis augustissimi Christiani Fre-
derick Hafniae, 1837, p. 1—100.
Bielz E. A. Fauna der Land- und Siisswasser-Mollusken Siebenbiirgens. 2 Aufl., Hermann-
stadt, 1867, S. 1—216.
Biggs H. E. J. Mollusca of the Iranien Plateau-III. — J. conchol., 1971, v. 27, p. 211 —
220.
Bigot L-. Recherches sur les groupements de gasteropodes terrestres: la constitution des
«grappes». — Vie et milieu, 1967—1968, v. 18, N 1, p. 1—27.
Binot D. Histologie, histochimie, cytologie de quelques formations glandulaires du te-
gument dlOncidiella celtica (Cuv.) (Gasteropode, Pulmone). — Cahiers biol. ma-
rine, 1965, v. 6, p. 325—346.
Bishara S., Hassan M., Kalliny A. Studies on some land snails injuries to agriculture
in U. A. R. — Rev. zool. et Jwt. Afric., 1968, v. 77, N 3—4, p. 239—252.
Bleakney J. S. Behavior of calciphilic Cepaea hortensis on acidic island off Nova Scotia. —
Nautilus, 1966, v. 79, N 4, p. 131—134.
Blinn W. C. Water in the mantle cavity of land snails. — Physiol. Zool. (Chicago), 1964,
v. 37, p. 329—337.
ЛИТЕРАТУРА
89
Block М. В. Epiphragms: some observation. — J. conchol., 1971, v. 26 (1), N 6, p. 388—
409.
Boettger, C. Ein systematisches Verzeichnis der beschalten Landschnecken Deutschlands,
Osterreich-Ungarns und der Schweiz. — Nachrichtenbl. dtsch. malak. Ges., 1911,
Bd 43 (1), S. 17—25.
Boettger C. Nochmals der Genotyp der Landschneckengattung Trichia Hartmann. —
Zool. Anz., 1928, Bd 77 (1/2), S. 1—4.
Boettger C., Wenz W. Zur Systematik der zu den Helicidenfamilien Campylaeinae und
Helicinae gehorigen tertiaren Landschnecken. — Arch. Moll., 1921, Bd 53, S. 1 —
55.
Boettger 0. Kaukasische Mollusken gesammelt von Herrn Hans Leder. — Jahrb. dtsch.
malak. Ges., 1880a, Bd 7, S. 109—150.
Boettger 0. Armenische und transcaucasische Mollusken aus einer Sendung des Hrn. Dr.
G. Sievers. — Jahrb. dtsch. malak. Ges., 1880b, Bd 7, S. 151—161.
Boettger 0. Diagnoses molluscorum novorum ab ill. Hans Leder in regione caspia Talysch
dicta lectorum. — Jahrb. dtsch. malak. Ges., 1880c, Bd 7, S. 379—383.
Boettger 0., Sechstes Verzeichniss transkaukasischer, armenischer und nordpersischer Mol-
lusken. — Jahrb. dtsch. malak. Ges., 1881a, Bd 8, S. 167—261.
Boettger 0. Diagnoses molluscorum novorum Transcaucasiae, Armeniae et Persiae. —
Nachrichtenbl. dtsch. malak. Ges., 1881b, Bd 13, S. 117—129.
Boettger 0. Siebentes Verzeichniss von Mollusken der. Kaukasuslander. — Jahrb. dtsch.
malak. Ges., 1883, Bd 10, S. 135—198.
Boettger 0. Liste der von Herrn 0. Retowski in Abchasien gesammelten Binnenmollu-
sken. — Ber. Senckenb. Nat. Ges., 1884, S. 144—155.
Boettger 0. Neuntes Verzeichniss (IX) von Mollusken der Kaukasuslander. — Jahrb.
dtsch. malak. Ges., 1886, Bd 8, S. 121—156.
Boettger 0. Die Binnenmollusken Transcaspiens und Chorossans. — Zool. Jahrb., 1889,
Abt. 3, Bd 4, S. 925—992.
Bonavita A. Conditions determinant la production de 1’epiphragme chez les gasteropodes
helicellines. — C. r. Acad, sci., 1965, v. 260, N 14, p. 4093—4094.
Bonavita D. Condition ecologiques de la formation de 1’epiphragme chez quelques he-
licides de Provence. — Vie et milieu, 1964, v. 15, N 3, p. 721—755.
Bornchen M. Untersuchungen zur Sekretion der fingerformigen Drusen von Helix pomatia
f L. — Z. Zellforsch., 1967, Bd 78, N 3, S. 402—426.
Bourguignat J.-R. Amenites malacologiques. Paris, 1856, t. I, p. 1—255; 1856—1860,
t. II, p. 1—216.
Bourguignat J.-R. Mollusques nouveaux, litigeux ou peu connus (4e decade). — Rev.
Mag. Zool., 1864, t. (2) 16, p. 193—212.
Brown A. Elimination of foreign particles by the snail, Helix aspersa. — Nature, 1967,
v. 213, № 5081, p. 1154—1155.
Brown T. Illustrations of the recent conchology of Great Britain and Ireland, with the
description and localities of all the species, marine, land and fresh water. 2nd ed.
London, 1844, 59 Taf.
Brusina S. Zur Retting unserer Mollusken-Fauna. — Nachrichtenbl. dtsch. malak. Ges.,
1904, Bd 36, S. 157—168.
Burch J. B. Land molluscs of the Southern Appalachians. — In: The Distributional History
of the Biota of the Southern Appalachians, Blacksburg, Virginia, 1969, p. 247—
264.
Biittner K., Ehrmann P. Beitragi zur Molluskenfauna des Ussuri-Gebietes. — Arch. Moll.,
1927, Bd 59, S. 217—250.
Cadarin M., Beladeen V. Contribufii la cunoa^terea morfologici sistemului nervos central
$i a neurosecrefiei la Xerophila obvia Kimakovicz (Helicidae). — Studia Univ.
Babes-Rolyai. Ser. biol., 1968, t. 13, N 2, p. 87—95.
Cameron R. A. D. The distribution of Helicodonta obvoluta (Mull.) in Britain. — J. con-
chol., 1972, v. 27, N 5/6, p. 363—369.
Campion M. The structure and function of the cutaneous glands in Helix aspersa. — Quart.
J. microscop. Sci., 1961, v. 102, p. 195—216.
Carney W. Mortality and apertural orientation in Allogona ptychophora during winter
hibernation in Montana. — Nautilus, 1966, v. 79, N 4, p. 134—136.
Chatfield J. E. The life history of the helicid snail Monacha cantiana (Montagu), with
reference also to M. carthusiana (Muller). — Proc. Malacol. Soc. London, 1968,
v. 38, N 3, p. 233—245.
Clench W. J., Turner R. D. Monographs of the Genera Papustyla, Forcartia and Meliobba
(Papuininae: Camaenidae). — J. Malacol. Soc. Austral., 1962, v. 6, p. 3—33.
Clench W. J., Turner R. D. Monographs of the Genera Megalacron and Rhytidoconcha
(Pdpuininae: Camaenidae). — J. Malacol. Soc. Austral., 1964, v. 8, p. 36—71.
Clench W. J., Turner R. D. Monographs of the Genus Rhynchotrochus (Papuininae: Ca-
maenidae). — J. Malacol. Soc. Austral., 1966, v. 9, p. 59—95.
90
ВВЕДЕНИЕ
Clench W. J., Turner В. D. Monographs of the Genus Letitia (Papuininae: Camaenidae). —
J. Malacol. Soc. Austral., 1968, v. 11, p. 32—49.
Clessin S. Galizische Arten und Varietaten. — Malakol. Bl., N. F., 1879, Bd I, N 12,
S. 42—48.
Clessin S. Mollusken aus Taurien. — Malakol. Bl., N. F., 1881, Bd III, S. 136—141.
Clessin S., Anhang zur Molluskenfauna der Krina. — Malakol. BL, N. F., 1883, Bd IV,
S. 37—52.
Clessin S. Deutsche Excursions-Molluskenfauna. 2 Aufl. Verl. Bauer und Raspe. Nurn-
berg, 1884, S. 1—658.
Cockerell T. D. A. Land Mollusca from the Maritime Province of Siberia. — Ann. and
Mag. Natur. History, ser. 9, 1924, v. 13, p. 579—583.
Cockerell T. D. A. Some helicoid snails from Manchuria. — Ann. and Mag. Natur. Hi-
story, ser. 9, 1926, v. 17, p. 226—228.
Cockerell T. D. A. Helicoid snails from Turkestan. — Ann. and Mag. Natur. History,
ser. 1928, v. 1, p. 361—366.
Cristofori J. de, Jan G. Catalogue in IV sectiones divisus rerum naturalium in museo Jo-
seph! de Cristofori et Georgii Jan. . . complectens adumbrationem oryctognosiae
et geognosiae atque prodromum faunae et florae Italiae superiores. Sectio II. Mi-
lano, 1832.
Croockewit H. W. E. Malakologische Resultate einer Reise nach der asiatische Turkei.
1 Einfiihrung. — Basteria, 1953, N 1—2, p. 17—28.
Da Costa E. M. The British Conchology. London, 1778.
Deblock R., Hoestland H. Donnees biologiques le Gastropoda littoral Theba pisana Mul-
ler aux limites septentrionales de «son extension». — C. r. Acad, sci., 1967, v. 265,
N 12, p. 893—896.
Draparnaud J.-P.-R. Tableau des mollusques terrestres et fluviatiles de la France. Pa-
ris, 1801, p. 1—116.
Draparnaud J.-P.-R. Histoire naturelie des mollusques terrestres et fluviatiles de la
France. Paris, 1805, p. 1—164.
Drozdowski A. Vergleichende Studien uber die Lungenmorphologie bestimmter Schne-
ckenarten (Gastropoda, Pulmonata). — Zool. pol., 1970, Bd 20, N 2, S. 217—256.
Drozdowski A., Burak I., Martinska D. Interdependence between the size of the specimen
and the construction of the radula in Helix pomatia L. and Cepaea nemoralis (L.). —
Studia Soc. sci. Torunensis, 1966, v. 8, N 1, p. 1—19.
Dubois de Montpereux. Voyage autour Caucase, chez les Tcherkesses et les Abkhases, en
Colchide, en Georgie, en Armenie et en Crimee. Paris, Gide, 1839, t. 2; 1839, t. 3;
1840, t. 4.
Dybowski W. Studien uber die Binnenmollusken des Amur-Gebietes. — Nachrichtenbl.
dtsch. malak. Ges., 1901, Bd 33, S. 129—144.
Eakin R. M., Brandenburger J. L. Differentiation in the eye of a pulmonate snail Helix
aspersa. — J. Ultrastruct. Res., 1967, v. 18, p. 391—421.
Ehrmann P. Mollusken.— In: Tierwelt Mitteleuropas. Leipzig, 1956, Bd II, N 1, S. 1—264.
Fernandez J. Nervous system of the snail Helix aspersa. I. Structure and histochemistry
of ganglionic sheath and neuroglia. — J. Compar. Neurol., 1966, v. 127, p. 157—
- 181.
Ferussac J. B. L. Tableaux systematiques des animaux mollusques classes en families
naturelles, dans lesquels en a etabli la concordance de tous les systemes; suivis
d’un prodrome general pour tous les mollusques terrestres ou fluviatiles, vivants
ou fossiles. Paris, 1821.
Fitzinger L. Systematisches Verzeichniss der im Erzherzogthume Oesterreich vorkommen-
den Weichtier. — In: Beitrage zur Landeskunde Oesterreich’s. 1833, Bd 3, S. 88—
122.
Flasar I. Der innere Bau der Befruchtungstasche bei Oxychilus draparnaudi (Beck) und
die Geschichte ihrer Entdeckung und Erforschung bei anderen Pulmonaten. —
Vest. Ceskosl. spolec. zool., 1967, Bd 31, N 2, S. 150—158.
Forcart L. Beitrage zur Kenntnis der Heliciden-Gattung Perforatella Schluter und deren
Untergattung Monachoides Gude & Woodward. — Verhandl. naturforsch. Ges.
Basel, 1946, Bd 57, S. 55—61.
Forcart L. Die systematische Stellung von Helix lanuginosa Boissy, und die Taxionomie
der Gattung Hygromia Risso. — Arch. Moll., 1954, Bd 83, N 4/6, S. 159—162.
Forcart L. Trichia Hartmann, 1840 — nomenklatorisch giiltig. — Arch. Moll., 1958,
Bd 84, N 4/6, S. 153—154.
Forcart L. New research on Trichia hispida (Linnaeus) and related forms. — Proc. I Eu-
rop. malac. congr., 1965, p. 79—93.
Forcart L. Systematische Stellung und Unterteilung der Gattung Sphincterochila Ancey. —
Arch. Moll., 1972, Bd 102, N 4/6, S. 147—164.
Frick W. Der Kalziumstoffwechsel bei Helix pomatia unter dem Einfluss wechselnder
Kohlensaureatmospharen. — Mitt. Zool. Museum Berlin, 1965, Bd 41, N 1,
S. 95—120.
ЛИТЕРАТУРА
91
Fuchs A., Kaufel Er. IV Land- und Siisswassermollusken aus Griechenland und von den
Inseln der Agaischen Meeres. In: Werner’s «Ergebnisse einer zoologischen Studien-
und Sounnelreise nach Griechenland, namentlich nach den Inseln des Agaischen
Meeres». — Sitzungsber. Akad. Wiss. Wien. Math.-naturwiss. KI. Abt. I. 1934,
Bd 143, H. 1 — 2, S. 71—90.
Fuchs A., Kaufel Fr. Anatomische und systematische Untersuchungen an Land- und
Siisswasserschnecken aus Griechenland und von den Inseln des Agaischen Mee-
res. — Arch. Naturg., N. F., 1936, Bd 5, N 4, S. 541—662.
Gaal I. Die sarmatische Gastropodenfauna von Rakosd im Komitat Hunyad. — Mitt,
a. d. Jb. ungar-geol. Anst., 1911, Bd 18 (1), S. 1—112.
Gabe M. La neurosecretion chez les mollusques et ses rapports avec la reproduction. —
Arch. anat. microsc. et morphol. exptl., 1965, t. 54, N 1, p. 371—385.
Garcia san Nicolas E. Estudios sobre la biologia, la anatomia у la sistematica del genero
Iberus Montfort, 1810. — Bol. R. soc. esp. hist, nat., 1957, p. 1—196.
Germain L. Mollusques Terrestres et Fluviatiles d’Asie-Mineure. Paris, 1936.
Gerstfeldt G. Ueber Land- und Siisswassermollusken Sibiriens und des Amur-Gebietes. —
Mem. Savants etrangers, 1859, Bd 9, S. 507—548.
Ghose K. Ch. Morphogenesis of the pedal gland in the giant land snail Achatina fulica
Bowdich. — Current Sci., 1963, v. 32, p. 167—168.
Ghose K. Ch. The structure of the heart of Achatina fulica and Macrochlamys indica and
pace-auricle in A. fulica, with a discussion on the same in gastropods. — Philip-
pine J. Sci., 1964, v. 93, N 2, p. 219—229.
Girod A. Distribuzione ed ecologia di Drepanostoma nautiliforme Porro, Helicodonta ob-
voluta (Muller), Helicodonta angigyra. (Rossmaessler) {Gastropoda, Pulmonata). —
Arch. Moll., 1968, v. 98, N 3/4, p. 121—133.
Gittenberger E. Zur systematischen Stellung von Helix lanuginosa Boissy, mit Neubeschrei-
bung eines Subgenus. — Bol. Soc. Hist. Nat. Baleares, 1968a, Bd 14, S. 63—68.
Gittenberger E. Zur systematik der in die Gattung Trissexodon Pilsbry {Helicidae, Heli-
codontinae) Gerechneten Arten. — Zool. meded., 1968b, Bd 43, N 13, S. 165—
172.
Gittenberger E. Eine' neue Art der Gattung Helicopsis {Gastropoda, Helicidae, Helicellinae)
aus Niederosterreich. — Basteria, 1969, Bd 33, N 1—4, S. 63—68.
Gittenberger E., Backhuys W., Ripken Th. E. J. De landslakken van Nederland. Konink-
lijke Neder. — Natuurhist. Ver., Amsterdam-Zuid, 1970, p. 1—177.
Grossu A. V. Gastropoda Pulmonata. Fauna Rep. Pop. Romine. — Mollusca, 1955, v. 3,
f. 1, p. 1—518.
Gude G. K. The Helicoid Land Shells of Asia. Corrections and Additions. — J. Malac.,
1904, v. 11, N 4, p. 93—94.
GudeG. K., Woodward В. B. On Helicella, Ferussac. — Proc, malacol. Soc. London, 1921,
v. 14, p. 174—190.
Haas F. Malakologische Bemerkungen und Neubeschreibungen. — Senckenbergiana,
1936, Bd 18, № 3/4, S. 143-154.
Habe T. Anatomical Studies on Japanese Land Snails. — Venus, 1955, v. 18, N 4,
p. 221-234.
Habe T. A New North Korean Land Snail, Trishoplita motonoi sp. nov. — Venus, 1963,
v.'22, N 3, p. 237—240.
Harry H. The anatomy of Chilina fluctuosa, reexamined, with prolegomena of the phylo-
geny of the higher limnic Basommatophora. — Malacologia, 1964, v. 1, N 3,
p. 355—385.
Hartmann J. D. W. Erd- und Siisswasser-Gastropoden der Schweiz. Mit Zugabe einiger
merkwiirdigen exotischen Arten. St. Gallen, 1840—1844, S. 1—37, 38—60, 117—
148, 149—156.
Held F. Notizen fiber Weichtiere Bayerns. — Isis (Oken), 1837, v. 4, p. 303—309; v. 12,
p. 901—919.
Hermann H. T. Optic guidance of locomotor behavior in the land snail, Otala lactea. —
Vision Res., 1968, v. 8, p. 601—612.
Hesse P. Helix frequens Mousson {Helicidae, Mollusca). — Изв. Кавк, муз., 1914, т. 6,
с. 253—270.
Hesse Р. Das Genus Levantina Kob. — Nachrichtenbl. dtsch. malak. Ges., 1918a, Bd 50,
S. 40-47.
Hesse P. Die' Subfamilie Helicodontinae. — Nachrichtenbl. dtsch. malak. Ges., 1918b,
Bd 50, S, 99—110.
Hesse P. Beitrage zur naheren Kenntnis der Subfamilie Fruticicolinae. — Arch. Moll.,
1921, Bd 53, S. 55—83.
Hesse P. Beitrage zur genaueren Kenntnis der Subfamilie Helicellinae. — Arch. Moll.,
1926, Bd 58, S. 113—141.
Hesse P. Zur Anatomie und Systematik palaearktischer Stylommatophoren. — Zoologica,
1931, Bd 81, S. 1—118.
93
ВВЕДЕНИЕ
Hesse Р. Zur Anatomie und Systematik palaearktischer Stylommatophoren. — Zoologica,
1934, Bd 85, S. 1—59.
Horne F. Accumulation of urea by a pulmonate snail during aestivation. — Comp. Bio-
chem. and Physiol., 1911, v. 38, N 3, p. 565—570.
Hudec V. Zur Definition der Unterscheidungsmerkmale am Geschlechtsorgan der Schnecke
Perforatella dibothrion (Kim.) und einiger verwandter Arten. — Biologia, Bra-
tislava, 1963, Bd 18, N 5, S. 348—360.
Hudec V. Systematische Stellung und Verbreitung von Trlchia bakowskii in der Tschecho-
slowakei. — Biologia, Bratislava, 1965, Bd 20, N 4, S. 245—259.
Hudec V. Zur Problematik des Vorkommens der Schnecken Helicopsis striata (Mull.) und
H. hungarica (Soos & Wagner) im Karpatenbecken. — Biologia, Bratislava, 1966,
Bd 21, N 3, S. 161—176.
Hudec V. Poznamky к anatomii. nekterych plzd z Madarska. — Casopis Nar. muz., odd.
prirod., 1970, V. 137, N 3/4, p. 33—43.
Hudec V. Poznamky к anatomii nekterych druhil plzft z Krymu. — Casopis Nar. muz.,
odd. prirod., 1912a, v. 141, N 1/2, p. 73—91.
Hudec V. Dodatek к anatomii nekterych druhil plzh z Krymu. — Casopis Nar. muz.,
odd. prirod., 1912b, v. 141, N 1/2, p. 101—104.
Hudec V. Bemerkungen zur Anatomie einiger Schneckenarten (Gastropoda) aus Ruma-
nian. — Ann. Zool. PAN, 1972c, Bd 29, N 10, S. 323—344.
Hudec V. Neue Erkenntnisse fiber kaukasische Fruticocampylaea-Arten und einige ver-
wandte oder konchyologisch ahnliche Formen (Mollusca, Helicidae). — Malak.
Abh. Staatl. Mus. Tierk. Dresden, 1912d, Bd 3, N 2, S. 251—275.
Hudec V., Lezhawa G. I. Kalitinaia n. subgen. des Genus Xerosecta Monterosato, 1892
(Helicidae, Helicellinae). — Arch. Moll., 1961a, Bd 96, N 3/6, S. 131 —137.
Hudec. V., Lezhawa G. I. Bemerkungen zur Erforschung der Landmollusken der Grusini-
schen Sozialistischen Sowietrepublik. — Sbor. Narodn. musea Praze, 1961b,
Bd 23B, N 3, S. 69—100.
Hudec V., Lezhawa G. I. Drei neue Heliciden aus der Grusinischen SSR. — Arch. Moll.,
1969a, Bd 99, N 1—2, S. 41—48.
Hudec V., Lezhawa G. I. Bemerkungen zur Erforschung der Landmollusken der Grusini-
schen Sozialistischen Sowietrepublik (II). — Sbor. Narodn. musea Praze, 1969b,
Bd 25B, N 3, S. 93—155.
Hudec V., Lezhawa G. I. Uber konchyologisch sehr ahnliche kaukasische Heliciden (Mol-
lusca, Pulmonata). — Malak. Abh. Staatl. Mus. Tierk. Dresden, 1910, Bd 3, N 1,
S. 15-31.
Hudec V., Biedel A. Neue Erkenntnisse fiber Chilanodon gerstfeldti (W. Dybowski) (Ga-
stropoda, Helicidae). — Arch. Moll., 1961, Bd 96, N 3/6, S. 124—130.
Hudec V., Vasatko J. Beitrag zur Molluskenfauna Bulgariens. — Acta Sc. Nat. Brno, 1911,
n. s., t. V, Fasc. 2, S. 1—38.
Hunter W. R. Physiological Aspects of Ecology in Nonmarine Molluscs. — In: Milbur,
Jonge. Physiol, of Mollusca. Vol. 1. 1964, p. 108—109.
Hutton Th. Notices of some land- and freshwater shells occuring in Afghanistan. —
J. Asiat. Soc. Benghal, N. S., 1849—1850, v. 18, N 2, p. 649—661.
Ihering H. Morphologic und Systematik des Genitalapparatus von Helix. — Z. wiss. Zool.,
1892, Bd 54, H. 1/2, S. 386—433; H. 3, S. 425—520.
Ihering H. System und Verbreitung der Heliciden. — Verhandl. zool.-hot. Ges. Wien,
1909, Bd 59, N 10, S. 420—455.
Jacobi A. Japanische beschalte Pulmonaten. Anatomische Untersuchungen des im Zo-
ologischen Museum der Kaiserlichen Universitat in Tokyo enthaltenen Materials.
I. Pulmonaten. — J. Coll. Sci. imp. Univ. Tokyo, 1898, v. 12 (1), p. 1 — 102.
Jeffreys J. G. A synopsis of the testaceous pneumonobranchous mollusca of Great Bri-
tain. — Trans. , Linn. Soc. London, 1830, v. 16 (2), p. 323—392.
Jong-Brink M. de. Histochemical and electron microscope observations on the reprodu-
ctive tract of Biomphalaria glabrata (Australorbis glabratus), intermediate host
of Schistosoma mansoni. — Z. Zellforsch., 1969, v. 102, N 4, p. 507—542.
Isarankura K., Runham N. W. Studies on the replacement of the Gastropod radula. —
Malacologia, 1968, v. 7 (1), p. 71—91.
Kaiser P. Bau, Entwicklung und Regeneration des Haarkleides von Trichia hispida (Lin-
naeus) zugleich ein Beispiel fur eine einfache Musterbildung im Tierreich. — Arch.
Moll., 1966, Bd 95, N 3/4, S. 111—122.
Kaleniczenko J. Revision des Escargots (Helices) russes enumeres par I. A. Krynicki. —
Bull. Soc. Nat. Moscou, 1853, v. 26, p. 68—94.
Kilias R. Weinbergschnecken. Berlin, 1960, S. 1—94.
Kilias R. Stamm Mollusca — Weichtiere. — In: Urania Tierreich. Wirbellose Tiere 1.1967,
S. 318—507.
Kimakowicz M. Beitrag zur Molluskenfauna Siebenbiirgens. — Verb. u. Mitt. Sieben-
bfirgens. Ver. Naturw. Hermannstadt, 1883—1884, S. 33—34.
ЛИТЕРАТУРА
93
Klika В. Verzeichniss der von Dr. V. Vavra wahrend seiner Kaukasus-Reise gesamelten
Binnenconchylien. — Spp. Abdr. Sitzungsb. Bohm. Ges. Wiss. Math.-naturw.
KI., 1893, S. 1—7.
Knipper H. Systematische, anatomische, okologische und tiergeographische Studien an
siidosteuropaischen Heliciden (Moll. Pulin.). — Arch. Naturg., N. F., 1939,
Bd 8, N 3/4, S. 327—517.
Kobelt W. Catalog der im europaischen Faunengebiet lebenden Binnenconchylien mit
besonderer Beriicksichtigung der in Rossmassler’s Sammlung enthaltenen Arten.
Cassel, 1871, S. 1—150.
Kobelt W. Zusatze und Berichtigungen zu meinem Catalog der im europaischen Faunen-
gebiet lebenden Binnenconchylien. — Jahrb. dtsch. malak. Ges., 1877, Bd 4,
S. 14-45.
Kothbauer H. Beitrage zur Fortpflanzengsbiologie und postembryonalen Entwicklung der
Weinbergschnecke (Helix pomatia L.). — Sitzungsber. Osterr. Akad. Wiss. Math.-
naturwiss. KI. Abt. I, 1972, Bd 180, N 1—4, S. .65—86.
.Kothbauer H., Schnitzler S. Haemagglutinine aus Cepaea hortensis (Miill.), C. nemoralis
(L.) und C. vindobonensis (Pfeiff.) (Helicidae, Gastropoda): Ihre Verwertbarkeit
fur die Artsystematik. — Z. zool. Syst. E'volut.-forsch., 1972, Bd 10, S. 133—
137.
Krynicki I. A. Novae species aut minus cognitae e Chondri, Bulimi peristomae helicisque
generibus praecipue Rossiae meridionalis. — Bull. Soc; Nat. Moscou, 1833, v. 6,
p. 391—436.
Krynicki I. A. Helices proprie dictae hucusque in limitibus Imperii Rossici observatae. —
Bull. Soc. Nat. Moscou, 1836, v. 9, p. 147—214.
Krynicki I. A. Conchylia. — Bull. Soc. Nat. Moscou, 1837, v. 10, p. 50—64.
Kuhlmann D. Neurosekretion bei Heliciden (Gastropoda). — Z. Zellforsch., 1963, v. 60,
N 6, p. 909—932.
Kuroda T. A Catalogue of the Non-Marine Mollusks of Japan, including the Okinawa and
Ogasawara Islands. — Malac. Soc. Jap., Tokyo, 1963, p. 1—71.
Lehmann R. Die lebenden Schnecken und Muscheln der Umgegend Stettins und im Pom-
mern mit besonderer Beriicksichtigung ihres anatomischen Baues. Cassel, 1873,
S. 1—238.
Lind H. Hibernating behavior of Helix pomatia L. (Gastropoda, Pulmonata). — Vid. medd.
Dansk naturhistor. foren. Knbenhavn, 1968, v. 131, p. 129—151.
Lind H. The functional significance of the spermatophore and the fate of spermatozoa
in the genital tract of Helix pomatia (Gastropoda: Stylommatophora). — J. Zool.,
London, 1973, v. 169, p. 39—64.
Lindholm W. A. Bemerkungen fiber Schnecken von Irkutsk (Sibirien). — Nachrichtenbl.
dtsch. malak. Ges., 1912, Bd 44, S. 62—68.
Lindholm W. A. Miszellen zur Malakozoologie des Russischen Reiches, I—XIII. —
Ежегод. Зоол. музея, 1913a, S. 151—167.
Lindholm W. A. Beschreibung neuer Arten und Formen aus dem Kaukasus-Gebiete. —
Nachrichtenbl. dtsch. malak. Ges., 1913b, Bd 45, S. 17—23, 62—69.
Lindholm W. A. Neue Heliciden aus dem Kaukasus-Gebiete. — Nachrichtenbl. dtsch.
malak. Ges., 1913c, Bd 45, S. 137—144.
Lindholm W. A. Ein Beitrag zur Kenntnis der Molluskenfauna der Krim. — Arch. Moll.,
1926, Bd 58, S. 161—177.
Lindholm W. A. Zur Systematik und Nomenklatur einiger Heliciden und ihrer Verwand-
ten. - Arch. Moll., 1927a, Bd 59, N 2, S. 116-138.
Lindholm W. A. Nachtrag zu meinem Heliciden-aufsatz in Heft 2 des Archivs fur 1927. —
Arch. Moll., 1927b, Bd 59, N 5, S. 316.
Lindholm W. A. Diagnosen neuer Landschnecken aus dem Ferghana-Gebiete. — Zool.
Anz., 19.27c, Bd 74, N 7/9, S. 97-102, 195-202.
Lindholm W. A. Neue Landschnecken aus den Russischen Zentralasien. — Arch. Naturg.,
1927d, Bd 92, H. 2, Abt. 2, S. 257—269.
Lindholm W. A. Zur Synonimie einiger kaukasischer Heliciden. — Arch. Moll., 1928,
Bd 60, S. 206—208.
Lindholm W. A. Die gezahnten Heliciden des Kaukasus. — Arch. Moll., 1929, Bd 59,
N 4/5, S. 205—211.
Linnaeus C. Systema naturae, ed. 10, reformata 1, Holmiae, 1758.
Lozek V. Klic ceskoslovenskych mekkysd. — Slov. Akad. Vied. Bratislava, 1956, p. 1 —
437.
Lozek V. Quartarmollusken der Tschechoslowakei. Praha, 1964, p. 1—374.
Lozek V. Stepni plzi Chondrula tridens (Muller) a Helicella obvia (Hartmann) v jiznich
Cechach. — Sb. Jihocesk. muz. Cesk. Budejov. Prir. vedy, 1970, t. 10, N 2,
p. 73—79.
Machin J. The evaporation of water from Helix aspersa. I. The nature of the evaporating
surface. — J. Exp. Biol., 1964a, v. 41, p. 759—769.
94
ВВЕДЕНИЕ
Machin J. The evaporation of water from Helix aspersa. II. Measurement of air flow and
the diffusion of water vapour. — J. Exp. Biol., 1964b, v. 41, p. 771—781.
Machin J. The evaporation of water from Helix aspersa. III. The application of evapora-
tion formulae. — J. Exp. Biol., 1964c, v. 41, p. 783—792.
Machin J. The evaporation of water from Helix aspersa. IV. Loss from mantle of the ina-
ctive snail. — J. Exp. Biol., 1966, v. 45, N 2, p. 269—278.
Machin J. Structural adaptation for reducing water-loss in three species of terrestrial
snail. — J. Zool., 1967, v. 152, N 1, p. 55—65.
Mandahl-Barth G. Recherches systematiques sur quelques especes du genre Geomitra (Heli-
cidae'). — Vid. medd. Dansk naturhistor. foren. Knbenhavn, 1937—1938, v. 101,
p. 137—152.
Martens E. Aufzahlung der von Dr. Alexander Brandt in Russisch-Armenien gesammelten
Mollusken. — Bull. Acad. St.-Petersb., 1880, Bd 24, S. 142—158.
Martens E. Conchologische Mittheilungen als Fortsetzung der Novitates conchologicae.
1 B., Kassel, 1881, S. ,1—101.
Martens E. Ueber Centralasiatische Mollusken. — Mem. Acad. Sci. St.-Petersb., 7 ser.,
1882a, Bd 30, N 11, S. 1—65.
Martens E. Ueber von Herrn Apollo Kuschakewitz gesammelte centralasiatische Land-
und Siisswasserschnecken. — Sitzungsber. Ges. naturf. Fr., Berlin, 1882b, S. 103—
107.
Martens E. Centralasiatische Landschnecken. — Sitzungsber. Ges. naturf. Fr., Berlin,
1885, S. 17—18.
Matzke M. Gastropoden am Vogelherd zwischen Ilmenau und Stutzerbach (Thur.). —
Malak. Abh. Staatl. Mus. Tierk. Dresden, 1968, Bd 2, N 2, S. 279—288.
Middendorff A. Reise in den ausserten Norden und Osten Sibiriens. 1851, Bd II, N 1,
Mollusken, S. 163—464.
Miller W. B. Greggelix, a new genus of autochtonous land snails (Helminthoglyptidae)
from Baja California. — Nautilus, 1972a, v. 85, N 4, p. 128—135.
Miller W. B. The anatomy of Speleodiscoides spirellum A. G. Smith, 1957. — Occasion,
papers Calif. Acad. Sci., 1972b, v. 97, p. 1—9.
Mollendorff O. Binnen-Mollusken aus Westchina und Centralasien. — Ежегод. Зоол. Му-
зея, 1899, S. 46—144.
Monterosato T. A. Molluschi terrestri delle isole adiacenti alia Sicilia. — Atti Accad. sci.
lett. e belle Arti, 1892, t. (3) 2, p. 1—33.
Moquin-Tandon A. Histoire naturelie des Mollusques terrestres et fluviatiles de France,
part 2. Paris, 1855, p. 1—646.
Mortillet G. Descriptions de quelques coquilles nouvelles d’Armenie, et considerations
malacostatiques. — Mem. Inst. Nat. Geneve, 1854, t. II, p. 1—15.
Mousson A. Coquilles terrestres et fluviatiles recueillies dans 1’Orient par M. le Dr. Alex.
Schlafli. — Vierteljahrschr. Nat. Ges. Zurich, 1863, Bd 8, p. 275—320, 368—426.
Mousson A. Coquilles recueillies par M. le Dr. Sievers dans la Russie,meridionale et asia-
tique. — J. conchyliol., 1873, t. 21, p. 193—230. ,
Mousson A. Coquilles recueillies par M. le Dr. Sievers dans la Russie asiatique. — J. con-
chyliol., 1876, t. 24, p. 137—148.
Mousson A. Coquilles terrestres et fluviatiles recueillies par M. Louis Graeser dans le
bassin de Г Amour. — J. conchyliol., 1887, t. 35, p. 10—32.
Muller О. E. Vermium terrestrium et fluviatilium sen animalium infusoriorum, helmin-
ticorum et testaceorum, non marinorum succincta historia, 2. Havniae et Lipsiae,
1774, p. 1—214.
Nopp H., Farahat A. Z. Temperatur und Regulation des Zellenstoffwechels bei. winter-
schlafenden Helix pomatia L. — Anz. Osterr. Akad. Wiss. math.-naturwiss. KI.,
1968, Bd 104, N 1—14, S. 171—175.
Ortiz de Zarate A. Observaciones anatomicas у posicion sistematica de varios helicidos
espaboles. V. Genero Oestophora Hesse, 1907. — Bol. R. Soc. esp. hist. nat. (B),
1962, t. 60, p. 81—104.
Pfeiffer C. Systematische Anordnung und Beschreibung deutscher Land- und Wasser-
Schnecken mit besonderer Riicksicht auf die bischer in Hessen gefundenen Arten. —
Naturg. Deutsch. Moll., 1828, Bd III, S. 1—84.
Pfeiffer L. Symbolae ad Historiam Heliceorum. 1846, v. 3, p. 1 — 100.
Pfeiffer L. Monographia Heliceorum viventium sistens descriptiones systematicas et
criticas omnium huius familiae generum et specierum hodie cognitarum. Lipsiae,
1848—1877, Bd 3, S. 1—711 (1853); Bd 4, S. 1—920 (1859); Bd 7, S. 1-674.
Pfeiffer L. Notiz uber serbische und sibirische Schnecken. — Z. Malak., 1853, S. 185—
189.
Pilsbry H. (in Tryon-Pilsbry). Manual of Conchology. 1886—1931, v. 1—28.
Pilsbry H. On the Zoological Position of Partula and Achatinella. — Proc. Acad. Natur.
Sci. Philadelphia, 1900, p. 561—567.
Pilsbry H. Non-marine Mollusca of Patagonia. Reports of the Princeton Univ. Exped.
to Patagonia, 1896—1899. V. 3, part V. Stuttgart, 1911, p. 513—633.
ЛИТЕРАТУРА
95
Pilsbry Н. Review of the Land Mollusca of Korea. — Proc. Acad. Natur. Sci. Philadelphia,
1926, v. 78, p. 453-475.
Pilsbry H. Review of Japanese land Mollusca. I. — Proc. Acad. Natur. Sci. Philadelphia,
1927, v. 79, p. 13—20.
Pilsbry H. The structure and affinities of Humboldtiana and related Helicid genera of
Mexico and Texas. — Proc. Acad. Natur. Sci. Philadelphia, 1928, v. 79, p. 162—
192.
Pilsbry H. Land Mollusca of North America (North of Mexico). V. I, part 1. Philadelphia,
1939, p. 1—573.
Pilsbry H. Land Mollusca of North America (North of Mexico). V. I, part 2. Philadelphia,
1940, p. 575-994.
Pilsbry H., Hirase. Descriptions of new Japanese Land Shells. — Nautilus, 1904, v. 17,
p. 114—126.
Pohunkovd H. The Ultrastructure of the Lung of the Snail Helix pomatia. — Folia morphol.,
1967, v. 15, N 3, p. 250—257.
Polinski W. Recherches sur 1’anatomie et la systematique des Xerophilines de la Pologne
et de la Transsylvanie. — Prace zool. Panstw. Mus. Przyrod., Warszawa, 1922,
t. 1, p. 166—184.
Polidski W. Anatomisch-systematische und zoogeographische Studien iiber Heliciden Po-
lens. — Bull. Acad. Sci et de Lett. ser. B, Krakow, 1924, p. 131—279.
Politiski W. Sur certains problemes du developpement morphologique et zoogeographique
de la faune des Alpes et des Karpates illustres par 1’etude detaillee des Helici-
des du groupe Perforatella auct. — Ann. Mus. Zool. Pol., 1929, t. 7, N 4,
p. 137—229.
Pomeroy D. E. Dormancy in the land snail, Helicella virgata (Pulmonata: Helicidae). —
Austral. J. Zool., 1968, v. 18, N 5, p. 857—869.
Pomeroy D. E. Some aspects of the ecology of land snail, Helicella virgata, in South Aust-
ralia. — Austral. J. Zool., 191)9, v. 19, p. 495—514.
Quattrini D. Osservazioni sulla ultrastruttura dei dotti escretori delle ghiandole salivari
di Helix aspersa Miiller {Mollusca Gastropoda Pulmonata). — Caryologia, 1967,
t. 20, N 2, p. 191—206.
Radwin G. Technique for extraction and mounting of gastropod radulae. — Veliger, 1969,
v. 12, N 1, p. 143—144.
Rainer M. Chromosomenuntersuchungen an Gastropoden (Stylommatophoren). — Ma-
lacologia, 1967, v. 5, N 3, p. 341—373.
Regondaud J. Developpement de la cavite pulmonaire et de la cavite palleale chez Lym-
naea stagnalis. — C. r. Acad, sci., Paris, 1961, t. 252, p. 179—181.
Renzoni A. Observations on the tentacles of Vaginulus borellianus Colosi {Mollusca: Ga-
stropoda: Soleolifera). — Veliger, 1969, v. 12, N 2, p. 176—181.
Retowski O. Liste der von mir auf meiner Reise von Konstantinopel nach Batum gesam-
melten Binnenmollusken. — Ber. Senckenb. Nat. Ges., 1889, S. 225—265.
Retowski 0. Materialen zur Kenntnis der Molluskenfauna des Kaukasus. — Изв. Кавк,
муз., 1914, т. 6, вып. 4, с. 271—347.
Rigby J. Е. Alimentary and reproductive system of Oxychilus cellarius. — Proc. Zool.
Soc. London, 1963, v. 141 (2), p. 311—359.
Risso A. Histoire naturelie des principals productions de 1’Europe meridionale et parti-
culierement de celles des environs de Nice et des Alpes Maritimes. Paris, 1826,
p. 1—439.
Rogers D. C. The fine structure of sensory neurons and their processes in the optic tentac-
les of Helix aspersa. — Z. mikrosk.-anat. Forsch., 1971, v. 84, № 1, p. 52—64.
Rosen О. B. Contribution a la faune malacologique terrestre du Turkestan (description de
deux especes nouvelles). — Feuille Jeunes Natur., 1897, p. 170.
Rosen О. B. Die Mollusken Ciskaukasiens und speciell des Kuban-Gebietes. — Ежегод.
Зоол. муз., 1911, т. XVI, с. 86—142.
Rosen О. В. Katalog der Schalentragenden Mollusken des Kaukasus. — Изв. Кавк, муз.,
1914, т. 6, с. 141-252.
Rossmdssler Е. A. Iconographie der Land- und Siisswasser-Mollusken, Bd 1—3. Fortge-
setzt von W. Kobelt; Bd 4—7 und N. F.; Bd 1—23 und Suppl. — B. Erschien bis
1920. 1835—1859.
Runham N. W. Rate of replacement of the molluscan radula. — Nature, 1962, v. 194
(4832), p. 992-993.
Runham N. W: A study of the replacement mechanism of the pulmonate radula. — Quart. .
J. Microscop. Sci., 1963, v. 104, p. 271—277.
' Sanchis C. A., Zambrano D. The Structure of the Central Nervous System of a Pulmonate
Mollusc {Cryptomphalus aspersa). I. Ultrastructure of the Connective Epineural
Sheath. — Z. Zellforsch., 1969, v. 94, N 1, p. 62—71.
Schileyko A. A. The system of the group Trichia s. lat. {Pulmonata: Helicoidea: Hygromi-
idae). — Malacologia, 1977, v. 17, N 1, p. 1—56.
Schlesch H. Ueber sibirische Heliciden. — Arch. Moll., 1928, Bd 60, S. 276—283.
96
ВВЕДЕНИЕ
Schluter F. Kurzgefasstes systematisches Verzeichniss meiner Conchyliensammlung nebst
Andeutung aider bis jetzt von mir bei Halle gefundenen Land- und Flussconchylien.
Halle, 1838, Bd 1—7, S. 1—40.
Schmidt H. Abhangigkeit der Darmgestaltung der Landpulmonaten von phylogenetischen
und okologischen Faktoren sowie von der Gehauseform. — Arch. Naturg., N. F.,
1936, Bd 5, S. 18—140.
Schrenck L. Reise und Forschungen im Amur-Lande in den Jahren 1854—1856. II. Mol-
lusken des Amur-Landes und des Nordjapanischen Meeres. St.-Petersburg, 1867,
S. 260—976.
Schmalbach G., Lickfeld K. G., Hahn M. Der mikromorphologische Aufbau des Linsenauges
der Weinbergschnecke (Helix pomatia L.). — Protoplasma (Wien), 1963, Bd 56,
S. 242—273.
Semper C. Reisen im Archipel der Philippinen, 2. Theil 3: Landmollusken (1—7). Wiesba-
den, 1870, S. 1—327.
Siemaschko J. Beitrag zur Kenntnis der Konchylien Russlands. — Bull. Soc. Nat. Mos-
cou, 1847, Bd 20, S. 93—131.
Sihler H. Das Liebesspiel der Weinbergschnecken. — Aus Heimat, 1954, Bd 62, N 1,
S. 16—19.
Slosarski A. Materyjaly do Fauny malakozoologicznej Krolestwa Polskiego. — Odb.
z. Pam. Fizjograf., 1881, v. 1, p. 1—64.
Smith B. J. The secretions o£ the reproductive tract of the garden slug, Arion ater. — Ann.
New York Acad. Sci., 1965, v. 118, Art. 24, p. 997—1014.
Salem A. Malacological Applications of Scanning Electron Microscopy. II. Radular Stru-
cture and Functioning. — Veliger, 1972a, v. 14 (4), p. 327—336.
Solem A. Molluscs from prehistoric sites in Afghanistan. — Trans. Amer. Philos. Soc.,
1972b, v. 62 (4), p. 57-84.
Solem A. Convergence in Pulmonate Radulae. — Veliger, 1973, v. 15 (3), p. 165—171.
Stephens G. J., Stephens G. C. Photoperiodic stimulation of egg laying in the land snail
Helix aspersa. — Nature, 1966, v. 212, N 5070, p. 1582.
Sturany R., Wagner A. J. Uber schalentragende Landmollusken aus Albanien und
Nachbargebieten. — Denkschr. k. Akad. Wiss. Wien, Math.-nat. KI., 1915, Bd 91,
S. 19—138.
Suzuki N. Behavioural and electrical responses of the land snail, Ezohelix flexibilis (Ful-
ton), to odours. — J. Fac. Sci. Hokkaido Univ., Ser. VI, 1967, v. 16, N 2, p. 174—
185. '
Talmadge R. R. Notes on coastal land snails. — Nautilus, 1967, v. 80, N 3, p. 87—89.
Taylor J. W. Monograph of the land and freshwater Mollusca of the British isles. Part III.
London, 1914, p. 1—160.
Taylor D. W., Sohl N. F. An outline of gastropod classification. — Malacologia, 1962,
v. 1, N 1, p. 7—32.
Tercafs R. R. Un nouveau mollusque terrestre pour la faune cavernicole beige Helicigona
lapicida L. (gaster. styl'l.). — Soc. roy. beige etudes geol. et archeol. «cherch. Wal-
lonie (Bull.)», 1958—1960, t. 17, p. 170—172.
Trappmann W. Die Muskulatur von Helix pomatia L. — Z. wiss. Zool. A, 1916, Bd 105,
N 4, S. 489—585.
Turton W. A manuel of the land and freshwater shells of British Islands, arranged accor-
ding to the modern systems of classifications: and described from perfect speci-
mens in the author’s cabinet: with coloured plates of every species. London, 1831.
Ulicny J. Uber Helix Clessini sp. n. — Malak. BL, 1885, Bd VII, N. F., S. 1—8.
Valovirta I. Land molluscs in relation to acidity on hyperite hills in Central Finland. —
Ann. zool. fennica, 1968, v. 5, N 3, p. 245—253.
Van Bruggen A. Additional notes on South African introduced molluscs, including a spe-
cies new to the list. — Ann. Natal. Mus., 1967, v. 18, N 3, p. 501—504.
Van Mol J.-J. Notes anatomiques sur les Helicarionidae (Gasteropodes pulmones). I.
Etudes des genres Macrochlamys, Parmarion et Xesta. — Ann. Soc. roy. zool. Belg.,
1968, t. 98, N 3, p. 189—215.
Van Mol J.-J. Notes anatomiques sur les Bulimulidae (Mollusques, Gasteropodes, Pul-
mones). — Ann. Soc. roy. zool. Belg., 1971, t. 101, N 3, p. 183—226.
Vappula N. Pests of cultivated plants in Finland. — Acta entomol. fennica, 1965, v. 19,
p. 1—239.
Verdcourt B. On the systematic position of some East African Helicidae I. — Arch. Moll.,
1969, v. 99, N 3—4, p. 175—185. '
Walden H. Historical and taxonomical aspects of the land Gastropoda in the North At-
lantic region.—In: North Atlantic Biota and their History, Pergamon Press, 1963,
p. 153—171.
Warburg M. R. On the water economy of some Australian land-snails. — Proc. Malac.
Soc. London, 1965, v. 36, N 5, p. 297—305.
Wiireborn I. Environmental factors influencing the distribution of land molluscs of an
oligotrophic area in southern Sweden.—Oikos, 1970, v. 21, N 2, p. 285—291?
ЛИТЕРАТУРА
97
Webb G. R. The mating-Anatomy Technique as applied to Polygyrid Landsnails. — Amer.
Naturalist, 1947, v. 81, N 797, p. 134—147.
Weber A. Ueber Binnenmollusken aus Ost-Turkestan, Kaschmir und West-Tibet. — Zool.
Jahrb., Abt. 3, Syst., Okol. u. Geogr., 1910, Bd 29, S. 299—312.
Weber A. Wissenschaftliche Ergebnisse der Reise von Prof. Dr.G. Merzbacher im zentra-
len und ostlichen Thian-Schan 1907/8. III. Mollusken. — Abhandl. Konig. Baye-
rischen Akad. Wiss. Math.-phys. KI., 1913, XXVI, Bd 5, S. 1—18.
Wenz W., Zilch A. Euthyneura. In: Handbuch d. Palaozoologie. Gastropoda, Th. 2. Ber-
lin, 1959—1960, S. 1—834.
Westerlund C. A. Neue Binnenmollusken aus Sibiriens. — Nachrichtenbl. dtsch. malak.
Ges., 1876, Bd 8, S. 97—104.
Westerlund C. A. Sibiriens Land- och Sotvatten-Mollusken. — Svenska vetensk. Acad.,
N. F., 1877, t. 14, N 12, p. 1—111.
Westerlund C. A. Fauna der in der palaarctischen Region lebenden Binnenconchylien.
II. Gen. Helix. Berlin, 1889, S. 1—473.
Westerlund C. A. Neue centralasiatische Mollusken. — Ежегод. Зоол. муз., 1896, т. 1,
с. 181—198.
Westerlund С. A. Beitrage zur Molluskenfauna Russlands. — Ежегод. Зоол. муз., 1897,
т. 2, с. 117—143.
Westerlund С. A. Novum Specilegium Malacologicum. — Ежегод. Зоол. муз., 1898, т. 3,
с. 155—183.
Westerlund С. A. Malacologische Bemerkungen. — Nachrichtenbl. dtsch. malak. Ges.,
1902, Bd 34, S. 35—47.
Wiegmann F. Binnen-Mollusken aus Westchina und Centralasien. — Ежегод. Зоол. муз.,
1900, ч. 1, с. 1—219.
Wurtz С. В. The American Camaenidae (Mollusca: Pulmonata). — Proc. Acad. Natur.
Soc. Philadelphia, 1955, v. 107, p. 99—143.
Yen T. C. Review and summary of Tertiary and Quaternary non marine mollusks of
China. — Proc. Acad. Natur. Soc. Philadelphia, 1943, v. 95, p. 267—309.
Yom-Tov Y. The biology of two desert snails Trochoidea (Xerocrassa) seetzeni and Sphin-
cterochila boissieri. — Isr. J. Zool., 1971, v. 20, N 3, p. 231—248.
Yom-Tov Y., Galun M. Note on feeding habits of the desert snails Sphincterochila bois-
sieri Charpentier and Trochoidea (Xerocrassa) seetzeni Charpentier. — Veliger,
1970, v. 14, N 1, p. 86—88.
Zilch A., Jaeckel S. G. A. Mollusken.—In: Die Tierwelt Mitteleuropas. Erganzung
zu II Bd., Lief. 1. Leipzig, 1962, S. 1—294.
7 А. А. Шилейко
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕНТНЫХ ВИДОВ НАДСЕМЕЙСТВА
HELICOIDEA ФАУНЫ СССР ПО РАКОВИНЕ
1 (4). При рассматривании снизу базальный край устья выглядит слегка изогнутым,
в виде лука; края устья без зубов, резко отогнуты под почти прямым углом;
устье округло-треугольное или в виде широкой изогнутой щели.
2 (3). Раковина плоская, базально-палатальный край устья оттянут вправо . . . ..
............................................. Helicodonta obvoluta (стр. 115).
3 (2). Раковина прижатая, базально-палатальный край устья почти параллелен
париетальному.............................Lindholmiola corcyrensis (стр. 116).
4 (1). Раковина иная.
5 (30). Устье раковины с зубами. Большой диаметр не более 20 мм.
6 (15). Париетальная стенка устья с 1—3 зубами.
7 (12). Париетальный зуб-один, в виде радиальной складки.
8 (11). На базально-палатальном крае устья имеются 2 зуба.
9 (10). Большой диаметр раковины не менее 9 мм. Вид обитает на Карпатах . . ..
..........................................Isognomostoma personatum (стр. 315).
10 (9). Большой диаметр не превышает 8 мм. Вид обитает в Восточной Сибири . . .
...........................................Triodopsis subpersonatum (стр. 347).
11 (8). На базально-палатальном крае устья имеется мощная губа, резко обрываю-
щаяся справа ..................................Paedhoplita laminata (стр. 302).
12 (7). Париетальных зубов 2—3.
13 (14). Париетальные зубы в виде двух сближенных параллельных бугорков . . .
...........................................Paedhoplita kirgisensis (стр. ,304).
14 (13). Париетальные зубы представлены сочетанием бугорка и подкововидной
пластинки, которая изредка распадается на два удлиненных бугорка ....
...........................................Paedhoplita lindholmi (стр. 303).
15 (6). Париетальная стенка устья лишена зубов; в крайнем случае они исчезающе-
слабые. На базально-палатальном крае имеются 2 почти одинаковых зуба;
изредка ближе к колумеллярному краю может присутствовать еще один малень-
кий зуб.
16 (17). Периферия раковины явственно угловатая. Вид обитает в Средней Азии
............................................... Odontotrema diplodon (стр. 167).
17 (16). Периферия раковины округлая или с очень легким углом. Распространение
иное.
18 (23). Виды обитают на Кавказе.
19 (20). Раковина почти шаровидная, тонкостенная.............,...............
........................................Karabaghia bituberosa (стр. 262).
20 (19). Раковина прижатая, твердостенная.
21 (22). Большой диаметр раковины 9—10 мм . . Diodontella stschukini (стр. 203).
22 .(21). Большой диаметр раковины 5—7 мм . . . Diodontella nubigena (стр. 203).
23 (18). Виды обитают в Европе или на Алтае.
24 (29). Раковина плоская или кубаревидная; в последнем случае вершина слегка
выступающая. Виды обитают в европейской части СССР.
25 (26). Раковина плоская........................Causa holosericum (стр. 317).
26 (25). Раковина кубаревидная.
27 (28). Большой диаметр раковины не превышает 9 мм..........................
...........................................Perforatella bidens (стр. 231).
28 (27). Большой диаметр раковины не менее 10 мм.............................
........................................Perforatella dibothrion (стр. 232).
29 (24). Раковина кубаревидная, вершина сглаженная. Вид обитает на Алтае . . .
...............................................Chilanodon bicallosa (стр. 228).
30 (5). Устье без зубов; на колумеллярном или базальном крае может быть утолщение
в виде наплыва. Большой диаметр раковины может превышать 20 мм.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО РАКОВИНЕ
'99
31 (36). На колумеллярном или базальном крае устья имеется зубообразное утолще-
ние, наплыв.
32 (35). Утолщение располагается на колумеллярном крае устья. Пупок широкий
или узкий, но не проколовидный.
33 (34). Раковина прижатая, пупок развернутый, перспективный..................
....................................... Bradybaena dieckmanni (стр. 137).
34 (33). Раковина шаровидная, пупок узкий, не Перспективный
. . ..................................... Bradybaena arcasiana (стр. 139).
35 (32). Утолщение располагается на базальном крае устья, пупок проколовйдный
............................................... Chilanodon gerstfeldti (стр. 229).
36 (31). Края устья лишены бугорков и наплывов.
37 (50). Губа ограничена базальным краем устья, часто резко обрывается при переходе
в палатальный край.
38 (45). Устье округлое или полулунное. Виды обитают в Европе.
39 (42). Раковина прижатая.
40 (41). Пупок цилиндрический, умеренно перспективный, ширина его составляет
1/6 большого диаметра ................................Trichia hispida (стр. 205).
41 (40). Пупок развернутый, перспективный, ширина его составляет 1/3 большого
диаметра .............................................Trichia concinna (стр. 207).
42 (39). Раковина кубаревидная.
43 (44). Завиток конический, высота раковины 5.5—6.0, большой диаметр 8—10 мм
......................................................Trichia bielzi (стр. 209).
44 (43). Завиток куполовидный, высота раковины 4—4.5, большой диаметр 6.5—
7.0 мм ....................................Edentiella bakowskii (стр. 210).
45 (38). Устье в виде узкой или широкой щели. Виды обитают в Средней Азии.
46 (47). Последний оборот с острым углом по периферии. Раковина имеет линзовидную
форму..........................................Angiomphalia lentina (стр. 300).
47 (46). Угол по периферии плавно закруглен и не придает раковине линзовидной
формы.
48 (49). Ширина пупка составляет большого диаметра............................
............................................. Archaica labianix (стр. 261).
49 (48). Ширина пупка составляет 1/12—1/j4 большого диаметра..................
...........................................Angiomphalia buamica (стр. 301).
50 (37). Губа, если имеется, на всех стенках устья (кроме париетальной) развита
примерно в равной степени или постепенно ослабляется от колумеллярного края
к ангулярной области.
51 (62). Раковина с ясным углом или килем по периферии, линзовидная, сверху и
снизу выпуклость раковины примерно одинаковая. Обороты в профиле имеют
резко угловатую или почковидную форму. Поверхность раковины без чешуй-
чатой скульптуры.
52 (53). Вид обитает в Средней Азии...........................................
.........................Bradybaena plectotropis smsp. plectotropis (стр. 150).
53 (52). Виды населяют Кавказ или европейскую часть СССР.
54 (57). Радиальная ребристость отсутствует. Виды обитают на западе европейской
части .СССР.
55 (56). Раковина твердостенная, с зернистой скульптурой. Устье цельное или почти
цельное; в последнем случае имеется ясная париетальная мозоль...................
............................................ Helicigona lapicida (стр. 313).
56 (55). Раковина тонкостенная, с тонкой частой спиральной скульптурой. Край
устья на париетальной стенке прерван.................Drobacia banatica (стр. 317).
57 (54). На раковине имеется более или менее ясная радиальная ребристость. Виды
встречаются на Кавказе.
58 (59). Раковина с резкими ребрами, ее периферия с грубым зазубренным килем.
Вид населяет низменные степные участки..........................................
.......................................Xerosecta sehelkovnikovi (стр. 254).
59 (58). Раковина со сглаженными ребрами, ее периферия угловатая или с гладким
не зазубренным килем. Виды обитают в горах.
60 (61). Палатальный край устья с тупым углом. Высота завитка равна высоте устья
или немного меньше. Поверхность раковины между радиальными ребрами
тонкозернистая ..................................Platytheba mingrelica (стр. 271).
61 (60). Палатальный край устья с острым углом и желобком. Высота завитка равна
половине высоты устья или еще меньше. Поверхность раковины между радиаль-
ными ребрами без зернистой скульптуры. . Platytheba prometheus (стр. 273).
62 (51). Киль или угол по периферии отсутствуют; если имеются, то не придают
раковине линзовидной формы. На поверхности раковины могут присутствовать
чешуйки.
63 (64). По периферии имеется притупленный угол. На поверхности располагаются
редкие, сильно сглаженные ребра. Окраска зеленовато-бурая или оливковая.
Большой диаметр 11—12.5 мм...................Caucasigena reminiscenta (стр. 199).
7»
100
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ '
64 (63). Признаки раковины иные.
65 (82). Поверхность раковины с волосками, придающими раковине характерный
«шерстистый» облик (длина волоска не менее 0.3 мм), или с радиальными мор-
щинками, на которых часто имеются кожистые шипики или чешуйки. Если
волоски утрачены, на поверхности раковины остаются ясные рубцы.
66 (71). Раковина с морщинками и кожистыми чешуйками или шипиками.
67 (68), Раковина с ясными треугольными шипиками, расположенными пятью спи-
ральными рядами. Вид обитает на Алтае . . Monachoides aculeata (стр. 238).
68 (67). Раковина покрыта радиально ориентированными чешуйками.
69 (70). Пупок широкий, развернутый, перспективный. Вид обитает в Тарбагатае
,.................................................Ponsadenia hirsute (стр. 119).
70 (69). Пупок очень узкий, цилиндрический. Вид обитает в Сибири.............
.......................................Monachoides stuxbergi (стр. 237).
71 (66). Раковина покрыта волосками.
72 (73). Высота завитка примерно равна высоте устья. Раковина хрупкая, без губы
или с очень слабой губой. Пупок проколовидный. Окраска от темно-роговой до
темно-бурой. Высота раковины 4.3—6.5, большой диаметр 6.0—9.0, малый диа-
метр 5.7—8.5 мм-.............................Pseudotrichia rubiginosa (стр. 238).
73 (72). Раковина иная.
74 (77). Большой диаметр не менее 15 мм. Виды обитают на Кавказе.
75 (76). Пупок узкий, неперспективный, частично прикрыт отворотом колумелляр-
ного края............................... Kokotschashvilia holotricha (стр. 188).
76 (75). Пупок неширокий (сквозь него виден целиком последний оборот), не прикрыт
или чуть прикрыт отворотом колумеллярного края................................
.........................................Euomphalia aristata (стр. 266).
77 (74). Большой диаметр не превышает 13 мм. Виды встречаются на Карпатах или
в Средней Азии.
78 (79). Волоски на раковине длинные (длина волоска 0.6—1 мм). Раковина хрупкая,
губа развита слабо или отсутствует. Вид обитает на Карпатах...................
.............................................Trichia villosula (стр. 208).
79 (78). Волоски на раковине умеренно длинные (длина волоска не более 0.4 мм).
Раковина не хрупкая, губа выражена более или менее ясно. Виды обитают
в Средней Азии.
80 (81). Раковина с широким развернутым пупком, сквозь который видны все пре-
дыдущие обороты, сильно уплощенная . . . Nanaja illuminate (стр. 165).
81 (80). Раковина с умеренно широким цилиндрическим пупком, слабо прижатая или
кубаревидная......................................Leucozonella caria (стр. 177).
82 (65). Волоски или чешуйки на поверхности раковины отсутствуют либо развиты
слабо и не придают раковине «шерстистого» облика. Если волоски имеются,
длина их не превышает 0.2 мМ. В случае их утраты рубцов не остается или они
исчезающе слабые.
83 (84). Раковина с двумя килями — один, более острый, проходит по периферии,
другой — ближе к шву, придает оборотам ступенчатую форму. Радиальные
ребра отсутствуют......................... Bradybaena cavimargo (стр. 161).
84 (83). Раковина не имеет двух килей; если же имеются кили такого типа, то раковина
с радиальными ребрами.
85 (120). Колумеллярный край устья расширен и полностью закрывает пупок; в край-
нем случае между отворотом колумеллярного края и базальной стенкой рако-
вины остается щель.
86 (87). Большой диаметр раковины не превышает 16 мм..........................
......................................... Monachoides vicina (стр. 235).
87 (86). Большой диаметр раковины не менее 18 мм.
88 (89). Раковина белая, одноцветная, почти плоская, толстостенная............
.........................................Levantina djulfensis (стр. 328).
89 (88). Раковина не белая, кубаревидная или шаровидная.
90 (95). Поверхность раковины покрыта выраженной в разной степени вермикулят-
ной скульптурой.
91 (92). Имеются ясные спиральные бороздки. Большой диаметр не превышает 25 мм
.......................................... . . . . Arianta arbustorum (стр. 310).
92 (91). Спиральные бороздки отсутствуют. Большой диаметр не менее 27 мм.
93 (94). Высота раковины приблизительно равна ее большому диаметру............
.......................................Cryptomphalus aspersa (стр. 324).
94 (93). Высота раковины заметно меньше ее большого диаметра.................
.........................................Eobania vermiculata (стр. 327).
95 (90). Вермикулятная скульптура отсутствует.
96 (109). Завиток конический или куполовидный; раковина шаровидная или шаро-
видно-кубаревидная, обороты относительно высокие. Колумеллярный край
устья высокий и почти отвесный, отворот его наверху широкий, книзу резко
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО РАКОВИНЕ
101
сужается и не переходит в базальный край. При 4—4.5 оборотах высота и'боль-
шой диаметр раковины не менее 25 мм.
97 (98). Поверхность раковины с микроскопической зернистой скульптурой . . . .
............................................................Helix buchi (стр. 343).
98 (97). Зернистая скульптура отсутствует.
99 (100). На раковине имеются темные радиальные размытые полосы либо фон очень
темный, а по периферии проходит неясная светлая лента...........................
................................................Helix lucorum (стр. 342).
100 (99). Окраска одноцветная или имеются темные спиральные ленты.
101 (102). Пупок в виде довольно широкой щели . . . Helix christophi (стр. 340).
102 (101). Пупок в виде очень узкой щели или полностью запаян.
103 (104). Скульптура представлена густыми и ясными спиральными бороздками и
тонкой неравномерной радиальной исчерченностью .................................
.................................................Helix lutescens (стр. 345).
104 (103). Спиральные бороздки отсутствуют либо выражены местами очень слабо.
105 (106). Высота раковины 38—45, большой диаметр 39—47 мм. Вид обитает на западе
европейской части СССР и завезен в ряд городов средней полосы...................
................................................Helix pomatia (стр. 346).
106 (105). Высота раковины 25—36, большой диаметр 28—38 мм. Виды обитают
в Крыму и на Кавказе.
107 (108). Края устья белые. Радиальная исчерченность тонкая и густая . . . .
.................................................... Helix nordmanni (стр. 338).
108 (107). Края устья розовые или красновато-бурые. Радиальная исчерченность
грубая и неравномерная, местами имеются элементы решетчатой скульптуры
...................................................... Helix albescens (стр. 340).
109 (96). Завиток и раковина иной формы; если такой же, то меньших размеров.
Колумеллярный край низкий, пологий, отворот его плавно переходит в базаль-
ный край.
110 (115). Большой диаметр раковины не менее 25 мм. Виды распространены на Кав-
казе.
111 (114). Последний оборот с решетчатой скульптурой. Поверхность раковины грубо
исчерчена.
112 (113). Темные спиральные ленты разбиты на ряды пятен. Последний оборот
в 1.5 раза шире предпоследнего .... Caucasotachea lencoranea (стр. 332).
113 (112). Спиральные ленты почти сплошные. Последний оборот менее чем в 1.5 раза
шире предпоследнего..........................Caucasotachea atrolabiata (стр. 332).
114 (111). Решетчатая скульптура отсутствует. Раковина тонко радиально исчерчена
.............................................. Caucasotachea calligera (стр. 335).
115 (110). Большой диаметр не превышает 25 мм.1 Виды распространены в Европе и
Предкавказье.
116 (117). Раковина ребристо радиально исчерчена. Пупок в виде узкой щели . . .
..................................................Сераеа vindobonensis (стр. 323).
117 (116). Раковина тонко радиально исчерчена. Пупок запаян.
118 (119). Края устья снаружи и внутри коричневые, губа красновато-коричневая
......................................................Сераеа nemoralis (стр. 320).
119 (118). Края устья и губа белые....................Сераеа hortensis (стр. 321).
120 (85). Пупок не запаян, хотя может быть очень узким; в любом случае пространство
между колумеллярный краем и базальной стенкой раковины щелью назвать
нельзя.
121 (122). Раковина плоская, массивная, толстостенная, с узким, но открытым пуп-
ком, белая, с 4 спиральными рядами размытых слабых пятен. Устье почти цель-
ное. Большой диаметр не менее 27 мм .... Levantina escheriana (стр. 328).
122 (121). Раковина иная.
123 (124). Раковина одноцветно черная или темно-бурая. Вид обитает на Карпатах
.....................................................Arianta aethiops (стр. 313).
124 (123). Окраска иная; если темно-бурая, то вид обитает не на Карпатах.
125 (128). На раковине имеется рельефный рисунок в виде светлых штрихов и непра-
вильной формы пятен, выраженных в разной степени; кроме того, может при-
сутствовать светлая спиральная лента. Большой диаметр не менее 12 мм.
126 (127). Раковина от прижатой до почти плоской, края устья сильно сближены.
Пупок широкий, развернутый. Вид обитает на Кавказе..............................
......................................... Shileykoia daghestana (стр. 244).
127 (126). Раковина от кубаревидной до почти шаровидной, края устья не сближены,
пупок узкий. Вид обитает в Средней Азии . . . Bradybaena scythica (стр. 145).
128 (125). Рисунок на раковине иной; если такой же, то большой диаметр раковины
меньше 10 мм.
129 (134). Скульптура представлена резкими радиальными ребрами.
1 Исключение: большой диаметр раковин Сераеа nemoralis изредка достигает 28 мм.
102
СИСТЕМАТИЧЕСКАЯ. ЧАСТЬ
130 (131). Раковина серая или желтоватая, либо коричневая со слабой светлой лентой
на периферии. Большой диаметр не менее 14 мм..................................
.......................................Oscarboettgeria euages (стр. 273).
131 (130). Раковина иной окраски, большой диаметр не более 13 мм.
132 (133). Устье очень косое, с мощной резкой губой; места его прикрепления сильно
сближены. Вид встречается на Кавказе ...........................................
..................................... Caucasigena tschetschenica (стр. 194).
133 (132). Устье умеренно косое, с широкой расплывшейся губой; места его прикрепле-
ния слабо сближены. Вид обитает в Средней Азии................................
...................................... Leucozonella crassicosta (стр. 180).
134 (129). Ребра отсутствуют или слабые, сглаженные.
135 (136). Раковина с ширококуполовидным завитком, покрыта сглаженными свет-
лыми радиальными ребрышками, сильнее выраженными на верхней поверх-
ности раковины; между ребрышками имеется точечная скульптура. Вид обитает
в Средней Азии ..................................Bradybaena tzwetkovi (стр. 151).
136 (135). Раковина иная.
137 (168). Виды встречаются к востоку от Байкала.
138 (147). Раковина очень хрупкая, сильно просвечивающая, с очень узким не пер-
спективным пупком.
139 (142). Раковина прижатая, с коричневой спиральной лентой (изредка одноцвет-
ная).
140 (1'41). Края устья заметно отвернуты. Вид обитает на Сахалине............
...............................-............Bradybaena duiensis (стр. 137).
141 (140). Края устья почти не отвернуты. Вид обитает на о-вах зал. Петра Великого
.................................................Bradybaena diversita (стр. 140).
142 (139). Раковина от почти шаровидной до шаровидно-кубаревидной, со спиральной
темной лентой или одноцветная.
143 (144). Раковина одноцветная — беловатая или светло-роговая, завиток кониче-
ский. Большой диаметр не превышает 20 мм. Высота завитка приблизительно
равна высоте устья ...............................Bradybaena fragilis (стр. 141).
144 (143). Раковина роговая или зеленоватая, иногда со спиральной лентой, завиток
куполовидный. Большой диаметр не менее 21 мм. Высота завитка обычно меньше
высоты устья.
145 (146). Края устья вполне сформированной раковины чаще всего отвернуты.
Вид обитает в Приморском крае.....................Bradybaena ravida (стр. 144).
146 (145). Края устья вполне сформированной раковины не отвернуты. Вид обитает
на Курильских о-вах............................ Bradybaena vulcanica (стр. 142).
147 (138). Раковина довольно прочная, слабо просвечивающая или непросвечивающая.
Пупок более или менее перспективный.
148 (155). Пупок широкий, развернутый.
149 (150). Скульптура представлена правильной тонкой радиальной ребристостью
............................................Bradybaena ussuriensis (стр. 131).
150 (149). Скульптура в виде радиальной исчерченности.
151 (154). Устье раковины оттянуто вправо, большой диаметр не превышает 16 мм.
152 (153). Высота завитка не превышает половины высоты устья.................
.......................................Lindholmomneme kudiensis (стр. 225).
153 (152). Высота завитка почти равна высоте устья...........................
. . . .............................Lindholmomneme. no tophila (стр. 226).
154 (151). Устье не оттянуто вправо, большой диаметр не менее 19 мм..........
......................................Bradybaena middendorffi (стр. 130).
155 (148). Пупок не широкий или узкий, цилиндрический.
156 (161). Раковина прижатая; если кубаревидная, то большой диаметр меньше 17 мм.
157 (160). Спиральная скульптура имеется.
158 (159). Большой диаметр раковины не менее 19 мм...........................
......................................Bradybaena transbaicalia (стр. 128).
159 (158). Большой диаметр раковины не более 17 мм...........................
......................................... Bradybaena schrencki (стр. 127).
160 (157). Спиральная скульптура отсутствует.................................
..................'............Lindholmomneme nordenskioldi (стр. 227).
161 (156). Раковина от шаровидной до кубаревидной; в последнем случае большой
диаметр не менее 20 мм.
162 (163). Раковина шаровидная, последний оборот почти одной ширины с предпо-
следним. На колумеллярном крае обычно имеется утолщение.......................
...........................................Bradybaena arcasiana (стр. 139).
163 (162). Раковина иная.
164 (165). Раковина резко или ребристо радиально исчерчена. Вид обитает на мате-
рике ............................................ Bradybaena maacki (стр. 132).
1 65 (164). Раковина без резкой радиальной исчерченности. Виды обитают на Сахалине
и Курильских о-вах.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО РАКОВИНЕ
103
166 (167). Раковина одноцветная . . ..........Bradybaena strelkovi (стр. 135).
167 (166). Раковина не одноцветная: имеются большей частью размытые желтовато-
коричневые радиальные пятна и полосы и спиральные ленты........................
........................................... Bradybaena weyrichii (стр. 133).
168 (137). Виды обитают к западу от Байкала.
169 (174). Виды обитают на Алтае и в Сибири.
170 (173). Раковина со спиральной скульптурой. Чаще всего имеются 1 или 2 темные
ленты.
171 (172). Раковина одноцветная или с одной темной спиральной лентой............
............................................Bradybaena schrencki (стр. 127).
172 (171). Раковина с двумя темными лентами — выше и ниже периферии ....
................................................. Bradybaena helvola (стр. 157).
173 (170). Раковина без спиральной скульптуры и без темных лент.................
................................Lindholmomneme nordenskioldi (стр. 227).
174 (169). Виды распространены на Кавказе, в Средней Азии и в европейской части
СССР.
175 (200). На последнем обороте имеются вмятины, носящие характер закономерной
скульптуры. В Средней Азии виды отсутствуют.
176 (191). Если оптическая ось глаза совпадает с осью пупка, сквозь пупок видно
не более половины предпоследнего оборота.
177 (178). На базальной части раковины имеется несколько тонких спиральных лент,
выраженных иногда слабо и разбитых на ряды пятен...............................
............................................. Cernuella virgata (стр. 250).
178 (177). Спиральные ленты отсутствуют или имеются 1—2 ленты — выше и ниже
периферии.
179 (180). Края устья окрашены в буроватый или красноватый цвета................
............................................Monacha carthusiana (стр. 286).
180 (179). Края устья светлые.
181 (186). Имеются 2 спиральные коричневые ленты (у отдельных раковин одна или обе
ленты отсутствуют). Последний оборот близ устья несколько оттянут вправо,
благодаря чему пупок эксцентрический.
182 (183). Ширина устья незначительно превышает его высоту или равна ей . . .
....................................................Hesseola solidior (стр. 263).
183 (182). Ширина устья заметно превышает его высоту.
184 (185). Большой диаметр раковины не превышает 18 мм..........................
........................................ Caucasocressa pratensis (стр. 305).
185 (184). Большой диаметр раковины не менее 20 мм..............................
......................................... Caucasocressa joannis (стр. 306).
186 (181). Раковина без темных спиральных лент, последний оборот не оттянут вправо.
Пупок концентрический, очень узкий.
187 (190). Вмятины покрывают последний оборот почти сплошь.
188 (189). Большой диаметр не превышает 13 мм. Вид обитает на Кавказе . . . .
...................................................Monacha caucasicola (стр. 283).
189 (188). Большой диаметр не меньше 14 мм. Вид обитает в Крыму.................
............................................ Monacha fruticola (стр. 282).
190 (187). Вмятины разбросаны по поверхности последнего оборота.................
...................................... Monacha subcarthusiana (стр. 289).
191 (176). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно
больше половины предпоследнего оборота.
192 (197). Окраска известково-белая или светло-серая; обычно имеется несколько
тонких темных лент, которые присутствуют и на базальной поверхности рако-
вины.
193 (194). Вид обитает в юго-западных районах европейской части СССР . . . .
...................................................Helicella candicans (стр. 224).
194 (193). Виды обитают в Крыму и на Кавказе.
195 (196). Раковина слабо прижатая — до кубаревидной. Вид обитает в Крыму
................................................. Helicopsis paulhessei (стр. 216).
196 (195). Раковина прижатая, нередко почти плоская. Вид обитает на Кавказе
.................................................. Cernuella tiflisiana (стр. 252).
197 (192). Окраска от желтой до каштановой; если есть темные ленты, то не на базаль-
ной стенке, а близ периферии.
198 (199). Скульптура представлена сочетанием грубых вмятин, спиральных бороздок,
рубцов от волосков и грубой радиальной исчерченности. На периферии имеется
размытая светлая лента............................Euomphalia strigella (стр. 269).
199 (198). Скульптура в виде тонкой радиальной исчерченности; вмятины редкие и
слабые. Спиральные бороздки и следы от волосков отсутствуют. Раковина одно-
цветная или с темной лентой над периферией . . Faustina faustina (стр. 309).
104
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
200 (175), Вмятины отсутствуют или выражены крайне неопределенно и не имеют
характера закономерной скульптуры; если же вмятины имеются, то виды оби-
тают в Средней Азии.
201 (204). Раковина от слегка прижатой до шаровидной. Спиральные бороздки ясные,
радиальная скульптура в виде тонкой морщинистости или исчерчепности.
Окраска одноцветная или с одной темной лентой выше периферии. Высота устья
примерно равна его ширине. Пупок узкий, но открытый. В Средней Азии виды
отсутствуют.
202 (203). Завиток куполовидный. Раковина почти шаровидная: индекс отношения
высоты раковины к ее большому диаметру 0.8—0.9...................................
.............................................Bradybaena fruticum (стр. 125).
203 (202). Завиток конический. Раковина прижатая: индекс отношения высоты рако-
вины к ее большому диаметру 0.6—0.7 . . . Bradybaena schrencki (стр. 127).
204 (201). Признаки раковины иные.
205 (208). Поверхность раковины с зернистой скульптурой. Виды обитают на западе
европейской части СССР.
206 (207). Пупок широкий, развернутый, перспективный ............................
.............................................Urticicola umbrosa (стр. 240).
207 (206). Пупок узкий, цилиндрический, не перспективный.........................
............................................Monachoides incarnata (стр. 234).
208 (205). Зернистая скульптура отсутствует. Виды обитают не только на западе евро-
пейской части СССР; если же имеется зернистость, то виды распространены на
Кавказе.
209 (214). Скульптура в виде радиальной ребристости, выраженной примерно оди-
наково на верхней и нижней поверхностях.
210 (211). По периферии раковины проходит нитевидный киль,1 нередко грубый и
зазубренный. Вид обитает в Крыму .... Helicopsis retowskii (стр. 215).
211 (210). Киль отсутствует; в крайнем случае имеется лишь слабый угол на пери-
ферии. Виды не обитают в Крыму.
212 (213). Одна из нескольких спиральных лент проходит выше периферии, нижняя
граница этой ленты ясная, а верхняя неопределенная. Вид обитает в Копетдаге
................................................. . Helicopsis likharevi (стр. 212).
213 (212). Спиральные ленты выражены в разной степени; нередко раковина почти
одноцветно известково-белая. Если выше периферии имеется темная лента,
то верхняя и нижняя ее границы выражены примерно в равной степени. Вид
обитает в юго-западных районах европейской части СССР............................
.............................................. Helicopsis striata (стр. 213).
214 (209). Радиальная скульптура представлена морщинистостью или исчерченностью;
если имеются ребра, то на нижней стороне раковины они выражены гораздо
слабее, чем на верхней.
215 (216). Раковина хрупкая, кубаревидная, одноцветно желтоватая, с редкими тон-
кими волосками. Обороты сильно выпуклые, шов глубокий............................
......................................... Plicuteria lubomirskii (стр. 204).
216 (215). Раковина в той или иной степени прижатая, не хрупкая, белая, серая или
роговая, часто со спиральными лентами. Обороты умеренно выпуклые, шов но
глубокий.
217 (218). Раковина серая, реже светло-роговая, со светлой лентой по периферии
................................................ Stenomphalia raxergieri (стр. 280).
218 (217). Раковина иной окраски; если окраска светло-роговая, то светлая лента
очень слабая, размытая.
219 (224). Пупок развернутый, перспективный.
220 (221). По периферии раковины проходит киль . . . Helicopsis filimargo (стр. 217).
221 (220). Киль отсутствует.
222 (223); Раковина тонко и неясно радиально исчерчена. Вид обитает в Крыму,
а также в окрестностях Мелитополя и Новороссийска..............................-
...............................................Helicopsis dejecta (стр. 216).
223 (222). Раковина тонко, но глубоко и густо радиально исчерчена. Вид обитает
в бассейне р. Днестра .............................Helicopsis instabilis (стр. 217).
224 (219). Пупок в начале развернутый, затем резко сужается, поэтому сквозь него-
видно не более полутора оборотов.
225 (230). Раковина в различной степени прижатая, белая, с темными спиральными
__ лентами разной ширины, часто разбитыми на ряды пятен; эти ленты присут-
ствуют и на базальной части. Иногда ленты развиты так интенсивно, что значи-
тельно маскируют белый фон; порой ленты отсутствуют. Губа развита слабо,
края устья тонкие, острые.
1 У некоторых популяций киль отсутствует.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО РАНОВИНЕ
105
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
(227) . Сквозь пупок виден весь предпоследний оборот или немного больше . . .
...........................................Xeropicta derbentina (стр. 220).
(226). Сквозь пупок видна часть предпоследнего оборота.
(229) . Сквозь пупок просматривается не более 1/i предпоследнего оборота ... ...
............................................Xeropicta krynickii (стр. 219).
(228) . Сквозь пупок просматривается 1/2—3/4 предпоследнего оборота ........
.........................................Xeropicta candaharica (стр. 222).
(225). Раковина не обладает совокупностью перечисленных признаков.
(300). Виды обитают в Средней Азии и на Алтае.
(235). Виды обитают в Копетдаге.
(234). Раковина ребристо исчерчена, с темными спиральными лентами ....
.............................................Helicopsis likharevi (стр. 212).
(233). Раковина почти гладкая, одноцветная . . Hesseola transcaspia (стр. 265)..
(232). Виды обитают на Тянь-Шане, Памире и Алтае.
(241). Раковина прижатая, светлая, одноцветная, почти гладкая.
(240). Обороты в профиле округлые, без намека на угол.
(239) . Сквозь пупок видна часть предпоследнего оборота.....................
.........................................Ponsadenia semenovi (стр. 121).
(238) . Пупок широкий, перспективный, развернутый...........................
........................................Bradybaena fedtschenkoi (стр. 160).
(237) . Обороты в профиле с легким, но заметным углом.................., •
......................................... Bradybaena perlucens (стр. 151).
(236). Раковина иная.
(243). Окраска раковины зеленовато-оливковая, почти одноцветная ....
.............................................Bradybaena tomyris (стр. 162).
(242). Окраска иная.
(245). Выше периферии имеется темная спиральная лента, фон белый или серый;,
иногда присутствуют неясные темные радиальные линии. Пупок узкий, непер-
спективный ...............Bradybaena plectotropis smsp. phaeozona (стр. 150).
(244). Сочетание признаков иное.
(269). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно-
больше половины предпоследнего оборота, чаще не меньше, чем целый обо-
рот.
(252). Выше и ниже периферии проходят две темные ленты, разделенные светлым
промежутком.
(251). Фон раковины роговой или светло-каштановый, но не белый (хотя часто-
очень светлый).
(250). Ширина устья примерно равна его высоте. Пупок в целом цилиндрический
............................................. Bradybaena helvola (стр. 157).
(249). Ширина устья превышает его высоту. Устье слегка оттянуто вправо, в ре-
зультате чего обнажена значительная часть предпоследнего оборота и пупок
приобретает воронковидную форму .... Bradybaena dichrozona (стр. 153).
(248). Фон раковины известково-белый или светло-серый, иногда с очень легким-
голубоватым оттенком........................Bradybaena stoliczkana (стр. 155).
(247). Схема окраски иная.
(254). Раковина от кубаревидной до почти шаровидной, красновато-коричневая,
густо и неравномерно радиально исчерчена, с широкой молочно-белой лентой-
по периферии. Вид обитает на южном Алтае . . «Bradybaena» boevi (стр. 163).
(253). Сочетание признаков иное.
(256) . Периферия раковины угловатая, сверху к периферии примыкает выражен-
ная в разной степени темная лента..........................................
......................... Bradybaena plectotropis smsp. necopinatus (стр. 150).
(255) . Периферия округлая; если имеется угол, то темная лента отсутствует,
а сам угол несколько светлее фона.
(258) . Раковина от кубаревидной до почти шаровидной .......................
......................... Bradybaena plectotropis smsp. almaatini (стр. 150).
(257). Раковина прижатая.
(262). Радиальная скульптура в виде грубых морщин. Поверхность раковины
с волосками, которые обычно сохраняются пожизненно на ранних оборотах,
но отсутствуют на последнем. Если волоски опадают, рубцов от них почти не-
остается. Большой диаметр раковины 16—21 мм.
(261) . Стилофоров две пары; нижняя пара заключает стрелы...................
....................................... Leucozonella ferghanica (стр. 169).
(260) . Стилофоры рудиментарные, одна пара; стрел нет.........................
......................................... Angiomphalia copiosa (стр. 292).
(259). Радиальная скульптура в виде тонкой исчерченности. Раковина без волосков,
и рубцов от них; либо раковина с волосками, которые пожизненно не сохра-
няются, но рубцы от них видны на последнем обороте. Размеры различ-
ные.
106
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
.263 (264). Места прикрепления устья сильно сближены, устье сильно скошено, с ясной
губой. Последний оборот энергично опущен, рубцы от волосков отсутствуют
...........................................Bradybaena pseudoferghanica (стр. 158).
264 (263). Места прикрепления устья почти не сближены, устье умеренно косое, без
губы или со слабой губой. Последний оборот прямой или слегка опущен, рубцы
от волосков обычно более или менее ясно видны, изредка отсутствуют.
265,(266). Последний оборот к устыо не опущен, рубцы от волосков очень слабые
• или отсутствуют............................Leueozonella retteri (стр. 175).
266 (265). Последний оборот к устью слабо опущен, рубцы от волосков ясные.
267 (268). Раковина одноцветная, стилофоров две пары . . Nanaja cumulata (стр. 164).
268 (267). Раковина одноцветная или с очень неясными радиальными полосами, стило-
форов одна пара па одной стороне вагины . . Archaicaheptapotamica (стр. 257).
.269 (246). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно
не более половины предпоследнего оборота.
270 (277). Форма раковины от кубаревидной до шаровидной. Окраска либо одноцветно
бурая, либо красновато-коричневая с ясной светлой лентой на периферии, либо,
наконец, на светлом фоне располагаются 2—4 спиральные коричневые ленты;
в любом случае вокруг пупка имеется светлое поле. Последний оборот в профиле
плавно закруглен.
271 (272). Раковина красновато-коричневая с желтоватой лентой на периферии . . .
..............................................Bradybaena bilaticincta (стр. 154).
272 (271). Раковина одноцветно бурая или светлая с темными лентами.
273 (274). Спиральные бороздки резкие. Диаметр раковины 22—28, высота 19—
27 мм ....................................Ponsadenia duplocincta (стр. 120).
274 (273). Спиральные бороздки вялые, иногда почти не выражены. Большой диаметр
раковины 19—27, высота 15—21 мм.
275 (276). Раковина правозавитая . . . Bradybaena lantzi f. dextrorsa (стр. 147).
276 (275). Раковина левозавитая .... Bradybaena lantzi f. sinistrorsa (стр. 147).
277 (270). Сочетание признаков иное.
.278 (279). Последний оборот оттянут вправо, благодаря чему пупочная впадина до-
вольно большая, а отверстие пупка сдвинуто к ее краю. Окраска белая, одно-
цветная или с исчезающей слабой светло-коричневой лентой по периферии и
иногда с размытыми рыжеватыми поперечными полосами или пятнами ....
..................„...............................Archaica apollinis (стр. 259).
279 (278). Последний оборот вправо не оттянут, пупок точковидный, окраска носит
иной характер.
.280 (281). Ниже периферии раковина коричневая, выше — серая. ,В проходящем
свете нижняя часть раковины просвечивает сильнее ..............................
.................__.....................Leueozonella hypophaea (стр. 178).
281 (280). Окраска верхней и нижней сторон раковины примерно одинакова.
.282 (283). Раковина шаровидная, толстостенная, белая, верхние обороты немного
темнее последнего, нередко имеют голубоватый или розоватый оттенок. Края
устья утолщены, не отвернуты, кроме колумеллярного. Губа отсутствует или
очень расплывшаяся..............................Leueozonella caryodes (стр. 170).
.283 (282). Раковина иная.
284 (285). Раковина шаровидная, тонкостенная. На поверхности имеются волоски, <
сохраняющиеся пожизненно.......................Angiomphalia exasperate (стр. 295).
285 (284). Форма раковины различная. Волоски отсутствуют или имеются только на
не вполне сформированных раковинах.
, 286 (291). Раковина белая с размытыми, радиальными, неправильно расположенными
полосами светло-коричневой или рыжеватой окраски; эти полосы иногда бывают
выражены только на нижней или только на верхней стороне раковины. Эмбрио-
нальные обороты выделяются рыжеватой окраской.
287 (288). Обороты сильно уплощены............Angiomphalia seductilis (стр. 298).
288 (287). Обороты умеренно или сильно выпуклые.
.289 (290). Раковина с рубцами от волосков и с ясными спиральными бороздками
........................................ Angiomphalia caelestimontana (стр. 293).
290 (289). Раковина без рубцов от волосков, со слабыми спиральными бороздками
• •............................................Angiomphalia almalensis (стр. 297).
291 (286). Радиальные полосы отсутствуют, эмбриональные обороты окраской не
выделяются.
292 (297). Раковина шаровидная.
293 (294). Большой диаметр раковины не превышает 12 мм...........................
. ...... и............................. Leueozonella mesoleuca (стр. 172).
294 (293). Большой диаметр раковины не менее 14 мм.
295 (296). Стилофоров две пары, нижние заключают стрелы.........................
........................f................Leueozonella rubens (стр. 171).
296 (295). Стилофоров одна пара, рудиментарных, без стрел.......................
Angiomphalia regeliana (стр. 296).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО РАКОВИНЕ
107
297 (292). Раковина от кубаревидной до прижатой.
298 (299). Последний оборот угловатый, светлая лента по периферии четкая ....
. .......................................Leueozonella rufispira (стр. 173).
299 (298). Последний оборот не угловатый или с едва заметным углом, светлая лента
по периферии выражена в разной степени . . Leueozonella angulata (стр. 176).
300 (231). Виды обитают на Кавказе.
301 (324). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно
не более половины предпоследнего оборота.
302 (303). Раковина известково-белая, с 1—2 темными, сильно просвечивающими спит
ральными лентами; изредка раковина одноцветная. Большой диаметр 18—20,
высота раковины 11—12 мм........................Caucasigena abchasica (стр. 198).
-303 (302). Раковина иная.
304 (305). На поверхности раковины имеются ясные следы от волосков, ориентирован-
ные вдоль оборотов..............................Monacha samsunensis (стр. 288).
305 (304). Рубцов от волосков нет или они вялые и редкие.
306 (313). Поверхность раковины покрыта, более или менее ясно выраженной зерни-
стой скульптурой.
307 (308). Зернистость представлена короткими высокими резкими морщинками,
ориентированными поперек оборотов; у молодых экземпляров на каждой мор-
щинке располагается слегка изогнутая чешуйка; у взрослых особей чешуйки
дольше всего сохраняются на скатах пупка. Края устья отвернуты, розоватые
.................................................. Stenomphalia maiae (стр. 278).
'308 (307). Зернистость представлена округлыми гранулами, лишенными чешуек.
Края устья прямые или умеренно отвернуты, беловатые или коричневые.
309 (310). Устье заметно оттянуто вправо. Вид обитает в западной части Кавказа
.................................................... Circassina frutis (стр. 250).
310 (309). Устье не оттянуто вправо. Виды обитают не только на западе Кавказа.
311 (312). Зернистая скульптура нерезкая. Имеется один вагинальный придаток. Вид
обитает в Талыше..................................Monacha talyschana (стр. 285).
-312 (311). Если вид найден в Талыше, зернистая скульптура очень четкая; у предста-
вителей некоторых популяций из других районов Кавказа зернистая скульптура
может быть очень слабой. Вагинальных придатков два . . .........................
....................................... Stenomphalia pisiformis (стр. 278).
313 (306). Зернистая скульптура отсутствует.
314 (315). Раковина желтая или белая, шаровидная, просвечивающая, почти гладкая.
Губа просвечивает сквозь стенку раковины в виде желтоватой полоски. Большой
диаметр не превышает 12 мм....................... . Stenomphalia selecta (стр. 275).
315 (314). Раковина иная.
316 (319). Губа четкая, высокая, располагается отступя от края.
317 (318). Высота завитка примерно равна высоте устья. Окраска от серовато-роговой
до интенсивно каштановой..................Kokotschashvilia phaeolaema (стр. 190).
318 (317). Высота завитка заметно меньше высоты устья. Окраска буро-желтая . . .
................................................ Teberdinia zolotarevi (стр. 183).
319 (316). Губа расплывшаяся, низкая, расположена близ края устья.
320 (323). Последний оборот к устью опущен.
-321 (322). Раковина почти одноцветная, с неясной светлой лентой по периферии . . .
................................................Circassina circassica (стр. 247).
322 (321). Раковина с 1—2 темными спиральными лентами, которые изредка бывают
настолько широкими, что образуют фон раковины; в этом случае между ними
остается светлый промежуток. У отдельных раковин ленты отсутствуют; тогда
раковина одноцветная ...........................Circassina bojenae (стр. 249).
323 (320). Последний оборот к устью почти не опущен.............................
.................................... Caucasigena thalestris (стр. 201).
324 (301). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно
не меньше половины предпоследнего оборота, обычно больше.
325 (326). На верхней стороне раковины имеются широкие размытые светло-коричне-
вые или каштановые радиальные полосы и пятна во всю ширину оборотов . . .
.............................................Hygrohelicopsis darevskii (стр. 181).
326 (325). На верхней стороне раковины полос и пятен указанного типа нет.
327 (330). Скульптура в виде радиальных ребер,z одинаково хорошо выраженных на
верхней и нижней поверхностях раковины.
328 (329). Последний оборот в 1.5 раза шире предпоследнего, снизу умеренно вы-
' пуклый. Сквозь пупок видно не более одного оборота............................
............................................ Xerosecta crenimargo (стр. 254).
329 (328). Последний оборот менее чем в 1.5 раза шире предпоследнего, снизу явст-
венно выпуклый. Сквозь пупок просматривается 1.5 оборота........................
........................................Xerosecta arcadiana (стр. 255).
330 (327). Скульптура не ребристая; если же ребра имеются, то на верхней поверх-
ности раковины они развиты лучше, чем на нижней.
108 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
331 (334). Большой диаметр раковины не превышает 9 мм.
332 (333). Раковина прижатая, почти одноцветно коричневая, без темных лент. Пери-
ферия угловатая, к устью угол сглаживается......................................
.........................................Caucasigena armeniaca (стр. 192).
333 (332). Раковина почти кубаревидная, светлая, с темными лентами. Периферия
округлая ..........................................Xeropicta parableta (стр. 223).
334 (331). Большой диаметр раковины не менее 10 мм.
335 (338). Периферия раковины в разной степени угловатая.
336 (337). К светлой ленте, проходящей по периферии раковины, снизу прилегает
коричневая лента с четкими краями . . . Caucasigena rengarteni (стр. 195).
337 (336). Ниже светлой периферической ленты коричневая лента отсутствует . . ..
................................................. Shileykoia daghestana (стр. 244).
338 (335). Периферия раковины плавно закруглена.
339 (342). Края устья сильно сближены.
340 (341). Поперек оборотов располагаются развитые в различной степени молочно-
белые штрихи, фон раковины роговой или каштановый. По периферии часто
проходит светлая лента.......................... Shileykoia daghestana (стр. 244).
341 (340). Радиальные белые штрихи отсутствуют, фон раковины белый или светло-
серый. Обычно имеются 2 спиральные темные ленты — выше и ниже периферии
................................................ Caucasigena eichwaldi (стр. 196).
342 (339). Края устья не сближены или умеренно сближены.
343 (344). Завиток плавно и широко закруглен, вершина сглажена и выступает незна-
чительно, чаще всего совсем не выступает. Обороты слабо выпуклые . . . .
......................................... Caucasigena sehaposchnikovi (стр. 199).
344 (343). Завиток конический; если куполовидный, то вершина не сглажена. Обороты
умеренно или сильно выпуклые.
345 (346). Спиральная скульптура ясно выражена по всей длине последнего оборота
.........................................Fruticocampylaea narzanensis (стр. 241).
346 (345). Спиральная скульптура отсутствует или слабо развита; в последнем случае
по длине последнего оборота выражена неодинаково.
347 (354). Окраска очень светлая; иногда за счет сгущения фона выше и ниже перифе-
рии возникают 2 спиральные ленты, тогда лучше всего выражены верхняя гра-
ница нижней ленты и нижняя граница верхней ленты; в общем эти ленты вы-
ражены слабо, порой лишь угадываются. В других случаях спиральные ленты
четкие, с ясными границами.
348 (349). Спиральные ленты коричневые, четкие, с ясными границами..............
..................................... Fruticocampylaea kobiensis (стр. 241).
349 (348). Спиральные ленты сильно размыты и образованы за счет сгущения фона
выше и ниже периферии; иногда раковина одноцветно белая или серая.
350 (351). Большой диаметр раковины не менее 22 мм..............................
.........................................Kokotschashvilia tanta (стр. 186).
351 (350). Большой диаметр раковины не превышает 21.5 мм.
352 (353). Индекс отношения большого диаметра к высоте раковины составляет
1.5— 1.6 ............................Kokotschashvilia , makvalae (стр. 184).
353 (352). Индекс отношения большого диаметра к высоте раковины составляет
1.3— 1.4 ..........................Kokotschashvilia eberhardi (стр. 187).
354 (347). Окраска светло- или темно-роговая, на периферии имеется размытая светлая
лента ............................................Euomphalia appeliana (стр. 268).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ, ПОДСЕМЕЙСТВ, РОДОВ,
ПОДРОДОВ И НЕКОТОРЫХ ПОДВИДОВ ПО ПРИЗНАКАМ ВНУТРЕННЕГО
СТРОЕНИЯ И РАКОВИНЫ
.1 (26). Семеприемник снабжен отходящим от его протока дивертикулом (отростком),
выраженным в разной степени.............................Helicidae (стр. 307).
2 (13). Слизистые железы вагины одноветвистые или двуветвистые. Бич не закручен
в правильную спираль...................................Ariantinae (стр. 308).
3 (12). Слизистые железы одноветвистые.
4 (7). Раковина линзовидная.
5 (6). Устье цельное или почти цельное. Раковина твердостенная, с грубой частой
зернистой скульптурой..................................Helicigona (стр. 313).
6 (5). Устье не цельное. Раковина тонкостенная, с тонкой густой спиральной
скульптурой............................................Drobacia (стр. 317).
7 (4). Раковина не линзовидная.
8 (11). Раковина плоская или прижатая. В устье имеются зубы.
9 (10). На париетальной стенке устья имеется пластинка. Бич длинный........
.............................................. Isognomostoma (стр. 315).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО АНАТОМИИ
109
10 (9). Париетальная стенка устья без зубов. Бич очень короткий, рудиментарный
............................................................ Causa (стр. 317).
11 (8). Раковина кубаревидная или шаровидная. Устье раковины без зубов . . .
............................................................ Arianta (стр. 310).
12 (3). Слизистые железы двуветвистые......................Faustina (стр. 308).
13 (2). Минимальное число ветвей каждой железы 4; если железы простые или дву-
ветвистые, бич закручен в правильную спираль...............Helicinae (стр. 319).
14 (15). Слизистые железы простые или двуветвистые, бич скручен в цилиндрическую
спираль................................................ Levantina (стр. 327).
15 (14). Слизистые железы дихотомически ветвятся пе менее двух раз, бич в спираль
не закручен.
16 (25). Внутри пениса располагаются одна за другой две папиллы.
17 (18). Общее число ветвей каждой железы равно 4............................
.................................. Cepaea, подрод Cepaea s. str. (стр. 320).
18 (17). Общее число ветвей каждой слизистой железы больше 4.
19 (20). Общее число ветвей каждой железы равно 8............................
......................................Cepaea, подрод Octadenia (стр. 322).
20 (19). Общее число ветйей каждой железы больше 8.
21 (22). Бич в 2—3 раза короче дивертикула семеприемника . . . Eobania (стр. 326).
22 (21). Бич не более чем в 1.5 раза короче дивертикула семеприемника, либо диверти-
кул короче бича.
23 (24). Раковина от шаровидной до шаровидно-кубаревидной. Колумеллярный край
устья высокий и отвесный...................................... Helix (стр. 337).
24 (23). Раковина кубаревидная. Колумеллярный край устья очень низкий и пологий
.......................................................Caueasotachea (стр. 330).
25 (16). Пенис заключает только одну папиллу; ниже ее располагается мощный полу-
кольцевой валик ......................................Cryptomphalus (стр. 324).
26 (1). Проток семеприемника лишен дивертикула.
27 (128). На вагине располагаются стилофоры или придатки.
28 (121). Семеприемник относительно массивный, резервуар прилегает к спермови-
дукту и не связан с нижней стенкой мантийной полости..........................
.................................................. Hygromiidae (стр. 163).
29 (62). Стилофоров 2 пары или 1 пара, располагаются симметрично относительно
вагины. Стрел две ........................................Trichiinae (стр. 163).
30 (33). Стилофоров, по крайней мере при наружном осмотре вагины, 1 пара.
31 (32). Нижняя часть каждого стилофора заключает полость, широко сообщающуюся
с просветом вагины. Никаких следов внутренних стилофоров нет..................
.................................................... Helicella (стр. 223).
32 (31). В нижней части стилофоров полостей нет. При вскрытии вагины обнаружи-
ваются рудименты внутренних (верхних) стилофоров..............................
..............................................: Hygrohelicopsis (стр. 181).
33 (30). Стилофоров 2 пары.
34 (37). Устье раковины с зубами.
35 (36). Раковина почти одноцветная, коричневая, с волосками и четким углом на
периферии . . '..........................................Odontotrema (стр. 166).
36 (35). Раковина с двумя темными лентами, светлая, без волосков и с округлой или
очень слабо угловатой периферией.........................Diodontella (стр. 202).
37 (34). Устье раковины без зубов.
38 (45)- На поперечном срезе папиллы пениса видна система из 3—8 продольных щеле-
видпых полостей, разделенных септами. Семепроводящнй канал сдвинут к одной
из стенок папиллы.
39 (40). Раковина кубаревидная, с широким куполовидным завитком и редкими
волосками................................................Edentiella (стр. 209).
40 (39). Раковина от кубаревидной до прижатой, с коническим, сильно уплощенным
завитком, без волосков...................................Caucasigena (стр. 191).
41 (42). Бич и эпифаллус длинные, примерно равной длины. Бич длиннее пениса
.......................................Caucasigena, подрод Anoplitella (стр. 199).
42 (41). Бич и эпифаллус короткие, обычно бич короче эпифаллуса или равен ему. Бич
и пенис примерно равной длины или пенис немного длиннее.
43 (44). Внутренние стилофоры не меньше наружных. Пупок очень узкий, более чем
наполовину прикрыт отворотом колумеллярного края .........................
.................................Caucasigena, подрод Dioscuria (стр. 201).
44 (43). Внутренние стилофоры меньше наружных. Пупок широкий или узкий, не
прикрыт или слегка прикрыт отворотом колумеллярного края .....................
........................Caucasigena, подрод Caucasigena s. str. (стр. 192).
45 (38). Папилла пениса не заключает полостей указанного типа.
46 (47). На дистальной части пениса имеется хорошо развитый конический придаток
...........................................................Xeropicta (стр. 219).
47 (46). Пенис без придатка.
по
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
48 (49). Внутренние стенки вагины покрыты продольными рядами призматических,
складок..................................................Plicuteria (стр. 203).
49 (48). Внутренние стенки вагины гладкие или несут продольные валики, не разби-
тые на ряды складок.
50 (51). Валики внутри вагины петлеобразно охватывают выходные отверстия стило-
форов. Интрапапиллярные полости и папиллярная лакуна отсутствуют . . .
.............................................................Nanaja (стр. 164).,
51 (50). Валики внутри вагины часто образуют лопасти возле отверстий стилофоров,
но не охватывают их снизу, а продолжаются в клоаку. Обычно имеются интра-
папиллярные полости и папиллярная лакуна. Если же вагинальные валики
петлеобразно охватывают отверстия стилофоров, то имеется папиллярная лакуна.
52 (53). Пенис заметно меньше стилофоров, папиллярная лакуна имеется, интралапил-
лярной полости нет......................Leucozonella, подрод Narina (стр. 178).
53 (52). Пенис примерно одного размера со стилофорами или крупнее, имеет вид .
простой трубки, либо на поверхности этой трубки имеется продольная борозда,
либо эта борозда замкнута и образует интрапапиллярную полость; в последнем,
случае от борозды остается папиллярная лакуна.
54 (55). Ретрактор правого омматофора не образует перехлеста с половым ретрактором
.........................................................Helicopsis (стр. 211).
55 (54). Ретрактор правого омматофора перекрещивается с половым ретрактором и.
располагается между пенисом и вагиной.
56 (57). Стилофоры тесно сближены попарно и не прилегают к стенке вагины. На по-
верхности папиллы пениса имеется борозда, стенки папиллы заключают две
продольные полости. Раковина одноцветная, буровато-желтая, без волосков и
следов от них ...........................................Teberdinia (стр. 183).
57 (56). Признаки внутреннего строения и раковины иные.
58 (59). Семепроводящий канал фиксирован внутри папиллы двумя противолежащими.
продольными тяжами, интрапапиллярная полость парная. Резервуар семеприем-
ника по объему примерно равен вагине вместе со стилофорами. Раковина при-
жатая, у молодых особей с волосками, у взрослых волоски могут опадать.
Большой диаметр раковины не более 10 мм.....................Trichia (стр. 205).
59 (58). Совокупность признаков иная.
60 (61). Семепроводящий канал внутри папиллы пениса или припаян одной стороной,
к внутренней стенке папиллы, или фиксирован 1—2 тяжами. Резервуар семе-
приемника заметно больше вагины вместе со стилофорами. Раковина без светлой,
периферической ленты...............................Kokotschashvilia (стр. 184).
61 (60). Папилла пениса в виде простой трубки, или на поверхности ее имеется про-
дольная борозда, или имеется одна интрапапиллярная полость. Резервуар семе-
приемника примерно равен по объему вагине вместе со стилофорами. Раковина,
со светлой лентой по периферии, заметной тем лучше, чем темнее фон раковины
............................Leucozonella, подрод Leucozonella s. str. (стр. 168).
62 (29). Строение дистальной части женского отдела иное.
63 (92). Стилофоров 1 или 2, расположенных на одной стороне вагины; они не изме--
нены в придатки, нижний заключает стрелу.
64 (69). Стенки папиллы плотные, мускулизованные, без синусов и полостей . . .
........................................................Archaicinae (стр. 256).
65 (66). Семепроводящий канал расположен резко эксцентрично..................
......................................Archaica, подрод Ugama (стр. 260).
66 (65). Семепроводящий канал занимает более или менее центральное положе-
ние.
67 (68). Бич примерно вдвое короче эпифаллуса, между пенисом и эпифаллусом натя-
нуты мускульные ленты . . . Archaica, подрод Archaica s. str. (стр. 259).,
68 (67). Бич не короче эпифаллуса, пениальные мускульные ленты отсутствуют или..
развиты очень слабо.....................Archaica, подрод Euarchaica (стр. 257).
69 (64). Стенки папиллы заполнены рыхлой паренхимой, часто заключают синусы,
капилляры, или паренхима распадается на отдельные радиальные пучки . . .
........................................................Hygromiinae (стр. 225).
70 (77). Стилофор один, никаких следов второго стилофора нет.
71 (72). Бич очень длинный, почти вдвое превышает суммарную длину пениса п эпи-
фаллуса. Проток семеприемника также длинный, многократно извит . . .
.........................................................Urticicola (стр. 239).
72 (71). Длина бича не превышает суммарную длину пениса и эпифаллуса. Проток
семеприемника прямой или слегка извит.
73 (74). Устье раковины с двумя зубами.................Perforatella (стр. 231).
74 (73). Устье раковипы без зубов.
75 (76). Раковина покрыта волосками, поверхность ее без зернистой скульптуры
.................................................... Pseudotrichia (стр. 238).
76 (75). Раковина без волосков, часто с кожистыми шипами или чешуйками и с зер-
нистой скульптурой.....................................Monachoides (стр. 234).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПО АНАТОМИИ
ИГ
77 (70). Стилофоров два; внутренний (верхний) иногда развит очень слабо и обнаружи-
вается только при вскрытии вагины.
78 (79). Слизистые железы отсутствуют...........................................
...............................Circassina circassica akramowskii (стр. 249).
79 (78). Слизистые железы имеются. . . .
80 (83). Внутренние стилофоры при наружном осмотре вагины не видны. Бич руди-
. . ментарный. ' '
81 (82). Раковина кубаревидная. Последний оборот незначительно шире предпослед-
него. Устье раковины с зубами или с небольшим наплывом на базальном,
крае устья ................................................Chilanodon (стр. 228).
82 (81). Раковина прижатая. Последний оборот заметно шире предпоследнего. Устье
без зубов ............................................ Lindholmomneme (стр. 225).
83 (80). Внутренние стилофоры видны при наружном осмотре. Бич не рудиментарный.
84 (87). Ретрактор правого омматофора не образует перехлеста с половым ретрактором,
и располагается в стороне от гениталий.
85 (86). Нижняя часть вагины в той или иной степени вздута и заключает цилиндриче-
скую полость, сквозь которую выдвигается пенис при совокуплении. Раковина,
ребристая или ребристо исчерченная..........................Xerosecta (стр. 253).
86 (85). Нижняя часть вагины не вздута, пенис выдвигается через клоаку, минуя ва-
гину. Раковина тонко исчерчена..............................Cernuella (стр. 250).
87 (84). Ретрактор правого омматофора перекрещивается с половым ретрактором и.
располагается между пенисом и вагиной.
88 (89). Бич не длиннее эпифаллуса. Внутри вагины имеется 1 пара высоких продоль-
ных складок, которые, спускаясь к клоаке, охватывают отверстия стилофоров,
затем отчленяются от стенки вагины и вдаются в атриальную полость единым
сосочком, по поверхности которого проходит глубокий желобок......................
................................................... Shileykoia (стр. 244).
89 (88). Бич длиннее эпифаллуса. Вагина внутри гладкая или с системой складок,
спускающихся в клоаку, но не образующих сосочковидных выростов.
90 (91). На поперечном срезе папиллы пениса видно, что от паренхимы стенок остались
отдельные радиальные тяжи, фиксирующие семепроводящий канал. Фон рако-
вины роговой, зеленоватый или желтоватый...................Circassina (стр. 246).
91 (90). Стенки папиллы пениса заполнены паренхимой, в которой располагаются
более или менее многочисленные капилляры и синусы. Фон раковины белый
................................................... Fruticocampylaea (стр. 241).
92 (63). Стилофоры видоизменены в вагинальные придатки.
93 (98). Вагинальных придатков два, они представляют собой рудименты стилофоров
и имеют вид округлых или удлиненных мешочков . .Paedhoplitinae (стр. 291).
94 (95). На париетальной стенке раковины имеются зубы в виде бугорков и (или)
складок................................................. Paedhoplita (стр. 302).
95 (94). На париетальной стенке зубов нет или они исчезающе слабые.
96 (97). Раковина линзовидная или обороты в профиле явственно угловатые. В устье
на базальной стенке имеется мощная губа, которая справа резко обрывается
. . .....................................Angiomphalia, подрод Lentiga (стр. 300).
97 (96). Раковина не линзовидная, обороты в профиле округлые или слабо угловатые.
Губа, если имеется, слабая и справа ре обрывается..............................
...................... Angiomphalia, подрод Angiomphalia s. str. (стр. 292).
98 (93). Вагинальных придатков 2 или 1; они представляют собой видоизмененные (но
не рудиментарные!) стилофоры и состоят из утолщенной базальной части, сужен-
ного отдела и концевого вздутия; либо эти придатки имеют веретеновидную
форму, пронизывают покровы вагины и в полости вагины концы их сближены.
В последнем случае вагинальных придатков всегда 2..............................
..................................................Euomphaliinae (стр. 261).
99 (120). Вагинальные придатки не пронизывают стенок вагины и состоят из трех
отделов. Нередко придатки имеют значительную длину и теряются среди сли-
зистых желез, по концевое утолщение всегда помогает отличить придатки от
желез.
100 (103). Раковина линзовидпая ...........................Platytheha (стр. 271).
101 (102). Вагинальный придаток 1 ..........................................
........................... Platytheha, подрод Platytheha s. str. (стр. 273).
102 (101). Вагинальных придатков 2 . . . Platytheha, подрод Jasonella (стр. 271).
103 (100). Раковина не линзовидная.
104 (105). Устье раковины с двумя зубами...................Karahaghia (стр. 262).
105 (104). Устье раковины без зубов.
106 (107). Раковина ребристая ........................Oscarboettgeria (стр. 273).
107 (106). Раковина не ребристая. . . . .
108 (111). Вагинальный придаток 1...................... . . Monacha (стр. 281).
109 (110). Половой ретрактор отсутствует.......................................
...............................Monacha, подрод Monacha s. str. (стр. 286).
пг
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
110 (109). Половой ретрактор имеется . . . Monacha, подрод Paratheba (стр. 282).
111 (108). Вагинальных придатков 2.
112 (113). Имеется вторичный чехол пениса (чехол папиллы), фиксированный одним
краем на внутренней поверхности пениального чехла, а другим — на поверх-
ности папиллы .............................................Euomphalia (стр. 266).
113 (112). Вторичный чехол пениса отсутствует. При основании папиллы в стенке
эпифаллуса имеется кольцевое разрыхление тканей.................................
................................................. Stenomphalia (стр. 275).
114 (115). Раковина серая, реже светло-роговая, с ясной белой лентой по периферии.
Бич и эпифаллус длинные, бич слегка извит, цилиндрический.......................
.................................Stenomphalia, подрод Harmozica (стр. 280).
115 (114). Окраска раковины иная. Эпифаллус умеренной длины или короткий, бич
короткий, почти прямой, конический.
116 (119). Базальная часть вагинальных придатков резко вздута, концевые утолще-
ния ясно выражены.
117 (118). Длина бича примерно равна длине эпифаллуса. Раковина с более или менее
ясной зернистой скульптурой . . Stenomphalia, подрод Diplobursa (стр. 278).
118 (117). Бич вдвое короче эпифаллуса. Раковина без зернистой скульптуры . . .
.......................... Stenomphalia, подрод Stenomphalia s. str. (стр. 275).
119 (116). Базальная часть вагинальных придатков слабо или умеренно вздута, кон-
цевые утолщения выражены нечетко................................................
.................................Stenomphalia, подрод Batumica (стр. 276).
120 (99). Вагинальные придатки веретеновидные, пронизывают стенки вагины . . .
.............................................................Hesseola (стр. 263).
121 (28). Семеприемник небольшой, резервуар его прилегает к нижней части белковой
железы и «пристегнут» соединительнотканной связкой к дну легочной полости
в районе реноперикардиального комплекса.................Bradybaenidae (стр. 118).
122 (123). Слизистые железы впадают непосредственно в стилофор или в дополнитель-
ный мешок, расположенный на поверхности стилофора . . Bradybaena (стр. 123).
123 (122). Слизистые железы впадают либо в парные резервуары, прилегающие
к клоаке, либо в перемычку — «мостик», соединяющий апикальную часть стило-
фора с атриальной областью.................................Ponsadenia (стр. 119).
4242(125). Слизистые железы впадают в «мостик», соединяющий вершину стилофора
с атриальной областью. Раковина одноцветная, светлая, прижатая, с округлой
периферией ......................Ponsadenia, подрод Ponsadenia s. str. (стр. 121).
125 (124). Слизистые железы впадают в парные резервуары, прилегающие к клоаке.
Раковина либо сильно уплощенная и одноцветно бурая, либо почти шаровидная,
одноцветно бурая или светлая со спиральными темными лентами.
126 (127). Резервуары слизистых желез небольшие, тесно прилегают к клоаке, но не
спаяны с нею. Раковина уплощенная, обороты в профиле резко угловатые
.................................... Ponsadenia, подрод Tarbagataja (стр. 119).
127 (126). Резервуары слизистых желез обширные, спаяны с поверхностью клоаки.
Раковина шаровидная, обороты в профиле округлые.................................
.................................. Ponsadenia, подрод Mesasiata (стр. 120).
128 (27). Стилофоры и вагинальные придатки отсутствуют.
129 (136). Слизистые железы имеются.
130 (131). Слизистая железа одна, неразветвленная, в разной степени гофрированная.
Устье раковины в виде широкой щели..............................................
................ Helicodontidae, Lindholmiolinae, Lindholmiola (стр. 116).
131 (130). Слизистая железа не одна; если одна, то состоит из двух ветвей неодинако-
вой длины.
132 (133). Слизистая железа одна, состоит из двух ветвей неодинаковой длины . . .
..........................Helicodontidae, Helicodontinae, Helicodonta (стр. 114).
133 (132). Слизистых желез несколько.
134 (135). Пенис и эпифаллус с мощными мускульными лентами. Внутри папиллы
пениса кольцевой полости нет. Бич длиннее пениса и эпифаллуса, вместе взятых
. . . Hygromiidae, Hygromiinae, Circassina circassica stephaniae (стр. 247).
135 (134). Пенис и эпифаллус без мускульных лент или эти ленты слабые. Внутри
пениальной папиллы имеется кольцевая полость. Бич короче пениса и эпифал-
луса, вместе взятых.............................................................
.... Hygromiidae, Euomphaliinae, Monacha, подрод Metatheba (стр. 287).
136 (129). Слизистые железы отсутствуют или находятся в явно рудиментарном со-
стоянии.
137 (138). Устье раковины с тремя зубами........................................
..........................Polygyridae, Triodopsinae, Triodopsis (стр. 347).
138 (137). Устье раковины без зубов.
139 (142). Раковина белая или светло-серая, одноцветная или с 1—2 коричневыми спи-
ральными лентами.
УСЛОВНЫЕ СОКРАЩЕНИЯ
113
140 (141). Пенис сильно вздут, почти шаровидный, переходит в эпифаллус без резкого
перегиба. Семеприемник отходит от вагины на значительном расстоянии от
клоаки. Раковина одноцветная......................................................
............Hygromiidae, Euomphaliinae, Monacha, подрод Boemica (стр. 289).
141 (140). Пенис не вздут, веретеновидный, на границе его с эпифаллусом имеется
резкий перегиб. Семеприемник отходит очень низко, почти от клоаки. Раковина
со спиральными лентами, отдельные экземпляры одноцветные -........................
.................. Hygromiidae, Metafruticicolinae, Caucasocressa (стр. 304).
142 (139). Раковина роговая, желтоватая или зеленоватая, с размытой светлой лен-
той по периферии, которая иногда лишь угадывается.................................
. . . Hygromiidae, Hygromiinae, Circassina circassica simpla (стр. 247).
УСЛОВНЫЕ СОКРАЩЕНИЯ И ОБОЗНАЧЕНИЯ
В описаниях
При обозначении размеров: ВР — высота раковины, БД — большой диаметр
(ширина) раковины, МД — малый диаметр раковины. СЖ — слизистые железы; для
обозначения числа и характера ветвления применена формула СЖ=Л1 X N, где М —
число желез,- a. N — число ветвей каждой железы.
В рисунках к систематической части
Б — бич (флагеллум);
БЖ — белковая железа;
В — вагина;
ВБ — вагинальные бугорки;
ВВ — вагинальные валики;
ВЛ — лопасти, образуемые вагиналь-
ными валиками в местах вы-
хода протоков стилофоров;
ВЛМ — верхняя лопасть мантийного ва-
лика;
ВИТ — вагинальные мускульные тяжи;
ВП — вагинальные придатки;
ВПП — верхняя лопасть папиллы пе-
ниса;
ГП — гермафродитный проток;
ДВВ — дистальные окончания ваги-
нальных валиков;
ДМ — дополнительный мешок стило-
фора;
ДП — дистальная папилла пениса;
ДСП — дивертикул семеприемника;
ИП — интрапапиллярная полость;
КВВ — кольцевой вагинальный валик;
КЛ — клоака, половой атриум;
КП — кольцевая полость внутри эпи-
фаллуса, пениального чехла и
папиллы пениса;
КР — кольцевое разрыхление тканей
в основании папиллы пениса:
ЛЛМ — левая лопасть мантийного ва-
лика;
М — матка;
МД — мембрана, связывающая ди-
вертикул семеприемника со
спермовидуктом, мембрана ди-
вертикула;
НИ — надклоачный пузырь;
НПП — нижняя лопасть папиллы пе-
ниса;
НСМ — нижняя стенка мантийной по-
лости;
НЧП — наружный чехол пениса;
ОП — отверстие пениса;
ОСП — отверстие протока семеприем-
ника;
ОСТ — отверстие стилофора;
П — пенис;
ПБ — пениальный бугорок;
ПВ — пениальные валики (складки,
расположенные на внутренней
поверхности пениального
чехла);
ПГ — пениальный гребень;
ПЖ — пениальная железа;
ПЛ — папиллярная лакуна;
ПЛМ — правая лопасть мантийного ва-,
лика;
ПМЛ — пениальные мускульные ленты;
ПП — папилла пениса;
ПР — простата;
ПРП — проксимальная папилла пе-
ниса;
ПС — полость стрелы;
ПСМ — пениальная соединительноткан-
ная мембрана (натянута между
семепроводом, эпифаллусом и
пенисом);
ПСП — проток семеприемника;
Р — «рельсы», направляющие склад-
ки в эпифаллусе;
РП — ретрактор пениса, половой ре-
трактор;
РПО — ретрактор правого омматофора;
РСТ — рудиментарный стилофор;
С — семепровод;
СЖ — слизистые железы;
СИП — септы в интрапапиллярной по-
лости;
СОД — спермовидукт;
СП — семеприемник;
СПК — семепроводящий канал;
СТ — стилофор;
ТП — тяжи внутри папиллы пениса,
возникающие в результате диф-
ференцировки паренхимы, за-
полняющей стенки папиллы;
8 А. А. Шилейко
114
I. HELICODONTIDAE
Ц — цекум, слепой вырост на по- ЧПП — чехол папиллы пениса, вторич-
нисе; ный чехол пениса;
ЧВ — чехол ваГины; ЭП — эпифаллус;
ЧП — чехол пениса; Я — яйцевод.
I. Семейство HELICODONTIDAE Hesse, 1907
Раковина прижатая, нередко почти плоская, коричневатых тонов, одноцветная;
скульптура зернистая или радиальная (от слабой исчерченности до ребристости),
спиральная скульптура отсутствует; нередко на раковине имеются волоски. Пупок
перспективный, цилиндрический или развернутый; последний оборот чаще всего в той
или иной степени угловатый; обороты узкие, тесно навитые, шов глубокий. Устье тре-
угольное, ромбическое или полулунное, края его не лежат в одной плоскости и плавно,
но сильно отвернуты, образуя «манжету»; при рассматривании снизу базальный край
выгнут наподобие лука. На базально-палатальном крае нередко бывают зубы, распо-
ложенные несколько отступя от края или имеющие вид вздутий на отвороте устьевого
края.
Половой аппарат характеризуется большим разнообразием. Стилофор один, из-
редка стилофоров 2 или они отсутствуют; иногда стилофор видоизменен в булавовидный
придаток. Слизистых желез одна-две, они располагаются на вагине. Семеприемник
без дивертикула, «пристегнут» к спермовидукту тонкими связками и перепонками и
тесно прилегает к нему; резервуар семеприемника овальный, относительно массивный,
не достигает белковой железы, проток семеприемника цилиндрический. Пенис без бича
и папиллы, реже со своеобразной папиллой; формы, у которых имеется бич, принадле-
жат, вероятно, К’ другим семействам.
Распространение. Средиземноморские страны; наибольшее разнооб-
разие — в Юго-Западной Европе, отдельные виды встречаются в Средней Европе.
В ископаемом состоянии семейство известно начиная с верхнего палеоцена (Европа).
I. Подсемейство HELICODONTINAE Hesse, 1907
[Раковина плоская, устье треугольное, пупок развернутый.
Стилофоры отсутствуют, слизистая железа одна, состоит из двух ветвей, неравно-
значных как по размеру, таки функционально. Пенис с очень короткой своеобразной
папиллой; дистальнее папиллы располагается клапан, ниже которого лежит мощный
продольный валик с продольной бороздой.
Распространение. Средняя и Южная Европа. В ископаемом состоянии
подсемейство известно начиная с нижнего олигоцена (Европа).
В настоящее время известен только один рецентный род.
Род HELICODONTA Fdrussac, 1821
Ferussac, 1821 : 9 (Helix part.); Fit zinger, 1833 : 98 (TrigonostomaY,
Held, 1837 : 12 (Gonostoma); W e n z, Zilch, 1959—1960:692.
[Признаки подсемейства.
Типовой вид — Helix obvoluta Muller, 1774.
fl. Helicodonta aff. evoluta Gaal, 1911.
Gaal, 1911 : 73; Стеклов, 1966 : 224, табл. XII, 217—219.
Материал: 3 экз. из типового местонахождения.
Раковина почти плоская, составлена 5.5—6 плавно и медленно нарастающими
оборотами. Завиток едва возвышается или при нормальном положении раковины не
виден. Последний оборот округлый, снизу слегка уплощен, к устью слабо опущен.
Окраска, видимо, была одноцветной. Скульптура очень характерная: помимо умеренно-
грубой радиальной исчерченности имеются частые мелкие радиальные вмятинки,
в центре каждой из которых находится небольшой бугорок. Устье слегка треугольное,
резко скошено, полулунно вырезано предпоследним оборотом. Несколько отступя от
края имеется очень низкая расплывшаяся губа; края устья тонкие, от ангулярной
области до места прикрепления колумеллярного края постепенно все более отгиба-
ются. Пупок очень широкий, перспективный. Размеры: ВР 4—6, БД 13—18, МД
11—15 мм.
Распространение. Найден в сарматских отложениях Предкавказья
(р. Аргудан) и Венгрии.
HELICODONTA
115
2. Helicodonta obvoluta (Muller, 1774). Табл. I, 1', рис. 42, 43.
Muller, 1774 : 27 (Helix')-, Fitzinger, 1833 : 98 (Trigonostoma); Cles-
sin, 1884, 2 Aufl. : 133, fig. 66 (Trigonostoma}-, L о z e k, 1956 : 216, t. 35, 4; 1964 : 303,
t. 27, 2 abc; Gittenberger et al., 1970 : 102, fig. 177, 183, Kaart 72; Ш и л e й к o,
1971г : 10, рис. 2, 3.
Материал: более 20 экз. из 4 местонахождений в Чехословакии, Польше и ГДР.
Раковина плоская, состоит из 6 оборотов, разделенных глубоким швом.
Последний оборот в профиле плавно широко закруглен, без наклонности к образованию*
Рис. 42—43. Helicodonta obvoluta (Miill.), Задней, ЧССР, 9 VII 1962. 42 — внешний
вид полового аппарата; 43 — пенис, вскрыт. (По Шилейко, 1971г).
угла. Окраска одноцветная, роговая. Скульптура представлена неправильной ради-
альной испорченностью; поверхность раковины с довольно длинными, умеренно густыми,
волосками, сохраняющимися, как правило, пожизненно. Устье округло-треугольное,
слегка или умеренно скошено, при рассматривании снизу базальный край слегка изог-
нут в виде лука (по А. А. Стеклову — «в профиле край устья изгибается петлеобразно»).
Края устья отогнуты и расширены. Размеры: ВР 5—6, БД 10—14, МД 9—12.5 мм.
Внутреннее строение. Материал: 2 экз. из Задиела, ЧССР, 9 VII 1962,
сбор и определение В. Гудеца; 5 — из Карлштейнского заповедника, ЧССР, 24 V 1969,
сбор Я. Бухара, определение наше; 7 — из Букеберга у Зюдероде, ГДР, VI 1969,
сбор и определение Э. Клаусса.
Стенки вагины внутри гладкие или покрыты неясными складками. Слизистая же-
леза двуветвистая, ветви ее неравной длины, различны по гистологическому строению
и выполняют разные функции: более длинная ветвь — собственно железа, а более ко-
роткая служит резервуаром для секрета длинной ветви. Пенис и эпифаллус одеты
общим чехлом. Внутренние стенки пениса несут несколько мощных продольных ва-
ликов, ил и складок; один из них короче других, выражен более резко и снабжен про-
дольной бороздкой, служащей, по-видимому, для ориентации сперматофора. Прок-
симальна я часть этого валика охвачена подушкообразным утолщением, поверхность
которого покрыта многочисленными коническими углублениями. Напротив этого утол-
щения расположено второе утолщение седловидной формы, несущее конические пш-
8*
116
I. HELICODONTIDAE
пики, соответствующие углублениям на противоположном утолщении. Гистологи-
ческое строение шипиков освещено в нашей работе (Шилейко, 1971г). В совокупности
описанные валики образуют своеобразный клапан, способный полностью разъединять
дистальный и проксимальный отделы пениса. В глубине пениса, за клапаном, имеется
очень короткая двулопастная папилла, охватывающая выходное отверстие эпифаллуса.
Половой ретрактор крепится не к диафрагме, а является ветвью колумеллярного
мускульного комплекса, точнее, той его части, которая дает начало ретрактору пра-
вого омматофора.
Резервуар семеприемника прилегает к спермовидукту примерно по середине его
протяжения; резервуар семеприемника неясно отграничен от протока.
Распространение и местообитания. Европа до Пиренеев, а се-
вернее — до Атлантического побережья (Girod, 1968), и Великобритания (Cameron,
1972). В СССР не обнаружен, но не исключено его присутствие в западных районах
Белоруссии, близ границы с Польшей. Обитает в подстилке лиственных горных и рав-
нинных лесов. Вид известен из плиоцена и плейстоцена Западной Европы.
II. Подсемейство LINDHOLMIOLINAE Schileyko, subfam. nov.
Раковина прижатая, с более или менее ясно выраженным углом на периферии;
устье полулунной формы; пупок узкий или умеренно широкий, цилиндрический.
Стилофоры отсутствуют, слизистая железа одна, неразветвленная, в разной сте-
пени гофрирована. Пенис без бича и папиллы, эпифаллус многократно извит и скручен;
«го изгибы плотно окутаны соединительнотканным чехлом и обнаруживаются только
при вскрытии.
Распространение. Южная и Юго-Восточная Европа.
1. Род LINDHOLMIOLA Hesse, 1931
Hesse, 1931 : 50; W е n z, Zilch, 1959—1960 : 693.
Признаки подсемейства.
Распрострайение. Южная Европа, к западу до Италии, к востоку до
Балканского п-ова и южных Карпат. В ископаемом состоянии род известен из верхнего
плиоцена (восточное Средиземноморье) и верхнего миоцена—верхнего плиоцена
(Предкавказье).
Типовой вид — Helix lens Ferussac, 1821.
fl. Lindholmiola fraudulosa (Steklov, 1966). Табл. I, 2.
Стеклов, 1966 : 225, табл. XI, 210—216 (Caracollina).
M атериал: 8 экз. из двух местонахождений в неогеновых отложениях Предкавказья.
Раковина плоская, с почти не возвышающимся завитком. Оборотов 5.5,
уплощенных, плотно навитых, медленно нарастающих, разделенных умеренно глубо-
ким швом. Последний оборот в профиле округлый, перед устьем внезапно опущен,
в базальной части уплощен. Окраска одноцветная. Скульптура в виде очень тонкой
неправильной радиальной исчерченности и следов от волосков; кроме того, на всех
раковинах наблюдалась вторичная скульптура в виде многочисленных вмятин от
прилегавших песчинок. Устье полулунное, сильно скошено, с тупыми утолщенными
краями. Колумеллярный край резко отогнут и слегка прикрывает цилиндрический
пупок, сквозь который просматривается не менее 2.5—3 предыдущих оборотов. Ба-
зальная часть раковины к пупку образует характерную покатость, свойственную пред-
ставителям рода. Размеры: ВР 7—8, БД 15—18, МД 14—16.5 мм.
Распространение. Верхний миоцен—верхний плиоцен Предкавказья.
2. Lindholmiola corcyrensis (Ferussac, 1839). Табл. I, 3; рис. 44, 45.
Ferussac, 1839, Hist. Nat., I : 21, Taf. 69e, Abb. 1—5 (Helix); R о s s m a s-
s 1 e r, 1838, Iconogr., 2, fig. 538 (Helix contorta); 1904 : 180 [Helix (Caracollina)
corcyrensis Zgl. (Helix torso.) si Friv. (Helix girva)j; Sturany, Wagner, 1915 : 32,
Taf. 2, Abb. 7a; G г о s s u, 1955 : 443, fig. 250 (Caracollina); Шилейко, 1971г : 12,
рис. 2, Б, 4; H u d e c, 1972b : 337, Abb. 9, t. Ill, 7.
Материал; более 30 экз. из окрестностей Кишинева и Болгарии.
Раковина с очень низким, едва возвышающимся куполовидным завитком
и сглаженной вершиной. Оборотов 6—7, узких, медленно нарастающих. Периферия
с плавным, но явственным углом. Окраска роговая, одноцветная или ср слегка наме-
чающейся светлой полосой на периферии. Скульптура в виде неправильной радиаль-
ной исчерченности или слабой морщинистости. Имеются многочисленные длинные
волоски,' сохраняющиеся пожизненно и видные нередко даже у пустых раковин.
LINDHOLMIOLA
117
Устье узкое, полулунное, края его отогнуты и образуют губу коричневато-розовой
окраски, часто со слабым фиолетовым оттенком. Устье умеренно скошено, последний
оборот к устью внезапно опущен. Пупок довольно широкий, но не развернутый, ци-
линдрический; слегка поворачивая раковину, сквозь пупок можно видеть все или почти
все обороты. Размеры: ВР 4—6, БД 9—12, МД 8—11 мм.
Внутреннее строение. Материал: 1 экз. из долины р. Вычи, зап. Ро-
допы, Болгария, 22 VI 1969; 2 — из окр. Ивайловграда, вост. Родопы, Болгария,
20 VI 1969, сборы и определения И. М. Лихарева; 5 — из окр. Кишинева (Дурлеш-
тский лес), 9 XI 1968, сбор и определение наши.
Рис. 44—45. Lindholmiola corcyrensis (Fer.), зап. Родопы, Болгария, 22 VI 1969. 44 —
внешний вид полового аппарата; 45 — дистальный отдел полового аппарата, вскрыт.
(По Шилейко, 1971г).
Внутренние стенки вагины несут несколько продольных складок, расположенных
бессистемно. На передней стенке вагины имеется мощное утолщение, благодаря кото-
рому полость вагины сообщается с атриальной полостью посредством очень узкой
щели между задней стенкой вагины и утолщением, которое, таким образом, играет роль
клапана, способного изолировать полость клоаки от женского отдела. Слизистая же-
леза одна, неразветвленная, с рыхлыми стенками, образует несколько тесно спаянных
извивов (гофрированная) или имеет многочисленные пережимы. При основании железа
резко сужается и впадает в вагину очень тонким протоком. Степень развития и внешний
вид железы находятся в прямой зависимости от ее функционального состояния (на
рис. 44 изображен июньский экземпляр; у ноябрьских, также вполне взрослых, сли-
зистая железа гораздо короче, тоньше и не изогнута).
Пенис и эпифаллус охвачены общим чехлом. Внутренняя поверхность пениса пок-
рыта короткими продольными складками, пенис в проксимальной части слепо замкнут.
•Эпифаллус впадает несколько сбоку посредством очень маленького отверстия; проток
эпифаллуса очень резко многократно скручен, стенки его железистые; эпифаллус
спускается вдоль пениса ниже места сообщения этих органов.
. ;. Резервуар семеприемника' прилегает it спермовидукту немного ниже середины
последнего и имеет округлую или молотовидную форму.
118
II. BRADYBAENIDAE
Распространение и местообитания. Населяет лиственную
подстилку равнинных и горных лесов Балканского п-ова и Греческого архипелага;
единичные находки в Италии; в СССР сделана одна находка в окр. Кишинева. В иско-
паемом состоянии встречается начиная с плиоцена (Юго-Восточная Европа).
2. Род OESTOPHORA Hesse, 1907
Hesse, 1907, Nachrbl. deutsch. malak. Ges., 39 : 76; О r t i z de Z a r a t e,
1962 : 6; W e n z, Zilch, 1959—1960 : 691.
Дифференциальное конхологическое отличие от видов рода Lindholmiola заключа-
ется только в отсутствии волосков на поверхности раковины.
Имеется один массивный стилофор, слизистых желез две-три, гофрированных в раз-
ной степени. Пенис без папиллы; в пределах рода наблюдается постепенная редукция
бича вплоть до полного исчезновения.
Этот род мы провизорно включаем в подсемейство Lindholmiolinae на основании
конхологического сходства с номинативным родом, а также в связи с формальным сход-
ством в строении совокупительного аппарата. Внешний вид слизистых желез также
в общем сходен, наличие или отсутствие стилофора в данном случае не может служить
препятствием к объединению этих двух родов в одно подсемейство, так как стилофор
вообще часто подвержен редукции. Окончательный вывод о таксономическом статусе
рода можно будет сделать после изучения внутреннего строения дистальной части
мужского отдела у большинства видов рода.
Распространение. В настоящее время ареал рода занимает Юго-Запад-
ную Европу (Пиренейский п-ов) и Северо-Западную Африку. В ископаемом состоянии
род известен начиная с нижнего олигоцена (Европа).
Типовой вид — Helix lusitanica L. Pfeiffer, 1848.
(Oestophora schileykoi (Prysjazlinjuk, 1974). Табл. I, 4.
Присяжнюк, 1974 ; 319, рис. 26—э {Perforatella).
Материал: 2 экз. из понтийских отложений левобережья Дуная.
Раковина прижатая, с очень низким коническим завитком, составлена 6 мед-
ленно нарастающими оборотами. Последний оборот в профиле слегка угловатый,
к устью не опущен. Скульптура в виде очень тонкой сглаженной радиальной морщи-
нистости. Устье полулунное, края его отвернуты и чуть утолщены, при рассматрива-
нии снизу хорошо заметен изгиб базального края, напоминающий ненатянутый лук.
Этот признак характерен для семейства в целом и позволяет отличать этот вид от пред-
ставителей рода Perforatella, на которых он действительно во многом похож. В левой
стороне базального края и на базально-париетальном крае имеются два удлиненных
зуба; последний развит лучше. Снаружи этим зубам соответствуют вмятины. Пупок
цилиндрический, сверху затянут тонкой мозолью. Размеры: ВР 4, БД 8, МД 6.8 мм.
Распространение. Вид известен пока из типового местонахождения
в окр. с. Виноградовка на оз. Ялпух, левый берег Дуная, верхний миоцен (понт).
II. Семейство BRADYBAENIDAE Pilsbry, 1939
Раковина различных размеров, формы и окраски; наиболее характерным конхо-
логическим признаком является присутствие ясных спиральных бороздок на поверх-
ности раковины, но эти бороздки могут быть выражены неясно или изредка отсутство-
вать.
Стилофор первично один, в редких случаях отсутствует. Слизистые железы сгруп-
пированы в два пучка, впадают в стилофор, часто через дополнительный мешок. Се-
меприемник без дивертикула, резервуар относительно небольшой, шаровидный, резко
обособлен от протока, «пристегнут» соединительнотканной связкой к задней части
дна мантийной полости в области сердца и почки. Проток семеприемника конический,
с расширенным в той или иной степени основанием. Бич и папилла пениса первично
отсутствуют, но изредка формируются вторично; , в этом случае папилла формируется
за счет погружения эпифаллуса в просвет пениальной трубки.
В пределах семейства принято выделять несколько подсемейств; однако в настоя-
щее время нет возможности ни составить их дифференциальные диагнозы,, ни опреде-
лить их границы и объем. В связи с этим все виды фауны СССР, относящиеся к этому
семейству, здесь обозначены без указаний на подсемейственную принадлежность.
Распространение. Наибольшее разнообразие наблюдается в Юго-Вос-
точной Азии; кроме того, виды семейства распространены на Филиппинских о-вах,
в Японии, на Дальнем Востоке, в Центральной и Средней Азии; отдельные виды прони-
кают в Европу. Систематическое положение видов, обитающих в Африке и относимых
по традиции к данному семейству, нуждается в уточнении.
PONSADENIA
119
В ископаемом состоянии представители семейства встречаются начиная с плио-
цена (Восточная Азия). В палеоцене Европы обнаружено небольшое число видов рода
Coneulota, который принято относить к данному семейству. Такая трактовка систе-
матического положения этого рода вызывает сомнение; род должен быть отнесен к дру-
гому семейству. Вообще геологическое распространение видов семейства изучено плохо.
1. Род PONSADENIA Schileyko, gen. nov.
Раковина трех типов: либо маленькая, уплощенная, с угловатой периферией,
либо прижатая, средней величины или небольшая, с округлой периферией, либо боль-
шая, почти шаровидная, с округлой периферией.
Слизистые железы впадают либо в парные резервуары, прилегающие к клоаке и
соединенные с апикальной частью стилофоров протоком, «мостиком», либо они впадают
в сам проток близ его середины.
От рода Bradybaena отличается наличием «мостика», первично представляющего
собой проток резервуаров слизистых желез.
Распространение. Тянь-Шань, Джунгарский хр. и Тарбагатай.
Типовой вид — Helix semenovi Martens, 1864»
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА PONSADENIA
1 (2). Раковина шаровидная. Большой диаметр не менее 20 мм. (Подр. Mesasiaia)
...................................................... 2. Р. duplocincta.
2 (1). Раковина прижатая. Большой диаметр не превышает 16 мм.
3 (4). Последний оборот резко угловатый. (Подр. Tarbagataja) .... 1. Р. hirsuta.
4 (3). Последний оборот плавно закруглен. (Подр. Ponsadenia) . . 3. Р. semenovi.
1. Подрод TARBAGATAJA Schileyko, subgen. nov.
Раковина маленькая, с едва возвышающимся завитком, одноцветная, бурая,
с углом на периферии и радиальными морщинками, несущими кожистые чешуйки.
Небольшие резервуары слизистых желез располагаются на конце общего протока
и прилегают к клоаке.
От других подродов того же рода резко отличается скульптурой и формой рако-
вины и значительной обособленностью резервуаров слизистых желез от клоаки.
Монотипический подрод.
Типовой вид — Brabybaena hirsuta Matiokin, 1966.
1. Ponsadenia (Tarbagataja) hirsuta (Matiokin, 1966). Табл. I, 5.
M а т ё к и н, 1966 : 124, табл. 4, рис. 7 (Bradybaena)', 1972 : 131, рис. 4, Г, Д, Е
{Bradybaena).
Материал: 4 не вполне сформированные раковины из Тарбагатая.
Раковина сильно уплощенная, с низким завитком и ступенчатыми оборо-
тами; оборотов 5.75—6. Последний оборот в профиле угловатый, к устью энергично
опущен. Скульптура в виде нерезких радиальных морщин, некоторые из них несут
кожистые чешуйки, располагающиеся на расстоянии 1—2 мм друг от друга. Устье
косое. Пупок не очень широкий, но перспективный и развернутый. Размеры: ВР 4.5—5,
БД 10.6—-10.8 мм.
Внутреннее строение. Материал нами не исследован, описание при-
водится в соответствии с данными П. В. Матёкина (1966).
Бутылковидный стилофор впадает в вагину у места перехода последней в клоаку.
При наружном осмотре два куста слизистых желез кажутся впадающими в два малень-
ких резервуара, расположенных по бокам суженной части стилофора близ места впа-
дения ее в вагину. У некоторых экземпляров хорошо заметна резкая килевидная
складка, идущая спереди назад по поверхности стилофора; эта складка представляет
собой вытянутую в длинную трубку заднюю часть стилофора («мостик»), которая сог-
нута в колено и прилегает к более широкой передней половине стилофора. Отмеченные
выше два резервуара слизистых желез представляют собой апикальное разветвление
этой вытянутой задней части стилофора. Таким образом, то расположение резервуа-
ров и слизистых желез, которое видно при внешнем осмотре, представляется резуль-
татом изогнутости верхней части стилофора, в разветвленный конец которой посред-
ством двух резервуаров впадают слизистые железы. Эти последние закладываются двумя
самостоятельными зачатками по обеим сторонам задней части стилофора.
Пенис длинный, цилиндрический, крючковидно изогнут.
Распространение и местообитания. Вид обнаружен в Джун-
гарском хр. и в Тарбагатае. Обитает в задернованных осыпях.
120
II. BRADYBAENIDAE
2. Подрод MESASIATA Schileyko, subgen. nov.
Раковина довольно большая, почти шаровидная, с округлыми оборотами, одно-
цветная или с темными спиральными лентами. Скульптура в виде неравномерной мор-
щинистости и спиральных бороздок.
Резервуары слизистых желез располагаются на поверхности атриальной области
и соединены «мостиком» с вершиной стилофора; слизистые железы впадают несколь-
кими отдельными ветвями в каждый из резервуаров.
Рис. 46—48. Ponsadenia (Mesasiata) duplocincta (Mart.), окр. оз. Иссык-Куль,
12 VIII 1970. 46 — внешний вид полового аппарата; 47 — стилофор, виде двух про-
тивоположных сторон, вскрыт; звездочка — «мостик»; 48 — пенис, вскрыт. (Ориг.).
От других подродов этого рода отличается большой шаровидной раковиной и мас-
сивными резервуарами слизистых желез, тесно спаянными с клоакой.
Монотипический подрод;
Типовой вид — Helix duplocincta Martens, 1879.
2. Ponsadenia (Mesasiata) duplocincta (Martens, 1879). Табл. I, 6; рис. 46—48.
Martens, 1879, Sitzungsb. naturf. Fr. Berlin : 125 (Helix)-, 1882a : 4, 56, t. 1,
fig. 1—7; t. 5, fig. 1 — 7 (Helix)', 5, 59, t. 1, fig. 8; t. 5, fig. 8, 9 (Helix paricincta); W es-
ter 1 u n d, 1889 : 122 (Helix); 1898 : 163 [Helix (Pomatia) stenroosi]; Kobelt,
1899, Iconogr., N. F., VIII : 64, fig. 1474 (Helix); Weber, 1913 : 5, fig. 1 (Helix);
Лихарев, P а м мельмейер, 1952 : 397, рис. 331. [Eulota (Eulota)]; M a-
t ё к и н, 1966 : 120, табл. 4, рис. 4 (Bradybaena); 1972 : 125, рис. 3, А, Б, В (Brady-
. Ъаепа).
Материал: более 50 экз. из 5 местонахождений на^Тянь-Шане.
Раковина твердостенная, шаровидно-кубаревидная или шаровидная, с ко-
ническим или слегка куполовидным контуром завитка, высота которого немного меньше
или равна высоте устья. Оборотов 6, умеренно выпуклых, довольно медленно нарастаю-
щих. Эмбриональные обороты гладкие, тонкойе'р'нистыс. Последний оборот в 1.5 раза
PONSADENIA
121
шире предпоследнего, к устью плавно, без перегиба, опущен. Окраска чаще образована
желтым или желтовато-бурым фоном и несколькими коричневыми спиральными лен-
тами. Лент может быть две: выше и ниже периферии, причем верхняя имеется и у шва
на предыдущих одном-двух оборотах. Нередко выше и ниже этих лент лежит по одной
более широкой размытой полосе (именно такая цветовая морфа представлена на табл. 1,6').
Иногда по периферии проходит одна широкая лента, образованная в результате рас-
ширения и слияния двух основных лент; наконец, известны случаи отсутствия лент,
тогда раковина либо одноцветно желтоватая, либо буровато-каштановая со светлым
полем вокруг пупка. В свое время большинство цветовых вариантов получило латин-
ские наименования; поскольку эти варианты есть проявления популяционной или ин-
дивидуальной изменчивости, названия здесь не приводятся. Скульптура представлена
неравномерной радиальной морщинистостью и спиральными бороздками, которые
местами прерываются радиальными морщинами. Устье округлое, слабо скошено,
места его прикрепления широко расставлены. Края устья утолщены, изнутри окайм-
лены слабой светлой губой. Колумеллярный край в начале почти отвесный, широко
отвернут и наполовину прикрывает пупок, остальные края устья почти не отвернуты.
Пупок узкий, неперспективный.’ Размеры: ВР 19—26, БД 22—27, МД 20—24 мм.
Внутреннее с т р о е н и е. Материал: 7 экз. из южного берега оз. Ис-
сык-Куль, 12 VIII 1970, сбор Л. В. Славошевской, определение наше.
Нижняя часть спермовидукта образует резкий двойной изгиб с сомкнутыми сто-
ронами. Яйцевод примерно вдвое короче вагины. Слизистые железы слабо гофриро-
ваны, некоторые из них раздвоены, общее число желез непостоянно. Железы впадают
в резервуар, разделенный неполной перегородкой на две камеры; в каждую камеру
открывается по нескольку слизистых желез — каждая железа самостоятельным про-
током. От резервуара слизистых желез отходит единый проток, просвет которого слабо
дифференцирован: на поперечном разрезе он представлен несколькими каналами, не
имеющими собственных стенок (рис. 47). Нижняя часть протока спаяна со стенкой сти-
лофора, далее канал несколько обособляется и впадает в верхнюю часть стилофора
не апикально, но несколько сбоку, образуя «мостик». Стилофор имеет форму сапожка,
верхняя часть его светлая, остальная часть пигментирована. Стрела короткая, зани-
мает самую верхнюю часть стилофора. Ниже стрелы внутренние стенки стилофора пок-
рыты необыкновенно мощными продольными складками,' разделенными глубоко вре-
занными бороздами на отдельные бугорки прямоугольной формы.
Имеется чехол пениса, закрепленный только у клоаки и охватывающий воротнич-
ком дистальную часть длинного цилиндрического пениса. Внутренняя поверхность ди-
стальной части пениса несет несколько четких продольных складок, контуры которых ПО’
мере удаления от клоаки начинают приобретать зубчатые очертания, и в прокси-
мальной части складки превращаются в ряды высоких лепестковидных выростов, распо-
ложение которых близ границы с эпифаллусом становится беспорядочным.
Резервуар семеприемника вжат в основание белковой железы, причем проток семе-
приёмника притянут к спермовидукту, а резервуар — к нижней стенке мантийной
полости.
Распространение и местообитания. Вид распространен пят-
нами в Тянь-Шане: от тугайных лесов долин левых притоков р. Или до-
восточной части Киргизского хр., минуя северные склоны Заилийского и Киргиз-
ского хр.; местами на побережье оз. Иссык-Куль. Гигрофильный вид, обитает на лес-
ных склонах лугово-лесо-степного и субальпийского поясов, в траве.
3. Подрод PONSADENIA s. str.
Раковина прижатая, светлая, без цветного рисунка, с округлой периферией, не-
большая, тонко скульптурирована.
Слизистые железы впадают в «мостик», соединяющий атриальную область с вер-
шиной стилофора; наблюдается высокая степень интеграции тканей покровов,
клоаки с резервуарами слизистых желез, в результате чего последние как таковые не
выражены.
Монотипический подрод.
3. Ponsadenia (Ponsadenia) semenovi (Martens, 1864). Табл. I, 7; IX, 81; рис. 49—51.
Martens, 1864, Malak. Bl., XI : 115, t. 3, fig. 6—8 (Helix); Мартенс}
1874 : 16. табл. I, рис. 12 (Helix); Martens, 1882a : 9, t. 2, fig. 5—7 (Helix); Gude,
1904 : 93.(Cathaica hermanni); 1905, J. Conch., XII : t. 4, fig. 10 (Cathaica hermanni);
Cockerell, 1928 : 363 (Cathaica keiseri); Л ин д г о л ь м, 1931 : 54 [Cathaica
(Pliocathaica)]; Лихарев, Раммельмейер, 1952 : 404, рис. 335 [Eulota (?)];
М а т ё к и н, 1966 : 120, табл. 4, рис. 1 (Bradybaena); 1972 : 124, 3, Г, Д, Е (Bra-
dybaena).
49
Рис. 49—51. Ponsadenia (Ponsadenia) semenovi (Mart.), ущелье p. Нарын
выше Токтогула, 31 V 1972. 49 — внешний вид полового аппарата;
50 — дистальная часть женского отдела, вскрыта; 51 — пенис, вскрыт.
(Ориг.).
BRADYBAENA
123
Материал: более 100 экз. из 26 местонахождений на Тянь-Шане.
Раковина в разной степени прижатая с куполовидным контуром завитка и
слегка выступающей вершиной. Оборотов 5—5.5, сильно выпуклых, медленно нарастаю-
щих, разделенных глубоким швом. Эмбриональные обороты гладкие. Последний обо-
рот в профиле правильно закруглен на всем протяжении контура, к устью несильно
и очень плавно опущен. Окраска почти одноцветная, без цветовых лент: светло-серая,
коричневато-розовая (чаще всего), слегка голубоватая с серым оттенком; нижняя по-
верхность обычно немного светлее верхней. Скульптура в виде тонкой радиальной ис-
черченности, с отдельными, более заметными радиальными складками на последнем
обороте'; кроме того, у большинства экземпляров имеются вялые спиральные бороздки,
выраженные лишь местами на верхней поверхности последнего оборота. Устье почти
правильно круглое, немного скошено, края его внутри окаймлены расплывшейся
•светлой губой, не отвернуты, кроме колумеллярного. Последний отвернут наверху,
но не закрывает пупка, сквозь который можно просмотреть лишь часть предпослед-
него оборота. Размеры: ВР 6—10, БД 9—15, МД 8—13 мм.
Мало изменчивый вид, варьируют в основном абсолютные размеры.
Внутреннее строение. Материал: 3 экз. из Заилийского хр., Аксай,
12 V 1970, сбор К. К. Увалиевой; 2 — из ущелья р. Аламедин на Киргизском хр.,
14 VI 1972; 4 — из ущелья Беш-Таш на северном склоне Таласского хр. близ г. Таласа,
5 VI 1972; 22 — из ущелья р. Нарын выше Токтогула, 31 V 1972; сборы и определе-
ния наши.
Спермовидукт в дистальной части образует крутое колено, края которого сопри-
касаются друг с другом, дистальная часть женского отдела массивная. Слизистые
железы в виде двух слабо ветвящихся пучков, протоки которых объединяются непо-
средственно перед впадением в своеобразный «мостик» между апикальной частью сти-
лофора и атриальной областью. Внутри этого «мостика» имеется проток, который на-
верху открывается в полость стилофора, а внизу — непосредственно в губчатые ткани
стенок клоаки. Эти ткани — продукт объединения собственно стенок клоаки и резер-
вуаров слизистых желез.
Пенис длинный, в различной степени извит, иногда даже слегка скручен. Внутрен-
няя его поверхность несет несколько продольных, иногда слабо ветвящихся или аносто-
мозирующих друг с другом валиков, которые прослеживаются вплоть до семепровода
без существенного видоизменения; эпифаллус не выражен. Чехол пениса начинаясь от
клоаки тянется вдоль пениса и постепенно сходит на нет, не прирастая к поверхности
пениса.
Диаметры яйцевода и основания протока семеприемника почти равны. Резервуар
семеприемника сильно вжат в основание белковой железы и верхнюю часть спермови-
дукта.
Распространение и местообитания. Почти весь Тянь-Шань,
кроме самых западных районов (отсутствует на Пскемском и Угамском хр.), южный
Алтай (Увалиева, 1963). Ксерофильный вид, обитает на открытых глинистых или щеб-
нистых участках со скудной растительностью; в жаркое время и на зиму зарывается
в почву на глубину до 10—12 см.
2. Род BRADYBAENA Beck,' 1837
Beck, 1837 : 18; Held, 1837 : 914; Hartmann, 1843 : 179 (Eulota);
Martens, 1860, Helic., ed. 2 : 56 (Acusta)' Lindholm, 1927a : 120 (Frutici-
cola); P i 1 s b r y, 1927 : 13 (Karaftohelix)', Лихарев, Раммельмейер,
1952 : 387 (Eulota); W e n z, Zilch, 1959—1960 : 642.
Раковина большей частью средней величины, в различной степени прижатая;
часто имеются спиральные темные ленты. Пупок различной ширины, но никогда не
бывает полностью запаянным.
Слизистые железы двумя пучками впадают либо непосредственно в стилофор, либо
в дополнительный мешок, расположенный на поверхности стилофора.
Распространение. Средняя, Центральная и вся Восточная Азия, к се-
веру до Камчатки; Филиппины и Япония. Два вида обитают в Европе. В ископаемом
состоянии род известен начиная с верхнего плиоцена (Центральная Азия).
Типовой вид — Helix similaris Ferussac, 1822.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ, ПОДВИДОВ
И НЕКОТОРЫХ ФОРМ РОДА BRADYBAENA
1 (34). Виды обитают в европейской части СССР, в Сибири и на Дальнем Востоке.
2 (7). Раковина от слегка прижатой до шаровидной, тонкостенная или умеренно
твердостенная. Радиальная скульптура в виде тонкой морщинистости или исчер-
ченности. Высота устья примерно равна его ширине. Пупок узкий, но открытый.
124
II. BRADYBAENIDAE
3 (4). Раковина с 2 спиральными темными лентами..............24. Br. helvola.
4 (3). Раковина одноцветная или с одной лентой.
5 (6). Раковина почти шаровидная: индекс отношения высоты раковины к ее боль-
шому диаметру 0.8—0.9.........................................1. Br. fruticum.
6 (5). Раковина прижатая: индекс отношения высоты раковины к ее большому диа-
метру 0.6—0.7 ........................................2. Br. schrencki.
7 (2). Совокупность признаков иная.
8 (9). Раковина шаровидная, высота завитка значительно превышает высоту устья.
Последний оборот почти одной ширины с- предпоследним. На колумеллярном
крае часто имеется зубообразное утолщение.....................11. Br. arcasiana..
9 (8). Раковина иная.
10 (19). Раковина очень хрупкая, сильно просвечивающая, с очень узким не перспек-
тивным пупком.
11 (14). Большой диаметр раковины не менее 21 мм.
12 (13). Края устья взрослых экземпляров чаще всего отвернуты. Спиральная исчер-
ченность резкая. Вид обитает в Приморском крае..................15. Br. ravida.
13 (12). Края устья взрослых экземпляров не отвернуты. Спиральная исчерченность
вялая. Вид обитает на Курильских о-вах.......................14. Br. vulcanica.
14 (11). Большой диаметр раковины не превышает 20 мм.
15 (16). Губа имеется; края устья и губа красноватые. Вид обитает на Сахалине . . .
................................................................ 9. Br. duiensis.
16 (15). Губа отсутствует или крайне слабая, белая; края устья светлые. Виды оби-
тают в южной части Приморского края.
17 (18). Раковина от кубаревидной до шаровидной. Пупок не менее чем наполовину
прикрыт отворотом колумеллярного края устья, чаще всего щелевидный. . . .
................................................................13. Br. fragilis.
18 (17). Раковина прижатая. Пупок почти не( прикрыт отворотом колумеллярного
края, округлый..................................................12. Br. diversita.
19 (10). Раковина не хрупкая, плотная или слабо просвечивающая, пупок перспек-
тивный.
20 (21). Большой диаметр раковины не превышает 11 мм. На базальном крае устья
обычно имеется удлиненное вздутие. На скатах пупка присутствуют многочис-
ленные чешуйки или гранулы ..................................10. Br. dieckmanni.
21 (20). Большой диаметр раковины не менее 12 мм. Базальный край устья без взду-
тия. Чешуйки или грапулы на скатах пупка отсутствуют.
22 (25). Пупок широкий, развернутый.
23 (24). Радиальная скульптура представлена правильной ребристостью. Большой
диаметр 12—16 мм.............................................5. Br. ussuriensis.
24 (23). Радиальная скульптура в виде исчерченности, но не ребристости. Большой
диаметр 21—26 мм............................................4. Br. middendorffi.
25 (22). Пупок не широкий или узкий, цилиндрический.
26 (27). Раковина прижатая. Вид обитает в Забайкалье .... 3. Br. transbaicalia.
27 (26). Раковина от кубаревидной до шаровидной. Виды обитают на Дальнем Во-
стоке.
28 (29). Раковина резко или ребристо радиально исчерчена. Вид обитает в бассейне
Амура и в Приморском крае . ....................................6. Br. maacki.
29 (28). Раковина без резкой радиальной исчерченности. Виды обитают на Сахалине
и Курильских о-вах.
30 (31). Раковина одноцветная . . . . ..................... 8. Br. strelkovi.
31 (30). Раковина не одноцветная: имеются большей частью размытые радиальные
полосы и спиральные ленты желтовато-коричневых тонов.
32 (33). Рисунок на раковине интенсивный. Высота раковины 20—22, большой диа-
метр 26—29 мм. Подвид встречается на Курильских о-вах...........................
.............................................. 7а. Br. weyrichii bocageana.
33 (32). Спиральные ленты и особенно радиальные полосы умеренно развиты; послед-
ние иногда отсутствуют. Высота раковины 20—31, большой диаметр 31—38 мм.
Подвид обитает на Сахалине ...........................7. Br. weyrichii weyrichii.
34 (1). Виды обитают в Средней Азии и в восточном Казахстане.
35 (36). На поверхности раковины имеется более или менее развитая вермикулятная
скульптура. Раковина шаровидная, толстостенная..................16. Br. scythica.
36 (35). Вермикулятная скульптура отсутствует. Раковина различной формы, стенки
ее различной толщины.
37 (48). Периферия последнего оборота с килем или явственным углом.
38 (43). Обороты ступенчатые: верхняя часть их у шва уплощенная.
39 (40). На поверхности раковины имеются радиальные ребра z...................
............................... 18е. Br. plectotropis smsp. transilensis.
40 (39). Радиальная скульптура в виде тонкой исчерченности.
41 (42). Обороты резко ступенчатые, киль острый. Подвид обитает на Тянь-Шане
................................................. 27. Br. cavimargo cavimargo.
BRADYBAENA 12$
42 (41). Обороты не резко ступенчатые, киль притупленный. Подвид обитает в Тарба-
гатае ...........................................27а. Br. cavimargo tarbagataica.
43 (38). Обороты не ступенчатые.
44 (45). Раковина ребристая ...............18d. Br. plectotropis smsp. plectotropis.
45 (44). Раковина гладкая или грубо исчерченная, но не ребристая.
46 (47). Раковина почти гладкая, вершина выступающая, завиток имеет слегка вогну-
тые очертания .............................. 25. Br. pseudoferghanica.
47 (46). Раковина грубо радиально исчерчена, вершина не выступающая, завиток
куполовидный ...............................18с. Br. plectotropis smsp. necopinatus.
48 (37). Периферия последнего оборота без киля; угол, если имеется, очень легкий и
выражен лишь в самом начале последнего оборота.
49 (50). Раковина с широким куполовидным завитком и уплощенной базальной частью,
покрыта сглаженными светлыми радиальными ребрышками, выраженными на
верхней поверхности раковины, но почти исчезающими па базальной поверх-
ности. Между ребрышками имеется точечная скульптура . . 19. Br. tzwetkovi.
50 (49). Раковина иная.
51 (54). Раковйиа сильно уплощена, тонко скульптурирована, с широким разверну-
тым пупком, одноцветно белая или верхняя поверхность немного темнее ниж-
ней. Темные спиральные ленты полностью отсутствуют.
52 (53). Последний оборот в профиле правильно закруглен . . 26. Br. fedtschenkoi.
53 (52). Последний оборот в профиле слегка угловатый, к устью угол исчезает . . .
.............................................................20. Br. perlucens.
54 (51). Раковина иная.
55 (58). Пупок развернутый, сквозь него просматриваются все обороты, ширина его
составляет 1/i—1IS большого диаметра. Имеются 2 темные спиральные ленты;
одна почти совпадает с периферией или расположена чуть ниже ее, вторая про-
ходит выше первой.
56 (57). Основной фон раковины известково-белый или светло-серый, иногда с очень
легким голубоватым оттенком. Высота раковины не превышает 9 мм ....
. . •........................................................ 23. Br. stoliczkana.
57 (56). Основной фон раковины от светло-рогового до темно-каштанового. Высота
раковины не менее 11 мм.......................................21. Br. dichrozona.
58 (55). Пупок цилиндрический, сквозь него просматривается не более 1.5 оборота,
ширина его не превышает 1/7 большого диаметра. Темные спиральные ленты
отсутствуют; если же имеются, то расположение их иное.
59 (68). Пупок очень узкий, частично прикрыт отворотом колумеллярного края,
сквозь него просматривается не более половины предпоследнего оборота.
60 (61). Раковина почти одноцветная, от оливковой до буровато-желтой. Спиральная
исчерченность еле намечается . .................................28. Br. tomyris.
61 (60). Раковина иной окраски. Спиральная исчерченность выражена ясно, хотя бы
на нижней стороне раковины.
62 (63). Раковина шаровидная, красновато-коричневая, с белой или желтой лентой
па периферии и светлым полем вокруг пупка.....................22. Br. bilaticincta.
63 (62). Раковина от прижатой до кубаревидной, белая, желтоватая или светло-рого-
вая, одноцветная или с темными спиральными лентами.
64 (65). Фон раковины известково-белый, выше периферии имеется одна темная лента;
когда раковина выцветает, эта лента видна в проходящем свете в виде просвечи-
вающей полоски .............................18а. Br. plectotropis smsp. phaeozona.
65 (64). Фон раковины от желтовато-белого до желтого, имеются 3 темные ленты:
верхняя прилегает к шву, средняя, наиболее четкая, занимает периферию, ниж-
няя расположена на базальной поверхности.
66 (67), Раковина правозавитая .......................17. Br. lantzi f. dextrorsa.
67 (66). Раковина левозавитая ........................17. Br lantzi f. sinistrorsa.
68 (59). Пупок умеренно узкий, не прикрыт или слегка прикрыт отворотом колумел-
лярного края, сквозь него просматривается от 1 до 1.5 оборота.
69 (70). Поверхность раковины с двумя спиральными лентами: выше и ниже перифе-
рии ............................................................24. Br. helvola.
70 (69). Поверхность раковины с одной слабой спиральной лентой выше периферии
или одноцветная . ..........................18b. Br. plectotropis smsp. almaatini.
1. Bradybaena fruticum (Muller, 1774). Табл. I, 8; рис. 52, 53.
Muller, 1774 : 71 (Helix); W esterlund, 1889 : 152 (Helix)', Лихарев,
Раммельмейер, 1952 : 390, рис. 317 (Eulota (Eulota)]; Grossu, 1955 : 421,
fig. 234 (Eulota); Ehrmann, 1956 : 119, t. 4, fig. 58 (Eulota); Lozek, 1956 : 190,
t. 37, 1 (Fruticicola); Zilch, Jaeckel, 1962 : 170: Lozek, 1964 : 284, t. 22,
la, b, c, 30, 3; Gittenberger et al., 1970 : 92, fig. 147, 148, Kaart 86.
Рис. 52—53. Bradybaena fruticum (Miill.), Подмосковье, с. Ромашково,.
5 V 1958. 52 — внешний вид полового аппарата; 53 — дистальная часть-
полового аппарата, вскрыта. (Ориг.).
BRADYBAENA
12Г
Материал: более 100 экз) из 32 местонахождений в Подмосковье, Прибалтике,
Крыму, на Карпатах, Северном Кавказе, в ГДР и Венгрии.
Раковина почти шаровидная, с округлым контуром завитка, высота которого,
примерно равна высоте устья или несколько больше, тонкостенная, просвечивающая.
Оборотов 5—6, выпуклых, плавно нарастающих. Последний оборот сильно вздут,
широко и правильно закруглен, к устью слегка и очень плавно опущен, немного мене е
чем в 1.5 раза шире предпоследнего. Остальные обороты также плавно выпуклые,
разделены умеренно глубоким швом. Окраска разнообразная: серовато-белая, желтая,
буроватая, каштановая, красновато-роговая; нередко над периферией проходит узкая
коричневая полоса, которая прослеживается у шва на одном-двух предыдущих обо-
ротах. В проходящем свете плотность фона и полосы одинаковая. Эмбриональные обо-
роты гладкие, скульптура дефинитивных оборотов представлена тонкими неправиль-
ными сглаженными морщинами (местами эти морщины более резкие) и густыми, тон-
кими и неглубокими спиральными бороздками. Устье круглое, умеренно скошено,
края его тонкие, острые, слабо отвернуты; колумеллярный край сильно отвернут и
слегка перекрывает узкий цилиндрический пупок, сквозь который просматривается
обычно не более одного оборота. Размеры: ВР 16—17, БД 18—22, МД 15—18.5 мм.
Внутреннее строение. Материал: 6 экз. из Подмосковья, с. Ромаш-
ково, 5 V 1958, сбор наш; 5 — из Венгрии, с. Хевиз близ оз. Балатон, 30 VII 1970,
сбор Д. А. Сахарова, определения наши; 4 — из Кведлинбурга, ГДР, V 1969, сбор,
и определение Э. Клаусса.
Нижняя часть спермовидукта и яйцевод образуют двойной изгиб, края которого,
большей частью прилегают друг к другу. Немного .ниже середины вагины впадает,
небольшой стилофор удлиненной или шаровидной формы. Дополнительный мешок,
часто того же размера и той же формы, что и стилофор. Слизистые железы парные, каж-
дая из них представлена парой ветвей гроздевидной формы с ячеистыми стенками;
однако эти ветви имеют тенденцию сливаться, и у некоторых экземпляров железы
неразделенные. Железы впадают в дополнительный мешок не апикально, их проток
становится единым уже внутри дополнительного мешка и впадает в полость стилофора
ниже основания стрелы. Надклоачный пузырь отсутствует, но стенка вагины, проти-
волежащая пенису, утолщена и заполнена очень рыхлой тканью; стенки этого участка
нередко немного светлее окружающих участков. Внутренние стенки вагины несут не-
правильные продольные валики, некоторые из которых в проксимальной части развет-
вляются, уходя одной ветвью в семеприемник, другой — в яйцевод.
Пенис массивный и длинный, чехол пениса хорошо развит, охватывает около трети
длины пениса и фиксирован, как и у большинства других видов, только у клоаки.
Внутренняя поверхность пениса в дистальной части несет множество беспорядочных,
коротких валиков и складок, в проксимальной части эта система несколько упорядо-
чивается и приобретает косое направление: складки сходятся к одной линии под острым;
углом, направленным в сторону семепровода. На этой же линии располагается широ-
кий, слабо дифференцированный пениальный гребень (см. поперечный срез пениса,
на рис. 52). Короткий половой ретрактор крепится широким основанием к месту пере-
хода эпифаллуса в семепровод.
Резервуар семеприемника сравнительно с другими представителями семейства
довольно велик. •
Распространение и местообитания. Вид населяет преимуще-
ственно пойменные луга с высоким травостоем и влажные леса на большей части Ев-
ропы. На запад доходит местами до берегов Атлантического океана (Пиренеи, Фран-
ция, Бельгия, Нидерланды, юго-восточная часть Великобритании); Альпы, северная
Италия, Средняя Европа, Карпаты, Балканский п-ов, южная часть Скандинавского)
п-ова, европейская часть СССР с Крымом и Северным Кавказом, кроме самых север-
ных районов.
К. К. Увалиева (1963) отмечает присутствие вида на южном Алтае; не исключено,
что автор имела дело с В г. schrencki.
Многократно найден в плейстоцене и голоцене Европы и южного Урала (Zilch,
Jaeckel, 1962; Lozek, 1964; Крылова, 1970).
2. Bradybaena schrencki (Middendorff, 1851). Табл. I, 9; рис. 54, 55.
M i ddendorf f , 1851 : 302, t. 30, fig. 20—26 (HeZZa); Gerstf eldt,
1859 : 520 (Helix)', Schrenck, 1867 : 672 (Helix)', Dybowski, 1901 : 133;
(Helix)', Лихарев, Раммельмейер, 1952 : 392 [Eulota (Eulota)]; Zilch,
J a e c k e 1, 1962 : 170.
Материал: более 100 экз. из 5 местонахождений в Архангельской обл. и на Алтае.
Р а к о вина слегка прижатая, с коническими контурами завитка, высота ко-
торого немного меньше высоты устья или равна ей. Оборотов 5—6, выпуклых, плавно,
нарастающих. Последний оборот в 1.5 раза шире предпоследнего, правильно округлый,
к устью не опущенили едва опущен. Окраска желтая, разных оттенков, или серая, реже.
128
II. BRADYBAENIDAE
однотонная, обычно же с тонкой темной лентой, проходящей несколько выше перифе-
рии. Скульптура — тонкая радиальная исчерченность и ясные спиральные бороздки.
Устье круглое, слегка скошено, внутри с очень тонкой расплывшейся губой. Колумел-
лярный край немного отвернут, остальные края почти не отвернуты. Пупок цилиндри-
ческий, открытый, но слабо перспективный. Размеры: ВР 9—12, БД 14 —17, МД
-12—15 мм.
Рис. 54—55. ВradybaenaTschrencki (Midd.), окр. Усть-Каменогорска, 28 VIII 1969.
54 — внешний вид полового аппарата; 55 — пенис, вскрыт. (Ориг.).
Вид по раковине похож на предыдущий, отличается коническим контуром завитка,
менее вздутым последним оборотом и меньшими размерами.
Внутреннее строение. Материал: 3 экз. из окр. Телецкого оз., VI 1967,
сбор Г. М. Овчинникова, определение наше; 3 — из окр. Усть-Каменогорска, 28 VIII
1969, сбор И. М. Хохуткина, определение наше.
Строение полового аппарата не имеет видимых отличий от того, что наблюдается
у предыдущего вида, кроме одной существенной детали. Если у Br. fruticum участок
стенки вагины утолщен и разрыхлен, то у этого вида покровы того же участка отслаи-
ваются и под ними образуется заметная полость.
Распространение и местообитания. Северо-Восточная Ев-
ропа, Сибирь, Алтай, Камчатка. Населяет влажные лесные участки и открытые места
близ воды с высокой травой и кустарником.
3. Bradybaena transbaicalia Schileyko, sp. nov. Табл. I, 10; рис. 56, 57.
Голотип хранится в Зоологическом институте АН СССР.
Материал: 8 экз. из одного местонахождения в Забайкалье.
Раковина прижатая, высота устья всегда превышает высоту широко-купо-
ловидного завитка. Вершина закругленная, сглаженная или чуть выступающая.
Последний оборот сильно вздут, по периферии правильно широко закруглен, у самого
устья несильно, но явно опущен без резкого перегиба. Окраска белесовато-желтая,
BRADYBAENA
129
монотонная, или имеется узкая коричневая полоса выше периферии последнего обо-
рота. Радиальная скульптура в виде неравномерных сглаженных морщин, спиральная
представлена ясными, слегка волнистыми бороздками. Устье почти'правильно круг-
лое, немного скошено, изнутри окаймлено очень широкой, но сильно расплывшейся
светлой губой, просвечивающей сквозь стенку раковины. Колумеллярный край уме-
ренно отвернут, далее по направлению к ангулярной'области’отворот постепенно схо-
дит на нет. Пупок неширокий, цилиндрический, но сквозь него просматриваются все
обороты. Размеры: ВР 14—16, БД 20—22, МД 17—18.5 мм. Голотип: ВР 14.3, БД 21.5,
МД 17.8 мм.
j По раковине вид похож на Br. middendorffi-, отличается менее прижатой’ракови-
ной, более узким пупком и круглым устьем.
Рис. 56—57. Bradybaena transbaicalia sp. nov., Забайкалье, хр. Хамар-Дабан,
10—13 VIII 1971. Паратип. 58 — внешний вид полового аппарата; 57— пенис,
вскрыт. (Ориг.).
Внутреннее строение. Материал: 4 экз. из Забайкалья, хр. Хамар-
Дабан, правый берег р. Мамай, 10—13 VIII1971, сбор И. М. Хохуткина, определение
наше.
Нижняя часть спермовидукта и яйцевод образуют крутой двойной изгиб, яйце-
вод и вагина приблизительно одной длины. Слизистые железы гроздевидные, впадают
в небольшой шаровидный резервуар, расположенный немного ниже середины стило-
фора; стилофор булавовидный, удлиненный, изогнутый. Разрыхленный участок стенок
вагины в значительной степени обособлен и образует железистый «пояс», имеющий при
рассмотрении спереди характерную бабочкообразную форму. Этот пояс представляет
собой дериват соответствующего участка вагины Br. fruticum.
Пенис умеренно длинный, булавовидный, нижняя треть его покрыта плотным чех-
лом. Внутренняя поверхность пениса покрыта мелкими складками, сходящимися под
углом к одной линии, на которой в проксимальной части пениса располагается хо-
рошо выраженный короткий продольный гребень. Эпифаллус короткий, половой рет-
рактор мощный.
gsj Распространение и местообитания. Вид пока известен только
из типового местонахождения на хр. Хамар-Дабан; И. М. Лихарев и Е. С. Раммель-
9 А. А. Шилейко
130
II. BRADYBAENIDAE
мейер (1952, стр. 392) указывают в описании Br. schrencki, что наряду с типичными
экземплярами встречаются более крупные: «высота 13—15, ширина 18—23 мм (forma
major)». По всей вероятности, эти сведения относятся к данному виду. Вид найден
в высокотравье (заросли осота) на берегу реки.
4. Bradybaena middendorffi (Gerstfeldt, 1859). Табл. II, 11', рис. 58—61.
Gerstfeldt, 1859 : 521, fig. 29 (Helix)-, S chr e nek, 1867 : 679 (Helix)-,
Mousson, 1887 : 17, t. 1, fig. 4 [Helix (Camaena) graeseri]; Cockerel 1, 1924 : 579
(Eulota eulemnisca), 571 (Hygromia amatoris)', Лихарев, Раммельмейер,
1952:395, рис. 326, 327, 328 [Eulota (Eulota)].
Материал: более 40 экз. из 9 местонахождений в Приморском крае.
Раковина низкоконическая, с широким низким куполовидным, плавно
закругленным завитком, высота которого равна высоте устья или меньше; иногда
даже высота завитка равна половине высоты устья. Стенки раковины тонкие, просве-
Рис. 58—61. Bradybaena middendorffi (Gerstf.), окр. Владивостока, 30 VI 1971. 58 —
внешний вид полового аппарата; 59 — дистальная часть женского отдела, вскрыта;
60 — место впадения яйцевода в вагину, вскрыто; 61 — пенис, вскрыт. Зеездочка —
железистый пояс. (Ориг.).
чивающие, поверхность блестящая. Вершина не выступает, сглаженная. Оборотов
около 6, выпуклых, тесно навитых. Последний оборот в профиле округлый, в 1.5 раза
шире предпоследнего; к устью не опущен или опущен незначительно и очень плавно.
Окраска бледно-желтая или желтовато-серая, над периферией проходит узкая темная
полоса, которая иногда может отсутствовать (одноцветные экземпляры описаны под
названием Hyrgomia amatoris — см. синонимику). Скульптура в виде тонкой неравно-
мерной радиальной исчерченности и четких спиральных бороздок. Устье широко-
овальное, косое, сильно вырезано предпоследним оборотом, места его прикрепления
широко расставлены. Края устья широко отвернуты, кроме верхнего, с расплыв-
шейся светлой губой. Верхний край отходит от стенки предпоследнего оборота вна-
чале почти горизонтально, затем, образуя плавную широкую дугу, переходит в пала-
тальный и далее в базальный край. Колумеллярный край короткий и сильно отвернут,
но не прикрывает широко развернутого пупка. Размеры: ВР 12—14, БД 21—26,
МД 18—22 мм.
Внутреннее строение. Материал: 4 экз. из окр. Владивостока (парк
Академгородка), 30 VI 1971, сбор наш; 2 — с о-ва Большой Пелис в зал. Петра Ве-
ликого, 12 IX 1972, сбор Н. М. Литвиненко; определения наши.
BRADYBAENA
131
Нижняя часть спермовидукта образует крутой изгиб или даже петлю. Яйцевод
несколько короче вагины и толще ее. Слизистые железы мощные, ячеистые, каждая
из двух желез впадает в дополнительный мешок отдельным протоком. Эти протоки
сливаются в пределах дополнительного мешка в общий узкий канал и впадают в сти-
лофор при основании короткой стрелы, сидящей на мускульной подушке. Дополни-
тельный мешок имеет толстые волокнистые стенки и лишен полости как таковой.
Полость внутри стилофора также отсутствует или очень маленькая; по существу
весь стилофор не что иное, как мускульная, почти шаровидная подушка с располо-
женной на ней стрелой. Полость матки резко сужена четырьмя мощными продоль-
Рис. 62—63. Bradybaena ussuriensis (West.), окр. Владивостока, 15 IX 1971. 62 —
внешний вид полового аппарата; 63 — дистальная часть полового аппарата, вскрыта.
(Ориг.).
ными валиками, разделенными глубокими узкими бороздами. По мере продвижения
наверх эти валики распадаются на несколько более мелких.
Короткий чехол пениса занимает не более четверти длины этого отдела, крепится
только у клоаки. Дистальная часть внутренней поверхности пениса занята четкими
округлыми продольными валиками, разделенными широкими бороздами. В прокси-
мальной части имеются мелкие косые складки и морщинки, сходящиеся под острым
углом к одной продольной линии, которая упирается в очень четкий короткий округ-
лый гребень, расположенный у отверстия эпифаллуса. Эпифаллус очень короткий,
половой ретрактор также короткий и мощный. Основание протока семеприемника
расширено незначительно.
Распространение и местообитания. Приморский край и бас-
сейн среднего Амура. Обитает в долинных широколиственных и смешанных лесах,
близ воды, часто встречается в сырых оврагах.
5. Bradybaena ussuriensis (Westerlund, 1897). Табл. II, 12; рис. 62, 63.
Westerlund, 1897 : 122 [Helix (Campylaea) ]; Величковский, 1924 : 154
(Eulota graeseri var. ussuriensis); Лихарев, Раммельмейер, 1952 : 394,
рис. 325 [Eulota (Eulota)].
9*
132
II. BRADYBAENIDAE
Материал: более 100 экз. из 6 местонахождений в Приморском крае.
Раковина прижатая, с широким и низким завитком куполовидных или
конических очертаний. Оборотов 5.5—6, выпуклых, медленно нарастающих. Послед-
ний оборот в профиле округлый, менее чем в 1.5 раза шире предпоследнего, к устью
еле опущен. Окраска светло-желтая или серовато-желтая, одноцветная или с одной
узкой коричневой лентой над периферией; нередко встречаются популяции, раковина
представителей которых почти не пигментирована, бесцветная или беловатая. Скульп-
тура в виде тонкой радиальной ребристости и четких густых спиральных линий. Устье
округло-треугольное, слегка оттянуто вниз и вправо. Края устья отвернуты, острые,
внутри с расплывшейся губой; колумеллярный край отвернут сильнее прочих, но не
закрывает развернутого перспективного пупка. ВР 7—9, БД 12—16, МД 10—13 мм.
Лектотип: ВР 7.8, БД 12.6 мм.
Внутреннее строение. Материал: 5 экз. с о-ва Большой Пелис
в зал. Петра Великого, 12 IX 1971, сбор Н. М. Литвиненко; 4 — из с. Пейшула При-
морского края, 4 VI 1971; 12 — из окр. зал. Восток в Японском море, 17 VI 1971;
1 — из окр. Владивостока, 15 IX-1971; сборы и определения наши.
Нижняя часть спермовидукта образует один изгиб. Тонкая вагина в 3—4 раза
длиннее яйцевода. Слизистых желез две, каждая открывается в дополнительный
мешок собственным протоком. Дополнительный мешок небольшой, впадает примерно
посредине тонкого удлиненного стилофора. Внутренняя поверхность вагины почти
гладкая, покрыта небольшими складочками. Железистый пояс охватывает вагину
чуть выше основания стилофора и примыкает с обеих сторон к дополнительному мешку.
Степень развития железистого пояса стоит в непосредственной связи с половой актив-
ностью особи.
Пениальный чехол покрывает от одной трети до половины пениса. В дистальной
части пениальной трубки имеются продольные валики, разделенные извилистыми или
зигзагообразными бороздками; реже эти бороздки прямые. Проксимальная часть
пениса несет множество коротких беспорядочных извилистых бороздок. У одного из
исследованных экземпляров наблюдалось образование временной папиллы пениса
за счет погружения эпифаллуса в просвет пениса (рис. 63). Эпифаллус примерно впо-
ловину короче пениса.
Распространение и местообитания. Прибрежные районы
южного Приморья. Населяет лесные биотопы всех типов, держится в основном у воды,
часто в трещинах скальных выходов в лесу.
6. Bradybaena maacki (Gerstfeldt, 1859). Табл. II, 13; рис. 64—66.
Gerstfeldt, 1859 : 518, fig. 27 (Helix); Schrenck, 1867 : 666 (Helix);
Mousson, 1887 : 16 [Helix (Fruticicola)], 17 (Helix (Fruticicola) middendorffi, non
Gerstfeldt, 1859]; Лихарев, Раммельмейер, 1952 : 393, рис. 322, 323
[Eulota (Eulota)].
Материал: более 100 экз. из 20 местонахождений в Приморье и Приамурье.
Раковина толстостенная, прочная, слабо просвечивающая и слабо блестя-
щая, ширококубаревидная, иногда почти шаровидная, с тупой вершиной и широким
завитком, контуры которого от почти конических до правильно куполовидных. Вы-
сота завитка обычно превышает высоту устья, однако в некоторых популяциях пре-
обладают особи, у которых высота завитка равна или даже меньше высоты устья.
Оборотов 6—7, довольно выпуклых, плавно нарастающих. Последний оборот не-
сколько шире предпоследнего, в профиле округлый, к устью почти не опущен. Окраска
образована желтым, светло-желтым или бурым фоном и тремя широкими коричневыми
лентами: верхней, примыкающей к шву; средней, по периферии; нижней, самой ши-
рокой, на базальной поверхности. Наиболее четкие контуры имеет центральная полоса,
верхняя выражена несколько слабее, а нижняя имеет размытые границы и в целом
заметно светлее прочих. В проходящем свете ленты и фон имеют примерно одинако-
вую плотность. Иногда встречаются экземпляры, лишенные либо нижней, либо верх-
ней ленты, либо и той и другой. Скульптура эмбриональных оборотов в виде тонкой
радиальной исчерченности, на дефинитивных оборотах имеется довольно равномерная
тонкая радиальная ребристость и ясные спиральные бороздки. Устье округлое, косое,
внутри светлое, с ясно просвечивающими лентами. Места прикрепления устья широко
расставлены и связаны тонкой мозолью. Края устья широко отвернуты, острые. Колу-
меллярный край отвернут сильнее прочих, слегка перекрывает не очень узкий, ци-
линдрический неперспективный пупок. Размеры: ВР 18—30, БД 23—35, МД 20—31 мм.
Внутреннее строение. Материал: 12 экз. из Хабаровского края, берег
Петропавловского оз., 10—12 VI 1970, сбор М. Д. Грибова; 6 — из Приморского
края, окр. с. Пейшула, 6 VI 1971, сбор и определения наши.
Спермовидукт резко складчатый, в нижней части образует резкий изгиб. Слизи-
стых желез 2—7, сливающихся непосредственно при основании; однако в большинстве
•случаев можно заметить группировку их в 2 грозди. Дополнительный мешок имеет
толстые стенки, но проток слизистых желез в пределах мешка расширения не образует
BRADYBAENA
133
и впадает в стилофор при основании стрелы (у изображенного на рис. 66 экземпляра
стрела отсутствовала). Стилофор очень длинный^ дистальная часть образует резкий
изгиб, часто двойной. На внутренней стенке имеется мощная складка, которая под-
нимается от клоаки; в области основания стрелы образует крутой поворот и возвра-
щается вновь вниз. Железистый пояс охватывает вагину и основание стилофора сплош-
ным кольцом.
Рис. 64—66. Bradybaena maacki (Gerstf.), Хабаровский край, берег Петропавловского оз.,
10—12 VI 1970. 64 — внешний вид полового аппарата; 65 — место впадения яйцевода
в вагину, вскрыто; 66 — дистальная часть полового аппарата, вскрыта. Звездочка —
железистый пояс. (Ориг.).
Пенис при самом основании покрыт коротким чехлом, фиксированным только
у клоаки. Эпифаллус не выражен. Внутренняя поверхность дистальной части пениса
покрыта продольными складками, разделенными бороздами; далее эта система скла-
док превращается в характерный узор «елочку», наконец, в проксимальной части
эти структуры трансформируются во множество тонких извитых складочек, стяги-
вающихся к хорошо выраженному продольному гребню. Основание семеприемника
слабо расширено или не расширено.
Очень характерной внутренней поверхностью обладает яйцевод. Здесь имеется
сложный узор из причудливо изогнутых и слегка ветвящихся мощных бугорков и
складок, разделенных глубоко врезанными бороздками (рис. 65). При сглаживании
этого узора объем яйцевода может значительно увеличиваться. Резервуар семеприем-
ника тесно прилегает к нижней части белковой железы.
Распространение и местообитания. От бассейна нижнего и
среднего Амура на юг — весь Приморский край и северная часть п-ова Корея, тер-
ритория Маньчжурии (ист. обл.) и бассейн Янцзы. Населяет все типы лесных биото-
пов; наибольшей плотности достигает в широколиственных влажных лесах, где встре-
чается чаще всего на поверхности подстилки. Во время продолжительных дождей
поднимается на деревья.
7. Bradybaena weyrichii weyrichii (Schrenck, 1867). Табл. II, 14-, рис. 67, 68;
Schrenck, 1867:669, t. 26, fig. 11—13 [Helix)-, Adams, 1868:459—472
(Helix serotina); Fulton, 1905, Ann. Mag. Nat. Hist., ser. 7, XV ; 92 [Eulota (Euhadra)
fiscina]; Pilsbry, 1927 : 13—20, fig. 1, t. 1, fig. 4—8 [Eulota (Karaftohelix) fiscina]",
Лихарев, Раммельмейер, 1952 : 408 [Eulota (Karaftohelix)]-, Kuroda,
1963:54 [Fruticicola (Karaftohelix)].
134
II. BRADYBAENIDAE
Материал: более 100 экз. из двух местонахождений в южной части Сахалина.
Раковина шаровидная, с широким коническим или слегка куполовидным
завитком и узкой вершиной, сравнительно тонкостенная. Оборотов 6—6.5, слабо
выпуклых, плавно нарастающих. Последний оборот округлый, немного шире пред-
последнего, к устью очень слабо и очень плавно опущен. Окраска образована желтым
фоном (нередко с коричневатым оттенком) и коричневыми спиральными лентами. Чаще
всего имеются три ленты: под швом, на периферии и на базальной части. Перифери-
ческая лента четкая, нижняя граница верхней полосы обычно в той или иной степени
Рис. 67—68. Bradybaena weyrichii weyrichii (Schrenck), окр. Южно-Сахалинска,
24 VIII 1971. 67 — внешний вид полового аппарата; 68 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
размыта, нижняя лента наиболее широкая и наиболее светлая. В проходящем свете
фон слегка плотнее периферической ленты. Нередко верхняя и (или) нижняя лента
отсутствуют. Кроме этого, часто присутствуют редкие размытые редкие радиальные
каштановые или темно-коричневые полосы. Скульптура на эмбриональных оборотах
отсутствует, на дефинитивных она представлена густыми радиальными морщинками
и складками и ясными глубокими спиральными бороздками. Устье округлое, косое,
внутри с толстой валикообразной расплывшейся губой. Места прикрепления устья
широко расставлены и соединены тонкой мозолью. Края устья острые, часто коричне-
ватые, сильно отвернуты. Пупок узкий, наполовину или несколько больше прикрыт
отворотом колумеллярного края. ВР 20—37, БД 31—38, МД 25—33 мм.
Внутреннее строение. Материал: 14 экз. из окр. Южно-Сахалинска,
24 VIII 1971; 5 — из южной части о-ва Кунашир, склоны вулкана Менделеева, 29 VIII
1971; сборы и определения наши.
Спермовидукт в нижней части резких изгибов не образует, яйцевод плавно изо-
гнут, вагина длинная и очень тонкая. Слизистые железы необыкновенно мощные, не
ветвящиеся, гроздевидные, число их колеблется от 3 до 12. Поверхность желез много-
бугорчатая, стенки ячеистые. Все ветви сливаются в общий проток непосредственно
при впадении в дополнительный мешок; однако у большинства особей все же наблю-
далось объединение их в две группы. Полость дополнительного мешка обширная,
стенки тонкие. Выводной проток дополнительного мешка проходит вдоль мускульной
подушки, на вершине которой располагается стрела, и открывается при основании
стрелы. Полость стилофора обширная, внутри почти гладкая или с нерезкими про-
BRADYBAENA 135
цокшш утолщениями. Полость вагины резко сужена продольными складками,
часть которых идет в яйцевод, а часть — в проток семеприемника. Железистый пояс
значительно обособлен от стенок вагины, охватывает ее ниже основания стилофора.
Пенис длинный, нижняя четверть его охвачена чехлом, который фиксирован на
обоих концах, заключая под своими покровами кольцевую полость. Эпифаллус корот-
кий. Внутри дистальной части пениса расположены продольные валики, разделенные
узкими глубокими бороздами; по мере приближения к проксимальному отделу эти
валики становятся более тонкими, затем рассыпаются на мелкие складки и приобре-
тают косое направление, стягиваются к хорошо выраженному продольному гребню,
которому снаружи соответствует небольшая, но ясная ямка. Основание протока
семеприемника немного расширено.
Распространение и местообитания. Сахалин. Обитает во
влажных лесах, в зарослях гигантских трав, в сырых местах.
7а. Bradybaena weyrichii bocageana Crosse, 1864.1
Crosse, 1864, J. Conch., XII : 284; XIV : 58, pl. 1, fig. 6, {Helix)-, Adams,
1868 : 459—472 {Helix miranda); Pilsbry, Hirase, 1904 : 121 {Eulota chishimana);
P i 1 s b r y, 1927 : 17 {Eulota urupensis); Лихарев, Раммельмейер, 1952:
408 [Eulota ( Karaftohelix) weyrichii part.].
Материал: более 20 экз. с о-вов Кунашир и Шикотан (южные Курильские о-ва).
Конхологические отличия от номинативного подвида заключаются в более интен-
сивном развитии цветовых лент и полос, общем потемнении фона и более мелких раз-
мерах: ВР 20—23, БД 26—29, МД 23—24 мм.
Анатомических отличий не найдено.
Распространение и местообитания. Курильские о-ва и се-
верная часть о-ва Хоккайдо. Встречается в тех же типах биотопов, что и предыдущий
подвид, а также в зарослях низкорослого бамбука.
8. Bradybaena strelkovi (Licharev et Rammelmeyer, 1952). Табл. II, 15', рис. 69—71.
Лихарев, Раммельмейер, 1952 : 408, рис. 340 [Eulota (Karaftohelix) ].
Материал: 12 экз. из двух местонахождений на Сахалине и Курильских о-вах.
Раковина шаровидная, толстостенная, но просвечивающая. Завиток почти
конический, высота его слегка превышает высоту устья. Вершина узкая и притуп-
ленная. Оборотов 5.5—6, выпуклых, плавно нарастающих, разделенных глубоким
швом. Последний оборот в профиле правильно округлый, немного шире предпослед-
него, к устью не опущен. Окраска монотонная: зеленоваго-серая, оливковая или
золотистая; встречаются отдельные экземпляры с одной спиральной бесцветной про-
свечивающей в проходящем свете, лентой. Скульптура на эмбриональных оборотах
отсутствует, на дефинитивных представлена неравномерной радиальной морщинистой
исчерченностью и ясными густыми спиральными бороздками. Устье округлое, косое,
места его прикрепления широко расставлены, края устья острые, светлые, сильно
отвернуты, внутри с широкой невысокой губой. Колумеллярный край отвернут силь-
нее других и частично перекрывает узкий неперспективный пупок. Размеры: ВР 17—
20, БД 23—26, МД 19—22 мм. Голотип: ВР 18.5, БД 24.2, МД 21.0 мм.
Внутреннее строение. Материал: 3 экз. из южной части Сахалина,
Ракума, 1946, сбор А. А. Стрелкова, определение И. М. Лихарева.
Дистальная часть спермовидукта и яйцевод образуют двойной изгиб, яйцевод
длинный и толстый, вагина еще длиннее, тонкая, стилофор впадает очень низко —
у самой клоаки. Слизистых желез 5—7, некоторые слабо ветвятся, массивные, с ячеи-
стыми стенками. Общим протоком железы впадают в небольшой шаровидный допол-
нительный мешок, расположенный немного выше середины стилофора. Дополнитель-
ный мешок внутри с обширной полостью; эта полость сужается в тонкий проток,
впадающий в полость стилофора при основании стрелы, которое охвачено высоким
соединительнотканным воротничком. Стрела короткая, расположена на мускульной
подушке, составляющей вершину стилофора. Железистый пояс фиксирован на поверх-
ности вагины выше стилофора отдельными неоформленными тяжами.
Чехол пениса охватывает 1/3—1/4 часть пениальной трубки, фиксирован на обоих
концах и слабо отграничен от стенки пениса. Внутреннее строение пениса, как у преды-
дущего вида, пениальному гребню также соответствует наружное углубление. Эпи-
фаллус почти не выражен. Основание семеприемника слабо расширено.
1 Подвидовое название опубликовано годом раньше видового; оно не использо-
вано в качестве видового для сохранения стабильности номенклатуры (Международ-
ный кодекс зоологической номенклатуры, Преамбула).
Рис. 69—71. Bradybaena strelkovi (Lich. et Ramm.), южн. Сахалин, VII 1946. 69 — внеш-
ний вид полового аппарата; 70 — место впадения яйцевода в вагину, вскрыто; 71 —
дистальная часть полового аппарата, вскрыта. (Ориг.).
Рис. 72—-74. Bradybaena duiensis (West.), южн. Сахалин, VII 1946. 72 — внешний вид
полового аппарата; 73 — место впадения яйцевода в вагину, вскрыто; 74 — дисталь-
ная часть полового аппарата, вскрыта. (Ориг.).
BRADYBAENA
137
Распространение и местообитания.' Южный Сахалин, южные
Курильские острова (Итуруп, Шикотан; на Кунашире не найден) (Лихарев, 1957а).
Обитает в кустарниковых зарослях.
9. Bradybaena duiensis (Westerlund, 1897,). Табл. II, 16; рис. 72—74.
Westerlund, 1897 : 121 (Helix); Лихарев, Раммельмейер, 1952 :
407, рис. 338, 339 [Eulota (Karaftohelix)].
Материал: 3 экз. из одного местонахождения на Сахалине.
Раковина прижатая, с широким коническим завитком, высота которого
немного меньше высоты устья, хрупкая, тонкостенная, просвечивающая. Оборотов 5,
слабо выпуклых, плавно нарастающих. Последний оборот сильно вздут, в профиле
правильно округлый, в 1.5 раза шире предпоследнего, к устью плавно и несильно
опущен. Окраска красновато-зеленая или тускло-золотистая. Над периферией послед-
него оборота проходит узкая коричневая лента с четкими контурами; на предыдущих
оборотах она скрыта под швом. Кроме того, во второй половине последнего оборота
имеется намек на вторую спиральную ленту в базальной части значительно шире
первой. Скульптура — тонкие редкие сглаженные радиальные морщины и правиль-
ные, очень тонкие спиральные бороздки; сочетание этих скульптурных элементов
местами создает впечатление тонкой зернистости. Устье округлое или слабоовальное,
умеренно косое, внутри с расплывшейся красноватой или белой губой. Места при-
крепления устья широко расставлены и соединены тонкой мозолью. Края устья ост-
рые, красноватые, сильно отвернуты, колумеллярный край примерно наполовину
закрывает узкий неперспективный пупок. Размеры: ВР 11—12, БД 16—17, МД 13 мм.
Голотип: ВР 8, БД 11 мм (раковина не вполне сформированная).
Внутреннее строение. Материал: 2 экз. с о-ва Сахалин, Ракума,
VII 1946, сбор А. А. Стрелкова, определение И. М. Лихарева.
Спермовидукт резко складчатый, яйцевод плавно изогнут, очень толстый и длин-
ный. Вагина внизу тонкая, кверху утолщается. Слизистых желез 5—10, ветви желез
не обнаруживают наклонности к вторичному ветвлению, массивные, булавовидные,
с ячеистыми стенками. Единым протоком все ветви открываются в дополнительный
мешок, внутри которого имеется небольшая, но ясная полость. Стилофор длинный,
изогнутый, бугорчатый, впадает очень низко, почти в клоаку; дополнительный ме-
шок располагается близ апикальной части стилофора, узкий проток его впадает в по-
лость стилофора при основании стрелы, которая охвачена воротничком на большей
части протяжения. Верхняя часть стилофора, где располагается стрела, отделена от
дистальной цилиндрической части ясным кольцевым валиком. Чехол пениса фикси-
рован на обоих концах, занимает примерно треть длины пениса, на продольном и
поперечном срезах видна полость между стенками пениса и пениального чехла. Ва-
лики в дистальной части пениса довольно ровные, затем приобретают вид «елочки»
и сходятся к ясно выраженному гребню в проксимальной части. Эпифаллус короткий.
Основание протока семеприемника сильно расширено. Поскольку семеприемник
фиксирован как внизу, так и наверху, длина его протока (если последний не извит)
определяется тем, насколько вытянут спермовидукт; так как у этого вида спермови-
дукт резко складчатый, длина его в нерасправленном состоянии невелика, следова-
тельно, проток семеприемника также короткий.
Распространение и местообитания. Вид обнаружен в двух
местах на Сахалине: окрестности Дуэ и Южно-Сахалинска. Обитает в лиственных
лесах и кустарниковых зарослях близ воды.
10. Bradybaena dieckmannl (Mousson, 1887). Табл. II, 17; рис. 75—78.
Schrenck, 1867 : 678 (Helix hispida L.); Mousson, 1887 : 13, t. 1, fig. 2
[Helix (Trichia)]; W esterlund, 1897 : 121 [Helix (Trichia) eutheta]; В ii t t n e r,
Ehrmann, 1927 : 229, t. 12, fig. 1—2 (Fruticicola); Schlesch, 1928 : 280, t. 12,
fig. 18—22 (Trichia); Лихарев, Раммельмейер, 1952 : 396, рис. 329, 330
[Eulota (Eulota)].
Материал: более 40 экз. из 4 местонахождений в Приморском крае.
Раковина шелковисто поблескивающая, от прижатой до кубаревидной;
стенки раковины просвечивающие, но довольно крепкие. Завиток конический или
слегка куполовидный, высота его чаще всего немного превышает высоту устья. Оборо-
тов 5—5.5, плавно нарастающих, выпуклых, чуть ступенчатых. Последний оборот
в профиле округлый, в конце немного опущен, в базальной части слегка уплощен.
Окраска одноцветная: бледно-желтая, желтовато-серая, красновато-роговая, иногда
раковина почти бесцветная. Скульптура эмбриональных оборотов представлена тон-
кой зернистостью, на дефинитивных имеются тонкие радиальные густые бороздки
(тонкая исчерченность) и легкие спиральные бороздки. На скатах пупка имеются мно-
гочисленные чешуйки или гранулы. Устье округло-треугольное, оттянуто вниз и
138
II. BRADYBAENIDAE
вправо. Наружный край умеренно отогнут, базальный и колумеллярный края широ-
кие и отвернутые, с ясной светлой губой, которая почти всегда несет в левой части
зубообразное вздутие. Места прикрепления устья широко расставлены. Пупок очень
широкий, развернутый и перспективный. Размеры: ВР 5.5—7, БД 7—12, МД 5—9 мм.
Внутреннее строение. Материал: 4 экз. из окр. Владивостока (парк
Академгородка), 30 VI 1971; 3 — с побережья Амурского зал. близ ст. Океанская,
15 IX 1971, сборы наши; 4 — с о-ва Наумова в зал. Петра Великого, 20 VI 1973,
сбор Р. Я. Братчика; определения наши.
Начиная от белковой железы и кончая клоакой половые пути не образуют не
только колен, но даже сколько-нибудь заметных изгибов (материал фиксирован в рас-
Рис. 75—78. Bradybaena dieckmanni (Mouss.), окр. Владивостока, 30 VI 1971. 75 —
внешний вид полового аппарата; 7в — яйцевод, вскрыт; 77 — дистальная часть жен-
ского отдела, вскрыта; 78 — пенис, вскрыт. (Ориг.).
I
I
правлением состоянии). Яйцевод очень широкий, вагина тоже. Слизистых желез
две, они длинные, массивные, с ячеистыми стенками, не ветвящиеся. In situ железы
очень тесно прилегают друг к другу, к вагине и к спермовидукту. Стилофор короткий,
дополнительный мешок несколько длиннее, и создается впечатление, что мешок рас-
положен не на стилофоре, но вклинивается между стилофором и стенкой вагины. Все
придатки дистальной части женского отдела вместе с вагиной окутаны соединительно-
тканным чехлом, вероятно, частично гомологичным железистому поясу, который
у исследованных экземпляров не обнаружен. Дополнительный мешок тонкостенный
и заключает обширную полость, проток которой впадает в узкий проток стрелы; по-
следняя не располагается на мускульной подушке, но погружена в ее ткани. Внутренняя
поверхность вагины несет несколько продольных валиков.
Чехол пениса короткий и тонкий, занимает около трети длины пениальной трубки.
Внутренняя поверхность пениса покрыта очень тонкими неровными складочками,
сходящимися к средней линии под острым углом, но геометрическая правильность
узора многократно нарушается. Характерная внутренняя структура имеется и в яйце-
воде; здесь расположен комплекс удлиненных бугорков, тесно прилегающих друг
к другу. Вероятно, отсутствие расширения при основании семеприемника связано
с тем, что роль этого расширения принял на себя яйцевод, т. е. сюда поступает пакет
со спермиями.
Распространение и местообитания. От бассейна нижнего и
среднего Амура до зал. Петра Великого. На Сихоте-Алине не найден. Встречается
в очень влажных лесных биотопах, чаще всего в заболоченных участках, среди зарос-
лей трав.
BRADYBAENA
139
11. Bradybaena arcasiana (Crosse et Debeaux, 1863). Табл. II, 18; рис. 79—81.
Gerstfeldt, 1859 : 519 [Helix fruticum part.); Crosse, Debeaux, 1863,
J. Conch., XI : 386 [Helix); 1864, XII : 316, t. 12, fig. 4 [Helix); Schrenck, 1867 :
671 [Helix); Mousson, 1887 : 17 [Helix); Cockerell, 1926 : 227 [Eulota mu-
rensis); Pilsbry, 1926 : 461, t. 35, fig. 7 [Ganesella virgo); Лихарев, Рам-
м'ельмейер, 1952:391, рис. 318, 319 [Eulota (Eulota)].
Материал: более 30 экз. из 3 местонахождений в Приамурье и Приморье.
Раковина почти шаровидная, умеренно тонкостенная, просвечивающая,
и высоким коническим или слегка куполовидным завитком и острой вершиной. Высота
завитка значительно больше высоты устья. Оборотов 6—6.5, выпуклых, разделенных
Рис. 79—81. Bradybaena arcasiana (Cr. et Deb.), окр. Смоляниновб, Примор-
ский край, 21 VII 1973. 79 — внешний вид полового аппарата; 80 — место
впадения яйцевода в вагину, вскрыто- 81 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
глубоким швом. Первые обороты нарастают медленно, два последних — быстро.
Последний оборот одной ширины с предпоследним и немного выше его. Периферия
последнего оборота в районе устья заметно нависает над швом (см. табл. II, 18) за
счет опускания последнего оборота в последней трети. Окраска желтовато-белая или
светло-бурая, чаще всего одноцветная, изредка встречаются раковины с одной корич-
невой спиральной лентой по периферий. Эмбриональные обороты гладкие. Скульп-
тура дефинитивных оборотов представлена тонкой радиальной исчерченностью и
явственными спиральными бороздками. Устье округло-треугольное, косое, края его
светлые, отвернутые, с тонкой белой губой. Колумеллярный край отвернут сильно,
на нем располагается выраженное в разной степени удлиненное утолщение, наплыв.
Пупок не очень узкий, но неперспективный, цилиндрический. Размеры: ВР 13—17,
БД 14—19, МД 15—17.5 мм.
Внутреннее строение. Материал: 2 экз. из окр. Смоляниново При-
морского края, 21 VII 1973, сбор В. А. Костырко, определение наше.
Нижняя часть спермовидукта и яйцевод образуют 2—3 резких изгиба. Вагина
длинная и тонкая, ее длина в 4—6 раз превосходит ширину. Слизистых желез две,
они не ветвятся, имеют гроздевидный облик, независимо впадают в дополнительный
мешок, полость которого очень узка. Стилофор небольшой, более чем вдвое короче
вагины, при основании изогнут. Дополнительный мешок слит со стилофором всем
ш
II. BRADYBAENIDAE
своим широким основанием. Полость стилофора четко разделена на два отдела: прокси-
мальный, куда впадает проток дополнительного мешка и где располагается короткая
стрела, сидящая на вершине мускульной подушки, и дистальный, более обширный,
широко сообщающийся с полостью вагины и клоаки. Железистый пояс не обнаружен.
Чехол пениса фиксирован только у клоаки, занимает не более одной четверти
длины пениса. Внутренняя поверхность пениса в дистальной части несет редкие неяс-
ные продольные складки, в проксимальной наблюдается характерная картина: тонкие
редкие косые складки, которые, пересекаясь, образуют правильный сетчатый узор.
Эпифаллус короткий, отграничен от пениса легким пережимом. Резервуар семе-
приемника относительно очень маленький.
Распространение и местообитания. Спорадически — бассейн
Амура (окрестности Хабаровска, Благовещенска-на-Амуре), Раздольно-Ханкайская
низменность, восточный берег Уссурийского зал., о-в Фуругельма, территория Маньч-
журии (ист. обл.) и северная часть п-ова Корея. Обитает в умеренно влажных био-
топах, держится в лесной подстилке и на склонах холмов. Наибольшая плотность
в Амуро-Зейской равнине наблюдается в полосе перехода открытых участков в дре-
весно-кустарниковые заросли и составляет там от 1 до 30 экз./м2 (Крастин и др., 1969).
12. Bradybaena diversita Schileyko et Bratschik, sp. nov. Рис. 82—84.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 18 экз. с островов зал. Петра Великого.
Раковина очень хрупкая, тонкостенная, сильно просвечивающая, прижа-
тая, с коническим завитком, составленным 4.5—5 оборотами; первые обороты нара-
стают медленнее последнего, ширина которого в 1.5 раза превышает ширину пред-
последнего. К устью последний оборот плавно и незначительно опускается, в профиле-
правильно широко закруглен, без намека на образование угла. Окраска состоит из
желтоватого или светло-рогового фона и спиральной коричневой ленты над перифе-
рией; эта лента на более ранних оборотах видна у шва, по существу, шов идет по
этой ленте, но к устью опускается чуть ниже ее. Изредка раковина одноцветная.
Рис. 82. Bradybaena diversita sp. nov. Голотип. (Ориг.).
Эмбриональные обороты почти гладкие. На дефинитивных оборотах радиальная
скульптура в виде слабых, очень редких и очень расплывшихся морщинок; спираль-
ные бороздки более ясные и правильные. Устье слегка овальное, ширина его немного
превышает высоту, слабо вырезано, скошено. Губы нет. Верхний край устья почти
прямой, колумеллярный и базальный немного отогнуты. Пупок цилиндрический,
узкий, но перспективный. Размеры: ВР 7—10, БД 11—15, МД 9—12 мм. Голотип:
ВР 8, БД 12.5, МД 10.5 мм.
По конхологическим признакам вид ближе всего к Br. duiensis; отличается мень-
шими размерами, более прижатым завитком, отсутствием отворота края устья и губы
и более широким пупком. По признакам внутреннего строения ближе всего стоит
к Br. fragilis, отличается более массивным стилофором и дополнительным мешком,
отсутствием резкой складчатости спермовидукта, более длинным пенисом и прямым
яйцеводом. Сближает эти виды высокая степень обособленности пениального гребня
и наличие чрезвычайно короткой вагины.
Внутреннее строение. Материал: 4 экз. с о-ва Два Брата в зал. Петра
Великого, 13 VI 1973, сбор Р. Я. Братчика, определение наше.
Спермовидукт в дистальной части образует двойной, очень крутой изгиб, края кото-
рого плотно сомкнуты. Яйцевод длинный, вагина очень короткая, так что проток семе-
приемника отходит почти от клоаки. Слизистые, железы ветвятся, стенки их ячеистые;
обе слизистые железы впадают отдельными протоками почти апикально в большой
дополнительный мешок, резко суженный при основании. Полость дополнительного
мешка неширокая, но вполне ясно выражена; узким протоком она сообщается с по-
BRADYBAENA
141
лостью булавовидного стилофора. Железистый пояс не обнаружен, но стенки соответ-
ствующего участка вагины утолщены и разрыхлены.
Чехол пениса морщинистый, занимает 1/(1—1/3 длины пениальной трубки. Вну-
тренняя поверхность пеписа несет нерезкие короткие косые складки. Пениальный
Рис. 83—84. Bradybaena diversita sp. nov., о-в Два Брата в зал. Петра Великого,
13 VI 1973. Паратип. 83 — внешний вид полового аппарата; 84 — дистальная часть
полового аппарата, вскрыта. (Ориг.).
гребень морфологически уже не является «гребнем», поскольку он фиксирован только
при основании; имеет вид придатка, утонченного на конце. Основание семеприемника
не расширено.
Распространение и местообитания. Вид известен с о-вов Два
Брата, Матвеева, Де-Ливрона и Дурново в зал. Петра Великого. Найден среди кам-
ней во влажных условиях.
13. Bradybaena fragilis Pilsbry, 1926. Табл. II, 19; рис. 85, 86.
P i 1 s b г у, 1926 : 455, t. 33, fig. 2 (Eulota); Лихарев, Раммельмейер,
1952:392, рис. 320, 321 [Eulota (Eulota)].
Материал: более 30 экз. из 4 местообитаний на юге Приморья.
Раковина почти шаровидная, тонкостенная, полупрозрачная или сильно
просвечивающая, блестящая, очень хрупкая. Оборотов 5—5.5, выпуклых, плавно
и быстро нарастающих. Завиток строго конический. Последний оборот в профиле пра-
вильно и широко закруглен,- преобладает над остальными, к устью слабо и очень
плавно опущен. Окраска одноцветная, светло-роговая, серовато-зеленоватая щли ра-
ковина почти бесцветная. Радиальная скульптура в виде тонкой и редкой исчерчен-
ности; спиральные бороздки ясные и частые, но неглубокие. Эмбриональные обороты
имеют микроскопическую зернистую или чешуйчатую скульптуру. Устье большое,
круглое, скошенное. Места его прикрепления не сближены, тонкие, отвернутые;
142
II. BRADYBAENIDAE
колумеллярный край отвернут сильнее и перекрывает пупок, оставляя от него узкую
щель. Губа не выражена. Размеры: ВР 10—15, БД 13—19, МД 10—15 мм.
Внутреннее строение. Материал: 2 экз. из заповедника «Кедровая
Падь», 23 VII 1946, сбор и определение И. М. Лихарева; 6 — с о-ва Фальшивого (зал.
Петра Великого), 4 VII 1973, сбор и определение Р. Я. Братчика.
Спермовидукт по всей длине образует несколько резких изгибов; мощный яйце-
вод также изогнут, проток семеприемника отходит очень низко, поэтому вагина ко-
Рис. 85—86. Bradybaena fragilis (Pilsbry), о-в Фальшивый в зал. Петра Великого,
4 VI 1973. 83— внешний вид полового аппарата; 84 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
роткая. Стилофор располагается также очень низко, у самой клоаки. Слизистых же-
лез две, точнее, два куста, поскольку форма желез своеобразна: это комплекс ветвя-
щихся гофрированных трубок с тонкими неячеистыми стенками, тесно прилегающих
друг к другу (на рис. 85 слизистые железы слегка разъединены). Каждый куст впадает
в дополнительный мешок отдельным протоком, не образующим расширения в преде-
лах мешка; общий проток желез сообщается с полостью стилофора при основании
очень короткой стрелы, окутанной чехлом. Железистый пояс, характерный для мно-
гих дальневосточных видов, не обнаружен.
Чехол пениса занимает от х/4 до почти 1/3 пениальной трубки, фиксирован только
у клоаки. Внутренняя поверхность пениса покрыта тонкими спиральными складками,
проксимальный гребень превращен в извилистый тонкий придаток. Проток семепри-
емника при основании не расширен. Эпифаллус не выражен.
Распространение и местообитания. Юг Приморского края
и северная часть п-ова Корея. Обитает во влажных местах (в основном в лесах и ку-
старниках) близ морского побережья.
14. Bradybaena vulcanica Schileyko, sp. nov. Табл. II, 20; рис. 87, 88.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 26 экз. из двух местонахождений на о-ве Кунашир.
Раковина' шаровидная, тонкостенная, хрупкая, просвечивающая, с кони-
ческим завитком, составлена 5 выпуклыми оборотами. Высота завитка меньше вы-
соты устья; у некоторых экземпляров чуть больше половины его высоты. Последний
оборот сильно вздут, явно преобладает над остальными, возле, устья почти вдвое шире
предпоследнего, по периферии округлый, к устью не опущен. Окраска одноцветная,
желтая с зеленоватым оттенком; у вполне взрослых экземпляров окраска к устью
BRADYBAENA
143
часто сгущается и становится бурой. Скульптура сложная: самые заметные скульптур-
ные элементы — неравномерные радиальные морщины, которые у шва выражены
резче, чем на периферии, местами грубые складки. Имеются также вялые тонкие спи-
ральные бороздки, видимые только при увеличении. Затем под бинокуляром заметны
чередующиеся матовые и блестящие спиральные линии (увеличение 10—15 раз). При
30-кратном увеличении видна тончайшая косая струйчатость, более четко выражен-
ная в пределах блестящих спиральных линий. Наконец, на многих раковинах местами
имеются вмятины. Похоже, что эти вмятины возникают при регенерации поврежден-
Рис. 87—88. Bradybaena vulcanica sp. nov., южная часть о-ва Кунашир, 2 IX 1972.
Паратип. 87 — внешний вид полового аппарата; 88'— дистальная часть полового
аппарата, вскрыта. (Ориг.).
него края устья по мере дальнейшего роста раковины; то, что такие вмятины встре-
чаются часто, объясняется чрезвычайной хрупкостью устьевых краев. Устье отно-
сительно большое, круглое, причем иногда высота его немного превышает ширину,
а иногда наблюдается обратное соотношение. Устье умеренно скошено, края его не
утолщены и лишены губы; отвернута лишь верхняя часть колумеллярного края.
Пупок открытый, узкий, неперспективный, при рассмотрении строго снизу (если
оптическая ось глаза совмещена с осью пупка) почти полностью перекрыт. Размеры:
ВР 15—17.5, БД 20.5—22, МД 16—18 мм. Голотип: ВР 17.3, БД 21.0, МД 17.0 мм.
По конхологическим признакам вид очень похож на Br. ravida, отличается более
мелкими размерами и более сложной и богатой скульптурой.
Внутреннее строение. Материал: 6 экз. из южной части о-ва Куна-
шир, подножье вулкана Головнина, 2 IX 1971, сбор и определение наши.
Спермовидукт резко складчатый, на границе его с яйцеводом имеется крутой
изгиб. Длина яйцевода примерно равна длине вагины. Установить число слизистых
желез трудно, так как они плотно сжаты и вторично ветвятся. В целом они образуют
мощный куст; стенки желез бугорчатые. Проток слизистых желез немного снизу впа-
дает в дополнительный мешок, заключающий обширную полость. Проток дополни-
тельного мешка длинный, открывается в полость стилофора при основании короткой
стрелы, расположенной на вершине мускульной подушки. Стилофор удлиненный,
дополнительный мешок почти шаровидный, располагается выше его середины, осно-
144
II. BRADYBAENIDAE
ванне стилофора тонкое. Выше стилофора вагина охвачена железистым поясом напо-
добие воротничка.
Пенис недлинный, нижняя его треть охвачена чехлом, фиксированным на обоих
концах либо только у клоаки. Половой ретрактор длиннее, чем у большинства дру-
гих видов рода. Внутренняя поверхность дистальной части пениса покрыта плотно
расположенными продольными валиками, пересеченными поперечными бороздками,
которые образуют елочкообразный узор. В проксимальной части эти бороздки ста-
новятся более мелкими и стягиваются к короткому гребню. Эпифаллус не выражен.
Основание протока семеприемника расширено, но уже яйцевода.
Распространение и местообитания. Известен только с
о-ва Кунашир (южные Курильские о-ва). Встречен в очень сыром заболоченном лист-
венном лесу на берегу ручья; на поверхности подстилки и на стеблях и листьях ги-
гантской гречихи.
15. Bradybaena ravida (Benson, 1842). Табл. Ill, 21; IX, 82; рис. 89—92.
Benson, 1842, Ann. Mag. Nat. Hist., IX : 486 (Helix); Reeve, 1852, Conch.
Iconica, VII : 130, fig. 398 (Helix); Gerstfeldt, 1859 : 517 (Helix); 519, fig. 28
(Helix selskii); Schrenck, 1867 : 663 (Helix), 665, t. 26, fig. 7—10 (Helix selskii);
Рис. 89—92. Bradybaena ravida (Benson), с. Пейшула Приморского края, 3 VII 1971.
89 — внешний вид полового аппарата; 90 — место впадения яйцевода в вагину,
вскрыто; 91 — дистальная часть женского отдела, вскрыта; 92 — пенис, вскрыт.
(Ориг.).
М о u s s о п, 1887 : 15, t. 1, fig. 3 (Helix (Cryptomphalus) cincto-inflata]; M б 1 len-
der f f, 1899 : 74 [Eulota (Acusta)]; Dybowski, 1901 : 134 (Helix selskii); C o-
ckerell, 1924 : 579 (Helix weyrichii, non Schrenck, 1867); 1926: 227 (Helix lavrushini);
Biittner, Ehrmann, 1927 : 236 (Eulota); Лихарев, Раммельмейер,
1952 : 405, рис. 336, 337 [Eulota (Acusta), E, (Acusta) selskii).
Материал: более 80 экз. из 30 местонахождений в Приморском крае.
Раковина от шаровидной до шаровидно-кубаревидной, с тупой вершиной
и узким, почти коническим завитком, высота которого равна высоте устья или не-
сколько меньше, хрупкая, тонкостенная. Оборотов 5—6, выпуклых, стремительно
BRADYBAENA
145
нарастающих. Последний оборот сильно вздут, почти вдвое шире предпоследнего,
явно преобладает над остальными, к устью не опущен. Окраска светло-роговая, жел-
товатая, оливковая или коричневая. Нередко раковина одноцветная; в других слу-
чаях немного выше периферии располагается узкая коричневая лента; наконец, из-
редка наблюдается присутствие трех лент; помимо центральной одна лента под швом,
другая на базальной поверхности. Две последние более широкие, размытые и не-
ясные; в проходящем свете ленты не плотнее фона. Скульптура в виде радиальной ис-
черченности — от тонкой* до довольно грубой, которая к шву становится более резкой.
Кроме того, имеются четкие, глубоко врезанные спиральные бороздки, рассекающие
большую часть радиальных морщин, в результате чего поверхность раковины иногда
местами мелкобугорчатая. Устье мало скошено, округлое, большое. Края его острые,
не отвернуты или отвернуты слабо. Колумеллярный край (верхняя часть его) нави-
сает над пупком. Иногда имеется слабая расплывшаяся губа. Пупок узкий, непер-
спективный. Размеры: ВР 20—30, БД 23—35, МД 18—28 мм.
Внутреннее строение. Материал; 2 экз. с о-ва Путятин в зал. Петра
Великого, VII 1957, сбор Т. Джамусовой, определение И. М. Лихарева; 3 — из окр.
с. Конгауз Приморского края, 22 VII 1972; 4 — из с. Пейшула Приморского края,
3 VII 1971; 5 — из окр. Владивостока, 26 IX 1971; 4 — с побережья зал. Восток При-
морского края, 16 VII 1971, сборы и определения наши.
Спермовидукт резко складчатый, сжат в компактный комок. Яйцевод вначале
сужен, в нижней части расширен. Вагина длинная и тонкая; стилофор впадает по
существу в клоаку. Слизистых желез от 3 до 12, массивных, вторично не ветвящихся,
с ячеистыми стенками. Каждая железа имеет удлиненное основание — ножку; сли-
ваются непосредственно при впадении в дополнительный мешок, имеющий обширную
полость. Проток дополнительного мешка впадает в узкий проток, в котором распо-
лагается стрела; основание стрелы находится в глубоком «колодце», а не на вершине
подушки. Нижняя часть стилофора расширена. Дополнительный мешок очень длин-
ный, его вершина далеко выдается за вершину стилофора. Железистый пояс охва-
тывает вагину выше стилофора.
Чехол пениса очень короткий, охватывает примерно пятую часть длины пениса,
фиксирован только у клоаки. Пенис внутри от начала до конца покрыт косыми пра-
вильными мелкими складками, сходящимися под углом вдоль одной линии; в прокси-
мальной части на этой линии располагается короткий продольный гребень, дисталь-
ная часть которого явственно обособлена от внутренней поверхности пениальной
трубки. Эпифаллус очень короткий. Основание протока семеприемника не расши-
рено, проток строго цилиндрический, но зато расширен участок яйцевода, соседствую-
щий с ним. Резервуар семеприемника сравнительно с другими дальневосточными пред-
ставителями рода очень большой; если он не наполнен, стенки его спадаются (рис. 89),
в противном случае резервуар шаровидный.
Распространение и местообитания. От бассейна среднего и
нижнего Амура до Южного Китая, охватывает весь Приморский край и большую часть
территории п-ова Корея. Населяет влажные широколиственные леса, часто подни-
мается в кроны деревьев. Очень обычный вид на Дальнем Востоке, но плотных попу-
ляций не образует.
16. Bradybaena scythica (Westerlund, 1898). Табл. Ill, 22', рис. 93, 94.
Westerlund, 1898 : 160 [Helix (Eucampylaea) ]; Lindholm, 1927d : 263
{Fruticicola roxane)', Линдгольм, 1931 : 52, t. I, fig. 1—3 {F. roxane); Лиха-
рев, Раммельмейер, 1952:403 [Eulota (Leucozonella) roxane].
Материал: более 40 экз. из 5 местонахождений на Тянь-Шане.
Раковина твердостенная, от кубаревидной до шаровидной, высота завитка
сравнима с высотой устья: может быть немного выше или немного ниже. Очертания
завитка явственно куполовидные. Оборотов около 6, умеренно выпуклых, плавно на-
растающих, разделенных неглубоким неровным швом. Последний оборот немного
менее чем в 1.5 раза шире предпоследнего, в профиле почти правильно округлый,
снизу совершенно не уплощен, к устью слабо и плавно опущен. Окраска образована
серовато-роговым или желтоватым фоном, на котором лежит сеть развитых в разной
степени неправильных белых пятен и штрихов; на периферии имеется размытая свет-
лая (иногда молочно-белая) лента, выше и ниже которой основная окраска, сгущаясь,
образует две более темные размытые ленты. В проходящем свете светлые элементы
окраски плотнее темных. Имеются 4 типа скульптуры: радиальные сглаженные мор-
щины, спиральные бороздки, тончайшая косая струйчатость, заметная местами при
значительном увеличении, и легкие продольные вмятины на последнем обороте. Устье
округлое, косое, места его прикрепления почти не сближены и связаны тонкой мо-
золью. Палатальный и базальный края почти не отвернуты и слегка утолщены, обра-
зуя перистом. Колумеллярный край слегка отвернут и очень немного прикрывает
Ю А. А. Шилейко
146
II. BBADYBAENIDAE
пупок, сквозь который можно просмотреть не более 3/4 предпоследнего оборота. Раз-
меры: ВР 15—22, БД 19—26, МД 16—22 мм. Лектотип: ВР 15.3, БД 20.8 мм.
Внутреннее строение. Материал: 1 экз. из Ферганского хр., с. Кы-
зыл-Унгур, 9 VI 1961, сбор И. М. Лихарева; 1 — из окр. Таджикабада, хр. Петра I,
7 VII 1968, сбор 3. Иззатуллаева; 8 — из северной части Ферганского хр., левый
берег р. Нарын ниже Токтогула, 31 V 1972, сбор и определения наши.
Нижняя часть спермовидукта образует 2—3 очень крутых изгиба, иногда сжатых
в единый комок: такая картина характера для особей, у которых спермовидукт в дан-
ный момент гипертрофирован; если же последний находится в стадии покоя, изгибы
Рис. 93—94. Bradybaena scythica (West.), Ферганский хр., берег р. Нарын ниже Ток-
тогула, 31 V 1972. 93 — внешний вид полового аппарата; 94 — дистальная часть
полового аппарата, вскрыта. (Ориг.).
выражены слабее или отсутствуют. Яйцевод в 2—3 раза короче вагины. Слизистые
железы собраны в 2 пучка, вторичное ветвление отсутствует или выражено слабо;
каждая ветвь имеет ровные или в различной степени бугристые стенки, ветви сли-
ваются непосредственно при впадении в дополнительный мешок. Этот последний у всех
исследованных экземпляров хорошо развит, впадает в базальную часть небольшого
стилофора и значительно длиннее последнего. Проток слизистых желез в пределах
дополнительного мешка несколько расширяется и широко сообщается с полостью
стилофора; внутренняя поверхность последнего покрыта характерными, беспоря-
дочно ориентированными короткими прямыми и изогнутыми складками и бугорками,
которые могут местами группироваться в продольные валики. Такое строение позволяет
предполагать, что стенки стилофора могут сильно растягиваться. Железистый пояс,
локальные разрыхления стенок вагины, равно как и надклоачный пузырь, отсутствуют.
Чехол пениса ясно выражен, охватывает воротничком нижнюю треть короткого
пениса. В проксимальной части пенис утолщен, иногда весьма значительно; внутрен-
няя поверхность его несет несколько неправильных продольных валиков, которые
близ семепровода сминаются в короткие складки и бугорки. В максимальном выра-
жении эти структуры имеют вид фестончатых лент. Основание протока семеприем-
ника расширено. Короткие беспорядочные продольные складки, покрывающие стенки
вагины, к месту отхождения протока семеприемника приобретают упорядоченность
и превращаются в серию высоких параллельных валиков, часть которых направ-
ляется в яйцевод, а часть — в семеприемник.
Распространение и местообитания. Вид спорадически рас-
пространен по хр. Петра I и Ферганскому. Встречен под большими камнями на сухих
открытых склонах. Видимо, экологический спектр вида очень узок.
BRADYBAENA
147
17. Bradybaena lantzi (Lindholm, 1927). Табл. Ill, 23, 24; рис. 95, 96.
Lindholm, 1927d : 261 (Fruticicola); Цветков, 19416 : 287—302 (Fr.
lantzi dextrorsa, Fr. lantzi sinistrorsa, Fr, lantzi m. steppensis, m. Montana, m. silvestris);
Лихарев, Раммельмейер, 1952 : 399 [Eulota (Eulota)]; Матёкин,
1966 : 119, табл. 3, рис. 7—8; 1972 : 116, рис. 1, А—Б.
Материал: более 100 экз. из 27 местонахождений на Тянь-Шане.
Раковина от кубаревидной до шаровидной, твердостенная, завиток кони-
ческий или слегка куполовидный, с узкой вершиной. Оборотов 5—6, довольно выпук-
лых, медленно и плавно нарастающих. Последний оборот в профиле округлый, к устью
немного и очень плавно опущен, в 1.5 раза или немного более шире предпоследнего.
Окраска образована желтым фоном разных оттенков и 3 спиральными темными (чаще
Рис. 95—96. Bradybaena lantzi (Lindh.), окр. Таласа, 3 VI 1972. 95 — внешний вид
полового аппарата; 96 — дистальная часть полового аппарата, вскрыта. (Ориг.).
всего коричневыми) лентами, верхняя из которых прилегает ко шву, средняя лежит
над периферией последнего оборота, нижняя — на базальной части; пупок окружен
светлым полем. Верхняя и нижняя ленты в различной степени размыты, средняя
с более четкими контурами. Встречаются экземпляры без срединной ленты или вообще
без лент, причем в этом случае раковина может сохранять довольно интенсивную
окраску фона или может не иметь окраски (альбинотические особи). Наконец, у неко-
торых экологических морф ленты развиты настолько интенсивно, что в значительной
степени маскируют фон, и раковина в результате имеет в целом темную окраску.
Скульптура эмбриональных оборотов тонкозернистая, дефинитивные обороты с нерав-
номерными грубыми радиальными морщинами; обычно местами на последнем обороте
наблюдаются слабые редкие спиральные линии. Устье почти правильно-округлое,
косое, края его почти прямые, слегка утолщены. Губа слабая, очень расплывшаяся,
или она отсутствует. Колумеллярный край сильно отвернут и частично перекрывает
узкий неперспективный пупок. Размеры: ВР 13—21, БД 17—27, МД 14—23 мм.
Полиморфизм данного вида выражается помимо широкой изменчивости в форме,
размерах и окраске раковины в существовании наряду с нормальными правозавитыми
популяциями популяций, состоящих из левозавитых особей. А. В. Яблоков и
А. В. Валецкий (1971) провели полевое изучение взаимного распространения дек-
стральных и синистральных популяций в сравнении с аналогичной работой Б. Н. Цвет-
кова (1938). Выяснилось, что за 30 лет с лишним конфигурация ареалов левозавитых
популяций заметно изменилась. Сборы моллюсков в зоне контакта между поцуляциями
«правых» и «левых» морф позволяют предполагать, что их смешение может проис-
10*
148
II. BRADYBAENIDAE
ходить лишь в масштабах, соизмеримых с радиусом индивидуальной активности
животного.
Было бы крайне интересным провести специальное исследование по сравнению
этих морф с применением электрофоретических, цитологических и физиологических
методов. В частности, показано, что при голодании синистральные особи быстрее
теряют в весе и быстрее погибают, чем декстральные (Гаузе, Смарагдова, 1939).
Внутреннее строение. Материал: 7 экз. из ущелья р. Пскем на
западном Тянь-Шане, 20 V 1972; 6 — из пойменного лиственного леса у р. Талас,
выше г. Таласа, 3 VI 1972; 5 — из верховьев р. Чаткал, 25 V 1972; 2 — из Киргиз-
ского хр. над с. Луговое, 6 VI 1972; 3 — из Киргизского хр., ущелье Карабалты,
11 VI 1972; 3 — из Киргизского хр., ущелье р. Аламедин, 14 VI 1972, сборы наши';
3 — из Алма-Аты, 11 VIII 1969; 6 — из восточного Казахстана, оз. Иссык, 22 VIII 1969
(левозавитые особи), сборы И. М. Хохуткина; 2 — из Заилийского хр., 13 IV 1966,
сбор К. К. Увалиевой; определения наши.
Нижняя часть спермовидукта образует один крутой изгиб с тесно сомкнутыми
краями. Слизистые железы в виде одного или нескольких кустов, интенсивность вет-
вления которых очень различна. Каждая ветвь имеет тонкие бугристые стенки, все
ветви объединяются в общий проток, который впадает в нижнюю часть дополнитель-
ного мешка и вливается в полость последнего не апикально, но сбоку. Дополнитель-
ный мешок развит в различной степени, но всегда более или менее ясно выражен,
полость его сообщается с полостью стилофора примерно на уровне конца стрелы,
основание которой окружено невысоким воротничком. Клоака, вагина и основание
стилофора охвачены светлым, плотным надклоачным пузырем; нижняя часть стило-
фора, пронизывая надклоачный пузырь, впадает очень низко, по существу в клоаку.
Пенис недлинный, пениальный чехол короткий, обычно морщинистый. Наруж-
ные покровы пениса интенсивно пигментированы, причем следы пигмента обнару-
живаются даже у тех особей, раковина которых не пигментирована. Проксимальная
часть пениса образует легкое вздутие, дистальная — цилиндрическая или слегка
коническая. Внутренняя поверхность пениса близ клоаки несет правильные четкие
продольные валики, которые внезапно распадаются на ряды бугорков, причем по мере
продвижения к эпифаллусу упорядоченность в расположении бугорков нарушается.
Эпйфаллус тонкий, граница его отмечена резким прекращением пигментации. Осно-
вание протока семеприемника плавно расширено, проток конический.
Распространение и местообитания. Тянь-Шань; обнаружен
на Алайском, Ферганском, Пскемском, Киргизском, Заилийском, .Джунгарском,
Таласском хребтах и в Таласской котловине. Вид в целом гигромезофильный;
наибольшую плотность образует на высокотравных среднегорных лугах, по бере-
гам рек и ручьев, в узких тенистых ущельях. В подобных местах имеется как бы ядро
популяции, из которого отдельные особи мигрируют даже на открытые склоны, где,
однако, держатся в дневное время под камнями, в осыпях и в трещинах скал.
18. Bradybaena plectotropis (Martens, 1864). Табл. Ill, 25—29; рис. 97—99.
Martens, 1864, Malak. Bl., XI : 114, t. 3, fig. 3—5 (Helix); Мартенс, 1874 :
13, табл. 1, рис. 8, табл. 3, рис. 39 (Helix phaeozona), 17, табл. 1, рис. 11 (Helix); Mar-
tens, 1882а : 11, t. 2, fig. 10, 11 (Helix phaeozona), 14 (Helix); Westerlund, 1889 :
153 (Helix phaeozona); 1898 : 159 (Helix palmeni); W e b e r, 1913 : 10, t. 1, fig. 6 [Cath-
aica (Pseudiberus) siigoschuriana]; Lindholm, 1927c : 195 (Fruticicola phaeozona
pseudarianta), 200 [Cathaica (Pseudiberus) stschukini]; Скворцов, 1940 : 372 (Fruti-
cicola almaatini); Цветков, 1940 : 391 [Cathaica (Pseudiberus) transilensis]; 389
(Fruticicola skwbrtzowi); 1941a : 283 [Cathaica (Pseudiberus) scalaris], 284 [C. (Pseudi-
berus) saxatilis], 285 [C. (Pseudiberus) necopinatus]; Лихарев, Раммельмейер,
1952 : 383 [Cathaica (Pseudiberus) necopinatus], 383, рис.312 [C. (Pseudiberus) siigoschu-
riana], [C. (Pseudiberus) transilensis], 384, рис. 313 [C. (Pseudiberus) stschukini], 385,
рис. 314 [C. (Pseudiberus)], 399 [Eulota (Eulota) almaatini], 400, рис. 332 [£. (Eulota)
phaeozona], 400 [Я. (Eulota) skwortzowi]; M а т ё к и н, 1966 : 118—119, табл. 3,
рис. 9; 1972 : 120, рис. 1, В—3, 2.
Данный вид представляет собой крайнее выражение конхологического поли-
морфизма. Изменчивость здесь такова, что представляется правильным считать весь
комплекс конхологических форм надвидом, состоящим из нескольких полувидов
(semispecies, smsp.). Эти взгляды были развиты П. В. Матёкиным (1966, 1972), изу-
чившим обширный материал по данной группе.
В общем можно констатировать, что эти полувиды, несомненно, привязаны к опре-
деленным типам биотопов, хотя биологический смысл образования какой-то конхо-
логической формы не всегда понятен. В качестве примера можно сослаться на кон-
хблогический комплекс из ущелья р. Аламедин на Киргизском хр. Здесь обитают резко
ребристые тонкостенные формы, имеющие на периферии коричневой или бурой рако-
вины резкий киль; эта форма четко связана с каменистыми осыпями, которые местами
BRADYBAENA
14 9
Имеются как на открытых, так и на залесенных склонах. Далее, в кустарниках вдоль
реки и в узких боковых ущельях живут улитки с конической или даже почти шаро-
видной раковиной, с правильно-закругленной периферией, тонко скульптурирован-
ные, светлые или с темными полосами и лентами и даже почти бесцветные. Наконец,
в одном месте у подножья крутых скал у самой реки обнаружена небольшая популя-
ция, занимающая промежуточное положение; раковина сильно прижатая, умеренно
скульптурированная, с углом (но не килем) выше периферии. Соответственно здесь
имеются полувиды plectotropis, almaatini, necopinatus.
Необходимо отметить также, что если встать на путь практического различения
всех форм изменчивости этого конхологического комплекса и считать их разными,
Рис. 97—99. Bl-adybaena plectotropis smsp. phaeozona (Mart.), Ферганский хр., берег
р. Нарын выше Токтогула, 1 VI 1972. 97 — внешний вид полового аппарата; 98 —
место впадения яйцевода в вагину; 99 — дистальная часть полового аппарата, вскрыта.
(Ориг.).
пусть даже близкими видами, то придется описывать в качестве самостоятельных
видов несколько десятков, если не сотен, популяций, поскольку в большинстве слу-
чаев каждая популяция имеет статистически достоверные отличия от соседней.
Отступая от принятой схемы, приводим вначале описание внутреннего строения,
свойственного всем членам надвида, а затем рассмотрим конхологические особенности
каждого из полувидов.
Внутреннее строение. Материал: 24 экз. из ущелья р. Аламедин
на Киргизском хр., 14—25 VI 1972 (полувиды almaatini, necopinatus, plectotropis);
8 — из ущелья Беш-Таш на Таласском хр., 4 VI 1972 (полувид plectotropis); 10 —
из Ферганского хр., берег р. Нарын выше Токтогула (полувиды phaeozona и alma-
atini), 1 VI 1972; сборы наши; 3 — из Сусамырской долины, 14 IV 1970, сбор
Л. В. Славошевской (полувид almaatini); 5 — из Алма-Аты, берег р. Алматинки,
13 VIII 1969, сбор .И. М. Хохуткина (полувид almaatini); 4 — из ущелья Каскасу
на Заилийском хр., 15 V 1968, сбор К. К. Увалиевой (полувид phaeozona); 3 — с юж-
ного берега оз. Иссык-Куль, 1970, сбор Б. П. Ушакова (полувид plectotropis); 6 —
из Казахстана, берег Б. Алматинского оз., 18 VI 1969, сбор К. К. Увалиевой (полу-
вид transilensis); определения наши.
150
II. BRADYBAENIDAE
Спермовидукт в различной степени изогнут или почти прямой, яйцевод длинный.
'Слизистые железы в виде 1—4 кустов впадают в общий проток, длина которого под-
вержена значительной индивидуальной изменчивости. Проток слизистых желез впа-
дает обычно близ основания дополнительного мешка, который может быть развит
лучше или слабее, но присутствует всегда, плотно примыкая к стилофору. Дисталь-
ная часть всех отделов женской части покрыта надклоачным пузырем, причем край
его иногда охватывает и нижнюю часть протока слизистых желез (рис. 97), в резуль-
тате чего этот проток пронизывает покровы пузыря.
Пенис умеренно длинный, при основании охвачен коротким чехлом. Внутренняя
поверхность дистальной части пениса покрыта очень высокими продольными склад-
ками, в результате чего просвет пениса на разрезе имеет вид многолучевой звезды; выше
середины пениса эти складки распадаются на множество высоких тонких ворсинок.
Эпифаллус тонкий, постепенно переходит в пенис. Основание протока семеприемника
незначительно расширено.
Распространение и местообитания. Тянь-Шань, предгорья
Памира, Западный Китай (восточные склоны Тянь-Шаня) и северная Индия (Каш-
мир). Политопный вид, населяет практически все типы биотопов в пределах ареала.
В известной степени описываемые ниже полувиды — отражение биотопической из-
менчивости.
18а. Bradybaena plectotropis smsp. phaeozona (Martens, 1874). Табл. Ill, 25.
Материал: более 100 экз. из 43 местонахождений на Тянь-Шане.
Раковина более или менее прижатая, с куполовидным низким завитком,
высота которого большей частью меньше высоты устья. Оборотов 6—6.5, не уплощен-
ных, разделенных неглубоким швом. Окраска белая с желтоватым или розоватым
оттенком, над периферией проходит четкая коричневая или черная лента, которая
прослеживается у шва на более ранних оборотах. Скульптура в виде равномерной
радиальной складчатости или исчерченности и неясных спиральных бороздок. Устье
широкоовальное, скошенное, внутри белое; спиральная лента почти не просвечивает,
сквозь стенку раковины. Края устья отвернуты. Пупок узкий, цилиндрический,
почти не закрыт колумеллярным краем. ВР 10—12, БД 15—18, МД 12.5—15 мм.
18b. Bradybaena plectotropis smsp. almaatini (Skwortzow, 1940). Табл. Ill, 26.
Материал: более 100 экз. из 27 местонахождений на Тянь-Шане.
Раковина от конической до шаровидной, высота слегка куполовидного
завитка равна или превышает высоту устья. Оборотов 6—6.5, слабо выпуклых, плавно
нарастающих. Последний оборот округлый, к устью немного опущен. Окраска чаще
всего коричневато-желтая, интенсивность окраски по поверхности раковины обычно
неодинаковая, в результате чего возникают радиальные размытые полосы; нередко
раковина одноцветная. Кроме того, иногда имеется спиральная темная лента. Скульп-
тура в виде грубой неравномерной радиальной исчерченности и слабых спиральных
линий. Устье круглое, косое, внутри палевое, с широкой, сильно расплывшейся губой.
Пупок шире, чем у предыдущей формы, цилиндрический, почти перспективный, не при-
крыт или слегка прикрыт отворотом колумеллярного края. Размеры: ВР 17--19,
БД 21—23, МД 18—19 мм.
18с. Bradybaena plectotropis smsp. necopinatus (Tzvetkov, 1941). Табл. Ill, 27.
Материал: более 40 экз. из 2 местонахождений на Киргизском хр.
Раковина прижатая, высота куполовидного завитка меньше высоты устья.
Оборотов обычно 5, на последнем имеется легкий, но хорошо заметный угол несколько
выше периферии. Окраска серая с очень легким коричневатым оттенком или кремо-
вая, причем угол светлее фона. Выше периферии нередко имеется очень слабая спи-
ральная темная лента. Радиальная скульптура — от грубой радиальной исчерчен-
ности до легкой ребристости, причем на нижней поверхности раковины эта скульп-
тура выражена слабее, чем на верхней. Устье очень косое, овальное или четырех-
угольное, углы очень сильно закруглены. Края устья широко отвернуты, места их
прикрепления заметно сближены. Пупок довольно широкий, но не развернутый. Раз-
меры: ВР 7—9, БД 13—20, МД 10—16.5 мм.
18d. Bradybaena plectotropis smsp. plectotropis (Martens, 1864). Табл. Ill, 28.
Материал: более 100 экз. из 32 местонахождений на Тянь-Шане.
Раковина от низкоконической, прижатой, до почти конической, линзовид-
иая, высота завитка меньше высоты устья (нередко меньше половины высоты устья).
Оборотов 5—6.5. Последний оборот снизу более выпуклый, чем сверху, с резким ки-
лем или углом на периферии, который совпадает со средней линией последнего оборота
BRADYBAENA
151
или сдвинут вверх. Окраска в целом бурая, коричневая или роговая, но часто угады-
ваются спиральные, еще более темные ленты; киль окрашен немного светлее фона.
Скульптура в виде радиальной ребристости, более или менее резкой, причем на ба-
зальной части ребра выражены слабее. Последний оборот к устью сильно опущен,
устье резко скошено, овальное, места его прикрепления сближены и соединены мо-
золью. В общем здесь можно говорить о наклонности к образованию цельного устья.
Устьевые края широко отвернуты, губа слабая или отсутствует; пупок умеренно раз-
вернутый, перспективный. Размеры: ВР 7—14, БД 14—22, МД 12—19 мм.
18е. Bradybaena plectotropis smsp. transilensis (Tzvetkov, 1941).-
Материал: 12 экз. из 2 местонахождений на северо-восточном Тянь-Шане.
Раковина сильно прижатая, высота завитка приблизительно равна высоте
устья. Оборотов около 5, в профиле обороты ступенчатые, т. е. помимо перифериче-
ского резкого киля на верхней стороне оборотов имеется более или менее ясно выра-
женный сглаженный угол. Окраска в общем бурая, киль немного светлее фона. Скульп-
тура — резкие косые радиальные ребра, на базальной части раковины выражены сла-
бее. Края устья слабо сближены, в различной степени отвернуты. Пупок почти ци-
линдрический, узкий, частично прикрыт отворотом колумеллярного края. Размеры:
ВР 6—8.5, ВД 10—15, МД 8.5—12.5 мм.
19. Bradybaena tzwetkovi Uvalieva et Soboleva, 1973. Рис. 100.
Увалиева, Соболева, 1973:1717, рис. 1, 2.
Материал: более 20 экз. из одного местообитания в Алма-Атинской обл.
Раковина кубаревидная, с широко закругленным куполовидным завитком
и сглаженной вершиной; высота завитка значительно превышает высоту устья. Обо-
ротов 6—7, медленно нарастающих, разделенных глубоким швом. Последний оборот
немного шире предпоследнего, к устью незначительно
опущен, снизу слегка уплощен. Окраска образована
кремовым, коричневым, желтоватым или светло-серым
фоном и одной спиральной темной лентой, проходящей
немного выше периферии на последнем обороте, а на
более ранних оборотах — над швом, не касаясь его; у
некоторых экземпляров имеется еще одна слабая лента,
расположенная над первой. По периферии проходит
светлая лента, низ раковины одноцветный. Скульптура
в виде сглаженных радиальных ребрышек, которые
хорошо выражены на верхней поверхности раковины,
но почти пропадают на нижней. Между ребрышками
имеется микроскопическая пунктировка; такая же
пунктировка покрывает эмбриональные обороты. Устье
косое, широкополулунное, с белой губой; края устья
острые, прямые, только колумеллярный край у пупка
линдрический, узкий. Размеры: ВР 9—12, БД 10—12.
100. Bradybaena tzivet-
Uvalieva et Soboleva.
(Ориг.).
слегка отвернут. Пупок ци-
Голотип: ВР 9.5, БД 10.1 мм.
В Зоологическом музее Московского университета хранятся сборы этого вида,
сделанные В. Н. Цветковым и определенные им как Cathaica brodskii; это название
упоминается также в статье (Цветков, Цветкова, 1950), но не сопровождается описа-
нием; следовательно, название относится к категории nomen nudum.
Внутреннее строение. Материал нами не исследован; судя по перво-
описанию, вид близок к Вт. plectotropis-. имеется небольшой дополнительный мешок,
надклоачный пузырь и умеренно длинный пенис. Внутреннее строение полового аппа-
рата неизвестно.
Распространение и местообитания. Вид встречен на равнин-
ных и горных пастбищах Алма-Атинской обл.
Рис.
kovi
20. Bradybaena perlucens (Rosen, 1901). Табл. Ill, 30; рис. 101.
Westerlund, 1898 : 158 [Heliz (Eucampylaea) indigena, non Mabille, 1895];
Розен, 1901 : 9 [1/. (Eucampylaed) ]; Линдгольм, 1931 : 59 [Cathaica (Kaz~
nakoviella)]; Лихарев, Раммельмейер, 1952:380, рис. 309 [Cathaica
{Cathaica}[; Матёкин, 1966 : 121; Лихарев, Старобогатов, 1967 : 190
(Cathaicay, Матёкин, 1972 : 120, рис. 7.
Материал: более 100 экз. из 4 местонахождений на Тянь-Шане.
Раковина прижатая, с коническим контуром завитка и невыступающей
вершиной. Оборотов около 5, выпуклых, быстро нарастающих. Последний оборот
не менее чем в 1.5 раза шире предпоследнего, к устью очень плавно и очень слабо опу-
щен, в начале с легким углом, исчезающим к устью. Окраска раковины живых мол-
152
II. BRADYBAENIDAE
люсков серовато-роговая или очень светло-каштановая, сверху немного темнее, чем
на базальной части. Пустые раковины быстро теряют первоначальную окраску и ста-
новятся белыми. Эмбриональные обороты с более или менее ясной микроскульптурой
в виде зернышек, расположенных в шахматном порядке. Скульптура дефинитивных
оборотов представлена тонкой струйчатой исчерченностью и местами грубыми склад-
ками. Устье вполне сформированных экземпляров немного раструбообразно расши-
рено, почти круглое, сильно скошено. Края устья почти равномерно отвернуты; сла-
бее^ всего отвернута верхняя часть палатального края, сильнее всего — колумелляр-
ный край. Пупок широкий, перспективный, умеренно или сильно развернутый. Раз-
меры: ВР 7—12, ВД 17—22, МД 14—17 мм.
Рис. 101. Bradybaena perlucens (Rosen), запов. «Ромит», Гиссарский хр.,
31 III 1967. Внешний вид полового аппарата и вскрытый пенис.
(Ориг.).
Внутреннее строение. Материал: 2 экз. из запов. «Ромит»,
Гиссарский хр., 31 III 1967, сбор и определение 3. Иззатуллаева; 3 — из предгорий
Чаткальского хр. близ с. Ала-Бука, 28 V 1972, сбор и определения наши.
Нижняя часть спермовидукта и яйцевод очень круто дважды изогнуты. Слизистые-
железы в виде одного куста, общее число ветвей непостоянно, но у всех изученных
особей обнаружены 2 главные ветви. Стилофор длинный, круто изогнутый; между
краями изгиба вклинивается дополнительный мешок, очень тесно прилегающий к ди-
стальной части стилофора и связанный с ним тонкими соединительнотканными пере-
понками. Слизистые железы впадают в дополнительный мешок почти апикально, по-
лость последнего слегка расширена и сообщается со стилофором узким каналом 'на
уровне середины стрелы. Следов надклоачиого пузыря не обнаружено.
Пенис длинный и тонкий, при основании охвачен коротким чехлом; внутренние
стенки его дистальной части покрыты прямыми продольными складками, которые за-
тем становятся зигзагообразными и в проксимальной части образуют правильный
узор. Эпифаллус очень неясно отграничен от пениса. Основание протока семеприем-
ника несильно, но явственно расширено.
Распространение и местообитания. Южный Тянь-Шань
(хребты Петра I, Гиссарский, Зеравшанский, предгорья Чаткальского хр.) и окр. Файз-
абада (Афганистан). По-видимому, предгорья Чаткальского хр. являются северной
границей ареала. Ксерофильный вид, встречается на открытых сухих каменистых
склонах; в жаркое время уходит в почву. Особенно плотные скопления образует в ме-
стах, где подстилающие горные породы, покрытые многочисленными трещинами, вы-
ходят на поверхность в виде ровных плит.
BRADYBAENA
153
21. Bradybaena dichrozona (Martens, 1885). Табл. IV, 31; рис. 102—104.
Martens, 1885 : 17 (Helix); Westerlund, 1889 : 106 (Helix); Лиха-
рев, Раммельмейер, 1952 : 401 [Eulota (Eulota)]; Матёкин, 1966 : 119;
1972 : 131 (Bradybaena pauiovskii pars).
Материал: более 30 экз. из 3 местообитаний на Чаткальском хр.
Раковина прижатая, со слегка куполовидным контуром завитка, высота
которого меньше высоты устья; чаще всего 2/3—3/4 этой величины. Оборотов 6, вы-
пуклых, в профиле у шва чуть ступенчатых, на периферии и снизу широко и пра-
Рис. 102—104. Bradybaena dichrozona (Mart.), верховья р. Касансай на Чаткальском хр.,
28 V 1972. 102 — внешний вид полового аппарата; 103 — дистальная часть женского
отдела, вскрыта; 104 — пенис, вскрыт. (Ориг.).
вильно закругленных. Последний оборот в 1.5 раза или немного более чем в 1.5 раза
шире предпоследнего, к устью несильно, но явственно очень плавно опущен. Окраска
характерная: немного выше периферии проходит отчетливая светлая лента, окаймлен-
ная с обеих сторон темно-коричневыми лентами, ширина которых равна ширине цент-
ральной или немного меньше. Остальная часть поверхности каштановая, заметно
менее интенсивно окрашена, чем коричневые ленты. Центральная лента разрезана
вдоль швом более ранних оборотов; нижняя темная лента проходит строго по пери-
ферии. Нижняя поверхность раковины окрашена в общем заметно светлее верхней.
В проходящем свете самую большую плотность имеет светлая лента, коричневые про-
свечивают сильнее фона. С гибелью животного,окраска быстро блекнет; дольше всего
сохраняются темные ленты. Скульптура в виде слабой неравномерной радиальной
морщинистости, у крупных экземпляров морщинистость грубая. На некоторых ра-
ковинах местами имеются вмятины. Спиральные бороздки еле намечаются или от-
сутствуют. Устье умеренно косое, просвет его почти круглой формы, края устья отог-
нуты тем сильнее, чем дальше от ангулярной области. Внутри устья четко выражена
наружная окраска, но в несколько ослабленном виде. Пупок почти не прикрыт, раз-
вернутый, широкий, перспективный. Размеры: ВР 11—17, БД 19—28, МД 16—23 мм-
154
II. BRADYBAENIDAE
Внутреннее строение. Материал: 6 экз. из верховьев р. Касансай на
Чаткальском хр., 28 V 1972, сбор наш; 5 — из долины р. Ала-Бука-сай на Чаткаль-
ском хр., 20 V 1972, сбор В. В. Горохова; определения наши.
Нижняя часть спермовидукта образует резкий двойной изгиб. Имеется один куст
слизистых желез, состоящий из 3—8 ветвей, которые обнаруживают наклонность к вто-
ричному ветвлению; ветви бугристые, с многочисленными поперечными перетяжками
и складками. Единым протоком слизистые железы впадают в широкий дополнитель-
ный мешок; в пределах мешка этот проток образует небольшое расширение, далее про-
низывает, сужаясь, ткани мешка и впадает в полость стилофора на уровне острия
Рис. 105—106. Bradybaena bilaticincta (Mart.), Чаткальский хр., окр. оз. Сары-Челек,
22 X 1972. 105 — внешний вид полового аппарата; 106 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
стрелы. Полость вагины сообщается с полостью стилофора небольшим круглым от-
верстием, а с полостью клоаки — длинной узкой щелью, возникшей за счет мощного
развития поперечной складки на границе вагины и клоаки. Надклоачный пузырь хо-
рошо развит, охватывает кольцом всю дистальную часть женского отдела.
Пенис длинный, чехол пениса занимает не более 1/i длины его. Продольные чет-
кие складки внутри дистальной части пениса превращаются близ очень' короткого
эпифаллуса в систему из множества высоких тонких бугорков, ворсинок. Основание
протока семеприемника незначительно расширено.
Распространение и местообитания. Чаткальский хр., высоко-
горье и среднегорье; через хребет не переходит и в долине р. Чаткал не обнаружен.
Ниже 500 м над ур. м., видимо, не спускается. Обитает в крупнообломочных осыпях,
в трещинах скал, неподалеку от воды.
22. Bradybaena bilaticincta (Martens, 1882). Табл. IV, 32; рис. 105, 106.
Martens, 1882а : 5, t. 1, fig. 12, 13 (Helix paricincta var. bilaticincta); Lind-
holm, 1926 : 262 (Fruticicola saturata); Лихарев, Раммельмейер, 1952 :
398 [Eulota (Eulota) paricincta var. bilaticincta); M а т ё к и н, 1966:120, табл. 4,
рис. 6; 1972 : 128, рис. 5.
BRADYBAENA
155
Материал: 22 экз. из 3 местонахождений на Чаткальском хр.
Раковина шаровидная, умеренно твердостенная или тонкостенная, сверху
и снизу примерно одинаково выпуклая, со слегка куполовидным контуром завитка.
Оборотов 5.5, умеренно выпуклых. Последний оборот явственно преобладает над
остальными, в профиле правильно широко закруглен, в 1.5 раза шире предпослед-
него. Окраска каштановая или красновато-коричневая, по периферии проходит белая
или желтая лента, вокруг пупка имеется светлое желтоватое поле. Возле перифери-
ческой ленты на большинстве раковин можно видеть сгущения фона, в результате
чего возникают еще две полосы — выше и ниже периферии последнего оборота, т. е.
схема окраски по существу такая же, как и у предыдущего вида. В проходящем свете
светлая лента темнее фона. Скульптура — неравномерная радиальная, довольно
тонкая морщинистость и редкие спиральные бороздки, которые могут отсутствовать
на верхней поверхности раковины, но видны на нижней стороне. Устье круглое, уме-
ренно скошено и немного, но ясно плавно опущено. Внутри устье светло-коричневое,
светлая лента на периферии хорошо различима. Края устья простые или слегка утол-
щенные, прямые или немного отвернуты. Отворот колумеллярного края наполовину
или меньше, чем наполовину, прикрывает узкий цилиндрический пупок, сквозь кото-
рый можно, поворачивая раковину, увидеть не более половины предпоследнего обо-
рота. Размеры: ВР 13.5—18, БД 17—21, МД 15—18.5 мм.
Внутреннее строение. Материал: 7 экз. из Чаткальского хр., окр.
оз. Сары-Челек, 22 X 1972, сбор В. В. Горохова, определение наше.
Дистальная часть короткого спермовидукта образует двойной изгиб, края кото-
рого плотно сомкнуты. Яйцевод короткий и мощный, вагина длинная. Слизистые же-
лезы в виде куста из 5—9 ветвей, не обнаруживающих наклонности к вторичному вет-
влению и впадающих единым протоком в стилофор. У всех вскрытых экземпляров до-
полнительный мешок наружно почти не выражен, но на продольном разрезе соответ-
ствующего участка стенки стилофора видно небольшое вздутие, внутри которого про-
ток слизистых желез расширяется. П. В. Матёкин (1966, 1972) обращает внимание,
напротив, на сильное развитие дополнительного мешка, имеющего бутылковидную
форму. Очевидно, для этого вида (как и для многих других среднеазиатских предста-
вителей семейства) характерна высокая изменчивость по данному признаку. Выходное
отверстие протока слизистых желез находится на уровне середины стрелы. Между по-
лостями стилофора, верхнего и нижнего отделов вагины имеется широкий поперечный
валик. Внутренняя поверхность вагины с необыкновенно мощными продольными
складками. В противоположность тому, что наблюдается у других видов, у которых
продольные складки, начинаясь в пределах нижнего отдела вагины, частью уходят
в яйцевод, а частью — в проток семеприемника, у данного вида внутренние стенки
яйцевода несут серию самостоятельных, чрезвычайно мощных складок, не связанных
со складками вагины (рис. 106). Эти складки обнаруживаются обычно даже без вскры-
тия вагины. Надклоачного пузыря нет.
Пенис длинный, при основании охвачен пигментированным чехлом. Внутреннее
строение пениса своеобразно: начинаясь от верхнего края пениального чехла, вверх
проходит глубоко врезанная зигзагообразная борозда, окаймленная парой мощных
валиков; этот комплекс виден при наружном осмотре как продольное бугристое взду-
тие на пенисе. В проксимальной части борозда и валики исчезают, разбиваясь на
множество крупных бугорков неправильной формы. Основание короткого протока
семеприемника расширено, но уже яйцевода.
Распространение и местообитания. Ферганский и Чаткаль-
ский хребты. Обитает во влажных кустарниковых зарослях близ воды.
23. Bradybaena stoliczkana (Nevill, 1878). Табл. IV, 33, 34; рис. 107—111.
N е v i 1 1, 1878, Yark. Miss. : 3, t. 1, fig. 4—6 (Helix); Martens, 1882a : 14,
t. 2, fig. 16 (Helix); Westerlund, 1898:161 [Helix (Eucampylaea) opposita];
Mo llendorf f, 1899 : 125 (Cathaica); Лихарев, 1955 : 183, рис. 1, Д, E, 2, 3
(Eulota pavlovskii); Матёкин, 1966 : 119, табл. 4, рис. 8 (Bradybaena pavlovskii);
1972 : 131, рис. 6, А, Б, В (Br. pavlovskii pars).
Материал: более 30 экз. из 3 местонахождений на Тянь-Шане.
Раковина от прижатой до почти плоской, с завитком, большей частью не
превышающим половины высоты устья. Вершина слегка выступающая; чем более
плоская раковина, тем сильнее выступает вершина. Оборотов около 5, сильно выпук-
лых, разделенных глубоким швом. Последний оборот в профиле округлый, иногда
с легким углом выше периферии, снизу не уплощен, к устью слегка очень плавно опу-
щен, в 1.5—2 раза шире предпоследнего. Окраска серовато-белая или кремовая,
иногда с очень легким голубоватым оттенком; имеются две коричневые спиральные
ленты, проходящие выше и ниже периферии; ленты развиты в разной степени, иногда
очень широкие, чаще узкие или умеренной ширины, прерваны светлыми промежут-
ками. Верхняя лента прослеживается у шва на более ранних оборотах. Эмбриональные
Рис. 107—111. Bradybaena stoliczkana (Nevill). 107—108 — южный берег оз. Иссык-
Куль, в 20 км к югу от Каджисая, 27 VII 1972. 109—111 — южный берег оз. Иссык-
Куль, берег р. Ак-Терек, 10 VII 1970. 107 и 109 — внешний вид полового аппарата;
108 и 111 — дистальная часть полового аппарата, вскрыта; 110 — проксимальная
часть пениса, вскрыта; звездочка — пакет со спермиями. (Ориг.).
BRADYBAENA 157
обороты гладкие или со следами чешуйчатой скульптуры; на дефинитивных оборотах
имеется грубая радиальная исчерченность и морщинистость, и местами вмятины и ред-
кие спиральные бороздки. Устье округлое, косое, внутри белое или коричневатое.
Места прикрепления устья сближены и соединены мозолью, верхний край устья от
ангулярной области сначала слабо изгибается кверху, затем, плавно выгибаясь, пе-
реходит в палатальный край. Края устья слабо отвернуты. Ширина пупка зависит
от ширины последнего оборота и от того, насколько он оттянут вправо (ср. фото 33
и 34 на табл. IV); однако в любом случае пупок развернутый и перспективный. Раз-
меры: ВР 4.7-9, БД 10—17, МД 8-14 мм..
Внутреннее строение. Материал: 5 экз. с южного берега оз. Иссык-
Куль, берег р. Ак-Терек, 10 VII 1970, сбор Л. В. Славошевской; 4 — с южного берега
оз. Иссык-Куль, в 20 км к югу от Каджисая, 27 VII 1972, сбор К. К. Увалиевой, опре-
деления наши; 2 — из долины р. Кум-Тар, 2 VIII 1934, сбор Д. Н. Кашкарова, опре-
деление И. М. Лихарева (голотип и паратип Eulota pavlovskti).
Спермовидукт прямой или извитой, но в нижней части его изгиб всегда имеется.
Слизистые железы состоят из 4—11 ветвей, объединяющихся непосредственно при
впадении в дополнительный мешок или стилофор, причем у одних экземпляров же-
лезы объединяются в два пучка, у других такое объединение почти не выражено. До-
полнительный мешок выражен в различной степени или редуцирован, проток слизистых
желез. впадает близ острия стрелы, которая почти целиком погружена в ткани вер-
шины стилофора. Следов надклоачного пузыря не обнаружено.
Дистальная часть пениса расширена, далее следует сужение, и в проксимальной
части вновь имеется вздутие, внутри которого у одного из экземпляров был обнаружен
пакет со спермиями (рис. 110). Чехол пениса у экземпляров из Каджисая был пол-
ностью слит со стенками пениса и как таковой не был выражен; в материале из до-
лины р. Кум-Тар наблюдалась в общем та же картина, но внутри расширенных сте-
нок в нижней части пениса имелась четкая кольцевая полость; наконец, у особей из
Ак-Терека был нормально выражен характерный для большинства представителей
рода пениальный чехол, фиксированный только у клоаки. Внутри пенис покрыт про-
дольными валиками, не образующими сколько-нибудь характерного узора. Основание
протока семеприемника расширено, проток конической формы.
Распространение и местообитания. Центральный Тянь-
Шань. Вид спорадически распространен, встречается на сухих каменистых или щеб-
нистых склонах на значительной высоте над уровнем моря.
24. Bradybaena helvola (L. Pfeiffer, 1853). Табл. IV, 35, 36\ рис. 112, 113.
L. Pfeiffer, 1853 : 188 (Helix)-, Лихарев, Раммельмейер, 1952 :
392 (Eulota schrencki part.); M а т ё к и н, 1966 : 127, табл. 4, рис. 5 (Bradybaena апа-
choretica); Увалиева, 1967 : 217, рис. 6, 7 (Br. likharevi)- М ат ё кин, 1972 :
127, рис. 4, А, Б, В (Br. anachoreticd).
Материал: более 30 экз. из 3 местонахождений на Алтае и Тарбагатае.
Раковина от прижатой до кубаревидной, с коническим или куполовидным
завитком, высота которого равна или меньше высоты устья. Оборотов 5.5—6. Послед-
ний оборот с очень легким углом на периферии, который порой едва угадывается;
этот оборот к устью не опущен или чуть опущен, не более чем в 1.5 раза шире пред-
последнего. Окраска построена по той же схеме, что и у трех предыдущих видов:
фон желтоватый или светло-каштановый, по периферии светлая лента, окаймленная
двумя темными лентами; верх и низ раковины темнее периферической полосы. Однако
нередко встречаются раковины, у которых интенсивность окраски периферической
ленты и фона примерно одинакова. Устье в той или иной степени скошено, края его
не сближены и отвернуты, кроме верхнего. Пупок почти не прикрыт, цилиндрический,
умеренно перспективный. Размеры: ВР 10—17, ВД 15—23, МД 14—20 мм.
Внутреннее строение. Материал: 1 экз. из окр. Усть-Каменогорска,
26 VIII 1969, сбор И. М. Хохуткина; 3 — из восточного Казахстана, Большенарым-
ский р-н, IX 1962, сбор К. К. Увалиевой; 2 — из'Тарбагатая, ущ. Сегизбей-булак
близ р. Каракол, V 1938, сбор П. А. Юматова; определения наши.
Нижняя часть спермовидукта не изогнута, но яйцевод скручен. Вагина тонкая,
прямая, длинная. Слизистые железы многобугорчатые, ветвящиеся, объединены
в 2 пучка, впадают в самую нижнюю часть стилофора, пронизывая надклоачный пу-
зырь. Последний хорошо развит и окутывает клоаку, основание стилофора и дисталь-
ный отдел пениса. Дополнительный мешок отсутствует; проток слизистых желез про-
ходит в стенке стилофора снизу вверх и впадает при основании стрелы, целиком по-
груженной в ткани вершины стилофора. Стилофор бугорчатый, с неровной поверх-
ностью, внутренние стенки его дистальной части покрыты отдельными мощными бу-
горками.
Чехол пениса необыкновенно мощный и длинный, охватывает почти 3/4 длины
пениса. Внутренняя поверхность пениальной трубки покрыта продольными склад-
158
II. BRADYBAENIDAE
ками, которые в проксимальной части становятся зигзагообразными. Эпифаллус ко-
роткий. Проток семеприемника почти цилиндрический, но все же можно заметить,
что его основание чуть большего диаметра, чем верхняя часть.
Распространение и местообитания. Тарбагатай и Алтай.
Вид описан по сборам Киндерманна из «Сибири»; однако в прошлом веке понятие
«Сибирь» было столь же обширно и неопределенно, как понятие «Туркестан»; во вся-
Рис. 112—113. Bradybaena helvola (L. Pfr.), Тарбагатай, ущелье Сегизбей-булак,
V 1938. 112 — внешний вид полового аппарата; 113 — дистальная часть полового
< аппарата, вскрыта. (Ориг.).
ком случае, большая часть Алтая относилась к Сибири. Обитает на кустарниковых
и травянистых склонах, на почве. При понижении влажности уходит под камни и
в почву.
25. Bradybaena pseudoferghanica Schileyko, sp. nov. Табл. IV, 37; рис. 114, 115,
Голотип хранится в Зоологическом институте АН СССР.
Материал: 16 экз. из 3 местонахождений на Джунгарском и Киргизском хребтах.
Раковина в различной степени прижатая: завиток — от почти плоского до
конического. Вершина выступающая. Оборотов 5—6, очень слабо выпуклых. По-
следний оборот не более чем в 1.5 раза шире предпоследнего, к устью сильно, но плавно,
без перегиба, опущен. Периферия угловатая, к устью угловатость сильно сглажи-
вается. Окраска от белесоватой с грязно-желтыми или рыжеватыми радиальными ко-
сыми полосами, края которых сильно размыты, до одноцветно желтой. Скульптура
в виде радиальных морщин и тонких спиральных линий, причем последние иногда
очень слабо выражены. Кроме того, на эмбриональных оборотах имеется очень тон-
кая сглаженная зернистость. Устье сильно скошено, края его слегка отогнуты, ост-
рые, места их прикрепления сильно сближены. Копумеллярный край слегка нави-
сает над пупком, но не закрывает его. Пупок развернутый, умеренно широкий, перс-
пективный. Размеры: ВР 9—12, БД 18—22, МД 15—17 мм. Голотип: ВР 9.3, БД 12.7,
МД 16.5 мм.
На Тянь-Шане обитают 3 вида, относящихся к трем крупным группировкам, ра-
ковины которых различить трудно. Это помимо описанного вида Angiomphalia copiosa
1
Рис. 114—115. Bradybaena pseudoferghanica sp. nov., Джунгарский xp.
верховья p. Чарса-чай, 12—15 VI 1969. Паратип. 114 — внешний вид
,полового аппарата; 115 — дистальная часть полового аппарата, вскпыта.
(Ориг.).
160
II. BRADYBAENIDAE
sp. nov. и Leueozonella ferghanica (Lindholm). Изучение типового материала по опи-
санному Линдгольмом виду показало, что название ferghanica надо оставить за пред-
ставителем рода Leueozonella (Hygromiidae, Trichiinae). Что касается Br. pseudofergha-
nica, то он отличается конхологически наличием спиральных бороздок и более резко
выступающей вершиной. Под названием Br. ferghanica этот вид фигурирует в работах
П. В. Матёкина (1966, 1972).
Внутреннее строение. Материал: 1 экз. из Джунгарского хр., вер-
ховья р. Чарса-чай, 12—15 VI 1969, сбор И. С. Даревского; 2 — из Киргизского хр.,
близ с. Чардовар, 6 VI 1972, сбор и определения наши.
Спермовидукт и яйцевод крутых изгибов не образуют. Слизистые железы двумя
то более, то менее сближенными вторично ветвящимися кустами впадают в нижнюю
треть стилофора, пронизывая покровы надклоачного пузыря; на исследованном ма-
териале дополнительного мешка не обнаружено, но П. В. Матёкин (1972) указывает,
что в пределах одной популяции наблюдается сильная изменчивость степени выра-
женности дополнительного мешка вплоть до полного исчезновения. .Пройдя сквозь
надклоачный пузырь, протоки слизистых желез проникают в стенку стилофора и,
пройдя внутри нее некоторое расстояние, впадают в нижнюю часть (не выше сере-
дины) полости стилофора. Стрела занимает немного более половины длины стилофора
и окружена при основании небольшим воротничком.
Чехол пениса хорошо развит, охватывает немного больше 1/3 пениса. Характер-
ная особенность вида заключается в присутствии на пенисе небольшого слепого вы-
роста — цекума. Внутренняя поверхность пениса в нижней части несет редкие мощ-
ные продольные валики, выше цекума — тонко гофрированные продольные складки.
Эпифаллус ясно отграничен от пениса, цилиндрический. Основание протока семепри-
емника значительно расширено.
Распространение и местообитания. По данным П. В. Матё-
кина (1972), распространен от Тарбагатая до Ферганского и Алайского хребтов, за-
хватывая и южные отроги Таласского хр. На Заилийском хр., вероятно, отсутствует.
Ареал нуждается в уточнении. Обитает на каменистых склонах, поросших кустарни-
ком, и в крупнообломочных осыпях.
26. Bradybaena fedtschenkoi (Martens, 1874). Табл. IV, 39; рис. 116—119.
Мартенс, 1874 : 16, табл. 1, рис. 9 (Helix); Westerlund, 1889 : 342
(Helix); Kobelt, 1893, Iconogr., N. F., VI : 75, fig. 1086 (Helix sturanyana); Gude,
1904 : 93 [Cathaica (Eucathaica) sturanyi]; Линдгольм, 1931 : 58 [Cathaica (Cam-
pylocathaica)]; Лихарев, Раммельмейер, 1952 : 380 [Cathaica (Cathaica)];
Матёкин, 1966 : 121; 1972 : 134.
Материал: более 40 экз. из трех местонахождений на Ферганском, Зеравшанском
и Кураминском хребтах.
Раковина прижатая, высота завитка меньше высоты устья, завиток кони-
ческий или куполовидный, но вершина всегда несколько выступающая. Оборотов
около 5, сильно или умеренно выпуклых, медленно и плавно нарастающих. Послед-
ний оборот примерно в 1.5 раза шире предпоследнего, к устью чуть опущен у самого
конца, в профиле правильно округлый. Окраска верхней поверхности свежих рако-
вин светло-коричневая с желтоватым оттенком, нижняя поверхность окрашена свет-
лее, вокруг пупка почти белая. Скульптура — тонкая и неравномерная радиальная
исчерченность, нередко раковина почти гладкая. Устье округлое, умеренно или
сильно скошено, края его в различной степени отвернуты, но чаще почти прямые,
кроме колумеллярного; пупок от развернутого до развернуто-цилиндрического, но
он всегда полностью перспективный. Размеры: ВР 6—10, БД 12—20, МД 10—17 мм.
Внутреннее строение. Материал: 12 экз. с берега оз. Маргузор на
Зеравшанском хр., 8 VIII 1973, сбор Ю. А. Мешкова; 2 — из с. Бободархон на Кура-
минском хр., 22 V 1972, сбор А. Мухитдинова; определения наши.
Нижняя часть спермовидукта и яйцевод образуют 1—3 крутых изгиба. Слизи-
стые железы в виде одного куста, состоящего из нескольких мощных ветвей с бугри-
стыми стенками. Дополнительный мешок в значительной степени редуцирован, од-
нако присутствие его всегда угадывается: вокруг единого ствола слизистых желез
имеется заметная припухлость стенки стилофора. Этот последний развит хорошо,
вершина его несколько утолщена. Стрела полностью залегает в канале верхней
части стилофора. Вагина и основание стилофора охвачены плотным надклоачным пу-
зырем. Внутренние стенки вагины покрыты продольными мощными валиками, разби-
тыми, полностью или частично, на ряды бугорков. Полости стилофора и вагины со-
общаются с полостью клоаки щелевидным отверстием благодаря присутствию попе-
речного, хорошо развитого валика, резко сужающего просвет нижней части полости
вагины.
Пенис довольно длинный, чехол пениса, напротив, короткий, занимает не более
1/s длины пениальной трубки. Внутренняя поверхность последней несет узкие, но
высокие продольные складки, которые в проксимальной части становятся зигзаго-
BRADYBAENA
161
образными. Эпифаллус длинный и очень тонкий — такой же длины, как и пенис, или
немного короче, и почти такой же толщины, как семепровод. Основание протока семе-
приемника по сравнению с верхней частью этого протока резко расширено.
Рис. 116—118. Bradybaena fedtschenkoi (Mart.), Зеравшанский хр., берег оз. Маргузор,
8 VIII 1973. 116 — внешний вид полового аппарата; 117 — дистальная часть жен-
ского отдела, вскрыта; 118 — пенис, вскрыт. (Ориг.).
Распростри не ние и местообитания. Зеравшанский, Алай-
ский, Заалайский, возможно, Чаткальский хребты, вершина Дигмайского плато,
предгорья Кураминского и Туркестанского хребтов. Обитает под камнями, среди
гниющих растений и под корнями многолетних трав и кустарников (Мухитдинов, 1972).
27. Bradybaena cavimargo cavimargo (Martens, 1879). Табл. IV, 38; рис. 119.
Martens, 1879, Sitzungsb. naturf. Fr. Berlin : 126 (Helix)' 1882a : 15, t. 2,
fig. 17 (Helix); Westerlund, 1896:187 [Helix (Jacosta) aspasta]; M 6 1 len-
der f f, 1899 : 116, t. 7, fig. 5 [Cathaica iacosta]; Лихарев, Раммельмейер,
1952 : 385, рис. 315 [Cathaica (?)]; Матёкин, 1966 : 120, табл. 4, рис. 2, 3; 1972 :
132, рис. 6, Ж, 3.
Материал: 18 экз. из 3 местонахождений на Алайском, Киргизском и Заилий-
ском хребтах.
Раковина прижатая, высота завитка не превышает высоту устья, иногда
составляет половину этой величины. Оборотов 5—6, медленно нарастающих. Послед-
ний оборот не более чем в 1.5 раза шире предпоследнего, к устью заметно опущен,
вершина выступающая. Обороты в профиле имеют характерные очертания: верхняя
сторона каждого оборота у шва плоская и почти горизонтальная, затем плавно, но
быстро перегибается вниз, вновь становится почти горизонтальной и немного выше
периферии образует второй угол, после чего плавно дугообразно изгибается, пере-
ходя в базальную часть. Нижний угол более резкий и может приобретать килевидный
облик; у молодых экземпляров оба угла выражены лучше, чем у взрослых, так как
11 А. А. Шилейко
162
II. BBADYBAENIDAE
к устью они заметно сглаживаются. Окраска от светло-серой, почти белой, до светло-
каштановой, на верхней поверхности раковины окраска более интенсивная; по ниж-
Рис. 119. Bradybaena cavimargo са-
vimargo (Mart.), Заилийский . хр.,
пос. Джандосова, 2 VIII1971. Внеш-
ний вид полового аппарата не вполне
нему углу проходит узкая светлая лента.
Скульптура на эмбриональных оборотах от-
сутствует или представлена очень тонкой
неясной зернистостью, на дефинитивных име-
ется очень тонкая радиальная исчерченность,
а также спиральные бороздки, которые об-
наруживаются местами. Устье округлое,
косое, края его тонкие, острые; базальный
и колумеллярный края умеренно отвернуты.
Места прикрепления устья в той или иной
степени сближены. Пупок развернутый, уме-
ренно широкий, перспективный. Размеры:
ВР 5—7, ВД 11—15, МД 9—12.5 мм.
Внутреннее строение. При-
водимое описание составлено в основном
по данным П. В. Матёкина (1966, 1972) и до-
полнено собственными наблюдениями по од-
ному не вполне взрослому экземпляру (За-
илийский хр., Каскеленский р-н, пос. Джан-
досова, 2 VIII 1971, сбор К. К. Увалиевой,
определение наше).
Нижняя часть спермовидуктэ крутого
изгиба не образует. Вагина длинная, сти-
лофор закладывается довольно высоко, но у
взрослых особей впадает немного выше
клоаки. Дополнительный мешок втиснут
между стилофором и стенкой вагины. Сли-
зистые железы в виде 2—3 ветвей впадают в
дополнительный мешок апикально. Пенис
взрослой особи. (Ориг.). длинный, прямой, у нашего экземпляра в
нижней четверти пениса наблюдалось форми-
рование пениального чехла. Эпифаллус выражен плохо. Основание протока семе-
приемника умеренно расширено.
Распространение и местообитания. Центральный и восточ-
ный Тянь-Шань и Гиссаро-Алай. Обитает на степных склонах в почве и мелкообло-
мочных осыпях. Встречается отдельными изолированными популяциями.
27а. Bradybaena cavimargo tarbagataica Matiokin, 1966.
M а т ё к и н, 1966 : 120; 1972 : 134.
Материал нами не исследован.
Согласно первоописанию, отличается от номинативного подвида более плавными
обводами завитка; оба угла в значительной степени сглажены.
Распространение и местообитания. Тарбагатай. Условия
жизни те же, что и у номинативного подвида.
28. Bradybaena tomyris (Lindholm, 1927). Табл. IV, 40.'
Lindholm, 1927с : 196 (Fruticicola); Лихарев, Раммельмейер,
1952 : 406 [Eulota (Acusta.?)].
Материал: единственный типовой экземпляр из Ферганского хр.
Раковина кубаревидная, умеренно тонкостенная, слегка просвечивающая;
высота завитка немного меньше высоты устья. Завиток конический, вершина при-
тупленная. Оборотов немного более 5, слабо выпуклых, медленно нарастающих. По-
следний оборот в профиле правильно округлый, к устью несколько опущен. Окраска
почти одноцветная, но в пределах раковины наблюдаются переходы от оливкового
до буровато-желтого цветов. Скульптура в виде грубой и неравномерной радиальной
исчерченности. Устье круглое, скошено. Места его прикрепления широко расставлены
и связаны тонкой мозолью. Верхняя часть палатального края устья острая, палаталь-
ный и базальный края немного, а колумеллярный — сильно отвернуты. Пупок узкий,
цилиндрический, немного менее чем наполовину прикрыт отворотом колумеллярного
края. Размеры: ВР 17.5, БД 22.2, МД 18.9 мм.
Внутреннее строение неизвестно.
Распространение. Единственный экземпляр найден на Ферганском хр.
в долине р. Чичкан. И. М. Лихарев и Е. С. Раммельмейер (1952) полагают, что этот
BRADYBAENA
16'3
вад может принадлежать дальневосточной группе Acusta. Однако у соответствующих
дальневосточных видов раковина значительно более тонкостенная, обороты нара-
стают значительно быстрее. Кроме того, это предположение несостоятельно с зоогео-
графической точки зрения.
29. «Bradybaena» boevi Uvalieva, 1967.
Увалиева, 1967 : 216, рис. 4, 5.
Материал: одна сформированная, слегка дефектная раковина (голотип) и одна
раковина молодого экземпляра.
Раковина слегка прижатая, высота завитка меньше высоты устья. Оборо-
тов 5.5, выпуклых, медленно нарастающих, разделенных умеренно глубоким' шром.
Последний оборот менее чем в 1.5 раза шире предпоследнего, к устью незначительно
и плавно опущен. Окраска красновато-коричневая, по периферии проходит широкая
(1.2 мм) молочно-белая лента. Скульптура в виде густой неравномерной радиальной
исчерченности и вялых спиральных бороздок. Устье почти круглое, косое; края, его
тонкие, слегка отвернутые, губа отсутствует. Пупок цилиндрический, но перспектив-
ный, не прикрыт отворотом колумеллярного края. Размеры голотипа: ВР 12, БД
17.3 мм.
Внутреннее строение. Приводится по первоописанию.
Спермовидукт короткий, яйцевод, напротив, длинный, цилиндрический. Две
слизистые железы, одна из которых близ вершины раздвоена, имеют ячеистые стенки,
впадают в вагину выше стилофора единым протоком. Стилофор массивный, цилиндри-
ческий. Пенис вздут, переходит в короткий эпифаллус. Имеется длинный бич, слегка
утолщенный в центральной части. Чехол пениса отсутствует. Проток семеприемника
при основании не вздут, тонкий, заканчивается небольшим овальным резервуаром.
Внутреннее строение гениталий неизвестно.
Несмотря на несовершенство описания и рисунка, можно быть уверенным, что
данный вид не относится к роду Bradybaena и, скорее всего, не принадлежит к се-
мейству Bradybaenidae.
Поводом для отнесения вида к данному семейству послужило наличие у вида
слизистых желез с ячеистыми стенками. Однако много других, гораздо более серьез-
ных признаков сближают этот вид с представителями подсемейства Hygromiinae
семейства Hygromiidae. К таким признакам относятся: наличие бича; впадение сли-
зистых желез в вагину, а не в стилофор; отсутствие чехла пениса; цилиндрическая
форма протока семеприемника; к тому же неизвестен характер крепления резервуара
семеприемника — к спермовидукту или к нижней стенке мантийной полости.
Вид, несомненно, своеобразен, но решить его систематическую принадлежность
можно будет только после достаточно тщательного исследования.
Распространение и местообитания. Вид найден на южном
Алтае, к северу от с. Алтай, на лесистом склоне среди трав.
f
III. Семейство HYGROMIIDAE Tryon, 1866
Раковина первично небольших или средних размеров, коричневатых тонов, с раз-
мытой светлой лентой на периферии и нешироким цилиндрическим пупком. Однако
разнообразие формы раковины и других конхологических признаков очень велико.
Стилофоров первично две пары — верхняя и нижняя, но число их может сокра-
щаться до 2, 1 или они могут редуцироваться полностью. Иногда стилофоры видоизме-
нены в вагинальные придатки, которых может быть 2 или 1. Слизистые железы рас-
полагаются на вагине выше стилофора, с плотными гладкими стенками, ветвятся
в различной степени, изредка отсутствуют. Семеприемник без дивертикула, резервуар
относительно массивный, овальный, молотообразный или мешковидный, прилегает
к спермовидукту и со стенкой мантийной полости не связан. Проток семеприемника
цилиндрический, в нижней части чаще всего не расширен. Бич и папилла пениса
первично имеются, но способны редуцироваться или видоизменяться. Папилла об-
разована за счет разрастания тканей вокруг выходного отверстия эпифаллуса. .
Распространение. Большая часть территории Палеарктики, многие
виды завезены в страны Нового Света. Наибольшее число видов — в Южной Европе
и на Кавказе. В ископаемом состоянии представители семейства встречаются начиная
с верхнего олигоцена (Европа).
I. Подсемейство TRICHIINAE Zilch et Jaeckel, 1962
Раковина небольшая или средних размеров, пупок никогда не бывает полностью
запаянным, поверхность раковины часто с волосками. Скульптура разнообразная.
Стилофоров 4, расположенных попарно, симметрично относительно вагины.
Верхняя пара лишена стрел и иногда редуцирована до такой степени, что обнару-
164
III. HYGROMIIDAE
живается только при вскрытии. В исключительно редких случаях стилофоров одна
пара, но они все равно располагаются по обеим сторонам вагины. Папилла пениса
от простой трубки до сложного органа, в стенках которого может залегать от 1 до 9
интрапапиллярных полостей, сообщающихся с полостью чехла пениса посредством
папиллярной лакуны.
Распространение. Европа, Средиземноморье с Кавказом и Средняя
Азия. В ископаемом состоянии подсемейство известно начиная с верхнего олигоцена
(Европа).
1. Род NANAJA1 Schileyko, gen. nov.
Раковина прижатая, до почти плоской; светло-коричневая или светло-роговая.
Пупок развернутый, перспективный. На поверхности раковины имеются волоски,
которые с возрастом могут опадать.
Правая и левая пары стилофоров сближены друг с другом на одной стороне ва-
гины. Папилла пениса без интрапапиллярных полостей и без папиллярной лакуны.
Вагинальные валики охватывают петлеобразно выходные отверстия стилофоров.
Ретрактор правого омматофора образует перехлест с половым ретрактором.
Род близок к роду Leucozonella, но у представителей этого рода стилофоры не
сближены на одной из сторон, вагинальные валики, как правило, не охватывают вы-
ходных отверстий стилофоров.
Распространение. Западный Тянь-Шань.
Типовой вид —• Nanaja cumulata Schileyko, sp. nov.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА NANAJA
1 (2). Сквозь пупок просматривается последний оборот. Большой диаметр 14—17 мм
.........................................................1. N. cumulata.
2 (1). Сквозь пупок просматриваются все обороты. Большой диаметр 10—12 мм . . .
....................................................2. N. illuminata.
1. Nanaja cumulata Schileyko, sp. nov. Табл. V, 41; рис. 120—123.
Голотип хранится в коллекции Зоологического института АЦ СССР.
Материал: 17 экз. из одного местонахождения в долине р. Пскем.
Раковина в различной степени прижатая, высота завитка не превышает вы-
соту устья или меньше его, часто значительно. Вершина выступающая. Оборотов 6,
умеренно выпуклых. Последний оборот в 1.5 раза шире предпоследнего, в начале
угловатый, к концу оборота угол сглаживается; к устью последний оборот слабо и
плавно в самом конце опущен. Окраска раковины — почти белый фон и чередующиеся
светло-роговые и кремовые широкие радиальные полосы, причем границы их крайне
неопределенны и число их непостоянно. В случае интенсивного развития полос свет-,
лый фон плохо заметен. По углу проходит размытая светлая лента. Скульптура со-
стоит из неправильных радиальных морщин, очень тонкой спиральной исчерченности,
выраженной местами, и многочисленных рубцов от волосков. На раковинах взрослых
экземпляров волоски почти всегда отсутствуют. Устье заметно скошено, края его
острые, не отвернуты, утолщены, но губа, если она есть, развита только в базальной
части. Колумеллярный край слегка отвернут. Пупок умеренно развернутый в начале
и цилиндрический в глубине; сквозь него просматривается весь предпоследний оборот
или более. Размеры: ВР 6.8—9.3, БД 14.4—17.0, МД 12.3—14.1 мм. Голотип: ВР 8.7
БД 16.5, МД 14.0 мм.
Изменчивость вида касается в основном относительной высоты завитка и распре-
деления кремовых поперечных полос.
Вид по раковине трудно отличим от Archaica heptapotamica и характеризуется
в основном развитием радиальных цветных полос и более ясной угловатостью по-
следнего оборота.
Внутреннее строен и'е. Материал: 12 экз. из типовой серии — за-
падный Тянь-Шань, верховья р. Пскем, правый берег, выше с. Нанай, 21 V 1972,
.сбор и определение наши.
Спермовидукт в дистальной части образует резкий изгиб. Вагина тонкая. СЖ
.3x2—3. Стилофоры сближены, так что спереди вагина не видна, а сзади она обна-
руживается в виде канала, проходящего по задней стороне стилофоров. Места впа-
. дения слизистых желез спереди закрыты стилофорами.
Пенис удлиненный, ненамного превышает длину вагины, эпифаллус почти та-
йкой же длины, как и пенис, бич несколько длиннее. Папилла пениса не заключает
1 Название дано в соответствии с названием населенного пункта, наиболее близ-
кого к месту находки представителей рода,— с. Нанай.
NANAJA
165
интрапапиллярных полостей и заполнена рыхлой паренхиматозной тканью. Между
семепроводом, эпифаллусом и пенисом имеется тонкая прозрачная перепонка. Проток
семеприемника отходит непосредственно за местом впадения слизистых желез и пов-
торяет изгиб спермовидукта. Небольшой округлый резервуар заметно не достигает
белковой железы.
Распространение и местообитания. Вид встречен в крупно-
обломочных малоподвижных осыпях в долине р. Пскем на западном Тянь-Шане.
2. Nanaja illuminata Schileyko, sp. nov. Табл. V, 42‘, рис. 124—126.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 4 экз. из одного местонахождения в долине р. Пскем.
Раковина почти плоская, с едва возвышающимся завитком и сглаженной
или чуть выступающей вершиной. Оборотов 5.5, уплощенных, медленно нарастаю-
Рис. 120—123. Nanaja cumulata sp. nov., верховья p. Пскем, 21 V 1972. Паратип.
120 — внешний вид полового аппарата; 121 — пенис, вскрыт; 122 — вагина, вскрыта;
123 — дистальная часть женского отдела, вид с другой стороны. (Ориг.).
щих. Последний оборот снизу явственно уплощен, с углом выше средней линии, ко-
торый заметно сглаживается к устью. Последний оборот в 1.5 раза шире предпослед-
него, к устью еле опущен. Окраска роговая, по углу проходит размытая светлая лента.
Свежие и не вполне сформированные раковины покрыты густыми, умеренно длинными
волосками. Скульптура в виде равномерных радиальных морщинок, спиральные
бороздки отсутствуют. Кроме того, после отпадения волосков остаются хорошо за-
метные рубцы; в связи, с тем, что радиальная морщинистость хуже выражена на ниж-
ней поверхности раковины, следы от волосков здесь выражены яснее, чем на верхней
поверхности. Устье полулунно-овальное, ширина его заметно превышает высоту.
Края устья острые, базальный край чуть отвернут, колумеллярный заметно отвернут.
Губа отсутствует или выражена очень слабо. Пупок развернутый, перспективный,
сквозь него просматриваются все обороты. Размеры: ВР 5—5.5, БД 10.9—11.6, МД
9.7—10 мм. Голотип: ВР 5.0, БД 10.9, МД 9.8 мм.
166
III. HYGROMIIDAE
Вид близок к предыдущему, отличается меньшими размерами, более резко вы-
:,раженной угловатостью последнего оборота и уплощенной базальной частью послед-
него оборота.
В.н утреннее строение. Материал: 2 экз. из долины р. Пскем выше
с. Нанай, 21 V 1972, сбор и определение наши.
„г : /Спермовидукт и яйцевод изгибов не образуют. СЖ 4X1—2. Стилофоры массив-
ные, .располагаются так же, как у предыдущего вида. Нижний отдел вагины очень
короткий.
Рис. 124—126. Nanaja illuminata sp. nov., долина p. Пскем, 21 V 1972. Голотип.
124 — внешний вид полового аппарата; 125 — пенис, вскрыт; 126 — вагина,
вскрыта. (Ориг.).
Пенис очень небольшой, в стенках пениального чехла имеется пениальная же-
леза. Самый длинный участок дистальной части мужского отдела — эпифаллус; бич
заметно короче, пенис еще короче. Проток семеприемника крутых изгибов не обра-
зует, резервуар небольшой, прилегает примерно к средней части спермовидукта.
Распространение и местообитания. Вид известен только из
типового местообитания: верховья р. Пскем между Пскемским и Угамским хребтами
выще с. Нанай. Обнаружен в лесистом узком ущелье, в лиственном опаде среди кам-
ней у ручья.
2. Род ODONTOTREMA Lindholm, 1927
Ы n d h о 1 m, 1927с : 228; Лихарев, Раммельмейер, 1952 : 409;
Wenz, Zilch, 1959—1960 : 646; S с h i 1 е у к о, 1977:7.
Раковина прижатая, коричневая, небольшая. В устье имеется два бугорковидных
зуба на базальной стенке.
Стилофоры булавовидные, слабо спаянные между собой и не утолщенные в ба-
зальной части. Вагинальные валики образуют небольшие лопасти в местах выхода
стилофоров и продолжаются в клоаку. Интрапапиллярные полости и папиллярная
лакуна отсутствуют, но на поверхности пениса имеется продольная борозда. Ретрак-
тор правого омматофора образует перехлест с половым ретрактором.
Распространение. Чаткальский хр.
Монотипический род.
Типовой вид — Odontotrema dipl.odon Lindholm,. 1927.
ODONTOTREMA
167
Odontotrema diplodon Lindholm, 1927. Табл. V, 43; рис. 127—129.
Lindholm, 1927c : 268; Лихарев, Раммельмейер, 1952 : 409,
рис. 341; Матёкин, 1954:90 (Trichia); 1966:123 (Trichia); Schileyko,
1977 : 7, figs 5—9, Pl. I, 1.
Материал: более 50 экз. из 3 местообитаний на Чаткальском хр.
Раковина сверху и снизу почти одинаково выпуклая; высота куполовидного
завитка не превышает высоты устья. Оборотов 5 или немного больше. Последний обо-
рот немного шире предпоследнего, немного выше средней линии с тупым углом, кото-
рый к устью сглаживается; угловатость имеется также вокруг пупка. К устью пос-
Рис. 127—129. Odontotrema diplodon Lindh., Сары-Челекский запов., Чаткальский хр.,
V 1972. 127 — внешний вид полового аппарата; 128 — пенис, вскрыт; 129 — вагина,
вскрыта. (По Schileyko, 1977).
ледний оборот незначительно опущен. Окраска в общем коричневая, с красноватым
или серым оттенком. Скульптура — густая и грубая радиальная исчерченность;
имеются многочисленные загнутые волоски, на периферии эти волоски несколько длин-
нее, нередко они опадают. Устье слегка ромбическое или в виде широкой изогнутой
щели, косое, внутри коричневое, с губой. Зубов 2: бугорковидный базально-палаталь-
ный и маленький базально-колумеллярный, представляющий собой радиальное
утолщение губы. Последний иногда отсутствует или может быть очень слабо развитым.
Колумеллярный край устья слегка отвернут. Пупок умеренно широкий, цилиндри-
ческий, почти перспективный. Размеры: ВР 4—5.5, БД 7—10, МД 7—8.5 мм.
Внутреннее строение. Материал: 14 экз. из Сары-Челекского запо-
ведника, V и X 1972, сбор В. В. Горохова, определение наше.
Нижняя часть спермовидукта изгиба не образует. СЖ 4X1. Стилофоры тонкие
и длинные, внешние спаяны с внутренними при основании, внутренние с вагиной почти
не спаяны. Вагина длинная, веретеновидная; внутри вагины имеются слабые вагиналь-
ные валики.
Бич несколько короче эпифаллуса; последний дважды изогнут и фиксирован в та-
ком положении мускульными лентами, входящими в состав соединительнотканной
перепонки. Пенис относительно очень массивный, веретеновидный, внутренняя по-
верхность его несет ветвящиеся, слегка гофрированные продольные складки. Папилла
пениса в общем цилиндрическая, с глубокой продольной бороздой, дистально сходя-
щей на нет. Концевая часть папиллы отграничена неполной кольцевой бороздой, выра-
женной в различной степени. Кроме семепроводящего канала, папилла никаких
полостей не заключает. На поверхности пениса имеются слабые продольные
168
III. HYGROMIIDAE
соединительнотканные ленты. Проток семеприемника не имеет резких изгибов
и плавно переходит в резервуар удлиненно-овальной формы, немного не достигающий
нижнего края белковой железы и плотно вжатый в поверхность спермовидукта.
Распространение и местообитания. Северо-восточная часть
Чаткальского хр. Населяет сырую подстилку кустарников и лиственных лесов, часто
вблизи воды.
3. Род LEUCOZONELLA Lindholm, 1927
Lindholm, 1927с : 263; Лихарев, Раммельмейер, 1952 : 401
[Eulota (Leucozonella) ]; Wenz, Zilch, 1959—1960 : 642 [Bradybaena (Leucozonella)]-,
Schileyko, 1977 : 8.
Раковина от шаровидной до прижатой; в последнем случае она по периферии мо-
жет быть слегка угловатой. Окраска светло-серых, желтоватых, бурых или красно-
ватых тонов; по периферии проходит светлая лента, неясно выраженная только в том
случае, если фон светлый. Пупок от точковидного до развернутого, перспективного.
Стилофоры не сближены на одной стороне вагины, располагаются симметрично,
основания их не вздуты. Папилла пениса с продольной бороздой, развитой в разной
степени, либо борозда замыкается, образуя интрапапиллярную полость, которая со-
общается с полостью пениального чехла посредством папиллярной лакуны; либо, на-
конец, интрапапиллярная полость вторично исчезает, но остается папиллярная ла-
куна (подрод Narina). Ретрактор правого омматофора образует перехлест с половым
ретрактором.
Распространение. Средняя Азия. В ископаемом состоянии род изве-
стен начиная с плейстоцена.
Типовой вйд — Helix rubens Martens, 1874.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА LEUCOZONELLA
1 (2). Раковина покрыта радиальными ребрами. (Подр. Narina) . 10. L. crassicosta.
2 (1). Радиальные ребра отсутствуют.
3 (4). Раковина двуцветная: выше периферии серая, ниже периферии роговая.
(Подр. .Narina)..........................................9. L. hypophaea.
4 (3). Раковина одноцветная или со светлой лентой на периферии.
5 (10). Раковина шаровидная.
6 (7). Раковина толстостенная, белая, верхние обороты немного темнее послед-
него, обычно с голубоватым или розоватым оттенком. Края устья слегка утол-
щены. (Подр. Leucozonella)....................................2. L. caryodes.
7 (6). Раковина тонкостенная или умеренно толстостенная, от желтоватой до крас-
новато-коричневой, верхние обороты окраской не выделяются. Края устья не
утолщены.
8 (9). Большой диаметр не более 12 мм. (Подр. Leucozonella) . . 4. L. mesoleuca.
9 (8). Большой диаметр не менее 14 мм. (Подр. Leucozonella) .... 3. L. rubens.
10 (5). Раковина от кубаревидной до прижатой.
11 (12). Раковина покрыта волосками. Большой диаметр не превышает 10 мм. (Подр.
Leucozonella) .............................................8. L. caria. .
12 (11). Раковина без волосков; если же имеются тонкие редкие волоски, то большой
диаметр не менее 17 мм.
13 (16). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно
не более половины предпоследнего оборота.
14 (15). Раковина прижатая. Последний оборот в профиле угловатый, светлая лента
по периферии четкая. (Подр. -Leucozonella).....................5. L. rufispira.
15 (14). Раковина кубаревидная. Последний оборот в профиле не угловатый или
с едва заметным углом, светлая лента по периферии выражена в разной степени.
(Подр. Leucozonella) ....................................7. L. angulata.
16 (13). Если оптическая ось глаза совмещена с осью пупка, сквозь пупок видно не
менее целого оборота.
17 (18). Раковина тонко радиально исчерчена. (Подр. Leucozonella) . . 6. L. retteri.
18 (17). Радиальная скульптура представлена грубой неравномерной морщинистостью.
(Подр. Leucozonella) ............................1. L. ferghanica.
1. Подрод LEUCOZONELLA s. str.
Верхняя и нижняя части раковины окрашены примерно одинаково интенсивно
или нижняя сторона немного светлее верхней; поверхность раковины в различной
степени скульптурирована, но радиальных ребер не бывает.
Пенис довольно массивный; папилла пениса простая или по ее поверхности про-
ходит борозда, которая иногда замыкается почти на всем протяжении, кроме самой
проксимальной части; эта открытая часть борозды представляет собой папиллярную
лакуну.
LEUCOZONELLA
169
Распространение. Тянь-Шань и отроги Памира. В ископаемом состоянии
подрод известен начиная с плейстоцена.
1. Leucozonella (Leucozonella) ferghanica (Lindholm, 1927). Табл. V, 44; рис. 130—
132.
L i n d h о 1 m, 1927c : 264 (Cathaica); Лихарев, Раммельмейер,
1952 : 381 (Cathaica); Schileyko, 1977 : 8, figs 10—15, Pl. I, 2.
Материал: 22 экз. из 2 местонахождений на Чаткальском хр.
Раковина прижата в различной степени, твердостенная, со слегка куполо-
видным или коническим, несильно выдающимся завитком и выступающей вершиной.
Оборотов 5.5—6, медленно нарастающих, сверху слабо, снизу более сильно выпуклых.
Рис. 130—132. Leucozonella (Leucozonella) ferghanica (Lindh.), Сары-Челекский запов.,
Чаткальский хр., 6 VII 1966. 130 — внешний вид полового аппарата; 131 — вагина,
вскрыта; 132 — пенис, вскрыт. (По Schileyko, 1977).
Последний оборот в 1.5—2 раза шире предпоследнего, немного опущен, по периферии
округлый или с очень легким углом немного выше средней линии; к устью угол исче-
зает. Окраска от светло-желтой до светло-роговой, одноцветная или с неясными ради-
альными полосами; светлая лента по периферии порой лишь угадывается.
Эмбриональные обороты гладкие; дефинитивные покрыты грубой радиальной исчер-
ченностью или морщинистостью, причем на верхней поверхности раковины скульптура
грубее, чем на нижней. Спиральные бороздки очень тонкие, выражены местами. Устье
овальное, сильно скошено; края его тонкие и острые; несколько отступя от края часто
присутствует сильно расплывшаяся губа. Места прикрепления устья умеренно сбли-
жены. Пупок очень развернутый, вполне перспективный. Размеры: ВР 9.5—12, БД
17—22, МД 14-18 мм.
Внутреннее строение. Материал: 2 экз. из Чаткальского хр., берег
оз. Кыла-Куль в Сары-Челекском заповеднике, 6 VII 1966, сбор А. И. Янковской,
определение И. М. Лихарева; 7 — из окрестностей Сары-Челекского заповедника,
.22 X 1972, сбор В. В. Горохова, определение наше.
Яйцевод образует два очень крутых изгиба, края которых плотно сомкнуты.
Длина прямой части вагины от изгиба до стилофоров в 6—7 раз превышает ее ширину.
СЖ 2X3—4. Стилофоры располагаются в нижней части вагины, шаровидные, область
стилофоров отделена от нижней части вагины неясным пережимом.
Пенис сильно вздут, массивный, веретеновидный. Папилла пениса прямая или
изогнутая в разной степени; борозда на ее поверхности неглубокая, занимает прокси-
170
III. HYGROMIIDAE
мальную часть папиллы. Ретрактор пениса недлинный и мощный. Проток семеприем-
ника почти не извит, заканчивается небольшим резервуаром неопределенно округлой
формы, немного не доходящим до белковой железы.
Распространение и местообитания. Северо-восточные склоны
Чаткальского хр. и окрестности Намангана. Населяет подстилку горных лесов, обычно
близ воды; изредка встречается на открытых местах, но в пределах биотопов с постоян-
ными влажными укрытиями (крупнообломочные осыпи большой мощности).
2. Leucozonella (Leucozonella) caryodes (Westerland, 1896). Табл. V, 45; рис. 133—
136.
Westerland, 1896 : 186 [Helix [Eulota) rubens var. caryodes]-, Lindholm,
1927b : 197 (Fruticicola conospira); Лихарев, Раммельмейер, 1952 : 401
(Eulota rubens part.); Матёкин, 1954 : 90 (Trichia conospira)- 1966 : 123, табл. 5,
рис. 8 (Tr. conospira)', Schileyko, 1977 : 9, figs 16—20, Pl. I, 3.
Материал: более 100 экз. из 12 местонахождений на Тянь-Шане.
Раковина почти шаровидная, толстостенная, с высоким куполовидным за-
витком; иногда высота раковины даже слегка превышает ее ширину. Оборотов 6, уме-
ренно выпуклых, разделенных глубоким швом. Последний оборот менее чем в 1.5 раза
Рис. 133—136. Leucozonella (Leucozonella) caryodes (West.), предгорья Киргизского хр.,
15 VI 1972. 133 — внешний вид полового аппарата; 134 — мантийный валик; 135 —
пенис, вскрыт; 136 — поперечный срез эпифаллуса. (По Schileyko, 1977).
шире предпоследнего, к устью медленно спускается начиная с середины предпослед-
него оборота. Окраска характерная: общий фон раковины известково-белый или
грязно-серый, со слабой периферической лентой. Верхние 2—4 оборота
коричневатые, темно-серые, изредка даже почти черные или имеют красновато-фиоле-
товый оттенок. Скульптура в виде неправильных и грубых радиальных морщин и ред-
ких спиральных бороздок, которые могут быть выражены очень слабо. Устье округлое,
умеренно косое, края его не отвернуты, кроме колумеллярного, но часто заметно утол-
щены. Пупок очень узкий, неперспективный. Размеры: ВР 12—17, БД 12—18,
МД 10—44 мм.
Внутреннее строение. Материал: 2 экз. из верховьев р. Чаткал,
LEUCOZONELLA
171
.26 V 1972; 6 — из предгорий Киргизского хр. близ г. Фрунзе, 15 VI 1972; 6 — из
ущелья Беш-Таш на северном склоне Таласского хр. близ г. Таласа, 4 VI 1972; сборы
и определения наши.
Яйцевод образует выраженный в разной степени, иногда петлеобразный изгиб.
СЖ 3X2. Верхний и нижний отделы вагины примерно равной длины. Стилофоры не
шаровидные, но удлиненные, почти равной длины. Вагина тонкая, заключает внутри
мощные вагинальные валики, лопасти в месте выходных отверстий стилофоров- выра-
жены нерезко.
Рис. 137—142. Leucozonella {Leucozonella} rubens (Mart.), ущелье p. Аламедин, Киргиз-
ский хр., 12 VI 1972. 137 — внешний вид полового аппарата; 138 — пенис, вскрыт
{звездочка — придаток папиллы); 139 — мантийный валик; 140 — вагина, вскрыта;
141 — поперечный срез папиллы пениса; 142 — поперечный срез эпифаллуса. (По Schi-
leyko, 1977).
Пенис массивный, веретеновидный, по его поверхности обычно проходят тонкие
соединительнотканно-мускульные ленты. Папилла пениса на поверхности несет рез-
кую борозду, плоскость которой составляет острый угол с сагиттальной плоскостью
папиллы. Бич длинный, но немного короче тонкого извитого цилиндрического эпи-
фаллуса, отдельные участки которого связаны между собой короткими мускульными
связками. Ретрактор пениса часто охватывает петлеобразно эпифаллус. Проток семе-
приемника тонкий, слегка извитой, без резкой границы переходит в удлиненно-оваль-
ный резервуар, который несколько не достигает белковой железы.
Распространение и местообитания. Центральный и север-
ный Тянь-Шань. Населяет открытые склоны, но предпочитает участки с высоким тра-
востоем, где образует скопления с плотностью до 30 экз./м2. В сухое время года уходит
в почву на глубину до 10—12 см.
3. Leucozonella (Leucozonella) rubens (Martens, 1874). Табл. V, 46; рис. 137—142.
Мартенс, 1874 : 12, табл. 1, рис. 6 {Helix); Martens, 1881 : 12, t. 4,
fig. 1—5 {H. rubens var. finschiana, var. concolor, var. zeiliana); 1882a : 8, t. 2,
fig. 1—4 {Helix); Wester lund, 1889 : 129 {Helix}-, Линдгольм, 1931 : 53
{Fruticicola}; Лихарев, Раммельмейер, 1952 : 401, рис. 333 . [Eulota
{Leucozonella}]; Лихарев, 1955:182 {Trichia); Schileyko, 1977 : 9, figs
21—26, Pl. I, 4. -
173
III. HYGROMIIDAE
Раковина умеренно толстостенная, шаровидная, с коническим или слегка
куполовидным завитком. Оборотов 6, верхние заметно более уплощены, чем нижние,
верхние обороты нарастают медленно и плавно, нижние — более стремительно. По-
следний оборот по периферии плавно закруглен (у не вполне сформированных рако-
вин слегка угловатый), к устью немного опущен, причем имеется легкий перегиб.
Окраска от светло-серой до коричневой и почти красной, в последнем случае спираль-
ная белая лента выражена особенно четко; верх раковины большей частью несколько-
темнее, чем нижняя сторона. Скульптура в виде неравномерной радиальной исчерчен-
ности и редких спиральных бороздок. Устье округлое, слегка косое, места его прикреп-
ления не сближены. Немного отступя от края устья имеется сильно расплывшаяся
губа, края устья слабо отвернуты. Колумеллярный край отвернут сильнее и не более-
чем наполовину прикрывает узкий неперспективный пупок; нередко пупок почти не при-
крыт. Размеры: ВР 12—15, БД 15—20, МД 13—17 мм.
Раковина вида чрезвычайно похожа на раковину Angiomphalia regeliana (ср. фото.
46 на табл. V и фото 166 на табл. XVI); раковина последнего имеет более узкий пупок
и в этом отношении походит на раковину L. caryodes, но отличается от L. caryodes
более тонкими стенками и окраской. Тем не менее, принимая во внимание изменчивость,
раковин всех трех видов, достоверно их можно определить только после анатомирова-
ния.
Внутреннее строение. Материал: 12 экз. из Заилийского хр., Кок-
пек, 2 VI 1970, сбор К. К. Увалиевой; 5 — из ущелья р. Аламедин на Киргизском хр.,
12 VI 1972, сбор и определения наши.
Нижняя часть спермовидукта и яйцевод образуют двойной изгиб, выраженный
в различной степени. Наружные стилофоры значительно массивнее внутренних, форма,
стилофоров приближается к сферической; вагина в области стилофоров вздута, к кло-
аке становится тоньше. Вагинальные валики то более, то менее мощные, но лопастей
не образуют или эти лопасти очень слабые.
Пенис веретеновидный, папилла почти коническая, несет на поверхности узкую,
глубоко врезанную борозду (рис. 141). При основании папиллы имеется придаток,
имеющий вид бугорка, конического выроста, или, в максимальном выражении, в виде-
губчатой пластинки, накладывающейся на папиллу. Бич короткий, эпифаллус в 2.5—
4 раза длиннее бича; на поперечном разрезе эпифаллуса виден ряд продольных вали-
ков, дифференциация которых в два комплекса едва намечается (рис. 142). Между
пенисом и эпифаллусом натянута соединительнотканная перепонка, в состав которой
входят и мускульные ленты. Проток семеприемника тонкий, довольно длинный,
заканчивается овальным резервуаром, обычно достигающим нижнего края белковой,
железы.
Распространение и местообитания. Вид широко, но споради-
чески распространен в Средней Азии. Встречается на открытых и слабо залесенных
склонах; особенно плотные скопления образует в местах с мощным травостоем.
4. Leueozonella (Leueozonella) mesoleuca (Martens, 1882b). Табл. V, 47, 48;:
рис. 143, 144.-
Ma r t e n s, 1882b : 105 (Helix); Westerlund, 1889 : 51 (Helix); 1896 : 186
[Helix (Eulota) rubens var. limitata]; Линдгольм, 1931 : 53, t. 1, fig. 4—6 (Fru-
ticicola); 54, fig. 1 (subsp. globuliformis); Лихарев, Раммельмейер,
1952 : 402 [Eulota (Leueozonella)]; Матёкин, 1954 : 90 (Euomphalia mecoleuca);
Лихарев, 1955 : 182, рис. 1, В (Trichia); Матёкин, 1966 : 124, табл. 5,
рис. 6,6а (Trichia).
Материал: более 30 экз. из 5 местонахождений на Тянь-Шане.
Раковина от шаровидной до конической, с куполовидным или, реже, кони-
ческим контуром завитка, высота которого приблизительно равна высоте устья.
Обор.отов 4.5—5, слабо выпуклых. Последний оборот снизу закруглен, в конце несильно,
но явственно опущен, вначале нередко со слабым углом, который к устью сглажива-
ется. Окраска красновато-коричневая или светло-роговая, светлая спиральная лента
проходит по периферии последнего оборота и у шва на предыдущих оборотах. Скульп-
тура в виде тонкой радиальной исчерченности, спиральные бороздки отсутствуют.
Поверхность раковины изредка покрыта очень тонкими мелкими светлыми волосками,
которые легко утрачиваются. Устье почти круглое, умеренно косое, края устья тон-
кие, не отвернуты. Места прикрепления устья соединены тонкой мозолью; колумел-
лярный край слегка отвернут, не более чем наполовину прикрывает очень узкий пу-
пок; однако у отдельных экземпляров при рассматривании строго снизу (т. е. если оп-
тическая ось глаза совмещена с осью пупка) пупка почтй не видно, в этом случае он
имеет вид широкой щели. Размеры: ВР 6.5—8, БД 9—11, МД 8—10 мм.
Внутреннее строение. Материал: 5 экз. из Ферганской долины,
Араван, 27 V 1972, сбор В. В. Горохова; 4 — из ущелья р. Аламедин на Киргизском хр.,
15 VI 1972; сбор и определения наши.
LEUCOZONELLA
173
Дистальная часть спермовидукта образует плавный изгиб. Наружные стилофоры
несколько массивнее внутренних, шаровидные или слегка удлиненный, область сти-
лофоров расширена, далее вниз вагина плавно сужается. СЖ 3—4X2. Вагинальные
валики развиты слабо, лопасти почти не выражены.
Пенис веретеновидный, умеренно вздут. Папилла пениса короткая, на поверх-
ности имеет глубокую борозду, прикрытую придатком, имеющим вид пластинки. Бич
а 2—3 раза короче цилиндрического эпифаллуса. Мембрана, натянутая между семе-
Рис. 143—144. Leueozonella (Leueozonella) mesoleuca (Mart.),
ущелье p. Аламедин, Киргизский хр., 15' VI 1972. 143 — внеш-
ний вид полового аппарата; 144 — пенис, вскрыт (звездочка —
придаток папиллы). (Ориг.).
проводом, пенисом и эпифаллусом, тонкая, прозрачная, без мускульных пучков.
Проток семеприемника ровный, без изгибов; резервуар шаровидный или овальный,
не достигает нижнего края белковой железы.
Распространение и местообитания. Киргизский, Таласский,
Чаткальский, Ферганский, Алайский, Заалайский, Туркестанский и Кураминский
хребты. Обитает на открытых склонах, в траве и зарослях кустарника.
5. Leueozonella (Leueozonella) rufispira (Martens, 1874). Табл. V, 49, 50;
рис. 145—147.
Мартенс, 1874 : 9, табл. 1, рис. 7, табл. 3, рис. 38 (Helix); L. Pfeiffer,'
1876 : 234 (Helix); Martens, 1881 : 14, t. 4, fig. 18—20 (var. albidorsalis); 1882a : 10,
t. 2, fig. 8 (var. albidorsalis); Westerlund, 1889 : 50 (Helix); Розен, 1901 : 8
(Helix rufispira var. maracadensis, var. fuscior); Cockerell, 1928 : 364 (Cathaica);
Лихарев, Раммельмейер, 1952 : 402 [Eulota (Leueozonella)}; Матёкин,
1954 : 90 (Trichia); J a e c k e 1, 1956, Mitt. Zool. Mus. Berlin, 32, 2 : 349, fig. 8a—d
(Cathaica rufispira hispida); Матёкин, 1966 : 124, табл. 5, рис. 7 (Trichia); Лиха-
рев, Старобогато в, 1967 : 191, рис. 19 (Trichia); S о 1 е m, 1972b : 63
(Trichia); Schileyko, 1977 : 10, figs 27-31, Pl. П, 6.
174
III. HYGROMIIDAE
Материал: более 100 экз. из 7 местонахождений на Тянь-Шане.
Раковина прижатая, со слабо выдающимся коническим или слегка куполот
видным завитком, высота которого обычно меньше высоты устья, реже равна ей, но
не больше. Оборотов 5—5.5, слабо выпуклых. Последний оборот явственно угловатый,
но к устью угол сглаживается; к устью последний оборот несколько опущен. Окраска
сверху от светло-коричневой до красновато-роговой, внизу постепенно светлеет, вок-
руг пупка имеется светлое поле. Периферическая светлая лента ясно выражена; ее
оптическая плотность значительно выше, чем плотность остальной части раковины.
Скульптура в виде тонкой радиальной морщинистости и редких складок. Спиральная
Рис. 145—147. Leucozonella (Leucozonella) rufispira (Mart.), перевал Анзоб, Гиссар-
ский хр., 28 VII 1968. 145 — внешний вид полового аппарата; 146 — пенис, вскрыт;
147 — мантийный валик. (По Schileyko, 1977).
скульптура видна только на хорошо сохранившихся раковинах в виде тончайших реб-
рышек, заметных при увеличении не менее чем bz30 раз. Иногда раковина с волосками.
Устье округлое, косое, места его прикрепления умеренно сближены, края его тонкие,
иногда с широкой, сильно расплывшейся губой. Колумеллярный край слегка отвер-
нут, но не прикрывает узкого неперспективного пупка. Размеры: ВР 6—8, ВД 9—14,
МД 7.5—12 мм.
В ущелье Беш-Таш на северном склоне Таласского хр. и в долине р. Шинг на Зе-
равшанском хр. обнаружены популяции, заметно отличающиеся от типичных: края
устья сближены, устье более косое, периферический угол сильно сглажен, размеры
более крупные: ВР 8—12, БД 15—17, МД 13—14 мм; у большинства особей пупок бо-
лее узкий, чем в типичном случае (фото 50 на табл. V).
Поскольку анатомических отличий от нормальных особей не найдено, и поскольку
эти популяции изолированы, мы воздерживаемся пока от возведения описанной формы
в ранг вида; считать ее подвидом также нецелесообразно, так как форма найдена в двух
удаленных друг от друга точках. По-видимому, это экологическая морфа, свойствен-
ная относительно влажным биотопам.
Внутреннее строение. Материал: 4 экз. с перевала Анзоб на Гиссар-
ском хр., 28 VII 1968, сбор и определение 3. Иззатуллаева; 6 — с верховьев р. Чаткал,
27 V 1972; 12 — из ущелья Беш-Таш на Таласском хр., 3 VI 1972, сборы наши; 5 —
из долины р. Шинг на Зеравшанском хр., 6 VIII 1973, сбор 10. А. Мешкова; определе-
ния наши.
Нижняя часть спермовидукта и яйцевод образуют выраженный в разной степени
изгиб с сомкнутыми сторонами. СЖ 3—4X2. Стилофоры относительно очень массив-
LEUCOZONELLA
175
ные, сферические, внутренние лишь слегка меньше наружных. Длина вагины в 3—5 раз
превышает ее ширину, вагинальные валики ясные, с хорошо выраженными лопастями
в местах выходных отверстий стилофоров.
Пенис шаровидный или немного удлиненный, папилла веретеновидная, с очень
глубокой продольной бороздой, которая у некоторых особей в дистальной части начи-
нает замыкаться; кроме того, неполная кольцевая борозда наблюдается иногда в про-
ксимальной части папиллы. Вич слегка короче круто изогнутого и слегка скрученного
эпифаллуса; изгиб эпифаллуса притянут к пенису соединительнотканно-мускульными
лентами. Проток семеприемника тонкий, почти прямой, резервуар овальный, заметно
не доходит до нижнего края белковой железы.
Рис. 148—150. Leucozonella (Leucozonella) retteri (Rosen), ущелье Кондара, Гиссар-
ский хр., 2 VII 1967. 148 — внешний вид полового аппарата; 149 — мантийный ва-
лик; 150 — пенис, вскрыт. (По Schileyko, 1977).
Распространение и местообитания. Тянь-Шань и предгорья
Памира; северный Афганистан. Обитает в каменистых осыпях; более крупные раковины
формируются в лесных участках, имеющих достаточную влажность, на небольших
высотах. Самые мелкие популяции обитают в высокогорье. Солем (Solem, 1972b) от-
мечает присутствие вида в верхнечетвертичных отложениях Афганистана.
6. Leucozonella (Leucozonella) retteri (Rosen, 1897). Табл. VI, 51; рис. 148—150.
Rosen, 1897 : 170 (Helix); W esterlund, 1902 : 20 (Helix); Лихарев,
Раммельмейер, 1952 : 403, рис. 334 [Eulota (Leucozonella)]; Матёкин,
1954 : 90 (Trichia); Лихарев, 1955 : 182, рис. 1, Г (Trichia); Матёкин, 1966 : 131
(Tr. rufispira part.); Schileyko, 1977 : 11, figs 32—36, Pl. II, 7.
Материал: 18 экз. из двух местонахождений на южном Тянь-Шане.
Раковина схожа с таковой предыдущего вида, но хорошо отличается нали-
чием ясных спиральных бороздок как на верхней, так и на нижней поверхности рако-
вины, и развернутым перспективным пупком. Размеры: ВР 8—12, БД 15—19, МД
13—17 мм.
Внутреннее строение. Материал: 4 экз. из ущелья Кондара на Гис-
сарском хр., 2 VII 1967, сбор и определение 3. Иззатуллаева.
Нижняя часть спермовидукта прямая, яйцевод тоже прямой или слабо изогнут.
СЖ 4X1—2. Стилофоры массивные, вздутые, шаровидные. Область стилофоров снизу
отграничена ясной перетяжкой. Вагина длинная, узкая, цилиндрическая, ее длина
176
Ш. HYGROMIIDAE
превосходит ширину в 5—6 раз. Вагинальные валики выражены нерезко, и лишь в об-
ласти выходных отверстий стилофоров видоизменяются в ясные лопасти.
Пенис массивный, веретеновидный. Папилла пениса характерной формы: прокси-
мальная часть цилиндрическая, затем следует резкое расширение, заканчивающееся
конической дистальной частью. Семепроводящий канал одной стороной спаян с вну-
тренней стенкой папиллы, а с трех сторон окружен обширной интрапапиллярной поло-
стью. По внутренней поверхности пениального чехла проходит мощный гребневидный
валик. Длина бича примерно равна длине эпифаллуса; последний образует 1—2 из-
гиба. Между пенисом и эпифаллусом имеется соединительнотканная мембрана, по по-
Рис. 151 —154. Leucozonella {Leucozonella) angulata (West.), долинаp. Пскем, 17 V 1972.
151 — внешний вид полового аппарата; 152 — пенис, вскрыт; 153 — пенис другого
экземпляра, вскрыт; 151 — вагина, вскрыта. (Ориг.).
верхности пениса проходят тонкие тяжи. Проток семеприемника слабо изогнут или
почти прямой, овальный резервуар касается нижнего края белковой железы.
Распространение и местообитания. Гиссарский, Зеравшанский
и смежные мелкие хребты. Обитает на открытых каменистых склонах и в осыпях.
7. Leucozonella (Leucozonella) angulata (Westerlund, 1896). Табл. VI, 52; рис. 151 —
154.
Westerlund, 1896 : 186 {Helix rubens var. angulata); Лихарев, Рам-
мельмейер, 1952 : 401 [Eulota {Leucozonella) rubens f. angulata].
Материал: более 100 экз. из 4 местообитаний в окрестностях Самарканда и в ущелье
р. Пскем.
Раковина от слегка прижатой до кубаревидной, со слегка куполовидным
завитком. Оборотов около 6, умеренно выпуклых. Последний оборот приблизительно
в 1.5 раза шире предпоследнего, в самом конце несильно, но явственно опущен. Ок-
раска от кремовой до роговой; при посветлении фона меняется в основном интенсивность
окраски последнего оборота, остальные обороты невыцветших раковин сохраняют ро-
говую окраску. Периферическая светлая лента даже на светлых раковинах заметна
хорошо, в проходящем свете плотнее (темнее) фона. Скульптура в виде неправиль-
ной поперечной исчерченности или даже морщинистости, спиральные бороздки вя-
лые, выражены местами, иногда,отсутствуют. Устье от почти круглого до широкооваль-
LEUCOZONELLA
177
ного, умеренно вырезано предпоследним оборотом, края его острые, отвернут лишь
колумеллярный край и частично базальный. Губа располагается чуть отступя от края
устья, слабая, расплывшаяся; в верхней части палатального края губа почти исчезает.
Пупок точковидный, в различной степени прикрыт отворотом колумеллярного края.
Размеры: ВР 7.5—11.7, БД И—17, МД 9—14 мм. Лектотип: ВР 11.5, БД 13.5, МД 11 мм.
Раковина этого вида пластична и обнаруживает, большей частью четкую зависи-
мость от окружающих условий: в затененных и влажных местах обитают особи с бо-
лее интенсивно окрашенной раковиной, имеющей тонкие стенки, малые размеры и от-
носительно более высокий завиток. На открытых местах, напротив, встречаются
экземпляры больших размеров с уплощенной раковиной, более плотными стенками
и более светлой окраской; между этими типами наблюдаются всевозможные переходы
по разным признакам.
Видовое название в переводе означает «угловатая»; необходимо отметить, что ра-
ковина этого вида, угловатая по периферии разве только в сравнении с типичной
L. rubens, и была описана как форма названного вида; угол по периферии у L. angulata
иногда имеется в очень ослабленном виде в начале последнего оборота, в чем мы убе-
дились, ознакомившись с типовой серией, хранящейся в Зоологическом институте
АН СССР.
Внутреннее строение. Материал: 5 экз. из Самаркандской обл.,
окр. Чупан-Ата, IV 1954, сбор 3. И. Обуховой; 22 — из долины р. Пскем между Пскем-
ским и Угамским хребтами, 17 V 1972, сбор и определения наши.
На границе спермовидукта и яйцевода имеется резкий перегиб. СЖ 4X2. Стило-
форы небольшие, шаровидные, наружные и внутренние почти одинаковых размеров.
Вагина в области стилофоров образует характерное расширение. Вагина длинная,
иногда даже без ее вскрытия просматриваются винтообразно направленные мощные
вагинальные валики, просвечивающие сквозь стенки вагины. Область стилофоров
отграничена от вагины более или менее ясным перехватом.
Пенис веретеновидный, длинный, внутренняя его поверхность несет 1—3 мощных
валика. Форма папиллы пениса способна значительно изменяться: у одних особей па-
пилла веретеновидная, но часто дистальный ее конец «оплывает», образуя грибооб-
разной формы фигуру (рис. 152, 153). По поверхности папиллы в проксимальной части
проходит борозда, но далее ее края смыкаются, образуя интрапапиллярную
полость. Таким образом, эта борозда 'представляет собой папиллярную ла-
куну. Тонкий бич примерно той же длины, что и конический эпифаллус, немного рас-
ширенный в дистальной части. Половой ретрактор очень мощный и короткий. Мем-
брана между семепроводом, эпифаллусом и пенисом тонкая, прозрачная. Проток се-
меприемника изгибов не образует, заканчивается небольшим овальным резервуаром,
заметно не достигающим белковой железы.
Распространение и местообитания. Юго-западный Тянь-
Шань. Встречается во всех основных типах биотопов — от влажных пойменных леси-
стых участков до открытых каменистых склонов.
8. Leucozonella (Leucozonella) caria Schileyko, 1977. Табл. VI, 56; рис. 155—157.
Schileyko, 1977 : 12, figs 37—41, Pl. II, 8.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 12 экз. из одного местонахождения на Гиссарском хр.
Раковина в различной степени прижатая, с почти коническим или немного
куполовидным завитком, высота которого всегда меньше высоты устья. Оборотов
4.5—5, выпуклых, довольно медленно нарастающих, разделенных глубоким швом.
Последний оборот чуть менее чем в 1.5 раза шире предпоследнего, к устью почти не опу-
щен. Эмбриональные обороты блестящие, неясно отграничены от дефинитивных и той
же окраски. Основной фон раковины коричневато-роговой; по периферии имеется раз-
мытая светлая лента. Скульптура — тонкая и довольно правильная радиальная ис-
черченность; спиральные бороздки отсутствуют. Поверхность раковины покрыта до-
вольно длинными изогнутыми на конце волосками. Когда волоски отпадают, на их
месте остаются рубцы в виде радиально ориентированных коротких морщинок.
Устье умеренно косое, почти круглое; внутри него, несколько отступя от края, имеется
очень расплывшаяся, довольно широкая светлая губа, которая у вполне половозре-
лых животных занимает весь край устья, а не ограничивается только базальной частью.
Колумеллярный край слегка отвернут, но не прикрывает развернутого, умеренно ши-
рокого перспективного пупка. Размеры: ВР 4.2—6, БД 7.7—8.8, МД 6.5—7.4 мм.
Голотип: ВР 4.7, БД 7.8, МД 6.5 мм.
Внутреннее строение. Материал: 3 экз. из Гиссарского хр., окр. Ходжи-
Обигарм, 28 V 1968, сбор 3. Иззатуллаева, определение наше.
Спермовидукт и яйцевод изгибов не образуют. СЖ 3X2—3. Стилофоры относительно
очень массивные, удлиненные, наружные плотно прижаты к внутренним. Вагина пря-
мая или изогнутая. Вагинальных валиков всего две пары, в местах выходных отвер-
стий стилофоров образуют хорошо выраженные лопасти.
12 А- А. Шилейко
178
III. HYGROMIIDAE
Пенис удлиненный, между пенисом и эпифаллусом имеется кольцевое вздутие.
Папилла пениса небольшая, овальная, занимает не более половины длины пениса.
Интрапапиллярная полость охватывает семепроводящий канал, спаянный с внутрен-
ней стенкой папиллы одной стороной. Чехол пениса внутри покрыт многочисленными
мелкими продольными складочками. Бич тонкий, несколько короче эпифаллуса. Эпи-
фаллус образует изгиб. Проток семеприемника почти прямой, заканчивается неболь-.
шим овальным резервуаром, заметно не достигающим нижнего края белковой железы.
Распространение и местообитания. Гиссарский хр. Обитает
на открытых склонах.
2. Подрод NARINA 1 Schileyko, subgen. nov.
Верхняя часть раковины окрашена светлее нижней, либо окраска раковины равно-
мерная, но в этом случае имеются радиальные ребра.
Рис. 155—157. Leueozonella (Leueozonella) caria Schileyko, окр. Ходжи-Обигарм,
Гиссарский хр., 28 V 1968. Паратип. 155 — внешний вид полового аппарата;
156 — вагина, вскрыта; 157 — пенис, вскрыт. (По Schileyko, 1977).
Пенис очень маленький; папилла пениса лишена интрапапиллярной полости, но
папиллярная лакуна имеется. Ретрактор правого омматофора образует перехлест
с половым ретрактором.
От других подродов этого же рода отличается очень малыми размерами пениса
и своеобразным устройством пениальной папиллы: наличие лакуны при отсутствии
интрапапиллярной полости.
Распространение. Алайский, Чаткальский и Ферганский хребты.
Типовой вид — Fruticicola hypophaea Lindholm, 1927.
9. Leueozonella (Narina) hypophaea (Lindholm, 1927). Табл. VI, 53, 54; рис. 158—
160.
Lindholm, 1927c : 198 (Fruticicola, Fr. balgutensis); Лихарев, Рам-
мельмейер, 1952 : 403 [Eulota (?)].
Материал: более 70 экз. из 2 местонахождений на Ферганском хр.
Раковина от конической до почти шаровидной, изредка прижатая. Завиток
более или менее куполовидный, вершина невыступающая, узкая, сглаженная. По-
1 Название образовано от названия р. Нарын, по берегам которой местами
в изобилии встречается типовой вид подрода.
LEUCOZONELLA
179
верхность раковины блестящая. Оборотов 5.5—6, умеренно выпуклых, изредка почти
ступенчатых. Последний оборот в профиле округлый, к устью едва опущен. Окраска
верхней стороны раковины белая, реже коричневатая или роговая, ниже периферии
серовато-роговая, просвечивающая. Нередки случаи, когда отличия в окраске между
верхней и нижней сторонами раковины настолько невелики (фото 53 на табл. VI),
что их можно обнаружить лишь при внимательном рассмотрении, или отсутствуют
вовсе. В этом случае раковину надо рассматривать в сильном проходящем свете,
лучше — левую сторону: ниже периферии она имеет восковую полупрозрачность,
что позволяет уверенно определять свежие раковины. Скульптура в виде густой ра-
Рис. 158—160. Leueozonella (Narina) hypophaea (Lindh.), Ферганский хр.,
берег р. Нарын над Ташкумыром, 31 V 1972. 158 — внешний вид поло-
вого аппарата; 159 — пенис, вскрыт; 160 — вагина, вскрыта. (Ориг.).
диальной исчерченности и неясных спиральных бороздок. Устье округлое, косоег
внутри светлое, с белой, сильно расплывшейся губой, расположенной немного от-
ступя от края устья. Места прикрепления устья широко расставлены; колумелляр-
ный край отвернут, но не закрывает или слегка прикрывает очень узкий точковидный
пупок. Размеры:-ВР 9—11, БД 12—15, МД 11—13 мм.
Внутреннее строение. Материал: 8' экз. из Тянь-Шаня, Ферган-
ский хр., берег р. Нарын над Таш-Кумыром, 31 V 1972, сбор и определение наши.
Спермовидукт в нижней части почти прямой или слегка изогнут на границе
с очень коротким яйцеводом. СЖ 3X2, верхний отдел вагины очень короткий. Наруж-
ные стилофоры значительно массивнее внутренних, все 4 стилофора располагаются
не в одной вертикальной плоскости, но внутренние как бы «выжаты» наружными
из пространства между ними и вагиной; в результате внутренние стилофоры видны
только при рассмотрении гениталий в том положении, как изображено на рис. 158;.
если их повернуть на 180°, внутренние стилофоры видны не будут. Вагинальный
валики в числе 2 пар, в месте выходных отверстий стилофоров образуют ясные лопасти
и тут же обрываются, не продолжаясь в клоаку.
Пенис очень маленький, длина его заметно меньше длины нижней части вагины,,
чехол пениса внутри гладкий. Папилла пениса простая, в виде трубки; папил-
лярная лакуна выражена хорошо, семепроводящий канал широкий. Бич примерно
12*
180
111. HYGROMIIDAE
равен почти прямому, не скрученному цилиндрическому эпифаллусу. Мембрана между
дистальными протоками мужского отдела тонкая, прозрачная. Половой ретрактор
крепится к эпифаллусу заметно выше пениса. Проток семеприемника прямой, резер-
вуар овальный, не достигает нижнего края белковой железы.
Распространение и местообитания. Ферганский и Алайский
хребты. Встречается на очень сухих открытых каменистых склонах.
10. Leucozonella (Narina) crassicosta Schileyko, sp. nov. Табл. VI, 55; рис. 161 —
164.
Голотип хранится в Зоологическом институте АН СССР.
Материал: более 100 экз. из 3 местообитаний на Чаткальском и Кураминском
хребтах.
Раковина от сильно прижатой до почти конической, составлена 5.5 выпук-
лыми оборотами, разделенными неровным глубоким швом. Вершина выступающая.
Последний оборот слегка угловатый, к устью почти не опущен, менее чем в 1.5 раза
шире предпоследнего. Окраска роговая, иногда с красноватым оттенком; молодые
особи имеют, как правило, более ярко окрашенную раковину. С возрастом окраска
Светлеет, причем посветление начинается с пространства, окружающего пупок; у
у большинства раковин нижняя половина в целом немного светлее верхней. Если по-
Рис. 161 — 164. Leucozonella (Narina) crassicosta sp. nov., окр. с. АлатВука в предгорьях
'Чаткальского хр., 29 V 1972. 101 — голотип, внешний вид полового аппарата; 162 —
шенис, вскрыт; 163 — вагина, вскрыта; 164 — дистальная часть женского отдела
экземпляра из Кураминского хр., 23 V 1972. (Ориг.).
'местить раковину под край непрозрачного абажура настольной лампы, видно, что
;ниже периферии раковина имеет восковую полупрозрачность (этот признак виден
только на свежих раковинах). Кроме того, имеется светлая периферическая лента.
’Скульптура в виде грубых редких, неправильно расположенных радиальных ребер
и слабых, но очень частых спиральных бороздок, лучше выраженных в промежутках
между ребрами и заметных при значительном увеличении. Популяции, обитающие
;на Кураминском хр., имеют более слабую радиальную ребристость, чем чаткальские.
’-Устье округлое или широкоовальное, слегка скошено. Губа отсутствует. Пупок не
HYGROHELICOPSIS
181
развернутый, но открытый, сквозь него просматриваются 1.5—2 предыдущих обо-
рота. Размеры: ВР 6—8.5, БД 10—13.5, МД 8.5—11.5 мм. Голотип: ВР 7.0, БД 11.5,
МД 10.0 мм.
Размерами и формой раковины вид больше всего походит на Leucozonella rufi-
spira; отличается менее резким углом по периферии и ребристой скульптурой.
Внутреннее строение. Материал: 17 экз. из окр. с. Ала-Бука в пред-
горьях Чаткальского хр., 29 V 1972; 2 — из северной части Кураминского хр.,
23 V 1972; сборы и определения наши.
Очень короткий яйцевод образует большей частью слабый плавный изгиб.
СЖ 3—4X2. Наружные стилофоры значительно массивнее внутренних, • последние
сближены на одной стороне вагины. У отдельных экземпляров наблюдается такое
положение стилофоров, как показано на рис. 164, т. е. они располагаются относительно
вагины почти мутовчато. Вагина короткая, очень плавно сужена к клоаке. Вагиналь-
ные валики в числе 2 пар, сближены попарно, в местах выходных отверстий сти-
лофоров образуют ясные лопасти и тут же обрываются или петлеобразно охватывают
снизу упомянутые отверстия.
Пенис маленький, заметно меньше вагины в области стилофоров. Папилла пениса
не заполняет полностью пениального чехла. Семепроводящий канал обширный, па-
пиллярная лакуна выражена хорошо. Бич слегка короче петлеобразно изогнутого
эпифаллуса. Проток семеприемника прямой, резервуар овальный, не достигает бел-
ковой железы.
Распространение и местообитания, Вид найден в двух ме-
стах: на юго-восточных склонах Чаткальского хр., вдоль левого берега р. Касансай,
на открытых, сухих, каменистых склонах; в северной части восточных склонов Ку-
раминского хр., среди камней. Живые особи обнаружены глубоко в расселинах скал
и под большими камнями.
А. Мухитдинов (1972) указывает, что среди Leucozonella rufispira, собранных
на Кураминском хр., имеются экземпляры с ребристой скульптурой раковины.
Очевидно, автор имел дело с данным видом.
4. Род HYGROHELICOPSIS Schileyko, 1977
Schileyko, 1977 : 13.
Раковина прижатая, лиловато-каштановая, с широкими размытыми светлыми
поперечными пятнами, занимающими всю ширину оборотов. Пупок неширокий,
цилиндрический, но перспективный. Волоски на раковине отсутствуют.
Внутренняя пара стилофоров целиком скрыта под соединительнотканным чехлом.
Внутри папиллы пениса имеется парная интрапапиллярная полость. Ретрактор пра-
вого омматофора не образует перехлеста с половым ретрактором.
Род не обнаруживает непосредственной близости ни с одним из известных родов;
строение папиллы пениса сближает его с европейскими Trichia, но от последних род
резко отличается раковиной и строением стилофоров.
Распространение. Кавказ.
Монотипический род.
Типовой вид — Hygrohelicopsis darevskii Schileyko, 1977.
Hygrohelicopsis darevskii 1 Schileyko, 1977. Табл. VI, 57; рис. 165—167.
Schileyko, 1977 : 14, figs 42-46, Pl. Ill, 9.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 3 экз. из 2 местонахождений в ущелье р. Чегем на Северном Кавказе.
Раков и н а сильно уплощена, с ширококуполовидным завитком и сглажен-
ной невыступающей вершиной. Оборотов 5, умеренно быстро и плавно нарастающих.
Последний оборот особенно быстро нарастает в высоту, так что париетальная стенка
устья составляет довольно острый угол со средней линией оборота, к устью заметно,
но без резкого перегиба опущен. Окраска состоит из лиловато-каштанового фона
и размытых светло-каштановых пятен; поверхность раковины имеет шелковистый
блеск. Эмбриональные обороты неясно отграничены от дефинитивных, гладкие, светло-
роговые. Скульптура в виде неправильных радиальных сглаженных тонких морщи-
нок, местами заметны слабые спиральные бороздки. Радиальные морщинки резче
всего выражены на верхней поверхности последнего оборота, базальная часть более
гладкая. Устье широкоовальное, большое, немного скошено; края его простые, ба-
1 Вид назван именем Ильи Сергеевича Даревского.
182
111. HYGROMIIDAE
зальный край слегка отвернут. Отступя от края устья имеется мощная высокая
снежно-белая губа, которая просвечивает сквозь стенку последнего оборота в виде
широкой полосы светлее окружающего фона, с неясными границами. В постэмбрио-
генезе образуется не одна, а 2—3 губы; более ранние губы также просвечивают в
в виде светлых полос. Сквозь пупок видна большая часть оборотов. Размеры:
ВР 5.5—6, БД 10—10.5, МД 8.8—9.0 мм. Голотип: ВР 6, БД 10.5, МД 9 мм.
Раковина вида напоминает раковину Caucasigena schaposchnikovi; отличается
постоянным отсутствием темных лент, более узким пупком, наличием светлых пятен;
Рис. 165—167. Hygrohellcopsis darevskii Schileyko, Чегемское ущелье,
9—10 VIII 1965. Голотип. 165— внешний вид полового-аппарата; 166— стило-
форы, вскрыты; виден рудиментарный внутренний стилофор; 167 — пенис, вскрыт.
(По Schileyko, 1977).
кроме того, париетальная стенка устья у С. schaposchnikovi направлена почти парал-
лельно средней линии оборота, а не под углом к ней.
Внутреннее строение. Материал: 2 экз. из Чегемского ущелья на
Северном Кавказе, 9—10 VIII 1965, сбор И. С. Даревского, определение наше.
Спермовидукт складчатый, почти прямой, яйцевод резко изогнут; длина его
примерно равна длине вагины. СЖ 4X1—2. Стилофоры чрезвычайно массивные;
при вскрытии вагины и плотного соединительнотканного чехла, окутывающего сти-
лофоры, обнаруживаются рудиментарные, очень маленькие внутренние стилофоры,
имеющие, тем не менее, собственные протоки, впадающие в полость стрелы.
Пенис вздутый, шаровидный. Папилла пениса в проксимальной части цилиндри-
ческая, в дистальной — шаровидная, в целом булавовидной формы. Семепроводящий
канал охвачен с двух сторон парной интрапапиллярной полостью, в базальной части
папиллы имеется небольшая папиллярная лакуна. Бич тонкий, примерно вдвое длин-
нее эпифаллуса. Последний дважды резко изогнут и фиксирован в таком положении
хорошо развитыми тяжами. Проток семеприемника длинный, слабо изогнутый,
с объемистым округлым резервуаром, почти достигающим белковой железы.
Особенного внимания заслуживает то обстоятельство, что ретрактор правого
омматофора располагается рядом с дистальными участками гениталий, не перекре-
щиваясь с ними; этот признак всегда сопряжен с определенным конхологическим
обликом (см. вводную часть).Данный вид — едва ли не единственное исключение:
подобный конхологический тип обычно связан с наличием перехлеста ретракторов.
TEBERDINIA
183
Распространение и местообитания. Вид пока известен только
из ущелья р. Чегем (центральная часть Большого Кавказа). Обнаружен на откры-
том склоне под камнями, на высоте 2000—2500 м над ур. м.
5. Род TEBERDINIA Schileyko, 1977
Schileyko, 1977 :15.
Раковина слегка прижатая, желтовато-бурая, почти одноцветная. Пупок про-
коловидный, почти неперспективный. Раковина без волосков.
Обе пары стилофоров развиты примерно в равной степени, тесно попарно прижаты
друг к другу и обособлены от стенки вагины. Папилла пениса имеет на поверхности
Рис. 168—170. Teberdinia zolotarevi (Lindh.), Тебердинский запов., 24 VII 1958.
168 — внешний вид полового аппарата; 169 — вагина, вскрыта; 170 — пенис, вскрыт.
, (По Schileyko, 1977).
глубокую борозду; семепроводящий канал охвачен парной интрапапиллярной по-
лостью. Ретрактор правого омматофора образует перехлест с половым ретрактором.
От близких родов резко отличается конхологическими признаками и строением
пениальной папиллы; в меньшей степени — небольшими стилофорами, не прилега-
ющими к вагине.
Р а с п р о с т р а не ни е. Кавказ.
Монотипический род.
Типовой вид — Helix (Fruticocampylaea) zolotarevi Lindholm, 1913.
Teberdinia zolotarevi (Lindholm, 1913). Табл. VI, 58; рис. 168—170.
Lindholm, 1913b : 17 [Helix (Fruticocampylaea) ]; Rosen, 1914 : 230
[Helix (Fruticocampylaea)]; Schileyko, 1977 : 15, figs 47—51, Pl. Ill, 10.
Материал: 2 экз. из Тебердинского заповедника (Северный Кавказ).
Раковина с куполовидным завитком, высота которого немного меньше
высоты устья, умеренно твердостенная. Вершина сглаженная, не выступающая.
Базальная часть заметно вздута; оборотов 5.75, уплощенных, разделенных мелким
швом. Последний оборот чуть угловатый, к устью слабо опущен, меньше чем в 1.5
раза шире предпоследнего. Окраска почти однотонная, желтовато-бурая, местами
фон незначительно сгущается. Скульптура в виде сглаженных радиальных морщин
184
III. HYGROMIIDAE
и вялых спиральных бороздок. Устье скошенное, в базально-палатальной части чуть
оттянуто вправо, края его простые, слегка отвернутые, сильнее других отвернут
колумеллярный край. Отступя от края устья имеется мощная высокая белая губа,
просвечивающая на затылке в виде светлой полосы. Сквозь очень узкий пупок видно
не более половины предпоследнего оборота. Размеры: ВР 6.7—7.5, БД 11—11.5,
МД 10 мм. Лектотип: ВР 7.3, БД 11.5, МД 10.0 мм.
Внутреннее строение. Материал: 1 экз. из Тебердинского заповед-
ника, 24 VII 1958, сбор Л. Е. Аренса, определение наше.
Спермовидукт на границе с яйцеводом образует очень плавный изгиб. СЖ 3X2.
Стилофоры маленькие, оба левых и оба правых плотно прилегают друг к другу и на-
правлены немного в стороны от вагины. Вагинальные валики узкие, но высокие;
в месте выхода протоков стилофоров образуют округлые лопасти.
Пенис удлиненный, чехол его на внутренней поверхности несет небольшие ва-
лики. Папилла пениса недлинная, мешковидная, на конце притупленная, примерно
половину ее длины занимает глубокая продольная борозда. Одна из интрапапилляр-
ных полостей прилегает к семепроводящему каналу, другая заходит в продольную
лопасть, образованную упомянутой бороздой (рис. 170). Бич немного длиннее пря-
мого цилиндрического эпифаллуса; между пенисом и эпифаллусом имеется кольце-
вое вздутие. Проток семеприемника очень тонкий, плотно втиснут в поверхность
спермовидукта, заканчивается объемистым мешковидным резервуаром, не доходящим
до белковой железы.
Гессе (Hesse, 1931 : 11, t. 1, fig. 1) дает изображение полового аппарата Fru-
ticicola caucasicola, принадлежащего несомненно к старой группе Trichia s. lat.
Однако видовое название caucasicola должно быть закреплено за представителем
рода Monacha, в чем мы убедились, ознакомившись с оригинальным материалом,
хранящимся в Зоологическом институте АН СССР. Вероятно, Розен, предоставивший
Гессе материал для анатомирования, ошибочно определил вид; на самом деле это
был скорее всего экземпляр, относящийся к данному виду.
Распространение. Пока сделаны только две находки в типовом место-
нахождении. Характер местообитаний неизвестен.
6. Род KOKOTSCHASHVILIA Hudec et Lezhawa, 1969
Hudec, Lezhawa, 1969a : 47 [Trichia (Kokotschashvilia)}', 1969b : 127
[Tr. (Kokotschashvilia)}', Schileyko, 1977: 16.
Раковина от слегка прижатой до кубаревидной, светлая, каштановая или ко-
ричневая, без темного рисунка. Пупок от слабо развернутого до узкого, цилиндри-
ческого. Раковина без волосков или с хорошо развитыми волосками.
Стилофоры хорошо развиты, внутренние прижаты к вагине. Папилла пениса
без борозды на поверхности, интрапапиллярная полость парная или один из разде-
ляющих ее продольных тяжей редуцируется, в результате чего названная полость
становится единой. Ретрактор правого омматофора образует перехлест с половым
ретрактором.
Род имеет отдаленное сходство по анатомическим признакам с Trichia s. str.,
но отличается более крупной и более шаровидной раковиной и чрезвычайно мас-
сивным резервуаром семеприемника.
Распростране ни е. Кавказ.
Типовой вид — Helix holotricha Boettger, 1884.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА KOKOTSCHASHVILIA
1 (2). Раковина хрупкая, покрыта волосками ...............4. К. holotricha.
2 (1). Раковина не хрупкая, волоски на раковине отсутствуют.
3 (4). Высота завитка примерно равна высоте устья. Пупок очень узкий, полупри-
крытый ..................................................5. К. phaeolaema.
4 (3). Высота завитка меньше высоты устья. Пупок умеренно узкий, открытый.
5 (6). Большой диаметр не менее 22 мм.........................2. К. tanta.
6 (5). Большой диаметр не превышает 21.5 мм.
7 (8). Индекс отношения большого диаметра к высоте раковины составляет 1.5—1.6
........................................................ 1. К. makvalae.
8 (7). Индекс отношения большого диаметра к высоте раковины составляет 1.3—1.4
.........................................................3. К. eberhardi.
1. Kokotschashvilia makvalae (Hudec et Lezhawa, 1969). Табл. VI, 59; рис. 171—175.
Hudec, Lezhawa, 1969a : 41, Abb. 1, 5 [Trichia (Xerocampylaea)]; 1969b :
129, Abb. 25, t. XI, 27 [Trichia (Xerocampylaea)}; Schileyko, 1977 : 16, figs 52—56.,
KOKOTSCHASHVILIA
185
Материал: 2 экз. из 2 местонахождений на Главном Кавказском хр.
Раковина слабо прижатая, с почти коническим завитком, высота которого
немного меньше высоты устья. Оборотов около 6. Последний оборот в 1.5 раза шире
предпоследнего, к устью заметно опущен, по периферии и снизу плавно и широко
закруглен. Окраска одноцветно белая, эмбриональные обороты светло-коричневые
или серые. Скульптура представлена тремя элементами: тонкой и неравномерной
радиальной исчерченностью, тонкой и редкой зернистостью и развитыми в слабой
степени спиральными бороздками, которые могут отсутствовать. Устье почти круглое,
внутри белое, слегка скошено. Места его прикрепления слегка сближены, края устья
тонкие, отвернутые, особенно базально-колумеллярный. Несколько отступя от края
Рис. 171—175. Kokotschashvilia makvalae (Hudec et Lezhawa), Грузия, окр. с. Балда,
3V 1967.777 — внешний вид полового аппарата; 172 — вагина, вскрыта; 173— пенис,
вскрыт; 174 — поперечный срез папиллы пениса; 175 — поперечный срез эпифаллуса.
(По Schileyko, 1977).
имеется тонкая узкая белая губа. Пупок умеренно широкий, но не перспективный:
сквозь него просматривается не более одного оборота. Размеры: ВР 14—15, БД 19.5—
21.5, МД 17—18.5 мм.
Внутреннее строение. Материал: 1 экз. из юго-западных склонов
Большого Кавказа, с. Балда Гегечкорского р-на, 3 V 1967, сбор Г. И. Лежавы, опре-
деление В. Гудеца; 1 — из центральной части южных склонов Большого Кавказа,
с. Квайса (Кванси) Джавского р-на, 19 X 1956, сбор К. Городкова, определение наше.
Нижняя часть спермовидукта и яйцевод изгибов не образуют. СЖ 4X2—3. Сти-
лофоры умеренно массивные, внутренние немного меньше наружных. Внутри вагины
имеются 4 продольных валика, сближенных попарно и занимающих латеральное по-
ложение; в местах выходных отверстий стилофоров лопасти почти не выражены.
Пенис веретеновидный, слабо вздут, чехол пениса внутри гладкий. Папилла
пениса веретеновидная, семепроводящий канал фиксирован двумя продольными
тяжами, развитыми примерно в равной степени; вдоль линии прикрепления одного
из тяжей имеются 1—2 тонких капилляра. Бич примерно вдвое короче слабо изогну-
того эпифаллуса; пениальная мембрана имеется, но развита довольно слабо. Проток
семеприемника умеренно извит, заканчивается обширным резервуаром, почти каса-
ющимся белковой железы.
186
III. HYGROMIIDAE
Распространение и местообитания. Судя по тем двум точкам,
откуда известен вид, он распространен спорадически в высокогорье центральной
и западной частей южных склонов Большого Кавказа. Обитает на хорошо прогре-
ваемых склонах с густым травостоем.
2. Kokotschashvilia tanta Schileyko, 1977. Табл. VI, 60; рис. 176—178.
Schileyko, 1977 : 16, figs 57—61, Pl. Ill, И.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 7 экз. из одного местонахождения на западном Кавказе.
Раковина от прижатой до почти кубаревидной: высота завитка немного
больше или немного меньше высоты устья. Завиток конический, вершина слегка
Рис. 176—178. Kokotschashvilia tanta Schileyko, Грузия, окр. с. Лебарде,
5 VII 1962. Паратип. 176 — внешний вид полового аппарата; 177 — вагина,
вскрыта; 178 — пенис, вскрыт. (По Schileyko, 1977).
сглаженная. Оборотов 6.5—7, умеренно выпуклых. Последний оборот в профиле
округлый, не менее чем в 1.5 раза шире предпоследнего, у шва немного ступенчатый,
к устью слабо опущен. Свежие раковины светло-рогового цвета, который, слегка
сгущаясь выше и ниже периферии, образует неясные ленты, немного более темные,
чем промежуток между ними; этот промежуток в виде светлой ленты слабо просле-
живается у шва на предыдущих оборотах. Скульптура в виде неправильной тонкой
радиальной исчерченности, местами имеется неясная зернистость и спиральные бо-
роздки, лучше выраженные на периферии и на базальной части раковины. Устье
округлое или слегка оттянуто вправо и скошено. Чуть отступя от края имеется белая
губа, которой снаружи соответствует светлая полоса. Пупок эксцентрический,
сквозь него полностью виден предпоследний оборот. Размеры: ВР. 14.5—18, БД 22—
27, МД 18.5—22 мм. Голотип: ВР 16.8, БД 25.5, МД 21.0 мм.
Раковина похожа на раковину предыдущего вида, но отличается более темной
окраской, наличием более или менее ясных спиральных бороздок, большим числом
оборотов и более крупными размерами.
Внутреннее строение. Материал: 4 /экз. из западной части южных
склонов Большого Кавказа, с. Лебарде Гегечкорского р-на, 5 VII 1962, сбор
М. Г. Нацвлишвили, определение наше.
KOKOTSCHASHVILIA
187
Яйцевод образует плавный изгиб, тонкий, книзу расширяющийся. СЖ 4\2 —3.
Стилофоры относительно массивные, шаровидные, внутренние лишь немного меньше
наружных. Вагинальные валики очень мощные, образуют широкие длинные лопасти.
Пенис состоит из двух отделов: шаровидно-веретеновидного проксимального
и цилиндрического дистального; в первом из этих отделов помещается папилла.
Как и у предыдущего вида, семепроводящий канал охвачен парной интрапапиллярной
полостью, но у данного вида одна из перемычек тонкая, другая фактически отсут-
ствует, т. е. семепроводящий канал плотно спаян с внутренней стенкой папиллы
(ср. поперечные срезы через папиллу на рис. 174 и 178). В стенке папиллы вдоль се-
мепроводящего канала также имеются капилляры. Бич примерно вдвое короче эпи-
фаллуса, притянутого к пенису многочисленными мускульными лентами. Проток
семеприемника тонкий, слегка извитой, без резкой границы переходит в мешковид-
ный тонкостенный резервуар, стенки которого у всех изученных экземпляров нахо-
дились в спавшемся состоянии. То обстоятельство, что семеприемник оказался
пустым, связано со временем сбора (июнь).
Характерно, что у К. eberhardi и К. holotricha, также собранных в июне—июле,
семеприемники были пустыми, а у К. makvalae и К. phaeolaema, собранных в начале-
середине мая, этот орган был наполнен и имел шаровидную форму, а стенки его были
гладкими, без морщин.
Распространение и местообитания. Вид пока известен только
из типового местонахождения в высокогорье западного Кавказа (с. Лебарде). Об-
' наружен на альпийских лугах.
3. Kokotschashvilia eberhardi 1 Schileyko, 1977. Табл. VII, 6'7; рис. 179—181.
Schileyko, 1977:19, figs 67-70, Pl. Ill, 12.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: 12 экз. из центральной части Большого Кавказа, между с. Каз-
беги и Сиони.
Гаковина слегка прижатая или почти кубаревидная, с коническим или
чуть куполовидным контуром завитка, высота которого приблизительно равна вы-
соте устья. Оборотов 6. Последний оборот менее чем в 1.5 раза шире предпоследнего,
к устью немного опущен, в профиле правильно широко закруглен. Вершина сгла-
женная или слегка выступающая. Окраска характерная: основной фон раковины
светло-палевый, к шву окраска постепенно светлеет, выше и ниже периферии фон
может сгущаться, образуя две более темные ленты. Вместе с тем необходимо под-
черкнуть, что раковина в целом очень светлая, и отмеченные ленты выделяются
только в силу того что лежат на почти белом фоне. В проходящем свете перифериче-
ская светлая лента и пространство у шва темнее фона. Скульптура — неправильная
радиальная морщинистость, резче выраженная у шва. При увеличении видно, что
морщины у свежих раковин светлее фона, белые. Губа просвечивает в виде беловато-
желтой полоски. Местами имеются спиральные бороздки и зернистость, которая
гораздо заметнее на базальной поверхности раковины. Устье почти круглое, умеренно
вырезано предпоследним оборотом, места его прикрепления слегка сближены. Не-
сколько отступя от края имеется хорошо развитая белая губа. На протяжении жизни
моллюска образуются 2—4 губы; более ранние также просвечивают в виде светлых
полос. Пупок неширокий, но почти перспективный. Размеры: ВР 10—12, БД 14—
15.5, МД 12—13 мм. Голотип: ВР 10.3, БД 14.5, МД 12.2 мм.'
Вид близок к К. makvalae, отличается меньшими размерами, немного более вы-
сокой раковиной, желтоватой окраской и более широким пупком.
Внутреннее строение. Материал: 5 экз. из центрального Кавказа,
между с. Сиони и Казбеги, VII 1969, сбор Э. Клаусса, определение наше.
На границе спермовидукта и яйцевода имеется крутой, но’ нерезкий изгиб.
СЖ 4X3—4. Стилофоры очень массивные, область стилофоров отделена от нижнего
отдела вагины неясной перетяжкой. Помимо четырех основных валиков, сближенных
попарно и занимающих латеральное положение, имеются дополнительные более
мелкие складки. Основные валики образуют очень неясные лопасти.
Пенис массивный, в большей или меньшей степени изогнут. Пениальные мускуль-
ные ленты отсутствуют, имеется лишь тонкая прозрачная мембрана, натянутая между
пенисом, эпифаллусом и семепроводом. Внутренняя поверхность пениального чехла
гладкая. Папилла пениса объемистая, мешковидная, с выходным отверстием в виде
широкой щели и крупной папиллярной лакуной. Семепроводящий канал, спаян со
стенкой папиллы и окружен обширной интрапапиллярной полостью. Стенка папиллы,
противолежащая семепроводящему каналу, покрыта многочисленными выростами
и складками (рис. 181). Длина тонкого бича составляет приблизительно половину
1 Вид назван именем Эбергарда Клаусса.
188
III. HYGROMIIDAE
длины цилиндрического, слабо изогнутого эпифаллуса. Проток семеприемника почти
прямой, не отграничен от резервуара, имеющего очень тонкие стенки; поскольку
у изученных экземпляров семеприемник оказался пустым, стенки его спались и смор-
щились. Резервуар не достигает белковой железы.
Рис. 179—181. Kokotschashvilia eberhardi Schileyko, южные склоны центр. Кавказа,
между с. Сиони и с. Казбеги, VII 1968. Паратип. 179 — внешний вид полового ап-
парата; 180 — вагина, вскрыта; 181 — пенис, вскрыт. (По Schileyko, 1977).
Обращает на себя внимание интенсивная пигментация паллиального нерва..
Распространение и местообитания. Пока известна лишь
одна находка на центральном Кавказе. Моллюски найдены на открытом каменистом
склоне, на высоте 2500—3000 м над ур. м.
4. Kokotschashvilia holotricha (О. Boettger). Табл. VII, 62; рис. 182—186.
О. Boettger, 1884 : 151 {Helix); Westerlund, 1889 : 70 {Helix);
R e t.owsk i, -1914 : 283 [Helix (Latonia) ]; Rosen, 1914 : 163 {Helix); Hesse,
1931 : 5, t. 1, fig. 4 {F,rutlcicola); Лихарев, Раммельмейер, 1952 : 453
[Trichia (Trichia)]; П u'dec, L e z h a w a, 1969a : 47, Abb. 4, 8 [Trichia (Kokot-
schashvilia)]; 1969b : 127, Abb. 23, t. XII, 29 [Trichia (Kokotschashvilia)]; Schi-
leyko, 1977 : 18, figs 62—66.
KOKOTSCHA SHVILIA
189
Материал: 9 экз. из 5 местонахождений на западном Кавказе.
Раковина почти кубаревидная, тонкостенная, просвечивающая, завиток
конический, вершина почти не сглажена. Последний оборот заметно вздут, в 1.5 раза
шире предпоследнего, в профиле немного овальный, к устью еле опущен. Оборотов
5.5—6, слабо выпуклых, быстро нарастающих. Окраска одноцветно роговая или
коричневая, иногда видна исчезающе слабая лента по периферии. Скульптура в виде
частой и резкой радиальной исчерченности, поверхность раковины с густыми во-
лосками, сохраняющимися пожизненно. Устье немного овальное, косое, края его
тонкие, острые, слабо отвернуты, внутри с очень тонкой губой. Заметно отвернут
только колумеллярный край, частично прикрывающий очень узкий пупок. Размеры:
ВР 10-13, БД 15-18.6, МД 13—16 мм.
Рис. 182—186. Kokotschashvilia holotricha (Bttg.), с. Цебельда близ Сухуми, 15 VI 1968.
182 — внешний вид полового аппарата; 183 — вагина, вскрыта; 184 — пенис, вскрыт;
185 — поперечный срез папиллы пениса; 186 — поперечный срез эпифаллуса. (По
Schileyko, 1977).
Внутреннее строение. Материал: 2 экз. из с. Цебельда близ Сухуми,
15 VI 1968, сбор Г. И. Лежавы, определение В. Гудеца; 2 — из долины р. Техури
в юго-западной части Большого Кавказа, 1 V 1971, сбор и определение наши.
На границе спермовидукта и яйцевода имеется резкий изгиб. СЖ 4\2. Область
стилофоров отграничена от нижнего отдела вагины нерезким перехватом. Вагиналь-
ные валики сближены попарно, занимают латеральное положение. Внизу они круто
обрываются, так что нижняя часть вагины и клоака внутри гладкие или с неправиль-
ными мелкими складками. Нижние окончания вагинальных валиков связаны между
собой поперечным утолщением.
Пенис веретеновидный, внутренняя поверхность чехла пениса на границе с кло-
акой несет продольные складки. Папилла объемистая, семепроводящий канал свя-
зан со стенкой папиллы лишь одним продольным тяжом, т. е. интрапапиллярная
полость окружает семепроводящий канал практически со всех сторон. В стенке па-
пиллы никаких других полостей или капилляров не обнаружено. Бич слегка кони-
ческий, короткий, примерно вдвое короче эпйфаллуса, S-образный изгиб которого
обеспечивается пениальными мускульными лентами, развитыми у данного вида осо-
бенно хорошо; однако степень изогнутости эпифаллуса может меняться в значитель-
ных пределах. Проток семеприемника толстый, совершенно нечувствительно пере-
ходит в резервуар, не достигающий белковой железы.
190
III. HYGROMIIDAE
Распространение и местообитания. Черноморское побережье
Кавказа (Сочи, Гагра, Сухуми); на восток отходит на незначительное расстояние
от моря вдоль северных (Майкоп) и южных (бассейн р. Техури) склонов Большого-
Кавказа. Населяет горные леса.
189
Рис. 187—190. Kokotschashvilia phaeolaema (Bttg.), Чегемское ущелье, 14 V 1970,
187 — внешний вид полового аппарата; 188 — особь in copuli, пенис вывернут,
вид со стороны подошвы; 189 — вагина, вскрыта; 190 — пенис, вскрыт. (По Schi-
leyko, 1977).
5. Kokotschashvilia phaeolaema (О. Boettger, 1886). Табл. VII, 63; рис. 187—190,
О. Boettger, 1886 : 138, t. 3, fig. 4а—е [Helix (Fruticocampylaea)]; Lind-
holm, 1913 : 140 [Helix (Fruticocampylaea) phaeolaema var. tenuitesta]; Rosen,
1914 : 171 (Helix)-, К алитина, 1958 : 59, рис. 1 (Trichia)-, Hudec, 1972d : 259r
Abb. 7 [Trichia (Xerocampylaea)]-, Schileyko, 1977 : figs 71—76, Pl. Ill, 13.
CAUCASIGENA
191
Материал: более 100 экз. из 3 местонахождений в Дагестане и Чегемском ущелье
на Северном Кавказе.
Раковина от кубаревидной до почти шаровидной, высота завитка равна
высоте устья или превышает ее. Контур завитка от строго конического до слегка
куполовидного, вершина сглаженная и немного выступающая. Оборотов 6, округлых,
разделенных беловатым швом. Последний оборот очень плавно закруглен, в конце
внезапно и достаточно резко опущен. Окраска коричневая, светло-роговая, кашта-
новая или желтоватая, светлых спиральных лент нет или одна исчезающе слабая
лента по периферии; на базальной стенке раковины просвечивают 2—5 губ в виде
светлых радиальных полос, кнаружи от которых, тесно прилегая к ним, располага-
ется полоса такой же ширины и более темная, чем фон раковины. Скульптура в виде
радиальных складок и ясной спиральной исчерченности. Устье округлое, косое;
внутри него, отступя от края, лежит толстая белая губа. Края устья слабо отвернуты;
колумеллярный край отвернут сильнее, слегка или наполовину прикрывает узкий
неперспективный пупок. Размеры: ВР 9—12, БД 11—16.0, МД 10—13.5 мм.
Внутреннее строение. Материал: 2 экз. из Дагестана, Хунзах-
ский р-н, 26 VIII 1955, сбор Т. Хасанова, определение И. М. Лихарева; 17 — из Че-
гемского ущелья близ водопадов, 14 V 1970, сбор и определение наши.
Яйцевод образует крутой изгиб. СЖ 4X2—3. Стилофоры очень хорошо развиты,
шаровидные, вагина в области стилофоров вздута. Вагинальные валики небольшие,
но в местах выходных отверстий стилофоров образуют четкие удлиненные лопасти.
Пенис объемистый, мешковидный или шаровидный, чехол пениса. внутри глад-
кий, только у самой клоаки имеются небольшие продольные складки. Папилла сильно
вздута, шаровидной или грушевидной формы. Семепроводящий канал подвешен на
двух продольных противолежащих тяжах и окружен парной интрапапиллярной
полостью. В отличие от остальных видов рода стенки папиллы толстые и заключают
дополнительные интрапапиллярные полости в виде разбросанных в толще стенок
небольших синусов, в результате чего стенка иногда выглядит губчатой. Длина бича
от 1/2 до ®/4 длины эпифаллуса, последний дважды изогнут и притянут к пенису хо-
рошо развитой мембраной, в состав которой входят мускульные волокна. Семепри-
емник чрезвычайно массивный, верхний край его соприкасается с нижним краем
белковой железы.
Характерной особенностью вида является интенсивная пигментация ряда внут-
ренних органов: матки, дистального отдела кишечника, мезентерия и эпиневраль-
ной оболочки окологлоточного нервного кольца.
Распространение и местообитания. Центральная и вос-
точная части Большого Кавказа. Встречается в сырых затененных ущельях с обиль-
ной растительностью или в местах выхода грунтовых вод.
7. Род CAUCASIGENA Lindholm, 1927
Lindholm, 1927а : 123 (Fruticocampylaea, sect. Caucasigena)', W e n z,
Zilch, 1959—1960 : 686 [Xerocampylaea (Caucasigena)]-, Schileyko, 1977 : 21.
Раковина от почти плоской до слегка прижатой, первично белая с двумя спираль-
ными темными лентами или слегка зеленоватая, или чуть красноватая, одноцветная.
Пупок большей частью развернутый, перспективный, гораздо реже узкий. Раковина
без волосков, лишь изредка у отдельных экземпляров могут присутствовать отдель-
ные редкие волоски.
Стилофоры хорошо развиты, прижаты к вагине. Семепроводящий канал внутри
пениальной папиллы очень узкий; окружающая его интрапапиллярная; полость раз-
делена продольными септами на 3—9 продольных полостей. Папиллярная лакуна
развита хорошо; кроме нее, никаких борозд на поверхности папиллы нет. Ретрактор
правого омматофора образует перехлест с половым ретрактором.
Распространение. Кавказ. В ископаемом состоянии род известен из
верхнемиоценовых отложений (меотис) Предкавказья.
Типовой вид — Helix eichwaldi L. Pfeiffer, 1846.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕПТНЫХ ВИДОВ РОДА С A UCASIGENA
1 (2). Раковина без темных лент. Пупок очень узкий, не менее чем наполовину при-
крыт отворотом колумеллярного края устья ........... . 10. С. thalestris.
2 (1). Раковина с темными лентами. Пупок умеренно узкий; если прикрыт, то менее
чем наполовину.
3 (12). Сквозь пупок просматриваются все или почти все обороты.
4 (7). Последний оборот в профиле правильно закруглен.
5 (6). Пупок развернутый, края устья сильно сближены, угол наклона устья не
менее 45°...............................................6. С. eichwaldi.
192
III. HYGROMIIDAE
6 (5). Пупок цилиндрический, края устья почти не сближены, угол наклона устья
не более 60°..........................................9. С. schaposchnikovi.
7 (4). Последний оборот в профиле более или менее угловатый.
8 (9). Раковина без радиальных ребер и без радиальных светлых штрихов . . . .
.........................................................3. С. arineniaca.
9 (8). Поверхность раковины покрыта радиальными светлыми ребрами или штрихами.
10 (11). Раковина с очень резкими белыми ребрами, одинаково выраженными на верх-
ней и нижней сторонах раковины.......................... 4. С. tschetschenica.
11 (10). Раковина со светлыми радиальными штрихами или слабыми ребрами, лучше
выраженными на верхней стороне раковины....................5. С. rengarteni.
12 (3). Сквозь пупок просматривается не более одного оборота.
13 (14). Последний оборот в профиле плавно закруглен. Окраска белая, обычно
с 1—2 коричневыми спиральными лентами......................7. С. abchasica.
14 (13). Последний оборот в профиле угловатый. Окраска зеленовато-бурая или олив-
ковая, со светлой лентой на периферии...................8. С. reminiscenta.
1. Подрод CAUCASIGENA s. str.
Раковина более или менее уплощенная, светлая, с двумя спиральными темными
лентами выше и ниже периферии; если раковина почти одноцветно коричневая, эта
окраска возникает за счет сильного развития спиральных лент. Волоски на раковине
развиты слабо или отсутствуют. Пупок в той или иной степени развернутый; если ци-
линдрический, сквозь него все равно можно увидеть больше одного оборота.
Бич толстый, конический, не длиннее половины эпифаллуса, реже той же длины.
Внутренние стилофоры ненамного меньше наружных. Семепроводящий канал подве-
шен внутри папиллы.
Распространение. Кавказ. В ископаемом состоянии подрод известен
из меотических отложений Предкавказья.
ф 1. Caucasigena (Caucasigena) psedachica (Steklov, 1966). Табл. VII, 64.
Стеклов, 1966 : 226, табл. XI, 205, 206 (Tropidomphalus).
Материал: 6 экз. из одного местонахождения в Предкавказье.
Раковина сильно прижатая, с широким коническим завитком. Оборотов 6,
умеренно быстро нарастающих, разделенных неглубоким швом. Последний оборот
почти вдвое шире предпоследнего, к устью энергично опущен, снизу немного уплощен.
Окраска не сохранилась. Скульптура в виде радиальной неравномерной морщини-
стости и очень тонкой, но ясной спиральной струйчатости. Устье сильно скошено,
края его слегка сближены и широко отвернуты; немного отступя от края имеется слегка
расплывшаяся, но довольно высокая губа. Пупок широкий, развернутый. Размеры:
ВР около 10, БД около 17, МД около 15 мм; точнее размеры привести невозможно,
так как весь материал представлен мятыми раковинами.
Распространение. Акчагыл и апшерон Предкавказья.
ф 2. Caucasigena (Caucasigena) fortangensis (Steklov, 1966). Табл. VII, 65.
Стеклов, 1966 : 226, табл. XI, 207—209 (Tropidomphalus).
Материал: 6 экз. из одного местонахождения в Предкавказье.
Раковина уплощена, с широким коническим или чуть куполовидным завит-
ком. Оборотов 6, более медленно нарастающих, чем у предыдущего вида: ширина по-
следнего оборота примерно в 1.5 раза шире предпоследнего. Последний оборот к устью
резко опущен, в базальной части уплощен, по периферии, возможно, слегка угловатый;
точно сказать трудно, так как все экземпляры типовой серии смяты. Скульптура в виде
тонкой и неравномерной радиальной исчерченности и очень слабых спиральных бо-
роздок, которые видны лишь местами на последнем обороте. Устье резко скошено,
места его прикрепления широко расставлены, при рассмотрении снизу устье выгля-
дит почти правильно круглым. Края устья острые, чуть отвернуты, колумеллярный
край отвернут сильно и частично перекрывает круглый неширокий пупок. Размеры:
ВР 6—7, БД 14—15, МД 12—13 мм.
Распространение. Меотический песчаник на р. Фортанге (Предкав-
казье).
3. Caucasigena (Caucasigena) arineniaca (L. Pfeiffer, 1846). Табл. VII, 66; рис. 191 —
195.
L. Pfeiffer, 1846, Syst. Hist. Helic.,III : 77 (Helix); 1846, in Conch.-Cab.,
ed. II : 132, t. 17, fig. 23—25 (Helix); Siemaschko, 1847 : 120 (Helix ziegleri
CAUCASIGENA
193
var. airumla), 121 (H. menetriesli nom. nov. pro H. nivalis Menetr.); Westerlund,
1889 : 108 (Helix)', Hesse, 1931 : 10, t. 1, fig. 6 (Fruticicola)', Schileyko,
1977 : figs. 77-82, Pl. Ill, 14.
Материал: 4 экз. из одного местообитания в Армении.
Раковина прижатая, сверху и снизу примерно одинаково выпуклая, уме-
ренно твердостенная. Оборотов 5, плавно нарастающих, слегка уплощенных. Послед-
ний оборот с закругленным углом по периферии, немного ' шире предпоследнего,
Рис. 191—195. Caucasigena (Caucasigena) armeniaca (L. Pfr.), Армения, гора Тодар,
18 VII 1951. 191 — внешний вид полового аппарата; 192 — вагина, вскрыта (звездоч-
кой помечена стрела); 193 — пенис, вскрыт; 194 — поперечный срез папиллы пениса;
195 — поперечный срез эпифаллуса. (По Schileyko, 1977).
к устью слегка опущен. Окраска коричневато-роговая, образована за счет интенсив-
ного развития спиральных темных лент, между которыми на периферии остается свет-
лая лента, в проходящем свете просвечивающая сильнее фона. Скульптура очень ха-
рактерная: сглаженные редкие радиальные морщины, хорошо заметные только при
увеличении, и редкие спиральные кожистые ребрышки в виде тончайших нитей; на
первый взгляд они создают впечатление спиральных бороздок, столь обычных для пред-
ставителей рода, но при увеличении в 30—40 раз при косом концентрированном свете
видно, что это положительные элементы скульптуры, т. е. выпуклые, а не врезанные.
Кроме того, на раковине имеются тонкие редкие, слабо изогнутые волоски, которые
легко опадают. Устье округлое, сильно скошено; внутри него отступя от края имеется
мощная желтоватая или белая губа, просвечивающая сквозь стенку раковины. Пупок
в начале широкий, развернутый, но затем резко сужается и становится цилиндриче-
ским, хотя остается перспективным. Размеры: ВР 4—5, БД 7.7—9, МД 6.5—
7.5 мм.
Внутреннее строение. Материал: 2 экз. из Армении, Степанаванский
р-н, гора Тодар (Бзовдал), 18 VII 1951, .сбор и определение Н. Н. Акрамовского.
Спермовидукт и яйцевод прямые. СЖ 4X2. Стилофоры очень характерные: на-
ружные удлинены, верхние концы внутренних выдаются за концы наружных.
Центральные части внутренних стилофоров выгнуты медиально и могут почти соприка-
саться. Внутренняя пара стилофоров стиснута с обеих сторон и заключает две.полости
(рис. 192). Вагинальные валики развиты хорошо, но лопасти почти не выражены.
Пенис слегка вздут, веретеновидный, на его поверхности имеются слабо развитые
соединительнотканные ленты. Чехол пениса внутри гладкий, папилла булавовидная
13 А. А. Шилейко
194
III. HYGROMIIDAE
или веретеновидная, с хорошо заметной папиллярной лакуной. Семепроводящий ка-
нал подвешен к внутренней стенке папиллы двумя продольными тяжами; имеются
2—3 интрапапиллярные полости, разделенные продольными септами (рис. 194). Бич
и эпифаллус короткие, бич примерно вдвое короче эпифаллуса, толстый, конический.
Эпифаллус цилиндрический, слабо изогнут. Между дистальными протоками мужского
отдела натянута тонкая прозрачная мембрана. Проток семеприемника почти прямой,
тонкий, резервуар небольшой, округлый, прилегает к верхней части спермовидукта.
Рис. 196—200. Caucasigena (Caucasigena) tschetschenica (Ret.), среднее течение р. Фиг-
дон на Сев. Кавказе, 19 IX 1970.196 — внешний вид полового аппарата; 197 — вагина,
вскрыта (звездочкой помечена стрела); 198 — пенис, вскрыт; 199 — поперечный срез
папиллы пениса; 200 — поперечный срез эпифаллуса. (По Schileyko, 1977).
Распространение и местообитания. В связи с путаницей,
существующей в литературе, в настоящее время трудно очертить ареал вида. Так, все
отмеченные находки этого вида на Северном Кавказе относятся не к этому виду,
а к С. rengarteni (Лихарев, Раммельмейер, 1952; Калитина, 1958, 1967); послед-
ний ошибочно сведен в синонимы к «Trichia» armeniaca Лихаревым и Раммельмейер;
настоящая С. armeniaca в указанный определитель вообще не включена.
Вид наверняка имеется на Малом Кавказе — от Бзовдальского (Базумского) до
Мровдагского хр. — и в юго-восточной части Большого Кавказа; возможно, здесь вид
представлен подвидом menetriesii Siemaschko, 1847, но, не имея материала, мы не ре-
шаемся говорить о различиях между популяциями, обитающими в этих районах.
Высокогорный вид, обитает на скалах и в осыпях.
4. Caucasigena (Caucasigena) tschetschenica (Retowski, 1914). Табл. VII, 67',
рис. 196—200.
Retowski, 1914 : 292 [Helix (Fruticocampylaea)}', Schileyko, 1977 :
figs 83-87, Pl. Ill, 15.
Материал: более 100 экз. из 1 местонахождения в центральной части Северного
Кавказа.
CAUCASIGENA i&5
Раковина от плоской до слегка прижатой, но вершина в любом случае вы-
ступающая, причем она выступает тем сильнее, чем более прижат завиток. Оборотов 5,
умеренно выпуклых, плавно нарастающих. Последний оборот менее чем в 1.5 раза шире
предпоследнего, к устью резко опущен, на периферии с углом или даже килем. Ок-
раска фона белая или светло-серая; выше и ниже периферии проходят темные спираль-
ные ленты, просвечивающие в проходящем свете сильнее фона. Радиальная скульп-
тура представлена относительно очень мощными ребрами, окрашенными в белый цвет
и пересекающими спиральные темные ленты. В промежутках между ребрами имеются
тонкие спиральные бороздки. Устье очень косое, от почти круглого до широкооваль-
ного, внутри с мощной и высокой белой губой. Края устья, особенно базальный и па-
латальный, отвернуты, а места их прикрепления сближены. Пупок вначале развернут,-
затем быстро сужается. Размеры: ВР 3.2—4.8, ВД 6.5—9.8, МД 5.6—8 мм. Для ориги-
нальных экземпляров Ретовский указывает следующие размеры: ВР 4—4.5, БД 7.7—8мм.-
Внутреннее строение. Материал: 8 экз. из Куртатинского ущелья
на Северном Кавказе, среднее течение р. Фиагдон, 19 IX 1970, сбор и определение наши,-
Яйцевод образует плавный изгиб, спермовидукт прямой. СЖ 4x2. Стилофоры-
вытянутые, внутренние несколько обособлены от вагины, хорошо развиты, наружные
примерно той же величины. Вагинальные валики резкие, но тонкие и лопастей не об-
разуют.
Пенис веретеновидно-шаровидный, папилла относительно очень массивная, мешко-
видная. Семепроводящий канал крепится одним продольным тяжом, интрапапилляр-
ная полость разделена продольными септами на 3—5 отделов. Бич конический, тол-
стый, вдвое короче прямого эпифаллуса; между семепроводом, эпифаллусом и пени-
сом натянута прозрачная мембрана. Проток семеприемника без изгибов, заканчивается
удлиненно-овальным резервуаром, не достигающим белковой железы.
Распространение и местообитания. Вид найден в двух местах:
типовое местообитание — гора Боноз-мта в Чечено-Ингушетии; большая популяция
обнаружена в среднем течении р. Фиагдон на Северном Кавказе. Обитает на отвесных
гладких известковых скалах.
5. Caucasigena (Caucasigena) rengarteni (Lindholm, 1913). Табл. VII, 68; рис. 201,.
202.
Lindholm, 1913с : 142, 143 [Helix (Fruticocampylaea)]; [H. (Fr.) gerassimovi];:
Лихарев, Раммельмейер, 1952 : 456 [Trichia (Xerocampylaea) armeniaca.
part.]; Schileyko, 1977 : figs 88—91, Pl. IV, 16, 17.
Материал: более 100 экз. из 5 местонахождений на Северном Кавказе.
Раковина от почти плоской до умеренно прижатой. Оборотов 5—5.5, слабо-
выпуклых. Последний оборот в начале с ясным углом, к устью пропадающим, в конце-
резко опущен. Темные спиральные ленты развиты очень хорошо, широкие, фон белый
или светло-серый. В проходящем свете темные ленты просвечивают сильнее фона. По- '
верхность раковины покрыта многочисленными радиальными складками или реб-
рами, гораздо более ясно выраженными на верхней стороне раковины; в промежутках
между радиальными элементами скульптуры имеются спиральные бороздки. У неко-
торых экземпляров наблюдаются редкие жесткие короткие волоски на верхней повер-
хности раковины. Радиальные складки в тех местах, где их не пересекают темные ленты,
окрашены в белый цвет. Устье широкоовальное, очень косое, места его прикрепления
сильно сближены, порой наблюдается наклонность к образованию цельного устья..
Чуть отступя от края лежит толстая и высокая белая губа; базальный и палаталь-
ный края отвернуты. Пупок развернутый, перспективный. Размеры: ВР 5.8—8.2,
БД 10—17, МД 7.5—14 мм (в первоописании ВР 7, БД 14.5 мм).
Внутреннее строение. Материал: 12 экз. из среднего течения р. Че-
гем возле с. Хуштосырт, 14 V 1970; 8 — из верховий р. Чегем возле с. Верхний Чегем,
19 V 1970; 14 — из урочища Ара-Боран между Чегемским и Баксанским ущельями,.
17 V 1970; 3 — из среднего течения р. Фиагдон, 19 IX 1970; сборыи определения наши..
Спермовидукт и яйцевод почти прямые. СЖ 4Х2. Стилофоры массивные, почти
шаровидные. Вагинальные валики выражены в различной степени, но всегда обра-.
зуют удлиненные лопасти.
Пенис состоит из двух отделов: веретеновидно-шаровидного проксимального, где
располагается папилла, и узкого цилиндрического дистального. По поверхности пе-
ниального чехла проходят соединительнотканные ленты, в состав которых входят,
по-видимому, мускульные волокна. Проксимальный отдел пениса внутри гладкий,
с мелкими морщинами и складками, дистальный несет многочисленные продольные
морщины, которые переходят в складки внутри клоаки. Папилла пениса объемистая,
мешковидная, с выходным отверстием в виде широкой щели и большими папиллярными
лакунами. Семепроводящий канал фиксирован двумя или тремя продольными тяжами;
интрапапиллярная полость разделена септами на 4—6 отделов. Эпифаллус прямой
или изогнутый, связан с пенисом очень тонкой прозрачной мембраной. Проток семе-
13* =
196
III. HYGROMIIDAE
приемника прямой или очень слабо изогнут, резервуар овальный, заметно не достает
до белковой железы.
Вместе с описанием данного вида Линдгольм (Lindholm, 1913) приводит описа-
ние еще одного вида —• Helix (Fruticocampylaea) gerassimovi и подчеркивает отличия
этого вида от Helix armeniaca, но из текста неясно, чем, кроме размеров, отличаются
между собой новые виды. Мы имели возможность ознакомиться с типовыми сериями
по обоим видам Линдгольма (хранятся в Зоологическом институте АН СССР) и нашли,
что различия между ними укладываются в рамки внутрипопуляционной изменчивости;
большинство особей из урочища Ара-Воран подходят под диагноз Helix gerassimovi,
в большинстве серий из Чегемского ущелья наблюдались плавные переходы между ра-
Рис. 201—202. Caucasigena (Caucasigena) rengarteni (Lindh.), Чегемское ущелье, окр.
с. Хуштосырт, 14 V 1970. 201 — внешний вид полового аппарата; 202 — дистальная
часть полового аппарата, вскрыта. (По Schileyko, 1977).
ковинами, подходящими под диагнозы обоих видов; анатомических различий не най-
дено. В связи с этим название «Я. gerassimovi» — синоним Caucasigena rengarteni.
Распространение и местообитания. Северо-восточная часть
Главного Кавказского хр. В самой восточной части ареала вид граничит с видом, кон-
хологически очень похожим — Shlleykoia daghestana (Hygromiinae)'. Указания на при-
сутствие вида в западном Азербайджане и большей частью в Дагестане касаются именно
этого последнего вида. Вид встречается на затененных скалах и в сырых гротах, боль-
шей частью в узких ущельях.
6. Caucasigena (Caucasigena) eichwaldi (L. Pfeiffer, 1846). Табл. VII, 69, 70;
рис. 203, 204.
L. Pfeiffer, 1846:77 (Helix); 1846, in Conch.-Cab., ed. 11:132, t. 17,
fig. 20—22 (Helix); W esterlund, 1889 : 108 (Helix); Lindholm, 1913b : 18
[Helix (Fruticocampylaea) eichwaldi var. nivicola}; Hesse, 1931 : 9, t. 1, fig. 7 (Fruti-
cicola); Лихарев, Раммельмейер, 1952 : 455, рис. 382 [Trichia (Xero-
campylaea)}; Hudec, Lezhawa, 1967b : 88, Abb. 10, t. IV, 9 [Trichia (Xeroca-
mpylaea)}; Schileyko, 1977 : figs 92—95, Pl. IV, 18.
Материал: более 60 экз. из 4 местообитаний в Чегемском и Дарьяльском ущельях
на Северном Кавказе и в северо-западном Дагестане.
Раковина в разной степени прижатая, с куполовидным завитком и более
или менее выступающей вершиной. Оборотов 5.5, плавно нарастающих, выпуклых,
CAUCASIGENA
197
разделенных глубоким швом. Последний оборот по периферии правильно округлый,
без намека на угловатость, не менее чем в 1.5 раза шире предпоследнего (нередко почти
вдвое шире предпоследнего), к устью резко опущен. Основная окраска грязно-белая,
имеются две бурые или темно-коричневые ленты, которые в проходящем свете сильно
просвечивают. В северо-западном Дагестане иногда встречаются мелкие популяции,
у которых ленты слабые или отсутствуют, а фон имеет явственный розоватый оттенок.
Скульптура в виде радиальных морщин или очень сильно сглаженных ребрышек, лучше-
выраженных на верхней поверхности раковины. Спиральные бороздки слабые, иногда
Рис. 203—204. Caucasigena {Caucasigena} eichwaldi (L. Pfr.), Царьяльское ущелье,
окр. с. Чми, 8 V 1970. 203 — внешний вид полового аппарата; 204— дистальная
часть полового аппарата, вскрыта. (По Schileyko, 1977).
отсутствуют; на последнем обороте, как правило, наблюдаются отдельные малозамет-
ные вмятины. Устье коротко-овальное, сильно скошено; края его тонкие, острые,
сильно отвернуты, вследствие чего устье старых особей может иметь раструбообразную
форму. Внутри устья, отступя от края лежит узкая высокая белая губа. Места прикреп-
ления устья сильно сближены. Пупок развернутый, перспективный. Размеры: ВР 7—10,
БД 12—20, МД 10—16 мм.
Вид похож на предыдущий, но у него никогда не наблюдается ни киля, ни даже
наклонности к образованию угла по периферии.
Внутреннее строение. Материал: 2 экз. из Дагестана близ с. Целитль,
10 VII 1973, сбор И. М. Лихарева; 12 — из Дарьяльского ущелья близ с. Казбеги,
9 V 1970; 9 — из Дарьяльского ущелья близ с. Чми, 8 V 1970; сборы и определения
наши.
На границе спермовидукта и яйцевода имеется выраженный в различной степени
изгиб. СЖ 4X2. Стилофоры большие, чуть удлиненные или шаровидные. Вагинальные
валики образуют небольшие лопасти в местах выходных отверстий стилофоров.
Пенис очень резко подразделен на цилиндрический дистальный и вздутый прокси-
мальный отделы. Папилла пениса чрезвычайно массивная, с большими папиллярными
лакунами; семепроводящий канал фиксирован 2—3 продольными тяжами, интрапапил-
лярная полость разделена септами на 6—9 узких отделов. Длина бича не более поло-
вины длины эпифаллуса, обычно несколько меньше. Эпифаллус не образует крутых
198
III. HYGROMIIDAE
изгибов. Проток семеприемника при основании изогнут, резервуар соприкасается
с нижним краем белковой железы.
Поскольку два последних вида иногда очень похожи и несомненно близки, приво-
дим анатомические различия. У С. rengarteni слизистые железы впадают примерно на
уровне верхних концов стилофоров; яйцевод прямой; проток семеприемника почти
не изогнут; длина бича не меньше, а обычно несколько больше половины длины эпи-
фаллуса; интрапапиллярная полость разделена на 4—6 отделов. У С. eichwaldi сли-
зистые железы впадают значительно выше уровня верхних концов стилофоров; яйце-
вод и проток семеприемника изогнуты; длина бича обычно меньше половины длины
эпифаллуса; интрапапиллярная полость разделена на 6—9 отделов.
Рис. 205—206. Caucasigena (Caucasigena) abchasica (Lindh.), долина p. Ин-
гури, 10 IV 19.71. 205 — внешний вид полового аппарата; 206 — пенис,
вскрыт. (Ориг.).
Распространение и местообитания. Восточная и северо-во-
сточная части северных склонов Главного Кавказского хр. В районе Крестового пе-
ревала вид заходит на южные склоны, но далеко вниз не спускается. Обитает в ущель-
ях на скалах; в высокогорье — под камнями среди альпийских ксерофитов, где обра-
зует популяции, состоящие из карликовых особей.
7. Caucasigena (Caucasigena) abchasica (Lindholm, 1927). Табл. VIII, 71; рис. 205,
206.
Boettger, 1883 : 170, t. 4, fig. 3—5 (Helix pontica, non Fischer, 1866); Rosen,
1911 : 106 (H. narzanensis var. pontica); Lindholm, 1927a : ,133 (Helix); Hesse,
1931 : 10, t. 1, fig. 5 (Fruticicola); Лихарев, Раммельмейер, 1952 : 457
[Trichia (Xerocampylaea)]; Hudec, Lezhawa, 1969b : 128, fig. 24, t. XI, 28
[Trichia (Xerocampylaea)].
Материал: более 100 экз. из 2 местонахождений в юго-западной части Большого
Кавказа.
Раковина от слегка прижатой до почти кубаревидной, с коническим завит-
ком, составлена 5—5.5 выпуклыми, плавно нарастающими оборотами. Последний обо-
CAUCASIGENA
199
рот почти вдвое шире предпоследнего, по периферии плавно и широко закруглен,
снизу едва уплощен, к устью резко опущен. Вершина не сглажена. Окраска извест-
ково-белая, спиральные ленты узкие, пространство между ними шире любой из лент,
которые в проходящем свете очень сильно просвечивают. У отдельных экземпляров
одна или обе ленты отсутствуют. Радиальная скульптура в виде тонкой и неравномер-
ной исчерчеиности; спиральные бороздки очень ясные.. Устье коротко-овальное или
почти круглое, слабо вырезано предпоследним оборотом, внутри белое, места его при-
крепления слегка сближены, края тонкие, отвернутые, особенно базально-колумел-
лярный. Немного отступя от края устья лежит тонкая узкая белая губа. Пупок ци-
линдрический, узкий, сквозь него видно не более предпоследнего оборота. Размеры:
ВР 11—12, БД 18-20, МД 16—17 мм.
Внутреннее строение. Материал: 6 экз. из долины р.Ингури на юго-
западном Кавказе, 10 IV 1971; 7 — из долины р. Техури немного восточнее предыду-
щего местонахождения, 24 IV 1971; сборы и определения наши.
Дистальная часть спермовидукта и яйцевод почти прямые. СЖ 2—4 X 2. Стилофоры
относительно небольшие, наружные шаровидные, внутренние несколько вытянуты.
Вагина тонкая. Вагинальные валики и лопасти в местах выхода протоков стилофоров
развиты умеренно.
Пенис веретеновидный, папилла большая, веретеновидная или цилиндрическая.
Семепроводящий канал подвешен только на одном продольном тяже. Септа одна, реже
2—3, причем септы разбиты на отдельные поперечные «перекладины». На стороне па-
пиллы, противолежащей семепроводящему каналу, имеется ряд неполных поперечных
перегородок, отвечающих «перекладинам» септы (рис. 206). Сравнительно с другими
представителями подрода бич тонкий, не столь явственно конический, немного короче
круто изогнутого эпифаллуса. Между семепроводом, пенисом и эпифаллусом имеется
тонкая перепонка. Проток семеприемника близ резервуара образует 1—3 коротких
резких колена. Резервуар большой, обычно изогнут, не достигает нижнего края белко-
вой железы.
Распространение и местообитания. Северные и южные склоны
западной части Главного Кавказского хр. Обитает на открытых склонах и долинах
горных рек, где держится под камнями и на травянистых растениях.
8. Caucasigena (Caucasigena) reminiscenta Schileyko, sp. nov. Табл. VIII, 72.
Материал: 11 экз. из Кавказского заповедника.
Голотип хранится в коллекции Зоологического института АН СССР.
Раковина почти кубаревидная, с куполовидными очертаниями завитка и
слабо выступающей или сглаженной вершиной, сверху и снизу примерно одинаково
выпуклая. По периферии проходит тупой, но ясный угол, к устью сглаживающийся.
Оборотов 6, умеренно выпуклых, равномерно и плавно нарастающих. Последний обо-
рот ненамного шире предпоследнего, к устью опущен. Окраска зеленовато-бурая или
оливковая, местами образует сгущения; по углу проходит размытая светлая полоса,
вокруг пупка окраска чуть светлее. Скульптура очень характерная: верхняя часть ра-
ковины покрыта редкими, сильно сглаженными ребрами, которые на базальной части
выражены значительно слабее; часть ребер окрашена светлее фона. Кроме того, име-
ются тонкие редкие правильные спиральные бороздки. Устье умеренно вырезано,
овальное, заметно скошено; края его отвернуты, за исключением верхнего. Отступя
от края имеется белая, узкая, но высокая губа. Пупок цилиндрический, узкий, не-
перспективный. Размеры: ВР 6.4—7.5, БД 11.3—12.2, МД 9.5—10.4 мм. Голотип:
ВР 7.4, БД '12.0, МД 10.3 мм.
От других видов рода отличается окраской и скульптурой.
Внутреннее строение неизвестно.
Распространение и местообитания. Пока известна только
одна находка, сделанная И. М. Лихаревым в Кавказском заповеднике на горе Псеашко.
Вид обнаружен на альпийском лугу среди камней на высоте 2000 м над ур. м.
2. Подрод ANOPLITELLA Lindholm, 1929
Lindholm, 1929 : 209; W е n z, Zilch, 1959—1960 : 687.
Раковина прижатая, белая или слегка лиловатая, спиральные ленты развиты
в разной степени, но всегда имеются. Волосков на раковине нет. Пупок умеренно раз-
вернутый, но достаточно широкий и перспективный.
Бич длинный, тонкий, несколько длиннее эпифаллуса. Наружные стилофоры го-
раздо крупнее внутренних. Семепроводящий канал подвешен к стенке папиллы.
Распространение. Северный Кавказ.
Монотипический подрод.
Типовой вид — Helix schaposchnikovi Rosen, 1911.
гоо
III. HYGROMIIDAE
9. Caucasigena (Anoplitella) schaposchnikovi (Rosen, 1911). Табл. VIII, 73, 74;
рис. 207—210.
Rosen, 1911 : 113, t. 3, fig. 2 (Helix); 1914 : 172, t. 1, fig. 4 (Helix antoni. non ,
Pfeiffer, 1876); Lindholm, 1927a : 134 (H. antonivagneri); 1929 : 208 (H. schapos-
chnikovi var. balkariensis); Л и x a p ев, Раммельмейер, 1952 : 455, рис. 383
(Trichia (Xerocampylaea)]; Schileyko, 1977 : figs 96—100, Pl. IV, 19, 20.
Материал: более 100 экз. из 4 местонахождений в Чегемском ущелье на Северном
Кавказе.
Рис. 207—210. Caucasigena (Anoplitella) schaposchnikovi (Rosen), Чегемское ущелье,
15 V 1970. 207 — внешний вид полового аппарата; 208 — мантийный валик; 209 —
пенис, вскрыт; 210 — поперечный срез эпифаллуса. (По Schileyko, 1977).
Раковина уплощенная, с широко закругленным куполовидным завитком,
нередко почти плоская; вершина сглаженная. Оборотов 5.5—6, слабо выпуклых, мед-
ленно нарастающих. Последний оборот широко и правильно закруглен, снизу выпук-
лый, в 1.5 раза или немного больше шире предпоследнего, к устью слегка опущен.
Фон раковины почти белый, с рисунком из двух коричневых широких лент и лилова-
тых пятен, линий или штрихов. Обычно превалирует один из типов окраски, причем
чем резче выделяются спиральные ленты, тем слабее второй тип окраски, вплоть до
полного исчезновения. Имеет место также обратная зависимость. В проходящем свете
ленты просвечивают, а фон гораздо плотнее; светлые разводы на раковине также имеют
большую оптическую плотность. Скульптура состоит из тонкой радиальной неправиль-
ной исчерченности и слабых спиральных бороздок, выраженных местами. Устье ши-
рокбовальное, косое, края его почти не отвернуты; места прикрепления устья широко
расставлены. Несколько отступя от края лежит толстая белая губа. Раковина взрос-
лого моллюска имеет еще от 1 до 4 более ранних губ, которые просвечивают сквозь
стенку раковины в виде светлых полос. Пупок в начале немного развернутый, затем
цилиндрический, умеренно широкий, почти перспективный. Размеры: ВР 5—8.5,
БД 10—15.5, МД 8.5—13 мм.
Вид представлен' двумя- экологическими морфами, различающимися окраской и
рисунком на раковине и вместе не встречающимися (фото 73 и 74 на табл. VIII). По
остальным конхологическим признакам эти морфы не различаются, анатомических
различий также не найдено.
CAUCASIGENA
201
В тенистых ущельях при слабой инсоляции и повышенной влажности обитает морфа
с двумя яркими широкими лентами, верхняя из которых прослеживается на предыду-
щих оборотах. В местах с интенсивной инсоляцией раковина лиловатая, часто с кашта-
новым оттенком, иногда даже чуть розоватая. По этому фону разбросан узор описан-
ного выше типа. С такими экземплярами имел дело Линдгольм, описывая var. balka-
riensis.
'Внутреннее строение. Материал: 12 экз. из урочища Ара-Боран
между Чегемским и Баксанским ущельями, 17 V 1970; 9 — из Чегемского ущелья
близ с. Хуштосырт, 15 V 1970; сборы и определения наши.
Спермовидукт в нижней части прямой или образует слабый изгиб. СЖ 3—4X2.
Вагина постепенно расширяется сверху вниз, в области стилофоров вздута. Наружные
стилофоры прижаты к внутренним, которые втиснуты между наружными и стенкой
вагины. Вагинальные валики и лопасти развиты в различной степени, но всегда при-
сутствуют.
Пенис массивный, вытянутый, внутри гладкий. Папилла пениса мешковидная,
с широко щелевидным выходным отверстием и хорошо развитыми папиллярными ла-
кунами. Семепроводящий канал одной стороной спаян со стенкой папиллы, интрапапил-
лярная полость разделена на 4—6 отделов. Бич извитой, не короче эпифаллуса, обычно
несколько длиннее, эпифаллус в той или иной степени скручен, образуя 1—2 петли.
Между изгибами эпифаллуса и пенисом имеются хорошо развитые мускульные ленты.
Проток семеприемника длинный, тонкий, извитой, длина его превышает длину спермо-
видукта, но благодаря изгибам овальный небольшой резервуар прилегает к спермови-
дукту и лишь слегка касается верхним краем белковой железы.
Распространение и местообитания. Восточная и централь-
ная части северных склонов Большого Кавказа. Обитает на очень влажных затененных
скалах и в сырых гротах.
3. Подрод DIOSCURIA Lindholm, 1927
Lindholm, 1927а : 123, 129 (Trochulus, sect. Dioscuria); Hesse, 1931 : 8
[Fruticlcola (Dioscuria)]; Лихарев, Раммельмейер, 1952 : 453 [Trichia
(D ioscuria)]; Wenz, Zilch, 1959—1960 : 686 [Trichia (Dioscuria)]; Schileyko,
1977 : figs 101-105, Pl. V, 22.
Раковина почти кубаревидная, одноцветная, зеленоватая или с легким краснова-
тым оттенком. Волосков на, раковине нет. Пупок очень узкий, почти целиком закрыт
отворотом колумеллярного края.
Бич примерно вдвое короче эпифаллуса. Наружные стилофоры меньше внутрен-
них. Семепроводящий канал не обособлен от стенки папиллы.
Распространение. Западная половина Кавказского перешейка.
Монотипический подрод.
Типовой вид — Trochulus thalestris Lindholm, 1927.
10. Caucasigena (Dioscuria) thalestris (Lindholm, 1927). Табл. VIII, 75; рис. 211 —
213.'
M о u s s о n, 1863 : 368 (Helix flaveola, non Kaleniczenko, 1853); О. В oettger,
1881a : 205 „(Я. flaveola); 1883 : 158 (H. flaveola); Lindholm, 1927a : 123, 130-
(Trochulus); Hesse, 1931 : 8, t. 1, fig. 2 (Fruticicola); Лихарев, Раммель-
мейер, 1952 : 454, рис. 380, 381 [Trichia (Dioscuria)]; Schileyko, 1977 :
figs 101—105, Pl. V, 2’2.
Материал: 3 экз. из 2 местонахождений на западном Кавказе.
Раковина лишь слегка прижатая, тонкостенная, просвечивающая, с шелковистым
блеском. Завиток строго конический или слегка куполовидный; в последнем случае
вершина выступающая. Оборотов 5—5.5, стремительно нарастающих. Последний обо-
рот почти вдвое шире предпоследнего, сильно вздут, снизу выпуклый, к устью слабо'
опущен, по периферии округлый или с исчезающе слабым углом, который к концу
оборота исчезает совершенно. Окраска одноцветно серовато-зеленая или красновато-ро-
говая; по периферии фон несколько светлее, за счет чего возникает очень размытая
спиральная светлая лента, которая может и отсутствовать. Вокруг пупка имеется более
светлое, чем фон, поле. Радиальная скульптура в виде тонкой густой исчерченности,
местами имеются более резкие морщины; спиральные бороздки ясные и густые. Устье
круглое или широкоовальное, оттянуто вправо и немного вниз, края его широко рас-
ставлены и отвернуты, тонкие, внутри с тонкой белой губой. Пупок проколовидный,
при рассмотрении строго снизу почти полностью закрыт отворотом колумеллярного'
края. Размеры: ВР 10—13, БД 15—19, МД 12.5—16 мм.
Внутреннее строение. Материал: 1 не вполне взрослый экземпляр
из Нового Афона, 2 VII 1913, сбор Н. В. Насонова, определение И. М. Лихарева.
202
III. HYGROMIIDAE
Спермовидукт и яйцевод ясных изгибов не образуют. СЖ 3—4X1—2. Внутрен-
ние стилофоры вздуты, наружные апикально утончены, тесно прилегают к внутренним.
Вагинальные валики умеренно развиты, лопастей не образуют.
Пенис удлиненный, чехол пениса внутри гладкий, папилла длинная, цилиндри-
ческая, семепроводящий канал проходит в толще стенки папиллы, интрапапиллярная
полость разделена на 3 отдела. Бич более чем вдвое короче эпифаллуса, притянутого
к дистальной части пениса посредством мощной мускульной связки, состоящей из
нескольких аностомозирующих лент. Проток семеприемника неоднократно изогнут,
резервуар, по-видимому, немного не доходит до белковой железы.
Рис. 211—213. Caucasigena (Caucasigena) thalestris (Lindh.), Новый Афон, 2 VII 1913.
211 — внешний вид полового аппарата (не вполне взрослый экземпляр); 212 — ва-
гина, вскрыта; 213 — пенис, вскрыт. (По Schileyko, 1977).
Наши данные несколько расходятся со сведениями, помещенными в работе Гессе
>(Hesse, 1931). По Гессе, бич немного длиннее эпифаллуса, пениальные мускульные
ленты отсутствуют, а проток семеприемника очень короткий. Необходимо учесть, что
рисунки Гессе иногда оказываются не совсем точными; мускульные и соединительно-
тканные связки этот исследователь вообще не принимал во внимание и не изображал.
С другой стороны, нами исследован только один не вполне зрелый экземпляр, и такой
признак, как длина бича, может оказаться подверженным возрастной изменчивости
(хотя на других видах мы этого не замечали). То же касается длины и степени изви-
тости протока семеприемника.
Распространение и местообитания. Вид спорадически распро-
-странен по юго-западным склонам Большого Кавказа, Колхидской низменности и се-
веро-западным районам Малого Кавказа. Встречается в лесах.
8. Род DIODONTELLA Lindholm, 1929
Lindholm, 1929 : 206; Лихарев, Раммельмейер, 1952 : 458;
W е n z, Zilch, 1959—1960 : 687.
Раковина сильно прижатая, светлая, со спиральными лентами. Пупок неширокий,
щилиндрический, но почти перспективный. Раковина без волосков. Внутри устья име-
ются два мощных зуба.
Внутреннее строение неизвестно.
Распространение. Центральная часть Северного Кавказа.
Типовой вид — Diodontella stschukini Lindholm, 1929.
PLICUTERIA
203
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА DIODONTELLA
1 (2). Последний оборот в профиле округлый. Большой диаметр раковины 9—10 мм
.................................................. 1. D. stschukini.
2 (1). Последний оборот с легким тупым углом выше средней линии. Большой диаметр
раковины 5—7 мм.....................................2. D. nubigena.
1. Diodontella stschukini Lindholm, 1929. Табл. VIII, 76.
Lindholm, 1929 : 206, t. 11, fig. 1; Лихарев, Раммельмейер,
1952 : 458, рис. 384.
Материал: 5 экз. из 1 местонахождения на Северном Кавказе.
Раковина уплощена, иногда почти плоская, завиток куполовидный, широко
закруглен, вершина невыступающая. Оборотов 5—6, очень медленно и плавно нара-
стающих, сверху слабо, снизу сильно выпуклых, разделенных глубоким швом. По-
следний оборот по периферии почти правильно закруглен или немного выше периферии
имеется легкая наклонность к образованию угла; к устью опущен, немного шире пред-
последнего. Фон раковины белый, иногда с легким синеватым оттенком; выше и ниже
периферии имеются широкие бурые ленты, местами прерванные светлыми радиальными
полосками, соответствующими более ранним губам. Кроме того, по поверхности пос-
леднего оборота разбросаны легкие коричневатые пятна. Скульптура в виде неравно-
мерной радиальной исчерченности и местами морщинистости. Устье полулунно-оваль-
ное, с тонкими, почти прямыми краями. Внутри устья лежит мощная губа с массивными
зубами — палатальным и базальным; губа просвечивает сквозь стенку раковины в виде
широкой светлой полосы. Пупок цилиндрический, но перспективный. Размеры:
ВР 4.5—5.5, БД 9—10, МД 8—9 мм. Лектотип: ВР 5.5, БД 10, МД 9 мм.
Распространение и местообитания. Известна только одна
находка в Чегемском ущелье; указание Линдгольма на более точное местонахождение
(«известковые скалы близ вершины Черах-кенд») связано с недоразумением: вершины
с таким названием в ущелье р. Чегем нет. Следовательно, этого моллюска надо искать
в названном ущелье на известковых выходах на высоте около 1500 м над ур. м.; ве-
роятнее всего, в районе водопадов по левому берегу.
2. Diodontella nubigena Lindholm, 1929. Табл. VIII, 77.
Lindholm, 1929 : 210, t. 11, fig. 2; Лихарев, Раммельмей-
e p, 1952 : 458. z
Материал: 4 экз. из 1 местонахождения на Северном Кавказе.
Раковина уплощена, с куполовидным закругленным завитком и слегка
выступающей вершиной. Оборотов 4—5, очень медленно и плавно нарастающих, тесно
навитых, снизу сильнее выпуклых, чем сверху, разделенных глубоким швом. Послед-
ний оборот лишь немного шире предпоследнего, над серединой с очень легким, сильно
сглаженным углом, к устью слабо опущен. Поверхность раковины матовая, грязно-
белая или серая, с цветными пятнами и размытыми темными лентами. Скульптура сла-
бая, в виде радиальной неясной исчерченности. Устье полулунно-овальное, с тонкими
неотвернутыми краями. Внутри устья располагается губа с хорошо развитыми базаль-
ным и палатальным зубами. Места прикрепления устья не сближены. Пупок неширо-
кий, но почти перспективный. Размеры: ВР 2.8—4.3, БД 5.25—6.75, МД 4—5.5 мм.
Лектотип: ВР 4.0, БД 6.2, МД 5.0 мм.
Распростра не н и е. Известна только одна находка в верховьях р. Чегем
на высоте около 2000 м над ур. м.
9. Род PLICUTERIA Schileyko, 1977
Schileyko, 1977 : figs 106—111, Pl. V, 23.
Раковина почти кубаревидная, желтоватая или светло-серая, одноцветная. Пу-
пок проколовидный. Молодые особи с волосками на поверхности раковины. Устье
без зубов, края его очень хрупкие.
Стилофоры хорошо развиты, прижаты к вагине. Семепроводящий канал подвешен
внутри папиллы на двух противолежащих тяжах; таким образом, интрапапиллярная
полость парная. Кроме папиллярной лакуны на поверхности папиллы имеется про-
дольная борозда. Ретрактор правого омматофора образует перехлест с половым рет-
рактором.
От близких европейских родов отличается более высокой и светлой раковиной,
стремительно нарастающими оборотами и глубоким швом; вагинальные валики раз-
биты на ряд бугорков, бич значительно короче эпифаллуса, проток семеприемника
очень короткий, резервуар относительно очень массивный.
204
III. HYGROMIIDAE
Распространение. Средняя Европа.
Монотипический род.
Типовой вид — Helix lubomirskii Slosarski, 1881.
Plicuteria lubomirskii (Slosarski, 1881). Табл. VIII, 78; рис. 214—218.
Slosarski, 1881 : 29 [Helix (Fruticicola)]; 'Ulicny, 1885 : 1 (Helix Cles-
sini); Лихарев, Раммельмейер, 1952 : 452 [Trichia (Trichia)]; G г о s s u,
1955:461 [Tr. (Trichia)]; Ehrmann, 1956:130 (Fruticicola); Lozek, 1956 *
206, Obr. 57, t. 32,1 {Trichia}; Zilch, Jaeckel, 1962:185 [Tr. (Trichia)]',
Lozek, 1964 : 299, t. 25, 3a—c '[Tr. (Trichia)]; Schileyko, 1977 -
figs 106-111, Pl. V, 23.
Материал: 12 экз. из 4 местонахождений на Карпатах.
Раковина чуть прижатая, с почти строго коническим завитком и острой
вершиной, тонкостенная. Оборотов 4.5—5, выпуклых, разделенных глубоким швом.
Последний оборот в профиле правильно и широко закруглен, к устью плавно и слабо
0.5 см
Рис. 214—218. Plicuteria lubomirskii (Slos.), Оломоуц, 4GCP, 16 IV 1964. 214 — внеш-
ний вид полового аппарата; 215 — вагина, вскрыта {звездочка — место выхода про-
токов стилофоров); 216 — пенис, вскрыт; 217 — поперечный срез папиллы пениса
в дистальной части, 218 — то же, в проксимальной части. (По Schileyko, 1977).
опущен. У взрослых особей волоски на раковине бывают редко, раковина молоди
с мелкими редкими волосками. Окраска желтовато-белая, однотонная. Скульптура
очень слабая, представлена неравномерной радиальной исчерченностыо и местами
сглаженными складками. Устье от круглого до коротко-овального, края его легко
обламываются, очень слабо отвернуты, иногда с едва намеченной губой. Пупок очень
узкий,' цилиндрический, не перспективный, не прикрыт или слегка прикрыт отворо-
том колумеллярного края. Размеры: ВР 5.5—7, БД 7—9, МД 6.3—7.8 мм.
Внутреннее строение. Материал: 1 экз. из Оломоуца, Моравия, ЧССР,
16 IV 1964, сбор и определение В. Гудеца.
Нижняя часть спермовидукта и яйцевод образуют слабые изгибы. СЖ4Х2—3.
Наружные стилофоры несколько лучше развиты, чем внутренние, верхние концы
последних выдаются за верхние концы наружных. Вагина довольно длинная, цилинд-
рическая, внутреннее строение ее очень характерно и своеобразно: вагинальные про-
дольные валики, разбиты на правильные ряды бугорков призматической формы; ни-
каких лопастей вагинальные валики не образуют.
TRICHIA
205
Пенис веретеновидный, чехол пениса внутри гладкий. Папилла пениса несет
на поверхности две продольные резкие, но неглубокие борозды и несколько неполных
кольцевых бороздок. В проксимальной части папиллы семепроводящий канал сосед-
ствует с одной обширной интрапапиллярной полостью, а в дистальной располагается
между двумя полостями (рис. 217 и 218). Мужской отдел в целом вытянут, не скручен
и не изогнут. Длина конического бича примерно вдвое меньше длины толстого ци-
линдрического эпифаллуса, связанного с семепроводом тонкой перепонкой. Проток
семеприемника толстый, прямой и короткий. Резервуар неправильно грушевидной
формы, прилегает к спермовидукту примерно на середине его длины.
Распространение и местообитания. Горные области Средней
Европы: Судеты, Татры, Карпаты с Предкарпатьем. Населяет подстилку лиственных
лесов — на равнине и в горах.
10. Род TRICHIA Hartmann, 1840
Held, 1837 : 914 (Fruticicola part.J; Hartmann, 1840 : 41; Lindholm,
1927a : 122 (Trochulus); C. Boettger, 1928 : 1; Hesse, 1931 : 4 (Fruticicola);
Лихарев, Рам м-е л ь м e й e p, 1952 : 448; W е n z, Zilch, 1959—1960 : 684;
Schileyko, 1977 : figs 128—142, Pl. VI, 27—29.
Раковина в разной степени прижатая, коричневатых тонов, почти одноцветная.
Светлая периферическая лента имеется почти всегда, но выражена слабо. Пупок
различной ширины. Молодь с волосками на раковине, которые могут сохраняться
пожизненно или утрачиваться по достижении половой зрелости. Устье без зубов
или с одним зубообразным утолщением губы на базальном крае. Губа обычно выра-
жена только на базальной части, справа часто резко обрывается.
Семепроводящий канал окружен парной интрапапиллярной полостью, спаян
по двум линиям с внутренней стенкой папиллы или подвешен на двух тяжах. Сти-
лофоры развиты в разной степени, но внутренняя пара всегда ясно выражена, прижата
к вагине или несколько обособлена. Бич, если и короче эпифаллуса, то не намного.
Ретрактор правого омматофора образует перехлест с половым ретрактором.
Распространение. .Средняя и Восточная Европа. В ископаемом состо-
янии род известен начиная с верхнего олигоцена (Европа).
Типовой вид — Helix hispida Linnaeus. Принято считать, что типовым видом явля-
ется Tr. filicina (L. Pfr.); однако Форкарт (Forcart, 1958) показал, что Гартманн,
устанавливая род Trichia с типовым видом Tr. filicina, на самом деле имел в виду
Tr. hispida.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА TRICHIA
1 (2). Высота завитка примерно равна высоте устья или даже превышает ее. Пупок не
менее чем наполовину прикрыт отворотом колумеллярного края устья ....
........................................................ 4. Tr. bielzi.
2 (1). Высота завитка меньше высоты устья. Пупок открытый или слегка прикрыт.
3 (6). Губа ограничена базальным краем устья и представляет собой четкую складку,
часто внезапно оборванную при переходе в палатальный край устья. Длина
волосков на раковине не более 0.4 мм.
4 (5). Ширина пупка составляет х/4—х/3 большого диаметра .... 2. Tr. concinna.
5 (4). Ширина пупка составляет Vs большого диаметра......1. Tr. hispida.
6 (3). Губа очень слабая, на базальном крае не образует складки. Длина волосков на
раковине 0.6—1 мм..................................3. Tr. villosula.
1. Trichia hispida (Linnaeus, 1758). Табл. VIII, 79; рис. 219—221.
Linnaeus, 1758 : 771 (Helix); Muller, \.T№ : 62, 63 (Helix sericea non auct.=
= Trichia plebeja); Westerlund, 1889 : 35 (Helix); P i 1 s b r y, 1939 : 16, fig. 9
(Hygromia); Лихарев, Раммельмейер, 1952 : 450, рис. 377; G г о s s u
1955 : 461, fig. 262; E h г m a n n, 1956 : 130, t. 5, fig. 65; Lozek, 1956:208,
t, 32, 3; Z i 1 c h, J h e c k e 1, 1962 : 185; Lozek, 1964 : 300, t. 25, la—c; For-
cart, 1965 : 79—89, fig. 1, Pl. 21; Gittenberger et al., 1970 : 101, fig. 175,
180, 181, Kaart 71; Schileyko, 1977 : figs 132-137, PI. VI, 28.
Материал: более 100 экз. из 23 местонахождений в европейской части СССР и
;В Западной Европе.
Раковина от прижатой до почти кубаревидной, с широким завитком, очер-
тания которого всегда в той или иной степени куполовидные, а высота немного мень-
ше высоты устья. Оборотов 5—6. Последний оборот с легким закругленным
углом выше средней линии, менее чем в 1.5 раза шире предпоследнего, к устью слабо
опущен. Окраска серовато- или красновато-бурая, светлая лента по периферии сла-
бая. Скульптура в виде тонкой и неравномерной радиальной исчерченности; поверх-
206
III. HYGROMIIDAE
ность раковины покрыта тонкими изогнутыми волосками, которые у взрослых особей
обычно утрачиваются, и на их месте остаются рубцы, хорошо заметные при увели-
чении. Устье коротко-овальное, базальный и колумеллярный края слегка отвернуты.
Отступя от края имеется светлая прямая губа, которая при переходе в палатальный
край более или менее резко обрывается. Пупок в начале развернутый, затем резко
сужается и становится цилиндрическим, но остается перспективным. Размеры:
ВР 4.5—5.5, БД 7—9, МД 6—7.5 мм.
Внутреннее строение. Материал: 12 экз. из Москвы, Ленинские
горы, 12 VII 1970; 13 — из Московской обл., пос. Заветы Ильича близ р. Серебрянки,
Рис. 219—221. Trichia hispida (L.), Москва, Ленинские горы, 12 VII 1970. 219 — внеш-
ний вид полового аппарата; 220 — вагина, вскрыта; 221 — пенис, вскрыт. (По Schi-
leyko, 1977).
15 VIII 1970; 4 — из Ленинграда, сад Зоологического института АН СССР,
28 IX 1965; 5 — из Львова, Стрыйский парк, 25 IX 1969; сборы и определения наши;
4 — из Кведлинбурга, ГДР, сад в черте города, IV 1969, сбор и определение Э. Кла-
усса; 3 — из окр. Кёльна, ФРГ, 30 V 1963, сбор X. Нордзика, определение 3. Гу-
деца.
На границе спермовидукта и яйцевода имеется выраженный в разной степени
изгиб. СЖ 3—4x2; нередко наблюдается вторичное ветвление отдельных ветвей,
в результате чего их общее количество может достигать 12, либо, наоборот, имеет
место редукция, тогда общее число ветвей падает до 6—5 и даже до 4. Внутренние
стилофоры развиты примерно в той же степени, что и наружные, или немного мельче;
верхние их концы всегда выдаются за верхние концы наружных, и место впадения
слизистых желез всегда располагается выше верхних концов внутренних стилофоров.
Вагина довольно длинная, цилиндрическая, вагинальные валики узкие, возле вы-
ходных отверстий стилофоров образуют слабые лопасти.
Пенис массивный, веретеновидный, дистальная часть его узкая, цилиндрическая.
Чехол пениса внутри гладкий. Папилла пениса удлиненная, семепроводящий канал
спаян с двух сторон со стенкой папиллы или в разной степени обособлен; тогда име-
ются продольные тяжи. Длина бича равна длине эпифаллуса или несколько больше.
Эпифаллус почти прямой или изогнут, иногда образует петлю. Дистальные отделы'
мужских протоков соединены тонкой мембраной. Проток семеприемника в различной
степени извит, резервуар овальный, прилегает к верхней части спермовидукта.
TRICHIA
207
Распространение и местообитания. Средняя и Восточная
Европа: к западу до Атлантики, к северу до 60° с. ш. (но по Атлантическому берегу
Скандинавии поднимается почти до северного Полярного круга), к востоку местами-
до Урала (Круликовский, 1889), к югу в общем до 50° с. ш., но местами встречается
значительно южнее. Ареал нуждается в уточнении. Обитает в лесах различных ти-
пов, кроме чисто хвойных. В ископаемом состоянии вид известен с плейстоцена.
(Европа).
2. Trichia concinna (Jeffreys, 1830). Табл. VIII, 80; рис. 222—224.
J е f f г' е у s, 1830 : 336 (Helix); Taylor, 1914 : 17 (Hygromia hispida part.);.
F о г с a r t, 1965 : 79—93, fig. 2, map. 1, pl. 2; Schileyko, 1977 : figs 128—
131, Pl. VI, 27.
Рис. 222—224. Trichia concinna (Jeffr.), Рожнява, ЧССР, 15 VIII 1972. 222 — внеш-
ний вид полового аппарата; 223 — вагина, вскрыта; 224 — пенис, вскрыт. (По Schi-
leyko, 1977).
Материал: 7 экз. из 2 местонахождений в Чехословакии.
Раковина сильно прижатая, с широким, слабо куполовидным завитком.
Оборотов 5—6. Последний оборот снизу уплощен, с более ясной, чем у предыдущего-
вида, угловатостью выше средней линии, почти в 1.5 раза шире предпоследнего,
к устью несколько опущен. Окраска почти одноцветно бурая, периферическая свет-
лая лента едва заметна. Поверхность раковины с шелковистым блеском, который
обеспечивается тонкой радиальной исчерченностыо. Устье широкоовальное, края
его умеренно отогнуты, особенно нижний. На базальном крае имеется белая прямая
губа. Пупок развернутый, широкий, перспективный. Размеры: ВР 3.5—5, БД 7.0—
9.5, МД 6—8 мм.
Helix (Trichia) nothra Westerlund, 1897 : 122 не имеет видимых отличий от дан-
- ного вида, в чем мы убедились, ознакомившись с голотипом, хранящимся в Зооло-
гическом институте АН СССР. Вестерлунд получил этот экземпляр от Эйхвальда
через Мейера с этикеткой «Кавказ». Однако настоящие Trichia на Кавказе отсутствуют,
поэтому указание на типовое местонахождение, вероятно, ошибочно. Следовательно,
мы считаем Н. nothra синонимом Tr. concinna.
Внутреннее строение. Материал: 4 экз. из Рожнявы, Словакия,
15 VII 1962, сбор и определение В. Гудеца.
208
HI. HYGROMIIDAE
Спермовидукт и яйцевод почти без изгибов. СЖ4Х2—3. Стилофоры коротко-
булавовидные, тесно сближены и частично сросшиеся. Область стилофоров неясно
отграничена от нижней цилиндрической части вагины. Вагинальные валики ясные,
лопасти очень слабые или отсутствуют.
Пенис удлиненный, папилла пениса цилиндрическая или слегка веретеновидная,
с крупной папиллярной лакуной. Семепроводящий канал спаян с двух сторон с внут-
ренними стенками папиллы и окружен двумя интрапапиллярными полостями.
Бич почти в 1.5 раза длиннее цилиндрического, слабо изогнутого эпифаллуса. Семе-
провод, эпифаллус и пенис связаны тонкой прозрачной мембраной. Проток семе-
приемника не только не извит, но даже почти не изогнут. Резервуар мешковидный,
пе касается белковой железы.
Рис. 225—227. Trichia villosula (Rssm.), Бабице, ЧССР, 16 VIII 1968. 225 — внешний
вид полового аппарата; 226 — вагина, вскрыта; 227 — пенис, вскрыт. (По Schileyko,
1977).
Распространение и местообитания. Средняя Европа, вклю-
чая южную часть Великобритании и Ирландию. Не исключено, что на восток дохо-
дит до границы СССР. Обитает в опаде лиственных лесов. Вид обнаружен в четверо
тичных и плиоценовых отложениях Европы.
3. Trichia villosula (Rossmassler, 1838). Табл. X, 91; рис. 225—227.
RossmSssler, 1838, Iconogr., II, 7:1, fig. 421 (Helix villosa Drap. var.
villosula); L. Pfeiffer, 1846, in Conch.-Cab., ed. II : 418, t. 148, figs 11, 12
(H. Pietruskyana); Westerlund, 1889 : 53 (Helix); Лихарев, Раммель-
мейер, 1952 : 451; Ehrmann, 1956 : 131 (Fruticlcola); Lozek, 1956 : 208,
t. 32, 4; 1964 : 301, t. 24, 2a—c; Schileyko, 1977 : figs 138—142, Pl. VI, 29.
Материал: 12 экз. из 4 местонахождений в Карпатах.
Раковина слегка прижатая, с широко и плавно закругленным завитком
и почти невыступающей вершиной. Оборотов около 5, плавно нарастающих. По-
следний оборот в профиле плавно закруглен, снизу не уплощен, немного шире пред-
последнего, к устью почти не опущен. Окраска коричневая, светлая периферическая
лента исчезающе слабая. Скульптура в виде неравномерной радиальной исчерчен-
ности; раковина покрыта длинными изогнутыми волосками, сохраняющимися по-
жизненно. Устье овальное, края его хрупкие, без губы внутри или губа очень слабая.
EDENTIELLA
209
Пупок умеренно широкий, не развернут, цилиндрический, перспективный. Размеры:
ВР 4.8—6.5, БД 7.5—10, МД 6.5—9 мм.
Внутреннее строение. Материал: 3 экз. из Бабице, Моравия,
16 VIII 1968, сбор и определение В. Гудеца.
На границе спермовидукта и яйцевода имеется изгиб. СЖ 4X1—2. Стилофоры
сильно вытянутые, плотно прилегают друг к другу и вагине, концы внутренних вы-
даются за концы наружных и достигают места впадения слизистых желез. Вагина
длинная, слегка суживающаяся к клоаке. Вагинальные валики лопастей не образуют.
Пенис веретеновидный, слегка изогнутый, при впадении в клоаку резко сужен
и образует перегиб. Папилла пениса булавовидная, с обширной папиллярной лаку-
ной и широким щелевидным выходным отверстием. Внутри чехла пениса на границе
с клоакой имеются продольные складки. Семепроводящий канал в пределах папиллы
фиксирован двумя тонкими продольными тяжами. Бич несколько-короче эпифаллуса;
последний плавно изогнут и соединен с пенисом мускульной лентой. Семепровод,
эпифаллус и пенис связаны тонкой прозрачной мембраной. Проток семеприемника
тонкий, образует 1—2 колена, резервуар округлый, немного не доходит до белковой
железы.
Распространение и местообитания. Вид широко распростра-
нен по территории Карпат. Встречается во влажных местах, по берегам горных речек,
в кустах или в лесном опаде, поднимается до 1600 м над ур. м. Обнаружен в четвер-
тичных отложениях Европы. ,
4. Trichia bielzi (A. Schmidt, 1860). Табл. X, 92.
A. Schmidt, 1860, Verhandl. siebenb. Vereins : 199 (Helix)-, Westerlund,
1889 : 33 (Helix)-, Kobelt, 1892, Iconogr., N. F., V : 2, fig. 7-19 [Я. (Pertoratella) ];
Лихарев, Раммельмейер, 1952 : 451; G г о s s u, 1955 : 458, fig. 259;
Lozek, 1956 : 204, t. 31, 5; 1964 : 298, Abb. 68; Hudec, 1965:245—260,
Abb. 3; 1972 : 332, Abb. 6, t. 1, 4.
Материал: 9 экз. из 4 местонахождений на Карпатах.
Раковина почти кубаревидная, со слегка куполовидным завитком и слегка
заостренной выступающей вершиной. Оборотов 5.5—6.5, выпуклых, узких, тесно
навитых, разделенных глубоким швом. Последний оборот по периферии с намеком
на угловатость, к устью еле заметно опущен, незначительно шире предпоследнего.
Окраска от роговой до красновато-коричневой, периферическая светлая лента может
отсутствовать (на светлых раковинах). Если лента имеется, она имеет легкий зелено-
вато-желтый оттенок и прослеживается у шва на предыдущих оборотах. Поверхность
раковины со слабым шелковистым блеском, слабо и неправильно радиально исчерчена.
Имеются редкие короткие тонкие волоски, присутствующие в основном на последнем
обороте. Если волоски утрачиваются, на их месте остаются слабые рубцы в виде бу-
горков, которые плотнее всего располагаются на базальной поверхности. Устье ко-
ротко-овальное, косое, слабо вырезано предпоследним оборотом, высота его превы-
шает половину ширины. При рассмотрении снизу расстояние между нижним и верх-
ним краем устья примерно равно половине его ширины. Пупок очень узкий, не ме-
нее чем наполовину прикрыт отворотом колумеллярного края. Размеры: ВР 5—7,
БД 8—10, МД 7—9 мм.
Внутреннее строение нами не исследовано. Приводимое описание
составлено по данным Полинского [Polinski, 1928 (1929)].
Спермовидукт и яйцевод изгибов не образуют. СЖ 2X4. Стилофоры резко сдви-
нуты вниз относительно слизистых желез, небольшие, компактные, удлиненные,
внутренние и наружные примерно равной величины. Вагина довольно длинная.
Бич слегка длиннее эпифаллуса или равен ему. Пенис веретеновидный, слегка
изогнутый, примерно такой же длины, как эпифаллус. Проток семеприемника пря-
мой, не извит, заканчивается мешковидным резервуаром, слегка касающимся белко-
вой железы.
Внутреннее строение гениталий неизвестно.
Распространение и местообитания. Широко, но спорадически
распространен по территории Карпат и Предкарпатья. Обитает в подстилке горных
и предгорных лиственных лесов, на высотах 200—600 м над ур. м.
И. Род EDENTIELLA Polinski, 1929
Polin ski, 1929 : 200; Wenz, Zilch, 1959—1960 : 685; Schileyko,
1977 : figs 154-159, Pl. VI, 31.
Раковина кубаревидная, с куполовидным завитком, почти одноцветная, светлой
окраски. Пупок проколовидный. Волоски на раковине сохраняются пожизненно.
Устье без зубов, но с высокой узкой губой, выраженной только на базальной части.
14 X. А. Шилейко
210
III. HYGROMIIDAE
Внутри папиллы пениса имеется система полостей, разделенных септами, как
у кавказских Caucasigena. Стилофоры удлиненные. Бич значительно длиннее эпи-
фаллуса. Ретрактор правого омматофора образует перехлест с половым ретрактором.
Распространение. Средняя Европа. В ископаемом состоянии род из-
вестен начиная с плейстоцена (Европа).
Типовой вид — Helix edentula Draparnaud, 1805.
Edentiella bakowskii (Polinski, 1924). Табл. X, 93; рис. 228—231.
Polinski, 1924:196, fig. 72—74 [(Trichia bielzi var. bakowskii)}; Лиха-
рев, Раммельмейер, 1952:451 [Tr. (Trichia) bielzi var. bakowskii];
230
Рис. 228—231. Edentiella bakowskii (Pol.), Закарпатье, окр. Рахова, 14 LX 1969.
28 — внешний вид полового аппарата; 229 — пенис, вскрыт; 230 — вагина, вскрыта;
231 — мантийный валик. (По Schileyko, 1977).
L о z е к, 1956 : 203, Obr. 56, t. 31, 5 (Trichia); 1964 : 297, Abb. 64 [Tr. (Edentiella)];
Hudec, 1965 : 245—246, Abb. 1,4; Schileyko, 1977 : figs 154—159, Pl. VI, 31.
Материал: 5 экз. из одного местонахождения в Закарпатье.
Раковина с высоким закругленным завитком, высота которого превышает
высоту устья примерно в 1.5 раза, и со сглаженной невыступающей вершиной. Обо-
ротов 6—6.5, узких, медленно нарастающих. Последний оборот ненамного шире
предпоследнего, с легким намеком на угловатость по периферии, к устью незначи-
тельно опущен. Окраска светло-коричневая, светло-роговая или желтоватая, моно-
тонная; светлая лента по периферии обычно имеется, но сильно размыта. Скульптура
в виде очень тонкой радиальной исчерченности. На поверхности раковины имеются
тонкие редкие волоски, сохраняющиеся пожизненно; если они опадают, рубцов от
них почти не остается. Устье широко-полулунное, скошено. При рассматривании
снизу расстояние между верхним и нижним краями устья составляет самое боль-
шее V3 часть ширины устья. Губа выражена ясно, но только на базальном крае. Пупок
круглый, узкий, цилиндрический, немного прикрыт отворотом колумеллярного края.
Размеры: ВР 2.5, БД 6—7, МД 5.7—6.2 мм.
Внутреннее строение. Материал; 5 экз. из Закарпатья, окр. г. Ра-
хова, 14 IX 1969, сбор и определение наши.
Яйцевод при переходе в вагину резко сужен и образует плавный изгиб. Сли-
зистые железы хорошо развиты, длина их приблизительно равна длине вагины или
немного меньше. По нашим наблюдениям, слизистые железы не собраны в пучки,
HELICOPSIS
211 .
но располагаются мутовчато вокруг вагины на уровне основания протока семеприем-
пика, общее число ветвей 6—8. Гудец (Hudec, 1965) отмечает наличие двух желез,,
каждая из которых состоит из 4 ветвей; очевидно, может иметь место любой из этих
вариантов. Стилофоры удлиненные, заметно обособлены от вагины, наружная и внут-
ренняя пара развиты примерло в одинаковой степени. Вагинальные валики ясно
выражены, но лопастей пе образуют или слабо вздуты у мест выхода протоков стило-
форов.
Пенис слегка вздут, дистальная часть его цилиндрическая. Папилла пениса
правильно веретеповидная. Семепроводящий канал подвешен на одном продольном
тяже; противолежащая стенка папиллы заключает систему интрапапиллярных по-
лостей, разделенных септами (рис. 229), причем отдельные полости могут сливаться
друг с другом. Все полости сообщаются с просветом чехла пениса посредством па-
пиллярной лакуны. Бич в 1.5—2 раза длиннее толстого прямого или слегка изогнуто-
го эпифаллуса, который плавно, без резкой границы переходит в пенис. Мембрана
между семепроводом, эпифаллусом ипенисом отсутствует или выражена крайне слабо.
Проток семеприемника тонкий, очень плавно переходит в удлиненно-овальный ре-
зервуар, заметно не доходящий до нижнего края белковой железы. У одного из ис-
следованных экземпляров резервуар был заметно обособлен.
Распространение и местообитания. Черногора (Гуцуль-
ские Альпы — Закарпатье); спорадически — средняя часть западных Карпат, за-
падная часть восточных Карпат, южные и северные склоны Низких Татр. Встреча-
ется на высотах порядка 1000 м над ур. м. в поясе субальпийских еловых лесов, в по-
ясе буково-пихтовых лесов, преимущественно во влажных прохладных местах; по
долинам .и ущельям спускается на высоту 800—700 м, где населяет ольшаники и бу-
ковые леса. В ископаемом состоянии вид известен начиная с плейстоцена (Европа).
12. Род HELICOPSIS Fitzinger, 1833
Fitzinger, 1833 : 101; С 1 е s s i n, 1876 : 149 (Striatinella); Wester-
lund, 1889:105 (Striatella); Sturany, Wagner, 1915:69 (Martha);
Hesse, 1926 : 131 (Helicella (Helicopsis)]', Lindholm, 1927a : 136 [Helicella
(Helicopsis)]; Лихарев, Раммельмейер, 1952 : 422 [Helicella (Helicopsis)];
Wenz, Zilch, 1959—1960:671.
Раковина от кубаревидной до прижатой, светлая, с несколькими тёмными Дей-
тами, выраженными в различной степени. Пупок не проколовидный, может быть
цилиндрическим или развернутым. Волоски на раковине всегда-отсутствуют.
Семепроводящий канал в пределах папиллы фиксирован 1—2 продольными тя-
жами; кроме того, интрапапиллярная полость может быть разделена на 2—3 отдела.
Независимо от этих полостей формируется щелевидная полость в дистальной части
эпифаллуса, которая может заходить в стенки пениального чехла и сливаться с ин-
трап апил л ярными полостями. Стилофоры развиты в различной степени, .наружные
крупнее внутренних. Бич короче эпифаллуса. Ретрактор правого омматофора не'Ьб-
разует перехлеста с половым ретрактором. ж;
Распространение. Средняя и Южная Европа, Крым, Закавказье,
Копетдаг; Малая Азия, Северная Африка. В ископаемом состоянии род известён
из нижнего плиоцена Европы и верхнего плиоцена Предкавказья. " .;
Типовой вид — Helix striata Miiller, 1774. - '
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕНТНЫХ ВИДОВ
РОДА HELICOPSIS
1 (2). Верхняя поверхность раковины в целом темнее нижней. Вид обитает в Копет-
даге .................................................... 3. Н. likharevi.
2 (1). Интенсивность окраски верхней и нижней поверхностей раковины примерно
одинаковая. Виды обитают на Кавказе, в Крыму и в европейской части СССР.
3 (6). Периферия последнего оборота несет выраженный в различной степени киль.
4 (5). Поверхность раковины тонко радиально исчерчена, ширина пупка составляет
х/4 большого диаметра................................. . . . 9. Н. filimargo-.
5 (4). Поверхность раковины ребристая, ширина пупка составляет 1/.—1/„ большого
диаметра ............................................ 5. Н. retowskii.
6 (3). Киль отсутствует; в крайнем случае последний оборот в профиле может быть
слегка угловатым.
7 (8). Скульптура в виде неравномерно расположенных притупленных радиальных
ребрышек . -...................................... .........4. Н. striata;
8 (7). Поверхность раковины тонко исчерчена.
9 (10). Поверхность раковины покрыта глубокой и густой исчерченностыо. Вид
обитает в бассейне р. Днестра............................. 8. Н. instabilis.
14*
212
III. HYGROMIIDAE
10 (9). Исчерченность поверхности раковины неясная. Виды обитают в Крыму, в ок-
рестностях Мелитополя и Новороссийска.
11 (12). Завиток слегка куполовидный. Рисунок образован несколькими коричневыми
спиральными лентами, 2 из которых лежат выше периферии . 7. Н. paulhessei.
12 (11). Завиток конический. Рисунок образован спиральными темными лентами, 1 из
которых лежит выше периферии..................................6. Н. dejecta.
f 1. Helicopsis sunzhica (Steklov, 1966). Табл. X, 94.
Стеклов, 1966:220, табл. X, 186—190 (НеИееИа).
Материал: более 30 экз. из 1 местонахождения в Предкавказье.
Раковина почти кубаревидная, с коническим контуром завитка, высота
которого меньше высоты устья, и выступающей, но сглаженной вершиной. Оборотов 5,
правильно нарастающих, сильно выпуклых, слегка ступенчатых, разделенных глу-
боко врезанным швом. Последний оборот правильно закруглен, немного более чем
в 1.5 раза шире предпоследнего. В конце его, над устьем ступенчатость выражена
особенно ясно; к устью последний оборот совершенно не опущен. Окраска светлая,
одноцветная; рисунок не сохранился. Скульптура представлена неравномерными,
сильно сглаженными радиальными складками. Устье почти круглое, косое, места
его прикрепления умеренно сближены. Края устья простые, острые, почти не отвер-
нуты. Отступя от края устья располагается широкая, но невысокая массивная губа;
внутри последнего оборота иногда заметны еще 1—3 губы, расположенные на разном
расстоянии друг от друга. Пупок цилиндрический; степень его перспективности
установить трудно. ВР 6—8, БД 8.5—10.5, МД 7—8.7 мм.
' Распространение. Акчагыльский ярус верхнего плиоцена Предкав-
казья.
f 2. Helicopsis libidinosa (Steklov, 1966). Табл. X, 95.
Стеклов, 1966:221, табл. X, 191—192 (Ilelicella).
Материал: более 20 экз. из одного местонахождения в Предкавказье.
Раковина слегка прижатая, с чуть куполовидным контуром завитка, вы-
сота которого приблизительно равна высоте устья или немного меньше. Вершина
не притупленная. Оборотов 5, выпуклых, округлых, не ступенчатых. Последний
оборот в профиле округлый, не более чем в 1.5 раза шире предпоследнего, к устью
постепенно и едва заметно спускается. В самом начале последнего оборота имеется
слабая наклонность к образованию угла (смотреть справа), который вскоре совер-
шенно исчезает. Окраска состоит из светлого фона и спиральных лент, которые при
жизни животного были, вероятно, почти черными, но на ископаемых раковинах вы-
глядят слегка темнее фона и отличаются восковой полупрозрачностью. В наиболее
полном наборе имеется 5 лент: тонкая, очень неясная, несколько ниже шва; следу-
ющая, наиболее широкая, непосредственно над периферией; чуть ниже периферии —
очень слабая тонкая лента; наконец, на базальной части имеются еще две ленты.
Первая и третья ленты часто совсем отсутствуют. Ленты обычно в той или иной сте-
пени разбиты на ряд пятен. Скульптура в виде несильной, но частой радиальной
морщинистости. Устье округлое, косое, с тонкими прямыми краями. Немного от-
ступя от края имеется сильно расплывшаяся губа. Пупок не прикрыт, узкий, круглый,
цилиндрический, сквозь него можно просмотреть немногим более одного оборота.
Размеры: ВР 6—7, БД 7.5—9, МД 6.5—8.2 мм.
Распространение. Апшеронский ярус верхнего плиоцена Предкавказья.
3. Helicopsis likharevi1 Schileyko, sp. nov. Табл. X, 96', рис. 232, 233.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: более 20 экз. из 2 местонахождений на Копетдаге..
Раковина прижатая, снизу заметно выпуклая. Оборотов 4.5, умеренно
выпуклых. Последний оборот немного шире предпоследнего, по периферии с легким
намеком на угол, к устью не опущен. Окраска выше периферии в целом темнее, чем
в нижней части раковины, состоит из рогового и темно-серого цветов; периферия
всегда светлая, выше нее проходит темная лента, нижняя граница которой выражена
-четко, а верхняя граница очень неопределенная из-за того, что рогового цвета «языки»
поднимаются беспорядочно вверх, достигая порой шва. В базальной части имеется
несколько слабо выраженных темных спиральных лент. Радиальные ребра, составля-
ющие дефинитивную скульптуру (эмбриональные обороты гладкие), окрашены пре-
имущественно в светлые тона, и местами светлые ребра пересекают полностью или
частично темную ленту над периферией. Устье слегка скошено, край его слегка от-
вернут только в колумеллярной части, губа отсутствует. Пупок узкий, сквозь него
1 Вид назван именем Ильи Михайловича Лихарева.
HELICOPSIS
213
мЬжио, слегка поворачивая раковину, просмотреть весь предпоследний оборот. Раз-
меры: ВР 4.5—6, БД 7—8.5, МД 6—7.5 мм. Голотип: 'ВР 5.7, БД 8.3, МД 7.4 мм.
По раковине вид похож на Н. striata и недавно описанный европейский вид
Н. austriaca (Gittenberger, 1969). Отличается более медленным нарастанием оборотов
и менее перспективным пупком.
Внутреннее строение. Материал: 2 экз. из ущелья Игдеджин (Колет-
даг), 25 IV 1953; 4 — из ущелья Шехин-дере (Копетдаг), 27 IV 1953; сборы И. М.
Лихарева, определения наши.
Спермовидукт при переходе в яйцевод образует небольшой изгиб. СЖ 2x2.
Длина участка вагины между основанием протока семеприемника и основанием сли-
зистых желез крайне незначительна. Стилофоры почти шаровидные, внутренние
Рис. 232—233. Helicopsis likharevi sp. nov., Копетдаг, ущелье Шехин-
дере, 27 IV 1953. Паратип. 232 — внешний вид полового аппарата;
233 — пенис, вскрыт. (Ориг.).
меньше наружных. Вагинальные валики умеренно развиты, образуют нерезкие ло-
пасти возле выходных отверстий стилофоров.
Пенис заметно вздут, стенки чехла тонкие, внутренняя их поверхность гладкая;
папилла коническая. Семепроводящий канал имеет довольно толстые стенки и ‘ под-
вешен к внутренней стенке папиллы одним продольным тяжом, расположенным почти
по касательной к стенкам папиллы и семепроводящего канала (см. поперечный срез
на рис. 233). Просвет последнего щелевидный. Семепровод почти вплотную притянут
тонкой мембраной к пенису и эпйфаллусу и сопровождает эти протоки вплоть до
клоаки. Длина бича составляет около V3 длины цилиндрического, почти прямого
эпифаллуса. Семеприемник имеет короткий, почти прямой проток и овальный резер-
вуар, не доходящий до белковой железы.
Распространение и местообитания. Копетдаг. Встречается
на каменистых склонах, днем держится под камнями и в осыпях.
4. Helicopsis striata (Muller, 1774). Табл. X, 97; -рис. 234, 235.
Muller, 1774 : 38 (Helix)-, Westerlund, 1889 : 251 (Helix); Polinski,
1922 : 170 (Helicella cereoflava podolica); Hesse, 1934 : 25 [77. (Helicopsis)];
Скворцов, 1938 : 180 (Helicella eudakowi nom. nudum); Лихарев, Раммель-:
мейер, 1952 : 422, рис. 347, 348 [17. (Helicopsis) ]; Grossu, 1955 : 430, fig. 240
[H. (Helicopsis)}; Ehrmann, 1956:121, t. 4, fig. 59 [17. (Helicopsis) ]; Lozek,
1956 : 195, t. 30, 1 [H. (Helicopsis) ]; Z i 1 c h, J a e c k e 1, 1962 : 175 [Heliconsis
(Martha) ]; L о z e k, 1964 : 289, t. 32, 2a—с; H u d e c, 1966 : 161—176, Abb. 1- 3
u. 5—6; Gittenberger, 1969 : 63—68, fig. 4, 7, 8.
214
III. HYGROMIIDAE
Материал: более 30 экз. из 4 местонахождений в европейской части СССР й ЧССР.
Раковина прижатая, изредка почти кубаревидная, вершина несколько
сглаженная. Оборотов 4.5—5, выпуклых, плавно нарастающих. Последний оборот
в профиле округлый или с легким намеком на угловатость в начале, в 1.5—2 раза
шире предпоследнего, к устью очень слабо опущен. Эмбриональные обороты темные,
окраска остальных оборотов белая или желтовато-белая, изредка одноцветная, но
чаще с многочисленными (до 8) коричневыми или черными спиральными лентами,
из которых наиболее широкая располагается над периферией последнего оборота
Рис. 234—235. Helicopsis striata (Miill.), окр. Курска, 16 VIII 1968. 234 — _
внешний вид полового аппарата; 235 пенис, вскрыт. (Ориг.)..
и. у шва на предыдущих оборотах. Нижние полосы обычно разбиты на ряд пятен. В про-
ходящем свете ленты просвечивают сильнее фона. Скульптура состоит из неравномер-
ных . притупленных ребрышек, между которыми видны тонкие радиальные бороздки.
Устье округлое, слабо вырезано предпоследним оборотом; места его прикрепления
слегка сближены. Края устья острые, простые, только колумеллярный слегка рас-
ширен. Губа тонкая, белая. Пупок умеренно широкий, почти перспективный. Раз-
меры: ВР 5—8.5, БД 7—12, МД 6—10.5 мм.
Внутреннее строение. Материал: 5 экз. из окр. Курска, с. Бар-
каловка, 16 VIII 1968, сбор и определение В. А. Николаева.
На границе спермовидукта и яйцевода имеется нерезкий изгиб. СЖ 2—3x2—4,
либо слизистые железы располагаются вокруг вагины мутовчато. Стилофоры мощные,
шаровидные или слегка удлиненные. Вагинальные валики хорошо развиты, сбли-
жены попарно, каждая пара занимает латеральное положение, лопасти в местах
выходных отверстий стилофоров слабые.
Пенис в разной степени вздутый, удлиненный. В стенках пениального чехла при
гистологическом исследовании обнаружена пениальная железа, имеющаяся у не-
которых среднеазиатских представителей подсемейства. Папилла не имеет постоян-
ной длины за счет развития кольцевой полости в основании папиллы, причем эта
HELICOPSIS
215
полость сливается с настоящей интрапапиллярной полостью; семепроводящий канал
подвешен на 1—2 тяжах только в дистальной части. Просвет семепроводящего канала
в виде узкой щели. Бич в 2—3 раза короче прямого или изогнутого эпифаллуса.
Проток семеприемника короткий, толстый, объемистый резервуар прилегает к спер-
мовидукту чуть выше его середины.
Распространение и местообитания. Западная и Средняя
Европа; восточная граница ареала неясна, но местами вид встречается в Молдавии,
западной Украине, доходит до Курска и Воронежа. Обитает на сухи'х, хорошо про-
греваемых лугах. В ископаемом состоянии известен из плейстоцена Великобрита-
нии и неоднократно найден в голоцене Средней Европы.
Рис. 236—242. Helicopsis retowskii (Cl.), Крым, пойма р. Бельбек, 6 VIII 1962. 236 —
внешний вид полового аппарата; 237 — вагина, вскрыта; 238 — пенис, вскрыт; 239 —
поперечный срез эпифаллуса в дистальной части; 240 — то же, в проксимальной части;
241 — поперечный срез папиллы пениса в проксимальной части; 242 — то же, в ди-
стальной части. (Ориг.).
5. Helicopsis retowskii (Clessin, 1883). Табл. X, 98, 99; рис. 236—242.
. Clessin, 1883 : 47, t. 3, fig. 1 (Helicella); Westerlund, 1889 : 323
(Helix); Пузанов, 1925a : 71, фиг. 3 (Helicella); Lindholm, 1926 : 172;
170 (Helicella (Jacosta) gireiorum]; Hesse, 1934 : 26, t. 5, fig. 44 (Helicella); Лиха-
рев, Раммельмейер, 1952 : 426 [Helicella (Helicopsis), (H.) gireiorum.];
Шилейко, 1967b : 32, рис. 3, В [Helicella (Helicopsis)]; Hudec, 1972a : 89,
Abb. 18, 38.
Материал: более 50 экз. из 4 местонахождений в Крыму.
Раковина в различной степени прижатая. Завиток различной высоты, но
не превышает высоту устья, нередко меньше половины этой величины. Оборотов 5—5.5,
умеренно выпуклых, плавно нарастающих, разделенных глубоким швом. Последний
оборот не более чем в 1.5 раза шире предпоследнего, к устью в различной степени, но
всегда очень плавно опущен. Окраска грязно-белая или кремовая, обычно с рисунком,
который может вторично исчезать: чаще всего две ленты располагаются выше и ниже
периферии, а на нищней стороне раковины могут располагаться еще 2—4 узкие ленты.
Последний оборот в профиле округлый или с развитым в различной степени килем —
от тонкого нитевидного до мощного зазубренного. Скульптура — от равномерной тон-
216 III. HYGROMIIDAE
кой до очень грубой ребристости (var. milaschevitschi Retowski). Степень развития реб-
рышек и киля зависит от одних и тех же внешних факторов, т. е. меняется вся скульп-
тура в целом. Устье округлое, косое, места его прикрепления немного сближены.
Края устья острые, простые, иногда внутри с тонкой губой. Пупок умеренно развер-
нутый, эксцентричный, почти перспективный. Размеры: ВР 5—8, БД 8—13, МД 7—
11.5 мм.
Внутреннее строение. Материал: 2 экз. из Севастополя, Черная
Речка, 20 VII 1962; 6 — из поймы р. Бельбек близ с. Голубинка (Крым), 6 VIII 1962;
сборы и определения наши.
Яйцевод плавно изогнут. Слизистые железы не обнаруживают тенденции к груп-
пировке в пучки, располагаются мутовчато; общее число ветвей 7—12. Наружные сти-
лофоры очень мощные, почти шаровидные, внутренние удлиненные, значительно мень-
ших размеров. Участок вагины между правой и левой парами стилофоров вздут.
Вагинальные валики очень мощные, но лопастей не образуют.
Пенис небольшой, шаровидный, с толстостенным чехлом, в стенках которого
можно подозревать присутствие пениальной железы (гистологического исследова-
ния не проводилось). Папилла мешковидная, семепроводящий канал в разрезе
щелевидный,; фиксирован двумя продольными тяжами. Один из этих тяжей соеди-
няет канал со стенкой папиллы, второй крепится к септе; ниже септы находится
ряд перегородок или одна перегородка, разбивающие интрапапиллярную полость
на отделы (рис. 241—242). Интрапапиллярных лакун несколько, хорошо развитых.
Семеприемник состоит из прямого протока и грушевидной формы резервуара, гра-
ница между которыми достаточно ясная. Семепровод притянут к пенису и эпифал-
лусу тонкой прозрачной мембраной. В основании папиллы пениса имеется выра-
женная в различной степени кольцевая полость.
Распространение и местообитания. Южная часть Крыма;
преимущественно горный Крым, но выходит и в степной Крым, доходя, в частности,
до Бахчисарая. Обитает в сухих открытых биотопах (степные участки, сухие поймы,
в горной части — яйлы). Держится под камнями и в дерновинах травянистых растений.
В жаркое сухое время уходит в почву на глубину нескольких сантиметров.
6. Helicopsis dejecta (Cjistofori et Jan, 1831). Табл. X, 100.
Crist of ori, Jan, 1832 : (Helix); Krynicki, 1836 : 197 (H. arenosa);
С 1 e s s i n, 1881 : 138 (H. substriata); Westerlund, 1889 : 339 (H. obvia var.
arenosa; var. dejecta); Hesse, 1934 : 25, t. 5, fig. 42 (Helicella); Лихарев, Рам-
мельмейер, 1952 : 424 [Helicella (Helicopsis) ]; G г о s s u, 1955 : 433, fig. 241
[Helicella (Helicopsis)]; H u d e c, 1972a : 90, Abb. 19, 37.
Материал: 12 экз. из 3 местонахождений в Крыму.
Раковина слегка прижатая, с широким коническим завитком, высота кото-
рого сильно варьирует, но не превышает высоту устья. Оборотов 5—6. Последний
оборот в 1.5 раза или немного более превышает ширину предпоследнего, к устью не-
много и плавно опущен, по периферии правильно закруглен. Окраска белая, одно-
цветная или с темными лентами, из которых наиболее широкие проходят выше и ниже
периферии и в проходящем свете просвечивают сильнее фона. Скульптура в виде очень
тонкой, но четкой радиальной морщинистости, местами имеются более грубые морщины;
однако при рассматривании невооруженным глазом раковина выглядит гладкой.
Устье округлое, слабо вырезано предпоследним оборотом, косое, места его прикрепле-
ния умеренно сближены. Края устья тонкие, острые, простые, губа слабая, расплыв-
шаяся. Пупок развернутый, перспективный. Размеры: ВР 6—11, БД 9—18, МД 7—
15.5 мм.
Внутреннее строение. Нами не изучено, описание приводится в со-
ответствии с данными Гессе (Hesse, 1934).
Спермовидукт и яйцевод почти прямые. Слизистые железы располагаются венцом
вокруг вагины и не обнаруживают тенденции к образованию пучков. Стилофоры до-
вольно массивные, булавовидные, внутренние заметно меньше наружных. Вагиналь-
ные валики высокие, в числе 4, попарно, видимо, не сближены.
Пенис вздутый, веретеновидный. Бич не длиннее 1/3 длины цилиндрического эпи-
фаллуса, который может петлеобразно изгибаться. Проток семеприемника почти пря-
мой, резервуар удлиненный, заметно не достигает белковой железы.
Внутреннее строение мужского отдела неизвестно.
Распространение и местообитания. Южный берег Крыма,
окрестности Мелитополя и Новороссийска. Встречается на открытых местах, на степ-
ных участках.
7. Helicopsis paulhessei (Lindholm, 1936). Табл. XI, 101.
Hesse, 1934 : 23, t. 5, fig. 40 (Helicella gasprensis Lindholm nom. mus.); Линд-
гольм, 1936 : 439, рис. 1 (Helicella); Миха рев, Раммельмейер,
1952 : 425 [Helicella (Helicopsis)].
HELICOPSIS
217.
Материал: 3 экз. из 1 местонахождения в Крыму.
Раковина данного вида чрезвычайно положа на раковину предыдущего.
Отличия: завиток имеет куполовидные очертания; если на раковине есть темные ленты,
то 2 из них лежат выше периферии последнего оборота, остальные ниже периферии.
Последний оборот не более чем в 1.5 раза шире предпоследнего; высота завитка в сред-
нем равна примерно высоте устья; в скульптуре присутствуют короткие вмятины на
последнем обороте. Размеры: ВР 9—10, БД 13.5—15.8, МД 11.5—13.8 мм. Лектотип:
ВР 9.0, БД 14.2, МД 12.0 мм.
Внутреннее строение. Нами не исследовано, приводимое описание
составлено по данным Гессе (Hesse, 1934).
Спермовидукт и яйцевод изгиба не образуют. Слизистые железы собраны в 4 пучка,
но ветви при основании не сливаются, т. е. каждый пучок лишен общего протока;
общее число ветвей И—12. Внутренние стилофоры развиты слабо, у 2 экз. замечена
только одна пара стилофоров, заметно вздутых в базальной части.
Пенис почти шаровидный, бич чуть длиннее половины эпифаллуса. Проток семе-
приемника мощный и короткий, резервуар неправильно овальный.
Внутреннее строение гениталий неизвестно.
Распространение и местообитания. Крым (Гаспра). Найден
в парках, на светлых местах, в траве и на камнях.
8. Helicopsis instabilis (Rossmassler, 1838). Табл. XI, 102, 103.
Rossmassler, 1838, Iconogr., II, 7:33, fig. 518 {Helix)-, С 1 e s s i n,
1879 : 9, t. 1, fig. 6 [77. (Xerophila) instabilis var. bakowskyana]; P о linski, 1922 : 167 —
171, t. 13, fig. 4—8 (Helicella); 1924 : 174, t. 11, fig. 29—34 (Helicella); Hesse,
1934 : 25 [(Helicella (Helicop sis)]; Лихарев, Раммельмейер, 1952 : 423
[Helicella (Helicopsis)]; G г о s s u, 1955 :428, fig. 238 [Helicella (Helicopsis)];
Hudec, Vasatko, 1971 : 26, Abb. 13, Pl. IV, 1.
Материал: 8 экз. из 3 местонахождений на юго-западе европейской части СССР.
Раковина сильно прижата — до совсем плоской, твердостенная; оборотов
4.5—5.5, выпуклых, разделенных глубоким швом, плавно нарастающих. Последний
оборот округлый, почти вдвое шире предпоследнего, к устью слегка опущен. Окраска
известково-белая или сероватая, одноцветная или с 1—10 темными лентами, выражен-
ными в разной степени. Скульптура в виде радиальной исчерченности, чаще всего глу-
бокой и густой, и спиральных бороздок, выраженных местами на последнем обороте.
Устье круглое, косое, слабо вырезано предпоследним оборотом; края устья простые,
острые, с расплывшейся губой. Места прикрепления устья немного сближены. Пупок
развернутый, широкий, перспективный. Размеры: ВР 6—8, БД 14—18.5, МД 11.5—15 мм.
Почти плоские экземпляры известны под названием Н. instabilis var. jachnoi
Clessin, 1890. Гудец и Вазатко (Hudec, Vasatko, 1971) именуют эту форму Helicopsis
(Helicopsis) instabilis jachnoi, считая, по-видимому, подвидом. Однако названная форма
встречается, по всей видимости, вместе с номинативной и, следовательно, таксономи-
ческого статуса не имеет (при условии, что обе формы связаны промежуточными вари-
антами). Однако взаимное распространение обсуждаемых форм выяснено недостаточно.
Внутреннее строение. Нами не исследовано, описание составлено по
литературным данным (Polinski, 1922, 1924; Hudec, Vasatko, 1971).
Спермовидукт и яйцевод почти прямые. СЖ 4x2—3. Стилофоры массивные, почти
равной величины; вагина от области стилофоров постепенно сужается к клоаке.
Пенис вытянутый, заметно длиннее бича; бич в свою очередь втрое короче эпифал-
луса. Семепровод притянут тонкой мембраной к эпифаллусу и пенису. Проток семепри-
емника сравнительно с другими видами рода тонкий и длинный, резервуар удлиненно-
овальный, не доходит до белковой железы.
Распространение и местообитания. Бассейн р. Днестра.
Обитает на открытых прогреваемых местах.
9. Helicopsis filimargo (Krynicki, 1833). Табл. XI, 104; рис. 243, 244.
Krynicki, 1833 : 435 (Helix); R ossmassler, 1835, Iconogr., I, 1 : 68
(H. taurica); 1837, Iconogr., I, 5 : 33, fig. 358 (Helix); Westerlund, 1889 : 343
(Helix); Пузанов, 1925a : 68, рис. 3 (Xerophila (Jacosta)]; Hesse, 1934 : 25,
t. 5, fig. 41 (Helicella (Helicopsis) ]; Лихарев, Раммельмейер, 1952 : 427,
рис. 351 [Helicella (Helicopsis)].
Материал: 18 экз. из 1 местонахождения в Крыму.
Раковина сильно прижатая — до почти плоской, со слабо выпуклым,
слегка куполовидным завитком и незначительно выступающей вершиной. Оборотов
5—5.5, умеренно выпуклых, у шва слегка ступенчатых. Окраска белая, одноцветная
или с тонкими спиральными лентами, которые обычно выражены слабо. Скульптура
в виде тонкой, густой и неясной радиальной исчерченности; на последнем обороте
часто имеются неясные продольные вмятины. Начиная с третьего оборота по перифе-
218
III. HYGROMIIDAE
рии тянется белый, четко отграниченный киль, выступающий над швом. На послед-
нем обороте у мелких раковин с низким завитком киль простирается до самого устья,
на раковинах крупных и высоких киль к устью заметно сглаживается. Устье округлое,
слабо вырезано предпоследним оборотом; если киль доходит до устья, палатальный край
с углом. Края устья острые, простые, с белой расплывшейся губой. Места прикрепле-
ния устья умеренно сближены. Пупок развернутый, очень широкий, вполне перспектив-
ный. Размеры: ВР 6.5—9, БД 12—17, МД 10—14 мм.
Внутреннее строение. Материал: 3 экз. из Севастополя, Черная
Речка, 20 VII 1962, сбор и определение наши.
Рис. 243—244. Helicopsis filimargo (Kryn.), окр. Севастополя,
20 VII 1962. 243 — внешний вид полового аппарата; 244 — пенис,
вскрыт {звездочка — участок пениса, гомологичный пениальному
придатку видов рода Xeropicta). (Ориг.).
Нижняя часть спермовидукта образует плавный изгиб. Слизистые железы в числе
6—8 не сливаются в пучки, но обнаруживают наклонность к вторичному ветвлению.
Внутренняя пара стилофоров развита значительно слабее внешней, наружные стило-
форы булавовидной формы; вагина постепенно сужается к клоаке.
Пенис разделен на 2 отдела: веретеновидный проксимальный и узкий изогнутый
дистальный. Папилла занимает проксимальный отдел, почти цилиндрическая. Семе-
проводящий канал в разрезе имеет форму трехлучевой звезды и фиксирован двумя
противолежащими продольными тяжами. Стенка чехла пениса в базальной части рас-
слаивается, и образованная таким образом полость сливается с парной интрапапил-
лярной полостью. Внутри дистального отдела пениса наблюдается структура, очень на-
поминающая то, что имеется внутри придатка пениса у'представителей рода Xeropicta
(см. ниже): имеются 2—3 складки, покрытые поперечными бороздками. Эта структура
была известна и ранее и считалась раздражающим' тельцем. Бич немного короче эпи-
фаллуса, тонкий; эпифаллус плавно изогнут. Пенис, эпифаллус и семепровод соединены
прозрачной мембраной. Проток семеприемника вслед за яйцеводом плавно изогнут,
заканчивается очень массивным удлинённо-овальным резервуаром.
Распространение и местообитания. Горный Крым, окрест-
ности Одессы, вилайет Зонгулдак в Турции (Croockewit, 1953). Обитает на степных
участках под камнями.
XEROPICTA
219
13. Род XEROPICTA Monterosato, 1892
Monterosato, 1892 : 24 [Helicella (Xeropicta) ]; Hesse, 1926 : 133
[Helicella (Xeropicta)]; L i n cl h о 1 m, 1927a : 136 [Helicella (Xeropicta)]; Л и x a-
p e в, Раммельмейер, 1952 : 428 [Helicella (Xeropicta)]; W e n z, Zilch,
1959—1960 : 672.
Раковина в различной степени прижатая, белая, одноцветная или с темными спи-
ральными лентами. Пупок в начале развернутый, затем цилиндрический, сквозь него
видно не более полутора оборотов. Волоски на раковине всегда отсутствуют.
. Семепроводящий канал в пределах папиллы фиксирован двумя продольными тя-
жами; таким образом, интрапапиллярная полость парная, но сливается с кольцевой
полостью, имеющейся в дистальной части эпифаллуса. В базальной части пениса имеется
конический придаток. Стилофоры небольшие, в высокой степени обособлены — верх-
ние от нижних. Бич короче эпифаллуса.. Ретрактор правого омматофора не образует
перехлеста с половым ретрактором.
Распространение. Средняя Азия, Кавказ, Крым, местами вд(оль се-
верного берега Черного моря, Балканский п-ов, Ближний Восток, Северо-Восточная
Африка.
Типовой вид — Helix krynickii Krynicki, 1833.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА XEROPICTA
'.1 (2). Большой диаметр раковины при 4.5—5 оборотах не превышает 7 мм ....
........................................................... 4. X. parableta.
2 (1). Большой диаметр раковины при том же числе оборотов не менее 10 мм.
3 (4). Сквозь пупок виден весь предпоследний оборот.........2. X. derbentina.
4 (3). Сквозь пупок видна часть предпоследнего оборота.
5 (6). Сквозь пупок просматривается не более х/4 последнего оборота..........
...................................................... 1.Х. krynickii.
6 (5). Сквозь пупок просматривается 1/2—3/4 последнего оборота ..............
..........................................................3. X. candaharica.
1 . Xeropicta krynickii (Krynicki, 1833). Табл. XI, 106; рис. 245—247.
Krynicki, 1833 : 434 (Helix); Clessin, 1881 : 137 .(Helix theodosiae);
Kobelt, 1884, Iconogr., X. F„ Г : 48, fig. 139, 140 (Helix); Пузанов, 1925a : 72,
фиг. 3 [Xerophila (Heliomanes)]; 74, фиг. 3—7 [Xerophila (Heliomanes) Orianda];
1926 : 91 [Xerophila (Heliomanes)]; Hesse, 1934 : 30, t. 6, fig. 48 (Helicella); Лиха-
рев, Раммельмейер, 1952 : 428, рис. 352a, 353, 354 [Helicella (Xeropicta)];
Grossu, 1955 : 435, fig. 244 [Helicella (Xeropicta)]; HI и л-е й к о, 1968а : 337,
рис. la [Helicella (Xeropicta)]; Hudec, Lezhawa, 1969b : 124, Abb. 21, t. IX,
24 [Helicopsis (Xeropicta)]; Hudec, Vasadko, 1971:26, Abb. 14 [Helicopsis
(Xeropicta) ].
Материал: более 100 экз. из 15 местонахождений в Крыму, на Кавказе, в окр.
Одессы и в Болгарии.
Раковина с широким завитком куполовидных или, чаще, конических очер-
таний, высота которого не меньше половины высоты устья. Оборотов 5—6; верхние
обороты нарастают медленно и слабо выпуклые. Последний оборот сильно вздут,
выпуклый, в' профиле правильно закруглен, примерно вдвое шире предпоследнего,
к устью слабо опущен. Окраска белая, одноцветная или с рисунком из темных лент
и пятен. Число лент и степень их развитости сильно изменчивы, но наиболее постоянны
две ленты: во-первых, лента, проходящая над периферией последнего оборота и над
швом предыдущих оборотов и доходящая до эмбриональной раковины; во-вторых; одна
из лент на нижней стороне раковины. Скульптура в виде тонкой и неравномерной ради-
альной исчерченности и спиральных бороздок, выраженных в разной степени. Устье
относительно очень большое, округлое, косое, слабо вырезано предпоследним оборо-
том. Места прикрепления устья несколько сближены, края его тонкие, простые, иногда
с очень тонкой светлой губой. Пупок характерный: сквозь него видно не более половины
предпоследнего оборота,располагается эксцентрично на дне широкой пупочной впа-
дины. Размеры: ВР 7.5—11, БД 12—15, МД 9.5—12 мм.
Внутреннее строение. Материал: 6 экз. из Крыма, окр. с. Голу-
бинка, 8 VIII 1962; 4 — из Ленкоранской низменности, 20 VII 1964; сборы и опреде-
ления наши; 1 — из Болгарии, VI 1969, сбор и определение И. М. Лихарева.
Нижняя часть спермовидукта образует резкий изгиб. СЖ 4X2—3. Верхняя пара
стилофоров впадает почти на уровне верхних концов наружных, удлиненно-булаво-
видной формы, в значительной степени отграничена как от наружных (нижних) сти-
лофоров, так и от вагины. Нижние стилофоры ненамного крупнее внутренних и короче
220
III. HYGROMIIDAE
их. Вагина цилиндрическая; вагинальные валики тонкие, в местах выхода протокой
стилофоров лопастей не образуют.
Пенис веретеновидный, при вскрытии обнаруживает четкое разделение на два
отдела, разграниченных мощным кольцевым валиком, Папилла занимает проксималь-
ную часть, стенки которой внутри гладкие. Семепроводящий канал внутри папиллы
фиксирован двумя тяжами. Дистальная часть пениса несет мощный конический при-
даток, внутри которого, почти целиком заполняя его полость, имеется продольное утол-
щение. Внутри дистальной части пе.чиса при основании придатка имеется еще 1—3 бо-
Рис. 245—247. Xeropicta krynickii (Kryn.), Болгария, VI 1969. 245 — внешний вид
полового аппарата {звездочка — пениальный придаток); 246 — пенис, вскрыт; 247 —
вагина, вскрыта. (Ориг.).
лее коротких утолщения, валика, каждый из которых покрыт многочисленными попе-
речными бороздками, что обеспечивает возможность значительного растяжения этих
валиков при выворачивании пениса и придатка. Бич в 2—3 раза короче цилиндри-
ческого эпифаллуса, образующего чаще всего 1—3 крутых изгиба. Проток семеприем-
ника повторяет изгиб спермовидукта, заканчивается мешковидным резервуаром, нем-
ного не доходящим до белковой железы.
Распространение и местообитания. Северо-восточное Среди-
земноморье: Балканский п-ов, Греция, Малая Азия, Иран, Южный берег Крыма,
окрестности Одессы, Новороссийска, Анапы, Астрахани; Ленкоранская низменность,
откуда по долинам горных рек поднимается до нагорно-степной зоны Талыша; вероятно,
западная часть Копетдага. Населяет степные участки, преимущественно на морском
побережье. В засушливые периоды поднимается по стеблям растений, образуя скопле-
ния.
2 . Xeropicta derbentina (Krynicki, 1836). Табл. XI, 107\ рис. 248, 249.
Krynicki, 1836 : 192 {Helix)-, Kobelt, 1877, Iconogr., V : 98, fig. 1433—1438
{Helix)-, Westerlund, 1889 : 342 {Helix)-, Пузанов, 1925a : 68, фиг. 3 [Xerop-
hila (Helicella) ]; H e s s e, 1934 : 28, t. 6, fig. 47 [Helicella (Xeropicta) ]; Л и x a p e в,
XEROPICTA
221
Раммельмейер, 1952 : 429, рис. 352в, 355 [Я. (Xeropicta) ]; G г о s s u,
1955 : 434, fig. 243, 245b [Я. (Xeropicta) ]; Hudec, Lezhawa, 1967b : 81,
Abb. 7, t. 6, 14 [Helicopsis (Xeropicta) ]; Шилейко, 1968a : 338, рис. 1, Я, 2, 11, 13
[Helicella (Xeropicta)].
Материал: более 100 экз. из 23 местонахождений в Крыму, на Кавказе и в окр.
Одессы.
Раковина от сильно прижатой до почти плоской; высота завитка чаще всего
не превышает половину высоты устья. Оборотов 5—5.5, выпуклых, быстро и плавно
Рис. 248—249. Xeropicta derbentina (Kryn.), окр. Дербента, 17 VII 1967. 248 —
внешний вид полового аппарата {звездочка — пениальный придаток); 249 — пенис,
вскрыт. (Ориг.).
нарастающих. Последний оборот в профиле плавно закруглен, почти вдвое шире пред-
последнего, к устью незначительно и плавно опущен. Окраска известковотбелая, без
рисунка или с темными лентами, часть из которых разбита на ряды пятен. Особенно
характерно наличие ряда таких пятен, прилегающего снизу к шву. Скульптура в виде
тонкой радиальной исчерченности и вялых спиральных бороздок. Устье чаще всего
почти правильно круглое, косое, внутри с расплывшейся губой, места его прикреп-
ления слегка сближены. Края устья тонкие, острые, простые. Пупок эксцентрический,
сквозь него полностью виден один оборот, редко — полтора оборота. Размеры: ВР 8—12,
БД 15-20, МД 13-18 мм.
Внутреннее строение. Материал: 3 экз. из Рикотского перевала на -
Малом Кавказе (Грузия), 26 IV 1971; 4 — из окр. с. Ахты, горный Дагестан, 16 VII1967;
12 — из окр. Дербента, 17 VII 1967; 8 — из нагорно-степной зоны Талыша, 3 VII 1964;
сборы и определения наши.
Организация полового аппарата в существенных чертах очень сходна с предыду-
щим видом. Основные отли ия: бич значительно короче — его длина не превышает
1/i длины эпифаллуса; кольцевой валик на границе дистальной и проксимальной
частей пениса развит слабо или отсутствует.
222
III. HYGROMIIDAE
Распространение и местообита ни я.. Южный берег Крыма;
спорадически вид распространен по всему Кавказскому перешейку и Малой Азии.
Розен (Rosen, 1914) сообщает о присутствии в Закавказье (Уплис-Цихе) Helix joppensis-
A. Schmidt; Лихарев и Раммельмейер (1952) полагают, что Розен имел дело с Xeropicta
krynickii, однако X. joppensis больше похожа на X. derbentina, и Розен, скорее всего,
имел в виду этот вид. Вместе с тем не исключено, что названия derbentina и joppensis —
синонимы, так как различия между ними крайне неопределенны (см.: Kobelt, Iconogr.,
N. F., 5 : 99, t. 144, fig. 1439). В этом случае в ареал вида надо включить еще Сирию'
и Палестину. Населяет степи и открытые склоны, встречается обычно большими скоп-
лениями; во время засухи образует «гроздья» на стеблях растений.
Рис. 250—251. Xeropicta candaharica (L. Pfr.), окр. Душанбе, 23 X 1971.
250 — внешний вид полового аппарата (звездочка — пениальный придаток);
251 — пенис, вскрыт. (Ориг.).
3. Xeropicta .candaharica (L. Pfeiffer, 1846). Табл. XI, 108; рис. 250, 251.
L. Pfeiffer, 1846, Proc. Zool. Soc., XJV : 37 (Helix); Hutton, 1849—
1850 : 650 (Helix); Martens, 1871, Malak. BL, 18 : 66, t. 1, fig., 4, 5 (H. krynickii
Andrz.); Мартенс, 1874 : 15, табл. 1, рис. 10 (H. derbentina Andrz.); Martens,
1882a : 47 (H. derbentina); Boettger, 1889b : 946—948 (H. krynickii Andrz. part.);
Розен, 1894 : 56 (H. millepnnctata Bttg, part.); Cockerell, 1928 ; 362 (Heli-
cella); Линдгольм, 1931 : 60 (Helicella); Hesse, 1934 : 28 (Helicella( Xeropicta)-
derbentina Kryn. var. candaharica) ]; Абрикосов, Цветков, 1945 : 210 (Heli-
cella); Лихарев, Раммельмейер, 1952 : 430, рис. 352, Б (Helicella (Xero-
picta)]; Лихарев, Старобогато в, 1967 : 190 (Helicella).
Материал: более 100 экз. из 9 местонахождений в Средней Азии.
Раковина очень похожа на раковину предыдущего вида. Отличия: высота
завитка, хотя и подвержена значительной изменчивости, в общем почти равна высоте
устья или немного меньше; пупок менее перспективный, сквозь него просматривается
примерно половина предпоследнего оборота. Размеры: ВР 8—10, БД 14.5—16,
МД 12.5-13.5 мм.
Внутреннее строение. Материал: 10 экз. из предгорий Чаткальского
хр. , берег р. Касансай у пос. Ала-Бука, 29 V 1972, сбор наш; 4 из того же места,
15 X 1972, сбор В. В. Горохова; 8— из окр. Душанбе, 12—23 X1971, сбор Л. Е. Урсо-
вой; 2 — из окр. Ленинабада, 29 VII 1967, сбор 3. Иззатуллаева; 2 — из окр. Джам-
була, 28 VII 1971, сбор К. К. Увалиевой; определения наши.
HELICELLA
223
Нижняя часть спермовидукта образует довольно резкий изгиб, яйцевод также изог-
нут или прямой. СЖ 4x2. Верхние стилофоры удлиненно-булавовидные, заметно обо-
соблены. Наружные (нижние) стилофоры удлиненно-овальные, немного крупнее вну-
тренних и расположены заметно ниже последних. Вагинальные валики лопастей не
образуют.
Пенис не обнаруживает столь четкого разделения на два отдела, как у X. krynickii,
Папилла пениса почти цилиндрическая, интрапапиллярная полость единая, семепро-
водящий канал подвешен на одном продольном тяже. При основании папиллы в стенке
эпифаллуса имеется небольшая кольцевая полость. Придаток пениса короче, чем у дру-
гих видов рода, но внутреннее строение его такое же; только продольная складка выра-
жена слабее или заменена 2—3 более мелкими складками. Длина бича — около поло-
вины длины эпифаллуса или немного больше. Проток семеприемника относительно тон-
кий, резервуар прилегает к спермовидукту немного выше его середины.
Распространение и местообитания. Средняя Азия: восточная
часть Копетдага — до Ферганского хр., Чаткальский хр., Ферганская долина, запад-
ная часть Киргизского хр., северный Афганистан. Населяет ровные и холмистые степ-
ные и полупустынные участки, в сухую погоду образует скопления на стеблях трав.
Выше предгорий не поднимается.
4. Xeropicta parableta (Boettger, 1881). Табл. XI, 105.
О. Boettger, 1881а : 212, t. 8, fig. 15а—с (Helix)- Westerlund,
1889 : 321 (Helix perableta ex errore).
Материал: более 30 экз. из трех местонахождений в Закавказье и Турции.
Раковина немного прижатая,со слегка куполовидными очертаниями завитка,
высота которого меньше высоты устья. Оборотов 5, довольно выпуклых, шов глубокий.
Последний оборот немного менее чем в 1.5 раза шире предпоследнего, округлый,
к устью не опущен. Окраска кремовая, с темным рисунком, сильно просвечивающим
в проходящем свете. Рисунок состоит из коричневых спиральных лент, в разной сте-
пени размытых; местами эти ленты соединяются друг с другом посредством неправиль-
ных радиальных или косых полос разной ширины. Низ раковины окрашен в целом
светлее; по периферии часто проходит кремовая лента. Самая яркая из темных лент про-
ходит по базальной части. Скульптура—неправильная, но довольно частая радиальная
исчерченность. Пупок умеренно развернут, не очень широкий, но перспективный.
Устье слегка скошено, округлое, немного вырезано предпоследним оборотом, внутри
с умеренно развитой губой. Края устья острые, не отвернуты. Размеры: ВР 3—5,
БД 6—7.5, МД 5.5—6.5 мм.
Внутреннее строение. Материал: 9 экз. из нагорно-степной зоны
Талыша, окр. с. Кельяз, 30 V 1974, сбор Я. И. Старобогатова, определение наше.
Несмотря на значительные конхологические отличия от прочих видов рода, на-
стоящий вид по признакам внутренней организации полностью отвечает родовому
диагнозу. Отличия от других представителей рода заключаются в следующих
деталях.
Бич очень короткий, конический. Семеприемник с тонким протоком и обширным
резервуаром. Характерные структуры внутри придатка пениса и в дистальной части
пениальной трубки менее резко выражены.
Распространение и местообитания. Вид описан по большой
серии, найденной в выбросах р. Араке у Нахичевани; живые моллюски обнаружены
в траве у ручья в нагорно-степной зоне Талыша.
14. Род HELICELLA Ferussac, 1821
F ё г u s s а с, 1821 : 41; Fitzinger, 1833 : 100 (Oxychilus part.); Held,
1837 : 903 (Xerophila)-, Hesse, 1926 : 118; Lindholm, 1927a : 135; Лихарев,
Раммельмейер, 1952 : 418; Wenz, Zilch, 1959—1960 : 668.
Раковина прижатая, белая, обычно с темными спиральными лентами. Пупок ши-
рокий, развернутый, перспективный. Поверхность раковины без волосков.
Интрапапиллярная полость образована за счет развития кольцевой полости в ди-
стальной части эпифаллуса, которая заходит в папиллу, в стенки пениального чехла
и в стенки эпифаллуса. Стилофоров одна пара, гомологичная наружным стилофорам
остальных родов подсемейства; нижняя часть стилофоров вздута, заключает широкую
полость. Бич значительно короче эпифаллуса. Ретрактор правого омматофора не об-,
разует перехлеста с половым ретрактором.
Отсутствие настоящей интрапапиллярной полости и своеобразное устройство сти-
лофоров свидетельствует о значительной изолированности рода от остальных таксо-
нов Trichiirae; однако, не имея возможности судить о его филогенетических связях,
мы оставляем этот род пока в составе названного подсемейства.
824:
III. HYGROMIIDAE
Распространение. Южная и Средняя Европа. В ископаемом состоянии
род известен начиная с нижнего плиоцена в пределах современного ареала.
Типовой вид — Helix itala Linnaeus, 1758.
Helicella candicans (L. Pfeiffer, 1841). Табл. XI, 109, 110; рис. 252—254.
L. Pfeiffer, 1841, Wiegm. Arch., I : 220 (Helix); Hartmann, 1844 : 148,
t. 45, fig. 1—6 (Helix obvia); Westerlund, 1889 : 339 (H. obvia); Лихарев,
Рис. 252—254. Helicella candicans (L. Pfr.), Болгария, окр. Пловдива, 15 IX 1950.
252 — внешний вид полового аппарата; 253 — пенис, вскрыт; 254 — дистальная
часть женского отдела, вскрыта. (Ориг.).
Раммельмейер, 1952 : 422; G г о s s и, 1955 : 425, fig. 235, 236; Ehrmann,
1956 : 124, t. 5, fig. 63; Lozek, 1956 : 200, t. 29, 1; Z i 1 c h, J a e c k e 1, 1962 : 174;
Lozek, 1964 : 289, Abb. 64 (Helicella obvia).
Материал: более 80 экз. из 9 местонахождений в Молдавии, Болгарии, Чехосло-
вакии и Венгрии.
Раковина в той или иной степени уплощенная, высота завитка не меньше
половины высоты устья, часто эти величины почти равны. Оборотов 5—5.5, слабо вы-
пуклых; первые медленно, два последних быстрее нарастают. Последний оборот сверху
немного уплощен, в профиле плавно и широко закруглен, к устью постепенно и плавно
опускается. Спиральные ленты чаще всего имеются. 1—2 наиболее яркие ленты про-
ходят по верхней стороне каждого оборота; остальные тонкие и менее четкие распола-
гаются на базальной части раковины. Нередко ленты распадаются на ряд пятен; осо-
бенно это касается базальных лент. Скульптура тонкая, представлена радиальной лег-
кой исчерченностью, которая лучше выражена на верхней стороне раковины, а также
вялыми спиральными бороздками и вмятинами. Нередко, однако, попадаются отдель-
ные экземпляры, у которых радиальная исчерченность переходит в морщинистость.
Устье от круглого до широкоовального, скошено, края его острые, простые, внутри
с тонкой губой. Пупок развернутый, вполне перспективный. Размеры: ВР 7—8,
БД 13—16, МД 11—14 мм.
LINDHOLMOMNEME
225
Внутреннее строение. Материал: 5 экз. из Венгрии, окр. с. Балатон-
фюред на оз. Балатон, 15 VII 1970; 3 — из Венгрии, окр. г. Печ, VII 1970, сборы
Д. А. Сахарова; 2 — из центральной Чехии, 22 V 1967, соор Я. Бухара; 2 — из Бол-
гарии, окр. Пловдива, 15 IX 1950, сбор А. Риделя; определения наши.
Нижняя часть спермовидукта и очень короткий яйцевод почти прямые. СЖ
4x2—3. Стилофоры очень массивные, удлиненные, в основании заметно вздутые.
Это вздутие отвечает расположенной в нижней части стилофоров полости, которая по
существу представляет собой резкое расширение вагины. От верхней части стилофора
эта полость отграничена кольцевым козырьком. Кроме того, в стенке вагины возле
места прикрепления верхнего края стилофоров имеется кольцевая полость.
Пенис веретеновидно-цилиндрический, внутренние стенки его почти гладкие,
папилла цилиндрическая, полость занимает не более половины ее длины, обычно меньше.
Бич значительно короче прямого эпифаллуса, обычно втрое короче. Проток семеприем-
ника короткий и толстый, прямой. Резервуар прилегает к середине спермовидукта.
Распространение и местообитания. Средняя и Юго-Вбсточ-
ная Европа с западными районами Украины и с Молдавией. Населяет степные участки,
иногда встречается по опушкам лесов. Во время летней засухи образует гроздья на
стеблях трав.
II. Подсемейство HYGROMIINAE Tryon, 1866
Раковина средних или небольших размеров, различной формы, с узким или ши-
роким пупком, который изредка может быть полностью запаянным. Окраска разно-
образная, чаще всего с одной неясной светлой лентой по периферии.
Стилофоров 2, расположенных на одной стороне вагины; в пределах подсемейства
наблюдается редукция внутреннего стилофора вплоть до полного исчезновения. Все
стенки папиллы заполнены рыхлой тканью, имеющей тенденцию распадаться на от-
дельные комплексы. Известен один случай ступенчатой редукции стилофоров и сли-
зистых желез (Circassina circassica).
Распространение. Большая часть территории Палеарктики, включая
Сибирь, Алтай и Дальний Восток. В Средней Азии подсемейство представлено только
одним широко распространенным видом — Pseudotrichia rubiginosa. В ископаемом со-
стоянии представители подсемейства встречаются начиная с верхнего олигоцена (Ев-
ропа).
1. Род LINDHOLMOMNEME Haas, 1936
Haas, 1936 : 149, Лихарев, Раммельмейер, 1952 : 439 (Zenobiella
part.); Шилейко, 1970:1311.
Раковина прижатая, до почти плоской. Окраска желтая или серая, почти одно-
цветная. Пупок развернутый или цилиндрический, но всегда достаточно широкий и
перспективный. Иногда на поверхности раковины имеются короткие редкие золотистые
волоски. Устье без зубов.
Внутренний стилофор рудиментарный, располагается при основании основного,
обнаруживается только при вскрытии вагины. Бич рудиментарный, папилла пениса
длинная, гладкая, толстостенная, стенки ее равномерно заполнены тканью. Имеются
хорошо развитые мускульные ленты, стягивающие пенис и эпифаллус. Ретрактор
правого омматофора образует перехлест с половым ретрактором.
Распространение. Алтай, Сибирь, южная часть Дальнего Востока.
Типовой вид — Helix rhysota Westerlund, 1896 (syn. Helix nordenskioldi Wester-
lund, 1876).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
РОДА. LINDHOLMOMNEME
1 (2). Спиральная скульптура отсутствует............3. L. nordenskioldi.
2 (1). Спиральная скульптура имеется.
3 (4). Высота завитка не превышает половины высоты устья . . . 1. L. kudiensis.
4 (3). Высота завитка почти равна высоте устья.........2. L. no tophi 1а.
1. Lindholmomneme kudiensis (Cockerell, 1924). Табл. XII, 111.
Cockerell, 1924 : 581 (Hygromia)- Ши лей к о, 1970 : 1313, рис. 4.
Материал: 3 экз. из' 1 местонахождения в Приморье.
Раковина почти плоская, с притупленным завитком, вершина слегка высту-
пающая. Оборотов 5—6, округлых, последний оборот в самом начале с легким намеком
на угол, в 1.5 раза шире предпоследнего, к устью слабо опущен. Окраска желтовато-
15 А. А. Шилейко
226
III. HYGROMIIDAE
роговая, однотонная. Скульптура — тонкая и неравномерная радиальная испорчен-
ность и гораздо более тонкие спиральные бороздки. Устье слегка оттянуто вправо,
опущено и скошено, очень характерной формы (смотреть при нормальном положении
раковины): край его в ангулярной области пемпого, но ясно поднимается, палаталь-
ный образует плавную дугу и в месте соединения с базальным краем закруглен; ба-
зальный край почти прямой, направлен под острым углом к средней линии последнего
оборота; колумеллярный край чрезвычайно короткий, вертикальный. Губа лежит
отступя от края устья, резче выражена на базальном крае и нижней части палаталь-
ного. Края устья тонкие, слегка отвернуты в нижней части устья. Пупок очень ши-
рокий, развернутый. Размеры: ВР 6.8—7, БД 14.8—15, МД 12.3 мм.
Рис. 255—256. Lindholmomneme notophlla (Cock.) из окр. с. Черниговка Приморского
края, 17 V 1912. 255 — внешний вид полового аппарата; 256 — дистальная часть
полового аппарата, вскрыта. (Ориг.).
Внутреннее строение неизвестно.
Распространение и местообитания. Вид найден в двух местах:
бассейн р. Кудия (типовое местонахождение) и окр. бухты Рудная в Приморском крае.
Лесной вид.
2. Lindholmomneme notophila (Cockerell, 1924). Табл. XII, 112; рис. 255, 256.
Cockerell, 1924 : 581 (Hygromia); Ш и л е й к о, 1970 : 1311, рис. 3, 3—4.
Материал: 3 экз. из 2 местонахождений в Приморье.
Раковина с умеренно возвышающимся, широким завитком и широкой вы-
ступающей вершиной. Оборотов 5—6, в профиле округлых, последний оборот при-
мерно в 1.5 раза шире предпоследнего, к устью незначительно опущен. Окраска очень
бледно-коричневая, монотонная; встречаются особи с золотистым оттенком фона. Скуль-
птура в виде тонкой радиальной исчерченности и многочисленных тонких спиральных
бороздок. Устье широкое, не оттянуто вниз и в сторону. Края устья отвернуты, губа
тонкая, близ колумеллярного края расширенная. Устье косое. У свежих экземпляров
на поверхности раковины имеются короткие волоски. Пупок почти цилиндрический,
но широкий и перспективный. Размеры: ВР 6—9, БД 11—13.5, МД 10—12.3 мм.
LINDHOLMOMNEME
.227
Внутреннее строение. Материал: 2 экз. из с. Черниговка Примор-
ского края, 17 V 1912, сбор Емельянова, определение И. М. Лихарева.
Нижняя часть спермовидукта и яйцевод почти прямые. СЖ 2X3. Стилофор до-
вольно крупный, удлиненно-яйцевидный. Рудиментарный стилофор виден при вскры-
тии вагины и нижней части основного стилофора; особенно хорошо он заметен на про-
дольных срезах.
Пенис длинный, цилиндрический. Мускульные ленты крепятся к его нижнему и
верхнему концам. Папилла длинная, тонкая, семепроводящий канал в разрезе имеет
Рис. 257—258. Lindholmomneme nordenskioldi (West.) из окр. г. Прокопьевска Кеме-
ровской обл., VI 1961. 257 — внешний вид полового аппарата; 259 — дистальная'
часть полового аппарата, вскрыта. (Ориг.).
форму трех- или четырех лучевой звезды. Бич в несколько раз короче цилиндрического
эпифаллуса, тонкий. Между дистальными протоками мужского отдела натянута про-
зрачная мембрана. Проток семеприемника прямой, заканчивается удлиненным резер-
вуаром, прилегающим к верхней части спермовидукта.
Распространение и местообитания. Вид найден в трех пунк-
тах Приморского края: окр. Конгауза (типовое местонахождение) и в Черниговке;
близ побережья бухты Песчаной у Владивостока найден один не вполне взрослый эк-
земпляр. Вид обитает в лиственном лесу.
3. Lindholmomneme nordenskioldi (Westerlund, 1876). Табл. ХП, 113; рис. 257
258.
W е s t er In nd, 1876 : 97 [Helix (Eulota)}; 1877 : 32, t. 1, fig. 2 [Helix (Eulota)];
1889 : 51 (Helix); 1896 : 185 [Helix (Trichia) rhusota et var. altaica]; S chlesch,
1928 : 276—279, t. 12, fig. 3—10 [Euomphalla); Лихарев, Раммельмейер,
1952:440, рис. 365, 366 (JZenobiella); Г у д e ц, 1969:26, рис. 1, 3 А—В
(Chilanodori); Шил ейко, 1970 : 1311, рис. 3, 1—2.
15*
838
III. HYGROMIIDAE
Материал: более 40 экз. из 4 местонахождений на Алтае.
Раковина умеренно прижатая, с тупым куполовидным завитком и слегка
выступающей вершиной. Оборотов 6—6.5, выпуклых, медленно нарастающих. По-
следний оборот в профиле правильно закруглен, лишь иногда в самом начале с лег-
кой угловатостью, в 1.5 раза шире предпоследнего, к устью слабо или умеренно опу-
щен. Окраска серая или золотистая, с неясной светлой лентой по периферии. Скуль-
птура в виде, радиальной густой равномерной ребристой исчерченности, выраженной
в равной степени на верхней и нижней сторонах раковины. Кроме того, на раковине
имеются редкие короткие золотистые, легко опадающие волоски, оставляющие не-
большие редкие рубцы. Спиральная скульптура отсутствует. Устье округлое или ши-
рокоовальное, косое, обычно слегка оттянуто вправо; места прикрепления устья ши-
роко расставлены и связаны ясной прозрачной париетальной мозолью. Края устья
тонкие, хрупкие, в базальной части отвернуты. Губа занимает только базальную
часть устья, очень слабая или отсутствует. Пупок слабо развернутый, почти цилин-
дрический, умеренно широкий, но перспективный. Размеры: ВР 7—8.5, БД 12—15,
МД 10—13 мм.
Внутреннее строение. Материал: 8 экз. из окр. г. Прокопьевска
Кемеровской обл., VI 1961, сбор Ю. Б. Бызовой, определение наше.
Нижняя часть спермовидукта и яйцевод образуют 1—2 изгиба. СЖ 2—3x2—4.
Стилофор чрезвычайно массивный; рудиментарный стилофор впадает в нижнюю часть
основного. Оба стилофора вдаются в полость вагины единым сосочком.
Пенис длинный, прямой или изогнутый; пениальные мускульные ленты мощные.
Папилла пениса длинная, семепроводящий канал в разрезе имеет трехлучевую форму.
Бич очень короткий, эпифаллус почти не изогнут. Проток семеприемника почти пря-
мой, резервуар небольшой, вытянутый, прилегает к спермовидукту немного выше
его середины.
Распространение и местообитания. Алтай, Южная Сибирь,
бассейн Амура и Приморский край. Границы ареала нуждаются в уточнении. Оби-
татель лиственных и смешанных лесов.
2. Род CHILANODON Westerlund, 1897
Westerlund, 1897 : 119; Lindholm, 1927a : 124 (Altaicola); W e n z,
Zilch, 1959—1960 : 682 (Altaicola)- Hudec, Riedel, 1967 : 96; Шилейко,
1970 : 1309.
Раковина высокая, с широким куполовидным завитком. Окраска роговая или жел-
тая. Пупок проколовидный. Раковина без волосков. Устье с 1—2 зубами; если зуб 1,
он может быть выражен слабо и иметь вид удлиненного утолщения губы в базально-
колумеллярной области.
Внутренний стилофор рудиментарный, при наружном осмотре гениталий не виден.
Бич рудиментарный, папилла пениса длинная, стенки ее равномерно заполнены рых-
лой тканью. Пениальные ленты выражены в разной степени. Ретрактор правого омма-
тофора образует перехлест с половым ретрактором.
Распространение. Алтай, Прибайкалье и низовья р. Уссури.
Типовой вид — Helix gerstfeldti Dybowski, 1901.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CHILANODO
1 (2). В устье раковины имеется 2 хорошо развитых почти одинаковых бугорковидных
зуба ....................................................1. Ch. bicallosa.
2 (1). Зубы в устье отсутствуют; губа в базальной части иногда образует бугорковидное
утолщение ................z..............................2. Ch. gerstfeldti.
1. Chilanodon bicallosa (L. Pfeiffer, 1853). Табл. XII, 11Ц рис. 259, 260.
L. Pfeiffer, 1853:638 (Helix)- Westerlund, 1889:32 (Helix)-,
Kobelt, 1892, Iconogr., N. F., V : 1, fig. 117 (Perforatella)- Лихарев, Рам-
мелц мейер, 1952 : 444, рис. 371 (Perforatella)- Шилейко, 1970 : 1309,
рис. 2, 1—2.
Материал: 17 экз. из 2 местонахождений на Алтае.
Раковина ширококоническая, состоит из 7—7.5 выпуклых оборотов, которые
нарастают медленно как в высоту, так и в ширину. Последний оборот в профиле очень
слабо угловатый, снизу немного уплощен, к устью опущен. Окраска роговая с красно-
ватым или желтоватым оттенком; по периферии проходит размытая светлая лента,
которая обычно прослеживается у шва на более ранних оборотах. Скульптура в виде
тонкой радиальной исчерченности, спиральная скульптура отсутствует. Устье распо-
лагается под базальной поверхностью предпоследнего оборота, широко изогнуто-ще-
Левидное. На губе базального края имеются 2 массивных зуба в виде бугорков, которым
CHILANODON
229
снаружи соответствуют слабые вмятины. Один зуб расположен в базально-колумел-
лярной части устья, второй — в базально-палатальной, причем второй расположен
глубже первого. Края устья слегка отвернуты, плоскость его скошена. Точковидный
пупок в различной степени прикрыт отворотом колумеллярного края. Размеры: ВР 6—7,
БД 8-9.5, МД 7.7—9.2 мм.
Внутреннее строение. Материал: 6 экз. из окр. Прокопьевска Ке-
меровской обл., VI 1961, сбор Ю. Б. Бызовой, определение наше.
Рис. 259—260. Chilanodon blcallosa (L. Pfr.) из окр. г. Прокопьевска Кемеровской обл.,
VI 1961. 259— внешний вид полового аппарата; 260 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
Спермовидукт и яйцевод почти прямые. СЖ 2—3X1—3. Стилофор относительно
очень массивный, вместе с дополнительным вдается в полость вагины единым сосочком.
Внутренняя поверхность вагины несет несколько очень слабых складок и одну более
мощную складку, спускающуюся в клоаку.
Пенис тонкий, почти цилиндрический, связан с эпифаллусом хорошо развитыми
пениальными мускульными лентами. Папилла пениса тонкая, длинная. Эпифаллус
прямой или в различной степени изогнут. Проток семеприемника довольно тонкий,
прямой, резервуар не доходит до белковой железы.
Распространение и местообитания. Алтай. Населяет подстилку
лиственных лесов, близ воды.
2. Chilanodon gerstfeldti (Dybowski, 1901). Табл. XII, 115\ рис. 261, 262.
W esterlund, 1897 : 119 (Helix sibirica, non Pfeiffer, 1853); Dybowski,
1901 : 136, fig. 1—3 (Helix)', Lindholm, 1912 : 64; Лихарев, Раммель-
мейер, 1952 : 445, рис. 372 (Perforatella)-, Hudec, Riedel, 1967 : 125—130,
Abb. 1,2; Г у д e ц, 1969 : 26—29, рис. 2; Шилейко, 1970 : 1310, рис. 2, 3—4.
230
III. HYGROMIIDAE
Материал: 12 экз. из 2 местообитаний в Прибайкалье.
Раковина ширококоническая, тонкостенная, состоит из 6 умеренно выпук-
лых, узких, медленно нарастающих оборотов. Последний оборот снизу уплощен, вну-
тренняя часть его плавно спускается к пупочной впадине. Периферия слегка углова-
тая и немного сдвинута вниз относительно средней линии последнего оборота, к устью
почти не опущенного. Скульптура в виде тонкой радиальной исчерченности. Устье ко-
сое, почти целиком накрыто предпоследним оборотом, округло-треугольное, с губой,
которая на базально-колумеллярном крае утолщается, образуя удлиненное зубообраз-
Рис. 261—262. Chilanodon gerstfeldt! (Dyb.) из Прибайкалья, 12 VII 1954. 261—
внешний вид полового аппарата; 262 — дистальная часть полового аппарата, вскрыта.
(Ориг.).
ное вздутие. На стыке базального и. палатального краев имеется еще один очень ма-
ленький бугорковидный зуб, . развитый иногда очень слабо. Пупок очень узкий,
круглый, может быть слегка прикрытым. Размеры: ВР 5.5—6, БД 6.5—7.5,
МД 6—7 мм.
Внутреннее строение. Материал: 6 экз. из Прибайкалья, Большие
Коты, 12 VII 1954, сбор Я. И. Старобогатова, определение наше.
Спермовидукт и яйцевод почти прямые. СЖ 2—3x2—4. Стилофор длинный, почти
цилиндрический, открывается в вагину вместе с крошечным дополнительным стило-
фором на конце единого сосочка.
Пенис в дистальной части немного сужен, в проксимальной — утолщен, слегка
изогнут, пениальные ленты умеренно развиты. Папилла длинная, ее просвет на попе-
речном разрезе имеет форму прямой или изогнутой щели. Эпифаллус длинный, бич
относительно чрезвычайно короткий — развит слабее, чем у любого другого представи-
теля подсемейства. Проток семеприемника без резких изгибов, резервуар прилегает
к верхней половине спермовидукта, но белковой железы не касается.
Распространение и местообитания. Прибайкалье; одна на-
ходка сделана близ устья р. Уссури. Обитает в пойменных сырых кустарниковых за-
рослях.
PERFORATELLA
231
3. Род PERFORATELLA Schluter, 1838
Held, 1837 : 915 (Trochiscus, non Heyden, 1816); Beck, 1837 : 2 (Petasia,
non Stephens, 1828); Schluter, 1838 : 4; Lindholm, 1927a : 124; Hesse,
1931 :16; Лихарев, Раммельмейер, 1952 : 443; Wenz, Zilch, 1959—
1960:681; Шилейко, 1970:1313.
Раковина высокая, с несколько заостренным завитком (вершина не сглаженная).
Окраска роговая, коричневая или каштановая. Пупок чрезвычайно узкий, не менее
чем наполовину закрыт, иногда почти полностью запаян. Раковина без волосков.
Устье с двумя хорошо развитыми зубами.
Рис. 263—264. Perforatella bidens (Chenin.) из Московской обл., 6 V 1966. 263 — внеш-
ний вид полового аппарата; 264 — дистальная часть полового аппарата, вскрыта.
(Ориг.).
Внутренний стилофор отсутствует. Бич не короче эпифаллуса, папилла с довольно
плотными стенками. Имеется тонкий наружный чехол пениса. Вагина окружена чех-
лом. Ретрактор правого омматофора образует перехлест с половым ретрактором.
Распространение. Восточная Европа., В ископаемом состоянии род
известен начиная с верхнего плиоцена (Европа).
Типовой вид — Trochus bidens Chemnitz, 1786.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ЕИДОЕ РОДА PERFORATELLA
1 (2). Большой диаметр раковины 7—9 мм......................1. Р. bidens.
2 (1). Большой диаметр раковины 10—12 мм.................2. Р. dibothrion.
1. Perforatella bidens (Chemnitz, 1786). ТаЫ. XII, 116; рис. 263, 264.
Chemnitz, 1786, Syst. Conch.-Cab. : 50,1.122, fig. 1052 (Trochus); Michaud,
1831, Compl. Hist. nat. Moll. France : 12, t. 14, fig. 26 (Helix); Westerlund,
1889 : 31 (Helix); Ehrmann, 1956 : 133, t. 5, fig. 68; Лихарев, Раммель-
мейер, 1952 : 443, рис. 370; Lozek, 1956 : 213, t. 35, 1; Zilch, Jaeckel,
1962 : 178; Hudec, 1963 : 348-360, Abb. 2, 8; L о z e k, 1964 : 294, t. 23, 2a—c;
Шилейко, 1970 : 1315, рис. 5, 3—4.
232
III. HYGROMIIDAE
Материал: более 100 экз. из 19 местонахождений в средней полосе европейской
яасти СССР, в Польше и ГДР.
Раковина ширококоническая, блестящая, с острой вершиной, но завиток,
тем не менее, куполовидных очертаний, не конический. Оборотов 7, очень медленно
нарастающих. Последний оборот незначительно шире предпоследнего, в самом начале
с тупым углом, к устью исчезающим. Возле устья последний оборот внезапно и резко
опущен. Скульптура в виде тонкой и неравномерной радиальной исчерченности.
Окраска светло-роговая или коричневая, с двумя размытыми светлыми лентами: одна
проходит у шва, другая — по периферии. Устье очень узкое, с отвернутыми краями и
губой, на которой расположены два массивных зуба, которым снаружи соответствуют
ямки, причем более глубокая ямка отвечает базально-палатальному зубу. Пупок почти
полностью закрыт отворотом колумеллярного края. Размеры: ВР 5—6.5, БД 7—9,
МД 6—8 мм.
Внутреннее строение. Материал: 3 экз. из Гарца, ГДР, V 1969, сбор
и определение Э. Клаусса; 4 — из Плохоцина у Варшавы, И V 1950, сбор и определе-
ние А. Риделя; 9 — из с. Апаринки Московской обл., 6 V 1966; 7 — из с. Ромашково
Московской обл., 3 XI 1968, сборы и определения наши.
Спермовидукт и яйцевод почти прямые. СЖ 3X1—3. Стенки вагины железистые,
опоясаны чехлом, который либо полностью окутывает нижнюю часть вагины воротнич-
ком, либо не полностью срастается с наружной стенкой вагины снизу и с боков, остав-
ляя открытый сверху карман. Стилофор впадает в вагину удлиненным сосочком, на
конце которого открывается проток стрелы.
Пенис длинный, тонкий, цилиндрический; соответственную форму имеет и папилла,
просвет которой на поперечном разрезе имеет форму многолучевой звезды. Бич не-
сколько длиннее суммарной длины пениса и эпифаллуса. Пениальные .мускульные
ленты отсутствуют. Проток семеприемника не извит, объемистый мешковидный тон-
костенный резервуар прилегает к спермовидукту немного выше его середины.
Распространение и местообитания. К северу вид распростра-
нен до Ленинградской и Новгородской областей, к востоку местами до Волги, к югу
до Харьковской обл., к западу до восточных предгорий Альп. Известен из верхнего
плиоцена Европы. Лесной вид; наибольшей плотности достигает в сырых ольшаниках,
черноольховых топях и сырых оврагах.
2. Perforatella dibothrion (Kimakowicz,J1884). Табл. XII, 117', рис. 265, 266.
В i е 1 z, 1867 : 58 (Petasia bidens var. major); Kimakowicz, 1883—1884 : 47
[Fruticicola (Petasina)]; Polinski, 1924:171, fig. 23, 25, fig. 43, 47 (Petasina);
Gr ossu, 1955 : 447; Ehrmann, 1956 : 134; L о z.e k, 1956 : 213, t. '35, 2;
Полевина, 1959 : 67; Zilch, Jaeckel, 1962 : 179; Hudec, 1963 :
348—360, Abb. 1, 7; Lozek, 1964 : 295, t. 23, la—с; Ш и л e й к о, 1970:1313,
рис. 5, 1—2.
Материал: более 40 экз. из 5 местонахождений на Карпатах.
Форма раковины приближается к шаровидной, со слегка сглаженной вер-
шиной, очертания завитка от почти строго конических до куполовидных. Оборотов
6.5—7.25, умеренно выпуклых, медленно нарастающих, тесно навитых. Последний
оборот незначительно шире предпоследнего, к устью внезапно и резко опущен. Окраска
от темно-роговой до каштановой и красновато-коричневой, светлая лента на перифе-
рии хорошо заметна, лента возле шва отсутствует или очень слабая. Скульптура в виде
правильной и частой радиальной исчерченности, лучше выраженной на верхней сто-
роне раковины. Базально-колумеллярный и базально-палатальный зубы очень хорошо
развиты, снаружи им соответствуют ямки. Пупок не менее чем наполовину, нередко
и полностью прикрыт отворотом колумеллярного края. Размеры: ВР 7—10, БД 9—12,
МД 8—10.5 мм.
Внутреннее строение. Материал: 1 экз. из Задиела, Словакия, 9 III
1962, сбор и определение В. Гудеца; 2 — из Бишчеди (Польша), 10 VIII 1952, сбор и
определение А. Риделя; 2 — из окр. Ужгорода, 6 IX 1969; 3 — из окр. Рахова, 17 IX
1969, сборы и определения наши.
Спермовидукт и яйцевод прямые. СЖ 3X1—2; обычно одна из них одноветвистая и
в различной степени гофрирована. Вагина окружена рыхлым чехлом, верхняя часть
которого лежит на вагине свободно. Внутренняя полость вагины и клоаки неполностью
перегорожена вдоль мощным валиком; таким образом, имеются отдельные пути для
выдвигания пениса и принятия пениса партнера.
Пенис прямой или образует петлю. Папилла тонкая, длинная, цилиндрическая,
с плотными стенками. Семепроводящий канал на поперечном разрезе имеет форму трех-
лучевой звезды. Пениальные мускульные ленты отсутствуют. Бич тонкий, длинный,
равен половине суммарной длины пениса и эпифаллуса или немного больше. Эпифаллус
тонкий, цилиндрический, от пениса наружно не отграничен; граница между пенисом
Рис. 265—266. Perforatella dibothrion (Kim.) из окр. Ужгорода, 6 IX I960.
265— внешний вид-полового аппарата; 266— дистальная часть поло-
вого аппарата, вскрыта. (Ориг.).
234
III. HYGROMIIDAE
и эпифаллусом отмечена местом прикрепления полового ретрактора. Проток семеприем-
ника почти прямой, заканчивается неопределенно фасолевидной формы резервуаром,
заметно не доходящим до белковой железы.
Распространение и местообитания. Карпаты. Вид населяет
подстилку лиственных лесов, особенно в сырых оврагах и по берегам ручьев.
4. Род MONACHOIDES Gude et Woodward, 1921
Fitzinger, 1833 : 95 (Monacha part.); Hartmann, 1841 : 41 (Trichia
part.); Gude, Woodward, 1921 : 179 (Monachoides, Zenobiella); Hesse, 1931 :
13 (Monacha part.); Лихарев, Раммельмейер, 1952 : 439 (Zenobiella);
Wenz, Zilch, 1959—1960 : 680 (Zenobiella), 682; Шилейко, 1970 : 1315.
Раковина от слегка прижатой до кубаревидной, с выступающей вершиной. Ок-
раска от желтовато-роговой до темно-бурой. Пупок узкий, неперспективный, иногда
полностью запаян. Раковина без волосков, но чаще всего с зернистой скульптурой и
кожистыми шипиками или чешуйками. Устье без зубов.
Внутренний стилофор отсутствует. Бич развит в различной степени. Имеется на-
ружный чехол пениса. Ретрактор правого омматофора образует перехлест с половым
ретрактором.
Распространение. Большая часть территории Палеарктики (без Даль-
него Востока). Основная масса видов сосредоточена в Европе. В ископаемом состоянии
род известен начиная с верхнего олигоцена (Европа, Северная Африка).
Типовой вид — Helix incarnata Muller, 1774.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА MONACHOIDES
1 (4). Большой диаметр раковины не превышает 9 мм. Виды обитают в азиатской
части СССР.
2 (3). Поверхность раковины несет 4—5 спиральных рядов треугольных кожистых
шипов......................................................4. М. aculeata.
3 (2). Поверхность раковины покрыта слегка изогнутыми удлиненными чешуйками
..........................................................3. М. stiixbergi.
4 (1). Большой диаметр раковины не менее 11 мм. Виды обитают в европейской части
СССР.
5 (6). Пупок не более чем наполовину прикрыт отворотом колумеллярного края
устья ....................................,...............1. М. incarnata.
6 (5). Пупок запаян или от него остается лишь узкая щель......2. М. vicina.
1 . Monachoides incarnata (Muller, 1774). Табл. XII, 118; рис. 267, 268.
Muller, 1774 : 63 (Helix)-, Westerlund, 1889 : 97 (Helix)-, Лихарев,
Раммельмейер, 1952:442, рис. 368 (Zenobiella)-, G г о s s u, 1955:449,
fig. 252 (Zenobiella)-, Ehrmann, 1956 : 132, fig. 90 (Monacha)-, Lozek, 1956 :
211, t. 33, 3; Zilch, J a e c k e 1, 1962 : 179 [Perforatella (Monachoides)]; Hudec,
1963 : 348—360, Abb. 3, 9 [Perforatella (Monachoides)]-, Lozek, 1964 : 293, t. 24,
3a—c, t. 30, 2; Gittenberger et al., 1970 : 99, fig. 168—170, 173, Kaart 69
[Perforatella (Monachoides)]; Шилейко, 1970 : 1315, рис. 6, 3—4.
Материал: более 40 экз. из 6 местообитаний в Средней Европе.
Раковина кубаревидная, с широким, почти строго коническим завитком.
Оборотов 6—6.5, плавно нарастающих, слабо выпуклых. Последний оборот в самом
начале слегка угловатый, к устью внезапно и резко опущен; углу перегиба соответ-
ствует широкая размытая светлая полоса — просвечивающая губа. Окраска от светло-
желтой до красновато-роговой, со светлой размытой лентой на периферии, которая
порой едва заметна. Свежие раковины с шелковистым блеском, старые — матовые.
Поверхность раковины очень тонко радиально неясно исчерчена; кроме того, имеются
многочисленные тонкие гранулы ромбической формы. В совокупности гранулы обра-
зуют характерный узор в виде плотной и частой сеточки. У свежих раковин каждая
гранула несет одну тонкую изогнутую, радиально ориентированную чешуйку; однако
эти чешуйки легко утрачиваются. Устье сильно скошено, широкоовальное, немного
оттянуто вниз и в сторону. Губа светлая, с желтым или Красноватым оттенком. Пупок
неперспективный, узкий, иногда полностью открытый, чаще прикрыт, но не более чем
наполовину. Размеры ВР : 9—11, БД 12—16, МД 10—13 мм.
Вестерлунд (Westerlund, 1898: 180) отмечает присутствие в окрестностях Киева
средиземноморского вида «Helix» revelata Ferussac. Несомненно, Вестерлунд имел дело
с молодой особью М. incarnata, которая очень похожа на «Я.» revelata, на.что указывал
еще Мокен-Тандон (Moquin-Tandon, 1855 : 221).
Внутреннее строение. Материал: 4 экз. из Гарца, ГДР, V 1969,
сбор и определение Э. Клаусса; 3 — из Казимежа над Вислой, Польша, 6—19 VII
MONACHOIDES
235
1957, сбор и определение А. Риделя; 1 —из окр. Враца, Болгария, 4 VII 1969, сбор и
определение И. М. Лихарева; 2 — из Предкарпатья, окр. г. Сторожинец, с. Майдан,
15 VII 1970, сбор Л. А. Шилейко, определение наше.
Спермовидукт и яйцевод без изгибов. СЖ 2X3—5; отдельные ветви вторично вет-
вятся. Стилофор булавовидный, вдается в полость вагины сосочком. Вагинальные ва-
лики слабые. Вагина и клоака окружены перивагиналыюй полостью, расположенной
в разросшихся рыхлых тканях стенок вагины.
Рис. 267—268. Monachoides incarnata (Mull.) из Болгарии, окр. Вроца, 4 VII 1969.
267 — внешний вид полового аппарата; 268 — дистальная часть полового аппарата,
вскрыта. (Ориг.).
Пенис не вздут, с тонкостенным двойным чехлом. Папилла очень длинная, с рых-
лыми стенками. Семепроводящий канал на поперечном разрезе четырехлучевой. Бич
и эпифаллус примерно равной длины. Граница между пенисом и эпифаллусом внешне
отмечена только местом прикрепления полового ретрактора. Проток семеприемника
умеренной длины, мешковидный резервуар не достигает белковой железы.
Распространение и местообитания. Горные области Средней
и Западной Европы; в СССР встречается в Карпатах и Предкарпатье, на восток местами
доходит до Днепра. Населяет подстилку лиственных и смешанных лесов. Известен
из четвертичных отложений в пределах ареала.
2 . Monachoides vicina (Rossmassler, 1842). Табл. IX, S3; XII, 119; рис. 269, 270.
R о ssmassl er, 1842, Iconogr., II, 11 : 3, fig. 689 (ffeZis); W esterlund,
1889 : 98 {Helix); Лихарев, Раммельмейер, 1952 : 442, рис. 369 {Zenobiella);
G г о s s u , 1955 : 451, fig. 254 {Zenobiella); Ehrmann, 1956 : 133, t. 5, fig. 67
{Monacha); Lozek, 1956 : 212, t. 33, 2; Zilch, Jaeckel, 1962 : 180 [Perfora-
tella {Monachoides)]; Hudec, 1963 : 348—-360, Abb. 4, 10 [Perforatella {Monachoi-
des)]; Lozek, 1964 : 294, t. 24, 4a—с; Шилейко, 1970 : 1315, рис. 6, 1—2.
Материал: 22 экз. из 4 местонахождений на Карпатах.
Раковина в общем кубаревидная, со слегка куполовидным контуром завитка.
Оборотов 6—6.5, плавно нарастающих. Последний оборот в профиле закруглен, к устью
236
III. HYGROMIIDAE
внезапно и резко опущен. Окраска от серовато-желтой до красновато-роговой, с раз-
мытой светлой лентой на периферии. Скульптура в виде слабой и неравномерной ра-
диальной исчерченности и зернистости; каждая гранула имеет вид маленького ребры-
шка, ориентированного радиально, гранулы расположены приблизительно в шахмат-
ном порядке. Устье овальное, сильно скошено, места его прикрепления почти не сбли-
жены. Края устья отвернуты. Губа белая, тонкая, лежит близ края устья и часто (осо-
бенно на базальном крае) несет небольшие утолщения. Пупок полностью закрыт отво-
ротом колумеллярного края, от него лишь иногда остается узкая щель. Размеры:.
ВР 8.5-11.5, БД 12-15.5, МД 10.5-13 мм.
Рис. 269—270. Monachoides viclna (Rssm.) из Закарпатья, окр. Рахова, 14 IX 1969.
269 — внешний вид полового аппарата; 270 — дистальная часть полового аппарата,
вскрыта. (Ориг.).
Внутреннее строение. Материал: 2 экз. из Задиела, Словакия, 9 VII
1962, сбор и определение В. Гудеца; 6 — из Закарпатья, окр. г. Рахова, 14 XI 1969;.
сбор и определение наши.
Спермовидукт при переходе в яйцевод слабо изогнут или прямой. СЖ 3X2. Длина
стилофора примерно равна суммарной длине клоаки и вагины. Чехол вагины спаян
со стенками этого органа; в чехле имеется щель в месте впадения пениса. Стенки ва-
гины очень рыхлые, железистые, на продольном разрезе видно кольцевое разрыхле-
ние тканей.
Пенис длинный, тонкий, цилиндрический, папилла также очень длинная; полость
ее на поперечном разрезе имеет вид изогнутой щели, одна из сторон которой грубо за-
зубрена. Тонкий бич слегка короче цилиндрического эпифаллуса; на границе пениса
и эпифаллуса имеются 1—2 резких изгиба. Проток семеприемника прямой, резервуар
мешковидный, заметно не доходит до белковой железы.
Распространение и местообитания. Карпаты. Лесной вид,
часто встречается в подстилке залесенных сырых оврагов. Хвойных лесов избегает.
Вид известен из плейстоценовых и более поздних отложений Средней Европы.
MONACHOIDES
237
3 . Monachoides stuxbergi (Westerlund, 1876). Табл. IX, 84; XII, 120; рис. 271, 272.
Westerlund, 1876 : 98 (Helix); S c h 1 e s c h, 1928 : 279, t. 12, fig. 11—15;
Лихарев, Раммельмейер, 1952 : 439 (Zenobiella rubiginosa part.).
Материал: 14 экз. из 3 местонахождений в Сибири.
Раковина прижатая, округлая, с куполовидным завитком, высота которого
заметно меньше высоты устья, и с выступающей слегка вершиной. Последний оборот
в профиле правильно и широко закруглен, в 1.5 раза шире предпоследнего, к устью
не опущен. Окраска светло-роговая, светлая лента на периферии очень слабая. Скульп-
тура чрезвычайно характерная, состоит из удлиненных коротких радиальных скла-
~Рис. 271—272. Monachoides stuxbergi (West.) из побережья Енисея близ пос. Мирное,
.7 VIII 1972. 271 — внешний вид полового аппарата; 272 — дистальная часть полового
аппарата, вскрыта. (Ориг.).
дочек, на каждой из которых имеется изогнутая чешуйка; чешуйки сохраняются по-
жизненно и остаются даже иногда на пустых раковинах. Устье почти круглое, края
его хрупкие,' не отвернуты, внутри с развитой в различной степени губой. Пупок от-
крытый или слабо прикрыт отворотом колумеллярного края. Размеры: ВР 5—6, БД7—
9.5, МД 5.5—8 мм. Лектотип: ВР 6.0, БД 9.3 мм.
Внутреннее строение. Материал: 4 экз. из Туруханского р-на, берег
Енисея у пос. Мирное, 7 VIII 1972, сбор Ю. И. Чернова, определение наше.
Яйцевод образует крутой изгиб. СЖ 2X2—3. Стилофор хорошо развит, в разной
степени изогнут. Вагина относительно короткая, широкая.
Пенис относительно длинный, цилиндрический, изогнут и может образовывать
петлю; между его дистальным и проксимальным концами имеются слабые мускульные
ленты. Папилла пениса короткая, толстостенная, с многочисленными полостями в стен-
ках. Бич не очень короткий, но тонкий, с признаками дегенерации. Между пенисом,
эпифаллусом и семепроводом натянута тонкая перепонка. Проток семеприемника об-
разует 1—2 резких изгиба. Резервуар прилегает к спермовидукту и отличается харак-
терной формой (рис. 271) — с выемкой на верхнем крае, причем эта выемка развита
не у всех экземпляров одинаково, иногда выражена слабо.
Распространение и местообитания. Сделано на менее 15 на-
ходок по берегам Енисея — от устья Подкаменной Тунгуски до Дудинки. Типовое
местообитание — с. Хантайка на правом берегу Енисея. Населяет припойменную
влажную разнотравную тайгу, в сырую погоду поднимается в верхние горизонты тра-
вяного покрова.
238
III. HYGROMIIDAE
4. Monachoides aculeata (Uvalieva, 1964). Табл. XIII, 121.
Увалиева, 1964 : 201, рис. 1, 2, 3 (Zenobiella); Шилейко. 1970 : 1318
[Pseudotrichia (Stygius)].
Материал: 8 экз. из 1 местонахождения па Алтае.
Раковина прижатая, с широким коническим или куполовидным завитком и
выступающей вершиной. Оборотов 4—4.5, выпуклых, почти ступенчатых, разделенных
глубоким швом. Последний оборот с очень плавным, широко закругленным углом на
периферии, в 1.5 раза шире предпоследнего, к устью чуть опущен. Окраска от красно-
вато-роговой до темно-бурой, почти одноцветная. Скульптура в виде тонких радиальных
линий, придающих раковине жирный блеск; кроме того, имеются радиальные ребры-
шки с расположенными на них 4—5 рядами кожистых треугольных шипиков, причем
верхний ряд виден и на предпоследнем обороте. Сильнее всего развиты шипики цен-
трального ряда, проходящего по периферии; слабее всего — у шва и на базальной части
раковины. Устье широкоовальное, края его острые, простые, не отвернуты, за исклю-
чением верхней части колумеллярного края, которая наполовину или немного меньше
прикрывает пупок. Размеры: ВР 6—6.5, БД 7.3—8.5, МД 6.3—7.2 мм.
Внутреннее строение. Нами не исследовано. Краткое описание мор-
фологии гениталий приводится в соответствии с первоописанием.
Стилофор очень крупный, веретеновидный, утонченный при основании; это дает
возможность предполагать отсутствие рудиментарного стилофора.
Пенис длинный, по-видимому, лишен мускульных лент. Бич очень короткий,
рудиментарный.
Внутреннее строение гениталий неизвестно.
Распространение и местообитания. Алтай; вид обнаружен
в лиственничных лесах в окр. Верхнекатунска, на высоте 1500—2000 м над ур. м.
5. Род PSEUDOTRICHIA Licharev, 1949
Лихарев, 1949 : 417; Шилейко, 1970 : 1317.
Раковина слегка прижатая, со сглаженной вершиной. Окраска роговая, разных
оттенков. Пупок узкий, неперспективный. Раковина с хорошо развитыми волосками,
которые сохраняются пожизненно. Устье без зубов.
Внутренний стилофор отсутствует. Бич слегка короче эпифаллуса. Наружного
чехла пениса нет. Ретрактор правого омматофора образует перехлест с половым рет-
рактором.
Распространение. Палеарктика.
Монотипический род.
Типовой вид — Helix rubiginosa A. Schmidt, 1853.
Pseudotrichia rubiginosa (A. Schmidt, 1853). Табл. XIII, 122; рис. 273, 274.
R ossm assler, 1838, Iconogr., II, 7 : 2, fig. 428 (Helix sericea Miiller part.);
A. Schmidt, 1853, Zeitschr. Naturwiss., I : 3 (Helix}; Clessin, 1879, Jahrb.
d. Malak. Ges., VI : 321, t. 12, fig. 4 (H. granulata); Westerlund, 1889 : 58 (He-
lix); 1897 : 120 [Helix (Trichia) annexa, H. ' (Tr.) verna, H. (Tr.) czekanowskii]; Poli-
nski, 1924 : 193, fig. 75—77 (Fruticicola czarnohorica); Лихарев, 1949 : 417
[Tricheulota (Pseudotrichia) shadini]; Лихарев, Раммельмейер, 1952 :
439, рис. 364 (Zenobiella), 453 (Trichia czarnohorica); G г о s s u, 1955 : 450, fig. 253
(Zenobiella); Ehrmann, 1956 : 131, fig. 89 (Monacha); Lozek, 1956 : 209, Obr.
58, t. 34, 1 (Monachoides); Hudec, 1963 : 348—360, Abb. 511 [Perforatella (Mona-
choides?)]; Lozek, 1964 : 292, t. 25, 2a—c (Monachoides); Gi ttenb erger et
al., 1970:100, fig. 167, 174, Kaart 70 [Perforatella (Monachoide;)]; Шилейко,
1970 : 1318, рис. 8.
Материал: более 100 экз. из 46 местонахождений в различных участках ареала.
Раковина в различной степени, но не сильно прижатая, чаще всего высота
завитка примерно равна высоте устья. Очертания завитка куполовидные, вершина
не выступает. Оборотов 4.5—5, выпуклых. Последний оборот не более чем в 1.5 раза
шире предпоследнего, в профиле правильно и широко закруглен, к устью не опущен.
Окраска красновато- или темно-роговая, иногда бурая, со светлой лентой на перифе-
рии, которая обычно выражена очень слабо. Поверхность раковины неравномерно,
тонко и резко радиально исчерчена, густо покрыта изогнутыми и прямыми волосками.
Устье округлое, немного косое, края его острые, простые, лишь нижний край немного
отвернут. Губы обычно нет или она выражена очень слабо. Пупок слегка прикрыт
отворотом колумеллярного края. Размеры: ВР 4.3—6.5, БД 6—9, МД 5.7—8.5 мм.
Внутреннее строение. Материал: 9 экз. из Московской обл., пойма
р. Пахры, 4 IX 1959; 15 — из Москвы, Ленинские горы, 5 IV 1969; сборы и определе-
URTICICOLA
239
ния наши; 2 — из Казимежа над Вислой (Польша), 6—19 VII 1951, сбор и определе-
ние А. Риделя; 3 — из дельты Волги, остров Бухтовый, 15 VII 1968, сбор Я. И. Старо-
богатова, определение наше.
Спермовидукт и яйцевод почти прямые. СЖ 2X2. Стилофор относительно очень
массивный, длинный, булавовидный или веретеновидный. Вагинальные валики развиты
слабо, ясно выражены лишь возле выходного отверстия стилофора, имеющего вид длин-
ной щели.
Пенис тонкий, от почти прямого до изогнутого довольно резко; имеются слабые
пениальные мускульные ленты. Папилла пениса тонкая, длинная, цилиндрическая;
семепроводящий канал на поперечном разрезе имеет неправильную форму (рис. 274).
Рис. 273—274. Pseudotrichia rubiginosa (A. Schm.) из дельты Волги, 15 VII 1968.
273 — внешний вид полового аппарата; 274 — дистальная часть полового аппарата,
вскрыта. (Ориг.).
Бич тонкий, эпифаллус цилиндрический, обычно изогнут. Проток семеприемника
крутых изгибов не образует, резервуар удлиненно-овальный или округло-треуголь-
ный, примыкает вершиной к нижнему краю белковой железы или к верхней части спер-
мовидукта.
Распространение и местообитания. Почти вся Палеарктика:
большая часть территории Европы, местами Кавказ, Западная Сибирь, юг Восточной
Сибири, Дальний Восток, окрестности Самарканда и пойма р. Кара-Шоро на Чаткаль-
ском хребте. Влаголюбивый вид, встречается в поймах рек, в сырых оврагах, часто
у уреза воды. Изредка встречается даже в хвойных лесах.
6. Род URTICICOLA Lindholm, 1927
Lindholm, 1927а : 124; Wenz, Zilch, 1959—1960 : 681; Шилейко,
1970 : 1315.
Раковина уплощенная, со слегка выступающей вершиной. Окраска сероватая
или роговая; пупок широкий, развернутый. Раковина без волосков, но с зернистой
скульптурой. Устье без зубов.
Внутренний стилофор отсутствует. Бич много длиннее эпифаллуса; соответственно
проток семеприемника также очень длинный. Наружного чехла пениса нет. Ретрактор
правого омматофора перекрещивается с половым ретрактором.
240
III. HYGROMIIDAE
Распространение. Южная часть Средней Европы от восточных Альп
до восточных Карпат. В ископаемом состоянии род известен начиная с плейстоцена
(Средняя Европа, южная Англия).
Монотипический род.
Типовой вид — Helix umbrosa С. Pfeiffer, 1828.
Urticicola umbrosa (С. Pfeiffer,. 1828). Табл. XIII, 123; рис. 275, 276.
С. Pfeiffer, 1828 : 27, t. 6, fig. 7 (Helix); Westerlund, 1889 : 44 (He-
lix); 'Лихарев, Раммельмейер, 1952 : 441, рис. 367 (Zenobiella); G г о s-
Рис. 275—276. Urticicola umbrosa (C. Pfr.) из Казимежа над Вислой, Польша, 6—
19 VII 1951. 275 — внешний вид полового аппарата; 276 — дистальная часть поло-
вого аппарата, вскрыта. (Ориг.).
s п, 1955 : 452, fig. 255 (Zenobiella); Ehrmann, 1956 : 132, t. 5, fig. 66 (Monacha);
Lozek, 1956 : 210, t. 33, 1 (Monachoides); Hudec, 1963 : 348—360, Abb. 6, 12
(Zenobiella (Urticicola)}; Lozek, 1964:291, Abb. 66 [Zenobiella (Urticicola)];
in и л e й к o, 1970 : 1317, рис. 7.
Материал: более 30 экз. из 5 местонахождений на юго-западе европейской части
СССР, в Болгарии, Польше и Венгрии.
Раковина сильно прижатая, с широким завитком, контуры которого в боль-
шей или меньшей степени куполовидные, но вершина не сглаженная. Высота завитка
немного превышает высоту устья или равна ей. Оборотов 5—5.5, плавно нарастающих,
сверху слабо выпуклых. Последний оборот снизу чуть уплощенный, на периферии с ту-
пым, но ясным углом, к устью резко, но не внезапно опущен, немного меньше чем вдвое
шире предпоследнего. Окраска от серовато-желтой до красновато-роговой, со слабой
периферической светлой лентой. Поверхность раковины тонко и неравномерно ради-
ально исчерчена, покрыта тонкими гранулами, расположенными неупорядоченно;
гранулы округлые, лучше выражены в низу раковины. Устье сильно скошено, широко-
овальное, места его прикрепления сильно сближены. Края устья отвернуты, особенно
FRUTI COCA MP YLAEA
241
нижний, острые, внутри с тонкой губой. Пупок вполне перспективный. Размеры;
ВР 5.5-7, БД 10.5—14, МД 8.5-11.5 мм.
Внутреннее строение. Материал: 2 экз. из Казимежа над Вислой,
Польша, 6—19 VII 1951, сбор и определение А. Риделя; 1 — из северо-западной Вен-
грии, 5 VI 1966, сбор Л. Пинтера, определение наше.
Спермовидукт и яйцевод прямые. СЖ 3X2—4. Стилофор очень массивный, длин-
ный, цилиндрический. Клоака и нижний отдел вагины вздуты за счет интенсивного
развития циркумвагинального разрыхления их стенок.
Бич не менее чем втрое длиннее эпифаллуса, многократно извит и скручен. Эпи-
фаллус образует 1—3 петлеобразных изгиба и, постепенно утолщаясь, переходит
в пенис, верхний и нижний отдел которого притянуты друг к другу мускульными
лентами. Папилла пениса умеренной длины, просвет ее щелевидный или трехлучевой.
Проток семеприемника сильно извит, поэтому, несмотря на его значительную длину,
резервуар прилегает к спермовидукту примерно на уровне середины последнего.
Распространение и местообитания. Горные области Средней
и Юго-Восточной Европы; в СССР — Карпаты и Предкарпатье. Населяет горные
широколиственные леса, предпочитает участки с повышенной влажностью.
7. Род FRUTICOCAMPYLAEA Kobelt, 1871
К о b е 11, 1871 : 13; Lindholm, 1928 : 544, 545; Hesse, 1931 : 16;
Лихарев, Раммельмейер, 1952 : 446; Wen z, Zilch, 1959—1960 :
683; Шилейко, 1972в : 1131.
Раковина от почти кубаревидной до слегка прижатой, с узкой несглаженной вер-
шиной. Окраска белая или светло-серая, первично с двумя темными спиральными лен-
тами, которые могут редуцироваться. Пупок не очень узкий, но цилиндрический, слабо
перспективный. Раковина без волосков и без зернистой скульптуры. Устье без зу-
бов.
Внутренний стилофор довольно хорошо развит. Длина бича превышает длину эпи-
фаллуса. Наружного чехла пениса нет, пениальные мускульные ленты хорошо раз-
виты. Стенки папиллы пениса обнаруживают тенденцию к образованию синусов и ка-
пилляров. Ретрактор правого омматофора образует перехлест с половым ретрактором.
Распространение. Кавказ.
Типовой вид — Helix narzanensis Krynicki, 1836.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА FRUTICOCAMPYLAEA
1 (2). Сквозь пупок просматривается не более одного оборота. Обычно имеются 2 тем-
ные ленты, верхняя из которых имеет четкие границы, а нижняя — размытые
. . . . ;.............................................1. Fr. narzanensis.
2 (1). Сквозь пупок просматривается не менее полутора оборотов. Чаще всего имеются
2 темные ленты, выраженные одинаково четко...............2. Fr. kobiensis.
1. Fruticocampylaea narzanensis (Krynicki, 1836). Табл. IX, S5; XIII, 124;
рис. 277—279.
Krynicki, 1836 : 172 (Helix); В о e t t g e r, 1883 : 164, t. 6, fig. 4a—c
[H. (Fruticocampylaea) Narzanensis var. Suanetica], 165, t. 6, fig. 3a—b (var. macrom-
phala), 167, t. 6, fig. 2a—b (var. cyclothyra); Westerlund, 1889 : 104 (Helix);
Л и n д г о л ь м, 19226 : 361 (Fruticocampylaea schamyli); Лихарев, Рам-
мель м е й е р, 1952 : 446, рис. 373, 374; Шилейко, 1972в : 1132, рис. 3; Н и-
d е с, 1972 : 251—275, Abb. 1, 2, 8, 11 [Fruticocampylaea (Fruticocampylaea)].
Материал: более 100 экз. из 32 местонахождений на Кавказе и в Закавказье.
Раковина с острым, почти коническим завитком, высота которого может быть
как больше, так и меньше высоты устья, твердостенная. Оборотов 6, выпуклых, вна-
чале медленно, под конец быстро нарастающих. Последний оборот й 1.5 раза или не-
много более чем в 1.5 раза шире предпоследнего, снизу выпуклый, в профиле плавно
закруглен, к устью резко опущен. Обороты у шва часто бывают слегка ступенчатыми.
Окраска очень светлая, с рисунком из 2 коричневых спиральных лент, в проходящем
свете просвечивающих сильнее фона. Одна лента выражена хорошо — узкая, яркая, с от-
четливыми границами, лежит над периферией последнего оборота; другая лента, гораздо
шире и бледнее первой, располагается ниже периферии; иногда эта лента настолько ши-
рокая, что вся нижняя часть раковины окрашена в светло-коричневый цвет. Наконец,
известны случаи сильного развития обеих лент. Скульптура в виде тонкой неравномер-
ной радиальной исчерченности и четких редких, слегка волнистых спиральных бо-
роздок. Устье округлое или широкоовальное, косое. Края устья тонкие, отвернутые;
немного отступя от края лежит белая губа, развитая в различной степени, причем
у более мелких экземпляров губа выражена резче. Пупок вначале слегка развернутый,
16 А. А. Шилейко
242
HI. HYGROMIIDAE
затем цилиндрический; сквозь него просматривается не более 1.5 оборотов (чаще —
не более одного оборота). Размеры: ВР 10—15, БД 14—24, МД 12—20 мм.
Внутреннее строение. Материал: 15 экз. из Дарьяльского ущелья1
близ с. Казбеги, 9 V 1970; 5 — из Чегемского ущелья, урочище Ара-Боран, 17 V 1970;
3 — из окр. с. Ихрек на берегу р. Самур (Дагестан), 15 VII 1967; сборы наши; 7 —
из Армении, окр. с. Н. Ахта, 29 VI 1960, сбор Н. Н. Дроздова; определения наши-
Рис. 277—279. Fruticocampylaea narzanensis (Kryn.) из г. Орджо-
никидзе, Сев. Осетия, 5 V 1970. 277 — внешний вид полового
аппарата; 278 — вагина, вскрыта; 279 — пенис, вскрыт. (По Ши-
лейко, 1972в).
На границе спермовидукта и яйцевода имеется слабый, но заметный изгиб. СЖ
2—3X2—4. Стилофоры развиты примерно в равной степени, но стрела имеется только
в наружном. Вагина вместе со стилофорами окутана мощным соединительнотканным
чехлом, из которого высовываются лишь вершины стилофоров. Клоака обширная,
внутри с несколькими продольными складками, которые уходят наверх и образуют
лопасти у выходного отверстия стилофоров, располагающегося па вершине тонкого,
довольно длинного сосочка.
Пенис вместе с эпифаллусом образует ясный резкий S-образнып изгиб, постоян-
ство которого обеспечивается мускульными лентами; между пенисом, эпифаллусом и
семепроводом имеется прозрачная мембрана. Папилла пениса удлиненно-коническая,
просвет семепроводящего канала на поперечном разрезе имеет Т-образную форму.
Стенки папиллы рыхлые, в их толще имеется серия синусов в виде тонких продольных
каналов. Бич примерно в 1.5 раза длиннее пениса и эпифаллуса, вместе взятых. Резер-
вуар семеприемника объемистый, граница между резервуаром и протоком пеяспая.
FRUTICOCA MP YLAEA
243
Распространение и местообитания. Главный Кавказский хре-
бет, кроме самых западных районов; Закавказье. Отсутствует в Колхидской низмен-
ности, в Аджарии. Куро-Араксинской и Ленкоранской низменности и в Талыше.
Эвритопный горный вид, встречается как в лесу, так и на открытых склонах, по в до-
статочно влажных условиях. Наибольшую плотность образует на скальных выходах
в лесу и зарослях кустарников.
Рис. 280—282. F ruticocampylaea kobiensis (Bttg.) из окр. Па-
санаури, VII 1969. 280 — внешний вид полового аппарата;
281 — дистальная часть женского отдела, вскрыта; 282 — пе-
нис, вскрыт. (Ориг.).
2. Fruticocampylaea kobiensis (Boettger, 1883). Табл. XIII, 125; рис. 280—282.
Boettgcr, 1883 : 166 [Helix (Fruticocpmpylaea)]; R os'en, 1911 : 107
(Helix); 1914 : 169, t. 2, fig. 6a—c (Helix, H. kobiensis var. jluxa); Лихарев, Pa м-
мелъмейер, 1952 : 447 (Fruticocampylaea narzanensis var. kobiensis).
Материал: более 30 экз. из 3 местонахождений.
Раковина с заостренным завитком, контур которого очень непостоянен:
может быть как коническим, так и куполовидным; соответственно у раковин с кониче-
ским завитком обороты более выпуклые, иногда ступенчатые; оборотов 5—5.5. Послед-
ний оборот немного менее чем вдвое шире предпоследнего, в профиле правильно за-
круглен, к устью плавно опускается. Шов глубокий. Окраска состоит из белого или
светло-серого фона и двух узких четких коричневых лент, развитых в равной степени,
сильно просвечивающих в проходящем свете; иногда нижняя лента или обе ленты от-
сутствуют, Скульптура в виде шелковистой радиальной исчерченности и .ясных спи-
ральных бороздок. Устье косое, края его немного сближены, тонкие, отвернутые, внутри
16*
244
III. HYGROMIIDAE
с губой, причем внутри последнего оборота может быть 1—3 губы. Сквозь пупок про-
сматривается около полутора оборотов. Размеры: ВР 8—11, БД 11.5—18, МД 9.5—
16 мм.
Внутреннее строение. Материал: 6 экз. из окр. с Пасанаури, южные
склоны Главного Кавказского хр., VII 1969, сбор Э. Клаусса, определение наше.
Нижняя часть спермовидукта и яйцевод изгиба не образуют. Места прикрепления
слизистых желез сближены, общее число ветвей 6—8, наклонности к вторичному вет-
влению не наблюдается. Наружный стилофор развит лучше внутреннего, нижняя часть
стилофоров обособлена от вагины. Стилофоры вдаются, таким образом, не в полость ва-
гины, а в своеобразный карман, образованный ее стенками. Полость этого кармана
сообщается с полостью вагины посредством узкой щели между парой продольных ва-
ликов.
Пенис короткий, слегка вздутый, соединен с эпифаллусом 1—2 мускульными лен-
тами; между этими лентами и эпифаллусом имеется прозрачная мембрана. Папилла ци-
линдрическая или булавовидная, семепроводящий канал на поперечном разрезе щеле-
видный, в толще рыхлых стенок имеется несколько, продольных синусов. Бич при-
близительно равен суммарной длине эпифаллуса и пениса, в различной степени извит.
Проток семеприемника образует несколько плавных изгибов, заканчивается удли-
ненно-овальным резервуаром, немного не доходящим до белковой железы.
Распространение и местообитания. Центральный Кавказ.
Обитает на открытых склонах на высотах не менее 1000 м над ур. м.
8. Род SHILEYKOIA Hudec, 1969
Г у д е ц, 1969 : 29 [Circassina (Shileykoia)]; Ш и л е й к о, 1972в : 1132.
Раковина сильно уплощена — до совершенно плоской, со слегка выступающей
вершиной. Окраска белая или желтовато-роговая, с двумя широкими спиральными
лентами выше и ниже периферии. Пупок развернутый, перспективный. Раковина без
волосков и без зернистой скульптуры, устье без зубов.
Внутренний стилофор развит лишь немного слабее наружного; оба стидрфора чрез-
вычайно массивные, вагина вздута. Стилофоры вдаются в-полость вагины сосочком,
по поверхности которого проходит узкий глубокий желобок. Пениальные ленты очень
мощные. В стенках папиллы пениса имеется хорошо развитая система крупных сину-
сов. Ретрактор правого омматофора перекрещивается с половым ретрактором.
Распространение. Восточный Кавказ.
Монотипический род.
Типовой вид — Helix daghestana Kobelt, 1877.
Shileykoia daghestana (Kobelt, 1877). Табл. XIII, 126; рис. 283—287.
Kobelt, Icouogr., 1877, .N. F., 5 : 28, t. 126, fig. 1217 (Helix}; Boettger.
1880a : 130 [Helix (Frutico-Campylaea) Eichwaldi var. daghestana]; Westerlund,
1898 : 158 [Helix (Fruticocampylaea) aliostoma]; Лихарев, Раммельмейер,
1952 : 455 [Trichia (Xerocampylaea) eichwaldi var. daghestana]; Г у д e ц, 1969 : 29,
ЗГ—Е, 4 [Circassina (Shileykoia)]; Шилейко, 1972в : 1132, рис. 4 (Schileykoia
lithophila); Hudec, 1972d : 251—275, Abb. 3 [Fruticocampylaea (Shileykoia)].
Материал: более 60 экз. из 4 местонахождений в Дагестане.
Раковина от сильно прижатой до плоской; однако вершина не сглажена.
Оборотов 6, умеренно выпуклых, плавно нарастающих. Последний оборот примерно
в 1.5 раза шире предпоследнего, по периферии закруглен или с тупым углом. Окраска
от почти белой до коричневато-желтой, с двумя хорошо развитыми темными спираль-
ными лентами разной интенсивности — от коричневатых до красновато-коричневых
тонов. Между этими лентами имеется серая или молочно-белая лента. Иногда темные
ленты настолько широкие, что образуют по существу фон раковины. В случае интенсив-
ной окраски могут присутствовать белые радиальные штрихи. Скульптура в виде не-
правильной радиальной исчерченности и спиральных бороздок, местами имеются вмя-
тины; у крупных, интенсивно окрашенных раковин часто присутствуют элементы вер-
микулятной скульптуры. Устье косое, с отвернутыми краями. Губа располагается от-
ступя от края устья, белая, широкая, просвечивает сквозь стенку раковины в виде
белой полосы; кроме того, имеется еще 1—3 более ранних губ. Пупок очень широкий,
сквозь него видны все предыдущие обороты. Размеры: ВР 4—8.5, БД 8—19, МД 7—
16 мм.
Внутреннее строение. Материал: 2 экз. из Дагестана, с. Гольбарт
Лакского р-на, 25 VI 1963, сбор Рухлядева; 7 — из окр. с. Акуша, 25 VI 1967; 2 — из
окр. с. Ихрек в южном Дагестане, 14 VII 1967; сборы и определения наши.
Нижняя часть спермовидукта и яйцевод плавно изогнуты. СЖ 2X2—4. Объеми-
стая вагина вместе с основаниями стилофоров охвачена тонким соединительнотканным
чехлом. Имеется пара вагинальных валиков, проходящих по дистальной части стило-
SHILEYKOIA
245
форов, которая обращена внутрь вагины в виде длинного сосочка. Между валиками
располагается глубоко врезанный желобок, ведущий в яйцевод.
Пенис подразделяется на цилиндрическую проксимальную часть и веретено-
видную дистальную; папилла длинная, булавовидная, начинается в проксимальной
части. Семепроводящий канал на поперечном разрезе щелевидный, дифференциация
паренхиматозной ткани стенок папиллы привела к образованию очень крупных про
Рис. 283—287. Shileykoia daghestana (Kob.) из окр. с. Акуша, Да-
гестан, 25 VI 1967. 283 — внешний вид полового аппарата; 284 —
вагина, вскрыта; 285 — поперечный срез вагины по линии а—а'
(звездочкой помечена борозда, проходящая по дистальной части
стилофоров); 286 — пенис, вскрыт; 287 — поперечный срез эпифал-
луса. (По Шилейко, 1972в).
дольных синусов. Пениальные мускульные ленты чрезвычайно мощные, связывают ди-
стальную часть пениса с проксимальной и с нижним отделом эпифаллуса. Бич кониче-
ский, чуть длиннее эпифаллуса. Умеренно длинный мощный проток семеприемника без
резкой границы переходит в объемистый мешковидный резервуар, не доходящий до
белковой железы.
Распространение и местообитания. Горная часть Дагестана.
Обитает на влажных скалах и под камнями на открытых местах. -
246
III. HYGROMIIDAE
9. Род CIRCASSINA Hesse, 1921
Hesse, 1921:62; Lindholm, 1927a : 125; Hesse, 1931 : 26; Л и x a-
p e в. Раииел ьмсйе p, 1952 : 434; W e n z, Zilch, 1959—1960 : 689;
Hudec, L e z h a w a, 1970 : 16 (Hela\, HI и л e й к о, 1972в : 1134; Hudec,
1972d : 276 (Abchasohela uom. nov. pro Hela).
Рис. 288—290. Circassina circassica circassica (Mouss.) из Бакуриани,
VII 1953. 288 — внешний вид полового аппарата; 289 — вагина,
вскрыта; 290 — пенис, вскрыт, и два поперечных среза папиллы —
в дистальной и проксимальной части. (По Шилейко, 1972в).
Раковина почти шаровидная, с вздутым последним оборотом' и не сглаженной
вершиной. Окраска почти одноцветная — желтоватая или зеленоватая; иногда имеются
1—2 темные спиральные ленты. Пупок очень узкий. Раковина без волосков, устьевых
зубов нет.
Первично имеются два развитых примерно в равной степени стилофора; однако
в пределах одного вида наблюдается ступенчатая редукция стилофоров и слизистых
желез. Пениальные ленты хорошо развиты. От паренхимы, заполняющей стенки па-
пиллы пениса, остаются отдельные тяжи. Бич очень длинный. Ретрактор правого ом-
матофора образует перехлест с половым ретрактором.
Распространение. Кавказ.
Типовой вид — Helix circassica Mousson, 1863.
CIRCASSINA
247
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CIRCASSINA
1 (2). Поверхность раковины покрыта более или менее ясно выраженной зернистой
скульптурой ...................................................3. С. frutis.
2 (1). Зернистая скульптура отсутствует.
3 (4). Раковина зеленовато-серая, желтоватая или красновато-роговая, часто со слабой
светлой лентой по периферии ................................1. С. circassica.
4 (3). Окраска от светло-желтой или зеленоватой до серовато-роговой, с 1—2 темными
спиральными лентами, которые иногда бывают настолько широкими, что за-
нимают большую часть поверхности; в этом случае на периферии остается свет-
лая лента ....................................................2. С. bojenae.
1. Circassina circassica circassica (Mousson, 1863). Табл. XIII, 127; рис. 288—290.
M о u s s о n, 1863 : 370 (Helix); К о b e 1 1, 1877, Iconogr., V : 78, fig. 1386
«(Helix); Westerlund, 1889 : 73 (Helix); Hesse, 1931 : 26, t. 4, fig. 27; Л и-
харев, Раммельмейер, 1952 : 435, рис. 359; Шилейко, 1972в : 1134,
рис. 5.
Материал: более 30 экз. из 5 местонахождений в Закавказье.
Раковина вздутая, высота завитка примерно равна высоте устья. Завиток
конический или, чаще, слегка куполовидный. Оборотов 6—7, верхние обороты упло-
щены, последний выпуклый, вздутый, в профиле правильно и широко закруглен,
к устью опущен, немного менее чем в 1.5 раза шире предпоследнего. Окраска большей
частью зеленоватая, реже желтоватая или роговая; периферическая светлая лента
•отсутствует или выражена чрезвычайно слабо. Скульптура в виде тонкой радиальной
исчерченности, которая местами выражена резче. Устье широкоовальное, косое, слегка
оттянуто вниз и вправо. Края устья острые, отвернутые, внутри с тонкой губой. Пу-
пок наполовину прикрыт отворотом колумеллярного края. Размеры: ВР 14—19, БД
18—22, МД 15-19 мм.
Внутреннее строение. Материал: 3 экз. из Грузии, окр.. Бакуриани,
VII 1953, сбор и определение И. М. Лихарева.
Нижняя часть спермовидукта и яйцевод почти прямые. СЖ 1—2X1—4. Стило-
форы почти одинакового размера, впадают в полость вагины единым сосочком, распо-
ложенным между двумя лопастями. Внутри вагины имеются 1—2 мощных продольных
валика.
Пенис в общем веретеновидный; папилла булавовидная, нередко с поперечными
складками. Стенки ее заключают обширную полость, в центре которой проходит се-
мепроводящий канал, подвешенный и растянутый многочисленными радиальными тя-
жами. Дистальная часть мужского отдела S-образно изогнута и фиксирована в таком
положении мощными пениальными мускульными лентами. Семеприемник неправильно
овальной или грушевидной формы, проток прямой или в различной степени изогнут.
Распространение и местообитания. Центральная часть юж-
ных склонов Главного Кавказского хр. и Закавказье. Населяет сырые леса..
la. Circassina circassica* stephaniae (Hudec et Lezhawa, 1970). Рис. 291—296.
Hudec, Lezhawa, 1967b : 89, Abb. 11, t. V, 12 (Circassina circassica part.);
1970:16, Abb. 1, 10 (Hela stephaniae); Шилейко, 1972b : 1134, рис. 6, 1—5.
Материал: 16 экз. из 5 местонахождений в западной Грузии.
От номинативного подвида отличается в основном несколько более вздутым по-
следним оборотом.
Внутреннее строение. Материал: 2 экз. из с. Верхняя Цебельда близ
Сухуми, 21 VIII 1905; 2 — из окр. Сухуми, 17 VII 1905; сборы Калишевского; 4 —
из Батуми, Ботанический сад, 7 IV 1971, сбор и определения наши.
Оба стилофора полностью отсутствуют, но слизистые железы в виде 2 пучков име-
ются. Внутри вагины присутствует кольцевой валик на границе с клоакой, а также
продольные валики, разбитые более или менее ясно на отдельные складки.
Распространение и местообитания. Прибрежные районы за-
падной Грузии. Населяет леса и кустарниковые заросли.
16. Circassina circassica simpla Schileyko, 1972. Табл. XIII, 128; рис. 297—301.
Rosen, 1911 : 111, t. 3, fig. 1 (Helix aurea non Dillwyn, 1817); Шилейко,
1972b : 1135, рис. 6, 6—11; Hudec, 1972d : 267, Abb. 4, 9 [Metafruticicola (Meta- .
fruticicola) shileykoi nom. nov. ].
Материал: более 100 экз. из 4 местонахождений на Северном Кавказе.
Конхологических отличий от номинативного подвида нет.
CIRCASSINA
249
Внутреннее строение. Материал: 2 экз. из плато Бичи-сып под Эль-
брусом, 17 V 1965, сбор М. Акбаева; 2 — из окр. Нальчика, 2 V1963, сбор Л. В. Ши-
лейко; 18 — из окр. Орджоникидзе, 5 V 1970; 5 — из Чегемского ущелья близ с. Ниж-
ний Чегем, 17 IX 1970; сборы и определения паши.
Отсутствуют все придатки женского отдела: стилофоры и слизистые железы. Ва-
гина связана со стеикой тела многочисленными соединительнотканными тяжами, раз-
витыми в различной степени. Внутренняя поверхность вагины покрыта складками не-
правильной формы, но местами угадывается первичное распределение их в продольные
ряды (нарис. 298 соответствующее место отмечено фигурной скобкой).
Гудец (Hudec, 1972d) считает эту форму представителем рода Metafruticicola
на том формальном основании, что вагина здесь не несет никаких придатков. Однако
строение совокупительного аппарата свидетельствует о принадлежности к роду Circas-
sina (ср. с данными по представителям подсемейства Metafruticicolinae).
Гессе (Hesse, 1931) отмечал 2 типа строения гениталий у данного вида, изображая
особь с одними только слизистыми железами из Батуми и особь без придатков женского
отдела. В тексте не указывается, откуда происходил материал по второй из форм, по
сообщено, что он получен от Розена, который его и определил. Розен обычно пользо-
вался материалом с западной части Северного Кавказа.
Распространение и местообитания. Северный Кавказ, кроме
самых восточных районов. Встречается во влажных лиственных лесах и в тенистых
ущельях.
1в. Circassina circassica akramowskii 1 Schileyko, 1972.
Ш и л e й к о, 1972в : 1135.
Голотип хранится в коллекции Института зоологии АН Армянской ССР.
Материал: 3 экз. из одного местообитания в северной Армении.
Конхологических отличий от номинативного подвида не обнаружено.
Внутреннее строение. Нами не исследовано; однако Н. Н. Акрамов-
ский любезно прислал нам рисунок, из которого ясно, что подвид представляет собой
последнюю из возможных комбинаций: отсутствуют слизистые железы, но имеются
оба стилофора.
Распространение. Северная часть Армении.
2. Circassina bojenae Hudec et Lezhawa, 1969. Табл. XIII, 129.
Hudec, Lezhawa, 1969a : 44, Abb. 1, 6; 1969b : 132, Abb. 26—27, t. 6, 12.
Материал: более 60 экз. из 1 местонахождения на юго-западном Кавказе.
Раковина с широким завитком, высота которого немного меньше высоты
устья. Очертания завитка слегка куполовидные. Оборотов 6.5, по мере нарастания ста-
новятся все более выпуклыми. Последний оборот немного мепее чем в 1.5 раза шире
предпоследнего, в профиле плавно и широко закруглен, к устью плавно, но сильно опу-
щен. Окраска от светло-желтой или слегка зеленоватой до серовато-роговой; в перво-
описаиии указано, что рисунок образован двумя темно-коричневыми спиральными лен-
тами, проходящими выше и ниже периферии. В долине р. Техури, западнее типового
местонахождения (ущелье р. Чери, окр. с. Балда Гегечкорского р-на) нами обнаружено
множество раковин этого вида, среди которых наряду с типичными формами имеются
одноцветные раковины, экземпляры с одной только лентой выше периферии и раковины
с двумя лентами, настолько широкими, что они по существу образуют фон. Скульптура
в виде тонкой и густой радиальной исчерченности и вялых спиральных бороздок, вы-
раженных местами на последнем обороте. Устье широкоовалыюе, слегка оттянуто вправо,
косое, края его острые, отвернутые, внутри с тонкой губой. Пупок слегка прикрыт от-
воротом колумелл яркого края. Размеры: ВР 15—20, БД 20—27, МД 17—22.5 мм.
Внутреннее строение. Материал: 2 экз. из ущелья р. Техури, 29 IV
1971, сбор и определение наши.
Спермовидукт и яйцевод почти прямые. СЖ 2x4—8. Стилофоры массивные, не-
много оттянуты вбок, наружный немного больше внутреннего.
Бич примерно вдвое длиннее эпифаллуса, который в свою очередь такой же длины,
как пенис. Папилла пениса толстостенная, в стенках имеются многочисленные, в ос-
новном радиальные тяжи. Между пенисом и эпифаллусом имеются мускульные лепты,
обеспечивающие постоянный крутой изгиб этих отделов. Проток семеприемника при-
мерно втрое длиннее резервуара, довольно толстый, резервуар неправильно грушевид-
ной формы, прилегает к верхней части спермовидукта.
Распространение и местообитания. Долины горных рек в за-
падной части южных склонов Большого Кавказа. Встречается на открытых склонах
среди камней и в кустарниковых зарослях, в частности среди самшита.
1 Подвид назван именем Николая Николаевича Акрамовского.
250
TIL HYGROMIIDAE
3. Circassina frutis (L. Pfeiffer, 1859). Табл. XIII, 130.
L. Pfeiffer, 1859 : 252 (Helix); Boettger, 1884 : 150 (Я. pachnodes);
Westerlund, 1889 : 73 (H. schuberti Roth var. frutls); Rosen, 1911 : 100 (He-
lix); 1914 : 163 (II. schuberti), 164 (H. frutis, H. pachnodes); R e t о w s к i, 1914 : 285
[Helix (Theba)]; Hesse, 1931 : 27, t. 4, fig. 28—30; Лихарев, Раммель-
мейер, 1952 : 436, рис. 360; Hudec, Lezhawa, 1969b : 134, Abb. 28, t. 12,
30; 1970 : 15-31, Abb. 8, 12, 14.
Материал: 3 экз. из 2 местонахождений на западном Кавказе.
Раковина почти кубаревидная, с острым завитком, имеющим строго кониче-
ский контур. Оборотов 6, слабо выпуклых. Последний оборот умеренно вздут, менее
чем в 1.5 раза шире предпоследнего, к устью плавно, но сильно опущен. Окраска крас-
новато-роговая, светлая периферическая лента развита в различной степени. Скульпту-
ра в виде тонкой радиальной исчерченности и микроскопической зернистости, особенно
ясной на ранних оборотах. Устье круглое или широкоовальное, с тонкими отвернутыми
краями и тонкой губой. Пупок чрезвычайно узкий, частично прикрыт колумеллярный
краем. Размеры: ВР 10—12, БД 13—16, МД 11—13.7 мм.
Внутреннее строение. Нами не исследовано; приводимое описание
составлено по литературным данным (Hesse, 1931; Hudec, Lezhawa, 1970).
Спермовидукт и яйцевод почти прямые. СЖ 4X3. Стилофоры очень массивные,
разпица в размерах между ними несущественна.
Бич тонкий, длинный, примерно в 1.5 раза длиннее эпифаллуса. Пенис наружно
мало чем отличается от эпифаллуса, примерно той же длины. Между дистальными
протоками мужского отдела натянута тонкая прозрачная мембрана и мощные пениаль-
ные ленты. Проток семеприемника недлинный и толстый, резервуар грушевидный,
объемистый, прилегает к верхней части спермовидукта.
Распространение и местообитания. Западная часть Большого
Кавказа и прилегающие территории западной Грузии. Обитает во влажных осыпях
и лесной подстилке.
10. Род CERNUELLA Schliiter, 1838
8 с h 1 ii t е г, 1838 : 6; Brown, 1844 : 49 (Heliomanes, non Newman, 1840);
Monterosato, 1892 : 24 (Xerolauta part.); Hesse, 1926 : 120 [Helicella (Cer-
nuella)]; 1934 : 5 [Helicella (Cernuella)]; Лихарев, Раммельмейер, 1952:
420 [Helicella (Cernuella)]; Wenz, Zilch, 1959—1960:666.
Раковина различной формы, co сглаженной вершиной. Окраска белая или светло-
серая, с несколькими темными спиральными лентами. Пупок от цилиндрического до раз-
вернутого, но в любом случае открытый или прикрыт отворотом колумеллярного края
незначительно. Скульптура в виде тонкой исчерченности. Волосков и устьевых зу-
бов нет.
Внутренний стилофор заметно меньше наружного. Пениальные ленты отсутствуют.
Паренхима стенок папиллы пениса разрыхлена, содержит кольцевую интрапапилляр-
ную полость или пару продольных полостей. Бич различной длины. Ретрактор пра-
вого омматофора не образует перехлеста с половым ретрактором.
Распространение. Европа и восточное Средиземноморье с Малой Азией
и Кавказом. В ископаемом состоянии род известен начиная с нижнего плиоцена (Ев-
ропа, Северная Африка).
Типовой вид — Helix virgata Da Costa, 1778.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CERNUELLA
1 (2). Сквозь пупок видны все обороты. В профиле обороты ступенчатые. Губа белая
........................................................ 2. С. tiflisiana.
2 (1). Сквозь пупок просматривается не более одного оборота. В профиле обороты пра-
вильно закругленные. Губа бурая или красноватая .......... 1. С. virgata.
1. Cernuella virgata (Da Costa, 1778). Табл. XIV, 131; рис. 302, 303.
Da Costa, 1778 : 79, t. 4, fig. 8 (Helix); Draparnaud, 1801 : 73 (H. va-
riabilis); Westerlund, 1889 : 165 (Helix); Hesse, 1934 : 6 [Helicella (Cer-
nuella)]; Лихарев, Раммельмейер, 1952 : 420, рис. 344, 345 [Helicella
(Cernuella)]; Gittenberger et al., 1970:97, fig. 160, 164, Kaart 65; H u-
d e c, 1972b : 104, Abb. 4 [Cernuella (Cernuella) virgata variabilis].
Материал: более 40 экз. из 12 местонахождений в Крыму, Болгарии и Франции.
Р а к о в и и а от слегка прижатой до почти шаровидной, с широким завитком слегка
куполовидных очертаний, высота которого сильно варьирует. Оборотов 5—6, умеренно
CERNUELLA
251
выпуклых, плавно нарастающих. Последний оборот в профиле округлый, в 1.5 раза
(иногда почти вдвое) шире предпоследнего, к устью почти не опущен. Окраска белая
или серая, изредка с желтоватым оттенком, обычно с более или менее развитым рисун-
ком из темных спиральных лент или рядов пятен. Чаще всего над периферией послед-
него оборота и у шва предыдущих оборотов имеется одна широкая лента, а на базаль-
ной части раковины — многочисленные узкие ленты. Скульптура в виде очень тонкой
радиальной исчерченности. Устье умеренно косое, округлое, внутри часто буроватое
или темно-желтое. Края устья острые, не отвернуты, внутри с довольно ясной губой,
которая может быть белой или окрашенной в коричневатый или красновато-бурый
Рис. 302—303. Cernuella virgata (Da Costa) из Ялты, 14 IX 1969. 302 — внешний вид
полового аппарата; 303 — дистальная часть полового аппарата, вскрыта. (Ориг.).
цвет. Пупок не очень узкий, но не перспективный, сквозь него видно не более полутора
оборотов, чаще — не более одного оборота. Размеры: ВР 8—12, БД 12—16, МД 10—
13 мм.1
Внутреннее строение. Материал: 9 экз. из Крыма, окр. Ялты, 14 IX
1969, сбор М. А. Кравченко; 2 — из Франции, окр. г. Арля, VIII 1971, сбор Д. А. Са-
харова; определения наши.
Нижняя часть спермовидукта образует плавный изгиб. СЖ 2x4—12; большое
число ветвей связано с тем, что ветви первого порядка часто образуют куст. Клоака и
вагина охвачены рыхлым соединительнотканным чехлом. На границе этих отделов
имеется кольцевидное утолщение, внутренняя стенка вагины покрыта многочислен-
ными, неправильной формы складками и бугорками, за счет выравнивания которых
этот отдел способен сильно растягиваться. Стилофоры вдаются непосредственно в по-
лость вагины.
Пенис почти цилиндрический, наружно не отграничен от эпифаллуса. Папилла
длинная, с поперечными складками или, при максимальном растяжении, гладкая.
В стенках папиллы имеется парная полость в виде двух узких продольных щелей. Бич
в 2—2.5 раза короче слабо изогнутого эпифаллуса. Основание протока семеприемника
сильно расширено, резервуар очень объемистый, прилегает к спермовидукту на уровне
его середины или несколько выше.
1 Указания на большие размеры относятся к близкому виду — С. cespitum
(Drap.), обитающему в Средиземноморских странах.
252
III. HYGROMIIDAE
Распространение и местообитания. Средиземноморье, пре-
имущественно северное; атлантическое побережье Европы вплоть до Великобритании;
Южный берег Крыма и окрестности Новороссийска. Завезен в Австралию. Населяет
открытые участки вдоль морских побережий, часто образует скопления на стеблях
трав. Обнаружен в плейстоцене Европы.
2. Cernuella tiflisiana (Lindholm, 1913). Табл. XIV, 132; рис. 304, 305.
Lindholm. 1913b : 20 (Xerophila); Лихарев, Раммельмейер,
1952:430 [Helicella (?)]; Rosen, 1914:230 (Helicella crenlmargo var. obtustor
part.).
Рис. 304—305. Cernuella tiflisiana (Lindh.) из Дарьяльского ущелья, окр. с. Чми,
8 V 1970. 304 — внешний вид полового аппарата; 305 — дистальная часть по-
лового аппарата, вскрыта. (Ориг.),
Материал: более 40 экз. из 3 местонахождений на Северном Кавказе и в окр. Тби-
лиси.
Раковина сильно уплощена, иногда почти совсем плоская. Оборотов 4.5—5,
плавно и быстро нарастающих, разделенных глубоким швом, слегка ступенчатых.
Последний оборот по периферии правильно закруглен, почти вдвое шире предпослед-
него, к устью не опущен. Окраска белая, наиболее яркие и широкие темные ленты
проходят выше и ниже периферии; между ними располагаются 2 очень узкие ните-
видные ленты и на базальной части раковины 1—2 ленты разной ширины. Если ри-
сунок исчезает, то в последнюю очередь блекнут широкие ленты. Скульптура в виде
тонкой и густой радиальной исчерченности, на последнем обороте видны короткие
продольные штрихи. Устье круглое, слегка скошено, места его прикрепления слегка
сближены, края острые, прямые. Далеко отступя от края имеется сильно расплыв-
шаяся белая губа. Размеры: ВР 6—6.6, БД 10.5—13.3, МД 9—11.5 мм.
Внутреннее строение. Материал: 3 экз. из окр. с. Чми на правом
берегу Терека в Дарьяльском ущелье, 8 V 1970, сбор и определение наши.
Нижняя часть спермовидукта и яйцевод не образуют резких изгибов. СЖ 4—
5x2. Наружный стилофор заметно больше внутреннего, стилофоры единым сосочком
впадают в своеобразный карман вагины, сообщающийся с собственно полостью ва-
гины посредством узкой щели между двумя вагинальными валиками.
Пенис состоит из вздутого дистального отдела и удлиненного проксимального,
который можно толковать и как нижний отдел эпифаллуса, так как папилла зани-
мает только вздутую часть пениса. В этом случае надо говорить о том, что половой
XEROSECTA
253
ретрактор крепится не к границе пениса и эпифаллуса, как это обычно наблюдается,
а к эпифаллусу. Внутри папиллы имеется кольцевая интрапапиллярная полость.
Бич заметно длиннее эпифаллуса. Семеприемник овальный, прилегает к верхней ча-
сти спермовидукта, его проток резких изгибов не образует.
Распространение и местообитания. Вид обнаружен в трех
местах: в г. Тбилиси и его окрестностях и в Дарьяльском ущелье на Северном Кав-
казе. Обитает на открытых склонах, среди травы и камней.
11. Род XEROSECTA Monterosato, 1892
Mo nt er osa t о, 1892 : 21; Hudec, Lezhawa, 1967a : 135 [Xerosecta
Kalitinaia) part.].
Рис. 306—307. Xerosecta schelkovnikoin (Bog.) из Грузии, окр. Касии,
14 V 1963. 306 — внешний вид полового аппарата; 307 — пепис,
вскрыт. (Ориг.).
Раковина от плоской до слегка прижатой. Окраска белая, серая или светло-ро-
говая, иногда с неясными спиральными темными лентами. Пупок от развернутого до
умеренно узкого, не прикрыт отворотом колумеллярного края. Скульптура — ра-
диальная ребристость, тонкая или очень резкая.
Внутренний стилофор меньше наружного. Пениальные ленты слабые или отсут-
ствуют. В основании папиллы пениса имеется кольцевая полость, заходящая в стенки
папиллы, чехла пениса и эпифаллуса. Бич длиннее эпифаллуса. Ретрактор правого
омматофора не перекрещивается с половым ретрактором.
Распространение. Восточное Средиземноморье и Кавказ. В ископае-
мом состоянии род известен начиная с плиоцена (Северная Африка, Предкавказье,
Франция).
Типовой вид — Helix explanata Muller, 1774.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА XEROSECTA
4 (2). Раковина сильно уплощена. Пупок широкий, развернутый, сквозь пего видны
все обороты . .......................................1. X. schelkovnikovi.
254
III. HYGROMIIDAE
2 (1). Раковина в различной степени прижатая. Пупок узкий, сквозь него видно не
более полутора оборотов.
3 (4). Последний оборот в 1.5 раза шире предпоследнего, снизу умеренно выпуклый.
Сквозь пупок просматривается предпоследний оборот ... 2. X. crenimargo.
4 (3). Последний оборот менее чем в 1.5 раза шире предпоследнего, снизу явственно
выпуклый. Сквозь пупок просматривается полтора оборота . 3. X. arcadiana.
1. Xerosecta schelkovnikovi (Bogatschev, 1936). Табл. XIV, 133; рис. 306, 307.
Богачев, 1936:62, рис. 13, 14 [Helix (Jacosta)]; Hudec, Lezhawa,
1967a : 131 —137, Abb. 1, 3 [Xerosecta (Kalitinaia)]; 1967b : 83, Abb. 8, t. 3, 7 [Xero-
secta (Kalitinaia) ].
Материал: 15 экз. из 2 местонахождений в восточной Грузии.
Раковина сильно уплощенная, вплоть до совершенно плоской, состоит из
5 быстро нарастающих оборотов. Последний оборот примерно в 1.5 раза шире пред-
последнего, к устью не опущен. По периферии проходит чрезвычайно резкий ните-
видный зазубренный киль, четко выраженный вплоть до устья и прослеживающийся
на предыдущих оборотах; поэтому обороты в профиле имеют сердцевидную форму.
Окраска белая или светло-роговая, спиральные темные ленты отсутствуют или исче-
зающе слабые. Поверхность раковины покрыта острыми резкими ребрами, окрашен-
ными светлее фона и одинаково хорошо выраженными на верхней и нижней поверх-
ностях раковины. Устье округлое или широкоовальное, с резкой выемкой, соответ-
ствующей килю. Края устья почти не отвернуты. Пупок очень широкий, развернутый.
Размеры: ВР 5—7, БД 12—15, МД 9—12 мм.
Внутреннее строение. Материал: 2 экз. из Грузии, окр. Каспи,
14 V 1963, сбор и определение Г. И. Лежавы.
Нижняя часть спермовидукта изгиба не образует. СЖ 3X2—3. Наружный стило-
фор заметно больше внутреннего. Вагина со стороны, противолежащей стилофорам,
в разной степени вздута; полость ее разделена резким кольцевым утолщением на два
отдела. Верхний отдел расширен, и в это расширение впадает проток стилофоров,
открывающийся на конце притупленного сосочка. Нижний отдел вагины, слитый с кло-
акой, заключает узкий изогнутый канал, служащий для выдвижения папиллы пениса.
Пенис короткий, папилла цилиндрическая, ее просвет на поперечном разрезе
щелевидный. Имеются слабые пениальные ленты и тонкая мембрана, натянутая между
пенисом, эпифаллусом и семепроводом. Эпифаллус длинный, извитой, примерно та-
кой же длины, как и бич. Проток семеприемника длинный, извитой, поэтому неболь-
шой резервуар не достигает белковой железы.
Распространение и местообитания. Вид описан из ашперон-
ских отложений (верхний плиоцен) Куринской депрессии; позже был обнаружен
в рецентном состоянии близ р. Куры у Каспи (вост. Грузии) и у Самуха (зап.
Азербайджан). Обитает в полынной 'полупустыне, держится под камнями и на стеб-
лях полыни.
2. Xerosecta crenimargo (L. Pfeiffer, 1848). Табл. XIV, 134; рис. 308—311.
К г у n i с k i, 1837 : 53 (nom. nudum); L. Pfeiffer, 1848, I : lli,(Helix);
Kaleniczenko, 1853 : 87 (Helix}; M о u s s о n, 1876 (Helix obtusior); W es-
ter 1 u n d, 1889 : 321 (Helix); К о b e 1 t, 1893, Iconogr., N. F., VI : 70, fig. 1077
[Xerophila (Jacosta)]; Лихарев, P а м м e л ь м е й е р, 1952:425, рис. 350
[Helicella (Helicop sis)]; Калитина, 1958 : 166, рис. 2 (Helicella); Hudec, Lez-
hawa, 1967a : 131—137, Abb. 2, 4 [Xerosecta (Kalitinaia) ].
Материал: более 100 экз. из 23 местонахождений на Кавказе.
Раковина в различной степени прижатая, но никогда не бывает плоской;
чаще всего высота завитка немного меньше высоты устья. Оборотов 4.5—5, выпуклых,
у шва слегка ступенчатых, разделенных глубоким швом. Последний оборот к устью
не опущен, примерно в 1.5 раза шире предпоследнего. По периферии проходит ните-
видный киль, выраженный в разной степени, часто отсутствующий совсем. Окраска
серая или кремовая, иногда одноцветная, киль, если имеется, немного светлее фона.
Однако чаще имеются слабые темные ленты, самая широкая проходит над килем;
остальные, нитевидные, проходят по базальной части раковины. Скульптура в виде
густой ребристости, причем ребра могут быть резкими или сглаженными, невысокими.
Устье почти круглое, края его простые, места прикрепления сближены. Пупок уме-
ренно широкий, сквозь него просматривается предпоследний оборот. Размеры: ВР 5—
6.5, БД 8-11, МД 6.5—9.5 мм.
Внутреннее строение. Материал: 21 экз. из Дагестана, окр. с. Ка-
рата, 6 VII 1967; 10 — из Дарьяльского ущелья у с. Чми, 8 V 1970; 10 — из Чегем-
ского ущелья у с. Верхний Чегем, 23 IX 1970; сборы и определения наши.
На границе спермовидукта и яйцевода имеется более или менее резкий изгиб.
СЖ 4x2. Стилофоры развиты в различной степени: наружный заметно больше, у ста-
XEROSECTA
255
рых особей он гипертрофирован, нижний конец его обособлен от вагины (рис. 309).
Противолежащая стенка вагины в разной степени вздута. Внутреннее строение ва-
гины — как у предыдущего вида (рис. 311).
Пенис неправильно цилиндрический, местами вздутый. Кольцевая полость в ос-
новании папиллы пениса развита особенно хорошо, далеко вдается в стенки эпифал-
луса; в результате длина папиллы способна значительно изменяться. Бич несколько
длиннее эпифаллуса. Проток семеприемника длинный и довольно толстый, изогнут
или слегка извит, резервуар небольшой, прилегает к верхней части спермовидукта
или слегка касается нижней части белковой железы.
Рис. 308—311. Xerosecta crenimargo (L. Р1г.) из Чегемского ущелья, 23 XI 1970,-
308 — внешний вид полового аппарата; 309 — вагина со стилофорами другой особи,
’310 — пенис, вскрыт; 311 — дистальная часть полового аппарата, вскрыта. (Ориг.)>
Распространение и местообитания. Центральное и восточ-
ное Предкавказье, Дагестан, восточное Закавказье. Хаас (Haas, 1936) сообщает
о присутствии вида в Малой Азии и Копетдаге. Однако надо иметь в виду.'что
в Копетдаге обитает похожий вид — Helicopsls likharevi\ возможно, Хаас имел в виду
этот последний вид. Обитает на открытых степных участках, образуя порой зна-
чительные скопления. Во время летней засухи и на зиму уходит в почву на глубину
нескольких сантиметров. Вид известен из апшеронского яруса верхнего плиоцена
Предкавказья (Стеклов, 1966).
3. Xerosecta arcadiana (Schileyko, 1967). Табл. XIV, 135\ рис. 312, 313.
III и л е й к о, 1967в : 28, рис. 1, ЗА [Helicella (Cernuella)].
Голотип хранится в коллекции Зоологического музея Московского государствен-
ного университета.
Материал: более 50 экз. из 1 местонахождения в нагорно-степной части Талыша. .
Раковина слегка прижатая, со слегка выступающей вершиной; высота за-
витка немного меньше высоты устья. Оборотов 4.5—5, выпуклых, ступенчатых. По-
следний оборот к устью не опущен, менее чем в 1.5 раза шире предпоследнего, в про-
филе округлый, снизу явственно выпуклый. У отдельных экземпляров имеется сла-
бый намек на киль. Окраска белая, серая илп кремовая, у молодых особей на рако-
256
III. HYGROMIIDAE
вине заметны слабые спиральные ленты. Скульптура в виде частых округлых ра-
диальных ребрышек, окрашенных немного светлее, чем промежутки между ними.
Устье почти круглое, умеренно скошено, края простые, не отвернуты. Отступя от
края устья имеется сильно расплывшаяся светлая губа. Пупок круглый или слегка
овальный, сквозь него видно не более полутора оборотов. Размеры: ВР 4.5—6.3, БД
8—9.7, МД 7—8 мм.
Внутреннее строение. Материал: 16 экз. из окр. с. Космальян в Та-
лыше, 27 IX 1966-, сбор и определение наши.
Рис. 312—313. Xerosecta arcadiana Schileyko из Талыша, 27 IX 1966.
Паратип. 312 — внешний вид полового аппарата; 313 — пенис,
вскрыт. (Ориг.).
Нижняя часть спермовидукта в разной степени изогнута. СЖ 4x2—3. Наруж-
ный стилофор значительно крупнее внутреннего; стенка вагины, противолежащая
стилофорам, не обнаруживает заметного утолщения.
Пенис коротко веретеновидный, папилла различной длины, что связано с су-
ществованием недлинной, но обширной полости в дистальной части эпифаллуса, ко-
торая заходит далеко вглубь стенок папиллы. Бич не менее чем вдвое длиннее эпи-
фаллуса. Проток семеприемника многократно извит, причем изгибы его притянуты
к спермовидукту и сохраняют относительное постоянство. Резервуар семеприемника
прилегает к спермовидукту заметно ниже белковой железы.
Распространение и местообитания. Нагорно-степная зона
Талыша (Зуванд) и, вероятно, смежные районы Иранского нагорья. Встречается
на открытых склонах на стеблях травянистых растений и в мелкообломочных осыпях.
III. Подсемейство ARCHAICINAE Schileyko, subfam. nov.
Раковина средних размеров, прижатая, с куполовидным завитком, высота кото-
рого всегда меньше высоты устья. Пупок цилиндрический, умеренно широкий, в са-
мом начале может быть слегка развернутым. Окраска от голубовато-белой до корич-
ARCHAICA
257
невой, почти одноцветная, с легкой светлой лентой на периферии; спиральных тем-
ных лент никогда не бывает. '
Стилофоров 2, расположенных на одной стороне вагины, внутренний всегда зна-
чительно меньше наружного, но всегда имеется. Стенки папиллы плотные, без си-
нусов и капилляров.
Распространение. Западный Тянь-Шань и Алтайский хр.
Род ARCHAICA Schileyko, 1970
Шилейко, 1970 : 1307.
Признаки подсемейства.
Типовой вид — Helix apollinis Martens, 1882.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ARCHAICA
1 (4). Последний оборот в 1.5 раза шире предпоследнего. Губа слабо развита или
. отсутствует.
2 (3). Последний оборот в профиле угловатый. Сквозь пупок просматривается боль-
шая часть оборотов ............................ . 1. А. (Е.) heptapotamica.
3 (2). Последний оборот в профиле закруглен или слабо угловатый лишь в самом на-
чале. Сквозь пупок видно не более одного оборота . . . . 2. A. (A.) apollinis.
4 (1). Последний оборот незначительно шире предпоследнего. Губа мощная, валико-
образная, ограничена базальным краем устья.............3. A. (U.) labianix.
1. Подрод EUARCHAICA Schileyko, 1970
Шилейко, 1970 : 1309.
Раковина уплощенная, светло-роговая, выше средней линии последнего оборота
имеется закругленный угол. Нередко на раковине имеются короткие золотистые во-
лоски. Сквозь пупок видны все или почти все обороты. Губа слабая, расплывшаяся.
Наружный стилофор в верхней части не заострен. Стенки чехла пениса довольно
тонкие. Бич и эпифаллус примерно равной длины, проток семеприемника довольно
длинный. Папилла пениса не шаровидная, обычно со складками и вмятинами.
Распространение. Западный Тянь-Шань.
Монотипический подрод.
Типовой вид — Cathaica heptapotamica Lindholm, 1927.
1. Archaica (Euarchaica) heptapotamica (Lindholm, 1927). Табл. XIV, 136;
рис. 314—320.
Lindholm, 1927c : 199 (Cathaica tianshanica); 1927d : 265 (Cathaica); Л и-
харев, Раммельмейер, 1952 : 381, рис. 310 [Cathaica (Cathaica)]; Л и-
х а р е в, 1955 : 181, рис. 1А (Zenobiella); Матёкин, 1966 : 126, табл. 5, 1 (Ze-
nobiella tzwetkovi); Шилейко, 1970 : 1309, рис. 1, 3—4.
Материал: более 100 экз. из 9 местонахождений на западном Тянь-Шане.
Раковина сильно прижатая, высота завитка немного меньше высоты устья.
Контур завитка почти конический, реже слегка куполовидный.. Оборотов 5.5—6,
сверху слабее выпуклых, чем снизу. Последний оборот с ясным углом, который к устью
сглаживается. Вершина раковины выступающая. Окраска роговая, различной ин-
тенсивности; чем темнее раковина, тем резче выделяется светлая лента, идущая по
углу. Радиальная скульптура представлена тонкой исчерченностыо, местами имеются
более грубые штрихи; спиральные бороздки очень вялые, имеются местами на послед-
нем обороте. Устье коротко-овальное, очень косое, края устья не отвернуты, кроме
колумеллярного и прилежащей к нему части базального, места прикрепления устья
сближены. Сквозь пупок просматриваются все обороты. Размеры: ВР 6—10, БД 12—
17, МД 10.5—15 мм.
Внутреннее строение. Материал: 12 экз. из верховьев р. Пскем,
19 V 1972; 7 — из’верховьев р. Чаткал, 27 V 1972; 2 — из Ферганского хр. вдоль р. На-
рын над Ташкумыром, 31 V 1972; сборы и определения наши; 5 — из каньона Аксу
на Таласском хр., сбор Шапошникова, определение П. В. Матёкина; 3 — из окр.
г. Джамбула, 27 VI 1937, сбор и определение Б. Н. Цветкова.
Дистальная часть спермовидукта и яйцевод образуют изгиб, выраженный в раз-
личной степени. СЖ 3X2—3. Внутренний стилофор маленький, наружный массив-
ный, при основании более или менее вздут. Это вздутие соответствует вагинальному
валику, который петлеобразно охватывает выходное отверстие стилофора.
Пенис умеренно вздут, в дистальной части сужен. Пениальные мускульные ленты
отсутствуют или развиты слабо. Форма папиллы пениса очень разнообразна
17 А. А. Шилейко
Рис. 314—320. Archaica (Euarchaica) heptapotamica (Lindh.) из окр. Джамбула, 27 VI 1937. 314 — внешний вид полового
аппарата; 315 — вагина, вскрыта; 316 — пенис, вскрыт; 317 — пенис экземпляра из верховьев р. Чаткал; 318 — пенис эк-
земпляра из Таласского хр.; 319, 320 — пенис экземпляров из верховьев р. Пскем. (Ориг.).
ARCHAICA
259
(рис. 316—320), стенки папиллы плотные, семепроводящий канал на поперечном раз-
резе не щелевидный. Длина бича колеблется, но в среднем примерно равна длине
эпифаллуса, который может быть прямым или изогнутым в различной степени, но
не скрученным. Проток семеприемника тонкий и довольно длинный, заканчивается
небольшим мешковидным резервуаром, немного не доходящим до белковой железы.
Распространение и местообитания. От северо-западной ча-
сти Ферганского и центральной части Таласского хребтов — на запад до самых за-
падных отрогов Тянь-Шаня; запад-
ная часть Киргизского хр. На Ку-
раминском хр., вероятно, отсутст-
вует. Встречается преимущественно
в крупнообломочных осыпях с высо-
ким травостоем, т. е. в биотопах с
особенно богатым набором микроус-
ловий.
2. Подрод ARCHAICA s. str.
Шилейко, 1970 : 1308.
Раковина от прижатой до почти
плоской; если завиток возвышается,
контур его почти конический. Ок-
раска очень светлая, нередко с го-
лубоватым оттенком. Раковипа без
волосков. Пупок слабо перспектив-
ный. Губа слабая или отсутствует.
Апикальная часть наружного
стилофора более или менее явственно
' заострена. Стенки чехла пениса
очень мощные. Бич значительно ко-
роче эпифаллуса, проток семеприем-
ника короткий. Папилла пениса без
складок, почти шаровидная.
Распространение. Юго-за-
падный Тянь-Шань и Алтайский хр.
Монотипический подрод.
2. Archaica (Archaica) apollinis
(Martens, 1882). Табл. XIV, 137,138;
рис. 321, 322.
Martens, 1882b : 105 (Helix);
Westerlund, 1889 : 342 (Helix);
1896 : 187 [H. (Helicella) diaphora];
Линдгольм, 1931 : 56, t. 1, fig.
7—9 [Cathaica (Xerocathaica) ]; 58, t.
1, fig. 10—11 (subsp. nubila); Лиха-
рев, Раммельмейер, 1952 :
381 [Cathaica (Cathaica)}; Лиха-
рев, 1955 : 180, рис. 1, Б (Zeno-
biella); M а т ё к и н, 1966 : 122 (Ze-
nobiella); Шилейко, 1970 : 1308,
рис. 1, 1—2.
322
321
Рис. 321—322. Archaica (Archaica)
(Mart.) из урочища Янгарык на р.
Ферганский хр., 23 VI 1928. 321—
вид
apollinis
Гульче,
внешний
полового аппарата; 322 — пенис,
вскрыт. (По Шилейко, 1970).
Материал: 16 экз. из 2 местонахождений на Алайском и Ферганском хребтах.
Раковина с возвышающимся в различной степени завитком, высота кото-
рого в любом случае значительно меньше высоты устья. Оборотов 5, слабо выпуклых.
Последний оборот может быть в самом начале с легким намеком на угловатость, но
при рассматривании раковины в нормальном положении слева всегда правильно за-
круглен; к устью последний оборот очень слабо опущен. Окраска известково-белая
или голубовато-белая, одноцветная или с редкими размытыми, неправильно располо-
женными радиальными, очень светлыми желтоватыми или рыжеватыми полосами.
Кроме того, на некоторых раковинах видна очень тонкая светло-коричневая лента,
верхняя граница которой выражена лучше нижней. Радиальная скульптура пред-
ставлена очень тонкими поперечными морщинами и складками; спиральные бороздки
вялые, выражены местами. Устье округлое, косое, места его прикрепления сближены.
Пупок вначале развернутый, затем резко сужается, но ширина его варьирует; сквозь
пупок просматривается от 0.5 до 1.5 оборотов. Размеры: ВР 7—9, БД 10—16, МД 9—
14 мм.
17*
260
III. HIGROMIIDAE
Внутреннее строение. Материал: 2 экз. из урочища Янгарык на
р. Гульче (Алайский хребет), 23 VI 1928, сбор Рейхардта, определение Линдгольма;
4 — из долины р. Нарын немного ниже Токтогула, Ферганский хр., 31 V 1972, сбор
и определение наши.
Нижняя часть спермовидукта образует незначительный изгиб. Рудиментарный
стилофор плотно прилегает к основному, вершина которого имеет коническую форму.
Вагинальные валики тонкие и многочисленные, винтообразно проходят по внутрен-
ней поверхности вагины, развиты в разной степени.
Рис. 323—327. Archaica (Ugama) labianix sp. nov. из ущелья p. Пскем, 17—21 V 1972.
Паратипы. 323 — внешний вид полового аппарата; 324 — вагина, вскрыта; 325 —
пенис, вскрыт; 326, 327 — пенисы других особей, вскрыты. (Ориг.).
Пенис почти шаровидный, связан с эпифаллусом хорошо развитыми пениальными
мускульными лентами. Папилла пениса сильно вздута, в состоянии покоя также почти
шаровидная, при совокуплении становится цилиндрической. Стенки папиллы уме-
ренно разрыхлены. Бич короткий, длина его примерно вчетверо меньше суммарной
длины пениса и эпифаллуса. Проток семеприемника толстый, недлинный, в разной
степени скручен, заканчивается мешковидным резервуаром, прилегающим к спермо-
видукту не выше его середины.
Распространение и местообитания. Вид обнаружен на Фер-
ганском и Алайском хребтах. Населяет скалы и осыпи с умеренным увлажнением.
3. Подрод UGAMA Schileyko, subgen. nov.
Раковина уплощенная, с куполовидным завитком. Окраска интенсивно кашта-
новая или коричневая; волоски отсутствуют. Пупок по всей длине цилиндрический.
Губа мощная, занимает базальную часть раковины, справа обрывается.
Наружный стилофор наверху не конический. Стенки пениального чехла довольно
тонкие. Длина бича примерно равна длине эпифаллуса. Папилла пениса удлиненная,
с продольными складками или без них, уплощенная, семепроводящий канал резко
сдвинут к одной из стенок папиллы.
От других подродов этого рода резко отличается конхологическими признаками
и строением пениальной папиллы.
ARCHAICA
261
Распространение. Северо-западный Тянь-Шань.
Монотипический подрод.
Типовой вид — Archaica labianix Schileyko, sp. nov.
3. Archaica (Ugama) labianix Schileyko, sp. nov. Табл. XIV, 139', рис. 323—327.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: более 100 экз. из 4 местонахождений в долине р. Пскем.
Раковина от почти плоской до прижатой, с широко и плавно закруглен-
ным контуром завитка, составлена 6 узкими, тесно навитыми оборотами. Последний
оборот имеет ясный, но плавно закругленный угол выше средней линии, незначи-
тельно шире предпоследнего, снизу уплощен, к устью не опущен. Окраска от роговой
и каштановой до темно-коричневой, иногда почти черной; чаще всего красновато-
роговая. Верхняя поверхность раковины окрашена интенсивнее нижней. По углу
последнего оборота проходит размытая светлая полоса, выраженная чаще всего слабо.
Раковина покрыта шелковистой радиальной исчерченностыо и очень тонкими спи-
ральными бороздками, которые лучше всего видны на предпоследнем обороте. Устье
в виде широкой изогнутой щели, слегка скошено, базальный край его очень плавно
переходит в палатальный. Отступя от края устья располагается мощная высокая
снежно-белая губа (отсюда видовое название), которая справа немного вздувается,
а затем плавно, но быстро обрывается. В течение постэмбриогенеза формируется от 2
до 6 таких губ, которые просвечивают сквозь стенку раковины в виде широких свет-
лых размытых полос. Пупок неширокий; несмотря на то что он не. развернут, сквозь
него можно, слегка поворачивая раковину, просмотреть большую часть предыдущих
оборотов. Размеры: ВР 5—5.5, БД 9—10.5, МД 8—9 мм. Голотип: ВР 5.4, БД 10,
МД 9 мм.
По конхологическим признакам вид очень похож на Angiomphalia buamica-, от-
личается более широким пупком и более сглаженным утлом выше периферии.
Внутреннее строение. Материал: 20 экз. из верхнего течения
р. Пскем между Пскемским и Угамским хребтами, 17—21 V 1972, сбор и определение
наши.
Дистальный отдел спермовидукта и яйцевод почти прямые. СЖ 4X2. Внутренний
стилофор втиснут в пространство между стенкой вагины и вздутым наружным стило-
фором. Вагинальные валики мощные; кроме двух соседствующих валиков, между
которыми открывается отверстие стилофоров, имеется один утолщенный внизу ва-
лик, входящий в щель между двумя противолежащими.
Пенис маленький, шаровидный, пениальные мускульные ленты отсутствуют.
Папилла состоит из двух продольных отделов, первый из которых в сечении более
или менее округлый и содержит семепроводящий канал; к нему примыкает пластинка,
заполненная рыхлой тканью и часто согнутая вдоль в разной степени (см. поперечные
срезы на рис. 325—327). Видимо, при совокуплении выпрямляется и заполняется
жидкостью именно эта пластинка, будучи аналогом os penis некоторых млекопитаю-
щих. Бич несколько короче прямого цилиндрического эпифаллуса; между семепро-
водом, эпифаллусом и пенисом натянута тонкая мембрана. Проток семеприемника
длинный, заканчивается небольшим мешковидным резервуаром, немного не доходя-
щим до белковой железы.
Распространение и .местообитания. Вид широко распростра-
нен в верхнем течении р. Пскем, где является массовой формой и встречается почти
во всех типах биотопов. Предпочитает сырые участки с камнями и высоким травостоем,
где образует плотные скопления.
IV. Подсемейство EUOMPHALIINAE Schileyko, subfam. nov.
Раковина маленькая или средних размеров, от уплощенной линзовидной до почти
шаровидной, с закругленной периферией. Пупок различной ширины, но никогда не
бывает очень широким или полностью запаянным. Окраска различных цветов, чаще
роговая или желтоватая, обычно с размытой светлой лентой по периферии, изредка
с темными лентами.
Стилофоры видоизменены в вагинальные придатки, часто вздутые на концах.
Первично таких придатков 2, но нередко 1 редуцирован, значительно реже редуци-
рованы оба придатка. В основании папиллы пениса имеется кольцевое разрыхление
тканей, которое превращается в обширную полость. В случае интенсивного разви-
тия этой полости и расслоения стенок пениального чехла папилла пениса не выдви-
гается, а выворачивается.
Распространение. Средиземноморье, Европа и Кавказ. В ископаемом
состоянии представители подсемейства встречаются начиная с верхнего миоцена
(Предкавказье), верхнего плиоцена и плейстоцена (Европа).
362
III. HYGROMIIDAE
1. Род KARABAGHIA Lindholm, 1927
Lindholm, 1927a : 124 [Perforatella (Karabaghia) ]; Лихарев, Гам-
ме л ьме йе p, 1952 : 444 (Perforatella part.); Wenz, Zilch, 1959—1960 : 682.
Раковина маленькая, почти шаровидная, тонко скулыггурированная. Пупок
узкий, неперспективный. Волоски на раковине отсутствуют. В устье имеются
2 зуба — базальный и палатальный.
Вагинальные придатки располагаются рядом друг с другом. Чехол папиллы пе-
ниса тонкий, стенки его не расслаиваются. Кольцевой полости в мужском отделе
Рис. 328—331. Karabaghia bituberosa (Lindh.) из Армении, вершина Архашан,
9 VIII 1952. 328 — внешний вид полового аппарата; 329 — вагина, вскрыта; 330 —
пенис, вскрыт; 331 — поперечный срез папиллы пениса. (Ориг.).
нет, кольцевое разрыхление в основании папиллы выражено очень слабо. Ретрактор
правого омматофора образует перехлест с половым ретрактором, но лежит в стороне
от пениса и вагины.
Распространение. Закавказье.
Монотипический род.
Типовой вид — Perforatella bituberosa Lindholm, 1927.
Karabaghia bituberosa (Lindholm, 1927). Табл. XIV, 140; рис. 328—331.
Rosen, 1914 : 227 [Helix (Petasia) bifrons non Lowe, 1833]; Lindholm,
1927a : 124 [Perforatella (Karabaghia)]; 1929 : 205, t. 11, fig. 4 (Perforatella); Лиха-
рев, Рамме л ьмейер, 1952 : 444 (Perforatella); Акрамовский, 1971а :
110 [Euomphalia (Karabaghia)].
Материал: 6 экз. из 2 местонахождений в Армении.
Раковина ширококоническая, приближается к шаровидной, с явственно
куполовидным контуром завитка и невыступающей вершиной. Оборотов 5—5.5, мед-
ленно и плавно нарастающих. Последний оборот в самом начале с легким намеком
на угловатость, в базальной части вздут, к устью еле опущен или прямой. Окраска
темно-роговая или желтоватая, с неясной светлой лентой на периферии. Скульптура
в виде радиальной исчерченности, выраженной в разной степени, спиральные бо-
роздки отсутствуют. Устье округло-треугольное, умеренно косое; верхний край устья
прямой, остальные отогнуты. Вблизи края устья имеется небольшая губа, на кото-
рой сидят зубы, причем палатальный имеет форму бугорка, а базальный — форму
HESSEOLA
263
i
I
удлиненной складки. Палатальному зубу снаружи соответствует глубокая ямка.
Сквозь пупок просматривается менее одного оборота. Размеры: ВР 4—4.5, БД 6—6.5,
МД 5.2—5.5 мм.
Внутреннее строение. Материал: 3 экз. из Армении, Кировакан-
ский р-н, вершина Архашан над с. Гамзачиман, 9 VIII 1952, сбор и определение
Н. Н. Акрамовского.
Спермовидукт и яйцевод прямые. СЖ 4X1. Вторичного ветвления слизистых
желез не наблюдается. Вагинальные придатки длинные, сильно сближены на одной
стороне вагины. Вагинальные валики развиты слабо.
Пенис сильно вздут, заключает коническую массивную папиллу, стенки которой
равномерно заполнены рыхлой паренхимой. Семепроводящий канал в пределах па-
пиллы сохраняет примерно то же строение, что и в пределах эпифаллуса, т. е. На по-
перечном срезе видна пара главных валиков и несколько более мелких противолежа-
щих им. Пениальные мускульные ленты слабо развиты. Бич примерно вдвое короче
круто изогнутого эпифаллуса. Половой ретрактор длинный и тонкий, петлеобразно
охватывает ретрактор правого омматофора. Проток семеприемника толстый и длин-
ный, почти прямой, резервуар мешковидный, касается нижнего края белковой же-
лезы.
Распространение и местообитания. Центральная часть Ма-
лого Кавказа. Встречается под камнями и среди травы на субальпийских лугах.
2. Род HESSEOLA Lindholm, 1927
Lindholm, 1927а : 125 {Euomphalia, sect. Hesseola); Wenz, Zilch,
1959—1960:689 [Euomphalia (Hesseola)].
Раковина небольшая,, почти кубаревидная, радиальная скульптура грубая, спи-
ральная, тонкая. Пупок узкий, неперспективный. На раковине могут присутствовать
очень тонкие волоски. Устье без зубов.
Вагинальные придатки располагаются рядом друт с другом, пронизывают стенки
вагины и концы их внутри вагины сближены. Чехол пениса расслаивается, образо-
вавшаяся полость проникает и в стенки папиллы, в результате чего последняя теряет
постоянную длину и форму. Ретрактор правого омматофора образует перехлест с по-
ловым ретрактором, но с гениталиями не перекрещивается.
Распространение. Закавказье и Копетдаг.
Типовой вид — Helix (Fruticocampylaea) adshariensis Lindholm, 1913 (syn. Helix
solidior Mousson, 1873).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА HESSEOLA
1 (2). Раковина белая или светло-серая, обычно с 2 спиральными коричневыми лен-
тами — выше и ниже периферии. Вид обитает в Закавказье . . 1. Н. solidior.
2 (1). Раковина роговая или каштановая, со слабой светлой размытой лентой по
периферии. Вид обитает в Копетдаге ......................2. Н. transcaspia.
1. Hesseola solidior (Mousson, 1873). Табл. XV, 141; рис. 332—334.
Mousson, 1873 : 200 [Helix (Campylaea) Narzanensis var. solidior]; К о b e 11,
1877, Iconogr., V : 30, t. V, fig. 1225 (Helix); R ose n, 1911 : 110 (H.' christophori);
Lindholm, 1913c : 141 [Helix (Fruticocampylaea) adshariensis]; Rosen, 1914 :
173, t. 1, fig. 5a—с (H. christophori); Hesse, 1931 : 25, t. 4, fig. 26 (Euomphalia ad-
shariensis); Акр а м о в с к и Й, 1949 : 153 (Metafruticicola pratensis part.); Лиха-
рев, Раммельмейер, 1952 : 447 (Fruticocampylaea adshariensis); Hudec,
Lezhawa, 1967b : 86, Abb. 9, t. IV, 10 (Fruticocampylaea narzanensis part.); Hu-
dec, 1972d : 272, Abb. 5, 6, 10 (Hesseola adshariensis; H. adshariensis antontomashevici
n. subsp.).
Материал: более 40 экз. из 5 местонахождений в Закавказье.
Раковина от прижатой до почти шаровидной, с широким, слегка куполо-
видным завитком и несглаженной вершиной. Высота завитка примерно равна высоте
устья; может быть как ниже, так и выше высоты устья. Оборотов 6—6.5, довольно
выпуклых, плавно нарастающих, разделенных глубоким швом. Последний оборот
в профиле плавно закруглен, немного менее чем вдвое шире предпоследнего, к устью
опускается плавно, без резкого перегиба. Окраска белая или светло-серая, одноцвет-
ная или, чаще, с двумя узкими коричневыми лентами, расположенными выше и ниже
периферии. В проходящем свете ленты просвечивают сильнее фона; изредка границы
их размыты, но обычно обе ленты четкие. Радиальная скульптура в виде грубой ис-
черченности; спиральные бороздки тонкие и густые, иногда выражены местами. Кроме
того, иногда наблюдаются короткие спиральные штрихи и слабые неправильной
формы вмятины, разбросанные по всей поверхности последнего оборота. Молодые
264
HI. HYGROMIIDAE
экземпляры густо покрыты тонкими волосками, которые с возрастом бесследно ис-
чезают. Устье круглое или слегка эллиптическое, косое, места его прикрепления не-
много сближены. Края устья тонкие, отвернутые, кроме самого верхнего отдела,
Отступя от края лежит валикообразная, хорошо развитая губа. Пупок узкий, сквозь
него просматривается не более чем один оборот. Размеры: ВР 9—12, БД 13—15, МД
11—13 мм.
Внутреннее строение. Материал: 4 экз. из Армении, Даралагез,
с. Гнишик, 15 VII 1949, сбор и определение Н. Н. Акрамовского; 3 — из окр. оз. Се-
ван, 1902, сбор Е. А. Елачича и А. А. Климантовича, определение наше.
Рис. 332—334. Hesseola solicitor (Mouss.) из Армении, с. Гнишик, 15 VII 1949.
332 — внешний вид полового аппарата; 333 — пенис, вскрыт; 334 — вагина,
вскрыта. (Ориг.).
Дистальная часть спермовидукта и яйцевод прямые. СЖ 3x2—3. Вагинальные
придатки длинные, развиты в равной степени, их основания охвачены общим чехлом
вместе с прилежащим участком вагины. Стенки вагинальных придатков значительно
мускулизованы.
Пенис чрезвычайно короткий, поэтому половой ретрактор крепится близ клоаки.
В определенном смысле можно говорить об отсутствии папиллы пениса как таковой,
потому что при максимальном втягивании эпифаллуса она исчезает, что в свою оче-
редь связано с существованием хорошо развитой кольцевой полости. Пениальные
мускульные ленты очень хорошо развиты. Конической формы бич в 1.5 раза короче
толстого эпифаллуса, в нижней части которого, на границе с пенисом, имеется резкий
перегиб. Эпифаллус тонкостенный, с обширной внутренней полостью. Проток семе-
приемника вначале довольно тонкий, затем быстро утолщается и сохраняет увели-
ченный диаметр до самого резервуара; ясная граница между протоком и резервуаром
отсутствует.. Резервуар прилегает к верхней части спермовидукта.
Распространение и местообитания. Западная и центральная
части Малого Кавказа. Населяет открытые, хорошо увлажняемые участки с сочной
травой.
HESSEOLA
265
2. Hesseola transcaspia (Boettger, 1889). Табл. XV, 742; рис. 335, 336.
Boettger, 1889 : 944, t. 27, fig. 12 [Helix (Carthusiana) ]; К о b e 11, 1892,
Iconogr., N. F., V : 13, fig. 745 [Helix (Latonia) ]; Л и x a p e в, Раммельмейер,
1952 : 462 (Euomphalia)-, Матёкин, 1966 : 123, табл. 5, рис. 2 (Euomphalia).
Материал: более 20 экз. из 2 местонахождений на Копетдаге.
Раковина приближается к шаровидной, с куполовидным завитком, высота
которого приблизительно равна высоте устья. Оборотов 4.5—5.5, умеренно выпук-
Рис. 335—336. Hesseola transcaspia (Bttg.) из Копетдага, 18 IV 1953. 335 —
внешний вид полового аппарата; 336 — дистальные части полового аппарата,
вскрыты. (Ориг.).
>лых, плавно нарастающих, разделенных мелким швом. Последний оборот в самом
начале с наклонностью к образованию угла, но довольно быстро оборот становится
в профиле правильно округлым; ширина его в 1.5 раза превышает ширину предпо-
следнего, к устью последний оборот внезапно опущен. Окраска светло- или красно-
вато-роговая, почти монотонная, на периферии с очень неясной светлой лентой. По-
верхность раковины густо, но тонко радиально исчерчена и покрыта короткими во-
лосками; если волоски опадают, остаются ясные многочисленные округлые рубцы.
Устье косое, круглое, иногда немного оттянуто вправо, в разной степени вырезано
предпоследним оборотом. Края устья острые, в базально-колумеллярной части от-
вернуты. Немного отступая от края устья лежит четкая высокая белая губа, которая
снаружи просвечивает в виде желтой полоски. Пупок слегка прикрыт отворотом
колумеллярного края, сквозь него просматривается не более половины предпослед-
него оборота. Размеры: ВР 6—8, БД 9—13, МД 8—10.5 мм.
Внутреннее строение. Материал: 4 экз. из Копетдага, Кара-Кала,
18 IV 1953, сбор И. М. Лихарева, определение наше.
Яйцевод прямой или слабо изогнутый. СЖ 2X2—3. Вагинальные придатки вере-
теновидные, на концах несут утолщения, их основания вместе с нижней частью ели-
Z66
III. HYGROMIIDAE
зистых желез охвачены многослойным соединительнотканным чехлом. Вагина тон-
костенная.
Пенис очень короткий, строение папиллы характерно для рода. Бич в 1.5—2 раза
короче толстого эпифаллуса, между изгибами которого натянуты мускульные ленты.
Половой ретрактор очень тонкий и длинный. Проток семеприемника почти прямой,
довольно толстый, семеприемник объемистый, мешковидный, прилегает к верхней
части спермовидукта.
Распространение и местообитания. Центральная и западная
части Копетдага. Встречается в достаточно влажных биотопах с растительностью.
3. Род EUOMPHALIA Westerlund, 1889
Westerlund, 1889:31, 32; Kobelt, 1904, Iconogr., N. F., 11:131;
Lindholm, 1927a : 124, Hesse, 1931 :19; Лихарев, Раммельмей-
e p, 1952 : 459; Wenz, Zilch, 1959—1960 : 688.
Раковина средних размеров, слегка прижатая, радиальная скульптура — до-
вольно грубая исчерченность; большей частью имеются также спиральные бороздки.
Раковина с волосками, которые сохраняются пожизненно или по достижении половой
зрелости утрачиваются. Пупок умеренно широкий, слабо развернутый, в разной
степени перспективный.
Вагинальные придатки не сближены на одной стороне вагины, впадают в вагину,
не пронизывая ее стенок, очень длинные, с тонким стволом. Благодаря характерному
расслоению стенки пениального чехла возникает своеобразный чехол папиллы, ко-
торый обеспечивает лишь частичное выдвижение папиллы при спаривании — не
более чем на 2/3 ее длины. Ретрактор правого омматофора образует перехлест с по-
ловым ретрактором, относительно гениталий способен легко перемещаться, то на-
ходясь в состоянии перехлеста, то проходя рядом с ними. Однако половой ретрактор
может отсутствовать.
Распространение. Средняя Европа и Кавказ. В ископаемом состо-
янии род известен начиная с раннего плейстоцена (Европа).
Типовой вид — Helix strigella Draparnaud, 1805.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА EUOMPHALIA
1|(2). Сквозь пупок просматривается не более 3/4 предпоследнего оборота.
................................................... 2. Е. appeliana.
2 (1). Сквозь .пупок просматривается не менее полутора оборотов.
3 (4). Раковина взрослых экземпляров покрыта волосками (длина волоска 0.6—1 мм),
сохраняющимися пожизненно.................................1. Е. aristata.
4 (3). Раковина взрослых экземпляров лишена волосков; раковина молоди покрыта
волосками (длина волоска 0.3—0.4 мм) .....................3. Е. strigella.
1. Euomphalia aristata (Krynicki, 1836). Табл. XV,fI45; рис, 337—340.
Krynicki, 1836 : 179 (Helix)-, Kobelt, 1879, Iconogr, VI : 36, fig. 1625
(Helix)- W esterlund, 1889 : 91 (Helix)- Лихарев, Раммельмейер,
1952 : 461, рис. 387; Hudec, Lezhawa, 1967b : 91, Abb. 12, t. Ill, 8.
Материал: более 30 экз. из 6 местонахождений на Кавказе.
Раковина почти кубаревидная, с широким куполовидным завитком, который
изредка бывает почти коническим; высота его приблизительно равна высоте устья.
Вершина слегка заостренная. Оборотов 6, умеренно выпуклых, медленно нараста-
ющих, разделенных глубоко врезанным швом. Последний оборот в 1.5 раза шире
предпоследнего, в самом начале со слабым углом, вскоре сглаживающимся, к устью
внезапно опущен. Окраска светло- или темно-роговая, со светлой лентой, выражен-
ной в разной степени. Скульптура в виде резкой и неравномерной радиальной исчер-
ченности; кроме того, по поверхности раковины проходят очень тонкие, неровные,
частые спиральные струйчатые кожистые ребрышки. Эти ребрышки у старых потер-
тых раковин отсутствуют, и спиральная скульптура представлена бороздками. Ра-
ковина покрыта многочисленными изогнутыми волосками; когда волоски опадают,
на их месте остаются ясно заметные рубцы; впрочем, волоски остаются пожизненно
и утрачиваются только у старых раковин. Устье круглое или широкоовальное, ко-
сое, места его прикрепления заметно сближены. Края устья острые, отвернуты, осо-
бенно сильно отвернут базальный край. Внутри устья имеется широкая, но невысо-
кая губа. Пупок глубокий, цилиндрический, перспективный. Размеры: ВР 12—15,
БД 16—21, МД 13—17.5 мм.
Внутреннее строение. Материал: 1 экз. йз окр. Абастумани, Гру-
зия, 1 X 1962; 1 — из окр. Орджоникидзе, гора Сапицкая Будка, 19 V 1964 (не вполне.
337
Рис. 337—340. Euomphalia aristata (Kryn.) из окр. Абастумани, 1 X 1962.
337^— внешний вид полового аппарата; 338 — поперечный срез папиллы
пениса; 339 — особь in copuli; 340 — не вполне взрослый экземпляр из окр.
Орджоникидзе, 19 V 1964. (Ориг.).
268
III. HYGROMIIDAE
взрослый); 2 — из Аджарии, Имеретинский хр., близ Адигени, 21 VII 1973; сборы
И. М. Лихарева, определения наши.
Изгиб нижней части спермовидукта, ясно выраженный у молодого экземпляра,
у взрослых почти отсутствует. Слизистые железы хорошо развиты, располагаются
мутовкой вокруг вагины, однако закладываются они попарно; нередко имеет место
вторичное ветвление отдельных желез. Вагинальные придатки не скручены, конце-
вые утолщения ясные. Внутреннее строение этих утолщений своеобразно; внутрен-
ний эпителий отстает от покровных тканей, образуя продольные складки.
Рис. 341—342. Euomphalia appeliana (Mouss.) из ущелья
р. Бзыби, VIII 1913. 341 — внешний вид полового аппарата;
342 — поперечный срез папиллы пениса. (Ориг.).
Пенис длинный, почти цилиндрический, дистальная часть его слегка сужена.
Дистальная часть папиллы имеет чрезвычайно рыхлые стенки, внутри которых рас-
полагаются многочисленные радиальные тяжи. Просвет папиллы обширный, имеет
сложную форму, соответствующую сечению эпифаллуса (рис. 337). Пениаль-
ные ленты хорошо развиты. Бич немного длиннее цилиндрического эпифал-
луса. Резервуар семеприемника заметно не достает до белковой железы, про-
ток его почти прямой. У молодого экземпляра половой ретрактор не обнаружен.
Распространение и местообитания. Западные и централь-
ные склоны Северного Кавказа; спорадическое распространение в Закавказье. Оби-
тает в подстилке лиственных лесов.
2. Euomphalia appeliana (Mousson, 1876). Табл. XV, 144; рис. 341, 342.
Mousson, 1876 : 36, t. 2, fig. 3 (Helix); К о b e 1 1, 1877, Iconogr., V ; 29,
fig. 1218, 1219 (Helix); Boettger, 1883 : 167, t. 6, fig. 5 [Helix (Fruticocampylaea)
narzanensis var. appeliusi Mousson f. depressa]; Westerlund, 1889 : 104 (Helix
EUOMPHALIA
269
appeliusi}', Rosen, 1911 : 104 {Helix}', 1914 : 167 {Helix}', Hesse, 1931 : 23, t. 3,
fig. 22; Лихарев, Раммельмейер, 1952 : 464.
Материал: 5 экз. из 3 местонахождений на западном Кавказе.
Раковина почти кубаревидная, с притупленным округло-коническим за-
витком, высота которого примерно равна высоте устья или немного меньше: Оборо-
тов 6,'слабо выпуклых. Последний оборот сильно вздут, в профиле округлый, в самом
начале с очень слабым углом, в 1.5 раза шире предпоследнего, к устью немного опу-
щен. Вокруг пупка имеется более или менее развитый угол (смотреть снизу). Окраска
светло- или темно-роговая, со светлой лентой на периферии. Скульптура в виде не-
равномерной радиальной исчерченности и мелкой струйчатой зернистости. Волоски
на раковинах взрослых экземпляров отсутствуют. Устье почти круглое, косое, края
его острые, тонкие, верхний слабо, нижний сильно отвернут. Места прикрепления
устья соединены отчетливой мозолью. Пупок довольно широкий, открытый, но не
развернутый и не перспективный: сквозь него видно не более одного оборота. Размеры:
ВР 12—14, БД 17-20, МД 15-17.5 мм.
Внутреннее строение. Материал: 1 экз. из ущелья р. Бзыби (южный
склон западной части Большого Кавказа), VIII 1913, сбор К. А. Сатунина, опреде-
ление И. М. Лихарева.
Нижняя часть спермовидукта и яйцевод резко изогнуты. Слизистые железы
располагаются мутовкой вокруг вагины и не обнаруживают тенденции к группировке
в пучки. Общее число ветвей — 12. Стволы вагинальных придатков тонкие, конце-
вые утолщения ясные. Эпителий внутри утолщений собран в несколько широких
складок и местами отстает от покровной ткани.
Пенис длинный, на границе с эпифаллусом очень резко изогнут, связан с эпи-
фаллусом мощными пениальными лентами. Папилла пениса занимает небольшую
часть дистального участка мужского отдела. Имеется чехол папиллы пениса, обес-
печивающий выдвижение ее на ограниченное длиной этого чехла расстояние. Бич
короче эпифаллуса. Резкой границы между протоком и резервуаром семеприемника
нет. Возможно, это связано с наличием у данного экземпляра сперматофора в семе-
приемнике.
Распространение и местообитания. Подкововидный ареал
вокруг западного крыла Главного Кавказского хр. от Кисловодска до Новороссийска,
вдоль побережья Черного моря и территория Абхазской АССР. Обитает в подстилке
горных лиственных лесов. Редкий вид.
3. Euomphalia strigella (Drapamaud, 1801). Табл. IX, 86; XV, 145; рис. 343—347.
Draparnaud, 1801 : 81 {Helix); Westerlund, 1889 : 92. {Helix); 1897 :
117 [Helix (Trichia) podolica}; Лихарев, Раммельмейер, 1952:460,
рис. 386; G г о s s u, 1955 : 453, fig. 256; Ehrmann, 1956 : 134, t. 5, fig. 69;
Lozek, 1956 : 214, t. 35, 3; Zilch, J aeckel, 1962 : 187; Lozek, 1964 : 302,
t. 23, 3a—c.
Материал: более 100 экз. из -23 местонахождений в окр. Москвы, в Молдавии,
Белоруссии, ГДР, Венгрии.
Раковина почти кубаревидная, реже слегка прижатая, с острым, чуть куполо-
видным завитком и несглаженной вершиной. Оборотов 6, выпуклых, быстро нараста-
ющих. Последний оборот в профиле округлый, к устью внезапно опущен, примерно
в 1.5 раза шире предпоследнего. Окраска от почти желтой до красновато-роговой
или серой, со светлой лентой по периферии. Поверхность раковины резко и неравно-
мерно радиально исчерчена, местами со спиральными бороздками и штрихами; кроме
того, на последнем обороте имеются многочисленные мелкие неориентированные
вмятины, благодаря чему поверхность напоминает порой сильно смятую и затем вновь
разглаженную алюминиевую фольгу. У молодых экземпляров раковина покрыта
редкими мелкими волосками, которые с возрастом чаще всего полностью утрачи-
ваются, оставляя после себя рубцы. Устье почти круглое, косое, места его прикреп-
ления сближены, края устья острые, слабо отвернуты, нижний край отвернут
несколько сильнее прочих. Несколько отступя от края располагается белая
губа, иногда с красноватым оттенком, просвечивающая на затылке последнего обо-
рота в виде желтой полосы. Пупок открытый, сквозь него просматриваются все или
почти все обороты. Размеры: ВР 9—12, БД 13—19, МД 11—15 мм.
Внутреннее строение. Материал: 6 экз. из окр. Кведлинбурга, ГДР,
V 1969, сбор и определение Э. Клаусса; 3 — из окрестностей Балатонфюреда, Венг-
рия, 15 VII 1970, сбор Д. А. Сахарова; 2 — из сосняка в окр. Брянска, VI 1965, сбор
М. Т. Лаврова; 13 — из с. Ромашково Московской обл., 3 XI 1963; 10 — из с. Архан-.
вельского Московской обл., 5 VI 1971; сборы и определения наши.
Дистальная часть спермовидукта и яйцевод почти прямые или слабо изогнуты.
СЖ 4X2. Длина вагинальных придатков превышает суммарную длину верхнего и
нижнего отделов вагины. Внутри вагины, начинаясь от клоаки, тянется выражен-
W/7
Рис. 343—347. Euomphalia strigella (Drap.) из с. Архангельского Московской обл.,
5 VI 1971. 343 — внешний вид полового аппарата; 344 — пенис, вскрыт (схематизи-
ровано); 345 — вагина, вскрыта; 346, 347 •— особь in copuli, вид справа и снизу.
(Ориг.).
PLATYTHEBA
' 271
ный в различной степени валик, который, разветвляясь возле мест впадения придат-
ков, заходит обеими ветвями в стволы этих придатков.
Пенис цилиндрический, папилла пениса фиксирована вторичным чехлом ближе
к дистальному концу, за счет чего обеспечивается частичное выдвижение папиллы
при совокуплении. Конический бич почти вдвое короче эпифаллуса. Ретрактор пра-
вого омматофора образует перехлест с половым ретрактором, но тем не менее часто
не перекрещивается с пенисом и вагиной. Мешковидный резервуар семеприемника
располагается на уровне верхней части спермовидукта.
Распространение и местообитания. Средняя и Северная
Европа, к северу до 61° с. ш., к югу до Черного и Каспийского морей, к востоку ме-
стами до Волги и даже левый берег Волги (у Казани — Круликовский, 1889); еди-
ничные находки на Урале: бассейн р. Белой, окр. Свердловска. Населяет преиму-
щественно умеренно увлажняемые лиственные и смешанные леса; в южной части
ареала встречается также в хвойных лесах и кустарниковых зарослях. В ископаемом
состоянии этот вид и близкие формы многократно найдены в плейстоцене и голоцене
Европы.
4. Род PLATYTНЕВА Pilsbry, 1894
К о b е 1 t, 1871 : 12 [Carthusiana (Nummulina, non d'Orbigny, 1826)]; Pils-
bry, 1894, Man. Conch., IX : 268 (Theba, sect. Platytheba); Hesse, 1921:63
(Theba, sect. Platytheba)', Li nd ho Im, 1927a : 123 [Trochulus (Xerocampylaea,
sect. Caucasojixus) ], 125 [Circassina (Jasonella)],. 126 [Monacha (Monacha s. str., sect.
Platytheba) ]; Лихарев, Раммельмейер, 1952 : 436; W e n z, Zilch,
1959—1960:674 [Monacha (Platytheba)].
Раковина средних размеров, линзовидная, с выраженной в разной степени ра-
диальной ребристостью. Раковина без волосков, пупок неширокий, но открытый.
Вагинальных придатков 2 или 1, при основании не сближенных. Имеется вторич-
ный чехол пениса (чехол папиллы). Ретрактор правого омматофора не всегда образует
перехлест с половым ретрактором; последний может отсутствовать.
Распространение. Западный Кавказ, Сирия, Палестина.
Типовой вид — Helix nummus Ehrenberg, 1831.
Из 5 видов, составляющих данный род, анатомически изучены 2: Pl. nummus
и Pl. mingrelica; анатомические различия между ними того же порядка, что и между
подродами близкого рода Monacha, т. е. у Pl. nummus имеется один вагинальный
придаток, а у Pl.mingrelica — два, что служит основанием для отнесения этих видов
к разным подродам. Напомним, что для членов подсемейства Euomphaliinae вообще
характерна тенденция к утрате вагинальных придатков (и полового ретрактора),
поэтому такой признак, как отсутствие или наличие придатка, в данном случае имеет
ранг не выше подрода.
Внутреннее строение второго вида фауны СССР — Pl. prometheus — не изучено,
но конхологически он стоит ближе к Pl. nummus, чем к Pl. mingrelica, почему мы
и вклч-^аем его провизорно в подрод Platytheba s. str.,
ТАБЛИЦАЩЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА1РЕА TYTHEBA
1 (2). Высота завитка равна высоте устья или немного меньше. Поверхность раковины
между радиальными ребрами зернистая. Киль тупой. Палатальный край устья
с тупым углом.....................................1. Pl. (J.) mingrelica.
2 (1). Высота завитка равна 0.5 высоты устья или еще меньше. Поверхность раковины
между радиальными ребрами лишена зернистой скульптуры. Киль острый.
Палатальный край устья с острым углом и желобком......................
........................................... 2. Pl. (Pl.) prometheus.
1. Подрод JASONELLA Lindholm, 1927
Lindholm, 1927а : 125; Hudec, Lezhawa,’ 1969b : 142.
Высота завитка равна высоте устья или немного Меньше ее; поверхность рако-
вины между радиальными ребрами с зернистой скульптурой. Палатальный край
устья слегка угловатый.
Вагинальных придатков 2, половой ретрактор образует перехлест с ретрактором
правого омматофора.
Распространение. Западный Кавказ.
Типовой вид — Circassina mingrelica (Hesse, 1921).
1. Platytheba (Jasonella) mingrelica (Hesse, 1921). Табл. XV, 146; рис. 348, 349.
Mousson, 1861, Vierteljahrsschr. naturf. Ges. Zurich, VI : 29 (Helix jasonis,
non Mayer, 1856); 1863 : 372 (H. jasonis); К о b e 1 t, 1879, Iconogr., VI : 1, fig. 1529
(H. jasonis); W esterlund, 1889 :30 (H. jasonis); Rosen, 1914 :160 [Я. (Nummu-
272
III. HYGROMIIDAE
lina) jasonis]; Retowski, 1914 : 282 [Я. (Nummulina) jasonis}-, Hesse, 1921 : 67
(Circassina mingrelica nom. nov.); Линдгольм, 1922a : 312 (Helix argonautarum
nom. nov.); Lindholm, 1927a : 125 [Circassina (Jasonella) ]; Hesse, 1931 : 27,
t. 4, fig.31 [Circassina (Jasonella)}-, Лихарев, Раммельмейер, 1952:
437, рис. 361 (Platytheba)', Hudec, Lezhawa, 1969b : 142, Abb. 32, t. IX, 23.
Материал: более 60 экз. из 2 местонахождений на западном Кавказе.
Раковина с куполовидным завитком и выступающей вершиной, умеренно
тонкостенная. Оборотов 6, слабо выпуклых, плавно нарастающих, разделенных мел-
ким швом. Последний оборот немного менее чем в 1.5 раза шире предпоследнего,
снизу выпуклый, к устью слегка и плавно опущен. По периферии проходит тупой
Рис. 348—349. Platytheba (Jasonella) mingrelica (Hesse) из долины
р. Техури, 29 IV 1971. 348 — внешний вид полового аппарата;
349 — пенис, вскрыт (схематизировано). (Ориг.).
нитевидный киль, который в сочетании с угловатостью последнего оборота придает
раковине характерней линзовидный облик. Окраска почти одноцветная, серовато-
белая или серовато-роговая, иногда с зеленоватым оттенком. Скульптура в виде рав-
номерных радиальных сглаженных ребер, окрашенных светлее фона. Промежутки
между ребрами покрыты ясной зернистостью, причем ориентация гранул такова,
что можно предполагать возникновение их за счет распадения тонких спиральных
бороздок. Устье овальное, косое, с плавным углом, соответствующим килю. Края
устья тонкие, острые, базально-палатальный край отвернут; губа имеется только
на базальном крае, развита слабо. Пупок перспективный. Размеры; ВР 9—12,
БД 20—24, МД 17—21 мм. '
В нутреннее строение. Материал: 3 экз. из окр. с. Талери в долине
р. Техури (юго-западные склоны Большого Кавказа), 29 IV 1971, сбор иопределение
наши.
На границе спермовидукта и яйцевода имеется небольшой изгиб. СЖ 4X2. Ва-
гина внутри несет многочисленные продольные складки; выходные отверстия ваги-
нальных придатков окружены кольцевыми валиками. Концевые утолщения вагиналь-
ных придатков внутри имеют по 3 продольные складки, причем одна из них располо-
жена напротив двух других и направлена в просвет между ними.
Пенис сложен вдвое и фиксирован в таком положении мускульными лентами.
Папилла пениса охватывается вторичным чехлом примерно посредине длины, чуть
ближе к дистальному концу, благодаря чему этот орган при совокуплении наполовину
выдвигается, а затем выворачивается. Бич конический, длина его слегка превышает
OSCARBOETTGERIA
273
половину почти прямого цилиндрического эпифаллуса. Резервуар семеприемника
не достигает белковой железы. Проток семеприемника близ резервуара образует
расширение; насколько этот признак стойкий, сказать пока нельзя из-за недостаточ-
ного материала. Гудец и Лежава (Hudec, Lezhawa, 1969b, 1970) такого расширения
не отмечают.
Распространение и местообитания. Западная часть Север-
ного Кавказа и западная часть южных склонов Большого Кавказа. Обитает в мало-
подвижных круннообломочных осыпях, на склонах с кустарником и травостоем, а
также в трещинах скал.
2. Подрод PLATYTHEBA s. str.
Высота завитка примерно равна половине высоты устья или чуть больше; поверх-
ность раковины между радиальными ребрами с тонкими спиральными ребрышками,
без зернистой скульптуры. Палатальный край устья с острым углом и желобком.
Вагинальный придаток 1, половой ретрактор отсутствует.
Распространение. Западный Кавказ, не восточнее Тбилиси, приле-
гающие районы Турции, Сирия, Палестина.
2. Platytheba (Platytheba) prometheus (Boettger, 1883). Табл. XV, 147.
Boettger, 1883 : 159, t. 4, fig. 6 [Helix (Nummulina) ]; Westerlund,
1889 : 29 (Helix); Kobelt, 1890, Iconogr., N. F., IV : 13, fig. 525 (Helix}-, Rosen,
1914 : 159 [Helix (Nummulina) ]; Лихарев, Раммельмейер, 1952 : 438,
рис. 362.
Материал: 3 экз. из одного местонахождения в западной Грузии.
Раковина с завитком в виде очень низкого конуса, причем одна сторона
этого конуса может быть чуть выпуклой; раковина очень тонкостенная. Вершина
почти не выступает. Оборотов 5, плавно нарастающих, почти плоских. Последний
оборот в 1.5 раза шире предпоследнего, к устью опущен, снизу выпуклый и более
гладкий, чем сверху. Окраска светло-роговая, вокруг пупка имеется размытое свет-
лое поле. Скульптура в виде густых радиальных тонких ребер, окрашенных светлее
фона, и тончайших кожистых спиральных ребрышек, заметных в пришовной части
последнего оборота. Киль острый. Устье очень косое, овальное, угол и желобок на
палатальном крае соответствуют килю. Места прикрепления устья сближены и со-
единены тонкой мозолью, края его тонкие и острые, внутри с сильно расплывшейся
губой. Пупок умеренно перспективный. Размеры: ВР 7—8, БД 18—20, МД 16—18 мм.
Внутреннее строение неизвестно.
Распространение и местообитания. Вид распространен
местами в западной части южных склонов Большого Кавказа (Млеты, Цагери) и в
в северо-восточной Турции (Карская обл.). Редкий вид. Обитает на скалах.
5. Род OSCARBOETTGERIA Lindholm, 1927
Lindholm, 1927а : 125 (Euomphalia, sect. Oscarboettgeria); Лихарев,
Раммельмейер, 1952 : 462 (Euomphalia, sect. ' Oscarboettgeria}; Wenz,
Zilch, 1959—1960 : 688.
Раковина средних размеров, г, слегка прижатая, радиальная скульптура
в виде косых ребер. Волоски отсутствуют. Пупок умеренно перспективный.
Вагинальные придатки не сближены, впадают в вагину щелевидным отверстием.
Вторичный чехол пениса (чехол папиллы) крепится к самому концу папиллы, в ре-
зультате чего этот орган при совокуплении целиком выворачивается. Ретрактор пра-
вого омматофора проходит между' пенисом и вагиной и перекрещивается с половым
ретрактором.
Распространение. Кавказ.
Монотипический род.
Типовой вид — Helix euages Boettger, 1883.
Oscarboettgeria euages (Boettger, 1883). Табл. XV, 149; рис. 350—353.
Boettger, 1883 : 161, t. 4, fig. 2; t. 6, fig. 1 [Helix (Eulota) ]; Wester-
lund, 1889 : 154 (Helix}; Kobelt, 1890, Iconogr., N. F., IV : 14, fig. 526 [Helix
(Eulota)]; Rosen,' 1911 : 109, t. 2, fig. 8a—c (Helix gagriensis); Hesse, 1931 : 23,
t. 4, fig. 24 (Euomphalia); Лихарев, Раммельмейер, 1952 : 462, рис. 388
(Euomphalia); Hudec, Lezhawa, 1969b : 140, Abb. 31, t. 14, 33—34 [Euomp-
halia (Oscarboettgeria) euages euages, euages gagriensis].
18 А. А. Шилейко
374
III. HYGROMIIDAE
Материал: 18 экз. из 3 местонахождений на западном Кавказе.
Раковина в общем кубаревидная, с заостренным коническим или слегка
куполовидным завитком, тонкостенная, но прочная, просвечивающая. Оборотов
6—6.5, слабо выпуклых, медленно нарастающих, разделенных глубоким швом. По-
следний оборот в самом начале с ясным закругленным углом на периферии, сглажи-
вающимся к устью, снизу слабо выпуклый, к устью внезапно и энергично опущен.
Окраска белая (иногда с легким желтоватым оттенком) или коричневая, по перифе-
рии проходит светлая лента, которая у светлых раковин еще светлее фона, а у корич-
невых выделяется ясно и имеет часто желтоватый оттенок. Радиальные ребра до-
вольно грубые, промежутки между ними покрыты тонкими спиральными бороздками,
Рис. 350—353. Oscarboettgeria euages (Bttg.) из окр. Гагры, 19 IX 1966. 350 — внеш-
ний вид полового аппарата; 351 — пенис, вскрыт (схематизировано); 352 — вагина,
вскрыта; 353 — полный набор слизистых желез другой особи. (Ориг.).
которые имеют тенденцию распадаться на ряды гранул. Устье круглое или коротко-
овальное, косое, края его острые, отвернуты, внутри окаймлены тонкой губой, причем
у коричневых раковин губа имеет розовый оттенок. Места прикрепления устья слегка
сближены. Пупок умеренно перспективный, сквозь него видно не более полутора
оборотов, обычно меньше, иногда слегка прикрыт отворотом колумеллярного края.
Размеры: ВР 9—13, БД 14—16, МД 12—15.5 мм.
Внутреннее строение. Материал: 3 экз. из окр. Гагры, 19 IX 1966,
сбор и определение Г. И. Лежавы.
Нижняя часть спермовидукта и яйцевод образуют двойной изгиб. Слизистые
железы хорошо развиты, расположены мутовкой, но обычно можно заметить их кон-
грегацию в 5 пучков; за счет вторичного ветвления общее число ветвей может дости-
гать 16. Дифференциация вагинальных придатков на отделы выражена нерезко,
но концевые утолщения ясные.
Пенис короткий, резко изогнут посредине, оба колена его стянуты тонкими лен-
тами; кроме них имеется мощная пениальная мускульная лента, соединяющая че-
хол пениса с проксимальной частью эпифаллуса. Вторичный чехол пениса (чехол
папиллы) охватывает папиллу апикально, благодаря чему выдвижение этого органа
при- спаривании исключено. Семепроводящий канал на поперечном разрезе имеет
многочисленные узкие высокие продольные складки, на которых могут располагаться
складки второго порядка. Бич несколько короче эпифаллуса. Проток семеприемника
иногда образует резкие перегибы. Резервуар неправильно овальный, немного не до-
ходит до белковой железы.
STENOMPHALIA
275
Распространение и местообитания. Западная часть северных
и южных склонов Большого Кавказа. Окр. Трабзона (Турция). Обитает в кустар-
никовых зарослях.
6. Род STENOMPHALIA Lindholm, 1927
L i n d h о 1 m, 1927a : 125 {Euomphalia, sect. Micromphalia, non Ancey, 1882);
1927b : 316 {Euomphalia, sect. Stenomphalia nom. nov.); 1928 : 206 {Euomphalia, sect.
Stenomphalia); Лихарев, Раммельмейер; 1952 : 466 {Euomphalia, sect.
Stenomphalia)’, Wenz, Zilch, 1959—1960 : 688 [Euomphalia (Stenomphalia) ].
Раковина маленькая или средних размеров, кубаревидная или шаровидная,
по периферии закруглена, скульптура различная, но не грубая. У молодых особей
раковина иногда с волосками. Пупок узкий, слабо перспективный.
Вагинальные придатки не сближены, короткие, большей частью ясно виден
вздутый базальный отдел, более тонкий средний и вздутый апикальный. В основании
папиллы пениса имеется явственное кольцевое разрыхление тканей, вторичный чехол
пениса отсутствует. При совокуплении папилла не выворачивается, а выдвигается.
Ретрактор правого омматофора перекрещивается с половым ретрактором, который
имеется всегда.
Распространение. Кавказ и Закавказье с прилежащими районами
Турции и Ирана; Копетдаг, Афганистан, и Сирия. В ископаемом состоянии род из-
вестен начиная с верхнего плиоцена (Предкавказье).
Типовой вид — Helix selecta Klika, 1893.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА STENOMPHALIA
1 (6). Зернистая скульптура имеется, иногда выражена местами.
2 (3). Раковина серая, реже светло-роговая, с четкой светлой лентой на периферии
................................................. 4. S. (Н.) ravergieri.
3 (2). Раковина светло-коричневая, роговая или каштановая; светлая лепта на пери-
ферии отсутствует или слабая, с неясными границами.
4 (5). Края устья широко отвернуты, окраска интенсивно каштановая. Зернистость
чрезвычайно четкая, гранулы имеют вид коротких гребней . 2. S. (В.) maiae,
5 (4). Умеренно отвернуты колумеллярный и базальный края устья, окраска от светло-
коричневой до роговой. Зернистость выражена в разной степени, гранулы в виде
округлых бугорков ...................................3. S. (D.) pisiformis.
6 (1). Зернистая скульптура полностью отсутствует......1. S. (S.) selecta.
1. Подрод STENOMPHALIA s. str.
Раковина маленькая, почти шаровидная, желтоватая, со спиральной скульп-
турой. ’
Вагинальные придатки маленькие, умеренно вздуты при основании. Бич втрое ,
короче короткого эпифаллуса. Вагина внутри с неясными продольными складками.
Распространение. Кавказ и Малая Азия.
1. Stenomphalia (Stenomphalia) selecta (Klika, 1893). Табл. XV, 150; рис. 354—356.
Krynicki, 1837 : 52 {Helix globula nom. nudum, non Lea, 1837); Kaleni-
czenko, 1853 : 85 {H. globula); Mousson, 1863 : 319 (H. inflata, non Deshayes,
1830); К о b e 1 t, 1892, Iconogr., N. F., V : 8, fig. 732 [77. (Latonla) globula];
Klika, 1893 : 2, 4 [Helix (Theba)]; Rosen, 1911 : 100 [Helix (Latonia) globula,
var. папа Bttg.]; Lindholm, 1913c : 137 [Helix (Theba) tschorochensis]; Rosen,
1914 : 162 [Helix (Latonia) globula]; Л индго л ь м, 19226 : 359 {Theba chrysomallis);
Hesse. 1931 : 21, t. 3, fig. 17 {Euomphalia); Лихарев, Раммельмейер,
1952 : 466, рис. 391 {Euomphalia, sect. Stenomphalia).
Материал: более 100 экз. из 28 местонахождений па Кавказе и в Закавказье.
Раковина округлая, высота устья приблизительно равна высоте завитка;
тонкостенная, просвечивающая, блестящая. Оборотов 5—5.5, довольно' выпуклых,
но шов сравнительно мелкий. Последний оборот сильно вздут, в профиле округлый,
снизу выпуклый, к устью плавно и незначительно опущен, примерно в 1.5 раза шире
предпоследнего. Окраска одноцветная, желтая разных оттенков или почти бесцвет-
ная (пустые раковины быстро мутнеют и становятся белыми). Скульптура в виде очень
тонкой радиальной исчерченности и еще более тонких спиральных бороздок; нередко
присутствуют очень тонкие волоски, особенно у пе вполне сформированных раковин.
Наконец, у отдельных экземпляров иа последнем обороте имеются отдельные, очень
слабые вмятины, разбросанные по всей поверхности оборота. Устье округлое, косое,
довольно сильно вырезано предпоследним оборотом; края его простые, острые, внутри
18*
276
III. HYGROMIIDAE
с узкой, но высокой губой, которая снаружи просвечивает в виде желтой полоски-
Пупок проколовидный, в разной степени прикрыт отворотом колумеллярного края.
Размеры: ВР 7—8.5, БД 8—12, МД 7—10 мм.
Внутреннее строение. Материал: 2 экз. из Армении, берег Занга
близ оз. Севан, 24 V 1902, сбор Е. А. Елачича и А. А. Климантовича, определение
И. М. Лихарева; 2 — из окр. с. Коджори близ Тбилиси, 25 IV 1971; 2 — из Чегем-
ского ущелья на Северном Кавказе в окр. с. Лечинкай, 23 IX 1970; 4 — из Чегем-
ского ущелья в окр. с. Хуштосырт, 15 V 1970; сборы и определения наши.
Нижняя часть спермовидукта и яйцевод образуют S-образный изгиб. СЖ 4x1—2.
Вагинальные придатки небольшие, по форме напоминают кегли. Концевые вздутия
Рис. 354—356. Stenomphalia (Stenomphalia) selecta (Klika) из Чегемского ущелья,
окр. с. Хуштосырт, 15 V 1970. 354 — внешний вид полового аппарата; 355 — пенис,
вскрыт (схематизировано); 356 — вагина, вскрыта. (Ориг.).
полые, ствол придатка заполнен многочисленными поперечными складками, в связи
с чем проток придатка очень узкий и извилистый.
Пенис с тонкостенным чехлом, папилла также тонкостенная, с обширным каналом.
Бич очень короткий, конический. Эпифаллус и пенис дважды резко изогнуты и. фик-
сированы в таком положении мощными мускульными лентами, образующими вокруг
пениального чехла более или менее плотную оболочку. Семеприемник имеет тонкий
проток и мешковидный резервуар, не доходящий до белковой железы.
Распространение и местообитания. Северный Кавказ, За-
кавказье (в Талыше и Куро-Араксинской низменности отсутствует), Турция, Сирия
(Germain, 1936), северный Иран (Biggs, 1971). Встречается на открытых склонах,
обычно недалеко от воды; лесов и сухих степных участков избегает.
2. Подрод BATUMICA Schileyko, subgen. nov.
Раковина средних размеров, шаровидная, интенсивно каштановая, без спираль-
ной скульптуры, но с ясными гранулами на поверхности.
Вагинальные придатки небольшие, слабо вздуты при основании. Внутри верх-
ней части вагины имеется своеобразный клапан, состоящий из коротких валиков.
Бич втрое короче эпифаллуса.
От Других подродов рода отличается темной окраской, широко отвернутой губой
и чрезвычайно четкой зернистостью. Из признаков внутренней организации харак-
терен клапан внутри вагины.
357
Рис. 357—360. Stenomphalia (Batumica) maiae (Hudec et Lezhawa) из Батуми, 7 IV 1971.
357 — внешний вид полового аппарата (экземпляр афалличный); 358 — вагина того же
экземпляра, вскрыта (звездочкой помечена система продольных складок)',-359 — пе-
нис экземпляра из долины р. Мачахели-цкали; 360 — то же, вскрыто. (Ориг.).
278
III. HYGROMIIDAE
Распространение. Юго-западное Закавказье.
Монотипический подрод.
Типовой вид — Euomphalia maiae Hudec et Lezhawa, 1969.
2. Stenoniphalia (Batumica) maiae (Hudec et Lezhawa, 1969). Табл. IX, 87; XV, 148;
рис. 357—360.
Hudec, Lezhawa, 1969a : 45, Abb. 3, 7 [Euomphalia (Harmozica)];
1969b : 137, Abb. 30, t. V, 11 [E. (Harmozica)].
Материал: 4 экз. из 2 местонахождений в Аджарии.
Раковина с острым ширококоническим или куполовидным завитком, вы-
сота которого немного меньше высоты устья. Оборотов 6, выпуклых, плавно нараста-
ющих, разделенных глубоким швом. Последний оборот сильно вздут, округлый,
в самом начале с легким углом, немного более чем в 1.5 раза шире предпоследнего,
к устью немного опущен. Окраска темная, каштановая или коричневая; светлая
периферическая лента слабая. Радиальная скульптура в виде густой и грубой ис-
черченности; зернистость представлена удлиненными гребневидными гранулами,
ориентированными радиально. У молодых экземпляров каждая гранула несет изо-
гнутую чешуйку. Устье круглое, косое, внутри с толстой белой или чуть розоватой
губой, края устья тонкие, светлые, острые, сильно и почти равномерно отвернуты.
Не отвернут только прилежащий к стенке последнего оборота верхний край устья.
Пупок слегка прикрыт отворотом колумеллярного края. Размеры: ВР 10—16,
БД 13.5—19, МД 11—16 мм.
Внутреннее строение. Материал: 1 экз. из Аджарии, долина р.
Мачахели-цкали, в 1 км от устья, 23 VII 1973, сбор И. М. Лихарева; 2 — из Батуми,
Ботанический сад, 7 IV 1971, сбор и определения наши.
Нижняя часть спермовидукта и яйцевод дважды резко изогнуты. СЖ 2—4X2.
Вагинальные придатки почти цилиндрические. Чуть ниже основания протока се-
меприемника имеется система коротких складок, выраженных в разной степени,
которые в совокупности образуют подобие клапана. Ниже этого клапана распола-
гается пара противолежащих продольных складок, резко оборванных как снизу,
так и сверху.
Недлинный пенис согнут вдвое и фиксирован в таком положении короткими
лентами. Между семепроводом, эпифаллусом и пенисом натянута прозрачная мемб-
рана. Папилла пениса тонкостенная, разрыхление тканей в ее основании хорошо
видно на продольных срезах. Конический бич в 2—3 раза короче вздутого эпифаллуса.
Резервуар семеприемника очень массивный, тонкостенный, прилегает к верхней
части спермовидукта?
Распространение и местообитания. Вид обнаружен пока
в двух точках, расположенных неподалеку друг от друга: в Батумском ботаническом
саду и в устье р. Мачахели-цкали. Обитает среди влажной субтропической расти-
тельности.
3. Подрод DIPLOBURSA Schileyko, 1968
Ш и л е й к о, 1968а : 344 [Euomphalia (Diplobursa) ].
Раковина средних размеров, кубаревидная, коричневатых тонов, без спиральных
бороздок, с выраженной в различной степени зернистостью.
Основания вагинальных придатков очень сильно вздуты, отделены от апикаль-
ного утолщения резкой перетяжкой. Внутренняя поверхность вагины с неясными
складками. Бич в 1.5—2 раза короче эпифаллуса.
Распространение. Кавказ, Закавказье, северный Иран, северо-восточ-
ная Турция, частично Копетдаг и Афганистан.
Монотипический подрод.
Типовой вид — Helix pisiformis L. Pfeiffer, 1852.
3. Stenoniphalia (Diplobursa) pisiformis (L. Pfeiffer, 1852). Табл. XVI, 151;
рис. 361—363.
L. Pfeiffer, 1852 : 131 {Helix); Westerlund, 1889 : 68, 69 {Helix);
Mousson, 1873 : 197, t. 7, fig. 4 {Helix arpatschaiana), 197, t. 7, fig. 5 {H. pseudog-
lobula);. Boettger, 1883 : 162, t. 5, fig. 6a—d {Helix flavollmbata); К о b e 1 t,
1892, Iconogr., N. F., V : 7, fig. 731, 9; fig. 734 [Helix (Latonia) ]; R о s e n, 1914 : 162
[Helix (Latonia) Flavollmbata, H. (Latonia) pisiformis]; Hesse, 1931 : 21, t. 3,
fig. 19 {Euomphalia); Лихарев, Раммельмейер, 1952 : 463, рис. 389
(E. pisiformis, E. arpatschaiana), 465 {E. assadovi); Акр амовский, Азарян,
1964 : 69—75 {Euomphalia); HI и л e й к о, 1968 : 344, рис. 6А [Euomphalia (Diplo-
bursa)].
STENOMPHALIA
279
Материал: более 100 экз. из 14 местонахождений в различных районах Кавказа.
Форма раковины подвержена значительной изменчивости; в общем она куба-
ревидная, но нередко бывает прижатой или почти шаровидной. Завиток заостренный,
слегка куполовидный. Оборотов 5—5.5, довольно выпуклых, плавно нарастающих,,
разделенных глубоким швом. Последний оборот сильно вздут, в профиле округлый,
в 1.5—2 раза шире предпоследнего, к устью нерезко опущен. Окраска от светло- до.
темно-коричневой, реже серовато-роговая; светлая лента на периферии очень слабая,
иногда не выражена совсем. Скульптура в виде тонкой неравномерной радиальной ис-
черчеиности и зернистости, выраженной в различной степени — от очень четкой, видной
Рис. 361—363. Stenomphalia (Diplobursa) pisiformis (L. Pfr.) из Дарьяльского ущелья,
•окр. с. Чми, 8 V 1970. 361 — внешний вид полового аппарата; 362 — пенис, вскрыт;
363 — вагинальный придаток, вскрыт. (Ориг.).
иногда невооруженным глазом, до слабо заметной. Нередко совокупность гранул дает
впечатление радиальной струйчатости. Раковина молодых экземпляров покрыта тон-
кими мелкими волосками; с возрастом они обычно опадают, но изредка остаются по-
жизненно. Устье круглое или коротко-овальное, косое, края его острые, отвернутые,
тонкие. Внутри устья возле края располагается тонкая светлая губа. Пупок может
быть открытым или прикрытым колумеллярным краем почти наполовину. Размеры:
ВР 5-12, БД 8-17, МД 7-14 мм.
Внутреннее строение. Материал: 6 экз. из Талыша, • Зуванд, окр.
•с. Космальян, 4 VII 1964; 4 — из Дарьяльского ущелья, окр. с. Чми, 8 V 1970; 2 — из
Дагестана, окр. с. Тлярата, 14 VII 1967; сборы й определения наши; 2 — из окр. г.Шуши
Нагорно-Карабахской АО, VIII 1964, сбор Г.' М. Овчинникова, определение наше;
.2 — из Армении, Ноемберянский р-н, окр. с. Айрум, 8 IX 1964; 2 — из Армении, Ме-
гринский р-н, окр. с. Личк, 14 VIII 1947; сборы и определения Н. Н. Акрамовского.
Спермовидукт и яйцевод почти прямые. СЖ 3X2—3. Разделение вагинальных при-
датков выражено резче, чем у любого другого вида: ствол придатков резко вздут, кон-
цевые утолщения отделены ясной шейкой. Вокруг выходных отверстий придатков
имеются кольцевые валики, вероятно играющие роль сфинктеров.
Пенис резко изогнут и фиксирован в таком положении хорошо развитыми мускуль-
ными лентами. Чехол пениса очень тонкий, полупрозрачный, контуры папиллы видны
без вскрытия чехла. Папилла пениса тонкостенная, просвет ее обширный, края выход-
ного отверстия гофрированы. Бич ненамного короче эпифаллуса. Проток семеприем-
ника прямой или волнисто изогнут, резервуар прилегает к верхней части спермови-
дукта.
280 in. HYGROMIIDAE
Распространение и местообитания. Вид широко распростра-
нен по всему Кавказскому перешейку, поднимаясь в горы до 2000 м над ур. м. Встре-
чается также в северном Иране и северо-восточной Турции (Germain, 1936).
По всей вероятности, предположение Лихарева и Старобогатова (1967) о том, что
данный вид должен быть сведен в синонимы к Helix bactriana Hutton, 1850, справед-
ливо. Мы воздерживаемся от этого шага потому, что материал по Н. bactriana не ис-
следован достаточно подробно. В этом случае ареал вида расширяется за счет обшир-
ных территорий южного и восточного Копетддга и предгорий.
Эвритопный вид, встречается по опушкам лиственных лесов и в редколесье, в за-
рослях кустарников. В открытых ландшафтах держится под камнями и в осыпях.
4. Подрод HARMOZICA Lindholm, 1927
Lindholm, 1927а : 125 (Euomphalia, sect. Harmozica); Лихарев, Рам-
мельмейер, 1952 : 463 (Euomphalia, sect. Harmozica).
Раковина средних размеров, кубаревидная, серая, с четкой белой лентой на пери-
ферии. Спиральные бороздки отсутствуют, зернистость выражена ясно.
Основания вагинальных придатков не вздуты, но все три отдела придатков выра-
жены. Внутренняя поверхность вагины с четкими складками. Бич и эпифаллус при-
мерно одной длины.
Распространение. Кавказ.
Монотипический подрод.
Типовой вид — Helix ravergieri Ferussac, 1835.
4. Stenomphalia (Harmozica) ravergieri (Ferussac, 1835). Табл. XVI, 152, 153;
рис. 364—367.
Krynicki, 1833 : 431 (Helix limbata, non Draparnaud, 1805); Ferussac,
1835, Bull, zool., sect. 2 : 21 (Helix ravergiensis nom. err.); Krynicki, 1836 : 182
(H. ravergii); L. Pfeiffer, 1845, Proc. Zool. Soc. London, 13 : 131 (H. caucasica);
Mousson, 1876 : 140 (H. ravergieri var. transcaucasica); Kobelt, 1877, Iconogr.,
V : 29, fig. 1220—1222 (H. ravergii); Westerlund, 1889 : 107 (Helix); Rosen,
1911 : 103 (H. ravergieri et var. transcaucasia); 1914 : 166 (H. ravergieri et var. transcau-
casia, f. transcaucasica); R e t о w s k i, 1914:287 [Я. (Theba) ravergieri et var. transcau-
casia]; Hesse, 1931 : 22, t. 3, fig. 20 (Euomphalia); Лихарев, Раммельмей-
e p, 1952 : 464, рис. 390 (Euomphalia); Стеклов, 1966 : 223 (Euomphalia);
Hudec, Lezhawa, 1967b : 96, Abb. 13, t. V, 11 [Euomphalia (Haromozlca)];
Шилейко, 1968a : 344 (Euomphalia); Hudec, Lezhawa, 1969b : 136,
Abb. 29, t. XIII, 32 [Euomphalia (Harmozica) ravergieri transcaucasica].
Материал: более 50 экз. из 13 местонахождений на Кавказе.
Раковина в общем кубаревидная, но относительная высота завитка варьи-
рует. Вершина слегка заостренная, контур завитка большей частью конический. Обо-
ротов 6—7, в разной степени выпуклых, плавно нарастающих, разделенных неглубо-
ким, местами неровным швом. Последний оборот вздут, в профиле округлый, не более
чем в 1.5 раза шире предпоследнего, к устью плавно, без резкого перегиба, опущен.
Окраска характерная: на светло-сером или, реже, на светло-роговом фоне белые спи-
ральные ленты: узкая, с четкими границами — чуть выше периферии и широкая,
размытая — у шва, и видна на более ранних оборотах. Первая из лент очень постоян-
ная, вторая изредка отсутствует. Скульптура в виде тонкой радиальной исчерченности^
местами со светлыми складками или морщинами, которые у шва выражены резче;
кроме того, имеется четкая зернистость и у отдельных раковин местами — вялые спи-
ральные бороздки. На раковинах молоди иногда имеются редкие тонкие волоски.
Устье округлое, косое, внутри с узкой, но высокой губой, края его’острые, отвернуты,
места прикрепления устья несколько сближены. Пупок узкий, не более чем наполовину
закрыт отворотом колумеллярного края, может быть открытым полностью. Размеры:
ВР 8—13, БД 12—18, МД 10—15.5 мм.
Внутреннее строение. Материал: 5 экз. из окр. с. Черная Речка, близ-
Нальчика, 5 V 1963, сбор Л. В. Шилейко; 6 — из окр. с. Ардон в Северной Осетии,
19 IX 1970, сбор и определения наши.
Дистальная часть спермовидукта и яйцевод прямые или образуют пологий изгиб.
СЖ 4X2. Вагинальные придатки развиты в различной степени, иногда скручены
(рис. 367). Протоки придатков очень узкие, извилистые, места их выхода в полость-
вагины окружены кольцевыми валиками.
Пенис короткий, чехол его тонкий, полупрозрачный. Папилла пениса также тонко-
стенная, внутренняя поверхность семепроводящего канала снабжена очень высокими
продольными складками, причем на поперечных срезах видно, что некоторые из них
расщеплены вдоль (рис. 365). Бич и эцифаллус длинные, изогнутые или слегка скру-
ченные. Пенис связан с эпифаллусом очень мощными мускульными лентами. Семепри-
MONACHA
281
емник состоит из тонкого протока и мешковидного резервуара, доходящего до ниж-
него края белковой' железы.
Распространение и местообитания. Вид широко, но споради-
чески распространен по Северному Кавказу, территории Дагестана и Закавказью.
Рис. 364—367. Stenomphalia (Harmozica) ravergieri (Fer.) из окр.
с. Ардбн в Сев. Осетии, 19 IX 1970. 364 — внешний вид поло-
вого аппарата; 365 — пенис, вскрыт; 366 — вагина, вскрыта;
367 — вагинальный придаток другого экземпляра. (Ориг.).
Населяет опушки лиственных лесов, заросли кустарников и открытые ландшафты.
Иногда поднимается на стебли трав. Известен из верхнеплиоценовых отложений Пред-
кавказья.
7. Род MONACHA Fitzinger, 1833
Fitzinger, 1833 : 95; Kobelt, 1871 : 11 (Carthusiana)-, Lindholm,
1927a : 126; Hesse, 1931 : 32 (Theba non Risso, 1926); Лихарев, Раммель-
мейер, 1952 : 431 (Theba)-, W e n z, Zilch, 1959-1960:673.
Раковина средних размеров, от кубаревидной и даже почти шаровидной до силь-
но прижатой, в профиле плавно закруглена, скульптура различная, нередко с вмяти-
282
III. HYGROMIIDAE
нами на последнем обороте. На поверхности раковины могут присутствовать волоски.
Пупок очень узкий, слабо перспективный.
Вагинальный придаток один или придатки отсутствуют; слизистые железы и по-
ловой ретрактор также могут отсутствовать. Вторичного чехла пениса нет, но имеется
единая интрапапиллярная полость, заходящая также в стенки пениального чехла.
Папилла пениса при совокуплении не выдвигается, а выворачивается. Ретрактор
правого омматофора образует перехлест с половым ретрактором, но лежит в стороне
от гениталий.
Распространение. Средиземноморье, Европа и Кавказ. В ископаемом
состоянии род.известен из верхнего миоцена (Предкавказье), плиоцена и плейстоцена
(Европа), а также из четвертичных отложений (Европа).
Типовой вид — Helix carthusiana Muller, 1774.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕПТНЫХ ВИДОВ 'И ПОДВИДОВ
РОДА МОП АС НА
1 (10). На последнем обороте имеются более или менее ясно выраженные бессистемно
ориентированные вмятины.
2 (3). Края устья красновато-бурые..................5. М. (М.) carthusiana.
3 (2). Края устья белые или светло-роговые.
4 (7). Контуры завитка слегка вогнутые, высота его заметно меньше высоты устья.
Вмятины на последнем обороте редкие и слабые.
5 (6). Раковина без волосков и следов от них..............................
.............................. 8. М. (В.) subcarthusiana subcarthusiana.
6 5). Раковина покрыта очень мелкими волосками или рубцами от них.........
.............................. 8а. М. (В.) subcarthusiana scrobiculosa.
7 (4). Завиток с прямыми или немного выпуклыми контурами, высота его примерно
равна высоте устья или незначительно меньше ее. Вмятины на последнем обо-
роте выражены ясно.
8 (9). Большой диаметр раковины не менее 14 мм.......... 1. М. (Р.) fruticola.
9 (8). Большой диаметр раковины не более 13 мм......2. М. (Р.) caucasicola.
10 (1). Вмятины на последнем обороте отсутствуют.
11 (12). Раковина покрыта очень тонкими и короткими волосками; если волоски утра-
чены, остаются многочисленные мелкие рубцы. Пупок проколовидный, не менее
чем наполовину прикрыт отворотом колумеллярного края устья ..............
.............................................. 6. М. (М.) samsunensis.
12 (11). Поверхность раковины без волосков и без рубцов от них. Пупок узкий, но
не проколовидный, слегка прикрыт отворотом колумеллярного края устья
................................................ 3. М. (Р.) talyschana.
1. Подрод PARATHEBA Hesse, 1914
Hesse, 1914 : 268 [Theba (Paratheba)]; W е n z, Zilch, 1959—1960:674.
Раковина от слегка прижатой до шаровидной, окраска от светло-серой до каштано-
вой. Зернистая скульптура отсутствует или очень слабая.
Имеется один вагинальный придаток; слизистые железы и половой ретрактор
присутствуют.
Распространение. Крым, Кавказ, Малая Азия.
Типовой вид — Helix fruticf'la Krynicki, 1833.
1. Monacha (Paratheba) fruticola (Krynicki, 1833). Табл. XVI, 154; рис. 368—369.
Krynicki, 1833 : 429 (Helix); Westerlund, 1889 : 76 (Helix); П у з a-
нов, 1925a : 64, фиг. 1, 7; Hesse, 1931 : 41, t. 6, fig. 54 (Theba); Лихарев,
Раммельмейер, 1952 : 433, рис. 356 [Theba (Theba)]; Hudec, 1972a : 90,
Abb. 31, 36.
Материал: более 100 экз. из 8 местонахождений в Крыму и в окрестностях Одессы.
Раковина от конической до шаровидной, с коническим или слабо куполо-
видным завитком и не сглаженной вершиной. Оборотов 6.5—7, плавно нарастающих.
Последний оборот вздут, в 1.5 раза шире предпоследнего, к устью почти не опущен.
Окраска светло-роговая, желтоватая или белая, монотонная^ Поверхность верхних
оборотов тонко радиально исчерчена, на последнем обороте местами имеются резкие
морщины и многочисленные по-разному ориентированные вмятины («удары молотка»).
Устье круглое или широкоовальное, косое; края его слегка отвернуты, внутри с тон-
кой светлой губой, просвечивающей сквозь стенку раковины. Пупок почти неперспек-
тивный: сквозь него видна незначительная часть предпоследнего оборота. Размеры:
ВР 12—15, БД 14—20, МД 12—17 мм.
MONАСНА
383
Внутреннее строение. Материал: 8 экз. из Одессы, VIII 1959; 10 —
из Крыма, окр. с. Краснолесье близ Симферополя, 3 VIII 1962; сборы и определения
наши.
Спермовидукт и яйцевод не образуют резких изгибов. СЖ 2—3X1—3. Вагиналь-
ный придаток внешне сильно напоминает одну из ветвей слизистых желез, располага-
ется непосредственно под ними.
Рис. 368—369. Monacha (Paratheba) fruticola (Кгуп.) из окр.
с. Краснолесье близ Симферополя, 3 VIII 1962. 368 — внеш-
ний вид полового аппарата; 369 — пенис, вскрыт. (Ориг.).
Пенис сложен двое, оба колена его стянуты тонкими лентами. Дистальная часть
семепроводящего канала в пределах папиллы — в том месте, куда не доходит коль-
цевая полость, — сложной формы: продольные складки развиты настолько интенсивно,
что общая площадь просвета незначительна. Бич тонкий, червеобразный, извитой,
несколько длиннее толстого, почти прямого эпифаллуса. Половой ретрактор
тонкий и длинный. Граница между протоком и резервуаром семеприемника очень
неясная, проток довольно толстый, резервуар немного не доходит до белковой железы.
Распространение и местообитания. Крым и Малая Азия;
завезен в Одессу. Населяет иногда открытые участки, но чаще встречается в зарослях
кустарников и широколиственных песах.
2. Monacha (Paratheba) caucasicola (Lindholm, 1913). Табл. XVI, 155\ рис. 370, 371.
Lindholm, 1913c : 138 [Helix (Theba)}-, Лихарев, Раммельмейер,
1952 : 457 [Trichia (?)].
384
III. HYGROMIIDAE
Материал: более 60 экз. из 2 местонахождений на Северном Кавказе.
Раковина от слегка прижатой до почти шаровидной, с заостренным кони-
ческим завитком, тонкостенная, но не хрупкая, просвечивающая, слабо блестящая.
Оборотов около 5, довольно выпуклых, плавно нарастающих. Последний оборот вздут,
в профиле плавно и широко закруглен, к устью слабо опущен, примерно в 1.5 раза
шире предпоследнего. Окраска одноцветная, от серовато-желтой до почти белой. Скуль-
птура в виде тонкой неясной радиальной исчерченности, на последнем обороте имеются
Рис. 370—371. Monacha (Paratheba') caucasicola (Lindh.) из Чегемского ущелья,
18 V 1970. 370 — внешний вид полового аппарата; 371 — пенис, вскрыт. (Ориг.).
многочисленные мелкие вмятины. Спиральные бороздки отсутствуют или выражены
у шва в виде следов. Нередко поверхность раковины несет очень тонкие мелкие во-
лоски. Устье почти круглое, умеренно косое, края его тонкие и острые, нижний край
отвернут. Отступя от края устья расположена выраженная в различной степени свет-
лая губа, просвечивающая сквозь стенку раковины в виде желтоватой полосы. Пу-
пок узкий, но открытый или почти открытый. Размеры: ВР 7—9, БД 9—13, МД 7.5—
11 мм.
Внутренне е строение. Материал: 12 экз. из окр. Эльбруса, 19 IV 1965,
сбор М. Акбаева; 2 — из Чегемского ущелья, 18 V 1970; сбор и определения наши.
Спермовидукт и яйцевод ясных изгибов не образуют. СЖ 2X2; расщепление сли-
зистых желез наблюдается, выше их середины. Вагинальный придаток четко подразде-
ляется на сильно вздутую базальную часть и концевое вздутие, между которыми име-
ется тонкая шейка.
Пенис не длиннее половины эпифаллуса, дистальная часть пениального чехла
соединена с эпифаллусом тонкой лентой, которая иногда распадается на несколько еще
более тонких лент. Просвет семепроводящегб канала в пределах папиллы имеет слож-
ную форму, причем площадь его крайне мала (рис. 371). Вич немного длиннее половины
MONACHA
285
почти прямого цилиндрического эпифаллуса. Половой ретрактор очень тонкий и длин-
ный. Семеприемник состоит из довольно толстого протока и резервуара молотообраз-
ной формы, не касающегося белковой железы.
Распространение и местообитания. Центральная часть Глав-
ного Кавказского хр. и северные его склоны. Встречается на альпийских и субальпий-
ских лугах, в зарослях кустарников. Во время засухи закапывается в почву. Ниже
500 м над ур. м. не спускается.
Рис. 372—373. Monacha (Paratheba) talyschana (Mart.)
из окр. Ленкорани, 12 VI 1964. 372 — внешний вид полового
аппарата; 373 — пенис, вскрыт. (Ориг.).
3. Monacha (Paratheba) talyschana (Martens, 1880). Табл. XVI, 156; рис. 372, 373.
Martens, 1880 : 154 (Helix); 1881 : 7, t. 3, fig. 4—7 (H. Talischana); Rosen,
1914 : 163 (Helix (Latonia) ]; Ш и л e ft к о, 1968a: 341, рис. 5Б, 6Б [Theba ( P aratheba)
longlflagellata]; 343, рис. 5B, 6B; 343, рис. 5Г, 6Г [Theba (Paratheba) maxima].
Материал: более 80 экз. из 32 местонахождений в Талыше и Ленкоранской низмен-
ности.
Раковина от слегка прижатой до шаровидной, с куполовидными очертани-
ями завитка. Оборотов 6, разделенных довольно глубоким швом. Последний оборот
в профиле широко и правильно закруглен, к устью сильно, но плавно опущен. Окраска
роговая разных оттенков — от светлого, желтоватого, до интенсивно коричневато-ро-
гового цвета; периферическая светлая лента большей частью заметна. Радиальная скуль-
птура — от тонкой исчерченности до грубой морщинистости; нередко имеется также
тонкая зернистость, выраженная местами. Устье в разной степени оттянуто вправо,
округлое или овальное. Край устья тонкий, коричневато-белый, слегка отвернутый,
с ясной белой губой, которая видна снаружи в виде желтоватой полоски. Пупок проко-
ловидный, в разной степени прикрыт отворотом колумеллярного края. Размеры:
ВР 7.5—17, БД 11—17.5, МД 9-15 мм.
Размеры, окраска, скульптура и форма раковины сильно варьируют, что дало
нам повод К описанию еще двух видов (см. синонимику); однако более поздний просмотр
материалов привел нас к убеждению, что характер этой изменчивости не дает оснований
к дроблению видов и связан с условиями обитания в данном биотопе.
286
III. HYGROMIIDAE
Внутреннее строение. Материал:' 55 экз. из различных районов Та-
лыша и Ленкоранской низменности; VI—VII 1964, III—IV 1965, IX 1966 гг.; сборы
и определения наши.
Нижняя часть спермовидукта и яйцевод прямые. СЖ 2—3X2—3. Вагинальный
придаток конический, иногда сложен вдвое, с продольными складками внутри.'
Пенис наружно не отграничен от эпифаллуса, пениальные мускульные ленты очень,
слабые или отсутствуют. Папилла пениса различной формы, чаще всего в дистальной
части немного утолщена. Кольцевая полость умеренно развита. На границе пениса
и эпифаллуса или немного выше может иметь место перегиб, или эпифаллус прямой,
немного длиннее бича. Ретрактор пениса умеренно длинный. Проток семеприемника
почти прямой; резервуар может примыкать как к нижней, так и к верхней его части,
но никогда не касается белковой железы.
Распространение и местообитания. Ленкоранская низмен-
ность и зона горных лесов Талыша. Встречается почти во всех типах лесных биотопов;
иногда на лесных опушках и изредка — на открытых склонах под камнями. Выше
зоны лесов не поднимается.
2. Подрод MONACHA s. str.
Раковина прижатая, белая. Зернистая скульптура отсутствует; имеются либо рубцы
от волосков, либо многочисленные вмятины на последнем обороте.
Имеются слизистые железы и вагинальный придаток; половой ретрактор отсут-
ствует.
Распространение. Северная Африка; Южная, Юго-Восточная, частично
Средняя Европа; Малая Азия; Кавказ. В ископаемом состоянии представители рода
известны из верхнеолигоценовых слоев (Европа), верхнего миоцена (Предкавказье),
плиоцена и плейстоцена (Европа).
f 4. Monacha (Monacha) externa Steklov, 1966. Табл. XVI, 157.
С т e к л о в, 1966 : 222, табл. XI, 201.
Материал: 12 экз. из 4 местонахождений в Предкавказье.
Раковина слегка прижатая, тонкостенная, хрупкая, блестящая, с кони-
ческим контуром завитка, высота которого немного меньше высоты устья. Оборотов
5.5—6, слабо выпуклых, медленно и плавно нарастающих. Последний оборот в профиле
немного овальный, но без угла, не более чем в 1.5 раза шире предпоследнего, к устью
если и опускается, то совсем немного (ни одного экземпляра с целым устьем не найдено).
Окраска одноцветная, белая. Скульптура нерезкая, в виде радиальной, слегка косой
исчерченности, выраженной более ясно у шва, и многочисленных более или менее
продольно ориентированных рубцов — следов от волосков. Устье округлое, колу-
меллярный край полностью прикрывает пупок, оставляя от него узкую щель. Размеры:
ВР 8—10, БД 11—13, МД 9—10 мм.
Распространение. Верхний миоцен Предкавказья.
5. Monacha (Monacha) carthusiana (Muller, 1774). Табл. XVI, 158; рис. 374—376.
Muller, 1774 : 15 .(Helix, non II. carthusiana Draparnaud, 1801); RossmSs-
s 1 e r, 1837, Iconogr., I : 1, fig. 364; Westerlund, 1889 : 15 (Helix); Лиха-
рев, Раммельмейер, 1952:432, рис. 357 [Theba (Theba)]; Grossu,
1955 : 437, fig. 246 (Theba); Ehrmann, 1956 : 135, t. 5, fig. 70 (Theba); Lozek,
1956 : 215, t. 24, fig. 2; Z i 1 c h, J aeckel, 1962 : 176 [Monacha (Theba)]; Lozek,
1964 : 291, Abb. 65; Gittenberger et al., 1970 : 99, fig. 166, 172, Kaart 68;
Hudec, Vazatko, 1971 : 29, Abb. 16, 17.
Материал: более 80 экз. из 17 местонахождений на юге и юго-западе европейской
части СССР, Болгарии и Венгрии. .
Раковина с очень низким, но правильно коническим завитком (очертания его
иногда даже явственно вогнутые), высота которого меньше высоты устья. Оборотов
5.5—6, слабо выпуклых. Последний оборот вздут, почти вдвое шире предпоследнего,
округлый, к устью плавно опущен. Окраска одноцветная, от молочно-белой до желто-
ватой, иногда ‘светло-серая, края устья красновато-бурые. Скульптура, особенно на
последнем обороте, решетчатая, состоит из множества легких вмятин. Кроме того,
имеется тонкая неясная радиальная исчерченность. Устье косое, широкоовальное,
края его острые, слегка отвернуты, кроме верхнего. Несколько отступя от края лежит
оплывшая толстая губа, видная снаружи в виде светлой полосы. Пупок не более чем
наполовину прикрыт отворотом колумеллярного края, нередко почти открытый.
Размеры: ВР 6—10, БД 10—16, МД 8—13 мм.
Изредка встречаются популяции, состоящие из карликовых особей (Helix praerupta
Westerlund, 1889 : 85) : ВР 4—5, БД. 6—8 мм.
MONACHA
287
Внутреннее строение. Материал: 1 экз. из Болгарии, окр. г. Кюстан-
дил, 26 VI 1969, сбор и определение И. М. Лихарева; 3 — из Болгарии, окр. г. Варна,
17 VIII 1950, сбор А. Риделя; 3 — из Венгрии, окр. г. Печ, VII 1970; 2 — из Венгрии,
с. Тихань на оз. Балатон, 22 VII 1970, сборы Д. А. Сахарова; 7 — из Севастополя,
Черная Речка, 20 VII 1962; сбор и определения наши.
На границе яйцевода и спермовидукта имеется более или менее ясный изгиб.
СЖ 3—4X2—4. Вагинальный придаток отходит почти у клоаки, четко разделен па
вздутый дистальный и тонкий проксимальный отделы; концевое вздутие выражено
слабо или отсутствует.
Рис. 374—376. Monacha (Monacha) carthusiana (Mull.) из Болгарии, гор. Кюстандил,'
26 VI 1969. 374 — внешний вид полового аппарата; 375 — пенис, вскрыт; 376 — ва-
гина и вагинальный придаток, вскрыты. (Ориг.).
Пенис сильно вздут, массивный, удлиненно-овальной формы. Интрапапиллярная
полость хорошо развита, но в стенки чехла пениса не заходит; в стенки папиллы захо-
дит почти до самого конца. Просвет семепроводящего канала в пределах-папиллы слож-
ной формы, напоминает многозубцовую фрезу. Бич — от х/2 до 1/3 длины прямого
цилиндрического эпифаллуса. Между пенисом, эпифаллусом и семепроводом натянута
тонкая прозрачная мембрана. Проток семеприемника короткий, резервуар овальный,
прилегает к спермовидукту на уровне верхней и средней трети его или ниже. У июль-
ских экземпляров из Крыма обнаружены в резервуаре семеприемника сперматофоры.
Распространение и местообитания. Средиземноморье, Юж-
ная Европа с южной Украиной, Крым, Новороссийск. Обитает на открытых степных
участках, нередко образует скопления, иногда встречается вместе с видами рода
Xeropicta в составе гроздей на стеблях трав. В ископаемом состояния встречается на-
чиная с нижнего плиоцена (Европа).
3. Подрод МЕТАТНЕВА Hesse, 1914
Hesse, 1914 : 268, [Theba (Metatheba) ]; Лихарев, Раммельмейер,
1952 : 433 [Theba (Metatheba)]; W e n z, Zilch, 1959 — 1960:675.
Раковина слегка прижатая, светло-роговая. Зернистая скульптура отсутствует;
имеются тонкие короткие волоски, если они утрачиваются остаются мелкие рубцы.
288
III. HYGROMIIDAE
Вагинальные придатки отсутствуют, слизистые железы и половой ретрактор име-
ются.
Распространение. Западный Кавказ, северная часть Малой Азии.
Типовой вид — Helix samsunensis L. Pfeiffer, 1868..
6. Monacha (Metatheba) samsunensis (L. Pfeiffer, 1868). Табл. XVI, 159; рис. 377,
378.
Рис. 377—378. Monacha (Metatheba) samsunensis (L. Pfr.)
из окр. Гагры, 14 IX 1966. 377 — внешний вид полового
аппарата; 378 — пенис, вскрыт. (Ориг.).
М о u s s on, 1863 : 318 (Helix frequens var. obscura)-, L. Pfeiffer, 1868 : 480
(Helix); Westerlund, 1889:70 (Helix), 75 (H. bifaria); Hesse, 1914:257,
fig. 7—9 (Theba), 259 (Th. samsunensis perfrequens, Th. samsunensis obscura); Reto
ski, 1914:285 [Helix (Theba) samsunensis Pfr. (=bifaria West.) et var. perfrequens
Bttg. (=frequens Bttg.)]; Hesse, 1931 : 43, t. 6, fig. 55 [Theba (Metatheba) ]; Лиха-
рев, Раммельмейер, 1952:433, рис. 358 [Theba (Metatheba)}; Hudec,
Lezhawa, 1969b : 125, Abb. 22, t. 10, 25 [Metatheba (Metatheba) ].
Материал: 12 экз. из 2 местонахождений в западной Грузии.
Раковина в общем прижатая: высота завитка всегда несколько меньше вы-
соты устья, однако общая форма раковины иногда приближается к шаровидной. Обо-
ротов 6—6.5, плавно нарастающих, слабо выпуклых. Последнийоборот сильно вздут, пла-
вно закруглен, к устью чуть опущен, примерно в 1.5 раза шире предпоследнего. Окраска
светлая, роговая, одноцветная или с намеком на светлую ленту на периферии. Скуль-
птура в виде тонкой радиальной исчерченности; чаще всего поверхность раковины по-
MONACHA
289
крыта волосками, заметными обычно только при увеличении. Когда волоски утрачены,
на поверхности видны многочисленные мелкие рубцы, выраженные в разной степени.
У некоторых раковин местами заметны очень вялые спиральные бороздки. Устье
округлое, косое, нижний край его очень слабо отвернут, колумеллярный край
возле пупка отвернут более резко и не менее чем наполовину прикрывает чрезвычайно
узкий пупок. Размеры: ВР 9—11, БД 11—15, МД 9—12.5 мм.
Внутреннее строение. Материал: 4 экз. из окр. Гагры, 14 IX 1966,
сбор и определение Г. И. Лежавы.
Спермовидукт относительно очень короткий, прямой. Слизистые железы распола-
гаются в самой верхней части вагины; каждая из двух желез состоит из 2—6 ветвей;
на рис. 377 изображен экземпляр с наиболее развитыми железами. Вагина очень длин-
ная, цилиндрическая, внутри с очень мощными и широкими продольными, слегка
винтообразными складками,
Пенис очень вздутый, форма папиллы зависит от ее функционального состояния,
просвет ее на поперечном разрезе имеет форму креста. Бич несколько длиннее корот-
кого, круто изогнутого эпифаллуса, притянутого к пенису тонкими пениальными лен-
тами. Половой ретрактор недлинный, очень мощный. Семеприемник мешковидный,
его проток короткий, толстый и прямой.
Распространение и местообитания. Черноморское побережье
Кавказа от Новороссийска до Батуми и побережье Малой Азии; в южной части ареала
вид местами отходит от морского побережья. Встречается на открытых местах, дер-
жится под камнями и на стеблях растений.
4. Подрод BOEMICA 1 Schileyko, subgen. nov.
Раковина от кубаревидной до сильно прижатой, белая, иногда с желтоватым от-
тенком. Зернистая скульптура отсутствует, на раковине могут присутствовать мелкие
волоски.
Вагинальные придатки и слизистые железы отсутствуют или находятся в явно
рудиментарном состоянии; половой ретрактор имеется.
Распространение. Черноморское побережье Кавказа и Малой Азии.
В ископаемом состоянии представители подрода известны из верхнемиоценовых от-
ложений Предкавказья.
Типовой вид Helix subcarthusiana Lindholm, 1913.
f 7. Monacha (Boemica) praeorientalis Steklov, 1966. Табл. XVII, 160,
Стеклов, 1966 : 223, табл. XI, 202—204.
Материал: 12 экз. из 2 местонахождений в Предкавказье.
Раковина почти кубаревидная, со строго коническим завитком и заострен-
ной вершиной. Оборотов 5.5, быстро и правильно нарастающих. Последний оборот
несколько вздут, округлый, чуть больше чем в 1.5 раза шире предпоследнего, к устью
заметно опущен. Окраска светлая, монотонная. Все обороты, кроме последнего, не-
равномерно и слабо радиально исчерчены; последний оборот местами с грубыми мор-
щинами, резче выраженными близ устья. Кроме того, видны следы спиральных бороз-
док (на базальной части раковины, смотреть при косом свете). В первоописании отме-
чено присутствие густых, очень маленьких рубцов от волосков; вероятно, это не рубцы,
а вторичная скульптура, обусловленная давлением песчинок на поверхность раковины,
так как эти вдавления, во-первых, имеют беспорядочное и неравномерное расположе-
ние, во-вторых, они округлые, тогда.как рубцы имеют вид коротких штрихов; наконец,
указанные вмятины неодинаковой формы. Устье слегка овальное, немного оттянуто
вправо, косое. Все края устья, кроме самой верхней части палатального, отвернуты.
Губа хорошо выражена, узкая, но довольно высокая. Пупок частично прикрыт отво-
ротом колумеллярного края. Размеры: ВР 9—10, БД 11—13, МД 8—10 мм.
Распространение. Верхний миоцен Предкавказья.
8. Monacha (Boemica) subcarthusiana subcarthusiana Lindholm, 1913. Табл. XVII,
611; рис. 379—381.
Boettger, 1884 : 149 (Helix carascaloides, non Bourguignat, 1856); Rosen,
1911 : 101 (H. carthusiana carascaloides)', Lindholm, 1913c : 139 [Helix (Theba)]
Hesse, 1914:263 (Theba orientalis roseni); R e t 0 w s k i, 1914 : 285 [Helix (Theba
orientalis var. roseni) (=H. carascaloides Bttg. et West. non Bourg.) et var. subcarthusi-
ana]; Лихарев, Раммельмейер,' 1952:434 [Theba (Metatheba) orientalis
subcarthusiana].
1 Подрод назван именем Рюрика Львовича' Бём.е.
19 А. А. Шилейко
290
HI. HYGROMIIDAE
Материал: 18 экз. из 2 местонахождений на западном Кавказе.
Раковина сильно прижатая, с коническим, слабо выдающимся завитком,
высота которого не превышает высоту устья, обычно немного больше половины вы-
соты устья. Оборотов^, умеренно выпуклых, плавно нарастающих. Последний обо-
рот сильно вздут, снизу выпуклый, к устью опущен, почти вдвое шире предпоследнего.
Окраска одноцветная, обычно белая, изредка желтоватая. Скульптура в виде тонкой
и неравномерной радиальной исчерченности и слабых редких вмятин на последнем
Рис. 379—381. Monacha (Boemica) subcarthusiana subcarthusiana
Lindh. из окр. станции Лазаревской Краснодарского края,
6 V 1965. 379 — внешний вид полового аппарата; 380 — пенис,
вскрыт; 381 — поперечный срез папиллы пениса. (Ориг.).
обороте. Кроме того, могут присутствовать вялые спиральные бороздки. Устье обшир-
ное, округлое, косое, наружный край его слегка отвернут. Губа тонкая, светлая, про-
свечивает сквозь стенку раковины в виде белой полосы. Края устья белые, иногда ры-
жеватые. Пупок узкий, но вполне открытый. Размеры: ВР 8.5—11.5, БД 13—17,
МД 11—14 мм. Лектотип: ВР 8.8, БД 13.3, МД 11.4 мм.
Внутреннее строение. Материал: 6 экз. из окр. ст. Лазаревской Крас-
нодарского края, 6 V 1965, сбор Р. Л. Бёме; 4 — из окр. Макопсе Краснодарского края,
6 IX 1971, сбор И. М. Хохуткина; определения наши.
Спермовидукт и яйцевод почти прямые. Вагина также прямая, цилиндрическая.
Придаточные органы женского отдела отсутствуют; самое большее, что удалось обна-
ружить, — у одного экземпляра небольшой пузырек и одну тонкую слизистую железу
в верхней части вагины (рис. 379). Названный пузырек представляет собой, вероятно,
рудимент вагинального придатка.
ANGIOMPHALIA
291
Пенис овальный, вздутый, в верхней и нижней его частях имеются более или менее
резкие кольцевые перехваты. Чехол пениса, особенно в дистальной части, мощный,
толстостенный. Пениальная папилла вздута, слегка уплощена в аксиальном направле-
нии или шаровидная; кольцевая полость заполнена чрезвычайно рыхлой тканью.
Просвет семепроводящего канала имеет форму многолучевой звезды. Бич несколько
длиннее эпифаллуса; пениальные ленты отсутствуют. Проток семеприемника почти
прямой, заканчивается объемистым резервуаром, прилегающим примерно к середине
спермовидукта.
Распространение и местообитания. Подвид распространен
вдоль Черноморского побережья Кавказа от южной части Краснодарского края до
Колхидской низменности; южнее замещается другим подвидом. Встречается на лугах
с сочной травянистой растительностью и на лесных опушках; днем держится под кам-
нями.
8а. Monacha (Boemica) subcarthusiana scrobiculosa Lindholm, 1922.
Линдгольм, 19226:360 (Theba); Лихарев, P а и м e л ь м e й e p,_
1952 : 434 [Theba (Metatheba) orientalis var. scrobiculosa].
Материал: 7 экз. из 1 местонахождения в Аджарии.
Раковина данного подвида отличается присутствием на верхней стороне
очень мелких волосков (особенно у молодых особей) или, когда волоски утрачены,
мелких рубцов; кроме того, раковина относительно более высокая и меньших раз-
меров: ВР 8—9.5, БД 13—13.5, МД 11 мм. Лектотип: ВР 9.2, БД 13.2, МД 11.0 мм.
Распространение и местообитания. Аджария. Населяет,
те же биотопы, что и номинативный подвид.
V. Подсемейство PAEDHOPLITINAE Schileyho, subfam. nov.
Раковина маленькая или средних размеров, от линзовидной до шаровидной.
Пупок от очень узкого до перспективного, развернутого. Окраска от почти белой
до красновато-роговой или темно-коричневой, чаще со слабой светлой лентой на пе-
риферии; темные спиральные ленты всегда отсутствуют. На базальном и париеталь-
ном краях устья могут присутствовать пластинчатые или бугорковидные зубы.
Стилофоры в той или иной степени редуцированы, видоизменены в вагинальные
придатки, часто имеющие вид небольших мешочков; придатков всегда 2. Кольце-
вые разрыхления тканей, равно как и интрапапиллярные полости, отсутствуют.
Папилла пениса обычно толстостенная, семепроводящий канал узкий, стенки папиллы
заполнены рыхлой тканью. В стенках пениального чехла обычно имеется пениаль-
ная железа.
Распространение. Тянь-Шань. В ископаемом состоянии представители
подсемейства найдены в плиоценовых отложениях Таджикистана.
1. Род ANGIOMPHALIA Schileyko, gen. nov.
Раковина линзовидная или периферия округлая; иногда форма раковины почти
шаровидная. На париетальной стенке устья зубов нет или они развиты чрезвычайно
слабо. Окраска от почти белой до темно-роговой.
Вагинальные придатки развиты в различной степени. В стенке пениального-
чехла имеется железа, которая может быть одиночной или парной. Папилла пениса
очень массивная, бич и эпифаллус длинные.
Распространение. Тянь-Шань. В ископаемом состоянии род обнаружен,
в плиоцене Таджикистана.
Типовой вид — Helix regeliana Martens, 1882.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ANGIOMPHALIA
1 (6). Периферия раковины угловатая или с килем.
2 (5). Сквозь пупок видны все или почти Все обороты.'
3 (4). Киль резко выражен вплоть до устья. Резкая губа ограничена базальным краем
и при переходе в палатальный внезаИно обрывается .... 7. A. (L.) lentina.
4 (3). Киль отсутствует, последний оборот в начале угловатый, к устью угол исчезает.
Губа слабая, расплывшаяся, выражена вдоль всего края устья или отсутствует
................................................1. A. (A.) copiosa.
5 (2). Пупок проколовидный, сквозь Яего видно не бойее половины предпоследнего
оборота ........................................8. A. (L.) buamica.
6 (1). Периферия раковины закругленная, без угла К киля.
7 (8). Раковина от прижатой до почти шаровидной. Окраска от светло-серой до
19*
293
in. HYGROMIIDAE
почти красной, co светлой лентой на периферии; радиальные цветные пятна и
полосы отсутствуют. Большой диаметр раковины не менее 14 мм..................
.......................................................... A. (A.) regeliana.
8 (7). Раковина иная.
9 (10). Раковина шаровидная, тонкостенная, покрыта тонкими волосками, сохраняю-
щимися пожизненно........................................... 3. A. (A.) exasperate.
10 (9). Раковина прижатая, твердостенная, волоски отсутствуют или имеются только
на не вполне сформированных раковинах.
11 (12). Обороты сильно уплощены......................6. A. (A.) seductilis.
12 (11). Обороты умеренно или сильно выпуклые,
13 (14). Раковина с рубцами от волосков и с довольно ясными спиральными борозд-
ками ............................... . . . . ... . . . 2. A. (A.) caelestimontana.
14 (13). Раковина без рубцов от волосков и с вялыми спиральными бороздками . . .
.....................................................5. A. (A.) almalensis.
1. Подрод ANGIOMPHALIA s. str.
Раковина слегка прижатая, кубаревидная или шаровидная. Периферия
округлая; угол если и есть, то не придает раковине линзовидной формы. Ок-
раска раковины светлая.
Вагина очень длинная, за редким исключением цилиндрическая. Придатки рас-
полагаются то в нижней, то в верхней ее части.
Рис. 382—384. Angiomphalia {Angiomphalia') copiosa sp. nov. из верховьев p. Чат--
кал, 25 V 1972. Паратип. 382 — внешний вид полового аппарата; 383 — пенис,
. вскрыт; 384— вагина другого экземпляра. (Ориг.).
Распространение. Т янь-Шань. В ископаемом состоянии подрод обнаружен
в плиоцене Таджикистана.
1. Angiomphalia (Angiomphalia) copiosa Schileyko,' sp. nov. Табл. XVII, 162;
' рис. . 382;—384. ,
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: более’ 100 экз. из 4 местонахождений на Чаткальском'хр.
ANGIOMPHALIA
293
Раковина уплощенная — от почти плоской до сильно прижатой, со слегка
куполовидным контуром завитка. Оборотов 6, слабо выпуклых, разделенных уме-
ренно глубоким швом. Последний оборот немного выше средней линии, с закруглен-
ным углом, в 1.5 раза шире предпоследнего, к устью плавно опущен. Окраска одно-
цветная, серовато- или желтовато-роговая. Скульптура в виде радиальной грубой
морщинистости и очень тонких спиральных бороздок, заметных лишь при значитель-
ном увеличении. Кроме того, на поверхности раковины имеются короткие волоски,
которые с возрастом утрачиваются. Устье широкоовальное, иногда почти круглое,
колумеллярный край заметно отвернут, базальный слабо отвернут, палатальный
в нижней части очень слабо отвернут, наверху прямой. Губа отсутствует или выражена
крайне слабо. В отличие от остальных членов подрода пупок развернутый, широкий,,
перспективный: сквозь него просматривается не менее/2 оборотов. Размеры: ВР 8.2—
10.5, БД 16.5—19, МД 13—16 мм. Голотип: ВР 10.4, БД 18.6, МД 15.7 мм.
Раковина вида почти неотличима от раковины Leueozonella ferghanica; для
определения необходимо исследовать внешний вид дистальной части женского
отдела.
Внутреннее строение. Материал: 14 экз. из верховьев р. Чаткал,
25 V 1972, сбор наш; 8 — из Чаткальского хр., 5 IX 1972, сбор Р. Я. Бутылина; оп-
ределения наши.
Спермовидукт и яйцевод почти.прямые. СЖ 4Х2. Вагинальные придатки довольно
хорошо развиты; у одних особей они имеют правильно цилиндрическую форму,
у других при основании придатков имеются более или менее ясно выраженные взду-
тия (рис, 384), внутри которых находится проток, впадающий в полость придатка.
Внутри вагинальных придатков проходят продольные складки.
Пенис чрезвычайно массивный, вздутый, в стенке его чехла залегает одиночная
железа, которая на разрезе (без гистологического исследования) выглядит как ясно
выраженное разрыхление тканей. Папилла пениса также массивная, коротко булаво-
видной или неправильной формы, толстостенная, с узким протоком. Стенки папиллы
заполнены тканью, насыщенной множеством синусов и капилляров. Эпифаллус срав-
нительно с пенисом тонкий, цилиндрический, основание его охвачено расширенным
нижним концом полового ретрактора. Бич тонкий и длинный, длина его примерно
равна суммарной длине пениса и эпифаллуса. Резервуар семеприемника неясно от-
граничен от протока, прилегает к верхней части спермовидукта.
Распространение и местообитания. Вид обнаружен на Чат-
кальском хр.; в пойме р. Чаткал является массовой формой^ Обитает в узкой полосе
прибрежных зарослей ольхи, где держится в лиственном опаде и среди скоплений
опавших сучьев; изредка встречается и в хвойных лесах (территория Афлатунского
лесхоза).
2. Angiomphalia (Angiomphalia) caelestimontana (Tzvetkov, 1940). Табл. XVII,
163, 164; рис. 385—389.
Цветков, 1940 : 390 (Cathaica); Лихарев,- ,Р а м м е.л ь м е й е р,
1952 : 382, рис. 311 (Cathaica); Матёкин, 1954 : 90 (Euomphalia); 1966 : 124,
табл. 5, рис. 4 (Euomphalia). -
Материал: более 50 экз. из 7 местонахождений на Тянь-Шане.
Раковина от. кубаревидной до в различной степени прижатой, с более или
менее явственно куполовидным контуром завитка и выступающей вершиной. Обо-
ротов 6, умеренно выпуклых. Последний оборот округлый или с едва намечающимся
углом в самом начале, причем этот угол яснее выражен у более прижатых форм.
Окраска кремовая, с одной слабой белой лентой по периферии и беспорядочно рас-
положенными светло-коричневыми или рыжеватыми размытыми радиальными по-
лосами. Эмбриональные обороты светло-коричневые, ясно выделяются на светлом
фоне раковины. -Скульптура в виде неправильной радиальной исчерченности, в ба-
зальной части выражена несколько слабее. Местами имеются вялые спиральные бо-
роздки и слабые-вмятины. Устье умеренно косое, края .-, его слабо отвёрнуты,
кроме колумеллярного, слегка прикрывающего пупок, сквозь который просматри-
вается лишь небольшая часть предпоследнего оборота’.' Размеры: ВР 8—11, БД 13—
16, МД 11—14.5 мм. '
Внутреннее строение. Материал: 5 экз. из окр. с. Кочкорка в вер-
ховьях р. Чу, 19 VI 1970, сбор Л. В. Славошевской; 3 — из южного берега оз.
Иссык-Куль южнее Каржисая, 27 VII 1972; 8 — из Заилийского хр., ущелье Боз-
гуль, 6 VI 1968; сборы К. К. Увалиевой; 3 — из ущелья р. Аламедин на Киргиз-
ском хр., 15 VI 1972; сбор и определения наши.
Дистальная часть спермовидукта и яйцевод могут быть прямыми или образо-
вывать двойной резкий изгиб;- длина яйцевода подвержена весьма значительной из-
менчивости. Вагинальные придатки развиты лучше, чем у других видов подрода,
бутылковидные, с более или менее ясно выраженным концевым утолщением, распо-
Рис. 385—389. A ngiomphalia (Angtomphalia) caelestimontana (Tzv.). 385, 386 —
из с. Кочкорка в верховьях р. Чу, 19 VI 1970; 387—389— из Заилийского хр.,
ущелье Бозгуль, 6 VI 1968. 385, 387 — внешний вид полового аппарата; 386, 388 —
пенис, вскрыт; 389 — вагина, вскрыта. (Ориг.).
ANGIOMPHALIA
295
лагаются выше середины вагины. СЖ4Х2. Внутри вагина покрыта мощными склад-
ками, резко сужающими ее просвет.
Пенис вздут, пениальная железа ясно выделяется на поверхности.чехла пениса
в виде серого овального пятна. Чехол пениса толстостенный, внутри с хорошо раз-
витыми продольными складками. Папилла массивная, веретеновидная, с узким про-
светом. Эпифаллус в различной степени извит, немного длиннее тонкого бича. Про-
ток семеприемника в большей или меньшей мере извит, резервуар прилегает к верхней
части спермовидукта, но белковой железы не касается.
Рис. 390—392. Angiomphalia (Angiomphalia) exasperata sp. nov. из окр. Бол. Алма-
тинского оз., 13 IV 1967. Паратип. 390 — внешний вид полового аппарата; 391 —
пенис, вскрыт; 392 — особь in copuH. (Ориг.).
Как видно из приведенных описаний раковины и гениталий, а также из иллю-
страций, вид сильно изменчив; однако эта изменчивость носит, по-видимому, непре-
рывный характер, что мешает нам считать этот вид комплексом видов. На рис. 385
и 387 изображены крайние случаи вариантов внешнего строения гениталий; то же
касается и фото 163 и 164 на табл. XVII.
Распространение и местообитания. Районы хребтов Тянь-
Шаня, обрамляющие оз. Иссык-Куль с юга, запада и севера; дальше всего на запад
вид проходит, видимо, по Киргизскому хр. Высокогорный вид, обитает на высотах
не менее 1500 м над ур. м.; населяет открытые склоны с развитым травянистым покро-
вом и кустарником, скальные выходы, крупнообломочные малоподвижные осыпи.
3. Angiomphalia (Angiomphalia) exasperata Schileyko et Uvalieva, sp. nov.
Табл. XVII, 165-, рис. 390—392.
Голотип хранится в коллекции Зоологического института АН СССР.
Материал: более 30 экз. из 4 местонахождений на Тянь-Шане.
Раковина от кубаревидной до шаровидной, с куполовидными контурами
завитка, высота которого примерно равна высоте устья. Оборотов 6, выпуклых. По-
следний оборот в профиле округлый, в 1.5 раза шире предпоследнего, к устью почти
не опущен. Окраска светло-роговая, желтоватая или красновато-роговая, со светлой
неясной лентой на периферии, которая прослеживается у шва на предыдущих обо-
ротах. Скульптура представлена радиальной исчерченностью или слабой морщи-
нистостью; поверхность раковины покрыта редкими тонкими короткими прямыми
золотистыми волосками. Устье округлое, умеренно косое, места его прикрепления
широко расставлены. Отступя от края устья имеется светлая широкая расплывшаяся
896
III. HYGROMIIDAE
губа. Размеры: BP 5.5—‘6.5, БД 8—11, МД 7—9.5. мм. Голотип: ВР 6.5, БД 8.7,
МД 7.8 мм.
Внутреннее .строение. Материал: 2 экз. — из окр. Большого Ал-
матинского оз., 13 IV 1967; 2 — из Заилийского хр., ущелье Бозгуль, 16 V 1968;
3 — из восточной части хр. Кунгей, 2 VII 1971; сборы К. К. Увалиевой, определения
наши.
На границе спермовидукта и очень тонкого яйцевода имеется выраженный в
в разной степени изгиб. СЖ4Х1—2. Вагинальные придатки маленькие, фасолевидной
формы, резко суженные при основании, впадают непосредственно под слизистыми
железами, внутри этих придатков имеются продольные складки. Вагина почти ци-
линдрическая, довольно тонкая и длинная.
Рис. 393—397. Angiomphalia (Angiomphalia) regeliana (Mart.) из Таласской котловины,
5 VI 1972. 393 — внешний вид полового аппарата; 394 — пенис, вскрыт; 395 — ваги-
нальный придаток, продольный разрез; 396 — вагинальный придаток другой особи
(звездочкой помечен дополнительный рудиментарный придаток); 397 — то же, про-
дольный разрез. (Ориг.).
Пенис неправильно веретеновидной, формы, на его поверхности ясно видна пиг-
ментированная парная пениальная железа в виде коричневато-серых пятен. Папилла,
пениса цилиндрическая, длинная, семепроводящий канал щелевидный. Внутренняя,
поверхность чехла пениса без резких складок. Бич слегка короче прямого или слегка,
скрученного эпифаллуса. Проток семеприемника почти прямой, небольшой овальный
резервуар иногда достигает нижнего края белковой железы.
Распространение и местообитания. Восточная часть Кир-
гизского и хребты Заилийский и Кунгей. Обитает на высотах не менее 1500 м
над ур. м., населяет альпийские и субальпийские луга, держится на хорошо увлаж-
няемых участках с высоким травостоем, под камнями и на траве.
4. Angiomphdlia (Angiomphalia) regeliana ' (Martens, 1882). Табл. XVII, 166; '
рис. 393—397.
.Martens, 1882 : 9, t. 2. fig. 4 .(Helix rubens nar. Regeliana); Лихарев,
Раммельмейер, 1952 :. 402 [Eulota (Leucozonella) rubens var. regeliana];
Матёкин, 1954 : 90 (Euomphalia rubens); 1966 : 124, табл. 5, рис. 3 (E. rubens)-. . ,
ANGIOMPHALIA
297
Материал: более 100 экз. из 14 местонахождений на Тянь-Шане.
Раковина от слегка прижатой до шаровидной, завиток имеет ясно куполо-
видные очертания. Оборотов 6, плавно нарастающих. Последний оборот в профиле
правильно округлый, к устью слабо и очень плавно опущен, менее чем 1.5 раза шире
предпоследнего. Окраска от почти белой до темно-красной; чем интенсивнее окраска
раковины, тем резче выделяется белая периферическая лента и тем яснее ее границы.
Скульптура в виде неравномерной, местами грубой радиальной исчерченности и
тонких редких спиральных бороздок, выраженных в разной степени. Устье округ-
лое, слегка косое; края устья внутри с очень невысокой расплывшейся губой, слабо
отвернуты. Пупок обычно наполовину прикрыт отворотом колумеллярного края,
сквозь него видно не более половины предпоследнего оборота, обычно еще меньше.
Размеры: ВР 11—15, БД 14—20, МД 12—17 мм.
В связи с путаницей, имеющейся в литературе, необходимо дать некоторые по-
яснения. Мартенс (1874) описал из бассейна верхнего Зеравшана по сборам
А. П. Федченко Helix rubens; в 1882 г. тот же автор описывает 4 формы этого вида:
concolor, finschiana, zeiliana и regeliana, которые отличаются формой завитка и окрас-
кой раковины. В 1896 г. Вестерлунд добавляет еще 3- формы: caryodes, limitata и ап-
gulata.
Первые сведения по морфологии гениталий этого моллюска имеются в кратком со-
общении Матёкина (1954), где констатируется, что вид Мартейса должен быть отнесен
к роду Euomphalia, причем точное местонахождение исследованных животных не
указано, но отмечено, что материал собран на Тянь-Шане. В 1955 г. Лихарев, иссле-
довав материал из Джунгарского и Киргизского хребтов, относит вид Мартенса к роду
Trichia, причем как заключение Матёкина, так и заключение Лихарева справедливы —
эти авторы имели дело с разными видами, которые конхологически почти неразли-
чимы. Сравнение с описанием и1 рисунками Мартенса заставляет полагать, что, опи-
сывая Н. rubens, автор имел дело'с представителем Trichiinae. Что касается названия
формы из группы «Euomphalia», то дело затрудняется тем, что типовые серии всех
названных форм представлены только пустыми раковинами. Однако большой мате-
риал, бывший в нашем распоряжении, показывает, что Helix regeliana, отличающийся
более прижатой раковиной, дает непрерывный ряд к красным шаровидным раковинам.
В связи с этим мы возводим названную форму в ранг. вида.
Различия между раковинами «Trichia» rubens и «Euomphalia» regeliana сводятся
к тому, что первый вид имеет более конический завиток, а второй — более куполо-
видный. Вместе с тем необходимо еще раз. подчеркнуть, что для точного определения
необходимо иметь спиртовой материал.
Внутреннее строение. Материал: 1 экз. из кишлака Канют на
хр. Петра I, 1 VII 1968, сбор и определение 3. -Иззатуллаева; 12 — из окр. с. Луго-
вое, Киргизский хр., 6 VI 1972; 9 —из ущелья Карабалты, Киргизский хр.,
11 VI 1972; 3 — из Таласской котловины близ г. Таласа, 5 VI 1972; 28 — из ущелья
р. Аламедин на Киргизском хр., 14 VI 1972; сборы и определения наши.
Спермбвидукт при переходе в яйцевод и верхняя часть вагины образуют двой-
ной изгиб. СЖ 3—4X2. Вагинальные придатки маленькие, удлиненно-овальные.
При вскрытии этих придатков нередко обнаруживаются утолщения в базальной
части, заключающие тонкие протоки, — свидетельство, что каждый придаток имеет
парное происхождение (рис. 395, 396). Вагина толстостенная, внутри нее немного
винтообразно проходят 4 чрезвычайно мощных валика и несколько более мелких
складок.
Пенис вздут, пениальная железа одиночная, слабо развита. Поверхность папиллы
пениса покрыта радиальными складками, за счет выравнивания которых длина па-
пиллы может заметно увеличиваться; выходное отверстие папиллы располагается
не апикально, но немного сбоку, что связано с асимметричным положением семе-
проводящего канала (рис. 394). По поверхности папиллы могут проходить 1—2 бо-
розды. Бич и эпифаллус умеренно извиты, примерно равной длины. Проток семепри-
емника не извит, прямой или слабо изогнут. Резервуар немного не доходит до бел-
ковой железы.
Распространение и местообитания. Вид широко распростра-
нен по Тянь-Шаню и северным отрогам Памира; населяет различные биотопы,
но лесов избегает. Чаще встречается на открытых участках с густым и высоким траво-
стоем, где обитают популяции с красноватой шаровидной раковиной. В более сухих
условиях раковина светлее и более прижатая. Близкая форма обнаружена
А. А. Никоновым в плиоценовых отложениях вдоль левого берега р. Зармас-сай на
Гиссарском хр,
5. Angiomphalia (Angiomphalia) almalensis Schileyko et Uvalieva, sp. nov.
Табл. XVII, 167; рис. 398, 399. .
Голотип хранится в коллекции Зоологического института АН СССР,
Материал: 5 экз. из 1 местонахождения в Джамбульской обл.
298
III. HYGROMIIDAE
Раковина слегка прижатая, высота куполовидного завитка немного меньше
высоты устья. Оборотов 6, равномерно нарастающих, разделенных умеренно глубоким
швом. Последний оборот на периферии очень правильно и плавно закруглен. Окраска
одноцветная, известково-белая. Скульптура в виде грубой неправильной радиальной
исчерченности и редких вялых спиральных бороздок; первые дефинитивные обороты
морщинистые, эмбриональные обороты желтоватые, гладкие. Устье немного скошено,
ширина его немного превышает высоту. Губа расположена чуть отступя от края,
в ангулярной части сходит на нет. Пупок очень узкий, проколовидный, слегка при-
крыт отворотом колумеллярного края устья. Размеры: ВР 11.5—12.5, БД 16—17,
МД 14—14.5 мм. Голотип: ВР 12.2, БД 16.8, МД 14.4 мм.
Рис. 398—399. Angiomphalia (Angiomphalia) almalensis sp. nov. из долины p. Чу
близ с. Токмак, 25 V 1969. Паратип. 398 — внешний вид полового аппарата;
399 — пенис, вскрыт. (Ориг.).
Внутреннее строение. Материал: 3 экз. из долины р. Чу близ с. Ток-
мак, 25 V 1969, сбор К. К. Увалиевой, определение наше.
Спермовидукт при переходе в яйцевод образует крутой изгиб; такой же изгиб
имеется и при переходе яйцевода в вагину. СЖ2ХЗ—4; слизистые железы распо-
лагаются возле основания семеприемника. Вагина очень длинная, внутри с хорошо
развитыми продольными валиками. Вагинальные придатки маленькие, в виде мешоч-
ков, располагаются ниже середины вагины.
Пенис очень объемистый, веретеновидный. Пениальная железа почти не раз-
вита и обнаруживается лишь местами. Папилла пениса в разной степени уплощенная,
с узким протоком; Бич несколько длиннее эпифаллуса, оба этих отдела извиты, но
эпифаллус не обвивается вокруг полового ретрактора. Проток семеприемника тон-
кий,, прямой, резервуар овальный, прилегает немного выше середины к поверхности
спермовидукта.
Вид близок к A. seductilis; отличается одноцветной раковиной, более узким пуп-
ком и отсутствием угла на периферии. Анатомические различия сводятся к тому,
что у данного вида яйцевод много длиннее, а эпифаллус не охватывает половой ретрак-
тор.
Распространение и местообитания. Пока вид известен только
из типового местообитания в долине р. Чу. Найден на открытом склоне холма.
6. Angiomphalia (Angiomphalia) seductilis (Westerlund, 1898). Табл. XVII, 168;
рис. 400—402.
Westerlund, 1898:157 [Helix (Theba)].
ANGIOMPHALIA
299
Материал: 14 экз. из 2 местонахождений на Киргизском и Заилийском хребтах.
Раковина прижатая, с широким и низким коническим или немного куполо-
видным завитком. Оборотов 6, слабо выпуклых, плавно и медленно нарастающих.
Последний оборот снизу слегка уплощен, в начале с легким углом, к устью слабо
опущен. Окраска состоит из серовато-белого или желтоватого фона, светлой раз-
мытой периферической ленты и радиальных неясных рыжеватых или светло-корич-
невых полос. Скульптура в виде резкой радиальной исчерченности и тонких частых
спиральных бороздок. Устье слегка скошено, широкоовальное, внутри белое или
рыжеватое, с белой расплывшейся губой. Края устья прямые, острые, базальный
Рис. 400—402. Angiomphalia {Angiomphalia) seductilis (West.) из Киргизского хр.,
12 VII 1972. 400 — внешний вид полового аппарата; 401 — пенис, вскрыт; 402 —
вагина, вскрыта. (Ориг.).
и колумеллярный слегка отогнуты. Пупок узкий, но немного шире, чем у предыду-
щего вида. Размеры: ВР 11—12, БД 17—20, МД 14.5—17 мм. Лектотип: ВР 11.4,
БД 18.2 мм.
Внутреннее строение. Материал: 5 экз. из Киргизского хр., близ
входа в Буамское ущелье, 12 VII 1972, сбор и определение наши.
Нижняя часть спермовидукта почти прямая; яйцевод как таковой почти отсут-
ствует, так как слизистые железы (СЖ 2X2—3) располагаются тотчас под местом
отхождения протока семеприемника. Вагина чрезвычайно длинная, верхняя часть
ее в той или иной степени изогнута. Вагинальные придатки маленькие, располага-
ются ниже середины вагины'. Продольные валики внутри вагины резкие и ши-
рокие.
Пенис сильно вздут; пениальная железа умеренно развита, одиночная. Папилла
пениса мощная, стенки ее сравнительно плотные, но имеются тем не менее продоль-
ные капилляры. Бич и эпифаллус длинные, примерно равной длины, извитые и скру-
ченные, чрезвычайно мощный половой ретрактор охвачен петлей эпифаллуса (у од-
ного из экземпляров эпифаллуСа был также скручен, но не охватывал ретрактора).
Проток семеприемника изгибов не образует, резервуар заметно не достигает белковой
железы.
Распространение и местообитания. Заилийский и вос-
точная часть Киргизского хр. Обитает в горной степи на камнях и стеблях
растений, а также в трещинах скал.
300
III. HYGROMIIDAE
2. Подрод LENTIGA Schileyko, subgen. nov.
Раковина сильно уплощенная, .линзовидная или с закругленным углом выше
средней линии. Окраска бурая или коричневая. Губа на базальном крае устья вы-
сокая и мощная.
Вагина недлинная, в различной степени вздутая в центральной части. Придатки
располагаются выше середины вагины.
От номинативного подрода данный резко отличается окраской и формой рако-,
вины,, а также присутствием губы на базальном крае устья. Анатомические отличия
заключаются в значительно более сильном развитии вагинальных придатков.
Распространение. Ферганский, Чаткальский и Киргизский хребты.
Типовой вид — Helix lentina Martens, 1885.
Рис. 403—404. Angiomphalia (Lentiga) lentina' (Mart.) из Ферганского хр., среднее
течение р. Нарын выше Токтогула, 1 VI 1972. 403 — внешний вид полового аппарата;
404 — пенис, вскрыт. (Ориг.).
7. Angiomphalia (Lentiga) lentina (Martens, 1885). Табл. XVII, 169, 170; рис..
403, 404. ,
Mart e.n s, 1885 : 18 (Helix); Lindholm, 1927c : 201 (Plectotropis mesasiana);
Лихарев, Раммельмейер, 1952 387, рис. 316 (Aegista mesasiana), 412,
рис. 343 (Paedhoplita).
Материал: более ,100 экз. из 2 местонахождений на Ферганском и Чаткальском
хребтах.
.Раковина сильно уплощена, линзовидная, высота завитка равна высоте
устья или немного меньше. Оборотов 5—6, медленно нарастающих, плоских. По-
следний оборот с острым углом; если завиток относительно высокий, то угол придает
оборотам ступенчатый облик. Последний оборот сверху плоский, снизу выпуклый,
немного шире, предпоследнего оборота, к устью чуть опущен. Окраска одноцветная —
коричневая или темно-роговая. Скульптура в виде грубой радиальной морщинисто-
сти, сверху выраженной резче, чем на нижней поверхности. Устье очень косое, в
в виде широкой щели, внутри устья располагается губа в виде резкой высокой ба-
зальной складки, резко обрывающейся справа. Пупок умеренно развернут, Перспек-
тивный. Размеры; ВР 4.5—5, БД 12—13, МД 10.8—11.6 мм.
ANGIOMPHALIA
301
Plectotropis mesasiana описан Линдгольмом по не вполне сформированным рако-
винам этого вида (фото 170 на табл. XVII).
Внутреннее строение. Материал: 18 экз. из среднего течения р. На-
рып выше Токтогула (Ферганский хр.), 1 VI 1972, сбор и определение наши.
Дистальная часть спермовидукта и яйцевод прямые. СЖ4Х2—3. Вагина притя-
нута к внутренней поверхности покровов моллюска многочисленными тонкими со-
единительнотканными тяжами. Вагинальные придатки развиты хорошо, почти ци-
линдрические, истончающиеся при основании и закругленные на концах. Внутрен-
няя поверхность вагины покрыта чрезвычайно мощными и высокими продольными
складками, так что на поперечном срезе площадь просвета вагины незначительна.
Рис.'405—408. Angiomphalia (Lentiga) buamica (В. Tzv. et E. Tzv.) из ущелья p. Чу,
VIII 1965. 405 — внешний вид полового аппарата; 406 — пенис, вскрыт; 407 — вагина,
вскрыта; 408 — вагинальный придаток, продольный разрез (звездочкой помечен ру-
диментарный дополнительный проток). (Ориг.).
Пенис очень большой, в стенке пениального чехла залегает обширная, но плохо
заметная пениальная железа. Папилла пениса занимает почти весь просвет чехла,
толстостенная, овальная или булавовидная; стенки ее рыхлые, насыщены множеством
синусов. Семепроводящий канал в разрезе имеет форму трехлучевой звезды. Бич
не более чем в 1.5 раза длиннее эпифаллуса. Длинный половой ретрактор нижним рас-
ширенным концом охватывает дистальную часть эпифаллуса. Проток семеприемника
недлинный, плавно переходит в относительно очень большой резервуар; если резер-
вуар пуст, стенки его спадаются. Семеприемник значительно не доходит до нижнего
края белковой железы.
Распространение и местообитания. Северная часть Ферган-
ского и восточные склоны Чаткальского хребтов; встречается отдельными популя-
циями. Гигрофильный вид; живет у воды в осыпях и в подстилке пойменных кустар-
никовых зарослей.
8. Angiomphalia (Lentiga)' buamica (В. Tzvetkov et E. Tzvetkova, 1943).
Табл. XVIII, 171; рис. 405—408.
Цветков, Цветкова, 1943 : 118 (Paedhoplita); Лихарев, Рам-
мельмейер, 1952 : 410 (Paedhoplita); Матёкин, 1954 : 90 (Euomphalia);
1966 : 123 (Euomphalia).
Материал: 6 экз. из 1 местонахождения в восточной части Киргизского хр.
Раковина сильно прижатая, с куполовидными контурами широко закруг-
ленного притупленного завитка. Оборотов 6—6.5, очень тесно навитых, медленно
302
III. HYGROMIIDAE
нарастающих, сверху уплощенных, разделенных узким, глубоко врезанным швом.
Последний оборот чуть шире предпоследнего, к устью не опущен, с тупым углом
несколько выше средней линии, сохраняющимся до самого устья. Окраска корич-
нево-роговая, одноцветная или с намеком на светлую ленту по углу. Скульптура
в виде грубой густой радиальной исчерченности или морщинистости, которая на верх-
ней поверхности раковины выражена сильнее, чем на нижней. Устье косое, в виде
широкой изогнутой щели с острыми краями, которые в базально-колумеллярной
области умеренно отогнуты. На заметном расстоянии от базального края устья лежит
четкая складкообразная губа, которая плавно начинается от столбика и довольно
резко (но более плавно, чем у предыдущего вида) обрывается близ периферического
угла. Снаружи губа просвечивает в виде желтоватой полоски. Нередко на париеталь-
ной стенке напротив губы имеется еще слабый наплыв, который можно обнаружить,
проведя концом тонкой иглы по париетальной стенке. В глубине раковины имеются
еще 1—3 губы, также просвечивающие сквозь стенку раковины. Пупок очень узкий,
цилиндрический. Размеры: ВР 4.5—6, БД 9—10, МД 7.4—9 мм.
Внутреннее строение. Материал: 4 экз. из ущелья р. Чу, VIII 1965,
сбор С. Л. Перешкольника, определение наше.
Дистальная часть спермовидукта и яйцевод прямые или со слабым изгибом.
СЖ2—3X2. Вагина умеренной длины; вагинальные валики достаточно четкие, раз-
делены узкими глубокими бороздами. Вагинальные придатки хорошо развиты, на
концах слегка утолщены; при основании имеются выраженные в разной степени
вздутия.
Пенис почти шаровидный, в стенке пениального чехла имеется слегка пигмен-
тированная пениальная железа, выделяющаяся серой или коричневатой окраской.
Папилла пениса относительно очень массивная, вздутая, с мощными стенками,
в которых имеется небольшое число мелких капилляров. Внутренняя поверхность
чехла пениса в дистальной части покрыта резкими складками.
Распространение и местообитания. Известен пока из типового
местонахождения — в ущельях рек Чу и Арасан в восточной части Киргизского хр.
Обитает в осыпях и под камнями.
2. Род PAEDHOPLITA Lindholm, 1927
Lindholm, 1927d : 266; Цветков, Цветкова, 1943:121; Лиха-
рев, Р а м м ел ь м е й е р, 1952 : 409; Wen z, Zilch, 1959—1960 : 646.
Раковина сильно прижатая, с ясным, но не острым углом выше средней линии.
На париетальной стенке устья имеются хорошо развитые зубы в виде пластинок или
бугорков. Окраска коричневатых тонов.
Вагинальные придатки маленькие. Пениальная железа отсутствует. Папилла
пениса относительно очень маленькая, занимает небольшую часть объема пениаль-
ного чехла. Эпифаллус недлинный, несколько короче бича.
Распространение. Тянь-Шань.
Типовой вид — Paedhoplita laminata Lindholm, 1927.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА PAEDHOPLITA
1 (2). На париетальной стенке устья имеется 1 зуб в виде поперечной складки . . .
.......................................’............. 1. Р. laminata.
2 (1). На париетальной стенке устья имеется 2—3 зуба. .
3 (4). Один из париетальных зубов округлый, бугорковидный, второй имеет вид
подкововидной пластинки, распадающейся иногда на 2 удлиненных бугорка
.....................................................2. Р. lindholmi.
4 (3). Париетальные зубы представлены двумя мощными, тесно сближенными гребне-
видными бугорками, один из которых расположен впереди другого.......
.............................................. 3. Р. kirgisensis.
1. Paedhoplita laminata Lindholm, 1927. Табл. XVIII, 172.
Lindholm, 1927d : 266; Лихарев, Раммельмейер, 1952 : 410.
Материал: 5 экз. из 2 местонахождений на Киргизском хр.
Раковина уплощенная, с низким округлым завитком, высота которого
составляет не менее половины высоты устья; может быть и равна ей. Оборотов 5—6,
ступенчатых, очень медленно нарастающих. Угол на последнем обороте очень сильно
сдвинут к шву. Окраска одноцветная, роговая. Скульптура в виде радиальной ис-
черченности, более резкой на верхней поверхности раковины. У молодых экземпля-
ров на поверхности раковины имеются мелкие единичные волоски. Устье узкое, ще-
левидное, косое, места его прикрепления широко расставлены. Губа очень высокая,
занимает базальный край устья; сверху свисает париетальный зуб в виде короткого-
PAEDHOPLITA
303
округлого гребня. При рассмотрении снизу видно, что этот зуб направлен не ради-
ально, а под некоторым углом к радиусу раковины. В процессе роста моллюска об-
разуются несколько пар подобных зубов (т. е. базальных и париетальных складок),
причем более ранние постепенно резорбируются. Пупок шире, чем у других видов
рода, вначале немного развернутый, затем цилиндрический. Размеры: ВР 3.5—4.3,
БД 6.5—9, МД 6-8 мм.
Внутреннее строение неизвестно.
Распространение и местообитания. Восточная часть Кир-
гизского хр. Обнаружен в хвойном лесу на высотах свыше 2000 м над ур. м.
Рис. 409—411. Paedhoplita llndholmi В. Tzv. et Е. Tzv. из ущелья Карабалты, Кир-
гизский хр., 11 VI 1972. 409 — внешний вид полового аппарата; 410 — пенис,
вскрыт; 411 — вагина, вскрыта. (Ориг.).
2. Paedhoplita lindholmi В. Tzvetkov et Е. Tzvetkova, 1943. Табл. XVIII, 173,174',
рис. 409—411 и 25, А, Б.
Цветков, Цветкова, 1943 : 119; Лихарев, Раммельмейер,
1952:411; М а т ё к и н, 1954:90 (Euomphalia lindcholmi nom. err.);, 1966:123
(Euomphalia).
Материал: более 100 экз. из 4 местонахождений на Киргизском хр.
Раковина в различной степени уплощенная. Оборотов 6, очень медленно
нарастающих, слабо выпуклых. Последний оборот чуть шире предпоследнего, к устью
совершенно не опущен, выше периферии с явственным в разной степени сглаженным
углом, причем иногда последний оборот стремительно нарастает в высоту, в резуль-
тате чего нижняя часть раковины становится более выпуклой, чем верхняя (фото
173 на табл. XVIII). Окраска довольно темная, коричнево-роговая, реже серовато-
роговая; по углу проходит слабая светлая лента. Устье косое, узкое, в виде изогнутой
широкой щели. На базальной стенке раковины имеется высокая губа; напротив нее,
на париетальной стенке расположены два зуба: небольшой, бугорковидный, лежит
впереди, близ столбика; другой — в виде подкововидной пластинки правее первого
(рис. 25, А, Б). Пупок по всей длине цилиндрический, узкий. Размеры: ВР 3.5—5,
БД 7—9, МД 6.2—8 мм.
Внутреннее строение. Материал: 3 экз. из окр. с. Чардовар, Мали-
новая щель, Киргизский хр., 9 VI 1972; 12 — из ущелья Карабалты, Киргизский хр.,
И VI 1972; 24 — из ущелья р. Аламедин близ г. Фрунзе, 15 VI 1972; сборы и опре-
деления наши.
304
III. HYGROMIIDAE
Яйцевод у верхней и нижней границы дважды резко изогнут. СЖ 2X1—2.
Сразу же под местом впадения слизистых желез располагаются очень маленькие
округлые вагинальные придатки. Вагина очень длинная, почти цилиндрическая.
Вагинальные валики сближены попарно, занимая латеральное положение; между
парными валиками могут присутствовать еще несколько небольших продольных
складок.
Пенис длинный, внутренняя поверхность чехла пениса покрыта геометрически
правильными гофрированными продольными складками (рис. 410). Папилла пениса
занимает от х/3 до х/2 длины чехла, сжата с боков и несет продольные бороздки.
В толще стенок папиллы, занятой рыхлой тканью, имеется разное число капилляров;
просвет папиллы в виде узкой щели. Эпифаллус очень тонкий, слегка извит. Половой
ретрактор прикрепляется не к месту перехода пениса в эпифаллус, а непосредственно
к эпифаллусу. Проток семеприемника очень тонкий, почти прямой , с маленьким ре-
зервуаром, прилегающим к верхней половине спермовидукта.
Распространение и местообитания. Северные склоны восточ-
ной части Киргизского хр. и Заилийский Алатау (найден на склонах гор Чилик и
Сийрек).
Встречается в осыпях, в сырых местах; среди пышной травянистой растительности
образует особенно плотные скопления.
3. Paedhoplita kirgisensis Licharev et Rammelmeyer, 1952. Табл. XVIII, 175.
Лихарев, Раммельмейер, 1952 : 411, рис. 342.
Материал: 2 экз. из 1 местонахождения на Киргизском хр.
Раковина сильно прижатая,, высота завитка меньше высоты устья. Обо-
ротов 5—5.5, очень медленно нарастающих; угол на последнем обороте проходит
значительно выше средней линии. Последний оборот- очень незаметно спускается
к устью. Окраска одноцветная, роговая. Скульптура в виде радиальной исчерчен-
ности, которая юверху выражена резче,;чем на базальной части. Последний и пред-
последний обороты покрыты очень мелкими волосками. Устье относительно более
широкое, чем у других видов рода; внутри него лежат 3 складки: базальная (пред-
ставляющая собой губу, которая занимает только Нижнюю, часть устья) и 2 пари-
етальные — тесно сближенные, одна позади другой, короткие, округлые, гребне-
видные. Пупок развернут умеренно, перспективный. Размеры: ВР 4, БД 8, МД 7 мм.
Внутреннее строение неизвестно.
Распространение и местообитани я.. Единственная находка
(2 пустые раковины) сделана А. Н. Кириченко в июле 1910 г. на Киргизском хр.
в верховьях р. Аламедин, на высоте 2620 м над ур. м. В 1972 г. мы были в указанном
месте, но собрать дополнительные материалы не удалось. На высоте свыше 1800 м
в этом ущелье растительность скудная, местами попадаются отдельные группы елей;
в изобилии встречаются скальные выходы и осыпи, большей частью крупнообломоч-
ные. Видимо, в этих биотопах и обитает рассмотренный вид.
VI. Подсемейство METAFRUTIGIGOLINAE Schileyko, 1972
Раковина средних размеров, с округлой периферией. Пупок неширокий, в общем
цилиндрический. Окраска белая или светло-роговая, первично с двумя темными
лентами — выше и ниже периферии. Устье без зубов.
Все придаточные органы женского отдела отсутствуют. Вагина очень короткая—
проток семеприемника отходит почти от клоаки. В стенках папиллы пениса и пениаль-
ного чехла формируются кольцевые полости; папилла от незамкнутой до замкнутой.
Распространение. Восточное Средиземноморье, Кавказ.
Род CAUCASOCRESSA Hesse, 1931
Hesse, 1931 : 28 [Fruticocampylaea f Caucasocressa) ]; Лихарев, Рам-
мельмейер, 1952 : 466 (Metafruticicola part.); W e n z, . Zilch, 1959—
—I960:,690; Шилейко, 1972г : 38; 1972д : 18.
Раковина от прижатой до сильно уплощенной, белая, с двумя четкими корич-
невыми лентами, которые могут вторично исчезать. Поверхность раковины без во-
лосков. Скульптура представлена зернистостью и спиральными бороздками, причем
у одних популяций может преобладать один тип скульптуры, у других — другой.
Чехол пениса тонкостенный, папилла полностью замкнута, с обширным семе-
проводящим каналом. Стенки дистальной части эпифаллуса и частично папиллы
содержат хорошо развитую кольцевую полость.
Распространение. Юго-западный Кавказ с прилежащими районами
Турции.
Типовой вид — Helix joannis Mortillet, 1854.
CAUCASOCRESSA
305
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА С A UCASOCRESSA
1 (2). Большой диаметр раковины 20—28 мм, высота завитка примерно равна высоте
устья................................................. 2. С. joannis.
2 (1). Большой диаметр раковины 16—18 мм, высота завитка меньше высоты устья
....................................................... 1. С. pratensis.
1. Caucasocressa pratensis (L. Pfeiffer, 1845). Табл. XVIII, 776; рис. 412, 413.
L. Pfeiffer, 1845, Proc. Zool. Soc. London, 13 : 132 (Helix); Reeve, 1852,
Conch. Iconica, 7 : 279, t. 145, fig. 936 (Helix)-, M о r t i 11 e t, 1854 : 9, t. 1, fig. 10
(H. joannis var. minor)-, Mousson, 1863 : 314 (H. pratensis var. Bayeri, var. nymp-
haea); К о b e 1 t, Iconogr., N. F., 5 : 31, t. 127, fig. 1226 (H. pratensis var. depressa);
Retowski, 1914 : 290 [H. (Fruticocampylaea) pratensis typ. et f. unicingulata];
Линдгольм, 1928 : 539 [Fruticocampylaea (Caucasocressa) ]; Hesse, 1931 : 18,
t. 2, fig. 13 [Fruticocampylaea (Caucasocressa)]; Лихарев, Раммельмейер,
Рис. 412—413. Caucasocressa pratensis (L. Pfr.) из окр. Боржоми,
28 VI 1917. 412 — внешний вид полового аппарата; 413 — ди-
стальная часть полового аппарата, вскрыта. (По Шилейко, 1972д).
1952 : 467, рис. 393 (Metafruticicola); Hudec, Lezhawa, 1969b : 144, Abb. 33,
t. 9, 23 [Metafruticicola (Metafruticicola)]; Ш и л e й к о, 1972д : 13, рис. 3.
Материал: 7 экз. из 4 местонахождений в Закавказье.
Раковина сильно уплощена, с почти невыступающей вершиной и низким
широким, слабо куполовидным завитком, высота которого не превышает высоту устья.
Оборотов 6, вначале медленно, затем быстрее нарастающих, умеренно выпуклых.
20 А. А. Шилейко
306
III. HYGROMIIDAE
Последний оборот почти вдвое шире предпоследнего, к устью внезапно, с перегибом,
опущен. Окраска белая, реже светло-желтая, спиральные коричневые ленты четкие,
проходят выше и ниже периферии; верхняя лента видна на предыдущих оборотах
у шва и прослеживается до самой вершины. В проходящем свете лепты просвечивают
сильнее фона. Изредка нижняя лента или обе отсутствуют. Кроме того, раковина иногда
покрыта мелкими серыми просвечивающими точками. Скульптура — тонкая радиаль-
ная исчерченность, ясная тонкая зернистость и выраженные в различной степени спи-
ральные бороздки. Устье округлое, очень косое; края его с тонкой белой губой или без-
губы, отвернутые, тонкие, колумеллярный край у основания расширен. Пупок вна-
чале немного развернутый, открытый, часть предыдущих оборотов можно просмотреть,
поворачивая раковину. Размеры: ВР 8.5—10, БД 16—18, МД 13—14.5 мм.
Внутреннее строение. Материал: 1 экз. из окр. Боржоми, 17 VII 1966,
сбор и определение Г. И. Лежавы; 1 — из окр. Боржоми, 28 VI 1917, сбор А. Морд-
вилко, определение И. М. Лихарева.
Нижняя часть спермовидукта образует небольшой, но резкий перегиб. Вагина
короткая и тонкая, внутри несет правильные и тонкие продольные складки.
Пенис почти веретеновидный, поверхность папиллы покрыта кольцевыми борозд-
ками, за счет выравнивания которых этот орган может удлиняться. Семепроводящий
канал на поперечном разрезе имеет форму многолучевой неправильной звезды, так как
в пределах папиллы в общем сохраняются складки, присутствующие внутри эпифал-
луса. Кольцевая полость хорошо развита, но заходит лишь в нижнюю часть стенок
папиллы. На границе пениса и эпифаллуса имеется резкий перегиб, половой ретрактор-
крепится дистальнее перегиба. Эпифаллус примерно вдвое длиннее пениса, почти пря-
мой, не извит. Длина бича немного превышает суммарную длину пениса и эпифаллуса
или равна ей. Резервуар семеприемника тонкостенный, очень массивный, мешковид-
ный, граница между резервуаром и протоком почти отсутствует. Когда семеприемник
пустой, стенки его спадаются (рис. 412). Основание протока семеприемника немного-
расширено и покрыто изнутри четкими упорядоченными гофрированными складками.
Семеприемник не достигает белковой железы.
Распространение и местообитания. Горные хребты, окаймляю-
щие с запада и юго-запада Картлийскую котловину, и прилегающая часть северных
склонов Малого Кавказа. Обитает на открытых влажных склонах и в ущельях.
X
2. Caucasocressa joannis (Mortillet, 1854). Табл. XVIII, 177, 178; рис. 414, 415.
Mortillet, 1854 : 8, t. 1, fig. 9 (Helix); Mousson, 1863 : 315 (Helix
delabris); Kobelt, 1877, Iconogr., N. F., V : 31, t. 127, fig. 1229 (II. delabris);
Boettger. 1881a : 210 (Helix (Fruticocampylaea) var. Joannis]; Lindholm,
1913b : 18 [Helix (Fruticocampylaea) joannis var. andronakii]; R e t о w s k i, 1914 : 292
(Helix (Fruticocampylaea) joannis typ. et f. pellucidepunctata]; Rosen, 1914:170
(H. joannis, H. delabris); JI и x а рев, Раммельмейер, 1952 : 467 (Metajruti-
cicola pratensis var. joannis); Шилейко, 1972д : 15, рис. 4 (Caucasocressa delabris).
Материал: более 30 экз. из 4 местонахождений в юго-западном Закавказье и Тур-
ции.
Раковина слегка прижатая, с завитком конических или куполовидных очер-
таний; вершина обычно слегка выступающая. Оборотов 6, равномерно и быстро на-
растающих, умеренно выпуклых. Последний оборот менее чем вдвое шире предпослед-
него, к устью энергично, но-без резкого перегиба, опущен. Окраска белая или светло-
серая, без желтого оттенка. Спиральные лепты темно-коричневые, нередко границы их
неровные, но ясные; располагаются выше и ниже периферии. Скульптура — топкая
радиальная исчерченность и зернистость, выраженная в разной степени; иногда могут
присутствовать и спиральные бороздки. Устье большое, оттянуто вниз и в сторону,
косое, округлое. Губа очепь слабая, расположена близ края устья. Края устья несильно-
отвернуты, кроме колумеллярного. Пупок неперспективный, умеренно широкий.
Размеры: ВР 9—16, БД 20—28, МД 16—23 мм.
Внутреннее строение. Материал: 2 экз. из Турции, Артвппскип
округ, окр. Ломашена, IV 1909, сбор П. Нестерова, определение И. М. Лихарева;
4 — из Аджарии, долина р. Мачахели-цкали, 22 VII 1973, сбор И. М. Лихарева, оп-
ределение наше.
Спермовидукт при переходе в яйцевод образует резкий, но короткий изгиб. Ва-
гина прямая, короткая, тонкая, внутри покрыта небольшими продольными склад-
ками.
Пепис слегка вздут, вместе с эпифаллусом образует S-образный изгиб; имеются
небольшие пениальные ленты. Папилла короткая, с обширным каналом и топкими
стенками. На поперечном разрезе папиллы видно несколько высоких тонких валиков,
которые могут давать дополнительные ответвления (рис. 415). В основании папиллы
имеется хорошо развитая кольцевая полость, заходящая далеко в стопки эпифаллуса.
Бич не длиннее половины эпифаллуса. Резервуар семеприемника очень большой, меш-
CAUCASOCRESSA
307
ковидный, неясно отграничен от протока. Основание протока очень немного расширено,
внутренняя поверхность его несет несколько четких, мелко гофрированных продоль-
ных валиков; резервуар заметно не достает до нижнего края белковой железы.
Распространение и местообитания. Юго-западная часть
Малого Кавказа и прилегающие районы Турции. Населяет открытые склоны
в местах выхода грунтовых вод, с обильным травостоем; встречается также на мокрых
скалах, среди растительности.
Рис. 414—415. Caucasocressa joannis (Mort.) из сев. Турции,
окр. Ломашена, VI 1909. 414 — внешний вид полового ап-
парата; 415 — дистальная часть полового аппарата, вскрыта.
(По Шилейко, 1972д).
IV. Семейство HELICIDAE Rafinesque, 1815
Раковина средних размеров или большая, разнообразной формы и окраски. Пупок
от широко развернутого до полностью запаянного. Изредка па раковине могут присут-
ствовать волоски. Устье разнообразной формы, в исключительно редких случаях
с зубами.
Стилофор всегда 1, слизистых желез 2, расположенных на вагине по бокам осно-
вания стилофора, простых или разветвленных в разной степени. Семеприемник пер-
вично с дивертикулом, притянут соединительнотканными связками к задней части
дна мантийной полости; резервуар небольшой, шаровидный, прилегает к основанию
белковой железы. Проток семеприемника топкий, в нижней части пе расширен.
Бич длинный, папилла пениса образована за счет разрастания дна полости пениса
20*
308
IV. HELICIDAE
вокруг выходного отверстия эпифаллуса; часто дистальнее ее образуется еще одна
папилла за счет складок внутреннего пениального чехла. Имеется пениальный бугорок.
Распространение. Европа, Северная Африка, Малая Азия, Аравийский
п-ов, Крым, Кавказ. В ископаемом состоянии представители семейства встречаются
начиная с эоцена (Северная Африка, Европа).
I. Подсемейство ARIANTINAE Morch, 1864
Раковина обычно средних размеров, очень разнообразная по всем конхологиче-
ским признакам; в редких случаях может быть цилиндрической или иметь париеталь-
ный зуб.
Слизистые железы простые или двуветвистые. Дивертикул семеприемника развит
очень хорошо.. Папилла пениса одна, изредка может состоять из двух лопастей.
Рис. 416; Faustina faustina (Rssm.) из Закарпатья, окр.
с. Квасы, 16 IX 1969. Внешний вид полового аппарата.
(По Шилейко, 19716).
Распространение. Европа. В ископаемом состоянии представители
подсемейства встречаются начиная с эоцена (Европа).
1. Род FAUSTINA Kobelt, 1904
К о b е 1 t, 1904, Iconogr., N. F., 11 : 131; Wenz, Zilch, 1959—1960 : 701
[Campylaea (Faustina}]; Шилейко, 19716 : 992.
FAUSTINA
309
Раковина прижатая, с развернутым перспективным пупком; периферия округ-
лая, устье без зубов; волоски и зернистая скульптура отсутствуют.
Папилла пениса двулопастная; эпифаллус, пенис и бич примерно равной длины.
Слизистые железы двуветвистые.
Распространение. Средняя и Юго-Восточная Европа. В ископаемом со-
стоянии род встречается начиная с верхнего плиоцена (Европа).
Типовой вид — Helix faustina Rossmassler, 1835.
ЧП
Рис. 417—418. Faustina faustina (Rssm.) из Закарпатья,
окр. с. Квасы, 16 IX 1969. 417 — пенис, вскрыт; 418 —
особь in copuli. (По Шилейко, 19716).
Faustina faustina (Rossmassler, 1835); Табл. XVIII, 179, 180; рис. 416—418.
Rossmassler, 1835, Iconogr., I, 2 : 4, fig. 93 (Helix); Westerlund,
1889 : 146 (Helix); К о b e 1 t, 1902, Iconogr., N. F., 9 : 24, t. 249, fig. 1913 (Campy-
laea faustina var. associata Rssm.); Лихарев, Раммельмейер, . 1952 : 470,
рис. 395 [Helicigona (Campylaea)]; G г о s s u, 1955 : 477, fig. 273 [Campylaea (Faustina)'];
Ehrmann, 1956 : 141, t. 5, fig. 78 [Helicigona (Campylaea) faustina]; L d z e k,
1956 : 220, Obr. 61, t. 34, 3 [Helicigona (Faustina)]; Zilch, Jaeckel, 1962 : 193
[Helicigona (Faustina)]; Шилейко, 19716 : 993, рис. 3.
Материал: более 40 экз. из 3 местонахождений в Карпатах и Татрах.
Раковина в различной степени уплощенная, с куполовидным завитком-и вы-
ступающей или сглаженной вершиной. Высота завитка не больше высоты устья. Обо-
ротов 4.5—5.5, выпуклых, плавно нарастающих. Последний оборот в 1.5 раза Шире
предпоследнего,, к устью опущен с плавным перегибом. Окраска либо одноцветная,
лимонно-желтая, красноватая или светло-роговая, либо имеется выше периферии одна
темная лента. Выше и ниже ленты часто наблюдается небольшое посветление фона.
310
IV. HELICIDAE
Скульптура в виде тонких и неравномерных радиальных сглаженных морщин; спи-
ральные бороздки отсутствуют. Устье коротко-овальное, косое, места его прикрепле-
ния немного сближены, края сильно отвернуты, внутри слегка утолщены в светлую
губу. Пупок широкий, сквозь пего видны все предыдущие обороты. Размеры: ВР
10—12, БД 15—23, МД 12—18 мм.
Внутреннее строение. Материал: 1 экз. из Низких Татр, 13 VII 1964,
сбор и определение В. Гудеца; 2 — из окр. Ужгорода (Невицкий замок), 6 IX 1969;
12 — из окр. с. Квасы близ г. Рахова, 12—18 IX 1969; сборы и определения наши.
Дистальная часть спермовидукта и яйцевод образуют крутой S-образный изгиб.
Вагина в различной степени вздутая. Ветви слизистых желез равной длины, их верх-
ние концы могут достигать половины длины спермовидукта. Стилофор массивный, ко-
ротко булавовидный.
Пенис в проксимальной части вздут. Папилла пениса занимает весь объем пениаль-
ного чехла, состоит из двух'лопастей, причем одна из лопастей значительно массивнее
другой и несет выемку, в которую укладывается меньшая лопасть. При совокуплении
выдвигается нижняя часть эпифаллуса вместе с папиллой, лопасти которой раскры-
ваются (рис. 418). Бич конический; половой ретрактор крепится к дистальной части
эпифаллуса. Пениальный бугорок небольшой, но четко выражен, располагается в ниж-
нем отделе пениса внутри небольшого расширения. Дивертикул семеприемника отхо-
дит очень низко, диаметр его значительно превышает диаметр протока; последний
длиннее спермовидукта, извит.
Распространение и местообитания. Карпаты и сопредельные
горные области Средней Европы. Встречается в горных и предгорных лесах; значитель-
ные скопления моллюсков наблюдаются в районах скальных выходов в лесу и в мало-
подвижных осыпях.
2. Род ARIANTA Turton, 1831
Turton, 1831 : 35; Martens, 1860, in Albers Helic., ed. 2 : 127 (Arionta)-
Hesse, 1931 : 85; Лихарев, Pa м м e л ь м е й е р, 1952 : 470; Wenz,
Zilch, 1959—1960 : 703; Шилейко, 19716 : 993.
Раковина от слегка прижатой до шаровидной, с чрезвычайно узким, почти закры-
тым пупком; периферия округлая, устье без зубов; волоски и зернистая скульптура
отсутствуют, имеются ясные спиральные бороздки.
Папилла пениса с двумя глубокими бороздками, за счет исчезновения которых
объем папиллы может очень сильно увеличиваться. Бич длиннее пениса и эпифаллуса,
вместе взятых. Слизистые железы одноветвистые.
Распространение. Средняя и Северная Европа. В ископаемом состоя-
нии род известен начиная с верхнего миоцена (Европа).
Типовой вид — Helix arbustorum Linnaeus, 1758.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ARIANTA
1 (2). Раковина твердостенная. Окраска: по желтому, каштановому или коричневому
фону разбросаны светло-желтые четкие штрихи и пятна; выше периферии часто
располагается коричневая лента........................1. A. arbustorum.
2 (1). Раковина тонкостенная, хрупкая. Окраска одноцветная — от темно-бурой до
почти черной........................................2. A. aethiops petrii.
1. Arianta arbustorum (Linnaeus, 1758). Табл. XIX, 181', рис. 419—421.
Linn aeus, 1758 : 771 (Helix)-, Westerlund, 1889 :147 (Helix)-, Taylor,
1914 : 417, t. 34 [Helicigona (Arianta)]-, Лихарев, Раммельмейер, 1952 : 471,
рис. 397, 398; G г о s s u, 1955 : 482, fig. 275, 276; Ehrmann, 1956 : 141, fig. 11,92;
Lozek, 1956 : 221, t. 37, 2; Zilch, Jaeckel, 1962 : 194 [Helicigona (Arianta)]-,
Lozek, 1964 : 309, t. 30, 5; G i t t e n b e r g e r et al., 1970 : 103, fig. 179, 182,
Kaart 74; Шилейко, 19716 : 995, рис. 4.
Материал: более 50 экз. из 4 местонахождений на Карпатах и Гарце (ГДР).
Раковина от шаровидно-кубаревидной до шаровидной, с притупленным сгла-
женным завитком, имеющим куполовидные очертания. Высота завитка не менее высоты
устья, обычно несколько больше. Оборотов 5.5—6. Последний оборот в профиле очень
плавно и широко закруглен, к устью энергично, но без резкого перегиба опущен.
Интенсивность окраски значительно варьирует; обычно она состоит из желтого, кашта-
нового или коричневого фона и четких светло-желтых штрихов и пятен, ориентирован-
ных большей частью вдоль оборотов. В проходящем свете эти элементы плотнее фона.
Кроме того, над периферией имеется узкая коричневая лента, имеющая меньшую опти-
ческую плотность, чем фон раковины. Скульптура в виде неравномерной радиальной
исчерченности и весьма четких густых и тонких спиральных бороздок. Устье круглое,
Рис. 419—421. Arianta arbustorum (L.) из Закарпатья, окр. с. Квасы, 16 IX 1969.
419 — внешний вид полового аппарата; 420 — пенис, вскрыт; 421 — особь in copuli.
(По Шилейко, 19716, с дополнениями).
I
312
IV. HELICIDAE
косое, места его прикрепления широко расставлены. Края устья широко отвернуты,,
белые, острые, переходящие в толстую губу. Пупок почти полностью запаян отворотом
колумеллярного края; от него остается узкая щель. Нередко пупок закрыт полностью.
Размеры: ВР 12—23, БД 18—25, МД 16—22 мм.
Внутреннеестроение. Материал: 8 экз. из окр. с. Квасы близ г. Ра-
хова, Закарпатье, 16 IX 1969, сбор и определение наши; 4 — из Гарца, ГДР, V 1969,
сбор и определение Э. Клаусса.
Рис. 422. Arianta aethiops petrii Kim. из Закарпатья,
окр. с. Квасы, 16 IX 1969. Внешний вид полового аппарата
не вполне взрослого экземпляра. (Ориг.).
Нижняя часть спермовидукта и яйцевод образуют двойной изгиб. Слизистые же”
левы необычайно мощные, верхние их концы могут достигать белковой железы. Сти-
лофор сравнительно небольшой, удлиненный. Внутри вагины имеется система резких
продольных складок, сливающихся в плотный узел в самой верхней части вагины.
Пенис относительно короткий, вздут, внутренняя поверхность его покрыта пра-
вильными резкими кольцевыми складками, за счет выравнивания которых пенис может
удлиняться. Папилла пениса в общем цилиндрическая, ее просвет имеет приблизительно
форму двутавровой балки. Половой ретрактор крепится к эпифаллусу ниже его сере-
дины. Пениальный бугорок хорошо развит, при совокуплении выворачивается и распо-
лагается позади отверстия пениса. Дивертикул семеприемника отходит низко; мемб-
рана, соединяющая его с поверхностью матки, особенно хорошо развита.
Слизистые железы, внутренняя поверхность стилофора и папилла пениса интен-
сивно пигментированы.
Распространение и местообитания. Средняя и Северо-За-
падная Европа; в СССР — Карпаты, Калининградская обл., окр. Ленинграда, острова
и прибрежные районы Эстонии, одна находка в Латвии. Населяет преимущественно
влажные широколиственные леса. В ископаемом состоянии встречается начиная
с верхнего плиоцена; многократно обнаружен в плейстоцене и голоцене в пределах
современного ареала.
HELICIGONA
313
2. Arianta aethiops petrii Kimakowicz, 1890. Табл. XIX, 182; рис. 422.
К imako wicz, 1890 : 69 (Helix aethiops var. petrii); Лихарев, Раммель-
мейер, 1952 : 472, рис. 399; Grossu, 1955 : 484; Шилейко, 19716 : 995.
Материал: 2 экз. из 2 местонахождений на Карпатах.
Раковина почти шаровидная, тонкостенная, завиток имеет явственно купо-
ловидный контур, высота его может быть больше высоты устья и меньше ее. Оборотов 5,
выпуклых, плавно нарастающих. Последний оборот в 1.5 раза шире предпоследнего,
округлый, близ устья слегка опущен. Окраска — от зеленовато-черной до темно-бу-
рой, почти одноцветная, вершина немного светлее. Скульптура в виде неравномерной,
но в общем слабой радиальной исчерченности, поверх которой как бы накинута тон-
чайшая сеточка с длинными ячейками, ориентированными радиально. Кроме того,
местами имеются выраженные в разной степени спиральные бороздки. Устье округлое,
косое, умеренно вырезано предпоследним оборотом, края его тонкие, почти не отвер-
нуты. Губа тонкая, светлая. Пупок щелевидный, почти полностью закрыт отворотом
колумеллярного края. Размеры: ВР 13—15, БД 18—20, МД 16.5—18 мм.
Внутреннее строение. Материал: 1 не вполне взрослый экземпляр
из окр. с. Квасы близ г. Рахова (Закарпатье), 16 IX 1969, сбор и определение наши.
Внешне морфология гениталий и их внутреннее строение у изученного экземпляра
очень сходны с тем, что наблюдается у предыдущего вида; несколько особенностей, ха-
рактерных для данного экземпляра, есть, возможно, проявление возрастной измен-
чивости. К таким особенностям относятся: резко суженная дистальная часть пениса,
относительно очень массивный стилофор, тонкая и изогнутая вагина (изображение
внешней морфологии гениталий вполне взрослой особи — см.: Polinski, 1924).
Распространение и местообитания. Черногора (Гуцульские
Альпы); Черновицкая обл. и северо-восточная Румыния; номинативный подвид
в СССР не встречается, распространен в Карпатах и смежных горных областях Сред-
ней Европы. Обитает на полонинах и по опушкам лиственных лесов.
3. Род HELICIGONA Ferussac, 1821
Ferussac, 1821 : 27; Turton, 1831 : 66 (Chilotrema); Beck, 1837 : 24
(Campylaea); Лихарев, Раммельмейер, 1952 ; 468; Wenz, Zilch,
1959—1960 : 703; Ш и л e й к о, 19716:995.
Раковина линзовидная, с развернутым пупком; устье без зубов; поверхность ра-
ковины с ясной зернистой скульптурой.
Папилла пениса без бороздок, с рыхлыми стенками. Длина бича приблизительно
равна суммарной длине пениса и эпифаллуса. Слизистые железы одноветвистые.
Распространение. Средняя и Западная Европа. В ископаемом состоя-
нии род известен начиная с нижнего плиоцена (Средняя Европа).
Типовой вид — Helix lapicida Linnaeus, 1758.
Helicigona lapicida (Linnaeus, 1758). Табл. IX, 88; XIX, 183; рис. 423—425.
Linnaeus, 1758 : 768 (Helix); Westerlund, 1889 :28 (Helix); Лихарев,
Раммельмейер, 1952 : 469, рис. 394; Ehrmann, 1956 : 137, t. 6, fig. 79;
Lozek, 1956 : 217, t. 36, 1; 1964 : 305, Abb. 70, t. 30, 4; G i 11 e n b e r g e r et al.,
1970 : 103, fig. 178, 184, Kaart 73; Шилейко, 19716 : 995, рис. 5.
Материал: более 20 экз. из 4 местонахождений в Карпатах и в Гарце (ГДР).
Раковина сильно уплощена, с ширококуполовидным низким завитком и слабо
выступающей вершиной, твердостенная. Оборотов 5, плоских, плавно нарастающих,
разделенных мелким швом. Последний оборот немного шире предпоследнего, снизу
слабо выпуклый, к устью внезапно и энергично опущен. Окраска от светло-серой
до бурой, изредка почти однотонная, чаще с рисунком из продольных и-поперечных
пятен, которые местами сливаются й образуют две спиральные ленты — выше и ниже
киля. Скульптура представлена тонкой и неясной радиальной исчерченностью и чет-
кой зернистостью, которая обычно видна даже невооруженным глазом. Устье ко-
ротко-овальное, очень косое, с притупленным углом на середине палатального края,
соответствующим килю. Места прикрепления устья сильно сближены, порой устье
почти цельное. Края устья сильно отвернуты, светлые. Колумеллярный край слегка
нависает над перспективным пупком.' Размеры: ВР 7.5—8.5; БД 14—17, МД 12—14.5 мм.
Внутреннее строение. Материал: 3 экз. из Гарца у Рюбелянда,
ГДР, VI 1969, сбор и определение Э. Клаусса; 1 — из Загорани, центральная Чехия,
8 IX 1969; 1 — из Чешского Крумлова, западная Чехия, 19 V 1969; сборы и опреде-
ления Я. Бухара.
Дистальная часть спермовидукта и яйцевод образуют двойной изгиб. Стилофор
булавовидный, вагина тонкая, цилиндрическая.
Рис. 423—425. Helicigona lapicida (L.) из Чешского Крумлова, зап. Чехия, 19 V 1969. 423 — внешний вид полового аппа-
рата; 424 — пенис, вскрыт; 425 — продольный разрез пениса и папиллы. (По Шилейко, 19716, с дополнениями).
ISOGNOMOSTOMA
315
Дистальная часть пениса вздута и заключает пениальный бугорок; далее пенис
сужается и вновь вздувается в проксимальной части, где располагается папилла.
Внутренние стенки пениального чехла, особенно в проксимальной части, часто несут
кольцевые бороздки. Папилла пениса толстостеппая, причем самый наружный и
самый внутренний слои стенок плотные, а между ними располагается чрезвычайно
рыхлый волокнистый слой (рис. 425). Длина бича примерно равна суммарной длине
пениса и эпифаллуса или немного меньше. Половой ретрактор крепится к эпифаллусу
значительно отступя от пениса. Дивертикул семеприемника на середине длины может
•образовывать петлю.
Распространение и местообитания. Западные и средние
сбласти Европы, включая Калининградскую обл. и побережье Латвии. • Населяет
горные и холмистые местности, встречается в лесах или зарослях кустарников, а
также в пещерах (Tercafs, 1958—1960).
4. Род ISOGNOMOSTOMA Fitzinger, 1833
F i t z i n g е г, 1833 :97;Schliiter, 1838 : 4 (Plicostoma); Albers, 1850 : 95
i( Ulostoma); Hesse, 1931 : 88; Лихарев, Раммельмейер, 1952 : 472;
Wenz, Zilch, 1959—1960 : 707; Ш и л e й ко, 19716 ; 996.
Раковина прижатая, округлая, с почти запаянным пупком; периферия округлая.
В устье имеются 3 зуба, раковина покрыта волосками; имеется зернистая скульптура.
Папилла пениса без продольных борозд, толстостенная. Длина бича немного
превышает суммарную длину пениса и эпифаллуса. Слизистые железы одноветвистые.
Распространение. Пиренеи, Альпы, Карпаты, Судеты. В ископаемом
состоянии род известен начиная с верхнего плиоцена (Европа).
Монотипический род.
Типовой вид — Helix personatum Lamarck, 1792.
Isognomostoma personatum (Lamarck, 1792). Табл. XIX, 184; рис. 426—428.
Gmelin, 1788, in Linnaeus Syst. nat. : 3621 (Helix isognomostoma part.);
Lamarck, 1792, Journ. Hist, nat., II : 348, t. 42, fig. 1 (Helix); Kimakowicz,
1883 : 27 [Anchistoma (Triodopsis)]; С 1 e s s i n, 1885 : 135, fig. 68 (Triodopsis); W es-
ter 1 u n d, 1889 : 27 (Helix); Hesse, 1931 : 88, t. 12, fig. 96; Л и x a p e в, Рам-
мельмейер, 1952 : 473, рис. 400; G г о s s u, 1955 : 465, fig. '264; Ehrmann,
1956 : 143, fig. 93; L oz ek, 1956 : 222, t. 36, fig. 5; Z i 1 c h, J a e c k e 1, 1962 : 196
[Isognomostoma isognomostoma (Gmelin, 1780) Schroter, 1784]; Loiek, 1964 : 310,
t. 28, 2a—c [Isognomostoma isognomostoma (Schroter, 1784)]; Ш и л e й к о, 19716 :
996, рис. 6. •_____
Материал: более 100 экз. из 10 местонахождений в Закарпатье, Чехии и ГДР.
Раковина с широко закругленным куполовидным завитком и сглаженной
вершиной; высота завитка всегда меньше высоты устья. Оборотов 5, умеренно выпук-
лых. Последний оборот снизу сильно вздут, к устью резко опущен. Окраска однотон-
ная, красновато-роговая, коричневая или бурая. Скульптура в виде неравномерной
радиальной исчерченности и тонкой зернистости; у свежих раковин каждая гранула
несет одну маленькую треугольную чешуйку. Кроме того, имеются многочисленные
волоски, чаще остающиеся пожизненно и даже некоторое время после гибели живот-
ного. Устье закругленно треугольное, края его сильно отвернуты, образуя манжету.
Губа толстая, пластинчатая, несет бугорчатые базальный и палатальный зубы. Кроме
того, по линии париетальной мозоли имеется мощная отвесная пластинка, которая
" в случае максимального развития загораживает устье настолько, что просвет его при
нормальном положении раковины не виден. Непосредственно за краем устья снаружи
проходит глубокая кольцевая борозда. Пупок запаян или от него остается узкая щель.
Размеры: ВР 5.5—6.5, БД 9—13, МД 7.8—11.5 мм.
Внутреннее строение. Материал: 3 экз. из Гарца, Бодеталь, VI 1969,
сбор и определение Э. Клаусса; 3 — из Карлштейнского заповедника, Чехия, 24 IV 1967,
сбор Я. Бухара; 5 — из с. Дубки близ Ужгорода, 5 IX 1969; 12 — из с. Квасы близ
Рахова, 19 IX 1969; сборы и определения наши.
Нижняя часть спермовидукта образует двойной изгиб, выраженный в разной
степени, причем у большинства экземпляров здесь наблюдается несколько вздутий.
Яйцевод чрезвычайно короткий. Стилофор удлиненно-булавовидный, вагина довольно
длинная.
Пенис прямой или изогнутый, эпифаллус иногда охватывает основание полового
ретрактора (рис. 426). Пенис не обнаруживает разделения на отделы; папилла удли-
ненно-коническая. При рассмотрении особей, находящихся in copuli, видно, что на
конце дистальной части женского отдела имеется точечное отверстие стилофора и по-
лулунное отверстие, ведущее в проток семеприемника. Длина бича превышает сум-
марную длину пениса и эпифаллуса. Проток семеприемника значительно тоньше и длин-
нее дивертикула, резервуар прилегает к границе спермовидукта и белковой железы.
DROBACIA—CAUSA
317
Распространение и местообитания. Все основные горные мас-
сивы Восточной, Средней и Западной Европы, а также предгорья. В горы поднимается
до высоты 1800 м над ур. м. В СССР встречается на Карпатах и в Предкарпатье.
Населяет широколиственные, реже смешанные леса, где чаще всего встречается под
корой упавших деревьев и под корягами, а также в лиственном опаде.
5. Род DROBACIA Brusina, 1904
Brusina, 1904 : 160; Ehrmann, 1956 : 138 (Dobracia nom. err.); W e n z,
Zilch, 1959—1960 : 703.
Раковина линзовидная, с открытым цилиндрическим пупком. Устье без зубов,
волоски на раковине отсутствуют, имеются ясные спиральные бороздки; зернистости
нет.
Папилла пениса без продольных борозд, тонкостенная. Длина бича приблизи-
тельно равна суммарной длине пениса и эпифаллуса. Слизистые железы нерасщепчен-
ные.
Распространение. Карпаты. В ископаемом состоянии род известен из
верхнего миоцена Румынии.
Монотипический род.
Типовой вид — Helix banatica Rossmassler, 1838.
Drobacia banatica (Rossmassler, 1838). Табл. XIX, 185.
Rossmassler, 1838, Iconogr., I, 2, fig. 457 (Helix); С 1 e s s i n, 188/ : 140,
fig. 55 (Campylaea); Kimakowicz, 1890 : 52 [Campylaea (Eucampylaea)]; Geyer,
1927, Unsere Land- u. Susswasser-Mollusken : 88, t. 5, fig. 24—26 [Campylaea (Dro-
bacia)]; G г о s s u, 1955:467, fig. 265, 266 [Helicigona (Drobacia)]; Ehrmann,
1956 : 138 [Helicigona (Dobracia)]; Zilch, Jaeckel, 1962 : 190 [Helicigona
(Drobacia)]; L о i e k, 1964:305, t. 26, fig. 3; t. 27, fig. 3 [Helicigona (Drobacia)];
III и л e й к o, 19716 : 1000 [Helicigona (Drobacia) ].
Материал: 2 экз. из 2 местонахождений в Карпатах.
Раковина сильно прижатая, хрупкая, составлена 6 очень плоскими оборо-
тами, разделенными мелким швом. Последний оборот незначительно шире предпослед-
него, у взрослых экземпляров плавно, но энергично опущен к устью. Окраска одно-
цветная,. роговая или желтоватая. Скульптура в виде слабой радиальной морщини-
стости и четких спиральных бороздок, заметных даже при небольшом увеличении.
Устье широкоовальное, угол на палатальном крае почти не выражен. Нижний край
устья отогнут. Пупок неширокий, но перспективный. Размеры: ВР 14—18, БД 25—30,
МД 23—27 мм.
Внутреннее строение. Материал: 1 не вполне взрослый экз. из За-
карпатья, окр. Рахова, VI 1966, сбор В. М. Мамаева, определение наше.
Нижняя часть спермовидукта и свободный яйцевод образуют плавный двойной
изгиб. Стилофор относительно небольшой.
Пенис слегка вздут, половой ретрактор крепится к эпифаллусу. Папилла пениса
занимает проксимальную часть пениса, внутри Дистальной части помещается пениаль-
ный бугорок. Дивертикул семеприемника длиннее и толще протока, резервуар неболь-
шой, шаровидный.
Распространение и местообитания. Трансильвания, Банат,
восточная Венгрия; в СССР сделана одна находка в южном Закарпатье. Обитает в гор-
ных широколиственных лесах и зарослях кустарников, неподалеку от воды. В плейсто-
цене вид был широко распространен по Средней Европе.
6. Род CAUSA Schileyko, 1971
Ш и л е й к о, 19716 : 997.
Раковина почти плоская, с развернутым перспективным пупком. В устье имеются
2—3 бугорчатых зуба, на поверхности раковины присутствуют редкие волоски, кото-
рые легко утрачиваются. Спиральные бороздки отсутствуют, зернистость слабая.
Папилла пениса короткая, с резкими кольцевыми бороздками. Бич очень корот-
кий, рудиментарный. Слизистые железы одноветвистые.
Распространение. Средняя Европа. В ископаемом состоянии род из-
вестен начиная с плейстоцена (Средняя Европа).
Монотипический род.
Типовой вид — Helix holosericum Studer, 1820.
Causa holosericum (Studer, 1820). Табл. XIX, 186; рис. 429—431.
Studer, 1820, Naturwissensch. Anz.; 11 : 87 (Helix); Rossmassler,
1835, Iconogr., I : 69, t. 1, fig. 20 (Helix); С 1 e s s i n, 1884 : 134, fig. 67 (Trigono-
318
IV. HELICIDAE
stoma)-, 1887, Molluskenfauna Oesterr.-Ungarns u. Schweiz : 112 (Trlgonostoma)- Ehr-
mann, 1956 : 144 (Isognomostoma)- Lozek, 1956 : 223, t. 36, fig. 6 (Isognomostoma)-,
Zilch, Jaeckel, 1962 : 197 (Isognomostoma)-, Lozek, 1964 : 311, Abb. 73
(I sognomostoma)-, Шилейко, 19716 : 998, рис. 7.
Материал: 6 экз. из 3 местонахождений в Татрах.
Раковина сильно уплощена, с едва возвышающимся завитком; оборотов 6,
узких, довольно медленно нарастающих. Последний оборот немного шире предпослед-
него, к устью опущен. Окраска одноцветная, светло-роговая. Скульптура в виде тон-
кой и неравномерной радиальной исчерченности и редко!! тонкой зернистости. Волоски.
Рис. 429—431. Causa holosericum (Stud.) из Краковского ущелья в Татрах,
25 VII 1958. 429 — внешний вид полового аппарата; 430 — клоака, вскрыта;
431 — пенис, вскрыт. (По Шилейко, 19716).
на поверхности раковины короткие, плохо заметны. Устье заметно скошено, края его
окаймлены светлой губой, на которой располагаются палатальный зуб и 1—2 базаль-
ных, причем тот из них, который лежит ближе к колумеллярному краю, развит слабее
и может отсутствовать. Пупок широкий, сквозь него видны все предыдущие обороты.
Размеры: ВР 5—6, БД 10—12, МД 9—10.2 мм.
Внутреннее строение. Материал: 2 экз. из Польши, Татры, Краков-
ское ущелье, 25 VII 1958, сбор и определение И. М. Лихарева.
Нижняя часть спермовидукта и яйцевод образуют крутой двойной изгиб. Стило-
фор относительно небольшой, вместе с основаниями слизистых желез притянут к ва-
гине соединительнотканными лентами; такие же ленты охватывают и нижнюю часть
вагины.
Пенис четко подразделяется на 2 отдела: шаровидный проксимальный и тонкий
цилиндрический дистальный; отдельные пениальные лепты стягивают названные от-
делы и обеспечивают относительное постоянство петли, которую образует дистальный
отдел пениса. Пениальный бугорок располагается непосредственно у клоаки. Папилла
пениса занимает шаровидный отдел, короткая, с многочисленными кольцевыми склад-
ками, которым отвечают кольцевые желобки на внутренней поверхности чехла. Бич
во много раз короче эпифаллуса; половой ретрактор крепится к эпифаллусу сильно-
KLIKIA
319
отступя от пениса — примерно на середине его длины. Резервуар семеприемника объ-
емистый, тонкостенный.
Распространение и местообитания. Альпы, Татры, западные
Карпаты и Судеты. Не исключено, что в СССР обитает в западных районах Белоруссии.
Встречается в широколиственных горных и равнинных лесах. В ископаемом состоя-
нии известен начиная с раннего плейстоцена (северная Франция, Чехословакия,
Польша).
7. Род KLIKIA Pilsbry, 1894
Pilsbry, 1894, Man. Conch., II ser., v. 9 : 289; 0. Boettger, 1909,
Nachr. Bl. dtsch. malak. Ges., 41 : 99 (Klikaia); Wenz, Zilch, 1959—1960 : 706.
Раковина слегка прижатая, округлая, сдовольно широким, но наполовину закры-
тым пупком. Спиральная скульптура отсутствует, зернистость правильная, тонкая
и четкая.
Рис. 432. Klikia giengensis zamechovense subsp. nov. из нижне-
сарматских отложений на р. Говорка близ с. Замехов. Голотип.
(Ориг.).
Распространение. Род известен начиная с эоцена до среднего плиоцена
(Средняя Европа).
Типовой вид — Helix osculant Thomae, 1845. — Табл. XIX, 187.
f Klikia giengensis zamechovense Prysjazhnjuk et Schileyko, subsp. nov. Рис. 432.
Голотип хранится в Институте геологических наук АН УССР.
Материал: 10 экз. из пресноводного известняка, нижнесарматские отложения
па р. Говорка близ с. Замехов.
Раковина сильно прижатая, почти плоская, со слабо возвышающимся
завитком, 4.75 оборота. Первые 3 оборота выпуклые, разделены глубоким швом,
предпоследний и последний обороты уплощены. Верхняя часть последнего оборота
сверху почти ступенчатая; в профиле последний оборот с ясным, но плавно закруг-
ленным углом, проходящим выше средней линии, к устью слабо опущен. Скульптура
в виде тонких радиальных морщин и очень маленьких бугорков, расположенных
в шахматном порядке. Устье скошенное, с утолщенными и резко отогнутыми краями,
образующими манжету. Пупок довольно широкий, но почти не развернутый, цилин-
дрический, незначительно прикрыт отворотом колумеллярного края. Размеры го-
лотипа: ВР 5.1, БД 9.8, МД 8.1 мм.
Распространение. Волыпский горизонт нижнего сармата Подолии.
II. Подсемейство HELICINAE Rafinesque, 1815
Раковина средних размеров или большая, чаще всего кубаревидная или шаро-
видная, реже уплощенная. Париетальные зубы всегда отсутствуют, на базальной
и палатальной стенках устья изредка могут присутствовать зубы в виде бугорков
или пластинок. Кроме того, на колумеллярном крае иногда имеется наплыв в виде
радиально ориентированного утолщения.
Слизистые железы ветвятся в различной степени; минимальное число ветвей
каждой железы — 4. Дивертикул семеприемника развит в различной степени, из-
редка редуцирован. Папиллы пениса 2, реже 1.
Распространение. Европа, Средиземноморье, Кавказ. В ископаемом
состоянии представители подсемейства встречаются начиная со среднего миоцена.
320
IV. HELICIDAE
1. Род СЕР ДЕД Held, 1837
Turton, 1831 : 33 (Tachea, non Fleming, 1822); Held, 1837 : 910; Hesse,
1920, in Kobelt’s Iconogr., N. F., 23 : 79; Лихарев, P а м м e л ь м е й е р,
1952 : 475; W е n z, Zilch, 1959-1960:716.
Раковина кубаревидная, окраска первично состоит из белого, серого, желтого,
розового или каштанового фона и 5 темных лент, причем число лент может умень-
шаться вплоть до полного исчезновения их. Скульптура в виде радиальной исчер-
ченности или тонкой ребристости. Пупок полностью закрыт.
Слизистые железы дихотомически ветвятся 2 или 3 раза, общее число ветвей
каждой железы — 4 или 8. Длина бича превышает суммарную длину пениса и эпи-
фаллуса, дивертикул семеприемника короткий. Пенис содержит 2 папиллы.
Распространение. Европа и Северный Кавказ. В ископаемом состоянии
представители рода встречаются начиная со среднего миоцена (Европа). Указания
на более ранние находки (олигоцен) нуждаются в проверке; вероятно, их следует
относить к Ariantinae.
Типовой вид — Helix nemoralis Linnaeus, 1758.
1
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА СЕРАЕА
1 (2). Раковина ребристо радиально исчерчена. Пупок запаян не полностью, от него
остается узкая щель...............................3. С. (О.) vindobonensis.
2 (1). Раковина тонко и неравномерно радиально исчерчена. Пупок запаян полностью.
3 (4). Края устья и губа белые.........................2. С. (С.) hortensis.
4 (3). Края устья коричневые, губа красновато-коричневая . 1. С. (С.) nemoralis.
1. Подрод СЕРАЕА s. str.
Окраска раковины желтая, каштановая или розовая. Поверхность тонко ради-
ально исчерчена. Пупок полностью запаян.
Задний отдел пениса короче переднего, дивертикул семеприемника много короче
лежащего ниже участка протока семеприемника. Каждая слизистая железа дихото-
мически ветвится дважды, общее число ветвей каждой железы — 4. га = 22.
1. Сераеа (Сераеа) nemoralis (Linnaeus). Табл. XIX, 188; рис. 433-
Linnaeus, 1758 : 773 (Helix); Taylor, 1910, Mon. Brit. Moll., Ill :274, t. 26
(Helix); Hesse, 1920, in Kobelt’s Iconogr., N. F., 23 : 80, t. 642, fig. 1—6; Лиха-
рев, Раммельмейер, 1952 : 476, рис. 403, 404A; Ehrmann, 1956 : 146,
t. 7, fig. 84; t. 6, fig. 86a; L о z e k, 1956 : 225, t. .38, fig. 3; 1964 : 313, Abb. 74; G i t-
tenberger et al., 1970 : 106, fig. 187, 189, Kaart 76; Ш и л e й к о, 1972г : 95—
99, рис. 12В, Д.
Материал: более 40 экз. из 4 местонахождений в Европе.
Раковина со слегка куполовидным завитком, высота которого превышает
высоту устья. Оборотов 4.5—5.5, слабо выпуклых, плавно и быстро нарастающих.
Последний оборот немного менее чем вдвое шире предпоследнего, к устью плавно,
но энергично опущен. Наиболее широкие темные ленты, если они присутствуют, —
четвертая и пятая (считая сверху). В проходящем свете ленты просвечивают немного
сильнее фона. Скульптура в виде тонких неравномерных радиальных морщинок и
тончайшей струйчатости, ориентированной в целом спирально; эта струйчатость
видна лишь при значительном увеличении. Устье широкое, очень косое, верхний
край его прямой или образует очень пологую дугу. Края устья, отворот колумел-
лярного края и париетальная стенка имеют темную окраску — от коричневой до
почти черной; губа красновато-коричневая. Пупок полностью запаян. Размеры:
ВР 16-18, БД 21-23, МД 18-19 мм.
Внутреннее строение. Материал: 12 экз. из Кведлинбурга, ГДР,
V 1969, сбор и определение Э. Клаусса.
Нижняя часть спермовидукта и яйцевод образуют резкий S-образный изгиб.
Стилофор чрезвычайно мощный, удлиненно-булавовидный, слизистые железы впа-
дают чуть выше его основания. Вершина стилофора далеко выдается за место перехода
вагины в яйцевод.
Пенис длинный, при переходе в клоаку образует более или менее ясный изгиб.
Благодаря хорошо развитым интрапапиллярным полостям обе пениальные папиллы лег-
ко меняют длину и форму; когда между ними находится уже сформированный клубок из
скрученного сперматофора, папиллы как таковые практически исчезают (рис. 39).
В прямой связи со способностью сперматофора скручиваться в клубок внутри пениса
стоит редукция дивертикула семеприемника, длина которого примерно в 1.5 раза
меньше длины протока семеприемника выше места его бифуркации. С этим же обсто-
ятельством связано и наличие небольшого расширения при основании протока се-
СЕРАЕА
321
меприемника. Длина бича превышает суммарную длину пениса и эпифаллуса; на-
ружно граница между этими отделами выражена нерезко и отмечена местом прикреп-
ления полового ретрактора.
Распространение и местообитания. Преимущественно се-
верные и средние районы Европы; вид заходит также в Пиренеи и занимает бблыпую
часть территории Великобритании. В СССР встречается местами вдоль северо-
Рис. 433. Сераеа (Сераеа) nemoralis (L.) из Кведлинбурга, ГДР,
V 1969. Внешний вид полового аппарата. (Ориг.).
западных границ: Латвия, Калининградская обл., о-в Хийумаав Балтийском море.
Населяет преимущественно умеренно увлажняемые разреженные леса, опушки,
встречается также на влажных лугах, в полосе дюн и в кустарниковых зарослях.
В ископаемом состоянии встречается начиная с плиоцена (Европа).
2. Сераеа (Сераеа) hortensis (Muller, 1774). Табл. XIX, 189; рис. 434, 435.
М ii 1 1 е г, 1774:52 (Helix); Taylor, 1911, Mon. Brit. Moll., 111:326,
t. 28 (Helix); Hesse, 1920, in Kobelt's Iconogr., N. F. 23 : 86, t. 642, fig. 7—10;
P i 1 sb r y, 1939 : 6, fig. 24; Лихарев, Раммельмейер, 1952 : 477,
рис. 404Б; Ehrmann, 1956 : 147, t. 6, fig. 85—86; L о z e k, 1956 : 226, t. 38,
fig. 2; Z i 1 c h, J a e c k e I, 1962 : 201; L о z e k, 1964 : 313, t. 29, fig. 3a—c; G i t-
tenberger et al., 1970 : 105, fig. 186, 188, Kaart 75; Ш и л e й к о, 1972г : 95—
99, рис. 12А.
Материал: более 30 экз. из 2 местонахождений в Прибалтике и ГДР.
Раковина похожа на раковину предыдущего вида, но меньше, более шаро-
видная, с более куполовидным контуром завитка; устье более узкое, верхний край
21 А. А. Шилейко
322
IV. HELICIDAE
его на значительном протяжении параллелен колумеллярному; губа, края устья
и париетальная стенка белые. Размеры: ВР 15—16, БД 18—21, МД 15—17.5 мм.
Внутреннее строение. Материал: 9 экз. из Кведлинбурга, ГДР,
IV 1969, сбор и определение Э. Клаусса.
Внутреннее строение в существенных чертах сходно с таковым предыдущего вида.
Отличия: слизистые железы длиннее, стилофор значительно короче, верхний его конец
лишь слегка выдается за место перехода вагины в яйцевод; длина дивертикула в 2—
2.5 раза меньше длины протока семеприемника выше места его раздвоения.
Рис. 434—435. Сераеа (Сераеа) hortensis (Mull.) из Кведлинбурга, ГДР, IV 1969.
434 — внешний вид полового аппарата; 435 — пенис, вскрыт. (Ориг.).
Распространение и местообитания. Центральная и Северная
Европа с Исландией; в СССР — большая часть территории Прибалтики, единичные
находки в Витебской обл. Завезен в Северную Америку, где встречается местами
от Ньюфаундленда и Квебека до Нантакета. Населяет те же биотопы, что и С. пе-
moralis, часто эти два вида встречаются вместе. В ископаемом состоянии встречается
начиная с плиоцена (Европа).
2. Подрод OCTADENIA Schileyko, subgen. nov.
Окраска раковины белая, серая или желтоватая. Поверхность в виде тонкой
радиальной ребристости. Пупок запаян, но полость его сообщается с окружающей
средой посредством узкой щели между отворотом колумеллярного края и стенкой
раковины.
Задний отдел пениса примерно такой же длины, как и передний, дивертикул
семеприемника не короче лежащего ниже участка протока семеприемника. Каждая
слизистая' железа дихотомически ветвится трижды, общее число ветвей каждой же-
лезы — 8. га=25.
СЕРАЕА
323
От номинативного подрода отличается бблыпим числом ветвей слизистых желез;
меньшими размерами стилофора и числом хромосом. Конхологические отличия ка-
саются преимущественно скульптуры.
Распространение. Юго-Восточная Европа, Крым и Северный Кавказ.
В ископаемом состоянии подрод известен начиная с нижнего плейстоцена (Европа).
Типовой вид — Helix vindobonensis Ferussac, 1821.
3. Сераеа (Octadenia) vindobonensis (Ferussac, 1821). Табл. XIX, 190; рис. 436j
437.
Рис. 436—437. Сераеа (Octadenia) vindobonensis (Fer.) из Болгарии, г. Кюстандил,
28 VI 1969. 436 — внешний вид полового аппарата; 437 — пенис, вскрыт. (Ориг.).
Ferussac, 1821, Hist. Moll. : 21, t. 32A, fig 6—7 (Helix sylvatica var. Vindo-
bonensis); Rossmassler, 1835, Iconogr., I, 1 : 60, fig. 7 (H. austriaca); К r y-
n i c k i, 1837 : 51 (H. Arvensis); Westerland, 1889 : 444 (Helix); Hesse;
1920, in Kobelt's Iconogr., N. F., 23 : 96, t. 643, fig. 8—12; Лихарев, Рам-
мельмейер, 1952 : 478, рис. 405, 406; G г о s s u, 1955 : 494, fig. 282; Ehr-
mann, 1956 : 145, t. 6, fig. 82; Lozek, 1956 : 224, t. 38, fig. 1; Zilch, J ae-
c k e 1, 1962 : 198; Lozek, 1964 : 312, t. 29, fig. 2a—с; Шилейко, 1972г : 95—
99, рис. 12C, E.
Материал: более 100 экз. из 8 местонахождений в южных и юго-западных рай-
онах европейской части СССР и в Венгрии.
Раковина с более высоким, почти коническим завитком, нежели у видов
Сераеа s. str., составлена 5—5.5 выпуклыми оборотами. Последний оборот немного
шире предпоследнего, к устью сильно и круто, но не внезапно (без перегиба), опущен.
Окраска белая или серая, иногда с желтоватым или коричневым оттенком. Рисунок
почти всегда образован 5 лентами: первая и вторая ленты узкие и светлее остальных
(вторая часто очень узкая или отсутствует); остальные три ленты широкие; самая
широкая пятая лента)располагается на середине нижней стороны последнего оборота.
В проходящем свете фон плотнее лент. Скульптура в виде радиальной ребристой
исчерченности и тонкой микроскопической и неравномерной спиральной исчерчен?
ности, выраженной в разной степени. Устье округлое, слегка оттянуто вниз и вправо;
21*
324
IV. HELICIDAE
колумеллярный край более отвесный, чем у прочих представителей рода. Отворот
колумеллярного края полностью закрывает пупок и на некотором протяжении
плотно прилегает к нижней стенке последнего оборота, затем он резко отходит от
стенки раковины, так что под ним образуется узкая щель, которая, однако, не со-
ответствует пупку. Размеры: ВР 17—20, БД 20—24, МД 17—20 мм.
Внутреннее строение. Материал: 3 экз. из Венгрии, с. Тихань на
берегу оз. Балатон, 26 VII 1970, сбор Д. А. Сахарова; 3 — из Одессы, VIII 1959;
5 — из Ужгорода, 5 IX 1969; сборы и определения наши; 1 — из Болгарии, г. Кю-
стандил, 28 VI 1969, сбор и определение И. М. Лихарева.
Изгиб яйцевода плавный и широкий. Слизистые железы относительно очень
длинные. Стилофор сравнительно маленький, вершина его не доходит до границы
вагины и яйцевода.
Пенис небольшой, внутреннее его строение характерно для рода (рис. 437). Ди-
вертикул семеприемника развит значительно лучше, чем у других видов рода, и от-
ходит от протока ниже, чем у них.
Распространение, как у подрода. Населяет редколесье равнинных
территорий, лесов избегает.
2. Род CRYPTOMPHALUS Charpentier, 1837
Charpentier, 1837 :6; Moquin-Tandon, 1855 : 174; Лихарев,
Раммельмейер, 1952 : 488 [Helix (Cryptomphalus) ]; Wenz, Zilch, 1959 —
1960 : 722.
Раковина в общем шаровидная, окраска желтых или коричневых тонов; спираль-
ные ленты присутствуют всегда, но иногда слабо выражены и часто разбиты на ряды
пятен. Скульптура вермикулятная; пупок запаян.
Слизистые железы обильно и беспорядочно ветвятся. Бич и дивертикул семепри-
емнйка очень длинные, значительно длиннее протока семеприемника и суммарной
длины пениса и эпифаллуса. Папилла пениса одна; ниже ее располагается очень
мощная высокая полукольцевая складка.
Распространение. Западная и Южная Европа и Северная Африка.
В ископаемом состоянии род известен начиная с верхнего плиоцена (Южная Европа,
Северная Африка).
Монотипический род.
Типовой вид — Helix aspersa Muller, 1774,
Cryptomphalus aspersa (Miiller, 1774). Табл. IX, 89; XX, 191; рис. 438—440.
M ii 11 e г, 1774 : 59 (Helix)-, Moquin-Tandon, 1855 : 174, t. 13, fig. 14,
32; Westerlund, 1889:450 {Helix)-, Kobelt, 1906, Helic., VI : 96, t. 319,
fig. 14—20; t. 320, fig. 1—12; Hesse, 1920, in Kobelt's Iconogr., N. F., 23 : 144;
Pilsbry, 1939 : 4, fig. 1, 3b—g; Лихарев, Раммельмейер, 1952 : 488,
рис. 415, 416 [Helix (Cryptomphalus)]; Ehrmann, 1956:149 [Helix (Cryptom-
phalus) ]; Gittenberger et al., 1970 : 106, fig. 190, 192, Kaart 77 [Helix (Cryp-
tomphalus) ].
Материал: 18 экз. из Франции, Испании и Алжира.
Раковина шаровидная или шаровидно-кубаревидная, сравнительно тонко-
стенная, просвечивающая, состоит из 4.5 выпуклых, быстро нарастающих оборотов.
Последний оборот почти вдвое шире предпоследнего, сильно вздут, оттянут вниз
и вправо, к устью сильно, но очень плавно опущен. Окраска желтая или желтовато-
коричневая; имеется 5 спиральных лент, обычно разбитых на ряды пятен; вторая
и третья ленты часто слиты в одну. Поперек лент проходят выраженные в разной
степени желтые извилистые или зигзагообразные линии и пятна. Скульптура очень
характерная — сеть тонких морщин, окрашенных светлее фона; в проходящем свете,
однако, морщины темнее фона. Устье коротко-овальное, высота его немного превы-
шает ширину или равна ей. Края устья тонкие, расширенные и отвернутые, с тонкой
‘белой губой. Размеры: ВР 27—-35, БД 27—38, МД 22—32 мм.
Внутреннее строение. Материал: 3 экз. из Алжира, окр. г. Бужи
(=Беджайя), 10 II 1973, сбор Н. Н. Воронцова; 3 —из Франции, окр. г. Монпелье,
VIII 1971, сбор Д. А. Сахарова; определения наши.
Спермовидукт при переходе в яйцевод образует резкий изгиб. Общее число вет-
вей каждой слизистой железы может достигать 20. Стилофор необыкновенно мас-
сивный, короткобулавовидный. Вагина внутри с резкими продольными складками.
Проксимальная часть довольно короткого пениса вздутая, почти шаровидная,
дистальная — цилиндрическая. Проксимальная часть заключает короткую шаро-
видную папиллу, несущую на поверхности многочисленные бугорки; стенки папиллы
полостей не заключают. Отделы пениса сообщаются друг с другом посредством щели
между стенкой пениального чехла и мощной полукольцевой складкой, расположен-
Рис. 438—440. Cryptomphalus aspersa (Mull.) из окр. Монпелье, Франция,
VIII 1971. 438 — внешний вид полового аппарата; 439 — пенис, вскрыт; 440 —
продольный разрез пениса, схема (звездочкой помечена полукольцевая складка).
(Ориг.).
326
IV. HELICIDAE
ной немного ниже папиллы. Место раздвоения протока семеприемника находится
на уровне нижней части спермовидукта. Дивертикул семеприемника и бич резко
и многократно извиты и скручены.
Распространение и местообитания. Южные и, местами, цен-
тральные части Западной Европы и Северная Африка. Вид завезен на острова Ат-
лантики, в Южную Африку, Австралию, Новую Зеландию, на о-в Гаити, во многие
пункты Мексики, Чили, Аргентины, Северной Америки (Ю. Каролина, Луизиана,
Рис. 441—442. Eobania vermiculata (Miill.) из окр. Севастополя, 12 VIII 1962. 441 —
внешний вид полового аппарата; 442 — пенис, вскрыт. (Ориг.).
Калифорния). В СССР обнаружен один раз в окрестностях Феодосии. Населяет лист-
венные леса, реже — кустарниковые заросли. Известен из верхнего плиоцена Си-
цилии и Алжира.
3. Род EOBANIA Hesse, 1915
Hesse, 1915, in Kobelt's Iconorg., N. F., 23 : 25; Л и x a p ев, . P а м м e л ь-
мейер, 1952 : 474; Wen z, Zilch, 1959—1960 : 712.
Раковина слегка прижатая, округлая, основная окраска от почти белой до ко-
ричневатой; одноцветная или, чаще, с 4—5 спиральными лентами, которые могут
распадаться на ряды пятен. Скульптура вермикулятная, пупок запаян.
Слизистые железы в нижней или средней части раздваиваются, и в верхней части
эти стволы обильно и беспорядочно ветвятся. Длина бича приблизительно равна
суммарной длине пениса и эпифаллуса, дивертикул семеприемника значительно длин-
нее. Папилл пениса 2, проксимальная очень длинная.
Р аспространение. Средиземноморье. В ископаемом состоянии род
известен начиная с нижнего плиоцена (Южная Европа и Северная Африка).
Типовой вид — Helix vermiculata Muller, 1774.
По всей вероятности название Eobania является младшим синонимом названия
Otala Schumacher, 1817; если это предположение подтвердится, то типовым видом
нужно считать Helix lactea Muller, 1774.
LEVANTINA
327
Eobania vermiculata (Muller, 1774). Табл. XX, 192; рис. 441, 442.
Muller, 1774:20 (Helix); Rossmassler, 1837, Iconogr., I, 5:6,
fig. 301 (Helix); Moquin-Tandon, 1855 : 159, t. 12, fig. 25—29 [Helix (Otala) ];
W esterlund, 1889 : 411 (Helix); Hesse, 1915, in Kobelt's Iconogr., N. F.,
33 : 26, t. 634, fig. 3—11; P i I s b r y, 1939 : 11, fig. 6d [Otala (Eobania)]; Лиха-
рев, Раммельмейер, 1952 : 474, рис. 401, 402.
Материал: более 60 экз. из Крыма и Франции.
Раковина с закругленным куполовидным завитком и сглаженной верши-
ной; высота завитка обычно немного превышает высоту устья. Оборотов 5, умеренно
выпуклых, плавно нарастающих. Последний оборот вздут, в профиле правильно
закруглен, в 1.5—2 раза шире предпоследнего, к устью энергично, с плавным пере-
гибом, опущен. Окраска от серовато-белой до коричневато-желтой, одноцветная
или, чаще, с 4—5 спиральными темными лентами, которые часто сливаются друг
с другом или распадаются на ряды пятен. Первая (верхняя) лента самая широкая
и располагается у шва; вторая и третья ленты нередко слиты и находятся выше пери-
ферии; четвертая и пятая не обнаруживают наклонности к слиянию и лежат ниже
периферии. Поверхность раковины неясно радиально исчерчена и покрыта много-
численными белыми короткими морщинами, беспорядочно ориентированными и пере-
плетающимися друг с другом; в проходящем свете эти элементы вермикулятной
скульптуры темнее фона. Устье коротко-овальное, очень косое, внутри с беловатым
налетом. Края устья острые, сильно отвернуты, утолщенные, светлые. Размеры:
ВР 17—22, БД 27—32, МД 21—24 мм.
Внутреннее строение. Материал: 4 экз. из Франции, окр. г. Мон-
пелье, VIII 1971, сбор Д. А. Сахарова; 2 — из Крыма, окрестности Севастополя,
12 VIII 1962; 3 — из Крыма, Ялта, Никитский ботанический сад, 2 X 1964; сборы
и определения наши.
Нижняя часть спермовидукта и верхняя часть яйцевода образуют крутой двой-
ной изгиб. Нижняя часть слизистых желез резко сужена и образует резкий изгиб;
затем их диаметр резко увеличивается и каждая железа раздваивается более или
менее близко к основанию. Эти две основные ветви всегда выражены четко, и лишь
выше середины начинают распадаться на ветви второго и последующих порядков.
Стилофор относительно небольшой, овальный. Его основание вместе с клоакой и
нижней частью вагины охвачено общим чехлом.
Пенис образует обычно 2 вздутия: дистальное, соответствующее пениальному
бугорку, и проксимальное, где располагается проксимальная папилла. Обе папиллы
длинные, без интрапапиллярных полостей; проксимальная папилла в 2—3 раза длин-
нее дистальной. Внутренняя поверхность пениального чехла в области проксималь-
ной папиллы покрыта очень высокими узкими складками, выраженными неравномерно.
Эпифаллус цилиндрический, изогнут, в разрезе его просвет имеет форму четырех-
лучевой звезды, каждый луч которой на конце раздвоен. Проток семеприемника
раздвоен примерно на середине длины; дивертикул длинный, многократно и резко
извит и скручен. Проток семеприемника выше места бифуркации тонкий, почти пря-
мой. .
Распространение и местообитания. Вид имеет типичный
циркумсредиземноморский ареал; завезен в Северную Америку (Новый Орлеан).
В Крым также завезен. Населяет открытые участки близ морского побережья.
В ископаемом состоянии гид известен начиная с нижнего плиоцена ^Румыния, Ита-
лия, Алжир).
4. Род LEVANTINA Kobelt, 1871
К о b е 11, 1871 : 19; Hesse, 1908, in Kobelt's Iconogr., N. F., 14: 139;
1918a : 40—47; Лихарев, Раммельмейеп. 1952 : 479; W e n z. Zilch,
1959—1960 : 719.
Раковина сильно уплощенная, белая, почти одноцветная, спиральные ленты
выражены слабо. Скульптура слабая, представлена тонкой радиальной исчерчен-
ностью. Места прикрепления устья сильно сближены, иногда край устья цельный.
Пупок всегда широкий, развернутый, но благодаря интенсивному развитию колу-
меллярного края в разной степени прикрыт, нередко полностью запаян.
Слизистые железы слабо развиты, простые или двуветвистые. Бич очень длин-
ный, закручен в спираль; дивертикул семеприемника также длинный, извитой. Па-
пиллы пениса две, обе хорошо развиты.
Распространение. Северо-западная Ливия, Передняя Азия, юг Ар-
мении и Нахичеванская АССР.
Типовой вид — Helix spiriplana Olivier, 1821.
328
IV. HELICIDAE
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА LEVANTINA
1 (2). Пупок перспективный, открытый, иногда слегка прикрыт отворотом колумел-
лярного края............................................2. L. escheriana.
2 (1). Пупок запаян полностью; в крайнем случае от него остается щель..
..................................................1. L. djulfensis.
1. Levantina djulfensis (Dubois de Montpereux, 1840). Табл. XX, 193; рис. 443, 444.
Dubois de Montpereux, 1840 : 24 (Helix)-, Bourguignat, 1856,
II : 63, t. 12, fig. 7—9 (Helix)-, Mousson, 1863 : 373 (Helix)- К о b e 1 t, 1890,
Iconogr., N. F., IV : 75, fig. 651, 652 (H. urmiensis); Westerlund, 1897:140
(H. placida); 1898 : 162 (H. casta); Kobelt, 1906, Helic., VI : 5, t. 30, Tig. 4—6;
Hesse, 1908, in Kobelt's Iconogr., N. F., 14 : 149, t. 384, fig. 1, 2 (H. urmiensis);
Лихарев, Раммельмейер, 1952 : 479, рис. 407—408.
Материал: 7 экз. из 2 местонахождений в Армении.
Раковина сильно прижатая, с низким завитком и умеренно выступающей
вершиной, твердостенная, составлена 4—4.5 оборотами. Обороты выпуклые, стре-
мительно нарастающие, разделенные глубоким швом; последний оборот примерно
вдвое шире предпоследнего, к устью плавно, но очень сильно опущен. Окраска го-
лубовато- или рыжевато-белая, рисунок образован 4—5 рядами слабых светло-ко-
ричневых пятен с размытыми контурами. Широкие эмбриональные обороты покрыты
очень тонкой струйчатой радиальной морщинистостью; скульптура дефинитивных
оборотов представлена слабой радиальной неравномерной исчерченностью, иногда
местами присутствуют слабые спиральные бороздки. Устье очень косое, овальное,
внутри белое, оттянуто вправо, места его прикрепления сильно сближены; парие-
тальная мозоль выражена в разной степени, иногда даже слегка утолщена. Края
устья сильно отвернуты, с тонкой губой. Колумеллярный край сильно расширен
и полностью закрывает пупок; иногда между колумеллярным краем и нижней стен-
кой раковины остается узкая щель. Отворот колумеллярного края несколько вдав-
лен в пупочную впадину. Размеры: ВР 16—18, БД 29—35, МД 23—27 мм.
Внутреннее строение. Материал: 4 экз. из Армении, окр. с. Мегри,
28 IV 1947, сбор и определение Н. Н. Акрамовского.
В нижней части спермовидукта имеется плавный, но хорошо выраженный изгиб.
Яйцевод короткий, не менее чем втрое короче вагины. Обе слизистые железы виль-
чатые или одна из них не разветвляется. Стилофор булавовидно-цилиндрический,
короче вагины.
Пенис веретеновидный, дистальная часть его сужена и круто изогнута; здесь
располагается слабо развитый пениальный бугорок. Дистальная папилла больше
проксимальной, покрыта многочисленными кольцевыми складками, стенки ее не
заключают интрапапиллярных полостей; просвет папиллы в поперечном разрезе
имеет Х-образную форму. Проксимальная папилла заключает интрапапиллярную
полость, однако семепроводящий канал фиксирован несколькими тяжами, что пре-
пятствует свободному изменению формы и длины папиллы. Витки спирали, образу-
емой бичом, сомкнуты. Эпифаллус цилиндрический, в 1.5 раза длиннее пениса. Про-
ток семеприемника раздваивается довольно низко, дивертикул скручен, все его из-
гибы связаны между собой и притянуты к матке прозрачной мембраной. У одного
из исследованных экземпляров внутри дивертикула обнаружен сперматофор,
(рис. 443), хвостовая часть которого заворачивалась в собственно проток семепри-
емника.
Распространение и местообитания. Южная Армения (Мег-
ринский и Разданский р-ны), Нахичеванская АССР, северо-западный Иран, се-
верный Ирак и прилежащие к нему территории Турции. Обитает в аридных горных
участках, на скалах.
2. Levantina escheriana (Bourguignat, 1864). Табл. XX, 194; рис. 445, 446.
Bourguignat, 1864:105, t. 12, fig. 8—11 (Helix); Westerlund,
1889 : 395 (Helix); К о b e 1 1, 1906, Helic., VI : 6, t. 300, fig. 7—12; Лихарев,
Раммельмейер, 1952 : 481; Акрамовский, 1971a : НО.
Материал: 4 экз. из 2 местонахождений в Армении.
Вид похож на предыдущий. Конхологические отличия: имеются 2—4 спираль-
ные ленты, которые также разбиты на отдельные пятна; раковина более уплощенная;
радиальная скульптура более резкая; на базальной части последнего оборота присут-
ствуют вмятины; пупок открытый, иногда слегка прикрыт отворотом колумелляр-
ного края; наблюдается наклонность к образованию цельного устья. Размеры:
ВР 14—16, БД 29—35, МД 23—27 мм.
Внутреннее строение. Материал: 3 экз. из Армении, Азизбеков-
ский р-н, окр. с. Арени (Арна), 12 V 1953, сбор и определение Н. Н. Акрамовского.
Рис. 443—444. Levantina djulfensis (Dub. de Mont.) из Армении, окр. с. Мегри,
28 IV 1947. 443 — внешний вид полового аппарата; 444 — пенис, вскрыт. (Ориг.).
330
IV. HELICIDAE
Нижняя часть спермовидукта и яйцевод почти прямые. Обе слизистые железы
двуветвистые. Стилофор большой, овальный, длиннее вагины. Яйцевод в
в 1.5—2 раза короче вагины.
Пенис в дистальной части тонкий, изогнутый, далее имеет веретеновидную форму.
Дистальная папилла пениса короче проксимальной, в стенках ее имеется кольцевая
полость; просвет семепроводящего канала на поперечном срезе имеет форму полу-
месяца' с короткими выростами. Форма просвета канала в пределах проксимальной
Рис. 445—446. Levanttna escheriana (Bourg.) из Армении, окр. с. Арени, 12 V 1953.
445 — внешний вид полового аппарата; 446 — пенис, вскрыт. (Ориг.).
папиллы менее постоянна; здесь, как и у предыдущего вида, имеются продольные,
слабо дифференцированные тяжи, фиксирующие семепроводящий канал. Пениаль-
ный бугорок рудиментарный,’ имеет вид небольшого утолщения одной из продольных
складок. Эпифаллус немного длиннее пениса; строение бича и семеприемника — как
у предыдущего вида.
Распространение и местообитания. Южная Армения и На-
хичеванская АССР, северный Иран, Армянский Тавр в Турции. Населяет сухие
каньоны полупустынной зоны, в жаркое время уходит глубоко в трещины скал (Ак-
рамовский, 1949).
5. Род CAUCASOTACHEA С. Boettger, 1911
С. Boettger, 1911:130 (Tachea, sect. Caucasotachea); Hesse, 1920,
in Kobelt's Iconogr., N. F., 23 : 100; Лихарев, Раммельмейер,
1952:481; Wenz, Zilch, 1959—1960:720.
Раковина в общем кубаревидная, роговая, желтая или буровато-желтая, со спи-
ральными темными лентами, из которых самая широкая проходит по базальной части
раковины. Скульптура — от тонкой радиальной исчерченности до грубой, иногда
CAUCASOTACHEA
331
с вмятинами. Места прикрепления устья не обнаруживают тенденции к сближению.
Пупок всегда полностью запаян.
Слизистые железы развиты очень хорошо, многоветвистые. Бич длиннее пениса
и эпифаллуса, вместе взятых, в спираль не закручен; дивертикул семеприемника
также очень длинный; папиллы пениса две.
Распространение. Кавказ, северный Иран и северо-западная Турция.
.В ископаемом состоянии род известен начиная с миоцена (Малая Азия и Кавказ).
Типовой вид — Helix atrolabiata Krynicki, 1833.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕПТНЫХ ВИДОВ
РОДА CAUCASOTACHEA
1 (4). На поверхности раковины присутствуют элементы решетчатой скульптуры.
2 (3). Решетчатая скульптура покрывает весь последний оборот. Спиральные ленты
черные или светло-коричневые (иногда отсутствуют). Большой диаметр не пре-
вышает 34 мм...........................................4. С. lencoranea.
3 '“’). Решетчатая или вермикулятная скульптура на последнем обороте выражена
местами. Спиральные ленты зеленовато-черные. Большой диаметр не менее
35 мм..................................................5. С. atrolabiata.
4 (1). Элементы решетчатой скульптуры отсутствуют, скульптура в виде неравномер-
ной радиальной исчерченности и тончайшей спиральной струйчатости . . .
........................................................ 6. С. calligera.
f 1. Caucasotachea andrussovi Steklov, 1962. Табл. XX, 195.
Стеклов, 1962 : табл. I, фиг. 1, 2 (Galactochylus sp.); 1966 : 228, табл. ХП,
220— 223- табл. XIII, 229.
Материал: 7 экз. из 2 местонахождений в Предкавказье.
Раковина коническая, со слегка притупленным завитком. Оборотов 5 или
чуть больше, плотно навитых, слабо выпуклых, разделенных мелким швом. Послед-
ний оборот в профиле с углом, который к устью полностью сглаживается, в конце
резко и сильно опущен. Рисунок образован 4 темными спиральными лентами, из
которых 2 лежат ниже периферии (верхняя из них — самая широкая), а 2 — выше.
Скульптура сложная, легко стирающаяся, различима только при хорошей сохран-
ности. Первые 1.5 оборота почти гладкие, блестящие, покрыты микроскопическими
волнистыми или прерывистыми складочками; у нижнего шва эти складочки стано-
вятся более грубыми. Более поздние обороты тонко и неравномерно исчерчены и
покрыты микроскопической зернистой скульптурой, которая становится более четкой
по мере нарастания оборотов; зернистость возникает за счет пересечения очень тон-
ких радиальных линий и тончайших складок, направленных под острым углом
к шву. На двух последних оборотах микрозернистая скульптура теряет правиль-
ность и переходит в сложный скульптурный узор, сочетающий довольно правильные
гранулированные поля и участки с волнистой струйчатостью. Исчерченность послед-
него оборота грубее, на нем выделяются многочисленные штрихи, образующие серию
‘беспорядочно расположенных коротких вмятин и выступов. Устье большое, сильно
скошенное, округлое, с оплывшей губой; края устья плавно и широко отогнуты, ко-
лумеллярный край резко отогнут. Размеры: ВР 15.5—22, БД 23—26.5, МД 20—21.5 мм.
Распространение. Вид известен из верхнего миоцена (сармат) запад-
ного Предкавказья.
f 2. Caucasotachea fortangensis Steklov, 1966. Табл. XX, 196.
Стеклов, 1966 : 229, табл. XII, 227', табл. XIII, 232—234.
Материал: более 20 экз. из 1 местонахождения в Предкавказье.
Раковина коническая, с почти коническим завитком, высота которого почти
равна высоте устья. Оборотов 5—5.5, правильно нарастающих, умеренно выпуклых,
разделенных мелким швом. Последний оборот в профиле округлый, к устью резко
спущен. На поверхности многих раковин заметен рисунок из темных спиральных
лент: одна — две широкие ленты на базальной части, одна — тоже широкая, непо-
средственно под периферией, и три более узкие ленты над периферией. Скульптура
образована частыми радиальными складками, часто правильными и довольно резкими.
Устье сильно скошено, с расплывшейся губой, немного оттянуто вправо, края его
плавно и широко отвернуты. Колумеллярный край спаян с основанием раковины
и полностью закрывает пупок; на нем имеется небольшое вздутие. В максимальном
выражении это вздутие загнуто внутрь устья в виде своеобразной складки и нависает
над внутренней стенкой устья. Размеры: ВР 18—19, БД 25.7—28.5, МД 21—23 мм.
Распространение. Предкавказье: меотический песчаник на р. Фор-
танге и «слои с Helix» на р. Ярык-су и р. Сулак.
332
IV. HELICIDAE
f 3. Caucasotachea kubanica Steklov, 1966. Табл. XX, 197.
Стеклов, 1966 : 230, табл. XII, 228.
Материал: 10 экз. из 1 местонахождения в Предкавказье.
Вид очень похож на предыдущий. Отличия: раковина более тонкостенная, за-
виток относительно более высокий (высота его превышает высоту устья), отворот
края устья узкий, размеры мельче: ВР 13—14.3, БД 19—21, МД 17—18 мм.
Распространение. Караганские слои (песчаник на р. Псефирь, Кавказ).
4. Caucasotachea lencoranea (Mousson, 1863). Табл. XX, 198; рис. 447, 448.
Mousson, 1863 : 376 (Helix atrolabiata var. Lencoranea); К о b e 1 t, 1906,
Helic., VI : 64, t. 312, fig. 8, 10, 11; t. 314, fig. 4—9 [Helix (Tachea)]; Hesse, 1920,
in Kobelt's Iconogr., N. F., 23 : 107, t. 645, fig. 5—9; t. 646, fig. 10; Лихарев,
Раммельмейер, 1952:484; Шилейко, 1968a : 345, рис. 8, 9.
Материал: более 50 экз. из 19 пунктов в Ленкоранской низменности и Талыше.
Раковина от слегка прижатой до, кубаревидной, с притупленным, почти
коническим завитком, высота которого больше или равна высоте устья. Оборотов 5,
слабо выпуклых, плавно нарастающих. Последний оборот в профиле округлый,
снизу слегка уплощен, не более чем в 1.5 раза шире предпоследнего, к устью плавно,
без резкого перегиба, но сильно опущен. Окраска от желтовато-коричневой до почти
желтой, с 5 черными или светло-коричневыми лентами; нередко отдельные ленты
отсутствуют, но одноцветные раковины встречаются редко. Ленты в большей или
меньшей степени разбиты на ряды йятен многочисленными светлыми штрихами.
Скульптура представлена грубой радиальной исчерченностью и многочисленными
вмятинами, желтоватые края которых выделяются на более темном фоне. Устье
округлое, сильно скошено, внутри с синеватым оттенком и просвечивающими лен-
тами. Края устья и париетальная стенка черные или темно-коричневые. На колумел-
лярном крае иногда заметно продольное утолщение, окрашенное светлее, чем осталь-
ная поверхность края. Размеры: ВР 18—25, БД 27—34, МД 22—27 мм.
Внутреннее строение. Материал: 23 экз. из 14 пунктов лесной зоны
Талыша и Ленкоранской низменности, VI—VII 1964, III—IV 1965 и IX 1966, сборы
и определения наши.
Дистальная часть спермовидукта переходит в яйцевод без изгибов. Слизистые
железы в общем очень длинные, но длина их зависит от функционального состояния.
Каждая железа делится на 3 основных ствола, в свою очередь ветвящихся: централь-
ный ствол распадается чаще всего на две ветви, боковые стволы ветвятся более ин-
тенсивно, в результате общее число ветвей 9—14.
Пенис слабо вздут, почти цилиндрический, пениальный бугорок длинный, но
нечетко дифференцированный. Дистальная папилла развита лучше проксимальной,
интрапапиллярные полости отсутствуют. Семепрсводящий канал в пределах дисталь-
ной папиллы имеет на разрезе вид трехлучевой звезды. Наружный чехол пениса от-
слаивается только в районе, отвечающем обеим папиллам. Внутренняя поверхность
пениса между папиллами несет мощный продольный валик. Бич примерно в 1.5 раза
длиннее суммарной длины пениса и эпифаллуса. Проток семеприемника разветвля-
ется примерно на середине длины; дивертикул семеприемника многократно извит
и заметно длиннее участка протока выше места раздвоения.
Распространение и местообитания. Талыш и Ленкоранская
низменность (АзССР). Иран — лесная зона хребта Эльбурс (юго-западное побережье
Каспийского моря) и прилегающие участки низменности. Лесной вид, на открытых
местах не встречается. Обитает в подстилке, но нередко поднимается на деревья,
изредка даже в кроны.
5. Caucasotachea atrolabiata (Krynicki, 1833). Табл. XX, 199; рис. 449, 450.
Krynicki, 1833 : 425, t. 9 [Helix (Acave) ]; Retowski, 1889 : 261
(H. atrolabiata yar. intercedens); К о b e 1 1, 1906, Helic., VI : 63, t. 312, fig. 4—6
[Helix (Tachea) stauropolitana Schmidt]; Rosen, 1911 : 115 (H. stauropolitana); R e-
t о w s k i, 1914 : 296 [77. (Tachea) atrolabiata f. tricolor Bttg., maculata-fasciata Bttg.,
alternans, translens, decussata Bttg.]; Rosen, 1914: 177 (Helix); Рошковский, 1919:
134—136; Hesse, 1920, in Kobelt's Iconogr., N. F., 23 : 104, t. 644, fig. 6—7, 7a;
t. 645, fig. 1—4 (H. atrolabiata stauropolitana); Линдгольм, 1928 : 548; Лиха-
рев, Раммельмейер, 1952 : 483, рис. 411, 412А; Hudec, Lezhawa,
1969b : 147, Abb. 35, t. 15, fig. 35.
Материал: 18 экз. из 3 местонахождений в западной части Большого Кавказа.
Раковина кубаревидная, твердостенная, с притупленным, почти кониче-
ским завитком, высота которого сильно варьирует, но всегда больше высоты устья.
Рис. 447—448. Caucasotachea lencoranea (Mouss.)
из окр. Ленкорани, 9 VII 1964. 447 — внешний
вид полового аппарата; 448 — пенис, вскрыт.
(Ориг.).
334
IV. HELICIDAE
Оборотов 5, умеренно выпуклых, плавно нарастающих. Последний оборот менее-
чем в 1.5 раза шире предпоследнего, к устью опущен с ясным, но плавным перегибом.
Окраска коричневато-желтая, элементы скульптуры желтые; имеются 4 зеленовато-
черные ленты, которые в проходящем свете имеют коричневато-красный цвет. Самая
узкая лента проходит под швом, самая широкая — по базальной части раковины,
часто захватывая и область пупка. Остальные 2 ленты располагаются выше и ниже
середины последнего оборота. Ленты в разных комбинациях могут отсутствовать,
иногда раковина одноцветная. Скульптура в виде сильной и неравномерной радиаль-
Рис. 449—450. Caucasotachea atrolabiata (Кгуп.) из долины р. Ингури, 10 IV 1971
449 — внешний вид полового аппарата; 450 — пенис, вскрыт. (Ориг.).
ной исчерченности, местами имеется решетчатая скульптура; кроме того, наблюдаются*
элементы вермикулятной скульптуры. Устье коротко-овальное, косое. Палатальная,
стенка устья, края и губа коричневые или почти черные; края устья широко отвер-
нуты, палатальный край плавно^выгнут. Отворот колумеллярного края припаян
к нижней стенке раковины, но при переходе в базальный край заметно отходит, так
что под ним образуется узкая щель. На колумеллярном крае часто имеется продоль-
ное или косое вздутие. Размеры: ВР 21—36 (в отдельных случаях до 38), БД 35—42,.
МД 25—36 мм.
Внутреннее строение. Материал: 2 экз. из окр. г. Хосты, V 1964,.
сбор И. И. Малевича; 1 — из долины р. Убин (северо-западная часть Большого Кав-
каза); 2 — из долины р. Ингури выше с. Джвари (юго-западная часть Большого
Кавказа), 10 IV 1971; сборы и определения наши.
Нижний отдел спермовидукта и яйцевод плавно изогнуты, причем яйцевод слегка
охватывает основание протока семеприемника. Длина слизистых желез изменчива,
но в общем меньше, чем у предыдущего вида. Стилофор удлиненно-булавовидный.
Пенис цилиндрический или образует 2 небольших вздутия, отвечающих папиллам.
На границе пениса и клоаки имеется небольшой пениальный бугорок, выше которого
помещается кольцевое вздутие, играющее, по-видимому, роль сфинктера. Прокси-
мальная папилла интрапапиллярных полостей не заключает; в стенках дистальной,
папиллы среди рыхлой паренхимы разбросаны многочисленные продольные щеле-
STEKLOVIA 335 -
видные лакуны. Таким образом, длина обеих папилл может меняться только за счет
' выравнивания кольцевых складок, имеющихся в большом числе на их поверхностях.
Просвет дистальной папиллы обширный, неправильной формы. Стенки пениального
чехла в пространстве между папиллами очень рыхлые, волокнистые; внутренняя
поверхность чехла в этом районе складчатая, здесь же располагается небольшой,
выраженный в разной степени бугорок, на конце которого открывается небольшое
отверстие, посредством которого просвет пениса сообщается с рыхлой паренхимой
стенки пениса. Длина бича несколько превышает суммарную длину пениса и эпифал-
луса. Проток семеприемника в нижней части образует более или менее ясно выражен-
ную петлю. Дивертикул семеприемника длиннее и толще протока выше места раз-
двоения.
Распространение и местообитания. Подкововидный ареал
вида охватывает западное крыло Большого Кавказа с севера, запада и юга. Обитает
в лиственных и смешанных лесах; часто поднимается на стволы деревьев. Близкая
форма известна из четвертичных делювиальных отложений северного Ирана.
6. Caucasotachea calligera (Dubois de Montpereux, £1840). Табл. XX, 200;
рис. 451—455.
Dubois de Montpereux, 1840 : 87 (ffelia,); M о u s s о n, 1863 : 375
(H. calligera et var. pallasii}; Retowski, 1914:296 [ff. (Tachea) latrolabiata
I. nemoraloides Martens, 1874, pallasii Dub.]; Рошковский, 1919 : 134 (Tachea
argonautarum}; Hesse, 1920, in Kobelt's Iconogr., N. F., 23 : 101, t. 644, fig. 1—5,
7b, 8 (C, atrolabiata}; Линдгольм, 1928 : 548; Лихарев, Раммель-
мейер, 1952 ; 484; Hude с, Lezhawa, 1969b : 146, Abb. 34, t. 15, fig. 36.
Материал: более 100 экз. из 6 местонахождений в западной части Грузии.
Раковина от слегка прижатой до кубаревидной, с притупленным коническим
или слегка куполовидным завитком, умеренно твердостенная. Свежие раковины бле-
стят, как лакированные. Оборотов 5, умеренно выпуклых, плавно нарастающих. По-
следний оборот немного шире предпоследнего, к устью сильно, но плавно опущен.
Окраска светло-желтая, первично с 3 темными лентами разной ширины, которые ни-
когда не имеют зеленоватого оттенка. Одна из лент располагается выше, две — ниже
периферии; часто число полос уменьшается до 2 или 1, или раковина одноцветная.
Поверхность раковины без вмятин, скульптура представлена тонкой и неравномерной
радиальной исчерченностью и тончайшей спиральной струйчатостью. Устье широко-
овальное, сильно скошено, палатальная стенка его светлая; париетальная стенка,
края устья и губа коричневые разных оттенков, иногда почти черные. Края устья ши-
роко отвернуты, на колумеллярном крае имеется продольный наплыв, часто хорошо
развитый и окрашенный немного светлее, чем прилежащие участки устьевого края.
Размеры: ВР 22—25 (изредка до 28), БД 27—34, МД 23—29 мм.
Внутреннее строение. Материал: 1 экз. из Батуми, Ботанический
сад, 5 IV 1971; 3 — из окр. с. Хоби в Колхидской низменности; 8 IV 1971; 22 — из до-
лины р. Техури на южных склонах западной части Большого Кавказа, 28—30 IV 1971;
сборы и определения наши.
Нижняя часть спермовидукта и верхний отдел яйцевода образуют небольшой, но
резкий двойной изгиб. Ниже изгиба яйцевод прямой. Слизистые железы длинные, вет-
вятся менее интенсивно, чем у других видов рода: общее число ветвей каждой железы
редко достигает 10, обычно 6—8. Стилофор почти цилиндрический.
Пенис длинный, с неясными вздутиями. Пениальный бугорок небольшой, гребне-
видный, выше него имеется полукольцевое вздутие. Дистальная папилла немного длин-
нее проксимальной, стенки ее полостей не заключают, просвет ее на поперечном се-
чении имеет характерную форму, напоминающую сочетание буквы «С» и ее зеркального
отражения, спаянных выпуклыми сторонами. Стенки внутреннего пениального чехла
между папиллами утолщены, этот участок способен значительно удлиняться, при сок-
ращении его возникают многочисленные резкие кольцевые складки. Длина бича варьи-
рует в значительных пределах, но не меньше суммарной длины пениса и эпифаллуса.
У одного ив исследованных экземпляров обнаружен сперматофор, расположенный внут-
ри дивертикула, но хвостовая часть его заворачивалась вверх, в проток семеприемника.
Распространение и местообитания. Западная часть Кавказа:
южные склоны Главного Кавказского хр., Колхидская низменность, западные районы.
Малого Кавказа.Северо-западная Турция. Обитает в лесах и кустарниковых зарослях.
f 6. Род STEKLOVIA Schileyko, gen. nov.^
Раковина шаровидная, светлая, одноцветная или с одной тонкой темной лентой
выше периферии. Скульптура в виде радиальной морщинистости и спиральных бо- •
роздок на последнем обороте. Пупок полностью запаян.
Рис. 451—455. Caucasotachea calligera (Dub. de Mont.) из долины p. Техури,
29 IV 1971. 451 — внешний вид полового аппарата; 452 — пенис, вскрыт; 453—
455 — поперечные срезы через сперматофор: 453 — в верхней части дивертикула
семеприемника, 454 — в нижней части дивертикула семеприемника, 455 — в про-
токе семеприемника выше места раздвоения. (Ориг.).
HELIX 337
Род близок к роду Helix- отличается очень четкой спиральной скульптурой, ши-
роко отвернутыми устьевыми краями и тем, что отворот колумеллярного края вжат
в базальную часть раковины.
Распространение. Плиоцен Предкавказья.
Монотипический род.
Типовой вид — Caucasotachea (?) maslovae Steklov, 1966.
f Steklovia maslovae (Steklov, 1966). Табл. XXI, 201.}
Стеклов, 1966 : 230, табл. XIII, 230, 231 [Caucasotachea (?)].
Материал: 12 экз. из 2 местонахождений в Предкавказье. г"'
Раковина с куполовидным завитком, высота которого немного меньше вы-
соты устья. Оборотов 4.5—5,.довольно выпуклых; последний оборот явно преобладает
над остальными, в профиле широко закруглен, немного шире предпоследнего, к устью
резко и внезапно опущен. Эмбриональные обороты гладкие, дефинитивные покрыты ра-
диальными морщинами; эти морщины на последнем обороте более грубые и правильные.
В начале последнего оборота появляются густо расположенные спиральные бороздки,
которые начинаются отдельными штрихами. Устье относительно очень большое, резко
скошено, края его очень широко отвернуты. При переходе колумеллярного края в ба-
зальный имеется четкий закругленный угол. Колумеллярный край косой, с продоль-
ным наплывом. Пупок закрытый, причем отворот колумеллярного края в верхней части
вжат в базальную поверхность раковины. Размеры: ВР около 20, БД около 26, МД око-
ло 22 мм; поскольку материал представлен мятыми раковинами, точнее размеры ука-
зать невозможно.
Распространение. Верхний плиоцен Предкавказья (акчагыльские слои
в Пседахской балке; акчагыльские и апшеронские слои на р. Сунже у с. Алды).
7. Род HELIX Linnaeus, 1758
Linn aeus, 1758 : 768; Ferussac, 1821 : 27 (Helicogena); Beck, 1937 : 43
(Pomatia); Hesse, 1918, NachrbL d. malak. Ges., 50 : 38 (Lindholmta part.); 1920,
in Kobelt's Iconogr., N. F., 23 : 115; Лихарев, Раммельмейер, 1952 :
486; Wenz, Zilch, 1959—1960:721.
Раковина от шаровидной до шаровидно-кубаревидной, светлая, с темными спи-
ральными лентами, реже с темными радиальными полосами, изредка одноцветная.
Скульптура в виде радиальной исчерченности или морщинистости; часто имеются ред-
кие, но ясные спиральные бороздки и вмятины на последнем обороте. Пупок закрытый
или щелевидный.
Слизистые железы многоветвистые, но не длинные. Длина бича и, следовательно,
дивертикула семеприемника подвержена значительной изменчивости (иногда даже
в пределах одного вида). Папиллы пениса две.
Распространение. Европа, Кавказ, Малая Азия, Ближний Восток.
В ископаемом состоянии род известен начиная со среднего миоцена (Европа) и верхнего
миоцена (Предкавказье, Украина, Крым).
Типовой вид — Helix pomatia Linnaeus, 1758.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РЕЦЕПТНЫХ ВИДОВ РОДА HELIX
1 (4). Раковина шаровидная, завиток округлый, с относительно низкими оборотами:
Колумеллярный край устья низкий, пологий, отворот его плавно сужается и
переходит в базальный крап.
2 (3). Поверхность раковины тонко исчерчена. Четвертая сверху темная лента лежит
ниже места прикрепления палатального края устья или верхний край ее только
касается края устья. Пупок в виде узкой щели...........2. Н. nordmanni.
3 (2). Поверхность раковины грубо исчерчена. Четвертая сверху темная лента лежит
выше места прикрепления палатального края устья или нижний край ее касается
края устья. Пупок в виде широкой щели..................3. Н. christophi.
4 (1). Раковина шаровидно-кубаревидная, завиток конический, с относительно вы-
сокими оборотами. Колумеллярный край высокий, почти отвесный, отворот
его книзу резко сужается и не переходит на базальный край,
5 (6). Поверхность раковины с микроскопической зернистой скульптурой . . . .
..........................................................6. Н. buchi.
6 (5). Зернистая скульптура отсутствует.
7 (10). Виды обитают в Крыму и на Кавказе.
8 (9). Эмбриональные обороты широкие, не больше чем в ,1.5 раза Уже первого дефи-
нитивного оборота, образуют широкую округлую вершину . . 4. Н. albescens.
9 (8). Эмбриональные обороты узкие, примерно вдвое уже первого дефинитивного
оборота, образуют узкую, слегка притупленную вершину ; . . 5. Н. lucorum.
22 А. А. Шилейко
338
IV. HELICIDAE
10 (7). Виды обитают в средних, западных и юго-западных областях европейской
части СССР.
11 (12). Высота и большой диаметр раковины не превышают 34 мм. Окраска одноцвет-
ная, светлая, изредка намечаются темные спиральные ленты.......................
........................................................ 7. Н. lutescens.
12 (И). Высота и большой диаметр раковины не менее 37 мм. Окраска светлая, с более
или менее ясным рисунком из 4 темных спиральных лент .... 8. Н. pomatia.
f 1. Helix maeotica Steklov, 1966. Табл. XXI, 202.
Стеклов, 1966 : 231, табл. XIII, 235, 236; табл. XIV, 237, 238.
Материал: 8 экз. из 2 местонахождений в Предкавказье.
Раковина шаровидная, с коническим, очень невысоким завитком, высота
которого меньше высоты устья. Оборотов около 5. Последний оборот очень сильно вздут,
к устью плавно опущен. Поверхность оборотов неравномерно радиально исчерчена, на
последнем обороте исчерченность переходит в слабую морщинистость. Устье большое,
слабо скошено, слегка оттянуто вправо, слабо вырезано париетальной стенкой устья.
Края устья слабо отвернуты и утолщены, образуют перистом. Колумеллярный край
Рис. 456—457. Helix nordmanni Mouss. из окр. с. Бенара на берегу р. Абастуманки
12 VI 1949 . 456 — внешний вид полового аппарата; 457 — пенис, вскрыт. (Ориг.)-
отвернут резко и полностью закрывает пупок. Размеры: ВР около 30, БД около 30,
МД около 24 мм; материал представлен мятыми раковинами.
Распространение. Верхний миоцен Предкавказья (меотические отло-
жения на р. Гумс и р. Аргудан).
2. Helix nordmanni Mousson, 1854. Табл. XXI, 203; рис. 456 , 457.
Mousson, 1854, Mitt. Naturf. Ges. Zurich : 366; Kobelt, 1906, Helic.,
VI : 181, t. 343, fig. 5—12 [Helix (Helicogena)]; Rosen, 1911 : 115 (H. vulgaris-
part.); Retowski, 1914 : 299 [Helix (Pomatia) vulgaris part. ]; R о s e n, 1914 : 179
HELIX
339
[Helix (Pomatia) vulgaris part.]; Hesse, 1920, in Kobelt’s Iconogr., N. F., 23 : 113,
t. 646, fig. 16—20; Лихарев, Раммельмейер, 1952 : 485 [Caucasotachea
(Lindholmia) ]; Hudec, Lezhawa, 1969b : 150, Abb. 37, t. 8, fig. 21 [Helix
(Lindholmia) ].
Материал: 8 экз. из 3 местонахождений в Закавказье.
Ра к о в и и а шаровидная, твердостенная, с округло-коническим завитком и
широкой сглаженной вершиной. Оборотов 4—4.5, слабо выпуклых, вначале медленно,
затем быстро нарастающих. Эмбриональные обороты гладкие, широкие, блестящие,
Рис. 458—459. Helix christophi Bttg. из Турции, окр. Ломашена, VI 1909.
458 — внешний вид полового аппарата; 459 — пенис, вскрыт. (Ориг.).
светло-желтые или серые. Последний оборот сильно вздут, в профиле широко и очень
правильно закруглен, вдвое шире предпоследнего, к устью очень плавно опущен.
Окраска от желтовато-белой до оливковой, с 5 светло- или темно-коричневыми лен-
тами, три из которых располагаются выше и две — ниже периферии. Самая тонкая —
вторая лента, самые широкие — первая, третья и пятая. Четвертая лента проходит
ниже места прикрепления верхнего края устья или ее верхняя граница касается края
устья. Пятая лента значительно отстоит от пупка. Скульптура в виде неравномерной
густой радиальной исчерченности или морщинистости, которая к концу последнего
оборота становится грубой, и редких вялых спиральных бороздок, выраженных ме-
стами только на последнем обороте. Устье большое, почти круглое, высота его слегка
превышает ширину, слабо вырезано предпоследним оборотом. Внутри устье светлое,
причем цветовые ленты сильно просвечивают и видны при рассмотрении изнутри устья.
Места прикрепления устья широко расставлены и соединены тонкой прозрачной мо-
золью. Края устья прямые, почти не отвернуты, слегка утолщены, образуя перистом.
Колумеллярный край отвернут сильно и почти целиком закрывает пупок, в результате
чего последний имеет вид узкой щели. Размеры: ВР 25—26, БД 29—31, МД 24—26 мм.
Внутреннее строение. Материал: 1 экз. из окр. с. Бенара на правом
берегу р. Абастуманки, 12 VI 1949, сбор Е. С. Кирьяновой, определение И. М. Лиха-
рева; 2 — из окр. с. Вордзил в долине р. Куры, 19 VII 1973, сбор Я. И. Старобогатова,
определение наше.
22*
340
IV. HELICIDAE
Спермовидукт в нижней части и яйцевод образуют двойной изгиб. Стилофор не-
большой, но широкий. Слизистые железы образуют два мощных ветвящихся куста,
но длина ветвей невелика: до места раздвоения протока семеприемника они не
достают.
Пенис образует два хорошо различимых вздутия, внутри которых располагаются
папиллы. Обе папиллы хорошо развиты, стенки их волокнистые, полостей не заклю-
чают. Просвет дистальной папиллы очень узкий, округлый; просвет проксимальной
папиллы имеет сложную изрезанную форму. Пениальный бугорок хорошо развит.
Эпифаллус приблизительно такой же длины, как и пенис. Бич длиннее суммарной длины
пениса и эпифаллуса, но заметно короче дивертикула семеприемника. Проток семе-
приемника разветвляется низко: на границе первой и второй трети длины.
Распространение и местообитания. Южная Грузия и се-
веро-восточная Турция (Ардануч). Обитает в зоне степей, среди нагорных ксерофитов,
под кустами в каменистых осыпях. Во время летней засухи зарывается в почву.,
3. Helix christophi Boettger, 1881. Табл. XXI, 204; рис. 458, 459.
О. Boettger, 1881а : 217 (Helix)', Westerlund, 1889 : 456 (Н. chri-
stophori, non Bossen, 1914); К о b e 1 1, 1897, Helic., IV : 783, t. 216, fig. 3, 4 [Helix
(Pomatia)]; 1915, Iconogr., N. F., 20 : 25, fig. 2797, 2798 (H. christophi atrolabioides);
Hesse, 1920, in Kobelt's Iconogr., N. F., 23 : 110, t. 646, fig. 11—15; Лихарев,
Раммельмейер, 1952 : 486 [Caucasotachea (Lindholmia) ].
Материал: 2 экз. из Закавказья и Турции.
Раковина очень похожа на раковину предыдущего вида. Отличия: поверх-
ность раковины грубо исчерчена; четвертая спиральная лента располагается выше
места прикрепления верхнего края устья или касается его нижним краем; вторая лента
иногда отсутствует; пупок в виде широкой щели, причем колумеллярный край в этом
месте слегка завернут внутрь; губа и париетальная мозоль более массивные и расплыв-
шиеся; при том же числе оборотов раковина больше: ВР 26—30; БД 32—35, МД 26—
28 мм.
Внутреннее строение. Материал: 1 экз. из окр. Ломашена, Артвин-
ский округ, северо-восточная Турция, VI 1909, сбор П. Нестерова, определение
И. М. Лихарева.
Спермовидукт в нижней части изогнут. Стилофор удлиненный, вершина его слегка
шаровидно утолщена. Слизистые железы в виде обильно ветвящихся кустов, короткие,
их ветви не поднимаются выше изгиба спермовидукта.
Пенис образует два вздутия, внутри которых имеются хорошо развитые папиллы.
Просвет дистальной папиллы узкий, просвет проксимальной папиллы в виде широкой
щели, по сторонам которой имеются глубоко врезанные бороздки (рис. 459). Бич не-
много длиннее суммарной длины пениса и эпифаллуса, дивертикул семеприемника
многократно извит, еще длиннее, чём бич. Проток семеприемника раздваивается при-
мерно посредине длины.
Это вид вместе с предыдущим принято выделять отдельным подродом Lindholmia
внутри рода Caucasotachea на том основании, что бич и дивертикул семеприемника у них
отличаются большой длиной (что характерно для рода Caucasotachea). Однако как род,
Caucasotachea, так и род Helix имеют четкий конхологический диагноз и резко отлича-
ются друг от друга; с другой стороны, оба этих сопряженных признака в пределах рода
Helix подвержены значительной изменчивости. Следовательно, подрод Lindholmia,
если признать его существование, надо относить к роду Helix. Учитывая вместе с тем
изменчивость названных признаков, мы не считаем целесообразным выделение этого,
подрода (равно как и других подродов рода Helix).
Распространение и местообитания. Западная часть Малого
Кавказа и северо-восточная Турция (Артвин, Ардануч). Населяет открытые участки,
встречается в зарослях кустарников и в осыпях.
4. Helix albescens Rossmassler, 1839. Табл. XXI, 205; рис. 460, 461.
Rossmassler, 1839. Iconogr., 2, 3/4 : 10, t. 44, fig. 585, 586; t. 44, fig. 582:
(H. vulgaris non Da Costa, 1778); Bourguignat, 1860, Rev. Mag. Zool., 12 г
160 t. 8 fig. 1—3 (H. obtusalis); Martens, 1860, Helic. : 143 (H. obtusata); W este-
rlund, 1889 : 456 (H. obtusata); 1897 : 123 (H. tntermissa); К о b e 1 t, 1906, Helic.,
VI : 176, t. 340, fig. 5, 6, 11, 12; t. 342, fig. 7—10; Rosen, 1911 : 115 [Я. (Pomatia)
vulgaris et. f. alba, (lava, unicincta, bicincta, bifasciata, quadrifasclata, ballionis Ret.];,
Retowski, 1914 : 299 [H. (Pomatia) vulgaris]; Rosen, 1914 : 179 (H. vulgaris);.
Hesse, 1920, in Kobelt's Iconogr., N. F., 23 ; 176, t. 653, fig. 1—6 (H. vulgaris);
Лихарев, Раммельмейер, 1952 : 489, рис. 417 [Я. (Helicogena) vulgaris];
Grossu, 1955 : 492, fig. 281 (H. vulgaris); Hudec, Lezhawa, 1967b : 95,
Abb. 15, t. 7, fig. 17 [H., (Physospira) ]; H u d e c, 1972a : 91, Abb. 22.
HELIX
341
Материал: более 40 экз. из 22 местонахождений на Кавказе, в Крыму и в Болгарии.
Раковина шаровидно-кубаревидная, с тупым коническим завитком, высота
которого немного меньше высоты устья. Оборотов 4, быстро нарастающих; эмбриональ-
ные обороты гладкие, выпуклые, блестящие. Последний оборот вздут, к устью плавно,
но сильно опущен. Окраска образована серовато-белым или коричневатым фоном и
5 светло-коричневыми лентами; однако число и ширина лент сильно варьируют (либо
раковина одноцветная, либо число лент достигает 6). Скульптура в виде радиальной
грубой и неравномерной исчерченности или морщинистости; на последнем обороте
местами могут присутствовать вмятины. Устье почти круглое, умеренно косое, высота
Рис. 460—461. Helix albescens Rssm. из Болгарии, Пловдив, 9 VI 1967. 460 — внеш-
ний вид полового аппарата; 461 — пенис, вскрыт. (Ориг.).
его немного больше ширины. Края устья тупые, почти не отвернуты, за исключением
колумеллярного, обычно несколько утолщены, окрашены в розовый или красновато-
бурый цвет. Колумеллярный край высокий и отвесный, широко отвернут и закрывает
пупок полностью или оставляет узкую щель. Места прикрепления устья широко рас-
ставлены и соединены тонкой мозолью. Размеры: ВР 27—36, БД 30—38, МД 24—32 мм.
Внутреннее строение. Материал: 1 экз. из Болгарии, Пловдив, го-
родской парк, 9 VI 1967, сбор и определение И. М. Лихарева; 1 — из Тбилиси, 25 XII
1966; 4 — из Крыма, окр. с. Краснолесье, 1 VIII 1962; сборы и определения наши.
На границе спермовидукта и яйцевода имеется крутой изгиб. Слизистые железы
умеренно ветвятся, общее число концевых ветвей каждой железы редко превышает 10.
Стилофор массивный, его основание вместе с прилегающим участком вагины охвачено
общим чехлом.
Пенис цилиндрический, пениальный бугорок небольшой, но ясный. Папиллы пе-
ниса короткие, каждая из них заключает кольцевую полость. Стенки пениса в простран-
стве между папиллами утолщены, bojh книстые, рыхлые; по внутренней поверхности
этого отдела проходит мощный продольный валик. Пенис и эпифаллус образуют изгиб,
стороны которого притянуты друг к другу тонкими лентами. Длина бича подвержена
значительной изменчивости: у экземпляра из Болгарии эта величина значительно пре-
вышала суммарную длину пениса и эпифаллуса, у экземпляра из Тбилиси бич при-
мерно в 1.5 раза длиннее, у крымских экземпляров бич имел примерно такую же длину,
как пенис и эпифаллус, вместе взятые. Место раздвоения протока семеприемника рас-
342
IV. HELICIDAE
полагается значительно выше его середины, дивертикул короче протока выше места
бифуркации.
Распространение и местообитания. Ареал вида циркумпон-
тический: Турция, Румыния, Болгария; Одесса, Николаев, Херсон, Мелитополь,
горный и степной Крым, западная и центральная части Северного Кавказа, места-
ми — Дагестан, Грузия, отдельные пункты Армении. Политопный вид, избегает
только аридных открытых биотопов и влажных горных теснин. Встречается в кустар-
никовых зарослях, по опушкам лесов и на скальных выходах.
Рис. 462—463. Helix lucorum L. из Болгарии, Стара Планина, 1 VII 1969.
462 — внешний вид полового аппарата; 463 — пенис, вскрыт. (Ориг.).
5. Helix lucorum Linnaeus, 1758. Табл. XXI, 206; рис. 462, 463.
Linnaeus, 1758 : 773; Krynicki, 1833 : 423—425, t. 10 (H. lucorum var.
taurica); Westerlund, 1889 : 468; К о b e 1 t, 1906, Helic., VI : 208, t. 344, fig. 1;
t. 345; fig. 1—3; Rosen, 1914 : 181 (H. taurica Kryn. et mut.-martens! Bttg.); Hesse,
1920, in Kobelt's Iconogr., N. F., 23 : 197, t. 657, fig. 4—9; t. 658, fig. 1—7; Лиха-
рев, Раммельмейер, 1952: 490, рис. 418; G г о s s u, 1955 : 490, fig. 279;
Hud ec, Lezhawa, 1969b : 149, Abb. 36, t. 7, fig. 18.
Материал: более 30 экз. из 12 местонахождений на Кавказе, в Крыму и в Болгарии.
Раковина шаровидно-кубаревидная, со слегка притупленным коническим
завитком почти правильных конических очертаний, высота которого примерно равна
высоте устья. Оборотов 4—4.5, слабо выпуклых, плавно нарастающих, разделенных
неглубоким неровным швом. Эмбриональные обороты выпуклые, гладкие. Последний
оборот сильно вздут, особенно в ширину, к устью плавно опущен. Окраска в общем
светлая, но на раковине имеются многочисленные коричневые радиальные полосы
с размытыми контурами, которые в случае интенсивного развития придают раковине
коричневую, довольно темную окраску. Кроме того, чуть выше периферии проходит
светлая лента, часто плохо различимая; в том случае, когда радиальные полосы раз-
виты особенно интенсивно, раковина приобретает яркую темно-коричневую окраску
с четкой светлой лентой выше периферии. На верхних оборотах имеются темные ленты.
Скульптура в виде резких радиальных складок, несколько усиливающихся у шва, и
HELIX
343
ясных, но редких спиральных бороздок. Устье коротко-овальное, косое, внутри бле-
стящее; края устья тупые, слегка утолщенные, белые или светло-коричневые, не от-
вернуты. Отвернут только колумеллярный край, полностью или почти полностью за-
крывающий пупок. Размеры: ВР 40—50. БД 41—52, МД 35 —44 мм.
Внутреннее строение. Материал: 1 экз. из Болгарии, Стара Планты,
1 VII 1969, сбор и определение И-. М. Лихарева; 2— из Крыма, окр. с. Голубинка,
4 VIII 1962, сбор и определение наши.
На границе спермовидукта и яйцевода имеется плавный изгиб. Слизистые железы
интенсивно ветвятся, общее число ветвей каждой железы достигает 16. Стилофор ци-
линдрический, наверху плавно закругленный.
Пенис цилиндрический, с неясными перетяжками. Пениальный бугорок развит
довольно слабо. Дистальная пениальная папилла заключает кольцевую полость,
широко сообщающуюся с полостью, возникшей в результате расслоения внутреннего
чехла пениса. Проксимальная папилла имеет сплошные стенки, заполненные рыхлой
тканью. Бич примерно вдвое длиннее пениса и эпифаллуса, вместе взятых. Проток
семеприемника раздваивается высоко, длина дивертикула немного меньше длины про-
тока выше места раздвоения.
Распространение и местообитания. Вид имеет широкий ареал,,
охватывающий Апеннинский п-ов, Балканский п-ов, Малую Азию, Сирию, Иран,
горный Крым, Черноморское побережье Кавказа (к северу до Сочи), Колхидскую низ-
менность с окаймляющими ее горными хребтами и восточное Закавказье (Шемаха,
Талыш).
Форму taurica Krynicki, 1833 (syn. Н. radiora Rossmassler, 1838) принято считать
подвидом; однако анализ взаимного распространения номинативной формы и формы
taurica, которые различаются окраской, заставляют считать название, предложенное
Криницким, синонимом названия lucorum.
Политопный вид, населяет как открытые места, так и леса.
6. Helix buchi Dubois de Montpereux, 1839. Табл. IX, 90; XXI, 207; рис. 464, 465.
Dubois de Montpereux, 1839 : 243; L. Pfeiffer, 1853, III :
181; Westerlund, 1889 : 462; Kobelt, 1906, Helic., VI : 259, t. 366,
fig. 1—5, 7, 8; R e t о w s k i, 1914:301 [Helix (Pomatia) buchi f. minor Bttg.];
Rosen, 1914 : 180; Hesse, 1920, in Kobelt's Iconogr., N. P., 23 : 204, t. 656,
fig. 3—9; Лихарев, Раммел ьмейер, 1952 : 491, рис. 419; Н u d е с,
L е г h a w a, 1967b : 93, Abb. 14, t. 6, fig. 13, t. 7, 16; Стеклов, 1966 : 231, табл.
15, 239—241.
Материал: 5 экз. из 2 местонахождений в Закавказье.
Раковина шаровидно-кубаревидная, с закругленным завитком, высота
которого варьирует, но в целом примерно равна высоте устья. Оборотов 5.5, быстро-
нарастающих, слабо выпуклых; эмбриональные обороты тонкозернистые, выпуклые,
блестящие. Последний оборот сильно вздут, особенно в ширину, к устью заметно опу-
щен. Окраска образована светло-желтым или желтым фоном и 5 бледно-коричневыми
лентами, причем нередко вторая и третья ленты сливаются; самая узкая лента —
верхняя. Скульптура представлена неравномерной радиальной исчерченностью, ме-
стами морщинистостью, которая у шва усиливается; благодаря этому шов неровный,
зазубренный. Кроме того, имеется микроскопическая зернистость и иногда вялые ред-
кие спиральные бороздки. Устье большое, коротко-овальное, косое, места его при-
крепления широко расставлены и чаще всего связаны мозолью, которая может быть,
утолщенной. Края устья простые, за исключением сильно отвернутого колумеллярного
края, утолщенные, белые или серые или коричневые. Колумеллярный край высокий,
слегка наклонный, широко отвернут и полностью закрывает пупок, но не спаян со-
стенкой раковины, так что остается узкая, но хорошо заметная щель. Размеры: ВР 46—
57, БД 45-60, МД 27-50 мм.
Внутреннее строение. Материал: 1 экз. из Батуми, 1936, сбор-
Е. С. Кирьяновой, определение И. М. Лихарева.
Нижняя часть спермовидукта образует пологую дугу. Яйцевод чрезвычайно корот-
кий. Каждая слизистая железа состоит из очень мощного ствола, который распадается’
на 3 ствола второго порядка, каждый из них вскоре обильно ветвится; общее число вет-
вей — около 20.
Пенис почти цилиндрический, пениальный бугорок тонкий, но высокий. Внутрен-
няя поверхность чехла пениса от пениального бугорка до основания дистальной па-
пиллы покрыта множеством тонких продольных складок, причем складки эти в об-
ласти папиллы ниже, в результате чего папилла лежит в своеобразном футляре, точно-
повторяющем все нюансы ее очертаний. Внутри дистальной папиллы имеется кольце-
вая полость, сливающаяся с полостью, образованной путем расслоения стенок пениаль-
ного чехла в пространстве между папиллами (рис. 465). Проксимальная папилла имеет
рыхлые, но цельные стенки, значительно короче дистальной. Внутренняя поверхность-
Рис. 464—465. Helix buchi Dub. de Mont, из Батуми, 1936. 464 - внешний вид поло-
вого аппарата; 465 — пенис, вскрыт. (Ориг.).
HELIX
345
камеры, заключенной между папиллами, покрыта мощными, неправильно расположен-
ными валиками и бугорками; стенка пениса в этом отделе многослойная, чрезвычайно
рыхлая. Бич примерно втрое длиннее пениса и эпифаллуса, вместе взятых; между се-
мепроводом, пенисом и эпифаллусом натянута прозрачная мембрана. Проток семе-
приемника раздваивается на уровне границы между второй и верхней третью, дивер-
тикул примерно в 1.5 раза короче протока выше места раздвоения.
Распространение и местообитания. Юго-западный Кавказ.
Встречается как на открытых местах, так и в лесах. В Картлийской котловине часто
во фруктовых садах среди растительных остатков и среди влажной травы. Близкая
форма обнаружена Стекловым (1966) в верхнеплиоценовых слоях Предкавказья (мео-
тические отложения рек Гумс и Аргудан).
7. Helix lutescens Rossmassler, 1837. Табл. XXI, 208.
R ossmassl er, 1837, Iconogr., 1,5:4, fig. 292; Westerlund, 1889 :
455; Kobelt, 1906, Helic., VI : 148, t. 331, fig. 7—12; Hesse, 1920, in Kobelt’s
Iconogr., N. F., 23:219, t. 659, fig. 8, 9; Лихарев, Раммельмейер,
1952:492 [Helix (Helicogena)]; Grossu, 1955:491, fig. 280; Lozek, 1956:
228, t. 37, fig. 4; Zilch, Jaeckel, 1962 : 202; Lozek, 1964 : 315.
Материал: 9 экз. из 3 местонахождений в Одесской обл. и в Венгрии.
Раковина шаровидно-кубаревидная, с притупленным коническим завитком,
высота которого обычно несколько меньше высоты устья. Оборотов 4—4.5, слабо вы-
пуклых, плавно нарастающих. Эмбриональные обороты гладкие. Последний оборот
Рис. 466—467. Helix pomatia L. из Болгарии, окр. г. Смолян, 7 V 1969.
466 — внешний вид полового аппарата; 467 — пенис, вскрыт. (Ориг.).
вздут, к устью очень плавно опущен. Окраска белая, серая или желтовато-белая,
чаще одноцветная, иногда с очень неясными темными лентами, которые в проходя-
щем свете светлее фона. Скульптура в виде умеренной радиальной морщинистости
и спиральных бороздок. Устье коротко-овальное, почти круглое, скошено, внутри бе-
лое или слегка розоватое. Места прикрепления устья широко расставлены и связаны
тонкой мозолью. Края устья простые, острые, почти не отвернуты; сильно отвернут
колумеллярный край, полностью закрывающий пупок или оставляющий очень узкую
23 А. А. Шилейко
346
ПРИЛОЖЕНИЕ
щель. Колумеллярный край высокий и почти отвесный. Размеры: ВР 27—33, БД 24—
31, МД 22—28 мм; реже ВР 32—34, БД 32—34, МД 29 мм.
Распространение и местообитания. Бассейн Дуная. В СССР
встречается в западных районах Украины, включая Житомирскую, Винницкую, Одес-
скую и Николаевскую области. Обитает в зарослях кустарников и по опушкам ли-
ственных лесов. Обнаружен в плейстоценовых и четвертичных отложениях Средней
Европы.
8. Helix pomatia Linnaeus, 1758. Табл. XXI, 209‘, рис. 466, 467.
Linnaeus, 1758 : 771; Westerlund, 1889:462; К о Ь е 1 1, 1906, Helic.,.
VI : 241, t. 362, fig. 1—3, 6; Hesse, 1920, in Kobelt's Iconogr., N. F., 23 : 216,
t. 659, fig. 10—13; t. 660, fig. 7—8; Лихарев, Раммельмейер, 1952 г
493, рис. 420 [Helix (Helicogena)]-, Grossu, 1955:486, fig. 278; Ehrmann,
1956 : 148, fig. 95; Lozek, 1956 : 227, t. 37, fig. 3; Zilch, Jaeckel, 1962 :.
202; L о z,e k, 1964 : 314, t. 29, fig. la—c; Gittenberger et al., 1970 : 107,.
fig. 191, Kaart 78,
Материал: более 100 экз. из 20 местонахождений в европейской части СССР,.
ГДР, Венгрии и Болгарии.
Раковина шаровидно-кубаревидная, с притупленным коническим или слегка
куполовидным завитком, твердостенная, матовая. Высота завитка немного меньше-
высоты устья или равна ей. Оборотов 4.5—5, выпуклых, быстро нарастающих. Эм-
бриональные обороты узкие, гладкие.Последний оборот сильно вздут, к устью слабо-
опущен. Окраска от беловато-серой до желтовато-бурой, с рисунком из 4 спиральных
лент, цвет которых — от светло-коричневого до коричневого с фиолетовым оттенком.
Скульптура представлена неравномерной морщинистостью и тонкими, нередко лишь
слегка намечающимися спиральными бороздками. Устье большое, коротко-овальное,
слегка косое. Края устья прямые или немного отвернуты, слегка утолщены, внутри
с тонкой белой или розоватой губой. Колумеллярный край высокий, отвесный, сильно-
отвернут и почти полностью закрывает пупок, оставляя от него только щель. Размеры:
ВР 38—45, БД 37—47, МД 32—40 мм.
Внутр е. инее строение. Материал: 1 экз. из Болгарии, Родопы, окр..
г. Смолян, 7 V 1969, сбор и определение И. М. Лихарева; 5 — из Кведлинбурга, ГДР,
сбор и определение Э. Клаусса; 3 — из Венгрии, окр. оз. Балатон, VI 1970, сбор-
Д. А. Сахарова; 6 — из окр. Ужгорода, 6 IX 1969; 2 — из Московской обл., с. Ба-
лашиха, 2 X 1970; сборы и определения наши.
Спермовидукт переходит в яйцевод без изгиба или образует пологую плавную дугу.
Яйцевод длиннее очень короткой вагины. Стилофор цилиндрический или слегка пере-
жат посредине. Слизистые железы обильно ветвятся.
Пенис почти цилиндрический, с небольшим вздутием возле клоаки, отвечающим!
пениальному бугорку, развитому в разной степени. Дистальная папилла пениса за-
ключает кольцевую интрапапиллярную полость, широко сообщающуюся с полостью,
возникшей за счет расслоения стенок чехла выше дистальной папиллы. Проксималь-
ная папилла короткая, сосцевидная, с толстыми рыхлыми цельными стенками. Об-
ширное пространство между папиллами покрыто многочисленными мощными, бесси-
стемно расположенными валиками и складками. Бич примерно вдвое суммарной длины!
пениса и очень короткого эпифаллуса. Дивертикул семеприемника рудиментарный,
короче поротока выше места раздвоения в 2—4 раза.
Распространение и местообитания. Средняя и Юго-Восточная-
Европа; в СССР — вдоль западных границ на территории Белоруссии, Украины, Мол-
давии и Прибалтики; завезен в Ленинград, Москву, Киев, Курск. Интродуцирован!
в Америку (Буэнос-Айрес, Монтевидео). Обитает в лиственных лесах, в зарослях ку-
старников, часто близ населенных пунктов. Неоднократно найден в плейстоцене и го-
лоцене примерно в пределах современного ареала; похожая форма встречена Стекло-
вым (1966) в акчагыльских слоях Предкавказья (на р. Ярык-су).
ПРИЛОЖЕНИЕ
Ниже дается описание вида, относящегося к другому надсемейству, но конхоло-
гически сильно напоминающего одного из представителей ArianVnae, в связи с чем;
его и принято относить к названному подсемейству.
ДОПОЛНЕНИЕ
347
Надсемейство POLYGYROIDEA
Семейство POLYGYRIDAE Pilsbry, 1895
Подсемейство TRIODOPSINAE Pilsbry, 1895
Род TRIODOPSIS Rafinesque, 1819
Раковина небольшая, прижатая, по периферии более или менее закругленная
или с углом. Пупок узкий, нередко полностью запаян. В устье 2—3 зуба: парие-
тальная пластинка и 1 —2 противолежащих бугорка. Пенис массивный, мешковидный,
без бича и папиллы, с системой складок внутри. Придатков женского отдела нет,
как и у прочих представителей надсемейства. Семеприемник с очень коротким прото-
ком.
Распространение. Северная Америка.
Типовой вид — Triodopsis lunula Rafinesque, 1831 (syn. Helix tridentata Say,
1816).
Triodopsis subpersonatum (Middendorff, 1851). Табл. XXI, 210.
Middendorfi, 1851:306, t. 30, fig. 27—29 (Helix)- Westerlund,
1889 : 27 (Helix); Лихарев, Раммельмейер, 1952 : 473 (Isognomostoma).
Материал: 1 экз. из окр. Аяна на берегу Охотского моря.
Раковина прижатая, со слегка куполовидным завитком, высота которого,
немного меньше высоты устья. Оборотов 5.5, умеренно выпуклых, плавно нарастаю-
щих. Последний оборот в 1.5 раза шире предпоследнего, к устью незначительно опу- -
щен, немного выше средней линии с закругленным углом. Окраска одноцветно бурая..
Скульптура в виде сглаженных радиальных морщин; местами эти морщины превра--
щены в ряды неясно выраженных удлиненных бугорков. Кроме того, имеются крайне
тонкие спиральные бороздки, заметные при увеличении порядка 30 раз. Устье косое,
с отвернутыми краями, образующими нерезко выраженную манжету. В устье 3 зуба:
париетальная треугольная пластинка, которая при рассмотрении .снизу имеет У-
образную форму, и бугорковидные зубы — базальный и палатальный. Пупок узкий,
цилиндрический, слегка прикрыт отворотом колумеллярного края. Размеры: ВР 4,
БД 6.8, МД 6 мм.
Внутреннее строение неизвестно.
Распространение. Вид найден в двух местах: Миддендорф нашел его
в окрестностях Удского острога; кроме того, он получил от Вознесенского 2 экз. из
Аяна. Вероятно, вид обитает на хр. Джугджур, поскольку поиски его непосредственно
в окрестностях Аяна, проведенные нами в июле—августе 1971 г., ив окр. Чумикана,
проведенные Я. И. Старобогатовым в сентябре 1972 г., оказались безрезультатными.
ДОПОЛНЕНИЕ
Когда текст был уже в наборе, мы получили материал по новому для науки свое-
образному виду, описание которого приводится ниже.
Bradybaena capillata Schileyko et Bratschik, sp. nov. Рис. 468—471.
Материал: более 100 экз. из 10 местонахождений на островах зал. Петра Великого.
Голотип собран на о-ве Малом.
Голотип хранится в коллекции Зоологического института АН СССР.
Раковина от прижатой до низко кубаревидной, сравнительно тонкостенная,
слегка просвечивающая. Высота завитка приблизительно равна высоте устья. Обо-
ротов 5, умеренно выпуклых, последний оборот менее чем в 1.5 раза шире предпослед-
него, по периферии закругленный, к устью слабо и плавно, но отчетливо опущен. Ок-
раска одноцветная — от светло-роговой до бурой. Эмбриональные обороты почти глад-
кие, неясно гранулированные. Скульптура дефинитивных оборотов представлена часто
и неравномерно расположенными сглаженными морщинками и тонкой радиальной
струйчатостью; на базальной поверхности при сильном увеличении заметны редкие
мелкие неровные кожистые спиральные ребрышки. Наконец, поверхность раковины
несет видные невооруженным глазом изогнутые волоски. Устье в целом овальное, места
его прикрепления не сближены, колумеллярный край умеренно отвернут, базальный
отвернут слабее, палатальный край прямой. Отступя от края устья залегает широкая,
сильно расплывшаяся губа, выраженная на базальном крае, но плавно исчезающая
23*
348
ДОПОЛНЕНИЕ
близ палатального. Пупок широкий, развернутый, перспективный; на скатах пупка за-
метны плохо развитые чешуйки. Размеры: ВР 5—6, БД 6.5—8.5, МД 5.5—7.5 мм. Го-
лотип: ВР 5.0, БД 8.1, МД 6.8 мм.
По конхологическим признакам вид напоминает Trichia hispida', отличается более
широким пупком, наличием кожистых ребрышек на базальной части раковины и
струйчатости между радиальными морщинками.
Внутреннее строение. Материал: 1 экз. из о-ва Наумова, 20 VI
1973; 2 — из о-ва Малого, 15 VII 1973; 8 — из о-ва Стенина, 20 IX 1975; сборы
Р. Я. Братчика, определение наше.
Рис. 468—471. Bradybaena capillata, sp. nov. 468, 469 — раковина голотипа, о-в
Малый в зал. Петра Великого; 470 — внешний вид полового аппарата паратипа,
о-в Стенина, 20 IX 1975; 471 —дистальный отдел полового аппарата того же экз.,
вскрыт.
Нижняя часть спермовидукта образует изгибы, стянутые вместе в плотный узел;
.яйцевод и вагина приблизительно одной длины. Слизистые железы сильно развиты,
представляют собой резко гофрированные длинные трубочки числом 4—6, причем
каждая железа впадает в стилофор самостоятельно. Никаких следов железистого
пояса и резервуара стилофора не обнаружено. Верхняя полость стилофора, где рас-
полагается стрела, отделена от нижней полости хорошо развитым кольцевым утолще-
нием, причем разделение стилофора на две полости видно даже при наружном осмотре
. стилофора.
Пенис тонкий, умеренно длинный, основание его охвачено коротким чехлом, фик-
сированным только у клоаки. .Посредине пенис может быть утолщенным либо по всей
ДОПОЛНЕНИЕ 349
длине равномерно цилиндрическим. Внутренняя поверхность пениса покрыта очень
неясными косыми складочками, стягивающимися к продольному утолщению, но ясно
дифференцированный пениальный гребень не обнаружен. Пенис переходит в эпифал-
лус очень постепенно, а граница между последним и семепроводом наружно вообще
не выражена; возможно, половой ретрактор крепится именно в месте перехода семе-
провода в эпифаллус.
Распространение и местообитания. Вид обнаружен на 10
островах зал. Петра Великого (о-ва Пахтусова, Наумова, Малый, Дурново, Козлова,
Пелис, Матвеева, Стенина, Де-Ливрона, Клыкова). > Населяет прибрежные заросли
кустарников и осыпи.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ1
abchasica, Caucasigena 47, 69, 107
.abchasica, Caucasigena (Caucasigena) 192,
198*
abchasica, Fruticicola 198
abchasica, Helix 198
abchasica, Trichia (Xerocampylaea) 198
A bchasohela 246
Achatina 21
aculeata, Monachoides 47, 70, 100, 234,
238
aculeata, Pseudotrichia (Stygius) 238
aculeata, Zenobiella 238
Acusta 60, 123, 163
adshariensis, Euomphalia 263
•adshariensis, Fruticocampylaea 263
adshariensis, Helix (Fruticocampylaea) 263
adshariensis, Hesseola 263
aethiops, Arianta 49, 101, 313
airumla, Helix ziegleri 193
akramowskii, Circassina circassica 111,
249
alba, Helix (Pomatia) vulgaris 340
albescens, Helix 49, 101, 337, 340,
341*
albescens, Helix (Physospira) 340
albidorsalis, Helix rufispira 173
aliostoma, Helix (Fruticocampylaea) 244
almaatini, Bradybaena plectotropis 105,
125, 149, 150
almaatini, Eulota (Eulota) 148
almaatini, Fruticicola 148
almalensis, Angiomphalia 48, 106
almalensis, Angiomphalia (Angiomphalia)
292, 297, 298*
altaica, Helix (Trichia) rhysota 227
Altaicola 228
alternans, Helix (Tachea) atrolabiata 332
amatoris, Hygromia 130
Ammonitellidae 44, 52, 53*
anachoretica, Bradybaena 157
andronakii, Helix ( Fruticocampylaea) joan-
nis 306
andrussovi, Caucasotachea 49, 331
Angiomphalia 111, 291
Angiomphalia s. str. 76, 111, 292
angulata, Eulota (Leueozonella) rubens
176
angulata, Helix rubens 176, 297
angulata, Leueozonella 46, 64, 65, 66, 107,
177
angulata, Leueozonella (Leueozonella) 168,
176*
annexa, Helix (Trichia) 238
Anoplitella 65*, 109, 199
antoni, Helix 2б0
antontomashevici, Hesseola adsharien
263
antonwagneri, Helix 200
appeliana, Euomphalia 48, 73, 108, 266,
268*
appeliana, Helix 268
appeliusi, Helix 269
appeliusi, Helix (Fruticocampylaea) nar-
zanensis 268
apollinis, Archaica 41, 48, 106
apollinis, Archaica (Archaica) 259*
apollinis, Cathaica (Cathaica) 259
apollinis, Cathaica (Xerocathaica) 259
apollinis, Helix 257, 259
apollinis, Zenobiella 259
arbustorum, Arianta 17, 40, 49, 100, 310,
311*
arbustorum, Helicigona (Arianta) 310
arbustorum, Helix 310
arcadiana, Helicella (Cernuella) 255
arcadiana, Xerosecta 48, 107, 254, 255,
256*
arcasiana, Bradybaena 46, 99, 102, 124,
139*
arcasiana, Eulota (Eulota) 139
arcasiana, Helix 139
Archaica 18, 110, 257
Archaica s. str. 110, 259
Archaicinae 63, 65, 75, 76, 110, 256
arenosa, Helix 216
arenosa, Helix obvia 216
argonautarum, Helix 272
argonautarum, Tachea 335
Arianta 33, 109, 310
Ariantinae 40, 56, 76, 77, 78, 79, 80, 108,
308, 320, 346
Arionta 310
aristata, Euomphalia 16, 40, 48, 73, 100,
266, 267*
aristata, Helix 266
armeniaca, Caucasigena 47, 108
armeniaca, Caucasigena (Caucasigena) 192,
193*
armeniaca, Fruticicola 193
armeniaca, Helix 193, 196
armeniaca, Trichia 194
armeniaca, Trichia (Xerocampylaea) 195
arpatschaiana, Euomphalia 278
arpatschaiana, Helix 278
1 Курсивом выделены синонимы, жирным шрифтом — названия родов, подсе-
мейств и семейств Helicoidea-, жирные цифры обозначают страницы с описаниями,
звездочкой отмечены страницы с рисунками, касающимися названного таксона.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
351
Arvensis, Helix 323
Ashfordia 75
aspasta, Helix (Jacosta) 161
aspersa, Cryptomphalus 3, 17, 19, 36, 44,
49, 80, 100, 324, 325*
aspersa, Helix 324
aspersa, Helix (Cryptomphalus) 324
assadovi, Euomphalia 278
associata, Campylaea faustina 309
Atenia 57, 58
atrolabiata, Caucasotachea 49, 101, 331,
332, 334*, 335
atrolabiata, Helix 331, 332
atrolabiata, Helix (A cape) 332
atrolabioides, Helix christophi 340
Aulacopoda 50, 51, 55
<aurea, Helix 2Ё1
austriaca, Helicopsis 213
•austriaca, Helix 323
bactriana, Helix 280
bakowskii, Edentiella 47, 99, 210*
bakowskii, Trichia 210
bakowskii, Trichia bielzi 210
bakowskii, Trichia (Edentiella) 210
bakowskii, Trichia (Trichia) bielzi 210
bakowskyana, Helix (Xerophila) instabilis
217
balgutensis, Fruticicola 178
balkariensis, Helix schaposchnikovi 200,
201
balllonis, Helix (Pomatia) vulgaris 340
banatica, Campylaea 317
banatica, Campylaea (Drobacia) 317
banatica, Campylaea (Eucampylaea) 317
banatica, Drobacia 49, 99, 317
banatica, Helicigona (Dobracia) 317
banatica, Helicigona (Drobacia) 317
banatica, Helix 317
Batumica 74*, 112, 276
Bayeri, Helix pratensis 305
bicallosa, Chilanodon 47, 71, 98, 228,
229*
bicallosa, Helix 228
bicallosa, Perforatella 228
bicincta, Helix (Pomatia) vulgaris 340
bidens, Helix 231
bidens, Perforatella 47, 71, 98, 231*
bidens, Trochus 231
bielzi, Helix 209
bielzi, Helix (Perforatella) 209
bielzi, Trichia 47, 99, 205, 209
bifarla, Helix 288
bifasciata, Helix (Pomatia) vulgaris 340
bifrons, Helix (Petasia) 262
bilaticincta, Bradybaena 46, 61, 106, 125,
154*
bilaticincta, Eulota paricincta 154
bilaticincta, Helix paricincta 154
bituberosa, Euomphalia (Karabaghia) 262
bituberosa, Karabaghia 48, 72, 98, 262*
bitiberosa, Perforatella 262
bituberosa, Perforatella (Karabaghia) 262
bocageana, Bradybaena weyrichii 124,
135
bocageana, Helix 135
Boemica 75, 113, 289
boevi, «Bradybaena» 46, 105/ 163
boissieri, Sphincterochila 42
bojenae, Circassina 48, 69, 107, 247, 249
Bradybaena 18, 20, 52, 61, 62, 112, 118,
123 163
Bradybaenidae 4, 30, 33, 43, 44, 51, 52,
53*, 55, 56, 57, 58, 61, 62, 72, 112,
118, 163
Bradybaeninae 59
brodskii, Cathaica 151
-buamica, Angiomphalia 48, 76, 99, 261
buamica, Angiomphalia (Lentiga) 291,
. 301*
buamica, Euomphalia 301
buamica, Paedhoplita 301
buchi, Helix. 49, 101, 337, 343, 344*
Buliminopsis 12
Bulimuloidea
caelestimontana, Angiomphalia 48, 106
caelestimontana, Angiomphalia (Angiom-
phalia) 292, 293, 294*
caelestimontana, Cathaica 293
caelestimontana, Euomphalia 293
calligera, Caucasotachea 49, 101, 331, 335,
336*
calligera, Helix 335
Camaenidae 4, 25, 30, 44, 51, 52, 53*,
55, 56, 76
Campylaea 308, 313
Canariella 57, 58
candaharica, Helicella 222
candaharica, Helicella (Xeropicta) 222
candaharica, Helicella (Xeropicta) deroen-
tina 222
candaharica, Helix 222
candaharica, Xeropicta 40*, 47, 67, 74,105,
222*
candicans, Helicella 47, 103, 224*
candicans, Helix 224
candidissima, Sphincterochila 49
caperata, Helicella 34
capillata, Bradybaena 347
Caracollina 57, 58, 116
carascalensis, Hygromia (Pyrenaearia) 68
carascaloides, Helix 289
carascaloides, Helix carthusiana 289
cardiostoma, Cathaica 60 ' i
caria, Leueozonella 38, 46, 64, 66, 100
caria, Leueozonella (Leueozonella) 168,
177, 178* .
Carthusiana 271, 281
carthusiana, Helix 282, ’286
carthusiana, Monacha 24, 48, 74, 103
carthusiana, Monacha (Monacha) 282, 286,
287*
carthusiana, Monacha (Theba) 286
carthusiana, Theba 286
carthusiana, Theba (Theba) 286
caryodes, Helix rubens 297
caryodes, Helix (Eulota) rubens, 170
caryodes, Leueozonella 41, 46, 64, 66, 106,
172
caryodes, Leueozonella (Leueozonella) 168,
170*
casta, Helix 328 .. .
Cathaica 54, 60
caucasica, Helix 280
caucasicola, Fruticicola 184
352
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
caucasicola, Helix (Theba) 283
caucasicola, Monacha 48, 103, 184
caucasicola, Monacha (Paratheba) 282, 283,
284*
caucasicola, Trichia (?) 283
Caucasigena 18, 38, 64, 65, 66, 67, 109,
191, 210
Caucasigena s. str. 67, 109, 191, 192
Caucasocressa 16, 38, 40, 69, 76, 113, 304
Caucasofixus 271
Caucasotachea 77, 109, 330, 331, 340
Causa 109, 317
cavimargo, Bradybaena 46, 100
cavimargo, Bradybaena cavimargo 124,
161, 162*
cavimargo, Cathaica (?) 161
cavimargo, Helix 161
Сераеа 3, 79, 80, 109, 320
Сераеа s. str. 78, 109, 320, 323
Cernuella 69, 70, 71, 111, 250
cespitum, Cernuella 251
Chilanodon 23, 68, 70, 71, 111, 228
Chilotrema 313
chishimana, Eulota 135
christophi, Caucasotachea (Lindholmia) 340
christophi, Helix 49, 101, 337, 339, 340
christophi, Helix (Pomatta) 340
christophori, Helix 263, 340
chrysomallis, Theba 275
ciliata, Ciliella 56
cinctella, Hygromia (Hygromia) 68
cincto-inflata, Helix (Cryptomphalus) 144
circassica, Circassina 48, 62, 69, 107, 225;
247
circassica, Circassina circassica 246*, 247
circassica, Helix 246, 247
Circassina 69, 70*, 75, 111, 246, 249, 271
Clausiliidae 25
Clessini, Helix 204
Cochlicella 12
concinna, Helix 207
concinna, Trichia 47, 99, 205, 207*
concolor, Helix rubens 171, 297
Coneulota 119
conospira, Fruticicola 170
conospira, Trichia 170
contorta, Helix 116
copiosa, Angiomphalia 48, 63, 75, 76, 105,
158
copiosa, Angiomphalia (Angiomphalia) 291,
292*
corcyrensis, Caracollina 116
corcyrensis, Helix 116
corcyrensis, Helix (Caracollina) 116
corcyrensis, Lindholmiola 30, 32*, 36, 46,
, 57, 98, 116, 117*
crassicosta, Leucozonella 46, 66, 102
crassicosta, Leucozonella (Narina) 168,
180*
crenimargo, Helicella 254
crenimargo, Helicella (Helicopsis) 254
crenimargo, Helix 254
crenimargo, Xerophila (Jacosta) 254
crenimargo, Xerosecta 34, 48, 107, 254,
255*
crenimargo, Xerosecta (Kalitinaia) 254
Cryptomphalus 109, 324
cumulate, Nanaja 46, 62, 66, 75, 106, 164,
165*
cyclothyra, Helix (Frnticocampylaea) nar-
zanensis 241
Cylindrus 12
Cyrnotheba 75
czarnohorica, Fruticicola 238
czarnohorica, Trichia 238
czekanowskii, Helix (Trichia) 238
daghestana, Circassina (Shileykoia) 244
daghestana, Fruticocampylaea (Shileykoia)
244
daghestana, Helix 244 '
daghestana, Helix (Frutico-Campylaea)
Elchwaldi 244
daghestana, Shileykoia 17, 38, 48, 69, 101,
108, 196, 244, 245*
daghestana, Trichia (Xerocampylaea) eich-
waldi 244
danubialis, Trichia 64, 65, 66
darevskii, Hygrohelicopsis 41, 46, 107, 181,
182*
dealhatus, Bulimulus 42
decussata, Helix (Tachea) atrolabiata 332
dejecta, Helicella 216
dejecta, Helicella (Helicopsis) 216
dejecta, Helicopsis 47, 104, 212, 216
dejecta, Helix 216
dejecta, Helix obvia 216
delabris, Caucasocressa 306
delabris, Helix 306
depressa, Helix (Fruticocampylaea) narza-
nensis appeliusi 268
depressa, Helix pratensis 305
derbentina, Helicella (Xeropicta) 220, 221'
derbentina, Helicopsis (Xeropicta) 221
derbentina, Helix 220
derbentina, Xerophila (Helicella) 220
derbentina, Xeropicta 47, 68, 105, 220,
221*, 222
dextrorsa, Bradybaena lantzi 125
dextrorsa, Fruticicola lantzi 106, 147
diaphora, Helix (Helicella) 259
dibothrion, Fruticicola (Petasina) 232
dibothrion, Perforatella 47, 69, 71, 98, 231,
232, 233*
dibothrion, Petasina 232
dichrozona, Bradybaena 23*, 46, 61, 105,
125, 153*
dichrozona, Eulota (Eulota) 153
dichrozona, Helix 153
dieckmanni, Bradybaena 36, 46, 99, 124,
137, 138*
dieckmanni, Eulota (Eulota) 137
dieckmanni, Fruticicola 137
dieckmanni, Helix (Trichia) 137
dieckmanni, Trichia 137
Diodontella 41, 65*, 109, 202, 203
Dioscuria 65*, 109, 191, 201
Diplobursa 112, 278
diplodon, Odontotrema 46, 64, 66, 98, 166-,
167*
diplodon, Trichia 167
diversita, Bradybaena 46, 60, 102, 124;
140*, 141*
djulfensis, Helix 328
djulfensis, Levantina 49, 100, 328, 329*
Dobracia 317
АЛФАВИТНЫЙ УКАЗАТЕЛЬ 353
Drepanostoma 57
Drobacia 108, 317
duiensis, Bradybaena 46, 102, 124, 136*.
137, 140
duiensis, Eulota (Karaftohelix) 137
duiensis, Helix 137
duplocincta, Bradybaena 120
I duplocincta, Eulota (Eulota) 120
duplocincta, Helix 120
duplocincta, Ponsadenia 15, 46, 61, 106
duplocincta, Ponsadenia (Mesasiata) 119,
120*
Dupotetia 77
eberhardi, Kokotschashvilia 47, 65, 108,
184, 187, 188*
Edentiella 64, 65*, 66, 109, 209
edentula, Helix 210
eichwaldi, Caucasigena 47, 108
eichwaldi, Caucasigena (Caucasigena) 191,
196, 197*, 198
eichwaldi, Fruticicola 196
eichwaldi, Helix 196
eichwaldi, Helix (Fruticocampylaea) 196
l' eichwaldi, Trichia (Xerocampylaea) 196
I1 Elona 77
Eobania 109, 326
escheriana, Helix 328
escheriana, Levantina 49, 101, 328, 330*
euages, Euomphalia 273
euages, Euomphalia (Oscarboettgeria)
euages 273
euages, Helix 273
< euages, Helix (Eulota) 273
euages, Oscarboettgeria 48, 72, 102, 273,
274*
Euarchaica 110, 257
eudakowi, Helicella 213
eulemnisca, Eulota 130
Eulota 123
i Euomphalia 71, 72, 73*, 74*, 112, 263,
266, 273, 275, 280, 297
Euomphaliinae 33, 38, 63*, 65*, 68, 69,
71, 74, 75, 76, 111, 112, 113, 261,
271
Eurystrophe 77
eutheta, Helix (Trichia) 137
i Euthyneura 22
evoluta, Helicodonta 46, 114
4 exasperata, Angiomphalia 48, 75, 106
exasperata, Angiomphalia (Angiomphalia)
292, 295*
explanata, Helix 253
externa, Monacha 48
externa, Monacha (Monacha) 286
• fasciola, Cathaica 60
Faustina 78, 109, 308
faustina, Campylaea 309
faustina, Campylaea (Faustina) 309
faustina, Faustina 48, 103, 308*, 309*
faustina, Helicigona (Campylaea) 309
faustina, Helicigona (Faustina) 309
faustina, Helix 309
fedtschenkoi, Bradybaena 46, 105, 125,
•I 160, 161*
fedtschenkoi, Cathaica (Campylocathaica)
160
fedtschenkoi, Cathaica (Cathaica) 160
fedtschenkoi, Helix 160
ferghanica, Bradybaena 160
ferghanica, Cathaica 169
ferghanica, Leucozonella 46, 63, 64, 66,
75, 105, 160, 293
ferghanica, Leucozonella (Leucozonella)
168, 169*
filicina, Trichia 205
filimargo, Helicella (Helicopsis) 217
filimargo, Helicopsis 47, 67, 104, 211, 217,
218*
filimargo, Helix 217
filimargo, Xerophila (Jacosta) 217
finschiana, Helix rubens 171, 297
fiscina, Eulota (Euhadra) 133
fiscina, Eulota (Karaftohelix) 133
flava, Helix (Pomatia) vulgaris 340
flaveola, Helix 201
flavolimbata, Helix 278
flexibilis, Ezohelix 27
fluxa, Helix kobiensis 243
fortangensis, Caucasigena 47
fortangensis, Caucasigena (Caucasigena)
192
fortangensis, Caucasotachea 49, 331
fortangensis, Tropidomphalus 192
fragilis, Bradybaena 36, 46, 60, 102, 124,
140, 141, 142*
fragilis, Eulota (Eulota) 141
fraudulosa, Caracollina 116
fraudulosa, Lindholmiola 46, 116
frequens, Helix 288
freytagi, Metafruticicola 77
Fruticicola 123, 201, 205
Fruticocampylaea 70*, 111, 241, 304
fruticola, Helix 282
fruticola, Monacha 48, 103
fruticola, Monacha (Paratheba) 282, 283*
fruticola, Theba 282
fruticola, Theba (Theba) 282
fruticum, Bradybaena 28*, 44, 46, 50, 59,
60, 61, 104,- 124, 125, 126*, 128, 129,
139
fruticum, Eulota (Eulota) 125
fruticum, Fruticicola 125
fruticum, Helix 125
frutis, Circassina 48, 107, 247, 250
frutis, Helix 250
frutis, Helix schuberti 250
frutis, Helix (Theba) 250
fusca, Hygromia 68
fuscior, Helix rufispira 173
gagriensis, Euomphalia ^Oscarboettgeria)
euages 273
gagriensis, Helix 273
Galactochilus 77, 331
gasprensis, Helicella 216
Gasullia 58
Geomitrinae 55, 56
Geophila 13, 17
gerassimovi, Helix (Fruticocampylaea) 195,
196
gerstfeldti, Chilanodon 47, 99, 228, 229,
230*
gerstfeldti, Helix 228, 229
gerstfeldti, Perforatella 229
giengensis, Klikia 49
354
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
jgireiorum, Helicella (Helicopsis) 215
gireiorum, Helicella (Jacosta) 215
girva, Helix 110
globula, Helix 275
globula, Helix (Latonia) 275
.globuliformis, Fruticicola mesoleuca 172
Gonostoma 114
graeseri, Cathaica 60
.graeseri, Helix (Camaena) 130
granulata, Helix 238
Halolimnohelix 4
Haplohelix 4
Harmozica 112, 280
Hela 246
Helicella 67, 68, 109, 223
Helicelllnae 34, 38, 39*, 40, 73, 80
Helicidae 18, 30, 42, 44, 51, 53*, 54, 56,
76, 77, 78, 108, 307
Helicigona 108, 313
Helicinae 18, 20, 55, 77, 78, 79, 80, 109,
319
Helicodonta 57, 58, 112, 114
Helicodontidae 17, 30, 49, 51, 55, 56, 57*,
, 58, 112, 114
Helicodontinae 112, 114
Helicogena 337
Helicopsis 65*, 67, 68, 110, 211
Heliomanes 250
Helix 15, 18, 23, 33, 62, 77, 109, 114, 331,
337, 340
Helminthoglypta 80
Helminthoglyptidae 33, 44, 51, 53*, 54, 55,
56, 59
helvola, Bradybaena 46, 61, 62, 103, 105,
124, 125, 157, 158*
helvola, Helix 157
Hemicycla 77
heptapotamica, Archaica 48, 62, 75, 106,
164
heptapotamica, Archaica (Euarchaica) 257,
258*
heptapotamica, Cathaica 257
heptapotamica, Cathaica (Cathaica) 257
heptapotamica, Zenobiella 257
Hermanni, Cathaica 121
Hesseola 40, 69, 74», 112, 263
hirsuta, Bradybaena 119
hirsuta, Ponsadenia 46, 61, 100
hirsuta, Ponsadenia (Tarbagataja) 119
hispida, Cathaica rufispira 173
hispida, Helix 137, 205
hispida, Hygromia 205, 207
hispida, Trichia 21, 36*, 47, 99, 205,
206*
Holopoda 50
holosericum, Causa 49, 98, 317, 318*
holosericum, Helix 317
holosericum, Isognomostoma 318
holosericum, Trigonostoma 317, 318
holotricha, Fruticicola 188
holotricha, Helix 184, 188
holotricha, Helix (Latonia) 188
holotricha, Kokotschashvilia 47, 65, 67,
100, 184, 187, 188, 189*
holotricha, Trichia (Kokotschashvilia) 188
holotricha, Trichia (Trichia) 188
hortensis, Сераеа 20, 37, 49, 101
hortensis, Сераеа (Сераеа) 320, 321, 322*
hortensis, Helix 321
Humboldtiana 15
Humboldtianidae 44, 51, 53*, 54, 55, 56
Hygrohelicopsis 64, 65*, 66, 109, 181
Hygromia 68, 69, 70*, 71, 205
Hygromiidae 15, 17, 30, 34, 37, 38, 39, 40,
41, 49, 51, 53*, 54, 55, 56, 58, 62, 63*,
65*, 67, 69, 72, 75, 76, 77, 79, 80, 109,
112, 113, 160, 163
Hygromiinae 39*, 56, 57, 62, 63*, 65*,
68, 70*, 71, 73, 75, 110, 112, 113, 163,
196, 225
hypophaea, Eulota (?) 178
hypophaea, Fruticicola 178
hypophaea, Leucozonella 46, 106
hypophaea, Leucozonella (Narina) 168, 178,
179*
illuminata, Nanaja 46, 100, 164, 165,166*
incarnata, Helix 234
incarnata, Monacha 234
incarnata, Monachoides 47, 70, 104, 234,
235*
incarnata, Perforatella (Monachoides) 234
incarnata, Zenobiella 234
indigena, Helix (Eucampylaea) 151
inflata, Helix 275
instabilis, Helicella 217
instabilis, Helicella (Helicopsis) 217
instabilis, Helicopsis 47, 104, 211, 217
instabilis, Helix 217
intercedens, Helix atrolabiata 332
intermissa, Helix 340
Isabella, Pleurodonte 55
Isognomostoma 3, 18, 108, 315
isognomostoma, Helix 315
isognomostoma, Isognomostoma 315
isseli, Lejeania 74
itala, Helix 224
jachnoi, Helicopsis instabilis 217
Jasonella 111, 271
jasonis, Helix 271
jasonis, Helix (Nummulina) 272
joannis, Caucasocressa48, 76,103, 305, 306,
307*
joannis, Helix 304, 306
joannis, Helix (Fruticocampylaea) 306
joannis, Metafruticicola pratensis 306
j oppensis, Helix 222
j oppensis, Xeropicta 222
Kalitinaia 253
Karabaghia 23, 74*, 111, 262
Karaftohelix 60,' 123
ketseri, Cathaica 121
kirgisensis, Paedhoplita 48, 76, 98, 302,
304
Klikaia 319
Klikia 18, 77, 319
kobiensis, Fruticocampylaea 38, 48, 69,
108,241,243*
kobiensis, Fruticocampylaea narzanensis
243 '
kobiensis, Helix 243
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
355
kobiensis, Helix (Fruticocampylaea) 243
Kokotschashvilia 16, 41, 64, 65*, 110, 184
krynickii, Helicella 219
krynickii, Helicella (Xeropicta) 219
(krynickii, Helicopsis (Xeropicta) 219
(krynickii, Helix 219, 222
(krynickii, Xerophtla (Heliomanes) 219
(krynickii, Xeropicta 47, 68,105, 219, 220*,
223
(kubanica, Caucasotachea 49, 332
kudiensis, Hygromia 225
kudiensis, Lindholmomneme 47, 102, 225
labianix, Archaica 41, 48, 75, 76, 99,
261
(labianix, Archaica (Ugama) 257, 260*,
261
>lactea, Helix 326
lactea, Otala 27, 42
iLaeocathaica 41
laminata, Paedhoplita 17, 48, 76, 98, 302
lantzi, Bradybaena 15, 46, 61, 62, 147*
lantzi, Eulota (Eulota) 147
lantzi, Fruticicola 147
.’lanuginosa, Hygromia (Ganula) 68
lapicida, Helicigona 17, 40, 49, 99, 313,
314*
lapicida, Helix 313
.lavrushini, Helix 144
lencoranea, Caucasotachea 49, 101, 331,
332, 333*
'Lencoranea, Helix atrolabiata 332
lencoranea, Helix (Tachea) 332
lens, Helix 116
lens, Lindholmiola 57
Lentiga 300
lentina, Angiomphalia 48, 76, 99, 111
lentina, Angiomphalia (Lentiga) 291, 300*
lentina, Helix 300
lentina, P aedhoplita 300
(Leucozonella 65*, 110, 160, 164, 168
Leucozonella s. str. 110, 168
Levantina 78, 109, 327
libidinosa,’ Helicella 212
iibidinosa, Helicopsis 47, 212
.likharevi, Bradybaena 157
likharevi, Helicopsis 47, 68, 104, 105, 211,
212, 213*, 255
limbata, Helix 280
limbata, Hygromia (Riedelia) 68
.limitata, Helix (Eulota) rubens 172
lindcholmi, Euomphalia 303
lindholmi, Euomphalia 303
lindholmi, Paedhoplita 41*, 48, 76, 98,
302, 303*
Lindholmia 337, 340
Lindholmiola, 57, 58, 112, 116, 118
Lindholmiolinae 112, 116, 118
Lindholmomneme 56, 68, 70*, 111, 225'
lithophila, Schileykoia 244
longiflagellata, Theba (Paratheba) 285
lubomirskii, Fruticicola 204
lubomirskii, Helix (Fruticicola) 204
lubomirskii, Plicuteria 47, 67, . 104, 204*
lubomirskii, Trichia 204
lubomirskii, Trichia (Trichia) 204
lucorum, Helix 49, 101, 337, 342*, 343
lunula, Triodopsis 347
lusitanica, Helix 118
lutescens, Helix 49, 101, 398, 345
lutescens, Helix (Helicogena) 345
maacki, Bradybaena 43, 46, 102, 124, 132,
133*
maacki, Eulota (Eulota) 132
maacki, Helix 132
maacki, Helix (Fruticicola) 132
macgillivrayi, Rhynchotrochus 55
macromphala, Helix (Fruticocampylaea)
Narzanensis 241
maculata-fasciata, Helix (Tachea) atrola-
biata 332
maeotica, Helix 49, 338
maiae, Euomphalia 278
maiae, Euomphalia (Harmozica) 278
maiae, Stenomphalia 16, 48, 107
maiae, Stenomphalia (Batumica) 275,
'277*, 278
major, Rradybaena schrencki 130
major, Petasia bidens 232
makvalae, Kokotschashvilia 47, 65, 108,
184, 185*, 187
makvalae, Trichia (Xerocampylaea) 184
maracadensis, Helix rufispira 173
martensl, Helix taurica 342
Martha 211
maslovae, Caucasotachea (?) 337
maslovae, Steklovia 49, 337
Massylaea 77
Mastigophallus 57
Maurohelix 77
maxima, Theba (Paratheba) 285
mecoleuca, Euomphalia 172
memnonis, Monachoides 71
menetriesil, Helix 193, 194
mesasiana, Aegista 3O0
mesasiana, Plectotropis 300, 301
Mesasiata 112, 119, 120
mesoleuca, Eulota (Leucozonella) 172
mesoleuca, Fruticicola 172
mesoleuca, Helix 172
mesoleuca, Leucozonella 37, 46, 66, 106
mesoleuca, Leucozonella (Leucozonella)
168, 172, 173*
mesoleuca, Trichia 172
Metairuticicola 249, 304
Metafruticicolinae 63*, 65*, 76, 77, 113,
249, 304
Metatheba 75, 112, 287
Micromphalia 275
middendorffi, Bradybaena 46, 60, 102, 124,
129, 130*
middendorffi, Eulota (Eulota) 130
middendorffi, Helix 130
milaschevitschi, Helicopsis retowskii 216
millepunctata. Helix 222
mingrelica, Circassina 271
mingrelica, Circassina (Jasonella) 271
mingrelica, Platytheba 48, 99
mingrelica, Platytheba (Jasonella) 271,
272*
minor, Helix joannis 305
minor, Helix ’ (Pomatia) buchi 343
miranda, Helix 135
Monacha 16, 33, 40, 71, 72, 74*, 111, 112,
234, 271, 281 '
356
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
Monacha s. str. 38, 40, 41, 75, 111,
286
Monachoides 16, 68, 69, 70», 71, 110,
234
Monadenia 44
montana, Fruticicola lantzi 147
murensis, Eulota 139
ncma, Helix (Latonia) globula 275
Nanai a 64, 65*, 110, 164
Narina 64, 65*, 66, 110, 168, 178
narzanensis, Fruticocampylaea 22*, 28*,
31*, 44, 48, 69, 108, 241, 242*, 263
narzanensis, Fruticocampylaea (Frutico-
campylaea) 241
narzanensis, Helix 241
narzanensis, Helix (Fruticocampylaea) 241
necopinatus, Bradybaena plectotropis 105,
125, 149, 150
necopinatus, Cathaica (Pseudiberus) 148
nemoralis, Сераеа 49, 101
nemoralis, Сераеа (Сераеа) 320, 321*, 322
nemoralis, Helix 320
nemoraloides, Helix (Tachea) atrolabiata 335
Nesiohelix 54
nivalis, Helix 193
nivicola, Helix (Fruticocampylaea) eichwaldi
196
nordenskioldi, Chilanodon 227
nordenskioldi, Euomphalia 227
nordenskioldi, Helix 225
nordenskioldi, Helix (Eulota) 227
nordenskioldi, Lindholmomneme 47, 102,
103, 225, 227*
nordenskioldi, Zenobiella 227
nordmanni, Caucasotachea (Lindholmia) 339
nordmanni, Helix 49, 101, 337, 338*
nordmanni, Helix (Helicogena) 338
nordmanni, Helix (Lindholmia) 339
nothra, Helix (Trichia) 207
notophila, Hygromia 226
notophila, Lindholmomneme 47, 102, 225,
226*
nubigena, Diodontella 47, 98, 203
nubila, Cathaica (Xerocathaica) apollinis
259
Nummulina 271
nummus, Helix 271
nummus, Platytheba 271
nymphaea, Helix pratensis 305
obscura, Helix frequens 288
obscura, Theba samsunensis 288
obtusalis, Helix 340
obtusata, Helix 340
obtusior, Helicella crenimargo 252
obtusior, Helix 254
obtusus, Cylindrus 40
obvia, Helicella 224
obvia, Helix 216, 224
obvoluta, Helicodonta 46, 49, 98, 114, 115*
obvoluta, Helix 115
obvoluta, Trigonostoma 115
Octadenia 78, 109, 322
Odontotrema 65*, 109, 166
Oestophora 58, 118
Onchidiella 21
opposita, Helix (Eucampylaea) 155
Oreohelicidae 30, 33, 44, 51, 53*, 55, 56,
Oreohelix 17
Orianda, Xerophila (Heliomanes) 219.
Oscarboettgeria 74*, 111, 273
osculum, Helix 319
osculum, Klikia 18
Otala 326
Oxychilus 223
pachnodes, Helix 250
Paedhoplita 18, 41, 76, 111, 302
Paedhoplitinae 63*, 65*, 75, 76, 111, 291
pallasii, Helix (Tachea) atrolabiata 335
pallasii, Helix calligera 335
palmeni, Helix 148
Papuina 25
Papuininae 44, 52
parableta, Helix 223
parableta, Xeropicta 47, 108, 223
Paratheba 75, 112, 282
paricincta, Eulota (Eulota) 120
paricincta, Helix 120
paulhessei, Helicella 216
paulhessei, Helicella (Helicopsis) 216
paulhessei, Helicopsis 47, 103, 212, 216
pavlovskii, Bradybaena 153, 155
pavlovskii, Eulota 155, 157
pekinensis, Cathaica 60
pellita, Metafruticicola 76, 77
pelluctdepunctata, Helix (Fruticocampylaea)
joannis 306
perableta, Helix 223
Perforatella 68, 69, 70*, 71, 110, 118, 231,
262
perfrequens, Theba samsunensis 288
perfrequens, Helix (Theba) samsunensis
228
perlucens, Bradybaena 46, 105, 125, 151,
152*
perlucens, Cathaica (Cathaica) 151
perlucens, Cathaica (Kaznakoviella). 151
perlucens, Helix (Eucampylaea) 151
personatum, Anchistoma (Triodopsis) 315
personatum, Helix 31.5
personatum, Isognomostoma 18, 40, 49, 76,.
98, 315, 316*
personatum, Triodopsis 315
Petasia 231
petrii, Arianta aethiops 310, 312*, 313
petrii, Helix aethiops 313
phaeolaema, Helix 190
phaeolaema, Helix (Fruticocampylaea) 190"
phaeolaema, Kokotschashvilia 19*, 44, 47,
65, 107, 184, 187, 190*
phaeolaema, Trichia 190
phaeolaema, Trichia (Xerocampylaea) 190
phaeozona, Bradybaena plectotropis 105,
125, 149*, 150
phaeozona, Eulota (Eulota) 148
phaeozona, Helix 148
Pietruskyana, Helix 208
pisana, Theba 77, 80
pisiformis, Euomphalia 278
pisiformis, Euomphalia (Diplobursa) 278
pisiformis, Helix 278
pisiformis, Helix (Latonia) 278
pisiformis, Stenomphalia 48, 107, 278
pisiformis, Stenomphalia (Diplobursa) 275,.
278, 279*
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
357
placida, Helix 328
Platytheba 74*, 111, 271
Platytheba s. str. Ill, 271, 273
plebeja, Trichia 205
Plectopylis 52*
plectotropis, Bradybaena 25*, 46, 52, 61,
62, 148, 151
plectotropis, Bradybaena plectotropis 99,
125, 149, 150
plectotropis, Cathaica (Pseudiberus) 148
plectotropis, Helix 148
Pllcostoma 315
Plicuteria 64, 65*, 110, 203
podolica, Helicella cereoflava 213
podolica, Helix (Trichia) 269
Polygyridae 112, 347
Polygyroidea 347
polytyla, Laeocathaica 41*
Pomatia ЗУ1
pomatia, Helix 3, 29*, 43*, 49, 101, 337,
338, 345*, 346
pomatia, Helix (Helicogena) 346
Ponsadenia 58, 61, 62, 112, 119
Ponsadenia s. str. 112, 119, 121
pontica, Helix 198
pontica, Helix narzanensis 198
praeorientalis, Monacha 48
praeorientalis, Monacha (Boemica) 289
praerupta, Helix 286
pratensis, Caucasocressa 48, 76, 103, 305*
pratensis, Fruticocampylaea (Caucasocressa)
305
pratensis, Helix 305
pratensis, Helix (Fruticocampylaea) 305
pratensis, Metafruticicola 263, 305
pratensis, Metafruticicola (Metafruticicola)
305
prometheus, Helix 273
prometheus, Helix (Nummulina) 273
prometheus, Platytheba 48, 99
prometheus, Platytheba (Platytheba) 271,
273
Prosobranchia 20
psedachica, Caucasigena 47
psedachica, Caucasigena (Caucasigena) 192
psedachica, Tropidomphalus 192
pseudarianta, Fruticicola phaeozona 148
pseudoferghanica, Bradybaena 46, 61, 62,
106, 125, 158, 159*, 160
pseudoglobula, Helix 278 -
Pseudotachea 77
Pseudotrichia 69, 70*, 71, 110, 238
Pulmonata 13
Pyrochilus 3
quadrifasciata, Helix (Pomatia) vulgaris
340
radlosa, Helix 343
ravergiensis, Helix 280
ravergieri, Euomphalia 280
ravergieri, Euomphalia (Harmozica) 280
ravergieri, Helix 280
ravergieri, Stenomphalia 48, 104
ravergieri, Stenomphalia (Harmozica) 275,
280, 281*
ravergii, Helix 280
ravida, Bradybaena 20, 36, 46, 102, 124,
143, 144*
ravida, Eulota (Acusta) 144
ravida, Helix 144
regeliana, Angiomphalia 37, 42, 48, 106,
172
regeliana, Angiomphalia (Angiomphalia)
292, 296*
regeliana, Eulota (Leueozonella) rubens
296
regeliana, Euomphalia 297
regeliana, Helix 291, 297
Regeliana, Helix rubens 296, 297
reminiscenta, Caucasigena 47, 99
reminiscenta, Caucasigena (Caucasigena)
192, 199
rengarteni, Caucasigena 38, 47, 67, 108,
198
rengarteni, Caucasigena (Caucasigena) 192
195, 196*
rengarteni, Helix (Fruticocampylaea) 195
retowskii, Helicella 215
retowskii, Helicella (Helicopsis) 215
retowskii, Helicopsis 34, 35*, 47, 104, 211,
215*
retowskii, Helix 215
retteri, Eulota (Leueozonella) 175
retteri, Helix 175
retteri, Leueozonella 46, 64, 66, 106
. retteri, Leueozonella (Leueozonella) 168,
175*
retteri, Trichia 175
revelata, Helix 234
rhysota, Helix 225
rhysota, Helix (Trichia) 227
roseni, Theba orientalis 289
roxane, Eulota (Leueozonella) 145
roxane, Fruticicola 145
rubens, Eulota (Leueozonella) 170, 171
rubens, Euomphalia 269
rubens, Fruticicola 171
rubens, Helix 168, 171, 297
rubens, Leueozonella 12*, 37, 46, 64, 66,
106, 177
rubens, Leueozonella (Leueozonella) 168,
171*
rubens, Trichia 171, 297
rubiginosa, Helix 238
rubiginosa, Monacha 238
rubiginosa, Monachoides 238
rubiginosa, Perforatella (Monachoides) 238
rubiginosa, Pseudotrichia 16, 33, 36, 48,
49, 70, 71, 100, 225, 238, 239*
rubiginosa, Zenobiella 237, 238
rufispira, Cathaica 173
rufispira, Eulota (Leueozonella) 173
rufispira, Helix 173
rufispira, Leueozonella 46, 64, 66, 107,
181
rufispira, Leueozonella (Leueozonella) 173,
174*
rufispira, Trlchla 173, 175
rugata, Camaena 55
samsunensis, Helix 288
samsunensis, Helix (Theba) 288
samsunensis, Metatheba (Metatheba) 288
samsunensis, Monacha 48, 107
358
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
samsunensis, Monacha (Metatheba) 282,
288*
samsunensis, Theba 288
samsunensis, Theba (Metatheba) 288
saturata, Fruticicola 154
saxatilis, Cathaica (Pseudiberus) 148
scalaris, Cathaica (Pseudiberus) 148
schamyli, Fruticocampylaea 241
schaposchnikovi, Caucasigena 41*, 47, 108,
182
schaposchnikovi, Caucasigena (Anoplitella)
192, 199*
schaposchnikovi, Helix 199
schaposchnikovi, Trichia (Xerocampylaea)
199
schelkovnikovi, Helix (Jacosta) 254
schelkovnikovi, Xerosecta 48, 99, 253,
253*, 254
schelkovnikovi, Xerosecta (Kalitinaia) 254
schileykoi, Oestophora 46, 58, 118
schileykoi, Perforatella 118
schrencki, Bradybaena 44, 46, 50, 60, 102,
103, 104, 124, 127, 128*, 130
schrencki, Eulota (Eulota) 127, 157
schrencki, Helix 127
schuberti, Helix 250
scrobiculosa, Monacha (Boemica) subcart-
husiana 282, 291
scrobiculosa, Theba 291
scrobiculosa, Theba (Metatheba) orientalis
291
scythica, Bradybaena 17, 46, 61, 101, 124,
145, 146*
scythica, Helix (Eucampylaea) 145
seductilis, Angiomphalia 48, 106, 298
seductilis, Angiomphalia (Angiomphalia)
292, 298, 299*
seductilis, Helix (Theba) 298
seetzeni, Trochoidea 42
seetzeni, Xerocrassa 34
selecta, Euomphalia 275
selecta, Euomphalia (Stenomphalia) 275
selecta, Helix 275
selecta, Helix (Theba)
selecta, Stenomphalia 48, 107
selecta, Stenomphalia (Stenomphalia) 275,
276*
selskii, Eulota (Acusta) 144
selskii, Helix 144
semenovi, Bradybaena 121
semenovi, Cathaica (Pliocathaica) 121
semenovi, Eulota 121
semenovi, Helix 119, 121
semenovi, Ponsadenia 41, 46, 61, 62, 105,
119
semenovi, Ponsadenia (Ponsadenia) 121,
122*
Semifruticicola 38
serbica, Semifruticicola 68
sericea, Helix 205, 238
serotina, Helix 133
setosa, Liburnica 40
shadini, Tricheulota (Pseudotrichia) 238
shileykoi, Metafruticicola (Metafruticicola)
247
Shileykoia 70*, 111, 244
stbirica, Helix 229
Sigmurethra 12, 23
Silvestris, Fruticicola lantzi 147
similaris, Bradybaena 50
similaris, Helix 123
simpla, Circassina circassica 77, 113, 247,
248*
sinistrorsa, Bradybaena lantzi 106, 125,
147
sinistrorsa, Fruticicola lantzi 147
skwortzowi, Eulota (Eulota) 148
skwortzowi, Fruticicola 148
solidior, Helix 263
solidior, Helix (Campylaea) Narzanensis
263
solidior, Hesseola 38, 40, 48, 103, 263..
264*
Soosia 57, 58
Sphincterochila 12, 24, 42
Sphincterochilidae 49, 51, 53*, 55, 56
spiriplana, Helix 327
stauropolitana, Helix atrolabiata 332'
stauropolitana, Helix (Tachea) 332
Steklovia 335
Stenomphalia 72, 73, 74*, 112, 275
Stenomphalia s. str. 112, 275
stenroosi, Helix (Pomatia) 120
stephaniae, Circassina circassica 112, 247,.
248*
stephaniae, Hela 247
steppensis, Fruticicola lantzi 147
stoliczkana, Bradybaena 46, 60, 61, 105,
125, 155, 156*
stoliczkana, Cathaica 155
stoliczkana, Helix 155
strelkovi, Bradybaena 46, 103, 124, 135,
136*
strelkovi, Eulota (Karaftohelix) 135
striata, Helicella (Helicopsis) 213
striata, Helicopsis 30, 32*, 47, 104, 211,
213, 214*
striata, Helicopsis (Martha) 213
striata, Helix 211, 213
Strtatella 211
Striatinella 211
strigella, Euomphalia 40, 48, 73, 103, 266,
269, 270*
strigella, Helix 266, 269
striolata, Trichia 64, 65
stschukini, Cathaica (Pseudiberus) 148
stschukini, Diodontella 47, 98, 202, 203
sturanyana, Helix 160
sturanyi, Cathaica (Eucathaica) 160
stuxbergi, Helix 237
stuxbergi, Monachoides 36, 47, 49, 70, 100,
234, 237*
stuxbergi, Zenobiella rubiglnosa 237
Stylommatophora 13
Suanetica, Helix (Fruticocampylea) Nar-
zanensis 241
subcarthusiana, Helix (Theba) 289
subcarthusiana, Helix orientalis 289
subcarthusiana, Monacha 48, 74, 103
subcarthusiana, Monacha (Boemica) sub-
carthusiana 282, 289, 290*
subcarthusiana, Theba (Metatheba) orien-
talis 289
sublecta, Cretigena 76, 77
Suboestophora 58
subpersonatum, Helix 347
subpersonatum, Isognomostoma 347
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ' 359
subpersonatum, Triodopsis 49, 98, 347
sub striata, Helix 216
stigoschuriana, Cathaica (Pseudiberus) 148
sunzhica, Helicella 212
sunzhica, Helicopsis 47, 212
Tachea 320, 330
talischana, Helix 285
talyschana, Helix 285
talyschana, Helix (Latonia) 285
talyschana, Monacha 48, 107
talyschana, Monacha (Paratheba) 282,
285*
tanta, Kokotschashvilia 47, 65, 108, 184,
186*
tarbagataica, Bradybaena cavimargo 125,
162
Tarbagataja 112, 119
taurica, Helix 217
taurica. Helix lucorum 342, 343
Teberdinia 64, 65*, 110, 183
tenuitesta, Helix (Fruticocampylaea) phaeo-
laema 190
thalestris, Caucasigena 47, 107
thalestris, Caucasigena (Dioscuria) 191,
201, 202*
thalestris, Fruticicola 201
thalestris, Trichia (Dioscuria) 201
thalestris, Trochulus 201
Theba 77, 271, 281, 282, 287
theodosiae, Helix 219
tianshanica, Cathaica 257
tiflisiana, Cernuella 48, 103, 250, 252*
tiflisiana, Helicella (?) 252
tiflisiana, Xerophila 252
Tingitana 3
tomyris, Bradybaena 46, 105, 125, 162
tomyris, Eulota (Acusta ?) 162
tomyris, Fruticicola 162
torsa, Helix 116
transbaicalia, Bradybaena 46, 60, 102, 124,
128, 129*
transcaspia, Euomphalia 265
transcaspia, Helix (Carthusiana) 265
transcaspia, Helix (Latonia) 265
transcaspia, Hesseola 48, 74, 105; 263,
265*
transcaucasia, Helix ravergieri 280
transcaucasica, Euomphalia ( Harmozica)
ravergieri 280
transcaucasica, Helix ravergieri 280
transcaucasica, Helix (Theba) ravergieri
280 '
transient, Helix (Tachea) atrolabiata 332
transilensis, Bradybaena plectotropis 124,
148, 149, 151
transilensis, Cathaica (Pseudiberus) 148
transitans, Cathaica 60
transsylvanica, Hygromia 71
transsylvanica, Hygromia (Lozekia) 68,
69
Trichia 64, 65*, 66, 67, 110, 181, 184, 205,
207, 234, 297
Trichia s. str. 17, 68, 184
Trichiinae 33, 54, 55, 62, 63*, 64, 65*,
66, 67, 68, 69, 71, 72, 75, 76, 77, 109,
160, 163, 223, 297
tricolor, Helix (Tachea) atrolabiata 332
tridentata, Helix 347
Trigonostoma 114
Triodopsinae 112, 347
Triodopsis 3, 112, 347
Trishoplita 59
Trissexodon 57
Trochiscus 231
Trochoidea 42
Trochulus 201, 205, 271
tschetschenica, Caucasigena 47, 102
tschetschenica, Caucasigena (Caucasigena),
192, 194*
tschetschenica, Helix (Fruticocampylaea)’
194
tschorochensis, Helix (Theba) 275
turriplana, «Oestophora» 58
tzwetkovi, Bradybaena 46, 102, 125, 151*
tzwetkovi, Zenobiella 257
Ugama 110, 260
Ulostoma 315
umbrosa, Helix 240
umbrosa, Monacha 240
umbrosa, Monachoides 240
umbrosa, Urticicola 48, 104, 240*
umbrosa, Zenobiella 240
umbrosa, Zenobiella (Urticicola) 240
undata, Monachoides 71
unicincta, Helix (Pomatia) vulgaris 340
unicingulata, Helix (Fruticocampylaea) pra-
tensis 305
Urguesella 4
urmiensis, Helix 328
Urticicola 68, 69, 70*, 71, 110, 239-
urupensls, Eulota 135
ussuriensis, Bradybaena 46, 52, 102, 124,
131*
ussuriensis, Eulota (Eulota) 131
ussuriensis, Eulota graeseri 131
ussuriensis, Helix (Campylaea) 131
Vaginulidae 44
variabilis, Cernuella (Cernuella) virgata
250
variabilis, Helix 250
ventouxiana, Monachoides 70
vermiculata, Eobania 3, 17, 42, 44, 49,
100, 326*, 327
vermiculata, Helix 326, 327
vermiculata, Helix (Otala) 327
vermiculata, Otala (Eobania) 327
verna, Helix (Trichia) 238
vicina, Helix 235
vicina, Monacha 235
vicina, Monachoides 47, 70, 100, 234, 235,
236*
vicina, Perforatella (Monachoides) 235
vicina, Zenobiella 235
villosula, Fruticicola 208
villosula, Helix 208
villosula, Helix villosa 208
villosula, Trichia 16, 47, 100, 205, 208*
vindobonensis, Сераеа 49, 101
vindobonensis, Сераеа (Octadenia) 320,
323*
vindobonensis, Helix 323
Vindobonensis, Helix sylvatica 323
virgata, Cernuella 34, 48, 71, 103, 250,
251* ' ' "
зво
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
virgata, Helicella (Cernuella) 250
virgata, Helix 250
virgo, Ganesella 139
vulcanica, Bradybaena 20, 46, 102, 124,
142, 143*
pulgaris, Helix 338, 340
vulgaris, Helix (Helicogena) 340
vulgaris, Helix (Pomatia) 338, 339, 340
Weyrichii, Bradybaena 46, 60, 103
Weyrichii, Bradybaena weyrichii 124, 133,
134*, 135
weyrichii, Eulota (Karaftohelix) 133
weyrichii, Fruticicola (Karaftohelix) 133
weyrichii, Helix 133, 134
Xerocampylaea 67, 271
Xerocrassa 41
Xerolauta 250
Xerophila 223
Xeropicta 30, 33, 41, 65*, 67, 68, 69, 109,
218, 219, 287
Xerosecta 41, 69, 70*, 71, 111, 253
Xero tricha 38
Zachrysia 44
zamechovense, Klikia giengensis 319*
zeiliana, Helix rubens 171, 297
zelebori, Xerocampylaea 38, 67
Zenobiella 225, 234
ziegleri, Helix 193
zolotarevi, Helix (Fruticocampylaea) 183
zolotarevi, Teberdinia 41, 46, 107, 183*
Zonitidae 50
Zonitoides 21
ТАБЛИЦА I
1 — Helicodonta obvoluta (MiilL), Карлштеннскпй заповедник, ЧССР; 2 — Lindholmiola
fraudulosa (Steklov), меотические отложения па p. Фортапге., голотип; 3 — L. согсугеп-
sis (Fee.), окр. Кишинева; 4 — Oestophora schileykoi (Prysjazhnjuk), понтийские отло-
жения левобережья Дуная, окр. оз. Ялпух, паратип; 5 — Ponsadenia (Tarbagataja)
hirsuta (Matiokin), Тарбагатай; 6 — P. (Mesasiata) duplocincta (Mart.), южный берег
оз. Иссык-Куль; 7 — P. (Ponsadenia) semenovi (Mart.), ущелье Беш-Таш на Таласском
хр.; 8— Bradybaena fruticum. (MiilL), Куртатипское ущелье на Сев. Кавказе; 9—
В. schrencki (Midd.), бассейн р. Вытегры, паратпп; 10 — В. transbaicalia sp. nov.,
Забайкалье, хр. Хамар-Дабап, голотип.
I А. А. Шилейко
ТАБЛИЦА II
11 — Bradybaena middendorffi (Gerstf.), зал. Петра Великого, о-в Б. Пелис; 12 —
В. ussuriensis (West.), зал. Петра Великого, о-в Попова; 13 — В. maacki (Gerstf.),
с. Пейшула Приморского края; 11 — В. weyrichii weyrichii (Schrenck), окр. Южно-
Сахалинска; 15 — В. strelkovi Lich. et Ramm., южн. Сахалин; 16 — В. duiensis (West.),
южн. Сахалин; 17 — В. dieckmanni (Mouss.), окр. Владивостока; 18 — В. arcasiana
(Crosse et Deb.), окр. Смоляпиново Приморского края; 19 — В. fragilis (Pilsbry),
запов. «Кедровая Падь»; 20 — В. vulcanica sp. nov., южная часть о-ва Кунашир,
голотип.
ТАБЛИЦА III
21 — Bradybaena ravida (Benson), о-в Путятин; 22 — В. scylhica (West.), долина p. На-
рын; 23 — В. lanizi (Lindh.) L dextrorsa, бассейн p. Некем; 24 — В. lantzi (Lindh.)
f. sinistrorsa, вост. Казахстан, окр. оз. Иссътк; 25 — В. plectotropis smsp. phaeozona
(Mart.), ущелье Каскасу, Заилийскии хр.; 26 — В. plectotropis smsp. almaatini (Skv.),
долина p. Нарын; 27 — В. plectotropis smsp. necopinatus (Tzv.), ущелье p. Аламедин,
Киргизский хр.; 28 — В. plectotropis smsp. plectotropis (Mart.), южный берег оз. Иссык-
Куль; 29 — В. plectotropis smsp. transilensis (Tzv.), берег Бол. Алмаатинского озера;
30 — В. perlucens (Rosen), Гиссарский хр.
ТАБЛИЦА IV
31 — Bradybaena dichrozona (Mart.), Чаткальский хр.; 32 — В. bilaticincta (Mart.),
окр. оз. Сары-Челек; 33 — В. stoliczkana (Nevill), долина р. Кум-Тар; 34 — В. sto-
liczkana (Nevill), южный берег оз. Иссык-Куль, 20 км к югу от Каджисая; 35 — В. hel-
vola (L. Pfr.), Болыпенарымское в вост. Казахстане; 36 — В. helvola (L. Pfr.), Тарбага-
тай; 37 — В. pseudoferghanica sp. nov., Джунгарский хр., голотип; 38 — В. cavimargo
cavimargo (Mart.), ущелье Карабалты, Киргизский хр.; 39 — В. fedschenkoi (Mart.),
окр. с. Бободархон на Кураминском хр.; 40 — В. tomyris (Lindh.), долина р. Чич-
кан, Ферганский хр., голотип.
ТАБЛИЦА V
41 — Nanaja cumulata sp. nov., бассейн p. Иском, голотип; 42 — N. illuminata sp. nov.,
бассейн p. Некем, голотип; 43 — Odontotrema diplodon Ljndh., окр. оз. Сары-Челек; 44 —
Leucozonella (Leucozonella) ferghanica (Lindh.), берег оз. Кыла-Куль в Сары-Челекском
запов.; 45 — L. (L.) caryodes (West.), ущелье Беш-Таш, Таласский хр.; 46 — L. (L.) ru-
bens (Mart.), ущелье р. Аламедпп, Киргизский хр.; 47 — L. (L.) mesoleuca (Mart.),
ущелье р. Аламедин, Киргизский хр.; 48 — L. (L.) mesoleuca (Mart.), окр. Аравана,
Алайский хр.; 49 — L. (L.) rufispira (Mart.), перевал Апзоб, Гиссарский хр.; 50 -
L. (L.) rufispira (Mart.), ущелье Беш-Таш, Таласский хр.
ТАБЛИЦА VI
51 — Leucozonella (Leucozonella) retteri (Rosen), ущелье Коидара, Гиссарский xp.;
52 — L. (L.) angulata (West.), долина p. Пскем; 53 — L. (Narina) hypophaea (Lindh.),
долина p. Нарын, Ферганский xp., выше Таш-Кумыра; 54 — L. (N.) hypophaea (Lindh.),
долина p. Нарын, Ферганский xp., близ с. Кара-Коль; 55 — L. (N.) crassicosta sp. nov.,
окр. с. Ала-Бука в предгорьях Чаткальского хр., голотип; 56 — L. (Leucozonella)
caria Schileyko, окр. Ходжи-Обигарм, Гиссарский xp., голотип; 57 — Hygrohelicopsis
darevskii Schileyko, Чегемское ущелье, голотип; 58 — Teberdinia zolotarevt (Lindh.),
Тебердинский запов.; 59 — Kokotschaslwilia makvalae (Hudec et Lezhawa), центральная
часть южных склонов Большого Кавказа, окр. с. Квайса; 60 — К. tanta Schileyko,
Грузия, окр. с. Лебарде Гегечкорского р-на, голотип.
ТАБЛИЦА VII
61 — Kokotschaslwilia eberhardi Schileyko, южные склоны центр. Кавказа, между
с. Сиони и с. Казбеги, голотип; 62 — К. holotricha (Bttg.), долина р. Техури; 63 —
К. phaeolaema (Bttg.), Чегемское ущелье; 64 — Caucasigena (Caucasigena) psedachica
(Steklov), акчагыльские отложения в Пседахской балке, Предкавказье, голотип;
65 — С. (С.) fortangensis (Steklov), меотические отложения па р. Фортанге, Пред-
кавказье, голотип; 66 — С. (С.) armeniaca (L. PIT.), Армения, гора Тодар; 67 —
С. (С.) tschetschenica (Ret.), Куртатипское ущелье па Сев. Кавказе; 68 — С. (С.) геп-
garteni (Lindh.), окр. с. Актопрак, Чегемское ущелье; 69 — С. (С.) eichwaldi (L. Pfr.),
окр. с. Чип, Дарьяльское ущелье; 70 — С. (С.) eichwaldi (L. Pfr.), Куртатипское
ущелье на Сев. Кавказе.
ТАБЛИЦА VIII
'll — Caucasigena [Caucasigena) abchasica (Lindh.), долина p. Иигурп; 72 — С. (C.) re-
miniscenta sp. nov., Кавказский залов., голотип; 73 — C. (A noplilella) schaposchnikovi
(Rosen), Чегемское ущелье; 74 — С. (Л.) schaposchnikovi (Rosen), урочище Ара-боран
между Чегемским п Баксанским ущельями; 75 — С. (Dioscuria) thalestris (Lindh..),
Новый Афон; 76 — Diodontella stschukini Lindh., Чегемское ущелье, голотип; 77 —
D. nubigena Lindh., Чегемское ущелье, голотип; 78 — Plicuteria lubomirskii (Slos.),
Оломоуц, ЧССР; 79 — Trichia hispida (L.), Московская обл., Ромашкове; 80 — Т. con-
cinna (Joffr.), Рожпява, ЧССР.
ТАБЛИЦА IX
Скульптура: 81 — Ponsadenia (Ponsadenia) semenovi (Mart.); 82 — Bradybaena ravida
(Benson); 83 — Monachoides vicina (Rssm.); 84 — M. stuxbergi (West.); 85 — Fruti-
cocampylaea narzanensis (Kryn.); 86 — Euomphalia strigella (Drap.); 87 — Stenomphalia
(Batumica) maiae (Hudec et Lezhawa); 88 — Helicigona lapicida (L.); 89 — Cryptompha-
lus aspersa (Miill.); 90 — Helix buchi Dub. de Mont.
V2I
ТАБЛИЦА X
91 — Trichia villosula (Rssm.), Бабице, ЧССР; 92 — T. bielzi (A. Schm.), окр. Рахова,
Закарпатье; 93 — Edentiella bakowskii (Pol.), окр. Рахова, Закарпатье; 94 — Helicop-
sis sunzhica (Steklov), акчагыльскне отложения на р. Сунже у с. Алды, Предкавказье,
голотип; 95 — Н. libidinosa (Steklov), апшеронские отложения в Заманкульской балке,
Предкавказье, голотип; 96 — Н. likharevi sp. nov., Копетдаг, ущелье Игдеджин,
паратип; 97 — Н. striata (Miill.), окр. Курска; 98 — Н. retowskii (Cl.), окр. Бахчиса-
рая; 99 — Н. retowski (Cl.), окр. Севастополя; 100 — Н. dejecta (Cr. et Jan), окр. Ново-
российска.
ТАБЛИЦА XI
101 — Helicopsis paulhessei (Lindh.), Гаснра, Крым, паратиц; 102 — H. instabilis (Rssm.),
окр. Кишинева; ЮЗ — II. instabilis Г. jachnoi Cl., окр. Беляевкл в низовьях Днестра;
104 — II. filiniaipto (Кгуп.), окр. Севастополя; 105 — Xeropicta parableta (Bttg.), Тур-
ция, окр. оз. Ban; 106 — X. krynickii (Кгуп.), шиша p. Лепкорап-чай; 107 — X. derbentina
(Кгуп.), окр. Дербента; 108 — X. canclaliarica (L. BIT.), окр. Душанбе; 109 — Heli-
cella candicans (L. Pfr.), окр. Кишинева; НО — II. candicans (L. Pfr.), окр. Пловдива,
Болгария.
ТАБЛИЦА XII
111 — Lindholmomneme kudiensis (Cock.) из окр. бухты Рудная, Приморье; 112 —
L. notophila (Cock.) из окр. Владивостока (не вполне сформированная раковина); 113 —
L. nordenskioldi (West.) из окр. Прокопьевска Кемеровской обл.; 114 — Chilanodon
bicallosa (L. Pfr.) из окр. Прокопьевска Кемеровской обл.; 115 — Ch. gerstfeldti (Dyb.)
из Прибайкалья; 116 — Perforatella bidens (Chemn.) из с. Апаринки Московской обл.;
117 — Р. dibothrion (Kim.) из окр. Рахова; 118 — Monachoides incarnata (Miill.)
из окр. Сторожинец, Предкарпатье; 119 — М. vicina (Rssm.) из окр. Рахова; 120 —
М. stuxbergi (West.) на берегу Енисея у пос. Мирное.
ТАБЛИЦА XIII
1.21 — Monachoides aculeata (Uvalieva) из окр. Верхискатупска, Алтай; 122 — Pseudo-
trichia rubiginosa (A. Selim.) из дельты Волги; 123 — Urticicola umbrosa (L. Pfr.) из Ка-
зимежа над Вислой, Польша; 124 — F ruticocampylaea narzanensis (Kryn.) из с. Чми
в Дарьяльском ущелье; 125 — F. kobiensis (Bttg.) из окр. Пасаиаури; 126 — Shileykoia
daghestana (Kobelt) из окр. с. Акуша, Дагестан; 127 — Circassina circassica circassica
(Mouss.) из Бакуриани; 128 — С. circassica sitnpla Schileyko из Орджоникидзе; 129 —
С. bojenae Hudec et Lezhawa из долины p. Texypn; 130 — C. frutis (L. Pfr.) из окр. Сочи.
ТАБЛИЦА XIV
131 — Cernuella virgata (Da Costa) из Ялты; 132 — C. tijlisiana (Lindh.) из окр. С. Чми
в Дарьяльском ущелье; 133 — Xerosecta crenimargo (L. РГг.) из окр. с. Карата, Дагестан;
135 — X. arcadiana Schileyko из окр. с. Космальян, Талыш, паратип; 136 — Archaica
(Euarchaica) heptapotamica (Lindh.) из ущелья р. Пскем; 137 — A. (Archaica) apollinis
(Mart.) из урочища Япгарык па р. Гульче, Алайский хр.; 138— А. (Л.) apollinis
(Mart.) из долины р. Нарын ниже Токтогула, Ферганский хр.; 139 — A. (Vgama)
labianix sp. nov. из ущелья р. Пскем, голотип; 140 — К arabaghia bituberosa (Lindh.)
из окр. оз. Севан.
ТАБЛИЦА XV
141 — Hesseola solidior (Mouss.) из окр. оз. Севан; 142 — Н. transcaspia (Bttg.) из окр.
Кара-Кала, Копетдаг; 143 — Euomphalia aristata (Kryn.) из окр. Абастумани; 144 —
Е. appeliana (Mouss.) из окр. Нальчика; 145 — Е. strigella (Drap.) из с. Архангель-
ского Московской обл.; 146 — Platytheba (J asonella) mingrelica (Hesse) из долины р. Те-
хури; 147 — Р. (Platytheba) prometheus (Bttg.) из окр. Цагери; 148 — Stenomphalia
(В atumica) maiae (Hudec et Lezhawa) из Батуми, Ботанический сад; 149 — Oscarboettge-
ria euages (Bttg.) из окр. Гагры; 150 — Stenomphalia (Stenomphalia) selecta (Klika)»
из Чегемского ущелья.
ТАБЛИЦА XVI
151 — Stenomphalia (Diplobursa) pislformls (L. Pfr.) из с. Чмп в Дарьяльском ущелье;
152 — S. (Harmozica} ravergieri (For.) из окр. с. Ардон в Сев. Осетии; 153 — S. (Я.) aff.
revergieri (Fer.) из апшеронских отложений в окр. Нальчика; 154 — Monacha (Рага-
theba) fruticola (Kryn.) из Краснолесья близ Симферополя; 155 — М. (Р.) caucasicola
(Lindh.) с северных склонов Эльбруса; 156 — М. (Р.) talyschana (Mart.) из окр. Лен-
корани; 157 — М. (Monacha) externa Steklov из меотических отложений на р. Фор-
танге, Предкавказье, голотип; 158 — М. (М.) carthusiana (Miill.) окр. с. Голубинка,
Крым; 159 — М. (Metatheba) samsunensis (L. Pfr.) из окр. Гагры; 160 — М. (Boemica)
praeorientalis Steklov из акчагыльских отложений в Пседахской балке, Предкавказье,
голотип.
ТАБЛИЦА XVII
161 — Monacha (Boemica)^subcarthusiana subcarihiistana]M\v\A\\. из окр. Макопсе, зап.
Кавказ; 162 — Angiomphalia (Angiomphalia) copiosa. sp. nov. из верховьев p. Чаткал,
голотип; 163 — A. (A.) caelestimonlana. (Tzv.) из окр. с. Кочкорка в верховьях р. Чу;
164 — A. (A.) caelestimonlana (Tzv.) с южного берега оз. Иссык-Куль южнее Каржи-
сая; 165 — A. (A.) exasperata sp. nov. из окр. Бол. Алматинского оз., голотип; 166 —
А. (Л.) regeliana (Mart.) из ущелья Карабалты, Киргизский хр.; 167 — A. (A.) alma-
lensis sp. nov. ип долины р. Чу близ с. Толман, голотлл; 168 — А . (А.) seductilis (West.)
из Киргизского хр.; 169— A. (Lentiga) lentina (Mart.) из долины р. Нарын; 170—
A. (В.) lentina ,inv. из Сары-Челекского заповедника (=«Aegista mesasiana Lindh.»).
Ш А. А. ТЦплейко
ТАБЛИЦА XVIII
171 — Angiomphalia (Lentiga) buamica (В. Tzv. et E. Tzv.) из ущелья p. Чу; 172 —
Paedhoplita laminata Lindh. из восточной части Киргизского хр., лектотип; 173 —
Р. lindholmi В. Tzv. et Е. Tzv. из ущелья Карабалты, Киргизский хр.; 174 — Р. lind-
holmi В. Tzv. et Е. Tzv. из верховьев р. Аламедин на Киргизском хр.; 175 — Р. kir-
gisensis Lich. et Ramm, из верховьев p. Аламедин на Киргизском хр., голотип; 176 —
Caucasocressa pratensis (L. Pfr.) из окр. Боржоми; 177 — С. joannis (Mort.) из Аджарии,
долина р. Мачахелис-цкали; 178 — С. joannis (Mort.) из Турции, окр. Ломашена;
179 — Faustina faustina (Rssm.) из окр. Ужгорода; 180 — F. faustina (Rssm.) из окр. Ра-
хова.
ТАБЛИЦА XIX
181 — Arianta arbustoruin (L.) из окр. Рахова; 182 — A. aethiops peCii Kim. из окр. Ра-
хова; 183 — Helicigona lapicida (L.) из Чешского Крумлова, ЧССР; 184 — Isognomo-
stoma personatum (Lam.) из окр. Рахова; 185 — Drobacia banatica (Rssm.) из окр. Ра-
хова; 186 — Causa holosericum (Stud.) из Краковского ущелья в Татрах; 187 — Kli-
kia osculum Thomae из нижнего миоцена Чехословакии; 188 — Сераеа {Сераеа) пето-
ralis (L.) из Кведлинбурга, ГДР; 189 — С. (С.) hortensis (Miill.) из Кведлинбурга,
ГДР; 190 — С. (Octadenia.) vindobonensis (Fee.) из Кишинева.
ТАБЛИЦА XX
191 — Cryptoinphalus aspersa (Mull.) из окр. Монпелье, Франция; 192 — Eobania
vermiculata (Miill.) из Ялты; 193 — Levantina djulfensis (Dub. de Mont.) из Армении,
окр. с. Мегри; 194 — L. escheriana (Bourg.) из Армении, окр. с. Арени; 195 — Cauca-
sotachea andrussovi Steklov из среднесарматских отложений на р. Белой у Майкопа,
голотип; 196 — С. fortangensis Steklov из меотических отложений на р. Фортанге,
Предкавказье, голотип; 197 — С. kubanica Steklov из караганских отложений р. Псе-
фирь у станицы Костромской, Предкавказье, голотип; 198 — С. lencoranea (Mouss.)
из окр. Ленкорани; 199 — С. atrolabiata (Kryn.) из окр. Хосты; 200 — С. calligera
(Dub. de Mont.) из долины р. Техури.
ТАБЛИЦА XXI
201 — Steklovla maslovae (Steklov) из акчагыльских отложении р. Сунжа у с. Алды,
Предкавказье, голотип; 202 — Helix maeotica Steklov из меотических отложений
на р. Гумс, Предкавказье, голотип; 203 — Н. nordmanni Mouss. из окр. с. Бенара
на правом берегу р. Абастуманки; 204 — Н. christophi Bttg. из Турции, окр. Ло-
машена; 205 — Н, albescens Rssm. из с. Голубинка, Крым; 206 — Н. lucorum L.
из с. Голубинка, Крым; 207 — II. buchi Dub. de Mont, из Батуми; 208 — Н. lutescens
Rssm. из окр. г. Печ, Венгрия; 209 — Н. pomatia L. из Ужгорода; 210 — Triodopsis
subpersonatum (Midd.) из окр. Аяна на берегу Охотского моря, паратип.
СОДЕРЖАНИЕ
Стр.
Предисловие .......................................................... - 3
Систематический указатель видов.......................................... 5
Введение .............................................................' 12
Общая характеристика моллюсков надсемейства Helicoidea.................. 12
Морфология и анатомия представителей Helicoidea с элементами физиологии 13
Внешний вид и раковина................................................ 13
Мантия, кожа и эктодермальные железы.................................. 18
Реноперикардиальный аппарат и легкое.................................. 22
Пищеварительный тракт ............................................... 24
Мускулатура .......................................................... 26
Нервный аппарат и нейросекреция....................................... 27
Половой аппарат ...................................................... 29
Биология представителей Helicoidea ..................................... 33
Питание ................................... . . . ................. 33
Размножение, жизненный цикл и длительность жизни..................... 34
Влияние химического состава почв на распространение Helicoidea........ 36
Понятие о жизненных формах............................................ 37
Защита от потери воды и летняя спячка............................ . 40
Зимняя спячка ........................................................ 43
Ритуальные формы поведения............................................ 44
Географическое распространение Helicoidea............................... 44
Основные направления морфологического развития Helicoidea во времени
и в пространстве........................................................ 50
Общие замечания. Исходные группы...................................... 50
Семейство Helicodontidae ............................................. 57
Семейство Bradybaenidae.............................................. 58
Семейство Hygromiidae ,............................................... 62
Подсемейство Trichiinae .......................................... 63
Подсемейство Hygromiinae.......................................... 68
Подсемейство Euomphaliinae ........................................ 71
Подсемейства Archaicinae и Paedhoplitinae........................ 75
Подсемейство Metafruticicolinae ................................... 76
Семейство Helicidae.................................................. 77
Методы исследования .................................................... 80
Сбор материала и фиксация............................................. 80
Камеральная обработка ................................................ 82
Литература ............................................................. 84
Систематическая часть .................................................. 98
Таблица для определения рецентных видов надсемейства Helicoidea фауны
СССР по раковине........................................................ 98
Таблица для определения семейств, подсемейств, родов, подродов и некоторых
подвидов по признакам внутреннего строения и раковины.................. 108
Условные сокращения и обозначения........................................ ИЗ
I. Семейство Helicodontidae ........................................... 114
СОДЕРЖАНИЕ
383
I. Подсемейство Helicodontinae ......................................... 114
Род Helicodonta Fer.................................................. 114
II. Подсемейство Lindholmiolinae........................................ 116
1. Род Lindholmiola Hesse ........................................... 116
2. Род Oestophora Hesse.............................................. 118
II. Семейство Bradybaenidae............................................... 118
1. Род Ponsadenia gen. nov............ . . ........................ 119
2. Род Bradybaena Beck .............................................. 123
III.. Семейство Hygromiidae .............................................. 163
I. Подсемейство Trichiinae ............................................ 163
1. Род Nanaja gen. nov........................................... 164
2. Род Odontotrema Lindh. . . ..................................... 166
3. Род Leucozonella Lindh.......................................... 168
4. Род Hygrohelicopsis Schileyko .................................. 181
5. Род Teberdinia Schileyko ....................................... 183
6. Род Kokotschashvilia Hudec et Lezhawa........................... 184
7. Род Caucasigena Lindh........................................... 191
8. Род Diodontella Lindh........................................... 202
9. Род Plicuteria Schileyko ....................................... 203
10. Род Trichia Hartmann............................................. 205
11. Род Edentiella Polinski ......................................... 209
12. Род Helicopsis Fitzinger ........................................ 211
13. Род Xeropicta Monterosato........................................ 219
14. Род Helicella Fer................................................ 223
II. Подсемейство Hygromiinae ......................................... 225
1. Род Lindholmomneme Haas ........................................ 225
2. Род Chilanodon West............................................. 228
3. Род Perforatella Schliiter ..................................... 231
4. Род Monachoides Gude et Woodw................................... 234
5. Род Pseudotrichia Licharev ..................................... 238
6. Род Urticicola Lindh.......................................... 239.
7. Род Fruticocampylaea Kobelt..................................... 241
8. Род Shileykoia Hudec ........................................... 244
9. Род Circassina Hesse............................................ 246
10. Род Cefnuella Schluter.......................................... 250
11. Род Xerosecta Monterosato ...................................... 253
III. Подсемейство Archaicinae ......................................... 256
Род Archaica Schileyko .............................................. 257
IV. Подсемейство Euomphaliinae ....................................... 261
1. Род Karabaghia Lindh............................................. 262
2. Род Hesseola Lindh. . . . .................................. 263
3. Род Euomphalia West.............................................. 266
4. Род Platytheba Pilsbry........................................... 271
5. Род Oscarboettgeria Lindh....................................... 273
6. Род Stenomphalia Lindh........................................... 275
7. Род Monacha Fitzinger .......................................... 281
V. Подсемейство Paedhoplitinae......................................... 291
1. Род Angiomphalia gen. nov......................................... 291
2. Род Paedhoplita Lindh......................................... . 302
VI. Подсемейство Metafruticicolinae..................................... 304
Род Caucasocressa Hesse .......................................... 304
384
СОДЕРЖАНИЕ
IV. Семейство Helicidae............................................. 307
I. Подсемейство Ariantinae ..................................... 308
1. Род Faustina Kobelt ...................................... 308
2. Род Arianta Turton........................................ 310
3. Род Helicigona Fer........................................ 313
4. Род Isognomostoma Fitzinger............................... 315
5. Род Drobacia Brusina ....................................... 317
6. Род Causa Schileyko ................................. . . . 317
7. Род Klikia Pilsbry.......................................... 319
II. Подсемейство Helicinae....................................... 319
1. Род Сераеа Held............................................ 320
2. Род Cryptomphalus Charp................................... 324
3. Род Eobania Hesse......................................... 326
4. Род Levantina Kobelt...................................... 327
5. Род Caucasotachea C. Bttg................................. 330
6. Род Steklovia gen. nov.................................... 335
7. Род Helix L............................................... 337
Приложение ............................................................ 346
Надсемейство Polygyroidea ............................................. 347
Род Triodopsis Rafinesque...................................... 347
Дополнение............................................................. 347
Алфавитный указатель латинских названий................................ 350
Анатолий Алексеевич Ш и л е й к о
НАЗЕМНЫЕ МОЛЛЮСКИ НАДСЕМ. HELICOIDEA
В серии: Фауна СССР. Моллюски. Т. III, вып. 6
Утверждено к печати Зоологическим институтом Академии наук СССР
Редактор издательства Л. В. Шоренкова
Технический редактор Л. М. Семенова
Корректоры О. И. Буркова, С. И. Семиглазова
ИБ № 8507
Сдано в набор 24.08.77. Подписано к печати 01.03.78. М-08332. Формат 70 X 1О8'/16- Бумага типограф-
ская № 2. Гарнитура обыкновенная. Печать высокая. Печ. л. 24 = 33,6 усл. печ. л. Уч,-
изд. л. 41,56. Тираж 1400. Изд. Ms 6564/6826. Тип. зак. Ms 654. Цена 6 р. 80 к.
Ленинградское отделение издательства «Наука», 199164, Ленинград, Менделеевская линия, д. 1
1-я тип. издательства «Наука», 199034, Ленинград, 9 линия, д. 12